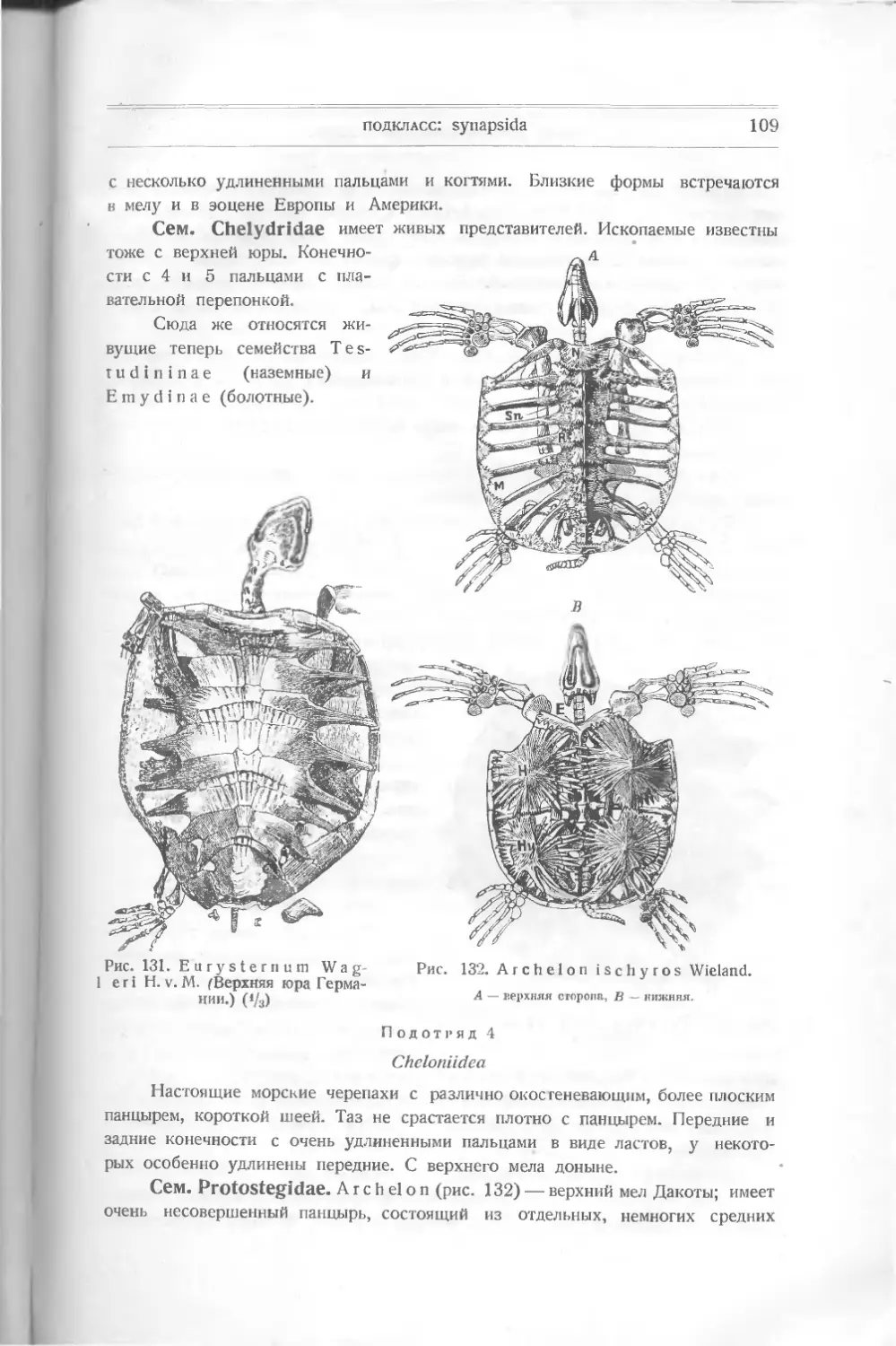
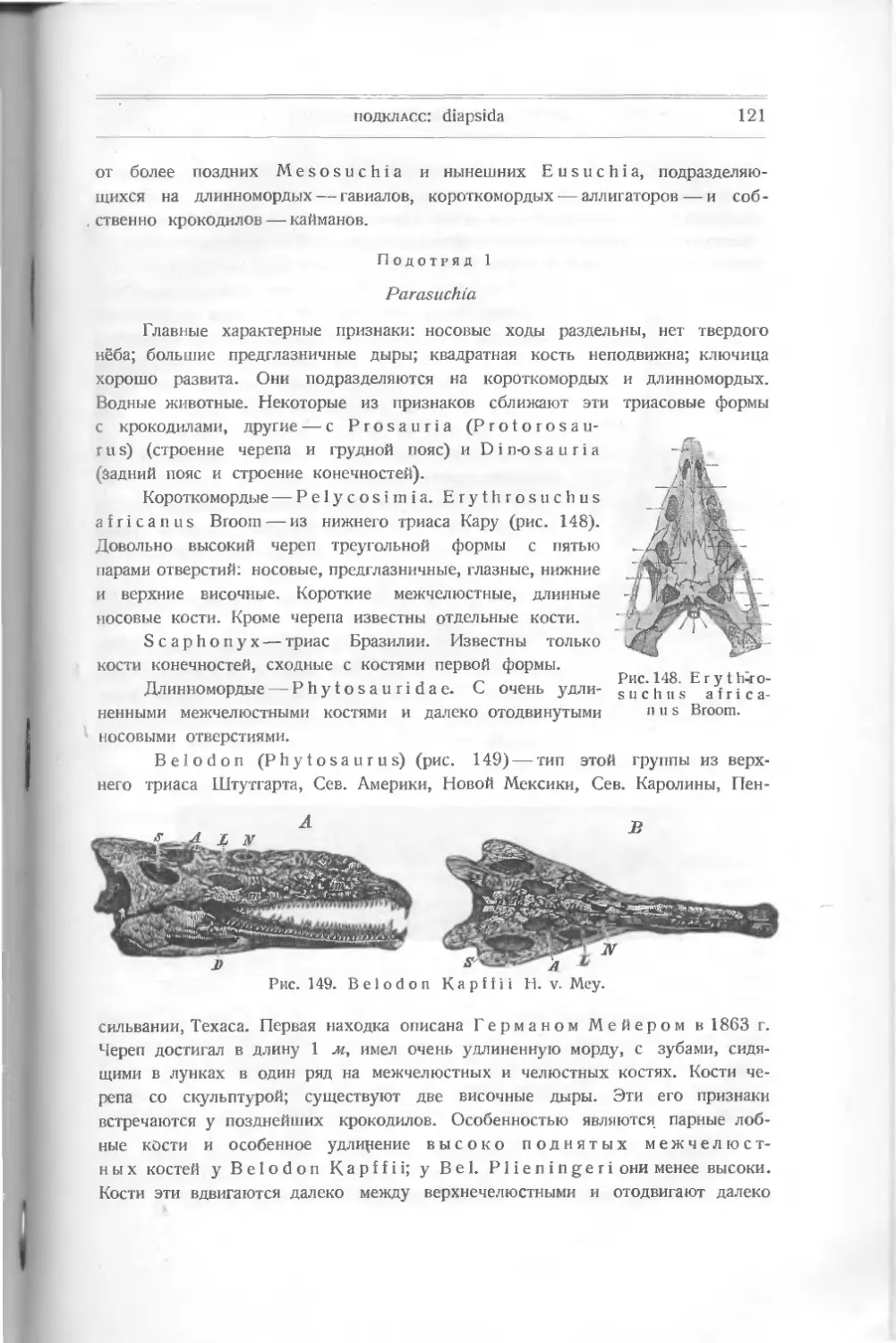
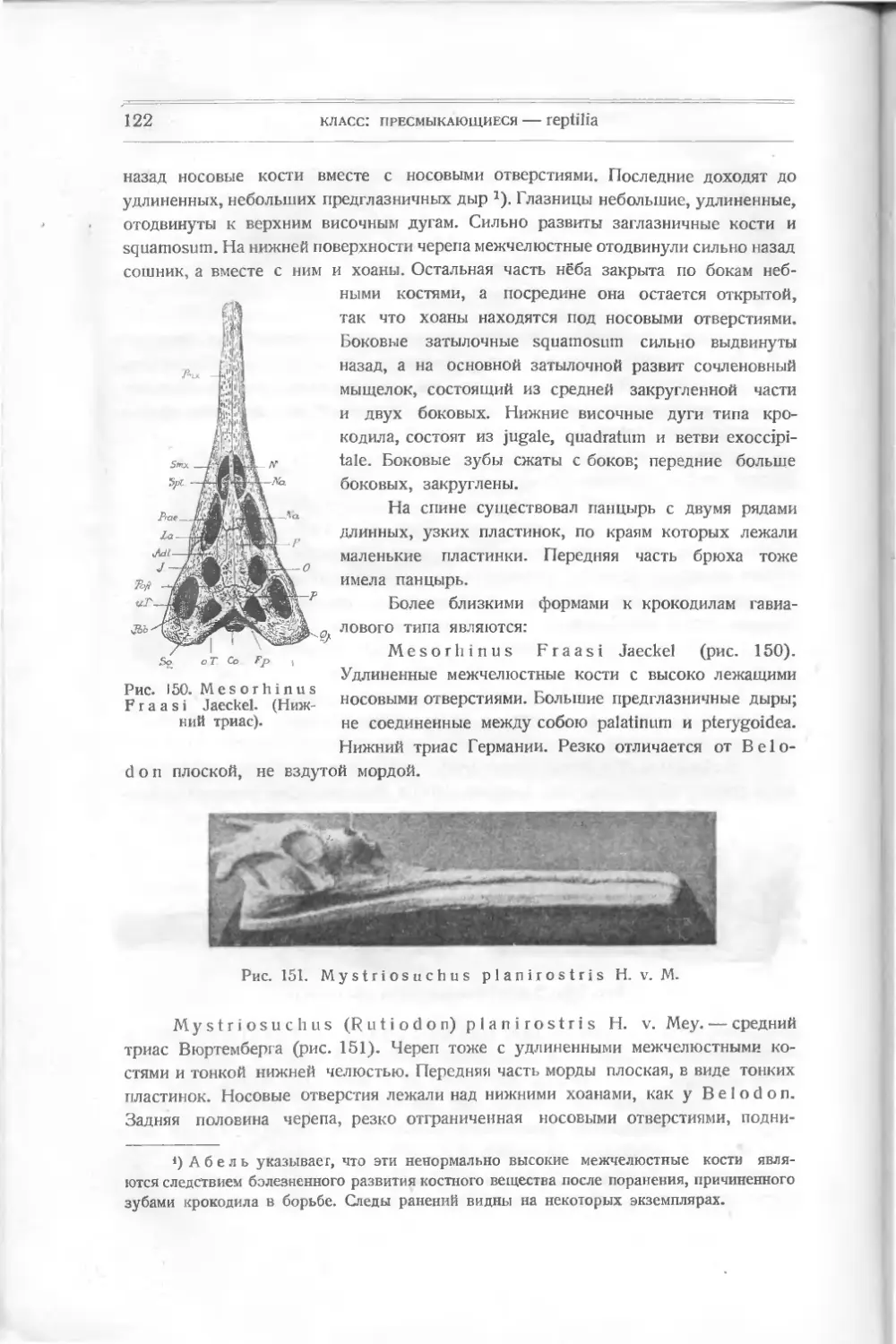
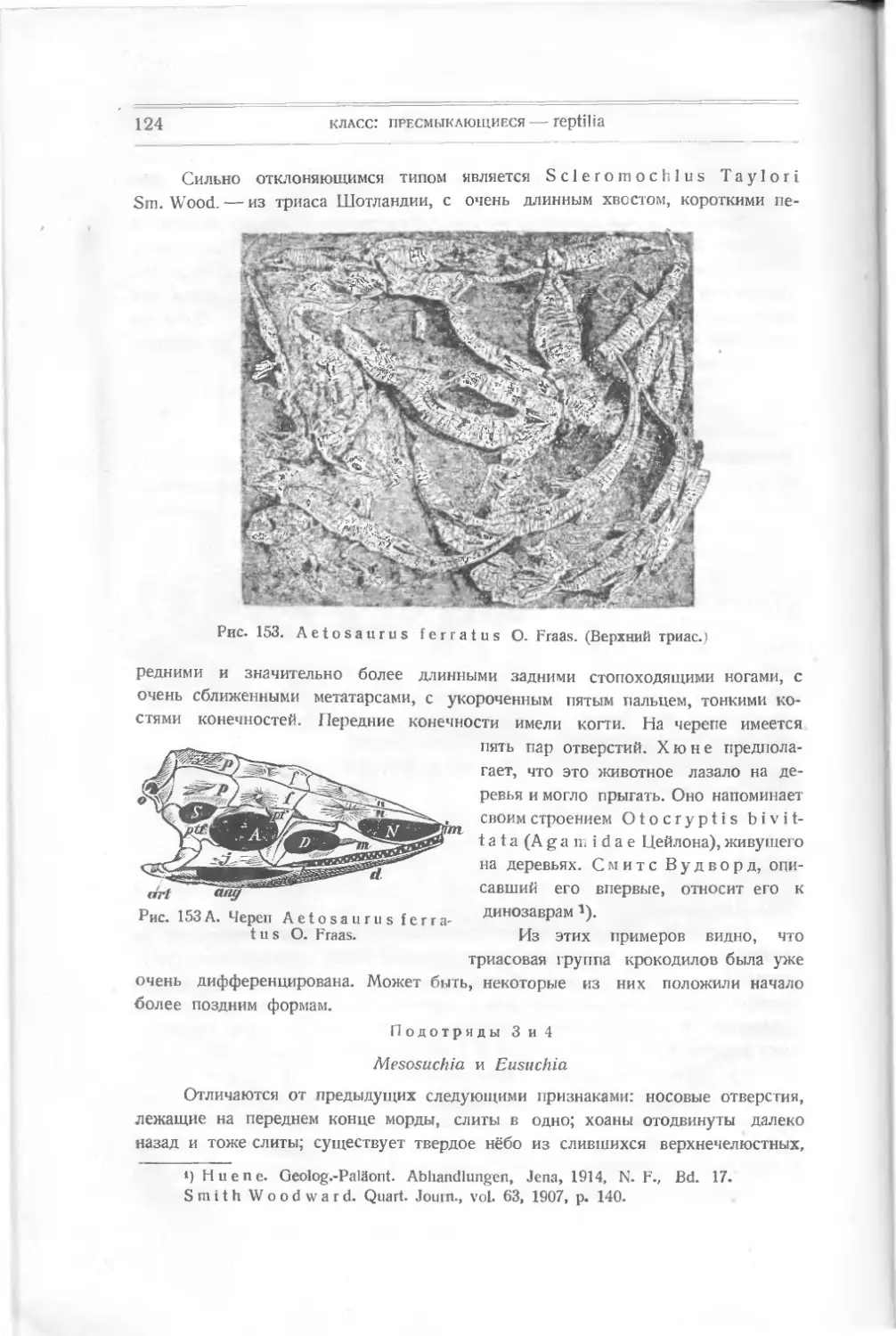
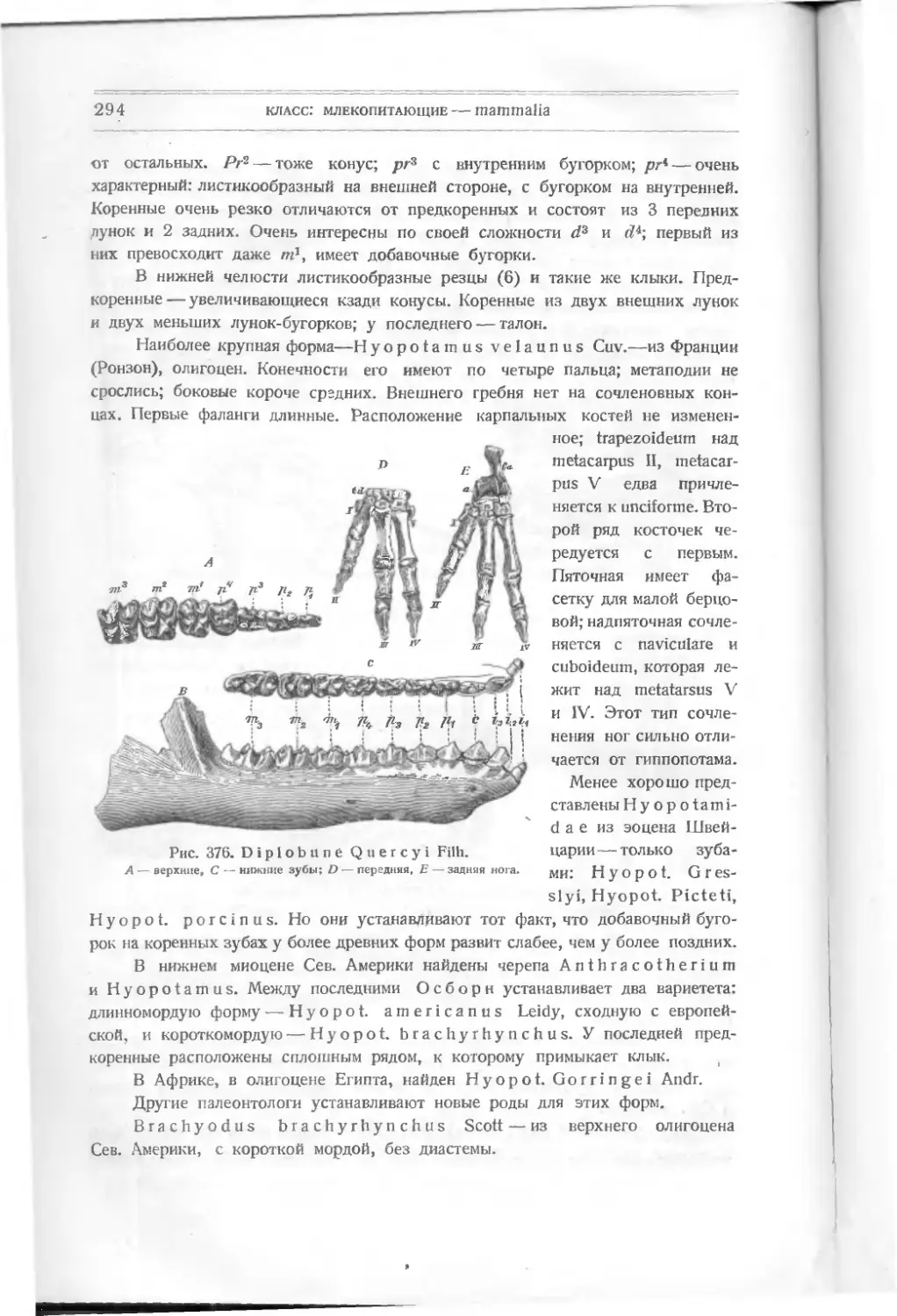
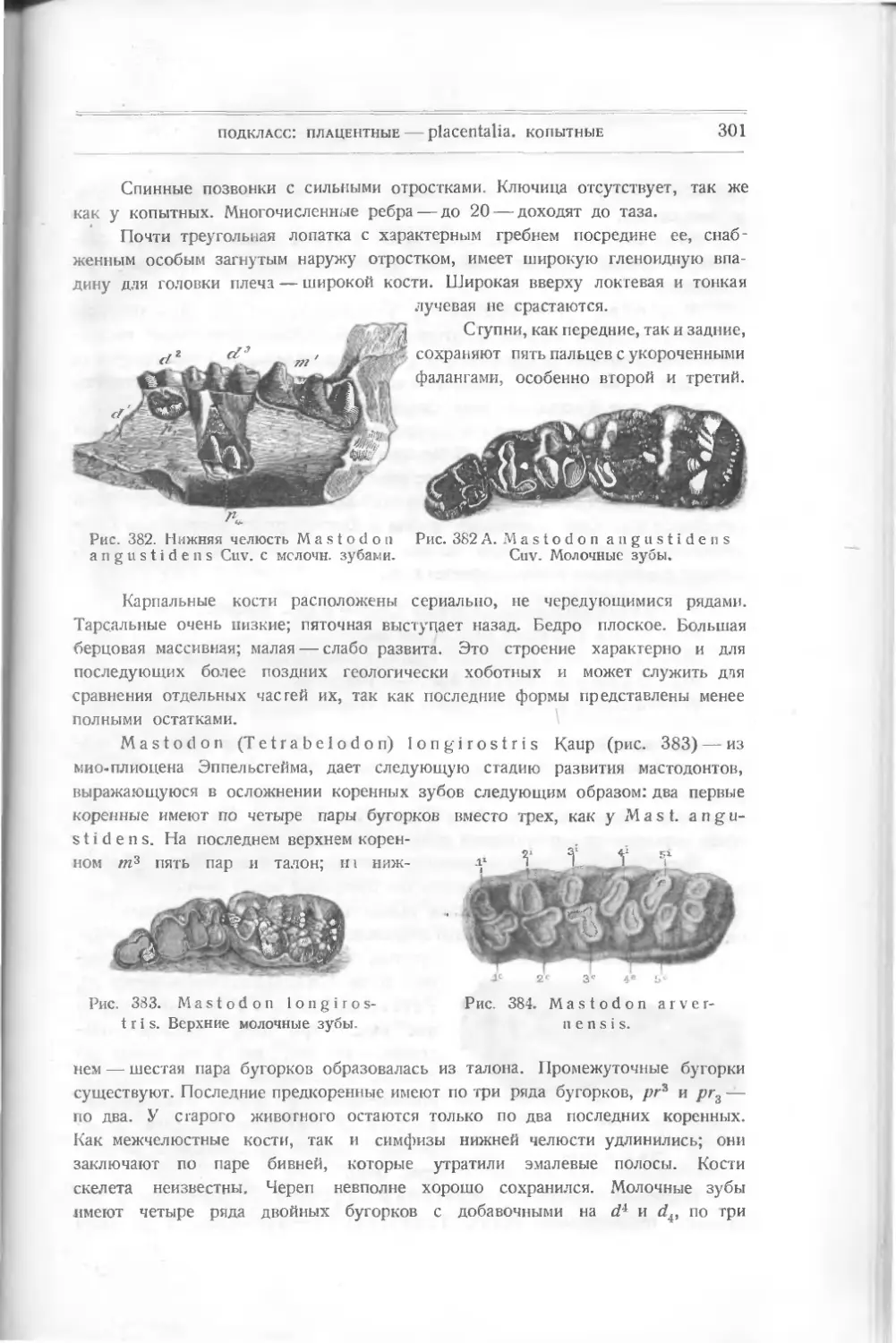
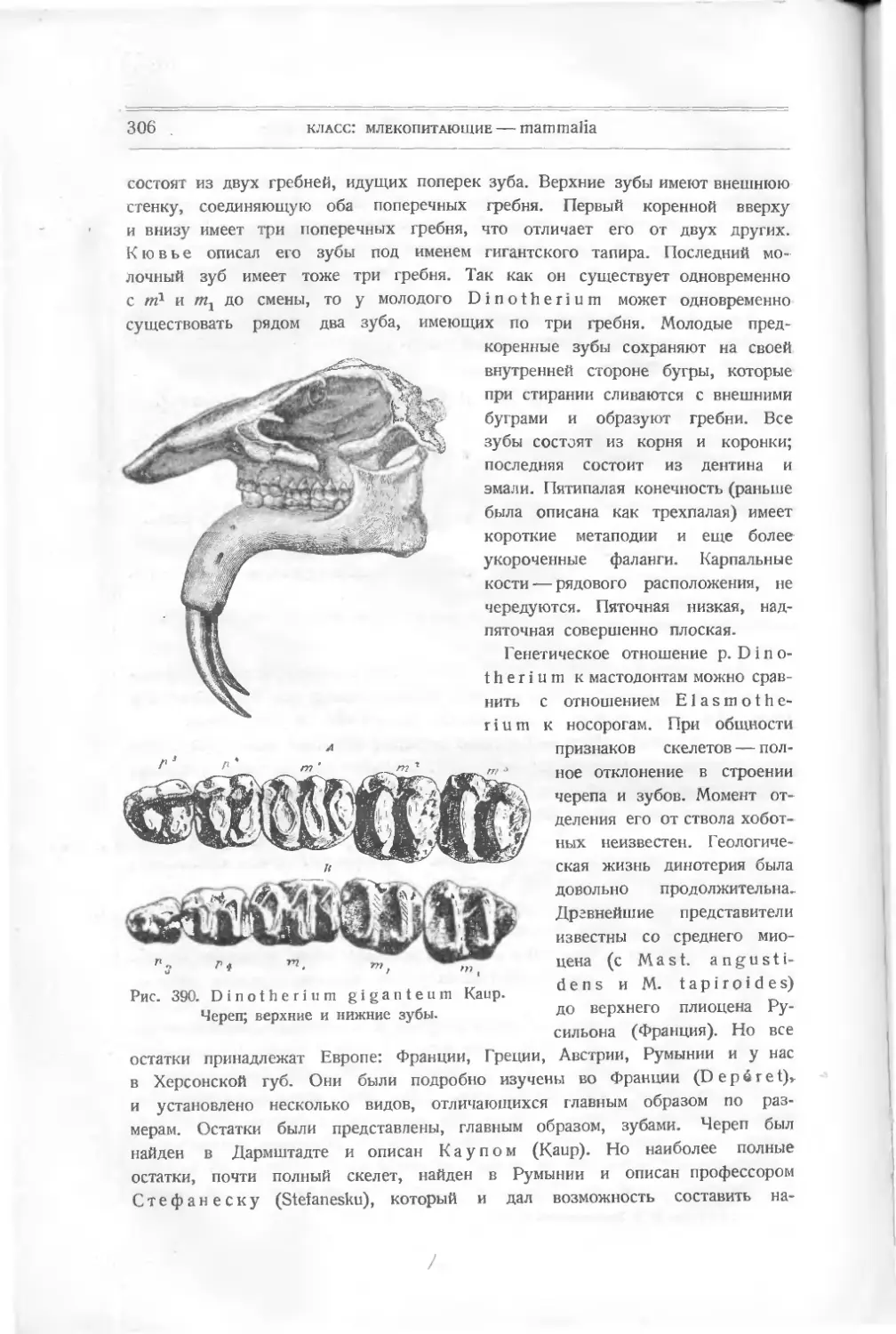
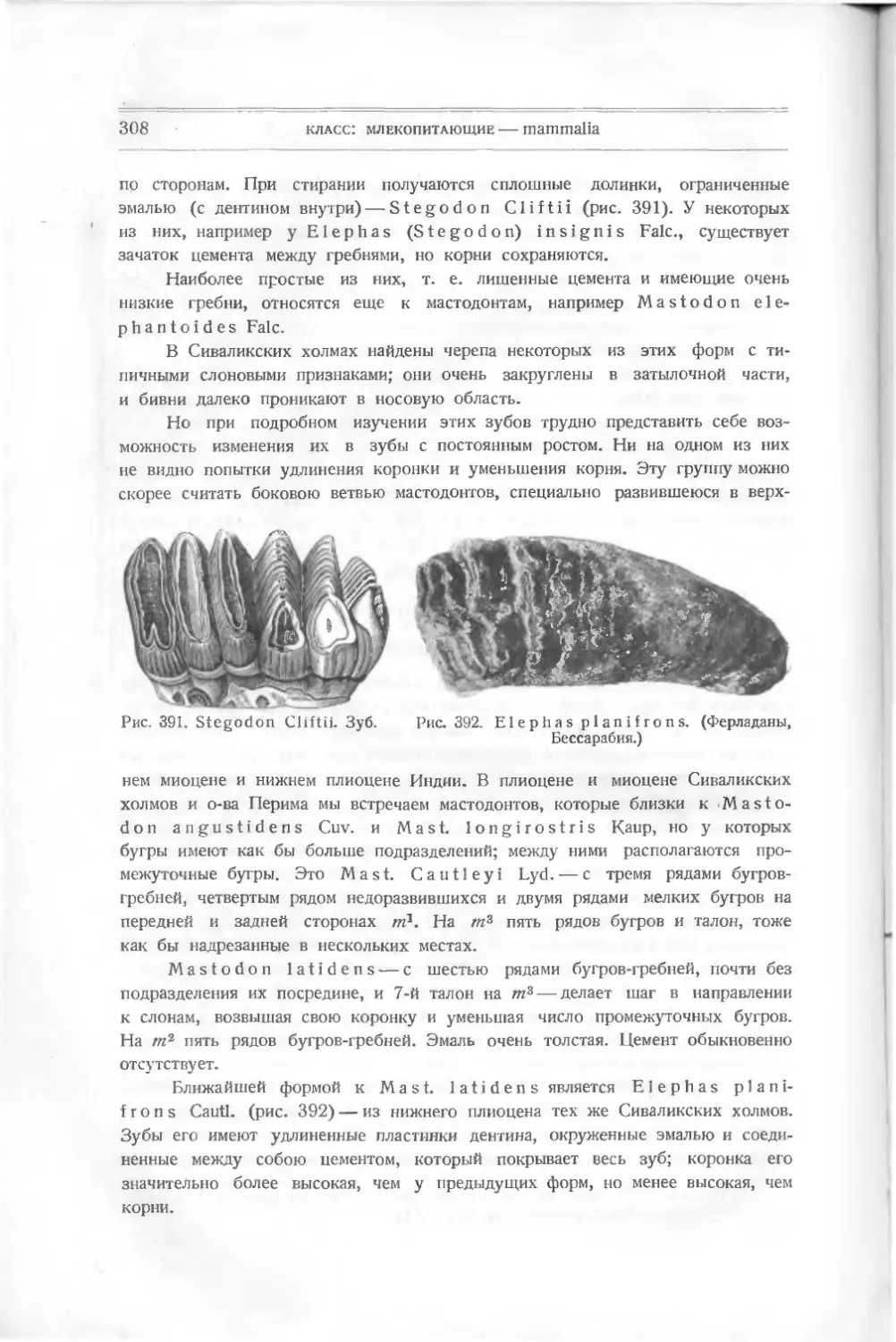
/
























































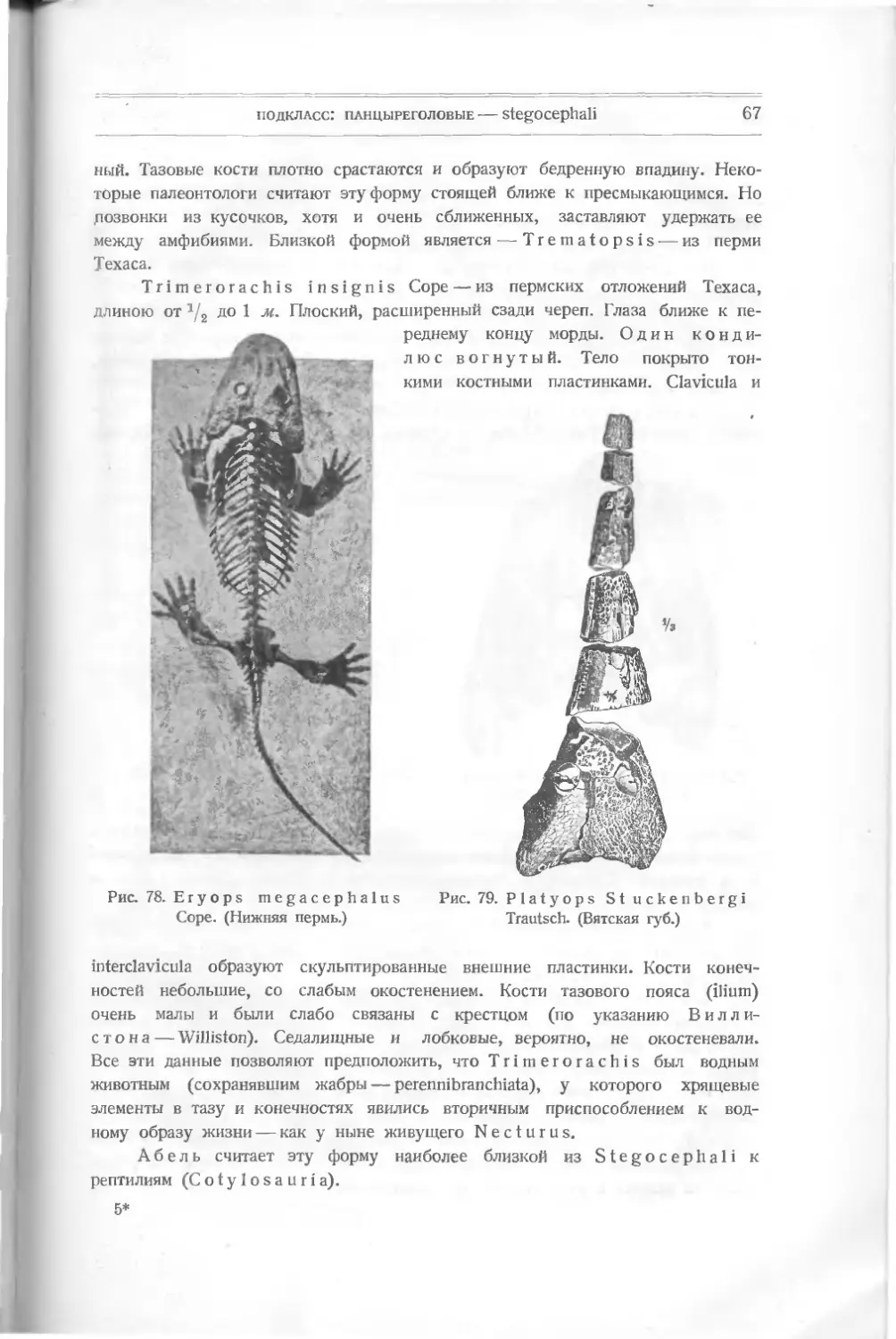
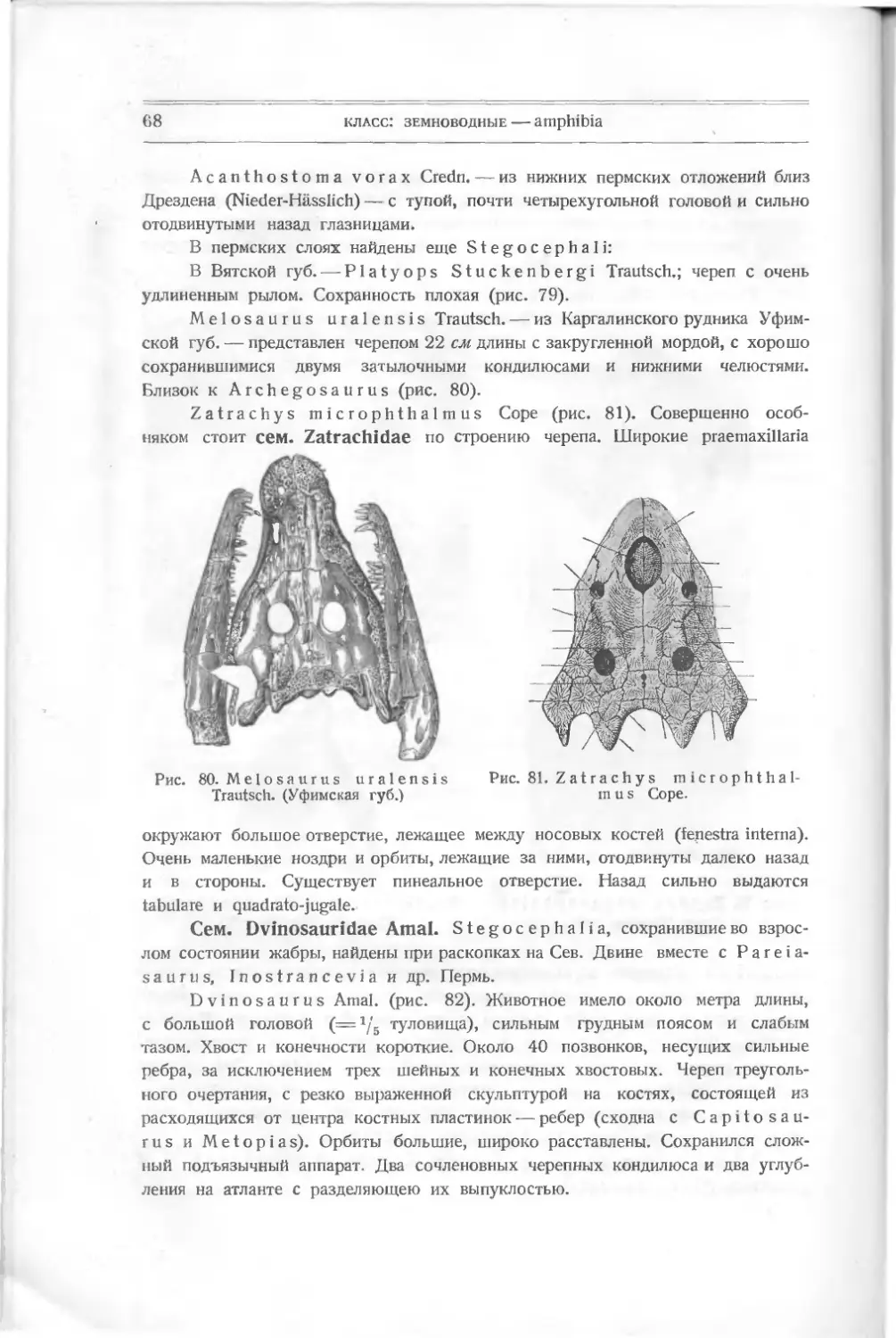


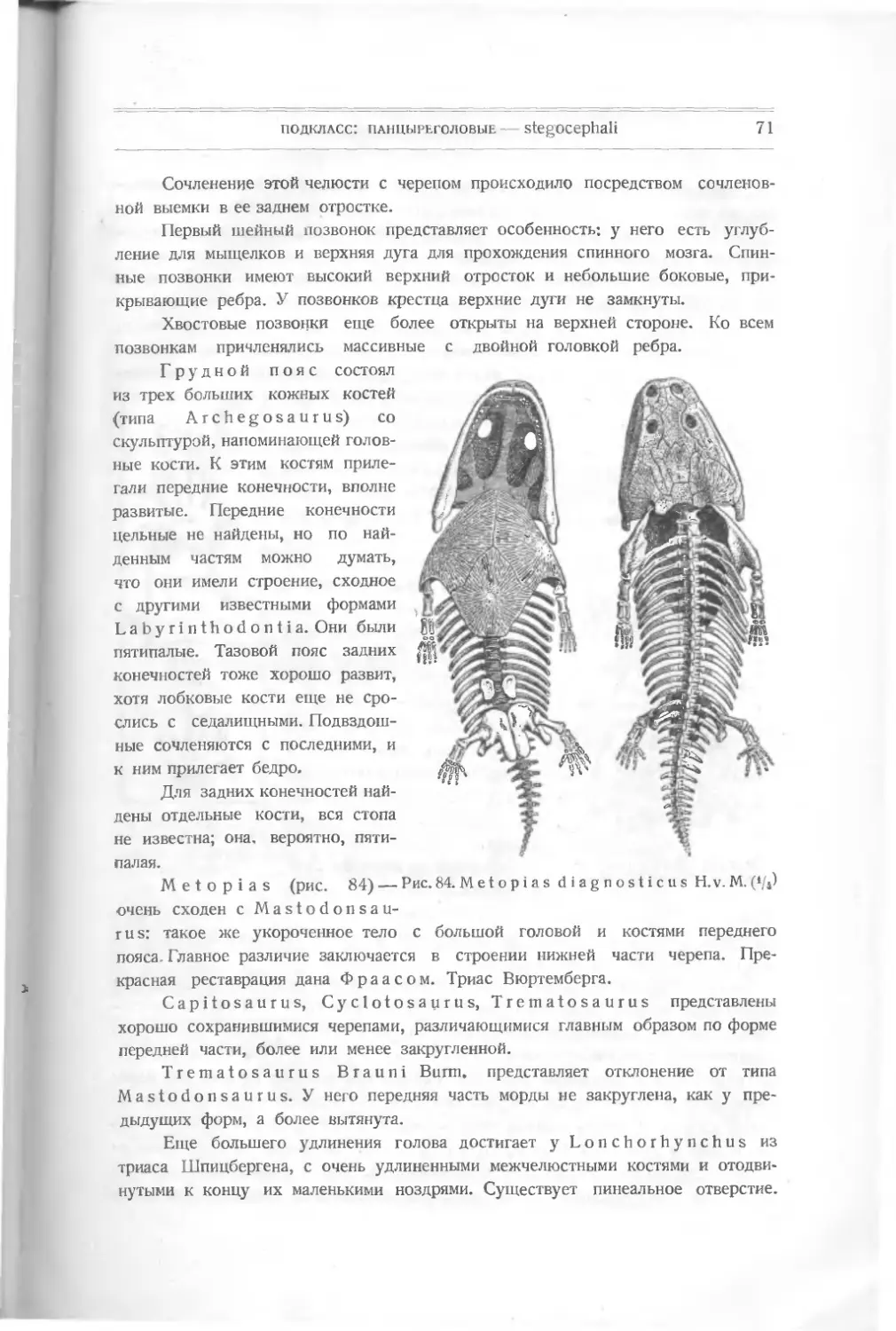
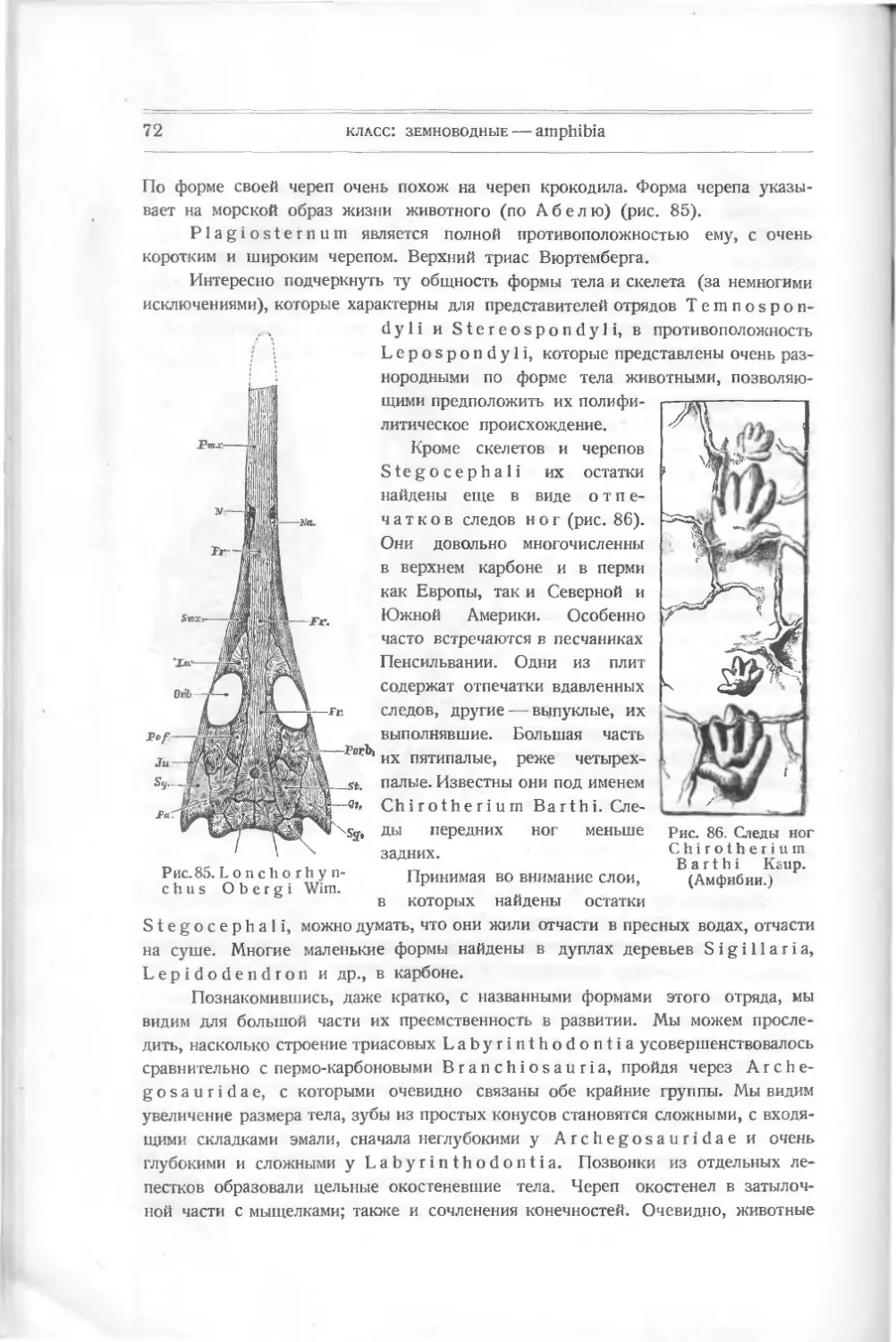

























































































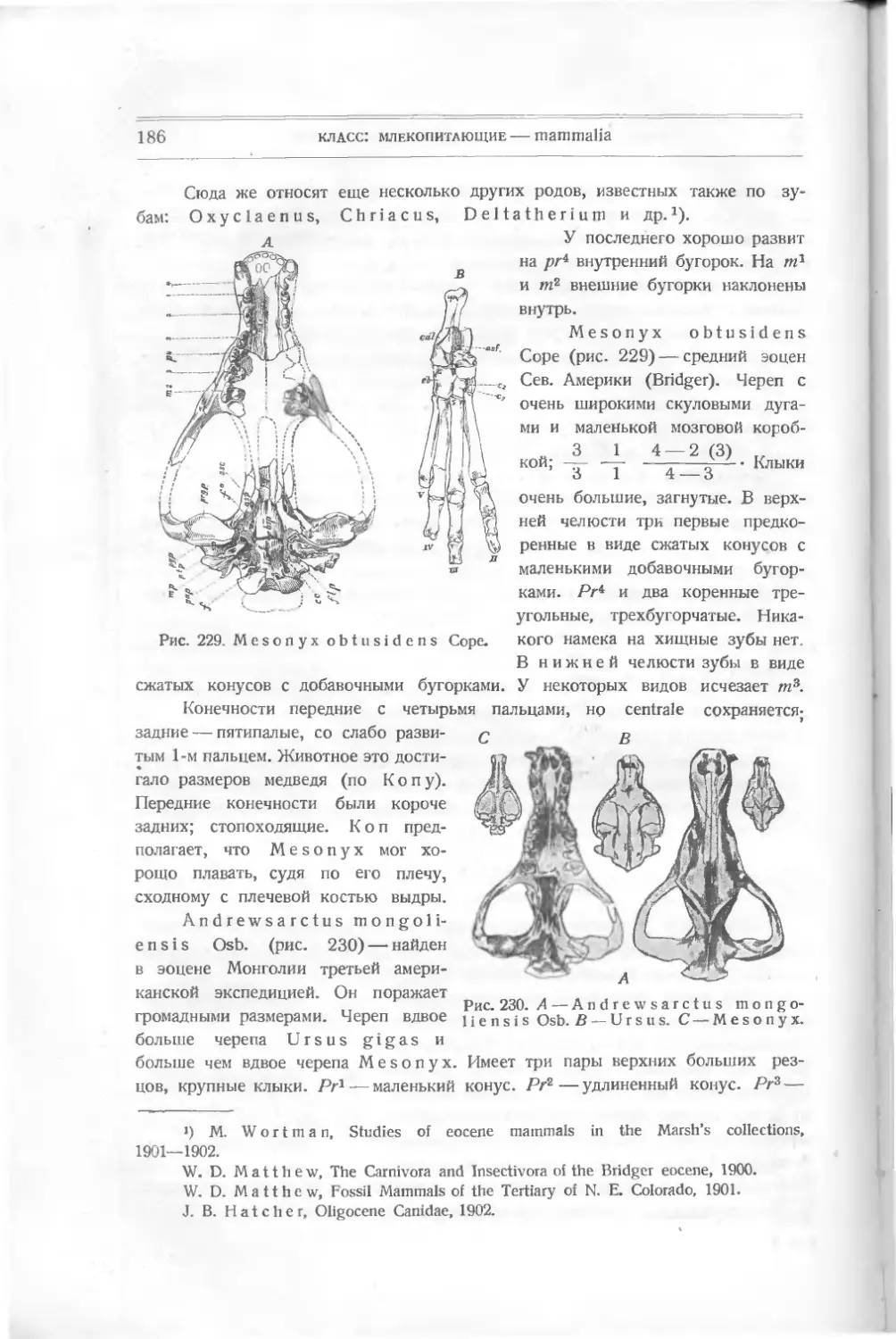


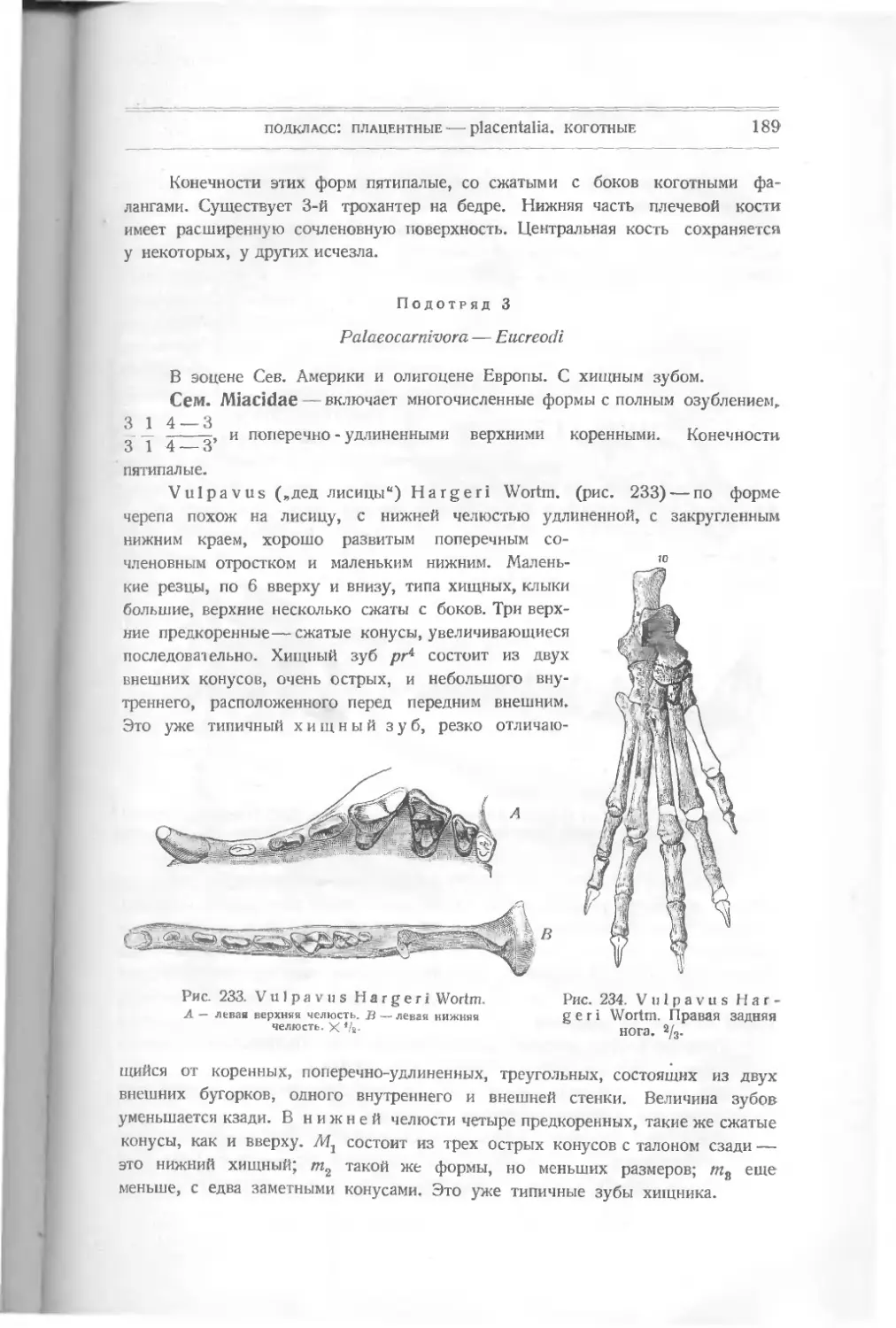
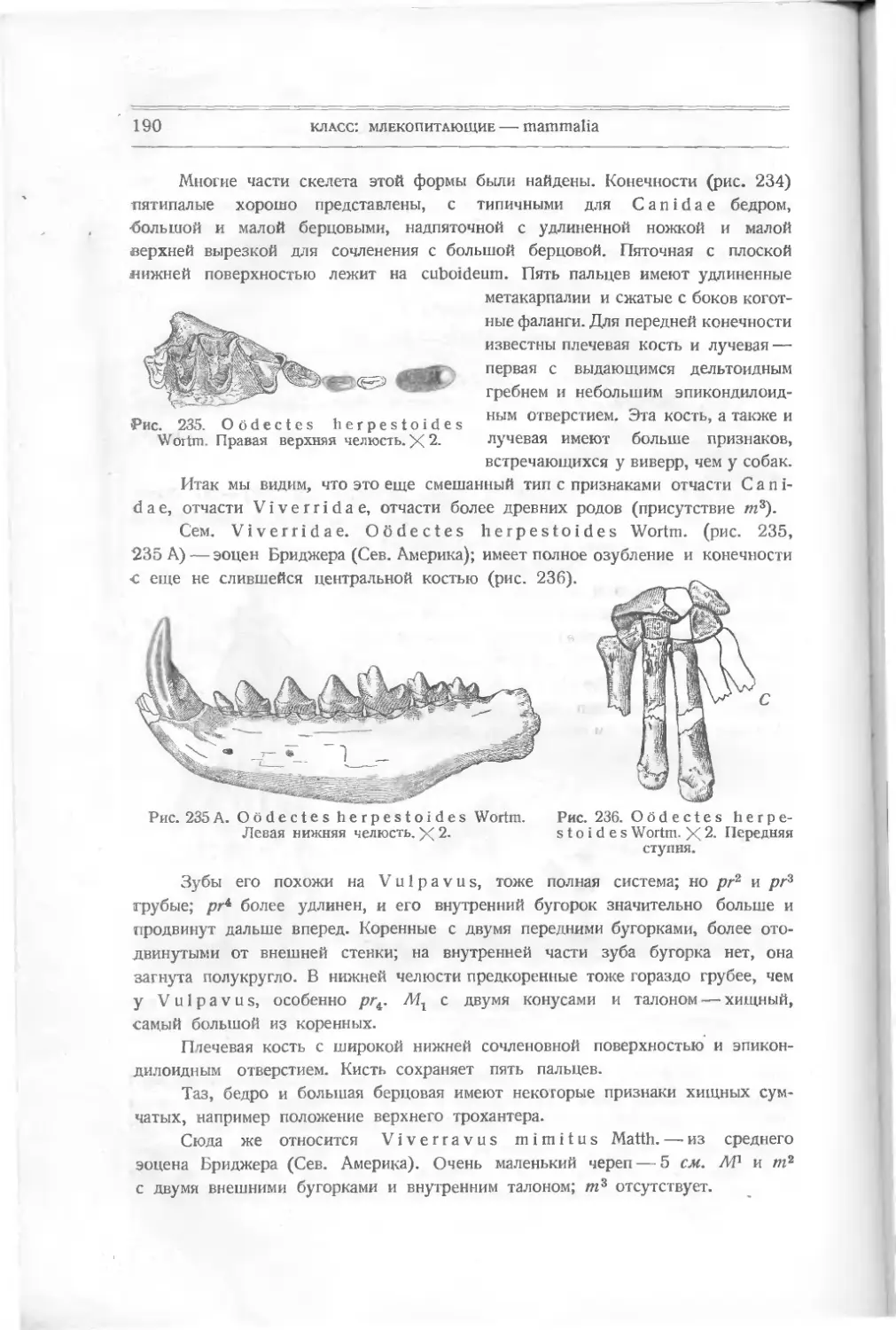
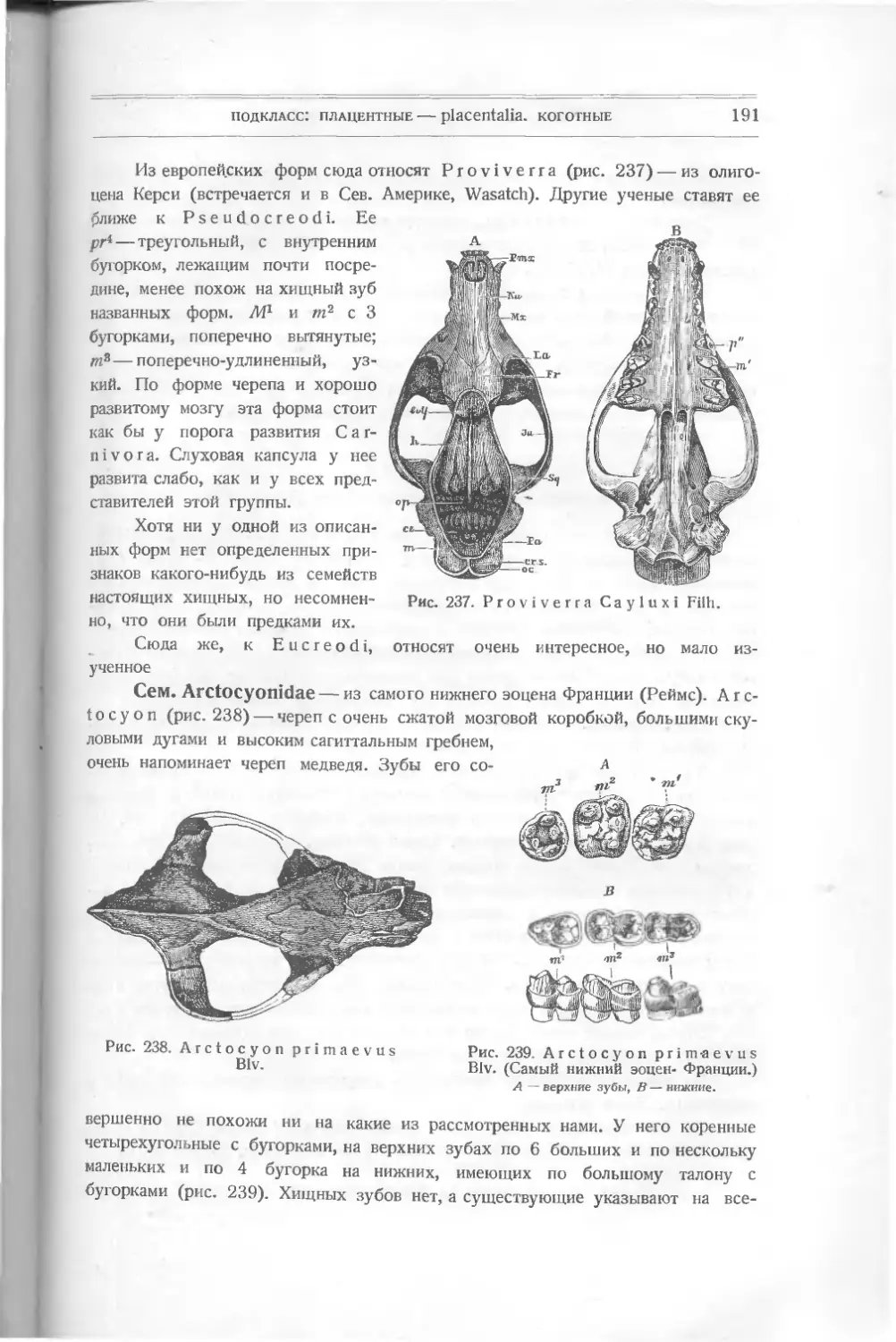
















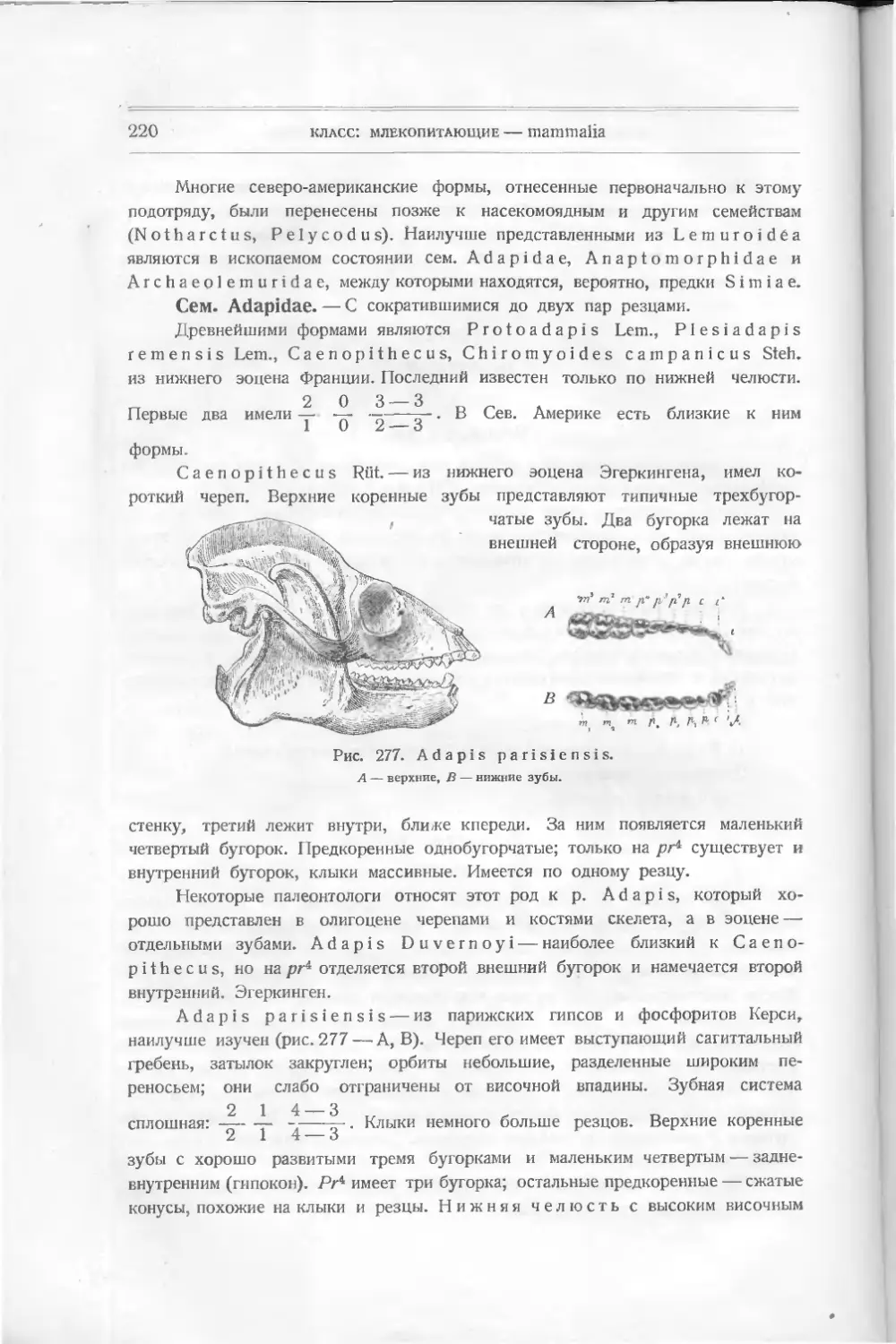


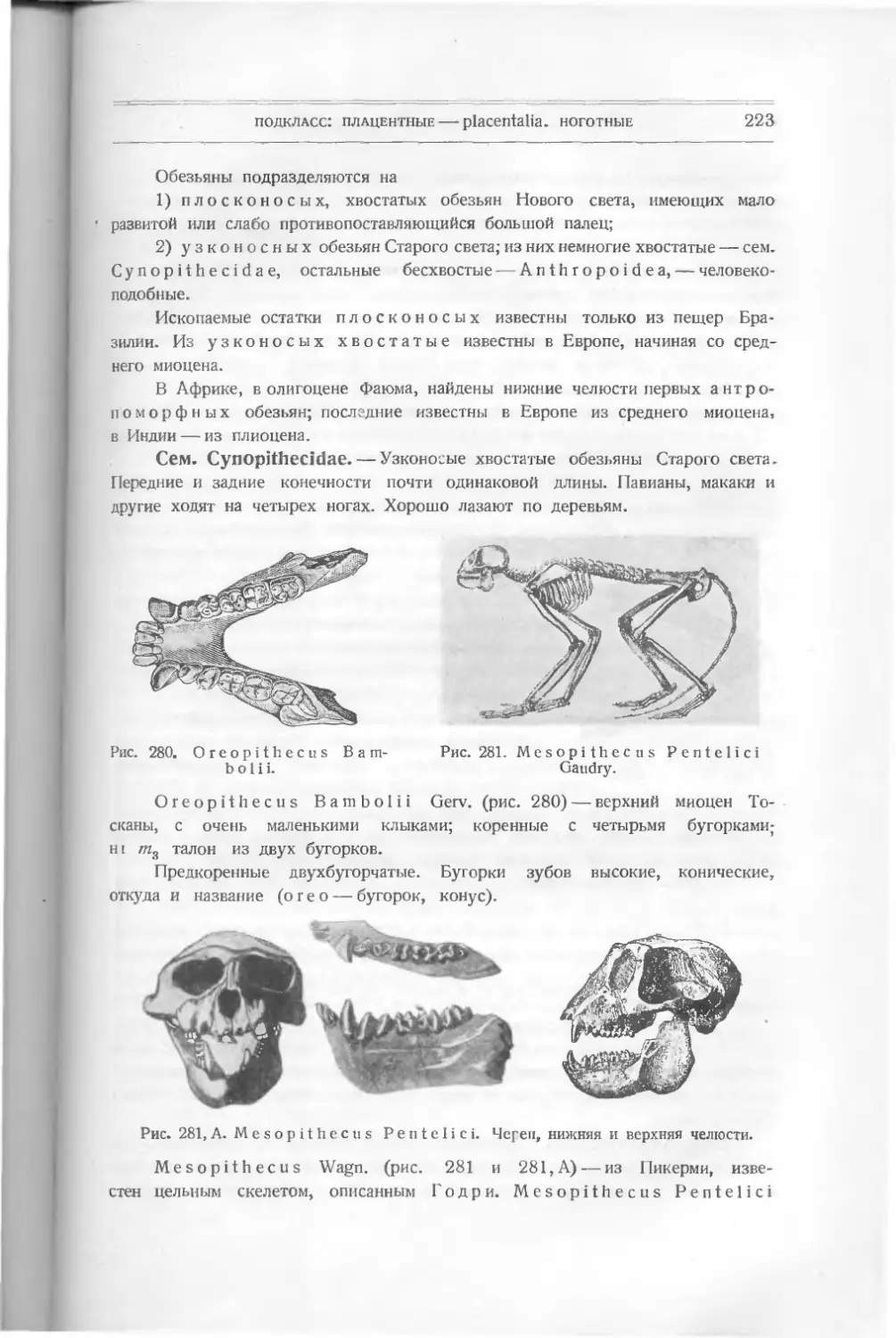


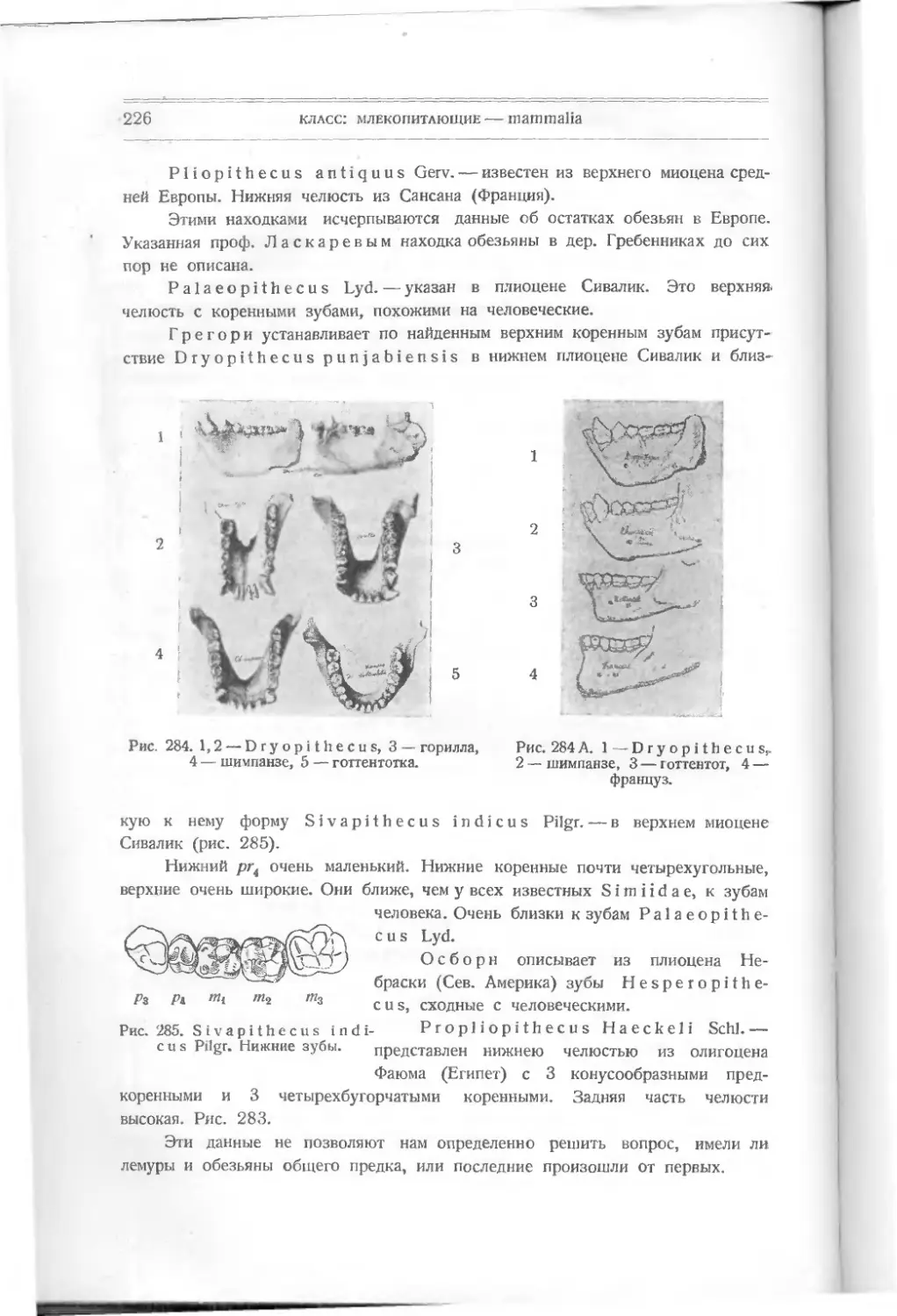








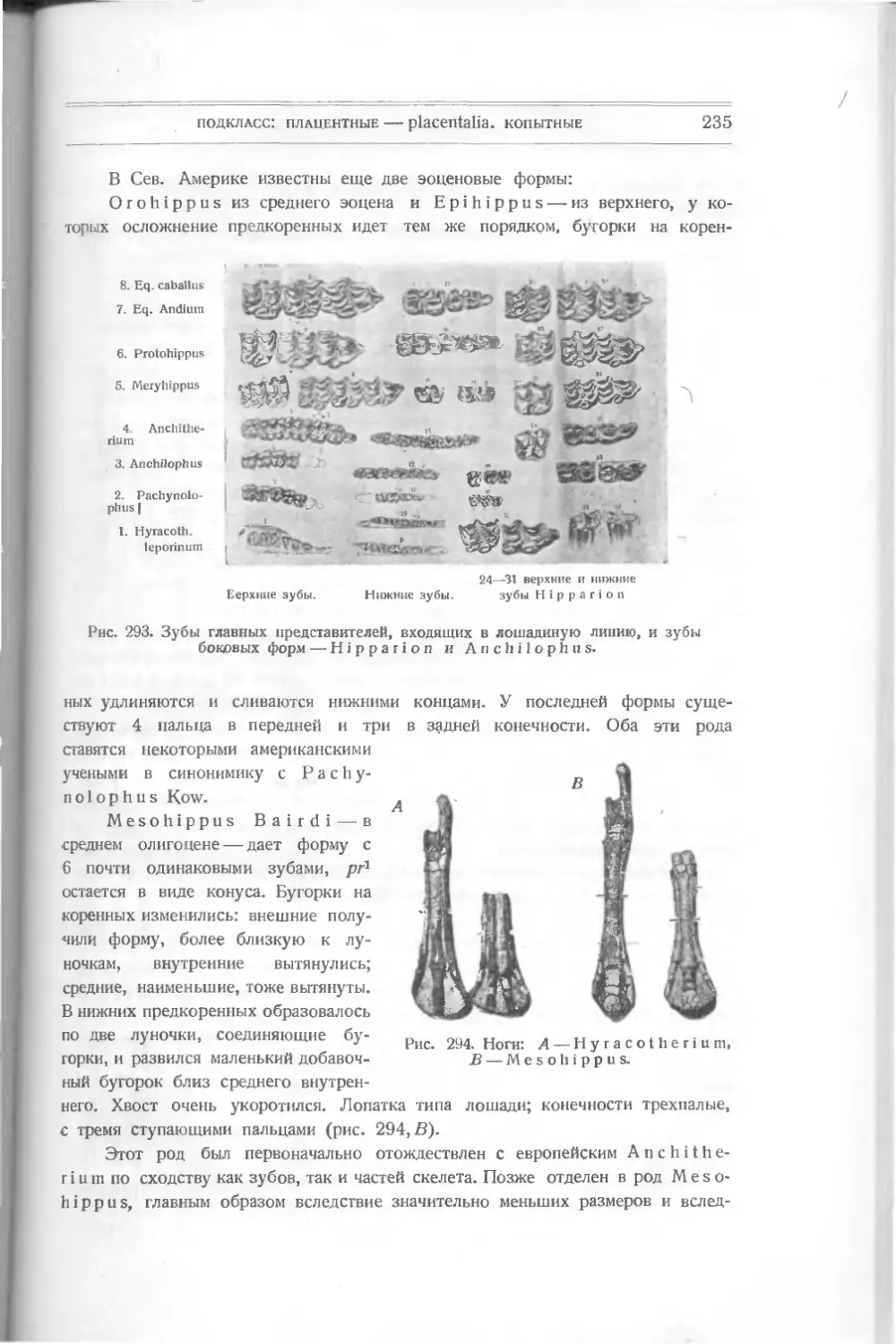










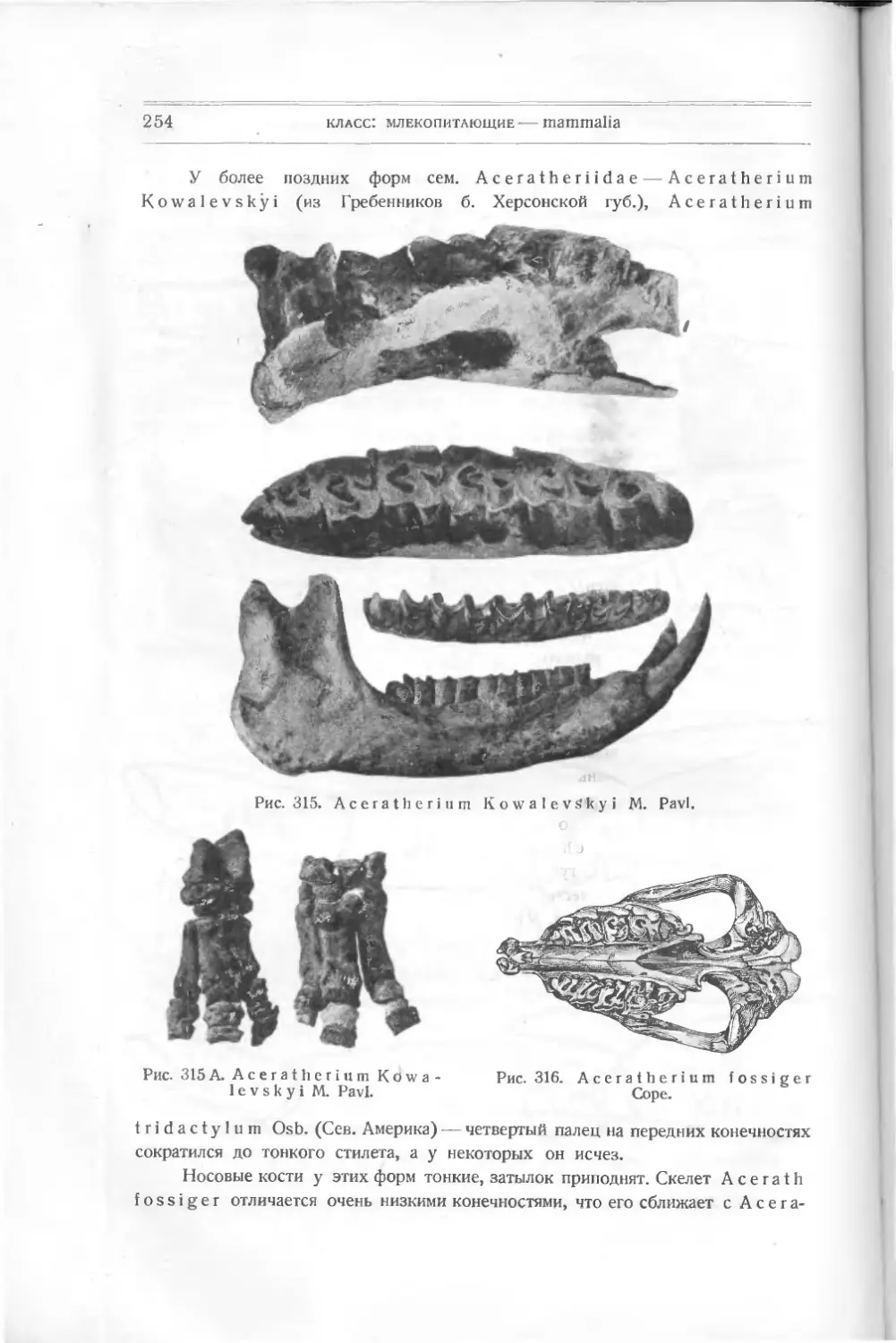

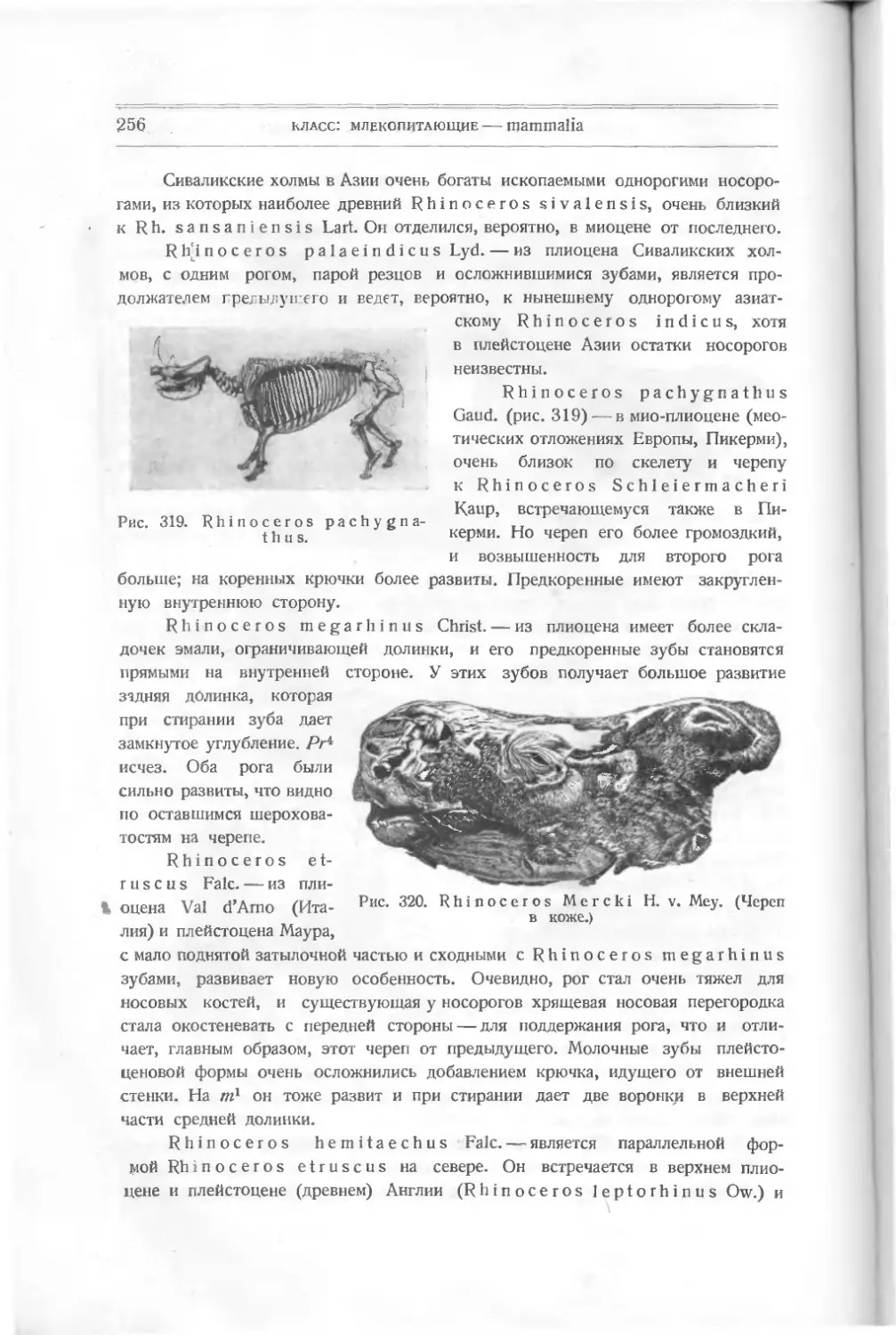



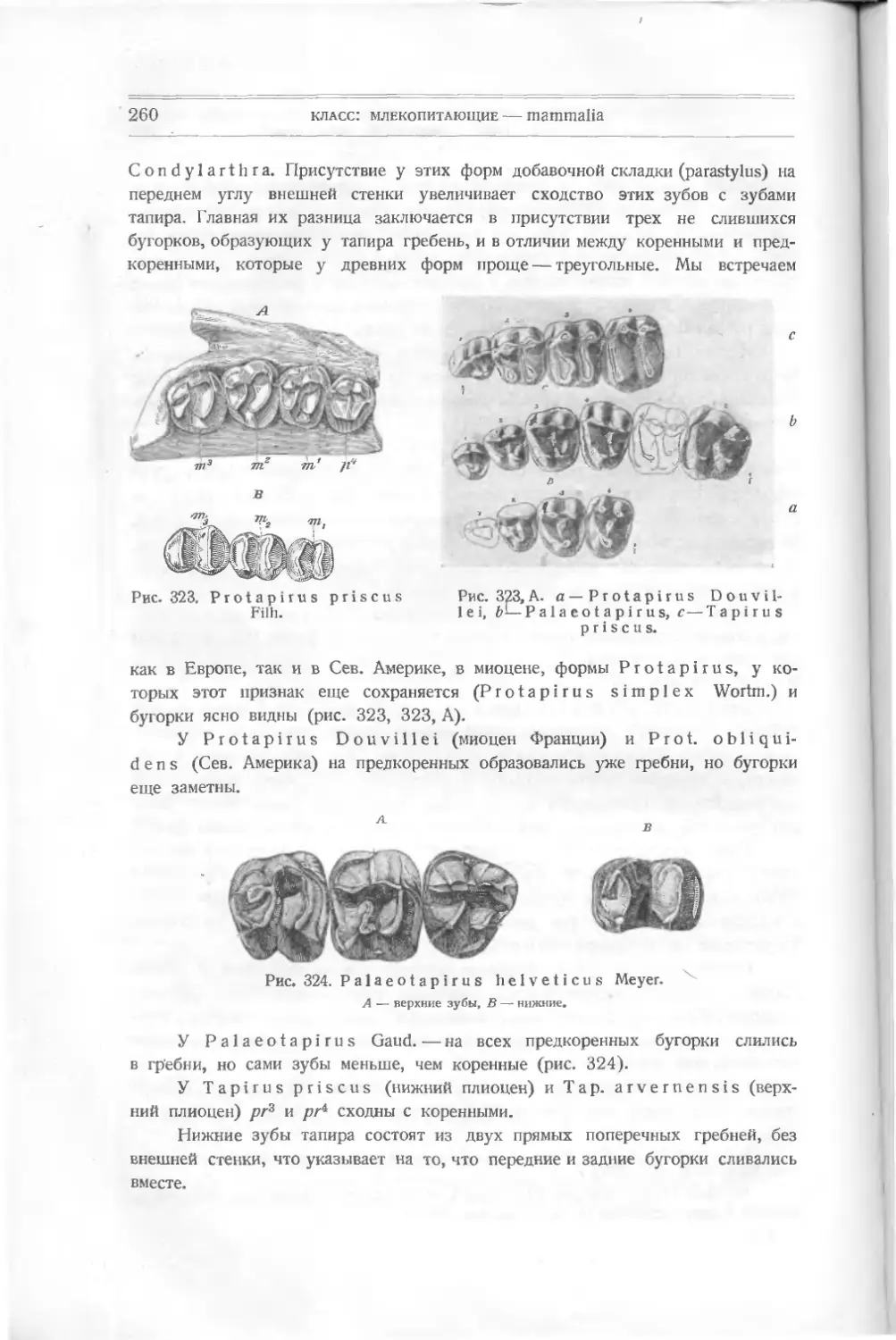


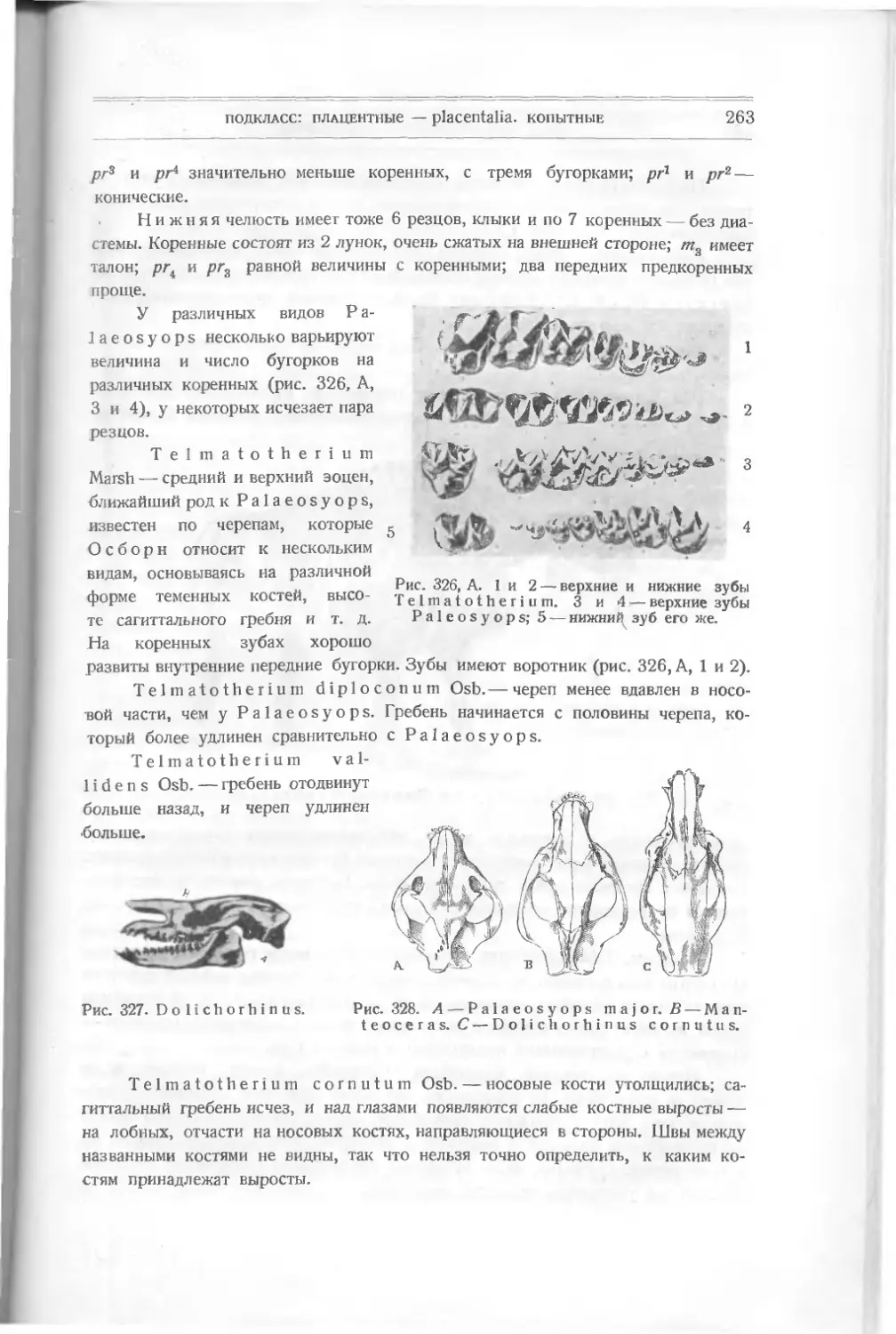








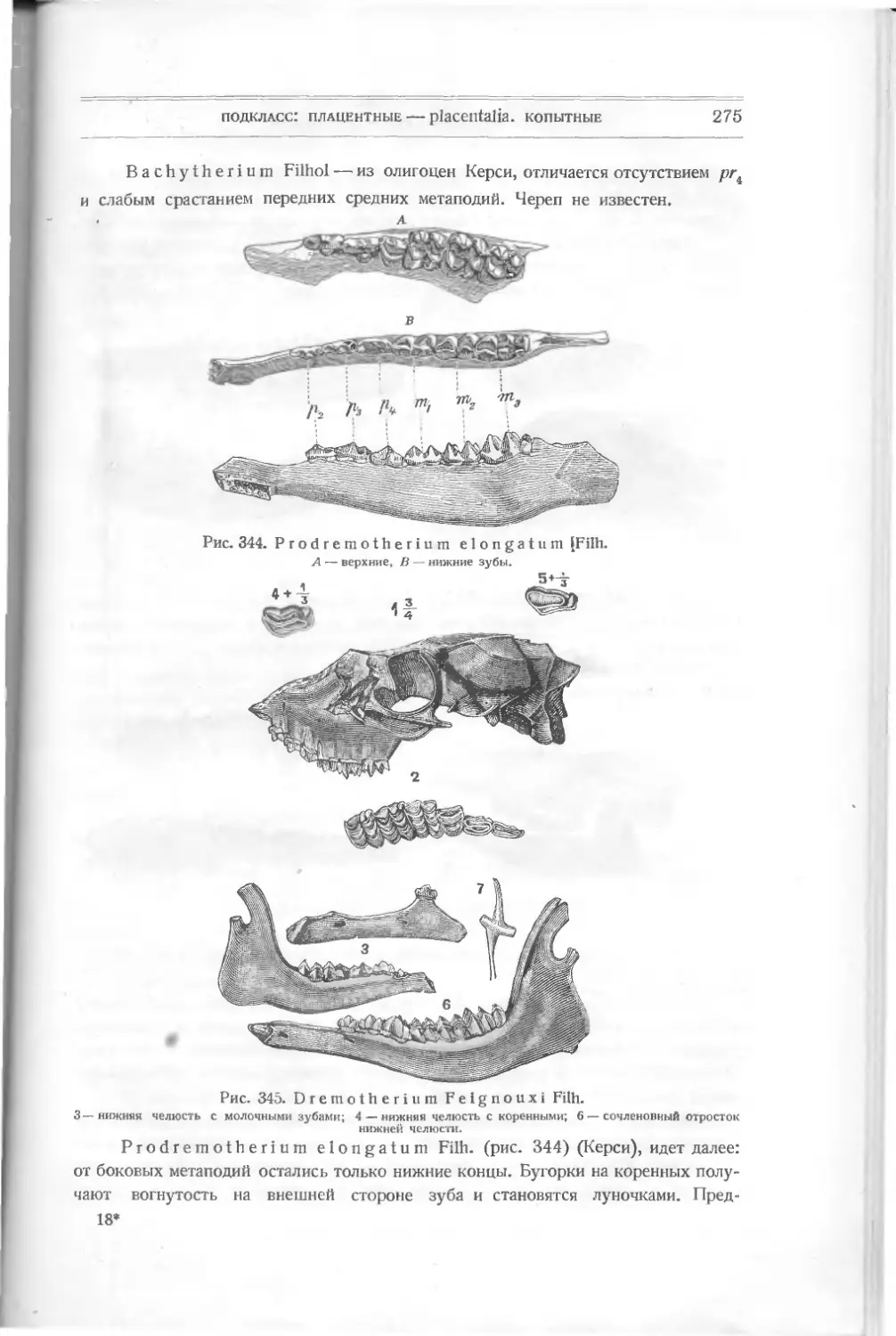

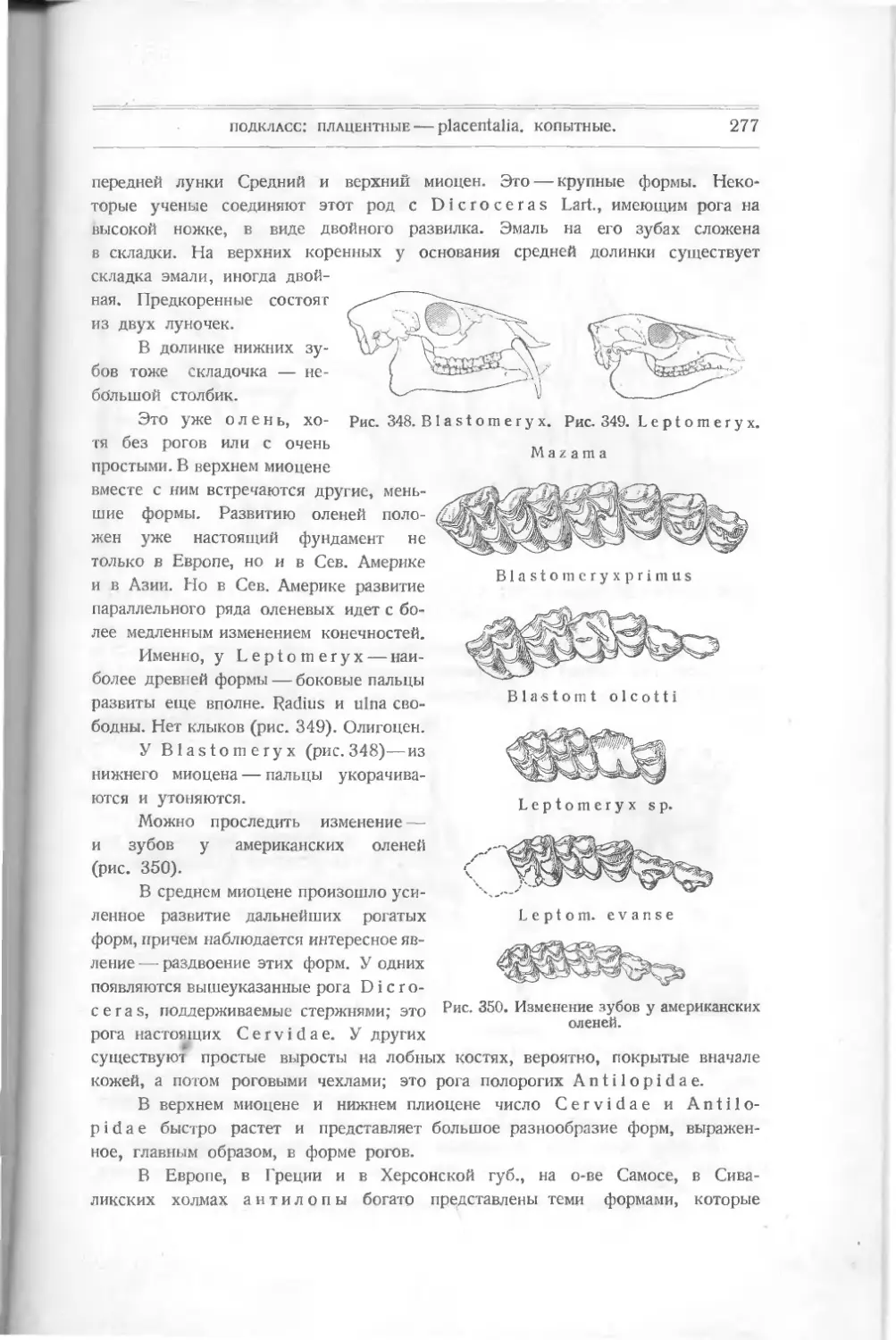


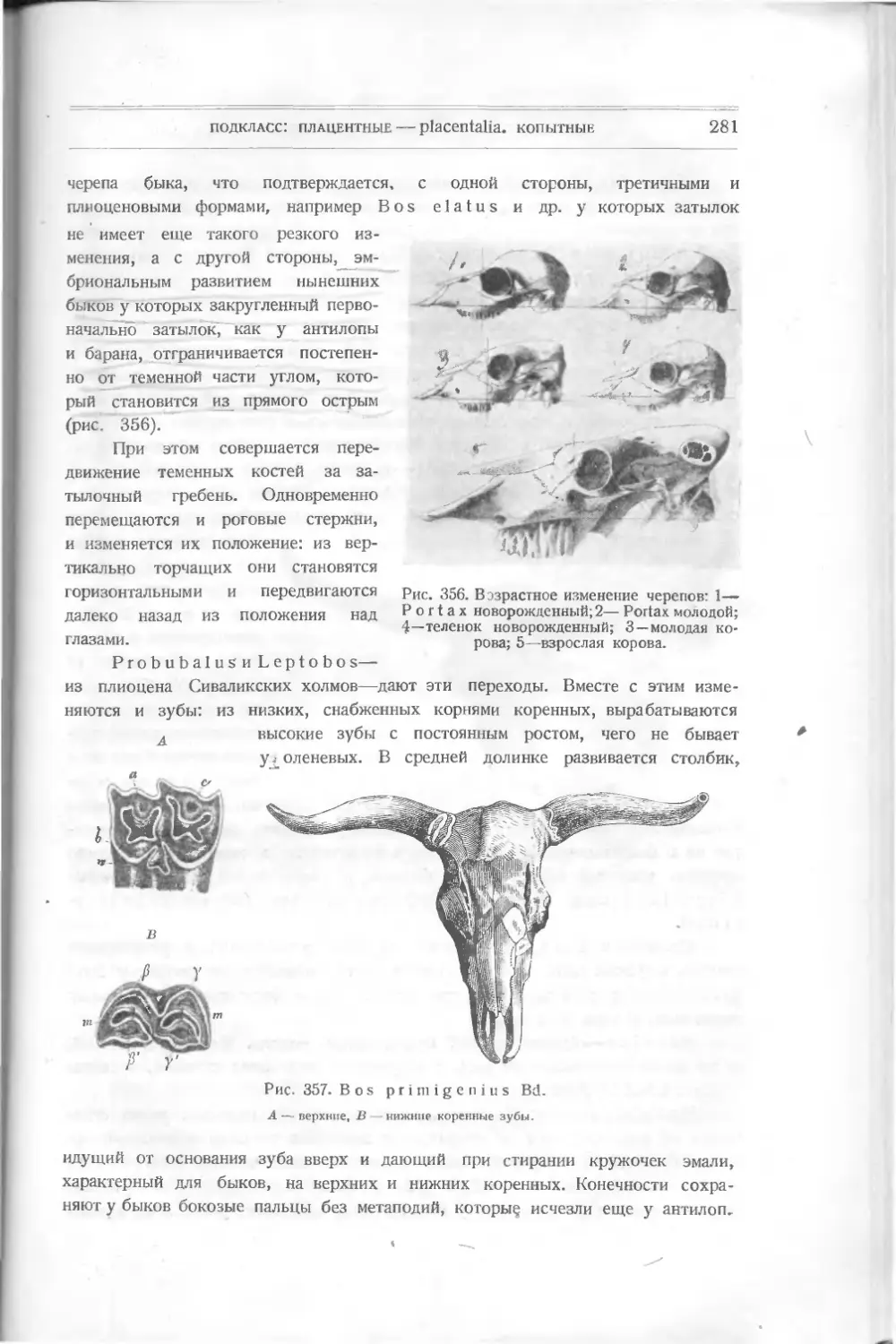

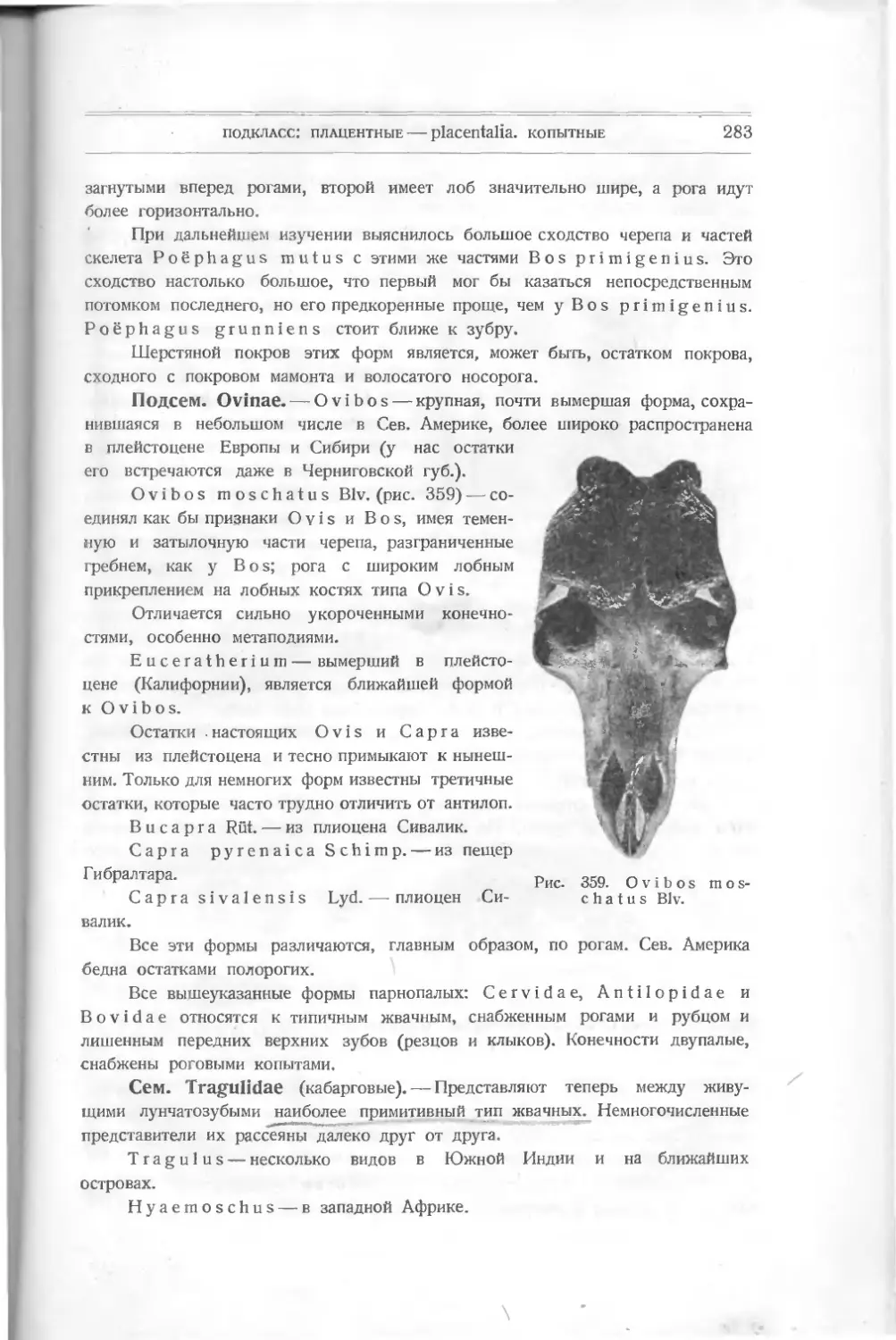














































































Текст
Проф. М. В. ПАВЛОВА
ПАЛЕОЗООЛОГИЯ
ЧАСТЬ ВТОРАН
V
Проф. М. В. ПАВЛОВА
ЧАСТЬ ВТОРАЯ
ПОЗВОНОЧНЫЕ
★
Научно-технической секцией Государственного ученого
совета рекомендовано в качестве пособия, для
высших учебных заведений
ГОСУДАРСТВЕННОЕ ИЗДАТЕЛЬСТВО
МОСКВА * ЛЕНИНГРАД
19 2 9
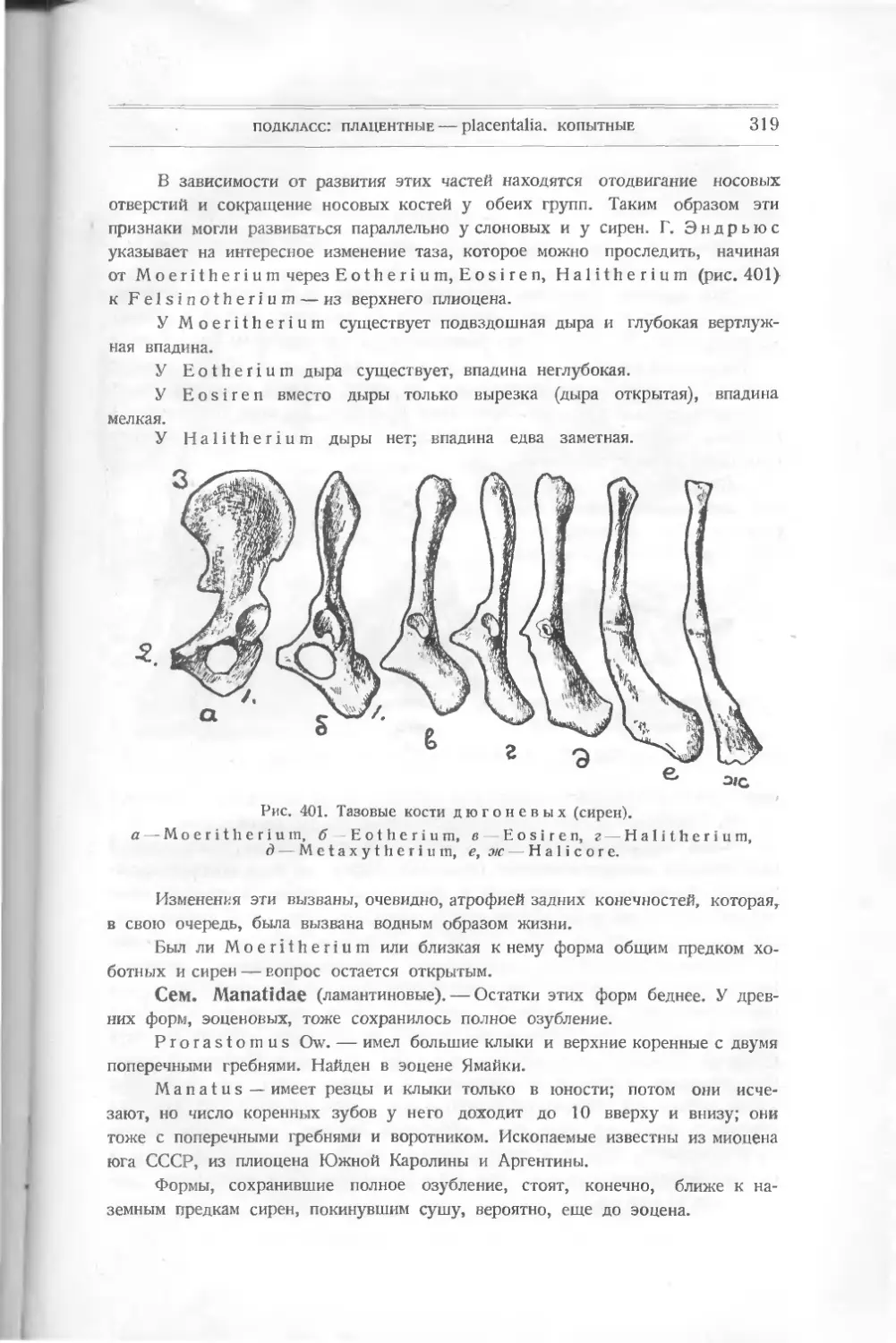
Приступая к изучению остатков ископаемых позвоночных, мы должны
прежде всего подчеркнуть их главные признаки, позволяющие объединить
весьма разных животных.
Конечно, прежде всего приходится указать на присутствие позвоноч-
ника, которому эти животные и обязаны своим названием. Это тот внутрен-
ний подвижной скелет, который расширен в своей передней части в мозговую
коробку, заключающую головной мозг; спинная удлиненная часть его защи-
щает спинной мозг, прикрывая его.
Вторым признаком являются двусторонняя симметрия животного
и спинное положение нервной системы.
Но костный скелет развит крайне различно у различных представителей
тех разнообразных отделов позвоночных, через которые шло развитие живот-
ных от низших рыб до высших форм — млекопитающих, известных как по
ныне живущим формам, так и по остаткам вымерших и исчезнувших форм.
Если мы не можем проследить перехода непосредственно от одной группы
к другой в различных отделах животных, то по ископаемым остаткам мы
знаем, что представители их появлялись постепенно, так как остатки эти
встречаются в последовательных слоях земной коры. В палеозое, как мы это
видели во введении к беспозвоночным, имеются только низшие рыбы, амфибии
и немногие рептилии; в мезозое рептилии достигают наибольшего развития и
разнообразия. Млекопитающие только появляются в этой эре, а развиваются и
достигают большого разнообразия в кенозое.
Но знаем ли мы, или можем ли надеяться открыть настоящих предков по-
звоночных? Некоторые зоологи указывают на сближение оболочников
(Tunicata) с позвоночными; другие ученые сближают с ними ланцетника
(Amphioxus). Некель (Jaeckel) высказывается за возможность развития позво-
ночных из форм, близких к скорпиону и древним паукообразным. Он указы-
вает на совпадение наибольшего развития гигантских паукообразных — Е и г у-
pteridae с появлением панцырных рыб и считает, что если эти рыбы произошли
от паукообразных, то это могло случиться именно в то время, когда послед-
ние достигли наибольшего развития и разнообразия. Палеонтология не дает до
сих пор указаний на первичную форму позвоночных, и трудно предположить,
чтобы последняя могла сохраниться, не имея твердого скелета. Предположение
Иекеля не находит подтверждения в строении названных животных. Вероятно,
что этой первоначальной формой является ланцетник. Но, конечно, не тот
II а в л о в а М. В. Палеозоология. Ч. II.
2
ВВЕДЕНИЕ
ланцетник, который живет теперь в море, зарываясь в песок (рис, 1), а его отда-
ленный предок, живший в море свободно. Одно можно сказать с уверенностью,
что первые формы позвоночных были водными животными и что
только позже, при изменениях в распределении морей и суши, они стали
изменять свой образ жизни, а соответственно этому и свою организацию,
переходя сначала в полусухопутных, а позже и в совершенно наземных
животных. Эти указания дает нам палеонтология. Однако И е к е л ь считает,,
что древнейшие рыбы жили на берегах рек и в пресных водах, переходя позже
и в моря1).
При изучении различных классов позвоночных были подмечены некоторые-
общие признаки, характеризующие различные моменты развития их. Так,,
например, американский ученый Коп (Соре) установил для рыб, как и для на-
земных позвоночных, что-
прогресс их развития’
не шел равномерно, а
происходил ритмиче-
ски: были смены усилен-
ного, беспокойного раз-
вития — настоящего-
застоя ппогпесса.
Рис. 1. Amphioxus lanceolatus (ныне живущий).
„ с — хорда, п — нервный ствол, д — жаберная часть кишечника.
прогресса и периоды сравнительного покоя,
во время которого животные изменялись, приспособляясь односторонне
к окружающей среде; они или развивались до громадных размеров, или
дегенерировали, но не давали высших прогрессивных форм. Так, например,
рептилии мелового периода при всем их богатстве не дали ни современных
рептилий, ни млекопитающих, которые получили начало гораздо раньше от
форм, менее специализировавшихся, с более общими признаками. Большая часть-
млекопитающих постепенно развивалась, проявив большое разнообразие в тре-
тичном периоде во время полного упадка рептилий. То же мы увидим при
изучении некоторых групп рыб и других позвоночных. Коп, американский
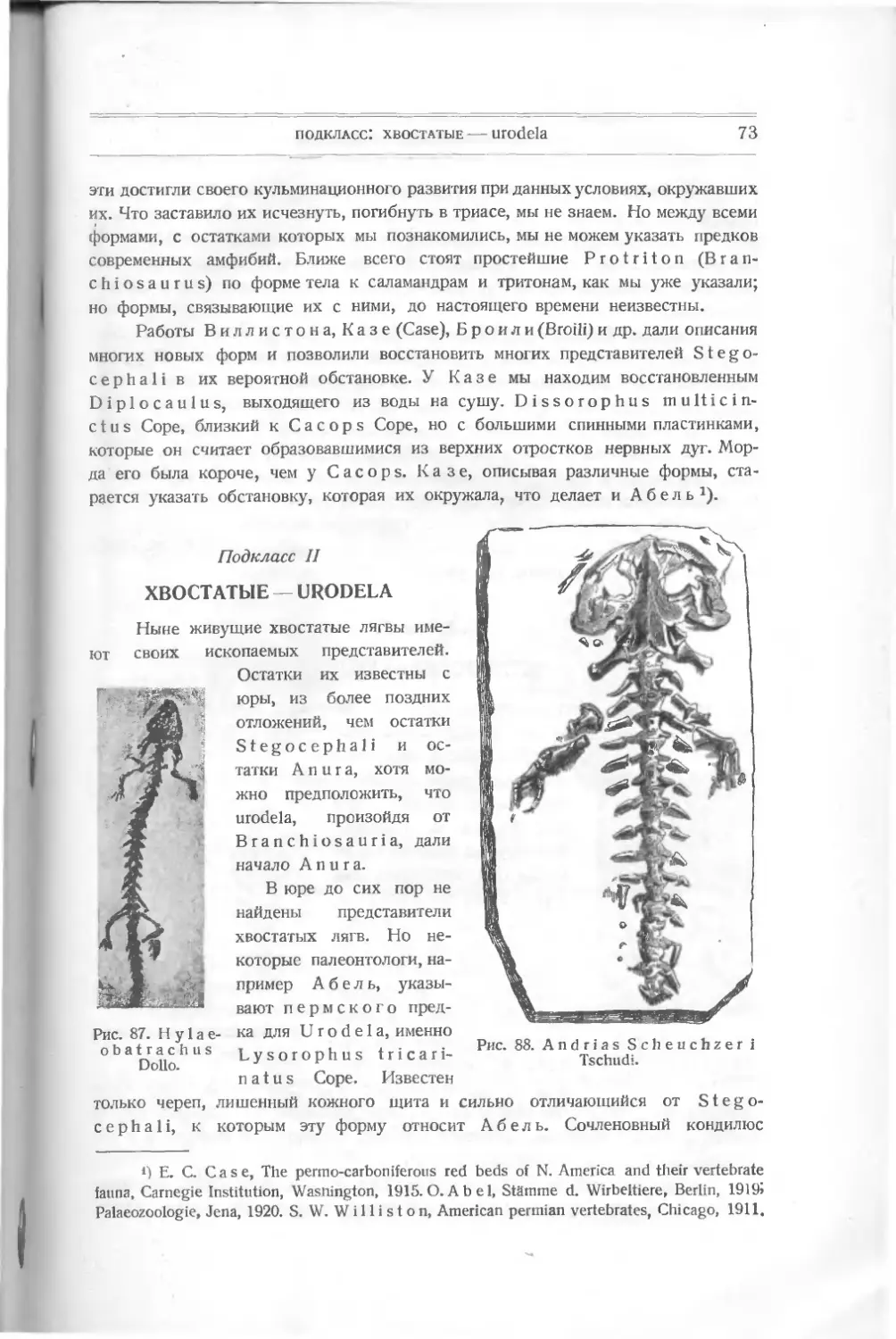
палеонтолог, первый подметил этот закон и назвал эти моменты существенного-
прогресса данной группы „точками выраженияили „моментами развития"
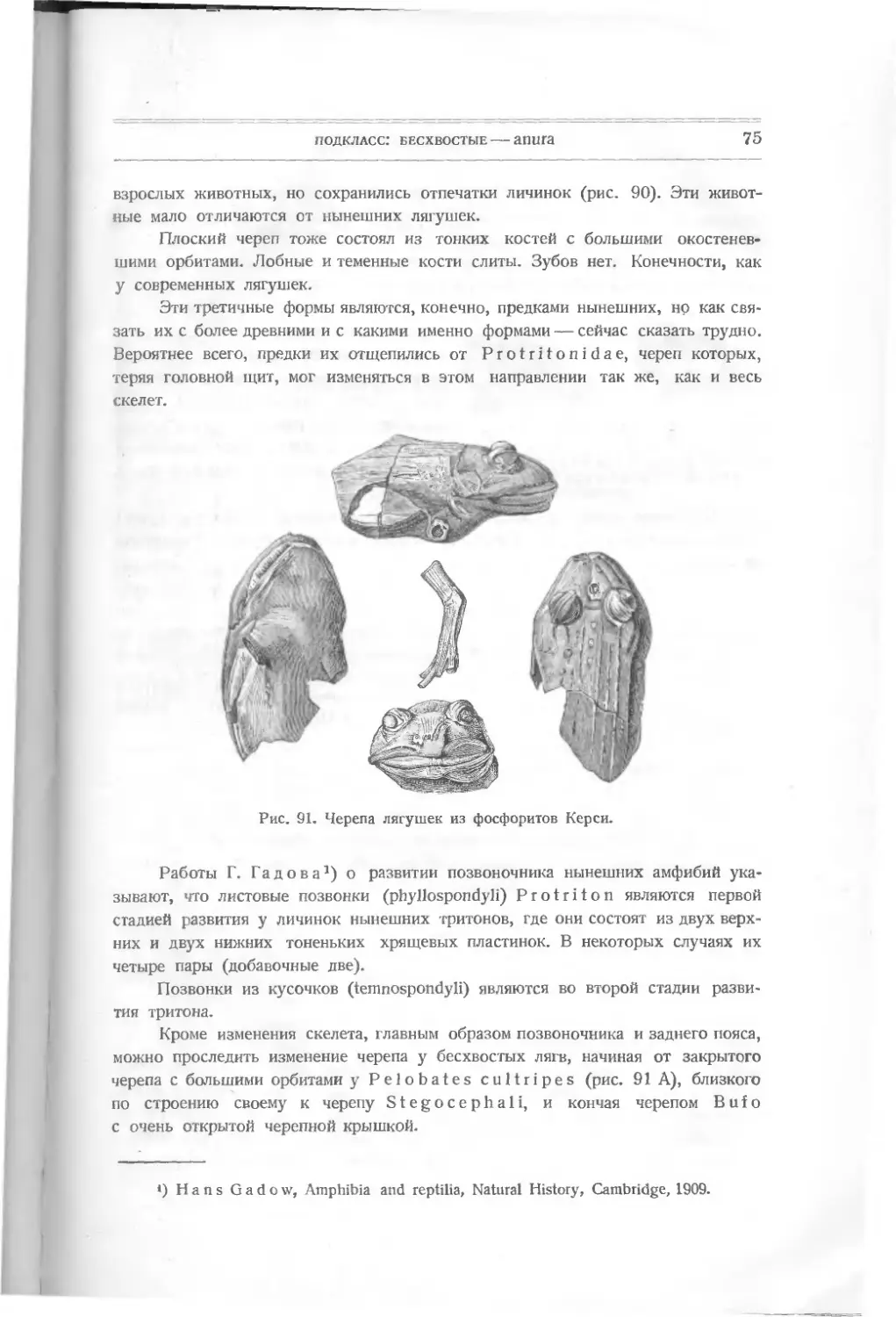
(expression points). Он видел в них проявление „поступательной силы"
(bathmic force). Эти моменты развития характеризуются окончательным установле-
нием некоторых важных признаков, которые были раньше несовершенными
или неустойчивыми. Закреплением их была достигнута возможность приспосо-
бления к разному образу жизни.
Нужно указать, что при изучении позвоночных приходится применять
методы, отличные от тех, которые применяются при изучении беспозвоночных.
Там мы имеем остатки в большинстве случаев в виде внешних покровов,
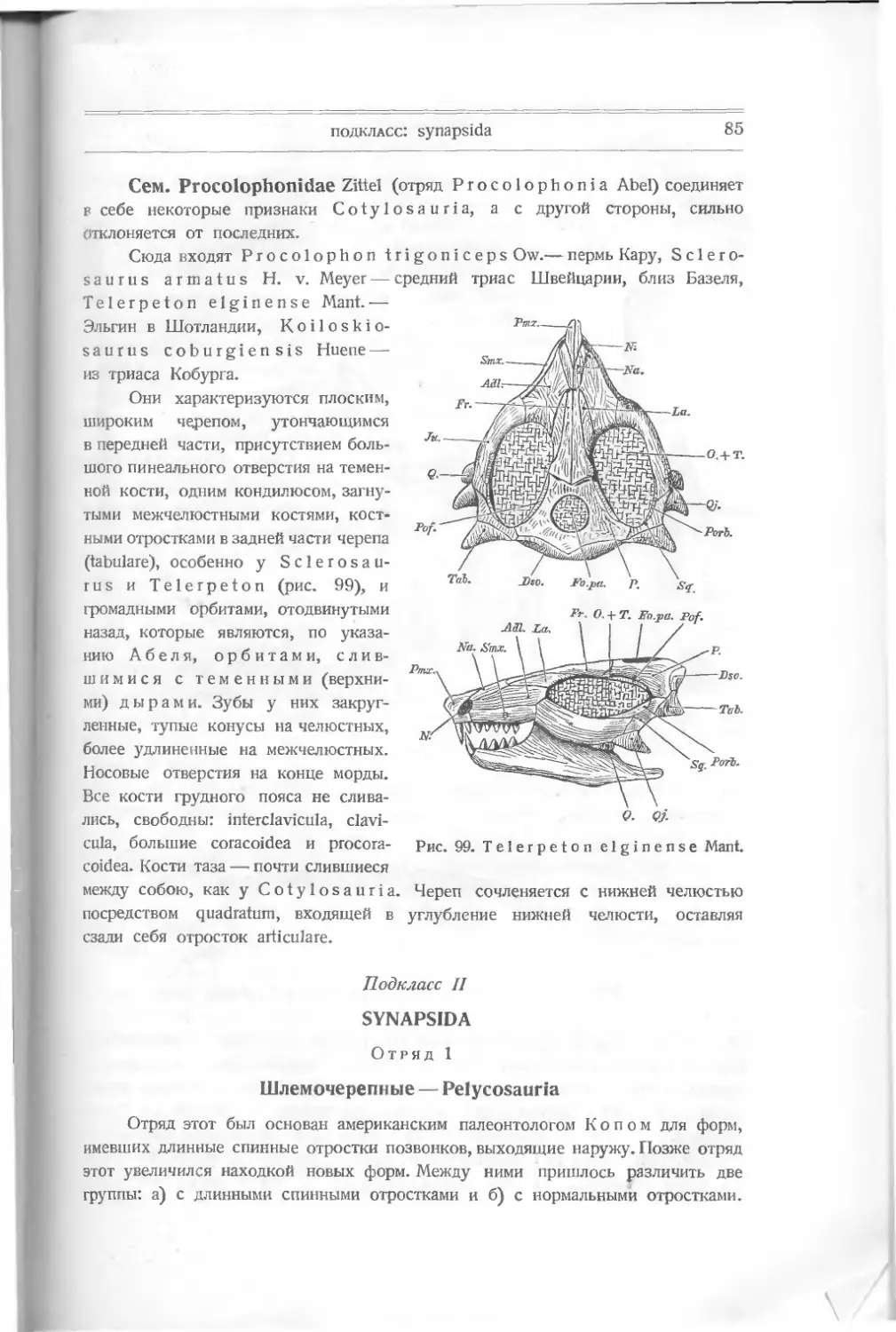
раковин или панцырных пластинок или же отпечатки мягких частей, и в боль-
шинстве случаев эти части дают или цельное животное, или хотя и не вполне
сохранившиеся, но характерные части его.
9 Otto Jaeckel, Die Wirbcltiere, 1911, S. 26.
ВВЕДЕНИЕ
3
У позвоночных, начиная с рыб и поднимаясь к высшим формам, мы
имеем настолько сложный и разнообразно построенный организм, несмотря на
сходство отдельных частей (скелета главным образом), что приходится счи-
таться с изучением не только отдельных типов животных, но и отдельных
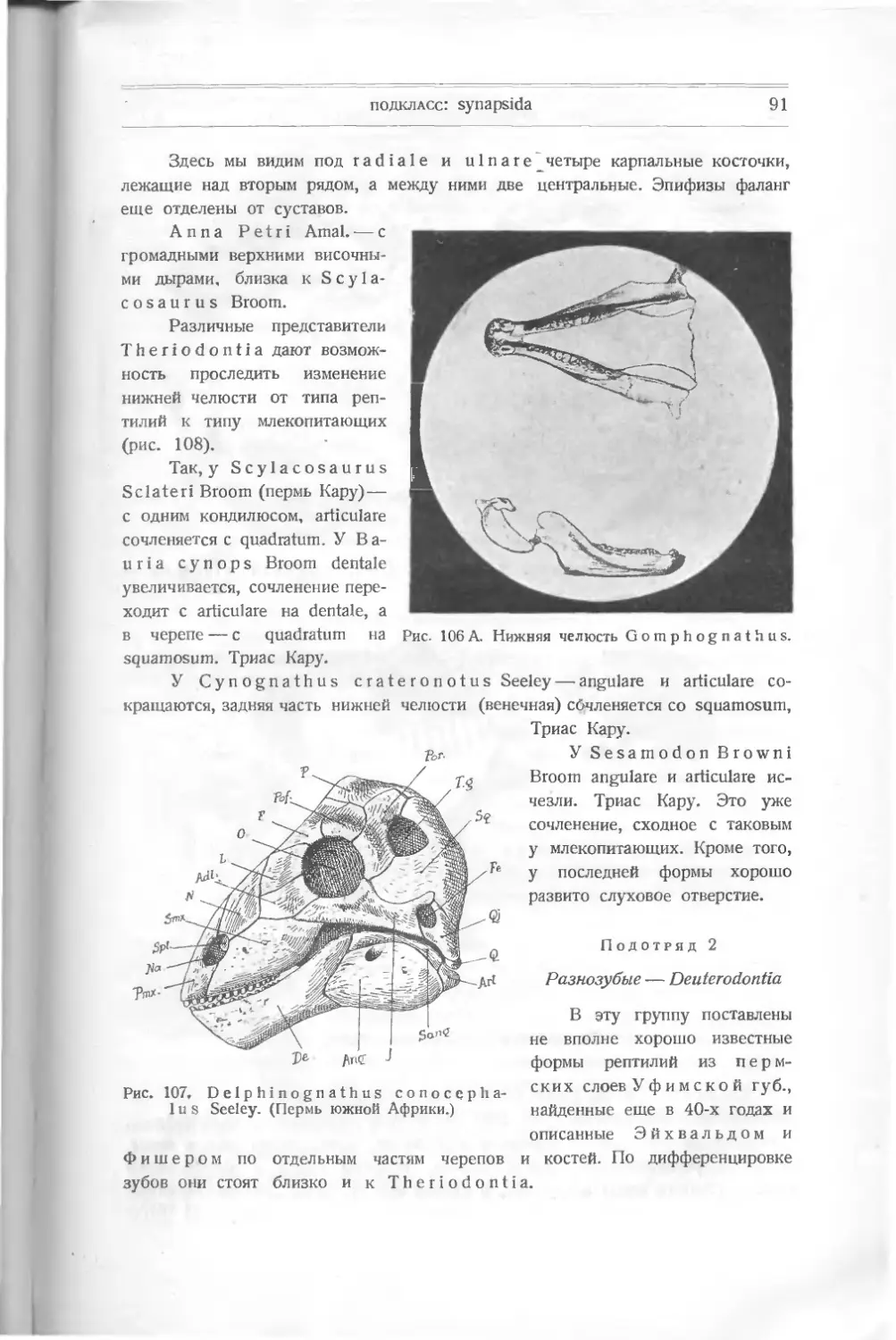
частей их — костей и зубов, — принимая во внимание, что многие из них кажутся
сходными при неглубоком знании их строения,
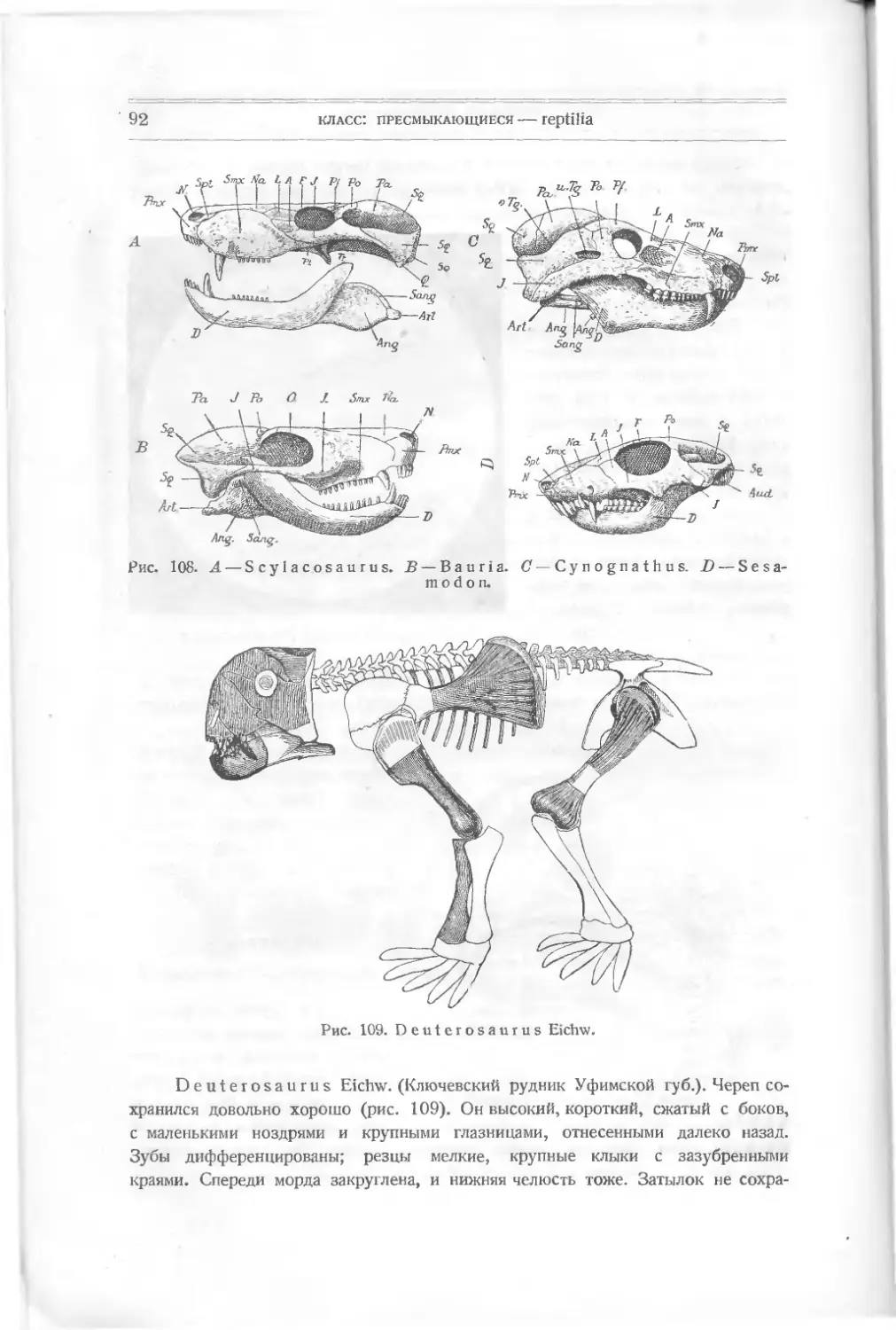
рептилий принимались за кости млекопитающих
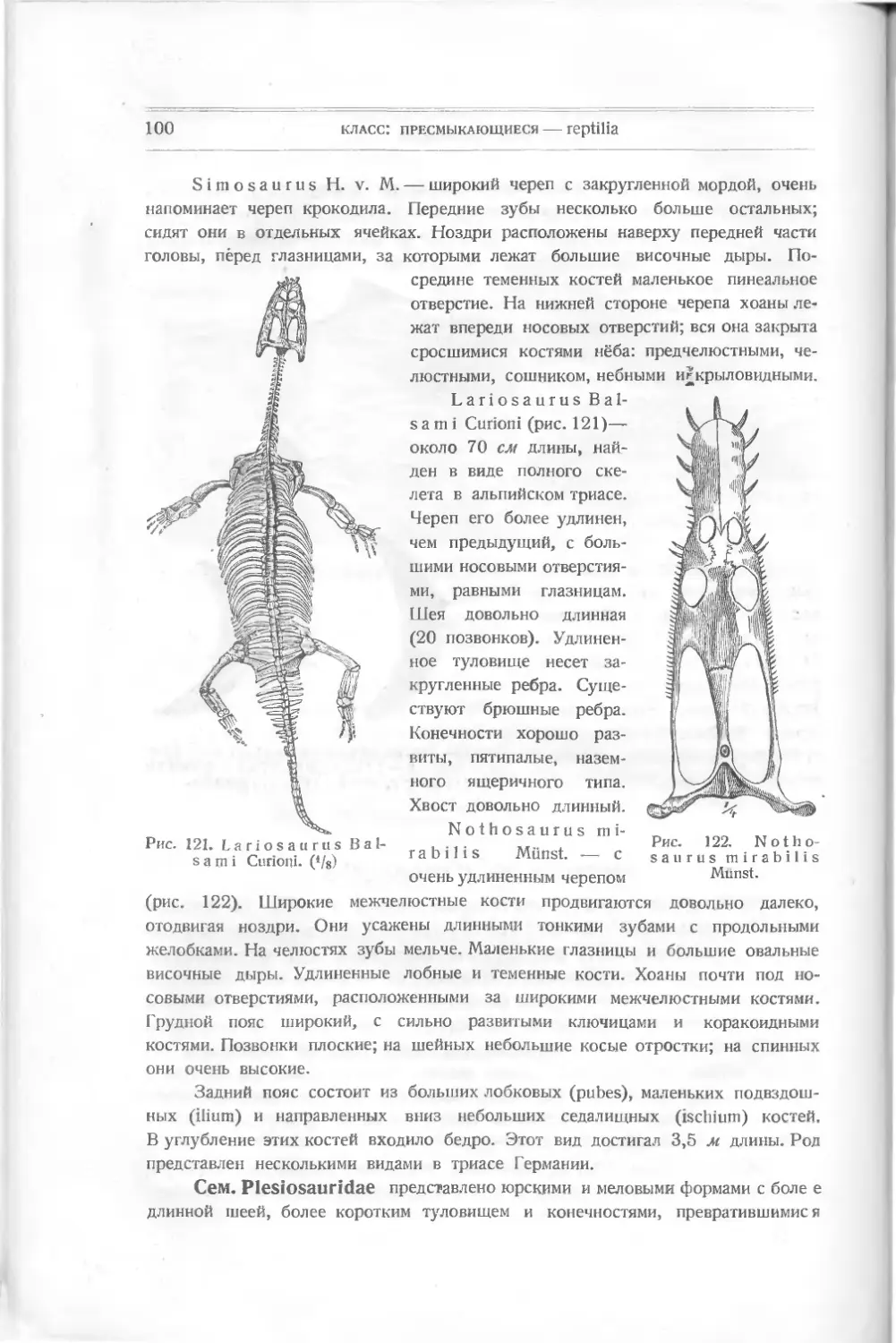
Нередко случалось, что кости
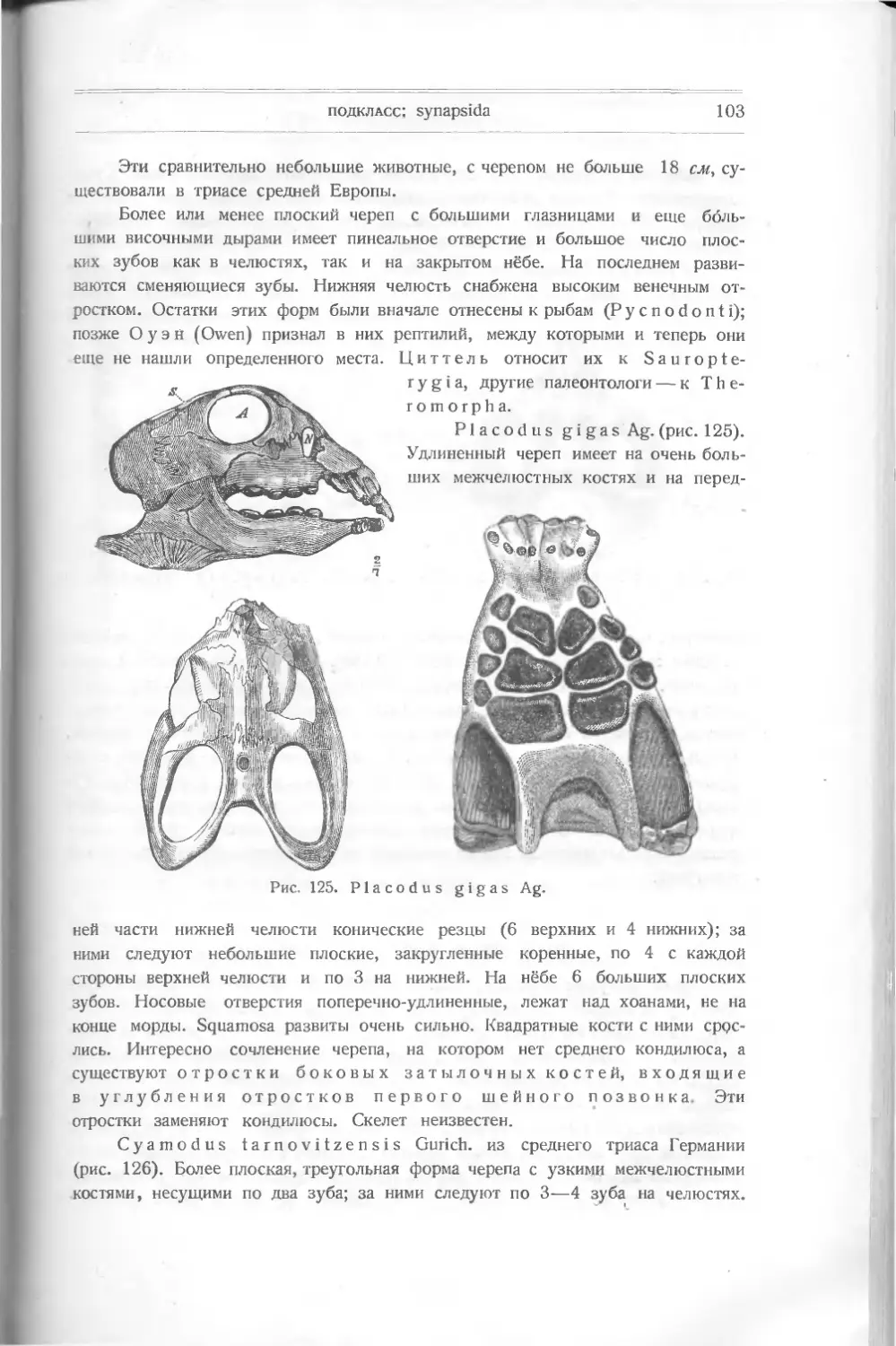
или птиц. Кювье (Cuvier)
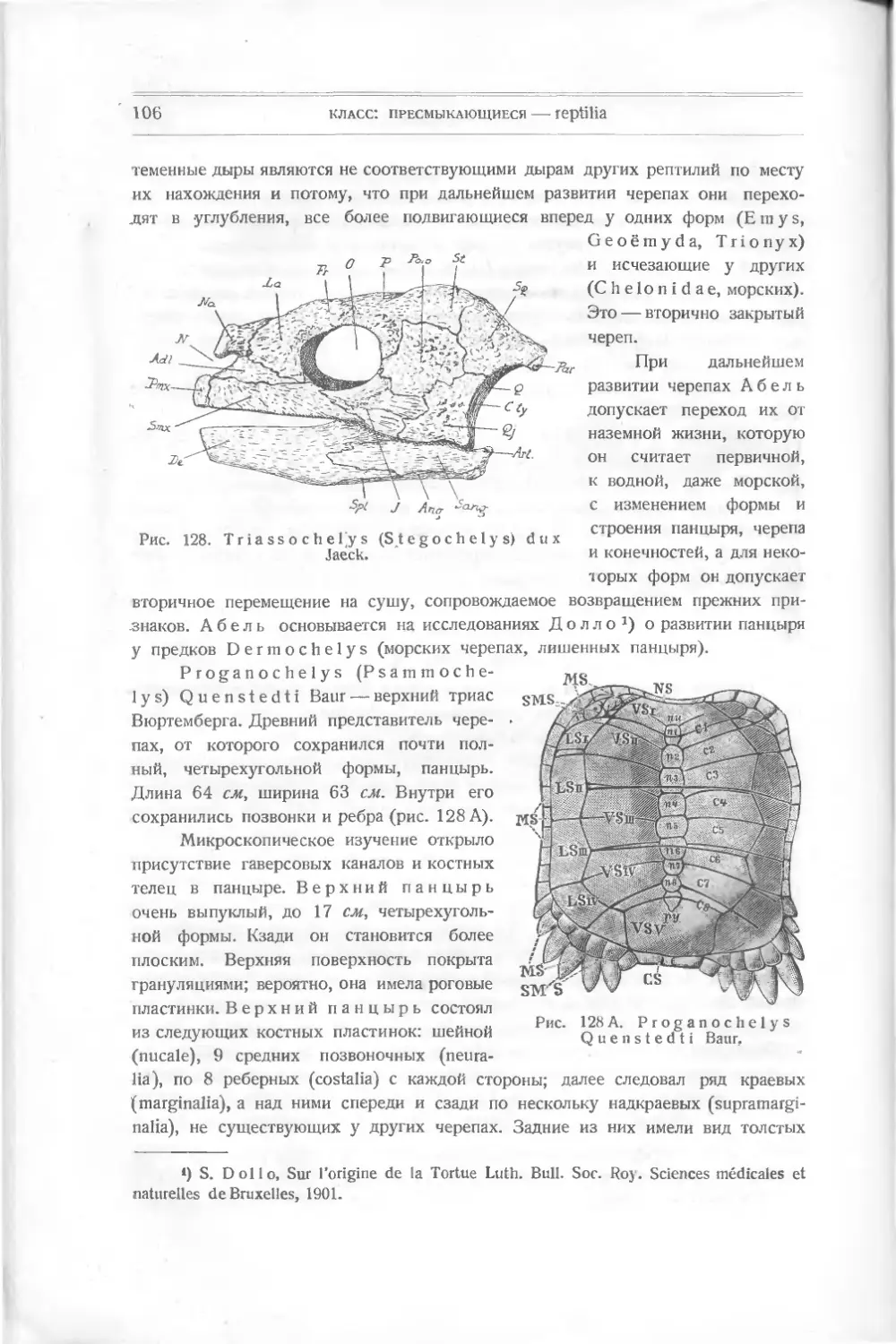
установил закон „соотношения органов" и мог по кусочку нижней че-
люсти, найденной в гипсах ^Монмартра, утверждать, что найденный там же
скелет принадлежит сумчатому, что и оправдалось. Но, во-первых, признак
Рис. 2. Схема развития
позвонка.
пс — нотохорда, sk.l — скелетород-
ное влагалище, sh. 1—оболочка хор-
ды; d.a— верхний, v.a — нижний
отростки.
строения нижней челюсти этой
группы млекопитающих слиш-
ком характерен для них, а во-
вторых, некоторые вымершие
рептилии и млекопитающие
соединяют в себе иногда такие
разнохарактерные признаки, что
только при большом знании
строения скелета можно опре-
делить принадлежность остатков
к тому или другому классу или
отряду.
Раньше чем изучать отдель-
ные типы позвоночных, нелиш-
ним будет дать здесь некото-
i
Рис. 3. Позвонки
акулы.
рые основные указания на развитие скелета, глав-
ным образом позвоночника и черепа. Для пред-
ставителей отдельных классов будут даны
соответствующие указания при
их изучении.
В своем простейшем виде внутренний скелет представляет эластиче-
ский стержень — хорду, состоящую из больших вакуольных клеток и
заключенную в чехол из соединительной ткани мезодермического происхо-
ждения — влагалище хорды. Эта так называемая нотохорда, или спинная
струна, проходит вдоль всего тела, несколько расширяясь в передней части
(будущей мозговой). Над нею лежит нервный ствол, который заключен
вместе с хордой в общий покров — скелетородное влагалище, дающий
начало твердым частям скелета — позвонкам (рис. 2). Развитие последних
бывает двух типов: 1) у основания первоначально образовавшихся отростков
позвонков отлагаются известковые частицы, образующие отдельные пластинки,
имеющие костное строение и сливающиеся в тела позвонков,— это у всех почти
позвоночных; 2) обызвествляются части влагалища хорды, образуя известко-
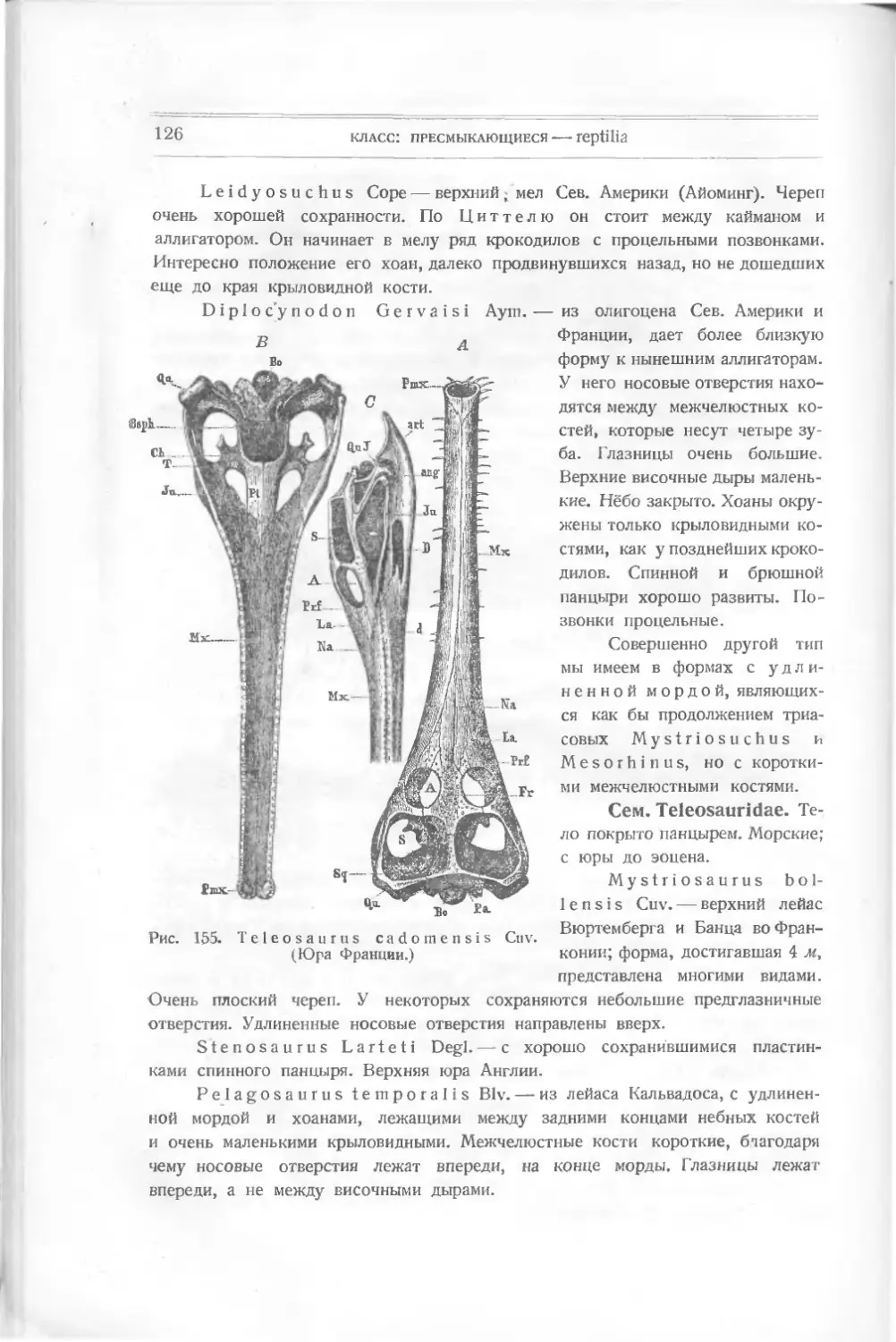
вые кольца без костных телец, — это у акуловых рыб. Первые позвонки
называются перихордальными, вторые — хордальными (рис. 3);
последние не имеют костного строения. По форме своей позвонки могут быть
4
ВВЕДЕНИЕ
Рис. 4. Зачаток черепа и висцеральных дуг
акулы.
Д’ — капсула обонятельная, А — зрительная, О —
слуховая.
различны: а) п л о с к и е, б) вогнутые с обеих сторон — амфицел ь ные, в) вы-
пуклые спереди и вогнутые сзади — опистоцельные, г) вогнутые спереди
и выпуклые сзади — процельные. Амфицельные характерны для рыб.
У высших позвоночных встреча-
ются у одних и тех же форм по-
звонки различных типов, кроме
амфицельных.
Первичный хрящевой череп у
рыб состоит из мозговой коробки
и трех
ной и
нижняя
первой
второй
капсул: слуховой, зритель-
обомятельной. Верхняя и
челюсти образуются
пары
пары
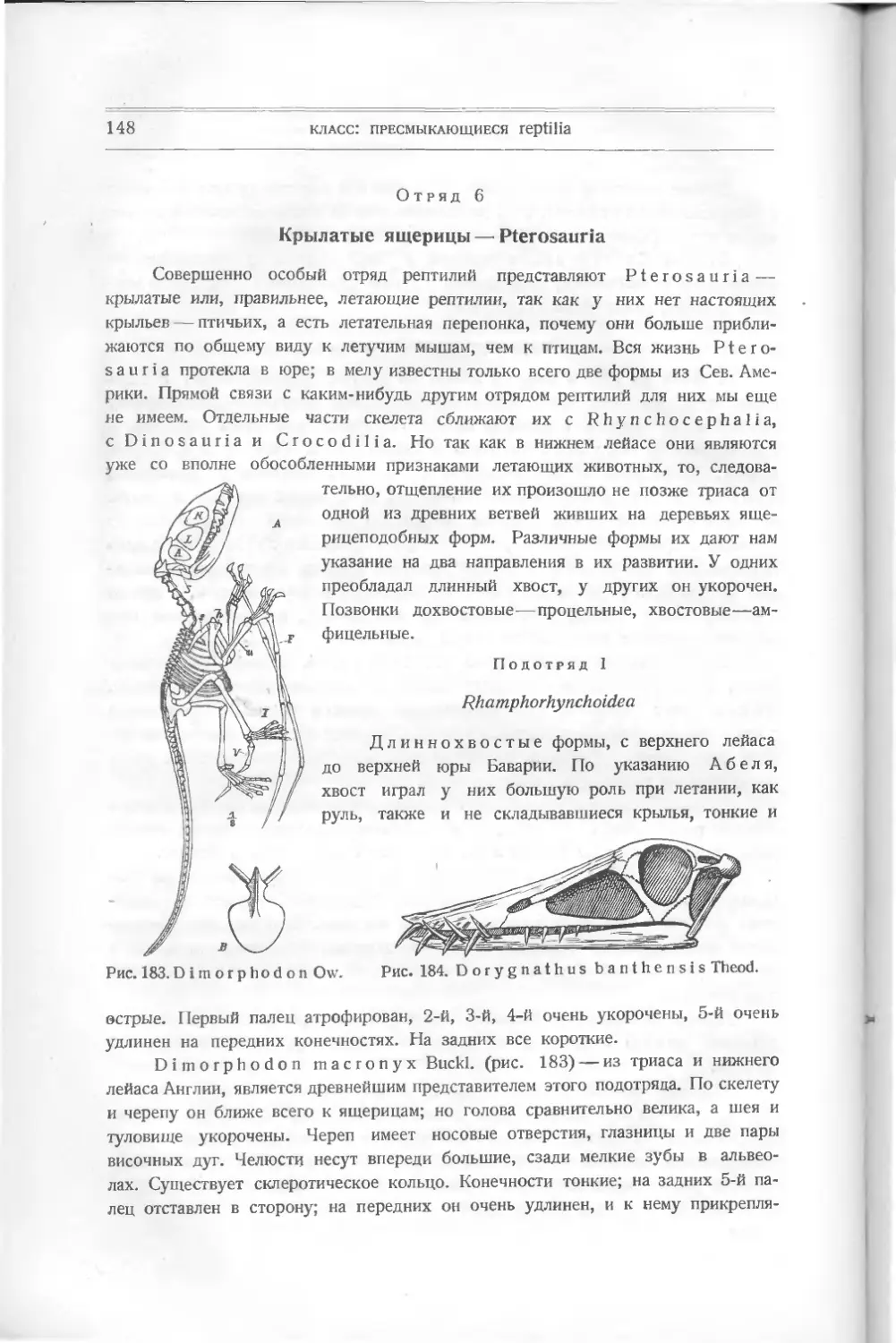
из
жаберных дуг; из
развиваются языко-
глоточная и подъязычная кости
(рис. 4). При дальнейшем изменении скелета хрящевые части окостеневают,
происходит слияние капсул, сращение верхней челюсти (palato-quadratum) с
черепными капсулами; сочленение с нижней челюстью происходит посред-
ством квадратной кости — задней части верхней челюсти, которая исчезает
у высших типов—млекопитающих. Кроме костей черепа, развившихся из
хрящевых частей, добавляются так называемые кожные кости. Они развива-
ются и у рыб. У высших форм они сливаются с хрящевыми костями, и их
трудно бывает различить.
Развитие органов движения очень различно у различных типов позвоноч-
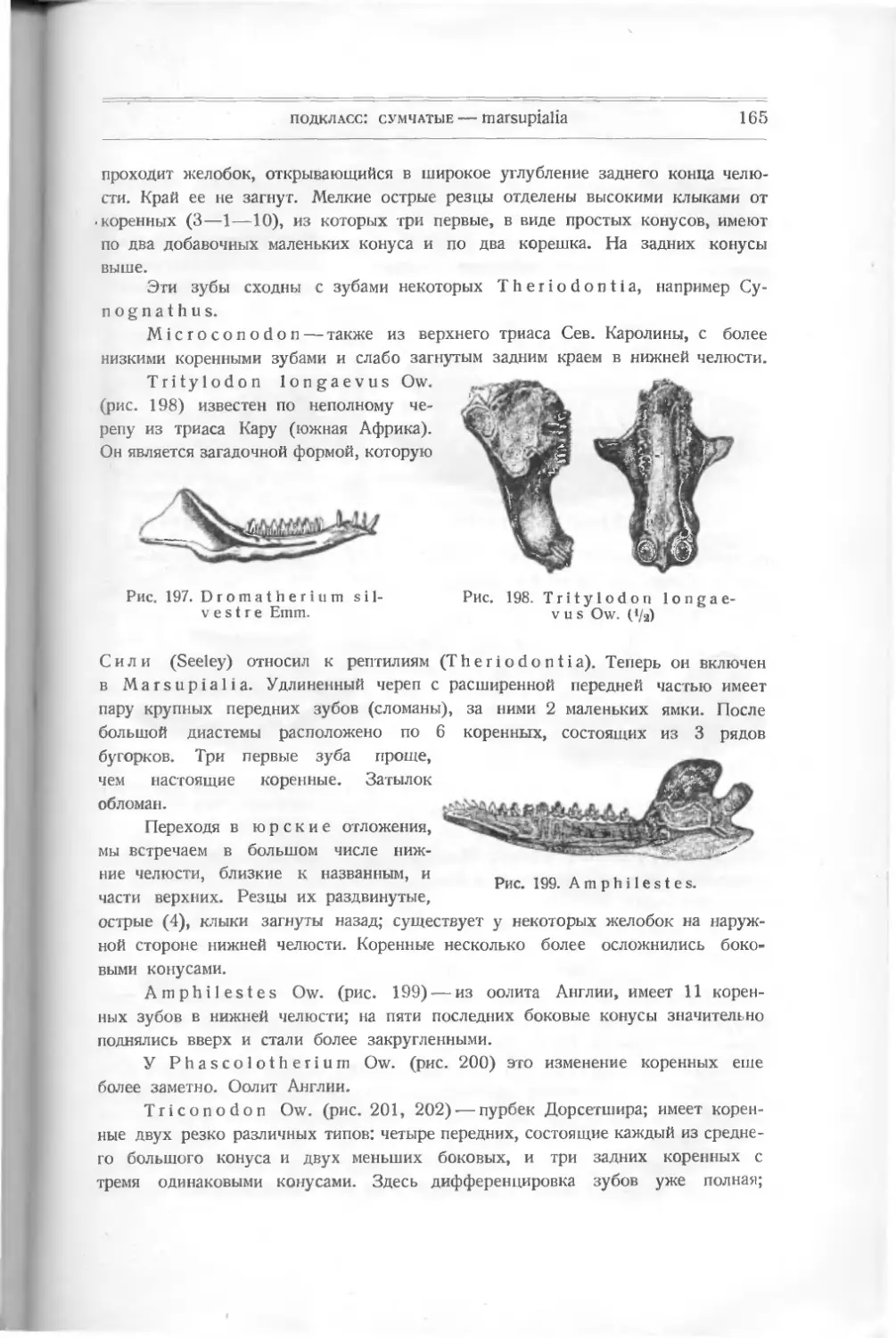
ных; о них придется говорить отдельно при изучении этих типов.
ОПИСАТЕЛЬНАЯ ЧАСТЬ
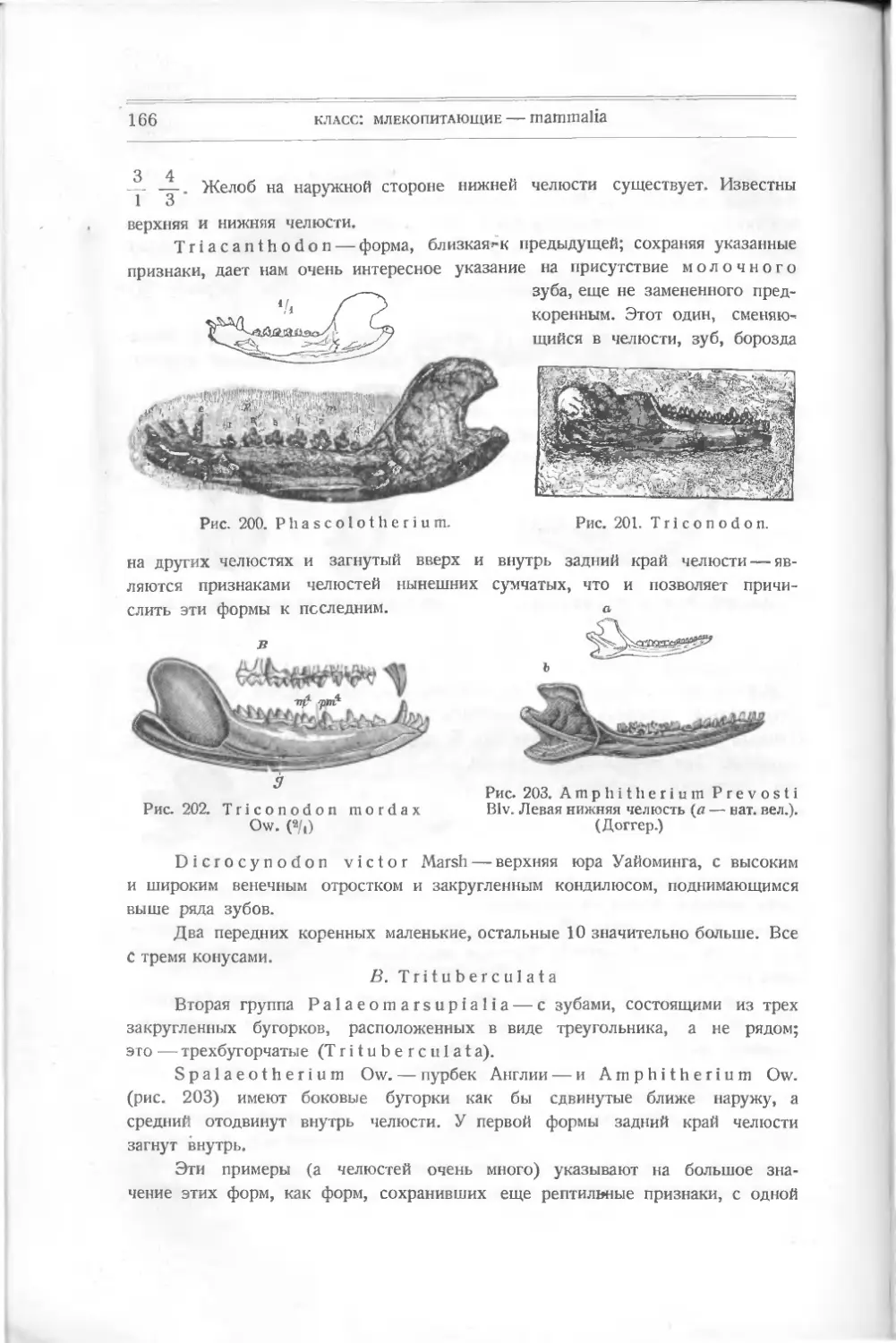
КЛАСС ПЕРВЫЙ
РЫБЫ - PISCES
Приступая к знакомству с ископаемыми остатками древних позвоночных,
мы начнем с простейших их представителей — р ы б, положивших начало всем
тем разнообразным типам позвоночных, которые существуют в виде более
совершенных, более поздних рыб, земноводных, пресмыкающихся, птиц и
млекопитающих.
Произнося слово рыб а, мы большею частью вызываем представление
о тех удлиненных животных, живущих в воде, которые снабжены внутрен-
ним скелетом, костистой головой, хорошо развитыми плавниками и тело ко-
торых покрыто чешуею. Это — костистые рыбы со скелетом: судак, селедка,
карась и т. п.
У некоторых людей к этому представлению присоединится мысль об
осетре, стерляди и других хрящевых рыбах, лишенных скелета и чешуи. Но
мало кто вспомнит, например, о миноге, змееобразной, мягкой рыбе, ли-
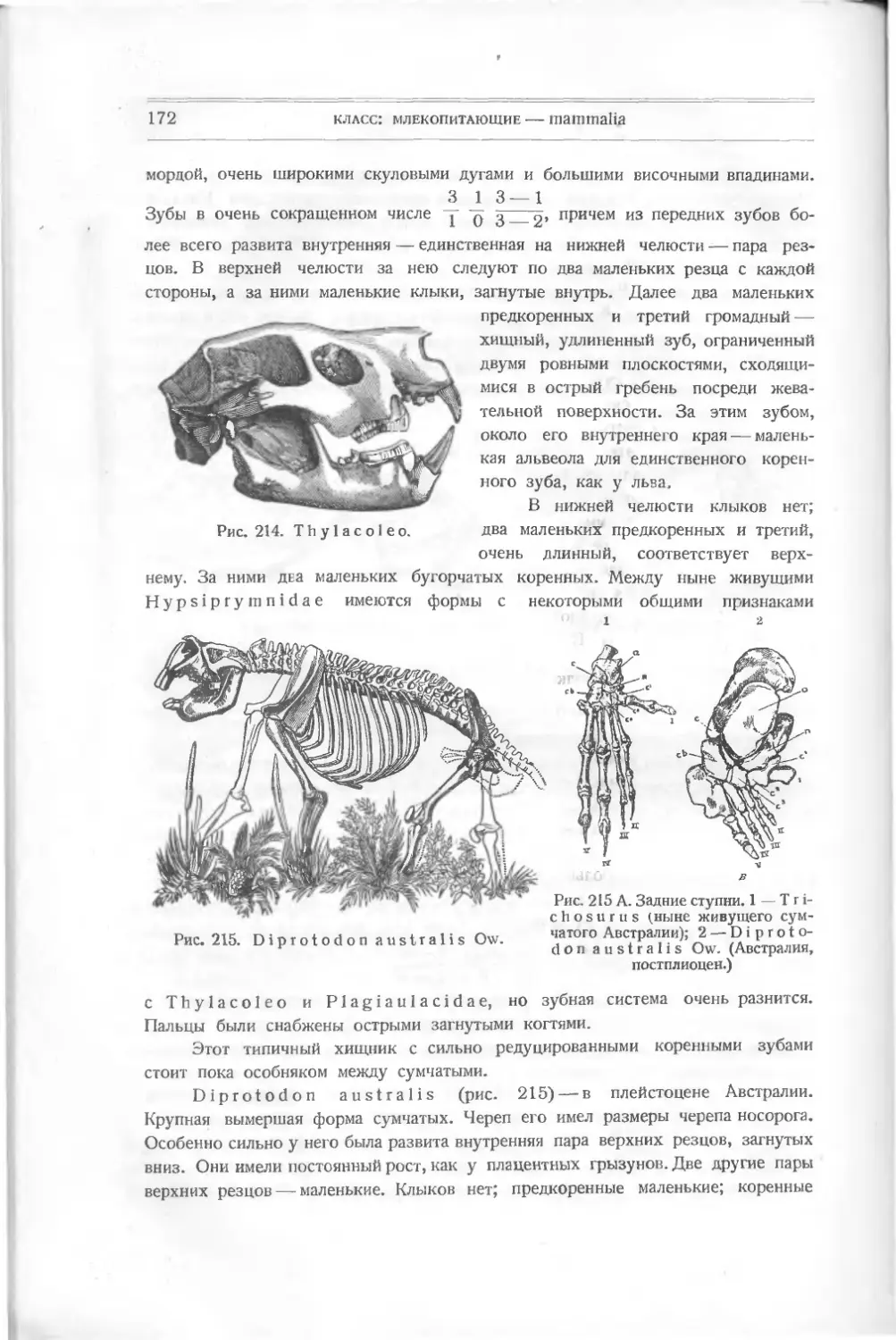
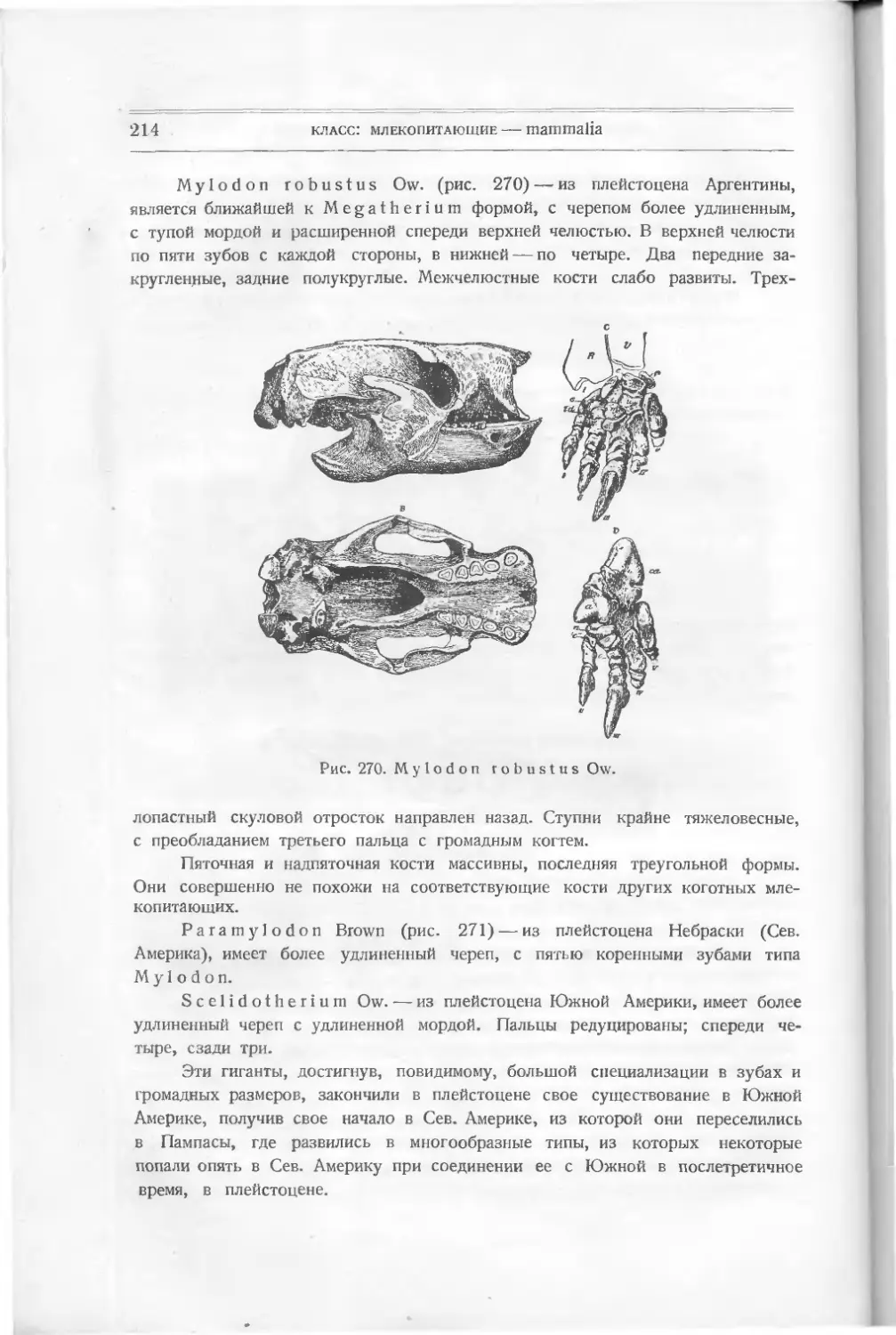
шенной не только костей, чешуи и плавников, но даже и челюстей. Изучение
развития ее открывает многие факты, объясняющие измененное строение других
рыб, хотя теперь некоторые зоологи отделяют миног (круглоротых) в отдель-
ный класс.
Кроме подклассов, к которым принадлежат упомянутые сейчас рыбы,
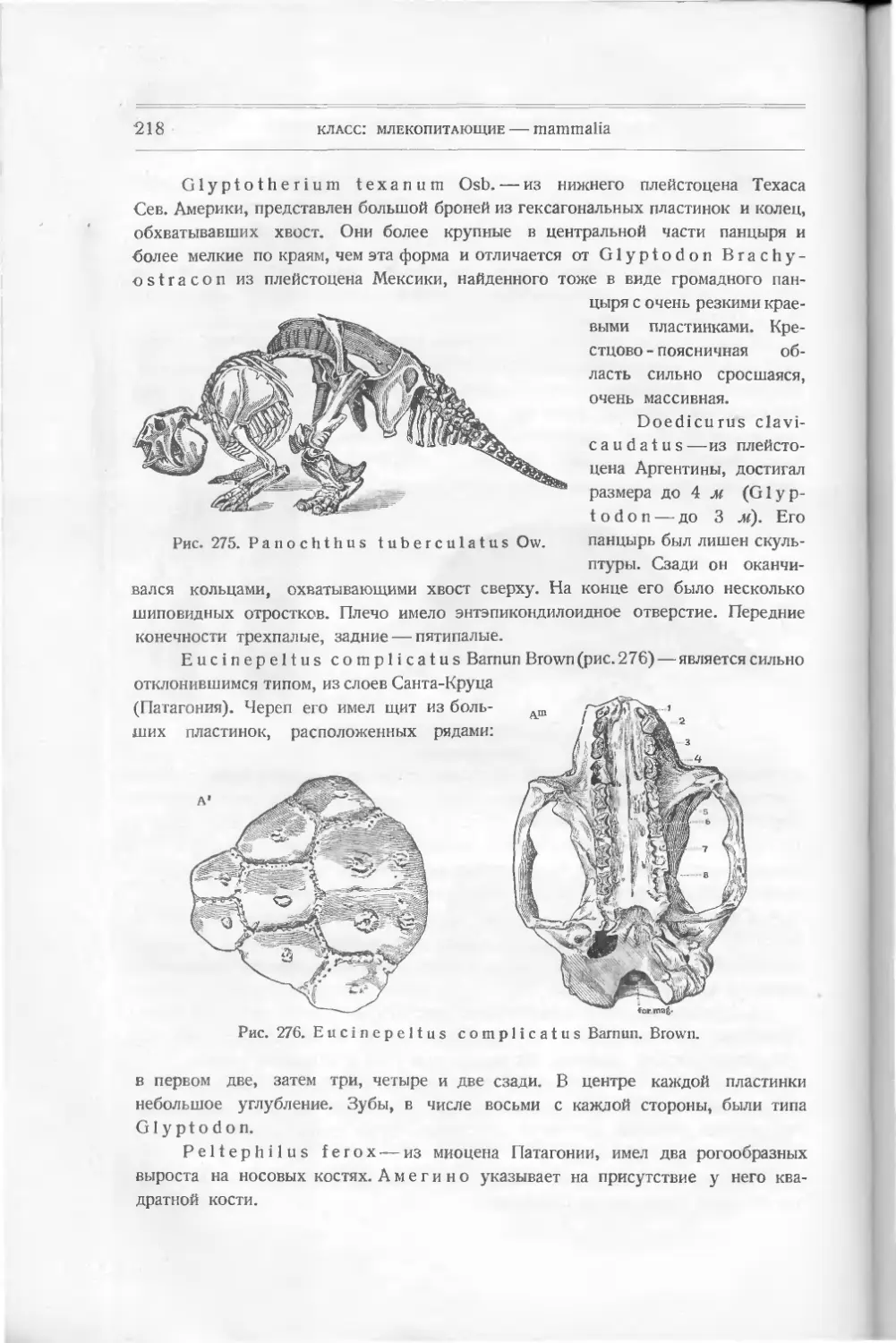
существуют еще и другие: двоякодышащие, акуловые, так что современные нам
формы класса рыб могут быть сгруппированы в ниже приводимые четыре
подкласса. Но кроме того существует еще подкласс вымерших рыб — Placo-
de г m i, панцырные, не вполне отвечающие признакам остальных рыб.
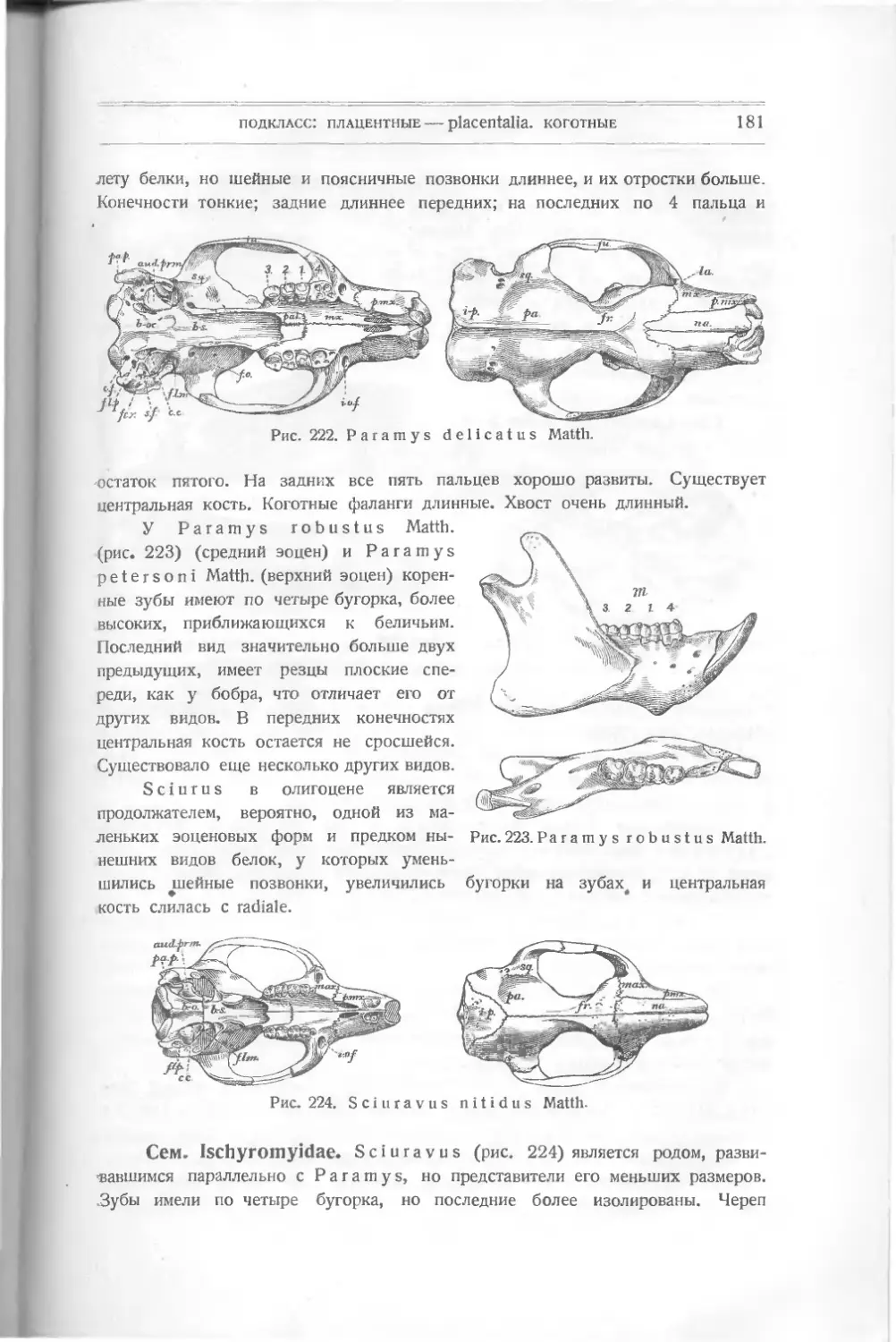
Ныне живущие рыбы разделяются на подклассы: . .
1. Круглоротые — Cyclostomi — миноги и другие близкие к ним.
2. Хрящеперые — Chondropterygii ]) (Elasmobranchii — пласти-
ножаберные)— акулы и скаты.
3. Химеры — Holocephali.
*) Я ввожу это название для Elasmobranch ii в соответствии с классифика-
цией по плавникам других рыб на Cross opterygii HActinopterygii, и заме-
няют слово „пластинчатожаберные" словом „пластиножаберные", т. е. состоящие из
пластинок, а не подобные пластинкам.
8
класс: рыбы — pisces
4. Совершенноротые — Teleostomi, которые подразделяются на
отряды:
Dipnoi — двоякодышащие.
Crossopterygii — кистеперые.
А с ti п о р t е г у g i i — лучеперые (куда входят и костистые).
Если мы сравним количественное отношение родов и видов ныне живу-
щих представителей различных подклассов и отрядов рыб, то узнаем, что
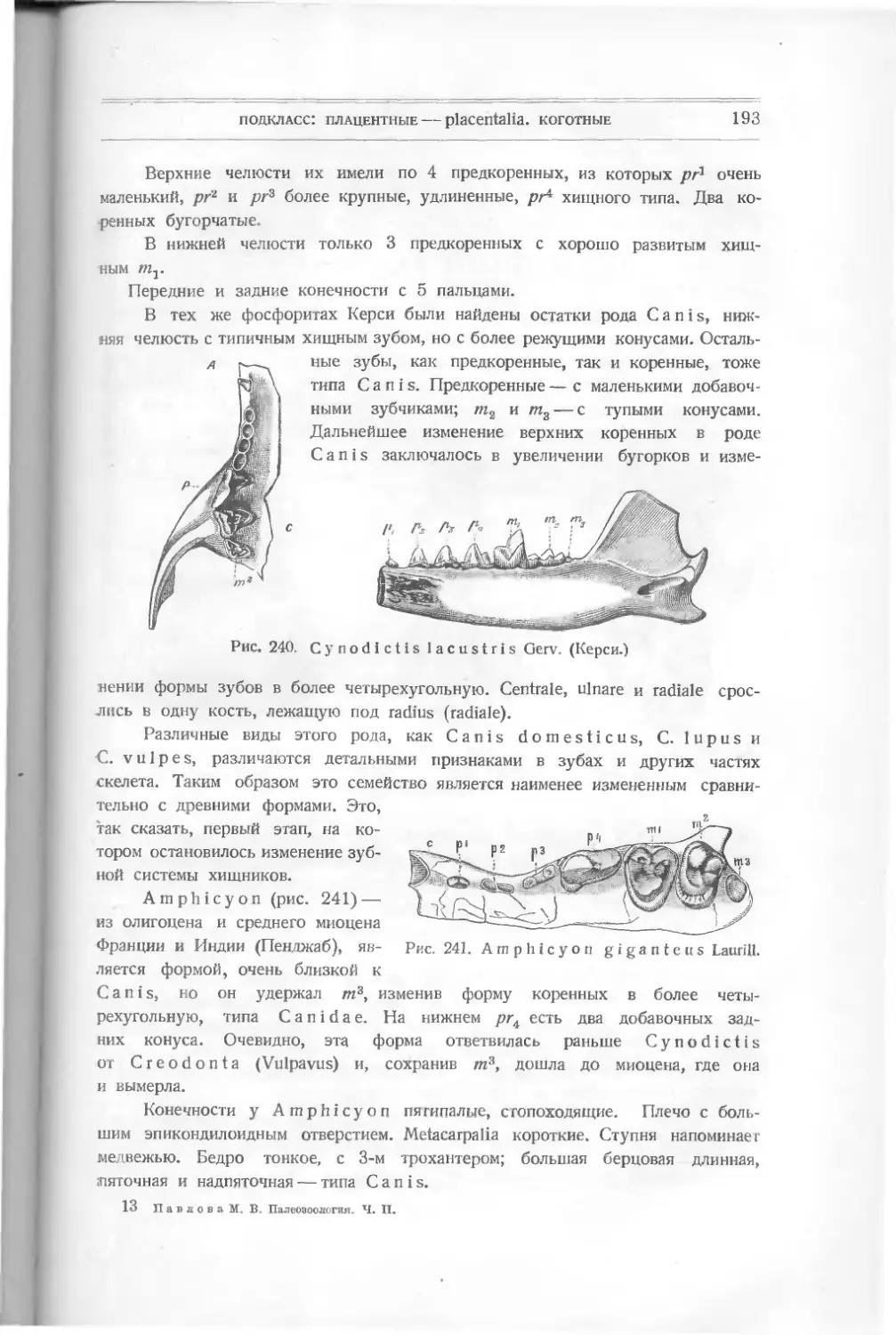
костистые рыбы, появившиеся позже всех других (в конце юрского пе-
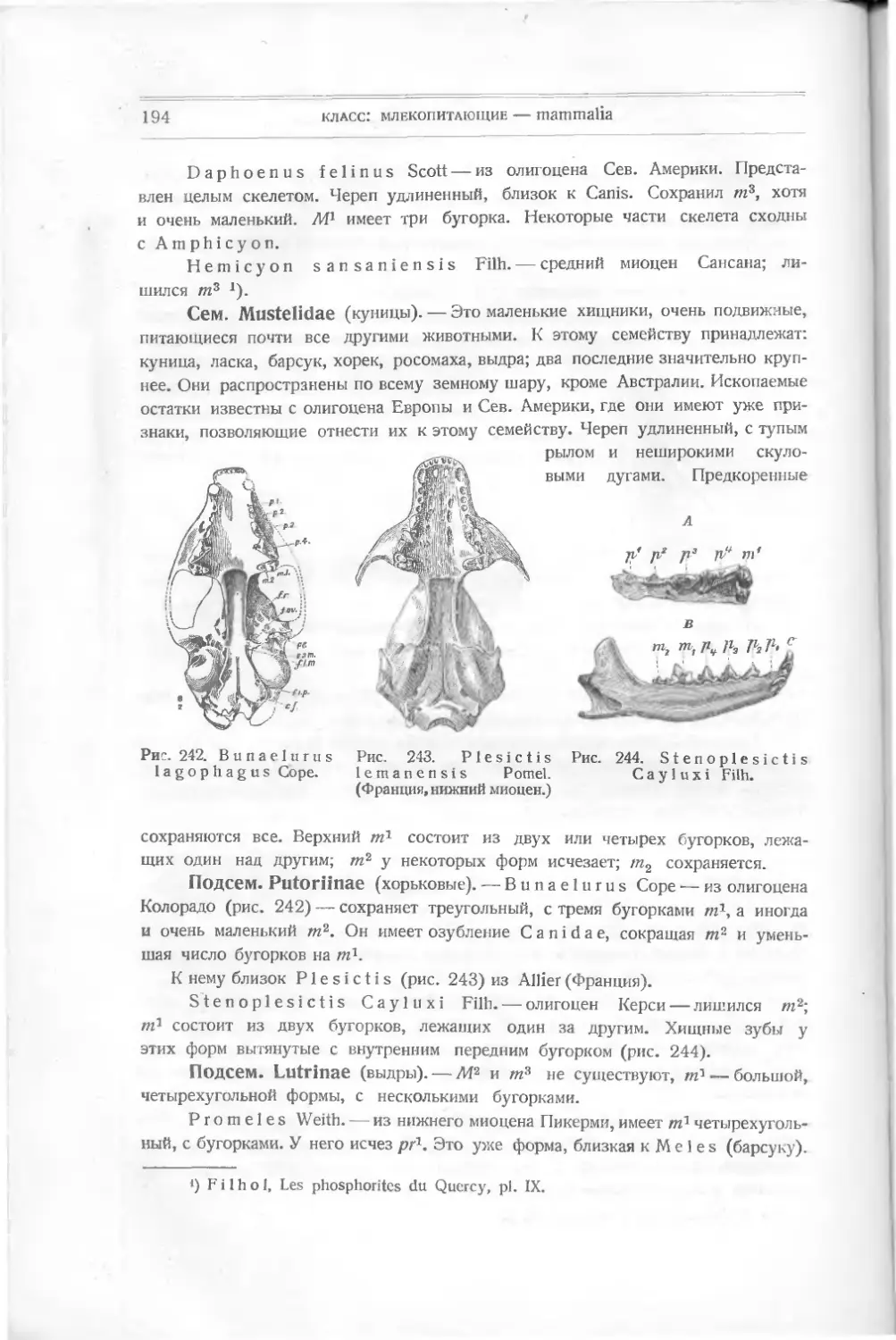
риода), составляют теперь 95 °/0 всех других. Действительно, в то время как
их роды, не говоря о видах, насчитываются сотнями, для других отрядов известно
не более 2—3 родов. Чем же вызвана такая разница? Чем можно объяснить
такое преобладание этих форм теперь, и существовало ли оно и раньше?
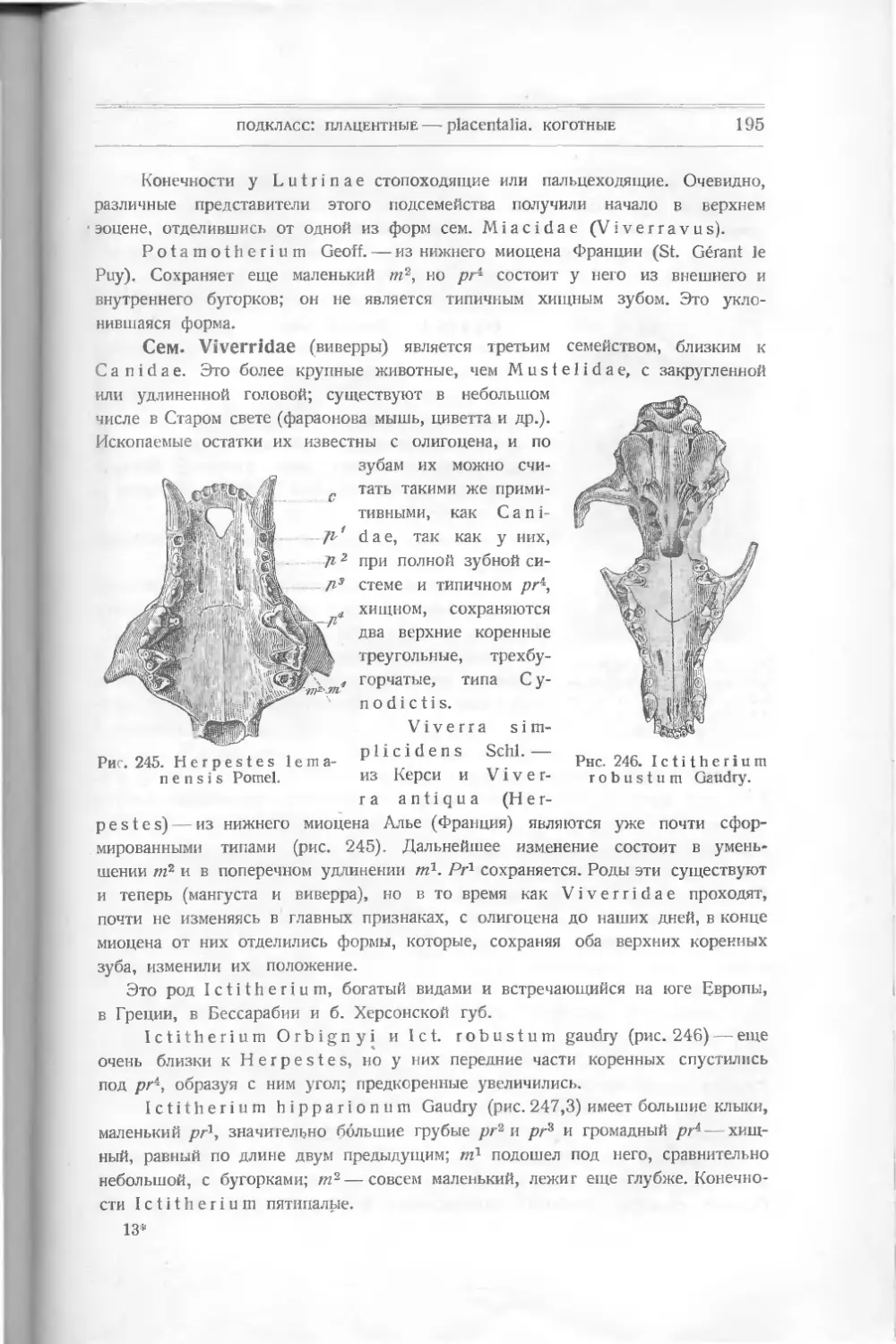
Мы уже упомянули об их отличительных признаках сравнительно с дру-
гими отрядами — о костном внутреннем скелете, о плавниках, о чешуе — и задаем
теперь вопрос, не в этих ли признаках скрываются их победа над другими
рыбами и возможность достижения широкого распространения и большого
разнообразия. Интересно проверить, каковы были их предки, что получили они
от них и что явилось в их организации последствием развития и изменения.
В силурийских слоях встречаются зубы, чешуи и плавниковые лучи
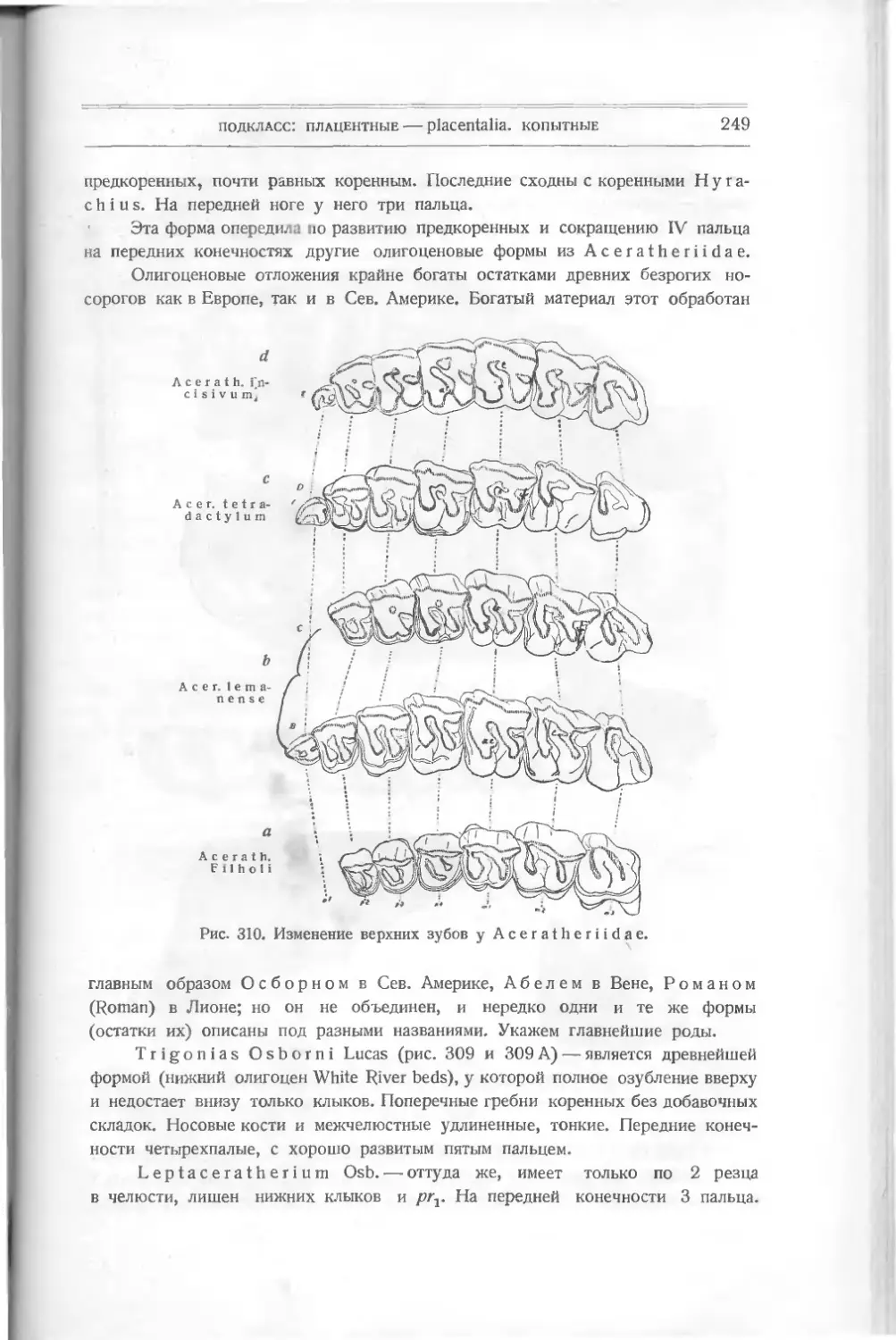
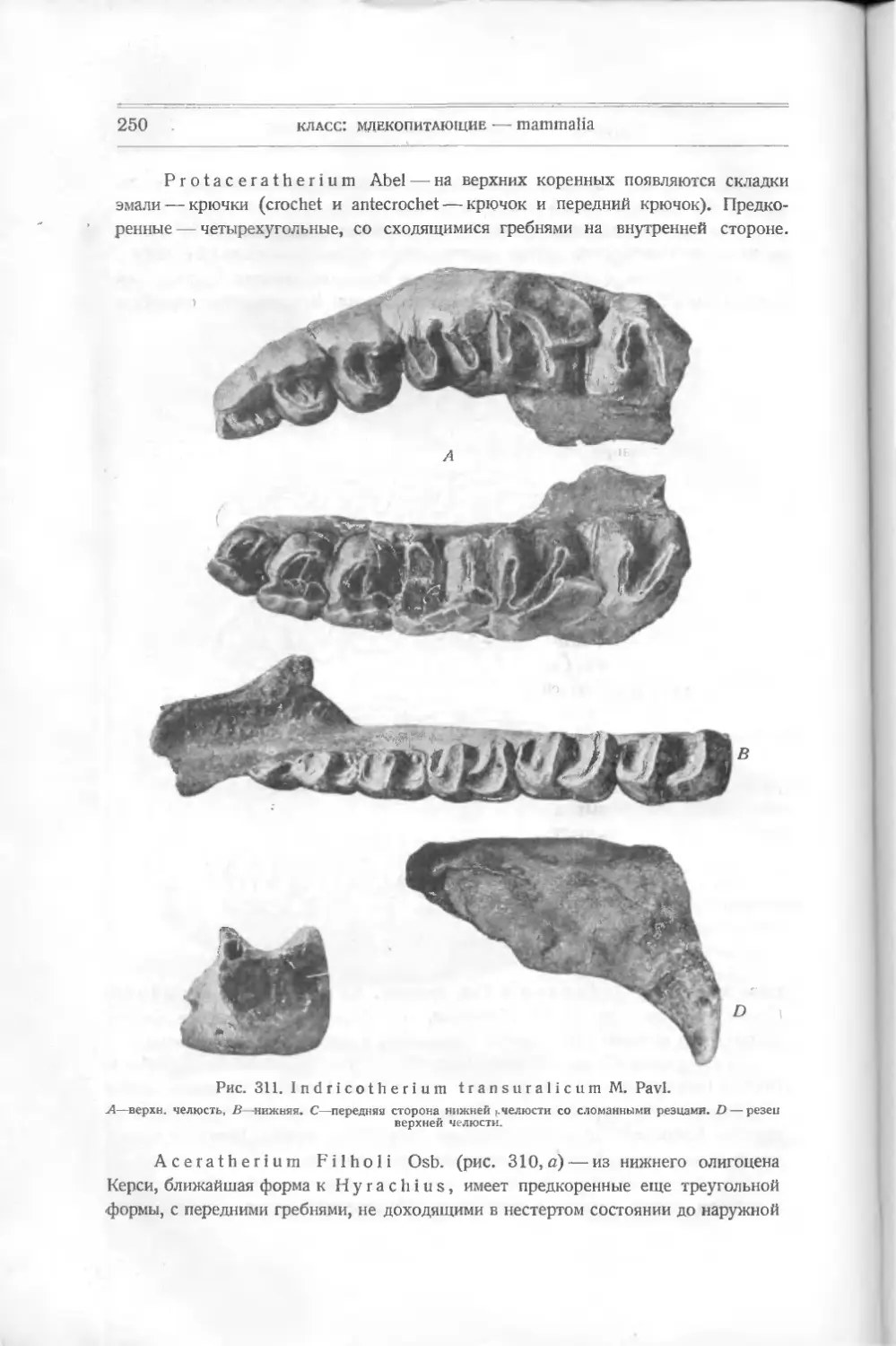
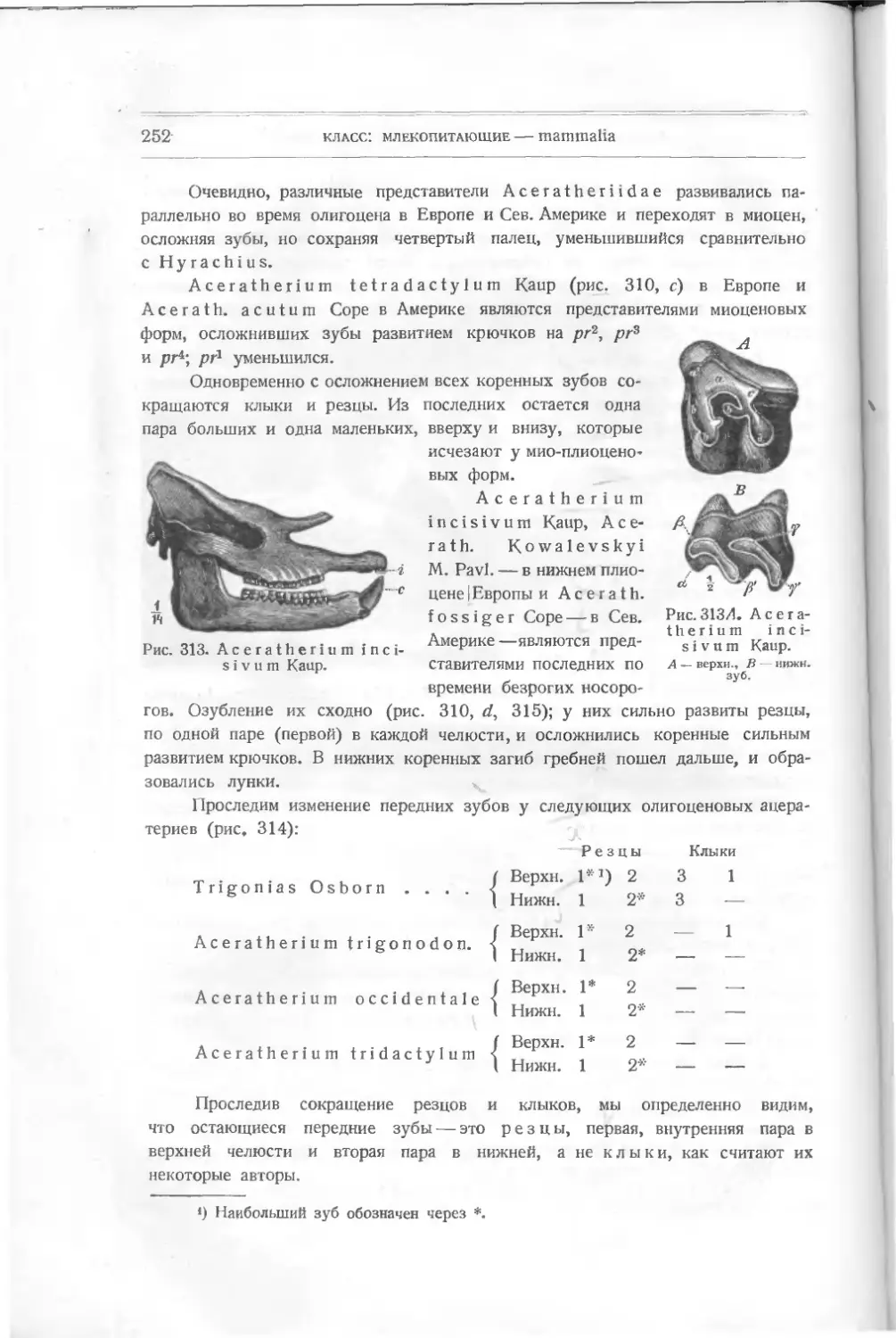
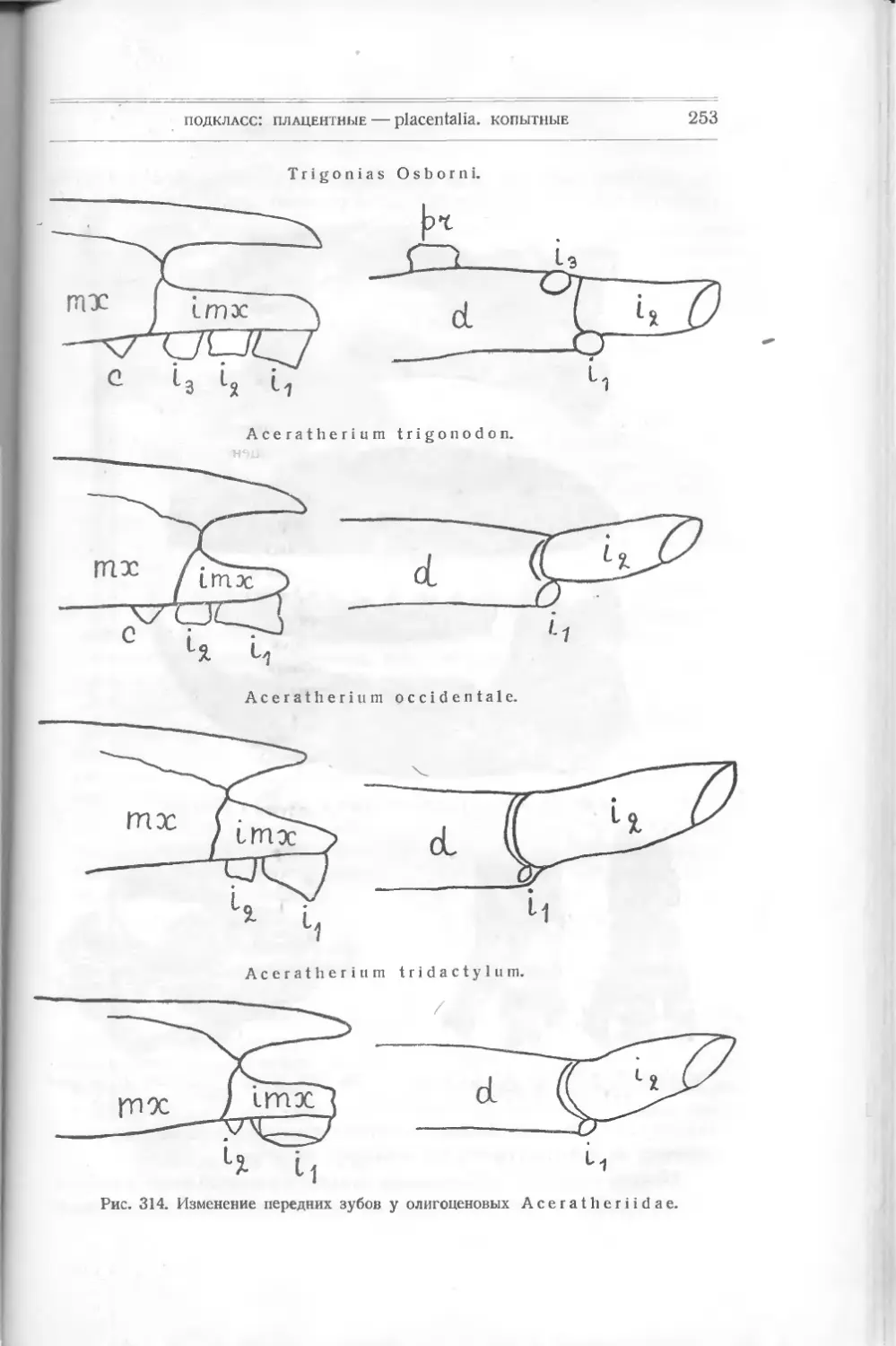
акул и других некостистых рыб, которые считались долгое время единственными
древними остатками этого класса. В 1899 г. английский ученый Траквайр
(Traquair) описал из верхне-силурийских и девонских слоев совершенно новые
формы животных, не существующие теперь и дающие основание считать их
близкими к предкам рыб и, может быть, положившими начало всей длинной
цепи позвоночных. Траквайр относит их уже к рыбам, и этого придержи-
ваются и другие палеонтологи — Кокен, Циттель (Koken, Zittel), хотя многие
признаки их не отвечают понятию о рыбах.
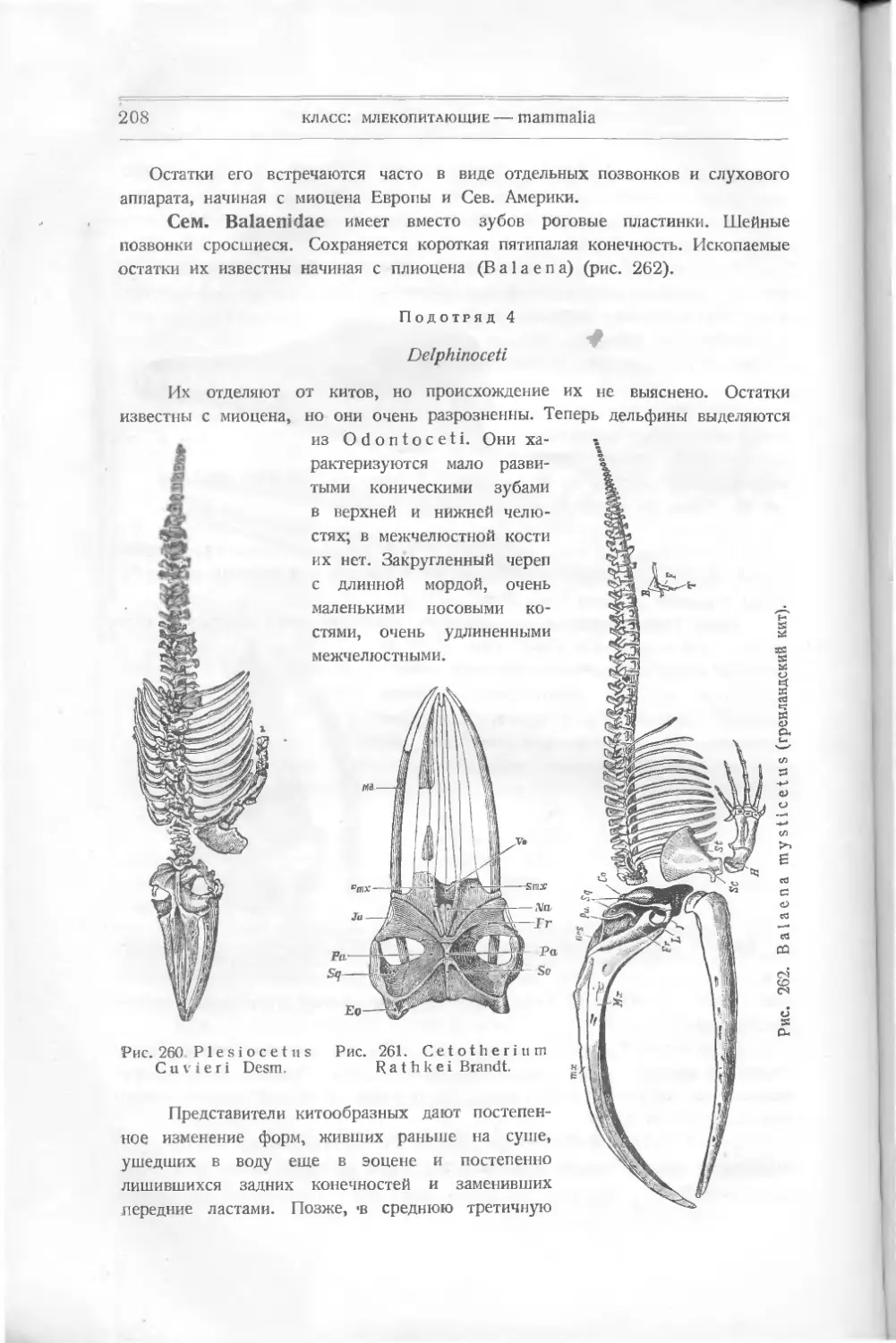
Их относят к подклассу панцырных — РI а с о d er m i, включающему дру-
гие, раньше найденные формы, тоже резко отличающиеся от настоящих. Раз-
личные палеонтологи классифицируют низших рыб различно. Так, Абель
совершенно отбрасывает общее название панцырных — Placodermi — и видит
в различных отрядах, входящих сюда, представителей подклассов. Ц и т т е л ь
сохраняет название Placodermi как подкласса. Так как в литературе встре-
чаются все эти названия, я считаю нужным дать их при классификации низ-
ших рыб.
Подкласс I
ПАНЦЫРНЫЕ — PLACODERMI
Панцирные характеризуются отсутствием парных плавников, отсутствием
внутреннего и развитием внешнего скелета, по которому их подразделяют на
несколько отрядов:
I. Heterostraci: Lanark!a, Thelodus, Drepanaspis,
Р t е г a s р i s.
подкласс: панцирные — placodermi
9
II. Osteostraci: Ateleaspis, Cephalaspis, Thyestes и др.
III. Antiarchi: Pterichthys, Asterolepis, Bothriolepis.
IV. An as pi da Traq.: Birknia, Lasanius, Pterolepis, Pha-
ryngolepis, Rhyncholepis.
Все они относятся к палеозойным формам.
Отряд 1
Разнощитковые — Heterostraci
Простейшей между ними формой является Lan ark ia Traq. — из верхнего
силура Шотландии (рис. 5). Это отпечаток маленького плоского тела длиною
в 3 см. Спереди оно расширено, имеет по бокам два выступа, напоминающие
плавники, за которыми тело суживается, удлиняется и
как бы раздвоенным хвостом. На переднем краю расши-
ренной части—голове — видны две черные точки —мо-
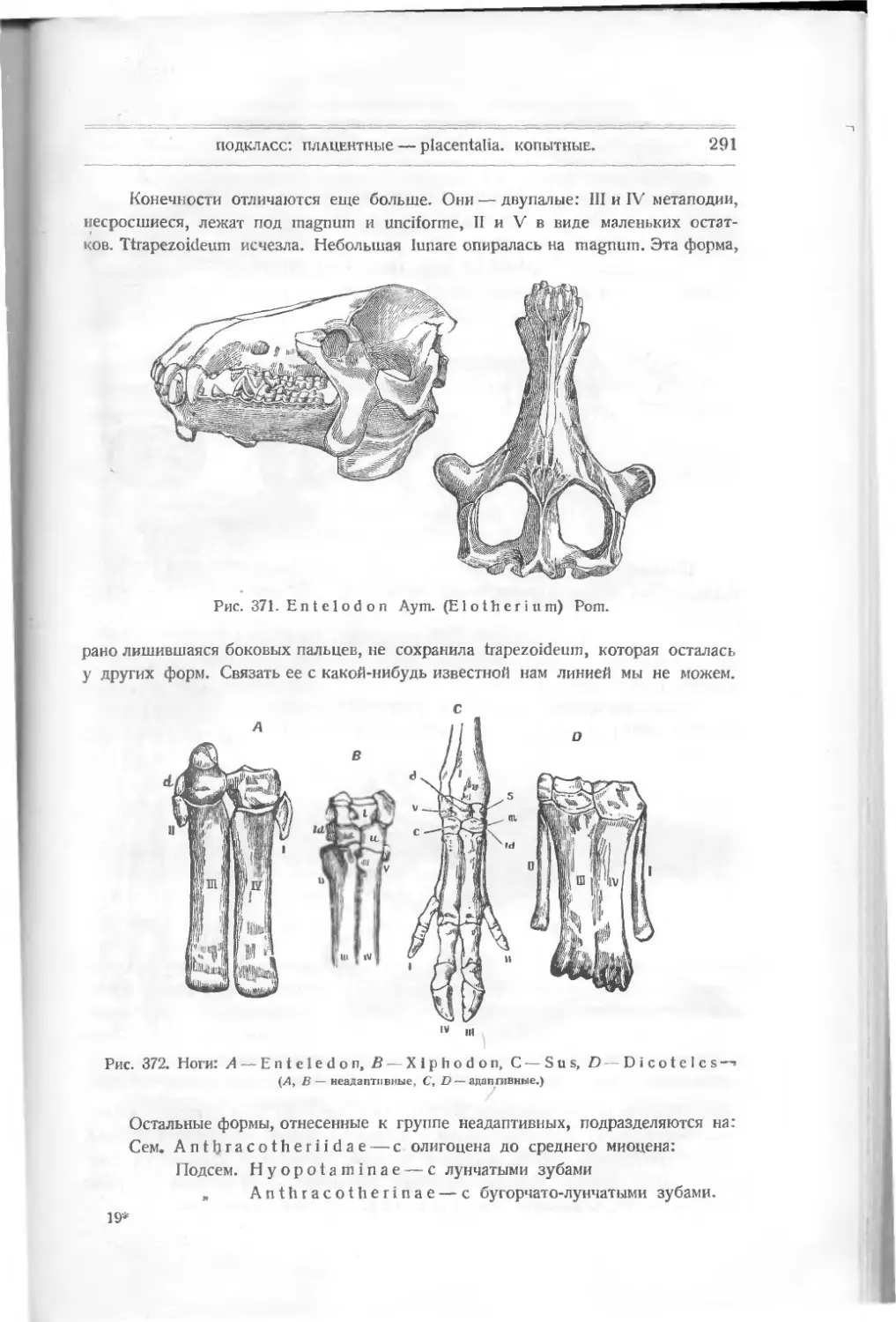
А
Рис. 5. Lanarkia
spinosa Traq.
жет быть, глаза. Все тело покрыто
мельчайшими полыми конусообраз-
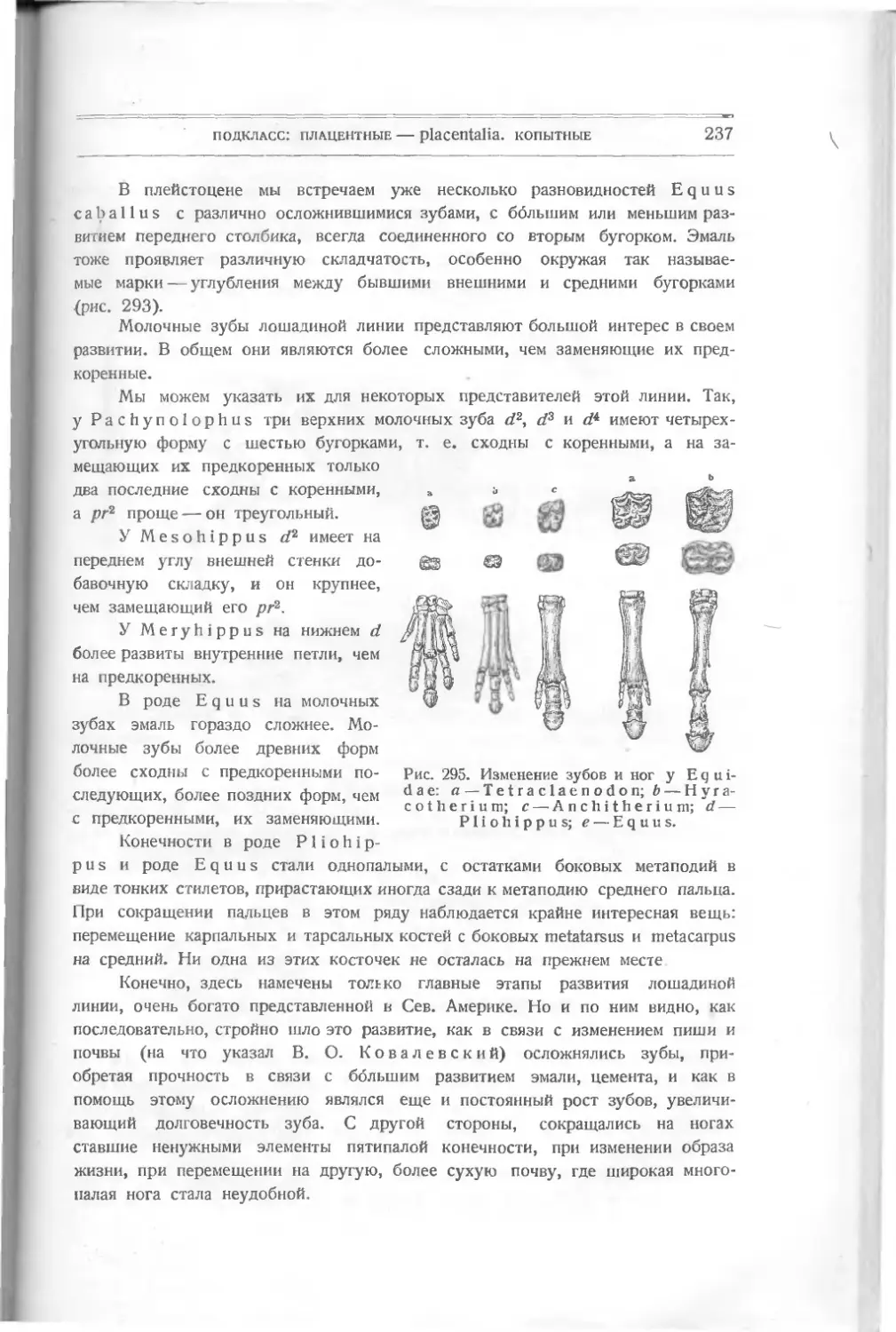
ными шипиками из дентина, без
отдельного основания. Это, очевидно,
первые защитительные органы, выде-
лявшиеся кожею животного.
Никаких намеков на внутренний
скелет еще нет. Установлено не-
сколько видов этого рода: L a n а г k i а
spinosa, Lanarkia horrid а.
Другая форма, найденная вместе
с Lanark! а,— Т h е 1 о d u s Traq. —
в верхнем силуре и в нижнем девоне
Шотландии,— похожа по форме тела
на первую, но шипики имеют основ-
ной край как бы отшнурованный от
остальной части — коронки. Снизу
оканчивается сзади
Рис. 6. Т h е 1 о d u's
s с о t i с u s Traq.
углубление, ведущее в пульпу. Форма
ши пиков бывает у некоторых видов
различная: круглая, овальная или
четырехугольная. Кроме того, посредине задней узкой
части тела заметна
складка, намекающая на спинной плавник.
Thelodus scoticus Traq. (рис.
похож на Lanarkia, отличаясь от нее
6) — из верхнего силура — очень
присутствием на спине складки —
плавника.
Thelodus Pagei Powrie (рис. 7) — из нижнего девона — имеет некото-
рые особенности, именно: 8 поперечных полосок, идущих по обеим сторонам
расширенной передней части тела и сходящихся к продольной складке. Может
быть, это зачатки жабер. Это очевидно форма, пошедшая вперед в своем
10
класс: рыбы — pisces
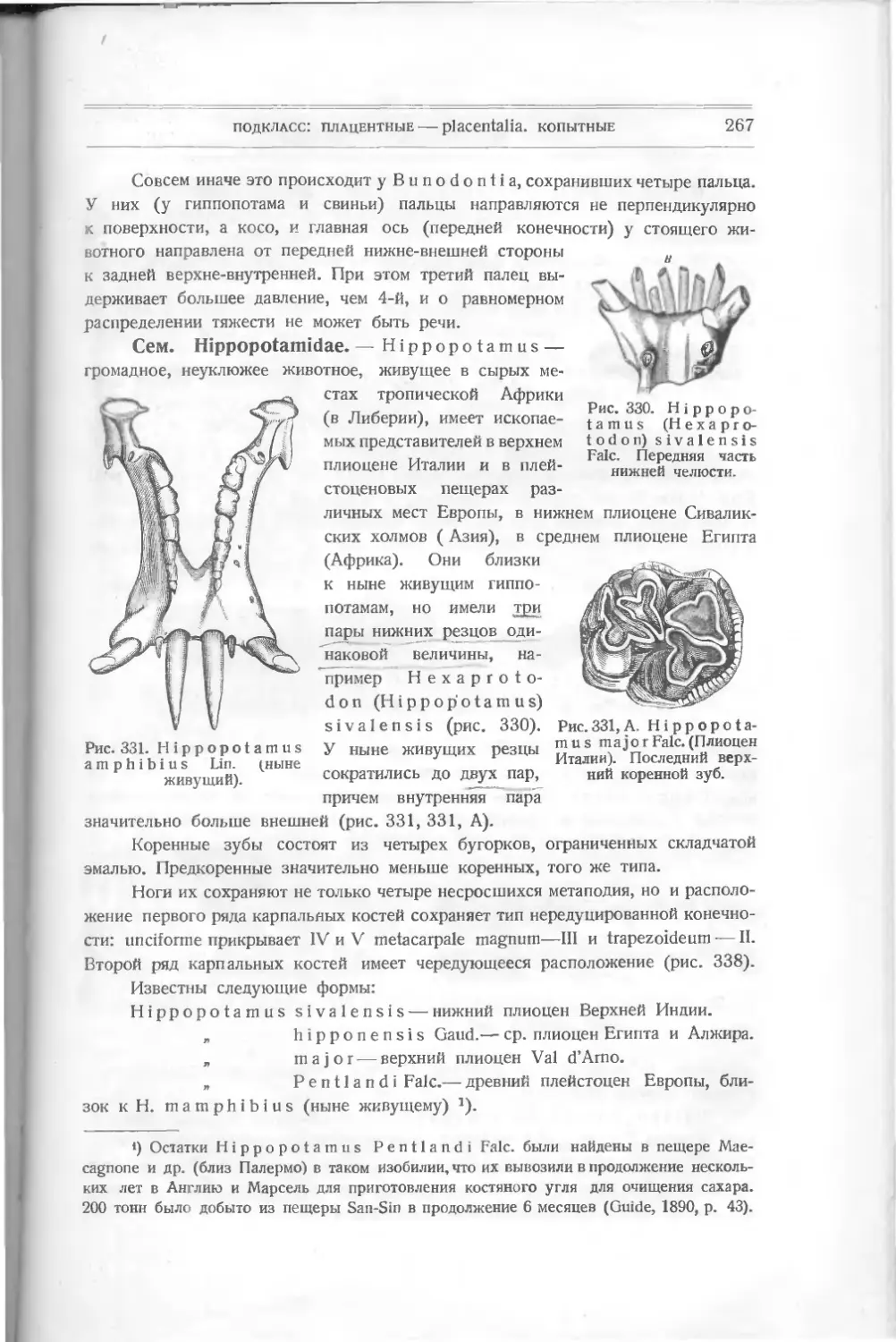
развитии как по внутренней организации (намек на жабры), так и по осложне-
Рис. 7. Thelodus Page!
Powrie.
В, С чешуйки.
девона Европы и Сев. Америки, имеет
нию защитительных органов (шипи-
ков) различной формы.
Drepanaspis Traq. (па-
лаш), из нижнего девона Рейнской
Пруссии, является как бы пере-
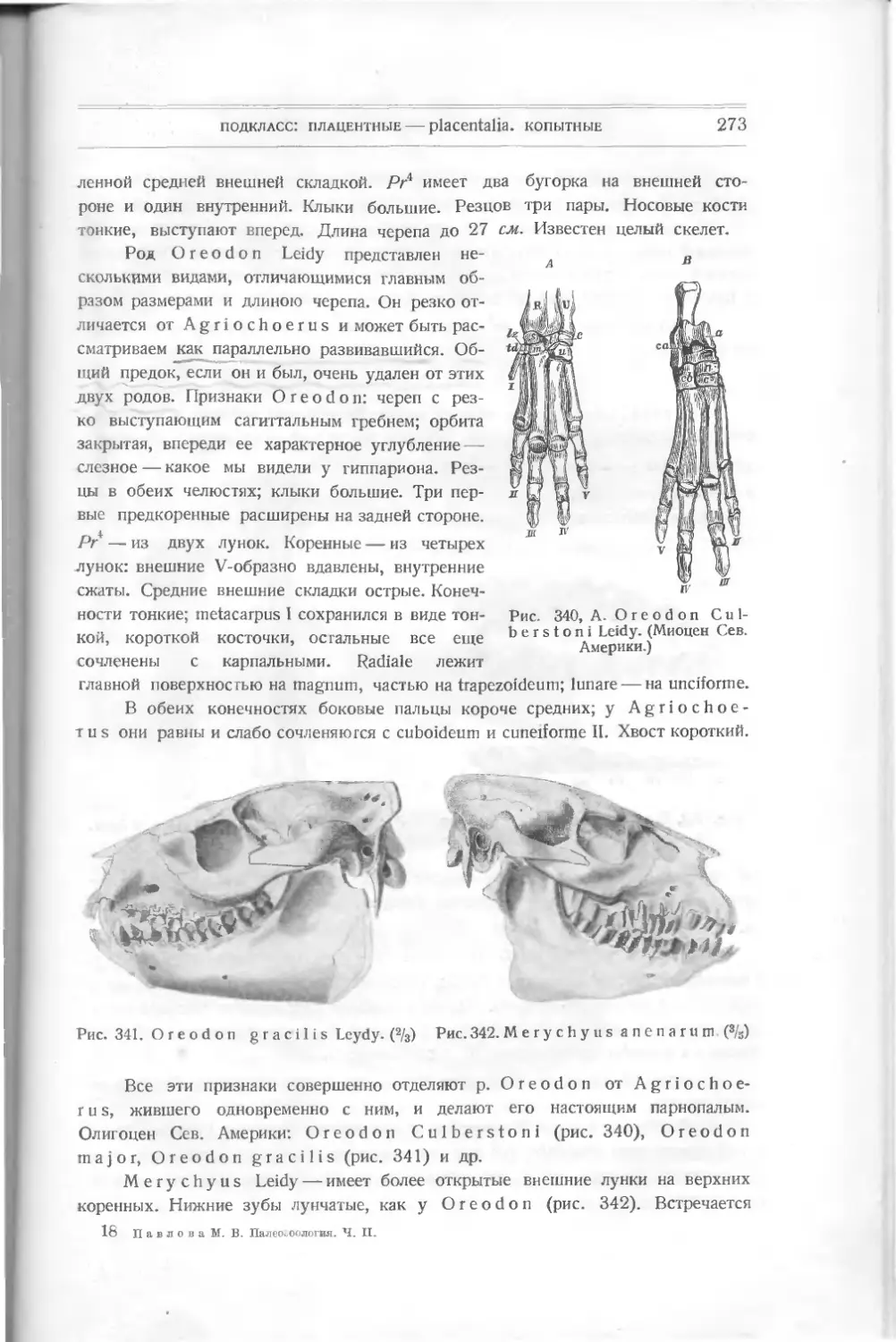
ходной формой к настоящим
панцирным рыбам. Он изучен
также Траквайром (рис. 8).
По форме тела он ближе к
Thelodus Pagel Powrie с рас-
ширенной передней частью тела и
суженной задней. Он достигал
60 см в длину. Покров его со-
стоял из маленьких и больших
щитков. Большие — спинной и
брюшной, два боковые верхние,
два нижние; два меньшие -— перед-
ние (носовые), около которых
намечается полоска — может быть,
ротовое отверстие. По бокам ши-
рокой части тела — глаза. Между
большими щитками лежат малень-
кие пластинки. Еще более мел-
кие покрывают узкую часть тела —
хвост и выступ его — плавник.
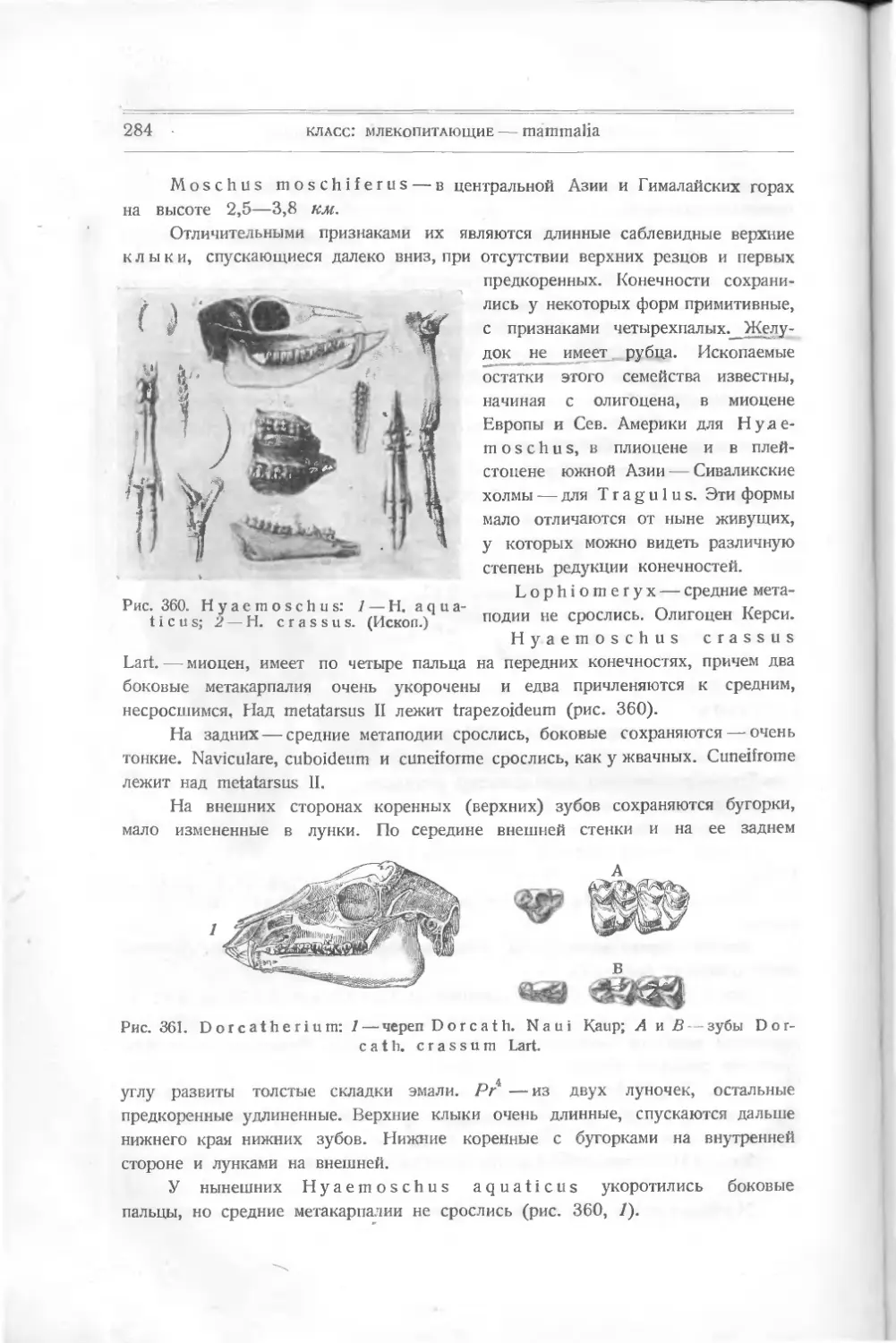
Вероятно, эти пластинки из ден-
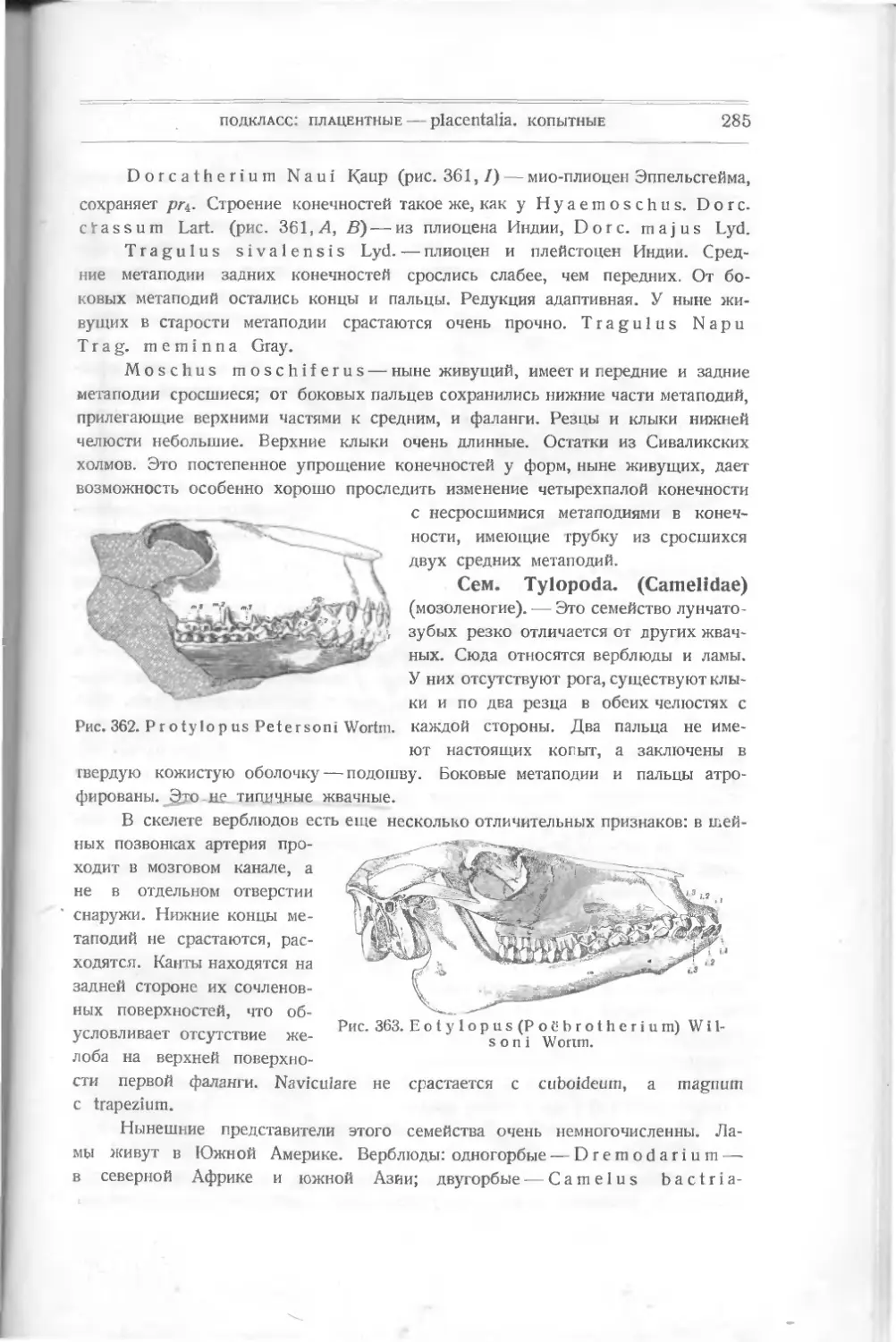
тина являются измененными щит-
ками и пластинками Thelodus,
слившимися вместе и расположен-
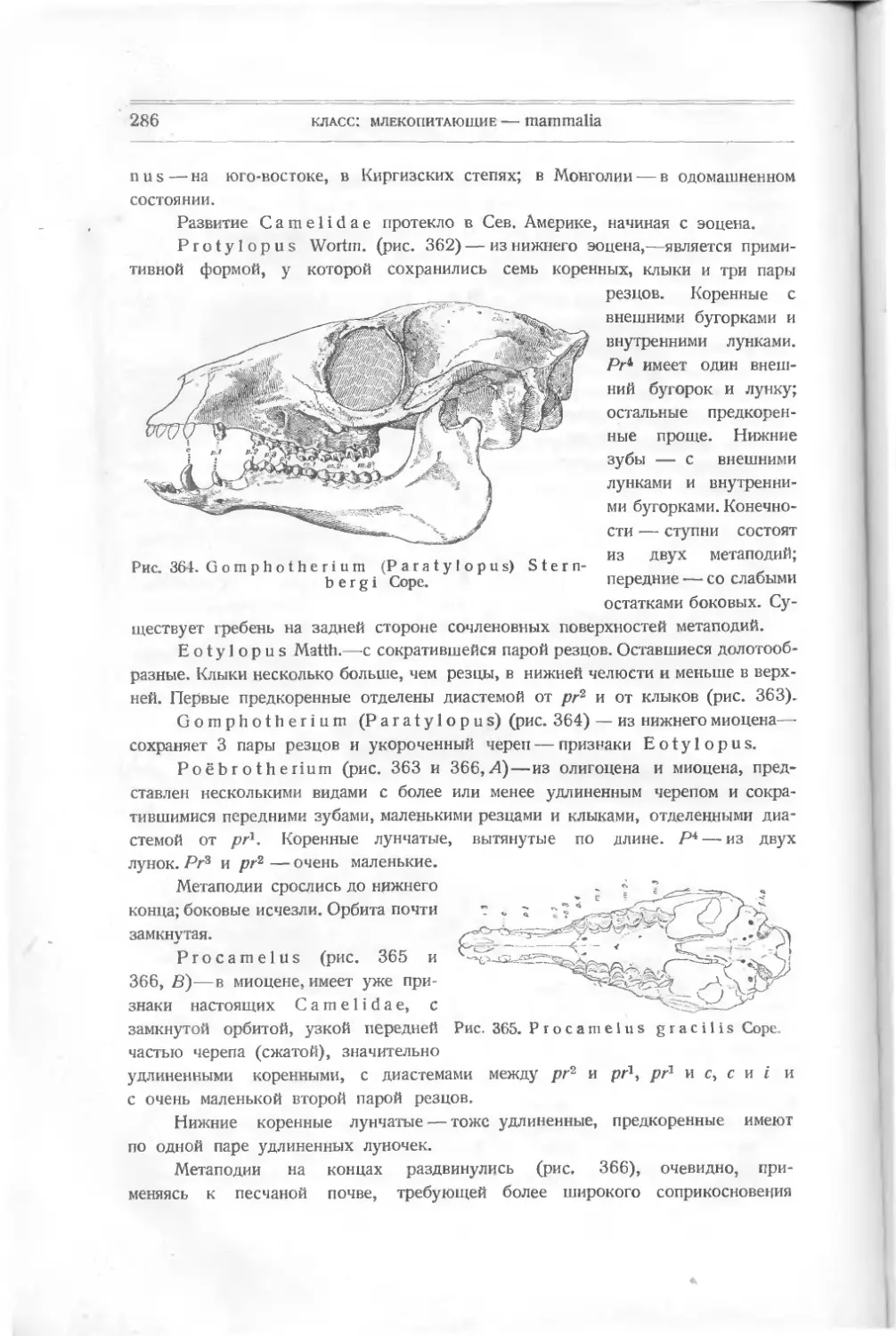
ными плотно друг возле друга.
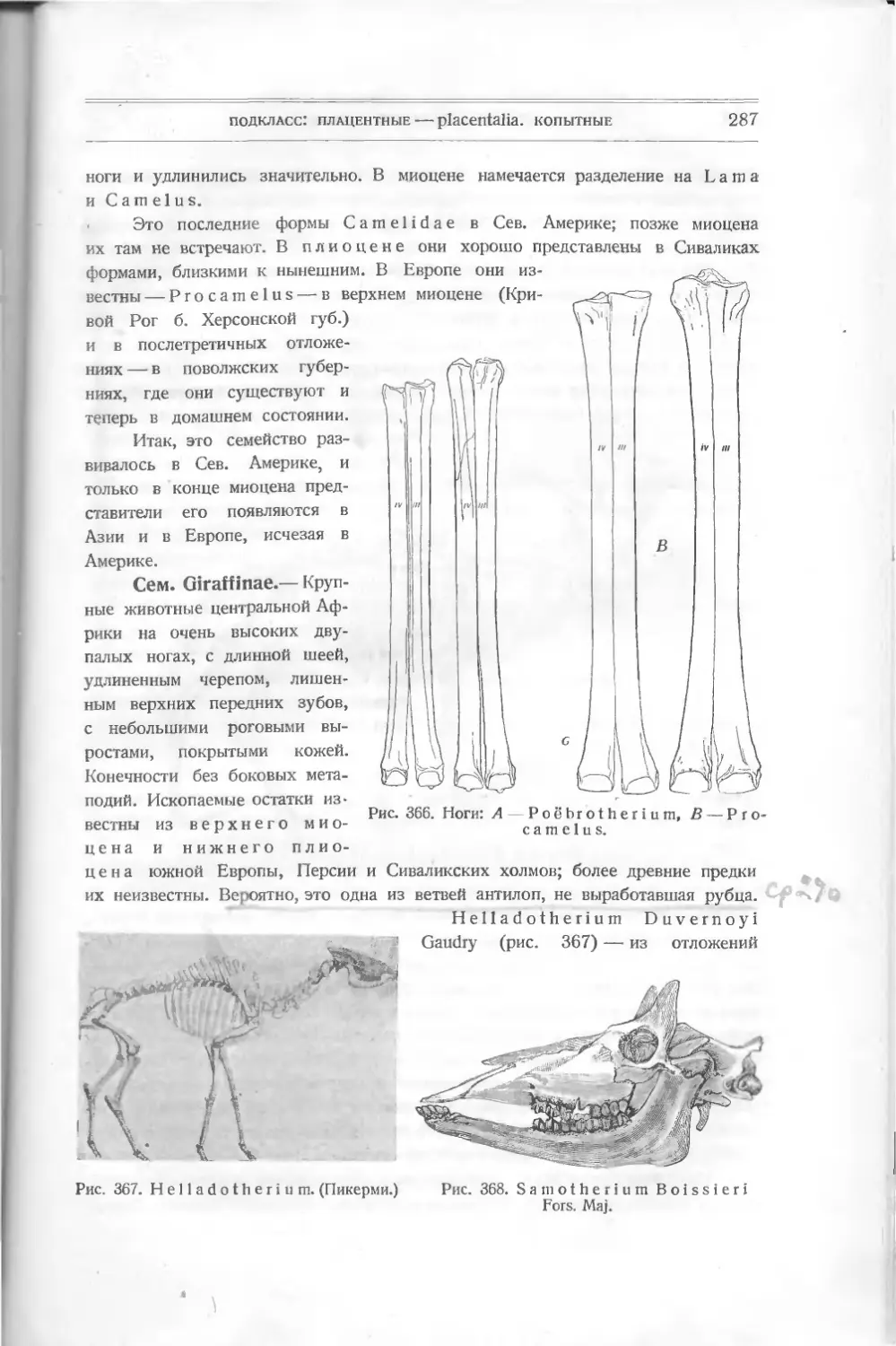
Pteraspis (крылатый щит)
(рис. 9), известный раньше,—то-
же из верхнего силура и нижнего
головной шит из восьми больших
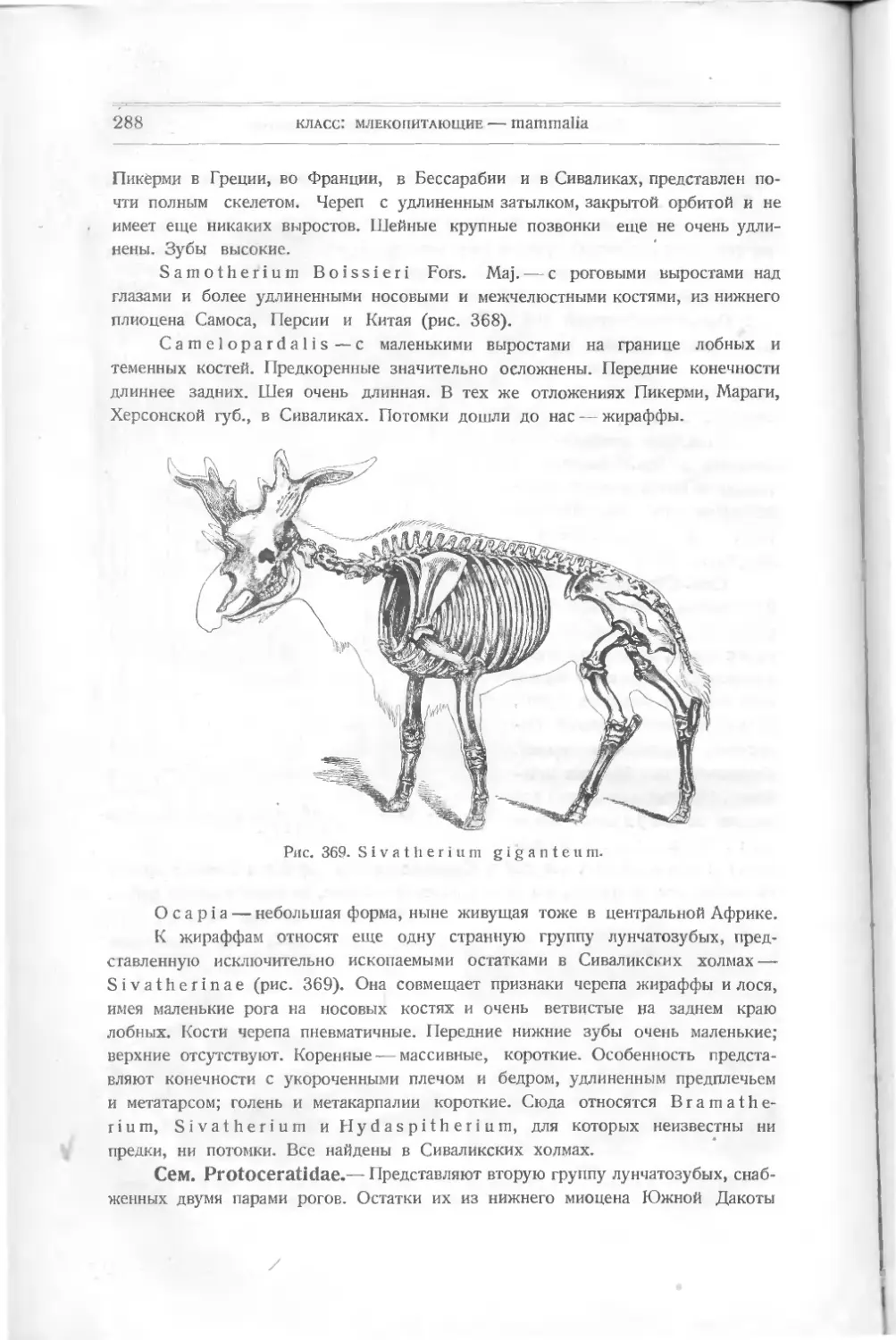
Рис. 8. Drepanaspis gemundenensis Traq.
— верхняя поверхность, 2? — нижняя; ж — ротовая пластинка, wid — сдиниая, mv — брюшная.
подкласс: панцырные—-placodermi
11
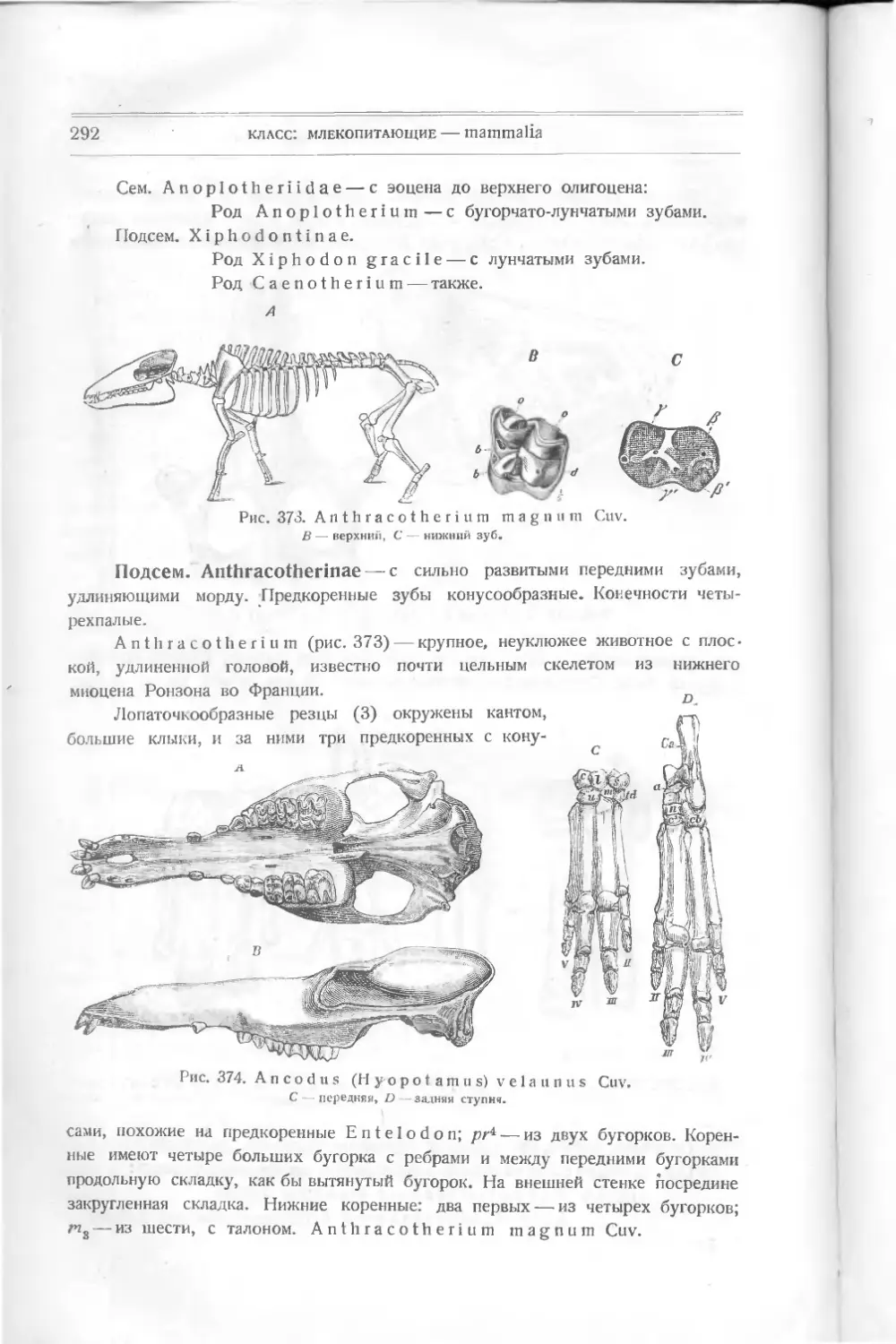
пластинок (маленьких между ними нет): спинной, брюшной, передней —
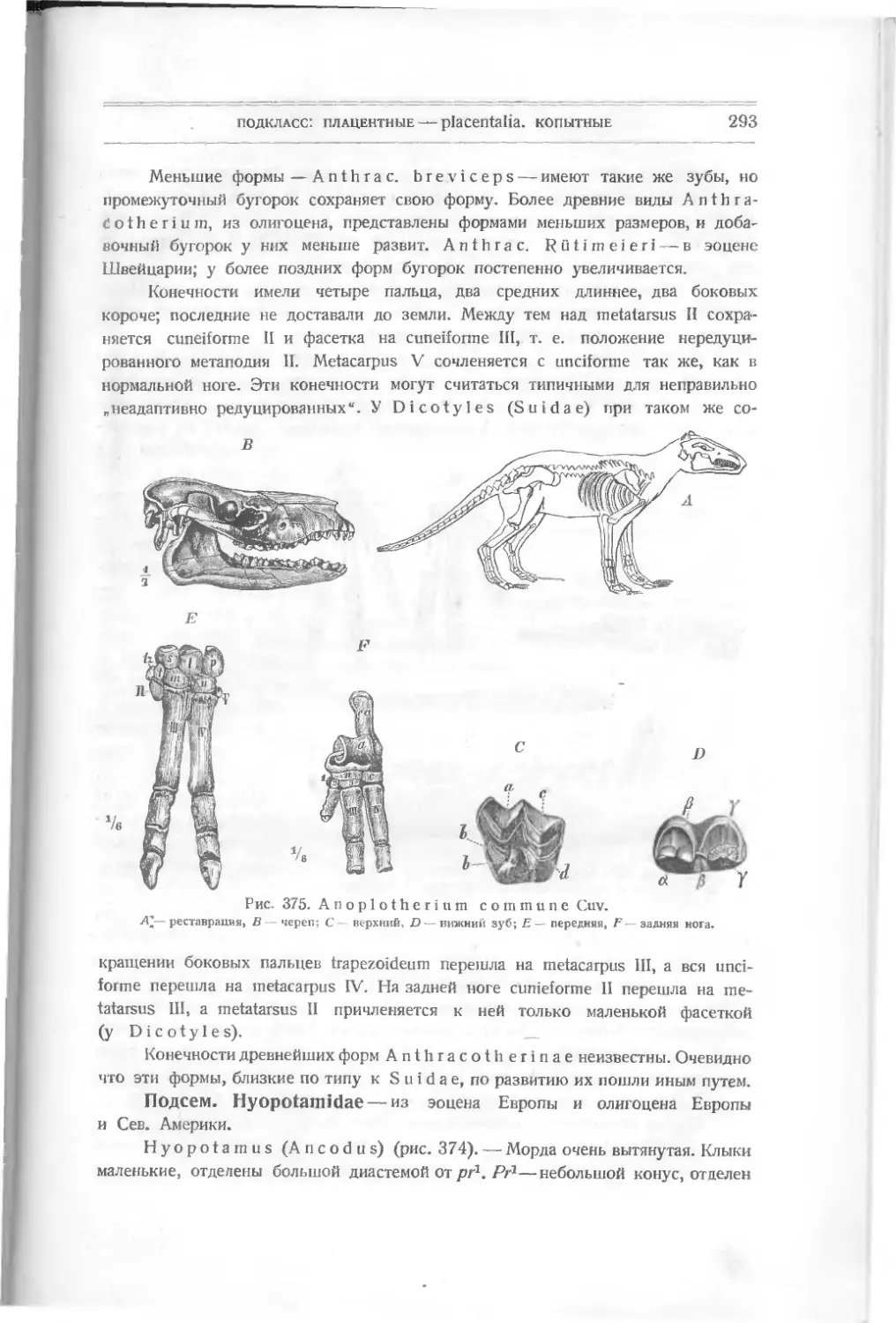
rostium, двух глазных, двух боковых и спинного шипа. Форма rostrum
бывает различна, более или менее удлинена у различных видов: Pte-
raspis Crouchi,
Pte г. Gosselati,
Pte r. rostratus.
Рот виден у всех
отчетливо, также и
глаза. Вся задняя
половина тела была
Рис. 9. Pteraspis rostratus Ag.
покрыта правильными ромбиками. Хвост удлинен. Строение пластинок слож-
нее, чем v Drepanaspis. Именно, каждая из них состоит из трех
слоев: 1) наружного, вазодентина, самою прочного, 2) пористого, толстого,
прорезанного каналами (в нем проходят сосуды) и 3) внутреннего перламутро-
вого, известкового, Это — значительное осложнение кожных покровов. Эти
формы представляют интерес, позволяя проследить, как мог оазвиваться твер-
дый покров их из маленьких шипиков, преобразующихся в пластинки
yLanarkia и Thelodus, в щиты у Pteraspis.
Отряд 2
Костнощитковые — Osteostraci
Голова покрыта большим костным щитом, тело-—поперечными мелкими
пластинками.
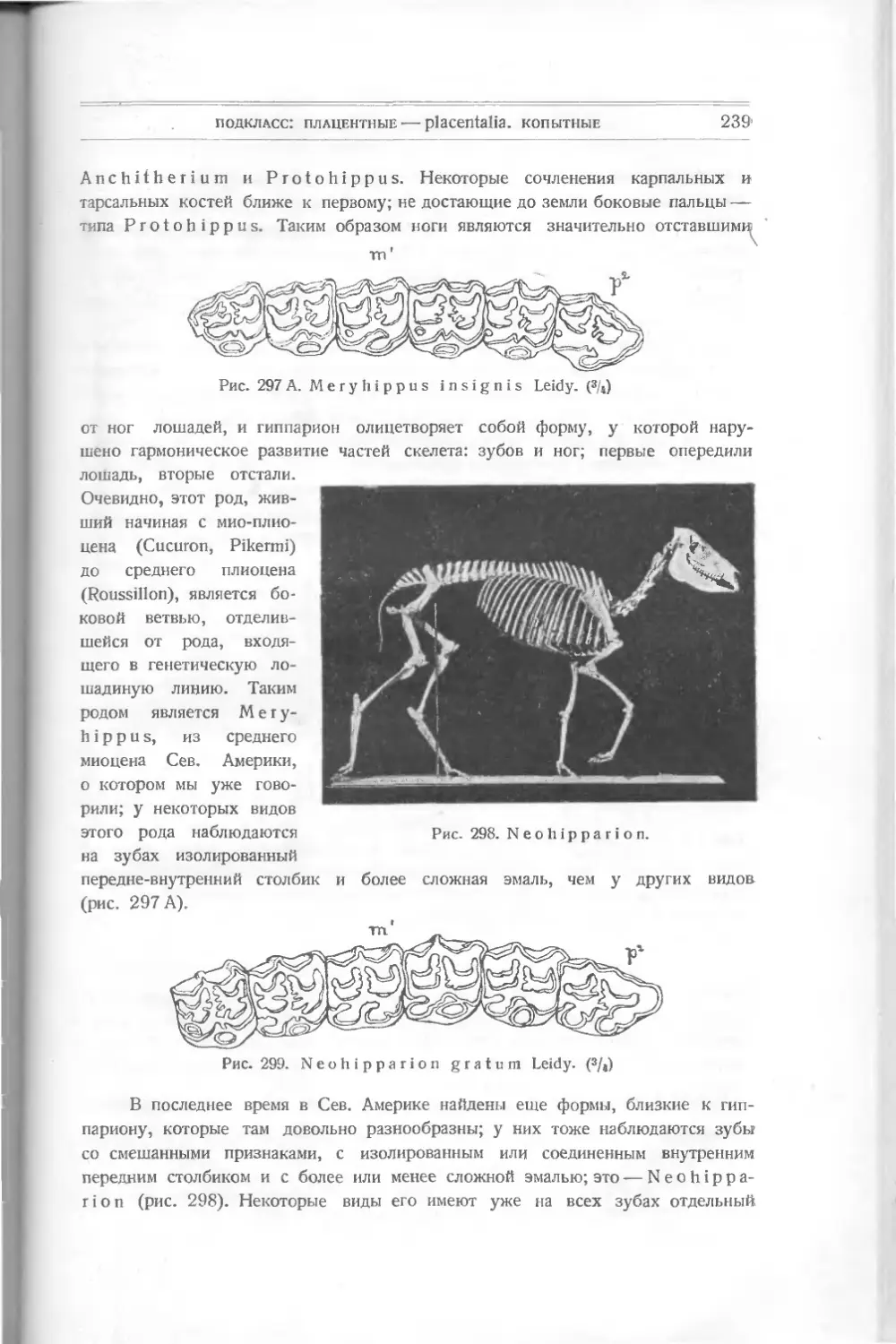
Сем. Cephalaspidae. По форме тела подходят ближе всего к The-
lodus Pagei, особенно более примитивные формы — например Ateleaspis
tesselata Traq. из верхнего силура Шотландии, у которого головной щит
состоит из маленьких полигональных трехслойных пластинок, образующих при
слиянии мозаику. Удлиненное тело покрыто мелкими пластинками, расположен-
ными в поперечные ряды. На задней половине тела имеется плавник посредине
тела и другой — на конце его.
Дальнейшее изменение в форме внешнего покрова представляет Cepha-
laspis (рис. 10). Головной щит его состоит из сросшихся пластинок, состоящих
Рис. 10. Cephalaspis Lyelli Ag.
также из 3 слоев, но в среднем
слое встречаются костные кана-
лы и тельца. На головном щите
его, посредине, видны несколько
отверстий, которым приписывается
различное назначение. Переднее,
круглое,— для непарного глаза, два парных под ним — глазные. Продолговатое
среднее— дыхательное; два маленьких за ним — слуховые. На краях щита
заметны щели, может быть, для прохождения щупалец. Конечно, все эти опре-
деления отверстий гадательны. (Ср. рис. 11 и 12.)
12
класс: рыбы — pisces
Головной щит Cephalaspis заканчивался по бокам двумя крыловид-
ными отростками. Поверхность щита покрыта бугорчатой скульптурой.
Этой английской нижне-девонской форме соответствуют найденные и
описанные Фр. Б. Шмидтом с о-ва Эзеля из верхнего силура: Thyestes
и Т г е ш a t a s р i s. От них найдены только щиты с характерными отверстиями,
расположенными как у Cephalaspis. Скульптура на них иная, также и
задний край. У Thyestes verrucosus Eichw. (рис. 11) сжатые бугорки,
расположенные рядами. На заднем краю, по бокам, находятся вырезки, обра-
зующие небольшие отростки; крыльев нет. Полный экземпляр Thyestes
найден в Англии, в нижнем девоне.
У Tretnataspis Schmidti Roh. (рис. 12) щит яйцевидный, без
скульптуры. Весь край слабо зазубрен. На нижней поверхности головного
щита, спереди, лежит ряд мелких отверстий, расположенных по кругу ]). Это
предполагаемые жаберные отверстия. В Норвегии найдена форма, близкая
к Cephalaspis: это —-Aceraspis с большим головным щитом. Слои,
в которых она найдена, относятся одними учеными к верхнему силуру, дру-
1 ими — к нижнему девону или считаются переходными слоями между этими
двумя системами.
Представители обоих этих отрядов, которые Абель соединяет
в один — Osteostraci, были морскими бентонными животными,
судя по их строению. Только Pteraspis мог иметь большую способность
к передвижению (большую подвижность).
Отряд 3
Крылатые рыбы — Antiarchi
Представители третьей группы имеют несколько пластинок на голове
и туловище, образующих щиты, и несколько сходны с Pteraspis. Отдель-
ные пластинки щитов соединяются между собою налегающими друг на друга
*) На нашем рисунке они изображены не все.
подкласс: панцырные — placodermi
13
краями. Пластинки усеяны звездчатыми бугорками. На средних и боковых пла-
стинках проходят каналы.
Существуют особые
придатки в виде как бы
передних конечностей,
состоящих из отдельных
суставов и помогавших
относятся: Asterolepis, Pterichthys,
Bothriolepis и другие девонские Placo-
dermi, входящие в семейство A s t е г о 1 е-
р i d а е.
Общая форма тела та же, что у пре-
дыдущих, т. е. рыбообразно удлиненная с
неправильным хвостовым плавником и дру-
гим— спинным.
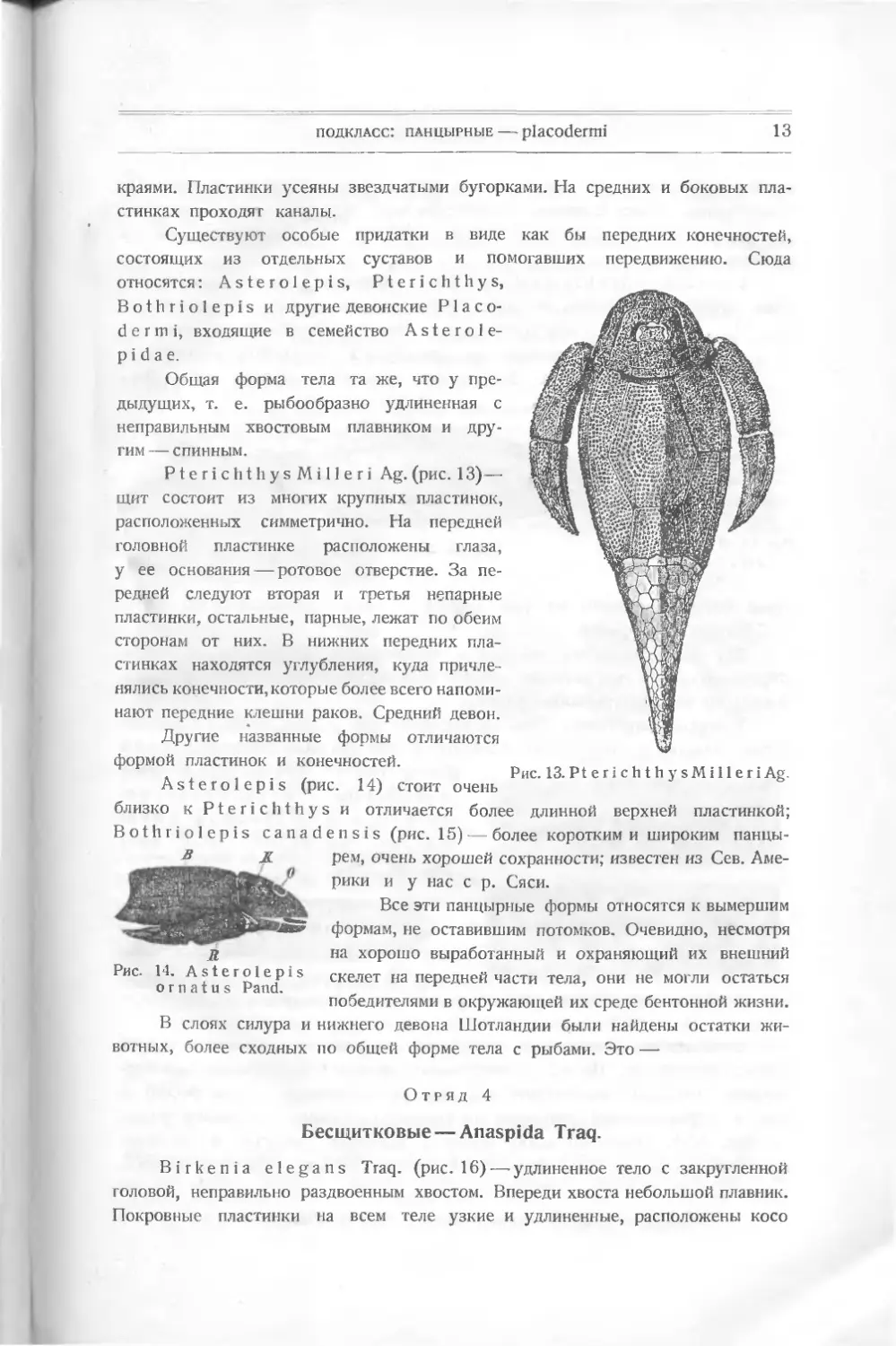
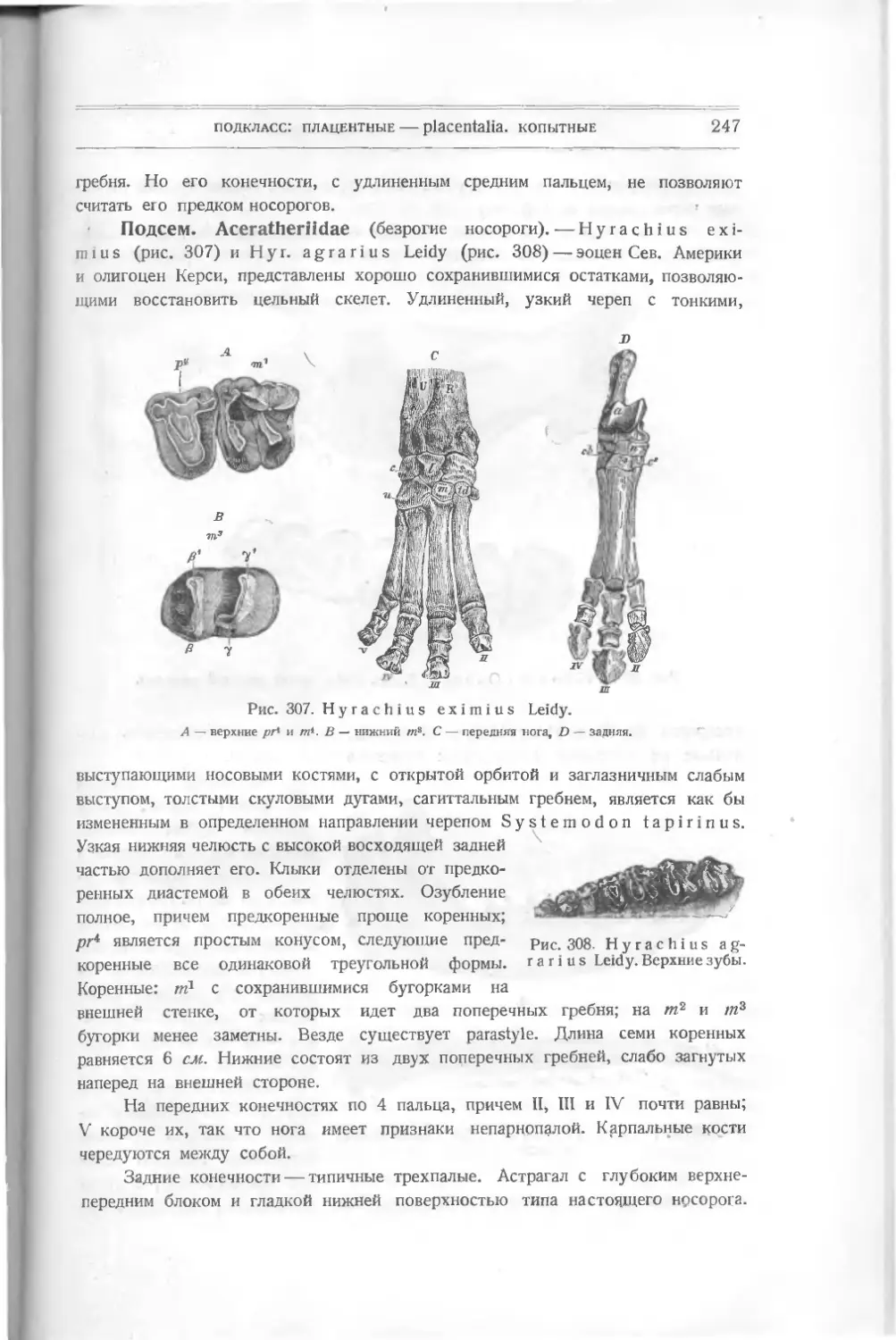
PterichthysMilleri Ag. (рис. 13)—
щит состоит из многих крупных пластинок,
расположенных симметрично. На передней
головной пластинке расположены глаза,
у ее основания — ротовое отверстие. За пе-
редней следуют вторая и третья непарные
пластинки, остальные, парные, лежат по обеим
сторонам от них. В нижних передних пла-
стинках находятся углубления, куда причле-
нялись конечности, которые более всего напоми-
нают передние клешни раков. Средний девон.
Другие названные формы отличаются
формой пластинок и конечностей.
Asterolepis (рис. 14) стоит очень
передвижению. Сюда
Рис. 13. Р t е г i с h t h у s М i 11 е г i Ag.
близко к Pterichthys и отличается более длинной верхней пластинкой;
Bothriolepis canadensis (рис. 15) — более коротким и широким панцы-
Я
Рис. 14. Asterolepis
о г п a t u s Pand.
В слоях силура и
вотных, более сходных
рем, очень хорошей сохранности; известен из Сев. Аме-
рики и у нас с р. Сяси.
Все эти панцырные формы относятся к вымершим
формам, не оставившим потомков. Очевидно, несмотря
на хорошо выработанный и охраняющий их внешний
скелет на передней части тела, они не могли остаться
победителями в окружающей их среде бентонной жизни,
нижнего девона Шотландии были найдены остатки жи-
по общей форме тела с рыбами. Это —
Отряд 4
Бесщитковые — Anaspida Traq.
Birkenia elegans Traq. (рис. 16) — удлиненное тело с закругленной
головой, неправильно раздвоенным хвостом. Впереди хвоста небольшой плавник.
Покровные пластинки на всем теле узкие и удлиненные, расположены косо
14
класс: рыбы — pisces
в несколько рядов. На голове мелкие шипики, расположенные группами. По
бокам головы, ближе к нижней поверхности тела, находятся восемь маленьких
круглых точек — может быть, жаберные отверстия. На спине находится ряд
крупных шипов.
Lasanius problematicus Traq. отличается более многочислен-
ными щитками-пластинками на спине. Форма тела такая же, с неправильным,
Рис. 15. Bothriolepis
canadensis. (*/2)
гегероцеркальным хвостом. Покровы этих форм от-
личаются от предыдущих отсутствием в них ден-
тина. Это только утолщения волокон кожи. Этот
Рис. 16. Birkenia elegans Traq.
отряд более изолирован от трех первых и более приближается по форме
тела и покровам к рыбам.
Так как остатки их найдены в пресноводных отложениях, то можно
предположить, что они населяли пресные воды верхне-силурийского или нижне-
деьонского периода (указание Абеля).
В верхне-силурийских или нижне-девонских слоях Норвегии найдены
формы, близкие к шотландским A n a s р i d а. Это три рода, описанные Кером
(Kiaer): Pterolepis nitidus, длина
11—13,5 см (рис. 17 и 17A); Pharyn go-
lepis oblongus, 19—20 cm\ Rhyn-
cholepis parvulus, 6—7 cm j)
(рис. 18, 19).
Рис. 17A. Pterolepis nitidus Kiaer.
Реставрация. (>/2)
Рис. 17. Р t е г о 1 е р i s n i t i d u s K'aer. (*/3)
Большинство экземпляров прекрасной сохранности; позволяют вполне
точное описание их. На их удлиненно-закругленном теле сохранились в нор-
мальном положении мельчайшие чешуйки, расположенные косыми рядами на
теле и определенными участками на голове, различными у разных родов.
На рис. 17 А, отчетливо видны точечные жаберные отверстия и глазницы.
На спине шипы — в один ряд. На брюшной стороне — анальный плавник.
*) Johan Kiaer, The Dovmtonian Fauna of Norway, 1924.
подкласс: круглоротые cyclostomi
15
Хвост дифицеркальный. (Траквайр принимал спинную сторону за
брюшную.)
Ротовое отверстие было снабжено дугообразным (arched) хрящом, который,
вероятно, можно сравнить с нижней челюстью настоящих рыб. Никаких сле-
дов внутреннего скелета не найдено — ни хрящевого, ни костного.
Особенностью этих Anasрidа является присутствие единствен-
ного носового отверстия, лежащего между передними краями глаз.
За ним лежит пинеальное отверстие. Это встречается еще только у
Cephalaspidae из Placodermi и у Cyclostomi. Этому признаку
Г е к е л ь (Haeckel) придает большое значение, что позволяет подразделять
Craniata на MonorrIrina и Diplorrhina1).
нем силуре.
Рис. 18. R h у п с h о 1 е р i s раг-
v u 1 u s Kiaer.
Anaspida были нектонпые, a Cepha-
laspidae— бентонные формы, очень рас-
пространенные в реках, озерах и эстуариях
северного атлантического континента, в верх-
Черепная крышка.
Рис. 19. а — Р t е г о 1 е р i s; b—Р h a i у n g o-
lepis; c —• R h у n c h о 1 e p i s.
Изучение панцирных рыб указывает, что, несмотря на видимое прогрес-
сивное развитие их наружного покрова, они вымерли, не оставив потомства
(может быть, исключая Anaspida). По своей организации они не могли
быстро передвигаться, плавать; кроме того, у них совершенно отсутствовал
внутренний скелет, и задняя половина тела оставалась мало защищенной твер-
дыми покровами, а между тем одновременно с ними жили крупные формы
беспозвоночных, как, например, Pterygotus между Gigantostraca, с ко-
торыми им, вероятно, приходилось бороться и от которых они погибали.
Может быть, здесь именно ярко выразился тот закон быстрого, но одно-
стороннего развития, в котором Коп видел серьезную причину для выми-
рания целых групп.
Подкласс II
КРУГЛО РОТЫ Е — CYCLOSTOMI
Этот подкласс является полною противоположностью панцырным рыбам,
так как лишен внешнего скелета. Немногочисленные представители, живущие
1) Haeckel, Systematische Phylogenie.
16
класс: рыбы — pisces
ныне, не имеют и внутреннего скелета. Это — миноги, мягкие, червеобразные
рыбы, покрытые кожей. Ископаемые остатки до недавнего времени не были
известны.
Но Траквайр нашел в слоях среднего девона Шотландии (Caithness)
форму, которую он
Рис. 20. Palaeospon-
dylus Gunni. Девон.
отнес KCyclostomi по форме рта. Это Palaeo-
spondylus, имеющий 3,5 см в длину. Эта форма
имеет расширенную переднюю часть — голову, за кото-
рой идет ряд известковых позвонков. Черепная ко-
робка разделяется на три области: переднюю с
лым
нюю
Сулу
ротовым отверстием, окруженным циррами,
с жаберными дугами и заднюю — слуховую
(рис. 20).
Позвонки имеют боковые отростки в средней
и нечто вроде плавниковых стержней в задней,
круг-
сред-
кал-
части
тела
что позволяет подразделить позвонки на три группы:
без отростков, с отростками и со стержнями.
Hypospondylus — из пермо-карбона Богемии—
вторая ископаемая форма, относимая к этому типу,
описанная Гекелем, имеет более сложную черепную
коробку, состоящую из отдельных пластинок, и жабер-
ные дуги. Весь позвоночник несет плавниковые под-
держки, на заднем конце его их два ряда. Некоторые
ученые считают эти формы личинками других животных: Palaeo-
spondylus — личинкой Coccosteus. Ни в одном из более поздних
отложений остатки Cyclostomi найдены не были. Нынешние представители
их считаются некоторыми учеными дегенерировавшими типами, пережитками
/ / / J /
Рис. 21. Скелет окуня.
очень первобытных форм, которые могут служить прототипом позвоноч-
ных, с едва зарождающимся внутренним скелетом, плавниками и головой,
снабженной ротовым отверстием, но без челюстей. Другие считают Cyclo-
s t о m i уклонившимися формами с развитым черепнохрящевым скелетом
(проф. Н. А. Северцов).
подкласс: круглоротые — cyclostomi
17
Чтобы понять строение и изменение скелета настоящих рыб, остатки
которых мы встречаем также начиная с силура и девона, мы должны позна-
комиться с общим строением Cyclostomi, являющихся как бы живыми
ископаемыми, сохранивших простейшее строение первичного скелета (позво-
ночника) предков позднейших форм.
У Bdellostoma скелет состоит из х о р д ы — спинной струны, за-
ключенной в оболочку, над которой протягивается спинной нерв, не-
сколько утолщенный спереди в головной части. Даже следов черепа не суще-
ствует. Дыхательные органы представлены жаберным мешком с отвер-
стиями наружу. Непарный спинной плавник, обра ювавшийся из
складки кожи, поднявшейся вдоль спины и переходящей на брюшную
сторону животного, поддерживается маленькими вертикальными кожными
складками.
У миноги (Petromyzon) в молодом возрасте признаки скелета почти
те же, что у Bdellostoma, но с возрастом он сильно изменяется. Хорда
сохраняется, но вокруг ее влагалища разви-
вается скелетородный слой, подни-
мающийся и над нервным стволом. В этом
слое — оболочке — отлагаются вертикальные
хрящевые отростки, обнимающие спинной
нерв. На передней части тела намечается
голова в виде хрящевой капсулы. В жабер-
Рис. 22. Позвонки ныне живущей
акулы.
ном мешке также появились хрящи, поддержи-
вающие его. Плавник идет по спине гораздо дальше к голове. Кожа голая, на
ней нет чешуи и каких-либо твердых образований в самой коже. По простоте
строения скелета минога является как бы следующей стадией после A m р h i о-
х u s (рис. 1), у которого нет ни жаберного аппарата, ни черепной капсулы.
Если мы сравним эти скелеты со скелетом костистой рыбы, например
окуня (рис. 21), то увидим громадную разницу. Вместо спинной струны протя-
гивается ряд соединенных между собой костных позвонков с отходящими от них
вверх и вниз отростками; первые из них обнимают спинной мозг (у миноги
они только намечались во влагалище хорды). Череп состоит из большого
числа костей, с причлененной нижней челюстью и мелкими зубами. Непарные
плавники, спинной, хвостовой и анальный, поддерживаются над телом хрящевыми
лучами, соединяющимися с лученосцами, идущими внутрь тела рыбы и в свою
очередь прилегающими к верхним остистым отросткам позвонков. Суще-
ствуют кроме того парные плавники, основная часть которых укрепляется
немногими костями в теле рыбы. Жабры закрыты жаберной крышкой. Тело
покрыто тонкими, прозрачными чешуйками.
Вероятно, эти признаки костистых рыб и дали им преимущество над дру-
гими отрядами. Каков же был путь, которым шло развитие костистых рыб, и
можно ли связать их скелет со столь отличным от них мягким скелетом миног?
Для решения этих вопросов обратимся к ископаемым остаткам более совер-
шенных рыб, живших начиная с палеозоя.
2 Па в л о в а М, В. Палеозоология, Я. II.
18
класс: рыбы — pisces
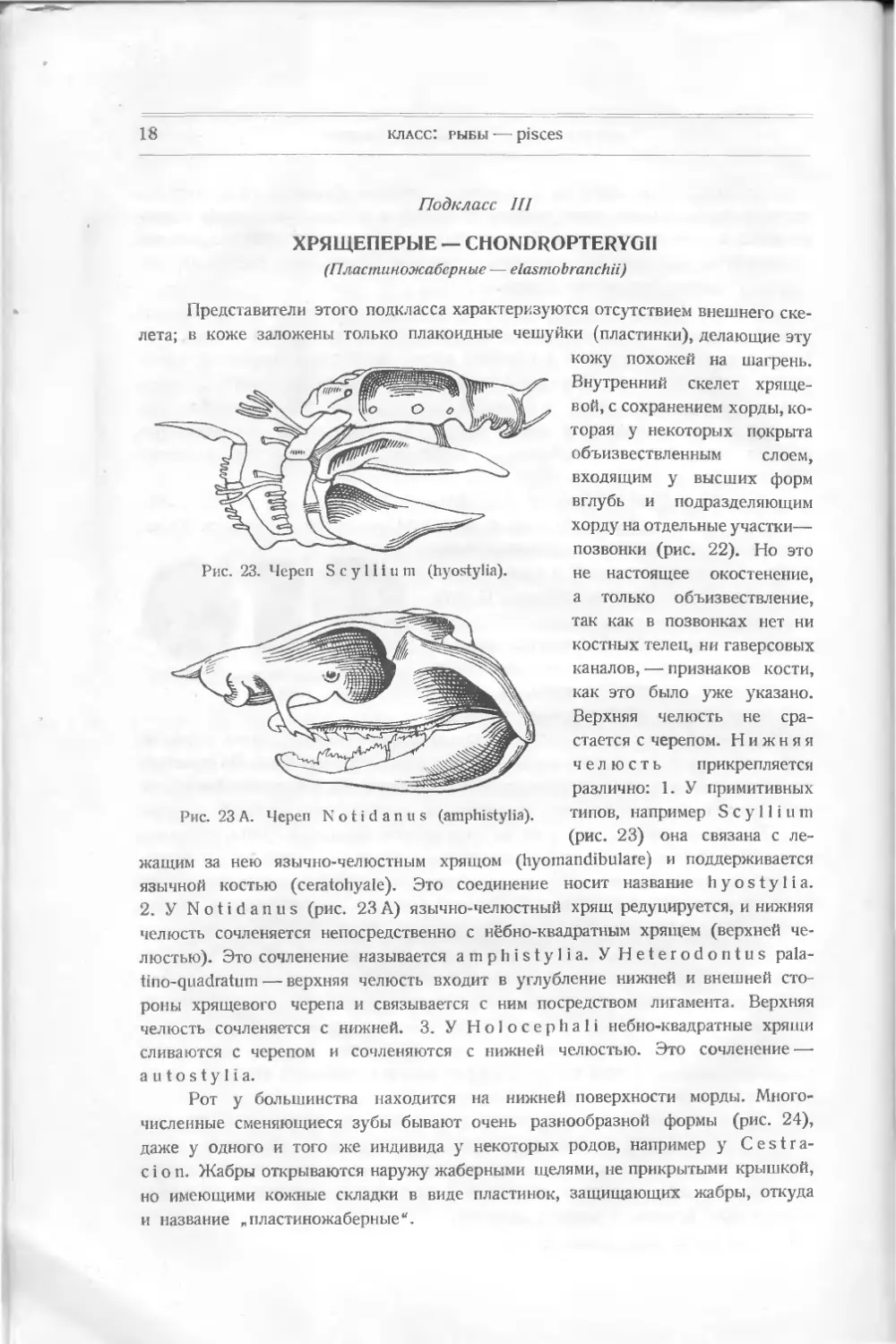
Подкласс III
ХРЯЩЕПЕРЫЕ — CHONDROPTERYGII
(Пластиножаберные — elasmobranchii)
Представители этого подкласса характеризуются отсутствием внешнего ске-
лета; в коже заложены только плакоидные чешуйки (пластинки), делающие эту
кожу похожей на шагрень.
Внутренний скелет хряще-
вой, с сохранением хорды, ко-
торая у некоторых покрыта
объизвествленным слоем,
входящим у высших форм
вглубь и подразделяющим
хорду на отдельные участки—
позвонки (рис. 22). Но это
не настоящее окостенение,
а только объизвествление,
так как в позвонках нет ни
костных телец, ни гаверсовых
каналов, — признаков кости,
как это было уже указано.
Верхняя челюсть не сра-
стается с черепом. Нижняя
челюсть прикрепляется
различно: 1. У примитивных
типов, например S с у 11 i u m
(рис. 23) она связана с ле-
жащим за нею язычно-челюстным хрящом (hyomandibulare) и поддерживается
язычной костью (ceratohyale). Это соединение носит название h у о s t у 1 i а.
2. У Notidanus (рис. 23 А) язычно-челюстный хрящ редуцируется, и нижняя
челюсть сочленяется непосредственно с нёбно-квадратным хрящем (верхней че-
люстью). Это сочленение называется amphistylia. yHeterodontus pala-
tino-quadratum — верхняя челюсть входит в углубление нижней и внешней сто-
роны хрящевого черепа и связывается с ним посредством лигамента. Верхняя
челюсть сочленяется с нижней. 3. У Holocephali небно-квадратные хрящи
сливаются с черепом и сочленяются с нижней челюстью. Это сочленение —
autostylia.
Рот у большинства находится на нижней поверхности морды. Много-
численные сменяющиеся зубы бывают очень разнообразной формы (рис. 24),
даже у одного и того же индивида у некоторых родов, например у С е s t г а-
с i о п. Жабры открываются наружу жаберными щелями, не прикрытыми крышкой,
но имеющими кожные складки в виде пластинок, защищающих жабры, откуда
и название „ пластиножаберные “.
подкласс: хрящецерые— chondropterygii
19
В парных плавниках первичный хрящевой скелет хорошо развит.
Юн состоит из основных пластинок (b a s а 1 i а), про-, мезо- и метаптеригий,
различно расположенных у различных родов. К ним прилегают у акул распо-
ложенное с одной стороны многочисленные хрящевые радиальные лучи
(radialia), несущие плавниковые роговые лучи кожного происхождения.
Рис. 24. Cochliodus
contortus Ag.
Задние и средние зубы.
А. В — Carchar о don. С — Galeocerdo. D — Notidanus. Е —
Oxyrrhina. F — Я у Ь о du s. G — А е г о d и s. Н, I — L a m и а.
Рис. 24 А. Зубы акул.
представлен теперь акулами (S е 1 а с h i i) и скатами (S q н а-
Этот подкласс
t i n i d а е), почти исключительно морскими обитателями-хищниками.
Ископаемые остатки их известны с силура, где они выражены отдель-
ными зубами и плавниковыми спицами.
О гряд 1
Proselachii Dod.
(Ichthyotomi Соре)
Парные плавники с сегментированной осью. Позво-
ночник совсем не расчленен, или слабо намечены под-
разделения влагалища хорды. Зубы с несколькими зубчи-
ками различной высоты. Древний палеозой (силур, девон).
Сем. Cladodontidae. Cladoselache (Сla-
do du s)— девон Сев. Америки; найден в нескольких
довольно хорошо сохранившихся экземплярах, что дало
возможность подробно изучить скелет и строение его
плавников (рис. 25). Раньше были известны только зубы
Cladoselache lamnoides
сквой), описанные под именем
кроме спинного, хвостового и
существовали парные, передние
их указывает на то, что они выработались из
служивших раньше для ползания (А б е л ь).
Поддерживались плавники хрящевыми
основными (basalia) и радиальными
На), дающими им широкое основание. Более
из Мячкова (под Мо-
Cladodus. У него,
анального плавников,
и задние; положение
органов,
Рис. 25. С1 a d о s е 1 а-
с h е F у 1 е г i Traq.
лучами,
(г a d i а-
подробно о строении ид будет
сказано позже.
2*
20
класс: рыбы — pisces
Сем. pieuracanthidae. Pleuracan thus (Xen acanthus) (пермь)
является наиболее интересным представителем палеозойных акул (рис. 26); он стоит
как бы между Cladoselache и Dipnoi и указывает на близкое отношение
этих двух групп, столь различных теперь, в более ранние геологические времена.
Длина его до 44 см. Через всю длину тела проходила хорда без следов окосте-
нения, вверху и внизу от которой располагались многочисленные объизвествлен-
ные отростки. Кожа была голая. Он сохранил цельным спинной плавник, перехо-
дящий в хвостовой и со слабым перерывом продолжающийся до анального.
Хвостовой плавник не раздвоен, закруглен. Основные и радиальные лучи,
поддерживающие спинной и хвостовой плавники, входят в кожу, но не связаны
с отростками влагалища хорды. Парные плавники достигли большого ра звития.
Передние прикреплялись плечевым поясом — в виде развилка. Основные лучи
хорошо сохранились; к ним прикрепляются с нижней стороны радиальные. На
верхней они слабо развиты. В задних плавниках основные пластинки приле-
Рис. 26. Р 1 е u г а с a n t h u s.
гают к хрящевому поясу, свободно лежащему в теле животного. Анальный
разделен на два маленьких плавника. Сохранились отпечатки пяти жаберных
щелей и хрящевого черепа с большими глазными впадинами, носовыми отвер-
стиями, нижней челюстью и длинной, тонкой спицей посредине заднего края
черепа, направляющейся назад. На черепе расположены неправильными ря-
дами кожные пластинки, сходные с пластинками Dipterus. Мелкие зубы
с тремя заостренными верхушками расположены в несколько рядов.
К этим палеозойным акулам стоит близко ныне живущая в Тихом океане
Chlamydoselache, открытая экспедицией „Челленджера“, сохраняющая
необъизвествленную хорду и зубы типа Pleuracanthus. Жабры прикрыты
складками кожи — приспособление к донной жизни и закапыванию в ил.
У плавающих акул существуют только жаберные щели.
Отряд 2
Настоящие акулы — Selachii
Сем. Hybodontidae. У большей части форм позвоночник расчленен.
Различные представители его существовали с нижнего карбона до мела.
Род Н у b о d u s (рис. 27). Удлиненное тело с двумя спинными плав-
никами, поддерживаемыми большими спицами. Парные небольшие. Хорда не
окостеневала. Верхние дуги длинные, они редуцированы в хвостовой области.
подкласс: хрящеперые — chondropterygii
21
Удлиненные зубы с несколькими добавочными конусами, число которых раз-
лично у различных видов. YHybodus р 1 i с a t i 1 i s из триаса три конуса
л два маленьких придатка. YHybodus retic-u 1 atus Ag. из нижнего
Рис. 27. Hybodus hauffianus Fraas.
лейаса средний из трех конусов очень сильно развит, а четвертый значительно
больше, чем у первого вида. У Hybodus polyprion из доггера пять боль-
ших конусов и два маленьких.
Интересна находка цельного Hybodus hauffianus Fraas в верхнем
лейасе (Гольг.маден — Вюртемберг), более метра длиною, с сохранившейся ко-
жей и отпечатками всех
ными дугами и нижней
челюстью. Сохранились
ребра, грудной пояс с
плавниками и желудок.
В последнем сохрани-
лись 250 рострумов ма-
леньких белемнитов,
длиною в 4 см. Этот
Hybodus хранится
в музее Штутгарта.
А с г о d u s из ни-
жнего лейаса сильно
отличается по зубам.
Это удлиненные кону-
сы, покрытые тонки-
ми складками эма-
ли, со слабо высту-
пающей вершинкой; зу-
бы одинаковые, рас-
положены в 6—7 ря-
хрящевых частей
черепа, с rostrum, с пятью жабер-
Рис. 28. Челюсть Cestracior Philippi Cuv.
(ныне живущей).
дов. Плавниковые шипы
существуют, как и у Hybodus.
Polyacrodus Jaeck., очень сходный по строению хрящевого скелета
с Hybodus, имеет зубы с более развитым дентином, чем у Acrodus.
В триасе и нижнем лейасе.
22
класс: рыбы — pisces
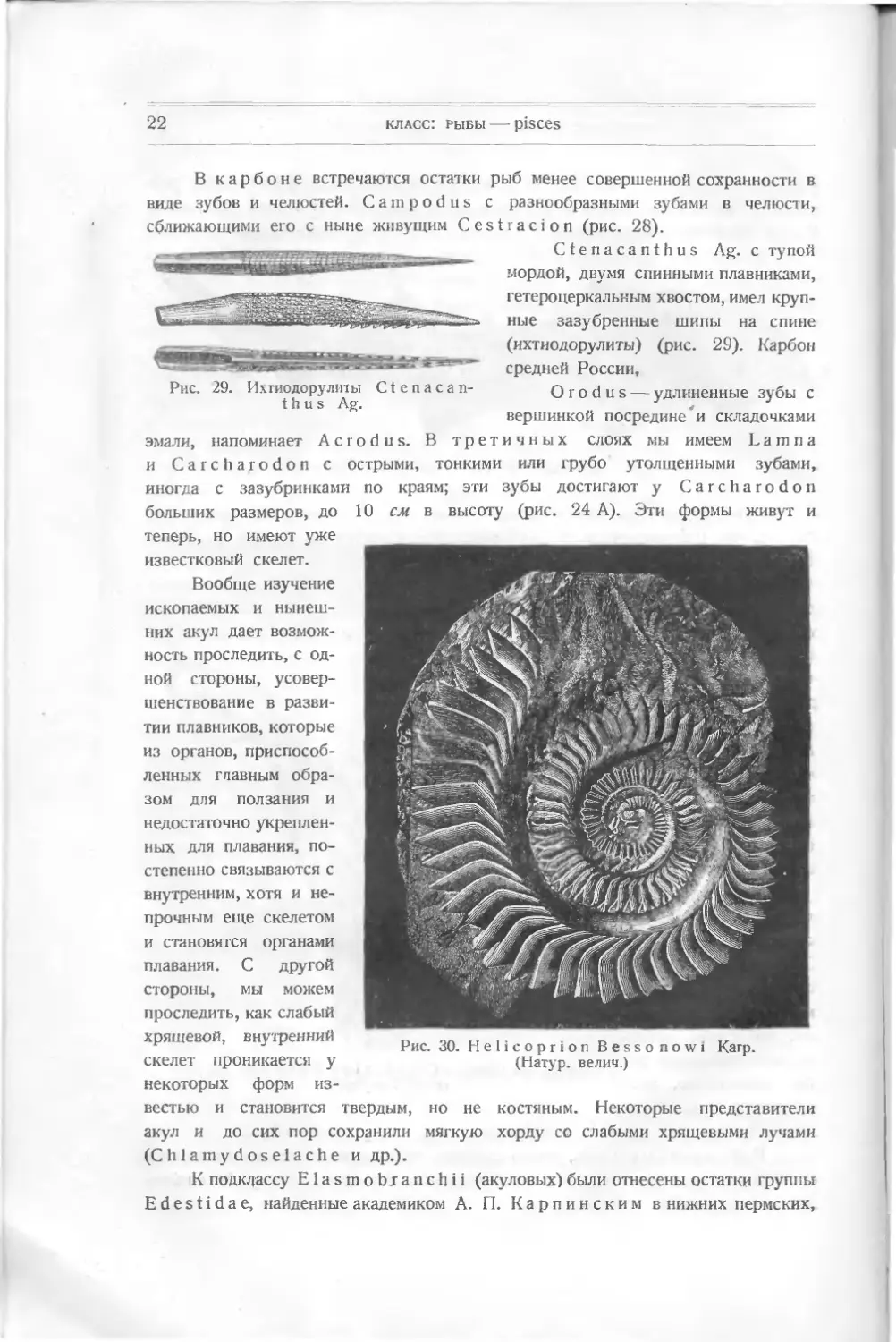
В карбоне встречаются остатки рыб менее совершенной сохранности в
виде зубов и челюстей. Campodus с разнообразными зубами в челюсти,
сближающими его с ныне живущим Cestracion (рис. 28).
Рис. 29. Ихтиодорулиты Ctenacan-
thus Ag.
Ctenacanthus Ag. с тупой
мордой, двумя спинными плавниками,
гетероцеркальным хвостом, имел круп-
ные зазубренные шипы на спине
(ихтиодорулиты) (рис. 29). Карбон
средней России,
О г о d н s — удлиненные зубы с
вершинкой посредине и складочками
эмали, напоминает Acrodus. В третичных слоях мы имеем L a m п а
и Carcharodon с острыми, тонкими или грубо утолщенными зубами,
иногда с зазубринками
больших размеров, до
по краям; эти зубы достигают у Carcharodon
10 см в высоту (рис. 24 А). Эти формы живут и
теперь, но имеют уже
известковый скелет.
Вообще изучение
ископаемых и нынеш-
них акул дает возмож-
ность проследить, с од-
ной стороны, усовер-
шенствование в разви-
тии плавников, которые
из ор*анов, приспособ-
ленных главным обра-
зом для ползания и
недостаточно укреплен-
ных для плавания, по-
степенно связываются с
внутренним, хотя и не-
прочным еще скелетом
и становятся органами
плавания. С другой
стороны, мы можем
проследить, как слабый
хрящевой, внутренний
скелет проникается у
некоторых форм из-
Рис. 30. Helicoprion Bessonowi Karp.
(Натур, велич.)
вестью и становится твердым, но не костяным. Некоторые представители
акул и до сих пор сохранили мягкую хорду со слабыми хрящевыми лучами
(Chlamydoselache и др.).
К подклассу Elasmobranchii (акуловых)были отнесены остатки группы
Edestidae, найденные академиком А. П. Карпинским в нижних пермских,
подкласс: хрящеперые — chondropterygii
23
артинских слоях. Эти остатки выражены у Helicoprion спиралью располо-
женными эмалевыми пластинками (зубами), уменьшающимися в размере внутрь
спирали (рис. 30).
По мнению акаде-
мика А. П. Карпин-
ского, это приросшие
зубы, которые при раз-
ворачивании спирали,
стираясь постепенно
снаружи ее, заменя-
Рис. 30A. Edestes crenulatus Hay.
лись другими; поэтому внутренние, более молодые —мелкие. Другие ученые тол-
куют иначе значение этих остатков, считая их за спинные придатки — органы за-
щиты, способные разворачиваться (Истман — Eastman).
Edestes (рис. 30 А) — другой род этой группы —
представлен органом, состоящим из удлиненных пласти-
нок с эмалевым зубом на каждой. Пластинки плотно
прилегали одна к другой — почти срастались. Их счи-
тают зубными пластинками.
cazo
Отряд 3
Скаты — Batoidei
пережитками
сравнительно
на морском
остатки их
Рис.31. Rhinobatis
m i r a b i 1 i s Wagn.
(Соленгофен, юра.)
Рис. 31 А. Р t у г h о-
d ifs
m a m i 11 а г i s
Ag-
Menaspis — найден
Скаты могут считаться наиболее изменившимися
древнего ствола акуловых. Они развились
поздно. Живут они
дне. Ископаемые
известны в виде
цельных отпечатков и зубов с
карбона, в мелу и в юре
(рис. 31). Главный представитель
в мелу Ptychodus — с плоски-
ми или выпуклыми эмалевыми
зубами, в виде пластинок с по-
перечными ребрышками (рис. 31 А).
Зубы были расположены на передней части челюсти.
в гиде отпечатка в верхне-пермских слоях Тюрингии. В соленгофенских
сланцах верхней юры найдены отпечатки цельных животных: S q u a t i n а,
Spatobatis, Rhinobatis.
Скелеты их мало отличаются от ныне живущих скатов.
В карбоне встречаются в большом разнообразии их зубы: Р s a m modus —
плоский; Petalodus — с закругленной коронкой, широким и длинным корнем;
Polyrhizodus—с бахромчатыми корнями (рис. 32) и большими ихтиодору-
литами (рис. 32 A). J a n е s s а — с мелкими мостовидными зубами и громадными
плавниками.
24
класс: рыбы—pisces
Отряд 4
Acanthodi Ag.
В девоне Англии найдены остатки представителей вымершей группы,
близкой к акуловым по строению скелета и плакоидных чешуй, но сильно
отличающихся по отсутствию зубов, веретенообразной форме тела и по строению
плавников. Они сохраняют хорду с зачаточными
перетяжками, намечающими многочисленные сегменты
(до 300) позвоночного столба. Хрящевой череп с
подвешенными челюстями и жаберными щелями без
кожных складок. В коже мелкие плакоидные пла-
стинки.
Сем. Acanthodidae — с парными плавниками,
снабженными спицами; число их и строение различны
у различных родов. Спинной плавник — один или два,
тоже со спицами.
С И m a t i u s (рис.
Рис. 32. Зуб Polyrhi-
zodus con cavus
Trautsch.
33) — два больших спинных
плавника, хвостовой —дифи-
церкальный и анальный. Пар-
ных несколько пар, причем
передняя и задняя пары наи-
большие, а между ними не-
сколько меньших. Такое
расположение указывает на
их образование из распав-
шихся на части боковых
Рис. 32 А. Ихтиодорулит
Polyrhizodus г os-
si с u s Inostr.
складок кожи, в которых развились местами роговые поддержки в виде спиц.
В роде Acanthodes мы наблюдаем при одном спинном плавнике не-
сколько очень различных типов парных плавников. У девонской формы А с a n t h.
Mitch elli и Ac. gracilis (пермь) существует только хвостовой, на месте
остальных сохранились только спицы.
У Acanthodes Wardi (рис. 34) — из верхнего карбона — имеются
плавники: спинной, анальный, хвостовой и две пары парных. Все они снабжены
впереди спицами. Верхняя часть хвостового плавника длиннее нижней, что делает
хвост неправильным — гетероцеркальным (признак девонских рыб). Эти плавники,
кроме хвостового, совершенно отличаются по своему строению от плавников
всех других рыб.
Подкласс IV
ХИМЕРЫ — HOLOCEPHALI
Ближайшие к акулам и скатам плакоидные рыбы. Они оставили мало сле-
дов своей жизни в древних слоях, которые и не могут дать указаний на их
развитие. Главными их отличиями от акул и скатов являются: жаберная
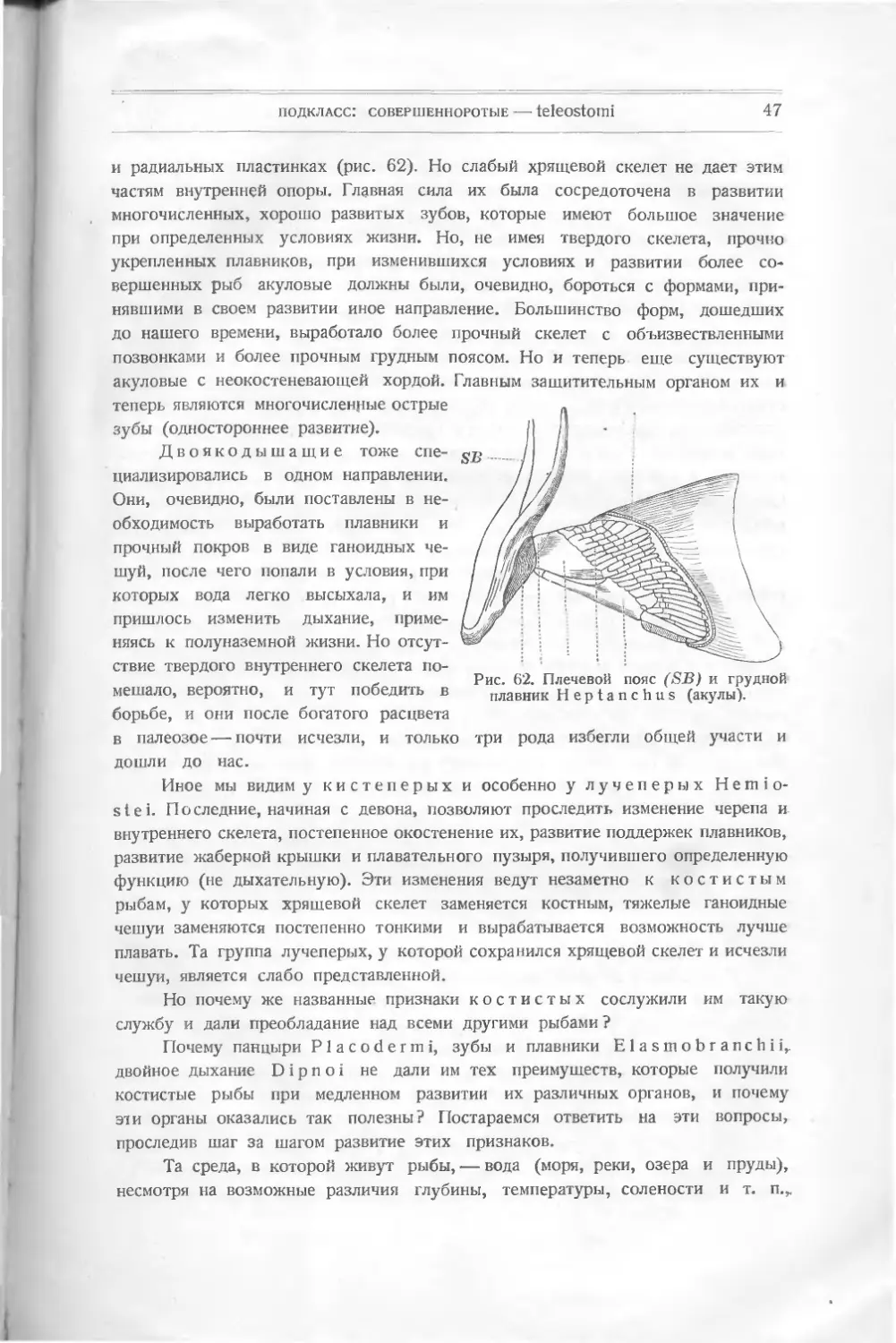
подкласс: arthrodira
25
крышка, зубы, приросшие к челюстям, и сочленение нижней челюсти не-
посредственно с черепом, без гиоидной дуги. Признаки эти указывают на зна-
чительное усовершенствование их организации.
Рис. 33. Изменение плавников у сем. Acanthodidae.
Подкласс V
ARTHRODIRA
Рядом с последним подклассом ставится еще группа девонских рыб, го-
лова и туловище которых покрыты панцырем, состоящим из больших щитов,
и напоминающих панцирных рыб. Но они резко отличаются от последних присут-
ствием хрящевого скелета с неокостеневшей хордой и с окостеневшими верх-
ними и нижними отростками. Существуют костные поддержки спинного и
брюшного плавников.
Некоторые палеонтологи сближают эту группу с Dipnoi, находя у не-
которых из них сходство в озублении, именно по 2—4 зуба в нижней челюсти.
26
класс: рыбы — pisces
Но существует большая разница как в расположении и образовании этих зубов,
так и в строении головных щигков и в присутствии грудно-брюшного панцыря.
Coccosteus decipiens Ag. может считаться главным представи-
телем этого подкласса, из нижнего девона Шотландии (рис, 34). Он
встречается также в девоне у нас, в Сев. Америке и в других странах1).
Рис. 33A. Acanthodes Wardi.
Голова, передняя
часть спины и грудь
были покрыты щитами,
состоящими из пласти-
нок, сходных с пла-
стинками щита A ste-
rol е р i s, и украшен-
ными звездочками.
Голова сочленя-
лась с двумя боковыми спинными пластинками, имевшими выдающиеся, закруглен-
ные головки, входившие в углубления головного щита и делавшие голову
подвижной. Нижняя челюсть сочленялась с черепом и несла по 2 боль-
ших прирастающих зуба. Такие же зубы существовали и на нёбе. В верхней
челюсти зубов не было.
Другой тип озубления представлен отдельными зубами — пластинками,
как у М у 1 о s t о ш а. Оба типа зубов указывают на возможность раздроблять
ими твердую пищу — раковины моллюсков и т. п.
Рис. 34. Coccosteus decipiens Ag. (Нижний девон.)
В грудном щите Смите Вудвард (Smith Woodward) и Некель
намечают у Coccosteus части грудного пояса: clavicula, interclavicula,
postclavicula. Покровы остальной части тела неизвестны. Хвост прямой, ди-
фицеркальный. Глаза окружены склеротинальным кольцом.
Dinichthys Newb., голова которого достигала 1 м, из верхнего де-
вона, имел на задней черепной пластинке выступ для сочленения с передним
щитком спинного панпыря.
9 О. Некель указывает две новых формы (Coccosteidca), найденные в де-
воне Вильдунгена с сохранившимся плавательным пузырем, внутреннее строение кото-
рого сходно с пузырем двоякодышащих рыб и отвечает функции легких (О. J а е с-
kel, Die Wirbeltiere 1911, S. 11).
подкласс: совершенноротые — teleostomi
27
Mylostoma — из девона Огио, имела четыре больших неправильно
треугольных передних зуба, которым соответствовали два зуба в нижней
челюсти.
Brachydirus — из верхнего девона Вильдунгена в Германии с остро-
конечной верхней челюстью, очень похож на Coccosteus. У Oxyos-
t е u s эта часть удлинена в виде рострума. Эта группа направляла, очевидно,
силы организма на выработку твердых внешних покровов, которые были ей
полезны при донном образе жизни, но при изменении условий оказались
неспособными защитить ее. Остатки их не известны позже девона, они най-
дены отчасти в морских, отчасти в пресноводных отложениях.
Подкласс VI
СОВЕРШЕННОРОТЫЕ — TELEOSTOMI
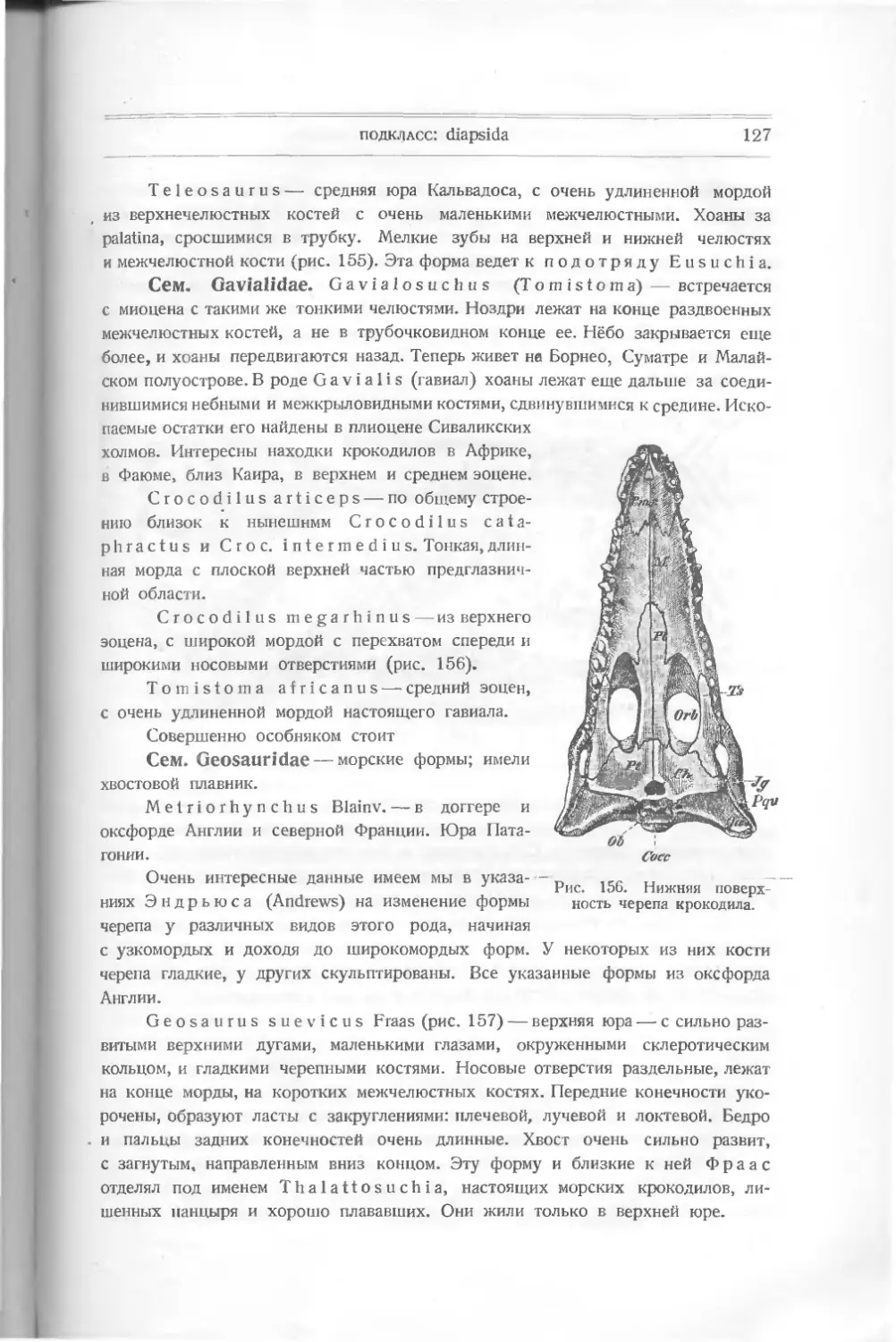
Переходя к знакомству с подклассом Teleostomi, мы встречаем новый
признак в покровах их — чешуи, которые и будут существовать почти у всех
остальных представителей класса, но будут иметь различный вид и строение.
Вот почему мы остановимся здесь, и раньше чем начнем изучать представите-
лей этого подкласса, мы познакомимся с различными видами кожных образо-
ваний у различных подклассов рыб.
К таким образованиям относятся, как мы уже видели, маленькие конусы
yLanarkia и Thelodus, пластинки и шипы различной формы у других
панцырных и у пластиножаберных. Мы видели, как отдельные маленькие конусы
заменялись пластинками, сначала изолированными, потом слившимися в щитки.
Все подобные образования возникают вокруг сосочков кожи, покрытых эпидер-
мисом, в слое которого отлагается в виде конусов или пластинок дентин,
покрытый эмалью и внутри прорезанный канальцами, несущими питание через
пульпу (см. рис. 7, С). Эти образования носят название плакоидных че-
шуй. Подними, в самой коже, отлагается базальная, или основная пла-
стинка, состоящая из фосфорнокислой извести — дентина. Подобное строение
чешуй соответствует строению зубов. У следующих групп рыб мы встречаем
настоящие чешуи, которые бывают толстые — ганоидные и тонкие — лептоидные.
Ганоидные состоят из костной основной пластинки, покрытой
блестящим слоем ганоина, похожего на эмаль, но не признанного всеми
учеными за таковую; некоторые видят в нем отложение, отвечающее дентину
плакоидных пластинок. Эта чешуя имеет иногда ромбическую форму
с загнутыми передними краями, входящими в желобки внутреннего края сосед-
ней чешуи; сочленяясь таким образом, чешуи остаются подвижными (Р о 1 у-
pterus). Другие ганоидные чешуйки могут быть закруглены; тогда они
налегают одна на другую, прикрывая задней половиной переднюю часть бли-
жайшей (И о 1 о р t у с h i u s). Чешуя обыкновенно покрывает все тело животного,
кроме головы, располагаясь косыми рядами. Ганоидная чешуя встречается глав-
ным образом у более древних чешуйчатых рыб.
28
класс: рыбы — pisces
Костистые рыбы, развившиеся позже, имеют тонкие лептоидные
чешуи, состоящие из основной, соединительной ткани, покрытой
блестящим слоем фосфорнокислой извести; эмаль отсутствует. Они
тонкие, упругие, блестящие, увеличиваются, нарастая с переднего края кон-
центрически; развиваются в особых кожистых мешечках и налегают друг на
друга черепицеобразно.. Различают чешуи закругленные, с ровными краями, и
другие — с мелкими зубчиками на заднем краю (неправильно называемые цикло-
идными и ктеноидными).
Большие пластины и щиты панцырных рыб состоят, как было указано,
из дентина и эмали или имеют костные тельца (головной щит Cephala-
spis). У осетровых большие спинные пластинки близки к ганоидным чешуям,
в которые они иногда переходят на хвосте.
Большие головные спицы — ихт и о д о р у л и т ы и плавниковые шипы
развиваются в зависимости от внутреннего хрящевого скелета; состоят они
из дентина, имеют пульпу и внедряются нижними концами в тело животного,
где и прикрепляются хрящевой пластинкой (головная спица Pleuracan-
t h u s). Плавниковые спицы имеют различное строение — роговое или костное.
В некоторых роговых шипах отлагается известь.
Зубы рыб в большинстве случаев состоят: коронка — из дентина, вазоден-
тина и эмали; корень — из вазо- или остеодентина. Они гомологичны плакоид-
ным чешуям. Головные кожные пластинки превращаются в настоящие черепные
кости у костистых рыб.
В подкласс совершенноротых — Teleostomi входят очень раз-
личные отряды рыб, объединенные общим признаком: окостеневшими, несущими
зубы челюстями, которые сочленяются с черепом посредством сочленовных
отростков. Скелет проходит различные стадии окостенения. Черепные кожные
кости принимают правильное расположение и входят в состав черепа. Жабер-
ная крышка окостеневает. Грудной пояс примыкает к заднему краю черепа.
Покрыты чешуями ганоидными или лептоидными. Они подразделяются на
Dipnoi, Crossopterygii и Actinopterygii1).
Представители их хорошо известны, начиная с девона. Все они имеют и
живых представителей. Вымерли только отдельные семейства.
Отряд 1
Двоякодышащие — Dipnoi
Эти рыбы, как показывает название, имеют двойное дыхание: кроме жабер
для водного образа жизни, они имеют воздушное дыхание, именно измененный
Ч Абель находит неправильным отнесение костистых к Actinopterygii,
указывая, что первые происходят из различных корней.
Я предлагаю данное подразделение Teleostomi, так как оно характеризуется
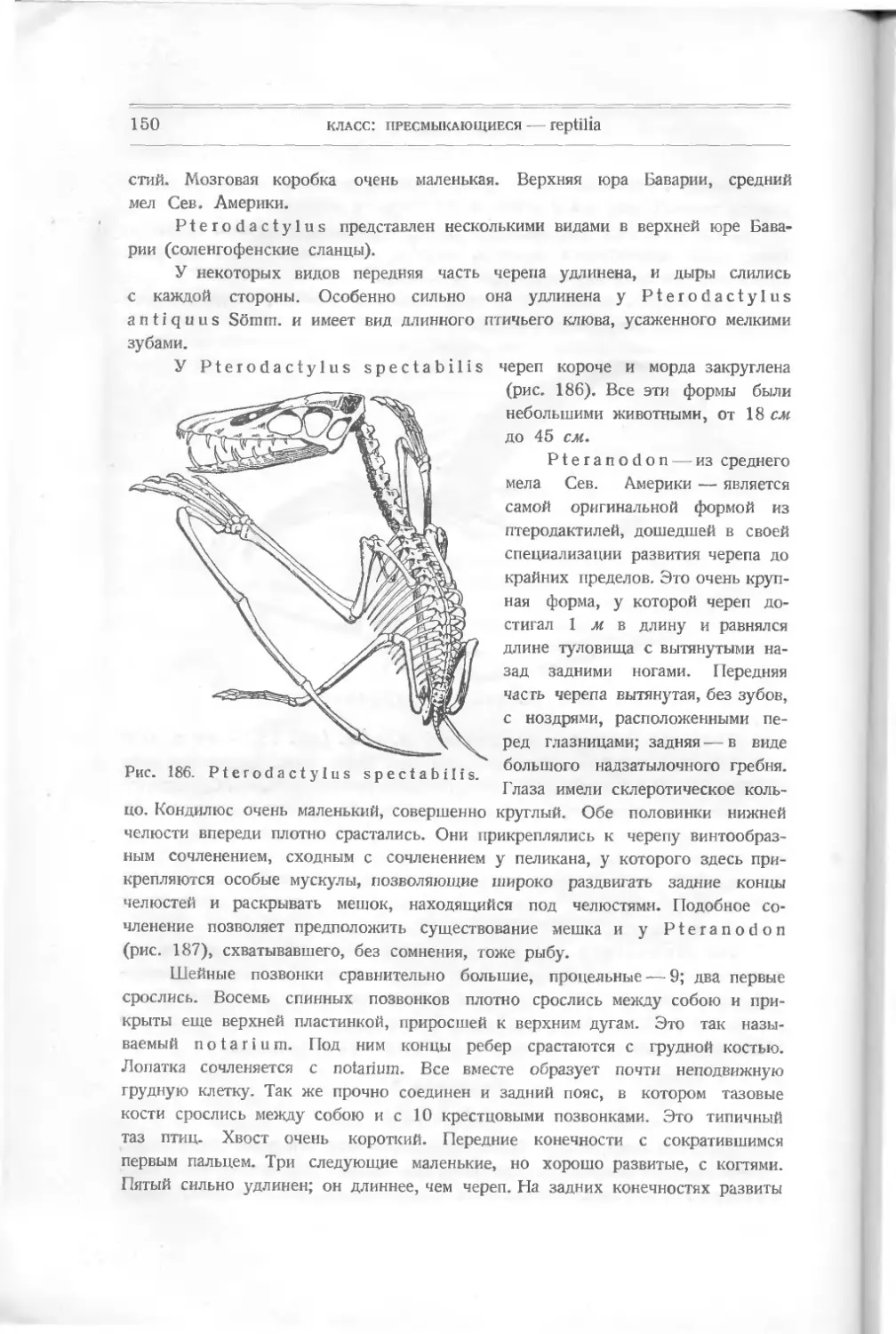
вполне определенными признаками.
Термин G а п о i d е i для обозначения части Actinopterygii, имеющих
ганоидные чешуи, считаю неудобным. Тогда нужно относить к ганоидным (по чешуе) и
Dipnoi и Crossopterygii.
подкласс: совершенноротые -— teleostomi
29
воздушный пузырь — прообраз легких. Характеризуются они хрящевым чере-
пом, покрытым кожными пластинками разных размеров у разных представите-
лей. Нёбно-квадратный хрящ сросся с черепом (верхняя челюсть). Сочленение
нижней челюсти типа autostylia.
Особенностью Dipnoi являются их зубы (рис. 35). В верхней и нижней
челюсти у большинства нет мелких конических зубов, а существуют две зуб-
ные пластинки: на нёбе и на внутренней стороне нижней челюсти, с гребнями
Рис. 35. Dipterus Valenciennesi
Ag- Нёбная часть с зубами. (Девон, Шот-
ландия.)
или бугорками. Это признак древних Dipnoi
(D i р t е г u s) и ныне живущего N е о с е г а-
todus. У Phaneropleuron и Sсайте-
Рис. 35А. Зуб Dipterus platyce*
р h а 1 u s Ag. (Девон, p. Сясь.)
nacia внешний ряд бугорков на пластинках функционирует как краевые
зубы. yUronemus на нёбе нет зубных пластинок, а только ряд отдель-
ных бугорков на краю его, сливающихся вместе у основания. Это как бы оста-
ток зубной пластинки, У ныне живущих Protopterus и Lepidosiren
Рис. З'т. Phaneropleuron Andersoni Huxley. (Девон.) (J/c)
пластинок совсем нет. Это различие зубной системы стоит а связи с образом
жизни животных — травоядным или хищным: острые зубы — у вторых, пло-
ские— у первых. Они изменялись при переходе от хищного к травоядному
образу жизни.
Жабры у Dipnoi открываются наружу одной щелью. Хорда не окосте-
невает. Ныне живущие формы представлены только тремя родами: Neocera-
todus, Lepidosiren и Protopterus. Главное отличие их от древних
форм заключается в присутствии мелких пластинок, покрывающих череп, и в со-
кращении парных плавников. Развитие воздушного дыхания могло быть вызвано
высыханием тех водных бассейнов, в которых жили их предки, и явилось
средством борьбы за жизнь. Это двойное дыхание сближает Dipnoi с пер-
выми наземными животными — земноводными или амфибиями, и они считались
раньше как бы переходными формами между этими последними и рыбами Но
изучение ископаемых остатков их показало, что, несмотря на сходство этого
и еще некоторых других признаков с признаками амфибий, Dipnoi не могли
30
класс: рыбы — pisces
быть их предками, так как в их скелете произошли изменения, не соответствующие
пути развития амфибий, — раньше, чем этот класс отчленился. Так, распо-
ложение костей черепной крышки у них не соответствует строению черепа
пермских амфибий. Зубы в виде сложных пластинок у Dipnoi не могли
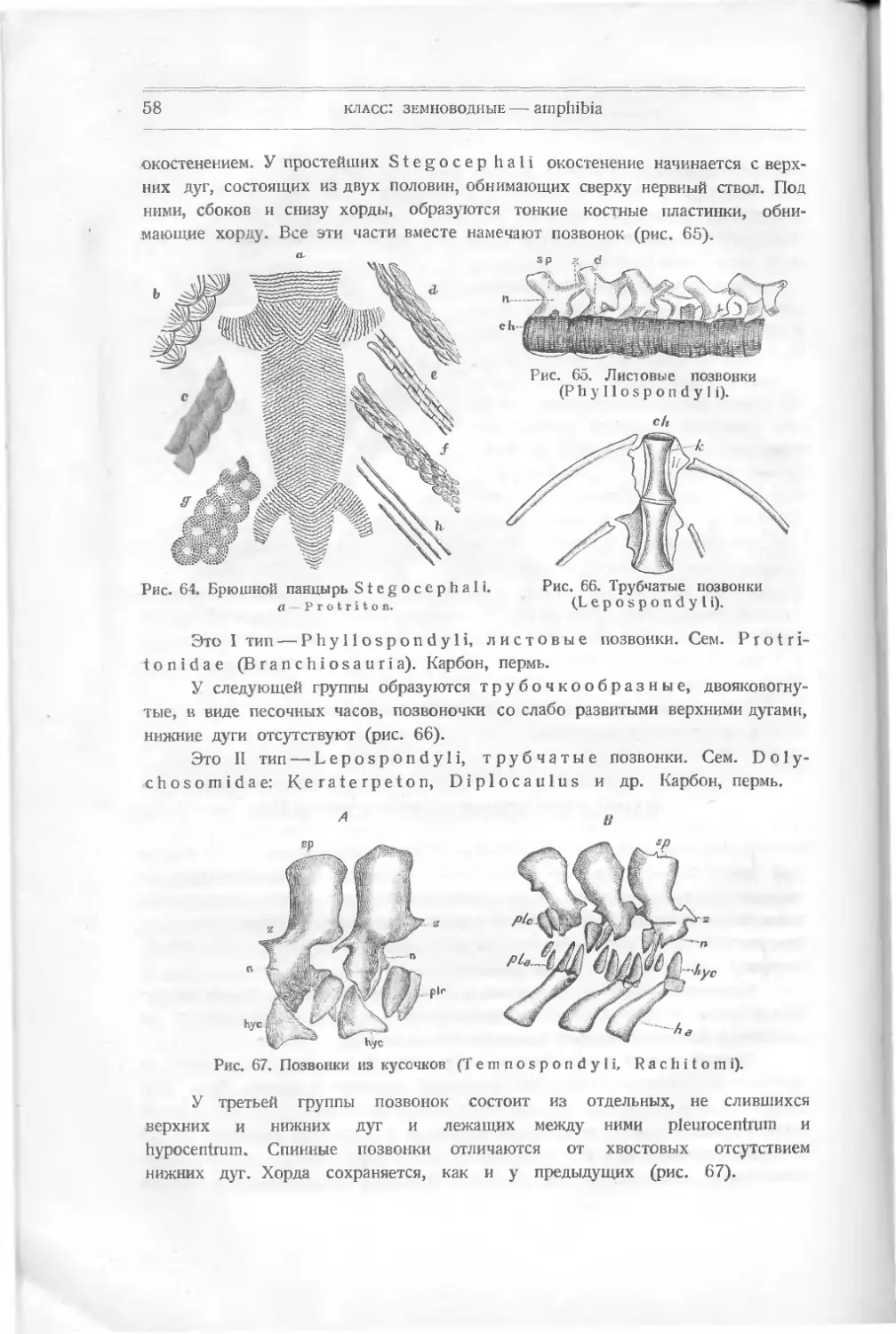
положить начало зубам щитоголовых амфибий — Stegocephali. Но мы
не имеем в палеонтологии никаких указаний на время появления у Dipnoi
воздушного дыхания
Ископаемые остатки Dipnoi встречаются главным образом в девоне,
который и является временем максимального их развития и специализации,
adf.
Рис. 37. Dipterus Valenciennesi Ag. (Девон.)
а нынешние формы являются только малочисленными
мимся с некоторыми представителями Dipnoi.
их остатками. Познако-
Рис. 37 A. Dipterus platycepha- Рис. 37 В. Dipterus Valencienne-
1 u s Ag. Черепная крышка. s * Ag.
Род Dipterus (рис. 37) представлен несколькими видами в нижнем
девоне Шотландии. Тело покрыто ганоидной чешуей с закругленными
кпаями. Хвост несимметричный. Парные плавники превращены в ласты, покры-
тые чешуей и отороченные плавниковыми лучами. Скелет их состоит из удли-
ненных basalia и radialia, несущих эти лучи. Двойной спинной плавник
с кожными поддержками лежит над анальным. Зубы не существуют на краю
челюстей; на нёбе две пластинки треугольной формы с зазубринками и две
пластинки на нижней челюсти, которая окостенела. Череп покрыт несколькими
рядами неправильно расположенных кожных пластинок (рис. 37 А, 37 В).
подкласс: совершенноротые — teleostomi
31
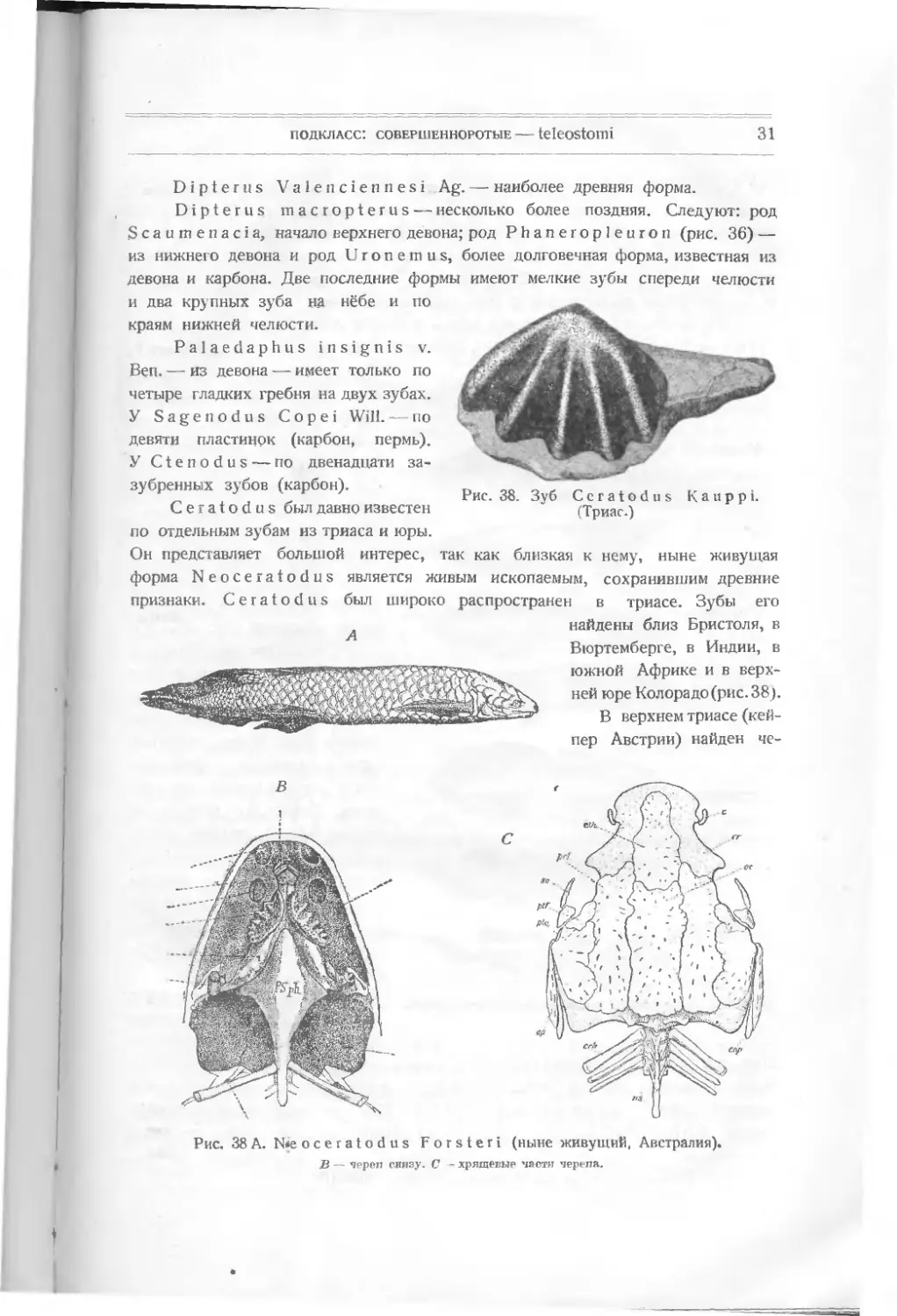
Dipterus Valenciennesi Ag. — наиболее древняя форма.
Dipterus macropterus — несколько более поздняя. Следуют: род
Scaumenacia, начало верхнего девона; род Phaneropleuron (рис. 36) —
из нижнего девона и род Uronemus, более долговечная форма, известная из
девона и карбона. Две последние формы имеют мелкие зубы спереди челюсти
и два крупных зуба на нёбе и по
краям нижней челюсти.
Palaedaphus insignis v.
Ben. — из девона — имеет только по
четыре гладких гребня на двух зубах.
У Sagenodus Copei Will. — по
девяти пластинок (карбон, пермь).
У Ctenodus — по двенадцати за-
зубренных зубов (карбон).
Ceratodus был давно известен
по отдельным зубам из триаса и юры.
Рис. 38. Зуб Ceratodus Kauppi.
(Триас.)
Он поедставляет большой интерес, так как близкая к нему, ныне живущая
форма Neoceratodus является живым ископаемым, сохранившим древние
признаки. Ceratodus был широко распространен в триасе. Зубы его
А найдены близ Бристоля, в
Вюртемберге, в Индии, в
южной Африке и в верх-
ней юре Колорадо (рис. 38).
В верхнем триасе (кей-
пер Австрии) найден че-
Рис. 38 А. №е oceratodus Forsteri (ныне живущий, Австралия),
В — чрреп снизу. С - хрящевые части черепа.
32
класс: рыбы — pisces
реп Ceratodus Sturi Tell., плохой сохранности, с таким же расположением
головных кожных пластинок, как у ныне живущей формы, но с большим
окостенением их. В раковинном известняке Баварии найдена хвостовая часть
Ceratodus, в которой роговые лучи хвостового плавника более прочно
построены, чем у нынешнего. Это указывает, что последний является дегенери-
ровавшим типом сравнительно с триасовым предком его.
Neoceratodus существует теперь только в Австралии. Это — N е о с е-
ratodus Forster! Giint. (рис. 38А). Он покрыт тоже круглой ганоидной чешуей.
Имеет спинно-хвостовой плавник
и совершенно не окостеневшую
хорду. Два большие зуба его
состоят из пяти гребней и ле-
жат позади маленьких зубов.
Они сходны с триасовыми зубами
Ceratodus Sturi Tell. Парные
плавники покрыты чешуей и име-
ют только ряд базальных лучей.
Череп покрыт двумя средними
пластинками и двумя рядами бо-
ковых пластинок (рис. 38 А, С).
Два другие ныне живущие
рода — Protopterus в тро-
пической Африке и L е р i d о-
siren в болотах области Ама-
зонки — еще более сократили
парные плавники, превратив-
шиеся у них в нитевидные при-
датки. Скелет их остался на
той же низкой ступени разви-
тия, как и у палеозойных
форм, с хрящевой, не окосте-
невшей хордой. У Lерido-
si г е п есть два маленьких
конических зуба в межчелюст-
ной кости.
Интересные данные нахо-
дим мы у Д о л л о г), изучав-
шего изменения плавников
Dipnoi, начиная с Dipterus Valenciennesi Ag. из нижней части
нижнего девона, проходя через Dipterus macropterus верхней части
нижнего девона, Scaumenacia curta Whit, нижней части верхнего
девона, Phaneropleuron Andersoni Huxl. верхней части верхнего
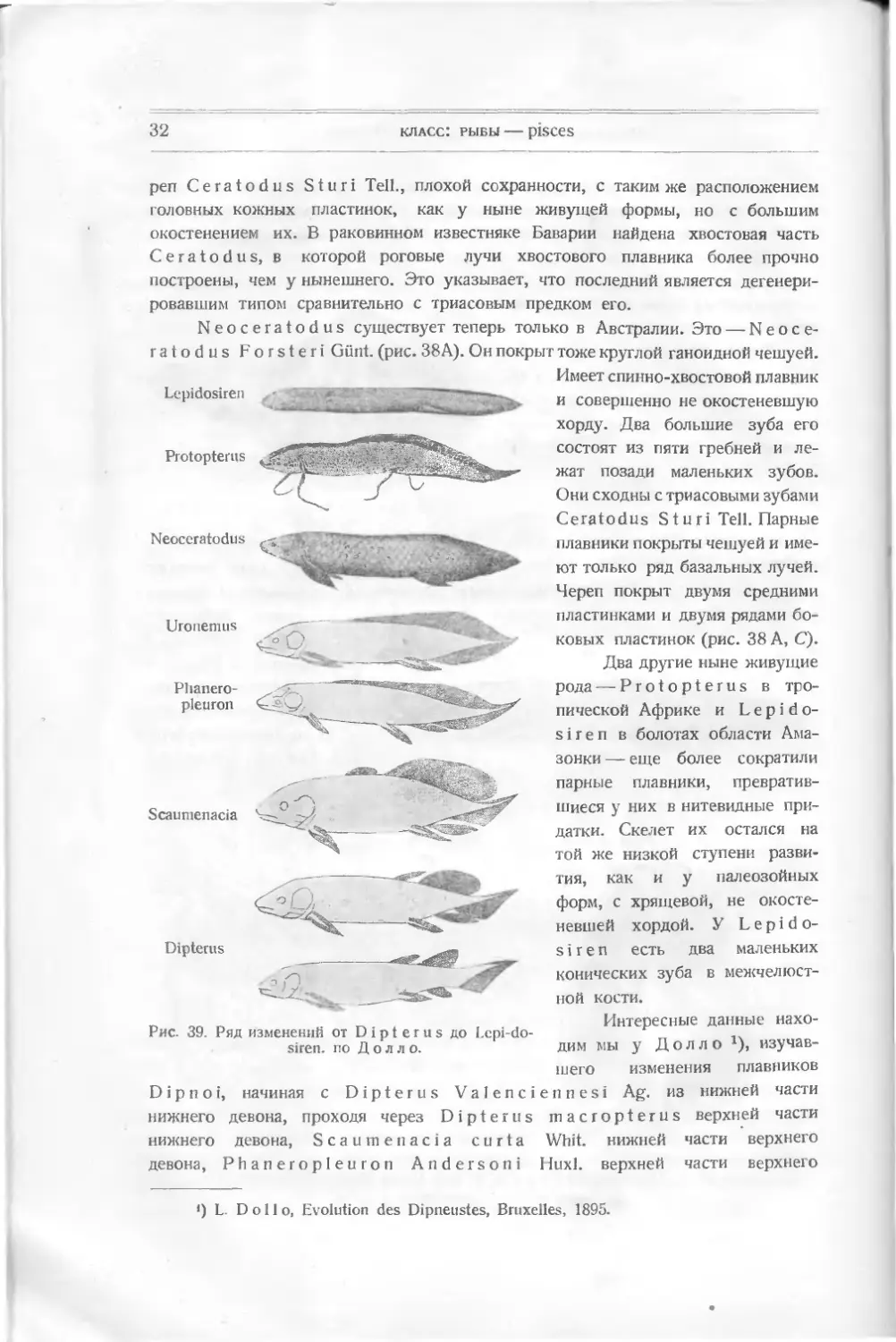
Рис. 39. Ряд изменений от Dipterus до Lepi-do-
siren. по Д о л л о.
<) L. D о 11 о, Evolution des Dipneustes, Bruxelles, 1895.
подкласс: совершенноротые — teleostomi
33
девона, Uronemus lobatus Ag. из нижнего карбона и заканчивая ныне
живущими: Neoceratodus Forsteri Glint., Protopterus annectens
<>w. и Lepidosiren paradoxa Fitz.
Мы видим (рис. 39), что древнейшая из этих рыб имеет форму настоя-
щей рыбы с гетероцеркальным хвостом, двумя спинными плавниками, неболь-
шим анальным и двумя парами парных плавников, имеющих вид ластов. Жа-
берная крышка широкая, покрыта ганоином; тело покрыто закругленными
ганоидными чешуйками с эмалью. У следующей формы, Dipterus m а с-
ropterus, хвост острее, задний спинной плавник больше. У S с а и me-
па с i а он еще более увеличивается, продвигаясь вперед. У следующих форм
он цельный и переходит в спинной. Тело рыбы становится веретенообразным
с дифицеркальным хвостом. Анальный плавник исчезает (сливается с хвосто-
вым). Исчезает также эмаль на жаберной крышке и на чешуе. Ласты утолщи-
лись у Uronemus и у Neoceratodus; у последнего сократилась жаберная
крышка, чешуя стала тоньше. У Protopterus чешуя стала мелкой, ласты уто-
нились. Наконец, у Lepidosiren они почти редуцировались; непарный плав-
ник уменьшился. На месте жаберной крышки — щель. Этот ряд наводит на
различные вопросы о прогрессивном или регрессивном изменении признаков,
так как морфологическое изменение их совпадает с геологическим положением
форм (кроме последних). Между тем гетероцеркальный хвост—признак более
совершенный, чем дифицеркальный, встречается здесь у более древних форм.
Также развитие жаберной крышки и разделенный на части непарный плав-
ник— все это говорит за регрессивное изменение форм. С этим согласен и
Дол л о, хотя это противоречит его теории „о невозвращении утерянных при-
знаков1'.
Он считает, что это изменение приобретенных признаков девонских D i-
pnoi было вызвано изменением условий жизни при переходе животных из чи-
стой воды в мутную (первое появление воздушного дыхания и уменьшение
жаберной крышки) и далее вынужденным пребыванием вне воды, в сырой
илистой местности, в какой и теперь они живут. Одновременно с этим умень-
шалась потребность быстрого движения, плавники стали сливаться. Очевидно,
мы имеем здесь регрессивное развитие, вызванное изменением
жизненных условий. Долл о, допуская это, видит в сплошных плавни-
ках позднейших форм другие элементы, чем у первичных форм со сплошным
плавником.
Protopterus и Lepidosiren развивались, вероятно, параллельно,
не проходя стадии один другого. Изучение эмбрионального развития их должно
дать ответ на этот вопрос.
Гёке л и (Huxley) указал в 1876 г. на родство Dipnoi с древнейшими
кистеперыми рыбами. Абель считает это родство очень близким. Он говорит,
что Dipnoi представляют собою непосредственно ответвившуюся от древних
совершенноротых ветвь (от Crossopterygii), развившуюся односторонне
благодаря жизни в илистых бассейнах, которые высыхали в жаркое время
года. Их плавательный пузырь превратился в легкие.
3 Павлова М. В. Палеозоология. Ч. II.
34
класс: рыбы — pisces
Как по покрову (ганоидные чешуи), так и по строению внутреннего
скелета и парных плавников и по числу спинных плавников этот древний
отряд сходен с отрядом Crossopterygii, к знакомству с которыми мы
и перейдем.
Отряд 2
Кистеперые — Crossopterygii
Представлен древними формами; они известны с девона и имеют только
два рода между ныне живущими рыбами: Polypterus (многопер) и С а 1 а-
moichthys в Ниле и бассейне тропической Африки, Сенегамбии. Представи-
тели этого отряда имеют два спинных плавника, кроме
Polypterus, у которого их целый ряд, и соединяют
Рис. 41. Зуб Rhizo-
dus Hibberti Ag.
Рис. 40. Osteolepis macrolepidotus Ag.
(Средний девон Шотландии.) (*/з)
в себе некоторые признаки акуловых, Dipnoi и лучеперых. Они удерживают
еще хрящевыми основные и радиальные части в закругленных или удлинен-
ных плавниках. Покрыты ганоидными чешуями, ромбическими или округлыми,
с эмалевым слоем. Ганоин только у Polypteridae.
Сем. Osteolepidae. Osteolepis macrolepidotus Ag. — средний
девон Шотландии (рис. 40). Удлиненная форма с дифицеркальным, направлен-
ным вверх хвостом, с короткими закругленными парными плавниками, ромбиче-
скими чешуйками. Кости черепной крышки, лежащие перед теменными (лобные),
сливаются в одну. На них существует пинеальное отверстие. Окостеневание
хорды начинается с хвостовой части, намечаются кольцевые позвонки.
Зубы мелкие, многочисленные, что сразу отличает Osteolepidae от Di-
pnoi, с которыми они имеют внешнее сходство.
Diplopterus Agassizii сходен с Osteolepis по форме тела и
расположению плавников, кроме хвостового, который не поднимается вверх, а
окружает прямой хвост. Тоже — девон Шотландии.
Сем. Rhizodontidae. Rhizodopsis sauroides Traq. и Rhizodus
Hibberti Ag. — из нижнего карбона, больших размеров, чем Osteolepis.
Покрыт закругленной чешуей. Грудные плавники удлинены. Головные кости
расположены симметрично, лобные не срослись. Гладкие, закругленные зубы.
Окостенение хорды, как у Osteolepis: кольцевые окостеневшие позвонки.
Особенностями рода Rhizodus являются плотное срастание межключич-
ной кости с ключицами и сжатые с боков зубы, дающие два острых края, а
у основания их входящие складки эмали (рис. 41).
подкласс: совершенноротые — teleostomi
35
Крайне интересным признаком является у некоторых Rhizodontidae,
например у Gyroptychius (рис. 41 А) и у Eusthenopteron Foordi,
окостенение базальных лучей во втором спинном и в анальном плавниках. Они
имеют по три основных луча, подходящих под широкий конец луча 2-го ряда,
который прилегает к
одному из спинных
отростков. Окостенев-
шие кольцевые по-
звонки находятся в
брюшной области (аб- рис. 41 A. Gyroptychius Traq.
доминальной).
Eusthenopteron имеет хвост не ровно закругляющийся, как у Gy-
roptychius, ас тремя лопастями. Окостеневающие базальные лучи спин-
ного и анального плавников могут быть приняты за начало заднего пояса.
Сем. Holoptychidae — с удлиненными передними плавниками. Н о 1 о-
ptychius Flemingi Ag. (рис. 42), представитель этого семейства, отли-
чается более толстым телом, тонким хвостом, очень удлиненными груд-
ными плавниками в виде ластов, как у Dipterus, с тремя рядами чешуй и
таким же, как у последнего, расположением и других плавников.
Рис. 42. Н о 1 о р t у с h i u s Flemingi Ag. (Девон.) (<|8)
A — чешуя; В — разрез зуба.
Чешуя очень крупная, закругленная, перекрывающаяся, с толстым слоем
ганоина. Голова вся покрыта кожными костями с бугорками ганоина. Лобные и
теменные кости раздельны, и нет пинеального отверстия ни на одной из них.
Крупные конусообразные зубы имеют много входящих складок эмали, что де-
лает их строение сложным. Раньше они были описаны под именем D е n d г о-
dus из девона Лифляндии. Мелкие зубы без складок. Окостенения хорды не
наблюдалось. Holoptychius является очень распространенной формой
в девоне Шотландии, Ирландии, Эстонии, в Канаде, в Соединенных Штатах,
в Бельгии и Богемии. Встречается и у нас. Некоторые виды достигали
больших размеров.
Glyptolepis macrolepidotus, из среднего девона Шотландии,
отличается от Holoptychius широкими, удлиненными передними плавни-
ками (с пятью рядами чешуй) и перемещенными близко к ним задними плав-
никами, тоже широкими.
Изучение остатков этого отряда представляет большой интерес в смысле
различной степени окостенения хорды, расположения черепных костей, формы
3*
36
класс: рыбы — pisces
плавников и строения зубов у различных родов. Ныне живущий Р о 1 у-
рterns, в Ниле и в Гамбии, сохраняет ромбические ганоидные чешуи.
Парные плавники его очень укорочены. На спине расположен целый ряд
маленьких плавников, как бы распавшихся из одного большого плавника.
Рис. 43. Undina penicillata Mstr.
(Верхняя юра.)
Внутренний скелет окосте-
нел, зубы с простой пульпой,
без складок эмали.
Особенностью Р о 1 у р t е-
r u s является отверстие
раздвоенного плава-
тельного пузыря, откры-
вающееся в глотку с брюшной
стороны, как у форм, дыша-
щих во щухом. У других рыб отверстие пузыря лежит со стороны пищевода.
Ископаемые остатки редки; встречаются в верхних третичных слоях.
К отряду Crossopterygii относят странную группу рыб, существо-
вавшую с перми до мела с почти неизмененными признаками древних
форм, не окостеневающей хордой.
Род Undina, представитель этой группы из верхней юры (рис. 43),
с плотно слившимися костями черепа, закругленным хвостом и большими лучами
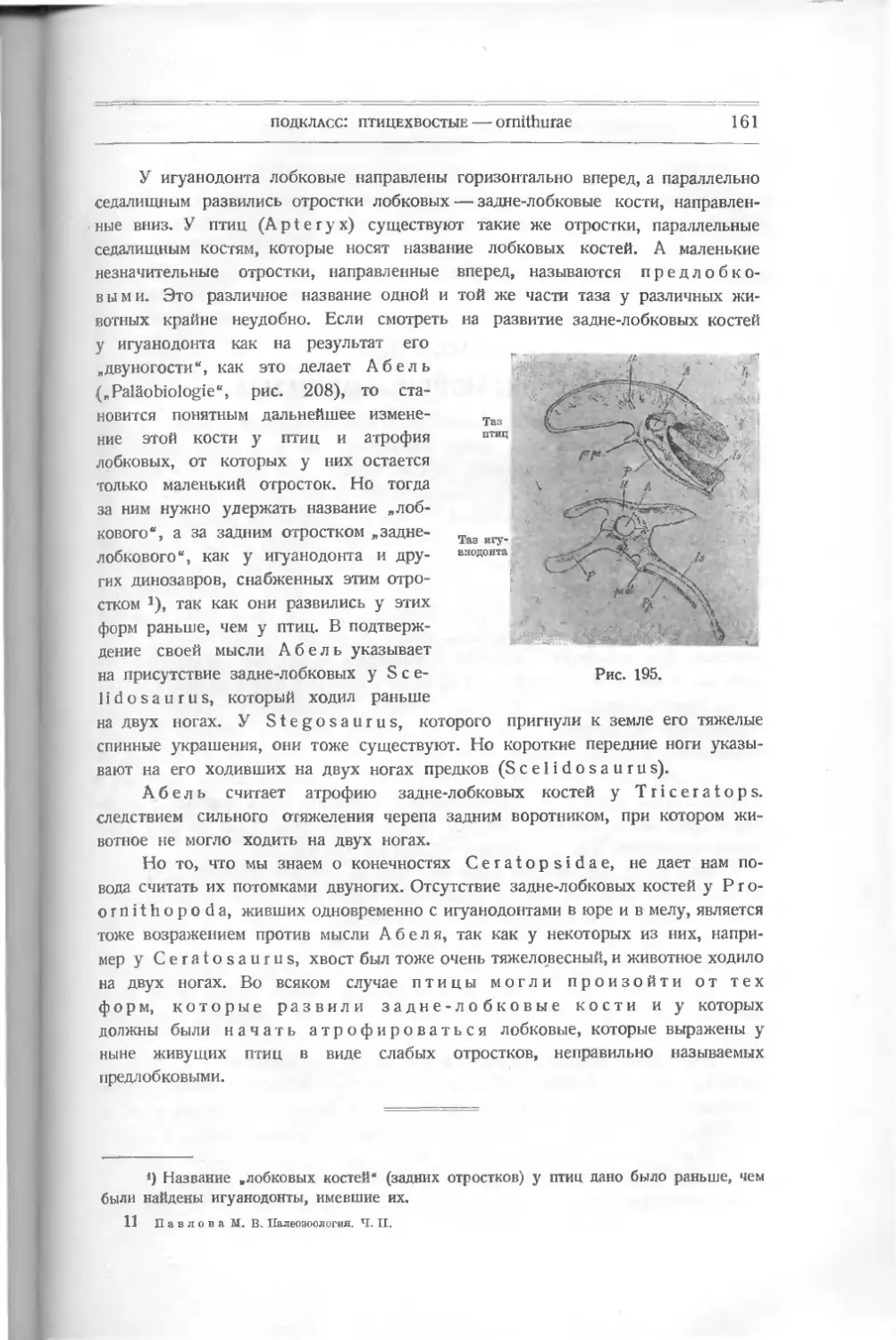
хвостового плавника, доходящими в теле до отростков не окостеневшей хорды.
Спинные, анальный и парные плавники не доходят до отростков хорды; осно-
вания их лежат в теле. У Undina сохранился окостеневший плавательный
пузырь Очевидно, это дегенерировавшая, рано отклонившаяся от первоначаль-
ного типа группа, вымершая в мелу.
Отряд 3
Лучеперые — Actinopterygii
Разделяются на Chondrostei, Hemiostei и Teleostei.
Первые и вторые (ганоидные других авторов) связаны с предыдущей
группой общностью покровов, имея ганоидные чешуи; но скелет Hemiostei
из хрящевого постепенно становится окостеневающим: образуются позвонки,
хотя все-таки сохраняется часть хорды. Парные плавники хорошо развиты.
Часто на переднем краю непарных, иногда и парных плавников развиваются
роговые шипы — ф у л ь к р ы. Сложный грудной пояс укрепляет передние
плавники. Хвост очень различной формы. Спинной плавник один. Teleostei
имеют окостеневший скелет. Остатки Actinopterygii известны с девона.
Абель подчеркивает существенное, по его мнению, отличие Actinopterygii
от Crossopterygii в расположении чешуй на хвосте. Он указывает на
образование хвостового плавника у Teleostei из хвостового и второго
анального плавников. Последний термин (analis secunda) не употреблялся
раньше при описании рыбьих плавников. Абель подтверждает это
рисунком хвоста молодого Lepidosteus. у которого хвостовой плавник
подкласс: совершенноротые — teleostomi
37
еще не слит с лежащим под ним вторым анальным, и другими рисунками,
на которых видно слияние этих плавников в один. Далее указывает
на то, что у Actinopterygii полоса, проходящая через чешую вдоль
тела рыбы, доходит до начала хвоста, на котором расположение чешуи
резко изменяется сравнительно с расположением на остальном теле, чего
не бывает ни у Dipnoi, ни у Crossopterygii. Этот признак (косое распо-
ложение чешуй хвоста) резко подчеркнут у Palaeoniscus macropomus
Ag. — пермь, Cheirolepis Traillii Traq.— девон, Amblypterus latus
Ag. — нижняя пермь, Coccolepis macropterus Traq. — вельд Бельгии
(Берниссар), Eurynotus crenatus Ag. — нижний карбон, Platysomus
striatus Ag. — пермь, Cheirodus granulosus Young, Dorypterus
Hofmanni Germ. — пермь. Абель пытается объяснить это изменение в рас-
положении чешуй на хвосте тем, что эти рыбы двигались не всем телом
змееобразно, а главным образом при содействии хвоста, вследствие большой
тяжести тела (особенно ганоидной чешуи). Но ведь их чешуя во всяком
случае была не легче чешуи Holoptychius.
Подотряд
Хрящевые — Chondrostei
Внутренний скелет и череп хрящевые. Голова покрыта костяным щитом.
Ганоидные чешуи на верхней части хвоста.
Сем. Chondrosteidae Sm. Woodw. Chondrosteus acipenseroi-
des — из нижнею лейаса. Удлиненное тело с почти правильным хвостом,
плавниками типа осетровых, с удлиненным черепом и ртом на его нижней
поверхности. Зубов нет; тело голое. Фулькры на верхней лопасти хвоста.
Внутренний скелет не вполне окостенел, но плавниковые поддержки доходят
до отростков хорды.
Рис. 44. А с i р е и s е г. Осетр.
Сем. Acipenseridae по строению внутреннего хрящевого скелета с едва
намеченным подразделением оболочки хорды, по отсутствию чешуи и развитию
только мельких кожных зерен и отдельных костяных пластинок вдоль спины,
на боках и на хрящевой голове считается некоторыми учеными дегенерирую-
щей группой (Acipenser — рис. 44) *).
*) Проф. А. Н. Северцов считает нынешних осетровых самостоятельным семей-
ством хрящевых рыб, а не дегенерирующим.
38
класс: рыбы — pisces
К подобному же типу боковых ветвей принадлежат, вероятно, два дру-
гих живущих рода Amia и Lepidosteus, мало изменившиеся с мезозоя
по внешности, но имеющие уже окостеневшие позвонки. Может быть, Amia
является потомком Megalurus верхней юры, с которым ее сближает форма
чешуй, закругленного хвоста, двойных, как бы рассеченных поперек, позвонков,
у которых задние половины несут дуги и отростки. Ископаемые ее остатки
известны из пресных вод третичных слоев Европы. Ныне она живет в
Сев. Америке.
У Lepidosteus тело покрыто ромбическими ганоидными чешуйками,
сочленившимися между собою. Позвоночный столб в различных стадиях раз-
вития. Современный Lepidosteus (южные штаты Сев. Америки) прибли-
жается по строению скелета KTeleostei и имеет опистоцельные по-
звонки, представляющие исключение между всеми рыбами.
П ОДОТРЯД
Полукостистые — Hemiostei
Древнейшим семейством является
Сем. Palaeoniscidae — с девона до юры; удлиненные формы со спин-
ным и анальным плавниками, неправильным хвостом и мелкими ромбическими
ганоидными чешуйками. Хорда сохраняется.
Cheirolepis Traillii Traq. (рис. 45) — из нижнего девона Шотлан-
дии; покрыт мельчайшей ромбической чешуей. Спинной плавник развит
слабо, придвинут к хвосту. Хвостовой плавник на нижней стороне хвоста.
Анальный и задние парные прикрепляются широкой поверхностью; передние
более сжаты. Плавники снабжены фулькрами. Черепная крышка сплошная:
пара маленьких темен-
ных, две боковые ви-
сочные составляют зад-
нюю часть черепа, вы-
тянутые лобные—сред-
нюю часть, широкая
носовая дополняет его
Рис. 45. Cheirolepis Traillii Traq.
спереди. Маленькие нижнечелюстные и далеко идущие назад верхнечелюст-
ные несут мелкие конические зубы. Глаза окружены костяным кольцом, за
ними лежат жаберные крышки. Этот тип черепа, характерный для всего
семейства, обнаруживает незначительные изменения у разных родов.
Другие виды Cheirolepis встречаются в верхнем девоне Сев. Америки
и у нас.
Palaeoniscus Freislebeni Blv. (рис. 46), ближайшая к первой
форма, очень распространена в пермских слоях Тюрингии (Германия) и Нор-
тумберланда (Англия); встречается и у нас. Известен еще с 1546 г. [Агрикола
(Agricola) и Геснер (Gesner)— 1565 г.]. Чешуя мелкая, ромбическая. Плавники
несколько уменьшились, как бы сжались. Хвост более правильный, чем у С h е i-
подкласс: совершеннонотые — teleostomi
39
folepis; верхняя часть его укоротилась, нижняя удлинилась. Впереди спин-
ного плавника ряд пластинок. Хорда остается неокостеневшей.
Elonichthys Robisoni Hibb. — нижний карбон. Форма очень близкая
KPalaeoniscus, но имеет более развитой спинной плавник, перед которым
расположено несколько круп-
ных закругленных пластинок.
Такие же пластинки лежат
за спинным плавником и
впереди анального. Зубы
расположены в два ряда на че-
люстях, внутренние крупнее.
Чешуя скульптированная.
Рис. 46. Palaeoniscus Freislebeni BIv.
Amblypterus — пермь. Близкая к предыдущей форма с крупными
черепными пластинками, большими плавниками, гладкой чешуей и такими же
пластинками на спине и на нижней поверхности, как у Elonichthys.
Coccolepis — по форме тела похож на предыдущие, но покрыт
крупной, тонкой, закругленной чешуей с бугорчатой поверхностью, далеко
накрывающей друг друга. Это — резкое отклонение в форме чешуи от целого
ряда названных форм семейства Palaeoniscidae. Нижний лейас Англии,
верхняя белая юра Германии, вельд Берниссара (Бельгия).
Сем. Caturidae — неполное окостенение позвонков. С a t и г и s (рис. 47)—
с нижнего лейаса до верхней юры — достиг большого прогресса в раз-
витии окостенения хорды и образования
лично развитых позвонков в разных частях
Рис. 47 А. Туловищные позвонки
Caturus furcatus Ag.
Рис. 47. Caturus elongatus Ag.
(Верхняя юра.)
Несколько видов рода
Caturus позволяют
проследить эти изменения и сра-
стание отростков с телами позвонков (рис. 47 А), внутри которых все-таки
остается еще хорда. Хвост более правильный, чем у предыдущих родов,
анальный плавник доходит
своим основанием до отрост-
ков позвонков. Чешуя у
одних форм ромбическая, у
других закругленная, почти
циклоидная.
Megalurus elegan-
tissimus Wagn. — из co-
Рис. 48. Megalurus elegantissimus Wagn.
(Верхняя юра, Соленгофен.)
ленгофенских сланцев верхней юры (рис. 48) — является более совершенной
40
класс: рыбы — pisces
с почти окостеневшим позвоночником, сильно измененным, закругленным
хвостом и удлиненным спинным плавником. Особенностью являются двойные
хвостовые позвонки, точно рассеченные поперек пополам. Чешуя закруглен-
plc
Рис. 48A. Eurycormus specio-
sus Wagn.
Туловищные позвонки. Хвостовые позвонки.
Oligopleurus
е s о с i и u s Thiol
ная. Зубы конические. Этот род является
самым совершенным из вымерших Н е-
m iostei. Связь его с другими фор-
мами еще не установлена. От Caturus
он сильно отличается формою чешуи
и прогрессивным развитием позвонков.
Двойные хвостовые позвонки и закруглен-
ный хвост встречаются у Eurycormus
(рис. 48А) и у ныне живущей Amia.
. — тоже из верхней юры и вельда —
с окостеневшей хордой и почти правильным хвостовым плавником,
закругленными тонкими чешуйками; он является как бы продолжением развития
Caturus. Встречается во Франции (Cerin) вместе с последним.
Это уже почти тип костистой рыбы, но с закругленной ганоидной
чешуей. К этой группе относится еще несколько родов; все они принадлежали
к хорошо плавающим в
открытом море, удлинен-
ным формам конца пале-
озоя и начала мезозоя,
до'«мела включительно.
Но одновременно с
ними 'существовали пред-
ставители другого образа
жизни, хотя они принад-
лежали также к Н e_m i-
о s t е i.
Это сем. Platyso-
Рис. 49. Eurynotus crenatus Ag.
midae, населявшее прибрежные, рифовые части моря. В противоположность
Palaeoniscidae, они имели плоское, короткое тело с удлиненным спинным
и анальным плавниками, уменьшенными парными и с коротким остро раздвоенным
хвостом.
Eurynotus crenatus Ag. (рис. 49) является как бы связующим
это семейство с сем. Palaeoniscidae — нижний карбон Эггенбурга. У него
тело укорочено. Хвост гетероцеркальный. Спинной плавник начинается со
средины спины, сначала очень высокий, потом переходит в короткие лучи.
Анальный — длинный, гетероцеркальный. Задняя пара плавников уменьшенная,
передняя — удлиненная. Зубы небольшие, тупо-конические.
PJatysomus striatus Ag. — в перми Англии. Плоская укоро-
ченная форма (рис. 50); сохранил еще маленькие задние плавники;
спинной и анальный—удлиненные, подходят к хвосту. Черепные кости более
слились.
подкласс: совершенноротые teleostomi
41
Cheirodus granulosus Young — карбон, значительно более высо-
кое тело, почти четырехугольное, с резкой границей начала спинного плавника;
анальный — удлиненный. Задняя пара исчезла; передняя маленькая. Хвост почти
правильный с узкими
лопастями. Хвост у
всех этих форм резко
отграничен особенным
расположением чешуи,
указанным выше.
Сем.Pycnodon-
tidae. Тело еще более
сжато, плавники раз-
виты слабо; хвост по
форме ближе к Р1 а-
tysomidae. Они то-
же покрыты ромбиче-
Рис. 50. Platysomus striatus Ag. (Англия, пермь.)
ской чешуей.
Окостенели дуги и отростки позвонков и ребра, переплетающиеся с под-
держками плавников. Фулькр нет. Особенность представляет озубление,
состоящее из нескольких рядов плоских, мостовидных зубов. От переднего
пояса известна только ключица.
Остатки Pycnodontidae известны с юры до эоцена. По строению
своему они являются как бы продолжателями Platysomidae с сильно изме-
нившимися зубами, в связи с изменением пищи, которую они перетирали.
Рис. 51. Gyrodus macrophthalmus Ag.
Зубы.
Gyrodus • macro-
phthalmus Ag. — с 5 ря-
дами бобообразных зубов
на нёбе (рис. 51). Средний
ряд крупнее остальных. В
верхней челюсти зубов нет;
в предчелюстной кости зубы
мелкие, режущие. В нижней
челюсти 4 ряда плоских
жевательных зубов. Из лито-
графского сланца Баварии.
Mesodon с большими спинным и анальным плавниками и ровным хво-
стовым. Хорда не окостеневает. Верхняя юра.
Microdon Wagneri Thioll. — более удлиненные клювообразные че-
люсти. Ряды крупных нижних зубов чередуются с маленькими зубами, распо-
ложенными неправильно. Внутренние части отростков отчасти охватывают
хорду в хвостовой части (рис. 52). Чешуя в задней половине тела очень тон-
кая. Верхняя юра.
Pycnodus Ag., близкий по строению зубов к предыдущим, имел
удлиненное тело. Спинной плавник гораздо длиннее анального. Чешуя на пе-
42
класс: рыбы — pisces
вредней части тела тонкая. По три ряда зубов на нёбе и в нижней челюсти.
Верхние и нижние дуги обхватывают хорду своими расширенными концами
{рис. 53). Верхний эоцен.
Рис. 52. Microdon Itieri Thioll.
Позвоночник.
Рис. 53. Pycnodus platessus
Ag. Два хвостовых позвонка.
При изучении этих семейств Hemiostei можно видеть связь между
ними и проследить изменение их строения в зависимости от изменения образа
жизни. Питались, вероятно, моллюсками.
Сгм. Stylodontidae. Характеризуются тонкими, иногда зазубренными
зубами, расположенными на краях челюсти. На небных костях зубы часто
крупные с вертикальной сме-
ной. Ромбические чешуи.
Semionotus Kapffi
Fraas — расширенное спереди
и суживающееся кзади тело
с косо срезанным хвостовым
плавником. Спинной плавник
довольно большой, впереди его
ряд заостренных чешуек. Пар-
ные плавники и анальный ма-
„ с, „ л • ,.. / г ленькие. Зубы все конические.
Рис. 54. Dapedius politus (pholidotus) J
Ag- Верхний триас Альп, Сев. Аме-
рики и южной Африки.
Dapedius politus (pholidotus) Ag. (рис. 54) — значительно более
широкая форма, с длинным спинным и анальным плавниками, с менее косо
•срезанным хвостом, маленькими парными. Внутренние поддержки плавников
доходят до концов длинных верхних и нижних отростков не окостеневшей
хорды. Глаза со склеротикальными пластинками. Покровные кости головы
с бугорками, покрытыми ганоином. Челюсти несут по краям столбикообразные
зубы, внутри — тоненькие, в виде щеточки. Хорошо представлен в нижнем и
верхнем лейасе средней Европы.
Lepidotus eivensis Blinv. (рис. 55) — крупная форма с толстой ром-
бической чешуей, большими спинным и анальным плавниками, слабо вырезан-
ным хвостом и резким изменением расположения чешуи у его основания. Зубы
тоже двух типов, как и у предыдущих. Передние — острые — для срезывания
кораллов (по мнению Абеля), задние — более плоские, закругленные—-для
перетирания пищи. Известен с верхнего триаса до верхней юры. Встречается
подкласс: совершенноротые — teleostomi
43
в юре восточной Индии и Соленгофена, в вельде Бельгии и восточной
Африки.
Lepidotus notopterus Ag. — более крупная форма.
Belonorhynchus gigas Sm. Woodw.— с удлиненной мордой, мел-
кими зубами. Голова покрыта кожными костями, более или менее сросшимися.
Зубы.
Рис. 55. Lepidotus ei vensis Blinv.
Хорда сохраняется. Парные плавники маленькие. ДиФицеркальный хвост с пря-
мым, ровным плавником. Спинной и анальный отодвинуты к хвосту. Ряд чешуй
идет вдоль спины и брюшной стороны. Хорошо представлен в триасе; отдель-
ными частями в лейасе.
Сем. Aspidorhynchidae представлено отклонившимися формами с очень
вытянутой мордой, с мелкими зубами
нижний мел.
Рис. 56 А. Кольцевые туловищные (а) и
хвостовые (b) позвонки Aspidorhynchus.
о верхний остистый отросток, и — нижний; w —
тело позвонка.
’Рис. 56. Aspidorhynchus acutiro-
s t г i s Ag. (Верхняя юра.)
pmd — praemandibula, pt — pterygoideum.
Aspidorhynchus (рис. 56 и 56 А) имеет тонкое, длиною до 1 м тело,
покрытое четырехугольными и удлиненно-ромбическими поперечными ганоид-
ными чешуями. Позвонки тонкие — кольцевые в туловище и с приросшими
дугами в хвосте. Особенностью является присутствие кости praemandibula в
нижней челюсти с мелкими зубами.
Подотряд
Костистые — Teleostei
Лучеперые Hemiostei были последними представителями рыб, по-
явившихся с палеозоя. Остатки костистых рыб, имеющих костный скелет и
покрытых тонкой чешуей, встречаются в мезозое с мела, что и дает возмож-
ность искать их предков между более ранними формами. Плавники устроены, как
у других Actinopterygii. Спинной — один. Кроме костного скелета, их
44
класс: рыбы — pisces
отличают от предыдущих форм: тонкая чешуя, лишенная ганоина, более легкая
и прозрачная, и присутствие межмускульных костей.
Сем. Leptolepidae является как бы переходным, соединяющим еще некото-
рые признаки Hemiostei с признаками костистых. С нижнего лейаса до мела.
Leptolepis — из соленгофенских сланцев (рис. 57). По общей форме
удлиненного тела, с правильно рассеченным хвостом, небольшими плавни-
Рис. 57. Leptolepis spratti-
f о г m i s Ag. (*/2)
шие нижние челюсти свободно
ками, он очень похож на Caturus
furcatus Ag. Но позвонки более око-
стенели, хотя хорда проходит еще через
их центры. Существуют тонкие ребра.
Верхние дуги не сливаются с позвон-
ками в брюшной области, а в хвостовой
они слиты, так же как и нижние. Боль-
подвешены. Зубы маленькие, конические.
Тело покрыто тонкими чешуйками, сохраняющими еще тонкий слой ганоина.
Непарные плавники и задние парные имеют роговые поддержки, но их
radialia и basalia не у всех доходят до внутреннего скелета. Грудные
плавники прикреплены посредством сложного пояса. Leptolepis spratti-
formis Ag., Lept. dubius Bl.
Как видно из описания, эта форма является как бы усовершенствован-
ным Caturus, сохраняя еще тонкий слой ганоина на чешуе, которая стала
много тоньше чешуи настоящих Hemiostei.
К этому семейству относятся еще немногие роды из верхней юры, от-
личающиеся некоторыми деталями строения: Thrissops, Tharsis и др.
Начиная с нижнего мела, мы имеем представителей настоящих ко-
стистых, еще близких к Leptolepidae.
Сем. Clupeidae. Удлиненное тело с раздвоенным хвостом покрыто ктено-
идными закругленными чешуями. В центре позвонка хорды не остается; по-
звонки совершенно окосте-
нели. Жаберная крышка
вполне развита. Один
спинной плавник. Сюда
относятся: С1 и р е а (рис.
58),Meletta, Pseudo-
b е г у х и др. типы.
Рис. 58. С1 u р е a v е п t г i с о s а Н. v. М. (Нижний миоцеи.)
Близкое к ним сем. Salmonidae, представленное тоже начиная с мела,
с трудом отличается от них по скелету, так как главный признак—.жи-
ровой плавник", лежащий за спинным, не сохраняется. Остатки ныне
живущего Mallodus встречаются в конкрециях ледниковой глины на
берегу Гренландии.
Rhinellus furcatus Ag. — из верхнего мела Сирии — представляет
отклонение от типа Leptolepidae; у него вытянутое тело и удлиненная
голова; плавники и хвост первого типа (рис. 59). Близ Бейрута остатки их нахо-
дят сотнями на плитах известняка, что указывает на их внезапную гибель.
подкласс: совершенноротые — teleostomi
45
Eurypholis — из тех же мест — вымершая форма, украшенная сильно
развитыми дермическими пластинками на голове, передней и боковых частях
туловища. Хвост раздвоенный.
Amphisyle Heinrichi Haeckel (рис. 60) — беззубая, с удлиненным
рылом. На спине костистый панцырь, оканчивающийся шипом. Олигоцен.
Рис. 59. Rhinellus furcatus Ag. (Верхний мел.)
Platax minor Pict, имеет тип рыбы с сильно укороченным вы-
соким телом, с громадными спинным и анальным плавниками типа Platyso-
m i d a e.
Beryx (рис. 61) имеет корот-
кое тело с крупными прозрачными
чешуями, большим спинным и широ-
ким раздвоенным хвостовым плав-
ником. В мелу.
Рис. 60. Amphisyle Heinrichi
Haeckel. (Олигоцен.)
Proteus molassus Cope — из сенона Канзаса — представляет
гиганта свыше 2 м в длину, с далеко Отодвинутым назад спинным
плавником,
Рис. 61.
правильно раздвоенным хвостом
В е г у х и его ктеноидная чешуйка.
и сильно развитым кост-
ным скелетом.
Из этих нескольких приме-
ров видно, что в мелу костистые
рыбы были представлены очень
разнообразными типами. Суще-
ствовали и Physostomi —
с пузырем, имеющим выходное
отверстие, и Physoclisti —
с пузырем без отверстия,—узкие
и широкие рыбы, имевшие предков между рыбами с хрящевым ске-
летом. В третичных слоях число разнообразных костистых рыб еще
более увеличивается, что продолжается и доныне, обусловливаясь различ-
ными условиями среды: глубиной воды, температурой ее, соленостью
и т. д. Но их коренные признаки остаются, за немногими исключе-
46
класс: рыбы—pisces
ниями, неизменными, что, невидимому, и дает им преимущество над всеми
другими отделами рыб J).
Резюмируя сказанное о строении различных групп рыб, мы посмотрим,
что именно и почему дало последней группе — костистым рыбам — пре-
имущество перед всеми другими, несмотря на то, что почти все подотряды, кроме
костистых, имели уже в девоне своих вполне обособившихся представителей.
Мы видели, что уже в силуре известны остатки твердых частей рыб:
зубы, спицы и даже целые отпечатки мягких рыб, например Lana-rkia. Зна-
чит, этот подкласс получил свое начало еще до силура, но, вероятно, отсутствие
внешнего твердого скелета способствовало их разрушению, и сохранились не-
многие экземпляры только с силура.
Мы застаем в д е в о н е уже разнообразных представителей обособившихся
подклассов, хотя еще и близких друг к другу.
Мы встречаем панцырных рыб с различно построенным внешним ске-
летом, начиная от зачатков его в виде маленьких пластинок до твердых пан-
цырей с боковыми придатками для передвижения—Pt erich thy s. Но вну-
тренний скелет у них отсутствует. Очевидно, этот подкласс пошел по ложному
пути развития, затратив слишком много материала организма на внешний пан-
цырь, не спасавший в борьбе с более подвижными хищниками, лучше снаб-
женными органами хватания, каковыми были тогда, например, громадные пауко-
образные Merostomata.
Представители отряда круглоротых, наоборот, слишком прене-
брегли внешним скелетом, и мы до сих пор не имеем указаний на их бли-
зость, или генетическую связь, с настоящими рыбами, хотя и имеем их пере-
житков в числе трех родов ныне существующих круглоротых (миног). Проф.
А. Н. Северцов выключает их даже из класса рыб, как было уже указано.
Две ископаемые формы, Palaeospondylus и Hypospondylus,
отнесенные условно сюда, имели уже внутренний костный скелет, совершенно
отсутствующий у нынешних форм.
У Elasmobranchii намечается — у более древних — и развивается
позже укрепление плавников; они получают поддержки в хрящевых основных
<) У нас известны некоторые представители следующих подклассов рыб:
П а н ц ы р н ы е — из девона Новгородской губ. (р. Сясь): Bothriolepis и др.
Пластиножаберные — зубы акул и скатов из каменноугольного известняка
и третичных слоев. Edestidae из пермо-карбона.
Д во!я к од ы ш а щи е — зубы Dipterus из прибалтийского девона, Cocco-
steus — Новгородская губ., р. Сясь.
Кистеперые — чешуи и обломки головных пластин Holoptychius из де-
вона Новгородской губ., р. Сясь.
Лучеперые — Paia eoniscus — пермский известняк и красный мергель Ка-
занской и Самарской губ., Platysomus — пермский известняк Оренбургской губ.
Костистые — Beryx, Clupea, Sparnodus (синяя глина около Киева}
и другие.
подкласс: совершенноротые — teleostomi
47
и радиальных пластинках (рис. 62). Но слабый хрящевой скелет не дает этим
частям внутренней опоры. Главная сила их была сосредоточена в развитии
многочисленных, хорошо развитых зубов, которые имеют большое значение
при определенных условиях жизни. Но, не имея твердого скелета, прочно
укрепленных плавников, при изменившихся условиях и развитии более со-
вершенных рыб акуловые должны были, очевидно, бороться с формами, при-
нявшими в своем развитии иное направление. Большинство форм, дошедших
до нашего времени, выработало более прочный скелет с объизвествленными
позвонками и более прочным грудным поясом. Но и теперь еще существуют
акуловые с неокостеневающей хордой. Главным защитительным органом их и
теперь являются многочисленные острые
зубы (одностороннее развитие).
Двоякодышащие тоже спе-
циализировались в одном направлении.
Они, очевидно, были поставлены в не-
обходимость выработать плавники и
прочный покров в виде ганоидных че-
шуй, после чего попали в условия, при
которых вода легко высыхала, и им
пришлось изменить дыхание, приме-
няясь к полуназемной жизни. Но отсут-
ствие твердого внутреннего скелета по-
мешало. вероятно, и тут победить в
борьбе, и они после богатого расцвета
Рис. 62. Плечевой пояс (SB) и грудной
плавник Heptanchus (акулы).
в палеозое — почти исчезли, и только
три рода избегли общей участи и
дошли до нас.
Иное мы видим у кистеперых и особенно у лучеперых Hemio-
stei. Последние, начиная с девона, позволяют проследить изменение черепа и
внутреннего скелета, постепенное окостенение их, развитие поддержек плавников,
развитие жаберной крышки и плавательного пузыря, получившего определенную
функцию (не дыхательную). Эти изменения ведут незаметно к костистым
рыбам, у которых хрящевой скелет заменяется костным, тяжелые ганоидные
чешуи заменяются постепенно тонкими и вырабатывается возможность лучше
плавать. Та группа лучеперых, у которой сохранился хрящевой скелет и исчезли
чешуи, является слабо представленной.
Но почему же названные признаки костистых сослужили им такую
службу и дали преобладание над всеми другими рыбами ?
Почему панцыри Placodermi, зубы и плавники Elasmobranchiir
двойное дыхание Dipnoi не дали им тех преимуществ, которые получили
костистые рыбы при медленном развитии их различных органов, и почему
эти органы оказались так полезны? Постараемся ответить на эти вопросы,
проследив шаг за шагом развитие этих признаков.
Та среда, в которой живут рыбы, — вода (моря, реки, озера и пруды),
несмотря на возможные различия глубины, температуры, солености и т. п.^
’ 48
класс: рыбы — pisces
представляет одинаковые механические условия для передвижения в ней. Эти
условия определяют типы строения различных организмов, живущих в этой
среде. При этом победа остается за теми, кто сумел наилучше приспособиться
к среде, развивая полезные органы.
Форма тела играет при этом значительную роль, и удлиненное, не-
сколько расширенное посредине, но не сплющенное — оно является наилучшим
рассекателем водной среды. Недаром наши суда, лодки, например, имеют
такую форму. Но в лодке нужен и руль и весла, без которых ей нельзя
правильно двигаться. У рыб роль руля исполняет хвост, роль весел — парные
плавники. Но у нее есть еще, кроме хвостового, другие непарные плавники:
спинной и анальный. Это — кили, удерживающие тело в устойчивом положении
при плавании. Но недостаточно иметь орудия движения; Нужно, чтобы они
были устойчивы и сами по себе и прочно соединены с движущимся телом,
составляя с ним одно целое, т. е. чтобы они были правильно координированы.
Следовательно, нужна прочная связь плавников, находящихся вне тела живот-
ного, с внутренней центральной его частью, которая, в свою очередь, должна
представлять подвижное, но устойчивое целое.
Просмотрим теперь последовательно развитие внутреннего скелета и ор-
ганов движения — плавников — у различных типов рыб. Начнем со скелета.
Припомним, что простейший позвоночник рыб представлен хордой, спин-
ной струной, состоящей из вакуольных клеток и заключенной в хрящевое вла-
галище, с лежащим над ним нервным тяжем, утолщающимся в передней части
тела. Оба они заключены в скелетородный слой, в котором закладываются
сначала отростки будущих позвонков, а позже и тела позвонков. У миноги
закладываются в верхней части этого влагалища хрящевые зачатки верх-
них отростков позвонков и поднимаются над спинным нервным тяжем на рав-
ном друг от друга расстоянии (сливаясь в дуги). Но никакого подразделения
хорды на отдельные участки — позвонки —• не замечается. Эта простейшая стадия
встречается также и у химер и у некоторых голых рыб, например у осет-
ровых, причем здесь образуются и нижние отростки, хрящевые, обни-
мающие кровеносный сосуд, образуя нижние дуги. Такая же хрящевая, не сег-
ментированная хорда встречается у древних акуловых и двоякодышащих. Из
нынешних подразделение хорды на отдельные участки намечено у Proto-
pterus; у Neoceratodus намечается начало окостенения частей дуг.
У некоторых из ныне живущих акуловых сохраняется хорда, но в ней
намечаются перетяжки на равном друг от друга расстоянии, где и начинается
отложение извести, разбивающее хорду постепенно на тела позвонков.
У других форм эти хрящевые участки — позвонки — пропитываются
больше известью'—объизвествляются и получают или форму удлиненную,
двояковогнутую с обоих концов, похожую на песочные часы, с остающейся
хордой внутри, или форму правильных плоских, более или менее толстых
кружков со слабой вогнутостью на концах. В разрезе позвонка можно видеть
последовательное образование и нарастание хряща, пропитанного известью. Но
никогда в нем нет костного вещества с костными тельцами. Отростки позвон-
подкласс: совершенноротые — teleostomi
49
ков у акуловых развиваются отдельно, не срастаясь с телами позвонков.
В ископаемом состоянии они редки. Сохранились у Hybodus.
У различных представителей кистеп ер ых строение и развитие по-
звонков различны. У девонского Holoptychius и близких к нему форм суще-
ствуют отростки позвонков, а тела их не развивались. У Rhizodontidae
позвонки существовали в виде окостеневших колец, но без отростков (девон
и карбон).
У лучеперых развитие позвонков достигает большого разнообразия.
Мы можем проследить на некоторых формах целый ряд их изменений по дан-
ным уже рисункам.
1. Так, рис. 52 дает нам часть позвоночника Mi crodon, на котором
верхние и нижние отростки образовали верхние и нижние части дуг, прикры-
вая часть хорды. Между ними она остается открытой, не’ замкнутой.
2. У Pycnodus (рис. 53) эти отростки пошли как бы навстречу друг
другу, покрывая хорду с боков, нарастая сверху и снизу; но тела позвон-
ков еще не образовались, их части являются как бы продолжением от-
ростков.
3. У Caturus (рис. 47А) заложены отдельные части тел позвонков:
вверху — pleurocentrum между верхними дугами, в виде овальных пластинок;
внизу — hypocentrum, пластинки треугольной формы над нижними дугами. Эти
пластинки, развиваясь постепенно, сближаются и сливаются вместе, образуя
кольца с приросшими к ним дугами.
Тела позвонков, вначале хрящевые, начинают позже постепенно окосте-
невать снаружи внутрь, оставляя еще долго в центре хорду, исчезающую по-
степенно, при переходе позвонка из хрящевого в костный. У Eurycormus
хвостовые позвонки раздвоены вертикально: одна из частей несет дуги, другая —
нет (рис. 48 А). На туловищных сохраняются еще pleurocentra и hypocentra, не
слившиеся с прилегающими к ним отростками. Внутри этих кольцевых позвон-
ков хорда сохраняется.
LeptolepisH Megalurus являются как бы завершителями перехода
ко все большему и большему окостенению тел позвонков и исчезновению
хорды у костистых; хрящевое вещество остается только между позвон-
ками. Так как окостенение идет к центру позвонка, то и форма его остается
в большинстве случаев двояковогнутой — амфицельной. Одновременно с этим
происходит срастание дуг с телами позвонков и обеспечивается ббльшая
крепость центральной части тела животного и охраняется драгоценный
орган — спинной мозг, заключенный в верхних дугах позвонков. В то же
время сохраняется подвижность органа благодаря отдельным позвонкам.
Что же происходит во время всех этих изменений хорды в передней ее
части? Мы уже видели у миноги незначительное ее расширение, захватываю-
щее переднюю часть нервного стержня — мозг — и образующее примитивный
хрящевой череп круглоротых.
У акул мы имеем более совершенную хрящевую головную кап-
сулу, подразделяющуюся на: 1) носовую часть; 2) глазную с орбитой;
4 Павлова М. В. Палеозоология. Ч. II.
50
класс: рыбы — pisces
3) затылочную, сочленяющуюся с pterygo-quadratum (верхней челюстью),
например у Notidanus. Нижняя челюсть подвешена к верхней. Это, так
сказать, тип черепа акул; хотя у некоторых из них кое-что изменяется в рас-
положении частей черепа, но верхняя и нижняя челюсти остаются отдельным
аппаратом, не срастающимся с черепной капсулой. Ротовое отверстие у боль-
шей части акул лежит не впереди морды, а снизу. Зубы многочисленны; запас-
ные расположены в несколько рядов.
У Dipnoi череп хрящевой с различным числом кожных костей, распо-
ложенных у некоторых из них не вполне симметрично. Верхние челюсти тоже
не срастаются с черепом.
У Crossopterygii происходит окостенение некоторых частей черепа —
хрящевых — и слитие кожных костей в меньшее число, с более правильным
расположением их. Челюсти окостеневают, и верхние срастаются с черепом,
что дает более прочное положение зубам и делает ненужными запасные
зубы акул.
У осетровых черепная капсула остается хрящевой, и сверх ее разви-
ваются кожные кости. Челюсти остаются примитивного типа.
У Actinopterygii, например у Palaeoniscidae, кожные кости
черепа покрыты ганоином, что увеличивает их прочность; число их здесь зна-
чительно. Это продолжение главной ветви развития рыб, которая идет через
Caturus и Leptolepis, с постепенным окостенением черепных и кожных
костей, к костистым рыбам, что сопровождается постепенным окостенением
позвоночника. Строение черепа и степень сложности костей у последнего от-
ряда очень различны, что и дает возможность подразделить его на много се-
мейств, но главный признак — полное окостенение позвонков — существует
почти у всех. Кроме хорошо сросшихся с черепом верхних челюстей, суще-
ствуют прочное сочленение нижних с черепом и хорошо развитая крышка,,
защищающая жабры.
Вот в общих чертах те изменения внутреннего скелета, которые мы
можем проследить, начиная от мягкой хорды круглоротых до костного позво-
ночника щуки, окуня и т. п., обусловливающего устойчивость тела костистой
рыбы и подвижность его.
Посмотрим теперь на органы передвижения у рыб и их изменение от
простейшей формы той же миноги до сложнейшей — костистых рыб. У рыб
существуют для передвижения плавники; как мы видели уже, они бывают
парные и непарные. Простейшей формой является непарный плавник, иду-
щий вдоль спины, переходящий на хвост и брюхо и останавливающийся1
у анального отверстия, впереди которого лежат парные плавники. Происхожде-
ние спинного плавника объясняется образованием складки кожи, тянувшейся
вдоль спины, перешедшей через хвост на брюхо. Парные расположены по
обеим сторонам тела. Происхождение их толкуется различно. Бальфур (Bal-
four) считает их самостоятельным образованием из таких же складок, как и
спинная. Г егенбауер ставил их развитие в зависимость от жабер-
ных складок.
подкласс: совершенноротые — teleostomi
51
Проф. Осборн (Osborn) признает развитие их из двух пар боковых
складок (у Cladoselaclie). Подробное изучение развития этих плавников
проф. Северцовым у осетровых вполне подтвердило это мнение.
В отряде Acanthodi было по одной длинной складке по обеим сторонам
тела или по две первоначальных, сливавшихся в одну.
Спинной плавник удлиненный развился раньше; он существует
у рыб, которые при движении изгибаются, и содействует устойчивости тела
в воде. Он встречается в этом виде у некоторых круглоротых — у миног, у
Protopterus, а в эмбриональном состоянии — у всех рыб.
Но для того чтобы эта первоначальная складка могла исполнять свое назна-
чение, не спадаясь, ей потребовалась поддержка, которая явилась первоначально
в виде тонких хрящевых лучей. По мере развития плавника целость его наруша-
лась, он распадался на передний и задний спинные, хвостовой и анальный. Под-
держки осложнились; у парных плавников развились два ряда лучей: основ-
ные и радиальные. Первые закладывались в коже и внедрялись в тело,
вторые направлялись в плавник и поддерживали его проксимальную часть. В ди-
стальной части плавника развивались роговые поддержки. Таким образом получи-
лись устойчивые непарные плавники различных акул, химер, двоякодышащих;
при этом основные и радиальные лучи перемещались, срастались, делая осно-
вания плавников более устойчивыми. Хвостовой плавник — руль—.имеет у
древних рыб неправильную форму, так как у них гетероцеркальный хвост;
при этом его верхний, более длинный конец сопровождает позвоночник, а ниж-
ний находится только на нижней стороне хвоста, например у акул. При даль-
нейшем развитии хвост делается более правильным, ровным, закругленным или
вырезанным; у костистых рыб хвостовой плавник расположен правильно, в конце
позвоночника в виде одной или двух лопастей.
Но эти плавники, как бы они ни были совершенно устроены сами по
себе, остаются у рыб, не имеющих твердого внутреннего скелета, без проч-
ной опоры, без связи с внутренней частью организма. Достаточно взглянуть
на известного нам Pleuracanthus, чтобы увидеть хорошо развитые спин-
ной и хвостовой плавники, имеющие хрящевые поддержки в теле, но не свя-
занные с отростками хорды. Последние проникнуты уже отчасти известью, но
хорда все еще сохраняется, тело позвонка еще не образовалось, и связи между
плавниками и внутренним скелетом нет. То же мы видим у Dipnoi и у
Crossopterygii, у которых плавники спинной и хвостовой разбились уже
на участки, отдельные плавники, с лучше развитыми поддержками, и только
у лучеперых устанавливается эта связь благодаря постепенному око-
стенению позвонков и срастанию их с дугами и отростками
(С a turns). Вполне же это укрепление плавников с внутренним скелетом
совершается у костистых рыб, с полным окостенением позвоночника, при
помощи основных и радиальных лучей. Тогда же развиваются фулькры у гано-
идных и лученосцы у костистых, которые в хвостовом плавнике сливаются
с остистыми отростками, и связь хвоста с позвоночником становится еще
прочнее.
4*
52
класс: рыбы—pisces
Парные плавники, весла рыб, еще более интересны в своем раз-
витии. Как мы указали, они появляются первоначально в виде двух или четы-
рех боковых кожных складок, идущих вдоль тела от жаберных отверстий до
анального отверстия. Они тоже распадаются на отдельные участки, со слабыми
внутренними вертикальными поддержками.
У древних пластиножаберных, у Climatius и Acanthodes (верхний
силур и девон) плавники эти были разбиты на отдельные (пять-шесть) малень-
кие плавнички, снабженные иногда впереди спицами. У некоторых остались
только спицы, лучи не сохранились в ископаемом состоянии. Нормально число
парных плавников — две пары: передняя — грудные, задняя — брюшные
плавники. Мы встречаем их в первичной стадии развития у интересной формы
древних акул Cladoselache Fyleri (Cladodus) Newb. (нижний девон
и карбон).
Грудные плавники, вытянутые в полуовалы, внедряются в тело одним рядом
основных хрящевых лучей. Над кожей в плавник входят радиальные хрящевые
лучи и кожные — роговые. Второй ряд базальных лучей продвигается глубже
внутрь тела (metapterygium) и дает начало лопатке (scapula), образующей
здесь первичный грудной пояс.
Брюшные плавники Cladoselache, более удлиненные по оси тела,
имеют почти такое же строение, с входящими основными хрящевыми лучами.
Второй ряд их тоже удлинен. Верхние его части (protopterygium) прилегают
к тазовым хрящам — зачатку тазового пояса. Средние спускаются вниз (meta-
pterygium) и заканчиваются у самцов длинным хрящом (pterygopodium)
(рис. 25).
Таким образом мы в этой древней акуловой рыбе видим начало обра-
зования связи передних — внешних — конечностей с внутренним скелетом. У
нынешнего Heptanchus эти отношения остаются почти такими же.
У Pleuracanthus, другой акулы — пермской, basalia удлинились
в самом плавнике, от них идут radialia к краям плавника. Внутри тела
basalia прилегают к раздвоенной ключице, подходящей к жаберным
дугам.
В сем. Acanthodidae, из девона и нижнего карбона, мы имеем
большое разнообразие поясов в соответствии с разнообразием грудных плав-
ников.
У Acanthodes Wardi (рис. 33) основные лучи несут кожистые
лучи, поддерживающие плавник. Радиальные лучи едва развиты в плавнике.
Нижний карбон.
У Diplocanthus striatus (нижний девон) базальный луч прилегает
к ключице, а к нему прикрепляются лучи плавника и т. д. Нижний пояс не-
известен.
У Dipnoi и Crossopterygii ластовидные плавники поддерживаются
тоже хрящевым поясом (рис. 62 В).
Все эти формы не употребляли еще своих парных плавников как орга-
нов плавания, они были большей частью донными, ползающими рыбами, упи-
подкласс: совершенноротые — teleostomi
53
рались плавниками или, поднимаясь немного в воде, балансировали при по-
мощи их.
У древних Actinopterygii грудной пояс состоит из двух половин,
разделенных на брюшной стороне. Части его соединены с костными пластин-
ками кожного скелета, что дает им значительную опору и устойчивость.
Переходя к костистым рыбам, мы видим осложнение
грудного пояса. В состав его входит уже целая группа кожных
костей (6 пар), посредством которых и укрепляются передние
а
Рис, 62 А. Г рудные плавники и пояса: а — Squatina (акула); b — Polypterus;
с — S а 1 m о.
плавники с одной стороны с черепом, а с другой — с жаберным скелетом,
что вполне упрочивает силу их движений
(рис. 62 А).
Брюшные плавники поддерживаются
задним поясом, развитым слабее переднего.
У многих костистых брюшные плавники
подвигаются вперед и прикрепляются под
грудными.
Достаточно посмотреть на скелет
окуня, чтобы оценить всю выгоду око-
стеневшего позвоночника, снабженного
Ъ с
Ъ а
Рис. 62 В. Грудной плавник N е о ce-
ratodus Forsteri.
длинными отростками, до которых доходят
поддержки непарных плавников и сложный передний пояс, прикрепляющий
передние плавники. Задние не имеют связи с внутренним скелетом (рис. 21).
Параллельно с изменением внутреннего скелета изменяется и внешний.
Тяжелые ганоидные чешуи (Dipnoi, Crossopterygii, Н е m i о st е i) защи-
щали тело рыб, но и отягощали его. Мы уже указали на утонение чешуй
и уменьшение слоя ганоина у Leptolepis и Oligopleurus. У настоящих
костистых они лишены ганоина, легки и прозрачны, что способствует лег-
кости тела, а следовательно и более свободному плаванию, что, в свою оче-
редь, позволяет легче избежать врага и схватывать добычу.
Из всех этих данных можно сделать следующий вывод.
В то время как одна группа рыб с хрящевым скелетом в различных
своих представителях развивала какой-нибудь один орган и, достигая этого,
побеждала, когда был полезен этот орган, другая группа рыб шла медленно
в своей эволюции, развивая равномерно различные части своего организма,
54
класс: рыбы — pisces
главным образом костный скелет, и достигла совершенства его, пройдя через
Palaeoniscidae и выработав настоящий тип костистых рыб, в котором
все органы находятся в соответствии. Это-то и дало им превос-
ходство и преобладание над всеми другими подклассами рыб. Теперь они за-
владели всеми водами земного шара в количестве 95 °/0. Малочисленные пред-
ставители Dipnoi и Crossopterygii зарываются в песок или ил.
Довольно многочисленные акулы держатся еще благодаря своим страш-
ным зубам; но сравнительно с костистыми, более безобидными, число их ни-
чтожно.
Немногие из последних остались донными с изменившимся скелетом;
большинство свободно плавает в морях и реках.
ТИП TETRAPODA JAECKEL
КЛАСС ВТОРОЙ
ЗЕМНОВОДНЫЕ - AMPHIBIA
Главным характерным признаком для этого класса является жаберное ды-
хание в юности (личиночное состояние) и легочное — во взрослом состоянии.
Напомню, что ныне живущие земноводные очень немногочисленны; все они
принадлежат к наземным жителям и жителям пресных вод. Это голые, холодно-
кровные животные. Сердце с двумя предсердиями и одним желудочком.
Ныне живущие амфибии подразделяются на:
1) Подкласс бесхвостых лягв — Anura, наиболее распространенным
представителем которых является лягушка.
2) Подкласс хвостатые — Urodela — более разнообразны. Сюда отно-
сятся: саламандры, тритоны и другие близкие формы, снабженные хвостом и
четырьмя конечностями.
3) Небольшой подкласс — змееобразные формы Gymnophiona — сем.
С о е с i 1 i а е, живущие в тропических странах Южной Америки, Ост-Индии
и Африки, лишены совершенно конечностей.
В отличие от рыб, в позвоночном столбе амфибий различают по-
звонки: шейные, грудные, крестцовые и хвостовые; у безногих лягв они со-
кращаются до трех отделов: шейные, грудные и хвостовые. Построены они
различно: есть амфицельные — двояковогнутые — как у рыб; процель-
ные— вогнутые впереди, выпуклые сзади; о п и с т о це л ь н ы е — выпуклые
спереди, вогнутые сзади. Закладываются они в виде четырех пар костных пла-
стинок: basidorsalia, basiventralia, interdorsalia, interventralia, которые сливаются
между собой различно, почему и получается различный тип позвонков, по
которым и установлена классификация ископаемых амфибий. У ныне живущих
амфибий позвонки более сходны между собою — они цельные.
Череп окостеневает только отчасти, т. е. немногие хрящевые части
первичного черепа превращаются в костно-хрящевые кости, остальные части
остаются хрящевыми всю жизнь (рис. 63). Кроме того, в строение черепа
входят кожные кости, т. е. местные окостенения кожи, как это было и у рыб.
Для некоторых вымерших будет указано особое строение черепа с добавоч-
ным костным щитом. Верхняя челюсть плотно срослась с черепом. Зубы рас-
положены или по краям челюстей, или также на сошнике.
Конечности. Наиболее отличительным признаком амфибий (и всех
других позвоночных-—Tetrapod a Jaeckel), отделяющим их от рыб, являются
конечности, состоящие из частей: плеча, предплечья и кисти —
56
класс: земноводные — amphibia
в передней конечности, бедра, голени и ступни — в задней. Пред-
полагаемое развитие их из передних (парных) плавников рыб до сих пор
не выяснено. Но различные ученые дают схемы их развития. Так, Г е г е н-
бауер считает скелет плавников древних акул той первичной формой,
из которой можно вывести конечности четвероногих.
Грудной пояс различен. Наиболее сложный у бесхвостых — Anura:,
sternum, clavicula, scapula, coracoideum. У Urodela (хвостатых) clavicula
отсутствует. Sternum отсутствует у Gymnophiona (безногих).
Также различно построен и задний пояс, в зависимости от развития
конечностей.У хвостатых он состоит из трех пар костей-пластинок: ischium —
седалищные, ilium — подвздошные и pubis — лобковые. У бесхвостых он
С л
Ртх Р^Х
Рис. 63. Череп лягушки.
А — сверху, В — сбоку, С — снизу.
сильно изменен в связи со способностью прыгать. Подвздошные кости
сильно удлинены и направлены горизонтально; они прикрепляются перед-
ним концом — лобковым — к крестцу. Седалищные — очень маленькие
кости (рис. 63 А).
Нам придется заниматься главным образом изучением хвостатых лягв.
Ископаемые остатки нынешних бесхвостых лягв известны только начиная
с юрских отложений, хвостатые — с нижне-меловых, и безногие совершенно
неизвестны. Но кроме того хвостатые представлены ископаемыми типами, жив-
шими в палеозое — в карбоне и перми, резко отличающимися от ныне живу-
щих. Это были первые обитатели суши из позвоночных, и они представляют
поэтому громадный интерес в смысле изменения органов дыхания и передви-
жения. Изменение это совершалось, конечно, в связи с изменением среды, в ко-
торой они жили, с высыханием лагун и вообще с увеличением континентов и
растительности. Эти изменения уже значительно выразились в девоне и пошли
еще дальше в карбоне.
Мы уже видели, что некоторые рыбы в девоне пытались приспособиться
к двойному дыханию — Dipnoi. Но мы не можем связать с ними ни одного
из земноводных, известных в ископаемом состоянии, которые являются очень
далеко ушедшими в своем развитии от рыб. Главным отличием являются
вполне сформированные конечности, снабженные пальцами. Раньше амфибий
подкласс: панцыреголовые — stegocephali
57
соединяли с пресмыкающимися в один класс, но присутствие двух затылоч-
ных отростков у первых и одного у вторых, различное дыхание и отличия
в строении черепа и позвонков заставили отделить их в особый класс.
У некоторых ископаемых рептилий, как мы увидим ниже, существовало тоже
по 2 затылочных отростка.
Мы уже указали на невозможность
связать генетически Dipnoi с Am-
phibia вследствие некоторых слиш-
ком специальных признаков Dipnoi
(зубы, головные кожные кости и т. п.).
Но можно указать на некоторое сход-
ство отдельных признаков древних зем-
новодных — Stegocephali с неко-
торыми рыбами из Crossopterygii,
именно с Rhizodopsis. Оно вы-
ражается в строении черепа, в при-
сутствии пинеального отверстия между
Рис. 63 А. Скелет современной
лягушки.
теменными костями, костных пластинок
вокруг глаз, в строении зубов, в неполном окостенении позвонков и т. д.
Напомним, что у Polypterus, современного представителя Crosso-
pterygii, имеется раздвоенный плавательный пузырь, открывающийся в глотку
с брюшной стороны. У амфибий и у всех наземных позвоночных в этом же
месте находится вход в гортань.
Все это заставляет ожидать найти предка амфибий между Crossopte-
rygii, а не между Dipnoi.
Подкласс 1
ПАНЦЫРЕГОЛОВЫЕ — STEGOCEPHALI
Ископаемые амфибии пермо-карбона и триаса, сгруппированные в отдель-
ный подкласс —панцыреголовые — Stegocephali, представляют не-
сколько типов, связанных между собою некоторыми общими признаками,
позволяющими их резко отличить от нынешних амфибий. Главный признак —
присутствие особого головного щита, состоящего из покровных костей,
которому они и обязаны своим названием.
Череп их состоит: а) из хрящевых костей, б) из кожных костей, вошед-
ших в состав черепа, и в) из покровных кожных костей, образовавшихся из
сросшихся маленьких зубных пластинок (по А б е л ю).
Другой признак у большинства — брюшной панцирь, состоящий из
роговых или костных пластинок различной величины и формы (рис. 64).
Окостенение черепа и позвонков различно у различных групп, так же как и форма
тела. Различная степень окостенения позвонков и слитие их основных частей
позволяют подразделить Stegocephali на четыре главные группы, начи-
ная с менее совершенного типа позвонков и доходя до позвонков с полным
58
класс: земноводные—amphibia
окостенением. У простейших Stegocep hali окостенение начинается с верх-
них дуг, состоящих из двух половин, обнимающих сверху нервный ствол. Под
ними, сбоков и снизу хорды, образуются тонкие костные пластинки, обни-
мающие хорду. Все эти части вместе намечают позвонок (рис. 65).
Рис. 64. Брюшной панцырь Stegocephali.
<1 — Protriton.
sp z d
Рис. 65. Листовые позвонки
(Р h у 11 о s р о n d у 1 i).
Рис. 66. Трубчатые позвонки
(Lepospondyli).
Это I тип — Phyllospondyli, листовые позвонки. Сем. Protri-
tonidae (Branchiosauria). Карбон, пермь.
У следующей группы образуются трубочкообразные, двояковогну-
тые, в виде песочных часов, позвоночки со слабо развитыми верхними дугами,
нижние дуги отсутствуют (рис. 66).
Это II тип — Lepospondyli, трубчатые позвонки. Сем. Dо 1 у-
chosomidae: Keraterpeton, Diplocaulus и др. Карбон, пермь.
Рис. 67. Позвонки из кусочков (Temnospondyli, Rachitomi).
У третьей группы позвонок состоит из отдельных, не слившихся
верхних и нижних дуг и лежащих между ними pleurocentrum и
hypocentrum. Спинные позвонки отличаются от хвостовых отсутствием
нижних дуг. Хорда сохраняется, как и у предыдущих (рис. 67).
подкласс: панцыреголовые — stegocephali
59
Это III тип — Temnospondyli, Rachitomi— позвонки из кусочков.
Сем. Archegosauridae: Actinodon, Dvinosauria Amal. и др.
Пермь, нижний триас.
Четвертая группа обнимает высших Stegocephali, имеющих сплош-
ные, плотные позвонки с верхними дугами и боковыми отростками.
Это IV тип — Stereospondyli, твердые позвонки (Labyrintho-
dontia). Сем. Mastodonsauridae: Metopias и др. Пермо-карбон,
верхний триас ’3).
Отряд 1
Phyllospondyli Credn.
(Protritonidae Gaudry, Branchiosauria Grtdn.)
Все вымершие палеозойные и триасовые Stegocephali принадлежат
к подклассу хвостатых амфибий. Простейшими между ними являются Protri-
tonidae Gaud.* 2) (Branchiosauria Credn.). По общей форме тела они
похожи на маленьких саламандр, имеют широкую
1 олову; по расположению костей черепа они сходны
с некоторыми формами из Rhizodontidae.
(Сравн. череп Rhizodopsis с черепом Pelo-
s a u г u s.)
Protriton (рис. 68). Глазницы, со склеро-
тическим кольцом, отодвинуты далеко назад; пи-
неальное отверстие существует; теменные и височ-
ные кости укорочены; в не окостеневшей затылочной
области отсутствуют затылочные мыщелки для сочле-
нения с первым шейным позвонком. Зубы распо-
ложены в один, ряд в верхней и нижней челюстях в
виде простых тонких конусов. Небольшое число их
находится и на сошнике. Хорда вполне сохра-
няется. Позвонки состоят, как мы сказали, из верх-
них дуг и двух тонких, редко сохраняющихся в
ископаемом состоянии листочков, обнимающих хорду
Рис. 68- Protriton
Gaudry (Branchio-
s a u г u s Credn.).
сверху и снизу.
Ребра снабжены одной сочленовной головкой. Кости скелета мало окосте-
нели. Брюшной панцырь развит хорошо, в виде тоненьких чешуек (см. рис. 64).
Protriton Gaudry (Branchiosaurus Credn.) — главный представитель
этого семейства — найден в громадном количестве в средних пермских слоях во
1) Подразделение по позвонкам на различные типы у разных авторов не оди-
наково. В „Палеозоологии" Абеля и его „Stamme" указано значительно большее число
групп, или, как он их называет, подотрядов амфибий, подразделяемых по позвонкам.
У Абеля и Циттеля, „Grundziige", 1923,два последние подотряда соединены в один—
Temnospondyli.
2) Удерживаю название Protritonidae Годри, как раньше установленное.
Это первые четвероногие — Eotetrapoda.
60
класс: земноводные —amphibia
Франции, близ Autun, и в Саксонии, близ Дрездена, где проф. К р е д н е р
(Credner) собрал их более 1 000 экземпляров. В пермских слоях Богемии они
тоже очень обильны и очень разнообразного возраста, начиная с личинок,
которые имели 4 пары жаберных дуг. Жабры исчезали у них по достижении
ими 6—7 см длины. Этому при-
Рис. 69. Protriton Thevenin различного
возраста.
сутствию жабер в детском возра-
сте они и обязаны своим назва-
нием — Protriton.Личинки име-
ют голое тело, которое с воз-
растом покрывается со стороны
брюха панцырем, состоящим из
мелких, закругленных на задней
стороне чешуек. Расположены они
чередующимися рядами, подсти-
лают грудь, брюхо, хвост и лапы;
все подвижны; таким образом, за-
щищая нижнюю поверхность живот-
ного, они в то же время не стес-
няли его движений. Взрослое живот-
ное достигало 10—15 см длины.
Конечности вполне сформиро-
ваны; передние имеют четыре, зад-
ние — пять пальцев. Прикрепля-
лись первые посредством хряще-
вого пояса, состоящего из ключиц,
лопаток, коракоидных и межклю-
чичных костей. Задний пояс со-
стоял из подвздошных и седалищ-
ных. Все кости с хрящевыми,
окончаниями.
Branchiosaurus salamandroides Credn. — из Богемии—считается
типом рода, длина его 6,5 см.
Branchiosaurus amblystomus Credn. — в Саксонии —
длина 12 см.
Protriton, описанный Го др и,-—из нижних пермских слоев О те на
(Autun) — отнесен позже к Branchiosaurus petrolei. Тевенен.
(Thevenin) дает целый ряд форм различного возраста Protriton из
Отёна1) (рис. 69).
Pelosaurus — -с более широкой головой и несколько укороченным
туловищем. На зубах появляются вертикальные складочки эмали. Раз-
<) Кроме того найдены в большом количестве остатки Р г о t г i t о п, до 60 отпечат-
ков, в угленосных слоях Комантри (Commentry) во Франции. Это вызвало вопрос о возра-
сте слоев: принадлежат ли они к карбону, как это принято считать, или к нижней перми?
Thevenin. Annales Paleontologiques, t. V.
подкласс: панцыреголовые — stegocephali
61
«еры больше, чем у Protriton, — до 20 см. Он же из Отёна описан
Тевененом.
Melanerpeton — до 13 см длины; грудной пояс осложнен добавочными
пластинками и удлиненной межключичной костью.
Отряд 2
Lepospondyli
(Pseudocentrophori Abel, Microsauridae Dawson)
Этот отряд представляет как бы отклонение от первоначального типа.
Сюда входят очень различные по внешнему виду формы с позвонками в виде
трубочек, вогнутых с обеих сторон.
Сем. Diplocaulidae. Удлиненное тело, около 1 м, похоже на тело
головастика с очень ма-
ленькими конечностями и
толстыми ребрами.
Diplocaulus Соре
(рис. 70) — с очень рас-
ширенной треугольной го-
ловой, с глазами на ее
передней поверхности, око-
стеневшей затылочной ча-
стью и плоскими конди-
люсами. Очень тонкий,
длинный хвост. Плечевая
кость имела энтепиконди-
лоидное отверстие для про-
хождения сосудов. Верх-
ний карбон и пермь Сев.
Америки. Громадная го-
лова при тонком длин-
ном теле затрудняет объ-
яснение движений этой
формы. Форма тела ука-
зывает на возможность
быстрого плавания и дви-
жения на суше, а тяже-
лая, балансирующая голо-
Рис. 70. Diplocaulus
magnicornis Соре.
Череп сверху и снизу.
ва на сближенных плоских кондилюсах не могла содействовать подоб-
ному движению.
Эта как бы исключительная форма, Diplocaulus, с резко раздвинутыми
задними концами черепа (tabularia), жившая в верхнем карбоне и перми, имела
своими предшественниками в карбоне формы, у которых череп дает как бы
62
класс: земноводные—amphibia
переходные типы от Diceratosaurus punctolineatus Соре (рис. 71),
у которого череп сжатый и tabularia направлены назад параллельно одна дру-
гой, через Batrachiderpeton
1 i п е a t u m Напе, et Atth., у кото-
рого tabularia раздвинуты и череп
принял более треугольную форму
(рис. 72), приближающуюся к Dip-
locaulus magnicornis Соре.
Рис. 71. Diceratosaurus punctolineatus Соре. (Сев. Америка,
Нижняя сторона черепа.
карбон.)
Верхняя сторона черепа.
Сем. Aistopodidae.
Змееобразное тело, конечности отсутствуют. Ребра
тонкие.
Dolichosoma — в карбоне Ирландии
и Богемии (рис. 73). Брюшной панцырь не
найден. Голова треугольной удлиненной фор-
мы с жаберными придатками у основания.
Длина животного 54 см. Число позвонков
доходило до 150. Позвонки с широкими верх-
ними дугами по форме напоминают песоч-
ные часы.
Сем. Microsaurfdae. По форме тела
ближе к саламандрам. Конечности передние
и задние пятипалые, хорошо развиты; хвост
длинный.
Scincosaurus crassus Fritsch.
(Keraterpeton). Г олова широкая, закруг-
ленная, снабжена у основания двумя выступами,
Рис. 72. Batrachiderpeton
1 i n е a t u m Hanc. et Atth.
направленными назад (рис. 74). Зубы мелкие, хвост длиннее тела, имеет до
75 позвонков, снабженных спинными дугами. Карпальные и тарсальные кости
подкласс: панцыреголовые — stegocephali
63
окостенели, как у Branchiosauria. Верхний карбон. Грудной щит не-
известен.
Эти формы, жившие одновременно в карбоне (Diplocaulus в перми),
представляют, очевидно, параллельные ветви, очень изменившиеся под влиянием
Рис. 73. Dolichosoma longissimum Fritsch.
(Верхний карбон.)
Рис. 74. S с i п с о s a U-
rus crassus Fritsch.
(Верхний карбон.)
изменения жизненных условий, но сохранившие общий тип позвонков. Пока
неизвестно, каковы могли быть эти изменения среды.
Отряд 3
Temnospondyli (Rachitomi Соре)
Более совершенный тип позвонков, состоящих из нескольких окостене-
вающих кусочков, обнимающих хорду, встречается в группе крупных, похо-
жих на крокодилов Stegocephali, достигавших длины до 15/2 м. Группа
эта является более объединенной многими общими признаками: формой тела,
формой черепа, большею частью треугольной, с большими глазницами, со вполне
окостеневшими костями скелета, с зубами, снабженными входящими складоч-
ками эмали.
Cochleosaurus — древнейший представитель из верхнего карбона
Богемии. Хорошо развиты кости черепа (рис. 75), который имеет наверху
отверстия, глазницы и хоаны; кондилюсы окостеневшие. Весь череп покрыт
костным щитом.
Loxomma — другая форма из верхнего карбона Англии. На ее головном
панцыре сильно выражены слизистые каналы (Schleimkanalen, Z it tel), значе-
ние которых точно не известно. Ее остатки указаны даже из нижнего карбона
64
класс: земноводные — amphibia
(менее совершенно сохранившиеся). Особенностью является расположение
зубов и хоан.
За передними зубами, в межчелюстной кости существует двойное отверстие.
За ним следует по зубу и отверстию для нижних зубов. Далее лежат носовые
отверстия на краю верхнечелюстных
костей. За ними отверстия для зубов
нижней челюсти и по большому зубу.
Затем следует промежуток, и опять по
зубу и отверстию. На самом краю меж-
челюстных и верхнечелюстных костей —
ряд мелких зубов. Нёбо совершенно
закрытое.
Сем. Archegosauridae. Аг-
chegosaurus DecheniH. v. М.
(рис. 76), давно известный из пермских
слоев Рейнской Пруссии, представляет
различную форму черепа в разное время
жизни. В молодости она треугольная,
с тупой мордой и орбитами, лежащи-
ми посредине черепной крышки; по сто-
Рис. 75. Cochleosaurus bohemicus
Fritsch.
Левая сторона черепа покрыта панцирем, с правой
он снят.
ронам лежат жабры. С возрастом жабры исчезают, а череп очень удлиняется,
орбиты кажутся передвинутыми ближе к затылку. Кондилюсы не окостеневают,
так что нет настоящего сочленения с атлантом. Зубы расположены по краям
верхней и нижней челюстей и по краям нёба. Передние конечности четырех-
палые, маленькие; задние — пятипалые, большие. Интересны кости грудного
пояса: episternum ромбической формы и треугольные ключицы, лежащие по
Рис. 76. А г с h е g о s a u г u s.
подкласс: панцыреголовые — stegocephali
65
сторонам его. Грудной щит состоял из тонких удлиненных чешуек, образо-
вавших пластинки. Черепа этой формы найдены в большом числе. Целый ске-
лет длиной в 1 м, прекрасной сохранности, найден в нижних пермских отло-
жениях Лебаха, близ Саарбрюкена
Actinodon Frossardi Gaudry — из нижних битуминозных сланцев
пермских слоев Отёна во Франции — известен в хорошей сохранности; най-
ден вместе с Protriton (рис. 77). По внешности это большая саламандра,
немного больше 70 см в длину, с широкой головою, имеющей на переднем
конце морды два носовых отверстия и почти
посредине головы глазницы. На заднем краю
черепа две вырезки, слуховые углубления
(Ohrenscblitze). Затылочная часть черепа око-
стенела, имеет два мыщелка, но поверхность
их вогнутая, что заставляет предполагать,
что их сочленовная поверхность состояла
из неокостеневшего хряша, который и не
сохранился. Зубы меткие, конусообразные,
Рис. 77 А. Грудной щит Actinodon.
Рис. 77. Actinodon.
расположены на верхней и нижней челюстях и на сошнике.
Позвонки состоят каждый из отдельных, не сросшихся частей; верхние
дуги развиты сильно. Каждая состоит из частей, прилегающих к basiventralia
и interventralia, охватывающих хорду снизу. Дуги окостенели не вполне.
К позвонкам причленяются ребра, из которых особенно сильно развиты ребра
верхней части туловища. Они сочленяются с боковыми отростками позвонков,
а не с телами их, которые, не будучи вполне окостеневшими, не могли дать
устойчивой опоры телу. Грудной и задний пояса, первый — типа Archego-
saurus, дают опору пятипалым конечностям. Длинные кости не имеют еще
окостеневших сочленовных поверхностей, как и у Protriton; внутри они
полые.
На экземпляре, описанном Г о д р и, сохранилась значительная часть груд-
ного щита, состоящего из отдельных удлиненных чешуек (3 см длины)
ганоидного типа, т. е. костных, очень твердых, покрытых блестящей эмалью
(с остеобластами) (рис. 77 А). Они были расположены косыми рядами, налегая
друг на друга и сходясь в центральной линии туловища внутренними концами.
Это общий тип панцыря Stegocephali. Годри указывает, что ни у од-
ного из мезозойных и кенозойных животных нет подобного щита. Только у
5 Па в л о в а М. В. Палеозоология. Ч. II.
66
класс: земноводные — amphibia
ныне живущей гаттерии, новозеландской рептилии, существуют подобные че-
шуйки на брюшной поверхности. Но этот признак не дает еще права сбли-
жать эти столь различные по строению других органов формы.
Вместе с указанными остатками Actinodon были найдены его копро-
литы (отбросы кишечника), которые по своей форме и по спиральной полоске,
обнимающей их. указывают на то, что кишечный канал этого животного имел
Рис. 77. В. Копролиты Actinodon.
спиральный клапан, какой существует
у ныне живущих акул. В некото-
рых из копролитов можно различить
чешуйки Palaeoniscus, которого
съедал Actinodon (рис. 77В).
Изучив подробно остатки этой
амфибии, Г о д р и восстанавливает
не только его внешность, но и образ
жизни. По зубам и черепу видно,
что это был хищник с маленькими
глазами и широкой пастью. Во
взрослом состоянии он дышал легки-
ми и жил более на суше, чем в
воде. Здесь он ползал на брюхе,
имея плоскую, как бы придавленную
форму тела, благодаря плоскому
тазу и широким ребрам.
При ползании тело его было очень легко подвижно благодаря гибкому
панцырю и позвоночнику. К массивным костям прикреплялись сильные спин-
ные мускулы, тоже облегчавшие движение.
Итак, мы видим, что из остатков скелета и отбросов кишечника перед
нами восстановилось цельное живое существо, позволяющее не только пред-
ставить себе его во плоти, но наблюдать даже, как оно жило, как бросалось
и хватало рыбёшку своей громадной пастью на берегу пермского озера. Эти
данные и заставили меня остановиться на этой форме несколько подробнее,
чем на других.
Интересно указать на то, что формы, близкие к Actinodon, были най-
дены в Германии — Archegosaurus Н. v. М.; в Индии—-Gondvanosau-
rus Lyd.; в Сев. Америке — Rhachitomus Lyd.; в Уфимской губ. — Zygo-
saurus Eichw. и Melosaurus Trautsch.; в Австралии и Южной Америке —
Bothriceps Huxley. Все это указывает на одновременное существование
близких между собой форм на расстоянии 7—8 тысяч километров друг
от друга.
Eryops Соре — из нижне-пермских отложений Техаса, найденный вна-
чале только в виде черепа и части позвоночника, известен теперь по цель-
ному скелету, восстановленному в музее Нью-Йорка (рис. 78). Позвонки типич-
ные, из кусочков — temnospondyli. Голова, закругленная спереди, очень
похожа на голову Actinodon. Передние конечности пятипалые. Хвост длин-
подкласс: панцыреголовые — stegocephali
67
ный. Тазовые кости плотно срастаются и образуют бедренную впадину. Неко-
торые палеонтологи считают эту форму стоящей ближе к пресмыкающимся. Но
цозвонки из кусочков, хотя и очень сближенных, заставляют удержать ее
между амфибиями. Близкой формой является — Trematopsis— из перми
Техаса.
Trimerorachis insignis Соре — из пермских отложений Техаса,
длиною от а/2 до 1 м. Плоский, расширенный сзади череп. Глаза ближе к пе-
реднему концу морды. Один конди-
люс вогнутый. Тело покрыто тон-
кими костными пластинками. Clavicula и
Рис. 78. Е г у о р s megacephalus Рис. 79. Platyops St uckenbergi
Cope. (Нижняя пермь.) Trautsch. (Вятская губ.)
interclavicula образуют скульптированные внешние пластинки. Кости конеч-
ностей небольшие, со слабым окостенением. Кости тазового пояса (ilium)
очень малы и были слабо связаны с крестцом (по указанию Вилли-
стона— Williston). Седалищные и лобковые, вероятно, не окостеневали.
Все эти данные позволяют предположить, что Trimerorachis был водным
животным (сохранявшим жабры — perennibranchiata), у которого хрящевые
элементы в тазу и конечностях явились вторичным приспособлением к вод-
ному образу жизни — как у ныне живущего Necturus.
Абель считает эту форму наиболее близкой из Stegocephali к
рептилиям (С о t у 1 о s a u г i а).
5*
68
класс: земноводные — amphibia
Acanthostoma vorax Credn. — из нижних пермских отложений близ
Дрездена (Nieder-Hasslich) — с тупой, почти четырехугольной головой и сильно
отодвинутыми назад глазницами.
В пермских слоях найдены еще Stegocephali:
В Вятской губ. — Platyops Stuckenbergi Trautsch.; череп с очень
удлиненным рылом. Сохранность плохая (рис. 79).
Melosaurus uralensis Trautsch. — из Каргалинского рудника Уфим-
ской губ. — представлен черепом 22 см длины с закругленной мордой, с хорошо
сохранившимися двумя затылочными кондилюсами и нижними челюстями.
Близок к Archegosaurus (рис. 80).
Zatrachys microphthalmus Соре (рис. 81). Совершенно особ-
няком стоит сем. Zatrachidae по строению черепа. Широкие praemaxillaria
Рис. 80. Melosaurus uralensis Рис. 81. Zatrachys microphthal-
Trautsch. (Уфимская губ.) mus Соре.
окружают большое отверстие, лежащее между носовых костей (fenestra interna).
Очень маленькие ноздри и орбиты, лежащие за ними, отодвинуты далеко назад
и в стороны. Существует пинеальное отверстие. Назад сильно выдаются
tabulare и quadrato-jugale.
Сем. Dvinosauridae Amal. Stegocephali а, сохранившие во взрос-
лом состоянии жабры, найдены при раскопках на Сев. Двине вместе с Р а г е i а-
saurus, Inostrancevian др. Пермь.
Dvinosaurus Amal. (рис. 82). Животное имело около метра длины,
с большой головой (—1/5 туловища), сильным грудным поясом и слабым
тазом. Хвост и конечности короткие. Около 40 позвонков, несущих сильные
ребра, за исключением трех шейных и конечных хвостовых. Череп треуголь-
ного очертания, с резко выраженной скульптурой на костях, состоящей из
расходящихся от центра костных пластинок — ребер (сходна с Capitosau-
rus и Metopias). Орбиты большие, широко расставлены. Сохранился слож-
ный подъязычный аппарат. Два сочленовных черепных кондилюса и два углуб-
ления на атланте с разделяющею их выпуклостью.
подкласс: панцыреголовые — stegocephali
69
Нижняя челюсть со сложным, примитивным числом костей. Хо-
рошо выражен бронхиальный апп
Lysorophus.
Амалицкий смотрит на эти формы
как на высший член Rhachitomi, как на
боковую прогрессивную ветвь преимущественно
водных форм, сохранивших жабры. Они соеди-
няют признаки пермских северо-американских
(Техас) примитивных форм с признаками
(прогрессивными) три асовых западно-евро-
пейских форм.
Установлено 3 вида по более или менее
расширенной сзади голоье: Dvinosaurus
primus, Dvin. secundus и Dvin. ter-
tius.
Discosaurus Netshajevi Riab.— в
Оренбургской губ.; найден почти полный скелет.
Характеризуется особенной формой грудных пла-
а р а т, сближающий ее с
Позвонки.
Рис. 82. Dvinosaurus Amal.
стинок в виде дисков.
Отряд 4
Stereospondyli (Labyrinthodontia)
Крупные Stegocephali с большой головой, коротким туловищем и
небольшим хвостом. Цельные позвонки, т. е. состоящие из вполне сросшихся
Рис. 83. Mastodonsaurus Jaeg.
Поперечный разрез зуба.
между собою частей. Иногда в
центре позвонка оставался след
хорды. Верхняя дуга имела
большой остистый отросток и
меньшие боковые.
Существовала эта группа с
нижнего карбона до верхнего
триаса включительно, имела
один общий признак в строении
зубов: именно конусообразные
зубы их имели входящую
эмаль, образующую внутри зу-
б а множество складок, делаю-
щих зуб в разрезе очень
сложным — лабиринтообразным,
откуда и название 1.abyrin-
thodontia (рис. 83). Зубы
эти являются как бы дальнейшим
осложнением зубов Arche-
70
класс: земноводные — amphibia
gosaurus. Эта сложность эмали давала зубу значительно большую плотность.
Они напоминают зубы Holoptychius. Весь скелет в этой группе тоже изме-
нился: он весь окостенел, и грудной панцырь стал ненужным и атрофировался.
Череп вполне костный, покрыт, так же как и у всех Stegocephali,
щитом из костных пластин со скульптурой. Затылочная, окостеневшая часть
несет выпуклые кондилюсы.
Зубы многочисленные, мелкие чередуются с несколькими крупными —
в челюстях, на нёбе и на сошнике.
Labyrinthodontia особенно хорошо представлены в триасовых отло-
жениях Германии: в Вюртемберге, Штутгарте и др. Остатки этой группы были
Рис. 83A. Mastodonsau-
rus Jaeg. (</20)
хорошо изучены и подразделены на несколько
родов, близких между собою. Лучше всех предста-
влены роды: Mastodonsaurus и Metopias.
Mastodonsaurus Jaeg. Хотя это живот-
ное и не было до сих пор найдено в виде цель-
ного скелета, но отдельные части его позволяют
составить себе о нем полное понятие.
Череп его имеет 110 см длины и поражает
плотностью строения. Кости его соединены плотно
между собою и покрыты щитом из окостеневших
кожных пластин. Их границы не совпадают с гра-
ницами черепных костей, и число их меньше по-
следних (рис. 83 А). На верхней поверхности череп
имеет три пары отверстий: 1) межчелюстные на
межчелюстных костях для входящих в них
двух больших зубов нижней челюсти; 2) носовые на границе носовых костей,
межчелюстных и челюстных; 3) глазницы, ограниченные лобной, предлобной,
скуловой, заднелобной и заднеглазничной костями. На заднем краю черепа нахо-
дится пара слуховых, не закрытых выемок и пинеальное отверстие между темен-
ными костями, функция которого не определена. Предполагают, что это оста-
ток исчезнувшего органа, может быть третьего глаза. Оно существует, как мы
указали, у всех Stegocephali, у некоторых Crossopterygii и у древ-
них R е р t i 1 i а. Кроме названных костей, в состав черепа входят слезные, ви-
сочные, скуловые и затылочные кости с мыщелками, очень выступающими и
очень сближенными между собою. На правой стороне черепа изображены кож-
ные кости со скульптурой, и видны их границы. На нижней поверхности че-
репа видны нёбные и нёбно-височные отверстия. Кости, входящие в состав ос-
нования черепа, видны на рисунке.
Верхние мелкие зубы многочисленны. Особенно поражают своими разме-
рами четыре зуба, лежащие с каждой стороны передней части сошника попар-
но, за рядом мелких зубов.
Нижняя челюсть состоит из двух сросшихся длинных костей—зуб-
ной и угловой, с большим числом мелких зубов и двумя очень большими
зубами, входящими в отверстия верхней челюсти.
подкласс: панцыреголовые — stegocephali
71
Сочленение этой челюсти с черепом происходило посредством сочленов-
ной выемки в ее заднем отростке.
Первый шейный позвонок представляет особенность: у него есть углуб-
ление для мыщелков и верхняя дуга для прохождения спинного мозга. Спин-
ные позвонки имеют высокий верхний отросток и небольшие боковые, при-
крывающие ребра. У позвонков крестца верхние дуги не замкнуты.
Хвостовые позвонки еще более открыты на верхней стороне. Ко всем
позвонкам причленялись
массивные с двойной головкой ребра.
Грудной пояс состоял
из грех больших кожных костей
(типа Archegosaurus) со
скульптурой, напоминающей голов-
ные кости. К этим костям приле-
гали передние конечности, вполне
развитые. Передние конечности
цельные не найдены, но по най-
денным частям можно думать,
что они имели строение, сходное
с другими известными формами
Labyrinthodontia. Они были
пятипалые. Тазовой пояс задних
конечностей тоже хорошо развит,
хотя лобковые кости еще не сро-
слись с седалищными. Подвздош-
ные сочленяются с последними, и
к ним прилегает бедро.
Для задних конечностей най-
дены отдельные кости, вся стопа
не известна; она. вероятно, пяти-
палая.
Metopias (рис. 84) — Рис. 84. Met о р i a s d i ag n о s t i c us H.v. M. (*/*)
очень сходен c Mastodonsau-
rus: такое же укороченное тело с большой головой и костями переднего
пояса Главное различие заключается в строении нижней части черепа. Пре-
красная реставрация дана Фраасом. Триас Вюртемберга.
Capitosaurus, Cyclotosaurus, Trematosaurus представлены
хорошо сохранившимися черепами, различающимися главным образом по форме
передней части, более или менее закругленной.
Trematosaurus Brauni Burm. представляет отклонение от типа
Mastodonsaurus. У него передняя часть морды не закруглена, как у пре-
дыдущих форм, а более вытянута.
Еще большего удлинения голова достигает yLonchorhynchus из
триаса Шпицбергена, с очень удлиненными межчелюстными костями и отодви-
нутыми к концу их маленькими ноздрями. Существует пинеальное отверстие.
72
класс: земноводные — amphibia
По форме своей череп очень похож на череп крокодила. Форма черепа указы-
вает на морской образ жизни животного (по Абелю) (рис. 85).
Plagiosternum является полной противоположностью ему, с очень
коротким и широким черепом. Верхний триас Вюртемберга.
Интересно подчеркнуть ту общность формы тела и скелета (за немногими
исключениями), которые характерны для представителей отрядов Temnospon-
dyli и Stereospondyli, в противоположность
Рис. 85. Lonchorhyn-
chus Obergi Wim.
Lepospondyli, которые представлены очень раз-
нородными по форме тела животными, позволяю-
щими предположить их полифи-
литическое происхождение.
Кроме скелетов и черепов
Stegocephali их остатки
найдены еще в виде отпе-
чатков следов ног (рис. 86).
Они довольно многочисленны
в верхнем карбоне и в перми
как Европы, так и Северной и
Южной Америки. Особенно
часто встречаются в песчаниках
Пенсильвании. Одни из плит
содержат отпечатки вдавленных
следов, другие — выпуклые, их
выполнявшие. Большая часть
их пятипалые, реже четырех-
палые. Известны они под именем
Chirotherium Barthi. Сле-
ды передних ног меньше
задних.
Принимая во внимание слои,
в которых найдены остатки
Рис. 86. Следы ног
Chirotherium
В art hi Каир.
(Амфибии.)
Stegocephali, можно думать, что они жили отчасти в пресных водах, отчасти
на суше. Многие маленькие формы найдены в дуплах деревьев S i g i 11 а г i а,
Lepidodendron и др., в карбоне.
Познакомившись, даже кратко, с названными формами этого отряда, мы
видим для большой части их преемственность в развитии. Мы можем просле-
дить, насколько строение триасовых Labyrinthodontia усовершенствовалось
сравнительно с пермо-карбоновыми Branchiosauria, пройдя через Arche-
gosauridae, с которыми очевидно связаны обе крайние группы. Мы видим
увеличение размера тела, зубы из простых конусов становятся сложными, с входя-
щими складками эмали, сначала неглубокими у Archegosauridaen очень
глубокими и сложными у Labyrinthodontia. Позвонки из отдельных ле-
пестков образовали цельные окостеневшие тела. Череп окостенел в затылоч-
ной части с мыщелками; также и сочленения конечностей. Очевидно, животные
подкласс: хвостатые— urodcla
73
эти достигли своего кульминационного развития при данных условиях, окружавших
их. Что заставило их исчезнуть, погибнуть в триасе, мы не знаем. Но между всеми
формами, с остатками которых мы познакомились, мы не можем указать предков
современных амфибий. Ближе всего стоят простейшие Protriton (Bran-
chiosaurus) по форме тела к саламандрам и тритонам, как мы уже указали;
но формы, связывающие их с ними, до настоящего времени неизвестны.
Работы Вилл и стон а, Казе (Case), Б р о и л и (Broili) и др. дали описания
многих новых форм и позволили восстановить многих представителей Stego-
cephali в их вероятной обстановке. У Казе мы находим восстановленным
Diplocaulus, выходящего из воды на сушу. Dissorophus multicin-
ctus Соре, близкий к Cacops Соре, но с большими спинными пластинками,
которые он считает образовавшимися из верхних отростков нервных дуг. Мор-
да его была короче, чем у Cacops. Казе, описывая различные формы, ста-
рается указать обстановку, которая их окружала, что делает и Абель1).
Подкласс II
ХВОСТАТЫЕ -URODELA
Ныне живущие хвостатые лягвы име-
ют своих ископаемых представителей.
Рис. 87. Н у 1 а е-
obatrachus
Dollo.
Остатки их известны с
юры, из более поздних
отложений, чем остатки
Stegocephali и ос-
татки Anura, хотя мо-
жно предположить, что
urodela, произойдя от
Branchiosauria, дали
начало Anura.
В юре до сих пор не
найдены представители
хвостатых лягв. Но не-
которые палеонтологи, на-
пример Абель, указы-
вают пермского пред-
ка для Urodela, именно
Lysorophus tricari-
na t u s Соре. Известен
Рис. 88. A n d г i a s S с h е и с h z е г i
Tschudi.
только череп, лишенный кожного щита и сильно отличающийся от Stego-
cephali, к которым эту форму относит Абель. Сочленовный кондилюс
*) Е. С. Case, The permo-carboniferous red beds of N. America and their vertebrate
fauna, Carnegie Institution, Wasnington, 1915. O. Abel, StSmme d. Wirbeltiere, Berlin, 19195
Palaeozoologie, Jena, 1920. S. W. W i 11 i s t о n, American permian vertebrates, Chicago, 1911.
74
класс: земноводные — amphibia
у него трехраздельный. Циттель („Grundzilge", 1923) относит Lysorophus
к Squamata (как и в 1911 г.).
В нижнем мелу (вельде) найден в пресноводных осадках скелет Н у 1 а е о-
batrachus Dollo (рис. 87), хвостатой лягвы, 10 см в длину, с тремя па-
рами окостеневших жаберных дуг. Последующие находки относятся к третич-
ным слоям.
В миоцене Швейцарии (Oeningen) найден скелет гигантской саламандры,
до Р/2 метра, близкой к ныне существующей японской саламандре Crypto-
branchus; это знаменитый скелет Andrias Scheuchzeri Tschudi (Ме-
galobranchus Tsch.) (рис. 88). Ископаемое это было признано первоначально
в 1771 г. за скелет человека и описано д-ром Шейхцером под именем „Ното
diluvii testis" (человек— свидетель потопа). В его широком, с большими глазни-
цами черепе кости сократились, слились больше, чем кости у Stegocepha-
1 i. Пинеального отверстия нет. Зубы маленькие, расположены на межчелюст-
ной, верхне- и нижнечелюстной костях, сошнике и на нёбе.
Andrias Tschudii Meyer —• меньший вид, найден в угольных слоях
верхнего олигоцена Богемии. Это уже настоящая саламандра.
Подкласс III
БЕСХВОСТЫЕ — ANURA
Ископаемые остатки лягушек известны, начиная с юры, в третичных
слоях. В нижнем эоцене они найдены в восточной Индии и Сев. Америке. Из
Рис. Я9. Р a J а е о-
batrachus Gau-
dry i, Vidal. (‘/О
передняя часть тела
верхнего эоцена, олигоцена
и нижнего миоцена остатки
известны во Фоанции, Верх-
ней Италии и Германии.
В Кембридже Испании найден
Видалем отпечаток ске-
лета Palaeobatrachus
G a u d г у i (рис. 89). Это на-
стоящая лягушка уже с ре-
дуцированным скелетом. В
олигоценовых фосфоритах
Франции (Керси) найдены
остатки лягушек Rana р i с-
tata F., совершенно осо-
бой сохранности (рис. 91),
Рис. 90. Ископаемые личинки
лягушки.
Это по большей части
с головой и лапами, превратившаяся в фосфорит. Это не
скелет, а точно окаменелое животное с кожей, сохранившей складки, и с вы-
пученными глазами, выражающими ужас.
Интересны остатки Palaeobatrachus grandipes Dieb., представ-
ленные в миоцене Германии несколькими видами. Найдены не только скелеты
подкласс: бесхвостые — anura
75
взрослых животных, но сохранились отпечатки личинок (рис. 90). Эти живот-
ные мало отличаются от нынешних лягушек.
Плоский череп тоже состоял из тонких костей с большими окостенев-
шими орбитами. Лобные и теменные кости слиты. Зубов нет. Конечности, как
у современных лягушек.
Эти третичные формы являются, конечно, предками нынешних, нр как свя-
зать их с более древними и с какими именно формами — сейчас сказать трудно.
Вероятнее всего, предки их отщепились от Protritonidae, череп которых,
теряя головной щит, мог изменяться в этом направлении так же, как и весь
скелет.
Рис. 91. Черепа лягушек из фосфоритов Керси.
Работы Г. Гадова1) о развитии позвоночника нынешних амфибий ука-
зывают, что листовые позвонки (phyllospondyli) Protriton являются первой
стадией развития у личинок нынешних тритонов, где они состоят из двух верх-
них и двух нижних тоненьких хрящевых пластинок. В некоторых случаях их
четыре пары (добавочные две).
Позвонки из кусочков (temnospondyli) являются во второй стадии разви-
тия тритона.
Кроме изменения скелета, главным образом позвоночника и заднего пояса,
можно проследить изменение черепа у бесхвостых лягв, начиная от закрытого
черепа с большими орбитами у Pelobates cultripes (рис. 91 А), близкого
по строению своему к черепу Stegocephali, и кончая черепом Вufо
с очень от крытой черепной крышкой.
<) Hans Gadow, Amphibia and reptilia, Natural History, Cambridge, 1909.
76
класс: земноводные — amphibia
Уже у Pelobates cultripes (рис. 91 А, А) мы замечаем, кроме
больших орбит, присутствие небольшого отверстия (fenestra — а) между задним
концом squamosa, quadratum и jugale.
yPelobates fuscus (рис. 91
отделяющая его от орбиты, становится
А в
а а
Рис. 91 А. А — Pelobates cultri-
pes. В — Pelobates fuscus.
Названные черепа все высокие, i
Rana, у которой орбита замкнутая. J
А, В) это отверстие увеличивается; кость,
уже: это — мост (Briicke) по Абелю.
У Ceratophrys Broili отвер-
стие увеличивается, мост утончается,
дыра становится сходной с височной
дырой, которую мы встретим у некото-
рых рептилий, но она совершенно дру-
гого происхождения.
yBufo agua мост нарушен —
произошло слияние височной дыры с
орбитой.
ie плоские, какие мы имеем у лягушки
r Urodela, например у М о 1 g е, орбита
не замкнута.
Ч
КЛАСС ТРЕТИЙ............
ПРЕСМЫКАЮЩИЕСЯ — REPTILIA
Почти все ныне живущие представители этого класса знакомы большин-
ству из нас: ящерицы, змеи, черепахи встречаются часто, и только кро-
кодилы, живущие в водах тропических стран, известны сравнительно меньше.
По внешнему виду последние стоят ближе всего к ящерицам.
Но в ископаемом состоянии мы встречаем не только остатки представи-
телей этих четырех подклассов пресмыкающихся, но еще нескольких более об-
ширных, совершенно исчезнувших, вымерших, несущих в себе гораздо более
разнообразия, чем ныне живущие. Сюда относятся большею частью крупные
животные, из которых одни жили на суше и ходили на четырех ногах, дру-
гие — только на задних, опираясь на хвост; еще иные жили в воде, напоминая по
форме тела то змей, то рыб, то ящериц. Наконец, некоторые из них подни-
мались в воздух при помощи выработанной перепонки между передними и зад-
ними конечностями, подобно летучим мышам.
Очевидно, животные этого класса, стараясь приспособиться к самым разно-
образным условиям жизни, достигали большого развития и разнообразия. В мелу
они достигают не виданного дотоле богатства форм, которое может сравниться
только с последовавшим в третичном периоде богатством млекопитающих. Но
в этом же меловом периоде они точно истратили всю силу развития, и большая
часть из них закончила свое существование. Некоторые отряды исчезли не бес-
следно: одни из них дали начало ныне живущим пресмыкающимся, другие —
высшим позвоночным—птицам, с одной стороны, и млекопитающим — с другой.
Чем же отличаются представители этого класса от предшествующих, и ка-
кие признаки обобщают столь различных представителей его?
Мы уже упомянули о внешнем сходстве крокодилов и ящериц; змеи ли-
шены конечностей, но по скелету похожи на два первые подкласса. Одни
только черепахи резко отличаются и от других рептилий и от амфибий.
Главными отличиями рептилий от последних являются: 1) легочное дыхание
в течение всей жизни, следовательно — отсутствие жабер, и 2) строение внутрен-
него скелета. В черепе увеличилось окостенение, что особенно заметно в верх-
них и основных затылочных костях, так как боковые окостеневали и у ам-
фибий. Эти кости окружают затылочную дыру; на основных затылочных костях
развивается один сочленовный отросток, condylus, сочленяющий череп с пер-
вым шейным позвонком, — вместо двух, как это было у амфибий и встречается
78
класс: пресмыкающиеся — reptilia
у немногих рептилий. Но у некоторых рептилий существуют и боковые конди-
люсы, сливающиеся с основным, в результате чего образуется тройной, сложный
затылочный отросток. Нижняя челюсть прикрепляется посредством квадратной
кости. Позвонки имеют более полное окостенение, чем у амфибий. Они бывают
амфицельные — двояковогнутые, или процельные — вогнутые спереди и
выпуклые сзади, или опистоцельные — выпуклые спереди и вогнутые сзади.
Конечности представляют много различий, от правильных пятипалых до
трехпалых, в зависимости от
образа жизни. Также
Рис.92. Sphenodon (Hatteria) punctatum Gray.
и пояса конечностей, пред-
ставляющие более или
менее цельный орган, осо-
бенно задний пояс — таз.
Остановимся несколь-
ко на строении черепа
рептилий, так как оно
положено Осборном
(Osborn) в основу клас-
сификации.
Черепа всех репти-
лий имеют в своем строе-
нии некоторые общие при-
знаки, например одни и те
же главные кости: межче-
люстные, носовые, лоб-
ные, теменные, затылоч-
ные, височно-скуловые и
верхнечелюстные. Неко-
торые из этих костей
образуют височные дуги, именно: squamosum — чешуйчатая, postfrontale —
залобная, postorbitale — заглазничная, quadrato-jugale — квадратно-скуловая и
jugale — скуловая; иногда supratemporale — надвисочная. Но эти кости дуг соеди-
няются между собою различно — в одну или в две дуги, что и дает два типа черепов.
У одних, например у гаттерии (рис. 92), если мы будем смотреть на
череп сбоку, мы увидим за лобными костями теменные, по обеим сторонам
которых лежат верхние височные отверстия, ограниченные parietalia — темен-
ными, squamosum, postorbitale и postfrontale. Это верхние височные дуги, огра-
ничивающие снизу верхние дыры. За ними следуют вторые, боковые височные
дыры, ограниченные снизу скуловыми костями—jugalia, образующими нижние
дуги, так что в черепе этого типа мы имеем две пары височных дыр, огра-
ниченных двумя парами височных дуг. Проф. Осборн назвал этот тип черепов
Diapsida — двойные дуги.
У другого типа — например у Gordonia из Dicynodontia — обе
дуги сближены (postorbitale, postfrontale, squamosum с jugale), и получиласч
одна пара дуг, ограничивающая одну пару дыр. Это Synapsida проф.
Осборна — слившиеся дуги (рис. 93).
класс: пресмыкающиеся — reptilia
79
При развитии этих двух типов может произойти исчезновение одной или
обеих дуг, что и ведет к новым типам; например, исчезновение верхней дуги
может привести к развитию черепа птиц, исчезновение обеих дуг — к черепу
млекопитающих. Величина и положение височных отверстий могут быть раз-
личны. Но между рептилиями есть еще одна группа, черепа которой предста-
вляют замкнутую коробку, без височных дыр, как у млекопитающих. Это Р а г е i а-
sauria или Cotylosauria, например Gordon!а (рис. 93). Проф. Осборн
относит их к Synapsida по развитию черепа.
У низших рептилий имеется одно нёбо, на
котором хоаны открываются щелями, разделенными
сошником (первичное нёбо), например у А ё t о s а и-
rus (древний триасовый крокодил). У других —
высших рептилий развивается второе нёбо, обра-
зующееся соединением в плоскости выростов верхне-
челюстных и небных костей, причем образуется
полость между двумя небами. Хоаны при этом
отодвигаются назад (ныне живущий крокодил).
Зубы существуют у всех рептилий, кроме
черепах. Они изменяются в зависимости от пищи
у хищных и травоядных. Наружный покров у боль-
шинства состоит из чешуй. У некоторых разви-
ваются кожные пластины. Некоторые остаются
голыми.
Принимая во внимание классификацию О с-
Рис. 93. G о г d о п i а.
бор на и внося в нее
Подкласс A n а р s i d а
некоторые изменения, мы
Подкласс Synapsida
имеем подклассы и отряды:
Подкласс D i а р s i d а
без дуг
Отряд Cotylosau-
r ia Соре (Р а г е i а -
s a u ria Seeley)
1 пара дуг
Отряд Pelycosauria
„ Theromorpha
„ Ichthyosauria
„ Plesiosauria
„ Placodontia
„ Testudinata
(часть)
2 пары дуг
Отряд Prosauria
(Rhynchocephalia)
„ Squamata.
„ Crocodilia
„ Dinosauria
„ Pterosauria
Мы переносим Ichthyosauria к Synapsida, как имеющих одну
видимую пару дуг, a Cotylosauria выделяем в новую группу — Апар-
s i d а, не имеющих дуг, с цельным котелкообразным черепом.
Присутствие или отсутствие височных дуг не может считаться признаком
более древним или более новым, так как между древнейшими формами мы
встречаем представителей тех и других отрядов:
§ ( Rhynchocephalia — Diapsida
S" । Theromorpha — Synapsida
и ( Cotylosauria—Anapsioa.
80
класс: пресмыкающиеся — reptilia
Ныне живущие относятся к D i а р s i d а, кроме черепах.
Но не все палеонтологи принимают эту классификацию; мы встречаем и
иную группировку, например по образу жизни животных — наземные, водные
и так далее.
Указать предка пресмыкающихся мы не можем, хотя на некоторые
формы из Stegocephali указывают как на промежуточные или, лучше
сказать, объединяющие эти два класса, и дают им название Proreptilia.
Так, Гадов считает Eryops и Trematops Сев. Америки за такие формы
благодаря срастающимся хвостовым позвонкам и несколько иному, более совер-
шенному составу и соединению между собою частей позвонков сравнительно
со Stegocephali.
Другие признаки, характерные для древних амфибий, — головной щит, не-
окостеневшие части позвонков и т. п. — заставляют других ученых считать Ery-
ops принадлежащим к классу амфибий. Абель считает Trimerorachis
из Stegocephali (с одним кондилюсом) очень близкой формой к пресмы-
кающимся.
Из рептилий наиболее близкой группой к амфибиям, по закрытому че-
репу, являются Anapsida (Cotylosauria Соре или Pareiasauria Seeley),
которые раньше были включены в отряд Theromorpha, но как по строению
черепа, так и по другим признакам должны быть выделены в отдельный отряд.
Подкласс I . . - .. , ..
ANAPSIDA PAVL.
Отряд
Котелкочерепные — Cotylosauria Соре
(Pareiasauria Seeley)
Древнейшая группа из пермо-карбона, верхне-пермских отложений и три-
аса. Крупные амфибиоподобные формы с совершенно закрытым черепом и
большею частью скульптированными черепными костями. Пинеальное отверстие
существует. Ноздри на конце морды — раздельные. Зубы конические, располо-
жены на нёбе, в углублении челюсти или в лунках. Одни Cotylosauria
относятся к protothecodonta, с зубами, сидящими в общем углублении, что
встречается и у Stegocephali, другие — к thecodonta, с зубами, сидящими
в отдельных альвеолах, что является высшей степенью специализации. Позвонки
амфицельные, глубоко вырезанные. Пятипалые конечности прикреплены срос-
шимися поясами. Этот последний признак является совершенно не соот-
ветствующим всем другим первичным признакам.
Сем. Pareiasauridae. Pareiasaurus Seeley является именно таким
коллективным типом, с головой треугольной формы или закругленной
спереди, напоминающей Labyrinthodontia; череп сплошной, швы не
заметны; кости очень шероховатые. Двойное носовое отверстие разде-
ляется снизу сошником (vomer), который, как и нёбо, покрыт двумя рядами
подкласс: anapsida pavl.
81
маленьких зубов (8 на нёбе). Нижняя челюсть имеет характерный, направлен-
ный вниз отросток почти посредине нижнего края ее. Затылочный condylus
очень большой. Позвонки имеют толстые отростки; крестцовых два. Кости таза
срослись и имеют углубление для головки бедра, характерное для млекопитаю-
щих. Грудной пояс состоит из широкой Т-образной межключичной кости,
к которой причленяются ключицы, лежащие задними концами на очень толстых
лопатках. Плечевая кость с энтэпикондилоидным отверстием; бедра очень укоро-
чены. Локтевая кость имеет высокий отросток для сочленения с плечом. Пальцы
снабжены когтями. Долго Pareiasaurus был известен только из перми
южной Африки (Кару). Но несколько лет тому назад были найдены в большом
числе его черепа и скелеты проф. В. П. Амалицким на Сев. Двине *).
Рис. 94. Pareiasaurus Karpinskii n.mal. (Сев.Двина, пермо-карбон.)
Pareiasaurus Karpinskii Amal., найденный цельным скелетом, за-
ключенным в песчаной линзе, имеет выпуклый, а не вогнутый позвоночник,
как это было раньше восстановлено Сили (Seeley) по африканскому экзем-
пляру. Вместе с этим скелетом найдены кожные звездчатые пластинки, покры-
вавшие частью тело животного. Длина скелета 2 м 45 с я, черепа—-48 см
(рис. 94).
Pareiasaurus tuberculatus Amal. Длина 2 м 60 см, с тремя
рядами кожных, звездообразных пластинок, расположенными по сторонам шеи.
Pareiasaurus horridus Amal.— с рогообразными отростками на щеках.
Pareiasaurus elegans — с небольшой сравнительно головой — 41 см-
Seymouria — американская и русская пермские формы, еще более по-
хожи по черепу на Stegocephali, с овальными ноздрями на конце морды,
овальными глазами посредине ее (рис. 95). На черепе панцыря не было; на нем
*) Diagnoses of the new formes of vertebrata and plants from the upper permian
on N. Dvina, изд. Росс, академии наук, 1922.
6 Павлова М. В. Палеозоология, ч. II.
82
класс: пресмыкающиеся — reptilia
Рис. 95. Seymouria bailorensis
Broili.
отчетливо видны швы между почти квадратными лобными и теменными костями.
Имеется пинеальное отверстие. Выемки для органов слуха открыты сзади черепа.
Конические зубы в челюстных и меж-
челюстных костях. На нёбе если и были
зубы, то небольшие. Ребра многочислен-
ные, начиная от шеи до конца хвоста; на
крестце они очень маленькие- Части таза
неплотно срослись, швы расходятся, но
бедро хорошо сочленялось, входя в верт-
лужную впадину. Оно имеет вырезку
на передней стороне и отросток посре-
дине задней для прикрепления мускулов
(3-й trochanter). Конечности пятипалые,
со многими фалангами. Эта форма еще
очень близка к Stegocephali, но
один затылочный кондилюс и строение
таза резко отличают ее от амфибий.
Виллистон считает ее примитивной
из Cotylosauria1). Академик Н. А.
Сушкин относит Seymouria к S t е-
gocephali.
В. П. Амалицким была найдена
на Сев. Двине, близ д. Соколки, форма,
очень близкая к Seymouria Will.,
которую он описал под именем Kot-
las siа (подрод рода Seymouria —
рис. 96). Длина ее 125 см, форма тела ящерицеобразная. Треугольный череп
суживается кпереди. Зубы конические, заостренные, очень маленькие. На задней
Рис. 96. Kotlassia Amal. (Seymouria Will.).
*) S. W. Williston, American permian vertebrates, Chicago.
S. W. Williston, Water reptiles of the past and present, 1914.
подкласс: anapsida pavl.
83
части крыловидной кости существуют шагреневые зубы, бугорчатые непра-
вильные шероховатости. Амалицкий установил этот подрод, главным образом,
вследствие большого расстояния, отделяющего его форму от американской формы.
Найден полный позвоночник с черепом, тазом и задней конечностью.
Рис. 97. Labidosaurus hamatus Соре. (Пермь.) (*/s)
Labidosaurus Соре — из перми Техаса, небольшое животное около
70 см в длину. Плоский череп с закругленной мордой и опускающейся вниз
межчелюстной костью в виде клюва напоминает Rhynchocephalia (рис. 97).
Кости черепа со скульптурой. Швы замкнуты. Зубы в межчелюстной (3) и челюст-
ных костях. Скелет менее массивен, чем у Pareiasaurus. Лопатка не захо-
дит на отростки позвонков. Плечевая кость очень выгнутая с энтэпикондилоид-
ным отверст Йем. Лучевая немного короче локтевой. Пять тонких пальцев. Кости
каждой половинки таза плотно срослись, образуя вертлужную впадину (рис. 97 А).
Задние конечности тоже с пятью тонкими пальцами. Хвост массивный. Позвонки
сохраняют хорду. Жил на берегу,
спускаясь в воду.
Limnoscelis paludis
Will. — из пермо-карбона Новой
Мексики (Сев. Америка), представ-
лен двумя скелетами, из которых
один почти полный, длиною до
22/2 м, сочленения в нем не нару-
шены. Череп плоский, почти тре-
угольный, с вытянутой мордой,
загнутыми вниз межчелюстными
Рис. 97A. Laoidosaurus hamatus Соре.
А—таз спереди, В—снизу.
костями, с тремя парами больших резцов. Зубы в челюстной кости уменьшаются
по направлению кзади. Швы черепа хорошо сохранились; существует пинеальное
отверстие. Нижние челюсти массивные, расширяющиеся кзади. Спинные по-
звонки имеют расширенные верхние отростки; тела позвонков вогнутые. Пояса
типа Pareiasaurus. Гленоидная впадина глубокая. Короткое плечо расши-
ряется книзу. Конечности пятипалые, с 3—5 фалангами. Это был хищник, что
видно по челюстям. По образу жизни это — земноводное животное, так же как
и Labidosaurus.
6*
84
класс: пресмыкающиеся — reptilia
Рис. 98. Elginia mirabilis Newt. (Верхняя
пермь.) (У3)
Даже краткое описание этих форм дает определенные указания на их
непосредственное родство со Stegocephali, при упорном сохранении неко-
торых примитивных признаков, как вогнутые позвонки, пинеальное отверстие,
массивная черепная коробка с костями со скульптурой, и на быстрое развитие
новых признаков, например один кондилюс и сросшиеся кости поясов.
Diadectes phaseolinus Соре — из перми Техаса и Новой Мексики —-
достигал 3 м длины, с закругленной мордой, с крепко построенным позво-
ночником, сильным хвостом и очень развитым грудным поясом и придаточными
пластинками на передних ребрах. Все это заставляет предполагать, что это было
роющее животное. Зубы передние режущие; задние (по 8) поперечно удлинен-
ные, что указывает на питание мягкой пищей: насекомыми, червями и т. п.
Diasparactus zenos Case —близкая к нему форма, указана в пермо-
карбоне Новой Мексики и является таким образом наиболее древней из С о-
tylosauria, а не Pareiasaurus, как считалось раньше.
Stephanospondylus
р u g п а х Stapp. — из перми
Саксонии — небольшое живот-
ное; череп напоминает по фор-
ме черепаший, с большим пи-
неальным отверстием и слухо-
выми углублениями.
Pantylus cordatus
Соре—пермь Техаса1). Широкая
голова со сжатой передней ча-
стью, крупными черепными
костями и глазами, расположенными впереди. Пинеального отверстия нет.
Мелкие зубы в челюстях и покрывают все нёбо.
Отклоняющейся от названных форм является Elginia mirabilis New-
ton (рис. 98). Верхняя пермь (Эльгин, Шотландия). Треугольный череп покрыт
костными пластинками, с большими буграми посредине. По краям черепа тон-
кие отростки; сзади два длинных рогообразных выступа. Носовые отверстия
на кончике морды. Громадные глазницы посредине черепа. Мелкие зубы с от-
шнурованной коронкой на челюстях. Абель сравнивает его с ныне живущими
Phrynosoma (Iguanidae) и молохом, но у них шипы роговые, а не
костные, как у Elginia 2).
Названные здесь формы составляют небольшую часть найденных и
изученных за последнее время Cotylosauria. Но и по ним уже видно,
насколько отряд этот был богато и разнообразно представлен в перми и как
было велико его распространение: на Сев. Двине, в Сев. Америке, в южной
Африке, в Саксонии.
<) R. В г о о m, On the Cotylosauria genus Pantylus Cope. Bull. Americ. Nat. Hist,
vol. 32, 1913.
2) 0. Abe 1, Die Stamme d. Wirbeltiere, 1919, S. 333—350.
подкласс: synapsida
85
Сем. Procolophonidae Zittel (отряд Procolophonia Abel) соединяет
в себе некоторые признаки Cotylosauria, а с другой стороны, сильно
отклоняется от последних.
Сюда входят Procolophon trigoniceps Ow.— пермь Кару, S с 1 е г о-
saurus armatus Н. v. Meyer — средний триас Швейцарии, близ Базеля,
Telerpeton elginense Mant. —
Эльгин в Шотландии, К о i 1 о s k i о-
saurus coburgiensis Huene —
из триаса Кобурга.
Они характеризуются плоским,
широким черепом, утончающимся
в передней части, присутствием боль-
шого пинеального отверстия на темен-
ной кости, одним кондилюсом, загну-
тыми межчелюстными костями, кост-
ными отростками в задней части черепа
(tabulare), особенно у Sclerosau-
rus и Telerpeton (рис. 99), и
громадными орбитами, отодвинутыми
назад, которые являются, по указа-
нию Абеля, орбитами, слив-
шимися с теменными (верхни-
ми) дырами. Зубы у них закруг-
ленные, тупые конусы на челюстных,
более удлиненные на межчелюстных.
Носовые отверстия на конце морды.
Все кости грудного пояса не слива-
лись, свободны: interclavicula, clavi-
cula, большие coracoidea и procora-
coidea. Кости таза — почти слившиеся
Рис. 99. Telerpeton elginense Mant.
между собою, как у Cotylosauria. Череп сочленяется с нижней челюстью
посредством quadratum, входящей в углубление нижней челюсти, оставляя
сзади себя отросток articulare.
Подкласс II
SYNAPSIDA
Отряд 1
Шлемочерепные — Pelycosauria
Отряд этот был основан американским палеонтологом Копом для форм,
имевших длинные спинные отростки позвонков, выходящие наружу. Позже отряд
этот увеличился находкой новых форм. Между ними пришлось различить две
группы: а) с длинными спинными отростками и б) с нормальными отростками.
86
класс: пресмыкающиеся — reptilia
Последние по форме тела близки к Cotylosauria. Изучение их проведено
главным образом Вилл истоном иКазе. .
Все формы этой группы найдены в перми и триасе, главным образом
Сев. Америки, но есть и европейские. Различной величины животные, от 70 см
Рис. 100. V а г а п о-
saurus acutiros-
trjs Broili.
до 1,5 м, ползали на брюхе. Имели одну верхнюю пару
височных дыр; у некоторых существовала маленькая
пара боковых дыр. Носовые отверстия были разделены.
Зубы крупные на челюстях и мелкие на нёбе. Пинеальное
отверстие. Коракоидная кость сливается с лопаткой у
взрослых форм.
Из группы, не имеющей отростков, выходящих
b
Рис. 100А. а — Varano-
saurus brevirostris
Will, b — Var. acutiro-
s t r i s Broili.
наружу, познакомимся с
Сем. Varanosauridae.
Представители его имели плос-
кий череп, сходный с черепом
нынешнего V а г a n u s.
Varanops и Varanо-
s a u г u s — полуназемные, бы-
стро двигавшиеся животные, из
нижней перми Техаса.
Varanosaurus brevi-
rostris Will. (рис. 100 А) —
череп с удлиненной мордой,
глазами, расположенными по-
средине черепа. Зубы (до 50)
мелкие, только спереди 2 боль-
ших. Спинные отростки низкие. Длинный
хвост. Пятипалые конечности
с тонкими пальцами. Последние фаланги удлинены. Животное достигало
Рис. 100В. Varanosaurus acutirostris Broili.
Р/2 метра длины. В грудном поясе сильно развита удлиненная межключичная
кость; существуют ключицы, тонкие и широкие лопатки. Плечо сильно рас-
ширено внизу, с эпикондилярным отверстием. Лучевая и локтевая кости почти
одинаковой длины. Тазовые кости не вполне срастались. Углубление для бедра
овальное. Лобковая и седалищная очень вытянуты, подвздошная с тонким от-
ростком. При соединении тазовых костей остается квадратное отверстие. Этот
таз резко отличается от таза Cotylosauria. У Varanosaurus ас и-
подкласс: synapsida
87
tirostris глаза — по бокам средины черепа. Было найдено 25 скелетов
Varanosaurus в Техасе.
Рис. 102. Clepsydrops natalis Cope.
Vara n о p s — близкая к Varanosaurus форма, с меньшим числом зубов.
Casea Broili Will. — другая ящерицеподобная форма с более корот-
кой, закругленной впереди мордой, большими глазницами по бокам черепа и
нижними дырами за ними (рис. 101). Один
большой кондилюс. Зубы—тупые конусы траво-
ядного типа, уменьшающиеся кзади. Нижняя
челюсть, очень низкая спереди, утолщается кзади.
Швов на черепе не видно. По строению скелет
Рис. 101. Casea Broili Will.
близок к Varanosaurus; конечности имеют такие же удлиненные, но более
тонкие коготные фаланги. Плечевая кость еще более расширена книзу, с эпи-
кондилярной дырой. Ребра сильно закруглены. Кости заднего пояса, соеди-
ненные швом, не вытянуты, как у Varanosaurus, а закруглены.
Сем. Clepsydropidae Соре — с выступающими остистыми отростками 1).
Ophiocodon — по об-
щему строению близок к преды-
дущим, но имеет более высокую
и узкую голову и спинные от-
ростки выше нормальных, но
над кожей не выступавшие.
Зубы — острые сжатые конусы
с режущими краями. Вероятно,
более наземное животное, чем
его современники в Техасе.
Пермь Новой Мексики.
Clepsydrops natalis
Соре (рис. 102) — с высокими
отростками, почти равными вы-
Рис. 103. Dimetrodon incisivus Соре.
соте тела на спине и уменьшающимися к шее и хвосту. Голова несколько
укорочена. Большой хищник. Пермь.
‘) Е. С. Case, The permo-carboniferous red beds of N. America and their verteb-
rate fauna, Washington, 1915.
S. W. Williston, American permian vertebrates, Chicago; id., Water reptiles of the
past and present, Chicago, 1914.
88
класс: пресмыкающиеся — reptilia
Dimetrodon incisivus Cope (рис. 103, 103 А) представляет следую-
щую ступень по развитию отростков и зубов. У него шлемообразный высокий че-
реп, сжатый с боков, с маленькими ноздрями спереди и глазницами, далеко
отстоящими назад. За ними лежали большие височные дыры. Межчелюстные кости
отграничивались от челюстных выемкой. В межчелюстных были большие кониче-
ские зубы, 3 пары; в челюстных второй зуб большой, загнутый, за ним многочи-
сленные меньшие. Они указывают на
Рис. 103А. Череп Dimetrodon.
хищность животного, жившего, вероятно,
на берегу озер. Конечности были пяти-
палые. Плечевые кости очень широкие
в нижней половине, с энтэпикондилоид-
ным отверстием. Высоко поднимавшиеся
спинные отростки придавали всему жи-
вотному устрашающий вид. Между от-
ростками была, по всей вероятности,
протянута кожистая перепонка. Хвост
не длинный. Сочленовные поверхности
костей хорошо развиты. Пермь Техаса.
Edaphosaurus cruciger Соре. Спинные отростки осложнились еще
поперечными придатками. Череп укорочен и более высок, чем у Dimetro-
don. Челюсти несут конические зубы; на нёбе перетирающие зубы. Пятипалые
конечности короче, чем у Dimetrodon. Это, очевидно, было ползающее
животное, отклонившееся в своем развитии от линии Clepsydrops и Di-
metrodon. Пермо-карбон Техаса.
Отряд 2
Звереподобные—Theromorpha Соре (Theromora Соре)
Этот отряд рептилий заключает в себе группы с очень различными при-
знаками, которые сближают их по внешнему сходству с зверями, откуда и
название. Они отнесены в подкласс Synapsida — с одной парой височных
дыр. Их жизнь прошла в перми и триасе, конце палеозоя и начале мезозоя.
Они были современниками древних Prosauria, Cotylosauria и Pely-
cosauria. Относительно классификации их далеко не все палеонтологи схо-
дятся. Мы принимаем разделение их на подотряды:
Theriodontia — зверезубые,
Deuterodontia — разнозуб ые,
Dicynodontia —двуклыковые.
Подотряд 1
Зверезубые — Theriodontia
Являются группой наземных хищников — пресмыкающихся с диффе-
ренцированными зубами, сходными иногда с зубами хищных млекопи-
тающих, откуда и название их — Theriodontia. Передние их зубы, в меж-
подкласс: synapsida
89
челюстной кости, — самые маленькие конусы — это резцы. По одному или по
два крайних в челюстных — большие конусы-—клыки, а остальные малень-
кие, иногда осложненные боковыми зубчиками — коренные. Все зубы с про-
стыми одиночными корнями. Дальнейшее отклонение от рептилий и приближе-
ние к млекопитающим заключается в затылочных кондилюсах, которые являются
здесь двойными или даже тройными (соединение оснований двух боковых, очень
Рис. 104. Inostrancevia.
сближенных, или с добавлением среднего, типичного для рептилий). На черепе
очень отчетливая одна пара височных дуг. Ноздри — на конце морды, хоаны —
посредине нёба. Зубов небных нет, или есть очень маленькие. Части грудного и
тазового поясов соединены швами. Позвонки амфицельные, являются здесь таким
же анахронизмом, как у некоторых пинеальное отверстие. Конечности пятипалые.
Нижняя челюсть с венечным отростком, сочленяющимся с черепом. Эта
группа, очевидно близкая к амфибиям
по двум кондилюсам, успела развить
в перми и триасе некоторые признаки
млекопитающих, после чего исчезла.
Мы не знаем представителей ее поз-
же триаса, после появления млеко-
питающих. Но какая ближайшая к этим
формам связала амфибий с млекопитаю-
щими — остается загадкой.
Inostrancevia Alexandri
Amal. (Сев. Двина) — большой перм-
ский хищник, у которого сильно раз-
виты верхние клыки, спускающиеся наружу до края нижней челюсти. За клы-
ками белые острые зубы (рис. 104, 104 А). Это — крупная форма, достигав-
шая 3 м длины. Плоский череп заканчивался спереди загнутыми вниз межче-
люстными костями с резцами. Носовые отверстия разделены на конце морды.
Глазницы отодвинуты далеко назад. За ними верхние височные дыры. Нижняя
челюсть сильно развита. Конечности пятипалые — звериные.
Другие виды Inostrancevia дают возможность проследить изменение
озубления !).
Рис. 104 A. Inostrancevia Ale-
xandri Amal. (Пермь Сев. Двины.)
*) П. А. Православлев, Gorgonopsidae. Северо-двинские раскопки
проф. В. П. Амалицкого, II, III, изд. Академии наук, 1927.
- 90
класс: пресмыкающиеся — reptilia
Cynognathus crateronotus Seeley (рис. 105) — тоже крупное жи-
вотное, до 2 м длины, с черепом в 40 см, похожим на предыдущий, но с более вы-
сокими теменными костями и сильно развитыми клыками, небольшими глазницами.
Коренные зубы (9) имеют боковые зубчики Четыре пары резцов в межчелюст-
ных костях — маленькие; в нижней челюсти — три пары (рис. 108 С). Найден
Рис. 105. Cynognathus crateronotus Seeley. (Триас южной Африки.)
цельный скелет: сохранились пояса и позвоночный столб из 29 позвонков;
они амфицельные.. Триас южной Африки (Кару).
Gomphognathus Seeley (Diademodon) (рис. 106, 106 А) до-
стигал еще большей специализации в зубах; тоже из триаса южной Африки.
Четыре пары верхних резцов, за ними по большому клыку; коренные (12)
притупленные, отделены от клыков небольшой диастемой. Величина их увели-
чивалась кзади; последние три уменьшались. Два затылочных кондилюса
раздельны. Нижняя челюсть с высоко поднимающимся венечным отростком,
имела 6 резцов, 2 больших клыка и по 12 коренных с каждой стороны.
Передняя часть морды отделяется впадиной от остальной части черепа. Брум
указывает на то, что у Gomphognathus Seeley и у Diademodon
В г о w n i была смена резцов, клыков и четырех предкоренных; коренные зубы были
снабжены бугорками: на больших зу-
бах— по четыре или по пяти, послед-
ний, маленький, имел два бугра: один —
большой, загнутый в виде конуса, дру-
гой — маленький, придаточный. Это уже
зубы типа млекопитающих.
Delphinognathus — с укоро-
ченным, высоким черепом, редуцирован*
ными зубами, однородными, не хищного
типа (рис. 107). Из пермских отложе-
ний южной Африки.
Dvinia prima Amal. близко примыкает к Gomphognathus. Пермь
Сев. Двины.
Несколько других родов стоят близко к вышеназванным: Lycosuchus —
пермь южной Африки, Galesaurus Ow. — триас южной Африки и др. Хотя
для форм этой группы не найдены конечности вместе с черепом или позво-
ночником, но сюда относят конечность, описанную под именем Therio-
d е s m u s Seeley, тоже из перми южной Африки (Кару),
Рис. 106. Верхняя и нижняя челюсти
Gomphognathus.
подкласс: synapsida
91
Здесь мы видим под radiale и ulnare четыре карпальные косточки,
лежащие над вторым рядом, а между ними две центральные. Эпифизы фаланг
еще отделены от суставов.
Anna Petri Amal.•— с
громадными верхними височны-
ми дырами, близка к S с у 1 а-
cosaurus Broom.
Различные представители
Theriodontia дают возмож-
ность проследить изменение
нижней челюсти от типа реп-
тилий к типу млекопитающих
(рис. 108).
Так, у Scylacosaurus
Sclateri Broom (пермь Кару) —
с одним кондилюсом, articulare
сочленяется с quadratum. У В а-
uria cynops Broom deutale
увеличивается, сочленение пере-
ходит с articulare на dentale, а
в черепе — с quadratum на Рис. 106А. Нижняя челюсть Gomphognathus.
souamosum. Триас Кару.
У Cynognathus crateronotus Seeley — angulare и articulare со-
кращаются, задняя часть нижней челюсти (венечная) сочленяется со squamosum,
рог.
Рис. 107, Delphinognathus conoceplia-
1 u s Seeley. (Пермь южной Африки.)
Триас Кару.
У Sesamodon Browni
Broom angulare и articulare ис-
чезли. Триас Кару. Это уже
сочленение, сходное с таковым
у млекопитающих. Кроме того,
у последней формы хорошо
развито слуховое отверстие.
Подотряд 2
Разнозубые — Deuterodontia
В эту группу поставлены
не вполне хорошо известные
формы рептилий из перм-
ских слоев У ф и м с ко й губ.,
найденные еще в 40-х годах и
описанные Эйхвальдом и
Фишером по отдельным частям черепов и костей. По дифференцировке
зубов они стоят близко и к Theriodontia.
92
класс: пресмыкающиеся — reptilia
Рис. 108. А— Scylacosaurus. В — Bauria. С— Cynognathus. D — Sesa-
me d о n.
Рис. 109. Deuterosaurus Eichw.
Deuterosaurus Eichw. (Ключевский рудник Уфимской губ.). Череп со-
хранился довольно хорошо (рис. 109). Он высокий, короткий, сжатый с боков,
с маленькими ноздрями и крупными глазницами, отнесенными далеко назад.
Зубы дифференцированы; р« зцы мелкие, крупные клыки с зазубренными
краями. Спереди морда закруглена, и нижняя челюсть тоже. Затылок не сохра-
подкласс: synapsida
93
нился. Височные дыры существуют. Лопатки очень расширены в верхнем
конце; другие кости пояса не
половине, в нижней тонкое,
также и бедро.
Rhophalodon Fisch,
(рис. ПО) имел более удли-
ненную морду с плоским вверху
черепом, маленькими глазница-
ми, снабженными склеротиче-
ским кольцом. Клыки большие;
коренные сжатые, острые. Ло-
патка почти четырехугольная,
без верхнего расширения. Длин-
сохранились. Плечо несколько шире в верхней
Рис. 110. Rhophalodon Fischer. (Пермь Уфим-
ской губ.)
ные кости конечностей не массивны. Ступни не известны ни у одной из этих
форм. Близкие формы к Deuterosaurus найдены в виде плохо сохра-
нившихся остатков при раскопках проф. Амалицким.
Подотряд 3
Двуклыковые — Dicynodontia
Как показывает название, эти формы характеризуются присутствием
только двух зубов — клыков — в верхней челюсти. Все остальные зубы
Рис. 111. Dicynodon
Trautscholdi Amal.
исчезли. Череп длиною ог 12 до 50 см, сжатый
с боков, имел большие височные дуги, глазницы,
расположенные по сторонам, ноздри спереди над за-
гнутыми межчелюстными костями. Существовало пи-
неальное отверстие. Пермь Сев. Двины и Кару (южная
Африка); нижний триас Шотландии.
Dicynodon pardiceps Ow.(южнаяАфрика).
Череп (рис. 112) с громадными клыками, направленными
вперед и вниз концами. Сильно развита squamosum,
передний отросток которой соединяется со скуловой,
а другой — нисходящий — с квадратной костью. Нёбо
вторичное, неполное, образовано из верхнечелюстных
и нёбных костей. Тонкий сошник вдвинут между
челюстными костями. Пятипалые конечности предста-
вляют интересные особенности. Плечо имеет дель-
тоидный гребень и энтэкондилоидное отверстие, суще-
ствующее у млекопитающих. Грудной пояс состоит
из сросшихся костей: удлиненной лопатки, имеющей
гребень — акромион, небольшой коракоидной и подкоракоидной.
В тазу кости тоже срослись неподвижно, и лобковые срослись еще
с 4 крестцовыми позвонками. Вертлужная впадина глубокая. Позвонки амфи-
94
класс: пресмыкающиеся — reptilia
цельные. Двойные сочленения ребер на шейных и ординарные на грудных напо-
минают Plesiosaurus. Отростки шейных позвонков похожи на крокодильи.
Dicynodon Trautscholdi Amal.— полный череп с нижней челюстью
(рис. 111) из пермских отложений Сев. Двины
отвечает по признакам черепу
Dicynodon pardiceps Ow.
Lystrosaurus latirostris
Ow. (рис. ИЗ). Известен полный скелет
с очень укороченными конечностями, что
указывает на водный образ жизни и
приспособления к ней, подобные при-
способлениям водных млекопитающих,
например морской коровы.
Lystrosaurus latirostrisOw.
Рис. 112. D i с у п о d о n р а г d i с е р s Ow. (тРиас КаРУ и Индии) — с менее вытя-
нутой передней частью черепа, чем у
Dicynodon, отодвинутыми вверх ноздрями, тройным кондилюсом. Глаза со
склеротическим кольцом. Клыки значительно меньше, чем у Dicynodon.
Кости конечностей на сочленовных поверхностях не вполне окостенели. Тип,
сохранивший древние признаки, но успевший сократить зубы (рис. 113).
Рис. ИЗ. Lystrosaurus latirostris Ow.
Oudenodon pusillus Jaeck.— более открытый и удлиненный череп
с овальным пинеальным отверстием между тонкими теменными костями. Височ-
ные дыры очень большие; глазницы также большие. Клыки зачаточные у одних
видов и отсутствуют у других (может быть самка). Кондилюсов два — отчет-
ливо видных. Передняя часть нёба закрыта. Пермь, триас южной Африки.
Oudenodon Venyukovi Amal. — череп из пермских отложений Сев.
Двины.
Gordonia Traquairi Newt, (пермь Сев. Двины и верхний триас Шот-
ландии) имеет череп еще более открытый благодаря большим височным дырам.
Сильно развита squamosum, сросшаяся с квадратной, сочленяющейся с нижней
челюстью. По форме черепа близка к Dicynodon, но клыки меньше. Длина
черепа 18 см.
Gordonia Аппае Amal. (рис. 114) и Gordonia rossica Amal. —
черепа из пермских отложений Сев. Двины, сходны с шотландскими.
подкласс: synapsida
95
Это сильно отклонившаяся группа от типа Theromorpha, сохранив-
шая пинеальное отверстие и развившая высшие признаки поясов и плеча. Ре-
дукция зубов, совершенно исключительная, позволяет проследить только исчез-
новение клыков у различных видов Oudenodon. Разнообразный кондилюс,
простой, тройной и двойной, является еще одной особенностью для Thero-
morpha.
Просматривая даже те немногие типичные формы, которые даны были
здесь для пермских рептилий, мы видим громадное разнообразие их
организации. Есть между ними типы, очень близкие к Stegocephali
(S е у m о u г i а), другие — близкие к нынешним ящерицам (Varanosaurus);
иные являют типы с признаками очень смешан-
ными (разных рептилий) — Protorosaurus; еще
иные — с признаками млекопитающих-хищных, —
Theriodontia, и самые удивительные — это те,
у которых признаки млекопитающих (таз) соеди-
няются с первичными признаками: пинеальным от-
верстием и сохранением остатков хорды в позвон-
ках,— Pareiasauridae.
Очевидно, мы застаем в конце пермского
периода кульминационные точки, до которых могли
дойти эти различные рептилии, после чего боль-
шинство совершенно исчезает — вымирает. Немно-
гие положили, вероятно, начало более поздним
формам, и только одна гаттерия является ныне
живущим представителем богатого пермского цар-
ства рептилий. Откуда появились они в перми?
До сих пор нет ответа. Отчего погибли, не вы-
Рис. 114. G о г d о n i а А п-
п а е Amal.
держав борьбы, несмотря на разнообразные приспособления организмов,— это
пока остается тоже неизвестным.
Прежде чем описывать остальные группы рептилий, интересно остано-
виться на упомянутых находках проф. В. П. Амалицкого: Pareiasaurus,
Seymouria, Inostrancevia и др., сделанных им на берегу Сев. Двины,
сходных с известными раньше из перми Сев. Америки и триаса южной Аф-
рики (Кару).
Работая в 1897 г. на берегу Сев. Двины, проф. Амалицкий на-
шел там остатки моллюсков Anthracosia и растений Glossopteris,
заставившие его отнести эти слои к пермским и предположить возможность
нахождения здесь рептилий, аналогичных найденным в южной Африке в таких же
слоях. Продолжительные раскопки склонов берега р. Сев. Двины дали бле-
стящие результаты, и в настоящее время более 50 скелетов найдено, и
многие из них восстановлены в музее Академии наук. Они указывают вместе
с растениями и моллюсками на одинаковую жизнь органического мира во время
пермского периода в столь отдаленных местностях, как Сев. Двина, провинция
Кару в южной Африке и Техас в Сев. Америке. Кости эти найдены в линзе
96
класс: пресмыкающиеся — reptilia
древнего речного песчаника, около деревень Ефимовской и Соколки, в склоне
речною берега; они были заключены в песчаниковые конкреции, как в футляры.
Некоторые кости были разрознены; в некоторых случаях сохранились целые ске-
леты, облепленные песчаником (рис. 115). Конкреции расположены в центре
Рис. 115. Конкреция, заключающая скелет Pa-
reiasaurus Amal.
сколько лет. Разнообразные формы из этой
песчаной линзы, и можно предпо-
ложить, что эти линзы, или чече-
вицы, являются остатками русла
древней реки, заполненного осад-
ками, и что попавшие в него
трупы животных расположились
вдоль русла.
Кроме названных рептилий
здесь были найдены остатки Ste-
gocephali (выше описанные
D v i п о s a u г i d а е).
Раскопки продолжались не-
коллекции обрабатываются еще и
теперь.
Отряд 3
Ichthyosauria
У ихтиозавров существует только одна пара височных дуг, и мы их от-
носим к Synapsida.
Морские животные. По общему строению тела они занимают особое по-
ложение и считаются происшедшими от наземных рептилий, ушедших в воду
и изменивших сообразно этому свой скелет. Раньше в них видели формы,
близкие к рыбам, откуда и название их; теперь их по внешнему виду и об-
разу жизни чаще всего сравнивают с китообразными млекопитающими — с боль-
шой головой, короткой шеей, вытянутой мордой, двумя парами .пастообразных
конечностей. Эта совершенно вымершая группа известна, начиная с триаса до
мела. Указать ее родство с какими-либо другими пресмыкающимися нельзя.
Общими признаками с плезиозаврами являются: сросшаяся с черепом квадратная
кость и ласты, но у ихтиозавров они имеют различное число рядов косточек. Общими
признаками отряда являются: рыбообразное тело с большой головой, вытя-
нутой мордой с многочисленными зубами, лежащими в общем желобе или
в альвеолах; глаза со склеротическим кольцом, очень укороченная шея. Хвост
с двумя лопастями; позвонки направлены в нижнюю лопасть. Тонкие
брюшные ребра. На спине кожный плавник. Тело голое. Позвонки слабо
амфицельные, почти платицельные. Дуги, не сросшиеся с телом позвонков.
Этот отряд подразделяется на два семейства: Mixosauridae nlchthyo-
s a u г i d а е.
Сем. Mixosauridae — формы известны из среднего триаса по отдель-
ным частям скелета, указывающим на более близкое их отношение к на-
земным животным. Atlas и axis не срослись. Зубы в альвеолах.
подкласс: synapsida
97
Mixosaurus (рис. 116) имел прямой остроконечный хвост. В передних
ластах было пять рядов косточек с удлиненными плечевой, лучевой и локтевой
ко< (ими. Пояс состоял из ключицы, лопатки, коракоидной и межключичной кос-
тей Задний пояс имел отдельные седалищные, лобковые и подвздошные кости.
Бедро, большая и малая берцовые, несколько удлиненные, и пять или три ряда
фаланг пальцев составляли конечность. Таз не скреплен с позвоночником.
Средний и верхний триас в Альпах, Шпицбергене и в южной Германии.
М е г г i a m i а (рис. 117, 3) — с очень редуцированными ластами, всего
три ряда фаланг. Плечо большое; локтевая и лучевая кости удлиненные, четырех-
угольные. Верхний
триас Калифорнии.
Mixo-
а t a v u s
Рис. 117. Передние ласты: 1, 1а— Ichthyosaurus Cony-
bear i. (Юра, мел.) 2. Mixosaurus cornulianus. (Триас.)
3. M-erriamia Zitteli. (Триас.)
Рис. 116.
s a u г u s
Quenst-
Сем. Ichthyosauridae. Юра и мел. Задние конечности сократились
больше; хвост двупопастный. Зубы сидят в общем желобе, у меловых форм
исчезают. Ноздри проходят по обе стороны сошника.
Очень распространенные формы в юре Англии, Германии, Франции,
Южной Америки, у нас и в мелу Европы, Индии, Австралии, Патагонии.
Род Ichthyosaurus представлен многочисленными видами. Найдены
целые скелеты животных различного возраста. Есть даже скелеты, за-
ключающие в себе скелеты зародышей. Морда различной ширины; большею
частью с очень тонкими межчелюстными костями, в которые входят носовые,
тоже очень тонкие. Лобные маленькие; на заднем конце их лежит пинеальное
отверстие, разделяя передние концы теменных костей. Зубы многочисленные
на внешнем краю челюстей, сидят в общем желобе. Они состоят из корня и
коронки, покрытой прочной эмалью, входящею внутрь складками. При стира-
нии они сменялись другими, сидящими за ними в желобе. Очень большие
глазницы со склеротическим кольцом. Квадратно-скуловая кость выдается по
краям черепа. Один кондилюс. Atlas и axis срастались. Грудной пояс хорошо
развит, как у первого семейства (рис. 119). Конечности представляют очень
7 Павлова М. В Палеозоология. Ч II.
98
класс: пресмыкающиеся — reptilia
большое разнообразие в числе рядов костей пальцев — от пяти до десяти
(рис. 117, 1 и 1 а), причем плечо и'^предплечье теряют свою удлиненную форму,
и часто локтевая и лучевая не отличимы от других костей ласта.
Ichthyosaurus communis — из лейаса, имеет от 6 до 8 рядов
фаланг.
Рис. 118. Ichthyosaurus quadriscissus Quenst. (S t е п о р t е г у g i u s).
(Верхний лейас.)
Ichthyosaurus quadriscissus (верхний, лейас) имеет по 5 рядов,,
как у Mixosaurus. Задние конечности развиты были слабее.
У Ichthyosaurus platydactylus (нижний мел) было восемь
рядов косточек, почти квадратных. Зубы значительно сократились.
Ichthyosaurus quadriscissus Qu.—длиной в 2,1 м, найден
в верхнем лейасе Вюртемберга в виде полного скелета с сохранившейся кожей.
el
Рис. 119. Ichthyosaurus communis Conyb.
Грудной пояс и ласт.
Род Ophthalmo-
saurus Seely(Baptanodon
Marsh) — верхняя юра Сев.
Америки. Пинеальное отвер-
стие очень маленькое. За-
тылочный кондилюс очень
выступает. Число зубов со-
кращено. Межчелюстные ко-
сти утолщены. Плечо сочле-
нялось тремя фасетками с
косточками предплечья.
Это разнообразное строе-
ние ластов вызывает вопрос
о целесообразности его, так
как увеличение числа рядов косточек не совпадает у форм одного и того же
геологического возраста. Интересное изменение наблюдается еще в строении
хвоста различных ихтиозавров.
У Mixosaurus триаса (см. рис. 120) прямой хвост, остроконечный.
Спинной плавник при этом расположен за задними ластами; хвостового нет.
подкласс: synapsida
99
У юрских форм молодых хвост имеет такую же форму, но у них спин-
ной плавник как бы подвинулся к хвосту (Ichthyosaurus quadriscis-
s u s). У взрослого эта лопасть увеличивается, так же как и нижняя; позвонки хвоста
у его основания загибаются вниз.
Наконец, у Ichthyosaurus
t г i g о п u s обе лопасти увеличи-
ваются, становятся одинаковыми;
позвонки хвоста очень утоняются
и сильно наклоняются вниз, хвост
как бы надламывается. Эти изме-
нения и надлом хвоста Абель
объясняет различием в условиях
движения животных, в зависимо-
сти от большей или меньшей глу-
бины водной среды.
Чаще всего остатки ихтио-
завров встречаются в виде пло-
ских позвонков, лишенных от-
ростков, например в Хорошове
под Москвой. Цельный плавник
Ophthalmosaurus найден на
берегу Москвы-реки в д. Мневни-
ках. Он был описан Траутшоль-
дом в 1879 г. На Волге, близ
г. Симбирска, также были найдены
остатки этого рода.
Рис. 120. Хвосты Ichthyosauri а: а—М i х о-
saurus; Ь — Ichthyosaurus quadri-
s с i s s u s; с, d — Ichth. trigonus.
Отряд 4
Plesiosauria Ow. (Sauropterygia)
Эта вымершая группа пресмыкающихся, жившая с триаса до верхнего
мела, объединялась еще недавно с Ichthyosauri а. Она характеризуется сле-
дующими признаками: имеет одну пару височных дыр и дуг, пинеальное от-
верстие, длинную шею, укороченное туловище, конечности в виде удлиненных
ластов, но с сохранением пяти рядов фаланг. Это-—морские, хорошо плававшие
животные, стоявшие ближе всего по образу жизни к Pythonornorpha.
Позвонки плоские, слабо амфицельные, с прирастающими отростками, без
остатка хорды. Грудной пояс хорошо развит, состоит из ключиц, лопаток и
коракоидных костей, соединенных вместе. Межключичная кость маленькая. Изучая
различные формы, можно просл< лить изменения их от форм, еще близких
к наземным ящерицам, до настоящих морских.
Сем. Nothosauridae. Представители его имеют удлиненные плечевую,
локтевую и лучевую кости, бедро и голень и настоящие пятипалые конечности-
Верхний триас.
7*
100
класс: пресмыкающиеся — reptilia
S i ni о s a u г u s H. v. M. — широкий череп с закругленной мордой, очень
напоминает череп крокодила. Передние зубы несколько больше остальных;
сидят они в отдельных ячейках. Ноздри расположены наверху передней части
головы, перед глазницами, за которыми лежат большие височные дыры. По-
Рис. 121. Lariosaurus Bal-
sa m i Curioni. (*/8)
средине теменных костей маленькое пинеальное
отверстие. На нижней стороне черепа хоаны ле-
жат впереди носовых отверстий; вся она закрыта
сросшимися костями нёба: предчелюстными, че-
люстными, сошником, небными игкрыловидными.
Lariosaurus Bal-
sa m i Curioni (рис. 121)—
около 70 см длины, най-
ден в виде полного ске-
лета в альпийском триасе.
Череп его более удлинен,
чем предыдущий, с боль-
шими носовыми отверстия-
ми, равными глазницам.
Шея довольно длинная
(20 позвонков). Удлинен-
ное туловище несет за-
кругленные ребра. Суще-
ствуют брюшные ребра.
Конечности хорошо раз-
виты, пятипалые, назем-
ного ящеричного типа.
Хвост довольно длинный.
Nothosaurus mi-
rabilis Miinst. — с
очень удлиненным черепом
Рис. 122. Notho-
satirus mirabilis
Munst.
(рис. 122). Широкие межчелюстные кости продвигаются довольно далеко,
отодвигая ноздри. Они усажены длинными тонкими зубами с продольными
желобками. На челюстях зубы мельче. Маленькие глазницы и большие овальные
височные дыры. Удлиненные лобные и теменные кости. Хоаны почти под но-
совыми отверстиями, расположенными за широкими межчелюстными костями.
Грудной пояс широкий, с сильно развитыми ключицами и коракоидными
костями. Позвонки плоские; на шейных небольшие косые отростки; на спинных
они очень высокие.
Задний пояс состоит из больших лобковых (pubes), маленьких подвздош-
ных (ilium) и направленных вниз небольших седалищных (ischium) костей,
В углубление этих костей входило бедро. Этот вид достигал 3,5 м длины. Род
представлен несколькими видами в триасе Германии.
Сем. Plesiosauridae представлено юрскими и меловыми формами с боле е
длинной шеей, более коротким туловищем и конечностями, превратившимися
подкласс: synapsida
101
в ласты с очень укороченными предплечьем и голенью и удлиненными фалангами.
Форма черепа, собственно передняя часть, различна у различных родов, с более
или менее вытянутой передней частью, как мы это видели и в сем. Norho-
sauridae.
dolichodermus Conyb. (Длиною 3 -и.) (Лейас.)
Рис. 123. Plesiosaurus
Рис. 123A. Plesiosaurus
Plesiosaurus (рис. 123) — с очень длинной шеей и маленьким чере-
пом с короткой мордой. Большие глазницы лежат по сторонам, на середине
черепа. Особенность представляет грудной пояс, состоящий из удлиненных ши-
роких коракоидных костей, образующих передний щит, приросших к нему
лопаток и маленьких треугольных ключиц. Задний пояс образует тоже щит
из сросшихся лобковых и седалищных костей. Подвздошные очень маленькие.
Форма и длина ластов у различных пред-
ставителей очень различны.
Plesiosaurus macrocepha-
1 us Buckl и Plesiosaurus Haw-
kin s i Ow. — из нижнего лейаса. Пос-
ледний известен почти полным скелетом,
около 1,5 м длиною (рис. 123 А).
Thaumatosaurus victor Н.
v. Meyer et Fraas—из верхнего лейаса—
имеет сжатую переднюю часть морды
при значительно большем черепе.
У Thaumatosaurus indi-
с u s (из верхней юры Индии) пере-
хват передней части морды более за-
метен и напоминает морду аллигатора.
Шея короткая.
Есть еще несколько юрских родов, представленных менее хорошо сохра-
нившимися остатками.
Peloneustes Lyd. достигал громадных размеров; голова около 1,5 м,
с короткой шеей и удлиненной симфизой нижней челюсти, напоминаю-
щей гавиала (Pliosaurus Ow.). Зубы в передней части черепа достигали
25 см.
102
класс: пресмыкающиеся — reptilia
Cryptocleidus oxoniensis Phil.—из верхней юры Англии—длиной
в 3,35 м. Короткомордый, с очень длинными ластами. Прекрасной сохранности
в музее Нью-Йорка.
Elasmosaurus Соре — из верхнего мела Сев. Америки — является
потомком короткомордых Plesiosauria с приподнятым затылком и большими
височными дугами. Скелет достигал 13 м (Филадельфийский музей). Очень боль-
шой длины достигали его ласты, всегда, однако, сохраняя пять рядов косточек (пять
пальцев). Коракоидные кости и лопатки не срастались, так же как и кости таза,
сходные с костями Plesiosaurus.
Trinacromerum Osborni Wil. (Dolichorhynchops Will.)-—
из верхнего мела Канзаса — представляет далеко отклоняющийся тип. Скелет
его более похож на скелет Ichthyosaurus, с удлиненной мордой, склеро-
тическим кольцом, поднятым затылком, коротким хвостом и удлиненными ла-
стами. Зубы мелкие и многочисленные (рис. 124).
Рис. 124. Trinacromerum Will. (Dolichorhynchops Osborni Will.).
Передний пояс — типа Plesiosaurus, в виде щита с громадными
коракоидными костями, маленькими лопатками и треугольными ключицами.
В заднем поясе очень удлинены седалищные кости; лобковые почти квадратные-
Передние и задние ласты почти одинаковые, с большим числом мелких фаланг,
расположенных в пять рядов.
Эти примеры указывают нам на то разнообразие, которого достигали
в юре и верхнем мелу плезиозавры с преобладанием двух типов: короткомордых
и длинномордых, В этом отношении их развитие сходно с развитием крокодилов.
Отчлениться они могли от пермских ящериц. Но при этом, уходя в воду, они
замкнули свои вторые височные дыры, сблизив обе дуги. У нас остатки их
известны, хотя очень неполно, больше в виде позвонков и частей конечностей
Elasmosaurus, Plesiosaurus, Pliosaurus, Thaumatosaurus и
др. Главные остатки этих форм обработаны проф. Н. Н. Боголюбовым’).
Отряд 5
Пластинчатозубые — Placodontia
Эта высоко специализировавшаяся группа представляет больщую особен-
ность в строении зубов, так что ее некоторые ученые называют А п о m а 1 о-
d о n t i а.
i) Н. Н. Боголюбов, Из истории плезиозавров в России, 1911, с 16 та-
блицами.
подкласс: synapsida
103
Эти сравнительно небольшие животные, с черепом не больше 18 см, су-
ществовали в триасе средней Европы.
Более или менее плоский череп с большими глазницами и еще боль-
шими височными дырами имеет пинеальное отверстие и большое число плос-
ких зубов как в челюстях, так и на закрытом нёбе. На последнем разви-
ваются сменяющиеся зубы. Нижняя челюсть снабжена высоким венечным от-
ростком. Остатки этих форм были вначале отнесены к рыбам (Pycnodonti);
позже Оуэн (Owen) признал в них рептилий, между которыми и теперь они
еще не нашли определенного места. Циттель относит их к Sauropte-
г у g i а, другие палеонтологи — к Т h е-
romorpha.
Placodus gigas Ag. (рис. 125).
Удлиненный череп имеет на очень боль-
ших межчелюстных костях и на перед-
Рис. 125. Placodus gigas Ag.
ней части нижней челюсти конические резцы (6 верхних и 4 нижних); за
ними следуют небольшие плоские, закругленные коренные, по 4 с каждой
стороны верхней челюсти и по 3 на нижней. На нёбе 6 больших плоских
зубов. Носовые отверстия поперечно-удлиненные, лежат над хоана ми, не на
конце морды. Squamosa развиты очень сильно. Квадратные кости с ними cpqc-
лись. Интересно сочленение черепа, на котором нет среднего кондилюса, а
существуют отростки боковых затылочных костей, входящие
в углубления отростков первого шейного позвонка. Эти
отростки заменяют кондилюсы. Скелет неизвестен.
Cyamodus tarnovitzensis Gurich. из среднего триаса Германии
(рис. 126). Более плоская, треугольная форма черепа с узкими межчелюстными
костями, несущими по два зуба; за ними следуют по 3—4 зуба на челюстях.
104
класс: пресмыкающиеся — reptilia
На нёбе два небольших зуба впереди и два крупных круглых сзади. Суще-
ствуют также боковые сочленовные кондилюсы. Глазницы маленькие, лежат за
ноздрями. Височные дыры очень большие. Ноздри над хоанами. Пинеальное
отверстие маленькое. Скелет неизвестен.
Placochelys placodonta Jaeck. (рис. 127)—верхний триас. По форме
черепа очень близок к Cyamodus, но межчелюстные кости более удлинены и
Рис. 126. Cyamodus tarnovitzen-
s i s Gurich.
заострены впереди, соответственно ниж-
ней челюсти. Зубов на этих частях нет;
Рис. 127. Placochelys placodonta
Jaeck.
вероятно, они были покрыты роговыми чехлами. В челюстях по 4 больших
плоских зуба, увеличивающихся кзади, на нёбе — 2. Нижняя челюсть с очень
большим, высоким венечным отростком. Существует средний и боковые конди-
люсы на основной и боковых затылочных костях. Животное было покрыто
щитом, состоящим из слившихся больших и малых закругленных пластинок.
Череп был тоже украшен пластинками. Конечности обращены в ласты с со-
хранившимся пятью пальцами, в которых число суставов увеличилось. По
внешнему виду это животное очень похоже на черепаху. Очевидно, это крайняя
форма той специализации, к которой направилась эта морская группа, пожи-
равшая и раздавливавшая своими плоскими зубами моллюсков и других морских
животных.
Отряд 6
Черепахи — Testudinata (Chelonia)
Ныне живущие черепахи, водные и наземные, являются благодаря покры-
вающему их панцырю, состоящему из брюшного щита и собственно спинного пан-
цыря, совершенно обособленным отрядом. Некоторые морские черепахи не имеют
панцыря и покрыты только кожей. Другими резкими признаками их являются
отсутствие зубов и роговой клюв, покрывающий переднюю часть верхней и
нижней челюстей. С некоторыми из них сходны по форме черепа из вымерших
пресмыкающихся Placodontia с исчезающими зубами, как Placochelys,
покрытый тоже панцырем, хотя и другого строения. Некоторые ученые считают
возможным видеть в последних формах предков черепах, но мы увидим, что
дали по этому вопросу новые находки ископаемых черепах в триасе.
подкласс: synapsida.
105
Нынешние черепахи живут — одни в морях, другие — в реках и на суше,
и различаются между собою как по строению панцыря, так и строением че-
репа и конечностей. При классификации их принимаются во внимание способ
втягивания головы и шеи внутрь панцыря и жизнь в различной среде.
Подотряд Amphichelydia (вымершие) — голова не втягивалась.
Подотряд Pleurodira — голова втягивается не по прямой линии, а бу-
дучи повернута на сторону. Пресноводные и наземные формы жарких стран.
Подотряд Crypto di га — голова втягивается по меридиональной линии.
Речные и сухопутные формы.
Подотряд Cheloniidea — шея не вполне втягивается. Морские
формы.
Подотряд Trionychia — шея как у С г у р t о d i г а; но формы со-
храняют очень древние признаки. Речные формы.
В связи с этими признаками стоит развитие шейных позвонков с силь-
ными боковыми отростками у Pleurodira и более слабыми у остальных.
Остатки ископаемых форм не всегда возможно определенно отнести к тому
или иному подотряду, так как они встречаются большею частью только в виде
панцырей. Древние формы выделяются в
Подотряд 1 '* : е ' :
, Amphichelydia ’) с - ‘f •...... -
Главные представители их найдены в триасе. Голова и шея не втягивались
под панцырь. В брюшном щите существовали добавочные средние пластинки,
mesoplastra, между передними и задними пластинками.
Triassochelys dux Jaeck. (рис. 128) — из верхнего триаса Гальбер-
штадта, недавно найденная и изученная И е к е л е м форма; представляет боль-
шой интерес по прекрасной сохранности. Верхний панцырь состоит из
12 neuralia, 9 costalia и 30 marginalia; они прикрывались четырьмя большими
средними пластинками и с каждой стороны—-боковыми и добавочными
(краевыми). Нижний щит состоял, главным образом, из сросшихся груд-
ных ребер. В плечевом поясе существует cleithrum,— косточка, подстилающая
лопатку и идущая от панцыря (от ребер) к щиту. Лопатка заканчивается
тонким отростком. Тазовые кости срослись между собою, с sacrum и с ниж-
ним щитом. Череп очень массивный, имеет задние височные дыры, ограни-
ченные снизу paroccipitale, с внутренней supraoccipitale, что указывает на не-
соответствие их с дырами (верхними или боковыми) других рептилий. Другой
особенностью являются зачаточные зубы по краям нижней и в верхней че-
люсти и маленькие на нёбе (на parasphenoidea, на vomer и на pterygoidea).
Абель указывает на Triassochelys Jaeck. как на ближайшую, древнейшую
форму к Cotylosauria, с закрытым черепом. Он указывает, что задние
4) К. Zittel, Grundziige der Paiaontologie, 1923, S. 300. Сюда относятся: Trias-
sochelys, Proganochelys, Proterochelys и др., включенные Абелем
в Cryptodir а.
' 106
класс: пресмыкающиеся — reptilia
теменные дыры являются не соответствующими дырам других рептилий по месту
их нахождения и потому, что при дальнейшем развитии черепах они перехо-
подвигающиеся вперед у одних форм (Е ш у s,
дят в углубления, все более
Рис. 128. Triassochelys (Stegochelys) dux
Jaeck.
Geoemyda, Trionyx)
и исчезающие у других
(С h е lo n i d а е, морских).
Это — вторично закрытый
череп.
При дальнейшем
развитии черепах Абель
допускает переход их от
наземной жизни, которую
он считает первичной,
к водной, даже морской,
с изменением формы и
строения панцыря, черепа
и конечностей, а для неко-
торых форм он допускает
вторичное перемещение на сушу, сопровождаемое
возвращением прежних при-
знаков. Абель основывается на исследованиях Д о л л о J) о развитии панцыря
у предков Dermochelys (морских черепах, лишенных панцыря).
Proganochelys (Psammoche-
lys) Quenstedti Baur — верхний триас
Вюртемберга. Древний представитель чере-
пах, от которого сохранился почти пол-
ный, четырехугольной формы, панцырь.
Длина 64 см, ширина 63 см. Внутри его
сохранились позвонки и ребра (рис. 128 А).
Микроскопическое изучение открыло
присутствие гаверсовых каналов и костных
телец в панцыре. Верхний панцырь
очень выпуклый, до 17 см, четырехуголь-
ной формы. Кзади он становится более
плоским. Верхняя поверхность покрыта
грануляциями; вероятно, она имела роговые
пластинки. Верхний панцырь состоял
„ „ Рис. 128A. Proganochelys
из следующих костных пластинок шейной Quenstedti Baur,
(nucale), 9 средних позвоночных (neura-
lia), по 8 реберных (costalia) с каждой стороны; далее следовал ряд краевых
(marginalia), а над ними спереди и сзади по нескольку надкраевых (supramargi-
nalia), не существующих у других черепах. Задние из них имели вид толстых
*) S. Doll о, Sur 1’origine de la Tortue Luth. Bull. Soc.
naturelles de Bruxelles, 1901.
Roy. Sciences medicales et
подкласс: synapsida
107
бугров и щитов. С внутренней стороны позвонки и ребра срастались с пан-
цырем. Тела позвонков очень тонкие.
Щит состоял из четырех частей: hyoplastron, myoplastron, hypoplastron
и xiphiplastron; он плотно срастался с верхним панцырем, но имел вырезы и не
был сплошным 11а внутренней поверхности видны места прикрепления тазовых
костей, а вверху приросшие части грудного пояса.
Proterochersis robusta Fraas (рис. 129) — тоже из верхнего триаса
близ Вюртемберга; найден панцырь и щит. Первый выпуклый, овальной формы
Спинной панцырь. Грудной щит.
Рис. 129. Proterochersis robusta Fraas.
с очень маленькими средними пластинками (neuralia), большими реберными
(costalia), маленькими краевыми (marginalia), без всяких придатков. На щите,
плотно соединенном на боках с панцырем, существуют промежуточные (me-
soplastron) пластинки. По размерам панцырь вдвое меньше, чем у Progano-
chelys. Абель считает Proterochersis древнейшим представителем
наземных Pleurodira.
Подотряд 2
Pleurodira
Имеют тоже тяжелый выпуклый панцырь, плотно срастающийся по сторонам
со щитом. Голова и шея втягиваются под панцырь боком. Шейные ребра
с сильными поперечными отростками. Таз срастается с панцырем. Это— пресно-
водные и наземные формы жарких стран. Ноги с 4—5 пальцами для хожде-
ния. С юры доныне: в Африке, на Мадагаскаре, в Бразилии (Ро d о с n е m i s).
Сем. Plesiochelydae. Plesiochelys solodurensis Riitim.—
верхняя юра (киммеридж) Швейцарии. Продолговатый панцырь соединен широко
со щитом. Средние пластинки тонкие. Близкая к нему форма — Platychelys
Wagn.—верхняя юра средней Европы. Известен по панцырю небольших раз-
меров (до 20 см), который имеет три ряда грубых пластинок, отороченных
ряд эм меньших, закругленных на задней стороне. Соединяется со щитом
неширокими перемычками.
108
класс: пресмыкающиеся — reptilia
Сем. Miolanidae — с единственным родом Miolania— не вполне хо-
рошо сохранившаяся форма из эоцена или верхнего мела Патагонии и плейсто-
цена Австралии. Череп Miolania argentina Ameg. совершенно закрытый,
украшен кзади несколькими парами рогообразных отростков (подобно Elginia
mirabilis). Глаза на конце морды, направлены вперед. Нижняя челюсть очень
толстая. Хвост имел костяной чехол (подобно G1 у р t о d о п). Эти признаки
сближают ее с ныне живущими Chelys fimbriata и отчасти с Plate-
m у s S р i х i i Южной Америки.
Подотряд 3
Cryptodira
Панцырь менее высокий, чем у Pleurodira, не вполне окостеневший;
таз не срастается с панцырем. Шейные позвонки не имеют больших отро-
стков; голова втягивается прямо. Конечности для хождения или плавания
(4—5 пальцев). Есть нынешние речные и сухопутные формы. Ископаемые —
начиная с верхней юры.
Сем. Thalassemydidae — вымершие. Thalassemys marina Fraas —
верхняя юра Швабии (рис. 130). На панцыре существуют по 4 маленьких боко-
Pijc. 130. Thalassemys
marina Е. Fraas. (Верхняя юра.)
вых фонтанели. Щит, открытый посредине, состоит из шести парных частей
(hyoplastron, hypoplastron, xiphiplastron), образующих как бы перекладины
прикрепленные по сторонам.
Eurysternum Wagleri Н. v. М.— верхняя юра, литографский сла-
нец Баварии (рис. 131). Эта форма тоже принадлежит к вымершему семейству,
соединявшему в себе признаки отчасти морских черепах — открытые фонта-
нели: одну — на панцыре и несколько по сторонам щита. Таким образом
щит соединялся отдельными перекладинами с панцырем, как у Cheloniidea
(морские). По форме отдельных частей щита, изогнутых hypoplastron, hyopla-
stron он ближе к болотным черепахам—Em у din а е. Конечности пятипалые
подкласс: synapsida
109
с несколько удлиненными пальцами и когтями. Близкие формы встречаются
в мелу и в эоцене Европы и Америки.
Сем. Chelydridae имеет живых представителей. Ископаемые известны
тоже с верхней юры. Конечно-
сти с 4 и 5 пальцами с пла-
вательной перепонкой.
Сюда же относятся жи-
вущие теперь семейства Tes-
r u d i n i n a e (наземные) и
Em у di па e (болотные).
Рис. 132. Archelon ischyros Wieland.
A — верхняя crop она, В — нижняя.
Рис. 131. Eurysternum Wag-
1 eri H. v.M. /Верхняя юра Герма-
нии.) (i/3)
Подотряд 4
Cheloniidea
Настоящие морские черепахи с различно окостеневающим, более плоским
панцырем, короткой шеей. Таз не срастается плотно с панцырем. Передние и
задние конечности с очень удлиненными пальцами в виде ластов, у некото-
рых особенно удлинены передние. С верхнего мела доныне.
Сем. Protostegidae. А г с h el о п (рис. 132) — верхний мел Дакоты; имеет
очень несовершенный панцырь, состоящий из отдельных, немногих средних
по
класс: пресмыкающиеся — reptilia
пластинок, доходящих до краев ребер; между ними остаются широкие свобод-
ные пространства. Щит состоит из четырех лучистых пластинок, тоже с про-
межутками. Эти пластинки были покрыты кожей. Шея короткая. Передние
пальцы длинные. Это типичная морская форма. У нее сильно изменен
череп. Он совершенно закрытый. Ложно-височные дыры наземных черепах
не существуют. Спереди сильно загнутые вниз межчелюстные кости в виде
клюва.
Protosphargis veronensis Cap.— верхний мел Вероны, с еще бо-
лее сократившимися, чем у Archelon, пластинками панцыря: hyoplastron,
hypoplastron и xiphiplastron.
Protostega Copei Wiel.-— очень близка к A r c h e 1 о п, из верхнего
мела Канзаса.
Caretta — ныне живущая в Средиземном море с чрезвычайно удлинен-
ными пальцами.
Toxochelys Bauri Wiel.— нижний мел Ганновера и верхний мел Кан-
iaca. С более открытыми фонтанелями, чем у Thalassemys, и особенными
костными придатками в виде бугорков на средних пластинках (нервных) пан-
цыря. Д о л л о считает их остатками эпитекального кожного покрова, указы-
вающими путь, по которому развились из наземных черепах Dermochelydae
(кожистые черепахи). Последних он считает формами, вторично перешедшими от
Рис. 133. Trionyx styria с u s Peters.
(Миоцен.)
прибрежного образа жизни к мор-
скому. В сем. Dermochelyidae
панцырь еще более редуцируется, и
преобладает кожный покров.
Некоторые формы из этого под-
отряда достигали громадных разме-
ров. Панцырь Protosphargis
(верхний мел) достигал 3 м.
Подотряд 5
Trionychia
Небольшая группа речных че-
репах с несовершенным панцырем.
Он состоит из среднего ряда пла-
стинок, покрывающих ребра, но не
доходящих до их концов, ребра
торчат из-под него. Щит имеет боль-
шие фонтанели. Весь костяной панцырь прикрыт кожей; роговых пластинок нет.
Щит и панцырь соединены слабо между собою. Остатки их известны с верхнего
мела; есть теперешние представители в жарких странах Азии, Африки и
Сев. Америки.
Тгюпух styriacus Pet. (рис. 133) — из миоцена.
Итак, в триасе мы имеем остатки наземно-пресноводных черепах. Начиная
с юры, мы встречаем Pleurodira и Cryptodir а. В мелу представлены
подкласс: diapsida
111
хорошо близкие к ныне живущим формы, морские и наземные, которые раз-
виваются параллельно. Морские — с более легким панцырем, ластообразными но-
гами и закрытым черепом. Речные и наземные •— с плотным панцырем, имеющим'
роговые пластинки, с ногами, снабженными когтями, и черепом с ложно-ви-
сочными впадинами. Это параллельное развитие могло бы позволить видеть-
в морских и наземных формах потомков общего предка. Но работы Д о л л о
и Абеля (вышеназванные) дают иные указания на развитие черепах. Они
считают наземные формы первичными, часть которых ушла в воду, и их ор-
ганы, приспособляясь к жизни в воде, сильно изменились. Это было бы про-
тиворечием высказанному Долло положению о „невозвратимости утерянных
признаков".
Подкласс 111
DIAPSIDA
Включает очень разнообразные отряды рептилий; мы начнем с древней-
ших, еще мало специализировавшихся.
* Отряд 1
Prosauria
(Rhynchocephalia Zittel) , »
У некоторых существуют клювообразно загнутые вниз межчелюстные кости,
откуда и название. Эти формы наземных хищников встречаются, начиная с н и ж-
них пермских слоев, в триасовых, в юре и в мелу, и один предста-
витель дошел до нашего времени. Это Hatteria, или Sphenodon, живущая
теперь в Новой Зеландии. Общими признаками для этого разнообразно
представленного отряда являются: позвонки амфицельные, часто с остатками
хорды; грудные ребра двуголовчатые; присутствуют брюшные ребра; хрящевая
грудная кость; конечности пятипалые; кожа покрыта роговыми чешуйками. Не-
которые формы соединяют как бы коллективные признаки различных групп.
Древнейшим представителем является Palaeohatteria (рис. 134) — из
нижне-пермских слоев близ Дрездена; напоминает своим скелетом ящерицу,
с длинным хвостом и пятипалыми конечностями. Сверху черепа мы видим
сильно развитые носовые кости, теменные с пинеальным отверстием, большие
глазницы с предглазничными и залобными костями и склеротическим кольцом.
За глазницами две пары височных дыр — верхние и боковые — с разграничи-
вающими их верхними и ограничивающими снизу нижними дугами. Это типич-
ная форма Diapsida Осборна. Снизу череп закрыт сошником, сходящимся
с расширенными верхними челюстями, образующими вместе второе нёбо. Зубы
мелкие, однорядные, расположены на межчелюстных и челюстных костях. Меж-
челюстные кости слабо загнуты вниз. Носовые отверстия не разделены. Квадратная
кость прикреплена неподвижно к squamosa. Позвонки двояковогнутые с остат-
ком хорды. Хвостовые позвонки срослись. и
112
класс: пресмыкающиеся — reptilia
Грудной пояс состоит из отдельных костей: удлиненной грудной (interclavi-
cula), к которой прилегают ключицы (clavicula); за ними лопатки (scapula)
с закругленными верхними краями и ниже их коракоидные (coracoideum). Плечо
без сочленовной головки, также и предплечье — они не окостеневали. Тазовый
пояс состоит из трех пар костей: сближенных между собой седалищных (ischium),
далеко расходящихся лобковых (pubes), соприкасающихся между собою под-
вздошных (ilium). Все соединялись хрящами. Бедро подходило к ямке между
этими костями и сочленялось, вероятно, хрящевой поверхностью, как и плечо.
Брюшные ребра хорошо развиты.
Рис. 134. Palaeo h atteria longicaudata Credn.
В — грудной пояс. С — таз. D — спинной позвонок. Е — хвостовой позвонок.
Крайне интересно сравнить эту нижне-пермскую ящерицеподобную
форму с вышеназванной ныне живущей гаттерией. Отличий мало. На черепе
те же кости с двумя парами височных дуг и височных дыр. Только дуги
у гаттерии более сильно развиты. Пинеальное отверстие сохранилось. Умень-
шилось число зубов на межчелюстных костях. Брюшные ребра сохранились.
Гаттерия — настоящее живое ископаемое. От ящериц эти формы отличаются
амфицельными позвонками, присутствием пинеального отверстия, соединением
симфиз нижней челюсти посредством связки; у ящериц они соединены швом.
Грудная кость—с хрящевым стержнем.
Очень близко к Palaeohatteria стоит Callibrachion Gaudry
Boule, найденный в пермских слоях Отёна вместе с Actinodon Gaudry.
Позвонки его плоские с большим остатком хорды. Череп не вполне хорошо
сохранился. Особенно хорошей сохранности передний пояс и передние ко-
нечности типа хищных ящериц. Длина 54 см.
подкласс: diapsida
113
няет
Pol уsphenodon Mtilleri Jack. — средний триас. С зубами, располо-
женными на нёбе и по краям челюстей. Самые маленькие — на крыловидной
кости в 4 ряда, два более длинных посредине и по 4 зуба по сторонам.
Эта триасовая форма является как бы измененной пермской Palaeohat-
ter 1 а.
Homaeosaurus Zittel — верхняя юра Соленгофена. Определенно сохра-
тип пермских Prosauria (рис. 135): амфицельные позвонки, пинеальное
отверстие, брюшные ребра. Она очень похожа на
нынешнюю гаттерию, но меньше ее — от 15 до 20 см.
В триасе мы встречаем формы, по которым собственно
и было установлено название Rhynchocephalia—
с межчелюстными костями, загнутыми вниз в виде
клюва.
Сем. Rhynchosauridae. Rhynchosaurus
а г t i с е р s Ow. (рис. 136). По общей форме тела и
черепа очень похож на Palaeo-
hatteria с двумя резко выра-
женными височными дырами и
дугами и большими глазницами.
Но у него нет пинеального отвер-
стия; межчелюстные кости не несут
зубов и загнуты вниз. Нижняя че-
люсть спереди раздвоена. Брюш-
ные ребра сильно развиты. Пояса
неизвестны. Длина до 27 см до
хвоста. Верхний триас.
Hyperodarpedo п—боль-
шое животное, свыше 2 м длины,
Рис. 136. Rhyn-
с широким черепом, большими chosaurus.
височными впадинами и резко
загнутыми вниз межчелюстными костями, входящими
между раздвоенными концами нижней челюсти. Мелкие
зубы на верхней челюсти и нёбе. Позвонки у этих
форм амфицельные; шейные у Hyperodarpedon —
опистоцельные. Верхний триас Шотландии (рис. 137).
Эта группа является отклонением от типа Palaeohatteria со слабо
загнутыми межчелюстными костями. Она потеряла древний признак — пинеаль-
ное отверстие, усовершенствовала шейные позвонки и вымерла в триасе. Мы
не встречаем позже ее потомков. В юре находим представителей других се-
мейств типа ящериц.
Sapheosaurus incisivus Jourd. (Sauranodon) — верхняя юра
Франции, по внешней форме близкая к Homaeosaurus, но резко отли-
чается по строению. Это — прогрессивная форма с процельными позвон-
ками, но лишенная зубов, имела загнутые вниз межчелюстные кости, маленькие
8 Па в.ю в а М. В. Палеозоология. Ч. I.
Рис.
г us
135. Homaeosau-
pulchellus Zittel.
(*/з)
114
класс: пресмыкающиеся — reptilia
височные впадины; нет пинеального отверстия. Брюшные ребра сохранились.
Очевидно, это продолжатель триасовых Rhynchocephalia.
Сем. Protorosauridae. Protorosaurus (рис. 138) из верхних перм-
ских слоев дает третий тип пермских ящериц. Он был описан в первый
раз в 1710 г. и с тех пор привлекал внимание многих ученых: Кювье,
Рис. 137. Hyperodarpedon Gordo п i Huxley. (*/4)
Г. ф. Мейера, Гексли и подробно был описан Сили в 1887 г. При-
знаки, характеризующие эту группу, до такой степени разнородны, коллективны,
так сказать, что ее с известным основанием относили то к одному, то к дру-
гому отряду пресмыкающихся. Ноздри расположены внизу морды, а не на
конце ее, как у форм, с которыми она имеет сходное строение черепа (крокодил).
Зубы расположены в челюстях и на нёбе в ямках, края которых под-
нимаются до коронок. Позвонки
двояковогнутые, вполне окосте-
невшие, с сильно развитыми спин-
ными отростками. Первый шейный
позвонок очень короткий, остальные
7 очень удлинены, с прилегающими
к ним ребрами. Они напоминают по-
звонки летающих рептилий и птиц.
Спинные позвонки (16) с очень
закругленной нижней стороной, снаб-
Рис. 138. Protorosaurus. жены хорошо развитыми закруглен-
ными ребрами, ограничивающими ши-
рокое туловище, указывающее на хорошее развитие внутренних органов.
Хвостовые позвонки с раздвоенными отростками.
Строение тазового пояса, поскольку он сохранился, приближается по
своей удлиненной форме к поясу летающих рептилий.
Бедро и берцовая кость имеют сочленовные поверхности типа Dino-
s a u г i а.
Передний пояс сохранился хуже, с большой coracoideum, соединяющей
признаки Plesiosauria и Dinosauria. Передние конечности значительно
подкласс: diapsida
115
короче задних, с хорошо развитыми пятью пальцами, имеющими 3—5 суста-
вов. Строение отдельных костей напоминает кости китообразных.
Из этого краткого описания видно, что Protorosaurus не может
считаться формой, близкой к какому-нибудь из ныне живущих пресмыкаю-
щихся. Это как бы коллективный тип, выработавший признаки, сближающие
его с различными формами, но, вероятно, не давший начала ни одному из по-
следующих типов. Мы не знаем предшествующую ему форму Развивался он
одновременно с другими пермскими Prosauria.
Champsosaurus Соре (Сев. Америка, Р/2 м длины) и Simaedo-
saurus Gerv. (Европа, 2г/2 м) являются меловыми и нижне-миоценовыми пред-
ставителями этого отряда, хотя и очень отклонившимися. Это, очевидно, полу-
водные пресмыкающиеся — с амфицельными позвонками и исчезнувшей хор-
дой. Шейные ребра двуголовчатые, спинные — простые. Череп сильно удлинен,
гавиалообразный, с височными дырами. Ноздри слившиеся. Зубы сидят неглубоко
в ячейках, имеют складки эмали. Грудной пояс массивный, конечности пятипа-
пые. Связать эту форму с другими Prosauria в настоящее время нельзя.
Ее и близкие к ней формы объединяют в подотряд Choristodira (Соре).
Отряд 2
Чешуйчатые — Squamata (Lepidosauria)
Начинают собою настоящих пресмыкающихся и заключают в себе змей и
ящериц, обобщенных признаками: процельными (вогнутыми спереди и выпу-
клыми сзади) позвонками и ребрами с одной головкой. Квадратная кость соеди-
нена с черепом только верхним
концом, имеется одна или две
пары височных дыр. Носовые
отверстия раздельны. Суще-
ствует в черепе особенная тон-
кая косточка —columella cranii,
упирающаяся одним концом в
pterygoideum, другим— в перед-
ний конец squamosum (рис. 139).
Кожа покрыта чешуйками или
Рис. 139. Череп Monitor niloticus (ныне
живущего).
кожными пластинками. Змеи живут в воде и на суше; ящерицы—животные
наземные.
Мы уже указали на сходсгво некоторых Prosauria с ящерицами и на
их отличия. Остатки настоящих змей и ящериц известны очень несовершенно.
Подотряд 1
Ящерицы — Lacertilia
Araeoscelis gracilis Will,
денная в перми Техаса и указывающая
признаки, не позволяющие поставить
8*
(рис. 140) — интересная форма, най-
на древность ящериц; однако она имеет
ее в предки позднейшим Lacertilia.
116
класс: пресмыкающиеся — reptilia
Найдено несколько почти цельных скелетов длиною 80 см, с очень длинным
хвостом. Конечности хорошо развитые, пятипалые, с очень удлиненным
четвертым пальцем на передних и задних конечностях. Главное отличие от
ящериц заключается в том, что
Рис. 140. Araeoscelis gracilis.
(Пермь Техаса.) (*/ю)
на месте нижней височной дыры
у Araeoscelis большая, тон-
кая костная пластинка, которая
является вторичным образованием
в черепе, закрывшим дыру, суще-
ствовавшую у ее предков, как это
наблюдается у некоторых черепах.
Существующая дыра Araeosce-
1 i s соответствует верхней дыре
Sphenodon. Абель считает
эту форму боковой ветвью, близ-
кой к Protorosaurus и Lepi-
d о s a u г i а.
Казе сближает ее с Clepsydropsidae и считает, судя по строению
ее тела, что она могла лазать по деревьям.
Незначительные остатки ящериц найдены в триасе Южной Америки —
Polygnatha; в верхней юре (в пурбеке) в очень неполном виде и
в нижнем мелу.
Macellodon — челюсти и кожные щиты. Верхняя юра.
Actaeosaurus Н. v. М.— из известняков
нижнего мела Истрии; известен скелет без черепа.
В третичных отложениях их много, но скелеты не-
полные. Iguana europaea Filh. — найдены ниж-
ние челюсти в олигоцене Франции. R h i п е u г а
Hatched Baur (рис. 141) — в олигоцене Сев. Аме- Hatched Baur,
рики — цельный череп с зубами. Глазницы не от-
делены от височных дыр. Все эти формы — уже близкие к нынешним, с про-
цельными позвонками, с симфизой, соединенной швом (в нижней челюсти),
с окостеневшим sternum. Установить непосредственную связь их с Prosauria
пока нельзя.
Подотряд 2
Змеи — Ophidia
Остатки змей до недавнею времени были известны только начиная с
нижних третичных отложений. Ими богат олигоцен Керси, а также пресно- ‘
водные миоценовые известняки Энингена, Штейнгейма и др. В них найдены
остатки ужей — Coluber, Ерарh is и др. В верхней третичной эпохе они
уже были богато представлены. Но в 1923 г. были описаны остатки змеи
Pachyopsis Woodwardi бароном Nopcsa как древнейшей змеи из нео-
подкласс: diapsida
117
кома (нижний мел) Герцеговины (рис. 142). Сохранился почти цельный скелет
с толстыми ребрами 3).
Коренная разница между двумя последними подотрядами — присутствие и
отсутствие конечностей — стоит в связи с другими особенностями скелета:
развитием ребер, отсутствием поясов у
змей — и вызывает интересный вопрос о
моменте разделения их. Но пока палеонто-
логия не имеет данных для решения этого
вопроса.
Отряд 3
Змееобразные — Pythonomorpha
Имеет представителей замечательно
хорошей сохранности. Найдены эти формы
в верхнем мелу Европы, Сев. Америки
и Новой Зеландии. Это—крупные змее-
образные с черепом ящерицы, с измененными в ласты конечностями, голые
морские хищники. Позвонки процельные, с хорошо развитыми верхними
отростками на шейных и спинных позвонках. На хвостовых развиты нижние
отростки. Особыми признаками отличается череп, в общем близкий по строе-
нию к черепу ящериц.
Большими размерами и формой, широкой вверху и суживающейся книзу,
-отличается квадратная кость. Орбита может быть замкнутая или открытая.
Существует пинеальное отверстие. Строение нижней челюсти указывает на ее
большую подвижность. Посреди ее длины существует особенное сочленение,
связывающее две ее половины и позволяющее расширять усиленно пасть. Это
строение сильно отличает ее от челюсти змей, у которых возможность широко
открывать пасть обусловливается особой подвижностью сочленения не только
квацратной кости, но и squamosum. Строение черепа у различных представите-
лей не одинаково, что будет указано при описании их.
Сочленовные отростки позвонков развиты хорошо в передней части ту-
ловища, в хвосте они иногда исчезают. Передний и задний пояса состоят из
окостеневших, но не сближенных вполне костей. Конечности очень укорочены,
но всегда в них можно различить кости пятипалой конечности. Это, очевидно,
боковая ветвь ящериц, отчленившаяся еще в триасе и развившая в меловую
эпоху свои особые признаки, превратившись в морских животных. В мелу же
они и вымерли. Наиболее полно изученными формами являются:
Thalattosaurus — из триаса Калифорнии.
Opetiosaurus — из неокома Далмации.
Mosasaurus — из мела Маастрихта, принятый сначала за кито-
образного.
<) Palaeontographica, Bd. LXV, 1923, Taf. VII, VIII.
118
класс: пресмыкающиеся — reptilia
Hainosaurus — из мела Бельгии.
Platecarpus t
Clidastes V — из мела Америки.
Tylosaurus I
Сем. Thalattosauridae Abel — триасовые представители Руth опо-
m о г р h а. Средний и верхний триас Калифорнии.
Thalattosaurus Alexandrae Мен. (рис. 143) — короткий череп с вы-
соко лежащими ноздрями, далеко входящими межчелюстными. Носовые кости
маленькие; лобные и теменные хорошо развиты, что делает черепную коробку
хорошо закрытой. Пинеальное отверстие маленькое. Склеротическое кольцо
массивное. Квадратная кость неподвижная, вырезана сзади. Зубы в челюстях и.
на нёбе. Позвонки амфицельные. Нижняя челюсть цельная. Резко отличается от
меловых представителей. Циттель относит это семейство к Rhynchoce-
phalia, Штейн м ан и Абель — к Р у t h'o п о ш о гр h а.
£ иР А
Рис. 143. Thalattosaurus Alexandrae
Merr. (Триас.
Рис. 144. 'Opetiosaurus
В u с с h i с h i Korn. (Неоком.)
Opetiosaurus Bucchichi Korn. (рис. 144)—из неокома о-ва Лезина
в Далмации — представляет как бы переходную форму oTVaranidae к Mosa-
saur i d а е. Он имел очень длинный хвост. Конечности еще мало укороченные —-
варанового типа, зубы на pterygoideum. Подвздошные кости соединялись с двумя
крестцовыми позвонками. Маленькая носовая кость лежит за удлиненными но-
совыми отверстиями варанового типа. Абель считает его предком мозазавров.
Сем. Mosasauridae. Mosasaurus Н. v. Meyer — мел; имеет тело
длинного крокодила, до 7 м. Череп с удлиненными межчелюстными и челюст-
ными костями, имеет одну пару височных дуг, ограничивающих верхние
впадины. Верхняя крышка черепа состоит из межчелюстных костей, подходящих
к тонким носовым, переходящим в лобные, сзади которых лежат теменные кости
с пинеальным отверстием и затылочные. По бокам черепа расположен ы верхне-
челюстные, сочленяющиеся с предлобными, ограничивающими глазницы, за ко-
торыми лежат залобные, образующие со squamosum и задней височной верхнюю
дугу. Между носовыми и верхнечелюстными костями лежат длинные носовые отвер-
подкласс: diapsida
119
стия. Большие конические зубы расположены в желобе верхней челюсти и
на
крыловидной кости. Число позвонков доходит до 130,
!з которых сто принадлежат хвосту. Все они вполне
окостенели и имеют спинные и боковые отростки,
развитые различно в различных частях туловища. Эта
форма была в первый раз найдена Германом
Мейером еще в 1780 г.; имеется ее череп, опи-
санный Кювье. Конечности не вполне известны.
Найдены отдельные части в Бельгии. Американские
формы сохранились более совершенно.
Platecarpus simus Marsh (рис. 146) — из
верхнего мела Канзаса. Сохранился весь скелет, имею-
щий около 150 позвонков. Верхние дуги хорошо сро-
слись с телами позвонков. Хорошо видно строение
поясов и конечностей с укороченными костями плеча и
предплечья, бедра и голени и с удлиненными фалан-
гами, метакарпами и метатарсами. Эти конечности
вполне отклоняются от конечностей Prosa uria и
стоят ближе к морским млекопитающим, ведущим
сходный с ними образ жизни. Череп — сходный с
предыдущим.
Tylosaurus — почти такой же хорошей со-
хранности, даже с хрящевыми частями. Так, например,
Рис. 145. Tylosaurus proriger Соре. (Верхний мел.)]
сохранились настоящие и ложные ребра; десять хря-
щевых ребер соединяются с грудной костью. Сохра-
нились кольца трахей и грудная хрящевая кость.
Рис. 146. Platecarpus coryphaeus. (Сев. Америка, мел.)
Длина достигала 8,5 м. Интересно сходство коракоид, без вырезки, с типич-
120
класс: пресмыкающиеся — reptilia
ными для ящериц коракоидами, например у V а г a n u s. Хвост загнут вниз,
и на нем и на спине существовал ряд плавников.
В черепе есть тоже некоторое сходство с Varanus. Все это дает по-
вод Осборну считать Tylosaurus древним отпрыском ящериц, приспо-
собившимся к жизни в море, к плаванию и сохранившим все-таки некоторые
признаки ящериц (рис. 145). Нижний сенон и верхний турон Сев. Америки.
Clidastes Соре (рис. 147) — очень близок к Platecarpus по
удлиненной форме черепа и форме тела. Лопатка сходна с лопа гкой Rhyncho-
с е р h а 1 i а.
Кости плеча и предплечья не только укорочены, но и очень плоские. Зубы
расположены, как у Mosasaurus. В туроне и нижнем сеноне Сев. Америки.
Рис. 147. Clidastes velox Marsh.
Большинство М о s а-
s a u г i d а е принадлежали к
страшным хищникам и были
снабжены большими острыми
зубами.
Но некоторые Р1 i о-
platecarpus из сенона
Маастрихта имели редуциро-
ванные зубы и питались.
вероятно, более мягкой пищей —-головоногими, подобно Орhthа 1 mо-
saurus из Ichthyosauri а.
Долл о описал мозазавра из верхнего мела Голландии — Gio bi de ns
Fraasi с еще более короткими и толстыми зубами, который, вероятно, пи-
тался иглокожими и двустворками и был хищник (durophag).
Эта удивительная меловая группа морских рептилий соединяет в себе
признаки змей и ящериц. Коп, много изучавший их, ставит Pythonomor-
р h а между змеями и ящерицами. Вымирание их в верхнем мелу кажется очень
странным, так как скелет их указывает на очень хорошее приспособление
к жизни в море, к плаванию и схватыванию добычи. Очевидно, это ветвь
ящериц, отчленившаяся от наземных форм, лишившаяся чешуи и пошедшая
в море, вероятно, в начале триаса.
Отряд 4
Крокодилы — Crocodilia
Недавно еще крокодилы относились к S q u a m a t а по их большому сход-
ству с ящерицами. Но изучение анатомических признаков заставило отделить
их в отдельный отряд.
Главными отличиями являются строение сердца и сросшаяся с черепом
квадратная кость.
Немногие ископаемые остатки известны из верхней перми; они встречаются
в триасе, юре и мелу. Старейшие триасовые формы, отличающиеся по некото-
рым признакам, отделяются под именем Parasuchia и Pseudosuchia
подкласс: diapsida
121
от более поздних Mesosuchia и нынешних Е u s u с h i а, подразделяю-
щихся на длинномордых — гавиалов, короткомордых — аллигаторов — и соб-
ственно крокодилов — кайманов.
Подотряд 1
Parasuchia
Главные характерные признаки: носовые ходы раздельны, нет твердого
нёба; большие предглазничные дыры; квадратная кость неподвижна; ключица
хорошо развита. Они подразделяются на короткомордых и длинномордых.
Водные животные. Некоторые из признаков сближают эти триасовые формы
с крокодилами, другие — с Prosauria (Protorosau-
r u s) (строение черепа и грудной пояс) и D i n-o s a u г i а
(Задний пояс и строение конечностей).
Короткомордые — Pelycosimia. Erythrosuchus
africanus Broom — из нижнего триаса Кару (рис. 148).
Довольно высокий череп треугольной формы с пятью
парами отверстий: носовые, предглазничные, глазные, нижние
и верхние височные. Короткие межчелюстные, длинные
носовые кости. Кроме черепа известны отдельные кости.
Scaphonyx — триас Бразилии. Известны только
кости конечностей, сходные с костями первой формы.
Длинномордые — Pnytosauridae. С очень удли- s u с h u s a f г i с а-
ненными межчелюстными костями и далеко отодвинутыми п 11 s Broom,
носовыми отверстиями.
Belodon (Phytosaurus) (рис. 149) — тип этой группы из верх-
него триаса Штутгарта, Сев. Америки, Новой Мексики, Сев. Каролины, Пен-
Рис. 149. Belodon Kapffii Н. v. Меу.
Сильвании, Техаса. Первая находка описана Германом Мейером в 1863 г.
Череп достигал в длину 1 м, имел очень удлиненную морду, с зубами, сидя-
щими в лунках в один ряд на межчелюстных и челюстных костях. Кости че-
репа со скульптурой; существуют две височные дыры. Эти его признаки
встречаются у позднейших крокодилов. Особенностью являются парные лоб-
ные кости и особенное удлинение высоко поднятых межчелюст-
ных костей у Belodon Kapffii; у Bel. Plieningeri они менее высоки.
Кости эти вдвигаются далеко между верхнечелюстными и отодвигают далеко
122
класс: пресмыкающиеся — reptilia
назад носовые кости вместе с носовыми отверстиями. Последние доходят до
удлиненных, небольших предглазничных дыр J). Глазницы небольшие, удлиненные,
отодвинуты к верхним височным дугам. Сильно развиты заглазничные кости и
squamosum. На нижней поверхности черепа межчелюстные отодвинули сильно назад
ним и хоаны. Остальная часть нёба закрыта по бокам неб-
сошник, а вместе с
Рис. 150. М es о г h i n u s
F г a a s i Jaeckel. (Ниж-
ний триас).
ними костями, а посредине она остается открытой,
так что хоаны находятся под носовыми отверстиями.
Боковые затылочные squamosum сильно выдвинуты
назад, а на основной затылочной развит сочленовный
мыщелок, состоящий из средней закругленной части
и двух боковых. Нижние височные дуги типа кро-
кодила, состоят из jugale, quadratum и ветви exoccipi-
tale. Боковые зубы сжаты с боков; передние больше
боковых, закруглены.
На спине существовал панцырь с двумя рядами
длинных, узких пластинок, по краям которых лежали
маленькие пластинки. Передняя часть брюха тоже
имела панцырь.
Более близкими формами к крокодилам гавиа-
лового типа являются:
Mesorhinus Fraasi Jaeckel (рис. 150).
Удлиненные межчелюстные кости с высоко лежащими
носовыми отверстиями. Большие предглазничные дыры;
не соединенные между собою palatinum и pterygoidea.
Нижний триас Германии. Резко отличается от Belo-
don плоской, не вздутой мордой.
Рис. 151. Mystriosuchus planirostris Н. v. М.
Mystriosuchus (Rutiodon) planirostris H. v. Mey. — средний
триас Вюртемберга (рис. 151). Череп тоже с удлиненными межчелюстными ко-
стями и тонкой нижней челюстью. Передняя часть морды плоская, в виде тонких
пластинок. Носовые отверстия лежали над нижними хоанами, как у В е 1 о d о п.
Задняя половина черепа, резко отграниченная носовыми отверстиями, подни-
<) Абель указывает, что эти ненормально высокие межчелюстные кости явля-
ются следствием болезненного развития костного вещества после поранения, причиненного
зубами крокодила в борьбе. Следы ранений видны на некоторых экземплярах.
подкласс: diapsida
123
малась высоко над передней — удлиненной. На ней расположены четыре пары
отверстий.
. Спинной панцырь состоял из четырех рядов пластинок; брюшной — из 30
пластинок. Жил Mystriosuchus, вероятно, на берегу и мог хорошо плавать.
Удлиненные морды этих животных с отодвинутыми далеко носовыми
отверстиями могли служить им для добывания пищи из рыхлой земли, как
это бывает у птиц с длинным клювом, по мнению Виллистона. Зубы их
указывают на хищный образ жизни, а не на растительный, как это предпо-
лагали раньше.
Подотряд 2
Pseudosuchia Zittel
Маленькие формы с коротким хвостом. Боковые носовые отверстия лежат
на переднем конце морды, между маленькими межчелюстными и большими но-
совыми костями. Позвонки амфицельные и платицельные. Передние конечности
короче задних. На спине
два ряда пластинок. Редко
в верхней перми, чаще
в среднем триасе.
Euparkeria ca-
pe n s i s Broom (рис.
152) — закругленная вы-
сокая морда с длинными
носовыми костями. Длин-
ный хвост с большими
Рис. 152. Euparkeria capensis Broom.
отростками. Триас Кару.
Ornithosuchus.
Короткий череп представляет особенности в своем строении: у него слиты
обе височные дыры и очень большие предглазничные дыры. Хоаны
отодвинуты далеко назад. Зубы слегка загнуты. Позвонки амфицельные. Под-
вздошные кости тонкие, длинные. Панцырные пластинки закруглены. Средний
триас Шотландии.
Aetosaurus ferratus Fraas (рис. 153, 153 А) — третий тип из тех же
отложений, наилучше изученный. Его треугольная голова оканчивалась острой,
короткой мордой со слабо развитой межчелюстной костью, вытесненной длинными
носовыми костями, за которыми лежат широкие лобные и теменные. Большие
удлиненные носовые отверстия находятся по сторонам на конце морды. За
ними следуют предглазничные дыры и глазницы. Две пары небольших височ-
ных дыр. Кости черепа гладкие. Немногочисленные зубы в глубоких альвео-
лах. Спина и хвост животного были покрыты прочными поперечными костными
пластинками. Хорошо развитые передние конечности короче задних. Все —
пятипалые и по строению более похожи на конечности ящериц, чем крокодила.
В триасе близ Штутгарта найдена целая плита в 70 см длиною более чем
с 24 индивидами различного возраста, хорошей сохранности (рис. 153).
124
класс: пресмыкающиеся — reptilia
Сильно отклоняющимся типом является Scleromochlus Taylori
Sm. Wood. — из триаса Шотландии, с очень длинным хвостом, короткими пе-
Рис. 153. Aetosaurus ferratus О. Fraas. (Верхний триас.)
редкими и значительно более длинными задними стопоходящими ногами, с
очень сближенными метатарсами, с укороченным пятым пальцем, тонкими ко-
стями конечностей. Передние конечности имели когти. На черепе имеется
пять пар отверстий. Хюне предпола-
гает, что это животное лазало на де-
ревья и могло прыгать. Оно напоминает
своим строением Otocryptis bivit-
t a t a (A g a m i d а е Цейлона), живущего
на деревьях. Смите Вудворд, опи-
савший его впервые, относит его к
динозаврам1).
Из этих примеров видно, что
триасовая группа крокодилов была уже
Может быть, некоторые из них положили начало
art апд
Рис. 153А. Череп Aetosaurus ferra-
tus О. Fraas.
очень дифференцирована,
более поздним формам.
Подотряды 3 и 4
Mesosuchia и Eusuchia
Отличаются от предыдущих следующими признаками: носовые отверстия,
лежащие на переднем конце морды, слиты в одно; хоаны отодвинуты далеко
назад и тоже слиты; существует твердое нёбо из слившихся верхнечелюстных,
*) Huene. Geolog.-Paiaont. Abliandlungen, Jena, 1914, N. F., Bd. 17.
Smith Woodward. Quart. Jomn., vol. 63, 1907, p. 140.
подкласс: diapsida
125
небных и крыловидных костей (у некоторых). Две пары височных дыр и дуг.
Предглазничные дыры постепенно исчезают. Все тело покрыто твердыми
роговыми пластинками. Задние ноги четырехпалые.
Начинаются Mesosuchia и Eusuchia с лейаса и доходят до нас в виде трех
типов: кайман и аллигатор — с закругленными широкими мордами и гавиал —
с тонкой, удлиненной.
Некоторые мезозойные и третичные формы позволяют нам проследить
прогрессивное изменение частей черепа в направлении к нынешним крокоди-
лам. Особенного внимания заслуживают перемещение небных костей дальше
назад и одновременное передвижение хоан к затылку. С другой стороны, про-
исходит укорачивание межчелюстных костей и передвижение носовых отвер-
стий на конец морды. Позвонки процельные или плоские.
Короткомордые Mesosuchia — юрские и меловые формы.
Рис. 154. Alligatorellus Beaumonti Jourd. (Верхняя юра.) (</2)
Alligatorellus — маленькая, до 22 см, юрская форма с короткой
закругленной спереди треугольной головой, большими глазницами и малень-
кими верхними дырами (рис. 154). Ноздри спереди разделены далеко прохо-
дящими тонкими носовыми костями. Хоаны ограничены небными и крыло-
видными костями. Позвонки амфицельные. Конечности пятипалые. Очень длин-
ный хвое г.
Эта форма связывает, может быть, триасового Aetosaurus с мело-
вым Notosuchus (сев. Патагония). Череп длиною в 18 см, с боль-
шими глазницами. Зубы верхней челюсти дифференцированы, в межчелюст-
ной кости три очень маленьких зуба и один большой; 7 зубов в верх-
ней челюсти. Нижняя челюсть у этой формы очень тонкая, с большой
дырой.
Berrtnissartia Dollo—вельд Бельгии, длиною до 1 м. Носовые отвер-
стия раздельны. По 20 зубов в челюсти. Передние конечности короче задних.
Спинной панцырь состоял более чем из двух рядов пластинок.
Libycosuchus brevirostrns v. Stromer — близкая к предыдущим
форма из сеномана Египта (Baharije).
126
класс: пресмыкающиеся — reptilia
Leidyosuchus Соре — верхний мел Сев. Америки (Айоминг). Череп
очень хорошей сохранности. По Циттелю он стоит между кайманом и
аллигатором. Он начинает в мелу ряд крокодилов с процельными позвонками.
Интересно положение его хоан, далеко продвинувшихся назад, но не дошедших
еще до края крыловидной кости.
Diploc’ynodon Gervaisi Aym.— из олигоцена Сев. Америки и
Франции, дает более близкую
форму к нынешним аллигаторам.
У него носовые отверстия нахо-
дятся между межчелюстных ко-
стей, которые несут четыре зу-
ба. Глазницы очень большие.
Верхние височные дыры малень-
кие. Нёбо закрыто. Хоаны окру-
жены только крыловидными ко-
стями, как у позднейших кроко-
дилов. Спинной и брюшной
панцыри хорошо развиты. По-
звонки процельные.
Совершенно другой тип
мы имеем в формах с удли-
ненной мордой, являющих-
ся как бы продолжением триа-
совых Mystriosuchus и
М е s о г h i n u s, но с коротки-
ми межчелюстными костями.
Сем. Teleosauridae. Те-
ло покрыто панцырем. Морские;
с юры до эоцена.
Mystriosaurus b о 1-
1 е n s i s Cuv. — верхний лейас
Вюртемберга и Банца во Фран-
конии; форма, достигавшая 4 м,
представлена многими видами.
Очень плоский череп. У некоторых сохраняются небольшие предглазничные
отверстия. Удлиненные носовые отверстия направлены вверх.
Stenosaurus Larteti Degl. —- с хорошо сохранившимися пластин-
ками спинного панцыря. Верхняя юра Англии.
Pelagosaurus temporalis Blv. — из лейаса Кальвадоса, с удлинен-
ной мордой и хоанами, лежащими между задними концами небных костей
и очень маленькими крыловидными. Межчелюстные кости короткие, б1агодаря
чему носовые отверстия лежат впереди, на конце морды. Глазницы лежат
впереди, а не между височными дырами.
подкласс: diapsida
127
Teleosaurus — средняя юра Кальвадоса, с очень удлиненной мордой
из верхнечелюстных костей с очень маленькими межчелюстными. Хоаны за
palatina, сросшимися в трубку. Мелкие зубы на верхней и нижней челюстях
и межчелюстной кости (рис. 155). Эта форма ведет к подотряду Eusuchia.
Сем. Gavialidae. Gavialosuchus (Tomistoma) — встречается
с миоцена с такими же тонкими челюстями. Ноздри лежат на конце раздвоенных
межчелюстных костей, а не в трубочковидном конце ее. Нёбо закрывается еще
более, и хоаны передвигаются назад. Теперь живет на Борнео, Суматре и Малай-
ском полуострове. В роде G a v i а 1 i s (гавиал) хоаны лежат еще дальше за соеди-
нившимися небными и межкрыловидными костями, сдвинувшимися к средине. Иско-
паемые остатки его найдены в плиоцене Сиваликских
холмов. Интересны находки крокодилов в Африке,
в Фаюме, близ Каира, в верхнем и среднем эоцене.
Crocodilus articeps — по общему строе-
нию близок к нынешним Crocodilus cata-
phractus и Croc, intermedins. Тонкая,длин-
ная морда с плоской верхней частью предглазнич-
ной области.
Crocodilus megarhinus—из верхнего
эоцена, с широкой мордой с перехватом спереди и
широкими носовыми отверстиями (рис. 156).
Tomistoma africanus —• средний эоцен,
с очень удлиненной мордой настоящего гавиала.
Совершенно особняком стоит
Сем. Geosauridae— морские формы; имели
хвостовой плавник.
Melriorhynchus Blainv. — в доггере и
Оксфорде Англии и северной Франции. Юра Пата-
гонии.
Соес
Очень интересные данные имеем мы в указа- - „ , .
Рис. 15b. Нижняя поверх-
ниях Эндрьюса (Andrews) на изменение формы ность черепа крокодила.
черепа у различных видов этого рода, начиная
с узкомордых и доходя до широкомордых форм. У некоторых из них кос гм
черепа гладкие, у других скульптированы. Все указанные формы из Оксфорда
Англии.
Geosaurus suevicus Fraas (рис. 157) — верхняя юра — с сильно раз-
витыми верхними дугами, маленькими глазами, окруженными склеротическим
кольцом, и гладкими черепными костями. Носовые отверстия раздельные, лежат
на конце морды, на коротких межчелюстных костях. Передние конечности уко-
рочены, образуют ласты с закруглениями: плечевой, лучевой и локтевой. Бедро
и пальцы задних конечностей очень длинные. Хвост очень сильно развит,
с загнутым, направленным вниз концом. Эту форму и близкие к ней Фраас
отделял под именем Thalattosuchia, настоящих морских крокодилов, ли-
шенных панцыря и хорошо плававших. Они жили только в верхней юре.
128
класс: пресмыкающиеся — reptilia
Итак, дифференцировка крокодилов произошла очень давно. Уже в
триасе мы встречаем два резко различных типа: с тупой мордой — Aetosau-
rus ис удлиненной — Mystriosuchus, и эти типы проходят через весь
мезозой, третичный период и доходят до нас, с той разницей, что у них со-
вершается постепенное изменение положения ноздрей и хоан и закрывается
нёбо. Эти изменения дают им возможность более продолжительного пребыва-
ния в воде. Они могут дышать, выставив длинную морду с ноздрями на
конце над водой и вбирать воздух, входящий в дыхательное горло через далеко
лежащие хоаны.
Рис. 157. Geosaurus suevicus Fraas. (Верхняя юра.) (*/в)
Изучение остатков крокодилов имеет большой интерес в смысле их гео-
графического распространения. Мы видели, как широко они были распростра-
нены в мезозое Европы и Сев. Америки и как они исчезли там в третичное
время. Крокодилы и аллигаторы существовали в Европе еще в миоцене. Ти-
пичный гавиал жил в плиоцене Индии и в эоцене Англии.
В Африке, их теперешнем отечестве, остатки известны только в эоцене.
Очевидно, при таком изменении их распространения большую роль должно
было играть изменение климата, охлаждение его в Европе и Сев. Америке,
заставившее эти формы перейти в Африку и на азиатские острова.
Отряд 5
Dinosauria
Под именем Dinosauria, „ужасных ящериц", объединяют в большую
группу очень разнообразных наземных пресмыкающихся. Общими признаками •
для них являются: отсутствие пинеального отверстия; оно существует только
у Diplodochus и Triceratops. Две пары височных дыр, ограниченных
костными дугами; две предглазничные дыры. Ключицы и эпистернум не из-
подкласс: diapsida
129
вестны. Лопатка очень большая, часто слитая с маленькой коракоидной костью.
Некоторые из этих признаков сближают их с крокодилами. Зубы в общем же-
лобке или в альвеолах.
Почти все животные, относимые сюда, принадлежат к вымершим, суще-
ствовавшим с триаса до верхнего мела. Их появление как бы совпадает
•с исчезновением Stegocephali.
Величина их, внешний вид и образ жизни очень различны. Некоторые из них
не больше кошки, другие достигали 18 л/ длины. Были хищники и травоядные.
Хотя все они снабжены хорошо развитыми четырьмя конечностями, но некоторые
двигались только на двух задних, опираясь на мощный хвост. Строение их позво-
ночника довольно разнообразно.
Встречаются позвонки амфицельные, платицельные и опистоцельные в раз-
личных частях позвоночника. Хотя мы не знаем их общего предка, но сохранение
некоторых древних признаков позволяет сблизить их с некоторыми R h у п-
chocephalia (Protorosaurus) и позволяет думать, что они отчленились
еще в перми, одновременно с крокодилами, от Rhynchocephalia, и раз-
вили к триасу свои специальные признаки.
Несмотря на большое разнообразие форм, Марш (Marsh), покойный
американский профессор, распределил их в три отряда1), основываясь на
строении их скелета, в связи с образом жизни:
1. Sauropoda (ящериценогие) — травоядные. Незначительные остатки
в триасе; главные представители в юре. Самые большие из динозавров. Bron-
tosaurus, Diplodocusn др.
2. Theropoda (звероногие) — хищные; в триасе, в мелу и в юре. Ап-
chisaurus — в триасе, Allosaurus, Ceratosaurus, Ornitholestes,
Compsognathus— в юре, Tyrannosaurus и др. — в юре и мелу.
Сили объединяет два первые отряда под именем Saurischia.
3. Praedentata — характеризуются присутствием особой кости в ниж-
ией челюсти—praedentale, входящей спереди между челюстными и соответ-
ствующей верхней межчелюстной кости, и присутствием задней лобковой (post-
pubes). Конечности различные, покровы также, почему Марш группирует их
еще в три подотряда (в юре и в мелу, травоядные):
a) Stegosauria — динозавры с костными пластинками на теле. S с е 1 i-
dosaurus, Stegosaurus, Р ala с ant h us — нижний мел.
6) Ceratopsidae — рогатые динозавры с громадным черепом, снабжен-
ным воротником и рогами. Ceratops, Triceratops и др.—верхний мел.
в) Ornithopoda — птиценогие. Iguanodon — нижний мел, Т г а с h о-
d о п и др. — верхний мел.
Изучение остатков динозавров, особенно обильных в Сев. Америке, при-
вело Хюне (Huene) к выводу, что они произошли не из одного корня,
как предполагалось раньше, а от двух различных представителей Pseudosu-
1) Здесь приведена классификация Марша, поэтому термины отряд и под-
отряд применены иначе, чем применяются теперь, также и название их.
9 Павлова М. В. Палеозоология. Ч. II.
130
класс: пресмыкающиеся — reptilia
chia (древних крокодилов): от первого развился отряд Saurischia Seeley»
включающий Theropoda и Sauropoda, от второго — Ornithischia
Seeley (Ornithopoda Marsh). Главным отличием между двумя отрядами
является присутствие у второго praedentale и postpubes.
Подотряд 1
Ящериценогие — Sauropoda
Травоядные ящериценогие динозавры — крупные животные, по внешнему
виду являются громадными ящерицеобразными животными, с маленькой головой
и наименее специализировавшейся организацией. Они ходили на 4 почти
равных ногах; были стопоходящими, с 5 пальцами и с очень длинным хвостом.
Сем. Atlantosauridae. Хорошо сохранившейся формой является Bronto-
saurus Marsh (рис. 158), найденный в верхней юре Уайоминга в Сев. Америке.
Рис. 158. Brontosaurus
excel sus Marsh. (Нижний мел.) (*/эо)
Л — 6-й шейный позвонок.
Длина его 18 м. Очень маленькая сравнительно с длиною тела голова с ко-
ническими зубами, расположенными на межчелюстной и передних частях че-
люстных костей. Тринадцать шейных позвонков представляют особенность —
присутствие удлиненных, раздвоенных боковых отростков; верхних отростков
нет. Позвонки эти выпуклы спереди, вогнуты' сзади — опистоцельные. Спинные
позвонки (10) с пустотами внутри, разделены слабыми перегородками внутри.
Спинные отростки у них большие; сочленовные поверхности позвонков гладкие.
Три крестцовых срастаются рано; два другие — позже.
В грудном поясе особенно выдаются длинные, расширенные книзу ло-
патки, слившиеся с коракоидными костями, прилегающими к двум окостеневшим
пластинкам грудной кости. Задний пояс состоит из трех пар не сросшихся,
но прилегающих друг к другу костей: ilium — широкие подвздошные, ischium —•
седалищные и pubes — лобковые удлиненные кости, направленные вперед. Сочле-
новные поверхности длинных костей не имеют окостеневших закруглений.
Стопа и кисть по массивности напоминают ноги хоботных млекопитающих.
Первый палец на ногах короткий; он снабжен большим когтем. Тяжеловес-
ные ноги представляют сгранный контраст с легким позвоночником и ма-
ленькой головой.
подкласс: diapsida
131
Diplodocus Marsh (рис. 159) найден в Сев. Америке, в тех же
слоях, в виде полного скелета и дает возможность изучить подробно строение
его, в общем сходное с Brontosaurus. Длина его превышает 20 м. Голова на-
поминает по общей форме лошадиную, с высоким затылком и закругленной перед-
ней частью (рис. 159). Очень большие слившиеся носовые отверстия лежат высоко
на черепе, между глазниц, за удлиненными носовыми костями; за ними лобные
короткие кости и теменные с пинеальным отверстием (единственный из S а и г о-
р о d а, имеющий их). Впереди глазниц, сильно отодвинутых назад, две пары пред-
глазничных дыр. Две задние очень удлиненные пары височных дыр ясно видны.
Затылок выдается далеко назад, прикрывая сочленовный кондилюс. Особен-
ность представляет расположение
зубов, по нескольку в одной ячей-
ке, на межчелюстной и передних
частях челюстных. Зубы стирались
постепенно, выдвигаясь один за
другим. Они имели цилиндриче-
скую форму с несколько сжатой
коронкой, покрытой тонкой эмалью.
Корни зубов длинные, полые вну-
три. Позвонки того же типа, как
и у Brontosaurus. Атлант
имеет углубление для сочленения
с черепом. Шейных—15, спин-
ных—10, все опистоцельные.
Крестцовых сросшихся три. Ноги
пятипалые, тяжеловесные, причем
удлиненная надпяточная кость,
астрагал, прикрывала три метатар-
сальных со срезанными, плоскими
Рис. 159. Diplodocus longus Marsh.
(Верхняя юра Колорадо.)
поверхностями. Первая фаланга имела тоже коготь, как и у Brontosaurus.
При изучении скелета этих форм приходят к выводу, что они жили в
болотах и в реках близ берегов и выходили на сушу. Формы эти были до недавнего
времени известны только в Сев. Америке. В 1922—1923 гг. третьей американ-
ской экспедицией в Монголии были найдены остатки близких к ним форм.
Благодаря щедрости американца Карнеги, сделавшего очень много
для развития науки в Америке, почти все главные музеи Европы обладают
прекрасными слепками скелета Diplodocus, что дает возможность изу-
чать его подробно. В Ленинграде слепок помещен в музее Академии наук.
При изучении Diplodocus различными учеными возник вопрос о том,
правильно ли восстановлен его скелет в Америке и на слепках в европейских
музеях. Г. Т о р н ь е (Tornier)п) определенно указывает, что строение конеч-
ностей и поясов не позволило бы этому животному ходить на вертикально
*) G. Tornier, Wie war der Diplodocus wirklich gebpaut, 1909.
9*
132
класс: пресмыкающиеся — reptilia
стоящих ногах, в том виде, как он восстановлен, потому что сочленовные по-
верхности его костей, так же, как и у ящериц, направлены в стороны. Сле-
довательно, ходить и бегать Diplodocus мог только так же, как это де-
лают ящерицы, т. е. с согнутыми в стороны ногами, припадая брюхом к земле.
Это противоречит взглядам на их водный образ жизни.
Строение позвонков шеи, по исследованию Т о р н ь е, указывает на ее
.вертикальное, согнутое в форме S, положение, а не на горизонтальное, какое
придают ему. Такое S-образное положение должно было давать животному,
жившему в прибрежных местах, возможность устремлять голову быстро вперед
.и схватывать добычу.
Хвост, очень тяжелый и длинный, при этом должен был тянуться по
земле. Таз не мог бы вынести его тяжести при его висячем положении.
Atlantosaurus Marsh. — Имеющиеся очень неполные остатки указы-
вают на строение скелета, близкое к Diplodocus.
Titanosaurus Lyd. — в вельде Западной Европы, в нижнем мелу Индии,
верхнем мелу Мадагаскара и Патагонии — является наиболее распространенной
географически и геологически формой. Американские формы отнесены Мар-
шем к юрским. В палеонтологии Ц и т т е л я они стоят в нижнем мелу.
Camaros aurus Соре (Morosaurus Marsh) отличается короткой за-
кругленной головой с громадными глазницами, носовыми отверстиями, распо-
ложенными высоко спереди, и височными дугами, что превращает все черепные
кости как бы в тонкие перекладины. Нижний мел Сев. Америки (Уайоминг)
и Англии. Morosaurus grandis Osb. — нижний мел Англии.
Gigantosaurus южной Африки. — В 1907 г. проф. Фраас из Штут-
гарта собрал интересную коллекцию остатков динозавров из верхне-меловых
отложений Тендагуры в южной Африке, а именно — костей конечностей не-
обыкновенно больших размеров:). При подробном изучении их и сравнении
с ранее известными остатками Фраас устанавливает их принадлежность к
Sauropoda и ставит новый род, Gigantosaurus, между Morosaurus
и Diplodocus, причем размеры этой формы превышали размеры последних.
Хвостовые позвонки были у него процельные, крестцовых четыре. Остатки
черепа совершенно неизвестны. Автор различает два вида: Gigantosaurus
africanus Fraas — с высокими задними ногами, как у Diplodocus,
и Gigantosaurus robustus Fraas — с более низкими и значительно более
грубыми задними конечностями. Число пальцев пять, с сильно развитыми пер-
выми и удлиненными коготными фалангами. Кости были найдены отчасти на
поверхности, не прикрытыми почвой, отчасти в отложениях, которые Фраас
сближает по петрографическим признакам с пресноводными юрскими отложе-
ниями Сев. Америки (Atlantosaurus - beds). Это сходство отложений верхней
юры и верхнего мела приводит автора к заключению, что во время их образо-
вания господствовали в Сев. Америке и в южной Африке одинаковые геоло-
гические условия.
’) Е. Fraas, Ostafrikanische Dinosaurier. Palaeontographica, Bd. 55, 1908.
подкласс: diapsida
133
Подотряд 2
3e,ep<iHi3Zue — Theropoda Marsh — (Pro-omithopoda— предптиценогие M. PavL)
Вторая группа динозавров откосится к хищным, и строение скелета их
соответствует этому образу жизни. Но при общем взгляде на скелет у неко-
торых форм, особенно если при реставрации его животное поставлено на четыре
ноги, его можно принять за Sauropoda:
небольшая голова, длинное тело с большим
длинным хвостом и четырьмя конечностями.
Однако при внимательном знакомстве разница
оказывается очень большой. Голова построена
гораздо легче, с большими дырами и тон-
кими костями. Пинеального отверстия нет.
Зубы конические, неравной величины, распо-
ложены в межчелюстных и в челюстных ко-
стях по всей их длине. Нижняя челюсть не
имеет венечного отростка и сочленяется по-
средством квадратной кости. Шейные по-
звонки опистоцельные. тонкие, с простыми
отростками. Спинные платицельные, реже
амфицельные. Конечности легкие, передние
А
Рис. 160. Anchisaurus major
Marsh.
короче задних, с хорошо развитыми тремя или четырьмя пальцами. Все строе-
ние скелета указывает, что эти животные ходили на задних ногах. Группа эта
хорошо представлена, начиная с триаса, в юре и в мелу Сев. Америки, Англии,
Рис. 161. Plateosaurus longiceps Jaeckel.
этого типа, свободно поднимавшееся при хождении
Франции, Германии, Па-
тагонии. Это были на-
стоящие обитатели су-
ши, бегающие и пры-
гающие за добычей.
А n с h isaurus
Marsh (рис. 160) — из
верхнего триаса Сев.
Америки, является древ-
нейшим из динозавров.
Это уже вполне сфор-
мировавшееся животное
на задние ноги, пальце-
ходящее.
Хвост, вероятно, тоже служил опорой. Голова его с большими пред-
глазничными дырами (3 пары) и глазницами напоминает птичью, а по хорошо
развитым височным дугам — Rhynchocephalia.
Позвонки, как и у всей этой группы, — полые, также и кости конечно-
стей, что делало весь скелет легким и облегчало движения животного. Перед-
ние конечности имели три хорошо развитые пальца и остатки четвертого и
134
класс: пресмыкающиеся — reptilia
пятого. Особенно хорошо развит первый палец. Из пальцев задних конечностей
четыре хорошо развитые, пятый — рудиментарный. Сочленовные поверхности
костей кисти и ступни развиты хорошо. Надпяточная кость, астрагал, подсти-
лает плотно большую берцовую, пяточная, кальканеум, •—малую берцовую, что
придает ноге вид птичьей.
Plateosaurus longiceps Jaeckel (рис. 161), недавно найденный в
И е к е л е м, представляет хорошо сохра-
нившийся скелет древнейшего в Европе
хищного динозавра, до 6 м длиною.
На передних конечностях сохранились
пять пальцев, хотя неравных разме-
ров. Три первые — длинные, с когтями,
два последние — маленькие. Задние ко-
верхнем триасе Германии и описанный
Allosaurus
Marsh. (Мел.)
Рис. 163.
agilis
А — задняя, В — передняя
нога.
Рис. 162. Ceratosaurus nasicornis Marsh.
нечности гораздо длиннее передних, с тремя длинными, снабженными когтями
пальцами.
Удлиненная морда имеет четыре пары больших отверстий по сторонам:
носовые, предглазничные, пазнччные и нижние височные; вверху—верхне-височ-
ные. Кондилюс выступает далеко назад.
Нижняя челюсть имеет посредине дыру, что отличает ее от челюсти
Anchisaurus и Allosaurus. Зубы заостренные, несколько загнутые.
Абель ставит эту форму у основания Theropoda. Остатки ее найдены
также во Франции (в триасе).
подкласс: diapsida
135
Ceratosaurus (рогатый) — верхняя юра Сев. Америки (рис. 162), имеет
рог на носу и два возвышения костей перед глазницами. Нижняя челюсть
имеет дыру посредине. Зубы в межчелюстной кости более мелкие, чем в
челюстях. Это еще более редуцировав-
шая свои конечности форма. У нее три
средние metatarsalia срослись
между собою наподобие птичьих. Та-
зовые кости тоже срослись между
•собою; подвздошные очень удлинились.
Передние конечности еще более укоро-
тились. Боковые пальцы утонились.
Это более специализировавшаяся форма
сравнительно с предшествующими.
Megalosaurus Buckl. — евро-
пейская юрская и меловая форма, до-
стигала размеров слона. Его бедро имело
в длину 1,1 м, лопатка—-55 см. Кроме
Рис. 164. Tyrannosaurus rex Osb.
(Верхний мел.) Вверху справа—малень-
кий Struthiomimus.
Европы его остатки известны из Индии, Мадагаскара, Австралии и Южной
Америки.
Veloceratosaurus — из Монголии, этого же типа.
Allosaurus Marsh (рис. 163) — верхняя юра Сев. Америки, дает даль-
Рис. 165. Podokesaurus holyokensis Talb.
лет найден не вполне цельный. Но отдельные
нейшую редукцию пальцев:
на передней конечности два
средние большие, два боко-
вые маленькие; на задней три
пальца, с преобладанием сред-
него. Пяточная и надпяточная
срослись с костями голени.
Удлиненная лопатка и приле-
гающая к ней коракоидная
примыкают к закругленной го-
ловке плеча. Тазовые общего
типа динозавров, образуют
дыру для бедренной головки.
Широкие подвздошные при-
легают к крестцу; седалищные
и лобковые направлены вниз.
Tyrannosaurusrex
Osb. (рис. 164) — меловое
чудовище Сев. Америки. Ске-
кости дают понятие о вели-
чине его. Большая берцовая имеет более метра в длину. Строение массивного
черепа напоминает Allosaurus — с шероховатостью над глазницами; но
кости более грубые, дыры меньше. Позвонки массивные, спинные и хвостовые.
136
класс: пресмыкающиеся — reptilia
Эта форма является продолжателем форм с грубым строением тяжеловесного
скелета, развивавшихся параллельно с гЬормами более легкого строения, кото-
рых Хюне отделяет под именем —
Рис. 166. Ornitholestes Hermann Osb. (Верхняя юра
Сев. Америки.)
Сем. Coelurosauridae Huene. — Podokesaurus (рис. 165) — из
триаса Сев. Америки, очень стройный скелет, с длинным хвостом, удлиненными
метатарсалиями; с равномерно развитыми тремя пальцами на передних и задних
конечностях.
Ornitholestes Osb. — верхняя юра Сев. Америки (рис. 166); длина
всего животного с хвостом — более
2 м. Скелет построен чрезвы-
чайно легко, кости и позвонки по-
лые. Череп с большими дырами.
Зубы: 4 — в межчелюстной кости,
10 — в верхней челюсти и 12 — в
нижней, занимают небольшое про-
странство. Главное отличие от
предыдущих форм заключается в
очень тонких конечностях, длин-
ном хвосте и в обилии ребер;
существуют шейные и хвостовые.
Это строение скелета с хватаю-
щими тонкими передними конеч-
ностями позволяет Осборну
Рис. 167 Compsognathus longipes предположить, что это животное
A. Wagn. (Юра.) могло с успехом гоняться за юр-
скими птицами.
Compsognathus Wagn. (рис. 167) — самый маленький из европейских
динозавров, величиною с курицу, имеет птичий череп, чрезвычайно открытый,
с зубами.
подкласс: diapsida
137
Весь скелет пневматичен. Шея длинная, соединялась с головой под
прямым углом. Передние конечности вдвое короче задних. На задних по
четгре развитых пальца; пяточная и надпяточная кости не срастаются с боль-
шой и малой берцовыми, только прилегают к ним. Эта форма как бы
отстала в развитии конечностей, но приблизилась к птицам по строению
черепа и костей.
Struthiomimus altus Osb. (рис. 168)—верхний мел Сев. Америки.
Беззубый маленький череп. На передних конечностях по три пальца
Рис. 168. Struthiomimus altus Osb.
с удлиненными последними фалангами, несколько загнутыми. Трехпалые задние
ноги с остатком пятого пальца и удлиненным третьим. Шейные позвонки ам-
фицельные, процельные и опистоцельные. Спинные опистоцельные. Суще-
ствуют грудные ребра. Напоминает по общему виду страуса; вероятно, вел
подобный ему образ жизни.
Ovirapror, —- беззубая форма, близкая к предыдущей, найдена в Мон-
голии (в мел}’). Предполагают, что он пожирал яйца Protoceratops,
близ которых найден его скелет.
По строению ног формы, обозначенные Маршем как Theropoda, стоят
гораздо ближе к птичьим, чем к звериным, как мы это видели; поэтому счи-
таю более правильным назвать их Pro-ornithopoda, в отличие от близких
к ним по ногам Ornithopoda J).
*) Марш установил название Theropoda по строению таза (лишенного задних
лобковых костей), сходного с звериным.
138
класс: пресмыкающиеся — reptilia
Особняком стоит Spinosaurus aegypticus Strom. — из нижнего мела
Египта (Gebel-el-Dist des Baharije Kessels), с высоко поднятыми спинными
отростками, достигавшими 1,80 м, указывающими на громадные размеры
животного. Шейные и спинные позвонки опистоцельные.
Подотряд 3
Praedentata Marsh (Qrnithischia Seeley)
Эта группа заключает в себе гораздо более разнообразные формы, чем
предыдущие. Все травоядные; они объединены двумя признаками: имеют в нижней
челюсти впереди вставленную добавочную кость, названную Маршем'— ргае-
dentale (предзубная); она отвечает верхней межчелюстной и не имеет зубов.
Вторым признаком являются особые выросты лобковых костей, направленные
Рис. 169. Hypsilopho don
F о х i Huxley. Передняя нога.
назад под седалищные, и носящие название post-
pubes — задних лобковых. Эти признаки отсут-
ствуют у предыдущих групп.
Жили представители этого подотряда» начи-
ная с юры, в мелу Европы и Сев. Америки. Prae-
dentata распадаются на резко разграниченные по
внешнему виду 3 семейства:
1. Ornithopodidae Marsh—птиценогие.
Большею частью это громадные животные, ходив-
шие на задних ногах, очень похожие на предыдущих
Pro-ornithopoda (Theropoda), являвшиеся
как бы параллельной ei( травоядной группой. Тело
у них голое. Corythosaurus, Camptosau-
rus, Iguanodon, Trachodon, Laosau-
rus и др.
2. Stegosauridae — ходившие на четы-
рех ногах, но передние укороченные. Они имели
различные костные придатки вдоль спины. По об-
щей форме тела они ближе всего к Sauropoda
Scelidosaurus, Stegosaurus и др.
3. Ceratopsidae — ходившие тоже на четырех ногах, но имевшие
рога и костный воротник на черепе. Eoceratops, Triceratops и др.
Сем. Ornithopodidae (Iguanodontidae). Hypsilophodon Foxi
Huxl. (рис. 169) — из вельда острова Уайта. Найдено несколько скелетов. Это
самый маленький и наименее специализировавшийся из Ornithopodidae.
Достигал 1 метра длины. Передние конечности короче задних, но все-таки
довольно длинные, имели 4 хорошо развитых пальца и остаток пятого. Ступня
значительно больше кисти, тоже с 4 хорошо развитыми пальцами, с преобла-
данием третьего и редуцированным пятым. Первый палец противопоставлялся. Все
пальцы имели когти. Интересно подчеркнуть существование зубов на межче-
люстной кости и склеротического кольца вокруг глаза: это, очевидно, при-
подкласс: diapsida
139
знаки, унаследованные от предков (Pseudosuchia, как предполагает
Абель для всех динозавров и птиц). Строение конечностей с противопоста-
вляющимся пальцем позволяет видеть в этой форме животных, лазавших по
деревьям, но перешедших к древесному образу жизни от наземного.
Род Iguanodon (рис. 170) известен из нижнего мела Англии в
многочисленных прекрасно сохранившихся ске-
долины Берниссара. Это громадные животные,
до верха
и в
незначительных остатках
летах из вельда Бельгии, из
достигавшие 18 м от хвоста
черепа; они передвигались на двух
задних трехпалых ногах, опираясь
иногда на хвост. Череп у них сра-
внительно небольшой, сжатый с бо-
ков. Большие носовые отверстия рас-
положены по сторонам
конца морды.
Рис. 170.А. Зубы. Iguanodon.
(Нижний мел.)
Рис. 170. Iguanodon Bernissar-
t е п s i s Bottt,
Глазницы небольшие; за ними громадные боковые височные дыры, ограничен-
ные сзади длинной квадратной костью, сочленяющейся с нижней челюстью.
Верхние дыры маленькие.
Верхняя и нижняя челюсти (кроме межчелюстной и предзубной костей)
имеют крупные листикообразные, покрытые эмалью зубы, расположенные на
внешней стороне в общем желобе (рис. 170, А). При стирании они заменяются
зубами следующего ряда.
Шейные позвонки не имеют ни отростков, ни ребер; они опистоцель-
ные. Спинные несут ребра и широкие спинные отростки. Хвост очень тяжело-
весный. Позвонки его с большими отростками. Длинная лопатка и небольшая
закругленная коракоидная кость прилегают к плечевой кости, сочленяющейся
с удлиненными костями предплечья. Грудная кость, из двух пластинок,
окостеневала. Четыре пальца передней конечности имеют но нескольку фаланг;
первый — в виде когтя. Таз имеет тонкие, удлиненные задне-лобковые кости,
идущие параллельно седалищным. Подвздошные очень удлинены (рис. 171).
Тазовые кости образуют впадину для головки бедра, на котором находится
3-й бугор (trochanter) для прикрепления мышц. Большая и малая берцовые кости
опираются на надпяточную и пяточную; последняя лежит над 4-м пальцем,
хорошо развитым, а надпяточная — над 3-м и 2-м; 5-го пальца нет совсем, от
1-го сохранился только небольшой остаток. Это строение ног имеет сходство
140
класс: пресмыкающиеся — reptilia
со строением птичьих ног и ног Pro-ornithopoda и указывает на боль-
шую редукцию их из пятипалых конечностей рептилий.
В 1917 г. найден скелет игуанодонта на о-ве Уайте, в вельде.
Интересное описание места нахождения Iguan.odon в Берниссарской
долине дает бельгийский ученый Дюпон1).
Вельдские отложения нижнего мела выполняют глубокую долину в 250 м,
на дне которой и жили игуанодонты. Отложения эти состоят из речного аллю-
вия вельдской эпохи. Долина была ограни-
чена стенами из каменноугольных известия- ‘'XSarlik'
Рис. 171. Тазовый поя'’ игуано-
донта.
Рис. 1Z2. Camptosaurus dispar
Marsh.
ков, внизу и на склонах которых находятся осыпи. Вельдские слои при-
крыты средне-меловыми — альбом.
В вельдскую эпоху в этой расщелине каменноугольных отложений про-
текала река, на берегу которой и жили игуанодонты, после смерти их ске-
леты затягивались илом, глиной. Эти слои с ископаемыми чередовались со
слоями без ископаемых. Все скелеты игуанодонтов, найденные здесь, принад-
лежат взрослым экземплярам.
Кроме этих животных здесь встречаются остатки крокодилов, черепах,
амфибий и рыб, а из растений — папоротников; но число этих форм незначи-
тельно сравнительно с игуанодонтами.
Большое число прекрасно собранных скелетов стоит в Брюссельском
музее. Некоторые хранятся в лежачем положении, в том виде, как они были
найдены. Кроме того найдены трехпалые следы — отпечатки ног игуано-
донтов, указывающие, что животное при хождении на хвост не опира-
лось, так как следов от него нет. На хвост они, вероятно, только опира-
лись, обгладывая листья. При ходьбе они балансировали. Длина отпечатка ноги
равняется 45 см.
*) Е. Dupont, Guide dans les collections, 1897.
подкласс: diapsida
141
Мы не знаем остатков игуанодонтов в Сев. Америке, но там хорошо
представлены другие формы этого семейства в юре и мелу, по которым
можно проследить изменение ног,
редукцию пальцев.
Camptosaurus Marsh—верх-
няя юра Сев. Америки (рис. 172)—по
форме скелета очень близок к игуа-
нодонту; череп отличается более пло-
ской формой и большими глазницами.
На передних конечностях пять паль-
Рис. 173. Psittacosaurus Osb.
цев; на задних — четыре, пятый неиз-
вестен, первый палец короткий. По-
следние фаланги удлинены. Незначительные остатки известны и в Европе.
Рис. 174. Trachodon amurensis Rfabinin.
Psittacosaurus Osb. — с более закругленной мордой, чем у игуано-
донта, и с загнутой вниз межчелюстной костью; найден в мелу Мош олии
142
класс: пресмыкающиеся — reptilia
третьей американской экспедицией полным скелетом. Он имел на всех ногах
по 4 пальца. Задние были значительно длиннее передних. Весь скелет построен
легко (рис. 173).
Laosaurus Marsh — юра Сев. Америки. Скелет менее тяжеловесный (мо-
жет быть—самка) с теми же признаками — отсутствие шейных ребер, большие
Рис. 175. Trachodon (мумия).
отростки на хвосте и т. д. Пять пальцев на руке хорошо развиты. На ногах три
удлиненных, тонких; 1-й утонен, 5-й редуцирован. Череп типа Camptosaurus.
Рис. 176. Saurolophus Osbor-
п i Brown. (Верхний мел.)
А — Prosaurolophus.
У Dryosaurus Marsh — юра Сев.
Америки, 1-й палец еще более редуцирован,
5-й исчез. Это тоже стройная форма.
Claosaurus Marsh — мел Сев. Аме-
рики— более тяжеловесная форма, подходя-
щая более к Iguanodon, но с плоским
черепом, имеет по той длинных тонких пальца
на передних конечностях, 1-й—укороченный.
На задних конечностях тоже по три пальца.
Надпяточная кость приросла к большой бер-
цовой. Задне-лобковая очень тонкая. Третий
trochanter развит хорошо. Лопатка изогну-
тая, с глубокой гленоидной впадиной. По не-
сколько расширенной передней части черепа
эта форма составляет как бы переход к
роду Trachodon mirabilis Leidy —
представляет большое отклонение в строе-
нии черепа, с расширенной передней ча-
стью в форме утиной головы. Мел Сев.
Америки.
Зубы, по форме близкие к зубам игуанодонта, расположены в общем желобе
в несколько чередующихся рядов, которые сменяются при стирании. Передние
конечности короткие, с 3 длинными пальцами, хорошо развитыми, и первым
сократившимся.
На ногах по три толстых пальца. Вся конечность тяжеловесная. На
концах пальцев копыта, а не когти.
подкласс: diapsida
143
Новая находка этой формы из верхнего мела Уайоминга — сохранившийся
скелет с покрывающей его кожей, в виде мумии — описаний Осборном
(рис. 175). Он указывает на присутствие в коже различной величины закругленных
пластинок, различно расположенных на теле. Под кожей хорошо сохранились
кости в их естественном соотношении одних к другим J).
Trachodon amurensis Riabinin был найден в Сибири, по р. Амуру,
в 60 км от г. Благовещенска, неполным
А. Н. Рябининым в Геологическом ко-
митете (рис. 174).
скелетом, восстановленным проф.
Рис. 177. Corythosaurus casuarius Brown. (Верхний мел.)
а — отпечаток, Ь — скелет, с — череп,
г
У Saurolophus Brown (рис. 176) часть носовых, предлобных и лоб-
ные кости вытянуты в кристу, торчащую над затылком.
Существует склеротическое кольцо, которого нет ни у одного из дино-
завров, кроме Hypsilophodon. Достигал 10 метров длины. Верхний мел
Сев. Америки.
Prosaurolophus Brown (рис. 176,Л) — криста менее вытянута. Верх-
ний мел Сев. Америки.
Corythosaurus casuarius Brown (рис. 177), недавно (191. г.) най-
денный и описанный Броуном, из мела Belly River Сев. Америки, дает совер-
шенно необычную форму черепа — с высоко поднятой, сжатой верхней частью,
напоминающей череп казуара, но сохранивший типичную для Trachodon
переднюю утинообразную часть морды. Найден с отпечатком всего животного.
Это перечисление различных близких между собою родов указывает на
большое разнообразие вторичных признаков, при строгом сохранении главных
признаков в семействе птиценогих (игуанодонтов).
Сем. Stegosauridae характеризуется присутствием больших костных
пластин и шипов, расположенных вдоль спины и хвоста. Формы юрские и
меловые Сев. Америки и Европы.
Скелет в общем сходен с Sauropoda; отличается небольшой относи-
тельно головой, присутствием шейных ребер, предзубной кости в нижней
*) Osborn, Memoirs Amer. Mus. Nat. History, june 1912.
144
класс: пресмыкающиеся — reptilia
челюсти и задне-лобковой в тазу; удлинением подвздошных. Сходными призна-
ками являются общая массивность скелета, сросшиеся сакральные позвонки и
пальцеходящие конечности. Позвонки амфицельные и процельные.
Scelidosaurus Ow. (рис. 178) — нижний лейас Англии, достигал 4,3 м
в длину. Позвонки амфицельные. Передние конечности имели четыре, задние —
Рис. 178. Scelidosaurus Ow.
три функционирующих паль-
ца. Очень крепкие ребра
доходили до крестца, к ко-
торому прилегали удлинен-
ные подвздошные кости. Зад-
не-лобковые массивные.
Пяточная и надпяточ-
ная кости неплотно приле-
гали к большой и малой
берцовым. Животное было
украшено на спине неболь-
шими, закругленными пла-
стинами, начиная от головы
до конца хвоста.
(рис. 179) — юра
стины на спине,
Уайоминга Сев. Америки,
до 12, расположенные в
Stegosaurus Marsh
имел большие треугольные пла-
один или два ряда. На конце
хвоста существовали четыре пары шипов,
развиты очень сильно. Череп по строению
но имеет предзубную кость. Многочислен-
ные зубы расположены в один ряд. Пе-
редние позвонки амфицельные, спинные —
платицельные.
Передние ноги значительно короче
задних. Лопатка с расширенным нижним
концом. Плечо, широкое вверху, сужи-
вается книзу. Локтевая высоко подни-
мается над лучевой.
Подвздошные кости, вытянутые по
Спинные отростки позвонков были
похож на череп Anchisaurus,
Рис. 179. Stegosaurus ungu-
latus Marsh.
позвоночнику, плотно прилегают к крестцу,
сросшемуся из четырех позвонков. Не вполне сходящиеся кости таза дают
углубление с небольшой дырой для головки бедра. Нижнее сочленение бедра
с большой берцовой—плоское. Пяточная и надпяточная кости подстилают
•большую и малую берцовые.
Palacanthus Hulke (рис. 180) — из вельда Англии — покрыт панцырем
из скульптированных кожных пластинок на крестцовой и задней части туловища.
Шея, спина и хвост покрыты шипами, как у Stegosaurus. Сюда же отно-
сится еще несколько других род ов, менее хорошо представленных ископаемыми
остатками.
подкласс: diapsida
145
Эти формы являются как бы измененными Ornithopodidae, опу-
стившимися на четыре ноги (передние короче задних). Они сохранили еще
задне-лобковые кости.
Сем. Ceratopsidae — отличаются резко от всех других динозавров своим
большим черепом с рогами и костным воротником, покрывающим затылок и
сросшиеся шейные позвонки. Форма воротника очень различна: он или сплош-
ной, или с более или ме-
нее большими дырами.
Рога бывают над глазами
и на носу. Обе пары ко-
нечностей почти одинако-
вой длины и имеют по пяти
пальцев на передних и по
три на задних, снабженных
копытообразными когтями.
Рис. 180. Palacanthus Hulke.
Крестец из сросшихся шести позвонков с плотно приросшим тазом — подвздош-
ными костями. Лобковые толстыезадне-лобковые в виде небольших отростков;
они почти неразвиты. Бедро значительно длиннее берцовых костей. Лопатка
талинная, сочленяется с костями коракоидной и плечевой,
которая короче ло-
Рис. 181 А. Зубы Tri-
ceratops.
Рис. 181. Triceratops flabellatus Marsh.
патки. Эти признаки установлены по скелету, описанному Маршем для рода
Triceratops из мела Сев. Америки, который и является типом (рис. 181).
Череп с очень маленькой мозговой полостью имеет спереди носовые от-
верстия. За носовыми костями лежат небольшие лобные; далее — залобные,
граничащие с теменными и с чешуйчатой (squamosum). Над глазницами
поднимаются большие рога, направленные вперед. На кончике носовых костей
маленький рог. У верхушки теменных находится пинеальное отверстие. Про-
должение теменных и чешуйчатые кости образуют широкий воротник с парой
височных небольших дыр. Нижняя челюсть короткая с предзубной костью —
клювообразной, как и межчелюстная. Край воротника имеет небольшие тре-
угольные придатки. Сзади воротник широкий, закругленный.
10 П а в л о в а М. В. Палеозоология. Ч. II.
146
класс: пресмыкающиеся —- reptilia
Верхние зубы представляют особенность: они имеют раздвоенный окрень,
что отличает их от зубов всех рептилий (рис. 181 А). Они развивались вер-
тикальными рядами, причем верхний стирающийся зуб заменялся новым.
Череп достигал 2 м длины.
S t er г h о 1 о ph u s — очень близкая к Triceratops форма, описана
Маршем по черепу. В последнее время найдено много новых форм в мелу
Сев. Америки, для которых основано более десяти новых родов, отличи-
тельные признаки которых
установлены по рогам, их положению и по
воротнику.
Monoclonius Brown (рис. 182) — имеет приподнятый вверх воротник,
с громадными дырами; он оторочен зубцами. Надглазные рога почти не раз-
Рис. 182. Monoclonius Brown.
виты; носовой очень большой, стоит
почти вертикально.
Anchiceratops Brown —
имеет очень тонкие, большие над-
глазные рога с значительно меньши-
ми дырами воротника, тоже припод-
нятого вверх.
Centrosaurus Lanibe — по-
чти с таким же воротником, но с
маленькими надглазными рогами и небольшим носовым, загнутым вперед.
Очень интересные находки сделаны американской экспедицией в Монго-
лии в 1923 г. Найдены более древние представители сем. Ceratopsidae-—
Protoceratops с менее развитыми воротником и рогами. Кроме того в тех
же меловых отложениях Монголии найдены яйца Protoceratops, и в не-
которых из них сохранились полуразвившиеся животные. Зоолог Эндрюс
(Andrews), участвовавший в экспедиции, устанавливает, на основании геологиче-
ских данных, картину жизни этих меловых динозавров, живших на берегу
озера и откладывавших яйца на песчаном берегу его, где из них развивались
детеныши под влиянием солнечной теплоты.
Недалеко от яиц были найдены скелет взрослого Protoceratops
(самки) и скелет Oviraptor, хищного динозавра, беззубого, вероятно, по-
жиравшего яйца (по мнению Эндрюса).
Л а м б е находит возможным установить три степени изменений этих форм:
1. От Eoceratops к Triceratops.
Надглазные рога увеличиваются, носовой остается маленьким; чешуйчатая
кость треугольная, широкая; теменное отверстие закрывается.
И. От Centrosaurus и Styracosaurus к Brachyceratops
(можно прибавить Monoclonius).
Надглазные рога остаются маленькими, носовой увеличивается; чешуйча-
тая кость остается небольшой; теменное отверстие уменьшается.
III. От Chasmosaurus к Torosaurus.
Надглазные рога увеличиваются, носовой уменьшается; чешуйчатая кость
удлиняется; теменное отверстие уменьшается.
подкласс: diapsida
147
Другие известные формы могут тоже войти в эти три группы. Во вторую
я включила Monoclonius. К сожалению, мы не имеем скелетов для этих
новых родов (кроме Monoclonius).
Остатки Ceratopsidae найдены в Сев. Америке с пресноводными
раковинами, с растениями, крокодилами, другими динозаврами и меловыми мле-
копитающими, описанными Маршем.
Распространение динозавров > к < . ;
К каким же общим выводам можем мы притти, познакомившись с глав-
ными представителями этого интереснейшего отряда рептилий — динозаврами?
Вся их жизнь протекла в мезозое, начиная с триаса, для очень немногих, и
заканчиваясь в мелу после богатого расцвета одних форм в юре, других
в мелу. Древнейшие из них являются, несомненно, отпрысками древнейших
Prosauria или крокодилов, но ближайших предков их мы еще не знаем.
Во всяком случае отщепление их произошло не позже конца пермского
периода; в триасе мы находим уже такие формы, как Anchisaurus, с ясно
выраженными признаками „двуногости“, которые имеются в юре у многочислен-
ных представителей травоядных и хищных, ходивших на двух ногах, и, с другой
стороны, имеем формы, утратившие эту особенность, но сохранившие еще
короткие передние ноги — Stegosauridae.
Более в стороне стоят меловые Ceratopsidae, ходившие на четырех
ногах, с остатком задне - лобковых костей и костным воротником черепа.
Абель хочет видеть в них возвращение двуногой формы к четвероногой
с остатком задне-лобковых костей. Очевидно, в конце мела условия жизни для
всей этой группы рептилий резко изменились, и она вымерла, несмотря на
разнообразное развитие и богатство форм.
Значение наземных позвоночных древнейших времен как пособия в истол-
ковании геологических факторов, и в особенности как проливающих свет на
разъединение и слияние континентов, оценивается все больше и больше.
Динозавры с их геологическим распределением почти через всю
мезозойную эру, с их почти мировым распространением, являются позвоноч-
ными вторичной эры, имеющими наибольшее значение. Это значение увеличи-
вается еще вследствие большого числа находок как их индивидуумов, так и
родов и удивительной способностью их приспособления к различным условиям
существования.
Динозавры вследствие большого географического распространения в про-
должение долгого существования являются, повидимому, очень гетерогенной
группой.
Недавние находки Protoceratops и других рептилий в мелу Монголии
указывают на их переселение отсюда в Сев. Америку, с одной стороны, и в за-
падную Азию — с другой, что дало там развитие близких к ним форм, с более
усилившимися признаками (воротник и рога у Ceratopsidae) и вызвало
большое разнообразие потомков. Причина полного исчезновения их в верхнем
мелу является и до сих пор загадкой.
10*
148
класс: пресмыкающиеся reptilia
Отряд 6
Крылатые ящерицы — Pterosauria
Совершенно особый отряд рептилий представляют Pterosauria —
крылатые или, правильнее, летающие рептилии, так как у них нет настоящих
крыльев — птичьих, а есть летательная перепонка, почему они больше прибли-
жаются по общему виду к летучим мышам, чем к птицам. Вся жизнь Ptero-
sauria протекла в юре; в мелу известны только всего две формы из Сев. Аме-
рики. Прямой связи с каким-нибудь другим отрядом рептилий для них мы еще
не имеем. Отдельные части скелета сближают их с Rhynchocephalia,
с Dinosauria и Crocodilia. Но так как в нижнем лейасе они являются
уже со вполне обособленными признаками летающих животных, то, следова-
тельно, отщепление их произошло не позже триаса от
одной из древних ветвей живших на деревьях яще-
рицеподобных форм. Различные формы их дают нам
указание на два направления в их развитии. У одних
преобладал длинный хвост, у других он укорочен.
Позвонки дохвостовые—процельные, хвостовые—ам-
фицельные.
Подотряд 1
Rhatnphorhynchoidea
Длиннохвостые формы, с верхнего лейаса
до верхней юры Баварии. По указанию Абеля,
хвост играл у них большую роль при летании, как
руль, также и не складывавшиеся крылья, тонкие и
Рис. 183.Dimorphodon Ow.
Рис. 184. Dorygnathus banthensis Theoa.
острые. Первый палец атрофирован, 2-й, 3-й, 4-й очень укорочены, 5-й очень
удлинен на передних конечностях. На задних все короткие.
Dimorphodon macronyx Buckl. (рис. 183) — из триаса и нижнего
лейаса Англии, является древнейшим представителем этого подотряда. По скелету
и черепу он ближе всего к ящерицам; но голова сравнительно велика, а шея и
туловище укорочены. Череп имеет носовые отверстия, глазницы и две пары
височных дуг. Челюсти несут впереди большие, сзади мелкие зубы в альвео-
лах. Существует склеротическое кольцо. Конечности тонкие; на задних 5-й па-
лец отставлен в сторону; на передних он очень удлинен, и к нему прикрепля-
подкласс: diapsida
149
лась, очевидно, перепонка одним краем, а другим — к спине и хвосту. Кости
скелета пневматичные, как у птицы, и таз прирос к крестцу.
Dorygnathus banthensis Theod. — верхний лейас сев. Германии,
Банц. Череп сравнительно большой, хвост и летательный палец короткие.
Очень большие предглазничные дыры (рис. 184).
Зубы острые, конические на praemaxillare и maxillare, уменьшающиеся
кзади, как у Rhamphorhynchus.
Рис. 185. Rhamphorhynchus.
Rhamphorhynchus Muenster! Н. v. М. (рис. 185) — из верхней
юры Баварии — такой же длиннохвостый тип. Череп тоже ящеричного типа,
с зубами в верхней и нижней челюстях, со склеротическим кольцом. Существуют
на черепе четыре пары дыр. У этой формы, найденной в соленгофенских сланцах
Баварии, сохранилась часть летательной перепонки. Она заканчивалась на пер-
вой половине хвоста. На конце его была еще небольшая добавочная лопасть.
На перепонке видны тонкие продольные струйки. Грудной пояс этих форм
состоял из грудной кости посредине и удлиненных лопаток и коракоидных;
ключица отсутствует.
Scaphognathus crassirostris Gold. — верхний лейас, верхняя юра
соленгофенских сланцев Баварии. Череп более высокий, несколько удлинен-
ный, не ящеричного типа, с четырьмя парами дыр. Зубы значительно более
редкие, чем у Rhamphorhynchus и Dimorphodon. Есть еще несколько
родов, менее хорошо представленных: Campylognathus и др.
Подотряд 2
Pterodactyloidea
Короткий хвост, крылья более широкие, причем 5-й удлиненный палец,
поддерживающий летательную перепонку, мог сгибаться, в противоположность
Rhamphorhynchus. Глазницы не вполне отделены от носовых отвер-
150
класс: пресмыкающиеся — reptilia
У Pterodactylus spectabilis
Рис. 186. Pterodactylus spectabilis.
цо. Кондилюс очень маленький, совершенно
стий. Мозговая коробка очень маленькая. Верхняя юра Баварии, средний
мел Сев. Америки.
Pterodactyl и s представлен несколькими видами в верхней юре Бава-
рии (соленгофенские сланцы).
У некоторых видов передняя часть черепа удлинена, и дыры слились
с каждой стороны. Особенно сильно она удлинена у Pterodactyl us
а п t i q u u s Somtn. и имеет вид длинного птичьего клюва, усаженного мелкими
зубами.
череп короче и морда закруглена
(рис. 186). Все эти формы были
небольшими животными, от 18 см
до 45 см.
Pteranodon — из среднего
мела Сев. Америки — является
самой оригинальной формой из
птеродактилей, дошедшей в своей
специализации развития черепа до
крайних пределов. Это очень круп-
ная форма, у которой череп до-
стигал 1 м в длину и равнялся
длине туловища с вытянутыми на-
зад задними ногами. Передняя
часть черепа вытянутая, без зубов,
с ноздрями, расположенными пе-
ред глазницами; задняя—в виде
большого надзатылочного гребня.
Глаза имели склеротическое коль-
круглый. Обе половинки нижней
челюсти впереди плотно срастались. Они прикреплялись к черепу винтообраз-
ным сочленением, сходным с сочленением у пеликана, у которого здесь при-
крепляются особые мускулы, позволяющие широко раздвигать задние концы
челюстей и раскрывать мешок, находящийся под челюстями. Подобное со-
членение позволяет предположить существование мешка и у Pteranodon
(рис. 187), схватывавшего, без сомнения, тоже рыбу.
Шейные позвонки сравнительно большие, процельные — 9; два первые
срослись. Восемь спинных позвонков плотно срослись между собою и при-
крыты еще верхней пластинкой, приросшей к верхним дугам. Это так назы-
ваемый п о t а г i и гл. Под ним концы ребер срастаются с грудной костью.
Лопатка сочленяется с notarium. Все вместе образует почти неподвижную
грудную клетку. Так же прочно соединен и задний пояс, в котором тазовые
кости срослись между собою и с 10 крестцовыми позвонками. Это типичный
таз птиц. Хвост очень короткий. Передние конечности с сократившимся
первым пальцем. Три следующие маленькие, но хорошо развитые, с когтями.
Пятый сильно удлинен; он длиннее, чем череп. На задних конечностях развиты
подкласс: diapsida
151
четыре пальца. Верхний ряд запястных костей слит с большой берцовой.
Расстояние между концами крыльев достигало 7 м.
Nyctosaurus gracilis Marsh —• с таким же удлиненным клювом, но
без заднего костного капюшона. Лопатка широкая, не сочленяется с notarium.
Хвост длинный, имел до 14 позвонков. Это менее специализировавшаяся форма
из верхнего мела Канзаса.
Хотя мы не в состоянии указать общего предка для рептилий и видели
между амфибиями близкие формы только для отдельных их отрядов, мы
можем, однако, набро-
сать, на основании гео-
логических и палеон-
тологических данных,
их развитие. Эти вы-
мершие животные в
большинстве случаев
были больших раз-
меров, но с малым
мозгом и холодной
кровью; они представ-
ляли большую плас-
тичность и разнооб-
разие в своем разви-
Рис. 187. Pteranodon Marsh.
тии и в механическом усовершенствовании, как указывает Осборн. Ка-
ждая из геологических эпох, в которой они жили, характеризуется опре-
деленными типами.
В перми Сев. Америки, Европы и Африки были исключительно назем-
ные и наземно-водные представители (Cotylosauria, Placodontia;
Pelycosauria, Pythonomorpha), исключительно специализированные.
Она совпадала с эпохой высыхания озер и лагун безводной Сев. Америки и с
отложением слоев песчаников дельтового и речного происхождения, в 2,7 км
мощностью, в южной Африке.
В триасе мы встречаем последовательно рептилий — представителей
наземной фауны — в Сев. Америке, Европе и Африке и первые указания на
существование морских форм в Сев. Америке.
В юре отложения больших внутренних континентальных морей дают
измененную рептильную морскую фауну (Plesiosauri a, Ichthyosauria)
в Европе и Сев. Америке.
Эволюция морских, пелагических рептилий продолжается с некоторым
перерывом в нижнем мелу и доходит до настоящей морской рептильной
фауны верхнего мела, где она и вымирает.
Между тем в триасовую эпоху в континентальных и речных отложениях
начинается эволюция как наземных динозавров, так и речных крокодилов,
л завершается эволюция наземных амфибий и прорептилий. Большой перерыв
•152 класс: пресмыкающиеся — reptilia
существует в истории наземной жизни до конца юрской эпохи и начала
меловой, когда начинается вторая стадия эволюции динозавров, открытая
в лагунных отложениях Северной и Южной Америки, давшая гигантских
Sauropoda.
Наконец, в верхнем мелу мы открываем обширные прибрежные отложения
сырых равнин, которые указывают на длительность существования наземных
рептилий в Сев. Америке, и Европе.
Изучая формы различных периодов, можно встретить вместе представите-
лей различного происхождения, но приспособившихся к жизни в одинаковых
условиях и приобревших сходные признаки 1).
<) Много интересных примеров указывает Осборн в ,The origin and evolution
oi life", London, 1918.
КЛАСС ЧЕТВЕРТЫЙ
ПТИЦЫ —AVES
Ныне живущие птицы, несмотря на большое разнообразие свое, объеди-
нены единым признаком, который всегда позволяет отличить их, а именно —
внешним покровом из перьев. В ископаемом состоянии перья сохраняются
только при особенно благоприятных условиях; следовательно, нужно знать
признаки скелета птиц, чтобы уметь определить принадлежащие птицам части
его, тем более, что некоторые из признаков скелета птиц встречались нам
уже у пресмыкающихся.
Первым важнейшим признаком является редукция передних конечностей,
доходящая иногда почти до полного их исчезновения. Срастание трех костей
плюсны; прирастание пяточной и надпяточной к большой и малой берцовым;
присутствие грудной кости и большее или меньшее развитие на ней грудного
гребня — киля; закрытый череп — без височных дыр, с большими орбитами
и одним сочленовным кондилюсом; неприросшая квадратная кость; отсутствие
зубов; таз, сросшийся с позвоночником; наконец, седлообразные сочленения
позвонков — дополняют признаки птиц. Многие из этих признаков варьируют
в зависимости от образа жизни птиц — более наземного или воздушного; но
в основе они остаются все теми же птичьими признаками. Желая отыскать
генетическую связь их с другими классами животных, приходится обратиться
к ближайшему классу — рептилиям и сравнить древнейшие ископаемые остатки
тех и других.
Мы видели уже, что маленький Compsognathus, между хищными
динозаврами, имел череп и конечности (задние) с признаком птичьих, хотя
кости плюсны здесь не вполне срослись.
У Ceratosaurus, напротив, срастание этих костей такое же, как у
страуса, но строение черепа резко отлично.
Pterodactylus имел большое сходство в строении таза с птичьим
(срастание его с крестцом), но его летательные органы являлись уже слишком
специализированным признаком, чтобы можно было искать в них предков
птиц. Все эти данные позволяют думать, что птицы произошли от одного из
наземных, лазающих по деревьям пресмыкающихся, вернее всего от неболь-
ших динозавров, изменивших задние конечности и сокративших передние
в попытке стать двуногими ]).
1) Н. Osborn считает, что птицы и динозавры произошли от общего предка
(в перми).
i
154
класс: птицы — aves
Долгое время ископаемые остатки птиц были известны только начиная
с мела. Но второй половине прошлого столетия были найдены два экземпляра
в верхних юрских слоях, настолько отличающихся от ранее известных бегающих
(R a t i t а е) и летающих (С а г i n a t а е), что для них установлен новый подкласс—
Подкласс I
ЯЩЕРЕЦОХВОСТЫЕ — SAURURAE
Род Archaeopteryx.—Найдены два экземпляра птицы, величиной с
грача, с отпечатками перьев, в верхне-юрских слоях Баварии. Один из них очень
хорошей сохранности; на нем отчетливо видна голова, позвоночник, конеч-
ности и хвост. Оуэн первый дал описание этой формы. Что было осо-
бенно поразительно в ней, это — непневматичные кости при существовании
перьев, хорошо развитые передние конечности и длинный хвост. Эти признаки
настолько не птичьи, что, будь эта форма найдена без перьев, ее, не колеб-
лясь, отнесли бы к рептилиям. Между тем благодаря оперению ее признали за
предка птиц Дамес (Dames) и др. ' t'йга
Дальнейшее изучение определенно указывало, что Archaeopteryx не
мог быть этим предком (профессор А. П. Павлов). Голова его по форме дей-
ствительно похожа на птичью; в ней отсутствуют височные дыры, существуют
предглазничные и удлиненные носовые отверстия; швы незаметны. Но суще-
ствуют склеротические кольца; челюсти обильно усажены зубами, сидящими
в альвеолах межчелюстных и челюстных костей. Позвонки амфицельные с сла-
быми отростками. Ребра маленькие на шейных и более крупные, одноголовча-
тые, на грудных позвонках. Передний пояс состоит из удлиненных частей:
лопатки, коракоидной кости и ключицы. Грудной кости нет. Передние конеч-
ности ящеричного типа, с удлиненным плечом, предплечьем и кистями, имею-
щими по три хорошо развитых пальца с когтями.
Задний пояс неполный; отчетливо видны только подвздошные кости. Задние
конечности длинные и тонкие, трехпалые, четвертый палец маленький. Пястные
кости неплотно сросшиеся между собою. Конечности эти, по указанию
Марша, вполне соответствуют конечностям Compsognathus.
Кроме всего указанного существуют перья, обильно одевающие хвост и
крылья и дающие основание отнести Archaeopteryx к птицам. Но этот
признак, при других вышеназванных, не достаточен для того, чтобы считать
археоптерикса предком нынешних птиц. В нем мы видим комбинацию при-
знаков древних рептилий — амфицельные позвонки, склеротическое кольцо, зубы
и пояса — с птичьими признаками: редукция пальцев до трех и перья, которые
не имели опоры для летания, так как пальцы передних конечностей еще на-
ходились вне крыла, служа органом хватания, чего нет у настоящих птиц.
Длинный хвост с перьями по обеим сторонам тоже мог только затруднять по-
лет, особенно при непневматичности костей, и едва ли мог быть при этих
условиях (перья) предшественником короткого птичьего хвоста. Вероятнее, что
укорачивание и сокращение хвостовых позвонков у предков птиц произошло
раньше развития перьев, так же как и дальнейшая редукция передних пальцев.
подкласс: птицехвостые — ornithurae
155
Поэтому правильнее будет смотреть на Archaeopteryx как на форму, ко-
торая слишком рано приобрела птичьи признаки — перья, не приспособив
к ним, однако, скелета, не выработав пневматичности в костях и не подготовив
конечностей для укрепления перьев при образовании настоящего крыла, даю-
щего возможность летать, а не перелетать; это еще затруднялось удлинен-
ным, тяжелым хвостом *). Это такая же боковая форма между птицами,
Рис. 188. Archaeopteryx Siemens! Dames. (Берлинский экземпляр.) (J/4)
какие мы будем встречать между млекопитающими. До сих пор это един-
ственный представитель Saururae 2).
Подкласс II
ПТИЦЕХВОСТЫЕ — ORNITHURAE
Сюда относятся все остальные птицы как ископаемые, так и нынешние.
Ископаемые представители известны, начиная с мела Сев. Америки,
на восточном склоне Скалистых гор. Они найдены вместе с мозазаврами,
плезиозаврами и другими рептилиями. Марш подразделил их на Odontol-
сае — с зубами в общем желобе, Odontormae — с зубами в отдельных
ячейках. 3-й отряд Anodontorni — беззубые.
По строению скелета и образу жизни птицехвостые подразделяются на
Ratif л е и Carinatae (бегающих и летающих).
*) Между ныне живущими птицами в Бразилии существует О р i s t h о с о m u s>
у птенцов которого во время долгого послеэмбрионального периода существуют неко-
торые признаки Archaeopteryx: передние конечности имеют когти. Хвост остается
более удлиненным, чем у других птиц, ио это указывает не на происхождение птиц
от Archaeopteryx, а на то, что и он и нынешние птицы произошли от форм, близ-
ких к динозаврам, или от общего с ними предка.
'-) Теперь некоторые авторы относят два экземпляра Archaeopteryx к двум
различным родам.
156
класс: птицы — aves
Отряд 1
Ratitae. Odontolcae. Marsh J).
Характеризуются сильной редукцией передних конечностей, отсутствием
киля на грудной кости. Зубы расположены в общем желобе.
Hesperornis (рис. 189) — мел Сев. Америки — крупная птица, имею-
щая высоту 1,05 м, обладала уже в своем скелете всеми признаками, типич-
ными для водяных, ныряю-
щих птиц. Череп напоминает
некоторых Pterosauria;
мозг — рептильного типа,
почти гладкий. Отличитель-
ными признаками являются
зубы, хорошо развитые в
обеих челюстях (кроме меж-
челюстной кости). Они рас-
положены в общем жело-
бе — 14 вверху, 33 вниз).
Между ними намечаются сла-
бые перегородки в виде
входящих выступов челюсти.
Каждый зуб состоит из ко-
ронки, загнутого конуса, по-
крытого эмалью, и более ши-
рокого корня, внутри кото-
рого развивается новый зуб.
Рис. 189. Hesperornis regalis Marsh. (*/6)- Укреплялись зубы в желобе
хрящом. Шея удлинена, со-
стоит из седлообразно сочленяющихся позвонков с верхними отростками, что дава-
ло шее прочность и гибкость. На нижней стороне позвонков тоже были отростки,
Ребра прикреплялись к позвонкам двойными головками, а спереди подходили
к широкой грудной кости, гладкой спереди. К ней прикреплялись: тонкая клю-
чица, более широкая коракоидная кость и длинная лопатка. От передней конеч-
ности осталась только тонкая плечевая кость, вставлявшаяся между лопаткой и
коракоидной и, очевидно, представлявшая весь остаток передней конечности.
В заднем поясе имеем удлиненные подвздошные кости, прикрывающие кре-
стец и большую часть хвостовых позвонков, приросших к ним. Седалищные и
лобковые, тонкие, длинные, направлены назад параллельно. Все эти кости обра-
зуют углубление для головки бедра, очень короткого. Голень удлинена; боль-
шая берцовая срослась с надпяточной — астрагалом. Три кости плюсны тоже
срослись между собою; наиболее длинный внешний палец, наиболее короткий —
*) Классификация птиц у различных авторов очень различная. Принимаю наименее
сложную.
подкласс : птицехвостые — ornithurae
157
второй. Таким образом тяжесть тела падала на внешний край конечностей, что
являлось полезным при плавательных движениях этих конечностей. Это описа-
н'ие указывает на полное приспособление этой настоящей меловой птицы
к плаванию, нырянию или хождению по берегу, одинаково с нынешними пти-
цами, ведущими такой же образ жизни. Эта форма очень далеко ушла от
рептильного типа, и только в виде воспоминания остались зубы и некоторые
признаки в черепе. Насколько далек от нее Archaeopteryx, насколько
нерационально его развит не сравнительно с настоящей, хотя и зубатой птицей
Hesperornis, видно из описания этой и следующей меловых зубатых
птиц.
Отряд 2
Garinatae. Odontormae. Marsh.
менее редуцированы. На грудной кости раз-
грудных мускулов. Тазовые кости
Передние конечности
вит киль — гребень для прикрепления
слабо развиты. Зубы в отдельных
ячейках.
Ichthyornis victor Marsh
(рис. 190) — типичная летающая птица
из верхнего мела Сев. Америки. Разме-
рами с голубя. В сильно вытянутом клюве
существуют зубы — в верхней и нижней
челюстях, кроме межчелюстной кости. Но
здесь они сидят в отдельных ячей-
ках. Передняя часть нижней челюсти,
заключающая зубы, очень похожа на
челюсть маленького мозазавра. Спереди
нижняя челюсть скреплена хрящом, как
и у Hesperornis. Позвонки амфи-
цельные. Передний пояс состоит из груд-
ной кости с килем. Ключица, коракоид-
ная кость и лопатка развиты почти равно-
мерно. Две последние служат для сочле-
нения с плечом, которое здесь хорошо
развито, так же как и предплечье. К ним
прикреплялись перья крыла; кисть редуци-
рована: она состоит из большого второго
пальца и меньших первого и третьего.
Тазовой пояс совершенно другого характера, чем у Hesperornis: дли-
на всех костей незначительна, подвздошные срастаются с 10 позвонками —
крестцовыми и хвостовыми. Тонкие ноги—с тремя хорошо развитыми и одним
маленьким пальцем. Эта птица определенно указывает, что в мелу тип летаю-
щих птиц был вполне хорошо выражен. Зубы являются единственным важным
отличительным признаком ее от позднейших птиц.
Рис. 190. Ichthyornis victor Marsh.
158
класс: птицы
aves
Gastornis— из нижних третичных слоев Англии и Франции, является
крупной бегающей птицей, она значительно крупнее Hesperornis. Боль-
шая берцовая кость имела 50 см длины. Череп — 36 см\ он ближе к черепу на-
стоящих птиц, но все-таки сохранил еще по одному большому зубу в челюсти
и состоит из несросшихся костей.
Отряд 3
Беззубые. — Anodontornae.
Сюда относятся все остальные птицы. В третичных слоях Англии и
Франции известны птицы, у которых зубов нет, но нижняя и верхняя челю-
Рис. 191. Odontopteryx toliapi-
с a s Ow.
сти имеют костные отростки, симу-
лирующие зубы (Odontopteryx
tо 1 i арiсus Ow.—рис. 191). Подоб-
ные отростки на челюстях встреча-
ются у наших гусей.
В миоцене Франции (St. Gerant
le Puy) встречаются птицы с настоя-
щей птичьей головой, без зубов, с
закрытыми швами и челюстями утки (Anas). Если подставим ее голову к ту-
ловищу Ichthyornis, то последний может быть принят за Anas. Оче-
Рис. 192. Prororhachus inflat us г. Amergh. (Патагония.)
видно, эти формы тесно связаны генетически, и Anas, лишившаяся зубов,
является как бы потомком этого мелового предка.
В третичных слоях (в миоцене) Патагонии найдены остатки колоссальной
болотной птицы из сем. пингвинов —
подкласс: птицехвостые — ornithurae
159
Prororhachus inflatus Amegh. (рис. 192) — череп величиною с ло-
шадиную голову, а вся высота птицы — 5 м. Позвонки были полые; кости
заключали в себе костный мозг. Череп очень сдавлен с боков, с очень длин-
ной межчелюстной костью. Носовые отверстия отодвинуты на средину черепа —
как у Belodon, они расположены по бокам. Сзади череп плоский. Суще-
ствуют небольшие височные дыры, сливающиеся с глазницами, а эти последние
с предглазничными. Квадратно-скуловая кость сочленяется с квадратной и отро-
стком верхне-затылочной. Это признаки
рептильного типа. Клюв подобен клюву
попугая. Нижняя челюсть имеет посре-
дине сквозную дыру. Сзади она сочле-
няется с квадратной, тоже рептильного
типа. Ноги трехпалые, с маленьким
4-м пальцем. Крылья короткие. Хвосто-
вые позвонки не срослись. Грудина не-
известна. Сильно отклонившийся тип
от ствола птиц. Штейнман (Stein-
mann) сближает его с крокодилом
(Belodon).
У Pelecyornis, близкой к пре-
дыдущей форме, существует небольшая
грудная кость. Рис. 193. dj at гут а.
Diatryma (рис. 193) — эоцен
Мексики и Gius — эоцен и миоцен Европы, относятся к этому же типу,
и указывают, что Prororhachus Патагонии имел близкие формы и в Ев-
ропе и в Сев. Америке
Из плейстоценовых слоев Мадагаскара известны остатки птиц сем. S t г и-
thionidae — страусовых.
Aepyornis— птица громадных размеров, со сравнительно маленькой
головой, с короткой, широкой грудной костью, с трехпалыми ногами. Большая
берцовая кость достигала 80 см длины. Целые скелеты, отдельные кости и яйца
найдены в болотистых отложениях северного Мадагаскара вместе с остатками
гиппопотамов, крупных лемуров (Megaladapis, Palaeopropithecus),
крупных черепах и крокодилов.
В юго-восточной области и в центре Мадагаскара найдены в ппибрежных
отложениях остатки яиц Aepyornis, образующих целые скопления. Яйца
эти в три раза больше яиц страуса.
Большое количество этих остатков в данных местах объясняется тем, что
птицы попадали сюда в сырое время года, так как только в сырых, болоти-
стых местах сохранялись вода и растительность, необходимые им для питания.
На многих костях птиц встречаются следы зубов крокодилов, увлекав-
ших их в воду ’).
*) L. М о n n i е г, Les Aepyornis. Annales de Paleontologie, Paris, 1913, t. VIII, f. 3—4.
160
класс: птицы—aves
напоминает таз динозавров, так же как
Рис. 194. Dinornis elephanto-
р u s Cw.
В послетретичных слоях Новой Зеландии найдены остатки гигантских бе-
гающих птиц из сем. Dinornithidae.
Dinornis elephantopus Ow. (рис. 194) до 3,6 м высоты, на очень тя-
желых ногах. Передние конечности совершенно атрофированы. Таз тяжеловесный,
и задние конечности. Бедро сравнитель-
но короткое. Четвертый палец исчез со-
вершенно. Метатарсы плотно срослись.
Плейстоцен Новой Зеландии.
Palapteryx имел четыре паль-
ца на задних ногах. Весьма вероятно,
что исчезновению Dinornis и близ-
кого к нему Palapteryx способство-
вал человек.
Эти чудовища стоят близко по строе-
нию скелета к ныне живущим в Новой Зе-
ландии маленьким киви (Apteryx), тоже
лишенным крыльев, но снабженным длин-
ным клювом, который помогает им оты-
скивать в сыром грунте червей. У D i-
п о г n i s клюв был острый, но короткий.
Ведут ли свое начало все ныне жи-
вущие бегающие птицы, очень разнооб-
разные, от Hesperornis — не уста-
новлено. При изучении ископаемых остат-
ков птиц, начиная с юрских —наиболее
древних из известных нам, мы видим, что
они имели сходство в различных ча-
стях скелета с различными пресмыкаю-
щимися, более всего с динозаврами. Но
все-таки это сходство не позволяет нам
указать их прямого предка между последними.
Можно думать, что он найдется в группе триасовых маленьких динозав-
ров, давших одновременно две ветви, ведущие к птицам:
а) летающим, хорошо развившим крылья, сохраняя нужные для прикреп-
ления их кости конечностей (передних), и
б) бегающим, с значительно более редуцированными передними конеч-
ностями. Новейшие исследования указывают, что различные представители
второй группы шли различными путями к этой редукции.
Интересно сравнить таз птиц с тазом Iguanodon и других динозав-
ров, имевших хорошо развитые задне-лобковые кости. Но начнем с форм, не
имевших этих костей. К таким принадлежат из динозавров Sauropoda, хо-
дившие на четырех ногах, и Pro-ornithopoda (Theropoda), ходившие
на двух. У тех и других лобковые удлиненные, направлены вниз и вперед,
а седалищные, такие же удлиненные, — вниз и назад (рис. 195).
подкласс: птицехвостые — ornithurae
161
вперед, называются предлооко-
той же части таза у различных жи-
на развитие задне-лобковых костей
Рис. 195.
У игуанодонта лобковые направлены горизонтально вперед, а параллельно
седалищным развились отростки лобковых — задне-лобковые кости, направлен-
ные вниз. У птиц (Apteryx) существуют такие же отростки, параллельные
седалищным костям, которые носят название лобковых костей. А маленькие
незначительные отростки, направленные
выми. Это различное название одной и
вотных крайне неудобно. Если смотреть
у игуанодонта как на результат его
„двуногости”, как это делает Абель
(„Palaobiologie”, рис. 208), то ста-
новится понятным дальнейшее измене-
ние этой кости у птиц и атрофия
лобковых, от которых у них остается
только маленький отросток. Но тогда
за ним нужно удержать название „лоб-
кового”, а за задним отростком „задне-
лобкового”, как у игуанодонта и дру-
гих динозавров, снабженных этим отро-
стком *), так как они развились у этих
форм раньше, чем у птиц. В подтверж-
дение своей мысли Абель указывает
на присутствие задне-лобковых у S с е-
lidosaurus, который ходил раньше
на двух ногах. У Stegosaurus, кот<
спинные украшения, они тоже существуют. Но короткие передние ноги указы-
вают на его ходивших на двух ногах предков (S с е 1 i d о s а и г и s).
Абель считает атрофию задне-лобковых костей у Triceratops,
следствием сильного отяжеления черепа задним воротником, при котором жи-
вотное не могло ходить на двух ногах.
Но то, что мы знаем о конечностях Ceratopsidae, не дает нам по-
вода считать их потомками двуногих. Отсутствие задне-лобковых костей у Р г о-
ornithopoda, живших одновременно с игуанодонтами в юре и в мелу, является
тоже возражением против мысли Абеля, так как у некоторых из них, напри-
мер у Ceratosaurus, хвост был тоже очень тяжеловесный,и животное ходило
на двух ногах. Во всяком случае птицы могли произойти от тех
форм, которые развили задне-лобковые кости и у которых
должны были начать атрофироваться лобковые, которые выражены у
ныне живущих птиц в виде слабых отростков, непоавильно называемых
предлобковыми.
пригнули к земле его тяжелые
<) Название „лобковых костей* (задних отростков) у птиц дано было раньше, чем
были найдены игуанодонты, имевшие их.
11 ПавловаМ. В. Палеозоология. Ч. II.
КЛАСС ПЯТЫЙ
МЛЕКОПИТАЮЩИЕ—MAMMALIA
Громадный класс млекопитающих животных, венчающих собою все пре-
дыдущие классы, может быть сразу охарактеризован рождением живых де-
тенышей и питанием их молоком матери. Следующими признаками являются
формы скелета: в черепе многие главные кости слиты между собою — лобная
состоит из предлобной, заднелобной и лобной, слившихся вместе; височные
дуги исчезают; отчасти они сливаются со скуловой костью, отчасти с височной
костью. Квадратная кость, характерная для низших позвоночных, включается в
сочленовный конец височной кости. Нёбо закрытое. Зубы дифференцированы,
и коренные имеют по два или по три корня. Существует смена зубов — молоч-
ные заменяются настоящими (резцы, клыки и четыре предкоренных у боль-
шинства). Два затылочных мыщелка развиваются на боковых затылочных ко-
стях, как у амфибий. Кости нижней челюсти срослись в одну с каждой стороны,
и обе половины срастаются спереди. Челюсти сочленяются с черепом посред-
ством венечного отростка. Тело у большей части животных покрыто шерстью.
Туловищные позвонки соединяются посредством межпозвоночных хрящей, не
существующих ни у птиц, ни у рептилий, шейные — посредством вогнутых и
выпуклых поверхностей. В грудном поясе коракоидная кость срослась с лопат-
кой в одну кость. Тазовые образуют цельный таз. Некоторые из этих изме-
нений встречались уже у рептилий — Theromorpha.
Этот класс представляет такое же, если не большее, разнообразие, как и
вымершие рептилии. Он так же преобладает в кенозое, как рептилии преобла-
дали в мезозое. Представители его многочисленны на суше, но существуют
живущие в воде и немногие, поднимающиеся в воздух.
Но ныне живущие рептилии так резко отличаются от млекопитающих,
что трудно даже представить себе генетическую связь между этими двумя
классами, если не быть подготовленным к этому изучением вымерших
форм.
Древнейшие ископаемые остатки млекопитающих известны из верхнего
триаса Швабии, Англии и Сев. Америки в виде отдельных зубов с двумя
корнями и немногих нижних челюстей. В юре их больше; по нижним челюстям
установлено 24 вида. В Сев. Америке эти остатки встречаются вместе с Atlan-
tosaurus. Все это очень маленькие формы, которые уничтожались хищными
рептилиями и может быть уничтожали их яйца. В мелу Сев. Америки остатки
млекопитающих также встречаются, но они незначительны. С началом тре-
подкласс: однопроходные — monotremata
163
т и ч н о й эры начинается их расцвет. При этом одни формы, развиваясь посте-
пенно, доходят до нашего времени, другие вымирают в разное время, не остав-
ляя потомков.
Классификация млекопитающих, данная разными учеными, основывается
на различных признаках их.
Мы примем подразделение их на три подкласса:
1. Однопроходных — Monotremata.
2. Бесплацентных — Marsupial!а.
3. Плацентных — Placentalia.
Сгруппируем их по форме конечностей так: коготные, копытные
и ноготные.
Эта группировка основана на форме конечностей, которые играют, можно
сказать, после мозга первостепенную роль в жизни животного и являются
в связи с зубной системой выразителями образа жизни.
Первая группа коготные — характеризуется конечностями, снабженными
когтями. Число пальцев большею частью пять; у некоторых они редуцируются,
но никогда не срастаются между собою. Хвост длинный. Зубы или только
конические, или конические и бугорчатые. Они включают:
а) однопроходных — Monotremata — несущих яйца: утконос
и ехидна, живущие в Австралии и на Вандименовой Земле,
б) бесплацентных — Marsupialia (сумчатых), тоже с очень
ограниченным географическим распространением:
Австралия, Южная Америка (Бразилия, Мексика).
Между плацентными мы встретим пред-
ставителей всех трех групп. К коготным отно-
сятся: хищные, грызуны, насекомоядные, неполно-
зубые, китообразные, с широким географическим
распространением. Остальные плацентные обни-
мают копытных и ноготных. Признак плацен-
та— общий для всех млекопитающих, кроме Monotremata и Marsu-
pialia.
Хотя плацента является признаком чисто зоологическим, не сохраняющимся
в ископаемом состоянии, но он связан с некоторыми изменениями скелета,
позволяющими различить этих животных и по ископаемым остаткам.
Рис. 196. Ornithorhyn-
chus. Коренные зубы ниж-
ней челюсти. (5/а)-
Подкласс I
ОДНОПРОХОДНЫЕ — MONOTREMATA
Ископаемые остатки этих форм очень ничтожны. Известны из послетре-
тичных отложений Австралии несколько костей скелета и обломки челюстей
с зубами, принадлежащие ехидне (Echidna) и утконосу (Ornithorhyn-
chus). Последний (рис. 196) имеет в молодости по два коренных зуба
в челюсти.
11*
164
класс: млекопитающие — mammalia
Между тем эти животные представляют громадный интерес, соединяя
в себе, с одной стороны, признаки рептилий — кладка яиц, с другой — очень
специализированные органы, как челюсти, лишенные зубов, молочные железы
без молока и т. д., и происхождение их возбуждает много вопросов.
Подкласс II
СУМЧАТЫЕ —MARSUPIALIA
Вышеназванные триасовые и юрские остатки зубов и челюстей,
иногда с рептильными признаками, не подходят вполне ни к одному из ныне
живущих отрядов. Они стоят ближе всего к некоторым сумчатым и могут
быть отделены под названием Palaeomarsupialia (древние сумчатые) как
отряд сумчатых. ,
Зубы эти бывают двух типов:
а) Передние в виде маленьких простых конусов, задние с двумя добавоч-
ными конусами: Cuspidata и Tricuspidata (Triconodontа). У неко-
торых из них конусы принимают вид 3 бугорков — зубы Trituberculata.
б) Зубы в виде пластинок с несколькими плоскими бугорками — это М u 1-
tituberculata (Allotheria Marsh).
Как простейшие и древнейшие формы, они были, вероятно, также ли-
шены плаценты.
Первые остатки принадлежали очень маленьким формам, жившим в Европе
и Сев. Америке.
Итак, мы подразделяем сумчатых на:
Отряд 1. Древние сумчатые — Palaeomarsupialia. Триасовые
и юрские формы, известные по отдельным зубам и челюстям.
Подотряд 1. Triconoconta: A. Tricuspidata, В. Trituber-
culata.
Подотряд 2. С. Multituberculata (Allotheria).
Отряд 2. Настоящие сумчатые — Marsupialia ver а.
Отряд!
Древние сумчатые — Palaeomarsupialia
Подотряд 1
Triconodonta
A. Tricuspidata
Кон усы расположены в один ряд
Dromatherium (рис. 197) — является наиболее древней верхне-триасо-
вой формой Сев. Америки (Сев. Каролина). Он представлен нижнею челюстью.
Длина ее 2,5 см. Нижний край выгнут посредине; по наружной стороне ее
подкласс: сумчатые — marsupialia
165
проходит желобок, открывающийся в широкое углубление заднего конца челю-
сти. Край ее не загнут. Мелкие острые резцы отделены высокими клыками от
коренных (3—1—10), из которых три первые, в виде простых конусов, имеют
по два добавочных маленьких конуса и по два корешка. На задних конусы
выше.
Эти зубы сходны с зубами некоторых Theriodontia, например Су-
nognathus.
Microconodon — также из верхнего триаса Сев. Каролины, с более
низкими коренными зубами и слабо загнутым задним краем в нижней челюсти.
Tritylodon longaevus Ow.
(рис. 198) известен по неполному че-
репу из триаса Кару (южная Африка).
Он является загадочной формой, которую
Рис. 198. Trirylodon longae-
vus Ow. (*/а)
Рис. 197. Dromatherium sil-
v е s t г е Emm.
Сили (Seeley) относил к рептилиям (Theriodontia). Теперь он включен
в Marsupialia. Удлиненный череп с расширенной передней частью имеет
пару крупных передних зубов (сломаны), за ними 2 маленьких ямки. После
большой диастемы расположено по 6 коренных, состоящих из 3 рядов
бугорков. Три первые зуба проще,
чем настоящие коренные. Затылок
обломан.
Переходя в юрские отложения,
мы встречаем в большом числе ниж-
ние челюсти, близкие к названным, и
части верхних. Резцы их раздвинутые,
Рис. 199. Amphilestes.
острые (4), клыки загнуты назад; существует у некоторых желобок на наруж-
ной стороне нижней челюсти. Коренные несколько более осложнились боко-
выми конусами.
Amphilestes Ow. (рис. 199) — из оолита Англии, имеет 11 корен-
ных зубов в нижней челюсти; на пяти последних боковые конусы значительно
поднялись вверх и стали более закругленными.
У Phascolotherium Ow. (рис. 200) это изменение коренных еще
более заметно. Оолит Англии.
Triconodon Ow. (рис. 201, 202)-—пурбек Дорсетшира; имеет корен-
ные двух резко различных типов: четыре передних, состоящие каждый из средне-
го большого конуса и двух меньших боковых, и три задних коренных с
тремя одинаковыми конусами. Здесь дифференцировка зубов уже полная;
166
класс: млекопитающие — mammalia
_ А. Желоб на наружной стороне нижней челюсти существует. Известны
верхняя и нижняя челюсти.
Triacanthodon — форма, близкая.-к предыдущей; сохраняя указанные
признаки, дает нам очень интересное указание на присутствие молочного
зуба, еще не замененного пред-
коренным. Этот один, сменяю-
щийся в челюсти, зуб, борозда
Рис. 200. Phascolotherium.
Рис. 201. Triconodon.
на других челюстях и загнутый вверх и внутрь задний край челюсти — яв-
ляются признаками челюстей нынешних сумчатых, что и позволяет причи-
слить эти формы к последним.
в
д
Рис. 202. Triconodon mordax
Ow. (8/t)
Рис. 203. Amphitherium Prevosti
Blv. Левая нижняя челюсть {а — нат. вел.).
(Доггер.)
Dicrocynodon victor Marsh — верхняя юра Уайоминга, с высоким
и широким венечным отростком и закругленным кондилюсом, поднимающимся
выше ряда зубов.
Два передних коренных маленькие, остальные 10 значительно больше. Все
С тремя конусами.
В. Trituberculata
Вторая группа Palaeomarsupialia — с зубами, состоящими из трех
закругленных бугорков, расположенных в виде треугольника, а не рядом;
это — трехбугорчатые (Trituberculata).
Spalaeotherium Ow. — пурбек Англии — и Amphitherium Ow.
(рис. 203) имеют боковые бугорки как бы сдвинутые ближе наружу, а
средний отодвинут внутрь челюсти. У первой формы задний край челюсти
загнут внутрь.
Эти примеры (а челюстей очень много) указывают на большое зна-
чение этих форм, как форм, сохранивших еще рептильные признаки, с одной
подкласс: сумчатые — marsupialia
167
стороны, а с другой, не только давших уже полную дифференцировку зубов,
но достигших уже формулы высших млекопитающих: 3 1 3 — 4 и осложнивших
задние коренные. Этот тип трехбугорчатых зубов принимается теперь за про-
тотип зубов высших млекопитающих.
Подотряд 2
С. Multituberculata (Masrh Alloiheria)
Второй тип зубов — многобугорчатые — резко отличается от первого мно-
гими бугорками на коренных зубах, расположенных рядами,
Microlestes (рис 204) — из триаса (близ Штутгарта) — представлен
несколькими зубами с удлиненной коронкой, разделенной
ком, по сторонам которого расположены бугорки (по 4).
Начиная с верхней юры известны следующие формы:
В о 1 о d о п (рис. 205) — известен по верхней челюсти
с 1 3 3 — 4; предкоренные имеют маленькие бугорки, непра-
вильно расположенные. На коренных бугорки лежат в два
ряда. Эти формы являются как бы переходными к мело-
вым Tripriodon Marsh и Meniscoessus Соре и тре-
тичным Poly mastodon — нижний эоцен Новой Мексики,
сверху желоб-
Рис. 204. Mic-
rolestes ап-
tiquus Plien.
Коренн. зуб. (,3/i ’
у которых имеется по три ряда бугорков на широких коренных. Интерес-
ным отклонением является в юре сем. Plagiaulacidae,—с сильно разви-
тыми резцами, вверху и внизу по два, и многобугорчатыми верхними корен-
Рис. 205 Bolodon crassidens Ow.
(А—ват. величина.)
ными. Нижние коренные сложены в
складки, подобно зубам грызунов.
Рис. 206. Plagiaulax minor
Ow. (а, Ь — увел., с — нат. вел.)
Plagiaulax Becclesi Falc. и Plag. minor Ow. (рис. 206) —
из пурбека Англии, с 6 нижними коренными зубами.
Некоторые из этих зубов (Microlestes) напоминают зачаточные зубы
Ornithorhynchus (утконоса), недавно открытые у этого загадочного млеко-
питающего, кладущего яйца и имеющего во взрослом состоянии роговой пло-
ский клюв. Это разнообразие зубов у древнейших форм указывает, что они
168
класс: млекопитающие — mammalia
произошли не одна от другой, а от еще более древних типов, может быть
нескольких. Наиболее примитивной может считаться форма, имеющая конусо-
образные рептильные зубы. Челюсти с коренными многобугорчатыми являются
уклонившимися боковыми ветвями. В мелу Сев. Америки мы встречаем остат-
ки Allotheria: Tripriodon Marsh и Meniscoessus Соре, но они
еще скуднее, чем в юре. Есть тут и обломки костей, для которых Марш
указывает древние признаки, как отдельную фасетку на лопатке для кора-
коидной и т. п. Но устанавливать какие-ни-
будь генетические отношения форм совер-
Рис. 208. Polymastodon taaense
Matth.
Рис. 207. Ptilodus gracilis Gidley.
(Нижний эоцен.) (2/3)
шенно невозможно в данное время по этим меловым остаткам. Между тем
именно меловая эпоха скрывает тайну развития млекопитающих, так как с
начала третичной эпохи мы встречаем уже обилие их.
Третичные формы:
Ptilodus gracilis Gidley — из нижнего эоцена Монтаны (рис. 207);
известен целый череп с очень укороченной нижней челюстью типа грызунов,
с большими резцами в нижней челюсти и 2 коренными, высоко поднятыми,
сжатыми с боков и снабженными вертикальными складочками. В верхней челюсти
резцы меньше; коренные многобугорчатые.
Череп сверху совершенно плоский. На нижней поверхности две большие
небные дыры.
Polymastodon attenuatus Соре (рис. 208) — самый нижний эоцен
Новой Мексики (Пуэрко). Размерами подходил к бобру. В обейх челюстях
имел по паре больших резцов типа грызунов. За верхними резцами, внутри челю-
сти, существовала пара маленьких резцов. Клыков не было. Следовала диастема, а
за нею по маленькому предкоренному и по два больших коренных в обеих челю-
стях. В верхней челюсти предкоренные зубы состояли из трех рядов бугорков,
задние — из двух рядов; в нижней — все зубы имели по два ряда бугорков.
Нижняя челюсть не имела загнутого заднего края.
подкласс: сумчатые — marsupialia
169
Зубы эти сходны по форме с зубами С i гп о 1 о гп у s Соре из мела Уайоминга,
найденными отдельно, и зубами Р t i 1 о d и s Gidley (верхней челюсти). Близких
форм к Р о 1 y-m a s t о d о п мы не знаем среди нынешних сумчатых.
Cimolomys — известны отдельные зубы из 2 и 3 рядов бугорков
(рис. 209).
Neoplagiaulax Lem. — нижний эоцен Франции. Близкая форма к
Plagiaulax, — у которой в нижней челюсти три передних коренных заме-
Рис. 210. Neoplagiaulax eocaenus.
Левая сторона. (Нижний эоцен.)
Рис. 209 Cimolomys nitidus
Marsh.
йены одним большим складчатым зубом. Верхние коренные из нескольких ря-
дов бугорков (рис. 210).
Отряд 2
Настоящие сумчатые — Marsupialia vera
Стоят особняком от других млекопитающих по отсутствию плаценты и
недоразвитию зародыша в утробе матери. Теперь они представлены в Австра-
лии и немногие в Мексике (D i d е 1 р h у s) и Бразилии (хищные). Отряд
этот представляет громадное разнообразие по образу жизни и строению ске-
лета. Есть между ними хищные, грызуны, лазающие и прыгающие. Характерным
органом являются марсупиальные кости, которые у самки поддерживают
сумку, в которой помещаются недоразвившиеся детеныши. Эти кости сочле-
няются с седалищными костями. Но так как они существуют у обоих полов,
то, очевидно, они развились не для поддержки мешка. Последние исследования
указывают на их соответствие с задне-лобковыми костями некоторых репти-
лий Другим характерным признаком сумчатых является замена только одного,
последнего, молочного зуба предкоренным.
На нижней челюсти характерен задний край, загнутый внутрь и наперед, и
желобок на внешней поверхности. Мозг маленький, почти гладкий у маленьких
форм. У более крупных существуют извилины, но простые.
Так как между нынешними сумчатыми встречаются формы, замещающие
представителей различных семейств плацентных, то их считали древнейшей груп-
пой, давшей начало этим семействам плацентных.
Мы имеем:
1) сумчатых хищных — Dasyuridae (сумчатые куницы),
2) сумчатых насекомоядных, как Myrmecobius (сумчатый
муоавьед),
170
класс: млекопитающие — mammalia
строго отграничивая их от сходных по
Рис. 211. Didelphys (ныне живущий).
3) сумчат-ых грызунов, как Phascolomys вомбат Ван-Димено-
вой Земли. Кенгуру напоминает даже копытных редукцией пальцев. На задних
конечностях они сокращаются до 2. Передние, очень укороченные, сохраняют
пять пальцев.
При дальнейшем изучении их остатков и остатков плацентных ока-
залось, что развитие тех и других шло параллельно и что сходные признаки
сумчатых и плацентных оказались результатом сходных условий жизни (кон-
вергенция признаков), а не генетического родства, так как другие признаки,
характерные для сумчатых, строго сохраняются и у ныне живущих форм,
им признакам плацентных коготных.
Кроме вышеописанных че-
люстей, которые относятся к сум-
чатым, ископаемые остатки по-
следних мало известны. Только
для сем. сумчатых крыс (Didel-
р h у d а е) они известны в верх-
нем эоцене, в гипсах Монмартра
близ Парижа и в Сев. Америке.
Это — Didelphys и Perathe-
rium (рис. 211 и 212). Позже
они встречаются в олигоцене Фран-
ции и Англии (это последний пе-
риод их жизни в Европе) и в
миоцене Колорадо. В послетре-
тичное время они существовали в
Средней и Южной Америке, где
живут и теперь (Мексика, Брази-
лия, Парагвай).
В Патагонии найден Рго-
thylacinus patagonicus —
в миоцене Santa Cruz. Он является
настоящим, представителем сумчатых хищников Dasyuridae и известен
цельным скелетом, с хищными зубами и длинным хвостом. Длина скелета —
140 см\ описан Синклером (W. С. Sinclair) (рис. 213).
В эоцене найдены отдельные кости и зубы. Другие остатки сем. Dasy-
uridae — сумчатых куниц — и Peramelidae — сумчатых барсуков —
известны только в послетретичных отложениях Австралии, их теперешнею
местожительства. Там же найдены остатки двух представителей совершенно
вымерших семейств сумчатых: Thylacoleo — хищника и Diprotodon —
гигантского грызуна.
Формы, близкие к Didelphydae, встречаются в юре (в пурбеке), напри-
мер Triacanthodon. Описанные Маршем остатки из мела Сев. Америки,
хотя и очень скудные, под именем Didelphodus близки к Didelphys.
Нас гоящий скелет, хотя и не полный, без черепа, был найден Кювье
Рис. 212. Didelphys fugax Соре (Рега-
t h е г i u гл).
а— верхняя, с, d—нижняя челюсть.
подкласс: сумчатые — marsupialia
171
в гипсах Монмартра. Встреченная им раньше здесь же нижняя челюсть имела
признак сумчатого — загнутый внутрь задний край, что и дало повод Кювье
отнести ее к Marsupialia и заставило думать, что в скелете, лежавшем
тут же, но еще не отпрепарированном, будут найдены сумчатые кости. При
расчистке скелета это оправдалось.
Величина всего скелета равнялась 16 см. К сожалению, череп не сохра-
нился. но нижняя челюсть имела признаки D i d е 1 р h у s, так же как и кости
скелета — особенно таз с марсупиальными хостяыи.
Под именем Peratherium описаны нижние челюсти, отличающиеся от
D i d е 1 р h у s большей величиною коренных зубов.
Рис. 213. Prothylacinus patagonicus Sincl. (Миоцен.)
В олигоцене Сев. Америки Коп нашел верхние и нижние челюсти с зубами,
которые отвечают формуле
5
нынешних Didelphydae: —
1 3 — 4
1 3 — 4
с про-
стыми конусообразными предкоренными.
Такие же зубы найдены в миоцене Германии и Франции; в более поздних отло-
жениях они не встречаются. Это семейство сумчатых представляет наибольший
интерес, так как существование его можно проследить с верхней юры (Т г i а с an-
thod о п) до миоцена в Европе и Сев. Америке, а в Южной Америке и доныне.
В то время как Didelphys до сих пор сохраняет пятипалые конеч-
ности с острыми когтями на четырех пальцах, и только пятый — тупой, закруг-
ленный, другой род — Macropus (кенгуру) — имеет четырехпалые задние ноги
с очень удлиненными пальцами, причем более всех пальцев развит четвертый.
Зубы у него тоже сократились:
_3
З-
0(1) 1—4
0 1 — 4’
К сожалению, мы до сих пор не имеем палеонтологических данных для
объяснения того, как происходили эти изменения. Кенгуру с раздутыми боками,
сильным хвостом, большими задними ногами и укороченными передними невольно
напоминают некоторых диназавров.
Thylacoleo Ow. (рис. 214) — из плейстоцена Австралии — назван
так Оуэном за сходство его черепа с черепом льва, которому он равен
и по размерам. От него найдены, кроме черепа с нижней челюстью, кости
передней конечности, таз и задняя нога; этих остатков достаточно, что-
бы составить понятие о цельном животном. Череп широкий сзади, с суженной
172
класс: млекопитающие — mammalia
мордой, очень широкими скуловыми дугами и большими височными впадинами.
3 1 3—1
Зубы в очень сокращенном числе у q g р, причем из передних зубов бо-
лее всего развита внутренняя — единственная на нижней челюсти — пара рез-
цов. В верхней челюсти за нею следуют по два маленьких резца с каждой
загнутые внутрь. Далее два маленьких
стороны, а за ними маленькие клыки,
Рис. 214. Thylacoleo.
предкоренных и третий громадный —
хищный, удлиненный зуб, ограниченный
двумя ровными плоскостями, сходящи-
мися в острый гребень посреди жева-
тельной поверхности. За этим зубом,
около его вну греннего края — малень-
кая альвеола для единственного корен-
ного зуба, как у льва,
В нижней челюсти клыков нет;
два маленьких предкоренных и третий,
очень длинный, соответствует верх-
нему. За ними два маленьких бугорчатых коренных. Между ныне живущими
Hypsiprymnidae имеются формы с некоторыми общими
признаками
2
Рис. 215 А. Задние ступни. 1 — Т г i-
chosurus (ныне живущего сум-
чатого Австралии); 2 — Diproto-
don australis Ow. (Австралия,
постплиоцен.)
Рис. 215. Diprotodon australis Ow.
с Thylacoleo и Plagiaulacidae, но зубная система очень разнится.
Пальцы были снабжены острыми загнутыми когтями.
Этот типичный хищник с сильно редуцированными коренными зубами
стоит пока особняком между сумчатыми.
Diprotodon australis (рис. 215) — в плейстоцене Австралии.
Крупная вымершая форма сумчатых. Череп его имел размеры черепа носорога.
Особенно сильно у него была развита внутренняя пара верхних резцов, загнутых
вниз. Они имели постоянный рост, как у плацентных грызунов. Две другие пары
верхних резцов — маленькие. Клыков нет; предкоренные маленькие; коренные
подкласс: сумчатые — marsupialia
173
сильно развиты, состоят из двух поперечных гребней каждый. В нижних челю-
стях пара больших с постоянным ростом резцов; предкоренные треугольной
формы; коренные с двумя гребнями. Весь скелет тяжеловесный с плотно при-
легающими друг к другу остистыми отростками. Особенностью передних ко-
нечностей является отсутствие олекранона, локтевого отростка. Плечевая
кость тонкая, так же как и бедро. Лопатка удлинена. Передние и задние
конечности сильно разнятся от конечностей других сумчатых. Пальцы очень
редуцированы, а тарсальные и метатарсальные кости сильно развиты. Может
быть, Diprotodon произошел от животного, лазавшего по деревьям, и изме-
нил так ненормально свои конечности, когда ему пришлось вследствие увели-
чения его размеров вести жизнь наземного животного.
Сопоставление этих двух постплиоценовых сумчатых подчеркивает раз-
нообразие тех специальных признаков, которые были выработаны представи-
телями сумчатых хищных и грызунов в зависимости от их питания. Очевидно,
это были крайние формы развития неизвестных линий, закончивших свою
жизнь в плейстоцене и не связанных непосредственно с линиями сумчатых,
дошедших до нас в виде представителей различных семейств.
Prothylacinus patagonicus Amegh. — из миоцена Санта-Круца.
Описан А м е г и н о (Ameghino) по остаткам, позволившим восстановить цельный
скелет. Это хищник из Dasyuridae, который может считаться предком
4 1 3 — 4
ныне живущих сумчатых волков (Thylacinus). Он имел у Пер-
вый палец сильно редуцирован. Форма эта является связующей между нынеш-
ним Thylacinus и эоценовыми предками, у которых был хорошо развит
первый палец, противопоставлявшийся, указывающий на возможность лазать
по деревьям. Устанавливается ряд форм, изменивших образ жизни и лишив-
шихся ставшего ненужным первого пальца (по указанию Абеля):
Amphiproviverra — с большим первым противопоставлявшимся паль-
цем (эоцен);
Cladosictis — большой палец укорочен (миоцен);
Prothylacinus — небольшой остаток от пальца (миоцен);
Thylacinus — палец исчез (ныне живущий). ‘ г-;
Зубы этих форм представляют тоже большой интерес при сравнении
с зубами ныне живущих.
Итак, Патагония дает нам наиболее интересные остатки этой обособлен-
ной группы млекопитающих — бесплацентных. !{ .. i; (
Раньше чем переходить к знакомству с плацентными, займемся вопросом
о развитии зубов млекопитающих (рис. 216).
Мы уже видели, что первичные формы триаса и юры имели зубы, очень
сходные с зубами Theriodontia. Это были простые конусы, по обеим сто-
ронам которых появились добавочные зубчики, сначала маленькие, потом до-
стигавшие размеров средних конусов, и зубы становились трехконусными.
Далее конусы притуплялись, закруглялись, и совершалось передвижение сред-
класс: млекопитающие — mammalia
174
«
него, причем наблюдалось его передвижение на внутренюю сторону в верхней
челюсти и на наружную — в нижней. Эти наблюдения и подробные исследова-
ния древнейших и позднейших зубов привели американских ученых, главным
образом Копа и Осборна, к возможности найти объяснение происхожде-
ния различных типов зубов млекопитающих от трехбугорчатого типа. Они
принимают за первоначальный тип „трехбугорчатый" зуб, причем на верхних
зубах первичный бугор — протокон — лежит на внутреннем краю зуба, а бо-
ковые бугорки — паракон (передний) и метакон (задний) — на наружной
стороне зуба. На нижних зубах, наоборот, сред-
ний протоконид — на наружной, а два боковые —
метаконид'* и параконид — на внутренней сторо-
не зуба.
Зубы с подобным расположением бугорков
встречаются и теперь у насекомоядных и хищных.
Дальнейшее изменение зубов, осложнение их, со-
вершается добавлением четвертого бугорка, появляю-
щегося сзади на внутренней стороне зуба, что
изменяет треугольную форму зуба в четырехуголь-
ную. Этот четвертый — добавочный — бугорок на-
зывается гипокон — на верхних и гипоконид — на
нижних зубах. Это типичная форма зубов для
Рис. 216. Развитие згбов
млекопитающих из 3-бугорча- Древних жвачных.
тых: а--нижние, Ь—верхние. Дальнейшим изменением являются добавоч-
ные бугры, развивающиеся между наружными и
внутренними буграми с каждой стороны. Это протоконюль — спереди и
метаконюль — сзади, для верхних зубов. На нижних добавочный является
на заднем конце, в виде складки или бугра — это талон. Эти зубы типичны
для древних непарнопалых. Случается, что два добавочные бугра развиваются
на верхних коренных раньше, чем развился четвертый бугор — гипокон, зубы
становятся пятибугорчатыми.
Такие встречаются у целой группы вымерших жвачных и у некоторых
насекомоядных.
Г р е б н и, характерные для многих зубов, являются, по мнению авторов
теории развития зубов млекопитающих из трехбугорчатых, следствием слияния
этих и добавочных бугров в гребни. Раньше на эти гребенчатые зубы (тапира,
например) смотрели как на примитивную форму. Конечно, не все зубы ныне
живущих млекопитающих, да и исчезнувших, можно подвести под это объ-
яснение их развития. Но, может быть, это является следствием того, что не
для всех сложных форм мы знаем первоначальную форму зуба, до момента ее
осложнения. Что касается названий бугорков, предложенных американскими
учеными, то они являются иногда сбивчивыми, так как не все ученые соглас-
ны признать в различных зубах аналогичные элементы. Некоторые зоологи
считают протокон за метакон других ученых, появляющийся, по их
мнению, раньше других бугров. Во всяком случае теория эта имеет в своем
подкласс: плацентные — placentalia, коготные
175
основании много данных быть принятой и приложимой при изучении ископае-
мых, так как у некоторых форм — особенно у копытных — мы можем про-
следить при изучении развития их осложнение трехбугорчатых предкоренных
и превращение их по форме в коренные.
Подкласс III .
ПЛАЦЕНТНЫЕ — PLACENTALIA
КОГОТНЫЕ
Отряд!
Насекомоядные — Insectivora
Простейшие из плацентных животных — большею частью маленькие пяти-
палые коготные животные с почти одинаковыми резцами, клыками и предко-
ренными; верхние коренные трех- или четырехбугорчатые, а нижние
с пятью острыми бугорками. Молочные зубы сменяются рано. Конечности ча-
сто роющие, пятипалые.
Из ныне живущих наиболее известны: еж, крот, землеройка в умеренном
поясе и сем. Centetidae на Мадагаскаре и Кубе.
До последнего времени остатки плацентных млекопитающих были из-
вестны только начиная с нижнего эоцена Сев. Америки и Европы. Выражены
они частями черепов и зубами. Костей скелета найдено мало. Но третья амери-
канская экспедиция открыла в 1924 г. в меловых отложениях Монголии
6 черепов плацентных млекопитающих с нижними челюстями. Слои были те
же, в которых найдены яйца и скелеты Protoceratops. Там же были най-
дены и остатки бесплацентных Multituberculata.
Животные, которым принадлежали шесть названных черепов, отнесены к
двум различным семействам и к трем новым родам:
Сем. Deltatherijdae, роды: Deltatherium, Hyotherium.
Сем. Zalambdodonta, р. Zalambdolestes.
Черепа их размерами не больше черепа крысы, с удлиненной мордой,
со слабо развитой мозговой капсулой (их изображений пока еще нет).
В первом семействе представлены предки хищных, близкие к Сгео-
d о n t а, во — втором предки насекомоядных.
Исследования Метью, Грегори и Элиа Смита определенно указывают,
что первичные хищные и насекомоядные жили на деревьях и что они были
предками других млекопитающих.
На первоначальную жизнь на деревьях указывает строение задних конечно-
стей у многих древних млекопитающих, и даже у человека в младенческом
возрасте — способность пальцев ноги схватывать.
Наиболее древними находками'считались Adapisorex и Adapisori-
culus Lem. — из самого нижнего эоцена Сернэ, близ Реймса. Это остатки
вымерших форм. Нижние челюсти имели 3 13 — 4; коренные были с тремя
бугорками.
176
класс: млекопитающие —mammalia
Palaeoryctes puercensis Matth. —.из самого нижнего эоцена (Тег-
rejon) Новой Мексики. Он представлен частью черепа с зубами
1 3 3 — 3
з ~з 3 и ни>кней челюстью, начиная с клыка. По типу зубов эту
форму сближают с представителями.
Сем. Centetidae (Zalambdodonta Gill., существующими на Мадага-
скаре и Кубе. Коренные зубы их характеризуются отсутствием внешних бугорков.
На предкоренных существует один внутренний бугорок и другой внешний по-
средине зуба, который считается двойным бугорком, из слившихся паракона и ме-
такона. На внешней стенке зуба находятся только складочки—метастиль (средняя)
и пара стиль (передняя).
Рис.217 Zalambdodonta: А — Palaeoryctes puercensis. (Нижний эоцен).
В — Didelphodus absarocae Соре.
Это тип зубов Zalambdodonta. Форма зуба удлиненнотреугольная.
Нижние коренные имеют острый, высокий сдавленный конус и добавочный
маленький задний. Pri такой же формы. Весь череп от 2 до 3 см.
Сем. Leptictidae. Palaeictops и Didelphodus, описанные
Копом из нижнего эоценаВазача и Уайоминга, очень близки к Centetidae,
но остатки их еще менее полны.
Нижние зубы Didelphodus осложнены маленьким ипередними зубчика-
ми. Matthew относит Didelphodus и Palaeictops Zalambdodonta.
Micropternodus Matth. — маленькие нижние челюсти из верхнего и
нижнего олигоцена Монтаны; очень похожи на челюсти, найденные в юре, с ко-
ренными, состоящими из среднего конуса с двумя маленькими боковыми.
У I с t о р s Leidy сохраняются три пары маленьких резцов, острые клыки
и такие же предкоренные. Это простейшая трехбугорчатая форма из олигоцена
Монтаны Сев. Америки.
Сем. Erinaceidae. Necrogymnurus Cayluxi (Cayluxothe-
r i u m) Filh. — из фосфоритов Керси, представлен хорошо сохранившимся чере-
пом с четырехбугорчатыми последними предкоренными и двумя коренными,
задний треугольный. Нижние коренные тоже из четырех маленьких бугорков.
Из олигоцена и миоцена Дакоты и миоцена Патагонии известны формы,
связывающие древних представителей с ныне живущими (на Мадагаскаре
подкласс: плацентные — placentalia, коготные
177
Кубе, в южной и западной Африке) формами М i с г о g а 1 е. По ним можно
проследить изменение зубов от простых конусов до трехбугорчатых, с малень-
кими добавочными.
Leptictis Leidy (рис. 218) — из нижнего миоцена Дакоты, вымершая
форма. Удлиненный череп имеет два продольных канта вверху. Резцы сокра-
щены до двух пар. Коренные трехбугорчатые.
Sorex pusillus Н. v. Меу.— из миоцена Франции (Grive St. Albin).
Очень хорошо представлены насекомоядные различных типов в отложениях
среднего эоцена Бриджера (Сев. Америка) ]).
Остатки ежей известны из олиго-
цена. G а 1 е г i х Pomel (Р а г a s о г е х Н. v.
Меу.) — с шестибугорчатыми коренными, яв-
ляется одной из тех форм, которые осложнили
свои зубы добавочными буграми. В Сев. Аме-
рике многие формы с подобными шестибу-
горчатыми зубами относились раньше к на-
секомоядным, позже они были переведены в
другие отряды. Остались немногие, напри-
мер Hyopsodus Matth. из среднего эо-
цена Уайоминга, который является предста-
вителем крупной вымершей формы, достиг-
шей уже в эоцене большей сложности зу-
Рис. 218. Leptictis Haydeni
Leidy. (Нижний миоцен.)
бов (шестибугорчатых коренных верхних и четырехбугорчатых нижних), чем
ныне живущие четырехбугорчатые насекомоядные.
Думаю, что эти формы с шестибугорчатыми зубами не могли принадле-
жать к насекомоядным. Им пришлось бы при дальнейшем развитии редуци-
ровать лишние бугры. Интересны находки в олигоцене и в нижнем миоцене
(Керси и Вейзенау) маленьких костей плеча (humerus) крота, единственной
части скелета.
Очевидно, развитие этого отряда шло медленно. Одни животные приспо-
соблялись в борьбе с другими к незатейливому образу жизни под землей,
направляя силы к рытью ходов, изменяя конечности, главным образом, и
оставляя зубы мало ушедшими от первоначального типа (крот). Другие выра-
батывали колючий покров (еж) или особые железы, выделяющие жидкость
неприятного запаха (вонючки).
Отряд 2
Летучие мыши — Chiroptera
Некоторые из родов (Vespertilio) принадлежат к насекомоядным, но
вместо того, чтобы зарываться в землю, они выработали перепонку, прикреп-
ляющуюся между удлиненными пальцами передних конечностей, и поднялись
в воздух, приспособившись в то же время к ночному образу жизни.
*) W. D. Matthew, The Carnivora and Insectivora of the Bridger basin, middle
eocene. Memoirs Amer. Mus. Nat. Hist., 1909, vol. IX.
12 Павлова M. В. Палеозоология. 4. И.
178
класс: млекопитающие — mammalia
По строению зубов они близки к древним насекомоядным. Их можно
подразделить на два типа:
М i с г о с h i г opt е г а — маленькие, питающиеся насекомыми, и
Mega chi г opt era крупные,— питающиеся плодами.
Ископаемые остатки с первых известны олигоцена (фосфориты Керси).
Они принадлежат к родам Rhinolophus и Pseudorhinolophus, близким
к нынешним. - .
Из Megachiroptera до сих пор известен только один скелет,
почти полный, из верхних лигнитов Monte Viale в верхней Италии. Это А г-
chaeopteropus transiens (летающая собака), описанный М е ши-
не л л и (Meschinelli). Это — крупное животное, до 40 см длиною, с хорошо сохра-
нившимися передними и задними конечностями.
Очевидно, Chiroptera отчленились от древнего типа с трехбугор-
чатыми зубами очень рано, может быть еще в мелу, и развивались параллельно
с настоящими насекомоядными, но в другом направлении, успев выработать
конечности для летания и изменяя зубы.
Сократился первый предкоренной, коренные сохранили треугольную форму.
Клыки очень развиты.
Отряд 3
Грызуны — Rodentia
В большинстве случаев это маленькие животные с сильно развитыми
резцами с постоянным ростом. Клыки отсутствуют. Коренные бывают в разном
числе, от 2 до 6, и различного строения.
Нижняя челюсть причленяется низкими, плоскими отростками, входящими
в углубления черепа и допускающими движение челюсти вперед и назад.
Отряд этот обособлен между нынешними млекопитающими и связан,
вероятно, генетически с древними, первичными насекомоядными. Как пища,
так и образ жизни разных семейств их различны, и в зависимости от этого
различно строение зубов и конечностей. Резцов бывает одна или редко две
пары. Коренные — или бугорчатые, например у мышей, с хорошо обособленными
корнями, или складчатые и пластинчатые с постоянным ростом,— у бобра, зайца.
Бегающие и лазающие по деревьям формы имеют хорошо развитые пяти-
палые конечности, приспособленные иногда к хватанию (белка). У форм,
живущих в воде, конечности изменены, снабжены перепонкой (бобр). Все эти
признаки принимаются во внимание при классификации, но главным.признаком
считается число резцов, по которым грызуны и разделяются на:
1. Simplicidentata — с од-
ной парой резцов в верхней челюсти:
1) бугорчатозубые (мыши,
белки),
2) складчатозубые (бобры и
полевки).
2. Duplicidentatа—с двумя
парами резцов в верхней челюсти
(заяц, пищуха). Коренные состоят
из пластинок; они призматической
формы. У молодых животных суще-
ствуют 3 пары резцов.
подкласс: плацентные — placentalia, коготные
179
Ископаемые остатки грызунов известны начиная с эоцена. Наиболее
древними являются семейства белок (Sciuridae) и сонь (Мyoxidae) — с
нижнего эоцена Сеь. Америки.
Бобры и зайцы — с нижнего миоцена Европы.
Мыши—с плиоцена.
Многие формы из ископаемых носят уже определенные, ясно выражен-
ные признаки, которые позволяют отнести их к которой-нибудь из названных
групп. Но есть формы, признаки ко-
торых не позволяют указать им опре-
деленное положение. Такой группой
является у
Сем. Tillodontia—из эоцена
Сев. Америки. Сейчас эти формы не
всеми признаются за грызунов, и
некоторые ученые относят их к
насекомоядным. Но, проследив из-
Рис. 219. Esthonyx Burmeisteri Соре.
менение их зубов, приходится отнести их к грызунам.
Esthonyx Burmeisteri Соре (рис. 219) — из нижнего эоцена Сев.
Америки (Wasatch), наиболее древняя форма; имеет две пары резцов (наруж-
ные — более крупные, с постоянным ростом), небольшие клыки и по шести
коренных, трехбугорчатых верхних и четырехбугорчатых нижних: —
1 3 — 3
1 3^3 ‘
Внутренняя пара нижних резцов очень маленькая, вторая пара отвечает по
Рис. 220. Tillotherium fodiens Marsh.
•маленькая, так же как и клыки. Коренные
3 — 3.
2^3 ’
размерам верхним. Другие
виды этого рода встреча-
ются в среднем эоцене Сер.
Америки (Bridger).
Tillotherium fo-
diens Marsh (рис. 220). —
В том же среднем эоцене
(Bridger) встречаем остатки
крупной формы, размером с
медведя, с удлиненным низ-
ким черепом, с одной парой
сильно развитых резцов (2-я
пара), покрытых снаружи
эмалью. Третья пара очень
верхние вытянутые поперек.
трехбугорчатые; нижние состоят из двух луночек. Сочленовная головка нижней
челюсти плоская.
Из скелета известны немногие кости: плечевая, бедро, малая берцовая,
надпяточная. Эти формы принадлежат к вымершей группе, рано сократившей
свои коренные зубы и резцы.
12* Д u Та 7 СЧХ ' «« ««МД* . 4 Д ...О
180
класс: млекопитающие—mammalia
Итак, хотя Т i 11 о d о п t i а имеют две пары резцов, но их нельзя отнести
к нынешним Duplicidentata, так как у них резцы, очень различные, рас-
положены рядом. Это — вымершая группа, не связанная с ныне живущими.
Подотряд 1
Simplicidentata
Сем. Sciuroidea — является наиболее древним и примитивным из гры-
зунов; оно появилось в эоцене Сев. Америки, прошлочерез олигоцен и нижний
миоцен и дало позднейших
потомков, дошедших до нас
в виде различных семейств.
Род Р а г a m у s Leidy
представлен несколькими ви-
дами в последовательных от-
ложениях эоцена, позволяю-
щими проследить изменение
этого рода. Это формы с пло-
ским черепом, с сильно загну-
тыми вниз верхними резцами,
лежащими в межчелюстных
костях. Носовые кости выдви-
нуты над ними. Коренные
отделены большим проме-
жутком, диастемой,
1 0 2 — 3 .
Т" 7Г 1 — 3
Рис. 221. Paramys delicatus Matth. Нижняячелюсуькороче верх-
ней, с высоким венечным от-
ростком и плоским сочленовным. Род этот очень близок, может быть
тождествен, с европейским Plesiarctomys Brav., менее хорошо пред-
ставленным ископаемыми остатками. Из ныне живущих он ближе всего
к р. Sciurus (белке).
Paramys primaevus — маленькое животное, встречающееся в
нижнем эоцене Вазача. Скелет не вполне сохранился. Зубы с низкими
бугорками.
Paramys delicatus Matth. (рис. 221, 222) — средний эоцен Брид-
жера. Коренные зубы имеют по три бугорка. Череп похож на Arctomys,
но с более короткой мордой и с небольшим нижнеглазничным отверстием
(foramen infraorbitale). Затылочный гребень невысокий. Глазницы не замкнуты.
Слуховая капсула не сохранилась; очевидно, она отпала, будучи не вполне
окостеневшей и не сросшейся с черепом. Нижняя челюсть с очень высоким
отростком. Зубы нижней челюсти с низкими бугорками. Скелет близок к ске-
подкласс: плацентные — placentalia, коготные
181
лету белки, но шейные и поясничные позвонки длиннее, и их отростки больше.
Конечности тонкие; задние длиннее передних; на последних по 4 пальца и
-остаток пятого. На задних все пять пальцев хорошо развиты. Существует
центральная кость. Коготные
фаланги длинные. Хвост очень длинный.
У Paramys robnstus Mattb.
(рис. 223) (средний эоцен) и Paramys
petersoni Mattb. (верхний эоцен) корен-
ные зубы имеют по четыре бугорка, более
высоких, приближающихся к беличьим.
Последний вид значительно больше двух
предыдущих, имеет резцы плоские спе-
реди, как у бобра, что отличает его от
других видов. В передних конечностях
центральная кость остается не сросшейся.
Существовало еще несколько других видов.
S с i u г u s в олигоцене является
продолжателем, вероятно, одной из ма-
леньких эоценовых форм и предком ны-
нешних видов белок, у которых умень-
шились шейные позвонки, увеличились
кость слилась с radiale.
Рис.223.Paramys robustus Mattb.
бугорки на зубах* и центральная
Рис. 224. Sciuravus nitidus Matth.
Сем. Ischyromyidae. Sciuravus (рис. 224) является родом, разви-
вавшимся параллельно с Paramys, но представители его меньших размеров.
Зубы имели по четыре бугорка, но последние более изолированы. Череп
182
класс: млекопитающие — mammalia
имеет более удлиненную переднюю часть; сагиттальный гребень отсутствует.
Слуховая капсула не вполне окостенела. Различные виды существовали в
продолжение всего эоцена в Сев. Америке.
Ischyromys Leidy (рис. 225) — в эоцене и олигоцене Южной Дакоты,,
больших размеров, чем предыдущий; слуховая капсула окостенела и сохрани-
лась. Зубы четырехбугорчатые, имеют входящие наружные и внутренние складки,
подразделяющие их на две части. Эта форма является как бы продолжением
развития Sciuravus в другом на-
правлении, с изменением зубов. По-
следующие формы относятся к:
Сем. Castoridae с родом S t е-
neofiber Н. v. М., встречающимся
Рис. 226. Castor europaeus Ow.
A — верхние, В—нижние.
Рис. 225. Ischyromys typus
Matth.
в олигоцене и нижнем миоцене Европы и Сев. Америки, у которого зубы
сохраняют еще корни, но складки на жевательной поверхности продвинулись
глубже, и бугорки стушевываются. Это изменение зубов продолжается у мио-
цено-плиоценового D i р о i d е s: зубы приобретают постоянный рост, теряют
корень, но поверхность их имеет не более трех складок.
Плиоценовые формы более сложны и дают переход к настоящим
Castorinae (рис. 226) — с многочисленными складками эмали, существую-
щими на всех 4 коренных зубах. Представители их известны с нижнего плей-
стоцена доныне — Castor europaeus.
Trogontherium — является вымершей формой, достигшей в плейсто-
цене очень больших размеров сравнительно с нынешними бобрами. Коренные
зубы уменьшаются кзади.
Итак, мы имеем две главные ветви, развившиеся из нижне-эоценового
Paramys с трехбугорчатыми коренными зубами. Американский ученый
д-р М е т ь ю, изучавший подробно развитие грызунов, указывает на развитие
других семейств грызунов от Paramys.
В олигоценовых слоях Египта (Фаюм) найдены челюсти маленьких гры-
зунов, близких к грызунам из нижнего олигоцена Европы (фосфориты Mouillac
во Франции). Это — Eomys Zitteli Schl. Сохранилась правая нижняя челюсть
с большим резцом, последним предкоренным и тремя коренными. Е'се снабжены
корнями. Предкоренной имеет три бугорка, коренные — по четыре, соединенных
между собою гребнями.
подкласс: плацентные — placentalia, коготные
183
Metaphiomys beadnelli Osb. — найден в более высоких слоях
тгх же отложений. Представлен тоже нижней челюстью. Коренные зубы слож-
нее. Они приближаются к Sciuroides Quercy Schl. Поверхность коронки
глубже подразделена на отдельные участки поперечными складками. Находки
эти представляют большой интерес, обобщая фауну столь отдаленных мест.
Epigaulus Hatcheri Gidley (рис. 227) и Ceratogaulus Matth.—
Совершенно особняком стоят эти грызуны, найденные в миоцене Канзаса.
Это чрезвычайно специализировавшиеся формы с двумя большими прямыми р о-
г, а м и на носовых костях,
над самыми глазами. Че-
реп сзади широкий, с вы-
ступающими назад кон-
дилюсами. Резцы развиты
сильно. Коренные сохра-
нились по одному боль-
шому в каждой челюсти,
с каждой стороны, с про-
дольными неправильными
складками эмали. Сзади
их находятся еще по две
Рис. 227. Epigaulus Hatcheri Gidley.
луночки от двух коренных. Очень сильно развитая лопатка и передние конеч-
ности с длинными коготными фалангами указывают на роющую функцию их.
Длина животного до хвоста 30 см. Это сложный тип с некоторыми призна-
ками бобра (роющие конечности); другие признаки сближают его более с
белками и с азиатскими формами Spalacinae.
Подотряд 2
Duplicidentata
Для сем. Leporidae Matthew считает родоначальником:
Palaeolagus — из олигоцена и миоцена Сев. Америки.Череп его сдав-
лен с боков. Задние ноги длиннее передних. Рг2 конусообразный; все осталь-
ные зубы пластинчатые. Вторая пара резцов лежит сзади первой и меньше ее.
Верхние резцы несут углубление на передней поверхности. Глазницы замкнутые.
Остатки рода Lepus известны из верхнего миоцена в Сев. Америке и
из нижнего миоцена Европы.
Интересно отметить то изменение черепа и особенно формы и сочлене-
ния нижней челюсти, которое произошло у грызунов вследствие развития
больших резцов с постоянным ростом. Нижняя челюсть их сравнительно с
черепом значительно короче, чем челюсти других коготных; она сочленяется
с верхней посредством закругленного отростка, входящего в гленоидную впа-
дину, далеко отодвинутую назад и имеющую удлиненную, по длине черепа,
поверхность, а не поперечную, как у других коготных: хищников, насекомояд-
184 класс: млекопитающие — mammalia
ных и др. Кроме того исчез постгленоидный отросток, что и позволило
нижней челюсти отодвинуться назад и зайти за скуловую дугу. Венечный
отросток нижней челюсти тоже значительно укорочен. Задняя половина челю-
сти расширена, и к ней прикрепляются мускулы, двигающие челюсть спереди
назад при грызении. Развитие этих особенностей Коп объясняет именно раз-
витием резцов и необходимостью при грызении заменить вертикальное движе-
ние челюстей, как у хищных, горизонтальным.
В связи с этим движением стоит и изменение коренных зубов и их
положения. У зайца зубы вдаются своими основаниями в угол глазничной
впадины.
Отряд 4
Хищные — Carnivora
Самыми распространенными и наилучше представленными по разнообразию
семейств, родов и видов между коготными являются хищные. Каждому хорошо
известны: кошки, собаки, волки, львы, тигры, медведи и многие другие хищ-
ники. Их объединяют многие общие признаки, позволяющие сразу отнести их
к этому отряду, и может быть, только медведи стоят несколько особняком. По
внешнему виду это формы с несколько удлиненной или закругленной головой,
пятипалыми или четырехпалыми конечностями, легко двигающиеся, снабженные
длинным хвостом. Зубная система их имеет признаки, указывающие на их
хищный образ жизни. За хорошо развитыми резцами следуют сильные, большие
клыки, предкоренные и коренные зубы. Последний предкоренной рг^ вверху и
первый коренной тг внизу развиты особенно сильно и носят название хищных
зубов. Все передние и предкоренные зубы состоят из заостренных конусов.
Коренные являются настоящими бугорчатыми зубами. Мозговая коробка и за-
ключающийся в ней мозг хорошо развиты. Особенностью передних конечностей
хищных является слияние трех костей запястья: centrale и ulnare слиты с radiale
в одну кость, на которую опирается лучевая.
Конечно, признаки эти, например число зубов, варьируют у различных
семейств, но тип их остается тот же.
Ныне живущие хищные разделяются на: С a n i d а е, М u s t е 1 i d а е, V i v е r-
ridae, Hyaenidae, Felidae и Ursidae.
Все они имеют ископаемых представителей начиная с олигоцена, позво-
ляющих в большинстве случаев проследить их развитие. В более древних слоях,
в эоцене, найдены остатки животных, тоже хищных, но имеющих несколько
иные признаки, что заставило выделить их в особый подотряд Creodonta.
Главное отличие их заключается в зубах: у них отсутствуют хищные зубы и
настоящие, сложнобугорчатые коренные; имеется большое число режущих
зубов или простых трехбугорчатых. В конечности карпальные кости не слиты,
и строение их — с далеко отходящим первым пальцем — указывает на иной
образ жизни — лазание по деревьям.
Первоначально найденные формы Creodonta были настолько далеки от
настоящих хищных, что их считали вымершей группой, не имеющей генетиче-
*
подкласс: плаценгные — placentalia, коготные
185
ской связи с более поздними формами. Но дальнейшие находки как в Европе,
так и в Сев. Америке позволили притти к другим выводам. Остатки этих древ-
них хищников оказались очень многочисленны и разнообразны и позволили
п< дразделить их на:
1-я группа. Примитивные Creodonta — еще близкие по озу-
блению к насекомоядным, например Tricentes. Это не сумевшие при-
способиться к окружающей среде формы, не давшие потомков.
Одни из них совсем не имели хищных зубов; это — Ас гео di: Meso-
nyx, Dissacus и др.
Другие имели ложные хищные зубы. Это — Pseudocreodi, сем.
Hyaenodontidae и Oxyaenidae: Hyaenodon, Pte г о don, Оху-
dепa, Patriofelis, Sinopa и др.
2-я группа. Paleocarnivora (прежде относились тоже к Creo-
donta). Это так называемые Eucreodi, имевшие хищные зубы, но раз-
лично устроенные конечности. У одних (сем. Arctocyonidae) radiale
срасталась с centrale — это вымершие формы. У других (сем. М i а с i d а е) ra-
dial» не срасталась с centrale.
Последние, изменяясь постепенно, дали потомство более поздних настоя-
щих хищников — Carnivora vera (3-я группа).
Итак, мы имеем три подотряда наземных хищных — Fissipedia:
.. „ . . (Acreodi,
1) Creodonta 2
(Pseudocreodi;
2) Palaeocarnivora — Eucreodi;
3) Carnivora vera.
Все они — наземные животные с нормально развитым скелетом. Но кроме
того’еще есть небольшая группа хищных, ушедших в воду и сильно изменив-
ших свой скелет; особенно сильно сократились конечности. Это — отряд
Pinnipedia — ластоногие, морские хищники.
Подотряд 1
ml ms «3
Creodonta Acreodi
Древнейшие формы из самого нижнего эоцена известны по зубам, при-
ближающимся к насекомоядным. Эти примитивные Creodonta, например Tri-
centes Соре — из Новой Мексики Сев. Америки
(рис. 228), имеют большие клыки и по три пред-
коренных и коренных. Рг1 отсутствует; рг2 в виде
маленького конуса; рг3 с внутренним бугорком; рг^
такой же, но более вытянут поперек. Верхние
коренные имеют по три бугорка и слабый воротник
на задней стороне. Нижние предкоренные — сжатые
конусы; коренные состоят из четырех маленьких
бугорков. Череп и кости скелета неизвестны.
Может быть, когда они будут найдены, положение этой формы изменится,
как это случалось с другими, отнесенными раньше к Creodonta.
Рис. 228. Tricentes sub-
trig о n u s Cope. (Самый
нижний эоцен.)
186
класс: млекопитающие — mammalia
Рис. 229. Mesonyx obtusidens Соре.
Рис. 230. А — Andrewsarctus mongo-
lie nsis Osb. В — Ursus. C — Mesonyx.
Имеет три пары верхних больших рез-
конус. РР—удлиненный конус. Рг3—
Сюда же относят еще несколько других родов, известных также по зу-
бам: Oxyclaenus, Chriacus, Delta therium и др.1).
У последнего хорошо развит
на рг4 внутренний бугорок. На пР
и т2 внешние бугорки наклонены
внутрь.
Mesonyx obtusidens
Соре (рис. 229) — средний эоцен
Сев. Америки (Bridger). Череп с
очень широкими скуловыми дуга-
ми и маленькой мозговой короб-
„ 3 1 4 — 2 (3) v
кой; — Клыки
О 1 тс -О
очень большие, загнутые. .В верх-
ней челюсти три первые предко-
ренные в виде сжатых конусов с
маленькими добавочными бугор-
ками. РР и два коренные тре-
угольные, трехбугорчатые. Ника-
кого намека на хищные зубы нет.
В нижней челюсти зубы в виде
сжатых конусов с добавочными бугорками. У некоторых видов исчезает т3.
Конечности передние с четырьмя пальцами, но centrale сохраняется-
задние — пятипалые, со слабо разви-
тым 1-м пальцем. Животное это дости-
гало размеров медведя (по Копу).
Передние конечности были короче
задних; стопоходящие. Коп пред-
полагает, что Mesonyx мог хо-
рошо плавать, судя по его плечу,
сходному с плечевой костью выдры.
Andrewsarctus mongoli-
е n s i s Osb. (рис. 230) — найден
в эоцене Монголии третьей амери-
канской экспедицией. Он поражает
громадными размерами. Череп вдвое
больше черепа Ursus gigas и
больше чем вдвое черепа Mesonyx.
цов, крупные клыки. РР — маленький
>) М. Wortman, Studies of eocene mammals in the Marsh’s collections,
1901—1902.
W. D. Matthew, The Carnivora and Insectivora of the Bridger eocene, 1900.
W. D. Matthew, Fossil Mammals of the Tertiary of N. E. Colorado, 1901.
J. B. Hatcher, Oligocene Canidae, 1902.
подкласс: плацентные — placentalia, коготные
187
трехбугорчатый с маленьким внутренним конусом. Рг4 — с более развитым
внутренним конусом. Коренные похожи на последний предкоренной. Очень
1онкие и широкие скуловые дуги и малая вместимость мозговой коробки харак-
терны для него. Осборн сближает его с Mesonyx.
Сюда же относятся Dissacus, Pachyaena и несколько других
родов.
Подотряд 2
Creodonta Pseudocreodi
Формы с несколько измененными зубами.
Hyaenodon — верхний эоцен и олигоцен Европы, Сев. Америки и Аф-
рики (Фаюм) — с сильно развитыми клыками;
2 1 4__2
7) г т---ч (Рис- 230- Два пеР-
& л. О
вне предкоренные сжаты по длине, как у Mesonyx, но значительно острее;
рг3 и рР снабжены внутренними бугорками. 1-й коренной меньше предкорен-
ных, т2 удлиненный, режущий. Эго ложно-хищный зуб. М3 не суще-
ствует. В нижней челюсти предко-
ренные сжатые, с острыми сред-
ними буграми; коренные удлинен-
ные, с добавочными острыми бу-
грами. Последний т3 состоит из
двух режущих гребней — ложно-
хищный зуб. Конечности со-
храняют пять пальцев, с отходя-
щим первым; centrale сохраняется.
По конечностям это более прими-
тивный тип, чем Mesonyx; по ко-
ренным зубам — ушедший дальше.
Этот род представлен очень
богато в фосфоритах Керси, во
Франции, причем размеры живот-
Рис. 231. Hyaenodon leptorhynchus
Laiser et Parieu. (Верхний эоцен.)
ных очень различны J). Американские формы более крупные. Вместе с Hyae-
nodon встречается в Европе, Африке и Сев. Америке другой представитель
Pseudocreodi, с зубами иной формы —
Pterodon (рис. 232) — с сильными клыками. Три предкоренные — ко-
нусы типа Hyaenodon; рг4 с внутренним бугорком, направленным вперед;
т1 и т2 — сильно развитые конусы с добавочными внутренними бугорками, на-
правленными внутрь и вперед, напоминают хищные зубы некоторых Carni-
vora; т3— маленький, поперечно удлиненный, тоже с внутренним бугром.
В нижней челюсти достигает наибольшей величины последний т3— ложно-
хищный; рг- нет. Конечности неизвестны.
*) Н. F i 1 h о 1, I es phosphorites du Quercy, Paris, 1877.
188
класс; млекопитающие — mammalia
Pterodon из Фаюма представлен хорошо сохранившейся нижней челю-
стью с тремя предкоренными и тремя коренными. Он отличается большими
размерами. На раздавленном черепе сохранились зубы от рг2 до т2 с характер-
ными признаками рода. Это Pterodon africanus Andr. (рис. 232).
Автор относит условно к этому роду несколько костей конечностей
(humerus, femur).
Anterodon — из Фаюма. Нижняя челюсть его длиннее челюсти Ptero-
don, со сходными зубами 4).
Из многочисленных С г е о d о n.t а можно указать еще на S i п о р а,
Oxyaena, Patriofelis и Limnocyon как на наиболее интересные формы.
Рис. 232. Pterodon.
Все они шли разными путями в выработке зубной системы и конечностей, но
не достигли, очевидно, прогрессивного развития и вымерли, не оставив
потомства.
Так, Limnocyon Сев. Америки как бы повторяет озубление Pterodon
Керси, с несколькими иначе направленными внутренними бугорками на рг4 и т1
(более к середине). Но его т2 имеет поперечно-удлиненную форму, более близ-
кую к т3 Pterodon. Последний отсутствует у Limnocyon.
Sinopa agilis Marsh стоит еще ближе по форме черепа, с узкой
сдавленной мозговой частью и вытянутой мордой, к Pterodon. Зубы его*
повторяют озубление последнего, но значительно меньших размеров. Рг* имеет
внутренний бугорок, как бы прототип хищного зуба. Коренные все тре-
угольные, удлиненные внутрь и наперед; т3 существует, сходный с другими
коренными, но меньше их.
Sinopa aethiopica — из Фаюма — представлена кусочком нижней
челюсти небольших размеров.
*) W. A nd ге ws, A descriptive catalogue of the tertiary vertebrata, London, 1906.
подкласс: плацентные-—placentalia, коготные
189
Конечности этих форм пятипалые, со сжатыми с боков коготными фа-
лангами. Существует 3-й трохантер на бедре. Нижняя часть плечевой кости
имеет расширенную сочленовную поверхность. Центральная кость сохраняется
у некоторых, у других исчезла.
Подотряд 3
Palacocarni-vora — Eucreodi
В эоцене Сев. Америки и олигоцене Европы. С хищным зубом.
Сем. Miacidae — включает многочисленные формы с полным озублением.
3 1 4 — 3
ЗТ 4 — 3’
и поперечно - удлиненными
верхними
коренными.
Конечности
пятипалые.
Vulpavus (ядед лисицы") Hargeri Wortm. (рис. 233) — по форме
черепа похож на лисицу, с нижней челюстью удлиненной, с закругленным
нижним краем, хорошо развитым поперечным со-
членовным отростком и маленьким нижним. Малень-
кие резцы, по 6 вверху и внизу, типа хищных, клыки
большие верхние несколько сжаты с боков. Три верх-
ние предкоренные— сжатые конусы, увеличивающиеся
последовательно. Хищный зуб рг^ состоит из двух
внешних конусов, очень острых, и небольшого вну-
треннего, расположенного перед передним внешним.
Sro уже типичный хищный зуб, резко отличаю-
Рис 233. Vulpavus Hargeri Wortm.
А — левая верхняя челюсть. В — левая нижняя
челюсть. X 44-
Рис. 234. Vulpavus Har-
geri Wortm. Правая задняя
нога. 2/3.
щийся от коренных, поперечно-удлиненных, треугольных, состоящих из двух
внешних бугорков, одного внутреннего и внешней стенки. Величина зубов
уменьшается кзади. В нижней челюсти четыре предкоренных, такие же сжатые
конусы, как и вверху. состоит из трех острых конусов с талоном сзади —
это нижний хищный; т2 такой же формы, но меньших размеров; ms еще
меньше, с едва заметными конусами. Это уже типичные зубы хищника.
190
класс: млекопитающие — mammalia
Многие части скелета этой формы были найдены. Конечности (рис. 234)
пятипалые хорошо представлены, с типичными для С a n i d а е бедром,
большой и малой берцовыми, надпяточной с удлиненной ножкой и малой
верхней вырезкой для сочленения с большой берцовой. Пяточная с плоской
лижней поверхностью лежит на cuboideum. Пять пальцев имеют удлиненные
метакарпалии и сжатые с боков когот-
Рис. 235. Oodectes herpestoides
Woi tm Правая верхняя челюсть. X 2.
ные фаланги. Для передней конечности
известны плечевая кость и лучевая —
первая с выдающимся дельтоидным
гребнем и небольшим эпикондилоид-
ным отверстием. Эта кость, а также и
лучевая имеют больше признаков,
встречающихся у виверр, чем у собак.
Итак мы видим, что это еще смешанный тип с признаками отчасти С a n i-
d а е, отчасти Viverridae, отчасти более древних родов (присутствие иг3).
Сем. Viverridae. Ofidectes herpestoides Wortm. (рис. 235,
235 A) — эоцен Бриджера (Сев. Америка); имеет полное озубление и конечности
с еще не слившейся центральной костью (рис. 236).
Рис. 236. Oodectes herpe-
stoides Wortm. X 2. Передняя
ступня.
Рис. 235A. Oodectes herpestoides Wortm.
Левая нижняя челюсть. X 2.
Зубы его похожи на V u 1 р a v и s, тоже полная система; но рг2 и рг3
грубые; ргл более удлинен, и его внутренний бугорок значительно больше и
продвинут дальше вперед. Коренные с двумя передними бугорками, более ото-
двинутыми от внешней стенки; на внутренней части зуба бугорка нет, она
загнута полукругло. В нижней челюсти предкоренные тоже гораздо грубее, чем
у Vulpavus, особенно рг±. с двумя конусами и талоном — хищный,
самый большой из коренных.
Плечевая кость с широкой нижней сочленовной поверхностью и эпикон-
дилоидным отверстием. Кисть сохраняет пять пальцев.
Таз, бедро и большая берцовая имеют некоторые признаки хищных сум-
чатых, например положение верхнего трохантера.
Сюда же относится Viverravus mimitus Matth. — из средгего
эоцена Бриджера (Сев. Америка). Очень маленький череп — 5 см. М1 и т2
с двумя внешними бугорками и внутренним талоном; т3 отсутствует.
подкласс: плацентные — placentalia, когогные
191
Рис. 237. Proviverra Cayluxi Filli.
Из европейских форм сюда относят Proviverra (рис. 237) — из олиго-
цена Керси (встречается и в Сев. Америке, Wasatch). Другие ученые ставят ее
ближе к Pseudocreodi. Ее
рг4 — треугольный, с внутренним
бугорком, лежащим почти посре-
дине, менее похож на хищный зуб
названных форм. /И1 и иг2 с 3
бугорками, поперечно вытянутые;
ms—поперечно-удлиненный, уз-
кий. По форме черепа и хорошо
развитому мозгу эта форма стоит
как бы у порога развития Car-
nivora. Слуховая капсула у нее
развита слабо, как и у всех пред-
ставителей этой группы.
Хотя ни у одной из описан-
ных форм нет определенных при-
знаков какого-нибудь из семейств
настоящих хищных, но несомнен-
но, что они были предками их.
Сюда же, к Eucreodi,
ученное
Сем. Arctocyonidae — из самого нижнего эоцена Франции (Реймс). Агс-
tocyon (рис. 238) — череп с очень сжатой мозговой коробкой, большими ску-
ловыми дугами и высоким сагиттальным гребнем,
очень напоминает череп медведя. Зубы
относят очень интересное, но мало из-
его со-
Рис. 238. Arctocyon primaevus
Blv.
Рис. 239. Arctocyon primaevus
Blv. (Самый нижний эоцен- Франции.)
А — верхние зубы, В — нижние.
вершенно не похожи ни на какие из рассмотренных нами. У него коренные
четырехугольные с бугорками, на верхних зубах по 6 больших и по нескольку
маленьких и по 4 бугорка на нижних, имеющих по большому талону с
бугорками (рис. 239). Хищных зубов нет, а существующие указывают на все-
192 класс: млекопитающие — mammalia
ядность животного и ближе всего стоят к зубам медведя. Кости скелета
неизвестны.
Другой род — Ana codon, известен только по зубам.
Может быть, дальнейшие находки укажут на это семейство как на древ-
нейших предков Ursidae.
Изучение более поздних форм, начиная с олигоцена, укажет нам на раз-
личное приспособление зубной системы хищных к различной пище. У них
специализация зубов выражается в упрощении коренных и усилении хищных
зубов, т. е. режущих, вместо бугорчатых — жующих. Конечности из пяти-
палых, стопоходящих или приспособленных к лазанию по деревьям, становятся
у некоторых четырехпалыми, пальцеходящими или хватающими.
Подотряд 4
Настоящие хищные—Carnivora vera (Fissipedia)
Мозг развит значительно больше, чем у Creodonta. Слуховые вздутия
окостенели. Хищные зубы существуют у всех, кроме медведей. Существуют
молочные: резцы, клыки и предкоренные, почему и можно отличить верхний
хищный—рг* — сменяющийся, а нижний хищный—т1 — несменяющийся. Молоч-
ные сложнее постоянных. В передней конечности срослись centrale, ulnare и
radiale. Надпяточная имеет значительное углубление вверху для большой берцо-
вой и выпуклость на нижнем конце для сочленения с naviculare. Плечевая кость
имеет две дыры на нижнем конце — для прохождения нервов и сосудов.
Сем. Canidae. — в олигоцене Европы и Сев. Америки мы встречаем пер-
вые формы, которые носят признаки этого семейства.
Cynodictis gregarius Сев. Америки известен почти полным скеле-
том. Череп его, слабо удлиненный, с широкими скуловыми дугами и широким
вместилищем мозга, снабженного извилинами, близок к черепу С a n i s. Из
трех резцов наибольший—крайний. Клыки большие, загнутые вниз. Рг1—ма-
ленький, рг2 и pH — острые конусы, сжатые по длине; рг* — длинный хищный
зуб с большим передним конусом и маленьким внутренним бугорком. /И1 со-
стоит из двух наружных, одного внутреннего бугорка, за которым лежит
небольшой кант, и двух маленьких промежуточных бугорков; т2—-меньше и
без промежуточных бугорков; wz3 не существует. Нижние предкоренные — сжатые
конусы, увеличиваются сзади. — хищный — из трех режущих конусов и за-
кругленного талона; т2 и т3—значительно меньше, и конусы тупее, чем у тА.
Это озубление очень близко к Vulpavus с той разницей, что корен-
ные более расширены внутри, и от3 исчез.
На передних и задних конечностях укоротились первые metacarpalia и
metatarsalia. Хвост длинный.
Из олигоцена Керси Фильоль описал более 17 видов Cynodictis
различных размеров — Cynodictis lacustris Filh. (рис. 240) и др.1).
*) Н. F i 1 h о 1, Les phosphorites du Quercy, Paris, 1877.
подкласс: плацентные — placentalia, коготные
193
Верхние челюсти их имели по 4 предкоренных, из которых рг1 очень
маленький, рг'1 и рг3 более крупные, удлиненные, рг* хищного типа. Два ко-
ренных бугорчатые.
В нижней челюсти только 3 предкоренных с хорошо развитым хищ-
ным тг
Передние и задние конечности с 5 пальцами.
В тех же фосфоритах Керси были найдены остатки рода С a n i s, ниж-
няя челюсть с типичным хищным зубом, но с более режущими конусами. Осталь-
ные зубы, как предкоренные, так и коренные, тоже
типа С а п i s. Предкоренные — с маленькими добавоч-
ными зубчиками; т2 и та— с тупыми конусами.
Дальнейшее изменение верхних коренных в
С а п i s заключалось в увеличении бугорков и
Г. Л;т
Рис. 240.
Cynodictis lacustris Gerv. (Керси.)
роде
изме-
нении формы зубов в более четырехугольную. Centrale, ulnare и radiale срос-
лись в одну кость, лежащую под radius (radiale).
Различные виды этого рода, как Canis domesticus, С. lupus и
С. vulpes, различаются детальными признаками в зубах и других частях
скелета. Таким образом это семейство является наименее измененным сравни-
тельно с древними формами. Это,
так сказать, первый эгап, на ко-
тором остановилось изменение зуб-
ной системы хищников.
Arnphicyon (рис. 241) —
из олигоцена и среднего миоцена
Франции и Индии (Пенджаб), яв-
ляется формой, очень близкой к
Canis, но он удержал т3, изменив форму коренных в более четы-
рехугольную, типа С а п i d а е. На нижнем рг4 есть два добавочных зад-
них конуса. Очевидно, эта форма ответвилась раньше Cynodictis
от Creodonta (Vulpavus) и, сохранив та, дошла до миоцена, где она
и вымерла.
Конечности у Arnphicyon пятипалые, стопоходящие. Плечо с боль-
шим эпикондилоидным отверстием. Metacarpalia короткие. Ступня напоминает
ме твежью. Бедро тонкое, с 3-м трохантером; большая берцовая длинная,
пяточная и надпяточная — типа Canis.
13
Павлова М. В. Палеозоология. Ч. П.
194
класс: млекопитающие — mammalia
Daphoenus felinus Scott — из олигоцена Сев. Америки. Предста-
влен целым скелетом. Череп удлиненный, близок к Canis. Сохранил иг3, хотя
и очень маленький. АР имеет три бугорка. Некоторые части скелета сходны
с Amphicyon.
Hemicyon sansaniensis Filh. — средний миоцен Сансана; ли-
шился иг3 !).
Ceiw. Mustelidae (куницы). — Это маленькие хищники, очень подвижные,
питающиеся почти все другими животными. К этому семейству принадлежат:
куница, ласка, барсук, хорек, росомаха, выдра; два последние значительно круп-
нее. Они распространены по всему земному шару, кроме Австралии. Ископаемые
остатки известны с олигоцена Европы и Сев. Америки, где они имеют уже при-
знаки, позволяющие отнести их к этому семейству. Череп удлиненный, с тупым
Рис. 242. Bunaelurus
lagophagus Cepe.
Рис. 243. Р 1 е s i с t i s
lemanensis Pomel.
(Франция, нижний миоцен.)
Рис. 244. Stenoplesictis
Cayluxi Filh.
сохраняются все. Верхний т1 состоит из двух или четырех бугорков, лежа-
щих один над другим; т2 у некоторых форм исчезает; т2 сохраняется.
Подсем. Putoriinae (хорьковые). — Bunaelurus Соре — из олигоцена
Колорадо (рис. 242) — сохраняет треугольный, с тремя бугорками иг1, а иногда
и очень маленький иг2. Он имеет озубление С a n i d а е, сокращая иг2 и умень-
шая число бугорков на иг1.
К нему близок Р1 е s i с t i s (рис. 243) из Allier (Франция).
Stenoplesictis Cayluxi Filh. — олигоцен Керси — лишился иг2;
иг1 состоит из двух бугорков, лежащих один за другим. Хищные зубы у
этих форм вытянутые с внутренним передним бугорком (рис. 244).
Подсем. Lutrinae (выдры). — М2 и ms не существуют, иг1 —большой,
четырехугольной формы, с несколькими бугорками.
Promeles Weith. — из нижнего миоцена Пикерми,имеет иг1 четырехуголь-
ный, с бугорками. У него исчез рг1. Это уже форма, близкая к М е 1 е s (барсуку).
*) Filhol, Les phosphorites du Quercy, pl. IX.
подкласс: плацентные — placentalia, коготные
195
Конечности у Lutrinae стопоходящие или пальцеходящие. Очевидно,
различные представители этого подсемейства получили начало в верхнем
эоцене, отделившись от одной из форм сем. Miacidae (Viverravus).
Potamotherium Geoff. — из нижнего миоцена Франции (St. Gerant le
Puy). Сохраняет еще маленький т2, но рг4 состоит у него из внешнего и
внутреннего бугорков; он не является типичным хищным зубом. Это укло-
нившаяся форма.
Сем. Viverridae (виверры) является третьим семейством, близким к
С a n i d а е. Это более крупные животные, чем Mustelidae, с закругленной
или удлиненной головой; существуют в небольшом
числе в Старом свете (фараонова мышь, циветта и др.).
Ископаемые остатки их известны с олигоцена, и по
Рис. 245. Herpestes lema-
il е n s i s Pomel.
зубам их можно счи-
тать такими же прими-
тивными, как С a n i-
dae, так как у них,
при полной зубной си-
стеме и типичном рР,
хищном, сохраняются
два верхние коренные
треугольные, трехбу-
горчатые, типа С у-
nodictis.
Viverra sim-
plicidens Schl. —
из Керси и Viver-
ra antiqua (Her-
Рнс. 246. Ictitherium
robustum Gaudry.
p e s t e s) - - из нижнего миоцена Алье (Франция) являются уже почти сфор-
мированными типами (рис. 245). Дальнейшее изменение состоит в умень-
шении т2 и в поперечном удлинении т1. Рг1 сохраняется. Роды эти существуют
и теперь (мангуста и виверра), но в то время как Viverridae проходят,
почти не изменяясь в главных признаках, с олигоцена до наших дней, в конце
миоцена от них отделились формы, которые, сохраняя оба верхних коренных
зуба, изменили их положение.
Это род Ictitherium, богатый видами и встречающийся на юге Европы,
в Греции, в Бессарабии и б. Херсонской губ.
Ictitherium Orbignyi и let. robustum gaudry (рис. 246)- еще
очень близки к Herpestes, но у них передние части коренных спустились
под рг4, образуя с ним угол; предкоренные увеличились.
Ictitherium hipparionum Gaudry (рис. 247,3) имеет большие клыки,
маленький рг1, значительно большие грубые рг2 и рг3 и громадный рг4 — хищ-
ный, равный по длине двум предыдущим; тг подошел под него, сравнительно
небольшой, с бугорками; т2— совсем маленький, лежит еще глубже. Конечно-
сти Ictitherium пятипалые.
13
196
класс: млекопитающие -— mammalia
Рис. 247. 1 — Ictitherium
Orbignyi. 2 — let. ro-
bust u m. 3 — let. hip-
pa r i о n u m. 4—H у a e n a
eximia.
Эти формы, исчезнув сами в плиоцене, ведут к новому семейству —
Сем. Hyaenidae. — Уже в том же мио-плиоцене Пикерми мы встречаем
форму, у которой рг4 имеет вид настоящего хищного зуба гиэны, но у ко-
торой за ним расположен т1 значительной величины; это — Hyaenictis
g / а е с a Gaudry. В нижней челюсти у него сохраняется маленький т2.
В тех же слоях находим остатки настоящей гиэны—
Hyaena eximia Gaudry (рис. 247,4) — с
очень сильными предкоренными и очень маленьким
wz1, лежащим под прямым углом с рг4.
В плиоцене Италии (долина Арно) гиэны до-
стигали больших размеров. Маленький тг сохраняется
в том же положении; сохраняется и рг1. Нижние
предкоренные становятся очень толстыми. Верхний
хищный зуб состоит из трех конусов наружных и
одного внутреннего.
У некоторых позднейших видов гиэн исчезают
рг1 и ргг
В послетретичных пещерах находили остатки
гиэн громадных размеров, превышающих нынешних:
Hyaena spelaea Goldf.
Итак, столь различные типы, как V i v е г г а и
Hyaena, оказываются имеющими общего предка и
связанными с ним постепенно изменявшимися фор-
мами, следы которых сохранили нам слои Пикерми в
Греции и соответствующие отложения в Бессарабии.
Сем. Felidae (кошки). — Иную редукцию в зубах мы видим
Felidae (кошек), являющихся между современными хищниками
кровожадными, совершенно отбросившими свои жевательные зубы и
нившими немногие режущие зубы.
Клыки их и предкоренные достигли
больших размеров. Морда большею
частью короткая, лоб закругленный,
что позволяет довольно легко опре-
делить черепа, относящиеся к этому
семейству.
Конечности передние пятипа-
лые, задние — четырехпалые; имеют
втяжные когти. Весь скелет пред-
ставляет соединение легкости и силы.
Остатки их известны тоже с
олигоцена Европы и Сев. Америки, Рис. 248. Machairodus.
где мы застаем уже редукцию пред-
коренных и коренных зубов одновременно, чего не было у предыдущих семейств.
Остатки кошачьих особенно многочисленны в Сев. Америке. Изучение их,
в сем.
самыми
сохра-
подкласс: плацентные — placentalia, коготные
197
начиная с древних отложений и проходя через последующие, позволило амери-
канским ученым установить двойную линию развития Felidae: одну, веду-
щую к нынешнему роду F е 1 i s, другую — к вымершим верхне-третичным и
послетретичным Machairodus (рис. 248).
У основания этих линий стоит род Proaelurus Filh. (Франция) —
с головой еще продолговатой. Он имеет все предкоренные зубы типа других
А В
Pseudaelurus Smilodon
Dinictis Haplophoneus
Рис. 249. Изменение озубления в сем. Felidae.
хищных, но в верхней челюсти у него сократился tri1, а в нижней — т&, суще-
ствующие у других современных ему хищников. Конечности длинные, тонкие, пяти-
палые, с отставленным первым пальцем; вероятно, животное лазало по деревьям.
Одна линия Felidae (рис. 249, А) начинается в Сев. Америке, в среднем оли-
гоцене, от D i n i с t i s, проходит через NimravusB верхнем олигоцене и Р s е и-
daelurus в среднем миоцене Европы, у которых постепенно сократились рг1,
увеличились рг3, рА в верхней челюсти, в нижней исчезли ргА и /п2, и
198
класс: млекопитающие — mammalia
форма головы более закруглилась. Эти формы привели к р. F е 1 i s, у которого
рг2 остался очень маленьким, рг3 и рг4 очень увеличились, т1 тоже остался малень-
ким, так что функционирующих осталось только по два зуба с каждой стороны.
Клыки увеличились и остались круглыми или слабо овальными, загнутыми вниз.
Рис. 249, А. Изменение озубления в сем. Felidae.
Параллельно с этой линией кошачьих шла другая, начинаясь также в олиго-
цене Сев. Америки от таких форм, как Haplophoneus — олигоцен Небраски
и Aelurictis— Керси, у которых сокращение зубов шло в том же порядке,
как и у выше названных параллельных им форм, но клыки увеличивались и
становились как бы сжатыми. В миоцене и плиоцене эти формы пол) чают
большое развитие; у них клыки, особенно верхние, достигают громадных раз-
меров, сжатие доходит до образования острых краев; это — кинжалы-зубы
(рис. 249, В).
подкласс: плацентные — placentalia, коготные
199
Род Machairodus, с исчезнувшим т2, в Европе и несколько соответ-
ствующих ему родов в Сев. Америке развивались прогрессивно во время всего
плиоцена. Из них S m i 1 о d о п Burm. достиг плейстоцена, где он и вы-
мер, несмотря на свое широкое распространение в Европе, Сев. и Южной
Америке. Очевидно, его клыки, полезные ему вначале, достигли таких размеров,
которые сделали их вредными. Его нижняя челюсть приобретает иное сочле-
нение, отбрасывающее ее назад, вследствие чрезмерного развития клыков. Весь
скелет Machairodus типичен для Felidae, с громадными чехлами над
коготными фалангамиJ).
Felis spelaea — вымерший пещерный лев, из настоящих кошачьих,
достигал больших размеров, чем ныне живущие Felidae.
Megalictis ferox Wortm. — из нижнего миоцена Южной Дакоты,
является интересной формой. Хотя автор относит eeKMustelidae, но как
по общей форме закругленной морды, так и
по редукции зубов он отклоняется от этого
семейства и приближается к Felidae. У
него исчез уже рг1, редуцирован рг2, рг3 —
небольшой, рг4 — со значительным внутрен-
ним бугром. Небольшой трехбугорча-
тый т1. Пятипалые конеч-
ности с довольно толстыми
metacarpalia и metatarsalia,
с расходящимися нижними
концами.
Этот краткий обзор
Carnivora указывает нам
определенно, что все эти
семейства развились из од-
Рис. 250. Ursus spelaeus. Череп и верхние
и нижние зубы.
ной или нескольких близ-
ких форм в верхнем эоце-
не и дали в позднейшем
развитии своем несколько стадий — этапов. На первой, с наименьшей редук-
цией зубов, остановилось сем. С а п i d а е, за ними пошло сем. Viverridae
с боковой ветвью, сильно сократившей коренные — сем. Hyaenidae. Сем.
Mustelidae занимает как бы промежуточную между ними стадию. И нако-
нец, сем. Felidae, достигшее наибольшего сокращения зубов, представляет
крайнюю стадию, причем из двух параллельно развертывавшихся линий одна
достигла до нас — род Felis, другая линия, с родом Machairodus, вы-
мерла в плейстоцене.
Многочисленные находки в Сев. Америке, вероятно, скоро пополнят недо-
стающие промежуточные формы.
*) W. D. Matthew, The phylogeny of the Felidae. Amer. Mus. of Natur. H.siory,
New York 1910, October.
200
класс: млекопитающие -— mammalia
Рис. 250, А. Ступни Ursus.
А — передняя, В — задняя.
Указанные здесь представители коготных составляют только незначитель-
ную часть описанных форм, найденных в Сев. Америке. Особенно богато пред-
ставлены там в прекрасно сохранившихся экземплярах Insectivora, Creo-
donta и Carnivora, во много раз превосходящие числом европейские формы.
Главные коллекции были собраны покойным профессором О. Маршем;
они обрабатываются несколькими палеонтологами еще и теперь.
Сем. Ursidae (медведи). — Это семейство стоит особняком от других
хищных как по общему тяжеловесному облику своему, так и по строению
зубов и конечностей — настоящих стопоходящих. Все коренные зубы медведя
коронки, снабженные низкими бугорками; режущих нет совер-
шенно. Из предкорениых сохранились очень
I J большие третий и четвертый (рис. 250,250, А).
I f Ископаемые остатки Ursidae извест-
ЙКГ; ны с верхнего плиоцена Европы. Ближайшей
формой к р. U г s u s является U г s a v u s, с
к тремя плоскими коренными, увеличивающи-
Л мися кзади; предкоренные — небольшие.
Г Ursus arvernensis — из плиоцена
Франции, с увеличенным третьим резцом и
большими клыками, имеет в верхней челюсти
три первые предкоренные очень маленькие, р1л — с двумя внешними бугорками
и внутренним воротником, два коренные с большими и маленькими бугорками;
яг3 исчез. В нижней челюсти яг3 сохранился.
Дальнейшее изменение состоит в исчезновении нижних и верхних пред-
коренных. При этом у Ursus spelaeus, послетретичного пещерного медведя,
внизу остается только рг4, у Ursus arctos — prs и рг4, а у нынешнего бу-
рого медведя существует и маленький рг2. Интересно подчеркнуть тот факт,
что до сих пор совершенно неизвестны остатки медведей в третичных отложе-
ниях Сев. Америки, несмотря на богатство этой страны остатками других хищных.
Из этого можно заключить, что медведи произошли из Евразии. Во время
ледникового периода они переселились из Европы в Сев. Америку (Ursus-
americanus, Ursus amplidens).
В плейстоцене Аляски, Калифорнии, Южной Каролины, Пенсильвании,
Мексики появляется род Arcthotherium Abel, который распространяется
и в Южной Америке. Остатки его были найдены на берегу Рио-Лаплаты,
в Пампасах.
Нижне-миоценовые и более древние предки медведей до сих пор точно
не известны. Раньше считали Amphicyon за такого предка, потом Н у а е п а г-
сtоs; но сравнительное изучение этих форм заставило видеть в Amphicyon
боковую ветвь С а n i d а е, сохранившую яг3, а в Hyaenarctos и других,
близких к нему формах членов сем. S u b и г s i Blainville. Из древних нижне-эоце-
новых Creodont а—A rctocyon имеет бугорчатые зубы типа всеядных, подхо-
дящие к зубам Ursus, как было упомянуто, и череп его очень близок к мед-
вежьему, со сжатой мозговой коробкой и широкими, тонкими скуловыми дугами.
подкласс: плацентные — placentalia, коготные
201
Ancodon ursidens Osb. — из верхнего или среднего эоцена Сев. Аме-
рики, известный по верхним и нижним коренным зубам, снабженным мелкими
бугорками, сближается Осборном с Arctocyon и Ursidae. Но остатки
этих форм очень неполны, и расстояние между ними и миоценовыми Ursidae
совершенно не выполнено. Остается ожидать новых находок, а пока вопрос
о генетическом отношении Ursidae к другим хищным остается открытым.
О т р я д 5
Тюленевые (ластоногие) — Pinnipedia
Хищники, ушедшие в воду с суши и изменившие соответственно этому
строение скелета.
Сюда относятся ныне живущие тюлени, моржи.
Теперь формы эти имеют очень простые, однообразные коренные зубы —
.небольшие конусы, и очень укороченные конечности, обращенные в ласты.
Укорочены, главным образом, кости плеча, предплечья, бедра и голени. Кисти,
как и ступни, сохраняют пять пальцев и все косточки карпальные и тар-
сальные.
Ископаемые остатки известны с миоцена; их встречают много в сармат-
ском известняке юга Бессарабии, и там мы застаем уже эти сократившиеся
в длине кости, легко позволяющие отличить их от соответствующих костей,
наземных хищников. Бедро и плечо являются почти четырехугольными костями,
с хорошо развитыми сочленовными головками для лопатки и таза. Лопатка
широкая, хорошо укрепляющая переднюю конечность (рис. 251, 2).
Недавно найдены остатки Phoca groenlandica на Мурмане в после-
ледниковых отложениях.
Остатки Phoca pontica из сармата Бессарабии описаны Траут-
шольдом (в 1850 г.) (рис. 251 (2) и Алексеевым (в 1920 г.).
Переселение тюленей в воду произошло, вероятно, в эоцене. Предков их
нужно искать между Creodonta.
Desmatophoca oregonensis Condon. — В миоценовых отложениях
берегов р. Орегона найден череп около 40 см длины, который как бы соеди-
няет признаки черепа Otaria и Phoca, но зубы его менее редуцированы
314—1
в своем строении. У него сохранились, хотя ине вполне, - -- —• От рр
есть альвеола, указывающая на один корень зуба. Рг2 сохранился на обеих
сторонах; он имеет два корня. Рг3 отсутствует, но в альвеоле заметно при-
сутствие двух корней. Рг4, соответствующий хищному зубу, имеет три корня
и два добавочных бугорка — внутренний и задний. Главный бугорок загнут
внутрь. Этот зуб представляет как бы переход от настоящего хищного зуба
наземных хищников (разрывающего мясо) к коническим зубам водных млеко-
питающих, более приспособленным к схватыванию скользкой рыбы. Томас Кон-
дон и Бортман (Wortman) видят в этой находке подтверждение взгляда
Рис. 251. 1 — Phoca groenlamlica Nilss. (Сев. ледов, океан.) 2— Phoca pon-
tic а Eichw: А, В — плечо; C, D — бедро. (Сармат.)
подкласс: плацентные — placentalia, коготные
203
о происхождении Pinnipedia от Creodonta и сближают зубы первых
с зубами Patriofelis1 *). Но у последнего еще не было настоящего
хищного зуба.
О т р я д 6
Китообразные — Cetacea
Громадные морские животные с сильно измененными передними конеч-
ностями и атрофированными задними, что сразу отличает их от другой группы мор-
ских млекопитающих — тюленей, морских хищных. Животные эти имеют боль-
шую удлиненную голову, короткую шею. Передние конечности очень укорочены,
грудной пояс сокращен; сохраняется лопатка. Задний пояс исчез; иногда суще-
ствуют незначительные остатки бедра. Существует хвостовой плавник, напра-
вляющий движения животного.
Ныне живущие китообразные разделяются на Odontoceti и Mys-
tacoc eti.
Odontoceti снабжены зубами, питаются рыбой. Зубы конические,
расположены в одной или обеих челюстях. Это — дельфины, кашалоты и т. п.
Mystacoceti — беззубые во взрослом состоянии; зубы только в эмбрио-
нальном состоянии. Верхняя челюсть и нёбо снабжены роговыми пластинками,
через которые вытекает вода, захваченная пастью, а мелкие животные остаются
в пасти и служат пищей чудовищу. Это — собственно киты.
В ископаемом состоянии известны более древние формы — Archaefl-
eet!. Из них одни — близкие к ныне живущим, другие — вымершие.
Подотряд 1
Древние киты — Archaeoceti
Под этим именем объединены древние китообразные, остатки которых
известны из эоцена Египта (Фаюм) и — одной формы — в Европе, Африке
и Южной Австралии (Z е u g 1 о d о п).
Protocetus atavus Fraas (рис. 252) — в среднем эоцене Фаюма. Жи-
вотное имело около 2 м длины и очень удлиненную, дельфинообразную морду.
Череп его, длиною 54 см, по строению сходен с черепом вымерших хищников
3 1 4 — 3
3 Т 4 — 3
Creodonta. Зубная
система
полная:
; коренные зубы — сжатые
с боков конусы, имели по три корня. Это первые шаги к редукции зубов
хищных наземных. Два первые предкоренные — простые конусы; рг3 и р/"1
имели добавочные задние зубчики, такие, как и у двух первых коренных,
спереди и сзади среднего конуса. Клыки имели два корня.
1) Th. Condon, A new fossil Pinniped from the miocene of the Oregon coast.
Bulletin Univers, of Oregon, vol. Ill, 1926, № 3.
204
класс: млекопитающие —- mammalia
Передние носовые отверстия лежали близко к концу морды, над клыками;
нижние отодвинуты далеко назад.
Кроме черепа известны позвонки, мало отличающиеся от позвонков на-
земных Creodonta.
Mesocetus (Eocetus) Fraas, найденный в тех же слоях Египта,
имеет несколько больший череп, до 1,1 м, того же строения. Поясничные
позвонки его более высокие.
Рис. 252. Protocetus atavus. (Египет, средний эоцен.)
Prozeuglodon atrox Andrew — из Фаюма, с молочными зубами,
значительно осложненными добавочными зубчиками по обеим сторонам глав-
Рис. 253. Z е u g 1 о-
d о n Isis Beadn.
(Длина 1 м.).
кого конуса 7И1 меньше, чем рг4. Средний эоцен.
Этот род по геологическому положению новее, чем
Eocetus; он занимает промежуточное место между
ним и выше лежащим Zeuglodon, который встре-
чается в ^Египте (Фаюм), в Европе (у нас и в Ан-
глии) и в Сев. Америке в верхней части среднего эо-
цена (рис. 253). У Zeuglodon Osiris And. первые
предкоренные — простые конусы, остальные имеют на
задней стороне добавочные конусы. На верхних коренных
конусы с обеих сторон, на нижних — только сзади.
У Zeugl. Isis Beadn. все предкоренные с добавочными
бугорками, как и коренные. Шейные позвонки очень уко-
роченные, но не сросшиеся; спинные — с высокими
отростками, плагицельные. Грудная кость состоит из
нескольких частей. Широкая лопатка срослась с корако-
идной, имеет небольшой
В передней конечности хорошо развитые, хотя и
локтевая и лучевая; кости кисти длинные.
Абель считает род Zeuglodon боковой
надсочленовный отросток.
укороченные: плечевая кость,
ветвью, вымершей в среднем
эоцене. Только Microzeuglodon,— форма, описанная из эоцена Кавказа
подкласс: плацентные — placentalia, коготные
205
Лейдекером по очень незначительным остаткам (кусочек нижней челюсти,
humerus и позвонок), поставлена им во главе дальнейшего развития китообразных.
Подотряд 2
Зубастые киты — Odontoceti
Формой, связующей Archaeoceti с Odontoceti (Squaloceti),
Абель ставит Agorophius pygmaeus (рис. 254) — из нижнего олигоцена
Южной Каролины. Здесь зубчики на
зубов меньше, чем у Zeuglodon, и
отсутствует сагиттальный гребень, сильно
развитый у последнего. Верхнечелюст-
ные кости покрывают своими концами
лобные. Межчелюстные доходят до но-
совых.
Squalodon (рис. 255)— из мио-
цена, представлен несколькими видами
с сильно удлиненными челюстями. Шей-
ные позвонки его еще более укорачи-
ваются и начинают срастаться; первый
шейный остается еще свободным. Число
зубов увеличивается, но они теряю г бо-
ковые зубчики, приближаются к кону-
сам дельфинов. У более поздних видов
зубы становятся простыми конусами и
легко выпадают.
Neosqualodon — миоцен Си-
передней и задней стороне коренных
Рис. 254. Agorophius pygmaeus
Abel. (*/6).
цилии, коренные зубы имеют большие острые зубчики на передней и зад-
ней стороне коронки. Десять последних коренных зубов имеют по два корня.
Рис. 255. Squalodon bariensis Jourd.
Тут наступило большое увеличение числа корней зубов сравнительно со
Squalodon, у которого только 7—8 зубов имеют по два корня. Челюсти
очень удлинены.
Prosqualodon Lyd.— из миоцена Аргентины. Зубы многочисленные,
зубчатые, некоторые коренные с тремя корнями. Передние конические. Абель
206
класс: млекопитающие — mammalia
считает эту форму боковой ветвью вследствие очень укороченной ее морды,
близкой к Agorophius.
Squalodon дал начало сем. Physeteridae (кашалотам) (рис. 256).
Переходной формой является Scaldicetus, найденный в миоцене и плиоцене
Европы, Сев. и Южной Америки (Scaldicetus patagonicus.) Зубы —
простые конусы, как резцы, так и коренные (22—24); они значительно проще,
чем у Squalodon; 2 корня слиты в один в каждом коренном зубе. Позже
исчезает эмаль на коронках, и зубы начинают редуцироваться сначала в меж-
челюстных костях (Prophyseter Dolloi из верхнего миоцена Антверпена),
Рис. 256. Physeter macrocephalus. (Длина животного —18 м.)
потом и в верхней челюсти. У ныне живущих кашалотов (Р h у s е t е г) только
на нижней челюсти остаются функционирующие зубы.
Первый шейный позвонок остается свободным, остальные сливаются до ше-
стого. Еижняя челюсть, длинная у Squalodon, постепенно укорачивается
у последующих форм. Череп не симметричный. Верхние челюсти имеют боль-
шое углубление в предносовой области, служащее для отложения жирового
вещества.
Сем. Acrodelphidae— является другою ветвью от Squalodon в
миоцене, с острыми зубами, ответвляющею от себя сем. Ziphiidae: Chone-
ziphius planirostris и Mioziphius belgicus Ab. — с двумя парами
больших передних зубов.
Последними представителями Acrodelphidae являются белуха, нарвал
и некоторые другие формы, неизвестные в ископаемом состоянии. В миоцене
Бельгии и Сев. Америки найдены остатки других форм из Acrodelphidae
(Ziphiodelphis Abel).
Сем. Eurynodelphidae — вторая ветвь от Acrodelphidae из верх-
него миоцена Бельгии и Сев. Америки: Eurynodelphis Cocheteuxi Abel.
Подотряд 3
Беззубые киты — Mystacocefi.
кЬепъ указывает на Patriocetus из верхнего миицена Лйнца (Ав-
стрия) как на промежуточную форму между Archaeoceti и Mystaccceti
подкласс: плацентные — placentalia, коготные
207
(рис. 257). Череп его, хорошо сохранившийся, указывает, что верхнечелюст-
ные кости подходят под лобные, как у китов. Зубы однородные, малень-
кие, со слабыми зазубринками на концах. Затылок сильно углублен. Морда
имеет треугольную форму.
Agriocetus austriacus является следующим представителем из самого
верхнего олигоцена Линца (Австрия) и связывает Patriocetus с Cetothe-
rium Rathkei— из миоцена
Бессарабии (по А б е л ю). Из-
вестна только задняя часть
черепа.
Для Mystacoceti яв-
ляется характерным присутствие
мелких зубов только в ранней
эмбриональной стадии, в чи-
сле 53. Функционирующих зу-
Рис. 257. Patriocetus Ehrlichi Abei.
бов у них нет.
Эти эмбриональные зубы являются остатками зубов Patriocetus, распав-
шихся на отдельные конусы: 7 зубов по 7 кснусов и 4 простых зуба = 53
зубам с каждой стороны (рис. 258).
Сем. Balaenopteridae. — Типичным представителем является Aulo-
cetus sammariensis Capell. (рис. 259), из
среднего миоцена Сан-Марино (Италия), с очень
изогнутыми наперед supraocciptale, напоми-
нающими затылок Zeuglodon, широкими
верхнечелюстными, узкими межчелюстными, уз-
кими и короткими носовыми костями. Темен-
Рис. 258. Схема подразделения зубов у Р а-
triocetus Abel.
Рис. 259. Aulocetus sammari-
е n s i s Capell.
ные сверху не видны. (Zittel, Grundziige, эту форму отнес KBalaeno-
р t е г i а а е).
Plesiocetus Cuvieri Desm. (рис. 260)—из плиоцена Италии (Monte
Pugnasco), известен почти полным скелетом —6,6 м. Грудная кость умень-
шилась; она состоит из одного куска. Кисть узкая. В нижней челюсти сохра-
нились ячейки от зубов.
Cetotherium Rathkei (рис. 261) — с очень крупным черепом и с груд-
ными плавниками. Позвонки спинные и хвостовые короткие. Ласты пятипалые.
Зубы исчезли.
208
класс: млекопитающие — mammalia
Остатки его встречаются часто в виде отдельных позвонков и слухового
аппарата, начиная с миоцена Европы и Сев. Америки.
Сем. Balaenidae имеет вместо зубов роговые пластинки. Шейные
позвонки сросшиеся. Сохраняется короткая пятипалая конечность. Ископаемые
остатки их известны начиная с плиоцена (В а 1 а е п а) (рис. 262).
Подотряд 4
Delphinoceti
Их отделяют от китов, но происхождение их не выяснено. Остатки
известны с миоцена,
но они очень разрозненны. Теперь дельфины выделяются
из Odontoceti. Они ха-
рактеризуются мало разви-
тыми коническими зубами
в верхней и нижней челю-
стях; в межчелюстной кости
их нет. Закругленный череп
с длинной мордой, очень
маленькими носовыми ко-
стями, очень удлиненными
межчелюстными.
Рис. 261. Cetotherium
R a t h k е i Brandt
Рис. 260 Plesiocetns
С u v i е г i Desm.
Представители китообразных дают постепен-
ное изменение форм, живших раньше на суше,
ушедших в воду еще в эоцене и постепенно
лишившихся задних конечностей и заменивших
передние ластами. Позже, 'в среднюю третичную
Рис. 262. Balaena mysticetus (гренландский кит).
подкласс: плацентные — placentalia, коготные
209
эпоху, стали изменяться и исчезать зубы —до замены их у некоторых групп
роговыми пластинками.
Некоторые ученые отделяют Archaeoceti от Cetacea и сближают
их с Pinnipedia — тюленями, считая обе группы происшедшими от древ-
них хищных — Creodonta1).
Отряд 7
Неполнозубые — Edentata
Название Edentata, беззубые, приходится заменить более правильным -—
неполнозубые, так как в этом классе только некоторые (муравьеды) лишены
зубов, другие животные имеют их в различном числе и различно расположен-
Рис. 263. 1 — ленивец. 2 — муравьед. 3 — армадилл. (Все три ныне живут в Южной Аме-
рике.) 4 — Mylodon. 5 — Hapalops. 6. Glyptodon. (Последние три—вымершие).
ными, но зубы лишены эмали. Весь отряд является теперь очень изолиро-
ванным как в смысле географического распространения, так и в смысле органи-
зации. Все живут в Южной Америке, кроме одной формы в южной Африке
(Orycteropus) и одной в Азии (Manis). Формы, живущие в Южной Аме-
рике, объединяются под именем Xenarthra, с простыми сочленениями зад-
них спинных и поясничных позвонков, а живущие в Старом свете—No-
marthra— имеют добавочные сочленовные поверхности на этих по-
звонках.
Эти ныне живущие неполнозубые как по образу жизни, так и по строе-
нию своему разделяются на три подотряда (рис. 263 (1—3) и 264 (1,3).
1. Bradypoda (Gravigrada) — ленивцы Южной Америки, покрытые
шерстью крупные животные, живущие в лесах, подвешиваясь на деревьях
своими трехпалыми конечностями и питаясь листьями и плодами. Характерен
1) Я остановилась несколько подробно на развитии китообразных, так как работы
Абеля дают много интересного об этом отряде хищников, изменившихся в водной
стихии и давших разнообразные группы.
14 Павлова М. В Палеозоология. 4. И. t
210
класс: млекопитающие — mammalia
их закругленный череп с очень укороченной лицевой частью. Резцы и клыки
почти всегда отсутствуют; коренные, от 3 до 4 с каждой стороны, цилиндриче-
ские. Характерна скуловая кость, спускающаяся на нижнюю челюсть, разделен-
ную сзади на три отростка: венечный, скуловой и нижний. Bradypus.
2, Vermilinguia — муравьеды, разделяются на: а) М у г m е с о р h a g а,
собственно муравьеды, покрытые шерстью, в южной Африке, и б) Manis —
ящеры, в южной Азии, покрытые роговыми чешуями J). Все живут в норах,
Передние ноги неполнозубых:
Рис. 264.
1 —Мyrmecophaga (нынеживущий).2—
М у I о d о п. (Плейстоцен.) 3 —-Bradypus
(ныне живущий). 4 — Hapalops (Миоцен.)
питаясь насекомыми и червями. Ко-
нечности у них пятипалые, с преобла-
данием среднего пальца. У Myrme-
с о р h a g а череп с очень вытян) тым
рылом, очень узкой нижней челюстью.
Зубов совсем нет; они существую,
только у одной формы — Orycte-
г о р u s и имеют совершенно особое
строение: они состоят из пентагональ-
ных призм из дентина, имеющих каж-
дая внутренний канал; общей пульпы
нет, зуб окружен тоненьким слоем
цемента. Живет Orycteropus в
южной Африке, Manis—-в Азии.
3. Dasypoda — броненосцы.
Животные, покрытые панцырем, со-
стоящим из костных пластинок, рас-
положенных рядами. Передние ряды
соединены подвижно, что позволяет
животному втягиваться под панцырь.
Передняя часть головы и хвост тоже
имеют покровы из пластинок. Череп
удлиненный, с узкой нижней челюстью. Между ними есть крупные формы — до
1,1 м и маленькие. Конечности короткие, с 4—5 пальцами и громадным когтем
на среднем пальце передних конечностей. Резцы имеет только Dasypus. Ко-
ренные — цилиндрические, бывают в различном числе. Питаются они главным об-
разом насекомыми. Живут в норах, выходят больше ночью. Южная Америка.
Ископаемые формы представляют большой интерес, указывая, с одной сто-
роны, на существование форм, близких к древним грызунам и исчезнувших со-
вершенно, а с другой стороны, указывая на совершенно иное распространение
неполнозубых в третичные времена сравнительно с нынешним. Ныне существу-
ющие неполнозубые имеют ископаемых представителей, начиная с мио-
цена. Кроме того существует эоценовая группа северо-американских форм,
которую можно считать предшественницей неполнозубых.
*) Некоторые применяют название „ящер” к рептилиям, что является большой
неправильностью.
подкласс: плацентные—placentalia, коготные
211
Подотряд 1
Ganodontia (Styhnodonta Wortm)
Рис. 265. Psittacotherium multi-
fragum Соре. (Нижний эоцен.) (*Д).
Известен начиная от нижнего эоцена Пуэрко до верхнего эоцена Бриджера;
он заключает формы, имеющие еще передние зубы: резцы и клыки.
Conoryctes comma Соре — самый нижний эоцен (Torrejon bed).
Удлиненный череп с сагиттальным гребнем. Нижняя челюсть высокая сзади.
Существуют зубы 2 1 4 — 3 вверху и внизу. Верхние коренные с тремя
бугорками; тг — очень маленький. Нижние коренные с пятью бугорками. Все
зубы с длинными корнями и низкими
коронками (Brachyodonta).
Hemiganus otari ides Co-
pe — из нижнего Пуэрко, имел уко-
роченную морду. В верхней челюсти
две пары маленьких резцов и пара боль-
ших клыков. Число верхних коренных
неизвестно, так как челюсть сломана
и передняя ее половина не сохранилась.
В нижней большие клыки, четыре пред-
коренных и три коренных, пара малень-
ких резцов. Нижняя челюсть круто
загнута вверх и имеет высокую расширенную заднюю часть, закругленную
вверху, характерную для Edentata. Сочленовный отросток тупой. Остатки
костей конечностей напоминают некоторых неполнозубых (М у 1 о d о п), также
и палец с характерной коготной фалангой.
Psittacotherium (рис. 265) — из верхнего Пуэрко,отделен от Hemi-
ganus толщей в 156 лг; по форме черепа и нижней челюсти близок к нему.
Но резцов у него одна пара вверху, внизу их нет, клыки меньше, корен-
ных по шести, они снабжены корнями. Верхние очень стерты — строение их
не видно. Нижние предкоренные имеют по два бугорка: наружный и внутрен-
ний; коренные — по четыре. Короткие массивные кости предплечья и кисти,
с громадными тремя последними фалангами — типа М у 1 о d о п. Также массив-
ный таз с сильно развитыми подвздошными костями. Этот род является
в ряду неполнозубых измененным потомком Hemiganus, по мнению
Бортмана.
Calamodon — из Вазача, нижний эоцен, известен только по нижней
челюсти, тоже массивной, широкой вверху и сзади и укороченной спереди,
имеет маленький резец с постоянным ростом, 4 предкоренных и 3 коренных с
корнями. Коронка имеет по 4 главных бугорка и одному добавочному. Поло-
жение этой формы не всеми признается одинаково. Некоторые относят ее к
Т i 11 о d о n t i a Marsh, древним вымершим i рызунам.
Stylinodon — эоцен Бриджера, с коротким, массивным черепом, без сагит-
тального гребня, с короткой широкой челюстью типа неполнозубых, сохраняет
14*
212
класс: млекопитающие — mammalia
еще клыкй; коренные цилиндрические, с постоянным ростом. Хорошо сохра-
нились: шейные позвонки, первый спинной, ребро, грудная кость, передняя
конечность с лопаткой, коротким плечом, предплечьем и кистью. Все эти
остатки позволяют отнести Stylinodon к предшественникам Edentata,
связанным с ними сходными признаками.
Подотряд 2
Ленивцы — Bradypoda (Xenarthra)
Существующие ныне только в лесах Южной Америки Bradypoda
представлены ископаемыми остатками в миоцене и плейстоцене Северной.
Средней и Южной Америки.
Рис. 266. Hapalops. Миоценовый предок ленивцев.
Сем.
Gravigradidae.— Hapalops Scott (рис.
266) — из миоцена Па-
тагонии, небольшое животное, около 1 м, сохранилось
в виде цельного скелета.
Рис<267. Megalonyx Jeffersoni Leidy.
По своим признакам он как бы со-
единяет в себе предка плейстоцено-
вых ленивцев и муравьедов.
Голова его слабо удлинена, но
не закруглена, как у настоящих ле-
нивцев, и не удлинена, как у му-
равьеда. Зубы его приспособлены к
перетиранию пищи, как у броненос-
цев; сохранились клыки. Конечности
снабжены большими когтями; согну-
тые передние конечности упирались
внешней стороной. Задние — стопохо-
дящие, как у муравьеда. Все пяти-
палые. Хвост мощный, с большими нижними отростками позвонков. Харак-
терна еще нижняя челюсть с тремя задними отростками. Это животное отча-
сти жило на деревьях, отчасти на земле, на что указывают его конечности.
подкласс: плацентные— placentalia, коготные
213
Hyperlestes garsonyanus Amegh. — из верхнего миоцена Санта-
Круца (Патагония). Тоже небольшое животное; череп равен 18 см, очень узкий,
'с заостренной верхней челюстью, клыками и четырьмя призматическими корен-
ными с каждой стороны. Эти оба рода имеют на бедре 3-й отросток —
трохантер.
Megalonyx Jeffersoni Leidy — из плиоцена Техаса и плейстоцена
Кентукки (Сев. Америка), сохраняет клыки (рис. 267). Морда его очень укоро-
Рис. 268. Megatherium am ericanum Blumb.
чена, череп тяжеловесный, с характерной нижней челюстью, с высоко под-
нятым венечным отростком и двумя другими менее высокими. Он составляет
переход к настоящим Gravigrada, гигантским травоядным ленивцам из
плиоцена и плейстоцена Америки, лишенным передних зубов.
Рис. 269. Megatherium americanum Blumb. Нижняя челюсть.
Сем. Megatheriinae.— Megatherium americanum Blumb. (рис. 268
и 269) найден в 1789 г. в плейстоцене Аргентины — почти полный скелет.
Укороченный череп его лишен передних зубов. Цилиндрические коренные, с
постоянным ростом, состоят как бы из двух пластин, сложенных вместе по
длине. На жевательной поверхности видны два продольных гребня с углубле-
нием между ними. Всего по четыре зуба с каждой стороны. Весь скелет очень
тяжеловесный.
Особенного внимания заслуживают ноги, снабженные громадными ког-
тями на средних пальцах, и толстый укороченный хвост. Скуловые кости
несут характерный, спускающийся на нижнюю челюсть отросток. Нижйий
край нижней челюсти сильно выгнут.
214
класс: млекопитающие — mammalia
Mylodon robustus Ow. (рис. 270) — из плейстоцена Аргентины,
является ближайшей к Megatherium формой, с черепом более удлиненным,
с тупой мордой и расширенной спереди верхней челюстью. В верхней челюсти
по пяти зубов с каждой стороны, в нижней — по четыре. Два передние за-
кругленные, задние полукруглые. Межчелюстные кости слабо развиты. Трех-
Рис. 270. Mylodon robustus Ow.
лопастный скуловой отросток направлен назад. Ступни крайне тяжеловесные,
с преобладанием третьего пальца с громадным когтем.
Пяточная и надпяточная кости массивны, последняя треугольной формы.
Они совершенно не похожи на соответствующие кости других коготных мле-
копитающих.
Paramylodon Brown (рис. 271) — из плейстоцена Небраски (Сев.
Америка), имеет более удлиненный череп, с пятью коренными зубами типа
Mylodon.
Scelidotherium Ow. — из плейстоцена Южной Америки, имеет более
удлиненный череп с удлиненной мордой. Пальцы редуцированы; спереди че-
тыре, сзади три.
Эти гиганты, достигнув, повидимому, большой специализации в зубах и
громадных размеров, закончили в плейстоцене свое существование в Южной
Америке, получив свое начало в Сев. Америке, из которой они переселились
в Пампасы, где развились в многообразные типы, из которых некоторые
попали опять в Сев. Америку при соединении ее с Южной в послетретичное
время, в плейстоцене.
подкласс: плацентные —- placentalia, коготные
215
Рис. 271. Paramylodon.
Подотряд 3
Муравьеды — Vermilinguia (Nomarthra)
Сем. Orycteropidae. — Беззубые, покрытые шерстью. Остатки их очень
немногочисленны, но заслуживают особенного внимания, так как встречаются
в Европе, начиная с олигоцена Франции (Керси), в меотических отложениях
Самоса, Пикерми и в б. Херсонской губ. (дер. Гребенники).
Orycteropus Gaudryi Fors. Maj. (рис. 272).—Был описан череп, най-
денный на о-ве Самосе в слоях Пикерми, мало отличающийся от современного.
Это было молодое животное, у которого существовали шесть коренных зубов и
альвеола 7-го. У взрослого исчезают 2-й, а иногда и 3-й предкоренные
(Andrews).
В тех же слоях Мараги (Персия) была указана находка зуба этого же
вида. В 1914 г. была найдена часть челюсти с тремя зубами этого же вида
в Херсонской губ., в дер. Гребенниках, в слоях Пикерми, которая установила
216
класс: млекопитающие — mammalia
присутствие этого зубатого муравьеда в средине третичного периода. Хотя
эта находка очень ничтожная, но строение зубов настолько характерно, что
позволяет определенно отнести ее к Orycteropus Gaudryi Fors. Maj.
По внешнему виду зубы похожи на зубы Megatherium, но они не сло-
жены из двух пластинок, а цельные, со срезанными сторонами покато
от середины. Особенностью является их строение, без общей пульпы. Жева-
Рис. 272. Orycteropus capensis Gm.
тельная поверхность зуба со-
стоит из маленьких призм
дентина с центральным кана-
лом каждая. Дальше вглубь
некоторые призмы и каналы
сливаются, стенки становятся
тоньше, каналы шире. Весь
зуб покрыт тонким слоем
цемента. Длина зуба 12 мм.
ширина 6 мм. Значение этой находки (из Гребенников) очень велико, так как
она расширяет площадь распространения этой южно-африканской формы в тре-
тичное время на европейском континенте.
Сем. Manidae— ящеры, покрытые чушуей. Остатки известны, начиная
с верхнего эоцена и олигоцена Франции (Керси): Necromanis, по указанию
Фильоля; из миоцена Баварии (Schlosser): Necrodasypus, Teuto
m a n i s и др. Род Manis известен из нижнего плиоцена Восточной Индии..
Все остатки очень незначительны.
Подотряд 4
Броненосцы—Dasypoda (Xenarthra)
Ныне существующие в Южной Америке Dasypoda (броненосцы) имеют
ископаемых представителей в миоцене и плейстоцене. Но есть и вымершие
формы, объединенные в особую
группу— Glyptodonta. Они ха-
рактеризуются присутствием верх-
него панцыря, похожего на пан-
цырь черепахи.
A. Dasypoda
Stegotherium (рис. 273)—
из миоцена Патагонии, представлен
Рис. 273. Stegotherium tesselatum
Amegh.
хорошо сохранившимся черепом, длиною в 13,5 см, с очень вытянутой мордой, без
скуловых отростков, с мелкими призматическими зубами: восемь в верхней че-
люсти, девять в нижней, с каждой стороны. В тех же отложениях найдены
менее значительные остатки рода Chlamydophorus, живущего ныне в
Южной Америке и имеющего подвижный панцырь из пластинок, располо-
женных рядами.
подкласс: плацентные—placentalia, коготные
217
Dasypus retusus Lyd. — из плейстоцена Аргентины, представлен
хорошо сохранившимся черепом с восемью зубами с каждой стороны.
В. Glyptodonta
Остатки этой вымершей группы броненосцев встречаются в третичных и,,
главным образом, послетретичных отложениях Америки вместе с остатками
Bradypoda (ленивцев).
Особенностью их является панцырь в виде черепашьего покрова, состоя-
щий из плотно сросшихся костных пластинок, большею частью гексагональных.
Он покрывал все животное кроме головы и хвоста. На лбу существовала крышка
из отдельных пластинок. Хвост был покрыт чехлом или
отдельными кольцами. Закругленный череп напоминает
череп Megatherium. Скуловой отросток направлен
вниз. Очень широкая лопатка; ребра сдвинуты наперед.
Рис. 274. Glyptodon ret с u at us
A, В — челюсти-
Особенностью является срастание отростков спинных позвонков между собою и
срастание позвонков: спинных, крестцовых и поясничных. Таз очень широкий;
верхняя часть хвоста с сильными отростками. Очевидно, все изменения скелета
стоят в тесной связи с образованием панцыря, со срастанием пластинок в один
неподвижный покров, требующий поддержки. Ноги пятипалые, с копытообраз-
ными последними фалангами. Зубы сложные и многочисленные.
Сем. Glyptodontidae.—-Со chi ops Amegh. — из миоцена Аргентины,
маленькое животное с мордой, не сильно укороченной, с нижней челюстью
невысокой и с простыми сравнительно зубами. Пластинки панцыря расположены
рядами и не вполне срослись.
Glyptodon reticulatus Ow. (рис. 274) — в плейстоцене Аргентины
громадные животные, около 3,2 м, более всего напоминающие черепаху.
Зубы имеют особое строение. Их много — от 7 до 8 с каждой стороны.
Каждый из них состоит из трех частей, как бы соединенных между
собою пластинок дентина, внутри которых проходит полоска вазодентина,
посылающая отростки в каждую пластинку. Зубы покрыты цементом. Рост
постоянный (рис. 274—А, В).
Рапосhthus tuberculatus Ow. (рис. 275) — ближайшая форма из
тех же слоев Аргентины и Бразилии.
218
класс: млекопитающие — mammalia
Glyptotherium t exanum Osb. — из нижнего плейстоцена Техаса
Сев. Америки, представлен большой броней из гексагональных пластинок и колец,
обхватывавших хвост. Они более крупные в центральной части панцыря и
более мелкие по краям, чем эта форма и отличается от Glyptodon Brachy-
ostracon из плейстоцена Мексики, найденного тоже в виде громадного пан-
цыря с очень резкими крае-
выми пластинками. Кре-
стцово - поясничная об-
ласть сильно сросшаяся,
очень массивная.
Doedicurus clavi-
caudatus —из плейсто-
цена Аргентины, достигал
размера до 4 м (Glyp-
todon — до 3 м). Его
Рис. 275. Panochthus tuberculatus Ow. панцырь был лишен скуль-
птуры. Сзади он оканчи-
вался кольцами, охватывающими хвост сверху. На конце его было несколько
шиповидных отростков. Плечо имело энтэпикондилоидное отверстие. Передние
конечности трехпалые, задние — пятипалые.
Eucinepeltus complicatus Barnun Brown (рис. 276) — является сильно
отклонившимся типом, из слоев Санта-Круца
Рис. 276. Eucinepeltus complicatus Barnun. Brown.
в первом две, затем три, четыре и две сзади. В центре каждой пластинки
небольшое углубление. Зубы, в числе восьми с каждой стороны, были типа
Glyptodon.
Peltephilus ferox — из миоцена Патагонии, имел два рогообразных
выроста на носовых костях. А м е г и н о указывает на присутствие у него ква-
дратной кости.
подкласс: плацентные — placentalia, ноготные
219
Эти постплиоценовые неполнозубые обеих Америк вымерли, достигнув
резкой специализации в выработке неподвижного тяжеловесного панцыря у
Glyptodonta и громадных размеров у Gravigrada. Менее специализиро-
ванные формы ленивцев и броненосцев продолжают свое существование, хотя
и в очень ограниченной области жарких стран.
2-я ГРУППА
НОГОТНЫЕ
Отряд 8
Primates Linne
К этому отряду относятся лемуры, или полуобезьяны, и обезьяны. Как бы
завершителем отряда является человек — Ното (подотряд В i m а п а).
Первый подотряд стоит близко к коготным по строению скелета и по
образу жизни; но у большинства форм на последних фалангах пальцев суще-
ствуют ногти, а не когти, и конечности их обладают хватательной способ-
ностью.
Лемуры и обезьяны живут на деревьях, питаются плодами, некоторые
из них всеядные; это — небольшие или крупные животные с круглой головой,
большими ушами и глазами, бесхвостые или с длинным хвостом; конечности
пятипалые с противопоставляющимся первым пальцем рук и ног, приспособлен-
ных к лазанию и хватанию.
Primates подразделяются на
1) Prosimiae (L е m и г о i d е а) (лемуры или полуобезьяны),
2) S i m i а е (обезьяны) и
3) В i m а п а (человек).
Подотряд 1
Полуобезьяны (лемуры) — Prosimiae
Тело и лицо покрыты шерстью; уши торчащие. На одних пальцах
когти, на других ногти. Скелет и зубы у лемуров более примитивного строе-
ния, чем у обезьян. Орбита хотя и замкнута, но не вполне изолирована;
она открывается в височную впадину. Слезный канал выходит вне орбиты.
Кости конечностей легкие, пустые, что облегчает лазанье по деревьям. Озуб-
ление полное. Мозг развит хорошо и снабжен извилинами, но они мало-
численнее, чем у обезьян. Ископаемые остатки лемуров известны с эоцена
Эгеркингена, в Швейцарии. По строению своему эти древние формы сильно
отличаются от лемуров, ныне живущих на Мадагаскаре, а отчасти в Африке и
южной Азии. Они стоят ближе к насекомоядным. Зубы их имеют по три
бугорка с небольшим добавочным четвертым. Пятипалые конечности сохраняют
центральную кость.
Но остатки эти очень несовершенны и редко состоят из цельных чере-
пов; цельные скелеты неизвестны.
220
класс: млекопитающие — mammalia
Многие северо-американские формы, отнесенные первоначально к этому
подотряду, были перенесены тзже к насекомоядным и другим семействам
(Notharctus, Pelycodus). Наилучше представленными из LemuroidCa
являются в ископаемом состоянии сем. Adapidae, Anaptomorphidae и
Archaeolemuridae, между которыми находятся, вероятно, предки S i m i а е.
Сем. Adapidae. — С сократившимися до двух пар резцами.
Древнейшими формами являются Protoadapis Lem., Plesiadapis
г е m е n s i s Lem., Caenopithecus, Chiromyoides c amp an i c us Steh.
из нижнего эоцена Франции. Последний известен только по нижней челюсти.
2 0 3______3
Первые два имели — — *—------—. В Сев. Америке есть близкие к ним
1 U Л - о
формы.
Caenopithecus Rut. — из нижнего эоцена Эгеркингена, имел ко-
роткий череп. Верхние коренные зубы представляют типичные трехбугор-
чатые зубы. Два бугорка лежат на
внешней стороне, образуя внешнюю
Рис. 277. Adapis parisiensis.
А — верхние, В — нижние зубы.
стенку, третий лежит внутри, бли ке кпереди. За ним появляется маленький
четвертый бугорок. Предкоренные однобугорчатые; только на рг* существует и
внутренний бугорок, клыки массивные. Имеется по одному резцу.
Некоторые палеонтологи относят этот род к р. Adapis, который хо-
рошо представлен в олигоцене черепами и костями скелета, а в эоцене —
отдельными зубами. Adapis Duvernoyi — наиболее близкий к Caeno-
pithecus, но на рг* отделяется второй внешний бугорок и намечается второй
внутренний. Эгеркинген.
Adapis parisiensis — из парижских гипсов и фосфоритов Керси,
наилучше изучен (рис. 277 — А, В). Череп его имеет выступающий сагиттальный
гребень, затылок закруглен; орбиты небольшие, разделенные широким пе-
реносьем; они слабо отграничены от височной впадины. Зубная система
2 1 4 — 3
сплошная: —-—— ---— .
2 1 4—3
Клыки немного больше резцов.
Верхние коренные
зубы с хорошо развитыми тремя бугорками и маленьким четвертым — задне-
внутренним (гипокон). Рг* имеет три бугорка; остальные предкоренные — сжатые
конусы, похожие на клыки и резцы. Нижняя челюсть с высоким височным
подкласс: плацентные — placentalia, ноготные
221
отростком. Коренные и последний предкоренной имеют по четыре маленьких бу-
горка, соединенных складочками эмали; остальные предкоренные конусообразные;
клыки больше верхних; резцы — маленькие лопаточки. Это — типичные зубы
для лемуров. Они, очевидно, очень близки к трехбугорчатым зубам насекомо-
ядных и отчетливо показывают осложнение зубов до шести бугорков раньше,
чем стать четырехбугорчатыми, как это предполагалось, да и теперь прини-
мается для некоторых зубов, относимых к лемурам — Ну op sod us, Pely-
с о d u s и др. из нижнего эоцена Сев. Америки.
Ad ар is magnus—тоже из Керси, но больших размеров, имеет зубы
несколько более сложные. Рг4 почти одинаков с коренными; рг3 и рг2 с двумя
внешними, хорошо развитыми бугорками и одним внутренним.
Из костей конечностей известны бедро и плечевая, почти одинаковой
длины; на бедре существует 3-й от-
росток — трохантер.
Астрагал с короткой закруг-
ленной ножкой и косой верхней
сочленовной поверхностью для Рис. 27й. Necrolemur.
большой берцовой.
A d а р i s с трехбугорчатыми зубами стоит у основания лемуров.
Megalad’apis— гигантская форма вымерших лемуров с о-ва Мадагаскара,
верхний плиоцен. Череп его достигал 25 см. Верхние коренные зубы трехбугорча-
тые, более простые, чем у большей части лемуров. Он является боковой ветвьюч).
Сем. Anaptomorphidae.— У них отсутствует сагиттальный гребень.
Necrolemur — из Эгеркин-
гена и Керси (рис. 278) — значи-
тельно меньших размеров, чем A d а-
а
Рис. 279. Anaptomorphus.
а — верхние, b — нижние зубы.
pis. Он имел удлиненный череп с большими глазницами, направленными вперед
2 1 з___з
и соединяющимися с височными впадинами. Зубы сокращены: — — ----г .
1 1 ) о — о
Коренные четырехбугорчатые. Малая берцовая кость срослась с нижним кон-
цом большой берцовой. Пяточная и надпяточная высокие, как у Т а г s i u s,
ныне живущей формы, сохранившей примитивные признаки.
Anaptomorphus homunculus Соре (рис. 279 — а, Ь) — из нижнего
эоцена Вазача (Сев. Америка), заслуживает особого внимания, изучен и описан
*) Forsyth Major, Megaladapis madagascariensis, 1894.
222
класс: млекопитающие — mammalia
одному внутреннему; нижние — четырехбугорчатые, удлиненные. Хотя
признаки — положение орбиты, слезное отверстие и другие — указы-
принадлежность этой древней формы к лемурообразным животным,
к Т а г s i u s, но другие признаки сближают ее уже с обезьянами:
Anaptomorphidae и к Pseudoloris. Ископаемые
Копом. Череп низкий, укороченный спереди, с закругленным затылком, с очень
большими глазницами. Зубы сокращены; число резцов неизвестно, клыки ма-
3__3
ленькие. Коренные -----; верхние поперечно-удлиненные, бугорчатые; по два
--
наружных бугорка на коренных и по одному бугорку на предкоренных, на
всех по
главные
вают на
близким
черепная часть развита значительно больше, чем лицевая, которая укорочена.
Глазницы более изолированы от височной впадины. Скелет неизвестен.
Pseudoloris parvulus Filh. — из фосфоритов Керси. Коренные
похожи на зубы Т а г s i u s.
Копи Шлоссер считают сем. Anaptomorphidae исходной груп-
пой для всех Anthropoidea, кроме S i m i i d a e.
Сем. Tarsiidae. — Ныне живущие являются очень примитивной группой
2 13____________________з
лемуров. Т а г s i u s имеет- — ---; верхние коренные трехбугорчатые, ниж-
113 — 3
ние пятибугорчатые. Зубы сходны с зубами лемуров из олигоцена Египта. По
строению плаценты Tar si us сближается с настоящими обезьянами. По зу-
бам он ближе к
остатки неизвестны. Живет в Восточной Индии. Выше олигоцена остатки ле-
муров не встречаются ни в Европе, ни в Сев. Америке. Очевидно, они пе-
реселились в конце олигоцена на Мадагаскар, Зондские о-ва и в Восточную
Индию, где живут изолированно и теперь. Из после третичных отложений Ма-
дагаскара, кроме Megaladapis, известны остатки лемура Р а 1 а е о р г о -
2 12___3
pithecus с зубной формулой: - - — ----• Другая форма из тех же отло-
Л U Л — о
жений — из вымершего сем. Archaeolemuridae, близка по строению зубов
к Cynopithecidae. Это А г с h а е о 1 е m u г Filh., с очень закругленным че-
репом, без сагиттального гребня, с большой
2 13 — 3
с глазами, направленными вперед-—----- .
2 0 о — о
коробки,
вместимостью мозговой
Клыки выдаются.
Подотряд 2.
Обезьяны — Simiae
Имеют лицо, не покрытое волосами, с менее выступающею переднею
частью, более закругленным затылком. Слезный канал открывается в глазницу,
совершенно изолированную от височной впадины. Конечности передние и зад-
2 12________________________________________________________________3
ние одинакового типа, с ногтями. Зубы в сокращенном числе: — — -----~ ;
Z 1 z — о
коренные четырехбугорчатые; предкоренные двухбугорчатые.
подкласс: плацентные — placentalia, ноготные
223
Обезьяны подразделяются на
1) плосконосых, хвостатых обезьян Нового света, имеющих мало
развитой или слабо противопоставляющийся большой палец;
2) узконосных обезьян Старого света; из них немногие хвостатые — сем.
Cynopithecidae, остальные бесхвостые — Anthropoidea, — человеко-
подобные.
Ископаемые остатки плосконосых известны только из пещер Бра-
зилии. Из узконосых хвостатые известны в Европе, начиная со сред-
него миоцена.
В Африке, в олигоцене Фаюма, найдены нижние челюсти первых антро-
поморфных обезьян; последние известны в Европе из среднего миоцена,
в Индии — из плиоцена.
Сем. Cynopithecidae. — Узконосые хвостатые обезьяны Старого света.
Передние и задние конечности почти одинаковой длины. Павианы, макаки и
другие ходят на четырех ногах. Хорошо лазают по деревьям.
Рис. 280. Oreopithecus В am-
bo 1 i i.
Рис. 281. Mesopithecus Pentelici
Gaudry.
Oreopithecus Bambolii Gerv. (рис. 280) — верхний миоцен То-
сканы, с очень маленькими клыками; коренные с четырьмя бугорками;
hi тя талон из двух бугорков.
Предкоренные двухбугорчатые. Бугорки зубов высокие, конические,
откуда и название (о г е о — бугорок, конус).
Рис. 281,A. Mesopithecus Pentelici. Череп, нижняя и верхняя челюсти.
Mesopithecus Wagn. (рис. 281 и 281,А) — из Пикерми, изве-
стен цельным скелетом, описанным Бодри. Mesopithecus Pentelici
224
класс: млекопитающие — mammalia
Gaudry—закругленная голова напоминает Anaptomorphus Соре, но заты-
лочная часть еще более укорочена и делает его похожим на Semnopithecus
юго-восточной Азии. По тяжелому скелету он ближе к Macacus. (Может быть,
эта форма и положила начало только что названным родам.) Скелет ее с очень
длинными конечностями, задние несколько длиннее передних; это указывает на
хорошо лазающее по деревьям животное. Хвост тоже длинный. Зубы пред-
ставляют как бы переход от заостренных бугорков Oreopithecus к более
плоским бугоркам настоящих Anthropoidea. Коренные резко отличаются от
предкоренных, состоя из четырех закругленных бугорков, предкоренные — из
двух. Клыки сильно развиты; резцы лопаточкообразные.
Semnopithecus известен из плиоцена Монпелье и Сивалик
(Semnop. palaeindicus Lyd.). Semnopithecus entellus — из
плейстоцена Индии — связывает плиоценовые формы с ныне живущими.
Libypithecus Markgrafi Strom.—из среднего плиоцена Вади
Натрун в Египте; представлен хорошо сохранившимся черепом, очень удли-
ненным, с сильно выраженным сагиттальным гребнем, расходящимися от него
двумя складками к внешним краям глазниц и сильно выраженными надглазнич-
__ ными дугами. Клыки (верхние) длинные.
ЦЦ Parapithecus Fraasi Schl. (рис. 282)—из
олигоцена Фаюма, представлен нижней челюстью с
очень расходящимися задними концами и суженной
передней частью. Резцы и клыки маленькие. Корен-
ные типичные четырехбугорчатые.
Вместе с этой челюстью найдена нижняя челюсть
с значительно закругленной передней частью. Это
^n^u^Fra^sVschlЬе Propliopithecus, близкий к европейскому
Pliopithecus, из миоцена (Гориах и Сансан),
которого уже относят к человекоподобным обезьянам, со сглаженными бугорками
на коренных и более сложными предкоренными. У последней формы имеется
на коренных базальный воротник, сходный с Hylobatus.
Из сем. Cebidae найдены в миоцене Патагонии представители трех
родов:
1) Homunculus patagonicus Amegh. — череп высоко поднятый
сзади, большие глазницы, направленные вперед; цельная нижняя челюсть
2 13______з
с — -г- --очень узкая впереди и высоко поднимающаяся сзади.
Л 1 «5 — <5
2) Anthropops perfectus Amegh. и
3) Eudiastatus lingulatus Amegh. Оба представлены неполными
нижними челюстями. Они имеют типичные признаки этого семейства и имеют
значение при изучении его, но не могут быть поставлены в предки обезьян
Старого света.
Сем. Anthropoidea. — Человекоподобные обезьяны Старого света. Ха-
рактеризуются возможностью вертикального положения тела, сильным разви-
тием клыков, сглаженными бугорками на коренных, сильно развитым мозгом,
подкласс: плацентные — placentalia, ноготные
225
отсутствием хвоста. Ныне живущие: оранг-утан, шимпанзе, горилла имеют в тре-
тичных слоях незначительное число более или менее близких форм. Зубная
О1О ______________1
формула: -- - ----
Z i z о
Сем. Sitniidae. — Dryopithecus J) Lartet (Gaudry) имел размеры
шимпанзе. Коренные зубы с четырьмя буграми, между которыми есть
маленькие, добавочные. Клыки развиты сильно. Известны две нижние че-
люсти с почти параллельно расположенными коренными зубами и часть плече-
вой кости.
Первая находка в 1856 г. (рис. 284) произвела большой переполох
между палеонтологами и антропологами, так как в первый раз были найдены
остатки человекоподобной обезьяны (они остались и
до сих пор единственными) в среднем миоцене
Гаронны.
Челюсть была очень массивная с торчащим
вверху клыком; рг3 был заостренный, pri — сжатый,
и два коренных с 4 главными бугорками и одним
добавочным. Л я рте, описавший ее, так резю-
мировал ее положение: „Эта новая ископаемая обезь-
яна относится к группе человекоподобных, но имеет
некоторые признаки высшего развития, чем оранг-утан, шимпанзе, горилла и гиб-
бон". Эти отличия заключались, по Л я рте, в строении зубов, направлении клы-
ков и сокращении передней части челюсти. В 1890 г. была найдена новая че-
люсть, тоже в среднем миоцене во Франции, которая привела Г о д р и к другому
выводу, так как направление клыка оказалось более косым, две пары резцов —
очень тонкие, рг3 — острые, а главное, подбородок очень косо срезан
назад, с сильным прогнатизмом (передняя часть первой челюсти была сломана).
Если сравним ширину челюсти (толщину кости, длину симфиза) Dryopi-
thecus Gaudry и других человекоподобных обезьян (гориллы, шимпанзе), то
окажется, что по всем этим признакам он стоит ниже их и бесконечно далеко
от самого примитивного нынешнего человека — готтентота (рис. 2842).
Очень узкое пространство между челюстями указывает на узкий, толстый
в корне язык, который отставал в своем развитии от языка гориллы и шим-
панзе и ни в каком случае не мог быть способен к членораздельной речи.
Прогнатизм у Dryopithecus больше, чем у шимпанзе (косая линия подбо-
родка) и ширина симфизы. Все это заставило Г о д р и поставить эту форму
ниже всех человекоподобных обезьян (рис. 284 и 284 А).
Г о д р и указывает, что очень молодые животные нынешних обезьян
имеют признаки более высоко организованных животных, чем Dryopithecus
во взрослом состоянии.
*) D г у о s — дуб. По соседству того места, где найдена первая челюсть
Dryopithecus, встречаются лигниты с остатками стволов деревьев, которые относят
к дубам, на которых, по предположению, и жил Dryopithecus.
15 Па в л о в а М. В. Палеозоология. Ч. II
226
класс: млекопитающие — mammalia
Pliopithecus antiquus Gerv. — известен из верхнего миоцена сред-
ней Европы. Нижняя челюсть из Сансана (Франция).
Этими находками исчерпываются данные об остатках обезьян в Европе.
Указанная проф. Ласкаревым находка обезьяны в дер. Гребенниках до сих
пир не описана.
Palaeopithecus Lyd. — указан в плиоцене Сивалик. Это верхняя»
челюсть с коренными зубами, похожими на человеческие.
Грегори устанавливает по найденным верхним коренным зубам присут-
ствие Dryopithecus punjabiensis в нижнем плиоцене Сивалик и близ-
Рис. 284. 1,2 — Dryopithecus, 3 — горилла,
4 — шимпанзе, 5 — готтентотка.
Рис. 284А. 1 — Dryopithecus^
2 — шимпанзе, 3 — готтентот, 4 —
француз.
верхние очень широкие. Они
Рз Pi «г т3
кую к нему форму Sivapithecus indicus Pilgr. — в верхнем миоцене
Сивалик (рис. 285).
Нижний prt очень маленький. Нижние коренные почти четырехугольные,
ближе, чем у всех известных S i m i i d a e, к зубам
человека. Очень близки к зубам Palaeopithe-
cus Lyd.
Осборн описывает из плиоцена Не-
браски (Сев. Америка) зубы Hesperopithe-
cus, сходные с человеческими.
Рис. 285. Sivapithecus i n d i- P г о pl i о p i t h e c u s Haeckeli Schl.—
cus Iilgr. Нижние зубы. представлен нижнею челюстью из олигоцена
Фаюма (Египет) с 3 конусообразными пред-
коренными и 3 четырехбугорчатыми коренными. Задняя часть челюсти
высокая. Рис. 283.
Эти данные не позволяют нам определенно решить вопрос, имели ли
лемуры и обезьяны общего предка, или последние произошли от первых.
подкласс: плацентные — placentalia, ноготные 227
Сем. Hominidae (человек). — Родство человека с обезьяной общеизве-
стно, но стоит посмотреть на скелет того и другой, чтобы сразу стало ясно
громадное расстояние между самыми близкими к человеку обезьянами, шимпанзе
или оранг-утаном, и современным человеком. Между отдельными костями ске-
лета конечностей есть сходство, например между костями передних конечно-
стей. Но достаточно взглянуть на череп обезьяны, на тяжеловесные кости
задних конечностей, более короткие, чем передние, с большим противопоста-
вляющимся пальцем, чтобы почти отказаться от сближения этих скелетов.
Особенно сильно отличается череп нынешнего человека с высоким лбом,
мало выдающимися краями глазниц, закругленным высоким затылком и нижней
челюстью с выступающим подбородком.
При этом сравнении нужно, однако, помнить, что мы имеем здесь дело
с двумя крайними типами — человекоподобная обезьяна современная и такой же
человек. Чтобы понять и оценить сходство или отличие различных признаков,
характеризующих высшие типы в различных линиях, нужно обратиться к древ-
нейшим представителям этих линий. Для обезьян мы знаем остатки более
древних форм; познакомимся с такими же остатками древнего человека — Ното
primigenius.
Еще в 1800 г. были впервые указаны остатки человека'—обломки
черепа, найденные близ Штутгарта (Cannstadt), которые впервые описал
Катрфаж (Quatrefages), французский ученый, в 1870 г. и отнес к древней-
шему человеку.
С тех пор много других находок было сделано, и между ними одна,
вызвавшая много научных споров. Это Pithecanthropus Dubois, найденный
в 1894 г. на о-ве Яве голландским ученым Дюбуа. Остатки состояли из че-
репной крышки, двух зубов и бедра, найденных в одних и тех же слоях, но
в различное время.
На Лейденском антропологическом конгрессе в 1894 г. из двенадцати
крупных авторитетов, решавших вопрос о зоологическом положении животного,
остатки которого были найдены, три высказались за принадлежность найден-
ных остатков ненормальному человеку, причем Вирхов стоял за то, что
это тип дегенерировавшего человека: болезнь его выразилась на костях бедра.
Три ученых стояли за принадлежность остатков обезьяне — неизвестной еще
до тех пор. Шесть — смотрели на эту форму как на промежуточную между
обезьяной и человеком.
Крышка черепа имеет очень удлиненную, низкую форму с совершенно
покатым лбом. Надбровные дуги очень развиты; при сравнении их с шимпанзе
и человеком они ближе к первому. Зубы похожи на зубы человека со сгладив-
шимися бугорками. Бедро очень длинное и слабо загнутое, с далеко высту-
пающей головкой и костными выростами на внутренней стороне верхней
части. Отложения с Pithecanthropus отнесены условно к верхнему
плиоцену.
Род Ното. — Следующие находки черепов, близких к человеку, отно-
сятся к р. Ното; они группируются в два типа: Homo primigenius и
15*
228
класс: млекопитающие — mammalia
Homo sapiens. Первый — с признаками, сближающими его с Pithecanth-
ropus, второй — ближе к современному человеку.
Homo primigenius.
Первый тип древнего человека заключает в себе два вида: Ното
neandertalensis и Homo heidelbergensis. К первому виду от-
носятся:
1) неандертальский человек, найден близ Эльберфельда, сначала
только в виде верхней части черепа и нескольких длинных костей скелета,
в 1858 г., потом в виде более полных черепов;
2) обломки черепов в С п и (Бельгия) (рис. 286) и
3) в пещере Крапины в Хорватии.
Все они характеризуются большой толщиною черепных костей, которую
проф. Вирхов объяснял в неандертальском черепе болезненным развитием,
утолщением, вызванным воспалением мозговых оболочек после поранения; на
черепе видны были следы ранений. Но после других находок этого же типа
выяснилось, что толщина черепных костей является
характерной для всего типа первичного чело-
века и лучше всего выражена в скелете и черепе,
найденных в пещере близ Шапель-о-Сен (Chapelle-
aux-Saints, Франция).
Проф. М. Буль (Boule) в 1911 г. дал по-
дробное описание цельного скелета с черепом,
хотя и раздавленным, но вполне хорошо вос-
становленным и дающим полное понятие о силь-
но выдающейся назад затылочной части черепа,
низком лбе, толстых надбровных дугах и т. д.
Слепок мозга дает почти гладкую поверхность со слабыми извилинами, кото-
рую можно сравнить только с мозгом обезьян и микроцефалов. Сравнение
черепа с шимпанзе и современным человеком тоже дает гораздо большее
сходство с первым, чем с последним.
Эта находка представляет большой интерес в том смысле, что положение
скелета указывает на его погребение. Это первый случай для Homo primige-
nius. Скелет найден в низкой пещере; он лежал в углублении, сделанном
в доломитовом известняке нижнего лейаса. На этом известняке и над ямой,
заключающей скелет, лежал слой толщиной от 30 до 40 см, состоящий из
обломков костей, кремневых орудий и кусков известняка. Из костей животных
можно было определить: носорога, лошадь, свинью, северного оленя, козу,
зубра, волка, лисицу, барсука и гиэну.
Наиболее древним остатком типа Homo primigenius считается
нижняя челюсть из мауэровских песков, близ Гейдельберга (вид Ното
heidelbergensis), найденная на глубине 24 м (рис. 287). Она мас-
сивнее всех других, с очень широкой задней половиной. Зубы типа чело-
веческих, коренные с 4—5 бугорками. Подбородок с выступающим верх-
ним краем.
Рис. 286 Череп неандерталь-
ского человека. (Спи, Бельгия.)
подкласс: плацентные — placentalia, ноготные
229
Интересно определить время, когда жили эти еще не вполне похожие
на нас человеческие существа.
Отложения, в которых были найдены остатки Pithecanthropus, были
первоначально отнесены к верхнему плиоцену. Дальнейшие исследования заста-
вили отнести их к нижнему плейстоцену.
Различные остатки Homo primigenius относятся к разным частям
плейстоцена, на что указывают древнейшие кремневые орудия, находимые
часто с этими остатками, и кости животных, живших одновременно с человеком.
По признакам челюсти и по нахождению с нею Rhinoceros Merckii
остатки из Гейдельберга древнее, чем неандертальский череп и чем человек
из Chapelle-aux-Saints Буля с указанны-
ми выше остатками животных. Вместо
Rhinoceros Merckii Булем указаны
Rhinoceros tichorhinus и мамонт.
Это средний плейстоцен с холодным клима-
том. Из орудий здесь встречаются только
каменные, хотя со следами отделки; сделан-
ных из кости нет.
Второй тип древнего человека —
Homo sapiens, более нам близкий,
был представлен несколькими расами:
Рис. 287. Нижняя челюсть Ното
heidelbergensis.
комбкапельской, галейхильской, брюнской, кроманьонской
и др. Он существовал и гораздо раньше эпохи мамонта и одновременно
с мамонтом. Ооудия его были вначале тоже из кремня, главным образом
ударные скребки и стрелы. Позже — орудия из кости. Черепа этого типа
значительно отличались от первого, хотя также преобладал удлиненный
затылок. Но лоб у них был более высокий, надбровные дуги мало выдава-
лись. Нижняя челюсть выступала вперед нижним краем. Черепные кости
тоньше.
Черепа эти были находимы: в пещерах Франции, в речных наносах
Англии и в лессе Моравии.
Eoanthropus Dawsoni Smith.
Среди остатков ископаемого человека особое место занимают череп и
нижняя челюсть из Piltdawn (Суссекс, Англия). Его сопровождали остатки
Mastodon arvernensis, плиоценовая форма. Эти остатки — во вто-
ричном залегании.
Остатки черепа: затылочная, теменная и височная кости, имеют форму,
близкую к современному человеку, с вполне закругленным затылком.
Нижняя челюсть, найденная отдельно и содержащая два зуба — wz2 и тг
с типичными 4 бугорками и пятым в виде талона, имеет на переднем нижнем
конце сломанной кости выступ, указывающий направление подбородка с вы-
ступающим верхним краем, т. е. типа обезьян. Этот палеолитический человек
представляет большую загадку, если приписать остатки черепа и нижнюю челюсть
одному и тому же существу, что многими оспаривается.
230
класс: млекопитающие — mammalia
Итак, все найденные до сих пор остатки человека в Европе относятся
к плейстоценовым и резко отграничены от человекоподобных обезьян.
По многим признакам наиболее совершенные обезьяны стоят ниже самого
первобытного человека.
В Южной Америке, в Лаплате, были указаны остатки человека в верхнем
плиоцене. Но там вообще определение возраста слоев далеко не точно, что
было указано при изучении различных млекопитающих. Нередко слои, отнесен-
ные первоначально к миоцену, приходилось переносить в плиоцен, и обратно.
В Бразилии остатки человека более поздних периодов известны в пещерах
вместе с глиптодоном, махайродоном и другими послетретичными формами.
3-я ГРУППА
КОПЫТНЫЕ
Первым признаком копытных, отличающим их от коготных, является
присутствие копыт — роговых чехлов на последних фалангах пальцев. Этот
признак, как будто незначительный на первый взгляд, сопровождается совсем
иным строением всего скелета и даже всего организма. Все животные, относя-
щиеся сюда, кроме сем. свиней, — травоядные. Значит, и органы питания будут
другие, главным образом зубная система. Органы передвижения тоже должны
измениться.
Нынешние копытные по числу пальцев подразделяются на две боль-
шие группы: 1) непарнопалых — Р е г i s s о d а с t у 1 a Ow. (М е s а х о п i a Marsh)
и 2) парнопалых — Artiodactyla (Paraxonia Marsh).
Представителями первых являются лошадь и носорог, вторых — бык и
свинья. Но есть формы, как, например, тапир, у которых на передней конечности
существует четыре пальца, а на задней три, что и затрудняет на первый взгляд
определение. Но если мы посмотрим на его переднюю ногу, то увидим, что
третий палец длиннее других, чего не бывает у парнопалых. Пятый — слабый,
едва причленяющийся. При этом очевидно, что центр тяжести тела проходит
через средину третьего пальца — как у лошади, а не между третьим и четвер-
тым, как у парнопалых. Тапир относится к непарнопалым, и ввиду таких слу-
чаев Марш и предложил названия: Mesaxonia — среднеосевые и Para-
xonia — бокоосевые.
Кроме этих типичных копытных к копытным в более широком смысле
относят: хоботных — слоновых, с пятью пальцами, титанотериев и некоторых
других. • ..........
Заключается ли разница конечностей только в числе пальцев, и все ли
конечности одной и другой групп развивались одинаково? Достаточно сравнить
ногу носорога, лошади, свиньи и быка, чтобы видеть разницу как в числе
пальцев, так и в плане строения конечностей.
Чтобы понять эту разницу, нам. нужно начать с простейшей пятипалой
конечности древнейшего копытного, близкой еще к конечности коготных, —
подкласс: плацентные — placentalia, копытные
231
Meniscotherium (Hyracops) (рис. 288). Здесь мы имеем по пяти паль-
це? на каждой конечности с наибольшим III; И и IV меньше его, но между
•собою равные, I и V еще меньше и тоже равные. Карпальные кости располо-
Рис. 288. Menis-
cotherium (Hy-
racops) socia-
1 i s. Кисть. (Ниж-
ний эоцен.)
жены рядами над метакарпальными, не чередуясь.
У носорога мы имеем по три пальца, недостает I и V; оставшиеся почти
не изменились. У свиньи сократился V; остальные почти равны между собою.
У лошади существует один, остальные исчезли, остались
верхние концы двух метаподий. У коровы тоже один, как
бы средний метаподий, но он состоит из двух сросшихся
вместе (III и IV) метаподий, что видно на разрезе кости,
называемой трубкой, каноном, на конце которого причле-
няются два пальца III и IV. От II и V остаются незначи-
тельные части. Является вопрос: были ли у предков этих
форм такие же ноги, или они развились из пятипалого
типа, изменяясь в различных направлениях? Изучая древ-
ние формы, мы можем проследить по ископаемым остаткам
.их действительные изменения. Мы увидим, что у них со-
вершалось сокращение метаподий и одновременно с этим
перемещение налегающих на верхнюю их поверхность кар-
пальных и тарсальных костей, которые передают тяжесть
всего тела на метаподий. Для экономии организма живот-
ного необходимо, чтобы каждая часть, получая питание, со-
вершала нужную работу и чтобы, в случае сокращения ка-
кой-нибудь части органа вследствие изменений условий
жизни, ближайшая к нему не оставалась бесполезной.
Изучая изменение конечностей и сокращение пальцев по ископаемым
формам, наш великий ученый В. О. Ковалевский установил важный
закон „правильной редукции" пальцев (адаптивной) у нынешних копытных и
противоположной ей „неправильной редукции", неадаптивной, которая под-
мечена им в целой группе вымерших парнопалых и существует у некоторых
непарнопалых.
При первой сокращение пальцев всегда сопровождается перемещением
освободившихся костей карпуса и тарсуса на остающиеся метаподий (мета-
карпалии и метатарсалии).
Кроме того иногда происходит срастание некоторых мелких костей.
При неправильной редукции названные кости, при сокращении пальцев, оста-
ются бесполезно на прежних местах, и нога лишается нужной связи между
верхней и нижней ее частями.
При изучении различных семейств копытных мы подробно познакомимся
с различным строением ног и изменением положения их косточек.
Зубы копытных резко отличаются от зубов коготных. Все коренные
состоят из бугорков или луночек: четыре бугорка или луночки у парнопалых,
шесть бугорков у непарнопалых, пять — у некоторых парнопалых. Клыки мало
отличаются от резцов. Режущих зубов нет.
232
класс: млекопитающие — mammalia
Отряд 9
Непарнопалые — Perissodactyla
Ныне живущие представлены немногочисленными семействами: это —
лошади, носороги и тапиры. Первые очень распространены во всех частях
света, вторые и последние существуют только в тропической Америке и
южной Азии.
Ископаемые представители известны для всех семейств. Но самый интерес-
ный материал дают остатки лошадиного семейства, которые позволяют вос-
становить генетическое развитие их, начиная с эоцена, где их древнейшие
представители носят еще не вполне определенные признаки и относятся к пер-
вичной группе копытных (Protungulata), которые являются здесь как бы
отвечающими по своему положению и по характеру признаков Palaeocarni-
vora, предшественникам различных хищных. Это Condylarthra (Protun-
gulata), у которых существуют еще некоторые признаки, сходные с Creo-
donta. Не будем останавливаться здесь на всех их представителях, так как
главные типы войдут в ос-
Рис. 289. Tetraclaenodon puercensis Scott.
А — верхние коренные, В — нижние. С—Задняя ступня.
нование линий разных ко-
пытных, другие еще не на-
шли своего определенного
положения среди древних
копытных.
Сем. Equidae. — Наи-
больший материал по иско-
паемым лошадям дала Сев.
Америка, и американские па-
леонтологи много сделали
для выяснения генетических
отношений отдельных форм.
Но первые указания были даны нашим ученым В. О. Ковалевским. Но-
вые формы, найденные после его работ, заставили многое изменить в сделан-
ных им выводах; но работы его сохраняют ценность по тому методу, которым
он шел в них, изучая каждую сочленовную фасетку и ее изменения *).
Некоторые ученые считают, что лошади развивались в Сев. Америке и
в Европу заходили только отдельные роды во время соединения обоих конти-
нентов.
Но так как многие роды общи для обеих частей света, хотя они и
не одинаково хорошо представлены, то мы дадим описание главных предста-
вителей вообще.
1) В. О. Ковалевский, Sur ГА n с h i t h e r i u m et sur 1’histoire paleonto-
logique des chevaux. Mem. Acad. St. Petersbourg, 1873; его же, Monographic der
Gattung Anthracotherium. Pala-ontographci, 1873, XXII.
подкласс: плацентные — placentalia, копытные
233
У основания сем. Equidae поставим Tetraclaenodon puercen-
sis Scott, один из видов p. Phenacodus (из Condylarthra), из ниж-
не'го эоцена Пуэрко (Сев. Америка) и Эгеркингена (Швейцария).
Различные авторы, отделив его от рода Phenacodus и оставив видо-
вое название puercensis, дают ему новые родовые названия: Protogonia
puercensis Соре, Euprotogonia р u ere en s i s Matth., Те t га с la е n о-
don puercensis Scott (рис. 289).
Phenacodus Wortmani Cope (рис. 290) не входит в эту линию.
Он имел зубы иного строения.
Рис. 290. Phenacodus Wortmani Соре.
По скелету и конечностям Tetraclaenodon puercensis Scott
еще далек от лошади. Но его пятипалые конечности имеют последние фа-
ланги закругленные, указывающие на присутствие копыт; I и V metatarsus
и metacarpus укорочены; os centrale исчезла; astragalus имеет внизу закру-
гленную сочленовную поверхность типа коготных (рис. 289, С). Хвост длинный
и тяжелый.
Череп по форме и озублению отличается от черепа всех коготных подъ-
емом к затылку. Особенно интересными являются зубы (рис. 289), снаб-
женные бугорками; рг1 и рг2 — простые конусы; рг3 имеет на наружной стороне
два слившихся бугорка и один внутренний; рг1 имеет два наружных, один
внутренний и два зачаточных добавочных между ними; его форма почти четырех-
угольная. Верхние коренные имеют по 6 бугорков: два наружных, два внутрен-
них и два промежуточных; они расположены в два ряда и несколько закруглены
спереди на т3, последние развиты слабо. Нижние коренные имеют по четыре
бугорка и маленькому придаточному. Это типичные буюрчатые зубы.
.234
класс: млекопитающие — mammalia
Р ютимейером
него эоцена Эгеркингена (Швейцария).
Hyracotherium — нижний
Поднимаясь немного выше в эоцене,
(Riitimeyer) описаны зубы
Рис. 291. Hyracotherium lepori-
n u m Ow.
В — верхний, С — нижний коренной.
Phenacodus из ниж-
и средний эоцен Европы и Сев. Америки,
встречаем форму, у которой внутренние
бугорки несколько сжаты и все
сближены между собою — на верх-
них коренных. Простейшей формой
является — Hyracotherium 1е-
р о г i n u m Ow. (рис. 291) — из
нижнего эоцена лондонской глины.
К сожалению, его скелет неизвестен.
По зубам он еще очень близок к
Tetraclaenodon puercensis.
Многочисленные виды этого рода,
найденные в Сев. Америке, указы-
вают на изменение бугорков, их
большее сжатие и сближение между
собою. У Hyracotherium in-
dex (Wasatch) мы имеем 2 бугорка
на рг2, на внешней стороне, на рг3 —
два на внешней, один внутренний
и один добавочный на передней
стороне. Это уже значительное осложнение предкоренных. Коренные зубы
с 6 сжатыми бугорками.
Конечности передние с 4 пальцами, задние — с тремя. При этом сохра-
нился еще trapezium — укороченный метаподий, причленяется к unciforme ма-
ленькой фасеткой.
В Америке описаны раз-
личные виды Hyracothe-
rium с осложняющимися зу-
•бами.
Е о h i р р u s Соре — ото-
ждествляется с Hyracothe-
rium leporinum Ow. Ну-
racotherium venticulum
Соре — известен почти цель-
ным скелетом (рис. 292). Перед-
няя конечность с 4 пальцами, задняя — с тремя (рис. 294, Л).
В. О. Ковалевский отделил от рода Hyracotherium форму
с более сложными предкоренными из среднего эоцена Франции под именем
Pachynolophus.
У него рг3 и рг'* по форме и числу бугорков приблизились к коренным;
j>r2 — трехбугорчатый, с маленьким 4-м бугорком. В нижней челюсти корен-
ные — с 4 бугорками. Скелет этой формы неизвестен.
подкласс: плацентные — placentalia, копытные
235
В Сев. Америке известны еще две эоценовые формы:
Orohippus из среднего эоцена и Epihippus — из верхнего, у ко-
торых осложнение предкоренных идет тем же порядком, бугорки на корён-
s. Eq. caballus
7. Eq. Andium
6. Protohippus
5. Meryhippus
Anchithe-
rium
3. Anchilophus
2. Pachynolo-
phus |
1. Hyracoth.
leporinum
Рис. 293. Зубы главных представителей, входящих в лошадиную линию, и зубы
боковых форм — Hipparion и Anchilophus.
них удлиняются и сливаются нижними концами. У последней формы суще-
ствуют 4 пальца в передней и три в задней конечности. Оба эти рода
ставятся некоторыми американскими
учеными в синонимику с Р а с h у-
nolophus Kow.
Mesohippus Bairdi—в
среднем олигоцене — дает форму с
6 почти одинаковыми зубами, рг1
остается в виде конуса. Бугорки на
коренных изменились: внешние полу-
чили форму, более близкую к лу-
ночкам, внутренние вытянулись;
средние, наименьшие, тоже вытянуты.
В нижних предкоренных образовалось
по две луночки, соединяющие бу-
горки, и развился маленький добавоч-
ный бугорок близ среднего внутрен-
него. Хвост очень укоротился. Лопатка типа лошади; конечности трехпалые,
с тремя ступающими пальцами (рис. 294, В).
Этот род был первоначально отождествлен с европейским А n с h i t h е-
r i u m по сходству как зубов, так и частей скелета. Позже отделен в род М е so-
hippus, главным образом вследствие значительно меньших размеров и вслед-
Рис. 294. Ноги: А — Hyracotherium,
В — Mesohippus.
236
класс: млекопитающие — mammalia
ствие находок в Америке Anchitherium, равного по размерам евро-
пейскому.
Anchitherium — средний миоцен Европы и Сев. Америки, дает по-
следующее развитие зубов с уменьшением рг1 и осложнением нижних
предкоренных большим развитием бугорка а1. Внизу 6 коренных одинакового
типа. Конечности трехпалые; все пальцы доставали до земли.
В миоцене Сев. Америки, кроме Anchitherium, известны несколько
форм меньших размеров, стоящих ближе к Mesohippus: это — Р а г a h i р-
pus с очень маленьким рг1 и Meryhippus с таким же рг1, но рг2 у него
удлинился, образуя выступ на передней стенке, что напоминает зубы лошади.
У некоторых видов последнего рода происходит уклонение рисунка зубов при
стирании их. Вместо того чтобы внутренний бугорок переднего ряда сливался
при стирании со средним, как это было у предыдущих форм и у некоторых
форм этого рода — Meryhippus campestris, здесь на некоторых зубах этот
бугорок остается изолированным, не слившимся со средним — у Meryhippus
i п s i gn i s.
Известно много остатков черепов этой формы, и найден почти полный
скелет, причем на передней конечности существуют остатки V пальца в виде
тонкой пластинки. Нам придется еще вернуться к этой форме.
Protohippus — в плиоцене Сев. Америки, представляет форму, у ко-
торой 6 коренных зубов имеют одинаковую поверхность. Внешние бугорки
изменились в лунки, следующие слились между собою по длине в виде за-
гнутых гребней. Зубы теряют корни и становятся гипсодонтными, с постоянным
ростом, хотя еще и невысокие. Развивается цемент на зубах. Нижние зубы
состоят из двух луночек, соединенных между собою особой петлей — из двух
стирающихся бугорков (а и а1).
Конечности трехпалые, но боковые пальцы не достают до земли и зна-
чительно передвинулись назад. Череп этой формы, да и весь скелет, стоит
очень близко к лошадиному.
Различные представители рода Pliohippus (Plioh. supremus,
Plioh. fossulatus и др.) дают ряд форм, связывающих род Protohip-
pus с родом Е q и и s.
Equus Stenonis — лз плиоцена Италии — и некоторые американ-
ские лошади дают нам последующую ступень в развитии лошадей. Глав-
ное изменение в зубах состоит в большем развитии зубов в высоту и в
преобладании переднего внутреннего бугорка (ставшего из бугорка стол-
биком, по мере роста зубов) над задним, очень уменьшившимся. Эмаль, огра-
ничивающая разные части зубной поверхности, изменилась. Складочки, появив-
шиеся уже на зубах Proto hippus, увеличились и осложнились у Plio-
hippus.
На нижних зубах петля средняя увеличилась, и образовалась другая —
задняя — полупетля. , -................................... £
Боковые пальцы значительно укоротились; фаланги исчезли, остались
только метаподии. Весь скелет — типа настоящей лошади.
подкласс: плацентные — placentalia, копытные
237
В плейстоцене мы встречаем уже несколько разновидностей Е q и и s
caballus с различно осложнившимися зубами, с большим или меньшим раз-
витием переднего столбика, всегда соединенного со вторым бугорком. Эмаль
тоже проявляет различную складчатость, особенно окружая так называе-
мые марки — углубления между бывшими внешними и средними бугорками
(рис. 293).
Молочные зубы лошадиной линии представляют большой интерес в своем
развитии. В общем они являются более сложными, чем заменяющие их пред-
коренные.
Мы можем указать их для некоторых представителей этой линии. Так
у Pachynolophus три верхних молочных зуба d2, d3 и d* имеют четырех
угольную форму с шестью бугорками, т. е. сходны с коренными, а на за
мещающих их предкоренных только
два последние сходны с коренными,
а рг2 проще — он треугольный.
У Mesohippus d2 имеет на
переднем углу внешней стенки до-
бавочную складку, и он крупнее,
чем замещающий его рг2.
У Meryhippus на нижнем d
более развиты внутренние петли, чем
на предкоренных.
В роде Е q и и s на молочных
зубах эмаль гораздо сложнее. Мо-
лочные зубы более древних форм
более сходны с предкоренными по-
следующих, более поздних форм, чем
с предкоренными, их заменяющими.
Рис. 295. Изменение зубов и ног у Equi-
dae: а — Tetraclaenodon; b — Нуга-
cotherium; с — Anchitherium; d—
Pliohippus; е — Equus.
Конечности в роде Pliohip-
pus и роде Equus стали однопалыми, с остатками боковых метаподий в
виде тонких стилетов, прирастающих иногда сзади к метаподию среднего пальца.
При сокращении пальцев в этом ряду наблюдается крайне интересная вещь:
перемещение карпальных и тарсальных костей с боковых metatarsus и metacarpus
на средний. Ни одна из этих косточек не осталась на прежнем месте
Конечно, здесь намечены только главные этапы развития лошадиной
линии, очень богато представленной в Сев. Америке. Но и по ним видно, как
последовательно, стройно шло это развитие, как в связи с изменением пиши и
почвы (на что указал В. О. Ковалевский) осложнялись зубы, при-
обретая прочность в связи с большим развитием эмали, цемента, и как в
помощь этому осложнению являлся еще и постоянный рост зубов, увеличи-
вающий долговечность зуба. С другой стороны, сокращались на ногах
ставшие ненужными элементы пятипалой конечности, при изменении образа
жизни, при перемещении на другую, более сухую почву, где широкая много-
палая нога стала неудобной.
238
класс: млекопитающие — mammalia
Кроме этих форм, входящих в генетическую линию лошадей, существуют
близкие к ним формы, но не могущие занять места в этой линии по некото-
рым признакам.
Род Hipparion. Одной из таких форм является гиппарион из меоти-
ческих слоев, еще сравнительно недавно считавшийся В. О. Ковалевским
Рис. 296. Hipparion gracile Каир.
и другими учеными непосредственным предшественником лошади. Но при
подробном сравнительно-анатомическом изучении его зубов и конечностей
с таковыми лошадиной линии оказалось невозможным оставить его на преж-
нем месте, несмотря на общее сходство скелета (рис. 296).
Зубы гиппариона (рис. 297) резко отличаются от зубов лошади значи-
тельно более сложными складками эмали и — на верхних зубах — передним
т'
Рис. 297. Hipparion gracile Каир. (3/4)
внутренним столбиком, совершенно изолированным от
среднего бугорка. Нижние зубы имеют тоже более сложную эмаль и до-
бавочные бугорки. Череп ближе к лошадиному, но отличается присутствием
особого предглазничного углубления, которое Годри назвал „лакримальной
впадиной".
Ноги его имеют по три пальца со всеми суставами, хотя боковые
пальцы уже не достают до земли. По ногам эта форма стоит как бы между
подкласс: плацентные — placentalia, копытные
239
Anchitherium и Protohippus. Некоторые сочленения карпальных и
тарсальных костей ближе к первому; не достающие до земли боковые пальцы —
типа Protohippus. Таким образом ноги являются значительно отставшим^
тп'
Рис. 297 A. Mery hippus insignis Leidy. (3/s)
от ног лошадей, и гиппарион олицетворяет собой форму, у которой нару-
шено гармоническое развитие частей скелета: зубов и ног; первые опередили
лошадь, вторые отстали
Очевидно, этот род, жив-
ший начиная с мио-плио-
цена (Cucuron, Pikermi)
до среднего плиоцена
(Roussillon), является бо-
ковой ветвью, отделив-
шейся от рода, входя-
щего в генетическую ло-
шадиную линию. Таким
родом является Мегу-
h i р р u s, из среднего
миоцена Сев. Америки,
о котором мы уже гово-
рили; у некоторых видов
этого рода наблюдаются
на зубах изолированный
передне-внутренний столбик и более сложная эмаль, чем у других видов
(рис. 297 А)
Рис. 298. Neohippar ion.
Рис. 299. Neohipparion gratuni Leidy. (3/4)
В последнее время в Сев. Америке найдены еще формы, близкие к гип-
париону, которые там довольно разнообразны; у них тоже наблюдаются зубы
со смешанными признаками, с изолированным или соединенным внутренним
передним столбиком и с более или менее сложной эмалью; это — Neohippa-
rion (рис. 298). Некоторые виды его имеют уже на всех зубах отдельный
240
класс: млекопитающие — mammalia
столбик и почти не отличаются от европейского гиппариона (рис. 299).
Очевидно, Neohipparion является формой, связывающей Meryhippus
с Hipparion, у которого все признаки изменившихся зубов укрепились и
отошли далеко от зубов лошади ]). Некоторые ученые как на предка гиппа-
риона указывали на средне-миоценовый Desmotippus Сев. Амёрики, —
форму, близкую к Anchitherium, у которого зубы еще имеют все бугорки
на коренных мало измененными, но эмаль более сложную.
Гиппарион существовал долго, почти до верхнего плиоцена, и имел боль-
шое географическое распространение: в Северной и Южной Америке установлено
много видов; на юге Европы, во Франции, Германии, в б. Херсонской губ., в Гре-
ции; в Азии — в Персии, в Сиваликских холмах указаны очень близкие формы.
В роде Е q и и s мы не встречаем ни отдельного столбика (разве у очень
старых животных с искаженной поверхностью зуба), ни столь сложной эмали.
Рис. 300. Palaeotherium me-
dium Cuv.
В палеонтологическом кабинете Московского
университета находится череп современной
лошади — не старой, у которой на двух зу-
бах передний столбик отделен (типа гиппа-
риона); это — шаг вперед (а не назад) в раз-
витии зубов лошади.
Молочные зубы лошадиной линии, всегда
более сложные, чем постоянные, как мы ука-
зали, никогда не имеют признаков зубов гип-
париона, а эти последние не имеют призна-
ков зубов лошади, а дают в молочных зу-
бах еще большую сложность.
Исчезновение гиппариона в начале плио-
цена, почти одновременное в различных
странах, кроме юга Франции (Roussillon),
должно быть объяснено, кроме неудачной
организации ног, еще каким-нибудь другим,
может быть, стихийным фактором. Последние измененные потомки его с уко-
роченными ногами известны из плиоцена южной Франции — Hipparion cur-
ium (Roussillon).
Сем. Palaeotheriidae. — Род Palaeotherium — из верхнего эоцена и
нижнего олигоцена Франции, является второй формой, близкой к лошадям, на что
еще указывал Кювье, но имевшей трехпалую конечность. Строение его скелета
и черепа — лошадиного типа (рис. 300). Но если рассмотрим зубы этой древней
формы, то мы не найдем им места в той линии, в когорую расположились изучен-
„ 3 14 — 3
ные нами формы. Их здесь — — ——--.
0 10 - 0
Но кроме рг1, все остальные предко-
ренные типа коренных, разве немного меньше размерами. Каждый из коренных
*) М. В. Павлова, Положение гиппариона в лошадиной линии по новым науч-
ным данным (Бюлл. Московского общества испытателей прирогы, 1925).
подкласс: плацентные — placentalia, копытные
241
состоит из двух продольных рядов бугорков, по три в ряд, причем внешние
приняли форму лунок, а внутренние стали овальными и загнулись назад внутрь
челюсти. Они соединены с внешними посредством маленьких, сжатых средних
бугорков, которые при стирании сливаются с внутренними. У некоторых ви-
дов зубы окружены воротником; у других он прерывается. Нижние коренные
состоят из двух луночек; добавочный бугорок отсутствует. Клыки хорошо
развиты. По форме зубов и по сложности предкоренных Palaeotherium
(рис. 301) отличается от современных ему Pachynolophus и Epihip-
pus лошадиной линии. Изменение бугорков шло у него иначе, чем у
этих форм.
Ноги его трехпалые, с маленькими остатками пятого metacarpus. Осталь-
ные метаподии длинные с короткими фалангами (рис. 302).
Род этот чрезвычайно богато представлен различными видами во Франции,
начиная со среднего эоцена в гипсах Парижа, в лигнитах Апта (верхний эоцен)
Рис. 301. Palaeotherium curium Cuv.
Рис. 302. Palaeothe-
rium medium Cuv.
и в фосфоритах Керси (олигоиен). Размеры различных видов очень различны.
Наибольший Palaeo th. magnum Cuv. достигал размеров лошади; наимень-
ший Palaeo th. (Paloplotherium) minus был не больше овцы. Виды
эти встречаются, как большие, так и маленькие, вместе в различных местностях
Франции. Кроме размеров они отличаются несколько и строением зубов,
большей или меньшей их сложностью, что вызвало отделение некоторых
форм в отдельный подрод.
Paloplotherium crassum Cuv. (рис. 303) — из грубого изве-
стняка Парижа (Anse), средний эоцен, является как бы предшественником
Palaeotherium. У него коренные построены по типу этого последнего из
6 удлинившихся бугорков. Предкоренные все проще и позволяют проследить
их осложнение от простого конуса рг1. Рг2 имеет два слившихся бугорка на
внешней стороне и один на внутренней. Рг3 имеет два бугорка на внешней
стороне, а на передней — промежуточный и внутренний бугорки слились; на
задней он имеет среднюю складку. Рг4 того же типа, но у него слились
16 п а в л о в а М. В. Палеозоология. Ч. II.
242
класс: млекопитающие — mammalia
внутренние бугорки в общую полукруглую складку. Осложняя предкоренные,
эта форма могла выработать зубы Palaeotherium1).
Но и для Р а 1 о р 1 о t h е г i u m мы не можем указать родственную форму
между предками лошади. Вероятнее всего, что она отделилась от кого-нибудь из
Condylarthra и повела свою отдельную линию, выразившуюся в большом
числе однородных форм, живших на очень ограниченном пространстве во Фрай-
ции в конце эоцена и в олигоцене, после чего их остатки не встречаются.
При этом одна ветвь изменяла предкоренные, и они становились похожими на
коренные, сохраняя рг\ Другая ветвь хотя и осложняла предкоренные, но они
оставались и в олигоцене проще, чем коренные; однако у них исчез рг1 (Pal.
minus, Pal. annectens).
В — верхний коренной, С — нижний.
Рис. 303. Palaeotherium crassum Cuv. (Верхний эоцен.)
Palaeotherium был указан В. О. Ковалевским в 1873 г.
как древнейший лошадиный предок. За ним следовали Anchitherium,
Hipparion и Equus.
Новые находки и новые сравнения форм привели к другим выводам и
заставили ввести в генетическую линию другие формы.
Интересно объяснение, которое дает Ковалевский способу уко-
рачивания пальцев трехпалой лошади, например Anchitherium, при
переходе к однопалой, с одним средним функционирующим пальцем, и к роду
Equus.
Три метаподия Anchitherium связаны между собою как общностью
развития, так и прилегающими к ним костями вверху и особенно внизу, где
их боковые стороны плотно соприкасаются друг с другом. При необходимо-
сти сокращения, укорачивания боковых пальцев, вследствие изменения режима,
организму трудно нарушить эти тесные отношения костей, и он обращает
свои силы на другое. Оставляя нижние концы II и IV метаподий на прежнем
месте и сохраняя их прежние размеры, начинает увеличивать, удлинять первые
фаланги III метаподия, и нога поднимается, а боковые пальцы становятся короче.
Когда они переставали вследствие этого функционировать, тогда получали меньше
<) В род Paloplotherium следует включить Pal. minus и Pal. a n-
nectens с зубами, более близкими к Palaeotherium, но имеющих предкорен-
ные более простые, чем коренные, и лишившихся рг*, как это указал Г о д р и
в „Enchainements".
подкласс: плацентные — placentalia, копытные
243
питания и начинали атрофироваться. В среднем пальце увеличивалась первая
фаланга, и развивался гребень на нижней сочленовной поверхности метаподия,
входящий в желоб первой фаланги и укрепляющий это сочленение, заменяя
боковые пальцы. Действительно, мы видим удлиненную первую фалангу у всех
однопалых лошадей, хотя бы и сохранивших еще остатки боковых пальцев.
Сем. Litopterna Aniegh. (гладкая пятка). — Представляет группу вы-
мерших форм пампасов Южной Америки, существовавших там с эоцена до
плейстоцена. Эти пальцеходящие непарнопалые по внешнему виду напоминают
формы лошадиные, но не по строению своих конечностей: в последних сохра-
няется сериальное расположение карпальных костей.
Рис. 304. Macrauchenia patagonica Ow.
Macrauchenia (рис. 304), известная целым скелетом, была давно
изучена Бурмейстером (Burmeister). Удлиненный череп ее имеет очень
короткие носовые кости и далеко назад отнесенные носовые отверстия, кото-
рые образуют большие дыры наверху черепа. Орбиты замкнуты. Озубление
полное. На внешней стенке зубов мы видим две глубокие лунки, от которых
идут, направляясь назад, стертые гребни, образовавшие (при стирании) замкну-
тые кружочки. Нижние зубы состоят из двух лунок. Шейные позвонки —
с плоскими сочленовными поверхностями.
Ноги у различных родов имеют по одному или по три пальца,
почти равной длины, причем расположение карпальных костей остается рядо-
вым, не смещенным; radiale над trapezoideum и trapezium, lunatum над magnum
и ulnare над unciforme, соприкасаясь с magnum. Сочленовные канты развиты
только на задней поверхности метаподий. На пяточной кости существует
фасетка для малой берцовой, как у парнопалых.
16*
244
класс: млекопитающие — mammalia
отверстиями в передней
Рис. 305.А — Theocodon
В — Thoatherium;
С— Diadiaphorus.
Theocodon — с 3 пальцами (рис. 305,А).
Diadiaphorus (рис. 305,С) — из верхнего миоцена Санта-Круца,
имеет очень укороченные боковые метаподии, с удлиненной первой фалангой
среднего пальца. Здесь radiale перешла отчасти на magnum, но над IV паль-
цем сохранился trapezoideum под radiale. Ulnare и unciforme лежат над IV
metacarpus.
В задней конечности calcaneum и cuboideum опираются на IV matatarsus,
не переходя на средний metatarsus, как это наблюдается у лошади.
Череп — укороченный, с короткими носовыми костями и с носовыми
части черепа, а не в верхней. Большие клыки
отделены от предкоренных диастемой. Предкоренные
несколько проще и меньше коренных, которые по
форме наружной стенки и гребней стоят ближе
всего к зубам Palaeotherium. Эта форма тоже
напоминает лошадиные, но ноги ее указывают
на неправильную редукцию пальцев относительно
запястья.
Thoatherium Amegh. (рис. 305, В) — из
верхнего миоцена Санта-Круца, дает еще более ре-
дуцированную конечность: это настоящая однопа-
лая нога лошади со слабыми остатками боковых
метаподий. Но карпальные кости у нее сохранили
рядовое расположение с unciforme и trapezoideum
над II и IV metacarpus. В задней ноге cuboideum и
calcaneum едва касаются III metatarsus, они лежат
над остатком IV metatarsus.
Нижняя челюсть, найденная вместе с Thoa-
therium, имеет 2 резца и 1 клык, небольшой,
отделенный маленькой диастемой от конусообразного рг1. Остальные зубы
имеют по две луночки.
Итак, все эти формы (есть еще несколько близких к ним) дают нам раз
личные типы непарнопалых конечностей, напоминающих лошадиные. Но при
близком знакомстве мы видим, что здесь редукция пальцев шла иным путем,
без перемещения соответствующих карпальных и тарсальных костей, т. е. тя-
жесть тела не распределялась правильно для передачи пальцам, как у форм
лошадиной линии, и Мэтью прав, называя эту группу патагонских непарно-
палых „ложными лошадьми" (Pseudo-Horse).
Интересно подчеркнуть, что названные формы Litopterna не стоят
в генетических отношениях между собою, да и по форме черепа они сильно
между собою различаются.
Но самым интересным является то, что формы с одним пальцем (Thoa-
therium) найдены в верхнем миоцене, с одним хорошо развитым и неболь-
шими боковыми (Diadiaphorus) — в миоцене и плиоцене, а с тремя хорошо
развитыми пальцами (Macrauchenia) — в плейстоцене.
подкласс: плацентные—.placentalia, копытные
245
Эти типы могут быть хорошей иллюстрацией „неадаптивной редукции"
непарнопалых конечностей, противоположной „адаптивной редукции" конечно-
стей форм лошадиной линии.
Каждая из этих форм отделялась от неизвестных предков и изменялась
по одному типу неадаптивной редукции в различных направлениях, и те из
них, которые раньше освободились от боковых пальцев, раньше и вымерли,
например Thoatherium. Macrauchenia с тремя пальцами дожила до
плейстоцена.
Сем. Rhinocerotidae (носороги). — Это семейство является полной
противоположностью лошадиным в смысле своего географического распростра-
нения. Насколько велико оно для первых, настолько сжато для вторых.
В настоящее время носороги известны только в немногих жарких странах
Азии и Африки. 4
Между ними различаются два типа:
1) Rhinoceros indie us — азиатский однорогий носорог, имеющий
резцы и покрытый шерстью. Близкий к нему Rhinoceros sondaicus,
тоже однорогий, живет на Зондских островах.
2) Rhinoceros bicornis — африканский, с двумя большими рогами,
лишен резцов. Шерсть только на ушах. Имеет длинную верхнюю губу, пи-
тается ветками. Близкий к нему по рогам Rhinoceros sim us отличается
короткой губой; питается травой.
Как бы промежуточными типами являются Rhinoceros lasiotis на
Суматре, с двумя небольшими рогами и резцами, покрыт шерстью, и Rhi-
noceros sumatrensis, с двумя маленькими рогами, тоже покрыт волосами,
самка без рогов. Последний как бы вариетет предыдущего.
Все они имеют трехпалые конечности, с преобладанием среднего пальца.
Ископаемые остатки носорогов известны со среднего эоцена Европы и
Сев. Америки, причем древние носороги дают третий тип — безрогих носоро-
гов — Aceratherium, большею частью снабженных четырьмя пальцами на
передних ногах.
В миоцене мы встречаем наибольшее их развитие в Сев. Америке и
в Европе и, кроме того, в Азии, где они появились в начале миоцена.
В плиоцене безрогие носороги исчезают в Сев. Америке; в Европе и
Азии продолжается широкое развитие изменившихся типов: однорогих и дву-
рогих носорогов.
Немногие формы существуют в плейстоцене, и еще меньшее количество
их известно теперь.
В Африке встречаются только незначительные остатки в плейстоцене и
ныне живущие двурогие формы. Ископаемые остатки носорогов из нижних
третичных отложений во многом отличаются от более поздних форм; у них
даже отсутствует главный признак — рог, но, тем не менее, другие признаки
скелета позволяют отнести их вполне определенно к этому семейству.
Обращаясь к группе первичных копытных (С о п d у 1 а г th г а), мы встре-
чаем там так же, как и для лошадей, формы, так сказать, „подготовляющие"
246
класс: млекопитающие — mammalia
развитие носорогов. К сожалению, остатки этих форм далеко не так полны,
как для лошадиной линии — Tetraclaenodon puercensis. Раньше чем
описывать эти формы, взглянем на скелет современного носорога и главным
образом на его конечности и озубление.
Ноги имеют по три пальца, с менее удлиненными метаподиями, чем
у лошади, и с укороченными тремя фалангами пальцев, с расширенной послед-
ней, копытной.
Зубы — верхние коренные более сложные, чем предкоренные, состоят
из внешнего гребня, образующего внешнюю стенку, от которой идут два по-
перечных гребня, снабженные различными выступами, направляющимися в сред-
нюю долинку, разделяющую их. Последний т3 имеет треугольную форму.
Нижние состоят из двух лунок. Клыков нет; резцы могут отсутствовать.
Изучая ископаемые формы, мы увидим развитие этих признаков.
Systemodon (из Со n dy lart hr а)—нижний эоцен Сев. Америки, Вазач.
Systemodon semihians Соре — имеет коренные зубы, состоящие
из двух больших внешних бугорков, двух меньших внутренних, слившихся
Рис. 306. Systemodon ta-
pir i n u s Cope.
с двумя маленькими промежуточными. Отличи-
тельной особенностью их от зубов Tetra-
claenodon puercensis являются значительно
большие первичные бугорки: протокон, метакон,
паракон и присутствие на переднем внешнем углу
зуба особой складки эмали (parastyle), которая
очень характерна для зубов носорогов и тапиров. Предкоренной рр — малень-
кий конус, стоит между клыком и рг2; остальные предкоренные имеют два
внешних бугорка и один внутренний; на рг2 он очень маленький. Нижние зубы
состоят из четырех бугорков, соединенных поперечными складками эмали. РР
отделен от рг2 диастемой.
Systemodon tapirinus Соре (рис. 306) — Вазач (Сев. Америка),
представлен раздавленным черепом с сагиттальным небольшим гребнем. Орбита
не замкнута, но ограничена сзади заорбитальным отростком. Носовые кости
узкие, доходящие почти до конца морды, так что носовое отверстие лежит
в конце ее (спереди). Нижняя челюсть узкая, с высокой задней частью. За
небольшими резцами находятся большие загнутые клыки. Предкоренные более
сложные, чем у System, semihians. Рг2 имеет большой внутренний бу-
горок; на рг3 он выражен еще сильнее, а на рР от него идет складка к зад-
нему наружному бугорку. Коренные верхние состоят из отчетливых внешних
бугорков, со складкой на переднем углу протокона, и двух гребней, идущих от
них из слившихся внутренних и промежуточных бугорков. На нижних коренных
по 4 бугорка. На стертых зубах бугорки не отличимы от поперечных гребней.
Вероятно, эти формы стояли у основания развития носорогов и ближайших
к ним сем. тапиров и лофиодонов. Подтверждение этому дадут находки более
полных остатков.
Heptodon Соре — очень близкая к Systemodon форма, у которой
бугорки на внешней стенке сгладились; зубы имеют по два поперечных
подкласс: плацентные — placentalia, копытные
247
гребня. Но его конечности, с удлиненным средним пальцем, не позволяют
считать его предком носорогов.
Подсем. Aceratheriidae (безрогие носороги). — Hyrachius exi-
mius (рис. 307) и Ну г. agrarius Leidy (рис. 308) — эоцен Сев. Америки
и олигоцен Керси, представлены хорошо сохранившимися остатками, позволяю-
щими восстановить цельный скелет. Удлиненный, узкий череп с тонкими,
п
Рис. 307. Hyrachius eximius Leidy.
А — верхние рг* и т\. В — нижний т*. С — передняя нога, D — задняя.
Рис. 308. Hyrachius ag-
rarius Leidy. Верхние зубы.
выступающими носовыми костями, с открытой орбитой и заглазничным слабым
выступом, толстыми скуловыми дугами, сагиттальным гребнем, является как бы
измененным в определенном направлении черепом Systemodon tapirinus.
Узкая нижняя челюсть с высокой восходящей задней
частью дополняет его. Клыки отделены от предко-
ренных диастемой в обеих челюстях. Озубление
полное, причем предкоренные проще коренных;
pi4 является простым конусом, следующие пред-
коренные все одинаковой треугольной формы.
Коренные: т1 с сохранившимися бугорками на
внешней стенке, от которых идет два поперечных гребня; на т2 и т2
бугорки менее заметны. Везде существует parastyle. Длина семи коренных
равняется 6 см. Нижние состоят из двух поперечных гребней, слабо загнутых
наперед на внешней стороне.
На передних конечностях по 4 пальца, причем И, III и IV почти равны;
V короче их, так что нога имеет признаки непарнопалой. Карпальные кости
чередуются между собой.
Задние конечности — типичные трехпалые. Астрагал с глубоким верхне-
передним блоком и гладкой нижней поверхностью типа настоящего носорога.
248
класс: млекопитающие — mammalia
В фосфоритах Франции Hyrachius представлен только зубами, осталь-
ные части скелета не найдены.
В Сев. Америке и в Европе, главным образом во Франции, известны из
олигоцена многочисленные остатки, главным образом черепов, разнообразных
Рис. 309. Trigonias Osborni Lucas. Зубы левой верхней челюсти.
носорогов из А с е га th eri i d a e — безрогих, позволяющие проследить раз-
личные их изменения и переход к более поздним формам.
Рис. 309 A. Trigonias Osborni Luc. Левая нижняя
челюсть.
Изменения касаются
как формы черепа, так и
строения зубов. Первые
заключаются в большем
или меньшем поднятии за-
тылочной части. Прогрессив-
ное изменение зубов состоит
в изменении формы предко-
ренных в направлении к ко-
ренным и в развитии доба-
вочных выступов — крюч-
ков — на поперечных греб-
нях верхних коренных.
Hyracodon pris-
с i d е n s — из олигоцена Ка-
нады, отличается прямыми
наружными стенками верх-
отходягцими от них гребнями и закругленными
них предкоренных, с двумя
внутренними сторонами. Три
подкласс: плацентные — placentalia, копытные
249
предкоренных, почти равных коренным. Последние сходны с коренными Нуга
chius. На передней ноге у него три пальца.
Эта форма опередила по развитию предкоренных и сокращению IV пальца
на передних конечностях другие олигоценовые формы из Aceratheriidae.
Олигоценовые отложения крайне богаты остатками древних безрогих но-
сорогов как в Европе, так и в Сев. Америке. Богатый материал этот обработан
d
А с е г a t h. i'n-
cisivumj
С
Acer, tetra-
dactylum
b
Acer. 1 e ma-
il e n s e
a
Acerath.
F11h о 1 i
Рис. 310. Изменение верхних зубов у Aceratheriidae.
главным образом Осборном в Сев. Америке, Абелем в Вене, Романом
(Roman) в Лионе; но он не объединен, и нередко одни и те же формы
(остатки их) описаны под разными названиями. Укажем главнейшие роды.
Trigonias Osborni Lucas (рис. 309 и 309А) — является древнейшей
формой (нижний олигоцен White River beds), у которой полное озубление вверху
и недостает внизу только клыков. Поперечные гребни коренных без добавочных
складок. Носовые кости и межчелюстные удлиненные, тонкие. Передние конеч-
ности четырехпалые, с хорошо развитым пятым пальцем.
Leptaceratherium Osb. — оттуда же, имеет только по 2 резца
в челюсти, лишен нижних клыков и pt\. На передней конечности 3 пальца.
250
класс: млекопитающие — mammalia
Protaceratherium Abel — на верхних коренных появляются складки
эмали — крючки (crochet и antecrochet — крючок и передний крючок). Предко-
ренные— четырехугольные, со сходящимися гребнями на внутренней стороне.
Рис. 311. Indricotherium transuralicum М. Pavl.
.А—верхи, челюсть, В—нижняя. С—передняя сторона нижней (.челюсти со сломанными резцами. D — резец
верхней челюсти.
Aceratherium Filholi Osb. (рис, 310, с) — из нижнего олигоцена
Керси, бллжайшая форма к Hyrachius, имеет предкоренные еще треугольной
формы, с передними гребнями, не доходящими в нестертом состоянии до наружной
подкласс: плацентные — placentalia, копытные
251
стенки зуба. Такую же форму верхних предкоренных мы встречаем у несколь-
ких американских форм, которые приводят нас к Acerath. occidentale
Leidy, у которого мы видим изменение предкоренных: все гребни дохо-
дят до внешней стенки на предкоренных; на тл развивается слабый вы-
ступ — передний крючок — на внутренней стороне переднего гребня, входящего
в среднюю долинку. Нижние коренные имеют два загнутые наперед гребня.
Затылок узкий, слабо поднятый. Зубы его находят и в Европе.
Acerath, lemanense (рис. 310, b) — из верхнего олигоцена Керси
имеет уменьшившийся рг1, остальные — четырехугольные, с двумя гребнями.
На /г1 — передний крючок; на т2.
появляется слабый крюч< к и на
заднем гребне, тоже направленный
в среднюю долинку.
Indricotherium Borys5).
В континентальных отложениях
Чалкар-Тениза, Тургайской обла-
сти, были найдены остатки крайне
интересной формы безрогого но-
сорога, изученного А. А. Б о р и-
сяком (1916—1917 гг.), давшим
ему название Indricotherium.
Рис. 312. Beludgitherium. (Сев. Америка.)
Остатки близкой формы изучены
мною. Indricotherium пора-
жает прежде всего своими размерами. Его III metacarpus имеет в длину 58 см.
Вся ступня с очень укороченными фалангами пальцев = 84 см. Зубы пре-
восходят вдвое зубы другиAceratheriidae.
Отложения, в которых найдены остатки Indricotherium, относятся
А. Борисяком к олигоцену.
Особенностью .этой формы являются передние зубы. У нее в верхней
челюсти существовало по одному большому резцу, в нижней — по два функцио-
нирующих и по два почти атрофированных.
Коренных сохранилось в обеих челюстях по 6; может быть, сохранился
и рг\ но челюсть обломана.
По строению коренных форма эта является примитивной. Зубы не имеют
крючка, но т3 треугольной правильной формы. Описаны два вида, различаю-
щиеся по зубам: Indricotherium turgaicum Вог. и Indricoth. tran-
suralicum M. Pavl. (рис. 311).
Близкие формы, может быть тождественные, найдены в Белуджистане
и в Монголии, которые Осборн описывает под именем Beludgitherium
(рис. 312), сближая его с Indricotherium.
*) А. А. Б о р и с я к, Остеология индрикотерия — Известия Российской академии
наук, 1917; Записки Академии наук, том 36.
М. Р a v 1 о w, Indricotherium transuralicum — Бюллетень Мо ск. обш.
испытателей природы, 1922.
252
класс: млекопитающие — mammalia
Очевидно, различные представители Aceratheriidae развивались па-
раллельно во время олигоцена в Европе и Сев. Америке и переходят в миоцен,
осложняя зубы, но сохраняя четвертый палец, уменьшившийся сравнительно
с Hyrachius.
Aceratherium tetr a d а с ty 1 u m Каир (рис. 310, с) в Европе и
Acerath. acutum Соре в Америке являются представителями миоценовых
форм, осложнивших зубы развитием крючков на рг2, рг3
и рг4; рг1 уменьшился.
Одновременно с осложнением всех коренных зубов со-
кращаются клыки и резцы. Из последних остается одна
пара больших и одна маленьких, вверху и внизу, которые
Рис. 313. Aceratherium inci-
si v u m Каир.
исчезают у мио-плиоцено-
вых форм.
Aceratherium
iпсisivum Каир, Ace-
rath. Kowalevskyi
M. Pavl. — в нижнем плио-
цене |Европы и Acerath.
fossiger Соре— в Сев.
Америке—являются пред-
ставителями последних по
времени безрогих носоро-
Рис. 313Л. Acera-
therium inci-
si v u m Каир.
А — верхи., В — нижн.
зуб.
гов. Озубление их сходно (рис. 310, d, 315); у них сильно развиты резцы,
по одной паре (первой) в каждой челюсти, и осложнились коренные сильным
развитием крючков. В нижних коренных загиб гребней пошел дальше, и обра-
зовались лунки. х
Проследим изменение передних зубов у следующих олигоценовых ацера-
териев (рис. 314):
Резцы Клыки
Trigonias Osborn . . . . < ( Верхи. pi) 2 3 1
| Нижн. 1 2* 3
Aceratherium trigonodon. < f Верхи. 1* 2 — 1
1 Нижн. 1 2* —
Aceratherium occidentale ' f Верхи. 1* 2 —
1 Нижн. 1 2* —
Aceratherium tridactylurn f Верхи. 1* 2 —
1 Нижн. 1 2* —
Проследив сокращение резцов и клыков, мы определенно видим,
что остающиеся передние зубы — это резцы, первая, внутренняя пара в
верхней челюсти и вторая пара в нижней, а не клыки, как считают их
некоторые авторы.
*) Наибольший зуб обозначен через *.
подкласс: плацентные — placentalia, копытные
253
Trigonias Osborni.
Aceratherium trigonodon.
Aceratherium tridactyl urn.
Рис. 314. Изменение передних зубов у олигоценовых Aceratheriidae.
254
класс: млекопитающие-—mammalia
У более поздних форм сем. Aceratheriidae — Aceratherium
Kowalevskyi (из Гребенников б. Херсонской губ.), Aceratherium
Рис. 315. Aceratherium Kowalevskyi М. Pavl.
Рис. 315 A. Aceratherium Ко w а - Рис. 316. Aceratherium fossiger
1 е v s к у i М. Pavl. Соре.
tridactylum Osb. (Сев. Америка) — четвертый палец на передних конечностях
сократился до тонкого стилета, а у некоторых он исчез.
Носовые кости у этих форм тонкие, затылок приподнят. Скелет А с е г a t h
fossiger отличается очень низкими конечностями, что его сближает с А с е г а-
подкласс: плацентные — placentalia, копытные
255
therium Kowalevskyi М. Pavl. (рис. 315); зубы этих форм тоже сходны
(рис. 316).
Подсем. Rhinocerinae. — Безрогие нижне-плиоценовые формы заканчи-
вают развитие носорогов в Сев. Америке, тогда как в Европе развиваются
настоящие Rhinocerotidae, с рогами, получившие начало в миоцене.
Рис. 317. Rhinoceros sansaniensis!
Lart.
В среднем миоцене одновременно с
Aceratherium tetradactylum
Каир встречается носорог, снабжен-
ный небольшим рогом—Rhinoceros
sansaniensis Lartet (рис. 317).
По коренным зубам и форме черепа
он близок к Aceratherium 1 е-
manense, но у него начинают
уменьшаться клыки и резцы, передняя
нога теряет четвертый палец, и начи-
нает вырабатываться тип настоящего
носорога.
В верхнем миоцене Rhinoceros Schleiermacheri Каир (рис. 318)
как бы продолжает развитие предыдущей формы. Передняя часть морды укора-
чивается, так как клыки и резцы исчезли. Носовые кости стали толще для
поддержания хорошо развившегося рога. На лобных замечается слабое возвы-
шение для зачатка второго рога;
затылок приподнимается. Коренные
зубы осложняются развитием крюч-
ков Ноги с хорошо развитыми тре-
мя пальцами с укороченными фалан-
гами. Остатки этих фирм найдены:
Rh. sansaniensis — во Фран-
ции, Rh. Schleiermacheri — в
Германии и в Херсонской губернии;
но целые скелеты неизвестны.
В осложнении зубов р. Rhi-
noceros наблюдается такая же
последовательность, как и у р.
Aceratherium (рис. 318).
Рис. 318. A — Rhinoceros sansaniensis,
В—Rhin. simorrensis, С—Rhin. Schlei-
ermacheri.
Два последние носорога могут
считаться родоначальниками всех
позднейших, которые сущее гвовали
в Европе и Азии в плиоцене и от которых развились ныне живущие азиатские
и африканские носороги, хотя точной связи между последними и выше назван-
ными сейчас указать нельзя.
Верхний миоцен может считаться временем расселения носорогов, так
как в этих отложениях остатки их известны и в Европе и в Азии. Что каса-
ется Африки, то до сих пор остатки их известны трлько в плейстоцене.
256
класс: млекопитающие — mammalia
Сиваликские холмы в Азии очень богаты ископаемыми однорогими носоро-
гами, из которых наиболее древний Rhinoceros sivalensis, очень близкий
к Rh. sansaniensis Lart. Он отделился, вероятно, в миоцене от последнего.
Rh'inoceros palaeindicus Lyd. — из плиоцена Сиваликских хол-
мов, с одним рогом, парой резцов и осложнившимися зубами, является про-
должателем предыдущего и ведет, вероятно, к нынешнему однорогому азиат-
Рис. 319. Rhinoceros pachygna-
thus.
скому Rhinoceros indicus, хотя
в плейстоцене Азии остатки носорогов
неизвестны.
Rhinoceros pachygnathus
Gaud. (рис. 319) — в мио-плиоцене (мео-
тических отложениях Европы, Пикерми),
очень близок по скелету и черепу
к Rhinoceros Sсh 1 еiегma chеri
Каир, встречающемуся также в Пи-
керми. Но череп его более громоздкий,
и возвышенность для второго рога
больше; на коренных крючки более развиты. Предкоренные имеют закруглен-
ную внутреннюю сторону.
Rhinoceros megarhinus Christ. — из плиоцена имеет более скла-
дочек эмали, ограничивающей долинки, и его предкоренные зубы становятся
прямыми на внутренней стороне. У этих зубов получает большое развитие
задняя долинка, которая
при стирании зуба дает
замкнутое углубление. Рг4
исчез. Оба рога были
сильно развиты, что видно
по оставшимся шерохова-
тостям на черепе.
Rhinoceros et-
r u s с u s Falc. — из пли-
1 опена Vai d’Arno СИта- Рис- 32°- Rhinoceros Mercki H. v. Меу. (Череп
k в коже.)
лия) и плейстоцена Маура,
с мало поднятой затылочной частью и сходными с Rhinoceros megarhinus
зубами, развивает новую особенность. Очевидно, рог стал очень тяжел для
носовых костей, и существующая у носорогов хрящевая носовая перегородка
стала окостеневать с передней стороны — для поддержания рога, что и отли-
чает, главным образом, этот череп от предыдущего. Молочные зубы плейсто-
ценовой формы очень осложнились добавлением крючка, идущего от внешней
стенки. На т1 он тоже развит и при стирании дает цве воронки в верхней
части средней долинки.
Rhinoceros hemitaechus Falc. — является параллельной фор-
мой Rhinoceros etruscus на севере. Он встречается в верхнем плио-
цене и плейстоцене (древнем) Англии (Rhinoceros leptorhinus Ow.) и
подкласс: плдцентныЕ — placentalia, копытные
257
Rhinoceros Merckei Meyer (рис. 320). У него существует тоже неполная
костная перегородка, поддерживающая носовые кости, как у Rhinoceros
etruscus Италии, но затылочная часть черепа поднята вверх значительно
выше.
На предкоренных увеличиваются складки эмали —на передней стороне
заднего гребня. На коренных (верхних) сильно развит крючок. Череп Rhino-
ceros Merckei (покрытый кожей), найденный в Сибири, был отнесен к
Rhinoceros tichorhinus, волосатому носорогу, встречающемуся там же.
Но присутствие полуперегородки, открытое в нем позже, заставило отнести
•его к Rh. Merckei Meyer.
У Rhinoceros tichorhinus (рис. 321) эта перегородка окостене-
вает и перегораживает носовую полость. В молодости окостенение ее
шло постепенно, и у ископаемых форм мы можем видеть различную
степень ее окостенения. При этом наблюдаются моменты в ее образова-
нии, когда она, окосте-
нев вся, еще не срослась
вполне с носовыми костя-
ми и может быть легко
выдвинута или выпасть са-
ма; в этих случаях череп
Рис. 321. Rhinoceros tichorhinus.
Khtichorhinus может быть принят зачереп, не имеющий перегородки и при-
надлежащий Rh. leptorhinus Cuv., жившему также в плейстоцене. Но
форма затылка и зубы у последнего отличаются: первый — меньшей высотой,
вторые — меньшей сложностью.
Зубы Rhinoceros tichorhinus (рис. 321,А) являются самыми слож-
ными. У него на коренных, кроме крючка (сг) и переднего крючка (асг),
отходящих от гребней во внутреннюю
долинку, развивается еще складка эма-
ли, идущая от внешней стенки тоже во
внутреннюю долинку (с/), что при сти-
рании зуба дает верхнюю часть долинки
в виде двух воронок, иногда глубоких.
Нижние коренные состоят из двух греб-
ней, сильно загнутых вперед на на-
ружной стороне. В некоторых зубах
найдены остатки пищи — хвоя и ива.
Верхние предкоренные меньше корен-
17 Па вдова М. В. Палеозоология. Ч. II.
Рис.321, А.Зубы Rlii п. tichorhinus.
А — верхний коренной, В — нижний корснног.
258
класс: млекопитающие — mammalia
ных, но тоже сложнее, чем у других носорогов» Полное отсутствие резцов
и клыков. Конечности имели по три хорошо сохранившихся пальца, с правиль-
ным перемещением карпальных костей.
Остатки этого носорога чрезвычайно широко распространены в Европе
(неизвестны только в Италии и Испании) и в северной Азии. Есть указание, что
тело его было покрыто шерстью, как у нынешних азиатских форм1). Его нельзя,
однако, поставить в генетическую связь ни с одним из ныне живущих видов
носорогов, вследствие сложности его зубов и костной носовой перегородки, не
существующей ни у одного из нынешних носорогов. Нужно искать предка ны-
нешних двурогих носорогов, Rhinoceros bicornis Африки, между потом-
ками Rhinoceros pachygnathus Gaudry из Пикерми.
Из этого краткого описания развития носорогов с эоцена до нашего
видно, что оно имело совершенно другое
времени
направление, чем развитие
лошадей. В то время как
Рис. 322. Elasmotherium sibiricum.
А — верхний зуб, В — нижний.
безрогих и рода Rhinoceros—с одним рогом.
последние развивались
стройным рядом с не-
сколькими небольшими
ответвлениями, развитие
носорогов шло как бы
кустообразно. До мио-
цена существовали без-
рогие носороги, но уже
в миоцене мы имеем па-
раллельное развитие и
рода Aceratherium—
В верхнем миоцене по-
являются двурогие, развиваются в Европе и расселяются в Азии. В конце плио-
цена мы имеем формы, у которых развивается костная полуперегородка, формы
с более плоским черепом — Rhinoceros etruscus — и другие с более
приподнятым затылком — Rhinoceros hemithacus. Наконец, в плейсто-
цене встречаются формы с полной костной перегородкой — в Европе и Азии.
В настоящее время в Азии и Африке живут двурогие и однорогие носороги
нескольких видов, представляя незначительные остатки богато представленного
в миоцене и плиоцене семейства.
Род Elasmotherium (рис. 322). — Род этот представлен черепами и
зубами, найденными главным образом в губерниях Самарской, Саратовской и
в Сибири, и костями конечностей. Череп отличается громадными размерами,
более метра, с громадным вздутием лобных костей, покрытым шероховатостью,
указывающею на присутствие большого рога. Носовые кости тонкие — оче-
видно, они не несли рогов. Нижняя челюсть узкая. Большую особенность
представляют зубы, сократившиеся до пяти коренных вверху и внизу, в виде
*) Большую редкость представляет сохранность остатков Rhinoceros
tichorhinus в виде мумий в нефтеносных слоях Старуни (Хорватия), с сохранив-
шейся сухой кожей, облегавшей кости в их естественном положении.
подкласс: плацентные — placentalia, копытные
259
удлиненных цилиндров, с постоянным ростом, с незакрытой пульпой. Они
состоят из дентина со складками эмали, образующими на верхних зубах внеш-
ний' гребень, от которого идут два поперечных гребня, как у носорогов. Но
эмаль, ограничивающая эти гребни, не гладкая, а вся сложена в глубокие
складочки, которые входят местами в среднюю долинку в виде крючков (более
удлиненных и сложных складок). Нижние зубы состоят из двух лунок. Все
зубы покрыты цементом. Резцов и клыков не было.
Конечности известны по отдельным костям плеча и предплечья,
бедра и голени и метаподий; все эти кости по форме своей вполне сходны
с костями носорога, но значительно больших размеров, даже чем самые круп-
ные носорожьи. Они были описаны Г о др и.
В 1913 г. остатки Е1 as m о th ег iu m — зубы — найдены на Кавказе
близ Ставрополя Кавказского и близ Баку. Все находки, кроме Е1 a s m о t h е-
rium caucasicum Borys., принадлежат одному виду — Elasmotherium
sibiricum. Вид А. Борисяка, хотя и представляет более простую форму,
но все-таки не дает указаний на генетическое отношение этого рода к носо-
рогам. До последнего времени Elasmotherium считался послетретичной
формой, хотя и не было на это точных указаний.
Теперь есть данные считать, по крайней мере, некоторых из них при-
надлежащими к третичной (самой верхней плиоценовой) фауне. В Баку найдены
два зуба в ракушечном известняке с Dreissensia rostriformis и
Dreis, polymorph а.
Enigmatherium stavropolitanum M. Pavl. — В Ставропольской
губернии найден зуб совершенно новой формы, вместе с прекрасно сохранив-
шимися зубами Elasmotherium sibiricum. Этот зуб соединяет в себе
некоторые признаки зубов носорога и эласмотерия. Он имеет корни, но на
его жевательной поверхности гребни имеют складчатую эмаль. Может быть,
это зуб одной из тех форм, через которые шел путь развития эласмотерия1).
Сем. Tapiridae является ближайшим к носорогам, отличается присут-
ствием четвертого пальца на передних конечностях, присутствием небольшого
хобота и иным строением зубов. В остальных частях скелета тапиры близки
к носорогам. Семейство это распадается на два подсемейства: а) собственно
Tapirinae и б) Lophiodontinae.
Первое представлено ископаемыми формами и ныне живущими в южной
Индии и Южной Америке; второе, вымершее рано, — эоценовыми формами
в Европе. Нынешние тапиры имеют ископаемых представителей, начиная с мио-
цена. Но древнейшими их предками могли быть или общие формы с предками
носорогов, или близкие к ним.
Прямые гребни верхних коренных зубов тапира, идущие от внешней
стенки, встречались нам уже у Heptodon и Systemodon, древних
9 А. Б о р и с я к, О зубнохМ аппарате Elasmotherium caucasicum Bor. —
Изв. Росс. акад, наук, 1914 г.
М. Павлова, Находки Elasmoth. sibiricum в Ставропольской губ. —
Записки Геолог, отделения О. Л. Е., Москва, 1915 г.
17*
260
класс: млекопитающие — mammalia
Condylarthra. Присутствие у этих форм добавочной складки (parastylus) на
переднем углу внешней стенки увеличивает сходство этих зубов с зубами
тапира. Главная их разница заключается в присутствии трех не слившихся
бугорков, образующих у тапира гребень, и в отличии между коренными и пред-
коренными, которые у древних форм проще — треугольные. Мы встречаем
Рис. 323. Protapirus priscus
Filh.
Рис. 323,А. я—Protapirus Douvil-
1 е i, b— Palaeotapirus, с— Tapirus
priscus.
как в Европе, так и в Сев. Америке, в миоцене, формы Protapirus, у ко-
торых этот признак еще сохраняется (Protapirus simplex Wortm.) и
бугорки ясно видны (рис. 323, 323, А).
У Protapirus Douvillei (миоцен Франции) и Prot. obliqui-
d е n s (Сев. Америка) на предкоренных образовались уже гребни, но бугорки
еще заметны.
в
Рис. 324. Palaeotapirus helveticus Meyer.
А — верхние зубы, В —• нижние.
У Palaeotapirus Gaud. — на всех предкоренных бугорки слились
в гребни, но сами зубы меньше, чем коренные (рис. 324).
У Tapirus priscus (нижний плиоцен) и Тар. arvernensis (верх-
ний плиоцен) рг3 и рг^ сходны с коренными.
Нижние зубы тапира состоят из двух прямых поперечных гребней, без
внешней стенки, что указывает на то, что передние и задние бугорки сливались
вместе.
подкласс: плацентные — placentalia, копытные
261
У Systemodon гребни имели слабые складки на внешней стороне.
Конечности древнего Heptodoп, как мы видели, имели уже в эоцене
четыре пальца на передней конечности и три на задней. И это строение ноги
сохранилось и у нынешних тапиров, с той разницей, что существовавшая
фибулярная фасетка на пяточной кости исчезла.
Сравнительно с носорогами тапиры представляют иное развитие. У отде-
лившихся от Condylarthra форм уже в начале миоцена начали сливаться
бугорки на зубах в гребни, а к концу миоцена остановилось развитие как
зубов, так и конечностей, сохранив консервативный тип признаков у немно-
гих дошедших до нас представителей, переселившихся в Азию и Южную
Америку, откуда известны их третичные остатки.
Подсем. Lophiodontinae представлено родом Lophiodon в эоцене
средней Европы, главным образом во Франции и Германии, несколькими
видами очень различных размеров. По скелету они близки к тапиру, с низким
черепом, с затылочным гребнем
ствовал небольшой хобот.
Узкая нижняя че-
люсть имеет высокий за-
гнутый венечный отро-
сток, широкий сочленов-
ный отросток и закруглен-
ный задний край.
Верхние коренные
зубы (6)— типа носоро-
га, с сильно выраженной 1/,
внешней стенкой и не-
сколько загнутыми попе-
и длинными носовыми костями; вероятно, суще-
Рис. 325. Lophiodon isselense Cuv.
речными гребнями, подразделений на бугорки нет. Передняя внешняя складка
(parastylus) развита больше, чем у тапира. Предкоренные проще коренных —
треугольные. Все зубы имеют сильно развитые воротники. Клык отделен
диастемой от рг1 и резцов (рис. 325).
Нижние коренные (6) с сильно загнутыми на внешней стороне гребнями.
На предкоренных гребни выражены менее ясно, чем на коренных.
Размеры лофиодонтов различны: от небольших — со свинью—до носорога.
Конечности Lophiodon leptorhinus наилучше изучены Dереге,
Передние — четырехпалые, с укороченным пятым метаподием, трехпалые задние
построены по типу ног Aceratherium tetradactylum, с укороченными
фалангами, что их отличает от ног современного тапира. Unciforme тоже отли-
чается от этой кости тапира: она не подстилает lunare, а только касается ее
одной фасеткой, как у Aceratherium.
Lophiod'on rhinoceroides Riit.— особенно крупный, с зубами, на-
поминающими носорога, но без крючков, с парас гилем. Как по строению зу-
бов, так и по скелету этот род близок к тапиру и носорогу и, вероятно,
имел общих с ними предков. Но почему-то жизнь его была коротка, при не-
262
класс: млекопитающие — mammalia
значительном географическом распространении в эоцене Франции и Германии.
В этом отношении он подобен роду Palaeotherium, представленному
в тех же слоях, но вымершему немного позже, в нижнем олигоцене.
Сем. Titanotheriidae. —- Под этим именем была объединена группа вымер-
ших млекопитающих, остатки которых найдены в олигоцене Сев. Америки. Как
показывает название, животные эти были больших размеров (титаны). Позже
к ним присоединили группу меньших форм из эоцена Сев. Америки, имевших
с ними некоторые сходные признаки, главным образом в строении зубов и
отчасти конечностей, и включили в отряд Perissodactyla, хотя они далеко
не отвечают характерным признакам этих форм. Объединяющими признаками
является полная зубная система, с уменьшением только числа резцов у позд-
нейших форм, строение коренных зубов и строение конечностей — трехпа-
лых задних и четырехпалых передних. Верхние коренные состоят из внешних
лунок и внутренних бугорков. Предкоренные проще — задний внутренний бу-
горок отсутствует. Семейство это естественно распадается на два подсемейства:
Рис. 326. Palaeosyops.
1) Palaeosyopsinae—
эоценовые тапирообразные формы и
2) Titanotheriinae — оли-
гоценовые, более крупные и тяжело-
весные формы, череп которых снаб-
жен двумя выростами — рогами, рас-
положенными рядом на передней
части головы.
Подсем. Palaeosyopsinae. —
Palaeosyops — из среднего эоцена
Сев. Америки — является наилучше
изученной формой; известен по почти
полному скелету (составленному из нескольких экземпляров) (рис. 326, 328, А).
По общему виду своему он больше всего напоминает скелет тапира, но
больших размеров.
Череп его отличается сильно вдавленными лобными костями и толстыми,
удлиненными носовыми костями, выдающимися дальше межчелюстных. Большие
височные впадины сильно сдавливают мозговую коробку, заключавшую не-
большой мозг. Глазницы не замкнуты, за ними лежит височный отросток. За-
тылочные мыщелки направлены назад.
Конечности высокие, с тонкими пальцеходящими ступнями. Передние —
с четырьмя почти равномерно развитыми пальцами, пятый немного меньше
остальных, задние — с тремя пальцами.
Озубление представляет особенности (рш. 326, А, 3, 4 и 5). В верхней
челюсти за шестью маленькими резцами следуют клыки и по семи коренных.
Коронки коренных несут на внешней стороне две лунки, очень углубленные, на
внутренней — бугорки: передний большой и задний (гипокон) различной
величины. На гп он или едва заметен, или не существует. На передней стороне
зубов существуют у некоторых видов промежуточные бугорки. Предкоренные
подкласс: плАЦЕнтные — placentalia, копытные
263
рг3 и рг4 значительно меньше коренных, с тремя бугорками; рг1 и рг2 —
конические.
. Нижняя челюсть имеет тоже 6 резцов, клыки и по 7 коренных — без диа-
стемы. Коренные состоят из 2 лунок, очень сжатых на внешней стороне; та имеет
лалон; рг4 и prs равной величины с коренными; два передних предкоренных
проще.
У различных видов Р а-
laeosyops несколько варьируют
величина и число бугорков на
различных коренных (рис. 326, А,
3 и 4), у некоторых исчезает пара
резцов.
Telmatotherium
Marsh — средний и верхний эоцен,
ближайший род к Palaeosyops,
известен по черепам, которые
Осборн относит к нескольким
1
3
Рис. 326, А. 1 и 2 — верхние и нижние зубы
Telmatotherium. 3 и 4 — верхние зубы
Paleosyops; 5 — нижни^ зуб его же.
видам, основываясь на различной
форме теменных костей, высо-
те сагиттального гребня и т. д.
На коренных зубах хорошо
развиты внутренние передние бугорки. Зубы имеют воротник (рис. 326, А, 1 и 2).
Telmatotherium diploconum Osb.— череп менее вдавлен в носо-
вой части, чем у Palaeosyops. Гребень начинается с половины черепа, ко-
торый более удлинен сравнительно
Telmatotherium v а 1-
1 i d е n s Osb. — гребень отодвинут
больше назад, и череп удлинен
•больше.
с Palaeosyops.
328. А — Palaeosyops major. В — Мап-
Рис.
teoceras.C—D ol i с h о г h i n u s cornutus.
Telmatotherium cornutum Osb. — носовые кости утолщились; са-
гиттальный гребень исчез, и над глазами появляются слабые костные выросты —
на лобных, отчасти на носовых костях, направляющиеся в стороны. Швы между
названными костями не видны, так что нельзя точно определить, к каким ко-
стям принадлежат выросты.
264
класс: млекопитающие — mammalia
Dolichorhinus — является представителем еще более вытянутого че-
репа с еще более грубыми носовыми костями и тоже зачаточными надглазными
выростами (рис. 327, 328, С).
Одновременно с этими формами, удлиняющими череп, существовали дру-
гие (в среднем и верхнем эоцене), сохраняющие короткий череп, например Мап-
te oceras Osb. и Limnohyops Marsh, и имеющие другие признаки, свой-
ственные Palaeosyopsinae.
Предки этих форм указаны в нижнем эоцене по менее хорошо сохранив-
шимся остаткам.
Род Lambdotherium Соре и отделенный от него род Е о t i t а-
п о р s Osb. имеют коренные зубы с неизменившимися внешними и внутрен-
Рис. 329. Titanotherium (Brontops)robustum Marsh.
ними бугорками и маленьким пятым, добавочным, между двумя передними-
Нижние коренные несут маленький добавочный бугорок на внутренней стороне,
сходный с бугорком зубов лошадиных форм. Передняя конечность, восстано-
вленная по нескольким обломкам, у Lambdotherium имеет четыре пальца
(метаподия).
Подсем. Titanatheriinae представлено в олигоцене (и нижнем миоцене—
Осборн) Сев. Америки. Их главный признак — присутствие костных выростов
впереди орбит, которые отличают их от Palaeosyopsinae. В остальном
скелет их имеет много общего с последними, но он тяжеловеснее; особенно —
конечности с укороченными предплечьем и голенью (рис. 329).
Пальцы на передней конечности сохранились в числе четырех, и не
только не последовало их сокращения, но пятый метаподий сравнялся со вто-
рым, и нога дает впечатление парнопалой. Только сочленение пятого пальца
очень маленькой фасеткой с unciforme и четвертым метаподием приближает ее
к ноге тапира. Очевидно, здесь произошло перемещение (сдвижение) unciforme
раньше, чем укоротился четвертый метаподий.
подкласс: плацентные — placentalia, копытные
265
Задняя конечность правильно трехпалая, с несросшимися cuneiforme111 и
cuneiforme11.
Озубление несколько изменилось. Резцы сократились: их по две пары
в обеих челюстях, и они очень маленькие. Клыки большие, прилегают
к ним. За ними следуют без диастемы семь коренных, имеющих на на-
ружной стороне по две лунки, более глубоких на настоящих коренных, чем
на предкоренных, и на внутренней стороне по два различно развитых бугорка.
На т и т2 передний бугорок больше заднего; на /и3 последний почти ис-
чез; на предкоренных существуют оба. Промежуточных бугорков нет. Нижние
зубы представляют тоже замкнутый ряд: 2 резца, 1 клык, 6 коренных; послед-
ние имеют по две лунки, — еще и талон. Итак, зубы и конечности Т i-t а-
notheriinae очень близки к этим же частям скелета Palaeosyopsinae.
Что касается главного их признака — рогов, то они долго служили раз-
общающим признаком, но с нахождением черепов различных видов Т е 1 m а-
totherium и др., у которых появляются зачаточные выросты над орбитами
на тех же местах, где они существуют и у Titanotheriinae, связь между
этими подсемействами может считаться установленной. Представители послед-
него очень многочисленны, и различные формы их были описаны различными
американскими учеными, причем нередко одно и то же животное получало
различные названия, даже родовые: Titanotherium— Brontops—Bron-
totherium — Menodus и т. д.
В 1896 г. Осборн соединил их всех под общим родовым названием
Titanotherium и расположил их в генетическом порядке, где можно ви-
деть увеличение рогов и сопутствующее ему уменьшение носовых костей. Позже
(в 1902 г.) при переработке этого материала с прибавлением нового, Осборн
разделяет опять р. Titanotherium и устанавливает четыре группы (phylum),
основываясь, главным образом, на длине черепа и величине рогов: Titano-
therium, Megacerops, Symborodon, Brontotherium.
Эти группы Titanotheriinae являются как бы продолжением четырех
групп Palaeosyopsinae, установленных им же: Palaeosyops, Telma-
totherium, Manteoceras, Dolichorinus.
У Titanotherium platyceras nTitanoth. ramosum выросты
достигают громадных размеров; череп кзади утончается, но зубная система
остается неизменной, так же как и строение конечностей.
Вместимость мозга у этих форм небольшая, но мозг имеет извилины и
сильвиеву борозду; большой мозг не прикрывает мозжечка.
Изучение этого подсемейства представляет большой интерес в смысле эво-
люции. Мы не можем указать сейчас ни одной формы, от которой, даже пред-
положительно, могли бы отчлениться древнейшие представители Titanothe-
riinae. Они являются в нижнем эоцене с готовыми признаками зубов и конеч-
ностей и развиваются вполне определенно, осложняя предкоренные, сокращая
зачаточные добавочные бугорки на коренных. Зубы у позднейших форм, состоящие
из двух лунок и двух бугорков, являются полной аномалией для непарнопалых. Они
напоминают зубы Anoplotherium. Передняя конечность тоже пытается удер-
266
класс: млекопитающие — mammalia
жать бесполезный уже ей пятый палец и дает тип неадаптивно редуцированной
конечности. С другой стороны, роговые выросты становятся, очевидно, лишним
бременем для организма, и это семейство, так богато и разнообразно предста-
вленное в эоцене и олигоцене, вымирает при непроизводительной затрате сил
организма на оказавшийся, вероятно, ненужным орган — сильно развитые рога.
Отряд 10
Парнопалые — Artiodactyla
Подразделяются на: 1) Bunodontia — бугорчатозубые и 2) Seleno-
dont i а — лунчатозубые.
Характерной чертой для тех и других является конечность с парным чи-
слом пальцев, в противоположность непарнопалым, причем у Bunodontia —
Suidae четыре метаподия остаются свободными. Вторым отличительным при-
знаком для сем. свиней являются бугорчатые зубы. У Selenodontia (у боль-
шей части) два средние метаподия срастаются в трубку, два боковые исчезают
(жвачные) или остаются (кабарговые). Зубы лунчатые. Большинство снабжено
рогами. Каждое из этих семейств имеет ископаемых представителей. Но кроме
этого в эоцене и олигоцене существовали парнопалые, которые имели конечно-
сти с сократившимися, но не сросшимися метаподиями, и у которых зубы но-
сили смешанный характер, состоя большею частью из лунок и из бугорков: это —
Bunoselenodontia. Эта вымершая группа, по исследованию В. О. Кова-
левского, относится вся к животным с „неадаптивно редуцированными ко-
нечностями". Но как бы ни изменялись конечности парнопалых, для них является
характерной и неизменной форма надпяточной кости — астрагала, имеющая
прямой блок на верхней и на нижней поверхностях, что его отличает от астра-
гала непарнопалых с косым блоком только на верхней стороне и с гладкой
поверхностью на нижней — для naviculare.
На пяточной кости (calcaneum) на передней стороне существует у парно-
палых фасетка для нижнего конца малой берцовой; у непарнопалых ее нет.
Подотряд 1
Бугорчатозубые — Bunodontia
В состав этого отряда входят сем. Suidae и сем. Н i р р о р о t a m i d а е.
Оба семейства сохраняют до сих пор признаки древних форм в строении зубов
и конечностей. Представители Suidae широко распространены во всех стра-
нах, тогда как гиппопотамы представлены только (одним родом) в тропической
Африке.
Интересное указание дает Абель на распределение тяжести тела на
пальцы у парнопалых и отличие между Bunodontia и Selenodontia
в этом смысле.
Он принимает, что у жвачных со сросшимися 3-м и 4-м метаподиями тя-
жесть тела передается равномерно на трубку, так, как она передается на 3-й
палец непарнопалых, например у лошади: в обоих случаях средние пальцы стоят
перпендикулярно к поверхности почвы.
f
подкласс: плацентные — placentalia, копытные
267
Совсем иначе это происходит у Bunodontia, сохранивших четыре пальца.
У них (у гиппопотама и свиньи) пальцы направляются не перпендикулярно
к поверхности, а косо, и главная ось (передней конечности) у стоящего жи-
вотного направлена от передней нижне-внешней стороны
к задней верхне-внутренней. При этом третий палец вы-
держивает большее давление, чем 4-й, и о равномерном
распределении тяжести не может быть речи.
Сем. Hippopotamidae. — Hippopotamus —
громадное, неуклюжее животное, живущее в сырых ме-
стах тропической Африки
(в Либерии), имеет ископае-
мых представителей в верхнем
плиоцене Италии и в плей-
стоценовых пещерах раз-
личных мест Европы, в нижнем плиоцене Сивалик-
ских холмов (Азия), в среднем плиоцене Египта
(Африка). Они близки
к ныне живущим гиппо-
потамам, но имели три
пары нижних резцов оди-
наковой величины, на-
пример Hexaproto-
don (Hippopotamus)
sivalensis (рис. 330).
У ныне живущих резцы
сократились до двух пар,
причем внутренняя пара
значительно больше внешней (рис. 331, 331, А).
Коренные зубы состоят из четырех бугорков, ограниченных складчатой
эмалью. Предкоренные значительно меньше коренных, того же типа.
Н 1 р р о р о-
(Н е х а р г о-
Рис. 330.
ta m u s
todon) sivalensis
Falc. Передняя часть
нижней челюсти.
Рис. 331. Hippopotamus
a m р h i b i u s Lin. (ныне
живущий).
Рис.331,A. Hippopota-
mus major Falc. (Плиоцен
Италии). Последний верх-
ний коренной зуб.
Ноги их сохраняют не только четыре несросшихся метаподия, но и располо-
жение первого ряда карпальных костей сохраняет тип нередуцированной конечно-
сти: unciforme прикрывает IV и V metacarpale magnum—III и trapezoideum •— II.
Второй ряд карпальных костей имеет чередующееся расположение (рис. 338).
Известны следующие формы:
Hippopotamus sivalensis — нижний плиоцен Верхней Индии.
„ hipponensis Gaud.— ср. плиоцен Египта и Алжира.
„ major — верхний плиоцен Vai d’Arno.
„ Pentlandi Falc.— древний плейстоцен Европы, бли-
зок кН. mamphibius (ныне живущему) *).
*) Остатки Hippopotamus Pentlandi Falc. были найдены в пещере Мае-
cagnone и др. (близ Палермо) в таком изобилии, что их вывозили в продолжение несколь-
ких лет в Англию и Марсель для приготовления костяного угля для очищения сахара.
200 тонн было добыто из пещеры San-Sin в продолжение 6 месяцев (Guide, 1890, р. 43).
268
класс: млекопитающие—mammalia
Hippopotamus sivajavanicus Dub.—древний плейстоцен Явы.
„ madagascarensis Grand. — из новейших отложе-
ний Мадагаскара.
Несколько карликовых форм известны из пещер острова Мальты, Кипра
и Крита, Hipp, minutus.
Гиппопотам может быть назван живым ископаемым: настолько у него со-
хранился древний тип конечностей и зубов. Более древ-
**** ' ние пРедки ег0 до сих п0Р неизвестны.
Сем. Suidae.— После гиппопотама свиньи явля-
Рис 332 Acotheru ются Jia-H6o iee примитивными формами из копытных по
lum. Нижние коренные, бугорчатым зубам и 4-палым конечностям. Ископаемые
остатки их (зубы), известные со среднего эоцена,
имеют признаки, изменение которых можно проследить до ныне живущего рода
S us. В еще более древних отложениях, в нижнем эоцене Эгеркингена
(в Швейцарии), найдены зубы еще более примитивного типа бугорчатых зубов.
Acotherulum Riit. имел коренные, состоящие из четырех одинаковых ма-
Рис- 333. Cebochoerus Riitimeyeri Stehlin.
леньких бугорков (рис. 332). Остатки его незначительны, так что только условно
можно поставить его у начала линии развития свиней.
Cebochoerus Gerv. — тоже из Эгеркингена, имеет по четыре главных
бугорка на верхних и нижних зубах и на первых еще добавочные бугорки,
Рис. 334. Palaeochoerus (Hyotheriuml Waterhausi Pom. (Миоцен.)
D—передняя конечность. и. в.
маленькие. Ргх— из двух удлиненных бугорков, остальные предкоренные пред-
ставляют конусы (рчс. 333).
Palaeochoerus Waterhausi Pom. — из нижнего миоцена — предста-
влен черепом с удлиненной передней частью и полным озублением. Четырех-
бугорчатые коренные имеют маленькие промежуточные бугорки. Рг1 — из трех
бугорков, рр —- из двух, остальные—простые конусы (рис. 334). Это — заметное
осложнение зубов сравнительно с Cebochoerus.
подкласс: плацентные — placentalia, копытные
269
У других видов Palaeochoerus осложнение идет дальше. У Р а 1. М е i s-
sneri Н. v. М. (рис. 335) появляется больше добавочных бугорков, воротник
складчатый на т и т . Также изменяются и нижние зубы, но три предко-
реннчх остаются конусами.
Hyotherium Soemmeringi Н. v. М. (рис. 336), из верхнего ми-
оцена, дает следующую стадию: число добавочных бугорков увеличивается,
главные выделяются меньше, и зубы становятся сходными с зубами современ-
ной свиньи — многобугорчатыми.
В миоплиоцене найдены остатки Sus erymanthius, в Пикерми и Гре-
бенниках, которая достигала очень больших размеров, имела сложные зубы и
особенность — над небольшими клыками на-
ружные костные выросты, которые не встре-
чаются у других Suidae (рис. 337). Это,
вероятно, боковая ветвь.
Ноги у S u i d а е изменяются параллельно
Рис. 336. Hyotherium Soemme-
ringi Н. v. Meyer. ‘/2 н- в-
Рис. 335. Palaeochoerus Meiss-
ner i Н. v. Meyer. ‘/2 н- в-
зубам (рис. 338). У Choerotherium (ближайшего к Cebochoerus) все
кости первого ряда карпуса лежат над метаподиями, metacarpes II касается
magnum. На передней стороне концов метаподий нет сочленовных гребней.
Unciforme над metacarpes IV и V. У Palaeochoerus metacarpes III оттеснил
metacarpes IV от magnum и сочленяется с trapezoideum; боковые пальцы стали
тоньше и короче. У Sus фасетка metacarpes III на trapezoideum стала боль-
ше, боковые пальцы утончились. У двух последних родов развились сочленов-
ные гребни на передних сторонах метаподий.
У Dicotyles (Sus Сев. Америки) вся trapezoideum перешла на
metacarpes III; metacarpes II только причленяется к III. Metacarpes V едва ка-
сается unciforme. Кости эти укоротились и утончились; они, очевидно, на пути
к исчезновению. Но кости корпуса, лежавшие раньше над ними, получили дру-
гую работу, и затрата организма на их питание окупается ею. В задних конеч-
ностях сохраняются в тарсусе три клинообразные кости: cuneitorme III — на
III metatarsus, cuneiforme II — на фасетках III и II; cuneiforme I— на другой,
фасетке IV. Они не срастаются. Dicoiyles встречается с верхнег, миоцена,
в плиоцене, плейстоцене и ныне живет в Америке.
Примитивные Suidae, общие в обоих полушариях в олигоцене, дали,
развиваясь, с миоцена в Старом свете нынешних S и йп а е, а в Америке
Dicotylinae, начиная с верхнего миоцена. Prosthenops, Platygonus
270
класс: млекопитающие-—mammalia
Mylogus, Dicot у les известны по черепам и зубам. У некоторых из ныне
живущих свиней, например у Phacochoerus, последний коренной стано-
вится очень сжатым и удлиненным. Он имеет до восьми рядов бугорков, и
длина его равняется длине остальных четырех коренных.
Вышеописанные ископаемые формы найдены в Европе, в Сиваликских
холмах Азии и в Сев. Америке.
Интересно замечание проф. О с б о пн а о развитии S u i d а е. Он указы-
вает, что тип, долго сохраняющий примитивные признаки, медленно развива-
Рис. 337. Sus erymanthius Gaudry.
А — верхние зубы. (Верхний миоцен Пмкерми и Гребенников.
ющийся. сохраняется долго, а его боковые ветви, быстро специализирующиеся
уклоняющиеся, обречены на вымирание. Он применяет это наблюдение к объ-
яснению устойчивости рода Sus по сравнению с боковыми вымершими (бо
лее односторонне совершенными) формами S u i d а е.
В нижнем эоцене Пуэрко (Сев. Америка) найдены остатки формы, отнесен-
ной к Condylarthra, с признаками, сближающими ее со свиньями; это —
Periptychus rhabdodon Соре. Это — пятипалое животное, с астра
галом без шейки, слабо выпуклым на нижнем конце.
подкласс: плацентные — placentalia, копытные
271
Коренные зубы его — четырехугольные с несколькими бугорками; они
меньше, чем закругленные предкоренные, состоящие из конуса с сильно раз-
витым воротником. Эти признаки зубов не позволяют считать Periptychus
предком Suidae.
Choerotherium.
Hippopotamus.
Рис. 338. Передние ноги.
Palaeochoerus.
Подотряд 2
Лунчатозубые — Selenodontia
Эти парнопалые характеризуются вполне определенным признаком — при-
сутствием четырех луночек на верхних и нижних коренных зубах. Сюда отно-
сятся все ныне живущие жвачные сем. Bovidae и Cervidae, объеди-
ненные еще несколькими признаками: присутствием рогов, сросшимися сред-
ними метаподиями на передних и задних конечностях и отсутствием передних
зубов — резцов и клыков в верхней челюсти. Это признаки, которые, очевидно,
стоят в связи со способностью отрыгать жвачку, с присутствием особого отдела
желудка — рубца (rumen), в котором хранится собранная пища (трава) и из
которого она попадает вторично в рот для пережевывания.
Некоторые лунчатозубые, сохранившие передние верхние зубы, не имеют
рогов;., они тоже жвачные и имеют четырехлунчатые зубы. Сюда относятся из
ныне живущих Tragulidae и Camelidae.
Ископаемые остатки Selenodontia известны с эоцена; между ними,
кроме форм, входящих в названные семейства, есть вымершие, например О г е-
odontidae, которые только по некоторым признакам могут быть отнесены
к четырехлунчатозубым; они имеют иногда добавочную пятую лунку.
Сем. Oreodontidae. — Небольшие животные с удлиненным или укоро-
ченным черепом, замкнутой или открытой орбитой, мало выдающимися носовыми
костями, глубокой впадиной впереди орбиты и полным озублением. Коренные —
четырехлунчатые, иногда с добавочной маленькой пятой лункой — бугорком.
Предкоренные проще коренных. Конечности пятипалые или четырехпалые
ТГ1
класс: млекопитающие — mammalia
с остатком первого пальца на передних ногах. С верхнего эоцена до плиоцена
в Сев. Америке.
Protoreodon — нижний эоцен. Наиболее древняя форма с тремя тре-
угольными предкоренными; рг со слабой внутренней лункой (рис. 339).
Рис. 339. Protoreodon parvus Scott
У близкой формы из верхнего эоцена, Protagriochoerus Scott, суще-
ствует добавочный бугорок на верхних коренных.
Нижние коренные у обеих форм имеют внутренние удлиненные бугор-
ки и внешние закругленные
лунки.
Agriochoerus Lei-
dy— является наиболее при-
митивной формой из оли-
гоцена, у которой сохра-
Верхняя челюсть.
Рис. 340. Orcodon Culbertsoni Scott. (г/3).
нился еще длинный хвост, пятипалые передние конечности С отходящим
первым пальцем, указывающие на то, что он еще лазал по деревьям. Задние
конечности четырехпалые. Орбиты открытые. Зубы четырехлунчатые; наружные
стенки их типа Hyopotamus — с глубокими V-образными лунками и закруг-
подкласс: плацентные — placentalia, копытные
273
ленной средней внешней складкой. Рг4 имеет два бугорка на внешней сто-
роне и один внутренний. Клыки большие. Резцов три пары. Носовые кости
тонкие, выступают вперед. Длина черепа до 27
Род О г е о d о n Leidy представлен не-
сколькими видами, отличающимися главным об-
разом размерами и длиною черепа. Он резко от-
личается от Agriochoerus и может быть рас-
сматриваем как параллельно развивавшийся. Об-
щий предок, если он и был, очень удален от этих
двух родов. Признаки О г е о d о п: череп с рез-
ко выступающим сагиттальным гребнем; орбита
закрытая, впереди ее характерное углубление —
слезное — какое мы видели у гиппариона. Рез-
цы в обеих челюстях; клыки большие. Три пер-
вые предкоренные расширены на задней стороне.
Рг — из двух лунок. Коренные — из четырех
лунок: внешние V-образно вдавлены, внутренние
сжаты. Средние внешние складки острые. Конеч-
ности тонкие; metacarpus I сохранился в виде тон-
кой, короткой косточки, ошальные все еще
сочленены с карпальными. Radiale лежит
см. Известен целый скелет.
А
в
Рис. 340, A. Oreodon С u 1-
b е г s t о n i Leidy. (Миоцен Сев.
Америки.)
главной поверхностью на magnum, частью на trapezoideum; lunare — на unciforme.
В обеих конечностях боковые пальцы короче средних; у Agriochoe-
tus они равны и слабо сочленяются с cuboideum и cune’forme II. Хвост короткий.
Рис. 341. Oreodon gracilis Leydy. (%)
Рис.342.Merychyus anenarum (3/s)
Все эти признаки совершенно отделяют р. Oreodon от Agriochoe-
rus, жившего одновременно с ним, и делают его настоящим парнопалым.
Олигоцен Сев. Америки: Oreodon Culberstoni (рис. 340), Oreodon
major, Oreodon gracilis (рис. 341) и др.
Merychyus Leidy — имеет более открытые внешние лунки на верхних
коренных. Нижние зубы лунчатые, как у Oreodon (рис. 342). Встречается
18 Па в л о в а М. В. Палеозоология. Ч. II.
*274
класс: м чекопитающие — mammalia
в среднем миоцене. В передней конечности metacarpus V вышел из-под
unciforme, metacarpus II отодвинулся из-под magnum и потерял фасетку для нее.
Возможно предположить, что представители этого рода состояли в гене-
тической связи с р. Oreodon и закончили в начале плиоцена (или конце
миоцена) сем. Oreodontidae. Agriochoerus потерял верхние резцы еще
в олигоцене, раньше, чем он сошел с дерева и изменил конечности.
Coloreodon имеет рг , почти равныйт1 (в нижнем миоцене, позже
не известен).
Жвачные (Ruminantia)
Tetraselenodon — в нижнем эоцене Эгеркингена найдены четырехлун-
чатые зубы очень маленькой формы, которые могут быть поставлены у основания
лунчатозубых — жвачных, d* — вполне типа гп',
i *
рг —трехбугорчатый.
Сем. Gelocinae. — Одновременно с Oreo-
dontidae существовала в олигоцене Европы дру-
Рис. 343, A. Gelocus communis Aym.
17 — передняя нога, 18 — задняя.
Рис. 343. Gelocus communis Aym.
А — верхние зубы, В — нижние.
гая группа парнопалых с не вполне еще лунчатыми зубами. Внешние
бугорки еще почти не изменились. Средняя внешняя складка существова-
ла. Рг1 исчез.
Gelocus Aymard (рис. 343, 343, А) — олигоцен Франции (Ронзон),
с низкими зубами; рг* — из двух лунок, остальные предкоренные — удлиненные;
коренные с 4 бугорками-лунками. Задние метаподий срослись; от боковых оста-
лись вверху и внизу тонкие косточки; средняя часть рассосалась. Naviculare
срослась с cuboideum; cuneiforme III с cuneiforme II.
В передней конечности средние метаподий очень сближены; боковые —
как у задних.
В. О. Ковалевский, подробно изучивший эту форму из Ронзона, видит
в ней предка всех жвачных, так как различные кости скелета Gelocus при
сравнении с костями последних указывают на их близкое сродство. Череп
не известен.
подкласс: плацентные — placentalia, копытные
275
Bachytherium Filhol — из олигоцен Керси, отличается отсутствием prt
и слабым срастанием передних средних метаподий. Череп не известен.
А
Рис. 344. Prodremotherium elongatum [Filh.
Рис. 345. Dremotherium Feignouxi Filh.
3—нижняя челюсть с молочными зубами; 4 — нижняя челюсть с коренными; 6 — сочленовный отросток
нижней челюсти.
Prodremotherium elongatum Filh. (рис. 344) (Керси), идет далее:
от боковых метаподий остались только нижние концы. Бугорки на коренных полу-
чают вогнутость на внешней стороне зуба и становятся луночками. Пред-
18*
276
класс: млекопитающие — mammalia
коренные рг2 и рг2 имеют по два бугорка и складку на внешней стенке и по бу-
горку на внутренней стороне. На нижних предкоренных появляются складочки,
направленные внутрь челюсти. Резцов верхних нет; клыки довольно большие.
Сем. Cervulidae. — Рогов еще нет, или встречаются небольшие развилки
у самцов. Зубы лунчатые; предкоренные осложнились: верхние — двухлунчатые,
нижние — с добавочными складочками. Верхние клыки существуют — большие.
Рис. 346. Amphitragulus lemanensis Filh.
Dremotherium (рис. 345) — очень близок к Prodremotherium,
в нижнем миоцене Оверни и Ульма. Боковые метаподий в виде нижних косто-
чек, как и у Prodremotherium; верхние концы приросли с задней стороны
к верхним концам средних метаподий,
и этим расширилась поверхность по- в
Рис. 347. Palaeomeryx Feignouxi Geoffr.
следних, передающая тяжесть тела средним пальцам. По строению ног он при-
близился к оленям.
Amphitragulus (рис. 346) — встречается вместе с предыдущей формой
в Оверни (нижний миоцен), отличается присутствием рг4, давно исчезнувшего
у других описанных форм. Рогов нет. На нижних зубах появляется складка —
„Palaeomeryx-Falte" Рютимейера. В ногах остаются нижние части метапо-
дий с фалангами. Метаподий шире, чем у Dremotherium.
Вероятно, эти две формы и положили начало: первая — Cervidae
(оленям), вторая —В о v i d а е (быкам).
Сем. Cervidae. Рога без чехлов. Palaeomeryx Н. v. Meyer
(рис. 347) — череп без рогов; нижние зубы со складкой на внешней стороне
подкласс: плацентные — placentalia, копытные.
277
зу-
не-
хо- Рис. 348. Blastomeryx. Рис. 349. Leptomeryx.
Maz a m а
передней лунки Средний и верхний миоцен. Это — крупные формы. Неко-
торые ученые соединяют этот род с Dicroceras Lart, имеющим рога на
высокой ножке, в виде двойного развилка. Эмаль на его зубах сложена
в складки. На верхних коренных у основания средней долинки существует
складка эмали, иногда двой-
ная. Предкоренные состоят
из двух луночек.
В долинке нижних
бов тоже складочка —
большой столбик.
Это уже олень,
тя без рогов или с очень
простыми. В верхнем миоцене
вместе с ним встречаются другие, мень-
шие формы. Развитию оленей поло-
жен уже настоящий фундамент не
только в Европе, но и в Сев. Америке
и в Азии. Но в Сев. Америке развитие
параллельного ряда оленевых идет с бо-
лее медленным изменением конечностей.
Именно, у Leptomeryx — наи-
более древней формы — боковые пальцы
развиты еще вполне. Radius и ulna сво-
бодны. Нет клыков (рис. 349). Олигоцен.
У Blastomeryx (рис. 348)—из
нижнего миоцена — пальцы укорачива-
Blastomeryxprimus
Blastomt olcotti
Leptomeryx sp.
Leptom. evanse
Рис. 350. Изменение зубов у американских
оленей.
ются и утоняются.
Можно проследить изменение —
и зубов у американских оленей
(рис. 350).
В среднем миоцене произошло уси-
ленное развитие дальнейших рогатых
форм, причем наблюдается интересное яв-
ление—-раздвоение этих форм. У одних
появляются вышеуказанные рога Dicro-
ceras, поддерживаемые стержнями; это
рога настоящих Cervidae. У других
существуют простые выросты на лобных костях, вероятно, покрытые вначале
кожей, а потом роговыми чехлами; это рога полорогих Antilopidae.
В верхнем миоцене и нижнем плиоцене число Cervidae и Antilo-
pidae быстро растет и представляет большое разнообразие форм, выражен-
ное, главным образом, в форме рогов.
В Европе, в Греции и в Херсонской губ., на о-ве Самосе, в Сива-
ликских холмах антилопы богато представлены теми формами, которые
278
класс: млекопитающие — mammalia
близки к нынешним африканским, с прямыми или загнутыми рогами, с глад-
кими или коленчатыми; позже меняется область их распространения, но при-
знаки остаются.
Рис. 351. Рога различных оленей: А, В - С е г v u 1 u s; С, D -Cervus elaphus;
F —D a m а. (Ныне живущие.)
ются на три главные группы, дошедшие
Рис. 352. Cervus elaphus.
Д — верхний коренной, В — нижний коренной;
С — передняя и D — задняя нога.
Олени изменяют рога в другом направлении: они начинают ветвиться и
в плиоцене дают много разнообразно разветвленных форм, которые распада-
: (рис. 351).
Cervus elaphus—бла-
городный олень, предки кото-
рого известны из плиоцена Per-
rier. Рога его с закругленными
стержнями и отростками, на-
правленными наружу. На но-
гах остаются фаланги боковых
пальцев (рис. 352). 7 м.
Cervus tarandus—
северный олень, с более ветви-
стыми, плоскими рогами; предки
известны только из плейстоцена
(рис. 353).
Наконец, Alces — лось,
с широкими, лопатообразными
рогами с небольшими отрост-
ками; предки найдены в меоти-
ческих слоях Гребенников — Alces meoticus М. Pavl.J).
В зубах произошла постепенная перемена; верхние передние исчезли,
коренные стали выше, нижние предкоренные осложнились добавочными
‘) М. Павлова. Ежегодник Палеонтолог, общества, 1926, т. IV.
подкласс: плацентные — placentalia, копытные
279
складочками. Лунки коренных остаются постоянными. У лося они осложняются
еще несколькими добавочными складками.
Procervus varia-
bills Alex. — P меотиче-
ских слоях Херсонской губ.
найдены очень разнообразно
представленные остатки оле-
ней (описаны А. Алексее-
в ы м) 1). Разнообразие вы-
ражено в рг зветвлении ро-
гов; некоторые из них за-
круглены, другие плоские;
число отростков и форма
их различны При этом
разнообразии название Pro-
cervus не вполне под-
ходит.
По сократившимся бо-
ковым пальцам олени раз-
Рис. 353. Cervus tarandus (var. Kloktun)
M. Pavl.
деляются на:
a) Plesiometacarpi — с остатками верхних частей боковых мета-
подий и фалангами II и V пальцев. Cervus elaphus.
б) Teleometacarpi — с остатками нижних концов боковых метаподий
с фалг нг 1ми. А1 с е s.
В плейстоцене встречаются гигантские формы оленей, близких к лосю,—
Рис. 354. Cervus euryceros Aldr.
Cervus euryceros (рис.
354) и C. latifrons — с гро-
мадными рогами, снабженными
большими надглазными отростка-
ми, достигшими, очевидно, тех
размеров, которые уже были
вредны для организма, и жи-
вотные эти вымерли. Более
простые формы существуют и
теперь. С исчезновением перед-
них зубов и развитием рогов
развился добавочный желудок — рубец, и животные стали жвачными.
Полорогие (Cavicornia.) Рога с роговыми чехлами.
Сем. Antilopidae. — Антилопы, живущие теперь главным обра-
зом в Африке, представлены ископаемыми остатками, начиная со сред-
него миоцена в Европе (Франция), роговыми стержнями с частями чере-
пов. В миоплиоцене (меотических слоях) в Пикерми, в Херсонской губ. и
1) А. Алексеев, Фауна позвоночных д. Ново-Елизаветовки, 1916.
280
класс: млекопитающие — mammalia
других местностях встречается большое количество их черепов и ча-
стей скелета; они подразделяются на несколько родов и видов, мало отли-
чаясь от ныне живущих. В послетретичных отложениях разных мест Европы
встречаются остатки Saiga tatarica, живущих теперь на востоке Европей-
ской части СССР.
Средний миоцен — Antilopa clavata Filh.
близкие KCervulinae j „ sansaniensis Filh.
I „ recticornis Filh. и др.
Мио-плиоцен —-Gazella deperdita Gew.
„ SchlosseriM. Pavl. (рис. 355)
(меотический ярус) „ brevicornis Wagn.
Tragoceras Wagn., Palaeoceras Wagn.
Palaeoryx Gaud, и др.
Плиоцен — Antilopa p al a e i n di c a Falc.
(Сивалики, Азия) Gazella porrecticornis Lyd.
(Алжир) Gazella atlantica и др.
Плейстоцен — Saiga tatarica J) (Европа).
Все эти формы характеризуются за-
кругленным сзади черепом, с сильно раз-
витыми теменными костями, рогами, рас-
положенными непосредственно над глазни-
цами и направленными вверх или назад,
отсутствием передних верхних зубов,
присутствием шести коренных, кроме
Saiga, у которой они сократились до.
пяти.
Сем. Bovidae.— Полорогие антило-
пы, появившиеся в миоцене и давшие боль-
шое разнообразие форм в плиоцене, резко
отличаются от домашнего скота — быков —
как формою черепа, так и разнообразием
рогов.
Ближе дру1их из Bovidae стоят
к антилопам козлы и бараны — О vis и
Capra, и очевидно, это те стадии, через
которые пришло .изменение черепа антилоп,
Рис. 355. Gazella Schlosseri вырабатывая тип Bovidae.
М' Pavl- Главное различие черепов состоит
в закругленной затылочной части антилоп
и баранов и косо срезанной — у быков.
Проф. Рютимейер, изучавший подробно эти различия, вполне опреде-
ленно ука «ал на постепенность этих изменений у древних форм при выработке
*) Н. Filhol. A. Gaudry, М. Pavlow — см. в списке лигературы.
подкласс: плацентные — placentalia, копытные
281
черепа быка, что подтверждается, с одной стороны, третичными и
плиоценовыми формами, например Bos elatus и др. у которых затылок
не имеет еще такого резкого из-
менения, а с другой стороны, эм-
бриональным развитием нынешних
быков у которых закругленный перво-
начально затылок, как у антилопы
и барана, отграничивается постепен-
но от теменной части углом, кото-
рый становится из прямого острым
(рис. 356).
При этом совершается пере-
движение теменных костей за за-
тылочный гребень. Одновременно
перемещаются и роговые стержни,
и изменяется их положение: из вер-
тикально торчащих они становятся
горизонтальными и передвигаются
далеко назад из положения над
глазами.
ProbubaluSH Leptobos—
из плиоцена Сиваликских холмов—дают эти переходы. Вместе с этим изме-
няются и зубы: из
Рис. 356. Возрастное изменение черепов: 1—
Р о г t а х новорожденный^— Portax молодой;
4—теленок новорожденный; 3—молодая ко-
рова; 5—взрослая корова.
низких, снабженных корнями коренных, вырабатываются
высокие зубы с постоянным ростом, чего не бывает
уз оленевых. В средней долинке развивается столбик,
Рис. 357. Bos primigenius Bd.
Л — верхние, В — нижние коренные зубы.
идущий от основания зуба вверх и дающий при стирании кружочек эмали,
характерный для быков, на верхних и нижних коренных. Конечности сохра-
няют у быков бокозые пальцы без метаподий, которые исчезли еще у антилоп..
282
класс: млекопитающие — mammalia
вых быков
Bovidae.—Ископаемые остатки плиоценовых и плейсгоцено-
очень разнообразны, что позволило Рютимейеру установить
три разновидности:
1. Bos primigenius (рис. 357) — первичный бык каменного века и
Bos trachoceros — наиболее к нему близкий,
2, Bos frontosus — бронзового века.
3. Bos brachyceros — свайных построек
Нынешний Bos taurus, домашний бык, считается потомком Bos pri-
migenius. Ближайшей к нему формой является Bison — зубр, немногие
представители которого существуют теперь в Сев. Америке и у нас на Кавказе
в диком состоянии и искусственно поддерживались в Беловежской пуще. (Те-
перь почти уничтожены с 1916 г.) Многочисленные остатки ископаемого би-
зона— Bison latifrons (рис. 358) — и других близких к нему видов с гро-
мадными рогами известны из верхнего плейстоцена Европы и Сев. Америки. Они
считались предками ныне живущих зубров, но при подробном изучении этих и
Рис. 358. Bison latifrons.
ныне живущих зубров
это трудно допустить,
особенно при сравне-
нии их рогов, В верх-
нем плиоцене и в ниж-
нем плейстоцене (в
песках МЛбаха, ти-
распольском гравии и в
других местностях) най-
дены черепа бизонов с
более короткими рога-
ми. При сравнительном
изучении этих черепов с длиннорогими можно установить параллельное разви-
тие их и факт вымирания длиннорогих в плейстоцене, а короткорогих можно
признать предками ныне живущих бизонов, и удержать для первых название
Bison latifrons, а для вторых Bi&onpriscus (Bison Schoeten-
s а с k i).
Отличается Bison latifrons от Bos primigenius укороченной
мордой, широким лбом и расходящимися почти горизонтально рогами; у Bos
primigenius рога закручиваются вперед. Зубы и кости скелета отличаются
очень мало у этих двух родов.
В u b а 1 и s — буйвол, третий представитель подсем. Bovidae, южный,
имеет особо расположенные рога с направляющимися вниз концами, и связан
с Probubalus Азии.
Poephagus—як Монголии, покрытый густой шерстью, резко отли-
чается по внешнему виду от предыдущих; некоторое сходство шерстяного по-
крова сближает его с зубром. Сравнительное изучение черепов Poephagus
m utus — дикого яка — и Poephagus grunniens — домашнего — указало
на очень важные различия между ними. Первый отличается узким лбом и резко
подкласс; плацентные — placentalia, копытные
283
широко распространена
Рис. 359. Ovibos mos-
chatus Blv.
загнутыми вперед рогами, второй имеет лоб значительно шире, а рога идут
более горизонтально.
При дальнейшем изучении выяснилось большое сходство черепа и частей
скелета Po6phagus mutus с этими же частями Bos primigenius. Это
сходство настолько большое, что первый мог бы казаться непосредственным
потомком последнего, но его предкоренные проще, чем у Bos primigenius.
Poephagus grunniens стоит ближе к зубру.
Шерстяной покров этих форм является, может быть, остатком покрова,
сходного с покровом мамонта и волосатого носорога.
Подсем. Ovinae. — Ovibos — крупная, почти вымершая форма, сохра-
нившаяся в небольшом числе в Сев. Америке, б(
в плейстоцене Европы и Сибири (у нас остатки
его встречаются даже в Черниговской губ.).
Ovibos moschatus Blv. (рис. 359) — со-
единял как бы признаки Оvis и Bos, имея темен-
ную и затылочную части черепа, разграниченные
гребнем, как у Bos; рога с широким лобным
прикреплением на лобных костях типа О vis.
Отличается сильно укороченными конечно-
стями, особенно метаподиями.
Euceratherium — вымерший в плейсто-
цене (Калифорнии), является ближайшей формой
к Ovibos.
Остатки .настоящих Ovis и Capra изве-
стны из плейстоцена и тесно примыкают к нынеш-
ним. Только для немногих форм известны третичные
остатки, которые часто трудно отличить от антилоп.
В и с а р г a Riit. — из плиоцена Сивалик.
Capra pyrenaica Schimp. — из пещер
Гибралтара.
Capra sivalensis Lyd. — плиоцен Си-
ва пик.
Все эти формы различаются, главным образом, по рогам. Сев. Америка
бедна остатками полорогих.
Все вышеуказанные формы парнопалых: Cervidae, Antilopidae и
Bovidae относятся к типичным жвачным, снабженным рогами и рубцом и
лишенным передних верхних зубов (резцов и клыков). Конечности двупалые,
снабжены роговыми копытами.
Сем. Tragulidae (кабарговые). — Представляют теперь между живу-
щими лунчатозубыми наиболее примитивный тип жвачных. Немногочисленные
представители их рассеяны далеко друг от друга.
Tragulus — несколько видов в Южной Индии и на ближайших
островах.
Hyaemoschus — в западной Африке.
284
класс: млекопитающие — mammalia
Moschus moschiferus — в центральной Азии и Гималайских горах
на высоте 2,5—3,8 км.
Отличительными признаками их являются длинные саблевидные верхние
клыки, спускающиеся далеко вниз, при
Рис. 360. Hyaemoschus: 1 — Н. aqua-
tic u s; 2 — Н. crassus. (Некой.)
отсутствии верхних резцов и первых
предкоренных. Конечности сохрани-
лись у некоторых форм примитивные,
с признаками четырехпалых.^Желу-
док не имеет _ рубца. Ископаемые
остатки этого семейства известны,
начиная с олигоцена, в миоцене
Европы и Сев. Америки для Hyae-
moschus, в плиоцене и в плей-
стоцене южной Азии — Сиваликские
холмы — для Т г a g u 1 u s. Эти формы
мало отличаются от ныне живущих,
у которых можно видеть различную
степень редукции конечностей.
Lophiomeryx — средние мета-
подии не срослись. Олигоцен Керси.
Hyaemoschus crassus
Lart. - - миоцен, имеет по четыре пальца на передних конечностях, причем два
боковые метакарпалия очень укорочены и едва причленяются к средним,
несросшимся, Над metatarsus II лежит trapezoideum (рис. 360).
На задних — средние метаподии срослись, боковые сохраняются—-очень
тонкие. Naviculare, cuboideum и cuneiforme срослись, как у жвачных. Cuneifrome
лежит над metatarsus II.
На внешних сторонах коренных (верхних) зубов сохраняются бугорки,
мало измененные в лунки. По середине внешней стенки и на ее
заднем
Рис. 361.
Dorcatherium: 1 — череп Dorcath. Naui
cath. crassum Lart.
Каир; А и В — зубы D о r-
углу развиты толстые складки эмали. Рг4 —из двух луночек, остальные
предкоренные удлиненные. Верхние клыки очень длинные, спускаются дальше
нижнего края нижних зубов. Нижние коренные с бугорками на внутренней
стороне и лунками на внешней.
У нынешних Hyaemoschus aquatic us укоротились боковые
пальцы, но средние метакарпалии не срослись (рис. 360, 7).
подкласс: плацентные — placentalia, копытные
285
Dorcatherium Naui Каир (рис. 361,/) — мио-плиоцен Эппельсгейма,
сохраняет рп. Строение конечностей такое же, как у Hyaemoschus. Dore,
classum Lart. (рис. 361,Л, В) — из плиоцена Индии, Dore, majus Lyd.
Tragulus sivalensis Lyd. — плиоцен и плейстоцен Индии. Сред-
ние метаподий задних конечностей срослись слабее, чем передних. От бо-
ковых метаподий остались концы и пальцы. Редукция адаптивная. У ныне жи-
вущих в старости метаподий срастаются очень прочно. Tragulus Napu
Trag. meminna Gray.
Moschus moschiferus — ныне живущий, имеет и передние и задние
метаподий сросшиеся; от боковых пальцев сохранились нижние части метаподий,
прилегающие верхними частями к средним, и фаланги. Резцы и клыки нижней
челюсти небольшие. Верхние клыки очень длинные. Остатки из Сиваликских
холмов. Это постепенное упрощение конечностей у форм, ныне живущих, дает
возможность особенно хорошо проследить изменение четырехпалой конечности
Рис. 362. Protylopus Petersoni Wortm.
с несросшимися метаподиями в конеч-
ности, имеющие трубку из сросшихся
двух средних метаподий.
Сем. Tylopoda. (Cam el idae)
(мозоленогие). — Это семейство лунчато-
зубых резко отличается от других жвач-
ных. Сюда относятся верблюды и ламы.
У них отсутствуют рога, существуют клы-
ки и по два резца в обеих челюстях с
каждой стороны. Два пальца не име-
ют настоящих копыт, а заключены в
твердую кожистую оболочку — подошву. Боковые метаподий и пальцы атро-
фированы. Это-щ* типичные жвачные.
В скелете верблюдов есть еще несколько отличительных признаков: в шей-
ных позвонках артерия про-
ходит в мозговом канале, а
не в отдельном отверстии
снаружи. Нижние концы ме-
таподий не срастаются, рас-
ходятся. Канты находятся на
задней стороне их сочленов-
ных поверхностей, что об-
условливает отсутствие же-
лоба на верхней поверхно-
Рис. 363. Eotylopus(Poebrotherium) Wil-
son i Wonm.
сти первой (Ьаланги. Naviculare не срастается с cuboideum, a magnum
с trapezium.
Нынешние представители этого семейства очень немногочисленны. Ла-
мы живут в Южной Америке. Верблюды: одногорбые — Dremodarium —
в северной Африке и южной Азии; двугорбые — Camelus bactria-
286
класс: млекопитающие — mammalia
n u s — на юго-востоке, в Киргизских степях; в Монголии — в одомашненном
состоянии.
Развитие С a m е 1 i d а е протекло в Сев. Америке, начиная с эоцена.
Protylopus Wortm. (рис. 362)—из нижнего эоцена,-—является прими-
тивной формой, у которой сохранились семь коренных, клыки и три пары
резцов. Коренные с
внешними бугорками и
внутренними лунками.
Рг1 имеет один внеш-
ний бугорок и лунку;
остальные предкорен-
ные проще. Нижние
зубы — с внешними
лунками и внутренни-
ми бугорками. Конечно-
сти — ступни состоят
из двух метаподий;
передние — со слабыми
остатками боковых. Су-
ществует гребень на задней стороне сочленовных поверхностей метаподий.
Eotylopus Matth.—-с сократившейся парой резцов.Оставшиеся долотооб-
разные. Клыки несколько больше, чем резцы, в нижней челюсти и меньше в верх-
ней. Первые предкоренные отделены диастемой от рг2 и от клыков (рис. 363).
Gomphotherium (Paratylopus) (рис. 364) — из нижнего миоцена—
сохраняет 3 пары резцов и укороченный череп—признаки Eotylopus.
Poebrotherium (рис. 363 и 366,А)—из олигоцена и миоцена, пред-
ставлен несколькими видами с более или менее удлиненным черепом и сокра-
тившимися передними зубами, маленькими резцами и клыками, отделенными диа-
стемой от рг1. Коренные лунчатые, вытянутые по длине. Р*— из двух
лунок. Рг3 и рг2 — очень маленькие.
Метаподий срослись до нижнего
конца; боковые исчезли. Орбита почти
замкнутая.
Procamelus (рис. 365 и
366, В)—в миоцене, имеет уже при-
знаки настоящих Camelidae, с
замкнутой орбитой, узкой передней Рис. 365. Procamelus gracilis Соре,
частью черепа (сжатой), значительно
удлиненными коренными, с диастемами между рг2 и рг1, рг1 и с, с и i и
с очень маленькой второй парой резцов.
Нижние коренные лунчатые — тоже удлиненные, предкоренные имеют
по одной паре удлиненных луночек.
Метаподий на концах раздвинулись (рис. 366), очевидно, при-
меняясь к песчаной почве, требующей более широкого соприкосновения
подкласс: плацентные — placentalia, копытные
287
ноги и удлинились значительно. В миоцене намечается разделение на Lama
и Camelus.
,<\Л
Рис. 367. Helladotherium. (Пикерми.)
Это последние формы Camelidae в Сев. Америке; позже миоцена
их там не встречают. В плиоцене они хорошо представлены в Сиваликах
формами, близкими к нынешним. В Европе они из-
вестны — Procamelus — в верхнем миоцене (Кри-
вой Рог б. Херсонской губ.)
и в послетретичных отложе-
ниях — в поволжских губер-
ниях, где они существуют и
теперь в домашнем состоянии.
Итак, это семейство раз-
вивалось в Сев. Америке, и
только в конце миоцена пред-
ставители его появляются в
Азии и в Европе, исчезая в
Америке.
Сем. Giraffinae.— Круп-
ные животные центральной Аф-
рики на очень высоких дву-
палых ногах, с длинной шеей,
удлиненным черепом, лишен-
ным верхних передних зубов,
с небольшими роговыми вы-
ростами, покрытыми кожей.
Конечности без боковых мета-
подий,
вестны
цена
цена
их неизвестны. Вероятно, это одна из
Рис. 366. Ноги: A - Poebrotherium, В — Pro-
cam e 1 u s.
Ископаемые остатки из-
из верхнего мио-
и нижнего плио-
южной Европы, Персии и Сиваликских холмов; более древние предки
ветвей антилоп, не выработавшая рубца.
Helladotherium Duvernoyi
Gaudry (рис. 367) — из отложений
Рис. 368. Sa mother! ti m Boissier!
Fors. Maj.
288
класс: млекопитающие — mammalia
Пикерми в Греции, во Франции, в Бессарабии и в Сиваликах, представлен по-
чти полным скелетом. Череп с удлиненным затылком, закрытой орбитой и не
имеет еще никаких выростов. Шейные крупные позвонки еще не очень удли-
нены. Зубы высокие.
Samotherium Boissieri Fors. Maj. — с роговыми выростами над
глазами и более удлиненными носовыми и межчелюстными костями, из нижнего
плиоцена Самоса, Персии и Китая (рис. 368).
Camelopardalis—с маленькими выростами на границе лобных и
теменных костей. Предкоренные значительно осложнены. Передние конечности
длиннее задних. Шея очень длинная. В тех же отложениях Пикерми, Мараги,
Херсонской губ., в Сиваликах. Потомки дошли до нас — жираффы.
Рис. 369. Sivatherium giganteum.
О с а р i а — небольшая форма, ныне живущая тоже в центральной Африке.
К жираффам относят еще одну странную группу лунчатозубых, пред-
ставленную исключительно ископаемыми остатками в Сиваликских холмах —
Sivatherinae (рис. 369). Она совмещает признаки черепа жираффы и лося,
имея маленькие рога на носовых костях и очень ветвистые на заднем краю
лобных. Кости черепа пневматичные. Передние нижние зубы очень маленькие;
верхние отсутствуют. Коренные — массивные, короткие. Особенность предста-
вляют конечности с укороченными плечом и бедром, удлиненным предплечьем
и метатарсом; голень и метакарпалии короткие. Сюда относятся Bramathe-
rium, Sivatherium и Hydaspitherium, для которых неизвестны ни
предки, ни потомки. Все найдены в Сиваликских холмах.
Сем. Protoceratidae.— Представляют вторую группу лунчатозубых, снаб-
женных двумя парами рогов. Остатки их из нижнего миоцена Южной Дакоты
подкласс: плацентные — placentalia, копытные
289
были найдены и изучены впервые проф. Маршем, и по ним установлена
зона Protoceras-beds, синхроничная с М1 оhiррus-beds в Орегоне.
, Protoceras cel er Marsh (рис. 370). — Черепа самцов снабжены двумя
парами рогов: одна — на верхних челюстных и другая — на теменных. У самок
одна пара на теменных костях. Длина черепа около 18 см, носовые кости
очень укорочены. Животное было величиною с овцу. В верхней челюсти
существуют довольно большие клыки, четыре предкоренные и три корен-
ные — конусообразные, рг* — из двух лунок, три коренные — типичные четырех-
лунчатые. В нижней челюсти три пары маленьких резцоь и клыки, уменьшаю-
щиеся кнаружи. Предкоренные отделены диастемой с обеих сторон; коренные —
типа жвачных. По зубам эта форма ближе всего стоит к Dremotherium и
Amphitragulus. Вместилище мозга было небольшое, но мозг имел глубокие
извилины. Передние конечности четырехпалые, с длинными боковыми пальцами,
но, вероятно, уже не достававшими до земли. Metacarpus II под trapezoideum,
за которой лежит trapezium под radiale. Metacarpus V причленен к unciforme
Редукция—-типа неадаптгвного.
Задняя нога со слабыми ос-
татками боковых метаподий. Средние
не срослись, как и в передних;
cuneitorme II не срослась с сипет
forme III. Нет канта на передней сто-
роне сочленовных поверхностей ме-
таподий. Если отдельные части ске-
Рис. 370. Protoceras celer Marsh.
лета и могут быть сравниваемы с некоторыми формами жвачных или вообще
лунчатозубых, то совокупность признаков, особенно череп, оставляет Proto-
ceras совершенно изолированным среди парнопалых. Присутствие двух пар
рогов одновременно с клыками и несросшиеся метаподий едва ли позволяют
отнести их к жвачным. Ни предки, ни потомки их не известны.
Интересно отметить строение молочных зубов у парнопалых. В работах
В. О. Ковалевского мы находим рисунки этих зубов как для адаптивных,
так и для невдаптивных форм и везде встречаем d* (последний, обозна-
ченный у него через d1) сходным с т1, a di — с тремя рядами бугорков,
так что оба эти зуба сложнее замещающих их предкоренных — р* и р4.
Сравнивая развитие, долговечность и распространение различных групп
парнопалых, мы видим, что преобладающее во всех отношениях значение
получили жвачные — Ruminantia, которые живут теперь как на Севере,
так и на Юге, как в холодных, так и в жарких странах. Чем же можно объ-
яснить такую их долговечность и преобладание над другими группами, которые
или вымерли рано, или если и живут, то на очень ограниченной площади
(Oreodontidae, Tragulidae)? Появление жвачных в конце эоцена и
начале олигоцена совпадает с расцветом неадаптивных форм: Anthracotheri-
idae, Anoplotheriidae и непарнопалых. Gelocus и его потомкам прихо-
дилось борогоСя за пищу с многочисленными травоядными современниками.
19 Па в л о в а М. В. Палеозоология. Ч. И.
290
класс: млекопитающие — mammalia
Вероятно, что неправильно редуцирующиеся конечности одной группы способ-
ствовали победе над ними форм, хотя и более слабых, но правильно разви-
вавшихся. Нужно, однако, думать, что главной причиной победы явилось изме-
нение пищеварительных органов, выработка' отдела желудка — рубца (rumen),,
который позволил лунчатозубым (начиная с миоцена) набирать пищу в запас и
на досуге, в тиши, без борьбы, переваривать ее. Но так как последний не
сохраняется в ископаемом состоянии, то, может быть, трудно указать, какие
формы из ископаемых можно считать обладавшими рубцом, и с какого вре-
мени он стал у них развиваться.
Те из нынешних форм, у которых сохраняются передние зубы и не раз-
виваются рога — Tragulidae, Camelidae — не имеют рубца. Те же, у
которых он существует, не имеют передних зубов, а имеют рога. Это дает
право считать, что ископаемые формы лунчатых, лишаясь передних зубов и.
вырабатывая рога, развивали одновременно рубец, получали возможность наби-
рать пищу в запас, чтб давало им большое преимущество над другими,
формами, не имевшими этой особенности организма. Средне-миоценовые
формы — антилопы и олени — были первыми из таковых, и дальнейшее разви-
тие их и их потомков продолжалось в верхнем миоцене, плиоцене, плейсто-
цене, и теперь они являются единственными из парнопалых, широко и разно-
образно представленными. Только некоторые из них, доведшие свои защити-
тельные органы — рога — до крайних пределов, например Cervus euryceros,.
Cervus latifrons, Cervus Sedgwicki, вымерли; также и бизоны
с громадными прямыми рогами — Bison latifrons.
Подотряд 3
Парнопалые с неадаптивной редукцией конечностей—Bunoselenodontla.
Entelodon (Elotherium) (рис. 371 и 372Л) — из олигоцена Фран-
ции (Ронзон) и Сев. Америки (Дакота) стоит особняком в этой группе, имея
настоящие бугорчатые зубы, что его сближает с Suidae. Но остальные части
скелета, особенно череп и конечности, резко отличаются. Череп крайне узкий
с очень малой вместимостью мозга и большими скуловыми впадинами, далеко-
отодвинутыми назад. Скуловые кости имеют широкий отросток, направляю-
щийся вниз и назад и доходящий до половины нижней челюсти. Орбита ма-
ленькая, закрытая. Полное озубление. Три маленьких резца, большие клыки.
рр — конусообразный, отделен диастемой; рг2 и рг3 — тоже конусы, но значи-
тельно большие; рг4 — четырехугольный, из двух бугорков. Коренные — из четы-
рех сливающихся бугорков с шероховатой поверхностью, имеют воротник, кант,
на внешней стороне; предкоренные окружены им. На нижних коренных — четыре
бугорка более разграничены. Предкоренные все конусообразные. Резцы с осо-
бым зубчатым воротником. Приближаясь несколько к свиным, зубы Entelodon
отличаются закругленной формой большими размерами и меньшим числом бу-
горков.
подкласс: плацентные — placentalia, копытные.
291
Конечности отличаются еще больше. Они — двупалые: III и IV метаподии,
несросшиеся, лежат под magnum и unciforme, II и V в виде маленьких остат-
ков. Ttrapezoideum исчезла. Небольшая lunare опиралась на magnum. Эта форма,
Рис. 371. Entelod on Aym. (El о th er i u m) Pom.
рано лишившаяся боковых пальцев, не сохранила trapezoideum, которая осталась
у других форм. Связать ее с какой-нибудь известной нам линией мы не можем.
Рис. 372. Ноги: А — Enteledon, В — Xiphodon, С — Sus, D Dicotelcs—>
(Л, В — неадаптивные, С, D — адапгивные.)
Остальные формы, отнесенные к группе неадаптивных, подразделяются на:
Сем. Anthracotheriidae — с олигоцена до среднего миоцена:
Подсем. Hyopotaminae — с лунчатыми зубами
„ Anthracotherinae — с бугорчато-лунчатыми зубами.
19*
292
класс: млекопитающие — mammalia
Сем. Anoplotheriidae — с эоцена до верхнего олигоцена:
Род Anoplotherium — с бугорчато-лунчатыми зубами.
Подсем. Xiphodontinae.
Род Xiphodon gracile — с лунчатыми зубами.
Род Caenotherium — также.
А
Рис. 373. Anthracotherium magnum Cuv.
В — верхний, С •— нижний зуб.
Подсем. Anthracotherinae — с сильно развитыми передними зубами,
удлиняющими морду. Предкоренные зубы конусообразные. Конечности четы-
рехпалые.
Anthracotherium (рис. 373) — крупное, неуклюжее животное с плос
кой, удлиненной головой, известно почти цельным скелетом нижнего
миоцена Ронзона во Франции.
Лопаточьиобразные резцы (3) окружены кантом,
большие клыки, и за ними три предкоренных с кону-
Рис. 374. А п с о d u s (Hyopotamus) v е 1 а и и u s
С — передний, D - задняя ступня.
сами, похожие на предкоренные Entelodon; рг4 — из двух бугорков. Корен-
ные имеют четыре больших бугорка с ребрами и между передними бугорками
продольную складку, как бы вытянутый бугорок. На внешней стенке посредине
закругленная складка. Нижние коренные: два первых — из четырех бугорков;
п3 — из шести, с талоном. Anthracotherium magnum Cuv.
подкласс: плацентные —placentalia, копытные
293
Меньшие формы — Anthrac. breviceps — имеют такие же зубы, но
промежуточный бугорок сохраняет свою форму. Более древние вилы A nth г а-
С о t h е г 1 и ш, из олигоцена, представлены формами меньших размеров, и доба-
вочный бугорок у них меньше развит. Anthrac. Riitimeieri—-в эоцене
Швейцарии; у более поздних форм бугорок постепенно увеличивается.
Конечности имели четыре пальца, два средних длиннее, два боковых
короче; последние не доставали до земли. Между тем над metatarsus II сохра-
няется cuneiforme II и фасетка на cuneiforme III, т. е. положение нередуци-
рованного метаподия II. Metacarpus V сочленяется с unciforme так же, как в
нормальной ноге. Эти конечности могут считаться типичными для неправильно
„неадаптивно редуцированных". У Dicotyles (Suidae) при таком же со-
Рис. 375. Anoplotherium commune Cuv.
Д}—реставрация, В— череп; С— верхний, D — нижний зуб; Е— передняя, F—задняя нога.
крашении боковых пальцев trapezoideum перешла на metacarpus III, а вся unci-
forme перешла на metacarpus IV. Ня задней ноге cunieforme II перешла на me-
tatarsus III, a metatarsus II причленяется к ней только маленькой фасеткой
(у Dicotyles).
Конечности древнейших форм Anthracotherinae неизвестны. Очевидно
что эти формы, близкие по типу к S u i d а е, по развитию их пошли иным путем.
Подсем. Hyopotamidae—из эоцена Европы и олигоцена Европы
и Сев. Америки.
Hyopotamus (Ancodus) (рис. 374). — Морда очень вытянутая. Клыки
маленькие, отделены большой диастемой от рг1. Рг1—небольшой конус, отделен
294
класс: млекопитающие — mammalia
от остальных. Рг1 — тоже конус; рг3 с внутренним бугорком; рг4— очень
характерный: листикообразный на внешней стороне, с бугорком на внутренней.
Коренные очень резко отличаются от предкоренных и состоят из 3 передних
лунок и 2 задних. Очень интересны по своей сложности rf3 и с?4; первый из
них превосходит даже тУ, имеет добавочные бугорки.
В нижней челюсти листикообразные резцы (6) и такие же клыки. Пред-
коренные— увеличивающиеся кзади конусы. Коренные из двух внешних лунок
и двух меньших лунок-бугорков; у последнего — талон.
Наиболее крупная форма—Нyopotamus velaunus Cuv.—из Франции
(Ронзон), олигоцен. Конечности его имеют по четыре пальца; метаподии не
срослись; боковые короче срэдних. Внешнего гребня нет на сочленовных кон-
цах. Первые фаланги длинные. Расположение карпальных костей не изменен-
Рис. 376. Diplobune Quercyi Filh.
А — верхние, С — нижние зубы; D — передняя, Е — задняя нога.
ное; trapezoideum над
metacarpus П, metacar-
pus V едва припле-
няется к unciforme. Вто-
рой ряд косточек че-
редуется с первым.
Пяточная имеет фа-
сетку для малой берцо-
вой; надпяточная сочле-
няется с naviculare и
cuboideum, которая ле-
жит над metatarsus V
и IV. Этот тип сочле-
нения ног сильно отли-
чается от гиппопотама.
Менее хорошо пред-
ставлены Hyopotami-
d а е из эоцена Швей-
царии — только зуба-
ми: Н у о р о t. G г es-
slyi, Hyopot. Picteti,
Hyopot. porcinus. Но они устанавливают тот факт, что добавочный буго-
рок на коренных зубах у более древних форм развит слабее, чем у более поздних.
В нижнем миоцене Сев. Америки найдены черепа Anthracotherium
и Hyopotamus. Между последними Осборн устанавливает два вариетета:
длинномордую форму -— Hyopot. americanus Leidy, сходную с европей-
ской, и короткомордую — Hyopot. brachyrhynchus. У последней пред-
коренные расположены сплошным рядом, к которому примыкает клык.
В Африке, в олигоцене Египта, найден Hyopot. Gorringei Andr.
Другие палеонтологи устанавливают новые роды для этих форм.
Brachyodus brachyrhynchus Scott — из верхнего олигоцена
Сеч. Америки, с короткой мордой, без диастемы.
подкласс: плацентные — placentalia, копытные
295
Bothriogenys Fraas i Schmidt — из нижнего олигоцена Египта
(Фаюм) с более удлиненной мордой и небольшой диастемой.
Ancodus leptorhinus Filh. — из олигоцена Франции (фосфориты
Керси), с очень удлиненной мордой и большими клыками.
Эта с голь распространенная форма имела в Англии (Hordwell) близкого
представителя с редуцированной конечностью, у которой сократились два бо-
ковые пальца.
Diplopus Ковалевского—найдены только кости конечностей вместе
•с настоящими Hyopotamus (четырехпалым); он представляет в расположении
костей конечностей сходство с Hyopotamus: вся cuneiforme III лежит над
metatarsus II, cuboideum— над IV.
Сем. Anoplotheriidae. — С короткой мордой; зубы без диастемы.
Конечности двух- или трехпалые.
Anoplotherium commune (рис. 375) — описан Кювье из гип-
сов Монмартра (нижний олигоцен), встречается в фосфоритах Керси и
в других местах. Плоский, короткий череп с беспрерывным озублением и ма-
ленькими резцами и клыками отличает его сразу от Hyopotamidae.
Коренные зубы состоят из двух внешних лунок, задней внутренней, переднего
внутреннего бугорка и промежуточного бугорка между передними — лункой
и бугорком. Форма лунок и бугорков изменяется на различных зубах, и иногда
трудно различить, который бугорок добавочный. Рг4— из двух луночек; остальные
предкоренные удлиненные, имеют по три складки на внешней стороне и не-
большие бугорки на внутренней. Нижние коренные состоят из луночек; пред-
коренные удлинены.
Конечности сохранили два средних несросшихся метаподия; от двух
боковых остались или незначительные части, или сохранился укороченный
II.
Но trapezium и trapezoi-
deum лежат над остат-
ком metacarpus И, кото-
рый сохранил даже
фасетку на magnum.
Все cuneiforme сохра-
нились.
У Diplobune
tridactylum, Dipl.
Quercyi (рис. 376)
и Dipl, bavaricum
Рис. 377. Xiphodon gracile Cuv.
(Anoplotherium
Cuv.) сохранились вторые пальцы с копытными фалангами (очень укороченные)
и остаток metacarpus I. Метаподий с таким же расположением карпальных и
тарсальных костей, как и у двупалой формы. Эго типичные неадаптивные
конечности Ковалевского.
Подсем. Xiphodontidae. — Имеют полное озубление с лунками на
коренных. Верхний олигоцен Европы. Маленькие животные.
296
класс: млекопитающие — mammalia
Xiphodon gracile Cuv. (рис. 377)—-с четырьмя правильными лун-
ками на коренных и маленькой добавочной полулуночкой на передней внутрен-
ней стороне; эта луночка иногда не развивается. Предкоренные удлиненные,
типа D. р 1 о b u п е, но более простые. Нижние коренные с внешними луноч-
ками и внутренними бугорками; рг4 такой же; prs и рг2 удлиненные.
Конечности тонкие — metacarpus III и IV не сросшиеся; trapezoideum —
над остатком IV. Редукция неадаптивная; животное вымерло в олигоцене.
Caenotherium (рис. 378) — из олигоцена Европы (Франция).
Коренные зубы лунчатые: три луночки в задней половине, две — в перед-
ней; все равные. Добавочная сзади; у всех других форм она была спереди.
РР — с двумя луночками; остальные предкоренные — удлиненные конусы. Ниж-
ние коренные состоят из двух луночек-бугорков; рг± с одной внешней луночкой
и двумя внутренними бугорками, остальные удлиненные. Весь череп равен 6 см.
Рис. 378. Caenotherium elongatum Filh.
Орбита замкнутая. Конечности четырехпалые, с trapezoideum над укороченным
metacarpus IV. Metatarsus II под cuneiforme II; cuboideum прикрывает metatarsus
IV и V, которые почти равны metatarsus III На передней конечности боковые
метаподий значительно короче средних.
Plesiomeryx Filh.— еще меньших размеров, с таким же черепом.,
зубами и конечностями. Керси.
Это главные представители с конечностями, неадаптивно редуцированными,
и с зубами, имеющими добавочный 5-й бугорок или луночку. Есть еще не-
сколько форм, представленных отдельными челюстями или зубами. Все они не
переходят нижнего миоцена и вымирают. Раньше некоторые из этих форм счи-
тались предками жвачных. Допускалось, чго исчезала пятая луночка, и зубы
становились четырехлунчатыми. Но тесная связь пятибугорчатых зубов или
пятилунчатых с неадаптивными конечностями, с одной стороны, развитие, а не
уменьшение, бугорков с геологическим возрастом животных и существование
одновременно с ними четыргхбугорчатых форм — с другой, заставляют отбро-
сить эту мысль о них как о предках жвачных и считать вымершей группой»
подкласс: плацентные — placentalia, копытные
297
получившей начало от эоценовых форм, у которых к 4 бугоркам добавлялся
пятый.
Pleuraspidotherium Lem. — из нижнего эоцена окрестностей
Реймса, с удлиненным, гладким черепом, бугорчатыми коренными, с добавочными
бугорками между двумя передними (иногда), с астрагалом, снабженным еще
шейкой, — может быть, станет у основания развития неадаптивных форм, если,
будут найдены связующие их формы.
ОтрядП.
Хоботные — Proboscidea
Ныне живущие хоботные представлены одним родом Е1 е р h a s — слонами,
живущими в тропических странах Азии и Африки и разделяющимися на два
вида: Elephas indicus и El. africanus.
Главные признаки слонов: удлиненный нос — хобот, два торчащих далеко
вперед в верхней челюсти громадных резца — бивни. Пятипалые конечности,
снабженные общим для всех пальцев копытом, что и дает повод причислять
их к копытным. По всем остальным признакам и по громадным своим разме-
рам слоны резко отличаются от последних.
Резко ограниченные теперь в своем географическом распространении,
слоны в третичную и особенно послетретичиую эпоху были расселены по всем
частям света, кроме Австралии. Для послетретичной формы—мамонта—труд-
нее указать местность в Европе, где он не существовал, чем места, где най-
дены его остатки. Эта форма — Elephas primigenius (первичный слон,
мамонт), являющаяся наиболее близкой к ныне живущим, считалась их предком.
Но подробное изучение строения зубов мамонта заставило отказаться от этого
и смотреть на него как на вымершую форму, особенно широко распространен-
ную на Севере, в противоположность нынешним — обитателям жарких стран,
так что „первичный слон“ является неподходящим названием.
Более древние остатки ископаемых слонов известны из различных отделов
верхних третичных слоев, причем вместе с этими формами и в более древних
слоях встречаются другие формы, хотя и близкие к слонам по общей организа-
ции и скелету, но отличающиеся резко по строению зубов: это — мастодонты.
Зубы слонов, коренные, состоят из пластинок дентина, покрытых эмалью
и соединенных между собой цементом. Они обладают постоянным ростом, и
корни появляются только у старых зубов. Из резцов сохраняется только одна
пара в верхних челюстях; клыков совсем нет.
Зубы мастодонтов имеют корни и коронки, состоящие или из закруглен-
ных бугорков — трех или четырех пар, или из трех или четырех пар гребней.
Коронка покрыта эмалью; цемента нет. Молочные зубы существуют у мастодон-
тов, у слонов они отсутствуют. Бивни имеются в верхних и в нижних челюстях.
Остатки мастодонтов, найденные в более древних отложениях, чем
остатки слонов, определенно указывают на более древнее существование масто-
донтов. Кювье установил для них название „мастодонт", т. е. „сосцевидные:
зубы", отделив их от слонов с пластинчатыми зубами.
298 •
класс: млекопитающие — mammalia
Подотряд! z
Mastodontidae
До недавнего времени древнейшей формой этой группы считался Masto-
don angustidens — из среднего миоцена Европы и Сев. Америки.
Но в последнее время были найдены остатки более древних форм
в Египте, близ Каира, в среднем и верхнем эоцене Фаюма, — в озерной области,
занимающей углубление в пустыне на запад от долины Нила, против не-
большого городка Васта, около 57 миль на юг от Каира. Это те флювиально-
морские отложения, из которых мы знаем уже остатки китообразных и гиопо-
тама. Эти находки подтвердили давнишнее предположение Осборна, что
предки хоботных должны быть найдены в Африке, откуда онн эмигрировали
в Евразию в конце эоцена или в начале миоцена, когда эти континенты
были соединены между собою. Факт этот имеет громадный интерес, так как
до этих находок „Африка считалась мрачным континентом для палеонтоло-
гии"— по словам Осборна.
Мастодонты разделяются по строению зубов на бугорчатозубых и гребне-
зубых. Все зубы снабжены корнями.
А. Бугорчатозубые мастодонты
Moeritherium lyonsi Andr. — считается наиболее древней формой
хоботных из среднего эоцена Фаюма. Он был величиною с тапира; череп
удлиненный, с плоской верхней поверхностью, совершенно
3 13____з 1
признаков хоботных (рис. 379, А). Зубы:- — ---йаиболее
2 v о — о
еще не имеет
развита вторая
пара резцов вверху и внизу. Коренные бугорчатые — из четырех бугорков. Пред-
коренные: рг2 (первый) — удлиненный, из нескольких бугорков; рг3 и рг4 —
трехбугорчатые; на т3 и т3 — сзади талоны. Кроме черепов от Moerithe-
rium известны позвонки, причем второй шейный имеет длинный зубовидный
отросток и напоминает этим свиной позвонок; тело его не укорочено, и голова
животного могла, вероятно, нагибаться до земли. Плечо узкое в своей верхней
части. Коракоидный отросток лопатки сильно развит и обхватывает головку
плеча с нижней стороны. Бедро больше подходит к слоновым, и у некоторых
видов оно как бы подходит к типу следующей формы — Moeritherium
trigonodon Andr. ; ;
Palaeomastodon (рис. 379,Z?) — из верхнего эоцена Фаюма, несколько
больших размеров, чем Moeritherium, по общему виду более похож на хобот-
ных, но имеет еще довольно длинную шею. Череп укорочен, нижняя челюсть удли-
нена. Носовые кости очень укорочены; существовал хобот. Из передних зубов
остались только резцы, по одной паре в челюстях; верхние длиннее нижних;
у тех и других на внешней стороне существует полоса эмали. Зубы эти
с постоянным ростом. Верхние бнвни несколько расходились и, очевидно,
пропускали между собою небольшой хобот.
подкласс: плацентные — placentalia, копытные
299
Предоренные:—рг2 удлиненный; рг3 и рг4—четырехбугорчатые; т1 и
имеют по шести бугорков, расположенных попарно; т3 — с добавочными
горками. Нижние — такие же, но более удлиненные; сократился рг2.
Найден молодой череп с молочными зубами, причем rf4 состоит
шести бугорков и похож на /п1; он сложнее, чем рг4. D3—значительно длиннее,
чем
т2
бу-
из
рг3, состоит из четырех бугорков. D2— не имеющий заместителя, маленький,
удлиненный. Межчелюстные
кости у молодого животно-
го более удлинены, чем у
взрослого (сравнительно с
черепом).
Первый шейный позво-
нок, atlas, большой, с глубо-
кими сочленовными впади-
нами для выступающих дале-
ко назад затылочных мыщел-
ков. Второй шейный позво-
нок, axis, имеет длинный зу-
бовидный отросток, похожий
на свиной.
/дз.
Рис. 379 A— Mocritherium. (Верхний эоцен Фаюма.) В -Palaeomastodon.
(Нижний олигоцен Фаюма.) С — Mastodon angustidens. (Миоыен Франции.) D—
Mastodon americanus. (Плейстоцен Сев. Америки.)
Передние конечности с широким плечом. Задние с плоским, высоким
бедром, с сильно развитым отростком — трохантером. Большая берцовая с рас-
ширенным вверху концом. Нижняя почти плоская поверхность большой берцовой
кости сочленяется с астрагалом. Все эти признаки сближают эти кости с ко-
стями хоботных. Пяточная и надпяточная указывают на животное с довольно
высокими ступнями. Пальцы неизвестны.
300
класс: млекопитающие — mammalia
Palaeomastodon представлен несколькими видами различной вели-
чины, из которых одни стоят ближе к Moeritherium по строению конечно-
стей, другие — дальше. У них
Рис. 380. Mastodon angustidens (ws).
замечается как бы переход жи-
вотных от водного образа жизни
к сухопутному, какой вели бо-
лее поздние хоботные: так сле-
дуют Moeritherium Andr.,
Palaeomastodon parvus,
Pal. beadnelli Andr.
Mastodon (T e t r a be-
lodon) angustidens Cuv. —
из среднего миоцена Европы и Сев. Америки, стоит близко к Palaeomasto-
don (рис. 379 С, 380 и 381) — особенно по строению зубов, но по размерам
своим он значительно больше его. Абель указывает на Tetrabelodon р у g-
m а е u s—из нижнего миоцена Алжира и нижнего миоцена Европы как на предше-
ственника Mast, angustidens. В Парижском музее восстановлен целый ске-
лет последнего из Симора (Франция) (рис. 381). Череп его укороченный, с закруг-
ленным лбом и затылком; межчелюстные кости короткие, несут пару длинных,
почти прямых бивней с полосою эмали. Передняя часть нижней челюсти очень
вытянута и несет пару более тонких и коротких бивней, чем верхние.
Верхние коренные зубы имеют по три ряда бугорков, как у Palaeo-
mastodon, т3—с четырьмя бугорками и слабым талоном. Два предкоренные
имеют по два бугорка.
Нижние зубы такие же,
но несколько уже. На та
задний талон подразде-
ляется на два небольших
бугорка. Между большими
бугорками на всех зубах
имеются маленькие, про-
межуточные, которые при
стирании сливаются с боль-
шими и осложняют их.
Молочные сложнее
предкоренных. Z)3 имеет
три ряда бугорков, как у
Palaeom astodon, плюс
добавочные (рис. 382).
Общее число коренных —
шесть вверху, пять внизу:
Рис. 331. Mastodon angustidens Cuv.
(В Парижском музее.)
3 — 3
2^3'
Шейные позвонки очень короткие, плоские,
с очень небольшими отростками, не позволявшими животному опускать низко
голову.
подкласс: плацентные — placentalia, копытные
301
Спинные позвонки с сильными отростками. Ключица отсутствует, так же
как у копытных. Многочисленные ребра — до 20 — доходят до таза.
Почти треугольная лопатка с характерным гребнем посредине ее, снаб-
женным особым загнутым наружу отростком, имеет широкую гленоидную впа-
дину для головки плеча — широкой кости. Широкая вверху локтевая и тонкая
Рис. 382. Нижняя челюсть Mastodon
angustidens Cuv. с мслочн. зубами.
лучевая не срастаются.
С гупни, как передние, так и задние,
сохраняют пять пальцев с укороченными
фалангами, особенно второй и третий.
Рис. 382 А. М a s t о d о n angustidens
Cuv. Молочные зубы.
Карпальные кости расположены сериально, не чередующимися рядами.
Тарсальные очень низкие; пяточная выступает назад. Бедро плоское. Большая
берцовая массивная; малая — слабо развита. Это строение характерно и для
последующих более поздних геологически хоботных и может служить дтя
сравнения отдельных частей их, так как последние формы представлены менее
полными остат ками.
Mastodon (Tetrabelodon) longirostris Каир (рис. 383) — из
мио-плиоцена Эппельсгейма, дает следующую стадию развития мастодонтов,
выражающуюся в осложнении коренных зубов следующим образом: два первые
коренные имеют по четыре пары бугорков
s t i d е n s. На последнем верхнем корен-
ном т'л пять пар и талон; hi ниж-
вместо трех, как у Mast, angti-
Рис. 383. Mastodon 1 о n g i г о s-
tris. Верхние молочные зубы.
Рис. 384. Mastodon arver-
п е n s i s.
нем — шестая пара бугорков образовалась из талона. Промежуточные бугорки
существуют. Последние предкоренные имеют по три ряда бугорков, рг3 и рг3 —
по два. У старого животного остаются только по два последних коренных.
Как межчелюстные кости, так и симфизы нижней челюсти удлинились; они
заключают по паре бивней, которые утратили эмалевые полосы. Кости
скелета неизвестны. Череп невполне хорошо сохранился. Молочные зубы
имеют четыре ряда двойных бугорков с добавочными на d4 и d4, по три
302
класс: млекопитающие — mammalia
ряда — на d3 и 6?3; d2 и d2 — небольшие, двухбугорчатые; последние зубы
не замещаются.
Этот вид очень богато представлен в музеях Западной Европы раз-
личными зубами, которые позволяют незаметно перейти к типичному для
плиоцена —
Mastodon (Tetrabelodon) arvernensis Croiz. Job. (Овернь)
(рис. 384) — с таким же числом бугорков на зубах. Осложняется только послед-
ний нижний коренной; но здесь бугорки обоих рядов не лежат один против другого,
как у двух первых видов, а чередуются между собою. Промежуточные остаются.
Это чередование бугорков не является внезапным: оно появляется и на некото-
рых зубах Mastodon longirostris и увеличивается постепенно, переходя
к Mastodon arvernensis. В Венском музее находится много зубов, кото-
рые в передней части сохраняют парное расположение бугорков, а в задней уже
происходит чередование бугорков, типичное для плиоценовой формы, Mast,
arvernensis. Она заканчивает собою в Европе группу мастодонтов с бу-
горчатыми зубами, в которую входят, кроме названных европейских форм,
близкие к ним, может быть, вариететы их:
Tetrabelodon angustidens Соре — в Сев. Америке.
Mastodon proavus Соре — там же.
Mastodon palaeindicus Lyd. — в Сиваликских холмах Индии.
Mastodon pyrenaicus Falc. — в Пиренеях.
Mastodon Pentelici Gaudry — в Пикерми.
Особенно богато представлена эта группа в Сиваликах Индии, где най-
дены хорошо сохранившиеся черепа с зубами и бивнями, например М a s t о-
donsivalensis в плиоцене. Для плейстоцена указаны отдельные зубы,
найденные в Буэнос-Айресе — Mastodon Humboldti. Но там не всегда
точно определяется геологический возраст отложений.
Интересно проследить изменение молочных зубов этих форм (Tetra-
belodon), подтверждающее генетические отношения между ними.
Для Moeritherium известен только один молочный — последний из
нцжней челюсти — d4. Хотя он плохо сохранился, но видно, что он имел четыре
бугорка, по два в ряде, т. е. форму корен-
ных зубов Moeritherium и форму
Palaeo mastodon. У последнего корен-
ные имеют три пары бугорков — trilo-
phodon — так же, как и последний мо-
лочный, предсказывая форму последнего
предкоренного у Mastodon angus-
tidens. У Palaeomastodon 5 ко-
ренных остаются до старости в челюсти
(рис. 385).
Рис. 385. Молочные зубы Palaeo-
mastodon.
Последний молочный у Mastodon angustidens имеет форму по-
следнего предкоренного Mast, longirostris — trilophodon, а у этого
подкласс: плацентные — placentalia, копытные
303
последнего он уже tetralophodon, как и коренные, и бугорки начинают сдви-
гаться, чередоваться, как у Mast, arvernensis.
У Mast, longirostris заменяется только один зуб с каждой стороны,
у Mast, arvernensis — тоже. Он имеет два ряда чередующихся бугорков
с большим числом добавочных.
Конечности у этой группы мастодонтов сохраняются без изменений,
пятипалыми, как и у более поздних хоботных.
Б. Гребнезубые мастодонты
Отклонение от бугорчатых зубов дает Mastodon tapiroides, встре-
чающийся вместе с Mast, angustidens в среднем миоцене Симура (Фран-
ция). У него бугорки сжаты с боков, что придает им форму гребней, особенно
при стирании, и на передней стороне зуба добавочные маленькие бугорки,
расположенные в косые ряды, идущие от вершины бугорка к подножию сосед-
него, У него по три гребня на двух первых коренных и по четыре на послед-
них. Эта форма, известная только по зубам, отделилась, вероятно, от Mast,
angustidens в начале его развития и положила начало новой группе, у
которой бугорки двух рядов сбились попарно и образовали гребни.
Mastodon turicensis (рис. 389, а) — из верхнего миоцена, стоит
близко к другой форме — Mast, tapiroides (рис. 389, b и 386); некоторыми
учеными эти формы даже отождествляются. У него тоже по три гребня на
коренных, кроме последних, где их по четыре, и слабый талон на т3. ho
здесь гребни почти цельные; они только немного подразделены посредине.
И здесь, как у бугорчатозубых, т3 сложнее, чем /я3. Ряды добавочных бугорков
выражены слабее. Нижние бивни значительно короче, чем у Mast, angusti-
dens. Число коренных не бывает одновременно больше трех. У старых жи-
вотных остаются два. В Европе и Сев. Аме-
рике; в последней — вариетет Mastodon
brevidens Соре.
Mastodon Borsoni Lart. (рис. 389, d
и 387). — Продолжает в плиоцене линию
гребнезубых мастодонтов. Число гребней
на т3 увеличилось: их пять. На /и3 пятый
в виде талона. Промежуточные бугорки почти
исчезли. Они сохраняются иногда на совер- рис. 386. Mastodon tap.roi des
шенно нестертых зубах. Остальные коренные Gaudry.
имеют по три гребня. Число эго остается
постоянным у гребнезубых мастодонтов. Громадные прямые бивни вверху,
небольшие — внизу. Встречается в Европе и Сев. Америке. Близ г. Николаева
найден почти целый скелет в 1860 г. Зубы хранятся в Академии наук.
Этот мастодонт имел громадные размеры и прямые бивни.
Mastodon oh i oti c u s = Ma s t. americanus (рис. 388). — На-
столько близок к Mast. Borsoni, что трудно отличить отдельные зубы.
На т3 четыре гребня и талон; на т3 — пять гребней; последний из них
304
класс: млекопитающие — mammalia
маленький и кроме того несет тало г. Гребни более массивны. На некоторых
зубах при стирании получается не овал, а углубление в виде листа клевера
добавочных
от остатков стершихся
бугорков.
Верхние бивни громадные, загну-
тые; нижние только зачаточные; у
взрослых их не бывает. Это главнее
отличие от Mastodon Borsoni. По-
следняя форма долго считалась при-
надлежащей только Европе, a Mast,
ohioticus — только Сев. Америке, в
Рис. 387. Mastodon Borsoni.
плейстоцене. Но на-
ходка почти полно-
го озубления масто-
донта в с. Песчана,
Подольской губер-
нии, и сравнение с
озублением масто-
донта из г. Николае-
ва позволили уста-
новить различие их
и других совместно
изученных форм и
признать присут-
ствие этих обоих
мастодонтов как в Европе, так
Mast. turicensis Schinz и
и в Сев. Америке. Они являются потомками
близок, может быть то-
ждествен, с Mast. Bor-
soni.
Молочные зубы этой
группы мало известны.
.Парте (Larte) указал
один d для Mast, turi-
censis (Sansan).
Таким образом мы
имеем для обеих групп
мастодонтов параллельные
фермы в Европе и в Сев.
Америке, тогда как в Азии,
в Сиваликах, очень богато
представлены только мас-
Mast, brevidens Cope. Последний очень
тодонты с бугорчатыми рис ggg. Mastodon americagus (ohioticus).
зубами, близкие к Mast.
з n g u s t i d е n s и М a s t. longirostris, настоящие же rpe6He3j бые не известны
подкласс: плацентные — placentalia, копытные
305
В группе гребнезубых мастодонтов мы видим такое же постепенное из-
менение, осложнение коренных зубов (рис. 389), как и в первой, с той раз-
ницей, что там зубы все осложнялись, добавляя сначала один и еще один
ряд бугорков до четырех. Здесь изменяется только последний коренной, до-
водя число гребней на та до шести. С увеличением этого зуба сокра-
щаются бивни.
Конечности остаются пятипалыми и у Mastodon ohioticus, от
которого найден в Сев. Америке целый скелет. В строении его скелета разницы
большой нет сравни-
тельно даже с Masto-
don angustidens.
(рис. 381).
Конечно, кроме
названных форм, связь
которых более опреде-
ленная, есть еще другие,
несколько отклоняю-
щиеся по форме черепа
или расположению бу-
горков или гребней на
М. arvern. и Borsoni.
М. longir. и americ.
Рис. 389. Изменение зубов мастодонтов (по Годри).
зубах.
Единственной формой, близкой к мастодонтам по скелету, но резко отличаю-
щейся от них по черепу и зубам, особенно бивням, является род Dinotherium
(рис, 390), связь которого с мастодонтами до сих пор не установлена.
Род Dinotherium. — Строение скелета с короткой шеей, массивными
конечностями повторяет скелет мастодонта. Преобладание локтевой кости над
лучевой, тонкий верхний конец последней; значительное преобладание большой
берцовой над малой, имеющей тоже верхний конец значительно тоньше, чем
нижний; пятипалая, не редуцированная конечность, все эти признаки вполне
совпадают с признаками хоботных вообще и затрудняют точное определение
отдельных костей этих форм.
Череп имеет совершенно определенные признаки, резко отличающие его
от всех других копытных. Плоская, удлиненная черепная крышка с несколько
поднятым затылком и длинными носовыми костями; очень маленькие орбиты
и громадные скуловые впадины, полное отсутствие верхних передних зубов —
вот его признаки.
Нижняя челюсть низкая, с круто загнутым вниз симфизом, заключает два
толстых, коротких бивня (резца), тоже загнутых вниз и назад. Это положение
бивней настолько странно, что при первом описании они были повернуты
вверх концами на сломанной челюсти. Такое направленное вниз положение
бивней встречается только у моржа, что давало повод считать и Dinothe-
rium тоже животным водным. Каждая челюсть динотерия заключает по
пяти коренных зубов, причем нижние несколько более удлинены, чем верхние.
По строению своему они ближе всего стоят к нижним зубам тапира, так как
20 Павлова М. В. Палеозоология. Ч II.
306
класс: млекопитающие — mammalia
состоят из двух гребней, идущих поперек зуба. Верхние зубы имеют внешнюю
стенку, соединяющую оба поперечных гребня. Первый коренной вверху
и внизу имеет три поперечных гребня, что отличает его от двух других.
Кювье описал его зубы под именем гигантского тапира. Последний мо-
лочный зуб имеет тоже три гребня. Так как он существует одновременно
с тУ и Шу до смены, то у молодого Dinotherium может одновременно
существовать рядом два зуба, имеющих по три гребня. Молодые пред-
коренные зубы сохраняют на своей
внутренней стороне бугры, которые
при стирании сливаются с внешними
буграми и образуют гребни. Все
зубы состоят из корня и коронки;
последняя состоит из дентина и
эмали. Пятипалая конечность (раньше
была описана как трехпалая) имеет
короткие метаподии и еще более
укороченные фаланги. Карпальные
кости — рядового расположения, не
чередуются. Пяточная низкая, над-
пяточная совершенно плоская.
Генетическое отношение р. Dino-
therium к мастодонтам можно срав-
нить с отношением Elasmothe-
rium к носорогам. При общности
л
Рис. 390. Dinotherium giganteum Каир.
Череп; верхние и нижние зубы.
признаков скелетов — пол-
ное отклонение в строении
черепа и зубов. ^Момент от-
деления его от ствола хобот-
ных неизвестен. Геологиче-
ская жизнь динотерия была
довольно продолжительна.
Древнейшие представители
известны со среднего мио-
цена (с Mast, angusti-
dens и М. tapiroides)
до верхнего плиоцена Ру-
сильона (Франция). Но все
остатки принадлежат Европе: Франции, Греции, Австрии, Румынии и у нас
в Херсонской губ. Они были подробно изучены во Франции (D е р ё г е t)r
и установлено несколько видов, отличающихся главным образом по раз-
мерам. Остатки были представлены, главным образом, зубами. Череп был
найден в Дармштадте и описан Каупом (Каир). Но наиболее полные
остатки, почти полный скелет, найден в Румынии и описан профессором
Стефанеску (Stefanesku), который и дал возможность составить на-
подкласс: плацентные — placentalia, копытные
307
стоящее представление об этом загадочном звере и установить его наземный
образ жизни. Эта форма названа Dinotherium gigantissimum Stef,
за ее размеры, много превышающие форму Каупа — Dinother. gigan-
teum1). Меньшие виды описаны под именами: Dinotherium Cuvier
Каир, Dinoth. medium Каир, Din. bavaricum H. v. Меу., Dino-
therium proavum Eichw., Dinotherium Koenigi Каир и др. Формы
эти, несмотря на свое разнообразие и многочисленность, представляют мало
отклонений от типа и всегда легко определяются по зубам.
Потомки, как и предки его, неизвестны. Абель указывает В ary th е-
rium grave Andr. — из верхнего эоцена Фаюма (Египет), известного по
нижней челюсти, сломанной спереди, и на Tetrabelodon dinotherioi-
d е s —-из верхнего плиоцена Канзаса, как на близкие формы к динотерию.
Последняя форма — с загибающимися вниз нижними челюстями (для бивней),
имеет коренные зубы типа Mast, arvernensis, почему и не может быть
близкой генетически к динотерию. У Barytherium прямая симфиза нижней
челюсти с очень узкими предкоренными и очень укороченное, с расширенным
нижним концом, плечо совершенно не подходят к типу динотерия.
Подотряд 2
Слоновые — Elephantidae
Сходство скелета мастодонтов со слоновыми и отличие их зубов
заставили Кювье, причислив их к хоботным, отделить под именем масто-
донтов, как мы это уже видели. Отличие слоновых, кроме существования
у них пластинчатых зубов с постоянным ростом (до старости), заключается
еще в полном отсутствии нижних бивней и отсутствии молочных зубов,
т. е. зубов, которые существуют у молодых животных на месте предкоренных
и которые заменяются выталкивающими их позже снизу предкоренными, как
это было у мастодонтов. У слонов все коренные зубы располагаются одни за
другими; их всего шесть, и появляются они постепенно, подвигаясь вперед по
мере стирания передних. У очень молодых форм существует три зуба, из ко-
торых первый очень маленький и рано выпадает. При постепенном стирании и
выпадении 2-го и 3-го постепенно выдвигаются сзади: четвертый, пятый и шестой.
У взрослых их бывает одновременно по два с каждой стороны, а в старости —
по одному, который появляется у современного слона на 40-м году жизни.
Мы не знаем точно форм, которые связывали бы мастодонтов со слонами.
Многими учеными принимаются за таковые слоны из группы Stegodonti-
dae Сиваликских холмов Индии.
Сем. Stegodontidae. — Эта довольно большая группа заключает формы,
у которых коренные зубы состоят из корня и коронки, сложенной из многих
гребней — не менее шести на передних зубах и до десяти на задних. Гребни
низкие, узкие, сжатые, со слабыми складочкообразными углублениями эмали
9 Остатки этого вида из окрестностей Кривого Рога описаны А. С. Серге-
евым — „Известия Геологического комитета", 1914.
20*
308
класс: млекопитающие — mammalia
пр сторонам. При стирании получаются сплошные долинки, ограниченные
эмалью (с дентином внутри) — Stegodon Cliftii (рис. 391). У некоторых
из них, например yElephas (Stegodon) insignis Falc., существует
зачаток цемента между гребнями, но корни сохраняются.
Наиболее простые из них, т. е. лишенные цемента и имеющие очень
низкие гребни, относятся еще к мастодонтам, например Mastodon е 1 е-
phantoides Falc.
В Сиваликских холмах найдены черепа некоторых из этих форм с ти-
пичными слоновыми признаками; они очень закруглены в затылочной части,
и бивни далеко проникают в носовую область.
Но при подробном изучении этих зубов трудно представить себе воз-
можность изменения их в зубы с постоянным ростом. Ни на одном из них
не видно попытки удлинения коронки и уменьшения корня. Эту группу можно
скорее считать боковою ветвью мастодонтов, специально развившеюся в верх-
Рис. 391. Stegodon Cliftii. Зуб.
Рис. 392. Elephas planifrons. (Ферладаны,
Бессарабия.)
нем миоцене и нижнем плиоцене Индии. В плиоцене и миоцене Сиваликских
холмов и о-ва Перима мы встречаем мастодонтов, которые близки к .Masto-
don angustidens Cuv. и Mast, longirostris Каир, но у которых
бугры имеют как бы больше подразделений; между ними располагаются про-
межуточные бугры. Это Mast. Cautleyi Lyd. — с тремя рядами бугров-
гребней, четвертым рядом недоразвившихся и двумя рядами мелких бугров на
передней и задней сторонах /и1. На /и3 пять рядов бугров и талон, тоже
как бы надрезанные в нескольких местах.
Mastodon latidens — с шестью рядами бугров-гребней, почти без
подразделения их посредине, и 7-й талон на та— делает шаг в направлении
к слонам, возвышая свою коронку и уменьшая число промежуточных бугров.
На т2 пять рядов бугров-гребней. Эмаль очень толстая. Цемент обыкновенно
отсутствует.
Ближайшей формой к Mast, latidens является Elephas plani-
frons Cautl. (рис. 392) — из нижнего плиоцена тех же Сиваликских холмов.
Зубы его имеют удлиненные пластинки дентина, окруженные эмалью и соеди-
ненные между собою цементом, который покрывает весь зуб; коронка его
значительно более высокая, чем у предыдущих форм, но менее высокая, чем
корни.
подкласс: плацентные — placentalia, копытные
309
Это уже настоящий слон с плоским лбом, одной парой верхних бивней
и зубами с цементом, но имеющими еще корни.
Остатки зубов и черепов из Сивалик очень многочисленны, и по ним
можно видеть различную степень перехода от бугров-гребней к толстым пла-
стинкам. На некоторых зубах сохраняется еще след средних бугров, дающих
при стирании форму ромба на середине пластинок. Связующим элементом
между этими формами
мастодонтов и слонами
является присутствие од-
ного молочного зуба с каж-
дой стороны, указанное
Фальконером (Fal-
coner), отсутствующего у
слонов.
Несколько лет тому
назад были найдены ос-
татки Elephas plani-
f г о n s Falc. (зуб) в де-
ревне Ферладанах, в Бес-
сарабии, в верхнем плио-
цене, а также и в Ав-
Рис. 393. Elephas meridionalis Nesti.
(В Парижском музее.)
стрии.
Дальнейшее измене-
ние ведет к нескольким
формам, хорошо представленным: так, в Европе — Elephas meridionalis
Nesti, в Сиваликах—Elephas hysudricus Falc., в Сев. Америке —
Elephas Columbi Osb.
Все эти слоны характеризуются громадными размерами, зубами с неболь-
шим числом толстых пластинок дентина, окруженных толстой эмалью и соеди-
ненных широкими слоями цемента; присутствием больших, не сильно загнутых,
бивней в верхней челюсти.
El. meridionalis Nesti — представлен хорошо сохранившимся скелетом
Durfort во Франции), черепами и отдельными костями в других странах
(рис. 393).
El. hysudricus Falc. — близкий к нему.
El. Columbi Osb. известен только по зубам.
Этими верхне-плиоценовыми формами заканчиваются третичные слоны.
Плейстоценовые слоны
Мы встречаем иногда в нижних слоях плейстоцена еще отдельные зубы
Elephas meridionalis, но чаще узкозубую разновидность его — Elephas
ant iquus. Зубы этой формы при почти одинаковом с El. meridionalis
числе пластинок имеют сравнительно узкую коронку.
310
класс: млекопитающие — mammalia
El. primigenius.
Прямым продолжателем EL mer i-
d i о n a 1 i s является слон из самых нижних
плейстоценовых отложений -— Elephas
Wusti М. Pavl. (El. trogontherii var.
meridionalis Pohl.), громадных раз-
меров, хорошо представленный в нижне-
ледниковом тираспольском гравии, в Тю-
рингии, в Мосбахе, близ Дармштадта и
d
El. trogontherii.
в других местах. Пол-
ный череп его не най-
ден, но находки зубов
и костей скелета мно-
гочисленны.
Зубы его с постоян-
ным ростом, по пяти
El. W ii s t i.
El. meridionalis.
El. a n t i q u u s.
Рис. 394. Изменение зубов у слонов. Верхние
коренные зубы.
в челюсти, развиваю-
щиеся постепенно, име-
ют корни только в старости
(рис. 394, с), состоят из от-
дельных пластинок, число
которых увеличивается по-
степенно к задним зубам.
Первый зуб неизвестен; мо-
жет быть, он очень рано
выпадал; второй имеет 6—7
пластинок; число их возра-
стает до 22 в последнем ко-
ренном. Пластинки, как и
эмаль, окружающая их, очень
толстые, но они разделяются
сравнительно тонким слоем
цемента, толщина которого
варьирует. На некоторых зу-
бах пластинки очень сбли-
жены. Интересно чрезвычай-
но большое сходство, даже
тождество зубов Е1. W 0-
sti Pavl., найденных в Ти-
располе, с зубами, описан-
ными Бюстом (Wrist) под
именем Elephas trogon-
therii Pohl, из таких же
отложений Тюрингии.
ii
в
Рис. 395. Elephas primigenius (мамонт). (Сибирь.)
В — Нижний коренной зуб.
Рис. 395A. Elephas primigenius (березовский мамонт). (Сибирь.)
312
класс: млекопитающие — mammalia
Кости скелета — значительно больших размеров, чем у всех других,
слонов.
Elephas imperator Osb. — Сев. Америка, чрезвычайно близок к по-
следним формам, имеет большие загнутые вверх и в сторону бивни. Лоб
узкий и плоский.
Дальнейшее изменение слонов в плейстоцене проявляется в зубах. Число-
их сохраняется прежнее, по 5 с каждой стороны в каждой челюсти. Увели-
чивается число пластинок в коренных зубах, и они утончаются. Эмаль стано-
вится тоньше, и у некоторых форм она образует маленькие складочки.
Elephas trogontherii Pohl. var. primigenius (рис. 394,ё) — счи-
тается характерным для межледниковых отложений Германии. Некоторые зубы
его тесно примыкают к зубам El. Wiisti Pavl.; некоторые ученые считают
их тождественными. Описанные Политом (Pohlig) зубы этой формы очень,
разнородны по числу пластинок.
El. trogontherii Pohl.—найденный в с. Троицком, под Москвой, по-
чти цельным скелетом, бесследно исчез. Часть нижней челюсти его оттуда же
находится в палеонтологическом кабинете Московского университета (рис. 394, d).
Почти цельный скелет его найден около г. Ярославля. Число зубных пласти-
нок больше, чем у Elephas Wiisti; они тоньше.
Elephas Columbi Osb. (Сев. Америка) — соответствует этой европей-
ской форме. В Азии остатки этих послетретичных форм неизвестны.
Elephas primigenius Blumb.—-мамонт (рис. 395 и'395 А), включает
все остальные формы более поздних слонов как в Европе, так и в Америке..
Еще недавно к нему относили теперь отделенного Политом Elephas tro-
gontherii Pohl. При дальнейшем более подробном изучении остатков Е1.
primigenius его придется еще подразделить на несколько разновидностей,
по степени осложнения зубов большим числом тонких пластинок и по строе-
нию их. Последние как по числу, так и по тонкости далеко опережают зубы
нынешних азиатских слонов. Другим признаком, удаляющим мамонта от ны-
нешних слонов, являются его громадные S-образно загнутые бивни и, наконец-
присутствие длинной шерсти, по крайней мере у форм, живших на Севере.
Нынешние слоны покрыты шерстью только в младенчестве. Эти признаки за-
ставляют смотреть на мамонта не как на предка нынешних слонов, а как на
конечную форму, завершающую линию плейстоценовых слонов (El. Wiisti —
El. trogontherii — El. primigenius).
Находки его чрезвычайно обильны у нас на Севере и в Сибири, где до-
бывают „мамонтовую кость"—его бивни, идущие на разные поделки. На Юге,
в Киевской, Полтавской губ., он является современником первобытного чело-
века, и остатки его сопровождают стоянки этого последнего. Такими являются
стоянки: Кирилловская в г. Киеве и Гонцовская в Полтавской губ.
Между этими формами мы находим одни — более близкие по зубам к Elephas.
trogonther i Pohl., другие — с более узкими пластинками.
Наибольший интерес представляют формы, найденные на крайнем севере
Сибири, в промерзлой почве, где они сохранились с мясом, шерстью, вну-
подкласс: плацентные — placentalia, копытные
313
тренностями, позволяющими вполне восстановить не только все животное, но
даже указать на его питание. Последние раскопки Герца и Волосео-
в и ч а указывают на присутствие перетерты» ветвей и игл хвойных деревьев
в желудке мамонта. Животные проваливались между льдами и замерзали, не
успев переварить проглоченную пищу, и сохранялись на многие тысячелетия.
Теперь они дают массу научных указаний на строение их мышц, внутренностей
и окружавшую их природу (рис. 395, 395 А, В, С, D).
Рис. 395 В. Нижняя челюсть с языком El. primigenius.
Стоянки с мамонтом в Европе и у нас на Юге дают понятие о культуре
его современника — человека. Найдены кремневые орудия и рисунки, выцарапан-
ные на кусках бивня. Это — изображения мамонта (рис. 396), оленя и жив-
ших одновременно с ними форм (киевская Кирилловская стоянка).
Каково же отношение нынешних слонов к ископаемым, если мамонт не
может считаться их предком?
Зубы нынешнего азиатского слона близко стоят по форме пластинок
к названным выше слонам и к Elephas primigenius, тогда как зубы
африканского слона имеют вместо овальных пластинок пластинки ромбиче-
ские (рис 397) и этим отличаются как от азиатского Elephas indicus,
так и от других слонов: от El. meridionalis до Е1. primigenius. Эти
различия заставляют искать для двух нынешних слонов различных предков.
314
класс: млекопитающие — mammalia
Elephas armeniacus — одна из разновидностей древних послетре-
тичных слонов, известная по зубам и нижней челюсти, стоящая близко к Е1.
Wiisti Pavl. и El. trogontherii Pohl.; имеет довольно широкие пла-
стинки с малыми промежутками цемента, причем пластинки задней поло-
вины зуба еще невполне слились и состоят из нескольких овалов. Эмаль слабо-
складчатая. Эти зубы очень близки к зубам Е1- i и d 1 с u s, и возможно, что
обладатель их, El. armeniacus, отчле-
Рис. 395 С. Нога мамонта с шерстью.
Рис. 395 D. Хобот мамонта.
плейстоцена и ушел в жаркую Индию. Между сиваликскими формами труд-
нее указать более близкую форму. Но остатки El. armeniacus еще
слишком неполны, чтобы можно было утверждать это его положение. В пале-
онтологическом музее Московского университета имеется его нижняя челюсть
с зубами.
Предком Elephas africanus, с ромбическими пластинками, многие
считают El. antiquus — древнего слона. Но другая форма, также из Сива-
лик, имеет эти признаки лучше выраженными. Это —
подкласс: плацентные — placentalia, копытные
315
Elephas priscus Falc. — с более ясно выраженными расширениями
пластинок посредине; он стоит ближе к Е1. africanus; некоторые ученые ото-
ждествляют его с европейским El. antiquus (Лейдеккер).
Вероятно, оба ныне живущих вида слона получили свое начало в самом
начале плейстоцена. Один ушел в Африку, другой остался в Азии, предоставив
европейский континент группе El. primigenius, которая претерпела позже
и ледниковый холод в Европе и Сибири и кончила полным вымиранием, чему
немало способствовал, помимо холода, и человек, каменные орудия которого
встречаются с костями мамонта в древних стоянках. Нынешние слоны тоже
жестоко истребляются при добывании их бивней и скоро, вероятно, вовсе
исчезнут.
Интересно подчеркнуть время существования слонов в Африке, в Азии и
в Европе. Африка была колыбелью предков хоботных и будет могилой послед-
них представителей слонов, не имея промежуточных форм. Азия со среднего
Рис. 396. Рисунок на кости. Мамонт.
Рис. 397. Elephas
africanus. Верхний зуб.
миоцена до нынешних времен дала массу разнообразных форм, которые были
почти так же богато представлены и в Европе, но только до исторических
времен.
При изучении развития хоботных приходится все время говорить об их
зубах, почти не касаясь других частей скелета, даже конечностей, этого глав-
ного показателя развития форм в других группах. Происходит это от того, что
конечности, как и весь скелет, сохраняются во всю долгую жизнь этого отряда
почти неизмененными. Уже у Mastodon angustidens из среднего мио-
цена мы застаем почти то же строение всего скелета и конечностей, в частно-
сти, которое имеем у ныне живущих слонов. Наблюдаются отдельные неболь-
шие только изменения.
До Mastodon angustidens мы не знаем полных скелетов, а потому
исходим от него.
Расположение карпальных и тарсальных костей у него рядовое (рис. 398),
которое и сохраняется у Elephas indicus, но несколько нарушено у Е1.
africanus, у которого происходит небольшое смещение мелких костей.
У Mastodon ohioticus lunare покрывает magnum и касается краями
trapezoideum и unciforme, т. е. несколько нарушает рядовое расположение. Unci-
forme имеет значительную фасетку на metacarpus III. У El. primigenius
происходит то же изменение; lunare более передвинута на trapezoideum, которую
316
класс: млекопитающие — mammalia
она отчасти покрывает. Это указано на скелете березовского молодого мамонта
академиком В. В. 3 а л е н с к и м. На скелете мамонта в Зоологическом музее
Московского университета, тоже сибирского, видно такое же расположение с до-
бавлением на Innate выступа на месте перехода ее с magnum на trapezoideum,
где как бы намечается фасетка. Это, очевидно, слабые попытки изменить пер-
вобытное строение ног. Изучение молодой конечности индийского слона дало
следующее: присутствие центральной кости в очень молодом возрасте, которая
сливается позже с radiale, но шов остается довольно долго — он исчезает только у
вполне взрослого слона; lunate не имеет фасетки на trapezoideum и сочленяется
только с magnum, a ulnare — cunciforme. Это присутствие centrale у молодого
слона очень характерно. Оно указывает на то, что первоначально расположение
карпальных костей было чередующееся, сериальное развилось позже.
Для распознавания отдельных костей скелета различных хоботных не
могут быть указаны определенные признаки. Для определения зубов слоновых
указывается число пластинок. Но для стертых зубов или зубов, не вышедших
еще вполне наружу из алвеол, это не всегда верный прием. Правильнее брать
для этого сравнения не всю жевательную поверхность зуба, а определенную
часть ее, например длиною в 10 см, отсчитать пластинки на ней и сравнить
с другой такой же поверхностью и с числом пластинок на ней. Такое срав-
нительное изучение зубов группы настоящих ископаемых слоновых дает сле-
дующую табличку, причем, конечно, берутся зубы, близкие по ширине.
Число пластинок на 10 сантиметров
максим миним.
Elephas primigenius 12'/3 10
и / Кирилловская | 12 9 I тип
| стоянка 1 8 . 7 II тип
Elephas trogontherii Pohl. var. primig. 7‘/. 6
Elephas Wiisti M. Pavl. var. merid. 7 5 V2
Тирасполь 7 5 ‘/2
Elephas meridionalis Nesti из Куяльника Pavl. 6 4 </2
Weithofer’a 6 4
Elephas antiquus Falc. Falconer’a 6 v2 5
(узкие зубы) 7 */2 5
Может ли быть указана теперь генетическая связь этой столь обособлен-
ной группы с какими-нибудь другими из известных отрядов млекопитающих?
Moeritherium, стоящий у основания хоботных, по указанию Эндрь-
ю с а, имел в коренных зубах некоторые признаки свиных, также и во втором
шейном позвонке; но его передние зубы сразу отличают его от этой группы:
преобладание второй пары резцов вверху и сокращение их в нижней челю-
сти до одной пары, а также и четырехбугорчатые предкоренные.
подкласс: плацентные — placentalia, копытные
317
Рис. 398. Е ephasef. anticusM. Pavl.
А — задняя, В — передняя ступня.
Интересно указание на некоторые обилие признаки, сближающие самого
большого из наземных животных, слона, с очень маленьким даманом — Нугах
(из Hyracidae), живущим в Абиссинии (Нугах abissinicus) и на юге
Африки (Нугах capensis). У него сохранилась центральная кость, несмотря
на значительное сокращение I и V пальцев и рядовое расположение карпаль-
ных костей; только unciforme не-
сколько передвинулась на meta-
carpus III. Существуют копытца на
пальцах. Передняя конечность —
четырехпалая, с остатком пер-
вого пальца; задняя — трехпа-
лая. Зубы его с корнями, носо-
рожьего типа, с внешней стенкой
и поперечными гребнями. Кроме
рядового расположения карпальных
костей и присутствия centrale, су-
ществующей также у очень моло-
к сем. Saghatheriidae, между
(рис. 399), как наиболее близкую
/3 1 4 —- 3\
полное озубление I — — —-— I с
\ О 1 х —— О /
Рис. 399. Mixohyrax niloticus
Schlosser. (</s)
дого слона, сходства нет.
Долго Нугах стоял совершенно изолированно среди ныне живущих
форм. Но в Египте, в Фаюме, найдены формы, отнесенные Эндрьюсом
которыми он указывает Mixohyrax
Форму к Нугах. У нее существует
преобладанием внутренней пары резцов
вверху и сплошным рядом корен-
ных, к которым прилегают клыки.
Коренные четырехбугорчатые. По-
чти вся группа Saghatheriidae
вымирает в олигоцене; немногие
формы переходят в нижний плио-
цен — Pliohyrax, найденный на
Самосе Форсит- Майором
(Forsyth Major).
Отряд 12
Морские коровы — Sirenia
Другая группа животных, с ко-
торою сближают слонов, кажется
еще более отдаленной от них.
Это сирены, морские коровы —-Sirenia, водные млекопитающие, пред-
ставленные ныне Manatus (ламантином) и Halicore (дюгонем), жи-
вущими в морях и в низовьях больших рек. У них передние конеч-
ности превращены в ласты и очень редуцированы, задние — по типу кито-
образных.
318
класс: млекопитающие — mammalia
Недавно еще существовал третий род Sirenia — Rliytina Stelleri
Cuv. (стеллерова корова Берингова пролива); но он был истреблен чело-
веком.
Сем. Halicoridae (дюгоневые). — В ископаемом состоянии остатки их
известны с эоцена в Европе, в Африке и Сев. Америке.
Eotherium — средний эоцен Египта. Передняя часть черепа не со-
хранилась. Молодое животное с dft и 3 коренными. У взрослого имеется
полное озубление.
Eosiren libyca Andr. (рис. 401, в) — средний эоцен Фаюма, с удлинен-
ным черепом, двумя передними бивнями, хорошо развитыми, и следами осталь-
ных почти исчезнувших передних зубов. Коренные четырехбугорчатые, с бо-
лее простыми предкоренными, близки по типу к Moeritherium.
Рис. 400. Halitherium Schinzi Каир.
Protosiren Fra-
asi Abel — верхний
эоцен Фаюма, как бы
связывает Eosiren с
олиго ценовым Hali-
therium Европы, у
которого морда укоро-
чена; осталась только
одна пара резцов, бивней. Коренные зубы получили добавочные бугорки; рг1
исчезли. Передние конечности укорочены; задние почти атрофированы.
Mesosiren Dolloi Abel — известны молочные зубы: верхние d2
представляют удлиненные конусы; d3 — треугольный; d4 — с четырьмя лунками и
двумя внутренними бугорками, похож на пг1. Средний эоцен верхней Италии.
1 0 2 — 3
Felsinotheri urn — из плиоцена Италии, имел --------------------.
U U £-----о
Halitherium (рис. 400) — средний олигоцен; эта сильно редуцирован-
ная форма очень близко стоит к Felsinotherium из верхнего плио-
цена и к ныне живущему дюгоню — Н а 1 i с о г е, который на первый
взгляд не имеет ничего общего с хоботными, а представляет им полную
противоположность. Но при сравнении находятся некоторые общие признаки,
как то:
1) удлиненные межчелюстные кости, вмещающие бивни;
2) слабо развитые носовые кости;
3) отодвинутые назад вследствие этого носовые отверстия;
4) положение крыловидных костей с сильно развитыми крыловидными
отростками, образующими чашкообразные выступы, поддерживающие альвеолы
коренных зубов.
Развитие этих признаков можно наблюдать у ископаемых форм; но
присутствие их можно объяснить зависимостью от развития других органов.
Так, удлинение межчелюстных — развитием бивней и у слонов и у сирен.
У ламантина и стеллеровой коровы зубы эти исчезли, а межчелюстные кости
стали еще большими и загнутыми.
подкласс: плацентные — placentalia, копытные
319
В зависимости от развития этих частей находятся отодвигание носовых
отверстий и сокращение носовых костей у обеих групп. Таким образом эти
признаки могли развиваться параллельно у слоновых и у сирен. Г. Эндрьюс
указывает на интересное изменение таза, которое можно проследить, начиная
от Moeritherium через Eotherium, Eosiren, Halitherium (рис. 401}
к Felsinotherium — из верхнего плиоцена.
У Moeritherium существует подвздошная дыра и глубокая вертлуж-
ная впадина.
У Eotherium дыра существует, впадина неглубокая.
У Eosiren вместо дыры только вырезка (дыра открытая), впадина
мелкая.
У Halitherium дыры нет; впадина едва заметная.
Рис. 401. Тазовые кости дюгоневых (сирен).
а — Moeritherium, б Eotherium, в Eosiren, г -Halitherium,
д — Metaxytherium, е, ж Hal i core.
Изменения эти вызваны, очевидно, атрофией задних конечностей, которая,
в свою очередь, была вызвана водным образом жизни.
Был ли Moeritherium или близкая к нему форма общим предком хо-
ботных и сирен — вопрос остается открытым.
Сем. Manatidae (ламантиновые). — Остатки этих форм беднее. У древ-
них форм, эоценовых, тоже сохранилось полное озубление.
Prorastomus Ow. — имел большие клыки и верхние коренные с двумя
поперечными гребнями. Найден в эоцене Ямайки.
Manatus — имеет резцы и клыки только в юности; потом они исче-
зают, но число коренных зубов у него доходит до 10 вверху и внизу; они
тоже с поперечными гребнями и воротником. Ископаемые известны из миоцена
юга СССР, из плиоцена Южной Каролины и Аргентины.
Формы, сохранившие полное озубление, стоят, конечно, ближе к на-
земным предкам сирен, покинувшим сушу, вероятно, еще до эоцена.
320
класс: млекопитающие —- mammalia
Отряд 13
Amblypoda
Обнимают собою группу вымерших эоценовых форм, живших в Сев. Аме-
рике и отчасти в Европе.
Это крупные, пятипалые, стопоходящие животные, снабженные копытами-
Они отличаются от настоящих копытных и от хоботных особым строением
черепа и зубов, по которым они распадаются на три семейства: более древние
1) Pantolambdidae и 2) Coryphodontidae и 3) более поздние
Dinoceratidae. Череп первых двух не имеет роговых придатков, череп
третьего имеет их три пары. Зубы тоже различные. По этой группировке и
признакам черепа они сходны с
стям они сходны с хоботными.
Coryphodon—из верх-
ней части нижнего эоцена Ев-
ропы и Сев.
давно известен
Titanotheriidae. По пятипалым конечно-
Америки, был
как древнейшая
го
Рис. 402. Pantolambda cavirictus Соре.
форма из Amblypoda; позже был найден в более низких частях нижнего
эоцена, Torrejon-bed Сев. Америки, представитель нового семейства —
Сем. Pantolambdidae Соре. — Pantolambda Соре, сохранивший
еще признаки хищного животного (рис. 402). Череп его имел высокий сагит-
тальный гребень; полное озубление с небольшими клыками. Пятипалые паль-
цеходящие конечности сохраняли центральную кость. Плечо имеет эпикондиляр-
ное отверстие, признак хищных; бедро с третьим отростком — trochanter;
надпяточная, с закругленной головкой внизу, сочленяется только с naviculare;
наверху имеется углубление для сочленения с большой берцовой. Изогнутый
первый метакарпус тоже является признаком хищных, так же как и длинный
хвост. Коренные зубы бугорчатые, причем предкоренные состоят из двух
бугорков: паракона и протокона; паракон стоит у основания V-образной лунки.
На собственно коренных от протокона и метакона идут две V-образные
лунки; внутренний бугорок (протокон) — как на предкоренных. На передней
стороне у некоторых зубов есть добавочный бугорок. Нижние зубы состоят
из двух луночек, сходных с луночками копытных. По этим признакам Осборн
находит возможным поставить род Pantolambda Соре у основания раз-
вития Amblypoda.
подкласс: плАцентныЕ—placentalia, копытные
321
Сем. Coryphodontidae Ow. — Крупные животные, размерами от та
нира до быка, представлены в верхней части нижнего эоцена Европы и Сев
Америки и очень характерны для этих слоев.
Coryphodon (рис. 403) по скелету ближе всего к тапиру, с удлинен
ными носовыми костями, глубокими скуловыми впадинами, составляющими про
должение открытых орбит. Верхняя часть черепа плоская. Озубление полное
Передняя часть морды перед клы-
ками как бы перетянутая. Листо
образная лопатка с гребнем посре-
дине. Конечности типа непарнопа-
лых, с одинаково развитыми локте-
вой и лучевой, большой и малой
берцовой, но с пятью пальцами.
Вместимость черепа очень мала, и
мозг гладкий, рептильного типа.
Полное озубление Coryphodon
(рис. 404) состоит из 6 небольших
Рис. 403. Coryphodon testis Соре.
резцов, значительно ббльших клыков, особенно верхних, и коренных, различ-
ных для верхней и нижней челюстей. Предкоренные верхние состоят из внеш-
них V-образных лунок и внутреннего бугорка — талона. Коренной пг1 состоит
из переднего гребня и внешне-задней лунки, которая до стирания имеет вид
косого гребня. При стирании этот гребень дает V-образную лунку, сходную
с лункой предкоренных. Размеры зубов увеличиваются кзади. Нижние пред-
коренные удлиненные, имеют широкие лунки и небольшой воротник на задней
Рис. 404. Coryphodon
hamatus Marsh. Нижние и верхние зубы.
стороне. На коренных лунки более сжаты на передней половине, на задней
образуется гребень. При сравнении с зубами Pantolambda видно сходство
предкоренных верхних; коренные очень различны.
Типичная ступня Coryphodon имеет сходство со слоновой. Все пять
пальцев хорошо развиты (рис. 405). Но сочлененье карпальных костей менее
сериальное; unciforme имеет фасетку для lunare, которая отделяет magnum от
ulnare. Metacarpus III подходит под magnum и unciforme одинаковыми фасетками.
21 Павлова М. В. Палеозоология. Ч. II
122
класс: млекопитающие — mammalia
В задней конечности naviculare лежит над тремя metatarsus (I, II и III). Астрагал
очень плоский, но у различных видов рода Coryphodon расположение этих
костей несколько варьирует и даже позволяет считать, что одни из Cory-
phodon, например С о г у ph о d о n h’amatus, были пальцеходящими, тогда
как другие, например Coryphodon
считает это изменение ноги вто-
ричным, вызванным иным образом
жизни, для последней формы —
жизнью в болотистых местностях.
Различные роды, установленные для
Coryphodontidae, объединены
Осборном в 1898 г. под общим
названием Coryphodon; виды раз-
личаются, главным образом, по строе-
нию и длине черепа и конечностей,
когда они известны. На некоторых
черепах Coryphodon встречается
слабое возвышение на лобных костях — над глазницами, например у Cory ph.
testis, и расширение краев теменных костей. На носовых костях возвыше-
ний нет (рис. 403).
I о b a t u s, — стопоходящими. Абель
Рис. 405. Coryphodon а и ах Соре.
Ступня левой ноги.
Нижняя челюсл
Рис. 1 Оо D i п о с е г a s m i г a b i 1 е (U i n t а 1 h е г i н т).
Сем. Dinoceratidae Marsh.— Характеризуется присутствием трех пар
рогов, причем первая - - маленькая - - лежит на носовых костях; вторая —
большая — на верхнечелюстных, над большими клыками, и третья — самая
подкласс: плацентные — placentalia, копытные
323
большая — представляет выросты на теменных костях. Кроме того, затылочные
кости расширяются с боков, и таким образом весь череп является сплошь уса-
женным роговыми выростами. Скелет более тяжеловесный, чем у Corypho-
don. Лопатка треугольной формы, похожа на слоновую.
Dinoceras mirabile Marsh (рис. 406) — наилучше изученная форма
из среднего эоцена Уайоминга. У него существуют все вышеуказанные признаки.
Характерной является еще нижняя челюсть с большим закругленным выступом
на передней части нижнего края, и межчелюстные раздвоенные кости, лишен-
ные зубов. Верхние саблевидные клыки опускаются на выступ нижней челюсти.
Нижние клыки небольшие. Зубы коренные, в числе шести вверху, кроме рг2,
имеют по два слившихся на внутренней стороне гребня, образующих глубокие
лунки. На настоящих коренных они сливаются не вполне, но все-таки сбли-
жены своими внутренними концами. Рг2—с одной широкой лункой, похож на
предкоренные Coryphodon (рис. 407, Л).
В.
А.
Рис. 407. Uintatherium (Dinoceras) mirabile. (Средний эоцен.)
Передняя
ступня.
Задняя
ступня.
Нижняя челюсть с желобовидной симфизой имеет тоже 6 зубов с греб-
нями, расположенными косо, со сходящимися внутрь концами. Вместимость
мозговой полости мала, и мозг совершенно гладкий, рептильного типа.
Конечности пятипалые являются в строении своем как бы следующей
ступенью за Coryphodon (рис. 407,В). Unciforme продвинута под lunare на
половину ее поверхности. Magnum имеет большую фасетку на metacarpus II, у
которого верхняя поверхность разделена на две фасетки. В задней конечности
cuneiforme III имеет фасетку на metatarsus II, чего не было у Coryphodon, а
naviculare причленяется к cuboideum. Астрагал еще более плоский, чем
у Coryphodon. При сравнении ног Dinoceras, Coryphodon и
Elephas, одинаковых как по числу пальцев, так и по форме их, можно
было бы принять, что это последовательное развитие ступней одной группы
животных, причем простейшей является Elephas с еще рядовым расположе-
нием костей; за ним следует Coryphodon со слабо сдвинутыми карпаль-
ными и тарсальными и, наконец, Dinoceras, у которого сериальное рас-
положение этих костей совершенно нарушено (рис. 408).
Кроме рода Dinoceras Marsh, сюда входят еще несколько других;
из них Uintatherium Leidy, более примитивный, сохраняет по четыре
нижних предкоренных. У Tinoceras укоротились шейные позвонки
сравнительно с Dinoceras. Число предкоренных у них одинаково.
21*
324
класс: млЕкопитлющие — mammalia
Главное отличие различных других родов основано на различии черепов
и величине рогов. У Uintatherium и Dinoceras череп короткий, клыки
направляются вниз, но число зубов различное.
У Loxolophodon Соре череп очень удлинен, клыки загнуты вниз
и назад. Известен цельный скелет.
Проф. Осборн описал в 1913 г. череп Bathyopsis из верхних
слоев нижнего эоцена, который он считает „идеальной стадией, предшеству-
юшей средне-эоценовым уинтатериям из отложений Бриджера". У этого жи-
вотного намечается роговое возвышение
на верхнечелюстных костях над
входящими в них клыками. Носо-
вые кости очень тонкие, никаких
намеков на возвышения на них
нет. Теменные рога едва наме-
чаются. (Нижняя челюсть его
была описана Копом в 1885 г.,
она отвечает челюсти, описанной
Осборном.)
У Elachoceras, ближай-
шей к нему формы, верхнечелюст-
ные и теменные возвышения уве-
личены, носовые кости обломаны;
неизвестно, были ли на них вы-
росты. На лобных, над глазницами,
незначительные костные складки,
которые Осборн тоже называет
Рис. 408. Передние и задние ступни. рогами. Но зубы у обеих форм
типа Dinoceras, резко отли-
чаются от типа Coryphodon, так что вряд ли можно считать эту форму
связующей указанные два типа. Она является только начальной для Dinoce-
r a t i i n a e, и связь между двумя большими семействами остается неустановлен-
ной. Между Titanotheriidae подсем. Palaeosyopsinae и Titano-
t h е г i i n a e, имеющие сходные зубы и отличающиеся тоже присутствием и
отсутствием роговых выростов, являются более связанными между собою.
Отряд 14
Южнокопытные — Notungulata
Новый отряд .млекопитающих, установленный Сант- Яго Рогом
(S. Jago Roth) для группы южно-американских вымерших копытных, за-
ключает некоторые формы, которые относились раньше к настоящим копытным,
гак как они снабжены копытами.
Животные, входящие сюда, очень разнообразны, и трудно указать
объединяющие их признаки.
подкласс: плацентные — placentalia, копытные
326
Они прошли довольно длинный геологический период — с начала эоцена
некоторые до миоцена, другие до плейстоцена. Строение зубов и конечностей
различно в различных семействах. Низкий, расширенный сзади череп является
характерным почти для всех. Размеры форм очень различны — от зайца до
носорога. Все травоядные; одни напоминают по озублению грызунов, другие —
неполнозубых. Наиболее хорошо представленными являются: <
Подотряд 1
Typotheria Zittel
С эоцена до плейстоцена, с пятипалыми конечностями; зубы без корней.
Аргентина.
Сем. Typotheriidae Amegh. — Наиболее длительная группа в геологи-
ческом отношении, с эоцена до плейстоцена. Озубление полное, передние
резцы увеличены. Диастемы или вовсе нет, или она небольшая.
Adapithecus secans Amegh. — наиболее древняя форма из верхнего
3 17
эоцена. Очень маленьких размеров. Весь зубной ряд - - - равен 3 см. Ко-
0 1/
ренные имеют внешнюю стенку с отходящими двумя гребнями, соединяю-
щимися на внутренней стороне. Три предкоренных с более закругленной
внутренней стороной. Рг\ клыки и резцы — удлиненные конусы. На нижних
коренных по две луночки.
Interatherium (Icochilus) Amegh. — величиною с кролика, миоцен
Санта-Круца, является типичной формой с низким черепом, расширенным в заты-
лочной части, с сагиттальным гребнем, с выдающимися надглазничными отро-
стками и очень толстыми скуловыми дугами. Носовые отверстия лежат в конце
морды. Нижняя челюсть широкая, толстая. Озубление полное и сплошное,
хотя клыки и боковые резцы Маленькие.
Строение зубов представляет некоторые особенности: предкоренные, почти
одинаковые с коренными, состоят из внешней стенки — двух слившихся бу-
горков — и идущих от нее двух поперечных гребней, оканчивающихся двумя
внутренними бугорками. РР конусо-
образные, так же, как и клыки и два д
боковых резца. Внутренняя пара резцов
развита больше; они долотообразные.
Нижние коренные состоят из двух
1ребней, а предкоренные — из луночки
и гребня; ргг — конус; нижние клыки
больше верхних; два резца маленькие,
передние развиты значительно. Зубы эти
более всего напоминают зубы древних
Рис. 409. Typotherium cristatum
грызунов. Serrl«
На передних конечностях сохра-
няются пять пальцев примитивной конечности с сериальным расположением
карпальных костей и центральной костью. На задней конечности I палец
326
класс: млекопитающие — mammalia
оттопырен, как у лазающих по деревьям животных. Последние фаланги узкие;
они более похожи на коготные, чем на копытные. Некоторые признаки длин-
ных костей конечностей тоже сближают эту форму с коготными. Большая и ма-
лая берцовые свободны. Плечо с незначительным эпикондилоидным отверстием
У Protypotherium australe Amegh., из верхнего миоцена Санта-
Круца, череп со сплошным озублением, менее массивен, чем у Interatherium.
Число пальцев четыре, V сократился и на передней и на задней конечности.
На передней укорочен и IV.
Typotherium cristatum Series (рйс. 409) — из древнего плей-
стоцена Пампасов (Буэнос-Айрес), величиною со свинью, имеет череп очень
похожий на Interatherium, но несколько удлиненный; затылочная часть
его также расширена; заглазничные выросты увеличены; скуловые дуги толстые.
Зубы сильно отличаются от Protypotherium. Вверху существуют
только передние резцы, покрытые эмалью спереди. Они очень широкие, сжа-
тые, стертые, отделены диастемой от пяти удлиненных коренных. Два перед-
них маленькие, состоят из внешней стенки и двух гребней, на задних их три.
Нижняя челюсть имеет по два резца и по четыре коренных. Все зубы приз-
матические; на некоторых существует цемент между гребней.
Передние конечности сохраняют пять пальцев; первый очень уменьшен
(у Protypotherium он почти исчез, а может быть, не сохранился?). Кар-
пальные кости чередуются; centrale слилась с radiale.
На задних конечностях первый палец, редуцирован больше, чем на перед-
них, что не встречалось у копытных.
К этому семейству относится еще несколько родов: Hegetotherium mi-
ra bile Sincl. — крупная форма из миоцена; Pachyrukos Moyani Amegh.—
гоже из миоцена, небольшое животное. В плейстоцене это семейство вымирает.
Подотряд 2
Toxodontia Amegh
Ноги трехпалые. Зубы с постоянным ростом.
Сем. Toxodontidae. — С миоцена Патагонии, в плейстоцене Аргентины и
Бразилии. Характеризуется большими размерами, с тапира и носорога, и трех-
палыми конечностями. Оно развивалось параллельно первому. Т о х о d о п
известен отдельными черепами и полным скелетом (рис. 410), в котором больше
всего бросаются в глаза укороченные передние конечности и сильно развитые
остистые отростки передних спинных позвонков. Черен укороченный, расши-
ренный сзади, носовые кости короткие с верхним носовым отверстием (у Т у-
potherium оно было на конце морды). Нижняя челюсть толстая, особенно
2 0 3-3 „
в задней половине. Зубная система неполная: — у --- Спереди две пары
резцов, клыков вверху нет; нижние резцы все сохранились, клыки маленькие-
Коренные верхние почти квадратные, с внешней стенкой и отходящими
от нее гребнями, число которых варьирует: 3—2 (рис. 410А); нижние
подкласс: плацентные — placentalia, копытные
327
более вытянутые, такого же строения. Нижние резцы направлены вперед, верхние
загнуты.
Кости плеча и предплечья укороченные, толстые; косое положение лу-
чевой, не срастающейся с локтевой, — как у хоботных; локтевая преобладает,
она имеет большой локтевой отросток.
Бедро длинное, почти вдвое длиннее голени. Пальцы — три, укороченные,
с толстыми, кривыми метаподиями и очень короткими фалангами—-типа хо-
ботных. Карпальные кости чередуются.
Рис. 410. Toxodon paraiiensis Ow
В задних конечностях кальканеум высокий, сочленяется с малой бер-
цовой, как у хоботных и у парнопалых. Астрагал закруглен внизу — имеет
небольшую шейку. Животное было пальцеходящим (рис. 410, А).
Adinotherium Amegh. — миоцен Санта-Круца, сохраняет вверху три
пары резцов, клык и рг1 очень маленькие. Коренные более сложные— больше
число гребней, отходящих от внешней стенки: их 3—5.
Morphippus imbricatus Amegh. — миоцен Патагонии.
Подотряд 3
Nesodontia Ow.
Миоцен Патагонии.
Сем. Nesodontidae — с сильно развитой парой резцов и трехпалыми
конечностями.
Nesodon Ow. (рис 411) -величиною с тапира, наичаще встречающаяся
форма в миоцене Патагонии (Санта-Круц). Конечности трехпалые. Строение
черепа — очень близкое к Toxodon Ow., но череп сохраняет полное озубле-
ние, причем его коренные сложнее, чем у Тox-odon. Особенно сильно развиты
две передние пары резцов вверху и боковая пара внизу. Клыки небольшие.
Коренные резко увеличиваются кзади, увеличивая число поперечных гребней.
Нижние коренные сжатые, удлиненные. У .некоторых форм появляется цемент
на коренных.
328
класс: млекопитающие — mammalia
Конечности настоящие трехпалые с копытными фалангами. Астра-
naviculare, имеет еще фасетку на cuneiforme HI; metatarsus
cuneiforme III
гал, сочленяясь с
III сочленяется с
Установить
d о n t i а трудно,
очень различное
и cuboideum. Ноги стопоходящие,
отношение между Toxodontia
живший одновременно с
строение зубов
генетическое
N es о d о и,
цене, имеет
2
Рис. 410A. Toxodon paranensis.
А — передняя, В — задняя ступня. С — верхняя челюсть.
и
и Neso-
Toxodon в мио-
конечностей. Toxodon
ушел очень далеко от него
в сокращении зубов. Ко-
нечности— пальцеходящие
у Toxodon и стопо-
ходящие у Nesodon
(рис. 410 А и 411).
Сем. Homalodon-
totheriidae Lyd. — Пол
ное озубление. Высоко лежащие носовые отверстия. Пятипалые конечности.
Homalodontotherium Huxley — из верхнего
нии, с полным озублением и пятипалой конечностью (рис.
наименее редуцированным типом. Зубы его тесно прилегают
Резцы и клыки небольшие. Коренные верхние почти квадратные,
стенку с двумя гребнями, отходящими
Lophiodon. Нижние коренные удли-
ненные, из двух гребней, идущих от
миоцена Патаго-
412), является
одни к другим,
имеют внешнюю
всего к зубам
нее. Они ближе
от
imbricatus Ow
Рис. 411. N е s о d о п
внешней стенки. Концы гребней подразделены. Носовые кости укорочены;
носовое отверстие очень большое. Орбиты почти закрыты. На передней ко-
нечности укорочен ] палец; остальные четыре почти равной длины.
Leontinia Gaudryi Amegh. — из нижнего миоцена Патагонии; такая
же крупная форма с очень утолщенными межчелюстными костями и высоко
лежащим носовым отверстием.
А б е л ь объясняет развитие этой оригинальной группы копытных, так же-
как и Litopterna, следующим образом
подкласс: плацентные — placentalia, копытные
329
Во время нижнего миоцена примитивная группа копытных перекочевала
из Сев. Америки в Южную. Вскоре после этого сообщение между двумя
Америками было чоервано, что и длилось до,плиоцена. В
в Южной Америке развивались потомки перекочевавшей
группы, давая разные типы, из которых некоторые были
совершенно несходны с северо-американскими и евро-
пейскими, другие же напоминали с первого взгляда
настоящих копытных. Вымирание их он объясняет вступле-
нием новых форм после соединения (в плиоцене) Сев.
Америки и Южной и пришествием в нее крупных хищни-
ков, которых не было в Южной Америке до тех пор
(кроме случайных хищников) и которые способствовали
уничтожению цветущей до тех пор фауны — копытных.
Подотряд 4.
Pyrotheria Amegh.
Распространение его ограничено олигоценом и ниж-
ним плиоценом Патагонии, стоит более изолированно
от других семейств. Полный череп неизвестен, но кости
и зубы указывают на крупное животное с укороченным
черепом и сокращенными зубами. 11ятипалые конечности.
это длительное время
Рис. 412. Н о m а 1 о-
dontotherium Se-
govia с Amegh.
Зубы с корнями1). Циттель (1923 г.) относит его к Amblypoda.
Pyrotherium Romeroi Amegh.-— наилучше изученное животное из
этого подотряда, было одним из самых больших южно-американских копытных
(рис. 413). Как в верхней,
так и в нижней челюсти су-
ществовало ТОЛиКО по одной
паре резцов. В верхней они
не сохранились, остались
только следы их. В нижней
они развиты сильно. Рг2—
удлиненный, с несколькими
неправильными бугорками; рг2
имеет спереди бугорок, сзади
гребень. Остальные коренные
состоят из двух гребней,
Рис. 413. Pyrotherium Romeroi Amegh. отороченных невысоким во-
ротником. Это — тип зубов
динотерия. Нижние челюсти очень расходятся кзади. Расстояние между двумя
рядами зубов узкое.
Первый молочный верхний зуб d- имеет хорошо развитые три бугорка,
d3 и di — по два гребня. Резцы состоят из цемента, эмали и особого вещества,
<) Работы: Ameghino, A. Gaudry, W. Sinclair.
330 класс: млекопитающие — mammalia
•свойственного бивням слонов,— „ivoire". Были и молочные резцы. Из костей ске-
лета известны: шейные позвонки, похожие на слоновые, такие же плоские, тонкие
и кости конечностей, которые представляют большую разницу в величине плеча
и предплечья. Последнее почти вдовое меньше первого, очень широкое вверху,
с сильно развитыми боковыми отростками. Нижняя сочленовная поверх-
ность плеча с лучевой — узкая. Последняя кость- занимает всю переднюю
поверхность предплечья; локтевая лежит за нею, с сильно развитым локтевым
отростком. Обе кости срослись. Это строение конечности указывает,
по Г о д р и, на ее роющую функцию, с большою подвижностью пальцев,
мускулы которых прикреплялись к сильно развитым боковым отросткам.
В задней конечности бедро тоже значительно выше голени. Оно плоское
с сильно выступающей головкой и плоской нижней сочленовной поверхностью.
Короткая большая берцовая сочленяется, не скрещиваясь, с малой берцовой,
как это было у Nesodon.
Плоский астрагал напоминает слоновый.
Сходство различных частей скелета с хоботными и зубов с зубами ди-
нотерия позволило Амегино и Лейдеккеру при первом описании находок
пиротерия отнести его к хоботным. Дальнейшее изучение заставило выделить
его в особое семейство вместе с некоторыми другими родами, менее хорошо
представленными.
Изучение главных типов сборной группы Notungulata показывает,
что между ними трудно установить генетические отношения, хотя некоторые
признаки и кажутся перешедшими от одного семейства к другому, например
сокращение передних зубов, осложнение коренных.
Подотряд Pyrotheria, наиболее древнее из Notungulata и исчез-
нувшее раньше других, настолько изолированы по зубам и форме длинных ко-
стей конечностей, что его представителей никак нельзя считать предками других
семейств. Скорее можно думать, что предок или предки этих форм Notungu-
lata скрыты в самых древних третичных слоях, а может быть, и в меловых,
а отдельные фермы являются параллельно развившимися и связанными только
в семейства, резко различные от всех копытных других стран1).
Другой такой же изолированной группой ископаемых Патагонии были
вышеописанные Litopterna, которые не находят сколько-нибудь близких
форм ни в какой другой стране.
Чем же можно объяснить такую изолированность третичных ископаемых
в Южной Америке, когда в то же время встречаются многие общие формы
третичных млекопитающих, особенно копытных, между формами Европы, Азии
и Сев. Америки: Equidae, Rhinocerotidae и др.?
Чтобы ответить на эти вопросы, мы обращаемся к выводам геологии,
основанным на изучении распределения материков в третичное время и до него,
в меловой период. Мы находим указания на то, что только к середине третич-
*) Амегино считает Pyrotherium предком Dinotherium. Annales del
Museo Nacional de Buenos Aires, 1906, p. 341.
подкласс: плацентные — placentalia, копытные
331
чого периода окончательно исчезла суша, соединявшая Африку с К )жаой
Америкой, которая была изолирована от Сев. Америки. Эта последняя в тре-
тичном периоде была соединена с Зап. Европой громадным материком, на ме-
'сте которого лежи-i теперь Атлантический океан (Атлантида). Имея перед собою
эти данные, можно объяснить развитие южно-американской фауны на том
большом континенте, который соединял ее в меловом периоде с Африкой и
который, опустившись под воду, оставил ее изолированной от Африки в тре-
тичном периоде. С другой стороны, она оставалась отделенной и от Сев. Аме-
рики, где развивались в это время формы, общие или близкие с европейскими;
разобщение Сев. Америки с Европой произошло в конце плиоцена. Эта изоли-
рованность континента Южной Америки" и должна была обусловить развитие
Рис. 414. Arsinoitherium Zitteli Beadn.
специальной фауны в третичном периоде, которая перешла отчасти и в после-
третичный. Мнение Абеля было изложено выше.
Общность некоторых отдельных признаков этих южно-американских форм
с европейско-азиатскими и северо-американскими указывает только на совпа-
дение (конвергенцию), а не на генетическую связь.
Отряд 15
Subungulata
Сем. Arsmoitheriidae Andr. — Arsinoitherium Zirteli Beadnell. - -
•олигоцен Африки (Фаюм) (рис. 414).
Громадное животное с парой больших роговых стержней, лежащих рядом
на носовых костях, и двумя меньшими рогами на лобных. Короткая морда
с высоко лежащими носовыми отверстиями, разделенными костной перего-
„ „ п, 3 1 4-3 „
родкой. I лазницы по бокам черепа открыты сзади. Зубы: — — --- • Резцы
о! 4 — о
и клыки не выдакнся. Нижняя челюсть узкая, с тонкими венечными отро-
3,32 класс: млекопитающие — mammalia
стками. Коренные зубы, состоящие из гребней, стираются ненормально. На пред-
коренных при стирании образуется внешняя стенка, от слияния внешних бугор-
ков. На коренных сливаются внутренние гребни и образуется внутренняя стенка,
а внешние концы гребней остаются свободными. Озубление сплошное.
Затылочные кондилюсы направлены назад. Бедро сжато спереди назад,,
имеет большой trochanter. Большая и малая берцовая — короткие. Лучевая толще,
чем локтевая. Плечо с большим дельтоидным гребнем. Надпяточная плоская,
со слабо выпуклой trochlea, сочленяется с cuboideum. Конечности пятипалые,
снабжены копытами. Передние напоминают конечности Uintatherium,
задние — Elephas. Расположение карпальных и тарсальных костей и мета-
подий, как у Elephas; отличается только астрагал, сочленяющийся с cubo-
ideum.
Известен только один род — Arsinoitherium Zitteli Beadnell, кото-
рого Ц ит те ль отнес к Subungulata. Форма эта стоит пока совер-
шенно особняком.
ИЗМЕНЕНИЕ КОНЕЧНОСТЕЙ У НЕКОТОРЫХ КОПЫТНЫХ И КОГОТНЫХ
Познакомившись с удивительной группой вымерших форм Not Un-
gulata, принадлежавших исключительно Южной Америке, мы вспоминаем, что
уже встречали между копытными той же страны группу Litopterna,
которая сильно разнилась от современных ей настоящих копытных Евразии
и Сев. Америки. Это различие вызывает желание более подробного сравнения
отдельных частей скелета у животных, во многом сходных между собою, но
таких различных, как Litopterna и Perissodactyla, с одной стороны,
и Notungulata и Elephantidae — с другой.
Без сомнения, сравнение конечностей в деталях, кроме указания числа
пальцев, может многое уяснить при изучении развития животных.
Проф. Го др и, в руках которого, в Естественно-историческом музее
Парижа, находился богатейший материал по ископаемым Патагонии, набросал
картину этого сравнения, и я считаю полезным передать ее здесь вкратце, что
послужит в то же время обзором различных типов конечностей.
Если мы сравним такие типы ног, как прямоходящие — Elephantidae
и Amblypoda, пальцеходящие — настоящие копытные, стопоходящие — из
хищных кошка, то сразу поймем, что между названными группами должна
быть большая разница вследствие различного употребления конечностей ’)•
У слона, передвигающего ноги, почти не сгибая их, они служат глав-
ным образом для поддержания тяжести его громадного тела. Это животное —
прямоходящее, все 4 конечности которого имеют одинаковую функцию медленно
передвигать тяжелое тело.
У лошади конечности служат для быстрого бега, унося далеко легкое
тело. Они касаются земли только концэдш п^дьцев — это пальцеходящее жи-
вотное. д
- и. ' мз -пйЮ лнжсщ
9 Albert Gaudry. Annales de Palfiontologie, Paris, 1906, 1908, 1909.
4s» /aFtf.
* U i.
подкласс: плацентные — placentalia, копытные
333
У кошки и многих других хищников функции передних и задних
конечностей различны. Передние служат не только для передвижения
тела, но главным образом, для хватания или рытья и, значит, требуют воз-
можности движения и в другом направлении (боковом). Кроме того, задние
конечности, которые во время исполнения передними не передвигающей, а иной
функции должны поддерживать всю тяжесть тела, не могут касаться земли одними
пальцами, а должны прочнее стоять на ней всей стопой—это стопоходящие.
Эти признаки являются нормальным выражением образа жизни наших евро-
азиатских и северо-американских форм и позволяют нам по строению их
конечностей составить понятие и о строении всего их организма. Посмотрим,
так ли это у форм южно-американских. Сравним плечевую и локтевую части
конечностей Dinoceras, Mastodon и Pyrotherium (рис. 1, 3 и 4
по Gaudry).
У первых двух обе части конечности почти одинаковой длины, причем
небольшая закругленная верхняя головка плеча входит в гленоидное углубление
лопатки, а нижняя поверхность (сочленовная) лежит почти вся на верхней
поверхности локтевой кости; лучевая лежит наискось.
У Pyrotherium локтевая очень уменьшена; всю переднюю поверх-
ность предплечья занимает лучевая кость, и тяжесть плеча распределяется на
radius и ulna. Плечо сочленяется с лопаткой очень широкой головкой, указы-
вающей на возможность подгибания передней конечности назад, когда живот-
ное ложилось. Локтевая и лучевая неподвижно срослись. Боковые отростки
на плечевой, служащие для прикрепления тяжей пальцев, сильно развиты.
Они отсутствуют у наших хоботных и развиты у носорогов. К сожале-
нию, число пальцев неизвестно у Pyrotherium. В строении задних ног
Pyrotherium представляет большое сходство с такими различными формами,
как Dinotherium и Dinoceras (сочленение бедра с большой и малой
берцовыми) (рис. 26—28 по Gaudry).
Сравним теперь пальцеходящих и возьмем из современных, например
лошадь, и южно-американского Proterotherium, величиной с козлика, имев-
шего один палец. У последнего сразу бросается в глаза присутствие острого
гребня (trochlea) на нижней внутренней стороне humerus, мешающего движению
изнутри наружу (de dedans еп dehors). Никогда не встречается подобный гре-
бень у наших бегающих копытных. У лошадей на этом месте — закругленная
сочленовная головка, позволяющая свободные движения вперед и назад.
Задние ноги Proterotherium в своих сочленениях представляю! еще
большее различие с лошадиными. У него боковые пальцы сократились почти
совсем. Metatarsus III закругленный; сократилась cuneiforme И, и naviculare
сочленяется только с cuneiforme III. Астрагал имеет шейку внизу, напоминает
кость хищника и отчасти парнопалого (без нижнего блока). Подобное сочле-
нение астрагала позволяло Proterotherium более сгибать ногу, чем это
может сделать лошадь, у которой астрагал неподвижно лежит на naviculare.
Судя по этому сочленению, задняя нога Proterotherium была как будто
более совершенна, чем у лошади.
334
класс: млекопитающие—mammalia
Интересно также сравнение стопоходящих передних конечностей с хватаю-
щими или роющими. Так, Nesodon, с тяжелым телом и черепом, с зубами,
похожими на носорожьи, имеет локтевое сочленение передних конечностей,
типичное для хищников и грызунов и близкое к человеческому. Нижний
конец его плеча лежит на скрещивающихся локтевой и лучевой костях. Но
локтевая развита в нижней части значительно больше, чем у хищных.
В задних конечностях, которые у стопоходящих должны давать большую
опору телу, чем у пальцеходящих (ближе к прямоходящим), нога так сгибается
в ступне, что дает прямой угол, и вся ступня касается земли. Naviculare лежит
впереди астрагала. У прямоходящих это сочленение неподвижно; astragalus при-
крывает более или менее naviculare. Гораздо труднее установить разницу между
ногой пальцеходящего и стопоходящего животного. Так, человек, вполне стопо-
ходящий, становится пальцеходящим при беге, танцах и при лежании. Но по-
верхность astragalus у стопоходящих (по которой двигается большая берцовая)
короче, чем у пальцеходящих, и длиннее, чем у прямоходящих.
У Neso-don astragalus представляет именно признаки пальцеходящего
животного: она узкая и сочленяется с большой берцовой, давая ей движение
сзади наперед. Эта узость astragalus дополняется широкой фасеткой на calcaneum
для' малой берцовой, что и делает ногу все-таки стопоходящей, т. е. имеющей
широкий упор в пяточном сочленении, но с иным соотношением костей, чем у сто-
походящих хищных. Чтобы хорошо понять положение astragalus в разнообразных
конечностях, нужно просмотреть табличку Годри (рис. 14—25, стр. 13).
После изучения конечностей, зная строение черепов и зубов у сравни-
ваемых форм, мы можем притти к выводу, что третичная и послетретичная
фауна млекопитающих Южной Америки, хотя и имеет некоторое сходство отдель-
ных форм с формами Евразии и Сев. Америки, представляет по общему харак-
теру совершенно особенную фауну, т. е. развившуюся в совершенно других
направлениях. Ее формы, хотя и могут быть отнесены к копытным или к не-
полнозубым, сходным по некоторым признакам с евразиатскими и северо-американ-
скими формами, все же представляют в каждой группе, даже семействе и роде,
такие отличия, которые вполне точно доказывают их независимое от наших
форм развитие, хотя древнейшие предки, может быть еще начала мелового
периода, и могли быть у них общими.
РАЗВИТИЕ РАЗЛИЧНЫХ ГРУПП КОПЫТНЫХ -
Резюмируя то, что известно о развитии копытных в широком смысле
этого слова, мы должны подчеркнуть очень разную судьбу разных групп их.
Указав древнейших предков для некоторых семейств, мы не можем указать
общего, еще более древнего предка для всех копытных. Он скрыт, вероятно,
в меловых отложениях.
Из предыдущего видно, что наиболее долговечными и широко распро-
страненными являются те из настоящих копытных, развитие которых шло
медленно и равномерно при изменении различных органов. На первом плане
пддкллсс: плацентные — placentalia, копытные
335
стоят в этом смысле лошади и жвачные. Свиньи хотя и сохранили первоначаль-
ное строение ног с четырьмя пальцами, но благодаря приспособлению ко всякой
пище одержали победу и дошли до нашего времени в разнообразных видах, широко
распространенных, чему немало способствовал и человек. Носороги и тапиры
занимают теперь очень незначительные географические площади в жарких
странах, будучи в третичную эпоху широко распространены. Их вымирание
шло постепенно в борьбе с более совершенными травоядными формами, кото-
рые могли переносить более холодный климат.
То же произошло и с хоботными, которые выработали длинные бивни и
громадные, устойчивые при перетирании пищи зубы, но сохранили конечности,
годные только для медленного передвижения и поддержания тела, а не изме-
нили их в органы более подвижные. Может быть, их громадные бивни и были
отчасти причиной их затруднения при борьбе с окружающей средой (в лесах).
Недолговечность большой группы парнопалых — с неадаптивной редукцией
конечностей, — не перешедшей верхнего олигоцена, объясняется невозможностью
бороться с правильно развившимися в это время жвачными и другими траво-
ядными.
Совсем другую картину представляют северо-американские титанотерииды
и амблиподы, устремившие очень рано силы своего организма не на правиль-
ное развитие различных частей его, а на слишком усиленное развитие таких
органов, как роговые выросты на черепе, достигавшие громадных размеров уже
к началу олигоцена, когда формы других линий только еще мало изменились,
развивая медленно различные органы и мозг.
Litopterna и Notungulata Южной Америки, хотя и дошли до плей-
стоцена (первые — с неправильно редуцированными конечностями), являются
загадочными формами, проследить развитие которых мы теперь не можем по
недостатку материалов. -r,t(
Вымирание палеотериев в олигоцене, после богатства и разнообразия видов
в эоцене, не может быть объяснено строением этих животных. Можно указать
только на слишком быстрое развитие, осложнение зубов и сокращение паль-
цев — сравнительно с медленным изменением этих органов, например, у лоша-
диной линии. Скорее можно предположить вымирание от каких-либо неизвест-
ных причин: болезнь или катастрофа геологического характера.
Конечно, эти недостатки организма — не единственные причины вы-
мирания групп животных. Но по данным примерам видно, насколько важную
роль они играли и должны были играть в переживании одних и вымирании
других форм.
Л>Г
— —.... 'Tisj-’C-o
' ГОМ
УКАЗАТЕЛЬ
К ПЕРВОЙ И ВТОРОЙ ЧАСТЯМ
Да" ! Ob
:/ U S.I ci С Л')
УКАЗАТЕЛЬ К ПЕРВОЙ И ВТОРОЙ ЧАСТЯМ.
Римские цифры указывают части, арабские — страницы, звездочки при
них — местонахождение рисунков.
А.
Acantherpestes I 203.
Acanthoceras I 293; —mammillare I 292;
— rotomagense I 292, 293*.
Acanthoceratidae I 292.
Acanthocladia I 92; — anceps I 91*.
Acanthodes II 24, 52; —gracilis II 24, 25*;
— Mitchelli II 24; — sulcatus II 25*;
— Wardi II 24, 26*. 52.
Acanthodi II 24, 51.
Acanthodidae II 24, 25*, 52.
Acanthodictya I 45.
Acanthograptus I 58;
— Granti I 57*"
Acanthosphaera I 34*, 35. '
Acanthostoma vorax II 68.
Acanthoteuthis speciosa I 307.
Acaste I 187*-
Aceraspis II 12.
Aceratheriidae II 247, 248, 249* 251, 252,
253*, 254.
Aceratherium II 245, 255, 258, 261; acutum
II, 252; — Filholi II, 249* 250; — fossiger
II, 252, 254*;—incisivum II, 249*, 252*;
— Kowalevskyi II, 252, 254*, 255; —lema-
nense II 249*, 251, 255; —occidenantale II
251, 252, 253*; — tetradactylum II 249*,
252, 255, 261; — tridactylum II 252, 253*.
254; — trigoaodon II 252, 253*.
Acetabularia I 54.
Acidaspis I 187*.
Acipenser II 37*.
Acipenseridae II 37.
Acmaea I 238; —Raincourti I 238*.
Acoeli I 307, 338.
Acotherulum II 268*.
Acreodi II 185.
Acrocidaris 1 161.
22*
Acrodelphidae II 206.
Acrodus II 19*, 21, 22.
Acrosalenia I 161, 162.
Acrotreta subconica I 101*.
Actaeon I 248; — perovskianus I 248*.
Actaeonella gigantea I 248; -voluta I 248.
Actaeonidae I 248.
Actaeonina 1248. . ..
Actaeosaurus II 116.
Actinocamax I 310, 315; — quadratus I 310*.
Actinoceras I 258; — abnorme I 258*.
Actinoconites I 306; — acutus I 308*.
Actinocrinus I 151; —stellaris I 151*.
Actinodon I 8; II 59, 65*, 66*, 112; — Fros-
sardi II 65.
Actinopora I 93.
Actinopterygii II 7, 8, 28, 36, 37, 43, 50, 53.
Adapidae 11 220.
Adapis II 220, 221; Duvernoyi II 220; — mag-
nus II 221; —parisiensis II 220*.
Adapisorex II 175.
Adapisoriculus II 175.
Adapithccus secans II 325. ,
Adinotherium II 327.
Adunata I 140, 150, 152, 153.
Aeger I 183, 184; —tipularius I 184*.
Aegoceras I 281; —capricornu I 281*.
Aegoceratidae I 281.
Aelgina I 192; —prisca I 192*.
Aelurictis II 198.
Aepyornis II 159.
Aetosaurus II 79, 125, 128; ferratus II 123,
124*.
Agamidae II 124.
Agassizocrinus I 145, 150; — dactylilormis 1
146*.
Agathammina I 24, 30; — milioloides 1 20*. >
Agathlceras uralicum I 272*.............../
Agelacrinidae 1 123. ....
340
УКАЗАТЕЛЬ
Agelacrinus I 123; — cincinnatensis I 123*;
Lebouri I 123; peleus I 123.
Aglaps na I 199.
Aglapsis I 199, 200.
Agnostus I 186, 187*, 188, 189, 194; —nudus
I 189*; —pisiformis I 189*.
Agoniatitidae I 266, 267.
Agorophius II 206; — pygmaeus II 205*.
Agriocetus austriacus II 207.
Agriochoerus II 272, 273, 274.
Ai’stopodidae II 62.
Alces II 278, 279; - - meoticus II 278.
Alcyonaria I 62, 64, 67, 68.
Alligatorellus II 125; — Beaumonti II 125*.
Allosaurus II 129, 134, 135; — agilis II 134*.
Allotheria II 164, 167, 168.
Alveolina I 31; —Bosel I 31*. : r -.’/.
Alveopora spongiosa I 72*. . t
Amalthaeidae I 279. "
Amaltheus I 278; — margaritatus I 279*; — spi-
natus I 263*.
Amblypoda II 320, 329, 332. -» :
Amblypterus II 39; —latus II 37.
Amia II 38, 40. „ ;
Amiella I 199. .
Amiskura sagittiformis I 119*.
Ammodiscus I 24, 30; — gordialis I 24*.
Ammonites angulatus I 39.
Ammonitidae I 255, 273.
Ammonoidea I 13, 254, 255, 263.
Amnicola I 249; — pseudoglobulus normalis I
250*.
Amnionites angulatus I 39.
Amphiastraea I 78; —gracilis I 78*.
Amphiastraeidae I 78. -
Amphibia I 13; II 55, 57.
Amphichelydia II 105.
Ampniclina I 111*.
Arnphicyon II 193, 194, 200; —giganteus II
193*.
Amphilestes II 165*.
Amphineura I 212, 253.
Amphioxus II 1, 17; — lanceolatus II 2*.
Amphip roviverra II 173.
Amphisyle Heinrichi II 45*.
Amphitherium II 166; —Prevosti 11 166*.
Amphitragulus II 276, 289; —lemanensis II
276*.
Amphiura I 169. '
Amplexus I 80; —wischerianus I 81*.
Ampullina patulla I 244.
Ampyx I 186, 187*, 190, 195; — nasutus I 190
— Volborthi I 190*.
Anacodon II 192.
Ananchytes I 166; — ovata I 166*.
Anapsida II 79, 80.
Anaptomorphidae II 220, 221, 222.
Anaptomorphus II 221*, 224; —homunculus
II 221.
Anarcestes I 266; — compressus I 267*; — ple-
bejus I 267*.
Anarcestidae I 266.
Anas II 158.
Anaspida II 9, 13, 14, 15.
Anaspides Tasmaniae I 182. ’ '
Anatina I 215.
Anatinidae I 216, 229.
Anchiceratops II 146.
Anchilophus II 235*.
Anchisaurus II 129, 133, 134, 144, 147;— ma-
jor II 133*.
Anchitherium II 235*, 236, 237*, 239, 240,
242.
Ancistropegmata I 102, 106. i Ь; о :
Ancodon ursidens II 201. - I <-.Lo.
Ancodus II 293; —leptorhinus II 295;
— velaunus II 292*. x
Ancyloceras I 293; — Matheroni I 293*.
Ancylopegmata I 102, 108.
Ancylus I 249. —
Andrewsarctus mongolicus II 186*.
Andrias Scheuchzeri II 73*. 74; — Tschudii II
74.
Anelasma rectangulum I 114*.
Angustisellata I 263, 264*, 275.
Anisomyaria I 230.
Anna Petri II 91.
Annelidae I 118. •
Anodonta I 222, 223. -
Anodontorni II 155,158. i
Anomalochladina I 41, 43.
Anomalocystidae I 130.
Anomalocystis I 130, 136; —disparilis I 131*.
Anomalodontia II 102.
Anoplotheiriidae II 289, 292, 295. -
Anoplotherium Г4; 265, 292, 295; — commune
II 293*, 295.
Anostraca I 176, 177.
Antedon I 137, 138*, 139 \ 140, 149*, 154;
— juticri I 149*; - -rosaceus I 149*.
Ante odon II 188.
Anthomorpha I 53.
Anthozoa I 36, 53, 54, 60, 61.
Anthracomarti I 206.
Anthracomartus I 207*; —trilobitus 1 206.
Anthracopalaemon I 183; gracilis I 183*.
УКАЗАТЕЛЬ
341
Anthracosia I 220, 222; II 95; — beaniana I
221*, —Venjukovi I 220, 221*.
Anthracosiidae 1 220.
Anthracotheriidae II 289, 291.
Anthracotheriinae II 291, 292, 293.
Anthracotherium II 292, 293, 294; — breviceps
II 293; —magnum II 292*; —Riitimeyeri II
293.
Anthropoidea II 222, 223, 224.
Anthropops perfectus II 224.
Antiarchi II 9, 12.
Antilopa clavata II 280; — palaeindica II 280;
— recticornis II 230; —sansaniensis II 280.
Antilopidae II 277, 279, 283.
Antira II 55, 56, 73, 74.
Apegmata I 102.
Apheocallites I 53.
Aphroditae I 120.
Apidae I 177.
Apiocrinilae I 147.
Apiocrinus Parkinsoni I 147*, 148.
Aptera I 208.
Apteryx II 160, 161.
Aptychus I 264*; — lamellostts I 265*.
Apus I 177, 189; —cancriformis I 178*.
Arachocystis I 130, 131, 136.
Arachnoidea I 203, 204.
Arachnomorpha I 185, 196.
Araeoscelis II 116; —gracilis II 115, 116*.
Araneae I 206.
Area I 214, 215*, 216*, — barbata I 219;
—diluvii I 219; — Noae I 219*.
Arcestes intuslabiatus I ТП*.
Arcestidae I 277.
Archaeoceti II 203, 205, 206, 209.
Archaeocidaris I 158*; —rossicus I 158*.
Archaeocyathidae I 52, 87.
Archaeocyathinae I 36, 51*, 52, 53, 54.
Archaeocyathus I 52*; —acutus I 51*.
Archaeofungia I 52.
Archaeolemur II 222.
Archaeolemuridae I 220, 222.
Archaeoniscus I 182; —Brodici I 182*.
Archaeopteropus transiens II 178.
Archaeopteryx II 154, 155, 157; — Siernensi
Il 155*.
Archaeosycon I 52.
Archegosauridae II 59, 64, 72.
Archegosaurus II 64*, 65, 66, 68, 69, 71, 72;
— Decheni II 64.
Archelon II 109, 110; —ischyros II 109*.
Archidesmus Macnicoli I 203.
Archijulus I 203.
Archimedes I 91; —indigae I 91*; —Worthe-
ni I 91*.
Archinocella I 238.
Archipolypoda I 203.
Arcidae I 219, 220.
Arctocyon II 191, 200, 201; — prlmaevus II
191*.
Arctocyonidae II 185, 191.
Arctomys II 180.
Arctotherium II 200.
Argiope decollate I 109*.
Argonauta I 254, 300, 301*; — argo I 300;
— hians I 300; — tuberculatum I 300.
Arietites I 280; — Bucklandi 1 280*.
Aristocystidae I 126, 136.
Aristocystis bohemicus I 126, 127*.
Arsinoitheriidae II 331.
Arsinoitherium Zitteli II 331*, 332. ,
Arihrodira II 25. '
Arthropoda I 174, 184, 196, 202.
Articulata I 97, 102, 138, 140, 147, 149, 154.
Artiodactyla II 230, 266.
Asaphidae I 192.
Asaphus I 186, 187*, 192, 195, — expansus 1
192*.
Ascoceras I 259; — bohemicum I 259*.
Ascones I 49.
Asellata I 263, 264*, 265, 266, 267.
Asidaspis I 194; —mira I 194*.
Aspidobranchia I 237, 239, 241, 248.
Aspidoceras I 290; — acanthicum I 290*; —per-
armatum I 290*; — Schilleri I 290.
Aspidoceratidae I 290.
Aspidorhynchidae II 43.
Aspidorhynchus II 43*; — acutirostris II 43*.
Aspidosoma I 169, 171; — petaloides I 170*.
Aspidura I 169; —loricata I 168*.
Assilina I 28*, 29.
Astarte I 224; — porrecta I 224*.
Astartidae I 224, 226.
Astenosoma I 159.
Asteracanthion rubens 1 169*.
Asteractinella I 46; — expansa I 46.
Asteroblastus 1 129, 134; —foveolatus 1 128,
-stellatus I 129*.
Astcroidea I 167, 169, 171.
Asterolepidae II 13.
Asterolepis II 9, 13, 26; - ornatus II 13*.
Asterozoa I 167, 171.
Astieria I 282.
Astraeidae I 76, 77.
Astraeospongia I 46; — meniscus I -46*.
Astraeospongidae I 46.
342
УКАЗАТЕЛЬ
Astrocoe iia I 75, 77.
Astroides calycularis I 63*.
Astrolites I 149.
Astropecten I 170.
Astrophiza I 20* 23.
Astrorhizidae I 23.
Astrosclera willeyana I 38*.
Astylocrinus I 145.
Astylospongia I 42; — praemorsa I 43*.
Atelaspis II 9; — tesselata II 11.
Atelostomata I 162, 165.
Athyris I 112.
Atlantosauridae II 130.
Atlantosaurus II 132, 162.
Atops I 187*.
Atractites I 306, 314; —applanatus I 306;
— quadratoides I 305, 308*.
Atremata I 98, 115.
Atrypa I 111, 112, 117; —reticularis I
111*.
Atrypidae I 102, 110, 111, 112. ........... ;
Attoides I 208.
Aturia I 263, 265; — zic-zac 1 262*.
Aucella I 231; — inflata I 231, 232*; — Key-
serlingi 1 231, 232*; —Lahuseni I 231*;
— mosquensis I 231*; — volgensis I 231.
Aulacobelus I 315.
Aulacoceras I 306, 314; — reticulatum I 306,
307*.
Aulacostephanus eudoxus I 292; — kirghi-
sensis I 292*; — pseudomutabilis I 292*.
Aulastraea I 178; —Schaferi I 78*.
Aul acetus sammariensis II 207*.
Atilocopium I 41; —aurantium I 41*.
Anlopora I 64, 67, 68, 92; — tubaeformis I
65*.
Auloroidea I 171.
Aves II 153.
Avicula I 215*, 216*, 231; — contorts I
231*.
Aviculidae I 230. 231.
Aviculopecten I 231.
Axinella I 40.
Axonolipa I 58, 59.
Axonophora I 56, 58.
Axophyllidae I 84, 87.
B. йА
Bachytherium II 275.
Bactrites I 266; —elegans I 266*.
Eaculitea I 297; — anceps I 297*; — Faujasi I
297.
Balaena II 208; — mysticetus II 208*.
Balaenidae II 208.
Balaenopteridae II 207.
Balanidae I 176, 179, 180.
Balanus I 180*.
Balatonites bogdoanus I 274*.
Baptanodon II 98.
Barroisia I 49, 54; — anastomans I 49*.
Barytherium II 307; —grave II 307.
Bathyopsis II 324.
Batocrinus pyriformis I 150.
Batoidei II 23.
Batrachiderpeton lineatum II 62*.
Bauria II 92*; —cynops II 91.
Bayanoteuthis I 310.
Bdellostoma II 17.
Belemnitella I 310, 315; — mucronata 1 307, 310*
Belemnitellidae I 305, 308, 310, 315.
Belemnites I 304*, 305*, 306*, 310; [—abso-
lutus I 308,309*, 313, 314, 315; —acuarius
I 304; —acutus I 308, 314; •- bipartitus I
309*, 310, 315; —breviaxis I 311*, 314;
— brevis I 314; — bruguerianus I 305*, 307;
— caniculatus I 308, 309*, 315; —clava-
tus I 314, 315; — dilatatus I 306, 309*;
— explanatus I 306, 308*; — giganteus
I 314; —hastatus I 308, 309*, 315; —kirghi-
sensis I 312*, 314; —lateralis I 312, 313*,
314; —latus I 309*, 310; —magnificus I
313*, 314; —mosquensis I 308, 309®, 311,
314, 315; — Oweni I 312, 313*, 314;—Pan-
der! I 308, 309*, 311, 314, 315; — paxillosus
I 303, 309*, 314; — russiensis I 308, 309*,
312, 314, 315; — semihastatus I 305; —sub-
extensus 1 311*, 312, 314; —subquadratus
I 312*, 314; — tripartitus I 310, 311*, 312,
314; — Rouilleri I 312*, 314.
Belemnitidae I 255, 300, 302, 305, 306, 315.
Belemnoidea I 255, 303, 314.
Belemnopsis canaliculatus I 308; - hastatus
I 308.
Belemnoteuthidae I 305, 315.
Belemnoteuthis I 304*, 305; — antiqua I 307*.
Belintirus I 201, 202; — reginae I 201*.
Bellerophon I 239, 240; — b carenus 1 239*;
— macrostoma I 239; — Urii I 239*.
Bellerophontidae I 239.
Beloceras inultilobatum I 268.
Belodon II 121, 122, 159; —Kapffii II 121*;
— Plieningeii II 121.
Belo.iorhynchus gigas II 43.
Beloptera I 303; — belemnitoidea I 303*.
Belosepia I 301, 304*; Blainvillei I 302, 303*
УКАЗАТЕЛЬ
343
Beloteuthis I 302; — Schiibleri I 303*.
Beltina I 198.
Beludgitherium II 251*.
Berenicea I 92, 93; — diluviana I 93*.
Bernissartia II 125.
Berriasella I 291; —Calisto I 291; —subriasa-
nensis I 291*. 292.
Beryx II 45*, 46.
Beyrichia I 179.
Billingsella I 105.
Biloculina I 31*.
Bimana II 219.
Bipartiti I 315.
Birkenia II 9; — elegans II 13, 14*.
Bison II 282; — latifrons II 282*, 290; — pris-
ons II 280; — Schoetensacki II 280.
Bithynia I 243.
Blastocystidae I 134.
Blastoidea I 121, 122, 126, 127, 129, 130, 131,
132*, 133, 134, 135, 136, 154.
Blastoidocrinus carcariaedeus I 134.
Blastomeryx II 277*; — Olcotti II 277*; —pri-
mus II 277*.
Blatta I 208; — abnormis I 208.
Bolodon II 167; — crassidens II 167*.
Bonorites I 269*.
Bos II 283; — brachyceros II 280; — elatus II
281; — frontosus II 282; —primigenius II
281*, 282, 283; —trachoceros II 282; —tau-
rus II 282,
Bothriceps II 66.
Bothriocidaris I 157;—Pahleni I 157*.
Bothriogenys Fraasi II 295.
Bothriolepis II 9, 13, 46; —canadensis II 13,
14*.
Bothriophyllum 1 81, 82*.
Botryocrinus I 138, 144.
Bouchardia rosea I 116*.
Bovidae II 271, 276, 280, 283.
Bovinae II 282.
Brachiopoda 1 12, 86, 89, 96, 97, 106, 194, 212,
213.
Brachyceratops II 146.
Brachydirus II 27.
Brachyodonta II 211.
Brachyodus brachyrhynchus II 294.
Bradyina I 26; —rotula I 26*.
Bradypoda II 209, 212, 217.
Bradypus II 210*.
Bramatherium II 288.
Branchiata I 175, 237, 238, 250.
Branchinecta I 176, 177; paludosa I 177*.
Branchiopoda I 176.
25*
Branchiosauria II 58, 59, 63, 72, 73.
Branchiosaurus II 59*, 73; amblystomus II 60;
petrolei II 60; salamandroides II 60.
Branchipus I 176.
Bronteus I 187* 192.
Brontops II 265; — robustus II 264*.
Brontosaurus II 129, 130, 131; —excelsus II
130*.
Brontotherium II 265.
Bryozoa I 89.
Bubalus II 282.
Bucapra II 283.
Buccinidae I 247.
Buccinum baccatum I 247*. : -ч
Buchiceras Ewaldi I 298.
Bufo II 75; — agua II 76.
Bugula I 89, 90; avicularis I 90*.
Bulimus I 250.
Bulla I 248. . .. 4
Bullidae I 248.
Bunaelurus II 194; —lagophagus II 194*.
Bunodontia II 266, 267.
Bunodus lunula I 201 *.
Bunoselenodontia II 266, 290.
Burgessia bella I 177*.
Buskia I 92, 94; tabulifera I 94*.
C.
Cacops II 73.
Cadoceras I 294, 299; — Elatmae I 294*;
Tchefkini I 296*.
Cadoceratidae I 294, 296.
Cadulus I 253.
Caenopithecus II 220.
Caenotherium II 292, 296; — elongatum II
296*.
Caetocrinus I 151.
Calamodon II 211. , . . -
Calamoichthys II 34.
Calcarina I 20*, 26.
Calceola I 87; —sandalina 1 86*, 87.
Calceolidac I 86, 87.
Calibrachion Gaudry II 112.
Callicrinus I 150.
Callocystidae I 126.
Callocystites Jcwetti 1 126*.
Calostylis I 78.
Calymene I 186, 187*, 193; Blumenbachi I
193*.
Calyptoblastea I 54, 56*- . (
Calyptrea I 242. . ,
Camarosaurus II 132.
344
УКАЗАТЕЛЬ
Camelidae II 271, 285, 286, 287, 290.
Camelopardalis II 288.
Camelus II 287; —bactrianus II 285.
Camerata I 140, 150, 152, 153, 154.
Campanuiaria I 56*, 89.
Campodus II 22.
Campophyllum I 80; — vermiculare I 81*.
Camptosaurus II 138, 141, 142; —dispar II
140'-.
Campylognathus II 149.
Canadia I 120; — setigera I 120*.
Canidae II 184, 190, 192, 193, 194, 195, 199,
200.
Caninia I 81, 83; — Schmidti I 82*.
Canis II 192, 193, 194; — domesticus II 193;
-lupus II 193; —vuipes II 193.
Caolinia I 251.
Capitosaurus II 68, 71. . . /...
Capra If 280, 283; — pyrenaica If 283; — si-
valensis II 283.
Caprinidae I 227.
Caprinula I 228.
Capulidae I 242, 248.
Capulus I 242; — hundaricus I 242*; — ru-
gosus I 242*.
Carbonicola I 220; —nova I 221*.
Carcharodon II 19*, 22.
Cardiidae I 223.
Cardioceras I 295, 299; —alternans I 295,
296*; — cordatum I 295*.
Cardita I 224; —Dunkeri I 214*; —planicosta
I 224*.
Cardium I 223; — edentulum I 223*; — edule
1 223*; — Fittoni I 223; — obsoletum I 223:
— porulosum I 223*.
Caretta К 110.
Carinatae II 154, 155, 157.
Carnivora II 184, 187, 191, 199, 200. ' '
Carnivora vera II 185, 192.
Carpocrinus I 153.
Carpoidea I 122, 130.
Carpospongia I 42; —globosa I 43*.
Caryocrinidae I 124.
Caryocrinus I 125; —- ornatus I 124*.
Caryophyllia I 76, 88; — cyiindracea I 76*.
Casea Broili II 87*.
Cassidulidae I 165.
Castor europaeus II 182*. ’ , ।
Casforidae II 182. f
Castorinae II 182.
Caturidae II 39.
Caturus П 39, 40, 44, 49, 50, 51; — elongatus
II 39*: —furcatus II 39*, 44.
Cavicornia II 279.
Cayluxotherium II 176.
Cebidae II 224.
Cebochoerus II 268,269; — Riitimeyeri II 268*,
Cellepora I 95.
Centetidae II 175, 176.
Centronella glansfagea I 109*.
Centrosaurus II 146.
Cephalaspidae II 11, 15.
CephalaspisI 13; 119,11, 12,28; —Lyelli II11*.
Cephalites I 48*.
Cephalopoda 1 212, 254, 255, 314.
Ceratiocaris I 181; —papilio I 181*.
Ceratites I 274, 275; —nodosus I 273, 274*.
Ceratitidae I 273, 275, 298.
Ceratodus II 31, 32; —Kauppi II 31*; —Sturi
II 32.
Ceratogaulus II 183.
Ceratophrys II 76.
Ceratops II 129. ‘
Ceratopsidae II 129, 138, 145, 146, 147, 161.
Ceratosaurus II 129, 135, 153, 161; —nasicor-
nis II 134*.
Ceratotrochus I 76; — duodecimcostatus I 76*.
Ceriopora astroides I 94*; —irregularis I 94;
— spongites I 94.
Cerithiidae I 246.
Cerithium I 246; giganteum I 246*; — marga-
ritaceum I 246*; — serratum I 246*.
Cermatia I 203.
Cervidae II 271, 276, 277, 283.
Cervulidae II 276, 280.
Cervulus II 278*.
Ccrvus elaphus II 278*, 279; — euryceros II
279*, 290; — latifrons II 279, 290; — Sed-
gwicki II 290; — tarandus II 278, 279*.
Cestracion II 18, 22; — Philippi II 21*.
Cetacea II 203, 209.
Cetotherium Rathkei II 207, 208.
Chaetetes I 65, 68; — Scheremetewi I 66*.
Chaetetidae I 65, 67.
Chaetognatha I 119,
Chaetopoda I 119.
Chama I 227; — exogyra I 227*.
Chamidae I 227. Cliamsosaurus II 115,
Chasmops I 187*.
Chasmosaurus II 146.
Cheilostomata I 94.
Cheirodus granulosus II 37, 41.
Cheirolcpis II 38; — Traillii II 37, 38*.
Cheirurus I 186, 187*, 193; — insignis I 194*..
Cheliter Hemprichii I 208.
Chelonia II 104.
УКАЗАТЕЛЬ
345
Chelonidae II 106.
Cheloniidea II 105, 108, 109.
Chelydridae II 109.
Chelys fimbriata II 108.
Chenendopora ‘I 44.
Chilopoda I 203, 204.
Chiromyoides campanictis II 220.
Chiroptera II 177, 178.
Chirotherium Barthi II 72*.
Chiton I 253; — priscus I 254*; — squamosus
I 254*.
Chitonidae I 212.
Chlamidophorus II 216.
Chlamydoselache II 20, 22.
Choerotherium II 269, 271*. .
Choia Carter! I 40. ,
Chondrostei II 36, 37.
Chondrosteidae II 37.
Chondrosteus acipenseroides II 37.
Chondropterygii II 7, 18.
Chonetes 1103,106; —sarcinulata 1104* —stria-
tella I 104*.
Choneziphius planirostris II 206.
Choristites Sowerby; I 114*.
Choristoceras I 274; — March! I 275*.
Choristodira II 115.
Chriacus II 186.
Cidaridae I 160, 161, 162.
Cidaris I 159, 160, 161, 162; — coronata I 160*;
— subvesiculosa I 160.
Cimolomys II 169; — nitidus II 169*.
Cirripedia I 176, 179.
Cistella I 110.
Cladiscites I 296; — tornatus I 278.
Cladoconus I 64, 67.
Cladodontidae II 19.
Cladodus II 19, 52.
Clagraptus I 57*.
Cladoselache I 13; II 19, 20, 51, 52; Fyleri II
19*, 52, — lamnoides II 19.
Cladosictis II 173. • :
Claosaurus II 142. ,
Clavella I 247. .
Clavirostridae I 305.
Clavularia I 65.
Clavulina Szabo! I 24*.
Cleodora I 251.
Clepsydropidae II 87, 116.
Clepsydrops II 88; —natilis II 87*:
Clidastes II 118, 120; — velox II 120*.
Climacograptus I 56; — kukersianus I 57*.
CHmacospongia I 39.
Climatius II 24, 52; —scutiger II 25*.
Clisiophyllium coniseptum I 81, 82*.
Clissiaphyllum I 53.
Clupea II 44, 46; —ventricosa II 44*.
Clupeidea II 44.
Clymenia I 265; — levigata I 265; — speciosa
I 265*, —undulata I 2b5*.
Clymeniidae I 255, 265, 266, 303.
Clypeaster I 156*, 164; — grandiflorus I 164*.
Clypeasteridae I 164, 165.
Cnemidium astroites I 44*.
Cocco.rinus I 138*.
Coccolepis II 39; —macropterus II 37.
Coccosteidea II 26.
Coccosteus II 16, 26, 27, 46; — decipiens 1126*
Coccoteuthis I 302.
Cochleosaurus II 63; — bohemicus II 64*.
Cochliodus contortus II 19*.
Cochloceras I 274.
Cochlops II 217.
Codaster I 134, 135; — trilobatum I 134*.
Codasteridae I 134.
Codonites I 134, 135; — steliiformis I 134.
Coeciliae II 55.
Coelenterata I 36, 89.
Coeloceras I 281; —subarmatum I 282*.
Coeloptychidae I 47.
Coeloptychium I 47; —subagaricoides I 47*.
Coelurosauridae II 136.
Coleoptera I 203, 209.
Collopegina I 41, 42.
Collyrites I 166.
Coloreodon II 274. ; t
Coluber II 116.
Comatula I 137.
Comatulidae I 138.
Compsognathus II 129, 136, 153, 154; — lon-
gipes II 136*.
Conchostraca I 176, 178.
Condylarthra II232, 242, 245, 246, 260, 261, 270
Congeria I 233; — subglobosa I 235*.
Conirostridae I 305.
Conocephalus I 190; •— Sulzeri I 191*.
Conoclypeus I 163.
Conocoryphe 1 187*, 190, 195; — Sulzeri 1191*.
Conodontae I 118*.
Conoryctes comma II 211.
Conularia I 252; — anomala I 252*; — quadri-
sulcata I 252*.
Corallium rubrtim I 62, 63*.
Corbicula fluminalis I 224*.
Cornuspira I 20*, 24, 30.
Cornuspiridae I 24, 31.
Corynoides I 56.
346
УКАЗАТЕЛЬ
Coryphodon II 320, 321,322,323,324*; —anax
II 322*; —hamatus II 321*, 322; —lobatus
II 322;—testis II 321*, 322.
Coryphodontidae II 320, 321, 322.
Corythosaurus II 138; — casuarius II 143*.
Coscinocyathidae I 52.
Coscinocyathus I 51*, 52*; —irregularis I 52.
Coscinopora I 48.
Coscinoporidae I 48.
Cosmoceras I 282, 283*, 293, 299,300; —eno-
datum I 283*; —Dunkani I 284*; —Jason I
283*; — ornatum I 284*.
Costata I 140, 142.
Cotylosauria II 67, 79, 80, 82, 84, 85, 86, 88,
.105, 151.
Crania I 101, 102; — ignabergensis I 101*.
Craniata II 15.
Craniidae I 101.
Craspedite.s kaschpnricus I 290; — nodiger I
290, 291*; — subditus I 290, 291*.
Craspedltidae I 290.
Craspedophylium I 53, 54.
Crassatllddae I 224.
Creodonta II 175, 184, 185, 187, 188, 192, 193,
200, 201, 203, 204, 209, 232.
Crepidula I 242; — aculeata I 242*.
Cribrospongia I 48; — concentrica I 48*.
Crinoidea I 130, 137, 140, 143, 149, 153, 154.
Crioceras I 293; Dusvalii I 293*.
Crioceratidae I 296.
Crisina I 92.
Cristellaria I 26; — rotulata var. Roemeri I 26*.
Crocodilia II 79, 120, 148.
Crocodilus articeps II127; — cataphractus II127;
— intermedins II 127; — megarhinus II 127.
Cromyocrinus I 145, 146*; — geminatus I
145*.
Crossopterygu II 7,8, 28, 33, 31, 36, 37, 50,
51, 52, 53, 54, 57, 70.
Crotalocrinus I 144, 147.
Crustacea I 175.
Cryptobranchus II 74.
Cryptocleidus oxoniensis II 102.
Cryptocrinidae I 122, 131.
Cryptocrinus I 131, 142; —cerasus I 131*.
Cryptocyclica I 13-<, 140, 147.
Cryptodira II 105; 108, 110. ,
Cryptostomata I 90, 91. . .. ;
Ctenacanthus II 22*. v ,
Ctenobranchia I 237, 241, 245. /
Ctenocrinus typus I 151*.
Ctenodus II 31.
Ctenophora I 36. ;
Cucullea I 219; —fibrosa I 219*. —hersilia I
219*.
Cupressocrinus crassus I 141*.
Curtonotus I 226.
Cuspidata I 164.
Cyamodus II104; —tarnovitzensis II 103, 104*.
Cyathaxonia I 76, 79*, 80, 86; — cornu I 80*.
Cyathaxonidae I 80.
Cyathocrinidae I 144, 154.
Cyathocrinus I 144, 154; — jawensis I 144*;
— longimanus I 144*; —ramosus I 144*.
Cyathocystites I 123.
Cyathophyllidae I 80, 83, 84, 87.
Cyathop'rylloides I 83.
Cyathophyllum I 83,86, 87;— caespitosurn 1 84;
— ceratites I 83*; — conicum I 83*; — he-
xagonum I 84*.
Cyathoseris I 74.
Cyathospongia 145. ?. г.
Cyclobranchia I 237,238. . .
Cyclolites I 74, 80; — undulata I 74*.
Cyclolobidae I 275, 277.
Cyclolobus I 276; — Stachei I 277*.
Cyclophthalmus senior I 205, 206*.
Cycloseris I 88.
Cyclosphaeroma trilobatum I 182*.
Cyclostoma I 237, 244.
Cyclostomata I 92, 94.
Cyclostomi II 7, 15, 16, 17.
Cyclostomidae I 244.
Cyclotosaurus II 71.
Cyclotus I 244.
Cylindrophytna I 43; — milleporatum I 43*.
Cynodictis II 192, 193, 195; — gregarius II192;
— lacustris П 192, 193*.
Cynognathus II 92*, 165; —crateronotus II
90* 91.
Cynopithecidae II 222, 223. : .
Cypellia I 48.
Cyphaspis I 193; Burmeisteri I 192*.
Cyphosoma I 161.
Cypridina I 176, 179. . .J
Cyprina I 224. if лЛ -- ;
Cypris I 179. ’• '
Cyrena semistriata I 224*; — subarata I 224*
Cyrenidae I 224.
Cyrtia I 112*, 114. ;
Cyrtina I 114.
Cyrtocalpis I 35; —urcolus I 34*.
Cyrtoceras I 259, 260; — corbulatum I 258
— Murchisoni I 258, 259*.
Cyrtolites I 240; —ornatus I 240*.
Cystasteroidea I 130.
УКАЗАТЕЛЬ
347
Cystechinoidea I 130.
Cystiphyllidae I 86, 87.
Cystiphyllum americanum I 86; —sihiriense I
85*, 86; — vesiculosum I 86.
Cystoblastidae I 126.
Cystoblastoidea I 130.
Cystoblastus Leuchtenbergi I 126*, 134.
Cystocidaridae I 157.
Cystocidaris I 129, 157; —pomum I 129*.
Cystocrinoidea I 130.
Cystoidea I 121, 122, 123, 128, 130, 131, 134,
135, 142, 143, 146, 152, 153, 154, 157.
Cytlierea I 215*, 224; —laevigata I 225*.
D.
Dactyloceras I 281, 284; — commune I 284*.
Dalmanites I 187*, 193; — Haussmann! I 193*.
Dama II 278*.
Dapedius politus (philodotus) II 42*.
Daphnia I 178.
Daphoenus felinus II 194.
Daraelites I 269*, 270*, 272, 278.
Dasypoda II 210, 216.
Dasypus II 210; —retusus II 217.
Dasyuridae II 169, 170, 173.
Davidsonia I 105, 106, 111.
Decapoda I 183, 255, 300, 301, 303.
Defrancia I 93.
Deiphon I 193, 195; — Forbesi I 194*.
Delphinoceti II 208.
Delphinognatnus II 90; — conocephalus II 91*.
Deltatheriidae II 175.
Deltatherium II 175, 186.
Dendrodus II 35.
Dendrograptus I 58.
Dendroidea I 58.
Dendrophyllia I 73*; — elegans I 73.
Dentalina I 26.
Dentalium I 212, 253*; - elephantium I 253*.
Dermochelyidae II 110.
Dermochelys II 106.
Deshayesia cochlearia I 244.
Desmatophoca oregonensis II 201.
Desmodonta I 214, 215*, 217, 229.
Desmothippus II 240.
Deuterodontia II 88,91.
Deuterosaurus II 92*, 93.
Diadectes phaseolinus II 84.
Diadetna I 159, 162.
Diademidae I 161.
Diademodon 1190; —Browni II 90.
Diadiaphorus II 244*.
Diapsida II 78, 79, 80, 111.
Diasparactus Zenos II 84.
Diatryma II 159*.
Dibranchiata I 254, 255, 299, 300.
Diceras arietinum I 227*.
Diceratidae I 223, 224.
Diceratosaurus punctolineatus II 62*.
Dicotyles II 269, 270, 291* 293.
Dicotylinae II 2S9.
Dicroceras II 277.
Dicrocynodon victor II 166.
Dictyoconites I 307; —reticulatus 1 307.
Dictyocyathidae I 52. U
Dictyocyathus I 53. . ‘
Dictyonema I 58; —flabelliformis I 57*.
Dictyonina 145, 46, 47. . „ г
Dictyospongia 145*. . .
Dictyospongidae I 45.
Dicyclica I 138, 140, 143, 147, 150, 152, 153.
Dicynodon II 94; — pardiceps II 93, 94*;
— Trautschojdi II 93*, 94.
Dicynodontia II 78, 88, 93.
Didelphodus II 170, 176; — absarocae II 176*.
Didelphydae II 170, 171.
Didelphys II 169, 170*, 171; — fugax II 170.
Didymograptus I 58; — informis I 56* .
Dielasma elongata I 109*.
Dimerocrinus I 153; — oligoptylus I 153*.
Dimetrodon II 88*; — incisivus II 87* 88.
Dimorphodon II 148*, 149; — macronyx II148.
Dimyaria I 216, 217, 229.
Dinichthys II 26. . ..--«г.
Dinictis II 197*. 198*.
Dinoceras II 323,324*, 333; — mirabile II 322*,
323*.
Dinoceratidae II 320, 322, 324.
Dinornis II 160; — elephantopus II 160*.
Dino.nithidae II 160.
Dinosauria II 79, 114, 121, 128, 148.
Dinotherium II 305, 306, 330, 333; —bavari-
cum II 307; — Cuvieri II 307; — giganteum
II 306*, 307; —g gantissimum II 307; — Koe-
nigi II 307; — medium II 307; - proavum
II 307.
Diphyp lyllum I 87.
Diplacanthus striatus II 52.
Dipleura I 187*.
Diplobune II 296; - - bavaricum II 295; —Quer-
cyi II 294*, 295; — tridaclylum II 295.
Diplocaulidae II 61.
Diplocaulus II 58, 61, 63, 73; — magnieornis
II 61*, 62. frhr.r.n
Diplocynodon Gervaisi II 126. .
348
УКАЗАТЕЛЬ
Diplodocus II 128, 129, 131, 132; —longus II
131*.
Diplograptus I 56, 57*, 58*.
Diplopoda I 203.
Dip'opora I 126.
Diplopterus Agassizi II 34.
Diplopus II 295.
Diplorrhina II 15.
Dipnoi II 8, 20, 25, 28, 29, 30, 32, 33, 34,
37, 47, 50. 51, 52, 53, 54, 56, 57.
Dipoides II 182.
Diprotodon II 170, 173; — australis II 172*.
Diptera I 208.
Dipterus II 20, 29, 30, 32*, 35, 46; —macro-
pterus II 31, 32, 33; — platycephalus II 29*,
30*; —Valenciennes! II 29*, 30*, 31, 32.
Discina I 100, 115*, 116; —elliptica I 100*;
— lamellosa I 100.
Discinidae I 100, 115.
Discoceras Danckelmanni I 261*; — teres I
261*.
Discoidea I 163; —cylindrica I 163*, — subu-
cula I 163*.
Discorbina I 26.
Discosaurus Netshajevi II 69.
Disparactus zenos II 84.
Dissacus II 185, 187.
Dissorophus multicinctus II 73.
Doedicurus clavicaudatus II 218.
Dokidocyathus I 53.
Dolichorhinus II 263*, 264, 265; — cornutus II
263*.
Dolichorhynchops II 102*.
Dolichosoma II 62; — longissimum II 63*.
Dol.chosomidae II 58.
Domatoceras I 262.
Dorcatherium II 284*; — crassum II 284*, 285;
— majus II 285; — Naui II 284*, 285.
Doricranites bogdoanus I 274*.
Doryderma I 43; —dichotomum I 43*.
Dorygnathus banthensis II 148*, 149.
Dorypterus Hofmanni II 37.
Dreissenia I 233; — polymorpha II 259; —ro-
striformis I 234*, II 259.
Dremodarium II 285.
Dremotherium II 276, 289; — Feignouxi II 275*.
Drepanaspis I 13; II 8, 10, 11; —gemunden-
sis II 10*.
Dromatherium II 164; —silvestre II 165*.
Dryopithecus II 225, 226*; — Larteti II 225;
— punjabensis II 226.
Dryosaurus II 142.
Duplicidentata II 178, 180, 183.
Duvalia I 306, 315.
Duvaliidae II 315.
Dvina prima II 90.
Dvinosauria II 59.
Dvinosauridae II 68, 96.,
Dvinosaurus II 68, 69*; — primus II 69; —se-
cundus II 69; — tertius II 69.
Dysodonta I 214, 215*, 217, 230.
E.
Ecardina I 98. ; .4^,
Eccyliomphalus I 240. • ''
Echidna II 163.
Echinidae I 155, 161.
Echinobrissus I 165.
Echinacaris I 181.
Echinoconus I 163; —Roemeri I 163*.
Echinocystites I 157.
Echinodermata I 121, 128, 130, 153.
Echinoencrinus I 126; — granatus I 125, 126*;
— striatus I 125, 126*.
Echinoidea I 157.
Echinolampas I 165.
Echinosphaerites I 124, 126, 135, 136;—auran-
tium I 124*; —infaustus I 124, 131.
Echinosphaeritidae I 124.
Echinus I 155*, 161.
Ectocochlia I 255.
Ectoprocta I 90.
Edaphosaurus cruciger II 88.
Edentata II 209, 211, 212.
Edestes II 23; — crenulatus II 23*.
Edestidae II 22, 46.
Edrioaster I 123; — Bigsbyi I 123*.
Eiffelia globosa I 46*’
Elachoceras II 324.
Elasmobranchii II 7, 18, 22, 46, 47.
Elasmosaurus II 102.
Elasmotherium II 258, 259, 306; — caucasicum
II 259; —sibiricum II 258*, 259.
Eldonia I 172*; — Ludwigi I 172*.
Eldonidae I 172.
Elephantidae II 307, 332. 1 ’
Elephas II 297, 323, 324*. 332; — africanus
II 297, 314, 315*, — antiquus I 309,
310*, 314, 315, 316, 317*; —armeniacus II
314; —Columbi II 309, 312; —hysudricus
II 309; — imperator II312;—indicus II 297,
313, 314, 315; —insignis II 308; —meridio-
nalis II 309*, 310*, 313, 316; —planifronsll
308*, 309; —primigenius II 297,310*, 311*,
312, 313*, 315, 316; — priscus II 315;
УКАЗАТЕЛЬ
349
Elephas trogontherii II 310*, 312, 314, 316;
— Wiisti II 310* 312, 314, 316.
Eleutherocrinus Cassedayi I 135*.
Eleutherozoa I 121, 155.
Elginia II 84; — mirabilis II 84*, 108.
Elliopsocephalus I 195.
Elonichthys II 39; — Robisoni II 39.
Elotherium II 290, 291*.
Emarginula exigua I 239*; — Miinsteri I 239;
— Schlotheimi I 239.
Emeradella Brocki I 200*.
Emydinae II 108, 109.
Emys II 106.
Enallohelia I 75.
Encrinasteridae I 171. .
Encrinurus I 187*.
Encrinus liliformis I 147*.
Endoceras I 257; — belemnitiforme I 258*, 259;
— bladius I 258*; — duplex I 257*.
Endocochlia I 255.
Endothyra I 26; — Bowmanni 1 26*.
Enigmatherium stavropolitanum II 259.
Entalophora I 92; — virgula I 92*.
Enteletes hemiplicata 1108*; — Lamarcki I 108*.
Entelodon II 290, 291*, 292.
Entomis I 179; — serrato-striata I 179*.
Entomostraca I 175, 176.
Entoprocta I 90.
Eoanthropus Dawsoni II 229.
Eobuthus I 202. : ..
Eoceratops II 138, 146.
Eocetus II 204.
Eocidaris I 160.
Eohippus II 234.
Eomys Zltteli II 182.
Eophrynus I 206; — Prestwichii I 207*.
Eoscorpius I 205, 208.
Eosiren II 318, 319*; —libyca II 318.
Eotetrapoda II 59.
Eotherium II 318, 319*.
Eotitanops II 264.
Eotylopus II 286; — Wilson! Il 285*.
Epaphis II 116.
Epigaulus Hatcheri II 183*.
Epihippus II 235, 241. 1
Equidae II 232, 233, 237*, 330.
Equus II 236, 237*, 240, 242; — Andium II
235*; — caballus II 235*, 237; — Stenonis II
236.
Erinaceidae II 176.
Eryma I 184; —ieptodactylina I 184*.
Eryon I 184; —propinquus I 183, 184*.
Eryops II 66, 80; —megacephalus II 67*.
Erythrosuchtis africanus II 121*.
Eschara I 95.
Estheria I 178*; — Hisingeri I 178*.
Estonyx Burmeisteri II 179*.
Estonioceras imperfectum I 261*.
Etherigia I 48.
Ethmophyllum I 51.
Euarmigera I 203, 204*.
Eubelemnitidae I 314.
Eucalyptocrinus I 149, 150; —rosaceus I 150,
151*.
Euceratherium II 283.
Eucinepeltus complicatus II 218*. , , ,.(1
Eucreodi II 185, 189, 191. ' r,
Eudiastatus lingulatus II 224. tnotq
Euechinoidea I 157, 159.
Eugerion I 209; — Boeckingi I 210*.
Euomphalidae I 240.
Euomphalus I 210; — tabulatus I 241*. :
Euparkeria capensis II 123*.
Euphoberia I 203. -.o
Euprotogonia puercensis II 233. ,
Eupsammia trochiformis I 73*. t . р-э
Eupsammidae I 72.
Eurycormus II 40, 49; —speciosus II 40*.
Eurynodelphidae II 206. „
Eurynodelphis Cocheteuxi II 206.
Eurynotus crenatus II 37, 40*.
Eurypholis II 45.
Eurypteridae I 198, 199; II 1.
Eurypterus I 196*. 197, 198; —Fischeri I
197*.
Euryptychites I 286.
Eurysternum Wagleri II 108, 109*.
Eusiphonella I 50.
Eusmilia I 78.
Eusthenopteron II 35; Foordi II 35.
Eusuchia II 121, 124, 125, 127.
Eutaxicladina I 41, 42.
Exogyra I 232; — columba I 232, 233*; —vir-
gula I 232, 233*.
Expleta I 79, 80.
Extrasiphonata 1 255, 265, 266.
F.
Fascicularia aurantium I 93*; — incrassata I
93*.
Favia I 77; — caryophylloides I 77*.
Favosites I 65, 66, 67*; —gothlandica I 66*.
Felidae II 184, 196, 197*, 198*, 199.
FelisII 197*, 198*, 199; — spelaea II 199.
Felsinotherium II 318,319. , л '
350
УКАЗАТЕЛЬ
Fenestella 1 91, 92; —oculata I 91*; —sara-
neana I 91*.
Fenestralia I 91. : .
Fibularia I 164.
Fissipedia II 185, 192.
Fissurella I 239; — graeca I 239*.
Fissurellidae I 239.
Fistulata I 140, 142, 143, 144, 150.
Flexibilia I 140, 153.
Foraminifera I 19, 20.
Forbesiocrinus I 153; —incurvus I 153*.
Fungia I 74.
Fungidae I 71, 74, 80, 86, 88.
Fusidae I 247.
Fusulina I 31, 32, 33, 55; — cylindrica 1 31,
32*; — longissima I 32*.
Fusulinella I 20*, 31, 32, 33; — Bockii I 32*;
— Brady I 32; — Struvei I 25, 32.
Fusuiinidae I 25, 33*.
Fusus I 237, 247; —ociculatus I 247*; —lon-
gaevus I 247*.
G.
Gadila I 253. • . ji/
Galeocerdo II 19*. '.i . . i
Galerix II 177.
Galesaurus II 90.
Gampsonyx I 183; —fimbriatus I 183*.
Ganodontia II 211.
Ganoidei II 28.
Gastigerina I 27*.
Gastornis II 158.
Gastrioceras I 268; —Jossae I 268*, 270*.
Gastrocoeli I 307, 308.
Gastropoda I 136, 212, 235, 241, 246, 251,
253.
Gaudryina I 24.
Gavialidae II 127.
Gavialis II 127.
Gavialosuchus II 127.
Gazella atlantica II 280; — brevicornis II 280.
deperdita II 280; — palaeindica II 280;
— porrecticornis II280; — Schlosser! II 280*.
Gelocina II 274.
Gelocus II 274, 289; — communis II 274*.
Geocoma 1 169; — carinata I 168*.
Geodites I 40.
Geogmyda II 106.
Geosauridae II 127. •
Geosaurus suevicus II 127, 128*.
Geoteuthis bollensis I 302, 303*.
Gephyrea I 119, 120. . ‘ J
Gephyroceras uchtense I 267, 268*......
Gephyroceratidae I 266, 267.
Geralinura I 205; —bohemica I 207*; — carbo-
naria I 207*.
Geraphrynus I 207; — carbonarius I 207*.
Gervillia I 230.
Gigantosaurus II 132; — africanus II 132; —ro-
bustus II 132.
Gigantostraca I 12, 196, 197, 198, 199, 202; Il
15.
Giraffinae II 287.
Gius II 159.
Glassia I 111.
Globidens Fraasi II 120.
Globigerina I 22, 25*, 27*.
Globigerinidae I 27.
Glossopteris II 95.
Glyphioceras diadema I 268*; — sphaericum I
268*.
Glyphioceratidae I 266, 268.
Glyptodon II 108, 209*, 218; — brachyostracon.
II 218; — reticulatus II 217*.
Glyptodonta II 216, 217, 219.
Glyptodontidae II 217.
Glyptolepis macrolepidotus II 35.
Glyptosphaerites 1 127; — Leuchtenbergi 1
128*.
Glyptosphaeritidae I 127.
Glyptostomata I 160, 161.
Glyptotherium texanum II 218.
Gnathostomata I 162.
Gomphoceras I 258; —bohemicum I 258*.
Gomphocyathus I 53*.
Gomphognathus I 14; II 90*, 91*.
Gomphotherium II 286; — Sternbergi II 286*..
Gondvanosaurus II 66.
Goniaster I 170; —Parkinsoni I 170*.
Goniatitae I 13.
Goniatitidae I 265, 266, 275.
Gonioclymenia speciosa I 265.
Goniomya I 229.
Goniophyllum I 87; —pyramidale I 86*.
Gordonia II 78,79*;—Annae 1194, 95*; —ros-
sica II 94; — Traquairi II 94.
Gorgonidae I 62.
Grammisia I 217; — hamiltonensis I 218*.
Granatocrinidae I 133.
Granatocrinus I 133, 135; —derbiensis I 133,
134*; — Norwoodi I 133*, 134*.
Graphularia I 62; — dcsertorum I 62*.
Graptolitha I 57*. . .
Graptolitidae I 56.
Gravigrada II 209, 213, 219.
Gravigradidae II 212.
УКАЗАТЕЛЬ
351
Gromia I 20*, 23.
Griinewaldtia I 112; —latilingius I 112*.
Gryphaea I 232; — arcuata I 233*.
Gymnoblastea I 54.
Gymnophionia II 55, 56.
Gyraulus I 249; — trochiformis sulcatus I 251*.
Gyroceras I 260; — alatum I 260*.
Gyrodus macrophthalmus II 41*.
Gyroptychius II 35*.
H.
Hainosaurus II 118.
Halicore II 317, 318, 319*. . .
Halicoridae II 318. . ...
Haliotis I 239, 241.
Halitherium II 318, 319*; - Schinzi II 318*.
Hallia I 83.
Halysites I 65; —catenularia I 65*.
Hamites I 297; —rotundatus I 297*.
Hapalops II 209*, 210* 212.
Haplocrinidae I 154.
Haplocrinus I 138*; — mespiliformis I 142*.
Haplophoneus II 197*, 198*.
Haplophragmium I 24; — Humboldti I 24*.
Haptopoda I 207.
Harpes I 186, 187*, 190, 195; —beningnensis
I 190*.
Harpoceras I 280, 281; — bifrons I 280;
— opalinum I 280*; — serpentium I 280;
— thouarsense I 280*.
Harpocerotidae I 280. . .
Haselia I 40; — palmata 1 40*.
Hatteria II 111; — punctata II 78*.
Hegetotherium mirabile II 326.
Heliastraea conoidea I 77, 78*.
Helicidae I 250.
Helicopegmata I 102, 110.
Helicoprion II 23; — Bessonowi II 22*.
Heliocrinus I 124; —balticus I 124*.
Heliodiscus I 35.
Heliolites I 63, 68; — interstinctus I 64*.
Heliolitidae I 63.
Heliopora I 63; — caerula I 64*; — Partschi I
63*.
Helioporidae I 63.
Helix I 242, 250; —fruticum I 250*.
Helladotherium II 287*; — Duvernoyi II 287.
Helmersenia I 101*.
Hemiaspis I 201; — limuloides I 201*.
Hemiaster I 167.
Hemicidaris I 161; —crenularis I 161*
Hemicosmites piriformis I 125*.
Hemicyon sansaniensis II 194.
Hemiganus II 211; —otariides II 211.
Hemiostei II 36, 38, 40, 42, 43, 44, 47, 53
Hemiptera I 208, 209.
Heptanchus II 47*, 52.
Heptodon II 246, 259, 261.
Hercoceras I 261; —mirum I 261*.
Herpestes II 195; —lemanensis II 195*.
Hesperopithecus II 226.
Hesperornis II 156, 157, 158, 160; —regalis II
156*.
Heterodonta I 215* 217, 218, 220, 224.
Heterodontus II 18. . . ;;!,ЕгСч .
Heterophyllia I 78. , . .
Heteropora I 93; —pustulosa I 93*. «gp .
Heterostraci II 8, 9.
Hexacorallia I 61, 62, 63, 64, 68, 69, 70, 71,
73, 74, 77, 78, 79, 80, 86.
Hexacrinus elongatus I 150.
Hexactinellidae I 44, 50. . • I . .1
Hexapoda I 255, 300.
Hexaprotodon sivalensis II 267*.
Hexax onae I 38.
Hildoceras I 280.
Hipparion II 235*, 238, 240, 242; — curtum II
240; — gracile II 238*.
Hipponyx I 242. < . .
Hippopotamidae II 266, 267.
Hippopotamus II 267, 271*; —amphibius II
267;—hipponensis II 267; — madaga-
scarensis II 268; — major II 267*; — minu-
tus II 268, Pentlandi II 267; — sivalensis II
267*; — sivajavanicus II 268.
Hippothoa I 95.
Hippurites I 228; — cornu vaccinum I 228*;
— radiosus I 228*.
Hirudina I 119. ; i.
Holaster I 166.
Holasteridae I 165.
Holectypus I 163; —depressus I 163; — orifi-
catus I 163*.
Holocephali II 7, 18, 24. :
Holoptychidae II 35.
Holoptychius II 27, 35, 37, 46, 49, 70; — Fle-
ming! II 35*. .
Holoscus I 45.
Holostamata I 160, 237, 242.
Holothuridae I 172.
Holothuroidea I 171.
Holtcnia Carpenteri I 37*.
Homaeosaurus II 113; — pulchellus II 113*.
Homalodontotheriidae II 328.
Homalodontotherium II 328; — Segoviae II329*.
352
УКАЗАТЕЛЬ
Hominidae II 227.
Homo II 219, 227; —heidelbergensis II 228*,
229*; — neandertalensis II 228*; — primi-
genius II227, 228, 229; —sapiens II 228, 229.
Homunculus patagonicus II 224.
Hoplites I 291; — auritus I 300; — Deshayesi I
292*; — Engersi I 292; — Raulini 1292,293*.
Hoplitidae I 291, 292.
Hungarites tropicus I 273.
Hyaemoschus II 283, 284*, 285; —aquaticus
II 284*; — crassus II 284*.
Hyaena II196; —eximia II196*; —spelaea II196.
Hyaenarctos II 200.
Hyaenictis graeca II 196. -
Hyaenidae II 184, 196, 199.
Hyaenodon II 185; 187; —leptorhynchus II
187*.
Hyaenodontidae II 185. , . , ; .
Hyalonema 145.
Hyalostelia I 45.
Hyboclypeus I 165, 166; —gibberulus I 165*.
Hybodontidae II 20.
Hybodus II 19*, 20, 21, 49; —hauffianus II
21*;—plicatilis II 21;—polyprion II 21;
— reticulatus II 21. : • ;
Hydaspitherium II 288.
Hydnoceras I 45; — bathensis I 45*.
Hydractinla I 54, 55; — echinata I 54; — in-
crustata I 54.
Hydriocrinus I 145; —pusillus I 146*.
Hydrocorallinae I 54, 55.
Hydromedusae I 54.
Hydrophoridae I 122, 123,
Hydrozoa I 36, 54, 55, 56, 89.
Hylaeobatrachus II 73*, 74.
Hylobatus II 224.
Hymenocaris I 184; —perfecta I 181; —ver-
micauda I 181*. -
Hymenoptera I 208.
Hyocrinus I 138; 143; 147; — bethellianus I
142*.
Hyolithes I 252; — elegans I 252*; — maxi-
mus I 252.
Hyopotaminae II 291, 293, 294, 295.
Hyopotamus II 272, 293, 294, 295; — america-
nus II 294; — brachyrhynchus II 294;
— Gorringei II 294; — Gresslyi II 294;
— Picteti II 294; — porcinus II 294; — ve-
launus II 292 s, 294.
Hyopsodus II 177, 221.
Hyotherium II 175; — Soemmeringi II 269*;
— Waterhousi II 268*.
Hyperlestes garsonyanus II 213.
Hyperodarpedon II 113; —Gordoni II 114*.
Hypoparia I 186, 187*, 188, 189.
Hypospondylus II 16, 46.
Hypsilophodon II 143; — Foxi II 138*.
Hopsiprymnidae II 172.
Hyrachius II 248, 249, 250; 252; — agrarius
II 247*; — eximius II 247*.
Hyracidae II 317.
Hyracodon priscidens II 248.
Hyracops II 231; —socialis II 231*.
Hyracotherium II 234, 235*, 237*; — index II
234; — leporinum II 234*, 235*; — venticu-
lum 234*.
Нугах II 317;—abissinicus II 317; —capen-
sis II 317.
‘ I.
Ibergiceras I 269*, 272.
Ichthyornis II 158; —victor II 157*.
Ichtyosauria II 79, 96, 99*, 120, 151.
Ichthyosauridae II 96, 97.
Ichthyosaurus II 97. 102; — communis II 98*;
— Conybeari II 97*; — platydactylus II 98;
— quadriscissus II 98*, 99; — trigonus II 99*.
Ichthyotomi II 19.
Icochilus II 325.
Ictitherium II195; —hipparionuui II 195, 196*;
— Orbignyi II 195,196*; — robustum II195*,
196*.
Ictops II 176,
Idmouea I 92, 94.
Iguana europaea II 116.
Iguanidae II 84.
Iguanodon II 129, 138, 139* 140* 142, 160;
— bernissartensis II 139*.
Iguanodontidae II 138.
Illaenus I 187*, 192; —Daemani I 192*;
— tauricornis I 195.
Imperforata I 20, 21, 71, 74,
Inadunata I 140, 143.
Inarticulata I 97, 98, 100, 102.
Indoceras Ismaeli I 298.
Indricotherium II 251; — transuralicum II 250*.
251; —turgaicum II 251.
Inexpleta I 79, 80.
Infradepressi I 314.
Infrasiphonata I 255, 265.
Inoceramus I 215, 233; —concentricus I 234*.
Inostrancevia II 68, 89*, 95; — Alexandri II
89*.
Insecta I 203, 208. . . : .
Insectivora II 175, 200. :
УКАЗАТЕЛЬ
353
ilntegripalliata I 217, 220, 224, 225, 227.
Interatherium II 325, 326.
Isastraea I 86; — helianthoides I 77*.
Ischadites Murchisoni I 51.
Ischyromyidae II 181.
Ischyromys II 182; —typus II 182*.
Isoarca I 219.
Isocardia I 224; — multicostata I 223*.
Isocrinus I 149,154.
Isodonta I 215*, 217, 230.
Isopoda I 181, 184, 186.
Itiria cabonetiana I 236*.
J.
Janessa II 23.
Jerea I 41, 42*: —kurskensis I 41; - - pyri-
formis I 41, 42*.
Jereica I 44*.
Julus I 203.
K.
Karpinskya 1 112; —conjugula 1 112*.
Keplerites I 282, 283, 299; — enodatum I 283*;
— gowerianum I 283*.
Keraterpeton II 58, 62.
Keyserlingia I 101; — Buchi I 101*.
Koiloskiosaurus coburgiensis II 85.
Koninckina I 111; —Leonhardi I 111*.
Koninckinidae I 102, 110, 111.
Kotlassia II 82*.
Kutorganacea I 100.
L.
Labelia 1 55.
Labidosaurus II 83; - hamatus II 83*.
Labyrinthodontia II 59, 69, 70, 71, 72, 80.
Lacertilia II 115.
Lagena I 25; — semistriata I 26*.
Lagenidae I 25.
Laggania cambria I 172,* 173.
Lagynis I 20, 23.
Lama II 287.
Lambdotherium II 264.
Lamellibranchiate I 212, 213, 214, 215, 236, 253.
Lamna II 19*, 22.
Lanarkia I 13; II 8, 9, 11, 27, 46; --horrida II
9; — spinosa II 9*.
Larsaurus II 138, 142.
Lapworthura I 169; — Miltoni I 168*, 171.
Lariosaurus Balsami II 100*.
Larviformia I 140, 142, 147, 154.
Lasanius II 9; — problematicus II 14.
Latirus I 236*, 237.
-'43 Павлова, M. В. Палеозоология. Ч. II.
ati “ llata I 263, 264*, 266, 267; 275.
Latzelia I 204; — primordialis I 204*.
Lacanites I 269*.
Lecanocrinus I 147, 153.
Leda I 219; — Deshayesi I 218*, 219.
Leidyosuchus II 126.
Leioceras I 280.
Lemuroidea II 219, 220.
Leontinia Gaudryi II 328.
Lepadidae I 176, 179.
Lepas I 179; — anatifera I 180*.
Leperditia I 179; — Hisingeri I 179*.
Lepidestes I 158; formosus I 159*; —laevis I
159*.
Lepidocentrus I 158; — rhenanus I 158*.
Lepidodendron II 72.
Lepidoptera I 208.
Lepidosauria II 115, 116.
Lepidosiren II 29, 32*, 33; paradoxa II 33.
Lepidosteus II 36, 38.
Lepidotus I 88; — eivensis II 42, 43*; - no-
topterus II 43.
Leporidae II 183.
Lepospondyli II 58*, 61, 72.
Lepralia I 95; — coccinea I 95*.
Leptaceratherium II 249.
Leptaena I 103, 106, 116; —rhomboidalis
105*; — sericea I 105*; — transversa I 105*.
Leptictidae II 176.
Leptictis II 177; -- Haydeni II 177*.
Leptobos II 281.
Leptolepidae II 44.
Leptolepis II 44, 49, 50, 53; — dubius II 44
— sprattiformis II 44*.
Leptomeryx II 277*; — evanse II 277*.
Lepus II 183.
Leucones I 49.
Leucosolenia I 49; - - lacunosa 1.38*.
Libellula I 210*.
Libycosuchus brevirostris II 125.
Libypithecus Markgrafi II 224.
Lichas I 18/*; —Boltoni I 195*.
Lichenodon I 136.
Lichenoides I 153; —prisons 1 125*.
Lichenopora tabulifera I 94*.
Lillia I 280.
Lima 1 233; —consobrina I 2311.
Limidae I 233.
Limnaea I 249.
Limnaeidae I 249.
Limnaeus I 249, 250; — pachygaster I 249*.
Limnocyon- II 188. . .
Limnohyops II 264.
.354
УКАЗАТЕЛЬ
Limnoscelis paludis II 83.
Lirriulava I 199.
Limulus I 185, 196, 200, 201, 202; Decheni I
202;—polyphemus I 201s; —Walchi 1201*,
202.
Lingula I 12,-98, 99, 115*, 116; — anatina I
99*; — Beani 1 99*; —tenuissima I 99*.
Lingulella I 99; —Davisii I 99*.
Lingulidae I 98, 115.
Linthia I 167.
Litharaea I 72, 75.
Lithistidae I 40, 41, 44, 46, 48, 49, 50.
Lithomantis I 209; — Woodwardi I 210*.
Lithostrotion I 84; — rossicum I 84*.
Litopterna II 243, 244, 328, 330, 332, 335.
Littorina I 243, 244*.
Littorinidae I 243.
Lituites I 13, 260: — lituus I 260*.
Lituola I 24, 25*; — Humboldti I 24*.
Lituolidae I 23, 25.
Lobites I 275; — delphinocephahis I 275*.
Loftusia persica I 55.
Loligo I 254.
Lonchorhynchus II 71; — Obergi II 72*.
Lonsdaleia floriformis I 84, 85*.
Lophiodon II 261, 328; — isselense II 261*;
-leptorhinus II 261; —rhinoceroides II261.
Lophiodontinae II 259, 261.
Lophiomeryx II 284.
Loriolaster I 170.
Louisella pedunculate I 173*.
Loxolophodon II 324.
Loxomma II 63.
Lucina I 224.
Ludwigia I 280.
Lunulites I 95; — cretacee I 95*.
Lutraria I 215*, 225.
Lutrinae I 194, 195.
Lycosuchus II 90'
Lyria I 247.
Lyrodesma I 226.
Lyrodesmidae I 226.
Lysorophus II 69, 74; — tricarinatus II 73.
Lyssacina I 45.
Lystrosaurus latirostris II 94*.
Lytoceras I 296, 297; — Liebigi I 296*.
Lytoceratidae I 278, 296.
M.
Macacus II 224.
Macellodon II 116.
Machairodus II 196*, 197*, 198*, 199.
Mackensia I 173; — costalis I 173*.
Macrauchenia II 243, 244, 245; —patagonica
II 243*.
Macrodon I 220; — hirsonensis I 221*.
Macropus II 171.
Macroscaphites I 296, 297.
Macrura I 183.
Mactra I 215, 225; —podolica 1 226?; — pon-
derosa I 225*.
Mactridae I 216.
Madrepora rudis I 72; — Solanderi I 72*.
Madreporaria I 75.
Madreporidae I 72.
Magas pumilus I 116*.
Magasella Cumingi I 116*.
Magellania I 117; —flavescens I 96*, 97*
116*; — venosa I 116*.
Malacostraca I 176, 180, 181.
Mallodus II 44.
Mammalia II 162.
Manatidae II 319.
Manatus II 317, 319.
Manidae II 216.
Manis II 209, 210, 216.
Manteoceras II 263*, 264, 265.
Manticoceras Backlundi I 268; — Pattersoni h
267*, uchtensis I 267, 268*.
Mantis I 209.
Marginella I 247.
Marrella I 189; — splendens I 188*
Marsipocrinus I 152; —magnificus I 152*.
Marsupialia II 163, 164, 165, 171.
Marsupialia vera II 164, 169.
Marsupites I 146; —testudinatus I 146*.
Mastigoteuthis I 301, 302*.
Mastodon II 333; — americanus II 299*, 303,.
304*, 305*; — Andium II 305*; angustidens
II 298, 299*, 300*, 301*, 302, 303, 304,305*,
306, 308, 315; —arvernensis II 229, 301*,
302, 303, 305*, 307; — Borsoni II 303, 304*,
305*; — brevidens II 303, 304; — Cautleyi
II 308; — elephantoides II 308; — Hum-
boldti II 302; - latidens II 308; — longi-
rostris II 301* 302, 303, 304, 305*. 308;
— ohioticus II 303, 304*, 305, 315; — pa-
laeindicus II 302; — Pentelici II 302; — pro-
avus II 302; — pyrenaicus II 302; — siva-
lensis II 302; —tapiroides II 303*, 305*,
306; - turicensis II 303, 304, 305*.
Mastodonsauridae II 59.
Mastodonsaurus II 69*, 70 s, 71
Mastodontidae II 298.
Mazama II 277*.
Meandroptychidac I 48.
УКАЗАТЕЛЬ
355
Meandroptychium I 48.
Mecochirus longimanus I 183*.
Medlicottia I 269*, 271*, 272.
Medusina costata I 60.
Medusites I 60; — Lindstroemi I 60*.
Meekoceras I 269*; — Hodgsoni I 273; — si-
biricum I 273*.
Megabelus I 314.
Megacerops II 265.
Megachiroptera II 178.
Megaladapis II 159, 221, 222.
Megalaspis I 190, 195; —extenuatus I 191*.
Megalictis ferox II 199.
Megalithisma foraminosa I 43* .
Megalobranchus II 74.
Megalodon I 223; — cucullatus I 223*.
Megalodontidae I 223.
Megalonyx Jefferson! II 212*. 213.
Megalosaurus II 135.
Megalurus II 38, 49; — elegantissimus II 39*.
Megamorina I 41, 43.
Meganeura Monyi I 209, 210*.
Megaphyllites I 276; — insectns I 276*.
Megateuthis paxillosus I 308.
Megatheriinae II 213.
Megatherium II 214, 216, 217; —americanum
II 213*.
Megathyridae I 102, 110.
Megathyris I 110; — decollate I 109*.
Megerlina lamarckiana I 116*.
Melania I 245.
Melanerpeton II 61.
Melaniidae I 245.
Melanopsis I 245.
Meles II 194.
Meletta II 44.
Melocrinidae I 151.
Melocrinus abconicus I 151; —typus I 151*.
Melonites I 159.
Melosaurus II 66. — uralensis II 68*.
Membranipora lapidosa I 94, 95*.
Menaspis II 23.
MeniscoSssus II 167, 168.
Meniscotherium II 231; —socialis II 231 .
Menodus II 265.
Menophyllum I 79.
Meristina I 114.
Merostomata I 176, 184, 196*, 198, 199, 200,
202, 205; II 46.
Merriamia II 97; — Zitteli II 97*.
Merychyus II273; — anenarum II 273-'.
Meryhippus II 235*, 236, 237, 239, 240; — cam-
pestris II 236; — insignis II 236, 239*.
Mesacanthodes Mitchelli II 25*.
Mesaxonia II 230.
Mesites I 128; —Pusirewskii I 128*.
Mesoblastus lineatus I 134*.
Mesocetus II 204.
Mesocystidae I 128.
Mesocystis I 128; — Pusirewkii I 128*.
Mesodon II 41.
Mesohippus II 235*, 236, 237; — Bairdi II 235.
Mesonyx II 185, 186*, 187; — obtusidens II
186*.
Mesopithecus II 223; — Pentelici II 223*.
Mesorhinus II 126; —Fraasi II 122*.
Mesosiren Dolloi II 318.
Mesosuchia II 121, 124, 125.
Metapaster I 170.
Metaphiomys Beadnelli II 183.
Metaxytherium II 319*.
Metazoa 119, 36.
Metopias II 59, 68, 70, 71; —diagnosticus I
71*.
Metriorhynchus II 127.
Miacidae II 185, 189, 195.
Michelinia I 66, 68; — lenticularis I 67; — ros-
sica I 68*.
Micraster I 166, 167; —cor testudinarium I
166*.
Microchiroptera II 178.
Microconodon II 165.
Microcyclus I 83.
Microdiscus I 187*.
Microdon II 49; — Itieri II42*, — Wagneri II 41.
Microgale II 177.
Microlestes II 167;—antiquus II 167*.
Micromitra superba 1100.
Micropternodus II 176.
Microsauridae II 61, 62.
Microseris I 86; — hemisphaerica I 74, 75*.
Microzeuglodon II 204.
Mijalia I 208.
Miliola I 20, 21, 31*.
Miliolidae I 24, 30, 31*.
Millepora I 55, 68; nodosa I 55*.
Milleporidae I 54.
Mimoceras 1 266; — coinprcssnin I 2665
Miohippus II 289.
Miolania II 108; —argentine II 108.
Miolanidae II 108.
Mioziphiits belgiciis II 206.
Mitra I 247.
Mitrocystis I 136; —mitra I 130*-
Mixohyrax II 317; —niloticus II 317*.
Mixosauridae II 96.
356
УКАЗАТЕЛЬ
Mixosaurus II 97, 98, 99*; — atavus II 97*;
— cornulianus II 97*.
Modiola I 233; — imbricata I 234*.
Moeritherium II 298, 299* 300, 302, 316, 318,
319*; Lyonsi II 298; — trigonodon II 298.
Molaria spinifera I 199, 200*.
Molge II 76.
Mollisonia gracilis I 188*.
Mollusca I 212.
Molluscoidea I 89, 118.
Moltkia I 62; — Isis I 62*.
Monactinellidae I 39.
Monaxonae I 38.
Monitor niloticus I 115*.
Monoclonius II 146*, 147.
Monocyclica I 138, 140, 147, 150.
Monograptus I 56, 57*.
Monomyaria I 216, 217, 230.
Monophyllites I 276; — Timonyi I 276*.
Monopleura I 227.
Monorrhina II 15.
Monotremata II 163.
Monticulipora I 67, 68; — Panderi I 67, 68*;
— petropolitana I 67, 68*; — ramosa I 68-
Montlivaultia I 76*, 86.
Morosaurus II 132; — grandis II 132.
Morphippus imbricatus II 327.
Mosasauridae II 118, 120.
Mosasaurus II 117, 118, 120.
Moschus moschiferus II 284; 285.
Multitub ere ulata II 164, 167, 175.
Murchisonia I 240.
Murex I 237; — Sedgwicki I 247; — spinicosta
I 247*; — tricarinatus I 247.
Muricidae I 247.
Mussa I 71*.
Mustelidae II 184, 194, 195, 199.
Mutela I 222.
Myidae I 215, 229.
Mylodon II 209* 210* 211, 214; robustus
II 214*.
Mylogus II 270.
Mylostoma II 26, 27.
Myophoria I 226; — decussata I 226; — lae-
vigata I 226*.
Myoxidae II 179.
Myriapoda I 203, 204,
Myrmecobius II 169.
Myrmecophaga II 210*.
Mystacoceti II 203, 206, 207.
Mystriosaurus bollensis II 126.
Mystriosuchus II 123, 126, 128; — planirostris
II 122*.
Mytilidae I 216, 233.
Mytilus I 214, 233.
Myzostamidae I 119.
N.
Najadites I 220; —compressa I 221*; - fabae-
formis I 221*.
Nannobelus I 314.
Nassellaria I 34, 35; — cyrtidea I 34*.
Natica I 236, 243, 250; — bulbiformis I 244i
— hantoniensis I 244*; — millepunctata I
244*; —patula I 244; —Willemeti I 24.4.
Naticidae I 241, 244.
Nautilidae I 13, 254, 255, 257, 259, 261, 263,
265.
Nautiloidea I 255.
Nautilus I 13, 254, 255, 256*, 257, 261, 262,
263; — bidorsatus I 256*; — cornutus 1 262*;
— danicus I 262*; — franconicus I 262;
— intermedins I 262*; — Konincki I 262;
— mosquensis I 261, 262*; — pompilius I
255*, 256*.
Nebalia I 181*.
Necrodasypus II 216.
Necrogymnurus Cayluxi II 176.
Necrolemur II 221*.
Necromanis II 216.
Necturus II 67.
Neobelemnitidae I 315.
Neoceratodus II 29, 31, 32*, 33, 48; — Forsteri
II 31*. 32, 33, 53*.
Neohipparion II 239*, 240; — gratum 11 239*.
Neolimulus I 201; — falcatus I 201*.
Neolenus I 193; —serratus I 187, 188*.
Neoplagiaulax II 169; —eocaenus II 169*;
Neusqualodon II 205.
Neotremata I 100, 115.
Nephthya I 62; — cretacea I 62*.
Nereites cambrensis I 118.
Nerinea I 236*, 245, 246; — dilatata I 245*;
— trinodosa I 245*.
Nerineidae I 245.
Nerita I 237, 241; Plutonis I 241*.
Neritidae I 241.
Neritina I 241, 244; —fluviatis I 242; —gra-
teloupana I 242*; —simulans I 242*.
Neritopsidae I 241.
Nesodon II 327, 328, 330, 334; — imbricatus
II 328*.
Nesodontia II 327, 328.
Nesodontidae II 327.
Neuroptera I 208, 209, 210.
Nimravus II 197*, 198*.
УКАЗАТЕЛЬ
357
Nodosaria I 25*,' 26, 28; — bacillum I 26*;
— guttifera I 26*.
Nomarthra II 209, 215.
Nonionina I 26. -
Norites I 269* 276.
Notharctus II 220.
Nothosauridae II 99, 101.
Nothosaurus mirabilis II 100*.
Notidanus II 18*, 19*, 50.
Notocoeli I 308, 310.
Notostraca I 176, 177.
Notosuchus II 125.
Notungulata II 324, 330, 332, 335.
Nubecularia novorossica I 32.
Nucleocrinus I 135; —Verneuli I 133.
Nucleolites 1 165.
Nucleospira I 112*.
Nucula I 214, 218, 219; — plancentina I 218*.
Nuculidae I 218, 220.
Nummulites I 28*, 29*. 30, 31, 33; —distans
I 28*.
Nummulitidae I 28.
Nyctosaurus gracilis II 151.
O.
Obollela I 99, 115*, 116.
Obolidae I 99, 100, 115.
Obolus I 99, 100; — Apollinis I 99*; — silu-
ricus I 99*.
Ocapia II 288.
Octocorallia I 62, 79.
Octopoda I 255, 300.
Octopus I 254, 300*.
Oculina I 55.
Oculinidae 1 75.
Odontoceti II 203, 205, 208.
Odontolcae II 155, 156.
Odontopteryx toliapicus II 158*.
Odontormae II 155, 157.
Ogygia corndensis I 195.
Olcostephanus hoplitoides I 286.
Oldhamia I 106.
Olenellus I 190; — Mickwitzi I 191*.
Olenidae I 190.
Olenus I 186, 187*, 190; —truncatus I 191*.
Olgodon I 220.
Oligopleurus II 53; — esocinus II 40.
Ommatostrephidae I 302.
Omphalotrochus discus I 240; —globosus I 240.
Omphyma I 81, 87, 106;— subturbinata I 83*;
turbinata 1 83*.
Oodectes herpestoides II 190*.
Opabinia regalis I 176*.
Operculina I 28.
Opetiosaurus II117;—Bucchichi II 118*.
Ophidia II 116.
Ophidioceras I 260.
Ophiocodon II 87.
Ophiurasteridae I 171.
Ophiuroidea 1 167, 168, 170, 171.
Ophthalmosaurus II 98, 99, 120.
Opisthobranchia I 237, 248.
Opisthocomus II 155.
Opisthoparia I 186, 187*, 190.
Oppelia I 281; —Ilexuosa I 281; —steraspis
I 281; —• tenuilobata I 281*.
Oppelinae I 280.
Orbicula I 115*.
Orbiculoides I 116.
Orbitoides I 29; —papyracea I 29*.
Orbitolina I 24, 28, 31; —lenticularis 1 24*.
Orbitolites I 30.
Orbitremites I 133; — Norwoodi I 133*.
Orbulina I 22, 27*.
Oreodon II 273, 274; — Culberstoni II 272*,
273*; —gracilis II 273*; —major II 273.
Oreodontidae II 271, 274, 289.
Oreopithecus II 224; — Bambolii II 223*.
Ornithischia II 130, 138.
Ornitholestes II 129, 136; —Hermanni II
136*.
Ornithopoda II 129, 130, 137.
Ornithopodidae II 138, 145.
Ornithorhynchus II 163*, 167.
Ornithosuchus II 123.
Ornithurae II 155.
Orodus II 22.
Orohippus II 235.
Orophocrinus I 131, 132, 134; — fusiformis I
132*; — verus I 134*.
Orthidae I 115.
Orthis I 105, 106; - calligramma I 105*;
— plana I 105*.
Orthoceras I 257, 258, 266; — dubium I 257*;
robustum I 258; —sericum I 257; —timi-
durn I 257*.
Ortoccraiidae I 13, 255, 257.
Orthoptera 1 208.
Orycteropidae II 215.
Orycteropus II 209, 210; - capensis II 216*;
— Gaudryi II 215, 216.
Osteolepidae II 34.
Osteolepis II 34; — macrolepidotus II 34*;
Osteostraci II 9, 11, 12.
Ostracoda I 176, 178, 179. . ‘
358
УКАЗАТЕЛЬ
Ostrea I 214, 216*, 232; —choroshovensis I
232, 233*; —digitalina I 232*.
Ostreidae I 231.
Otaria II 201.
Otoceras tropicum 1 273; — Woodwardi I 274.
Otocryptis bivittata II 124.
Ottoia I 120; — minor I 120*.
Ovibos II 283; — moschatus II 283*.
Ovinae II 283.
Oviraptor II 137, 146.
Ovis II 230, 283.
Oxyaena II 185, 188.
Oxyaenidae II 185.
Oxyclaenus II 186.
O.xynoticeras I 278,279; —catenulatum I 279*;
— fulgens I 279*; oxynotum I 279*.
Oxyosteus II 27.
Oxyrrhina II 19*.
P.
Pachyaena II 187.
Pachynolophus II 234, 235*, 237, 241.
Pachyodonta I 215, 217, 227.
Pachyopsis Woodwardi II 116, 117*.
Pachyrukhos Moyani II 326.
Pachyteuthis acutus I 308.
Palacanthus II 129, 144, 145*.
Palaeacmaea I 238.
Palaeaster I 170; —eucharis I 170*.
Palaeechinoidea I 157.
Palaeechinus I 158.
Palaeedaphus insignia II 31.
Palaeictops II 176.
Palaeobatrachus Gaudryi II 74*; —grandipes
II 74.
Palaeocampa I 204; — anthrax I 204*.
Palaeocaris I 183; — typus I 182*.
Palaeocarnivora II 185, 189, 232.
Palaeoceras II 280.
Palaeochoerus II 269, 271*; —Meissneri II
269*; - Waterhousi II 268*.
Palaeoconcha I 216, 217, 220, 225, 229.
Palaeocyclus porpita I 80.
Palaeocypris I 179; —Edwardi I 179*.
Palaeodictyoptera I 208.
Palaeodiscus I 157; —ferox I 157.
Palaeohatteria II 111, 112, 113; -longicau-
data II 112*.
Palaeolagus II 183.
Palaeomarsupialia II 164, 166.
Palaeomastodon II 298, 299*, 300, 302*;
— Beadnelli II 300; — parvus II 300.
Palaeomeryx II 276; —Feignouxi II 276*.
Palaeomutela I 220; — golovkinskiana 1 221*,
— irregularis I 221*; — Keyserlingi I 221*.
Palaeoniscidae II 38, 39, 40, 50, 54.
Palaeoniscus I 8; II 39, 46, 66; - - Freislebeni
II 38, 39*; — macropomus II 37.
Palaeophonus nuncius I 204, 205*.
Palaeopisthacanthus I 208.
Palaeopithecus II 226.
Palaeopropithecus II 159, 222.
Palaeoryctes puercensis II 176*.
Palaeoryx II 280.
Palaeospondylus II 16, 46; — Gunni II 16*.
Palaeostraca I 176, 184, 265.
Palaeosyops II 262*, 263*. 265; — major II 263*.
Palaeosyopsinae II 262, 264, 265, 324.
Palaeotapirus II 260*; — helveticus II 260*.
Palaeotheriidae II 240.
Palaeotherium I 4; II 240, 241, 242, 244, 262;
— crassum II 242*; — curtum II 241*;
— magnum II 241; — medium II 240*, 241*;
minus II 241.
Palapteryx II 160.
Palasterina I 171; — primaeva I 170, 171*.
Paloplotherium II 242; — annectens II 242;
— crassum II 241; —minus II 241, 242.
Paludina I 6, 243, 244; — casaretto I 243*;
— craiovensis I 243*; — Laskarevi I 243*;
— leiostraca I 243*.
Paludinidae I 243*.
Panochthus tuberculatus 11 217, 218*.
Panopaea I 214, 229; — peregrina I 229*.
Pantolambda II 320, 321; - cavirictus II 320*;
Pantolambdidae II 320.
Pantylus cordatus II 84.
Paradoxides I 186, 190; — bohemicus I 185*.
Parahippus II 236.
Paramylodon II 214, 215*.
Paramys II 180, 181, 182; —delicatus 11 180*,
181*; — Petersoni II 181; —primaevus 11
180; — robustus II 181*.
Parapithecus Fraasi II 224*.
Paraprolecanites I 269*, 272.
Parapronorites 1 269*, 271*, 272.
Parasmilia I 75; — centralis I 75.
Parasorex II 177.
Parasuchia II 120, 121.
Paratylopus II 286; - - Sternbcrgi 11 286*.
Paraxonia II 230.
Pareiasauria II 79, 80.
Pareiasauridae II 80, 95.
Pareiasaurus I 14; II 68, 80, 81, 83, 84, 95,
96*; — elegans II 81; —horridus II .81;
— Karpinskii II 81*; —tuberculatus II 81.
УКАЗАТЕЛЬ
359
Patella I 238, 239, 242, 248: - nitida I 238*;
—Raincourti I 238*.
Pa'tellidae I 238, 239.
Paterina labradorica I 99; — superba I 99,
100*.
Patlerina I 115*, 116.
Patriocetus II 206, 207*; — Ehrlich! II 207*.
Patriofelis II 185, 188, 203.
Pavlovia I 284, 285*.
Pecten I 214, 216, 231; — fibrosus I 232*.
Pectinidae I 230, 231.
Pectunculus I 219; — obovatus I 219*.
Pedipalpa I 205.
Pelagosaurus temporalis II 126.
Pelagothuria Ludwigi I 172; —natatrix I 172*.
Pelecyornis II 159.
Pelecypoda I 212,213.
Pelmatozoa I 121, 122, 135.
Pelobates cultripes II 75, 76*; — fuscus II 76*.
P.eloneustes II 101.
Pelosaurus II 59, 60.
Peltephilus ferox II 218.
Pelycodus II 220, 221.
Pelycosauria II 79, 85, 88, 151.
iPelycosimia II 121.
Penaeus Meyeri J 183.
Peneroplidae I 28.
Pennatulidae I 62.
Pentacrinidae I 148, 154.
Pentacrinus I 137, 138, 139, 149, 154; — ca-
put Medusae I 147*; —subangularis I 148*,
149.
Pentameridae I 102, 107.
Pentamerus I 112; —conchidium I 108; —ga-
leatus I 107, 108*.
Pentephylium I 135.
Pentremites I 132, 133*, 134, 135, 136; —ele-
gans I 132; —florealis I 132, 133*; —Go-
doni I 132*; —pyriformis I 132, 133*, —sul-
catus I 133*.
Pentremitidae I 132
Peramelidae II 170.
Peratherium II 170*, 171.
Perforata I 21, 68, 71, 73, 74, 75.
Peripatus I 174, 175*, 204.
Periptychus II 271; — rhabdodon II 270.
Perisiphonidae I 56*.
Perisphinctes I 281, 284, 287, 291; colubri-
nus I 284, 285*; — dorsoplanus I 285*;
— Martell! I 284, 291; — Pavlovi I 284;
— polyplocus I 285; — scopinensis I 285;
—submutatus I 285*; —Tiziani I 285, 286*.
Perisphinctidae Г 284, 290, 291.
Perissodactyla II 230, 232, 262, 332.
Permia I 87.
Perna I 232; — Sandbergeri I 234*.
Pernidae I 232.
Peronidella I 50; - - cylindrica I 50; — duraosa
I 50*.
Petalaxis I 84, 85®.
Petalia I 209.
Petalodus II 23.
Petromyzon II 17..
Peytoia Nathorsti I 61*.
Phacochoerus II 270.
Phacopidae I 193.
Phacops I 186, 187*, 193, — fecundus I 193*;
— latifrons I 193*.
Phaenoschisma I 133, 135.
Phalangiotarbi I 207.
Phaneroplettron II 29, 31, 32*; — Andersoni II
29*, 32.
Pharetrones I 38, 49, 50, 92.
Pharyngolepis II 9, 15*; — oblougus И 14.
Phascolomys II 170.
Phascolotherium II 165, 166*.
Phenacodus II 233, 234; - Wortman! II
233*.
Phillipsastraea I 84, 86; — radiata I 85*.
Phillipsia I 185, 186, 192, 193, 194, - gem-
muliforma I 192*.
Phoca II 201; —groenlandica II 201, 202*;
— pontica II 201, 202*.
Pholadidae 1 229.
Pholadomya I 214, 229; —canaliculate 1229*
— Candida I 229.
Pholadomyidae I 216, 229.
Pholas I 214, 229; — Lovesquei I 229*.
Phragmoceras I 259, — ventricosum I 259*.
Phragmoteuthis I 306; — bisinuata I 307*.
Phryganidae I 210.
Phrynosoma II 84.
Phyllocaridae I 176, 180, 181, 184, 187.
Phylloceras I 277; — heterophyllum I 277*.
Phylloceratidae I 276, 277, 278.
Phyllopoda I 176, 178, 181, 181.
Phyllospondyli II 58*, 59, 75.
Physa 1 250.
Physeter II 206; macrocephalus II 206*.
Physeteridae II 206.
Physoclisti II 45.
Physostomi II 45.
Phytosauridae II 121.
Phytosaurus II 121.
Piesetrobelus I 314.
Pinecoceras I 278; — parma Г 278*.
2в‘
360
УКАЗАТЕЛЬ
Pinacoceratidae I 278.
Pinna I 215, 232; —lanceolata I 234*.
Pinnipedia II 185, 201, 203, 209.
Pirittocrinus transitor I 144.
Pisces II 7.
Pithecanthropus II 227, 229.
Placentalia II 163, 175.
Placochelys II 104; — placodonta И 104*.
Placocystidae I 130.
Placocystis I 136; —forbesianus I 130.
Placodermi II 7, 8, 13, 15, 47.
Placodontia II 79, 102, 104, 151.
Placodus gigas II 103*.
Placoparia I 187*.
Placophyllia I 78.
Plagiaulacidae II 167, 172.
Plagiaulax II 169; —Becclesi II 167; — minor
II 167*.
Plagiodon I 222.
Plagioptychus I 227; —Anguilloni I 228*.
Plagiosternum II 72.
Planctoceras I 259.
Planorbis I 249, 250, 251; — corneus I 249*;
— Kleini I 249; — oxystoma I 249*, 251*;
— planorbiformis I 249, 251*; —scalaris I
249; —trochiformis I 249, 251*; —trochi—
formis elegans I 250*; — trochiformis pla
norbiformis I 251*; — trochiformis sulcatus
I 251*; —trochiformis trochiformis I 151*;
— turbiformis I 249.
Platax minor II 45.
Platecarpus II 118,120; — coryphaeus II 119*;
— simus II 119.
Platemys Spixii II 108.
Plateosaurus longiceps II 133*, 134.
Platyceras I 242; —infundibuliformis I 242;
— neritoides I 242*.
Platychelys II 107.
Platycrinidae I 150.
Platycrinus I 137, 138*, 150, 153; —triginti
dactylus I 150*.
Platygonus II 269.
Platylenticeras catenulatum I 279; —fulgens
I 279.
Platylenticeridae I 279.
Platyops Stuckenbergi II 67*, 68.
Platyostoma I 242; — niagarensis I 242*.
Platysomidae II 40, 41, 45.
Platysomus II 46;';—striatus II 37, 40, 41*.
Plecanium I 24, 25*.
Plectambonites I 105*.
Plesiadapis remensis II 220.
Plesiarctomys^II 180.
Plesictis II 194; —lemanensis II 194,
Plesiocetus Cuvieri II 207, 208*.
Plesiochelydae II 107.
Plesiochelys solodurensis II 107.
Plesiomeryx II 296.
Plesiometacarpi II 279.
Plesiosauria II 79, 99, 102, 114, 151.
Plesiosauridae II 100.
Plesiosaurus II 94, 101, 102; — dolichodermus..
II 101*; — Hawkinsi I 101*; — macrocepha-
lus II 101.
Plesiosiro I 207.
Plesioteuthis I 305; •— priscus I 302.
Pieuracanthidae II 20.
Pleuracanthus II 20*, 28, 51, 52.
Pleuraspidotherium II 297.
Pleurocystidae I 129.
Pleurocystis I 129*, 130, 136.
Pleurodictyum americanum I 66; — problema-
ticum I 66, 67*, 68, — stylophorum I 66(.
67.
Pleurodira II 105, 107, 108, 110.
Pleurojulus I 203.
Pleuromyidae I 229.
Pleurophyllum I 48*.
Pleurostoma I 48; — stellatum I 48*.
Pleurotomaria I 240; —Buchi I 240*.
Pleurotomariidae I 240.
Plicatula I 230; — pectinoides I 230*.
Pliohippus II 236, 237*; — fossulatus II 236;.
— supremus II 236.
Pliohyrax II 317.
Pliopithecus II 224; — antiquus II 226-
Plioplatecarpus II 120.
Pliosaurus II 101, 102.
Podocnemis II 107.
Podokesaurus II 136; —holyokensis II 135*..
Poebrotherium II 286, 287*; — Wilsoni II 285*.
Pogphagus II 282; —grunniens II 282, 283;.
— mutus II 282, 283.
Polyacrodus II 21.
Polycoelia I 80; —profunda 1 81*.
Polygnatha II 116.
Polymastodon II 167, 169; —attenuatus II
168*; — taaense II 168*.
Polypora I 91; — Goldfussi 1 91*.
Polupteridae II 34.
Ploypterus II 27, 34, 36, 53*, 57.
Polyptychites I 284, £86, 287; - Beani I'
286*; — gravesiformis I 286; — latissimus Г.
286; — Keyserlingi I 286, 287*; — ramuli-
costa I 286.
Polyptychitidae I 285, 286, 287-
УКАЗАТЕЛЬ
361
Polyrhizodus II 23; —concavus II 24*; — ros-
sicus II 24*.
Polysphenodon Miilleri II 113.
Polystomelia I 30*.
Polytremacis I 63.
Polyxenis I 203.
Popanoceras I 276; —multistriatum I 277*.
Porambonitidae I 102, 108.
Porcellia I 240; — Puzosi I 240*.
Porcelliidae I 240.
Porifera I 75.
Porites I 72; --incrustans I 72; — ramosa I
72*.
Poritidae I 72.
Porocrinus I 143; — conicus I 143*; —radia-
tus I 143*.
Porosphaera I 50.
Porospongia I 48. ,
Portax II 281*.
Potamites I 246.
Potamotherium II 195.
Poteriocrinus I 144, 145; --multiplex I 145*.
Praedentata II 129, 138.
Praecardiurn I 217; —fidens I 218*.
Prestwichia rotundata I 201*.
Primates II 219.
Primitia I 179.
Pristiograptus 1 59*.
Proaelurus II 197.
Proboscides II 297.
Probubalus II 281, 282.
Procalosorna minor I 210.
Procamelus II 286, 287* — gracilis II 286*.
Procervus II 279; — variabilis II 279.
Procolophon trigoniceps II 85.
Procolophonia II 85.
Procolophonidae II 85.
Prodaraelites I 272.
Prodremotherium II 276; — elongatum II 275*.
Productella I 103.
Productidae I 102, 110, 111.
Productus 1 102, 103, 106; — Cancrini I 103,
104*; —giganteus 1 103*, 111; —horridus
I 103, 104*; —- semireticulatus I 103*.
Proetidae I 192.
Proetus I 186, 187*, 192.
Proganochelys II 105, 107. — Quenstedti II
106*.
Progonoblattina 1 208; — helvetica I 209*.
Prolecanites I 269* 271, 272; —lunulicosta I
270*.
Prolecanitidae I 269, 275.
Promedlicottia I 272.
Promeles II 194.
Pronorites I 269*, 271, 272; — cyclolobus 1
270*.
Pronoritidae I 273.
Pro-ornithopoda II 133, 137, 138, 140, 160,.
161.
Proparia I 186, 187*, 193.
Prophyseter Dolloi II 206.
Propinacoceras I 269*,
Propliopithecus II 224, 225*; —Haeckeli II.
226.
Propteticus I 209.
Prorastomus II 319.
Proreptilia II 60.
Prororachus II 159; — inflatus II 158*, 159-
Prosauria II 79, 88, 111, 113, 115, 116, 119,
121, 147.
Prosaurolophus II 142*, 143.
Proscorpius I 204; - Osborni I 204, 205*.
Proselachii II 19.
Prosimiae II 219.
Prosobranchia I 237, 238, 248.
Prosqualodon II 205.
Prosthenops II 269.
Protaceratherium II 250.
Protagriochoerus II 272.
Protapirus II 260; — Douvillei II 260*; — ob-
liquideus II 260; — priscxis II 260*; — sim-
plex II 260.
Protaster I 168, 171; — Miltoni I 168*.
Proteroblastus Schmidti I 128*.
Proterochelys II 105.
Proterochersis II 107; —robusta II 107*.
Proterotherium II 333.
Proteus molassus II 45.
Proth\lacinus II 173; —patagonicus II 170„.
171*, 173.
Protoadapis II 220.
Protobelemnitidae I 314.
Protocaris I 177, 184; —Marshii I 177*.
Protoceras II 289; — celer II 289*.
Protoceratidae II 288.
ProtocMatops II 137, 146, 147, 175.
Protocetus atavus II 203, 204*.
Protocrinidae 1 127.
Prot >crinites oviformis I 127, 128*.
Protocrinus I 136.
Protogonia puercensis 11 233.
Protohippus II 235*, 236, 239.
Protolimulus I 202.
Protolycosa I 206; — anthracophila I 207»..
Protopharetra I 53*.
Protophasma Dumasii I 209*.
•362
УКАЗАТЕЛЬ
Protopterus II29,32% 33,48,51; — annectens II
33.
Protoreodon II 272; — parvus II 272*.
Protorosauridae II 114.
Prot rosaurus II 95, 114*, 115, 116, 121, 129.
Protoschizodon I 226.
Protosiren Fraasi II 318.
Protosphargis II 110; —veronensis II 110.
Protospongia I 45; — fenestrata I 45*.
Prot ispongidae I 45.
Prostega Copei II 110.
Protostegidae II 109.
Protozoa I 19, 36, 37.
Protrachyceras Archelaus I 275.
Protremata I 115.
Protriton II 58*, 59*, 60% 61, 65, 73, 75.
Protritonidae II 58, 59, 75.
Protungulata II 232.
Protylopus II 286; — Petersoni II 285*.
Protypotherium II 326; — australe II 326.
Proviverra II 191; —- Cayluxi II 191*.
Prozeuglodon atrox II 204.
Psammechinus I 156*.
Psammochelys Quenstedti II 106.
Psatnmodus II 23.
Pseudaelurus II 197*.
Pseudobe us I 315.
Pseudoberyx II 44.
Pseudocaninia I 81, 82*.
Pseudocentrophori II 61.
Pseudocreodi II 185, 187, 191.
Pseudol'oris II 222; — parvulus II 222.
Pseudomon otis I 231.
Pseudorhinolophus II 178.
Pseudosuchia II 120, 123, 129, 139.
Psiloceras I 280, 281, 284; —planorbis I 281*.
Psittacosaurus II 141*.
Psittacotheriuni II 211; —multifragum II 211*.
Pteranodon II 150, 151*.
Pteraspis II 8, 10, 11, 12; — Crouchi II 11;
— Gosselati II 11; — rostratus II 11*.
Pterichthys I 13 II 9, 13, 46; — Milleri II 13*.
Pterinea I 231.
Pterocodon I 35.
Pterodactyloidea II 149.
Pterodactylus II 150, 153; — antiques 11 150;
— spectabilis II 150*.
Pterodon II 185, 187, 188*; —africanus II-188-
Pterolepis II 9, 15*; —nitidus II 14*.
Pteropoda I 237, 251, 252.
Pterosauria II 79, 148, 156.
Pterygotus I 197, 198; II 15; II — auglicus I
197*; — bilobus I 197.
Ptilodus II 169; — gracilis II 168*.
Ptychites gibbus I 278*.
Ptychitidae I 278, 281.
Ptychodus II 23; —mammillaris II 23*.
Ptychoparia I 187*, —cordilleria I 187.
Pulmonata I 237, 244, 248.
Pulvinulina I 22.
Pupa I 250; —diliolum I 250*.
Putorlinae II 194.
Pycnodonti II 103.
Pycnodontidae II 41.
Pycnodus II 41, 49; —platessus II 42*.
Pygaster I 162, 163, 165; — umbrella I 163*.
Pygope diphya I 109, 110*.
Pygurus I 165; — royerianus I 165*.
Pyrotheria II 329, 330.
Pyrotherium II 330, 333; —Romeroi II 329*.
Pythonomorpha II 99, 117, 118, 120, 151.
Q.
Ouenstedticeras 1294,299; — Lamberti 1 295*;
Leachi I 294; — Mologae I 294; — rybinski-
anum I 294.
Quinqueloculina I 31*.
R.
Radiolaria I 19, 33.
Radiolites 1 228.
Radix I 249.
Rana II 76; — pictata II 74.
Ratitae II 154, 155, 156.
Receptaculidae I 50.
Receptaculites I 50; —Neptuni I 50*; —or-
bis I 50*.
Reptilia II 70, 77.
Requienia I 227; — ammonia I 227*.
Retiolites I 57*, 59;—Geinitzi I 59*.
Retzia I 114; —triganella I 114*.
Rhachitomi II 58*, 59, 63, 69.
Rhachitomus II 66.
Riacophyllltes I 277; —neojurensis 1 277;
tortisulcatus I 277.
Rliamphorhynclioidea II 148.
Rhampliorhynchus 11 149*; Muenstcri 11
149.
Rhaphidonema 1 50.
Rhaphistoma I 238.
Rheophax I 20*, 23, 25, 26.
Rhinellus furcatus II 44, 45*.
Rhineura Hatcher! II 116*.
Rhinobatis II 23; — mirabilis II 23*.
УКАЗАТЕЛЬ
363
Rhinocerinae II 255.
Rhinoceros II 255, 258; — bicornis II 245,
258; — etruscus II 256, 257, 258; — hemi-
taechus II 256, 258; — indicus II 245, 256;
- lasiotis II 245; —leptorhinus II 256, 257;
— megarhinus II 256; — Merckii II 229,
256*, 257; — pachygnathus II 256% 258;
— palaeindicus II 256; — sansaniensis II
255*, 256; — Schleiermacheri II 255*, 256;
— simorrensis II 255*; — simus II 245; —si-
valensis II 256; — sondaicus II 245;- — sumat-
rensis II 245; — tichorhuius II229, 257*, 258.
Rhinocerotidae II 245, 255, 330.
Rhinolophus II 178.
Rh’pidocrinus I 152; —• crenatus I 152*.
Rhizodontidae II 34, 35, 49, 59.
Rhizodopsis II 57, 59; — sauroidcs II 34.
Rhizodus II 34; — Hibberti II 34*.
Rhizomorina I 41, 44.
Rhizophyllum I 86.
Rhizopoda I 19, 20.
Rhizostomites 161; —lithographicus I 61*.
Rliombifera I 124, 143.
Rhopalobelus I 315.
Rrophalodon II 93*.
Rhynchocephalia I 13; II 79, 83, 111, 113,
114, 118, 120, 129, 133, 148,
Rhyncholepis If 9, 15*; — parvulus II 14,
15*;
Rhynchonella I 98, 106, 108, 111, 112* 117*.
cuboides I 107*; —Fischeri I 107*; —li-
vonica I 107; —loxiae I 107; —Meyendorfi
I 107*; — octoplicata I 106*, 107; — oxy-
optycha I 106*, 107; —Panderi I 107:
— personata 1107; — pleurodon I 107; —prin-
ceps I 107; — psitracea I 98*; — pugnus I
107; —varians I 107; —Wilsoni I 107.
Riynchonellidae I 102, 106, 110, 112.
Rhynchosauridae II 113.
Rhynchosaurus II 113*, —articeps 113.
Rhytina Stelleri II 318.
Richthofenia I 106; —lavreciaua I 106*.
Richthofeniidae I 102, 106.
Rimula I 239.
Rissoidae I 243.
Rodentia II 178.
Rotalia I 21*, 28; — Akneriana I 26; —so-
lida 1 26.
Rudistae I 215, 228.
Rugosa I 53, 78, 79, 87, 106.
Ruminantia II 274, 289.
Rustellacea I 100.
Rutiodon planirostris II 122.
S.
Saccamina I 20, 23, 26; — Carteri I 23, 24*.
Saccocoma I 143, 154; —pectinata 1 142*,
143; — tenella I 143
Sagatheriidae II 317.
Sageceras I 271, 272.
Sagenocrinus expansus I 153.
Sagenodus Copei II 31.
Sagillaria II 72.
Saiga II 280; —tatarica II 280.
Salena I 162; —petalifera I 162*.
Salenidae I 161, 162.
Sahno II 53*.
Salmonidae II 44.
Samotherium Boissieri II 287*, 288.
Sao 1 194; —hirsute I 189*.
Sapheosaurus incisivus II 113.
Sauranodon II 113.
Saurischia 11 129, 130.
Saurolophus II 143; —Osborni 11 142*.
Sauropoda II 129, 130, 131, 132, 133, 138,
143, 152, 160.
Sauropterygia 11 99, 103.
Saururae II 154, 155.
Saxicava arctica I 213*.
Scalaria I 245.
Scalariidae I 245.
Scaldicetus II 206; — patagonicus II 206.
Scalpellum I 179.
Scaphites aequalis I 294*; - compressus I
300; - - Conradi I 300; — spiniger I 264*,
293.
Scaphognathus crassirostris II 149.
Scaphonyx II 121.
Scaphopoda I 212, 252, 253.
Scarabaeides deperditus I 210*.
Scautnenacia II 29, 31, 32*, 33; —curta II 32.
Scelidosaurus II 129, 138, 144*. 161.
Scelidotherium II 214,
Scenella I 238.
Schiz.odonta I 215*, 217, 225.
Schizodus I 226, 230; - obscnrus I 227*.
Schizopoda I 183.
Schloenbachia I 296.
Schuchertia I 171: - stellata 1 170, 171*.
Schwagerina 1 31, 32; — princeps I 33*.
Scincosaurus crassus 11 62, 63*.
Sciuravus II 181, 182; — nitidus II 181*.
Sciuridae II 179.
Sciuroidea II 180,
Sciuroides Quercy II 183. 4
Sciurus II 180, 181.
364
УКАЗАТЕЛЬ
Sclerotnochlus Taylori II 124.
Sclerosaurus II 85; — armatus II 85.
Seorpionidae I 204.
Scurria I 238; — nitida I 238 *.
Scutella I 164; —subrotundata I 164*.
Scutoplan.es I 173.
Scylacosaurus II 91, 92*; — Sclateri II 91.
Scyllium II 18*.
Scyphocrinus I 154.
Scyphozoa I 36, 60.
Selachii II 19, 20.
Selenaria I 90, 95.
Selenodontia II 266, 271.
Seloris I 182; — paradoxa I 182*.
Semionotus Kapffi II 42.
Semnopithecus II 224; — entellus II 224: —pa-
laeindicus II 224.
Sppia I 254, 302, 304*; — oilicinalis I 302*.
Sepioidea I 255, 301, 302, 305.
Serpula I 118, 246; —convolute I 118; —gor-
dialis I 118; —Umax I 118; —sciabs I 118;
—spirulaea I 118.
Serpulidae I 118*, 246, 253.
Sertularia I 56.
Sertularidae I 56*.
Sesamodon II 92*; — Browni II 91.
Seymouria II 81, 82*, 95; —bailorensis II 82*.
Sicanites I 269*, 271.
Sidneva I 199; —inexpectans I 199*.
Siliquaria I 247.
Sipiaedosaurus II 115.
Simbirkskites I 287, 299; — coronatiformis j
288*;— Decheni I 289*; — discofalcatus I
290; — elatus I 288*, 289; — inverselobatus
I 289*; — progrediens I 289*; — versicolor
I 287*, 289, 290.
Simiae II 219, 220, 222.
Simiidae II 222, 225, 226.
Simosaurus II 100.
Simplicidentata II 178, 180.
Sinopa II 185, 188; —agilis II 188; —aethio-
pica II 188.
Sinupalliata I 217, 220, 224, 229.
Siphonaria I 248; — crassicostata I 248*.
Siphonariidae I 248.
Siphonia I 41, 42; —pirum I 42; —tulipa I
42 s.
Siphonostomata I 237, 242, 245.
Siphonotreta I 101; —verrucosa I 101*.
Siphonotretidae I 100, 115.
Sirenia II 317, 318.
Sismondae Bellardi I 300, 301*.
SivapiAecus indicus II 226*.
Sivatheriinae II 288.
Sivatherium II 288; —giganteum II 288*.
Sinimonia I 197, 198*.
Smilodon II 197*, 198*, 199.
Solarium I 236*.
Solecurtus I 225.
Solen I 225; — vagina I 226*.
Solenidae I 216, 218.
Solenomya I 218; — vetusta I 218*.
Solenopsis I 218, 225.
Sorex pusillus II 177.
Spalacinae II 183.
Spalacotherium II 166.
Sparnodus II 46.
Spatangidae I 166, 167.
Spatangopsis I 60*.
Spatangus I 166; —Desmaresti I 167*.
Spatobatis II 23.
Sphaeronites globosus I 127*.
Sphaerospongia tesselata I 51.
Sphenodon II 111, 116; —punctatus II 78*.
Spinosaurus aegypticus II 138.
Spiractinella I 46.
Spirialis I 251.
Spirifer I 112, 114, 117; —Archiaci I 113; —
fasciger I 114*; —Kleini I 113*; —macro-
pterus I 113*; —mosquensis I 103*, 113*,.
114; —rectangulus I 114; —speciosus I
113*; —Strangwayi I 114; —striatus I 114;
— superbus I 113; —supramosquensis I
114; —Trautscholdi I 114; —trigonalis
114; —undulatus I 113.
Spiriferidae I 102, 110, 112, 113.
Spiriferina I 117.
Spirigera I 112, 114; — ambigua I 114*.
Spirillina I 20, 24.
Spiroceras I 293.
Spirocyathidae I 53.
Spirocyathus I 53; — atlanticus I 51*.
Spiroloculina I 31*.
Spiropora I 92, 94; — verticellata I 92*.
Spirula I 254, 303, 304*.
Spirulidae I 303, 315.
Spirulirostra I 304*; — Bellardi I 303.
Spondylidae I 230.
Spondylus I 215*, —spinosus I 230*; —te-
nuispina I 230.
Spongia I 36, 38.
Spongilla I 40.
Spumellaria I 35; — discoidea I 34*; — sphae-
roidea I 34*.
Spyridiocrinus I 152.
Squaloceti II 205.
УКАЗАТЕЛЬ
365
Squalodon II 205, 206; — bariensis II 205*.
Squamata II 74, 79, 115, 120.
Squatina II 23, 53*.
Squatinidae II 19.
Stauria I 79; — astraeiformis I 84*.
Staurocephalus 1 194,195; —Murchison! I 194*.
Stauroderma I 48.
Staurosphaera I 34*.
Stegocephali I 13; II 30, 57, 58*, 59, 63, 65,
67, 68, 69, 70, 72, 73, 74, 75, 80, 81, 82,
84, 95, 96, 129.
Stegochelys dux II 106*.
Stegodon Cliftii II 308*; — insignis II 308.
Stegodontidae II 307.
Stegosauria II 129.
Stegosauridae II 138, 143, 147.
Stegosaurus II 129, 138, 144, 161; —ungu-
latus II 144*.
Stegotherium II 216; —tesselatum II 216*.
Stellispongia I 49, 94; — varlabilis I 49*.
Stelmatopora I 90.
Stemmatocrinus cornuus I 144*.
Steneofiber II 182.
Stenoplesictis Cayluxi II 194*.
Stenopterygius II 98*.
Stenosaurus Larteti II 126.
Stephanoceras I 282, 286, 295; •— Braikenridgi
I 282; — coronatum 1 282*, 286; - hymphri-
esianum I 282*.
Stephanoceratidae I 282, 294.
Stephanocrlnus I 142, 154; —angulatus 1142*.
Stephanophyllia I 74; — elegans I 72, 73*
Stephanospondylus pugnax II 84.
• Stereospondyli II 59, 69, 72.
Sterrholophus II146.
Stomatopora I 92, 93; - - dichotomy I 92*;
— dichotomoides I 95.
Stomechinus I 161; —denudatus I 161*.
Strabops Thacheri I 198.
Streptelasma I 81; — europaeutn I 82*.
Stringocephalidae I 102, 109.
Stringocephalus I 109; — Burtini I 110*.
Stromatopora I 55, 94; — eifeliensis I 55;
— polyostiolata I 55*; — tuberculata I 55.
Stromatoporella I 55.
Stromatoporidae I 55.
Strophalosia I 103;—horyescens I 101*.
Strophomena I 103, 105, 106; - rliomboidalis
I 104*.
Strophomenidae I 102, 103, 115.
Struthiomimus II 135*; —altus II 137*.
Struthionidae II 159.
:Stylaster I 55.
Stylina Delabechei I 77, 78*.
Stylinodon II 211, 212.
Stylinodonta II 211.
Styiiola I 251, 252; — clavulus I 252*; — recta
I 252*; — striatula I 252*.
Stylodontidae II 42.
Stylonurus I 198.
Stylophora I 75; — subreticulata I 75*.
Stylophoridae I 75.
Styracosaurus II 146.
Styracoteuthis orientalis I 310*.
Subpronorites I 272.
Subungulata II 331, 332.
Subursi II 200.
Suidae II 266, 268, 269, 270, 271, 290. 293.
Suinae II 269.
Suprasulcatl I 315.
Sus II 21’8, 269, 270, 291*; — erymantheus II
269, 270*.
Sycones I 49, 54.
Symbatocrinus I 138*.
Symborodon II 265.
Synapsida II 78, 79. 85, 88, 96.
Synaptidae I 173.
Synaptula I 173; —hydrifortnis I 173*.
Syncarida I 182.
Syringocnetna I 53*.
Syringocnetnidae I 53.
Syringopora I 64, 65; — gracilis I 65*; — pa-
rallels I 65*.
Systemodon II 246, 259, 261; — semihianus II
246; —tapirinus II 246*. 247.
T.
Tabulata I 62, 64, 68, 92, 94.
Tapes I 224; — gregaria I 225*.
Tapiridae II 259.
Tapirinae II 259.
Tapirus arvernensis II 260; — priscus II
260*.
Tarsiidae II 222.
Tarsius II 221, 222.
Taxocrinus I 153; -- niultibrachiatus I 153*.
Taxodonta 1 214, 215*, 217, 218.
Teleometacarpi II 279.
Teieosauridac II 126.
Teleosaurus II 127; - cadomeusis 11 126*.
Teleostei II 36, 38, 43.
Teleostomi II 8, 27, 28.
Telerpeton II 85; — elginense II 85*.
Tellina I 225; —planati I 226*.
Tellinidae I 225.
366
УКАЗАТЕЛЬ
Telmatotherium II 263*, 265; —cornutum II
263; — diploconutn II263; — vallidens II 263.
Telotremata I 115.
Temnoptychites I 286, 287*, 290.
Temnospondyli II 58*. 59, 63, 66, 72, 75.
Tentaculites I 252; — acuarius I 252*; —orna-
tus I 252*.
Terebella lapilloides I 118.
Terebratella dorsata I 109*; — rubicunda I
116*.
Terebratula I 98*, 108, 109, 117; —biplicata
I 109*; — chilensis I 109; —coarctata 1109;
elongata I 109*;—fischeriana I 108*;—glans-
fadea I 109*; — gracilis I 109*; — obesa I
108*; — substriata I 109; - vitrea I 109;
— vulgaris I 108*, 109*.
Terebratulidae I 102, 105, 108, 110, 112, 116*,
117.
Terebratulina I 109; - gracilis I 109*.
Teredina I 229; — personate I 229*.
Teredo I 229.
Testicardines I 102.
Testudinata II 79, 104.
Testudininae II 109.
Tethyopsis Steinmanni I 40, 41*.
Tetrabelodon II 302, 307, 310; —angustidens
Il 300, 302; — arvernensis II 302; — dino-
therioides II 307; longirostris II 301; —pyg-
maeus II 300.
Tetrabranchiata I 254, 255, 257.
Tetracidaris I 160. Tetracladina I 41, 44. Tctraclaenodon II 237*; — puercensis II 232*,
233, 234, 246. Tetracorallia I 62, 69, 70, 71, 76, 77, 78, 79,
80, 86. Tetractinellidae 1 40, 41. Tetragraptus I 58; —Bigsbyi I 59; — fruti-
cosus I 57*, 59. Tetrapoda II 55. Tetraselenodon II 2'4. Tetraxoniae I 38, 40. Teuthoidea I 301. Teutomanis II 216. Textularia I 24, 28; carinata I 24*.
Textularidae I 24, 28. Thaecocystis I 123. Thalassemydidae II 108. Thalassemys II 110; —marina II 108*
Thalattosauridae II 118.
Thalattosaurus II 117; —Alexandrae II118*.
Thalattosuchia II 127.
Thamnastraea I 74, 84, 86; — prolifera I 74*.
Thamnastraeidae I 74, 75.
Tharsis II 44.
Thaumatosaurus II 102; — indicus II IOIl
victor II 101.
Thecidea I 105; —digitata I 105; —mediter-
ranea 1105; —papillata I 105, 106*; — ver-
micularis I 105.
Thecididae I 102, 105.
Thecocystidae I 123.
Thecocystis sacculus I 123*.
Thecoidea I 122.
Thecosmilia 176; —trichotoma I 76*.
Thecospira I 111, 112*; —Haidingeri I 111*.
Thelodus I 13; II 8, 9, 10, 11, 27; — Pagei II
9, 10*, 11; —scoticus II 9*.
Theocodon II 244*.
Theonea aurantium I 93*.
Theriodesmus II 90.
Theriodontia II 88, 91, 95, 165, 173.
Theromora II 88.
Theromorpha I 13, 14; II 79, 80, 88, 95, 103,
162.
Theropoda II 129, 130, 133, 134, 137, 138,
160.
Thoatherium II 244*, 245.
Thomisus I 208.
Thrissops II 44.
Thyestes II 9, 12; —verrucosus II 12*.
Thylacinus II 173.
Thylacoleo II 170, 171, 172*.
Tiarechinus I 159; — princeps I 159*.
Tillodontia II 179, 180, 211.
Tillotherium fodiens II 179*.
Timanites acutus I 267, 268*.
Tinoceras II 323.
Tirolites I 274; — cassianus I 273; — Smiria—
gini I 273*.
Tissotia Fourneli I 298*.
Titanophasma I 209.
Titanosaurus II 132.
Titanotheriidae II 262, 320, 324.
Titanotheriinae II 262, 264, 265, 324.
Titanotherium II 265; — platyceras II 265;.
— ratnosum II 265; — robustum II 264*.
Tomistoma 11 127; — africanus II 127.
Tornoceras simplex I 267*.
Torosaurus II 146.
Toxastcr I 166.
Toxochelys Bauri II 110.
Toxodon II 326, 327, 328; — paranensis IL
327*, 328*.
Toxodontia II 326, 328.
Toxodontidae II 326.
УКАЗАТЕЛЬ
367
Tracheata I 175, 202, 203.
Trachodon II 129, 138, 142*, 143; — amu-
rensis II 141*, 143; —mirabilis II 142.
Trachyceras Archelaus I 275; — austriacum I
275.
Trachyteuthis I 302.
Trachytidae I 275.
Tragoceras II 280.
Tragulidae II 271, 283, 289, 290.
Tragulus II283,284; — meminna II285; — Napu
II 285; — sivalensis II 285.
Tremataspis II 12; — Schmidti II 12*.
Trematocystis I 127*.
Trematops II 80.
Trematopsis II 67.
Trematosaurus II 71; —Brauni II 71.
Triacanthodon II 166, 170, 171.
Triacrinidae I 154.
Triacrinus altes I 141; —astus 1 141*; —fla-
gellifer I 141*.
Triarthrus I 186.
Triassochelys II 105; — dux II 105, 106*.
Triaxonae I 38.
Tricentes II 185; — subtrigonus II 185*.
Triceratops II 128, 129, 138, 145*, 146, 161;
— flabellatus II 145*.
Trichosurus II 172.
Triconodon II 165, 166*; —mordax II 166*,
Triconodonta И 164.
Tricuspidata II 164.
Trigonia I 215*, 225, 226; — costata I 226*.
Trigonias Osborni II 248*, 249, 252, 253*.
Trigoniidae I 225.
Trigonoscorpio I 208.
Trilobitae I 184, 185, 202.
Triloculina I 31*.
Trimerella Lindstroemi I 100*.
Tritnerellidae I 100.
Tritnerorachis II 67, 80; —insignis II 67.
Trinacromerum II 102*; — Osborni II 102.
Trinucleus I 186,187*, 189, 195; —Goldfussi I
189*.
Trionychia II 105, 110.
Trionyx II 106; —styriacus II 110.
Tripriodon II 167, 168.
Triquetra I 222.
Triton I 247.
Tritonidae I 247.
Tritubercuiata II 164, 166.
Tritylodon longaevus II 165*.
Troastocrinus I 135; —pyriformis I 133.
Trochammina I 20*, 24, 26.
Trochidae I 240, 243.
Trochoceras I 260; — optatum I 260*.
Trochocyathus I 76; — mitratus I 76*.
Trochocystis I 130; —bohemicus I 131*.
Trocholites dcpressus I 251*.
Trochosmilidae I 75.
Trochus I 240; —podolicus I 241*.
Trogontherium II 182.
Tropites subbullatus I 275*.
Tropitidae I 275.
Trucifelis II 197*.
Truncatula I 92; — repens I 93*.
Tryblidium 1238; —reticulatum I 238*'
Tubicola I 118.
Tubipora I 65; — mu аса I 63*; — purpurea 1.
63, 64, 65.
Tubulariae I 54.
Tubularidae I 5:.
Tubuliporidae I 92.
Tunicata II 1.
Tuponla lineata I 40.
Turbinidae I 240, 243.
Turbinolia I 75, 80, 86; — Bowerbanki J 75*.
Turbinolidae I 75, 76*.
Turbo I 240; — puschianus I 241*.
Turbonitella I 243.
Turrilites I 297; — catenatus I 297*; — poly-
plocum I 297.
Turritella I 245, —imbricataria I 245*; —te-
rebra (turris) I 245*.
Turritehidae I 245, 246.
Tylopoda II 285.
Tylosaurus II 118,119,120; — proriger II119*.
Typotheria II 325.
Typotheriidae II 325.
Typotherium II 326; — cristatum II 325*, 326-
Tyrannosaurus II 129; —rex II 135*.
U.
Udenodon II 95; — pusillus II 94; —Venju-
kovi II 94.
Uintacrinus socialis I 149*.
Uintatherium II 323, 324, 332; —mirabile II •
322*, 323*.
Umbrellidae I 248.
Uncites 1 114. i ,
Undina П 36; penicillata II 36*.
Ungula I 99.
Ungulella I 99.
Unio I 222, 223; - atavus I 222*; — flabella-
l.us I 222*.
Unionidae I 220.
Uraster rubens I 169*.
36$
УКАЗАТЕЛЬ
Urodela II 55, 56, 73, 76.
Uronetnus II 29, 31, 32*, 33; — lobatus II 33.
Ursavus II 200.
Ursidae II 184, 192, 200, 201.
Ursus II 186*, 200*; — americanus II 200;
— amplidens II 200; — arctos II 200; —arver-
nensis II 200; — gigas II 186; — spelaeus II
199*, 200.
V
Varanidae II 118.
Varanops II 86, 87.
Varanosauridae II 86.
Varanosaurus II 86, 87, 95; — acutirostris II
86*; — brevirostris II 86*.
Varanus II 86, 120.
Vauxia I 47; — dignata I 46*.
Vauxinidae I 47.
Veloceratosaurus II 135.
Venericardia I 224; —planicosta I 224*.
Veneridae I 224.
Ventriculites I 47; — pedester I 47*.
Ve.driculitidae I 47.
Venus I 215, 224; — Haidingeri I 214*.
Vermes I 118.
Vermetidae I 246.
Vermetus arenarius I 247; — Intordus I 247;
lutnbricalis I 246*.
Vermilinguia II 210, 215.
Verruculina I 44*.
Vespertilio II 177.
Vincularia virgo I 95*..
Virgatites I 284, 285; —Pallasi I 285, 286*;
virgatus I 286*.
Viverra II 196; — antique II 195; — sitnpli-
cidens II 195.
Viverravus II 195; —mimitus II 190.
Viverridae II 184, 190, 195, 199.
Vivipara I 243.
Vlasta I 217; — bohemica I 218*.
Voluta I 247.
Volutidae I 247.
Vulpavus II 190, 192, 193; — Hargeri II 189*.
W.
Waagenoceras 1 276.
Waldheimia I 117; —flavescens I 96*, 97*,
109; —fischeriana I 108*.
Waptia I 181, 187; —fieldensis I 181*.
Willemoesia I 184.
Wiwaxia I 120.
Woodocrinus I 137*, 145.
Worthenella cambria I 119*.
X.
Xenacanthus II 20.
Xenarthra II 209, 212, 216.
Xenodiscus nivalis I 273; — Schmidti I 273*.
Xiphodon I 4; II 291*; — gracile II 292, 295*.
296.
Xiphodontinae II 292, 295, 296.
Xiphosura I 196, 197, 200, .202.
Xylobius I 203; — mazonus ( 204*.
Y.
Yoldia I 218; — actica I 218*.
Z.
Zalambdodonta II 175, 176*.
Zalambdolestes II 175.
Zaphrentidae I 79, 80, 83, 87.
Zaphrentis I 81, 83. 86; —intermedia I 82*.
Zatrachidae II 68.
Zatrachys microphthalmus II 68*.
Zeuglodon II 203, 204, 205, 207; — Isis II
204*; — Osiris II 204.
Ziphiidae II 206.
Ziphiodelphis II 206.
Zygosaurus II 66.
Zygospira I 111.
Zynodictyum I 34*-
Zyocrinus I 135.
ОГЛАВЛЕНИЕ
Введение
Стр.
1
. ОПИСАТЕЛЬНАЯ ЧАСТЬ.
Класс п е р в ы й. Рыбы — Pisces ....................... 7
Подкласс!. Placodermi............................................ 8
Отряд 1. Heterostraci....................................... 9
„ 2. Osteostraci’:'.................................... 11
„ 3. Antiarcha........................................• 12
„ 4. Anaspida.......................................... 13
Подкласс II. Cyclostomi......................................... 15
Подкласс III. Chondropterygii (Elastnobranchii)................ 18
Отряд 1. Proselachii (Ichthyotomi)......................... 19
„ 2. Seiachii.......................................... • 20
„ 3. Batoidei.......................................... 23
„ 4. Acanthodi......................................... 24
Подкласс IV. Holocephali ........................................ —
ПодклассУ. Arthrodira........................................... 25
Подкласс VI. Teleostomi......................................... 27
Отряд 1. Dipnoi.......................................... 28
„ 2. Crossopterygii.................................... 34
„ 3. Actinopterygii.....................•.............. 36
Подотряд Chondrostei.................................. 37
„ Hemiostei....................................... 38
„ Teleostei....................................... 43
ТИП TETRAPOD A.
Класс второй. Земноводные — Amphibia................................... 55
Подкласс!. Stegocephali .......................................... 57
Отряд 1. Phyllospondyli ..................................... 59
» 2. Lepospondyli....................................... 61
„ 3. Temnospondyli....................................... 63
, 4. Stereospondyli...................................... 69
Подкласс II. Urodela.............................................. 73
Подкласс III. Anura............................................... 74
Класс третий. Пресмыкающиеся — Reptilia................................. П
Подкласс!. Ana psida.............................................. 80
Отряд Cotylosauria (Pareiasauria)............................. —
Подкласс II. Synapsida............................................ 85
Отряд 1. Pelycosauria......................................... —
„ 2. Theromorpha (Theromora)............................. 88
VI
ОГЛАВЛЕНИЕ
Стр.
Подотряд 1. Theriodontia...................................... 88
„ 2. Deuterodontia..............•..................... 91
„ 3. Dicynodontia..................................... 93
Отряд 3. Ichthyosauria............................................ 96
„ 4. Plesiosauria (Sauropterigia).............................. 99
„ 5. Placodontia............................................. 102
, 6. Testudinata...............................................104
Подотряд 1. Amphichelydiae..............v...................*.105
„ 2. Pleurodira......................................107
„ 3. Cryptodira..................................... 108
„ 4. Cheloniidea.....................................109
5. Trionychia....................................... 110
Подкласс Diapsida.........................................................Ill
Отряд 1. Prosauria (Rhynchocephalia).............'.................. —
„ 2. Squamata (Lepidosauria).....................................115
Подотряд 1. Lacertilia......................................... —
2. Ophidia......................................... 116
Отряд 3. Pytonomorpha ..............................................117
4. Crocodilia................................................120
Подотряд 1. Parasuchia........................................ 121
, 2. Pseudosuchia.....................................123
„ 3. Mesosuchia и 4. Eusuchia..........................124
Отряд 5. Dinosauria................................................. 128
Подотряд 1. Sauropoda................................... .... 130
„ 2. Theropoda (Pro-ornithopoda)......................133
„ 3. Praedentata (Ornithischia).......................138
Распространение динозавров...............................................147
Отряд 6. Pterosauria............................................. 148
Подотряд 1. Rhamphorhynchoidea................................. —
„ 2. Pterodactyloidea................................ 149
Класс ч e т в е р т ы й. Птицы — Aves..........................................153
Подкласс I. Saururae.................................................... 154
„ П. Ornithurae.................................................. 155
Отряд 1. Ratitae (Odontolcae)......................................156
2. Carinatae (Odontormae)................................... 157
„ 3. Anodontornae..............................................158
Класс пятый. Млекопитающие — Mammalia........................................ 162
Подкласс!. Monotremata................................................. 163
„ II. Marsupialia...................................(..............164
Отряд 1. Palaeomarsupialia.................. . .................... —
Подотряд 1. Triconodonta...................................... —
A. Tricuspidata....................................... —
В Trituberculata........................................166
Подотряд 2. Multituberculata (Allotheria).....................167
Отряд 2. Marsupialia vera ........................................ 169
Подкласс III. Pla c ent alia.............................................175
К or о T H Ы E.
Отряд 1. Insectivora.............................................. 175
» 2. Chiroptera............................................... 177
, 3. Rodentia................................................. 178
ОГЛАВЛЕНИЕ
VII
Стр.
Подотряд 1. Simplicidentata ..................................180
„ 2. Duplicidentata....................................183
Отряд 4. Carnivora....................................................184
Подотряд 1. Creodonta. Acreodi................................185
„ 2. Creodonta. Pseudocreodi ..........................187
„ 3. Palaeocarnivora (Eucreodi) ....'..................189
4. Carnivora vera (Fissipedia) ., . . 192
Отряд 5. Pinnipedia............................................... 201
„ 6. Cetacea....................................................203
Подотряд 1. Archaeoceti........................................ —
, 2. Odontoceti........................................205
„ 3. Mistacoceti..................................... 206
4. Delphinoceti.................................... 208
Отряд 7. Edentata.....................................................209
Подотряд 1. Ganodontia (Stylinodontia)........................211
„ 2. Bradypoda.........................................212
„ 3. Vermilinguia......................................215
4. Dasypoda..........................................216
A. Dasypoda.....................’.........................—
B. Glyptodonta..........................................217
H о г о т н ы Е.
Отряд 8. Primates.....................................................219
Подотряд 1. Piosimiae.......................................... —
„ 2. Simiae.............................................222
Копытные.
Отряд 9. Perissodactyla...............................................232
, 10. Artiodactyla.........................................266
Подотряд 1. Bunodontia ......................................... —
„ 2. Selenodontia.......................................271
Ruminantia ..............................................274
Подотряд 3. Bunoselenodontia.................................. 290
Отряд 11. Proboscidea.................................................297
Подотряд 1. Mastodontidae......................................298
А. Бугорчатозубые мастодонты ............................ —
В. Гребнезубые мастодонты...............................303
Подотряд 2. Elephantidae.......................................307
Плейстоценовые слоны.....................................309
Отряд 12. Sirenia.....................................................317
„ 13. Amblypoda..........................................320
» 14. Notungulata.............................................. 324
Подотряд 1. Typotheria.............................• • .... 325
„ 2. Toxodontia........................................326
„ 3. Nesodontia ..................................... 327
„ 4. Pyrotheria........................................329
Отряд 15. Subungulata............................................. 331
Изменение конечностей у некоторых копытных и ко-
го т н ы ................................................................332
Развитие различных групп копытных...............................334
Указатель к / и II частям................................................ 337
ОТПЕЧАТАНО!
в 1-й Образцовой типографии
Гиза. Москва, Пятницкая, 71.
Главлит.А-6315. Н.ЗО. Гиз 18465.
Заказ № 2034. Тир. 3 000 экз.