/






























































































































































































































Автор: Колаковский Э.
Теги: пищевая промышленность в целом производство и консервирование пищевых продуктов пищевое производство промышленность кулинария пищевая промышленность
ISBN: 5-10-002019-9
Год: 1991




Текст
E.KofakowskiЭ.КолаковскийТехнологияРЫБНОГОфаршаTechnologiafarszOwrybnych
Э.КолаковскийТехнологияРЫБНОГОфаршаEdward KofakowskiTechnologiafarszowrybnych
Edward KofakowskiTechnologia
farszdw
rybnychParistwowe Widawnictwo
Naukowe • Warszawa
1986
Э.КолаковскийТехнологияРЫБНОГОфаршаПеревод с польского В. Е. ТишинаПод редакцией кандидата технических наук
Л. И. БорисочкинойфМосква
ВО “Агропромиздат"
ББК 36.94
К60
УДК 664.95.65Редактор Л, J1. КожинаКолаковский Э.К60 Технология рыбного фарша /Пер. с польск. В. Е. Тишина;Под ред. J1. И. Борисочкиной. — М.: Агропромиздат, 1991. —
220 с.: ил.ISBN 5-10-002019-9В книге обобщены результаты исследований и технология производ¬
ства рыбного фарша во многих странах мира. Приведены характеристика
рыбного сырья, методы его первичной обработки, указаны вспомогатель¬
ные материалы, применяемые при производстве рыбного фарша. Описаны
физико-химические изменения, происходящие в фарше под влиянием
различных факторов. Рассмотрены способы получения фарша разных ви¬
дов, требования к качеству фарша, его устойчивости при хранении, приве¬
дены примеры использования.Для технологов, ихтиологов и других специалистов рыбной промыш¬
ленности.Кз_-Г ,24„,ISBN 5 —10—002019—9 (СССР) © Copyright by Panstwowe Wydawnictwo
ISBN 83-01-05952-4 (Республика Naukowe, Warzawa 1986
Польша) © Перевод на русский язык ВО ’’Агро¬промиздат”, 1991
ОТ РЕДАКЦИИС целью рационального использования океанических
рыб, которых трудно реализовать в привычном для населе¬
ния виде, сначала в Японии, а затем в других странах была
разработана перспективная технология изготовления из них
рыбного фарша, который служит полуфабрикатом для про¬
изводства различных кулинарных изделий (колбас, соси¬
сок, котлет, пельменей и др.). Фарш можно готовить из та¬
ких видов рыб, как минтай, сайра, хек, путассу, налим, мар¬
лин, пристипома, а также треска, терпуг, ставрида, камбала,
морской окунь, тунец, акулы и др. Однако особо важное
значение придается использованию маломерных нежирных
рыб, что позволяет направлять дополнительные массы
сырья на выработку пищевых продуктов вместо использо¬
вания его на кормовые цели.В отечественной рыбной промышленности в последние
годы также эффективно развивается рыбокулинарное про¬
изводство, однако при переработке на фарш мелких рыб
промышленность испытывает значительные трудности. Ис¬
следования, проведенные в нашей стране и касающиеся тех¬
нологии приготовления рыбного фарша, не решили многих
вопросов в этой области. В связи с этим книга польского
ученого Эдварда Колаковского ’’Технология рыбного
фарша”, посвященная комплексу проблем фаршевого про¬
изводства, представляется весьма ценной и достойно вос¬
полняющей пробел в области теории рыбного фарша. В ней
компетентно и с привлечением большого научного материа¬
ла (в том числе и результатов исследований отечественных
специалистов-технологов) рассматриваются сырье и вспомо¬5
гательные материалы, применяемые для приготовления
фарша, анализируются изменения белков, липидов и других
компонентов мяса рыбы в процессе приготовления фарша и
при его хранении.Большое внимание в книге уделено влиянию на качест¬
во фарша первичной обработки сырья, способов его хране¬
ния, посмертных изменений, степени измельчения мяса
рыбы. Подробно рассмотрена технология приготовления
рыбного фарша, влияние на его качество промывок водой,
стабилизация фарша с использованием разнообразных до¬
бавок, биохимическая модификация и др.Приведены технологические процессы производства
пастеризованного, соленого, сушеного и ферментированно¬
го фаршей, а также требования к их качеству и срокам хра¬
нения.Есть основания надеяться, что предлагаемая книга ока¬
жется полезной для научных и практических работников
рыбной отрасли, а изложенные в ней сведения будут спо¬
собствовать повышению качества рыбного фарша и кули¬
нарных изделий, вырабатываемых на его основе.Редакция выражает благодарность канд. техн. наук
Н. И. Рехиной за помощь, оказанную при подготовке ру¬
кописи к печати.
ПРЕДИСЛОВИЕТехнология рыбного фарша относится к новейшим процессам пе¬
реработки рыбы, открывающим новые возможности в области рацио¬
нального использования морского животного сырья, особенно малоцен¬
ной рыбы. В мировой технической литературе до сих пор отсутствуют
обобщающие работы на эту тему. Настоящая книга заполняет этот про¬
бел, охватывая весь комплекс проблем, связанных с технологией рыб¬
ного фарша. В первых разделах книги рассмотрены теоретические осно¬
вы технологии измельченного мяса рыбы, при этом особое внимание
уделяется происходящим в нем физико-химическим изменениям и спо¬
собам их замедления или предотвращения, а также способы получения
пищевых продуктов из измельченного мяса рыбы. На основе дополни¬
тельных теоретических исследований изложены новейшие достижения
в области получения стабилизированного, соленого, сушеного, фермен¬
тированного и других видов рыбного фарша, приведены технологиче¬
ские схемы процессов, описаны производственные линии, а также маши¬
ны и устройства. Остальные разделы посвящены описанию условий хра¬
нения рыбного фарша, а также способов обеспечения его устойчивости.Автор
1. ОБЩИЕ СВЕДЕНИЯ1.1. МИРОВОЕ ПРОИЗВОДСТВО РЫБНОГО ФАРШАПроцесс получения рыбного фарша состоит из трех основных эта¬
пов: предварительной разделки рыбы на тушку, куски или филе; отде¬
ления мяса от несъедобных частей рыбы (крупных и мелких костей,
кожи и т. п.) с помощью сепараторов, во время этой операции мышечная
ткань измельчается;, переработки измельченного мяса для обеспечения
его устойчивости при хранении и изменения функциональных свойств.Таким образом, рыбный фарш — это измельченное мясо рыбы, от¬
деленное механическим способом от несъедобных частей, подвергнутое
дополнительной технологической обработке с целью увеличения срока
хранения, сформированное в блоки и замороженное.Промышленное производство мороженого рыбного фарша возникло
в Японии в 60-х годах. Это был фарш типа сурими, полученный из много¬
кратно промытого питьевой водой мяса, перемешанного затем с солью
и другими добавками. На специфику технологии фарша основное влия¬
ние оказали традиционные требования японского рынка, предпочитаю¬
щего продукты с характерной резинистой консистенцией — камабоко,
чикува и др. Вначале производство фарша осуществлялось главным
образом на береговых предприятиях, а с 1967 г. его стали производить
также и в море (табл. 1). Основным сырьем для производства фарша
служит минтай, из которого вырабатывается свыше 90 % сурими.Таблица 1. Производство фарша типа сурими в Японии
в 1965-1980 гг. (в тыс. т)Производствофарша196519671969197119731975197719791980На берегу244593179205200234162106В мореВсего8323984103196180359218423187387162396190352183'289Помимо особенностей, связанных с традиционными требованиями
японского рынка при производстве продуктов типа паст*, много труд¬* Пастообразные продукты включают главным образом камабоко, чикува,
ханпен, сатсума-агэ и некоторые рыбные колбасные изделия.8
ностей вызывает экономическая сторона производства сурими, в основ¬
ном это большой расход питьевой воды и значительные потери мы¬
шечных белков во время промывки фарша. Именно поэтому европей¬
ские страны долгое время не проявляли заинтересованности в развитии
производства рыбного фарша. Лишь модификация технологии, заклю¬
чающаяся в уменьшении расхода питьевой воды, и разработка новых
способов производства, исключающих промывку, а также быстрое раз¬
витие механизации процессов отделения съедобной части рыбы методом
сепарации вызвали в конце 60-х годов значительный интерес к техноло¬
гии производства рыбного фарша во многих научно-исследовательских
центрах и на промышленных предприятиях [188] Важным стимулом
развития этого производства в мире была растущая доля в морских
уловах так называемых малоценных рыб, а также повышение цен на тра¬
диционное рыбное сырье.К важнейшим производителям рыбного фарша в настоящее время
относятся Япония, Советский Союз, Канада, Дания, Исландия, Норвегия,
Южная Африка, Великобритания и Польша.В Польше первые попытки получения двукратно промытого пить¬
евой водой фарша были предприняты Институтом морского рыболов¬
ства в Гдыне [124, 253] . Непромытый фарш с введением смеси нату¬
ральных добавок был впервые получен сотрудниками Технологическо¬
го института пищевых морепродуктов Сельскохозяйственной академии
в Щецине в 1972 г. на траулере ’’Щитно” производственного объедине¬
ния ’’Одра” в Свиноустье [226]1.2. ПРЕИМУЩЕСТВА ПЕРЕРАБОТКИ РЫБЫ НА ФАРШВыход съедобной части — важнейший критерий оценки способа пе¬
реработки рыбы. В этом смысле производство рыбного фарша можно
считать наиболее рациональным и современным способом переработки
рыбного сырья. При этом выход съедобной части рыбы достигает 40—
60 %, тогда как при филетировании он составляет всего 28—33 %
(табл. 2).Применение сепараторов дает возможность получать съедобное
мясо из отходов от филетирования рыбы (табл. 3), включающих хреб¬
товую кость с прирезями мяса, обрезки, кусочки теши рыбы, что по¬
зволяет значительно рациональнее использовать рыбное сырье.К преимуществам производства рыбного фарша как способа комп¬
лексной переработки водного сырья относятся также:возможность обработки разных видов рыбы, ракообразных и мол¬
люсков, в том числе и непригодных для филетирования на механизи¬
рованных линиях;сохранение на судне только мяса рыбы, что позволяет коренным об¬
разом улучшить экономические показатели использования холодильных
камер. Несъедобные части рыбы, которые при обычных методах пер-9
Таблица 2. Выход фарша и филе из морской рыбыВыход (в % массы нераз-Рыбаделанной рыбы)ИсточникфаршфилеПикша54,832,1[386]Мерланг55,333,0Хек46,133,7Морской налим40,232,7Морской петух42,0-Аргентина62,9-Путассу54,433,5Бычок (зеленая нототения)44,033,0Тихоокеанский хек44,730,0[68]Терпуг48,135,0Gadus macrocephalus40,428,0(тихоокеанская треска)Sebastodes flavidus41,729,0(морской окунь, морской ерш)Sebastodes pinninger40,528,0(оранжевый окунь)Parophrys vetulus (речная камбала)47,333,0Таблица 3. Выход съедобных частей из отходов от филетирования[209, 324]РыбаВыход фарша из отходов (без головы
и внутренностей) от филетирования,
% массынеразделаннойрыбыотходовLimanda ferruginea1647(желто хвостая камбала)Sebastes marinus19-2031,2-38,6(морской окунь)Gadus morhua12-1959-66(треска)Melanogrammus aegleflnus1756(пикша)Pollachius vixens (сайда)18-2060-72,2Merluccius bilinearis1755(серебристый хек)Anarhichas lipus (зубатка)2066Brosmebrosme (менек)227010
вичной обработки входят в состав продукта, при производстве фарша пе¬
рерабатываются на рыбную муку непосредственно на судне;снижение трудоемкости обработки рыбы благодаря большим воз¬
можностям механизации и автоматизации процессов производства
фарша;простота получения из фарша различных современных рыбных
продуктов.Для производства рыбного фарша и получения из него различных
продуктов можно применять непрерывнодействующие линии с высо¬
ким уровнем автоматизации. Это облегчает соблюдение требуемых
режимов технологического процесса и улучшает санитарные условия
производства. Большое значение имеет также возможность упрощения
процесса переработки фарша благодаря использованию его в качестве
сырья. Примером этого может служить исключение операции предвари¬
тельного размораживания при производстве гамбургеров и крокетов
[241].1.3. НАПРАВЛЕНИЯ ИСПОЛЬЗОВАНИЯ РЫБНОГО ФАРШАСамой простой формой использования рыбного фарша является его
применение в качестве облагороженного рыбного сырья для приготов¬
ления различных блюд на предприятиях общественного питания и в до¬
машних условиях. Более всего для этой цели пригоден мороженый ста¬
билизированный фарш из рыбы или ракообразных, выпускаемый в роз¬
ничной упаковке (в брикетах и картонных коробках). Например, фирма
’’Уэгменс Фуд Маркетс” (США) выпускает рыбный фарш в упаковках
по 250 г. На упаковке напечатаны рецепты блюд, которые можно при¬
готовить из этого фарша.Популярным направлением использования рыбного фарша, особен¬
но в США и странах Западной Европы, является производство рыбных
продуктов, изготовленных методом распиливания мороженых блоков
(порции, рыбные палочки и др.). Преимущество этого метода — отсут¬
ствие необходимости повторного замораживания пищевого продукта,
а также дополнительного измельчения фарша, как при переработке
размороженного фарша. Для переработки на такие продукты больше
всего подходит стабилизированный фарш с добавками, увеличивающими
его водоудерживающую бпособность и предотвращающими денатура¬
цию белков, или вкусовыми приправами, а также фарш, промытый
водой и отделенный на сепараторе [208]. Может быть также использо¬
ван нестабилизированный фарш из тощей рыбы, однако качество полу¬
чаемого продукта в этом случае хуже, главным образом из-за быстро из¬
меняющейся консистенции [280] . Для переработки на порции идет
фарш из крупноизмельченного мяса, цвет на разрезе светлый, равномер¬
ного оттенка.В некоторых странах измельченное мясо рыбы применяется для ла¬11
минирования (соединения отдельных пластин) мороженого рыбного
филе с целью получения монолитных блоков, предназначенных для рас¬
пиливания на порции. Количество добавляемого фарша не превышает10 % (Великобритания) или 12—14 % (Норвегия) общей массы блока
[208]Еще одним направлением использования рыбного фарша является
производство из него формованных продуктов. В этом варианте к из¬
мельченному мясу добавляют различные наполнители, изменяющие в
требуемом направлении реологические свойства, вкус и запах продук¬
та. Для получения дополнительного эффекта — улучшения вкуса и со¬
вершенствования технологии — измельченное мясо рыбы можно смеши¬
вать с мясом креветок, устриц и других беспозвоночных [22, 369]
Технология формованных изделий из измельченного мяса рыбы предус¬
матривает применение различных видов добавок и способов обработ¬
ки, в результате чего получаются такие продукты, как рыбные гам¬
бургеры, крокеты, фрикадельки, тефтели и др. [225] Как правило,
продукты выпускаются в мороженом виде, хотя весьма часто реализу¬
ются и как готовые блюда, кулинарные изделия, а также используют¬
ся для выработки консервов. Для производства формованных продук¬
тов наиболее пригодны крупноизмельченный, стабилизированный и пас¬
теризованный фарш, а также их смеси. Фарш может быть переработан
после предварительного размораживания либо без размораживания
с использованием соответствующих дробилок для измельчения моро¬
женых рыбных блоков [228] На многих предприятиях по производст¬
ву формованных продуктов используют мясо рыбы, отделенное меха¬
ническим способом от тушек свежей или размороженной рыбы.Изготовление рыбных колбасных изделий и продуктов типа кама¬
боко, чикува и др. — одно из старейших направлений использования рыб¬
ного фарша. Однако для приготовления этих изделий фарш должен от¬
вечать довольно строгим требованиям, особенно с точки зрения раство¬
римости белков, поэтому не всякий рыбный фарш пригоден для про¬
изводства таких изделий. Этим требованиям наиболее полно удовлет¬
воряет промытый сырой фарш типа сурими [361] и стабилизированный
фарш из рыбы средней жирности, в котором не происходит значитель¬
ного уменьшения растворимости белков во время хранения в заморо¬
женном виде. Минимальная растворимость белков в 4 %-ном растворе
хлористого натрия при производстве рыбных колбас но польской тех¬
нологии [225] составляет 6 г белка на 100 г 4 %-ного раствора хлорис¬
того натрия. Фарш с более низкой растворимостью белков должен
быть улучшен добавлением соответствующих препаратов.Для производства паст в основном пригодны все виды фарша.
Кроме того, применение различных добавок, а также разнообразные
методы хранения делают эту группу изделий весьма перспективной с
точки зрения разнообразия ассортимента. В качестве примеров можно
привести рыбоовощные мороженые или охлажденные пасты [228] ,
12
пастеризованные пасты [369] , заменители мяса лосося [401] , кремы,
майонезы [86,131] и др.Фарш может применяться для производства разнообразных кули¬
нарных изделий - различных готовых блюд, горячих и холодных за¬
кусок и т. п. [299] Важную роль в этом играет высокая степень готов¬
ности фарша для переработки — нет необходимости в первичной обра¬
ботке рыбы, отсутствуют отходы, фарш легко соединяется с различны¬
ми вспомогательными ингредиентами. Растет интерес к фаршу также
как к сырью для производства консервов. Разработанные к настоящему
времени продукты свидетельствуют о значительных возможностях ис¬
пользования фарша для производства консервов [154]., но необходи¬
мы дальнейшие исследования с целью улучшения консистенции стерили¬
зованного фарша.Рыбный фарш дает возможность развивать производство рыбо-муч-
ных изделий. Кроме традиционных пирожков и других изделий, состоя¬
щих из двух частей (теста и начинки), рыбный фарш использовали как
сырье для производства рыбных макарон [29, 387] , рыбной соломки
[6] , рыбо-кукурузных, рыбо-картофельных хлопьев [326, 256] и дру¬
гих продуктов.Самостоятельным направлением становится использование рыб¬
ного фарша в качестве заменителя белка в пищевых продуктах. Это
дает возможность реализовать такой фарш, который с точки зрения не¬
которых свойств обладает меньшей технологической пригодностью- для
переработки. Например, жесткая консистенция фарша из некоторых
тресковых рыб ограничивает его применение для производства моро¬
женых рыбных порций и рыбных палочек. Вместе с тем фарш из этих
рыб обладает слабо выраженными рыбным запахом и вкусом, что яв¬
ляется положительным свойством при применении такого фарша в ка¬
честве заменителя мяса при производстве различных мясных продук¬
тов, например сосисок, котлет, начинки для пирожков и др. Показано,
что замена 10—15 % измельченной говядины фаршем из минтая в паро¬
вых котлетах существенно не ухудшает вкус и консистенцию этих
продуктов [428]Использование фарша, приготовленного из съедобных отходов от
филетирования (хребтовые кости, обрезки мяса и т. п.), а также из так
называемой второй фракции (содержащей много красного мяса), огра¬
ниченно из-за их кровянистого цвета и нестабильности при хранении,
зато такой фарш может быть использован в качестве добавки в мясные
изделия, главным образом из говядины. Оптимальное содержание
рыбного фарша в мясо-рыбных гамбургерах, паштетах и готовых блю¬
дах составляет 25—50 % в зависимости от вида рыбы [206, 210, 211]
Мясо-рыбные продукты дешевле продуктов из чистого мяса животных,
но не уступают им по вкусу и пищевой ценности.На основе изучения важнейших направлений использования рыбного
фарша можно утверждать, что они весьма разнообразны и зависят как13
от вида фарша и степени его переработки, так и от привычек и традиций
питания населения каждой страны. Подавляющая часть изготовляемого
рыбного фарша используется как высокоценное сырье в рыбообраба¬
тывающем производстве. Это открывает новые возможности в рацио¬
нальном использовании морского пищевого сырья, а также механиза¬
ции и автоматизации технологических процессов. К сожалению, попу¬
ляризация изделий из фарша и продвижение их на рынок сбыта не поспе¬
вают за развитием техники и технологии.Другая существенная проблема - необходимость холодильной цепи
и поддержания высокого санитарного уровня производства и реализа¬
ции. Часто нерешенность этой проблемы служит причиной отказа от но¬
вых технологий, а следовательно, и от технологических и экономиче¬
ских преимуществ. Внедрение новых изделий из рыбного фарша требует
тщательной отделки конечного продукта. Кроме вкусовых достоинств,
существенную роль играют такие факторы, как форма, величина пор¬
ции, способ хранения продукта, тип и качество упаковки и др.
2. ОБЩАЯ ХАРАКТЕРИСТИКА ТКАНЕЙ,
ВХОДЯЩИХ В СОСТАВ РЫБНОГО ФАРШАОсновным компонентом рыбного фарша являются скелетные мыш¬
цы туловища, хвостового стебля и плавников. Присутствие других
тканей в фарше считается загрязнением, попадающим в мясо во время
его отделения от несъедобных частей. Это в основном кусочки кожи,
мелкие кости позвоночника, плавников, частицы почек, плавательного
пузыря, включения крови.2.1. МЫШЕЧНАЯ ТКАНЬ (МЯСО)2.1.1. Состав мышцТело рыбы в основном составляют мышцы туловища: спинные и
боковые.Большая боковая мышца (musculus lateralis magnus) расположена
обычно вдоль боковой линии. Горизонтальная перегородка из соедини¬
тельной ткани делит ее на надосевую, или спинную (m.dorsolateralis),
и подосевую, или брюшную (m.ventrolateralis), части. Мышца состоит
из многочисленных миомеров, разделенных между собой миосептами
из соединительной ткани. Число миомеров соответствует числу позвон¬
ков, а их размеры изменяются по длине рыбы (рис. 1)*. Цвет большой
боковой мышцы белый, так как ее волокна не содержат миоглобина,
поэтому обычно ее называют белой мышцей.Боковая подкожная мышца (m.rectus lateralis) распо¬
лагается под кожей вдоль боковой линии, заполняя желобок между
краями спинной и брюшной мышц (рис. 3). В противоположность бе¬
лой мышце боковая подкожная мышца характеризуется высоким со¬
держанием миоглобина, находящегося в саркоплазме, который придает
мышце красный цвет. Отсюда название — красная, или темная, мышца.К красным мышцам относятся также другие подкожные мышцы,
которые не играют такой важной роли в технологии рыбного фарша, как
боковая подкожная мышца.* С увеличением длины рыбы ширина миомеров возрастает (рис. 2) .15
Рис. 1. Расположение миомеров туловищных мышц трески Gadus morhus и скумб¬
рии Scomber scombrus [ 34]:а - вид сбоку; б - вид со стороны спинки; в - в-ид снизу; 1 - миомеры; 2 - мио-
септы; 3 - горизонтальная межмышечная перегородка; 4 - большая боковая
мышца (спинная часть); 5 - большая боковая мыщца (брюшная часть)• .!• '
•1 • •1 • •I. •г • г► ••25 37,5 50 62,5 75 87,5 100Полная длина рыбы, смРис. 2. Зависимость между полной длиной трески и шириной миотома № 12 (счи¬
тая от головы) большой боковой мышцы [283]Рис. 3. Поперечный разрез по туловищу в приголовке (а), середине (б), у хвос¬
та (в) отдельных видов рыб:1 - верхняя хребтовая мышца; 2 - вертикальная перегородка; 3 - сердцевина по¬
звоночника; 4 позвонок; 5 - большая боковая мышца (спинная часть); 6 -
горизонтальная перегородка; 7 - красная боковая мышца; 8 - большая боковая
мышца (брюшная часть); 9 - аорта; 10 - почка
С точки зрения выполняемой двигательной функции относитель¬
ная масса красного мяса (мышц) зависит от степени подвижности
рыбы в водной среде. Больше всего красных мышц содержится в теле
пелагических рыб, а меньше всего - в теле рыб, ведущих малоподвиж¬
ный образ жизни (табл. 4).Таблица 4. Содержание красных мышц в мясе рыбы (в % общего
количества мышечной ткани) [126]СемействоЧислоиссле¬дован¬ныхвидовСодержа¬
ние крас¬
ного мяса
в мыш¬
цах, %СемействоЧислоиссле¬дован¬ныхвидовСодержа¬
ние крас¬
ного мяса
в мыш¬
цах, %Scombridae226,1Bothidae27,2(Скумбриевые)(Ромбовые)Clupeidae519,8Lophiidae17,2(Сельдевые)(Удилыциковые)Carangidae118,3Triglidae27,0(Ставридовые)(Морские петухи)Sparidae115,7Cottidae26,2(Спаровые, мор¬(Рогатковые)ские караси)Centrolophidae15,9Cyclopteridae114,8(Центролофовые)(Пинагоров ые)Scorpaenidae45,5Mugilidae114,5(Скорпеновые)(Кефалевые)Anarhichadidae35,1Squaloidae114,3(Зубатковые)(Катранообразные)Lumpenidae15,0Ammodytidae112,9(Лумпеновые)(Песчанковые)Macrouridae34,5Pomatomidae112,7(Макрурусовые)(Л уфарев ые)Gobiidae14,5Gadidae1410,6(Бычковые)(Тресковые)Labridae3,5Pholididae111,1(Губанов ые)(Маслюков ые)Trachichthyidae13,5Scyliorhinidae210,2(Большеголовые)(Кошачьи акулы)Congridae12,8Argentinidae29,4(Морские угри)(Аргентиновые)Trachinidae12,4Zeidae19,3(Морские дракон¬(Солнечниковые)чики)Pleuronectidae78,9Chimaeridae10,6(Камбаловые)(Химеровые)Anguillidae18,8Caproidae10,5(Угреобразные)(Кап ров ые)Soleidae28,1Trichiuridae10,5(Солеевые, косо¬(Вол о со хво стовые)ротые)Rajidae30Osmeridae27,4(Скатов ые)(Корюшковые)18
ПродолжениеЧислоСодержа¬ЧислоСодержа¬иссле¬ние крас¬иссле¬ние крас¬Семействодован¬ного мясаСемействодован¬ного мясаныхв мыш¬ныхв мыш¬видовцах, %видовцах, %Aeopiidae 1 0 Bramidae 1 О(Лисьи акулы) (Морские лещи)Callionymidae 1 0 Berycidae 1 О(Морские мыши) (Бериксовые)У тресковых рыб, из которых в основном изготовляется фарш,
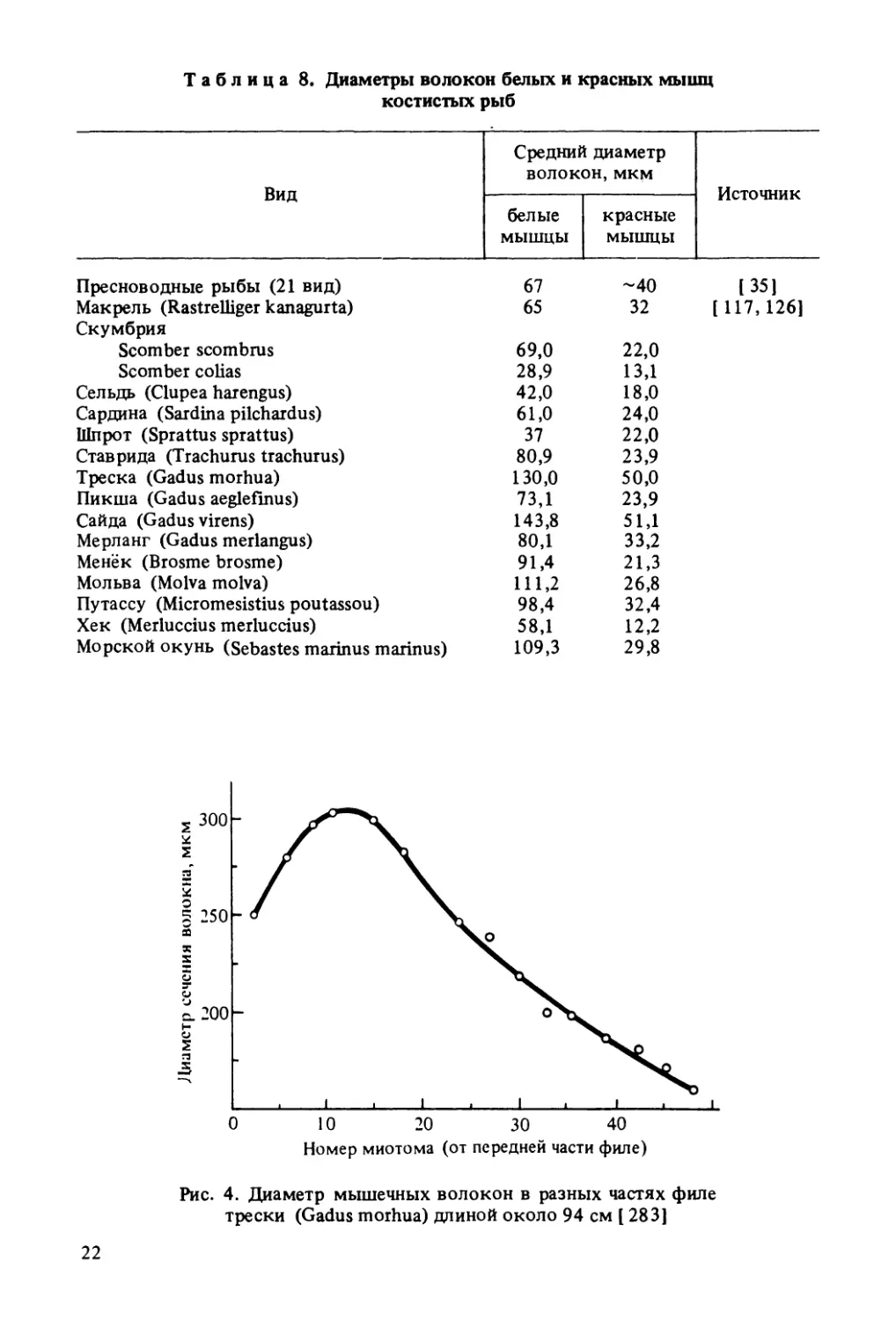
среднее содержание красных мышц составляет примерно 10 %, в том
числе у менька и мольвы не превышает 2 %, хека - около 5,5 %, мер¬
ланга, пикши, сайды — 12 %, у трески — 17 %, а у морского налимчи-
ка Phycis blennoides (Brunnich) достигает 25 % [126]Красное мясо значительно отличается от белого по химическому
составу и ферментативной активности. Это связано главным образом
с кислородным характером обмена веществ в красных мышцах. В то
время как в белых мышцах метаболические процессы идут в бескис¬
лородных условиях, а энергия образуется главным образом в резуль¬
тате распада гликогена, в красных мышцах главным источником энер¬
гии являются липиды [141, 284] . Большинство ферментов, содержащих¬
ся в красных мышцах, характеризуется более высокой активностью,
чем в белых мышцах. Это относится и к деметилазе триметиламино-
ксида [461] , фосфатазе, фосфорилазе, аминопептидазе [335] , аргина¬
зе [308], холинэстеразе [377] и липооксидазе [366] Из гидролаз
особого внимания заслуживает липаза, активность которой в красных
мышцах примерно в 5 раз выше, чем в белых (табл. 5).Таблица 5. Содержание жира и активность липазы в красных
и белых мышцах макрели Rastrelliger kanagurta [117]МышцыСодержание жира, %Активность липазы,
10“6 л СС>2/ (мг белка ч)Красные 5,75±0,62 255,15±21,55Белые 1,71±0,32 55,59±8,21В красных мышцах содержится меньше воды (примерно на 19 %),
белка (на 22 %) и золы (примерно на 12 %), чем в белых мышцах, зато
заметно больше липидов. В общем можно принять, что содержание липи¬
дов в красных мышцах в 2—5 раз выше, чем в белых (табл. 6), хотя у не¬
которых видов рыб, особенно тощих, это различие значительно больше.
Например, у хека (Merluccius capensis, М.paradoxus) осеннего вылова в
красном мясе липидов содержится в 20 раз больше, чем в белом [495]19
Таблица 6. Содержание (в %) воды, белка, жира и золы в.красных (Кр)
и белых (Б) мышцах некоторых видов рыб [ 40]ВидМышцыВодаБелокN•6,25ЛипидыЗолаСельдь (Clupea harengus)Кр57,815,528,2'1,30Б74,022,013,01,40Сайда (Pollachius virens)Кр77,818,62,51,13Б78,419,90,51,27Скумбрия (Scomber scombrus)Кр54,214,929,71,18Б65,521,213,11,24Палтус (HippoglossusКр62,011,327,00,82hippoglossus)Б77,414,57,01,10В красном мясе содержится также больше веществ, ускоряющихокисление липидов - миоглобина, гемоглобина, цитохрома С (табл. 7).Таблица 7. Содержание (в мг%) хромопротеидов в красном (Кр)
и белом (Б) мясе некоторых видов рыб [ 309]ВидГемоглобин +
+ миоглобинМиоглобинЦитохром СКрБКрБКрБCololabis sajra5103627141,82,0(сайра, скумбрещука)
Katsuwonus vagans21851606503120,01,5(полосатый тунец)
Makaira mazara102014510012,30,1(голубой марлин)
Makaira mitsukurii115025660014,40,8(полосатый марлин)
Neothunnus macropterus2510140111508,41J(желтоперый тунец)
Scomber japonicus98010390013,00,1(скумбрия)Seriola quinqueradiata40012150217,20,9(желто хвост)
Thunnus orientalis257230512221007,50,3(голубой тунец'*Содержание миоглобина в красном мясе возрастает от головы к
хвосту так, что в хвостовых участках мышц миоглобина содержится
в 1,5—2 раза больше, чем в приголовных [144, 145] . В мышцах некото¬
рых видов рыб, например сардины, содержится термоустойчивый мио-
глобин, выдерживающий даже пятиминутное нагревание при темпера¬
туре 100 °С без заметных признаков денатурации [503] В процессе
самоокисления 1 моль миоглобина красного мяса рыбы поглощает
в среднем 0,34 моля кислорода [310] Высвобождающийся в резуль¬
тате изменения pH или термического разрушения миоглобина этот кис-20
пород может участвовать в различных окислительных процессах, про¬
исходящих в фарше. Переход оксимиоглобина в метмиоглобин, а так¬
же окисление жира являются основными причинами изменения цвета
(побурения) красного мяса, в то время как в белом мясе побурение
является главным образом результатом реакции Майяра [393]В красном мясе содержится также значительно больше железа, не
входящего в состав гема, чем в белом [284] На долю такого железа
приходится 56-75 % общего количества железа в красном мясе [106]В связи с этим, несмотря на незначительное различие в составе жирных
кислот и даже более высокое содержание полиеновых кислот (С _6,
С20_5» ^22-5^ в белом мясе [490, 495] , красное мясо обычно более
подвержено окислительным изменениям жира, чем белое [123, 195,
2741Кроме того, причиной большей подверженности липидов красного
мяса окислению может быть высокое содержание свободного гистидина
[8], поскольку эта аминокислота оказывает выраженное каталитиче¬
ское действие [397]2.1.2. Гистологическое строение скелетных мышцФизические свойства рыбного фарша, а также экстрагируемость
миофибриллярных белков в процессе перемешивания в воде в значи¬
тельной степени определяются строением скелетных мышц, и особен¬
но длиной и диаметром мышечных волокон, наличием в мышцах крове¬
носных сосудов, содержанием и строением соединительных тканей.Мышечные волокна имеют форму валиков, закруглен¬
ных с обоих концов и прикрепленных к перегородкам из соединитель¬
ной ткани. Волокна расположены в миомере таким образом, что они
слегка наклонены по отношению к продольной оси тела, при этом слои,
близкие к поверхности, состоят из сильно скошенных волокон, более
длинных, чем в глубоких слоях.Размеры мышечных волокон колеблются в широких пределах
в зависимости от вида и возраста рыбы, вида мышц и локализации
миомера в мышцах. Диаметр волокон туловищных мышц может состав¬
лять от 10 до 380 мкм, а длина от нескольких сотен микрон до 14 мм
[125, 287] Белые мышцы состоят из толстых мышечных волокон с
небольшим количеством саркоплазмы, но с многочисленными воло¬
концами. Красные мышцы состоят из тонких мышечных волокон
(табл. 8) с большим количеством саркоплазмы. Самые длинные и са¬
мые широкие волокна находятся в 11 —14-м миомере филе, считая
от головы, самые короткие и тонкие — в миомерах, расположенных
в хвостовой части филе (рис. 4).От положения мышцы зависит также и наличие в ней кровеносных
сосудов. Например, на 1 мм2 туловищных мышц окуня приходится
около 167 волокон (диаметр 27 и 74 мкм) и 76 капилляров, тогда как21
Таблица 8. Диаметры волокон белых и красных мышц
костистых рыбВидСредний диаметр
волокон, мкмИсточникбелыемышцыкрасныемышцыПресноводные рыбы (21 вид)67-40(35]Макрель (Rastrelliger kanaguita)6532[117,126]СкумбрияScomber scombrus69,022,0Scomber colias28,913,1Сельдь (Clupea harengus)42,018,0Сардина (Sardina pilchardus)61,024,0Шпрот (Sprattus sprattus)3722,0Ставрида (Trachurus trachurus)80,923,9Треска (Gadus morhua)130,050,0Пикша (Gadus aeglefmus)73,123,9Сайда (Gadus virens)143,851,1Мерланг (Gadus merlangus)80,133,2Менёк (Brosme brosme)91,421,3Мольва (Molva molva)111,226,8Путассу (Micromesistius poutassou)98,432,4Хек (Merluccius merluccius)58,112,2Морской окунь (Sebastes marinus marinus)109,329,8Номер миотома (от передней части филе)Рис. 4. Диаметр мышечных волокон в разных частях филе
трески (Gadus morhua) длиной около 94 см [ 283]22
в хвостовой части их число возрастает до 281 волокна (диаметр 24 и
63 мкм) и 139 капилляров на 1 мм2 [141]Зависимость между средней длиной мышечного волокна рыбы и ее
объемом можно описать уравнением [426]D = —23,731og V + 37,85; (г = 0,844, Р < 0,001),где D — средняя длина мышечного волокна в поперечном сечении филе
на расстоянии 2/3 его длины, начиная от головы, мкм; V — объем ры¬
бы, см3Т7 1 hwl
V — — 7Т 3 4’где И — максимальная высота рыбы, см; w — максимальная ширина
рыбы, см; / — полная длина рыбы, см.2.1.3. Структура мышечного волокнаМышечное волокно состоит из пучка расположенных параллельно
волнистых волоконец, окруженных поверхностной оболочкой - сарко¬
леммой, а также жидкостью, заполняющей пространство между воло¬
концами — саркоплазмой. Кроме волоконец, в саркоплазме имеются
различные трубчатые или пузырьковые образования (аппарат Гольджи,
митохондрии, лизосомы, саркоплазматическая сетка) и большое число
ядер, поэтому мышечное волокно называют не клеткой, а клеточным
комплексом или плазмодием [125] Исключение составляют полосатые
мышцы гребешка, волокна которых имеют только одно ядро (Morrison
and Odens, 1968). «Волоконца состоят из саркомеров, охватывающих весь тем¬
ный отрезок и прилегающие к нему с обоих концов светлые отрезки.
Границей саркомера являются тонкие пластинки (оболочки) Z (рис. 5).
Длина саркомера зависит от вида мышечного волокна. Большие воло¬
конца белых мышц состоят из саркомеров длиной около 1,82 мкм (за
исключением рыб семейства Anguilla — 1,98 мкм и Cobitis - 2,04 мкм,
а волоконца красных мышц - из саркомеров длиной около 2,04 мкм
(угорь - 2,47 мкм) [141] У некоторых рыб (например, миноги) дли¬
на саркомера достигает 4 мкм [201]Темный отрезок саркомера состоит в основном из миозина, а свет¬
лый — из актина. Актин присутствует в виде тонких нитей (длина при¬
мерно 2 мкм, диаметр 5 мкм), называемых миофиламентами, которые
одним концом плотно сидят в пластинке Z, а второй конец их свобод¬
но размещается между более короткими и более толстыми миофила¬
ментами миозина (длина примерно 1,5 мкм, диаметр 10 мкм). Поверх¬
ность тонких миофиламентов гладкая, толстые миофиламенты имеют
мелкие кольца, расположенные на расстоянии 6—7 мкм друг от друга23
£<&фуУgy$>Нити миозина
(более толстые)-^Мышечные волокна5Мышечное волокнот-| pi—I 1 МышечноеИ III I волоконце(миофибрилл)Саркомер\Нити актина
(тонкие)Рис. 5. Схема структуры скелетной мышцы на разном уров¬
не ее строения(см. рис. 6). Сокращение мышцы происходит путем втягивания тонких
нитей между толстыми, что приводит к изменению формы мышцы
без изменения ее массы.2.2. СОЕДИНИТЕЛЬНАЯ ТКАНЬ МЯСАЭто название включает различные ткани, образующие волокнистое
вещество, расположенное под кожей, между миомерами (миосепты)
в межклеточном пространстве, а также в сарколемме. Соединительная24
ткань состоит из бесструктурного желеобразного, так называемого
’’подпорного” вещества, а также различных волокнистых и клеточных
образований. Жировая ткань, присутствующая главным образом в под¬
кожном слое, а также в миосептах, — это измененная соединительная
ткань. Волокнистые образования состоят в основном из коллагеновых,
а также из немногочисленных эластичных волокон. Они существуют
в виде пучков и единичных волокон, образующих трехмерную сетчатую
структуру. Благодаря этим свойствам соединительная ткань создает как
бы поддерживающую решетку, связывающую и защищающую отдельные
пучки мышц.Прочность соединительной ткани на растяжение меньше, чем проч¬
ность соединительной ткани мяса животных, что обусловлено невысоким
содержанием эластина и коллагена в элементах волокон. Коллагеновые
волокна легко поддаются сгибанию и скручиванию, однако сопротивля¬
ются растяжению - их удлинение при разрыве составляет всего несколь¬
ко процентов. Пучок коллагеновых волокон сечением 1 мм2 выдержи¬
вает нагрузку до 6 кг [466] . Под действием температуры выше 40 °С*
коллаген сокращается так, что по достижении температуры 60—70 °С
волокна укорачиваются на У4-У3 своей первоначальной длины. При
более продолжительном действии указанной температуры большая часть
коллагена переходит в желатин. Разбавленные кислоты и основания вы¬
зывают набухание коллагеновых волокон, их распад на отдельные цепи
тропоколлагена и даже на отдельные полипептидные цепочки [383]
Продукты термической или химической денатурации коллагена легко
поддаются воздействию протеолитических ферментов.К особенностям волокон эластина относится значительная растяжи¬
мость (свыше 50 %) и возможность возврата к первоначальной длине,
а также высокая устойчивость к действию' химических веществ и темпе¬
ратуры. Эластин, нагретый в воде до температуры 137 °С, не растворя¬
ется и не образует клейкой взвеси. Присутствие эластина играет важную
роль в снижении крошливости мяса и сохранении его плотности, осо¬
бенно в процессе термической обработки. В мясе рыбы содержание
эластина низкое, таким образом можно принять, что основным компо¬
нентом соединительной ткани является коллаген. Так, в коже мор¬
ских рыб содержание эластина составляет 2,3-4,0 %, а коллагена 9,6-17,1 %, что соответствует 8,6-12,7 и 35-54,4 % в пересчете на сухую
массу [473]2.3. КОЖАПрисутствие кусочков кожи в фарше ухудшает его цвет, а также
снижает возможный срок холодильного хранения.* Температура сокращения коллагена (7^) рыбы может колебаться в преде¬
лах от 33 до 58 °С. Например, для коллагена кожи рыб холодных морей Ts сос¬
тавляет 33-52 °С, рыб теплых морей - 49-58 °С [ 272].25
В крже рыбы имеются слизистые железы, выделяющие гликопро¬
теид муцин, который при смешивании с водой образует слизь. После
смерти рыбы это вещество становится идеальной питательной средой для
микроорганизмов и основным источником заражения мышечной ткани.В ухудшении цвета фарша основную роль играют окрашенные клет¬
ки, присутствующие в эпидермисе или в самой коже.В меланофорах содержатся зерна меланина коричнево¬
го или черного цвета. В этих зернах красящее вещество агрегировано на
сетчатой белково-фосфолипидной основе. Меланин - полимер продуктов
окисления тирозина, нерастворимый в воде и кислотах. Образование его
из тирозина катализируется ферментом тирозиназой (фенолоксидазой),
весьма распространенным в мире животных и растений [287]Гуанин — продукт распада нуклеотидов - содержится в гуано-
форах в виде бесцветных кристаллов длиной 2-20 мкм. Гуанофоры
обычно расположены слоями на наиболее обширных участках поверх¬
ности кожи.Липофоры чаще всего содержат каротиноидное красящее ве¬
щество липохром. Рыба получает его, поедая водоросли или рако¬
образных. Липохром двухцветный — желтый в ксантофорах и
красный в эритрофорах. Оба красящих вещества легко вымыва¬
ются органическими растворителями, применяемыми для экстракции
липидов. Состав каротиноидов может быть различным в зависимости от
вида рыбы. Основным каротиноидом в коже морских рыб является
тунаксантин (30—65 % общего содержания каротиноидов), а в ко¬
же пресноводных рыб — л ю т е и н (20—40 %). Имеются также зеак-
сантин, цинтиаксантин, криптоксантин, изокрипто ксантин, астацен,
L-дорадецин, 0-каротин [305, 306, 307] В коже некоторых видов рыб
(например, скорпеновых) в значительном количестве содержится розо¬
во-красное красящее вещество астаксантин, придающее этим видам
рыб характерный цвет [477] . Желтый краситель, нерастворимый в спир¬
те (этиловом), присутствует в аллофорах. Он относится к группе фла-
винов и обусловливает флуоресценцию рыб.Общее содержание комплекса красителей в коже рыбы 1,3—45 мг%
[372].В живой рыбе красящие вещества связаны с белками, липидами или
нуклеотидами. Разрыв этих связей в результате денатурации белков,
гидролиза липидов или других физико-химических изменений приводит
к высвобождению красящих веществ, которые в зависимости от хими¬
ческих свойств могут образовывать взвеси в воде или растворяться
в липидах, что вызывает изменение окраски и подкожное пожелтение
мышечной ткани.К весьма характерным следует отнести изменения астаксантина
(3,3'-диокси-4,4'-дикето-/3-каротин), который в результате энолизации
приобретает свойства кислоты и образует соли. Однако эти соли неус¬
тойчивы и при доступе воздуха в результате окисления превращаются
26
в астацен (3,3'-4,4'-тетракето-0-каротин) — пигмент интенсивного чер¬
ного цвета [271]Окисление каротиноидов в рыбном фарше может происходить под
действием ферментов, например липооксидазы, содержащейся в боль¬
ших количествах в темном мясе [366] , или других факторов нефермен¬
тативного характера (кислород, свет, продукты окисления жира, ионы
металлов и др.) [269] Эти реакции часто идут с постоянной скоростью.
Окисление под воздействием кислорода начинается с ненасыщенных
связей в ионах. Каротиноиды в этой фазе отбирают из окружающей
среды кислород и действуют как антиокислители. В следующей фазе
процесс окисления достигает полиеновых связей, и каротиноид может
действовать как вещество, ускоряющее окисление (прооксидант)
[44] . Ксантофилы более устойчивы к окислению, чем каротины.В коже тощей рыбы (треска, путассу) содержится в среднем от 0,2
до 3,9 % жира [473] Зато в коже жирной рыбы содержание жира зна¬
чительно выше и составляет: в ставриде — 13,3, сардине — 45,4, тихооке¬
анской скумбрии — 48,2 % [468] , в атлантической скумбрии 20—32,6 %
весной и 39,8—54,9 % осенью [195]Состав липидов кожи колеблется в зависимости от сезона добычи
рыбы, в определенные периоды содержание фосфолипидов может быть
высоким (свыше 50 %). Кожа тощей рыбы характеризуется обычно
более высоким содержанием фосфолипидов (свыше 50 %). В коже то¬
щей рыбы содержится больше фосфолипидов и стеролов, чем в коже
жирной рыбы. Полиеновых жирных кислот в коже рыбы часто содер¬
жится больше, чем в мясе. Например, содержание полиеновых кислот
с 4—6 двойными связями (в процентах к общему содержанию насыщен¬
ных кислот) в коже тихоокеанской скумбрии составляет 0,93, а в мыш¬
цах 0,74-0,79 [469] В коже содержится также значительно больше
железа (примерно 17,5 мкг%) и цинка (19,7 мкг%), чем в мясе (соот¬
ветственно 10,8 и 4,0 мкг%). Содержание в коже остальных тяжелых ме¬
таллов близко к их содержанию в мясе [194]Приведенные выше различия позволяют сделать вывод, что остатки
кожи в мороженом рыбном фарше окисляются быстрее, чем мясо. Из
данных, представленных в табл. 9, следует, что примерно после 3 нед
хранения при температуре —5 °С степень окисления гомогената кожи
в 3 раза превышала степень окисления в пересчете на содержание жира
и более чем в 8 раз в пересчете на содержание ткани кожи по сравнению
с тканью мяса.Вместе с тем одинаковая окисляемость жира, экстрагированного из
свежих тканей, может свидетельствовать о наличии в коже специальных
веществ, ускоряющих окисление (прооксидантов). Кстати, эти предпо¬
ложения нашли в дальнейшем подтверждение [196]Скорость окисления липидов кожи в процессе хранения мороженой
рыбы значительно выше скорости окисления других тканей. В коже та¬
кой рыбы, как тихоокеанская скумбрия, сардина и ставрида, после ме-27
Таблица 9. Подверженность гомогенатов различных тканей
тихоокеанской скумбрии и липидов гомогенатов окислению жира
после 22 сут хранения при температуре -5 ° С [ 468]Вид тканиСодер¬жаниелипи¬дов,Подверженность окислению в пере¬
счете на 1 кг ткани*ТБЧ после хра¬
нения в течение
2 ч при 37 °СТБЧ при хранении
в течение 24 ч при
температуре -5 °С
в присутствии
ионов медиТБЧ после
22 сут хра¬
нения при
-5 ° С (в пере¬
счете на 1 кг
ткани)Мышца целиком11,99,32813Красная мышца25,2436869Внутренности32,611011296Кожа35,51860112ТБЧ - тиобарбитуровое число.* Сразу после приготовления препарата.сячного хранения при температуре -5 °С ТБЧ в 13—19 раз выше, чем
вообще в мясе, и в 4—12 раз выше, чем в красном мясе [469]Присутствие даже в небольших количествах измельченной кожи в
рыбном фарше ускоряет его окисление, так как наряду с кожей в нем
содержится быстроокисляющийся подкожный жир, а следовательно, ка-
ротиноиды и другие прооксиданты [218] , несмотря на то что некото¬
рые компоненты кожи (например, меланины) обладают антиокислитель-
ными свойствами [218]2.4. КРОВЬКровь состоит из плазмы и кровеобразующих элементов, к кото¬
рым относятся красные и белые кровяные тельца и тромбоциты. Кровь
служит носителем многих белковых, небелковых, липидных и мине¬
ральных веществ, а также ферментов. Способность поглощения и выде¬
ления кровью кислорода объясняется содержанием в ней гемоглобина,
присутствующего в красных кровяных тельцах (эритроцитах) .Содержание гемоглобина в крови рыбы, ведущей подвижный образ
жизни, составляет около 10 %, а у рыбы, ведущей малоподвижный об¬
раз жизни, — около 6,5 % [100]Главным источником загрязнения рыбного фарша кровью является
передний отрезок аорты, расположе ной под позвоночником между
почками. В процессе прохождения рыбы через сепаратор сгустки крови,
находящиеся в аорте, выдавливаются в фарш. В мышцах красные кро¬
вяные тельца присутствуют главным образом в соединительнотканевых
оболочках, разделяющих пу^ки мышечных волокон. Гемолиз красных28
о-^э 1 з—о 2 •—• 3Рис. 6. Влияние добавления оксигемоглобина на изменения ТБЧ (/4) и поглощение
кислорода (В) в гомогенатах мышц форели (3 в.ч. мяса на 1 в.ч. воды) в процессе
хранения при температуре -20 °С 144711 - контрольная проба; 2 - проба с добавлением 2,7 мг НЬ02-Ре/кг; 3 - проба
с добавлением 6,2 мг НЬ02-Г;е/кгкровяных телец, происходящий во время хранения рыбы перед перера¬
боткой, изготовлением фарша и его замораживанием, вызывает высво¬
бождение гемоглобина и переход его в окружающую среду (фарш).
Одновременно происходят химические изменения гемоглобина, в част¬
ности его окисление (присоединение кислорода без окисления железа)
и переход в оксигемоглобин. Один грамм гемоглобина присоединяет
около 1,94*мг атмосферного кислорода [385] Присутствие в фарше
оксигемоглобина увеличивает его окисляемость (рис. 6).Ослабление связей гемоглобина с кислородом, например в резуль¬
тате снижения pH мяса в посмертный период или термического разло¬
жения белка, приводит к высвобождению кислорода гемоглобина, вслед¬
ствие чего он становится катализатором окислительных процессов
в фарше.2.5. КОСТИЧасти скелета, попадающие в рыбный фарш, - это главным образом
кости, кусочки ребер, колючих отростков позвонков и костистые нарос¬
ты на коже (жучки), реже кусочки позвоночника, которые легче всего
Удалить механическим способом.29
Все эти элементы нежелательны с точки зрения безопасности по¬
требления продуктов, но их влияние на химические свойства фарша не¬
велико. Наиболее опасны мелкие кости, присутствующие в мясе, еди¬
ничные, имеющие игло- или Y-образную форму.В костях рыбы содержится много минеральных веществ, особенно
фосфора и кальция - соответственно 1,1-2,4 и 1,5-5,7 % [513] , одна¬
ко эти элементы встречаются главным образом в виде нерастворимых
в воде солей, в результате чего они практически не попадают в мышеч¬
ную ткань в процессе приготовления фарша.2.6. ПОЧКИПочки рыбы располагаются под осевым скелетом (позвоночником)
в виде длинных парных органов вдоль всей полости тела. Почки состоят
из нефронов, пронизанных многочисленными разветвляющимися крове¬
носными сосудами. Кроме того, в почках имеется каналец для образо¬
вания и отведения мочи [125] В почках снулой рыбы содержится боль¬
шое количество гемоглобина и мочевины. Из тканей рыбного фарша
почки имеют наибольшую проокислительную активность. Например,
проокислительная активность тканей почек превышает таковую мяса
спины более чем в 50 раз [175] Кроме того, высокая активность дс
метилазы наряду с высоким содержанием ТМАО в почках (примерно
850Miifo) [285] делают ткани почки наиболее нежелательной примесью
в рыбном фарше.
3. ИЗМЕНЕНИЯ, ПРОИСХОДЯЩИЕ
В ИЗМЕЛЬЧЕННОМ МЯСЕ РЫБЫ3.1. ФАКТОРЫ, УСКОРЯЮЩИЕ
НЕЖЕЛАТЕЛЬНЫЕ ИЗМЕНЕНИЯКроме типичных явлений, вызывающих нежелательные изменения
мороженого рыбного сырья, рыбный фарш подвержен действию допол¬
нительных факторов, которые значительно интенсифицируют эти изме¬
нения. К ним прежде всего относятся: разрушение первоначальной струк¬
туры мяса (измельчение); содержание в массе измельченного мяса тка¬
ней, катализирующих нежелательные процессы в фарше, например жи¬
ра, красных мышц, почек, крови, кожи и др.; попадание в фарш некото¬
рого количества воздуха в процессе отделения мяса от костей и пере¬
мешивания.Процесс механического отделения мяса вызывает значительное
разрушение его первоначальной структуры и ускорение многих хими¬
ческих реакций, вероятно, в результате высвобождения ферментов из
клеточных структур, а также контакта ферментов с субстратами. В свя¬
зи с этим при хранении мороженый фарш быстрее подвергается небла¬
гоприятным изменениям, чем филе. Однако эти изменения зависят от
вида рыбы и ее химического состава. В жирной рыбе большое значение
имеет содержание подкожного жира. Например, фарш из очень жир¬
ной сельди (свыше 20 % жира) окисляется значительно быстрее, чем не-
разделанная рыба, а скорость окисления фарша из тощей сельди (около
5 % жира) такая же, как неразделанной рыбы. По степени окисления
фарш из скумбрии с содержанием жира 17 % почти не отличается от ту¬
шек даже после 3,5 мес хранения при температуре -28 °С [218] Одна¬
ко в фарше любого типа из жирной рыбы содержится больше белково¬
липидных комплексов, чем в сырье других видов обработки [218]У тощей рыбы измельчение мяса приводит главным образом к уве¬
личению содержания продуктов распада ТМАО (табл. 10) и связанному
с ним уменьшению растворимости белков. Кроме того, липиды быстрее
окисляются в фарше из тощей рыбы, чем в филе [491] , однако это
различие не оказывает существенного влияния на запах продукта. Ос¬
новной причиной ухудшения качества мороженого фарша из тощей
рыбы являются изменения консистенции, и особенно сочности после
тепловой обработки, а также вкуса (табл. 11)31
Таблица 10. Изменение содержания диметиламина и муравьиного альдегида
в фарше и филе из хека (Merluccius merluccius) и трески (Gadus macrocephalus)
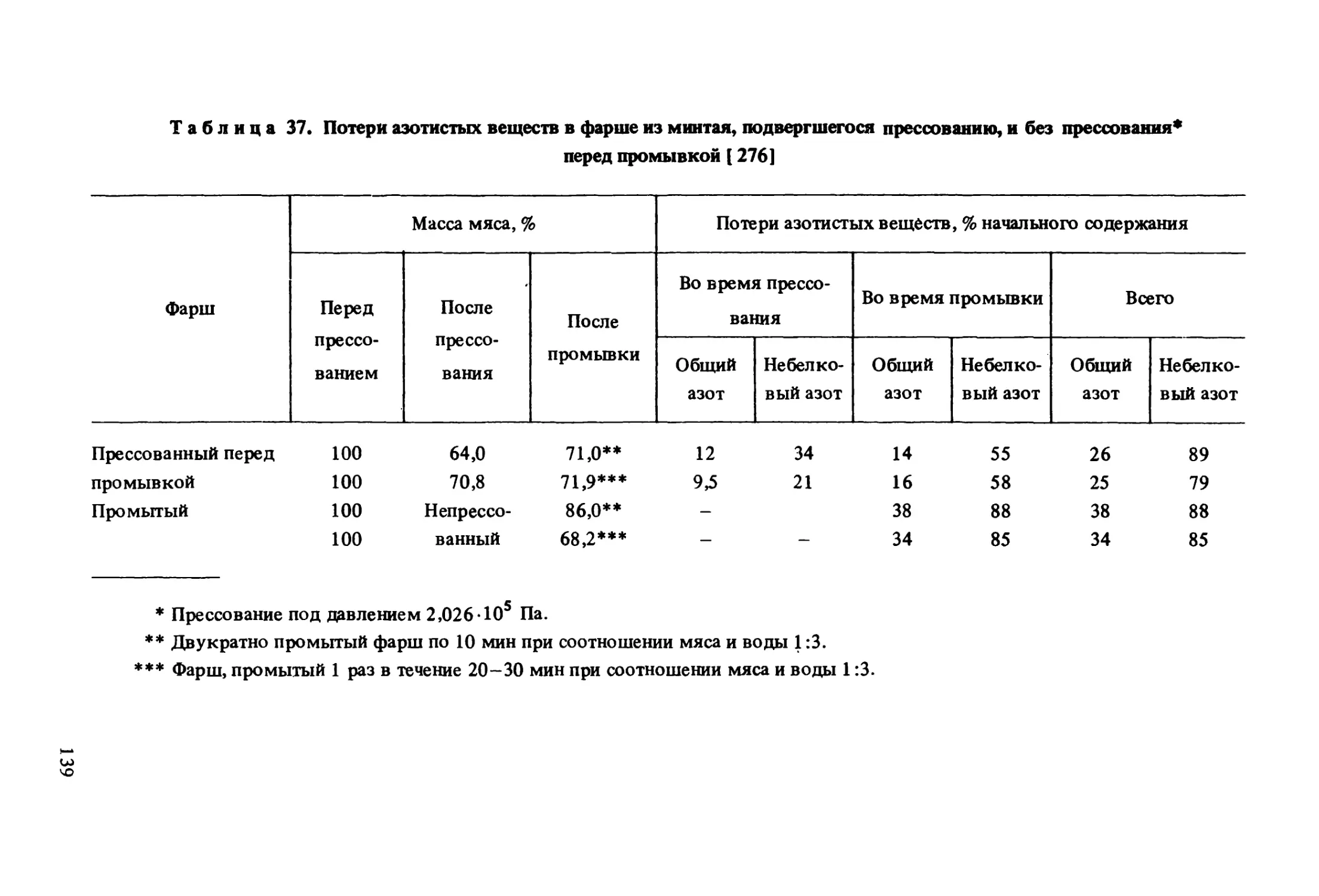
в процессе хранения прй температуре -26° С [68]РыбаПродуктПродолжительность хранения, мес12Диметиламин,мкг/гХекФиле7,47,813,821,025,10,98551,581Фарш22,031,038,250,864,00,99253,460ТрескаФиле8,715,323,028,331,10,98741,927Фарш36,741,759,863,265,50,94602,637Муравьиный альдегид, мкг/гХекФиле9,719,021,626,831,80,98391,733Фарш22,024,626,136,947,20,94252,090ТрескаФиле14,519,822,323,834,80,94481,487Фарш20,536,936,945,481,10,91004,323Примечание.Фарш приготовленна сепараторе(YamigayaFish Separator), диа-метр отверстий барабана 4 мм. Ь, г — коэффициенты в уравнении регрессии.Таблица 11. Средние* результаты органолептической оценки** порций***,
вырезанных из мороженых блоков фарша или филе, хранившихся
при температуре -26 ° С [ 68]РыбаПродуктПродолжительность хранения, мес12ХекФилеКонсистенция**
7,79 7,507,667,047,09Фарш7,437,297,375,856,71ТрескаФиле7,928,138,057,466,92Фарш7,777,787,435,786,09ХекФилеСочность**
6,50 7,395,686,947,21Фарш6,506,975,966,437,14ТрескаФиле6,737,337,047,427,42Фарш5,226,205,085,576,45ХекФиле7,23Вкус**7,767,437,477,16Фарш7,566,906,606,176,12ТрескаФиле7,917,877,507,657,76Фарш7,017,226,936,676,3032
ПродолжениеРыбаПродуктПродолжительность хранения, мес12Общая привлекательность**ХекФиле5,776,756,136,326,50Фарш5,455,794,964,775,03ТрескаФиле6,5 37,146,636,766,73Фарш5,175,925,395,025,13* п = 48; ** по 9-балльной шкале (9 - высшая оценка, 1 - низшая); *** пор¬
ции, панированные влажной и сухой панировочной смесью и обжаренные во фри¬
тюре.Увеличение степени измельчения фарша, например в результате
его повторной сепарации в стрейнере или перемешивания без добавле¬
ния стабилизаторов, ускоряет неблагоприятные изменения реологиче¬
ских свойств в процессе его хранения в мороженом виде. Это заметно
отражается на качестве готовых изделий (табл. 12). Прежде всего воз¬
растают резинистость и волокнистость и снижается водоудерживающая
способность фарша.Таблица 12. Влияние степени измельчения мяса морского окуня
на консистенцию обжаренных порций, приготовленных из мороженых
блоков фарша, хранившегося 12 мес при температуре -18 °С [ 322]Консистенция* обжаренныхХарактеристика фаршапорций из фарша, хранивше¬
гося (в мес)06912мерно 7 мм)Тонкоизмельченный фарш (после стрейнера, да
метр частиц примерно 1,4 мм)Смесь грубо- и тонкоиз мельче иного фарша (1:1)4,64,64,44,43,93,43,63,74,24,23,64,0* Шкала оценки консистенции: 5 - очень хорошая; 4 - хорошая; 3 - доста¬точно хорошая (удовлетворительная) ; 2 - на грани допустимого для потребления;1 - плохая.33
Самой низкой водоудерживающей способностью обладает средне-
измельченный фарш с частицами размером 1,5—3 мм. При размере час¬
тиц более или менее 1 мм водоудерживающая способность возрастает
[30] , что в первом случае обусловлено уменьшением степени повреж¬
дения первоначальной структуры мышечной ткани, а во втором - воз¬
растающей долей фракции белков, растворимых в воде и набухающих в
процессе связывания воды.Наибольшие физические и химические изменения в измельченном
мясе рыбы происходят в начальный период хранения (обычно до 1 мес)
[510].Механическое разрушение мяса во врзмя сепарации вызывает также
некоторые изменения его химического состава, как правило, неблаго¬
приятные с точки зрения сохранения качества фарша. Например, пока¬
зано [290], что в фарше из Catostomus commersoni содержится меньше
фосфолипидов (0,534 мг%) и больше карбонильных соединений
(0,89 мкмоль/г), чем в филе из той же рыбы (соответственно 0,370
и 0,61). Это объясняется, по-видимому, тем, что в процессе сепарации
подкожный жир отделяется от кожи тщательнее, чем во время филетиро-
вания. Кроме того, при сепарации из измельченного мяса удаляется не¬
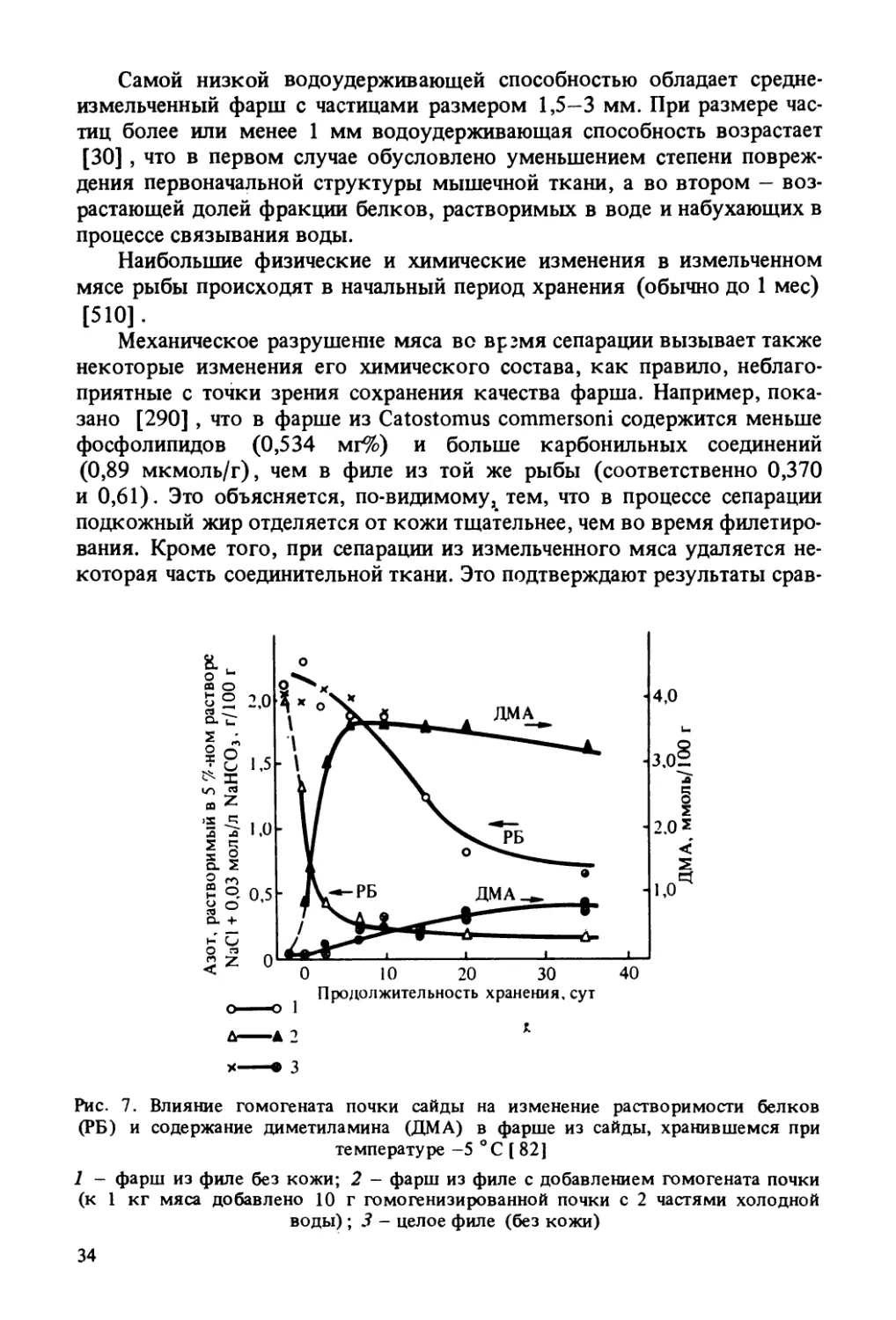
которая часть соединительной ткани. Это подтверждают результаты срав-Рис. 7. Влияние гомогената почки сайды на изменение растворимости белков
(РБ) и содержание диметиламина (ДМА) в фарше из сайды, хранившемся притемпературе -5 °С [ 82)1 - фарш из филе без кожи; 2 - фарш из филе с добавлением гомогената почки
(к 1 кг мяса добавлено 10 г гомогенизированной почки с 2 частями холодной
воды) ; 3 - целое филе (без кожи)34
Продолжительность хранения, суто 1
• 2
• 3Рис. 8. Изменение содержания клеточного сока, образовавшегося под давлением,
и свободно вытекающего из измельченного мяса трески, которое хранилось притемпературе —5 ° С [ 82] :1 - измельченное филе; 2 - измельченное филе с добавлением крови (16,5 см3/кг
мяса) ; 3 - фарш из отходов от филетироъаниянительных исследований [494] химического состава мяса, отделенного
механически и вручную. В мясе, отделенном механически, содержится
значительно больше жира и меньше - белков стромы. Однако сущест¬
венного различия в содержании миофибриллярного белка и золы не ус¬
тановлено.Присутствие тканей, катализирующих нежелательные физико-хи-
мические изменения в фарше, чаще всего связывают с недостаточно тща¬
тельным обескровливанием рыбы и неполным удалением почек во вре¬
мя первичйой обработки. При чрезмерном прижатии ленты к барабану
сепаратора в фарш попадает значительное количество красного мяса
и кусочков кожи. Отрицательное влияние присутствия этих тканей в
фарше на качество и сроки его возможного хранения отмечено во мно¬
гих работах. Некоторые из таких данных представлены на рис. 7-10.
В общем можно утверждать, что все перечисленные выше ткани уско¬
ряют окислительные и гидролитические процессы, происходящие в жире,
а также деметилирование ТМАО и уменьшение растворимости белков в
мороженом рыбном фарше. Их активность заметнее проявляется по
мере увеличения срока хранения (обычно после одного месяца хране-35
Продолжительность хранения, сутА А 2Рис. 9. Изменение растворимости белков (снижается) и содержания диметиламина
(возрастает) в фарше из трески, хранившемся при температуре -5° С [82]:1 - фарш из филе; 2 - фарш из отходов филетирования (с почкой) ; 3 - фарш из
филе с добавлением крови (16,5 см3/кг мяса)ния) и возрастает с увеличением содержания этих тканей в фарше. В фар¬
ше из жирной рыбы, содержащем кровь, ткани почек, темное мясо и ко¬
жу, особенно интенсифицируются процессы окисления жира. При оди¬
наковом содержании в фарше (1 %) сильнее всего действуют почки и
кровь, слабее кожа. В количествах, практически встречающихся в фар¬
ше, кожа играет роль основного катализатора окислительных процес¬
сов жира, что, вероятно, связано с наличием подкожного жира.В фарше из тощей рыбы кровь, почки, кожа и темное мясо катали¬
зируют главным образом процессы гидролиза жира и разложение ТМАО
до диметиламина (ДМА) и свободных жирных кислот. Наибольшей ак¬
тивностью обладают почки и кровь, а самой низкой — кожа, независимо
от содержания этих тканей в фарше [244] Остатки в фарше названных
тканей снижают не только продолжительность его хранения, но и тех¬
нологическую пригодность. Изделия из фарша, полученного из тушек,
менее приемлемы, чем из фарша, выработанного из обесшкуренного фи¬
ле^ приемлемость изделий, изготовленных из фарша, полученного из
отходов от филетирования, оказалась лишь удовлетворительной
(табл. 13).36
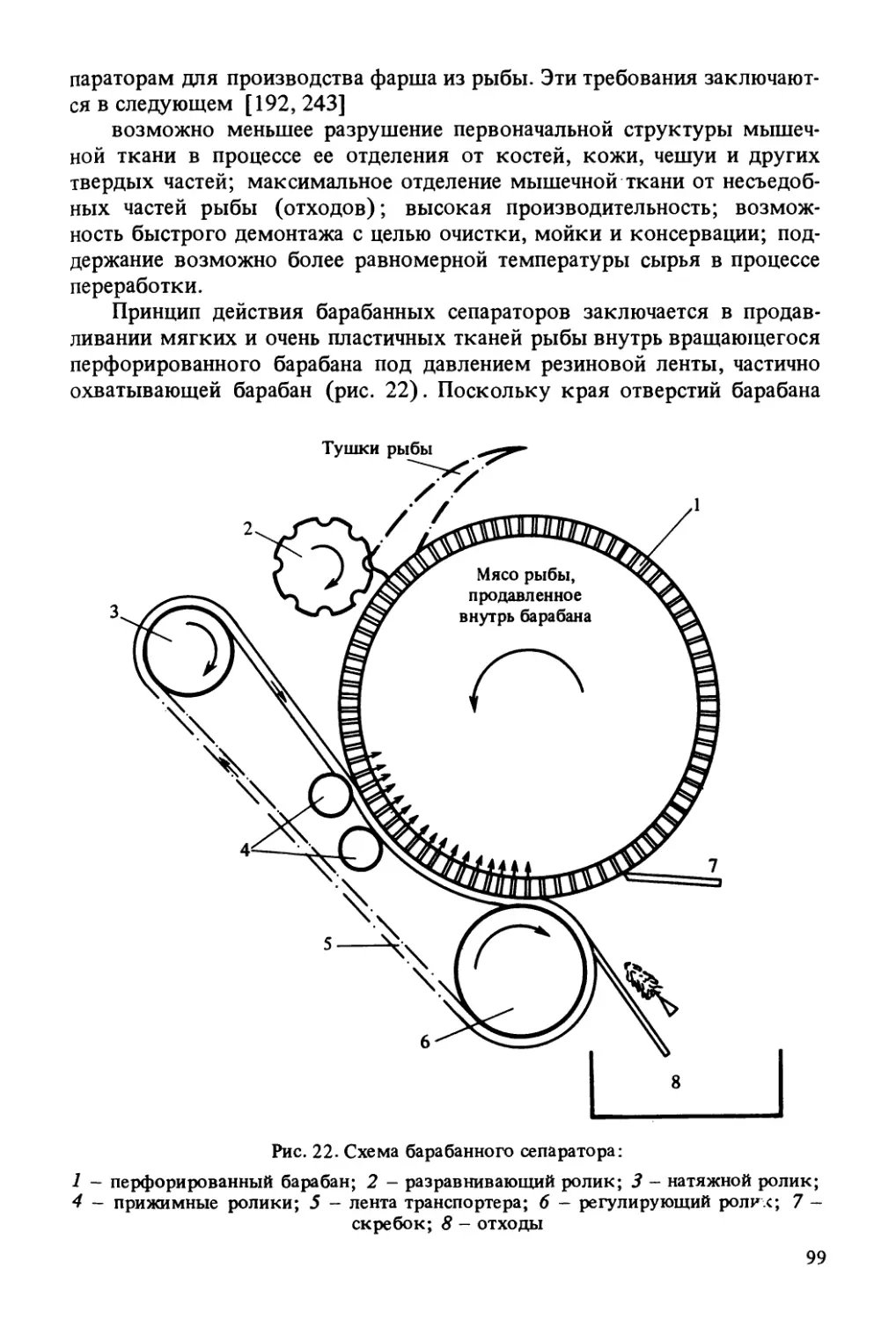
Такой пористый, очень липкий и пластичный материал, как фарш,
весьма подвержен воздействию воздуха. Воздух проникает в фарш
главным образом в момент сепарирования мяса и во время его переме¬
шивания со стабилизирующими добавками. Отделять воздух от фарша
с помощью предпрессовки не рекомендуется, так как возможно разру-Исследованная концентрация, %Рис. 10. Влияние добавления крови, почки, красных мышц и кожи на окислитель¬
ные и гидролитические изменения жира в измельченном мясе сельди, хранившем¬
ся в течение 10 сут (д) и 70 сут (б) при температуре -15 °С [ 82]:1 - LN (мг 0/100 г жира) - перекисное число; 2 - LK (мг КОН/ г жира) - кис¬
лотное число почка i исследованные концентрации: 0; 0,1; 0,2; 0,3...1 %
кр. кровь )КМ - красные мышцы 1 исследованные концентрации:К - кожа / 0, 1, 2, 3...10 %Ъ1
Таблица 13. Сравнение приемлемости рыбных порций из филе и фарша,
полученного из филе, тушек и отходов от филетирования некоторых видов рыбы[22]РыбаПродолжи¬
тельность хра¬
нения при тем¬
пературе
-18 °СОбщая приемлемость (1-9 баллов)ЦелоефилеФарш изфилебезкожифилескожейприпозвоноч-
ного мяса и об-
резей от филе¬
тированиятушекТихоокеанский06,985,624,324,‘344,68хек125,785,204,163,624,42Терпуг05,924,604,463,385,5412-----Скорпена06,805,665,824,084,78126,064,564,883,584,46Морской язык05,664,724,322,663,44126,124,904,522,743,80шение его крупнозернистой структуры. Только в атмосфере инертных
газов фарш можно освобождать от воздуха без заметного ухудшения
его консистенции.3.2. БЕЛКИ МЫШЦ И ИХ ИЗМЕНЕНИЯС точки зрения пищевой ценности и технологической пригодности
белок является наиболее ценным компонентом рыбного фарша. Изме¬
нения белков непосредственно отражаются на функциональных свойст¬
вах рыбного фарша (например, консистенции, способности связывать
воду, эмульгировать жир) и его биологической ценности. Сведения об
изменениях белка совершенно необходимы для получения рыбного
фарша высокого качества.3.2.1. Характеристика белков мяса рыбыБелки мяса рыбы состоят из многих фракций, различающихся раст¬
воримостью, pH изоэлектрической точки, молекулярной массой и темпе¬
ратурой коагуляции (табл. 14). Около 25 % общего количества белков
плохо растворяется в водопроводной воде (например, во время промыв¬
ки фарша). Кроме обычных альбуминов (миоген, миоальбумин), к этой
группе относится также глобулин х, который в деионизированной (дис¬
тиллированной) воде не растворяется, однако при смешивании фарша
с водой он ведет себя подобно альбуминам, поэтому минеральные соли,
содержащиеся в тканевом соке мяса рыбы и в водопроводной воде,
создают достаточно высокую ионную силу для его растворения. Белки38
саркоплазмы — наиболее сложная и разнообразная фракция, ко¬
торую с помощью электрофореза можно разделить на несколько фрак¬
ций, иногда более десяти [81, 223] Таким образом белки саркоплазмы
состоят из нескольких десятков белков, имеющих разные свойства.
В состав фракции белков саркоплазмы входит около 100 различных
ферментов, катализирующих, кроме всего прочего, реакции гликолити-
ческого цикла, гидролиз белков и полипептидов, ферментативное раз¬
ложение ТМАО и многие другие. Эта группа белков содержит, кроме
того, хромопротеиды крови и мышц (гемоглобин и миоглобин), а так¬
же цитохромы.Таблица 14. Состав и характеристика белков мяса костистых рыбСодержа¬Молеку-pH изо-ТемператураРастворимостьФракцияние (в %лярнаяэлектри-коагуляции,(необходимаябелковобщегомасса,ческой°Сионная силасодержа¬ЛГ-103точкираствора*)ния бел¬ков)А. Белки сар¬20-3018-4654-8,540-56 'коплазмы10-1581-1506,5-6,7Растворимые вмиоген, деионизирован¬миоальбу-2-2,53-3,5ной водеминмиоглобин0,5-2,016,8глобулин X10-15Около 1605,2Менее 0,05Б. Миофибрил-60-7043-5304,7-5,535-55Около 0,55лярные белкимиозин30-40500-5305,4-5,540-45(0,35-1,0)актин15-2543-1304,7тропо мио¬2-2,5525,1зинтропонин18,3-40,2В. Белки сое¬2-4**,динительной8-11*3тканиколлаген320-3507,0-7,844,6-5 3,7*4Нерастворимыеэластин60-100135*5в нейтральныхконектин61,7-67,0*°солевых и раз-(ретикулин)бавленных раст-ворах щелочей* При pH 5,2; ** костистая рыба; *3 хрящевая рыба; *4 температура сокра¬
щения; *5 пограничная температура термической стабильности; *° температура
желатинизации.По аминокислотному составу отдельные компоненты белков сарко¬
плазмы значительно различаются между собой [251] , однако в общем
эти белки относятся к полноценным.39
Самую многочисленную группу, исключительно ценную в биологи¬
ческом отношении, но наиболее подверженную изменениям в процессе
хранения мороженого фарша, составляют миофибриллярные
белки. К ним относятся: миозин, актин, тропомиозин и тропонин.
Миозин и актин, составляющие в сумме около 50 % всех белков мышеч¬
ной ткани, являются основными белками, способными сокращаться.
Миозин обладает свойствами фермента, называемого АТФазой, т. е.
способен катализировать реакцию гидролиза АТФ. Высокое содержание
сульфгидрильных групп (SH) делает этот белок высокоактивным и
чувствительным компонентом, вызывающим денатурацию [264] Мио¬
зин особо чувствителен к присутствию ионов тяжелых металлов и тем¬
пературе, а его денатурация сопровождается уменьшением раствори¬
мости. Трипсин легко расщепляет миозин на меромиозины, отделяя от
него легкие боковые цепи с молекулярной массой 17500—26500. Мио¬
зин темных мышц имеет две боковые цепи, а миозин белых — три [492]
Актин не обладает свойствами фермента, хотя в состоянии присоеди¬
нять молекулы АТФ, и может существовать в двух формах: глобуляр¬
ной - G-актин и фибриллярной - F-актин. Последний прочно связан с
АТФ и ионами кальция. Под действием физиологической ионной силы
молекулы G-актина полимеризуются с образованием четырехрядной
структуры F-актина. Эта реакция сопровождается распадом АТФ [320]В добываемом рыбном сырье большая часть миозина присутствует
в виде комплекса — актомиозина. Показано [24] , что в мясе
пресноводной рыбы, подвергнутой анализу сразу после обезглавлива¬
ния, содержание растворимого актомиозина составляет всего 0,76—2,8 %,
а после 1—4 сут хранения возрастает до 25—65 % общего количества
растворимых белков. Повторное снижение содержания актомиозина при
дальнейшем хранении рыбы (после 5 сут) обусловлено денатурацион-
ными изменениями. Отношение миозина к актину в актомиозине колеб¬
лется от 2 до 4 [66] Актомиозин, в котором больше лецитина, харак¬
теризуется также более высоким содержанием миозина [438, 439]В растворах, ионная сила которых ниже 0,35, актомиозин разрушается,
поэтому один из методов отделения актомиозина от миозина заклю¬
чается в диализе вытяжки этих белков в буферном растворе с ионной
силой 0,25 при pH 7,1 [173]Тропомиозин и тропонин выполняют функции белков, регулирую¬
щих сокращение мышц. Первый характеризуется заметно асимметрич¬
ным строением, состоит из двух частей — спиралей, оплетающих G-ак¬
тин, и высокоустойчив к денатурации. В комплексе с тропомиозином
проявляется глобулярный белок тропонин. Молекулы тропонина состо¬
ят из трех частей с молекулярной массой 40200, 23500 и 18300. Часть
с молекулярной массой 23500 прекращает активность АТФазы незави¬
симо от концентрации ионов Са2+ и называется ингибирующим факто¬
ром TN-I. Самая легкая часть (TN-C) имеет сильное сродство с ионами
кальция и прекращает активность АТФазы только в отсутствие этих
40
ионов. Самая тяжелая составная часть тропонина (TN-B или TN-T)
обнаруживает сильное сродство как с тропомиозином, так и с актином,
и поэтому называется связующим фактором [320] Теория сокраще¬
ния мышц и участие мйофибриллярных белков в процессе сокраще¬
ния и релаксации саркомеров весьма подробно описаны в литературе
[413]Третью группу мышечных белков составляют белки соедини¬
тельной ткани, называемые также белками стромы. Они
состоят главным образом из коллагена, а также эластина и конектина
[144] В мышцах костистых рыб содержится около 3 % этих белков, а в
мышцах хрящевых — около 10 %, тогда как в мясе убойного скота их
содержание составляет около 15 % общего количества белка. Белки
стромы характеризуются значительной химической и термической ста¬
бильностью [431] Они нерастворимы в солевых растворах, разбав¬
ленных щелочах и кислотах, за исключением раствора разбавленной ук¬
сусной кислоты (0,2 моль/л), в котором растворяется коллаген, а точ¬
нее, его элементарные частицы (тропоколлаген).По аминокислотному составу эти белки заметно отличаются от дру¬
гих мышечных белков. Глицин составляет около */з всех аминокис¬
лот. Специфичным для этих белков является высокое содержание Прони¬
на (примерно 11 %), гидрооксипролина (3,5—14 %), наличие гидроокси-
лизина, отсутствие триптофана и очень низкое содержание цистина, цис-
теина и тирозина [144, 203, 478] . С этой точки зрения названные белки
нельзя признать полноценными. Зато они выполняют в тканях весьма
важные опорно-связывающие функции.3.2.2. Денатурация белковДенатурация белков вызывает изменения нативного строения их мо¬
лекул без нарушения структуры первой степени.В белке все полярные группы с дипольным моментом взаимно
компенсированы или подвергаются сольватации* частицами воды, а не¬
полярные группы по закону энтропии втягиваются внутрь макрочасти¬
цы. На этом основании можно утверждать, что внутри частицы белка
обладают свойствами восков, а снаружи — свойствами мыл. Под воздей¬
ствием денатурационных факторов ’’восковая” часть белка распрямляет¬
ся или переходит в менее упорядоченную структуру. Нативная форма
глобулярных белков обладает минимальным количеством свободной
энергии и стабилизирована прежде всего действием энтропии.* Сольватация — воздействие частиц растворителя на ионы или частицы раст¬
воренного вещества в результате взаимного притяжения полярных частичек раст¬
ворителя и ионов или полярных частичек растворенного вещества. Сольватация
в водных растворах называется гидратацией.41
Нативная формаРис. 11. Схема денатурации белка (Whitaker and Tannenbaum, 1977)При переходе от нативного состояния в денатурированное каждая
макрочастица белка должна пройти через активное состояние (порог
активизации), связанное с определенной энергетической величиной.
Из активного состояния макрочастица белка может перейти в нативное
или денатурированное состояние в зависимости от дальнейшего дейст¬
вия химических факторов. Переход белка из нативной формы в денату¬
рированную сопровождается изменением количества свободной энер¬
гии - термодинамического потенциала AF, который является суммой
изменений отдельных процессов [105] . При этом свободная энергия
AF складывается из изменений энтропии белка при распрямлении спи¬
ралей (изменение структуры) А^ол/ и из энеРгии переноса боковых це¬
пей аминокислот из внутренних слоев частицы в водную фазу AfuПри необратимом процессе прирост энтропии системы превышает
количество тепла AQ, полученного при нагревании до температуры Т,Д5 >-р-В изолированной системе, в которой происходят необратимые про¬
цессы (AQ = 0 и AS > 0), энтропия возрастает. При распрямлении мо¬
лекулы белка, которое сопровождается возрастанием степени свободы
элементов пептидной цепи, когда неподвижная система превращается
в гибкую, свободно висящую нить (рис. 11), энтропия значительно воз¬
растает.К основным признакам денатурации белка относятся: ухудшение
растворимости (главным образом, глобулярных белков); уменьшение
липкости растворов белков; снижение способности связывать воду;
уменьшение активности АТФазы миозина или актомиозина.42
3.2.3. Факторы, вызывающие денатурацию белкаДенатурация белка в рыбном фарше может быть результатом вы¬
мораживания воды (главным образом, в мороженом сыром фарше),
тепловой обработки (в вареном фарше), действия кислот (в подкис¬
ленном фарше), поваренной соли (в соленом фарше) и других соеди¬
нений, обладающих способностью образовать сетевидные связи в белке
(например, муравьиного альдегида).3.2.3.1. Действие тепловой обработкиПовышение температуры настолько резко ускоряет процесс денату¬
рации, что создается впечатление спонтанной денатурации при опреде¬
ленной температуре для данного белка. Отсюда следует, что денатурация
имеет очень высокий температурный коэффициент (около 600). Внут¬
ренняя энергия белков во время денатурации возрастает до 210—
295 кДж/моль. Для актомиозина эта величина составляет около
230 кДж/моль независимо от вида рыбы [421]Подвижные фракции белка рыбы начинают денатурировать уже при
температуре 25—30 °С. Например, миозин скумбрии примерно в
1000 раз менее стабилен при температуре 30 °С, чем при температуре0 °С, при этом миозин красных мышц в 3,3-3,7 раза более термостаби¬
лен, чем миозин белых мышц [492]Денатурация мышечного белка рыбы происходит в две стадии.
Первая стадия идет при температуре 30—60 °С, с отчетливо выраженным
прямым отрезком кривой денатурации в диапазоне температур 35—
55 °С, при этом в процесс вовлекается 65—98 % всего белка. Вторая ста¬
дия начинается при температуре выше 62 °С и заканчивается при дости¬
жении температуры 78—80 °С, при этом расходуется остальной белок,
превращающийся в термостабильную фракцию, в состав которой входят
главным образом ферменты, миоглобин и некоторые сложные белки.
Таким образом можно принять, что при нагревании до температуры
80 ° С денатурации подвергается практически все 100 % мышечного бел¬
ка. Образуется характерный нерастворимый осадок (гель), а в бульоне
остаются только полипептиды и пептиды — основные продукты термиче¬
ского разложения белков, фракция коллагена, превратившегося в же¬
латин, гистоны и протамины. Скорость денатурации белка рыбы связана
экспоненциальной зависимостью с ростом температуры и логарифмиче¬
ской - с продолжительностью нагревания [293]Следовательно, при подборе условий тепловой обработки необхо¬
димо принимать во внимание ее продолжительность после достижения
температуры нагревания. Для достижения полного эффекта денатурации
продолжительность обработки должна составлять не менее 5-7 мин.Белки стромы подвержены несколько иным изменениям под дейст¬
вием тепловой обработки, чем миофибриллярные и саркоплазматиче-43
ские белки. Вначале происходит сокращение и набухание коллагеновых
волокон при температуре, зависящей от вида рыбы и содержания ами¬
нокислот, а затем наступает термическое разложение волокон в присут¬
ствии воды до образования желеобразного глютина, который по мере
дальнейшего нагревания переходит в нежелирующие продукты разложе¬
ния (желатозы). Температура сокращения и разложения коллагена рыбы
ниже аналогичной температуры для коллагена теплокровных животных.
Это объясняется главным образом более низким содержанием аминокис¬
лот, а следовательно, и меньшим числом поперечных связей в коллагене
рыбы. Если коллаген говядины после нагревания в течение 2,5 мин под¬
вергается желатинизации примерно на 10 %, то коллаген рыбы при тех
же условиях нагревания разрушается на 50-60 и до 75 % [478]Зависимость между разложением коллагена и продолжительностью
нагревания для разной температуры представлена в виде логарифмиче¬
ской функции (рис. 12). При температуре 60—100 °С после очень быст¬
рого разложения на первой минуте начинается вторая стадия (b =
= —0,072 -г-0,112) и заканчивается через 5 мин. Дальнейшее разложе¬
ние происходит медленно (Ъ = —0,014 н--0,026). Так, даже после дли¬
тельного нагревания при температуре 60, 80 и 100 °С определенное коли¬
чество коллагена мяса рыбы (в среднем 30, 25 и 20 % общего содержа-Продолжительность нагревания, %Рис. 12. Влияние температуры и продолжительности нагревания на желатинизацию
коллагена мяса рыбы [ 478]44
ния коллагена) не желирует. После стерилизации мяса рыбы при низкой
температуре (ниже 115 °С) количество нежелатинизированного колла¬
гена составляет 10—15 %. При высокотемпературной стерилизации (свы¬
ше 121 °С) желатинизации подвергается практически весь коллаген.
Добавление соли заметно снижает скорость желирования коллагена.
Например, при добавлении 2,5 % соли после 30-минутного нагревания
при температуре 80 ° С желирует только 40 % коллагена, тогда как без
добавления соли — около 75 % [478]Повышение кислотности мяса ускоряет разложение коллагена в
процессе нагревания, особенно в начальной фазе (до 5—10 мин). Напри¬
мер, после 5-минутного нагревания при температуре 80 °С в мясе скумб¬
рии при естественном pH желированию Подвергается 45—50 % коллаге¬
на, а после подкисления до pH 5,0 свыше 70 % [478] Таким образом,
добавление хлористого натрия, и подкисление перед тепловой обработ¬
кой могут применяться для улучшения реологических свойств фарша.
Если коллаген разлагается только до желатина, то после охлаждения
последний может связывать частицы фарша и улучшать его водоудер¬
живающую способность. При далеко зашедшем разложении желатина
(до желатоз) улучшение реологических свойств фарша становится
уже невозможным.3.2.3.2. Влияние электролитовКаждый электролит, добавленный в фарш в соответствующем
количестве, способствует коагуляции белков. Это действие объясняется
частичным или полным освобождением белковых молекул от их элект¬
рического заряда. Заряды коллоидных частиц, образующиеся на поверх¬
ности молекул (адсорбционный слой), нейтрализуются ионами проти¬
воположного знака. Так, катионы снижают отрицательные, а анионы —
положительные заряды коллоидных частиц. Быстрая коагуляция проис¬
ходит в изоэлектрической точке. Коагуляция гидрофобных коллоидов
обычно идет интенсивно уже при небольших концентрациях ионов, но
коагуляция гидрофильных коллоидов с молекулами, покрытыми
слоями молекул воды, т. е. при более высоких концентрациях элект¬
ролита, проходит медленнее. При повышении концентрации электроли¬
тов, пока величина электрокинетического потенциала частиц меньше
критической, происходит медленная коагуляция белков, поскольку
чем выше заряд иона, тем сильнее его коагулирующее действие.Последовательность коагуляции ионов описывается лиотропными
рядами Хофмейстера. Например, способность одно- и двухосновных
катионов вызывать коагуляцию коллоидов возрастает в следующей
последовательности:С < R; < К+ < Na+ < Li+;BA2 < Sr2+ < Ca2* < Mg2+ ;45
анионов:CNS" < Г < NO3 < Вг" < СГВ сложных растворах измельченного мяса рыбы действие соли не
так однозначно, как в модельных растворах. Неорганические соли
способны изменять и стабилизировать нативную структуру мышеч¬
ных белков, повышать липкость фарша, не изменяя его пластичности,
или образовывать эластичный гель, особенно если фарш смешан с опре¬
деленным количеством соли (табл. 15).Таблица 15. Влияние некоторых солей на реологические свойства
рыбного фарша [ 318]АнионКатионLi+Na+NH$к+Mg2+Са2+Си2+Ва2+Fe3+А13+Цитрат____SO4-----+ОООАцетат--+О+СГ+++++ООоООВг"++NO3+++++ооГ+++CNS”++++Оcol'+-+- вместо эластичного геля образуется густая паста
+ образуется эластичный гель
о белки коагулируютПри низких концентрациях (0,05—0,2 моль/л) большинство солей,
особенно сернокислых, увеличивает желирующую способность фарша
в соответствии с количеством экстрагированного белка. При средних
концентрациях (0,2-1,0 моль/л) только некоторые соли (например,
NaCl, КС1, KNO3) способствуют образованию эластичного геля в фарше,
в то время как под действием остальных солей (например, KJ, K2S04,
KCNS) белок в той или иной степени денатурирует. При высоких кон¬
центрациях (3,0 моль/л) все соли оказывают явно выраженное денату¬
рирующее воздействие на белок [319]Высаливание может привести к разрушению твердых или жидких
соединений белка.Под действием кислот или оснований происходит ионизация групп,
расположенных в неполярной внутренней части микрочастицы белка.
Ионизированные группы притягивают гидратационную оболочку, кото¬
рая таким образом перестает соответствовать первоначальной структуре46
молекулы белка, и равновесие нарушается в сторону денатурированной
формы, не имеющей характерной складчатой пространственной струк¬
туры.3.2.3.3, Вымораживание водыВ результате отвода тепла от поверхности блока рыбного фарша на¬
чинается образование льда, в то время как более глубокие слои, еще не
достигшие криоскопической температуры, остаются незамороженными.
Перемещение границы льдообразования связано с образованием кон¬
центрированного пересыщенного раствора. Концентрация низкомоле¬
кулярных и диссоциированных веществ в тканевых растворах обычно
бывает выше 0,13 моль/л. Зависимость между концентрацией раствора
в насыщенном слое С на расстоянии х от границы льдообразования и
концентрацией раствора на границе раздела С0 можно представить в ви¬
де следующего уравнения [72]С = Со-е-™,где п — коэффициент, характеризующий уменьшение концентрации по
мере удаления от границы льдообразования.Если представить зависимость в полулогарифмической форме,
коэффициент п примет видXФорма кристаллов энергетически неустойчива и в конце концов
изменяется тем быстрее, чем выше температура хранения. Процесс
перекристаллизации может полностью остановиться лишь при эвтекти¬
ческой температуре, которая для пищевых продуктов составляет при¬
мерно —65 °С [71] Однако исследования последних лет показали,
что определенная часть воды, содержащейся в продуктах, не замерзает
даже при очень низких температурах, а процесс образования льда закан¬
чивается лишь в момент снижения температуры до -30 °С.В мясе рыбы количество связанной воды в среднем составляет
0,35 кг на 1 кг сухой массы [378] , что соответствует 6—10% общего
ее содержания. Доля незамороженной воды в мясе рыбы возрастает по
мере повышения температуры замораживания. В мясе рыбы, заморо¬
женной до температуры —30, -20, -10 и -5 °С, остается соответст¬
венно 8—10, 9,5-11, 13—16 и 20—23 % незамороженной воды. Темпера¬
тура замораживания, определяющая непосредственно его скорость,
в значительной степени влияет также на количество и состав незаморо¬
женной воды. При медленном замораживании, когда скорость диффузии
растворенного вещества больше скорости образования кристаллов,
концентрация минеральных веществ и белков в незамороженной жидкой47
фракции значительно возрастает. При этом создаются благоприятные
условия воздействия соли на бейки. Это подтверждают результаты ис¬
следований [47] , показавшие, что растворимость миофибриллярных
белков форели в КС1 при концентрации 0,45 моль/л снижается быстрее
при температуре —10 °С, чем при —20 и —30 °С. В измельченном мясе
рыбы эти изменения происходят быстрее, чем в ткани с неразрушенной
клеточной структурой.Охта и Танака [356] показали, что в жидкой фракции, отделенной
с помощью центрифуги из замороженного мяса рыбы, концентрация
минеральных веществ в 2—3 раза выше, чем в соке незамороженного
фарша, несмотря на то что качественный состав этих веществ в обеих
жидких фракциях сходен. По концентрации ионы, присутствующие в не¬
замороженной фракции сока из свежей рыбы, можно выстроить в поряд¬
ке убывания содержания: К > Р > Na > Cl > Mg > Са. При этом концент¬
рация иона фосфора более чем в 4 раза превышает концентрацию нат¬
рия, почти в 10 раз концентрацию магния и более чем в 22 раза концент¬
рацию кальция. В процессе замораживания и холодильного хранения
концентрация отдельных ионов значительно изменяется. Кроме биохи¬
мических процессов (например, увеличение содержания ионов фосфора
в результате ферментативного дефосфорилирования нуклеотидов и фос-
фосахаров), основной причиной изменения концентрации ионов и увели¬
чения чувствительности мышечных белков к денатурации в процессе
замораживания фарша становятся такие физико-химические явления,
как ассоциация ионов, уменьшение толщины слоя диффузионной воды,
разрушение сольватационной оболочки.Ассоциация ионов. В соответствии с теорией сильных электролитов
Дебай—Хюкеля часть ионов может подвергаться ассоциации. В концент¬
рированных растворах электролитов в результате действия сил притя¬
жения между ионами вблизи каждого иона располагается больше ионов
противоположного заряда, чем одноименно заряженных ионов. Явление
ассоциации ионов приводит к образованию достаточно устойчивых пар
или троек ионов (+—, + — + или — + —), которые, однако, не об¬
разуют не диссоциированных химических молекул. Чем выше концент¬
рация раствора, тем заметнее указанные явления. Таким образом, в кон¬
центрированных растворах сильных электролитов ионов кажется мень¬
ше, чем их действительно находится в растворе. С этой точки зрения для
определения эффективной (мнимой) концентрации вещества в соответ¬
ствии с его действием используется понятие активности а:а = fc,где с — действительная концентрация вещества; /— коэффициент актив¬
ности, корректирующий концентрацию вещества до величины, соответ¬
ствующей его действию. Этот коэффициент стремится к единице (f =
= 1) по мере уменьшения концентрации раствора. Например, для кон¬48
центрации раствора КС1 0,1 моль/л / = 0,769, для концентрации
0,0001 моль/л / = 0,982.Из приведенной зависимости следует, что в разбавленных растворах
электролитов коэффициент активности такой же, как и во всех раство¬
рах одинаковой ионной силы*. В свою очередь, в соответствии с законом
Дебай—Хюкеля возрастание ионной силы приводит к уменьшению сред¬
него коэффициента активности (у±) электролита, диссоциирующего на
ионы с валентностью z+ hz"log 7* = -0,5091 (z+.z-)VTЭто уравнение позволяет рассчитывать средние коэффициенты ак¬
тивности электролитов в растворах концентрацией не выше 0,01 моль/л.
Уравнение можно также использовать для расчета активности отдельных
ионов, если вместо произведения валентностей подставить квадрат ва¬
лентности данного иона, например Z2+ Для растворов концентрацией
выше 0,1 моль/л величина среднего коэффициента активности ионов
данного электролита (fm±) может быть определена по уравнению Де-
бай-Хюке ля-Б рёнстеда:+ А _ + z — yJT- W *с|-где А и В — коэффициенты, зависящие от вида растворителя и темпе¬
ратуры. Для воды температурой 25 °С А = 0,509,1? = 0,330; а — эффек¬
тивный диаметр ионов (минимальное расстояние, на которое могут сбли¬
зиться разноименные ионы) зависит от вида электролита, а = 0,2-=-1,1 нм;
с — эмпирический коэффициент; fm*, fm~ — коэффициенты активности
катиона и аниона.В результате ассоциации ионов pH незамороженной жидкой фракции
изменяется. Изменения pH пищевых продуктов в процессе заморажи¬
вания [483, 485] обусловлены в основном следующими изменениями,
происходящими в незамороженной жидкой фракции: ростом концент¬
рации растворенных веществ вследствие образования кристаллов льда,
ассоциацией ионов в результате возрастания концентрации электроли¬
тов, взаимодействием белков с ионами растворенных веществ.В мясе мороженой рыбы pH составляет обычно 1,0—1,5, при этом
минимальный pH достигается после 1-2 мес хранения при температуре* Величины средних коэффициентов активности электролитов зависят не толь¬
ко от концентрации данного электролита, но и от концентрации всех остальных
ионов в растворе. Для учета этого фактора используется понятие ионной силы
раствора/, определяемой уравнением: / = /22CfZ f (С/ - концентрацияz-го иона;
Zz* - валентность /-го иона). Например, для раствора концентрацией 0,1 моль/л
NaCl и 0,2 моль/л Na2SC>4 ионная сила составит/ = i (Na+-12 + (Cl") I2 + (SOj-) -22 = i (0,5 + 0,1 + 0,8) = 0,7 моль/л.49
-18 °С, после чего pH снова возрастает. Однако даже при более длитель¬
ном хранении pH мороженого мяса рыбы несколько ниже (на 0,2-0,3)
первоначального [484] Этими изменениями в основном объясняется
снижение растворимости белков и связанное с этим изменение консис¬
тенции и водоудерживающей способности мяса рыбы. Келли [196]
установил влияние pH на снижение растворимости белков мяса трески,
хранившейся при температуре -20 °С. Для трех значений pH мяса рыбы
зависимость между растворимостью белков S (в % растворимого азота
от общего азота) и продолжительностью хранения х (в мес) он описал
следующими уравнениями регрессии:pH >6,7 S = 85,00- 0,74*;pH 6,4 -6,7 5 = 89,75 - 1,780х;pH < 6,4 S =86,8.5 - 2,38д:.Как видно из этих уравнений, величина отрицательного коэффици¬
ента значительно возрастает при уменьшении pH.Уменьшение толщины диффузионного слоя. Согласно теории сильныхэлектролитов Дебай—Хюкеля толщина диффузионного слоя —-— за¬
висит от ионной силы раствора_1 = 10004 кТ *Р 8 тгЩе2 }где е - величина элементарного заряда; е — диэлектрическая проницае¬
мость; к — постоянная Больцмана; Т — абсолютная температура (К);
/ - ионная сила раствора, NQ — число Авогадро.Уменьшение толщины диффузионного слоя, который чаще всего
становится внешней оболочкой мицеллы (рис. 13), способствует как
коагуляции белков, так и ассоциации ионов. Эти процессы происходят
во время концентрирования раствора соли и растворимых белков в неза¬
мороженной фракции. Полностью диффузионный слой исчезает, когда
величина электрокинетического потенциала £ = 0. Тогда заряд гранулы
равен сумме зарядов адсорбционной оболочки. Точка, в которой усло¬
вия коагуляции оптимальны, так как система наименее устойчива, на¬
зывается изоэлектрической. В действительности коагуляция коллоидов
происходит уже при значениях £, меньших критических и составляющих
примерно 25-30 мВ [420] Таким образом, возрастание ионной силы
в процессе вымораживания воды вызывает уменьшение толщины диффу¬
зионного слоя и тем самым способствует коагуляции белков и ассоциа¬
ции ионов.Удаление сольватационной оболочки. Для протекания процесса
коагуляции белков необходимо удаление сольватационной (гидрата-
ционной) оболочки. Неорганические ионы, вызывающие изменение
водных структур, окружающих молекулу белка, могут уничтожить на¬
тивную структуру [489, 490] В зависимости от степени неупорядо-50
© © @© ° ©© ©©© ©© © ©© © ©Рис. 13. Схема строения отрицательной (д) и положительной (б) мицеллы кол¬
лоида:1 - поли молекулярное ядро; 2 - абсорбционная оболочка (внутренний слой
ионов); 3 — диффузионная оболочка (внешний слой ионов)ченности (энтропии) окружающего водного слоя все анионы и катионы
можно разделить на структурообразующие, которые приводят к обра¬
зованию сильного электростатического поля и создают плотный абсорб¬
ционный слой, и разрушающие водные структуры, образующие толстый
диффузионный слой [247, 337] Одновалентные ионы (кроме F" и
Li”) увеличивают энтропию системы и разрушают упорядоченные
водные структуры, в то время как двухвалентные (Mg2+, Са2+, Ва2+,
SO4") и трехвалентные (POj”) ионы ее уменьшают [489] . Ионы, кото¬
рые не влияют на водные структуры (например, ацетата, тетраметила¬
мина) , не изменяют нативной структуры белков [247, 489] .3.2.3.4. Действие продуктов окисления
липидов на белкиСвободные жирные кислоты. В начале 60-х годов считали, что дена¬
турацию белка мяса рыбы в процессе ее холодильного хранения вызы¬
вают в основном свободные жирные кислоты [10, 12, 93] Показано,
что оптимальные условия взаимодействия свободных жирных кислот
с миофибриллярными белками возникают в растворах соли с ионной
силой / = 0,5 при pH 7,2. Такая концентрация соли обнаруживается в
клеточном соке трески, охлажденной до температуры —1,5 С. Концент¬
рация соли в незамороженной жидкой фракции в процессе заморажива¬
ния считается основным фактором, ускоряющим взаимодействие сво¬
бодных жирных кислот с белками [13] . Неодинаковое снижение раство¬51
римости белков отдельных видов мороженой рыбы в процессе хране¬
ния пытались объяснить отношением содержания свободных жирных
кислот к содержанию нейтральных жиров, играющих в этом случае за¬
щитную роль по отношению к белкам [365]Предпринята попытка установить графическую зависимость между
отношением содержания свободных жирных кислот к содержанию нейт
ральных жиров и количеством нерастворимого актомиозина в мясе мо¬
роженой рыбы. Хотя для значительной части видов промысловых рыб
существование такой зависимости подтверждалось (например, в мясе
рыбы, у которой соотношение свободных жирных кислот и нейтраль¬
ных жиров составляло 2, после 4-5 мес хранения содержалось около
4,5 % нерастворимого актомиозина) [365] , для некоторых рыб, осо¬
бенно тех, в мясе которых содержалось менее 80 мг% нейтральных жи¬
ров, эта зависимость не подтверждалась. Ошибка здесь, по-видимому,
заключается в том, что нейтральные липиды нельзя считать единствен¬
ным веществом, предотвращающим денатурацию белка.Модельные исследования гомогенатов мяса рыбы [12] показали,
что свободные жирные кислоты образуются главным образом в про¬
цессе гидролитических изменений фосфолипидов саркоплазменной
сетки, расположенной вблизи миофибриллярных белков. Оставшиеся
неразложившимися фосфолипиды также способны защищать белки
мяса от действия свободных жирных кислот. В связи с этим предложен¬
ные зависимости между содержанием нерастворимого белка и отноше¬
нием содержания свободных жирных кислот к общему содержанию
липидов за вычетом содержания свободных жирных кислот в мясе пред¬
ставляются правомерными, так как они равным образом подтверждают
защитное действие нейтральных жиров на мышечные белки. Дополни¬
тельными доводами в пользу неоднозначности зависимости между соот¬
ношением содержания свободных жирных кислот и нейтральных жиров
и растворимостью белков являются также быстрая денатурация мы¬
шечных белков некоторых жирных рыб с высоким содержанием нейт¬
ральных жиров [140] и интенсивное снижение растворимости белков
в мороженом мясе тощей хрящевой рыбы (например, Raja maculata,
Scyllium canicula) на том этапе процесса холодильного хранения, когда
содержание накопленных свободных жирных кислот в мясе еще очень
незначительно [364]Несмотря на приведенные условия, свободные жирные кислоты
считались одним из основных факторов денатурации мышечных белков
рыбы. Под действием свободных жирных кислот растворимость белка
быстро снижается. Связывание белком свободных жирных кислот, осо¬
бенно с длинной углеродной цепью (Ci6-Ci8), а частично и с короткой
настолько сильное, что практически их нельзя экстрагировать органиче¬
скими растворителями [137] Целые миофибриллы мяса рыбы легче
вступают во взаимодействие со свободными жирными кислотами, чем
отдельные миофибриллярные белки. Из мышечных белков наиболее
52
быстрое уменьшение растворимости под действием свободных жирных
кислот наблюдалось у тропомиозина. В присутствии муравьиного альде¬
гида в мясе рыбы белок быстро денатурирует под действием свободных
жирных кислот [58]Лецитин. Тагучи и Икеда [438] показали, что в препаратах актомио¬
зина, независимо от степени их очистки, содержится постоянное, точно
определенное количество лецитина, который составляет большую часть
липидов, связанных с актомиозином рыбы [438] Отдельные виды рыб
различаются по содержанию лецитина, связанного с актомиозином, при
этом актомиозин, в котором содержание лецитина выше, содержит
также больше миозина.Лецитин, по-видимому, свободно связан с актомиозином и легко
экстрагируется смесью этилового спирта и этилового эфира (спирто¬
эфирной смесью 3:1), особенно из замороженных препаратов актомио¬
зина [438], или выделяется центрифугированием при высокой частоте
вращения [436]Лецитин сохраняет АТФазную активность актомиозина и тяжелого
миозина В или действует в направлении, противоположном действию
холинфосфогидролазы, которая нейтрализует активность АТФазы
[439] Введение лецитина восстанавливает активность АТФазы, предва¬
рительно дезактивированной с помощью холинфосфогидролазы.Наличие лецитина в очищенных препаратах миозина было подверг¬
нуто сомнению [363] , поэтому его воздействие на белки мышц нельзя
считать полностью доказанным. Если допустить, что лецитин является
постоянным спутником миозина, то его гидролитические и окисли¬
тельные изменения приводили бы к снижению растворимости белков
мяса рыбы. Вместе с тем стабилизация ’’актомиозинового” лецитина
(например, путем добавления альфа-токоферола) или введение новой
порции лецитина в восстановленной (неокисленной) форме предот¬
вращало бы чрезмерное снижение растворимости белков рыбного
фарша.Защитное действие лецитина на мышечные белки можно также объ¬
яснить тем, что он предохраняет саркомеры миозина и актина от воз¬
действия слишком высокого содержания кальция, который препятст¬
вует их структурным изменениям. Доказано, что однослойные оболоч¬
ки лецитина способны связывать ионы кальция в физиологических диа^
пазонах pH [202]Взаимодействие окисленных липидов с аминокислотами и бел¬
ками. В мороженом рыбном фарше взаимодействие окисленных липи¬
дов с азотистыми соединениями может быть причиной не только умень¬
шения растворимости белков, но и снижения их пищевой ценности и
неферментативного побурения. Механизм взаимодействия липидов с
белками очень сложен и многообразен, доказательством чего, кстати,
является наличие в липидно-белковых комплексах химических связей
различных типов. Наибольшую роль играют, вероятно, ионные и гидро¬53
фобные связи, а в значительно обводненных системах также силы Ван-
дер-Ваальса [186, 375, 403] Реакция липидов с белками может приво¬
дить также к образованию липидно-белковых комплексов и связей ти¬
па белок—белок, катализированных изменениями липидов. Некоторые
исследователи считают, что образовывать комплексы с белками спо¬
собны только окисленные липиды [258, 341, 336] , другие полагают,
что окисление липидов лишь облегчает их реакцию с белком, но белки
могут взаимодействовать и с неокисленными липидами, например при
перемешивании рыбного фарша [218]Взаимодействие аминокислот и белков с первичными продуктами
окисления липидов (гидроперекисями) не вызывает изменения органо¬
лептических характеристик продукта, однако биологическая ценность
его снижается в результате разрушения некоторых аминокислот. На¬
пример. цистеин окисляется до цистина, затем до окиси сульфата, суль-
фана и сульфоновой кислоты [375] Подобным образом метионин
окисляется до окиси сульфата и сульфона. Индоловое кольцо трипто¬
фана легко повреждается в присутствии окисленных липидов [91]Свободные радикалы жира могут легко отрывать водород сульф-
гидрильных групп белков, что ведет к их агрегации [392] и снижению
растворимости белков. Связь липидов с белками может осуществлять¬
ся также посредством мостиков из ионов металлов, например Са2 + ,
Mg2+, что подчеркивает значение ионной силы в процессе взаимодейст¬
вия [115, 53] Таким образом, сложный механизм взаимодействия ли¬
пидов с белками обусловлен состоянием как белка, так и липидов, а
также другими факторами.При температуре ниже точки денатурации белка актин легче реаги¬
рует с липидами, чем миозин. Например, при температуре 4 °С актин
связывает около 33 % нейтральных липидов и примерно 48 % полярных
липидов, тогда как миозин связывает только следы липидов. Вместе с
тем при температуре выше точки тепловой денатурации белков способ¬
ность актина связывать липиды незначительно изменяется - возраста¬
ет для нейтральных липидов и снижается для фосфолипидов. Способ¬
ность к связыванию у миозина заметно возрастает. Например, миозин,
нагретый в течение 15 мин при температуре 50 °С, связывает около18,2 % полярных липидов и 32,3 % нейтральных липидов, а нагретый
в течение 30 с при температуре 70 °С - 70,4 % нейтральных липидов по
отношению к его массе [404] С липидами реагируют также белки сар¬
коплазмы, хотя и значительно слабее, чем миофибриллярные белки,
часто только в результате замораживания рыбного фарша [218] Таким
образом, главную роль во взаимодействиях с липидами в мороженом
рыбном фарше играют миофибриллярные белки, при этом заморажива¬
ние является фактором, заметно катализирующим процесс.Реакции вторичных продуктов окисления липидов (альдегиды,
кетоны) с аминокислотами и белками весьма сходны с реакциями
Майяра, так как способны вызывать неферментативное побурение пи-
54
щевых продуктов. От продуктов этих реакций в значительной степени
зависят органолептические свойства рыбного фарша, особенно запах.
В результате реакции альдегидов со свободными аминокислотами об¬
разуются основания Шиффа, а с аминокислотами, входящими в состав
белка, — разнообразные сетчатые связи [74, 487]Бутткус [46] изучал реакцию малонового альдегида с миозином и
обнаружил, что в ней принимают участие боковые цепи молекулы белка.
При хранении мороженого фарша наиболее активными с малоновым
альдегидом были остатки аминокислот: тирозина, лизина и аргинина.
Малоновый альдегид легче всего реагирует со свободными еаминными
группами лизина, при этом образуются поперечные связи у молекул
белка [59] В результате взаимодействия окисленного сложного мети¬
лового эфира линоленовой кислоты с небелковыми азотистыми вещест¬
вами мяса скумбрии образуются окрашенные мономеры и полимеры
[332] Ниже на примере взаимодействия актомиозина форели, хранив¬
шейся при температуре —20 °С с капроновой кислотой и пуриновым ос¬
нованием, схематично изображен механизм вторичных реакций продук¬
тов окисления липидов с белками [447] (см. схему на стр. 56).Как показано на схеме, пропионовый альдегид реагирует с груп¬
пами SH развернутых молекул актомиозина, образуя тиоацеталь, в ре¬
зультате чего белок становится нерастворимым. Капроновая кислота
соединяется с актомиозином водородными или гидрофобными связями.
Эта реакция непосредственно не вызывает уменьшения растворимости
актомиозина. Однако, поскольку образованные связи не уравновеши¬
вают силу ковалентных связей, образующихся в актомиозине в процессе
холодильного хранения, растворимость белка также снижается [450]
Биологическая ценность белков в результате их взаимодействия
со вторичными продуктами окисления липидов во время холодильного
хранения рыбы и рыбных продуктов снижается, при этом теряются ли¬
зин, триптофан и метионин [234, 260]К другим факторам, влияющим на реакцию окисленных липидов с
аминокислотами и белками, относятся прежде всего pH, концентрация
соли, температура, доступ атмосферного кислорода, света и др. Несмот¬
ря на многочисленные исследования в этой области, результаты неодно¬
значны. Например, в модельных системах эта реакция быстрее всего про¬
ходит при pH, близком к изоэлектрической точке белка [474] , а в мо¬
роженом рыбном фарше — при естественном pH мышечной ткани [218]В модельных смесях миозина с липидами, нагретых в течение 15 мин при
температуре 100 °С, снижение pH от 7 до 4-5 не оказывало существенно¬
го влияния на связывание полярных липидов, зато вызывало снижение
количества связанных нейтральных липидов примерно на 9 %. Актин наи¬
более активно связывает нейтральные липиды при pH 5,5 и 3, а полярные
липиды - при pH ниже 5 [403]Перемешивание и измельчение рыбного фарша облегчает взаимодей¬
ствие липидов с белками, по-видимому, в результате увеличения площа-55
| Жирная кислота
• Водородные или
гидрофобные связиди контакта между реагирующими субстратами [218] .В некоторых слу¬
чаях процесс окисления жира в мороженом рыбном фарше может даже
приостановиться.3.2.4. Изменения белков и консистенция фаршаНаибольшее влияние на консистенцию рыбного фарша оказывают
изменения миофибриллярных белков. Изменения коллагена, несмотря
на высокую водоудерживающую способность желатина (табл. 16), иг¬
рают второстепенную роль в образовании консистенции, поскольку
мясо рыбы бедно коллагеном. В мышечной ткани без костей и кожи
содержание коллагена составляет всего 0,3-0,6 %, тогда как в говядинеI сорта 30 % (по сырой массе) [478]Из миофибриллярных белков миозин обладает самой высокой спо¬
собностью образовывать эластичный гель и эмульгировать жир в нагре-
56
Таблица 16. Сравнение водоудерживающей способности геля
гомогената мышц, миозина и белков стромы мяса рыбы [317]Вид геляpHВо до удерживающая способность,
г воды на 1 г белкаГомогенат мышц4,58,35,56,68,512,3Миозин6,18,7Белки стромы4,518,46,915,18,821,7тых гомогенатах мяса рыбы [112, 333, 395, 475] Это возможно бла¬
годаря специфическому строению молекулы миозина и наличию в нем
многочисленных групп SH, из которых большинство высокоактивно
[204]. Миозин (главным образом, концевые группы молекул) спо¬
собен агрегироваться, а также образовывать трехмерную сетку попереч¬
ных связей, что объясняется распрямлением двойной сетчатой сердце¬
вины молекулы [165, 191] . Группы SH окисляются в процессе нагре¬
вания или под действием других факторов, что приводит к образованию
в белке сильных ковалентных связей типа S-S. При нагревании актомио¬
зина карпа от 20 °С до 80 °С в течение 2 ч число активных (доступных)
групп SH, расположенных внутри молекул белка, а также окисленных
групп SH заметно возрастало [169] Имеется много данных, свидетель¬
ствующих о том, что образование геля связано не только с окислением
групп SH до SS, но и появлением в белке связей типа SH—SS. Например,
введение цчстеина, по-видимому, повышает способность фарша жели-
ровать после термической обработки. То же самое относится и к цисти-
ну, который в этих условиях редуцируется до цистеина. Редуцирующие
соединения, например нитрит натрия, тиосульфат натрия, кислый суль¬
фат натрия и другие, вероятно, действуют на фарш после тепловой об¬
работки как окислители, например КВг03 [168]Способность миозина к желированию зависит от ряда факторов,
важнейшими из которых являютря температура и продолжительность
тепловой обработки, отношение миозина к актину в белке, концентра¬
ция белка, наличие добавок, активность протеолитических ферментов
и др.Необходимая для образования геля продолжительность нагревания
миозина при температуре 60 ° С составляет около 10 мин, при этом
даже двухчасовое нагревание не ухудшает его реологических свойств
[167]. Полученный при этих условиях белковый гель отличается зна¬
чительной эластичностью и высокой водосвязывающей способностью,
несмотря на то что образующий его белок находится в нерастворимом
(денатурированном) состоянии. Консистенция такого геля нежная, по¬
скольку структура его состоит в основном из распрямленных цепей57
миозина (актомиозина), а число образовавшихся групп SS не слишком
велико. По мере увеличения числа связей S-S в геле его плотность и
твердость возрастают, что сопровождается частичной потерей питатель¬
ной ценности белка, особенно переваримости, поскольку связи типа
S-S относительно устойчивы к действию протеолитических ферментов
[318]. В процессе образования геля из миозина и актомиозина можно
выделить два заметных температурных оптимума. Гелеобразование
быстрее всего происходит в фарше, оставленном при температуре 30-
35 0С [480] или нагретом при температуре 60—70 °С [165] В резуль¬
тате слишком высокой температуры и продолжительного нагревания мо¬
жет произойти вторичная деградация образованного геля (белкового
полимера), при этом консистенция фарша ухудшается. При нагревании
актомиозина карпа при температуре 80 °С неблагоприятные изменения,
выражающиеся в снижении содержания активных групп SH в геле, об¬
наруживаются уже через 30 мин [169] В процессе нагревания тонкоиз-
мельченного мяса скумбрии при температуре 90 °С [348] уже спустя
25 мин количество абсорбированной воды уменьшалось, а количество
свободной воды в геле увеличивалось. Это объясняется структурными
изменениями белков, а именно образованием новых водородных мости¬
ков между полипептидными цепочками. Во время нагревания при темпе¬
ратуре 100—120 °С тяжелые цепочки меромиозина распадаются на лег¬
кие, так что уже через 60 мин нагревания при 120 ° С тяжелые цепочки
не обнаруживаются. Увеличение продолжительности нагревания свыше
5 ч при данной температуре приводит также к быстрому росту количе¬
ства растворимых в 5 %-ной хлористоводородной кислоте продуктов
разложения миофибриллярных белков [216] Доказательством разло¬
жения SH-групп в процессе нагревания мяса при температуре 80-120 °С
является образование сероводорода [136]Увеличение отношения миозина к актину в белке увеличивает проч¬
ность его геля после тепловой обработки. Максимальная прочность ге¬
ля (550 Н/м2) достигается при молярном отношении миозина к актину,
равном 2,7:1, что в пересчете на массу составляет 15:1 [164] . Это озна¬
чает, что наиболее благоприятные условия для гелеобразования созда¬
ются, когда 15—20 % белков присутствуют в виде актомиозина, а осталь¬
ные в свободном виде. Актомиозин в этом случае действует как связы¬
вающий и структурообразующий фактор, кроме того, улучшаются
механические свойства геля. Связывающее и структурообразующее дей¬
ствие актомиозина зависит от содержания F-актина: чем его больше,
тем оно сильнее. F-актин легко и быстро коагулирует при температуре
около 60 °С, тогда как G-актин не разрушается даже при 80 0 С [436]
Прочный и эластичный гель, который после нагревания фарша
способен связывать его отдельные частицы, может образовывать толь¬
ко растворимая неденатурированная фракция миофибриллярных бел¬
ков. Несмотря на то что существующая зависимость между раствори¬
мостью белков и структурой фарша неоднозначна [55], в общем можно
58
принять, что степень растворимости миофибриллярных белков и их кон¬
центрация в фарше определяют его консистенцию после тепловой обра¬
ботки. В этом свете особое значение приобретает предотвращение денату¬
рации белков в процессе хранения. Для получения достаточно плотного
и однородного геля необходима экстракция миофибриллярных белков
из мышечных волокон в межволокнистом пространстве, как это делает¬
ся при изготовлении фарша типа сурими, предназначенного для изготов¬
ления рыбной ветчины и колбас. Это достигается увеличением ионной
силы жидкой фракции фарша до уровня, при котором растворимость
миозина и актомиозина максимальна, т. е. I = 0,3 -г 0,4 [165] Макси¬
мальное по сравнению с естественным снижение pH мяса рыбы увели¬
чивает механическую прочность белкового геля после термической об¬
работки, однако при этом возрастают его твердость и хрупкость при од¬
новременном снижении эластичности. Это результат синерезиса, вызы¬
вающего ’’скручивание” белковых цепей путем удаления части связан¬
ной воды. Максимальная прочность актомиозинового геля (после нагре¬
вания при температуре 80 С в течение 60 мин) наблюдается при pH 6,3
[167], а миозинового геля (после нагревания при температуре 65 0 С
в течение 25 мин) - при pH 6,0 [165, 396]Дальнейшее понижение pH, особенно ниже ь,0, значительно ослаб¬
ляет гелеобразующую способность миофибриллярных белков после
тепловой обработки и уменьшает водоудерживающую способность
геля [317,396]Действие добавок на консистенцию подвергаемого тепловой обра¬
ботке рыбного фарша проявляется по-разному. Некоторые соединения,
понижающие температуру криоскопической точки (например, глута-
мат натрия, ЭДТК), соединения, диссоциирующие актомиозин (три-
фосфат натрия, гексаметафосфат), и другие (например, цитрат натрия)
ускоряют гелеобразование и увеличивают прочность белкового геля,
в то время как сахара и спиртосахара не обладают такими свойствами
[351].Отрицательное влияние протеолитических ферментов на консистен¬
цию фарша становится заметным при очень медленном повышении тем¬
пературы в процессе его хранения. Активизировавшиеся в таких усло¬
виях протеолитические ферменты успевают вызвать гидролиз белка до
их полной инактивации, которая происходит при температуре 80-85 °С.
Протеолитические изменения белков во время нагревания рыбного
фарша касаются главным образом тропомиозина и миозина. Тропомио-
зин рыбы отличается высокой температуроустойчивостью [56] и весьма
подвержен протеолитическому разложению [95, 267] Протеолитиче-
ское разложение миозина в процессе нагревания измельченного мяса
рыбы при температуре ниже 70 ° С приводит к уменьшению содержания
тяжелых цепочек миозина и образованию легких цепочек [55] , в резуль¬
тате чего консистенция фарша ослабляется.Фермент, катализирующий разложение миофибриллярных белков59
во время нагревания, обнаруживается в белках саркоплазмы. Установле¬
но, что оптимальная температура протеолитической активности этого
фермента 60 °С, а оптимальный pH 8,0—8,5 [54]Исследование фрагментов миозина, полученных в результате его
разложения химотрипсином, показало [396] , что гелеобразующая спо¬
собность головок молекул миозина весьма низка и составляет всего
45±3,5 Н/м2 при температуре 50 °С, гелеобразующая способность серд¬
цевины молекулы — 105,0±2,0 Н/м2, в то время как для целой молеку¬
лы миозина эта величина достигает 180,0±5,0 Н/м2Таким образом, для получения полного гелеобразующего эффекта
необходимо предотвратить разрушение миозина, что при тепловой об¬
работке фарша достигается его быстрым нагреванием до необходимой
температуры.Химически чистый триметиламиноксид относительно устойчив к
действию кислот, оснований и температуры. В растворе ТМАО с pH 3,0—9,0, выдержанном в течение 7 ч при температуре 107 °С в стеклянном
сосуде, не было обнаружено никаких химических изменений [391]В рыбном сырье разложение ТМАО — одна из основных причин сниже¬
ния качества. Особенно это относится к измельченному мясу, посколь¬
ку скорость разложения ТМАО в нем обычно в 2—3 раза выше, чем в
филе или тушках.По характеру изменений при разложении TMAQ в мясе рыбы мож¬
но выделить три типа реакций:восстановление ТМАО до ТМА, вызванное в основном психрофиль-
ными грамотрицательными бактериями (Achromobacter, Enterobacter,
Pseudomonas и др.), при температуре выше криоскопической точки, оп¬
тимальной в диапазоне 25-35 °С.разложение ТМАО до диметиламина (ДМА) и муравьиного альде¬
гида (МА) в результате активности эндогенных ферментов (деметила-
зы), присутствующих во фракции белков саркоплазмы ткани рыбы, в
охлажденном, подмороженном и замороженном мясе рыбы, и особен¬
но интенсивно в диапазоне температур от 3 до —5 °С, но также и при
более низкой температуре (до —40 °С)3.3. РАСПАД ТРИМЕТИЛАМИНОКСИДА (ТМАО)
В МЯСЕ РЫБЫСН3 фермент,I восстановительфермент,восстановитель60
СНзIферментн3с\Н3С - N = О
INH + НСНО;СН3Н3Стермическое разложение ТМАО, в основном до ДМА, МА и ГМА
в результате нагревания мяса рыбы при температуре выше 60 °С в при¬
сутствии естественных катализаторов, например гемопротеидов (мио-
глобина, гемоглобина), некоторых аминокислот (например, цистеина),
ионов металлов (Fe2+ и др.)* Характерной чертой этой реакции являет¬
ся относительно высокая доля ДМА в продуктах термического разло¬
жения ТМАО. Даже в очень свежем сырье, подвергшемся тепловой об¬
работке, отношение ДМА:ТМА100 составляет более 50 %, тогда как
в первой реакции продуктом восстановления ТМАО является почти ис¬
ключительно ТМА. Интенсивность разложения в значительной степени
зависит от вида рыбы, а особенно от доли белого и красного мяса в
мышцах. В красных мышцах это разложение, как правило, идет значи¬
тельно быстрее, чем в белых, что обусловлено прежде всего различием
в содержании катализирующих веществ.В мороженом фарше из сырой рыбы происходит главным образом
ферментативное разложение ТМАО до ДМА и МА.Фермент, катализирующий это разложение, представляет собой
белковую фракцию экстракта тканей рыбы, которую удается выделить
методом высаливания сернокислым аммонием при концентрации, соот¬
ветствующей 20—40 % полного насыщения раствора. Специфическая ак¬
тивность полученного таким образом фермента составляет для белого
мяса минтая 0,3—0,8, для красного мяса — 1,6—7,3 ДМА или МА на 1 мг
белка. В тканях внутренних органов она значительно выше, например
в почках активность фермента составляет 190—2000, в пилорических
придатках — 49—210, в желчном пузыре она в 450—1200 раз выше, чем
в белых мышцах (табл. 17) .Дальнейшая очистка фермента путем молекулярной фильтрации на
декстриновых гелях и колоночной хроматографии на геле Sephadex
DEAE А-25 позволили получить препарат со специфической активностью
примерно в 48 раз выше, чем в сыром экстракте тканей. Максимальная
активность наблюдается при pH 7,1-7,2, при этом как снижение pH, так
и его повышение вызывает резкое уменьшение активности, при pH 3
и pH 10 активность почти равна 0. Наибольшее снижение активности в
результате подкисления наблюдается в диапазоне pH 7,1—5,8 и 4,2-3,0
для очищенного фермента и 7,1—6,0 для неочищенного. Подщелачивание3.3.2. Характеристика фермента,
разлагающего ТМАО до ДМА и МА61
Таблица 17. Сравнение специфической и относительной активности
фермента, разлагающего ТМАО до ДМА и МА в различных
тканях минтая [461]Специфическая активностьОтносительнаяактивность***Ткани и органыN ДМА на 1 мг
белка*МА на 1 мг
белка**ДМАМАБелая мышца0,5 (0,3-0,8)0,61,01,0Красная мышца4,1 (1,6-7,3)3,5(1,9-5,1)8,25,8Сердце26,6(5,6-122,0)127,2 (41,2-213,2)5 3,2212,0Печень7,6(0,3-15,5)0,8 (0,7-1,0)15,21,3Пилорические72,1 (24,3-105,2)54,1 (39,7-68,5)144,290,2придаткиКишки156,4 (85,0-284,0)157,1 (137,3-176,9)312,8261,8Желудок93,6 (28,5-115,0)174,5 (152,5-196,5)187,2290,8Желчный пузырь405,0 (225,0-610,0)732,7 (696,7-768,7)810,01221,2Селезенка89,4 (33,7-131,0)129,3(121,6-136,9)178,8215,5Почка300,5 (99,8-820,0)129,4(105,6-153,1)601,0215,7Яичники1,7 (0,2-3,6)1,93,43,2Семенники1,3 (0,2-2,8)0,72,61,1* Среднее из 10 определений; **среднее из двух определений; *** по сравне¬
нию со специфической активностью белой мышцы, принятой за 1.pH рнРис. 14. Влияние pH на активность фермента, разлагающего триметиламиноксид
(ТМАО) до диметиламина (ДМА) и муравьиного альдегида (МА) в мясе мин¬
тая [ 461]а - фермент, экстрагированный фосфатным буферным раствором концентрацией
0,05 моль/л (pH 7,2) из пилорических придатков и очищенный методом высалива¬
ния [фракция 15-30 % (NH4)2S04, молекулярной фильтрации на Sephadex G-200
и двукратной колонковой хроматографии на DEAE Sephadex А-25; б - фермент
водной вытяжки мышечной ткани, хранившейся 2,5 мес при температуре -18 °С62
Рис. 15. Влияние температуры и срока хранения на стабильность фермента, разла¬
гающего ТМАО до ДМА [ 461] :а - фермент, изолированный из пилорических придатков -минтая и очищенный
путем высаливания, фильтрации на декстриновом геле и. колонковой хроматогра¬
фии на Sephadex А-25; б - водная вытяжка из мяса карпа (неочищенный фермент)среды с pH 7,1 до 8,0 вызывает снижение активности более чем в 3 раза
независимо от степени очистки (рис. 14).Очищенный фермент относительно чувствителен к нагреванию: уже
при 20 ° С обнаруживается снижение стабильности, а после 30-минутного
нагревания при 40 °С остается лишь 12 % первоначальной активности.
Однако термическая инактивация фермента в мышечной ткани проходит
значительно труднее. Для полной инактивации фермента необходимо
нагревание в течение 30 мин при температуре выше 70 °С, хотя уже при
температуре 50—60 °С наблюдается заметное снижение активности
(рис. 15). Было показано [502] , что нагревание при 100 °С в течение5 мин вызывает полную инактивацию ферментов, разлагающих ТМАО
в пилорических придатках минтая. При нагревании этих ферментов
в буферном растворе при pH 5,0 скорость инактивации возрастала,
достигая максимума уже при температуре выше 60 ° С [462]К относительно стабильным относятся ферменты, восстанавливаю¬
щие ТМАО в почках. Для полной их инактивации в буферном растворе
при pH 7,1 необходимо нагревание при температуре выше 75 °С.63
3.3.3. Реакция муравьиного альдегида с белкамиРеакция муравьиного альдегида с белками мышц вызывает сни¬
жение их растворимости, агрегацию и связанные с этим нежелательные
изменения мышечной ткани. Муравьиный альдегид реагирует с боль¬
шинством функциональных групп белков, особенно легко с €-аминовой
группой лизина и гидроксилизина [415] В результате реакции образу¬
ется одна формальдегидная связь и гидрооксиметиленовое производное:R - NH2 + СН20 R — NH - СН2ОНВ этих соединениях формальдегид связан относительно слабо, одна¬
ко блокирование отдельных функциональных групп уменьшает способ¬
ность белков связывать воду и ухудшает их растворимость. Например,
в мороженом филе минтая заметное уменьшение растворимости белков
в растворе КС1 концентрацией 0,6 моль/л наступает уже после накопле¬
ния в мясе 2,2-2,5 % азота диметиламина [461]Реакции второго типа, которые происходят в процессе длительного
хранения мяса рыбы, приводят к образованию стабильных связей второ¬
го порядка между муравьиным альдегидом и соседними цепочками мо¬
лекул белка или даже внутри одной и той же молекулы белка при по¬
мощи метиленовых мостиков:2R - NH2 + СН20 -> R1 - NH - СН2 - NH - R2 - Н2В этом случае можно заключить, что действие муравьиного альдегида
направлено на образование сетчатой структуры мышечных белков.
Образование поперечных связей внутри молекул и между молекулами
белка приводит к его агрегации и образованию структуры геля [65]В реакциях с муравьиным альдегидом участвуют в основном мио¬
фибриллярные белки, хотя в них задействованы и саркоплазматические
белки [414] Из миофибриллярных белков легче всего реагируют
тропомиозин и тяжелые цепочки миозина, которые полностью утрачи¬
вают растворимость уже после добавления в измельченное мясо рыбы
0,04 % МА, зато актин и компонент с молекулярной массой 37400 более
стабильны. Эти соединения даже после добавления 0,4 % муравьиного
альдегида остаются еще частично неденатурированными [57]Блокирование некоторых функциональных групп белков муравьи¬
ным альдегидом может приводить, помимо уменьшения растворимости,
к снижению пищевой ценности, особенно с точки зрения доступности
лизина и переваримости. Например, добавление в измельченное мясо
трески 1 % МА вызывает 50 %-ное уменьшение содержания доступного
лизина в белке [379] Однако в мороженом фарше, в котором содержа¬
ние муравьиного альдегида, как правило, не превышает 0,015 %, изме¬
нения не столь существенны.64
3.3.4. Факторы, влияющие на скорость
разложения ТМАО в мясе рыбы3.3.4.1. Содержание ТМАО в мясе водных
промысловых животныхОсобенно много ТМАО содержится в рыбах с белым мясом, нижним
расположением рта, морских моллюсках и ракообразных (табл. 18). Он
обнаружен также во внутренних органах этих животных. Например, в ик¬
ре сельди содержится 22-139, в молоках 158-514 мг% ТМАО. В пресно¬
водной рыбе ТМАО совсем не содержится или содержится в значительно
меньшем количестве, чем в морской (обычно менее 50 мг%). Содержа¬
ние ТМАО в мясе морской рыбы зависит главным образом от ее вида,
размера, района промысла и сезона вылова. Рыба с белым мясом, напри¬
мер минтай и другие, характеризуется высоким содержанием ТМАО
в белых мышцах, а у рыб с темным мясом, например скумбрии, сарди¬
ны, тунца, содержание ТМАО в красном мясе больше, чем в белом
(табл. 19). Сезонные колебания ТМАО в мясе рыбы связаны в основ¬
ном с температурой воды; максимум приходится на зимний период.
В рыбе, выловленной в арктических районах, содержится больше ТМАО,
чем в рыбе умеренных и тропических вод. Содержание ТМАО в мясе
рыбы увеличивается с возрастом.В общем можно принять, что мышечные белки водных животных
с высоким содержанием триметиламиноксида подвергаются болееТаблица 18. Содержание ТМАО в мясе некоторых рыб,
моллюсков и ракообразных [,285, 391, 441]РыбаТМАО, мг%РыбаТМАО, мг%ДиапазонСред¬нееДиапазонСред¬нееМалая акула_1155Мерланг225Катран710-1035927Морская камбала120-360200Скат250-1359870Скумбрия96-198149Минтай470-785657Сырть-92,3Треска100-1080420Щука14-10049Тихоокеанский397Форель0-9948кальмарУгорь-36,9Креветка318-397332Окунь-26Сайда213-525314Линь-24,9Морской окунь310Карась-21,2Ершоватка160-570310Густера-9,1Пикша102-750308Форель0 94,5Палтус270Осетр0Сельдь108-32423865
Таблица 19. Содержание азота ТМАО, азота ТМА, азота ДМА в белом (Б) и красном (Кр)мясе некоторых видов рыбы [461]РыбаN ТМАО, мг%N ТМА, мг%N ДМА, мг%БKpБKpБKpЯпонская скумбрия12,921,41,13,50,51,4Scomber japonicus(5,3-22,5)(12,4-27,4)(0,1-2,8)(0,7-12,1)(0,1-1,1)(0,4-2,1)Тихоокеанская сардина3,026,60,10,700,6Sardinops melanostica(1,9-3,4)(23,9-30,4)(0-0,2)(0,4-0,8)(0,4-0,8)Бонито2,518,00,2502,1Katsuwonus pelamis(1,9-3,0)(15,6-20,4)(0,1-0,3)(4,2-5,6)(1,9-2,3)Альбакор3,013,20,17,00,14,8Thunnus albacora(3,0-3,1)(10,0-16,5)(6,6-7,3)(0-0,1)(4,7-4,9)Сайра10,811Д0,11,00,20,5Cololabis sajra(9,6-13,4)(8,6-15,5)(0-0,2)(0,5-1,4)(0-0,4)(0,2-0,7)Японская ставрида61,340,70,41,50,20,7Trachurus japonicus(54,9-67,9)(39,4-42,6)^0,2-0,5)(1,0-2,2)(0,1-0,5)(0,6-0,8)Минтай111,957,30,41,30,55,3Theragra chalcogramma(104,8-118,2)(54,4-61,4)(0,2-0,6)(1,0-1,5)(0,4-0,6)(2,7-6,8)Угольная рыба111,247,00,10,40,11,1Anoplopoma fimbria(105,0-117,5)(45,0-49,0)(0,1-0,2)(0,3-0,6)(0,1-0,2)(1,0-1,2)
интенсивным химическим изменениям в процессе холодильного хране¬
ния, чем белки животных с низким содержанием ТМАО в мясе.Разложение ТМАО до ДМА и МА особенно быстро идет в мясе трес¬
ковых рыб, некоторых рыб родов Scopeliformes и Macruriformes, а также
в мясе моллюсков и ракообразных, что обусловлено значительным со¬
держанием в нем триметиламиноксида и высокой активностью демети-
лазы в сырье.В мясе тресковых рыб самая высокая скорость восстановления
ТМАО в процессе холодильного хранения (при —5 °С) наблюдается
у хека, средняя у трески, минтая и менька, а самая низкая — у пикши
[51, 52] Удаление красного мяса из филе этой рыбы перед заморажи¬
ванием вызывает заметное снижение и даже полное прекращение увели¬
чения содержания ДМА. Образования ДМА в мясе палтуса, морского
окуня и зубана не установлено. В жирной рыбе содержание МА в про¬
цессе холодильного хранения увеличивается значительно медленнее, чем
в тощей, что можно легко подтвердить на примерах сельди и балтий¬
ской трески. Эта зависимость, хотя и неоднозначно, сохраняется также
и для некоторых тощих рыб с содержанием липидов менее 5 %. Рехина и
Рамбеза [389] изучали скорость увеличения содержания муравьиного
альдегида в мороженом стабилизированном фарше, а также содержание
липидов в мясе и разделили всю исследованную рыбу на три группы
(табл. 20).Таблица 20. Скорость увеличения содержания муравьиного альдегида (МА)
я стабилизированном мороженом фарше в процессе хранения
и содержание липидов в мясе [ 389]ГруппаСреднее содержание МА в стабилизирован¬
ном фарше* после 6 мес хранения
при температуре -20 °С (мг%)Содержа¬
ние липидов
в мясе, %РыбаI120,3-0,7Мерланг, менё'кII70,3-0,7Сайда, полярнаятресочка1Д-2,8Хек3,7-4,7Морской карась1,1-2,8СеребрянкаIII0,5-1,33,4-4,7Красный мор-ской петух* С добавлением 1,5 % поваренной соли, 1 % сахарозы и 1 % цитрата натрия.Из таблицы видно, что для большинства исследованных видов рыб
существует обратно пропорциональная зависимость между скоростью
увеличения МА в процессе замораживания и холодильного хранения и
содержанием жира в мясе.67
3.3.4.2. Влияние температуры замораживанияВо время хранения рыбы при температуре ниже —30 °С взаимодей¬
ствие муравьиного альдегида с белком происходит очень медленно и не
создает особых трудностей. Если же рыбу хранить при более высокой
температуре, что обычно встречается на практике, то разложение ТМАО
происходит достаточно быстро (рис. 16). Разложение триметиламинокси-
да - одна из главных причин уменьшения растворимости белков и свя¬
занных с ним неблагоприятных изменений консистенции.С повышением температуры замораживания активность ферментов,
вызывающих разложение ТМАО до муравьиного альдегида, увеличива-Рис. 16. Влияние продолжительности замораживания на образование диметиламина
(ДМА) и муравьиного альдегида (МА) в мясе минтая [4611ВремяРис. 17. Влияние температуры ра скорость образования диметиламина в инкуби¬
рованных водных вытяжках мяса минтая [ 461 ]68
ется. Наибольшая активность ферментов наблюдается в начальный пе¬
риод замораживания при температуре от —3 до —7 °С (рис. 17). Это
указывает на важную роль скорости кристаллизации льда во время за¬
мораживания.3.3.4.3. Влияние катализаторов и ингибиторовНаиболее сильными катализаторами процессов разложения ТМАО
в мясе рыбы служат миоглобин и гемоглобин. Они активизируют как
ферментативное, так и химическое разложение ТМАО в процессе на¬
гревания мяса. Гематин благодаря содержанию трехвалентного железа
также оказывает каталитическое действие.Редуцирующие вещества, как правило, ускоряют разложение ТМАО,
при этом некоторые из них, например аскорбиновая кислота, NaHS03
и другие, оказывают очень сильное действие, тогда как FeCl2, SnCl2
и некоторые другие - слабое. Большинство неорганических окислите¬
лей ингибируют образование ДМА и МА в мороженой рыбе. Таким об¬
разом, этот ряд можно представить следующим образом:SnCl2,KMn04, FeCl3, KBr03, Н202 [461]Из свободных аминокислот заметное каталитическое действие ока¬
зывает цистеин, остальные в этой роли имеют небольшое значение.В фарше, передержанном в незамороженном состоянии, опосредо¬
ванным бактериальным катализатором разложения ТМАО является хло¬
ристый натрий в концентрации 2—3 %. Концентрация хлористого натрия
выше 9 % полностью прекращает бактериальное разложение ТМАО
[354]3.4. ЛИПИДЫ И ИХ ИЗМЕНЕНИЯВыше рассматривалось действие продуктов гидролиза и окисления
липидов на мышечные белки. Теперь остановимся на наиболее важных
проблемах, касающихся изменений липидов с точки зрения технологии
производства рыбного фарша, при этом особое внимание уделим фак¬
торам, определяющим химические изменения жиров и образование
продуктов окисления липидов, имеющих запах.3.4.1. Подверженность липидов рыбы окислениюВысокая окисляемость липидов обусловлена прежде всего высоким
содержанием ненасыщенных жирных кислот, незначительным содержа¬
нием естественных антиокислителей, наличием в мясе рыбы естествен¬
ных прооксидантов, а также активностью тканевых ферментов, катали¬
зирующих процессы гидролиза и окисления липидов.В липидах мяса рыбы содержится в среднем на 32,5 % больше нена-69
сыщенных жирных кислот, чем в жире млекопитающих. Кроме того,
они характеризуются высокой степенью ненасыщенности жирных кис¬
лот. В то время как в жире убойного скота содержатся лишь следы поли¬
еновых жирных кислот с тремя или более двойными связями, в липидах
мяса рыбы это содержание довольно значительно. Например, в сельдевых
содержание полиеновых жирных кислот составляет 13—42, тунцовых —
30-40, треске - 24—61 % общего количества жирных кислот [393]В группе полиеновых кислот значительную долю составляют кислоты
с пятью и шестью двойными связями. В липидах балтийской сельди, со¬
держащих 19,33 % полиеновых жирных кислот, на долю гексаеновых
кислот приходится 6,31, пентаеновых — 6,00, тетраеновых — 4,32 и
триеновых - 2,7 % [224] В пересчете на сырую массу содержание поли¬
еновых кислот колеблется в диапазоне 0,2—7,4 (в среднем 1,4 %), со¬
держание докозагексаеновой кислоты (С22_б) 0,07—2,6 % (в среднем
0,65 %). Особенно высоко содержание полиеновых жирных кислот в
жирной рыбе (сардина, сельдь, скумбрия, лосось). Оно достигает 4 %,
из которых около 3,5 % приходится на кислоту С22_6* В мясе тощей
рыбы (тресковые, камбаловые) содержание полиеновых жирных кис¬
лот и докозагексаеновой кислоты ниже и составляет соответственно
около 0,37 и 0,15 % [424]В жире тощей рыбы содержится довольно много фосфолипидов. На¬
пример, в липидах трески их содержание составляет 64—91,4 %, в липи¬
дах шпрота — всего 4,4—10 % [393] Поскольку фосфолипиды харак¬
теризуются высоким содержанием полиеновых кислот, особенно гекса¬
еновых и пентаеновых, а также значительной подверженностью гидро¬
лизу, даже низкое содержание жира в мясе тощей рыбы играет сущест¬
венную роль в формировании функциональных свойств фарша, особенно
растворимости белков.По сравнению с другими жирами сходной степени ненасыщенности
(например, растительными) рыбные жиры бедны токоферолом, который
является их основным естественным антиокислителем. Среднее содер¬
жание токоферола в жирах мяса рыбы составляет 2—15 мг% [41, 432],
тогда как в растительных жирах — чаще всего 40-120 мг% [26] . Кроме
того, в жире рыбы практически содержится только а-токоферол [101] ,
в то время как в растительных жирах присутствуют все остальные изо¬
меры токоферола, характеризующиеся наиболее высокой антиокисли-
тельной активностью. Чем жирнее рыба, тем меньше содержание в ней
токоферола [142] , что с точки зрения технологии неблагоприятно.Из естественных прооксидантов, содержащихся в мясе рыбы, прежде
всего следует упомянуть гемопротеиды миоглобин и гемоглобин [405]
и негемовое железо. Особенно богата ими жидкая фракция гомогената
красных мышц. В процессе сепарации и перемешивания измельченного
мяса происходит распределение этих катализаторов окисления липидов
во всей массе фарша, что ускоряет процесс окисления липидов. Проокис-
лительная активность гемопротеидов и неорганического железа выше70
при кислой реакции (pH ниже 6,0), чем при естественном pH мяса рыбы
[106, 406]В мясе рыбы содержится много ферментов, вызывающих изменения
жира в процессе холодильного хранения. К ним относятся липазы, фос-
фолипазы и др. Тканевые ферменты особенно ответственны за гидро¬
литическое расщепление жира в фарше.Активность липаз и фосфолипаз как гидролаз неразрывно связана
с количеством свободной воды [138], которое уменьшается в резуль¬
тате вымораживания. Однако прямой зависимости между активностью
гидролитических ферментов, разлагающих липиды, и температурой за¬
мораживания фарша нет, так как замораживание (размораживание)
заметно повышает активность ферментов. Это, вероятно, результат вы¬
свобождения ферментов из клеточных органелл, поврежденных в ре¬
зультате кристаллизации льда и облегчения контакта фермента с суб¬
стратом. Больше всего продуктов гидролиза липидов образуется в
фарше, хранящемся при температуре от —5 до -7 °С. По мере сни¬
жения температуры скорость гидролиза уменьшается, так что при тем¬
пературе ниже —20 °С она значительно ограничена, а при более низкой
температуре —30 °С гидролиз практически полностью прекращается.
Поскольку липазы и фосфолипазы характеризуются высокой устойчи¬
востью при замораживании, то даже во время хранения при низкой
температуре после определенного периода происходит значительное на¬
копление свободных жирных кислот. В мясе тощей рыбы (треска)
основным источником свободных жирных кислот являются фосфоли¬
пиды, в меньшей степени — триглицериды и эфиры холестерина, а в мясе
жирной рыбы (сельдь, шпрот) - триглицериды [427]Ацетилгидролаза фосфолипидов вызывает деацилирование лецити¬
на (холинфосфатид) до лизолецитина [177] , который впоследствии
разлагается ацетилгидролазой лизолецитина до глицерофосфохолина и
свободных жирных кислот [512] Не исключается также непосредст¬
венное разложение лецитина до глицерофосфохолина в присутствии
ацетилгидролазы лизолецитина [176]Оптимальная активность липаз и фосфолипаз обнаруживается в диа¬
пазоне pH от 7,0 до 8,0, а значения pH ниже 5 значительно ограничивают
гидролиз триглицеридов и фосфолипидов [177, 374] Тепловая обработ¬
ка фарша при 60—80 °С перед замораживанием значительно замедляет
гидролитические реакции липидов [268]3.4.2. Продукты окисления липидов,
обладающие запахомОкисление липидов - одна из основных причин появления непри¬
ятных запахов мороженого фарша, особенно из жирной рыбы и рыбы
средней жирности. Можно предположить, что, как и при замораживании
других пищевых продуктов, содержащих жир, ухудшение запаха мо-71
роженого рыбного фарша вызывается главным образом низкомолеку¬
лярными соединениями, образующимися в процессе окисления ненасы¬
щенных жирных КИСЛОТ (прежде всего С20:5>С22:6 И С18 :1) ПРИ содей¬
ствии гемопротеидов, мышечных оксидаз и других каталитических фак¬
торов, например света. Важнейшие из низкомолекулярных соединений
карбонильные соединения, особенно такие алифатические альдегиды,
как л-алкен из ряда С2—Ci2, я-алк-2-ен из ряда С4—Сц и л-алк-2,4-дви-
$н из ряда С9—С12. Кетоны в летучих карбонильных соединениях пред¬
ставлены только очень незначительной фракцией или вообще отсутст¬
вуют [252].Алифатические альдегиды имеют очень низкие пороговые значения
органолептического восприятия. Запахи обнаруживаются при пороговых
концентрациях от 0,01 до нескольких десятков мкг/г. Для сравнения
можно показать, что пороговые концентрации для всех жирных кислот
составляют 5-15 мг/г, для кислот C8-Ci2 20—60 мг/г [265]Приведенные выше данные показывают, что для заметного ухуд¬
шения запаха мороженого фарша достаточно окисления или другого
процесса расщепления даже очень незначительных количеств липидов.В летучей фракции окисленного жира сардины идентифицировано
43 соединения, среди которых преобладали уксусная кислота, пропио-
новый альдегид, пропионовая кислота, этиловый спирт, бутиловый
альдегид, амиловый альдегид и амилен [331]Нежелательный запах мороженой трески, появляющийся в процес¬
се хранения, обусловлен прежде всего образованием энантового альдеги¬
да (цис-4-гептаноальдегида), а радужной форели — пропионового аль¬
дегида и капроновой кислоты [450] Кроме того, пропионовый альде¬
гид — основной компонент летучих веществ окисленного жира скумб¬
рии [325]При хранении мороженой рыбы в ней образуются также этиловый и
метиловый альдегиды, ацетон и пропионовая кислота. В то время как
количество этих соединений возрастает, содержание в мясе рыбы кис¬
лот С2о: 5 > С22 :б ? С is: 1, наличие которых можно установить лишь после
1 мес хранения рыбы при температуре —20 °С, уменьшается.В вареном фарше состав пахучих веществ еще сложнее, чем в моро¬
женом. Кроме множества продуктов термического окисления липидов
(альдегиды, кетоны, спирты, эфиры, углеводороды и циклические со¬
единения), к этой группе относятся еще продукты разложения ТМАО
(триметиламин, диметиламин и монометиламин), а также продукты де-
карбоксилирования аминокислот: л-пропиламин, и-бутиламин [214] ,
сероводород [508] и др.В соленом фарше так же, как и в солено-сушеной мороженой рыбе,
кроме альдегидов и летучих оснований, могут присутствовать летучие
жирные кислоты, но их содержание чаще всего очень незначительно
[155,497]72
3.5. НЕФЕРМЕНТАТИВНОЕ ПОБУРЕНИЕСреди различных типов неферментативного побурения наибольшее
значение в технологии изготовления и хранения рыбного фарша имеют
реакции, происходящие между редуцирующими сахарами и аминокис¬
лотами, пептидами, белками и аминами, продуктами окисления липи¬
дов (особенно карбонильными пахучими соединениями) и аминокис¬
лотами, пептидами, белками и аминами, а также окисление гемопроте¬
идов (гемоглобина, миоглобина).В первых двух случаях явления побурения связаны с наличием ами-
новых соединений и карбоксильных групп, поэтому этот тип нефер¬
ментативного побурения можно назвать карбонилоаминовым. Эта реак¬
ция называется реакцией Майяра [151, 152, 517]Реакция Майяра представляет собой комплекс различных, часто
взаимосвязанных химических процессов, предвестниками которых
являются редуцирующие моносахара и аминокислоты, находящиеся в
свободном или связанном состоянии, первичные и вторичные амины, а
также аммиак. Зиемба [516] показал, что третичные амины вызы¬
вают побурение так же, как и компоненты коптильного дыма безамин-
ное побурение. Для более полной характеристики реакции Майяра ее
можно разделить на этапы, ограничившись реакцией между аминокис¬
лотами и редуцирующими сахарами [312]В начальной стадии (этап I) редуцирующий сахар, например глюко¬
за, реагирует с аминным соединением, например глицином, с образова¬
нием- моноглюкозилглицина, который, будучи неустойчивым соедине¬
нием, подвергается затем перегруппировке Амадори и превращается
в монофруктозилглицин (этап II). На III этапе монофруктозилглицин
(МФГ) может реагировать со следующей молекулой глюкозы, после
перегруппировки образуя дифрукто зил глицин (ДФГ).перегруп¬пировкаАмадориЭтап IIГлюкозаН ОН6ОН1 6
ch2-nh-ch2-cooh НОСЭтап III
^ 1 Глюкоза2 Перегруппировка
АмадориМонофруктозил¬
глицин (МФГ)73
ДФГ-рН5,5 CHj-NH—CHj—СООН_ IС =0Iно—НС—он
НС—он
сн,он<j:HOс=оIсн2
НС—он
не—онСНа0НДругиекарбонильныесоединенияО О
II II
СН3—С—С—Нft °СН3—С—Й—СНз БиацетилСН f
ОНft-С—СНзСНОиккIнс-онIсн2онМетилглиоксальГндрооксиацетон2-фуриловый альдегидЭти соединения в дальнейшем разлагаются до карбонильных или
других (например, гетероцикличных веществ). На последующей ста¬
дии могут происходить, различные вторичные реакции, которые при¬
водят к образованию полимеров темного цвета, а также характерных
пахучих веществ. Одной из важнейших и наиболее известных реакций
разложения аминокислот является разложение аминокислот по Штре-
керу, которая идет по следующей схеме [400]R уО1 , ^Н - С - NH2 + R-C-C-R" -*R-C + R' - СН - С -I II II \ I IIСООН 0 0 Н NH2 о- R" + С02Образовавшиеся в результате этой реакции альдегиды становятся
основным источником пахучих соединений. Они могут снова реагиро¬
вать с аминными соединениями с образованием альдиминов. На заклю¬
чительной стадии из альдимина образуются азотистые полимеры и сопо¬
лимеры, называемые меланоидами. Продукты образуются на начальной
стадии реакции, бесцветны, обнаруживают абсорбцию в области ультра¬
фиолетового спектра излучения. На промежуточной стадии образуются
бесцветные или желтого цвета продукты, интенсивно абсорбирующие
ультрафиолетовое излучение при длине волны 375 нм.74
Очень важным фактором, влияющим на интенсивность реакции
Майяра, является также количество и качественный состав субстрата.
Скорость реакции прежде всего зависит от температуры, времени и pH.
Можно принять, что повышение температуры на 10 ° С вызывает четы¬
рехкратное увеличение скорости реакции [118] Логарифм оптической
плотности, характеризующий окрашивание смеси глюкозы с глицином,
связан прямой зависимостью с квадратом времени [99] Скорость ре¬
акции значительно возрастает, когда pH смеси становится выше изо-
электрической точки аминокислот [509]В фарше, хранившемся при требуемой низкой температуре не боль¬
ше 6 мес, реакции Майяра приводят только к образованию бесцветных
продуктов, поэтому при оценке качества, основанной на органолептиче¬
ских показателях, они не имеют большого практического значения, хотя
и оказывают некоторое влияние на питательную ценность белка [234]
Влияние реакции Майяра на качество сырого мороженого фарша прояв¬
ляется только при более длительном сроке хранения (побурение поверх¬
ности блоков), а также в фарше, подвергшемся Перед замораживанием
тепловой обработке (варке, сушке и т. п.). В таком фарше нефермен¬
тативное побурение обусловлено термическим разложением ТМАО до
ДМА, которое особенно быстро идет при aw < 1 [423] Рыбный фарш
с добавлением крахмала, сахара и белковых препаратов, а также фарш
и коагуляты из ракообразных более подвержены реакции Майяра, чем
мясо без добавок.Простые аминные соединения (например, аминокислоты) реагиру¬
ют быстрее, чем сложные (например, белки). При нагревании с глюко¬
зой среди 17 аминокислот наиболее активны лизин, орнитин, далее трип¬
тофан и аргинин; глутаминовая кислота практически инертна; цистин
проявляет ингибирующие свойства [315]Среди сахаров пентозы более активны, чем гексозы [470] Однако
глюкоза, присутствующая в мясе рыбы в больших количествах, оказы¬
вается важнее для побурения, чем рибоза. Важная роль отводится глю-
козо-6-фосфату, вызывающему более темное окрашивание с глицином,
чем глюкоза [384] Допускается, что в реакцию с аминокислотами
вступает полуацетальная форма сахаров с открытыми цепями в молеку¬
лах, доля которой в молекуле глюкозы составляет 1,0, в молекуле
ксилозы - 16,7, а рибозы - 354,0.Ионы металла (Cu2+, Fe2+), карбоновые кислоты (яблочная, лимон¬
ная) и их соли, а также некоторые фосфаты (например, Na2HP04) вы¬
ступают как активаторы побурения. Муравьиный альдегид ингибирует
побурение модельных систем [511], но катализирует побурение мяса
крабов с добавлением углеводов (Нагасава, 1959). Атмосферный кис¬
лород катализирует побурение, а энергия света оказывается ингибирую¬
щим фактором. В этом случае побурение является результатом реак¬
ции аминов с редуцирующими сахарами [315] .Активными ингибиторами реакции Майяра являются вещества,75
мгновенно реагирующие с карбонильными соединениями, такие, как
диоксид серы, кислый сернокислый натрий, гидроксиламин и семикар-
базид [99] , и в меньшей степени некоторые аминокислоты (например,
цистеин), хлористый кальций и тринатрийфосфаты [511]Изменение pH фарша в щелочную сторону (до pH 10,6±0,1), а еще
сильнее в кислотную ускоряет-реакцию Майяра. Частичное обезвожива¬
ние фарша вызывает ускорение реакции Майяра.Результатом реакции Майяра являются не только видимые хромо¬
форы, но и ухудшение вкуса и запаха, а также снижение питательной цен¬
ности мороженого рыбного фарша. Образование запаха связано прежде
всего с разложением аминокислот по Штрекеру, обусловленным при¬
сутствием дегидроредуктазы, которая образуется при помощи сахаров.
Если образованные альдегиды летучи, они являются источником запаха
(см. раздел 3.4.2). Разложение метионина ведет к образованию в фарше
летучих сернистых соединений, которые редко изменяют его органолеп¬
тические свойства. В процессе тепловой обработки фарша происходят
карбонильно-аминовые реакции, продукты которых придают ему горь¬
кий вкус [179] . Интенсивность этих реакций зависит от интенсивности
некоторых, например автолитических, реакций в мясе, состава экстраги¬
руемых веществ и других факторов.При неферментативных реакциях побурения рыбного фарша проис¬
ходят значительные изменения гемоглобина и миоглобина, что обус¬
ловлено высокой активностью образующихся красящих веществ. В то
время как оксигемоглобин млекопитающих стабилен при pH до 4,
оксигемоглобин рыбы в большинстве случаев (около 80 %) превращает¬
ся в дезоксигемоглобин уже при снижении pH с 6,8 до 6,4. Окисление
гемоглобина трески при температуре 15 °С при pH 7,0—6,4 происходит
в 30—50 раз быстрее, чем окисление гемоглобина млекопитающих
[197] . Оксигемоглобин, встречающийся главным образом в красных
мышцах, легко переходит в коричневый метмиоглобин. Протеканию
описанных выше реакций способствуют недостаточное охлаждение рыбы
перед переработкой на фарш (слишком резкое снижение pH во время
посмертного окоченения), неправильное обескровливание и доступ
атмосферного кислорода. Кроме побурения и других изменений окрас¬
ки, эти реакции могут вызывать появление в фарше нежелательного за¬
паха. Это особенно заметно при взаимодействии окисленных гемопроте¬
идов с сероводородом, продуктами которого являются сульфогемогло-
бин, сульфомиоглобин и другие пигменты зеленого цвета [215] Эти
изменения можно эффективно предотвращать, ограничив доступ атмос¬
ферного кислорода в фарш в процессе его хранения и размораживания.3.6. ИЗМЕНЕНИЯ НУКЛЕОТИДОВНуклеотиды, содержащие группу ОН в положении ”G” кольца пу¬
ринового основания, а также в положении ”5” рибозы или дезоксири-
7S
Гипоксантин+Рибозо - 1 - фосфатАТФАТФазаАДФМиокиназаАденоэинмоно- АМФдезаминаза^ Инози|)монофосфат 5'нуклеотид^^
фосфат (АМФ)Гидролаза нуклеоэидаГипоксантин
+ рибозаРис. 18. Схема разложения АТФ в мышечной ткани рыбыбозы, соединенной эфирной связью с фосфорной кислотой, потенциаль¬
но определяют вкус продукта. Самый важный нуклеотид, который при¬
дает желаемый вкус мясу рыбы—инозин-5-монофосфат (ИМФ),- обра¬
зуется в процессе разложения аденозин-5-трифосфата (АТФ), а точнее, в ре¬
зультате гидролитического дезаминирования АМФ (рис. 18). Дезаминаза
АМФ наиболее активна при pH 6,5-7,0 и температуре 38 °С, однако вы¬
держивание в течение 30 мин при температуре 50 °С не вызывает ее
полной инактивации (Хидака и Саито, 1960). Этот фермент, встречаю¬
щийся во фракции миофибриллярных белков и особенно активный
в скелетных мышцах, ведет к быстрому накоплению ИМФ в мясе рыбы.
Содержание ИМФ в период пикового аккумулирования может достигать6 мкмоль на 1 г мышечной ткани. Идеальным с точки зрения техноло¬
гии было бы поддержание сырья в таком состоянии в течение всего
срока его хранения. К сожалению, присутствующие в мясе рыбы фос-
фомоноэстеразы снижают его вкусовые свойства. Разложение ИМФ
не только приводит к утрате желаемых вкусовых свойств мяса рыбы, но
и очень часто вызывает образование веществ, дополнительно ухудшаю¬
щих вкус. Инозин, образующийся из инозинмонофосфата, не имеет
вкуса, а образующийся в результате его ферментативного гидролиза
гипоксантин придает продукту характерный горький вкус.Скорость дефосфорилирования ИМФ в мясе рыбы значительно
выше, чем в мясе теплокровных животных. Содержание ИМФ в мясе
морской промысловой рыбы обычно достигает максимума за период
°т нескольких часов до нескольких суток после вылова, а уже на вторые
сутки с момента смерти рыбы уменьшается примерно на 50 %, тогда как
в мясе убойного скота — лишь примерно после двух недель хранения
[419].77
Разложение ИМФ идет до расходования субстрата или до потери ак¬
тивности 5-нуклеозидазы, которая при температуре О °С стабильна по
крайней мере в течение 6 лет, но после 10 мин нагревания при темпе¬
ратуре 42 ° С активность снижается примерно наполовину, а максимум
ее обнаруживается при pH 7,6 [457] Ионы Мп2+ в меньшей степени
Mg2+ активизируют 5-мононуклеотидазу, однако этилендиаминтетра-
уксусная кислота (ЭДТА) и пирофосфаты (Мп4Р207) уже при концент¬
рации 1-10 мкмоль/л оказывают ингибирующее действие на фермент.Скорость разложения нуклеотидов зависит главным образом от то¬
го, как долго рыба сохраняла активность в орудиях лова, от ее вида,
температуры во время обработки и хранения мороженого фарша,
pH и многих других факторов. В рыбе, долго агонизировавшей в ору¬
диях лова, содержится значительно меньше АТФ и ИМФ, чем в необес¬
силенной (табл. 21). В зависимости от продолжительности хранения
рыбы во льду, необходимой для достижения уровня разложения нук¬
леотидов, соответствующего К = 20 %*, рыб можно разделить на три
группы [97] достигающие этих величин спустя 1-2 сут хранения
(минтай, дальневосточная треска), спустя 3—6 сут (ставрида, скумб¬
рия) и спустя 7—14 сут (Pagrus major, Mylio macrocephalus, Paralichthys
olivaceus, P. indicus, Bembrus japonicus, Saurida undosquamis, Platycepha-
lus indicus и др.).Таблица 21. Влияние продолжительности нахождения рыбы в орудии
лова на содержание нуклеотидов в ее мясе (в мкмоль/г) [ 129,182]НуклеотидТреска (Gadus callarias)Морской язык (Parophrys vetulus)неутомленнаяобессиленнаянеутомленныйобессиленныйАТФ5,340,263,912,15АДФ0,580,430,230,94АМФ0,690,570,080,17ИМФ1,265,860,732,11Из этих данных следует, что для тресковых, являющихся основным
сырьем для производства фарша, характерно очень быстрое разложение
нуклеотидных соединений в мясе, что связано с высокой активностью
5-нуклеотидазы.* Коэффициент К, определяющий подверженность мяса рыбы разложению
нуклеотидных соединений, определяется по уравнениюИнозин + ГипоксантинК = ,АТФ + АДФ + АМФ + ИМФ + Инозин + Гипоксантингде перечисленные сокращения означают содержание отдельных соединений в мясе.
Для рыбы сразу после обезглавливания К равен 3,5±1,9 %, а для мяса I сорта обыч¬
но превышает 20 % [ 97].78
Для болеё полной характеристики изменений вкуса в результате раз¬
ложения нуклеотидных соединений необходимо определение состава
образующихся при этом продуктов, особенно инозина и гипоксантина.
Сырье водного происхождения можно разделить на инозин- и гипоксан-
тинобразующее. Гипоксантинобразующая рыба когда-то получила на¬
звание ’’щелочной”, поскольку инозинмонофосфатная фосфогидролаза
обнаруживает максимальную активность при щелочном pH. В свою оче¬
редь, инозинобразующую рыбу называют ’’кислотной”. К инозинобра-
зующей рыбе относятся тунцы и палтусы, некоторые виды макрели,
скумбрии, ставриды, японский анчоус, тихоокеанская сельдь, марлин,
а также креветки. Содержание гипоксантина в мясе перечисленных ви¬
дов рыб настолько низкое, что после длительного хранения во льду,
когда рыба уже непригодна в пищу, оно не превышает 1,5 мкмоль/г
[149]. С этой точки зрения такая морская рыба напоминает пресно¬
водную, в которой содержание гипоксантина после 2 нед хранения не
превышает 1 мкмоль/г [87] . К гипоксантинобразующим рыбам отно¬
сятся прежде всего некоторые камбаловые (Lepidosetta mochigarei,
Pleuronectes pallasi, Platichthus stellatus, Paralichthus olivaceus, Limanda
aspera, L. herzensteini, Eopsetta grigorjewi, морские окуни (Sebastes tac-
zanowskii, S. inermis, S. schlegeli), головоногие моллюски. В морских оку¬
нях и камбалах, хранившихся 2—5 сут во льду, содержание гипоксанти¬
на в мясе составляло 4,5—6,5 мкмоль/г.Большинство тресковых, так же как и лососевые, палтусы и угре¬
вые, относятся к группе со средней скоростью накопления гипоксанти¬
на в мясе. Когда они становятся непригодными в пищу, содержание
гипоксантина в мясе достигает 2—3 мкмоль/г J45, 149] . Однако в мясе
тресковых ИМФ очень быстро разлагается. Следовательно, ухудшение
вкуса их мяса связано не столько с накоплением гипоксантина, сколько
с разложением основного стимулятора желаемого вкуса мяса рыбы, ка¬
ким является ИМФ, особенно если принять во внимание, что мясо трес¬
ковых относительно бедно другими вкусовыми соединениями, например
свободными аминокислотами. Так, в течение первых двух суток хране¬
ния во льду первоначальное содержание ИМФ в мясе минтая снижается
почти на 75 %, тогда как у морского карася всего на 10—20 % [98]Понижение температуры только частично тормозит дефосфорили-
рование и дезаминирование 5-рибонуклеотидов, так как большинство
ферментов, катализирующих их разложение, относительно устойчиво
к низким температурам. Например, фосфогидролаза 5-рибонуклеоти-
да, катализирующая разложение ИМФ и инозина, сохраняет активность
до температуры —20 °С, а фосфотрансферазы, катализирующие дефос-
форилирование нуклеотидов, остаются активными даже при темпера¬
туре —30 ° С [180] . При температуре 0-^-26 °С энергия активации ИМФ
в мясе меч-рыбы (Xiphias gladius) составляет 110,5 кДж независимо от
вида мышц, а энергия инактивации гипоксантина — 126 и 140 кДж со¬
ответственно для всего мяса и только для красных мышц [94].79
В мясе рыбы, хранившейся при температуре выше криоскопиче-
ской точки, постоянная скорости реакции к, вызывающей разложение
нуклеотидов, возрастает экспоненциально температуре. Например,
в мясе карпа, хранившегося при температуре 0—25 °С, эту зависимость
можно описать уравнением: к = 0,0053f2 + 0,0455f + 0,318 (t — темпе¬
ратура, °С) [357] В красных мышцах рыбы разложение ИМФ проис¬
ходит значительно быстрее, чем в белых мышцах [108] , однако ско¬
рость образования гипоксантина в мышцах обоих видов весьма сходна
(табл. 22).Таблица 22. Влияние температуры на скорость образования
гипоксантина в мясе (белые и красные мышцы) и отдельно
в красных мышцах скумбрии (Scomber scombrus) [108]Температура,°СПродолжительность
хранения, сутУравнение регрессииВсе мышцы (а)Красные мышцы (б)00-15 (а)у = 0,09+0,104*у = 0,18* + 0,110-20 (б)50-15 (а)у — 0,16 + 0,15*у = 0,28* + 0,200-20 (б)100-20 (а,б)у= 0,81* -0,02у = 0,92* - 0,34150-20 (а,б)у= 1,10* -0,34у = 0,88* + 0,36В большинстве случаев значительного различия в скорости разло¬
жения ИМФ в свежей (незамороженной) и размороженной рыбе не на¬
блюдалось [198]. Однако в тех случаях, где оно имелось, это свиде¬
тельствовало об интенсивном влиянии замораживания и разморажива¬
ния на скорость разложения нуклеотидов в мясе рыбы. Например, за¬
мораживание и размораживание хека вызывает увеличение содержа¬
ния гипоксантина на 0,328 мкмоль/г. Примерно столько же гипоксан¬
тина образуется после 3—4 сут хранения рыбы во льду [45]Разложение нуклеотидов происходит относительно быстро и в мо¬
роженом сырье. Это подтверждают приведенные ниже уравнения ре¬
грессии, характеризующие скорость разложения инозинмонофосфата
(ИМФ) и образования гипоксантина (Ги) в мясе мороженой меч-рыбы,
хранившемся при трех различных температурах [94]ИМФ - 18 °С у = 3,58 - 0,029л: (Р = 0,005; п = 8) ;ИМФ — 26 °С у = 4,54 — 0,021* (^ = 0,05; п = 4)Ги — 8 °С у = 0,46 + 0,0316л: (Р= 0,001; п = 7)Ги - 18 °С у = 0,32 + 0,0064л: (Р= 0,001; п = 8) ;Ги - 26 °С у = 0,30 + 0,0007* (Р пренебрежимо мало),где у — содержание компонента, мкмоль/г, х — продолжительность хра¬
нения, нед.80
На основании этих данных можно в общем виде принять, что для
практически полного предотвращения нежелательных изменений нукле¬
отидов в мороженом мясе рыбы желательно хранить его при температуре
не выше -25 °С. Хранение рыбы при более высокой температуре да¬
же в течение короткого времени (например, в результате недоста¬
точно быстрого охлаждения после вылова, медленного заморажива¬
ния или домораживания) ведет к быстрому разложению нуклеотидов и
накоплению в мясе продуктов их распада. Например, при температуре
-8 °С содержание гипоксантина в мясе меч-рыбы увеличивается пример¬
но в 4 раза быстрее, чем при -18 °С. Это означает, что уже спустя месяц
хранения при такой температуре содержание гипоксантина практически
в 3 раза превысит его минимальное содержание (0,2 мкмоль/г), уста¬
новленное Джоунзом [178] , при котором органолептически обнаружи¬
вается привкус горечи в мясе. Даже если считать, что содержание гипок¬
сантина по Джоунзу слишком незначительно1, не вызывает сомнения
установленная многими исследователями высокая корреляция между
содержанием ИМФ и гипоксантина в мясе и вкусом его после тепловой
обработки (табл. 23).Таблица 23. Зависимость между содержанием инозинмонофосфата (ИМФ)
и гипоксантина в мясе и вкусом его после тепловой обработки*Вид рыбыСпособхране¬ниясырьях - содержание инозин¬
монофосфата, мкмоль/г- содержание ги¬
поксантина,* мкмоль/гу - результат органолептической оценки
вкуса мяса по lQO-балльной шкале**Ис¬точ¬никАтлантиче- Хранение у = 41,5 + 7,5* у = 81-17,7*ская скумб- во льду Р < 0,001, п= 105 Р< 0,001 [108]рия (Scorn- п = 105ber scombrus)Меч-рыба Замора- .у = 22,4+18,5* ^=139-114*(Xiphias жив а ни е Р = 0,05 п = 97 Р = 0,05 п = 97gladius) -8° С [94]Замора- у= (-21,1) +26,8* у= 111 - 115*жив а ни е Р = 0,05 п = 97 Р= 0,05-18 °С п =91* Кусочки мяса толщиной около 2 см, завернутые в алюминиевую фольгу,
нагревали 12-15 мин в кипящей воде.**100 баллов - максимальная оценка; 40 баллов - продукт в пищу непри¬
годен.1 Показано [ 108, 422], что вкус горечи в мясе рыбы четко обнаруживается
при более высоком содержании гипоксантина - 1,2-3,0 мкмоль/г.81
Степень разложения АТФ в мясе рыбы в значительной степени обус¬
ловливает качество фарша, изготовленного из этой рыбы. Высокая
корреляция между содержанием инозинмонофосфата и гипоксантина
в мясе минтая и качеством сурими, полученным из этой рыбы, подтверж¬
дено Фудзии и др. [111] . Фарш хорошего качества получали только из
рыбы, содержание ИМФ в которой было не ниже 10—16 %, а гипоксан¬
тина — не выше 8—10 % общего содержания АТФ и продуктов ее разло¬
жения. Для этого рыба должна была храниться во льду не более 3—4 сут..В мороженом рыбном фарше нуклеотиды изменяются быстрее,
чем в филе или неразделанной рыбе, так как механическое повреждение
первичной структуры ускоряет ферментативные процессы. В вареном
фарше дополнительным фактором, вызывающим качественные и коли¬
чественные изменения нуклеотидов, служит тепловая обработка. Если
процесс варки проходит с нарушением технологического режима, потери
ИМФ в результате термической активности 5-нуклеотидазы, максимум
которой приходится на температуру около 40 °С, могут быть значи¬
тельными [419] . При соблюдении режима (например, если фарш нагре¬
вается острым паром в куттере-смесителе) содержание ИМФ снижается
обычно не более чем до 70 % первоначального его содержания в сыром
мясе.Подкисление фарша тормозит разложение ИМФ, так как при pH 4,0—
5,0 5-нуклеотидаза подвергается значительному разрушению в противо¬
положность слабощелочной среде (pH 7,6), в которой ее активность
максимальна [457]
4. ОСНОВНОЕ СЫРЬЕ И ЕГО ОБРАБОТКА4.1. РЫБНОЕ СЫРЬЕ ДЛЯ ПРОИЗВОДСТВА ФАРШАПо данным ФАО [131] , около 40 млн т морских биологических
ресурсов ежегодно не используются из-за отсутствия соответствую¬
щих методов первичной обработки. Эти ресурсы, включающие более
1000 видов рыб, ракообразных и морских головоногих моллюсков,
являются потенциальным сырьем для производства рыбного фарша.
До сих пор лишь незначительная часть биологических ресурсов нахо¬
дит практическое применение при производстве рыбного фарша. На¬
пример, в 1976 г. всего 46 видов рыб и ракообразных перерабатыва¬
лось на фарш [294] Можно предположить, что это число будет с каж¬
дым годом возрастать по мере освоения новых промысловых районов.В Японии среди нескольких десятков видов рыб, перерабатываемых
на фарш, ведущее место занимает минтай (Theragra chalcogramma),
а также такие виды, как морские окуни, хек, тихоокеанская треска, тер¬
пуги, марлин, Chloropthalmus albatrosis [43, 362,446] .В СССР на фарш перерабатываются главным образом малоценные
виды морских и пресноводных рыб, которые из-за низкого качества
мяса, малых размеров и других причин имеют низкую технологическую
пригодность и не находят широкого спроса. На фарш перерабатывают в
основном путассу, сайду, минтая, аргентину, карася, речного окуня и др.[103, 386]. Разработана также технология производства фарша из
сабли-рыбы и ставриды [83, 85]. Предпринимались попытки изготов¬
лять фарш из кильки и анчоуса [416] .В Канаде для производства фарша вначале предназначалась в ос¬
новном пресноводная рыба, которая не пользуется спросом из-за боль¬
шого количества костей. В настоящее время для получения фарша ис¬
пользуется более десяти различных видов пресноводных и морских
рыб, а также омары и крабы. Наибольшее промысловое значение имеют
треска, пикша, хек, менёк и сайда. Фарш производится, как правило,
из рыбы с белым мясом, блоки которого изготовляются из одного или
нескольких видов этих рыб [37, 280).В Скандинавских странах, а также в Великобритании получило рас¬
пространение производство фарша из отходов от филетирования и даже
из целого филе некоторых видов рыб с белым мясом. В зависимости83
от степени подготовки сырья получаются продукты разных классов
качества, различающиеся цветом, консистенцией, сроком возможного
хранения, а также направлением использования.4.2. ВЛИЯНИЕ ПОСМЕРТНЫХ ИЗМЕНЕНИЙНа качество мороженого рыбного фарша в значительной степени
влияют быстрота переработки сырья и стадия посмертных изменений
мышечной ткани в момент замораживания, которые зависят в основном
от продолжительности агонии рыбы в орудиях лова перед переработкой
ее на фарш и температуры хранения после вылова, особенно в период
наступления посмертного окоченения.Активность рыбы в момент траления или хранения на борту судна
вызывает быстрый распад АТФ и посмертные изменения в бескисло¬
родных условиях. Эти процессы сопровождаются увеличением скорости
образования молочной кислоты в результате распада гликогена. По¬
скольку скорость образования молочной кислоты в утомленной рыбе
в 2—3 раза выше, чем в неутомленной [27, 31, 425] , pH мяса в период
посмертного окоченения значительно понижается. В фарше, получен¬
ном из утомленной рыбы, растворимость миофибриллярных белков во
время хранения и после размораживания снижается быстрее, чем в фар¬
ше из отдохнувшей рыбы. Особенно заметно растворимость миофибрил¬
лярных белков снижается, если pH мышечной ткаг:и становится ниже6,0. Например, после 1 ч хранения при температуре 24- 25 °С pH мышеч¬
ной ткани агонизировавшей в эксперименте скумбрии перед обезглав¬
ливанием составлял 5,75 и в ней содержалось всего 25—40 % нераствори¬
мых миофибриллярных белков [249]Утомление рыбы не оказывает существенного, влияния на раствори¬
мость белков саркоплазмы. Снижение растворимости миофибриллярных
белков тем больше, чем выше температура хранения рыбы во время на¬
ступления посмертного окоченения. С повышением температуры pH
мышечной ткани снижается, а сокращение миомеров увеличивается.
Критической считается температура 17 ° С [181] У рыбы, хранившейся
при более высокой температуре, мышцы так сильно и резко сокраща¬
ются во время посмертного окоченения, что создающееся при этом на¬
пряжение вызывает разрыв ткани, соединяющей отдельные миомеры,
в результате чего первоначальная структура мяса в значительной сте¬
пени разрушается. Если посмертное окоченение происходит после размо¬
раживания, то изменения в тканях еще сильнее, поскольку, кроме ин¬
тенсификации гликогенолиза, замораживание вызывает механическое
повреждение мышечных тканей [283, 506] и снижает прочность соеди¬
нительной ткани.Консистенция фарша, изготовленного из рыбы в стадии посмерт¬
ного окоченения и быстро размороженного, изменяется больше, чем
фарша, размороженного медленно [283] После тепловой обработки
84
такой фарш теряет много клеточного сока и приобретает жесткую
волокнистую консистенцию.Во время агонии рыбы молочная кислота, диффундируя из мышц
в кровь, вызывает изменение окислительно-восстановительного потен¬
циала. Химическое сродство кислорода к гемоглобину уменьшается
[31] , в результате чего избыток его освобождается в мышцах. Кстати,
этот кислород может играть роль окислителя липидов в начальный
период хранения рыбного сырья.Консистенция фарша, изготовленного из рыбы, находящейся в ста¬
дии после посмертного окоченения, меньше изменяется во время замо¬
раживания, чем из рыбы в стадии посмертного окоченения или до его на¬
ступления. Таким образом, с технологической точки зрения степень све¬
жести, в которой должен находиться замороженный фарш, зависит от
предусмотренного заранее срока его хранения. Фарш, предназначенный
для кратковременного хранения в мороженом виде (до 3 мес), должен
быть изготовлен из рыбы, прошедшей посмертное окоченение или из ры¬
бы, находящейся в стадии его окончания. Фарш, предназначенный для
более продолжительного хранения в мороженом виде, как и фарш, пе¬
рерабатываемый без предварительного размораживания, следует изго¬
товлять из рыбы в стадии до наступления посмертного окоченения.
Медленное прохождение стадии посмертного окоченения во время хра¬
нения мороженого фарша обеспечит высокое качество продукта без
опасности прохождения посмертного окоченения во время разморажи¬
вания.Сикорский [413] сообщил, что даже при хранении мороженой ры¬
бы при низкой температуре полное дефосфорилирование АТФ в ткани
также происходит достаточно долго и после размораживания процессы
посмертного окоченения не проявляются. В мороженом измельченном
мясе дефосфорилирование происходит быстрее, чем в филе, так как
изменение структуры в результате измельчения ткани усиливает актив¬
ность АТФазы [114]4.3. ВЛИЯНИЕ СПОСОБА ХРАНЕНИЯ РЫБЫ
ПЕРЕД ПЕРЕРАБОТКОЙ НА КАЧЕСТВО ФАРШАФарш самого высокого качества получают из рыбы, охлажденной
немедленно после вылова льдом, смесью льда и воды или морской во¬
дой. Если посмертные изменения у рыбы (например, сардины) проис¬
ходят быстро, даже короткая задержка на палубе судна без соответст¬
вующей защиты вызывает заметное ухудшение качества мяса, и особен¬
но его вкуса и способности к образованию эластичного геля во время
тепловой обработки (табл. 24, 25)Если рыбу предполагается направить на переработку после прохож¬
дения посмертного окоченения, следует стремиться к тому, чтобы тем¬
пература во время прохождения посмертного окоченения была как мож-85
Таблица 24. Влияние способа и продолжительности хранения сардины на изменение содержания
продуктов разложения АТФ и растворимость белков в мясе, а также на свойства камабоко,
приготовленного из этого мяса [163]Способу храненияПродол¬житель¬ностьхране¬ния,сутПродукты разложе¬
ния АТФ, относитель¬
ные % (от АТФ)гипо¬ксан¬тинРастворимостьмышечныхбелковино¬ИМФв во¬в 3,75 %-номpHпроч¬коли¬тест назинде,раствореностьчествопереги¬мг%NaCl,геля,сока,бание*азотамг% азотаг-см%Свойства камабокоРезультат орга¬
нолептической
оценки по 10-
балльной шкалеПересыпание льдом сра¬003,896,5102314337,09140436,9АА8,0зу после вылова, через11,05,793,396214087,2073936,0АА5,72 ч хранения в воде, ох¬21,611,886,68924617,1349940,2А5,2лажденной льдом33,212,484,57673347,2656439,9А4,5На палубе в течение 1,5 ч00,55,194,596212127,0876937,5А6,1при температуре 13-13,812,383,97564127,1940348,1В-С4,214 ° С, затем охлаждение25,317,077,78653247,1642547,2D4,0небольшим количеством
льда в течение 4 ч с по¬
следующим хранением в
воде, охлажденной льдом* Согласно 5-балльной шкале, разработанной Нисия [ 344], для ломтика камабоко толщиной 3 мм и диаметром 3 см опре¬
деляют: АА (5 баллов) - отсутствие трещин при складывании вчетверо; А (4 балла) - трещины при складывании вчетверо, от¬
сутствие трещин при складывании вдвое; В (3 балла) и С (4 балла) - частичное и полное разламывание при сложении пополам;
D (1 балл) распадается на кусочки при нажатии пальцем.
Таблица 25. Влияние способа и продолжительности хранения рыбы перед переработкой на качество фарша (сурими),
а также продолжительности хранения мороженого фарша на свойства камабоко, приготовленного из этого фарша [163]Способ храненияПродолжительностьхранениярыбы до
обработки,
сутфарша (сури¬
ми) при
-40° ССвойства камабоко, изготовленного из фарша (сурими)pHПроч¬
ность
геля,
г-смКоличество клеточ¬
ного сока, выделив
шегося под давле¬
нием, %Тест на
переги¬
бание*Результат органо¬
лептической оцен¬
ки (по 10-балль¬
ной шкале)Пересыпание льдом сразу пос¬06,92109036,0АА8,2ле вылова, через 2 ч хранения12 нед7,0960439,2А6,2в воде, охлажденной льдом26,9536950,1-4,337,1025448,4-4,007,0087336 ЛАА-17,1552045,А-24 мес7,0236350,2--37,1528851,1--На палубе в течение 1,5 ч при06,9073340,0А6,2температуре 13-14 °С, затем12 нед6,9825854,23,8охлаждение небольшим коли¬26,9723459,13,2чеством льда в течение 4 ч с по¬06,9853344,7АА-следующим хранением в воде,14 мес7,ю'26557,5-охлажденной льдом27,0522161,4* Объяснения см. в табл. 24.
но ниже. Такую рыбу рекомендуется хранить при температуре О °С
[181]Выбор способа охлаждения рыбы зависит от ее вида. Для хранения
рыбы, у которой пигменты кожи легко переходят в мясо, не следует
применять в качестве охлаждающей среды морскую воду. Если таких
явлений не наблюдается, морская вода является лучшей охлаждающей
средой, чем лед. Показано [109] , что хранение хека в охлажденной
морской воде при температуре ±1 °С перед обработкой увеличивает
срок хранения мороженого фарша по меньшей мере на 1 нед по срав¬
нению с фаршем, полученным из рыбы, хранившейся во льду. В сарди¬
не, хранившейся в морской воде со льдом при 5 °С, К возрастает с 10
до 40 после 7 сут, а при хранении только во льду - после 3 сут [332]Качество рыбы, предназначенной для переработки на фарш, можно
значительно улучшить, применяя охлажденную морскую воду, насыщен¬
ную диоксидом углерода [25, 63] , или (при охлаждении жирной рыбы),
морскую воду с добавлением антиокислителей и синергических ве¬
ществ, например фосфорной, лимонной кислот, глутамата натрия в ко¬
личествах соответственно: 0,38; 0,048 и 0,48 % [132].Даже при соблюдении требуемых условий хранения рыбы качество
изготовленного фарша снижается экспоненциально продолжительности
хранения его в замороженном состоянии. Кроме микробиологических
причин, основной причиной снижения качества является уменьшение
растворимости белков [63] и связанные с этим изменения консистен¬
ции фарша [259] По данным Пелиховского и Нодзиньского [373] ,
для получения мороженых продуктов высокого качества и соответст¬
вующей стойкости при хранении рыбу перед замораживанием не следует
хранить в охлажденной морской воде более суток. Хранение жирной ры¬
бы в охлажденной морской воде в течение 2 сут значительно снижает
срок ее возможного хранения в замороженном виде.4.4. ФАКТОРЫ, ОПРЕДЕЛЯЮЩИЕ ТЕХНОЛОГИЧЕСКУЮ
ПРИГОДНОСТЬ РЫБНОГО СЫРЬЯТехнологическая пригодность рыбного сырья для переработки на
фарш зависит от ряда биологических, технологических и технических
факторов. Накопленные к настоящему времени данные не позволяют
однозначно определить и классифицировать эти факторы, чтобы они
полностью могли характеризовать пищевую ценность сырья, поэтому
изложенные ниже сведения должны прежде всего помочь в выборе
сырья для производства фарша, а также способа его облагораживания
и стабилизации. На основании полученных ранее данных для производ¬
ства фарша предназначаются, как правило, менее ценные виды рыбы и
съедобные отходы от филетирования, что позволяет рациональнее ис¬
пользовать доступное рыбное сырье в существующих технико-техноло¬
гических условиях.88
К факторам, определяющим технологическую пригодность рыбы
для производства фарша, относятся прежде всего следующие: способ и
продолжительность хранения рыбного сырья перед его переработкой на
фарш; химический состав мяса, и особенно наличие веществ, катализи¬
рующих нежелательные изменения в фарше; качество и тщательность
первичной обработки, а также способ сепарирования измельченного мя¬
са; морфометрические особенности рыбы, тесно связанные с ее пригод¬
ностью для механической первичной обработки; наличие паразитов и др.На фарш необходимо направлять свежую рыбу очень хорошего ка¬
чества и только в исключительных случаях использовать для этих целей
мороженую рыбу. Продолжительность хранения рыбы после вылова
должна быть возможна более короткой. Сырой непромытый фарш вы¬
сокого качества получается из рыбы, переработанной до наступления
посмертного окоченения. Например, хранение зауриды во льду в тече¬
ние 3 сут ухудшает консистенцию фарша, и особенно его способность
образовывать эластичный гель после тепловой обработки примерно на
50 % [252]. Быстрое ухудшение вкуса мяса рыбы объясняется главным
образом распадом 5-рибонуклеотидов (см. раздел 3.6), а частично также
накоплением продуктов обмена веществ микроорганизмов (амины,
аммиак и др.). При выработке промытого фарша (сурими) тощую рыбу
(в зависимости от вида) можно хранить от нескольких до десяти и бо¬
лее суток в охлажденном виде (от -1 до 3,3 °С) без заметного ухудше¬
ния ее технологической пригодности. Однако жирную рыбу с высоким
содержанием красного мяса следует перерабатывать на фарш сразу после
вылова. Показано [162] , что даже хранение в течение одних суток во
льду или охлажденной морской воде снижает качество сурими главным
образом из-за появления резкого рыбного запаха, ухудшения цвета и
снижения способности фарша образовывать гель.Допустимое время хранения охлажденной тощей рыбы до перера¬
ботки на фарш зависит в основном от посмертных изменений SH- и
SS-групп, а также содержания миофибриллярных белков, особенно мио¬
зина, в мясе. Если содержание миозиновых белков высоко, а число SH-
групп снизилось незначительно, то даже после относительно длитель¬
ного хранения рыбы в охлажденном виде получаемый продукт облада¬
ет хорошей консистенцией и в общем высоким качеством. Например,
камабоко, изготовленное из непромытого мяса тихоокеанской трески,
хранившейся сутки во льду, обладает еще хорошей эластичностью, но
уже только приемлемой консистенцией, камабоко из рыбы, хранившей¬
ся 5 сут, — очень слабой консистенцией и в общем приемлемым качест¬
вом, тогда как камабоко, изготовленное из промытого фарша (сури¬
ми), характеризуется хорошим качеством даже после 9 сут хранения
рыбы во льду.Из тихоокеанского хека, речной камбалы и катрана получают обыч¬
но фарш с низкой гелеобразующей способностью после тепловой обра¬
ботки независимо цт продолжительности хранения рыбы во льду и ме¬89
тода улучшения качества измельченного мяса (с промывкой, без про¬
мывки) . Из мольвы можно получать хороший промытый фарш даже пос¬
ле 10-суточного хранения ее во льду [257]Симидзу [408], сравнивая пригодность девяти видов морских и
пресноводных рыб для производства камабоко, разделил их на виды с
высокой гелеобразующей способностью после тепловой обработки
(консистенция - выше 6 баллов по 10-балльной шкале) и низкой (ниже
5 баллов). После промывки измельченного мяса водой консистенция
камабоко улучшалась у исследованных видов рыб в среднем на 2—
3 балла (табл. 26).Замораживание рыбы перед переработкой на фарш весьма замет¬
но снижает ее технологическую пригодность, особенно если она храни¬
лась довольно длительное время. Даже у рыб с высокой гелеобразую¬
щей способностью это свойство теряется на 50—60 % после 2 нед хране¬
ния в мороженом виде [259] Скорость замораживания не оказывает
столь существенного влияния на снижение технологической пригодности,
как продолжительность и температура хранения. Если эти условия не
отвечают рекомендациям Международного института холода [316],
то быстро развивающиеся физико-химические изменения приводят к
заметному снижению технологической пригодности сырья.Изменения химического состава, как одного из факторов, опреде¬
ляющих технологическую пригодность рыбы, происходят в основном в
скелетных мышцах. Однако нельзя игнорировать и те ткани, которые,
попадая в измельченное мясо, существенно изменяют его химический и
ферментный состав (например, почки, кожа, кровь и др.). Рассматривая
в комплексе технологическую пригодность рыбы для производства
фарша, необходимо иметь в виду следующие факторы: отношение со¬
держания миофибриллярных белков к содержанию белков саркоплаз¬
мы; подверженность мышечных белков денатурации во время хранения
в замороженном виде; соотношение содержания полярных и нейтраль¬
ных липидов; наличие гемопротеидов, присутствующих главным обра¬
зом в красных мышцах, крови и почках; активность тканевых фермен¬
тов, в основном деметилазы, липазы и протеаз.Эти вопросы уже рассматривались в разделах 2 и 3. Ниже изложены
некоторые дополнения.Тощая рыба обладает, как правило, более высокой технологиче¬
ской пригодностью для производства фарша, чем жирная и рыба средней
жирностй после короткого периода хранения в мороженом виде (1—
3 мес). Однако быстро развивающиеся денатурационные изменения в
тощей рыбе могут досрочно привести к снижению технологической при¬
годности фарша прежде всего из-за уменьшения водоудерживающей
способности и ухудшения его консистенции. В фарше, изготовленном
из такой рыбы, содержится большое количество несвязанной жидкой
фракции, а после тепловой обработки он приобретает волокнистую и
твердую консистенцию. Соответствующая защита жирной рыбы или90
Таблица 26. Гелеобразующая способность пасты из промытого (П) и непромытого (Н) мяса
некоторых видов рыб после нагревания до 50 ° С [ 408]Гелео бразу кнцая
способностьВид рыбыСпособобра¬боткиТемпература тепловой обработки пасты50 °С90 °СПрочность
геля* SE,
г/см2Органолептическая
оценка консистенции
по 10-балльной шка¬
ле**Прочность
геля* SE,
г/см2Органолептическая
оценка консистенции
по 10-балльной шка¬
ле**ВысокаяБелый горбыльН57685457Aigyrosomus aigentatusП106496388Scombrops boopsН----П1003107749Рыба-ящерицаН36163576Saurida elongataП81897609СтавридаН45074476Trachurus japonicusП74896448НизкаяКарасьН622631Carassius cuvieriП211531КарпН422241Cuprinus carpioП15651084Кунья акулаН60602Mustelus manazoП1712576* Определена как произведение S-Е, где S - граница прочности геля в тесте по релаксации (г/см2), Е - удлинение разры¬
ва Л L/L.** 9—10 — очень плотная; 7-8 - плотная; 5-6 - средней плотности; 3-4 - крошащаяся, ломкая; 1-2 - очень крошаща¬
яся, ломкая; 0 - рассыпчатая.
рыбы средней жирности от самоокисления липидов, например путем
использования газонепроницаемой упаковки и низкой температуры
хранения, делает эту рыбу равным, а иногда даже лучшим по качеству
сырьем для производства фарша по сравнению с тощей рыбой. Основ¬
ное преимущество жирной и среднежирной рыбы — относительно мед¬
ленное снижение растворимости мышечных белков во время хранения
в мороженом виде. Однако после кратковременного хранения в моро¬
женом виде (например, сразу после замораживания) содержание раст¬
воримых мышечных белков, в основном миофибриллярных, в этой рыбе
меньше, чем в тощей.Содержание миофибриллярных белков в фарше связано прямоли¬
нейной зависимостью с эластичностью и отрицательной линейной зави¬
симостью с количеством отжатого сока в продуктах после их тепловой
обработки (рис. 19), именно поэтому из рыбы с более высоким отно¬
шением миофибриллярных белков к белкам саркоплазмы в мясе
получается более эластичный фарш после тепловой обработки. Из рыбы с
высоким содержанием белков саркоплазмы получаются твердые и ма¬
лоэластичные гели, поскольку эти белки после коагуляции плотно за¬
полняют межволоконное пространство и делают структуру мяса непод¬
вижной [442] Несмотря на то что содержание миофибриллярных
белков почти во всех рыбах одинаково, содержание белков саркоплазмы
значительно колеблется в зависимости от вида. В общем можно принять,
что рыба, ведущая подвижный образ жизни, характеризуется более вы¬
соким содержанием белков саркоплазмы в мясе. Например, процентное
отношение подвергающихся термической коагуляции белков саркоплаз¬
мы к актомиозину возрастает у перечисленных ниже видов рыб в сле¬
дующем порядке: каменный окунь (31,2 %), карась (33,9 %), карп
(42,9 %), заурида (45 %), ставрида (48,9 %), круглая сельдь (52,7 %),
барракуда (60,1 %), скумбрия (73,0%) [410]Заметное влияние фракции растворимых в воде белков на снижение
гелеобразующей способности после тепловой обработки (в пастах типа* 2
8 £и '2О W ** UU DUUБелок, растворимый в 3,75 %-ном растворе
хлористого натрия, мгЫ/100 гРис. 19. Зависимость мевду растворимостью белков промытого фарша (сурими)
и свойствами изготовленного из него камабоко [163]92
камабоко) и на возрастание денатурационных изменений белков во
время замораживания подтверждено сравнительным анализом 12 видов
морских рыб [349] .Жирная рыба характеризуется не только более высоким содержани¬
ем азотистых веществ, растворимых в воде, но и более высокой раст¬
воримостью саркоплазматических белков по мере увеличения ионной
силы раствора, которое наблюдается во время перемешивания измель¬
ченного мяса. Растворимость белков саркоплазмы жирной рыбы (на¬
пример, скумбрии, сардины) логарифмически возрастает с увеличением
ионной силы раствора от 0 до 0,15, тогда как белки саркоплазмы тощей
рыбы (например, минтая) в этом диапазоне ионной силы характеризуют¬
ся меньшим или большим снижением растворимости (рис. 20). Это раз¬
личие объясняется в основном наличием фракции белков саркоплазмы,
элюационная способность которой к = 0,2—0,3 на геле Sephadex G-200
[409].При производстве фарша из мороженой рыбы ее технологическая
пригодность определяется главным образом степенью развития денатура¬
ционных изменений мышечных белков (у тощей рыбы) и степенью
окисления липидов (у жирной рыбы и рыбы средней жирности). Эти
изменения, как известно, зависят в основном от продолжительности
и температуры хранения в мороженом виде, а также специфических
свойств мяса рыбы. Значительное влияние на технологическую пригод¬
ность рыбного сырья оказывает вид изготовляемого фарша: для непро¬
мытого фарша качество сырья важнее, чем для промытого. Вместе с тем
промытый соленый фарш (каэн-сурими, см. раздел 6), приготовленный
из сырья низкого качества, быстрее теряет стабильность при хранении,
чем фарш без добавления хлористого натрия (муэн-сурими) [170]Применяемые методы первичной обработки, а также сепарации
измельченного мяса должны гарантировать максимальное удаление та¬
ких тканей, как почки, кровь, кожа, красное мясо, которые являются
основным источником хромопротеидов гидролитических ферментов.
Даже незначительное количество этих тканей может заметно снизить ка¬
чество и возможный срок хранения измельченного мяса, несмотря на то
что главный компонент фарша (белые мышцы) характеризуется значи¬
тельной стабильностью. Например, в почках рыбы активность демети-
лазы в 300 раз, а щелочной протеиназы в несколько тысяч раз выше,
чем в мышечной ткани [430, 461] Ферменты почки также более устой¬
чивы к действию высокой температуры.Морфометрические характеристики рыбы имеют решающее значе¬
ние при определении ее пригодности для первичной механической обра¬
ботки и выхода измельченного мяса. Крупная рыба веретенообразной
формы требует дополнительного разделения тушки на части перед се¬
парированием мяса, а также удаления передней части позвоночника.
Для рыбы с большим количеством различных костистых образований
на коже (жучек) необходимо применение сепараторов с меньшим диа-93
Растворимость белков,100SardinopsmelanostictaV-I I «10050010050EopsettagrigorijewiSauridaundosquamis_i i iScomberjaponicusBranchiostegusjaponicusCarassiuslongsdorfii0,1 0,2 0,3
Ионная силаCoryphaenahippurusNemip terns
virgatusTheragracholcogrammal i »0,1 0,2 0,3Рис. 20. Влияние ионной силы раствора на растворимость белков саркоплазмы мяса
18 видов костистых рыб [ 409 ]
метром отверстий в барабане или дополнительных устройств для очист¬
ки измельченного мяса (стрейнеров). Эти вопросы более подробно рас¬
смотрены в разделе 4.6.Важной проблемой является наличие паразитов в некоторых видах
рыбы, служащих сырьем для изготовления фарша (например, путассу
и тихоокеанский хек). Большое количество паразитов, даже безвредных
для здоровья, может сделать рыбу непригодной для переработки на
фарш. Это особенно относится к паразитам, визуально обнаруживаемым
в мясе рыбы, например спорообразующие микроорганизмы рода Kudoa,
которые образуют характерные цисты. Мышечная ткань, сильно заражен¬
ная этими паразитами, приобретает мажущуюся консистенцию. Проте¬
кание процессов ферментативного разложения тканей обусловлено дей¬
ствием протеолитических ферментов Kudoa, а также собственных проте-
олитических ферментов и лизосомных ферментов рыбы [496] . Если па¬
разиты сосредоточены в органах, не используемых в пищу (например,
внутренности), а также в теше, что характерно для нематод родов Ani-
sakis и Terranova, при разделке рыбы тешу необходимо удалять [273]
Наиболее выгодный с точки зрения технологии вид разделки рыбы
перед переработкой на фарш — тушка без теши и переднего отрезка по¬
звоночника (от плечевого пояса до анального отверстия).4.5. ПЕРВИЧНАЯ ОБРАБОТКА РЫБЫ И ЕЕ ВЛИЯНИЕ
НА КАЧЕСТВО ФАРШАДайер [92] выделил пять типов сырого фарша в зависимости от ви¬
да первичной обработки рыбы: фарш из потрошеной обезглавленной
рыбы с удаленной брюшной пленкой; фарш из обезглавленной потро¬
шеной рыбы без удаления брюшной пленки; фарш из отходов от фи-
летирования рыбы так называемым резом 2V; фарш из отборных от¬
ходов от филетирования со шкурой - теши и др.; фарш из очень мелкой
рыбы или целых ракообразных.К этим типам фарша следует добавить еще фарш из филе или теши,
а также фарш из обезглавленной потрошеной рыбы с удалением части
позвоночника одновременно с почками и плавательным пузырем.Такое разнообразие типов фарша в зависимости от вида первичной
обработки приводит к тому, что качество и срок возможного хранения
конечного продукта могут довольно значительно варьировать, даже если
фарш изготовлен из одной и той же рыбы. Особенно это относится к не¬
промытому фаршу.Самый лучший фарш можно получить из филе с удаленной брюш¬
ной пленкой. Однако направлять такое сырье на фарш нецелесообразно,
поскольку само по себе филе пользуется большим спросом, поэтому ос¬
новным сырьем для производства товарного рыбного фарша следует
считать тушки и некоторые отходы от филетирования.Степень зачистки тушки оказывает большое влияние на качество95
и технологическую пригодность фарша. В общем можно принять, что
более тщательная зачистка тушки перед переработкой снижает интен¬
сивность неблагоприятных изменений в мороженом фарше. Однако этот
эффект по-разному проявляется для тощей и жирной рыбы. В фарше из
тощей рыбы, например трески, изготовленном из тушки с удаленной
передней частью позвоночника одновременно с почками и частью аорты,
увеличение содержания муравьиного альдегида (МА) и снижение раст¬
воримости белков в процессе хранения происходит значительно мед¬
леннее. Однако окисление жира, по-видимому, не зависит от степени
зачистки тушки. В фарше из жирной рыбы зачистка тушки заметно
замедляет процесс окисления липидов и уменьшает степень их гидро¬
литических изменений, но на растворимость мышечных белков зачист¬
ка тушки существенного влияния не оказывает (Колаковский и др.,
1982). Таким образом, удаление почек, сгустков артериальной крови и
брюшной пленки предохраняет фарш из тощей рыбы прежде всего от
быстрого снижения растворимости белков, уменьшения его эмульги¬
рующей и стабилизирующей способности, а фарш из жирной рыбы -
от быстрого сокращения срока возможного хранения в результате
окисления жира.Один из весьма эффективных способов разделки - вырезание пе¬
редней части позвоночника от головы до анального отверстия. Полу¬
ченный в результате такой разделки продукт представляет собой час¬
тично распластанную тушку с удаленной передней частью позвоночника
вместе с костистыми отростками, почками, плавательным пузырем и
частью спинной аорты. Такую разделку производят филетирующие
машины фирмы ’’Баадер” (например, Баадер 33) или соответствующие
приставки (Баадер 82) [102] С помощью рыборазделочных машин
производственного объединения ’’Дальмор” типа DP2 можно получить
тушку с подрезанными тешами — так называемый ’’американский”
рез. Такой способ разделки может быть рекомендован, если теша зара¬
жена паразитами [88]К недостаткам описанных выше способов первичной обработки
рыбы относится то, что в отходах, особенно при позвоночнике, оста¬
ется значительное количество мышечной ткани, поэтому более рацио¬
нальными являются такие способы обработки, которые обеспечивают
получение соответствующим образом зачищенной тушки без выреза¬
ния переднего отрезка позвоночника. На рис. 21 представлена схема
машины фирмы ’’ТРИО Инжиниринг”, предназначенной для обработки
неразделанной или потрошеной рыбы для получения тушки с удалени¬
ем почки, плавательного пузыря и брюшной пленки без частичного
удаления позвоночника или теши. Высокая степень зачисткет тушки
достигается благодаря применению в машине специальных устройств
для дочистки, а также регулированием направления струй воды. Маши¬
на специально предназначена для обработки трески массой 300—1500 г
и хека массой 150—500 г с производительностью соответственно 30—5096
laшнпн-н2a 2За 4%-$-ф4а 5Рис. 21. Схема установки фирмы ”ТРИО Инжиниринг А/S” для переработки нераз-
деланной или потрошеной рыбы на тушки с удалением почки, плавательного пузы¬
ря и брюшной пленки с оставлением позвоночника и теши:А — машина для обезглавливания и потрошения; Б — машина для зачистки тушки;
1 — головоотсекающее устройство; 1а - подающее устройство; 2 - нож, разрезаю¬
щий брюшную полость; 2а - вспомогательный стационарный нож; За - комплект
щеток для зачистки тушки снаружи; 3, 4, 5 - набор фрез для зачистки брюшной
полости; 4а, 6а - сопла для подачи струй воды под давлением; 6 - нож для разре¬
зания хвостового стебляи около 150 рыб в минуту. Расход воды составляет около 200 л/мин.В США фирма ”Jla Пайн Б рос” (г. Гладстон) выпускают машину
(Модель 22) для обезглавливания и потрошения мелкого хека (массой
63—380 г), предназначенного для производства фарша [314] . Модель 22
позволяет при относительно высокой производительности (около
60 рыб в минуту) перерабатывать хек на тушки, распластанные в хвосто¬
вой части с полным удалением внутренностей, брюшной пленки и плава¬
тельного пузыря. Выход фарша из полученного таким образом полуфаб¬
риката высокий, при обработке хека массой 200—250 г он достигает
50 % целой рыбы (табл. 27).Для мелкой рыбы (килька, анчоус, шпрот и др.) стандартная меха¬
ническая обработка может оказаться нерентабельной, поэтому все чаще
применяют упрощенные методы обработки рыбы (особенно малоцен-97
Таблица 27. Пооперационный выход рыбы при переработке хека
(Merluccius bilinearis) на машине фирмы ”JIa Пайн” (Модель 22)
и сепараторе ’’Бибун” в зависимости от размера [314]ПродуктВыход (в %) в зависимости
от массы рыбы, гСредний выход, %ДО125125-190190-250свыше250от целой
рыбыот обезглав¬
ленной рыбыНеразделанная рыба100100Обезглавленная ры¬70,070,5баПолностью зачищен¬49,852,5ная тушкаФарш43,447,8100100100-70,568,569,910053,751,151,974,350,340,045,465,0ной) перед изготовлением фарша. Например, способ, предложенный
Скачковым [416] , предусматривает разрезание промытой неразделан-
ной мелкой рыбы на кусочки длиной около 1 см и нагревание получен¬
ной массы паром при температуре 100-105 °С в течение 0,5 с или в во¬
де при температуре 90-100 °С в течение 1 с. Процесс ошпаривания не¬
обходим для лучшего отделения внутренностей и кожи на последую¬
щих этапах обработки. Кроме того, это позволяет удалить сгустки кро¬
ви, а также частично извлечь подкожный и мышечный жир. Во время об¬
работки содержание подкожного жира снижается примерно на 30 % для
анчоуса и более чем на 10 % для кильки.Немедленно после тепловой обработки кусочки рыбы охлаждают и
подвергают интенсивному перемешиванию с водой для удаления внут¬
ренностей, сгустков крови, чешуи и частично кожи. Промытые кусочки
рыбы направляют на центрифугирование или прессование для удаления
избытка воды и далее на сепаратор. Описанный способ имеет опреде*-
ленные недостатки, к которым относятся потеря белковых веществ во
время обработки, а также возможность загрязнения фарша бактериями
пищеварительного тракта рыбы.4.6. СЕПАРИРОВАНИЕ МЯСА4.6.1. Сепараторы и способы сепарированияОтделение (одновременно с частичным измельчением) мышечной
ткани рыбы от несъедобных твердых частей производится с помощью
сепараторов, которые раньше называли устройствами для отделения
костей. Среди более десятка моделей сепараторов, выпускаемых раз¬
личными зарубежными фирмами [188, 192, 210, 211] , наиболее пригод¬
ными для переработки рыбы на фарш оказались барабанные и шнековые
установки, которые лучше отвечают требованиям, предъявляемым к се-
98
параторам для производства фарша из рыбы. Эти требования заключают¬
ся в следующем [192,243]возможно меньшее разрушение первоначальной структуры мышеч¬
ной ткани в процессе ее отделения от костей, кожи, чешуи и других
твердых частей; максимальное отделение мышечной ткани от несъедоб¬
ных частей рыбы (отходов); высокая производительность; возмож¬
ность быстрого демонтажа с целью очистки, мойки и консервации; под¬
держание возможно более равномерной температуры сырья в процессе
переработки.Принцип действия барабанных сепараторов заключается в продав-
ливании мягких и очень пластичных тканей рыбы внутрь вращающегося
перфорированного барабана под давлением резиновой ленты, частично
охватывающей барабан (рис. 22). Поскольку края отверстий барабанаРис. 22. Схема барабанного сепаратора:1 - перфорированный барабан; 2 - разравнивающий ролик; 3 - натяжной ролик;
4 - прижимные ролики; 5 - лента транспортера; 6 - регулирующий роли с; 7 -скребок; 8 - отходы99
острые, а прижимающее усилие ленты возрастает постепенно, можно
говорить не только о раздавливании и проталкивании, но и о частичном
вырезании кусочков мышечной ткани с более или менее сохранившейся
первичной структурой. Эти кусочки имеют форму, близкую к цилиндру,
а их длина зависит от диаметра отверстий в барабане. Таким образом,
сырье, которое оказывается между барабаном и лентой, разделяется на
две фракции: измельченное мясо проходит через перфорацию внутрь ба¬
рабана, а твердые части (отходы) остаются на его поверхности. Измель¬
ченное мясо удаляется из барабана вмонтированным в него неподвиж¬
ным шнеком, а отходы снимаются с поверхности скребком. Поскольку
в результате трения рабочих элементов их температура может возрас¬
тать, в некоторых сепараторах на резиновую ленту позади барабана на¬
правляется струя воды, что, помимо охлаждения, облегчает очистку
ленты. При подаче крупных тушек или недостаточно размороженной
рыбы в сепараторе может произойти проскальзывание сырья, что вызы¬
вает уменьшение пропускной способности установки. Установка снаб¬
жена специальным подающим валиком, который благодаря клиновид¬
ным выступам препятствует проскальзыванию рыбы во время сепара¬
ции и, кроме того, регулирует толщину слоя сырья.К барабанным сепараторам можно отнести также установки типа
’’Паоли”, обычно работающие в комплекте с волчком, в котором про¬
исходит раздавливание сырья перед сепарированием, поэтому в рыбной
промышленности такие сепараторы находят ограниченное применение.Шнековые сепараторы изготовляются на базе шнекового пресса,
в котором сжатие сырья происходит в результате уменьшения простран¬
ства между перфорированным кожухом и шнеком (рис. 23). Вращаю¬
щийся шнек вызывает перемещение сырья вдоль рабочей части установ¬
ки с одновременным выдавливанием частиц мяса наружу. Отходы, ко¬
торые не смогли пройти через отверстия решетки (кожуха), выталки¬
ваются сквозь концевое отверстие, закрытое подпружиненной задвиж-100Рис. 23. Схема сепаратора шнекового типа
Рис. 24. Схема установки для дополнительной очистки фарша (стрейнер)кой. Установки такой конструкции применяются для переработки на
фарш сырья, предварительно измельченного на волчке (рис. 24), или ча¬
ще для более полной очистки фарша как вторичные сепараторы (стрей-
неры). Диаметр отверстий в перфорированном кожухе шнековых сепа¬
раторов обычно составляет 1,2—2,6 мм.4.6.2. Влияние величины давления ленты барабана
на выход мяса и содержание в нем костейВеличина давления ленты на барабан заметно влияет на содержание
костей в измельченном мясе, при этом большое значение имеют вид
рыбы и длина тушки: крупные тушки дают мясо с меньшим содержа¬
нием костей, чем мелкие.Вопреки предположениям зависимость между величиной давления
ленты на барабан и выходом мяса не является линейной. Вначале уве¬
личение давления до 3—4 (по 7-балльной шкале) вызывает экспонен¬
циальное увеличение выхода мяса (чем мельче тушка, тем больше вы¬
ход) . Самый высокий выход достигается при давлении, равном 4—5 бал¬
лам независимо от вида рыбы (рис. 25). Увеличение давления ленты от
4—5 до 7. баллов вызывает резкое увеличение содержания костей в мясе.
Так, при максимальном давлении это содержание оказывается в 2—4 ра¬
за выше, чем при оптимальном (рис. 26). Это увеличение происходит в
основном за счет самых мелких костей (0— 1 мм). Можно предположить,
что под действием высокого давления ленты отламываются части костей.
Чрезмерное давление ленты на барабан во время сепарации вызывает
также переход в фарш кусочков кожи и практически всего количества
подкожных мышц, что в конечном счете приводит к ухудшению цвета
фарша. В случае жирной рыбы, наблюдается, кроме того, незначительное
уменьшение содержания воды и равномерное увеличение содержания
липидов в фарше, вероятно, в результате перехода в него большей час¬
ти подкожного жира. В фарше из тощей рыбы существенного различия101
Давление ленты на барабан, баллыРис. 25. Зависимость выхода мяса из тушек рыбы от давления ленты на барабан
сепаратора. Давление измеряется по 7-балльной шкале: 7 баллов - максимальное
давление; 1 балл - минимальное давление. Давление регулируется поворотом
ручки, регулирующей натяжение ленты:1 - путассу (длина 20 см, масса 62,5 г); 2 - ставрида крупная (длина 22 см,
масса 125 г); 3 - ставрида мелкая (длина 15 см, масса 52,6 г*Давление ленты на i
1Рис. 26. Зависимость содер¬
жания костей в измельчен¬
ном мясе от давления ленты
на барабан:а - длина кости 3 мм; б -
длина костей < 3 мм: 1 -
путассу (длина 20 см, мас¬
са 62,5 г); 2 - ставрида
мелкая (длина 16 см, масса
52,6 г)
химического состава в зависимости от величины давления ленты на
барабан не наблюдается. В процессе изготовления фарша из тушки оп¬
тимальная величина давления ленты на барабан для мелкой рыбы долж¬
на составлять 3—4, а для крупной — 4—5 баллов по 7-балльной шкале
(на примере DX16).Выход фарша зависит от таких факторов, как форма рыбы, доля
мышц, наличие жучек на коже, способ хранения сырья и др. Рыбы вере¬
тенообразной формы с гладкой поверхностью, например хек, мерланг,
путассу и другие, обычно дают высокий (более 80 % массы тушки) вы¬
ход фарша (табл. 28). Меньший выход фарша характерен для костля¬
вых, имеющих мало мышц, жучки на коже рыб (ставрида и морской
окунь). Установлено, что из размороженной рыбы выход фарша боль¬
ше (табл. 29), чем из охлажденной (см. табл. 28). Это объясняется,
вероятно, более легким отделением несъедобных частей от разморо¬
женного мяса, первичная структура которого нарушена заморажива¬
нием.Дополнительное влияние на увеличение выхода 'фарша из мороже¬
ного мяса рыбы может оказывать также поглощение мясом воды во вре¬
мя размораживания струями воды. Большое значение имеет температу¬
ра сырья во время сепарирования. Очень низкая температура (ниже— 1 С) вызывает проскальзывание рыбы между барабаном и лентой,
что приводит к неполному отделению съедобных частей. Вместе с тем
слишком высокая температура может быть причиной значительныхТаблица 28. Выход фарша из рыбы, охлажденной льдом [ 209]4.6.3. Влияние вида рыбы и способа
хранения сырья на выход фаршаРыбаЛатинское названиеВыход, %Тихоокеанский хек
Мерланг
Красный хек
Короткоиглый морской
окуньТихоокеанская треска
ТерпугМорской окуньБелый горбыльТихоокеанская сельдьАнчоусСкумбрияКамбалаТихоокеанская речная кам¬
балаMerluccius productus
Merluccius dilineares
Urophycis chuss
Sebastes brevispinis88,486,083,080,6Gadus macrocephalus
Ophiodon elongatus
Sebastes marinus
Micropogon endulatus
Clupea harengus pallasi
Engraulis mordax
Scomber scombrus
Parophrys vetul
Platichthys stellatus69.477.069.475.069.592.665.084.1
79,4103
Таблица 29. Выход фарша из размороженных тушеки филе рыбы [ 226]РыбаВыход фарша (в %)из тушкииз филеСкумбрия90,693,2(90,3-91,1)(92,4-95,2)Сардина—95,1(93,8-96,2)Ставрида81,1(80,7-82,3)Минтай82,9(81,8-83,4)-Хек86,591,8(83,1-88,5)(91,5-92,3)потерь тканевого сока во время сепарирования мяса. Максимальный
выход фарша получают из рыбы, размороженной до температуры
—0,5-г1 °С. При этом получается фарш с относительно низкой темпе¬
ратурой, так как во время сепарации температура продукта повышает¬
ся всего на 1 °С.
5. СЫРЬЕ И ВСПОМОГАТЕЛЬНЫЕ МАТЕРИАЛЫ5.1. ДОБАВКИДобавки чаще всего применяются в качестве стабилизаторов из¬
мельченного мяса рыбы, реже — как вкусоароматические, связующие
вещества и наполнители. Иногда эти функции выполняются одновре¬
менно. Добавки вносятся как в сырое измельченное мясо (например,
стабилизированный фарш), так и в мясо, подвергнутое технологиче¬
ской обработке, — промытое, вареное и т. п.В качестве добавок в основном применяют олигосахара, полифос¬
фаты, глутамат натрия, антиокислители и натуральные растительные
приправы. Из других добавок следует упомянуть некоторые неоргани¬
ческие соли, гидроколлоиды и эмульгаторы.5.1.1. СахараВ рыбных фаршах сахара применяются главным образом как добав¬
ки, предотвращающие денатурацию миофибриллярных белков в про¬
цессе замораживания. В литературе можно встретить упоминания об
антиокислительном действии как самих сахаров [262] , так и продук¬
тов их взаимодействия с аминокислотами [ 166] или пептидами [278].Механизм защитного действия сахаров по отношению к белкам изу¬
чен не полностью, хотя известно, что действие их зависит главным обра¬
зом от содержания в молекуле групп ОН”, молекулярной массы и
пространственной структуры молекулы сахара [147,158, 367] . Наиболь¬
шее значение среди упомянутых факторов приписывают гигроскопич¬
ности сахаров, обусловленной наличием многочисленных групп ОН“,
благодаря которым они могут соединяться с боковыми цепями моле¬
кул белка, увеличивая гидратацию. Образующиеся таким образом
усиленные и упорядоченные водные оболочки стабилизируют строение
белков и препятствуют взаимодействию боковых цепей во время вы¬
мораживания воды [150] , которое Коннелл [64] считает основной
причиной денатурации белков мяса рыбы во время замораживания.
Антиденатурационное действие сахаров объясняют также тем, что в
процессе замораживания воды прекращается миграция ионов в мясе.
Увеличение концентрация сахаров в растворе вызывает снижение элект-105
ропроводности. При концентрации сахаров 8—10 % миграция ионов
электролита замедляется настолько, что актомиозин мяса рыбы, поме¬
щенного в такой раствор, во время хранения при температуре —20 °С
практически не подвергается денатурации [347].Антиденатурационное действие сахаров в мороженом рыбном фар¬
ше зависит главным образом от его pH, концентрации и вида добавляе¬
мого сахара. Наиболее сильное действие сахаров обнаруживается в нейт¬
ральной среде. Изменение pH фарша на единицу в кислотном направле¬
нии вызывает большее снижение антиденатурационной активности са¬
хара, чем такое же изменение pH в щелочную сторону [18].Антиденатурационная активность сахаров возрастает логарифми¬
чески до равновесного состояния системы сахар—актомиозин, при
этом наиболее эффективно в диапазоне 0—5 %, хотя для полного пре¬
кращения денатурации актомиозина требуется концентрация сахара
в фарше, в 20 раз превышающая концентрацию в нем актомиозина
[16, 18]. Для практических целей можно принять, что добавление са¬
харов в фарш в количестве, троекратно превышающем содержание
актомиозина, значительно замедляет его денатурацию во время хране¬
ния мороженого фарша [17], но добавление сахаров в количестве
0,3 % содержания актомиозина существенно не влияет на процесс де¬
натурации.Оптимальная концентрация сахаров в фарше достаточно высока
и составляет 3—10 % в зависимости от вида фарша и сахара. Так, в мо¬
роженом фарше сурими она составляет 8—10 %, в мороженом муэн-су-
рими — около 8 % [352] , в фарше, предназначенном для производст¬
ва камабоко (перемешанном с 2,5 % поваренной соли), — около 10 %
[157], а в фарше сублимационной сушки — 3—10 % сырой массы пе¬
ред замораживанием [301]. В фарше, в который добавлялись элект¬
ролиты, которые могут оказывать денатурирующее действие на мы¬
шечные белки во время замораживания (например, поваренная соль,
полифосфаты и т. п.), количество добавленных сахаров должно быть
соответственно выше, тогда как в промытом фарше, освобожденном
от большинства электролитов, оно может быть более низким [18,125]В зависимости от эффективности замедления денатурации мышеч¬
ных белков рыбы сахара можно расположить следующим образом:
трисахариды > дисахариды > моносахариды [185] . Из моносахаридов
наиболее сильное действие оказывают глюкоза, фруктоза, манноза, га¬
лактоза, а самое слабое и даже негативное — рибоза и ксилоза. Из диса¬
харидов явно антиденатурирующее действие оказывают сахароза, лак¬
тоза, трегалоза и мелибиоза. Активность трисахаридов (например,
рафинозы) примерно в 2—3 раза выше, чем дисахаридов (сахарозы).
Заметного различия между редуцирующими и нередуцирующими саха¬
рами не установлено. Спиртосахара обычно обладают несколько более
слабым защитным действием, чем соответствующие им сахара, при этом
эффективность циклических спиртосахаров (например, инозита) сход-106
на с эффективностью алифатических сахаров (например, сорбата) и в
обычных случаях увеличивается пропорционально размеру молекулы
[185, 302]. Это подтверждает вывод о том, что антиденатурационная
эффективность сахаров или спиртосахаров зависит в основном от содер¬
жания групп ОН“ и массы молекул.5.1.2. ПолифосфатыПолифосфаты известны главным образом как вещества, увеличи¬
вающие водоудерживающую способность белков мяса убойного скота
и рыбы [122, 134, 328] . Они также оказывают антиокислительное дей¬
ствие [39, 381], задерживают денатурацию миофибриллярных белков
мяса рыбы во время замораживания, особенно повторного [279, 288,
358] , улучшают структурно-механические свойства рыбного фарша
[50] и даже замедляют прорастание спор некоторых микроорганиз¬
мов [479] .Такое широкое действие полифосфатов, часто определяемое как
фосфатный эффект, обусловлено влиянием этих добавок на диссоциа¬
цию актомиозина, растворимость миофибриллярных белков, набухание
белков, оставшихся в нерастворимой фракции, связыванием ионов
двухвалентных металлов и др. [229] Действие полифосфатов зависит
от вида использованного фосфата и его содержания в фарше. Для рыб¬
ной промышленности наиболее пригодны тринатрийфосфат (Na3P04),
пирофосфат натрия (Na4P207) и калия (К4Р207), а также стекловид¬
ный метафосфат натрия. С точки зрения взаимодополнения свойств
этих солей рекомендуется применять полифосфаты в виде соответст¬
вующих смесей, например смеси Хамина*.Оптимальное количество полифосфатов, вносимых в измельченное
мясо рыбы, составляет 0,15—0,30, хотя способность связывать воду в
фарше обнаруживается также в пределах более высокой концентрации
(примерно до 1 %), а максимум антиокислительной активности поли¬
фосфатов приходится скорее на более низкие концентрации (0,02-
ОД %). Избыток полифосфатов не рекомендуется с токсикологиче¬
ской точки зрения. Показано, что при содержании полифосфатов в ко¬
личестве 1 % у молодых крыс обнаружено патологическое изменение
почек [73]. Избыточное содержание полифосфатов в пищевых про¬
дуктах может вызывать нарушения минерального состава вследствие
связывания кальция, железа, меди и магния. Допустимая суточная
доза (ДСД) фосфора для человека составляет 70 мг на 1 кг массы те¬
ла [339]* Препарат Хамина голландского производства имеет следующий состав:
натрийфосфат щелочной - 29,4 %, пирофосфат натрия - 4,6 %, более конденсиро¬
ванные фосфаты - 23,9 %, pH 1 %-ного раствора Хамина составляет 8,4.107
5.1.3. Глутамат натрияСчитается, что это вполне безопасная с точки зрения токсикологии
добавка [96] В технологии продуктов морского происхождения при¬
меняется глутамат натрия в основном как стимулятор вкусовых свойств
рыбных изделий и измельченного мяса рыбы [229] Антиденатурацион-
ная активность глутамата натрия в отношении миофибриллярных белков
мяса рыбы [350, 486] позволила использовать глутамат в качестве ан-
тиденатуранта и в технологии рыбного фарша [352] . Для получения
заметного антиденатурационного эффекта содержание глутамата натрия
в рыбном фарше должно составлять примерно 0,42 % [350] , хотя для
измельченного мяса карпа сублимационной сушки удовлетворитель¬
ные результаты получены при концентрации 3 % по сырой массе перед
замораживанием [300]В модельных исследованиях, проведенных на растворе актомиози¬
на, выделенного из мяса рыбы (карп, ставрида), антиденатурационная
активность глутамата натрия при концентрации 0,3 моль/л была по мень¬
шей мере столь же высока, как и активность глюкозы при концентрации
1 моль/л [350] .5.1.4. АнтиокислителиПрименение антиокислителей особенно необходимо для фарша из
жирной рыбы. Наиболее эффективны синтетические антиокислители —
бутилгидроксианизол (БОА), бутилгидрокситолуол (БОТ) пропил-
галлат, этилендиаминтетрауксусная кислота (ЭДТК) и их производные.Согласно теории отрицательного катализа эти соединения обладают
редуцирующими свойствами (разложение перекисей жира) и, кроме
того, способны восстанавливаться, благодаря чему даже в малой кон¬
центрации (например, 0,01 %) могут в течение нескольких месяцев ус¬
пешно предотвращать окисление жира в мороженом рыбном фарше.Оптимальная концентрация синтетических антиокислителей в моро¬
женом рыбном фарше составляет 0,01—0,05 %, хотя достаточная актив¬
ность достигается и при более низких концентрациях. Показано [458] ,
что добавление 0,06 % БОА или БОТ позволяет увеличить срок хранения
мороженого фарша из морского окуня при температуре —18 °С с 3 до
9-12 мес.Польское законодательство, касающееся пищевых продуктов, не
позволяет применять синтетические антиокислители при хранении рыб¬
ных продуктов. Единственными допущенными стандартами веществами
антиокислительного действия являются а-токоферол и аскорбиновая
кислота, применение которых требует, однако, разрешения органов
здравоохранения.Поскольку антиокислительная активность токоферола и токотрие-
нола выше, чем активность а-токоферола [402, 467] , токоферол чаще108
всего применяется в виде концентрата смеси токоферолов. Эффектив¬
ная концентрация токоферола для мороженого рыбного фарша состав¬
ляет 0,02—0,05 % [221] , а для глазури — 0,15-0,05 % [451] Недостат¬
ком токоферола как добавки является трудность его введения в моро¬
женый рыбный фарш, так как он растворяется только в жире и спирте.
Таких проблем не возникает в случае применения аскорбиновой кисло¬
ты, характеризующейся хорошей растворимостью в воде (1 г кислоты в
3,5 см3 воды и 1 г ее натриевой соли в 2 см3). Аскорбиновая кислота
не относится к антиокислителям, скорее это синергическое вещество,
а ее антиокислительная активность объясняется сильным редуцирующим
действием. Обнаружено, что для фарща из жирной рыбы добавление
0,05 % аскорбиновой кислоты оказалось более эффективным, чем вне¬
сение 0,02 % а-токоферола [235] Индукционный период жира с до¬
бавлением этих антиокислителей составлял соответственно 6 и 5 мес,
тогда как в контрольной пробе фарша (без добавок) - всего 1,5 мес.
Подтверждена также способность натриевой соли D-изомера аскорбино¬
вой кислоты увеличивать срок хранения фарша из чукучана Catostomus
sp., хранившегося при температуре -4 °С [161] .Из других веществ, обладающих антиокислительными свойствами,
все большее значение приобретают естественные антиокислители, при¬
меняемые в виде различных концентратов и препаратов растительного
и животного происхождения. Эти добавки описаны в книге Э. Колаков-
ского ’’Технология рыбных продуктов” (Варшава, 1984).5.1.5. Натуральные приправыПриправы, широко известные как вкусоароматические добавки,
при соответствующем подборе могут играть в рыбном фарше также роль
антиокислителей [237] В зависимости от антиокислительной актив¬
ности (на примере мороженого рыбного фарша из балтийской сельди,
хранившегося при температуре —20 °С), приправы можно разделить
на три группы [230, 238] приправы с выраженным антиокислитель-
ным действием, например свежий и сушеный чеснок, зеленый и черный
перец, можжевельник, тмин, розмарин, гвоздика и др.; приправы со сла¬
бым антиокислительным действием - молотый красный перец, мускат¬
ный орех; приправы с проокислительным действием, например лавро¬
вый лист, майоран, кориандр.Антиокислительная активность приправ зависит в основном от сос¬
тава и содержания присутствующих в них фенольных соединений, в ос¬
новном флавоноидов, кислот и других веществ [60] Содержание
эфирных масел существенно не влияет на антиокислительную актив¬
ность приправ, а иногда эти вещества действуют как проокислители
[368] Антиокислительная активность приправ, как и большинства
фенольных антиокислителей, зависит от их количества. Высокие (вы¬
ше 0,4 %) концентрации приправ неэффективны и даже могут спо¬109
собствовать окислению. Наиболее эффективное возрастание антиокис-
лительной активности наблюдается по мере увеличения количества при¬
правы с 0,05 до 0,3 % [239] При составлении смесей из нескольких
приправ, которые по отношению друг к другу могут оказывать синер¬
гическое действие, оптимальное содержание каждой приправы состав¬
ляет примерно 0,1 % массы фарша [221] Натуральные приправы (на¬
пример, кверцетин) оказывают заметное синергическое действие на ас-’
корбиновую кислоту, токоферол и флавины [219]Нагревание молотых натуральных приправ при температуре 110—
160 °С повышает их антиокислительную активность, поэтому для ста¬
билизации мороженого рыбного фарша применяют стерилизованные
приправы [220]. Исследования показали, что сушка черного перца
(влажность 12 %) в течение 40-60 мин сухим горячим воздухом при
температуре 110, 135 и 165 °С позволяет значительно снизить количе¬
ство спор бактерий (на 99,8 %). Дополнительное искусственное увлаж¬
нение черного перца до влажности 24,5 % значительно ускоряет уничто¬
жение спор (снижение численности на 99,9 % после нагревания в тече¬
ние 20 мин при температуре 160 °С), что, однако, вызывает потерю ха¬
рактерного аромата перца [183]Молотые натуральные приправы обладают значительной способ¬
ностью к набуханию и связыванию воды (1—2 г на 1 г приправы) [118] ,
благодаря чему могут повышать водоудерживающую способность
фарша.5.1.6. Гидролизаты, изоляты
и белковые концентратыПрименяются главным образом как средства, повышающие способ¬
ность фарша связывать воду, естественные антиокислители или как ве¬
щества, препятствующие взаимодействию (синерезису и агрегации)
коллоидных структур. Примером гидролизата желатина является пре¬
парат Addi-Fro 71, предложенный шведской фирмой ’’Эстрако АБ”.
Его добавляют в фарш в количестве 3—6 %. В Институте технологии про¬
дуктов морского происхождения Сельскохозяйственной академии в
Щецине установлено, что антиокислительная активность гидролизатов
желатина в значительной степени зависит от молекулярной массы и сос¬
тава пептидов. Наиболее эффективны в качестве антиокислителей препа¬
раты желатина, гидролизованного в растворе уксусной кислоты концент¬
рацией 1 моль/л в течение 90 мин при слабом кипении [418] . Не исклю¬
чено, что гидролизаты желатина оказывают также антиденатурационное
действие и замедляют протеолитические процессы в мороженом рыбном
фарше, однако это требует подтверждения.Из изолятов и белковых концентратов чаще всего применяют расти¬
тельные препараты (Promin D, Promic Р-6), соевую муку, клейковину,
реже препараты животного происхождения (казеин, порошковое моло¬110
ко и др.). В невысокой концентрации (до 3 %) эти препараты могут быть
использованы для улучшения реологических свойств фарша, главным
образом сырого непромытого, а также вареного и соленого. Однако при
более высокой концентрации (4—10 %) большинство белковых препара¬
тов вызывает изменение органолептических свойств фарша, в основном
вкуса и запаха [3, 61, 456] В промытом фарше (сурими) добавление
свыше 3 % изолята сои (Promic Р-6) вызывает заметное ухудшение не
только органолептических свойств, но и снижение технологической при¬
годности. Камабоко, изготовленное из такого фарша, характеризуется
значительной потерей сока под давлением и сниженной прочностью
геля [19]Влияние добавки белковых препаратов на консистенцию в значи¬
тельной степени зависит от их воздействия на pH фарша. Щелочные бел¬
ки несколько улучшают, а кислые белки заметно снижают способность
фарша образовывать эластичный гель после тепловой обработки [504]5.1.7. ПолисахаридыПолисахариды как высокомолекулярные соединения весьма гид¬
рофильны, имеют характерную структуру и могут быть использованы
в технологии рыбного фарша в основном как средства, связывающие
свободную воду, импрегнирующие (пропитывающие), препятствующие
синерезису белковых соединений [229] Кроме того, полисахариды
способствуют формированию требуемой консистенции фарша, особен¬
но если технологический процесс предусматривает тепловую обработку
(вареный фарш), поскольку они устойчивы при хранении и к действию
высокой температуры. Более того, их способность связывать воду под
влиянием нагревания увеличивается.К важнейшим полисахаридам, применяемым в качестве добавки
к рыбному фаршу, относятся: альгинат натрия [189] , натриевый глици-
нат целлюлозы [338], амилопектин [445] , карбоксиметилцеллюлоза
[3], каррагенин [329] и камеди - главным образом рожкового дере¬
ва и бобов рожкового дерева [229] Оптимальная концентрация этих
добавок относительно низкая и не превышает 0,5 %, а часто составляет
менее 0,3 %. Передозировка полисахаридов может вызвать нежелатель¬
ные изменения фарша, способствовать созданию липкой и мажущейся
консистенции. Например, в мороженом фарше из ставриды такие явле¬
ния наблюдаются при добавке 0,3 % альгината натрия* [221] В качест¬
ве импрегнирующего средства полисахариды могут действовать как ан¬
тиокислители липидов мороженого рыбного фарша [482]* Protanal SF-120, изготовляемый норвежской фирмой ”Протан и Фаген-
тун А/С” в Драммене.111
5.1.8. ЭмульгаторыЭто поверхностно-активные вещества, способствующие дисперги¬
рованию и эмульгированию жира, повышающие липкость и замедляющие
денатурацию белков. Вносятся в основном в фарш, предназначенный для
изготовления рыбной ветчины и колбас [9, 158, 184]Из синтетических эмульгаторов чаще всего используют продукты
обезвоживания спиртов с шестью гидроксильными группами (сорби-
ты)*, соединенные эфирными связями с жирными кислотами или
окисью этилена (торговые названия Span, Tween, Arlacel [70]). Эмуль¬
гаторы типа Tween являются полисорбатами, в которых количество
окиси этилена строго определено, например Полисорбат 80 (полиокси-
этилен-20-монооцейнат** сорбита) и Полисорбат 60 (полиоксиэтилен-
-20-моностеарат сорбита) и др.Несмотря на очень ценные эмульгирующие свойства, эти соединения
не совсем безопасны с токсикологической точки зрения и в некоторых
странах их использование в качестве добавок запрещено. Например,
эфиры жирных кислот вызывают значительное увеличение массы почек
у крыс уже при содержании их в корме в количестве 2,5 %, а при кон¬
центрации 5—10 % происходят изменение печени и задержка роста [159]
Эфиры полиоксиэтилена и стеариновой кислоты вызывают у подопыт¬
ных животных опухоли мочевого пузыря [339] Не вызывает опасений
сам сорбит, который по калорийности не отличается от глюкозы, и его
использование разрешается, особенно в диетических продуктах для
больных диабетом [339].Из естественных эмульгаторов чаще всего применяется лецитин в
количестве 0,2—1,5 % [456], который из-за относительно высокой
чувствительности к концентрации солей и pH (Каратани, 1979), а также
значительного содержания жира (30—40 %) не так эффективен, как син¬
тетические добавки.5.1.9. Другие добавкиК другим добавкам, применяемым в технологии производства фар¬
ша и заслуживающим внимания, относятся прежде всего хлористый
натрий и цитрат натрия. Поваренная соль, добавленная в количестве* Раствор сорбита, нагреваемый в присутствии кислотного катализатора,
подвергается внутреннему обезвоживанию и переходит в циклическое соедине¬
ние с эфирными связями и четырьмя гидроксильными группами, сорбитан, ко¬
торый в дальнейшем обезвоживается и переходит в сорбит - соединение, содер¬
жащее две гидроксильные группы.** Монооцейнат — соединение с 8-членным кольцом в структурной фор¬
муле. - Прим. пер.112
2,5—4 %, действует как растворитель миофибриллярных белков (на¬
пример, при производстве сурими), а в количестве, превышающем
20 %, — как денатурат и консервант (например, в соленом и солено-су-
шеном фарше). Кроме того, соль замедляет гидролиз и окисление липи¬
дов. Не рекомендуется добавлять в фарш поваренную соль в количе¬
стве менее 1 %, поскольку в этом случае она действует как проокисли¬
тель [335] . Цитрат натрия применяется в основном как добавка, предот¬
вращающая желирование мороженого фарша в процессе его хранения
[360] , поэтому его рекомендуется применять особенно для такого
фарша (например, каэн-сурими), в состав которого, помимо других до¬
бавок, входят соли щелочных металлов с лиотропными анионами (на¬
пример, хлористый натрий), вызывающие желирование [315]5.2. УПАКОВКАТребования, предъявляемые к упаковке мороженого рыбного фар¬
ша, в основном идентичны требованиям, предъявляемым к упаковке
рыбных продуктов [240] Фарш, однако, еще более подвержен нежела¬
тельным изменениям, вызванным плохой упаковкой, чем рыба и моро¬
женые рыбные продукты, поэтому особенно высокие требования предъ¬
являются к таким свойствам упаковки, как газопроницаемость (в слу¬
чае фарша из жирной рыбы и рыбы средней жирности) и механическая
прочность. Эти требования весьма однозначно определяют выбор вида
упаковки, хотя с точки зрения ассортимента это может быть комбини¬
рованная упаковка типа пленка + коробка из гофрированного или ли¬
того картона или однослойная упаковка типа коробки из литого кар¬
тона.Для фарша из тощей рыбы особенно рекомендуется упаковка (па¬
кет) из полиэтилена низкой плотности (высокого давления). Это плен¬
ка характеризуется весьма низкой проницаемостью для водяного пара,
устойчивостью к действию низкой температуры (до —40 °С), а также вы¬
сокой эластичностью и механической прочностью. В Польше производят
несколько разновидностей полиэтилена, из которых полиэтилен I —
020/60 допущен органами здравоохранения к применению в пищевой
промышленности.При упаковке фарша из жирной и среднежирной рыбы следует
прежде всего применять комплексные (многослойные) пленки, полу¬
чаемые соединением слоев различных пластмасс со свойствами, допол¬
няющими друг друга. Примером самой простой и самой тонкой комп¬
лексной пленки является полиэтилен-целлофан. Из других комплексных
пленок заслуживает внимания полиэтилен-полиамид. Из однослойных
пленок для жирной рыбы можно применять полиэтилен низкого давле¬
ния (высокой плотности), который характеризуется средней газопрони¬
цаемостью и низкой проницаемостью для водяного пара. Дополнитель¬
ным преимуществом этой пленки, так называемой ’’бумагоподобной”,113
является устойчивость к действию высокой температуры (до 110 °С),
что позволяет разогревать фарш прямо в упаковке.Для упаковки мороженого фарша могут применяться монолитные
или сборные коробки. Различают коробки из пластмасс и из картона.
Примером коробок из пластмасс могут служить упаковки типа ”Поли-
онда”. Они изготовляются из полиолефиновых пластмасс, в частности
этилен-пропиленовых сополимеров. Эта пластмасса обладает очень хо-
ропдщи механическими свойствами, высокими показателями паро- и
водонепроницаемости [233]. Коробки изготовляются также из литого
или гофрированного картона, ламинированного пластмассовой пленкой,
например полиэтиленом, полиэтиленом, модифицированным микровос-
ками,и др. Одним из лучших ламинированных материалов, предназна¬
ченных для изготовления коробок, является смесь сополимера этилена
и винилацетата или диацетата гликоля и винил ацетата (Elvax) [62] .
6. ТЕХНОЛОГИЯ ПРОИЗВОДСТВА РЫБНОГО ФАРШАВ зависимости от применяемых способов обработки выпускается
рыбный фарш следующих видов: стабилизированный, промытый водой,
вареный или готовый к употреблению, соленый, сушеный и ферменти¬
рованный. Представленную классификацию следует понимать скорее
как условную, поскольку существует фарш с признаками, отвечающи¬
ми одновременно нескольким его видам.Технология изготовления фарша в значительной степени определя¬
ет его физические и химические свойства, от которых, в свою очередь,
зависят способы его дальнейшей переработки.6.1. СТАБИЛИЗИРОВАННЫЙ ФАРШСреди разных видов рыбного фарша наибольший интерес с техно¬
логической и экономической точек зрения представляет стабилизирован¬
ный фарш. Это объясняется в основном тем, что по химическому сос¬
таву и свойствам он лишь незначительно отличается от измельченного
мяса рыбы.Стабилизированный фарш можно изготовлять из промытого фарша,
полученного из тушек рыбы, или фарша, разделенного на фракции по
содержанию темных мышц. Иногда используют фарш, полученный из
отходов от филетирования, содержащий большое количество крови.
Разделение фарша из тушек на две фракции применяется главным об¬
разом при использовании рыбы, содержащей значительное количество
красного мяса. Фракционирование фарша осуществляется путем обра¬
ботки тушек или филе в два приема - сначала на барабане со слабо на¬
тянутой лентой, затем ’’отходы” от первого сепарирования пропускают¬
ся через сепаратор с туго натянутой лентой (рис. 27). Первая фракция
фарша (60—70 % съедобных частей) [67] состоит в основном из белых
мышц и не содержит кусочков кожи или подкожного красного мяса.
Содержание крови в этой фракции низкое. Это позволяет получать
светлый фарш, характеризующийся хорошей консистенцией и устойчи¬
востью при хранении. Вторая фракция состоит в основном из темного
мяса и оставшегося белого. В состав этой фракции входит значительное115
количество кусочков кожи, почек и мелких костей. Если такой фарш
предназначен для производства порционных изделий, кости должны
быть измельчены или растерты. Для этого фарш второй фракции про¬
пускают через специальное измельчительное устройство (стрейнер)
с мелкими отверстиями в решетке или используют коллоидную мельни¬
цу (см. рис. 27). Цвет второй фракции обычно красно-серый или бурый.
Иногда перед обработкой на стрейнере фарш промывают водой. Как с
точки зрения технологической пригодности, так и из-за срока возмож¬
ного хранения фарш второй фракции уступает фаршу первой фракции
или целому фаршу. Это различие особенно заметно при применении оди¬
накового способа стабилизации фарша.116
Цель стабилизации - увеличить продолжительность хранения фарша.
Существует много способов стабилизации рыбного фарша; основные
из них: добавление стабилизирующих веществ и создание внутренних
стабилизирующих систем в результате биохимических или физических
модификаций измельченного мяса.6.1.1. Фарш с добавлением стабилизирующих вещертвГлавным стабилизирующим фактором в таком фарше являются
химические вещества естественного происхождения или искусственные,
которые консервируют мясо рыбы, но существенно не изменяют его
основных исходных свойств.Эти вещества можно разделить на пять основных групп:
повышающие водоудерживающую способность фарша и таким об¬
разом улучшающие его консистенцию, например полифосфаты, натрие¬
вые и калийные соли альгиновой кислоты, карбоксиметилцеллюлоза,
обычный и модифицированный крахмал, поваренная соль, белковые
препараты и др.;антиденатуранты, например ди- и трисахара, некоторые карбониль¬
ные кислоты, аминокислоты, пептиды и др.;синтетические и натуральные антиокислители и эмульгаторы, такие,
как ВОА, БОТ,Tween 80, Span 80, приправы и экстракты приправ, ас¬
корбиновая кислота, токоферолы, полифенильные соединения и др.;вкусовые добавки: глутамат натрия, поваренная соль, сахар, при¬
правы и др.;синтетические и натуральные красители: Амарант, Эритрозин, Пур¬
пур Понсо 3R, молотый красный перец, каротиноидные препараты и др.Из перечисленных добавок наибольшее значение для улучшения
сохранности рыбного фарша имеют антиокислители и антиденатуранты.
Применение антиокислителей особенно необходимо в фарше из жирной
рыбы и рыбы средней жирности, поскольку основная причина ухудше¬
ния его качества во время морозильного хранения - окисление жира.
Показано [438] , что снижение концентрации антиокислителей (БОА,
БОТ) из смеси добавок, применяемых для стабилизации фарша, почти
до уровня контрольного образца (фарш без добавок) сокращало срок
хранения фарша из зеленухи. Применение антиденатурантов имеет
большое значение, особенно* для увеличения стойкости при хранении
фарша из тощей рыбы, например из трески.Для хранения незамороженного фарша при температуре ниже точки
кристаллизации воды (примерно —3 °С) в него вводятся добавки, сни¬
жающие криоскопическую точку, например хлористый натрий,, этило¬
вый спирт, многоосновные спирты и др. [507] . Механизм действия этих
веществ описан в работе Мацумото [304]Большинство применяемых добавок относится к нескольким груп¬
пам одновременно, так как благодаря своим специфическим свойствам117
могут выполнять одновременно несколько стабилизирующих функций.
Примером может служить глутамат натрия - широко известный стиму¬
лятор вкусовых свойств, который в технологии рыбного фарша при¬
меняется главным образом как антиденатурант (см. раздел 5).На практике в измельченное мясо рыбы чаще всего вносят одновре¬
менно несколько добавок с дополняющими друг друга функциональ¬
ными свойствами. В табл. 30 приведены некоторые стабилизирующие
добавки, применяемые при производстве рыбного фарша. Из этих дан¬
ных следует, что количество этих добавок обычно не превышает 10 %,
а часто значительно ниже (0,2—5 %). В наибольших количествах добав¬
ляют сахара (5-8 %), в минимальных — приправы, эмульгаторы и анти¬
окислители (0,006—0,15 %).Стабилизирующие добавки могут быть внесены в фарш в процессе
осторожного перемешивания или после предварительной гомогенизации
их с небольшим количеством измельченного мяса. Перемешивание фар¬
ша с добавками производится в фаршемешалках с низкой частотой вра¬
щения в течение возможно короткого времени (3—5 мин), чтобы не выз¬
вать чрезмерного разрушения структуры мяса. Чаши фаршемешалок
могут быть полукруглой формы, с ленточными или лопастными рабочи¬
ми перемешивающими органами [429] . На рис. 28 представлена линия
производства стабилизированного рыбного фарша на судах проектов
В-417 и В-418. Полученное после сепарирования измельченное мясо рыбы
накапливается в бункере, откуда через отверстие в дне под действием
силы тяжести подается в фаршемешалку ленточно-лопастного типа.
В процессе перемешивания в измельченное мясо вносятся добавки из до¬
затора, размещенного над смесителем. Другой способ внесения добавок
применяется в США при производстве модифицированных блоков мо¬
роженого фарша, предназначенного для изготовления рыбных палочек.
К 2,5 кг измельченного мяса рыбы добавляют 1 кг поваренной соли,1 кг сахара, 0,3 кг глутамата натрия, 0,15 кг трифосфата натрия и 5 кг
воды, после чего смесь гомогенизируют (например, в куттере-миксе-
ре) до получения однородной густой массы. Полученный таким обра¬
зом продукт перемешивают со 100 кг измельченного мяса в фаршеме¬
шалке (примерно 50 об/мин) в течение 2—5 мин [211] .Стабилизирующие препараты могут быть также приготовлены в
виде смеси без мяса рыбы. Это обеспечивает более равномерное их рас¬
пределение в фарше, а также позволяет выявить дополнительные функ¬
циональные свойства препарата. Так, один из таких препаратов получа¬
ют путем эмульгирования растопленного при температуре 100 ° С сор-
битола с моноглицеридами и выдержки их при комнатной температуре
до застывания, затем твердую массу измельчают в порошок. Препарат
добавляют в измельченное мясо в количестве 4,3 %, его применяют так¬
же в смеси с другими добавками [158] Еще один стабилизирующий
препарат применяют в виде эмульсии [463] Его приготовление заклю¬
чается в гомогенизации одной весовой части растопленных при темпе-118
Таблица 30. Некоторые добавки (в %), применяемые при производстве рыбного фаршаДобавкаВариант рецептурыIIIII IVVI VII VIII IXXIAddi-Fro 71*Апьгинат натрияАмилопектинАскорбат натрияБОАБОТХлористый натрий
Сахарная пудра
Цитрат натрия
ЭДТКТринатрийфосфат
Натрийгликолят целлюлозы
Глутамат натрия
Hamina S**Лимонная кислота
Мономиристинат изосорбита
Мономиристинат сорбита
Мономиристинат сорбитола
Моноолеат сорбата
Растительное масло
Натуральный перец
Перец зеленый
Пирофосфат натрия
Полисорбат 80
Prebest TPJH
Сахароза
Сорбитол
Сушеный чеснок
Полифосфаты
ВодаИсточник0,05440,3
[ 338][104]0,0060,0061,00,150,30,007510,007515[458]0,20,0170,0150,130,020,0170,83ОД0,2 0,02-3,00,2 0,20,15 0,150,1 0,1 ОД ОДОДОД ОД ОД ОД[445] [184] [9] [237] [237] [237] [237]
[221] [221] [221] [221]ОД[189]
ПродолжениеДобавкаВариант рецептурыXIIXIIIXIVXVXVIXVIIXVIIIXIXXXAddi-Fro 71*Альгинат натрияАмилопектинАскорбат натрияБОАБОТХлористый натрий
Сахарная пудра
Цитрат натрия
ЭДТКТринатрийфосфат
Натрийгликолят целлюлозы
Глутамат натрия
Hamina S**Лимонная кислота
Мономиристинат изосорбита
Мономиристинат сорбита
Мономиристинат сорбитола
Моноолеат сорбата
Растительное масло
Натуральный перец
Пе^ец зеленый
Пирофосфат натрия
Полисорбат 80
Prebest ТР***Сахароза
Сорбитол
Сушеный чеснок
Полифосфаты
ВодаИсточник0,1ОД0,5ОД0,250,5[161] [161] [161]1,00,150,31,05[322]1,50,50,31,01.51.5
0,41,00,010,010,50,51,03,01,50,41,51,51,5[261] [390] [Морал, 1972] [386] [ 386]* Гидролизат желатина; ** смесь: тринатрийфосфат (32,9 %), пирофосфат (21,4 %), ортофосфат (10,4 %); *** смесь: Dсорбитол (87 %) , моноплицериды (6,5 %), полифосфаты (6,5 %).
Рис. 28. Схема линии производства стабилизированного фарша на судне типа В-417[193]:1 - головоотсекающая и рыборазделочная машина; 2 - рыборезка; 3 - элеватор;4 - горизонтальные ленточные транспортеры; 5 - моечная машина; 6 - сепаратор;7 - бункер для отходов; 8 - бункер для сырья; 9 - мешалка; 10 - емкость для
стабилизированного фарша (с насосом); 11 - дозатор добавок; 12 - стол упа¬
ковкиратуре 60 ° С моноглицеридов с тремя весовыми частями 53 %-ного раст¬
вора сахарозы, подогреве до более высокой температуры и охлаждении
смеси до комнатной температуры. Полученную таким образом эмульсию
добавляют в измельченное мясо в количестве 1 % вместе с сорбитолом
(5 %) и полифосфатом (0,2 %).К группе стабилизированного фарша можно отнести также смеси
типа паст, из которых после формирования в блоки и замораживания
изготовляют порционные блюда. В их состав, кроме стабилизирующих
и консервирующих добавок, входят также вкусовые, ароматические и
красящие вещества, вода, жир, соль и др. В сумме количество всех до¬
бавок может достигать 40 % рыбной массы (табл. 31). По консистен¬121
ции пасты могут быть крупно- или тонко измельченными типа колбас¬
ного фарша. После стерилизации продукт готов к употреблению.Таблица 31. Состав модифицированного фарша (рыбной пасты),
приготовленного на основе измельченного мяса рыбы (в %) [ 369]Тип смесиКомпонентбазоваяс запахом
морского
гребешкас зе¬
леным
лукомобработан¬
ная коптиль¬
ной жид¬
костьюИзмельченное мясо65,065,065,065,0Масло (растительное)18,018,018,018,0Вода8,08,07,76,45Модифицированный крахмал
(фосфатный)5,05,05,05,0Кукурузный крахмал2,02,02,02,0Поваренная соль1Д1,11,21,2Глутамат натрия0,50,50,50,5Трифосфат натрия0,150,150,150,15Сорбат калия0,100,100,100,10Риботид*0,050,050,050,05Span 800,350,350,350,35Tween 800,350,350,350,35Папаин0,0030,0030,0030,003Ароматизирующий препарат**-0,30--Сахар-0,0065--Диметил сульфат-2,10--Зеленый лук сублимированный--0,25-Розовый пищевой краситель---0,0022Красный перец молотый-- *-0,06Томат-паста---0,25Коптильная жидкость
Charsol Н-10———0,22Экстракт приправ № 2700---0,85Ароматический препарат, приго----0,10товленный из мяса* Смесь равных ьесовых частей динатриевых солей инозиновой и гуанило-
вой кислот.** Имитация запаха морского гребешка.6.1.2. Фарш, стабилизированный с помощью
биохимической модификацииВ фарше, относящемся к этой группе, увеличение срока хранения
достигается в результате биохимических реакций, вызванных физиче¬
скими, химическими и микробиологическими воздействиями и про¬
цессами в мясе. В механизме реакции стабилизации непосредственное122
участие принимают продукты из субстрата или мяса рыбы, причем само
мясо не подвергается существенным изменениям при переработке.
Этот вид биохимической модификации фарша заключается в том, что
в измельченном мясе рыбы активизируются или вызываются реакции
образования натуральных консервирующих продуктов или продуктов,
вызывающих замедление и даже прекращение неблагоприятных изме¬
нений в мясе. Коротко этот метод можно назвать ’’направленным само-
консервированием мяса рыбы” В соответствии с этим методом для уве¬
личения срока хранения рыбного фарша его механически перемешивают
перед замораживанием, добавляют воду в процессе замораживания,
понижают кислотность измельченного мяса, используют протеолитиче-
ские ферменты и т. д.6.1.2.1. Стабилизация путем перемешивания
измельченного мясаПеремешивание свежеприготовленного измельченного мяса рыбы
в течение 2—10 мин (в зависимости от вида) при температуре 5—15 °С
с интенсивностью, позволяющей получить липкость примерно 100—
150 сП при Dr=48,6 приводит к образованию естественных стабилизиру¬
ющих систем, которые в принципе увеличивают срок хранения мороже¬
ного рыбного фарша [220] . Этот эффект обусловлен в основном взаи¬
модействием некоторых азотистых веществ с жировыми, в результате
чего процесс автоокисления жира замедляется. Показано [218], что су¬
ществует тесная зависимость между содержанием аминного азота и окис¬
лением жира рыбного фарша. Установлено, что из многих азотистых
веществ в реакции с жиром участвуют главным образом щелочные и се¬
росодержащие аминокислоты, а также некоторые пептиды. Эти про¬
дукты могут образовываться в измельченном мясе под влиянием пере¬
мешивания, которое облегчает также связывание вновь образованных и
существовавших в мясе азотистых веществ с жиром, в результате чего
предотвращается окисление жира. Катализирующее влияние перемеши¬
вания на скорость гликолитических реакций в измельченном мясе трес¬
ки, хранившейся при температуре, — 3-6 °С, подтвердили Наумэн и Дай¬
ер [353]. Описанный выше способ удлинения срока хранения моро¬
женого рыбного фарша особо эффективен для жирной рыбы, у которой
изменения липидов являются главной причиной снижения качества.
Перемешивание, особенно если оно производится под вакуумом, спо¬
собствует не только протеканию естественных реакций стабилизации,
но и частичному эмульгированию жира, который благодаря этому ста¬
новится менее подверженным окислению. Показано, что изменение
перекисного числа и ТБЧ в перемешанном фарше из сельди, хранившем¬
ся в течение 9 мес при температуре —27±2 °С, происходит в арифмети¬
ческой прогрессии, а в неперемешанном — в геометрической [230]В фарше из тощей рыбы (балтийской трески) перемешивание, по-123
Неперемешанный фаршПеремешанный фаршПродолжительность хранения, месРис. 29. Изменения ТБЧ и содержания муравьиного альдегида в перемешанном и
неперемешанном фарше из балтийской трески разной степени первичной обра¬
ботки [ 246]мимо замедления окислительных процессов липидов, значительно задер¬
живает увеличение содержания муравьиного альдегида (МА) на на¬
чальном этапе хранения мороженого фарша (1—3 мес), т. е. в период,
когда в неперемешанном фарше содержание МА увеличивается очень
быстро. Эффективность влияния перемешивания зависит, однако, от
степени зачистки рыбных тушек перед переработкой на фарш и опре¬
деляется тщательностью удаления при разделке естественных катали¬
заторов нежелательных химических реакций, которыми являются поч¬
ки, кровь и плавательный пузырь (рис. 29).Установление оптимальных условий перемешивания рыбного фарша
с целью увеличения его стойкости при хранении является предметом124
исследований, проводимых в Институте технологии пищевых продуктов
морского происхождения Сельскохозяйственной академии в Щецине.
Полученные к настоящему времени результаты свидетельствуют, что
слишком интенсивное перемешивание фарша (куттерование) нежела¬
тельно, поскольку оно интенсифицирует гидролиз жира [217] и ухуд¬
шает консистенцию фарша.6.1.2.2. Стабилизация путем перемешивания
измельченного мяса с водойПоказано [263, 264, 265] , что вода, добавленная в модельные сис¬
темы пищевых продуктов, действует как антиокислитель, вероятно,
в результате разведения субстратов и катализаторов химических реак¬
ций, а также образования естественных защитных оболочек. Критиче¬
ский предел активности воды (0W)> при котором скорость окисления
жира рыбы в белково-жировых эмульсиях наиболее высока, находится
в пределах 0,32-0,75 [515] При дальнейшем возрастании величины
aw окислительные процессы в жире замедляются. Приведенные выше
теоретические основы могут служить предпосылкой для разработки
технологии стабилизации фарша путем добавления воды. Проведенные
до настоящего времени исследования с использованием фарша из тощей
рыбы (путассу) не дают, однако, достаточных оснований для однознач¬
ного решения вопроса о применении этого метода. Растворимость белков!
в измельченном мясе мороженой рыбы, куттерованном в течение 10 мин
с добавлением 23 % воды и хранившемся затем при температуре —20 °С,
значительно уменьшалась и консистенция быстрее ухудшалась, чем в
мясе, полученном сепарированием из немороженых тушек на установке
Баадер 694 без добавления воды [38] . Вероятно, однако, что эти разли¬
чия (не в пользу первого метода) были вызваны в значительной степени
разным режимом хранения рыбного сырья перед переработкой на фарш.
Кроме того, метод стабилизации водой дал бы, по-видимому, лучшие
результаты, если бы фарш был приготовлен из жирной рыбы или рыбы
средней жирности, которая менее чувствительна к денатурации белков.
Добавление воды оказывает катализирующее влияние на протеолитиче-
ские процессы и может вызвать образование естественных антиокисли¬
телей в мясе (растворенные аминокислоты, пептиды и др.) подобно
тому, как это происходит в процессе перемешивания фарша без добав¬
ления воды.6.1.2.3. Стабилизация путем понижения кислотности
измельченного мясаСтабилизация рыбного фарша путем понижения кислотности осно¬
вана на том, что pH влияет на активность деметилазы, разлагающей
ТМАО до ДМА и МА (см. раздел 3). Следовательно, изменяя pH фарша125
в кислотном и щелочном диапазоне, можно значительно снизить актив¬
ность ферментов и тем самым уменьшить количество образующегося в
мясе формальдегида, который, как известно, вызывает неблагоприятные
изменения в белках. Однако понижение pH в кислотном диапазоне вызы¬
вает денатурацию белков и ухудшает функциональные свойства фарша,
который из-за этого имеет ограниченное применение при производстве
рыбных продуктов. Зато повышение pH до 8 снижает активность деме-
тилазы почти так же эффективно, как и подкисление до pH 6, но не вы¬
зывает уменьшения растворимости мышечных белков. Напротив, такой
фарш во время хранения в мороженом виде характеризуется значитель¬
но более высокой растворимостью белков по сравнению с фаршем, име¬
ющим естественный pH [250] , что согласуется с широко известной за¬
висимостью между pH и растворимостью белков мяса рыбы [412] .Показано [143] , что повышение pH мяса сардины до 7,6 предохра¬
няло его от денатурации миофибриллярных белков во время посмерт¬
ного хранения при температуре 26 °С более эффективно, чем хране¬
ние при 5 °С.Скорость термической инактивации СА2+ АТФазы при pH < 6,6
была в 2-7 раз выше, чем при pH 7,0. Таким образом, понижение
кислотности может быть простым и эффективным способом стабилиза¬
ции фарша, особенно из тощей рыбы. Такой способ, предложенный
Костухом и др. [254] , заключается в понижении кислотности до pH
фарша 7,0—7,2 с одновременным восстановлением триметиламинокси-
да при температуре 0—4 °С для уменьшения скорости образования
формальдегида.При стабилизации рыбного фарша этим методом следует избегать
слишком высокой щелочности из-за возможности рацемизации при этом
L-аминокислот до D-аминокислот, а также образованием лизиналанина
(ЛАЛ) — соединения с токсическими свойствами. Заметное образова¬
ние Л АЛ начинается при pH около 9 [110] , при этом нагревание белков
выше температуры их денатурации многократно ускоряет эти реак¬
ции [27]6.1.2.4. Стабилизация фарша путем применения
протеолитических ферментовИзвестны также способы биохимической модификации рыбного
фарша, осйованные на применении ферментов растительного (например,
папаин, фицин), животного (трипсин, виокиназа) и грибкового проис¬
хождения (опизин и теризин). Их добавляют в основном с целью частич¬
ного гидролиза мяса и улучшения таким способом его функциональных
свойств [48]Показано (Быкова, 1971), что добавление фицина в количестве
0,05—0,1 % значительно увеличивает водоудерживающую способность
фарша из трески и содержание растворимого белка. Хотя исследования126
проводились с фаршем из мороженой рыбы, было доказано, что приме¬
нение протеолитических ферментов открывает новые возможности при
производстве измельченного мяса рыбы.6.1.3. Фарш, стабилизированный с помощью
физической модификацииНекоторые новые и немногочисленные разновидности рыбного
фарша позволяют применять методы стабилизации на основе физиче¬
ских процессов, не вызывающих существенных биохимических измене¬
ний в мясе. Примером этого может служить фарш, насыщенный инерт¬
ным газом, чаще всего диоксидом углерода или азотом. Газ вводится
в массу во время интенсивного перемешивания в герметичном куттере.
Процесс обычно заключается в предварительном перемешивании массы
при пониженном давлении с целью удаления воздуха. Затем в куттер по¬
дается инертный газ до достижения определенного давления смеси газа
с фаршем. В зависимости от давления газа и интенсивности перемеши¬
вания фарш насыщается газом, благодаря чему его структура становится
пористой [240]. Это позволяет не только улучшать стойкость фар¬
ша при хранении, но и придавать ему желаемые реологические
свойства.Наибольшая эффективность процесса насыщения рыбной массы
углекислым газом достигается при использовании кратковременного
(1—3 мин) куттерования фарша под давлением (около 196 кПа) [266]Дополнительным преимуществом использования диоксида углеро¬
да вместо других газов (например, азота) для стабилизации фарша явля¬
ется то, что этот газ, частично реагируя с аммиаком и триметиламином
[321], улучшает органолептические свойства фарша.6.2. ПРОМЫТЫЙ ФАРШОсновным процессом, улучшающим качество и стойкость фарша при
хранении, является промывка измельченного мяса водой для удаления
веществ, прямо или косвенно вызывающих неблагоприятные химиче¬
ские и физические изменения в процессе хранения мороженого фарша —
гемопротеидов, небелковых азотистых веществ, например ТМАО, фер¬
ментов саркоплазмы. Дополнительным результатом промывки является
увеличение содержания в фарше миофибриллярных белков и улучшение
его реологических свойств — эластичности и плотности после терми¬
ческой обработки. Увеличение содержания миофибриллярных белков,
однако, происходит за счет уменьшения содержания белков саркоплаз¬
мы, которые из многократно промытого фарша вымываются практи¬
чески полностью. С экономической точки зрения производство промы¬
того фарша менее оправданно, чем производство стабилизированного
фарша, так как, кроме нежелательных компонентов, во время промыв-127
ки удаляются также высокоценные пищевые компоненты. Кроме того,
после промывки фарш обычно стабилизируют соответствующими добав¬
ками.6.2.1. Промывка как процесс, улучшающий
свойства фаршаПромывка оказывает благоприятное воздействие на свойства фарша,
особенно на его цвет, запах и консистенцию. Улучшение цвета является
главным образом результатом удаления следов крови, тканевого сока,
а также определенного количества каротиноидов. Это влияние особенно
очевидно в фарше из тощей рыбы, а также полученном из отходов от
филетирования, содержащих определенное количество тканей почек.
Зависимость между содержанием крови и цветом фарша из трески
(Gadus morhua), определенная в системе координат X, Y, Z, описывает¬
ся следующим уравнением регрессии [120]Содержание крови (в %) = -0,082^ + 2,65 (г = -0,92; Р< 0,1),где X — цвет фарша (показатель Гарднера X).Для жирной рыбы с высоким содержанием в ткани красных мышц
промывка не дает таких очевидных результатов с точки зрения улучше¬
ния цвета, так как жир препятствует вымыванию, а присутствующие
в жирной рыбе пигменты находятся главным образом во внутрикле¬
точном пространстве.Эффективное влияние промывки на улучшение запаха фарша обус¬
ловлено тем, что многие вещества, придающие ему запах, относятся к
растворимым в воде. Однако изменение качества мороженого фарша
в процессе хранения определяется не только абсолютным содержанием
пахучих соединений, но прежде всего удалением соединений, которые
превращаются в вещества, имеющие запах и катализирующие образова¬
ние запахов. Например, в результате однократной промывки фарша
ставриды пятикратным объемом воды в течение 10 мин удаляется
82—95 % ТМАО, 53—70 % липидов и 50—75 % минеральных веществ
[459] Особенно эффективно вымываются низкомолекулярные компо¬
ненты азотистых веществ — аммиак, амины, ТМАО, свободные амино¬
кислоты и другие, а также вещества липидного характера — свободные
жирные кислоты, альдегиды и др. Влияние промывки на улучшение ста¬
бильности рыбного фарша очевидно, особенно после длительного хра¬
нения мороженого фарша (рис. 30). Более высокое качество промыто¬
го фарша с точки зрения запаха прежде всего объясняется ограничением
процессов окисления жира. Однако проблема запаха рыбного фарша ос¬
тается весьма сложной.Имеется много данных, свидетельствующих, что рыбный запах воз¬
никает в результате реакции между продуктами окисления жира и три-
метиламином. Реакции такого типа могут идти в фарше, в котором128
Продолжительность хранения, месРис. 30. Сравнение стабильности запаха промытого и непромытого фарша из мор¬
ского окуня, хранившегося при температуре -18 °С. Оценка производится по
5-балльной шкале (5 - органолептические признаки окисления отсутствуют; 4 -
слабый запах окисления; 3 — незначительный запах окисления; 2 - запах окис¬
ления средней интенсивности; 1 - сильный запах окисления); римские цифры
рядом с кривыми означают номер исследования [ 322]имеются жир и следы или большие количества таких третичных азотис¬
тых соединений, как триметиламин, триметиламиноксид, а также холин,
который может образовываться в результате самоокисления лецитина.Эффективное улучшение реологических свойств фарша, например
способности к образованию геля, эластичности и других, связывается
прежде всего с увеличением содержания миофибриллярных белков и
удалением белков саркоплазмы. Последние, как известно, замедляют
процесс образования эластичного геля в фарше во время тепловой об¬
работки. Добавление в промытый фарш водных вытяжек мышечной
ткани снижает его эластичность после тепловой обработки. Эти изме¬
нения исчезают при добавлении водных вытяжек, из которых удален
белок [359]Консистенция промытого рыбного фарша в значительной степени
зависит от содержания воды в конечном продукте. Слишком тщатель¬
ное удаление избытка воды ухудшает его консистенцию. Самую луч¬
шую консистенцию имеет фарш, в котором содержание влаги после
промывки и удаления избытка воды несколько выше, чем до промыв¬
ки (табл. 32).Показано [371] , что увеличение содержания воды на 4,5 % в промы¬
том модифицированном фарше существенно не влияет на консистенцию,
но уменьшение содержания воды на 1,6 % по сравнению с первоначаль¬
ным вызывает ее заметное ухудшение.129
Таблица 32. Влияние содержания воды на консистенцию
промытого фарша из мороженого морского окуня (Sebastodes sp.),
хранившегося в течение 12 мес при температуре —18 °С [ 322]Продолжительностьцентрифугирования,мин**Содер¬жаниеводы,%**Оценка консистенции фарша по 5-балльной
шкале* в зависимости от срока хранения
(в мес)0369120 (контрольная проба -79,54,03,84,03,73,7без промывки)284,04,24,04,23,83,91081,94,24,04,03,84,03077,93,83,73,53,33,4* Шкала оценки консистенции: 5 - очень хорошая, 4 - хорошая, 3 - удовлет¬
ворительная, 2 - терпимая, 1 - плохая.** После однократной промывки фарша водой (4 в.ч. воды + 1 в.ч. фарша).6.2.2. Оптимизация операций промывки6.2.2.1. Теория промывкиУдаление растворимых в воде компонентов измельченного мяса
рыбы основано на массообмене между двумя фазами - дисперсионной
средой (водой) и дисперсной фазой (мясом) во время их перемешива¬
ния. Из разных теорий, объясняющих механизм перехода массы через
межфазную поверхность из одной фазы в другую [429] , наиболее вер¬
ной представляется теория пенетрации. Она рассматривает процесс пе¬
рехода массы в дисперсионную среду как ряд непрерывных процессов
диффузии компонента от межфазной поверхности в жидкость, сопри¬
касающуюся с этой поверхностью и проникающую в глубь дисперсион¬
ной среды. Межфазная поверхность не является статичной, постоянной
величиной как в теории пограничных слоев, а образуется со стороны
жидкости из элементов, каждый из которых в течение короткого, но
одинакового времени находится в контакте с дисперсной фазой. Со
стороны жидкости постоянный ламинарный слой отсутствует, а турбу¬
лентность достигает самой межфазной поверхности. Движение массы
осуществляется в условиях непрерывной диффузии частиц от межфаз¬
ной поверхности к элементу жидкости в течение времени контакта т.
Этот процесс описывается уравнением непрерывной диффузии (так
называемый закон Фика):
ъсл Ь*СА
где г — время диффузии; х — расстояние до межфазной поверхности;
Dji - коэффициент диффузии; Сд — концентрация компонента А,
киломоль А/м3 или кг/м3Принимая, что концентрация компонента Л для элемента жидкости
на межфазной поверхности Сд2 и на значительном удалении от этой по¬
верхности Сд2 величины постоянные, что допустимо, так как время кон¬
такта незначительно, получаем для решения уравнения следующие ко¬
нечные условия:1) для* = 0 т> 0 СА = Сд2 равновесная концентрация;2) для* = оо т > о Сл = CAq;3) для* > 0 г = 0 Q = Сдо начальная концентрация.На основании второго закона Фика выведено уравнение [494] ,
определяющее степень экстракции белков саркоплазмы из белков
мяса минтая в зависимости от времени и пути (расстояния) диффузии.
Исходной точкой этих исследований была разработка формулы, по¬
зволяющей вычислить снижение концентрации белка R на модели слоя -
прямоугольной пластинке или плоскости, параллельной направлению
волокон в растворе саркоплазмы и погруженной в дистиллированную
воду:с — С„ о 00 1 л^ =~Г = “~2 тг — -ехр [-(2д2+1)2тг2-7-] ,со~Ч тг п=0 (2п +1) ’ l2где R — степень экстракции белка с учетом величины снижения концент¬
рации (от 1 до 0); С — средняя концентрация белка в пробе по истече¬
нии времени экстракции; С0 — начальная концентрация белка в пробе
(для t = 0) в момент начала процесса диффузии (принимается, что
С0 постоянна для каждой точки исследуемого слоя); Cs — концентра¬
ция белка в растворителе (воде); t - время (продолжительность) диф¬
фузии; D — постоянная диффузии; L — длина пути диффузии, равная
половине толщины пластинки; п — числа по порядку лг = 1,2,3 00.Отбросив все члены суммы, за исключением первого, или приняв
« = 0, получаемLПоказано, что скорость массообмена зависит от расположения во¬
локон в модельном слое. Для волокон, расположенных параллельно по¬
верхности слоя, применяется функция t/L1»5, а для перпендикулярно
расположенных волокон — функция t/L2. Несмотря на это различие, кри¬
вые экстракции растворимых в воде белков имеют сходную форму для
обоих упомянутых выше случаев и соответственно описываются зависи¬
мостями t/L1*5 и 0,28t/L2. Степень экстракции белков саркоплазмы
(уменьшение логарифма величины сниженной концентрации К) возрас¬
тает логарифмически в зависимости от времени экстракции: чем ближе131
Продолжительность диффузии, чРис. 31. Влияние продолжительности диффузии (h) на снижение концентрации
белков саркоплазмы (R) в мясе минтая разной степени измельчения [ 394]:а - направление переноса массы параллельно расположению мышечных волокон;
б - направление переноса массы перпендикулярно направлению мышечных во¬
локоннаправление диффузии к направлению мясных волокон (рис. 31), тем
быстрее происходит экстракция. Однако добиваться слишком большо¬
го снижения концентрации миофибриллярных белков в мясе при одно¬
кратной промывке (например, для частиц диаметром 5 мкм R = 0,1)
не рекомендуется по технологическим соображениям, так как время
диффузии при этом слишком велико (t > 21 ч), зато при меньшем сни¬
жении концентрации (например, при R = 0,5) это время непропорцио¬
нально уменьшается (t < 2 ч). Таким образом, применение многократ¬
ной промывки даже при меньшем снижении концентрации белков сарко¬
плазмы для получения конечного результата может оказаться выгоднее
и эффективнее, чем более желательная однократная промывка.Прямолинейная зависимость между логарифмом длины пути диф¬
фузии (L) и логарифмом времени диффузии (t) для отдельных уровней
R (рис. 32) иллюстрирует, в свою очередь, весьма существенное влияние132
бРасстояние диффузии, смРис. 32. Зависимость между расстоянием (L) и продолжительностью диффузии
(Л) на четырех выбранных уровнях концентрации саркоплазменных белков (Л)[394]:а, б - как на рис. 31размера частиц мяса на скорость вымывания веществ, растворимых
в воде.Указанные выше зависимости в основном подтверждают теорию
массообмена в условиях неустойчивой диффузии, в соответствии с ко¬
торой скорость переноса массы пропорциональна прежде всего раз¬
ности концентраций компонента и площади межфазной поверхности
[429]В принудительных системах турбулентное движение весьма затруд¬
няет теоретическое определение коэффициентов массообмена, так кзк
конечный результат зависит от многих факторов. Кроме таких основных
параметров диффузии, как разность концентраций вымываемого компо¬
нента дисперсной фазы и дисперсионной среды, площадь межфазной по¬
верхности и время диффузии, на практике действуют также факторы,
определяющие условия перемешивания, например тип мешалки, диа¬
метр емкости, частота вращения мешалки, и реологические свойства
смеси — липкость и др. В связи с этим для полной оптимизации про¬
цесса промывки на практике чаще всего бывает необходимо проведе¬
ние экспериментальных исследований. Для обобщения выводов этих ис¬
следований используются критериальные уравнения типа:Sh = BRewSc",133
где Sh - число Шервуда; Re — число Рейнольдса; Sc — число Шмидта;
В, т, п — постоянные коэффициенты.Традиционный вид числа Рейнольдса для мешалок, которые чаще
всего применяются в исследованиях такого рода,где п — окружная скорость концов лопаток мешалки (п = ndn), d -
диаметр мешалки; q - плотность смеси; т? - липкость смеси.При неполном взвешивании частиц измельченного мяса в воде
рост числа Рейнольдса вызывает значительное увеличение интенсив¬
ности массообмена, так как масса частиц, принимающих участие в про¬
цессе, возрастает. Зато если все частицы мяса взвешены, увеличение ин¬
тенсивности перемешивания вызывает уже значительно меньшую интен¬
сификацию процесса промывки, поэтому применение мешалки с более
высокой скоростью вращения, чем это необходимо для взвешивания
частиц мяса в воде, нецелесообразно. По этим же соображениям нера¬
ционально размещать в емкости перегородки, так как это вызывает
значительное возрастание мощности перемешивания и мало влияет на
интенсивность массообмена [429]6.2.2.2. Влияние pH воды или раствораПодкисление воды до pH, близкого к изоэлектрической точке
белка, улучшает эффективность удаления небелковых азотистых ве¬
ществ из мяса рыбы при значительном снижении потерь белка во время
промывки. Например, при промывке одной весовой части измельчен¬
ного мяса трески двумя весовыми частями воды при pH 5,0 снижение
содержания ТМАО увеличивается примерно на 20 %, зато потери белка
уменьшаются на 34 % по сравнению с промывкой при pH 6,5 (табл. 33).
Осветление мяса по мере снижения pH воды свидетельствует о том,
что подкисление облегчает также экстракцию гемопротеидных соеди¬
нений, в основном гемоглобина и миоглобина. То же самое наблюда¬
лось во время промывки свиного мяса водой при pH от 7,0 до 4,0 [471]
Следует, однако, отметить, что слишком сильное подкисление воды для
промывки иногда приводит к частичной денатурации белка мяса и ухуд¬
шает консистенцию фарша, особенно при pH воды ниже 5,5. Оптималь¬
ный pH промывной воды для фарша из рыбы составляет примерно 5,5.Промывка мяса водой с добавлением солей щелочных металлов и
слабых кислот (например, соды Na2C03, NaHC03) применяется в основ¬
ном с целью повышения желирующей способности фарша после тепло¬
вой обработки при производстве камабоко. Этот процесс заметно пред¬
отвращает снижение желирующей способности промытого фарша в про¬
цессе его хранения в замороженном виде [200] , а частично также окис¬
ление липидов, если фарш изготовлен из жирной рыбы [444] .134
Таблица 33. Влияние pH воды, применяемой для промывки*,
на содержание азотистых веществ в фарше из трески (Gadus morhua),
а также цвет фарша и содержание в нем воды [ 120]pH водыСреднее снижение содержания,
% содержания в непромытом
фаршеТМАО,мкмоль/кгБелок,г/кгСодержание
воды в про¬
мытом фарше,Цвет промытого
фарша (показа¬
тель X Гардне¬
ра)7,030,212,686,635,56,528,110,885,836,76,027,79,5986,238,25,530,67,3884,742,65,033,87,1782,049,0* 1 в.ч. измельченного мяса с частицами диаметром около 5 мм плюс 2 в.ч.
воды при температуре 20 °С. Продолжительность промывки при осторожном
перемешивании около 5 мин.Чаще всего применяются растворы NaHC03 концентрацией пример¬
но 0,5 % или смеси равных количеств NaHC03 и Na2C03 [330] При
применении растворов более высокой концентрации (например, 1 %-ного
раствора NaHC03 при pH 8,0) требуется дополнительная промывка
фарша водой после промывки раствором соды [248] Недостаток
промывки с применением солей щелочных металлов — меньшая эф¬
фективное удаления из измельченного мяса небелковых азотистых
веществ, например летучих оснований и ТМАО, чем при промывке
в кислой среде.6.2.2.3. Влияние количества воды
и кратности промывокПри однократной промывке фарша небольшим количеством воды
(от 2 до 5 в. ч. воды на 1 ч. фарша) величина потерь обычно больше за¬
висит от продолжительности промывки, чем от соотношения фарша и
воды. В свою очередь, при более высоком соотношении воды и фарша
(например, 10:1) влияние времени промывки в диапазоне 10—30 мин
на снижение содержания минеральных солей, белка и липидов оказы¬
вается несущественным (табл. 34).Увеличение числа промывок значительно увеличивает потери белка
саркоплазмы и уменьшает выход фарша (табл. 35). Это связано с боль¬
шими потерями массы мяса (главным образом, в виде взвесей) во вре¬
мя отделения воды от промытого фарша, особенно если для этой цели
применяется прессование.135
Таблица 34. Влияние соотношения мяса и воды и продолжительности
промывки на содержание пищевых компонентов в фарше из ставриды [459]Весовое
соотно¬
шение мя¬
са и водыПродол¬
житель¬
ность про¬
мывки,
минСодер¬
жание
воды в
фарше,%Поте¬
ри мас¬
сы мя¬
са, %Белок
N х 6,25 %Липиды,%Минераль¬
ные соли, %Со¬
де р-
жа-
ниеПо¬териСо-дер-жа-ниеПо¬териСо-дер-жа-ниеПо¬тери1:21079,5331,5618,0521,290,7670,870,5759,763074,2431,6117,0326,821,4545,340,5164,521079,0132,4617,6525,911,2653,610,5264,661:33078,4030,7818,1724,051,1657,460,4768,19578,8927,4917,9519,570,4880,990,6751,401:51072,2327,4917,5123,931,0460,330,5859,213078,3829,8518,0423,651,7335,760,4569,161:101078,4628,2617,0623,540,6674,850,4667,653079,4131,8217,5524,200,7372,340,4866,43Таблица 35. Влияние числа промывок на содержание водорастворимых
белков и выход фарша из хека и минтая [ 43]Вид обработкиСуммарноевремяпромывки,Выход фарша, %
нераз деланной
рыбыСодержание водо¬
растворимых бел¬
ков, %минХекМинтайХекМинтайБез промывки_59,641,7100100С промывкойоднократной1040,931,745,377,4”2039,4-69,7-двукратной1034,9-21,8-”1535,4-28,8-2033,931,320,354,53034,2-10,9-трехкратной1530,5-36,4-”3030,530,811,413,8четыре хкратной2026,528,020,336,1Многократная промьюка способствует также впитыванию большого
количества воды частицами фарша, поэтому ее отделение затрудняется,
а полученный продукт характеризуется повышенным содержанием влаги
(табл. 36). В фарше типа сурими, изготовленном по классической техно¬
логии, которая предусматривает применение больших количеств воды
по сравнению с количеством мяса (например, 7:1), а также многократ¬
ную (4—6 раз) промывку, содержание воды в конечном продукте пре¬
вышает 86 %. Следует, однако, отметить, что в таких условиях практи¬
чески полностью вымываются вещества, растворимые в воде, а также
Таблица 36. Влияние одно- и двукратной промывки фарша из морского
петуха на потери питательных веществ мяса (соотношение мяса и воды
в процессе промывки 1:3) [ 84]Содержание, %НебелковыйФаршВодаЖирБелокМинераль¬
ные солиазот, % об¬
щего содер¬
жанияАВАВАВазотаБез промывки73,22,48,621,580,22,59,32,9С однократной
промывкой70,41,25,817,183,01,88,71,1С двукратной
промывкой83,30,53,113,785,01,48,40А - по сырой массе.
В - по сухой массевесь тканевый жир. Удаление жира происходит главным образом во вре¬
мя прессования или центрифугирования промытого мяса.6.2.2.4. Влияние продолжительности промывкиПродолжительность промывки измельченного мяса рыбы должна
обеспечить максимальное удаление растворимых веществ при возможно
меньшей потере массы мяса. Однако оптимизация этих факторов затруд¬
нена из-за неблагоприятного влияния продолжительности промывки на
содержание растворимых веществ и потери мяса. Данные для примера
представлены на рис. 33.При короткой промывке (около 5 мин) потеря массы мяса относи¬
тельно невелика (около 27,5 %), тогда как количество извлекаемых ве¬
ществ, растворимых в воде, довольно значительно и составляет для
минеральных солей свыше 50 %, а для ТМАО свыше 80 %. В диапазоне
продолжительности промывки от 5 до 30 мин количество удаленных
минеральных солей возрастает логарифмически, а удаление ТМАО пря¬
мо пропорционально продолжительности промывки. Это означает, что
увеличение времени промывки с 5 до 10-15 мин весьма эффективно
влияет на вымывание из мяса минеральных солей, не вызывая сущест¬
венного уменьшения массы фарша. Увеличение продолжительности про¬
мывки с 15 до 30 мин значительно улучшает эффективность удаления
из мяса триметиламиноксида при значительном возрастании потерь
массы фарша. Дальнейшее увеличение длительности промывки обычно
приводит к большим потерям массы фарша, но не влияет заметно на эф¬
фективность удаления веществ, растворенных в воде.Большие потери массы фарша при более длительной промывке137
Продолжительность промывки, минРис. 33. Влияние продолжительности промывки на динамику вымывания раство¬
римых в воде веществ из измельченного мяса ставриды (Trachurus trachurus) и
уменьшение его массы; весовое отношение воды к мясу 5:1, перемешивание мед¬
ленное [459]объясняются прежде всего образованием взвесей, трудноотделимых от
воды как декантацией, так и центрифугированием. Это является резуль¬
татом чрезмерного набухания мяса, которое прямо зависит от свежести
сырья. Оптимальная продолжительность промывки измельченного мяса
рыбы при тщательном перемешивании с 2—5 в.ч. воды составляет около
10 мин. Если для отделения воды применяется декантация, к этому вре¬
мени необходимо прибавить 8—10 мин на отстаивание смеси и 4—5 мин
на слив жидкой фракции [388] . При других методах отделения избытка
воды добавочное время составляет от нескольких до десятков минут.Фарш, полученный из менее свежей или мороженой рыбы, обладает
меньшей влагоудерживающей способностью и требует более продолжи¬
тельной промывки.6.2.2.5. Влияние дополнительных процессов
предварительной обработки мяса рыбыИзмельченное мясо перед промывкой можно обрабатывать различ¬
ными способами, например прессованием, варкой и др. В результате
такой обработки потери белка заметно уменьшаются, при этом эффек-138
Таблица 37. Потери азотистых веществ в фарше из минтая, подвергшегося прессованию, и без прессования*перед промывкой [ 276]ФаршМасса мяса, %Потери азотистых веществ, % начального содержанияПередпрессо¬ваниемПослепрессо¬ванияПослепромывкиВо время прессо¬
ванияВо время промывкиВсегоОбщийазотНебелко¬
вый азотОбщийазотНебелко¬
вый азотОбщийазотНебелко¬
вый азотПрессованный перед10064,071,0**123414552689промывкой10070,871,9***9,52116582579Промытый100Непрессо¬86,0**-38883888100ванный68,2***34853485* Прессование под давлением 2,026 105 Па.** Двукратно промытый фарш по 10 мин при соотношении мяса и воды 1:3.*** Фарш, промытый 1 раз в течение 20-30 мин при соотношении мяса и воды 1:3.
тивность удаления небелковых азотистых веществ, минеральных солей
и жира не снижается.Например, мясо, поверхность которого обработана горячим паром,
более подвержено вымыванию липидов, чем полностью сырое, поэтому
содержание липидов в таком мясе уменьшается пропорционально уве¬
личению соотношения воды и мяса, тогда как величина потерь белка
практически не изменяется (табл. 37). Положительное влияние пред¬
варительного прессования на уменьшение потерь белка обусловлено в
основном тем, что растворимость небелкового азота в предпрессовом
соке выше, чем растворимость общего азота (табл. 38).Таблица 38. Потери питательных веществ во время промывки
фарша, полученного из мелкой рыбы методом Скачкова* [172]Соотношение фарша
и воды во время
промывкиВодаБелокА ВА ВБез промывки 80,50±1,34 100 16,29±1,03 100
(контрольная проба)1:2 81 Д2±1,16 100,77 15,84±0,99 97,24
1:4 82,26±0,69 102,19 14,57±0,56 89,44
1:6 81,93±0,20 101,78 14,92±0,10 91,59ПродолжениеСоотношение фарша
и воды во время
промывкиЛипидыКалорийность,кДж/кгА ВБез промывки2,64 ±0,46100,00355(контрольная проба)1:21,53±0,2157,953221:41,28±0,3948,482921:61,01 ±0,2238,26287А - содержание, % массы фарша; В - % массы контрольной пробы (непромы¬
тый фарш) - принято за 100 %.* Измельчение пробы на кусочки, поверхностное нагревание (ошпаривание)
паром, отделение внутренностей, приготовление фарша.6.2.3. Группы и характеристика промытого фаршаВ зависимости от способа промывки фарш можно разделить на про¬
мытый после сепаратора и промытый в сепараторе; в зависимости от
кратности промывки измельченного мяса на фарш, промытый 1 раз,
2 раза и многократно. Как правило, промытый фарш, изготовляемый
в производственных условиях, дополнительно стабилизируют различ¬
ными добавками, поэтому фарш можно также разделить на стабилизи-
140
рованный (модифицированный) и промытый нестабилизированный.
Типичным примером промытого после сепаратора и стабилизирован¬
ного фарша является сурими, производимый в больших количествах
в Японии. Как пример промытого в сепараторе и стабилизированного
фарша можно привести полуфабрикат, производимый в США при изго¬
товлении рыбных палочек [206]6.2.3.1. Фарш, промытый после сепаратораПромывка производится в специальных установках, которые пред¬
ставляют собой емкость с округлым дном, снабженную горизонтальной
низкооборотной мешалкой. Типичным примером промытого фарша
является сурими.Способ изготовления сурими, разработанный Нисия и др. [345,
346] и запатентованный в Японии [343] , позволяет получить рыбный
фарш, характеризующийся высокой стабильностью в процессе моро¬
зильного хранения и способностью образовывать эластичный и плотный
гель после тепловой обработки. Эти свойства достигаются, во-первых,
многократной промывкой мяса водой для удаления веществ, катализи¬
рующих нежелательные изменения в фарше, и белков саркоплазмы,
что вызывает концентрирование миофибриллярных белков, характе¬
ризующихся высокой гелеобразующей способностью после тепловой
обработки, и, во-вторых, перемешиванием или куттерованием промы¬
того фарша со стабилизирующими добавками с целью тщательного рас¬
пределения, а также частичного выделения миофибриллярных белков
из мышечных волокон (твердая фаза) в окружающую жидкую среду.Приготовленный таким образом мороженый фарш характеризуется
высокой растворимостью белков даже после длительного хранения.
Миофибриллярные белки, находящиеся в основном в растворе стабили¬
зирующих добавок (антиденатурантов), легко экстрагируются из фарша
в процессе его переработки, поэтому фарш типа сурими можно назы¬
вать препаратом мяса рыбы. Его используют в основном для производ¬
ства продукта типа камабоко, который обладает характерной резинис-
той консистенцией.Сырье. На производство сурими идет в основном рыба со свет¬
лым мясом, низким содержанием жира, например минтай (Theragra
chalcogramma), горбыли (Agryrosomus argentatus, Nibea mitsukuri, Nibea
schlegelli и др.), зауриды (Saurida elongata, S. argynophanes и др.), мар¬
лин (Maccira ampla), путассу (Macromesistius poutassou, M. australis),
камбалы и др. Основным критерием пригодности рыбы для производ¬
ства сурими является способность мяса рыбы образовывать прочный
и эластичный гель после тепловой обработки. Это особенно важно при
изготовлении камабоко, поэтому в течение длительного времени основ¬
ным сырьем для производства сурими были горбыли, так как эта рыба
хорошо хранится во льду, а изготовленный из нее фарш не теряет спо¬141
собности гелеобразования. Например, установлено, что горбыль приго¬
ден для приготовления сурими даже после 3—4 нед хранения во льду,
тогда как для другой рыбы это время ограничивается несколькими
сутками [362] Примером может служить минтай, который с точки
зрения относительно быстрой потери способности образовывать гель
пригоден для производства сурими лишь в течение нескольких суток
после вылова.Несмотря на то что сурими можно изготовлять и из мороженой
рыбы, для этого, однако, лучше всого подходит свежая (немороженая)
рыба высокого качества. В последние годы для производства сурими
все шире используется жирная рыба с темным мясом — сардина, скумб¬
рия и др. Поскольку процесс промывки позволяет удалить значительное
количество тканевого жира и пигментов, полученный продукт по цвету
в основном сходен с сурими из тощей рыбы. Однако технологическая
пригодность рыбы с темным мясом значительно ниже, чем рыбы с белым
мясом, так как полученный из нее фарш характеризуется весьма слабой
способностью образовывать эластичный гель после тепловой обработки.
Это вызвано главным образом более значительным снижением pH в по¬
смертный период и содержанием большого количества темных мышц
в этом сырье [162] Кроме того, резкий рыбный запах и непродолжи¬
тельный срок хранения фарша свидетельствуют о том, что рыбу с тем¬
ным мясом невыгодно использовать в качестве сырья для производства
сурими.Хранение и первичная обработка сырья. Сырь¬
ем для сурими чаще всего служит неразделанная рыба, хранившаяся
во льду или в охлажденной морской воде. После сортировки рыбу обез¬
главливают и потрошат обычно вручную, что позволяет собирать непо¬
врежденные гонады. У крупной рыбы удаляют также переднюю часть по¬
звоночника для улучшения отделения мяса на сепараторе и обеспечения
хорошего качества фарша, так как одновременно с позвоночником уда¬
ляются также почки и большая часть брюшной пленки.Если первичная обработка механизирована, рыбу обычно разделы¬
вают на тушку с удалением почки, брюшной пленки и теши с оставлени¬
ем позвоночника или на филе с удалением внутренностей вместе с позво¬
ночником. Полученную после первичной обработки (разделки) тушку
или филе дважды промывают и оставляют для стекания воды.Отделение (сепарирование) мяса. Для отделения
мяса чаще всего применяют барабанные сепараторы типа ’’Бибун” с диа¬
метром отверстий в барабане 5-7 мм. При производстве сурими из не¬
которых видов рыбы (например, ставриды) применяют двухступенча¬
тую установку для отделения мяса на сепараторе (рис. 34) со слабым и
сильным натяжением ленты на барабане. Это позволяет повысить выход
съедобной части и уменьшить количество воды, необходимой для про¬
мывки фарша.П ромывка. Отделенное мясо немедленно промывают пресной142
sfzJ]/!2 _z—ТраулерОхлаждение рыбы
в воде со льдомРучное
обезглавливание
и потрошениеперемешивание с промывкой
при температуре Ю °СПеремешивание
измельченного
мяса с добавкамиЗамораживаниеРис. 34. Технологическая схема производства суримиводой при весовом соотношении рыбы и воды от 1:2 до 1:10, чаще всего
от 1:5 до 1:7. Количество потребляемой воды зависит прежде всего от
вида и степени свежести рыбы. Промывку повторяют от 2 до 6 раз, чаще
всего 3—4 раза. Первая промывка продолжается около 20 мин, последу¬
ющие — не более 10 мин, чтобы не допустить чрезмерного набухания
мяса. Температура смеси в процессе промывки не должна превышать
10 °С.Емкость для промывки фарша имеет дно полукруглой формы и
снабжена низкооборотной мешалкой (10—16 об/мин) лопастного типа.
Вода и фарш подаются в емкость и отводятся из нее по трубопроводам.143
При периодической промывке фарша порядок операций следующий
[89] в начале цикла промывки открывают вентиль на трубопроводе
подачи фарша, одновременно включается мешалка с таймером и начи¬
нается заливка емкости пресной водой, затем подачу фарша и воды
прекращают, содержимое емкости перемешивают в течение опреде¬
ленного времени. Мешалка выключается автоматически, что позволяет
смеси отстаиваться. Отстой сливают (декантируют), а оставшийся фарш
подается насосом в отцеживающее устройство, чаще всего вращающий¬
ся сетчатый барабан. После передачи фарша в следующую емкость цикл
промывки повторяется. Многократную промывку можно производить
без вращающегося барабана для отделения влаги, с использованием
только декантации, т. е. поочередно заливая и сливая воду после отстаи¬
вания. В этом случае промывку производят в одной емкости. Примене¬
ние вращающегося сетчатого барабана для отделения воды от фарша
между отдельными промывками позволяет уменьшить расход воды или
кратность промывок без снижения эффективности процесса.Отделение избытка воды и измельчение. В полу¬
ченном после промывки и слива воды мясе обычно содержится 92-94 %
воды, т. е. на 15—20 % больше, чем перед промывкой [361] Это до¬
полнительное количество воды, однако достаточно прочно связано с мя¬
сом и трудно отделяется. Для удаления избытка воды применяют шнеко¬
вые прессы (рис. 35) или специальные центрифуги.После выгрузки из устройств для отделения от фарша избытка во¬
ды в некоторых случаях он оказывается так сильно спрессованным,
что требует измельчения, которое производят на волчке.Очистка промытого мяса на стрейнере. Целью
этой операции является удаление из фарша небольших костей, кусочков
кожи, брюшной пленки и темного мяса.Вход измельченного мяса и водыРис. 35. Схема шнекового пресса для удаления избытка воды из промытого фарша1349]1 - перфорированный цилиндр; 2 - шнек; 3 - винт для регулирования давления;4 - вал144
Размер отверстий в сетке стрейнера составляет 1,4— 1,6 мм для фар¬
ша первого (А) класса качества и 2,3—2,8 мм для фарша второго (В)
класса качества.Перемешивание с добавками. Очищенное мясо пере¬
мешивается с добавками в куттере или низкооборотной мешалке.
Чаще всего добавляют 5—10 % сахаров и 0,2—0,4 % полифосфатов.
Если продукт должен быть соленым (каэн-сурими), то к фаршу добав¬
ляют 2,5 % хлористого натрия и 5 % сахарозы, после чего его перемеши¬
вают около 20 мин до получения клейкой массы. В сурими без добавле¬
ния соли (муэн-сурими) в качестве постоянной добавки используют по¬
лифосфаты (смесь равных частей полифосфата натрия и фосфата нат¬
рия) , а состав остальных добавок зависит от качества рыбы, используе¬
мой для получения фарша, и его назначения (табл. 39). Сахарозу и сор¬
бит добавляют в сурими, предназначенное для производства камабоко
высокого качества, требующее получения светлого продукта. Например,
в рейто-сурими добавляют 8-10 % сахарозы. Глюкозу добавляют в су¬
рими более низкого качества, используемое в основном для производ¬
ства колбас, так как побурение фарша, вызванное реакцией глюкозы
с аминокислотами, не влияет отрицательно на цвет продукта. К фаршу
с очень высокой способностью к гелеобразованию во время изготовле¬
ния и хранения в мороженом виде рекомендуют добавлять 1,5 % лимон¬
нокислого натрия [360]. При механическом перемешивании фарша с
добавками его температура не должна превышать 10 °С.Упаковка и замораживание. Готовый фарш фасуют
в пакеты из полиэтиленовой пленки, помещают в металлические формыТаблица 39. Стандарты на сурими из минтая (Theragra
chalcogramma), установленные в 1965 г. Хоккайдской ассоциацией
сурими [ 323 ]Макси¬
маль¬
ное со¬
держа¬
ние
воды,
%Добавки, %Добавка крах¬
мала при пе¬
реработке су¬
рими на кама¬
боко с целью
проведения
теста на элас¬
тичность, %КласскачестваХарактеристикасырьясаха¬розасор¬битглю¬козаполи-фос¬фатыА специ¬
альныйВ стадии посмертного
окоченения79-5-0,20а'После окончания по¬
смертного окоченения805——0,23ВПосле хранения рыбы
во льду в течение 1-
3 сут8250,27сПосле 3-4 сут хране¬
ния во льду83——50,210145
и замораживают в горизонтально-плиточных морозильных аппаратах при
температуре ниже —35 °С. Для фасовки фарша в пакеты применяется
специальное устройство прямоугольной формы, расположенное у вы¬
ходного сопла. Устройство формует блоки массой 10 кг, толщиной
55 и шириной 354 мм. Фарш нагнетается непосредственно в пакет, поме¬
щенный в форму, которая двигается по ленточному транспортеру. После
замораживания блоки фарша укладывают в картонные ящики и хранят
при температуре от —20 до -28 °С.Выход продукта. Этот показатель зависит от вида и каче¬
ства сырья, а также от кратности промывки. Чаще всего выход колеб¬
лется в пределах 20—25 % массы неразделанной рыбы. Выход мяса на
первом сепараторе составляет 40—50 % массы неразделанной рыбы.
На втором сепараторе можно получить до 10 % мяса из отходов, полу¬
ченных после первого сепарирования (см. рис. 34).Примеры японских линий для производства сурими. Сурими может
быть изготовлено на берегу из свежего или мороженого сырья, а также
в море на борту промыслового судна, где в качестве сырья используют
исключительно свежую рыбу.На рис. 36 представлена линия японской фирмы ”Тайё Ейгё”, предна¬
значенная для производства сурими на береговых предприятиях. Произ¬
водительность линии составляет около 10 т готового фарша в сутки,’
что в пересчете на сырье составляет 30—40 т неразделанной рыбы. Линия
предназначена для переработки трески, сайды и морского окуня. Линию
обслуживают 29 человек, из которых 6 занимаются разделкой рыбы на
двойное филе с кожей, 10 — сепарацией мяса, а остальные 13 — промыв¬
кой фарша, отделением избытка влаги, очисткой и упаковкой. Эти дан¬
ные свидетельствуют о высокой трудоемкости производства сурими. Да¬
же при снижении числа промывок до двух оно значительно более трудо¬
емко, чем производство обесшкуренного филе при помощи разделоч¬
ных машин [253]На рис. 37 представлена схема линии производства сурими из пела¬
гических рыб, например японской скумбрии (Pneumatophorus japonicus),
сардины (Sardinops melanostica) и др. Эта линия, устанавливаемая в ос¬
новном на береговых предприятиях, интересна тем, что в ней имеется
специальная установка для выделения и сбора жира. Это танк вмести¬
мостью 1,2 м3, снабженный уловителем жира, горизонтальной мешал¬
кой и установленным над дном горизонтальным шнековым транспорте¬
ром. Фарш подается в установку и выгружается из нее системой насосов
непрерывного действия. Жироотделитель установлен в начале линии,
между накопительной емкостью, служащей для сбора измельченного мя¬
са после сепарации, и винтовым прессом. Применение установки для
сбора жира позволяет извлекать около 30 % тканевого жира, остальной
жир теряется во время промывки, удаления избытка воды и очистки
мяса, в результате чего конечный продукт содержит всего 3 % жира
[351].146
Рис. 36. Схема линии производства сурими на береговых предприятиях фирмы ”Тайё Ейгё (Япония) [ 351]:1 - бункер для свежей рыбы; 2 - головоотсекающая машина; 3 — машина для разделки морского окуня; 4 - машина для раз¬
делки трески; 5 - бункер для тушек; 6 - транспортер для отходов; 7, 8 - филетирующие машины; 9 - барабанная мойка;
10 — машина для производства фарша из филе; 11 - емкость для смешивания фарша с водой (1:1); 12 — насос для перекачи¬
вания фарша; 13 - емкость для промывки; 14 - шнековый пресс; 15 - насос для подпрессовой воды; 16 - емкость для под-
прессовой воды; 17 - центрифуга для отделения подпрессовой воды; 18 - шнековый транспортер; 19 - волчок; 20 - устрой¬
ство для очистки фарша; 21 - шнековый транспортер; 22 - охлаждаемый смеситель; 23 - устройство для фасовки фарша;
24 - транспортер для перемещения фасованных порций в морозилку; 25 - контактная морозилка
Рис. 37. Сх&ма линии производства сурими из пелагических рыб [351] :1 - сепараторы; 2, 5 - шнековые прессы; 3 - емкости для промывки фарша; 4 -
сепаратор жира; 6 - мешалки; 7 - фасовочные машиныСледующая характерная особенность этой линии — установка стрей-
нера непосредственно у места последней промывки мяса. Таким обра¬
зом, в стрейнер попадает мясо с большим количеством воды, что об¬
легчает его очистку. Под этой операцией понимается в основном уда¬
ление из фарша кусочков темного мяса, остатков кожи, костей и чешуи.
Из стрейнера очищенное мясо подается в винтовой пресс, а оттуда непо¬
средственно на перемешивание и фасовку.Для судовых условий японская фирма ’’Отака” разработала линию
производства промытого фарша производительностью около 20 т сырья
(неразделанная рыба) в сутки (рис. 38). При числе занятых 6-8 чел.
в смену расход пресной воды составляет 24 т/сут.Технологический процесс осуществляется следующим образом
[89] . Неразделанная рыба подается ковшовым элеватором в барабан¬
ную мойку 1 (см. рис. 38), откуда ленточным транспортером 2, а за¬
тем элеватором 3 и транспортером 4 направляется в буферный накопи¬
тель 5. Из накопителя рыба загружается вручную в две—четыре филети-
рующие машины 6. Полученное филе (двойное) подается транспорте¬
ром 7 под струями воды на транспортер 8, где производится его ручная
зачистка. Далее филе поступает в сепаратор 9. Отходы удаляются из148
сепаратора транспортером 11, а измельченное мясо транспортером 10
подается в емкость 12, где перемешивается с водой и далее в виде пуль¬
пы с помощью насоса 13 подается в емкость 14, где происходит про¬
мывка. После окончания промывки пульпа перекачивается насосом 15
в накопитель 16, откуда под действием собственной силы тяжести попа¬
дает в расположенную ниже установку 17 для первичного центрифуги¬
рования избытка воды. Частично обезвоженное мясо направляется в
шнековый пресс 18, где происходит окончательное удаление избытка
воды до треб'^мого содержания ее в фарше. Отжатая масса из пресса
шнековым транспортером 19 подается на волчок 20. Измельченное мясо
взвешивают на весах 21 заранее заданными порциями, которые перено¬
сятся в куттер 22, куда вносят также полный набор добавок. Тонкоиз-
мельченное (растертое) мясо направляется по желобу в машину 23,
формующую блоки и фасующую фарш в пакеты.Другие варианты производства фарша, промытого после сепарации.
В 1965—1975 гг. большой интерес к производству промытого фарша
возник также во многих странах Европы и в Северной Америке. В ре-149
зультате были разработаны упрощенные методы производства промы¬
того фарша, которые позволяют получить продукт, более близкий
требованиям европейского потребителя, чем японское сурими. Неко¬
торые из них описаны ниже.В СССР фарш производят в основном на промысловых судах.
В НИКИМРП были разработаны и изготовлены сепараторы серии ’’Фарш”
для производства фарша, которые могут быть установлены в каждой
технологической линии. Исследования, проведенные во ВНИРО и других
научно-исследовательских организациях [386, 388] , позволили решить
многие технологические проблемы, касающиеся прежде всего способов
промывки фарша.Испытаны следующие варианты: трехкратная комбинированная
промывка: первая в течение 2 мин, вторая в течение 3 мин в пресной
воде, третья в течение 3 мин в 1 %-ном растворе поваренной соли; трех¬
кратная обычная промывка пресной водой, как описано выше; дву¬
кратная комбинированная промывка: первая в пресной воде при соот¬
ношении воды и мяса 3:1 в течение 10 мин. После 5-минутного отстаи¬
вания вторая промывка в 1 %-ном растворе поваренной соли в течение
5 мин; обычная двукратная промывка в пресной воде при соотношении
мяса и воды 1:3, 1:4, 1:10, 1:12; комбинированная двукратная промыв¬
ка: сначала морской водой в течение 15 мин, затем пресной водой в те¬
чение 5 мин при соотношении мяса и воды 1:3, после чего 5-минутное
отстаивание фарша перед центрифугированием.Оказалось, что оптимальной является двукратная промывка при
осторожном перемешивании мяса с 3—4 в.ч. воды [330] . Установлено,
что увеличение отношения воды к мясу даже до 12:1 не улучшает замет¬
но качества фарша и его стойкости при хранении. Не дало существен¬
ных результатов и применение на последнем этапе промывки 1 %-ным
раствором поваренной соли. Использование морской воды на первом
этапе промывки признано недопустимым, так как она оставляет в
промытом фарше соединения, придающие продуктам, изготовленным
из такого фарша, горький вкус. Установлено, что применяемый способ
промывки и отделения избытка воды должен гарантировать содержа¬
ние растворимых в воде азотистых веществ менее 20 % их первоначаль¬
ного содержания, а содержание воды в конечном продукте не должно
превышать 86 %.Практическим результатом исследований, проведенных в СССР,
было начало производства промытого фарша на нескольких рыболов¬
ных судах на рубеже 60-х и 70-х годов. В качестве примера можно при¬
вести линию производства промытого фарша из минтая на судне ’’Севе¬
родвинск” (рис. 39). Рыбу разделывали на двойное филе на машинах
LW-520 с зачисткой брюшной пленки вручную. Выход филе составлял
27-30 % массы неразделанной рыбы. После промывки и стечки филе
пропускали через машину для изготовления фарша типа ’’Неопресс”,
снабженную барабаном с диаметром отверстий 4 мм. Полученный фарш
150
C=F)f? U vLIEРис. 39. Схема линии производства промытого фарша на плавбазе ’’Северо¬
двинск” [7]:1 - рыбомоечная машина; 2, 5 - сетчатый элеватор; 3 - стол; 4 - филетировоч-
ные машины; 6 - сепаратор; 7 - шнековый транспортер; 8, 11 - промывочные
танки; 12, 26 - накопительные танки; 10,13 - ротационные сита; 14,16 - шнеко¬
вые транспортеры; 18 - наполнительная машина; 15 - фильтр для рыбного мяса;
17 - куттер; 19, 21 - рольганги; 20 - фасовочный стол; 22 - контактные моро¬
зилки; %3 — гидропривод; 24, 25 - насосы для перекачивания пульпы (смесь
фарша и воды); 28 - насос с фильтром; 29 - расходный водяной танк; 30 - во-
доохладитель; 31 - циркуляционный насос; 32 - насосы для подачи воды; 33 -
щит автоматики; 35—37 — транспортеры для отходов; 38 - установка для про¬
мывки ротационных сит и шнекового пресса
дважды промывали пресной водой при весовом соотношении фарша и
воды 1:3. Промывка осуществлялась поочередно в двух 600-литровых
емкостях. Емкости снабжены лопастными мешалками, вращающимися
с частотой 24 об/мин. После первой и второй промывки фарш отжимали
на вращающемся барабанном сите и направляли в сборник вмести¬
мостью 1500 л. Для предотвращения желирования фарш в сборнике
постоянно перемешивался мешалкой с частотой вращения 24 об/мин.
После второй промывки и отделения избытка воды на вращающемся
сите фарш с содержанием воды 93—95 % подается в пресс для оконча¬
тельного обезвоживания. Очистку промытого фарша от остатков кожи и
костей производили на стрейнере с барабаном (диаметр отверстий2,4 мм). Перемешивание промытого мяса с добавлением 0,4 % поли¬
фосфатов и 1,5 % сахарозы осуществляли на быстроходном куттере с
частотой вращения ножевого вала 1500 об/мин, вместимость чаши кут-
тера 150 л. Содержание воды в фарше после перемешивания его с добав¬
ками составляло около 83 %. Затем продукт механически дозировали
по 10 кг в пакеты из полиэтилена и укладывали в морозильные формы.
Для увеличения срока хранения фарша и уменьшения отклонений массы
блока от установленной применяется дополнительное устройство для
частичного удаления воздуха из пакета во время дозирования фарша.
Замораживание производится в горизонтально-плиточных морозиль¬
ных установках при температуре —35 °С. Время, необходимое для дос¬
тижения температуры —18 °С в центре блока, составляет 2—5 ч [7].В Польше разработкой технологии получения промытого фарша за¬
нимались научные сотрудники Морского института рыболовства в Гды¬
не. Лучшие результаты получены при перемешивании фарша в течение
10 мин с двумя частями питьевой воды, охлажденной так, чтобы темпе¬
ратура смеси во время промывки не превышала 6 °С. Для стабилизации
промытого и очищенного на стрейнере фарша предложены два вариан¬
та добавок: альгинат натрия + сорбит и смесь полифосфатов типа Hami-
па [124]6.2.3.2. Фарш, промытый в сепаратореФарш, относящийся к этой группе, промывается в барабане сепара¬
тора в процессе отделения мяса от несъедобных частей. В принципе мож¬
но говорить в этом случае о прямом совмещении операций сеиарации и
промывки. Для этого служат специальные устройства, снабженные при¬
ставкой для орошения барабана водой, а также дополнительным бараба¬
ном, выполняющим роль сита, отцеживающего промытый рыбный фарш.
Рекомендуемый диаметр отверстий в барабане сепаратора составляет
5—7 мм, а в отцеживающем барабане — 0,3—0,5 мм. Воду через пристав¬
ки направляют на барабан, она попадает внутрь барабана через отверстия
и перемешивается с фаршем, таким образом осуществляется его про¬
мывка. Отделение воды от промытого фарша происходит во время про-
152
хождения смеси через барабан-сиго, длина которого составляет 90—
108 см. Описанный способ промывки имеет определенные преимущества,
хотя и относится к менее эффективным. Промывка фарша непосред¬
ственно после отделения мяса от костей и кожи позволяет удалить зна¬
чительное количество крови, так как в этот момент она еще не образует
сгустков и легко растворима в воде. Зато другие вещества мяса — бел¬
ки саркоплазмы, жир и минеральные соли — удаляются только частич¬
но, поскольку вымывающее действие воды ограничено в основном по¬
верхностным слоем отдельных частиц фарша. Установлено, что при ис¬
пользовании двух-трех весовых частей воды во время промывки этим
способом удаляется лишь 10 % компонентов мяса (по сухой массе)
[211]. Промывка в сепараторе должна применяться в основном
для улучшения цвета фарша, изготовляемого из рыбы со светлым
мясом.Применение для промывки в сепараторе щелочного раствора саха¬
ров и (или) аминокислот позволяет соединить процесс промывки мяса
с его модификацией и стабилизацией. Полученный таким способом
мороженый фарш характеризуется повышенной способностью к обра¬
зованию эластичного геля после тепловой обработки даже после дли¬
тельного хранения [199].6.3. ПАСТЕРИЗОВАННЫЙ ФАРШВ основе технологии пастеризации рыбного фарша лежит прежде
всего использование тепловой обработки для инактивации фермен¬
тов, вызывающих неблагоприятные изменения в мороженом сыром мя¬
се; денатурация нативных мышечных белков для перевода их в менее
активную форму; уменьшение активности воды а в результате частич¬
ного обезвоживания фарша (термическое удаление клеточного сока);
частичное удаление с клеточным соком нежелательных химических со¬
единений — минеральных солей, триметиламиноксида, летучих основа¬
ний, свободных жирных кислот и т. п.; частичное отбеливание фарша
в результате разложения натуральных красящих веществ мяса — мио-
глобина, гемоглобина и др.По этим же причинам физико-химические изменения, происходя¬
щие в пастеризованном рыбном фарше во время хранения в мороже¬
ном виде, значительно менее выражены, чем в сыром. Это особенно ка¬
сается реологических свойств фарша, а также изменений продуктов фер¬
ментативного разложения триметиламиноксида во время хранения
мороженого фарша. Плотность вареного фарша лишь незначительно
увеличивается в процессе хранения (рис. 40), а его эластичность даже
имеет тенденцию уменьшаться (рис. 41), тогда как в сыром фарше эти
изменения весьма заметны и свидетельствуют о неблагоприятном явле¬
нии — желатинизации.153
Продолжительность хранения, месо 1
02• 3©4Рис. 40. Влияние хранения при температуре - 24 °С
на плотность сырого и вареного фарша из трески
[421]:1 - сырой фарш; 2 - сырой фарш, очищенный на
стрейнере; 3 — вареный фарш; 4 - вареный очищен¬
ный фаршО1 * 1 • 1 1 I I L1 2 3 4 5 6 7 8
Продолжительность хранения, месФ 1О 2
• 304Рис. 41. Влияние продолжительности хранения при тем¬
пературе -24 °С на эластичность сырого и вареного фар¬
ша [421]:1—4 — то же, что и на рис. 40
£ 1 2 3 4 5 6 7^ Продолжительность хранения, мес9 102• 3©4
® 5Рис. 42. Изменение летучих аммонийных оснований в сыром и вареном фарше
трески, хранившемся при температуре -24 ° С [ 421 ] :1-4 - то же, что и на рис. 40; 5 - обычный вареный фарш с добавлением 0,8 %
хлористого натрияИзменения содержания летучих оснований в пастеризованном моро¬
женом фарше минимальны по сравнению с изменениями в сыром фарше,
особенно после длительного хранения (рис. 42).6.3.1. Оптимизация технологии6.3.1.1, Выбор оптимальной температуры
и продолжительности тепловой обработкиКак уже отмечалось, одна из главных целей тепловой обработки
фарша — удаление излишка воды (клеточного сока) в результате теп¬
ловой денатурации мышечного белка. Эта цель легко достижима, так
как мышечные белки рыбы весьма чувствительны к тепловой обра¬
ботке, что обусловлено свойствами их основного компонента — мио¬
фибриллярных белков. Уже при температуре 30-37 °С происходит
50 %-ное снижение растворимости миофибриллярного белка мяса ры¬
бы, при этом у рыбы со светлым мясом это снижение происходит быст¬
рее, чем у рыбы с темным мясом. Повышение кислотности мяса до
pH 5,5 вызывает снижение температуры денатурации половины мио¬
фибриллярных белков до 11 ° С для минтая и до 20—24 ° С для сардины155
Рис. 43. Термостабильность миофибриллярных белков различных видов рыбы[249]:1 - минтай; 2 — сардина; 3 - скумбрия; 4 - тунеци скумбрии (рис. 43). Нагревание рыбного фарша до 40 ° С завершает
первый этап денатурационных изменений мышечных белков, доказа¬
тельством чего является практически полная инактивация Mg2+ АТФазы
миозина [440]Нагревание фарша до 40-60 °С приводит к образованию в белке
новых поперечных связей, в основном бисульфидного типа, а часто
также эфирных, причем эти связи стабильны и не нарушаются даже пос¬
ле добавления кислоты или щелочи [135] При температуре 60 °С,
соответствующей коагуляции миогена, измельченное мясо рыбы об¬
ладает минимальной способностью связывать воду, что определяется
по количеству сока, выделившегося при тепловой обработке, или по
уменьшению массы [407] . Теоретически можно было бы принять эту
температуру за оптимальную, если бы не тот факт, что она не гаранти¬
рует полной инактивации некоторых термоустойчивых ферментов мы¬
шечной ткани, а также необходимого снижения численности микроор¬
ганизмов в фарше. К термоустойчивым ферментам относятся в основ¬
ном деметилаза почек, щелочная протеаза мышц, щелочная фосфатаза
и др. Остаточная активность щелочной фосфатазы в мясе рыбы, прогре¬
том при температуре 60 °С в течение 30 мин, составляет 12—31 % мак¬
симальной активностью [121] Температурный оптимум активности156
щелочной протеазы мышц рыбы составляет 60 °С, а практически пол¬
ная ее инактивация происходит лишь при температуре 85 ° С [55] Эта
термоустойчивая протеаза в значительной степени содействует ослаб¬
лению консистенции пастеризованного фарша, особенно если его нагре¬
вают обычными методами, которые характеризуются медленным повы¬
шением температуры в зоне высокой активности щелочной протеазы.
Установлена высокая корреляция между консистенцией пастеризован¬
ного фарша и активностью щелочной протеазы в сыром мясе рыбы
[270]Условия тепловой обработки, достаточные для денатурации боль¬
шинства мышечных белков (нагревание до 78—80 °С и поддержание
этой температуры в течение 5—7 мин, см. раздел 3.2.3.1), не могут
считаться оптимальными для тепловой обработки измельченного мяса
рыбы. Эти противоречия еще более углубляются, если в измельченное
мясо включена ткань почек или пищеварительные ферменты, проник¬
шие в мышцы из пищеварительного тракта во время хранения рыбы
перед разделкой. Для полной инактивации деметилазы почек необхо¬
димы температуры выше 80 ° С [281, 433] Термостабильный комп¬
лекс протеолитических ферментов пищеварительного тракта инактиви¬
руется лишь при температуре 65—70 °С в зависимости от вида рыбы
[434]Таким образом, нагревание измельченного мяса до недостаточно
высокой температуры (60—70 °С) приводит к ослаблению консистен¬
ции фарша в результате ферментативного гидролиза мышечных белков,
главным образом миозина и тропомиозина. Этот гидролиз вызывают
прежде всего протеолитические ферменты, присутствующие в сарко¬
плазме мышц. Вместе с тем быстрое нагревание измельченного мяса
до температуры около 85 ° С позволяет получить фарш плотной и элас¬
тичной консистенции [54, 452]Применение ’’высокотемпературной” пастеризации необходимо так¬
же с бактериологической точки зрения. Даже при соблюдении соответ¬
ствующих санитарных условий в процессе обработки измельченное мясо
рыбы оказывается зараженным значительным количеством аэробных
гетеротрофных микроорганизмов родов Pseudomonas, Moraxella, Arthro-
bacter, Flavobacterium, Miccococcus [255, 514]В некоторых случаях невозможно также избежать присутствия ана¬
эробных болезнетворных бактерий, например Clostridium botulinum
тип Е и других, которые при благоприятных условиях могут образовы¬
вать токсины. В связи с этим, хотя при нагревании измельченного мяса
рыбы до 70 °С и выдерживании при такой температуре в течение 2—
4 мин уничтожается практически вся вегетативная флора (табл. 40),
на практике применяют значительно более жесткие режимы тепловой
обработки. Чаще всего применяется так называемый ’’ботулиновый ми¬
нимум”, предусматривающий нагревание мышечной ткани до 82,2 °С
в течение 30 мин [4]157
Таблица 40. Зависимость между продолжительностью варки, температурой
в центре батонов фарша и снижением числа вегетативных бактериальныхклеток [ 297}Продолжительность
варки, минТемпература в геометриче¬
ском центре батона, ° СОбщее число микроорга¬
низмов в 1 г фарша0-1,3-10"404411045054110360653670671580765908201008426.3.1.2. Влияние степени измельчения мяса
перед тепловой обработкойСпособность сырого мяса, полученного в результате сепарирования,
удерживать тканевый сок после тепловой обработки довольно низка,
что обусловлено его ’’зернистой” (пористой) структурой и низкой
липкостью. Низкая водоудерживающая способность мяса проявляется
в относительно большой потере сока в результате варки, для жирной
рыбы и рыбы средней жирности она составляет обычно 15—25 %, а для
тощей рыбы — 20—40 %.Рис. 44. Изменения массы фарша (а) и количества тканевого сока (б) в процессе
варки куттерованного и некуттерованного фарша из балтийской сельди при
температуре 102,5±0,5 °С [ 230] :1 - некуттерованный фарш; 2 - куттерованный фарш158
Рис. 45. Изменение содержания растворимого белка в 5 %-ном растворе хлористо¬
го натрия в куттерованном и некуттерованном фарше из балтийской сельди в
процессе варки при температуре 102,5±0,5 °С [ 230]Для увеличения водоудерживающей способности пастеризованного
фарша измельченное сырое мясо перед тепловой обработкой дополни¬
тельно измельчают, перемешивают в мешалке или куттере-смесителе
(с добавлением соли или без него) либо пропускают через волчок или
стрейнер. Эффективность этого процесса зависит от степени измельче¬
ния мяса и условий тепловой обработки [295] Слишком тонкое из¬
мельчение мяса дает фарш резинистой консистенции, особенно если в
процессе тепловой обработки образующийся гель не разрушается меха¬
нически. Если температура и продолжительность тепловой обработки
слишком велики, влияние измельчения уменьшается. Например, потери
массы фарша из балтийской сельди в результате измельчения в куттере-
смесителе в течение 1 мин при 1420 об/мин и нагревания паром при тем¬
пературе 102,5± 1 °С более 30 мин такие же, как и фарша, не подвергав¬
шегося куттерованию, тогда как при обработке паром не более 20 мин
потери массы куттерованного фарша были примерно на 50 % ниже, чем
некуттерованного (рис. 44).Измельчение мяса вызывает улучшение его теплопроводности,
благодаря чему в куттерованном фарше требуемый уровень термиче¬
ской денатурации белка достигается быстрее, чем в некуттерованном
(рис. 45), при этом в нем сохраняется значительно больше связанной
воды (см. рис. 44). Кроме того, куттерованный фарш после тепловой
обработки более плотный и менее жесткий, чем некуттерованный [230]159
6.3:1.3. Влияние добавки солиСоль, добавленная в измельченное мясо в концентрации, превышаю¬
щей 1 %, вызывает частичное растворение миофибриллярных белков в
процессе перемешивания, в результате чего липкость фарша повышается
(рис. 46), а следовательно, увеличивается его водоудерживающая спо¬
собность (рис. 47). Оптимальная концентрация соли в фарше 1,5-3 %,
однако по вкусу более 1,5 % ее обычно не требуется [296] . Добавление
соли уменьшает количество сока, вытекающего во время пастеризации
фарша, примерно на 40 % по сравнению с фаршем без добавки соли
[299]Пастеризованный мороженый рыбный фарш может быть изготовлен
из свежего или мороженого сырья. Однако продукт более высокого ка¬
чества получается из свежей рыбы, переработанной непосредственно
после вылова. На рис. 48 представлена универсальная схема техноло¬
гического процесса производства мороженого пастеризованного рыбного
фарша. Она предусматривает два варианта: тепловую обработку рыбы
перед сепарированием мяса и тепловую обработку измельченного мяса.В первом варианте рыбу, разделанную на тушку, нагревают острым
паром при температуре 95£5 °С в паровой камере периодического дей¬
ствия или в варильнике непрерывного действия в течение 10—20 мин.
Вареную рыбу загружают в сепаратор для отделения кожи и костей.
Полученную массу пропускают через винтовой пресс с целью удаления
избытка воды так, чтобы ее содержание в конечном продукте не пре¬
вышало 68 %. После предварительного охлаждения рыбной массы до
10—12 °С ее загружают в формы и замораживают до достижения внутри
блока температуры-18 ° С [6]На рис. 49 представлена разработанная в СССР и испытанная на судах
типа БМРТ линия производства вареного фарша, предназначенная в ос¬
новном для переработки минтая. Испытания, проведенные в условиях
промысла, показали, что при изготовлении фарша из вареной рыбы
потери массы значительны. Самые большие потери наблюдаются во вре-Рис. 46. Влияние содержания соли на липкость пас¬
теризованного рыбного фарша [ 299]160
Рис. 47. Влияние содержания соли на водоудерживающую способность пастеризо¬
ванного рыбного фарша [ 299] :1 - выделение сока, %; 2 - содержание NaCl, %Рис. 48. Технологическая схема процесса производства пастеризованного мороже¬
ного рыбного фарша
Рис. 49. Схема линии производства мороженого пастеризованного фарша из минтаяна БМРТ [ 6]:1 - рыборазделочная машина; 2 - транспортеры с рабочими местами для дополни¬
тельной зачистки тушек от брюшной пленки; 3 - ковшовый элеватор; 4 - бара¬
банная мойка; 5 - бланширователь; 6 - стол для подготовки сырья к варке; 7 -
транспортер; 8 - сепаратор ”Фарш-4”; 9 - шнек для измельчения мяса; 10 — шне¬
ковый пресс; 11 - вакуум-наполнительная машинамя варки тушек (6-8 %) и отделения избытка влаги из вареного мяса
(6-7 %), зато потери при охлаждении и замораживании фарша состав¬
ляют всего 1 — 1,5 %. Из-за этих потерь выход готового продукта до¬
вольно низок: для минтая он составляет всего 17—18 % массы неразде-
ланной рыбы [6] Это компенсируется, однако, относительно высокой
стабильностью вареного фарша в мороженом виде (см. раздел 7) и по¬
вышенным содержанием сухих веществ в продукте.По второму варианту сырьем для производства фарша служит сы¬
рая рыба, разделанная на тушку. Полученное в результате сепарирования
измельченное мясо дополнительно измельчается и очищается в стрейне-
ре или волчке, а затем перемешивается с солью (1,5 %) и другими вку¬
совыми или стабилизирующими добавками. В зависимости от вкуса по¬
требителя в фарш могут быть добавлены также другие компоненты,
например свиное сало, растительное масло, сливочное масло, маргарин
и др. Для улучшения цвета, а частично также вкуса и реологических
162
свойств фарша в СССР в него добавляют 15—25 % пасты ’’Океан” из ан¬
тарктического криля [298] . Фарш смешивается с добавками в кутте-
рах или куттерах-смесителях в течение соответственно 3—5 и 1—2 мин.
Температура фарша после куттерования не должна превышать 10—12 °С.
Если поддержание такой температуры невозможно, в измельченную
массу следует добавить 5—8 % измельченного льда [299] Полученный
после куттерования фарш пастообразной консистенции фасуется в ме-
талличесще формы вместимостью примерно 10 кг с крышкой или в син¬
тетическую оболочку (батоны массой около 2 кг) и пастеризуется.Нагревать фарш необходимо быстро: температура 80 ° С в геометри¬
ческом центре блока или батона должна достигаться не более чем за
40 мин. Это осуществляется нагреванием блоков паром температурой
около 98 ° С или погружением батонов с фаршем в горячую воду (95—
98 °С), подогреваемую паром, который подается через жиклеры. На
практике продолжительность тепловой обработки колеблется от 1 до1,5 ч в зависимости от геометрических размеров блока фарша и спосо¬
ба его нагревания [297, 299] .Очень быстрое нагревание фарша может быть достигнуто в куттере-
смесителе, приспособленном для работы под давлением и вакуумом.
В этих аппаратах фарш перемешивается и обрабатывается острым па¬
ром, который подается прямо в герметически закрытую ванну. Про¬
должительность подогрева фарша до 85 ° С составляет всего 3—5 мин
в зависимости от его первоначальной температуры и давления подводи¬
мого пара. Предварительное удаление воздуха из фарша перед тепловой
обработкой позволяет использовать некоторые естественные добавки
(например, приправы) не только как вещества, формирующие вкусо¬
ароматические или физические свойства фарша, но и как консервирую-
ще-стабилизирующие [227] Кроме того, тепловая обработка измельчен¬
ного мяса, происходящая одновременно с перемешиванием, препятст¬
вует чрезмерному желированию продукта в процессе морозильного хра¬
нения. После отделения избытка воды (например, на барабанном сите)
получают фарш однородной консистенции.После тепловой обработки фарш немедленно охлаждают до 8—11 °С.
Для охлаждения батонов применяют комбинированный метод: сначала
батоны охлаждают струями воды до 10—11 °С для достижения в центре
батона температуры 30—32 °С, а затем воздухом, охлажденным до 3—
4 °С; для охлаждения блоков используют воздух температурой —10-г
—30 °С [297] В куттере-смесителе фарш охлаждают путем отвода тепла
испарения воды при пониженном давлении, а также с помощью водяной
рубашки, в которую подается охлажденная вода.Замораживание батонов фарша чаще всего производят блоками в
горизонтальных плиточных или туннельных морозильных установках
с принудительной циркуляцией воздуха. Оптимальная температура
замораживания ниже —30 °С, а температура хранения не должна отли¬
чаться более чем на Уз от температуры замораживания.163
Продолжительность хранения, месРис. 50. Изменение водоудерживающей способности пастеризованного мороженого
фарша в процессе хранения при различной температуре [ 299]:1 - температура замораживания -8 °С, хранения -8 °С; 2 температура замора¬
живания -8 °С, хранения -23 °С; 3 - температура замораживания -35 °С, хране¬
ния -8 °С; 4 - температура замораживания -35 °С, хранения -23 °СВареный фарш весьма чувствителен к отрицательным последстви¬
ям рекристаллизации в процессе хранения, поскольку содержащаяся
в нем вода связана относительно слабо. Последствия рекристаллизации
проявляются в виде увеличения потерь клеточного сока при разморажи¬
вании (рис. 50) и ухудшения консистенции.6.4. СОЛЕНЫЙ ФАРШ6.4.1. Теоретические основы ускоренного посолаИзвестно, что поваренная соль и мышечные белки обладают высо¬
ким сродством к воде. При низкой концентрации соли молекулы белка
абсорбируют на поверхности ионы хлора, набухают и их способность
связывать воду увеличивается [135] Вода перемещается из раствора
соли (тузлука) в мясо до достижения равновесия. Фарш в этом случае
становится более липким, при органолептической оценке его консистен¬
ция часто определяется как ’’мажущаяся”. При высокой концентрации
ионов Na+ и СГ, когда количество доступной свободной воды уменьша¬
ется, между белками и солью обнаруживается конкуренция за оставшу¬
юся свободную воду. Поскольку ионы электролита связаны с водой
сильнее, чем частицы белка, вода перемещается в обратном направлении:
с частиц мяса в окружающую жидкую фазу (тузлук). Происходит осаж¬
дение белков из раствора, называемое высаливанием [135], коагуля¬
цией [472] или денатурацией [69]164
Если небольшой кусочек мяса рыбы погрузить в раствор хлорис¬
того натрия заданной концентрации, то по истечении определенного вре¬
мени достигается состояние равновесия, причем относительное содержа¬
ние соли в двух этих фазах (мясо—тузлук) будет зависеть от начальной
концентрации тузлука. При концентрации соли до 1,5 моль/л (около8,5 %) соотношение содержания соли в мясе и тузлуке возрастает, тогда
как при концентрации выше 2 моль/л (около 11,5 %) отношение содер¬
жания соли в мясе к содержанию ее в тузлуке снижается и при концент¬
рации 5,7 моль/л (около 33 %) достигает минимума (рис. 51).Указанная выше зависимость была установлена Дель Валье и Ни¬
керсоном [79] при разработке способа ускоренного посола измель¬
ченного мяса рыбы с целью его частичного обезвоживания в результате
денатурации мышечных белков.Соль выполняет при этом двойную функцию: коагулирует мышеч¬
ные белки и связывает отнятую воду, а также консервирует продукт,
предохраняя его от порчи.Однако чтобы денатурирующее действие соли было эффективным,
ее концентрация в измельченном мясе должна быть соответствующей.
Высокая концентрация соли необходима с микробиологической точки
зрения, а также для максимальной инактивации тканевых ферментов.
Минимальная активность воды для большинства бактерий составляет
0,90-0,96, для дрожжей - 0,88, для плесеней — 0,75, что приблизи¬
тельно соответствует следующим концентрациям водного раствора
NaCl при температуре 25 °С: 1,2-2,8 %, 3,55 % и 6,83 % [413] ОднакоКонцентрация NaCl, моль/лРис. 51. Изменение коэффициента равновесия соли в мясе меч-рыбы (кусочки мяса
диаметром 3,8 см и толщиной 0,6 см погружали в раствор хлористого натрия раз¬
личной концентрации на 48 ч при Температуре 25 °С до достижения состояния рав¬
новесия. Соотношение объемов мяса и тузлука 1:1) I 78]165
галофильные микроорганизмы могут развиваться при концентрациях
соли 15-20 %, а некоторые даже при концентрации 20-25 % [160,488]
Для полного прекращения восстановления ТМАО до ТМА в соленом
мясе трески, хранившемся при 25 °С, необходима 16 %-ная концентра¬
ция NaCl, достаточная также для полной инактивации протеолитиче-
ских ферментов.6.4.2. Способы ускоренного посола
и их характеристикаСпособы ускоренного посола, разработанные в 1968—1972 гг., пер¬
воначально использовали для посола рыбного филе. Исходным сырьем
служило филе без кожи, на последующих этапах технологического про¬
цесса его измельчали, а конечным продуктом был солено-сушеный фарш.
Среди разных способов ускоренного посола можно выделить два базо¬
вых: метод Дель Валье - Никерсона [75, 79] и метод Мендельсона -
Андерсона [11] Несмотря на значительное сходство технологии
(рис. 52), способы несколько различаются по характеру некоторых опе¬
раций, возможности механизации производственных процессов, себе¬
стоимости производства, а также по качеству готового продукта.По методу Дель Валье — Никерсона просаливание филе происходит
в процессе измельчения на волчке с добавлением 20—100 % соли. Это
позволяет значительно увеличить площадь поверхности мяса, сопри¬
касающейся с солью. Однако такой ’’сухой” способ посола вызывает
значительное просаливание фарша и насыщение его воздухом, что при¬
водит к окислению содержащегося в фарше жира. При сушке на солн¬
це, которая в условиях тропиков продолжается от 3 до 6 сут [80] , окис¬
ление жира происходит практически в течение всего процесса. Получен¬
ный фарш обладает характерным прогорклым запахом жира и не всегда
приемлем для населения, непривычного к продуктам такого рода.
Из-за высокого содержания соли (50—60 %) продукт перед потреблени¬
ем необходимо отваривать в воде в течение 10 мин. Для варки, отмочки
и ’’освежения” фарша требуется большое количество воды [313]Количество потребляемой при этом соли велико, поскольку она
добавляется в избыточном количестве: обычно 70-100 % массы сырого
мяса [80] При использовании крупнозернистой соли проникновение
ее в мясо в начальный период посола — процесс относительно медлен¬
ный, поскольку соль пржде всего должна раствориться, поэтому после
измельчения филе с солью полученная смесь подвергается дальнейше¬
му перемешиванию в волчке или в мешалке в течение 2—5 мин с целью
тщательного распределения соли и денатурации белков. Для рыбы с вы¬
соким содержанием жира (например, сардина) продолжительность пере¬
мешивания может достигать 20 мин. Способ относительно прост и быстр,
не требует холодильной цепи. Срок хранения готового продукта в усло¬
виях тропиков составляет 2 года.166
Рис. 52. Технологическая схема производства соленого фарша из обесшкуренного
рыбного филе по методу Дель Валье - Никерсона (а) и Андерсона - Мендель¬
сона (б)Способ Андерсона — Мендельсона [11] основан на мокром посоле.
Филе измельчается в куттере в течение 30 с (2* 15 с) и смешивается с туз¬
луком в соотношении 1:1, что соответствует концентрации соли около
25 %. Полученную смесь выдерживают при комнатной температуре в
течение 15 мин при осторожном перемешивании для полного осаждения
белков. Дальнейшая обработка сходна с описанной выше, но получен¬
ный после прессования фарш высушивается в вакуум-сушилке при 50 ° С
в течение 4 ч, а затем упаковывается под вакуумом в пакеты из много¬
слойной пленки. Срок хранения полученного таким способом продукта
при комнатной температуре составляет 11 мес [313] . Количество конеч-167
ного продукта относительно высокое, так как уже во время посола
мясо защищено от доступа воздуха тузлуком, а в процессе хранения —
герметичной упаковкой.Следует, однако, отметить, что способ Андерсона — Мендельсона
до сих пор не внедрен в промышленном масштабе, тогда как метод
Дель Валье — Никерсона уже в начале 70-х годов был испытан в полу-
производственных условиях на рыбообрабатывающем предприятии
в г. Прогресо (Мексика). В табл. 41 приведен выход полуфабриката и
готового продукта из некоторых видов рыб, полученный в полупроиз-
водственных условиях.Таблица 41. Выход полуфабриката и готового продукта
в процессе экспериментально-промышленного производства
солено-сушеного фарша из некоторых видов рыбы [ 80]ПоказательВид рыбыРагап-thiasfurci-ferEucinosto-
mus gula
Diapterus
plumieriEphine-phelusmonoAemulumespurusВыход филе, % неразделанной рыбы41,946,032,628,6Количество внесенной соли, %75757575Выход измельченного соленого мяса,. /о.от массы филе170173172172от массы неразделанной рыбы71,079,856,049,4Выход фарша после прессования*, %от массы измельченного солено¬72,472,370,873,8го мясаот массы целой рыбы51,957,839,836,4Выход солено-сушеного фарша, %**от прессованного фарша91,289,480,489,6от целой рыбы47,451,132,032,6от филе114,0112,098,0114,0* Измельченное соленое мясо, отжатое на гидравлическом прессе под дав¬
лением около 13,5 МПа в течение 1,25 мин.** Фарш, сформованный в виде лепешек массой примерно 470 г и высушен¬
ный на солнце до влажности 17 %.Канадские исследователи [499, 500, 501, 32, 33] использовали
метод Дель Валье — Никерсона для получения фарша из мяса рыбы, из¬
мельченного методом сепарирования, а также значительно оптимизиро¬
вали технологический процесс, главным образом с точки зрения сниже¬
ния содержания соли в конечном продукте.168
6.4.3. Оптимизация ускоренного посола
измельченного мяса6.4.3. 1. Влияние добавки солиКоличество соли, необходимое для правильного высаливания бел¬
ков и обезвоживания измельченного мяса, зависит от многих факторов:
вида рыбы и химического состава ее мяса, скорости перемешивания
массы в процессе посола, температуры, степени измельчения мяса, разме¬
ра кристаллов соли и др. Больше всего на количество вносимой соли
влияет водоудерживающая способность мяса.Показано [77] , что оптимальное количество вносимой соли прямо
пропорционально водоудерживающей способности мышечной ткани.Количество соли (%) = 1,48 (WHC) - 74,87, где WHC - водоудер¬
живающая способность мышечной ткани, определенная путем спрессо¬
вывания 2 г мяса между двумя взвешенными листамй фильтровальной
бумаги под давлением свыше 140 атм [15] ._ jqq Масса воды, отжатой из пробы
Масса пробыРыба с низкой водоудерживающей способностью мяса, например
корифена, требует меньшего количества соли, чем рыба с высокой во¬
доудерживающей способностью мяса, например бонито (более 40 %).
Мясо, недостаточно насыщенное лью, превращается в густую, лип¬
кую массу, трудноотдающую воду в процессе прессования. Вместе с
тем, если соли слишком много, фарш после прессования становится
хрупким и ломким.Интенсивное перемешивание измельченного мяса с солью (при
куттеровании, гомогенизации) ускоряет процесс посола так, что ко¬
личество соли может быть ограничено до минимума, обеспечивающего
максимальное насыщение жидкой фазы мяса при возможно меньшем
избытке соли, остающейся в мясе в виде кристаллов. Например, для из¬
мельченного мяса тощей рыбы с содержанием воды 80—83 % минималь¬
ная добавка соли, гарантирующая олное высаливание белков, состав¬
ляет 25 % [501]При производстве фарша из жирной рыбы необходимо добавлять
много соли, поскольку присутствующий в мясе жир не только затрудня¬
ет ее проникновение в мышечные волокна, но и защищает мышечные
белки, снижая их способность к денатурации. Например, в измельченном
мясе балтийского шпрота, содержащем 14,5 % жира, количество выде¬
ленной центрифугированием жидкой фракции (тузлука) возрастает ло¬
гарифмически с увеличением содержания соли в смеси до 40 %, после
чего начинает выделяться вода (рис. 53). Жир затрудняет также отделе¬
ние тузлука от фарша. Содержание соли в конечном продукте возраста-169
0I 1 • 1 1 U.10 20 30 40 50Содержание хлористого натрия в смеси, %Рис. 53. Влияние содержания хлористого натрия в измельченном мясе балтийского
шпрота, куттерованном с солью в течение 20, 40, 60 и 120 с на количество отцент-
рифугированного тузлука [ 245]ет экспоненциально с увеличением содержания соли в смеси и тем быст¬
рее, чем дольше длится посол [245]Таким образом, оптимальная добавка соли (около 40 %), обеспе¬
чивающая максимальное выделение воды из измельченного мяса жирной
рыбы, вызывает также его чрезмерное просаливание. Например, содер¬
жание соли в мясе шпрота, куттерованного в течение 60 с с добавлением
40 % поваренной соли и центрифугированного для отделения избытка
соли и воды (тузлука), достигает 50 % и выше, поэтому на практике
количество добавляемой соли ниже того, которое позволяет добиться
максимального удаления воды из измельченного мяса жирной рыбы.
Для измельченного мяса шпрота рекомендуется добавлять 30 % соли
при продолжительности куттерования 30-40 с, что позволяет получить
фарш с содержанием соли менее 20 % и сухой массы более 60 % [245]6.4.3.2. Влияние температуры посолаПовышение температуры мясо-солевой смеси ускоряет денатурацию
белков в процессе посола. Денатурация белков, определяемая по коли¬
честву тузлука, которое можно отделить от мяса, быстрее всего проис¬
ходит в диапазоне относительно низких температур (10—30 °С), кото¬
рые оказывают денатурирующее действие только на миофибриллярные170
рис. 54. Влияние температуры
посола на количество выделен¬
ного тузлука и содержание во¬
ды в отжатом фарше из трески
(весовое соотношение мяса и
соли 3:1, продолжительность
посола 3 мин, отжим в шнеко%7060вом прессе) [501]50403020 • • Содержание воды в отжатом фарше, %о Количество отжатого тузлука10 20 30 40 50
Температура посола, °Сбелки (см. рис. 54). Зависимость между массовым соотношением жид¬
кой фракции (W) и плотной фракции (NSS) в соленом фарше трески
после прессования и температурой посола (Т) определяется степенным
уравнением регрессии [501]где Т — температура посола в диапазоне 10—80 °С.Это свидетельствует о незначительном влиянии более высокой тем¬
пературы (40—80 °С) на процесс просаливания фарша и подчеркивает
решающую роль соли как денатурирующего фактора мышечных белков.Оптимальная температура посода измельченного мяса рыбы для по¬
лучения соленого фарша составляет 35-38 °С [500]Потери белков, происходящие в процессе ускоренного посола из¬
мельченного мяса рыбы, обусловлены прежде всего тем, что определен¬
ная часть трудноосаждаемых белков растворяется в тузлуке, а в про¬
цессе их механического отделения часть взвеси переходит в жидкость.
Величина потерь зависит в основном от концентрации соли в смеси,
а также от температуры и продолжительности посола. Например, при
посоле измельченного мяса трески при температуре 35 °С количество6А.З.З. Потери белков171
белков, растворимых в тузлуке (у), уменьшается экспоненциально рос¬
ту процентного содержания соли в смеси (х) [501]у (мг/г тузлука) = 77,45е-0»0757*Увеличение температуры смеси от комнатной до 38 °С вызывает
более чем двукратное уменьшение количества белков, растворенных в
тузлуке.Если посол продолжается более 30 с, потери общего белка (N-6,25)
обычно не превышают 30 % массы сырого мяса, из которых примерно
V3 приходится на небелковый азот [245] Содержание общего белка
в тузлуке колеблется от 0,1 до 1,5 % в зависимости от вида рыбы
(табл. 42) и составляет в среднем около 1 %. О величине потерь белка
свидетельствует также количество тузлука, отделенного от фарша.Таблица 42. Сравнение содержания воды, соли и белка
в быстропосоленном рыбном фарше перед прессованием и после него,
а также в подпрессовой жидкости (посол по методу Дель Валье - Никерсона)[77]РыбаФарш после переме¬
шивания с солью
перед прессованиемФарш после прес¬
сования под дав¬
лением 18 МПаПредпрессоваяжидкостьСодержание, %ВодаNaClБелокN•6,25ВодаNaClБе¬локВодаN•6,25NaClБелокN•6,25Испанская мак¬
рельScomberomorussierra51,533,818,337,043,532,575,337,40,1Кефаль
Mugil cephalus55,729,614,638,737,030,472,939,40,4Полосатый тунецKatsuwonuspelamis49,432,218,843,335,626,970,936,50,3КарпCyprinus carpio55,829,122,848,231,331,773,836,81,5КорифенаCoryphaenahippurus54,524,017,749,522,716,767,940,30,1Для уменьшения потерь белка до минимума следует прежде всего
стремиться к полному осаждению белка в процессе высаливания.172
6.4.4. Схема и описание процесса изготовления
соленого фарша из измельченного мяса рыбыНа рис. 55 представлена обобщенная схема изготовления соленого
фарша из измельченного мяса рыбы, полученного методом сепариро¬
вания.Наиболее пригодна для производства соленого фарша тощая рыба
с белым мясом — треска, сайда, путассу, минтай и др. Можно использо¬
вать также рыбу средней жирности (южноафриканскую сардину, жел¬
тую нототению, жирного хека) и жирную рыбу после нереста, когда
она не годится для производства обычных продуктов. Способ и продол-fttc. 55. Технологическая схема производства соленого фарша из измельченного
мяса рыбы, полученного сепарированием173
жительность хранения рыбы перед переработкой не играют такой су¬
щественной роли, как при производстве стабилизированного фарша.
Для переработки на соленый фарш может быть использована свежая
рыба даже II класса качества и мороженая тощая рыба, хранившаяся
более длительный период (6—12 мес), если непосредственно после раз¬
мораживания ее направляют в посол. Следует избегать направления на
переработку рыбы, не достигшей стадии посмертного окоченения, по¬
скольку ее мясо обычно пенится в процессе перемешивания с солью,
что затягивает его просаливание.Оптимальный диаметр частиц измельченного мяса с точки зрения
скорости его просаливания составляет 3-7 мм. Однако если для пере¬
мешивания мяса с солью применяют *уттёры или куттеры-смесители,
величина частиц и способ получения измельченного мяса не имеют су¬
щественного значения.Для посола измельченного мяса следует применять мелкозернис¬
тую соль, так как она быстрее растворяется. Продолжительность и ско¬
рость перемешивания мяса с солью должны быть подобраны так, чтобы
просоленное мясо было крупно- или среднеизмельченным, а не пасто¬
образным. Внешним признаком окончания просаливания считается по¬
явление свободного тузлука.Отделение избытка воды и соли от соленого измельченного мяса
производится на центрифугах, гидравлических и шнековых прессах.
Применение шнековых прессов позволяет сделать процесс изготовления
соленого фарша непрерывным. Прессы должны иметь барабан с отвер¬
стиями диаметром примерно 0,8 мм, частота вращения шнека — 30-
50 об/мин. Однако в фарше, отжатом на шнековом прессе, содержится
больше воды (48—50 %), чем в центрифугированном (34 %), поэтому
при производстве солено-сушеного фарша чаще применяют центрифуги
и гидравлические прессы [500] К их недостаткам относятся прерывис¬
тость работы и необходимость применения дополнительного оборудова¬
ния (волчка) для измельчения полученного ”жома” перед сушкой.
Фарш, полученный на шнековом прессе, имеет крупнозернистую
настолько рыхлую структуру, что измельчения перед сушкой не
требуется.Для предохранения от порчи полученный фарш можно высушивать,
охлаждать или замораживать, либо перерабатывать непосредственно
после тепловой обработки на другие продукты, например паштеты. Суш¬
ку следует проводить быстро, лучше всего при пониженном давлении до
достижения влажности продукта 18—22 %. Не рекомендуется, чтобы сни¬
жение воды в фарше было ниже 15 %, так как это вызывает ускорение
неблагоприятных химических изменений в результате частичного разру¬
шения мономолекулярного водного слоя. Высушенный продукт следу¬
ет упаковывать в паро-газонепроницаемую пленку. Рекомендуется при¬
менять пакеты из комбинированной пленки с заваркой их под вакуумом
или в атмосфере инертного газа.174
Если сырой фарш (после отжатая тузлука) хранится при понижен¬
ной температуре, период хранения можно использовать для созревания
продукта, благодаря чему он приобретает органолептические свойства,
близкие к свойствам соленых продуктов. Продолжительность созре¬
вания соленого фарша составляет 3—4 сут при температуре 18—25 °С
и 2—3 нед при температуре 3—5 °С [500]Для увеличения срока хранения охлажденного фарша применяются
консерванты, задерживающие развитие галофильных бактерий. Чаще
всего применяют сорбиновую кислоту, добавляя ее непосредственно
в фарш в количестве около 0,3 % или одновременно с водой при про¬
мывке.6.5. ПРОЧИЕ ВИДЫ ФАРШАКроме упомянутых способов изготовления рыбного фарша, из¬
вестны и другие, основанные чаще всего на соединении нескольких опе¬
раций тепловой обработки или на применении биотехнологии. Такой
фарш используется в основном в качестве белковых препаратов, вку¬
соароматических добавок, загустителей, стабилизаторов, а также пище¬
вых продуктов, готовых к употреблению. К продуктам такого типа
относятся прежде всего сушеный и ферментированный фарш.6.5.1. Сушеный фаршСпособ изготовления сушеного фарша основан на предварительной
варке или пропаривании мяса рыбы для удаления избытка воды с по¬
следующим высушиванием полученной массы либо сушке подверга¬
ется непосредственно сырое мясо. Сырьем для производства сушеного
фарша обычно служит тощая рыба с белым мясом, которая в процессе
сушки не окисляется так быстро, как жирная рыба. Например, по техно¬
логии, разработанной во ВНИРО (СССР), сырьем для производства фар¬
ша служит обезглавленный хек, замороженный блоками. Рыбу размора¬
живают до температуры 0-г-1 °С, потрошат и режут на куски длиной 6 см,
после чего варят в кипящей воде в течение 10 мин при весовом соотно¬
шении рыбы и воды 1:2. После стекания воды вареную рыбу пропуска¬
ют через сепаратор с отверстиями барабана 4-5 мм и сушат до влаж¬
ности менее 10 %. Сушку производят горячим воздухом (40 °С), про¬
пускаемым через слой фарша со скоростью 2—4 см/с. Для увеличения
поверхности испарения фарш, уложенный слоями, непрерывно пере¬
мешивают. Повышение температуры сушки сверх 40 ° С заметно умень¬
шает растворимость белка и ухудшает функциональные свойства про¬
дукта [286]Согласно технологии, разработанной в Научно-исследовательском
институте рыбной промышленности (FIRI) в ЮАР, сырьем для произ¬
водства сушеного фарша служит филе хека. Филе режут на куски дли¬175
ной 15 см и варят в течение 15 мин, после чего заворачивают в полотно
и прессуют в течение 10 мин под давлением около 60 кПа, при этом
масса мяса уменьшается примерно наполовину.Отжатую массу измельчают в течение нескольких секунд в обыч¬
ном куттере и направляют на вальцовую сушилку. Иногда к отжатому
мясу добавляют вещества, предотвращающие нежелательные измене¬
ния в процессе сушки - БОТ (0,05 %), пиросульфат натрия и др.
[20]В США разработан метод сушки сырого фарша, по которому измель¬
ченное мясо тощей рыбы (тихоокеанского хека), предварительно пере¬
мешанное с хлористым натрием (0,5 %), высушивается в вальцовой су¬
шилке. Температура поверхности валков сушилки 150 °С, продолжи¬
тельность нахождения слоя фарша (0,8-0,13 мм) на барабане около
20 с. Содержание воды в конечном продукте 5-8 %. Это позволяет при
соответствующей упаковке хранить продукт при комнатной температуре
минимум 3 мес без значительного ухудшения качества. Для получения
качественного продукта необходимо, чтобы измельченное мясо рыбы
содержало не более 2 % жира. В противном случае жир, налипая на бара¬
бан, ухудшает условия сушки, из-за чего консистенция массы становится
клейкой [428]Исследования по усовершенствованию технологии сушеного фарша
направлены в основном на улучшение функциональных свойств готового
продукта. Важнейшей задачей является предотвращение чрезмерной де¬
натурации белка в процессе тепловой обработки, вследствие чего
продукт становится слишком твердым и в дальнейшем плохо впитывает
воду. Для этого предлагается варить рыбу в 50 %-ном растворе сорбита
[327], уменьшая продолжительность обработки до 10 мин. Показано
[28] , что гидроколлоиды и мука злаковых эффективно улучшают ре-
гидратационные свойства сушеного фарша. Особенно этому способству¬
ют такие добавки, которые в процессе тепловой обработки (сушки)
сохраняют высокую способность связывать соду, например альгинат
натрия, модифицированные соевые волокна и др.Большие надежды связывают с сублимационной сушкой измельчен¬
ного мяса рыбы, которая позволяет значительно уменьшить потерю раст¬
воримости белка по сравнению с обычной сушкой [174]6.5.2. Ферментированный фаршВ основе производства ферментированного фарша лежит использо¬
вание стартовых культур микроорганизмов для преобразования углево¬
дов в молочную кислоту, снижающую pH среды с целью создания опти¬
мальных условий для активности протеолитических ферментов тканей
в кислотном диапазоне pH, способствующем созреванию мяса, э также
хранения продукта путем подавления микроорганизмов, неспособных к
росту в кислой среде (например, гнилостных бактерий).176
Поскольку мясо рыбы бедно углеводами, они вносятся извне в виде
различных добавок или их смесей. В этих добавках сахара могут нахо¬
диться в форме полисахаридов (например, пшеничная мука, отруби, ва¬
реный картофель, крахмал и др.) или олигосахаров (сахароза, мальто¬
за и др.). Независимо от вида источника углеводов конечным продук¬
том их гидролиза должны быть простые сахара, которые в бескислород¬
ных условиях превращаются микроорганизмами в молочную кислоту
(молочнокислое брожение). От вида источника глюкозы зависит выбор
стартовых штаммов, в качестве которых чаще всего используют молоч¬
нокислые бактерии (Lactobacillus brevis, L. plantarum, L. cellobiosus
и др.) или плесени (Aspergillus oryzae, A. flavus и др.) [289] . Молочно¬
кислые бактерии не способны гидролизовать нативный крахмал в проти¬
воположность плесеням, которые легко гидролизуют его до фермента¬
тивных сахаров [213]. В группу молочнокислых бактерий входят гомо-
ферментные штаммы, образующие из сахаров практически чистую мо¬
лочную кислоту, и гетероферментные штаммы, которые, кроме молоч¬
ной кислоты, образуют в разных количествах такие побочные продукты,
как уксусная кислота, этиловый спирт и др. Таким образом, правиль¬
ный выбор стартовых штаммов лежит в основе технологии ферментиро¬
ванного фарша, хотя известны также способы получения такого фарша
без добавления инокулята, путем использования лишь бактериальной
флоры рыбы.Регулирование активности ферментов в процессе ферментации дос¬
тигается с помощью температуры, а также хлористого натрия, кото¬
рый одновременно играет роль консерванта. В фарше, изготовленном из
вареного или пропаренного мяса рыбы, процесс ферментирования проис¬
ходит обычно без добавления поваренной соли, иногда' ее вводят лишь
в готовый продукт. Ниже в качестве примеров описаны некоторые
способы изготовления ферментированного фарша.Ферментирование с добавлением поваренной соли [453]. Измель¬
ченное мясо рыбы (75 %), пшеничные отруби (10 %), соль (14,5 %)
и препарат Aspergillus oryzae (0,5 %) после тщательного перемешивания
оставляют на 2 нед при температуре 25—30 °С для созревания. Первые
признаки ферментации (размягчение мяса) появляются после 4 сут;
через 5 сут появляется характерный запах, а между 7-ми и 13-ми сут фер¬
ментация проходит с самой высокой скоростью, что проявляется в за¬
метном увеличении содержания несвязанной жидкой фракции [454].
Кислотность смеси возрастает с pH 6,0—6,5 в начале до 4,5 в конце про¬
цесса. Созревший полуфабрикат подогревают до кипения для преры¬
вания ферментативной активности, охлаждают и разделяют с помощью
пресса или центрифуги на жидкую и плотную фракции. Плотная фрак¬
ция после перемешивания или гомогенизации до однородной массы
(пасты) превращается в готовый продукт, выход которого составляет
45—50 % массы использованного сырья.Ферментация без добавления поваренной соли [148] Измельченное177
мясо рыбы пастеризуют при температуре 80 °С в течение 20 мин с целью
инактивации протеолитических ферментов, быстро охлаждают до тем¬
пературы 30 °С, добавляют воду (25 %) , сахарозу (около 7,5 %), препа¬
рат молочнокислых бактерий (0,5 %), содержащий штаммы Lactobacillus
helveticus и L. acidophilus, азотнокислый калий (0,02 %) как средство,
препятствующее маслянокислому брожению, и хлористый кальций
(0,02 %) в качестве вещества, улучшающего консистенцию фарша, после
чего все тщательно перемешивают и инкубируют при температуре 30° С
в течение 24 ч. В конце этого периода pH смеси становится ниже 4,5.
Затем смесь отжимают на сыроваренном прессе для отделения несвязан¬
ной жидкой фракции, которая составляет 40—50 % первоначальной мас¬
сы компонентов, полученные блоки фарша погружают в 20 %-ный раст¬
вор соли на 1,5 ч, подсушивают и упаковывают нод вакуумом в пленоч¬
ные пакеты. Срок хранения продукта при комнатной температуре сос¬
тавляет около 5 мес.
7. ТРЕБОВАНИЯ К КАЧЕСТВУ
И СРОК ХРАНЕНИЯ ФАРША7.1. ТРЕБОВАНИЯ К КАЧЕСТВУК качеству фарша предъявляются органолептические, физик о-хими-
ческие и микробиологические требования. Из органолептических
свойств наибольшее значение имеют запах, вкус и консистенция фарша.
Физико-химические показатели, все еще недостаточно изученные, чаще
всего используют для контроля за технологическими процессами (со¬
держание применяемых добавок, например поваренной соли в соленом
фарше), а также при оценке показателей, характеризующих функцио¬
нальные свойства фарша при выработке из него некоторых видов про¬
дукции (например, растворимость белков фарцщ, предназначенного для
производства рыбных колбас). Микробиологические требования каса¬
ются в основном защиты человека от болезнетворных бактерий, а также
санитарного контроля за процессом производства.Требования к качеству значительно различаются в ^висимости от
вида фарша. При оценке качества мороженого сырого фарша наиболь¬
шее внимание уделяется запаху, вкусу и консистенции. Оценка этих
свойств обычно производится по пробе, подвергнутой модельной тепло¬
вой обработке, хотя для увеличения точности оценки рекомендуется
также исследовать сырой фарш непосредственно после его разморажи¬
вания. Модельная тепловая обработка заключается в варке или запека¬
нии фарша в установленных условиях. Например, пробу фарша массой
около 200 г заворачивают в алюминиевую фольгу и отваривают на пару
или запекают в духовом шкафу при температуре около 205 °С, после
чего нагревают до тех пор, пока температура в геометрическом центре
образца не достигнет 70 °С.Запах и вкус фарша должны быть естественными, присущими
данному виду рыбы. Не допускается присутствия посторонних вкуса
и запаха, запахов окислившегося жира или свидетельствующих о бак¬
териальной порче фарша. В фарше самого низкого качества допус¬
кают, однако, ослабление естественных запаха и вкуса мяса или их
ухудшение, вызванное хранением мороженого фарша. В стандартах
США, которые предусматривают деление стабилизированного фарша
на три класса качества, запах и вкус являются основными критерия¬
ми оценки, тогда как остальные показатели, включая и консистенцию,179
трактуются как равнозначные и выражаются суммарно в виде числа
физических дефектов незначительной, средней и высокой интенсив¬
ности (табл. 43).Таблица 43. Критерии оценки качества рыбного фарша
согласно стандартам США [ 498]КласскачестваВкус и запахМаксимальное число физических
дефектовНезначи¬тельныеСредниеЗначитель¬ныеАХорошие300ВДостаточно хорошие510СУ дов л етво рител ьные731Консистенция фарша должна быть характерной для пра¬
вильно вымешанного крупноизмельченного мяса с хорошо различимыми
частицами. Клеточный сок в фарше должен быть хорошо связан, фарш
должен быть сочным, плотным, но невязким. Такие свойства, как губ-
чатость или резинистость, после размораживания свидетельствуют о да¬
леко зашедших денатурационных изменениях белков, в частности об их
агрегации и ассоциации. Об этом же говорит наличие большого коли¬
чества несвязанной жидкой фракции (клеточного сока), которая отде¬
ляется при небольшом нажатии пальцем, а при снятии нагрузки частич¬
но вновь уходит в фарш. После тепловой обработки в таком фарше об¬
наруживается большая потеря массы, а при пробе ”на зуб” он оказыва¬
ется твердым, вязким и сухим. Нежелательна также мажущаяся или
слишком хрупкая (неплотная) консистенция, что является прежде
всего следствием чрезмерной протеолитической активности, вызванной
обычно ферментами, попавшими в мясо из внутренностей. В фарше
низшего класса качества допускается слегка резинистая, кашеобраз¬
ная или несколько суховатая консистенция. Для нестабилизированно-
го фарша эти критерии могут быть более смягченными. Остальные тре¬
бования к качеству касаются различных дефектов, часть из которых
может быть определена в мороженом виде, а часть - после разморажи¬
вания фарша. К дефектам, оцениваемым на мороженых блоках фарша
(после извлечения их из упаковки), относятся: пожелтение и высуши¬
вание поверхности, повреждения боковых сторон и углов, неправильная
форма, неоднородность размеров и массы блоков. К основным дефек¬
там, которые оцениваются после размораживания блоков, относятся:
наличие костей и неоднородность цвета в массе продукта. В табл. 44
представлена более подробная идентификация этих дефектов в соответ¬
ствии со стандартами США.Для определения количества костей в фарше рекомендуется ис¬
пользовать метод Ямамото и Уонга [507] , заключающийся в разведении
180
Таблица 44. Физические дефекты мороженого фарша
согласно стандартам США [ 498]Дефект качестваСтепень интенсивности дефекта1. Неоднородность
размеров2. Неоднородность
массы3. Наличие желтых пя¬
тен на поверхности4. Высушивание
верхности5. Повреждение граней
и углов блока6. Неправильная фор¬
ма блокаА. Оценка мороженого фаршаДва или более отклонения от принятой длины, ширины
или высоты блока (в каждом размере может быть оценен
только один дефект)Н (незначительный) - до ±3 мм
С (средний) - 3-i9,5 мм (±)3 (значительный) - более ±9,5 мм
Н - не более 57 г
3 - более 57 гПожелтение или побурение поверхности мороженого бло¬
ка, вызванное хранением:Н - незначительное
С - среднее
3 - значительноСублимация лы** из верхних слоев блока, в виде ’’хо¬
лодного ожога”, на поверхности, превышающей 5 % по¬
верхности блокаН - легко удаляется (соскабливается)С - требует применения ножа или другого предмета для
удаления3 - трудно удаляется из-за значительного проникновения
ожоговых пятен в глубь блокаНеправильная форма или механические повреждения
отдельных граней или углов блока. Повреждение грани
в двух местах или угла во всех трех плоскостях принима¬
ется за одно ’’полное” повреждение.Число ’’Полных” повреждений:Н - 2
С- 33-4 или болееНаличие воздушных или ледяных карманов либо углуб¬
лений, видимых повреждений на поверхности или на раз¬
резе блока. За единицу дефекта принимается карман
объемом 10 -2,5 1,5 см3
Н - 1-3 единицы
3 - свышё 3 единицБ. Оценка размороженного фарша7. Неоднородность Видимые кусочки кожи, брюшной пленки, чешуя или
цвета в массе про- сгустки крови; число очагов с поверхностью 0,4 см2 на
дукта 1кг фарша:Н - Ю-гЗО
С-31^60
3 - более 608. Наличие костей Острые и твердые кости или их кусочки длиной свыше6 мм; их число на 2,5 кг фарща:Н - 1-й; С - 3-i4; 3 - свыше 4, но не более 10181
ПродолжениеДефект качестваСтепень интенсивного дефектаВ. Оценка фарша после тепловой обработки9. Дефекты консистен- Н - фарш несколько губчатый или резинистый, однако
ции без признаков мажущейся консистенцииС - среднегубчатый или среднерезинистый фарш3 - фарш сильно губчатый, резинистый, сухой или оченьмажущийсяпробы фарша раствором мочевины концентрацией 3 моля/л + 0,02 моля/л
едкого натра при температуре 35 ±5 °С в течение примерно 16 ч.При оценке промытого фарша, кроме упомянутых выше характе¬
ристик и критериев качества, применяются еще и другие, специфические
для этой группы продукты. В случае сурими это содержание воды и спо¬
собность образовывать эластичный гель после тепловой обработки.
Эластичность камабоко, изготовленного из сурими, определяется пене¬
трометром или органолептическим методом, при котором образец
складывается пополам и вчетверо. Кроме того, в качестве вспомогатель¬
ных показателей при лабораторных испытаниях определяют тиобарбиту-
ровое число (ТБЧ) и растворимость белков в растворе хлористого калия
концентрацией 0,6 моль/л [42]Чтобы отличить соленый сурими от несоленого, определяют содер¬
жание хлористого натрия или общее содержание золы (табл. 45).Таблица 45. Сравнение основного химического состава
соленого и несоленого сурими из минтая [ 303]Каэн-сурими, содер¬
жащий 2,5 % соли
и 5 % сахараМуэн-сурими, содер¬
жащий 0,2 % полифос¬
фатов и 5 % сахараВодаЖирОбщий белок
ЗолаУглеводы74,00,413,62,39,776,60,414,40,77,9Новейшие исследования в области оценки качества промытого фар¬
ша ведутся в направлении разработки объективных показателей, харак¬
теризующих качество фарша, а особенно его технологическую пригод¬
ность. Эти показатели включают также содержание и функциональные
свойства тех белков, которые более всего влияют на желаемые показа¬
тели фарша как сырья для переработки актомиозинов. Критерии оценки
технологической пригодности сурими по содержанию актомиозина и ак¬
тивности Са2+ — АТФазы представлены в табл. 46.182
Таблица 46. Содержание актомиозина и активность Са2* - АТФазы
в мороженом сурими в зависимости от класса качества [ 190]КласскачестваСодержание
актомиозина,
мг/10 г
суримиАктивность Са2+ - АТФазыспецифическая
мкмоль неорг.
Р/ (мин-мг акто¬
миозина)общая
мкмоль неорг.Р/
(мин 10 г сурими)SA10180,306312 100В9010,311280 80В*8350,290242 77А8090,295239 76При оценке качества несырого (вареного, соленого, ферментирован¬
ного и т. д.) фарша большое внимание уделяется также химическому
составу и пищевой ценности белка. Происходящие в таком фарше с от¬
носительно большой скоростью процессы неферментативного побурения
обычно ведут к значительному снижению биологической ценности белка.
Это особенно касается солено-сушеного фарша, который хранится при
температуре окружающей среды (табл. 47) . В ферментированном фарше
основным показателем качества является pH, который должен быть ни-Таблица 47. Сравнение основного химического состава,
пищевой ценности, численности микроорганизмов в свежеприготовленном
солено-сушеном фарше из Nicholsina usta и после длительного хранения[76]Содержание, %PER (белковая эф¬
фективность)Численность
бактерий в
1 г продуктарезуль¬
тат, по¬
лучен¬
ный в
иссле¬
довани¬
яхрезультат
пересчета
при усло¬
вии, что
PER казеи¬
на ра¬
вна 2,5Фаршводыбел¬касолиNPUоб¬щаягало-филовСвежеприготов¬ленный12,930,956,23,6±0,42,789,9±6,1900012000После 18 месхра-7,931,061,42,8 ±0,42,177,9± 4,1бнения в условиях
тропикова - фарш, спрессованный под давлением около 14 МПа и отформованный в
виде лепешек массой около 0,5 кг, затем высушенный на солнце в течение 4 сут
(температура 33-37 °С, относительная влажность 65-75 %); б - менее 30 при
разведении 1:10.183
же 4,5, а также содержание продуктов гидролиза белка. Принимается,
что в достаточно созревшем продукте отношение содержания небелко¬
вого азота к общему азоту должно превышать 45 % [454] . Порча фарша
происходит, когда содержание аммиака возрастает до 90 мг% [455]Микробиологическая оценка рыбного фарша заключается в опреде¬
лении присутствия в нем болезнетворных и индикаторных бактерий,
а также в сравнении полученных результатов с нормативами, установ¬
ленными стандартами. Некоторые бактерии вызывают пищевые отравле¬
ния в результате интоксикации ядами (токсинами), которые образуют¬
ся вне клеток (например, золотистый стафилококк Staphylococcus aure¬
us), в результате действия других возникает токсикоинфекция, третьи
бактерии не образуют токсинов, переходящих в продукт, но размножа¬
ются в пищеварительном тракте человека, что также приводит к отравле¬
нию (например, кишечные палочки рода Salmonella, дизентерийные па¬
лочки рода Shigella). В последнее время при исследовании рыбного фар¬
ша на наличие болезнетворных бактерий предлагают также включать
определение мезофильной анаэробной палочки Vibrio parahaemolyticus,
поскольку она повсеместно распространена в морской среде [493, 514]
Из индикаторных микроорганизмов исследуют кишечную палочку
(Escherichia coli), которая присутствует в пищеварительном тракте чело¬
века и животных. Наличие этих микроорганизмов в фарше свидетельст¬
вует о несоблюдении санитарно-гигиенических правил производства,
и прежде всего личной гигиены работников. Степень обсемененности
фарша в процессе производства можно также определить по общей чис¬
ленности психрофильных бактерий. В процессе производства фарша
обсемененность обычно многократно возрастает: в 1000 раз в сепариро¬
ванном мясе (нестабилизированный фарш) и в 10 000 раз в промытом
фарше [435] Увеличение обсемененности сепарированного мяса проис¬
ходит главным образом в результате плохого ухода за оборудованием, а
промытого фарша — при перекачивании фарша через систему трубопро¬
водов, а также во время отделения от фарша избытка воды. В стабили¬
зированном фарше дополнительными источниками бактериальной об¬
семененности могут быть добавки, например приправы. Пользуясь
показателем общей численности микроорганизмов, следует помнить,
что микроорганизмы, чувствительные к действию низкой температу¬
ры, гибнут в процессе замораживания и холодильного хранения. Если
обсемененность свежеприготовленного фарша составляет 104 — 10s
клеток на 1 г, то, например, после двухмесячного хранения она не
должна превышать 103-104 клеток на 1 г [435]Летальное действие замораживания тем сильнее, чем ниже темпе¬
ратура хранения замороженного фарша [116], но даже длительное
хранение в замороженном виде не приводит к полному уничтожению
изначально присутствующей микрофлоры [514]Это свидетельствует о необходимости постоянного микробиоло¬
гического контроля мороженого рыбного фарша. Важнейшей целью
184
этого контроля является предотвращение обсеменения фарша болезне¬
творными бактериями из-за несоблюдения определенных технологи¬
ческих и санитарных требований.Принятые в Польше микробиологические требования для стаби¬
лизированного рыбного фарша не допускают наличия в нем палочек
группы Salmonella в 20 г, палочек группы Escherichia coli и грамполо-
жительных стафилококков в 0,01 г продукта [518]7.2. СРОК ХРАНЕНИЯ ФАРШАСрок хранения рыбного фарша зависит прежде всего от вида рыбы,
способа консервирования (стабилизации), вида и качества упаковки,
температуры хранения.Влияние этих факторов на изменение качества фарша при хранении
зависит от его вида. В сыром мороженом фарше указанные факторы
определяют практически все изменения компонентов мяса рыбы. В пас¬
теризованном фарше эти факторы касаются главным образом окисления
липидов и взаимодействия продуктов их окисления с белками. В соле¬
ном и ферментированном фарше, кроме этих изменений, большое влия¬
ние на срок хранения фарша оказывает неферментативное побурение.Влияние вида рыбы на срок хранения фарша определяется свойст¬
вами измельченного мяса, а точнее, его подверженностью нежелатель¬
ным физико-химическим изменениям. Для фарша из жирной рыбы
эти изменения прежде всего касаются окисления липидов. Таким об¬
разом, качество фарша из жирной рыбы снижается в основном в ре¬
зультате появления запаха и вкуса окисленного жира. В нестабилизи-
рованном мороженом фарше из жирной рыбы запах появляется после
2—4 мес хранения в зависимости от вида рыбы и температуры хране¬
ния (табл. 48). Появление заметного запаха и вкуса окисленного жира
практически означает полную непригодность фарша для потребления,
что для большинства жирных и среднежирных рыб наступает после
4—5, а для сардины — после 2,5—3 мес хранения.Введение в фарш антиокислителей значительно удлиняет срок его
хранения. Например, хорошее качество мороженого фарша из ставри¬
ды, скумбрии и салаки с добавками соответствующих смесей естест¬
венных приправ сохраняется в течение 5—6 мес хранения при темпера¬
туре —28 °С, а полностью непригодным для употребления он становит¬
ся примерно после 9 мес хранения [220, 221] Добавление антиокис¬
лителей и искусственных синергических веществ удлиняет срок хране¬
ния мороженого фарша из рыбы средней жирности более чем в 3 раза
по сравнению с нестабилизированным фаршем (табл. 49).Качество мороженого фарша из тощей рыбы ухудшается в основ¬
ном в результате снижения водоудерживающей способности и ухудше¬
ния консистенции. Эти изменения обусловлены главным образом денату¬
рацией мышечных белков. В общем можно принять, что срок хранения185
Таблица 48. Время хранения измельченного мяса из жирной рыбы
н рыбы средней жирности до момента появления заметных органолептических
признаков окисления жираРыбаСодер¬
жание
жира в
мясе, %Темпера¬
тура хра¬
нения,
°СПродолжительность хранения
(мес) до момента органолеп¬
тического обнаружения
признаков окисленияначальныепризнакизаметные запах
и вкус окислен¬
ного жираИсточникЖирная сардина2,5-281,5-22,5-3[ 220,221]Тощая сардина23[220,221]Ставрида-283-3,54-5[220,221]Скумбрия-283-44-5[220,221]Балтийская-28ок. 45-5,5[220,221]сельдьКилька8,1-181,5-20[417]Тюлька17,7-1820[417]Хамса5,3-1830[417]Катран8,7-182-30[417]Таблица 49. Влияние стабилизирующих добавок на возможный срок хранения
мороженого фарша из каменного окуня (Sebastes flavidus) [ 458]ПартияПрименяемые добавкиПродолжитель¬
ность хранения,
месСтепень окисле¬
ния по 10-балль¬
ной шкале**АВсе добавки*, включая антиокислите-010,0ли БОА и БОТ310,0610,097,5127,0ВВсе добавки, за исключением антиокис-010,0лителей34,0СБез добавок010,034.0* Состав применяемых добавок, %: вода - 5, рафинированное растительное
масло - 1, соль - 1, сахар - 1, трифосфат натрия - 0,15, глутамат натрия - 0,3,
БОА и БОТ по 0,006. Полисорбат 80 и сорбат моноолеиновой кислоты - по 0,075.** При степени окисления ниже 6 баллов фарш к дальнейшему хранению не¬
пригоден.мороженого фарша из тощей рыбы прямо пропорционален стабильности
(устойчивости к денатурации) миофибриллярных белков при замора¬
живании. Фарш из минтая, трески, путассу и других рыб, для которых
характерно быстрое снижение растворимости миофибриллярных белков
186
в процессе морозильного хранения, быстро теряет технологическую при¬
годность. Если фарш не стабилизирован, срок его хранения ограничи¬
вается 2-3 мес. После этого как водоудерживающая способность, так
и способность к эмульгированию жира обычно настолько низки, что
фарш после размораживания практически непригоден для производства
продуктов, изготовляемых на основе эмульсии (сосиски, паштеты).
Однако такой фарш можно использовать для производства продуктов,
порционируемых путем распиливания или формованных из измельчен¬
ного мороженого мяса, минуя операцию размораживания [241]В последнем случае срок хранения увеличивается до 5 мес. Неста-
билизированный фарш из рыбы, мышечные белки которой характери¬
зуются высокой устойчивостью к денатурации при замораживании
(например, аргентина), может храниться даже до 9 мес. Следует, одна¬
ко, отметить, что органолептическая опенка этого фарша со второй
половины срока хранения низкая, часто он едва приемлем, отмечается
значительная потеря сока и относительно твердая, волокнистая кон¬
систенция после термической обработки.Эффективное предотвращение денатурации белков в измельченном
мясе тощей рыбы при замораживании способствовало бы удлинению воз¬
можного срока хранения фарша. Это достигается с помощью антиденату-
рантов и средств, повышающих во до удерживающую способность мяса
(см. раздел 5), а также применением низких температур заморажива¬
ния и хранения. Например, в измельченном мясе минтая, замороженном
при температуре —40 °С и хранившемся при той же температуре, раст¬
воримость мышечных белков снижается незначительно [114], но хране¬
ние при более высокой температуре (выше —20 °С) ускоряет денатура¬
цию белков. В стабилизированном фарше из сайды, замороженном при
температуре —40 °С и хранившемся при -30 °С, активность Са2+ -
АТФазы после 25 мес хранения снизилась всего на 15 %, при этом все
свойства, делающие его исключительно пригодным для производства
камабоко, сохранились [113] . Фарш из других видов тощей рыбы не
выдерживает столь длительного хранения. В общем можно принять,
что максимальный срок хранения стабилизированного фарша состав¬
ляет 9-12 мес.Использование герметичных упаковок для улучшения качества и
удлинения срока хранения мороженого фарша представляется наибо¬
лее целесообразным, если он изготовлен из рыбы средней жирности
(содержание жира в мясе 2-10 %). Консистенция фарша из такой рыбы
не столь быстро изменяется в процессе хранения, как фарша из тощей
рыбы (см. рис. 56). Вместе с тем окисление жира так явно не влияет на
снижение его качества, как в случае жирной рыбы. Однако основным
фактором снижения качества этого фарша в процессе хранения являет¬
ся окисление липидов (рис. 57). Поскольку содержание естественных
проокислителей в мясе среднежирной рыбы невысоко, окисление жира
происходит главным образом в наружных слоях блока мороженого187
Продолжительность хранения, месРис. 56. Влияние морозильного хранения при температуре -20 °С на изменение
консистенции измельченного мяса рыбы разной жирности [ 334]. Содержаниежира в фарше (в %) :1 - из тихоокеанской трески - 0,6; 2 - минтая 2+ Sebastes babcocki (смесь 1:1) -
2,2; 3 - минтая - 0,8; 4 - Sebastes alutus - 2,2; 5 - Sebastes babcocki - 2,2; 6 -
Sebastolobus alascanus - 2,0; 7 - катрана - 8,7; 8 - Atheresthes stomias - 5,5фарша (рис. 58), что свидетельствует о значительном влиянии доступа
воздуха на эти изменения. Проникновение кислорода через слой замо¬
роженного фарша происходит быстрее, чем через слой льда [107] , поэ¬
тому глазирование фарша и применение газонепроницаемых упаковок
значительно замедляют скорость окислительных изменений липидов.
Динамика окислительных изменений липидов в наружных слоях фар¬
ша, упакованного в газонепроницаемую пленку, подобна динамике их
изменений во внутренних слоях (см. рис. 58). Отсюда следует, что
основной причиной окисления липидов в мороженом фарше из средне¬
жирной и тощей рыбы в процессе хранения является кислород воздуха,
а не эндогенный кислород, абсорбированный измельченным мясом во
время его изготовления или выделенный соединениями, которые его
связывают, например миоглобином и гемоглобином.В промытом фарше, который характеризуется низким содержанием
жира и естественных проокислителей, основным фактором, определяю¬
щим возможный срок хранения, являются денатурационные изменения
белков. Для обеспечения возможно более длительного хранения такого
фарша желательно поддержание как можно более низкой температуры
хранения. Например, при температуре -20 °С срок хранения сурими из
188
минтая составляет 12—13 мес, а при -10 °С — всего 2—3 мес [171]
На срок хранения фарша влияют качественный и количественный сос¬
тав стабилизирующих добавок. Например, соленый сурими (каэн-сури-
ми) лучше сохраняет высокую пригодность. к обработке в начальный
период морозильного хранения (до 4—5 мес при температуре —20 °С),
чем несоленый (муэн-сурими), однако по истечении этого периодаа бПродолжительность хранения, месРис. 57. Влияние продолжительности хранения при температуре -20 °С на изме¬
нение ТБЧ в мороженых блоках измельченного мяса рыбы с разным содержани¬
ем жира [334]:а - внешняя часть блока (слой толщиной 3 мм); 6 внутренняя часть блока
(около 30 мм от поверхности); остальные обозначения как на рис. 56189
12345678 12345678Продолжительность хранения, месРис. 58. Изменение ТБЧ в отдельных слоях блоков мороженого измельченного
мяса серебристого хека, хранившихся при температуре -6,7 °С в обычной и газо¬
непроницаемой упаковке [ 277] :а - картонные провощенные коробки; б - такие же коробки, завернутые в лами¬
нированную пленку типа Marvelseal 1312 с низкой проницаемостью для кислорода.
(Блоки размерами 25,4 X 15,3 X 5,7 см, массой 23 кг распиливали перед отбором
проб ленточной пилой на куски толщиной 0,5 см, которые укладывали параллель¬
но наибольшей поверхности блока, при этом образовалось по пять слоев: 1 - на¬
рушенный слой; 2-4 - средние слои; 5 - слой, расположенный в середине блока)качество его быстро ухудшается в основном из-за гелеобразования.
Изделия типа камабоко i несоленого сурими по качеству уступают
изделиям из соленого сурими, однако несоленый фарш хранится доль¬
ше, минимум 10 мес, при температуре —20 °С [360]Мороженый пастеризованный фарш также может храниться доволь¬
но долго. Для фарша, изготовленного из свежей рыбы, срок хранения
при температуре —21 ± 1 °С составляет 7-8 мес, а из мороженого сырья -
не более 4 мес [154] Чем дольше хранится рыба до переработки на
фарш, тем короче возможный срок хранения пастеризованного моро¬
женого фарша. Стабилизирующая активность добавок в пастеризо¬
ванном фарше ниже, чем в сыром, поэтому чаще всего они представляют
собой вкусоароматические вещества [299] Дальнейшее удлинение
190
срока хранения пастеризованного фарша прежде всего требует разра¬
ботки новых способов улучшения консистенции. Кроме механических
(перемешивание) и биотехнологических (применение протеолитических
ферментов) процессов, большие возможности в этой области открывает
применение соответствующим образом подобранных гидроколлоидов
(например, гуммиарабики Locust, Guar и др.).Длительный срок хранения ферментированного фарша обусловлен
не только низким pH продукта, но и, вероятно, образованием антибио¬
тиков в процессе ферментации [146] Однако эти вопросы требуют
дальнейшего изучения.Срок хранения сушеного и солено-сушеного фарша зависит в основ¬
ном от условий сушки, вида и способа упаковки. Задача заключается
в поддержании низкой активности воды (aw), предотвращающей раз¬
витие микроорганизмов, и ограничении доступа атмосферного кисло¬
рода к продукту. Если эти условия выполняются, срок хранения фарша
при комнатной температуре достигает 12 мес. Для солено-сушеного
фарша из жирной рыбы часто необходимо применять антиокислители.Итак, возможный срок хранения рыбного фарша не является таким
однозначным критерием, как в случае изделий, у которых как состав,
так и условия хранения жестко предопределены. Предотвращая или ог¬
раничивая действие внутренних (биологических) и внешних (техно¬
логических) факторов, обусловливающих нежелательные физико-хими-
ческие изменения, происходящие в фарше при хранении, срок его хране¬
ния можно значительно удлинить.
ЛИТЕРАТУРА1. Addison R, F., Ackman R. G.: Separation of triglycerides and free fatty acids on
Sephadex LH-20-Лш/. Bioch., 28;515-522, 19692. Addison R. F. i in.: Distribution of fatty acids in cod flesh lipids. J. Fish. Res.
Bd Can., 25; 2083-2090, 19683. Akiba М., Motohiro Т., Tanikawa E.: Preventing denaturation of the proteins in
frozen fish muscle and fillets. I. Effects of additives on the quality of frozen minced
fish muscle. J. Food Technol., 2; 69-78,19674. Alderman G. G., King G. J., Sugiyama H.: Factors in survival of Clostridium bo-
tulinum type E spores through the fish smoking process, f. Mila Food Technol.,
35; 163-166, 19725. Aleksiejewa Т. I., Bazilewicz W. I., Wangesowa W. A, Gotowaniec W. A., Gle-
bowa L. N.: Utuczszenie technofogii proizwodstwa ’’Sofomki iz rybnogo tjesta”.
Rybn. Chozj. nr 2, 69-71, 19746. Aleksiejewa Т. I., Bojcowa Т. М.: Wozmoznyj wariant proizwodstwa wareno-
morozenogo farsza iz mintaja na sudah tipa BMRT. Rybn. Chozj. nr 10, 53, 54,19747. Aleksiejewa Т. I., Pupikin A. Ja.: Linija po proizwodstwu piszczewogo farsza iz
mintaja na PS ’’Sewierodoneck”. Rybn. Chozj. nr 8, 54-57, 19738. Amano K., Bito М.: Consequence of free amino acids generated from decom¬
posing fish muscle.Bull. Jap. Soc. Sci. Fish., 15(12); 10-16, 19519. Amano H., Yoshida C., Nakamura A.: Cryoprotectant for frozen ground fish.
Jpn. Kokai Tokkyo Koho 80 07, 017 (Cl. A23 LI1325), 18 Jan. 1980, Appl.
.78/78, 241, 28 Jan. 1978. Uprawniony z patentu: Kao Soap Co., Ltd., Japonia198010. Anderson M. L., King F. J., Steinberg M. A.: Effect of linolenic, linoleic and
oleic acids on measurement of protein extrability from cod skeletal muscle using
the solubility test. J. Food Sci., 28; 286-288, 196311. Anderson M. L., Mendelsohn J. М.: A rapid salt-curing technique. J. Food Sci.,
37; 627-628, 197212. Anderson M. L., Steinberg M. A.: Effect of lipid content on protein-sodium lino-
lenate interaction in fish muscle homogenates. J. Food Sci., 29; 327-330, 196413. Anderson M. L., Steinberg M. A., King F. J.: Some physical effects of freezing fish
muscle and their relation to protein-fatty acid interaction. In: The Technology
of Fish Utilization, ed.by R. Kreuzer, pp. 105-110. FAO, Fishing News (Books)
Ltd., London 196514. Anonimowo: Why fish fingers top the market. Fishing News Intern., 14(6); 30-34,197515. AOAC.: Official Methods of Analysis, (9th ed.) Assoc. Agric. Chemists, Washing¬
ton 196016. Arai K., Fukuda М.: Studies of musculatur proteins of fish. XII. Effect of tempe¬
rature on denaturation of actomyosin ATPase from carp muscle. Bull. Jap. Soc.
Sci. Fish., 39(6); 625-631, 1973192
17. Arai К., Takahashi H., Saito A.: Studies on musculatur proteins of fish. III. In¬
hibition by sorbitol and sucrose on the denaturation of carp actomyosin during
frozen storage. Bull. Jap. Soc. Sci. Fish., 36(3); 232-236, 197018. Arai K., Takahashi H.: Studies on musculatur proteins of fish. XI. Effect of free¬
zing on denaturation of actomyosin ATPase from carp muscle. Bull. Jap. Soc.
Sci. Fish., 39(5); 533-541, 197319. Areche N., Fuji Y.: Effects of vegetable protein added to frozen surimi on the pro¬
perties of its kamaboko. Bull. Tokai Reg. Res. Lab. No. 100, 1-7, 197920. Atkinson A.: Storage tests on dried hake mince. An. Rep. FIRI, 30; 71-73, 197621. Aymard C., Cuq J. L., Cheftel J. C.: Formation of lysino-alanine and lanthiamine
in various food proteins, heated at neutral or alkaline pH. Food Chem. No 3,
1-5, 197822. Babbitt J. K., Crawford D. L.: Quality and utilization of minced deboned fish
muscle. Second Technical Seminar on Mechanical Recovery and Utilization of
Fish Flesh. Boston, June 12-13, pp. 32-43, ed. by E. R. Martin National Fishe¬
ries Institute, Washington 197423. Babbitt J. K., Law D. K., Crawford D. L.: Acceptance of a fish-shrimp portion
utilizing machine-separated minced fish flesh. J. Food Sci., 39; 1130-1131, 197424. Baliga B. R., Mooijani M. N., Lahiry N. L.: Changes with pyrophosphate, conta¬
ining buffer and precipitation of protein fraction at //2=0,225 during storage of
fresh-water fish. Food Technol., 16; 84-86, 196225. Barnett H. J., Nelson R. W., Hunter P. J., Bauer S., Groninger H.: Studies on the
use carbon dioxide dissolved in refrigerated brine for the preservation of whole
fish. Fishery Bulletin. 69(2); 433-442, 197126. Bauemfeind J. C., Desai I. D.: The tocopherol content of food and influencing
factors. Critical Rev. in Food Sci. and Nutrition, 8(4); 337-382, 197727. Beamish F. W. H.: Glycogen and lactic acid concentrations in Atlantic cod (Gadus
morhua) in relation to exercise. J. Fish. Res. Bd Can., 25(5); 837-851, 196828. Bello R. A., Pigott G. М.: A new approach to utilizing minced fish flesh in dried
products. J. Food Sci, 44(2); 355-359, 197929. Bhatia D. S., Moorjani M. N., Iyengar J. R., Visweswaraiah K.: Fish macaroni.
Food Science, 8; 1-3, 195930. Bietova Z. I.: Wlijanje stiepieni izmiefczenija farsza na jego w&gouderziwajuszczu
sposobnost. Trudy WNIRO, 90; 34-37, 197431. Black E. C., Manning G. Т., Hayashi K.: Changes in levels of hemoglobin, oxigen,
carbon dioxide, pyruvate and lactate in venous blood of rainbow trout (Salmo
gairdneri) during and following serve muscular activity. J. Fish. Res. Bd Can.,
23(6); 783-795, 196632. Blight E. G.: Salt minced fish. Ed. by Fisheries and Marine Service Environment
Canada, Halifax N. S. (broszura), 1-15, 197633. Blight E. G.: A note on salt minced fish. In: Handling, Processing and Marketing
of Tropical Fish, pp. 291-292. Tropical Product Inst., London 197734. Blin P. C., Balea Т., Prudhomme М.: Une nouvelle methode d’identification des
poissons: la diagnose myologique. Recu de Medicine Vet.,79; 483-501, 195335. Boddeke R., Slijper J., van der Stelt A.: Histological characteristics of the body
musculature of fishes in connection with their mode of life. Koninkl. Nederl.
Akademie van Wetenshappen, Proc. C., 62; 576-588, 195936. Bolee effektiwnoje ispolzowanie meticich widow siga pri pomoszczi predwari-
telnoj oczistki i farszirowki ryby. Prospekt firmy Trio Engineering Ltd A/S37. Bond R. М.: Background paper on minced fish. FA О Fisheries Circular No. C332,
Codex Fisheries Committee, Rome, Italy, 1-34, 197538. Borderias A. J., Moral A., Garcia-Matamaras E.: Storage properties of blue whi¬
ting after mincing by different methods. Intern. J. Refrig., 3(1); 42-46, 198039. Boyd J. W., Southcott B. A.: Effect of polyphosphates and other saltes on drip
loss and oxidative rancidity of frozen fish. J. Fish. Res. Bd Can., 22(1); 53-67,
40. Braekkan О. R., Lambertsen G., Myklestad H.: Alpha-tocopherol in some of
fishes. Fisk. Dir. Skr.f Serie Teknologiske unders kelsel, 3(8); 1-42, 195941. Braekkan O. R., Lambertsen G., Myklestad H.: Alpha-tocopherol in some orga¬
nisms and fish oils. Fiskeridirektorates Skrifter, 4(8); 1-11, 196342. Bruun A., Olsen К. B.: Studierejse til Japan august 1971. Fiskeri-fiskeriindustri
og anvendelse af fiskeprodukter i Japan. Fiskeriministeriets Fors Ogslaboratorium.
Denmark 197143. Budenko M. S., Kuzmiczewa G. М., Rambeza E. F.: Isledowanije po utoczeniju
niekotorih rezimow zagotowki piszczego rybnogo farsza. Trudy Atlant NIRO
nr 54, 25-31, 197344. Bunnell R. H., Bauemfeind J. C.: Chemistry, uses and properties of carotenoids
in foods. Food Technol., 16(7); 36-43, 196245. Burt J. R., Simmonds С. K.: Hypoxanthine as an indicator of freshness in iced
cape hake before freezing and after thawing. In: Fish inspection and quality cont¬
rol pp. 196-200. FAO, Published by Fishing News (Book) Ltd., London 197146. Buttkus H.: The reaction of myosin with malonaldehyde. J. Food Sci., 35; 558—
560, 196747. Buttkus H.: Accelerated denaturation of myosin in frozen solution. J. Food Sci.,
35; 558-564, 197048. Bykova V. М.: Faktory wlilajuszczije na wlagouderziwojuszczuju sposobnost
mjasa ryby i teplokrownych ziwotnych. Mirow. Rybot. nr 11, 65-67, 196949. Bykova V. М.: Puti uluszczenia kaczestwa farsza iz mrozonoj ryby. Ryb. Chozj.
nr 2, 48-51, 197250. Bykova V. М.: Wlijanje niekotorych dobawok к rybnom farszu na jego struktu-
ralno-mechaniczeskie swoistwa. Trydy'WNIRO, 95; 44—48, 197451. Castell С. H.: Metal-catalyzed lipid oxidation and changes of protein in fish. J. Am.
Oil. Chem. Soc., 48; 645-649, 1971a52. Castell С. H., Smith B., Noal W.: Production of dimethylamine in muscle of seve¬
ral species of gadoid fish during storage, especially in relation to presence of dark
muscle. Fish. Res. Bd Can., 28; 1-5, 1971b53. Chapman D.: Physical studies of lipid-lipid and lipid-protein interactions. Lipids,
4(4); 251-256, 196954. Cheng C. S., Hammann D. D., Webb N. B.: Effect of thermal processing on min¬
ced fish gel texture. J. Food Sci, 44(4); 1080-1086, 1979a55. Cheng C. S., Hammann D. D., Webb N. B., Sidwell V.: Effects of species and
storage time on minced fish gel texture. J. Food Sci., 44; 1087-1092, 1979b56. Cheng C. S., Parrish F. C. Jr.: Heat-induced changes in myofibrillar proteins of
bovine longissimus muscle. J. Food Sci, 44; 23-26, 1979c57. Childs E. A.: Interaction of formaldehyde with muscle in vitro. J. Food Sci., 38;
1009-1011, 197358. Childs E. A.: Interaction of free fatty acids with fish muscle in vitro. J. Fish. Res.
Bd Can., 31(1); 111-116, 197459. Chio S., Tappel A. L.: Inactivation of ribonuclease and other enzymes by peroxi-
ding lipids and by melonaldehyde. Biochemistry, 8(7); 2827-2832, 196960. Chipault J. R., Mizino G. R., Hawkins J. М., Lundberg W. О.: The antioxidaht
properties of natural spices. Food Res., 17; 46-55, 195261. Christians O.: Untersuchung iiber die Verarbeitung kleiner Fische und Beifang
mit'Hilfe von Gratenseparatoren zu Fischfarce. Food Res., 80-82, 196862. Chrzanowski S.: Krotki przegl^d opakowan z kartonu i tektury do ryb i produk-
tow rybnych ze szczegolnym uwzgl^dnieniem opakowan do asortymentow mro-
zonych. Zesz. Centr. Lab. Przem. Rybnego, 19(13); 24-30, 1971>3. Collins J., Reppond K. D., Bullard D. A.: Black rockfish, Sebastes melanops:
changes in physical, chemical and sensory properties when held in ice and in car¬
bon dioxide modified refrigerated seawater. Fish. Bull., 77(4); 865-870, 1980194
64. Connel J. J.:' Studies on the proteins of fish skeletal muscle. 7. Denaturation and
aggregation o;f cod myosin. Biochem. J., 75; 530-538, 196065. Connel J. J.: The role of formaldehyde as a protein crosslinking agent acting
during the frozen storage of cod. J. Sci Food Agric., 26; 1925-1929, 197566. Connel J. Ji, Howgate P. E.: Studies on the proteins of fish skeletal muscle. 6. Ami¬
no acid composition of cod myofibrillar proteins. Biochem. J., 71; 83-86, 195967. Crawford D. L., Law D. K., Babbitt J. K.: Yield and acceptability of machine
separated minced flesh from marine food fish. J. Food Sci., 37; 551-553, 197268. Crawford D. L., Law D. K., Babbitt J. K., McGill L. A.: Comapative stability and
desirability of frozen pacific hake fillet and minced flesh blocks. J. Food Sci.,
44(2); 363-367, 197969. Crean P. B.: The light pickle salting of cod. J. Fish. Res. Bd Can., 18(5); 833-
844, 196170. Cyganska J., Witwicka J.: Emulgatory i emulsje w przemysle spozywczym. WPLiS,
Warszawa 196771. Czizow G. B.: Procesy cieplne w technology chfodniczej produktow zywnoscio-
wych. WNT, Warszawa 197472. Czizow G. B., Czuranow O. A.: Tworzenie krysztatow lodu w produktach zywnos-
ciowych przy zamrazaniu. Materiaiy z konferencji NOT, SITPSpoz., Warszawa 197073. Datta P. K., Frazer A. C., Sharrat М., Sammons H. G.: Biological effects of food
additives. II - Sodium pyrophosphate. J. Sci. Food Agric., 13(11); 556-566, 196274. Davidkova E., Svadlenka I., Rosmus J.: Interaction of malonaldehyde with collagen.
I. The reaction condition. Z. Lebensm. Unters.-Forsch., 153; 13 — 16, 197375. Del Valle F. R.: Un metodo nuevo para la conservacion rapida у barata del pescado.
Technologia de Alimentos, 7(1); 12-16, 197276. Del Valle F. R., Bourges H., Haas R., Gaona H.: Proximate composition, protein
quality and microbial counts of quick-salted, freshly made and stored fish cakes.
J. Food Sci, 41; 975-976, 197677. Del Valle F. R., Gonzales-Inigo J. L.: A quick-salting process for fish. 2. Behavior
of different species with respect to the process. Food Technol., 22(9); 85-88,
1968a78. Del Valle F. R., Nickerson J. T. R.: Studies on salting and drying fish. I. Equili¬
brium considerations in salting. J. Food Sci, 32; 173-179, 196779. Del Valle F. R., Nickerson J. T. R.: A quick-salting process for fish. 1. Evaluation
of the process. Food Technol., 22(8); 104-106, 1968b80. Del Valle F. R., Padilla М., Ruz A., Rodriguez R.: Pilot plant production of and
large scale acceptance trials with quick-salted fish cakes. J. Food Sci., 38(2); 246-250, 197881. Dingle J. R., Eagles D. E., Neelin J. М.: Electrophoretic studies of proteins extrac-
table from post-rigor cod at low ionic strenght. J. Fish. Res. Bd Can., 12(1); 75-83,
195582. Dingle J. R., Hines J. A.: Protein instability in minced flesh from fillets and frames
of several commercial Atlantic fishes during storage at -5°C. J. Fish. Res. Bd Can.,
32; 775-783, 197583. Dolbisz G. A.: Ispolzowanie atlanticzeskoj ryby - sabli dla prigotowlenija piszcze-
wogo morozennogo farsza. Rybn. Chozj. nr 2, 67-69, 197384. Dolbisz G. A.: Ispolzowanie morskogo pietuha dla prigotowlenija piszczewogo mo¬
rozennogo farsza. Rybn. Chozj. nr 2, 70-72, 197485. Dolbisz G. A., Noskova W. J.: Piszczewoj farsz iz atlantyczeskoj stawridy. Rybn.
Chozj. nr 10, 66-68, 197186. Dreosti G. М., Doesburg J. J., Atkinson A., Lamprecht E.: Fish chesse spread. FIRI
Annual Report nr 2, 56-57, 196787. Dugal L. C.: Hypoxanthine in iced freshwater fish. J. Fish. Res. Bd Can., 24(11);
2229-2239, 1967195
88. Dutkiewicz D.: Maszyny i urz^dzenia do obrobki nowych gatunk<5w ryb i bezkr^-
gowcow. W: Zagospodarowanie nowych surowcow rybnych. Materiaty konferencyjne
Sekcji Przemyshi Rybnego SITPSpoz. i Zjednoczenia Gospodarki Rybnej, ss. 57 -72,
Szczecin - 6 listopada 197889. Ditkiewicz D., Wotoszyk W.: Maszyny i urz^lzenia do produkcji farszow. W: Stu-
dium do programu rozwoju produkcji farszow rybnych w Polsce. Opracowanie
zbiorowe pod kierownictwem E. Kordyla, Osrodek Wydawniczy MIR w Gdyni,
59-92, 197090. Dworschak E.: Nonenzymatic browing and its effect on protein nutrition. CRC
Critical Rev. Food. Sci. and Nutr., 13(1); 1-38, 198091. Dworschak E., Czuczy^P.: The changes of some essential amino-acids during sto¬
rage in infant formulae based on the one hand glucose on the other. I. Studies in
aqueous solutions. A eta Pharm. Hung., 45; 40-45, 197592. Dyer W. J.: Stability of minced fish an important factor in standards formulation.
In: Second Technical Seminar on Mechanical Recovery and Utilization of Fish Mesh.
Massachusets, Boston, June 12-13, ed. byR. E. Martin, Director Science and Tech¬
nology, National Fischeries Institute, Washington, D.C. 197493. Dyer W. J., Dingle J. R.: Fish proteins with special reference of freezing. In: Fish
as Food, vol. 1 (ed. by G. Borgstrom), pp. 275-327. Academic Press, New York
and London 196194. Dyer W. J., Hiltz D. J.: Nucleotide degradation of frozen swordfish muscle. J. Fish.
Res. Bd Can., 26(6); 1597-1603, 196995. Ebashi S., Kodoma A.: Native tropomyosin-like action of tropomin on trypsin-
treated myosin B. J. Biochem., 60; 733-735, 196696. Ebert A. G.: The dietary administration of L-monosodium glutamate, DL-mono-
sodium glutamate and L-glutamic acid to rats. Toxicol. Lett., 3(2); 71-78, 197997. Ehira S.: Biochemical study on the freshness of fish. Bull. Tokai Reg. Fish. Lab. No.88, Dec., 1-123, 197698. Ehira S., Uchiyama H.: Formation of inosine and hypoxanthine in fish muscle du¬
ring ice storage. Bull Tokai Reg. Fish. Res. Lab. No. 75, Oct., 63-73, 197399. Ellis G. P.: The Maillard reaction. Advances Carbohydrate Chem., 14; 63-68, 1959100. Engel D. W., Davis E. М.: Relationship between activity and blood composition in
certain marine teleosts. Copeia No 3, 586-587, 1967101. Engelhardt F. R., Geraci J. R., Walker B. L.: Tocopherol composition of frozen
Atlantic herring (Clupea harengus harengus) tissues and oil. J. Fish. Res. Bd Can.,
32(6); 807-809, 1975102. Erichsen L. B.: Baader. In: Second Technical Seminar on Mechanical Recovery and
Utilization of Fish Flesh. Massachusets, Boston, June 12-13, ed. by R. E. Martin,
Director Science and Technology, National Fisheries Institute, Washington, D.C.
1974103. Ertel L. J.: Technologija prigotowlenija piszczewogo farsza iz mintaja. Rybn. Chozj.
nr 7, 46-48, 1968104. Extraco A. B.; Summary of investigations made by the Swedish Institute for Food
Preservation Research, Gothenburg and Research Laboratory for the Norwegian
Canning Industry, Stavanger (prospekt firmowy) 1978105. Fasold Hj Budowa biatek. PWN, Warszawa 1977106. Fischer J., Deng J. C.: Catalysis of lipid oxidation: a study of mullet (Mugil cepha-
lus) dark flesh and emulsion model system. J. Food Sci., 42(3); 610-614, 1977107. Flink J. М., Goodhart М.: Transport of oxygen through ice and frozen minced fish.
J. FoodProc. Pres., 2(3); 229-248, 1979108. Fraser D. J., Pitts D. P., Dyer W. J.: Nucleotide degradation and organoleptic qua¬
lity in fresh and thawed mackerel muscle held at and above ice temperature. J. Fish.
Res. Bd Can., 25(2); 239-253, 1968109. Frazer H. D., Smith L. B.4 Lemon D. W., Dyer W. J.: Deteriorative changes du¬
ring frozen storage in fillets and minced flesh of silver hake processed from round196
fish held in ice and refrigerated sea water. J. Fish. Res. Bd Can., 33(11); 2560-
2567, 1976110. Friedman М., Zahnley J. C., Masters P. М.: Relationship between in vitro diges-
tability of casein and its content of lysino-alanine and D-amino acids. J. Food
Sci., 46; 127-131, 1981111. Fujii Y., Watanabe K., Maruyama Y.: Relation between the ATP-breakdowns in
ice stored Alaska Pollock meat and the quality of ’’frozen surimi”. Bull. Tokai
Reg. Fish. Res. Lab. No. 75, Oct., 7-14, 1973112. Fukazawa Т., Hashimoto Y., Yasui Т.: Effect of some proteins on the binding
quality of an experimental sausage. J. Food Sci., 26; 541-543, 1961113. Fukuda Y., Hasegawa Y.: Processing of pollack meat. 1. Preseivability of frozen,
ground pollack meat. Aomori - ken Suisanbutsu Kako Kenkyusho Shiken Kentyu
Hokoku (publ. 1976), 50-51, 1975 (wg Chemical Abstracts, 90; 1979, 476,
90:532800114. Fukuda Y., Kakehata K., Arai K.: Denaturation of myofibrillar protein in deep-sea
fish by freezing and storage. Bull. Japan. Soc. Sci. Fish., 47(5); 663-672, 1981115. Fullington J. G.: Lipid-protein interaction. Bakers Digest, 43(6); 34-39, 1969116. Ganowiak Z.: Zagadnienia bakteriologiczne zwijzane z ocenj sanitarno-higie-
niczn^ ryb mrozonych. Studia i Materiafy MIR w Gdyni, Seria D, nr 6, 1-77,1972117. George J. C.: A histophysiological study of the red and white muscles of the
mackerel. Am. Midi. Nat., 69; 487 -494, 1962118. Gerhardt J.: Quell- und Emulgiervermogen von Gewiirzen. Fleischwirtschaft,
49(3); 341-345, 1969119. Giffe J. W., Urbin М. C., Fox J. B., Landmann W. A., Seidler A. J., Sliwinski R. A.:Chemia tkanki mi^sniowej. W: Nauka о miqsie i produktach miesnych, praca
zbiorowa, ss. 61-109. WPLiS, Warszawa 1969120. Gill T. A., Dingle J. R., Smith-Lall B., Stanley D. W.: Improved utilization of fish
protein - Quality enhancement of mechanically deboned fish. J. Inst. Can. Sci.
Technol. Aliment., 12(4); 200-202, 1979121. Gjelman A. G., Nj eh am kin B. L.: Sezonnaja izmjencziwost aktiwnosti i tempera-
tumy charakteristik szxzelocznoj fosfatazy sudaka i leszcza kurskogo zaliwa.
Trudy Atlant NIRO nr 64, 31-35, 1976122. Gordon A.: Polyphosphate treatment of fish. Food Manuf. nr 7, 57-58, 1971123. Gordon M. S.: Oxygen consumption of red and white muscle from tuna fishes.
Science, 87; 159-161, 1968124. Goralczyk A., Pielichowski J., Zalewski J., Salmonowicz J., Kochanowski J.,
Dutkiewicz D., Woioszek W., Jutkiewicz L., Duszenko J.: Produkcja mrozonych
farszow rybnych oraz przygotowanie jej do wdrozenia na statkach rybackich.
Opracowanie wykonane na zlecenie Centrum Techniki Okr|towej, MIR Gdynia,
listopad (maszynopis) 1975125. Grabowska J., Sikorski Z.: Technological quality of minced fish preserved by
freezing and additives. Acta Alimentaria, 2(3); 319-326, 1973126. Greer-Walker N., Pull G. A.: A survey of red and white muscle in marine fish.
J. Fish. Biol, 7; 295-300, 1975 # f127. Grodzinski Z.: Ukfad mi^sniowy. W: Anatomia porownawcxa krpgowcow, pod
red. H. Szarskiego, ss. 281-304. PWN, Warszawa 1980128. Grodzinski Z.: Anatomia i embriologia ryb, swyd. 3. PWRiL, Warszawa 1981129. Guardia E. J., Dollar A.: The influence of ante-mortem factors and gamma irra¬
diation on the degradation of 5 ribonucleotides in the muscle of English Sole
(Parophrys vetulus). J. Food Sci.,. 30; 223-227, 1965130. Guinan J. A.: New product concepts for minced fish. U. S. Dept, of Commerce
News, No. 3, 1-2, 1974131. Gulland J. A. (ed.): The fish resources in the ocean. Fishing News (Books) Ltd.,
London 1971197
132. Gutierrez A. L.: Additive for preserving fish by preventing oxidation. Span. Pat.
470, 798 (Cl. A23B), 01 Feb. 1979. Appl. 15Jun 1978 (Gufix S.A.) 1978133. Hamm R.: Biochemistry of meat hydration. Advances in Food Res., 10; 356-443,
1960134. Hamm R.: Phosphate in meat products. Dtsch. Lebensm.-Rundsch., 76(8); 263-269, 1980135. Hamm R., Deatherage E. E.: Changes in dehydration solubility and changes of
muscle proteins during heating of meat. Food Res., 25(5); 587-610, 1960136. Hamm R., Hofmann R.: Changes in the sulphydryl and disulphide sroups in beef
muscle proteins, during heating. Nature, 18(5003); 1269-1271, 1965137. Hamosh М., Atia R., Shapiro B.: Fatty acid uptake and esterification by fish
muscle./. Food Sci., 31(2); 146-150, 1966138. Hanaoka K., Toyomizu М.: Acceleration of phospholipid decomposition in fish
muscle by freezing. Bull. Japan. Soc. Sci. Fish., 45(4); 465-468, 1979139. Hanson S. W. F., Olley J.: Apparent losses in lipid recovery from tissue homoge-
nates. Biochem. J., 89; 101P-102P, 1963140. Hanson S. W. F., Olley J.: Observations on the relationship between lipids and pro¬
tein deterioration. In: The Technology of Fish Utilization, ed. by R. Kreuzer,
pp. 111-115. FAO, Fishing News (Books) Ltd., London 1965141. Harder W.; Anatomy of fishes. E. Schweizerbart’sche Verlagsbuchhandlung (Na-
gele und Obermiller), Stuttgart 1975142. Hardy R., Mackie P.: Seasonal variation in some of the lipid components of sprats
(Sprattus sprattus). J. Sci. Food Agric., 20(4); 193-198, 1969143. Hashimoto A., Arai K.: The effects of pH and temperature on the stability of
myofibrillar Ca-ATPase from some fish species. Bull. Japan. Soc. Sci. Fish., 44(12);
1389-1393, 1978144. Hashimoto K., Watabe S., Kono М., Shiro K.: Muscle protein composition of sar¬
dine and mackerel. Bull Japan. Soc. Sci. Fish., 45(11); 1435-1441, 1979a145. Hashimoto K., Yamaguchi K., Takeda N., Ogawa K.: Distribution of hemoprotein
in the muscle of mackerel and sardine. Bull. Jap. Soc. Sci. Fish., 45(10), 1331 —
1333, 1979b146. Hamulu G.: Biologisk konservering of livsmedel. Swedish Institute for Eood Pre¬
servation Research, Report No. 282, 1970147. Haurowitz F.: Chemistry and Biology of Proteins, p. 187. Akademic Press Inc.,
New York 1950148. Herborg L., Johansen S.: Fish cheese: the preservation of minced fish by fermen¬
tation. In: Proceedings of the Conference on the handling, processing and mar¬
keting of tropical fish, pp. 253-255. Tropical Products Institute, London 1977149. Hiltz D. F., Dyer W. J., Nowlan S., Dingle J. R.: Variation of biochemical quality
indices by biological and technological factors. In: Fish inspection and quality
control, pp. 191-195, FAO, Published by Fishing News (Books) Ltd., London1971150. Hironaka Y., Hayashi S., Ooshiro Z.: Interaction between protein and sugar during
frozen storage. Л/em. Fac. Fish. Kagoshima Univ., 25(1); 101-105, 1976151. Hodge J. E.: The chemistry of browning reactions. J. Agr. Food Chem., 1; 928-
943, 1953152. Horubafe A.: Podstawy przechowalnictwa zywnosci. PWN, Warszawa 1975153. Hotta М.: The Japanese surimi market. FAO Infofish Marketing Digest, No. 1,
17-20, 1982154. Hwan E. A.: Soweirszenstwowanije technotogii proizwodstwa rybnego farsza i
jego ispolzowanije. Obzornaja informacfa, Seria: Obrabotka ryby i morieproduk-
tow, wyp. 5, 1-38, 1978155. Iida H., Nakamura K., Tokunaga Т.: Changes in volatile components formed in
salted and dried sardine during frozen storage. I. Search for origin of volatile
components. Bull Tokai Reg, Fish. Res. Lab. No. 98, 77-85, 1979198
156. Ikeuchi Т., Simidu W.: Study on cold storage of brayed fish meat for the mate¬
rial of kamaboko. II. Effects of saccharides and others on setting of brayed fish
meat. Bull. Japan. Soc. Sci. Fish., 29(2); 157-160, 1963a157. Ikeuchi Т., Simizu W.: Study on cold storage of brayed fish meat for the mate¬
rial of kamaboko. III. Effects saccharose concentration on setting of brayed fish
meat. Bull Japan. Soc. Sci. Fish., 29(3); 258-262, 1963b158. Inamine S., Matsuda Т., Ueno R.: High quality frozen ground fish. Jpn. Kokai
Tokkyo Koho, 79, 154, 560 (Cl A24B4/06), 05 Dec. 1979, Appl 78/59, 967,
22 May 1978, Uprawniony z patentu: Kabushihi Kaisha Ueno Seiyaku Oyo Ken-
kyujo, Japonia 1979159. Ingram A. J., Butterworth K. R., Gaunt I. F., Grasso P., Gangolli S. D.: Short¬
term toxicity study on sorbitan mono-oleate (Span 80) in rats. Food Co'smet.
Toxicol., 16(6); 535-542, 1978160. Ingram М., KitcheU A. G.: Salt as a preservative for food. J. Food Technol., 2;
1-15, 1967161. Iredale D. G., York R. K.: Effects of chemical additives on extending the shelf
life of frozen minced sucker (Catostomidae) flesh. J. Fish. Res. Bd Can., 34; 420-425, 1977162. Ishikawa S., Nakamura K., Fujii Y.: Fish jelly product (kamaboko) and frozen
minced meat (frozen surimi) made of sardine. I. Freshness and handling of the
fish material affecting on its kamaboko forming ability. Bull Tokai Reg. Res.
Lab. No. 90, 56-66, 1977163. Ishikawa S., Nakamura K., Fujii Y., Yam an о G., Sugiyama Т., Shinozaki K., To-
bita K., Yamagushi Y.: Fish jelly product (kamaboko) and frozen minced meat
(frozen surimi) made of sardine. III. Influence on the treatment methods for
materials just after catch on the kamaboko forming ability of sardine meat. Bull
Tokai. Reg. Fish. Res. Lab. No. 99, 31-42, 1979164. Ishioroshi М., Samejima K., Arai K., Yusui Т.: Effect of blocking myosin-actin
interaction in heat-induced gelation of myosin in the presence of actin. Agric.
Biol. Chem., 44(9); 2185-2194, 1980165. Ishioroshi М., Samejima K., Yasui Т.: Heat-induced gelation of myosin: factors
of pH and salt concentrations. J. Food Sci., 44(5); 1280-1284, 1979166. Itoh H., Kawashima K., Chibota I.: Antioxidant activity of browning products
of triose sugar and amino acid. Agr. Biol. Chem., 39(1); 283-284, 1976167. Itoh H., Yoshinaka R., Ikeda S.: Gel forming ability of carp actomyosin. Bull
Japan. Soc. Sci. Fish., 45 (1); 73-77, 1979a168. Itoh H., Yoshinaka R., Ikeda S.: Effects of inorganic reducing agents on the gell
formation of fish meat heating. Bull Japan. Soc. Sci. Fish., 45(4); 455-458,
1979b169. Itoh H., Yoshinaka R., Ikeda S.: Behavior of the sulfhydryl groups of carp acto¬
myosin by heating. Bull Japan. Soc. Sci. Fish., 45(8), 1019-1022, 1979c170. Iwata К., Kanna K., Umemoto S., Okada М.: Study of the quality of frozen sto¬
red Alaska pollack surimi. I. The influence of freshness of the material and chan¬
ges in storage temperature. Bull. Japan. Soc. Fish., 36; 626-633, 1970171. Iwata K., Okada М.: Protein denaturation in stored frozen Alaska pollack muscle.I. Protein extractability and kamaboko forming ability of frozen surimi. Bull
Japan. Soc. Sci. Fish., 37(12); 1191-1198. 1971172. Ivanowa E. E., Petriczenko L. K., Szulzenko L. V., Skaczkov V. P.: Piszczewoj
farsz iz melkoj presnowodnoj ryby. Rybn. Chozj. nr 12, 55-56, 1980173. Jebsen J. W.: Proteins in fish muscle. In: Fish in Nutririon, ed. by E. Heen and
R. Kreuzer. Fishing News (Books), Ltd., London 1962174. Jensen М. H.: Chemical and textural changes resulting from freeze drying of min¬
ced cod flesh. Lebensm.-Wiss.u.-Technol, 12; 243-345, 1979175. Jonas R. E. E., Bilinsld E.: Utilization of lipids by fish. III. Fatty acid oxidation
by various tissues from sockeye salmon (Oncorhynchus nerka). J. Fish. Res. Bd
Can., 21(3); 653-656, 1964 199
176. Jonas R. E. E., Bilinski E.: Glycerylphsphorylcholine and related compounds in
rainbow trout muscle stored.at -4°C. J. Fish. Res. Bd Can., 24; 273-280, 1967a177. Jonas R. E. E., Bilinski E.: Phospholipase A activity in rainbow trout muscle.
/. Fish. Res. BdCan., 24(12); 2555-2562, 1967b178. Jones N. R.: Fish flavors, pp. 61-81. Proc. Flavor Chem. Symp. Campbell Soup
Co., Camden, New Jersey 1961179. Jones N. R.: Enzyme systems in fish muscle related to flavour. Abstracts of 1 th.
Intern. Congress of Food Science, p. 8 (takze: J. Sci. FoodAgric., 12; 167, 1962)180. Jones N. R.: Interconversions of flavorous nucleotide catabolites in chilled and
frozen fish. Proc. XI Int. Congr. Re frig. Munich, Paper IV-5, 917-922, 1963181. Jones N. R.: Problems associated with freezing very fresh fish. In: Fish handling
and preservation. Organisation for Economic Co-operation and Development,
Paris, rozdz. II, 31—56, 1965182. Jones N. R., Murray J.: The acid-soluble nucleotides of codling (Gadus callaris)
muscle. Biochem. J., 77; 567-575, 1960183. Kaczan-Glegola H., Lewinska I.: Wptyw wybranych warunkow termicznej stery-
lizacji suchej na stopien zniszczenia spor Bacillus sp. w pieprzu czarnym (Piper
nigrum). Praca magisterska pod kier. E. Ceronika, AR w Szczecinie 1981184. Kaneyama Т., Tomiyasu K.: Preservation of frozen fish meat paste with Prebest
TP433 New Food Ind., 21(12); 49-55, 1979185. Kanna K., Sakuraba М., Aihara Y.: Denaturation of fish muscle protein by dehyd¬
ration. V. Preventive effect of sugars and sugar-alcohols on protein denaturation
by dehydration. Bull. Tokai Reg. Fish. Res. Lab. nr 11, 69-76, 1974186. Karel М.: Protein-lipid interactions in biosystems. J. Food Sci., 38; 756-763,1973187. Kamicki Z.: Maszyny do produkcji farszu. Separatory. Komunikaty Zaktadow
Technologii i Mechanizacji Morskiego Instytutu Rybackiego w Gdyni, nr 5, 1975.188. Kamicki Z.: Stan i kierunki rozwoju produkcji farszow rybnych. TGM, nr 6,
334-337, 1975189. Kamicki Z., Kordyl E., Gora A., Salmonowicz J.: Sposob otrzymywania farszow
rybnych. Pat. Tymczasowy PRL nr 14624, zgtoszony 14.01.1912 (P. 152891),
Kl. 53c, 3/03; opis patentowy opublikowano 30.04.1915; uprawniony z patentu:
Morski Instytut Ryhacki, Glynia (Polska) 1972190. Kato N.. Uchiyama H.: A simplified method for determination of actomyosin as
total Ca +-ATPase activity for fish flesh quality. Bull. Tokai Reg. Fish. Lab. No.18, 33-39, 1974191. Kawakami H., Morita J., Takahashi K., Yasui Т.: Thermal denaturation of myo¬
sin, heavy meromyosin and subfragments. J. Biochem., 70; 635-639, 1971192. Kawka Т.: Urzq^dzenia do odkostniania mi^sa rybnego. Wyd. Uczelniane Akademii
Rolniczej w Szczecinie, nr 26, 1-32, 1980193. Kawka Т., Kolbiarz A., Ptociak J.: Linia technologiczna do produkcji stabilizo-
wanego farszu rybnego na statku B-417 i B-418. W: Badania nad opracowaniem,
doskonaleniem i wdrozeniem technologii przetworSw z farszow rybnych pod
kptem produkcji wielkoprzemyslowej\ cz. 1. Technologia produkcji farszow ryb¬
nych. Praca wykonana na zlecenie PPDiUR ’’Odra” w Swinoujsciu, Akademia
Rolnicza w Szczecinie (maszynopis) 1978194. Ke P. J., Ackman R. G.: Metal - catalyzed oxidation in mackerel skin and meat
lipids. J. Amer. Oil. Chem. Soc., 53(10); 636-640, 1976195. Ke P. J., Ackman R. G., Linke B. A., Nash D. М.: Kifferential lipid oxidation in
various parts of frozen mackerel. J. Food. Techn., 12; 37-47, 1977196. Kelly К. C.: Factors affecting the texture of frozen fish. In: Freezing and Irra¬
diation of Fish, pp. 339-342, ed. by R. Kreuzer. Fishing News (Books) Ltd.,
London 1969197. Kelly К. C., Little W. Т.: Brown discoloration in pre-rigor cut fish fillets. J. Food
Techn., 1; 121-129, 1966200
198. Kemp В., Spinelli J.: Comparative rates of IMP degradation in unfrozen and frozen
and thawed (slacked) fish. J. Food Sci., 24; 132-135, 1969199. Kibun Co., Lt<L: Fish deboning. Jpn. Kokai Tokkyo Koho 8081569 (Cl.
A23L1/325), 19 June 1980, Appl. 78/152/05, 12 Dec. 1978, 1980a200. Kibun Co., Ltd.: Ground fish preparation for fish paste production. Jpn. Kokai-
Tokkyo Koho 8081570 (Cl. A23L1/325), 19 June 1980, Appl. 78/152, 706,
12 Dec. 1978, 1980b201. Kilarski W.: The fine structure of striated muscles in teleosts. Z. fur Zellforschung,
79; 562-580, 1967202. Kimizuka H., Koketsu K.: Binding of calcium ion to lecithin film. Nature, 196;
995-996, 1962203. Kimura S., Kubota М.: Studies on elastoidin. IV. Hydrothermal stability of elas-
toidin. Bull Japan. Soc. Sci. Fish., 34(11); 1036-1039, 1968204. Kimura J., Murozuka Т., Arai K.: Comparative studies on fishes. Bull Japan.
Soc. Sci. Fish., 43(3); 315-321, 1977205. King F. J.: A list of companies offering meat-bone separators to the seafood in¬
dustry. Technical Note No. 6, National Marine Fisheries Servide, Glucester, Massa-
chusets, 1973a206. King F. J.: Improving the supply of minced blocks for the fish stick trade: a prog¬
ress report Marine Fisheries Rev. Marine Fish Rev., 35(8); 26-32, 1973b207. King F. J.: Acceptability of main dishes (entrees) based on mixtures of ground
beef with ground fish obtained from underused sources. J. Milk and Food Techn.,
36(10); 504-508, 1973c208. King F. J.: Past, present and future uses of minced fish. Marine Fish. Rev., 39(4);
1-4, 1977209. King F. J., Carver J. H.: How to use nearly all the ocean’s food. Commer. Fish.
Rev., 32(12), 12-21, 1970210. King F. J., Flick G. J.: Beefish and hamburger patties rate equally in taste prefe¬
rence tests. Food Product Develop., 7/9(11); 31-33, 1973a211. King F. J., Flick G. J.: Beefish patties. Marine Fish. Rev'., 35(7); 31-33, 1973b212. King F. J., Heiligman F., Wierbicki E.: Solubilized fish muscle as a food binding
material. Marine Fish. Rev., 36(1); 18-20, 1974213. Ко S. D.: Tape fermentation. Appl. Microbiol, 23; 976-978, 1972214. Koizumi Ch., Cao К. Т., Non aka J.: Undesirable odor of cooked sardine meat.
Bull Japan. Soc. Sci. Fish., 45(10); 1307-1312, 1979215. Koizumi Ch., Nonaka J.: Green discoloration of frozen rock sole. J. Tokyo Univ.
Fish., 64(2); 95-99, 1978216. Kokuryo H., Seki N.: Degradation of fish myofibrillar proteins by heating at high
temperatures. Bull Japan. Soc. Sci. Fish., 46(4); 493-498, 1980217. Kotakowska A.: PortSwnanie zmian oksydacyjnych i hydrolitycznych thiszczu w
mrozonych farszach rybnych, rybach calych i filetach ze sledzia baJtyckiego skfa-
dowanych w temperaturze 253K. Biul. Centr. Lab. Chtodnictwa, 10(1); 51-58,
1976218. Kotakowska A.: Wptyw wybranych czynnikow technologicznych na jefczenie
ryb mrozonych, Rozprawy Nr 67, Wyd. AR w Szczecinie 1979219. Kotakowska A., Gajowiecki L., Kotakowski E.: Zastosowanie kwercetyny do
przediuzania trwatosci mrozonych farsxow rybnych. VI Sesja Nauk. Kom. Techn
i Chemii Zywn PAN, Olsztyn 24-26.01.1975, Streszczenie doniesien, s. 160220. Kotakowska A., Kotakowski E., Tchorzewski S.: Sposob wytwarzania mrozonego
farszu rybnego. Pat. PRL 105400, sgtoszony 22.12.1976 (P. 194677), Cl. A23B4 06, uprawniony z patentu: Akademia Rolnicza, Szczecin (Polska) 1976221. Kotakowska A., Kotakowski E., Szybowicz Z., Deutry J., Wianecki М.: Badania
nad opracowaniem, doskonaleniem i wdrozeniem technologii przetworow z far-
szow rybnych pod kqtem produkcji wielkoprzemysfowej, cz. 1., Technologia far-
szow rybnych. Praca wykonana na zlec. PPDiUR ” Odra” w Swinoujsciu. Aka¬
demia Rolnicza, Szczecin (maszynopis) 1978
222. Kotakowska A., Lachowicz K., Gajowiecki L., Kotakowski E.: Spos6b przedtu-
zania trwatosci mrozonych farszow rybnych. Zastrzezenie patentowe P. 189749,
19.05.1976, Akademia Rolnicza w Szczecinie 1976223. Kotakowski E.: Elektroforeza biafek sarkoplazmy mi^sa sledzia na folii z octanu
celulozy. Zesz. Nauk. WSR w Szczecinie, nr 35, 211-225, 1971a224. Kotakowski E.: Przyczyny szybkiego psucia si^ ryb. W: Konserwacje surowca
rybnego. Referaty z konferencji szkoleniowej NOT, 1-17, Szczecin 1971b225. Kotakowski E.: Nowa technologia kieibas rybnych. Przem. Spoz., 30; 409-411,
1976a226. Kotakowski E.: Sagadnienia technologiczne produkcji farszow rybnych. Przem.
Spoz., 30(1); 23-24, 1976b227. Kotakowski E.: Niektore zapdnienia technologiczne^ w produkcji farszow ryb¬
nych. W: Technologia farszow rybnych jako surowcow dla przetworstwa. Mate-
riafy konferencyjne Sekcji Przem. Rybnego SITPSpoz. i Zjednoczenia Gospldarki
Rybnej, ss. 1-37, Szczecin, 27 czerwca 1977228. Kotakowski E.: Pasty rybno-warzywne. Przegl. Gastron, nr 2, 9-11, 1981229. Kotakowski E.: Technologia mrozonych przetworow rybnych. Wyd. Morskie,
Gdansk (w druku)230. Kotakowski E., Borzychowski J.: Zmiany fizykochemiczne w kutrowanym i
niekutrowanym farszu ze sledzia baltyckiego podczas parowania (w przygoto-
waniu do druku)231. Kotakowski E., Chero L. H., Sypniewska G.: Wptyw mieszania na zmiany fizy¬
kochemiczne w mrozonych farszach rybnych (w druku)232. Kotakowski E., Chero L. H., Sypniewska G.: Wpfyw stopnia opraeienia ryb na
zmiany fizykochemiczne w mrozonym farszu sktadowanym w temperaturze -27±
±2° С (w przygotowaniu do druku)233. Kotakowski E., Chrzanowski S., Orwat G.: Opakowania z tworzyw sztucznych w
przemy&le rybnym. Wyd. Morskie, Gdansk 1976234. Kotakowski E., Fik М., Karminska S.: Investigations into changes in the protein
nutritive value of frozen fish sausages produced from fresh and frozen minced
meat In: Freezing and storage of fish, poultry and meat, pp. 59-63. Issued by
HR Annexe 1972-3, Paris 1972a f235. Kotakowski E., Gajowiecki L., Kotakowska A., Safacki М., Leszczynska М.: Wpiyw
niektorych antyoksydantow i surowcow pomocniczych na trwatasc mrozonych
farszow z ryb ttustuch. Materiaty z V Dorocznej Sesfi Naukowej Komitetu Tech¬
nologii i Chemii Zywnosci PAN, s. 92. Gdansk 24-26 stycznia 1974236. Kotakowski E., Kaminski L., Safacki M.:Porownanie wydajrosci farszu z wyb-
ranych gatunkow ryb morskich na farszownicy typu Selo-Bibun. Przem. Spoz.,
30(2); 60-61, 1976237. Kotakowski E., Kotakowska A., Gajowiecki L.: Sposob wytwarzania mrozonego
farszu rybnego. Pat PRL Nr 82390, zgioszony: 07.10.1972 (P. 158155), G.
A23B4/06, uprawniony z patentu: Akademia Rolnicza, Szczecin (Polska) 1972b238. Kotakowski E., Kotakowska A., Rakus S.: Stabilizing properties of natural spi¬
ces on the fat of frozen minced herring flesh. In: Freezing and storage of fish,
poultry and meat, pp. 65-72. Issued by HR, Annexe 1972-2, Paris 1972c.239. Kotakowski E., Kotakowska A., Szybowicz Z.: Wptyw st^zenia czosnku na jego
wtasciwosci konserwacyjne w mrozonych kietbasach rybnych. VI Sesja naukowa
Komitetu Technologii i Chemii Zywnosci PAN, s. 163. Olsztyn 24-26 stycznia
1975a240. Kotakowski E., Lachowicz K.: Sposob wytwarzania farszu szczegolnie rybnego.
Zastrzezenie patentowe P. 194676, 22.12.1976. Akademia Rolnicza w Szczecinie1976241. Kofakowski E., Lachowicz K., Kaminski L.: Sposob wytwarzania wyrobow for-
mowanych z mrozonych farszow rybnych. Pat. PRL P. 221519. Data zgloszenia
25.01.1980. Akademia Rolnicza w Szczecinie 1980202
242. Kotakowski E., Lachowicz K., Szybowicz Z., Satacki М., Kotakowska A. i in.:
Technologia przetworow z farszow rybnych. Praca wykonana na zlecenie PPDiUR
’’Odra” w Swinoujsciu, AR Szczecin (maszynopis) 1975b243. Kotakowski E., Lachowicz K., Swierczynska Z., Mozdyniewicz S., Ogrodnik S.,
Jakacki W.: Sposob wytwarzania past rybnych. Pat. PRL 109114, zgtoszono
22.10.1977 (P. 201924) Cl. A23L1/325,1977244. Kotakowski E., Maliszewski A.: Wptyw dodatku krwi, nerki, miQsnia czerwonego
lub skory na jetczenie tiuszczu w rozdrobnionym miqsie ryb sktadowanym w tem-
peraturze -15° С (w druku)245. Kotakowski E., Pinedo Sanchez R. E.: Ustalenie optymalnych parametrow szyb-
kiego solenia rozdrobnionego mi$sa szprota battyckiego (w druku)246.' Kotakowski E., Silva G. R. A., Sypniewska G.: Porownanie zmian fizykochemicz-
nych w niemieszanych i mieszanych farszach z dorsza baltyckiego sktadowanych w
temperaturze -28° С (w przygotowaniu do druku)247. Kotodziejska J.: Wptyw soli K, Na, Ca i Mg na zamrazalnicze zmiany miofibry-
lamych bialek xyb. Praca doktorska pod kier. Z. E. Sikorskiego. Politechnika
Gdanska 1980248. Komatsu K., Nozaki H., Kato N., Nakajima H.: Ground fish - crustacean producti.
Jpn. Kokai Tokkyo Koho 7955, 753 (Cl. A23L1/325), 04 May 1979, Appl
77/120,125, 07 Oct. 1977, 1979249. Konagaya S., Konagaya Т.: Denaturation at moderate temperatures of myofib¬
rillar protein of red-meat fish: a possible cause of Yaka-Niku. Bull. Tokai Reg.
Fish. Res. Lab. nr 96, 61-14, 1978250. Konopczynski W.: Wptyw wybranych dodatkow na wfasciwosci reologiczne rozd¬
robnionego mi^sa z dorsza i sledzia battyckiego. Praca magisterska pod rier.
E. Kotakowskiego, Akademia Rolnicza w Szczecinie (maszynopis) 1982251. Konosu S., Hamoir G., Pechere J. F.: Carp myogens of white and red muscles.
Properties and amino acid composition of the main low-molecular weight compo¬
nents of white muscle. Biochem. J., 96; 98-112, 1965252. Kopecky A., Kocova P.: Zluknuti tuku z organoleptiskego hleoliska. Prum Pot-
ravian, 18(1); 49-51, 1967253. Kordyl E. (kier. oprac. zesp.): Studium do programu rozwoju produkcji farszow
rybnych wPdlsce. Osrodek Wydawniczy MIR, Glynia 1970254. Kostuch S., Sadowska М., Sikorski Z. E.: Sposob przygotowania farszu z mi$sa
ryb chudych. Zastrzezenie patentowe, P. 187186T, 12.02.1976. Politechnika
Gdanska 1976255. Kowalska A., Tchorzewska L.: Bakteriologiczna ocena farszow rybnych wjjpro-
dukowanych na statkach przetwomiach PPDiUR ’’Odra” Swinoujscie. Praca ma-
gisterska pod kier. E. Ceronika, Akademia Rolnicza, Szczecin 1980256. Krishnaswamy M. A., Lahiry N. L., Kadkol S. B., Rao S. V., Revankar G. D.,
Chandra К. C. S.: A nutritional evaluation of fish-potato flake. Food Technol.,
19(4); 197-200, 1965257. Kudo G., Okada М., Miyauchi D.: Gel forming capacity of washed and unwashed
flesh of some pacific coast species of fish. Marine Fish. Rev., 35(12); 10-15, 1973258. Kummerow F. A.: Formation of lipid-protein complexes. Nurt. Rev., 24(5); 147-149, 1966259. Kurokawa Т.: Kamaboko-forming ability of frozen and ice stored lizard fish.
Bull. Japan Soc. Sci. Fish., 45(12); 1551-1555, 1979260. Kuusi Т., Nikkfla О. E., Salolainen K.: Formation of malonaldehyde in frozen
Baltic herring and its influence on the changes in proteins. Z. Lebensm. Unters.-
Forsch., 159; 285-286, 1975261. Kuzmiczewa G. М., Bidenko M. S.: Issledowanije kaczestwa tonkoizmiefczennogo
farsza treski с razlicznymi stabilizinguszczimi dobawkami. Trudy Atlant NIRO,
nr 64, 78-83^1976262. Kyzlink V., Curda D.: Einfluss der Saccharose und Zuganglichkeit des Sauerstofs
auf den Oxydationsverlauf der L-Ascorbins'aure im flussigen Medium. Z. Lebensm.-
Unters., 263-273, 1969263. Labuza T. P., Chon H. E.: Decrease of linoleate oxidation rate due to water at
intermediate water activity./. Food Sci., 39; 112-113, 1974264. Labuza T. P., Heidelbaugh N. D., Silver М., Karel М.: Oxidation at intermediate
moisture contents. J. Am. Oil Chem. Soc., 48; 86-90, 1971a265. Labuza T. P., Silver М., Cohn М., Heidelbaugh N. D., Karel М.: Metal-catalyzed
oxidation in the presence of water in foods. /. Am. Oil. Chem. Soc., 48; 527-531,
1971b266. Lachowicz K.: Wykorzystanie gazowego dwutlenku w^gla do ksztaltowania teksty-
ry przetworow z rozdrobnionego mi^sa ryb. Praca doktorska pod kier. E. Kofa-
kowskiego, Akademia Rolnicza, Szczecin (maszynopis) 1980267. Laki K.: The composition of contractile muscle proteins. /. Cell, and Comp. Phy¬
siol49; 249-254, 1957268. Lall B. S., Manzer A. R., Hiltz D. F.: Preheat treatment for improvement of fro¬
zen storage stability at -10° С in fillets and minced flesh of silver hake (Merlucius
bilinearis). J. Fish. Res. Bd Can., 32(8); 1450-1454, 1975269. Land D. G.: Stability of plant pigments. In: Recent advances in food science,
vol. 2, ed. by J. Hawthorn and J. M. Leitch, pp. 50-56. Butterworths, London
1962270. Lanier Т. C., Lin T. S., Hamman D. D., Thomas F. B.: Effects of alcaline protease
in minced fish on texture of heat processed gels. J. Food Sci, 46(6); 1643-1645,
1981271. Larry D., Salwin H.: A new carotenoid pigment in shrimp. /. Am. Oil Assoc.
Chem., 49(3); 681-693, 1966272. Lasek W.: Kolagen, chemia i wykorzystanie. WNT, Warszawa 1978273. Laskowski J., WojtasW.: Problemy jakosciowe zwi^zane z nowymi surowcami.
W: Zagospodarowanie nowych surowcow rybnych. Mat. konf. Sekcji Przem.
Rybnego SITPSpoz. iZjedn. Gosp. Rybnej, ss. 73-90, Szczecin, 6 listopada 1973274. Lee С. М., Toledo R. Т.: Degradation of fish muscle during mechanical deboning
and storage with emphasis on lipid oxidation. J. Food Sci, 42(6); 1946-1649,1977275. Leszczynska М.: Wptyw dodatkow na zmiany substancji tluszczowych w mro¬
zonych farszach ze sledzia i makreli. Praca magisterska pod kier. E. Kotakows-
kiego, Akademia Rolnicza w Szczecinie (maszynopis) 1973276. Levanidov I. P., Ertel i. Ja.: О wzmoznosti sokraszczenija roshoda wody pri
proizwodstvije osobogo morozenogo farsza. Rybn. Chozj. nr 2, 72-73, 1974277. Licciardello J. J., Ravesi E. М., Allsup M. G.: Minced whiting block oxidation
studies show need for hish density packaging. Quick Frozen Foods, No. 5, 58-59,
76-77, 83, 1977278. Lingnert H., Eriksson С. E.: Antioxidative Maillard reaction products. II. Pro¬
ducts from sugars and peptides of protein hydrolysates. /. Food Proc. and Pre-
serv., 4(3); 173-181, 1980279. Linko R. R., Nikkila О. E.: Inhibition of the denaturation by salt of myosin in bal-
tic herring. J. Food Sci., 26(6); 606-610, 1961280. Linton О. М.: General direction and trends in Canada on the labeling of products
prepared from mechanically recovered fish flesh. In: Second Technical Seminar
on Mechanical Recovery and Utilization of Fish Flesh., ed.by R. E. Martin,
pp. 249-256. Boston, Massachusetts, June 12-13, 1974. National Fisheries Ins¬
titute, Washington, D. C. 1974281. Lisovaja W. P., Njehamkina N. P.: О proniknowjenii szczetocznych peptygidrolaz
w myszczecznuju tkan stawrydy i skumbrii. Trudy Atlant NIRO, nr 64, 20-25,
1976282. Love R. М.: Studies on the Noth Sea cod. I. Muscle cell dimensions. /. Sci Food
Agric9(4); 193-198, 1958204
283. Love R. М.: Protein denaturation in frozen fish. VII. Effect of the onset and
rigor mortis on denaturation. J. Fish. Res. Bd Can., 13; 534-545, 1962284. Love R. М.: The chemical biology of fishes. Academic Press, London and New
York 1970285. Love R. М., Lovern J. A., Jones N. R.: The chemical composition of fish tissues.
Her Majesty's Stationary Office, Special Report No. 69, 1-62, London 1959286. Lagunov L. L., Kriwoszeina L. I.: Razrabotka sposoba potuczenija wareno-sus-
zenogo rybnogo farsza. Rybn. Chozj. nr 10, 70-72, 1973287. Lukiewicz S.: The biological role of melanin. I. New concepts and methodical app¬
roaches. Folia Hist, et Cyt., 10(1); 93-108, 1972288. MacCallum W. A., Shiej H. S., Chalker D. A., Dyer W. J., Idler D. R.: Polyphosp¬
hate treatment of frozen cod. 2. Effect /in drip, yield, lipid hydrolysis and pro¬
tein extractability in twice-frozen Newfoundland summer trap and fall cod. J. Fish.
Res. Bd Can., 21(3); 539-548, 1964289. Mackie I. М., Hardy R., Hobbs G.: Fermented fish products. FAO Fish. Reports
No. 100, 1-59, 1971290. Mai J., Kinsella J. E.: Lipid composition of dark and white muscle from white
sucker (Catostomus commersoni). J. Food Sci., 44; 1101-1105, 1979291. Mai J., Kinsella J. E.: Composition of lipids and proteins of deboned minced and
filleted white sucke (Catostomus commersoni). J. Food Biochem., 3(4), 229-239, 1980292. Makinodan V., Ikeda S.: Studies on fish muscle protease. I. On the existence of
two kinds of proteinase active in acid and slightly alkaline pH range. Bull. Jap.
Soc. Sci. Fish., 35(7); 672-676, 1969293. Markowski B., Sokotowska A., Wilska S.: Kinetyka zmian tkanki rybnej podczas
obrobki cieplnej i zmiany sokow tkankowych. Zesz. CLPR w Gdyni, 19(14);
1-92, 1971294. Martin R. E.: Mechanicallv-deboned fish flesh. Food Technol., 30(9); 64-70,
1976295. Mastow A. М., Mastowa G. B., Prudowskaja E. Ja.: Wlijanije stepjeni izmiefczenija
rybnogo farsza na jego wjazkost i wtagoudierziwajuszczuju sposobnost. Ekspress-
Informacja CNIITEIRH. Obrabotka ryby i moreproduktow, nr 4, 5-10, 1973a296. Mastow A. М., Mastowa G. B., Prudowskaja E. Ja.: Wlijanije procentogo soder-
zanija soli na strukturalno-mehaniczeskije pokazateli i wfagouderziwajuszczuju
sposobnost farsza. Ekspress-Informacja CNIITEIRH, Obrabotka ryby i morepro¬
duktow, nr 4, 11-16, 1973b297. Mastowa G. W., Krytow В. K., Skomorowskaja I. R., Prudowskaja E. Ja.: 3. Wareno-
morozenyj rybnyj farsz. Ekspress Informacja, Obrabotka ryby i moreproduktow,
3(1); 4-8, 1975b298. Mastowa G. W., Prizrenowa I. I., Prudowskaja E. Ja., Skomorowskaja I. R., Ser-
gejewa D. B.: Issledowanije sposobow tieptowoj obrabotki pri proizwodstwje
wareno-morozenogo rybnogo farsza. Ekspress-Informacja, 3(10); 2-8, 1975a299. Mastowa G. W., Skomorowskaja I. P., Prudowskaja E. Ja., Krytow В. K.: Proiz-
wodstwo wareno-morozenogo rybnogo farsza. Piszcz. Promyszl, Moskwa 1978300. Matsuda Y.: Influence of sodium glutamate on the protein denaturation of lyophi-
lized carp myofibrils during storage. Bull. Jap. Soc. Sci. Fish , 45(6); 733-736,
1979a301. Matsuda Y.: Influence of sorbitol on the protein denaturation of lyophilized carp
myofibrils during storage. Bull. Jap. Soc. Sci. Fish., 45(5); 581-584, 1979b302. Matsuda Y.: Influence of sugars on denaturation of lyophilized carp myofibrilar
protein during storage. Bull. Jap. Soc. Sci. Fish., 45(6); 737-743, 1979c303. Matsuda Y.: Influence of storage temperatura on the Kamaboko-fQfming ability
of lyophilized Alaska pollack ’’surimi”. Bull. Jap. Soc. Sci. Fish., 45(4); 511-515,
1979d304. Matsumoto J. J.: Denaturation of fish muscle proteins during frozen storage.205
In: Proteins at low temperatures. Advances in Chemistry, Series No. 180, 205-
224,1979305. Matsuno Т., Higashi E., Akita Т.: Caro ten oid pigments in Gobies and five related
fishes. Bull Jap. Soc. Sci. Fish., 39(2); 159-163, 1973306. Matsuno Т., Nogata S., Chiba K.: Comparative biochemical studies of carotenoids
in fishes - V. Comparative studies of carotenoids between fresh water origin and
sea water origin striped millets. Bull. Gap. Soc. Sci. Fish., 41 (4); 459-464, 1975307. Matsuno Т., Nogata S., Sato Y., Watanabe Т.: Comparative biochemical studies
of carotenoids in fishes - II. Carotenoids of horse mackerel, swellfishes, porcupi¬
ne fishes and striped mullet. Bull. Soc. Sci. Fish., 40(6); 579-584, 1974308. Matsuura F., Baba H., Mori Т.: Chemical studies on the red muscle (’’Chiai”) of
fishes - I. Oceurence of arginase in the red muscle of fishes. Bull. Jap. Soc. Sci.
Fish., 19; 893-898, 1953309. Matsuura F., Hashimoto K.: Chemical studies on the red muscle (’’Chiai”) of
fishes - II. Determinations of the content of hemoglobin, myoglobin and cytoch¬
rome С in the muscles of fishes. Bull Jap. Soc. Sci. Fish., 20; 308-312, 1954310. Matsuura F., Hashimoto K., Yamaguchi K.: Oxygen absorption in autoxidation
of fish myoglobins. Bull Jap. Soc. Sci. Fish., 29(11); 1005-1010, 1963.311. McGill A. S., Hardy R., Guntrone F. D.: Further analysis of the volatile compo¬
nents of frozen cold stored cod and the influence of these flavour. J. Sci. Food
Agric., 28; 200-205, 1977312. McWeeny D. J., Knowles М. E., Hearne J. F.: The chemistry of non-enzymic brow¬
ning in foods and its control by sulphites. J. Sci. Food Agric., 25; 735-746, 1974313. Mendelson J. М.: Rapid techniques for salt-curing fish. A review. J. Food Sci.,
39; 125-127, 1974314. Mendelson J. М., Connors T. J., Callan J. G.: A machine for heading and eviscera¬
ting small fish. Marine Fish. Rev., 39(2); 11-18, 1977^315. Merkuze Z.: Wptyw metali na brunatnienie roztworow glikozy z lizyn§. Roczniki
PZH, 14(1); 65-70, 1963316. Mi^dzynarodowy Instytut Chtodnictwa. Zalecenia do produkcji i obrotu mrozonej
zywnosci (tytufr oryg.: Recommendations for the processing and handling of fro¬
zen foods), tlum. A. St^pkowski. Wyd. Czasopism Technicznych NOT, Warszawa1972317. Migita М., Mizuno F.: Water holding capacity of ’’myosin” gel. Bull Jap. Soc. Sci.
Fish., 32(12); 1043-1046, 1966318. Migita М., Okada М.: Setting phenomenon of fish muscle. I. Influence of electro¬
lytes. Bull Jap. Soc. Sci. Fish., 18(3); 25-31, 1952a319. Migita М., Okada М.: The setting phenomenon of the muscle. II. Influence exerted
by alkaline salts. Bull Jap. Soc. Sci. Fish., 18(4); 159-170, 1952b320. Minakowski W.: Biochemia krggowcow. PWN, Warszawa 1981321. Miwa K., Tokunaga Т., Iida H.: Studies on protecting methods of occurrence of
bad odors and their removing methods in fish processing factories. III. Deodori-
zation of ammonia and trimethylamine by carbon dioxide treatment and water
washing combination method. Bull. TodaiReg. Fish. Res. Lab., 87; 7-13, 1976322. Miyauchi D.: Storage studies on minced rockfish and the use of carp in minced
products. In: Second Technical Seminar on Mechanical Recovery and Utilization
of Fish Flesh, ed.by R. E. Martin. Boston, Massachusetts, June 12-13, National
Fisheries Institute, Washington, D. C. 1974323. Miyauchi D., Kudo G., Patashnik М.: Surimi - a semi-processed wet fish prote¬
in. Marine Fish. Rev., 35(2); 7-9, 1973324. Miyauchi D., Streinberg М.: Machine separation of edible flesh from fish.. Fishery
Industrial Res., 6(4); 165-171, 1970325. Mizushima Y., Takama K., Zama K.: Lipid oxidation at low intermediate mois¬
ture content. II. Corelation between common lipid oxidation measurements and206
volatile compounds. Hokkaido Daigaku Suisangakubu Kenhyu Ino, 30(2); 172-179, 1979326. Mooijani W. N., Balakiishnan R., Kxishnaswamy M. A., Lahiry N. L.: Fish protein
enriched corn starch. Research and Ind., 7; 307-309, 1962327. Motosugi М., Dohi S., Ishikawa М.: Improvement of processes for boiling and
drying of shipjack. VII. Improvement of the texture of shipjack fish. Shizuoka-
ken Kogyo Shikenjo Hokoku, No. 23, 73-78, 1979328. Murray С. K.: Polyphosphates dipa for fish. Torry Research Station Advisory
Note No. 31, 1967329. Nachenius R. J., Atkinson A.: Comminuted fish material for canning. Annual
Report of Fishing Industry Research Institute (FIRI), rozdz. 2.4, 9-11, 1966330. Nakajama H., Kato N.: Ground shark meat production. Jpn. Kokai Tokkyo Ko-
ho 79, 105, 272 (CL A23L1/325), 18 Aug. 1979, Appl. 78/9, 259, 01 Feb. 1978,
1979331. Nakamura K., Iida М., Tokunaga Т.: Separation and identification of odor in
oxidized sardine sil. Bull. Jap. Soc. Sci. Fish., 46(3); 355-360, 1980332. Nakamura М., Nishi K., Watanabe Т., Tamoto K.: Utilization and processing
sardine (Sardinops melanosticta). I. Changes in the freshness of sardine during
iced and water-iced storage. Hokucuishi Geppo, 34(12); 9-14, 1977333. Nakayama Т., Sato Y.: Relationship between binding quality of meat and myo-
fibryllar proteins. 3. Contribution of myosin A and actin to Theological proper¬
ties of heated minced-meat gel. J. Texture Stukies, 2; 75-79, 1971334. Nakayama Т., Yamamoto М.: Physical, chemical and sensory evaluations of fro-
zen-stored deboned (minced) fish flesh. J. Food Sci., 42(4); 900-905, 1977335. Nambudiry D. D.: Lipid oxidation in fatty fish: the affect of salt content in the
meat. J. Food Sci. Tech., 17(4); 176-178, 1980336. Narayan K. A., Sugai М., Kummerow F. A.: Complex formation between oxi¬
dized lipids and egg albumin. J. Am. Oil Chem. Soc., 41; 254-259, 1964337. Nemethy G.: The structure of and aqueous solutions. In: Low Temperature Bio¬
logy of Foodstuffs, edi by Hawthorn J. and Rolfe E. J., pp. 1-25. Pergamon
Press, Oxford, 1968338. Nikken Chemicals Co., Ltd. Frozen ground fish stabilizer. Jpn. Kokai Tokkyo
Koho 80 58, 058 (Cl. A23B4/08), 30 Apr. 1980, Appl. 78/130, 843, 24 Oct.
1978, 1980339. Nikonorow М.: Substancje obce dodawane celowo do zywnosci i zanieczyszczenia
techniczne. WPLiS, Warszawa 1966340. Nikonorow М.: Toksykologia zywnosci. PZWL, Warszawa 1979341. Nishida Т., Kummerow F. A.: Interaction of serum lipoproteins with the hydro¬
peroxide of methyl linoleate. J. Lipid. Res., 1; 450-455, 1960342. Nishimoto J., Koreeda N.: Protein denaturation and the changes of gel-forming
capacity in the rinsed muscle during frozen storage. Nipoon Suisan Gakkaishi,
45(8); 989-993, 1979343. Nishiya K.: Manufacture of frozen surimi. Japan Patent. No. 306857, 1963a344. Nishiya K.: Process of frozen Alasca pollock surimi. In: Special Report of Sym¬
posium on Cold Storage of Fish Meat. Jap. Soc. Sci. Fish., 211-222, 1963b345. Nishiya K., Tamoto K., Tanaka O., Takeda F., Fukumi Т.: Studies on freezing
of surimi (fish paste). III. Month. Rept. Hokkaido Muncpl. Fish. Lab., 17(9),
1959346. Nishiya K., Tamoto K., Tanaka O., Takeda F., Fukimi Т.: Studies of freezing of
surimi (fish paste) V. Month. Rept. Hokkaido Muncpl. Fish. Lab., 19(4); 1961347. Niwa E., Mori B., Miyake М.: Retardative mechanism of protein denaturation
by addition of saccharides during cold storage of minced fish meat (surimi). I. Bull.
Jap. Soc. Sci. Fish., 39(1); 61-67, 1973348. Niwa E., Nakayama Т., Hamada I.: State of water in fish flesh determined by near
IR technique. Nippon Suisan Gakkaishi, 46(9); 1147-1150, 1980
349. Noguchi S.: Recent advances in the utilizing and processing techniques in the Japa¬
nese fishery. The 5th International Ocean Development Conference, September
25-29, Tokyo, Preprints (I) Session C2, 43-55, 1978350. Noguchi S., Matsumoto J. J.: Studies on the control of the denaturation of the
fish muscle proteins during of frozen storag. I. Preventive effect of Na-glutamate.
Bull Jap. Soc. Sci Fish., 36(19); 1078-1087, 1970351. Noguchi S., Matsumoto J. J.: Modification of Thermal and Non-Thermal Gelation
of Carp Actomyosin by Additive Compounds. Bull. Jap. Soc. Sci Fish., 44(3);
273-278, 1978352. Noguchi S., Shinoda E., Matsumoto J. J.: Studies on the control of denaturation
of fish muscle proteins during frozen storage. V. Technological application of
cryoprotective substances on the frozen minced fish meat. Bull Jap. Soc. Sci
Fish., 41(7); 779-786, 1975353. Nowlan S. S., Dyer W. J.: Effect of mincing on glycolitic activity on prerigor
Atlantic cod (Gadus morhua) muscle stored in ice or frozen. J. Fish. Res. Bd
Can., 31(4): 473-476, 1974354. Nozawa E., Ishida Y., Kadota H.: Combined effect of NaCl and temperature on
TMA production by some bacteria isolated from chilled salted fish. Bull Jap. Soc.
Sci Fish., 45(11); 1395-1399, 1979355. Ogata Т., Mori М.: A histochemical study of hydrolytic enzymes in muscle fibres
of various animals. J. Histochem. Cytochem., 23; 1465-1467, 1963356. Ohta F., Tanaka K.: Some properties of the liquid protion in the frozen fish moscle
fluid. Bull Jap. Soc. Sci Fish., 44(1); 59-62, 1978357. Ohta F., Hirahara Т.: Rate of degradation of nucleotides in cooking-stored carp
muscle.Mem. Fac. Fish. Kagoshima Univ., 26; 97-102, 1977358. Ohta F., Yamada Т.: Denaturation of carp actomyosin by freezing in phosphates
buffer. Bull Jap. Soc. Sci Fish., 44(1); 63-66, 1978359. Okada М.: Effect of washing on the jelly forming ability of fish meat. Bull Jap.
Soc. Sci Fish., 30(3); 255-261, 1964360. Okada М.: Effect of sodium citrate on the keeping quality of frozen surimi (fro¬
zen fish paste). In: Freezing and Irradiation of Fish, ed. R. Kreuzer, pp. 312-315.
Fashing News (Books), Ltd., London 1969361. Okada М.: Fish paste products. In: Utilization of Marine Products, pp. 73-80.
Overseas Technical Cooperation Agency, Tokyo 1972362. Okada М., Noguchi E.: Trends in the utilization of Alaska pollack in Japan. In:
Fishery products, ed.by R. Kreuzer, pp. 189-193. FAO, Fishing News (Books),
Ltd., London 1974363. Olcott H. S., Richards E. G., Van der Venn J., Medwadowski B. F.: Absence of
lipid residues in purified myosin. Bull. Jap. Soc. Sci. Fish., 34(12); 1112-1113,1968364. Olley J., Pirie R., Watson H.: Lipase and phospholipase activity in fish skeletal
muscle and its relationship to protein denaturation. J. Sci Food Agric., 10; 501-516, 1962365. Olley J., Stephen E., Farner J., Robertson L: A critical look at two objective
tests for cold storage deterioration. J. Food Techn., 2; 207-216, 1967366. Ono Т., Nagayama F.: Studies on the lipoxidase in fish tissues. II. Lipoxidase
action of liver and dark muscle. Bull Jap. Sci Fish., 23(4); 211-214, 1957367. Ooshiro Z., Hironaka Y., Hayashi S.: Preventive effect of sugars on denatura¬
tion of fish protein during frozen storage. Mem. Fac. Fish., Kagoshima Univ.,
25(1); 91-99, 1976368. Palitzsch A., Schulze H., Lotter G., Steichele A.: Untersuchungen iiber die Wir-
kung von Naturgewiirzen, Gewiirzekstrakten, atherischen Oelen, Extraktiens-
riickstanden und synthetischen Antioxydantein auf den Abbau von Schweinefett
und Modell-Lipiden. III. Mitteilung: Gewiirzextrakte, bei der Extraktion anfa-208
llende wasserdampffliitige und nichtfliichtige Extraktbestandteile sowie Extrak-
tionsriickstande. Meischmrtschaft, 54(1); 63-68, 1974369. Patashnik М., Kudo G., Miyauchi D.: Smooth, white spread from separated fish
flesh forms a base for flavoured dips, snack items. Food Product Development, No.
7/8, 307-310, 1973370. Patashnik М., Kudo G., Miyauchi D.: Bone particle content of some minced fish
muscle products. J. Food Sci., 39(3), 588-591, 1974371. Patashnik М., Miyauchi D., Kudo G.: Objective evaluation of texture of minced
black rockfish (Sebaste sp.) during frozen storage. J. Food Sci., 41; 609-611,1976372. Paweljewa L. GM Wfasowa W. W.: Ob izmienienii karotenoidow i produktow okis-
lenija w zirie kaspiskoj kilki pri jejo zamoroziwanii i chotodilnom chranienii.
Trudy WNIRO, 88; 5—13, 1972373. Pielichowski J., Nodzynski J.: Wpfyw wst^pnego schtadzania £ledzi i ostrobokow
w ozi^bionej wodzie morskiej i lodzie na jakosc produktow mrozonych. Prace
MIR w Gkyni, seria B, 17; 161-186, 1978374. Podeszewski Z., Stodolnik K.: An influence of sodium chloride on the degree of
triglycerides of fish oil hydrolysis in relation to the temperature of storage. Papers
of the XIV International Congress of Refrigeration, Moskwa 1975375. Pokorny J., Janicek G.: Wechselwirkung zwischen Proteinen und oxydierten Li-
piden. Nahrung, 19; 911-914, 1975376. Pokorny J., Janicek G., Davidek J.: Determination of the interaction products
of proteins with lipids. Zesz. Probl. Post. NaukRol., 167; 155-170, 1975377. Рога A. E., Sildan N., Wittenberger C.: Muscle cholineesteraze in fish. Stud. Univ.
Babes-Bolyai, Ser. Biol, nr 1, 85-87, 1965^378. Postolski J., Gruda Z.: Zamrazanie zywnosci. WNT, Warszawa 1974379. Poulter R. G., Lawrie R. A.: Studies on fish miscle protein: nutritional consequen¬
ces of adding low concentrations of formaldehyde and/or linoleic acid to cod
muscle. Lebensm.-Wiss.u. Technol, 12; 47-51, 1979380. Rambeza E. F., Rjehina N. I.: Wlijanije himiczeskogo sostawa miasa ryby na kac-
zestwo i sroki hranienija piszczewogo morozenogo'rybnogo farsza. Rybn. Chozj.
nr3, 66-68, 1980381. Ramsey М. B., Watts В. М.: The antioxidant effect of sodium tripolyphosphate
and vegetable extracts on cooked meat and fish. Food Technol., 17(8); 102-105, 1963382. Regenstein J. М., Regenstein С. E.: Minced fish: the economical, versatile and
nutritional meat. International Institute of Refrigeration, Commissions C2, Dl,
D2, D3, Boston (USA) - 1981-4, lc-9c383. Reich G.: Kolagen. Zarys metod, wyniki i kierunki badania. WNT, Warszawa 1970384. Reynolds Т. М.: Chemistry of nonenzymatic browning. I. The reaction between
aldoses and amines. Adv. Food Res., 12; 1-46, 1963385. Riggs A.: Studies of the hemoglobins of Amazonian fishes: on review: Comp.
Biochem. Physiol., 62A; 257-272, 1979386. Rjehina N. I. (red.): Technotogija rybnych farszej iz matomiernych ryb. WNIRO,
Naucznoj otczet po temje 38, ss. 1-94, 1972387. Rjehina N. I., Agapowa S. A., Pjatnickaja I. N., Worobiewa N. A.: Ispolzowanije
morozenogo farsza iz mintaja как bieflcogo obogatitelja piszczewyych produktow.
Trudy WNIRO, 73; 72-76, 1977388. Rjehina N. I., Lewanidow I. I., Bidjenko M. S., Puklicz G. I.: Proizwodstwo ryb¬
nogo piszczego farsza na sudah. IV. Nauczno-Techniczeskaja Konferencja po
Razwitju Ftota Rybnoj Promyszlennosti i Promyszlennogo Rybotowstwa Socja-
listiczeskich Stran. Dokiad No. 4.12, Leningrad 1972389. Rjehina N. I., Rambeza E. F.: Wlijanije formaldegida na denaturacjonnyje izmieni-
enija morozenogo promytego rybnogo farsza pri hranienii. Rybn. Chozj. Nr 4,
68-71, 1981
390. Rodin E. W.: Hotodilnaja technotogija rybnych produktow. Piszcz. Promyszl.,
150-157, Moskwa 1976391. Ronold O. A., Jakobsen F.: Trimethylamine oxide in marine products. J. Soc.
Chem. Ind., 66, 160-166, 1947392. Roubal W. Т., Tapp el A. L.: Damage of proteins, enzymes and amino acids by
peroxidizing Lipids. Arch Biochem. Biophys., 113; 5-8, 1966393. Rzawskaja F. W.: Ziry ryb i morskich mlekopitajuszczih. Piszcz. Promyszl., Mosk¬
wa 1976394. Sakai М., Miki М.: Extraction rate of water-soluble protein from several-sized
fish flesh. Nippon Shokuhin Kogyo Gakkaishi, 25(1); 616-621, 1978395. Samejima K., Hashimoto Y. Y., Yasui Y., Fukazawa Т.: Heat gelling properties
of myosin, actin, actomyosin and myosin subunits in saline model system. J. Food
Set, 34; 242-246, 1969396. Samejima K., Ishioroshi М., Yasui Т.: Relative roles of the heed and tail portions
of the molecule in heat-induced gelation of myosin. J. Food Sci., 46(5); 1412-
1418, 1981397. Saunders D. H., Coleman J. E., Hampson J. W., Wells P. A., Riemenschnei-
der R. W.: Autoxidation of fatty materials in emulsion. J. Pro-oxidant effect of
histidine and trace metals on the oxidation of linoleate esters. J. Am. Oil Chem.
Soc., 39; 434-439, 1962398. Sawant P. L., Magar N. G.: Studies on frozen fish. II. Some chemical changes occu-,
ring during frozen storage. Food Technol.,15(8); 347-350, 1961399. Savolainen K.: Formation of formaldehyde in Baltic herring and cod and related
changes in the fish muscle proteins during frozen storage. Scand. Refrigeration,
4(4a); 141-143, 1975400. Schonberg A., Moubacher R.: Strecker degradation of a-amino acid. Chem. Rev.,
50; 261-277, 1952401. Schubring R., Tiilsner М.: Verfahren zur Herstellung eines seelachsanlichen Salz-
fischerzeugnisses aus Farsch. Lebensmittelindustrie, 25(4); 1-7, 1978402. Seher A., Ivanow S. A.: Natiirliche Antioxydantien. I. Die antiohydative Wirkung
der Tocotrienole. Fette, Seifen, Anstrichmittel, 75(10); 606-608, 1973403. Shenouda S. Y. K., Pigott G. М.: Lipid-protein interaction during aqueous extrac¬
tion of fish protein. Actin-lipid interaction. J. Food Sci., 40; 523-532, 1975404. Shenouda S. Y. K., Pigott G. М.: Fish miofibryllar protein and lipid interaction
in aqueous media at detected by izotope labelling, sucrose gradient centrifugation,
polyacrylamide electrophoresis and electron paramagnetic resonance. Work suppor¬
ted by National Sea Grant Programm, 1—30, 1977405. Shibata N., Kinumaki Т.: Research of prooxidants existing in fish meat. Bull.
Tokai Reg. Fish. Res. Lab. No. 95, 91-95, 1978a406. Shibata N., Kinumaki Т.: Effect of iron compound on the oxidation of fish oils.
Bull. Tokai Reg. Fish. Res. Lab. No. 95, 97-103, 1978b407. Shimidu W., Takeda K.: Studies on the muscle of the aquatic animals. 10. Hyd¬
ration and dehydration of fish muscle. Bull. Jap. Soc. Sci. Fish., 25(10); 18-20,
1959408. Shimizu Y.: Species variations in gel-forming ability of fish meat paste. Bull. Jap.
Soc. Sci. Fish., 40(2); 175-179, 1974409. Shimizu Y., Karata S., Nishioka F.: Extractability of proteins for fish skeletal
muscle at low ionic strengths Bull. Jap. Soc. Sci. Fish., 42(9); 1025-1031, 1976410. Shimizu Y., Nishioka F.: Species variations in heat coagulation properties of fish
actomyosin-sarkoplasmatic proteins systems. Bull. Jap. Soc. Sci. Fish., 40(3);
267-270, 1974411. Siebert G.: Enzymes of marine fish muscle and their role in fish spoilage. In: Fish
in Nutrition, eds. E. Heen and R. Kreuzer, pp. 80-82. FAO, Fishing News (Books)
Ltd., London 1962210
412. Sikorski Z. E.: Technologia zywnosci pochodzenia morskiego. WNT, Waiszawa
1971413. Sikorski Z. E.: Technologia zywnosci pochodzenia morskiego, wyd. 2. WNT,
Warszawa 1980414. Sikorski Z. E., Kostuch S., Kotodziejska J.: Zur Frage der Proteindenaturierung
im gefrorenen Fischfleisch. Die Nahrung, 19(9/10); 997-1003, 1975415. Sikorski Z., Olley J., Kostuch S.: Protein changes in frozen fish. Critical Rev. in
Food Sci. and Nutrition, 8(1); 97-128, 1976416. Skaczkow W. P.: Razrabotka tiechnotogii proizwodstwa siszczego farsza iz mato-
miernych ryb. Rybn. Chozj. nr 9, 56-58, 1974417. Skaczkow W. P., Judina O. P.: Chranienije piszczeso farsza iz mieikich ryb. Rybn.
Chozj. nr 6, 54-55, 1975418. Skorupska I., Grudzien J.: WpJyw dodatku hydrolizatow zelatyny na zmiany
fizydochemiczne mrozonego farszu ze sledzia baftyckiego. Praca magisterska
pod kier. E. Kotakowskiego, Akademia Rolnicza w Szczecinie (maszynopis) 1981419. Slowik М.: Przemiany 5-rybonukleotydow w tkance miqsniowej zwierz^t rzeznychi ryb. Gosp. Miqsna nr 3, 9-11, 1970420. Sobczyk L., Kisza A.: Chemia fizyczna dla przyrodnikow. PWN, Warszawa 1975421. Sorensen Т.: Effect of frozen storage on the functional properties of separated
fish. In: The production and utilization of mechanically recovered fish flesh (min¬
ced meat), ed.Keay J. N., pp. 56-65. Conference proceedings, April 7-8, Torry
Research Station, Aberdeen 1976422. Spinelli J.: Effect of hypoxanthine of the flavor of fresh and low-dose irradiated
Petrale sole fillets. J. Food Sci., 30; 1063-1067, 1065423. Spinelli J., Koury B.: Nonenzymatic formation of dimethylamine in dried fishery
products./. Agric. Food Chem., 27(5); 1104-1108, 1979424. Stansby М. E.: Polyunsaturates and fat in fish flesh. J. Am. Dil Assoc., 63(12);
625-630, 1973425. Stevens E. D., Black E. C.: The effect of intermittent exercise on carbohydrate
metabolism in rainbow trout, Salmo gairdneri. J. Fish. Res. Bd Can., 23(4); 471-485, 1966426. Stickland N. C.: Relationship between size of muscle fibers and body dimensions
in number of teleosts. Experientia, 31(11); 1279-1281, 1975427. Stodolnik L., Podeszewski Z.: Lipidy przemystowych ryb baityckich. Cz. IV.
Zmiany frakcji lipidowych w czasie sktadowania ryb mrozonych. Bromet. Chem.
Toksykol., 8(1); 9-21, 1975428. Streinberg M. A., Spinelli J., Miyauchi D.: Minced fish as an ingredient in food
combinations. In: Proceedings of the Conference on the handling, processing and
marketing of tropical fish, pp. 245-248. Tropical Products Institute, London1977429. Str$k F: Mieszanie i mieszalniki. WNT, Warszawa 1981430. Su H., Lin T. S., Lanier Т. C.: Contribution of retained organ tissues to the alka¬
line protease content of mechanically separated Atlantic cioakes (Micropogon
undulatus), J. Food Sci., 46(6); 1650-1653, 1981431. Subba Rao G. N.: Physico-chemical investigations on the muscle proteins of some
commercial species of fish caught in North-East Scottish waters. Thesis for the deg¬
ree of Ph. D., Aberdeen, Part II, Section C, 272-305, 1948432. Suggi K., Kinumaki Т.: Distribution of vitamin E in a few species of fish. Bull.
Jap. Soc. Sci. Fish., 34(5); 420-428, 1968433. Svensson S.: Stabilization of fish mince from gadoid by pretreatment of the fish.
SIK Service Serie Nr 655, 1-29, 1979434. Szendeijuk W. I., Njekrasowa G. Т.: Termostabilnost kompleksa proteolityczes-
kich fermentow piszczewaritelnogo trakta ryb pri jestestwiennom znaczenii pH.
Trudy Atlant NIRO nr 54, 13-16, 1973211
435. Szkolnikow S. S.: Mikrobiotogiczeskije issledowanija zamorozennogo farsza iz
mintaja w processe jego izgotowlenija i hranienija. Trudy WNIRO, t. 95,. 38-43,1974436. Taguchi Т.: Lecithin residues in carp muscle myosin В and its ATPase activity.
Bull Jap. Soc. Sci. Fish., 38(9); 969-972,1972437. Taguchi Т.: Activation mechanism of carp muscle myosin В ATPase by lecithin.
Bull. Jap. Soc. Sci. Fish., 40(1); 63-66, 1974438. Taguchi Т., Ikeda S.: Studies on the properties of fish actomyosin. III. Lecithin
found the actomyosin from fish muscle. Bull. Jap. Soc. Sci. Fish., 34(4); 339-343,
1968a439. Taguchi Т., Ikeda S.: Studies on the properties of fish actomyosin. IV. Effect of
lecithin on the ATPase activity of actomyosin from fish muscle. Bull Jap. Soc.
Sci. Fish., 34(5); 414-441, 1968b440. Taguchi Т., Kikuchi K., Tanaka М., Suzuki K.: Heat coagulation of fish actin. Bull
Jap. Soc. Sci. Fish., 45(5); 639-642, 1979441. Takagi М., Murayama H., Endo S.: Trimethylamine and trimethylamine oxide
contens of fish and marine invertebrates. Bull. Jap. Soc. Sci. Fish., 18(2); 261-267, 1967442. Takahashi Т.: On toushness of boiled fish meat. In: Fish in Nutrition, ed. by
E. Heen and R. Kreuzer, pp. 224-225, FAO, Fishing News (Books) Ltd., London
1962443. Takahashi H., Kaneko H.: Changes in the lipids of marine product during treat¬
ment processing and storage. 1. Lipid composition before and after washing of
’’otoshimi” meat from Japanese sardine. Hokusuishi Geppo, 35(10); 1-5, 1978444. Takahashi H., Teruhiko S.: Changes in the lipids of marine product during treat¬
ment processing and storage. 2. Changes in the lipids of ’’otoshimi” and ’’suri¬
mi” from sardine during frozen storage. Hokusuishi Geppo, 36(9); 188-198,
1979445. Takahashi K., Matsuura K.: Prevention of actomyosin denaturation in frozen
ground fish. Ipn. Kokai Tokkyo Koho 7998, 361 (Cl A23B4/08), 03 Aug. 1979,
Appl 78/5, 707, 21 Jan. 1978. Uprawniony z patentu. Nishida Shigeru (Japonia)446. Takahata К.: О proizwodstwie rublennogo i rastiertogo mjasa ryby (surimi) i sosi-
sok iz ryby. Materiaty reklamowe firmy Kanematsu-Gosho Ltd. (thim. z j^z. japon-
skiego), Leningrad, 1968447. Takama K.: Insolubilization of rainbow trout actomyosin during storage at -20 °C.II. Mechanism of propanal or caproic acid reaction with actomyosin to induce
insolubilization. Bull. Jap. Soc. Sci. Fish., 40(6); 589-593. 1974a448. Takama K.: Changes in the flesh lipids of fish during frozen storage. V. Autoxida-
tion of triglycerides and lecithin during cold storage. Hokkaido Daigaku Suisan
Gakubu Kenkyulho, 25(2); 154-161, 1974b449. Takama K.: Changes in the flesh lipids of fish during frozen storage. V. Accele¬
rative substances of lipid oxidation in the muscle of the rainbow trout. Bull. Fac.
Fish. Hokkaido Univ., 25(3); 256-263, 1974c450. Takama K., Zama K., Igaiashi H.: Changes in the flesh lipids of fish during frozen
storage. III. Relation between rancidity in fish flesh and protein extrability. Bull
Jap. Soc. Sci. Fish., 38(6); 607-612, 1972451. Tanaka K.: Experiment of natural tocopherol used for frozen fish as an antioxi¬
dant. Refrigeration, 48(548); 11-16, 1973452. Tanikawa E.: Marine products in Japan Koseisha-Koseikaku Company, Tokyo 1971453. Tanikawa E., Akiba М., Akiba S.: Studies on the manufacture of Gyomiso (fer¬
mented fish paste). I. Experiments on the manufacture of Gyomiso. Bull Jap.
Soc. Sci. Fish., 15(11); 689-695, 1950a454. Tanikawa E., Akiba М., Akiba S.: Studies on the manufacture of Gyomiso (fer¬
mented fish paste). II. The chemical changes of the fish meat protein during the
ripening of Gyomiso. Bull Jap. Soc. Sci. Fish., 15(11); 695-696, 1950b212
455.456.457.458.459.460.461.462.463.464.465.466.467.468.469.470.471.472.473.474.Tanikawa E., Akiba М., Akiba S.: Studies on the manufacture of Gyomiso (fer¬
mented fish paste). III. The relation berween the amount of salt added and ri¬
pening velocity and the creation of ammonia during the ripening of Gyomiso.
Bull Jap. Soc. Sci. Fish., 15(11); 696-702, 1950cTanikawa E., Motohiro Т., Akiba М.: Development of fish products with parti¬
cular reference to frozen minced fish muscle (surimi). In: Freezing and Irradia¬
tion of Fish, ed. by R. Kreuzer, pp. 304-311. FAO, Fishing News (Books) Ltd.,
London 1969Tarr H. L. A., Gardner L. J., Ingram P.: Pacific cod muscle 5-nucleotidase. J. Food
Sci., 34(6); 637-640, 1969Teeny F. М., Miyauchi D.: Preparation and utilization of frozen blocks of min¬
ced black rockfish muscle. J. Milk and Food Technol, 35(7); 414-417, 1972
Tejada М., Borderias A. J., Moral A.: Effects of washing of horse mackerel (Trachu¬
rus trachurus L.) minces on the removal o£ substances detrimental to preservation
in cold storage. I. I. R. Commisions C2, Dl, D2, D3 - Boston (USA), No. 4, 1-5,1981Toguchi Т., Kikuchi K., Ogumi М., Tanaka М., Suzuki K.: Heat changes of myosin
В Mo2+-ATPase and ’’setting” of fish meat paste. Bull Jap. Soc. Sci. Fish., 44(12);
1363-1366, 1978Tokunaga Т.: Biochemical and food scientific study on trimethylamine oxide and
its related substances in marine fishes. Bull Tokai Reg. Fish. Res. Lab. No. 101,
1-129, 1980Tomioka K., Ogushi I., Endo K.: Studies on dimethylamine in foods. II. Enzymic
formation of dimethylamine from trimethylamine oxide. Bull. Jap. Soc. Sci. Fish.,
40; 1021-1026, 1974Tomita Т., Mizukoshi М., Kanda Т., Shimomura Т., Tamiyasu K.: Quality impro¬
vement of meat paste product. Jpn. Kokai Tokkyo Koho 79 28, 464 (Cl. A23L1
325), 17 Sep. 1979, Appl 75/8, 511, 20. Jan. 1975. Uprawniony z patentu: Kao
Soap Co., Ltd., Japonia 1979Tomlinson N., Geiger S. E., Dollinger E.: Chalkiness in halibut in relation to muscle
pHfcnd protein denaturation. J. Fish. Res. Bd Can., 22; 653-663, 1965
Tomlinson N., Geiger S. E., Dollinger E.: Chalkiness in halibut in relation to muscle
pH and protein denaturation. J. Fish. Res. Bd Can., 23; 673-680, 1966
Towamicki R.: Histologia ogolna i szxzegofowa ryb. WSR, Szczecin (skrypt do uzyt-
ku wewn^trznego) 1965Toyama K., Shinazu М.: The protection of marine product from their deterioration
due to the oxidation of oil. XIII. Applicability of concentrated natural tocopherol
mixture. Bull. Jap. Soc. Sci. Fish., 38(5); 487-495, 1972Toyomizu М., Hanaoka K.: Lipid oxidation of the minced ordinary muscle of fish
during storage at -5 °C and susceptibility to lipid oxidation. Bull Jap. Soc. Sci.
Fish., 45(8); 1007-1010, 1980aToyomizu М., Hanaoka K., Nakamura Т.: Lipid oxidation in the skin during storage
of whole fish at -5 °C and the method to predict the susceptibility of fish to lipid
oxidation. Nippon surian Gakkaishi, 46(8); 1011-1017, 1980b
Toyomizu М., Yamazaki Т., Komori Y.: Studies on discoloration of fishery pro¬
ducts. III. Participation of ribose in rusting of fish muscle in model dehydrated sys¬
tems. Bull Jap. Soc. Sci. Fish., 34(2); 143-145, 1968Trautman J. C.: Effect of temperature and pH on the soluble proteins of ham.
J. Food Sci., 31; 409-414, 1966Tressler D. K., Lemon J. McW.: Marine products of commerce. Reinhold Publishing
Company, New York, 1951Treszczewa W. I., Sotomatina L. F.: Chimiczeskij sostaw kozi okeaniczeskih ryb.
Rybn. Chozj. nr 11, 67-68, 1978Trumbetas J., Fioriti J. A., Sims R. J.: Study of lipid-protein interaction using
pulsed NMR. J. Amer. Chem. Soc., 56(10); 890-893, 1979213
475. Tsai R., Cassens R. G., Briskey E. J.: The emulsifying properties of purified muscle
proteins. J. Food Sci., 37; 286-289, 1972476. Tsuchiya Т., Tsuchiya Y., Nonomura Y., Matsumoto J.: Prevention of freeze dena¬
turation of carp actomyosin by sodium glutamate. J. Biochem., 77(4); 853-862,1975477. Tsukuda М., Katahara Т.: Composition of the esterified fatty acids of astaxanthin
diester in the skin of seven red fishes. Bull. Tokai Reg. Fish. Res. Lab. No. 77, 89-95, 1974478. Tiilsher М., El-Bedawey F.: Der warmebedingte Kollageabbau bei der Fishverar-
beitung. Lebensm.-Ind. nr 4, 175-179, 1980479. Uchiyama H., Amano K.: The softening spoilage of fish sausage. V. Effect of so¬
dium pyrophosphate and sorbic acid on the growth of spore of Baccillus sirculans.
Bull Jap. Soc. Sci. Fish., 25(7-9); 531-537, 1959480. Ueda Т., Shimizu Y., Simidu W.: Species difference in fish muscles. I. The gel-for-
ming ability of heated ground muscles. Bull. Jap. Soc. Sci. Fish., 34(4); 357-361,
1968a481. Ueda Т., Shimizu Y., Simidu W.: Species fifference in fish actomyosin. III. The ve¬
locity and mechanism of heat-denaturing reaction. Bull. Jap. Soc. Sci. Fish., 34(4);
351-356, 1968b482. Valencia Q. A.: Wptyw hydrokoloid6w na wfasciwosci fizyczne farszow rybnych.
Praca doktorska pod kier. E. Kotakowskiego, Akademia Rolnicza w Szczecinie
(maszynopis) 1982483. van den Berg L.: The effect of addition of sodium and potassium chloride to the
reciprocal system: KH2P04-Na2HP04-H20 on pH and composition during free¬
zing. Arch. Biochem. Biophys., 84; 305-310, 1959484. van den Berg L.: Changes in pH a some frozen foods during storage. Food Technol.,
15(9); 434-437, 1961485. van den Berg L., Rose D.: Effect of freezing on the pH and composition of sodium
and potassium phosphate solutions: the reciprocal system KH2P04-NaHP04-H20.
Arch. Biochem. Biophys., 81; 319, 1959486. Varga S. G., Sims P., Michalik P., Regier L. W.: Growth and control of halophilic
microorganisms in salt minced fish. Can. Inst. Food Sci. Technol J., 1978487. Velisek J., Davidek J., Pokorny K., Grundova K., Janicek G.: Reactions of glyoxal
with glycine. II. The influence of reaction conditions on the course of reaction.
Z. Lebensm. Unters.-Forsch., 149; 323-329, 1972488. Venkataraman R., Sreenivasan A.: Salt tolerance of marine bacteria. Food Research,
19(3); 311-313, 1954489. von Hippel P. H., Schleich Т.: In: Structure and stability of biological macromole¬
cules, ed.by Timasheff S. N. and Fasman G. D., p. 417. Marcel Decker, New York1969490. von Hippel P. H., WangK.-Y.: On the conformational stability of slobular proteins.
The effects of various electrolytes and nonelectrolytes on the thermal ribonuclease
transition. J. Biol Chem., 240; 3909-3923, 1965491. Wada S., Koizumi Ch., Nonaka J.: A comparison of lipid deterioration in minced
and sliced cod flesh during storage at 0°C. Bull Jap. Soc. Sci. Fish., 43(5); 595 —
599, 1977492. Watabe S., Hashimoto K.: Myosins from white and dark muscle of mackerel. Some
physico-chemical and enzymatic properties. J. Biochem., 87; 1491-1499,1980493. Waters М. E., Garrett III E. S.: Hazards assessment of minced fish flesh. In: Second
Technical Seminar on Mechanical Recovery and Utilization of Fish Flesh, ed. by
R. E. Martin, pp. 217-236. Boston, Massachusetts, National Fisheries Institute,
Washington, D. C. June 12-13, 1974494. Webb N. B., Hardy E. R., Giddings G. G., Howell A. J.: Influence of mechanical
separation upon proximate composition, functional properties and textural cha¬214
racteristics of frozen Atlantic croaker muscle tissue. J. Food Sci, 41; 1277-1281,1976495. Wessels J. P. H., Spark A. A.: The fatty acid composition of the lipids from two
species of hake./. Sci Fd Agric., 24; 1259-1370, 1973496. Widera L., Pyszczuk B.: Histopatologiczne zmiany tkanki mi^sniowej i t^cznej
morszczuka wywofane inwazjjj. pasozytow Kudoa. Zesz. Centr. Lab. Przemyshi
Rybnego, 23(26); 27-33, 1975497. Wierzchowski J., Severin М.: Kwasy lotne z par^ wodn^ jako podstawa do oceny
przydatnosci spozywczej ryb. Roczniki PZH. nr 3, 301-305, 1962498. Wildlife and Fisheries. United States standards for grades of frozen minced fish
blocks. Code of Federal Regulations, 1979499. Wojtowicz М. B., Fierheller M. G., Regier L. W.: Observations on salting minced
lean fish. Paper presented at 20 Annual Conference of A.F. Т., Newport, R.I. 1975500. Wojtowicz М. B., Fierheller M. G., Regier L. W.: Making ’’instant” salt minced fish.
New Series Circular (Fesheries and Oceans Canada) nr 68, 1:13, 1978501. Wojtowicz М. B., Fierheller M. G., Legendre R., Regier L. W.: A technique for
salting lean minced fish. Department of Fisheries and the Enviroment, Canada,
Fisheries and Marine Service Technical Report nr 731, 1-15, 1977502. Yamada K., Amano K.: Studies on the biological formation of formaldehyde and
dimethylamine in fish and shellfish. Bull Jap. Soc. Sci Fish., 31; 60-64, 1965503. Yamaguchi K., Takeda N., Ogawa K., Hashimoto K.: Properties of mackerel and
sardine myoglobins. Bull Jap. Soc. Sci. Fish., 45(10), 1335-1339, 1979504. Yamamoto A., Nagao K.: Studies on the gel formation of ’’surimi” added with
plant proteins. Teikoku Gakuen Kiyo No. 5, 11-18, 1979505. Yamamoto Т.: Low temperature storage of ground fish. Jpn. Kokai Tokkyo К oho
79,140, 766 (Cl. A2384/08), 01 Nov. 1979, Appl 78/45, 931, 20 Apr. 1978,1979506. Yamanaka М., Nakagawasai Т., Kikachi Т., Amano K.: Studies on the construction
of carp muscle. I. Remarkable differences between rigor mortis and thaw rigor.
Nippon Suisan Gahhaishi, 44(10); 1123-1126, 1978507. Yamatoto М., Wong J.: Simple chemical method for isolating bone fragments in
minced fish flesh./. Food Sci, 39(6); 1259-1269, 1974508. Yamazawa М.: Studies on formation of foul odors by heating of frozen surimi.
Part 1. Behkviour of hydrogen sulfide volatile bases and formaldehyde. Aichi-ken
Shokuhin Kogyo Shikensho Nempo, 18; 56-62, 1977509. Yang R., Shin D. B.: A study on the amino-carbonyl reaction. Hanguk Sikp'um
Kwahakhoe Chi Korea, 12(2); 86-96, 1980510. Yoshimura K., Kubo S., Soma Т., Murakami K.: On the frozen ’’Otoshimi” (Crushed
fish meat). Bull. Jap. Soc. Sci Fish., 25(8); 34-41, 1960511. Yu М. H., Wu М. Т., Wang D. J., Salunkhe D. K.: Nonenzymatic browning in syn¬
thetic systems containing ascorbic acid, amino acids, organic acids and inorganic
salts. Can. Inst. Sci. Technol. J., 7(4); 279-283, 1974512. Yurkowski М., Brockerhoff H.: Lysolecithinase of cod muscle. /. Fish. Res. Bd Can.,
22; 643-652, 1965513. Zajcew W. P., Kizeweter J. W., Lagunow L. L., Makarowa T. J., Minder L. P., Pod-
sewafow W. N.: Technotogija rybnych produktow. Piszczewafa promyszl, Moskwa
1965514. Zaleski S. J.: Mikrobiologia zywnosci pochodzenia morskiego. Wyd. Morskie, Gdansk1978515. Zama K., Takama K., Mizushima Y.: Effect of metal salts and antioxidants an the
oxidation of fish lipids during storage under the conditions of low and intermediate
moistures./. Food. Proc. Pres., 3; 249-257, 1979516. Ziemba Z.: Proba wyjasnienia powstawania zabarwienia produktu w^dzonego. Praca
doktorska pod kier. J. Tilgnera, Politechnika Gdanska 1964517. Ziemba Z.: Reakcje brunatnienia w zywnosci. Przem. Spoz., 20(7); 40-51, 1966518. ZN-79(ZGR-09111). Norma zakiadowa. Surowce rybne mrozone. Farsze rybne.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬААктиваторы побурения 75
Активность антиокислительная 10?6110- воды 48- проокислительная 70- сахара антиденатурационная 106
Актомиозин 40Альбумины 38
Альдегид муравьиной 64, 67
Альдегиды алифатические 72
Антиокислители 108, 109, 110
Астаксантин 26
Ассоциация ионов 48649ББактерии болезнетворные анаэробные
157- молочнокислые 177Белки миофибриллярные 40, 54, 56,84, 92- саркоплазмы 39- стромы 41ВВетчина и колбаса рыбные 112
Вещества стабилизирующие 117
Виды рыбного фарша 115
Влияние температуры посола 170
Вода свободная 71- связанная 47
216Волокна мышечные 21, 23
Выход продукта 146- фарша 103ГГемоглобин крови 28, 29
Гидролиз 71
Гидроперекиси 54
Гипоксантин 79
Глутамат натрия 108
Гуанин 26ДДенатурация белка 42, 43
Дефекты мороженого фарша физи¬
ческие 181Дефосфорилирование 85
Диметиламин (ДМА) 36, 60
Добавки вкусовые 1053Замораживание батонов фарша 163- рыбы 90Запах и вкус фарша 179
ИИзделия кулинарные из рыбного фар¬
ша 13Источники бактериальной обсеменен-
ности фарша 184
кнКамабоко 12
Кислоты гексаеновые 70- докозагексаеновые 70- полиеновые жирные 70- свободные жирные 51
Компоненты, добавляемые в рыбный
фарш 162Консистенция фарша 180
Концентрация соли в фарше 160
Кости 29, 30Критерии оценки сурими 182
Куттер-смеситель 159,163
ЛЛецитин 53Линии производства сурими японские
146,148Линии производства промытого фарша
из минтая на судне ’’Северодвинск”
150,152 вареного фарша 160, 162Липкость фарша 15&Липофора 26ММашины рыборазделочные 96, 97
Меланофоры 26
Меланоиды 74Метод Андерсона - Мендельсона 166,
167- Дель Валье - Никерсона 166- сушки сырого фарша 176
Минимум ботулиновый 157
Миозин 40Модификация биохимическая 122
Мышцы красные 15, 19
Муцин 26Наличие паразитов в рыбе 95
Натрий хлористый 112, 113ООболочки синтетические для фасовки
фарша 163- сольватационная 50Обработка тепловая модельная 179- фарша тепловая 155, 156, 157
Окисление липидов 71
Оптимизация ускоренного посола 169
Отделение избытка воды и измель¬
чение 144- (сепарирование) мяса при производ¬
стве сурими 142Отравления пищевые 184Оценка качества несырого фарша 183 промытого фарша 182- рыбного фарша микробиологическая184Очистка промытого мяса на стрейнере
147ППалочки группы Salmonella и Esche¬
richia coli 185- кишечные 184Паста ’’Океан” из антарктического
криля 163Пастеризация высокотемпературная
157Паштеты рыбные 174
Перемешивание измельченного мяса123, 124- с добавками 145
Побурение неферментативное 73
Полисахариды 14217
Полифосфаты 107
Посмертные изменения 84, 85
Посол ускоренный 164
Потери белков 171, 172
Прессы гидравлические 174- шнековые 174Пригодность рыбы технологическая 89
Приправы 109- вносимые в фарш 163
Продолжительность промывки 137,
138Промывка 142
РРесурсы биологические 83
Реакция Майяра 73, 75, 76
Рыба жирная 93- тощая 90- средней жирности 92ССепаратор типа ’’Бибун” 142
Сепараторы 98, 99,100- серии ’’Фарш” 150
Слой диффузионный 50
Созревание соленого фарша 175
Соотношение воды и фарша при про¬
мывке 135Способность гелеобразующая 92
Способ разделки мелкой рыбы 97, 98
Способы охлаждения 88- ускоренного посола измельченного
мяса рыбы 166Способность водоудерживающая 158,
159Средства, повышающие способность
фарша связывать воду, 110
Срок хранения фарша 185Стабилизаторы измельченного мяса
105Стабилизация измельченного мяса 125,
126Стандарты США 179,180
Степень зачистки тушки 95- обсемененности фарша 184
Стафилококки грамположительные185Стрейнер 144
Сурими 8Сушка сублимационная 176Схема производства соленого фарша173, 174Сырье для производства сурими 141гТехнология ферментированного фарша
177Токотриенол 108
Токоферол 70
сетокоферол 70Триметиламиноксид 31, 35, 60, 65, 67
УУпаковка 187- и замораживание 145, 146- мороженого фарша 113,114ФФарш вареный 164- куттерованный 159- пастеризованный 153, 154, 155- промытый 127 в сепараторе 152, 153- несоленый (муэн-сурими) 93 после сепаратора (сурими) 141 соленый (каэн-сурими) 93- рыбный 83218
мороженый пастеризованный 160- солено-сушеный 183- соленый 72, 164-стабилизированный 117, 118, 121,122, 125,127- сушеный 175, 176- ферментированный 176, 177
Ферментирование без добавления пова¬
ренной соли 177, 178- с добавлением поваренной соли 177
Ферменты в мясе рыбы 71- протеолитические 126- содержащиеся в красных мышцах 19
Формы металлические для фасовки
фарша 163XХранение и первичная обработка сырьяпри производстве сурими 142
ЦЦентрифуги 174
Цитрат натрия 112, 113ЧЧикува 12Число промывок 135,136- тиобарбитуровое (ТБЧ) 182ШШтаммы стартовые 177
ЭЭмульгаторы 112
Эффективность белковая 183
СОДЕРЖАНИЕОт редакции 5Предисловие . 71. Общие сведения 81.1. Мировое производство рыбного фарша 81.2. Преимущества переработки рыбы на фарш 91.3. Направления использования рыбного фарша 112. Общая характеристика тканей, входящих в состав рыбного фарша 152.1. Мышечная ткань (мясо) 152.2. Соединительная ткань мяса . 242.3. Кожа 252.4. Кровь 282.5. Кости 292.6. Почки 303. Изменения, происходящие в измельченном мясе рыбы 313.1. Факторы, ускоряющие нежелательные изменения 313.2. Белки мышц и их изменения .... 383.3. Распад триметиламиноксида (ТМАО) в мясе рыбы. 603.4. Липиды и их изменения 693.5. Неферментативное побурение 733.6. Изменения нуклеотидов 764. Основное сырье и его обработка 834.1. Рыбное сырье для производства фарша 834.2. Влияние посмертных изменений 844.3. Влияние способа хранения рыбы перед переработкой на качество
фарша 854.4. Факторы, определяющие технологическую пригодность рыбного
сырья 884.5. Первичная обработка рыбы и ее влияние на качество фарша 954.6. Сепарирование мяса 985. Сырье и вспомогательные материалы 1055.1. Добавки 1055.2. Упаковка 1136. Технология производства рыбного фарша 1156.1. Стабилизированный фарш 1156.2. Промытый фарш . 1276.3. Пастеризованный фарш 1536.4. Соленый фарш 1646.5. Прочие виды фарша 1757 Требования к качеству и срок хранения фарша 1797.1. Требования к качеству 1797.2. Срок хранения фарша 185Литература 192Предметный указатель 216
Колаковский ЭдвардТЕХНОЛОГИЯ РЫБНОГО ФАРШАЗав. редакцией В. Е. Машковский
Художник А. Ф. Серебряков
Художественный редактор К. Е. Мацегорин
Технический редактор Й. Г. Гоголевская
Корректор Н. Я. ТумановаИБ № 7041Сдано в набор 17.07.90. Подписано в печать 17.10.90. Формат 60 X 881 /i6.
Бумага кн.-журн. Гарнитура Пресс-Роман. Печать офсетная. Уел. печ. л. 13,72.
Уел. кр.-отт. 13,96. Уч.-изд. л. 15,26. Изд. № 176. Тираж 1600 экз. Заказ №4032Цена 4 р. 40 к.Ордена Трудового Красного Знамени ВО ’’Агропромиздат”, 107807, ГСП-6,
Москва, Б-78, ул. Садовая-Спасская, 18.Московская типография № 9 НПО ’’Всесоюзная книжная палата” Государствен¬
ного комитета СССР по печати. 109033, Москва, Волочаевская, 40.
В 1991 ГОДУВ ВО ’’АГРОПРОМИЗДАТ”ВЫХОДИТ В СВЕТ КНИГАЛьюис М., ТаулерК., НилссонЛ и др.
Новые технологии в молочной промышленности. Т. 1. Пер.
с англ. Нидерланды, 1986, 28 л.В книге на основе прогрессивных технологий, разрабо¬
танных в последнее время, обсуждаются вопросы тепловой
обработки молока, сепарирования и переработки сливок;
получение сливочного масла и выработка бутербродных ма¬
сел; сушка молока и молочных продуктов; использование
молочных сьюоротки и казеина. Большое внимание уделяет¬
ся автоматизации технологических процессов и контролю
за качеством выпускаемых продуктов.Для научных работников, инженеров-конструкторов и
технологов молочной промышленности.
В 1991 ГОДУВ ВО ’’АГРОПРОМИЗДАТ”ВЫХОДИТ В СВЕТ КНИГАЛьюис М., Т а у л е р К., Н и л с с о н Л. и др.
Новые технологии в молочной промышленности. Т. 2. Пер.
с англ. Нидерланды, 1986, 28 л.В книге на основе прогрессивных технологий, разрабо¬
танных в последнее время, обсуждаются вопросы производ¬
ства кисломолочных продуктов, твердых и мягких сыров,
особенности выпуска замороженных молочных продуктов.
Особое внимание уделено современной лабораторной прак¬
тике в молочном деле.Для научных работников, инженеров-конструкторов и
технологов молочной промышленности.
В 1991 ГОДУВ. ВО "АГРОПРОМИЗДАТ”ВЫХОДИТ В СВЕТ КНИГАБартельмес Д. Гидробиологические основы рыб¬
ного хозяйства во внутренних водоемах. Пер. с нем. ФРГ,
1981,20 л.В книге изложены вопросы структуры и функции озер¬
ных экосистем во взаимодействии с составом ихтиофауны.
Показана связь биопродукции с первичной и вторичной про¬
дукцией и составом рыбного населения. Рассмотрены прин¬
ципы улучшения рыбных запасов и перспективы управления
рыбным хозяйством.Для специалистов рыбного хозяйства.
4 p. 40 к.Рыбный фарш — это измельченное мясо рыбы, отделен¬
ное механическим способом от несъедобных частей, под¬
вергнутое дополнительной технологической обработке для
увеличения срока хранения, сформированное в блоки и за¬
мороженное.Технология рыбного фарша относится к новейшим
процессам переработки рыбы, которые открывают новые
возможности в области рационального использования жи¬
вотного сырья, особенно малоценной рыбы.Рыбный фарш служит полуфабрикатом для выработки
различных кулинарных изделий (колбас, сосисок, котлет,
пельменей и др.) .