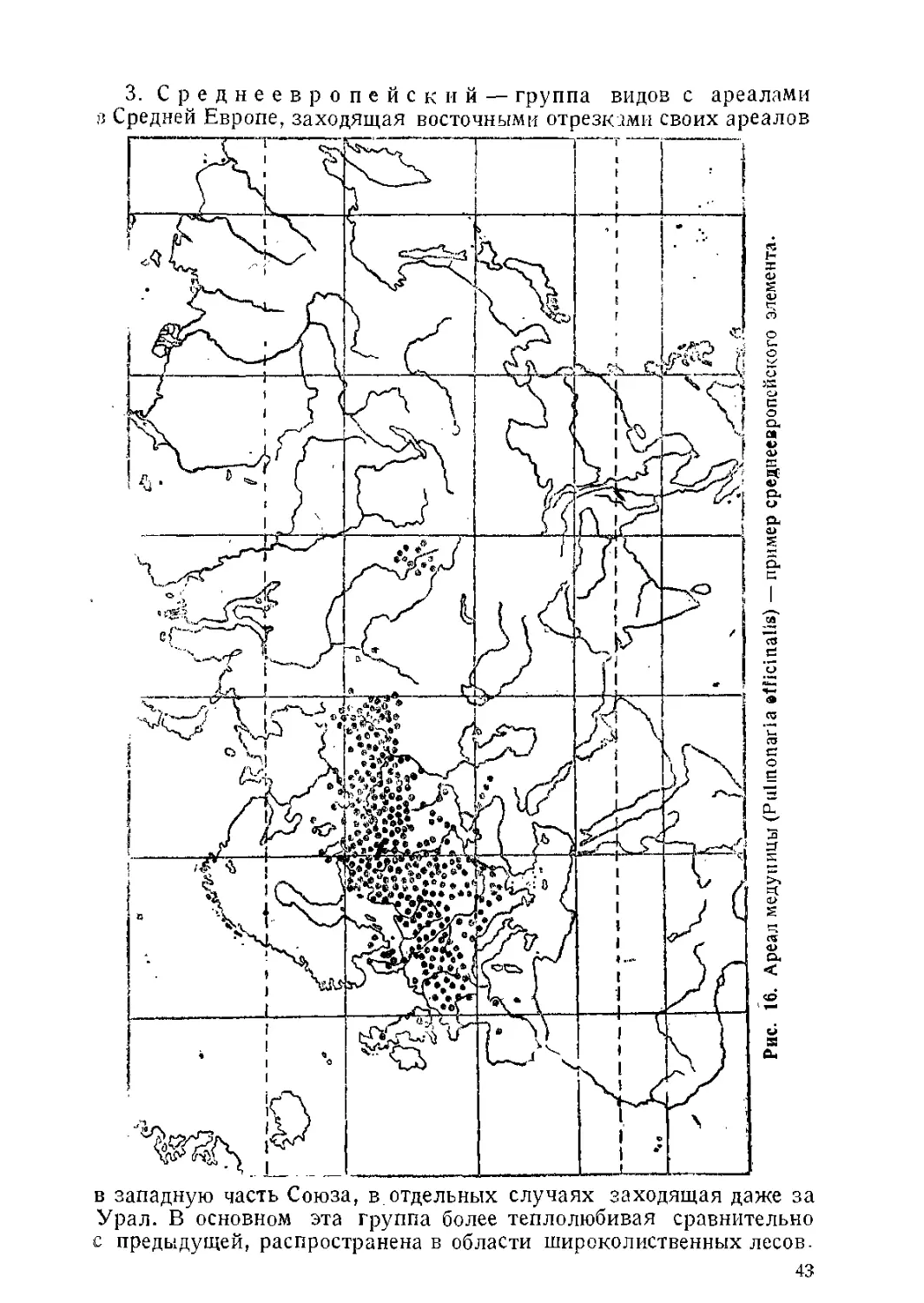
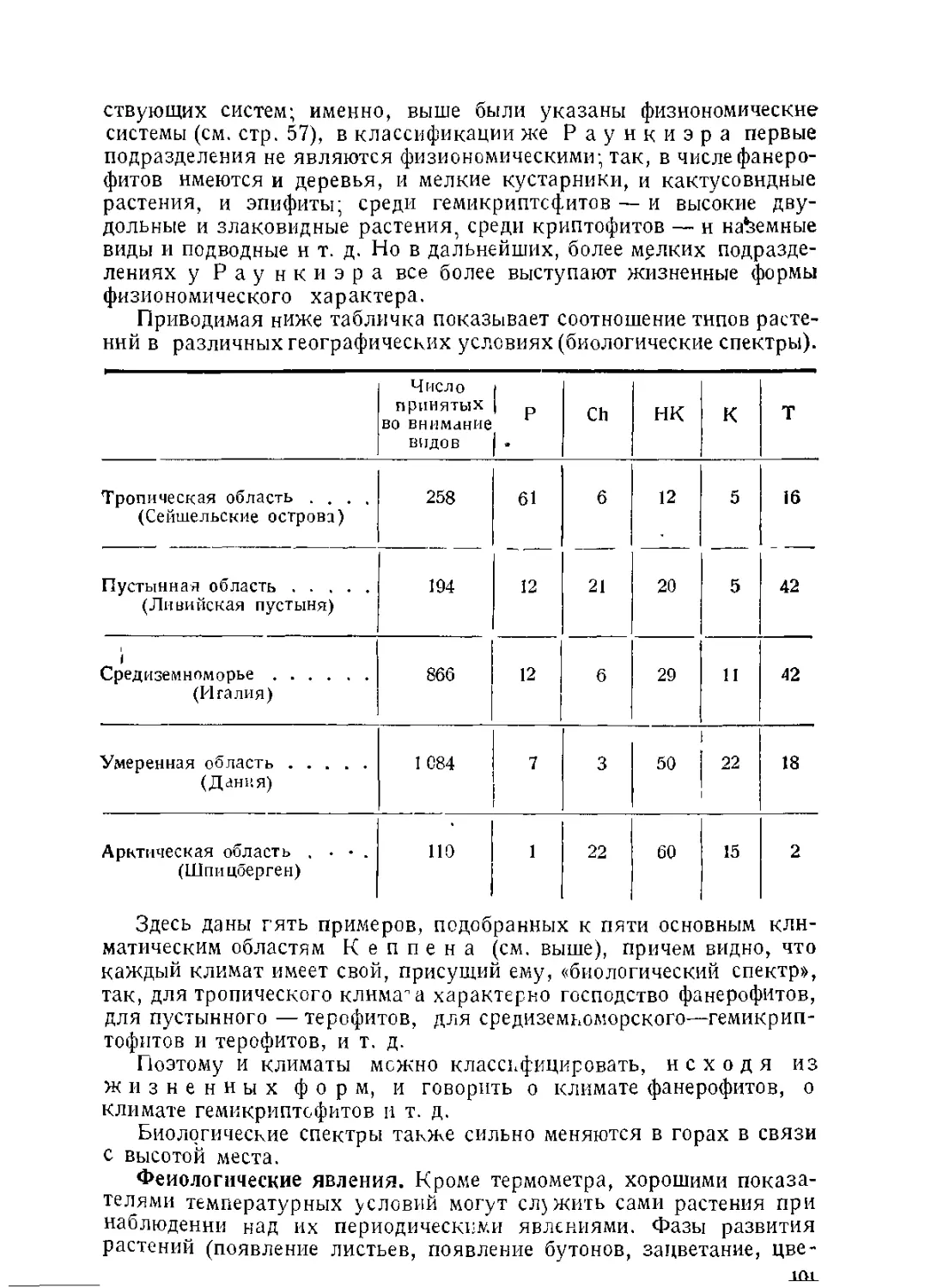
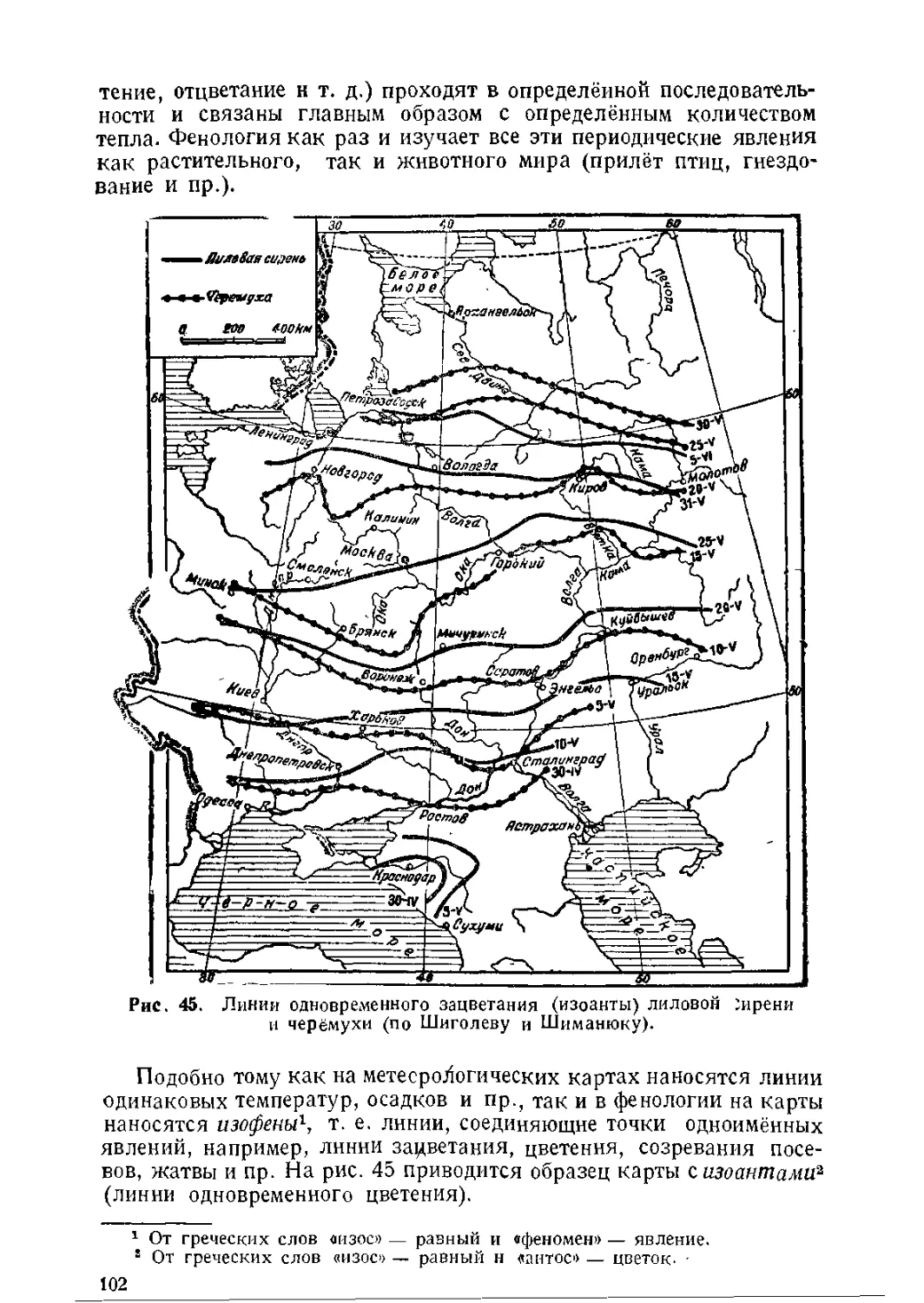
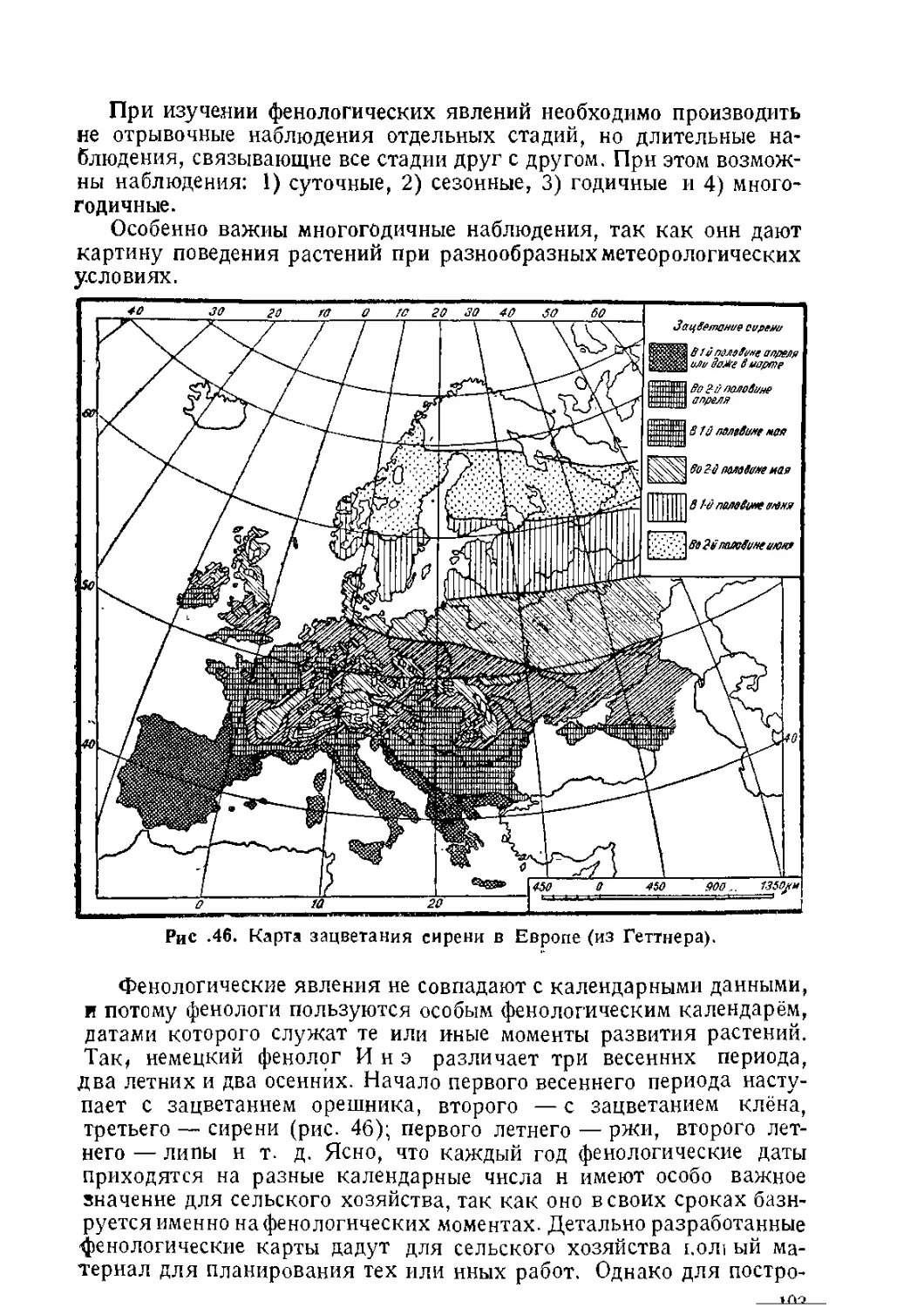
/

































































































































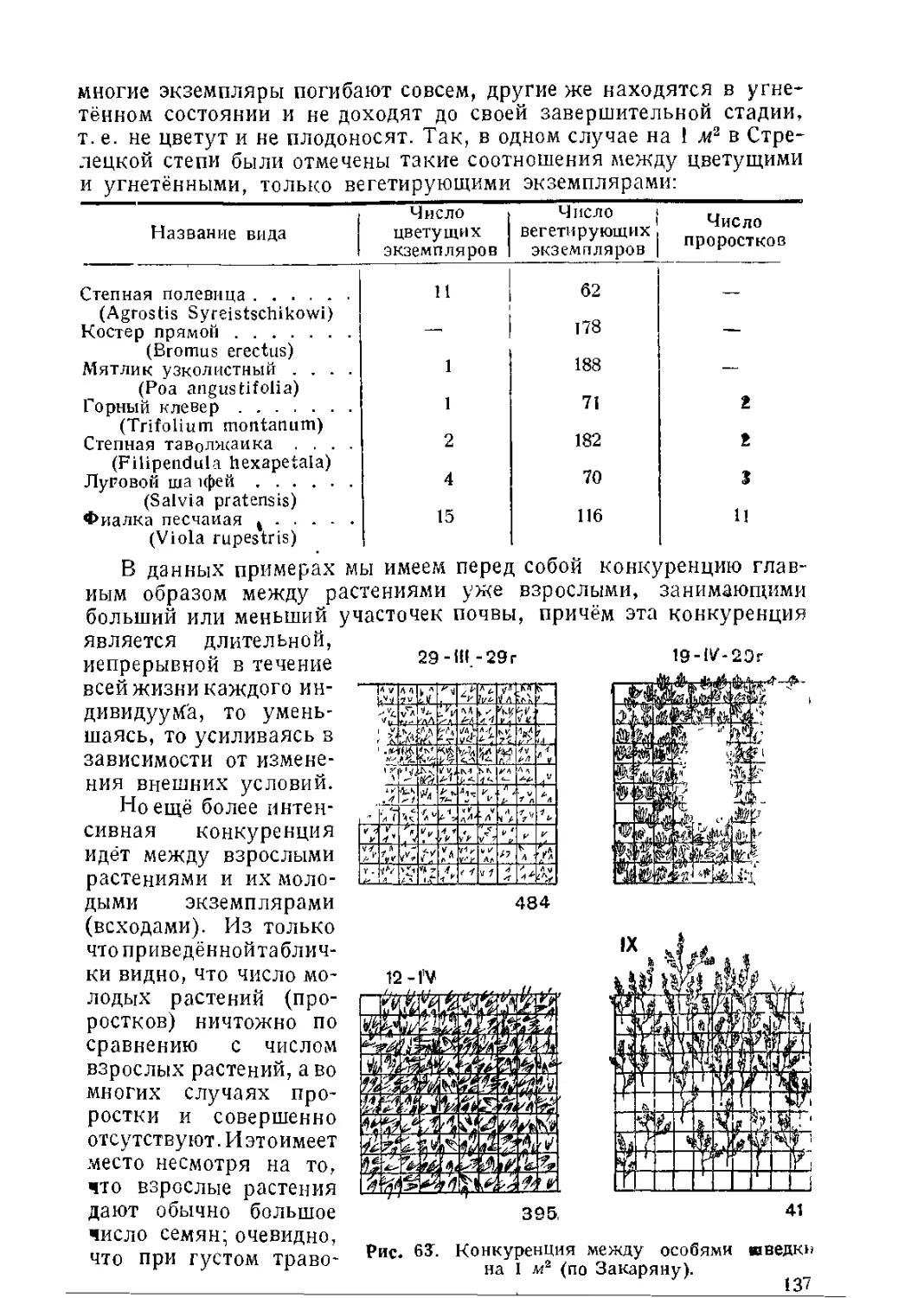

















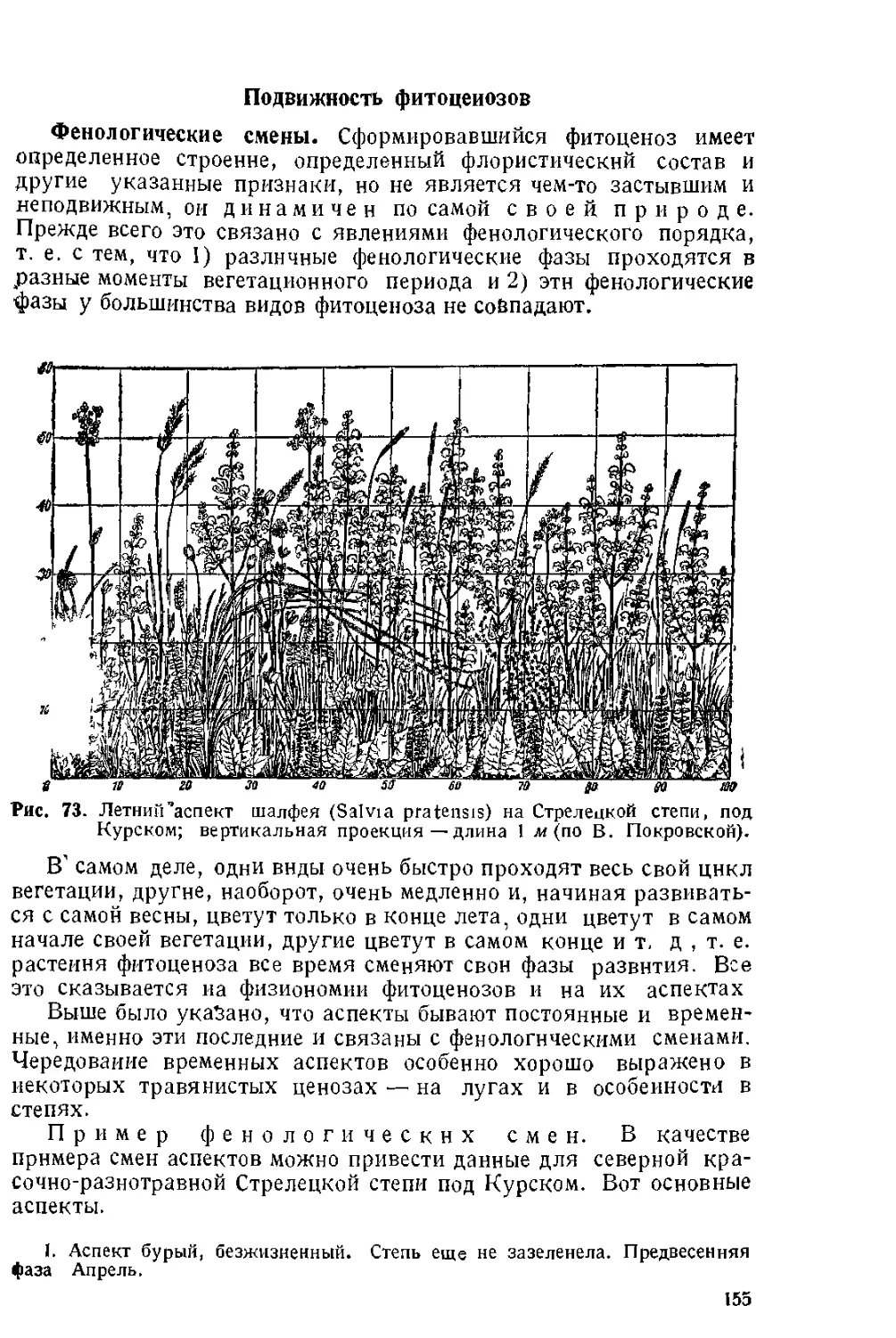












































































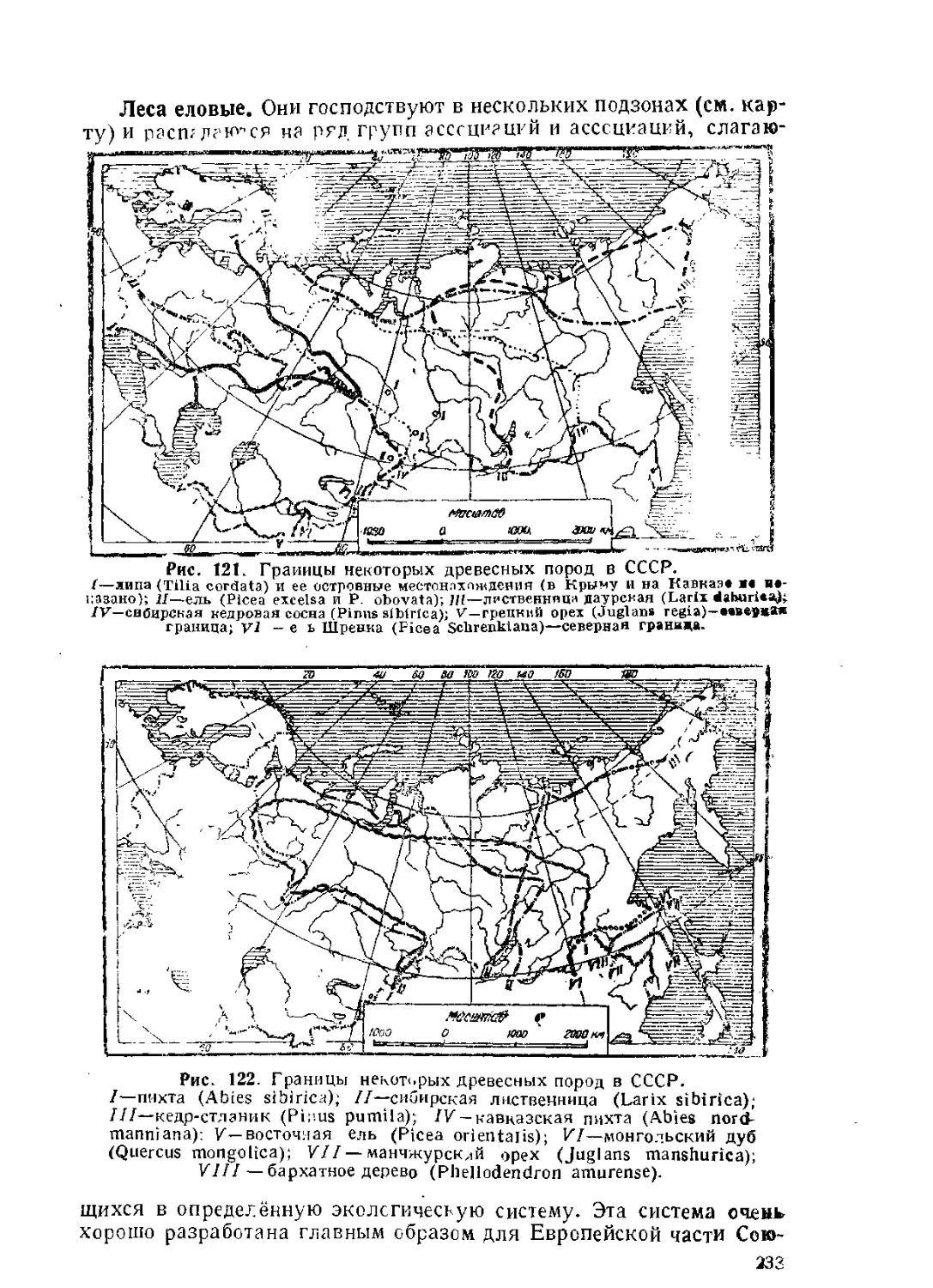






















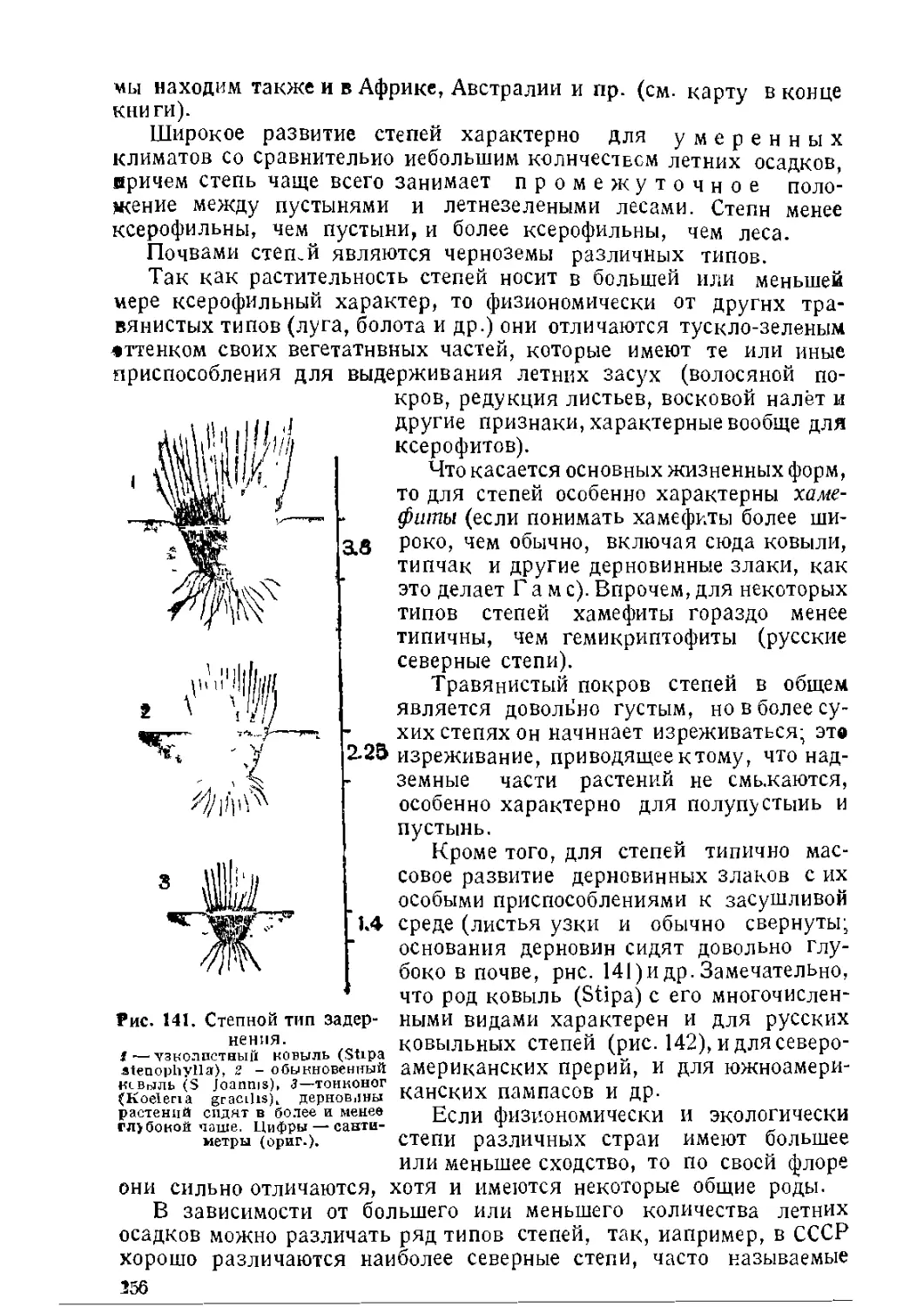








































































































































































































Текст
Книга является руководством по фитогеографии, экологии
и фитоценологии для студентов пединститутов и универ-
ситетов. Особенно много внимания в книге уделено вопросам
экологии и фитоценологии. Руководства с подобным содер-
жанием на русском языке не имеется.
ПРЕДИСЛОВИЕ К ПЕРВОМУ ИЗДАНИЮ
Данное руководство соответствует курсу «География растений» в програм-
мах пединститутов. Оно распадается на пять основных отделов: I. Учение об
ареале, II. Учение об экологических факторах, ]Ц. Учение о фитоценозах*
IV. Основные типы растительного покрова, V. Флористические обтает . В
сущности, к географии растений надо отнести лишь отделы I и V, по св ему
объему самые небольшие. Отдел II всецело относится к экологии а III и IV —
к фитоценологии. Таким образом, если дать название курсу, вполне отвечаю-
щее его содержанию, то это будет: «Основные элементы географии растений,
экологии и фитоценологии >. В курсе затрагиваются три основные проблемы:
проблемы местообитания, пространства и смен, касающиеся как отдельных
видов, так и фитоценозов, т. е. весь тот круг вопросов, который Р ю б е л ь
считает возможным объединить как геоботанику.
Мы не склонны объединять все указанные дисциплины в одну науку, думаем,
правильнее считать геоботанику синонимом фитоценологии; если же действи-
тельно в данной книге и в данном курсе объединяются все эти дисциплины, то
делается это по соображениям практического порядка: курс должен дать сту-
дентам элементы различных дисциплин, ио, несомненно, связанных друг
с другом, и притом студентам пединститутов. В университетах подобный курс
распадается на ряд самостоятельных курсов, как «Экология растений», «Фи-
тогеография», «Фитоценология».
Наибольшее внимание уделяется в курсе вопросам экологии (отдел II) и
основным типам растительного покрова (отдел IV), они составляют две трети
всей книги. Это имеет своё основание в том, что моменты экологии совершенно
необходимы для более глубокого понимания растительности и потому, что все
процессыздесь динамичны по своему существу. Описание же основных типов рас-
тительности крайне важно для студента-географа, так как здесь он знакомится
с главнейшими ландшафтами. Мы пытались построить эту главу о растительных
типах тоже иа экологическом принципе.
Не все основные типы растительности описаны одинаково полно; более полно
описаны типы, развитые в пределах СССР или важные в общегеографическом
отношении.
Важный последний отдел о флористических областях не мог быть развит
так, как он этого заслуживает, главным образом потому, что этот отдел трвйует
от студента хороших знаний по флоре и систематике, чего, конечно, у студентов
географических факультетов нет, так как никаких ботанических курсов, кро-
ме данного, им не читается. Поэтому даже и в том виде, в каком отдел о флори-
стических областях представлен в книге, он будет студенту труден, но будет
служить известным справочником в дальнейшей его педагогической работе.
3
Кстати замечу, что отсутствие курса морфологии и систематики растений в пла-
нах географических факультетов ничем оправдано быть не может.
Хотя генетический принцип везде пронизывает географические главы книги,
одиако особой главы, посвящённой геологической истории растений, здесь
нет (так как нет её и в программах). Основная причина, — это перегруженность
программы и особенное усиление экологического принципа. Думается, однако,
что эта глава должна быть.
Трудность написания книги была очень велика: надо было выбрать из огром-
ного материала наиболее важное и представить его возможно проще и понятнее.
Думаю, что научность изложения от этого нисколько не пострадала.
Так как для того, чтобы получить представление о ландшафтах, требуются
физиономические представления, то необходимо было снабдить книгу ландшафт-
ными рисунками, фотографиями, картами и пр.
Мы думаем, что книга даёт материал не только для студентов пединститутов,
она будет полезна и для студентов университетов, особенно для почвенно-
географических факультетов; кроме того, она будет полезна и вообще для нату-
ралистов.
В заключение считаю необходимым выразить свою признательность всем
лицам, оказавшим мне содействие при написании этой книги.
Проф. В. Алехин
Москва, 28 /1 1937 г.
ПРЕДИСЛОВИЕ КО ВТОРОМУ ИЗДАНИЮ
При подготовке 2-го издания «Географии растений» я принял во внимание
ряд указаний, которые были сделаны различными лицами, и в соответствии с
этим внёс некоторые дополнения в текст. Из более крупных дополнении нужно
указать следующие: 1) добавлена небольшая глава «Краткий очерк истории
фитогеографии, экологии н фитоценологии» и два приложения: «Происхож-
дение культурных растений» и «Систематический справочник семейств растений
с краткими характеристиками»; 2) значительно расширен материал по раститель-
ности СССР; 3) также расширен материал, касающийся роля человека и его
воздействия на растительность.
Прибавление характеристик всех семейств, имеющихся в тексте,
мне кажется, является очень важным, так как даёт возможность учащимся
получить некоторые представления о географическом распространении, а так-
же об основных морфологических и систематических чертах приведенных
семейств.
Учебник в основном написан по соответствующей программе, поэтому из
ряда пожеланий различных лиц я не мог принять во внимание следующие:
1) дать отдельную главу по растительности СССР, 2) дать характеристики рас-
тительности отдельных континентов и 3) объединить флористические подразде-
ления земной поверхности с подразделениями растительного покрова.
Что касается объединения флористических подразделений и подразделе-
ний растительности, то я думаю всё же, что это две различные стороны дела
и что раздельное их представление может дать более ясное понимание наблю-
дающихся явлений.
Содержание книги несколько шире ее названия. Название, отвечающее
содержанию, — это «Основы фитогеографии, экологии и фитоценологии». Так,
мне кажется, должен называться курс, читаемый в педвузах.
Число рисунков и карт в настоящем издании значительно увеличено. Не-
которые рисунки заменены новыми. Вместо двух карт даются три: карта рас-
тительности СССР и две мировых карты растительности (по Брокмаи-Ерошу н
БСАМ).
Проф. В. Алехин
13 октября 1943 г.
ВВЕДЕНИЕ
Ботаника —наука о растениях —изучает растения и их со-
четания с различных сторон. При этом можно наметить несколько
направлений. f
Прежде всего ботаника занимается изучением растительных
форм и расчленения растений; это основа всего ботанического зна-
ния, так как познание всякого предмета начинается с изучения
его внешней формы. Здесь мы имеем морфологическое направление.
Но, кроме внешнего расчленения растений (внешняя морфология),
мы имеем дело также и с внутренним расчленением, изучение ко-
торого представляет предмет внутренней морфологии, или, как
чаще говорят, анатомии растений.
Вторым направлением является физиологическое. Физиология
растений имеет задачей изучение жизненных процессов в растении
(поступление веществ, передвижение их в растении, рост, движение
и пр<).
Третье направление, экологическое, изучает отношение растений
к среде («экология» —от греческих слов «ойксс» —дом, жилище
и «логос» —учение, т. е. учение о «домашнем обиходе» растения).
Так как внешний вид растения во многих случаях зависит от усло-
вий окружающей среды, то „экология стремится изучить раститель-
ные формы и связать их с такими элементами среды, как темпера-
тура, влажность, свет и т. п.
Четвёртое направление в ботанике — географическое. Здесь
основное внимание уделяется изучению размещения видов на зем-
ной поверхности; виды распределяются крайне разнообразно: одни
имеют более широкое распространение, другие встречаются редко
и только в определённых условиях. Изучить всё это разнообразие
и его причины (и в настоящем и в прошлом) —является одной из
основных задач географии растений.
Пятым направлением будет генетическое, изучающее вопросы про-
исхождения, генезиса, изменчивости и наследственности растений.
Наконец шестым направлением является систематическое, ко-
торое ставит своей задачей дать обзор всех растительных видов,
населяющих земную поверхность, а также живших в прежние гео-
логические времена. Этот обзор имеет целью расположить виды
так, чтобы возможно ближе отразить филогенети ческие,
родственные, отношения; в основе системы видов должно лежать
представление об эволюционном развитии растительного мира.
Указанное разнообразие направлений в изучении растений
представляет в то же время одно неразрывное целое
и не может быть строго разграничено. В ряде случаев экология
сливается с физиологией, морфология незаметно переходит в эко-
логию и т. д.
Ботаника имеет предметом своего изучения не только раститель-
ные виды, но также и те сочетания растений, из которых
состоит растительный покров земной поверхности. Такие объеди-
нения растений, как леса, болота, степи и пр., представляют бота-
нические единицы особого рода и называются фитоценозами (от
греческих слов «фитон» — растение и «койнос» — общий), являясь
объектом изучения особого раздела ботаники — фитоценологии.
Фитоценология изучает фито ценозы с различных точек зрения,
совпадающих с теми, которые имеют место при изучении отдельных
растений; так, можно изучать морфологию, экологию, географию
фитоценозов и пр.
Таким образом, ботаника распадается на два раздела, из ко-
торых один имеет своим объектом изучение видов растений, а дру-
гой — фитоценозов. И тот и другой подразделяются на ряд отделов
(направлений).
В дальнейшем основное внимание будет уделено таким отделам
ботаники видов, как география и экология растений, и разделу фито-
ценологии; остановимся на этом несколько подробнее.
География растений представляет отдел ботаники, изучающий,
как уже было указано, распространение растений по земной поверх-
ности и законы этого распространения. География растений имеет
дело не только с современным распространением растений, она за-
нимается также вопросами о прежнем распространении расте-
ний в различные геологические периоды и эры; таким образом, мы
можем говорить о географии растений мелового периода, каменно-
угольного периода и др. В последних случаях география растений
основывается исключительно на ископаемых остатках растений, их
отпечатках и пр., которых, конечно, всегда имеется недостаточное
количество; поэтому наши знания о распространении растений
в прошлые геологические времена можно считать несомненно
неполными.
Однако не следует думать, что современное распространение ра-
стений известно нам с исчерпывающей полнотой: ведь очень большое
число видов растений остаётся до сих пор неизвестным науке; это
с очевидностью следует из того, что каждый год открываются и опи-
сываются всё новые и новые виды, относящиеся как к низшим, так
и к высшим растениям. Что же касается распространения уже из-
вестных растений, то н здесь их географизм во многих случаях оста-
ётся далеко ещё невыясненным; в ряде случаев вновь открываемые
пункты нахождения тех или других видов в корне меняют наши
представления о территории, занятой этими видами.
Но, как было указано, растительные виды обычно сгруппиро-
вываются в особые сочетания—фитоценозы; поэтому наряду с
географией видов мы имеем дело и с географией фито ценозов (осо-
бый отдел фитоценологии).
Изучение географии фитоценозов связано с ещё большими труд-
ностями по сравнению с географией видов, так как здесь материала
значительно меньше; что же касается фито ценозов в геологической
истории земли, то в этом отношении наши сведения крайне скудны.
При изучении географизма видов и фитоценозов можно ограни-
читься лишь изучением плсщади распространения, границ, сопо-
ставлением флор и растительности различных континентов.
Объектами при этом могут служить не только виды, но и роды,
семейства и т. д., а также такие фитоценологические единицы, как
ассоциации, формации, классы формаций и пр.
Так как в основе подобных исследований лежит установление
площади размещения вида или фитоценоза (или других единиц),
так называемого ареала (от латинского слова «ареа» — площадь),
то направление географии, которое этим занимается, можно на-
звать ареалологическим и можно говорить об ареалологической гео-
графии (видов и фитоценозов).
Однако мы не можем ограничиваться только формальным уста-
новлением ареалов и их границ, — нас интересуют причины, вы-
звавшие конфигурацию тех или иных ареалов, соотношения ареа-
лов и пр. Эти причины в основном сводятся к двум: 1) причины
экологические и 2) причины генетического порядка.
Действительно, в очень многих случаях площади ареалов тесно
связаны с известными условиями существования и в первую оче-
редь с климатом и почвами, и потому экологическая география ра-
стений имеет своей задачей установить эту связь и объяснить ареалы
условиями современной обстановки1.
Однако в большинстве случаев очень трудно объяснить совре-
менное размещение видов (и фитоценозов), исходя только из дей-
ствующих в настоящее время причин, приходится принимать во
внимание также те условия, которые имели место в истории данной
страны или местности. Таким образом, здесь возникает вопрос о
происхождении, генезисе и эволюции современных ареалов и флор.
Всеми этими вопросами и занимается генетическая география.
Таким образом получается следующая схема:
География видов География фи то ценозов
Ареалологическая Ареалологическая
Экологическая Экологическая
Генетическая Генетическая
Ареалологическая география видов обычно называется флори-
стической; генетическая география иногда называется историче-
ской. Данную схему подотделов фитогеографии можно применить
к любому геологическому периоду при изучении размещения ра-
стений этого периода, причем подотделы генетической географии
связывают флоры и растительность ближайших друг к другу пери-
одов.
1 Н ельзя относить к экологической географии растений, как это очень часто
делают, изучение отношений между растениями п окружающей их средой, —
это является предметом экологии растений; надо помнить,что воснове географии
всегда должно лежать представление об ареале.
8
Приведенное подразделение географии видов и фито ценозов
нужно лишь для лучшего понимания самого предмета, но ни в ка-
ком случае не предполагает самостоятельного, отдельного суще-
ствования указанных подотделов. Наоборот,—и это вполне
естественно,—последние так тесно связаны между собой, что один
подотдел почти незаметно сливается с другими.
Что же касается экологии, то ее мы определяем как направление
ботаники, изучающее отношения растений к окружающей внешней
среде и, следовательно, влияния этой последней на растительный
организм. Но подобные влияния имеют место также и между сре-
дой и сочетаниями растений —фитоценозами. Поэтому и здесь мы
имеем два отдела: в одном объектом является растение, в
другом — фитоценоз. Первый отдел называется аутэколосией (отдел
ботаники видов), второй —синэкологией (отдел фитоценологии), при-
чём, конечно, границы между этими отделами достаточно условны.
Наконец, фитоценология, как уже было сказано, имееет дело с
фитоценозами, рассматривая их разносторонне, со всех воз-
можных точек зрения. Подчинить фитоценологию экологии,
как это делают в большинстве случаев американские авторы,
нельзя, так как, наоборот, — синэкология представляет часть
фитоценологии, а аутэкология касается лишь отдельных растений.
Исходя из тех же оснований, нельзя подчинить фитоценологию
фитогеографии, так как география фитоценсзов есть только часть
фитоценологии, а география видов не может относить-
ся к фитоценологии, так как является отделом ботаники видов. По-
следнее обстоятельство особенно необходимо учитывать, так как
включение фитоценологии в географию вообще имело место не толь-
ко у более старых русских авторов, когда фитоценология только
лишь зарождалась, но это имеет место и в самое новейшее время,
чем, несомненно, затемняется сущность рассматриваемых отделов.
В дальнейшем изложении будут даны основы: 1) географии
растений, 2) экологии растений и 3) фитоценологии, причём в по-
следнем разделе особое внимание посвящено географии фитоцено-
зов.
КРАТКИЙ ОЧЕРК ИСТОРИИ ФИТОГЕОГРАФИИ, ЭКОЛОГИИ
И ФИТОЦЕНОЛОГИИ
Фитогеографию часто называют молодой отраслью ботаники;
это и верно и неверно. Верно потоку, что фитогеография сравни-
тельно недавно, и много позже других ботанических отделов (на-
пример морфологии и систематики), завоевала всеобщее призна-
ние" и вошла в число предметов университртского’преподавания.
Неверно потому, что начало фитогеографии относится к глубокой
древности.
Первые географические идеи зародились в древней Греции, за
несколько столетий до нашей эры. Однако греки не выходили за
пределы современной средиземноморской (флористической) под-
области и потому не имели материала для сравнительных геогра-
фических представлений. Лишь поход Александра Маке-
донского в Персию и Индию (334—323 гг. до н. э.) вывел гре-
ков за пределы средиземноморской области и познакомил их с
растительностью других стран—с центральноазиатскими степя-
ми-пустынями и с тропическими лесами. Стали возможными бо-
танико-географические сопоставления. Первые работы в этом на-
правлении принадлежат Теофрасту (ученику знаменитого
мыс. ителя Аристотеля), который обработал материалы, со-
бранные во время похода Александра (он сравнил растения
различных стран, указал на влияние климата и почвы, отметил
своеобразие мангровой растительности, различал растения, при-
сущие горам, долинам и т. д.).
Однако в дальнейшем идеи фитогеографии не получили своего
развития ни в древнем Риме, ни в средние века, невидимому, по-
тому, что не были поняты и опередили своё время (как это часто
бывало в истории различных наук).
Лишь через 2000 лет фитогеография возрождается вновь, при-
чём основание её как науки можно отнести к 1807 г., когда Алек-
сандр Гумбольдт опубликовал «Идеи о географии ра-
стений». Гумбольдта справедливо называют «отцом гео-
граф и ра стен ий».
Правда, можно назвать немало ботаников конца XVIII столе-
тия, предшественников Гумбольдта, высказывавших те или
иные фИТогеографические мысли и положения. Так, Галлер
в 1768 г. дал описание горных поясов в Швейцарии и даже сравни-
вал их с широтными зонами; француз С у л а в и (Soulavie) в
ш_
1779 г. установил для юга Франции пять климатических поясов
в горах и привёл ряд экологических и фенологических наблюдений.
Наконец, нужно особенно отметить Вильденова, который
в 1792 г. опубликовал свои «Основы травоведения», где он затро-
нул ряд вопросов распространения растений (влияние климата,
сглаживающее влияние водной среды, особенности в распростра-
нении растений гор и долин и т. д.) и коснулся причин историче-
ского порядка, указав на возможность в прошлом соединения мате-
риков и островов, ныне разъединённых. В работах Виль-
денова уже намечались трн главные линии географии ра-
стений; флористическая, экологическая и генетическая (истори-
ческая).
Однако заслуга создания новой науки —географии растений —
принадлежит всё же Гумбольдту (1769—1859), любимому
ученику Вильденова, У всех предшественников Гум-
больдта недоставало географического кругозора, Гум-
больдт же получил в результате путешествий личное знаком-
ство с различными частями земного шара (знаменитое путешествие
в Америку, где он пробыл 5 лет, путешествие по Сибири на Алтай,
к Каспийскому морю и др.). Гумбольдт не являлся только
учёным-ботаником, в его работах мы находим глубоко-комплексное
понимание явлений, он положил основы и ряда других дисциплин
(климатология, метеорология и др.). В работах Гумбольдта ещё
в большей мере, чем уВильденова, мы находим материалы
по трем направлениям географии растений: флористическому,
экологическому и генетическому. Особо нужно отметить то, что
Гумбольдт впервые ввёл в науку представление о физионо-
мических формах растений, которые в дальнейшем получили столь
большое значение в экологии. Гумбольдт, кроме т< го, впер-
вые упо' р„бил слово «ассоциация», обозначая этим группы различ-
ных рас ений, а в своих ландшафтных описаниях он инстинктивно
исходил из формаций (хотя не употреблял этого термина).
Физиономические представления Гумбольдта были восприняты
некоторыми ботаниками-путешественниками (М арци у с —1824;
Лунд — 1835; Me йе н —1834 и др.), особенно Марциусом, ко-
торый дал замечательные картины природы Бразилии; однако
большинством современников они поняты не были и возродились
вновь лишь в работах Кернера (1863) и Гризебаха (1872).
Из других авторов начала прошлого столетия следует отме-
тить Августа Пирама' де Кандоля, Робер-
та Броуна (путешествие в Конго и в Австралию), Вален-
б е р г а (Лапландия, Карпаты) и др.
В результате работ указанных авторов явилась возможность
дать обобщение некоторых фактов и идей в фитогеографии, что
сделал датчанин С к о у (Schouw) в 1822 г. в своих «Основах об-
щей географии растений». Если в работах более ранних авторов
наме.чалось, как было уже указано, подразделение географии ра-
стений на три основных направления, то С-к о у впервые дал
ясные формулировки для этих направлений, обозначив их особыми
названиями. «Основы?) С к о у были первой сводкой, руковод-
ством по фитогеографии, причём особое внимание он уделил вопро-
сам экологической и флористической географии (не генетиче-
ской).
Что касается генетической географии растений, то для середи-
ны прошлого столетия нужно указать имена таких выдающихся
исследователей, как Форбс, Гукер и Альфонс де
Кандоль. Первый, находясь под сильным влиянием работ
знаменитого геолога Л я й е л л я, опубликовал работу о про-
исхождении фауны и флоры Британских островов (1840), в кото-
рой показывал, что большая часть видов этих островов проникла
сюда в результате миграции. Гукер в своих многочисленных
работах открыл новое направление в генетической географии,
связанное с происхождением целых флор, но не отдельных видов,
как это имело место до него. Эта широта географического анализа
сделалась возможной лишь в связи с тем, что Гукер совершил
ряд путешествий и получил знакомство с флорой почти всего-зем-
ного шара (его работы по флоре Австралии, Новой Зеландии, Тас-
мании, гГалапагосским островам, Азорским островам и т. д.).
В 1855 г. появился замечательный труд Альфонса де
Кандоля «Ботаническая география» («Geographic botanique
raisonnee») в двух томах; здесь впервые мы находим сводку всех
имевшихся знаний в области генетической географии, почему по
сграведливости А. де Кандоля можно считать основате-
лем этого отдела географии растений. Стало совершенно ясно, что
одних климатических условий для объяснения современного рас-
пространения растений недостаточно, так как распространение
растений в значительной мере обусловлено прежде существовав-
шими условиями. Труд А. де Кандоля — это вторая свод-
ка по географии растений после сводки С к оу (1822).
Однако дальнейшее развитие географии растений, и особенно
генетической географии, сковывалось теми воззрениями (постоян-
ство, неизменяемость видов, вера в творческие акты и Др.), которые
существовали в то время. Внимательное изучение природы обна-
руживало факты и явления, несовместимые с господствующими
взглядами.
Дарвин в 1859 F-- в своей книге «Происхождение видов» раз-
бил все имевшиеся противоречия, доказав эволюционное развитие
всего, органического мира. Работы Дарвина произвели пол-
ный переворот в биологии и в частности в географии растений. Зна-
менитое кругосветное путешествие на корабле «Бигль» было ре-
шающим для принятия Дарвин ы м идеи об эволюции расти-
тельного и животного мира. Как известно, по Дарвину имеется
три фактора эволюции: первый фактор — свойства организма,
второй—внешняя среда, третий —взаимоотношения между ра-
стениями (конкуренция). Последний фактор имеет особо важное
значение; что же касается влияния среды, то оно обычно сильно пе-
реоценивается. Установление Дарвин ы м важности конкуренции
видов следует особо отметить, и Дарвина можно считать одним
из основоположников той отрасли ботаники, которая называется в
настоящее время фитоценологией (см. ниже) и занимается изуче-
12
нием взаимоотношений между растительными видами в раститель-
ных сочетаниях (фитоценозах).
Дальнейший шаг вперёд в развитии географии растений —
это работа Энглера, особенно его труд, вышедший в 1879—
1882 гг., «Опыт истории развития растительного мира, в особен-
ности флористических областей, с третичного периода». Проблема,
.затронутая Энглером, крайне сложна, но тем не менее он
ее блестяще разрешил. Одним из новых моментов в работе Эн-
глера следует считать то, что для систематических целей (для
выяснения родственных отношений между видами) был использо-
ван географизм (ареалы) этих видов, что впоследствии прочно во-
шло в систематику.
Идеи Энглера нашли многочисленных последователей.
У нас нужно особенно отметить Д. И. Л и т в и н о в а; его^рабо-
та «Геоботанические1 заметки о флоре Европейской России» (1890)
поставила вопрос о некоторых особенностях флоры Европейской
России, которые можно было объяснить, лишь принимая во внима-
ние историю (с третичного времени) нашей равнины. Другие ра-
боты Литвинова продолжали развивать и углублять его
мысли. Дальнейшее развитие эти идеи получили в работах Па-
ч о с к ог о «Основные черты в развитии флоры юго-западной
России» (1910) и др. и в ряде исследований Козо-Полянско-
г о в Курской и Воронежской областях (его научно-популярная
книжка «В стране живых ископаемых» — 1931), а также в работах
Лавренко и др.
Таковы основные этапы генетической географии растений; од-
нако необходимо-ещё указать на то, что вопросы миграций расте-
ний, вопросы, связанные со сходством или различием флор мате-
риков, островов и пр., получили особое освещение в результате
выдвинутой Вегенером (1912) гипотезы перемещения
материков в горизонтальном направлении (в русском пере-
воде книга Вегенера вышла в 1925 г., — «Происхождение
материков и океанор»). Проверка гипотезы Вегенера на бо-
таническом материале (работы Ирмшера и др.) показала
приемлемость этой гипотезы, хотя многие факты в некотором от-
ношении и трудно объяснимы. Гипотеза Вегенера имеет
преимущество перед другими гипотезами в том отношении, что она
исходит из непосредственного (в прежнее время) соприкосновения
материков, устраняя гипотетическую связь суши, так называемые
мосты, якобы существовавшие между ныне имеющимися матери-
ками.
Возвращаясь к развитию географии растений в прошлом сто-
летии, можно видеть, что элементы экологии имели место уже у
самых первых авторов. Так, С к о у, в своей вышеназванной
книге, рассматривает экологические1 2 3 факторы, как тепло, влаж-
1 Термин «геоботаника» Литвинов понимал в ином смысле, чем обычно; это—
отдеп ботаники, изучающий связь растений с историей страны.
3 Нужно заметить, что термины «экологические», «экология» в то время не
существовали. Отдел биологии, изучающий влияние внешних условий на орга-
низмы, впервые назвал экологией лишь Геккель в 1866 г.
13
ность, свет и др., в том же порядке, как это делается и в настоящее
время. Помимо этого, С к о у подразделяет растения ио их место-
обитанию на песчаные, известковые, болотные, скальные и т. д.,
а в зависимости от господства тех или иных видов он употребляет
такие названия, как ericeta, fageta, querceta и др., однако пред-
ставления о «сообществах» у С к о у, невидимому, не было.
Приуроченность растений к определённым условиям существо-
вания особенно интересовала ботаников того времени. Известна дис-
куссия, которая возникла по вопросу о том, какие свойства почвы
являются определяющими — физические (Турман, 1843) или
химические (Унгер, 1836). Но особенное развитие экологи-
ческое направление получило лишь в самом конце прошлого сто-
летия, и особенная заслуга в этом отношении принадлежит датско-
му ботанику Варм ингу (1895). Книга Варминга «Эко-
логическая география растений» послужила мощным толчком для
дальнейших экологических исследований и не потеряла своего зна-
чения и до последнего времени1. Она даёт многочисленные сведе-
ния по экологии растений, однако почти исключительно морфолого-
анатомического характера.
Это был первый этап в развитии экологии, этап описатель-
н ы й. Но уже и в то время наметилось другое направление в
экологии. Так, в 1898 г. появился капитальный труд Ш и м п е р а
(на немецком языке) под названием «География растений на фи-
зиологической основе», где сделана попытка не ограничиваться
лишь4 констатацией морфолого-анатомических признаков растений,
но и объяснять их причинами физиологического характера. Так
как для физиологического объяснения требуется соответствующий
эксперимент, постановка опытной проверки,то экология прибли-
жается к наукам экспериментального направления, в частности,
к физиологии растений. Однако это только приближение, и даже
в 3-м издании книги Шимпера в 1935 г. (теперь: Ш и м п ер-
Фабер) в предисловии указано, что изложение географии ра-
стений с физиологическим объяснением наблюдаемых фактов —
дело будущего. Тем не менее, это 3-е издание содержит много ма-
териалов в данном направлении2.
Физиологическое направление в экологии растений особенно
сильно выражено в книге Лундегорда «Влияние климата и
почвы на жизнь растений» (1936, перевод с немецкого).
Необходимо отметить, что большие успехи в области экологии
достигнуты в Америке. Учение об «индикаторах»—показателях
различных почв, различных климатических условий и др. получи-
1 Книга Варминга была переведена на русский язык два раза. Первое изда-
ние под названием «Экологическая география растений» было напечатано в
Москве (1901 г.), второе издание вышло в свет в Петербурге в 1902 г. под на-
званием «Распределение растений в зависимости от внешних условий (экологи-
ческая география растений)». Последнее издание имеет дополнительную главу,
написанную Танфильевым, «Главнейшие черты растительности России», являю-
щуюся первой подробной сводкой по растительности России (с картой расти-
тельности).
1 К сожалению, книга Шимпера до енх пор не переведена на русский язык.
14
ло там особенное развитие, главным образом благодаря работам
Клементса («Растения-индикаторы» — Plant indicators,
1920). Большое значение имеет также книга Уивера и
Клементса «Экология растений» (1929, 2-е издание — 1939),
дающая полное представление об успехах и методах американской
экологии, причём экология понимается американцами (а также
англичанами) очень широко, включая сюда и фитоценологию (см.
ниже).
В СССР экологическое направление в ботанике стало разви-
ваться главным образом лишь в советское время. Проблемы засу-
хоустойчивости, морозостойкости, продвижения кулыур на север,
освоения пустынь и т. д. самым тесным образом связаны с позна-
нием экологических свойств растений.
Как уже было указано, экология выделилась из экологической
географии в конце прошлого столетия, но Экологическая геогра-
фия в первое время (например в книге Варминга) включала
в себя и другую отрасль ботаники, ставшую теперь самостоятель-
ной, — фитоценологию.
Краткая история фитоценологии такова. Несомненно, что та-
кие единицы растительности, как лес, болото, луг, степь и т. д.,
были знакомы и первобытному человеку. Однако ботаники чуть
ли не до конца прошлого столетия из-за деревьев, в букваль-
ном смысле слова, не видели леса и «сообществе» (фито-
ценоз) не было предметом их изучения. Однако столь наглядный
признак фито ценозов, как «физио комичность», обратил на себя
внимание ещё Гумбольдта (см. выше), и он дал описания
физиономических ландшафтных картин, хотя и не являющихся
строго очерченными единицами растительности, но очень близ-
ких к формациям ряда последующих авторов. В сущности,
Гумбольдт, как кажется, был первым ботаником, исходив-
шим при'описаниях растительности из самой растительности, но
не из «местообитания». На местообитаниях основывались не толь-
ко предшественники Гумбольдта, но также и многие позд-
нейшие авторы (С ко у, Хеер н др.). Более прочная осно-
ва фитоценологических представлений была заложена лишь в се-
редине прошлого века работами Гризебаха (1838—1845 и
др.), Лоренца (1858) иКернера (1863). Первый продол-
жил физиономическое направление Гумбольдта, дал бо-
лее детальную систему основных жизненных форм и впервые пра-
вильно определил единицу растительности, названную им фор-
ма ц и е й (впрочем, объём этой единицы не был у него вполне
определён). Необходимо отметить классическую работу Гризе-
баха «Растительность земного шара» в двух томах (вышла в
1872 г., русский перевод в 1874 — 1877 гг.), которая явилась первой
попыткой дать описание растительности земного шара в связи с
климатическими особенностями на основе жизненных форм и расти-
тельных формаций. Это сочинение не потеряло значения и до
настоящего времени.
Но особенно важны для истории фитоценологии работы Ло-
ренца и Кернера. Работа первого автора относится к
15
изучению предальпийских болот Зальцбурга, работа Кернера
посвящена растительности придунайских стран. Лоренц в
своих описаниях не только базируется на растительных естествен-
ных единицах, но делает попытку дать классификацию этих еди-
ниц, устанавливая восходящий ряд последних. Эго первая попытка
в данном направлении. Кроме того, уЛоренца мы имеем ряд
данных по анализу травостоя.
Основная заслуга Кернера в том, что он впервые подхо-
дит к познанию структуры растительных ценозов —его формации
слагаются из особых сочетаний растений (Bestande), которые рас-
полагаются ярусами, т. е. формация состоит из наслоения
ярусов один на другой. Помимо этого, Кернер дал прекрас-
ные картины подвижности формаций в течение вегетационного
периода (фенологические смены). Кернер в своих описаниях
исходит из формаций, имеющих закономерное сложение, и впер-
вые говорит со о со бо й ботанической дисциплине, изучающей
растительные формации, которую он предлагает назвать «Pflan-
zenphysionormk».
Большое значение в развитии фитоценологии имели работы
скандинавских исследователей: фон Поста (1851), X у л ь т а
(1881), а в дальнейшем Се р на Идера, Однако в своё время
работы первых двух авторов почти не оказали влияния на совре-
менников, так как были написаны на мало доступном шведском язы-
ке. П о с т, невидимому, впервые применил метод «пробных пло-
щадок», X у л ь т дал ряд методов аналитического изучения ра-
стительности. Сернандер — известный болотовед, изучая бо-
лота и их четвертичную историю, ввёл представление о сменах
растительности; он является основателем упсальской (г. Упсала)
шкоты фито це но логов, занимающей сейчас видное положение в
Европе (в настоящее время глава школы дю-Р и э).
В середине прошлого столетия появились работы Дарвина.
Как указано было выше, Дарвин особое значение придавал
взаимоотношениям (конкуренции) между растениями, фактору,
лежащему в самой основе растительных ценозов. Однако эти идеи
Дарвина в то время не получили дальнейшего развития, так
к; к учение о растительных сочетаниях находилось тогда в самом
зачаточном состоянии.
Для конца прошлого столетия необходимо указать уже упомя-
нутую выше книгу Варминга «Экологическая география ра-
стений». Эта книга дала толчок к мощному развитию экологии, но
содержит также очень большой материал по «сообществам» расте-
ний всего немного шара, причём характеристики «сообществ» да-
ны в ней главным образом с экологической точки зрения, считая,
что между определённой растительностью и местообита-
нием существует непременная связь.
В конце прошлого столетия и в начале нынешнего очень бол; -
шое значение для развития фитоценологии имели работы Шре-
тера и Ф л а о. Первый был основателем швейцарской школ, i
фитоценологов, второй—французской (в Монпелье). Эти школы
существуют и в настоящее время, главой швейцарской (цюрихской;
16
сейчас является Р ю б е л ь, главой школы в Монпелье — Браун-
Блан к э. Большая заслуга Шретера и Фл а о со-
стоит в том, что они много труда посвятили установлению основных
фито це но логических единиц, так как до того времени обычно фи-
гурировала довольно неопределённая «формация». Свои предло-
жения они внесли на Международный ботанический конгресс в
Брюсселе в 1910 г., где вообще впервые обсуждались вопросы фи-
тоценологического характера. С тех пор упрочились две единицы
растительности: ассоциация (основная) и более крупная —
ф.о р м’а ц и я.
Несколько слов о развитии фитоценологии в Северной Америке.
Первые работы фитоценологического характера появились там
лишь в самые последние годы прошлого столетия и, как кажется,
под влиянием европейских исследователей (особенно под влиянием
«Экологической географии» В а р м и н г а). Особенности амери-
канских работ заключаются во всестороннем применении метода
квадратов (в Америке этот метод упрочился намного раньше, чем
в Европу) и в динамическом изучении растительности. В послед-
нем отношении изучались смены (так называемые сукцессии) ра-
стительности: каждая сукцессия представляет определённую се-
рию, распадающуюся на ряд стадий и завершающуюся стадией,
названной климаксом. Учение о климаксах особенно ха-
рактерно для американской фитоценологии и до настоящего вре-
мени. Отметим Клементса, как наиболее выдающегося пред-
ставителя американской фитоценологии и экологии.
Что касается истории фитоценологии в России, то сё возник-
новение здесь относится к самому концу прошлого столетия. Осо-
бенно много в этом отношении сделал П а ч о с к и й, который в
1891 г. указал, что отдел ботаники, изучающий сочетания растений,
представляет особую дисциплину и имеет особый объект ис-
следования, именно — растительную формацию. Эту новую на-
уку П а ч о с к и й назвал «флорологией» = (современной фитоцено-
логии). Первыми же авторами в России, приблизившим, ся к обла-
сти фитоценологических представлений, были Коржинский
к Краснов (1888).
Для конца прошлого столетия и начала нынешнего следует ука-
зать ряд выдающихся исследователей, называвшихся в то время
«ботанико-географами», которые изучали растительность России,
имея объектом изучения растительную формацию. К их числу от-
носятся Танфильев, Краснов, Крылов и Вы-
соцкий1. Особенно много сделала в смысле укрепления и раз-
вития фитоценологических представлений казанская школа ис-
следователей: кроме указанного выше К о р ж и н с к о г о, на-
зовём Гор дягина, который первым в России стал применять
при описаниях пробные площадки. 1 2
1 Однако нельзя не указать, некоторые „отал к и ; ого времени ие по"
нимали идей фиюиы ^логпп, наир ivcp, Ьекегив графил растений»»
2 Географин растени
17
отдел i
УЧЕНИЕ ОБ АРЕАЛЕ
Ареал
Каждый растительный вид находится на земной поверхности
обычно в огромном количестве особей, распределённых на пло-
щади, которая представляет так называемый «ареал» вида. Ареал —
это площадь (область) обитания вида. Без познания а реалов, конечно,
совершенно невозможно делать какие-либо заключения о геегра-
физме тех или иных видов, и поэтому ареал является основны.ч
объектом в географии растений.
Кроме размещения отдельных видов, мы можем изучать разме-
щение растительных ценозов, и таким образом мы имеем дело:
I) с ареалами видов (также родов, семейств и пр.) и 2) с ареалами
фитоценозов (их различных таксономических единиц).
Размеры ареала. Площадь ареала, определённая для каждого
данного вида или ценоза, может сильно варьировать для различных
объектов, и амплитуда в этом отношении очень велика: от поверх-
ности почти всей суши до одного ограниченного участка на этой
поверхности. В первом случае это будет растение, распространён-
ное крайне широко (так называемый космополит), во втором —
растение исключительно редкое.
Между этими двумя крайностями наблюдаются все переходы,
но вообще широко распространённых видов значительно меньше,
чем видов с более ограниченным распространением.
Растения-космополиты. Строго говоря, растений-космополитов
в буквальном смысле слова, т. е. таких, которые были бы распро-
странены везде, где возможна растительная жизнь, мы не знаем:
да и трудно представить себе растение, которое могло бы одинако-
во произрастать и в т^ ндре,и в тропическом лесу, и в пустыне, к ьа
болотах и т. д. Под космополитами обычно понилг ют такие растения,
которые широко распространены на всех континентах, но, конеч-
но, в ряде областей и районов могут и отсутствовать. Наибольшее
число космополитов относится к водным растениям, что объясняет-
ся большей однородностью условий водной среды и более легкими
способами расселения. Из числа этих водных растений можно на-
звать тростник (Phragmites communis), частуху (Alisma plantago
aquatica), некоторые ьиды рдеста (Potamogeton) и др. Встречают-
ся также космополиты и среди растений влажных местообитаний,
например лужайник (Limosella aquatica).
20
Космополитами являются также некоторые сорные растения,
связанные в своём распространении с деятельностью человека:
мокрица (Stellaria media), крестовник (Senecio vulgaris), осот (Son-
chus oleraceus), крапива (Urtica urens и U. ioica), одуванчик (Ta-
raxacum vulgare) и др.
Гораздо реже космополитами являются растения сухих мест,
так как здесь разнообразие условий существования крайне вели-
ко; сюда можно отнести папоротник-орляк (Pteridium aquilinum),
папоротник Polypodium vulgare и некоторые другие виды; однако
эти виды далеко не являются абсолютными космополи-
тами, и например Polypodium vulgare отсутствует на мно-
гих территориях (в большей части Европейской части СССР).
Широко распространённые виды иногда называют эврихорными
(от греческих слов: «эврис» — широкий и «хорос» — пространство),
а виды, ограниченные в их распространении, — стенохорными
(от греческих слов: «стенос» — узкий и «хорос» — пространство).
Последние решительно преобладают среди цветковых растений,
и по де Кандолю имеется лишь около 0,001 % цветковых,
распространённых на 1/4 земной поверхности, остальные распростра-
нены на меньшей территории.
Если растения-космополиты очень редки, то космополитных
фитоценозов нет совсем, так как фитоценозы состоят из ряда ви-
дов, которые в этом случае почти все должны были бы бытькосмопо-
литами. Исключение могут составить лишь те фитоценозы, где
господствует только один вид при ничтожном участии других,
как, например, в фитоценозе тростника. Но вообще, как правило,
фитоценозы являются в своём распределении стеыохорными, даже
в таких крупных подразделениях, как степи, пустыни, тундры и
т. д. В своих же более мелких подразделениях фитоценозы обна-
руживают ещё большую территориальную ограниченнссть.
Как крайний предел стенохории надо считать такие случаи,
когда вид или фитоценоз ограничен в своём распространении лишь
одной точкой на земной поверхности, одним пунктом. Так, например,
эльдарская сосна (Pinus eldarica) растёт в Грузии лишь на площа-
ди около 50 га, сосна Станкевича (Pinus Stankeviczi) известна для
двух пунктов Крыма (Судак и мыс Айя), камчатская пихта (Abies
gracilis) известна лишь на восточном берегу Камчатки (одна роща)
и т. д. Для травянистых растений можно было бы привести немало
подобных примеров, причем, как правило, такие стенохорные виды
особенно многочисленны в горных странах, а на равнинах обычно
отсутствуют. Укажем несколько примеров: лишь на мелах юго-во-
стока Европейской части встречается ряд меловых видов — Lina-
ria cretacea, Silene cretacea, Artemisia hololeuca и др.; замеча-
тельный каучуконос тау-сагыз (Scorzonera tau-saghiz) встречается
лишь в горах сыр-дарьинского Кара-Тау и в Туркестанском хребте;
известны виды, встречающиеся лишь на одной какой-либо горе, и пр.
Все указанные виды относятся к числу эндемико в1,— это
не только виды, ограниченные лишь одним пунктом, но также и
1 9 н д ем и к — от греческих слов: «эн» — внутри и «демос» — народ.
21
все те виды, которые вообще имеют более или менее ограниченный
ареал (рис. 1). Так, можно говорить об эндемиках Алтая, Кавка-
за нт. д., но, понимая эндемики более широко, можно говорить
даже об эндемиках целого материка или его части, например Ав-
стралии, Южной Америки и пр. (рис. 208, 211).
Рис. 1. Ареал Daphne cneorum; заштрихованная пло-
щадь— более или менее сплошное распространение,
точки — отдельные местонахождения (по Гаевскому).
Небольшой участок ареала на востоке относится к
очень близкому виду D. Julia
Кроме эндемичных видов, имеются и эндемичные ценозы. В одних
случаях эндемичные ценозы своим существованием тесно свя-
заны с эндемичными видами, входящими в их состав, особенно в
случае преобладания этих видов, в других —такой связи может
не быть. Так, рощн эльдарской сосны, сосны Станкевича, камчат-
ской пихты, фитоценозы с присутствием тау-сагыза и др. являют-
ся эндемичными в связи с наличием в них эндемичных видов, но
такие фитоценозы, как разнотравные степи Европейской части
СССР, «осиновые кусты» области Донского ледникового языка и
др., тоже эндемичны, хотя виды, составляющие их, являются ши-
роко распространенными: так, например, в «осиновых кустах» го-
сподствуют осина и ряд обычных кустарников и травянистых ра-
стений.
22
О причинах эндемизма и о значении эндемиков для целей гео-
графии растений будет сказано дальше.
Размещение индивидуумов по площади ареала. Нельзя пред-
ставлять себе, что индивидуумы того или иного вида или отдельные
участки фитоценозов занимают весь свой ареал сплошь, но индиви-
дуумы одних видов встречаются более часто, других менее ча-
С1о. В этом отношении в природе наблюдается неограниченное
Рис. 2. Распространение альдрованды (Aldrovandia vesiculosa) в
Европе; пример прерывистого распространения (по Хечи с допол-
нениями Гамса и Алехина).
разнообразие, но вообще можно различать два типа растений:
эвритопные и стенотопные («топос» по-гречески—место, о значении
слов «эврис» и «стенос» см. выше); первые —очень распространён-
ные виды, мало разборчивые к местам своего произрастания, имею-
щие широкую «экологическою амплитуду»: вторые — связаны
в своем произрастании с достаточно узко ограниченными условия-
ми (например известковые, кислые, торфяные, песчаные почвы,
избыток воды и т. п.).
К числу свритопных относятся, например, наши обычные зла-
ки— пырей (Agropyrum repens), костер безостный (Bromus inermis),
мятлик луговой (Роа pratensis); к числу стенотопных—клейкая оль-
ха, связанная обычно с поймами рек и отсутствующая на водораз-
делах; ясень, избирающий наиболее богатые почвенные условия,
подбел (Petasites spurius) — лишь по песчаным отмелям рек, ряд
23
растений — лишь на меловых обнажениях, другие — лишь на
сфагновых моховых болотах (Sphagnum — особый мох) и пр.
Вообще же, нет ни одного вида, который занимал бы свой аре-
ал сплошь, наоборот, обычно большие площади ареала бы-
вают совершенно лишены индивидуумов данного вида (рис. 2).
В этом отношении имеется ряд различий, но если ареал обуслов-
лен климатически, т. е. если данный вид за пределами
ареала не может существовать по условиям климата, то чаще всега
наполнение индивидуумами будет наиболее густым в центральных
частях ареала и менее густым в частях периферических, так как *
последних вид находит менее благоприятные условия существо-
вания (например дуб).
Кроме различий в экологических амплитудах, у разных видов
в наполнении ареалов индивидуумами играют роль тацдсе и неко-
торые свойства видов: способность к обильному размножению,
способность к энергичному распространению, способность давать,
сплошные насаждения и др.; так например, в подзоне дубрав дуб
дает сплошные насаждения, а клён обычно сюда вкраплен отдель-
ными экземплярами или группами, лещина (орешник) в дубравах
дает сплошные заросли, а европейский бересклет зд.сь же растет
отдельными особями. В некоторых типах степей типчак (Гс-
stuca sulcata) представлен щеткой из бесчисленного числа экзем-
пляров, а ряд других видов обычно разбросан по степи отдельными,
редкими экземплярами. Некоторым видам свойственно вообще
очень редкое, спорадическое распространение,такие растения та-
ежной области, как подъельник (Epipogon aphyllum) или калипс®
(Calypso bulbosa), в пределах своего ареала встречаются иногда
настолько редко, что их с трудом можно найти.
Методические указания. При установлении ареала нередко от-
мечают крайние периферические точки нахождения данного вида,
соединяют их друг с другом Л1 нией, в результате чего получается
замкнутая площадь разнообразных конфигураций. Однако при по-
добной методике получается слишком обобщенное и Hej едъ о лож-
ное представление об ареале, пригодное, впрочем, для общих це-
лей. Более правильно нанесение на карту лишь отдельных пунктов
нахождений растения по всему ареалу (рис. 1, 2 и 3).
Типы ареалов
Основные типы ареалов. Независимо от своих размеров, ареалы
по своим очертаниям имеют различные конфигурации и различ-
ные типы строения. Прежде всего различают сплошные и несплош-
ные ареалы; последние называются также разъединенными или
дизъюнктными (от латинского слова «дизъюнктус» — не находя-
щийся в связи).
Сплошные ареалы представляют одну целостную площадь оби-
тания вида; при разъединенных ареалах ареал распадается на две,
три и более частей; иногда в пределах ареада имеются лишь отдель-
ные пункты, удаленные друг от друга, где встречается данное рас-
тение (рис 11 "десь ареал как бы разорван на известное числе
24
небольших площадей или даже пунктов. Примером такого ареала
может служить водное насекомоядное растение Aldrovandia vesicu-
losa (рис. 2), имеющее Островные местонахождения в Европе (Юж-
ная Франция, Италия и другие места Западной Европы), в СССР
(Белоруссия, Украину, Воронежская и Курская области, низовья
Волги, Амурская область), в Японии и в Восточной Австралии.
Другим примером может служить папоротник Cystopteris sudeti-
са, известный для скалистых мест Судетских и Карпатских тор-
ге о го ла во ав _ мо_____teo__iso ня .
------------ -------L. IvAl _ _ ХрИ л _j______~ -4— — —L 4— k
а? о ж ы so во vo ср ио iso igg j
Рис. 3. Местонахождения Arctous alpina в Евразии—пример прерывиетвг»
ареала; растение..встречается также и в Северной Америке (по Бушу).
на русской равнине этот папоротник найден в ряде отдельных пунк-
тов, особенно в восточных частях, а также на Урале и в Сибири.
Хорошим примером видов с разъединенными ареалами явля-
ются виды альпийские, или высокогорные: сюда относятся многие
виды камнеломок (Saxifraga), горечавок (Gentiana) и др., встречаю-
щиеся на горах Европы, на Кавказе и пр., а в промежуточных
равнинных местах совершенно отсутствующие (рис. 3). Разъеди-
ненные ареалы обусловлены причинами исторического характера
(см. ниже).
Совершенно ясно, что если бы обвести крайние точки всех этих
местонахождений, то о распределении сида внутри ареала
нельзя было бы получить никакого представления, поэтому пра-
вильнее в подобных случаях обозначать местонахождения точками.
Особом типом ареалов являются ленточные, где отдельные
участки территории, занятые каким-либо видом, вытянуты в виде
лент (рис. 4). Сюда относятся виды, распространенные в поймах
рек, по их террасам или по скалистым берегам; например подбел
(Petasites spurius) встречается вдоль пойм по песчаным наносам,
по песчаным террасам рек встречается смолевка (Silene parvifl»-
га) н т. д.
Сплошные ареалы нередко на своих границах переходят в лен-
точные; в качестве примера укажем на некоторые степные расте-
2
ния (Gypophila paniculata, Centaurea marschalliana и др.), про-
израстающие в степной полосе на различных местообитаниях, на
севере же, на границе своего ареала, они встречаются лишь на
песках вдоль рек, давая ленточные полосы. Ряд степных видов по
каньонам р. Днестра продвигается-на запад (рис. 5). Ленточные
полосы дает также на севере дуб, произрастающий здесь лишь в
поймах рек.
Рис 4. Распростпапение сибирской лиственницы на сегеро-востоке Евро-
пейской части СССР Пример ареала, б&столщего из ленточных полос вдоль
рек (поперечная штриховка); крестики — отдельные местонахождения, тре-
угольники — места ископаемых находок (по А Ильинскому)
Ареалы, внутри которых отдельные местонахождения не пре-
вышают расстояний, доступных растениям при их обычных спосо-
бах распространения, можно вообще называть связанными.
Причины различных конфигураций и границ ареалов. Выше
были указа1 ы некоторые причины границ ареалов Основные при-
чины таковы.
1. Климатические. Вид занимает данную территорию
потому, что она по своим климатическим особенностям соответ-
ствует его потребностям в тепле, влажности и пр. За границами
ареала вид уже произрастать не может в силу неблагоприятных
климатических условий Таковы, например, на нашей равнине
2б
северные и южные границы многих растений лесов, степей, сфаг-
новых болот и т. д.
2. Эдафические, или почвенно-грунтовые. Здесь от-
сутствие соответствующих местообитаний делает невозможным бо-
лее широкое расселение вида, хотя климатические условия этому
не мешают. Зак, некоторые меловые растения юго-востока Евро-
Рис. 5. Ленточное распространение некоторых видов в Западной
Украине, как следствие эдафических условий (каньоны р. Днестра)
и миграции на запад по долине р. Днестра (по Гаевскому).
пейской части СССР при более широком распространении мелового
субстрата имели бы и более широкие ареалы, сосна на русской
равнине могла бы дальше продвигаться к югу при присутствии
здесь песчаных пространств, многие растения гранитов искал мог-
ли бы на нашей равнине иметь более широкое распространение
(рис. 5).
3. Механические Препятствием к расселению растений
служат не климатические или эдафические условия, а такие, во
многих случаях непреодолимые, препятствия, как моря, океаны,
горные цепи и пр В частности флора удаленных островов обычно
крайне сроеобрг^'Ш, ”то стоит в связи с невозможностью заноса
27
семян через водные пространства (впрочем, здесь действуют и дру-
гие причины — см. ниже).
4. Биотические. Препятствием к расселению растений
являются. 1) конкурирующие виды, 2) растительные ценозы, соз-
дающие особые условия, не позволяющие проникать в них расте-
ниям других условий существования, так, на русской равнине
преьяювг ем для расселен* я к щв<ру нека орых стегных рас-
тений служат обширные леса, и действительно, разреживание ле-
Рис. 6 Современное распространение семейства магнолиевых (в узком
смысле) — пример разъединенного ареала (по Хетчинсону).
сов и их вырубка открывают пути для продвижения степных ра-
стер ий далее современной их северней границы, 3) отсутствие тех
или иных животных (например опылителей) и др.
5. Исторические. Эти причины уже не связаны с со-
временными условняли существования на земной поверхности.
Растения, населяющие землю, за немногими исключениями видов,
молодых по своему происхождению, возникли в прошлые геологи-
ческ! е времена, нередко при иных климатических условиях, при
ином распределении суши и воды, нередко при ином строении зем-
ной поверхности. Естественно, что ареалы подобных видов совер-
шенно не могут быть поняты исходя из современных отношений,
но эти ареалы играют исключительную роль в географии растений
(рис. 6 и 9)
6. Во многих случаях границы ареалов всецело связаны сче-
тов е к о м, человек в результате своей хозяйственной деятель-
ности уничтожает местонахождения тех или иных видов, так, в
Европейской части СССР вследствие распашки степей уничтоже-
ны наиболее северные местонахождения ряда степных видов. В
других случаях человек способствует расширению ареаг ов — рас-
пространение сорных растений, порубки лесов дают возможность
растениям открытых мест расширять свои ареалы.
28
Вообще же надо сказать, что объяснение границ ареалов дей-
ствием какого-либо одного фактора представляется в большинстве
случаев очень трудным и даже невозможным, так как в природе,
как правило, мы имеем совместное действие факторов.
Устойчивость ареалов. Можно представить себе, что один вид
достиг во всех направлениях своих климатических границ, другой
находится в процессе расселения, значит, ареал первого будет уже
сформировавшимся, второго — находящимся в процессе форми-
рования. Ареалы первого рода нередко называют стабильными,
второго — лабильными Стабильность ареалов надо относить, од-
нако, лишь к определенному моменту и понимать несколько
условно, так как абсолютной неподвижности в природе вообще
быть не может.
Стабильность ареалов может быть обусловлена различными,
только что указанными причинами.
Причины лабильности ареалов:
1. Вид не достиг еще своих климатических границ
2. Вид не достиг своих эдафических границ.
3. Вид в результате своего угасания сокращает площадь своего ареала
4 Вид изменяет ареат в связи с геологическими причинами (опускание,
поднятие суши и т. д )
5. Вид, в резучьтате воздействия человека, сокращает свой ареал, нли,
наоборот, расширяет.
Возможны и другие причины Лабильности ареалов.
Таким образом видно, что лабильность ареалов может быть
прогрессивной (расширение ареала) и регрессивной (сокращение
ареала).
Формирование ареалов
Миграции. При формировании ареала вопрос прежде всего
идет о расселении того или иного вида го территории. Каждое
растенье обладает известньми средствам^ .для сьоего распростра-
1еьия или использует для этого различные внешние агенты Эти-
ми агентами являются вода, ветер, животные и человек, которые
разносят зачатки или части растений, вообще называемые диаспо-
рами. С другой стороны, некоторые растения и вне зависимости
от внешних агентов могут расселяться на большие пространства
(разбрасывание семян самими растениями, передвижение при по-
мощи корневищ и укореняющихся побегов и пр ). Более подробно
о способах расселения растений будет изложено в отделе II.
Необходим, несомненно, огромный промежуток времени для
того, чтобы вид заселил какой-либо континент, а тем более всю
семную поверхность При этом приходится думать о длительных
и сложных миграциях (от латинского слова «миграцио» — пере-
селение). Если в настоящее время, как уже было указано, на зем-
ной поверхности нет космополитов в абсолютном смысле, тоь как
свидетельствуют палеоботанические данные, в более ранние гео-
лшические т^срыоды, напр! мер в каменноугольный и юрский пе-
риоды, имело место очень большое сходство флор континентов, в
настоящее время далеко удаленных один от другого.
29
Невозможность для растений преодоления обширных водных
пространств, разъединяю!! их континенты, привела к представле-
нию о существовании мостов, соединявших когда-то эти континен-
ты (теория мостов)1. По этим мостам или перемычкам суши и могли
расселяться растения (и животные); в дальнейшем мосты опусти-
лись. Теория мостов еще сравнительно недавно занимала в науке
солидное положение, так как иначе объяснить сходство во флоре,
например, Австралии с Южной Африкой или Африки с Индией бы-
ло бы очень трудно.
В новейшее время, благодаря появлению теории Вегенера,
вопрос с миграциями значительно упрощается. По Вегенеру,
земля в древние геологические времена и ещё в каменноугольный
Рис. 7. Карта земной поверхности и положение поносов (кружки
с® стрелками) в каменноугольный период. Заштрихованы океани-
ческие впадины, точками обозначены морские пространства (по
Кёппе н-Вегенер у).
период представляла одну сплошную глыбу (рис. 7), на которой
расселение растений могло происходить беспрепятственно. В даль-
нейшем эта сплошная глыба распалась на части, которые разо-
шлись в стороны, в результате чего образовались отдельные, ино-
гда очень далеко отстоящие континенты. Таким образом получи-
лась не миграция растений, а миграция участков суши, населён-
ных этими растениями.
Не надо думать, однако, что теория мсстсв для'понимания рас-
пространения растений совершенно отпадает; она сохраняется
для мостов меньших протяжений. Так например, теория мостов
с успехом может быть применена для объяснения сходства во фло-
ре Крыма, с одной стороны, и Малой Азии и Балканского полу-
острова, — с другой, также Кавказа и Балканского полуострова
и пр.
Теория миграций, как мы видим, выдвигает предположение,
что вид возникает в определённом пункте, а затем отсюда распро-
30
страняется; таким образом, основное положение таково, что аре-
ал при своём возникновении представ-
ляет очейь ограниченную площадь, а за-
тем (п.:и благоприятных условиях) даёт концентричесю е площади,
всё более и более обширные.
Но в вопросе формирования ареала имеется и другая точка
зрения, а именно, что вид в момент своего возникновения уже име-
ет ареал, представляющий часть ареала материнского вида
(т. е. вида родона аальиого). На этой части ареала состав биоти-
пов (см. стр. 64) начинает меняться в зависимости от изменений
условий существования или в связи с появлением мутаций и др.
В результате в отдельных частях ареала появляются новые формы
а даже виды. Пачоский, развивший эту теорию, приводит
эяд примеров: так, ель, занимая обширный ареал в Северной Ев-
ропе и Азии, не является однородной по всему этому ареалу; в
Европе она представлена европейской елью (Picea excelsa), а в
^зин (а также на северо-востоке Европейской части СССР) — си-
бирской (Р- obovata), в сущности очень мало отличающимися
яруг от друга; кроме того, на Урале ив некоторых других местах
встречаются формы ели, являющиеся. как бы промежуточными
лежду двумя указанными. Возникшие'близкие морфологические
рормы затем конкурировали друг сругом; в результате в одних
тастях ареала (в Европе) наиболее жизнестойкой оказалась форм ,
зазыв емая европейской елью, в других частях (Сибирь)более
лриспособленной оказалась ель сибирская. В некоторых частях
1реала, например на Урале, пока ещё ни одна из форм не получи-
та преобладающего значения, и потому здесь мы и находим ряд
|эорм, хотя и очень близких, но всё же отличающихся морфологи-
гески. Можно привести примеры, когда и этого ещё нет, но когда
эодоначальный вид уже начинает диференцироваться на мелкие
:диьицы; так, в южно русских степях обычным растением является
сасатиц— ирис (Iris pumila), но различные экземпляры его, при
тождестве морфологического строения, имеют различные окраски
тветка —лиловую, голубую, желтую, белую и др. (так называе-
мый полихроизм в окраске). Каждый экземпляр всегда имеет оп-
>еделённую окраску, и этот признак является наследственным.
5дссь, таким образом, по всему своему ареалу вид распался на
[ьетные расы, причём, вероятно, в дальнейшем различные расы
упрочатся в различных частях ареала и произойдёт диференциа-
ция ареалов.
Приблизительно такую же картину дает южностепной тюльпан
(Tulipa Schrenkii) с жёлтыми и красными цветками. Особенно ин-
тересно чересполосное чередование некоторых очень близких ви-
дов (Anchusa officinalis и A. ochroleuca на западе Украины), -что
является уже шагом вперёд в деле расчленения в начале общего
ареала.
В общем можно думать, что формирование ареалов происходит
двумя путями: 1) путём расширения и 2) путём распадения родо-
начальных ареалов; эти два пути не исключают один другой и
нередко имеют место одновременно.
31
Центры ареалов. Часто говорят о «центрах» ареалов, употре-
бляя этот термин в различных смыслах. Можно представить себе
такие центры: 1) центр геометрический; 2) центр, где возник вид
и откуда он стал мигрировать; 3) центр, где вид наиболее распро-
странён и обилен. Однако эти центры обычно между собой не сов-
падают. Центр геометрический и центр возникновения могут сов-
падать лишь в том случае, если вид возник недавно в одном пункте
и может, в силу благоприятных условий, распространяться во все
стороны, т. е. это может иметь место у молодых ареалов.
Ареалы старые, особенно дошедшие от прежних геологических
времён, претерпели, конечно, ряд изменений в своей территории,
и центр возникновения вида обычно находился вне современных
границ ареала, т. е. 'вид, возникший в каком-либо пункте,
в дальнейшем проделал длинный путь миграций, упрочился где-
либо в отдалённой стране и совершенно вымер или был уничтожен
в местах своего первоначального ареала. Искать в подобных слу-
чаях центр возникновения можно лишь на основании палеобота-
нических данных.
Центр ареала геометрический (или современный) имеет вообще
очень небольшое значение, кроме тех случаев, когда ареал обуслов-
лен климатически; в таких случаях современный центр часто сов-
падает с центром обилия, гб<, как уже было указано, климатически
обусловленные ареалы на своих границах имеют значительно мень-
шее обилие индивидуумов того или другого вида. Так, многие
степные виды нашей равнины и к югу и к северу встречаются всё
реже н реже; например, ковыли (Stipa Lessingiana, S. stenophyl-
!a и др-), из двудольных — Filipendula hexapetala, Adonis verna-
lis, Gypsophila paniculata и ряд других.
Разъединение (дизъюнкция) ареалов
Причины разъединений. Выше было указано, что разъединение
.ареалов обусловливается причинами исторического характера и
нередко очень далёкое отстояние частей ареала не может быть объ-
яснено современными условиями. Причины возникновения разъе-
динённых ареалов могут быть довольно различны.
1. Один сплошной ареал мог разделиться на части вследствие
изменения климатических условий в некоторых его частях и от-
мирания- там вида.
2. Разъединение могло произойти в результате миграций ра-
стений и отмирания их в пределах прежнего ареала; так, в ледни-
ковый период в Европе древняя альпийская флора спустилась с ,
гор и была зажата между покрытыми льдом Альпами и краем на-
двинувшегося с севера великого ледйика. По окончании леднико-
вого периода и с повышением температуры эта флора, с одной, сто-
роны, стала подниматься иа горы и дала современную альпийскую
флору, а с другой стороны, вслед за отступающим ледником, стала
продвигаться к северу и дала элементы арктической флоры. П слу-
чилось разъединение ареалов, так как на среднеевропейских рав-
нинах альпийская флора совершенно вымерла.
32
3'. Разъединение могло произойти в результате вытеснения в
некоторых частях ареала одной расы другими близкими расами
без изменения климатических условий (см. вышеприведённые при-
меры формирования ареала ели и чересполосных ареалов).
4. Нередко разъединения вызваны опусканием участков суши под
уровень морец и океанов. Такое объяснение разъединения ареалов
ещё недавно широко применялось в географии растений (теория
мостов — см. выше), так как иначе объяснить многочисленные
разъединения на^отдалённых континентах представлялось невоз-
можным.
5. Разъединение ареалов произошло в результате разъеди-
нения или расхождения континентов и участков су-
ши. Этот способ разъединений Ьбъясняет сравнительно недавно
(1915) возникшая теория Вегенера. По этой теории современные
континенты являются разошедшимися частями некогда сплошно-
го континента. Эта теория превосходно объясняет большое сход-
ство флоры Европы и Северной Америки, Южной Америки и Аф-
рики, Южиой Африки и Австралии и др. Теория Вегенера
будет в дальнейшем пояснена рядом примеров.
6. Возможно скачкообразно еч расселение растений
с образованием особых отдалённых частей ареала. Это может иметь
место д.г я споровых растений с их крайне лёгкими спорами.
7. В образовании разъединённых ареалов мог участвовать и
человек, в иных случаях преднамеренно. Так, агава (Agave ameri-
сапа), кроме мексиканского ареала, имеет ещё и «искусственный»
ареал в средиземноморских странах. Сюда же относятся многие
сорные растения, попавшие в Европу и сильно распространив-
шиеся, например мелколепестник канадский (Erigeron canadensis)
из Северной Америки и др.
Кроме перечисленных причин разъединений, существуют и
некоторые другие, но они имеют гораздо меиьшее значение.
При разъединениях ареалов вопрос идет не только о разъеди-
нениях видов; мы можем говорить о разъединениях родов, се-
мейств, порядков и других систематических единиц.
Примеры разъединений. Так как разъединения играют очень
большую роль в географии растений н в филогенетической система-
тике, то необходимо привести здесь некоторые примеры их.
Разъединения находятся на различных континентах:
I . Евразийско-североамериканское разъединение — ви-
ды или другие систематические единицы одну часть ареала имеют в Европе н
Азии, а другую в Северной Америке, будучи разъединены океаном. Примеры:
все семейство Salicaceae (ивовые) — Евразия (некоторые виды в Северной Африке)
и Северная Америка; роды лиственница, пихта, ель (Евразия и Северная Амери-
ка); род вереск (Caliuna); род Ribes (рис. 8) н др.; что касается видов, то можно
привести очень большое количество примеров.
2 . Североатлантическое — одна част ареала находится в
Европе, другая — в Северной Америке, разъединённые северной частью Ат-
лантического океана. Виды: плаун (Lycopodium inundatum), осока желтая
(Carex flava), лобелия (Lobelid Dortmanna) и др.
3 . Северотихоокеанское — одна часть ареала в Азии (пре-
имущественно в восточной части), другая— в Северной Америке, разъединён-
ные Тихим океаном. Роды* катальпа (Catalpa), хвойный род Тоггеуа, замеча-
3 География растений ^3
тельное по форме своих листьев тюльпанное дерево (Linoaendron — рис. 9),
все семейство Calycanthaceae и др
4 Пантропическое — в пределах всего пояса тропиков н су б-
тропиков — Азия, Африка и Америка семейство замин ых (из саговников)
Zamiaceae
Рис 8 Ареал рода смородины (Ribes)— пример евразпйско-американ-
ского разьединения (по Хетчиисону)
5 Палеотропическое — части ареалов в пределах тропиков Ст?
рого Света — Африка, Азия Сюда относятся разъединения между Восточной
Африкой и Индией, Африкой и Мадагаскаром и некоторые другие Роды Атч-
sophylia (рис 10), Nepenthes и дг
100 120 140 150 №0 150 140 120 100 80 60
Рис 9 Дреал тюльпанного дерева (Liriodendron) В Северной Аме-
рике— L tulipifera, в Китае — L chinensis (очень близки! bi д) (по
Ферма льду)
6 Неотроп и ческое — части ареалов находятся в пределах тропи-
ков Нового Светт
7 Южнотихоокеанское — части ареалов находятся в Южной
Америке, на островах Тихого океана, в Новой Зеландии, Австралии Род Dn-
mys (рис 11)
8 Южноатлантическо е—части ареалов, разъединенные южной
частью Атлантического океана, находятся в Южной Америке, Ад>рике и
на Мадагаскаре Род Ravenala — дерево путешественников Мадагаскар —
Гвиана
34
9 А и i a p кт и чес к о е — части ареалов находятся в южных частях
Южной Америки, Южной Африки, Новой Зеландии, Австралии и на Антарк
гическом материке (на последнем в ископаемом состоянии) Род Nothofagus —
южный бук
Г яс 10. Ареал рода Anisophylla из семейства мангровых (Rhizophora-
сеае) — пргшер палеотропического разъединения (по Хетчинсону).
Рис 11. Ареал рода Dnmys —пример южнотихоокеанского разъеди-
нения (то Хетчинсону).
Не указывая других типов разъединения между мате-
риками, коснемся некоторых разъединений более частного
порядка, именно, разъединений в пределах континентов
Подобных примеров можно было бы привести чрезвычайно много
чцесь мы приведем лишь некоторые
3* 35
1. С р е д и з ? я и )иорски е разъгди нения - очень аразно-
збразны и могут быть разделены на ряд подтипов, касающ 1хся различных ча-
стей Средиземноморской области, именно; а) Италия — Балканский полуостров,
Рис. 12. Ареал Symphytum tauricum — пример одного
аз средиземноморских разъединений: Крым — Кав-
каз—Малая Азия—Балканы (по Гаевскому).
Рис- 13. Пример австралийского разъединения
ареал рода Jacksonia (из семейства бобовых) (по
’ Хетчинсону).
б) Балканский полуостров — Крым, в) северные части Средиземноморья—ост-
рова Средиземного моря и др. Сюда примыкают разъединения, близкие по
существу и территориально, но захватывающие, в пределах того м.-е Сретб) 7''''”-
поморья, различные континенты: а) Крым — Малая Азия — Закавказье, б) се-
верные части Средиземноморья — Северная Африка и др.( рис. 12).
Некоторые примеры разъединения: Крым—Малая Азия — западные среди-
земноморские страны: земляничное дерево (Arbutus andrachne), крымская сос-
на (Pinus pailasiana) н др. (рис. 120).
2. Австралийские разъединения — очень интересны,
причём одни части ареалов лежат в Западной Австралии, другие — в Восточной;
разъединяющей территорией является Центральная Австралия (рис. 13).
Что касается территории СССР, то здесь можно установить
самые разнообразные тиг.ы разъединений как в горных странах
(Крым — Кавказ, Кавказ — среднеазиатские горы и др.), так и
на равнинных территориях. Укажем лишь несколько примеров:
J) Leontice altaica — юго-западная Украина — Алтай и некоторые соседние
территории; 2) Daphne altaica — Алтай и сопредельные части Курской, Воро-
нежской и Харьковской областей (в последнем случае очень близкий вид Daphne
Sophia); 3) род Cymbaria — небольшой ареал на юге Европейской части СССР,
а затем на юге Сибири, Монголии, Манчжурии.
Наблюдаются и более близкие разъединения: так, Cymbaria borysthenica,
имеющая небольшой ареал на нижнем Днепре, другой, еще более ограниченный
ареал имеет в лрисальских степях. Целый ряд разъединений будет отмечен в
дальнейшем изложении.
Реликтовые ереаг.ы и реликты. Как было указано раньше, одни
ареалы увеличиваются в своей площади, другие, наоборот, умень-
шаются. Если площадь ареала сокращается до небольших преде-
лов, то перед нами реликтовый ареал, а данный вид — реликт.
Реликтовые ареалы наиболее очевидным образом могут быть до-
казаны при помощи ископаемых остатков. Так, реликтовость аре-
алов саговников (Cycadales) может быть бесспорно доказана палео-
ботаническими данными; также со всей очевидностью это устанав-
ливается для гинкго (Ginkgo biloba) — последнего остатка
некогда (в мезозое) хорошо представленного класса Gingkoales;
помимо культуры гинкго встречается в диком состоянии лишь в
Западном Китае. Также реликтовыми необходимо считать ареалы
многих хвойных (Sequoia, Taxodium и др.), многочисленные остатки
которых мы находим в ископаемом виде. Однако, если ископаемые
остатки хорошо сохраняются у древесных видов, то у травянистых
нередко не находят никаких ископаемых следов. В подобных слу-
чаях хорошим критерием является положение данного вида в фи-
логенетической системе; сама по себе редкость вида ещё не
доказывает его реликтовссти, но редкость, соединённая с изоли-
рованным положением в системе, является решающим фактором.
Так, для гинкго решающим является его вполне изолированное
положение в системе, даже если бы отсутствовали какие-либо иско-
паемые остатки.
Нужно заметить, что реликты (от латинского слова «релик-
тус» — оставленный), т. е. растения, оставшиеся от прошлых гео-
логических времён, не всегда имеют ограниченный, реликтовый,
ареал, но нередко распространены довольно широко; так, напри-
мер, многие виды и роды семейства вересковых (Azalea, Ledum,
Andromeda, Cassandra и др.) являются, несомненно, третичными
реликтами, но и в настоящее время широко распространены. Та-
ким образом реликтовые виды ие всегда имеют реликтовые ареалы.
37
Реликты и их возраст. Возраст реликтов может быть весьма
различен: имеются реликты мезозойские, третичные, ледниковые
и послеледниковые; несомненно, значение реликтов тем больше,
чем больший возраст они имеют.
На территории СССР имеются реликты третичные и более близ-
ких к нам времён. Третичные реликты сохранились на тех терри-
ториях, которые не подвергались оледенению в ледниковый период
и где растения могли всё время оставаться на своих местах. Сюда
относятся в первую очередь западное побережье Кавказа, область
Талыша на юго-востоке Кавказа, Дальневосточный край, отчасти
Крым. Некоторые авторы считают, что третичные элементы могли
сохраниться и на тех частях русской равнины, которые не были
покрыты ледником: на возвышенностях Среднерусской, Подоль-
ской, Приволжской и др.
Ледниковые реликты, по крайней мере на терри-г
тории Европейской части СССР, сохранились в очень незначитель-
ном количестве, так как большая часть этой территории была по-
крыта мощным ледником; значит, ледниковые реликты могли со-
храниться на месте лишь там, где ледникового покрова не
было. Сюда можно отнести, например, находки на болотах Средне-
русской возвышенности таких растений, как багульник (Ledum),
клюква (Oxycoccos palustris), брусника (Vaccinium vitis idaea) и
др. Такие виды, как низкорослая берёзка (Betula папа), во-
дяника (Empetrum nigrum, рис^ 136), пушица (Trichophorum alpi-
num) и многие другие, считаемые реликтами для ряда мест Запад-
ной Европы, у нас имеют несколько иное значение, так как все
современные местонахождения этих видов находятся в области
оледенения — все эти виды переселились сюда из других мест в
послеледниковое время.
Из числа реликтов послеледникового возраста нужно указать на
реликты ксеротермические, т. е. оставшиеся и остающиеся н а ме-
сте со времени тёплого и сухого периода (ксеротермического)—'
одного из периодов, бывших после ледникового. Реликты этого
периода представлены видами степного характера; их можно встре-
тить и севернее степной полосы в области лесов в прибалтийских
странах, в Московской, Калининской областях, даже под Ленин-
градом и пр.; эти степные виды, как можно предполагать, остались
здесь со времени ксеротермического периода.
Явление эндемизма
Эндемизм и эндемические виды (эндемики). Эндемиками назы-
вают такие виды, которые занимают ограниченный ареал в преде-
лах какой-либо области, и в других частях земной поверхности
не встречаются. Это вид, присущий исключительно данной стране
или части страны. Так, можно говорить об эндемиках Кавказа
или Крыма, об эндемиках океанических островов и т. д. Эндемизм
может быть более широкий и более узкий — можно говорить об
эндемике Кавказа, распространённом в различных его районах,
но также и об эндемике какого-либо одного района. Имеется ие-
3*
лало эндемиков, ограниченных в своём распространении крайне
1ИЧТ0ЖИЫМ районом и даже одним пунктом. Выше были указаны
некоторые примеры — эльдарская сосна, сосна Станкевича и др.
Эндемизм имеет двоякую природу: или ограниченность ареала
есть результат сокращения некогда более обширного ареала, или
же ограниченный ареал говорит о недавнем возникновении вида,
не успевщего ещё распространиться более широко.1 В первом слу-
чае мы имеем эндемизм реликтовый, или консервативный, во вто-
ром — прогрессивный. Таким образом, возраст эндемиков может
быть крайне различен, и эндемики могут быть очень моло-
дыми по своему происхождению..
Древние эндемики, конечно, являются реликтами, но эти два
понятия всё же не совпадают, так как реликты могут иметь более
широкое распространение и не быть присущими только одной ка-
кой-либо области.
Отношение числа эндемиков к общей флоре какой-либо страны
может быть весьма различным: так, процент эндемиков в ряде
равнинных областей СССР равен 0, тогда как на Новой Зеландии
он равен 72, на острове св. Елеиы — 85.
Островные флоры. Эндемизм находится в прямой зависимости
от древности данной территории и от и з о л я ц и и её
от соседних территорий, так как только при этих условиях могла
сохраниться древняя флора, не смешиваясь с флорами соседних
стран и развиваясь самостоятельно.
Классическим примером эндемизма являются флоры островов—
островные флоры. Однако богатый эндемизм обнаруживают далеко
не все острова; острова, недавно отделившиеся от суши, или океа-
нические острова молодого происхождения, обы .но совершенно
не имеют эндемиков, подтверждением чего являются молодые ост-
рова кораллового ''происхождения (Бермудские, Полинезийские
и др.). Только те острова, которые являются очень древними, име-
ют флору с резко выраженным эндемизмом. Сюда относятся как
материковые острова, давно потерявшие связь с материками (Но-
вая Зеландия, Новая Каледония, Мадагаскар и др.), так и океа-
нические острова древнего происхождения. Хорошим примером
последних являются острова Гавайские, Галапагосские, св. Еле-
ны, Канарские и ряд других. Процент эндемичных видов здесь
колеблется от 50 до 85. Здесь сохранились, с одной стороны, мно-
гочисленные древние виды, вымершие на соседних континентах
(эндемики консервативные), а с другой стороны, возникли и но-
вые виды, не распространившиеся на соседние территории вслед-
ствие изолированного положения островов (эндемики прогрес-
сивные).
Горные страны и эндемизм. Подобно островам, и горные страны
имеют ряд моментов, благоприятных для сохранения древних эле-
ментов. Здесь прежде всего имеется крайнее разнообразие условий
существования (разнообразные климатические условия, разно-
образие горных пород и почв), что сильно отражается на б о г а т-
с т в е самой флоры- Разнообразие условий ведёт к тому, что ви-
ды всегда находят для себя наиболее благоприятные условия, в
39
то время как на равнинах, даже при незначительных изменениях
климата, отдельные виды могут исчезнуть; в некоторых случаях
наступающие новые виды могут даже совершенно вытеснить ста-
рые. В горах эти последние всегда найдут для себя то или другое
подходящее убежище.
Особенно богаты эндемиками горные страны, древние геоло-
гически, так как при этом большую роль играет изолированность
их. Горная цепь и окружающая её равнина в данном случае могут
быть уподоблены острову, затерявшемуся среди океана.
В качестве примера горных стран приведем Кавказ. Из 5767 видов, известных
там в настоящее время (невидимому, Кавказ имеет не менее бОООвндов), эндемн-
ки составляют 1153 вида, или 19,8% всей флоры. По отдельным естественным
районам Кавказа эндемики распределяются неодинаково: особенно богаты энде-
миками горные (высокогорные) районы, по сравнению с районами низменными
(степными л пустынными). Так, в районе Большого Кавказа имеется 369 эндеми-
ков, а в северных предгорьях, степях и в пустынных равнинах по р. Куре со-
ответственно 35,18 и 42. Еще более показательны относительные числа в процентах
от общего числа видов каждого района. Что касается эндемичных ро-
дов, то их для Кавказа насчитывается десять. Эндемики Кавказа явля-
ются или консервативными или прогрессивными.
Эндемизм прогрессивный и викарирукщие виды. 'Нередко в
горных областях, в результате изоляции отдельных районов, тот
или иной вид разделяется на ряд более мелких видов, каждый из
Рис. 14. Ареалы викарирующих видов цикла Arabis petraea. Распростра-
нение каждого вида на карточке обозначено особым значком или штриховкой
и географически почти всегда обособлено одно от другого (гну Бушу).
которых занимает отдельный район. Один ареал родительского
вида разбивается, таким образом, на ряд более мелких ареалов.
Подобные виды и их ареалы называются викарирующими, или
замещающими (от-латинского слова «викариус» — заменяющий,
замещающий). Классическим примером являются викарирующие
виды рода горе чавки (Gentiana) в Швейцарских Альпах (Ветт-
ш те й н). Можно привести и другие примеры (рис. 14).
40
Вновь возникающие, таким образом, виды являются, конечно,
эндемиками для данного района,*но уже эндемиками прогрессивными.
Однако викарирующие виды возникают не только в условиях
пространственной изоляции, они могут возникать и в пределах
одного п того же района. В этом случае изолирующим фактором
будет разнородность почвенно-грунтовых условий. Так
если в каком-либо районе имеются чернозёмные, песчаные i
известковые почвы, то вполне возможно возникновение из одною
родительского вида соответствующих рас или молодых видов
приуроченных к данным почвам.
Подобный процесс может иметь место и на равнинах, и русские
ботаники уже давно указывали
нередких в степной полосе.
Черноземные виды
Thymus Marschallianus
(чабрец степной)
Koeleria gracilis
келерия степная)
Festuca sulcata
(типчак)
Silene densiflora
(смолевка)
на «замещающие пары» видов,
Песчаные" виды
Thymus odoratissimus
(чабрец песчаный)
Koeleria glauca
(келерия сизая)
Festuca duriuscula
(овсяница)
Silene parviflora
(смолевка мелкоцветная)
Можно было бы привести и другие многочисленные примеры
викарирующих видов.
В последнее время на основании материала южной части на-
шей степной полосы наметилось очень большее число викарирую-
щих видов с почвенной изоляцией. Так, например, установлен ряд
мелких видов чабреца, связанных с различными субстратами (мел,
гранит, пески и др.), но и в пределах одного и того же субстрата
возможно возникновение викарирующих видов; установлено, что
песчаный вид василька (Centaurea margaritacea) даёт эндемиче-
ские более мелкие виды, различные в пределах речных систем Буга,
Днепра и Дона (по их песчаным берегам), и более того, в пределах
каждой из указанных речных систем песчаные виды борового лево-
бережья отличны от песчаных видов правобережья, носящего бо-
лее южный характер. В результате установлено более 10 эндемич-
ных видов. Вообще последние данные по пескам юга степной поло-
сы обнаружили энергичный процесс видообразования, имеющий
место именно на песках.
Элементы флоры
Изучая ареалы растений, легко можно видеть, что онн до край-
ности разнообразны в своих конфигурациях; можно почти навер-
ное сказать, что нет двух ареалов, которые были бы тождественны
в своих границах. Но, с другой стороны, одни ареалы имеют боль-
шое сходство в своём расположении и очертаниях, другие же со-
вершенно исключают друг друга. Можно сгруппировать ареалы
со сходным восновных чертах размещением на земной
поверхности. Группы видов с более или менее совпадающим раз-
мещением их ареалов представляют определённые элементы фло-
ры, причём каждая группа — это особый элемент.
В данном случае, мы говорим о географическом элементе, так
хак здесь для сгруппирования видов критерием служит лишь гео-
графическое размещение; но иногда анализируют флору с точки
зрения элементов генетических или исторических-^ при этом генети-
ческий элемент — это группа видов, имеющих общую родину, а
исторический виды, пришедшие на данную территорию в одно
и то же время. Установление элементов двух последних категорий
нередко представляет очень большие трудности.
Рис. 15. Местонахождения крупки (Draba g'acialis) — арктический элемент;
распространена также в арктической Америке (по Бушу).
Географический элемент флоры устанавливается, как сказано,
на основе современных ареалов, независимо от их ге
зиса, но он является отправным моментом для дальнейшего анали-
за флоры с точки зрения её происхождения.
Главнейшие элементы флоры СССР. Подразделение флоры
СССР на географические элементы крайне сложно, к тому же для
очень многих видов мы не располагаем достаточными данными;
поэтому лишь для примера можно указать главнейшие географи-
ческие элементы флоры:
1. А р кт и че с к и й — группа видов, ареалы которых находятся
на крайнем севере, в зоне материковых тундр и на арктических ост-
ровах. Он распадается на ряд более дробных элементов, например,
западно-арктический и восточно-арктический. С другой стороны,
некоторые арктические виды имеют части своих ареалов (дизъюнк-
ции) на Кавказе, Алтае н’др., поэтому можно говорить об
элементах арктическо-кавказском, арктическо-альпийском и др.
(рис. 15).
2. Северный (или бореальный) — группа видов
с ареалами, расположенными в основном в пределах северных ча-
стей лесной области, именно в области хвойных лесов. Здесь тоже
имеются дальнейшие подразделения: евробореальный — лишь в
Европейской части, сиббореальный — в Сибири и др.
42
3. Среднеевропейский — группа видов с ареалами
а Средней Европе, заходящая восточными отрезками своих ареалов
в западную часть Союза, в.отдельных случаях заходящая даже за
Урал. В основном эта группа более теплолюбивая сравнительно
с предыдущей, распространена в области широколиственных лесов.
43
Примеры: обыкновенный дуб — доходит до Урала (рис. 111),
остролистный клён (а также клёны полевой и татарский), ясень,
граб, бук (рис. 108), зимний дуб (Quercus petraea), такие травя-
нистые виды, присущие широколиственным лесам, как копытень,
петров крест (Lathraea squamaria), медуница (Pulmonaria offici-
nalis — рис. 16) и др,
4. Атлантический — почти отсутствует на территории
СССР, так как в этот элемент входят виды атлантических" при-
40 ' 30 да Ю О Ю 20 30 Л? (W ~
Рис. 47. Ареал'горицвета (Adonis vernalis) — пример понтического элемента
В Западной Европе — отдельные местонахождения (точки) (по Штернеру с
изменениями Алехина).
брежных частей Европы (рис. 135): всё же некоторые виды более
продвигаются на восток; из растении, растущих на вашей тер-
ритории, можно указать лобелию (Lobelia Dortmanna), восковник
(Myrica gale).
5. Понтический — группа видов, главным образом, юж-
норусских степей (рис. 17), но встречающихся также в румынских
и венгерских степях (если виды встречаются в основном в венгер-
ских степях, то это паннонский элемент). Сюда относятся много-
численные виды наших степных пространств: горицвет (Adonis
vernalis), чистец (Stachys recta), фиолетовый коровяк (Verbascum
phoeniceum), жёлтая скабиоза (Scabiosa ochroleuca), степная виш-
ня (Prunus fruticosa), ракитник (Cytisus ruthcnicus) и т. п. Пан-
нонский элемент у нас представлен очень слабо.
44
6. Сарматский — объединяет виды, занимающие терри-
торию между южными степями и хвЬйными лесами на севере, не
заходя далеко на запад, за западные границы Союза. Это виды
менее теплолюбивые сравнительно с понтическими. Несколько
примеров: горошек (Vicia pisiformis), фиалка холмовая (Viola
collina), астрагал песчаный (Astragalus arenarius)1.
7. Средиземноморский — группа видов, распро-
странённых в сухих областях, окружающих Средиземное море,
и на востоке произрастающих на побережьях Чёрного моря — в
Р __________ 30
Рис. 18. Ареал самшита (Buxtis sempervirens) — пример средиземно-
морского элемента. Точки —современные местонахождения, треуголь-
ники-местонахождения, уничтоженные культурой (по Христу).
Крыму и на Кавказе (здесь также в прикаспийских районах). Де-
ревья и кустарники с вечнозелеными кожистыми листьями и сухо-
любивые травы. Примеры: земляничное дерево (Arbutus andrach-
пе), самшит (Buxus sempervirens) (рис. 18), сумах (Rhus coriaria),
дикий жасмин (Jasminum fruticans) и т. д. Некоторые авторы в сре-
диземноморский элемент'включают также переднеазиатский и цен-
тральноазиатский элементы.
8. Переднеазиатский. Сюда относятся виды, имею-
щие ареал в странах Передней Азии — от границ Ирана на восто-
1 В последнее время некоторыми русскими авторами сарматский элемент трак-
туется в ином смысле: под названием этого элемента объединяют виды, охваты-
вающие своими ареалами главным образом северный Казахстан (и отчасти юж-
ную Сибирь): это в общем степные виды. Лучше, во избежание путаницы, гово-
рить здесь о севера казахстанском элементе.
45
ке до берегов Средиземного моря. В основном — это растения сухих
горных стран. Распадается на ряд элементов более узкого значе-
ния, из которых отметим иранский, в общем совпадающий с Иран-
ским наюрьем и заходящий в наши пределы в Закавказье.
9. Центральноазиатский — приурочен к Сред-
ней Азии, к её крупным горным цепям (Тянь-Шань, Памиро-Алай,
Тарбагатай, Алтай). Он очень сложен и распадается на ряд более
дробных элементов.
10. Т у р а н с к и й — объединяет группу видов, ареалы ко-
торых в основном связаны с пустынями Туранской низменности
Рис. 19. Ареал белой полыни (Artemisia terrae albae) —
туранскнн элемент (по Е, Коровину).
Средней Азии. Элемент пустынного характера. В основных чертах
это арало-каспийский элемент некоторых авторов, который, однако,
обычно понимается несколько шире. Типично туранским элемен-
том является группа среднеазиатских пустынных полыней (Arte-
misia) (рис. 19). <
11. Манчжурский — группа видов, имеющая основную
площадь ареала в Манчжурии и заходящая в южные части Даль-
невосточного края. Ряд деревьев и кустарников широколиствен-
ного типа: манчжурский орех (Juglans manshurica— рис. 118),
манчжурская аралня (Aralia manshurica), бархатное дерево (Phel-
lodendron amurense — рис. 119), разнолистная лещина (Corylus
he ter ophy 1 la) и др.
Элементы флоры Кавказа. Специально для Кавказа можно ука-
зать еще некоторые географические элементы, пространственно
более ограниченные:
12. Кавказский—состоит из видов, связанных своими ареалами
с Большим Кавказом; сюда входят кавказские эндемики (лесные и альпий-
ские).
13. Колхидский — группа видов, имеющих свой ареал в Колхид-
ской провинции Кавказа, т. е. в западном Закавказье (Аджария, Абхазия в
более северное побережье). Виды лесные, известняковые, горно - луговые
46
Большинство составляют древ'ше по своему генезису элементы (третичнйе).
понтический дуб (Quercus pontica), рододендрон (Rhododendron Smirnowii),
берёза (Betula Medwedewji) и др.
14. Гирканский— виды, занимающие крайний юго-восток Кавказа,
но главная масса видов сосредоточена за границей—в северном Иране. Древ-
ние третичные элементы (главным образом лесные виды): Parrotia persica,
гледичия (Gleditschia caspia), шелковая акация (Albizzia julibrissiп), род
Danae и др.
Приведённый перечень географических элементов СССР дале-
ко неполон, к тому же анализ нашей флоры в этом отношении еще
не завершен.
В заключение нужно указать, что географические элемент! t
на какой-либо территории не исключают друг друга, но любая
флора состоит из нескольких элементов, так как ареалы последних
перекрывают друг друга.
Для примера дадим анализ флоры Большого Кавказа (по Гросс-
гейму): всего здесь насчитывается 1 261 вид, которые ^распадаются
на такие элементы, выраженные в процентах:
Колхидский................................12,7
Бореальный (северный).....................38,2
Понтический............................... 5(б
Средиземноморский.........................13,2
Переднеазиатский.......................... 9,7
Тура некий................................ 0,1
Кавказский (собственно)...................20,0
Пришлые виды ............................. 0,5
Данный пример представляет «спектр географических элемен-
тов» и показывает роль тех или иных элементов в сложении флоры;
так, можно видеть, что на Большом Кавказе, центре видообразо-
вания кавказских видов, последних имеется 20%,’вся же осталь-
ная флора имеет свои центры на других территориях.
Число видов в различных частях земной поверхности
В настоящее время число видов на земной поверхности, неви-
димому, надо считать не менее 500 ООО; из этого числа покрытосе-
менные составляют около 160 000, грибы — около 200 000. По всей
вероятности, при более полном изучении флоры число покрыто-
семенных увеличится до 200 000 видов.
Установление более точных цифр связано с рядом затруднений:
1) флора ещё недостаточно изучена, особенно это касается низших
организмов; 2) представление о систематическом объёме вида не-
сколько субъективно, что приобретает особенное значение в послед-
нее Бремя, когда наблюдается усиленное дробление крупных ви-
дов на более мелкие (один крупный вид может распадаться на не-
сколько десятков более мелких); поэте му систематическое значение
исчисляемых видов равноценно, и раздробление ещё неподраз-
делёкных видов приведёт, конечно, к новым цифровым данным.
Закономерности в распределении видов. Распределение видов
по различным странам и территориям земного шара крайне неодно-
родно, но подчинено известным закономерностям. Однако все приво-
димые цифровые данные имеют ряд указанных выше недостатков,
47
:юда же присоединяется ещё различие в площадях территорий,
сравниваемых между собой. Тем не менее с подобными дефекта-
ми числа видов являются вполне показательными. Приведём неко-
торые цифровые данные для различных частей СССР (цифры
относятся лишь к цветковым растениям):
I. Северные территории
Число видов
Земля ненцев............................. 342
Полуостро г Канин............................265
Новая Земля..................................200
Земля Франца-Иосифа...........................37
Таймырский полуостров....................... 200
Остров Диксон . 104
Между нижним течением Лены и Хатанг........450
Земля чукчей (Анадырский округ)..............180
Таким образом, в северных районах число видов не поднимает-
ся выше 500, а собственно в тундрах не доходит даже до 300'.
//. Средние территории
Число видов
Восточная по швина Калининской области . . . 930 (приблизительно)
Московская область . . . . ,..................1 100
Южная половина Горьковской области..............I 100
Тамбовская область .........................1 150
Орловская область............................. 1 120
Курская область.................................I 300
Здесь цифры колеблются около 900—1 ЗОЭ, причем наибольшее
количества видов приходится на самую южную Курскую область.
/У/; Южные территории
Число видов
Херсонщина .............................около 1 500
Область среднего н нижнего Допа........около 1 600
Крым.........-...............................2 010
Кавказ....................................... 5 767
Средняя Азия (горная часть!................ .6 000
Средняя Азия (пустынная часть)................600
В южных частях число видов ещё увеличивается и достигает
очень больших цифр на Кавказе и в горных частях Средней Азии.
На основании приведённых цифр, хотя они и не являются аб-
солютно сравнимыми, можно установить несколько общих законо-
мерностей:
1. Число видов определённо повышается при движении к ю г у,
что является несомненным следствием улучшения климатических
условий.
2. Число видов на равнинах резко падает с увеличением с у -
хости воздуха, например в пустынях Средней Азии. Это име-
ет место везде при сравнении областей с морским и сильно конти-
нентальным климатом.
3. Горные страны (даже если они сухие) имеют исключительное
богатство видов. Причины были указаны выше: разнообра-
зие климатических, грунтовых, почвенных условий, роль гор,
как убежищ для сохранения древних элементов флоры и др.
48
4. Последнее обстоятельство, что горы могут служить убежи-
щем древних флор, приводит к установлению еще того факта, что
древние территории флористически богаче территорий,
геологически более молодых.
Богатство видами более древних участков суши свойственно
не только горным странам, но также и территориям более или ме-
нее равнинного характера. Можно привести такие цифры для юго-
западной Украины (по Пачоскому), где даётся количество видов,
свойственных лишь той или иной части территории:
Число ВИДОВ
Лесостепь подольского типа.....................224
Причерноморские степи ..................... . . 129
Полесье ........................................94
Первое число почти вдвое превышает второе и третье, так как
область подольской лесостепи представляет более древнюю сушу
сравнительно с другими областями.
Выводы, к которым мы пришли в результате» некоторого анализа
флор Союза, вполне приложимы к тем отношениям, которые име-
ют место вообще на земной поверхности.
Прилагаемая карта (по Вульфу) даёт представление о раз-
мещении флор с различным количеством видов (рис. 20).
Особенного внимания заслуживают огромные количества ви-
дов, характерные для стран тропических и субтропических.
Вот некоторые данные для ряда стран:
I. Азия
Число видов
Индия......................................21 СЯ0О
Остров Борнео...............................11 000
Филиппинские острова........................10 000
II. Африка
Тропическая Африка..........................13 000
Капская Земля...............................12 000
Сахара...................................... 300
III. Америка
Южная Мексика.....................................7 000
Куба..............................................7 000
Бразилия....................................40 000
Обращает на себя внимание исключительное богатство видами
тропических стран, особенно Бразилии. Здесь имеются две основ-
ные причины: 1) крайне благоприятные климатические условия и
2)ненарушаемое существование раститель-
ности с древнейших в р е' м ё н.
Конечно, простое сопоставление всех этих цифр, как уже ука-
зывалось, не может быть сделано; так, цифры для Бразилии (40000)
и Курской области (1 300) несравнимы, так как площади их крайне
неодинаковы; однако всё ^ке приблизительные соотношения полу-
• 49
4 География растений
чить можно, если сравнить Бразилию с СССР; так, весь Союз име-
ет лишь около 14 000 видов, при достаточно дробном их понимании.
Площадь же нашего Союза — одна шестая часть земной поверх-
ности — в 21/2 раза больше Бразилии. Таким образом, богатство
видами Бразилии выступает крайне ярко. То же нужно сказать
и относительно Индии, тропической Африки и др.
Соотношения систематических единиц, флористические спектры
и пропорции. Чтобы дать флористическую характеристику какой-
либо страны, нужно иметь данные: 1) о числе видов, 2) о числе
эндемиков, 3) о географических элементах и их соотношениях.
Необходимы также и некоторые другие данные, прежде всего —
соотношения числа видов таких систематических единиц, как се-
мейства.
Так, например, для арктической области указывается такой
нисходящ! й ряд семейств по степени уменьшения в них числа ви-
дов: осоковые (Сурегасеае), злаки (Gramineae), крестоцветные
(Cruciferae), гвоздичные (Caryophyllaceae), лютиковые (Ranuncu-
laceae), камнеломковые (Saxifragaceae), вересковые (Ericaceae) и
сложноцветные (Compositae). На первом месте, как видим, стоят
осоковые и злаки, на одном из последних (приведены не все семей-
ства) — сложноцветные. .
Для Кавказа порядок семейств получается совершенно иной
(по Гроссгейму):
Число о.
ВИДОВ '°
Сложноцветные............................ 774
Бобовые............................ . . 542
Злаки.....................................420
Крестоцветные.............................323
Зонтичные.................................300
Гвоздичные................................283
Лилейные................................. 258
Розоцветные.............................. 237
Губоцветные...............................232
13,3
9,9
7,3
5,5
5,2
5,0
4,4
4,1
4,0 и т. д.
Даже из сопоставления двух приведенных примеров видно,
что господствующие семейства в различных областях различны
так, на Кавказе выдающуюся роль играют сложноцветные, в аркти-
ческих странах они находятся на последних местах; на Кавказе
Большую роль играют бобовые, зонтичные, лилейные и др., в ар-
ктических же стр< нах эти семейства не имеют никакого значения,
и т. д.
Представляя флору страны в процентных соотношениях всех
имеющихся там семейств, мы получаем «флористический спектр».
Можно дать спектры и более крупных систематических единиц
(порядки и др.).
Иногда составляют для разных стран сравнительные соотношения числа ви-
дов, родов и семейств (так называемые пропорции).
Семейства Р оды В и ды
Средняя Европа...................
Западная Австралия ..............
Кергеленекие острова ............
120 800 3 500
98 618 3 700
14 18 21
51
4*
Соответствующие пропорции будут:
1:6,6: 29,2
1:6,0: 37,0
I : 1,3 : 1,5
т. е. на каждое семейство в первом случае приходится 6,6 рода и 29,2 вида,
а в последнем — 1,3 рода и 1,5 вида. Причины бедности видами в пределах
родов и семейств на островах могут быть двоякого рода; это или результат
молодости островов и недостаточности времени для поселения других видов,
или же это показывает (на отдалённых океанических островах) древность
флоры, в которой сохранились лить немногие виды в различных семействах;
для решения вопооса необходимы данные относительно эндемизма этих видов.
Родовой коэфициент. Он представляет отношение числа видов к числу
родов на определенной территории. Эго отношение может быть выражено
двояким образом: или взять за основу число видов, приходящихся в сред-
нем иа один род, или же каждый раз исходить из 100 видов и указывать, по
скольким родам эти виды распределяются. Так, если флора какого-либо
участка содержит 106 видов, распределяющихся по 99 родам, то по первому
способу каждый род будет иметь в среднем 1,18 вада (106: 90),
по второму же способу нужно составить уравнение 103:90= 100 : х, т. е.
если на 106 видов приходится 90 родов, то сколько же родов придётся на
100 видов? В данном примере х — 85; таким образом, 100 видов распреде-
ляются по 85 родам.
Последний способ более удобен и нагляден, в то время как первый
да£т по большей части дроби, а самые числа колеблются между 1 и 6, т. е.
в очень небольших пределах.
Отношение числа родов к числу видов обычно обозначают как родовой
коэфициент-, в указанном примере он равен 85%. Впервые указанные отно-
шения быт и выведены Гумбольдтом.
Интересна связь родового коэфицченга с величиной исследуемой терри-
тории. В приводимых ниже примерах даются территории, уменьшаю-
щиеся по своей площади (по Жаккару).
Вся флора Швейцарии...................на
Область Триента .......................»
Область массива Вильдгори . . ........»
Только альпийские луга (нескольких областей)»
Альпийские луга (только Юры)...........»
Только один луг на Юре.................»
100 видов 26 родов коэф.
100 » 45 » >
100 » 50 » »>
109 » 57 » »
‘00 » 60 > »
100 » 85 » »
26%
45%
50%
57%
60%
85%
В данном примере коэфициент повышается от 26 до 85%, а с другой
стороны, соответствующие площади уменьшаются в своих размерах (вся
Швейцария — и один луг). Так как ясно, что чем больше по площади тер-
ритория, тем внешние (экологические) условия разнообразнее, то отсюда
следует такая закономерность' родовой коэфициент обратно пропор-
ционален разнообразию экологических условий.
Можно предпотагать, чтс участки, наиболее однородные по своим эко-
логическим устовням, будут иметь родовой коэфициент равным 10,%. г. е.
каждый род будет представлен одним видом. И действительно, на площад-
ках'в 109 м2 в лесах и иа болотах крэфициент нередко составляет 100%,
в степях же, при огромном богатстве флористического состава, он не под-
нимается выше 80%.
Родовой коэфициент очень высок также на древ я их (океанических и мате-
риковых островах), где господствуют м о и о т и п н ы е роды (т. е- роды
с одним видом). Здесь, однако, основная причина—вымирание видов н
сохранение лишь немногих.
ОТДЕЛ II
УЧЕНИЕ О ВЗАИМОСВЯЗЯХ МЕЖДУ РАСТЕНИЯМИ
И СРЕДОЙ (ЭКОЛОГИЯ)
Изменчивость растений
Растительные организмы крайне сильно зависят от тех внешних
условий, которые их окружают; пределы растительной жизни
достаточно широки: растения могут существовать и на далёком се-
ьере при крайне низких зимних температурах (до—60° и более);
могут расти и в сухих знойных пустынях, где температуры на по-
верхности почв достигают 80°; могут существовать в горячих источ-
никах и в холодных северных морях, в глубинах морей и на высо-
чайших рсрах. Можно думать, что на земной поверхности нет вообще
мест, где не могли бы существовать растения; если, однако, >их нет
на далёком севере и юге, то этому мешают вечные снега и льды, но
каждый участочек суши, освобождающийся на короткое полярное
лето ото льда, имеет те или иные виды растений.
Раз мы имеем такую широкую амплитуду внешних условий, то
естественно, что растительные организмы должны были к этим усло-
виям соответственно приспособляться, и, конечно, растения тропи-
ков, полярных стран, водных глубин или засушливых пустынь
должны физио комически и экологически представлять совершенно
разные типы.
Но и в пределах одного и того же вида на-
блюдается обычно очень значительная изменчивость, так как различ-
ные особи одного и того же вида могут произрастать в достаточно
различных условиях: одно растение растёт на более сухом месте,
другое — на более влажном, одно растёт на полнсмсолнечном свету,
Другое —в тени; более плодородные почвы способствуют более
пышному росту, более тощие дают угнетённые экземпляры, и т. д.
Повседневные наблюдения убеждают нас в этом: комнатные рас-
тения, выросшие на окнах, обращённых на юг и на север, очень
сильно отличаются по своей внешности; унавоженная грядка в саду
является условие^ для получения хороших овощей, для развития
пышно цветущих растений и пр.
В ряде случаев можно наблюдать, что даже о д и н и тот
ж е экземпляр перестраивается в своём развитии и внешнем виде
при изменении внешних условий. Так, комнатные растения
при перестановке нх со светлого окна на тёмное, будут сильно
53
реагировать на изменение условий, меняясь в своих внешних фор-
мах. То же имеет место и в тех случаях, когда высыхает водоем, и
растение водоема оказывается в совершенно других условиях. Все
время внешняя среда влияет на растения формирую ще.
В природе можно подобрать ряд особей одного и того же вида,
выросших в различных внешних условиях и настолько не похожих
друг на друга по своим вегетативным частям, что их легко можно
принять за различные виды, в то время как все это результат воз-
действия окружающих условий; эти воздействия очень разнообразны
и многочисленны.
Тот отдел ботаники, который изучает взаимоотношения между
растениями и внешней средой, называется экологией. Морфолог
изучает и классифицирует внешние формы растений и закономерно-
сти их внешнего и внутреннего сложения; эколог ставит эти формы
в связь с внешними условиями, стараясь их объяснить, исходя из
последних.
Однако эколог, как было уже указано выше, изучает ие только
связь отдельных растений со средой (аутэкология), объектом иссле-
дования может быть установление связи и взаимоотношений между
средой и сочетаниями растений, таких, как леса, луга,
степи, болота и т. д. (синэкология), которые тоже обнаруживают
очень тесную связь с внеш-
ними условиями, представ-
ляя вполне закономерные
сочетания растений (подроб-
нее о сочетаниях растений
см. ниже). Однако практи-
чески эти два отдела разъ-
единить очень трудно.
Среди признаков расте-
ний можно различать при-
знаки двоякого рода: одни,
Рис 21 Изменение формы листьев в зайи- которые представляют так
самости от условий среды — надводные и называемые систематические
подводлье листья—у американского вод- признаки, признаки наслед-
ного растения Cabomba (из Хетчидсопа) ственные, обычно не изме-
няющиеся при различных
внешних воздействиях (например форма венчика, число тычи-
нок, форма пестика, плода и т. п.), и другие —очень пластич-
ные, чутко улавливающие изменение внешних условий [величина
и форма листьев (рис. 21), интенсивность опушения, длина междо-
узлий, форма крон деревьев (рис. 22) ит. п] Первые называются
обычно организационными, вторые—приспособительными. Если эко-
логическое значение первых чаще всего познается путем наблюде-
ний и рассуждений, то вторые могут служить объектом для экспери-
ментирования. Граница между признаками первого и второго родов
достаточно условна.
Значение экспериментальной экологии огромно —здесь имеется
возможность изменять по нашему желанию растительные формы и
направлять их по заранее избранному плану. Разрешение вопросов
54
луговодства, лесоводства,
связанных с экологи-
ческими проблемами,
а также вопросов
подбора засухоустой-
чивых и морозостойких
форм, акклиматизации
и продвижения культур
в тундры и в пустыни и
многих других важней-
ших проблем, в част-
ности создания новых,
практически ценных
сортов растений, не-
мыслимо без глубоких
экологических иссле-
дований (в соединении
с работами генетиче-
скими, морфологиче-
скими и др.).
полеводства и других) теснейшим образом
Рис. 22 Сосны, выросшие на открытом месте
(справа) и в лесу (слева) близ Тимирязевской
с.-х академии, под Москвой (ориг.).
Жизненные формы
Если систематика имеет дело с растительными видами как основ-
ными единицами, то для экологии систематическое родство не имеет
главного значения, — экология имеет дело с жизненными
формами. Жизненная форма—это результат длительного
приспособления растения к местным условиям существования,
выраженный в его внешнем облике.
Так как влияние одних и тех же внешних условий на растения
совершенно различных систематических групп может дать один и
тот же внешний эффект^ то, следовательно, жизненная форма не
имеет ничего общего с систематическим родством. Если десять
тычинок—почти неизменный систематический признак бобовых,
совершенно не зависящий от тех условий, где эти представители
бобовых живут (теплый, холодный климат, сырые, сухие места
и т. д ), то по своим жизненным формам бобовые, в зависимости от
различия внешних условий, в которых они находятся, представляют
очень большое разнообразие: они могут быть деревьями, кустарни-
ками, лианами, травами; эти же последние в свою очередь распа-
даются иа жизненные формы большего или меньшего значения.
Особенно разнообразны жизненные формы среди травянистых
растений Растения степей по своему облику ничего общего не имеют
с растениями водных местообитаний и болот; так, например, в степях
растения весьма различных семейств дают нередко такую характер-
ную жизненную форму, как «пеоекати-поле», имеющую почти пра-
вильную форму шара; погруженные в воду растения (например
рдесты, зостера и др ) дают длинные тесьмовидные или расчленен-
ные на узкие многочисленные участки листья (уруть, роголистник
и др.). Общеизвестна кактусовидная жизненная форма (рис. 23),
55
имеющая сочный стебель, лишённый нормальных листьев и покры-
тый колючками (метаморфоз листьев) — это форма пустынь; но
интересно, что ту же кактусовидную форму в соответствующих
сходных условиях выработали не только представители семейства
кактусовых, но и представители других семейств, совершенно не-
родственных (например молочайные, ласточковые и др.). Лишь
вовремя цветения можно видеть, что столь сходные
внешне растения относятся
к совершенно различным се-
мействам.
Лишь очень редко жиз-
ненные формы связаны с
систематическими подразде-
лениями; сюда относится
семейство нимфейных, все
представители которого яв-
ляются водными расте-
ниями и имеют характер-
ные округлые плавающие
листья (наши жёлтая кув-
шинка , белая водяная ли-
лия, тропическая виктория-
регия и др.).
Уже давно ботаники де-
лали попытки выделить
среди огромного разнообра-
зия растительных видов
некоторые основные «расти-
тельные формы» (иногда
называемые также основ-
ными формами»), исходя
прежде всего из физионо-
мического (ландшафт-
Фис. 23. Кактусовидная жизненная форма
У растений различных семейств:
bchmocereus Blankii-^верхний слева (сем. нан-
ЧД совых); Heurnia Penzigii—в середине (сем.
ласточниковых); Echldnopsis ccreiformis—ншнний
слева (то же); Euphorbia beaumieranar—справа
(Сем. молочайных) (по Веттштейну и др.).
ного) момента. Так, ещё в самом начале прошлого столетия «отец
географии растений» Александр Гумбольдт впервые
установил 19 форм растений, физиономически очень отличных друг
от друга. Вот некоторые из них:
I. Форма пальм-4 высокий тонкий ствол с пучком перистых или
пальчатых листьев (рис.
2. Форма ба и а — невысокий ствол с большими расщепленными
листьями, как у банановых (рис. 24).
3. Форма мимоз— деревья чаще всего с зонтиковидной кроной и
многократно-перистыми листьями.
4-Форма хвойных деревьев.
5. Форма кактусов и ди а я — округлый пли цилиндрические
сочный стебель с колючками, без листьев (рис. 23).
6. Ф о р м а л и а н — деревянистый тонкий ствол, взбирающийся на другие
растения и пользующийся ими как подпорками (рис. 82).
7. Форма лавровых деревьев — сравнительно невысокие де-
ревья с широколанцетными вечнозелёными кожистыми листьями.
8. Форма злаков и дная — травянистые растения с узкими длин-
ными линейными листьями (злаки, осоки, ситники и др.)
Из приведенных примеров видно, что здесь не играет роли при-
надлежность растений к тем или иным систематическим группам;
так, лианы могут относиться
к самым разнообразным се-
мействам, также лавровые, как-
тусовидные и т. д.
Системы жизненных форм.
После Гумбольдта более раз-
работанную систему жизнен-
ных форм дал Гризебах (1872)—
он устанавливает уже ,54 фор-
мы (в дальнейшем—60), сгруп-
пировывая их в семь основ-
ных типов: деревянистые рас-
тения, суккуленты, лианы,эпи-
фиты, травы, злаковидные
травы, клеточные растения.
Деревянистые растения делят-*
ся в свою очередь на ряд жиз-
ненных форм —деревья, ку- ?
стар ники и др., травы—на
травы с олиственными стебля-
ми, с безлистными стеблями и
бесстебельные травы с розеткой
прикорневых листьев (папорот-
ники) и т. д.
В дальнейшем многие авто-
ры давали системы жизненных
форм (X у л ьт, В а р м и н г,
Друде, Раункиэр, Гаме,
Дю-Риэ и др.), детализируя их
в большей или меньшей степени.
В настоящее время имеется
много систем , жизненных
форм, причём число этих форм
доходит у некоторых авторов
до 55 (например система Друде) с целым рядом подразделений.
Не касаясь жизненных форм всего СССР, укажем лишь основ-
ные формы, характерные главным образом для равнин Европейской
и Азиатской частей Союза.
Рис. 24. ФизионТОшческая форма
«банана»—дерево путешественников
(Ravenala madagascariensis).
А. Жизненные формы с деревянекицима*стеблями
1. Деревья. Многолетние растения с деревянеющими многолетними
надземными частями; ясно выражен ствол с ветвями, на определённой высоте
образующими крон}(. Обычно выше 2 м:
а) летнезелёные деревья, с более или менее широкими, опадающи-
ми на зиму листьями; большинство наших деревьев;
б) вечнозелёные деревья с более или менее широкими листьями;
встречаются лишь в более южных частях: Крым, Кавказ и др. (лавр, зем-
ляничное дерево);
в) вечнозелёные деревья с игловидными листьями-хвоями; наши
хвойные деревья:
57
2. Кустарники. Многолетние растения с дерев имеющими над-
земными частями; ствола нет, так как ветвление начинается у самой земли, в
результате чего образуется несколько более или менее тонких стволиков. В
лесах образуют ярус подлеска. Высота (чаще всего) от 0,8 до 3 м. Распадаются
на такие же группы, как и деревья:
а) летнезелёные — большинство наших кустарников (орешник, кру-
шина, бересклет, калина и т. п.);
б) вечнозеленые лиственные—в Крыму и на Кавказе (веч-
нозелёная крушина, лавровишня, рододендрон и пр.); на равнинах СССР не
встречаются;
в) вечнозелёные х в-о й н ые — виды можжевельника.
3. Кустарнички, Как кустарники, но значительно меньших разме-
ров, обычно не выше 0 ,5 м. Располагаются в травянистом ярусе и при описаниях
нередко находятся в списках «травянистого покрова» (что, конечно, неверно).
Распадаются на те же группы, что и предыдущие основные растительные
формы:
а) летиезел ё и ы е — эта форма реж других (черника, голубика);
б) вечнозелёные лиственные с более или менее широкими
листьями (большинство кустарничков наших сфагновых болот: брусника, ан-
дромеда, Кассандра, багульник и пр.);
в) вечнозелёные с игловидными или чешуйчатыми
листьями (из наших растений — вереск); сюда относятся растения верес-
кового или эрикоидного («Erica» — вереск — род, имеющий многочисленные
виды) типа.
В последних подразделениях можно различать ещё прямостоячие формы от
ползучих; так, в подразделении «а» ползучими формами будут некоторые по-
лярные ивы, в подразделении «б» — клюква с её ползучими стеблями и вечно-
зелёными листьями.
Б. Жизненные формы переходного типа — между деревяиеющими и травя-
нистыми или имеющие представителей обоих типов
Если предыдущая группа заключала жизненные формы с деревяиеющими
многолетними стеблями (стволами) — деревья и кустарники, если
следующая заключает формы с травянистыми однолетними стеблями — т р а-
в ы, то имеется ряд жизненных форм, которые, строго говоря, нельзя отнести
ни к той, ни к другой категории. Так, например, виды грушанок имеют зиму-
ющие надземные стебли, а некоторые даже и вечнозелёные листья; у некоторых
суккулентов перезимовывают все надземные части. Подобные формы, конечно,
не являются травами в обычном смысле. Особый тип представляют полукустар-
ники, — очень хорошо выраженный переходный тип. В данную же группу мы
относим лианы, как жизненную форму, представленную деревянистыми и тра-
вянистыми растениями (может быть, правильнее лианы выделить в совершенно
особую группу).
1. Полукустарники. По внешнему виду очень похожи на кустар-
нички и отличаются тем, что если у кустарничков все надземные стеблевые части
перезимовывают, то у полукустарников перезимовывают лишь нижние части
стеблей, а верхние отмирают каждый год осенью; таким образом, растение в
своих нижних частях деревянистое, а в верхних— травянистое, являясь про-
межуточной формой между кустарником и травой и вполне оправдывая своё
название. Неправильно (что иногда делают) относить к полукустарникам н е-
большие кустарники, названные выше кустарничками; это в сущности
тоже кустарники, но лишь небольших размеров.
К полукустарникам относится много растений более южных мест (южные
степи, полупустыни, пустыни). Из более известных растений: чабрец (богоро-
дицкая травка)/ виды полыней (Artemisia) и др.
Следует отметить, что нередко к полукустарникам причисляют чернику
(отнесённую нами к кустарничкам); однако у черники наблюдается отмирание
не верхушек стеблей, но иногда стебли отмирают от самого основания.
2. Р а с т е н и я-п о д у ш к и. Подобные растительные формы полу-
чаются в результате образования многочисленных побегов и их повторных
ветвлений; так как рост побегов очень ограничен, то получаются приземистые,
очень плотные подушки (о подробностях см. ниже). Побеги, несущие листья,
многолетние, несущие цветки к зиме отмирают. Растительная форма, харак-
терная для альпийских высот, тундр, пустынь (рис. 149 и 206).
3. Суккуленты1. Растения имеют своеобразный облик, так как их
стебли или листья очень сочны и утолщены вследствие накопления в этнх-орга-
нах большого количества воды. Среди суккулентов можно отличить две
жизненные формы: 1) листовые суккуленты и 2) стеблевые суккуленты; приме-
ром первых является молодило (Sempervivum), вторых — солерос (Salicornia
herbacea). Одни суккуленты в надземных частях многолетни, другие — одно-
летни, третьи — являются однолетниками.
4. Вечнозелёные травы. Внешний облик трав, ио и листья и
стебли многолетни. Хорошим примером являются грушанки (Pirola rotundifo-
!ia, Ramischia secunda, Chimaphila umbellata и др.).
5. Зимнезелёные травы. Эта жизненная форма ещё ближе к
настоящим травам; здесь перезимовывают стебли (или часть стеблей), являю-
щиеся многолетними; листья на вегетативных побегах живут обычно два се-
зона, иа цветущих — к зиме отмирают. Пример — зеленчук (Galeobdolon
luteum). ~-
6. Лианы. Имеют длинный тонкий стебель, нуждающийся для своей
опоры в соседних растениях. Особенно обильно представлены лианы в тропи-
ческих лесах н там имеют деревянистый многолетний стебель. У нас деревяни-
стые лианы встречаются лишь в более южных местах (например, плющ в Крыму
и на Кавказе и др.). Из травянистых лиан СССР назовём хмель и вьюнок (Са-
lystegia). Так как лианы могут быть и деревянистыми н травянистыми, то мы и
их помещаем в промежуточную группу.
В. Жизненные формы с травянистыми стеблями
Надземные стеблевые части и листья этих растений на зиму обычно отмирают.
Среди травянистых растенпй*надо различать наземные, эпифитные н водные,
сильно отличающиеся по своему внешнему облику. По продолжительности своей
жизни они подразделяются на многолетние, двулетние и однолетние. Многолет-
ние растения — наиболее обширная группа травянистых растений, они могут
существовать очень продолжительное время и, начиная с известного возраста,
в большинстве случаев каждый год цветут и плодоносят. Это растения поликар-
пические (от греческих слов «полис»— многий и «карпос»— плод), т. е. дающие
плоды много раз. Однако некоторые многолетники плодоносят один раз в
жизни.
Что касается двулетних и однолетних растений, то они являются исклю-
чительно монокарпическими (от греческих слов «монос» — один и «карпос» —
плод) и, следовательно, плодоносят только один раз в жизни. В сущности,
однолетники живут гораздо меньше года, всего иеско-лько месяцер или недель, и
в это время цветут, плодоносят, а затем нацело отмирают, и зимний период про-
водят в виде семян. Двулетники для завершения плодоношения требуют двух
вегетационных периодов — первое лето они, в большинстве случаев, дают лишь
прикорневую розетку листьев, совершенно ие развивают стебля, но запасают
в своих подземных органах питательные вещества (примеры — морковь,
редька и др.), на второе лето цветут, плодоносят и затем отмирают-
I. Наземные растения (прикрепляются к почве)
В этой группе имеется наибольшее количество жизненных форм. Из их числа
триводятся лишь некоторые:
1. Высокие двудольные травы. Иногда до 2 м, с высоким
стеблем, более или менее равномерно усаженным листьями. Многолетники или
двулетники. Иван-чай, осот (Cirsium), будяк (Onopotdon) н др.
2. Травы средней высоты. Листья более или менее равномерно
определены на стеблях. Эги травы очень разнообразного характера, но
>бычно с более или менее широкими листьями двудольного типа.
3. Злаковидные травы. Травы с длинными, узкими листьями, как
/ злаков. Кроме злаков, сюда относятся также осоковые растения и некоторые
фугие (рис. 25).
1 Суккулент — от латинских слов «суккус» — влага и «лентусо — про-
юлжительный.
59
4. Ситин ковн дные травы. Травы с безлистными зелеными
прутьевидными стеблями. Некоторые виды ситников, камышей и др.
5. П а п р о т н и к в ы е т р вы. Миогол'тники : многократнорас-
сеченными листьями,
♦
Рис. 25. Злаковидная
жизненная форма —
кобрезия (Cobresia
Bellardu) из сем. осо-
ковых (по Е. Коро-
вину)-
прпкреп чяются, свободно
выходящими из горизонтального (по большей части)
корневища. Нередко листья папоротника очень
велики, например папоротника «страусов перо»
(Strnthiopteris).
6 Розеточные травы. Листья лишь
в прикорневой розетке, стебель очень сильно уко-
рочен. Виды одуванчика, подорожника и др.
7. Травы типа «перекати-поле».
Сильно ветвистые стебли или соцветия, дающие в
общем форму шара. Многие растения степей.
Однолетники по своей жизненной форме
могут относиться к злаковидным (однолетние злаки)
к розеточным (крупка весенняя), к перекати-поле
и др
Двулетники в первый год жизни отно-
сятся к розеточным травам во второй — к высоким
травам, к перекати-поле и др.
По чая система жизненных форм наземных
трав еще не разработана.
II. Эпифитные растения (прикрепляются
к коре или листьям других растений)
В нашей флоре иет эпифитных растений, отно-
сящихся к цветковым растениям, реже имеются
эпифиты - папоротники (в Крыму, на Кавказе, в
Уссурийском крае) Вообще эпифиты крайне харак-
терны для тропических лесов с их обилием влаги,,
они пользуются деревьями лишь ^ак местом при-
крепления, не заимствуя от них никаких питатель-
ных веществ. Как особую группу эпифитов состав-
ляют эпифиллы (от греческих слов «эпи»— на и
«филлон» — чист), поселяющиеся на листьях де-
ревьев.
III. Водные растения (прикрепляются
ко дну водоемов или совсем не прикрепляются)
1. Плавающие растения с листьями,,
плавающими на поверхности воды, причем одни
растения прикрепляются ко дну водоемов (желтые
кувшинки, белая водяная лилия), другие же не
плавая (виды рясок, лягушатник— Hydrocliai is
morsus ranae).
2. Погруженные растения — они целиком погружены в воду и
выставляют над водой лишь свои соцветия во время цветения. Некоторые же
виды цветут под водой. Сюда относятся многочисленные рдесты, уруть, рого-
листник и г. п (рис. 26)
Иногда как особую группу жизненных форм указывают болотные растеният
погруженные в воду лишь своими нижними частями и высоко выносящие над
водой свои стебли и соцветий. Однако эти растения легко могут разместиться по
вышеприведенным группам, например прибрежник (Lythrum) можно отнести
к высоким травам, осоки — к злаковидным, камыши (Scirpus) — к ситниковид-
ным и т д , тем более, что большинство болотных растении может расти
п без воды на почве.
Г. Жизненные формы мхов и низших растений
Выше были приведены жизненные формы лишь наиболее обыкновенные
(полный перечень был бы очень велик) и к тому же относящиеся только к выс-
шим растениям. Что касается мхов и низших растений (лишайники, водоросли!,
60
грибы), то здесь тоже имеется большое разнообразие жизненных форм, обусло-
вленных прежде всего средой; так, одни нэ этих растений живут на суше, дру-
,ие — в водной среде, третьи — прикрепляются к живым растениям (эпифиты),
еще некоторые являются сапрофитами и паразитами.
Рис. 26. Жизненные формы водных погруженных растений:
1—рдест (Potamogeton), 2—другой вид рдеста, 3—уруть (Мупор-
hyllum), 4—роголистник (GeratophyHutn) (по Сыреищ икону).
Некоторые из жизненных форм мхов н низших растений играют большую
роль в физиономии, в ландшафте территорий и потому должны быть указаны.
1. Жизненная форма типа кукушкина льна (Polytnchum). Довольно высо-
кий жесткий стебель с многочисленными узкими листьями. В лесах п на боло-
тах виды кукушкина льна образуют иногда сплошные ковры.
2. Жизненная форма типа гипсовых мхов. Стелющиеся зеленые мхи с мел-
кими чешуйчатыми листьями. Образуют в некоторых типах леса сплошные
ковры.
3. Жизненная форма типа сфагновых мхов (Sphagnum). Мхи торфяных
болот белого (также бурого, красного и др) цвета, сильно гигроскопичные,
покрывающие болота сп тошным покровом и способствующие накоплению торфа.
4. Жизненная форма кустистых лишайников типа оленьего лишайника
(Cladonia). Небольшие кустики белого оленьего лишайника, сплошь покры-
вающие почву сухих сосновых лесов, а иа севере дающие сплошной покров
кладониевых лишайниковых тундр.
5.-Многочисленные жизненные формы эпифитов. Эпифитами могут быть
---ГЧ.Т.Л 111 .г,тгИ_ПППЛППГ ПИ
При установлении жизненных форм решающим фактором, не-
сомненно, должен быть фактор экологический (но не систематический)
Так, например, жизненные формы «дерево с опадающей листвой»
«растение-подушка», «лиана» и другие могут относиться к совершен-
но различным семействам. Тем не менее все же фактор системати-
ческий иногда сказывается; так, «папоротниковые травы», «злако-
видные травы» и другие—эти жизненные формы не выходят за
пределы соответствующих (хотя и крупных) систематических под-
разделений.
Вообще вряд ли возможна физиономическая система
жизненных форм, построенная только на экологических принципах,
так как нередко внешняя форма в известней степени связана
с систематическим положением, например «хвойные» деревья, «зла-
ковидные» травы и др. Поэтому возможны как преимущественно
физиономические системы жизненных форм, так и экологические;
в последних при основных подразделениях играют роль именно
экологические факторы. Пример экологической системы будет дан
ниже (см. стр. 98).
Мы остановились довольно долго на физиономических жизненных
формах потому, что они являются элементами ландшафтаили даже
определяют ландшафт и вследствие этого для общего знакомства
с растительным покрове^ важнее, чем систематические единицы
(например зона хвойных лесов, злаковые степи, район
широколиственных лесов и т. д.).
Вопросы генезиса жизненных форм. Очень интересны генети-
ческие взаимоотношения различных жизненных форм.
Не устанавливаясь на данном вопросе во всей его полноте, выясним
лишь генетические отношения между «деревами» и «травами» у по-
крытосеменных растений.
Какая жизненная форма из этих двух должна считаться санов-
ной и какая производной? Некоторые цифры дадут нам материал
для соответствующих выводов; прежде всего несколько цифр по-
кажут процент травянистых видов в ряде областей умеренного и
тропического типа из всего числа видов в данной стране.
Умеренные области
Число всех видов1 Число видов трав %
СССР 14 704 12 588 86
Франция . . 3 424 3492 89
Германия . . 1 117 947 85
Северные части США . . . .... 2 662 2 089 78
Южные части США . . . . 2 266 1 666 74
Норвегия 857 741 86
Швейцария 1899 1 726 91
1 Число видов в этой таблице и в следующих в большинстве случаев меньше
соответствующих цифр в отделе «Флористические области земного шара», где
даются цифры согласно новейшим данным. Для наших целей, однако, это зна-
чения не имеет.
62
Тропические области
Число всех 'видов Число видов трав %
Тропическая Африка . . . . 8 577 3 560 42
Британская Индия 10 454 4 344 42
Цейлон 1 793 670 37
Ява ... 3 183 867 27
Малайский полуостров . . . 3 252 553 17
Долина Амазонки 2 209 265 12
Из таблицы совершенно ясно видно, что в северных стенах в
ряде случаев процент трав во много раз больше, чем в тропи-
ческих.
Процент травянистых видов в отдельных семействах для умерен-
ных и тропических стран очень различен:
Семейства Число видов трав в разных странах в %
Северная умеренная зона Бразилия Индия Тропиче- ская Африка
Амарантусовые 100 79 44 84
Сложноцветные 100 39 91 80
Молочайные 95 14 10 29
Фиалковые 100 6 52 IS
Бобовые 90 22 38 41
Вербеновые 100 20 4 7
Как видно, большинство приведённых семейств в умеренных
областях состоит исключительно из травянистых видов, а в тропи-
ческих областях процент травянистых видов в некоторых семействах
ничтожен. Многие тропические семейства (магнолиевые, аноновые,,
эбеновые и др.) совершенно не имеют травянистых представителей,
и вообще более половины всех семейств двудольных не
имеет травянистых видов.
Нет сомнения, что тропическая флора—наиболее древняя,
развивающаяся беспрепятственно с отдаленных времен третичной
эры. Она представлена главным образом древесными видами. Вы-
работка последних связана с теплым вегетационным периодом,
продолжающимся круглый год.
Травянистые же формы покрытосеменных растений предста-
вляют редуцированные формы, возникшие в результате похолодания
климата и сильного сокращения вегетационного периода. Все эти
соображения географического порядка подтверждаются филогене-
тическими и палеоботаническими данными, (покрытосеменные прои-
зошли от голосеменных, среди которых травы отсутствуют; наиболее
древние порядки покрытосеменных состоят лишь из древесных
форм и т. д.).
Приведем ещё несколько фактов, касающихся соотношений
деревьев и трав на древних океанических и материковых островах;
здесь нередко эндемические роды совершенно не имеют травянистых
представителей.
Острова Фиджи
Число видов трав %
Всего видов .... 563 81 14
Не эндемичных видов Эндемичных видов не эндемичных 292 76 26
родов 251 5 2
Видов эндемичных родов 10 Острова Галапагосские —
Всего видов 446 270 61
Не эндемичных видов . Эндемичных видов не эндемичных 220 177 80
родов 215 93 43
Видов эндемичных родов 11 — —
Острова Гавайские
Всего видов 582 138 24
Не эндемичных видов Эндемичных видов не эндемичных 82 62 76
родов 257 55 21
Видов эндемичных родов ...... 243 21 8,5
Что касается травянистых видов покрытосеменных растений,
то можно не сомневаться, что формы наземные являются более
древними; водные формы произошли от наземных путём приспосо-
бления к водной среде; вторичными в большинстве случаев явля-
ются и однолетники; они представляют приспособление к засушли-
вым условиям и к короткому влажному периоду (особенно харак-
терны для пустынь). Таким образом, для покрытосеменных растений
эволюция жизненных форм в основном шла таким путем:
_ травы многолетние >> травы водные
Деревья - кустарники-н£земнь1е <травы одно-
летние 1
Биотипы и экотипы
Если каждый вид даёт определённую жизненную форму, то
более детальное изучение показывает, что внутри самого вида на-
блюдаются более мелкие экологические единицы. Виды в современ-
ном представлении состоят из очень большого числа так называемых
биотипов, причём под последними понимается совокупность инди-
видуумов, генетически одинаковых. В различных условиях не все
би'типы одинаково жизненны; и в результате конкуренции оста-
ются в данной обстановке лишь биотипы, наиболее отвечающие
данным условиям, что, конечно, сказывается и на их внешнем облике.
Совокупности подобных биотипов Турессон не так давно на-
звал экотипами, понимая под ними мелкие наследственные
единицы, экологически соответствующие данной среде.
В последнее вре\1я экотипы подразделяют на климатипы, эда-
фотипы, ценотипы. Первые возникли под влиянием климата,
1 В иных случаях однолетники в дальнейшей эволюции дали снова много-
летние формы/
64
Рис. 27. Изменение внешнего облика растений
в связи с изменением внешних условий. Нежник
(Heliantliemum vulgare):
J—иа равнине' 2 и 3 —в горах (Т и 2 имеют одинако-
вый масштаб, 3 — увеличение вдвое). По Бопнье.
вторые — почв, третьи — под воздействием какой-либо раститель-
ной группировки, например, ценотипы леса, луга и т. дА
Впрочем, выделять эти категории экотипов нередко довольно
трудно, так как обычно действует не один какой-нибудь фактор, а
совокупность их.
Необходимо иметь в виду, что внешний облик растений может
иметь двойную природу: или это экотип, т. е. наследственная форма,
аомонирующая с усло-
вия ми среды; ил и же
перед нами модифика-
ция, т. е. формеК, тоже
изменённая под влия-
нием данной среды, но
наследственно не обу-
словленная. Экотип при
изменении внешних ус-
ловий хотя несколько
и изменяется, но со-
храняет свои основные
черты, свои морфоло-
гические признаки, мо-
дификация же нередко
совершенно изменяет
свой облик. Узнать в
природе, имеем ли мы
в том или другом слу-
чае эхотип или же моди-
фикацию, в большин-
стве случаев очень труд-
но, для этого необходим
эксперимент, т. е.
пересадка и культура
избранных растений в
различных условиях.
Очень большое зна-
чение имели в своё вре-
мя опыты Б о н н ь е
над перенесением неко-
торых видов растений
нз равнинных условий
в горные (рис. 27),
причём непосредственно
стений равнин (высокий рост, крупные листья, удлинённые
междоузлия и ряд анатомических особенностей) в горах совер-
шенно изменился (низкий рост, более мелкие листья, сосредо-
точенные близ основания стебля, укороченные междоузлия и пр.).
1 Учение Турессона об экотипах дало основание упрекать его в
антидарвинизме, так как у него отсутствуют указания на возникновение в при-
роде новых биотипов. Однако нельзя сомневаться в том, что под воздействием
внешних условий постоянно идет процесс возникновения новых биотипов.
5 География растений 65
обнаружилось, что внешний облик
Чтобы материал опыта был совершенно однороден, Боннье один
и тот же экземпляр делил на части и э® части сажал в разных
условиях. В данных опытах Боннье i мел^дело, как видно из ре-
зультатов, с модификациями (не наследственными формами).
Однако в дальнейшем Турессон показал, что опыты Боннье
представляют лишь частный случай и не могут быть обобщены,
так как в одних случаях данные Боннье подтверждаются, в дру-
гих — нет. И в последнем случае растения при пересадке в иные
Рис. 28. Сивец (Succisa рте и /за)—пример экотипов (два левых экземпляра)
и модификации (правый экземпляр). Первые удержали в культуре свою
внешнюю форму, второй—из низкорослой формы сделался высокорослым
(по Турессоиу).
условия сохраняют свои основные морфологические признаки, свой
внешний облик: н зкоросгые растения гор остаются низкорослыми
п при всех других условиях и т. д. Очень наглядны эксперименты
Турессона над кульбабоЙ (Leontodon autumnalis), прибереж-
ником (Lythrum salicaria), сивцом (Succisa praemorsa—рис. 28)
И Др.
Работы Поплавской в Крыму привели к выделению ряда экотипов:
астра (Aster а тле 11 us) даёт собственно горный экотип в горах лесистой местности
(oect, montanus) и альпийский экотип на Яйле (oect. alpinus); последний имеет
более низкий рост, компактное соцветие, более раннее цветение и пр. Трёх-
летняя культура этих двух экотипов в одинаковых условиях показала, что все
их особенности являются стойкими. Подобные же экотипы были установлены у
девясила (Inula ensifolia): экотип горный (oect. montanus) и экотип альпийский
(oect. alpinus), у золотой розги (Solidago virga a urea) и др.
Так как внешне очень сходные формы могут иметь различную в смысле на-
следования признаков природу, то целесообразно это отмечать и в соответству-
ющих названиях растений, указывая сущность той или иной формы; так, напри-
мер, горная форма наследственного порядка должна обозначаться как oect.
montanus, а ненаследственная, как phoen. montanus (модификация, или фе-
нотип).
Убедиться в том, является ли данная форма экотипом или же
это простая м. дификация1, вызванная влиянием окружающих усло-
1 В вопросе о природе модификаций мнения учёных расходятся; большин-
ство считает их ненаследственнымн в противоположность Лысенко и его после-
дователям.
66
внй, как уже указано, возможно лишь путём соответствующих
культур; не только облик растений — его жизненная форма —
может 1 меть либо наследственную, либо приспособительную при-
роду, но также это относится и котдельным призна кам
растений; так, в иных случаях сильно опушённое растение в
культурах может терять своё опушение, в других случаях опушение
остаётся при всех условиях эксперимента; свёрнутые листья у одних
злаков остаются свёрнутыми и при различных условиях культуры,
у других же развертываются и становятся плоскими н т. д.
Здесь широко применяется эксперимент, и экология становится
экспериментальной наукой. Нельзя не указать,
что ещё недавно экология была наукой чисто описательной, но
при этом объяснение какого-либо признака, какой-либо раститель-
ной формы являлось чисто умозрительным, или спекулятивным.
Только на основе эксперимента можно судить о сущности того или
иного приспособления. Поэтому нужно с большой осторожностью
подходить к толкованию приспособлений и признаков растений.
Среда растений
Так как внешние условия (среда) накладывают определённый
отпечаток на растительный организм, то, изучая растительные ор-
ганизмы, совершенно необходимо изучать эту среду.
Средой нужно считать совокупность всех факто-
ров, оказывающих влияние на растение или на растительный
ценоз. Этих факторов немало, и в дальнейшем они будут указаны.
Однако не все факторы, находящиеся в окружающей среде, необхо-
димы для прохождения вполне нормального цикла развития растений:
так, например, в почве находится очень много химических элементов,
из которых, однако, для растительных организмов нужны лишь
некоторые; с другой стороны, в почве может быть много влаги,
гораздо больше, чем нужно для данного растения; также может
быть избыток света, кислорода и т. д.
Поэтому для нормально растущего растения обычно среда пре-
доставляет: 1) необходимые факторы нередко в количествах больших,
чем растению нужно, и 2) ряд факторов, для него ненужных и даже
вредных; вредными могут стать и необходимые факторы в случае их
избытка (например избыток света и др.). Отсюда следует, что лишь
часть среды безусловно необходима для растения; эту совер-
шенно необходимую часть среды можно назвать условиями суще-
ствования (или средой в узком смысле). Если условий суще-
ствования нет налицо, — растение отсутствует даже при крайне
благоприятных условиях среды вообще; так, в тропическом
климате многие растения полярных стран расти не могут х.
Каждое растение и каждый участок растительности занимают
на земной поверхности определённый географический пункт
{местонахождение). Но растение, произрастая в данном пункте,
1 Часто понятие «условия существования*» употребляют в общем смысле,
в смысле вообще внешних условий,
5* 67
связано с его экологическими условиями; совокупность всех эколо-
гических свойств пункта (участка) представляет местообитание1.
Основные факторы среды. Среда, как сказано, слагается из ряда
отдельных факторов, объединяемых в группы или факторы круп-
ного порядка. Факторы эти следующие:
1. Климатический. Основной фактор, накладывающий
свой отпечаток на распределение и характер как отдельных видов,
так и фитоценозов. Климат в основных чертах определяет расти-
тельность.
2. Э д а ф и ч е с к и й. Здесь играют роль почвы, подпочвы,
грунты. Если климат может давать определённый отпечаток целой
стране, то внутри неё почвенно-грунтовые условия могут вносить
те или иные местные изменения; так, например, в области еловых ле-
сов, соответствующих определённому климату, могут быть вкрап-
лены участки сосновых лесов, если встречаются песчаные простран-
ства; в области степей на меловой почве может развиваться совер-
шенно своеобразная растительность и т. д.
3. Орографический, или фактор рельефа. Рельеф
страны или рельеф данного участка крайне резко сказывается на рас-
тительности, и растительность ровных мест, склонов, повышений
или впадин будет сильно отличаться; здесь играет роль также сте-
пень наклона участка и пр.
4. Биотический. Это группа факторов, охватывающих
влияние органической природы — животных и растений. Живот-
ные влияют в результате пастьбы, распространения семян и плодов;
насекомые опыляют растения; роющие животные разрыхляют землю
и пр. Растения влияют на растения, конкурируя друг с другом,
сожительствуя или создавая особую среду-
5. Влияние человека. Влияние человека на расти-
тельность может быть прямым и косвенным; оно очень разносторон-
не и, несомненно, должно быть выделено в особый фактор.
б. Исторический. Всеми перечисленными факторами
не всегда можно объяснить присутствие растения на данном место-
обитании (а также и строение или состав растительных сочетаний),
так как растительный организм сложился в результате длительной
истории и нередко имеет те или иные пережитки прошлого. Также и
растительность данного участка может содержать формы, уцелев-
шие от прошлые геологических времён.
Однако выделяя исторический фактор в особую категорию, нужно
указать на то, что и другие факторы нельзя рассматривать в отрыве
ст истории; поэтому исторический фактор не равноценен остальным.
Каждый из приведённых факторов распадается на целый ряд
факторов более мелкого порядка. Так, климатический фактор объ-
единяет в себе влияние воздуха, света, тепла, влажности и пр.:
\Некоторые авторы (Сукачев) предлагают пользоваться несколько
ином терминологией: вместо местообитания (в указанном смысле) говорить
«условия местопроизрастания), а под «условиями местообитания» понимать
совокупность факторов, входящих в состав среды-на всем протяжении ареала
вида или ассоциации. Таким образом «местообитание» — совокупность мест
всех обитаний вида (ассоциации) и их условий.
68
эдафический — физические и химические свойства почвы, подпочвы,
материнской горной породы и т. п.
Среди факторов всех категорий можно различать факторы двоя-
кого рода: во-первых, такие, которые непосредственно,
прямо действуют на растения или ценозы, и во-вторых, такие,
которые действуют лишь косвенно.
Так, например, из эдафических факторов химические свойства
почвы действуют на растения непосредственно, а материнская под-
стилающая порода —косвенно, поскольку она влияет на формиро-
вание почвы, её режим и т. д.
Из биотических факторов пастьба, сенокошение, распространение
семян —примеры прямого воздействия, но такой фактор, как ис-
кусственное осушение местности, влияет на растения лишь кос-
венно, поскольку при этом сначала создаётся новый почвенный
режим, а уже последний начинает оказывать влияние на растения.
Таким образом, все вообще факторы можно делить на прямо-
действующие и косвеннодействующие.
К прямодействующим факторам относятся физические и хими-
ческие свойства среды: вода, соли, свет, тепло и воздух; из
биотических факторов — сенокошение, пожары, перенесение семян,
поедание скотомой т. п.
К косвеннодействующим факторам нужно отнести: общий климат
страны, общие условия рельефа, материнскую горную породу, почву
я пр., из биотических—осушку местности, отношение и влияние
f одних ценозов на другие (нгпример^леса на соседнюю степь) и пр.
? Косвенные факторы иногда называются знтопическими, и их сово-
’ купность даёт так называемую «энтопию».
Инсгда к подразделению факторов подходят иначе и делят нх на
первичные и вторичные (Т. Ф р и з). Первые обусловлены действием
неорганической среды (их называют также экзогенными), а вторые
являются результатом влияния растительных организмов (эндоген-
ные). Например, свет на открытых местах —это первичный фактор,
а в лесу —свет, проходя через древесный полог, изменяясь в своих
свойствах, является уже вторичным фактором. Вторичные факторы
называют также «биологически измененными» (Сукачев).
Необходимо учитывать, что в природе имеет место совокуп-
ное действие факторов, и потому изучение влияния отдельных
факторов на растение нли фитоценоз представляет очень большие
трудности. Если же факторы при изложении рассматриваются от-
дельно, то это делается, конечно, для лучшего уяснения. В дальней-
шем мы будем придерживаться подразделения факторов на клима-
тические, эдафические и другие, хотя это деление не может не вы-
звать ряда возражений. Так, например, воздух находится, кроме
атмосферы, и в почве и, значит, в последнем случае должен отно-
ситься к эдафическому фактору; то же самое можно сказать о воде,
температуре и пр.; свет в лесу, как биотически изменённый фактор,
межно рассматривать в числе биотических факторов и т. д.
Краткое рассмотрение факторов удобнее всего начать с’климати-
ческих, как наиболее существенных и важных. Их можно подразде-
1 Тал’ ВОЗДуХ, Т'^^О,
69
Климатические факторы
1. В о з д у. х
Воздух, как известно, состоит из 4/5 частей азота, кислорода
и, кроме того, ничтожных количеств угольной кислоты, некоторых
других газов (аргона, гелия) и водяных паров. Кроме того, в воздухе
могут находиться те или другие примеси.
Воздух для жизни большинства наземных растений имеет исклю-
чительное значение, так как кислород необходим для дыхания, а
углекислый газ —для фотосинтеза. Зелёные наземные растения
без воздуха существовать вообще не могут.
И тем не менее воздух как фактор экологический имеет сравнитель-
но с другими климатическими факторами очень небольшое значение.
Здесь особенно ясно то различие, которое Существует между факто-
ром вообще и его экологической ролью; точно так же
становится ясным различие между задачами физиологии и экологии
растений.
Задачей физиологии растений в данном случае является изучение
нормальных процессов дыхания и ассимиляции углекислого газа.
И если бы везде на земной поверхности эти процессы протекали
одинаково, то экологу здесь было бы делать нечего, так как он изу-
чает соотношения организмов с местными условиями. Состав
воздуха очень однороден, или во всяком случае он изменяется по
сравнению с такими факторами, как вода и температура, в очень
слабой степени. Поэтому последние и имеют огромное экологическое
значение по сравнению с воздухом.
Переходя к рассмотрению экологического значения воздуха и его
составных частей, остановимся сначала на кислороде и углекислом
газе (азот, как известно, значения не имеет).
Кислород. Он всегда имеется в воздухе в совершенно до-
статочном для дыхания количестве и потому экологического значе-
ния не имеет, но в воде (реки, озера) и в почвах содержание его
падает. К тому же в те органы растений, которые находятся в
воде, кислород проникает лишь через оболочки клеток путём
осмотического газообмена (так как здесь устьиц нет), что происхо-
дит крайне медленно; ирн этом кислород поступает медленнее, чем
углекислый газ. В болотистых почвах содержание кислорода иногда
настолько мало,что растения должны перестраивать свою морфо-
лого;анатомическую систему. Так, корни и корневища ряда бо-
лотных видов развивают особую ткань с полостями для наполнения
их воздухом (так называемая аэренхима), н эти полости соединены
с надземным воздухом. Если воздухоносная ткань находится на
корнях снаружи, то эти корни бывают белого цвета.
Подводные органы приспосабливаются главным образом путём
увеличения наружной поверхности. Это до-
стигается несколькими способами (рис. 26): 1) путём рассечения
листьев на многочисленные доли: уруть (Myriophyllum), роголист-
ник (Ceratophyllum), лютик (Ranunculus); 2) путём сильного удли-
нения листьев — тесьмовидные листья ряда рдестов (листья подоб-
70
кого рода имеют и другое значение — они хорошо приспособлены
к текучей воде). У некоторых растений пластинки листьев стано-
вятся очень тонкими, что облегчает проникновение кис-
лорода. Очень интересно мадагаскарское растение Aponogeton с его
продырявленными листьями, состоящими почти из одних жилок
(рис. 39).
Особого упоминания заслуживают «дыхательные» корни неко-
торых болотных и мангровых (см. ниже, стр. 190) растений, которые
обладают отрицательным геотропизмом и вследствие этого растут
не вниз, а вверх, поднимаясь над поверхностью болота (рис. 52).
Воздух входит через особые отверстия в воздушные полости дыха-
тельных корней и далее проходит в подземные части корней. Кроме
многочисленных мангровых растений, дыхательные корни имеют
некоторые пальмы, панданусы, сахарный тростник и др.
Углекислый газ. Он имеет исключительное значение для зелёных
хлорофиллоносных растений, так как всё органическое вещество
растений строится за счёг усвоения (ассимиляции) СОа в процессе
фотосинтеза; последний же, как известно, осуществляется в природе
за счёт солнечной энергии и происходит исключительно на свету.
Химическое выражение процесса при фотосинтезе таково:
6СО2 6Н2О = Се Н12 Ов 4- 6О2,
т. е. в результате получается сахар и свободный кислород.
Углекислый газ содержится в воздухе в ничтожных количествах —
0,03%, или 0.57 мг на литр воздуха (в среднем), но потребление его
растениями в процессе фотосинтеза за год составляет около T/3S
всего запаса углекислого газа в атмосфере (около 2100 биллионов
килограммов), т. е. весь запас СО2 был бы потреблён'растениями
всего лишь в 35 лет, если бы не было источников восстановления
этих запасов.
Источники восстановления запасов СО2: горение, дыхание
животных и растений, выделение СО2 в источниках и при вулкани-
ческих извержениях и деятельность почвенных микроорганизмов.
Очень большое экологическое значение имеет последний источник,
так называемое «почвенное дыхание», во-первых, потому, что интен-
сивность его очень велика, а во-вторых, потому, что это создаёт
благоприятные условия для фотосинтеза растений нижних ярусов
фитоценозов, как например травянистых растений лесов, приуро-
ченных к нижним слоям атмосферы, богатым СО2.
По Люидегорду, в одном лесу содержание СО3 в при-
почвенном слое было больше, чем в свободной атмосфере, на 154%.
а на высоте груди на 47—70%. Выделение же СО3 на площади в 1 га
за 1 час даёт такие соотношения:
Песчаная почва — 2,0 кз Лесная почва букового леса — 15,4—22,0 кг
Глинистая почва— 3,97 кг Лесная почва ольшаннпка— 11,7—23,4 кг
Интересно, что растения тенистых местообитаний, т. е. там, где
происходит энергичное почвенное дыхание, имеют устьица почти
исключительно на нижней стороне листьев, что, быть может, является
приспособлением для лучшего улавливания СО2, выделяющегося
из почвы.
71
Содержание СО2 в воздухе не является таким постоянным, как
предполагалось раньше, оно подвержено весьма значительным
колебаниям; другими словами, СО3 может играть роль экологи-
ческого фактора.
х Примеси воздуха. Очень большое значение для жизни растений
имеют те или иные примеси к нормальному составу воздуха. Эти
примеси находятся в воздухе, особенно городов и промышленных
центров, где в воздух выделяются значительные количества вред-
ных газов. Особенно вредной является сернистая кислота.
Уже давно известно, что в больших городах на стволах деревьев
отсутствуют лишайники, являющиеся показателями чистого воздуха.
Так же вреден городской воздух для хвойных (особенно для пихты),
причём происходит отмирание верхушек деревьев, неправильное
ветвление, отмирание хвои и пр. Даже ничтожные количества сер-
нистой кислоты, по некоторым данным даже 1/1 000 000 объёма,
вызывают те или иные вредные влияния.
В посчеднее время постройка заводов и возникновение больших промыш-
ленных центров влечет за собой усиление этих вредных влияний (у нас в Союзе,
ряде мест Германии и др.). Можно наблюдать не только повреждения отдель-
ных деревьев, но даже гибель больших площадей леса. По Рейсу затрону-
тая влиянием вредных газов площадь лесов Германии составляем 90 000 га,
из которых 9 000 га леса совершенно погибли.
Различные деревья неодинаково реагируют на вредные газы, причем можн»
установить такую шкалу чувствительности, начиная с. более чувствительных:
Хвойные1 пихта, ель, сосна, лиственница.
Лиственные: бук, дуб, клен, липа, ясень, груша, вяз.
В некоторых местах СССР, вблизи химических заводов можно набтюдать-
полную гибель сосновых участков, в то время как береза не показывает ника-
ких признаков угнетения. В Москве в последние годы очень плохо развиваются
хвойные и в ботаническом саду у них наблюдаются все признаки отмирания,,
тогда как лет 25—30 назад они росли достаточно хорошо.
В связи с этим в городах и фабричных поселках встаёт очень важ-
ный вопрос о подборе ассортимента древесных растений, устойчи-
вых против вредного действия газов.
Вредное действие дымовых газов очень сложно. Прежде всеге
они вызывают нарушение фотосинтеза, а также влияют на дыхание,
транспирацию, отток ассимилятор и др. •
В последнее время Красинский дал ряд новых материа*
лов по газоустойчивости растений, причём он различает три вида
газоустончивости:
1. Биологическую —растения быстро восстанавливают
повреждённые газом органы (американский клён, бузина и др.).
2. Морфолого-анатомическую — растения имеют те
или иные структурные особенности, ограничивающие газообмен и
потому затрудняющие проникновение газов в листовые ткани (ряд
суккулентов, многие растения с жёсткими листьями —азалии,
буксус и др.).
3. Физиологическую — это наиболее сложный вид га-
зоустсйчивости, Опыты показывают, что здесь особую роль играет
окисляемссть клеточного содержимого: газоустойчивость тем выше,
чем меньше окисляемость. Так как окисляемость связана с система-
тическим положением видош, то и газоустойчивость связывается с
72
определёнными родами и семействами (очень газоустойчивы жимо-
лостные, паслёновые, кипарисовые и др.).
Движение воздуха. Большое значение для растений имеет также
движение в о з д у хГа (ветер). Действие его может быть пря-
мое и косвенное. При прямом влиянии ветер может так или иначе
механически повреждать растения, а при косвенном — создавать
лучшую или худшую обстановку для произрастания. Так, напри-
мер, морские ветры, дующие с моря, приносят иногда очень много
влаги, а дующие из глубины континентов действуют иссушающе,
уничтожая иногда растительность. Юго-восточные суховеи в области
степей Европейской части СССР хорошо известны. Даже по берегам
морей и океанов может расстилаться пустыня (например, Сахара),
если сухие тёплые ветры постоянно дуют с континента на море и
морской влажный воздух не может попасть на материк. Но иногда
прибрежные пустыни могут быть и при морских ветрах- так, на при-
ме р, по берегу Тихого океана в Южной Америке длинной полосой
тянутся пустыни; здесь с океана дует холодный ветер; попадая in
теплый материк, он нагревается, но вместе с тем становится очень
сухи м, что и влечёт за собой образование пустыни.
Так как чем ближе к поверхности земли, тем сила ветра стано-
вится меньше (вследствие трения), то, естественно, наибольшему
воздействию ветра подвержены древесные растения. Большой из-
вестностью пользуются флагообразные формы крон деревьев, явля-
ющиеся результатом полного отмирания ветвей с наветренной
стороны. Особенно хорошо можно наблюдать такие формы по бере-
гам морей, на высоких горах и пр. Причины отмиранья ветвей с
одной стороны кроны при очень сильных ветрах могут быть чист©
механического свойства, но чаще всего это объясняется иссушающим
действием ветра. Под действием ветра ветви испаряют влаги
во много раз больше, чем обычно, в результате чего клетки растения
теряют воду в таком количестве, что их жизнедеятельность стано-
вится невозможной. Особенно влияет на растения иссушающее
действие ветра в тех случаях, когда растение находится на сухих
почвах или на почвах холодных, из которых вода может извлекаться
лишь с большим трудом. Последнее явление как раз имеет место
на северных пределах леса, и иссушающее действие ветра — основ-
ная причина северной границы леса. По Ч и л ь м а н у, здесь
действуют иссушающие зимние ветры, и растение погибает, испа-
ряя больше, чем может само получить из мерзлой земли. Город-
ков же считает, что особенное несоответствие между поступаю-
щей и испаряемой водой имеет место летом, когда растение испаряет
очень много, но получать из почвы много воды не может, так как
почва слишком холодна вследствие пр] су стгия вечной мерзлоты,
и, таким образом, несоответствие в расходе и приходе воды имеет
место в продолжение круглого года.
В гораздо меньшей степени действие ветра сказывается на более
низких деревьях и кустарниках. Однако и здесь при наличии осо-
бенно сильных ветров получаются ветровые формы. Так, в Крыму,
близ «Ласточкина гнезда», можно видеть очень интересные ветро-
вые формы можжевельника (Juniperus excelsa) — получаются низ-
7Л
кие кустарники н даже почти стланики с ветвями, обращёнными в
противоположную от господствующих ветров сторону ('рис 291
ЛаЗЛ"ЧНЫе ДСРСВЬЯ и даже различные виды одного и того же
рода неодинаково страдают от ветров. Так, менее страдают- горная
сосна, береза и особенно тутовое дерево; наиболее подвержены
воздействию ветра: обыкновенная сосна, лиственница и особенно
чувствительны некоторые вечнозеленые деревья, как лавр, олив-
ковос дерево, мирта, а из летнезелёных - тополь, ивы, боярышник
среди хвойных ,акже некоторые являются очень стойкими против
действия ветра, например: американская ель (Picea canadensis),
Рис. 29. Влияние сильных прибрежных ветров на изменение облика мож-
жевельника (Juniperus excelsa) у (-Ласточкина гнездам в Крыму. Нормально
этот можжевельник—деревцо правильной формы (ориг.).
крючковатая сосна (Pinus uncinata), почему их и сажают на мор-
ских побережьях.
Сильные порывы ветра могут или повреждать деревья или даже
вырывать их с корнем. Из наших деревьев особенно страдает ель,
обладающая поверхностной корневой системой, и можно нередко
наблюдать огромные ели, вывороченные со всеми корнями (рис. 30);
сосна, имеющая стержневой корень, прочно укреплена в почве, —
у неё ломается ствол (рис. 31), а в случае сильных резких порывов
ветра может оторваться вся крона вместе с верхней частью ствола.
Дубы очень сильно противостоят ветру, и приходилось видеть, что сильные
порывы ветра только рвут листья на части, деревья же остаются неповрежден-
н ыми.
Ветровалы иногда захватывают большие площади и создают в наших север-
ных лесах, удаленных от жилья, почти непроходимые завалы из стволов упав-
ших деревьев. Особенно сильные бывают ветровалы в более разреженных на-
саждениях, так как в сомкнутых насаждениях ветер встречает большую сипу
сопротивления.
Напомним, чтов 1904 г, в Москве ураганом была уничтожена Анненгофская
сосновая роща (в Лефортове), от которой теперь не осталось никакого следа.
Положительное действие ветра. Кроме вредного воздействия на
растения, ветер вообще является крайне важным фактором: он
способствует опылению у ветроопыляемых видов, разносит споры,
семена, плоды, части растений и даже целые растения (перекати-
поле).
74
Интересно обратить внимание на то. что ветроопыляемые ра-
стения, почти как правило, связаны с участками или территориями,
свободными для действия ветра; так, например, можно указать на
луга, нередко покрытые сплошь злаковым покровом (злаки опы-
ляются ветром), на осоковые болота, на злаковые степи и т. д. Боль-
шинство наших деревьев, образующих леса, тоже опыляется ветром
(сосна, ель, береза, дуб, осина и пр.), внутри же леса ветро-
опыляемых растений очень мало. Имеются также указания, что на
океанических островах, где обычны сильные ветры, до трети всех
видов являются ветроопыляемыми.
Те части растений, которые разносятся ветром, или очень легки (споры,
некоторые семена, например, семена орхидеи Goodyera repens, весят 0,000002 г,
Stanhopea ocuhta— 0,000003 г, Gymnadenia conopea—0,000008 г и др,), или
снабжены летучкаРми, крыльями и другими приспособлениями. Растения,
дающие «'перекати-поле», при высыхании становятся очень легкими, предоставляя
75
довольно большую ударную поверхность (многочисленные стебли, сухие листья
и др ) для ветра (см. ниже).
Хотя имеются указания на то, что зачатки’растений могут'пере-
носиться на сотни километров, однако ветер в большинстве случаев
является агентом скорее - местного значения.
В связи с последним положением можно обратить внимание на ряд исследо"
ваний, касающихся заселения растениями острова Кракатау Как известно,
этот небольшой остров, лежащий в Зондском архипелаге между Суматрои и Явой
(В 37 км ©т первой и в 40 км от второй), в 1883 г сделался жертвой вулканического-
Рис. 31. Разрушения в лесу от тайфуна близ Киото, в Японии
(лес из Pinas densiflora)
*
извержения, причем, как предполагал Треп б, вся растительность его была
уничтожена. Посетив остров через три года (в 1886 г ), Т р е п б обнаружил
там 22 вида цветковых растений и И видов папоротников и сделал заключение,,
что при заселении островов растениями гпрают роль ветер и морские течения,
а также птицы. Что касается ветра, то вывод отсюда был такой, что расстояние
в 40 км не является пределом для разнесения зачатков растений, т е , другими
словами, ветер является агентом не только местного значе-
ч и я. Этот вывод сыграл большую роль в географин растений и на нем обычно
стали базироваться Дальнейшая дискуссия проблемы Кракатау обнаружила,
что, поводимому, не вся первоначальная растительность острова бы >а уничто-
жена извержением, и, значит, современная флора острЬва могла развиться не-
зависимо от ветра и морских течении (Б а к е р) Однако, новейшие данные
©пять говорят о том, что значительная часть флоры была занесена с соседних
•стровов. Но вообще вся проблема Кракатау не может решить вопроса о заносе
на большие расстояния и о роли ветра как агента широкого
значения (здесь расстояние всего 40 км).
2. Свет
Свет играет исключительную роль в жизни зеленых растений,
так как процесс фотосинтеза идет только в присутствии света: без
света ж и ° н ь зеленых растений н ° з м о ч п
/6
В результате фотосинтеза растения вырабатывают органические
вещества и пополняют запасы потенциальной энергии в природе,
и потому без зеленых растений невозможно
существование человека и животных.
Однако анализ исключительной роли света в указанном смысле
принадлежит физиологии растений, точно так же, как и рассмот-
рение важного вопроса о роли света в универсальном у растений
процессе испарения (транспирация) Это в основном проблемы фи-
зиологические, а не экологические.
Но свет становится экологическим фактором, как только мы
начинаем изучать его действие не вообще, ав конкретных
географических, топографических и фитоценологических условиях^.
Особенно большую роль он играет при взаимоотношениях растений
в их сочетаниях (фитоценозах)
При отсутствии или недостатке света растения, как известно,
становятся этиолированными, что сказывается в сильном удлине-
нии междоузлий и, наоборот, в уменьшении листовых пластинок.
Однако подобное формирующее воздействие недостаток света оказы-
вает на растения двудольные, тогда как у однодольных (злаки)
междоузлия укорачиваются, а листовая пластинка даже стано-
вится шире. В природной обстановке явления этиолирования в
большей или меньшей степеги наблюдаются в сомкнутых насажде-
ниях, в фитоценозах, где соседние растения затеняют друг друга.
Достаточно посмотреть на экземпляры одного и того же вида, вырос-
шие на свободном месте или же в густом древостое или травостое,
чтобы убедиться в этом. Так, деревья в лесу имеют тонкий цилин-
дрический ствол, очищенный от сучьев, и сравнительно небольшую
крону (рис. 32), тогда как деревья того же вида, выросшие на сво-
боде, имеют более толстый, приближающийся к коническому, ствол
с широкой развесистой кроной Также и травянистые растения в
травостое как бы тянутся кверху, имея длинные стебли и черешки
и располагая свои листья в верхних частях стебля.
Что касается настоящего этиолирования с отсутствием хлоро-
фил а, то оно редко наблюдается в природе, например, в глубине
пещер, куда птицы заносят семена и где семена могут про-
растать.
На различных точках земного шара — на экваторе, в полярных,
умеренных странах — сила света различна; различна также и
продолжительность освещения за отдельные сутки. Сила света
также различна на уровне моря и на высотах.
Свет, падающий, на любое растение, слагается из прямого сол-
нечного света, сильно измененного при прохождении атмосферы,
и из рассеянного, или диффузного, отраженного от неба и облаков,
кроме того, можно говорить в некоторых случаях еще о «нижнем
свете» (Unterlicht), отраженном от почвы; он имеет особенное зна-
чение для растительности по берегам озер, рек и морских берегов,
где отражение от водной поверхности может быть очень сильным,
именно «нижний свет» может составлять до х/в «верхнего света».
«Нижний свет» также имеет очень большое значение для растений,
растущих на л еловых обнажениях, ослепительно блестящих в сол-
77
нечные дни (он влияет здесь и на архитектуру растений — строение
листьев н пр.).
Силу света измеряют □'•злечными способами, но наиболее простым является
способ Бунзена — Роско э, основанный на почернении хлористо-сере-
бряной бугмаги под действ/геи химических лучей солнца. Че.м .медленнее проис-
ходит почернение, тел! сила света, естественно, меньше. Сравнение производят
78
с «нормальным тоном>>, который получается от нанесения на бумагу I части
чистой сажи и 1 000 частей окиси цинка. Если почернение хлорнсто-серебряной
бумаги до нормального тона происходит в течение 1 секунды, то мы имеем силу
света в I ^унзеновскую единицу; если для этого потребуется 2 секунды, то сила
света равна бунзеновской единицы и т. д.
Солнечные и теневые растения. Все растения по своей потреб-
ности в свете делятся на солнечные (гелиофиты) и теневые (сциофиты).
Солнечные и теневые растения отличаются друг от друга по
своим листьям и морфологически и анатомически; так, у первых
листья обычно мельче, имеется хорошо развитая столбчатая парен-
хима, хлорофилловые зёрна более мелки; у вторых—. истья шире,,
преимущественно развита губчатая паренхима с крупными меж-
клетными пространствами, хлорофилловые зерна крупны.
Солнечные и'теневые растения различно относятся к свету.
Очень слабый свет нередко не может быть использован растением
при фотосинтезе, но с увеличением света возрастает и энергия фото-
синтеза; однако это возрастание происходит не беспредельно, и у
ряда солнечных видов максимальная ассимиляция достигается при
г/3 яркости дневного света, у тенев х — при \119 света (например
у кислицы). При более же ярюм свете интенсивность а симиляции
стоит на одном и том же уровне.
Что касается низшего предела света при фотосинтезе, при кото-
ром растения могут существовать, то, естественно, у теневых рас-
тений он ниже, чем у солнечных; так, у растений наших лесов он
лежит около 1/90 полного дневного света, а в тропических лесах
(в более благоприятных условиях)—даже около 1/1?0. При более
низких пределах уже будет так называемая мёртвая лесная
тень.
Однако фотосинтез может иметь место н при меньшей интенсив-
ности света, но в этих случаях накопление органического вещества
идёт медленнее, чем её потеря в процессе дыхания. Та точка, при
которой ассимиляция СО3 при фотосинтезе уравновешивает потерю
органического вещества при дыхании, называется компенсационной
точкой. Таким образом, если растение длительно пребывает при
интенсивности света, или соответствующей компенсационной точке
или находящейся ниже её, то оно голодает и отмирает.
В таких условиях нередко находятся листья в глубине кроны
у деревьев и, возможно, прикорневые листья многих травянистых
растений, в результате чего они отмирают.
В летнее время многие растения самых нижних ярусов леса жи-
вут при интенсивности света ниже компенсационной точки; они
определённо голодают, но живут или за счет весенних и осенних
запасов, накапливаемых в то время, когда интенсивность света
лежит выше компенсационной точки (вследствие отсутствия листьев
на деревьях), или же за счет отдельных кратковременных вспышек,
ассимиляционной деятельности, когда на листья теневого растения
попадают солнечные блики. Естественно, что эти расте-
ния растут крайне медленно.
Потребность в свете у различных расте.1 мй (Lichtgenuss). О и
выражается в бунзеновских единицах так: L равна числу се-
79
к у н Д -п р и полном дневном свете, делённому
на число секунд в условиях данного место-
обитания. Это — относительная сила света;
для растений, растущих на полном свету, £=-]-= 1, в затенённых
же местах L всегда меньше единицы: т/2, х/3 и т. д. В тенистых лесах
относительная сила света очень мала: так, по Леммермайеру,
в глубине букового леса в середине лета она составляла
всего у*.
Естественно, что в лесах и в нижних ярусах других фитоценозе
могут произрастать лишь очень теневыносливые растения.
Вопрос о большей или меньшей теневыносливости древесных пород давно
интересует лесоводов, так как это обстоятельство имеет очень важное значение
для смен древесных пород, причём, естественно, породы более теневыносливые
вытесняют менее теневыносливые. Была установлена такая шкала пород, на-
чиная с наименее теневыносливых: лиственница, береза, осина, сосна, дуб,
ясень, клён, ель, пихта, бук, тисс, самшит. При составлении подобных шкал
по 1ьзуются методами как объективного, так и субъективного порядка. В послед-
нем случае принимают во внимание густоту облиствления (чем гуще листва,
тем теневыносливее порода), очищаемость от сучьев (чем менее ствол очищается
от сучьев, тем порода теневыносливее), нзреживание с возрастом насаждения
(чем меньше с возрастом остаётся экземпляров на единицу площади, тем порода
менее теневынослива) и т. д. Светлые лиственничные'или березовые леса, конеч-
но, представляют полную противоположность темным еловым и особенно пих-
товым и буковым лесам.
Методы объективного учёта теневыносливости основаны на измерении
латпсадной и губчатой ткани в листьях (С у р о ж), на явлениях этиолирова-
ния побегов (Т у р с к и и), на различной чувствительности хлорофиллонос-
ного аппарата к свету (Л ю б и м е н к о). При этом при применении послед-
нее метода оказалось, что отрезки листьев, находящиеся в пробирке и поме-
шенные в особые ящики с боковым отверстием, начинали усвоение СО2 при
весьма различной величине этого отверстия; бук — при 4 см2, пихта и липа —
при 9 см2, сосна — при 49 см2, берёза — при 64 см2, а лиственница и белая
акация — при 100 см2,
В отношении потребности в свете можно установить три группы
растений:
1. Растущие при полном свете (L=l). Сюда относятся почти все
однолетники, растения степей, пустынь..
2. Растущие при полном свете (L -1), но могущие переносить и
некоторое затенение (L < 1). Многие растения опушек и растения
светлых насаждений.
3. Растения, не переносящие полного освещения (L всегда мень-
ше единицы). Это главным образом растения лесов.
Потребность в свете не является величиной постоянной для дан-
ного вида: она меняется с возрастом (например проростки и подрост
у деревьев более теневыносливы, чем более взрослые экземпляры),
с изменением времени года (весной потребность в свете больше,
чем летом), с изменением широты места (на севере L увеличивается),
с изменением почвы (на плохих почвах L увеличивается) и т. д.
Большая потребность в свете в высоких широтах и на горах,
несомненно, связана с тем, что свет в данном случае замещает недо-
стающее количество тепла, На верхней границе леса на освещенные
места выходят не только отдельные виды, но и отдельные ярусы леса.
80
Кроме всего сказанного, свет оказывает на растения иформи*
ру ю щ ее действие. Это формирующее действие света сказы-
вается не только во внешне-морфологических признаках (свет во-
обще задерживает рост, и стебли растений и другие части на свету
являются укороченными), но также и в признаках анатомического
строения: большое различие в развитии губчатой и палисадной
ткани в зависимости от силы освещения, на что было указано выше.
Направляющее действие света. Особенно наглядно направляющее
действие света: стебли направляются к свету, и листья нередко
становятся перпендикулярно к его лучам. У ряда растений наблю-
дается мозаичное расположение листьев (рис. 33). Укажем ещё
на обычное сорное растение — мальву (Malva rotundifolia). листья
которой крайне чувствительны к свету, причём листовые пластинки
в течение всего дня стоят перпендикулярно к свету; утром они
обращены на восток, в полдень — на юг, вечером — на запад.
Движения соцветий-корзинок обыкновенного подсолнечника всем
известны — отсюда и название этого растения.
Очень интересны движения, зависящие от влияния света, которые
наблюдаются в цветках растений и вызывают раскрывание и за-
крывание венчиков цветков. Для раскрывания различных цветков не-
обходгша различная интенсивность света, и поэтому одни виды раскрываются
очень рано утром, другие — позже, третьи — еще позже. Это явление послужи-
ло основанием для составления цветочных часов, ив ботанических
садах подобные часы нередко устраивались (особенно в прежнее время); по ним
можно определить время дня-
3 География растений 81
Лепестки венчиков некоторых видов в высшей степени чувствительны к той
или иной интенсивности света, Интересно указать на те наблюдения, которые
были произведены летом 1936 г. во время солнечного затмения. Так, в Стрелец-
кой степи, под Курском, цветки некоторых растении раскрылись рано утром до
начала затмения, с первых фаз затмения они стали медленно закрываться до
момента наибольшей фазы, а затем так же медленно стали раскрываться. Кривая
движения лепестков венчика вполне совпала с кривой затмения, т. е. с кривой
интенсивности света, Особенно наглядно все эти отношения можно было на-
блюдать у дикого льна (Linum perenne), причем нужно отметить, что самое
затмение было неполным и день был не вполне ясный, и тем не менее чувствитель-
ность растения проявилась в высшей степени.
шении австралийские
Рис. 34. Компасное расте-
ние — чатук (Lactuca sca-
riola ; слева—вид растения
с за ада на восток, справа—
с юга на север.
Имеется немало растений, которые ставят свои листья ребром
или под различными углами к свету. Очень известны в этом отно-
алипты, леса из которых почти не дают
тени (см. ниже стр. 2084 У нас в степях
имеется ряд растений с вертикально рас-
положенными листьями: Serratula сеп-
tauroides, Inula ensifolia, Centaurea trincr-
via, Chondrilla juncea и др. Эти растения
представляют переход к так называемым
компасным растениям, листья которых:
узкой стороной направлены на юг — се-
вер, а широкой—на восток и на запад,
благодаря чему они избегают интенсив-
ного освещения среди дня.
Из наших растений здесь можно указать
на дикий салат-, атук (Lactuca scariola),
растущий на сорных местах и по дорогам
(рис. 34). Особенно показательны экзем-
пляры, растущие на сильно освещенных
местах, тогда как в тени компасность
нередко почти совсем не выражена. Ряд
хороших компасных растении имеется в
Америке (Silphium).
Свет как ботанико-географический
фактор. Очень большое значение имеет
свет как фактор ботанико - географиче-
ского порядка. Более продолжительное,
хотя и более слабое, освещение в высоких
широтах вызывает ряд крайне интересных
явлений. Здесь для вызревания многих
растений требуется менъШИй срок, чем на юге: например в
северной Гердшнии ячменЬ оканчивает свое развитие в 100 дней,
в северной Норвегии — в 89 дней, а в Якутске даже в 80 дней
и менее. Далее, при разнице в один градус широты (111 км)
в апреле цветение растений в Скандинавии дает разницу в 4 дня
(т. е. растения, находящиеся к северу на 111 км, запаздывают на
4 дня), в мае запаздывание составляет 2 дня, в июне—Р/2 Дня, в
июле — V2 Дня> а в августе никакой разницы не наблюдаемся,
так как более продолжительное освещение летом сглаживает тем-
пературные отличия.
82
Очень большое влияние на развитие растений оказывает соотно-
шение между продолжительностью дня и ночи (фотопериодизм),
что было установлено Гарнером и Аллардом в Ь23 г.
В настоящее время различают растения длинного дня (растения
северных широт) и короткого дня (растения тропические и суб-
тропические). Так называемые географические опытные посевы
совершенно совпадают с теорией фотопериодизма: так, продолжи-
тельность от всходов до цветения у растений длинного дня была
такова (в днях):
Мак (Papaver somniferum)
Рыжик (Camelina glabrata)
Овес шатнловский № 18 .
Мурман Архангельск Краснодар
35 45 87
29 28 56
36 41 81
К растениям длинного дня относятся также режь, ячмень и др.
Продолжительность времени от всходов до цветения у растений
короткого дня (в днях):
Каменная степь СеверньЙ _
( Ворсеежская обл ) Кавказ 1?шкент
Клещевина (Ricinus communis) 70 54 49
Хлопчатник (Gossypium hir-
sutum)........................ 122 76 42
Соя (Soya hispida)............. 123 82 44
Сюда относятся еще бобы, рис, сорго, просо, табак и др.
Анализ указываемых явлений приводит к вопросу о происхожде-
нии культурных растений: очевидно, что растения короткого дня
принадлежат по своему происхождению к тропическим и субтро-
пическим областям, а расте ия длинного дня — к умеренным.
В последнее время Лысенко дал иное толкование явлениям
фотопериодизма, связывая их с учением стадийности в развитии
растений, именно со «световой стадиев» (следующей за стадией
«яровизации»). Эта световая стадия связана с определен-
ным количеством света. После прохождения этой
стадии все дальнейшие стадии развития могут проходить при раз-
личных условиях фотопериодизма. Подобное понимание фотопе-
риодизма открывает большие возможности в продвижении растений
короткого дня на север, а длинного дня— на юг, при этом растение
должно получить определенное количество света во время световой
стадии.
Если же поставить вопрос, кладет ли свет предел для распро-
странения на земле растений, то надо сказать, что на земной по-
верхности нет ни слишком кмнсгс, ьи слишком светлого места,
где бы не могли существовать растения- но, конечно, в темноте
эти растения без хлорофилла.
3. Вода
Физиологическое значение воды огромно; если
те илн иные растения (низшие) могут существовать без воздуха,
а также без света, то без воды они существовать не могут; вода
содержится во всех частях растения и составляет нередко более
6* 83
90% всего веса растения. Все основные жизненные функции растения
требуют присутствия воды.
Как известно, минеральные питательные вещества почвы доступны для расте-
ния лишь в водном растворе, всасываемом корнями, фотосинтез требует
обязательного присутствия воды (см формулу на стр 71). передвижение
веществ по растению опять-таки происходит в водном растворе, самое
передвижение растворов и поднятие их по стеблю тесно связано с процессом
испарения (транспирации) воды растением (главным образом через тистья),
ктеточный сок состоит главным образом из воды, протоплазма без воды
существовать не может (Содержит около 80% воды ) и т д, Особое значение
воды состоит в том, что она приводит протоплазму, состоящую из гидрофиль-
ных котлоидов (вбирающих воду и разбухающих), в особое жизненное
состоя ние.
Таким образом, вода находится во всех частях растения. Но
находится ли вода везде и в природе9 Вода в природе находится
везде, но количество ее подвержено очень большим коле-
баниям: от местообитаний, перенасыщенных водой (болота), мы
находим все переходы к местам с ничтожным количеством воды,
даже недостаточным для жизни растений (пустыни). В некоторых
местах воды избыток, но она находится в состоянии, недоступном
растениям (ледяные пространства Арктики и Антарктики).
Все это необычайное разнообразие в количествах воды
как в широком географическом смысле (тропики и умеренные страны,
примо] ские страны и континентальные и др.), так н в топографи-
ческом (склоны, понижения, водоразделы и т. д) приводит к тому,
что вода, кроме своего исключительного физиологического значения,
играет роль первостепенного экологического фактора.
Различают три категории растений в связи с их отношением к
воде, гигрофипы, мезофиты и ксерофиты. Первые — это растения,
произрастающие в условиях избыточного увлажнения (водные
растения, растения болот и пр ), последние — растения, растущие
при недостатке воды (растения пустынь, степей), мезофиты — ра-
стения средних условий увлажнения (растения лугов, влажных
лесов и др )
Недостаток воды может иметь две основных причины: сухость
физическую и сухость физио логическую В последнем случае вода
может иметься в избытке, но растение не может воспользоваться
этой водой вследствие ряда причин* или почва слишком богата cons- ми
(солончаки), или почва богата гуминовыми кислотами (торфяные
болота), или температура почвы слишком низка (тундры и вообще
полярные страны). Во всех этих случаях растения не могут извле-
кать достаточное количество воды, и почвы, физически мокрые,
являются физиологически сухими.
Растения трех указанных категорий размещаются как по гео-
графическим областям (ксерофиты — в области пустынь, гигрофи-
ты — обитатели влажных тропиков и пр ), так и в пределах не-
больших участков в связи с различной влажностью (так, например,
вверху по склонам — ксерофиты, в средней части — мезофиты, а у
основания, на болоте—гигрофиты).
Внешность растения, его облик прежде всего
отражает условия годного па кима, причем недостаток воды особенно
:'4
разнообразно сказывается на строении растения. Вообще же вода —
фактор, оказывающий формирующее действие.
Ксерофиты и их признаки. Ксерофиты по своему облику отли-
чаются от гигрофитов, и еще издали можно видеть, какой тип расти-
тельности мы имеем перед собой, у ксерофитов зелень тусклая,
иногда седая (степь), тогда как гигрофиты выделяются своей яркой
зеленью (влажный луг).
Ксерофитами еще недавно считали растения, растущие на сухих
почвах в оолее или менее засушливых условиях и приспособленные
к наименьшему испарению воды, это, однако, нельзя считать верным,
так как в настоящее время мы знаем, что многие ксерофиты испа-
ряют достаточно много воды (см. стр. 87). Обычно главнейшими
признаками ксероморфной структуры ? вляются такие:
1. Сильное уменьшение (редукция) поверхности. При
этом листья становятся узкими или свертываются (виды ковылей и
вообще многие злаки). В других случаях вместе с уменьшением
поверхности они становятся жесткими (склерофиллы), иногда ли-
стья почти совсем редуцируются, и роль листьев берет на себя
стебель [кузмичева трава, испанский дрок (рис. 35), казуарина
(рис. 213) и др ]
2. Сильно утолщаются наружные стенки эпидермиса, вслед-
ствие чего, конечно, сильно понижается транспирация.
3. На поверхности растений, особенно на листьях, находится
более или менее выраженный волосяной покров из волосков
различного строения. Большинство степных растений имеет подоб-
ное защитное приспособление; некоторые из них являются совер-
шенно серебристыми или войлочными (Salvia aethiopis, Convolvulus
lineatus и Др.). Ниже указывается замечательное серебряное дерево
с юга Африки (стр. 374).
4. На поверхности растения находится восковой налёт,
и растение приобретает сизый цвет; часто видовые названия рас-
тений обозначаются, как giaucus (сизый): Asperuia giauca, Koeieria
glauca и пр. Кроме того, хороший пример подобных растений —
Centaurea ruthenica.
Интересна пальма (Kiopstockia), где восковой налёт имеет до
5 мм толщины.
5. Присутствие водосодержа
Рис. 36. Различные типы устьиц на листах
ксерофитов:
I—погруженное устьице у Amherstia nobilis; 2—то
же у Hakea suaveoleus, 3—восковое кольцо во-
круг устрица у молочая (Euphorbia tirukalli), 4—
уе.ьице с извилистым каналом у пальмы Nipa fru-
ticans (по Габерланду).
щей ткани или отдель-
ных клеток, богатых во-
дой. Вследствие этого
р асте н и я дел аются соч-
ными, приобретают ха-
рактерный внешний вид
и называются суккулен-
тами. Суккуленты могут
быть стеблевыми, когда
сочным является стебель
(кактусы, молочай и др.)
и листовыми, когда тол-
стыми и сочными стано-
вятся листья (агава, алоэ;
из наших растений: мо-
лодило — Sempervivum,
очиток — Sedum).
6. Особое строение
устьиц. Здесь у ксеро-
фитов наблюдается ряд
изменений в их строении,
сводящихся к уменьшению
испарения, причем устоН-
ца погружаются ниже
поверхности1! листа или над устьицем образуется восковое коль-
цо, или между замыкающими клетками уст ица имеется ориги-
нальный извилистый канал, или происходит закупорка уст&иц
и т. п. (рис. 36).
Кроме того, можно указать и на ряд других приспособлений,
как, например, уменьшение межклетных пространств,
увеличение склеренхимы и пр.
Но кроме морфолого-анатомических признаков ксерофитам при-
сущ и ряд физиологических признаков. Особенное значение имеет
повышенное осмотическое давление в клетках, которое
усиливает сосущую деятельность корня. Благодаря этому растения
могут извлекать из почвы большие количества воды и получать воду
86
тогда, когда она недоступна растениям без повышенного осмотиче-
ского давления.
Кроме того, в клетках вырабатываются особые гидрофильные
коллоиды, обладающие способностью очень сильно удерживать воду
и, таким образом, сохранять водные запасы при усиленной транспи-
рации.
Ксероморфная структура и вообще ксероморфные признаки могут эначиЭ
тельно варьировать даже в пределах различных частей одного и того же индиви-
дуума и в течение различных периодов его жизни.
Обычно первые стадии развития наземных растений проходят при более
(благоприятном водном режиме, чем последующие, и потому в первых стадиях
Рис. 37. Иллюстрация закона Заленского: т—участок нижнего листа,
2—верхнего: на каждом участке на левой половине нанесены жилки
листа, на правой—дано расположение устьиц (на нижнем эпидермисе).
растение обнаруживает признаки мезофитов, а во взрослом состоянии признаки
ксерофитов. Закономерности, которые при этом наблюдаются, представляют со-
держание закона Заленского (Заленский — русский исследователь,
изучивший эти явления). Именно, если проанализировать чистья одного и того
же индивидуума, расположенные на различных частях стебля, то наибол низко
расположенные листья по морфолого-анатомическим признакам очень сильно
•тличаются от верхних, связываясь с ними всеми переходами. При этом нижние
листья всегда бывают более крупными, имеют более крупные клетки, более
крупные, но редко расположенные устьица, сравнительно слабо развитую
сетэ жилок (сосудисто-волокнистых пучков); верхние листья имеют менее круп-
ные пластинки, менее крупные, часто расположенные устьица, сильно развитую
сеть жилок (рис. 37).
Что верхние листья более засухоустойчивы, показывает простое наблюде-
ние растения во время засушливого периода: нижние листья желтеют и опа-
дают. верхние же остаются жизнедеятельными.
В резучьтате этих наблюдений мы приходим к непонятному на первый взгляд
выводу, что ксерофиты (в данном случае листья с ксероморфной структурой)
испаряют си чьи ее, чем не ксерофиты первые имеют больше устьиц
и больше жилок на определенную площадь листа.
Исследования последнего времени дали нам более глубокое и
правильное понимание явлений ксёрофитнзма и они же обнаружили,
что многие ксерофиты испаряют воды больше, чем не ксерофиты,
что совершенно как бы не вяжется с представлением о ксерофитах
вообще. Так, в развитие мыслей Заленского, в исследованиях
Келлера брались различные виды одного и того же
р ® д а, но произрастающие в различных условиях сухости, и
87
сравнительно исследовались на указанные выше анатомические
признаки, причём обнаружились такие данные (в каждом случае
даны два наблюдения — I и II):
Общая длина Число устьиц 1 Величина кле-
Asperula жилок на I ел!3 на 1 мм2 ток эпидермиса
поверхности нижней по- верхней по-
(ясменник) листа, верхностн верхностн
в мм листа листа в мм2
A. odorata
(леснсй вид. наименее ксеро- I 316 63 0,0057
фнльный) II 308 49,3 0,0064
A. tinctoria
(вид опушек, северных сте- I 581 97,7 —
пей, средний по ксерофчль-
ности) II 703 150,3 —
A. glauca
(вид более южных степей, Г 1075 385 0,0051
наиболее ксерофильный) | II 1206 499,2 0,0038
Что касается интенсивности транспирации, то
были получены такие относительные величины за д ва^сле дующих
один за другим часа (I и 11) на солнечном свету:
I II
Asperula odorata ...... 54 54
A. tinctoria ...........81 67
A. glauca .............100 100
Отсюда ясно, что ксерофит Asperula glauca испаряет больше,
чем не ксерофиты.
По М а к с и м о в у, у ксерофитов имеются такие с т р у к*
т у р н ы е признаки:
1) уменьшение размера всех клеток, в том числе и клеток устьиц; 2) утол-
щение клеточной оболочки; 3) развитие палисадной ткани; 4) более густая сеть
жилок; 5) увеличение числа устьиц.
К числу же признаков физиологических относятся: 1) усиленная
транспирация; 2) усиленная ассимиляция; 3) повышенное осмотическое давле-
ние; 4) повышенная способность переносить завядание.
То, что у ксерофитов возможна усиленная транспирация, объ-
ясняется тем, что у них обычно хорошо развита корне-
вая с и с т е м а, и нередко корни опускаются очень глубоко,
что даёт им возможность получать воду из глубоких слоёв почвы.
Кроме того, ксероморфная структура всегда связана с высокой
концентрацией клеточного сока и повышенным осмотиче-
ским давлением. В результате имеет место более энергичное
всасывание из почвы воды, чем у не ксерофитов.
Вообще группа ксерофитов (в прежнем понимании) неодно-
родна; одни из них, как оказалось, имеют сильную транспи-
рацию, в то время как другие соответствуют их прежнему толко-
ванию, имея слабую транспирацию и другие классические при-
знаки ксерофитов (уменьшенная поверхность, запасы воды, вос-
ковой налёт и др.). Это—суккуленты. Интересно, что слабая транс-
пирация влечёт за собой уменьшенный газообмен с атмосферой и,
следовательно, слабое поступление СО2, результатом чего является
слабый рост; кроме того, дыхание здесь не доходит до образо-
вания СО2 и Н2О, а лишь до щавелевой, яблочной и других органи-
ческих кислот, которые в процессе ассимиляции опять превраща-
ются в углеводы.
Если наличие защитных приспособлений суккулентов понятно,
то для других ксерофитов требует пояснения их двойственность:
с одной стороны, сильное испарение, а с другой, — ксероморфное
строение. Однако ксерофиты каждый день испытыгают недостаток
воды, так как в полуденные часы испарение достигает очень больших
величин, и бот тогда все защитные приспособления препятствуют
завяданию растений (устьица закрываются, волосяной покров
уменьшает испарение и пр,). Вообще же у ксерофитов водный режим,
подвержен резким колебаниям, и защитные приспособле-
ния выполняют своё назначение во время превышения транспирации
над поступлением воды из почвы.
Для ксерофитов, кроме того, характерна их способность в ы-
де ржи в а т ь завядание.
Вопросы ксерофитнзма тесно связаны с вопросами засухо-
устойчивости и имеют огромное практическое значение,
при чём засухоустойчивость это, во-первых, способность пере-
носить засуху и, во-вторых, способность легко опра-
вляться после завядания.
Засухоустойчивые расы растений илн находят в природе, или
же их выводят путём селекции. Природные ресурсы в этом отноше-
нии далеко не исчерпаны.
В засушливых районах (Средняя Азия, юго-восток Европей-
ской части н др.) борьба с засухой может итти в двух направле-
ниях: I) выработка засухоустойчивых рас и 2) выработка рас ско-
роспелых, у которых период от цветения до созревания протекает
в короткое время, до наступления засухи. Последние нн в коем
случае не могут считаться ксерофитами; это — растения, убегаю-
щие, образно выражаясь, от засухи.
Поэтому так называемые эфемеры, т. е. те растения, которые
очень быстро проходят весь свой цикл вегетации (вплоть до образо-
вания семян) весной, до наступления засухи, конечно, не могут
относиться к ксерофитам. Эфемеры обычны в степях и пустынях.
Это или однолетники или лукоьично-кл; бневые растения с крайне
непродолжительным циклом развития (тюльпаны, гиацинты, кро-
кусы); из однолетников — Draba verna, виды Alyssum и др- Ьсё
это растения, убегающие от засухи, и нх ни в коем случае
нельзя считать ксерофитами, как это иногда неправильно делают.
Гигрофиты и их признаки. Растения гигрофиты не испытывают
недостатка в воде и потому не имеют никаких приспособлений к за-
щите от излишней транспирации. Наоборот, они имеют : 1) большие
тонкие листья, 2) удлинённые стебли, 3) слабо развитые корьевые
системы, так как вода имеется в избытке.
Кроме этих общих признаков, у ряда растений имеются
некоторые специальные приспособления:
89
1) сеть межклетников, усиливающих газообмен;
2) развитие особых органов — гидатод, при помощи которых
происходит выделение избытков воды в капельно-жидком состоянии;
число гидатод может быть довольно велико, например, у Gono-
caryon pyriforme в среднем на
I л/л/2 верхней поверхности
листа находится 55 гидатод, на
нижней—58 (рис. 38);
3) «капельные острия» на
листьях растений тропических
лесов, по которым вода стекает,
как по жолобу (например фи-
кус— Ficus religiosa);
4) сосочки на листьях (на-
пример Begonia imperialis), что,
увеличивая поверхность, спо-
собствует более быстрому испа-
рению избытка воды;
5) строение устьиц (напри-
мер у ряски — Lemna minor)
показывает полное отсутствие
приспособлений к затруднению
газообмена.
Гидрофиты и их признаки.
В группе гигрофитов нужно вы-
делять особо гидрофиты, т. е.
растения собственно водные,
растущие в воде нли погружён-
во время цветения выставляют
Рис. 38. Некоторые приспособления
гигрофильных растений'
1—лист фикуса (Ficus religiosa) с капель-
ным острием; 2 —гидатода на листе Gono-
♦агуоп pyrlforme— в 'Д в разрезе. 3 - то
же/ вид сверху; 4 — сосочки на поверх-
ности листа бегонии (Begonia imperialis);
по Шталю (1, 4) и Габерланду (2, 3).
ные в неё; некоторые из них лишь
над водой свои цветки (виды рдестов, водные лютики, валлисне-
рия), а другие даже цветут под водой (наяды, роголистник и др.).
Некоторые приспособительные
признаки гидрофитов'.
1) у величение поверхности
листьев (тесьмовидные илн рас-
сеченные листья), так как газо-
обмен затруднен вследствие умень-
шения количества кислорода в
воде и вследствие того, что он
совершается лишь через клеточ-
ные оболочки. Кроме того, данное
строение листьев более приспо-
соблено для текущей воды. У
мадагаскарского растения Аропо- Рис 39. мадагаскарское водное рас-
geton последнее приспосооление темне Aponogeton fenestralis (ориг).
достигается иным путем (про-
дырявленные листья) (рис. 39);
2) слабое развитие кутикулы, так как нет необходимости затруд-
нять транспирацию;
3) почти полное отсутствие устьиц;
90
4) периферическое положение хлорофиллоносного аппарата, так
как света в воде недостаточно;
5) расположение сосудисто-волокнистых пучков в центральном
тяже (защита от растяжения растения текущей водой);
6) при спокойной воде полная редукция механической ткани.
Почти все эти признаки приспособительного характера и могут
наблюдаться у ряда земноводных видов, когда они попадают в вод-
ную среду и должны соответственно перестроить свою морфологию
(известный пример — водный лютик).
Вода и распространение
растений. Вода имеет боль-
шое значение при рас-
пространении се-
мян и плодов, мень-
шее — при о п ы л е и и и.
При этом семена и плоды
имеют различные приспо-
собления к плаванию: осо-
бые плавательные пузыри
или плавательные ткани
(например у плодов кокосо-
вой пальмы); в некоторых
случаях воздухом запол-
няется лишь промежуток
между стенкой плода и се-
менем. Известны многочис-
ленные примеры, когда семе-
на и плоды плавали в мор-
ской воде до 3—4 месяцев и
не теряли своей всхожести.
Среди наших растений
хорошо плавают плоды неко-
торых болотных и прибреж-
ных осок, заключенные в осо-
Рис. 40. Валлиснерия (Vallisneria spiralis):
1—женское растение со спирально скрученными
цветоножками; 2 — мужское растение с отры-
вающимися и плавающими мужскими цветками.
бые вздутые мешочки, напол-
ненные воздухом; так же пла-
вают семена болотных ирисов
.(плавательные пузыри). У
одного клевера (Trifolium fra-
gifcfum) чашечка развивается в виде лодочки; у некоторых видов
щавеля роль лодочки играет высохший околоцветник и т. д.
Что касается роли воды как опылителя, то вода переносит пыльцу
у некоторых водных растений (например наяда и др.), причём в
этих случаях пыльцевые зёрна имеют обычнощид нитей, что обеспечи-
вает большую вероятность опыления.'
У некоторых растений вода в процессе опыления используется
у ещё иным способом: так, например, у совершенно погружённого
растения валлиснерни (Vallisneria spiralis), обычного в наших ак-
вариумах, лишь цветение и опыление происходят над водой; при
этом мужские цветки отрываются от своих цветоножек и всплывают
91
на поверхность воды, а женские цветки выступают из воды и нх
рыльца находятся на одном уровне с тычинками свободноплаваю-
щих мужских цветков. Благодаря движению воды мужские цветки
приходят в соприкосновение с женскими (рис. 40).
В распределении воды на земной поверх-
ности не наблюдается такой правильности, как в распределении
света и тепла, и на небольшой территории, в связи с разнообразием
топографических условий, можно найти как ксерофитные и гигро-
фитные, так и промежуточные мезофитные местообитания. Особен-
ное значение для растительности имеет не абсолютное количество
атмосферных осадков, а распределение их по временам года
(см. отдел IV), а кроме того, характер осадков: ливни или
продолжительные, но слабые дожди и т. п., — всё это в комбина-
ции с тепловым режимом создаёт разнообразные климатические
условия. /
Если приведённые выше примеры указывают на роль воды как
климатического фактора, то всё же нельзя не видеть, что содержание
воды в атмосфере очень тесно сочетается с содержанием воды в
почве; так, гигрофиты обычно растут на почтах с большим содер-
жанием воды, ксерофиты — наоборот. Влажная или же сухая поч-
ва — это уже факторы эдафического порядка. Таким образом,
подразделение факторов на климатические и эдафические довольно
условно, и подчас трудно бывает сказать, какая вода играет
большую роль — атмосферная или почвенная.
4. Тепло
Тепло — это один из важнейших факторов физиологического
порядка. Каждая функция растения требует того или другого ко-
личества тепла, причём наиболее интенсивно растительная жизнь
протекает при достаточно высоких температурах (н достаточных
количествах влажности) в тропических странах. Нельзя сомневаться,
что растительность умеренных и холодных стран возникла в резуль-
тате соответствующего понижения температуры.
Поскольку суммы тепла в различных точках земной поверхности
в настоящее время крайне различны, тепло выступает в качестве
очень важного фактора экологического значения, определяющего в
основных чертах распределение растений на поверхности земного
шара. Смена тепла от полюсов к экватору влечет за собой изменение
растительности и распределение последней по зонам: тундры, леса,
степи, пустыни, тропические леса, причём температурные условия
сочетаются с водным режимом. Зонами же мы называем широкие
полосы растительности (н соответственно почв), обусловленные
климатом.
Классификация климатов. Имеется несколько попыток клас-
сифицировать климаты.
Наиболее удачной нужно признать классификацию Кёппена.
Он различает 5 основных климатов:
1. Тропический климат с обилием дождей.
Климат, не имеющий холодного времени. Иногда имеется одни
92
период засухи. Характерная растительность этого климата —
влажнотропический лес (периода засухи нет, осадков более 2 м),
саванны и саванновые леса (засуха не менее двух месяцев).
2. Сухой климат. Он зависит от недостатка осадков илн
от их неравномерного выпадения. Сюда относятся пустыни без
зимних морозов и пустыни и степи с зимними морозами; первые,
например, характерны для Сахары, вторые — для Средней Азии.
3. Умеренно-тёплый климат. Средняя температура
самого тёплого месяца выше 22°. Зима более или менее холодная.
Примером может служить средиземноморский климат и ряд других,
4. Субарктический климат, или умеренно-
холодный. Лето умеренно-тёплое со средней температурой
10—22° и более илн менее обильными дождями; зима холодная со
снежным покровом. Растительность—главным образом хвойные леса
(область тайгн).
5. Снежный климат. Июльская изотерма ниже 10°, про-
должительная холодная зима. Тундры и ледяные пустыни Арктики
и высоких гор. Сюда Же относится и Антарктика.
При характеристиках климатов очень большое значение имеет
температура, ьо, естественно, также и другие климатические эле-
менты, как количество влаги, распределение тепла и влаги по от-
дельным временам года и т. п.
Очень близкую классификацию даёт де-Мартонн, но он различает
В групп климатов:
1. Тёплый климат без периода сухости.
2. Тёплый климат с сухим периодом.
3. Умеренно-тёплый климат без морозного периода.
4. Умеренный климат с морозным периодом.
5. Тёплый климат пустынь.
6. Холодный климат пустынь.
7. Холодный климат с умеренно-тёплым летом.
8. Холодный климат без тёплого времени года.
Иной принцип при классификации климатов положил в основу
Р а у н к и эр. Он взял за основу само растение, его отношение
к неблагоприятному времени года. Об этой классификации будет
сказано ниже после обсуждения вопроса о жизненных формах,
установленных Р ау нкиэром (см. стр. 28).
Несмотря на то, что тепло — фактор первостепенного порядка,
нельзя по внешнему облику растения сказать, выросло ли оно при
избытке тепла или при недостатке, в то время как на основании
предыдущего всегда совершенно ясно — имело ли растение избыток
или недостаток воды. Тепло, таким образом, не является
фактором формирующего порядка. Мы почти не знаем
внешних приспособительных признаков, вызываемых холодом или
избытком тепла. По внешнему виду никогда нельзя' определить,
сможет лн данное растение перезимовать в зелёных листьях, или
листья прн первом морозе отомрут. Защита от излишней транспи-
рации видна непосредственно, защита же от тепла и холода внешне
ничем не выражается. Так, например, такие нежные растения, как
мокрица (Stellaria media), мятлик (Роа annua), анютины глазки
(Viola tricolor), прекрасно у нас перезимовывают, не замерзая, в
93
то время как многочисленные растения с более жёсткими лис
и стеблями (например осоки, хвощи и др.) та зиму теряют свои
надземные части. Нет никаких сомнений в том, что здесь особую
роль играют свойства плазмы, различные у различных растений.
/Можно сказать, что на земной поверхности почти нет такого
места, где бы не могли существовать растения из-за отсутствия теп-
ла; если в некоторых полярных странах нет растений, то это зависит
от того, что там нет земли, свободной от снега и льда, но и там на
каждом освободившемся от льда участке земли развиваются хотя
растения.
Три кардинальные точки. Из
бы на короткий срок немногие
Рис. 41. Ложечная трава (Cochlearia
arctica) (по Чильману).
(минимальная температура —68°)
вает более 200 видов.
физиологии растений известно,
что для каждого растительного
вида нужно различать три
основные кардинальные точки:
верхнюю, нижнюю и опти-
мальную.
Верхняя точка—это верхний
температурный предел, ниж-
няя— нижний предел, а опти-
мальная точка — это точка
наиболее благоприятной тем-
пературы. При- этом для раз-
личных функций (для асси-
миляции, дыхания, роста нт. д.)
и д: я различных С1а;ий раз-
вития растеш й положение этих
точек различно.
Жизненная амплитуда ра-
стений весьма разнообразна.
В то время как некоторые тро-
пические растения повреждают-
ся от холода при -ф 2° или
даже при -ф 5°, растения се-
вера свободно выдерживают
очень низкие температуры и,
например, в Верхоянске (Во-
сточная Сибирь) при средней
температуре декабря в —46,3°,
января —50,1°, февраля —44,5е
растут леса, и флора насчиты-
Давно известно, что ложечная трава (Cochlearia arctica, рис. 41)
на северных побережьях Сибири в листьях и бутонах переносит
суровую зиму с температурой до — 46° и весной продолжает своё
развитие (Ч ильма н). Так же переносят зиму наши растения
(маргаритка — Bellis perennis, мокрица — Stellaria media, анютины
лазки — Viola tricolor и др.), выходя из-под снега с прошлогод-
ними зелёными листьями и не успевшими осенью распуститься бу-
тонами. Многие наши травянистые растения не теряют иа зиму
94
листьев, являясь зимнезелёными (например, зеленчук — Galeob-
dolon luteum, копытень — Asarum europaeum).
Наблюдения и опыты показывают, что одна из причин гибели
от холода — это потеря клетками воды; у замёрзших растений в-
межклетниках наблюдаются ледяные кристаллы. Вообще части
растений, бедные водой, гораздо лучше переносят низкие темпера-
туры; например сухие семена могут выносить до—80°, а набухшие
отмирают при более высокой температуре. Однако, например, мо-
крица (Stellaria media) в своих вегетативных частях более богата
водой, чем многие другие растения, отмирающие на зиму.
Давно отмечено, что гораздо вреднее, чем низкие температуры,
оттаивание, особенно, если оно идёт быстро. Но и здесь
нет общих правил; в Альпах, на Алтае и на других горах можно
наблюдать, что венчики раскрывшихся цветков ночью совершенно
промерзают и становятся хрупкими, как стекло, утром же оттаи-
вают, не обнаруживая никаких повреждений. Никаких приспособ-
лений (внешних) от холода здесь не имеется.
Зимой в большинстве случаев для растений вредна не низкая температура,
а сильное испарение под влиянием сухих зимних ветров. В резуль-
тате испарения клетки растений теряют воду в количестве, превышающем
необходимый минимум н вследствие этого отмирают. Естественно, что отмирают
ветви и части растений, выступающие над снегом, и чем выше растения, тем
больше они подвергаются риску замерзнуть; поэтому во многих местах высота
деревьев и кустарников соответствует глубине снежного покрова. Интересны
опыты, показывающие, что различные деревья испаряют в зимнее время раз-
лично; так, по Иванову, транспирация на 1 сма поверхности в относитель-
ных величинах (причём транспирация 1 сл*а годовалого побега лиственницы
принимается за единицу) составляет:
лиственница ... 1 сибирский кедр . 1,9 ель сибирская . . 0,8
пихта сибирская . 0,3 сосна 0,6 ель европейская . 1,5
Как видно, лиственница, теряющая на зиму хвою, испаряет своими годо-
валыми побегами больше, чем хвоя хвойных пород (кроме ели европейской).
Ещё больше, чем лиственница, испаряют зимой деревья, лишённые зимой листь-
ев: клен— 1,45, ольха серая—1,86, бук—2,91, граб—4,58.
Из сказанного становится понятно, почему северные границы лиственных
пород не идут так далеко на север, как хвойных.
Отсутствие ряда лиственных пород в Сибири тоже зависит от силы транс-
пирации; так, по Г о р дя г и н у, ветви дуба дают максимальные суточные
потери воды (к дубу близок ясень), вследствие чего он и отсутствует в Сибири.
Морозостойкость. Изучение проблемы морозостой-
кости растений крайне важно как теоретически/ так и практи-
чески. Основная цель — получение морозостойких сортов, не вы-
мерзающих при низких температурах.
Причины вымерзания, невидимому, достаточно сложны и еще
не выяснены в полной степени. Высказывалось мнение о том, что
причиной вымерзания является обезвоживание клеток — в меж-
клетниках образуются ледяные кристаллы; последние сосут воду из
прилегающих клеток; однако ряд опытов не подтверждает того, что
непосредственной причиной вымерзания является обезвоживание.
Также не может считаться правильным мнение об отравлении
протоплазмы вредными, сильно концентрированными веществами
(соли, дубильные вещества и др.) при повышенной концентрации
клеточного сока.
95
Высказывалось также мнение, что причиной вымерзания явля-
ются кристаллы льда, образующиеся в межклетниках и механи-
чески производящие разрушение клеток. В последнее время при-
ходят к убеждению в том, что при вымерзании имеет место совокуп-
ное влияние ряда неблагоприятных воздействий на клетку, причём
наибольшую роль играют два фактора: обезвоживание и механи-
ческое давление ледяных кристаллов на обезвоженные клетки-
Действительно, одно образование кристаллов льда в межклетниках
не вызывает гибели клеток, но в сильно обезвоженных клетках
поверхностный слой протоплазмы легко повреждается от давления
льда, ледяные кристаллы проникают в протоплазму, что приводит
к сильному изменению её строения и к замерзанию.
Однако можно думать, что у различных растений процесс вымер-
зания может видоизменяться.
Так как вымерзание связано со свёртыванием протоплазмы, то
пси усилении морозостойкости вопрос идёт о химических измене-
ниях протоплазмы, направленных к увеличению её способности
сопротивляться свёртыванию, об уменьшении воды в клетках и
накоплении некоторых веществ (например, сахаров), усиливающих
морозостойкость.
Для некоторых растений установлено, что морозостойкие сорта имеют в сво-
их клетках большее количество растворимых углеводов-сахаров, чем неморозо-
стойкие; при этом можно искусственно повышать морозостойкость, вводя в клетки
те илн иные вещества.
При вымерзании играют роль и условия оттаивания. Имеются указа-
ния, что гибель растений нередко имеет место при быстром оттаивании.
Однако, как показывают более новые данные, гибель растений от оттаивания
происходит в природной обстановке очень редко, но при оттаивании обычно
усиливаются повреждения, полученные растением при замерзании. Вообще
же п здесь отмирание клетки происходит в результате механического поврежде-
ния протоплазмы, так как обезвоженный протопласт при быстром, оттаивании
также быстро изменяет свой объём.
Некоторые сорта растений, например, некоторые юж но а мери канские сорта
картофеля, обладают очень высокой холодостойкостью—свойством, наслед-
ственно закреплённым. Однако ряд опытов показывает, что возможно и искус-
ственное з а к а л и в а и и е растений против холодов, искусственное вы-
ведение морозостойких сортов.
Многолетние растения должны иметь приспособления для того,
чтобы пережить зимний холодный период. Для этого они проходят
сложный процесс закаливания, клеточное содержимое претерпевает
ряд изменений. Растения должны подготовиться 'к зиме. Действи-
тельно, если ветки сосны, берёзы и др. вымерзают (в холодильник: х)
летом прн —8°, то зимой она выносит морозы до —40° и ниже; ози-
мые ,в сентябре погибают в холодильниках при —7°, а зимой выно-
сят —20° и ниже и т. д. У озимых растений при закаливании нако-
пляется большое количество сахара, у многолетних растений сначала
накапливается крахмал, который затем переходит в защитные ве-
щества (сахар и др.), у многих древесных растений крахмал пере-
ходит не только в сахар, но и в масла. Эти защитные вещества не
замерзают при очень сильных морозах или совсем неспособны за-
мерзать (масла), что даёт возможность растениям выдерживать низ-
кие температуры.
Рис. 42. Сольданелла (Soldanella alpina)
пробивает снег (по Мартонну).
Верхняя кардинальная точка тоже является раз-
ичной у разных растений. Так как высокая температура связана с
ысыханием, то лучше всего её выдерживают органы с небольшим
содержанием воды (семена, споры — до 100—130°). Вообще же
на земной поверхности мало мест, где бы не могли жить растения
из-за высокой температуры (например кратеры, фумаролы вулканов
и пр.), так как даже в горячих источниках (до 80°) находятся бак-
терии и сине-зелёные водоросли. Для большинства цветковых рас-
тений губительны действующие некоторое время температуры
40—50°.
Нередко можно наблюдать вредное действие высоких температур
на деревья, оказавшиеся после порубок на сильно освещённых
местах: камбий на стороне, обращённой к солнцу, отмирает, и по-
лучаются как бы ожоги коры.
Так как поверхностные
слои почвы нагреваются го-
раздо сильнее, чем воздух
(в пустынях даже до 80—84°),
то, естественно, от сильного
нагревания должны страдать
нижние части растений, со-
прикасающиеся с почвой.
Поэтому у многих ксеро-
фильиых злаков при основа-
нии развиваются так назы-
ваемые туники из остатков
листьев; остатки нижних
листьев сохраняются также
у многих ксерофильных дву-
дольных, защищая осног ания
стеблей (в иных случаях
остатки нижних листьев у
растений высокогорий могут
защищать их от холода).
Существенной защитой от
сильного нагревания расте-
ний служит постоянная транспирация, отнимающая тепло от са-
мого растения, вследствие чего части растения всегда имеют более
низкую температуру сравнительно с окружающей средой.
Оптимальная точка — это наиболее благоприятная
температура для жизни растения. Нужно различать оптимум гар-
монический, абсолютный и экологический. Если гармонический оп-
тимум наиболее благоприятен для той или иной функции, то абсо-
лютный — это та температурная точка, при которой данная функция
проявляется наиболее энергично; например дыхание при повышении
температуры идёт всё более и более интенсивно, достигая абсолютно-
го оптимума, но этот последний не является наиболее благоприятным
(гармоническим), который лежит значительно ниже. Экологиче-
ский же оптимум — это совокупность гармонических лптимямов.
Нужно указать на то, что различные функции растения (про-
растание, рост, ассимиляция, дыхание и пр.) имеют различные
кардинальные точки, так, например, нижняя точка для дыхания
— 10°, а для ассимиляции (Picea excelsa)—40°.
Очень интересны нижние точки для роста, которые для большин-
ства растений лежат выше 0°, но у ряда альпийских растений
(Soldanella — рис. 42, Crocus н др.) рост идет при температурах,
близких к точке замерзания. То же наблюдается и у наших ве-
сенних эфемеров (Anemone, Corydalis, Ficaria и др.), способных
к подснежному развитию в зимние месяцы, благодаря чему
они и зацветают так рано весной (рис. 43).
Рис. 43. Подснежное развитие. Молодые стебли'пролески (Мег-
cunahs perenms) с бутонами и (слева) молодые листья чистяка
(Ficaria ranunculoides). Все разрыто из-под снега в начале апреля
(по Кожевникову).
Можно думать, что эти растения являются реликтами ледни-
кового периода, сохранившими свойственный им ритм развития
и до настоящего времени.
Система жизненных форм Раункиэра
Выше было указано, что положение перезимовывающих орга-
нов растений на различном уровне над землей имеет очень
большое значение при сильных зимних ветрах. Положение этих
органов в связи с условиями неблагоприятного времени года
(холод или засуха) н было принято Раункиэром в основу
его классификации жизненных форм. Ра у н к и эр различает
пять основных типов (рис 44)
I. Фанерофиты (Р)Почки возобновления находятся
1 От греческих слов «фанерос» — явный, открытый и «фнтон» — растение.
98
Рис. 44. Жизненные формы по Рауикиэру-
1- фанерофит,
и 2ь—подтипы хамофитов; За,
растений зачерненные на
Зь, Зс—подтипы гемикриптофитов, 4а, 4ь—подтипы криптофитов, 3—терофит,
рисунке, перезимовывают, незачерненные— отмирают на зиму.
Част^
высоко над поверхностью почвы и или совсем не защищены почеч-
ными чешуями (влажные тропические леса), илн же защищены
ими. Сюда относятся главным образом деревья и кустарники, но
также и стеблевые суккуленты, эпифиты и некоторые другие. Фане-
рофитами особенно богаты влажные тропические и субтропические
страны.
Этот большой тип распадается на несколько подтипов, из ко-
торых укажем:
мегафанерофит ы1—очень высокие деревья, гыше 30
мезофанерофиты — белее низкие деревья—от 8 до 30 м;
микрофанерофиты — по большей части кустарники;
на нофа н*е рофит ы1 — мелкие к\старники.
Первые два подтипа обозначаются условно ММ, микрофанеро-
фиты— М, нанофанерофьты — N.
II. Хамефиты (Ch)1 2. Почки находятся сравнительно
невысоко над земной поверхностью и защищены чешуями и снегом.
Мелкие кустарнички и полукустарники (брусника, черника, вереск
и др.). Характерны для холодных поясов. Растения не образуют
каждый год нижние части надземных побегов (короткий вегетацион-
ный период). Распадаются на ряд подтипов.
III. Гемикриптофиты (НК)3. Почки возобновления
гемикриптофитов лежат на уровне поверхности почвы, т. е. на
зиму отмирают все надземные части. Они распадаются на под-
типы и сюда относится большая часть травянистых растений уме-
ренных широт.
IV. Криптофиты (К)4. Отмирают на зиму не только
все надземные части, но н часть цодземного стебля, и потому
почки возобновления находятся на известной глубине под землей
(луковичные, клубневые, корневищные растения) или под водой.
Все криптофиты — травянистые растения. Много криптофитов на-
ходится в степях.
Криптофиты обычно подразделгются на геофиты (G)5 — сухо-
путные растения, гелофиты6 (болотные растентя) и гидрофиты
(водные растения); почки возобновления последних двух 1 азван-
ьых групп находятся под водой (условно две последние группы
обозначаются НН).
V. Т е р о ф и т ы (Т) 7. Всё растение на зиму отмирает целиком
и перезимовывают лишь семена. Однолетники. Особенно характерны
для пустынь с их коротким периодом влаги.
Классификация жизненных ферм Раункиэра сильно
отличается в своих основных подразделениях от других соответ-
1 От греческих слов «мегас» — большой, «мезос» — средний, «микрос» —
маленький и от латинского слова «нанус» — очень маленький.
а От греческих слов лхамай» — над землей и «фитои» — растение.
а От греческих слов «геми» — наполовину, «крипто» — скрываю, «фитон»—
растение.
4 От греческих слов «крипто» — скрываю и «фитон» — растение.
6 От греческих слов «ге» — земля и «фитон» — растение.
в От греческих .слов «гелос» — болото и «фитон» — растение.
7 От греческих слов «терос»—лето н «фитон»—растение.
*00
ствующих систем; именно, выше были указаны физиономические
системы (см. стр. 57), в классификации же Р а у и к и э р а первые
подразделения не являются физиономическими; так, в числе фанеро-
фитов имеются и деревья, и мелкие кустарники, и кактусовндные
растения, и эпифиты; среди гемикриптофитов — и высокие дву-
дольные и злаковидные растения, среди криптофитов — и наземные
виды и подводные н т. д. Но в дальнейших, более мелких подразде-
лениях у Р а у н к и э р а все более выступают жизненные формы
физиономического характера.
Приводимая ниже табличка показывает соотношение типов расте-
ний в различных географических условиях (биологические спектры).
Число принятых во внимание ВИДОВ 1 р * Ch нк К Т
Тропическая область .... (Сейшельские острова) 258 61 6 12 5 16
Пустынная область (Ливийская пустыня) 194 12 21 20 5 42
i Средиземноморье (Италия) 866 12 6 29 11 42
Умеренная область (Дания) 1 С84 7 3 50 22 18
Арктическая область . • • . (Шпи цберген) ПО 1 22 60 15 2
Здесь даны гять примеров, подобранных к пяти основным кли-
матическим областям К е п п е н а (см. выше), причем видно, что
каждый климат имеет свой, присущий ему, «биологический спектр»,
так, для тропического климата характерно господство фанерофитов,
для пустынного —терофитов, для средиземноморского—гемикрип-
тофитов и терофитов, и т. д.
Поэтому и климаты можно класскфидировать, исходя из
жизненных форм, и говорить о климате фанерофитов, о
климате гемикриптофитов и т. д.
Биологические спектры также сильно меняются в горах в связи
с высотой места.
Фенологические явления. Кроме термометра, хорошими показа-
телями температурных условий могут сложить сами растения при
наблюдении над их периодическими явлениями. Фазы развития
растений (появление листьев, появление бутонов, зацветание, две-
тение, отцветание н т. д.) проходят в определённой последователь-
ности и связаны главным образом с определённым количеством
тепла. Фенология как раз и изучает все эти периодические явления
как растительного, так и животного мира (прилёт птиц, гнездо-
вание и пр.).
Подобно тому как на метеорологических картах наносятся линии
одинаковых температур, осадков и пр., так и в фенологии на карты
наносятся изофены1, т. е. линии, соединяющие точки одноимённых
явлений, например, линии зацветания, цветения, созревания посе-
вов, жатвы и пр. На рис. 45 приводится образец карты с изоантами2
(линии одновременного цветения).
1 От греческих слов «изос» — равный и «феномен» — явление.
! От греческих слов «изос» — равный н «антос» — цветок. -
102
При изучении фенологических явлений необходимо производить
не отрывочные наблюдения отдельных стадий, но длительные на-
блюдения, связывающие все стадии Друг с другом. Прн этом возмож-
ны наблюдения: 1) суточные, 2) сезонные, 3) годичные и 4) много-
годичные.
Особенно важны многогодичные наблюдения, так как онн дают
картину поведения растений при разнообразных метеорологических
Рис .46. Карта зацветания сирени в Европе (из Геттнера).
Фенологические явления не совпадают с календарными данными,
и потому фенологи пользуются особым фенологическим календарём,
датами которого служат те или иные моменты развития растений.
Так, немецкий фенолог И н э различает три весенних периода,
два летних и два осенних. Начало первого весеннего периода насту-
пает с зацветанием орешника, второго — с зацветанием клёна,
третьего — сирени (рис. 46); первого летнего — ржи, второго лет-
него — липы н т. д. Ясно, что каждый год фенологические даты
приходятся на разные календарные числа н имеют особо важное
значение для сельского хозяйства, так как оно в своих сроках бази-
руется именно на фенологических моментах. Детально разработанные
фенологические карты дадут для сельского хозяйства i.oni ый ма-
териал для планирования тех или иных работ. Однако для постро-
15Q0
аосю
2500
2000
1500
VOOO
500
ения карт требуется массовый материал для очень
большого числа точек.
На севере фенологические стадии очень связаны с продолжи-
тельностью освещения (дл! ниый день), на это уже было указано
(см. стр. 8а). Здесь со(Тветствующие фенологические явления
летом наступают неск лько раньше, чем весной, и если весной 1*
широте, давал на севере запаздывание цвете-
ни' е а 4 дня, то цветение ржи запаздывает
на 3 д; я, а жатва ржи лишь на2,4 дня в сред-
нем. Наоборот, вгорах запаздывание усили-
вается: для цветения ржи на каждые 100 м
подъем запаздывание составляет 4 дня, а для
жатвы—5—7 дней; впрочем, здесь влияют
другие факторы-
Интересна схема Шрётера-Гамса
(рис> 47), показывающаях наступление раз-
личных фенологических периодов на различ-
ных высотах, из которой видно, что с вы-
сотой: 1) сокращается число периодов; 2) ве-
сенние периоды запаздывают; 3) осенние
наступают раньше: на высотах выше 2 000 м
весна прямо переходит в осень; 4) на скло~
иах условия более благоприятны, чем у
оснований гор.
Фе! ©логические явления, как было вы-
яснено, протекают в определенные сроки, и
каждое из них наступает при определенной
сумме тепла (сумма средних нлн максималь-
ных суточных температур, начиная от весен-
него исходного дня). Однако, если указан-
ными суммами тепла иногда и пользуются,
все же необходимо заметить, что э о явление
очень сложное, так как низкие температуры
могут иметь иногда исключительное значение
сравнительно с более высокими, что доказа-
но, например, яровизацией.
Фенологические наблюдения позволяют
изучать ритмы развития у растений, они
в очень многих случаях устанавливают,
что Э1и ритмы не совпадают с температур-
ными и вообще с климатическими условиями
места произрастания этих растений. Впро-
чем, об этом будет сказано ниже (см. стр. 133).
ходя из учения о стадийно-
мы можем изменять
Рис 47. Схема изме-
нения фенолог ве-
ских времен roia с
высотой в горах. За-
штр: хованная часть
схемы соответствует
периоду зимнего по-
коя. ЯСНО ВцДНО
(особенно слева) бо-
дес благоприятное
по iO/кечце склонов
по сравнению с по-
до ивой горы.
1 — начало весны; 2—
первая половина ррсны,
3 — вт( п 1я по obi на вес-
ны 4-перв 'Я половила
лет! 5—вторая полови
на .ни, 6 —начало осе-
ни, 7 — о( ень S— повл-
ияй осень Ц- фры с 1ов 1
и а ОнСАНье — высо а в
метр ах, цифры сверху—
месяцы ода (по Гансу
с изменениями).
Яровизация И
с т и е развитии растений (Л ы с е н к о),
обычную длительность тех или иных фенологических фаз. Имен-
но, первая-стадия, названная стадией яровизации, требует опре-
деленной температуры, причем эта температура очень различна
для различных видов и даже сортов.Яровизация дает возможность
104
С
заставить озимую пшеницу вызревать в первый же год, поздне-
спелые сорта пшениц превращает в раннеспелые, заставляет хлоп-
чатник созревать раньше и т. д.
ри этом недостающий в природных условиях фактор (повышен-
ная или пониженная температура н пр.) дается семенам, тронув-
шимся в рост. Семена соответствующим образом проращиваются,
зародыши начинают показываться из оболочки, и это--момент
применения яровизации апример семена твердых пшениц выдер-
живаются 10—15 дней при температуре!—5°). Затем семена слег-
ка просушиваются и сеются.
Трудно даже предвидеть все последствия открытия процесса
яровизации — человек получил в нем могучее средство управления
природными явлениями.
Несомненно, яровизация сыграет огромную роль и в аккли-
матизации растений, понимая под акклиматизацией приспособлен
ьие растений при переносе их в новые условия существования.
Акклиматизация и натурализац 1Я. Если при акклиматизации
растение переносится в страну с иными условиями, чем имеющиеся
на его родине, то при натурализации условия являются тожде-
ственными, однако на практике очень трудно разграничить два этих
понятия, тем более, что трудно представить себе две страны, со-
вершенно тождественные во всех отношениях. Поэтому целесооб-
разнее все соответствующие явления рассматривать как аккли-
матизацию то более, то менее глубокую. Акклиматизация невоз-
можна без детального познания экологических свойств растений,
потому что здесь необходим тот или иной сдвиг экологического
105
оптимума. Так как гармонические оптимумы различны для разных
функций, то становится понятным, почему при акклиматизации
растение иногда вегетирует, но ие приносит цветков; так, многие
растения умеренной зоны (наши плодовые деревья, тмин, синяк —
Echium vulgare и др.) подтропиками не цветут, так как для разви-
тия цветков требуется более низкая температура, чем для вегета-
тивных органов.
Вопросы акклиматизации всегда играли очень большую роль в сельском
хозяйстве, в парковом, садовом деле и т. п. Трудно, например, представить
себе Крым без кипарисов, магнолий, павловний, лавров и т. п., ио все это —
результат акклиматизации (рис. 48/. В настоящее время в зеленом строительстве,
в лесном деле и в сельском хозяйстве вопросы введения (интродукции) новых
пород приобретают исключительное значение.
Эдафические факторы
Для большинства растений почва совершенно необходима, как
для их п р и к р е п л е н и я, так и для п и т а и и я. Не при-
крепляются к почве лишь свободиоживущие растения, взвешенные
в воде, паразиты, эпифиты и некоторые другие, прикрепляющие-
ся к другим организмам.
Высшие растения при-
крепляются к почве на-
столько прочно, что вы-
звать их с корнями очень
трудно, а в большинстве
случаев просто невозмож-
но. Выше уже было ука-
зано, что даже ураганные
ветры только ломают не-
которые деревья, но ие в
состоянии выдернуть их
с корнями. Так обстоит
дело у многолетних ра-
стений с более или менее
мощной корневой систе-
мой. Лишь однолетние
растения не имеют необ-
ходимости в сильном раз-
витии корневой системы
и потому легко вы-
дергиваются с кор-
нем из земли.
Рис. 49. Баниан (Ficus bengalensis). Для лучшего прикрепления
к почве растения имеют раз-
личные приспособления; так, у растений болотных мест тропических областей
нередки так называемые ходульные корни, отходящие косо от нижних
частей стебля и укрепляющие растения в недостаточно прочных почвах (на-
пример, панданусы, мангровые деревья и др.). Среди наших растений некоторое
подобие ходульных корней мы встречаем у ольхи.У баниана имеются столбо-
видные, поддерживающие крону корни (придаточные, рис. 49). Целый ряд типов
укрепляющих корней известен у деревьев тропических и субтропических лесов,
106
Интересны корни втягивающие. Они втягивают на известную глуби-
ну в почву луковицы, клубни и корневища многих наших растений (луки,
пролески, орхидеи, ирисы и др.). Эти корни не дают также розеточным расте-
ниям подниматься выше и все время притягивают их к земле. Втягивая в землю,
втягивающие корни, конечно, более прочно прикрепляют растения.
Значение почвы в питании растения общеизвестно. Все
зольные элементы, необходимые для растений, получаются именно
из почвы; сюда относятся: фосфор, сера, калий, кальций, же-
лезо, магиий. Поглощаются элементы в виде растворимых в воде
солей и всасываются корнями, вернее — корневыми волосками и
самыми тонкими корешками.
Как известно, поступление воды (и содержащихся в ней солей) в клетки
корневых волосков обусловлено разностью осмотических давлений внешнего
(почвенного) раствора и клеточного содержимого. Чем выше осмотическое да-
вление в клетках, тем с большей силой идет всасывание воды из окружающей
почвы; при равенстве давления всасывание может совсем прекратиться. При
всасывании вода проходит через клеточную оболочку, представ тяющую мелко-
порнстую пленку, и через постенный слой протоплазмы, обладающий полупро-
ницаемыми свойствами (т. е. способностью пропускать воду и некоторые мине-
ральные соли, но почти не пропускать органических веществ).
Детальное рассмотрение вопросов питания растений относится
к области физ: ологии растений, поэтому перейдем к почве как
экологическому фактору эдафического порядка.
Эдафические факторы для растений имеют крайне важное зна-
чение, однако они проявляются на фоне обшеклиматических усло-
вий, так как в одной и той же климатической зоне растительность
может быть очень разнообразной в связи с разнообразием эдафи-
ческих условий. Растения могут быть прикреплены к голому суб-
страту (неизмененная материнская порода), но могут расти и на
почве, если понимать под последней лишь те дневные или близкие
к ним горизонты материнской породы, которые изменены под воз-
действием климата, животных и растительных организмов (почва,
кроме того, тесно связана с рельефом и отражает историю данной
страны). При таком толковании почвы (Докучаев) ие всякий
субстрат является почвой, ио почва всегда содержит остатки ра-
стительного и животного происхождения (гумус). Вообще же почва
без деятельности живых организмов образоваться не может.
Экологическое значение почв зависит от их крайнего разнообра-
зия '
Так как почва, с одной стороны, имеет определенную структуру,
а с другой стороны, содержит определенные химические соедине-
ния, то, естественно, можно говорить о физических и хи-
мических свойствах почв. К физическим свойствам относят-
ся: механический состав, влагоемкость, водопроницаемость, ка-
пиллярность, структура, плотность, окраска почв и т. п.; к хими-
ческим — валовой состав почвы, гумус почвы, реакция почвы,
солевой режим и пр.
Вопрос о том, какие свойства почвы — химические или физи-
ческие — имеют большее значение, являлся, и иногда является и
теперь, предметом споров, причем представителем первого направ-
ления был У и г е р (1836), а второго — Турман (1843).
Однако надо думать, что подобное рассмотрение вопроса односто-
107
роине, и нельзя сомневаться в том, что химические и физические
свойства почв находятся в тесном сочетании между собой.
Растения-иидикатсры. Уже давно было подмечено, что одни ра-
стения связаны с определёнными почвами, тогда как другие не
являются показательными в этом отношении. Ещё Унгер под-
разделил растения на: 1) почвопостоянные, 2) почвопредпочитаю-
щие. 3) индиферентные.
Дальнейшие наблюдения показали, с одной стороны, правиль-
ность нередкой приуроченности растений к известной почве, а с
другой стороны, указали на необходимую осторожность в этом
вопросе, так как в различных местах почвоприурочениость может
изменяться. Особенное развитие учение о почвоприуроченности
растений получило в Америке (К л е м е и т с) — учение об «ин-
дикаторах». Можно говорить сб индикаторах чернозёмных почв,
песчаных, солончаковых и др., об индикаторах почв избыточно
увлажнённых, среднеувлажнённых и др., об индикаторах почв
каменистых и рыхлых и т. д. Установление индикаторов имеет
большое практическое значение, так как растения при этом могут
сл\жить непосредственными указателями эдафических условий
без применения длительных почвенных исследований. Приведём
лишь некоторые примеры индикаторных растений.
Эвтрофные растения (т греческих слов «эу» —
хорсшо и «трофэ» — питание), всегда связанные* с богатыми пом-
вамп. Сюда относятся многочисленные растения черноземных степ-
ных пространств, а также широколиственных лесов (сныть, про-
леска — Mercurialis, ясменник и др-), тучных лугов и пр.
Олиготрофные растения (ст греческих слов
«олкгсс» — небольшой и «трофэ» — питание)'. Они довольствуют-
ся для своей жизни почвами, бедными питательными веществами,
и растут на тощих оподзоленных суходолах, на верховых болотах
и др. Сюда относятся: белоус (Nardus stricta), вереск (Calluna vul-
garis), росянка, сфагновые мхи верховых болот и пр. Верховые
(сфагновые) болота нередко называют олиготрофными. Быть мо-
жет, насекомоядная росянка (Drosera) своим насекомоядным обра-
зом жизни возмещает отсутствие питательных веществ в субстрате.
Нитратные растения. Они указывают на богат-
ство нитратов в почве, что обычно имеет место вблизи жилья че-
ловека. Хорошим примером является обыкновенная крапива,
всегда сопровождающая человека. Присутствие крапивы в лесу
часто указывает на место прежнего жилья. Дикое произраст ние
крапивы в олыпанниках связано с богатыми почвами этих угодий
и с обилием здесь птиц.
Кальцифильные растения- По вопросу о связи растений с из-
вестковыми почвами имеется обширная литература. Различают
растения кальцифильные («предпочитающие» кальций), кальци-
фобные (избегающие кальция) и индиферентные. К кальцифилам
относится большинство степных растейпй (степные почвы богаты
известью); такие растения, как Gypsophyla altissima, Astragalus
austriacus, Centaurea ruthenica и др., сразу указывают на повышен-
ное вскипание потвы с НС1.
108
Но особенно обильно кальцифилы развиваются на известковых обнажениях
и иа обнажениях чистого мела. Последние крайне типичны для юго-востока
Европейской части СССР по нижним течениям Дона, Донца, Волги и других
рек. Здесь встречается много видов с видовыми названиями «меловые»: Linaria
cretacea, Scrophularia cretacea, Silene cretacea и т. д. Целый ряд эндемиков
связан с этими мелами.
Это в общем оригинальная ^меловая флора», сыгравшая в русской ботани-
ческой географии большую роль. Она дала повод для научной дискуссии, про-
должавшейся длительное время в конце прошлого и в начале этого столетия.
Одни авторы (Литвинов, Пачоский и др.) считали её древней,
реликтовой, другие (особенно Т а л и е в) связывали её присутствие с деятель-
ностью человека, считая, что меловые обнажения явились в результате усилен-
ного выпаса. Однако нет никаких оснований связывать меловые обнажения
исключительно с деятельностью человека, к тому же меловая флора богата
своеобразными эндемиками и рядом реликтовых видов. В последнее время
«реликтовая теория» получила ещё ряд новых фактов (К о з о-П о л я иски й).
В связи с кальцифильными растениями особенно усиленно об-
суждался вопрос о том, какие свойства почв являются определяю-
щими — химические или физические. Сторонники физической тео-
рии указывали на то, что кальцифилы являются ксерофитами, а
кальцифобы — гигрофитами; это верно лишь отчасти, так как
многие кальцифилы произрастают на субстратах различной влаж-
ности, ио обязательно я^держзших известь (Orchis militaris,
Gentiana cruciata, Brunella grandiflora и др.).
С другой стороны, можно привести примеры искусственных
культур типичных кальцифобов на почвах, богатых известью;
например, сфагнум культивировался на мелу, вереск — на извест-
ковой почве и пр. Очевидно, кальций как таковой, не играет
уж такой решающей роли.
Вопрос о калышфилах усложняется ещё тем, что их индикатор-
ное значение очень относительно, и хороший инди-
катор известковых почв одной местности может в другой местности
переходить, и на другие почвы. Так, такой хороший кальцифил,
как Cypripedium calceolus, можно встретить и на болотистых ме-
стах. Лиственница в Баварии и Зальцбурге растет на известковых
почвах, в Швейцарии и Тироле —на почвах, бедных известью, а в
Карпатах — на различных почвах. Можно привести немало и
других подобных примеров.
Шим пер, основываясь на этих фактах, думал, что подобное
явление можно объяснить тем, что здесь мы имеем у одного и
того же вида различные физиологические и экологические
формы: то кальцифильные, то кальцифобные, то индиферентные.
В других случаях весьма большую роль играет в природе кон-
куренция между видами, так как при устранении конкуренции не-
которые меловые виды становятся вида^й индиферентными; так,
например, в ботанических садах они могут произрастать на
самых различных почвах, в природе же произрастают на извест-
ковых и меловых почвах потому, что не встречают там конку-
ренции среди других видов.
В разных частях своего ареала кальцифильные виды обнаружи-
вают неодинаковое отношение к почвенным условиям: в централь-
ных частях ареала, там, где виды находятся в наиболее благо-
109
приятных условиях существования, они более или менее безраз-
личны к почвам, а в периферических частях ареала становятся
все более и более почвопостоянными, избирая известковые почвы.
Так, многочисленные степные растения, встречающиеся в области
степей как в самой степи, так и на склонах, на лугах и пр., на сво-
ей северной границе встречаются исключительно иа известковых
южных склонах.
Реакция среды. В вопросе о кальцифилах, как показали иссле-
дования последних лет, большое значение имеет реакция среды,
которая может быть кислой, щелочной и нейтральной. Еще.
Пауль показал, что для сфагновых мхов губительна ие из-
весть, а щелочная реакция СаС03, так как эти мхи могут хороша
расти иа гипсе (CaSO4).
Реакция среды зависит от количественных соотношений свободных ионов
водорода Н* и гидроксильных ионов ОН' (НаО ОН' + Н‘), причем если в
избытке находятся свободные ноны водорода, то среда кислая, при избытке
свободных ионов гидроксила — среда щелочная, а при одинаковой концентра-
ции водородных и гидроксильных ионов среда является нейтральной. В 1 л-
воды при 18°С количество диссоциированной воды равно 10-14 грамм-молеку-
аамилиЮ-7 грамм-ионам Н (0,0000001г) и 10~7 грамм-ионам ОН' (0,0000001г).
Для чистой воды существует такая закономерность
[Н ]х [ОН'] =10“7 1(Г-’=Я=КГМ-
Так как произведение из числа иоиов Н’ и ОН' является величиной по-
стоянной (К), то, естественно, при увеличении числа иоиов Н' число иоиов
ОН' будет уменьшаться. Предположим, что число ионов Н‘ стало больше
10-", тогда величина 10-7 должна сделаться больше, но 10-"-= --р, и, значит,
при увеличении этой дроби показатель знаменателя должен сделаться меньше,
апример нли в (10-6 или 10—5).
Показатель степени для ионов водорода условно обозначается как pH и в
последнем примере pH для нейтральной жидкости равнялся 7, а прн увеличе-
нии числа ионов водорода получил значение 6 или 5. Таким образом, при уси-
лении кислой реакции pH уменьшается (pH = 7, рН=6, pH =5 .. ), а при ослаб-
лении кислой реакции pH увеличивается (рН=8, pH=9 ..), т. е. в кислой среде
мы имеем значение pH <7, а в щелочной—pH >7.
По отношению к реакции среды можно различать три группы
растений:
оксифильные (кислые)1 pH < . . . . 6,7;
нейтрофильные pH = . 6,7—7,0э
базифйльные (щелочные) pH > . . . . 7,0.
К числу оксифильных растений относятся такие виды, как
черника, душистый колосок, ландыш, щучка (Deschampsia caes-
pitosa), молиния (Molima coerulea) и другие растения кислых лугов
и болот (рис. 50), к числу нейтрофильных — трясунка, овсяница
луговая и др., к числу базифильных —многочисленные растения
степей, полупустынь, пустынь и др.ЛК нейтрофильным или очень
слабо базифильным относятся рассмотренные выше кальцифиль-
ные растения, и вопрос о кальцифилах тесно связывается с вопро-
сами реакции среды. В самом деле, СаСО3 представляет соединение
сильного основания со слабой кислотой и реагирует, как слабая
1 Кислые растения называют также ацидофильными (от латинскою сповг
«ацидус» — кислый).
ПО
щелочь; там, где кальций находится в избытке, он нейтрализует
гумусовые кислоты, образующиеся прн гниении растений, и почва
здесь реагирует или слабо щелочио, или слабо нейтрально. Поэто-
му растения, связанные с нейтральной или слабо щелочной реак-
цией, и являются «кальцифилами».
Значение pH ие является строго постоянным для определенных
видов, ио может колебаться в известных пределах. В разных поч-
венных типах и в различных областях pH дает значительные ко-
лебания. Так, в умеренных климатах преобладают кислые почвы
Рис. 50. Зависимость развития от крестовника (Senecio silvaticus) и pH поч-
вы; цифры—значения pH (по Ольсеиу).
(например в области распространения бука рН=6,5—4,5), в тёп-
лых климатах — нейтральные почвы, а в жарких (аридных) об-
ластях — щелочные, со значением pH ^>7. Вообще же, на-
сколько известно, пределы pH колеблются от 2,9 до II.
Наши культурные растения почти все являются или нейтрофиль-
ными, или базифильными, поэтому при внесении минеральных
удобрений нужно остерегаться того, чтобы не сделать почву кислой.
Солончаковые растения — галофиты. Галофильная раститель-
ность местами занимает обширные территории, имея широкое бо-
танико-географическое значение. У нас огромные пространства
Средней Азии, а также юго-востока Европейской части СССР имеют
засоленные почвы и галофильиую растительность; в Средней Азии
встречается особый тип пустынь — солончаковый. Солончаковая
растительность встречается также везде по берегам морей и океа-
нов, а также иа месте высохших морских водоемов. В Западной
Европе можно указать на солончаковые луга Бельгии и Голлан-
дии, заливаемые морскими приливами. Очень характерны заросли
по берегам морей и рек в тропических странах, состоящие из свое-
образных «мангровых» деревьев и кустарников (см. стр. 188).
Галофиты связаны с почвами, засоленными легко раствори-
мыми солями натрия, кальция и магния (хлористые и сернокислые
соли), подобные почвы называются солончаковыми и солончака-
ми (последние очень сильно засолены). Они встречаются лишь там,
111
где осгдков немного и где легко растворимые соли не могут сильно
выщелачиваться или там, где эти соли вновь накапливаются (на-
пример в результате морских приливов).
Солончаки нужно отличать от солонцов- первые обычно являют-
ся влажными или сырыми и у них засоление идёт от самой поверх-
ности почвы; солонцы — сухи, и засоление начинается глубже,
Кроме того, они имеют
Рис. 51. Солерос (Salicor-
nia herbacea).
определённую структуру (столбчатые со-
лонцы). Галофиты свойственны лишь
солончакам, на солонцах типич-
ные галофиты отсутствуют, и здесь
характерны ксерофиты (впрочем, расти-
тельность солонцов изучена мало).
Так как вообще растения с трудом
переносят большое содержание солей в
почве и, например, 2—3% NaCl дей-
ствует, как яд, то, очевидно, на засо-
ленных почвах (по берегам морей засо-
лённость может доходить до 2.7—3,2%)
могут произрастать лишь особо приспо-
собленные растения.
Интересно, что галофиты могут расти
и на обыкновенной садовой почве, хотя
они здесь накопляют солей больше, чем
негалофиты. Если в золе обыкновенных
растений NaCl меньше 5%, то в золе
галофитов процент этот зн житель но
выше: у Aster tripolium—49,9% (сте-
бель), у Chenopodium maritimum—47,08%
и т. д.
Если галофитизм есть приспособление, то тем
не менее он во многих случаях связан с опреде-
ленными семейства ми, так как многие семейства
почти исключительно состоят из галофитов —
Chenopodiaceae, Plumbaginaceae, Frankeniaceae,
тогда как в других семействах и даже в группах
семейств почти совершенно нет галофильных ви-
дов (Orchidaceae, Amentiflorae, Urticaceae и др.).
Очевидно, галофитизм связан с известными кон-
ституцнонными особенностями, чего совершенно
нет У кальцифидов, у различных ксерофитов
и пр.
Галофиты очень характерны по своим внешним признакам и в
большинстве случаев морфологически схожи с суккулентами, имея
мясистые стебли и листья, сильную редукцию поверхности и пр.
Различают (Келлер) для вашего юго-востока несколько основных типов
галофитов:
1) тип солероса (Saltcornia herbacea)— сочные растения с сильно редуциро-
ванными листьями и цилиндрическими мясистыми члениками стеблей (рис. 51);
2) тип шведки (Suaeda maritima) — мясистые растения, листья тоже сочные,
ие редуцированные;
3)фтип кокпека (Atriplex сапа, также A. vemicifera) — здесь очень ори-
гинальна наружная водоносная ткань из крупных пузыревидных волосков,
развитая иа поверхности листьев;
112
4) тип немясистых солянок (виды кермека— Statice, Tamarix, Frankenia и
Др.); У последних на вегетативных органах имеются оригинальные железки,
выделяющие капельки* горько-солёного раствора, вследствие чего в клетках
находится значительно меньше легко растворимых солей.
Имеются и другие типы.
Все галофиты, кроме наружных морфологических черт, имеют
ряд особенностей анатомического и физиологического свойства. Во-
обще у галофитов, в. связи с повышенной концентрацией почвенных
растворов, наблюдается повышенная концентрация клеточ-
ного сока и высокое осмотическое давление
(иногда до 150 атмосфер). Без этих свойств корни растений гало-
фитов ие были бы в состоянии всасывать из почвы необходимые
им минеральные вещества.
Опыты и наблюдения показывают, что суккулеитиость гало-
фитов — прямое следствие повышенного осмотического давления,
причём даже растения иегалофиты иа заселённых почвах дают
суккулентные формы.
Галофиты являются очень хорошими индикаторами засолён-
ных почв. Различают облигатные и факультативные галофиты.
К первым, связанным исключительно с засолёнными почвами,
относятся: солерос (Salicornia herbacea), солончаковая астра (As-
ter tripolium), кермек (Statice Gmelini), шведка (Suaeda maritima)
и др.; ко вторым — Salsola kali, Kochia arenaria, Scirpus mariti-
1Ши т. п.; последние могут расти иа заселённых и незаселённых
почвах. Облигатные галофиты, как, например, солерос, ие только
растут на засоленных почвах, но именно здесь развиваются наибо-
лее пышно (Келлер). Значит, в этом случае нельзя смотреть
на приуроченность к засоленным почвам как на средство освобо-
диться от конкурентных видов, не способных произрастать иа этих
засолённых почвах. Однако для многих галофитов произрастание
на засолённых почвах связано с явлениями конкуренции: здесь
они встречают меньше конкурентов. Связь большинства галофи-
тов с солончаками объясняется недостаточным водоснабжением
их на других почвах (солонцы, чернозёмы) — они интенсивно
транспирируют и имеют маломощную корневую систему.
Физические свойства почв. Очень важным фактором для про-
израстания растений являются физические особенности почв. Ме-
ханический состав почв может быть очень различен: ои зависит
от тех пород, на которых образовалась почва (почвы щебневатые,
песчаные, глинистые и др.). Иногда почвы как таковой иет, и ра-
стения поселяются на голых скалах.
Для выяснения механического состава производят механи-
ческий анализ почв, причём частицы (зёрна) почвы подраз-
деляются на группы {фракции) по их величине.
Фракции почвы в мм
Камни ..........................
(крупный ..................
средний..................
мелкий ..................
8 География растений
более 10 в диаметре
от 10 до 7 >>
» 7 » 5 »
й 5 >> 3 а
113
(крупный .................
средний .................
мелкий ..................
(крупная .................
средняя .................
мелкая ..................
Ил (глина)......................
от 3 ДО 1 в диаметре
л 1 ?> 0.5 л
0,5 » 0,05 »
» 0,05 » 0,01 »
>> 0,01 •> 0,005 >
» 0,005 » 0,001 »
меиее 0.001 »
Хрящ н крупный песок называют скелетом почвы. Хрящ,
и песок всех фракций относят к крупноземам, пыль и ил — к мел-
коз'ё мам.
Присутствие в почвах тех или иных частиц в различных соот-
ношениях приводит к разностям почв по их механическому составу..
Нужно отличать классификации почв по их механическому
составу от классификации по почвообразовательному процессу
(генетическая классификация). Почвообразовательный процесс мо-
жет итти по подзолистому (в лесной зоне), чернозёмному (в степ-
ной зоне), болотному и другим типам, но, например, чернозёмный
тип почвообразования может иметь место на почвах глинистого,
песчаного или другого механического состава; то же относится к
подзолистому типу и др. Поэтому глинистые почвы могут быть
чернозёмными, подзолистыми, засолёнными и др.
От величины частиц (зерен) почвы зависят такие важные свой-
ства почв, как проницаемость, водоподъемная
способность (капиллярность) и влагоёмкост ь. Раз-
личают полную и относительную влагоёмкость.
Полная влагоёмкость — это наибольшее количество воды, ко-
торое та или другая почва может в себя принять; однако не вся эта
вода удерживается почвой, и часть её уходит в нижиие слои. Остав-
шееся количество воды представляет относительную влагоёмкость,
Для растений важна, конечно, последняя. Чем почва более мелко-
земмста и чем в ней больше капилляров, тем больше относитель-
ная влагоёмкость; в суглинистом чернозёме удерживается 59% воды,
(от веса сухой почвы), в суглинке —43%, в супесчаной почве -
23%.
Для растений, всасывающих воду, особенное значение имеет
то количество воды, которое доступно растению («физиологически
доступная влага»). В разных почвах эти количества весьма различи
ны. Количество влаги, физиологически недоступной, составляет в
чернозёмных почвах 8—8,5% общего объёма почвы, в глинистой
почве — 6,5—7%, в песке — 2—2,5%.
Таким образом, в песчаных почвах почвенная влага исполь-
зуется в гораздо большей степени.
Что карается проницаемости, то крупнозернистые почвы очень
проницаемы, а мелкозернистые (глины), наоборот, проницаемы
слабо, вследствие чего в соответствующих условиях (избыток во-
ды, понижение в рельефе и др.) и происходит заболачивание. Ка-
пиллярное поднятие тоже связано с величиной зерна, и оио тем
больше, чем мельче зёрна' почвы; наблюдения показывают, что
после дождя дороги высыхают гораздо скорее, чем места по сторо-
нам дорог; также известно, что на полях так называемый «чёрный
пар» очень сильно предохраняет почвы от высыхания; в первом ука-
занном случае капиллярность усиливается в результате уплот-
нения дороги, а во втором — ослабляется, так как при распашке
мелкозернистая структура почв переходит в комковатую.
В самой тесной зависимости от механического состава нахо-
дится воздушный и водный режимы почв. При до-
статочном количестве воздуха в почве (а следовательно, и кислор»-
Рис. 52. Мангровое дерево (Sonneratia alba) в Сиаме, с воздушными
корнями—пневматофорами, выходящими вертикально из ила. На зад-
нем плане-ходульные корни других мангровых деревьев (изе‘Мартонна).
да) мы имеем почвы с хорошей аэрацией, в противном случае все
процессы в почве протекают в условиях анаэробиоза. Плохая аэра-
ция почвы приводит к закисанию почвы, к накоплению в ней ра-
стительных остатков (залежи торфа) и вызывает у растений ряд
анатомических и морфологических особенностей, например воз-
душные полости в подземных органах, выступание корней иад по-
верхностью земли (рис. 52) и др. (см. стр. 7 1).
В связи с механическим составом почв можно наметить неко-
торые экологические типы растений (растения - индикаторы).
Псаммофиты (от греческих слов «псамма» — песок, «фитои» —
растение) или песколюбы — растения, обитающие на песчаном
субстрате (песчаные берега рек, песчаные пустыни и др.). Очень
много видов связано исключительно с песками, что нередко отра-
жен® и в их названиях: гвоздика песчаная (Dianthus arenarius),
астрагал песчаный (Astragalus arenarius), кохия песчаная (Kochia
arenaria) и пр. Многие виды являются викарирующими иа песках
и иа других местообитаниях: Silene densiflora — в степи, S. parvi-
flora — иа песках, Jurinea arachnoidea — в степи, J. cyanoides —
на песках и др. Псаммофиты представлены жизненными формами',
выработавшимися в борьбе с сильной трансцирацией (особенно в
пустынях) и с подвижностью самого субстрата. Именно, они име-
ют ряд ксероморфных особенностей — редукция листьев, афиль-
иые формы и др. Что касается подвижности субстрата, то защитой
от засыпания песком является способность перемещения почек
возобновления, причём они всё время перемещаются кверху; в
других случаях защитой служат длинные горизонтальные* корни
и корневища, скрепляющие песок. Большая часть псаммофитов
имеет семена и плоды, распространяющиеся прр помощи ветра —
оии сиабжеиы летательными аппаратами, или сильно вздуты, или
представляют упругие шарики и т. д.; всё это делает их очень лёг-
кими, оии остаются иа поверхности или близ поверхности, не за-
рываясь глубоко в песок.
Наибольшее разнообразие псаммофитов мы находим в песчаных
пустынях (см. стр. 315).
На каменистых и щебенистых местообитаниях имеются особые
экологические типы: литофиты и хасмофиты.
Литофиты (от греческого слова «литое» — камень и «фитои» —
растение). Это растения, растущие непосредственно на камнях
и скалах; в этих условиях могут расти лишь лишайники и водо-
росли. Из лишайников это прежде всего так называемые накипные
(корковые) — они окрашивают скалы в различные цвета: чёрный,
бурый, красный и др. Поселяясь на голом камне, накипные лишай-
ники в результате своей жизнедеятельности делают поверхйость
субстрата шероховатой и дают возможность селиться и листоватым
лишайникам.
При дальнейшем разрушении камней образуются трещинки и
поселяются мхи, а затем и некоторые цветковые. Здесь очень хо-
рошо выражена смена (сукцессия) растительности на первоначаль-
но голом субстрате: лишайники — мхи — цветковые растения.
Хасмофиты (от греческих слов «хазма» — щель, расщелина
и «фитон» — растение). Это растения щебенистых и каменистых
местообитаний, растущие в расщелинах скал (скальные растения).
Они имеют обычно длинные крупные корни, которые проникают
глубоко в щели между камнями и щебнем, где скопляются частицы
мелкозёма. По своей жизненной форме — это подушки, а также
дернины. Наряду с ксерофитами здесь можно вс!ретить и мезо-
фитов, так как условия увлажнения иа данных местообитаниях
очень различны. Хасмофиты — это дальнейшая стадия развития
растительности (сравнительно с литофитами) иа камнях и скалах.
Структура почв. Частицы почвы — глинистые, пылеватые, пес-
чаные, обычно связываются в комочки больших или Меньших раз-
меров. Если почва состоит из комочков, то оиа называется струк-
ту р н о й, в противном случае — пылеватой (распа-
дается иа отдельные частицы). Величина комочков почвы коле&
лется от 0,25 до 10 мм (реже выходит за эти пределы).
Структурные почвы обладают признаками, полезными для ра-
стений, именно, между комочками почвы находятся более или ме-
нее крупные щели, куда свободно проходит воздух и куда поступа-
ет дождевая вода. Бесструктурная почва скважностью ие облада-
ет, дождь почти в неё не проникает, воздух может лишь находить-
ся в капиллярных порах.
Причины структурности различны — оии могут быть физико-
химическими или биологическими. Среди первых укажем присут-
ствие извести, которая способствует образованию комочков. Био-
логические причины: жизнедеятельность дождевкх червей в поч-
ве, которые пропускают через свой пищеварительный канал ча-
стицы почвы и склеивают их пищеварительными соками; влияние
корневых систем, которые пронизывают почву и разбивают её иа
отдельности, причём корни, кроме того, оплетают и связывают
частицы почвы.
Так как структурные почвы благоприятны для роста растений,
то роль человека, очевидно, должна быть направлена к созданию
и поддержанию структурных почв: неумелая обработка почвы ве-
дёт к разрушению структуры и к распылению почвы. Для созда-
ния почвенной структуры лучшими средствами являются: посев
трав злаковых и бобовых, известкование, запашка в почву навоза
и др.
Человек в результате своей деятельности может как'ухудшать
качества почв/так и улучшать их. Каждая почва может при соответ-
ствующих на неё воздействиях превратиться из плохой в хорошую.
Недаром говорится, что «нет плохих почв, а есть плохие хозяе-
ва». При улучшении качеств почв можно воздействовать как на
поверхностные слои (пахотные), так и иа более глубокие; так, в
подзолистых почвах, занимающих в СССР огромные территории,
пахотные слои бедны питательными веществами, которые выне-
сены в более глубокие слои (на глубину в 40—60—80 см). Вывора-
чивание этих слоёв наружу сразу обогатит поверхностные слои и
сделает их плодородными. В данном направлении ведутся в СССР
опыты, дающие очень хорошие результаты.
Орографические факторы
Орографические факторы играют очень большую роль при
расчленении растительного покрова. К иим относятся: высота
над уровнем моря, рельеф местности, наклон склоибв (угол накло-
на), направление склона по отношению к тем или иным странам
горизонта (экспозиция)и др.
О влиянии высот уже ие раз говорилось выше: опыты
Б о н и ь е с пересадкой растений из долин в горы, формирование
особых высотных экотипов, изменение числа фенологических вре-
мён года и пр.
Высота над уровнем моря в высокогорных районах сказывает-
ся очень резко на характере растительности и на основных лаид-
117
«афтах. Так, иа высоких горах наблюдается смеиа поясов
растительности в связи с изменением климатических условий;
если, например, высокие горы находятся в области пустынь (на-
пример в Средней Азии), то, поднимаясь в горы, мы будем иметь
такую последовательную смену поясов (снизу вверх): пояс степей,
пояс лесов, пояс безлесных альпийских пространств.
Влияние рельефа. В пределах каждого пояса растительность
сильно варьирует в связи с различными особенностями рельефа,
и вообще наибольшая пестрота растительного покрова связана
Рис. 53. Примеры комплексов и экологических рядов; бук-
вами обозначены различные ассоциации; направление стре-
лок указывает увеличение влажности (ориг.).
именно с горами. Влияние орографического фактора особеннв хо-
рошо можно наблюдать в горах. '
Однако и в равнинных местностях рельеф оказывает очень боль-
шое влияние на растительность- Так, например, в степях можно
различать как макрорельеф (общее расчленение), так и мезо- и
микрорельеф. Если в степях макрорельефом будет основной ба-
лочный рельеф, то западины на степи будут относиться к мезорелье-
фу, а кочки в западинах или холмики роющих животных иа ров-
ной степи — к микрорельефу.
В связи с рельефом растения и фитоцеиозы располагаются по
так называемому экологическому ряду, чем ниже по рельефу, тем
влажность будет большей, и растения дают ряд по усилению гигро-
фитности (высотный экологический ряд). Это особенно хо-
рошо можно видеть по склонам балок, в степных западинах, на
лугах с их меняющимся рельефом и т. п.
На степных склонах можно различать по крайней мере три хо-
рошо выраженные полосы растительности: верхнюю часть, сред-
нюю и иижиюю, —это различные ассоциации. В западйиах на
степях или лугах, если они по своему очертанию являются более
или менее округлыми, можно наблюдать ряд концентрических поя-
сов, обычно очень хорошо отграниченных Друг от друга. Подобных
поясов может быть 5—7 и более (рис. 53). Здесь иа небольшой пло-
118
щади имеется несколько ассоциаций, каждая из которых занимает
незначительную площадь («комплекс ассоциаций?*).
При исследовании подобных западин и вообще при исследова-
нии растительного покрова с частыми сменами одних ассоциаций
другими метод пробных площадок (см. стр. 165) мало применим,
и основным методом должен являться метод экологических рядов,
где главное внимание уделяется сравнительному изуче-
нию ассоциаций или растений данного конкретного ряда. При,
этом особенно ясно обнаруживается экологическая сущность того
или другого вида. Так, одни виды встречаются только в верхних
частях ряда и, значит, являются наиболее ксерофильиыми, другие —
Рис. 54. Схема правила лредварен11я,Северное (влаголюби-
вое) растение (/) переходит на юге на северные склоны
и дно балок, южное (2) — при движении к северу перехо-
дит на лучше нагреваемые южные склоны (ориг.).
только в нижних частях (наименее ксерофильиые), третьи прохо-
дят более или менее равномерно вдоль всего ряда (растения с ши-
рокой экологической амплитудой) и т. д.
Иногда ничтожного повышения или понижения рельефа до-
статочно, чтобы растительность изменилась довольно резко, что
особенно часто наблюдается в более южных засушливых местах,
где для растений важна каждая капля воды; на севере микрорель-
еф отражается на растительности менее резко.
I Экспозиция склонов. Огромное значение имеют угол на-
клона и экспозиция склонов. В последнем отношении
наиболее резкие противоположности имеют место между север-
ными и южными склонами, что особенно хорошо можно наблюдать
иа наших степных безлесных пространствах. Растительность юж-
ных склонов обычно имеет более южный, более ксерофильиый ха-
рактер сравнительно с растительностью ровных,так называемых
плакорных пространств. Это объясняется тем, что южные склоны
более нагреваются, так как солнечные лучи падают на них под
большим углом. На северных склонах отношения как раз противо-
положные, и их растительность носит более северный характер
сравнительно с плакорными местами.
Так, в наших северных степях на открытых южных склонах
можно наблюдать массовое развитие шалфея поникшего (Salvia
nutans), ковыля - тырсы (Stipa capillata) и других видов, в пла-
корных условиях не встречающихся здесь, ио растущих в более
южных степях. На северных склонах северных степей мы находим
ряд видов (Koeieria Delavignei, Festuca rubra и др.), встречающих-
ся в плакорных условиях лишь в наиболее северных степях.
В смысле уменьшения ксерофитности намечается такой ряд:
южные склоны —плакорные места —северные склоны. Склоны
других экспозиций (западные, восточные) носят промежуточный
характер. Так как плакорная растительность наиболее отвечает
119
климатическим условиям страны, растительность южных склонов
несёт элементы более южных плакорных пространств, а север-
Рис. 55. Тропические цветки с длинной
трубочкой венчика, опыляемые ноч-
ными бабочками, имеющими хоботок,
равный длине трубочки венчика. Сле-
ва—Oxyanthus hirsutus, справа—Ехо-
stema flonbundum (уменьшено иа
(по Шимперу).
от основной линии северо-
нений.
иых склонов—элементы более
северных плакорных мест, то
очевидно, что, изучая расти-
тельность этих частей рельефа,
мы можем представлять себе и
растительность более южных и
более северных плакорныхтер-
риторий. Здесь можно говорить
о «правиле предварения»: «пла-
корный вид или плакорная
растительность предваряется
на юге или на севере в соот-
ветствующих условиях место-
обитания». Таким образом,
изучая растительность дан-
ного пункта, мы можем со-
ставить себе представление о
более широком географиче-
ском распределении растений
(рис. 54).
Особенно хорошо, как уже
указано, явления предварения
можно наблюдать в степях, в
открытых безлесных местно-
стях; в лесной зоне травяни-
стая растительность находится
под сильным воздействием дре-
весного яруса, создающего осо-
бые микроклиматические усло-
вия.
Совершенно ясно, что пра-
вило предварения хорошо
выражено лишь при известной
сравнимости более южных и
более северных территорий и
при относительной ненару-
шеиности естественного расти-
тельного покрова. Сильное
влияние человека, островные
выходы тех или других гор-
ных пород и т. п. вносят су-
щественные изменения в за-
кономерные явления предва-
рения; реликтовые растения
1 и эндемики могут также
давать некоторые отклонения
южных климатических нзме-
120
Биотические факторы
Биотические факторы касаются взаимоотношений между ра-
стениями и средой органической; растение и животный мир, рас-
тение и растение (все факторы, рассмотренные выше, касались
взаимоотношений между растениями и неорганической средой).
Биотические отношения очень многочисленны и подчас сложны
и здесь затрагиваются лишь в самых общих чертах. Влияние
человека целесообразнее выделять из числа биотических
факторов и рассматривать особо (см. стр. 127).
Растения и животный
мир. Здесь связи могут
быть весьма разнообраз-
ны:
1. Животные яв-
ляются агентами при
опылении цвет-
ков (главным образом
насекомые, реже птицы и
млекопитающие) (рис. 55).
Способы опыления цветков
очень разнообразны отсут-
ствие опыляющего животного
в иных случаях приводит к
невозможности размножения
и существования того или
иного вида, и отсутствие этого
вида в данной местности, при
благоприятных климатических
и эдафпческих условиях, мо-
жет быть объяснено лишь
биотическим фактором. Давно
известен пример с красным
клевером, не Приносящим се-
мян в тех странах, где нет
опылителей - шмелей.
2. Животные игра-
ют важную роль при
распространении
семян п плодов.
При этом роль животных
может быть различна:
а) семена и плоды у ряда растений снабжены зацепками, крючочками и пр.,
которыми онп прицепляются к животным и могут переноситься послед-
ними на большие расстояния (рис. 56);
б) сочные плоды поедаются животными (особенно птицами), и таким обра-
зом разносятся семена;
в) нередко семена имеют особые мясистые придатки н растаскиваются мура-
вьями, поедающими эти придатки;
г) иногда животные намеренно растаскивают плоды; например белки
разносят лесные орехи, жолуди и др.
Если процесс распространения семян и плодов животными вообще
можно назвать зоохорией1, то в первом случае мы имеем эпи зоохорию3, в» 1 2
1 Зоохория — от греческих слов «зооно — животное и «хорео» — иду,
распространяюсь.
2 От греческого слова «эпи’> — на.
121
втором эндозоохорию1, в третьем — мирмекохорию1 * 3 и в четвертом — син-
зоохорню8.
К числу эпизоохорных плодов относятся плоды липучки (Echniosper mum),
гравилата (Geum), репейника (Agrimonia), чериокореня (Cynoglossum) и др.
Эидозоохорные плоды имеют смородина, барбарис, вишня, бузина рябина,
черника, малина и т. д, ’
К числу мирмекохорных растений относятся: чистяк, хохлатки, чистотел,
гусиные луки (рис. 57), различные виды фиалок и пр.
Примеры спнзоохорных плодов уже были указаны.
Рис. 57, Гусиный лук (Gagea erubescens);
при созревании семян растение падает,
семена высыпаются иа землю и растаски-
ваются муравьями.
Интересно, что все мирме-
кохорные растения относятся
к растениям травянистого по-
крова леса, большинство эндо-
зоохорных—к кустарникам ле-
са, где гнездятся птицы, аэпи-
зо о хорные — в большинстве
случаев растения открытых
мест. Если вспомнить, что пло-
ды и семена деревьев разно-
сятся главным образом вет-
ром, то мы имеем в лесу очень
тесную связь между его яруса-
ми и определенными способами
распространения ^плодов.
3. Животные в
результате вь-паса ока-
зывают влияние на
травостой. Это влия-
ние сказывается в таких
направлениях:
а) скот угнетает более сла-
бые элементы травостоя;
б) способствует разраста-
нию растений, чем-либо за-
щищенных (растения колючие, пахучие, с млечным соком н пр.).
в) засоряет сорными растениями хорошие луга, степи и пр.;
г; втаптывает семена растении в почву, способствуя лучшему прорастанию.
4. Животные своей деятельностью изменяют характер
и свойства почвы, создавая для растений иные экологические
условия. Вот несколько примеров:
а) животные утаптывают почву, изменяя ее физическую структуру; при
этом в почвах нередко усиливается капиллярность, и растительный покров ста-
новится более ксерофитным;
б) степные роющие животные (сурки, суслики и др.) перерывают почву,
выносят на поверхность иижние горизонты почвы ит. д.: растительность «сур-
чин» резко выделяется иа общем фоне степи;
в) земтяные черви, пронизывая почву своими ходами, способствуют рыхле-
нию н, следовательно, аэрации почвы; имеются указания, что до известной
г убины каждая частица почвы перерабатывается земляными червями в течение
немногих лет;
г) мураввд также перемешивают и разрыхляют почву;
д) животные удобряют почву и делают её более плодородной.
5. Животные могут служить пищей растениям
(насекомоядные растения), для чего у растений вырабатываются
сложные приспособления.
1 От греческого слова «эндо» — внутри.
3 От греческого слова «мирмекс» — муравей.
3 От греческого слова «син» — вместе.
122
Среди нашей флоры насекомоядными растениями являются
виды росянки (Drosera, рис. 159), обычно растущие на моховых
болотах, виды пузырчатки (Utricularia) в водоёмах и иа болотах,
жирянка (Pinguicula) иа болотах; среди чужеземных растений —
североамериканские Sarracenia (рис. 161) и Dionaea (мухоловка
рис. 162), тропические непентесы и др.
/6. Животные вступают с растениями иногда в сложные
взаимоотношения, например, удивительные взаимо-
отношения между растениями и муравьями в тропических
странах.
Эти взаимоотношения настолько
своеобразны и интересны, что за-
служи ва ют то го, что бы на ии х
остановиться. Муравьи в тропиках
играют исключительную роль и
наносят очень большой вред мест-
ной и культурной растительности;
так, в тропической Америке, там,
где встречаются некоторые виды
муравьёв, совершенно невозможны
культуры кофе, апельсинов, роз и
других растений, потому что так
называемые муравьи-листорезы уни-
чтожают иа растениях все листья
(рис. 58).
Огромные полчища муравьёв,
нападают на листья деревьев и
вырезают в них небольшие кусоч-
ки, которые на спинах уносят в
свои гнёзда. Массы муравьёв обра-
зуют зелёные (от кусочков листьев)
движущиеся потоки. В своих гнёз-
дах муравьи пережевывают кусочки
Рис. 58. Справа-лист, край ко-
торого изрезан муравьями-листо-
резами (Atta discigera), слева—
муравьи.листорезы, нагруженные
кусг.чкамн листа, сползают по
стеблю растения (по А. Меллеру).
листьев, выплевывают их, и полу-
чившаяся мягкая масса служит для
посадки мицелия гриба Rozites gan-
gylophora; гриб всё время поддер-
живается в вегетативном состоянии
и даёт особые образования, напоми-
нающие кольраби и служащие муравьям пищей (муравьиные «сады»).
Если в данном прй.мере растения терпят ущерб, муравьи же
получают пользу, то в других случаях взаимоотношений между
растениями и муравьями имеет место обоюдная польза, и эти от-
ношения могут быть названы симбиотическими (от греческих
слов «син» — совместно и «биос» — жизнь).
Здесь муравьи поселяются на растениях—или, правильнее,
в растениях —и получают от последних пищу, но в свою очередь
приносят растениям пользу, защищая их от других муравьёв,
именно, от муравьёв-листорезов (явление мирмекофила и)1.
1 От греческих слов «мирмекс» — муравей и «филео-> — люблю.
123
Имеется ряд типов подобных симбиотических взаимоотноше-
ний в тропических областях Виды рода Acacia имеют полые тон-
костенные колючки при основании листьев, которые муравьи
пробуравливают, где и поселяются. Пищей муравьям служат осо-
бые яйцевидные или грушевидные тельца на концах листочков
перистых листьев, богатые белковыми веществами («белтовские
тельца» — по имени Белта, открывшего явление мирмеко-
Рис. 59. Цекропия (Сес-
ropia cinerea)— верхняя
часть стебля, внизу сте-
бель вскрыт, чтобы по-
казать полость стебля,
где поселяются муравьи
Пр I основании черешков
мюллеровские тельца
(слева — в увеличенном
виде).
Рис 60 Мирмекодия (Myrmecodia) На верхнем ри-
сунке клубень разрезан вдоль, чтобы показать поло-
сти, где поселяются муравьи (по ьВеттштейну)
филин); кроме того, при основании черешков листьев находятся
нектарники, выделяющие сахаристую жидкость
У видов Cecropia (особенно С adenopus) муравьи поселяются
в полых стволиках, пробуравливая особые утонченные места, за-
метные на стебле в виде округлых углублений Пищей мурцвьялг
счужат особые тельца, находящиеся при основании черешков
(«мюллеровские тельца» — по имени открывшего ихМюллера,
рис 59)
Можно установить целый ряд градаций в смысле все большего
приспособления растений к защищающим их муравьям; так, если
у Acacia муравьи пробуравливали стенки колючек, то у Cecropia
124
Рис 61 Эпифитная орхидея (Jonopsis panicu-
lata) на ветке апельсинного дерева в Бразилии
(из Шимпера).
существуют уже особые углу бления на стебле, а у других видов
(Humboldia laurifoha, Ficus inaequahsu др ) на стеблях находятся
готовые отверстия.
Особый тип мирмекофильиых растений представляют эпифит-
ные виды Myrmecodia и Hydnophytum на Малайском архипелаге,
у которых полости находятся не в междоузлиях стебля, как у пре-
дыдущих видов, а имеются многочисленные сообщающиеся полости
в клубнях (рис 60)
Кроме мирмекофильиых растений с полостями в органах стеб-
левою типа, имеется немало мирмекофильиых растений с мура-
вьями, живущими в полостях, связанных с листьями
Влияние растений друг на друга Выше было указано, что мно-
гие пришлые, иноземные растения не уживаются в дикой природе;
это один из примеров того, что борьба за существование между
видами, конкуренция
из-за места, пищи, све-
та и пр. приводит к
гибели одних и к вы-
живанию других
Связи между расте-
ниями и влияние их
друг иа друга могут
быть весьма разнооб-
разны Среди этих яв-
лений нужно разли-
чать, во-первых, те свя-
зи, когда лишь од н~а
сторона получает
пользу, другая же по
большей части прямой
ущерб, а во-вторых,
связи, при которых
обе стороны полу-
чают пользу в большей
или меньшей степени
Вторые связи обычно
называют с и м б и о т и-
ческими, хотя неко-
торые авторы симбио-
связг
т
зом называют
всякого рода ,
применяют это
нятие очень
р о к о
е
по-
ш и-
I Связи, дающие одностороннюю пользу
I Растения паразиты— органически связаны со своим «хозяином* и всю
вою пищу получают от него
2 Эпифиты — пользуются другими растениями только как местом при-
репления и поселения (рнс 61), причем особый тип эпифитов — эпифиллы
рис. 84) — поселяются на листьях (тропические страны)
125
3. Лианы — пользуются другими растениями как подпорой, благодаря чему
и не затрачивают много материала на постройку своих стеблей (рис. 82); сюда
же относятся лазящие растения, снабженные усиками и пр.
II. Связи, дающие обоюдную пользу (симбиоз)
L Связи между бобовыми и бактериями. На корнях бобовых образуются
клубеньки в результате проникновения бактерий>из почвы в корни. Бактерии
становятся паразитами клеток корней, но в то же время усваивают свободный
азот воздуха и в виде нитратов предоставляют его бобовому растению. Извест-
но, какое важное значение имеет данный вид симбиоза для сельского хозяйства.
2. Связи между высшими растениями и грибами (зидотрофная микориза).
Гриб в виде гиф поселяется в клетках коры некоторых деревьев, кустарников и
травянистых растений н здесь паразитирует; высшие растения переваривают и
усваивают грибные гнфы. В ряде случаев (некоторые орхидные) высшее расте-
ние без микоризы не может существовать, н даже семена не прорастают при
отсутствии гиф гриба.
3. Связи между водорослями и грибами. Симбиоз между некоторыми видами
этих организмов приводит к формированию особых, н морфологически н эколо-
гически, организмов, называемых лишайниками. Морфологически лишайники
не имеют ничего общего с грибами или водорослями, а экологически они явля-
ются обитателями обычно сухих, иногда исключительно сухих местообитаний
(сухие голые скалы), в то время как грибы — растения мезофитных, а водо-
росли — гигрофитных условий. Физиологические взаимоотношения между водо-
рослями и грибами в лишайниках довольно сложны; грибы получают от водо-
рослей, содержащих хлорофилл, органическую пищу, но в свою очередь дают
им воду и растворённые в ней вещества.
Однако чаще всего взаимоотношения между растениями таковы,
что они, не находясь между собой в непосредственной связи, так или
иначе влияют друг на друга: конкурируют из-за питательных ве-
ществ почвы, из-за света, из-за места на данном участке и пр. Мно-
гие из подобных связей будут рассмотрены ниже в отделе фитоце-
нологии, относясь уже к синэкологии.
Синузии. В растительном покрове имеются сочетания видов,
более тесно или менее тесно связанных между собой; некоторые
виды обычно существуют совместно, относясь к тождественным
или близким жизненным формам, имея однородные ритмы разви-
тия и создавая как бы определённую жизненную среду. Совокуп-
ность таких видов составляет синузию. Эти последние можно
считать как бы экологическими единицами в растительных ассоци-
ациях.
Можно различать несколько' категорий синузий по степени их экологиче-
ской связанности.
Синузия iso порядка — совокупность особей одного и того же вида, обще-
ственно растущего. Примеры: заросль тростника, камыша, совокупность сосен
в сосновом лесу и т. п.
Синузия 2-го порядка — совокупность особей различных видов, но принад-
лежащих к одной и той же жизненной форме. Примеры: совокупность широко-
лиственных деревьев наших дубрав (дуб, липа, клён и др.), совокупность широ-
колиственных дубравных злаков н осок и т. п.
Синузия 3-го порядка — совокупность особей, принадлежащих к различным,
но экологически равноценным жизненным формам. Пример: совокупность степ-
ных ксерофитов, относящихся, однако, к различным жизненным формам, имею-
щим различные приспособления (опушенные виды с листьями, виды с редуци-
рованными листьями, виды с жёсткими, стоящими ребром листьями и др.).
Синузия 4-го порядка — совокупность особей, относящихся к жизненным
формам, экологически не равноценным, но принадлежащих всё же к одной я
той жг жизненной форме боле общего значения. Примеры: различны ярусы
126
растительных ассоциаций; так, например, древесный ярус может состоять из раз-
личных «деревьев-) (жизненная форма общего порядка), но относящихся к
«лиственным», и «хвойным» (экологически ие равноценные жизненные формы).
При установлении синузии можно принимать во внимание и фенологические
моменты (ритмы развития), но здесь мы этого касаться не будем.
Влияние человека
Человек является одним из наиболее важных факторов, влияю-
щих на растения и на растительные сочетания. Влияние человека
на окружающую его природу имело место, конечно, с самых от-
далённых времен, но особенно усилилось за последнее время.
Это влияние может быть бессознательным и сознательным. Когда
первобытный человек собирал в большом количестве в лесу или в
другом каком-либо ценозе те или другие полезные ему растения,
то он этим самым изменял соотношения обилия различных видов;
когда он выжигал или стравливал степи, то растительный покров
претерпевал большие изменения и т. д. —всё это примеры бес-
сознательного влияния. Бессознательное влияние человека могло
приводить к положительным для человека результатам, но часто
влекло за собой отрицательные изменения растительности. С другой
стороны, бессознательный отбор, имевший место у первобытного че-
ловека, привёл, по Дарвину, к формированию ряда культурных
растений (см. приложение «Культурные растения»). Что касается
сознательного влияния человека на растения, то оно в большин-
стве случаев приводит к положительным результатам, так как
искусственный отбор является средством к улучшению тех или иных
видов и к созданию новых культурных растений. В результате
сознательного влияния мы можем улучшать также растительные
ценозы и конструировать новые.
Влияние человека в данное время на растительный покров так
разносторонне и глубоко, что, пожалуй, с трудом можно найти
место на земной поверхности с совершенно неизмененной расти-
тельностью. Многие типы растительности являются полукуль-
ту р н ы м и или даже возникшими всецело в результате дея-
тельности человека. Так, наши северные суходольные луга возник-
ли на месте уничтоженных лесов, заливные луга в поймах рек
занимают места вырубленных пойменных лесов, наши обширные
березняки и осинники —результат многократных порубок перво-
бытных лесов и т. д.
Но если изменены первоначальные ценозы и даже возникли
новые, то не меньшие изменения произошли в первоначальной
флоре, которая, с одной стороны, обогащается новыми видами,
прежде в данной флоре не содержавшимися, а с другой —лишает-
ся некоторых видов. Так, например, какой-нибудь степной район
может совершенно лишиться большого количества степных видов
вследствие распашки в этом районе степей; в ближайших окрест-
ностях городов замечается постепенное исчезновение более редких
видов; если некоторые виды ещё не исчезли, то обилие их (число
экземпляров) резко падает. Поднимается вопрос об охране
редких и интересных растений, что у нас составляет задачу Всерос-
127
сийсксго общества охраны природы (и за границей имеются со-
ответствующие организации).
Синантропные растения. Обогащение флоры человеком может
быть сознательным и бессознательным. До последнего времени
имело место главным образом последнее. Целый ряд видов расте-
ний, помимо воли человека, сопровождал его при его расселении,
следуя за ним. Сюда относятся сорные растения, иначе называе-
мые синантропными (т. е. связанными с человеком, от греческих
слов «син»—вместе, «антропос»—человек). Среди сорных ра-
стений можно различать несколько групп: археофиты—сорня-
ки, существовавшие ещё в доисторические времена, неофиты —
сорняки, более нового времени, для которых можно указать ино-
гда даже год их появления. Как археофиты, так и неофиты явля-
ются растениями пришлыми, отсутствующими в местной флоре,
однако среди сорных растений имеется группа апофитов, —
это местные растения, но легко переходящие иа культурные и дру-
гие местообитания, создаваемые человеком.
Из числа археофитов назовём ряд растений, известных для
поселений человека каменного века: куколь (Agrostemma Githa-
go), белая марь (Chenopodium album), дымянка (Fumaria offici-
nalis), ярутка (Thlaspi arvense), лопух (Arctium lappa), посевной
василёк (Centaurea cyanus), лебеда (Atriplex hastata), посевной
костёр (Bromus secalinus) и многие другие.
Из неофитов, занесённых в Европу, можно указать следующие:
водяная чума (Elodea canadensis)—появилась в 1836 г. (из Се-
верной Америки), мелколепестник (Erigeron canadensis)—по-
явился в XVIII столетии (из Северной Америки), энотера (Oeno-
thera biennis)—в начале XVII столетия, мимулус (Mimulus lu-
teus) —в 1850 г., недотрога мелкоцветная (Impatiens parviflora)—
в середине прошлого столетия и др.
К числу апофитов можно отнести немало растений местной
флоры, легко становящихся сорняками. После распашки степей
ряд степных видов начинает произрастать в посевах, в огородах,
на иасыпях, у дорог и т. п. Так, в посевах встречаются нередко
в гораздо большем обилии, чем на целинных степях, такие степные
виды: резак (Falcaria vulgaris), льнянка (Linaria Biebersteinii),
чина клубненосная (Lathyrus tuberosus), будяк (Carduus hamulo-
sus), шалфей (Salvia nemorosa) и др.; у дорог очень обычны: василёк
(Centaurea diffusa), австрийская и полевая полыни (Artemisia
austriaca и A. campestris), люцерна жёлтая (Medicago fa lea ta).
типчак (Festuca sulcata) и др.
К группе апофитов относится ряд видов, из которых одни бо-
лее, а другие менее легко переходят иа культурные местообитания
и являются сорными то в большей, то в меньшей степени. Эта
группа постепенными переходами связана с пришлыми сорными
растениями.
Большинство пришлых сорных растений поселяется на куль-
турных почвах или на местах, лишённых первоначальной расти-
тельности (у поселений, при дорогах, на искусственных обнажени-
ях и др.). Те виды, которые приурочены к посевам, называются
128
сегетальными (от латинского слова «сегеталис»—пашенный),
а произрастающие на мусорных местах, у жилья —рудеральными
(от латинского слова «рудералис»—мусорный, сориый). К
числу первых нужно отнести: посевной василёк, куколь, посев-
ной костер, тысячеголовник (Vaccaria parviflora) и др., к числу
вторых—лопух (Arctium lappa), белену (Нуoscyamus niger), виды
будяка (Carduus), ряд крестоцветных (особенно Sisymbrium Loese-
Ш) и др. Некоторые виды растут почти исключительно вдоль до-
рог, как, например, спорыш (Polygonum aviculare), на юге также
якорцы (Tribulus terrestris) и др.
Способы заноса пришлых растений могут быть очень разнообраз-
ны; семена заносятся при посредстве транспорта (железные дороги,
судоходство и др.), вместе с товарами, с зерновым и семенным ма-
териалом и т. д. Войны и связанные с ними передвижения людей
и транспорта имеют исключительное значение в этом отношении.
Пришлые растения, связанные почти всегда с человеком, ред-
ко переходят в дикую природу, в естественные фитоценозы. Здесь
они встречают, очевидно, очень сильную конкуренцию со стороны
дикорастущих растений и быстро погибают. Лишь в редких случаях
они входят в состав местных фитоценсзов и даже вытесняют тузем-
ные видЫ; так, например, указанная выше водяная чума почти
повсеместно встречается в речках и ручьях в Европе. Энотера
встречается по песчаным берегам рек и вообще по песчаным местам,
производя впечатление дикорастущего растения.
Большинство пришлых растений поселяется на местах с раз-
реженной растительностью, на насыпях и др.; в случае зарастания
этих местообитаний сплошным растительным покровом пришлые
растения быстро исчезают, не выдерживая конкуренции с мест-
ными растениями.
Подобно пришлым, и культивируемые человеком растения пе-
реходят в дикую природу лишь в виде исключения. Так, в знаме-
нитом саду Разумовского под Москвой культивировалось в начале
XIX столетия не менее 8 000 видов, однако они не перешли в дикую
природу, если не считать одного недавно найденного близ Горенок
американского растения (Sisyrinchium angustifolium); в Никитском
ботаническом саду близ Ялты за все время его существования
культивировалось не менее 10 000 видов, из которых одичало не бо-
лее 1% видов (Станков); в ботаническом саду Демидова,
существовавшем в конце XVIII столетия в Москве на месте совре-
менного Нескучного сада, насчитывалось, не считая дикорастущих,
около 2 000 видов, из которых сейчас встречаются лишь Impatiens
parviflora и коноплевидная крапива (Urtica cannabina). Москов-
ский ботанический сад за всё время своего существования (более
200 лет) совершенно не обогатил местную флеру, если не считать
недотроги мелкоцветной.
Лишь в очень редких случаях растения из ботанических садов
проникают в местную природу; так, недотрога мелкоцветная ме-
стами встречается в большом обилии и, например, в Нескучном
саду в Москве, иногда развивается сплошь, вытесняя все другие
растения; по берегам рек в западной части Европейской территории
Географая растений
СССР местами даёт заросли аир (Acorns calamus), культивировав-
шийся как лекарственное растение. Одиако примеров подобного
рода очень мало.
Сознательное обогащение флоры человеком. Что касается со-
знательного обогащения флоры, то здесь вопрос идет о внедрении
в естественные ценозы растений, полезных в том или ином отноше-
нии. Уже давно луговодами применялся прием подсева ряд? зла-
ков и других растений в травостои естественных лугов с целью
повышения их качества. То же имело место в лесоводстве —под-
садка соответствующих древесных пород и закультивирование
полей и открытых площадей. В последнее время делаются попытки
внедрения (интродукции) новых древесных пород, отсутствовавших
в местной флоре, главным образом североамериканских или даль-
невосточных.
Однако роль человека не заканчивается только обогащением
флоры и внедрением видов растений, отсутствующих в данной
флоре. Человек идёт дальше, создавая новые сорта и виды и делая
попытки создать новые растительные сочетания (фитоценозы).
Прсисхождение новых видов может иметь место как результат
бессознательной или же сознательней деятельности человека.
О возникновении ряда культурных растений путем бессознатель-
ного отбора сказано ниже (см. стр. 270), сейчас же укажем на то,
что новые виды могут возникать помимо воли человека,
только лишь как следствие его хозяйственной деятельности. Сюда
относятся замечательные новые виды сорных растений, тесно свя-
занные с определёнными культурами и выработавшиеся в послед-
них; так, в посевах ржи выработались особые виды, в посевах
льна —другие, в иных условиях совершенно не встречающиеся.
В посевах льна выработался даже ряд видов, названных видовым
названием «linicolae», т. е. произрастающие во льне: рыжик льня-
ной (Camel ina irniccla), плевел льняной (Lclium liniccla), горец
льняной (Polygonum linicola) и др.; плоды (или семена) всех этих
видов по размеру сходны с семенами льна, при обработке не отде-
ляются от последних и переходят таким образом в посевной мате-
риал. Это—результат длительного естественного отбора, шедшего
под косвенным воздействием человека. В посевах ржи встречаем
особый подвид погремка (Alectcrolophus m? jor var. apterns), который
отличается от других подвидов, растущих на лугах, и пр.: данный
подвид называется apterus, т. е. лишённый крыла, так как у него
семя не имеет широкой крылатой оторочки, как у подвида в есте-
ственных ценозах. У этого последнего семена развеваются ветром
(крылатое семя!), что было бы нецелесообразно для семян сорного
подвида, примешивающихся к зернам ржи. Имеется и ещё ряд
растений, выработавшихся в условиях различных культур.
Косвенным путем человек способствовал появлению новых
видов или рас растений своими ежегодными покосами в
травянистых ценозах; так, на лугах появились ранние и псздиие
расы, т. е. цветущие и плодоносящие до покоса и после покоса
(сезонный диморфизм —см. ниже, стр. 270).
Однако несравненно большее значение имеет сознательная дея-
130
тельность человека в формировании новых видов, полезных в том
или ином отношении. Здесь мы имеем в виду «культурные растения»,
которые известны лишь в культуре и в дикой природе отсутствуют.
О происхождении культурных растений и о тех методах, которые
применял и применяем человек при их формировании, см. в прило-
жении «Культурные растения». ч
Человек и создание новых растений. Исключительно важным
вопросом является вопрос опеределке природы ра-
стений. Вопрос этот во всю ширь поставлен только в СССР
в связи с общим строительством. Правда, еще до революций Ми-
чурин провел ряд работ по скрещиванию между растениями
с целью переделки их природы, но в то время работы Мичурина
почти игнорировались. Мичурина можно смело назвать
выдающимся экспериментатором, новатором в данном направлении;
он разработал новые методы и успешно их применял (более под-
робно см. на стр. 384). Прямым последователем Мичурина
является Лысенко, но если Мичурин производил свои
исследования над деревянистыми плодо-ягодными растениями, то
Лысенко свои работы ведёт над травянистыми растениями
полевых культур (пшеница, рожь, картофель и др.). Замечательны
работы Лысенко над превращением озимых сортов в яровые,
исходя из учения о стадийности в развитии растений; это—под-
линная переделка природы этих растений. Было выяснено, что
озимые и яровые сорта представляют по свойству ози-
мо с т и один непрерывный ряд, причём для них различны лишь
условия в первой стадии развития — в стадии яровизации (разли-
чия в температуре и в продолжительности действия низких темпе-
ратур). Изменение этих условий влечёт за собой и изменение
свойств озимости. Не менее интересны работы, связанные с выро-
ждением картофеля на юге: на юге урожай картофеля был очень
низким,, клубни получались мелкие и вообще картофель подвер-
гался различным болезням вырождения. Лысенко выяснил,
опять-таки исходя из учения о стадийности, что время образования
и роста клубней картофеля очень неблагоприятно, так как в это
время почва бывает суха и температура (как в почве, так и в воз-
духе) слишком высока. В результате клубни получаются очень
мелкими. Для получения более хороших урожаев ранних
сортов необходимо перенести развитие картофеля вменее жар-
кий и более влажный период (хотя бы на полмесяца
раньше), а для ускорения развития нужна предпосевная
специальная обработка клубней. Эта обработка состоит в том, что
клубни проращивают при 12—15° в течение 10—15 дней, а затем
выдерживают при той же температуре на свету 25—30 дней. Эт;от
процесс Лысенко и называет я р о в и з а ци-е й.
Что касается вырождения поздних сорте в картофеля на юге,
то это связано с тем, что его посадка производится в слишком жар-
кое время, и температурные условия для клубнеобразования не-
благоприятны; поэтому необходима более поздняя посадка, чтобы
клубнесбэазование пришлось на время с более низкими темпера-
турами после спадения летней жары, Таким образом, каждая
9* 131
стадия развития будет проходить при наиболее благоприятных
условиях. Это яркий пример управления развитием рас-
тения, для чего необходимо глубокое знание экологии растения.
Переделка растений возможна как путём подбора соответствую-
щих условий, так и путём взаимодействий двух и более растений
между собой (см. работы и методы Мичурина, Держа-
вина, Цицина и др. на стр. 385). Получение ржаио-пше-
ничных гибридов, пшенично-пырейных многолетних гибридов и
др.—это огромный шаг в переделке растений.
Современная жизнь полна смелых попыток по перестройке ра-
стений на пользу человека. Сюда относятся также работы К и з То-
рина по продвижению плодовых деревьев в Сибирь и в
северные районы нашей страны. Основная мысль здесь та, что толь-
ко припочвенная зона в 25—30 см высотой является наи-
более благоприятной для выращивания в суровых климатических
условиях нежных плодовых пород; выше 30 см при суровых зимах
имеет место отмирание стволов и ветвей.
Как следствие отсюда —плодовые деревья должны быть не
выше указанной зоны и иметь стелющуюся форму: получается
особый тип сада —ба хчево-стелющийся (плоды иногда
непосредственно лежат на земле), причём главный приём —при-
гибание дерева и ветвей к земле. Подобный тип сада даёт воз-
можность продвижения на север плодовых пород на 7—8° широты
сравнительно с прежними северными границами (на 700—800 км).
Последний пример показывает, какое значение имеет соответ-
ствующее воспитание растения. Индивлдуальный под-
ход к растению с учётом всех его потребностей является основой’
стахановского движения при полевых культурах растений. Мы
знаем, какие успехи достигнуты при этом на свекловичных полях
и на многочисленных других культурах.
В общем, успехи в СССР по переделке растений, по созданию
новых сортов, по улучшению имеющихся сортов и др. огромны,
и, нет сомнения, не остановятся на достигнутом.
Исторические факторы
Так как современная флора сложилась в результате длитель-
ного исторического процесса, то, очевидно, многие факты совре-
менной действительности не могут быть объяснены современными
климатическими, эдафическими и другими факторами. Растения
имеют неодинаковый возраст, и в то время как одни являются
сверстниками и даже более молодыми сравнительно с человекол!
(примеры новых видов в посевах), другие являются пережитками
весьма отдалённых времён. Некоторые виды, несомненно, являют-
ся последними вымирающими остатками (реликтами) древних ти-
пов. И во многих случаях присутствие этих растений обусловлено
историческими причинами. Многие примеры этого приведены в
отделе «Флористические области».
Длительная и сложная эволюция многих существующих теперь
видов не могла не отразиться и на внешнем облике и на внутрен-
132
них свойствах растений. Поэтому попытки объяснить морфологи-
ческие и физиологические особенности растений только современ-
ными условиями существования нельзя считать правильными.
В этом отношении очень интересно проанализировать ритмы
развития растений: эти ритмы сохраняются и в том случае, если
растения культивируются в совершенно других климатических
условиях. Так, например, известно, что наши дубы, буки, плодо-
вые деревья и другие растения в более теплых областях всё же
сбрасывают свои листья на время, соответствующее нашей зиме,
хотя это не оправдывается условиями температуры. Присматри-
ваясь к нашим растениям, можно видеть, что ритмы развития их
очень различны. Вот некоторые примеры.
1. Растения вегетируют все тёплое время, ио зима .для них
представляет время вынужденного покоя, тогда как в более тёп-
лом климате Западной Европы они вегетируют круглый год (мар-
гаритка, Glechoma hederaceuin и т. д.).
2. Растения развиваются с весны, в дальнейшем цветут и пло-
доносят, и вегетация идёт до осени. Йз деревьев—липа, из кустар-
ников — бересклет, калина; очень многие травянистые растения.
3. Растения цветут и вегетируют ранней весной, но вегетация
очень скоро совершенно прекращается (например в июне). Многие
весенние травянистые растения: ветреницы, хохлатки, гусиные
луки, подснежники (Galanthus) и др. — растения-эфемеры.
4. Весной появляются листья, летом имеется период покоя,
осенью же начинается цветение. Некоторые шафраны (Crocus),
безвременник (Colchicum).
Если ограничиться этими примерами, то можно совершенно
ясно видеть, что виды первой и второй группы гармонируют бо-
лее и менее в своих ритмах с ритмом климата Средней Европы,
виды же третьей и четвёртой групп находятся в полной дисгармо-
нии. В самом деле, растения третьей группы используют лишь очень
небольшой отрезок вегетационного периода, более холодный и,
казалось бы, менее благоприятный, а растения четвёртой группы
используют весну и осень, наиболее же благоприятное время (ле-
то) проводят в состоянии покоя.
Эти соотношения дают основания считать, что если оба ритма
(развития и климата) идут параллельно, то перед нами растения,
выработавшиеся в данном климате, в противном случае можно
предполагать, что растение выработалось при иных климатических
условиях и имеет родину, соответствующую им. Так, ритм рас-
тений четвёртой группы соответствует климатическому ритму
средиземноморскому, а второй группы — пр и ле д-
виковому, высокогорному. Последние растения, сохранив
сво,й ритм развития с ледникового времени, в настоящее время
заняли в общем климатическом ритме соответствующее место.
Однако не только отдельные виды, но и растительные ценозы
очень часто не гармонируют в своих ритмах с ритмами климатиче-
скими. Причины могут быть различные — или это отражение исто-
рии, или это связано с влиянием человека. Так, на лугах уко-
роченный ритм развития можно связать с ежегодными покосами
133
и уничтожением растений, более поздно цветущих; иа сфагновых
болотах не используется полностью климатический ритм, и в летние
месяцы там почти отсутствуют цветущие виды (кроме росянок и
других).
Кроме того, следует подчеркнуть, что многие наши фитоцеиозы
не насыщены видами с точки зрения их гармонии с современным
климатом, т. е. они находятся ещё в состоянии приспособления.
Более глубокое изучение циклов развития видов и ценозов по-
зволяет подойти более основательно к истории флоры и раститель-
ности.
Совокупное влияние факторов
В природе обычно имеет место совместное влияние различных
факторов, причём выделить отдельно тот или иной фактор иногда
крайне трудно. Если выше каждый фактор рассматривался от-
дельно, то это было сделано для удобства из. сжения.
В настоящее время нельзя считать правильным известный закон
минимума Либиха, который говорит о том, что решающим
является тот фактор, который находится в минимуме, т. е. в слу-
чае недостатка или отсутствия одного необходимого элемента,
при наличии всех остальных, почва становится бесплодной для всех
тех растений, для жизни которых этот один элемент является не-
обходимым. Таким образом, урожай повышается или понижается
в точной пропорции с увеличением или уменьшением
.минеральных веществ, вносимых в почву.
Нам теперь известно, что этот закон имеет лишь относительное
значение, и закон относительности говорит лишь об относительном
действии фактора, причём это относительное действие тем больше,
чем более данный фактор находится в минимуме сравнительно с
остальными факторами; при увеличивающейся интенсивности фак-
тора относительное действие уменьшается и приближается к нулю
в области максимума. Таким образом, экологически дей-
с твующий фактор, как правило, фактор в минимуме. Однако
и остальные факторы тоже не лишены некоторого значения, так
как неблагоприятное действие фактора в минимуме другие факто-
ры люгут так или иначе компенсировать при тех или иных
сочетаниях.
Нужно особо подчеркнуть значение ограничивающего
фактора (Блэкман), под которым понимается фактор
недостаточный, останавливающий и сильно задерживающий тот
или иной процесс (например рост растения). При увеличении огра-
ничивающего фактора данный процесс возобновляется, и растение
получает возможность произрастать в данной обстановке.
Замещение факторов. Большое затруднение при определении
и разграничении действия ряда факторов вносит явление заме-
щения факторов. Это явление состоит в том, что один
фактор может замещаться другим в полной мере или отчасти. Это
замещение факторов позволяет растениям произрастать при
разных внешних условиях в случае возможной
замены одних факторов другими. Если ограничиться тремя кате-
134
горними факторов (климатические, эдафические, биотические \ то
можно говорить о таких случаях замещения:
1. Климатические факторы замещаются другими климатическими.
2. Эдафические факторы замещаются другими эдафическими.
3. Биотические факторы замещаются другими биотическими.
4. Климатические — эдафическими и наоборот.
5. Климатические — биотическими и наоборот.
6. Эдафические — биотическими и наоборот.
Некоторые примеры по этим шести случаям пояснят сказанное.
1. Определенное количество осадков может замещаться влажностью воздуха.
Тепло может замещаться светом. Примеры были указаны в разделе «Свет».
-2. Физическая сухость оказывает то же действие, что и физиологическая
сухость. Засоленная почва действует, как сухая почва.
3. Выпас может приводить к тем же результатам, что и покос, уничтожая
ряд растений. Вырубка может замещаться конкуренцией видов; при вырубке
соснового леса нередко появляется ель, но ель также сменяет сосиу в резуль-
тате конкуренции как более теневыносливая порода.
4. Более северный влажный климат на юге замещается влажной почвой, и
северные виды на юг продвигаются по долинам рек. С>хой климат замещается
на севере известняковой или песчаной почвой, и южные растения в более север-
ных местах произрастают на известняках или на песках.
5. Влажный воздух замещается защитой деревьев: вереск на востоке при
более сухом воздухе растет лишь в лесах, а на Балтийском побережье с влажным
воздухом — на открытых местах (см. также стр. 247). Вечнозеленые виды в более
континентальном климате растут лишь в подлеске (рис. 62).
Рис. 62. Иллюстрация замещения факторов: ве(чнозелёные виды идут в
более континентальном климате в подлесок, где деревья как бы замеща-
ют более влажный климат (замещение климатического фактора биоти-
ческим; по Рюбелю).
6. Плодородная почва замещается унавоживанием. Ослабление конкуренции
между видами замещается засолёнными почвами, так как иа засоленных почвах
могут существовать виды менее конкурентно сильные. Выпас, уплотняя почву,
создаёт повышенную капиллярность, и травостой становится более ксеро-
фпльным.
Эти немногие примеры показывают, что внутренние причины при одном и
том же внешнем эффекте могут быть весьма различны.
Всё сказанное делает изучение экологических условий очень
трудным и должно предостерегать от априорных экологических
заключений. В большинстве случаев необходим эксперимент.
1 Биотические факторы в данном месте понимаются более широко, включая
и влияние человека.
О^ДЕЛ III
УЧЕНИЕ О ФИТОЦЕНОЗАХ
Растительные организмы — индивидуумы, почти как правило,
произрастают в природе не изолированно, вне связи друг с другом,
но дают определенные сочетания, причём эти сочетания могут со-
стоять из индивидуумов одного и того же вида или из индивиду-
умов различных видов. Чаще всего мы встречаем сочетания раз-
личных видов, причём последние представлены большим или
меньшим числом экземпляров.
Конкуренция между растениями. В иных случаях индивидуумов
на участке может быть немного, в других же, при особо благопри-
ятных условиях существования, число индивидуумов может быть
очень значительным. Так, в окрестностях Ташкента в эфемеровой
пустыне (см. стр. 313) на 1 м2 было зарегистрировано более 5000 экз.
растений, относящихся к 47 видам. Большое число экземпляров
объясняется тем, что большинство растений являлось мелкими
однолетниками, но и при почти полном отсутствии однолетников
число экземпляров может быть велико; в Стрелецкой степи под
Курском на 1 м2 нами было отмечено 1939 экз., принадлежащих
к 77 видам (Роа angustifolia —189 экз., Bromus erectlis - 178, Fi-
lipendula hexapetala—184, Viola rupestris—131, Veronica incana—1,
Leucanthemum vulgaris — 1 и т. д.).
Такое исключительно большое число экземпляров представляет
явление более редкое, обычно «экземплярная насыщенность», т. е.
число экземпляров на определённую площадь, значительно меньше;
так, в южных степях и особенно в пустынях растительный покров
сильно разрежен, ина Lit можно найти лишь несколько растений.
В особо крайних условиях существования встречаются участки,
совершенно лишённые высших или каких бы то ни было растений.
При сильной насыщенности территории между отдельными инди-
видуумами возникает конкуренция из-за света, питательных ве-
ществ в почве, воды и т. п. Нужно различать конкуренцию между
индивидуумами одного и того же вида и между индивидуумами
различных видов (внутривидовая и межвидовая
конкуренция).
Чем ближе по своим жизненным потребностям те или иные инди-
видуумы, тем больше между ними конкуренция и наоборот. Ещё
Дарвин указывал, что борьба между видами и особями может
быть то более, то менее л£е стоксп. В результате конкуренции
13в
многие экземпляры погибают совсем, другие же находятся в угне-
тённом состоянии и не доходят до своей завершительной стадии,
т. е. не цветут и не плодоносят. Так, в одном случае на 1 м2 в Стре-
лецкой степи были отмечены такие соотношения между цветущими
и угнетёнными, только вегетирующими экземплярами:
Название вида Число цветущих экземпляров Число вегетирующих экземпляров Число проростков
Степная полевица (Agrostis Syreistschikowi) 11 62 —
Костер прямой (Bromus erectus) 178 —
Мятлик узколистный .... (Роа angustifolia) 1 188 —
Горный клевер (Trifolium montanum) 1 71 2
Степная таволжаика .... (Filipendula hexapetala) 2 182 2
Луговой ща 1фей (Salvia pratensis) 4 70 3
Фиалка песчаиая t (Viola rupestris) 15 116 1!
В данных примерах мы имеем перед собой
конкуренцию глав-
ным образом между растениями уже взрослыми, занимающими
больший или меньший участочек почвы, причём эта конкуренция
является длительной,
непрерывной в течение
всей жизни каждого ин-
дивидуума, то умень-
шаясь, то усиливаясь в
зависимости от измене-
ния внешних условий.
Но ещё более интен-
сивная конкуренция
идёт между взрослыми
растениями и их моло-
дыми экземплярами
(всходами). Из только
чтоприведённойтаблич-
ки видно, что число мо-
лодых растений (про-
ростков) ничтожно по
сравнению с числом
взрослых растений, а во
многих случаях про-
ростки и совершенно
отсутствуют. Иэтоимеет
место несмотря на то,
что взрослые растения
дают обычно большое
12-l'V
395,
Рис. 63L Конкуренция между особями шведки
на I м2 (по Закаряну).
19 - IV-29г
число семян;очевидно,
что при густом траво-
стое семена или ие прорастают, или же погибают в
стадии проростков в результате конкуренции с более старыми
экземплярами.
Интересны данные о количестве выживающих экземпляров в
природе, что видно на рис. 63 (наблюдения производились в Му-
ганской степи, в Закавказье, на небольших площадках в 1 лг).
Наблюдения производились на «чистых» площадках, где никаких
других растений не было. Особенно много экземпляров погибло
на плошадках со шведкой блестящей (Suaeda splendens), так как
её всходы, благодаря небольшим размерам, растут очень густо;
сразу выделяются более сильные и более слабые экземпляры, по-
следние скоро погибают, и в дальнейшем идёт конкуренция между
более сильными. Вот цифры уменьшения числа экземпляров:
29 марта.............. 484 экз. 19 апреля............. 395 экз.
5 апреля............. 467 » 26 » 391 »
12 395 » сентябрь.............. 41 >
Таким образом, полный жизненный цикл завершили лишь
несколько более 8% экземпляров. Но и это имеет место лишь в том
случае, если семена попадают в подходящую экологическую обста-
новку; в противном случае все семена и проростки могут погибнуть.
Закономерные, сложившиеся сочетания растений— фитоценозы
Как ни велика конкуренция между экземплярами растений,
но так или иначе на определенном участке подбирается известный
комплекс видов. Является ли этот комплекс случайным, состоит
ли он из растительных видов любого сочетания? Наоборот, наблю-
дения показывают, что в каждом отдельном случае мы имеем вполне
определённые, постоянно повторяющиеся сочетания растительных
видов. На севере мы имеем одни комбинации видов, на юге — дру-
гие; в пустынях эти комбинации и сами виды, их слагающие, бу-
дут совершенно иными, чем в степях, на лугах, в лесах и т. д. Эти
комбинации растений отличаются не только своим различным ви-
довым составом, они отличаются и своим внешним видом, своей
физиономией. Последняя настолько бросается в глаза,
что люди, с ботаникой ничего общего не имеющие, давно устано-
вили такие понятия, как степь, луг, болото, лес и т. п.
Но такое, например, физиономическое объединение, как «лес»,
распадается на ряд более мелких подразделений, как леса хвойные,
лиственные вечнозелёные, лиственные с опадающей листвой и пр.;
эти подразделения распадаются ещё дальше; так, например, хвой-
ные леса распадаются на еловые, сосновые, пихтовые, листвен-
ничные и пр., достаточно хорошо физиономически различающиеся;
возможны и дальнейшие подразделения, с которыми мы познако-
мимся дальше. Сказанное о лесах применимо также и к другим
широким физиономическим объединениям, как болото, степь и пр.
Все эти объединения достаточно постоянны и, что особенно
важно, повторяются в сходных экологических условиях;
так, например, на песчаных почвах обычно развиваются сосновые
леса, на глинистых — еловые, на деградированных чернозёмах —
дубовые и т. п. Эта повторяемость указывает на то, что здесь перед
138
нами не случайное собрание видов, а некоторый комплекс видов,
сжившихся между собой и обладающих определённым флористи-
ческим составом и определённой физиономией. Подобное сочетание
растительных видов называется фитоценозом (фитоценоз от грече-
ских слов «фитон» — растение и «койнос»— общий); это основной
объект фитоценологии (геоботаники).
Фитоценоз (согласно 'предложению Брюссельского ботани-
ческого конгресса в 1910 г.) не представляет понятия таксономи-
ческого (систематического)' значения, как, например, в систематике
вид или род, это — растительные объединения как более крупного,
так и более мелкого объёма. Так, фитоценозом мы можем назвать
как лес вообще, так и лес хвойный и подразделения последнего, а
также и данный конкретный участок. Фитоценоз — это общее по-
нятие г.
Основные проблемы при изучении фитоценозов. При изучении
фитоценозов можно ставить себе ряд основных проблем, совпадаю-
щих с теми проблемами, которые имеют место при изучении видов.
Эти проблемы таковы:
1. Проблема морфологии, изучающая сложение и структурные
признаки фитоценозов; подобно виду, каждый фитоценоз; имеет
определённое строение, различное у различных фитоцеиозов: вод-
ные фитоценозы построены иначе, чем наземные; травянистые —
иначе, чем древесные; северные леса построены много проще, чем
южные, тропические, и т. д. С основными структурными призна-
ками мы познакомимся дальше.
2. Проблема экологии, изучающая связь и взаимоотношения
между фитоценозами, с одной стороны, и окружающей средой,
с другой, так как каждый фитоценоз находится в самой тесной свя-
зи со своей средой (местообитанием). Если морфология водных фи-
тоцеиозов сильно отличается ог морфологии наземных (например
заросли кувшинок и водяных лилий в реках и злаковые заросли
на лугах), то эта морфология связана с различными условиями су-
ществования, и экология стремится объяснить указанную связь,
объяснить экологически имеющиеся структурные особенности. Наи-
более ценным, однако, объяснение может быть лишь в том случае,
если наши априорные предположения оправдываются при соответ-
ствующем эксперименте. Как экология видов, так и экология фито-
ценозов в последнее время-всё более и более становится экспери-
ментальной наукой. 7
3. Проблема смен, или эволюции фитоценозов. Фитоценозы
как известные закономерные комплексы растений возникли в резуль-
тате длинного эволюционного процесса, но и дойдя до своего бо-
лее совершенного состояния, не остаются в покое — они эволю-
ционируют всё дальше и дальше, хотя и не так заметно и быстро.
Но, кроме того, в природе мы везде наблюдаем как разрушение
уже сложившихся фитоценозов, так и все стадии их восстановле-
ния, когда, например, на обнаженном почему-либо месте поселя-
1 Однако некоторые геоботаники (особенно ленинградские) фитоценозом
называют только конкретные участки растительных сочетаний (см. ниже).
139
ются растения и постепенно складываются в более или меиее устой-
чивое целое. Одним словом, если проблема морфологии — это в
основном проблема статики фитоцеиозов, то проблема смен — это
проблема динамики.
4. Проблема географии фитоценозов, или распределения их на
земной поверхности География фитоценозов имеет дело с их аре-
алами, с распределением последних по земной поверхности и с вы-
яснением основных закономерностей этого распределения Геогра-
фия фитоценозов тесно связана с учением о ландшафтах
5 Проблема классификации (систематики) фитоценозов. Она
ставит себе целые расчленение растительного покрова на единицы
различных таксономических значений и составление определен-
ной системы этих единиц». Только таким путем можно разобраться
в огромном количестве фитоценозов, имеющихся на земной поверх-
ности, иначе они представляют материал, совершенно не поддаю-
щийся обозрению и сопоставлению
В дальнейшем мы более подробно остановимся на основных
намеченных проблемах, теперь же можно дать такое определение
фитоценологии это отдел ботаники, имеющий объектом своего изу-
чения фитоценозы (растительные ценозы), познаваемые со всех
возможных точек зрения (с точки зрения морфологии, экологии,
географии и т д ) Нередко данный отдел ботаники называют гео-
ботаникой.
Термин «геоботаника) был впервые употреблен немецким ботаником Гри-
зебахом в 1866 г в смысле географии растений, приставка ге (по-гречески
«ге» — зе тля) к стову ботаника должна была указать, что этот отдел изучает
растения в их распределении по з е м л е, по земному шару В конце
прошлого столетия некоторые русские ботаники под геоботаникой понимали
•тде i htvkh, устанавливающий связь между растениями и историей и жпзны®
почв (например, Краснов), Литвинов же назвал геоботаникой т®
нлправ jhi е, которое имеет целью связать распределение современных видов с
поошлои геологической историей страды И в настоящее время имеются различ-
ные толкования этого термина Так, например, швейцарский ботаник Рюбель
включает в геоботанику три основных проблемы проблему пространства, место
•битачия и изменчивости, причем эти проблемы касаются не только фитоценозов,
но и отдельных видов (таким образом, сюда включается география видов, эко-
огия видов и изменчивость видов) Имеются еще и другие толкования геобота
н ги Л? чше или совсем не употреблять этого термина, пли же пользоваться им
как синонимом фитоценологии, что мы и будем делать
Формирование фитоцеиозов
Каждый фитоценоз, достаточно сформировавшийся, имеет,
как уже было указано, определенный флористический состав и
определенную структуру Целесообразнее, однако, прежде выяс-
нения морфологии (структуры) фитоценозов остановиться сфор-
мировании их, начиная с самых начальных стадий.
Каждое обнаженное место может представлять
большие или меньшие удобства для поселения растений и форми-
рования фитоценозов. При этом нужно различать обнаженные
места двоякого рода во-первых, те, на которых прежде не было
растительности и которые, следовательно, не содержат никаких
растительных остатков, ни живых (семена, остатки корней и пр ),
ни мертвых (гумус отмерших растений), — это первичные обнажен-
ий
ные места, во-вторых, нужно отличать обнаженные места, которые
можно назвать вторичными, так как иа подобных местах раститель-
ность уже существовала (и, быть может, не раз), но была тем или
иным способом уничтожена, на таких местах сохраняются в земле
некоторые живые части и во всяком случае большее или меньшее
количество гумуса
Первичные обнаженные места могут возникать различными пу-
тями они могут образоваться путем оползней, обвалов, в результате
деятельности текучей воды (эрозионные процессы — рис 64), ледни-
ков, уничтожающих при своем движении всякую растительность,
Рис 64 Обнаженные места в Небраске (Северная Америка)—«плохие земли»,
результат водной эрозии (по Мак-Дуголлу).
а также от действия ветра, особенно в областях с песчаным суб-
стратом Огромные обнаженные территории могут возникать в ре-
зультате поднятия дна морей над уровнем воды Возможны и мно-
гие другие пути
Вторичные обнаженные места тоже возникают различными
путями, так, пожары или сильная засуха могут нацело уничто-
жить имевшуюся растительность, возможно также вымокание
растительности от затопления водой и пр.
С развитием культуры обнаженные места появляются в резуль-
тате деятельности человека, особенно часто возникают вторичные
обнаженные места—распашка степей, корчовка лесов, после чего
часто дело А оходит до черного пара Однако деятельность человека,
приводящая к возникновению «полезных» обнаженных мест, мо-
жет иногда по недосмотру способствовать появлению вредных
обнаженных мест как вторичного, так и первичного характера
В качестве вторичных можно привести места лесных пожарищ,
примерами первичных могут служить эрозионные процессы в ре-
зультате распашки степей, уничтожения лесов и пр , способствую-
щие образованию оврагов, обнажений с голыми склонами и т д
141
Сукцессии. Рассмотрим несколько примеров формирования ра-
стительности на обнажённом месте, которое особенно хорошо мож-
но наблюдать по берегам рек с песчаными отмелями; места эти
первоначально лишены каких-либо растений, представляя пере-
мытый песок, в понижениях—с иловатыми частицами. Постепенно
здесь поселяются те или иные виды, прорастая из семян, заноси-
мых водой, птицами, ветром и другими агентами. В числе первых
видов здесь поселяются некоторые ситники (Junciis), некоторые
растения из семейства маревых (Chenopodiaceae), череда (Bidens)
и др. Все эти растения разбросаны отдельными экземплярами на
голом субстрате, и в их взаимном расположении не наблюдается
никакой правильности, так как занос семян происходит совершенно
случайно.
Когда некоторое количество экземпляров поселилось на опреде-
лённой территории, то снабжение семенами данного участка уже
обеспечено и, кроме того, многие растения способны размножать-
ся вегетативно. Вместо отдельных экземпляров появляются группы
или скопления экземпляров, иногда называемые «колониями».
Кроме того, внедряются и новые виды и новые экземпляры, несом-
кнутого ковра растительности ещё нет, и нельзя установить каких-
либо закономерностей в создавшейся растительной группиров-
к е. Термином «группировка» можно обозначать подобные не-
определённые и часто случайные комплексы растений.
Через некоторое время к числу растений-пионеров присоеди-
няется подбел (Petasites), имеющий крупные лопуховидные листья-и
длинные корневища; при дальнейшем своём развитии он даёт почти
сплошные заросли; остальные растения ютятся под его листьями.
В дальнейшей эволюции между экземплярами подбела посе-
ляются некоторые виды ив (особенно Salix triandra), и через из-
вестное количество времени получаются густые заросли ивняков.
Подбел теперь находится в угнетённом состоянии, но зато появ-
ляется ряд новых видов, особенно некоторые злаки с ползучими
корневищами, как костёр безостый (Bromus inermis), пырей обык-
новенный (Agropyrum repens), а также некоторые двудольные.
Если подобная эво'люция растительности наблюдается в лес-
ной зоне, то ивняк со временем сменяется лесом. Господетвующей
древесной породой является здесь дуб (в средних областях Европей-
ской части СССР); в более северных частях место дубазаним етель.
К дубу примешиваются вяз, осина, а также некоторые кустарники:
калина, крушина, шиповник и др. В травянистом покрове появля-
ются лесные растения; иногда в большом количестве развивается
ландыш.
В дальнейшем эволюция (при постоянстве климатических усло-
вий) не идёт дальше дубового леса, но и не останавливается, так
как лес как таковой претерпевает те или иные изменения внутри
себя, однако менее заметные и не выходящие за пределы данного
типа. Тип дубового леса можно поэтому считать «заключительной»
стадией описанного процесса.
Рассмотренный процесс формирования растительного покрова,
начавшийся на обнажённом месте и завершившийся устойчивым
142
в данных условиях дубовым лесом, представляет так называемую
сукцессию (иногда её называют просто «сменой»); она состоит, как
мы видели, из ряда стадии, причём нужно различать начальные,
промежуточные и заключительную стадии. Совокупность стадий
представляет серию.
При постоянстве климатических условий необходимость сук-
цессии в растительном покрове обусловлена следующими, постоян-
но действующими причинами:
1. Всегда имеется избыток семян и плодов, приносимых на дан-
ную территорию.
2. Всегда между населяющими растениями идёт конкуренция,
и биологически более приспособленные вытесняют более слабые.
3. Сама среда (местообитание) постоянно меняется в результате
жизнедеятельности растений (отложение гумуса и пр.); возникают
новые условия^ более пригодные для появления новых видов.
В вышеприведённом примере ход сукцессии осложняется так-
же тем, что здесь действует ещё одна причина, направляющая и
ускоряющая описанную эволюцию. Именно, рассматриваемая сук-
цессия связана с берегом реки, где ежегодно весной происходит
заливание полыми водами реки и отложение того или иного
материала (песок, глина, ил и пр.), несомого этими водами. Здесь
к указанным выше причинам, связанным с жизнью
самого фитоценоза, прибавляется причина внешнего характе-
ра, извне направляющая ход сукцессии.
На данном примере хорошо видно, что сукцессии растительного
покрова могут зависеть от двух причин — внутренних и внешних.
В первом случае сукцессии называются эндодинамическими, во
втором — экзодинамдческыми. В нашем случае сукцессия имеет
смешанный характер.
Приведём ещё один пример сукцессии, болёе простой, связанной
лишь с внутренними процессами (эндодинамической). Перед нами
распаханная степь, доведённая до абсолютно чистого чёрного пара.
Однако .это — вторичное обнажённое место, так как чернозём рас-
паханной стели очень богат гумусом, и кроме того, в нём имеется
обычно немало семян, остатков корней, корневищ и т. п. И в самом
деле, на чёрном пару, если его не поддерживать в чистом состоянии,
сейчас же появляются всходы различных растений, а также ростки
от имеющихся в почве корневищ. Иногда быстро появляются кур-
тины розового осота (Cirsiujn arvense), отрастающие от глубоко
расположенных подземных органов этого растения.
В общем пар довольно скоро сплошь покрывается растениями,
в распределении которых не замечается никакой правильности. За-
селение пара идёт и за счет зачатков растений, находящихся в
почве, и за счёт семян, приносимых со стороны. Особое внимание
обращает на себя в этой первой стадии зарастания обилие и мощ-
ное развитие однолетников и двулетников. Это стадия так называ-
емых бурьянов, достигающих нередко роста человека.
В дальнейшем начинают появляться растения многолетние,
особенно снабжённые длинными корневищами; куртины этих ра-
стений разрастаются во все стороны и, смыкаясь, иногда образуют
143
сплошной ковёр. Наиболее типичен для данной стадии пырей (Ag-
ropyrum repens); иногда обильно развивается зубровка (Hierochloa
odorata). Эта стадия по своему флористическому составу и внешнему
виду совершенно не похожа на предыдущую н может быть назва-
на корневищной стадией.
Через некоторое время корневищные растения в связи с уплот-
нением почвы начинают изреживаться, и здесь поселяются степ-
ные двудольные со стержневыми корнями, а также злаки дерно-
винного типа, из которых особенно характерен типчак (Festuca
sulcata); начинают поселяться также такие типичные степные зла-
ки, как ковыли. Одним словом, на данной стадии характерные
черты степной растительности все более и более усиливаются.
Постепенно появляются все новые и новые типично степные
растения; крупные дерновины ковыля образуют равномерный по-
кров, представляя уже сформировавшуюся ковыльную степь. Эта
стадия является «заключительной».
Данная сукцессия представляет пример эндо динамической сме-
ны (конкуренция растений влияние растений на поч-
ву и пр.); однако немалую роль здесь играет также и изменение
физической структуры почвы (почва становится всё менее рыхлой,
капиллярность усиливается и пр.), т. е. и здесь причины экзоди-
намического характера все же присутствуют.
Рассмотренные сукцессии могут быть названы ещё сингенети-
ческими (сингенез от греческих слов «син» — с, совместно и «гене-
зис» — происхождение); здесь перед нами проходит процесс фор-
мирования или восстановления нарушенного фитоценоза.
При изучении начальных стадий сукцессий всякого рода (как
на первичных, так и вторичных обнажённых местах) обращает на
себя внимание то обстоятельство, что растительный покров прохо-
дит несколько стадий в своем формировании, а именно:
1. Отдельные растения беспорядочно разбросаны по всему уча-
стку и нередко на таком расстоянии, что совершенно не оказыва-
ют какого-либо влияния друг на друга.
2. Травянистый покров уже сомкнулся, растения, конечно,
уже влияют друг на друга, но бросается в глаза случайность ком-
бинаций видов, не видно какой-либо выработанной системы отно-
шений. Здесь можно различать два основных варианта:
а) растительный покров односоставный, т. е. состоящий из од-
ного доминирующего вида, почему-либо быстро занявшего терри-
торию и сильно размножившегося; например сплошные заросли
пырея на залежн, костра безостого на овражных выносах почв,
заросли щавелька (Rumex acetosella) на паровых полях и пр.;
б) растительный покров разносоставный, т. е. состоящий из
нескольких видов, но расположенных группами, пятнами отдель-
ных видов, быстро размножившихся и занявших определенные
участки территории. Подобную картину можно наблюдать на за-
лежах. заросших пашнях, лесосеках и т. п.
3. Травостой не только сомкнулся, но каждое растение нашло
себе соответствующее место среди других. В травянистых фитоце-
нозах обычно наблюдается мозаичное распределение видов с опре-
144
делёнными количественными отношениями экземпляров, В древес-
ных фитоценозах особенно хорошо выражен подбор видов, имею-
щих различную высоту и располагающихся своими верхними ча-
стями в различных горизонтах.
Указанные. стадии сингенетической сукцессии могут так или
иначе видоизменяться, но основным в этом процессе является то, что
растительный комплекс сформировывается, получает известную мор-
фологическую структуру и закономерные отношения между видами.
ПЬдобный комплекс видов мы и называем фитоценозом, разу-
мея под этим понятием нечто структурно сложившееся, и в таком
случае первые стадии сингенетического процесса можно назвать
только «группировкой» растений; здесь всё ещё случайно, и даже
влияние одних растений на другие может отсутствовать. Естествен-
но, что группировки вполне постепенно переходят
в фитоценозы. Некоторые авторы не подразделяют сочетания ра-
стений на группировки и фитоценозы и называют фитоценозами
все сочетания растений, в которых имеются влияния растений
друг на друга.
Морфология фитоценозов
Сложившийся фитоценоз име?т, как уже указывалось, опреде-
лённые структурные признаки и определенную физиономию. Та-
кие фитоценозы, как ковыльная степь, разнотравная степь, сос-
новый лес с вересковым покровом, сосновый лес с белым наземным
лишайниковым покровом, еловый лес с черникой и т. п., имеют
вполне определённую физиономию и нередко резко отличаются друг
от друга. Надо, однако, заметить, что у одних фитоценозов физионо-
мия остаётся почти неизменной в течение всего вегетационного
периода, у других она претерпевает ряд изменений.
Физиономия фитоценоза даёт так называемый асп ект (внешний
вид). Поэтому фито ценозы бывают с постоянными и меняющимися
аспектами (о последних будет сказано ниже).
Физиономия и аспекты крайне важны для познания фитоцено-
зов, подобно тому как внешняя морфология определяет собой тот
или иной растительный вид. Отграничение фитоценозов в природе
прежде всего основывается на их физиономии.
Физиономия фитоценозов является следствием: 1) флористи-
ческого состава, 2) количественных отношений между индивиду-
умами имеющихся видов, 3) распределения видов и некоторых
других признаков.
Флористический состав (набор видов, входящих в состав фито-
ценоза). Он может быть очень разнообразен и ни в какой степени не
стоит в связи с систематическими отношениями этих видов. В ос-
новном здесь играют роль моменты экологического значения. Так
как было указано, что каждый фитоценоз связан с определёнными
внешними условиями, то можно было бы думать, что в фитоцено-
зах подбираются виды одинаковой и близкой экологии. Одиако это
совсем не так. Возьмём для примера какой-либо фитоценоз леса;
он состоит из сочетания очень различных жизненных форм: дере-
вья, кустарники, травы злаковидного типа, травы широколистного
10 География растений 145
типа, мхи, лишайники и др. Совершенно ясно, что жизненные по-
требности и экология этих растений крайне различны. То же име-
ет место в большей или меньшей степени и в любом фитоценозе.
Таким образом, виды фитоценоза в основном экологически
разноценны-
Но как возникает подобная экологическая равноценность видов
на местообитании, имеющем в общем определенную экологию? Мож-
но привести такой упрощенный,
пример; иа какую-либо неболь-
шую площадку попало несколь-
ко семян солнечного вида, ко-
торые дали взрослые растения;
таких растений, предположим,
на данной площадке может раз-
меститься 10 экземпляров, Оче-
видно, что другие виды с по-
добным же отношением к свету
на площадке поселяться не мо-
гут, однако могут поселиться
такие виды, которые являются
более теневыносливыми и могут
разместиться под тенью первого
вида. Далее можно представить
себе какой-либо третий вид,
обладающий еще большей тене-
выносливостью и поселившийся
под тенью первых двух видов
(рис, 65). В данном простом
примере имеются, таким обра-
зом, три различных вида, ра-
стущих на одном и том же
участке и экологически разно-
ценных (в смысле теневыносли-
вости). Но можно представить
и другие сочетания видов с
различной экологией, но пре-
красно уживающихся на одной
и той же площади, например,
одно растение имеет более по-
верхностную корневую систе-
му, другое—более глубокую,
третье — еще более глубокую,
Рис. 65. Схемы взаимного расположения и соответственно с этим эти три
видов, экологически разноценных j растения предъявляют различ-
сверху-особи толььо одного вида; в середине- тпрппвяиий к впяге мп-
осоои двх* видов, внизу-особи^трех^. видов ные треоования к влаге, ми-
(ориг.) неральному питанию и пр., и,
значит, вполне могут существовать совместно.
В результате на определенной (однородной) площади подбирает-
ся известный набор видов (флористический состав), находящихся
между собой в тех или иных взаимоотношениях, причем конкурен-
146
ция между ними несколько ослаблена тем, что они предъявляю!
к среде неодинаковые требования (в смысле света, мине-
рального питания и пр.).
Однако среди флористического состава могут находиться и ви-
ды, имеющие близкие экологические потребности, —виды эколо-
гически равноценные, — и тогда конкурентные отношения между
ними выражены, конечно, более сильно. Но не нужно забывать,
что многие виды в фитоценозе находятся в очень большом числе
индивидуумов, и так как последние экологически рав-
ноценны, то и конкуренция среди них должна достигать
сильного напряжения.
Итак, в фитоценозах подбираются растения: 1) экологически
разноценные (большинство видов) и 2) экологически равноценные
(некоторые виды и индивидуумы одних и тех же видов).
Как уже было указано, в сложившихся фитоценозах флористи-
ческий состав является достаточно определенным, а потому нужно
предполагать, что растения с указанными различными экологи-
ческими потребностями должны находиться в определенных уста-
новившихся отношениях. Другими словами, фитоценоз должен
обладать известной структурой.
Ярусность. Ярусное расчленение — это один нз структурных
признаков сложившихся фитоценозов: растения различной высоты
(л соответственно различных экологических свойств) располагают
свои верхние части на разных уровнях, отчасти перекрывая друг
друга. В различных фитоценозах бывает различное число ярусов,
Рис. 66. Схемы ярусности в лесу слева—трехъярусный, справа—пятиярус-
ный фитоценоз (ориг ).
так, например, в сосновых лесах наблюдаются такие структуры
(рис. 66):
1. Сосна — в 1-м ярусе, олений лишайник — во 2-м ярусе (двухъ-
ярусный ценоз).
2, Сосна — в 1-м ярусе, черника — во 2-мярусе, зеленыймох —
в 3-м ярусе (трехъярусный ценоз).
3. Сосна — в 1-м ярусе, орешник — во 2-м ярусе, черника —
в 3-м ярусе, зеленый мох — в 4-м ярусе (четырехъярусный ценоз).
10* 147
4. Сосна — в I-м ярусе, орешник — во 2-м ярусе, черника в
3-м ярусе, кислица — в 4-м ярусе, зеленый мох — в 5-м ярусе
(пятиярусный ценоз).
Возможны и более многоярусные цжозы, в приведенных приме-
рах в каждом ярусе указан господствующий вид (доминант-
ный). В одних случаях ярусное сложение выражено очень отчет-
ливо (леса, болота), в других — менее хорошо (некоторые типы
лугов, степей, тропические леса и др ). Последнее имеет место в
фитоценозах с богатым флористическим составом, когда подбира-
ются виды весьма разнообразной высоты. Вообще же число ярусов
увеличивается. I) в связи с увеличением числа видов, 2) в связи
с улучшением условий существования и 3) в связи с возрастом фи-
тоценоза. Наиболее древние фитоценозы, например, в тропиках,
существующие с давних времен (с третичного периода), имеют
сложное ярусное строение, так как многочисленные виды могли
приспособиться к условиям совместного существования. Эволю-
ция фитоценозов, можно думать, идет в направлении усложнения
ярусного строения.
Что каса'ется размещения различных видов по ярусам, то оче-
видно, что виды (также индивидуумы), экологически равноценные,
располагаются в одном и том же ярусе, а экологически разноцен-
ные — в различных. Так, в приведенных примерах такие виды,
как сосна, орешник, черника, зеленые мхи и др., являются различ-
ными жизненными формами, экологически равноценными, и рас-
пределяются по разным ярусам.
Выше было указано, что растительные виды могут в большем
количестве разместиться на данном участке, если '.корневые систе-
мы их будут располагаться не на одинаковой глубине, так дей-
ствительно и обстоит дело в природе, т. е. в фитоценозах, помимо
надземной ярусности, имеется также и подземная ярус-
ность. В степях, например, можно различать в типе три подземных
яруса: верхний, наименее глубокий, в котором находятся корни
однолетних растений, луковицы, клубнн н другие, неглубоко рас-
положенные корневые системы, средний, состоящий главным об-
разом из корней злаков (ковыли и др.), и наиболее глубокий, в
котором располагаются корни стержнекорневых двудольных.
Располагаясь на различных глубинных уровнях, корневые сис-
темы в стегях как бы не конкурируют друг с другом из-за необхо-
димых питательных веществ, но это не совсем так, и если кон-
куренция за минеральное питание ослаблена, то конкуренция за
воду достаточно сильна: верхние корневые системы перехватыва-
ют нередко почти всю влагу, вообще недостаточную в степях.
На лугах с моховым покровом можно хорошо наблюдать че-
тыре яруса корневых систем. Наиболее высоко лежащие корневые
системы расположены в самом моховом ковре и при сдирании по-
следнего легко удаляются, несколько глубже, но в самом поверх-
ностном слое почвы, размещаются более глубокие корневые системы;
еще глубже находятся корневища злаков и двудольных, наконец,
глубже всего опускаются корни некоторых двудольных.
148
Не приводя ещё других примеров (рис. 67), можно вообще от-
метить, что благодаря подземной ярусности на одном и том же
местообитании могут совместно расти столь различные типы ра-
стительности, как ксерофиты, мезофиты и даже гигрофиты или,
с другой стороны, галофиты и негалофиты. Дело в том, что у ксеро-
фитов корневые системы могут находиться в верхних иссушенных
горизонтах почвы, а у мезофитов и гигрофитов — доходить до уров-
Рис 67 Ярусность надземная и подземная в травянистом покрове
хвойного леса; хорошо выделяется ярус кислицы, видно и несколько
подземных ярусов (по КуЙяла).
ня грунтовых вод. Вот почему в степной и полупустынной обста-
новке степные растения могут находиться бок-о-бок с таким при-
брежно-водным растением, как тростник. То же имеет место и
для галофитов и негалофитов, так как верхние горизонты могут
быть засолены, а нижние — нет.
Из сказанного достаточно ясно, что в результате ярусности
двоякого рода (надземной и подземной): I) на определенном уча-
стке фитоценоза может поселиться иногда очень большое число
видов и 2) эти виды могут относиться к экологически разнородным
жизненным формам.
Если в типе фитоценозы многоярусны, то реже они бывают
одноярусными. Так как в последнем случае нередко фитоценоз
состоит лишь из одного растительного вида, то, естественно, этотвид
представлен большим количеством экземпляров, образуя так на-
зываемые «чистые заросли». Причины образования и существова-
ния подобных чистых зарослей могут быть различны, в ряде слу-
чаев они характерны для крайних условий существования:
для местообитаний очень влажных (заросли тростника по берегам
рек, заросли камыша в водоемах), для местообитаний сильно засо-
ленных (заросли солянок) и др. В этих случаях конкуренты от-
сутствуют вследствие неблагоприятных условий. В других
149
случаях чистые заросли — это одна из стадии сингенетического
процесса, когда какой-либо вид временно развивается в большом
числе экземпляров (например заросли пырея на залежах и др.).
Возможны и другие случаи существования чистых зарослей.
В общем нужно различать фитоценозы простые (чистые зарос-
ли) и сложные (из многих видов); с другой стороны, — одноярус-
ные и многоярусные.
Чем больше в фитоценозе ярусов, тем ои является более на-
сыщенным, более совершенным. Различают фитоценозы более
я р у с н о насыщенные и менее яруснонасы-
щенные. Более сложившиеся, более старые фнтоценозы яв-
ляются более ярусно насыщенными.
Растительные ценозы в большинстве случаев имеют сомкнутый
растительный покров, и отдельные экземпляры растений располо-
жены близко один от другого, оказывая на своих соседей большее
или меньшее влияние (рис. 68). Подобные фитоценозы нередко
называют закрытыми. Но иногда, например в полупустынях, пу-
стынях и пр., между отдельными экземплярами находятся свобод-
ные промежутки — это так называемые открытые фитоценозы,
где напервый взгляд отсутствует влияние растений друг на друга.
Однако в последних случаях фитоценоз, открытый в своих над-
земных частях, является закрытым в подземных частях вследствие
взаимного влияния корневых органов, т. е. является по существу
опять-таки закрытым. И можно думать, что вообще открытых фито-
ценозов не может быть, раз одним из признаков фито-
ценоза является влияние растений друг на друга; даже открытые
внешне фитоценозы могут быть закрытыми по существу, причем
большее количество особей на данном участке уже поселиться не
может (рнс. 69). Если же в природе и существуют открытые (н над
150
и под землёй) группировки растений, то это или случайные собра-
ния растений или начальные (пионерные) стадии при формирова-
нии фитоцеиозов. Фитоценоз по своей сути является закрытым.
Обилие. Обил и е — это количе-
ство индивидуумов того или
другого вида; оно зависит от ряда
причин: I) от свойств данного вида
(одни виды обычно встречаются отдель-
ными экземплярами, а другие в боль-
ших количествах), 2)отконкуррентных
отношений с другими видами и от под-
бора этих последних, 3) от экологиче-
ских условий, в которых данный фито-
ценоз находится. Имеются и еще неко-
торые другие причины. Экологические
условия для обилия тех или иных
видов важны потому, что один и тот же
вид в разных условиях развивается то
более, то менее обильно. ч
В иных случаях в фнтоценозах виды
в лице их индивидуумов разбросаны
диффузно, образуя во время цветения
пестрый ковер, в других — в том
или ином ярусе известные виды домини-
руют. Выше были приведены примеры
3
Рис. 69. Схема закрытых фи-
тоценозов различных типов:
1—фитоценоз, закрытый в своих
надземных и подземных частях;
2—закрытый лишь в подъемных
частях, 3—открытый и в надзем-
ных и в подземных частях, не
закрытый по существу (орпг ).
доминирования: чистые заросли, раз-
личные фитоценозы сосновых лесов.
Доминирующие растения называются
доминантами — они всецело господ-
ствуют. Так, в еловом лесу в его дре-
весном ярусе ель может быть доминантом без примеси каких-либо
других видов; в других случаях к доминанту может присоеди-
няться другой вид, играющий важную роль в сложении яруса,
но с меньшим обилием, — это будет субдоминант.
Доминанты могут находиться во всех ярусах фитоценоза: так,
четырехъярусный фитоценоз (ель — орешник — черника — зелё-
ный мох) имеет четыре доминанта, каждый из которых господству-
ет в соответствующем ярусе. Доминанты играют исключительную
роль при составлении названий фитоценозов (см. ниже).
Можно различать доминанты постоянные и временные, первые
остаются таковыми в течение всего вегетационного периода или
даже всего года (вечнозеленые растения), временные же домини-
руют нередко лишь в продолжение небольшого отргезка периода
вегетации, сменяясь другими, тоже временными доминантами.
Примеры: кратковременное цветение тюльпанов в южных степях,
незабудок в северных степях, голубых подснежников (Scilla сег-
пиа) в дубравах, белых ветрениц (Anemone nemorosa) в наших за-
падных лесах и т. п. (рис, 70).
Фитоценологическая роль домина нтов в каком-либо фитоцено-
зе различна и, конечно, доминанты верхнего яруса леса имеют
151
большее значение в экологии фитоценоза, чем доминанты нижних
ярусов. Поэтому доминант, определяющий строй и, в известной
мере, флористический состав, можно называть эдификатором (ла-
Рис. 70. Временный (весенний) доминант—анемон (Anemone nemorosa) в лесах.
Смоленской области (фото Т. Т. Трофимова).
тинское слово кодификатор»—строитель) фитоценоза. Так, эди-
фикатором в буковом лесу будет бук (доминант верхнего яруса),
в еловом лесу — ель, в ковыльных степях — ковыль и т. д.| V
Методические указания к уче° м
обилия. Учет обилия ведется в полев й
обстановке илн субъективно (на глазо-
мер), илн объективными методами. Так
как обилие зависит от числа экзем-
пляров, то при объективных методах
производится подсчет на определенной
площади (например на I м2) числа
экземпляров каждого вида илн, в слу-
чае трудности отграничения отдельных
экземпляров (у корневищных растений),
числа стеблей. При более точных
исследованиях наносят на бумаге поло-
жение каждого экземпляра, например,
на 1 м2 (так называемые квадраты
обил и я). При субъективной оценке
обилия определяют на глаз количествен-
ные отношения экземпляров, пользуясь
различными шкалами обили я.
Наиболее проста и очень употребительна
четырехбальиая шкала обилия Друде с
аки ми условными обозначениями:
Рис. 71 Схема травостоя, квалифи-
цируемого по Друде как soc., но
имеющего на одной и той же площади
очень различное число экземпляров—
смыкание надземных частей не свя-
зано непосредственно с обилием
(ориг-).
152
'эос. (sociales) — растения смыкаются своими надземными частями, *€ря-
Эуют фон (фоновые растения) — «ф»;
сор. (copiosae) — растения встречаются обильно, т. е. в очень большом
или большом числе экземпляров — «об1);
ар. (sparsae) — растения встречаются в небольшом количестве, рассеянно,
изредка— «изр.»;
sol. (solitariae) — растения встречаются редкими экземплярами, редко—«р».
Более правильно, однако, считать приведенную шкалу трёхбальной, так как
ступень soc. не даёт представления о числе экземпляров, а имеет дело о разме-
рами их и с характером надземных частей (рис. 71).
Поскольку рассмотренные признаки (флористический состав,
ярусность, обилие) познаются в результате изучения опреде-
лённых конкретных участков, то они и назы-
ваются «аналитическими», так как при этом производится анализ
растительного покрова1.
Среди подобных аналитических признаков можно указать еще
покрытие и жизненность.
Покрытие. Обилие ещё не даёт представления о фитоценологи-
ческой роли отдельных видов, так как при одном и том же числе
экземпляров на какой-либо площади роль различных видов может
быть весьма различна. Покрытие имеет дело с внешним ви-
дом растения, с характером его надземных органов.
Методические указания. Покрытие учитывают, наблюдая растительный ковер-
сверху вниз и замечая, какую часть почвы в процентах на определенной площадке
занимает тот или иной вид своими надземными частями (какую он дает проек-
цию). Наблюдение производят или на глаз или пользуясь особой сеточкой с
делениями (сеточка Раменского), наставляя ее над травостоем и опре-
деляя вполне точно процент покрытия.
Жизненность. Из приведённых выше примеров конкурентных
отношений между видами можно было видеть, что в сложившихся
фитсценозах лишь небольшое число экземпляров какого-либо вида
проходит полный цикл своего развития и обсеменяется, а боль-
шинство экземпляров остается лишь в вегетативном состоянии;
таким образом, даже в пределах одного фитоценоза «жизненность»
разных экземпляров различна. Ещё более различна жизненность
в разных фитсценозах, в различных экологических условиях, при-
чём в одних фитсценозах какой-либо вид плодоносит обильно, в
других лишь вегетирует, в третьих даже и вегетирует слабо. Чем
лучше в данном фитоценозе развивается тот или иной вид, тем,
очевидно, условия существования для него благоприятнее, поэтому
жизненность даёт указание на приспособленность
видов к соответствующим фитоценозам.
Методические указания. При отметках в полевой обстановке пользуются
тремя степенями жизненности: 3—вид проходит полный цикл развития; 2—вид
в данных условиях лишь вегетирует; 1—вид вегетирует слабо. Эти отметки де-
лают на основании наблюдений над несколькими (многими) экземплярами к
выводят общее заключение.
1 В сущности эти признаки лишь условно могут быть названы анали-
тическими, так как при их изучении анализ всё время сочетается с синтезом.
153
2Q
25 50 75 100
Рис. 72. Аспекты (весенние) на Стрелецкой степи под Курском в вертикальных проекциях (длина 1 лг) Верхняя проекция—
ранняя весна, аспект прострела (Pulsatilla patens), нижняя — аспект горицвета (Adonis vernalis), конец апреля
(п» В- Покровской)
Подвижность фитоцеиозов
Фенологические смены. Сформировавшийся фитоценоз имеет
определенное строение, определенный флористический состав и
другие указанные признаки, но не является чем-то застывшим и
неподвижным, ои динамичен по самой своей природе.
Прежде всего это связано с явлениями фенологического порядка,
т. е. с тем, что I) различные фенологические фазы проходятся в
разные моменты вегетационного периода и 2) этн фенологические
фазы у большинства видов фитоценоза не совпадают.
Рис. 73. Летний "аспект шалфея (Salvia pratensis) на Стрелецкой степи, под
Курском; вертикальная проекция—длина 1 л* (по В. Покровской).
В1 самом деле, одни виды очень быстро проходят весь свой цикл
вегетации, другие, наоборот, очень медленно и, начиная развивать-
ся с самой весны, цветут только в конце лета, одни цветут в самом
начале своей вегетации, другие цветут в самом конце и т, д , т. е.
растения фитоценоза все время сменяют свои фазы развития. Все
это сказывается иа физиономии фитоцеиозов и на их аспектах
Выше было указано, что аспекты бывают постоянные и времен-
ные, именно эти последние и связаны с фенологическими сменами.
Чередование временных аспектов особенно хорошо выражено в
некоторых травянистых ценозах — на лугах и в особенности в
степях.
Пример фенологических смен. В качестве
примера смен аспектов можно привести данные для северной кра-
сочно-разнотравной Стрелецкой степи под Курском. Вот основные
аспекты.
I. Аспект бурый, безжизненный. Степь еще не зазеленела. Предвесенняя
фаза Апрель.
155
2. Аспект бурый, но с лиловыми н светложелтыми пятнами. Фаза цветения
прострела (Pulsatilla patens) — лиловые цветки и осоки (Carex hutnilis) —
светложелтые пыльники. Конец апреля — начало мая (рис. 72).
3. Аспект с золотисто-желтыми цветками адониса (Adonis vernalis) и неж-
ноголубыми гиацинтами (Hyacinthella leucophaea). Фаза цветения адониса и.
гиацинта. Сплошного зеленого ковра еще нет. Первая половина мая (рис. 72.),
4. Аспект — бледножелтый от цветущих баранчиков (Primula otticinalis).
Имеется уже зеленый ковер от начинающих вегетировать многочисленных
растений. Фаза баранчика. Середина мая.
5. Аспект с белыми пятнами анемона (Anemone Silvestris), сочевичняка
(Orobus albus) и с лиловыми куртинами касатика-ириса (Iris aphylla). Фаза ане-
Рис. 74. Позднелетний аспект с дельфиниумом (Delphinium cuneatum)* вер-
тикальная проекция—длина 1 м (по В. Покровской).
мона и сочевичника. Степь уже совсем зеленая. 20-е числа мая.
6. Аспект голубой от массы незабудок (Myosotis silvatica); желтые пятна
крестовника (Senecio campester). Фаза незабудок. Конец мая — начало июня.
7. Аспект лиловый от изобилия цветков лугового шалфея (Salvia pratensis),
яркожелтые пятна козлобородника (Tragopogon orientalis). Фаза шалфея
(рнс.73). Первая половина июня.
156
3. Аспект в основном белый от белых цветков поповника (Leucanthemum
vulgare), горного клевера (Trifolium montanum) и степной таволжанкн (Fili-
pendula hexape tai а). Вторая половина июня.
9. Аспект розовато-желтый от розового эспарцета (Onobrychis arenaria) и
желтого подмаренника (Galium verum). Начало июля.
10. Аспект буроватый от отцветающих - растении, на этом фоне выделяются
крупные синие соцветия дельфиниума (Delphinium cuneatum). Средина июля
(рис. 74).
11. Аспект еще более бурый и тусклый. Местами выделяются темнокрасные
соцветия чемерицы (Veratrum nigrum). Конец июля— начало августа.
12. Аспект совсем бурый; вновь зацветающих растений нет; все растения
стоят почти высохшие, бурые. С августа до снега.
В приведённой смене аспектов указаны лишь основные виды,
и вся эта смена для наглядност! сильно упрощена. Кроме яркого
примера фенологических смен, здесь ещё раз можно убедиться в
том, что виды, составляющие фитоценоз, экологически крайне
разнообразны, так как совершенно ясно, что виды, дающие аспект
ранней весной, и виды, достигающие своего полного развития в
конце лета, требуют для своей жизнедеятельности совершенно
различных условий.
С другой стороны, на данном примере можно видеть, что набор
видов с различными фенологическими циклами даёт возможность
всем этим видам разместиться на небольшой территории, тогда
как если бы они все развивались одновременно, совместное суще-
ствование такого же числа видов было бы фактически
невозможно.
По аналогии с рассмотренными выше явлениями ярусности
(надземной и подземной), приводящими к тому, что на данной пло-
щади размещается большое число видов, здесь можно говорить
тоже о ярусности, но не в пространстве, а во
в р е м е и и.
Смены в связи с метеорологическими условиями. Если феноло-
гические смены наблюдаются и констатируются на протяжении
одного вегетационного периода, то смены, связанные с метеорологи-
ческими условиями, обнаруживаются из сопоставления р .ститель-
ности двух и более вегетационных периодов (различное
количество осадков, различные температурные условия и др.).
В чем же сказываются смены метеорологического характера?
Прежде всего в самом характере травостоя, причем в засушливые
годы травостой бывает низкорослым и значительно более разре-
женным. Некоторые виды, которые имели степень жизненности 3,
т. е. развивались вполне нормально, при сильной засухе могут
иметь степени жизненности 2 или I. Некоторые виды могут в иные
годы совсем выпадать из травостоя. В результате всего этого и
физиономия фитоценозов и самые аснскты претерпевают различные
изменения; так, ярко выраженный аспект одного года на другой
год может быть представлен очень слабо или даже совсем выпасть.
Поэтому нужно различать аспекты постоянные (константные), пов-
торяющиеся из года в год, и непостоянные (неконстантные), в не-
которые годы выпадающие. Естественно, что наиболее типичны
для фитоценозов аспекты постоянные.
157
Сукцессии, их различные типы. Только что рассмотренные
смены свидетельствуют о том, что фитоценоз по самой своей природе
подвижен. Однако все эти смены не приводят к нарушению и из-
менению основных свойств и признаков фитоценоза; последний
остается как таковой в своем более или менее ненарушенном виде.
Но имеется ряд смен другого порядка, которые приводят к к о-
ренной перестройке иногда всех признаков фитоцеио-
зов. Чтобы смены последнего рода более ясно отличать от смен,
выше рассмотренных, целесообразно говорить здесь о сукцес-
сиях1, тем более, что это слово означает «продвижение вперед».
Каждая сукцессия всегда начинается на обнаженном месте и заканчивается
стадией, наиболее устойчивой при данных физико-географических условиях.
Так как обнаженные места бывают первичными н вторичными, то п сукцессии
могут быть первичными и вторичными Примеры сукцессий того и другого рода
бытн приведены выше первичная сукцессия—формирование растительности
на песчаной отмели реки, вторичная — восстановление степной растительности
иа распаханном месте (вторично обнаженное место). Каждая сукцессия имеет
ряд последовательных стадий, число которых_в различных случаях различно
Рис. 75. Смена поясов растительности в мелких водоемах
т—пояс погребенным растении (распадается на несколько роясов), 2—пояс растений с
птавающцмн листьями, 3—пояс камыша, 4—пояс тростника, 5 —пояс осоь (но Сукачеву).
(у вторичных сукцессий число стадий обычно меньше). Сукцессии в лице их
серий сильно различаются в зависимости от характера обнаженного места, на
котором сукцессии имеют свое начало, поэтому отличают песчаные, глинистые,
каменистые, водные и другие серии. Водные серии, например, очень наглядно
представ тены при зарастании речек или водоемов в результате отложения
остатков растений (корневища, стебли н пр.) дно водоема повышается, и одна
раститетьность сменяется другой (рис. 75, более подробно см. ниже).
Наиболее совершенную стадию серин называют «заключитель-
ной», и так как эта заключительная стадия обычно находится в
соответствии с климатическими условиями, то американцы (а за-
1 Сукцессия от латинского слова «сукцессио»—преемннчество, вступление
иа чье-либо место, продвижение вперед. Отсюда видно, что смысл
слова ссукцессия» не позволяет говорить о ('фенологических сукцессиях», повто-
ряющихся каждый год.
158
тем и другие) называют ее «климаксом». Так, в лесной зоне климак-
сом будут являться лесные фитоценозы, в степных —f степные и
т. д.1. Заключительные фитоценозы иногда называют кореннымиг
так как онн представляют коренной растительный покров, не на-
рушенный человеком, развившийся в соответствии с общеклимати-
ческими условиями,
Сукцессии рассмотренного характера обусловлены
главным образом развитием самой расти; егь нести; их вообще
можно Нс звать фитогенными; по Сукачеву — зто сингенетиче-
ские сукцессии. Известны еще н другие фьпогенные сукцессии
(см., например, рис. 76).
Совершенно особо стоят сукцессии, которые так или иначе н а-
рушают или изменяют ход сукцессий, начавшихся
на обнаженном месте. Эти изменения вызываются внешними
причинами, причем можно различать такие сукцессии:
1. Клнматогенные, обусловленные изменением климата (их нужно
отличать от рассмотренных выше смен, зависящих от метеорологических колеба-
ний из года в юд и не влекущих за собой изменений климата в целом).
2. Эдафогенные, обусловленные изменением почвенно-грунтовых
условий, вне зависимости от деятельности человека (например изменение расти-
тельности в результате естественного заболачивания местности, естественной
осушки, наносов рекой и пр ).
3. Зоогенные, вызванные влиянием животных (выпас, влияние рою-
щих животных, влияние тех или иных насекомых-вредителей и пр.).
4. Ан тропогенные, вызванные воздействием человека (искусствен-
ное орошение, осушка, воздействие путем порубок, покосов и т. п.).
По большей части эти сукцессии проявляются в смешанном виде так, на-
пример, антропогенные сукцессии в одних случаях сочетаются с эдафогеннымнл
в других — с фитогенными и др.
Рис. 76. Смена сосны елью (схема)—несколько стадий (ориг.).
Все эти внешние влияния могут действовать или на заключи-
тельные стадии или же на те илн иные промежуточные стадии, изме-
няя таким образсм «нормальный ход» первичных сукцессий. Сукцес-
1 Учение о климаксах в том виде, как оно часто трактуется в Америке и за
границей вообще, носит метафизический характер, так как стадия климакса
представляется как бы завершительной, к которой стремятся все фитоценозы
какой-либо территории. Однако даже при не меняющихся внешних условиях
фитоценоз не остается вполне неподвижным в силу целого ряда изменений,
имеющих место внутри самого фитоценоза. Поэтому, говоря о «заключительной»
стадии нельзя приписывать ей абсолютного значения.
Разработка учения о сукцессиях и климаксе принадлежит американскому
ботанику Клементсу.
159
сии, зависящие от внешних воздействий, Сукачев называет
экзодинамическими сукцессиями, в противоположность эндодинами-
ческим, где внешние влияния играют небольшую роль (в первичных
сукцессиях).
Можно говорить о прогрессивных и регрессивных сукцессиях.
Первые обычно ведут к усложнению фитоценозов (усложняется
структура, увеличивается органическая масса, и др.), вторые при-
водят к упрощению и деградации фитоценоза. При нормальном
ходе сукцессий мы имеем обычно прогрессивный процесс; зоогенные,
.антропогенные и другие влияния обычно ведут к регрессу.
Особо следует ещё отметить сукцессии ценогенетические; здесь
дело идёт о формировании фитоценозов как таковых в
истории земли и растительного покрова. Так, ценогенетическая
сукцессия, например дубового леса, сводится к возникновению в
истории земли этого типа и дальнейшей его эволюции. Подобные
сукцессии Сукачев называет филоценогенетическими, так как в
связи с развитием ценозов шел и ф 1логенез видев.
Изучение сукцессий совершенно ясно показывает, что расти-
тельный покров в целом находится в постоянном движении и что
даже хорошо сложившиеся фнтоценозы не остаются неизменными
во всех своих деталях, хотя и изменяются достаточно медленно.
Одни сукцессии могут протекать довольно быстро и регистри-
роваться путём непосредственных наблюдений из года в год, дру-
гие же требуют слишком длительных сроков и устанавливаются
сопоставлением ряда фактов. Так, сукцессии на болотах устанав-
ливаются путём изучения торфяных слоёв, отложившихся в отда-
лённые времена, климатические сукцессии устанавливаются на
основании сопоставления исторических фактов, и т. д.
Таким образом подвижность фитоценозов зависит от ряда при-
чин. Эти причины следующие: I) смены фенологические, 2) смены
метеорологические, 3) сукцессии как внутреннего, так и внешнего
характера. Кроме того, всё время имеет место сукцессия ценоге-
нетическая.
Если ко всему сказанному добавить, что глубокие климатиче-
ские изменения в геологической истории приводили к «вековым»
сменам, обычно очень сильно или почти нацело перестраивавшим
растительность, то станет совершенно ясным, что движение
лежит в основе фитоценоза.
Были попытки представить фитоценоз как «равновесную си-
стему», прилагая к нему так называемую теорию подвижного равно-
весия. При этом принималось, что фитоценоз в своих изменениях
колеблется около какого-то среднего состояния, выходя из равно-
весия лишь под действием внешних причин. А отсюда
следует, что фитоценозу как таковому движение непрнсуще. Подобное
толкование фитоценоза противоречит диалектическому пониманию
природы.
Устойчивость фитоценозов
В природе наблюдается очень большая устойчивость сложив-
шихся «коренных» фитоценозов, которая состоит в том, что послед-
160
ние, будучи нарушены, более или менее полно и скоро возвращаются
к своему исходному типу. Так, если распахать степь с её сложив-
шимися фитсценозами, то через известное время она восстана-
вливается во всех своих основных признаках (стадии вос-
становления были указаны выше); точно так же, если вырубить в
лесостепной полосе дубовый лес, то он рано или поздно восстана-
вливается, проходя ряд стадий с другими древесными породами
(особенно с осиной). Число подобных примеров может быть очень
велико.
Эта устойчивость как бы не согласуется с той большой подвиж-
ностью. которая лежит в природе фитоценоза. Однако это не так.
В самом деле, как было сказано выше, фитоценоз представляет
подчас очень сложное целое с выработавшимися отношениями вну-
три себя, находящееся в соответствии с окружающими условиями
существования. А раз это так, то подобный комплекс видов должен
обладать известной устойчивостью, во всяком случае большей,
чем какой-либо комплекс более случайного состава.
Обычно устойчивость фитоценозов и их способность восстанавли-
ваться является всё же относительной; так, восстановление какого-
либо фитоценоза идёт в общем до основного типа и хотя,
например, распаханная степь восстанавливается, но точное по-
вторение имевшихся отношений, конечно, невозможно. Для вос-
становления, кроме того, совершенно необходимо лишь частичное,
в смысле площади, нарушение данного фитоценоза, так как если
фитоценоз нарушен на всём его ареале, то отпадает
целый ряд условий, необходимых для всякого восстановления рас-
тительности (уничтожаются виды, затрудняется перенос семян и пр.).
Вот почему нередко исчезают бесследно реликтовые участки тех
или иных древних фитоценозов.
Систематика фитоцеиозов
Ассоциация. Подобно тому как в систематике растений имеется
ряд систематических (таксономических) единиц: вид, рсд, семейство,
порядок, класс и др., так и в фитоценологии для того, чтобы сделать
обозримым всё огромное разнообразие фитоценозов, нужна какая-
то система с определёнными таксономическими единицами. Всё
время употреблявшийся в предыдущем термин «фитоценоз», как было
выяснено, не имеет такс комического значения, а имеет широкое,
общее значение. Основной систематической единицей, по пред-
ставлениям большинства'геоботаников, является в фитоценологии
ассоциация. Ассоциация — это наиболее мелкая, хоро-
шо улавливаемая ф и з и о н о м и ше с к и, единица
растительного покрова. Так, например, на заливном лугу доста-
точно отчётливо вырисовываются различные физиономически одно-
родные участки: пёстрый участок с обилием цветущих двудольных;
более однотонный с господством лисохвоста; яркозелёный с пре-
обладанием осок и т. д. Всё это отдельные ассоциации: разнотравная
ассоциация, лисохвостовая, осоковая и др., однако представленные
на данном лугу лишь своими конкретными участками.
В самом деле, подобные же участки — лисохвостовые, осоковые
„ 16!
И'Географии растении
и др. — можно встретить н на том же лугу и на других лугах в
сходных условиях существования; вообще ассоциация в природе
представлена многими (иногда очень многочисленными) участками,,
причем совокупность последних и даёт ассоциацию. Отсюда видно,
что объектом полевого изучения является прежде всего конкрет-
ный участок ассоциации. Ещё некоторые примеры ассоциа-
ций — в сосновом лесу можно различить такие: сосновый лес
с ярусом из брусники, сосновый лес с черникой, сосновый лес с лишай-
никовым покровом, сосновый лес с господством кислицы и др. Все
эти ассоциации хорошо физиономически различаются.
Ассоциации (вернее, нх участки) в природе отграничиваются
в одних случаях очень отчётливо, иногда даже крайне резко, так
что границы между ними можно проводить очень хорошо, в других
же случаях границы довольно условны. Здесь наблюдается то же,
что и в систематике видов, где различают «хорошие)) и «плохие» виды.
Причина резкости границ участков ассоциаций во многих случаях
зависит от конкурентных отношений доминирующих видов.
Вряд лн целесообразно вместо термина «участок ассоциации»
употреблять «фитоценоз», как это делает Сукачёв, а за ним
ряд ленинградских геоботаников, причем в этом случае ассоциация
слагается из конкретных фитоценозов. Но употребление термина
«фитоценоз» и в общем смысле (как вообще закономерное сочетание
расте[ ин) и в конкретном (как участок гсссциации) мсж<.т приводить
к путанице; кроме того, это противоречит указаниям Брюссельского
конгресса и довольно установившемуся смыслу этого термина у нас
и за границей.
Основные таксономические единицы. Однако, если ассоциация
является наиболее мелкой, хорошо различаемой физио-
номически, таксономической единицей в фитоценологии, то она
распадается на еще более мелкие единицы, так называемые социа-
ц и и, по терминологии шведских ботаников; но последние ф и з ио-
ном и чески, не улавливаются или улавливаются с
трудом. Так, например, ассоциация соснового леса с черникой
может распадаться на социации в связи с различиями в моховом
покрове: в одних случаях моховой покров состоит из мха Pleurozium
Schrebtri, в других —13 мха Hylccomium prolifcrum. В обоих
случаях общее строение остаётся одинаковым, меняется лишь
нижний, физиономически не играющий роли, ярус:
сосна — черника — мох (Pleurozium);
сосна — черника — мох (Hylocomium).
Точно так же в ассоциации с господством лисохвоста можно
выделить социации в результате замещения одних второстепенных
видов другими. То же щноси.ся и к другим асе циаьиям. Соцнация-
ми можно пользоваться при более детальных исследованиях.
Если ассоциации.распадаются на более мелкие единицы — со-
циации. то, с другой стороны, ассоциации соединяются в более круп-
ные таксономические единицы — в «группы ассоциаций»; последние
соединяются в формации, группы формаций, классы формаций и,
наконец, в типы растительности. Тип растительности — это наиболее
крупное подразделение растительного покрова земного шара.
162
Классификация Брокмаи-Ероша и Рюбеля. В последнее время
чаще всего различают четы> е основных типа растительности: Lignosa
(от латинского слова *lignosus» —деревянистый) — древесно-кус-
тарниковый тип: Herbosa (от латинского слева «herbosus» — травя-
нистый)— травянистый, Deserta (от латинского слова «deserta» —
пустыня) — пустынный и Errantia (от латинсксго слова «errans» —
блуждающий) —тип, состоящий из организмов, не прикрепленных
к субстрату, взвешенных в водной, почвенной или воздушной среде.
Первые три типа объединяют фитоценозы с растениями, прикреп-
лёнными к субстрату, причем Herbosa включает в себя травянистые
фитоценсзы с более или менее сплошным травостоем, a Deserta —
фитоценозы с сильно разреженным травостоем, не смыкающимся
над землей.
Эти основные типы (I, И, III, IV) дальше распадаются (по Брок-
ман-Ерошу и Рюбелю) на классы формаций (I, 2, 3...) и группы
формаций (а, Ь, с...):
I. Lignosa — древесно-кустарниковый тип.
1. Pluvnlignosa («плюьиум» 1—дожди)—влажные (дождевые) тропиче-
ские древесные и кустарниковые фитоценозы.
a) Pluviisilvae («плювиум» — дождь, «сильва» — лес)— влажнотропические
леса; 4
б) Pluviifruticeta («фрутицес» — кустарники) — влажнотропические ку-
старниковые фитоценозы.
2. Laurilignosa («лаурус» — лавр)—древесные и кустарниковые фитоце-
нозы лаврового типа.
a) Laurisilvae («сильва» — лес) — леса лаврового типа;
b) Laurifiuticeta («фрутнцес» — кустарник) — кустарниковые фнтоценозы
лаврового типа.
3. Duriligr.osa («дурус» — жесткий) — древесные и кустарниковые жестко-
листные (вечнозеленые) фиюценозы.
a) Duiisilvae— леса жестколистного типа;
b) Durifruticeta — кустарниковые фнтоценозы жестколистного типа.
4. Ericiligncsa («эрика» — вереск) — деревянистые фнтоценозы с листьями
верескового типа (мелкие, вечнозеленые).
a) Ericifruticeta — кустарниковые фитоценозы с листьями верескового типа
(леса подобного типа в природе отсутствуют).
5. Aestiligncsa («эстас» — лето) — древесные и кустарниковые фитоценозы
с листвой в летнее время года.
a) Aestisilvae — «летние» леса, т. е. с листвой в летнее время года;
b) Aestifruticeta — кустарниковые фитоценсзы с листвой в летнее время года.
6. Hiemihgncsa («гпемс» — ы л а) — древесные и кустарниковые фнтоценозы
с листвой в зимнее дождливое время года.
а) Н ietnisilvae — леса с листвой в зимнее дождливое время года (летом —
засуха);
Ъ) Hiemifruticcta — кустарниковые фнтоценозы с листвой в зимнее дождли-
вое время года. *
7, AcicuJilignosa («ацикулум» — игла,хвоя), — древесные и кустарниковые
фнтоценозы с листьями в форме игл илн чешуй.
a) Aciculisilvae — леса хвойные;
b) Acicul।fruticeta — кустарниковые хвойные фитоценозы.
II. Herbosa — травянистый тип («гербсзус» — травянистый).
8. Ternherbosa («терра» — земля, суша) — наземные травянистые фнтв-
ценозы.
a) Duriherbosa («дурус»— жесткий) — травянистые фнтоценозы с жёсткими
растениями, например степи;
1 Все названия ет латинских еле*.
16S
b) Sempervirentiherbosa («семпер» — всегда, «виревс» —зеленеющий) — веч-
нозелёные травянистые фитоценозы.
с) Altherbosa («альтус» — высокий) — травянистые фитоценозы с высокими
травами — субальпийские луга, заливные луга.
9. Aquiherbosa («аква»—вода) — травянистые фитоценозы, растущие
воде или на сильно влажных местах.
a) Emersiherbosa («эмерсус» — выдающийся из воды)— фитоценозы, выда-
ющиеся из воды; прибрежная растительность болота;
b) Submersiherbosa («субмерсус» — погружённый)— фитоценозы, погру-
жённые в воду;
с) Sphagniherbosa (Sphagnum— особый род болотного мха, типичный для
верховых болот) — мохово-сфагновые болота верхового типа.
III. Deserta («дезерта» — пустыня)— пустынный тип.
a) Siccideserta («сиккус»— сухой)— пустыни, вызванные сухостью кли-
мата — сухие пустыни;
b) Fr i go г i deserta («фригус»— холод)— пустыни, вызванные холодным кли-
матом — холодные пустыни;
с) Litorideserta («литус»— берег)— прибрежные засолённые пустыни по
берегам морей;
d) Mobilideserta («мобилис» — подвижный) — «подвижные» пустыни на пес-
чаных, подвижных от ветра, ме.стах;
е) Rupidesertt («рупес» — камень) — каменистые пустыни;
f) Saxideserta («саксум» — скала) — сюда относится растительность скал,
а также стволов деревьев; этот класс представле водорослями, лишаиниками
и мхами, поселяющимися на голом камне или на коре деревьев и не требующими
для своего поселения продуктов разрушения скал или почвы.
Класс «каменистые пусты ин» представлен группировками высших растении,
укореняющихся и требующих продуктов разрушения.
IV. Errantia («эрранс» — блуждающий) — тип, состоящий из группировок
низших организмов, ие прикрепленных < субстрату, «блуждающих».
a) Aquerrantia («аква»»— вода)— группировки организмов, живущих в
толпы воды, та с называемый «фитопланктон»;
b) Soierrantia («соль»— почва> — группировки организмов, находящихся
в толще почвы, образующие фнтоэдафои («эдафон» — по-гречески почва),
с) Aererranti («аэр> — воздух) — группировки, находящиеся в воздушно!
толще, так называемый фитоаэрон.
В данной классификации г, если рассматривать её снизу вверх,
более простые сочетания растений, по мере движения вверх, все
более и белее усложняются; так, межно видеть такие ступени:
1. Низшие организмы, не прикрепляющиеся к субстрату, на-
ходящиеся в очень слабой связи друг с другом (Errantia).
2. Тоже низшие организмы, но некоторые уже более организо-
ванные (лишайники, мхн), более тесно связанные между собой, но
не имеющие корневых систем и корневой конкуренции. Одноярусные
группировки (Saxideserta).
3. Г1о большей части уже расчленённые на ярусы группировки
и фитоценозы нз высших растений с корневыми системами и корне-
вой конкуренцией. Однако растительный пскров обычно разрежен,
не сомкнут (Deserta, за вычетом Saxideserta).
4. Многоярусные и одноярусные фитоценозы с сомкнутым тра-
вянистым покровом, связанные с влажными почвами, бедные фло-
ристически (Aquiherbosa).
1 В последней редакции (1933 г.) Рюбель увеличил число классов фор-
маций до 28, возведя все группы фор.маций данной классификации в ранг клас-
сов формаций.
164
5. Многоярусные, обычно богатые флористически, фитоценозы,
связанные с сухими почвами (Terriherbosa).
6. Иногда очень сложные фитоценозы с добавлением к травяни-
стым ярусам ещё кустарниковых и древесных (Lignosa). Эта ступень
может быть ещё разделена на ряд усложняющихся ступеней.
Вся классификация построена на ф и з и о н о м и ч е с к и-
эколог и ческой основе.
Пример классификации для'лесов. В приведенной классифи-
кации даны три высшие таксономические единицы: типы, к л а с с ы
и группы формаций; более низкие единицы отсутствуют.
Если же взять (для примера) древесно-кустарниковый тип, и при-
вести и высшие и низшие таксономические единицы, то получится
такой нисходящий ряд:
1. Т и п растительности. Lignosa —древесно-кустарниковый тип.
2. Класс форм ций. Aciculilignosa — древесные и кустарниковье
фитоцеиозы с листьям! в виде хвои или игл (все хвойные леса + все хвойные
кустарниковые фитоценозы).
3. Группа фо? наций. Aciculisiiva е—все хвойные леса (сосновые Н-
е.ювые + пихтовые и т. д.).
4. Формация. Pineta silvestris — все сосновы леса из обыкновенной
сосны; другой формацией будет Piceeta excelsae— еловьв леса из европейской
ели и др.
5, Группа ассоциаций. Сосновый лес распадается hi ряд групп
ассоциаций, из которых одна характерна наземным покровом m зеленого мха
(Hylocomium) — группа hylocomiosa (Pineta silvestris hylocomiosa— бор-зеле-
номошник); другая — наземным покровом из оленьего лишайника (Cladonia) —
группа ciadinosa (Pineta silvestris cladinosa — бор-беломошник); третья —
присутствием кустарникового подлеска и ряда растений дубравного типа —
группа nemorosa (Pineta silvestris nemorosa) и т. д.
6. Ассоциация. Бор-зеленомошник распадается на целый ряд ассо-
циаций: бор-черничник—Pinetum myrtillosum (черника—Vaccinium myrtillus);
iop-брусничиик с ярусом из брусники; бор-кис л и чин к с кислицей (Oxalis асе-
tosella) и др. Все эти ассоциации связываются общностью наземного покрова
из зелёного мха.
Ассоциация — наиболее обычный объект полевого исследования раститель-
ного покрова.
7. С о ц и а ц и я. Ассоциация бора-черннчника распадается в зависимости
вт того, какой зелёный мох находится в наземном покрове, на социации: сосна—
черника — Hylocomium proliferum (одна социация); сосна— черника— Pleu-
rozium Schreberi — другая социация.
Отдельные участки социации об . чио не занимают в природе таких больших
площадей, как участки ассоциаций, ио дают нередко небольшие пятна.
Методические указания. Объектом полевых исследований при
изучении растительного покрова являются конкретные участки социаций или
ассоциаций. Более крупные единицы получаются в результате объединения
близких ассоциаций, групп ассоциаций и пр. Могут исследоваться естест-
венные участки социаций или ассоциаций, но при более точных исследо-
ваниях выделяют в пределах естественных участков искусственные пробные
площадки определенных размеров в 1 м2, в 100 м2 или в лесах до 1 га
(метод пробных участков). На этих площадках учитывают основные аналитиче-
ские признаки: флористический состав, ярусность, обилие, жизненность и др.
В начале же анализа делается физиономическое описание. Описание раститель-
ности дается на фоне основных условий существования (рельеф, геологическое
строение, почва и др.). В результате каждого описания соответствующей социа-
ция или ассоциации дается название.
Составление названий социаций и ассоциаций. Основные пра-
вила (приёмы) составления названий таковы. Для социации не-
105
обходимым является выделение ярусов и доминант каждого яруса и
расположение в одну строчку этих доминант, начиная с верхнего
яруса, например:
Pinus Silvestris — Vaccinium myrtillus — Pleurozium Schreberi
(сосна) (черника) (мох)
При этом доминанты различных ярусов соединяются знаком
тире. Если же в одном и том же ярусе имеется несколько доминант,
то они соединяются знаком плюс.
При составлении названий ассоциаций можно пользоваться теми
же приемами, например:
Pinus — Ledum palustre — Sphagnum (различные виды).
Однако для ассоциаций часто применяют другой способ, а
именно, берут два (или три) вида, наиболее важных (по доминиро-
ванию и обилию) в данной ассоциации, и к главному из них при-
бавляют окончание — е t u т, а ко второму по значению — окон-
чание о s u m (во всех случаях указанные окончания присоеди-
няются к корню слова).
Например, сосновый лес (сосна — Pinus) с господством брус-
ники (Vaccinium) будет иметь название Pinetum vacciniosum. Ду-
бовый лес (дуб — Quercus) с подлеском из липы (липа — Tilia)
будет иметь название Quercetum tiliosum и т. д. В более сложных
случаях можно иметь такое название; Piceeto-Pinetum vaccinioso-
oxalidosum.T. е. елово-сосновый лес с господством брусники и ки-
слицы.
Некоторые авторы пользуются и ещё другими способами.
Фитоценозы и история страны
Уже не раз указывалось, что фитоценозы тесно связаны с соответ-
ствующими условиями существования; одн ко понять фитоценоз,
исходя только из этих последних, не представляется возможным.
В самом деле, можно представить себе на земной поверхности ряд
участков, находящихся в крайне близких условиях существования,
но флора п растительность которых будет очень сильно разли-
чаться (например, наши и североамериканские степи, наши и амери-
канские широколиственные леса и т. д.). Лишь физиономия
этих соответствующих фитоценозов будет очень сходной в силу поч-
ти тождественности экологических условий. Подобные фитоценозы
получили название изоценозов.
Причиной, почему при сходных условиях имеются различные
наборы видов, является несовпадение их ареалов. Последние же в
очень многих случаях связаны с историей тех или иных участков
суши. Так, если две территории издавна были разъединены друг от
друга, то, естественно, их флора должна самым коренным образом
различаться. Мы приходим, таким образом, к установлению того
факта, что без знания и с т о р и и территории понять фито-
ценоз в его флористическом сложении невозможно.
При анализе фитоценозов нередко можно видеть, что одни фло-
ристические элементы являются более новыми, другие же более
древними. Это происходит от того, что фитоценоз складывается в
166
своей ценогении постепенно, ас другой стороны, здесь не-
редко мы имеем наследие далекого прошлого. Так, если
какая-либо территория издавна существует как суша, то в совре-
менной растительности могут находиться такие элементы, которые
сохранились от прежних В[ смен, нередко с иными кли-
матическими условиями. Другими словами, можно предположить,
что некоторые современные фито ценозы представляют ряд наслоений,
ряд пережитков прошлых отношений. Эти пережитки обычно мало
гармонируют с современными экологическими условиями и ведут
к той или иной дисгармонии.
Таким образом, ясно, что одной современной среды для пони-
мания фитоценозов недостаточно. Если же сопоставить все приве-
денные выше соображения, то в заключение можно сказать, что под
фитоценозом нужно понимать вполне закономерные (не случайные)
сочетания видов, сложившиеся: 1) в результате взаимоотношений
между видами, 2) приспособлений к условиям данной среды и
3) связанные с историей данной территории и её флоры; как след-
ствие—они являются достаточно устойчивым и.
Если так понимать фитоценоз, то можно ли считать фитоценоза-
ми посевы культурных растений, сочетания сорных растений и др.?
Очевидно нет, так как подобные сочетания прежде всего неустойчи-
вы (например посевы, предоставленные самим себе, сейчас же сме-
няются другими группировками), случайны, обнаруживая все при-
знаки примитивных растительных сочетаний. Чтобы отличать эти
примитивные сочетания от сложившихся фитоценозов, их целесо-
образно называть просто «группировками» (сюда же относятся
первые стадии сингенеза—см. выше). Группировки могут быть
естественными и искусственными. Таким обра-
зом, сочетания растений подразделяются на группировки
п ф и т о ц е н о з ы.
Фитоценология, как учение о сочетаниях растений, изучает как
сложившиеся фитоценозы, так и группировки, так как послед-
ние обычно являются первыми стадиями при формировании фито-
ценозов и постепенно в них переходят.
Некоторые авторы все сочетания растений, если имеются
между последними те или иные взаимовлияния, называют фитоце-
нозами (включая сюда посевы и др.); однако в этом случае объеди-
няются сочетания, качественно очень различные.
ОТДЕЛ IV
ОСНОВНЫЕ ТИПЫ РАСТИТЕЛЬНОГО ПОКРОВА
Схема идеального континента
В основном растительность земной поверхности зависит от обще-
климатических условий и соответственно с этим обнаруживает те
или иные изменения. Там, где имеются большие пространства суши,
можно наблюдать постепенное изменение растительюсти с севера
на юг, причем отчетливо вырисовываются растительные зоны, про-
тягивающиеся в основном с запада на восток и сменяющиеся с севера
на юг. Это — полосы растительности широкого фитоценологическогл
значения (порядка класса формаций), зависящие главным
образом от климата. Эти зоны-полосы определяют ландшафты
соответствующих территорий.
Особенно хорошо зоны выражены в СССР на его обширных
равнинных пространствах, причем с севера ща юг здесь имеются
та и. основные зоны: тундра — лес — степь — пустыня-, каждая из
них в свою очередь распадается на подзоны и связывается пере-
ходным полосами, как лесотунора, лесостепь, полупустыня.
При меньших пространствах суши и изрезанности её морями,
заливами и пр. указанная зональность сильно нарушается, а в иных
случаях, в силу особых климатических условий, зоны занимают
не широтное протяжение, а меридиональное, как, например, неко-
торые зоны в Северной Америке.
Если определённые закономерности ярко бросаются в глаза на
кар ах растительности земного шара, то ещё лучшее выражение
они получили бы в том случае, если бы земная поверхность пред-
ставляла сплошную сушу. Брокман-Ерош даёт схему
распределения растительности на подобном «идеальном» континенте,
который представляет сплошную сушу, граничащую на западе по
прямой линии с морем (рис. 77). При этом в распределении расти-
тельности отразились следующие условия: 1) уменьшение коли-
чества тепла от экватора к полюсу; 2) постоянные западные влаж-
ные ветры (дующие с моря на материк) в умеренных широтах; 3) су-
хие ветры (пассаты), дующие с материка на море, по обе стороны
экватора; 4) уменьшение количества осадков от берега моря в
глубь материка.
Свою схему Б р о к м а н-Е р о ш строит главным образом для
северного полушария, так как южное, занятое преимущественна
161
водными пространствами, менее показательно в этом отношении,
но в общем подчиняется тем же закономерностям.
Эти основные закономерности видны из прилагаемой схемы.
Здесь прежде всего имеет место чередование лесных и безлесных
SlCClOCSEftrt
> у Q о Y е о v о
Рис. 77. Схема распределения основных классов и
групп формаций растительности на идеальном кон-
тиненте (по Брокмаи-Ерошу).
широтных полос: 1) на экваторе и по сторонам его—тропические и
субтропические леса' 2) далее к северу - полоса пустынь и степей.
3) опять лесная полоса (очень широкая), 4) безлесная северная
полоса —тундра. В связи с этим в северном полушарии имеются
три границы леса, отделяющие лесные полосы от безлес-
ных.
В основном на данной схеме имеется 10 классов н групп форма-
ций (они имеют ландшафтное значение), распределяющихся так:
1- Тропическая лесная полоса распадается на
три класса формаций: Pluviilignosa, Laurilignosa, Hiemilignosa.
При этом первый —влажнотропические леса, которые развиваются
•собенно в западных частях, выклиниваясь к востоку, так как
влаги в этом направлении становится всё меньше. Эти леса окай-
16 s
«мляют лавровые леса с кожистыми лакированными вечнозелёными
листьями, требующие меньшего количества тепла и влаги. В ещё
менее влажном климате и при летнем засушливом периоде разви-
ваются зимнезелёные леса; как видно на схеме, они идут в глубь
континента. Зимнезелёные леса включают в себя саванновые
леса и саванны. Последние представляют переход к следующей
(к северу) полосе.
2. Безлесная полоса пустынь и степей. Она
с 'Стоит из сухих пустынь (Siccideserta), от самого берега моря иду-
щих в глубь материка, сменяющихся на севере в более континен-
тальных частях степями (Duriherbosa). То обстоятельство, что сухие
пустынй доходят до самого моря, обьясняется постоянно дующими
здесь пассатами (влажные западные ветры, дующие с моря, здесь
отсутствуют). В схеме Б р о к м а н-Е р о ш а сухие пустыни в
большинстве случаев прямо переходят в леса; при большей
детализации пустыни должны, невидимому, везде смыкаться с ле-
сами через полосы Duriherbosa.
3. Умеренная лесная полоса. Эта полоса содер-
жит ряд классов формаци I: Aesti lignosa (летнезелёные леса), Acicu-
11 lignosa (хвойные леса), Durilignosa (жестколистные леса) и Lauri-
lignosa (лавровые леса). Большие пространства заняты летне-
зелёными п хвойными лесами. Первые развиваются в достаточно
мягком приморском климате, но с уже довольно холодным зимним
периодом, поэтому они на зиму сбрасывают листья; вторые распо-
лагаются далее к востоку в более континентальных условиях, при-
чём, несмотря на то, что не сбрасывают на зиму хвою, испаряют
меньше, чем летнезелёные. Переходным типом являются жестко-
листные леса, расположенные между лесами летнезелёными и сухи-
ми пустынями и развивающиеся при достаточном количестве тепла,
но при недостатке влаги. Они вечнозелёны, но имеют обычно узкие
листья, часто стоящие ребром; это — леса Средиземноморья. Что
касается лавровых лесов, то они занимают в данной полосе умерен-
ных лесов лишь очень небольшую территорию. Они находятся в том
месте, где пассаты начинают сменяться западными морскими ветрами
и где, следовательно, при ещё достаточном количестве тепла влаги
больше. Можно предполагать, что при отсутствии пассатов лавровые
леса с\ бтропкческие и умеренные смыкались бы.
4. Безлесная северная полоса. Она находится
к северу от полярной границы леса и содержит две группы форма-
ций; Sempervirentherbcsa и Frigcrideserta; первая занимает узкую
полосу, приуроченную к приморскому климату, и состоит из низко-
рослых растений, переносящих в зелёном состоянии неблагоприят-
ное время года под зимним снежным покровом, а вторая — холод-
ные пустыни, лишённые деревьев.
ВзаимсотношенЙя растительных типов в приморском климате
более сложны, чем в континентальном, вдали от моря. В последнем
случае ясно выражены те зоны, которые были указаны выше: тунд-
ры— леса —степи — пустыни; под экватором имеется ещё одна
юна — Hiemilignosa, т. е. зона саванновых лесов и саванн.
ПО
Сравнивая распределение растительных типов на идеальном
континенте с тем распределением, которое существует на земной
поверхности, можно видеть, что наибольшее сходство наблюдается
в приатлантических частях Старого Света.
В конце книги даны две карты растительности земного шара:
одна составлена Б р о к м а н-Е р о ш е м, вторая — дана по
ЕСАМ. На карте Б р о к м а н-Е р о ш а имеется всего 10 под-
разделений, совпадающих с теми, которые даны для идеального
континента. Карта БСАМ значительно сложнее, число подразделе-
ний здесь 24; некоторые обозначения на обеих картах одинаковы
(летнезелёные леса, степи, пустыни), другие — сильно различаются.
Так, на карте БСАМ отсутствуют такие обозначения, как зимне-
зелёные и лавровые леса, вместо них даны тропические и субтропи-
ческие леса различных типов, саванны и ду.; имеются и многие
Другие различия; к оме того, границы даже одноимённых классов
формаций не всегда совпадают. Мы намеренно приводим эти две
карты, во-первых, для того, чтобы показать,что взгляды различных
авторов в вопросе подразделения растительности неодинаковы,
и во-вторых, думаем, что сопоставление этих различных карт по-
служит для лучшего уяснения материала.
В дальнейшем мы остановимся на распределении и характеристи-
ках осн . ных классов формации. Они дают совершенно особые
ландшафты и физиономически резко отличаются друг от
друга. Если на флористических подразделениях и картах основное
внимание уделяется флористическим элементами и их генезису,
то подразделения карт фитоценологических прежде всего физио-
ном и чны.
Помимо классов и групп формаций, имеющихся на схеме иде-
ального континента БрокманЕроша, будут указаны неко-
торые другие подразделения, которые отсутствуют на данной схеме
(из-за масшта а), но которые важны в том или ином отношении.
Все эти подразделения физиономически очень различны и дают
особые, хорошо различающиеся, ландшафты. В ряде случаев мы
отступаем от классификации Б р о к м а н-Е р о ш а.
I. КЛАСС ФОРМАЦИЙ — ВЛАЖНОТРОПИЧЕСКИЕ ДРЕВЕСНЫЕ И КУС-
ТАРНИКОВЫЕ ФИТОЦЕНОЗЫ (PLUVII1 IGNOSA)
1. Группа формаций— влажнотропические леса (Pluviisilvae)
Общая характеристика. Влажнотропические или «дождевые»
леса — это тропический тип древесной растительности, связанный с
наиболее благоприятными для растительности условиями суще-
ствования на земной поверхности: избыток тепла и влаги, коти, ые
к то'гу ж распределены более или менее равномерно на протяже-
нии всего года. Дождевые леса вечнозелёны, с крупными, по большей
части блестящими листьями, с почками, не защищёнными почечны-
ми чешуями, с массой эпифитов, эпифиллов и лиан (рис. 78).
Влажнотропические леса занимают большие пространства по обеим сторо-
нам экватора и не переходят к северу н к югу за пределы тропиков: они встре-
чаются в Африке (область р. Конго, область Великих озер, восточный берег
171
Мадагаскара и ряд островов)' в Америке (долина Амазонки, Гвиана, восточный
верег Центральной Америки, большая часть Малых Антильских островов),
в Австралии (Новая Гвинея, Тихоокеанские острова, небольшие участки в
самой Австралии), в Азии (Филиппинские, Молуккские, Зондские острова,
кроме восточной части Явы, южная часть полуострова Малакки).
Климатические условия всех этих областей более или менее
•днообразны, и количество осадков, почти как правило, более 200
см, доходя до 400 см и более (в горах Гавайских островов — до
17?
1200 см). В местах с более или менее равномерным распределением
осадков развиваются вечнозелёные леса, там же, где дождливый
период сменяется сухим, развиваются так называемые муссоновые
леса, или зимнезелёные (см. стр. 1S8).
Влажность воздуха очень велика, достигает 90%.
Температура круглый год колеблется в пределах 25—30е, и во-
обще колебания между температурой наиболее тёплого и наиболее
холодного месяца составляют 1—6°, самый же холодный месяц
имеет среднюю температуру выше 18°. Вот несколько примеров
климатических элементов для некоторых пунктов:
Батавия (на о- Яве) . . .
Борнео ................
Устье Нигера...........
Ямайка.................
78
72
84
74
26 0 26,5 25,4 180 35
26.9 27,7 26 1 323 49
25 5 26,6 24,4 366 63
19,6 21,2 18,3 282 51
3
10
24
8
Максимальные температуры редко превышают 35—36°.
Интенсивность света, в противоположность прежним предста-
влениям, в тропических странах совсем не высока и нередко ниже,
чем в более высоких широтах. Причина этсго — особое состояние
атмосферы и прежде всего ее богатство водяными парами.
Ф и з и о н о м и ч е с к и тропические леса сильно отличаются
от лесов умеренных климатов (рис. 79). Прежде всего они очень
богаты видами, и с трудом можно найти два рядом стоящих дерева,
относящихся к одному и тому же виду: ио бога с.ву ] идов они на-
поминают северные разнотравные степи (см. стр. 260). Известно
указание В а р м и н г а, что он в Бразилии на площади в 3 кв.
мили отметил около 400 видов деревьев (однако иногда тропические
леса состоят из немвспх ендов деревьев и даже из одного).
Фитоценологическая структура. Разнообразие древесных пород
o6ycA0BJ ивает особую структуру тропических лесов:
верхняя граница древесных крон ье представляет прямой горизон-
тальной линии, а неправильную, зубчатую; это зависит ст
того, что высота деревьев различных видов различна. При взгляде
на лес сверху (например с горы) опять-таки резко бросается в глаза
очень неровная поверхность лесных крон, а также самая окраска
листвы леса. Она никогда не бывает однотонной, как, например,
в наших дубовых и других лесах, но представляет пеструю
мозаику различных тонов зелёного, бурого, жёлтого, оливко-
173
вого и других оттенков; кроме того, на этом мозаичном фоне там и
сям вкраплены яркие пяша то красного, то белого цье1а —это
цветут те или иные деревья.
Фитоцено логическая структура ipc пьчсыи х
лесов достаточно разнообразна: в г нь х а учиг х сьи со-
в
з
।1 мы без того [ а, " ак как по д; е (к i j i а ны < бра-
hv стволов, ветвей и j i стьы , в дру rv х с у ч<. ях
174
в лесу господствует глуоокая тень и под деревьями почти нет под-
леска и травянистого покрова.
Деревья в лесу располагаются в несколько ярусов
(4—5), тогда как в наших лесах деревья образуют 1—2 яруса.
В результате —сложность фитоценологического строения.
Что касается самих деревьев, то они поражают строй-
ностью и высотой своих С1ВСЛОВ, почти как правило, очень слабо
ветвящихся, причем ветвление не идет датьше ветвей 3—4-го
порядка (в нашг х лесах до 5—8 порядков) Mhoi i е деревья совсем
не ветвятся, например пальмы, древовидные папоротники, некоторые
Рис. 80 Досчовидные корни у Ficus variegata (в центре)
и у F. cordifoha (счева)—разтичные типы (по Фаберу).
двудольные. Изучить систематический состав деревьев крайне труд^
но, так как различать цветки и лгстья в высоко расположенной
кроне, спутанной лианами* и эгифитами, совершенно невозможно;
приходится сснсБЬваться на угьЕпл х иногда пледах, поваленных
деревьях и т п Hai более же надежный способ—это определение
по коре и анатсА ичееккм признакам ствола
Для дерегьев характерен высокий стр о йн ый ств'о л—
они как бы стремятся обогнать в росте другие и вынести свои кроны
кверху Вообще устремление ввер х —одна из характер-
ных черт тропического леса Однако высокие стволы нуждаются в-
укреплении своего основания, для ^его служ т развивающиеся у
некоторых видов в нижних частях ствола так называемые доско-
лидные корни, которые в виде крыльев или контрфорсов отходят
от ствола (рис 80) Впрочем, роль досчатых корней еще недостаточ-
но выяснена В качестве примеров деревьев с особенно характер-
ными подобными корнями следует указать на некоторые виды фи-
кусов, Parkia africana и многие другие.
17»
Листья на деревьях большие, вечнозелёные, обычно
жесткие, кожистые, нередко имеют консистенцию жести и в избытке
содержат кремнезём (в тропической почве кремнезёмная кислота
находится в растворённом виде). Обычно листья блестят, как бы
покрыты лаком, и очень редко покрыты волосками. Все эти призна-
ки указывают на ксе ром о р фн о е строение листьев, что
кажется на первый взгляд странным для растений влажного тропи-
ческого климата. Однако в жаркое время дня листья очень сильно
гранспирируют, и наступает несоответствие между поступлением и
испарением (ток воды должен подняться по очень высокому стволу^.
Сильной транспирации способствует также и большая сила ветра на
высоте древесных крон. Блестящая поверхность листьев отражает
лучи слишком интенсивного света, а с другой стороны, она
служит приспособлением для быстрого скатывания q листа дожде-
вой воды.
Кора тропических деревьев слабо развита, гладкая, иногда
блестящая, в иных случаях имеет даже зелёный цвет вследствие
просвечивания хлорофилла.
Что касается систематического состава деревьев, то в него входят
многочисленные представители двудольных тропических семейств,
также встречаются пальмы, древовидные папоротники, бамбуки.
Однако пальмы не играют той роли, как это иногда представляют.
Располагаясь в несколько ярусов, деревья имеют различную высоту
и в своих нижних ярусах смыкаются с ярусом кустарников.
Кустарники тропического леса крайне разнообразны и
принадлежат главным образом к двудольным. Подобно деревьям,
«ни слабо ветвя ся и этим отличаются от кустарников наших лесов.
Листья их по большей части крупны, нежны, редко — кожисты.
В противоположность л. стьям высоких деревьев они не имеют
приспособлений ксерофи. ьного характера, так как транспирация
в глубине леса очень невелика.
Травянистый покров довольно разнообразен, но в
разных условиях развит различно; так, в наиболее тёмных участ-
ках леса он почти совершенно отсутствует, на опушках же и на
более освещенных местах достигает оольшого великолепия (рис. 81).
Иногда в последних случаях травы поднимаются выше роста чело-
века. Особенно нужно отметить виды семейства имонрных (Zingi-
beraceae), выделяющиеся иногда своими оригинальными и краси-
выми соцветиями.
Но обращает внимание на себя то, что травянистый покров
менее разнообразен сравнительно с древесным, и что нередко один
какой-либо вид, без примеси других, покрывает большие участки,
чего обычно в древесных ярусах не бывает. Получается картина,
в б р а т и а я тому, что наблюдаемся в наших лесах. И в травя-
нистом покрове травы обычно слабо разветвлены, имеют удлинён-
ные осн, розетки отсутствуют.
Очень важную роль в травянистом покрове играют споровые
растения: се лаги не л ли, плауны и особенно папо-
ротники, причём последние представлены многочисленными
видами. Нельзя не указать на папоротники семейства Нушепо-
176
phyllaceae, у которых листья крайне нежный, кроме жилок, неред-
ко состоят нз одного слоя клеток. Это указывает на необычайную
Нис. 81. Травянистый покров в тропическом лесу в Мексике. Хорошо выде-
ляются крупные листья Anthurium и стебли Selaginella с мелкими листьями
(похожи на ветви елок) (по Карстену).
влажность воздуха среди напочвенной растительности тропических
лесов, прн которой только и возможно существование таких нежных
листьев: они могут впитывать воду всей своей поверхностью. Как у
12 География растеши' 177
крайних гигрофитов, корни здесь развиты чрезвычайно слабо и
почти' совсем отсутствуют. Злаки обычно представлены очень
слабо.
Если вспомнить, что деревья верхних ярусов имеют нередко как
бы жестяные, жесткие листья ксероморфной структуры, а с дру-
гой стороны, листья папоротников в глубине леса имеют пластинки
из одного слоя клеток, то совершенно очевидна та огромная
амплитуда в условиях существования, которая iimUt место
в различных ярусах тропических лесов. Представление о том, что
все растения влажных тропических лесов являются ярко выражен-
ными гигрофитами, неправильно: фитоклимат здесь крайне разно-
образен.
Однако указанные черты тропического леса еще не дают о нем
полного представления, мы пока говорили лишь об «ярусной» рас-
тительности, но именно во влажном тропическом лесу необычайно
171
пышное развитие получает так называемая «внеярусная» раститель-
ность. Эта растительность представлена лианами и эпифитами.
Обилие лиан и эпифитов — одна из самых характерных черт влажно-
тропических лесов, обращающих ш себя внимание путешественников.
Лианы. Они крайне разнообразны и относятся к весьма различ-
ным семействам (рис. 82). Эта жизненная форма, стремящаяся к
свету, как н деревья, обладает тем преимуществом, что не разви-
вает толстого стебля и может быстро взбираться на самые высокие
Деревья. Приспособления, при помощи которых лианы взбираются
по деревьям, различны: они или перебрасываются с ветки на ветку,
задерживаясь своими колючками, или развивают особые придаточ-
ные корни-прицепки или усики; в иных случаях они обвивают стволы
и ветки деревьев. Всползая по
деревьям, они свешиваются фе-
стонами, петлями разных раз-
меров и форм, иногда тяжесть
лиан настолько велика, что
ветви обламываются, и лианы
падают на землю, затем взби-
Рис 83. Мох (Barbella javanica)—
эпифилл—свисает с листьев во
влажнотропическом лесу, на о.
Яве (по Фаберу).
179
раются на другие деревья и т. д. Длина пальм-ротангов (виды рода
Calamus) может достигать 300 м\ это — одни из самых длинных рас-
тений. Вообще же пальмы-лианы среди лиан наиболее характерны
и часты. Очень важную роль играют также лианы из семейства
ароидных (Philodendron и др.). Упомянем также лианы из родов
фикуса, перца (Piper), ванили (Vanilla), Bauhinia с плоскими вол-
нистыми стеблями и пр.
Эпифиты. Эпифиты не менее, чем лианы, характерны для влаж-
нотропических лесов. Они крайне разнообразны как системати-
чески, так и экологически. Огромное количество их относится к'
папоротникам и орхидным. Эпифитами усажены стволы и ветви
иногда в таком огромном количестве, что получаются как бы особые
воздушные сады. Эпифиты лучше, чем какая-либо другая жизнен-
ная форма тропических лесов, способны добраться до света, по-
селившись в самых верхних частях древесных крон. Эпифитами
могут быть не только травянистые виды, но также и кустарники и
деревца. Еще лучше приспособляются эпифиллы (рис. 83 и 84):
они поселяются на самых листьях (к эпифиллам относятся не только
мхи ” чо тп’огд.т и цветковые пдстения).
30
Естественно, что экологически эпифиты крайне различны, так
как виды внутренних тенистых частей леса сильно отличаются от
видов, поселяющихся на вершинах деревьев: если первые явля-
ются гигрофитами, то среди последних мы имеем резко выраженные
ксерофиты (сочные листья, особые клубни, запасающие воду, и т. д.).
Некоторые эпифиты запасают воду в особых вместилищах: знамени-
тая дисхидия (D.ischidia, рис. 85), бромелин и др.
Из сказанного можно заключить, что различные ярусы тро-
пических лесов имеют крайне отличающиеся фитоклиматические
условия и что такие факторы, как свет и влага1, имеют здесь
исключительное значение.
Обилие влаги в нижних частях леса связано с целым рядом
приспособлений для удаления излишней влаги (см. стр. 90): капель-
ные острия у листьев, гидатоды на листьях, бархатистая поверх-
ность листьев и др. (рис. 38). Обращает на себя внимание также п
то, что среди травянистых растений многие имеют на листьях при-
хотливые р и с у н к и из пятен и полос различных цветов (белого,
красного и др.), как, например, у бегоний (они культивируются
в оранжереях)- другие растения отливают металлияески-синим цве-
том. Экспериментальных объяснений этим явлениям ещё нет.
Как уже указывалось, количество света в различных ярусах
различно; до почвы иногда доходит лишь 1/140 света, причём пре-
делом для цветковых здесь является1/^—Чмо-Но места с ничтож-
ным количеством света на небольших участках сменяются и более
светлыми (даже до 1/9). Интересно, что это связано с солнечными
бликами и с отражением от блестящих лакированных листьев, а
кроме того, с тем явлением, о котором говорилось выше, именно —
с зубчатой крышей леса: получается разорванность крон
и возможность проникновения света.
Вечнозелёность тропического леса и периодичность роста. Пред-
ставление о том, что вечнозелёные деревья имеют'длительно суще-
ствующие листья и непрерывный рост, неправильно. Обычно деревья
сбрасывают свою листву по частям (1—б раз в год), через известные
промежутки времени (независимо от времени года), и никогда не
стоят без листьев. Интересно, что сбрасывание более старых листьев
у различных экземпляров одного и того же вида происходит
в разные сроки.
Несмотря на крайне благоприятные в течение круглого года
условия, вечнозеленые, подобно другим деревьям, имеют период
покоя. Указания на то, что многие тропические деревья растут
беспрерывно, невидимому, неправильны. Очень оригинальна «ин-
дивидуальность» ветвей, когда на отдельных ветках или на отдель-
ных системах ветвей распускаются новые листья независимо от
других ветвей. Распускание листьев у многих деревьев дает красоч-
ные картины: у одних — молодые листья яркокрасные, у других —
буро-красные, у иных — белые.
Так каквнешние условия влажнотропического леса очень постоян-
ны, то период покоя связан, невидимому, с внутренними причинами.
Сравнительная бедность цветами тропических лесов. Мощность
вегетации трошщеску^ лесов, поражают1?а наблюдателя, находится
181
в известном контрасте с бедностью или невзрачностью цветков, о
чем говорят все исследователи этих лесов (Уоллес и др.)- Очень
многие деревья имеют либо самоопыляющиеся цветки, либо опы-
ляемые ветром. Пышное развитие цветков мы находим лишь на
опушках или в осветленных местах, а также при рассеченном релье-
фе (овраги, долины). Иногда деревья или кустарники цветут яркими
цветками, но они обычно растут отдельными экземплярами (не
образуют, как у нас, скоплений) и теряются среди тонов зеленого
цвета, выделяясь лишь отдельными пятнами. Собрания
прекрасных орхидей, ароидных и других в наших оранжереях
совсем не дают представления о тропическом лесе, так как все эти
растения не дают там таких красочных скоплений. Возможно, что
сравнительно небольшое число красиво цветущих, опыляемых жи-
вотными, цветков зависит от недостатка этих животных. Трудн©
представить себе, сколько нужно было бы насекомых для опыления
многочисленных растений тропического леса.
Подобно листопаду и развертыванию новых листьев, время
цветения не приурочено к определенному периоду, а распределено
по всему году, что также является причиной бедности цветами
тропических лесог. Но многие виды цветут круглый год, однако
беспрерывное цветение нужно понимать в том смысле, что всегда
можно встретить цветущие экземпляры; и здесь характерна инди-
видуальность отдельных растений и их ветвей: в то время как одни
деревья или даже отдельные ветви цветут, на других в то же время
уже имеются гк еды.
Среди ряда астений с кратковременным цветением
имеет место залючательное явление: на пространстве многих квад-
ратных километров все экземпляры какого-либо вида расцветают
в один и тот же день (некоторые виды кофейного дерева,
некоторые бамбуки, пальмы и др.).
Сравнительная бедность цветками тропических лесов отчасти и
субъективное представление — цветки на высоких деревьях снизу
не видны.
Каулифлория. Наконец в связи с цветением нужно указать на
характерное явление у тропических деревьев, на каулифлории.;, т. е
возникновение цветков прямо на стволе; позже стволы бывают уве-
шаны плодами (рис. 86). Видов с каулифлорией насчитывается в
тропиках более 1000, из их числа назовем дерево какао, хлебное
дерево (Artocarpus), хурму (Diospyros), многочисленные фикусы
(до 50 видов в Азии) и т. п. Физиономически это явление очень ори-
гинально. Причины каулифлории недостаточно ясны; так, одни
авторы связывают это явление с опылением при помощи бабочек,
летающих в глубине леса; другие видят в каулифлории примитивное
явление, когда генеративные и вегетативные (ассимилирующие)
части на растении пространственно разъединены, что подтверждается
и некоторыми палеоботаническими данными. Тогда расположение
цветков среди листьев кроны, например у деревьев наших лесов,
нужно считать вторичным явлением.
В целом влажнотропический лес необычайно сложен; там гос-
подствует крайнее разнообразие жизненных форм и эти последние
заполняют всевозможные места. Поэтому насыщенность леса как
флористически, так и фитоценологически очень велика. Это —
результат необычайно длительной эволюции.
"Обращает на себя внимание полная противополож-
но с т ь нашему лесу; тогда как в наших лесах флористическое
богатство и разнообразие жизненных форм свойственны нижним
травянистым ярусам, в тропических лесах — самым верхним.
Краткий обзор тропических лесов в различных странах. Тро-
пические леса в Азии. Особенно хорошо изучены леса
на о. Яве и, в частности, лес у Tjibodas, который хотя и находится
иа высоте 1 500 м, но содержит Бесхарактерные черты равнинного
тропического леса. Его флористический состав: деревьев—165 видов,
кустарников и трав 350, лиан 65 видов. Деревья не образуют ярусов,
а располагаются как бы беспорядочно, но этот «беспорядок» зави-
сит от того, что между ярусами размещаются еще дополнительные
внеярусные виды. В лесах Явы встречаются в качестве эпифитов
такие замечательные мирмекофильные растения, как Myrmecodia
(рис. 60) и Hydnophytum (см. стр- 125).
Малезийские тропические леса Азии, в противоположность
Америке, очень бедны растениями с красивыми крупными цвет-
ками. Здесь господствуют папоротники и растения с невзрачными
цветками: кроме того, очень сильно развито вегетативное размноже-
ние. Но бедность цветками зависит еще от того, что вообще цветение
распределено на весь год, и нет скопления цветущих видов, как
в наших лесах, в тёплые месяцы.
Особый колорит яванским лесам придают древовидные папорот-
ники Alsophila (до 25 м) и более низкорослые Cyathea и Dicksonia
(рис. 87>.
184
На Филиппинах господствуют леса главным образом из деревьев,
семейства Dipterocarpaceae (до 40—50 я высоты).
Тропические леса в Австралии. Развиты лишь
на Новой Гвинее и на восточном берегу континента. Они очень схо-
жи с лесами малезийского типа, имея, впрочем, и некоторые австра-
лийские элементы.
Тропические леса в Африке. Развиты по реке Конго и
в низовьях р. Нигера, по побережью Гвинейского залива, отчасти
на Мадагаскаре. По своей территории они уступают лишь лесам по
Амазонке, — это область okoj о 600 000 кмг, сплошь покрытая ле-
сом, флористически очень богатым и разнообразным, с очень большим,
количеством эндемиков (о некоторых флористических элементах
см. стр. 357).
Тропические леса в Америке. Они занимают всю
область р. Амазонки с ее притоками, все Атлантическое побережье
до Южного тропика и др.; леса по Амазонке наиболее обширны по
своей территории,они знамениты тем, что здесь впервые натуралисты
(Гу мбольдт) познакомились с влажнотропическими лесами.
Гумбольдт назвал эти леса «гилеей» («гилейсн» — по-гречески
лес) Ч
Некоторые авторы считают
эти леса наиболее роскош-
ными из всех тропических ле-
сов, но они вообще очень
схожи экологически ифизио-
номически с соответствую-
щими лесами Малезии. Раз-
личают два основных типа:
«игапо» -влажнстрсйический
лес на более низких местах,
временами заливаемый рекой,
и «эте»—на более высоких
.местах, уже не заливаемых.
Более богат флористически
последний, заключая много
эндемических видов.
Особенно обращает на
себя внимание обилие
в г и л е е эпифитов,
причем характерны эпифиты
из американского семейства
бромелиевых^ последние по-
разительны по своим фор-
мам и по яркости окраски
цветков. Бромелин относятся
к «цистерновым» эпифитам
(рис. 88), так как тесно со-
прикасающиеся в розетке листья образуют воронку, где скопляется
1 Иногда гилеями называют вообще влажнотропические леса и других кон-
тинентов.
185
года (до J л), минеральные и органические вещества, служащие
для питания и всасываемые основаниями листьев. Так же харак-
терны в качестве эпифитов кактусы (особенно виды Rhipsalis).
Некоторые авторы указывают ещё на одну черту гилеи: эпифиты
из семейств Агасеае и Clusiaceae дают многочисленные воздушные
иеразветв ленные корни, которые отвесно спускаются вниз, укре-
пляются в почве и натягиваются, как проволока.
Очень важным по своему значению является каучуконосное
дерево хевея (Hevea brasiliensis).
Не приводя списков других наиболее характерных древесных видов гилеи
(Bertholletia excelsa, Сагуосаг villosum, Dipteryx odorata и др.), укажем, что
дтя берегов рек особенно типично изобилие различных видов с ( ’
Phytelephas, AstocariumH др.); многие пальмы растет и на бол
местах (Euterpe—рис. 79, Attalea и др.).
В области гилеи имеется целый ряд мирмекофильны р ,
т. е. «сожительствующих» с муравьями (виды рода Cecropia, re-
plans и др.). Интересны «муравьи-листорезы», муравьи, разводя-
щие «сады», и пр. (см. стр. 122).
Наконец, нужно добавить, что здесь, в тихих заводях м онки
и её притоков (а также р. Ориноко), встречается культивируемая в
наших оранжереях виктория-регия (Victoria regia), растение из
семейства нимфейных (рис. 89).
Субтропические и умереино-субтрояические леса. Кроме влажно-
тропических лесов, нужно отличать еще леса субтропические и
86
умеренно-субтропические. Если, как выше было указано, тропи-
ческие леса не выходят за пределы тропиков, то субтропические
развиваются за пределами последних, именно, в Южной Бразилии,
в Северной Мексике, на Флориде, а также в Натале (Африка), в
Квинслэнде н Новом Южном Уэльсе (Австралия). Близкий к суб-
тропическим лесам тип представляют горные влажнотропи-
ческие леса, которые покрывают склоны гор (на высотах 1 200—
1 500 м) в пределах тропической области, например восточные
склоны Анд в средних частях Южной Америки, южные склоны
Гималаев и пр.
Субтропические и горные леса незаметно переходят в тропи-
ческие и отличаются от последних рядом отрицательных признаков:
разнообразие видов, особенно древесных, не так велико; количество
эпифитов и деревянистых лиан сильно уменьшается; резко падает
число видов с каулифлорией и с досковидными корнями. С другой
стороны, усиливается роль мхов и папоротников, особенно
древовидных. Последние местами даже могут образовывать
особые папоротниковые леса, как, например, в Новой Зеландии,
на горах Антильских островов; в других случаях они образуют
более низкие ярусы леса.
Умеренно-субтропические леса при большом
количестве осадков развиты в тепло-умеренных областях (особенно
в Японии, в Юго-восточном Китае, в Южном Чили, на Новой Зе-
ландии и др-). В этих лесах ещё более усиливаются отрицательные
признаки субтропических лесов.
Некоторыми авторами умеренно-субтропические леса и часть
субтропических относятся к «лавровым» лесам (Laurilignosa), рас-
сматриваемым нами ниже.
Влажнотропический лес и человек. Девственность тропиче-
ских лесов не нужно переоценивать; местами эти леса сохра-
нили свой примитивный характер, особенно там, где они зани-
мают большие площади и мало доступны (в бассейне р. Ама-
зонки, в Центральной Африке); однако в ряде мест они или
совсем уничтожены, или носят вторичный характер. Так, почти
уничтожены влажнотропические леса на Западном Малабарском
побережье в Индии, на Цейлоне, в Индо-Китае, быстро уничто-
жаются на некоторых Зондских островах (Ява, Суматра) и т. д.
Можно различать экстенсивные и интенсивные формы воздействия
человека и, соответственно, экстенсивные и интенсивные культуры.
К экстенсивным воздействиям относятся так называемая система
банту в экваториальной Африке, система рай (rays) в Индо-Китае
и др. Здесь единственным орудием земледелия является мотыга.
Во время короткого, более сухого, периода рубится подлесок и более
мелкие деревья, которые затем сжигаются: зола служит удобрением,
другие удобрения (от животных) не применяются. Более крупные
деревья уничтожают, раскладывая у их оснований костры, но не-
которые деревья остаются и в дальнейшем играют роль семенников.
После расчистки леса разводят различные культурные растения,
особенно бананы, маниок, кукурузу и др. После нескольких лет
культуры подобные плантации забрасываются и они начинают
187
зарастать дикими растениями. Последние очень быстро завладевают
своей прежней территорией, что объясняется очень благоприятными
условиями для роста в тропических областях, В первый год особен-
но быстро завладевают территорией лианы, затем появляются дре-
весные растения, давая свою поросль от корней, оставшихся в поч-
ве. Некоторые деревья с мягкой древесиной растут чрезвычайно
быстро. Во всяком случае через десять лет восстанавливается лес
до 15—25 ж высотой с густым подлеском; однако лес этот нужно
называть вторичным, так как он очень сильно отличается от
исходного, перви чного леса. Основное отличие — это фло-
ристическая бедность, зависящая от того, что многие виды были
совершенно уничтожены и медленно заносятся из окружающих
пространств. Имеются данные, что восстановление первичного ха-
рактера леса идет настолько медленно, что даже нескольких сот лет
для этого недостаточно — лес остается беден древесными породами.
Местами тропические' леса хищнически разрушаются тем, что
производится выборочная рубка тех или других ценных древесных
пород (хинные деревья в Южной Америке, деревья с ценной дре-
весиной и др.). В иных случаях леса потеряли почти все свои ценные
в практическом отношении породы.
Как уже было указано, большие пространства влажнотропиче-
ских лесов совершенно уничтожены. Их место заняли культуры
тропических полезных растений. Это или древесные растения:
дерево какао, кофейное дерево, хинное дерево, кокосовая пальма,
ряд каучуконосов и др., или злаки — особенно рис, реже кукуруза.
Во многих местах юго-западной Азии (Суматра, Индо-Китай
и др.) после тех или иных культур территории зарастают злаком
Imperata arundinacea, называемым аланг-аланг, или диким сахар-
ным тростником (Sacharum spontaneum). Образуемые им заросли
настолько густы, что не дают возможности быстро восстановиться
первоначальному влажнотропическому лесу.
2, Группа формаций—влажнотропические кустарниковые фитоце-
иозы (Pluviifruticeta) илн мангровая растительность
Общая характеристика. Этот оригинальный тип растительности
связан с тропическими странами, именно с их морскими плоскими
берегами, защищенными от волн и ветров (заливы, лагуны и др-),
и подвержен действиям прилива и отлива.
Наибольшее развитие данный тип растительности имеет под
экватором, hj он продвигаемся и на север и на юг, почти не выходя,
однако, из пределов тропиков: на севере доходит до южной Кали-
форнии, Флориды, Бермудских островов, Персидского залива,
Красного моря, островов Лючу, на юге — до Южной Бразилии,
Эквадора, Новой Зеландии, Нового Южного Уэльса и Наталя.
Мангровая растительность представлена вечнозелеными кустар-
никами, кустарниковидными деревцами или небольшими деревья-
ми (некоторые, однако, до 30 м высоты). Но в общем она имеет ха-
рактер густых кустарниковых зарослей, почему и относится рядом
авторов к влажнотропическим кустарниковым фитоценозам (Р1п-
viifruticeta)
188
Нужно отличать две области мангров — восточную (Азия, Ав-
стралия, Микронезия и восточный берег Африки), и западную (за-
падный берег Африки и Америки); первая богата мангровыми
кустарниками идеревьями, вторая—бедна. Однако, вообще говоря,
флористический состав беден; в восточной области 22 вида, в за-
падной—всего 4 вида.
Все эти виды распределены по 8 семействам: из них наиболее
важны семейства Rhizophoraceae (9 видов) (роды Rhizophora и Вги-
Рис. 90. Общий вид мангровых зарослей во впемя отлива—видны сплетения
ходульных корней (по Шантцу).
guiera), Verbenaceae (род Avicennia), Sonneratiaceae (род Sonneratia).
Мангровые заросли крайне однообразны по своему составу,
что зависит от того, что плоды видов, входящих в их состав, хорошо
переносятся морскими течениями.
Если смотреть на мангровые заросли с моря во время прилива,
то кроны деревьев как бы выходят из воды, во время же отлива
обнажаются стволы и целая система особых корней — ходульных и
дыхательных. Эти корни особенно типичны для манГровых зарослей.
Ходульные корни. Они укрепляют деревья в илистой почве,
особенно характерной для мангровых побережий (рис. 90), хотя
мангры иногда могут расти и на других субстратах. Ил обычно
имеет голубовато-черный цвет и очень беден кислородом, во время
прилива насыщен водой и представляет мало устойчивый субстрат.
Вот почему мангровые растения и образуют так хорошо и обильно
пгвю"? vn^i'-пьные корни.
189
Впрочем, подобные корни развиты лишь у некоторых видов
главным образом у тех, которые образуют в зарослях внешний
(по отношению к морю) пояс. Здесь нужно отметить два вида
Rhizophora — Rh. mucronata и Rh stylosa.
Илистый субстрат особенно благоприятен для размножения
мангровых, В этом отношении замечательна опягь-таки Rhizophbra
mucronata, сна принадлежит к числу живородящих растений,
г е таких, у которых зародыш без периода покоя прорастает в
плоде, находящемся еще на дереве. Здесь зародыш имеет очень длин-
ное (до 60 ел/) подсемядольное колено булавовидной формы, затем
он отрывается от плода и, вследствие имеющегося утолщения в ниж-
ней части, падает вертикально вниз, зарываясь в ил и давая новое
растение Живородящими являются и многие другие виды.
Дыхательные корни (пней в матофоры). У некоторых видов
мангровых имеются дыхательные корни различного рода (рис. 91)
Последние совершенно необходимы, так как ил, как сказано выше,,
почти не содержит кислорода. У одних видов корни змеевидные
ползучие, и их верхний утолщенный край снабжен дыхательными
отверстиями (Xylocarpus granatum), у других — корни коленчато
выступают из ила (Bruguiera gymnorhiza), у третьих вследствие
отрицательного геотропизма корни растут вверх и выступают из
ила, как молодые стебли спаржи (Avicenma, Sonneratia, рис. 52)
у видов Rhizophora, имеющих ходульные корни, нет особых дыха-
190
тельных корней, но функцию последних играют выступающие hs
воды части ходульных корней Все указанные образования при-
способлены для передачи кислорода в подземные части.
Мангровые растения—галофиты. Так как растения мангровых
зарослей растут по берегам морей и заливаются морской водой во
время приливов, то естественно, что они должны относиться к га-
ло фитам. Однако представление о том, что они —факультатив-
ные галофиты, неправильно, почти все они являются галофитами
облигатными в большей или меньшей степени, и в связи с
этим находится распределение видов в прибрежных зарослях, одни
занимают внешний пояс (Rhizophora mucronata или Sonneratia).
другие располагаются дальше от берега, третьи занимают самое
внутреннее положение (рис. 92).
Рис 92 Схематический профиль на морском побережье у Танга (восточная
Африка)—ясно видна смена поясов растительности*
/-пояс, «пшенный растительности, 2—пояс с г сподством Avicenma 3-пояс Cenops
/—поя Rhizophora 5—пояс Sonneratia Верхняя гоьиаонгальная линия (пунктирная)
ысшии уровень прилива, средняя—нормапг ныи прилив, нижняя^ЦП-нлэшии уровень
отлива ; 1
Мангровые растения являются галофитами. Листья их в большей
или меньшей мере суккулентны Однако, как показали новейшие
исследования, мангры испаряют воды больше, чем многие гигрофил -
ные деревья, это связано с очень большим осмотическим давлением
в клетках корней и^истьев
Интересно, что осмотическое давление во время прилива и в»
время отлива различно так, во время прилива в листьях и корнях
Avicennia marina давление достигает 163,2 и 96,6 ат, при отливе—
соответствующие цифры составляли 82 и 50,3 Таким образом, в»
время прилива, когда мангровые заросли заливаются соленой
водой, осмотическое давление повышается почти вдвое. Эта регу-
лировка осмотического давления — интереснейшее приспособ-
ление мангровых Естественно, что при такой большой засасываю-
щей силе клеток и возможна сильная транспирация Однако какую
же роль играет суккулентность у листьев мангровых? Здесь, неви-
димому, избыток воды идет на транспирацию в то время, когда
растение перестраивается в начале прилива с низкого осмотической
давления на высокое.
Однако новейшие исследования не подтверждают приведен-
ных данных, по эь м исследованиям осмотическое давление у мангро-
вых значительно меньше у Sonneratia alba в среднем 32 ат, у
191
Rhizophora mucronata 33 ат, у Avicenma marina 34,5—62,0 am,
вообще же осмотическое давление довольно устойчиво и не меняется
во время прилива и отлива, но оно выше, чем в почве, н этого пре-
вышения достаточно для успешного водного снабжения, суккулент-
ность листьев не стоит в связи с отдачей воды во время изменения
осмотического давления (последнее не имеет места), и листья не могут
спадаться, так как стенки водоносной ткани достаточно крепки.
Не замечается также разницы в концентрации почвенного раствора
со время прилива и отлива 3ai адочным является водное снабжение
(02
зародыша у живородящих видов —здесь осмотическое давление
лежит не только ниже, чеад у материнского растения, но ниже поч-
венного раствора.
Вместе с водой в мангровые растения поступает много поварен-
ной соли Для удаления последней *на листьях имеются особые
железки, при этом листья иногда почти сплошь покрыты кри-
сталлами соли
Дальше от моря, на менее соленом субстрате в устьях рек, мангры
уступают господство низкорослой пальме — Nipa fruticans, иногда
покрывающей огромные пространства (формация «нипа»). Эта фор-
мация поднимается вверх по рекам в области действия морских
приливов (рис. 93).
В западной области мангровых, как сказано, имеется всего лишь
4 вида мангровых- внешние части заро л. й занимает Rhizophora
mangle, самые внутренние — Lagunculana racemosa, промежуточное
положение занимают два вида Av^cenma.
Эпифитов в мангровых зарослях очень мало. В западной области
можно указать на бромелин и лишайники.
И. КЛАСС ФОРМАЦИ “'—ДРЕВЕСНЫЕ И КУСТАРНИКОВЫЕ ФИТОЦЕНО-
ЗЫ ЛАВРОВОГО ТИЛА (LAURlLlGhOS )
1. Группа формаций—лавровые леса (Laurisilvae)
Особенно больших площадей эта группа фоэмащ й не занимает
и распространена на Канарских островах, на Мадейре, в Порту-
галии. в Северной Америке (на Тихоокеанском побережье и на
Флориде), в Южгой Америке (в Чили и в Патагонии), в Японии,
по южному склону Гималаев, в Новой Зеландии и в некоторых
других странах.
Общая характеристика. Леса лаврового типа вечнозелены и могут
состоять из деревьев двудольных и голосеменных. Двудольные имеют
очень характерный тип листа, по большей части пластинка крупная,
овальная, всегда блестящая, кожистая и лишенная волосков, по
отношению к свету всегда стоит перпендикулярно. Ти-
пичным примером подобного листа является лавровый (отсюда и
название). Кроме того, листья имеют свежезеленую окраску и часто
относительно крупные межклетники Указанные признаки говорят о
близости лавровых лесов к тропическому лесу, но имеется ряд
существенньх отличий, почки на концах побегов уже одеты почеч-
ныл ! -пшуяли (однако растения подлеска почечной чешуй могут
не пиль, так как в глубине леса флтоклимат более благоприятный),
отсутствует ряд признаков влажнотропического леса, нет каули-
флории, нет обилия эпифитов, деревья лишены досков дных кор-
ней, на листьях отсутствуют капельные острия и т д
Леса подобного типа по своей внешности несколько схожи с жест-
колистными вечнозелеными лесами, но резко различаются своими
листьями в жестколистных лесах они никогда не стоят перпенди-
кулярно к свету, никогда не блестят (о других признаках этих
лесов см. на стр. 202).
Леса лаврового типа могут состоять не только из двудольных
деревьев, но также из хвойных, именно из тех родов, которые эко-
193
логически, а также до известной степени и физиономически, на-
поминают лавровые леса из двудольных. Сюда относятся две группы
хвойных, морфологически отличающиеся одна от другой, во-первых,
это хвойные с плоской блестящей хвоей, расположенной
на ветвяхтак, что получается
впечатление перистого листа
двудольных. Прекрасным
примером является северо-
американское красное дерево
(Sequoia sempervirens, рис
94) с «ложноперистыми» ве-
точками (очень похожи на
перистые листья), стоящими
перпендикулярно к свету1.
Хуже данный тип выражен
у тисса (Taxus и Cephalo-
taxus)
Другая группа хвойных
имеет тесно сидгщие че-
шуйчатые хв и, такчтоветка
в целой! представляет как бы
один длинный Л1 ст, который
к тому же стоит более или
менее перпендикулярно к
свету,таковы американские
ту и (Thuja occidentals, Т. he-
terophyПа и др ).
Правда, «лавровыми» та-
кие олиственные веточки
хвойных можно назвать лишь
с большой долей приближен-
ности, но леса из указанных
Рис 94 Лес”из красного дерева (Sequoia хвойных занимают террито-
semperxirens) в Калифорнии (из Мак- риИ, климатически
Дуголла). крайне сходные с
теми, где развиваются типичные лавровые двудольные.
Климат этих лесов в основном близок к субтропическому, тем-
пература более или менее равномерна в течение целого года (так,
например, в Калифорнии у С.-Франциско средняя температура
января 10°, а июля 14°), равномерно распределение осадков, боль-
шая влажность воздуха- По сравнению с тропическим лесом, тем-
пература здесь несколько ниже и сезонные колебания более зна-
чительны.
Положение листьев, перпендикулярное к свету, обусловлено,
невидимому, большой облачностью и большой влажностью атмос-
феры, что влечет за собой ослабление интенсивности света, и харак-
терно главным образом для растений подлеска.
1 Сходный по морфологии своих листьев болотный кипарис (Taxodium
distichum) ие может быть отнесен сюда, так как он является летнезеленым.
194
Двудольные «лавровые» леса. Прекрасным примером лесов
лаврового типа являются лавровые леса на Канарских островах;
они располагаются здесь на высоте 400—1 300 м в поясе очень вы-
сокой облачности. В них господствует очень сильная влажность и
интенсивность света сильно понижена, особенно в глубине леса.
Доминирующее дерево —Канарский лавр (Lantus canariensis) с ши-
рокими кожистыми листьями. Многие виды подлеска тоже имеют
крупные вечнозеленые листья. Очень обильны папоротники и мхи,
они растут как на почве, так и на деревьях в качестве эпифитов.
Особенно красив эпифит — папоротник Davalha canariensis. Сход-
ные леса встречаются и на о. Мадейре.
Двудольные «лавровые» леса в Америке имеют место в средней
и северной Флориде, в Чили, в Патагонии и др. В Чили они пред-
ставлены вечнозеленым буком (Nothofagus betuloides) с листьями
лаврового типа. Обилие круглый год дождей, хотя и при более
низких температурах, обусловливает их присутствие. И здесь очень
обильны мхи, папоротники (особенно Hymcnophyllaceae). Южнее
леса становятся лишь летнезелеными, но подлесок остается вечно-
зеленым (см. стр. 220).
«Лавровые» леса из хвойных дере ьев. Они особенно характерны
для Тихоокеанского побережья Северной Америки. В iiiraie Ьа-
шингтон, в его западных частях, климат очень мягкий и очень оо-
гатый осадками (100—250 см осадков), крайне большая облачность
и насыщенность воздуха водяными парами; зАе-ьпо западным скло-
нам Каскадных гор развиваются вечнозеленые леса из гш антских
деревьев в среднем около 1 м толщиной и в очень густом насаждении
(расстояние между деревьями 2—4 м). В нижних частях склонов
гор леса состоят из цуги (Isuga heterophylla), туи (ihuja gigantca)
с их плоско распростертыми ветвями, улавливающими свет, сюда же
примешивается дугласова пихта (Pseudotsuga) и др. В более высоких
частях гор господствуют другие хвойные, пихты, белая сосна, а
также Chamaccypabs nootkatensis. Эти леса отличаются очень
Полиной растительностью.
Замечательные леса находятся на калифорнийском побережье
при исключительно однообразной температуре в течение всего года
(см. выше). Верхний ярус образован великолепными экземплярами
красного дерева (Sequoia sempervnens) с красивыми горизонтальными
ветвями, напоминающими сложные листья, и блестящими, как листья
лавров. В нижних ярусах ряд вечнозеленых, земляничное дерево
(Arbutus), магония с блестящими листьями (Mahonia nervosa) и др.
2. Группа формаций—кустарниковые фитоценозы лаврового типа
(Laurifruticeta)
Они могут также состоять из двудольных или из хвойных, морфо-
логия и экология их листьев такова же, как у «лавровых» деревьев.
Область распространения их шире, чем у лесов лаврового типа,
что объясняется тем,что кустарники лаврового типа нередко входят
в лесные фитоценозы в качестве подлеска, причем верх-
ние ярусы подобных лесов могут состоять из деревьев с опадающей
листвой или из хвойных деревьев умеренных широт. Здесь наблю-
13* 195
дается явление замещения факторов: равномерный океанический
климат замещается климатом леса (фитоклимат), более влажным и
более равномерным, чем в окружающих безлесных пространствах.
При таких условиях лавровые кустарники могут проникать в иные
климатические области, в области лесов с опадающими листьями
или хвойных лесов, причем последними их представителями можно
считать нашу бруснику и другие вересковые с блестящими широ-
кими листьями.
Отдельные примеры лавровых кустарниковых фитоцеиозов. В
Средиземноморье мы находим заросли из благородного лавра (Laurus
nobil's), столь распространённого там в культуре. Однако в есте-
ственном состоянии лавр — горное растение более влажного клима-
та (в горах Малой Азии, в Далмации), растущее нередко в подлеске
(в Далмации).
Очень характерны на Кавказе заросли лавровишни (Primus
laurocerasus) н рододендрона (Rhododendron caucas cum). Первая
имеет листья ярко выраженного лаврового типа (крупные, блестя-
щие) и встречается на известных высотах в качестве подлеска боко-
вых лесов или лесов нордманновой пихты. Рододендрон же образует
обширные густые заросли на верхней границе лесов в субальпийском
поясе, но поднимается и выше, до 2 900 м (на Главном хребте и в
западном Закавказье) (рис. 95), интересно, что в западных частях,
196
в условиях большей влажностй, он встречается на склонах вс-ех
экспозиций, а далее на восток переходит на более влажные север-
ные склоны. Под рододендроном находим водянику, чернику, папо-
ротники, плауны и др. Характерны почвы — темноцветные, торфяни-
стые, в результате плохого проветривания и неполного разложения.
Рододендроны вообще характерны для горных высот. Так, на
Кавказе мы встречаем еще заросли Rhododendron ponticum; для
гор Центральной Европы крайне типичны Rh. hirsutum, Rh. fer-
rugineum; рододендроны пышно развиты также в Гималаях и др.
Хотя заросли рододендронов обычно развиваются в условиях,
сильно отличных от типичного климата лавровых растений, но
глубокий снежный покров умеряет действие зимних холодов.
Выделение лесов и кустарниковых фитоценозов лаврового типа (Р ю б е л ь,
Бро к ча н-Е р о ш) не является общепринятым. Чаще всего фнтоценозы
^энного типа распределяют среди других растительных типов. Так, собственно
тавровые леса (с господством Laurus canariensis) относят к субтропическим ле-
сам, леса вечнозеленых буков, леса юго-восточного Китая н др. — к умеренно-
субтропическим лесам, хвойные леса лаврового типа — к хвойным лесам вооб-
ще, заросли рододендронов— к верещатникам. Однако, целый ряд общих черт,
указанных выше, оправдывает, невидимому, выделение особого класса форма-
ций Lauri lignosa.
Ill, КЛАСС ФОРМАЦИЙ—ЗИМНЕЗЕЛЕНЫЕ ДРЕВЕСНЫЕ И КУСТАРНИ-
КОВЫЕ ФНТОЦЕНОЗЫ (HIEM1LIGNOSA)
Две группы формаций: 1) зиинезеленые леса—Hiemisilvae и 2) зим-
иезеленые кустарниковые фнтоценозы—Hlemifruticeta)
Зимнезеленые леса распространены почти исключительно в
пределах тропиков и характерны для тех территорий тропического
пояса, где выражен континентальный климат или где периодически
дуют то сухие, то влажные ветры (муссоны), С тропическими лесами
они связаны переходами, но отличаются тем, что во время летней
засухи теряют свои листья, быстро развивающиеся вновь с наступле-
нием дождливого периода.
Данный класс формаций довольно разнообразен и представляет
в сущности ряд классов, зависящих в своем развитии от количества
осадков в дождливое время года. При большом количестве осадков
развиваются муссоновые леса, наиболее приближающиеся к тропи-
ческим. при меньшем количестве — саванновые леса, при еще мень-
шем — леса сменяются фитоценозами колючих кустарников или же
переходят в саванны
В частности для муссоновых лесов количество осадков соста-
вляет около 150 см\ температурные различия между средними тем-
пературами самого теплого и самого холодного месяцев доходят до
8° (во влажнотропических лесах не превышают 1—6°). Имеется
ясно выраженный сухой период, однако почва не становится слишком
сухой. При меньших количествах осадков (90—150 см), с разницами
средних температур в 14° и периодом засухи в 4—6 месяцев разви-
ваются саванновые леса, или саванны: при еще более крайних усло-
виях развивается «колючее редколесье» (или колючие кустарни-
ковый фитоценозы).
197
На карте Брокман-Ероша все эти подразделения объединены в
«зимнезелёные леса», на других картах (например на карте БСАМ)
они представлены отдельно.
Муссоновые леса. Особенно развиты в Юго-восточной Азии,
именно на ряде восточных островов Зондского архипелага (Тимор,
Целебес, восточная часть Явы), в Индостане (Декан) и в Индо-
Китае (Бирма). В основном они очень схожи с тропическими лесами,
но в период засухи, с июня по октябрь, большинство’ деревьев
лишено листьев. Господствующим деревом является тиковое дерево
с очень крупными листьями (Tectona grandis — из сем. вербеновых);
оно образует сплошные насаждения, достигающие 20—25 м высоты
(иногда 40 м). Это дерево даёт прекрасный материал для постройки
судов.
Во время Засухи деревья стоят голыми, и сухие листья шуршат
под ногами; большинство сопровождающих деревьев тоже лишено
листьев, но некоторые деревья являются вечнозелёными. Кустар-
ники вообще многочисленны. Травы во время засухи по большей
части стоят сухими, а однолетники в это время совершенно отсут-
ствуют.
Наиболее засушливые месяцы в восточной части Явы — август
и сентябрь. В ноябре, с началом муссоновых дождей, тиковое дерево
начинает покрываться листьями, которые в молодом состоянии бы-
вают красными. В это время цветёт большинство кустарников,
несколько позже цветут травы. Месяцы, менее богатые цвету-
щими видами, — апрель, май, июнь. Интересно, что и во время
сухих месяцев имеются цветущие растения; так, в июле и
августе цветёт дерево' Butea monosperma (сем. мотыльковых) с
крупными огненно-красными цветками; в это же время цветут ама-
риллисовые.
Вообще же в тиковых лесах цветов гораздо больше, чем во влаж-
нотропических, так как здесь больше света и суше.
Но богатство жизненными формами здесь меньше; лианы хотя
местами и часты, но всё же их меньше, и они часто теряют свои
листья в засушливый период; эпифиты тоже менее обильны и при-
урочены главным образом к кронам деревьев; деревья менее высоки
и лишены досковидных корней.
Там, где засуха продолжается 4—6 месяцев, где почва сильно
высыхает и где в дождливое время выпадает меньше осадков, раз-
виваются леса ксерофильного типа. Они очень разнообразны и
флористически и фитоценологически и связаны с континентальными
областями тропиков. В частности они занимают большие простран-
ства в Африке, Южной Америке, меньше — в Австралии и Азии
(Индостан). Их можно подразделить на два основных т и-
п а: саванновые и колючие ксерофильные леса (или кустарники).
Саванновые леса. Основные признаки саванновых лесов таковы:
деревья стоят довольно редко — гораздо реже, чем в муссоновых
лесах, по большей части низкорослы, часто корявы; листья не дают
сплошного покрова, и крона дает много света; часто листья бывают
перистыми и имеют в иных случаях расширенные зелёные черешки;
последние ассимилируют и в то время, когда листочки перистых
листьев осыпались. Встречаются также и некоторые вечнозелёные
деревья (пальмы, хвойные). Подлесок в саванновых лесах или со-
вершенно отсутствует, или же состоит из ксерофильных кустарни-
ков, Но особенно развит травянистый покров из злаков и двудоль-
ных — его пышное развитие связано с достаточным количеством
света в лесу, Лианы редки, эпифиты встречаются чаще. В сухое
время года деревья стоят совершенно голые, трава бурая и сухая.
Саванновые леса тесно связаны с саваннами, где деревья раз-
бросаны по травянистому покрову лишь отдельными редкими эк-
земплярами. Саванны описываются ниже как травянистые жест-
колистные фитоценозы (см. стр. 249).
Колючие ксерофильные леса («колючее редколесье») и колючие
кустарниковые фитоцеиоэы развиваются при ещё более континен-
тальном климате и при большей продолжительности засухи. Это —
наиболее ксерофильные группы деревянистых формаций тропиков.
В большинстве случаев это невысокие деревца или кустарники с
листвой, опадающей во время засушливого периода. Листья часто
перистые и нередко очень колючие. Особенно характерны бебсвые:
виды акаций, Prosopis, Parkinsonia и др.
Часто здесь встречаются тонкоствольные лианы, нередки эпифи-
ты, но травянистый покров беден, в частности^ злаками. Особенно
характерно в этих лесах то, что здесь всегда богато представлены
колючие растения (отсюда их название).
5 Подобные фитоценозы особенно развиты в Африке, Америке,
Австралии и т. д., местами занимая обширные площади. В некото-
рых местах они разрежены и при дальнейшем ухудшении климата
в сторону засушливости сменяются пустынями.
Краткий^обзор ксерофильных лесов в разных странах., Ксеро-
фильные леса в Африке. Как саванновые леса, так и колю-
чие фитоценозы в тропических областях Африки крайне разнообраз-
ны. Первые указываются для Восточной Африки, для области Банг-
веоло, Южного Судана, Абиссинии, Северной Родезии и пр. Деревья
относятся главным образом к бобовым (Acacia, Berlinia, Erythrina
и Др.) и имеют до 20 м высоты. Особенно интересны саванновые
леса типа «плодовых садов», названные так потому, что по внешнему
виду они удивительно похожи на наши яблоневые сады, заросшие
травой, с низкорослыми сильно ветвистыми деревьями, вроде наших
Яблонь, Подобные «сады» имеются в Восточной Африке, в Сомали
и т. д. (рис, 96),
Для ксерофильных лесов характерны деревья с перистыми
листьями; особенно многочисленны акации; часты также виды и
с тройчатыми листьями, Лианы представлены большим количеством
видов. Травянистый покров богат видами лишь на более осветлён-
ных местах. Вообще же эти фитоценозы в Африке крайне разно-
образны, Интересно указать на формацию кактусовидных молочаев
в Восточной Африке.
Ксерофильные леса в Америке, В тропических
частях Америки очень широко представлены все типы ксерофиль-
ных лесов, связанные, с одной стороны, с влажнотропическими
леса?»» с другой, — с саваннами и пустынями.
199
Встречаются также саванновые леса типа «плодовых садов»
(в Венецуэле, Восточном Перу и др ) И здесь характерны деревья
из бобсвых (Cassia), иногда встречаются и кактусы (американская
особенность).
Крайне интересным типом ксерофильных лесов является так
называемая «каатинга» Бразилии, занимающая обширные простран-
ства между кампосами (см. стр. 254) на юге и влажнотропическими
лесами Амазонки и ее притоков на севере. Каатинга состоит из це-
лого ряда типов, причем можно различать каатинги, состоящие или
из кустарников, или из деревьев в кустарниковой каатинге заросли
могут быть очень густы Деревья и кустарники обычно теряют
листья во время засухи Особенно характерно здесь обилие кактусов
и растений с шипами и колючками. В древесной каатинге на деревьях
можно встретить многочисленные эпифиты, особенно из американ-
ского семейства бромелиевых и в частности замечателыую Tillan-
dsia usneoides («луизианский мох»), свешивающуюся длинными ни-
тями с ветвей, встречается также эпифит — ваниль (Vanilla). Среди
деревьев преобладают представители бобовых, сами деревья часто
низкорослы, с широкой распростертой кроной, немало колючих
молочайных, некоторые из которых древовидны (Manihot dichotoma
дает кау 4ук), об обилии кактусовых (тоже нередко древовидных —
Cercus до 10 м) уже было сказано. На почве — почти исключительн©
мелкие кактусы и бромелин.
201
Замечательным элементом каатинги является дерево из семейства
Bombacaceae— Cavanillesia arborea (рнс. 97), толстый ствол кото-
рой имеет несколько метров в поперечнике, сужен кверху и книзу
и очень похож на редьку. В стволе запасается большое количество
воды (борьба с засухой)
Не останавливаясь на ряде других американских типов колючих
лесов и кустарников, укажем на своеобразные, очень распространен-
ные в Северной Америке (Новая Мексика, Техас, отчасти Нижняя
Калифорния) фитоценозы типа сухих саванн с господством «мескито»
(так называется Prosopis glandulosa, а также и другие виды этого
рода). Это — небольшие деревья или кустарники. Здесь же встре-
чаются кактусы (Cereus giganteus) и др. Заросли мескито предста-
вляют уже переход к полупустыням,
IV. КЛАСС ФОРМАЦИЙ—ЖЕСТКОЛИСТНЫЕ ДРЕВЕСНЫЕ И КУСТАРНИ-
КОВЫЕ ФИТОЦЕНОЗЫ (uURILIGNOSA)
Две группы формаций: 1) жестколистные леса (Duris lvae)n 2) жест-
колистные кустарниковые фнтоценозы (Durifruticeta)
Общая характеристика. Класс формаций жестколистных лесов
и кустарников тесно связан с определенными климатическими усло-
виями и у же внешне указывает на значительную сухость климата—
растения производят впечатление ксерофитов. Очень характерны
листья, которые обычно так хорошо отражают внешние условия
су шествования, они жестки, хорошо снабжены механической тканью
(склерофильные листья), вечнсзелены, пластинка обычно невелика
и часто покрыта волосками, чтобы уменьшить нагревание пластин-
ки, они стоят не перпендикулярно к солнечным лучам, а косо,
поэтому лучи скользят по листьям.
К данному классу относятся не только растения с жесткими
листьями. У некоторых видов на стеблях листья совершенно ре-
дуцируются, и стебли становятся зелеными — получаются жизнен-
ною формы с прутьевидными жесткими стеблями. Хорошим приме-
ром последних может служить испанский дрок ( ис 35).
Листья, в противоположность лавровому типу, нико'да не быва-
ют блестящими, но тусклые или серо-зеленые, часто матовые от смо-
листых выделений из особых железок В листьях имеется ряд механи-
ческих приспособлений, чтобы они не спадались во время засушливо-
го периода устьица обычно погружены ниже поверхности, и ряд при-
способлений задерживает транспирацию через устьица. Очень рас-
пространено образование колючек, нередко колючими бывают листья.
Жестколистные растения в большинстве случаев красиво цветут,
причем преобладает желтая окраска цветков. Многие виды вы-
деляют эфирные масла, что вместо с пахучими цветками придает
зарослям жестколистных растений сильный своеобразный аромат.
Почки защищены довольно слабо, некоторые виды (например
оливковое дерево) совершенно лишены почечных чешуй.
Классическим местом развития жестколистных лесов и кус-
тарниковых фитоценозов является Средиземноморская
о б л а с т ь, т. е. северные и южные берега Средиземного моря
(большая часть Пиренейского и Апеннинского полуострова, южная
802
часть Балканского и др., южный берег Крыма относится сюда же).
Вторая территория сильного развития жестколистных лесов —
Рис, 98. Прохождение январской изотермы в 4° и июльской в 20° (по Рюбелю).
Рис. 99. Границы распространения (жирная линия) маслины (Olea euro-
раеа) в Средиземноморье (по Рюбелю).
Калифорния, она простирается также на север вдоль гор южного
Орегона. Большие площади заняты данным типом в Австралии
20»
(на юге Виктории и в Юго-западной части Австралии). Кроме этих
трех основных территорий, жестколистный тип развит в Южной
Африке (Капская область) и в Чили между 40 и 50° ю. ш. Таким
образом, жестколистный тип в большей или меньшей степени развит
во всех пяти частях света.
Климатические условия областей жестколистных лесов очень
характерны. В Средиземноморской области лето очень сухое и жар-
кое (средняя температура самого жаркого месяца 22—28°, осадков
в июле от 2 до 23 мм), зима — не холодная (средняя температура
января 5—12°) и с большим количеством осадков. Годовое количест-
во осадков оО—75 см, но большая часть их выпадает зимой. В летние
месяцы небо безоблачно и беспрерывно сияет солнце, зимой, несмот-
ря на осадки, очень много солнечных дней. Область Средиземно-
морской жестколистной растительности хорошо очерчивает изитерма
января в 4° и на крайнем западе изотерма июля в 20°. — только к
югу от этих изотерм лежит средиземноморский климат (рис. 98),
часто называемый климатом маслины (оливкового дерева) (рис. 99).
Неблагоприятным временем года, как видно из сказанного, явля-
ется сухое и жаркое лето, поэтому растения должны выраба-
тывать ксерофильные приспособления, к числу которых относятся и
жесткие листья и прутьевидные стебли; с другой стороны, весной
здесь характерны многочисленные луковичнье и клубневые эфемеры
(о флористической характеристике Средиземноморья см. стр. 341).
Во время летней жары жестколистные растения находятся,
как показывают исследования, как бы в состоянии покоя и ассими-
лируют очень слабо, Ассимиляционная деятельность имеет место
главным образом весной.
Климатические условия Кал! форнии и ех областей Австралии,
где развт ты жестколистные леса, очень схожи с указанными для
Средиземноморья.
Рассматриваемый класс'формаций включает две группы форма-
ций: же стколистные леса (Durisilvae) и же с т к о л и , i к ы е
кус; ар hi кизые ф! то це н озы (Dunfruticeta). Особенно разви-
ты последние, что именно и характерно для данного клас-
с а, тогда как во всех других деревянистых классах (тропические
леса, летнезеленые, хвойные и пр.) на первый план выступают
деревья, соответствующие же кустарниковые формации имеют под-
чиненную роль. Одной из причин этого в Средиземноморье является
влияние человека, так как вырубки лесов, распашки и
пр. имели здесь место с отдаленнейших времен. И в настоящее время
воздействие человека на дикую природу очень велико. Имеется
не мало районов, где не только уничтожены леса, но и почвы изме-
нены так, что восстановление леса невозможно: лишенные нормаль-
ного растительного покрова, почвы смыты, развеяны ветром и на по-
верхности находится голый камень.
Однако сильное развитие кустарниковых формаций, возможно,
вызывается и климатическими причинами. В самом деле, жестко-
листные леса в их естественном состоянии не дают пышного разви-
тия, и деревья здесь не достигают таких больших размеров, как это
имеет место и в тропических и в умеренных лесах. Причина кроется
204
в том, что теплое время года не совпадает с
дождливым. Поэтому области жестколистных лесов — это
области угасания леса и его деградации в кустарниковые формации.
Можно думать, что и в докультурное время кустарниковые формации
были здесь развиты, хотя, конечно, в меньшей степени, чем в настоя-
щее время.
Рассмотрим основные растительные формации четырех главных
областей жестколистных лесов и кустарниковых фитоценозов.
Жестколистные формации в Средиземноморье. Жестколистные
леса состоят из вечнозеленых дубов — каменного дуба (Quercus
ilex), в западной части также пробкового дуба (Q. suber). Последний
особенно распространен в Португалии, Испании, Марокко и Алжире
(менее часто на Корсике и в Италии), поселяясь на основных поро-
дах, вто время как каменный дуб характерен для почв известковых.
В Крыму эти два дуба 5* диком состоянии не встречаются, но культи-
вируются в западной части Ю кного берега. Эпифиты из высших
растений почти совершенно отсутствуют, лиан очень мало (однако
имеется плющ, Tamus communis, Smilax и др ). Эти веч! озсленые
дубовые леса (особенно из пробкового дуба) достаточно светлы, и
потому в них обычно хорошо развит подлесок и травянистый покров;
подлесок состоит из многих видов, характерных и для маквиса
(см. ниже), т. е. кустарниковых вечнозеленых зарослей.
Из вечнозеленых деревьев и кустарников дубовых лесов укажем:
земляничное дерево (Arbutus untdo), вереск (Erica arborea), мирту
(Myrtus communis), виды ладанника (Cistus) и др.
205
Что касается маслины, этого характернейшего дерева Среди-
земноморья, то дикорастущих лесов маслины в настоящее время не
имеется, а в культурном состоянии она играет исключительную
роль в общем ландшафте страны (рис. 100).
Несравненно большую роль, чем леса, в Средиземноморье играют
кустарниковые фитоценозы, представленные весьма различными
формациями и носящие некоторые народные названия, так, разли-
чают маквис (корсиканское название), гаригу (юг Франции), томил-
ляры (Испания), фригану (Греция) и др.
Маквис распространен почти по всему Средиземноморью, но
осе б. н. о предпочитает более влажные условия, поэтому более
развит в западных частях и особенно на Корсике. Он состоит
из кустарников со склерофильным и эрикоидными листьями,
а также из прутьевидных форм, в среднем вы.о а кустарников
колеблется от Р/2до 4 м. В зависимости от ряда условий в маквисе
преобладают то одни, то другие виды. Наиболее пышную раститель-
ность имеют маквисы с преобладанием земляничного дерева (Arbutus
unedo) до о—8 м высоты, некоторые деревья (Ph Пугеа) достигают
даже 11 м, из других деревьев отметим мастичное дерево (Pistach
lentiscus), мирту (Myrtus communis). Это — уже почти лес. Другую
группу маквисов составляют заросли с господством ладанник!
(Cistus), еще укажем на маквис из олеандра (Ncrium oleander):
последний характерен для берегов ручьев и речек. В Крыму маквис
отсутствует.
Г а рига распространена в Средиземноморье больше, чем преды-
дущий тип она состоит из низкорослых вечнозеленых кустарников
и полукустарников не выше 1 м.
креме того, она беднее видами,
причем кустарники обычно не
образуют сплошных зарослей.
Есе это позволяет думать, ч^о
гарига в большинстве случаев—
это по сравнению с маквисом
дальнейшая при воздействии
человека регрессивная стадия.
Впрочем весенняя эфемеровая
флора гариги очень богата.
Можно различать целый ряд фор-
маций гариги в связи с условиями
существования, причем в разных слу-
чаях господствуют различные виды.
Очень распространена гарига с гос-
подством. кустарникового дуба (Quer-
cus coccifera) (рис 101) с колючими
жесткими листьями (по-провансаль-
ски он называется Garou ha—отсюда
слово гарига) Среди других видов
типичны для гариги такие растения,
как чабрец (Thymus), розмарин (Rosmarinus), дафме (Daphne gnidium), дрок
(Genista scoparn) и др Другим типом гариги являются заросли карликовой
пальмы (Chamaerops hurmhs), единственной пальмы (дикорастущей) Европы, ее
заросли встречаются в южной Испании, иа Балеарских островах, в Сици-
лия, в Алж н''г‘ ч Марокко
206
Т оми л л я р ы — особый тип кустарниковых формаций с гос-
подством ароматичных и обычно сильно опушенных растений, хотя
«тсмилляры» (от растения Thymus) — испанское народное название,
но этот термин обычно распространяется на аналогичные формации
всего Средиземноморья. Среди ряда формаций укажем томилляры
с господством Thymus, с розмарином (Rosmarinus), с лавандой
(Lavandula). Все они очень ароматичны, что ^лужит защитой от
уничтожения скотом.
На востоке имеется своеобразный тип кустарниковых формаций
— ф р и г а н а (на Балканском полуострове, особенно в Греции,
на Крите, в Малой Азии). Здесь наиболее типичны вечнозеленые
кустарники с колючками и шипами, к которым при-
соединяются ароматичные губоцветные и прутьевидные кустарники.
Колючие кустарники нередко имеют шаровидную форму (Poterium
spinosum, Euphorbia acanthiothamnos, Genista acanthoclada и др ).
Фригана предпочитает сухие южные склоны, В Крыму фригана
отсутствует.
При еще более усиленном влиянии человека кустарниковые
формации деградируют все более и более, и, наконец, мы находим
пространства, почти совершенно лишенные растительности (антро-
погенная пустыня). Можно установить такой ряд деградации в связи
с влиянием человека: жестколистный лес — маквис — гарига —
каменистый выгон —пустыня.
В СССР, именно в Крыму, все эти вечнозеленые кустарниковые
формации отсутствуют (Станков), хотя в русской литературе
нередко указывался для Южного берега Крыма маквис (Реманн.
А г г е е н к о и др). Правда, в Крыму имеются некоторые вечно-
зеленые элементы, но они или не ооразуют кустарниковых зарослей
или же совсем не характерны для средиземноморского маквиса
(например Ruscus ponticus — рис. 191). Имеющиеся же кустарни-
ковые заросли состоят из кустарников с опадающими листьями и
ближе всего подходят к балканскому шибляку (см. стр. 229). Во-
обще же н_, жно заметить, что такие названия формаций, как «мак-
вис», «шибляк» и др., очень мало определенны с ботанической (и
особенно фитоценологической) стороны.
Обзор жестколистных формаций в других странах. Жестко-
листные леса в Америке распространены в южном
Орегоне и в нижней Калифорнии (Северная Америка), в климати-
ческих условиях, очень близких к средиземноморским. И здесь
также имеются и леса и кустарниковые заросли, и здесь господствую-
щими деревьями являются вечнозеленые дубы, число которых
больше, чем в Европе. Помимо дубов, имеются и Другие вечнозе-
леные (в частности американский вид Arbutus M^nziesn). Основные
ассоциации. Qtkruis d^nsiflora —Arbutus, Q agrifoha —Arbutus и др.
По калифорнийскому побережью обильно развиты кустарниковые
формации, чрезвычайно похожие на маквис. Этот тип растительности
называется здесь чапараль. Последний состоит из .многочислен-
ных вечнозеленых кустарников некоторые виды дубов и ряд
видов, напоминающих средиземноморские. Очень характерным р *с-
тением является розоцветное Adenostoma fasciculatum, своим внеш-
207
ним видом похожее иа вереск (Erica). Очень богаты видами то-
локнянка (Arctostaphylos) и род из крушиновых — Ceanothu
(рис. 102).
Небольшой район вечнозеленых жестколистных кустарников
имеется, кроме того, в среднем Чили (Южная Америка) на высотах
1 000—2 000 м В более благоприятных условиях над кустарниками
Рис 102 Жестколистная флора Калифорнии (в чапарале)
I—icknostonia fasciculatum, 2—Ceanothus cunffttus, з—Quercus dumosa, 4—Ce anothus
papHIosus (из Шимпера)
возвышаются деревья Физиономически и систематически заросли
кустарников похожи на калифорнийские
Жестколистные леса в Австралии Развиты
главным образом на юго-западе, а также в некоторых юго восточных
частях Здесь жестколистные леса играют гораздо большую роль,
чем в других аналог !чных областях, причем они занимают до трети
всей юго-западной провинции. Главное господствующее дерево
здесь эвкалипт с рядом видов Основную роль играют Еиса-
ly plus marginata, Е di versicolor, Е redunca, дающие одно-
составные леса из того или другого вида, примесь других деревьев
лишь в виде исключения Леса эвкалиптов очень оригинальны
высокие, стройные деревья до 60—70 м, со стволами, лишенными
ветвей, до 40—50 м высоты, хотя деревья стоят довольно часто, но в
лесу очено светло, так как листья эвкалиптов расположены ребром
последнее способствуют пышному развитию вечнозеленого подлеска,
который состоит из многочисленных видов, узнать которые по внеш-
нему ваду без цветов почти невозмо/кно — настолько схожи веге-
тативные части Особенно обильны виды из семейств бобовых и
20S
протейных (Proteaceae). Эпифитов и лиан нет. Вообще физиоиоми-
чески эвкалиптовые леса Австралии очень отличаются от вечно-
зеленых дубовых лесов Средиземноморья и Калифорнии, предста-
вляя все переходы к эвкалиптовым лесам саваннового типа. Эти
пост Д1 и^ занимают обширные пространства на северо-востоке.
Господствующими деревьями являются эвкалипты (ряд видов),
образующие светлые насаждения благодаря положению своих ли-
стьев (рис 103)
Рис. 103 Саванновый лес из эвкалиптов в Квинслэн де—Австралия
(из Ши и пера)
Из других деревьев очень типичны характерные для Австралии
акации, часто с зонтиковидной кроной, а также казуарины. Травя-
нистый покров разнообразен и меняется в своих аспектах в зависи-
мости от времени года
Жестколистные кустарниковые формации
в Австралии крайне разнообразны и напоминают средиземно-
морск! й маквис, они не превышают 1—2 м в высоту и носят местное
название скрэб Растительность в иных случаях очень близка к под-
леску эвкалиптовых лесов Листья скрэба жестки, стоят часто реб-
ром, иногда типа вересковых или хвойных, серо-зеленого цвета,
тусклые, часто покрытые восковым или смолистым налетом Иногда
вместо гистьев филлодии (например у акаций), ряд растений имеет
колючки (рис 104) Сравнительно со средиземноморскими кустарни-
ками растения скрэба имеют некоторые особенности крайне харак-
терны изолатеральные листья (т. е. листья, у которых верхняя
14 География пастеяий 2(К
Рис 104 Жестколистная флора в Австралии
1 Banksia serrata, 2—Dryandra mucronulata 3—Chori-
zema triangularis, 4 —Myoporum tuberculatum, 5 —
Acacia flonbjnda, 6—Hakea saligna (из Шимпера)
и нижи я я стороны анатомически построены одинаково) и раз-
личные приспособления к погружению устьиц и их защите
Скрэб занимает в юго-западных,центральных и некоторых других
частяхАвстралии огром-
ные пространства и фи-
зиономически представ-
ляет крайне унылую кар-
тину, при взгляде с воз-
вышенности — это бес-
конечное серо-зеленое
море густых кустарни-
ковых зарослей, заблу-
диться в которых ничего
ие стоит. На первый
взгляд совершенно не-
возможно р^ зобраться
в видовом составе —так
похожи вегетативные
органы Лишь во время
цветения можно видеть,
насколько скрэб богат
видами.
Господствуют семей-
ства: бобовых (целый
ряд эндемичных родов,
очень многочисленны
акации), миртовых (ку -
старниковые эвкалип-
ты, Melaleuca), протей-
ных и др , кроме того,
характерны прутьевид-
ные казуарины (рис
213) и др Травянистых
растений очень мало
При наблюдениях
над скрэбсм в течение
целого года можно
установить ряд аспек-
тов, сильно отличаю-
щихся Друг от друга
Описанная картина без-
жизненного серо-зеленого скрэба относится к марту — апрелю,
когда вегетация почти прекращается из-за засушливого вре-
мени. В мае скрэб* пестрит разнообразными цветками крас-
ного, розового, желтого, белого цветов. В конце июля темпе-
ратура достигает своего минимума, а количество осадков — макси-
мума (как раз в противоположность Средиземноморью), и в это время
заросли цветущих аканий представляются желтыми, а остальные
р стения дают пестрый ковер В августе — то же причудливое
смешение различных красок.
Южноафриканская жестколистная расти-
тельность. Здесь, в Капской области, жестколистные кустар-
никовые заросли распространены очень сильно Господствуют виды с
листьями эрикоидного и игольча-
того типэв из различных семейств
(рис. 105). Вересковые представ-
лены особенно обильно (более 400
видов), также бобовые, рутовые,
крушиновые, протейные и др.
Так как местами в Капской
области воздух очень влажен, то
вересковые заросли приближа-
ются к приаглантическим западно-
европейским «верещатникам» (см.
стр. 247), с которыми и среди-
земноморский маквис связан пе-
реходами.
Жестколистные леса и человек
Уже при описании различных
типов этих лесов и жестколист-
ных кустарниковых фитоценозов
была указана исключительная
роль человека при формировании
некоторых из них. Это особенно
относится к жестколистным форма-
циям Средиземноморской области.
Это та область, к которой отно-
сятся столь хорошо известные сло-
ва Энгельса: «людям, которые
в Месопотамии, в Греции, вМалой
Азии и в других местах выкорче-
вывали леса, чтобы добыть таким
путем пахотную землю, и не сни-
лось, что они этим положили нача-
Рис. 105 Жестколистная флора в
Капской области
1 —Gnidia pmifolia, 2— Leucadendr^e
Levisancus. 3 — Coleonema album; 4 —.
Agathosma capitatum (из Шимпера).
ло нынешнему опустошению этих стран, лишив их вместе с лесами
центров собирания и хранения влаги Когда альпийские итальянцы
вырубили на южном склоне гор хвойные леса, так заботливо охра-
няемые на северном, они не предвидели, что этим подрезывают
корни скотоводства в их области, еще меньше они предвидели, что
этим лишают свои горные источники воды на большую часть года,
с тем еще эффектом, что тем более бешеные потоки они будут изли-
зать в долину в период дождей» («Диалектика природы»).
Восстановлению лесов в Средиземноморье препятствует не толь-
ко то, что их уничтожение имеет длительную историю, насчитываю-
щую тысячи лет, а также и то, что сами климатические условия не
являются благоприятными для быстрого их восстановления (сухость
климата). Те кустарниковые фитоценозы, которые столь характер-
ны для Средиземноморья, являются в большинстве случаев дери-
ватами леса, но кустарники сами в результате выпаса деградируют
и превращаются в заросли колючих низкорослых растений (гарига,
томилляры и др.). В дальнейшем связный растительный покров
нарушается, почва смывается или развеивается ветрами и обнажается
каменистый субстрат, на котором уже не могут расти растения унич-
тоженных ^формаций. В результате получается пустыня; для есте-
ственного восстановления прежней растительности нужны тысяче-
летия и отсутствие воздействия человека, что, конечно, не может
иметь места. Так обстоит дело на обширных территориях Среди-
земноморья, в СССР же — в Крыму на его Южном берегу. Необхо-
димая задача — не восстанавливать леса в их прежнем виде, а
производить облесение новым ассортиментом древесных пород,
Сообразно новым создавшимся условиям.
V. КЛАСС ФОРМАЦИЙ —ЛЕТНЕЗЕЛЁНЫЕ ДРЕВЕСНЫЕ И
КУСТАРНИКОВЫЕ ФИТОЦЕНОЗЫ (AESTILIGNOSA)
Летнезеленые леса одеты листвой лишь в благоприятное время
года, осенью листья опадают, и в холодное время деревы стоят
без листьев. Эго повторяется из года в год. Полчки на деревьях, как
Рис. 106. Границы некоторых древесных пород в Европей-
ской части СССР.
212
правило, одеты почечными чешуями — защита на неблагоприятное
время. К этому же классу относятся и кустарниковые формации
летнезеленого типа (Aestifruticeta).
Летнезеленые леса и кустарниковые фитоценозы распространены
почти исключительно в северном полушарии, в его умеренных
частях, находящихся под влиянием океанического климата. В час-
тях более континентальных летнезеленые леса переходят в иголь-
чато-хвойные; это особенно хорошо можно видеть в умеренной зоне
Евразии, где летнезеленые леса, столь типичные для Западной
Европы, выклиниваются в Европейской части СССР (рис. 106), и
Азиатская часть в своих более континентальных частях лишена
совершенно этих лесов.
1. Группа формаций — летнезелёиые лега (Aestisilvae)
Общая характеристика. Летнезеленые леса распространены в
Западной Европе в ее умеренных частях, на юг. (главным обра-
зом) Европейской части СССР (узкая их полоса тянется и в Запад-
ной Сибири между тайгой и степями), в Крыму и на Кавказе, иа
Дальнем Востоке, в Манчжурии и на востоке Китая, на Камчатке,
Сахалине, на северных Японских островах, в Северной Америке,
в приатлантических штатах. В Южной Америке летнезеленые леса
развиты в Патагонии, на других же континентах южного полушария
ие представлены, не имея там подходящих условий.
Климатические условия территорий, занятых летнезелёиыми
лесами, таковы: по крайней мере четыре летних месяца имеют тем-
пературы выше 10*; средняя температура самого теплого месяца
®т 13 до 22 —23*, самого Холодного до — 6° (но в наиболее
континентальных частях СССР до—10—12*). Максимум осадков
падает на теплое время года; месяц, наиболее богатый осадками,
имеет около 100—130 л/л/ осадков (в континентальных частях СССР
менее 100 л/л/).
Благоприятные условия во время вегетации (теп-
ло + осадки) дают возможность пышного развития растительности
мезофильного типа (с некоторыми исключениями, см. ниже), листья
на деревьях имеют широкие пластинки (бук, дуб, клеи и др.), по-
этому подобные леса называются широколиственными. Очень немно-
гие летнезелеиые деревья имеют небольшие листовые пластинки (бе-
рёза, осина), и леса из этих деревьев называются мелколиственными.
Листья вообще яркозеленые и так как они приурочены к летнему
влажному периоду, то имеют довольно нежную консистенцию.
Редко листья очень крупны (платан, конский каштан) или пе-
ристы (ясень, рябина, орех — Jug Ians — рис. 118). Вообще размеры
и форма листьев варьируют очень слабо сравнительно с деревьями
тропических лесов. Опушение листьев обычно отсутствует.
Листья не приспособлены к зимнему неблагоприятному вре-
мени и на зиму опадают; в этом видят защиту деревьев от излиш-
него зимнего испарения. Действительно, почва зимой очень холодна,
являясь физиологически сухой. Кроме того, стволы и ветви защи-
щены толстой корой, а почки плотными чешуями, нередко смоли-
стыми. Все эт© — защита от зимнего испарения.
Летом и зимой летиезелёные леса, естественно, физиономически
йчеиь несхожи. Летом эти леса тоже дают различные, физиономиче-
ские картины в зависимости от пород, входящих в их состав (на-
пример тёмные тенистые буковые леса и осветлённые берёзовые
леса), или от других причин; так, светлые опушки предоставляют
все условия для пышного развития крон деревьев и для более пол-
ного развития кустарников, отчего опушечная стена леса является
очень густой; в глубине леса совершенно другая картина: высокие
стволы с верхушечной кроной, более слабо развитой подлесок и пр.
Обращает внимание обильное ветвление и, как следствие этого,
хорошо развитая крона; бук даёт ветви 8-го порядка (в тропическом
лесу ветвление редко идёт выше 5-го порядка).
Интересное отличие от тропических лесов состоит в том, что
деревья верхнего яруса имеют в общем одну и ту же высоту, и если
посмотреть на лес сверху, то он представляется ровной зелёной
поверхностью. Это зависит от очень небольшого числа видов де-
ревьев, слагающих верхний ярус, причём нередко здесь имеется
лишь один вид, занимающий очень большие пространства, например
обширные чисто буковые леса, чисто дубовые леса и т. п. Впрочем,
иногда летиезелёные леса более богаты древесными породами.
В своём фитоценологическом сложении летиезелёные леса в
ряде случаев обнаруживают довольно сильно развитую ярусность;
так, можно различать два древесных яруса (один — более высокий,
Другой — более низкий), кустарниковый, несколько травянистых^
зависящих от трав различных высот (до трёх ярусов) и напочвенный
ярус (состоящий из мхов, лишайников, водорослей). В других же
случаях ярусное сложение выражено слабо, а в очень тенистых
буковых лесах иногда летом отсутствует всякая растительность, и
вся почва покрыта плотным покровом из старых спавших листьев.
Ряд признаков отличает летиезелёные леса от тропических;
здесь отсутствуют илн почти отсутствуют: I) лианы, 2) эпифиты,
3) каулифлория, 4) досковидиыё корни-подпорки у стволов.
В летнезелёных лесах недостаточно тепла и влажности для
быстрого роста лиан и, кроме того, зимний период очень неблаго-
приятен для длинных тонких стволов. Все же некоторое число
лиан имеется: хмель, плющ, ломонос, дикий виноград, смилакс и
другие. В наших лесах встречается лишь хмель, в Крыму,
на Кавказе, на Дальнем Востоке и в Северной Америке число лиаи
возрастает.
Эпифиты, относящиеся к цветковым растениям, вообще отсутст-
вуют; можно лишь в виде исключения указать иа несколько эпифи-
тов из цветковых растений в лесах Японии или Северной Америки
(в последнем случае луизианский «мох» — Tillandsia). Но эпифиты,
относящиеся к мхам, лишайникам, водорослям, представлены в боль-
шом количестве видов (это также имеет место и в хвойных лесах):
они поселяются иа коре деревьев, особенно у их оснований. Впрочем,
нужно заметить, что, говоря об эпифитах, обычно имеют в виду
эпифиты, относящиеся к высшим растениям (креме мхов).
Каулифлория имеется в виде исключения, а досковидные корни
в таком виде, как в тропических лесах, совершенно отсутствуют,
имеются лишь некоторые намёки на них, особенно на влажных
почвах, у клейкой ольхи, бука и некоторых других.
Очень характерен световой режим. Здесь наблюдается два
световых максимума: один весной, когда деревья ещё не оделись
листвой, другой — осенью, когда листва леса начинает изрежи-
ваться, но вегетация ещё продолжается (рис. 107). В летние месяцы
.. ~-------.--,---i —г —|
Quercus robur ..........
Carpinus betulus . . .
Corylus avellana . . .
Anemone nemorosa . . .
Xsrurn. maculatuiii . .
Sc ilia nutans ....
Ficaria verna,..........
Ranunculus auricomus .
Primula acaulis ....
A jug a reptans ........
<raleobdolon luteum .
Hlechoma hederacea . .
Рис. 107. Продолжительность вегетации ряда растений дубового леса (в Англии);
видно, что эфемеры (Ficaria, Scilla и др.) начинают вегетировать задолго до
распускания листьев у дуба. (Quercus). Чёрные полосы—вегетация. Буквы
внизу—начальные буквы месяцев (по Салисбери).
интенсивность света очень сильно падает, особенно в лесах из таких
теневыносливых пород, как, например, бук. Таким образом, кривая
света имеет в основном две вершины.
Подобный световой режим резко отзывается иа травянистой
растительности леса. Ранней весной для летиезелёных лесов крайне
характерен резкий взрыв весенней растительности, со-
стоящей главным образом из эфемеров-многолетииков. Они очень
быстро зацветают и затем так же быстро заканчивают свою вегета-
цию, принося плоды и семена; к июню они совершенно выходят из
травянистого покрова, скрываясь под землёй в виде корневищ,
луковиц, клубней и пр. Взрыв весенних эфемеров обычно очень
красочен, так как цветки их ярко окрашены, а число экзем-
пляров так велико, что травянистый покров кажется совершенно
сплошным; летом, как сказано, от него не остаётся никакого следа.
Интересно, однако, что иногда в буковых лесах во время весеннего
светового максимума совершенно отсутствуют эфемеры (см. стр. 219).
В летние месяцы число цветущих растений травянистого покрова
сравнительно невелико, и красочных аспектов почти совсем нет.
Лишь на опушках и на лесных полянкахмы находим пёстрые ковры
цветов. Позднелетиее изреживание леса не сказывается у нас на тра-
вянистой растительности, так как к этому времени все растения уже
начинают заканчивать евою вегетацию. Однако в Западной Европе
замечается осенью некоторое усиление вегетации (см. рис. 107).
Большинство деревьев цветёт рано весной до распускания ли-
215
стьев и является ветроопыляемыми 1 поэтому цветки их невзрачны и
часто собраны в сережки Цветение до распускания листьев связано
с лучшим обеспечением опыления Только небольшое число видов
опыляется насекомыми, из европейских деревьев здесь нужно отме-
тить клен, цветущий весной, и липу, цветущую среди лета и своим
цветением намечающую переход ко второй половине лета Цветки
лнпы Содержат нектар и сильно пахнут, что обеспечивает опыление
Деревьев, находящихся даже в глубине лесных массивов
Рис 108 Ареал 6yKa(Fagus sdvatica) в Средней Европе Места с редкой штри-
ховкой—редкая встречаемость, белые места внутри ареала—отсутствие бука.
Точки вне ареала—искусственные посчдкн В Крыму—Fagus taunca на Кавка-
зе—F on entails (по Леммермайеру с изменениями Алехина).
Большая часть деревьев имеет семена или плоды, снабженные
крыльями или вообще приспособлениями для перенесения ветром,
что, конечно очень важно для завоевания лесом новых территорий
У растений кустарниковых и травянистых ярусов опыление и
рассеивание плодов происходит иным способом Впрочем, об этом
было сказано выше (стр 122)
Наиболее распространенные формации лет-
незеленых лесов следующие
Буковые леса. Формация буковых лесов — одна из наиболее
важных для Западной Европы, однако бук избегает как слишком
влажного морского климата (Ирландия, запад Англии), так и более
континентального (рис 108) На русской равнине бук встречается
216
лишь на западе Украины, доходя на восток до б. пограничной
р Збруч (в одном районе и на левом берегу этой речки, рис. НО).
В более северных частях Западной Европы буковые леса занима-
ют равнины, а на юге переходят в горы, занимая определенный
пояс, ниже которого находятся дубовые или каштановые леса (по-
следние на почвах, лишенных извести) В Крыму и на Кавказе бу-
ковые леса тоже занимают определенный высотный пояс
Хотя буковые леса Европы состоят из различных видов бука
(в Западной Европе и в Западной Украине — Fagus sJvatica, в
Крыму — F taurica, представ-
ляющ1 й переход между евро-
пейским и кавказским буком,
на Кавказе — F onentahs/,
но фитоценологически они очень
схожи, бук — основной и един-
ственный доминант, опред ляю-
Щ1Й все остальные отношения
в силу своей исключительной
теневыносливости Листья бука
выдерживают до х/80силы света,
это чрезвычайно низкая вели-
чина, так как такое теневое ра-
стение, как Hepatica triloba,
начинает отмирать при 1/37 Силы
света В результате, буковые
леса часто лишены подлеска и
летних травянистых растений
Особенно характерно пышное
развитие весенних эфемеров
Anemone nemorosa, Scilla
bifolia, Galanthus mvahs
109 Белый (западный) подснеж-
ник (Galanthus nivalis)
(рис 109), виды D.ntaria
Для Западной Европы Рис
наиболее характерны так е
ассоциации Fag^tum asperulosum odoratae, F alliosum ursini, F ca-
ricosum albae, F bromosum ramosi и др В равнинных условиях
наиболее характерна первая ассоциация с Anemone nemorosa ранней
весной Интересно, что в подлеске нередко встречаются вечно-
зеленые виды тисс (Taxus baccata), остролист (Ilex aquifolium),
а плющ иногда покрывает почти сплошь почву, это — результат
замещения факторов в глубине леса фитоклимат замещает собой
более мягкий океанический климат.
В СССР (кроме Крыма и Кавказа) буковые леса встречались
до воссоединения Западной Украины лишь в Проскуровском округе
на самом запад. государственной границы, занимая чистыми
насаждениями пространство всего лишь около 400 га Здесь устано-
влены ассоциации Fagetum asperulosum, F alhosum urs,ni, F athy-
riosum filix feminae и др Однако это лишь восточный форпост
1 Однако некоторые авторы (Вульф) полагают, что в Крыму имеются как
типичные F siivitica и F onentahs, так к переходный между ними F taurica.
217
Ряс. НО Росточная граница бука в Западной Украине,
течки—отдельные местонахождения восточной границы
Сплошного распространения (по Шаферу)
буковых лесов, которые в западных областях Украины играют
очень большую роль (рис НО) Здесь бук образует леса как по
склонам Карпат, так и на равнинных местах в Подольском Ополье
По склонам Карпат бук дает хорошо выраженный пояс чистых
буковых лесор в пределах 500—1 200 м, но отдельные деревья под-
нимаются и значительно выше Важным спутником бука является
европейская пихта (Abies alba)—дерево, до присоединения западных
областей не встречавшееся в наших пределах (рис 128) Отметим
также тисс (Taxus
baccata),enb(Picea
excelsa) Буковые
леса Ополья этих
спутников не име
ют Если в арпат
ских б> ковых ле
сах очень важную
роль играет пихта,
то в Ополье не
меньшую роль
играет граб (Саг
pinus Betulus) Из
наиболее харак
терных травяни
стых видов буко
вых лесов отметим
ясменник (Aspe
rula odorata), ве-
рсникх горную
(Veronica monta
па), зубянку (Den
*ап bulbifera),
хохлатку (Coryda
hs cava), лук-че
ремшу (Allium
ursinum)
Помимо пихты,
в буковых лесах
Западной Украины
встречается немало видов, отсутствовавших до того в СССР из кустар-
ников—жимолость (Lonicera nigra), из трав: Aposeris foetid a, Digitalis
purpurea, Helleborus purpurascens, Lysimachia nemorum и т д
Для Крыма очень характерны буковые леса с весенним эфемером
зубянкой (Dentaria quinquefoha) ассоциация Fagetum dentariosum,
это прекрасные высокоствольные леса, но лишенные других де-
ревьев и подлеска Летом травянистый покров почти отсутствует
Выше 1 100 м бук становится искривленным и растет куста-
ми — Fagetum subalpinum.
Особо большие площади занимают буковые леса на Кавказе В
связи с высотой их ассоциации меняются, в районе Сочи —Крас-
ная Поляна установлен такой ряд (К о ж е в и и ков)
2 1S
Высота над уровнем моря
Fagetum laurocerasorunj. —...........,............. 200 и
(подлесок из лавровишни . Laurocerasus)
F trachystemonosum ............................... 1 000 м
(травянистый покров развит очень слабо, обильнее других
Trachystemon orientals)
F fihcosum ........................................ 1 880 м
(травянистый покров из папоротников)
F. subalpinum .... ..................... 1 980 м
(полное отсутствие эфемеров, лежащий главный ствол бука).
Своеобразная особенность последней ассоциации — лежащий
главный ствол бука с саблевидно изогнутыми ветвями связан с
Рис til. Ареал дуба (Quercus robur) Точечная граница на востоке—
восточная граница ясеня (Fraxinus excelsior) (по Рубнеру и др )
глубокими^снежными навалами (больше 8 л/т) Здесь наблюдает-
ся «подснежное» развитие бука он развивает листья и цветет в то
время, когда лежит еще снежный покров Естественно, что эфеме-
ры не находят здесь подходящей экологической обстановки: без-
листной светлой стадии лес не имеет
Буковые леса Северной Америки Они раз-
виты в восточных штатах США и на юго-востоке Канады Здесь
господствует американский бук (Fagus americana), ио доминиро-
вание разделяет с иим также сахарный клен (Acer saccharum)
Эти леса особенно замечательны своей осеиией раскраской, так как
кроме сахарного клена красивого по своей раскраске, встречают-
219
ся и некоторые другие виды- Осенью — все тона красных и жёлтых
оттенков. Американские буковые леса менее тенисты, и здесь можи®
встретить много травянистых видов; леса богаты также и деревья-
ми — более 40 видов. Встречаются и лианы: «дикий виноград»
(Parthenocissus quinquefolia), обычное культивируемое у нас ра-
стение, и несколько видов настоящего дикого винограда (Vitis).
Дубовые леса. Они распространены по всей Западной Европе,
в Европейской части СССР, иа Дальнем Востоке, в Северной Аме-
рике и т. д. В Европе три лесообразующих вида дуба: зимний дуб
(Quercus petraea), летний черешчатый дуб (Q. robur, рис. 111) и
пушистый дуб (Q. pubescens), каждый из которых обычно встре-
чается в чистых насаждениях. Они идут на запад (Ирландия, за-
падная Англия) и иа восток (Европейская часть СССР) дальше,
чем буковые леса. В Западной Европе дубовые леса раньше (на-
пример в средние века) были распространены гораздо шире, так
как искусственно поддерживались человеком из-за желудей, слу-
живших кормом для свиней. По*явление в Европе картофеля резке
отразилось на откормке свиней и одновременно на судьбе дубовых
лесов: последние естественным путём быстро стали заменяться
буковыми, как гораздо более теневыносливыми.
В горах дубовые леса обычно образуют более низкий пояс срав-
нительно с буковыми. Что касается размещения дубовых лесов с
господством черешчатого и зимнего дуба, то первые предпочитают
более тяжёлые и сырые почвы, а вторые — более лёгкие и сухие
и не заходят так далеко на север.
Так как дуб является менее теневыносливым деревом, чем бук,
то света в глубь дуоового леса проникает гораздо больше, и потому
там может хорошо развиваться подлесок и травянистый покров.
В наиболее благоприятных случаях можно различать в дубовом
лесу до 7 ярусов.
По своему флористическому составу и в своих ассоциациях
дубовые леса очень различны с запада на восток. Так, в Ирландии
можно встретить леса из зимнего дуба, где субдоминаитом является
вечнозеленый остролист (Ilex aquifolium), и где имеется ещё це-
лый ряд вечнозелёных растения (земляничное дерево — Arbutus
unedo, рододендрон, тисс), некоторые папоротники субтропических
лесов (Hymenophyllum, Osmunda) — всё это совместно с черникой,
кислицей, золотой розгой и т. д. Интересно, кроме того, что неко-
торые виды поселяются как эпифиты на стволах дубов. Подобный
лес напоминает некоторые леса на острове Яве.
Это — крайний западный океанический тип; другой крайностью
могут служить дубовые леса юга Европейской части СССР, назы-
ваемые дубравами. Они образуют особую подзону широколиствен-
ных лесов, но как исключения распространены и в зоне хвойных
лесов и в степной. К востоку от Урала дубовые леса (и дуб) не встре-
чаются по условиям климатического характера.
Наши дубравы основной породой имеют черешчатый дуб (Quer-
cus robur), к которому присоединяются другие широколиственные
породы: клёны (остролистный, полевой, татарский), ясень, липа,
ильм. Иногда дуб встречается в чистых насаждениях. Для под-
220
леска очень характерен орешник. В травянистом покрове —мио-
равное широкотравье»: сныть, медуница, ясменник, зеленчук,
копытень (рис. 112), колокольчики и др.; кроме того, ландыш, со-
ломонова печать (рис. 113, 114); злаки и осоки также имеют широ-
221
кие пластинки (Bromus Benekeni, Brachypodium silvaticum, Ca-
rex pilosa и др.). Развитие плоских широких пластинок, по всей
Рис. ИЗ. Ландыш (Convaliaria majalis)
вероятности, связано с боль-
шой влажностью, с понижен-
ным испарением и со слабым
освещением. Это имеет место и
в буковых лесах.
Для дубрав очень характе-
рен раниевесенний аспект эфе-
меров: лазоревые подснежники
(Scilla cernua), жёлтые вет-
"реницы (Anemone ranunculoi-
des) и чистяк (Ficaria ranun-
culoides), розовые зубяики
(Dentaria), лиловые хохлатки
(Corydalis solida) и др- Столь
раннее и быстрое развитие этих
видов зависит от того, что они.
обладают подснежным
развитием, т. е. развиваются
ещё тогда, когда снег покры-
вает почву; это возможно пото-
му, что температура почвы здесь
около 0° (рис. 115).
Дубравы в Европейской части СССР слагаются из целого, ряда
ассоциаций, но наиболее распространены такие: Quercetum aego-
podiosum (снытевая дубрава), Q. galeobdolosum (зеленчуковая
дубрава)— в западных районах; Q. caricosum pilosae (осоковая
дубрава) — на более сухих местах. Во всех этих ассоциациях под-
лесок из орешника-
Рис. 115. Листья подснежника (Scilla cernua) ранней весной пре-
вил ись из-под снега; на некоторых экземплярах у нее имеются
бутоны (по Келлеру).
Подзона сплошных дубрав в СССР (см. карту в конце книги)1
тянется довольно узкой полосой, однако дубравы встречаются и в
подзоне смешанных лесов, а также и в степной зоне. В послед-
нем случае участки дубрав, комбинируясь с участками степей,
дают «лесостепь» (в более северных частях), или же дубовые лески
связаны исключительно с балками (в более южных частях) —
«байрачные» лески. Здесь иа водоразделах всецело господствует
степь. Взаимоотношения между лесом и степью довольно сложны:
они изучались рядом исследователей (см. стр. 265).
В Северной Америке дубовые леса тоже занимают
более континентальные районы сравнительно с буковыми; в приат-
лантических штатах они расположены далее в глубь страны и
соприкасаются с областью прерий при посредстве переходной
полосы лесостепи (см. ниже). Дубовые леса состоят в отличие
от европейских, из нескольких видов дубов. Впрочем,
североамериканские широколиственные леса чаще состоят из
большого числа других, кроме дубов, широколиственных пород,
чем ещё больше отличаются от европейских (рис. 116): мно-
гочисленные виды клёнов, (Juglans), гикори (СагузА
тюльпанное дерево из магнолиевых (Liriodendron tulipifera)
(рис 117); реже — платаны, Celtis, Ostrya и др Нередки лианы.
О причинах гораздо большего богатства лесов Северной Америки
сравнительно с европейскими см. ниже (стр 350)
Широколиственные леса востока Азии. Богатые широколист-
венные леса находятся также в восточном Китае, Япстии н в юж-
ных частях Дальнего Востока (китайско-японская флористическая
подобласть) Здесь они непосредственно переходят на юге в субтро-
224
пическ! е леса, и на севере — в хвойные и, таким образом, можно
наблюдать все стадии этих переходов
Северная часть подобласти вклю
1ает в себя Мднчи^рию, Приамурье и При-
уссуриискин край, Северный Китаи и Север-
ную и Среднюю Японию Здесь господствуют
хвойные и широколиственные породы, послед
иле с листвой, опадающей зимой (летнезеле
ные), из хвойных — особенно виды сосны, а в
более южных частях также криптомерия (Cry р-
tomena japomca), цефалотаксус (Cephalota-
xus drupacea) и др Широколиственные по-
роды представлены многочисленными видами,
и чеса северных частей китаиско японской под
области гораздо богаче европейских Здесь
встречаются виды дуба (в Приамурье Quercus
mangohca), виды ореха (Juglans), кленов и
целый ряд родов, отсутствующих в Европе
софора (Sophora ррошса), гледичия (Gledi
tschia sinensis), павловния (Pawlownia impe-
rials), аилант (Ailanthus glanduiosa), магно
лня (Magnolia kobus—Япония), камелия (Ca-
meha j iponica —- Япония) и много других
Перечисленные растения в Европе культиви-
руются в парках (у нас в Крыму и иа Кавка-
зе) являясь прекрасными декоративными
растениями
Рис 117 Тюлшанное дереве
(Liriodendron chmense)—ветвь
с цветками (вид близкий к
сев американскому)
В этих летнезеленых лесах обычно очень богат подлесок, состоящий из
видов родов Hydrangea, Lomcera (жимолость), Syringa (сирень), Rhodo
dendron (рододендрон), Li gustrum (бирючина) и др Большинство этих родов
представлено многочисленными видами,
Рис 118 Представитель да jib не г осточных
шигоко чнетвен ых лесов — манчжур кий
орех (Juglans manshurica) Ветвь с сопве
тнями и плод (по В Л Комарову).
отсутствующими в Европе
Помимо обильного подлеска
хорошо представлены, особенно в
более южных частях, лианы и эпи
фиты, к северу постепенно совсем
выпадающие
Подобласть в своих наи-
более северных частях захо-
дит и в наши пределы, но
флора является здесь иаибо
лее обедненной, хотя все же
на Дальнем Востоке мы на-
ходим два дуба (Quercus
mongo hca и Q ckntata),
ясени (Frax nus man^hunca,
F rhynchophy Па), липы (Ti-
ha munshurica, T amurensis),
несколько видов Ulnius (вя-
зы), граб (Carp nuS cordata),
орех манчжурский (Juglans
manshurica) (рис 118), бар-
хатное дерево (Fhelloden-
dren amurtnse) (рис 119) и
другие — одним словом, на-
бор видов, более богатый,
чем в Европе
15 ГелгпаФпя растений
225
называют субтропической провинцией.
Рис. 119. Представитель дальневосточных
широколиственных лесов— бархатное де-
рево (Phellodendron amurense). Ветвь с
плодами (по В. Л. Комарову).
Южная часть китайско-японской подобласти занимает южную Япо-
нию, южный Китай, Юннань, Сечуань и Гималаи. Здесь, в силу благоприят-
ных климатических условий и древности самой территории, флора особенно
богата, являясь субтропической, поэтому эту южную часть подобласти обычно
е - _ , Кроме обычных, широколиственных
пород (дубы, клены и пр,), здесь
уже встречаем пальмы (Trachy-
carpusexcelsa, Livistona chinen-
sis), саговники (Cycas revoluta),
южные хвойные (Podocarpus, Cun-
ninghamia, Glyptastrobus и пр/Ь
такие роды, как Piper, Camphora,
Aralia, Melia и т. п, Все это со-
здает ландшафт субтропического
характера.
Широколиственные леса
южного полушария. Как уже
указывалось, они встреча-
ютсялншь в Патагонии и на
Огненной Земле, доходя на
севере до 45ст. Основным де-
реном является южный бук—
Nothofagus; встречаются в
подлеске некоторые вечно-
зелёные виды (например
барбарис). Эти леса, особен-
но зимой, похожи на леса
северного полушария, когда
они стоят без листьев и за-
сыпаны снегом, а глубина
снежного покрова в некото-
рые зимы доходит здесь до
1— Р/3 М.
Мелколиственные леса (бе-
резовые и осиновые). Сюда
относятся леса из различных
видов берёзы (Betu 1а) и осины
(Populus tremula). В большинстве случаев их нужно считать вто-
ря чными, возникшими в результате порубок широколиственных
и хвойных лесов. Таковы, например, березняки и осинники, зани-
мающие в СССР очень большие пространства почти в любой об-
ласти (например в Московской). В других случаях, березняки воз-
никают на заброшенных пашнях (сюда заносятся семена), но и
здесь являются временным типом, так как в дальнейшем сменяются
коренными, менее светолюбивыми породами (главным образом
елью). Однако в ряде случаев берёзовые леса являются пер втл ч-
и ы м и, длительно существующими; например своеобразные ле-
са из каменной берёзы (Betula Ermani) в периферических частях
Камчатки (см. карту в конце книги), берёзовые леса и колки берё-
зы в западносибирских степях (здесь мы имеем берёзовую лесо-
степь), березняки на Кольском полуострове (В. tortuosa). Что ка-
сается осинников, то они в качестве первичных насаждений
встречаются значительно реже, Можно указать иа «осиновые кус-
ты», своеобразные осиновые колки, разбросанные по северным
степям (в понижениях, западинах) в области Донского ледникового
языка, Известным аналогом «осиновых кустов» являются «берё-
зовые колки» Западной Сибири, тоже разбросанные по степям.
Леса пойменные и болотистые
Выше были описаны леса, развитые иа водораздельных прост-
ранствах в пределах соответствующих зон (леса зонального типа).
Однако леса могут спускаться в речные долины, могут развивать-
ся на пониженных болотистых местах, давая здесь как бы включе-
ния в основной зональный тип — леса интразонального типа.
Они связаны с орографическими и эдафическими условиями и от-
носятся к различным классам формаций: к летиезелёным, к иголь-
чатохвойным/ к влажнотропическим и др- Особое значение имеют
у нас летнезелёиые (о влажнотропических болотистых фитоцено-
зах см. «Мангровые заросли») (стр. 188). Их можно подразделить
на две группы: леса в речных длинах на почвах с проточной
водой — пойменные (Fluviisilvae) и леса на заболоченных почвах,
с недостатком кислорода — болотистые (Раludisilvae).
Пойменные леса. Оии связаны с ежегодным залива-
нием полыми водами и отложением минеральных и органических
веществ; поэтому почвы здесь богаты питательными веществами и
содержат кислород в достаточном количестве. В результате выруб-
ки этих лесов возникают поёмные луга (см. стр. 268), При отсут-
ствии покоса и выпаса луга опять покрываются лесами,
Пойменные леса не связаны с лесной зоной, но по долинам рек
развиваются и в других зонах — в степной и даже в пустынной.
Это — эдафически обусловленные формации, развивающиеся при
достаточном количестве воды в почве. В безлесных областях леса
растут вдоль берегов рек и называются галл^рейиыми
лесами; они выделяются зелёными полосками на фоне безлесных
равнин. В Средней Азии пойменные леса называются тугайными
(тугаи).
Пойменные леса довольно разиобразны, так как в долинах
рек рельеф и почвы меняются иногда даже иа небольших расстояни-
ях. Но здесь встречаются почти исключительно лиственные леса.
По самым берегам рек очень характерны заросли ив, а также то-
поля, дальше от реки — дубовые леса. Последние в СССР очень
распространены в южных поймах, но и на севере дуб идёт по доли-
нам дальше на север, чем по водоразделам; можно предполагать,
что еще недавно дубовые леса в долинах северных рек были более
широко распространены, так как иа дне рек и в берегах находят
дубовые стволы, называемые черным дуйом. Но наряду с дубом в
северных поймах встречаются и хвойные — ель, а восточнее —
пихта.
Пойменные леса в Северной Америке тоже состоят главным
образом из лиственных пород (тополя, вязы, клены, дубы и др.)
и, как и^водораздельные леса, значительно богаче древесными по-
родами, чем в Европе.
мт
Пойменные леса тропиков изучены очень мало; во многих слу-
чаях тропический лес доходит до рек почти в неизмененном состоя-
нии, в других случаях по берегам находятся обширные заросли
бамбуков нли леса особого характера (см. выше тропические
леса по р- Амазонке).
Леса болотистые (лесные болота). Они развиваются
нри близком стоянии грунтовых вод, как правило, в пониженных
местах и представляют облесенные низинные болота. Эти послед-
ние богаты питате льными веществами и поэтому называются звтроф-
ными\ они могут быть травянистыми (см. стр. 285), лесными, ре-
же кустарниковыми. Различают целый ряд типов лесных болот;
для Европейской части СССР можно указать березовые низинные
болота, ольховые и др.
Очень распространенным» типом является березово-осоково-коч-
карное болото с подлеском из ив, крушины и пр. В травянистом
иокрове два вида крупнодерновиниых осок (Carex caespitosa и С. ра-
radoxa), образующих крупнее койки-
Крайне характерным типом лесных низинных болот являются
ольховые болота из клейкой ольхи (Alnus glutinosa), так назы-
ваемые олыианники. Приурочены они чаще всего к тем понижениям
речных долин, которые примыкают к коренным берегам (притер-
расные понижения). Здесь на очень богатых почвах при избыточ-
ном увлажнении и развиваются эти ольшаиники с очень типичным
набором растений как подлеска, так и травянистого покрова, а
основном почти ие меняющимся на больших пространствах. В под-
леске — всегда крушина, некоторые ивы, чёрная смородина, ка-
лина; в травянистом покрове — некоторые осоки, горько-сладкий
паслён, таволжанка вязолнстиая, обыкновенный вербейник и др-
всегда встречаются папоротники Dryopteris thelypteris и D- spinu-
losa и хмель, В более северных местах в древостое появляется ель
на востоке и пихта. Ольхи обычно сидят на высоких кочках (между
ними — вода) и снабжены нередко как бы ходульными Корнями,
укрепляющими деревья в неустойчивой почве; однако здесь имеет-
ся лишь отдалённый намек на соответствующие отношения в ман-
гровых лесах; как в СССР, так и в Западной Европе ольшаиники
удивительно сходны. Можно смотреть на них, как на очень древ-
ние, прочно сложившиеся фитоценозы.
Под тропиками лесные болота описаны во многих
местах; деревья принадлежат к различным тропическим семей-
ствам, Нередки лазящие пальмы. У деревьев обычно развиты
укрепляющие досковидные корни и дыхательные корни (пневма-
тофоры). Травянистый покров скуден, но торфообразование очень
сильно.
2. Группа формаций —летнезелёные кустарниковые фитоценозы
(Aest if ruticeta)
Во многих случаях они обусловлены биотически (например
•решииковые заросли после вырубки леса, дубовые кустарники в
результате постоянного обкусывания скотом и т. д,), ио нередко
являются естественными (заросли различных рп-’пц цн Salice-
tum — по берегам рек, в горах и пр., заросли зелёной ольхи —
Alnetum viridis в субальпийском поясе и т. д.). Остановимся на
нескольких примерах и прежде всего на кустарниковых фитоценв-
зах средиземноморской области.
Шибляк1. Эю—заросли летнезеленых кустарников
•чень различного флористического состава, развившиеся обычив
на месте уничтоженных лесов и распространенные от Испании иа
восток, но особенно в Югославии, Болгарии, Румынии, в СССР —
в Крыму и на Кавказе.
Очень распространен шибляк с господством кустарникового
пушистого дуба (Quercus pubtscens), но несомненно он связан с
вырубксй лесов из пушистого дуба и с дальнейшим стравливанием-
Другие примеры: шибляк с господством держн-дерева (Paliuretum
australis), сирени (Syringetum vulgaris) — Болгария, Сербия, бар-
бариса (Eerberidetum vulgaris) и т. д.
В Крыму на Южном берегу очень распространены кустарнико-
вые заросли типа шибляка. Можно различить ряд формаций: Quer-
cetum pubescentis — заросли кустарниковидного пушистого ду-
ба (очень сильно распространены), Carpinetum duinensis — низко-
рослые заросли граба, Paliuretum australis — заросли держи-
дерева, Cotinetum ccggygriae — заросли сумаха-желтинника, Rh»-
retum coriariae — заросли ксжевенного сумаха и др. Нет сомне-
ния в том, что крымский шибляк в большинстве случаев — резуль-
тат разрушительной деятельности человека;особенно это относит-
ся к низкорослым дубовым н грабовым зарослям, возникшим на
месте порубок дубовых лесов, при их дальнейшем стравливани*-
Продолжакщееся стравливание приводит к полному уничто-
жению растительности — «биотическая» пустыня. Подобную пус-
тыню можно видеть в ряде мест Южного берега Крыма, напри-
мер в окрестностях Симеиза и др. Несомненно, восстановление
леса, создание новых лесных насаждений — первоочередная за-
дача на Южном берегу Крыма.
Заросли держн-дерева (Paliuretum australis) распространены
также в Новорссснйскем’райсне на Кавказе и тоже являютсяслед-
ствием уничтожения первобытной природы.
Дерезняки. В наших степях мы находим нередко заросли стен-
ных кустарников, близкие к шибляку. Обычно в степ-
ной литературе эти заросли называю ся дерезняками, так как иног-
да состоят из зарослей колючего кустарника дерезы (Caragana
frutex). В других случаях господствуют другие кустарники: тори
(Prunus spinosa) — терновники, степная вишня (Prunus fruticosa)—
вишарники, бобовник (Amygdalus папа), таволжанка (Spiraea
crenifolia и S, bypericifoНа) и др. Чаще эти кустарники образуют
чистые заросли того или другого вида, в которых произрастает
много степных растений и некоторые высокорослые виды, свой-
ственные собственно дерезнякам. На дерезняки некоторые авторы
смотрят как на авангард лесной растительности при её надвигании
на степь.
Слово сербского происхождения.
22»
Дерезняки в большинстве случаев представляют естественные
формации и, невидимому, в первобытных степях занимали большие
пространства, Однако большая их часть распахана. В настоящее
время дерезняки в северных степях обычно приурочены к южным
склонам: в южных степях дерезняки связаны с понижениями, луч-
ше увлажняемыми. Терновники и вишарники иногда местными
жителями называются «садами», так .как они в урожайные годы
дают массу плодов (тёрн и дикая вишня).
Летиезелёные кустарниковые фнтоценозы тундровой зоны. Они
развиты главным образом в самых южных частях тундровой зоны,
образуя даже особую подзону кустарниковых тундр. Эти кустар-
никовые фнтоценозы состоят из низкорослой берёзки (Betula папа,
к востоку от р. Енисея — В- exilis) и из различных видов ив (Sa-
lix). Ивняки местами очень густы и даже трудно проходимы (в
южной части тундр до 3 м высотой, обычно же около 1 з<). Более
подробно об этих кустарниковых фитоценозах будет сказано ниже
(см, стр, 301), Данный тип растительности, по мнению Город-
к о в а ведёт своё происхождение с конца третичного периода.
Летнезелёные леса и человек. Летнезелёные леса в настоящее
время, конечно, не занимают таких сплошных площадей, как
показано на карте (см. в конце книги). Огромные площади обез-
лесены, и леса разбросаны лишь отдельными участками, нередко
клочками. Территории, занятые летнезелёнымн лесами, — это
как раз места с сильно развитой культурой (Западная Европа,
США и др,). В СССР под этими лесами очень плодородные почвы—
деградированные чернозёмы, занятые в настоящее время полевыми
культурами (пшеница, рожь). После порубки летнезелёиых широ-
колиственных пород они обычно сменяются мелколиственными,
более светолюбивыми, как временным типом, а затем постепенно
могут вновь восстановиться. Так, например, дубовые леса в Евро-
пейской части СССР после порубки обычно сменяются осиной
(мелколиственная порода). При повторных порубках получаются
сплошные осинники. В других случаях при порубках и дальнейшем
стравливании скотом, который ежегодно объедает молодые кусты,
получаются кустарниковые заросли дуба, называвшиеся в б. Орлов-
ской губ. «площадями». Местные жители даже думали, что это
особый кустарниковый дуб.
Для изучения дубрав и для постановки опытов по их рекон-
струкции организован госзаповедник «Тульские засеки» (около
7-000 га) близ г.Крапивны. Небольшой широколиственный заповед-
ник «Лес на Ворскле» (1 200 га) находится на юге Курской области.
VI. КЛАСС ФОРМАЦИЙ — ИГОЛЬЧАТОХВОЙНЫЕ ДРЕВЕСНЫЕ
И КУСТАРНИКОВЫЕ ФИТОЦЕНОЗЫ (ACICULILIGNOSA)
1. Группа формаций—игольчатохвойиые леса (Aciculisilvae)
Общая характеристика. Формации данного класса состоят из
хвойных деревьев или кустарников, имеющих листья в виде более
Ъли менее удлинённых игл с развитой по большей части механи-
ческой тканью (реже — листья чешуйчатые). В этот класс форма-
ций нельзя включить вообще все леса с господством хвойных, так
как среди последних имеется целый ряд родов с широкими листья-
ми (Agatliis, Podocarpus) или с листьями, расположенными так,
что они напоминают листья сложны# (Sequoia sempervirens) и др-
Леса из «хвойных» последнего рода, как уже было сказано,
относятся к формациям жестколистных лесов, или лавровых,
Игольчатохвойиые леса развиты исключительно в северном
полушарии в умеренных широтах (в субарктической области)-
Здесь они занимают огромные пространства в северной Евразии
и Северной Америке, иа своей северной границе образуя северную
границу леса вообще.
Климатические условия области этих лесов в основных чертах
таковы. Самый тёплый месяц имеет среднюю температуру 10 — 19°,
самый холодный 4- 3 и —52° (в Сибири). Только 1—4 месяца имеют
температуру выше 10°, н вообще вегетационный период короток.
Климат подобного рода имеет место также и в горах, почему
игольчатохвойные леса развиты на северных равнинах, а также на
известной высоте в горах.
Сравнительно с летнезелёными лесами игольчатохвойные, как
видно на схеме идеального континента и иа карте земного шара,
занимают более северные и более континентальные территории.
Кроме этого, они занимают более плохие почвы (например песча-
ные в области летнезелёных) и более бедные кислородом (болота),
В общем Ош», таким образом, менее требовательны и климатически
и эдафически. Почвы подзоны хвойных лесов относятся к подзоли-
стому типу с более или менее резко выраженным подзолистым бе-
лесым горизонтом (горизонт вымывания) и нередко с конкрециями
ортштейна (рудяка) в более глубоком горизонте вмывания.
Южное полушарие не имеет территории соответственного клима-
та и потому, как уже сказано, игольчатохвойиые там отсутствуют,
но зато там имеются хвойные, относящиеся к другим формациям.
Игольчатохвойиые леса представляют очень характерный ланд-
шафт, несколько отличающийся от госпсдствэ тех или иных пород;
ель, пихта дают конусообразные, иногда узко остроконечные кроны
(сибирская пихта), сосны — округлые. Лиственницы отличаются
тем, что их хвоя опадает на зиму, и зимой лиственничный лес похож
на лес из летнезелёных деревьев Ч
Еловые н пихтовые леса, вследствие своей большей теневынос-
ливости сравнительно с сосновыми и лиственничными, являются
более тёмными и мрачными, чем последние, которые более разре-
жены и светлы. Поэтому иногда говорят о тёмнохвойных (рис. 120)
и сеетлохвойных лесах.
Нужно заметить, что хвойные геологически являются вообще
очень древней группой, и потому говорить о жизненных формах
хвойных, как отражающих современные условия, можно лишь с
большой осторожностью.
Обращает на себя внимание в северных игольчатохвойных лесах
то обстоятельство, что они обычно, особенно в Евразии, состоят лишь
1 Быть может, лиственничные леса правильнее относить к летнезелёным.
231
из о л но г о какого-либо вида деревьев; например обширные леса
из одной ели, из одной сосны, лиственницы и т. п. Флористическая
бедность древесного яруса здесь исключительна. Границы основных
пород даны на рис. 121 и 122.
232
Леса еловые. Они господствуют в нескольких подзонах (см. кар-
ту) и расгкдгюпся на ряд, групп ассоциаций и ассоциаций, слагаю-
Рис. 121. Границы некоторых древесных пород в СССР.
С—яипа (Tilia cordata) и ее островные меетонахпжденпя (в Крыму п на Кавказ* < вы-
казано); Я—ель (Picea excelsa и Р. obovata); Ш—лиственница даурская (Larix daburicaj;
ТУ—сибирская кедровая сосна (Pinns sib(rfca); V—грецкий орех (Juglans regia)-MBe#»a*
граница; у7 - е ь Шренка (Picea Schrenkiana)—северная граница.
Рис. 122. Границы некоторых древесных пород в СССР.
/—пихта (Abies sibirica); II—сибирская лиственница (Larix sibirica);
III—кедр-стланик (Pi;ius pumila); /У-кавказская пихта (Abies nor<^
manniana): У—восточная ель (Picea orientaiis); VI—монгольский дуб
(Quercus mongolica); VII — манчжурский орех (Juglans manshurica);
VIII —бархатное дерево (Pheliodendron amurense).
щихся в определённую экологическую систему. Эта система очень
хорошо разработана главным образом для Европейской части Сою-
233
за. Гак как принципы этой системы вполне приложимы и к другим
хвойным породам, то остановимся на них более подробно.
Можно установить пять
основных групп ассо-
циаций еловых лесов, при-
уроченных к определенным
местообитаниям (по Сука-
чеву):
1. Ельник и-з е л е -
номошники (Piceeta liy-
Iccomicsa). Это по большей
iaciH трехъярусные фито це-
нозы -в первом ярусе ель,
во втором — брусника, чер-
ника или кислица (рис. 123),
в третьем — гипновые з е-
левые мхи. Почвы более
или менее богатые, хорошо
дренированные.
2. Ельник и-д о л го-
мо ш и и к и (Piceeta po-
ly trichosa), В иижнем ярусе
мох кукушкин лен (Poly-
trichii’Ti), называемый также
тслгнм мхом. Два или три
яруса. Почвы сходные, но
менее дренированные, след-
Рис 123. Ельник-кисличник под Москвой ствием чего является уже
('фого А. п, Шиманюка). легкое заболачивание: при-
сутствие кукушкина льна — первый .показатель заболачивания.
3 Ельники сфагновые (Piceeta sphagnosa). В нижием ярусе
сфагновые мхи Два или три яруса. Почвы еще более заболо-
ченные. равнинные. Сфагновый мох при заболачивании—следую-
щая ступень за кукушкиным льном,
4. Ельники болотно-травяные (Piceeta uligi-
noso-herbosa). Здесь более или менее высокий травянистый покров.
Развиваются на дне логов, по речкам и пр- при сильной заболочен-
ности, но при проточности вод.
о. Ельники сложные, или ельники с дуб-
равными элементами (Piceeta composita, или Р. пе-
morosa). Здесь фитоценологическая структура более сложна
появляется очень хорошо выраженный ярус кустарников (орешник,
кустарниковидная липа и др.). В древесном ярусе может присут-
ствовать дуб, в травянистом — нередко имеется немало растений,
связанных обычно с дубравами (ясменник, сныть, медуница и т, п.).
Почвы богатые, нередко с известковой подпочвой. Данный тип
представляет как бы известный переход к лесам широколиствен-
ным (дубравы).
Взаимные соотношения указанных групп можно представить
я такой схеме:
234
?. sphagnosa
I___.___. -
P. polytrichosa
P. hylocomiosa
P. nemorosa
P. uliginoso-
herbosa
В схеме центральной группой является Piceeta hyh-
comiosa, находящаяся в средних условиях существования, налево
идет заболачивание, направо — улучшаются почвенные условия,
вниз — увеличивается влажность, но проточность воды остается.
Рис ’"124. Профиль, показывающий положение различных типов и ассоциаций
елового леса 'в рельефе.
1—сльник-бруснич тик; s—ельник-кисличник, 3—ельник-черничник, 4—ельник травяной;
5—ельник-дочгомолнин, 6-сфагновое болото с низкорослой сосной (ориг.)
Рис. 125. Вегетативное размножение при помощи подземных побегав
и корневищ у растений елового леса.
1 -вороний глаз (Pans quadnfoha); 2— седмичник (Trientalis europaea), 3—
Кислица (Oxalis acetosell а) (по Сукачеву),
В пределах каждой нз пяти групп имеется значительное
число ассоциаций например, в пределах центральной группы:
235
ельники-брусничники, ельники-кисличинки, ельиики-чериичникк
н Др- распределяются закономерно по профилю (рис. 124).
Экологические особенности еловых ле-
сов н вообще темнохвсйнь,х (пихта, сибирский кедр), Оии до-
статочно своеобразны, целый ряд видов тесно связан с еловыми
лесами, являясь его «спутниками» или «верными» видами: литнея,
•дноцветковая гру шавка, альпийская цг.рцея, 'орхидные —
Gocdyera, Epipogon, Calypso и, др,
Приведем некоторые экологические черты еловых лесов: 1) очень
сильно развито вегетативное размножение, так как большое зате-
Рмс 126, Профиль, показывающий положение в рельефе различных типов и
ассоциаций соснового леса.
/—бор-беломошник; S—бор-брусничник, 3—бор-1в ю-чнин; бор-долгомошнин, 3 —
сфагновое болото с низкорослой сосной, 2 и 3—ассоциации группы б оров-зелен ом ож-
инков (ори г.).
нение мало благоприятствует появлению цветков (рис. 125);
2) очень характерна белая окраска цветков, что делает их более
заметными; 3) много видов вечнозеленых или зимнезелёных исполь-
зующих длительно, без перерыва, недостаточное количество света;
4) отсутствуют. весенние эфемеры, очень характерные в летне-
зеленых лесах, так как весенней светлой фазы здесь нет; 5) скуд-
ное освещение способствовало переходу ряда видов иа сапрофити-
ческое питание.
Ель имеет огромное географическое распространение, в Европе
•на представлена европейской елью (Picea excelsa), а иа северо-
востоке Европейской части СССР и в Сибири — сибирской елью
(Р. obovata): различия между ними не так велики К Другая очень
распространенная порода в СССР — это сибирская пихта (Abies si-
birica). Она вместе с елью и сибирским кедром (сибирский «кедр» —
это не кедр, а особый вид сосны — Pinus sibirica) образует в за-
падносибирской низменности обширные, несколько заболоченные
леса, представляющие так называемую тайгу, Вообще тайга
в западносибирском смысле имеет три основных признака: силь-
ную затененность, заболоченность и отсутствие элементов широ-
колиственных лесов. Однако тайгу можно понимать и более ши-
1 На сев.-зап. Европы (Карелия, Финляндия н др.) нужно отметить еме
финскую ель (Picea fennica.j.
роко и относить к ней северные хвойные леса Европейской части
СССР, лишенные широколиственных элементов, и лиственничные
леса Восточной Сибири (лиственничная тайга).
Леса сосновые. В Евраз! и могут быть представлены той же
схемой, что и еловые (см. выше), однако с той разницей, что здесь,
кроме пяти основных групп ассоциаций, большое значение имеет
шестая, занимающая наиболее сухие положения и в схеме
располагающаяся выше группы hylocomiosa. Это — Pineta cladi-
nos, — боры-беломошники (или лишайниковые боры с лишайни-
ком Cladonia), располагающиеся на бедных сухих почвах. Вообще
же сосновые леса (Pineta) развиваются иа песчаных почвах, пред-
ставляя включения в ту или иную подзону. На приложенном про-
филе видно взаимное расположение различных типов сосновых
лесов (рис. 12b).
Экологические особенности сосновых лесов из обыкновенной
сосны (ninus silvestris) связаны с осветлёиностью этих лесов, с
бедностью их субстрата и, по большей части, сухостью. В сухих
типах преобладают ксерофиты, а на юге в сосцовые леса заходят
многочисленные степные элементы вместе с ковылём.
Еловые и сосновые леса и их взаимоотношения. Обычно в при-
роде эти леса занимают различные местообитания; ель занимает
глинистые почвы, сосна — песчаные, но так как ель способна про-
израстать и иа песчаных почвах, то здесь возникают конкурентные
отношения между этими двумя породами. Исход борьбы обычно
бывает благоприятен для ели, так как она, сравнительно с сосной,
значительно более теневынослива и затеняет сосну, которая по-
степенно погибает. Можно в природе наблюдать все стадии смены
сосновых лесов еловыми (см, выше, рис. 76). Лишь иа самых бедных
и сухих почвах (в типе Pineta cladinosa) сосна может ие бояться
конкуренции ели. Но возникает вопрос; раз ель почти иа всех место-
обитаниях может вытеснить сосну, то как объяснить существование
обширных сосновых лесов, до сих пор ещё не вытесненных елью?
Объяснением служат лесные пожары (как естественные от молнии,
так и от неосторожного обращения с огнём), которые в сосновых
лесах представляют обычное явление и нередко захватывают ог-
ромные территории. При этих пожарах прежде всего страдает
ель, имея поверхностную корневую систему — она таким обра-
зом выпадает из древостоя. Сосна имеет более глубокую корневую
систему и менее страдает от пожаров. В дальнейшем ель может опять
поселиться в сосновом лесу, но новый пожар опять её уничтожает.
При отсутствии пожаров сосновые леса, можно думать, в большей
своей части сменились бы еловыми.
Хвойные леса Центральной и Восточной Сибири. Они си льне
отличаются от европейских и западносибирских — здесь очень
большие территории занимают леса лиственничные .из
сибирской лиственницы (Larix sibirica) в бассейне р. Енисея и из
даурской лиственницы (L. dahurica) в бассейне средней Лены.
И здесь можно отличать различные типы: более сухие, более влаж-
ные, более заболоченные и т, д., т. е. приведенная выше схема
приложима и в данном случае.
2ЭТ
Везде в области лиственничных лесов встречается сосна, кото-
рая дает лесные типы, аналогичные тем, которые дает лиственни-
ца, ио лишь с сосновым древесным ярусом. Как показывают наблю-
дения, сосна вытесняет лиственницу, однако это зависит не от боль-
шей теневыносливости первой (в этом отношении эти две породы
почти не различаются), а от их различного отношения к пожарам.
Лиственница от пожаров сильнее страдает, чем сосна, и выпадает
из древостоя
Обобщенная схема типов хвойных лесов в СССР. Соотношения
хвойных пород и их типов могут быть представлены в обобщен-
ной схеме (рис,
127), построенной по
указанному выше
принципу (см. еловые
леса), причем по вер-
тикальной линии уси-
ливается влажность
от типа Ck(cladinosa)
до (J. h. (uliginoso-
herbosa), направо от
центральной точки
Ox. (oxalidosa—кис-
личники, централь-
ная ассоциация груп-
пы йу1осопио5а)идут
более богатые типы'
TiL (tiliosa с липо-
вым подлеском) и Q
(quercetosa — с ду-
Рис. 127. Обобщенная схема типов лесов (по Сукачеву). бОм), налево —забо-
лачивающиеся тнпьк Pl. (polytrichosa) и Sph. (sphagnosa). Разме-
щения на схеме типов направо или налево, конечно, условны
Экологическая амплитуда каждого лесного типа представлена
на схеме кругом или замкнутой линией, более или менее прибли-
жающейся к кругу, причем видно, что для пихтовых лесов
(Abiegneta) она меньше всех других пород, а у сосновых лесов
(Pineta) — наибольшая; видно также, что еловые леса (Piceeta),
леса сибирской лиственницы (Lariceta sibiricae), сибирского кедра
(Cembreta)1 ие заходят на слишком сухие местообитания, наиболее
влажные местообитания пригодны лишь для еловых лесов и т. д.
Хвойные леса Западной Европы. Наиболее характерные х в о й-
н-ы е породы Западной Европы (кроме обыкновен-
ной сосны цели) — это европейская лиственница (Larix europaea)
и пихта (Abies alba), произрастающие в Альпах и иа Карпатах
(в СССР — на восточных Карпатах); равнины заняты главным об-
разом летнезелеными лесами. Пояс лиственничных лесов лежит в
пределах 1 200—2 300 м, это светлые леса, вследствие чего в них
сильно развит травянистый покров (тип Lancetum pratosum).
1 Сибирский кедр—Pinus sibirica, иначе называется Р. cembra ssp sibirica
Д_2£2_
Рис. 128 Ареалы европейских и малоазиатских пихт. В центре ^вропы - Abies alba, па юге Пиренейск.
полуострова—A. pinsapo, на Сицилии—A. nebrodensis, в Греции—A. cephalonica, в средних частях Балкан-
«кого полуострова—A. Borissii regis, на Кавказе—A. ^nordmanniana, на севере Малой Азин—A. bornmullenana,
г 9 юге М^лой^Азии —A. cihcica (по Рубнеру).
В Пиренеях — леса из Pinus uncinata, в горах южной Европы—
из Pinus nigra; в Греции — белая европейская пихта замещается
другим видом — Abies cephalonica (рис. 128).
Хвойные леса Северной Америки, Эти леса занимают в Северной
Америке очень большие территории: большую часть полуострова
Лабрадор, значительные части Аляски и широкую полосу, свя-
зывающую эти два полуострова, Вдоль берегов хвойные леса далеко
продвигаются на юг; на западе вдоль тихоокеанского побережья,
особенно по горным цепям до самой Центральной Америки (Скали-
стые горы, Каскадные, Сиерра-Невада и др ), иа востоке —по при-
атлаитическсй прибрежной равнине до Мексиканского залива.
По сравнению с европейскими лесами здесь имеется очень боль-
шое разнообразие хвойных пород, именно, ряд особых
родов /Tsuga, Pseudotsuga, Sequoia, Thuja), т кроме того, миого-
2W
численные виды сосны, а также ели, пихты, лиственницы, можже-
вельники и пр. Основной причиной этого богатства нужно считать
благоприятные условия миграций во время ледникового периода:
все эти породы могли свободно отступить иа юг, а затем снова вер-
нуться после отступания ледника-
Как и в Европе, в бо-
лее холодных частях Се-
верной Америки хвоиные
широко распространены
на равнинных террито-
риях, в более же южных
частях они переходят в
горы, занимая опреде-
ленный пояс.
Разнообразие хвой-
ных пород и местных
условий делают трудным
дать в кратком изложе-
нии более или менее пол-
ное представление о наб-
людающихся отношени-
ях. Но в основном мы
имеем следующее Наибо-
лее далеко идет на север
лиственница (Larix ame-
псапа), на севере Канады
преобладающим деревом
хвойных лесов* является
сосна Pinus Banksiana,
к которой присоединя-
ются некоторые ели,
бальзамическая пихта
(Abies balsamea), лист-
венница н некоторые
мелколиственные (осина,
береза) На Аляске леса
ситхинскон ели (Picea
sitchensis), цуги (Tsuga
heterophyПа и Т. пкг-
tensiana), аляскинского
riTnutkaensis)13 В^бъас- рис' 1301 Лес из мам0НТ0Ва дерева (Sequoia gigan-
ти Великих озер и р св. tea> в Сиерра-Невдда в Калифорнии (по Упхару).
Лаврентия леса состоят
главным образом из веймутовой сосны (Pinus strobus, рис, 129), к которой
присоединяются Pinus resinosa,туя (Thuja occidentilis), цуга (Tsuga occiden-
tals). Вдоль восточного побережья хвойные леса далеко спускаются к югу н,
например, в штатах Вирджиния л Каролина леса состоят главным образом из
сосны Pinus rigida (также Р. taeda). z
Особенно развиты хвойные леса на западном побережье Северной Америки,
причем в различных районах они представлены особыми видами так для Бри-
танской Колумбии наиболее характерны дугласова пихта (Pseudotsuga Dougla-
бп), туя, Pinus Murreyana, в Скалистых горах—ряд видов сосны, в горах Сиер-
ра-Невада — желтая сосна (Р. ponderosa), гигантская секвойя или мамонтово
дерево (Sequoia gigantea) (рис. 130) и др.
Ряд дополнительных данных указан в отделе «Флористические области»
(см. ниже).
Средиземноморские хвойные леса. Они развиты в странах, не-
посредственно окружающих Средиземное море, и главным образом
состоят из видов сосны — алеппская сосна (Finns halepensis) и
др. Для Крыма очень характерны леса из крымской сосны (Р. pal-
16 ^ографня растений 241
lasiana, рис. 131), занимающие южные склоны Яйлы Кроме сос-
новых лесов, леса из кипариса (Cupressus sempervirens) на Крите
и в Малой Азии и из древовидного можжевельника (Juniperus
excelsa) — на южном берегу Крыма
Надо упомянуть еще о кедровых лесах из настоящего
кедра (Cedrus), подобные леса встречаются на Атласе в Север-
ной Африке (атласский кедр—рис. 189), в Передней Азии, в частно-
сти в горах Ливана (ливанский кедр), и в Гималайских горах, в
Афганистане и Белуджистане (гималайский кедр). Все они зани-
мают высоты выше 1 000 м, доходя (в Гималаях) до 3 200 м.
2 Группа формаций—хвойные, кустарниковые фитоценозы
(Aciculif ruticeta)
Данная группа формаций не имеет широкого распространения:
относящиеся к ней формации чаще всего приурочены к особь м
климатическим и эдафическим условиям (субальпийский пояс
и Др.). Отметим низкорослые заросли можжевельника (Juniperus
depressa) на горах Крыма, заросли кедровника (Pinus pumila) в
подгольцовом поясе в Восточной Сибири, заросли казацкого мож-
жевельника (Juniperus sabina) на мелах юго-востока Европейской
части СССР и др.
Леса пойменные и болотистые (хвойные). В поймах рек Евро-
пейской части СССР особенно распространены, как уже было ука-
зано, леса летнезеленые (см. выше). Однако в более север-
ных частях лесной зоны в поймах встречается ель, а на северо-во-
стоке также и сибирская пихта, очень характерны на севере ель-
ники по долинам мелких лесных ручьёв и речек (так называемый
тип Piceetum rivulare), подобные ельники имеют народное название
согра (в некоторых местах — шохра). Другим типом ельников
являются ключевые ельники (Piceetum fontina 1е) в местах выхода
ключей, обычно — это начальная стадия заболачивания.
Однако ель не выносит чрезмерного избытка влаги и особенно
заболачивания^ Сосна в этом отношении менее прихотлива, и мы
встречаем на низинных болотах, кроме березняков н ольшанников
(стр.228) также н сосновые насаждения. Но особенно характерна
сосна на моховых болотах (ель в этих условиях совершенно отсут-
ствует), здесь она образует цельй ряд форм и в наиболее типичных
случаях представлена редкими низкорослыми экземплярами
(стр. 290).
В пой'мениых лесах Северной Америки, так же как н в Европе,
господствуют летиезелёные породы, ьо в более кжгьх местах
встречаются н хвойные'. цуга, туя и виргинский можжевельник.
На болотах в Северней Америке господствующими породами яв-
ляются те или другие хвойные, к которьм присоединяются листвен-
ные. особенно распространен белый кипарис (Clit maecyparis thyoi-
dcs). В более южных местах господствует болотный кипарис (Taxo-
dium distichum), креме того, магнолия (iVrgnclia grandifleva), не-
которые сосны, на Флориде особенно характерен болотвьй кипарис
со своими дыхателгньми корнями, высту пак ними из болотной
почвы вследствие недостатка там кислорода, Деревья болотвог»
ле* _
кипариса имеют причудливый вид (рис. 132), так как они не разви-
вают такой правильной кроны, как другие хвойные, и обычно гу-
сто покрыты свисающим вниз луизианским «мхом» (Tillandsia).
Кроме болотного кипариса, встречается ряд других деревьев: Nys-
sa biflora, красный клен и др., а также и одна пальма (Sabal Adan-
sonii). Болота с болотным кипарисом, кроме Флориды, встреча-
ются и на севере до Вирджинии и по МиссисСипи до Миссури и Ил-
линойса. Самая южная часть Флориды занята обширными боло-
тами с господством злака Cladium effusum, на более повышенных
местах — болотный кива рис и некоторые сосны,
Хвойные леса и человек
Хвойные леса, занимающие в Евразии oiротные территории
(см, в конце книги карту растительности), не являются сплошны-
ми, как это представляется по карте. Обширные пространства
(особенно в Европейской части) давно распаханы или заняты по-
селениями. Интересно при э ом отметить, что в первую очередь
выкорчевываются и распахиваются те типы леса, которые занимают
наиболее богатые почвы;так, в Приветлужье прежде всего раскор-
чевываются леса елово-пихтовые с богатым травянистым покровом,
или же занятые ассоциацией ельника-кисличника и др. Но кро-
ме уничтожения лесов, огромные изменения в составе лесов вы-
званы порубками и п*астьбой в лесах скота.
Порубка ведет в большинстве случаев к полной смене одних
пород другими, причём обычно более теневыносливые древесные
породы сменяются менее теневыносливыми (в связи с осветлением
места после порубки). Так еловые и сосновые леса Европейской
части сменяются осиной и берёзой, причём эти породы
занимают в настоящее время огромные пространства (осиновые
и березовые леса), Однако эти леса в р е м е и и о г о типа, так как в
дальнейшем они возвращаются к своему исходному типу (ельники и
сосняки). В иных случаях после порубок возникают сероолыианни-
ки (из Alnus incana). Местами, особенно в центральных областях,
мелколиственные породы играют такую большую роль, что с тру-
дом можно найти исходный (коренной) тип леса. Восстановление
исходного типа сильно затрудняется пастьбой в лесу скота, уничто-
жающего всходы деревьев. В результате —временные типы могут
существовать неопределённо долгое время.
Вопрос об охране лесов, с одной стороны, а с другой —
об их реконструкции и о создании лесов, более ценных
с производственной стороны, представляет в данное время крайне
важную задачу. Можно подобрать ряд пород, быстро растущих,
дающих большое количество ценной древесины. Североамериканские
и дальневосточные породы особенно пригодны в этом отношении,
В заключение укажем ряд заповедников в области хвойных
лесов: центрально-лесной в Калининской области (35 000 га), Пе-
чорско-Илычский в бассейне рр. Печоры и Илыча (около 1 м ли-
га), Кондо-Сосвенский в Западной Сибири (около 800 000 га) и др.
Особо нужно отметить замечательный заповедник Беловежская
Пуща, расположенный в западной части Белоруссии. Эго —
•грешный лесной массив в 130 000 га, состоящий из многочис-
ленных типов еловых, сосновых, дубовых, грабовых и ольхо-
вых л сов; массив этот имеет в общем хорошую сохранность, так
как он ещё до войнй 1914 г. охранялся, как место царской охоты.
Здесь ещё в 1914 г. насчитывалось 727 зубров, в 1915 г. — 127,
а в 1921 г. был убит последний зубр (часть зубров была вывезена
в Германию). В последнее время велись работы по восстановлению
зубра. Во флоре Беловежской Пущи мы имеемтакие породы, как пих-
та (Abies alba), сидячецветььй дуб (Qucrcus petraea), тисс (Taxus
baccata); все сии имеют здесь свои северовосточьые крайние пункты.
V1J. КЛАСС ФОРМАЦИЙ — ДЕРЕВЯНИСТЫЕ ФИТОЦЕНОЗЫ С ЛИСТЬЯ-
МИ ВЕРЕСКОВОГО ТИПА (ERIC1LIONOSA)
Группа формаций—кустарниковые фьтсцеисзы верескового типа
(вересковые пустошн, верещатники— Ericifruticeta)
Общая характеристика. Верещатники представляют физиоио-
мически очень своеобразный тип растительности: это пространства,
покрытые вечнозелёными кустарниками и кустарничками, относя-
щимися почти исключительно к семейству вересковых, и обуслов-
ленные как климатически, так и эдафически.
Наибольшего развития они достигают в западной и североза-
падной (приатлантической) Европе, именно в северной Фран-
ции, Бельгии, Голландии,
Ирландии, Великобрита-
нии, ;Дании и северной
Германии;
режья Балтийского моря
они доходят до Эстонской
ССР- 'Кроме того, имеют
распространение на западе
Скандинавии. Вне Европы
почти не развиты, за
исключением некоторых
местностей Северной Аме-
рики, Африки (Капская
область), Фалклэндских
островов и других.
Кроме умеренных обла-
стей, близкие к вересковым
пустошам формации встре-
чаются в Арктике, Евразии
и Северной Америке, а
кроме того, иа высотах в
вдоль поое-
альпийском поясе гор. Рие. 133. Растения с листьями эрикоидног*
Физиономически дан- w я „ ™па-
1—обыкновенный вереск (CalJuna Tulgarts); 2—еврс-
НЗЯ Группа формации «ейский вереск (Erica tetralix) (по Вагнеру),
отличается тем, что вхо-
дящие в ее состав вечнозелёные кустарники имеют листья
эрикоцдного типа (от слов Erica — вереск); они характерны тем,
что имеют игловидную удлиненную форму (рис. 133), зависящую
246
чаще всего от того, что листья завернуты своими краями наружу
н свернуты так, что снизу остается лишь одна щель, часто эта щель
закрыта волосками, у нашего вереска листья игловидные трехгран-
ные Впрочем, в ряде формаций, входящих в этот класс, имеются
господствующие виды с широкими листьями (толокнянка — Arcto-
staphylos)
Хотя большинство видов, как сказано, относится к семейству
вересковых, но здесь встречаются также представители
Empetraceae (водяника — Empetrum), Epacridaceae (в южном полу-
шарии), миртовых (Myrtaceae), бобовых (дрок — Genista, Ulex) и др
Развитие вересковых формаций связано с определенными кли-
матическими и эдафическими условиями Кли-
мат холодно-умеренный зимы относительно мягки, а лето про-
хладно, годовые амплитуды невелики (10—16°) Очень характерно
равномерное распределение осадков в течение всего года,
а также большая влажность воздуха, сюда присоединяется
еще большая облачность В общем—это прохладный и влажный оке-
анический климат
Почвы верещатников очень бедны, так как соли выщелачиваются
обильными осадками в более глубокие горизонты По большей ча-
сти почвы сильно песчанисты и сильно оподзолены Очень характе-
рен на известной глубине (иногда 30 си) горизонт ортш.ейна (ру-
дяка), образовавшегося за счет вымывания гумусовых и других ве-
ществ из более высоких горизонтов и выпадения их в более глубоких
Для вересковых формаций характерно, кроме того, образование
грубого гумуса за счет многочисленных отпадающих листьев
Внешний вид вересковых кустарников очень своеобразен, имея
большое число признаков ксероморфного строения узкие листья
с завернутыми краями, с погруженными устьицами, с волосками
в желобке листа и пр. Однако считать вересковидные кустарники
ксерофитами в обычном смысле и нельзя по ряду причин Преж-
де всего они испаряют очень много, что совершенно необходимо,
так как иначе они не могли бы брать из бедной минеральными ве-
ществами почвы достаточное количество солей Способность к испа-
рению у них должна быть особенно велика вследствие того, что они
наиболее обильно развиваются в областях с очень влажным воздухом
Однако для понимания ксероморфности строения необходимо
иметь в виду то обстоятельство, что основные территории, занятые
верещатниками, представляют области очень сильных ветров Эти
ветры, особенно осенью, зимой и ранней весной, когда почва доста-
точно холодна, действуют иссушающе Мелкие эрикоидные листья,
во-первых, менее испаряют, чем крупные листья, а во вторых, круп-
ные листья могли бы более страдать от разрывов Здесь можно го-
ворить об анемоморфных (анемос по-гречески — ветер) листьях
Перед растениями верещатников стоит такая задача нужно ис-
парято много, так как почва бедна, но крупные листья непригодны,
так как господствуют си ьные ветры Отсюда — образование листьев
эрикоидного типа, но в очень большом количестве, имеются данные,
что на одном экземпляре вереска (Calluna) может находиться до
75 000 листьев.
Характерно, что вересковые формации ограничиваются приат-
лантическими областями и не идут на восток, в области более конти-
нентального характера Однако на востоке вересковидные растения
(в частности вереск) образуют в лесах особый ярус,
вне лесов встречаются реже, и обычно как вторичное явление после
порубки лесов Здесь прекрасный пример замещения факторов —
климатические факторы замещаются биотическими в лесах вереск
находит фитоклимат, схожий с морским
Имеются в литературе мнения, что вересковые формации приат-
лантических стран являются не коренными, а связанными с деятель-
ностью человека и развились на месте вырубленных лесов, посколь-
ку преобладают песчаные почвы, можно думать о сосновых лесах.
Однако трудно представить себе, чтобы данный тип растительности,
обусловленный так хорошо климатически н эдафически, был вторич-
ным на всей своей территории, по видимому, будучи естественным,
он расширил свою территорию за счет вырубленных лесов
Описываемый тип растительности мы называем «верещатниками»,
или «вересковоьми пустошами» Немецкое название «Heiden» не пе-
реводится более удачно, чем «пустоши», однако последний термин не
дает правильного представления о типе
Верещатники прнатлантические Наиболее распространенным
кустарником эрикоидных формаций является обыкновенный вереск
(Callima vulgaris, рис 133, 134), из которого главным образом состоят
Рис 134. Ареал вереска (Calluna vulgaris), встречается также в Северной
Америке и Гренландии (по Бушу)
верещатники Голландии, северо-западной Германии, Великобрита-
нии и западных берегов Швеции В конце лета цветущие верещат-
ники очень красивы, и красочные лиловые пространства мало
отвечают термину «пустошь» Ряд сопутствующих растений (про-
стрел — Pulsatilla, дрок — Genista,золотая розга — Solidago и др )
позволяет наметить особые ассоциации На местах более влажных
и при более влажном климате появляется другой род вересковых —
Епти < самый распространенный вид Епса hlralix, рис 133 и 135).
247
Наиболее богаты видами Erica верещатники Великобритании и Ир-
ландии. Bg время цветения верещатники из Erica дают Фиолетовые
и пурпуровые аспекты. Если верещатники состоят из других родов,,
то и физиономически они сильно отличаются; так, заросли ULx
Galii в И;'ланд! и во вр°мя цветения имеют золотистый аспект.
Рнс- 135. Ареал европейского вереска (Erica tetralix); штриховка—
сплошное распространение, точки—отдельные местонахождения (по
Гранлунду).
Описанные приатлантические верещатники наиболее типичны
для данного класса формаций, так как в них обыч о господствуют
различные верески. Однако верещатники можно понимать и в более
широком смысле и относить сюда заросли и других вечнозелёных
(а иногда и не вечнозелёных) кустарничков (водяника, голубика,
толокнянка и др.). При этом нужно различать горно-альпийские и
арктические «верещатники».
Верещатники горно-альпийские н арктические. Первые особенно
хорошо выражены в Скандинавии. Здесь выше границы леса мы
встречаем не субальпийские н альпийские луга, как в Альпах и.
в других южных горных цепях, а кустарниковые заросли, альпий-
ские же луга выражены очень слабо. В последнее время эти вереско-
вые заросли подробно описаны скандинавскими ботаниками; можно
в основном различать несколько главных типов с господством таких
видов: водяники (Empetrum nigrum рис. 136). низкорослой берёзки
(Betula папа), черники, голубики, Phyllodoce coerulea, Cassiope
tetragona, куропаточьей травы (Dryas) и некоторых других. Однако
во всех этих типах мхи н лишайники играют исключительную роль.
Эти типы на севере смыкаются с тундрой, представляя в сущности
нагорную тундру. На больших высотах в Альпах и Карпатах
имеются формации L dselcuria procumbens и Vaccinium, очень схо-
жие с арктическими.
Горно-альпийские верещатники, особенно на более северных го-
рах (в Скандинавии, также на Урале, в горах Восточной Сибири
др.\ очень приближаются к арктическим кустарнико-
вым формациям, развитым и в тундрах и на арктических
островах (Гренландия, Шпицберген и т. д.). Те же Empetrum, Vac-
cinium, Cassiope, Arctostaphylos,. Dryas, Betula папа и другие соп-
ровождаемые многочисленными лишайниками и мхами. Здесь име-
ются различные кустарничковые формации тундр, о чем
более подробно будет сказано ниже
Взрещатники внеевропейских стран. Типичные верещатники
вида—один в северном полушарии, другой—в южном (по Хетчинсону).
Яровых кустарничковых формаций, заросли, напоминающие ириат-
лантические "верещатники, встречаются в Новом Брауншвейге. В
Южной Америке, в южной Патагонии, в Чили, на Фалклэндских
островах встречаются заросли водяники, но не чёрной (Empetrum
nigrum), как в северных странах, а красной (Е. rubrum — рис. 136).
Особенное разнообразие и обилие вересковых характерно для Кап-
ской области, где имеется более 450 видов рода Erica; здесь влаж-
ность воздуха и облачность более способствуют процветанию этих
растений, чем количество осадков. Вопрос о том, куда отнести «ве-
рещатники» Капской области — к верещатникам собственно или к
жестколистным кустарниковым фитоценозам (Dur ifruticeta) не-
ясен; невидимому, там нужно отличать и те и другие.
VIII. КЛАСС ФОРМАЦИЙ — ТРАВЯНИСТЫЕ ЗИМНЕЗЕЛЕНЫЕ КСЕРО-
ФИЛЬНЫЕ ФИТОЦЕНОЗЫ (HIEMIDURIHERBOSA)
Саванны
Мы выделяем саванны в особый класс формаций. Брокман-
Е р о ш на своей карте включает саванны в класс зимнезелёных
24S
лесов, лто, конечно, искусственно. Р ю б е л ь включает саванны
в класс Durilierbosa вместе со степями, что также не может быть
•правдано. В параллель зимнезеленым и летнезеленым лесам нам
представляется целесообразным выделить травянистые классы фор-
маций — зимнезеленые ксерофильные фитоценозы—с в ниы (Hiemi-
durilierbosa) и летнезеленые ксерофильные фитоценозы — степи
(Aestidunh^rbosa), и те и другие тесно связываются с соответ-
ствующими классами древесных формаций.
Общая характеристика. Саванны — это тропический тип
травянистой растительности ксерофильного характера; от
ртепей, развитых в умеренных областях, они отличаются прежде всего
тем, что не лишены совершенно деревьев, как наиболее типичные сте-
пи, но здесь по травянистому покрову разбросаны деревья отдель-
ными более или менее ред»<ими экземплярами. Интересно отметить,
что присутствие деревьев очень характерно для всех тропи-
ческих фармаций вплоть до тропических пустынь, что, очевид-
но, связано с большим количеством тепла. Но, конечно, в тропиче-
ских травянистых пространствах и в пустынях выработались особые
типы ксерофильных деревьев.
Саванны распространены под тропиками очень широко: в тропи-
ческой Африке (особенно в восточной части), в Южной Америке, в
Гвиане и по р. Ориноко (так называемые льяносы), а также в Брази-
лии (каиписы) кроме того, в Австралии и Передней Индии.
Климат саванн характеризуется значительным количеством осад-
ков, около 90—150 см, выпадающих, однако, неравномерно,
но приуроченных к дождливому сезону, который каждый год че-
редуется с периодом засушливым, почти лишенным дождей (про-
должительность его 4—6 месяцев), так, например, в бразильских
к ьмпосах из общего числа осадков в 150 см в течение мая — августа
выпадает лишь 10 см, в саваннах Западной Африки при сумме осад-
ков около 100 см в течение июля — сентября осадков выпадает
лиШо около 1 см. В ю же время температурные условия меняются
Очень незначительно, и все время температура достаточно высока,
для первого примера средняя температура дождливых месяцев около
18—21°, а засушливых 14—15°, для второго примера соответствую-
щие цифры таковы: 20—24° и 18—20°.
Саванны, как сказано, очень характерны тем, что везде разбро-
саны одиночные, реже группами, деревья ксерофильного типа —
ландшафт чрезвычайно оригинальный и для умеренных широт не
известный. Экология этих ксерофильных деревьев до известной сте-
пени может напоминать лишь то, что мы имеем в Средиземноморье.
Основные черты ксерофильных деревьев гаковы: они
низкорослы, суковаты, часто с неправильным стволом, но имеются
и деревья высокие и стройные очень значительных и даже гигантских
размеров. Что касается более низкорослых деревьев, то они более
всего похожи на наши яблоневые деревья (см. выше саванновые леса
типа плодовых садов, стр. 199).
Ксерофильные деревья имеют ряд приспособлений для перене-
сения засушливого периода: прежде всего они сбрасывают свои жест-
кие листья. Очень часто листья перистые, листочки у них по-
250
движим и могут принимать положение, лучше защищающее их от
солнечных лучей. Вечнозеленые листья очень сильно опушены и
имеют консистенцию жести. Интересно, что почки очень хорошо
защищены почечными чешуями.
Одна из наиболее характерных черт деревьев саванн — это их
очень своеобразная крона, она плоская и имеет форму зонтика,
сверху иногда как бы обрезана. Разбросанные по саванне подоб-
ные деревья создают оригинальный ландш'афт (рис. 137). В более
редких случаях крона состоит из ряда плоских ярусов, удаленных
один от другого. Зонтиковидная форма присуща деревьям раз-
личных семейств, — другими словами, это определенная жиз-
ненная форма, связанная с условиями саванн. Можно думать, что
подобная форма является защитным приспособлением от сильных
ветров и их механического н иссушающего действия: i лавн ям о сказом
действию ветра здесь подвергается узкий край древесной кроны.
Обращает на себя внимание имеющаяся у многих деревьев очень
толстая корка, покрывающая кору: это еще одно приспособление
против засухи.
Одной из наиболее интересных черт саванн является присутст-
вие там деревьев, иногда исключительных размеров, что
имеет место, как мы видели выше, и в ксерофильных лесах. Подоб-
ные деревья запасают в стволах огромное количество воды для за-
сушливого периода.
Чго касается кустарников, то они не менее ксероморфны,
чем деревья,особенно бросается в глаза сильное развитие подземных
частей в ущерб надземным. Подземные части служат для запасания
воды.
Лианы на деревьях редки, интересно, однако, отметить, что встре-
чаются кустарники из таких семейств и родов, которые вообще со-
держат лишь формы типа лиан. Можно предполагать, что здесь
251
мы имеем возврат к исходному типу. Эпифиты почти отсутствуют
(однако встречаются некоторые эпифнты-фикусы).
Травянистый покров саванн обычно не' является
сплошным, причем между дерновинами растений видна красная поч-
ва латеритового типа, реже—травостой сплошной. В большинстве
случаев преобладают злаки до метра высотой, однако иногда они
достигают и большей величины (до 3 л/, а еще реже — н более) При
сомкнутости травяного покрова передвижение в саваннах очень
Рис 138. Баобаб (Aaansoma digitata) в африканской саванне, справа и слева
зонтиковидные акации (по*Буссе).
затруднено и почти невозможно. Злаки обнаруживают ксероморф-
ную структуру — листья узки и прямостоячи, у двудольных листья
по большей части невелики, жестки и нередко совсем редуцируют-
ся. Интересно, что некоторые двудольные имеют клубни, в которых
запасается также и вода (у нас, как известно, это свойственно глав-
ным образом однодольным)
Так как высокий травянистый покров в виде сухих остатков сохра-
няется и на следующий год, то саванны в общем имеют буроватый
тон и лишены ярких зеленых и цветистых тонов наших степей и лу-
гов. Луковичные почти отсутствуют Лишь в более разреженных
типах саванн цветистость усиливается. Зеленым ковром покрыва-
ется саванна лишь в случае уничтожения ее растительности пожа-
ром.
Травянистая растительность саванн связана с рельефом местно-
сти, особенно роскошна она на более пониженных местах — выше
роста человека, на повышенных же местах она значительно ниже.
В долинах растут леса.
353
Африканские саванны. Они занимают большие пространства,
чем в какой-либо другой части света, особенное развитие они имеют
® восточной Африке н к югу от Сахары. Все сказанное выше поило-
^кимо и к африканским саваннам Травянистый покров состоит глав-
ным образом нз высоких злаков (Andropogon, Panicum, Chlora и
др ), двудольные играют подчиненную роль. Из деревьев очень
характерны зонтиковидные акации, а особенно знаменитый баобаб—
«хлебное дерево обезьян» (Adansonia digitata — рис 138). Это —
гигантское дерево (до 25 л<) с необычайно толстым стволом (до 9,5 м
в поперечнике). Интересно, что баобаб — это дерево исключительно
253
саванн; оно не выносит присутствия других деревьев. В древесине
имеются большие скопления воды, что объясняет непонятное на пер-
вый взгляд Присутствие баобаба в климате саванн. Баобаб может
достигать возраста в 4 5 тысяч лет. Свое название он получил бла-
годаря своим плодам, сочная мякоть которых сьедобна, сам же плод
имеет вид большого длин-
ного огурца. Из других
Деревьев в африканских
саваннах встречаются не-
которые пальмы (рис. 139);
«т^етиг, креме тиГъ, дре-
вовидные молочаи.
Южноамериканские са-
ванны. Как уже было ска-
зано, в Южной Америке
саванны развиты в двух
областях, — к северу и к
югу от влажнотропических
лесов р. Амазонки. Как
льяносы (в Венецуэле
н Гвиане),так и кампосы
(в центральных частях
Бразилии) представляют
по большей части ландшаф-
ты паркового характера,
хотя местами имеются про-
странства, совершенно ли-
шенные деревьев. На более
влажных местах встреча-
ются леса, по берегам
рек—галлерейного типа.
В льяносах дожди на-
чинают выпадать лишь с
апреля, зимой же долго
держится засуха. Благо-
даря обильным дождям
льяносы покрываются
роскошной растительно-
стью из злаков (Panicum,
Paspalum, Hymmachne) и ряда двудольных. Интересен ландшафт
с пальмой (Copernicia tectorum), стволы которой нередко одеты це-
лой сетью корней эпифита-фикуса.
Кампосы занимают главным образом водоразделы между Амазон-
кой и Ла-Платой. Деревья низкорослый корявы, очень много кустар-
ников, из трав преобладают злаки (Paspalum, Panicum, Andropogon,
Aristida), а также многие осоковые- из двудольных — бобовые и
сложноцветные (Eupatorium, Vernonia). Для деревьев характерна
зонтиковидная крона.
Австралийские саванны. Занимают значительные пространства
во внутренних частях Австралии (на карте Брокман-Ероша они
обозначены степями). Господствующими деревьями являются эвка-
липты и замечательные древовидные лилейные, так называемые
«травяные деревья» (Xanthori lioea — рис 140).
Саванны (также саванновые леса) и человек
Первоначальное воздействие человека на эти типы растительности
сводилось к влиянию выпаса домашнего скота. Естественно, что скот
производил изменения в первобытной растительности — уничтоже-
ние более хороших в кормовом отношении растений, большее или
меньшее уничтожение мертвого покрова из отмерших частей расте-
ний и др. Саванновые леса нередко превращаются во вто-
ричные саванны вследствие уничтожения деревьев человеком для
своих нужд, а саванны могут становиться совершенно лишенными
деревьев. Уничтожение деревь в могло иметь место или в резуль-
тате порубок или же всл.дствне выжигания сухой травы, к торое
постоянно применялось пастушескими племенами (и теперь еще при-
меняется) для улучшения травостоя, так как накапливающий^!
мертвый покров сильно подавляет молодые растущие части растений
(это — примитивная пастушеская система). 1
При большей оседлости племен выпасание скота на травянистых
пространствах комбинируется с залежной системой хозяйства; са-
ванновые леса и саванны освобождаются от древесных растений и
земля обрабатывается мотыгой. После нескольких лет культуры
земля забрасывается в залежь, а затем снова через известное время
обрабатывается (суданская система агрикультуры — в Судане в
Африке).
На Мадагаскаре саванновые леса после их выжигания н после
ряда лет культур превращаются в кустарниковые заросли (так
называемая савска — Savoka), несколько схожие с средиземно-
морским маквисом (см. стр. 2С6). Эти (заросли очень бедны флори-
стически, что и вообще типично для растительности вторичного,
характера. Дальнейшее выжигание приводит обычно к образованию
травянистых пространств характера степей, которые также дегра-
дируют все больше и больше. Гюд конец может получиться нечто
вроде пустыни.
IX. КЛАСС ФОРМАЦИЙ—ТРАВЯНИСТЫЕ ЛЕТНЕЗЕЛЕНЫЕ КСЕРО-
ФИЛЬНЫЕ ФИТОЦЕНОЗЫ—СТЕПИ (AESTIDURIHERBOSA)
Общая характеристика. Для этого класса формаций наиболее
типичны степи—травяньстье, лишенные деревьев, фитоценсзы
большего иЙи меньшего ксерофильного характера. Наибольшие
пространства степи завил акт в СИР, протягиваясь широкой поло-
сой по Европейской и Азиатской частям с запада на восток до
р. Оби и далее на восток уже в пределах Китая (в наших пределах к
востоку от р. Оби лишь отдельными участками), кроме того, в Север-
ной Америке (здесь степи называют^ прериями). Небольшие участки
степей («пушты») находятся в Дунайской низменности, в Венгрии;
довольно обширные пространства степей в Южной Америке (Арген-
тина и Уругвай)— так называемые «пампасы» (или «п м. ь»). Степи
255
аз
2-23
1.4
мы находим также и в Африке, Австралии и пр. (см. карту в конце
кии ги).
Широкое развитие степей характерно для умеренных
климатов со сравнительно небольшим количеством летних осадков,
нричем степь чаще всего занимает промежуточное поло-
жение между пустынями и летнезелеными лесами. Степи менее
ксерофильны, чем пустыни, и более ксерофильны, чем леса.
Почвами степ.й являются черноземы различных типов.
Так как растительность степей носит в большей или меньшей
мере ксерофильный характер, то физиономически от других тра-
вянистых типов (луга, болота и др.) они отличаются тускло-зеленым
оттенком своих вегетативных частей, которые имеют те или иные
приспособления для выдерживания летних засух (волосяной по-
кров, редукция листьев, восковой налёт и
другие признаки,характерные вообще для
ксерофитов).
Что касается основных жизненных форм,
то для степей особенно характерны хаме-
фиты (если понимать хамефиты более ши-
роко, чем обычно, включая сюда ковыли,
типчак и другие дерновинные злаки, как
это делает Гаме).Впрочем, для некоторых
типов степей хамефиты гораздо менее
типичны, чем гемикриптофиты (русские
северные степи).
Травянистый покров степей в общем
является довольно густым, но в более су-
хих степях он начинает изреживаться; этв
изреживание, приводящеектому, что над-
земные части растений не смыкаются,
особенно характерно для полупустынь и
пустынь.
Кроме того, для степей типично мас-
совое развитие дерновинных злаков с их
особыми приспособлениями к засушливой
среде (листья узки и обычно свернуты;
основания дерновин сидят довольно глу-
боко в почве, рнс. 141)идр.Замечательно,
что род ковыль (Stipa) с его многочислен-
ными видами характерен и для русских
ковыльных степей (рис. 142), и для северо-
американских прерий, и для южноамери-
канских пампасов и др-
Если физиономически и экологически
степи различных стран имеют большее
или меньшее сходство, то по своей флоре
они сильно отличаются, хотя и имеются некоторые общие роды.
В зависимости от большего или меньшего количества летних
осадков можно различать ряд типов степей, так, например, в СССР
хорошо различаются наиболее северные степи, часто называемые
задер-
Рис. 141. Степной тип
нения.
1—узколистный ковыль (Stipa
atenophylla), 2 - обыкновенный
ксвыль (S Joannis), 3—тонконог
(Koeieria gracilis), дерновины
растений сидят в более и менее
глубокой паше. Цифры — санти-
метры (ориг.).
356
луговыми, и более южные,— ковыльные, переходящие в степи
полупустынного характера. В Северной Америке различают более
влажные высок"злаковые и менее влажные низкозлаковые прерии.
Степи СССР. Очень трудно и даже невозможно дать одну общую
ландшафтную картину для степей, даже если остановиться иа ка-
ком-либо одном типе, так как одна из наиболее характерных черт
степной растительности вообще — это сильная динамичность
растительного покрова в смысле фенологических смен- В самом
деле, флора степи складывается из растений, не совпадающих в своих
фенологических фазах и, следовательно, физиономически выделя-
ющихся в самые различные моменты вегетационного периода. Выше
(см. стр. 155) мы познакомились с очень ярким примером подобного
рода и видели, что на Стрелецкой степи под Курском можно разли-
чать до 12 красочных фаз, сменяющихся одна другой. Эти фазы
настолько красочны, что степь приобретает то голубую, то фиоле-
товую, то белую, то желтую окраску в связи с массовым развитием
тех или иных господствующих видов. Значит, говорить об общей
физиономической картине степи нельзя.
Подобно северным степям (Стрелецкая степь относится к север-
ным степям), такую же-смену аспектов дают и более южные ковыль-
ные степи. Так, например, для ковыльных степей Днепропетров-
17 ГеогваФия растений 257
щины с обилием красочных двудольных растений можно установить
такие основные красочные аспекты.
I. Ранней весной появляются многочисленные луковичные, а также клубне-
вые растения: желтые тюльпаны, бледноголубые гиацинты, золотистые гусиные
луки, снежнобелые птицемлечники, белые крокусы и др. Кроме того, цветут
такие корневищные растения, как яркозолотистые адонисы, бледнофиолето-
вые фиалки п др. Перед нами либо пестрый ковер этих нежных ранневесенних
видов, либо участки с преобладанием тех или иных из указанных растений.
Рис. 143. Ареал Centaurea orientalis—один из примеров
общности флористических элементов русских и венгерских
степей (по Гаевскому)
Почти все эти растения — эфемеры, очень быстро выходящие из степного .тра-
востоя и «убегающие» от летней засухи.
2. В мае степь совершенно преображается: это время пышного развития зла-
ков, в частности ковыля (Stipa Lessingiana). Степь напоминает седое волную-
щееся море, когда травостой колышется от легкого ветра. Большую роль играют
и другие злаки; типчак (Festuca sulcata), тонконог (Koeleria gracilis), костёр
(Bromus erectus). Немало также красочного разнотравья: фиолетовые бабки
(Salvia nutans), красноватые юринен (Jurinea arachnoidea), белые тринии
(Trinia Henningii) и многие другие. Эго— ковыльно-красочно-разнотравная
фаза. Однако, ковыль довольно скоро сбрасывает свои серебристые ости, п потому
неправильно думать, что ковыльные степи всегда серебрятся ковылем.
3. В июне зацветает целый ряд других двудольных видов, например, желтый
василек (Centaurea orientalis, рис. 143), злаки почти все уже отцвели.' В данную
фазу очень характерны растения, образующие «перекати-поле» (см. ниже) —
своеобразная жизненная форма, тесно связанная со степным простором. В это
время степь пестрит разноцветными шарами этих растений: белые шары качима
(Gypsophila paniculata), фиолетовые — кермека (Statice tatarica, рис. 144),
красно-фиолетовые— травы-ветра (Phlomis pungens), голубые— котовика
(Nepeta ucrainica).
4. В июле степь снова затягивается пеленой ковыля, но другого вида—имен-
но, ковыля-тырсы (Stipa capillata). Ости его волосовидные (не перистые), и степь
в это время отливает золотисто-зелёным оттенком этих остей. И в это время
цветет ряд двудольных, но в случае пышного развития тырсы они почти тонут в
море густого травостоя.
5. В августе число цветущих видов сильно падает, и общая физиономия степи
определяется многочисленными засохшими или засыхающими растениями.
Но все же некоторые виды только что зацветают: степная астра (Linosyris villosa),
полынь (Artemisia austriaca), одуванчик (Taraxacum serotinum). Появляются не-
которые однолетники; в случае влажной погоды, например, очень обильна
жёлтая очанка (Odontites lutea).
6. В сентябре почти нет цветущих растений, некоторые еще отцветают, но в
случае дождей степь дает некоторое усиление вегетации: появляются молодые
листья у дерновинных злаков, у двудольных при основании стеблей появляются
небольшие побеги и т. д. В таком состоянии степь уходит под снег.
Необходимо указать, что степям несвойственно явление, кото-
рое считают часто характерным для степей, — именно они не
имеют летнего замирания растительности. Это особенно хо-
рошо можно видеть из фенологической смены цветущих и зацве-
Как видно, после достижения максимума в конце мая н начале
нюня число цветущих видов всё время медленно падает и
падения числа цветущих видов именно в середине лета не замечается.
Лишь в наиболее южных степях, в причерноморских (например в
Аскания-Нова), наблюдается в середине лета некоторое выгорание
растительности, а затем новый взрыв с сильным развитием ковыля-
тырсы. Ярко же выраженный летний перерыв вегетации мы находим
лишь в полупустынях и пустынях.
Вообще в русских степях наблюдается целый ряд закономерных
изменений в растительности степей при движении с севера на юг.
1. Травостой всё более и более разреживается и густой траво-
стой северных степей сменяется сильно разреженным с пустыми
промежутками между растениями.
2. Красочность степей сильно уменьшается, так как число
двудольных к югу падает.
3. На севере—господство многолетников, к югу — значительно
усиливается роль однолетников, причём последние развиваются в
пустых промежутках между многолетними растениями. Обилие
однолетников особенно характерно для пустынь.
4. Роль широколистных ’ злаков (косдёр — Brcmus erectus,
овёс—Avenastrum pubescens и др.) к югу сильно падает. Их место за-
ступают злаки узколистные (койылн, типчак й др.).
5. Сменяется целый ряд ковылей: Stipa Joannis — S. steno-
phylla — S. Lessingiana — S. ucrainica; последний ' ковыль — наи-
более южный. В более южных степях обилие ковылей особенно велико-
6. Видовая насыщенность (т. е. число видов на определенную
площадь) показывает такие цифры для 1 л!2: на орловских (наиболее
северных) степях около 30 видов, на курских — около 50 и более
(в отдельных случаях—80), на старобельских—25, на асканий-
ских—12.
В общем ксерофитизация степей к югу всё более усиливается, и
степи постепенно переходят в полупустыни.
Рис. 144. Кермек (Statice tatarica). Ветви соцветия при созре-
вании семян загибаются кнаружи, потучается шаровидное обра-
зование, которое затем отрывается от корня (оригЛ.
Особенно характерной жизненной формой для степей является,
как уже было сказано, перекати-поле. Эта жизненная форма выра-
боталась независимо в самых разнообразных семействах, но осо-
бенно, конечно, среди двудольных. Наибольшее число представи-
телей перекати-поля встречается в средних частях степной зоны.
В основном мы здесь имеем растения шаровидной формы, что дости-
гается обильным ветвлением или другими способами (рис. 144).
При созревании семян растения многолетние отделяются от своих
подземных частей, однолетние — выдергиваются с корнями н,
будучи очень легкими, начинают перекатываться по степи, разбра-
сывая свои семена. Отделение от корневищ или корней происходит
чаще всего в результате того, ыто стебель внизу становится чрез-
вычайно хрупким и переламывается даже от небольшого толчка;
в иных случаях отделение происходит от перегнивания стебля позд-
ней осенью или зимой (например тслстые стебли зонтичных). Инте-
ресно, что при разбрасывании семян во время перекатывания, в ряде
случаев выработались приспособления, ведущие к тому, что семена
разбрасываются постепенно, вследствие чего они рассеиваются на
большой площади (например отверстия коробочек становятся более
узкими нли развиваются волоски, затрудняющие высыпание
семяи и др.).
Можно предполагать, как выяснилось в са^лое последнее время,
что роль растений перекати-поле сводится не только к тому, чтобы
рассеивать по степи свои семена. Имеются виды (например Hippo-
marathrum crispum и др.), у которых роль перекати-поле играют
многочисленные листья прикорневой розетки, которые отрываются
от корня и перекатываются, стебель же с плодами остается на месте.
Здесь, невидимому, имеет место освобождение растения от своих
многочисленных остатков, которые, накопляясь из года в год,
затрудняли бы дальнейший рост растений. Получается как бы
с а м о о ч истк а растений. У тех же видов, у которых отрываю-
щиеся части содержат семена илн плоды, отдет ением от корня сразу
решаются две задачи: рассеиваются семена и происходитсамоочистка
растения. Последнее особенно важно потому, что обычно перекати-
поле развивает большое число стеблей (а также нередко листьев),
которые давали бы, при отсутствии отрывания, за ряд лет мощный
мертвый покров, тормозящий весенний рост растений.
Вопросы классификации русских степей достаточно разработаны.
В самых общих чертах основные типы степей таковы:
I. Северные от е п и. Красочное разнотравье северной®
типа (Salvia pratensis, Filipendula hexapetala, Anemone Silvestris
и др.) с широколистными злаками (костер — Bromus erectus,
дикие овсы — Aveiiastrum pubescens, A. Schellianum, полевица —
Agrostis SyreistschikcMi). Из ковылей — ковыль обыкновенный —
Stipa Joannis и узколистный — S. stenopliylla.
II. Южные степи. Ковыльно-типчаковые степи с южным
разнотравьем и узколистными злаками (ковыль, типчак, келерия—
Koeleria gracilis). Из ковылей — ковыль Лессинга—S. Lessingiana,
ковыль-тырса— S. capillata, на востоке — S. rubens.
1. Большое количество разнотравья (к р а с о ч ны й к о в ы ль-
н и к). Более северный вариант.
2. Небольшое количество разнотравья (бес*красочный
ковыльник)- Более южный вариант. На западе большую роль
играет ковыль украинский (Stipa ucrainica).
Стспн Европейской части СССР переходят за Урал и идут сплош-
ной полосой до р. Оби, далее же на востоке в пределах СССР раз-
биваются на отдельные острова. Этн сибирские степи
имеют много аналогичных черт со степями европейскими, однак®
рельеф их значительно менее расчленен, вследствие чего имеются
обширные пониженные равнинные пространства, где большую роль
играют «займища», т. е. травяные болота с тростником, и где широк®
распространены солончаки и солонцы В результате мы имеем слож-
ность экологических отношений, и степные виды сочетаются с рас-
тениями более влажных местообитаний и с солончаковыми.
26*
И флористически сибирские степи сильно отличаются от евро-
пейских, они беднее видами, и ряд растений европейских степей
здесь отсутствует. С другой стороны, появляются некоторые сибир-
ские виды, а далее на восток и виды монгольского происхождения
В Европейской части СССР степи почти распаханы, в Азиат-
ской — еще занимают очень большие территории. О влиянии вы-
паса на степной травостой см. стр. 267.
Венгерские пушты. Значительные пространства были заняты
степями (онн почти целиком распаханы) в Придунайской
низменности и Венгрии, здесь степи называются «пуштами» и зани-
мают главным образом пространство между Дунаем и Тиссой. Они
довольно схожи с южнорусскими степями. Почвами венгерских
степей являются главным образом песчанистые черноземы.
Можно различать несколько степных формаций с господство/л
ковылей (Stipa Joannis, S. capillata), с господством злака Polhnia
gry Hus и др.
Растительность начинает развиваться с самой ранней весны, и аспекты сме-
няются до конца лета Первые растения расцветают в последних числах марта
степь покрывается тысячами золотистых звездочек гусиного лука (Gagea pusi Па)
и многими однолетниками (Draba verna, Veronica verna, Saxifraga tndactyhs
i др) В апреле цветут лютик (Ranunculus pedatus), прострел (Pulsatilla
pratensis), барвинок (Vinca herbacea), лапчатка (Potentilla pratensis) M а й —
это наиболее пышный расцвет растительности, это — пестрый ковер цветов,
в эго время цветут ковыли и другие злаки, а также многочисленные двудольные
виды В июне степь блекнет, но еще многие растения зацветают вновь В
июле чис то цветущих видов совсем невелико (Galium veruni, Asperula cynan-
chica, Onobrych'S arenana и др.)
Нередки участки с господством однолетнего костра (Bromus
tectorum), но это — результат усиленного выпаса.
Североамериканские прерии. Очень большие пространства в
Северной Америке заняты степями, называемыми прериями. Об-
ласть прерий с севера на юг протягивается от 60° в Канаде до 30°
в Техасе, а с востока на запад — от 89° в Иллинойсе до 107° в Уайо-
минге. Наиоолее важны для американских прерий три рода злаков
Stipa, Agropyrum и Bouteloua, которые встречаются почти везде
и присутствуют в любой ассоциации. Из этих злаков последний в
евразийских степях не встречается.
В прериях — пять основных типов’ настоящая пре-
рия, луговая прерия, смешанная прерия, низкозлаковая прерия,
дерновинно-злаковая прерия.
(.Настоящая прерия Она занимает срединное по-
ложение в области прерий и развита в штатах: Северная и Южная
Дакота, Небраска, Канзас, протягиваясь,таким образом, в долгот-
ном направлении с севера иа юг, при этом долгота 100° является
как бы осью этого типа прерий. Почвой служит структурный глу-
бокий чернозем, увлажняемый на большую глубину, вследствие
чего здесь много глубоко укореняющихся видов, подразделяющихся
на три яруса по положению их корневых систем: на глубине 1 м,
1,66 м и глубже.
Данный тип степи называют типом Stipa —Koeleria. Из
злаков доминируют: Stipa spartea, Andropogon scoparius, Koele-
262
ria gracilis, Bouteloua racemosa. Можно наметить ряд аспектов.
Весной развиваются многие виды, имеющие, однако, невзрач-
ные цветки, особенно обильна Antennaria campestris, на более вы-
соких местах цветистость несколько больше. Летом цветистость
значительно повышается. Psora lea flonbunda, Erigeron ramosus,
Achillea millefolium и т д В конце лета и осени: астры,
золотая розга, Helianthus, Liatris и др. Особенно характерны для
осени сложноцветные.
Период вегетации продолжается около 100 дней, и прерия за-
сыхает лишь осенью, летнего перерыва вегетации от засухи здесь
нет. Травостой вначале около 2U см высотой, летом доходит до 50 см,
причем наиболее высокие растения достигают 80—120 см.
2. Луговая прерия (или тип Andropogon). Распростра-
нена к востоку от настоящей прерии, главным образом к востоку
от Миссури (штаты Айова, Сев Миссури, Иллинойс, Индиана и др.).
Это более влажный тип, количество осадков здесь до 100 см (в настоя-
щей прерии около 75 ел/) Ландшафтные отличия от настоящей прерии
таковы, в балках лес (главным образом дуб), травостой более высо-
кий и сомкнутый. Наиболее характерны виды рода Andropogon
(A. furcatus, A. nutans, A scoparius, A saccharoides), также Stipa,
Bouteloua, Koeleria, Agropyrum Есть основание думать, что до
культуры здесь был лес. В настоящее время луговая прерия к во-
стоку при посредстве промеж т чной полосы (лесостепь) переходит
в область широколиственных лесов (см. выше).
3. Смешанная прерия (или тип Stipa — Bouteloua).
Распространение к западу от настоящей степи (штаты Мон-
263
тана, Уайоминг, отчасти Северная и Южная Дакота, Колорадо и
др., на севере проникает в Альберту и Саскачеван). В основном
это — переходный тип между высокозлаковыми прериями (настоя-
щая прерия) и ннзкозлаковыми (см. ниже). Из высоких злаков здесь
Stipa, Andropogon, Agropyrum, Bouteloua racemesa; из низких —
Bouteloua gracilis и Buchloe dactyloides. Получается двухъярусное
сложение: высокие злаки, а в промежутках между ними низкие
злаки (рис. 145). Большинство видов имеет глубокие корни, как и *
настоящей прерин. Иногда высокие злаки выпадают из травостоя,
Рис. 146. Американское степное растение «трава бизонов» (Buchlog dactyloide*^.
Слева—женский экземпляр, справа—мужской (из Шимпера).
остаются лишь низкие, и тогда прерпя получает характер низкв-
злаковой; это является результатом неумеренного выпаса.
4. Дернов инн о-з лаковая прерия (или тип Agro-
pyrum — Stipa). Этот тип прерин характерен для песчаных холмов
в области смешанной и настоящей прерии и очень сходен со сме-
шанной прерией, лишь дерновины злаков располагаются более
разреженно. Тип идёт ещё далее на запад до восточных частей шта-
тов Вашингтона и Орегона и связан со смешанной прерией всеми
переходами.
Все четыре рассмотренных типа прерий могут быть объединены
как высокозлаковые; следующий тип будет состоять из
низких злаков.
5. ' Низкозлаковая прерия (илн тип Euchloe —
Bouteloua). Связана с «Великой равниной». Распространена иа
большой территории от северо-запада Небраски и запада Как аса
до северо-запада Техаса и Аризоны, но также заходит на юго-восток
Юты и юго-запад Колорадо. Небольшое количество осадков (окол®
40 см) вызывает сухость почвы, которая усиливается ещё тем, что
около трети и более выпадающих осадков стекает по поверхности.
264
не попадая вглубь. Это происходит от того, что почва здесь очень
мелкозерниста (чернозём). В результате почва не промачивается
глубже 65 см, и господствуют мелко укореняющиеся растения.
Эта прерия монотонна и уже в июне высыхает (продолжительность
вегетации около 60 дней). Господствуют два низких злака — Buch-
loe dactyloides (трава бизонов, рис. 146) и Bouteloua gracilis (трава
грама), причём они занимают до 75—90% всего травянистого ковра
и так переплетают почву своими корнями, что почти не допускают
других растений. Травостой или сплошной или несколько разрежен,
высотой от 5 до 8 см.
Взаимоотношения степи и леса. Степь и лес, граничащие друг
с другом, дают очень причудливую линию контакта; в СССР она то
поднимается на север, то опускается на юг. С другой стороны, остров-
ки лесов проникают далеко на юг в степную область как по водораз-
делам, так и особенно по поймам рек и по балкам; но и обратно, ост-
ровки степей (или вообще степной растительности) вкраплены среди
лесов лесной области (острова степей в Горьковской области, в При-
уралье и др.). Эта область контакта леса и степи уже давно получила
название лесостепи (иногда говорят даже об особой лесо-
степной зоне). Чем обусловлена граница между лесом и степью?
Что за причина безлесья степей? И 'были ли степи безлесны до по-
явления человека? Все эти вопросы были поставлены в конце прош-
лого столетия и сыграли огромную роль в развитии русской ботани-
ческой географии, как науки. В разрешении их приняли участие все
виднейшие представители этой отрасли знания. И вопрос решался
по разному. Так, одни исследователи решающее значение в без-
лесьи степей видели в климатических условиях (Бэр, Шимпер,
отчасти Высоцкий и др.), другие выдвигали на первый план
почвенно-грунтовые условия, в особенности засолённость почв
(Т а н ф и л ь е в), третьи выдвигали физические свойства почв,
особенно их мелкозёмистость и плохую проницаемость для воды
(Косты ч е в и др-); ещё иные решающую роль отводили кон-
курентным отношением между лесом и степью, считая лес более
сильной биологически формацией (К о р ж и н с к и й), или смот-
рели на степь, как на определённое звено в генезисе растительного
покрова (три стадии развития: пустыня—степь — лес; Пачо-
с к и й). Имелись ещё и другие точки зрения.
Как видим, для объяснения безлесья указывались причины
климатические, эоафические, биотические и генетические.
Однако совершенно невозможно решать вопрос о безлесье сте-
пей вообще, так как сами степи, как было выяснено выше, далеко
неоднородны как по своему географическому положению, так и по
растительности. Еопрос о южных степях может решаться иначе,
чем для северных степей. Кроме того, несколько односторонне
искать причину безлесья лишь в сд! ой какой-либо группе факто-
ров — в природе мы имеем совместное действие факторов.
Что касается южнорусских степей и настоящих и низкозла-
ковых прерий Северной Америки, то нельзя сомневаться в том,
что их существование обусловлено общей природной
обстановкой (сухой континентальный климат, почвы богатые
2Й5
солями и др.); эти степи непосредственно переходят в полупусты-
ни и пустыни (при ещё более неблагоприятных условиях). Во-
обще же степь — это совершенно необходимое природное звено
между лесом, с одной стороны, и пустыней — с другой. Лес не
может, конечно, прямо переходить в пустыню, необходим проме-
жуточный тип растительности, и этим типом является степь.
Те части степной зоны, которые находятся в непосредственной
близости к лесной зоне (наши северные степи) во многих случаях
носят как бы переходный характер и потому называются л е с о-
степью; здесь участки степи и леса находятся в сложных взаимо-
отношениях. В более южных частях леса приурочены к балкам и
местам большего увлажнения. К северу лесных участков становится
всё больше и больше, и степь в конце концов почти сходит на нет—
от неё остаются лишь остепнённые поляны. ' Так медленно и посте-
пенно совершается переход от степи к лесу.
В подзоне северной лесостепи в распределении участков степин
леса играют роль уже не общеклиматические условия, а те или
другие местные условия; микроклимат, небольшие колебания рель-
ефа и др.
Таким образом, степь — это такое же естественное природное
явление, как лес, пустыня и др.; можно привести ряд свидетельств
различного рода (почвенные, исторические, биотические), которые
доказывают, что степи на юге СССР существовали и в доистори-
ческое время, т. е. не были связаны с деятельностью человека (уни-
чтожение лесов). Иногда говорят об «отвечном» существовании южно-
русских степей; однако это нужно понимать условно, т.е. вне связи
с деятельностью человека. Но мы знаем, что в третичное время и в
начале послеледникового в связи с иными климатическими условия-
ми на месте наших степей были леса.
Пампасы Южной Америки. Занимают обширные равнины на юге
Южной Америки от Патагонии (на юге) и почти до области Чако
на севере; на востоке они доходят до берегов Атлантического океана,
а на западе —до предгорных частей Кордильер. В общем пампасы
занимают большую часть Аргентины и Уругвая.
Почвой является чернозём, близкий к европейскому. Подпоч-
вой служит лёсс. По ландшафту пампасы, подобно степям, пред-
ставляют необозримые пространства, покрытые травянистой расти-
тельностью. В общем растительность здесь ксероморфного характера,
и злаки имеют буро-зелёную или сероватую окраску. Весной при
известном количестве осадков пампасы яркозелёные.
Вегетация начинается с октября, и целый ряд злаков и двудольных расте-
ний дает постепенно сменяющиеся аспекты. Из злаков первым зацветает Роа,
а затем цветут Stipa, Aristida/ Melica, Bromus, Briza; в ноябре— Ischnemum,
Festuca, Koeleria и др. В декабре — январе появляются виды Panicum. Paspa-
lum, Andropogon и др. Это разгар лета. Многие злаки цветут еще позже (февраль—
март), намечая уже начало осени.
Кроме злаков, развиваются многочисленные двудольные; весной — красивые
портулаковые (Portulacaceae), многие гвоздичные ^(Silene, Arenaria), мотыльковые
(Lupinus, Desmodium, Vicia и др.), вербеновые и т. д. Особенно красива красная
вербена (Verbena chamaedryfolia). Из сложноцветных весной встречается лишь
крестовник (Senecio).
266
С начала лета появляются жёлтые и голубые цветки ирисовых — прекрасные
Cypella и Alophia, многие дербенниковые (Lythraceae), миртовые (Myrtaceae),
паслёновые (Solanaceae), а также многочисленные сложноцветные, которые
для этого периода особенно характерны; из них для южноамериканской флоры
типичны сложноцветные с двугубым венчиком (у сложноцветных Старого Света
двугубых венчиков не встречается, только лишь одногубые). В это время цветут
высокие зонтичные и другие.
В конце лета особенно много сложноцветных, большинство родов отличны
от европейских. Вообще же обилие сложноцветных — это характерная особен-
ность пампасов.
Данное краткое описание относится к пампасам Уругвая.
Местами развиваются болота, физиономически напоминающие
наши болота, в более западных частях начинают попадаться засо-
лённые участки.
Пампасы, как места, благоприятные по своим климатическим
условиям, уже давно используются как пастбища и распахиваются,
так что во многих местах от первобытной растительности ничего не
осталось. Часто большие территории занимают сорняки, пришед-
шие сюда вместе с европейцами; особенно известен в этом отношении
кардон (Cynara cardunculus), являющийся просто бичом страь.ы.
Южноафриканские степи в верхних частях Оранжевой р ки
представляют травянис.ые n;oCTj анства степного характера, н в
д угих местах более напоминают полупустыни с более разреженн м
травостоем. Близка к степям и «пустыня» Калахари (см. стр. 317).
А з ст р а л и й с к и е степи правильнее относить к саваннам
(см. стр. 525).
Степи и человек
Степи издавца связаны с человеком, именно, ещё в доистори-
ческие времена. Кочевые первобытные племена использовали степь
как обширные кормовые угодья. В истории русского и украинского
народов степи играли выдающуюся роль. Для военных целей степь
нередко на больших пространствах выжигалась (степные «палы»),
чтобы лишить наступающего неприятеля подножного корма; выжи-
галась также и для улучшения травостоя (при этом уничтожались
отмершие остатки травостоя прошлых лет). В настоящее время степи
и прерии почти целиком распаханы, однако, например, в Западной
Сибири осталось еще немало целинных пространств,!, е. участков
никогда не пахавшейся степи. Степи распаханы потому, что они
занимают наиболее плодородные почвы — чернозёмы различных
типов; сейчас территория бывших степей занята главным образом
зерновыми культурами (пшеница, ячмень и др.), в более южных
областях — кукурузой и подсолнечником. Там, где степи ещё не
распаханы, они используются в качестве прекрасных кормовых
угодий (животноводческие совхозы, конные заводы и др.). Степное
сено лучше поедается скотом, чем сеяное, так как оно по составу
более разнородно, содержит ряд предпочитаемых животными расте-
ний и пр. Так как степные участки во многих случаях очень сильно
выпасаются, то естественный травостой претерпевает ряд измене-
ний — менее устойчивые по отношению к выпасу растения исчезают,
количество несъедобных растений становится больше и т. д. Уста-
новлен ряд стадий изменения растительности степей при выпасе,
соответственно умеренному, сильному и чрезмерному выпасу.
287
Умеренный выпас не вредит пастбищу, наоборот, — скот, вытапты-
вая и поедая прошлогодние остатки, способствует более хорошему
росту травы весной. При чрезмерном выпасе пастбище монет стать
совершенно непригодным для дальнейшего выпаса — разрастаются
в массах несъедобные растения, обнажается голая почьа и т. Д-
Определение нормальной нагрузки скота на определенную площадь
выпаса — основная задача спецвалнстсв-кормсьнвов.
Продолжающаяся распашка степей может привести к полному
уничтожению степных фитоценозов, степного ландшафта, столь
связанного с историческим прошлым русского народа. Поэтому
вполне своевременна организация степных запсвеониксв, которые
сохраняют образцы дикой природы, позволяют решить ряд важных,
еще не выясненных вопросов — вопросы образования чернозема,
сложения и возникновения степных фитоценозов и их истерии и
т. д. Заповедники служат также резервуарами многочисленных по-
лезных растений; главнейшие степные заповедники: знаменитый
заповедник «Аскания-Нова» в причерноморских степях (в настоя-
щее время 1 400 га), «Центрально-Черноземный» в Курской области,
(около 3 500 ас), «Поперечеиская степь» близ Пензы (около 2Ь0 га
«Наурзумский» в северном Казахстане (300 000 га) и другие.
X. КЛАСС ФОРМАЦИЙ—ТРАВЯНИСТЫЕ МЕЗОФИЛЬНЫЕ ФИТОЦЕНОЗЫ
С БОЛЕЕ ИЛИ МЕНЕЕ ВЫСОКИМ ТРАВОСТОЕМ—ЛУГА (PRATOHERBOaA)1
Фитоценозы этого класса формаций отличаются от фитоцеиозов
предыдущего класса (Aestiduriheibcsa) тем, что слагающие их виды
«мезоморфный (белее или менее широкие листья, соответствующее
анатомическое строение и др.) в противоположность степям с их
ксероморфным обликом и строением растений. В укреплении на-
земных частей растений наряду с механической тканью играет
роль тургор (у растений степей главная роль принадлежит механи-
ческой ткани).
Данный класс формаций объединяет фитеценозы лугов соб-
етвенно—травянистые фитоценозы мезофильного характера. Так
как мезофильные условия особенно благоприятствуют произраста-
нию леса, то в лесных областях Еврспы, Азии и Северной Америки
луга в большинстве случаев являются етордчьыли образованиями,
возникшими после вырубки лесов и поддерживаемыми в состоянии
луга постоянными покосами (косьба уничтожает всходы древесных
пород).
Луга чаще всего можно встретить в поймах рек (так называемые
поемные, или заливные, луга), в северных частях лесной области
на водоразделах (суходольные, или материксеые, луга), на горах
выше границы леса — субальпийские п альпийские луга. Впрочем,
1 По классификации Б рок ма н-Е р о ш а и Рюбе ля фор,мации
данного класса относятся отчасти к классу Altherbosa, отчасти к Semperviren-
tiherbosa. В частности к последнему классу относятся и заливные луга, однак»
если в Западной Европе в связи с мягкими зимами достаточно хорошо выражена
ееечнозеленость» лугов, то в более континентальном климате это не имеет места.
Вообще же высота травостоя и общая его мезофильность являются признаками,
более существенными для объединения соответствующих формаци
268
альпийские луга правильнее относить к особому классу формаций
<см. ниже).
Растительный покров лугов обычно очень пышный (за исключе-
нием суходольных лугов на бедных подзолистых почвах), с густым
и довольно высоким травостоем, причём почва сплошь задернована.
По своим жизненным формам луга характеризуются преобладанием
гемикриптофитов} другие жизненные формы не играют почти ни-
какой роли. В более мягком климате, например в Западной Европе,
луга остаются зелёными и на зиму, и в их состав входит значительное
число хамефитов- в областях же с суровыми зимами и с глубоким
снежным покровом хамефиты отсутствуют.
Луга заливные. Они занимают в поймах рек огромные простран-
ства, нередко достигая десятка и более километров в поперечнике
(многие сибирские реки, Волга и др.). Некоторые авторы полагали,
что такие факторы, как ледоход, обусловили безлесье пойм (Г р е б-
н е р), другие связывают безлесье пойм с продолжительным зали-
ванием их полой водой, что вредно отзывается на росте древесных
пород. Но можно считать достаточно установленным, что все эти
факторы не могут иметь решающего значения. Лишь в отдельных
случаях в низовьях некоторых рек весеннее затопление настолько
продолжительно, что, несомненно, препятствует росту деревьев.
Если так обстоит дело в лесных областях, то в степных областях
в иных случаях рост деревьев затруднён сильной засолённости*
пойменных почв. В большинстве же случаев и здесь, в условиях
повышенного увлажнения, леса хорошо развиваются, образуя так
называемые «галлерейные» леса, тянущиеся длинной лентой по
обоим берегам рек.
Флористический состав заливных лугов довольно разнообразен,
хотя флористически они значительно менее богаты, чем другие
травянистые типы, например, степи; это зависит, повидимому, от
молодого возраста лугов.
Так как заливные части речных долин (поймы) очень разнооб-
разны по своим экологическим условиям, то и растительность их
расчленяется на ряд сильно отличающихся фитоценозов. На более
сухих местах преобладают многочисленные двудольные растения, на
местах среднего увлажнения чаще всего господствуют злаки, на
наиболее влажных местах — главным образом осоки и некоторые
кислые злаки-
Из злаков некоторые дают почти сплошные заросли и очень
характерны для лугов: лисохвост (Alopecurus pratensis), мятлики
(Роа palushis, Р. pratensis), полевица (Agrostis alba), овсяница
луговая (Festuca pratensis), канареечник (Phalaris arundinacea),
щучка (Detchampsia caespitosa); по более южным рекам —
бекманця (Beckmannia eruciformis), лисохвост русский (Alopecurus
ventricosus) и др- Из луговых осок укажем Carex gracilis, С. Go-
odenoughii, С. vesicaria, в более северных частях — С. aquatilis;
впрочем, осоки особенно характерны для луговых болот.
Многочисленное разнотравье представлено разнообразными се-
мействами; бобовые менее обильны и приурочены преимущественно
к более сухим местообитаниям.
269
Луга дают очень разнообразные физиономические картины;
это зависит прежде всего от разнообразия рельефа, и, глядя на
луг с высокого берега реки, можно хорошо различать пятна, полосы
и вообще отдельные участки различных оттенков и цветов — зелё-
ные, розовые, жёлтые и другие, что зависит от физиономии различ-
ных фитоценозов в данный момент в связи с цветением тех или иных
господствующих видов. Но разнообразие физиономических картин
зависит также и от того, что здесь, подобно степям, наблюдаются
довольно хорошо выраженные, хотя и в меньшей степени, чем
в степях, фенологические смены. При этом можно различать извест-
ное число фаз.
Так как луга каждый год скашиваются, а в более тёплых и влаж-
ных частях Западной Европы покос иногда производится до трёх
раз в течение вегетационного периода, то здесь должно наблюдаться
влияние этого биотического фактора на растительность. И действи-
тельно, имеется ряд указаний на приспособления растений в данном,
направлении; так, наприм.р, некоторые растения цветут и плодоно-
сят до покоса, и, следовательно, от покоса не страдают, другие пло-
доносят во время первого покоса, третьи.— во время второго и т. д.
Естественно, что те растения, которые не успели дать семена ко
времени покоса, должны постепенно исчезнуть с луга.
Одним словом, покос гесемненно оказывает определённое
отбирающее действие (В е т т ш т е й н). На лугах можно
прекрасно наблюдать явление сезонного диморфизма, когда поли-
морфный вид разбился на более ранние н более поздние формы,
тогда как средние формы были покосом нацело уничтожены. В
этом отношении известны такие роды, как очанка (Euphrasia), по-
гремок (Alectorolophus) и другие, виды которых имеют весенние и
позднелетние расы.
Заливные луга отличаются среди других формаций ещё и
" тем, что ежегодно они затопляются весенними полыми водами.
Это имеет очень важное значение, так как полые воды несут
с собой взвешенный в воде материал (илистые, глинистые, пес-
чаные частицы), который и отлагается на пойме. Благодаря отло-
жению Э1Их частиц почвы пойм становятся очень плодородными.
Самый процесс отложения частиц называется 'седиментацией (л
латинско, о слов i «седиментум» — осадок), а почвы называются
аллювиальными (от латинского слова «аллюу.ио»— наносная земля).
Луга суходольные, или материковые. Они развиваются на во-
доразделах, в лесной зоне, на месте сведённых лесов и потому яв-
ляются вторичными. Южные суходольные луга, развивающиеся
южнее границы леса,—это уже степи, т. е. природное явление,
в своём возникновении не связанное с человеком (см. выше). Можно
различать целый ряд типов материковых лугов в связи с почвами,
условиями увлажнения и пр. Разнообразие материковых луг> в зави-
сит также от возраста, от их бывших распашек и т. д. Травостой
обычно низкий, почва замшелая, покрытая мохом, продуктивность
лугов невелика. Можно различать то злаковые травостои, то раз-
нотравные. Из злаков наиболее обычны: белоус (Nardus stricta),
дающий «белоусники», полевица (Agrostis vulgaris), душистый ко-
почти полная за-
а также рослый
лосок (Anthoxanthum), щучка (Deschampsia caespitosa) и др. Все эти
виды олиготрофны и указывают на бедность почв.
Луга субальпийские. Развиваются в горных странах, как раз
выше верхнего предела леса, на высоте 1 500—2 500 м (на Кавказе
и в Средней Азии). Впрочем, высота их расположения зависит от
ряда причин: от географического положения, от экспозиции скло-
нов, от условий влажности и т. д. Занимая особый горный пояс,
они выше переходят в альпийские луга (см. ниже).
Для субальпийских лугов характерна
дернованность почвы и полное покрытие,
травостой.
Субальпийские луга дают
сложные сочетания злаков и
двудольных растений, причём
последние физиономически на-
кладывают на луга тот4 или
иной отпечаток- Во время своего
цветения они пестрят различ-
ными красками. Для субаль-
пийских лугов Средней Азии
особенно характерны герани
(Geranium collinum), а также
белые анемоны (рис. 147), оран-
жевые купальницы, розовые
астры и т. д.
На Кавказе (в Юго-Осетии,
по данным Бушей) субаль-
пийские луга представлены це-
лым рядом ассоциаций и очень
пестры по своему составу. Из
злаков доминируют: костё р (Вго-
mus variegatus), душистый ко-
nocoK(Anthoxanthum odoratum),
овёс (frisetum flavescens), по-
левица (Agrostis рlanifоlia)и др.;
из бобовых—особенно клевера
(Trifolium ambiguum и Т- Рис. 147. Анемон (Anemone narcissiflora).
canescens), из разнотравья —
лютиковые, горечавки, сложноцветные и др. По Г р о с с г е и м у,
на Кавказе можно различать три основных типа этих лугов: влажные,
мезофильные и сухие; в первых двух типах преобладают двудольные
(рис. 148), в последнем—злаки (особенно Bromus variegatus и
Brachypodium pinnatum).
Вопрос о происхождении субальпийских лугов недостаточно
ясен; многие авторы склоняются к тому, что эти луга развилис
вторично в результате искусственного понижения верхней границы
леса. И, действительно, элементы субальпийских лугов встречаются
везде на полянках н опушках на верхней границе леса.
Другие авторы (особенно Буш) настаивают на том, что час
субальпийских лугов первична, т.е.они никогда не были под лесом,
271
субальпийские луга богаты видами, и некоторые виды являются
эндемичными, например, Cbaerophyllum confusum Pyrethrum Bu-
schianum.
Высокотравие. В западных частях Кавказа (Колхида, Юго-
Ос.тт ) в пределах субальпийского пояса ршвито т^к называемое
«высокотравил , причем некоторые виды достигают 2—2,5 мвысоты.
Этот тип содержит ряд третичных реликтов: лилии (LiHum Szovi-
tsianum и др.), борщевики (Heracleum ponticum и др.), коло-
кольчик (Campanula lactiflora), ряд сложноцветных — телекия (Tele-
kia speciosa), девясил (Inula magnifica) и др. Кроме того, характер-
ны: валериана (Valeriana alliariaefolia), крестовики (Senecio nemo-
rensis и др ), цефалярия (Cephalaria tatarica), борец (Aconitum
nasutum), дельфиниумы (Delphinium) н др.
Можно различить две основные формации высокотрав’я:
1) высокотравие, совершенно не имеющее нижних ярусов и состоя-
щее из перечисленных и других высокорослых видов, и 2) высоко-
травие, имеющее три яруса, причем второй ярус состоит из ряда
лесных видов (папоротники, грушанки, кислица и др.), третий
(нижний) состоит из мхов. Обе эти формации связаны с близким
стоянием грунтовых вод, особенно близка грунтовая вода во
второй формации.
Высокотравие — тип почти всегда вторичный (на месте бывших
участков леса).
272
Луга и человек
Луговые формации, пожалуй, больше, чем какие-либо другие,
связаны с человеком, человек не только своей деятельностью влияет
на изменения травянистого покрова, но луга, повидимому, в боль-
шинстве случаев обязаны человеку и самим своим существованием.
Это относится и к заливным лугам, и к суходольным, и в значитель-
ной мере к субальпийским. Это полукультурные форма-
ции, возникшие пос^е вырубки лесов. В настоящее время эти угодья
очень сильно эксплоатируются, представляя важные кормовые
площади, воздействие человека сказывается в двух направлениях —
покосы и пастьба скота. О влиянии покоса было уже ска-
зано. Что касается пастьбы, то она в зависимости от ее интенсивно-
сти приводит к различным последствиям, кроме того, нужно прини-
мать во внимание условия существования того или другого луга,
время выпаса и др- Так, например, весенний выпас на влажных
заливных лугах может приводить к кочковатости луга, что, конечно,
нежелательно, усиленный выпас на сухих лугах может приводить к
еще большей их ксерофитизации вследствие уплотнения почвы
(усиление капиллярности).
Некоторые типы лугов (особенно заливных) представляют крайне
ценные угодья по составу своего травостоя, другие же нуждаются
в тех или иных улучшениях, как, например, подсев соответствую-
щих трав, боронование и т. д. Особенно плохие травостои дают север-
ные суходольные (материковые) луга — здесь необходимы коренные
перестройки их фитоценозов.
В последнее время некоторые заливные луга распахиваются для
посевов хлебных злаков и для огородных культур, однако распашка
их дает неблагоприятные последствия (почвы сносятся полыми
водами и др ).
Субальпийские луга главным образом выкашиваются, хотя в
иных случаях служат пастбищами. Иногда они очень засорены в
результате неумеренного выпаса, сильно распространена буквица
крупноцветная (Betonica grand if lora), виды щавелей (Rumex), виды
манжетки (Alchimi 11а) и др. В иных случаях очень сильно развивает-
ся белоус (Nardus stricta), почти не трогаемый скотом. Засорение
лугов нередко связано с деятельностью грызунов-землероев, раз-
рыхляющих субстрат (появляются крупные щавели, чемерица —
Veratrum Lobelianum и др.).
Для улучшения суб ль. ийскик лугов необходимо принимать соот-
ветствующие меры; так, для Юго-Осетии рекомендуются такие меро-
приятия (Б у ш). 1) запрещение пастьбы ранней весной и поздней
осенью (в это время почва пропитана водой и пои пастьбе дерн легко
разрушается)^ 2) борьба с землероями, засоряющими луга, 3) раннее
выкашивание —в конце цветения, до высыхания травостоя, в этом
случае сорняки еще не обсеменились, 4) регулирование пастьбы.
Что касается распашки и боронования лугов, то на склонах (даже
в 15°) они не допустимы, так как ведут к снесению почвенного слоя
и к явлениям эрозии, но распашка на ровных местах иногда Дает
хорошие результаты и наблюдались случаи, когда распаханные
[R География растений 273
белоусники сменялись более хорошим злаковым травостоем. Не-
обходимо также дальнейшее развитие сенокошения, так как при этом
дёрн на сильно измененных выпасом участках может восстанавли-
ваться.
Значение лугов как основной кормовой площади — огромно,
и потому их всестороннее изучение крайне необходимо.
К сожалению, специально луговых заповедников в СССР нет,
но в некоторых заповедниках наряду с другими растительными
типами имеются и луга (например Печорско-Илычский заповедник,
луга по рр. Печоре и Илычу и др.); кроме того, имеется три вы-
хухолевых заповедника в поймах рр. Оки, Клязьмы и Хопра; &
двух первых луга занимают основную площадь (выхухоль придер-
живается .лу говых водоемов).
Субальпийские луга имеются в ряде горных заповедников, осо-
бенно же хорошо представлены в Кавказском заповеднике (взапад-
ной части Главного Кавказского хребта).
XI. КЛАСС ФОРМАЦИЙ — ТРАВЯНИСТЫЕ АЛЬПИЙСКО-ТУНДРОВЫЕ
ФИТОЦЕНОЗЫ С НИЗКИМ ТРАВОСТОЕМ (FRIGIDOHUMILIHERBOSA>
Сюда мы относим травянистые фитоценозы, характерные для
высокогорных условий (альпийские луга, альпийские лужайки),
а также для тундровой зоны (дерновинные тундры, луговинные
тундры), Для всех этих фитоценозов типичен низкий травостой при
сплошном обычно задернении почвы. Б р о к м а н-Е рот относит
алопийские луга к классу формаций Sempervirentiherbosa, а указан-
ные формации тундр — к классу тундровых формаций вообще
(Frigorideserta). Однако «вечнозеленость» не является присущей
только этому классу, так как и в степях, например, у ряда растений
имеются под снегом зеленые прикорневые листья и молодые нобли.
Что касается отнесения тундровой растительности к «холодной
пустыне», то это надо считать неправильным, хотя бы потому, что^в
тундре прежде всего имеется сплошной травяной (или моховой)
покров, которого пустыня совершенно лишена.
Растения, относящиеся к данному классу формаций, выделяют
в экологическую группу психрофитов, т. е. растений холодной
почвы (физиологическая сухость).
Альпийские луга. На горах, поднимающихся выше границы
леса, на верхнем пределе растительности выше пояса субальпий-
ских лугов развиваются альпийские луга, называемые в Западной
Европе — маттами (Matten). Альпийская растительность по своей
экологии и по своим признакам очень сходна с растительностью
арктических областей. Общим прежде всего является короткий
вегетационный период; в результате —господство многолетников,
среди которых особенную роль играют хамефиты и гемикрипто-
фиты; однолетники почти совершенно отсутствуют.
Подобное соотношение жизненных форм вполне согласуется с
условиями существования, так как короткого лета недостаточно
для прохождения полных жизненных циклов, как у однолетних
растений; хамефиты имеют то преимущество, что у них остаются
274
нижние части стеблей, в иных случаях перезимовывают и листья,
и растения затрачивают материал главным образом на развитие
генеративных органов.
Целый ряд признаков условий существования общих для альпий-
ских лугов и тундр, однако имеется немало и отличий; так,
в альпийских условиях:
]) отсутствует вечная мерзлота;
2) обычно выпадает большое количество осадков;
3) имеются особенности в составе света, так как в высокогорных
областях свет менее изменён, чем в более низких, где при прохож-
Рис. 149. Жизненная форма «растение-подушка »'(Draba
bryoides) — Северный Кавказ (высота около 2000 лт). Сле-
ва—общий вид, справа—продольный разрез (ориг.).
дении через слой атмосферы особенно поглощаются коротковолновые
лучи;
4) очень важное значение имеет в горах световой режим, так как
здесь интенсивность света значительно большая, чем на уровне
моря, а кроме того, в южных горах продолжительность дня зна-
чительно меньше, чем в тундрах, поэтому растения тундр могут
быть отнесены к растениям длинного дня, а альпийских лугов — к
растениям короткого дня.
Если сопоставить высокогорные условия тропических гор с
условиями тундр, то здесь имеется ещё одно важное отличие, а имен-
но, если в тундрах имеется хотя и короткий, но относительно
теплый (10° и выше) вегетационный период, то в горах (например
Анды) круглый год наблюдается одинаковая низкая
температура, всего несколько градусов выше 0°, а на больших
высотах температура не поднимается выше 0° и, значит, исключает
растительную жизнь.
Наиболее характерные признаки альпийских растений: 1) низкий
рост (около 10—15 см), 2) прикорневое расположение листьев,
образующих прикорневую розетку, 3) сильная укорененность меж-
доузлий, в результате чегостебель очень сильно укорочен;4) нередко
встречается жизненная форма «растение-подушка» (рис. 149); 5) у
некоторых видов имеются лишь безлистные цветочные стрелки;
6) сильно развито вегетативное размножение, так как образование
семян не всегда обеспечено в случае ранних холодов; 7) яркая
окраска цветков, как результат интенсивного освещения и как
приспособление к обеспечению опыления, 8) более крупные, чем на
равнинах, размеры цветков.
Рис. 150. Альпийские растения на хребте Сиерра-Невада в Испании.
1—Dianthus brachianthus; 2 — irtamisia granatensis 3—Linam nevadensis, 4—
'^Ranunculus„acetosellaeformis, 5—Viola^nevadensis (из Шлмпера).
Обращает на себя внимание”еще одна очень интересная черта
альпийских растений — это крайняя устойчивость про-
тив замерзания. Даже и летом, во время вегетации, вслед-
ствие сильного излучения, температура иногда ночью опускается
ниже 0°, и венчики некоторых цветков замерзают и становятся
хрупкими, как стекло, но под действием лучей солнца быстро от-
таивают, и цветки продолжают цвести.
Альпийские растения обычно образуют очень низкорослый
травостой, сплошь покрывающий почву. На больших высотах он
изреживается и. наконец, совершенно сходит на нет
276
Растительный ковёр состоит из растений, обнаруживающих
те или иные признаки ксерофитов (рис. 150): узкие свернутые листья
у злаков и осок, восковой налет и различные виды опушения, сочные
листья н стебли (суккуленты) и т. д. Особенно нужно отметить
жизненные формы подушковидных растений (рис. 151), розеточные
формы, прижимающиеся к почве, шпалерные формы и др.
Рис. 151. Схематические разрезы через различные типы рас-
тений-подушек.
j—Andro>ace helve tic а; 2-бииы Saxifraea, J-Silene aeauhs, 4—ее более
старый экаемнллр J— Androsace alpina, *—Carex firma—мвлодой экземп-
ляр (по Хаури).
На горах центральной и южной Европы шатты» играют исклю-
чительную роль и представляют более или менее густой дерн из
злаков (особенно Роа, Festuca, Agrostis, Sesleria, Trisetum — рис.
152), осок (Carex firma, C. curvula, C. sempervirens) и других осоко-
вых (Elyna). Сюда присоединяются многочисленные двудольные.
Разнообразие условий существования приводит к преобладанию
тех или иных растений и к формированию ряда фитоценозов. Особен-
ную известность получила в последнее время ассоциация с господ-
ством Carex curvula, исследованная рядом швейцарских авторов.
Она имеет широкое распространение в западных, центральных и
восточных Альпах на высотах 2 400—3 000 м, спускаясь до 2 100 м
277
и поднимаясь местами до 3 300 м. Излюбленные местообитания —
это ровные или слабо покатые пространства на основных горных
породах. Carex curvula даё1 толстый слой гумуса с реакцией рН =
= 4,2— 5,4, на котором поселяются и другие сопровождающие расте-
ния. Из последних наиболее типичны: Chrysanthemum alpinum,
Sesleria disticha, Роа alpina, Avenastrum versicolor, Polygonum vivi-
parum и др. Большая часть видов ассоциаций перезимовывает под
снежным покровом с зелеными вегетативными частями, поэтому швей-
царские авторы относят её, равно как и вообще «матты», к классу
Sempervirentiherbosa. Из жизненных форм здесь господствуют
гемикриптофиты, на втором месте стоят хамефиты.
Другие хорошо исследованные ассоциации швейцарских Альп—
это Caricetum firmae и Elynetum myosuroides. Интересны взаимо-
отношения этих ассоциаций и ассоциации Caricetum curvulae.
Последняя является наиболее кислой, a Caricetum firmae, наоборот,
базифильной при оптимуме рН=7,0—7,5. Elynetum занимает среднее
положение при рН=5,5—6,2.
В СССР высокогорные луга очень хорошо развиты на горах
Средней Азии и на Кавказе. В основном можно различать два наи-
более хорошо развитых типа среди целого ряда имеющих более
ограниченное значение; во-первых, осоко-злаковые луга, где господ-
ствуют различные виды осоковых и злаков, а во-вторых, альпий-
ские «лужайки» или альпийские «ковры», где физиономическую и
основную роль играют двудольные.
В Средней Азии особенно характерны для первого типа к об-
рез и е в ые луга из двух видов кобрезий (Cobresia Bellardii и
С. schoenoides—из семейства осоковых); это — низкорослые дер-
278
невинные растения обычного вида осок (рис. 25), которые почти
сплошь задерновывают почву и имеют свои многочисленные корни
в верхних слоях почвы. Лишь немногие виды растений могут ужи-
ваться с плотным дерном кобрезий. В общем же —это очень одно-
тонные пространства.
На Кавказе характерны альпийские луга с осокой Carex tristis,
к которой присоединяются С. Huetiana, С. oreophila и другие мелкие
осоки. И здесь иногда дерн, образованный этими осоками, является
настолько густым, что другие растения с трудом проникают в него.
Луга с С. tristis широко распространены как на Большом, так и на
Малом Кавказском хребте.
Отметим также луга с господством кобрезий (Cobresia Bellardii,
С schoenoides и др.).
Западноевропейские луга с Carex curvula, кавказские и средне-
азиатские с кобрезиями можно считать географически замещающими
типами, так как по своему сложению и по своей физиономии они
очень близки.
Альпийские лужайки (ковры). Однотонные и бедные флористически
осоковые луга резко отличаются от альпийских «лужаек» с их оби-
лием видов, имеющих в большинстве случаев ярко окрашенные
цветки. Если говорят о красочности альпийских лугов, то при этом
имеют в виду не монотонные осоковые луга, а пестрые лужайки с
господством двудольных. Разнообразие видов нередко очень велико,
и контрастирующие окраски различных цветков дают сложный ковер
красок. Особенно в первый раз поражают миниатюрные размеры
растений и, с Другой стороны, — относительно крупные и ярко
окрашенные цветки: вот маленькая горечавка (Gentiana) всего в
несколько сантиметров, с одиночным темносиним цветком, вот коло-
кольчик (Campanula), такой же высоты тоже с одним крупным
цветком, а спустишься ниже к опушке леса — и горечавки, и коло-
кольчики представлены
высокими видами с много-
численными цветками.
Все лето альпийские
лужайки пестреют разно-
образными цветками [не-
забудки, маки, лютики,
колокольчики, горечавки
(рис. 153), васильки, астры
ит. д.], и тому, кто хотя
бы раз побывал на этих
высотах, трудно забыть
Рис. 153. Горечавка — типичное альпийское контрасты цветов и интен-
растение (Gentiana acaulis). сивность окраски. Осенью
развиваются в иных местах
шафраны-крокусы различных окрасок — яркожелтые, синие, белые
(рис. 154).
Флористически альпийские лужайки различных гор, конечно,
сильно отличаются, но некоторые роды распространены повсеместно:
279
Gentiana, Primula, Pedicularis, Myosotis и др. Широко известен
эдельвейс (Leontopodium — рис. 155).
В более влажных частях Главного Кавказского хребта, по Б у ш уг
нет резкого различия между осоковыми лугами и «коврами», так как
красочные двудольные виды в большей илн меньшей степени пред-
ставлены в обоих типах.
Рис. 154. Группа крокусов (Crocus Scharojani) на Кавказе (сентябрь),
Домбай—склоны г, Мусатчери (около 2 000 м) (фото В. Г. Сочеванова).
В северных горных странах и на северо-востоке Азин описан-
ная растительность постепенно сменяется моховыми, кустарнико-
выми и лишайниковыми фитоценозами, представляющими нагорные
тундры-
В горных цепях Северной Америки альпийская ра-
стительность физиономически очень близка к растительности,,
описанной для Европы. Ив Южной Америке, в Андах,
например, в районе «пуны» встречаются хорошо развитые матты,
но в несколько изменённом виде: здесь злаки и осоки отступают
на задний план, но очень сильно развиты растения-подушки и розе-
точные формы, приближающиеся к подушковидным. Также при-
сутствуют и шпалерные фбрмы.
Снеговые ложбинки. Растительность снеговых ложбинок (Schnee-
talchen) европейских Альп заслуживает упоминания потому, что
это первый растительный ценоз, более или менее хорошо охаракте-
ризованный в ботанической литературе (Освальд Хеер,
1834), а во-вторых, потому, что он широко распространен в Запад-
ной Европе. Снеговые ложбинки находятся в альпийской зоне
280
Альп. Это — небольшие понижения или ложбинки, где зимой ско-
пляется больше всего снега; летом он тает и сильно увлажняет
почву, на которой развивается своеобразный растительный покров.
Такие ложбинки имеют много влаги круглый год, низкую темпера-
туру и короткий вегетационный период. Они характерны не только
для запада Европы и для Скандинавских гор, но встречаются и у
нас на Кавказе и на^Алтае.^В более высоких частях гор снеговые
Рис. 155. Эдельвейс (Leontopodium alpinuni) на альпийских лугах бл. Тянь-
шанской обсерватории — 3572 м над ур. моря (по фото Е. Е. Сочеванова).
ложбинки покрыты снегом круглый год н, конечно, лишены расти-
тельности.
Основным фитоценозом снеговых ложбинок для Альп является
Salicetum herbaceae, где господствует приземистая ива (Salix her-
bacea), имеющая шпалерную форму. Из наиболее характерных видов
укажем: Cardamine alpina, Gnaphalium pusilium, Soldanella pusilla,
Veronica alpina, Sibbaldia procumbens, Ranunculus alpestris
(рис.156) и ряд других. Вообще же флористический состав ложбинок
не богат.
Интересно, что все растения снеговых ложбинок, кроме Salix
herbacea, являются зимнезелёными, так как они всё время под
снегом имеют зелёные листочки и побеги и обнаруживают рост,
хотя, конечно, и очень слабый. Это обусловливается тем, что тем-
пература почвы под снегом около 0° и даже выше.
Некоторые виды могут расти под снегом н более интенсивно;
так, знаменитая альпийская сольданелла (Soldanella pusilia) закла-
дывает под снегом стебель и бутоны и выставляет свои цветки над
281
снегом,^пробивая снеговой покров в несколько сантиметров (см.
На Кавказе нет сольданеллы, но аналогичные явления можно
наблюдать у видов гусиного лука (Gagea), первоцвета (Primula)
и др., пробивающих снеговой покров.
Лее это дало Брокма н-Е рошу основания для отнесения
писываемого фитоценоза к Sempervirentiherbosa
Если в Альпах, Пиренеях, Карпатах, в горах Шотландии, на
Кавказе растительность снеговых ложбинок действительно связана
с ^ложбинка ми и понижениями, то в Скандинавии в альпийской
•бласти аналогичная растительность занимает огромные простран-
ства не только в пониженных, но и на ровных местах и склонах.
Поэтому говорить здесь о растительности «снеговых ложбинок» уже
не приходится.
Но интересно, что флористически Salicetum herbaceae Сканди-
навии очень схож с среднеевропейским и представлен большинством
тех же видов; отсутствуют лишь настоящие «альпийцы», как Sol-
danella, Chrysanthemum alpinum и др.
‘Тундровые луга (луговинные тундры). Растительность тундро-
вой зоны слагается из ряда формаций, относящихся к разным клас-
сам и группам формаций (Aestifruticeta, Humiliherbosa, Eubryosa,
Lichenosa и др,). Одни формации являются зональными, другие —
интразональными, к последним относятся тундровые луга, являю-
щиеся лишь включениями в основной зональный тип (о зональных
типах см. стр. 301).
2в2
Тундровые луга обычно связаны *с отрицательными формами
рельефа, с лощинами, оврагами и др., где скопляется и долго
держится снег, причем здесь имеется избыточное увлажнение
проточными водами и более глубоко оттаивающая почва. В резуль-
тате этих условий развивается травянистый покров, а мхи
и лишайники отступают на второй план, так как для последних
эти условия неблагоприятны.
Травянистый покров не высок и довольно разнообразен по фло-
ристическому составу, в одних случаях господствует разнотравие,
в других—злаки. Разнообразие травянистого покрова зависит
также и от фенологических смен в течение вегетационного периода
(ряд аспектов).
Луговинные тундры (тундровые луга) особенно часто
встречаются в европейских и западносибирских тундрах, централь-
носибирские же тундры с малым количеством зимних осадков для
них неблагоприятны. Нередко они развиваются по самому краю
снежных сугробов в оврагах и выделяются цветистыми, пестрыми
коврами на фоне однотонной вообще тундры.
Из злаков господствующими видами являются: виды щучки
(Deschampsia alpina и D. flexuosa), лисохвост (Alopecurus pratensis),
виды мятлика (Роа alpigena, Р. alpina, Р. arctica), душистый коло-
сок (Anthoxanthum odoratum) и др.; среди разнотравия укажем:
зиббальдия (Sibbaldia procumbens), фиалка (Viola biflora), лютики
(Ranunculus borealis, R. nivalis, R. pygmaea), незабудка (Myosotis
alpestris) и др. В различных частях тундровой зоны господствуют
то те, то другие из перечисленных видов.
Вот красочная картина луговинной тундры на севере
полуострова Канина (Андреев): ранней весной (в конце июня
и в начале июля) цветут горечавки (Gentiana verna и G. tenella),
потом появляются зеленоватые метелки чемерицы (Veratrum Lo-
belianum), на склонах белые цветки дерена (Cornus suecica), розовые
пятна мытника (Pedicularis sudetica, Р. vertici I lata), желтые цветки
лапчатки (Potentilla alpestris), купальницы (Trollius europaeus),
фиалки (Viola biflora), голубые соцветия незабудки (Myosotis alpes-
tris), астрагала (Astragalus alpinus) и др.
Дерновиниые тундры. Подобно луговинной тундре они являются
интразональным типом и включены отдельными участками в основ-
ной фон тундры. Развиваются на склонах и имеют более ксерофиль-
ную растительность, сравнительно с луговинной тундрой, так как на
склонах меньше задерживается снега и они освобождаются от нег®
с ранней весны. Господствуют цветковые растения. Травостой низкий,
около 15—20 слц в южных частях тундры несколько выше. Между
дерновинными и луговинными тундрами имеется вполне определен-
ная связь: первые располагаются на склонах (особенно в верхних
частях), вторые — при основании склонов. Из цветковых растений
наиболее характерны: дриада (Dryas octopetala) и овсяница (Festuca
supina). Общее число видов не менее 60 (особенно разнотрав-
ные виды и некоторые кустарнички); лишайниково-моховой покров
развит довольно хорошо.
Другие типы тундр будут рассмотрены ниже (см. стр. 301).
2вв
Рассмотренные типы растительности (альпийские, тундровые луга)
некоторые авторы относят к пустошным лугам (Шен ников),
«днако вряд ли альпийские лужайки с их сложным ковром красок
и тундровые разнотравные ковры могут иметь эпитет «пустош-
ные». Да и вообще термин «пустошные» луга не может считаться
удачным.
[Альпийские и тундровые луга и человек
Альпийские луга используются человеком с самых отдалённых
времен, так как население горных стран занимается главным образом
скотоводством, н альпийские (и отчасти субальпийские) луга явля-
ются главным местом для выпаса. Сюда скот выгоняется на всё
время вегетационного периода (так называемые летние пастбища).
Так как норм выпаса до последнего времени не существовало, то,
понятно, выпас мог коренным образом изменять естественный по-
кров. Это и действительно имеет место в очень многих случаях,
и альпийские (а также субальпийские) луга иногда представляются
сильно изменёнными. Так, появляются вторичные ассоциации
белоуса, растения, очень мало съедобного; в других случаях по-
являются обширные заросли колючих чертополохов (Cirsium и др:),
некоторых несъедобных и ядовитых растений и т- д. Помимо изме-
нения в отрицательную сторону травостоя, при усиленном выпасе
имеется ряд побочных явлений: обнажается иа склонах подпочва,
образуются осыпи и др.
Исследования летних пастбищ Азербайджана (Г россгейм
с сотрудниками) показали, что основными мерами улучшения
этих пастбищ являются: 1) строгое соблюдение норм выпаса; факти-
чески же выпас производится обычно сверх всяких норм. Однако
рядом наблюдений установлено, что альпийские пастбища менее
страдают от чрезмерного выпаса сравнительно с субальпийскими
(это стоит в связи с жизненными формами — подушки, розетки и
др.,—более устойчивыми к скотобою); это свойство пастбищ издав-
на учтено скотоводами: 2) очистка пастбищ от колючих растений и
сорняков; 3) постановка опытной работы по изучению местных трав
и слагающих их фитоценозов.
Подобная опытная работа уже проводилась в ряде мест. Так,
Г россгей м с сотрудниками работали на высокогорных лу-
гах Азербайджана; в Юго-Осетии горно-луговой стационар на
высоте 2 200 Л1 над уровнем моря работает, начиная с 1934 г.
(Буш) и др.
' Стационарные длительные наблюдения над альпийскими лугами
особенно хорошо могут быть поставлены (и уже поставлены) в за-
поведниках. В СССР имеется ряд горных заповедников, в которых
альпийские луга занимают большие площади. Именно на Кав-
казе: Кавказский заповедник (в западной части Главного Кавказ-
ского хребта — около 400 000 га), Тебердинский (бассейн р. Теберды
на северном склоне Главного хребта — около 60 000 га), Гуниб-
ский (в нагорном Дагестане); в Средней Азии: Алма-Атинский
близ г. Алма-Ата (около 400 000 га), Аксу-Джебаглы-Су (в Талас-
ском Ала-Тау — около 80 000 га) и др.
284
•Луговинные и дер и овинные тундры имеют
важное хозяйственное значение, так как здесь олени находят сочные
травянистые корма как летом, так и осенью. Так как обычно по
соседству с луговинными тундрами находятся скопления снега, то
летом сюда привлекает оленей и прохлада от снегов.
XII. КЛАСС ФОРМАЦИЙ — ТРАВЯНИСТЫЕ ГИГРОФИЛЬНЫЕ ФИТОЦЕ-
НОЗЫ. ТРАВЯНЫЕ БОЛОТА (EMERSIHERBOSA).
Общая характеристика. Данный класс формаций объединяет
травянистые фитоценозы, связанные с сильным или даже постоянно
избыточным увлажнением. Господствуют гигрофиты как с высоким,
так и с низким травостоем. Одни авторы относят этот класс к боло-
там (травяным), другие — к болотистым лугам. Повидимому более
правильно первое, так как фитоценозы данного класса очень тесно
связаны с другими типами болот.
Для болот вообще характерно обилие, влаги, а так как влага
естественно собирается в отрицательных формах рельефа (котло-
вины, понижения, речные долины и т. д.), то здесь и развиваются
болота. Другими словами, болота обычно являются как бы в к л ю-
ч е н и я м и в основной фон растительности, покрывающей водо-
разделы (интразональный тип растительности). Однако это имеет
место лишь в тех случаях, когда водоразделы достаточно сухи (на-
пример в степной области, в южных частях лесной); в случае же
достаточного количества влаги болота могут возникать и непосред-
ственно на водоразделах, что наблюдается в северных частях лесиой
области, в лесотундре и пр.
Дать определение болота не всегда легко, так как сюда отно-
сятся, как ниже будет указано, очень разнообразные типы расти-
тельности, но вообще можно сказать, что это есть тип гигрофильной
растительности, не погружённой в воду (или во всяком случае
возвышающейся над уровнем воды своими вегетативными частями).
С болотами часто связывают представление об отложении торфа;
это верно в очень многих случаях, но не всегда, поэтому вводить в
определение болота признак торфообразования не представляется
возможным.
Болота представляют очень широкое понятие и могут быть рас-
пределены по различным классам формаций; так, можно различать:
травяные болота (они относятся к Emersiherbosa); моховые болота
(относятся к Sphagniherbosa); болота, покрытые лесом (лесные болота);
эти последние могут быть покрыты березой и другими деревьями
с опадающей листвой (AestisiJvae), сосной (Aciculisilvae), вечно-
зелёными мангровыми деревьями — мангровые болота (PJuviifru-
tice ta) и т. д. Одним словом, сюда относятся фитоценозы очень
разнообразной фитоценологической структуры: начиная от моховых,
слагающихся главным образом из мхов, и кончая сложными дре-
весными (лесные болота были описаны выше, см. стр. 228).
Ниже рассматриваются лишь травяные болота, они относятся
к так называемым низинным болотам (низинные болота, кроме
того, могут быть лесными с господством ольхи, берёзы, сосны —
см. выше).
285
Низинные травяные болота. Они нередко представляют обширные
пространства осок и злаков; разнотравие выражено слабо. Харак-
терные черты подобных травяных болот:
1) почти все растения многолетни;
2) очень многие имеют корневища, что даёт возможность этим
растениям образовывать обширные заросли: виды осок, пушицы,
тростник, рогоз и т. д_;
3) ряд растений имеет ксероморфный облик, что, невидимому,
связано с присутствием гумусовых кислот, вызывающих физиоло-
гическую сухость;
4) болотные почвы ^едны кислородом; так как все поры почвы
заполнены водой, то в корнях многих растений развиваются воз-
духоносные полости, куда воздух поступает из наружной атмосферы;
5) болотные почвы, как правило, кислые со значением pH
чаще всего между 4 и 6.
Можно указать ряд типов травяных болот, но особенно рас-
пространены у нас осоковые болота с различными видами осок; к
осскам нередко присоединяются мхи, образуя гапново-осоковые бо-
лота (здесь большую роль играют гипновые мхи Drepanocladus,
Calliergcn и пр.) и сфагаово-гипново-осоковые болота. -Местами, осо-
бенно в плавнях южнорусских рек, огромные территории занимают
тростниковые и рогозовые (Typha angustifolia) болота.
В травяных болотах основную роль играют однодольные растения;
это г меет место в Европе, Северной Америке (Phragmites, Dulichium,
Tjpha и др.), Африке (на верхнем Ниле Cyperus — известный па-
пирус), в трошках (бамбуки, ароидные и др.); на травяных болотах
Цейлона, креме осоковых и ситниковых, физиономическую роль
играют виды Ericcaulon: цветки у них собраны в соцветия, крайне
похожие на соцветия сложноцветных—как бы «сложноцветные»
среди однодогьгых.
Сукцессии на болотах. Низинные болота находятся в пониженных
частях рельефа ив своём водном питании непосредственно связаны с
'рунтовыми водами, богатыми минеральными в/ществами, поэтому
эти болота называются также эвтрофными.
Низинные (эвтрофные) болота постепенно повышают свой уровень.
Это зависит от того, что остатки растений этих болот в бедной кисло-
родом почве не могут вполне разлагаться, почему и происходит их
накопление. Мало-помалу накопляется торф, и субстратом растений
является уже не минеральная почва, а торфяной слой. Естествен-
но, что при этом меняются условия существования для растений,
и травянистый покров претерпевает те или иные изменения.
При увеличении мощности торфяного слоя, который неизменно
нарастает всё больше и больше, условия существования становятся
всё более и более отличающимися от первоначальных (рис. 157).
Здесь наблюдается последовательная смена раститель-
ных ассоциаций, которая в основном сводится к тому, что рас-
тительность становится всё более и более олиготрофной. Это зави-
сит от того, что грунтовые воды, богатые минеральными веществами,
всё менее и менее принимают участие в питании растений,
так как уровень торфяной массы повышается. Наконец поверхностный
286
уровень торфа и растения, на нём находящиеся, отрываются от
грунтовых вод, и последние уже почти ие принимают участия в пи-
тании растений: болото переходит в стадию атмосферного питания,
где в растения поступают лишь те минеральные вещества, которые
находятся в атмосферной (дождевой) воде. Это уже болота олиго-
трофного типа, т. е. сущертвующие при небольшом количестве
минеральных веществ на бедных субстратах, — болота сфагновые
Рис. 157. Генетическая схема болотных формаций:
питания сменяются боло1ами грунтового питания,
атлюсферного питания. На схеме видна, кроме того,
ассоциаций (по Аболпну).
болота озёрно-речного
а последние — болотами
последовательная смена
(господствующий мох — сфагнум), или верховые (по своему положе-
нию), относящиеся к классу формаций Sphagniherbpsa.
Целый ряд ассоциаций сменяет друг друга при переходе ни-
зинных болот в верховые, причём промежуточная стадия развития
болота носит название болот переходного типа. Так как условия
питания здесь становятся хуже, то появляются некоторые мало тре-
бовательные сфагновые мхи, некоторые менее требовательные осоки
и другие растения.
При этом получается такой генетический ряд: Submersiherbosa
(погружённая растительность) —Emersiherbosa (низинные болота) —
Sphagniherbosa (сфагновые болота). Здесь, таким образом, перед нами
один из очень показательных примеров сукцессий (см. выше, стр. 157).
Ход сукцессии можно хорошо представить себе, если рассмотреть
те процессы, которые протекают при зарастании более или менее
мелководных озёр.
Здесь в ряде случаев резко выступает распределение раститель-
ности поясами от более глубоких частей водоёма к более мелким.
Основные пояса таковы (рис. 75).
1. Пояс микрофитов (от греческих слов «микрос»—малый икфитон»—
растение). Это наиболее глубокий пояс, и растения, его составляющие, принад-
лежат к споровым растениям (зеленые водоросли, сине-зеленые водоросли,
диатомовые п др.), среди которых встречаются и довольно крупные водоросли,
но все растения погружены в воду.
2. Пояс макрофитов (от греческого слова «макрос» — большой).
Здесь уже более крупные растения, образующие менее глубокий пояс, причем
уже не только споровые растения (харовые водоросли, некоторые мхи), но и ряд
цветковых растений — особенно виды рдестов, роголистник (Cera tophyHum) и
2в7
др. Все растения и здесь погруженные, некоторые и цветут под водой, другие
же выставляют цветки над водой.
3. Пояс широколистных рдестов. Ещё на меньших глуби-
нах (3-5 м) располагаются широколистные рдесты (Potimogeton perfoliatus,
Р. lucens п др.), уруть (Myriophyllum) и пр., но все эти виды, тоже погружен-
ные в воду, выставляют над водой во время цветения свои соцветия.
4. Пояс водных лилий. Ещё менее глубокий пояс, растения
чооторого/не только выставляют над водой цветки, но у которых ц листья погру-
жены не целиком—они плавают, т. е.
верхняя их сторона соприкасается с
воздухом, а нижняя—с водой. Сюда
относятся водяная лилия (Nymphaea),
(рис. 158), кувшинка (Nuphar),плава-
ющий рдест (Poti moge ton па tans).
5. П о я с камышей. Здесь
мы находим растения, уже высоко
возвышающиеся над водой, погру-
женные лишь своими нижними ча-
стями (глубина не больше 2 м). Осо-
бенно типичен камыш (Scirpus licus-
tris), образующий нередко сплошные
заросли.
б. Пояс тростника. Иногда
очень резко отделяется от предыду-
щего и состоит из густых зарослей
тростника (Phragmites communis).
7. Пояс мелководных
растений. В пределах этого
мелководного пояса наблюдается
большое разнообразие видов, так как
здесь для растений условия более
благоприятны, чем при более глубо-
ких водоемах, где часто имеются
лишь 1—2 вида, которые в данном
поясе доминируют потому, что дру-
гих видов с соответствующей эколо-
гией в данной флоре не имеется. Для
мелководного пояса характерны осо-
ки (Carex gracilis, С. rostrata и др.),
сусак (Butomus), частуха (Ahsma),
цикута (Cicuta), ирис (Iris pseuda-
corus), прпбережник (Lythrum salicaria) н ряд других.
Растительность этого последнего пояса постепенно переходит в
наземную растительность на берегу водоёма, представляющую уже
нередко травяное болото. Перечисленные пояса растительности
нередко дают указанный ряд, в других случаях тот или иной пояс
выпадает или бывает выражен не так отчётливо.
Надводные пояса физиономически очень хорошо отличимы и не-
редко достаточно резко отграничены один от другого. Растительность
подводных поясов представляет так называемый бентос (стр. 293).
Данный экологический ряд является в то же самое время и гене-
тическим, так как каждый пояс в своем существовании не является
долговечным, но сменяется соседним, менее глубоководным. Это
зависит от того, что каждый пояс в результате отложения своих
органических остатков повышает уровень дна водоёма и дает воз-
можность поселяться видам соседнего пояса, расположенного на
менее глубоких местах. Таким образом, всё время происходит н а-
ступание поясов от края водоёма вглубь:
288
пояс мелководных растений наступает на пояс тростника, этот
последний на пояс камыша и т. д., пока весь водоём не будет за-
торфован остатками растений (корневища, стебли, листья и т. д.).
После заторфовывания всего водоёма (различают торфы, в зави-
симости от их происхождения, различного характера: осоковый, трост-
никовый, кэмышёвый и др.) образуется травяное болото. Здесь перед
нами пример сукцессии прогрессивного порядка, так как и флори-
стическая сложность и фито цен о логическая всё время повышается.
Однако болота возникают не только описанным способом — для
образования болот вообще необходим избыток воды, и потому воз-
можны и другие способы их образования.
Основные способы образования болот та-
ковы:
1. Заторфовываиие водоёмов (рек, озёр и др.). Кроме описанного
способа, возможны ещё и другие.
2. Заболачивание суши- в лесах, на лугах, на пожарищах
в лесу и др.
3. Заболачивание в местах выхода ключей внизу склонов и на
самых склонах.
Однако не все указанные способы образования болот приводят
именно к травяным болотам; так заболачивание в лесах приводит
к лесным болотам, на местах выхода ключей —к мохово - гип-
новым и др.
Болотоведы все указанные фитоценозы объединяют под назва-
нием болот, но если исходить при классификации растительности
из фитрценологических и физиономических представлений, то пра-
вильнее болота распределить по различным классам формаций,
как это и сделано в предыдущем изложении.
XIII. КЛАСС ФОРМАЦИЙ — СФАГНОВЫЕ ОЛИГОТРОФНЫЕ ФИТОЦЕ-
НОЗЫ — СФАГНОВЫЕ (ВЕРХОВЫЕ) БэЛОТА ' (SPHAGN.HEfiBOSA)
Этот класс формаций связан главным образом с лесными областя-
ми, так как для свсего развития требует известного количества
осадков л тепла.' В степьых областях и в тундрах сфагновые болота
почти не имеют подходящих условий для своего развития. Таким
образом, эти болота связаны с определённым климатом.
Общая характеристика. Растительный покров на сфагновых
болотах настолько своеобразен,что они вряд ли могут быть отнесены
к типу травянистых фитоцеиозов (Herbosa), куда их относит Брок-
мац-Ерош (см. стр. 163), но представляет совершенно особый тип.
Прежде всего здесь господствуют сфагновые мхи, всё время своей
верхушкой нарастающие всё выше и выше. Уже это одно создает
особый экологический режим. Здесь могут по-
селяться лишь те растения, которые обладают способностью пере-
мещать свои точки роста на более высокие уровни, чтобы не быть
погребёнными в сфагновой толще, к тому же растения олиготроф-
ного типа. Хорошо известный пример в этом отношении — росянка
(рис. 159).
Кроме различных видов сфагнума (Sphagnum medium, S. fuscum,
S. recurvum и др.) здесь находим ряд растений, крайне характер! ых
География растений 293
для болот данного типа, а именно, вересковые кустарнички: андро
меда, Кассандра, багульник, голубика, черника, брусника, клюква,
такие многолетники, "
как
36 мм
1905
шеихцерия, ринхсспора, морсшка, пушица
(Enophorum vag па turn), осоки (особенно
Со rex hmosa), в бог ее северных местах
сюда пргсседгняются водяника (Empetrum
n grom) нгзкорослая березка (Betula папа)
и некоторые другие
Все эти виды могут быть доминантами
и, кслб!ькруясь друг с другом, дают
очень большое число ассоциацгй. Если
добавить к приведенному переселению
видов шье несколько менее обильных
(виды росянок, некоторые осоки, марьян-
ник — Melampyrum pratense), то этим
почти исчерпывается весь флористический
состав сфагновых болот
В облесен! ых болотах сосна — един-
ственная древесная порода, но она очень
низкоросла (около 2—3 к), с мелкими шиш-
ками и оиенькороткой хвоей Замечательна
разг обидь есть сосны, найденная в При-
балтике, куста рникового типа,
т е разветвлен! ая с самого основания; вся
он а к тему же нередко погребена в торфе,
и над поверхностью сфагнума выставля-
ются лишь веточки длиной в несколько
сантиметров (рис 160)
Растнтель! ость сфагновых болот ' яв-
ляется олиготрофней и скафильноЙ, pH
здесь не поды-мается выше 5
Обращает на себя внимание ксероморф-
ность большинства видов сфагновых болот
(восковой налет, волосяной покров, кожи-
стссть листьев, пегружеы е усты-ц и т д ),
но вопрос о причинах ксероморфнзма еще
и е вг олне ясен Повидг м ому, здесь причи-
ны различны у очень рано разв! вающн хся
весной растений i сероморф! зм может
вызывать холодная вода, у ычнезеленых
от 31Мнего испарения; но не ьзя не
Рис 159 Стебетек росян-
ки (Drosera) нарастающий
сеоеи верхушкой, видеч
прирост сте^етька за гри
года (этот прирост опреде-
ляется приростом сфагно-
вой тотщ(, по Бертч^)
ксероморфизм — защита
обратить также внимания на то, что летом во время засух поверх-
ность сфагновых болот настолько пересыхает, что здесь в это время
влаги вряд ли больше, чем на каком-л! бо i сключ! только ксеро-
фит! ом мсстосбчтаьь и
Основные типы сфагновых болот Сфэгно«ье болота и*еют ряд тигсв Наи-
более характесны дая Западной Европы болота, имеющие выпуклую
форму, т е Han6oRee повышекнье в своих центра аьпых частях (имеют форму
часового стек аа) Это зависит оа ряда причин оогееси ьнос напастанпе сфа< нума
в центре, так как по периферии боаее эвтрофные условия для него менее благо-
приятны, кроме того, си аьная гигроскопичность Сфагнума, вследств! пгеь
260
Рнс 160 Особая скорма болотной сосны (Pinus silvestris f pumila)
Пунктир на рисунке—поверхность болота (по Аболнну)
Рис. 162 Насекомоядное растение—
венерина мухоловка (Dionaea) (по
Кернеру).
сфагновый массив впитывает воду, как губка Встречаются болота и п л or
кой формы и еще некоторых других типов. Сфагновые болота могут быть н е
облесенные и облесенные.
Рельеф выпуклых болот довольно разнообразен, здесь различают крае-
вые скаты, радиальные ложбины, краевое канавообразиое
понижение на месте перехода болота в минеральный берег. Кроме того,
на поверхности болота обычно хорошо диференцироваиы кочки н понижения
между ними — мочажины.
Собственно верховые болота выпуклой формы характерны для
более влажного климата Западной Европы, облесенные болота
встречаются в более континен-
тальном климате—на востоке
Европы и в Сибири.
В Северной Аме-
рике сфагновые болота
развиты главным образом на
северо-востоке; в централь-
ных частях, невидимому,
отсутствуют., По своей ра-
стительности ог и очень
сходны с европейскими, н©
вереск на них отсутствует,
наоборот, водяника (Empet-
rum nigrum) очень распро-
странена. Характерным дере-
вом на болотах является ли-
ственница (Lar х laricina).
В Южной Америке
сфагновые болота 'Источа-
ются в АнДах, а также на
Огненной Земле. Различные
типы их указываются и для
Новой Зеландии.
Интересно добавить, что
сфагновые болота, очень бед-
ные питательными веще-
ствами, отличаются от других типов растительности пргсутствием
насекомоядных растений: роды росянка (Drosera) и жи-
рянка (Pirgaicula); в Северной Америке, кроме того, Sarracenia
(рис. 161) и Dionaea (рис. 162).
Травяные и сфагновые болота и человек
Растительность болот не представляет особой ценности; неко-
торые типы болот, например некоторые осоковые, имеют с кормовой
точки зрения отрицательный характер. Поэтому вопросы улучшения
кормовой ценности болот обычно связаны с их мелиорацией, именно
с осушением. В результате осушения уровень грунтовых вод пони-
жается, и тогда на болотах получают возможность селиться более
ценные растения. Местами болота sai в мают обширные территории
(например Пинские болота на Западе Белоруссии), и первоочередной
задачей является их осушение, к чему уже и приступлено.
292
Верховые болота совсем не имеют кормовой ценности, но имеют
значение как основное местопроизрастание некоторых кустарничков,
дающих съедобные плоды (например клюква, морошка и др.), кото-
рые имеют большое хозяйственное значение. Интересен вопрос о
селекции клюквы и о культурах се более ценных сортов.
большое значение болот, именно торфяных, состоит в том, что
©ни дают нередко очень мошные залежи торфа, а последний соста-
вляет значительный процент нашего топливного баланса и является
источником электроэнергии для некоторых наших электростанций.
Торф идет также на удобрение, на подстилку для скота, иа изгото-
вление изоляционных плит, ряда химических веществ (креозот,
нарафин, метиловый спирт и др.)
Специально болотных заповедников у нас нет, но в ряде заповед-
ников имеются крупные болотные массивы: в Белорусском—в вер-
ховьях рр. Березины и Западной Двины, в Центрально-Лесном—в
Калининской области и др. Обширные низинные болота имеются в
Астраханском заповеднике в дельте р. Волги.
XIV и XV. КЛАССЫ ФОРМАЦИЙ - ФИТОЦЕНОЗЫ ВОДНЫХ РАСТЕНИЙ,
ПОГРУЖЕННЫХ В ВОДУ, ПЛАВАЮЩИХ НА ПОВЕРХНОСТИ ВОДЫ ИЛИ
ВЗВЕШЕННЫХ В ВОДЕ (ayBMERSlHERBOSA И AQVERRAHTIA)
Растения, обитающие в водной среде (в морях, реках, сзерах и
яр ), по своему отношению к этой среде-могут быть крайне различны;
один из них плавают на поверхности воды, не будучи совсем
прикреплены; другие почти всеми частями погружены в воду,
прикрепляясь к диу водоемов, третьи — взвешены в толще
воды и могут в ней или пассивно или активно перемещаться.
Первые организмы в своей совокупности представляют плейстон
(плавающие организмы), вторые — бентос (подводные прикреплен-
ные), третьи — планктон (взвешенные в воде организмы).
Нужно отметить, что планктон, бентос и плейстон представляют
совокупность и растительных и животных организмов Если иметь
в виду только растения, то тогда говорят о фитопланктоне, фито-
вентосе и др. В дальнейшем мы будем касаться лишь растительных
©ргаиизмов.
Высшие растения больше всего представлены в бентосе, меньше
в плейстоне и совершенно отсутствуют в планктоне.
Бентос можно объединить вместе с плейстоном в один класс
формаций Submersi herbosa.
Бентос. Он состоит из очень большого количества фитоценозов
Одни растения (высшие растения) прикрепляются ко дну своими кор-
нями, или ризоидами (ризобентос), другие (водоросли) прикрепля-
ются особыми выростами на своих талломах (гаптобентос).
Водоросли в бентосе морей и океанов имеют исключительное
значение; из цветковых здесь встречается главным образом «морская
трава» (Zostera), образующая нередко обширные подводные заросли
(рис 163). В зависимости от глубины различают несколько поясов,
а именно.
1) литоральный пояс — внутри границ приливов и
©тливов; господствуют °елрные и бурые воторосли, из последних
293
особенно фукусовые; в тропиках — главным образом красные и
зелёные водоросли;
2) сублиторальный пояс — от нижней границы от-
лива до глубины 40 лг; зеленые водоросли уменьшаются в количестве,
красные играют всё большую
и большую роль; в этом поясе
находятся морские цветковые
растения;
3) элиторальный по-
яс—глубже 40 м; здесь почти
исключительно красные всдо-
рссл , доводящие до 100 м у
даже более.
В пресноводном беи ссе на
более мелких мьаах исключи-
тельную роль играют цветковые
растения, которые массами за-
полняют иногда речки, заводи,
пр^ды и пр Что касается водо-
рослей, то это главным образом
зеленые водоросли, р звизаю-
щиеся до глубины в 2—3 л:
впрочем, в озерах с очень
прозрачной водой (например
Бойка л) водоросли могут спус-
каться до 20—2 (Ж
Выше было уже указано (см.
стр. 287), что водная раститель-
ность в водоёмах распреде-
ляется поясами, и что в резуль-
тате своей жизнедеятельности
Рис. 163. «Морские травы»- слева—
Zostera manna, справа — Posidonia
oceanica (по Щнмперу).
она приводит к заторфовыванию озер, прудов и пр.
Некоторые экологические особенности водных растений были
указаны выше (см. стр. 70).
Вообще же нужно отметить, что так как условия водной среды
довольно однообразны, то и водная растительность не имеет такого
разнообразия форм, приспособлений и пр., как наземная раститель-
ность. Удобство же расселения при посредстве воды приводит к
большому флористическому однообразию водных фитоценозов.
Особо следует здесь отметить своеобразный фитоценоз, имеющий
место в тропических областях и развивающийся на скалах в быстро-
текущей воде (водопады, быстрые потоки и др.). Растения этого
фитоценоза принадлежат к семейству Podostemonaceae и по внеш-
нему виду совершенно сходны с печёночными - мхами, представляя
плоские или нитевидные зелёные образования, прикрепляющиеся к
подводным скалам особыми прицепками—видоизменёнными корня-
ми (онс. 164).
Плейстон. Играет значительно меньшую роль, чем бентос. Среди
низших растений укажем огромные скопления оторвавшихся бурых
водорослей Sargassum в Атлантическом океане («саргассовое море»).
294
Среди высших растений имеется ряд видов, плавающих на по-
’рхности воды, главным образом в более защищенных местах (в
з .ливах, прудах, зарослях крупных прибрежных растений). Так,
<. стоячих водоемах у нас обыч! ы виды рясок, сплошь покрывающие
' "!ёныч покровом всю поверхность воды; южнее встречается саль-
ния (водный папоротник) и др.
Рис. 164. Представители семейства Podostemonaceae.
J—О «ж one fiexuosa, 2—Podostemon olivaceus, 3—Tnsticha hypnoides (по Туласнэ),
Планктон (Aquerrantia). Он состоит только из низших растений
(бактерии, жгутиковые, водоросли), а также из низших животных.
Все эти организмы или взвешены в воде и перемещаются пассивно,
или же могут передвигаться самостоятельно (например при помощи
жгутиков). Различают планктоны соленых и пресных водоемов, а
также загрязненных водоемов. Планктон играет исключительную
роль в питании более крупных животных (например рыб).
Разнообразие планктона очень велико в связи с географизмом,
.а также на различных глубинах и в различное время года.
Водная растительность и человек
Уже было сказано, что планктон представляет основную пищу
для <рыб, и потому для рыбного хозяйства исключительное значение
HAieer состав планктона и его динамика в течение вегетационного
периода. Поэтому и составу планктона и его динамике уделяется
особое внимание.
Что касается высшей растительности водоёмов, то она, особенно
в последнее время, привлекает к себе внимание в связи с устрой-
ством крупных водных вместилищ на ряде наших рек (канал Волга-
Москва с рядом водовместилищ—«Московское море» и др., Куйбы-
шевская плотина с образованием крупных водоемов и другие гидро-
технические сооружения). Здесь особое значение имеют вопросы
естественного зарастания вновь образуемых водоемов, направлен-
295
ность этого зарастания, установление соответствующих сукцессий.
Некоторые водные растения обладают способностью быстро за-
нимать большие площади, очень интенсивно размножаясь корне-
вищами. Классическим примером крайне быстрого вегетативного
размножения является элодея (Elodea canadensis), называемая «во-
дяной чумой». Это американское растение местами так размножилось
яутём вегетативного разрастания, что заполнило некоторые каналы
и водоёмы, мешая рыбной ловле, а иногда даже судоходству (от-
сюда и название растения).
Так как в ряде случаев зарастание наносит вред водоёмам, те
вопрос может итти о выработке мероприятий, препятствующих
этому зарастанию, а в других случаях зарастание должно
регулироваться самим человеком. В результате — управление
водными фитоценозами.
XVI и XVII, КЛАССЫ ФОРМАЦИЙ — МОХОВЫЕ И ЛИШАЙНИКОВЫЕ
ТУНДРОВЫЕ ФИТОНЕНОЗЫ С ГОСПОДСТВОМ ЗЕЛЕНЫХ МХОВ И ЛИ-
ШАЙНИКОВ (EUBRYOSA И LICHENOSA)
Моховые и лишайниковые фитоценозы имеют очень широкое
распространение в зоне тундр и особенно в той её подзоне, которая
называется подзоной лишайниково-моховых тундр. Эта подзона
является наиболее типичной для тундры, занимая срединное поло-
жение между более южными и более северными подзонами (о рас-
членении тундры на подзоны см. ниже, стр. 301).
Моховые тундры. Б подзоне лишайниково-моховых тундр моховые
тундры занимают плоские повышенные пространства, являясь зо-
нальным типом. Моховые тундры часто заболочены, а на востоке бы-
вают кочкарными. Помимо плоских пространств, занимают также от-
крытые склоны холмов; на субстратах грубых, песчаных, вместо мохо-
вых развиваются лишайниковые или лишайниково-моховые тундры.
Моховая дернина в моховых тундрах состоит из таких мхов:
Aulacomnium turg-dum, Camptothecium trichoides, Dicranum elon-
gatum, Hylocomium proliferum, Polytrichum strictum, Ptilidjum
ciliare и др. Все эти мхи переплетаются между собой так тесно,
что выделить дернинку одного какого-либо вида очень трудно; это,
невидимому, связано с сильным освещением, так как в лесах, где мхи
занимают нижний ярус и где они находятся в сильном затенении,
мы имеем коврики тех и других видов мхов без примеси
других видов. Над моховым покровом в моховых тундрах мы на-
ходим разреженный травянисто-кустарничковый ярус из ряда трав
(мятлик — Роа arctica, осока —Carex rigida, живородящий горец—
Polygonum viviparum и др.) и кустарничков (куропаточья трава —
Dryas punctata, виды шпалерных ив — Salix ро laris, S. reticulata,
голубика — Vaccinium uligmosum, брусника — Vaccinium vitis
idaea и др.).
Моховые тундры распространены в западной и центральной ча-
стях тундровой зоны до р. Лены, восточнее же представлены очень
слабо (здесь особенно развиты кочкарные тундры с кочками, образо-
ванными пушицей — Eriophorum vagina turn и кочкообразующей
осокой — Carex Soczavaeana.
Моховые тундры с кустарничковым ярусом. Они отличаются
моховых тем, что хорошо выражен ярус из низкорослых кустар-
ничков; из последних укажем низкорослую берёзку (Betula папа,
к востоку от р. Енисея — В. exilis), ряд ив (Salix glauca, S. pul-
ch га н др.), вересковые кустарнички (Ledum pa lustre — багульник.
Vaccinium uliginosum — голубика). Мохово-кустарничковые тундры
наиболее распространены в подзоне кустарниковых тундр, причём
высота и густота кустарничкового яруса связаны с глубиной снеж-
ного покрова, так как последний защищает кустарнички от силь-
ных зимних ветров, иссушающих и повреждающих выступающие над
поверхностью снега побеги. На открытых пространствах лишай-
никовс-моховой подзоны с её сильными ветрами мохово-кустарнич-
ковые тундры занимают более защищённые от ветров места, куда
сносится снег.
Лишайниковые тундры. Если моховые и кустарничковые тундры
представляют зональный тип растительности, то лишайниковые
тундры связаны с песчаными грунтами, являясь интразональным
типом. Моховые тундры — это как бы напочвенный покров хвойных
лесов, лишённый на севере древесного яруса, лишайниковые тундры
представляют лишайниковый ярус сосновых и лиственничных
лесов.
Лишайниковые тундры не остаются однообразными по раститель-
ности, но меняются в зависимости от географического положения.
Именно, в западных частях (до р. Енисея) преобладают ягельные
(род С lad onia) тундры, в центральных частях (центральная конти-
нентальная Сибирь) — алекториевые (род Alectoria), на крайнем
востоке — опять ягельные тундры. Ягельные тундры Связаны с
большим Снежным покровом на западе и на востоке.
В ягельных тундрах господствуют в напочвенном покрове различ-
ные виды ягелей-кладоний (Cladonia Silvatica, С. rangJerina,
С. uncial is) и др., в более южных частях также С. alpestris. Нсскольк©
торфянистая дернина этих тундр имеет 8— 10см в толщину.
В травяно-кустарничковом разреженном ярусе наиболее характерны
такие травы: некоторые злаки (Arctagrostis latifclia, Hiercchloa
alpina), осока (Carex rigida), ожига (Luzula confusa), мытник (Pedi-
cular! s hirsuta) и др.; из кустарничков преобладают: альпийская
толокнянка (Arctcus alpina), водяника, брусника, голубика, низко-
рослая берёзка, багульник (Ledum decumbens) и др.; на восток©
флористический состав несколько меняется.
Ягельные тундры не остаются однородными не только в связи с
географическим положением, но и с условиями местообитания —
©ни или приближаются л о растительности к моховым (на более
тяжёлых грунтах) или к болотистым тундрам с торфяным горизон-
том (на заболоченных песках).
В центрально-сибирских тундрах, как уже сказано, развиты
не ягельные, а алекториевые тундры; для ягелей здесь неблагоприят-
ны: 1) малоснежные зимы и 2) сухость лета (отсутствие защитных
приспособлений от сухости воздуха). Кроме алекторий (Alectoria
©chroleuca преимущественно, также A. nigricans) характерны также
цетрарии (Cetraria cucullata и С. nivalis); отметим ещё Bryopogon
897
Jivorgcns и др. Ягели в а лекториевых тундрах встречаются лишь как
примесь. Кустарничково-травяной ярус еще более разрежен-
Так как ягельные тундры дают прекрасные корма для оленей, те
их растительность, в связи с неумеренным выпасом, сильно стра-
дает. Мы можем наблюдать в природе все стадии деградации туьдр
в результате выпаса (вторичные тундры). Алекториевые тундры
И’леют значительно меньшую хозяйственную ценность, так как
а лектории поедаются оленями слабо.
Как указано выше, моховые и лишайниковые тундры осо-
бенно характерны для наиболее типичной туьдровой подзоны—
лиш йниково-моховой. Выше были отмечены ещё и такие типы:
кустарниковая тундра (относится к Aest fruticcta), луговинная и
дерновинная (относятся к Fjrgidohumihherbosa). Имеются и ещё
другие типы.
Таким образом «тундра» как зона слагается из большого числа
формаций, относящихся к различным классам формаций, что связан®
со значительной протяженностью тундры с юга на север (ряд подзон),
с эдафнчсскими и орографическими условиями.
Краткая характеристика тундровой зоны. Ч~обы дать пред-
ставление птунДре в целом, ниже приводится краткое общее опи-
сание тундровой зоны.
Тундрами заняты огромные пространства на со г ере Евразии,
на севере Америки, включая все северные острова. Именно тундры
занимают северное побережье ~ Скандинавии и Koj ьского полу-
острова, Канин полуостров, пространство между устьями рр. Пе-
чоры и Оби, включая весь полуостров Ямал, все пространство
между устьями рр. Оби, Енисея и Лены, включая Таймыр, далее
к востоку прибрежную полосу северней 70° северней шпроты, почти
весь ЧукскктЙ полуостров. В Северней Америке тундры встре-
чаются и на белее южных широтах, местами заходя на юг за 60°
северной широты и развиваясь, таким образом, на широте Ленин-
града и даже Москвы. В южном полушарии тундры почти нераз-
виты, так как там нет условий для их широкого развития. Тундры—
фптоценс логи чески достаточно разнообразны. Так, встречаются
пространства, покрытые мхами, лишайниками, осоками, разно-
травными видами, кустарничками, ивняками, на юге — сфагно-
выми болотами. Поэтому здесь нужно говорить о различных
классах формаций. В целом' же тундра образует кли-
матическую зону и имеет характерный ландшафт.
Основной признак тундры—это её безлесье?Деревья появля-
ются лишь на южной границе тундр, образуя подзону лесотундры,
которая далее на юг переходит уже в леса. Однако в тундре пре-
обладают не травянистые типы растительности, а мохово-
лишайниковые и кустарниковые.
Растительность тундр находится в крайне неблагоприятных
условиях: длинная и суровая зима, короткое и холодное лето, при-
чём средняя температура самого тёплого месяца не выше 10°, малое
количество атмосферных осадков (не более 25 см), причём зимой
мало снега (всего около 10% годичных осадков), очень сильные вет-
ры. К особенностям тундр нужно прибавить продолжительное сол-
298
ночное сияние летом почти в течение круглых суток, а в более вы-
coki х широтах —и кр)г;ые с^тки.
Почва т) ндр очень холодна и на известней глубине (за некоторы-
ми исключениям! ) находится вечная мерзлота, при-
чём летом почва оттавьает jn шь до глубины в 15—20 см.
Флористически тундры бедны сравните; ьно с бот ее южными
растительными титамн; особенно мало видов на северных островах:
Новая 3V мл я—200 видов. Земля Франца-Иосифа—всего 38 видев.
Флора тундр не имеет особых, свойственных тотько ей семейств.
Наиболее характерны: вересковые, ивовые, осоковые, злаки, юти-
ковые, крестоцветное, розоцветные.
Д; я Антарктики особенно характерны розоцветные (род Асгепа),
зонтичные (Az rdla), крестоцветные (Pnngler) и гвоздичные (Lyal-
lia). Очень характерны также мхи и лишайники.
Вегетационный период короток—всего 2—2% месяца. На Шпиц-
бергене 86 цветковых видов по своему цветению распределяются так:
Июнь | Июль
11—20 | 21—30 | 1—10 11—20 21-31
Число ВИДОВ 8 | 16 1 29 25 8
При продвижении сельскохозяйственных культур в тундровую зо-
ну особенно неблагоприятна hi зкая температура почвы—здесь необ-
ходимо выведение мсро"о тонких сортов, также сортов длинного-д ня.
Жизненные формы тундр. Неблагоприятные, условия тундры
приводят к гыр*быке у цветковых растений жизненных форм,
близких в общем к формам альпийских высот.
29»
Рее 168. Шпалерная форма сетчатой ивы
(Salix reticulata) (по Шнмперу).
1. Здесь господство хамефитов и гемикриптофитов. Эти формы
о л у чают преимущества при коротком периоде вегетации; терофитов,
т. о. однолетников, нет, за единичными исключениями.
2. Очень много вечно-
зелёных — кустарнички
брусничного и верескового
типов. Они имеют готовый
аппарат для ассимиляции.
3. Многие растения
очень низкорослы.
4. Нередки шпалерные
формы — целый ряд ив
(Salix polari's, S. herbacea,
S. reticulata —рис. 165 и
166); растения как бы
прижимаются к поверх-
ности почвы, которая на-
гревается сильнее воздуха
(иногда до 30° и выше).
5. Типична форма растений-подушек (Draba alpina, Eritrichium
villosum и др.).
6. Корни -растений в большинстве случаев находятся в самых
поверхностных слоях почвы, которая менее холодна.
Имеются ещё и другие особенности тундровой флоры, причём
надо отметить удивительную способность многих растений перено-
сить всё холодное зимнее время, не теряя вегетативных органов и
даже цветков. Классическим примером является ложечная трава
(Cochlearia arctica), выдержавшая температуру—46° (см. рис. 41).
У большинства подобных растений не имеется каких-либо внешних
защитных приспособлений от холода и это, невидимому, обуслов-
ливается осмотическим давлением и свойствами клеточных кол-
лоидов.
У арктических растений обращает на себя внимание крайне
медленный рост, что приводит к низко рос лости и к мик рофнлли*
(к мелколистности). Если сравнить виды, растущие и в тундрах
и в более южных областях, то это выступает особенно наглядно.
Рост в толщину так незначителен, что у можжевельника, стволик
которого имел 83 мм толщины, было насчитано 544 годичных колец.
Давно уже было обращено внимание на ксероморфный облик
арктических растений и на целый ряд приспособлений, характерных
для ксерофитов (редукция поверхности, опушение, завёртывание
краёв листьев и т. д.). Обычно это связывают с физиологической
сухостью, зависящей от низких почвенных температур. Впрочем,
экспериментов в этом направлении почти нет.
Растительность тундр СССР. Она слагается из целого ряда ти-
пов. Нередко тундру неправильно представляют себе как обширные
заболоченные пространства, покрытые мхами и мелкими кустар-
ничками, Но кроме моховых туйдр, как уже было указано выше,
имеются ещё и другие. Разнообразие тундр зависит от различия в
субстрате, в рельефе и в географическом положении.
зн
В направлении с юга иа север можно установить такие подзоны
тундр (не считая лесотундровой подзоны, промежуточной между
лесом и тундрой): куста рниковую^ лишай ни ко во-моховую, аркти-
ческую. Ещё севернее — исключительно на островах Ледовитсг©
океана — находится уже снеговая зона с арктическими
пустынями.
Кроме зональных подразделений (с юга на север), нужно раз-'
дичать ряд подразделений (провинций) с запада на восток (всего 12,
самая западная провинция Кольского полуострова, самая восточ-
ная — Чукотско-Анадырских гор).
Лесотундровая подзона. Здесь наблюдается чере-
дование безлесных участков с участками древесной растительности.
Первые — с типично тундровой растительностью, вторые — пред-
ставлены своеобразными формациями криволесья и редколесья.
Очень характерны такие жизненные формы, как стланики и полу-
етланики (более подробно о лесотундре см. ниже, стр. 303).
Кустарниковая подзона уже совершенно безлесна.
Кустарники распространены не только по склонам холмов, но и на
ровных местах. Возможность их существования обусловлена доволь-
йо значительными зимними осадками и более слабыми ветрами:
©ни находятся под снегом, и ветры не уничтожают молодые побеги;
именно последнее и имеет место в более северных тундрах, где ветры
очень сильны и, действуя иссушающе на молодые побеги, приводят
их к гибели. По своему сложению кустарниковые туч дры трех-'
ярусны: кустарниковый ярус— кустарничково-травян'стый — мо-
хово-лишайниковый. Из кустарников особенно характерны: низко-
рослая берёзка (Betula папа), к востоку от р. Енисея близкий заме-
щающий вид — В. cxilis, виды ив (Salix phylicifolia, S. lapponum,
S. glauca, S. lanata и ряд других), багульник и др. На песчаных
почвах — почти исключительно низкорослая берёзка. К востоку
от Лены видовой состав иь меняется и особенно обильны: S: lix baica-
lensis, S. pulchra; также появляется ольховник, (Alnus frut.cosa).
В кустарничк* во*травянисл м ярусе: брусника, водяника, не-
которые осоки, Festuca supina и др.
В лишайн) ксво-мсховсм ярусе—многие виды мхов (Aulacom-
Mium, Dicranum, Poly trichum, Ну loco mi uni prchfu'um и др.) и
лишайнт ки: Cetraria cucullata, кладонии (Cladonia gracilis, C. Sil-
va tica), Pcltigcra и др.
Фитоценозы кустарниковой тундры нужно отнести к группе
формаций Aest frut.ceta.
К северу от кустарниковых тундр располагается подзона ли-
шайнико-моховых тундр. Это—наиболее типичная тундра.
Моховая тундра здесь покрывает все плоские, обычно заболочен-
ные, пространства, а также склогы холмов, причём на песчаных
почвах её замещает ль шт йы ксвая тур дра. В данной подзоне
кустарники спускаются на склоны и постепенно редеют к северу.
Фитоценозы моховых и лишайниковых тундр относятся к классам
формаций Eubryosa и LicJiencsa (стр. *296).
Ещё более северная подзона — это арктическая тундра.
Здесь уже совершенно нет кустарниковых ассоциаций; моховые
301
тундры замещаются полигональными f особенно там, где господствуют
сильные зимние ветры. Б местах с менее сильными ветрами (централь-
ная Сибирь) полигоналы ые тундры развиты менее, а господствуют
сухие дриадовые (рис. 167) и моховые. Полигональные тундры
связаны с сильными зимними
ветрами и очень незначительным
снежным покровом. Напочвен-
ный покров деградирует, и поч-
ва растрескивается на много-
угольные (полигональные) от-
дельности, причём раститель-
ность удерживается лищь п©
трегциш ам и ложбин кам, между
-которыми — голая почва. П©
трещинкам и ложбинкам ютятся
ундровые мхи, лишайники и
екоторые цветковые растения,
апример лисохвост (Alopecurus
Ipinus), осока (Carex г gida),
1ЮТГК (Ranunculus nivalis), щуч-
ка (Dcschampsia arctea) и др.
На местах, особенно подвер-
женных ветрам, растительность
иногда почти отсутствует или
растительный покров разорван-
1 ый,что характерно для севернее
расположенной (на островах)
снеговой зоны. Здесь находятся
настоящие холодные пустыни
(Fngondeserta — ь см. ниже,
стр. 320).
Рассмотренные типы тундр имеют огромные пространственные
протяжения, но существует ряд типов тундр, или ограниченных
небольшими участками, или встречающихся в разных подзонах.
В южных частях, особенно в лесотундре, очень развита бугристая
тундра, связанная с замерзанием воды в верхних слоях торфа и,
следовательно, с расширением объёма и с соответственным вспучи-
ванием поверхности. Б северных частях преобладает пятнистая
тундра с голыми участками почвы; вопрос происхождения сё вы-
зывает у разных авторов разногласия, и, невидимому, в разных
случаях оно различно. Б понижениях находятся осоковые тундры,
представляющие в сущности низинные болота, на склонах в
верхних ч стях, где снег сдувается, развивается более ксерофиль-
ьый 'равяы-клыйпокров из Кstuca supina, Dryas octopetala и др.
(дерно инные тундры), а вщ зу склонов, где за. ёжч вается снег,—
луговинные тундры с большим количеством разютрав}ых видов.
На ьыхедах гор} ых пород разв! ваются скалисто-щебнесые тучдры,
имеющие сильно разрежен ый растпелы ын г.скрсв; здесь особенно
типична жизненная форма растений-подушек. Имеются ещё и дру-
гие типы тундр,
302
Болота с господством сфагнумов (верховые болота) на южной
•крайне тундр начинают к северу cvei яться другими типами. Для
сфап умов вссбще неблагоприятен высокий урсвеьь вечной мер-
злоты и неглубокий снежный покров. Bit, ечающиеся в тундре
сфагнумы относятся к нимнным сфагнумам, развивающимся в
пониженных местах.
Очень большие пространства, особенно в северо-восточных ча-
стях Сибири, занимают нагорные т у и д р ы, развитые на
гольцах. В основном господствуют лишайниковые тунд-
ры (доминируют Cladonia alpestris или Alcctoria ochrclcuca и Bryo-
pogon diverge ns) ‘с разрежем ым травяно-кустарничковым ярусом
(брусника, голубика, Cassiope tetregena, Ltzula ccnfusa и др.).
Лишайниковые нагорные тутдры (гольцовый пояс) ниже сме-
няются горно-тундровыми кустарниковыми фитоценозами (подголь-
цовый пояс). В восточной Ст 61 ри и на Дальнем Востоке мы имеем
заросли Pinus pumila, Alnus frut>.ccsa, Betula Middcndcrfii.
Нижняя граница этих поясов, в связи с географическим положе-
нием, находится на различной высоте: от 100 м на островах Ледо-
витого океана до 2000 на Алтае.
Тундры Северной Америки, насколько известно,
в основном схожи с тундрами Евразии. В наиболее северных частях
находятся каменистые тундры со скудной растительностью из мхов,
лпшайнгксв и немнеп х цветковых растений. Очень распространён-
иым в Северной Амер! ке типом тундр являются ягельные тундры,
покрывающте обширт ые пространства. Флористически североаме-
рнканскге тундры довольно схожи с евроазийскими.
Взаимоотношения тундры и леса. Тут дры не переходят непосред-
ственно в леса, но, как уже было указано, на юге тундровой зоны
имеется особая лесотундровая подзона. Эта подзона носит как бы
переходный характер, но по ряду признаков её лучше относить
всё же к тундрам. Лесотундра начинается на северной границе
лесной зоны и граничит с тундрой там, где с водоразделов исчезает
древесная растительность. Лесотундра — это чередование участков
безлесг ых с ti пичко туьдровей растительностью, участков со свое-
образной древесной растительностью и лесотундровых болот.
Древесные участки водоразделов представлены своеобразными фор-
мациями криволесья и ре колесья (в долинах леса главным образом
таёжного Tina) —они характер}ы лишь для подзоны лесотундр;
почти I-сключительн о свойствен г ы псслсдь ей и крупнобугргстые бо-
лота (бугры в среднем до4л1 высоты, иногда до 8 м). Редколесье очень
широко распростра! сн.о (ель, береза, лиственница), деревья здесь
имеют 3-—8 м высоты и сильно разрежены (на 1 га всего 300—500
экземпляров). Характерной чертой является также присутствие
таких Ж! з;еы ых форм, как стланика и полустланики (некоторые
ветви поднимаются вверх), отсутствующих в лесной зоне. Криво-
лесье развито особенно блл з морск! х пебсрежгй, а также в условиях
ropi ых лесотундр (i-anpi мер га север} см Урале и др.).
Древсс| ые нгсаддеьгя гесотугдр отт г чаются от сбытых лесов
рядом npt знаков: 1) деревья oaei ь угнетены, 2) древесный ярус
сильно разрежен, 3) i нжи-ie ярусы состоят из светолюбивых
30?
лишайников и кустарников (вследствие достаточного освещения).
Наиболее далеко на север идут такие породы: берёза (Betula tortuosa)
на Кольском полуострове, ель. и сибирская лиственница между
р, Печорой и Уралом, сибирская лиственница между Уралом и
Хатангой, а восточнее—даурская лиственница.
Вопрос о причинах северной границы древесных пород в связи
с безлесьем тундры решался различными авторами различно: не-
достаток тепла (Г р и з е б а х), холодные влажные ветры (М и д-
д е н д о р ф), низкие температуры и заболоченность (С ом мь е,
Т а н ф и л ь е в), иссушающее действие зимних ветров (Чиль-
м а н). Как было указано выше, более всего вероятна теория
Городкова об иссушающем действии ветров особенно во
время вегетационного периода.
Несомненно, что лесотундровое редколесье находится в тесной
связи с неблагоприятными для роста деревьев условиями. Но в чём
состоит основная причина редкого стояния деревьев? Многие авторы
(Визнер, М о р оз ов) указывали набольшую потребность в свете у
растений севера и вместе с тем на ослабленную там силу света; однако
активность света в Арктике выше, чем в средних широтах. Много
данных за то, что причина разреженности древостоя—в холодной поч-
ве, лежащей над вечной мерзлотой: корни не углубляются, а распро-
страняются вширь, что влечёт за собой корневую конкуренцию и
отмирание известного числа экземпляров (А. Г р и г о р ь. е в).
Что же касается самой северной границы древесных насажде-
ний, то здесь находятся лишь отдельные группы дере-
вьев или отдельные экземпляры, расположенные
очень далеко друг от друга. Корневая конкуренция в подобных
случаях уже не может иметь места. Можно предполагать, что подоб-
ное групповое расположение вызвано’ различной глубиной оттаи-
вания почвы на различных, даже oyienb близко лежащих участках.
Лесотундру в последнее время подразделяют на две подзоны:
северную лесотундру и южную. Последняя имеет облесен-
ность 20—30%, и в лесных насаждениях обычны сочетания несколь-
ких древесных пород; в северной лесотундре древесные участки
состоят из одной какой-либо породы, облесенность водоразделов
очень мала, леса главным образом в долинах рек.
Интересен вопрос о динамике северной границы леса. Почти
несомненно, что тундра в ксеротермический период (один из перио-
дов послеледникового времени) была покрыта лесом — остатки
пней, погребённые лесные почвы, мошные остатки мёрзлого торфа,
теперь там не образующегося, и др. Ряд авторов полагает, что и -в
настоящее время продолжается наступание тундры на лес,, однако
имеются и противсполсжьые данные — отступание в некоторых
местах границы вечной мерзлоты на север, прогрессивное распро-
странение на север по долинам рек ряда древесных пород и др.
Тундры и человек
Растительность тундр издавна испытывает воздействие со ето-
роны человека — оленеводство исключительно присуще тундровой
зоне. В иных случаях воздействие выпаса столь значительно (в слу-
304
чае присутствия больших стад и длительного их пребывания на
определённой территории), что растительные ассоциации коренным
образом видоизменяются. Поэтому нужно различать тундры пер-
вичные и вторичные, причём вторые обязаны своим
существованьем чрезмерному выпасу. И здесь, конечно, можно
уставов! ть различь ые градации в интенсивности выпаса — уме-
ренный, силщый, чрезмерный. По времени выпаса в тундрах разли-
чают два т1 па пастбн щ: летние и з и м и и е. Первые содер-
жат главным образом растения, которые иа зиму отмирают
в своих надземь ых частях (осоки, злаки, разнотравные виды
и др.), или сбрасывают листья (различные виды ив). Зимние
пастбища — это преимущественно лишайниковые, с различными
видами лишайников. Хорошими летними пастбищами служат ив-
няки, луговинная тундра, осоковые заросли, кустарниковые тундры
и т. д.
Летом олени совершенно не нуждаются в лишайниках, послед-
ние нужны лишь тогда, когда травянистая растительность отмирает.
Среди лишайников первое место занимают виды ягеля — Cladonia,
нередко называемого «олений моха (однако это не «мох», а лишай-
ник). Уже это название указывает на важность этого лишайника
для корма оленей. Лишайниковые тундры обычно связаны с песча-
ными грунтами и вообще широко распространены в более север-
ных частях тундровой зоны, но уже в подзоне арктических тундр
они постепенно деградируют. Причина последнего явления, по-
мимо неблагоприятных климатических условий, также неумерен-
ный выпас. Особенно вреден выпас лишайниковой тундры ле-
том, когда от выпаса лишайники страдают гораздо больше, чем
зимой.
Если от сильного выпаса страдают вообще все типы тундр, то
особенно он сказывается на лишайниковых тундрах, что связано
с тем, что npvpocf лишайников после скусывания происходит очень
медленно; при этом прирост по направлению к северу всё падает;
например прирост кустистых лишайников даёт в среднем такие
цифры: в лесной зо} е 4—6 лш в лето, в лесотундре 3—4 лглг, в без-
лесных подзонах 2—3 мм, в арктической подзоне i—2 мм. Ничтож-
ные цифры прироста говорят сами за себя и дают указания отно-
сительно рационального использования лишайниковых пастбищ:
установление числа голов оленей на определённую площадь, дли-
тельность пребывания стада в одном месте и т. д.
Кладониевые (ягельные) тундры, как выше было указано, раз-
виты в западных частях тундры до р. Енисея, а затем на крайнем
востоке (здесь более снежная зима и влажное лето), в центральной
же части зоны господствуют алекториевые и цетрариевые тундры
(лишайники Alectoria и Cetraria). Хозяйственная ценность послед-
них гораздо ниже, так как эти лишайники поедаются оленями не-
охотно.
Заповедники в тундровой зоне пока отсутствуют, если ие счи-
тать таёжно-тундрового заповедника на Кольском полу-
острове — Лапландский заповедник, иначе называемый «Чуна-
тундра» (200 000 га),
2Ф География растений 305
Пустыни (Deserta)
Пустыни занимают территории, наименее благоприятные для
существования растений. Здесь, конечно, отсутствуют леса, но и
травянистая растительность не может образовать сомкнутого
покрова; последний или разрежен («открытые» фнтсцсксзы), или же
совершенно отсутствует. Таким образом, влияния растений друг га
друга или нет совсем, или же оно менее значительно, чем в друп х
типах растительности.
Можно различать прежде всего два класса формаций: сухие
пустыни (Siccideserta) в сухом климате и холодные пустыни (Frgo-
rideserta). Последние развиваются в арктических областях (в снего-
вой зоне) и в высокогорных условиях выше альпийских лугов, где
субстрат является физиологически сухим.
Но отсутствие сомкнутого растительного покрова может быть
вызвано и рядом других условий, вследствие чего можно различать
и другие классы формаций пустынь,Вообще же пустыни могут быть
таковы:
1) физически сухие (пустыни тёплого и сухого климата);
2) физиологически сухие (пустыни холодного климата);
3) пустыни, вызванные крайне засолёнными почвами (эдафи-
ческая причина) — по берегам морей, на местах высохших морских
водоемов, на солонцах и т. д. (Litoridcserta);
4) пустыни, обусловленные подвижностью субстрата; например
песчаные пространства, перевеваемые ветром (Mobilideserta);
5) пустыни, вызванные неблагоприятными свойствами субстрата,
когда на камнях и скалах могут поселяться растения лишь по тре-
щинам, или же мхи и лишайники прямо на голом камне (Saxide-
serta и Rupideserta);
6) нужно особо отметить пустыни, вызванные биотическими
факторами, именно деятельностью человека и животных. Здесь,
при усиленном воздействии на растительный покров любого сложе-
ния, могут возникать (в результате выпаса, распашки, вытаптыва-
ния и т. д.) сильно разреженные ценозы и даже пространства, совер-
шенно лишенные растительности.
Подобное понимание пустынь гораздо шире обычного представ-
ления. Ниже даётся описание пустынь лишь двух первых типов,
имеющих наибольшее географическое значение и занимающих об-
ширные территории.
XVIII. КЛАСС ФОРМАЦИЙ — ФИТОЦЕНОЗЫ С РАЗРЕЖЕННОЙ РАСТИ-
ТЕЛЬНОСТЬЮ, ОБУСЛОВЛЕННОЙ СУХИМ И ТЕПЛЫМ КЛИМАТОМ —
СУХИЕ ПУСТЫНИ (&ICCIDESERTA)
Географическое распространение сухих пустынь. Области, зани-
маем, ые сухими пустынями, гаходятся в основном вне тропиков,
и в схеме (см. схему идеального континента) протягиваются к илу
и к северу от тропической полосы лесов. Особенно хорошо выра-
жена полоса пустынь в северном полушарии Старого Света—сна
тянется здесь от самого Атлантического океана (в Северной Африке),
через Аравию и Центральную Азию почти до Тихого OKeaia.
Здесь находятся -:г'' наиболее обширных пустыни — Сахара'в Аф-
306
рике и Гоби в Азии; кроме того, нашц обширные центрально-азиат-
ские пустыни.
В Северной Америке пустыни менее обплщы; здесь с;.и лежгт
между 28—38° с°вср: о?' широты и 105—120° жгзднсй долготы,
главным образом в пг? Lx Юта, Пдаго, Ор^'ои и Н^-жда, отчасти
в Аризоне, Калифорнии и Мека ь н
В южном полушарии гшбслпле ко 1лсшадп тхсгь-ди рас-
положены в центральных частях Австралии, менее значите: ьные—
в южных частях Африки. В Южной Америке пустыни занимают
узкую полосу вдоль западного побережья в пределах 5—25° южной
широты, а также западные части Аргентины.
Пустыни в основном — области с очень ьсбольшим количеством
осадков, не превышающим 30 см, а во мцопмх местах осадков вы-
падает гораздо меньше: в Мерве — 15,5 см'т в Бухаре—13,5 см,
в Репетеке —10,3 см, и можно указать немало пунктов, где осадков
даже менее 10 ом: Кашгар (в Центральной Азии) — 5 см, Каир—3 см
п др., в некоторых же местах Чилийской пустыни —всего 0,5 см.
Бывают годы, когда в некоторых пустынях количество осадков так
ничтожно, что совершенно не поддаётся намерению.
Кроме небольшого количества осадков, для пустынь характер-
ны: 1) ничтожная влажность воздуха, 2) очеьь высокие температуры,
3) резкие температурные контрасты, 4) сильная инсоляция, обусло-
вливаемая небольшой облачностью, 5) часто сильные и порывистые
ветры.
Все эти условия крайне неблагоприятны для растительности н
вызывают необходимость чрезмерного испарения. Вообше разрьв
между количеством осадков и испарением очень велик, и нередко
испарение в 10 и более раз превышает выпадающие осадки.
Так, например, в алжирской Сахаре это превышение может
достигать 50—60 раз! В Ливийской же пустыне испарение соста-
вляет около 4 000 мм в год, а дождь бывает один раз в 4—5 лет.
Ясно, что за счет атмосферной влаги растения пустынь жить i е
могут; основные источники их водоснабжения — это грунтовая
вода и влага почвы, получающаяся в резулынте конденсации водя-
ных паров из воздуха.
Не менее неблагоприятны для растительности и эдафичсские
условия. Обычно в почве пустынь имеется недостаток воды и бед-
ность в органических веществах; кроме тоге, почвы сильно засо-
лены, так как ничтожное количество осадков недостаточно для
выщелачивания почвенных солей.
Жизненные формы пустынь. Растительность пустынь состоит из
различных жизненных форм, но в основном можно выделить две
группы, совершенно различно приспособившиеся к жизни в обста-
новке пустынь: одна группа состоит из видов, приспособив-
ш и х с я к перенесению неблагоприятных условий пустыни (рас-
тения ксерофиты), другая — из видов, не переносящих засухи и
заканчивающих свою вегетацию до наступления ее. Первые пред-
ставлены многолетниками (особенно полукустарниками и травами),
вторые—по большей части однолетниками.
20* Qfi7
Пустынные многолетники. Они , обладают Це-
лым рядом приспособлений, так как должны энергично вбирать
воду из сухой почвы или медленно испарять её.
Чго касается всасывания веды из почвы, то здесь имеет место
или повышенное осмотическое давление в корнях, или особые
устройства корневых систем. Осмотическое давление очень велико,
и для растений египетских пустынь приводятся очень большие
цифры в 19—50 ат (для листьев), сосущая сила корней ешё более
Рис. 168. Схема распределения корней песчаного злака Aristida
Karel ini (в плане). Корни распростр -няются в горизонт i льном на-
правлении (каждая клетка равна 1 лА Цифры—количество метров).
высока (до 100—160 от), что, конечно, позволяет всасывать даже
небольшие ко л и чеса ва влаги.
Корневые системы или очень сильно развиты вглубь
и тогда достигают грунтовых вед, причём здесь интересен необычай-
но быстрый рост корней, в первое время в прямой ущерб надземным
частям; но очень часто грунтовые воды растениям недоступны (они
лежат на глубине 100 м и более), и тогда корневые системы разви-
ваются вширь; последнее наблюдается у трав и кустарников
(рис. 168); виды Callig-num в Кара-Кумах имеют боковые корни
длиной в 10—15 м и эю не представляет исключения. У некоторых
видов (полыни, эфедра и др.) можно наблюдать диференциацию
корней: приповерхностные корни эфемерны и всасывают весеннюю
влагу, а более глубокие функционируют в остальное время веге-
тационного периода, пронизывая почву по всем направлениям.
Уменьшение транспирации у пустынных расте-
ний достигается различными способами, но в основном является
результатом редукции испаряющей поверхности (листьев). Вообще
имеются такие формы приспособлений;
1. Развиваются афилльные формы (от греческих слов «филлон»—
лист и «а» —пристав! а, означающая отрицание). При этом листья
чрезвычайно редуцируются и в крайних случаях их #аже с трудом
можно обнаружить (примеры: саксаулы, виды Calligonum в средне-
азиатских пустынях, Casuarina—в пустынях Австралии и т. п,).
Получаются безлистные фермы, у которых в фотосинтезе основ
ную роль играют зелёные стебли.
2. Нередки гетерофильные фермы (ст греческого слова «гетерос»—
разный, различный), у которых весенние листья отличаются от
летних, более приспособленных к засухе.
3. Нередки растения с жёсткими листьми и с толстой кожи-
цей, защищающей от излишнего испарения.
4. Ряд растений пустынь накопляет в своих частях запасы во-
ды* В отдельных случаях вода содержится в волосках, имеющих
вид пузырьков, густо покрывающих листья (например у полуку-
старника кокпека —Atrip lex сапа), но чаще запасы боды содер-
жатся в тканях стебля и листьев (суккуленты). Суккуленты очень
типичны для некоторых пустынь земного шара: кактусы, молочаи,
мезембриантемумыит. п. У кактуса (Cereusgigantcus) ь 15 жьь.сстсй
было обнаружено 2 000—3 000 л воды (Северная Америка). Количе-
ство воды у некоторых суккутентов составляет до 96% веса.
5. Ряд растений имеет защитные покровы (волоски, Бесковой
налёт и т. п*), о чём не раз говорилось при описании ксерсфг льных
растений.
Имеется у пустынных растений и ещё ряд характерных призна-
ков; укажем лишь на одно свойство пустынных растенкй, именно
на усиленную склеротизацию вегетативных частей. В зависимости
от сухости среды у растений развиваются твёрдые механические
элементы: листья и пазушные побеги превращаются в кслючки.
Вообще колючие растения характерны для пустынь.
В засол ё н и ы х пустынях растения, кроме крайней сухо-
сти, должны выносить и исключительно высокое засоление почв.
Здесь возможны два способа: или растения повышают до- вы-
сокой степени своё осмотическое давление, или они обладают спо-
собностью выделять наружу избыток солей. И тот и
другой способы имеют место в природе. Некоторые виды выделяют
соли через особые поры на листьях (в наших среднеазиатских пу-
стынях: гребенщик — Тапи rix, кер мок — Stc tice и др.). Такие же
растения, как солерос (Salicornia herbacea), имеют высокое осмоти-
ческое'' давление.
Весенние эфемеры. Вторая группа растений, харак-
терная для некоторых пустынь, — это однолетники или же видь:
многолетние, но быстро проходящие весной весь свой цикл веге-
тации. Выше мы видели, что в биологическом спектре пустынь осо-
бенно выделяются терофиты. Однолетние эфемеры устроены очень
просто—это низкорослые сднсстебельные формы 30—40 см вы-
соты, а в очень засушливые годы лишь 1—2 см. Они относятся к
различным семействам. Многолетние эфемеры в своих надземных
частях схожи с однолетними, но подземные их части должны выдер-
жав 1ть летнюю засуху; поэтому последние представлены большей
частью клубнями и луковицами. Даже злаки, обычно не образую-
щие луковичек, дают их при сснсвавки стеблей.
Характерно для эфемеров их необычайно быстрое развитие вес-
ной: пусты ни по крыва ются и ногда с пло шным ковром цве тущих
309
растений. Вегетация эфемеров про-
должается около 2—2г/2 месяцев, ио
ряд видов весь свой жизненный цикл
проходит недели в три. Некоторые
пустыни особенно богаты весенними
эфемерами и потому называются эфе-
меровыми (например у нас в Сред-
ней Азии—рис. 169).
Различные типы пустынь. Нужно
оговориться, что не везде для пустынь
характерно присутствие весенних эфе-
меров,—это прежде всего связано со
временем выпадения осадков* Осад-
ки могут выпадать в значительном
количестве весной (тогда развиваются
эфемеры), ио могут небольшими пор-
циями распределяться и по всему
периоду вегетации (в этих случаях
эфемеров почти нет).
Этот характер^выпадения осадков
позволяет говорить о пустынях раз-
личных т и п о в: одни с весен-
ними осадками (соленые пустыни
Туркестана, пустыни Месопотамии,
Северной Африки, пустыня Карру
в Южной Африке и др ), другие с
осадками, равномерно распределен-
ными по вегетационному периоду
(южный Казахстан, центральноазиат-
ские пустыни, цеитральноавстралий-
ские, североамериканские в области
«Великого бассейна» и др.). Однако
во всех этих случаях сумма осадков
остается более или менее однообраз-
ной и не превышает 30 см в год.
По своему субстрату, и, соответ-
ственно, растительности пустыни
очень разнообразны: г л и и истые,
песчаные, каменистые,
солончаковые и др. В даль-
нейшем мы остановимся на главных
пустынях земной поверхности.
Сахара. Она занимает огромное
пространство в 9 млн. яле2 и пред-
ставляет сложную систему более
повышенных и более пониженных
гористых пространств. По субстрату
здесь можно выделить такие типы
пустынь: скалистые, каменистые (хря-
щевые) и песчаные. Пустыню пересе-
ло
кают сухие русла — «вади». Лишь наиболее глубокие долины и
места выхода ключей, «эазисы», дают резкий контраст с пустын-
ными бесплодными пространствами.
В различных частях Сахары выпадение осадков по времени раз-
лично: в алжирской Сахаре максимум приходится на весну и осень,
а в центральных частях ее, в ливийской, египетско-аравийской,
правильности не наблюдается, да к тому же осадки крайне скудны.
Рис. 170. Группа финиковых пальм (Phoenix dactylifera) — Египет
(из Веттштейна).
Крайние пределы температуры в тени от 7,5° до 52,4°; особен-
но сильно нагреваются почвы, наблюдались температуры в 69,8—
74,4е в течение нескольких дней подряд.
Флора Сахары местами крайне скудна или даже совершенно
отсутствует. Более богаты растениями лишь те места, где имеется
вода, «оазисы». Наиболее характерным растением является здесь
финиковая пальма (Phoenix dactyl f_ra, рис. 170), крометого, несколь-
ко видов акаций (Acacia), называемых гумми-акацизми, так как
они дают клейкие вещества. В более южных частях (к юго-востоку)
появляется пальма Hyphaene thebaica.
Обращает на себя внимание сильное развитие однолетников и
галофитов; наоборот, луковичные растения, подушкообразные, ред-
ки, а также редки и сильно суккулентные виды, очень ха-
рактерные для южноафриканских и американских пустынь Неви-
димому, это обстоятельство, помимо причин генетического характе-
ра, связано с небольшим количеством осадков—в сущности здесь
нет воды, которую суккуленты могли бы запасать.
Для песчаных пустынь очень характерен злак Aristida pungens,
с сильно развитыми поверхностными корнями до 20 м длиной, улав-
ливающими влагу с большой площади.
311
В центральных частях Сахары, на больших протяжениях отсут-
ствуют какие-либо растеш я.
Аравийская пустыня Она также бесплодна, кроме с ы сев, где
растет опять та же финиковая пальма, акации, а также Bcrassus,
3!2
дающий аравийский бальзам. Интересны некоторые однолетники,
при созревании приобретающие шарообразную форму, обламываю-
щиеся и перекатывающиеся от ветра, так называемые «перекати-
поле»; сюда относятся виды родов Anastatica, Odontospermum —
«иерихонские розы». Интересен также съедобный лишайник (Leca-
nora esculenta), который легко переносится ветром, так как он к
земле не прикрепляется. Обычен также пустынный злак Anstida,
некоторые полукустарники, некоторые сочные растения (суккулен-
ты)— все они носят сильно ксероморфиый отпечаток.
Наиболее обширные пространства в Азин занимают центрально-
азиатские и арало-каспийско-туранскне пустыни (последние у нас
часто называются среднеазиатскими — они находятся в пределах
СССР)
Среднеазиатские пустыни. Они очень разнообразны, по положе-
нию и климату распадаются на северные (южноказахстанские) и
южные. Границей между ними является линия, проходящая не-
сколько южнее Аральского моря и Балхаша. В северных пустынях
«садки распределены более или менее равномерно, а в южьых хо-
рошо выражен весенигй максимум; в результате в последнем
случае обильное развитие эфемеров.
По своей растительност! , которая тесно связана с субс ратом,
среднеазиатские пустыни можно подразделить на полисные (г; и-
и истые), солянковые (солончаковые), кустарниковые
(п е е ч а н ы е) и др
Для глинистых северных пустынь (рис. 171) особен-
но характерны полыни (Artemisia) и различные виды из семейства
солянковых. Из полыней исключительную роль играет Artemisia
terrae albae, которая комбинируется с рядом видов лебедовых и
дает особые типы пустынь: полынь — биюргун (Anabasis salsa),,
полынь — кскпек (Atriplex сапа), полынь — черный саксаул (Halo-
xylon aphyllum)/ др Это—крайне монотонные полынные и со-
лянково-полынные пустыни с господствам полукустарников и других
ксерофитов *
Черный саксаул — это небольшое деревцо, совершенно безлист-
ное; оно образует заросли, производящие впечатление небольших
лесков. По своему внешнему виду — это типичный ксерофит.
Полный контраст представляют глинистые пустыни южных
частей Средней Азш ; это — своеобразные эфемеровые густыми.
Раст! тыльный' покров госит мезофильный хар i тер и весной на-
столько густ, что представляет сплошную дернину, и
его аожно сравнить с растительностью на лугах; гочва сов^с-
шенно гропнтана въгой Лсюм все эф.меры обмирают и вы-
ветрив ются; поверхность по^вы становится почти совершенно
roj ой и сухой
Основу дернины в эфемеровой пустыне образуют осока (Сагех
pachystyhs) и луковичный мятлик (Pea bulbcsa), к которым присо-
единяются эфемеры многолетние и особенно однолетние. Последних
не менее 80% всех видов
В последнее время некоторые авторы (Ш е н н и к о в) относят
эфемеровые пустыни к лугам. Однако луга характеризуются
31’
вегетацией в течение всего тёплого времени года и эфемеры для них
совершенно чужды.
Солянковые (солончаковые) пустыни. Эют тип
связан с речными террасами Сыр-Дарьи, Амур-Дарьи, Мургаба
л др. и вообще с пониженными местами. Засоление главным обра-
зом за счет хлоридов (особенно поваренной соли). Растительность
состоит из галофитов, главным образом из суккулентов; особенно
'34
распространены различные солянки (Halonemum strobilaceum,
Halostachys caspia, Salicornia herbacea и др.). Встречается ряд
растений, у которых особые железки иа листьях выделяют солевые
растворы, образующие при высыхании кристаллы и корочки солей
(Frankenia, Tamarix, Reaumuria и др.).
Нередки пространства, совершенно лишенные какой бы то ни
было растительности (мёртвые пространства).
Однако солянковые пустыни менее монотонны, чем полынные,
глинистое. Растительность даёт ряд меяющнихся аспектов: весной —
яркозелеиая, летом—жёлтая, осенью—розовых, кроваво-красных
и фиолетовых оттенков. Осенью показывается и молодая зелень
новых отпрысков.
Кустарниковые (песчаные) пустыни (рис. 172).
Большие территории Средней Азии заняты песками: северные
Кара-Кумы, Му юн-Кумы, Прибалхашские пески, Кызыл-Кумы,
южные Кара-Кумы и др. В противоположность другим типам пустынь
растительность песчаных пустынь отнюдь не является скудной,
так как здесь имеется достаточно благоприятный водный
режим: атмосферная влага быстро проникает в песок; капил-
лярность очень слаба; пески обладают способностью сгущать водя-
ные пары в почве во влагу.
Поэтому на песках существуют не только травянистые растения,
но также кустарника и деревья. Из кустарников Calligonum (до
30 видов), Ammodendron (5 видов), Halimodendron, Tamarix и др.
Замечательным деревом пустынь является белый саксаул (Haloxylon
persicum). В апреле и мае, когда цветут кустарники, эти пустыни,
по выражению некоторых исследователей, представляют сове р-
ш е и и ы й с а д.
Кроме особого водного режима, песчаные пустыни имеют ещё
одну важную особенность — их субстрат родвижен и под действием
ветра перемещается, образуя бугры (барханы). Растения должны
обладать способностью бороться с этой подвижностью субстрата,
иначе они будут засыпаны песком. Здесь имеется ряд приспособле-
ний для перемещения растений как в горизонталь-
ном, так ив вертикальном направлении. В первом слу-
чае образуются длинные горизонтальные шнуровидные корни, во
втором — растение перемещается все выше и выше, и новые корни
возникают на верхних частях стебля. В зависимости от степени
подвижности песков различается и растительность.
Меиьшче пространства занимают другие типы пустынь, напри-
мер. камен иста я (гипсовая). Основной ее признак—большая
или меньшая каменистость и сильная (до 50%) гипсоносность. В се-
верных каменистых пустынях особенно распространена полынно-
боялычная пустыня (полынь—Sa Isola arbuscula, также S. la rici foil a).
Полынь Artemisia terrae albae на юге сменяется A. herba alba. Ос-
новной ландшафт южных пустынь — полынно-солянковый. Гипсо-
ьосность грунтов очень резко сказывается на своеобразии флоры—
здесь очень много эндемиков.
Центрально-азиатские пустыни. Они занимают огромные про-
странства от Памира и Тянь-Шаня до Большого Хингана на про-
315
тяжении более 4 000 км и представляют возвышенную страну, ле-
жащую на высоте 1 000—1 500 м. Эта страна почти со всех сторон
ограничена горами и имеет исключительно континентальный климат
(в Яркэнде средние температуры самого теплого и самого холодного
месяца 4-27,6° и —6,9°, в Урге +17,5е и —26,2°).
В общем растительность очень схожа с северными аралокаспий-
сними пустынями, так как осадков здесь мало и они выпадают более
или менее равномерно. Очень характерны полыни; наиболее хорошо
представлены семейства лебедовые, крестоцветные, свннча^ковые,
сложноцветные.
Восточная часть центральноазиатских пустынь — Гоби, или
Шамо, —тоже очень бедна видами; и здесь на огромных простран-
ствах господствует погынь. Далее к востску пустыня принимает
характер, приближающейся к степи.
Южноафриканские и американские пустыни. Для большинства
из них особенно характерно обглге крупных суккулентов: в Старом
Свете — молочаи, в Новом — кактусы.
Рис. 173 Растения-суккуленты южноафриканских пустынь
1 и г—Crassula p\ramiclalis (2—। ид сбо< v, 2—вил сверху;, 3—С column an s3 rf-Mesem
bnanthemunij.Bcilu3i , 5—М calcareum (по Веттштеину). растения а, 4 и 5 очень похонн
на камни—*расгенил-качни»
Особенно богаты кактусами мексиканские пустыни; кроме как-
тусов встречаются также юкки и агавы (средн последних i звестная
Agave amencana, дающая национальный мексиканский напиток).
Кактусы различны по своей величине и форме: то шарообразны, то
в виде лепешек, то в виде цилиндров или канделябров. Высота их
316
нередко превышает рост человека, в иных случаях достигает 10 М
и больше. Кроме кактусов, встречаются различные колючие кустар-
ники-
Совершенно отлична пустыня Великого бассей-
на в юго-западной части США. Здесь господствует полынь —
Artemisi^ tndentata; растение имеет серо-зеленый цвет и высоту
до 1 м; его экземпляры расположены так, что между ними можно
свободно проходить. Другие виды физиономически не играют ни-
какой роли, и пустыня имеет крайне однообразный вид.
Среди южноафриканских пустынь по своему резко пустынному
климату особенно выделяется пустыня Н а м и б, тянущаяся
по западному берегу Африки к северу от устья Оранжевой реки.
Здесь растительность чрезвычайно скудна; cpe/yi жизненных
форм господствуют суккуленты, особенно род Mesembrianthemum
(рис. 173). Но после редких дождей пустыня может покрываться
пестрым ковром красных, желтых и белых цветков (рис. 174).
|В местах с более или менее близким стоянием грунтовых вод раз-
вивается замечательное растение Welwitschia mirabilis (рис, 199);
другое растение Acanthosicyos horrida (см. стр. 359) имеет корни
до 15 м в глубину. Эти два вида являются хорошими показателями
относительной близости грунтовых вод.
Далее на восток, в глубине страны, лежит так называемая п у-
стыня Калахари; в большей своей части она представляет
травянистые, хотя и разреженные пространства с рядом древесных
форм (ряд акаций и др.). Особенно обращает на себя внимание бо-
гатство родов и видов злаков.
317
Южноафриканская пустыня Карру по оби-
лию суккулентов подобна мексиканским пустыням и полупустыням,
но здесь совершенно другие растения. Среди растительности Карр)
можно отличить такие основные жизненные формы:
1) листовые суккуленты — особенно мезембриантемумы (не-
обычайное разнообразие видов), также многочисленные алоэ, из
которых некоторые древовидны, виды рода Crassula и многие другие1:
2) стеблевые суккуленты — многочисленные молочаи, некоторые
из которых очень напоминают по внешности американские кактусы
и являются древовидными, а также стапелии и др.;
3) низкорослые кустарники, нередко снабженные колючками;
4) растения с подземными, запасающими воду, органами.
Австралийские пустыни. Они занимают значительные простран-
ства в центральных частях материка. Наиболее бедна осадкам и
область, называемая Эремеей (местами 13—15 см в год). Раститель-
ность находится в связи с субстратом; сильно засоленные почвы
богаты листовыми суккулентами из лебедовых, но здесь же встре-
чаются кустарники австралийского типа: акации, казуарины, эвка-
липты и др. В песчаных местах иа обширных территориях развита
так называемая с п и и и ф е к с о в а я пустыня (или «Spimfex») с го-
сподством злаков (однако доминируют виды рода Triodia, лншь на
востоке род Spinifex); в более пониженных местах казуарины, эвка-
липты п замечательные «травяные деревья»—Xanthorrhoea (рис. 254).
Нужно отметить, что для австралийских засушливых областей за-
мечательно то, что у растений очень редки стеблевые колючки и
шипы; сами листья становятся очень жесткими и оканчиваются
колючим острием, т. е, колючесть достигается иными путями:
злаки типа «Spinifex» тоже имеют такие же лнст'ья.
Пустыни н человек
Пустыни, несомненно, с давних времён использовались челове-
ком-кочевнг ком, причем верблюд и овца —это наиболее приспособ-
ленные к кормам пустыни животные, Животноводство было основ-
ной базой при использовании природных ресурсов пустыни. Хотя
пустыни по своим климатическим условиям и пригодны для земле-
делия, но недостаток воды является основным препятствием к раз-
витию последнего. Там, где имеется вода, вполне возможны раз-
личные культуры и рост деревьев; об оазисах Сахары было уже
сказано. В большинстве случаев растительность оазисов является
культурней: финиковые пальмы, оливки, фиговые деревья и др ,
различные травянистые культуры: хлебные злаки, ячмень, сорго,
пенициллярия и др., а также кормовые растения.
Для СССР orpc.vr.ee значение имеет проблема освое-
ния среднеазиатских пустынь. При этом, конечно, должны быть
1 Некоторые виды Mesembrian themum, Crasstila и др. очень похожи п
своим листьям на камни. Это—один из примеров мимикрии (подражанж
среди растений. Очевидно, подобные растения могут избегнуть поеданю
травоядными животными (животные принимают их за камни)
318
тщательно проанализированы местные условия и в возможно полней
степени изучена экология как местных растений, так и вводимых
в культуру. Так как основной ограничивающий фактор в пустыне—
это недостаток воды, то освоение пустыни должно быть связано с ее
орошением или с более рациональным использованием имеющихся
в наличности запасов воды. Наибольший эффект при освоении nj-
стыии должно иметь, конечно, орошение. В этом отношении пред-
приняты и предпринимаются у нас грандиозные работы: замечатель-
ный Ферганский канал, проблема орошения полупустынных прост-
ранств в низовьях р. Волги и др. В условиях орошенной пустыни
возможны самые разнообразные культуры: различные плодовые
деревья (персики, абрикосы, гранаты, грецкие орехи, яблоки, гру-
ши и др.), такие южные культуры, как виноград, арбузы, дыни,
также южные волокнистые, масличные, каучуконосы и др. (хлоп-
чатник, клещевина, гвайюла и т>д.); укажем еще рис и другие зла-
ки. Однако нельзя оросить, пользуясь имевшимися водами, всю
пустыню, нужно рационально использовать местные запасы влаги,
использовать зимние и весенние осадки. При этом необходим подбор
таких растений, которые проходят цикл своей вегетации быстро,
до наступления летних засушливых месяцев. Здесь необходим под-
бор растений скороспелых — «убегающих» от засухи. Кроме того,
возможны такие засухоустойчивые культуры, которые имеют мощ-
ную корневую систему, опускающуюся до грунтовых вод.
Имеются данные о хороших результатах в отношении таких куль-
тур: арбузы, сорго, просо, гаолян и др. При этом интересно указать,
что для сохранения почвенной влаги, помимо поверхностного рыхле-
ния почвы, применяется покрытие почвы бумажной «мульчей» или
битумной эмульсией (получается на поверхности почвы пленка в
1,5—2 мм толщиной, задерживающая испарение). Таким образом,
земледелие вполне возможно при соответствующих агротехнических
мероприятиях и на неполивных территориях (так называемое «богар-
ное» земледелие).
Однако, помимо поливного и богарного земледелия, пустыни
Средней Азии представляют обширную арену для животноводства.
Здесь вопрос стоит в направлении улучшения имеющихся пастбищ и
в создании наиболее рациональных кормовых угодий. Для этого не-
обходим подбор соответствующих растений—более ценных с кормо-
вой стороны, вегетирующих более продолжительное время и т. д.
Разные типы пустынь (глинистые,.песчаные и др.) имеют, конеч-
но, свои специфические особенности при их освоении. Так, в част-
ности, песчаные пустыни особенно нуждаются в закреплении песков,
которые, как правило, разбиты у каждого поселка или колодца-
И в этих пустынях имеется немало растений, вполне пригодных для
этой цели (например колосняк — Elymus gigantlieus). Очень важна
в песчаных пустынях проблема саксаулового хозяйства — саксаул
дает прекрасное топливо, уголь и др.; здесь задача состоит в восста-
новлении уничтоженных саксаульников и заращении новых площа-
дей. Богатство песчаных пустынь различными жизненными формами
растений дает большие перспективы для их хозяйственною исполь-
зования.
319
XIX. КЛАСС ФОРМАЦИЙ — ФИТОЦЕНСРЫ с разреженной расти-
тельностью, ОБУСЛОВЛЕННОЙ ХОЛОДНЫМ КЛИМАТОМ-ХОЛОДНЫЕ
ПУСТЫНИ (PK1GORIDESE«1 А)
Пустыни, обусловленные холодом, — это, во-первых, пустыни
высокогорные, а во-вторых, арктические и антарктические.
Тепло находится здесь в минимуме. Еб. ьшую часть года гос-
подствует зимний покой. Здесь нельзя говорить о весне или осени,
так как период вегетации очень короток. Растения прижимаются
к земле, так как она нагревается лучше, чем воздух.
Растительность находится почти у пределов жизни, она очень
скудна флористически, так как лишь немногие виды могут выдер-
живать такие крайние условия; с другой стороны, сплошной расти-
тельный покров здесь отсутствует, и лишь немногие более благо-
приятные местообитания несут как бы оазисы растительности,
в остальном —это почти безжизненные пустыни с редкими отдель-
ными экземплярами.
Высокогорные холодные пустыни. Они лежат выше альпий-
ских ковров и лужаек (см. выше). Обнаженные скалы, каменистые
россыпи, каменистые морены и т. д. —все это места, мало пригодные
для поселения растений (рис. 175). Высший предел цветковых ра-
стений доходит до очень значительных высот; так, в Альпах неко-
торые виды доходят до высоты в 4 000—4 270 м, в Андах — до 4 730 м;
в Тибете на высоте 5 000—5 362 м имеется до 50 видов, а в Гимала-
320
ях Arenaria musciformis была найдена на высоте 6 220 м. Однако на
крайних высотах чаще встречаются лишайники; так, иа вершине
Килиманджаро в 6 010 м находятся лишь одни лишайники.
Жизненные формы высокоальпийских растений таковы же, как
у альпийских, но рост растений еще более подавлен. Особенно
хорошо представлены формы подушек, розеток и шпалерные
формы.
В Альпах необычайно красочно на коренных породах сочетание
подушек темноголубых ге циан (Gentiana bavarica subacaulis),
нежнорозового прол ом ника (And rosace alpina — рис. 176) и небесно-
голубой Eritrichium папиш На известковых породах встречаются
другие виды. В качестве характерного высокогорного растения
нужно отметить приледииковый лютик (Ranunculus glacialis).
Вообще большинство высокоальпийских видов хотя и низкорос-
лы, но крайне изящны и красочны.
Примером холодных пустынь является Памир, находящийся
на высоте 3 000— 4 000 м и представляющий пустынные плоскогория.
Осадков здесь выпадает 58 мм в год (в среднем), а в иные годы до
21 мм\ они выпадают главным образом летом; зимой же снежный по-
кров, который мог бы защищать растительность от холода, отсут-
ствует. Средняя годовая температура — 1°. Зимой температура
падает до — 46,7°, а летом может подниматься до 30°; в период ве-
гетации температура может понижаться до 0° и ниже. Температура
почвы на ее поверхности иногда доходит до 33,5°, и вообще роль поч-
вы в смысле теплового режима растений очень велика
21 География растений 321
Подобная экологическая обстановка крайне неблагоприятна дгя
произрастания растений: они прижимаются к почве, находя здесг
более благоприятную среду. Основные приспособления в данном
направлении таковы: ряд растений дает подушковидные формы
(Acantholimon, Oxytropis), другие растения стелятся по земле и
разрастаются в стороны, некоторые дают характерные «гсдьмины
кольца»1 (Astragalus Muschketovi, некоторые злаки, а также харак-
терное памирское растение-полукустарник терсскен — Eurotia се-
рне. 177. Ведьм :чо кольцо, образованное Saussurea sp бл Тянь-панской обсер-
ватории на высоте 3672 м (по фото Е. Е. Сочеванова).
ratoidcs); ряд растений окутан при ссн сванки остатками отмерших
листьев, крайне интересна скученность растений при нсключгтегьюй
разреженности травянистого покрова; так, гногда в подушке какого-
либо вида включено несколько других более мелких bi-дов (до 12);
растения не только прижимаются к почве, но и друг к другу.
Среди 500В1дов Памира около 70 видов являются одюлетниками
Это явление характерно для сухих пустынь, но недостаточно понят-
но для условий Памира.
На Памире имеются такие типы пустьць: каменистые, такырио-
солончаковые и песчаьые. Первые занимают нанболыше простран-
ства и особенно характерны. Ландшафт? ым растер! ем здесь являет-
ся распростертый полукустары к терсскен, но он разбросан отдель-
ными экземплярами, и на пространстве ь 100 л*2 имеется сбычро не
более 10 экземпляров, т. е. растительность является совершенно
1 «Ведьмины кольца» образуются у растений, растущих плотной дернови-
ной, от того, что центральные части дерновины отмирают, а периферические
части разрастаются в стороны — получается кольцо, диаметр которого при
дальнейшем разрастании все увеличивается (рис. 177).
322
открытой (вообще же в этом типе пустыни встречается всего 4 более
или менее постоянных вида).
Арктические холодные пустыни. Они развиты в «снеговой» зо-
не, нежащей севернее зоны тундр, исключительно на островах Ле-
довитого океана (но южные части Новой Земли еще относятся к тун-
дре). На более южных и на равнинных островах ледники отсутствуют
или развиты очень слабо, и здесь находятся каменистые россыпи,
пустынные пространства с полигональными почвами, там и сям от-
дельные клочки тундровых формаций и т. д. На гористых островах
в более северных частях Ледовитого океана лишь узкие прибрежные
полосы, обычно низменные, пригодны для растительности. Здесь
лишь в на! более благоприятных условиях можно найти клочки арк-
тических лугов и травяно-гипноьых болот; до 70% площади, сво-
бод! ей от льдов, занято щебенчатыми пустынями, каменистыми рос-
сыпями, полигональными почвами; здесь можно встретить лишь ци-
чтажн ые клочки ра стительн ости.
Сильнейшие ветры, сдувающие с поверхности весь снег, явля-
ются одним из факторов, влияющих на отсутствие связного расти-
тельного покрова.
Вообще здесь замечательны контрасты между зелеными, иногда
пестрыми от цветов участочками растите л внести и лишенными ра-
стительности ледяными пустынями. На Шпицбергене наиболее раз-
вита растительность в глубине фиордов, далеко вдающихся в глубь
островов; также более богата растительность южных, лучше на-
греваемых склонов, где можно говорить о небольших «садиках»
с прекрасно цветущими растениями, особенно если оии удобрены
благодаря присутствию многочисленных птиц: иебесноголубые Ре-
21 ’ 328
lemonium hutnile, бледиожёлтые маки Papa ver radicatum (рис. 178)
и др. На Земле Франца Иосифа растительность ещё более скудна.
Но даже на нунатаках и на ледяных полях внутренней Гренлан-
дии все же встречаются некоторые растения; например на нунатаках
можно встретить довольно значительное число высших растений,
а на льдах — некоторые водоросли. Так, водоросль Anabaena Nor-
denskioeldi окрашивает в пурпурно-бурый цвет значительные про-
странства ледниковых полей внутренней Гренландии.
Вообще можно думать, что низкие температурные условия ни-
где на земной поверхности не ставят препятствий для существова-
ния растений.
Холодные пустыни и человек
Освоение этих пустынь представляет, конечно, большие труд-
ности и в ряде случаев является вопросом будущего. Однако в не-
которых случаях в этой области сделаны очень большие успехи, в
частности в освоении пустынь высокогорного Памира. Здесь освое-
ние идет в таких направлениях: введение важных культурных ра-
стений, введение в культуру местных растений, создание новых ра-
стительных форм, наиболее отвечающих местным условиям. В пер-
вом направлении вводятся зерновые культуры (пшеница, ячмень,
рожь и др.), огородные (капуста, лук, морковь, картофель и др.),
кормовые (люцерна, клевер, чина, эспарцет и др.) и другие культуры,
отсутствовавшие до сих пор на Памире. Опыты в отношении ряда
указанных растений проходят вполне успешно. Что касается мест-
ных растительных ресурсов, то здесь испытываются некоторые дико-
растущие злаки, бобовые и др., имеющие кормовое значение. Соз-
дание новых растительных форм базируется на отдалённой гибри-
дизации между видами и родами. Таковы основные, уже осущест-
вляемые, пути в деле освоения Памира.
ОТДЕЛ V
ФЛОРИСТИЧЕСКИЕ ОБЛАСТИ И ПОДОБЛАСТИ
В данном отделе даются лишь самые основные флористические
подразделения и то в возможно кратком изложении, так как
указания различных семейств, да к тому же в большинстве случаев
неизвестных для начинающих, обычно дают очень мало. Для
углубления познаний флористических особенностей земной поверх-
ности необходимы солидные сведения по систематике растений.
Некоторую помощь могут оказать краткие характеристики всех
семейств, упомянутых в тексте, которые даны в приложении (см.
стр. 401).
В основе флористических подразделений лежит генетический
принцип, т. е. общность происхождения. Чем меньше нарушений
испытывала данная область (в смысле климата, в смысле изменения
соотношений воды и суши и пр.), тем более целостна и однородна ее
флора. Для установления генезиса, конечно, особенно важны палео-
ботанические данные, но так как их по большей части не имеется,
то основой для флорютических подразделений служит современ-
ное распределение (ареалы) систематических единиц.
Флористический пр г нцип может в ряде случаев давать пред-
ставление и о ландшафтах; так, если сказать, что на данной терри-
тории господствует порядок хвойных или, в другом случае, семейство
злаков, то получаются соответствующие физиономические представ-
ления о хвойных лесах или о злаковых лугах или степях. Но так как
в большинстве случаев виды в пределах семейств могут представ-
лять и деревья, и кустарники, и травы, то семейства не могут дать
достаточных физиономических представлений.
Основными единицами при характеристике флори-
стических подразделений берут обычно семейства, изучая их аре-
алы. При этом сказывается, что одни семейства являются чисто
тропическими, отсутствуя во внетропических странах (например
семейства пальмовых, ано новых, мимозовых, эбеновых и т. п.),
другие встречаются исключительно во внетропических странах
(ивовые, первоцветные и др.); некоторые семейства встречаются
исключительно в Америке (кактусовые, канновые, бромелиевые),
в Австралии (цефалотовые —семейство насекомоядных и пр.) и т. д.
То же, но в еще большей степени, можно сказать и относительно
родов.
На флористических картах устанавливают обычно такие под-
325
разделения: области (иногда называемые «царствами»), подобласти,
провинции, округа. В настоящее время наиболее принято большин-
ством ботаников деление земной поверхности на шесть обла-
стей голарктическую, палеотропическую, неотропическую, австра-
лийскую , капскую и антарктическую (рис. 179). Седьмой является
океаническая область (иногда различают несколько океанических
областей).
I. Голарктическая область
Общая характериетш а. Наиболее крупная по территории область;
заш мает больше половины всей суши, а именно всю Европу с се-
вер ,ыми островами, почти всю Азию (за исключением Индостана,
Индокитая, Филиппинских островов, Малайского архипелага и
других прилегающих сюда островов), северную половину Африки
(до южной граш цы Сахары), почти всю Северную Америку (исклю-
чая большею часть Мексики) с Гренландией и другими северными
островами Это — в общем внетропическая область всего север-
ного полушария. Область представляет почти сплошной пояс,
далеко переходящий за полярный круг как в Евразии, так и в Аме-
рике. Все другие области не имеют такого поясного расположения,
но ограничены территориями в пределах некоторых градусов дол-
готы .
В основных своих чертах флора голарктической области, несмо-
тря на разъединен! е Атлантическим и Тихим океанами, настолько
близка, что нет никаких оснований предполагать различный ее
генез* с и сел неваться в существовании некогда соединения матери-
ков Евраз! и и Северной Америки. Подобное соединение, согласно
теории Вегенера, существовало до четвертичного периода,
когда произошло горизонтальное перемещение этих двух материков.
Однако огромная территория голарктической области н весьма
различные климатические условия в различных ее частях (Япония—
Новая Земля—Сахара—прерии Северной Америки и т д ), ко-
нечно, сказываются на современной флоре всех этих частей, но фло-
ра всей этей территории, как сказано, имеет общий генезис и проис-
ходит от арктотргтичной флоры1. Эта флора была богаче современ-
ной и имела много общих черт с современными флорами юга Северной
Америки и Восточной Азии; сна была представлена многочисленными
видами деревьев, как, например, секвойи, болотный кипарис (Та-
xodium distichum — рис. 132), криптомерия и другие хвойные,
тюльпанное дерево, магнолия, катальпа, кельрейтерия, орехи (Jug-
lans), многочисленные дубы, клены, тополя и пр.
В настоящее время в голарктической области богато представле-
ны семейства умеренных широт, именно буковые (Fagaceae), бере-
зовые (Betulaceae), ивовые (Sabcaceae), лютиковые (Ranunculaceae),
гвоздичные (Caryophyllaceae), маревые (Chenopodiaceae), крестоцвет-
ные (Cruciferae), розоцветцые (Rosaceae), зонтичные (Unibelhferae), ио-
1 Арктотретнчнои называется та флора третичного периода, которая была
распространена на территории современной голарктической области и в боль-
шей своей части была уничтожена ледником.
ричниковые (Scrophulariaceae), первоцветные (Primulaceae), сложно-
цветные (Compositae), осоковые (Сурегасеае), злаки (Gramineae) и др
Отсутствуют или почти отсутствуют тропические семейства па-
леотропической и неотропической областей, относящиеся к папорот-
никовым (Marattiaceae, Gleichen'aceae, Cyatheaceae и др.), голосе-
менным (Cycadaceae, Zamiaceae) и покрытосеменным (Cactaceae,
Myristicaceae, Anonaeeae, Nepenthaceae, Gesneriaceae, Bignoniaceae,
Cannaceae, Palmae и многие другие).
Огромное протяжение области,—на юг местами до 10° с. ш.
(Сахара), а на север до пределов возможного существования назем-
ных растений,—и большое разнообразие климатических условий вы-
зывает расчленение области на ряд подобластей; разные авторы ;а-
ют несколько иные подразделения, но наиболее удобным будет де-
ление на такие подобласти: арктическую, евро-сибир-
с к у щ, китайско-японскую, понт и че с к о-це п-
тральноазиатскую, средиземноморскую, севе-
роафр и канско -индийскую, ма ка р о н е з и й с к у ю,
североамериканскую атлантическую, севе-
роамериканскую подобласть прерий, северо-
американскую пацифическую — всего 10 подобла-
стей. Эти подобласти подразделяются на провинции, некоторые гз
последних будут указаны дальше.
Даем самую краткую характеристику указанных подобластей.
1. Арктическая подобласть. Занимает наиболее северные части
области и протягивается циркумполярно (циркум — латинское слово
«вокруг»); кроме того, занимает наиболее высокие части Скандина-
вии, Великобритании, Северного Урала и др. Здесь совершенно отсут-
ствуют все древесные семейства, характерные для голарктики. Типич-
ны кустарники— ивы и березка (Salix, ВеЫа)икустарнички из се-
мейства вересковых (Епсасеае) — Vaccimum, Cass.ope, Phyllcdcce.
Характерна куропаточья трава — дриада (Dryas, из семейства ро-
зоцветных). Наиболее хорошо представлены травянистые растения
таких семейств: осоковые (особенно род Carex), злаки (особенно роды
Роа, Agrostis), лютиковые, крестоцветные (особенно роды Draba,
Cochleana, Вгауа), розоцветные (Potentilla). Характерны различные
виды мхов и лишайников, которые развиваются настолько обильно,
что дают ландшафтные картины: моховые и лишайниковые тундры.
Однако обилие цветковых тоже местами дает ландшафтные типы,
дерновинные и луговинные тундры.
Флористически подобласть, как уже было отмечено выше, бед-
на. Вот некоторые цифры, касающиеся арктической Америки: Грен-
ландия— 390 видов, арктический Архипелаг—204 вида, аркти-
ческая часть Северной Америки —450 видов.
В части подобласти, относящейся к СССР, из ив характерны та-
кие виды: Salix polaris, S. reticulata, S. lapponum, S. glauca, S. rep-
tans, S. lanata, S pulchra и др. К востоку от реки Енисея — S. Ьа-
icalensis; одни ивы стелются по земле, образуя шпалерные формы,
другие дают сплошные заросли до 1 м высоты (на юге до 3 м). Березка
Betula папа, а на востоке от р. Енисея близко родственная В exi-
lis стелются по земле и образуют «ерники», не превышающие 1 м.
28
Из мхов характерны: Aulacomnium palustre, A. turgidum, Camp-
tothecium trichoides, Dicranum elongatum, Drepanccladus uncina-
tus, Hylocomium proliferum, Poly trichum strictum и некоторые сфа1-
нумы (S. Girgensohnii, S. squarrosum и др.).
Из лишайников особенно типичны: исландский лишайник (Се-
trana islandica), разные виды оленьего лишайника или ягеля (С1а-
donia rangifenna и др.), Peltigera aphtosa, Stereocaulon alpinurn,
иа севере средней части Сибири —Alectoria ochroleuca и т. д.
Дерновинные тундры богаты злаками, некоторые из которых
имеют дерновины: белоус (Nardus stricta), щучка (Deschampsia
flexucsa), овсяница (Festuca brevjfolia), несколько видов мятликов
(Роа alpina, Р. alpigena, Р. arctica) и др. На луговинках имеются
некоторые яркие двудольные: незабудка (Myosotis alpestris), лю-
тик (Ranunculus borealis), фиалка (Viola bi flora), некоторые крупки
(Draba glaciahs, D. hirta), валериана (Valeriana capitata) и др.
2. Евро-сибирская подобласть. Это—наибольшая по своей площа-
ди подобласть, занимающая всю среднюю Европу и Азию; большая
часть этой области находится в пределах СССР. В Европе она захо-
дит в северные части Пиренейского полуострова, занимает большую
часть Италии и северные части Балканского полуострова. В ланд-
шафтном отношении — это главным образом область лесов, область
массового развития древесных видов из семейств, наиболее
типичных для голарктической области в целом; из хвойных роды:
ель, пихта, сосна, лиственница (рис. 180); из покрытосеменных
роды: дуб, бук, клен, граб и др.
32*
На такой обширной территории намечается целый ряд флористи-
ческих провинций, причем в основном хвойные породы занимают
Сибирь и северные части подобласти в Европе (север Европейской
части СССР, Финляндию, Скандинавский полусстрсв). Леса же
лиственные —правильнее, широколиственные (основные древесные
породы: дуб, бук, клён, ясень, липа и др.), — развиты главным
«бразом в провинциях Западной Европы.
Подобласть занимает в СССР обширные территории, протяги-
ваясь от западных границ до Тихого океана и Охотского моря. Фло-
ристически она богаче, чем арктическая подобласть, но все ,ье для
•тдельных районов число видов колеблется между 800—1200.
Вот некоторые примеры:
Число видов Число ВИДОВ
Прибалтийская часть Лениград- Московская область . . л 1 100
екой области........... около 900 Полесье.................>> 1290
Восточная половина Калинин- Якутия .... . . » 1 190
ской области ..............около 930 Камчатка . . » 828
Северную границу подобласти (и вообще северную границу ле-
сов) образуют: на Кольском полуострове — северный вид березы
(Betula tortuosa), между Печорой и Уралом — ель и сибирская лист-
венница, в Сибири дз Хатанг^ — сибирская лиственница, восточ-
нее— даурская лиственница. На юге подобласти имеются такие
древесные породы: в Европейской части—дуб, в Западной Си-
бири— берёза, за Енисеем — берёза, сосна и лиственница, за
Байкалом главным образом даурская лиственница. Таким образом,
в Европейской части южная и северная границы лесов образо-
330
Рис. 182. Лес из восточной ели (Picea
orientahs) по берегу р. Аманаус (северный
склон Большого Кавказского хребта)
(фото В. Г. Сочеванова).
ваны очень различными экологически древесными породами, а на
востоке Сибири даурская лиственница проходит с самого севера
до самого юга.
Наиболее широко распространёнными породами в лесной области
СССР являются ель, обыкновенная сосна и сибирская пихта (рис.
121, 122). Ель не однородна на всём пространстве Европейской и
Азиатской частей, но представляет два очень близких вида: европей-
ская ель (Picea excelsa — рис. 181) и сибирская ель (Р. obovata).
Первая — в Европейской части (также в Западной Европе), кроме
северо-востока и Предуралья1, вторая — к востоку от территории,
занятой европейской елью. В Европейской части из хвойных к запа-
ду от р. Мезени встречаются лишь европейский ель и сосна, к восто-
ку от неё начинают появляться сибирские породы: лиственница,
сибирская ель и пихта и (ближе к Уралу) кедровая сосна.Таким об*/
разом, намечаются различные флористические провинции (см. в
конце карту растительности СССР). За Уралом в Западной Сибири
господствуют сибирские хвой-
ные породы: сибирская ель,
сибирская пихта, сибирсю я
лиственница, кедровая сосна
(особая западно-сибирш ая
провинция). К востоку от р.
Енисея, в области правь х
его притоков—провинция с
господством сибирской лист-
венницы, а в бассейне р. Jlei ы
и других рек, впадающих в
Северный Ледовитый океан,—
провинция с господстюм
даурской лиственницы. На
Охотском побережье и oct -
бен но на Дальнем Восто-
ке — аянская ель (Picea р-
zoensis), кедровник (Pinus
р и mi la) и др.
Всех хвойных в СССР1 2 3
имеется 61 вид, из которые
сосна имеет 14 видов, ель-
10, пихта — 8' лиственни-
ца— 6, можжевельник (Ju
niperus) — 21, микробиота
(Micro bi eta) — 1, тисс — 2.
Среди елей, кроме указан-
ных, отметим: на Кавказе —
восточную ель (Picea оrienta-
ils, рис. 182), в Средней
1 На северо-западе Европы (Карелия, Финляндия и др.) выделяют
особый вид ели — Picea fennica.
2 В нижеследующем перечне древесных пород имеется в виду весь СССР, а
не только евро-сибирская подобласть.
331
Азии — ель Шренка (Р. Schrenkiana) и тяньшанскую ель (Р. tian-
schanica—,рис. 183), на Дальнем Востоке—корейскую ель (Р. ко-
raiensis), на Сахалине—ель Глена (Р. Glehni), на Камчатке — ель
камчатскую (Р. kamtchatkensis); последние три имеют очень огра-
ниченное распространение.
Что касается di хт, то, кроме сибирской, укажем еще такие:
белую пихту (Abies alba) на западе Белоруссии и Украины, нордм^н-
нову пихту (A. nordmanniana) на Кавказе (рнс. 184), пихту Семено-
ва (A. Semenovi) в Средней Азии и несколько пихт на Дальнем Во-
стоке.
Из сосен большие пространства занимают обыкновенная сосна
и меньшие — кедровая сосна; остальные ограничены зьачительно
меньшими территориями. На Кавказе 6 видов сосны, из которых не-
которые занимают очень небольшие площади. Так, зльдарская сос-
на (Pinus eldanca) в Грузии ограничена площадью всего в 50 га,
пицундская сосна (Р. pithyusa) встречается лишь га Черноморском
побережье от Анапы до мыса Пицунда. Обе являются эндемиками.
В Крыму 3 вида сосен (см. ньже). На Дальнем Востоке —
4 вида, из которых большое распространение имеет кедровник, г ли
кедровый стланик (Р. punnla)—кустарник, идущий на запад до
Байкала и верхнего течения р. Лены. На Карпатах Р. cembra
и Р. mughus. Интересно, что в Средней Азин нет ни одного вида
сосны.
3.3 2
рис. 184 Лес из кавказской пихты (Abies
nordnianniana) по р- Домбай-Ульгеи иа
Северном Кавказе, 1 750 м над ур. м.
(фото В. Г, Сочеванова).
Среди лиственниц наибольшие плошади занимает сибирская
^Larix sibirica) и даурская лиственницы (L. dahurica); в западной
Украине (на Карпатах) про-
израстает европейская лист-
венница (L. europaea).
Основные хвойные по-
роды, занимающие обшир-
ные равнинные пространства
СССР, в более южных ча
стях подобласти сменяютс у
широколиственными, ОД1 акс
не на всей своей южной гра
нице, но только в Европы -
ской части Союза и на Даль-
нем Востоке.
В средних частях Евро-
пейской территории широ-
колиственные породы, при-
мешиваясь к хвойным, обр< -
зуют «смешанные леса»,
далее на юге — леса искш
чительно широколиственно
типа. Основной породой пос-
ледних является дуб (Quer-
cus robur); к нему прпмеп и-
вается остролистный ю ен
(Acer platanoides), ясень
(Fraxinus excelsior), ильм
(Ulmus scabra) и др. На
западе и юго-западе Украины
сюда присоединяется еще ряд
других западноевропейских
широколиственных видов (деревьев, кустарников и трав). По направ-
лению к востоку дубовые леса беднеют, ясень не идет восточнее
Волги, а дуб доходит лишь до Урала, и в Сибири дубовые леса от-
сутствуют. Широколиственные породы вновь появляются на Даль-
нем Востоке уже в пределах китайско-японской подобласти (см.
стр. 336). В Сибири из всех широколиственных пород встречается
лишь липа (в западносибирской низменности, в предгорьях Алтая
и еще в нескольких пунктах).
Имеющиеся данные говорят о том, что в третичное время широко-
лиственные породы Европейской части и Дальнего Востока смыка-
лись, и потому липа на Алтае должна считаться реликтом.
На пространстве Европейской части Союза можно очень хорошо
наблюдать большое влияние на юго-западе западноевропейских
элементов, а на северо-востоке—сибирских. Первые все более и
более теряют свое значение к востоку и северо-востоку, а послед-
ние — к западу и юго-западу. Помимо древесных пород, то же на^-
блюдается и для ряда кустарников и для многочисленных травяй|г
стих видов
Широколиственные породы в СССР богаче всего представлены
на Кавказе и на Дальнем Востоке.
Дуб имеет в СССР 19 видев, из которых 17 встречаются на Кав-
казе, а 2 —на Дальнем Всстске. Салым распространенный вгдем,
как указано, является Quercus robur, на юго-западе Украины
к нему присоединяются Q. petraea и Q. pubescens, ко всем этим
видам на Кавказе присоединяется еще 14 видов (Q. macranthcra,
Q. Hartwisiana, Q. ibenca, Q. pentea и др.).
Кленов в СССР 24 вида, причем на Кавказе 10 видов (Acer Traut-
vetten, A. hyrcanum, A. ibericvm и др.), па Дальнем Всстске 7 ви-
дов (клен желтый — A. ukurunduense, клен приречный — A. gin-
nala, клен зеленокорый — A. tegmentesum, клен маньчи%урск1 й—
A. manshuricum и др.); в Средней Ал и —7 видов. В Европейской
части, кроме остролистного клена, встречается еще полевей ктен
(A. campestre), татарский (A. tatancum) и на юго-западе Украины
явор (A. pseudoplatanus).
Среди лгп наиболее распространена обыкновенная липа (Tilia
cordata). На юго-западе Украины встречаются еще липа серебри-
стая (Т. argentea) и лп га к[.угнол стьая (Т. granchfcha). На Кав-
казе 7 видов, на Дальнем Всстске—липа маньчжурская (Т. manshu-
rica) и липа амурская (Т. amurcnsis).
Ясени представлены мегышм чп слом видев. Креме обыкновен-
ного ясеня, в Крыму и на Кавказе растет еще Frax>nus схусагра;
несколько видев встречается еде в Средней ?зиь; ьа Дальнем Во-
стоке 2 вида (F. manshunca и F. rhynehephy 11а).
Род Ulmus представлен 12 видам! ; в Европейской части иль;»
(Ulmus s^abra), вяз (U. laevs), бересты (U. fchacea и U, subcrcsa)
и др., 5 ыд^в в Крыму и на Кавказе, другие в Средней Азии,
в Восточней Сибйри и на Дальнем Всстске.
Граб имеет в СССР всего 4 вида. На юго-западе Европейской va-
сти (в Белоруссии и на западе Украп гы) встречается сбыкневеы ый
граб — СаГ| inus betulus, на Кавказе — 3 ei да. На Дальнем Востоке
«ссбый вид — С. cordata.
Бук в Европейской части встречается лппьь в верхнем Придне-
стровье на западе Украины и в Бессарабии (Fcgus silvat са),ьа Кав-
казе — F. orientals, в Крыму переходный вид между этпд.и двумя —
F. taunca.
Интересно отметить, что ряд приведенных листвеьп ых пород
совершенно отсутствует в Средней Азии, а именно: дуб, липа, граб,
бук.
Хвойные и широколиственные породы после порубок и гежаров
обычно сменяются более светолюб! еым1 породами, особенно бе-
резой и осиной, и эти последы е на больших территориях замени-
ли в настоящее время основные породы.
Объединение в одну подобласть территорий с хвойными и широ-
колиственными породами обусловлено геьезьсом фтс[:ы — лоса
последнего типа в третичное время занимали гораздо большие тер-
ритории, покрывая и юг Сибири и смыкая западноевропейские леса
с дальневосточными.
334
Что касается географических элемент о’в флоры, тс
здесь основную роль играют элементы бореальный, сред-
неевропейский и атла н»и чески й. Последний наи-
более полно представлен в приатлантических часп х подобласти
(особенно во Франции, Англии, отчасти в Прибалтийской провин-
ции): Myrca, Genista anghca, вереск—Erica cmerea, Ulex и др.
В наиболее южных частях подобласти в Западной Е р пе (на
Пиренейском и Апеннинском полуостровах) встречаются уже такие
южные виды, как благородный каштан и вечнозеленый дуб. Кроме
того, в южных частях имеет место проникновение с редиз ем-
номорского элемента.
Вследствие большего количества осадков во многих частях под-
области (сссбенно на севере Европейской части СССР й в Западной
Сибири) обильно представлены болота, в общем с крайне однород-
ным флористнчесю м составом; на низинных болотах — тростник
(Phragmites), виды осск (Carex) и др., на моховых болотах—виды
мха (Skhcgnum), осоки и ряд кустарничков из семейства вересковых
(багульник, андромеда, Кассандра, голубика). На севере, кроме то-
го, морошка (Rubus chamaemorus— рис. 185), поленика (R. arcti-
cus), водяника (Empetrum nigrum).
Довольно неоднородная флора подобласти, помимо разнообра-
зия современных климатических условий, является наследием лед-
никового периода с его оледенением, уничтои<ением арктотретичной
флоры на обширных территориях и с дальнейшими миграциями.
365
Естественная растительность в более южных частях подобласти
подверглась очень большим изменениям под влиянием культуры.
Это особенно сказалось в Западной Европе, где местами совсем не
сохранилось дикой растительности. Вместо естественной раститель-
ности широко распространены многочисленные культуры, в север-
ных частях — главным образом рожь, овёс и картофель, в более
южных —пшеница, ячмень, кукуруза, а в наиболее южных частях
в западной Европе и на Кавказе также табак, виноград и благород-
ный каптан.
Особо следует указать на провинцию евро-сибирских
высоких гор, которая отдельными округами представлена
в Пиренеях, Альпах, на Апеннинах, Балканах, Кавказе, Тянь-
Шане, на Гималаях и др. Флора здесь резко отличается от флоры
равнин и представляет смесь различных элементов — арктического
и средиземноморского, арктического и центральноазиатского и др.
В разных горных странах комбинации элементов, конечно, различ-
ны. Особый отпечаток придаёт резко выраженный эндемизм (как
консервативный, так и прогрессивный).
На Кавказе, как и вообще на высоких горах, различают альпий-
скую и субальпийскую флору; первая представлена низкорослыми
травами, вторая — высокорослыми. Среди последних такие высокие
травы, как кавказская скабиоза (Scabiosa caucasica), живокость
(Delphinium), водосбор (Aquilegia), валериана (Valeriana), телеки*
(Telekia speciosa), крестовник (Senecio) и др. Собственно альпийская
флора состоит из низкорослых видов таких родов, как колокольчик,
горечавка (Gentiana), первоцвет (Primula), незабудка (Myosotis),
фиалка (Viola) и др. Имеется много эндемиков (о некоторых других
особенностях см. выше).
Китайско-японская подобласть. Занимает югс-восток Азии и
включает Японию, южную часть Сахалина, Уссурийский край и
Приамурье, Маньчжурию, Корею, юго-восточную часть Китая, север-
ную часть Индо-Китая и восточные Гималаи. На юге она доходит в
некоторых частях до тропика Рака и переходит в палеотропическую
область.
Подобласть очень богата осадками: в год выпадает не менее 50
с,«, а местами даже более 150 см. Дождливое время года — лето.
На юге зимы очень мягки, на севере же они достаточно суровы.
Подобласть имеет довольно однообразную флору, чю стоит в
связи с историческими причинами: вся эта страна не подвергалась
оледенению в ледниковый период и, таким образом, растительность
могла свободно развиваться и сохранять свои прежние флористи-
ческие особенности. В последнем отношении данная подобласть име-
ет известное сходство с североамериканскими подобластями (см. ни-
же) и отличается от рассмотренной евро-сибирской. Во всяком слу-
чае флора китайско-японской подобласти является прямой преем-
ницей флоры третичной и очень богата.
Однако протяжённость подобласти приблизительно на 30° ши-
роты обусловливает значительные флористические особенности в раз-
личных её частях, особенно в северных (в пределах СССР) и наибо-
лее южных (об основных древесных породах см. выше, стр. 224).
336
185. Жень-шень (Panax ginseng);
Ряс.
растение отцвело—плоды (по Кёрнеру).
Более других разнообразна и богата флора некоторых районов
Гималаев — много вечнозелёных деревьев: магнолии, древовидные
рододендроны, камелии. Это одно из наиболее интересных мест на
земной поверхности. Здесь сталкиваются западные средиземномор-
ские элементы и восточноазиатские (Cornelia, Aucuba, Aralia, Deut-
zia и др.); имеются американские роды. Кроме того, характерны
древовидные папоротники, а также кедр гималайский (Cedrus deo-
dara).
Травянистая флора подобласти также очень богата. Нужно упо-
мянуть знаменитый жень-
шень (Panax ginseng — рис.
186), который издавна ис-
пользуется китайцами в ме-
дицине.
Здесь, в восточной Азии,
мы имеем крайне посте-
пенный переход от
субтропической флоры лесов
к умеренной, и леса субтро-
пические непосредственно
смыкаются с лесами умерен-
ными (уже голарктической
подобласти). В данном отно-
шении— это одно из харак-
терных мест земного щара.
Нельзя не напомнить,
что в юго-западном Китае
ещё произрастает в диком
виде такое «живое ископае-
мое)). как гинкго (Ginkgo
biloba), единственный ныне
живущий представитель из
вымершего порядка Ginkgoa-
les (рис. 187).
В пределы СССР китай-
ско-японская подобласть за-
ходит лишь в южные час-
ти Дальнего Востока. Выше
уже говорилось о богатстве
Дальнего Востока широко-
лиственными породами и о
том, что здесь имеет место
реликтовая флора третичного
времени. Кроме кр> пных
деревьев, здесь немало деревьев второй величины и кустарников,
образующих подлесок.
Монгольский дуб, маньчжурский ясень, бархатное дерево, амур-
ская липа и др. уже были указаны. В подлеске — амурская сирень
(Syringa amurensis), маньчжурская лещина (Corylus manshurica),
(ралия (Aralia manshurica), маакия (Maackia amurensis — сем.
Геогр*фтя раств»*
рис. 187. Гинкго (Ginkgo biloba)—«жи-
вое ископаемое»; ветвь с мясистыми
семенами и с листьями, имеющими ха-
рактерное дихотомическое жилкование.
мотыльковых) и многие другие. Очень характерны лианы, отсут-
ствующие в широколиственных лесах Европы: амурский виноград
(Vitis amurensis), лимонник (Schizandra chinensis — сем. магно-
лиевых), актинидия, или «киш-
миш» (Actinidia kolomicta), а
в Юишо-уссурийском крае и
ещё другой очень крупный
вид —A. arguta. В числе эпи-
фитов — папоротник Polypodi-
um lineare. Всё это (не говоря
о многочисленных характерных
травянистых видах) говорит
о древности этой флоры. До-
бавим, что на гольцах Сихотэ-
Алиня встречается эндемичный
род Microbiota (М. decussata).
Численность флоры такова:
Китай — около 20 000 видов,
Япония — более 5 500, Ко-
рея — более 2 000, Маньчжу-
рия — 1 660.
В настоящее время во мно-
гих местах, особенно в Китае и
Японии, обширные пространст-
ва лишены своей естественной
растительности и заняты куль-
турными площадями. Кроме
более северных культур, здесь
возделывается особенно рис и
чайный куст; чайные культуры, как известно, здесь имеют большую
древность.
Китайско-японская подобласть важна для нас как один из оча-
гов разнообразия культурных растений; подобных очагов в настоя-
щее время установлено восемь, находящихся в разных частях земной
поверхности. В нашей подобласти находится «восточноазиатский»
очаг в юго-западных горных частях Китая; здесь, как установлено,
имеют свою родину такие культурные растения, как просо, гречи-
ха, некоторые клубнеплоды и корнеплоды, ряд плодовых деревьев—
абрикос, персик, слива, некоторые цитрусовые, многие лекарствен-
ные растения. Япония и Китай — родина многочисленных декора-
тивных растений.
4. Понтическо-цеитральноазиатская подобласть. Занимает боль-
шую территорию, простирающуюся полосой с востока Европы по-
чти через всю Азию на протяжении 100° долготы (приблизительно
от 30 до 130° по Гриничу). На западе она занимает равнины Вен-
грии и юга Европейской части СССР; в Азии — степные простран-
ства западной Сибири, внутренние части малоазиатского полуостро-
ва, Иран, среднеазиатские части СССР, Монголию и Тибет.
В общем для подобласти характерно почти полнее отсутствие
лесов и преимущественное развитие травянистых и полукустарнн-
338
новых ценозов. Флористически она выделяется присутствием не-
которых семейств и родов. Типичны маревые (Chenopcdiaceae), та-
марисковые (Tamaricaceae), парнолнстиые (Zygophyllaceae), а из ро-
дов двудольных: астрагал (Astragalus), смолевка (Silene), калли-
гонум (Calligonum), солодка (Glycyrrhiza), кузгния (Ccusinia), ва-
силек (Centaurea), из однодольных: ковыль (Stipa), лук (Allium),
тюльпан (Tulipa), петушки (Iris) и др.
В общем все эти растения являются сухолюбивыми (ксерофи-
тами) в соответствии с климатически ми условиями. Осадков здесь
выпадает иногда крайне мало, характерна также большая ампли-
туда между крайними температурами. Господствует флора пустынь,
полупустынь и степей.
В пределах подобласти имеется несколько географических эле-
ментов; понтический, североказахстанский,
ту ра некий, переднеазиатский и др., что указы-
вает на флористическую её неоднородность и на возможность под-
разделения на флористические провинции. Последних также имеется
несколько.
Провинции, растительность которых представлена степями, вы-
деляются своим сравнительно богатым флористическим составом.
Так, в Херсонщине насчитывается до 1 500видов, в Донском крае —
1 600 видов, в тд время как пустынные части Средней Азии имеют
около 600, Тибет—700 видов. Впрочем, в Малой Азии имеется не
менее 7 000 видов (однако нужно иметь в виду, что периферические
части Малой Азии входят уже в средиземноморскую подобласть).
Понтическая или южнорусская степная провинция (преобладает
понтический элемент) в связи с разнообразием климатических и
почвенных условий неоднородна по флоре на всей своей территории.
Однако такие роды, как ковыль (Stipa), овсяница (Festuca), костёр
(Bromus), келерия (Koeleria), а из двудольных —шаг фей (Salvia),
василёк (Centaurea), серпуха (Serratula), вероника (Veronica), адо-
нис (Adonis), астрагал (Astragalus) и др., представлены по всей
провинции.
Что касается видов, то лишь сравнительно небольшое число
их распространено по всей провинции (Festuca sulcata, Stipa capil-
ia ta, Koeleria gracilis и др.), большинство же тяготеет то к более
северным, то к более южным частям провинции. Так, из ковылей,
столь характерных вообще для данной провинции, в более северных
частях особенно развиты S. Joannis и S. stenephyПа, в более
южных — S. Lessingiana и S. ucrainica, за Волгой и в Западной
Сибири — S. rubens. Для более северных частей провинций укажем
ещё такие виды: шалфей луговой — SGvia pratensis, таволжанку —
Filipendula hexapetala, румянку' — Echium rubrum, мытник —
Pedicularis comosa, незабудку—Myosctis silvatica, ряд клеверов
и т. д. В более южных частях появляются многие луковичные расте-
ш я, представленные в северных частях очень слабо, именно виды
тюльпанов (Tulipa), луков (Allium), птицемлечников (Ornithcgalum),
гусятников (G.'gea) и др.; из двудольных для более южных
ч стей ”'рактсрны роды: астрагал (много видов), кермек, шал-
фей и
22* 330
Помимо флористического богатства отдельных частей провинции,
велика и видовая насыщенность, т. е. число видов на определённую
площадь. Так, в северных степях на 1 лР иногда находится до 80
видов; в южных степях число их постепенно падает (в степях Аска-
нии-Нова всего 12 видев).
Провинция западно-сибирских степей близка к южнорусской,
но отличается рядом особенностей. Прежде всего, ряд степных
видов (европейских) здесь выпадает (Sjlvia nut ins, Stichys recta.
Echium rubrum, Hyacinthella leucophaea, TrifJium m^ntanum и
т. д.), но, с другой стороны, значительную роль играют сибирские
элементы, отсутствующие или редкие в южнорусской провинции
(виды полыней, ковыль Stipa rubens, Castilleja pallida, Seseli hypo-
marathrum, обильное развитие Ave nj strum deser to rum и т. д.). В при-
енисейских степях признаки отличия ещё более усиливаются. Фло-
ристическое богатство западносибирских степей сравнительно с
европейскими менее велико.
Туранская провинция, или провинция среднеазиатских пустынь
(преобладает тура некий элемент), имеет ряд характерных родов
как древесных и кустарниковых, так и травянистых; все они носят
обычно отпечаток крайней ксероморфности. Деревья —саксаул
(белый саксаул — Haloxylon persicum — на песках, чёрный сак-
саул — Н. aphyllum — на глинистых местах). В глинистых пусты-
нях крайне типичен ряд полыней (Artemisia), анабазис (Anabasis),
лебеда (Atriplex); в солончаковых пустынях—семейства: солян-
ковые (S ilsclaceae), кермековые (Plumlaginaceae), тамарисковые
(Tamaricaceae), гвоздичные (Caryophyllaceae) и др.; в песчаных
пустынях, кроме саксаула, каллигенум (Calligcnum дет 30 видов),
AmiiKdendron, астрагал (до 20 видов) и многие другие.
Особую провинцию представляет Памир, плоскогорье на
высоте 3 000—4 000 лк Эго — холодная пустыня с сухим и холодным
климатом. Здесь насчитывается около 500 видов растений. Очень
характерны такие виды: Acantholimon diapensioides из семейства
Plumbaginaceae (в виде подушки), полынь —Artemisia Scorniako-
vii; особенную роль в ландшафте играет терескен — Eurotia ceratoi-
des —призёмистый полукустарник; из злаков очень характерен
восточный ковыль (Stipa orientulis), а также мятлик (Роа attenu.ita)
и типчак. Из бобовых нужно отметить род Oxytropis, а из других
двудольных—семейство крестоцветных, некоторые роды которого
играют очень большую роль. Видовая насыщенность ничтожна;
на каменистых почвах нередко можно встретить I—4 вида, на
более мелкозёмистых — не более 10 видов.
На увлажнённых почвах встречаются различные осоки (Carex —
ряд видов), кобрезия (С bresia), из злаков: Festuca, Роа, Alopecurus
mucromtus, Hcrdeum secaL’num и ряд других.
Особая провинция—пустыня Гоби; она имеет целый ряд флори-
стических особенностей. Очень разнообразна флора армянско-
иранской провинции, куда входит центральная Малая Азия, Арме-
ния и Иран (преобладает переднеазиатский элемент). Эго сильно
расчленённая страна с разнообразными экологическими условиями,
где всё же преобладают условия, благоприятные для ксерофильной
340
флоры. Так как периферия Малой Азии относится уже к средиземно-
морской подобласти, то во флоре заметно влияние средиземноморского
элемента: древовидный можжевельник (Juniperus excelsa), кевовое
дерево (Pistacia mutica) и др.
Понтическо-центральноазиатская подобласть во Многих местах
в настоящее время совершенно лишена дикой растительности, осо-
бенно в степных провинциях, а также местами в Малой Азии; в
последней культурные воздействия имели место с отдалённейших
времён истории человека. ‘
Рассматриваемая подобласть сыграла огромную роль в куль-
турной истории человека; она явилась также родиной многочислен-
ных растений, введённых человеком в культуру. На юге подобласти
имеются два основных оч^га происхождения этих растений: один -
среднеазиатский в горах Средней Азии, другой —переднеазиатский
в гористых странах Малей Азии и Ирана. Первый центр дал ряд
пшениц (мягкая, карликовая и др.), основные бобовые—горох,
чечевицу, конские бобы, чину, ряд масличных растений, хлопчатник-
гузу и др. Второй (переднеазиатский) дал целых девять видов пше-
ницы, режь, ряд плодовых деревьев —алычу, черешню, миндаль,
айву и др., кроме того, виноград. Из сказанного совершенно ясно
значение флоры Передней и Средней Азии. Кроме Азии, пшеница
(твёрдая) получила своё начато в Абиссинии (см. ниже).
5. Средиземноморская подобласть. Занимает все страны, лежащие
по берегам Средиземного меря: большую часть Пиренейского полу-
острова, южную полосу Франции, прибрежные части Апеннинского
полуострова, прибрежные части Балканского полуострова (всю
почти Грецию), южный берег Крыма, большую часть Кавказа,
побережье М и ой Азии, Сирию, Палестину, побережья Египта,
Алжира и Марокко; кроме того, все острова Средиземного моря
(Бтлеарские, Сардинию, Корсику, Сицилию и др.). Таким обра-
зом, подобласть образует кольцо вокруг всего Сре-
диземного моря, причем можно отличать северное,
более расчленённое полукольцо и южное —слабо расчленённое.
Подобласть характеризуется сухим и жарким летом и мягкой
зимой с большим или меньшим количеством осадков; к востоку
зимы становятся более суровыми. Флора является в общем довольно
однородной; господствует географический средиземномор-
ской элемент. Имеются также и арктотретичные элементы, но
можно думать, что современная флора произошла из тропической
в результате всё увеличивающейся сухости. Вообще для подобласти
характерно: 1) присутствие многих вечнозелёных видов, 2) вегета-
ция в течение всего года (зимой различные цветущие растения),
3) присутствие многочисленных южных культурных растений.
По ряду фактов, по тем связям, которые имеются у средиземно-
морской флоры даже с очень отдаленными странами, можно думать,
что эта флора развивалась спокойно с давних времен (связи с Кап-
ской областью, Абиссинией, Азией, даже с Австралией).
Флористически подобласть характеризуется рядом сосен (Pinus
halepensis, Р. pinaster, Р. pinea) (пис. 188), такими вечиозел ё-
н ы ми, как вечнозеленые дубы (Quercus ilex, Q. coccifera—рнс. 101),
341
маслина (Olea europaea), лавр, мирта (Myrtus communis), олеандр
(Nerium oleander), виды Ruscus (иглица) с жёсткими кладодиями
(кладодий—это листовидный плоский стебель), земляничное де-
рево (Arbutus unedo). Немало деревьев и с опадающими листьями:
благородный каштан (Castanea vesca), платан (Platanus orientalis),
характерный кустарник V(tex agnus castus и др.
КакАи вообще в засушливых областях, здесь весной развивается
много луковичных и клубневых (большинство из них весенние
эфемеры) из семейств лилейных (тюльпаны, Narthecium), ирисовых
(шафраны — Crocus, ирисы), амариллисовых (нарциссы, подснеж-
ники — Gilanthus —рис. 109). Летом характерны многочисленные
губоцветные, богатые эфирными маслами, зонтичные, сложно-
цветные и др.
Помимо лесов с хвойными и вечнозелёными деревьями, осо-
бенно по сухим местам берегов морей, развиваются своеобразные
заросли ксерофильных кустарников, называемые «маквис», «гарига»
и др. (более подробно см. выше, стр. 206).
Как ни однородна средиземноморская подобласть, но она рас-
падается на две провинции: западно-средиземноморскую и восточно-
средиземноморскую; первая занимает все западное Средиземноморье
к западу от Апеннинского полуострова, включая западное побе-
режье последнего, вторая—восточное Средиземноморье к востоку
от Апеннинского полуострова.
Для западной провинции характерны деревья: Pipits pinaster,
вечнозелёный дуб (Querdus ilex), пальма (Chamaerops humilis) —
342
это единственная пальма в Европе (юг Испании, Си-
цилия, Балеарские острова; также Сев. Африка); из кустарников —
виды ладанника (Cistus), вереска (Erica), нежника (Helianfhemum);
из трав—виды льнянки (Linaria) и др. В горных лесах Атласа (Север-
ная Африка) атласский кедр (Cedrus atlantica —рис. 189) и нуми-
дийская пихта (Abies numidica).
Ряс. 185. Группа атласских кедров в горах Атласа (на севере Африки)
(из Шимпера).
Восточная провинция лишена указанных характерных деревьев
западней провинции; приведённые кустарники представлены значи-
тельно меньшим числом видов, зато в большем количестве появля-
ются роды, характерные для степей: Astragalus, Teucrium, Phlomis,
Allium, Crocus и др. Отметим такие хвойные, как пихты (рис.
128) — Abies cephalonica (Греция) и A. cilicica (Малая Азия), кипа-
рис (Cupressus), сосны (Pinus pallasiana и Р. рейсе), для горных
лесов Ливана особый вид кедра —ливанский кедр (Cedrus Libani).
На Южном берегу Крыма, тоже относящегося к средиземномор-
ской подобласти, обнаруживается угасание средиземноморских черт.
Здесь средиземноморская флора развита от мыса Айя (на западе)
и тянется до Алушты и немного восточнее, занимая южный склон гор
приблизительно до высоты 300—320 м. В этом поясе господствую-
щие породы (особенно в прошлом): древовидный можжевельник
(Juniperus excelsa) и пушистый дуб (Quercus р u be see ns). Встречается
также крымская сосна (Pinus pallasiana —рис. 190), господствую-
щая, однако, в более высоком поясе, а кроме того, кевовое (терпен-
тиновое) дерево —Pistacia mutica. Из растений подлеска: дикий
343
Рис. 191. Иглица (Ruscus)—часть стеб-
[Я с листовидными кладодиями (плоские
стебли) и с цветкам^ на них.
жасмин (Jasminum fruticans),
ладанник (Cistus), иглица (Rm-
scus—рис. 191), сумах (Rhus
canaria), скомпия (Cotinus cog-
g}gna) и др. Вечнозеленые,
столь характерные для Среди-
земноморья, почти отсутству-
ют; можно назвать лишь зем-
ляничное дерево (Arbutus
!ndr chne—рис. 190), иглицу
(Ruschs pcnticus), ладанник
(Cistus tJ uncus) и лиану —
nj ющ (HecLra helix). Кроме
крымской сосны, занимающей
пояс выше 250 м, интересна
эндемичная сосна Станкевича
(Pmus Stankeviczi), встречаю-
щаяся лишь на мысе Айя и б л иг
Судака. Она близко родственна
кавказской пицундской сосне.
Средиземноморский харак-
тер флоры Южного берега Кры-
ма усиливается еще присутстви-
ем ряда понтических элементов
(средиземноморского происхож-
дения).
344
Ряд крымских элементов встречается и на Кавказе в Новорос-
сийском районе, но непосредственной связи между этими двумя
районами нет, и, как можно думать, в прошлом связь была мере:
понтическую сушу, соединявшую указанные районы в третичное
время.
Особенно хорошо средиземноморская флора в СССР представлена
на Кавказе — на Черноморском побережье и на западном берегу
Каспийского моря.
О богатстве Кавказа древесными видами уже было сказано
Особенно богаты леса Колхиды в западном Закавказье (низмен-
ность р. Риона и прилегающее побережье) и Талыша (Ленко-
ранская низменность на юго-востоке Кавказа). Здесь встреча-
ются реликтовые, третичные виды, и в этом отношении данные
районы можно сравнить с Уссурийским краем, Японией и вос-
точными штатами Северной Америки, сохранившими элементы тре-
тичной флоры.
Для Колхиды укажем такие деревья: лапину (Pterocarya fraxi
niftha из сем. ореховых), каштан (Gstanea vesca), дзелкву (Ztlkova
сгепЛа), гранатник (Punica gran^tum) и др. Из вечнозеленых расте-
ний. остролист (Ilex aquif hum), рододендрон (Rhododendron pon-
ticum), иглицы (виды Ruscus), самшит (Buxus sempervirens). Вечно-
зеленая лиана-плющ (Hedera colchica), а также и ряд других
лиан говорят о близости к субтропикам На это же указывает па-
поротник Pclypodium vulgc re, встречающийся здесь уже в виде
эпифита. Отметим еще очень крупный папоротник на болотах —
Osmund.i reghlis.
Ф,.ора Талыша несколько отлична, но носит тот же третичный
характер. Очень характерно эндемичное железное дерево (Рагго-
tia persica); также встречаем лапину, дзелкву, вечнозеленые кустар-
ники — Ruscus и D.in. e, много лиан. Интересно, что в числе эпифитов
встречаются некоторые цветковые растения (не только лишайники,
мхи и папоротники).
Некоторые авторы причисляют Кавказ вместе с Балканами,
Апеннинами и др. к евро-сибирсксй подобласти. ч
Влияние человека сказалось с особой силой на дикой флоре
средиземноморских стран, так как последние явились ареной и
культурной и разр¥шительной деятельности человека с древнейших.
времен человеческой истории (Египет, Греция, Рим). Многие тер-
ритории целиком заполнены культурными растениями. Возможность
культур во многих местах усиливается искусственным орошением
с гор.
Главнейшими культурами являются: пшеница, маис, табак,
виноград, различные овощи, местами рис и хлопчатник; из деревь-
ев: маслина, фиговые деревья (инжир), цитрусовые, гранаты,
фисташки. Многочисленны культивируемые декоративные растения:
пальмы, кедры, акации, эвкалипты, тюльпанное дерево, кипарисы и
мноте другие; они придают Средиземноморью особый колорит. Тан_
в Крыму искусственные насаждения на южном берегу создаю!
ландшафты, совершенно не свойственные его дикой природе (кипа-
рисы — рис. 192, магнолии, платаны и т. п.).
Средиземноморье является не только колыбелью человеческой
ку»туры, здесь находится и один из очагов происхождения культур-
ных растений — «средиземноморский» очаг: здесь имеют свою родину
маслина, рожковое дерево (Cent nia siliqua), целый ряд овощных
растений и др. Масли-
н а настолько характерна
тля Средиземноморья, что
его иногда называют «стра-
ной маслины» (см. выше,
ареал маслины —рис. 99).
6. Североафриканско-ннднй-
ская пустынная подобласть.
Эго подобласть пустынь или
полупустынь; она занимает
асю Северную Африку к
:еверу от 15—18° с. ш., за
исключением присредиземно-
морской полосы, почти весь
Аравийский полуостров, Ме-
сопотамию, прибрежные ча-
сти Ирана и Индии, пустыни
по р. Инду и на Пенджабе.
В эту подобласть входит
величайшая пустыня —Са-
хара, а также Нубийская и
Аравийская. Пустынность
подобласти вызвана крайне
неблагоприятными климати-
ческими условиями; местами
количество осадков достигает
mo л „ , « ЛЛ -Два 20 см в год, а средние
Ряс. 192. Аллея пирамидальных кипарисов ’ г
(Cupressus sempervirens var. pyiamidalis) в температуры ИЮЛЯ ДОХОДЯТ
Карасане (Южный берег Крыма) (фото ДО 36° и даже более. Очевид-
В. В. Алехина). но, что при таких климати-
ческих условиях флора долж-
н1 быть крайне скудной; так, для Центральной Сахары приводится
. ишь 300 видов, а в некоторых её районах число видов падает до
2—3 десятков. Для Аравии в целом приводится более 2000 видов,
но число видов пустынных частей Аравии, несомненно, очень невели-
ко (точных данных4 нет).
Сахара—геологически молодая территория, ещё в третичное
время была под морем, а затем заселилась и с юга и с севера, являясь
до известной степени как бы переходом между Голарктикой и Южной
Африкой. Об основных чертах флоры подобласти было сказано
выше.
Культуры в данной подобласти развиты лишь воазисах и
по долинам рек, особенно по р. Нилу в Египте и по рр. Тигру и
Ефрату в Месопотамии. По р. Нилу возделывают многочисленные
1 ультурные растения —рис, сахарный тростник, хлопчатник и
многие другие. В оазисах — финиковая пальма (рис. 170),
346
акации, в Аравии—Borassus, кофе (Coffea arabica —рис. 193); Acacia
arabica даёт гуммиарабик.
По р. Нилу необходимо указать «священный» лотос египтян
(Nymphaea lotos). Индийский «священный» лотос относится к другому
роду Nelumbiuna — N. nuciferuna и распространен в Индии, на
Цейлоне, в Малайском архипелаге, в Иране и др. Интересно нахож-
Рис. 193. Ветвь кофейного дерева (Coffea arabica), с цвет-
ками и плодами (из^Веттштейна)
дение его в СССР —в низовьях р*р. Куры и Аракса, в дельте
р. Волги (рис. 194) и на Дальнем Востоке.
Если средиземноморскую подобласть можно назвать страной
маслины, то данную подобласть можно назвать страной фин id-
le о в о й пальмы. Однако в настоящее время финиковая пальма
в диком состоянии неизвестна (впрочем, вопрос этот ещё недостаточно
выяснен), н можно думать, что она произошла из близких видов
(аравийско-индийского или абиссинского). Один экземпляр пальмы
дает в год 100—250 кг плодов.
7. Макаронезийская переходная подобласть1. Эта подобласть
имеет очень небольшую территорию, занимает острова Атлантиче-
ского океана на северо-западе Африки: Канарские, Азорские, Зе-
леного Мыса и Мадейру. По флоре — это переходная ступень между
средиземноморской и африканско-тропической флорой. Основанием
для выделения этой флоры в особую подобласть является то, что
хотя она обнаруживает многие связи с третичной флорой Европы,
но имеет вполне самостоятельное развитие. Последнее сказывается
Макаронезия от греческих слов «макарос»—счастливый и «незсс'>— ветров.
347
Рис. 195. Канарская финиковая пальма
(Phoenix Jubae) на о Тенерифе (фоте
О Лаутербаха из Друде).
«’’резком эндемизме (консервативном и прогрессивном), который
характерен для осн ровов^ подобласти.
Многие виды и роды являются средиземноморскими: земля-
ничное дерево, лавр, Tamils; с другой стороны, присутствуют неко-
торые тропические афри-
канские роды. Очень типич-
ны ка на река я финикова я
пальма (Phoenix Jubae —
рис. 195) и знаменитое дра-
коновое дерево из лилейных
(Dracaena draco — рис 196).
Длительная изолированность
сказалась в выработке свое-
образных морфологических
фоом у таких столь обычных
в Голирктике редев, как мо-
лочай (Euphorbia), осот (Son-
chus), румянка (Echium),
живучка (Sempervivum).
В равнинных местах ха-
рактерны небольшие кустар-
ники и суккуленты, в го-
рах — вечнозеленые леса с
господством Канарского лав-
ра (Laurus canariensis), с
рядом вечнозеленых видов в
подлеске, с многочисленными
папоротниками и мхами.
Это — леса лаврового типа
(см выше). Выше — леса из
особой Канарской сссны (Pi-
nus canariensis), а еще выше --различные кустарники (Cistus,
Daphne и пр ).
Макаронезия в общем представляет очень хороший пример
древних островов материкового типа со своеобразной флорой, бо-
гатой эндемнками. Упомянем Cytisus nubigens (ракитник) — энде-
мик, ограниченный в своем распространении лишь Тенерифским
ником.
Что касается числа видов, то на Канарских островах насчиты-
вается 1 552 вида, на острове Мадейре—648 (без папоротников)
на Азорских островах- 478 (то> е без папоротников).
Многсч!сленгы культурные растения: обычные зерновые злаки,
маис, виноград, сахарный тростник, табак, ананасы, банаьы, фини-
ковые пальмы, также кофейные деревья и чайные кусты. Укажем
еще опунции.
8 СевероЕмерикансная атлантическая подобласть. Она занимав1
всю восточную полов! ну Северней Америки по течению и к востоку
от Миссгссипи, почти всю Флориду и Лабрадор, всю южную Канаду
и более южные части Аляски. Таким образом, подобласть, занимая
304
Рис 196 Драконовое дерево (Dracaena draco)
• . Тенерифе, справа — отдельньи цветок
основную территорию в атлантической части Америки, простирается
полосой до Тихого океана, имея сходство в этом отношении с евро-
сибирской подобластью, тоже простирающейся от Атлантического
до Тихого океана
На данной тер-
ритории господству-
ют древесные породы,
» ив смысле ланд-
шафта леса из
этих пород очень
сходные лесами евро-
сибирской подобла-
сти, но число древес-
ных пород гораздо
больше, нэпом!
ная в этом отношении
китайск о-японскую
подобласть Роды те
же, что и в евро-си-
бирской провинции
дуб, бук, клен, бере-
за, тополь, ива, из
хвойных—сосна, ель,
пихта, лиственница
(все они, однако,
представлены други
мп видами); но сюда
на юге присоединя-
ются деревья новых
семейств ореховые
(Juglandaceae), маг-
нолиевые, лавровые
Целый ряд новых ро-
дов хвойных Taxo-
dium (на самом юге),
Tstiga, Chamaecy pa-
ns Различные про-
винции области характеризуются присутствием тех или других
из указанных пород (см ниже) Однако сходство флор евро-
сибирской и атлантической подобластей настолью велько, что нет
никаких сомнений в их общем генезисе; по теории Вегенера, как уже
было указано, расхождение материков Евразии л Америки произош-
ло уже в самом конце третичного периода
Что касается большего богатства С Америки видами, в первую
очередь древесными, то это — следствие ледникового периода в
связи с орсграф! чески ми условиями, так, при наступании с севера
ледниковых масс расц дельность должна была отступить к югу ь
области с более благоприятными условиями При этом в Европе
ее отступанию к югу препятствовали горы (Альпы и др ), распо-
ложенные по большей части в широтном направлении; в Америке
350
горных цепей подобного расположения не имелось, наоборот, гор-
ные цепи имели меридиональное протяжение Естественно, чт» в
последнем случае растения могли свободно передвинуться на юг, а
затем, после отступания ледника, вернуться вновь на более северные
территории, в Европе же отступившая растительность по большей
части погибла Таким образом, в Америке сохранились многие виды
аркготретичной флоры
Рассматриваем я подобласть да леконе однородна по своей флере
ее самые северные части (на Аляске и в Канаде) находятся за поляр-
ным кругом, а самые южные (берега Мексиканского залива и север-
ная Флорида) подходят почти к тропику Рака; поэтому можно на-
метить ряд провинций, из которых северные соприкасаются с тунд-
рой, а южные i меют субтропическую флору.
В основном, подобно тому, как это имеет место в евро-сибирскоь
подобласти, и здесь северные части заняты хвойными лесами, а юж-
ные — широколиственными
Что касается числа видов, то имеются данные для ряда районов
Канады и Соединенных Штатов, именно.
Числе
< видов
Канада — северо-западная часть . . 40®
» — Манитоба, Саскачеван, Ачьберта 2 300
» — Квебек, Онтэрио 3 000
Соединеннее Штаты — центральные и северо-восточные штать! 4 885
» > — юго восточные штаты . 6 680
Сопоставление цифр с соответствуют7 ми Ц1 фрами для Европы
г оказывает значительно большее флористическое богатство Америки
(даже если соответствующее территории не совсем сравнгмы); так,
юго-восточные штаты имеют число видов значительно большее,
чем весь Кавказ, северные (хвойные) районы Канады имеют 2—3
тысячи видов
Основные провинции подобласти Провин-
ция Канады и А теки — это господство хвойных деревьев; здесь
главную роль играют те же роды, что в Евразии —сосна, ель,
.оственница, но представ пенные совершенно другими вида-
ми В Евразии господствует обыкновенная и сибирская ель (Picea
cxcelsa и Р obovata), здесь — черная ель, а ближе к Аляске —
белая ель (Р n gra и Р alba), эти деревья занимают огромное тер-
ритории Сосны, занимавши е главным образом песчаные и камени-
стые почвы, тоже представлены двумя основными и замещающими
видами Pintis banksiana, а на Аляске — Р тшгауапа Наша сосна —
Р silvestns — отсутствует Нужно указать еще па очень распростра-
ненную лиственницу (особая американская лиственница — Larix
amencana), также на пихту (Abies balsamea) Белая ель и амери-
канская лиственница идут дальше всего на север, доходя до самой
тундры К господствующим хвойным примешиваются некоторые
лиственные, тоже параллельные евро-сибирским — береза и тополь,
представленные такими видами: Betula papynfera, Populus tremuloi-
des, P bals . uiifera и др
Кустарники и травы в лесах очень разнообразны, t меется ряд сход-
ных видов с евро-сибирскими (морошка, багульник, голубика и др )
351
tt~ > Q /V 80 Ы) 4S
™ ГУМТ-я МИШИН ЧМ« ШГТ-Л« И- ПН № -
Fhc. 197. Ареал семейства Sarracenia-
ceaek ограниченный лишь Америкой.
г t > г — ареалы различные родов, еходя-
квх в вто семейство 2 — ареал рода Sarra
ееша (по Хетчинсон,).
Провинция Великих озер тоже характерна господством хвойных
пород, но уже других видов: веймутова сосна (Pinus strobus) и другие
сосны, канадская «ель» (Tsuga canadensis), американский можжевель-
ник (Juniperus occidentabs)и некоторые виды елей и пихты. Листвен-
ные представлены березами, кленами и др. Из кленов характерны
сахарный и пенсильванский
клен (Acer saccharum и А. реп-
silvamcum). На болотах на-
секомоядная саррацения (Sarra-
сеша) — все семейство Sarrace-
niaceae только в теплых ча-
стях Америки (рис. 161 и 197)
ПровинцияМиссиссипи и Ал-
леган расположена к югу от
предыдущей и занимает терри
торию от средних частей бассей-
на р. Миссиссипи до Атланти-
ческого океана. Это область
широколиственных лесов. Ланд-
шафта о она опять-таки сход-
на с соответствующими обла-
стями Европы, но флористиче-
ски, и прежде всего в том, что
касается древесных пород, очень
отличается от них. Роды же
почти одни и те же. Интересен
дубе многочисленными видами,
виды кленов и пр. Роды, от-
сутствующие в Европе, здесь
таковы: тюльпанное дерево (Liriodendron), гикори (Сагуа) с двумя
очень распространенными видами (С. ovata и С. cordiformis), на
самом юге — магнолия. Характерен орех (Juglans).
Провинция южноатлантических штатов. Здесь растут уже
бамбуки, бромелии, некоторые пальмы (Sabal). Крайне типичен
болотный кипарис (Taxcdium distichum). Упомянем оригинальный
«луизианский мох» (Tillandsia usneoidts —из семейства броме-
лиевых), совершенно лишенный корней и поселяющийся эпифитно
на деревьях (рис. 132); при обилии атмосферной влажности он
может расти даже на телеграфных проволоках. Осадки здесь
достигают 12—14 см в год, и преобладают теплые югозападные вет-
ры — в общем некоторая аналогия с климатом Китая и Японии
Особо нужно отметить растительность прибрежной песчаной рав-
нины от Техаса до Нью-Джерсея. Это господство сосны (Pinus rigida,
Р. taeda, Р. palustris). Южная часть Флориды относится к нео-
тропической области (см. стр. 368).
В рассматриваемой подобласти первоначальная флора местами
почти уничтожена (особенно в Соединенных Штатах); чю касается
культурных растений, то здесь культивируются почти все расте-
ния, о которых было упомянуто при описании подобластей Европы
и Азии. Огромные площади заняты пшеницей и папе ом, а также
352
хлопчатником. Если Европа дала Америке многочисленные куль-
турные растения, то Америка в свою очередь дала целый ряд де-
ревьев, кустарников и травянистых растений, разводимых у нас с
теми или иными целями. Декоративными растениями являются:
веймутова сосна (Pinus strcbus), североамериканский орех (Jug-
la ns cinerea), белая акация (Robinia pseudacacia), тополя — канад-
ский (Pop ulus canadensis) и бальзамический (Р. ba Isa mite га),
американский клён (Acer negundo) и др. Белая акация, кроме того,
играет большую роль при степном лесоразведении, как сильно за-
сухоустойчивая порода; гледичия (Gleditschia triacanthos) широко
используется на юге при устройстве колючих изгородей.
9. Подобласть североамериканских степей (прерии). Занимает внут-
ренние части Северной Америки, простираясь полосой с северо-
запада (приблизительно от 60°) на юг о-ю г о-в о с т о к поч-
ти до тропика Рака. Своеобразие климатических условий, завися-
щее от распределения суши и моря, от направления горных цепей,
ветров и пр., приводит к тому, что простирание этой про-
вкнции с травянистой флорой совершенно отличается от соответ-
ствующей травянистой провинции Евразии, где климатические
условия последовательно сменяются с севера на юг, почему и степ-
ная полоса простирается с запада на восток
Число видов довольно велико: для области прерий и равнин
центральных частей Северной Америки указывается около 4000 ви-
дов. Хотя это число включает в себя и некоторые соседние к степям
территории, но отсюда видно, что европейские степи флористи-
чески несколько беднее.
Присутствие травянистых пространств связано с континен-
тальным климатом: небольшое количество осадков (50—70сти), вы-
падающих главным образом летом, высокие летние температуры
и сильное испарение, зимы холодные и с очень небольшим количест-
вом осадков.
Хотя американские степи физиономически схожи с русскими,
но флористически они очень различны; большинство главных родов
представлено особыми американскими видами. Из злаков особен-
но характерны: «трава грама» (Bouteloua hirsuta), трава бизонов
(Buchloe dactyloides — рис. 146), виды бородача (Andropogcn),
тонконога (Koeleria) и др.; последний род типичен и для русских
степей. Двудольные представлены главным образом такими семей-
ствами: бобовые, сложноцветные, норичниковые; из бобовых роды
Desmodium, Amorpha, отсутствующие в Европе, из сложноцветных
крайне характерны такие роды, как астра (Aster), рудбекия (Rud-
beckia), ленок (Coreopsis), сильфиум (Silphium), полынь (Arte-
misia), золотая розга (Solidago). Многие растения прерий введены
в культуру как декоративные растения —астры, рудбекия, ленок
и др
К югу прерии переходят в сухие пустынные области Мексики и
Калифорнии.
Прерии в настоящее время почти целиком распаханы, пред-
ставляя необозримые поля пшеницы и маиса.
23 География растений.
353
10. Североамериканская пацнфическая подобласть. Занимает за-
падное побережье Северной Америки к югу от Аляски до Калифор-
нии, в глубь страны доходит до Скалистых гор. Таким образом,
подобласть климатически неоднородна: прибрежные места имеют
умеренный морской климат, а восточнее располагаются обширные
возвышенности между Сиеррой-Невад>й и Скалистыми горами с
растительностью пустынного и полупустынного характера.
Число видов в различных частях подобласти от 3 000
до 4 000.
Отличительная черта прибрежных и горных частей подобласти —
обилие хвойных пород; в этом отношении тихоокеанское побережье
занимает первое место на земной поверхности. Число видев исчис-
ляется несколькими десятками (вспомним, что в
средних частях евро-сибирской подобласти в пределах нашей Евро-
пейской части лишь два хвойных дерева —ель и сосна). Кроме
многочисленных видов, особенно сосны, а также пихты и ели, встре-
чается много родов, совершенно отсутствующих в Европе.
Имеются значительные различия в древесных породах более
северных и более южных частей подобласти. Для первых нужно
указать такие виды: Picea sitchonsis, Tsuga m^rtnsian?, Thuja
gig.ntea (туя), дугласова пихта (Pstuok tsuga D.ugLsii), Chamae-
cyparis nutk .Lnsis, многочисленные виды сосны, также пихты и
ели. Для более южных частей особенно нужно отметить два вида
секвойи: секвойя вечнозелёная (S.quJa semporvirens —рис. 94) и
секвойя гигантская (S. gige ntea—pre. 130). Первая особенно
типична для Калифсрш йского побережья, вторая —для горных
лесов Сиерры-Невады. Гигантская секвойя (маментово дерево)—
это исполинское дерево, достигающее огромной высоты —до 150 м
и до 16 м в поперечнике; это — одно из величайших деревьев мира.
Вечнозелёная секвойя, иначе называемая красным деревом, не
достигает указанной высоты, но имеет ещё большую толщину. Эти
два великолепных дерева везде в соответствующих клгматических
условиях введены в культуру (у нас, например, в Крыму).
Упомянем ещё для Мексика неких Кордильер необычайно кра-
сивую сосну Монтезумы (Pinus Montezumae) с длинными спускаю-
щимися хвоями.
Местами в подобласти, в более сухих районах на каменистых
почвах, развиты заросли кустарников, на юге вечнозелёных, пред-
ставляющие так называемый «чапараль» (chappar.. 1). Здесь имеются
многочисленные американские роды, кроме того, некоторые виды
дуба и пр.
Совершенно особый флористический район представляет об-
ласть североамериканского плоскогорья между Скалистыми горами
и Сиеррсй-Невадсй. Это—обширная полупустыня и пустыня с
рядом высыхающих, некогда обширных озёр; последнем остатком
является Большое солёное озеро. Мостами осадки не превышают
11 Стив год; летние температуры очень высоки. Hr i бои ее характерным
растением является полынь (Artemisia trick nt t.). В самых южных
частях встречаются такие роды, как кактус-цереус (Cercus), юкка
(Yucca) и др. Здесь уже переход к мексиканским пустыням,
354
Вообще флористически подобласть довольно сильно отличается
ет атлантических частей Америки и имеет на юге некоторые связи
с неотрспическсй областью (например кактусы).
Флора подсбластн дала очень много видов, введённых в Европе
в культуру. Выли уже указаны такие красивые деревья, как ги-
гантская секвойя (иначе называемая мамонтовым деревом, или
веллингтонией), вечнозелёная секвойя, дугласова пихта, сосна
Монтезумы и др. Из кустарников упомянем магонию (Ma honia
aquifJium) с блестящими вечнозелёными листьями, многочислен-
ные травянистые растения, разводимые в цветниках,—лобелия,
годецгя, ленок (Coreopsis) и ряд других.
В заключение укажем, что на территории СССР находятся в
большей или меньшей их части такие подобласти Голарктгки; арк-
тическая, евро-сибирская, китайско-японская,понтическо-централь-
ноазиатская и средиземноморская.
П. Палеотропическая область
Общая характеристика. Область занимает Африку к югу от
тропика Рака (за исключением небольшой юго-западной части,
принадлежащей к Капской области—см. ниже), юго-западную
часть Аравии, Индостан, Индо-Китай с южными частями Китая,
Рис. 198. Ареал семейства пальм, ограниченного тропическими странами
показаны северная и южная границы (по Друде).
Филиппины, МалаЙскт й архтпелаг, Новую Гвинею, Новую Зе-
ландию и острова Тихого океана (Ф1 джт, Со мог, Санды.чевы и др.).
По размерам зант маемой площадь эта область является второй
после голарктической, включая части Африки, Азии, Австралии
и ряд островов Т» хого океана.
Так как рассматриваемая область находится в тропическом
поясе, то, естественно, здесь господствуют тропические семейства,
из которых укажем лишь некоторые: Menispertnaceae, Begoniaceae,
Sapindaceae, Myrtaceae, Gesneriaceae, Bignaniaceae, Palmae (рис.
198). Однако все эги семейства встречаются как в палеотропиче-
ской, так и в неотропической областях; но представлены в них
обычно различными родами.
Собственно палеотропическими семействами нужно считать не-
пентесовые (Nepenthaceae—рис. 203), панданусовые (Pandancr
сеае), двукрылоплодные (Dipterocarpaceae). Многие семейства, имея
общетропическое распространение, в Старом Свете представлены
значительно большим числом видов; сюда среди других относятся:
аноновые (Апапасеае), эбеновые (ЕЬепасеае), ласточниковые (As-
clepiadдсеае), тыквенные (Cucurbitaceae). В то же время, ряд не-
отропических семейств совершенно отсутствует: бромелиевые (Вго
meliaceae), канновые (Саппасеае), маркгравиевые (Marcgraviaceae)
и некоторые другие; почти отсутствует семейство кактусовых (Сас-
taceae), столь типичное для тропиков Нового Света. Нужно отме-
тить очень сильное развитие мангровой флоры, особенно по бере-
гам Индийского океана, относящейся к ряду семейств (в неотро-
пической области представлена слабо). _____
Соединение в одну флористическую область Африки и южных
азиатских полуостровов, островов Малайского архипелага и пр.,
в настоящее время не связанных между собой, подтверждается как
палеоботаническими данными, так и теоретическими представления-
ми В е г е н е р а о разломах и дальнейшем расхождении частей
первоначально единого сплошного материка. Так, по Вегене-
р у, связь Африки через Мадагаскар с Индией существовала ещё
в самом начале третичной эры и лишь в эоцене произошло их рас-
хождение, а связь Индии с Австралией существовала лишь до юр-
ского периода мезозойской эры.
Таким образом, те дизъюнкции, о которых юворилось выше,
являются естественным следствием расхождения тех или иных тер-
риторий палеотропической суШи, не привлекая сюда обычно сом-
нительной теории «мостов». В общем значительная однородность
флоры данной области стоит в связи с общностью генезиса.
Флористически область очень богата, что вообще характерно
для тропиков. Вот некоторые цифры:
Число видов
(приблизительно)
Тропическая Африка ............................. 13 000
Капская земля ................................... 12 000
(впрочем, большая её часть относится к Капской флористической
области)
Мадагаскар ................................................ 5 500
Абиссиния ................................................1 650
Индия ..................................................21 000
Цейлон ................................................. 3 074
Малайский полуостров.................................... 6 920
Борнео ................................................. 11 000
Ява ..................................................... 5 000
Филиппины .............................................. 10 000
Новая Зелаидия ....................................... 1843
356
Вся палеотропическая область распадается на три основных
подобласти: I) индо-африканскую, 2) малезий-
с к у ю нЗ) ново-зеландскую. Можно выделить ешё
подобласть Сандвичевых островов и подобласть африканских ост-
ровов (о. Св. Елены и др.).
1- Индо-африканская подобласть. Это наиболее крупная подоб-
ласть палеотропической области, занимающая Среднюю и Южную
Африку с Мадагаскаром и переднюю Индию. Она расчленяется
на несколько провинций в связи с различием климатических усло-
вий. Так, если продвигаться в Африке с севера на юг, то из области
Сахары (голарктическая область) мы переходим в полосу степей,
далее, в бассейне р. Конго, сменяющихся влажнотропическим
лесом; ещё далее на юг тропический лес вновь сменяется саванна-
ми и степями, а на самом юге (особенно на юго-западе) саванны и
степи-переходят в пустынные пространства. Таким образом, суще-
ствует как бы симметричное распределение основных растительных
ландшафтов, что особенно хорошо выражено в западней половине
Африки,- пустыни —степи и саванны —тропические леса —сте-
пи н саванны—пустыни. j
Такое распределение ландшафтов, как уже говорилось, стоит
в связи с климатическими изменениями: так, тропические леса тре-
буют очень большого количества осадков (не менее 180 см в год),
причём они должны равномерно распределяться в течение всего
года, саванны же развиваются при меньшем количестве осадков
(меньше 150 Сти), причём осадки выпадают неравномерно и засушли-
вые периоды сменяются влажными. Небольшое количество
осадков и сильные засухи приводят к развитию полупустынь
и пустынь.
Необходимо добавить, что сухое время года и, следовательно,
замедление или прекращение вегетации, приходится в средних
частях Африки на летнее время, в одних местах — с мая по октябрь,
в других — с июня по август. В результате вегетация идёт глав-
ным образом зимой; в это время одеваются листвой деревья и кустар-
ники, летом же во время засухи листва опадает. Леса подобного
рода называются зимнезелёными (Hiemisilvae) в проти-
воположность нашим лиственным — летнезелёным (Aes-
tisilvae).
Наибольшие пространства занимают в данной подобласти са-
ванны н степи, меньшие пространства—тропический лес, ещё
меньшие —пустыни.
Подобласть распадается на ряд провинций; одни из них лесные
с флорой влажнотропических лесов, другие — степные с флорой
степей и отчасти пустынь.
Западне африканская провинция влажнотропических лесов за-
нимает главным образом бассейн р. Конго, а также южное побе-
режье Гвинейского залива.
Древесные породы здесь крайне разнообразны: роды семейств
Moriceae (например фикус —ряд видов), Апспасеае, Bombacaceae,
Leguminosae (Albizzia) и т. п. Нужно упомянуть ещё масличную
пальму — Elaeis guineensis, важное здесь полезное растение.
357
О влажнотропических лесах как о своеобразном типе лесов уже было
сказано выше (см. стр. 171).
Степи и саванны занимают в индо-африканской под-
области огромные пространства, главным образом в Африке, имен-
но по южной окраине Сахары, в Абиссинии, в южной части Афри-
ки поперёк всего кон1инен1а; кроме того, меньшие площщи на Ма-
дагаскаре и в передней Итдии. Местами—это открытые травя-
нистые пространства, но в большинстве случаев это или саванны,
или кустарниковые степи ксерофального характера. На южной гра-
нице Сахары (например в Судане) находятся саванны или <парко-
Рис. 199. Вельвичия (Welwjtschia mirabilis); направо — более стапын_экзем'
пляр, налево — более молодой с соцветиями (по Мартонну)-
вые степи», т. е. степи с разбросанными по ним деревьями (судан-
ская провинция парковых степей); из травянистых растений, до-
минирующих в степном ковре, особенно распространены роды Реп-
nisetum и Cymbopogon; из деревьев особенно характерны виды Aca-
cia, Borassus, Adansonia (A. digitata). Указанные древесные роды
вообще характерны для африканских саванн (рис. 138).
В Абиссинии и в прилежащих территориях (североафриканская
возвышенная и степная провинция) очень распространены кустар-
никовые степи из многих видов кустарников и небольших дере-
вьев (между прочим виды акации). На больших высотах — горные
луга из злаков Andropogon и Sporobolus с многочисленными одно-
дольными из лилейных, ирисовых, амариллисовых и двудольными.
Восточные части Африки относят к восточноафриканской степ-
ной провинции. Это главным образом степи и саванны с многочислен-
ными видами акации, алоэ и др.
358
Южные части Афэики (кроме крайнего юго-запада) представляют
южноафриканскую переходную провинцию, являющуюся по своим
флориС1Ическим элемешам переходом к капской флористической об-
ласти. Сюда относится бассейн р. Оранжевой, пустыни: Калахари,
Карру, Дшарлэнд и др. По своей растительности —это или степи
(Трансвааль и Оранжевая провинцгя), или пустыни. Дг я степей ха-
рактерен особенно злак Aristida brevjfJia и многие виды акации.
Для пустынных территорий с их более сухим климатом особен-
но типичны растения-суккуленты (роды Mesembrianthemum, мо-
лочай и алоэ — А1:ё), а также многочисленные виды акацги; из
злаков —роды Aristida, Pennisetum и др. Для пустынь Дамарлэн-
да (Намиб) нельзя не указать замечательного голосеменного расте-
ния вельвнчии (Wj wits: hi a mirabilis), короткий ствол которого
похож на обрубок с двумя длинными лентовидными листьями (рис.
199). Род вельвичия интересен в систематическом отношении — он
стоит в системе совершенно особняком (относится к порядку Gneta-
les), имея только один вид, и представляет прекрасный пример кон-
сервативного реликта и эндемк это — «живое ископаемое». Стоит
ещё указать на одно
очень характерное здесь
растение из тыквенных —
Acanthosicyos horrida,
длинные корни которого
имеют до 15 м (!) длины.
Переходный Характер
рассматриваемых южных
ч 1стей Африки сказывает-
ся в появлении некото-
рых элементов капской
флоры; из последних
укажем на виды рода
пеларгониума (Pelargoni-
u m),вереска (Erica),семей-
ство Proteaceae и др.
Флора острова Мада-
гаскар, несмотря на бли-
зость материка, доволь-
но оригинальна и пред-
ставляет хороший пример
флоры древних материко-
вых островов. Достаточно
сказать, что из 5 500 ви-
дов более 3 000 видов
являются эндемика ми.
Это указывает на изоли-
Рис 200 Дерево путешественников (Rave-
nala madagascariensis) (фото М. М. Местер-
гази).
рованность в развитии
флоры. Флора имеет различные связи: западная часть Мадагаскара
имеет большое сходство с Африкой, восточная —с Индией. Связь
с Индией хорошо устанавливается через Маска ренские и Сейшель-
ские острова. Эти острова с Мадагаскаром образуют провинцию
35*
Ряс. 201. Дынное дерево (Carica papaya) с
плодами (из Веттштейна).
восточноафриканских островов. Интересны связи Мадагаскара и с
весьма отдаленной Южной Америкой: только в за их двух местах встре-
чается известное дерево путешественников (Ravenala—рис. 200),
Персднеиндинекая провинция занимает Малаба рский берег, низ-
менность р. Ганга, западные Гималаи и другие. Главнымобразомтро-
пические леса. Можно назвать некоторые пальмы: Caryota urens,
Corypha umbraculifera, Phoenix Silvestris, а также баниан (Ficus
religiosa), акацию (Acacia catechu) и др. Характерны для данной
провинции джунгли — непроходимые, часто болотистые за-
росли бамбука и других растений, стволы которых тесно перепле-
тены лианами; среди лиан много видов лазящих пальм.
Многочисленные территории и ндо-а фрика некой подобласти яв-
ляются ещё «девственными», особенно тропические леса внутрен-
них частей страны, отчасти
саванны, но из года в год
культуры занимают всё
большие пространства. Мож-
но указать многочисленные
культурные или культиви-
руемые растения, например,
кофейное дерево (рис. 193),.
банан, рис, индиго, сахарный
тростник, маньок, бататы,
дынное (тыквенное) дерево
(рис. 201), каучуконосные,
пряные растения и др.
В данной подобласти нахо-
дятся два очага происхожде-
ния культурных растений: I)
абиссинский и 2) индомалай-
ский. Первый дал твёрдую
пшеницу, ячмень (однако эти
растения имеют здесь, неви-
димому, вторичный центр —
см. стр. 388 и 391), кофей-
ное дерево (Coffea arabica),
второй очаг (захватывающий
и Индокитай и отчасти Ма-
лайскую область) дал таьие
растения, как рис, многие
зерновые бобовые, сахарный
тростник и плодовые тропи-
ческие деревья: кокосовую
пальму, цитрусы, банани др.
2. Малезнйская подобласть.'Занимает весь Индо-Китай с Малак-
кой, Цейлон, Большие и Малые Зондские острова, Филиппинские
острова, Новую Гвинею и некоторые острова Полинезии (Новую
Каледонию, Фиджн, Товарищества, Самоа и др.), Иначе называет-
ся областью муссонов, так как здесь характерными ветрами явля-
ются юго-западные муссоны, приносящие обильные дожди. Коли-
360
чество осадков очень велико, во всяком случае ие менее 2 м в «год,,
но в некоторых случаях доходит до 14 м (!).
В основном почти вся территория занята высокими тропически-
ми деревьями, образующими типичный влажнотропический лес.
Число видов очень велико. Ниже приводятся некоторые цифры для
отдельных частей подобласти:
Числ»
видов
Малаккский полуостров ................................. 6 290
Борнео ................................................11 000
Ява ....................................................5 000
Филиппинские острова ..................................10 000
Цейлон ................................................ 3 074
Общее число видов подобласти более 15 000, причем эти виды
размещены очень плотно; так, имеются указания, что иногда на про
странстве 1кл1анахо'
дится до 2 000 видов,
т. е. столько видов,
сколько имеется во
всём Крыму или поч-
ти столько, сколько
во всей современной
Венгрии.
Господствуют раз-
личные тропические
семейства (Guttife гае,
Pandanaceae, Pipe-
raceae, Zingib. raceae,
La uraceae.Dipterccar-
paceae, Palmae, Ne-
penthaceae—рис. 202
и 203). Также сильно
представлено семей-
ство бобовых. Из
пальм, обильных на
Малайских остро-
вах, назовём: Ссгу-
pha gebrnga, Areca
catechu, Nipa, также
лазящая Calamus
(ротанг). Пальмы
многочисленны так- Рис. 202. Расте ние-кувшнн (Nepenthes raffleataea);
Же на Филиппинских слева — кувшин (верхняя часть листа), справа —
островах (кроме то- соцветие („з Веттштейна).
го, здесь встречаются
деревья из семейства Dipterocarpaceae, камфарное, гвоздичное,
тиковое — Tectoпа grandis). Очень характерны фикусы (Ficus).1;
На больших высотах, например в Гималаях, выше границы ле-
сов развиваются альпийские луга с рядом северных видов.
Растительность Новой Гвинеи носит до известной степени пе~
М’
Рис. 203. Ареал рода Nepenthes (растение-кув-
шнн), показывающий тесную связь между
„Мадагаскаром, Индо-1(нта.:м и Мт езийской
подобластью (по Хетчинсону),
подобласти (главный продукт
Рис. 204. Группа кокосовых пальм (Cocos
nucifera) на Сандвичевых (Гавайских)
островах (по фото М. М. Местергаэи).
реяодный характер, имея ряд элементов Австралийской области:
виды эвкалипта (Eucalyptus) и рода Melaleuca.
Чго касается Полинезийских островов (Новая Каледония, Фид-
жи, Самоа и др.), то для
них особенно характерен
род хвойных: подока р-
пус (Podocarpus) и агатис
(Ag. fhis, рис. 215). Эти
хвойные в отличие от на-
ших голарктических хвой-
ных имеют плоские, более
или менее широкие ли-
стья, похожие на листья
двудольных деревьев.
Богатая флора Мале-
зии включает очень боль-
шое число растений, по-
лезных для человека;
многие из них уже указа-
ны для предыдущей под-
области. Особенно нужнФ
•тметить широкое распространение кокосовой пальмы (рис. 204),
культивируемой везде на ос
кокосовой пальмы—жирное
кокосовое масло); также
нельзя не отметить бананы
(виды рода Musa), одно иа
важнейших пищевых расте-
ний Малезии. Многочислен-
ные растения дают пряности:
корицу (Cinn im mum), гвоз-
дику (Caryophyllus), мускат-
ный орех (Myristica), перец
(Piper) и т. п. Много расте-
ний, дающих красящие ве-
щества (индиго, куркума и
др.), каучук (виды фикуса)
и т. п. Многие деревья тро-
пических лесов дают ценную
древесину (например,тиковое
дерево —Tectona grandis).
Укажем, наконец, бамбук
(рис. 205).
В М алезии и Индии на-
ходится один нз очагов про-
исхождения культурных ра-
стений—индо-малайский (см.
выше), захватывающий Ин-
достан, Индо-Китай и часть
Малайских островов.
Ж
3. Новозеландская подобласть. Эта подобласть занимает Новую
Зеландию и несколько островов, расположенных к югу от неё; ино-
гда причисляется к Австралийской области, но флора её в значи-
тельно большей степени носит малезийский характер. Северные
части Новой Зеландии лежат в области муссонов, но и вообще эти
©строза получают много влаги,/вследствие чего создаются условия
для субтропических лесов.
Характерны вечнозелёные двудольные деревья, древовидные
напоротники, а также своеобразные хвойные. Из последних роды:
Dacrydium, Podocarpus, Aga this имеют не хвою, а более или ме-
нее широкие листовые пластинки; среди многочисленных древо-
видных папоротников —роды Cyathea, Dicksonia и др.; на горах—
южный бук (Nathofagus) и хвойное Lib о се dr us. Очень оригинальна
растение-подушка Raoulia eximia (рис. 206).
Обилие осадков приводит к образованию на горах ледников,
которые сползают далеко вниз в пояса, занятые лесом, причём мож-
но наблюдать любопытные близкие соседства ледников и пальмовых
деревьев.
Первобытная растительность во многих местах крайне наруше-
на, вместо неё образуются сплошные заросли папоротника-орляка
(Pteridium esculenturn, близкий к Р. aquilinum); очень много за-
носных акклиматизировавшихся растений, количество которых до-
ходит почти до 25% всей флоры.
Полезных растений здесь очень мало. Нужно упомянуть доволь-
но распространённое в Европе декоративное растение—новозе-
ландский лен (РЬогтшт tenax), дающий грубое волокно (рис. 207).
М8
Если ниже pacnoj ож^нные части Новой Зеландии носят вполне
малезийский характер, то её высокогорная флора имеет целый
Рис. 206. Деревянистое
растение-подушка (Ra-
•ulia eximia) на южном
острове Новой Зелан-
дии (по Дю-Риэ)
ряд черт, сходных с горами. В сеточной
Австралии, Тасмании и даже Южной Аме-
рики.
4. Подобласть Сандвичевых (Гавайских)
островов. Фтора этих островов носит впол-
не тропический характер и известна своим
выдающимся эндемизмом — по некоторым
данным, более 90% видов являются энде-
миками. Характерны многочисленные дре-
вовидные папоротники. Интересно, что из
этой группы островов геологически более
древние имеют и значительно более бога-
тую и с большим количеством эндемиков
флору.
5. Подобласть африканских островов. Иног-
да в качестве особой подобласти выделяют
несколько островов, лежащих к западу от
Африки —Св. Елены Тристан-да-Кунья, к
югу-востоку—Св. Павла и др. Эти острова
очень богаты эндемиками и показывают по
своей флоре родственные отношения как с
Африкой, так и с Америкой. Интересна
флора небольшого острова Св. Елены. Здесь
вся первоначальная флора содержит 45
видов цветковых (40% эндемиков) и 26 па-
поротников (20% эндемиков); пришлых же
видов в настоящее время насчитывается д*
400, т. е. почти в 6 раз больше. Это —один
из наиболее ярких примеров влияния че-
ловека на изменение состава флоры. На
острове Павла имеется всего 15 видов высших
растений.
III. Не отрои нче ска я область
Общая характеристика. Эта область занимает почти всю Мек-
сику, всю Центральную Америку с Антильскими островами, самую
южную оконечность Флориды и всю Южную Америку до 40° юж-
ной широты, т. е. за исключением южных её частей (относящихся
к антарктической области), кроме того, ряд островов Тихого океа-
на, примыкающих к американскому материку. Называется неотре-
пической потому, что расположена в тропиках Нового Света.
Здесь в основном, конечно, господствуют тропические
семейства; они в общем те же, что и в палеотропической об-
ласти (см. выше), но в большинстве случаев представлены другими
родами и видами. Однако имеется ряд чисто .американских се-
мейств: кактусовые (Cactaceae), бромелиевые (Brome liaceae), канио-
364
вые (Саппасеае), Xyndaceae, киианиевыс (Juhaniaceae), настурци-
ей не (Tropaeolaceae рис. 208) Некоторые палеотропические се-
мейства (см. выше) совершенно отсугивуют. Другие же, хотя встре-
чаются под тропиками и Старого и Нового Света, но в последнем
случае представлены бблыним количеством видов; сюда относятся
такие семейства: стр а ст но цветные (Passifloraceae), молочайные
(Euphorbiaceae), кермесовые
(PhytJaccaceae), никтагиновЫе
(Nyctaginaceae), пасленовые
(Solanaceae), миртовые (Myrta-
сеае) и др.
Большинство голарктиче-
ских семейств или отсутству-
ют, или имеются в небольшом
количестве видов; обычно они
представлены лишь на горах,
в климатических условиях,
более сходных с северными
областями. Однако некоторые
голарктические семейства хо-
рошо развиты и в неотропиче-
жой области: сложноцветные,
бобовые и злаки.
Рис. 2( Ареал семейства настурцие-
вых aeolaceae) (по Хетчинсону).
36&
Нужно добавить, что мангровая флора, столь богато представ’
ленная на побережьях Индийского океана, в Америке очеьь бедна
видами, хотя и характерна как таковая.
Флористически область очень богата, что вообще типично дер
тропических стран; вот некоторые цифры:
Числе
видов
Неверная Мексика ..........................................3 040
Южная Мексика .............................................7 000
Венецуэла ................................................ 6 800
Вразилия .................................................40 000
Чили ..............*..................................... 5 500
Парагвай . . .................................... 4 220
Уругвай ............................................ 2 234
Оетров Куба .........................................7 000
Неотропическая область распадается на три подобласти: 1) цен-
тральноамериканскую, с ксерофт льной флорой, 2) т ру-
ническую, 3) подобласть Анд.
1. Центральноамериканская подобласть. Эта подобласть за ни мает
иочти всю Мексику (кроме Гирных цепей с голарктической флорой)
и полуостров Калифорнию. Таким образом, здесь имеется очень
близкое соседство и возможность обмета флор голарктическей и
неотрспической. В частности, на мексиканских Кордильерах (про-
должающих к югу более северные горные цепи) нахсдятся сосново-
дубовые леса; из сосен—сосна Монтезумы (Pinus Mcntosumae), а так-
же и другие виды,кроме того дубы,ольхи и т. д.; в травянистом покро-
ве имеются такие голарктические роды, как подмаренник (Galium)r
грушанка (Piro 1а), валерьяна (Valeriana), крестсвни к (Stпесit) и др.
Бзльшая часть подобласти занята высоким и обширным плоско-
горьем о крайне засушливым климатом (сколе 60с.м осадков в год),
что влечет за собой господство элементов полупустыни, В мекси-
канских полупустынях и пустынях встречаются многочисленные
кактусы (роды: опунция—Opuntia, Cereus, Echinccactus, Me mil-
iaria), агавы (Agave), виды Dasylirion, различима виды семейств
брсмелиевых, а также юкки (ргс. 209).
В низменных местах, особенно по низменным побережьям,
условия благоприятны для развития тропической растите; внести;
здесь имеются древовидные папоротники, саговники (роды Diocn,
C^ritjzamia), пальмы, виды семейства банановых и пр. Из эпгфи-
тов типичны американские брсмелии.
Нельзя не упомянуть о присутствии здесь ряда третичных ви-
дов. Особенно замечателен таксодиум (Тахо clium muert па turn):
известны экземпляры очень большого возраста, один гз KCTOfbix
достигает 17 м в поперечнике. К числу «живых ископаемых» нужно
также отнести указанные выше роды саговников.
Добавим, что наиболее северный пункт мангровой растительно-
сти — южная оконечность Калифорнийского полуострова.
Мексика —один из очагов происхождения ряда вультурных
растений, именно «центральноамериканский». Из этих растений
укажем: маис, дерево какао, хлепчатнгк-упланд, амергваьсвие
тыквы, фасоли и пр. Нужно указать агаву (Agave amtricana), ина-
че называемую «столетником;?, дающую волокна (механическая
ткань гистьев) и особый напиток (сок срезанных молодых соцветий'?
а также ваниль (Vanilla planJoha) из семейства орхидных, плоды
которой употребляются как пряность.
В Мексике имеются многочисленные культуры как американ-
ских, так и внеамериканеких растений.
2. Тропическая подобласть. Занимает Центральную Америку (к
Ю1 у от Мексики), Атильские острова, южную часть Флориды и всю
Южную Америку до 40° ю. ш. Подразделяется на несколько провин-
ций, отличающихся своими климатическими условиями; наиболее
влажные провинции представляют влажнотропический лес, про-
винции с меньшим количеством влаги и неравномерным её распре-
делением представляют саванны и степи
Центральное положение в подобласти вдоль экватора занимает
яровинция типичных влажнотропических лесов, представляющих
колоссальный массив, бэлыш й, чем где-либо на земной поверхно-
сти (см. выше, стр. 185). Эш леса были названы Гумбольдтом «гилеей»
(«гилейон» по-гречески —лес).
Флористические провинции, охватывающие перешеек, соединяю-
щий Северную и Южную Америку, Антильские острова с южной
частью Флориды и самые северные части Южней Америки, пред-
ставлены в основном также влажнотропическими лесами с теми или
иными признаками отличия. Впрочем, во многих местах, особенно
на Антильских островах, первобытная растительность сохранилась
лишь небольшими участками, а обширные пространства заняты
культурами или нарушенной растительностью. Вообще здесь ха-
рактерно присутствие в лесах многочисленных пальм и особенно
древовидных папоротников (особенно виды родов Alsophila и Суа-
thea). Очень интересно отметить присутствие на горах и вообще на
более повышенных местах таких северных древесных пород, как дуб
и сосна, что указывает на известное смешение двух различных флор-
умеренных широт и тропической. На побережьях местами развиты
мангровые заросли. Вполне тропический характер имеет расти-
тельность и в южной части Флориды: около десятка пальм, саговник
(Zamia floridana), древовидные мареновые (Rubiaceae), целый ряд
тропических родов, отсутствующих в других частях США. Сильно
развиты болота со знаменитым болотным кипарисом (Taxodium di-
stichum — рис. 132).
Остальные территории данной подобласти заняты зимнезелёиы-
ми лесами, саваннами и степями, за исключением прибрежных ча-
стей и долин крупных рек, где развивается влажнотропический
лес несколько иного флористического состава. Можно выделить
несколько провинций, где представлены такие типы растительно-
сти как льяносы, каатинги, кймпосы и пампасы. Краткая характе-
ристика последних была дана выше.
3. -Подобласть Анд. Она тянется более или меиее широкой полосой
с севера иа юг вдоль всего южноамериканского материка, по его
западному побережью. Эта подобласть, захватывающая всю гигант-
скую цепь Анд, по своей флоре крайне разнообразна, так как, во-
первых, имеет очень большое протяжение с севера на юг, а во-вто-
рых, занимает различные горные ступени и горные страны.
В основных чертах можно сказать, что иа восточных склонах Аид
развиты влажнотропические леса, на западных склонах—расти-
тельность пустынного типа (за исключением побережья под тро-
368
никами и в Чили), на повышенных [местах —горные степи и высо-
когорная растительность тропических областей.
Влажнотропические леса, развитые главным образом на восточ-
ных склонах и переходящие непосредственно в гилеи по р. Амазон-
ке, под экватором переходят
и на западные склоны. Эти
леса в общем схожи с лесами
гилеи, здесь встречаются
представители почти тех же
тропических семейств. Леса
доходят до высот в 3 400 м
Особенно нужно отметить
очень характерное здесь хин-
ное дерево (Cinchona — рис.
210) и коку (Erythroxylon
coca); из листьев последнего
добывают кокаин.
Несколько южнее эквато-
ра и до 30° io. ш. все запад-
ное побережье занимают
очёнь сухие территории —
пустынные степи и иногда
пространства, почти совер-
шенно лишённые всякой
растительности. Имеющаяся
растительность представлена
жесткими травами, колю-
чими кустарниками.
Пустынные области встре-
чаются и в повышенных
горных частях подобласти;
так, к югу от тропика, меж-
ду цепями Кордильер на-
ходятся высокие равнины,
занятые пустынями (напри-
мер пустыня Атака-
Рис. 210. Ветвь хинного дерева (Cinchona
officinalis) с соцветием (по Кернеру).
ма); далее к югу высокие равнины между Кордильерами предста-
вляют высокогорную степь с колючими кустарниками (так назы-
ваемые пуны). Из трав здесь вид ковыля — Stipa ichu, из колючих
растений — акации, коллегия (Colletia), известная своими кладо-
диями, некоторые кактусы.
Побережье в Чили ие носит пустынного характера, а представ-
ляет вечнозелёные субтропические леса. Здесь встречается мыльное
дерево, дающее мыльную кору (Quillaja saponaria).
Что касается высокогорной флоры Анд, то она носит альпийский
характер с жизненными формами типа розеток и подушек. Замеча-
тельно, что здесь присутствуют многочисленные роды Голарктики,
как Alchimilla, Gentiana, Saxifraga, Viola, Astragalus и др., с дру-
гой же стороны, —ряд арктических родов: Acaena Azorella, Bo-
lax.
34 География растений
ЗГ.О
Несколько слов о флоре Галапагосских островов, известной бо-
гатством своих эндемиков: на 350 встречающихся здесь видев энде-
миков около половины. В основном здесь тропический лес; интерес-
ны мимозы и древовидные сложноцветные, многочисленные лиаиы
и эпифиты. Нужно отметить полнее отсутствие на этих островах
пальм.
В богатой флоре Центральной и Южней Америки имеются мно-
гочисленные полезные растения. Ряд деревьев даёт каучук; особен-
но важна в этом отношении бразильская Hevea bra si 1 iensis, также
Castilloa elnstica. Другие дают важные медицинские средства —
хинное дерево (Cinchona), кока (Ery thrcxylon coca); многочислен-
ные деревья дают ценную древесину: палисандровые деревья (Dal-
bergia и Machcrium), красное сандальное (Caos' Ipinia echinata)r
чёрное (Brya ebenus); многие деревья дают смету, луб и пр.; молоч-
ное дерево (Gclactcdendrc п) даёт сок, употреблясмьй как молоко;
костяная пальма употребляется для приготовления пуговиц (Phyte-
1 ер has macrocarpa; идут в дело семена), другие пальмы служат для
плетения шляп; Carludcvica —род, близкий к пальмам, даёт ма-
териал для панамских шляп; плоды некоторых деревьев употреб-
ляются на сосуды (Lecythis — горшечное дерево и др.). Упомянем
ещё два можжевельника (Juniperus virginiana и J. barbadensis),
древесина которых незаменима для карандашей и сигарных ящи-
ков. Различные травянистые растения также имеют практическое
применение; упомянем ваниль и т. д.
В Андах (главным образом в приэкваториальных частях) нахо-
дится один из очагов происхождения культурных растений; именно
здесь имеют с^ою родину многочисленные виды картофеля, табак,
ананас, земляной орех и др.
Очень трудно перечислить многочисленные культурные расте-
ния (некоторые из только что перечисленных уже введены в ьуль-
туру). Назовём: рис, сахарный тростник, табак, кофейное дерево,
дерево какао, бананы, ананасы, хлопчатник, маиьок (или тапиока),
виноград, агаву, в степных районах—маис, пшеницу, картофель
и т. д.
IV. Австралийская область
Эта область занимает всю Австралию и остров Тасманию. Осо-
бенно типично австралийская флора представлена в западней части
Австралии; северные и восточные части имеют ряд черт, сходных
с малезийской подобластью, поэтому некоторые авторы относят
разные части Австралии к разным флористическим областям. Од-
нако во всех частях Австралии так много своеобразных, чг сто ав-
стралийских флористических черт, что вполне целесообразно всю
Австралию выделять как особую область.
Флористически Австралия очень богата, общее число видов —
12 049, из которых около 9 086 видов являются эндемиками,
т. е. эндемики составляют не менее 75% флоры. Отсюда ясно
видно исключительное своеобразие австра;г йской флоры.
Ниже приводятся данные по числу видов для отдельных частей
Австралии:
370
Числе
ВИДОР
Западная Австралия ......................................4 384
Южная Австралия ........................................ 2 208
Провинция Виктория ......................................2 171
Новый Южный Уэльс ...................................... 3 773
Каинслэнд ........................... '................. 4 395
Северная Австралия ..................................... 2 248
Остров Тасмания ........................................ 1 127
В приведенных данных бросается в глаза флористическое бо-
гатство западных частей Австралии, с одной стороны, и восточных —
с другой; это объясняется и климатическими и генетическими при-
чинами. Кроме флористического богатства этих двух частей, ин-
тересно указать на их флористическую обособленность: по данным
некоторых авторов, эти две части Австралии имеют и е более
10% общих видов, что указывает на изолированное развитие флор
западней и восточной частей.
Особенно типично, как уже указано, собственно австралийские
черты флоры представлены в западней части. Очень характерны
семейства Prcteaceae, Resticnaceae, Gocdeniaceae, Myoperaceae,
Epacridaceae, Cephalotaceae (рис. 211) и др. Из семейства Prctea-
ceae характерны такие
роды, как Н; кеа, Вап-
ksia, Grevillea, Осо-
бенно нужно указать
на роды из семейства
миртовых — эвкалипт
(Eucalyptus)n Melaleuca
и из семейства бобо-
вых —акацию (Acacia)
с весьма многочислен-
ными видами.
Если северные и
восточные части Австра-
лии имеют значитель-
ное сходство с Мале-
Рис. 211. Ареал семейства Cephalotaceae,
строго ограниченного лишь юго-западом
Австралии (по Хетчинсону).
зией, то Западная Авст-
ралия более сходна с
Каплэндом и Южной
Америкой. И действительно, по Вегенеру, связь Австралии
с Индией и Цейлоном порвалась в юрском периоде, а с Южной
Америкой—лишь в третичную эру, когда Австралия отделилась
от Антарктического материка, при посредстве которого была
связь с Америкой.
Австралия представляет высокое плато, приподнятое по краям;
местами вдоль берегов проходят горные цепи; горы задерживают
влагу, и потому центральные части Австралии очень сухи, на об-
ширных территориях представляя пустыни. Лишь восточный бе-
рег находится в весьма благоприятных условиях (до 200см осадков),
почему здесь и развивается влажнотропический лес, ландшафтно
сильно схожий с лесами малезийской подобласти. Здесь очень мио-
'24‘f
37J
гочислениы пальмы и другие тропические деревья. Особо следует
отметить юго-восточный округ с древовидными папоротниками (А1-
sophila, Cyathea, Dicksonia) в смеси с эвкалиптами и протейными
(Proteaceae).
Вся остальная Австралия, в связи с большим или меньшим
количеством осадков, представляет саванны, степи, заросли ку-
старников или пустыни.
Для Австралии наиболее характерен вечнозеленый эндемичный
род из семейства миртовых —эвкалипт (рис. 212); он представлен
Рис. 212. Ареал рода эвкалипт (Eucalyptus) (по Берри).
более чем 400 видами, многие из которых растут насаждениями
Эвкалипты настолько типичны для Австралии, что обычно австра-
лийские ландшафты определяются именно эвкалиптами. Послед-
ние встречаются в самых различных фитоценозах (леса, саванны,
пустыни и др.), представляя разнообразные жизненные фермы: то
это величайшие в мире деревья, до 150 м высотой, то это более
низкие деревья, то это небольшие кустарники. Листья у эвкалип-
тов стоят ребром к солнцу, благодаря чему солнечные лучи прони-
кают до почвы; к тому же, эвкалиптовые леса ие бывают густыми,
давая светлые, редкие насаждения (рис. 103); поэтому в них мало
тени и хорошо развивается травянистый покров.
Другим очень сильно распространенным родом является акация.
она представлена тоже большим числом видов —до 400, встречаю-
щихся в самых различных условиях: в лесах, степях, пустынях.
Для Австралии характерны акации с филлодиями (листовидные
черешки).
Флористически Австралия может быть подразделена иа три про-
винции: 1) северо-восточную, 2) западную и 3) срединную, так назы-
ваемую «эремею».
372
О северо-восточной провинции, захватывающей северное и восточ-
ное побережье, уже было сказано: провинция эремея занимает наи-
большую территорию: всю Центральную Австралию с северо-запад-
ным побережьем и значительней частью южного. Это главным об-
разом саванны и степи, а во внутренних частях —пустыни. Особен-
но развиты ксерофкльиые кустарники, дающие густые, нередко
колючие, непроходимые заросли, называемые «скрэб»; последний
представлен различными типами и в высшей степени характерен
для описываемых территорий. Флористи-
чески он состоит из многочисленных видов
акации, эвкалиптов и некоторых других
растений. Для пустынных частей нужно
указать кустарниковые заросли из эвка-
липтов и видов казуарииы (Casuarina —
рис. 213), древнего рода с безлистными
ветвями.
Западная провинция (вернее юго-за-
падный угол) — это наиболее ориги-
нальная часть Австралии. Здесь
господствуют вечнозеленые жестколист-
ные леса,главным образом из различных
видов эвкалиптов с акациями, казуарк-
нами7и протейными. В саваннах, менее
распространенных, нельзя не отметить
оригинальное древовидное лилейное Хап-
therrhoea (рис. 140).
Интересно еще добавить, что на го-
рах Южной Австралии и в Тасмании,
наряду с австралийскими видами, име-
ются уже некоторые антарктические, как
южный бук (Nothcfagus). В Тасмании,
кроме того, многочисленные хвойные:
Dacrydium, Phy llccladus, Podocarpus
(рис. 214 и 215) и др.
К числу древних элементов Австра-
Рис, 213. Ветвь казуарииы
(Casuarina equisetifolia) с
цветками и плодами (п®
Энглеру).
лии относятся саговники; здесь имеется
два эндемических рода: Macrczamia
(15 видов) и монотипный род Bowenia
(1 вид), кроме того, несколько видов
Cycas.
Австралия очень бедна по
лезными расте-
ниями; например, здесь совершенно отсутствуют местные пло-
довые деревья. Однако некоторые эвкалипты, акации и др. упо-
1ребляются как декоративные растения. Некоторые виды эвка-
липтов высаживают на болотистых малярийных местах; это спо-
собствует уменьшению и прекращецию малярии (например у иас
на Кавказе), так как эвкалипты растут очень быстро и осушают
болота.
Ввезены и культивируются в Австралии очень многие культур-
ные растения: хлебные злаки (особенно пшеница), виноград, ко-
фейное дерево и т. д. Особого очага происхождения культурных
растений Австралия не имеет
с широкими листьями
I—Agathis loranthifoha; 2—Podocarpus purdi-
Рис 214. Своеобразное
«х^о ное’* с ьладодиями
(Phyllocladus rhomboidcs).
Отдезьная ветвь— видно,
что кладодии выходят из
пазух чешуевидных листьев
(го Культеру и Чембер-
лену).
cana. Ветви с листьями (по Культеру я Чем-
берлену).
V. Капская область
Капская область занимает лишь очень небольшую часть Юго-
западной Африки, но имеет все признаки самостоятельной области
Климат в общем напоминает средиземноморский. Флористически
область очень богата; точных цифр не имеется, ио во всяком случае
число видов нужно принять в несколько тысяч (флора Капской зем-
ли имеет около 12 000 видов, но это более обширная территория,
чем капская флористическая область).
Характерная черта — бедность древесными ви-
дам и; почти все деревья, встречающиеся здесь, свойственны тро-
пической Африке; господствуют вечнозеленые кустарники с жест-
кими листьями и многочисленные многолетники.
Очень характерен вечнозеленый род Erica (вереск) с несколь-
кими сотнями видов (более 450), семейство протейных (Proteaceae)
с различными родами (до 260 видов), ряд других семейств (например
Restionaceae, Oxalidaceae, Amaryllidaceae), многочисленные роды
ирисовых, такие роды, как Pelargonium, Helichrysum и др.
Из протейных нужно особо отметить замечательное серебряное
дерево (Leucadendron argenteum), имеющее серебристо-белые листья,
покрытые многочисленными прижатыми волосками.
Флора Капской области имеет многосторонние связи; замеча-
тельна связь ее с флорой Австралии —семейства Proteaceae, Re-
stionaceae, Epacridaceae и др.; число общих родов не менее 20 (между
прочим Drosera, Pelargonium, Me’sembrianthemum); одиако имеет-
374 _
ся и ряд различий: в Капской области отсутствуют акации
с филлодиями и, наоборот, в Австралии отсутствуют сочные мо-
лочаи, виды алоэ и др.
Связь с Южной Америкой очень слаба, хотя и там и здесь при-
сутствуют протейные и некоторые роды, например росянка.
Интересна флористическая связь Капской области с Голаркти-
кой. Не считая родов почти космополитных, как тростник, осока,
ситник и другие, можно насчитать до 50 родов, свойственных и
Капской области п Европе: лютик, лен, фиалка, манжетка, кра-
пива, роза, келерия, ковыль и т, д. Но, конечно, очень миогв
бореальных семейств здесь отсутствует.
Значительные связи, естественно, имеются с флорой более се-
зерйо расположенных частей Африки, но об этом было сказано
зыше при описании южноафриканской переходной провинции
см. стр. 359).
Не может быть сомнения в том, что Южная Африка находилась
з соединении с Австралией, Антарктическим материком и Южной
Америкой, По В еге и е ру, Африка отделилась от Антаркти-
ческого материка лишь в юрском периоде.
Культурные влияния местами не только сильно нарушили перво-
бытную флору, но даже создали новые ландшафты; так, выше было
сказано о почти полном отсутствии местных деревьев, одиако в на-
стоящее время вокруг поселений можно видеть группы различных
деревьев: здесь и дубы северного полушария и сосны из Среди-
земноморья, по берегам рек — ольха. Имеются и другие деревья.
Капская область дала, со своей стороны, целый ряд декора-
тивных растений, разводимых в Европе в садах, оранжереях и пр.
Сюда относятся многочисленные верески с довольно крупными,
очень различно у различных видов окрашенными цветками, раз-
личные виды пеларгонии (в общежитии называемых геранями),
красиво цветущие лилейные, амариллисовые, ирисовые и др.
Местных культурных растений очень немного, но ввезено много
растений из других стран: виноград, плодовые деревья и т. д.
VI. * Антарктическая область
Эта область занимает самую южную часть Южной Америки
(Патагонию и часть Чили), Огненную Землю, ряд антарктических
островов (Фальклэндские, Южная Георгия, Кергелены, Кэмбэлл,
Ауклэнд и др.), огромный Антарктический материк.
Флористически область очень бедна, особенно на юж-
ных островах, находящихся в очень неблагоприятных климати-
ческих условиях Приводим некоторые данные:
Число
видов
Огненная Земля . 615
Фальклэндские острова .................. 115 (только цветковые)
Аукчэнд 85
Кэмбэлл 61
Чэрзмские . . , .......... 5
375
Флора области обнаруживает 'целый ряд черт, роднящих ее
с флорами южных частей Новой Зеландии, Австралии и, в мень-
шей степени, Капской области; так, южный бук (Nothofagus)
встречается иа юге Южной Америки, в Новой Зеландии, в Австра-
лии, род Gunnera (семейство Halorrhagidaceae) — в Южной Аме-
рике, Новой Зеландии, Тасмании, южной Африке и др. (рис. 216
и 217); семейство Proteaceae — иа юге Южной Америки, в Австра-
лии, Южной Африке (и в некоторых других местах); можно
привести и другие примеры.
Рис. 216 Ареал рода Gunnera, указывающий на связи между Южной Амери-
кой, Африкбй, Зондским архипелагом, Тасманией н Новой Зеландиейj
(по Хетчинсону)-
Особенно характерен для области южиый бук (Nothofa-
gus), роды Асаепа (из розоцветных) и Azorella и Bolax (из
"ОНТИЧНЫХ).
На юге Южной Америки в западной части, с ее более влажным
климатом, господствуют леса, в восточной, более сухой, преобла-
дают пространства с растительностью степною характера. В лесах
встречается несколько видов южного бука, дримис из магнолие-
вых (Dnmys), многие хвойные Araucaria imbricata (образует даже
юса — рис 218), Libocedrus, Fitzroya, Saxegothaea; сюда же за-
ходят довольно многочисленные представители тропических се-
мейств из тропической Америки (лавровые, миртовые и др ); много
лиан, эпифитов, папоротников и мхов.
Болота Огненной-Земли крайне сходны с горными болотами Тас-
мании и Новой Зеландии На антарктических островах в связи
с холодным климатом флора очень бедна; имеется много болот и
торфяников с подушками Bolax, Azorella, Асаепа
На Антарктическом материке до сих пор найдено лишь одно
цветковое растение — Aira antarctica, доходящее до 62° ю ш ;
остальная флора состоит из мхов и лишайников
Ископаемые находки иа южных островах указывают на богатую-
древесную растительность, погибшую при оледенении Антаркти-
ческого материка.
376
Нет сомнения, что современная флора Антарктической области
представляет раздробленные и уцелевшие остатки флоры Антар-
ктического материка, связывавшего Америку, Африку и Австра-
лию (связь Антарктики с Южной Америкой продолжалась до чет-
вертичного периода)
VII. Океанические области
В основном здесь господствуют морские водоросли (споровые
растения), из цветковых — не более 30 видов (например род Zos-
tera — морская трава). Однородность среды обусловливает, ко-
нечно, и достаточно однородную флору.
Вопрос о флористических подразделениях океанов ещё недо-
статочно разработан, но во всяком случае можно различать три
екеанические области: северную (бореальную), тропическую и
южную (австральную).
Для тропической области характерны багряные водоросли и
многочисленные виды Saigassum (бурая водоросль), для северной
вбласти—такие роды, как фукус (Fucus), ламинария (Lamina-
ria), лессония (Lessonia) и другге, для южной области — макро-
цистис (Macrocystis), Durvillea и др.
Свободно жив}шие растения, не прикреплённые к морскому
ану, образуют фитопланктон, прикреплённые же около бере-
[ ов — фитобентос. Последний на разных глубинах очень разли-
чен (см. выше, стр. 293).
ПРИЛОЖЕНИЕ 7
КУЛЬТУРНЫЕ РАСТЕНИЯ И ИХ ПРОИСХОЖДЕНИЕ
Культурные и культивируемые растения. Культурными ра-
стениями обычно называют такие растения, которые возделывают-
ся (культивируются) человеком для тех или иных целей,
будучи полезными в том или другом отношении; в дикой
природе они не встречаются. Таким образом, культурные растения
всегда связаны с человеком и всегда прино-
сят ему пользу. Однако ие всякое полезное растение яв-
ляется культурным. В дикой природе имеется очень большое чи-
сло полезных человеку растений, которые используются челове-
ком непосредственно в дикой природе: человек собирает плоды ди-
ких растений (у нас в лесах—дикая малина, черника, брусника
и др.) и употребляет их в пищу; в других случаях человек соби-
рает те или иные части растений (цветки, листья, подземные части
и др) для лекарственных целей (валериана, адонис, наперстянка,
беладоина и т. д.); в иных случаях производится сбор диких ра-
стений потому, что оии дают дубильные, красящие вещества и т. д.
Все только что перечисленные полезные растения являются дико-
растущими и в своём существовании и происхождении со-
вершенно не связаны с человеком.
Однако человеку не выгодно то, что те или иные полезные ра-
стения находятся далеко от его жилья, или же, хотя и произра-
стают от него близко, но разбросаны отдельными редкими экзем-
плярами, что сильно затрудняет сбор их. Поэтому человек стре-
мится иметь все эти полезные растения непосредственно у своег»
жилья и в большом количестве: высевает около жилья лекарствен-
ные растения, переносит из леса кусты чёрной смородины, выса-
живает деревца рябины и др.
Первобытный человек сеял около своего жилья главным обра-
зом семена растений, которые были для него важны в пищ е-
в о м отношении, поддерживая их существование соответствую-
щим уходом; без работы со стороны человека эти растения исче-
зают. Такие растения имеют некоторую связь с человеком, ио всё
же их нельзя назвать культурными. Это — «культивируемые» ра-
стения, ио не культурные. Так, если на наших лугах мы высеваем
такие кормовые растения, как лисохвост, тимофеевку, костёр и
37»
др., собирая их семена в дикой природе, то мы лишь культивиру-
ем эти растения; если они лучше развиты, чем соответствующие ди-
кие экземпляры, то это зависит от внешних условий, например,
•т более разрыхленной и улучшенной почвы и др. Часто при-
знаки культивируемых растений являются модификациями,
т. е. ненаследственными.
Культурные растения не только культивируются человеком,
но обязаны ему большим или меньшим числом своих признаков,
наследственно закреплённых. Человек сам создаёт нуж-
ные для него сорта растений (также и породы животных).
Человек, создавая новые сорта растений или новые признаки
нх, делает это с какой-либо определённой целью, стре-
мясь получуть наиболее для себя полезное растение. Поэтому у
культурных растений обычны признаки, полезные для человека,
но н е полезные и нередко даже вредные для са-
мого растения. Это—характерная черта культурных растений.
Из числа этих признаков укажем некоторые:
1) растения иногда утрачивают приспособления для распро-
странения семян и плодов или эти приспособления становятся
менее ярко выраженными (например стержень колоса культурной
ржи не разламывается, у дикорастущих видов разламывается,
то же имеет место у культурного и дикого овса);,
2) культурные растения в иных случаях теряют различные
защитные приспособления от поедания их животными (колючки,
шипы и др.), становясь почти беззащитными;
3) размеры плодов иногда увеличиваются, и нередко плоды
становятся бессемянными; плоды при этом теряют для растений
всякое значение;
4) иногда цветки становятся махровыми, т. е. тычинки и пе-
стики превращаются в лепестки; цветок теряет своё значение, так
как семян он дать не может. В дикой природе растения с по-
добными признаками были бы обречены на более или менее
•ыстрсе исчезновение.
Возраст культурных растений. Возраст различных культур-
ных растений крайне различен. Относительно некоторых даже
трудно сказать, являются ли оии лишь культивируемыми или же
стали уже культурными. В дикой природе и до настоящего времени
имеются ещё безграничные запасы полезных, но ещё ие исполь-
зуемых растений. Можно ли назвать культурным растением не-
давно открытый в Средней Азии одуванчик кок-сагыз, являю-
щийся прекрасным каучуконосом и засеваемый у нас на многих ты-
сячах га? Невидимому, нет, так как возделываемый кок-сагыз — это
только возделываемое дикое растение. Но недалеко то время, когда
в культуре возникнут формы кок-сагыза, не имеющиеся в дикой
природе и ещё более богатые каучуком; они и послужат материа-
лом для выработки уже культурных растений.
В последние годы в СССР ведётся усиленная работа по пере-
смотру её флоры иа полезность. Обнаружено много новых
полезных растений, и, несомненно, в ближайшее время число куль-
турных растений сильно возрастёт.
380
Однако если многие культурные растения формируются у нас на
1 лазах, то, с другой стороны, известно немало растений, культура
которых относится к очень отдалённым временам. Таковы, напри-
мер, пшеница, ячмень, просо, рис и др., которые возделывались
еще за несколько тысячелетий до начала нашей эры; так, пшеница
возделывалась на равнинах Месопотамии ещё до основания Вави-
лона (6000 лет до нашей эры), жители Египта возделывали пше-
ницу за 4 000 лет, а в Китае культура её известна за 3 000 лет до
нашей эры (кроме того, культура ячменя, риса, проса и сои). Зёрна
пшеницы и ячменя обычны в свайных постройках в Швейцарии,
в Австрии и других частях Западной Европы, относимых к неолиту
(новый каменный век) —от 3 000 до 500 лет до нашей эры. Куль-
турные растения Америки не столь древни, хотя культуры куку-
рузы, табака и др. всё же имеют возраст не менее 2 000 лет.
Таким образом, одни культурные растения являются нашими
современниками, другие имеют уже возраст более 6 000 лет. Есте-
ственно, что в продолжение этих 6 000 лет возник целый ряд
новых культурных растений. Так, например, рожь упоминается
лишь римским автором Плинием (1 век нашей эры) и не была
известна ни в Египте, ии во время свайных построек; гречиха ста-
ла возделываться лишь в средние века, кофе стал известен в XV
веке, кормовая свёкла возникла в XVI веке, сахарная свёкла в
XVII веке и т. д.
Если вопрос о происхождении культурных растений, получен-
ных сравнительно недавно, вполне или достаточно ясен, то не так
обстоит дело с культурами древними, возникшими в доисториче-
ские времена. Культура пшеницы, например, возникла более 6 000
лет назад—можно ли точно указать её предка и восстановить
процесс окультуривания? Для решения этих вопросов возможны
два пути: ботанико-систематический и ботанико-географический.
Основные очаги происхождения культурных растений. Здесь
особенно важны исследования, произведённые в недавнее время
в СССР. Многочисленные экспедиции посетили важнейшие области
земного шара и произвели сбор коллекций возделываемых расте-
ний со всех мест их возделывания. Огромный собранный мате-
риал показал, что очаги культурных растений приурочены глав-
ным образом к горным странам (Гималаи, горы Абиссинии,
Кордильеры в Америке и др., в Старом Свете —полоса между
20 и 40° с. ш.), и древние земледельческие культуры в долинах
рр. Тигра, Евфрата, Инда, Нила и др. являются уже в т с-
р и ч и ы ми. Ив самом деле, долинные культуры, связанные
нередко с искусственным регулированием водного режима, немыс-
лимы без крупных государственных объединений и без значитель-
ных затрат человеческого труда, чего, конечно, при возникновении
человеческой культуры ие было. В горах же небольшие долинки
и склоны легко использовались для культуры даже немногочислен-
ными племенами, ие говоря о том, что горы представляли для
них естественные крепости.
В результате исследований последнего времени установлено 8
основных очагов (центров) происхождения культурных растений:
381
1. Во сто ч н о а з и а теки й. Он схватывает горный цен-
тральный и западный Китай и прилегающие к нему низменные
части. Это самый крупный очаг разнообразия культурных расте-
ний — 136 видов; среди них — просо, гречиха, некоторые клуб-
неплоды и корнеплоды, некоторые плодовые деревья —абрикос,
яерсик, слива, ряд цитрусовых, многие лекарственные растения
и Др-
II. Индо-малайский. Охватывает Индию (Гималаи
и их строги), горы Индо-Китая, часть островов Малайского архи-
пелага. Может быть подраздет ён иа индийский собственно и индо-
малайский (в узком смысле). Первый имеет 117 культурных видов,
второй —55 видов; среди них: рис, сахарный тростник, многие
зерновые бобовые, ряд плодовых тропических деревьев (кокосовая
пальма, цитрусы, банан и др.).
III. Среднеазиатский. Горные части Средней Азии
(Памир, Тянь-Шань и др.). Имеет 42 вида культурных растений: ряд
пшениц (мягкая и др.), основные бобовые (горох, чечевица, кон-
ские бобы и др.), хлопчатник-гуза, ряд масличных растет й и др.
IV. П е р е д н е а з и а т с к и й. Охватывает горные страны
Малой Азии, Закавказья, Ирана и др. Имеет 83 вида, из которых
укажем девять пшениц (в это число входят дикие пшеницы — одно-
зернянка и двузернянка), рожь, виноград, плодовые-деревья: гру-
ша, алыча, айва, миндаль и др., кормовые бобовые —эспарцеты,
люцерна и пр.
V. Средиземноморский. Охватывает страны, ле-
жащие по берегам Средиземного моря: полуострова Пиренейский,
Апеннинский, Балканский; побережья Малой Азии, Патентны,
Северней Африки, включая ньжнюю часть долины р. Игла. Имеет
34 культурных вида, из которых укажем маслину, рожковое де-
рево (Ceratcnia siliqua), различные свошные и др.
VI. Абиссинский. Занимает Абгссинскую горную стро-
ну в верховьях р. Нила. Имеет 38 культурных видов: твёрдую пше-
ницу, ячмень, кофейное дерево (Ccffea ага bip) и др.
VII. Центральноамерика некий. Занимает повы-
шенные страны южной Мексики и Центральной Америки. 49 куль-
турных видов, из которых наиболее важны: кукуруза, тыква,
хлепчатник-упланд, перец, дерево какао и др.
VIII. Южноамериканский. Находится в средней
части Кордильер (Перу, Эквадор, Боливия и др.). Имеет 62 куль-
турных вида, из которых особенно важны: картофель, табак, ана-
нас, земляной орех и др.
Таковы восемь основных очагов. Если же вссточисгзиатский
•чаг подразделить на центральискитайш-вй и на загаднекитай-
ский, индо-малайский подразделить на собственно индгйсы й и на
индо-малайский в более узком смысле и, кроме того, пртбавить
небольшой по числу культурных растений североамериканский
(подсолнечник и земляная груша), то число очагов увеличится до
эдини^дцати.
Из приведённых данных видно, что некоторые культурные ра-
стения имеют один определённый очаг (пентр) своего возникневе-
382
ния (например кукуруза, рожь, подсолнечнику другие возникли
в своих разных сортах в различных, иногда очень отдалённых
ечагах (например хлопчатник имеет два очага: Центральную Аме-
рику и Среднюю Азию, пшеница имеет три очага —среднеазиат-
ский, переднеазиатеккй и абиссинский, груша — восточиоазиат-
ский и переднеазиатский и т. д.). Из этого уже ясно, что нельзя
говорить об одном предке какого-либо культурного растения,
например пшеницы.
Интересно, что одни части земного шара дали очень много куль-
турных растений, другие не дали почти ничего. Так, Южная Азия
дала более 400 видев, т. е. д в е трети всего мирового разно-
образия культур; с другой стороны, две такие большие террито-
рии,как Австралия и Южная Африка, почти не дали никаких куль-
турных растений. Последнее обстоятельство объясняется, по Дар-
вину, не тем, что там не было местных растений, которые могли бы
послужить материалом для выведенья культурных видов, а тем,
что там не было туземных земледельческих племён, которые могли
бы рспсльзсвать этот матерь ал.
Происхождение культурных растений. На примере Австралии
мы только ч;о видели, что noj езные в том или другом отношении
дикие растения ие становятся культурными, если человек не ска-
зывает иа эти растения известного воздействия. При этом, конечно^
воздействия, которые оказывал человек в доисторические времена
и в настоящее время, очень различны.
По Дарвину, основным приёмом получения улучшен-
ных сортов растений у первобытного человека был бессозна-
тельный отбор. Этот отбор (как и всякий другой) мог иметь
место лишь при условии изменчивости растений, что дей-
ствительна и наблюдается в природе —ведь нельзя найти двух
•собей одного и того же вида, совершенно похожих друг на друга
во всех своих признаках. Бессознательный отбор у первобытного
человека состоял в том, что безотчётно отдавалось,предпочтение
особям какого-либо вида с теми или другими полезными для чело-
века признаками. Эю делалось при сборе пледов, семян и др., как
в местах их дикого произрастания, так и тогда, когда для удобства
сбора эти растения высаживались или высеивались непосредствен-
но близ жилищ. Так, например, при высеивании злаков зерновки
собирались, естественно, с особей, имеющих более крупные
зерновки, при посадке плодовых деревьев выбирались особи с более
крупными и более вкусными плодами и т. д.
Такой бессознательный отбор продолжался долгое время
и происходил очень медленно. Несомненно, он продолжался ты-
сячелетиями, шаг за шагом накоплялись мелкие полезные признаки,
что и привело к тому, что данное растение стало,сильно отли-
чаться ст своих предков. Вот почему многие культурные растения
уже в самом начале человеческой истории как бы совершение
готовы во всех своих основных признаках. Вот почему так
трудно и часто совершенно безуспешно искать непосредственных
предков культурных растеши—весь процесс формирования по-
следних совершался в далёкие доисторические времена.
3bS
Наглядным примером длительности бессознательного отбора
может служить культура груши. Нет сомнения, что плоды дикой
1 РУ ши служили пищей человеку с самой глубокой древности; ио,
по свидетельству П л н н и я, ещё и в Риме груши были очень
низкого качества.
Бессознательный отбор продолжался до сравнительно иедавнег»
времени, когда он уступил место отбору сознательному,
при котором человек уже ставил себе определённую цель и доби-
вался её ^методически и более или менее быстро. Таким образом,
время действия сознательного отбора, в противоположность от-
омру бессознательному, обычно очень невелико.
Метод сознательного отбора впервые был применён в середине
прошлого века Вильмореном для получения повышен-
ного содержания сахара в клубнях свекловицы; этот метод отбора
с целью улучшения какого-либо сорта в тех или иных его призна-
ках называется селекцией. Если сознательный, искусственный,
отбор, в связи со скрещиванием, является, по Да рви ну, главной
движущей силой при выработке культурных растений, то не-
малое значение имеют и некоторые другие методы, применяемые
главным образом за последнее время (см. ниже).
Чго касается скрещивания, то оно даёт возможность
быстро вводить в данное растение те или иные полезные нам при-
знаки, отсутствующие в нём. При скрещиваниях селекционер стре-
мится создать тот ассортимент признаксв, который для него наи-
более желателен. В своё время и Дарвин указывал на роль
скрещивания при выработке культурных растений.
- Не считая скрещивание (гибридизацию) исключительным ме-
тодом, всё же нужно отметить очень большую его роль в вопросе
происхождения культурных растений. Яркой иллюстрацией зна-
чения гибридизации служат работы Мичурина и его после-
дователей.
В своих работах по выведению новых сортов плодовых растений
М и ч у р и н, как известно, применил впервые целый ряд нс вых
методов: I) метод подбора пар растений-производителей, 2) метод
«посредника», 3) метод предварительного вегетативного сближения
и др.
Первый метод, важнейший в учении Ми чурина, состоит
в том, что при скрещивании (гибридизации) берутся пары растений-
производителей, очень удалённых по месту своего произ-
растания; так, например, брались такие пары: материнское ра-
стение— дикая уссурийская груша, отцовское—французский
культурный сорт бере-рояль; материнское растение —дикий ус-
сурийский виноград, отцовское — коринка греческая и т. д. Пре-
имущество скрещивания географически удаленных родителей со-
стоит в том, что избегается доминирование местиы^ неблагоприят-
ных признаков, наоборот, вводятся новые полезные 'признаки.
Если в этом методе гибридизация не выходила за пределы ви-
дов одного и того же рода, к тему же видев более или менее близких
между собой (близкие виды груши, винограда и др.), т° при втором
методе дело шло об отдалённой межвидовой’ гибридизации:
334
вишня—черёмуха, абрикос —слива и др. (все эти растения при-
надлежат к одному роду Prunus в широком его понимании) и даже
межродовой (яблоня —вишня,яблоня —абрикос), что до послед-
него времени считалось невозможным (потомство оставалось бесплод-
ным). Ми чу р и н, при невозможности непосредственного скре-
щивания подобных видов’; сначала выводил про м е ж у точ-
ную форму — «посредник», которую затем и скрещивал с дру-
гой родительской формой (получались указанные выше гибриды,
т. е. гибриды межродовые).
При методе предварительного вегетативного сближения
черенки гибридного растения в самом раннем его возрасте (1—2 года)
прививались в крону того взрослого растения, с которым было же-
лательно его скрестить; при этом подвой оказывает сильное воздей-
ствие на привитые ранневозрастные черенки.Это воздействие про-
должается до цветения привоя, и в дальнейшем скрещивание про-
исходит более легко. Таким путём были получены гибриды между
рябиной и грушей, айвой и грушей и др. Не касаясь ещё некоторых
других методов (например, метод применения смеси пыльцы раз-
личных видов для обеспечения отдалённых скрещиваний), можно
видеть, что для получения новых сортов растений Мичурин
на первое место ставил гибридизацию.
Большую роль гибридизации при получении новых форм
признаёт и Лысе нко. Он предложил в качестве приёма для
повышения урожайности и для улучшения свойств растений ис-
кусственно производить в иутрисо ртовое скрещивание у
са моопыл я ющи хся сортов (например, у пшениц). Пои
этом будут скрещиваться растения с несколько иными свойствами,
в то время как при самоопылении обновления не происходит; в
результате — более жизнеспособное потомство.
Замечательны работы Державина, Цицина и неко-
торых других по выведению многолетних сортов тех куль-
турных растений, которые до сих пор являлись лишь однолетни-
ми; таковы наши культуры ржи, пшеницы, подсолнечника, сорго,
такие кормовые'растеиия, как вика, чина и др. Преимущества мно-
голетних культур совершенно очевидны,' причём главное их пре-
имущество — это их большая урожайность сравнительно с одно-
летними, что стоит в зависимости от их более мощной корневой
системы, заполняющейся к осени запасными питательными веще-
ствами. Ц и ц и н путём скрещивания однолетней пше-
ницы (культурной) и многолетнего дикого пырея получил
многолетние пшенично-пырейные гибриды,
Державин получил многолетние сорта путём скре-
щивания культурных однолетних сортов ржи, подсолнечника,
сорго и др. с многолетними дикими видами тех же родов. Кроме
того, многолетнюю высокоурожайную пшеницу путём .гибриди-
зации пшениц с многолетней рожью (р ж а н о- п ш е н и ч н ы е
гибриды).
Нет сомнения, что и в дикой природе скрещивание играло боль-
шую роль при возникновении основных культурных растений.
А потому правильно ли искать одного предка того или дру-
25 География растений 385
того растения? Не правильнее .ци искать г р у п п у видев,
которая дала основной материал при скрещивании? Послед-
нее положение выдвигается некоторыми исследователями (К с-
м а р.о в).
Помимо искусственного отбора и гибридизации, большую роль
в получении новых видов и сортов растений некоторые авторы при-
писывают мутациям. Под мутациями понимают внезапно
появившиеся, но наследственно стойкие формы, в тех или иных
признаках резко отклоняющиеся от других особей того же вида
(К о р ж и н с к и й, де Фриз). Мутации отбираются и слу-
жат материалом для дальнейших селекционных работ. Мутации
очень нередки среди культивируемых растений, например формы
деревьев с пирамидальной кроной, формы с плакучими ветвями,
с сильно разрезными листьями, с махровыми цветками, с различ-
ной окраской цветков, с различной окраской плодов (например у
кукурузы) и т. п. Однако в происхождении большинства
наших культурных растений мутации, как показывают соответст-
вующие изыскания, особой роли не играли.
Средн методов получения новых свойств культурных расте-
ний нужно указать давно известный метод п р и в и вок, при-
меняемый главным образом на древесно-кустарниковых растени-
ях. Для нас особенно здесь важно влияние привоя (прививаемое
растение) на подвой (растение, на которое прививают). В целом
ряде случаев это влияние сказывается очень сильно; так, выше
оыло сказано о методе вегетативного сближения Мичурина.
Другим методом Мичурина в данном направлении является
метод «ментора», который состоит в том, что' здесь в крону молодо-
го тбридного растения прививаются черенки сортов, обладающих
теми или иными другими желательными свойствами, например
особой урожайностью, особой зимостойкостью, более развитой
корневой системой и др. Привой при этом оказывает соответствую-
щее влияние на подвой: повышается урожайность, зимостойкость
и пр. В других случаях мы знаем примеры влияния подвоя на при-
вой, например при прививке обыкновенной яблони на морозостой-
кой сибирской повышается морозостойкость первой; то же имеет
место при прививке мандарина на трехлистном померанце (Citrus
trifoliatus) и др,
Таковы основные методы получения новых сортов растений и
улучшения тех или иных признаков их. В ряде случаев при вы-
ведении новых сортов происходит некоторая перестройка
хромосомного аппарата, как основного носителя наследственности.
Новые формы получаются, кроме того, в результате увелы
ч е н и я числа хромосом, именно, формы с тройрым, четвер-
ным и др. набором хромосом (полиплоиды).
Эти полиплоиды отличаются рядом признаков как анатомиче-
ского, так и морфологического порядка. Но особенно важно при
этом увеличение в большинстве случаев размеров как все-
го растения, так и его клеток —явление гигантизма.
Интересно то, что формы с измененным числом хромосом по-
лучаются как в результате мутаций, так и некоторых искус-
386
стве иных воздействий: хлоре формирование, рентгенизация,
действие радия, обработка различными реактивами и др. Эти
достижения последнего времени открывают новые перспек-
тивы в дальнейшей работе по получению более совершенных
сортов культурных растений.
Из искусственных воздействий укажем на интересные новей-
шие работы по обработке алкалоидом колхицином семян
нли взрослых частей растений (колхицин получается из растений
рода Colchicum). При этом получаются триплсиды (тройное число
хромосом), тетраплоиды (четверное) и даже октоплсиды (восемь
наборов хромосом). Этот новый метод (1937) в настоящее время
очень усиленно разрабатывается. Не менее эффективно воздей-
ствие другого вещества —аценафтена.
Таковы методы получения новых видов и сортов растений во-
обще и культурных сортов в частности. Очень большое число сор-
тов возникает у нас на глазах; некоторые примеры этого были ука-
заны выше.
Число культурных и культивируемых растений очень велико
(20 000—30 000), из них около 1 000 видов являются более или ме-
нее широко распространёнными, не считая многочисленных де-
коративных растений.
Культурные растения подразделяют на группы, на основе тех
или других признаков. Так, их иногда делят на три группы:
1) хлебные растения (хлебные злаки, гречиха), 2) кормо-
вые растения (корнеплоды, злакй, бобовые и др.); 3) техниче-
ские растения —прядильные, масАичные, красильные и т. п.
Можно также делить по характеру продуктов, производи-
мых растениями: крахмалоносные, белковые, сахароносные, мас-
личные, дубильные, каучуконссы, красильные и пр. Иногда делят
по способу культуры: полевые, овощные, бахчевые, садовые и пр.
По времени введения в культуру растения полевой куль-
туры распадаются на две группы:
Первая группа. Основные древние культуры — пер-
вичные культуры; они известны лишь в культурном состоянии
(пшеница, кукуруза, хлопчатник, лён и т. п.).
Вторая группа, Культуры, возникшие из сорняков,
засорявших основные первичные культуры, — вторичные
культуры (рожь, овёс, некоторые виды картофеля, горчица и др.).
В дальнейшем мы остановимся лишь на некоторых более важ-
ных культурах и выясним их происхождение. Однако полной до-
стоверности в ряде случаев здесь быть не может, так как происхо-
ждение многих культур относится к глубочайшей древности.
I. Некоторые культурные растения умеренной полосы
Пшеница (Tri tic urn). В Новый Свет и в Австралию культура
пшеницы проникла, как достоверно известно, лишь в новые века,
поэтому предков её нужно искать в Евразии и в Африке. Однако
вопрос очень осложняется дем, что «пшеница»—это целый ряд
видов, сильно отличающихся между собой. Они отличаются как
25* 3«7
морфологически, так и своим набором хромосом. Так, отличают
14 видов пшениц, подразделяя их на три группы: 1) виды с 7 хро-
мосомами, так называемые однозернянки: Triticum monococcuni
(дикая) и Т. spontaneum (культурная); 2) виды с 14 хромосомами:
твёрдая пшеница (Т. durum), английская (Т. turgidum), полба эммер
(Т. dicoccum), дикая двузернянка (Т. dicoccoides) и др.; 3) ви-
ды с 21 хромосомой: мягкая пшеница (Т. vulgare), карликовая
(Т. compactum) и др. Эги виды в свою очередь распадаются на целый
ряд более мелких систематических единиц, число /Ке селекцион-
ных сортов крайне велико, почти безгранично. У мягких пшениц
установлено 166 признаков, у твёрдых 78 признаков, имеющихся
в различных комбинациях.
Пшеница—это основная первичная культура, возникшая в
доисторические времена (Передняя Азия, Египет, Китай и др.).
Есть указания, что культура пшеницы была известна за 50 000
лет до нашего временц. Вообще это—древнейшее культурное
растение. Говорить о происхождении пшеницы вообще нельзя,
нужно говорить о происхождении каждого вида пшеницы.
В настоящее время, по мнению некоторых ученых, очагом сор-
тового разнообразия большинства пшениц с 21 хромосомой (мно-
гочисленные формы мягких пшениц, сорта карликовой пшеницы
и др.) является территория между Гиндукушем и Гималаями (сред-
неазиатский очаг). Для твёрдых пшениц (14 хромосом) очагом сор-
тового разнообразия является Северная Африка, особенно Абис-
синия (абиссинский очаг). Для наибольшего числа сортов однозер-
нянки (с 7 хромосомами)и некоторых других видов очагом является
Малая Азия (также Крым) — переднеазиатский очаг.
Можно ли считать однозернянку за наиболее древний вид, дав-
ший начало всем остальным? Некоторые прежние авторы действи-
тельно придерживались этого мнения, однако, как выяснилось,
однозернянок нет на родине как мягких, так и твёрдых пшениц.
Вывод такой — разные виды имеют различные пути про-
исхождения. Однако несомненно то, что эволюция пшениц шла в
направлении увеличения числа хромосом (7—14—21), причём
элементарная группа с 7 хромосомами имеет дикорастущего пред-
ставителя (дикая однозернянка —Т. aegilopoides), группа 14-хро-
мосомная (твёрдые пшеницы) имеет также одного дикорастущего
поедставителя на всю эту большую группу (дикая двузернянка —
Т. dicoccoides), группа мягких пшениц (21 хромосома) —совер-
шенно не пмееет дикорастущих видов.
Имеются данные о том, что культурная однозернянка произо-
шла от однозернянки, дико растущей в восточной части Среди-
земноморья, и что культурная двузернянка произошла от дико-
растущей двузернянки (Сирия и Палестина), но это не решает
вопроса о происхождении других видов пшениц и особенно мяг-
ких. Если нет других близких дикорастущих предков, то нет ли
родов, близких пшенице, с которыми она могла бы давать гиб-
ридные формы? Оказывается, та£ие роды имеются: эгилопс (Aegi-
lops), пырей (Agropyrum), рожь (Secale); о ржано-пшеничных гиб-
ридах уже говорилось выше. Особенно близок к пшенице род эги-
388
л one, который имеет четыре вида и обладает очень большой измен-
чивостью, причём интересно то, что изменчивость этого растения
вполне параллельна изменчивости пшеницы, а имен-
но, имеются мягкие и твёрдые формы, остистые и безостые, с
различной окраской колоса и т. д. Между видами эгилопса и пше-
ницей имеются различные гибриды, что показывает их близкое
родство. Можно думать поэтому, что твёрдые пшеницы (кроме дву-
зернянки) имеют происхождение от видов рода эгилопс в ре-
зультате их скрещивания как между собой, так и с другими близ-
кими родами (Agropyrum и др.). Мягкие пшеницы произошли от
твёрдых пшениц. Кроме скрещиваний, большую роль при этом
играл сначала бессознательный, а затем сознательный отбор.
Дикорастущие однозернянки (Triticum aegilopoides), двузер-
нянка (Т. diccccoides) и виды рода Aegilops распространены в вос-
точном Средиземноморье или в Малой Азии, и потому можно ду-
мать, что именно здесь место возникновения ьультурных сортов
пшеницы. Что касается двух других очагов сортового разнообра-
зия, пшениц— абиссинского (твёрдые пшеницы) и среднеазиат-
ского (мягкие пшеницы), то не являются ли эти очаги вто-
ричными?
Имеются и другие мнения относительно происхождения пшениц.
Так, Фляксбергер думает, что пшеница возникла ещё
в третичное время и была сорным растением около поселений до-
исторического человека. Человек взял её в культуру такой, ка-
кой она была в природе. На это заключение наводит его тот факт,
что, по его исследованиям, зёрна пшеницы из свайных построек
Швейцарии очень сходны с различными формами современных
пшениц. Однако взгляд этот встречает целый ряд возражений,
да и кроме того невозможно допустить, чтобы культурные растения
оставались почти неизменёнными столь продолжительное время.
Культура пшеницы как в мировом масштабе, так и в пределах
СССР, по площади занимает первое место среди всех
других культур —всего около 150 млн. га. Обычно культура
пшеницы связывается с более тёплым климатом и с более богаты-
ми почвами. Однако селекция более морозостойких сорто.в и со-
ответствующий уход за посевами даёт возможность преодолевать
северные пределы культуры, бывшие в дореволюционное время.
Так, до революции пшеница возделывалась главным образом в
зоне степей и в подзоне широколиственных лесов, в настоящее же
время она культивируется даже в подзоне хесйтых лесов север-
ного типа (напргмер в Архангельской области). Вообще в лесней
полосе посевная площадь пшеницы увеличилась с 1913 по 1937 г.
в 9 раз, а в Московской области — в 278 раз.
Число сортов пшеницы, культивируемых у нас, очень велико.
Различают формы яровые и озимые, остистые и безостые и др. Боль-
шое значение имеют твёрдые яровые пшеницы, так как из них по-
лучают лучшие сорта крупчатки, лучшие сорта макарсн, манная
крупа и пр. Различные сорта этих пшениц таковы: арнаутка, ру-
банка, белотурка, гарновка и др. Среди мягких гшеьгц можно
указать белоколосую безостую (белоколоска, полтавка, улька,
гирка белая и др,), красноколосую безостую (красноколоска, гир-
ка), белоколосую остистую (русская, русак) и др.
Рожь (Secale cereale). Культура ржи сравнительно с пшеницей
очень молода. Зёрна ржи совершенно отсутствуют в
египетских пирамидах, в свайных. постройках Швейцарии и др.,
где обычно имеются зерна пшеницы и ячменя.
Впервые, как кажется, говорит о ржи римский писатель Пли-
ний. Рожь возникла в Старом Свете, так как в Америке во время её
открытия ржи не было. Интересно, что в Риме хлеб из ржи не особен-
но ценился; так, П л и н и й указывает, что ржаной хлеб годится толь-
ко для утоления голЬда и в высшей степени противен для желудка.
В настоящее время площадь Жультуры ржи составляет около
45 млн. га, т. е. в 3 с лишним раза меньше площади культуры пше-
ницы, и рожь стоит цо занимаемой площади на пятом месте. В СССР
рожь по площади занимает второе место (около 22 млн. га). Однако
сравнительно недавно рожь по площади была на первом месте. Это
объясняется тем, что рожь вообще менее требовательна к климату н
почве и могла давать известные урожаи даже при крайне отсталом
крестьянском „хозяйстве. Ещё недавно писалось, что рожь являет-
ся главнейшим хлебным злаком славянских народов, а также
немцев. Однако для СССР этого сказать уже нельзя.
Вопрос о происхождении ржи крайне интересен. Ещё недавно
предком ржи считали те или другие дикорастущие виды ржи, имею-
щиеся на Кавказе или на юге Европейской части СССР. Так, дикая
Sccale fragilis растёт близ Херсона, и, значит, культура ржи могла
возникнуть на юге СССР. Однако последние исследования застав-
ляют совсем иначе смотреть на данный.вопрос. Именно, они обнару-
жили: 1) крайнее разнообразие сортов ржи в Афганистане, Таджики-
стане, Иране, Армении и некоторых прилегающих странах, т. е.
там, где культура ржи имеет очень небольшое значение, 2) сорный
характер ржи в этих странах в культурах пшеницы и ячменя.
Рожь—это сорняк среди посевов. >
Рожь менее прихотливое растение по сравнению с пшеницей, и
потому при продвижении на север и в горы древних куль-
тур (пшеницы или ячменя), имевших рожь в качестве сорняка,
рожь, как менее прихотливое растение, брала верх над культур-
ными растениями, н эти последние совсем выпадали. Оставалась,,
таким образом, одна рожь, сделавшаяся новым культурным
растением. Это имело место в более северных сравнительно с Афга-
нистаном и Туркестаном странах, а именно на территории Евро-
пейской части СССР, в Сибири, в Германии.
Таким образом, рожь, у- это вторичная культура, причём
здесь мы имеем крайне интересный пример культурного растения,
получившегося в результате естественного отбора (а не искурствек-
ного, как это обычно для культурных растений).
Интересно ещё отметить, что сорная рожь переднеазнатского
очага очень разнообразна в своих формах, культурная же рожь
является довольно однородней, утерявши свое сортовое разно-
образие. Конечно, и культурная рожь имеет ряд сортов, но они раз-
личаются не так сильно'
390
Ячмень (Hordeum sativum). Пъ п лоща пи культуры ячмень в
мировом масштабе занимает шестое место (37,8 млн. га), в СССР —
четвёртое место; впереди его стоят пшеница, рожь, овёс (из мировых
I ультур, кроме того, кукуруза и рис).
Ячмень является как кормовым растением для скота, так и
употребляется для пивоварения, а также даёт муку и крупу. Это
вообще неприхотливое растение как в отношении климата, так и
почвы, поэтому оно идет очень далеко на север и высоко в горы;
также разводится на юге в степях, в Туркестане и др.
Hordeum sativum —сборное название нескольких видов н мно-
гочисленных сортов. Ячмень — это древняя, первичная куль-
тура, он был известен за 4 ООО лет до нашей эры. Найден в древней-
ших памятниках Вавилонии, в древнейших египетских гробницах
и т. д. Ячмени делят на два основных вида — двурядный
(Н. distichum) и многорядный (Н. polystichum), а последний
в свою очередь подразделяется на четырёхрядный и шестнрядный.
Некоторые авторы считают, что ячмени произошли от дикого
двурядного ячменя (Н. soontaneiim), который и сейчас растет в
диком состоянии в Средней и Передней Азии (Туркменистан, За-
кавказье, Палестина, Месопотамия и др.). Однако этот дикий ячмень
не имеет того сортового разнообразия, -каким отличается культур-
ный ячмень (этот последний даёт крайнее разнообразие сортов:
остистые, безостые, широколистные, широкочешуйчатые и т. д.).
Кроме того, как показали последние исследования, ячмени имеют
не один очаг сортового разнообразия, а два, именно китайский и
особенно абиссинский (имеет 18 эндемичных разновидностей, не
считая сортов), причём скрешивание некоторых видов из этих двух
очагов не даёт хороших результатов, что говорит об отдалённости
этих вит’гв. Однако, если считать первичны^ очагом культуры
ячменя Юпо-западную Азию, то указанные очаги сортового разно-
образия, быть может, являются вторичными и сильно разошедши-
мися в своих сортах.
Но имеются и другие точки зрения. Так, Ши ма н' считает
первичными не двурядные, а шестирядные ячмени (первые — это
редукционные типы), а Ларионов предками ячменей считает
виды рода Elymus (колосняк), у которых колоски сидят на оси
колоса по нескольку пучками, причём шестирядные и двурядные
ячмени ведут своё начало от общего предка (путём уменьшения
до двух числа колосков в пучке).
Овес (Avena sativa). Среди культур земного шара по своей пло-
шпди овёс занимает четвёртое место (более 55 млн. ад), в СССР овёс
съ пт на третьем месте по площади (около 18 млн. га). Культура
овса особенно развита в северных странах, но имеется также и в
южных странах, где овес главным образом возделывается в горах.
Овёс в своих признаках не является однородным, а даёт крайнее
разнообразие форм (формы метёлок, присутствие или отсутствие
с~тей, различный цвет колосковых чешуй, опушение зерновок и т. п.).
Ччсло форм очень велико.
Культура овса имеет значительно меньшую древность сравнитель-
но с пшеницей и ячменём. В древнем Египте она была неизвестна,
391
однако зёрна овса находят в свайных постройках Европы бронзовог»
века и в поселениях Германии, современных им. Римляне удивля-
лись, что германцы пекут хлеб из такого варварского растения.
Вероятным предком овса ещё недавно считали дикий овёс —
овсюг (A. fatua), который столь обычен у нас, особенно на юге, в
посевах культурного овса (у него более мелкие зерновки и ломкая
ось колосков). Однако мы теперь знаем, что имеется ряд видов
культурных овсов и, естественно, нужно говорить об истории каждог»
из них. Повидимому, их происхождение имело место в разных ме-
стах. Так, указывается пять центров происхождения культурного
овса: 1) Средиземноморье, 2) Абиссиния, 3) Северо-западная и Север-
ная Европа, 4) Азия ci Закавказья до Китая (здесь место происхожде-
ния нашего овса—A. sativa), 5) Китай. Каждый из этих центров имеет
свои особые культурные овсы, резко отличные друг от друга; так?
в Китае имеются овсы с голыми крупными зерновками (A. nuda),
в Абиссинии —совершенно своеобразные овсы (A. abissinica) и пр.
Каким же образом дикие овсы сделались культурными расте-
ниями? Здесь произошло то же, что выше было указано для ржи.
Именно, в Иране, при совершенном отсутствии овсяных культур,
овёс (A. sativa) встречается лишь в посевах полбы в качестве обыч-
ного сорняка. И в других местах культуры полбы овёс всегда нахо-
дится в этих культурах в виде сорняка (сама полба — одна из древ-
нейших культур). С переносом культуры полбы на север или
в горы, т. е. в более суровые условия, полба стала выпадать,
и овёс сделался самостоятельной „культурой. То же.
повидимому, имело место и в других центрах происхождения овсов,
но в каждом центре были свои особые сорные виды; овёс всюду
являлся вторичной культурой.
Просо обыкновенное (Panicum miliaceum). Среди мировых культур
занимает по площади восьмое место (около 25 млн, го), в СССР—
шестое. Сравнительно с пшеницей занимает несколько более южные
места (очень засухоустойчиво), в горы идёт менее высоко. Это —
древняя культура, по древности, повидимому, не уступающая
пшенице (древний Египет, свайные постройки и др.). В Америку
просо было привезено из Европы. Всего имеется около 50 сортов,
причём различия довольно невелики; сравнительно с другими куль-
турами (например с пшеницей) просо довольно однородно.
Предки проса неизвестны, так как дикое просо до сих пор не най-
дено. Большая однородность проса позволяет предполагать, чте
культурное просо имеет лишь одного предка.
Гречиха (Fagopyrum esculentum). Среди мировых культур зани-
мает по площади скромное место (находится в третьем десятке), в
культурах СССР —десятое. По сбору зерна СССР занимает первое
место. В некоторых западных странах (например Англия) гречиха
имеет лишь кормовое значение или запахивается в качестве зелёноге
удобрения. Гречиха малотребовательна к климату п почве, и потому
её культуры продвигаются далеко на север. Наша обычная гречиха
разводится в двух разновидностях: одна, с крылатыми плодами,
культивируется в восточных областях и в Сибири; другая с тупыми
рёбрами плода —на западе н в Западной Европе. Существование
392
этих двух разновидностей, невидимому, связано с тем, что гречиха
проникла в Европу двумя путями: в Россию в средние века, в Запад-
ную Европу из Турции в XV—XVI веках. Дикого предка нашей
гречихи указать нельзя, можно предполагать, что она была введена
в культуру в Гималаях, будучи раньше одним из местных сорняков.
Сорный характер гречихи подтверждается тем, что другой вид
гречихи — «дикуша» (F, tataricum) массами встречается в днксм
состоянии в Сибири, а местами является тягостным сорняком пше-
ницы. Этот вид тоже иногда культивируется, но особого распростра-
нения не получил. Итак, гречиха, можно думать, вторичная
культура.
Картофель (Solanum tuberosum). Наравне с зерновыми культу-
рами картофель занимает очень видное место; среди мировых культур
по площади находится на девятом месте, в СССР занимает пятое место.
Его называют то овощным растением, то «корнеплодом» (однако,
клубень картофеля не утолщённый корень, а подземный стебель).
Родина картофеля — Южная Америка, где его куль-
туры появились задолго до прихода туда европейцев. Первые сведе-
ния о картофеле относятся к 1553 г. В Европу картофель попал
прежде всего в Испанию (в XVI веке), а затем в Англию, Францию,
Германию; в последней в XVIII веке он упрочился. В России кар-
тофель появился в начале этого же столетия, но лишь за столом
богатых людей; в конце XVIII века правительством делались попыт-
ки распространить культуру картофеля среди населения, однако
эта мера успеха не имела вследствие инертности местных властей.
-Новые меры были предприняты у нас в 40-х годах прошлого столетия
после очень сильного неурожая в 1839 г.; с тех пор культура карто-
феля у нас упрочилась, хотя и не вполне безболезненно, так как
крестьяне подозревали правительство, столь решительно насажда-
ющее картофель, в каких-то замыслах, предпринятых во вред им.
Отсюда —ряд волнений («картофельные» бунты).
В Южной Америке культура картофеля, несомненно, восходит
к доисторическим временам. Предком культурного картофеля не-
редко считают дикорастущий в Чили Solanum tuberosum, однако
вероятнее всего ои находится там в одичалом виде. Но в Южной
Америке немало действительно дикорастущих видов картофеля с
подземными клубнями. Скорее всего картофель — это результат
скрещивания ряда видов. Последнее подтверждается частым бес-
плодием сортов картофеля —признак гибридов, а также необычай-
ным сортовым его разнообразием (более 1 000 сортов), так как труд-
но предположить, чтобы таксе разнообразие могло иметь место при
одомашнении одного дикорастущего предка. Були так смотреть
на дело, то Solanum tuberosum в дикой природе никогда не суще-
ствовал.
Интересен вопрос, связанный с вырождением картофеля в более
тёплых странах, о чём, однако, было сказано выше (см. стр. 131).
II. Некоторые более южные культуры
Кукурузу (мане —Zea mays), Играет исключительную роль
среди мировых культур, по площади занимает второе месте
39?
^первое —пшеница); в СССР по площади занимает восьмое место.
Происхождение —американское (это единственный американский
хлебный злак); в Европе первые указания относятся к 1500 г.
Ко времени открытия Америки кукуруза возделывалась там везде,
где позволяли это климатические условия; возникновение культуры
восходит, невидимому, к очень отдалённым временам. В настоящее
время кукуруза распространена широко во всех южных странах,
в Европе — в средиземноморских странах, у нас — в южных частях
Союза.
Сортовое разнообразие кукурузы крайне велико, известно более
300 разновидностей и несколько тысяч различных сортов. Кукуруза
обладает сгчень сильной изменчивостью в результате климатических
влияний, а также подвержена мутационным изменениям.
Кукуруза даёт муку, крупу, сахар, спирт, масло (из зародышей
зерновок), корм для скота и пр., поэтому при селекции кукурузы
преследуют те или иные цели.
Днкой кукурузы не обнаружено; по ряду-своих признаков она
~оит особняком среди злаков (женское соцветие —початок
с мясистой осью).
Вопрос о происхождении кукурузы связывается с мексиканским
злаком теосинте (Euchlaena mexicana), возделываемым главным
сбоазом в качестве кормового растения. Основное отличие его от
кукурузы то, что у него початка не образуется, а зёрна сидят на осн
не густо. Одни авторы считают, что кукуруза произошла от скрещи-
вания теосинте с каким-то другим дикорастущим злаком, другие —
от скрещивания между собой её отдельных рас и разновидностей
без привлечения доугих видов. Указывается ещё один возможный
путь — мутация. Именно, початок кукурузы можно рассматривать
как фасциацию (сращение) боковых двурядных ветвей соцветия
теосинте (фасциация же — оДнн из видов мутации).
Итак, кукуруза имеет предком теосинте и, вероятнее всего,
произошла от него путём мутации.
Подсолнечник (Helianthus annuus). В мировом масштабе под-
солнечник стоит по площади его культуры в самом конце второго
десятка, в СССР же —на седьмом месте. Это уже одно указывает
его, роль в нашем хозяйстве. Культуры подсолнечника —преи-
щественно на юге н юго-востоке Союза. Сплошные культуры на
бани известны всякому, проезжавшему по этим местам.
Подсолнечник имеет очень многочисленные применения., но
’вным образом известен как масличное растение, кроме
'э, как лакомство, корм для скота (жмыхи), топливо, удобрение,
' медонос и пр.
Подсолнечник — американское растение; в Европе стал культи-
ловаться с XVII века, в СССР же культура его имеет лишь столет-
й возраст. В Америке подсолнечник был в культуре ещё, пови-
димому, задолго до её открытия европейцами.
Подсолнечник и в настоящее время встречается в диком состоянии
в юго-западных штатах Северной Америки. Кроме того, там же
имеется ряд однолетникх и многолетних видов этого же рода. Нет
сомнения, что дикий Helianthus annuus дал начало культурному
’стеникк Однако, быть может, в возникновении последнего приняли
;астие и другие дикие виды подсолнечника, образуя помеси.
Наибольшее разнообразие сортов подсолнечник дает в СССР;
'о объясняется, невидимому, тем, что ввезенный материал был
знороден в своих формах, а кроме того, и тем, что именно в СССР
дсолнечник усиленно возделывается и, значит, здесь и йужно
кидать разнообразия форм.
Табак (Nicotiana). Когда европейцы открыли Америку, то
курение табака там было широко распространено, В Еврспу табак
был привезён в XVI веке (название Nicotiana в честь француза
Nicot, положившего начало возделыванию табака во Франции),
В настоящее время табак культивируется во всех частях света (в
Австралии культуры не обильны). Наилучший табак получается
и а Кубе (Северная Америка), на Суматре (Малайский архипелаг),
'жже в Турции (в Малой Азии сорта самсун и трапезонд) и на Фи-
мппинских островах (сорт манилла). Вообше табак для своего
развития нуждается в тёплом и влажном лете, В СССР высшие сорта
газделываются в Закавказье, в Крыму и в южных частях Европей-
ской части, низшие сорта — в средних черноземных областях.
Культурные табаки имеют крайнее разнообразие сортов и форм,
относящихся к нескольким видам; из последних наиболее известны
N, tabacum (сигарный или настоящий табак) и N. rustica (махорка).
В диком виде ни эти, ни другие культурные виды табаков в Америке
ге найдены; невидимому, они являются гибридами видов дикора-
стущего табака.
III. Некоторые тропические н субтропические культуры
Хлопчатник (Gossypium — несколько видов). Как среди мировых
культур, так и среди культур СССР хлопчатник занимает седьмое
м^сто и следует не посредстве нно за основными зерновыми культурами.
Одно это указывает на его важную роль. Это — сольно теплолю-
бивая культура, требующая средней годовой температуры 12—25°
и среднего годового количества осадков 120—450 лл. Хлопчатник—
главнейшее прядильное растение, даёт также важный побочный
продукт—хлопковое масло (из семян) и др. -
, Несомненно, хлопчатник очень древняя культура, но первые
указания на неё (для Индии) мы находим в V веке до нашей эры у
греческого историка Геродота, Из Индии хлопчатник распро-
странился и на восток и на запад, однако лишь с VIII века он полу-
”т<л более широкое распространение на севере Африки н на юге
иропы.
При открытии Америки обнаружили, что хлопчатник имеется
там, и давно разводится. Таким образом, оказалось, ито хлопчат-
ок имеет два, независимых друг от друга, очага своей культуры.
С"1 к американские, так и азиатские виды относятся
ктомужероду Gossypium, откуда ясна общность их происхожде-
ния, но культуры являются независимыми. Основной индийский
гид—G. herbace urn, в Аме пике—G. barba dense (на Малых Антиль-
ских островах), G. reruvianum (дико в Бразилии"1, G. hirsutum
^так называемый уплэнд — происхождение не ясно). Для амери-
395
канских видов типично присутствие 52 хромосом, для азиатских— 26,
что указывает на их отдаленность; последнее псдтверждгетсяи их
•чень плохим скрещиванием.
Помимо двух указанных очагов возникновения хлопчатника,
некоторые авторы (Зайцев) указывают еще третей —афри-
канский; в Африке действительно имеются дикие виды
(G. arboreum), но культура его, по всей вероятности, возникла
в недавнее время.
Для СССР культуры хлопчатника имеют огромнее значение, и
наша страна в настоящее время обеспечена своим хлопком. Это
является результатом улучшенных культур и продвижения их в
совершенно новые районы. Так, хлопчатник возделывается не
только в Средней Азии и Закавказье, но и в Европейской части
СССР (Украина, Ростовская, Сталинградская области и др.).
Это осеверение ещё более значительно, чем для пшеницы.
Стахановские методы обработки почвы и ухода за культурами дали
рекордные урожаи в отдельных случаях: 125 н даже 151 ц с га
(в Америке лишь до 60 ц с га). Кроме того, введена культура
египетского хлопчатника (помесь двухамериканских видов),
дающего длинные шелковистые волоски — наилучшее волокно.
Рис (Oryza sativa). Это одна из виднейших мировых культур;
по занимаемой площади стоит на третьем месте (пшеница, куку руза у
рис) —78 млн. га и является главным питательным продукт gal ье
менее чем для 800 млн. человек. Рис —тропическая и субтропи-
ческая болотная культура (поля покрывают водой на 12—18 см);
развита во всех пяти частях света. В СССР, из-за климатических
условий, развита лишь в немногих областях (Узбекистан, Закав-
казье, Дальний Восток) и стоит по плошади в третьем десятке.
Культура риса очень древняя, она возникла во всяком случае
за несколько тысячелетий до нашего времени и вероятнее всего в
Индо-Китае, отсюда распространилась на Яву и другие острова,
в Индию, Китай и др. (в Китае известна за 8000 лет д®
нашей эры). Древний Египет, Вавилон, Палестина риса не знали.
Европейцы впервые познакомились с ним во время похода Але-
ксандра Македонского в Индию (в 326 г. до нашей эры), однако вве-
дение этой культуры в Средиземноморье относится лишь к концуг
VII и началу V111 века, причём ввели ее арабы, покорившие сре-
диземноморские страны. В Америке рис стал культивироваться
лишь с конца XVII века.
Рис (культурный) очень богат сортовым разнообразием: только
в Индостане имеется более 2 ОСО сортов. Имеются формы с остями**
и без остей, с просвечивающими сухими зерновками и непросвечи-
вающими, с большим содержанием крахмала и с малым, с прямо-
стоячим стеблем и с плавающим и т. п. Всё это — результат и с-
ку с с т в е н н о го отбора, скрещивания, а также мутаций.
Известны ли предки риса? Многие авторы указывают, что рис и
до сих пор имеется в дикол! состоянии в Индии и а Индо-Китае,
однако другие предполагают, что это рис одичавший. В этой области
необходимы дальнейшие исследования.
Вообще же род Oryza имеет 17 видов, из которых 7 встречаются
396
в диком состоянии на юге Азии. Однако они сильно отличаются от
О. sativa (у него листья имеют сильно удлинённые язычки) и с ним
непосредственно не связаны.
Вряд ли правильно мнение Комарова, что О. sativa — это
результат случайного скрещивания двух или более диких видов
риса. Какие же это виды — неизвестно. Пока ещё лучше подождать
выяснения вопроса о характере произрастания «одичавшего» риса
в Индо-Китае.
В истории человечества рис сыграл огромную роль. Его куль-
тура дала материальные возможности для существования в при-
митивных государствах густого населения,
Чайный куст (Thea sinensis нли Camelia theifera).. Европейцы
познакомились с чаем очень недавно — он был ^первые ввезён
лишь в 1517 г.; однако широкое распространение получил значи-
тельно позже; в Англии—в конце XVIII века, в России — в
начале XIX в. (впервые в Россию был привезён в 1638 г.).
Родиной чайной культуры является Китай; чай стали разводить
в III веке нашей эры, но он был известен и много раньше как меди-
цинское растение.
Вопрос о происхождении чайного куста совершенно ясен — он
и в настоящее время встречается дикорастущим в виде деревца в
юго-западном Китае. Многочисленные культурные формы получились
в результате искусственного отбора и гибридизации.
Помимо китайского чайного куста, разводится ещё ассамский
(Т. assamica) с более крупными листьями, растущий в диком со-
стоянии в Индо-Китае.
Основные центры возделывания чая: Китай, Индия, Цейлон,
Ява. В СССР чай возделывается в Западном Закавказье, особенно
около Батуми, а также в Талыше (Прикаспийский район). Некоторые
попытки культуры чая делались в России еще в 40-х годах прошлого
столетия, но лишь в самом конце столетия на это дело было обращено
большое внимание. До революции площадь культур едва доходила
до I 000 га; в настоящее время площадь занимает около 50 000 га.
Делаются попытки по продвижению культуры чая на север (одно-
летние культуры). f
Кофейное дерево (Coffea arabica —рис. 193). Первое знакомство
с кофе как с продуктом, обладающим возбуждающими свойствами,
имело место в Абиссинии; отсюда кофе проник к арабам в начале
XVI столетия. Кофе как напиток очень быстро вошел в употребление
у арабов, а в дальнейшем и у турок. В Европе кофе появился в те-
чение XVI столетия и быстро получил широкое распространение.
Родина кофе — Абиссиния, здесь он встречается в диком виде.
Это — Coffea arabica. Имеются и другие дикорастущие виды, из
которых часто культивируется либерийский кофе — С. liberica
(родом из Либерии). Лучшим кофе считается арабский (именно его
культуры в Абиссинии и в южной Аравии, так называемый «мокко»).
Культура кофе почти исключительно развита в тропиках. Наи-
лучшие климатические ^условия 15—25° тепла и не менее 2200 мм
осадков в год. В настоящее время главным районом культуры являет-
ся Бразилия, кроме того, тропическая Африка, Индо-Китай, Ява и
mt
др. В СССР культура кофе возможна лишь в условиях очнеьхорощо
защищённого грунта.
В результате многолетней культуры кофе имеет ряд разновид-
ностей, однако он представляет пример культурного растения, мало
отличающегося от своего дикого предка.
Шоколадное дерево, дерево какао flheobroma cacao). В противо-
положность чаю и кофе, какао происходит из Америки. Впервые
европейцы узнали какао в 1510 г., в период завоевания испанцами
Мексики; однако туземцы культивировали дереьо, несомненно,
очень задолго до этого времени. Какао—это древнемексикансюе.
слово, равным образом, и шоколад («чоко латль», на языке дру-
гих древнемексиканских племен — «какао с водой»;.
Через несколько лет (1520 г.) какао было ввезено в Испанию^
и обычай пить шоколад распространился по всей Европе в течение
XVII и XVIII веков. В Россию шоколад проник в XVIII веке, но в
качестве напитка не получил и до последнего времени широкого
распространения; какао идет главным ооразом на выделку кон-
фект.
В настоящее вре^я какао широко культивируется во всех тропи-
ческих странах с равномерными осадками не меньше I 000 мм.
обычно же 2 000 мм и больше и со средней годовой температурой
23—26°. Первое место занимают культуры на Золотом берегу в Аф-
рике, второе — в Бразилии. Но плодоносить какао может и в оран-
жерейной культуре (плодоносит, например, в Ленинграде).
Культивируется главным образом вид i h. cacao, однако имеется
несколько других видов (родина всех их—экваториальная часть
Америки). '1 h. cacao очень изменчив и потому при] оден для куль-
ту ры. Целый ряд культурных форм получен при помощи искусст-
венного отбора, прививкой и окулировкой. Метод тибридизации не
трименим, так как цветки какао настолько нежны, что невозможно
тропзвести искусственное опыление. На примере какао мы также
видим культурное растение, мало отличающееся от своего предка.
Хинное дерево (Cinchona — ряд видов — рис. 210). Ьодьна
хинного дерева (сем. мареновых), как и шоколадного дерева, —
Америка, именно влажные восточные склоны Анд Южной Америки
между 10° с. ш. и 20° ю. ш., на высоте чаще от I ООО м до
2 400 ль
Впервые европейцы познакомились ' с действием хинной
к о р ы лишь в 1038 г. (местное население давно знало это действие,
но скрывало от европейцев). В течение нескольких лет новое лекар-
ство быстро распространилось по всей Европе.
Хинное дерево имеет несколько видов с различным процентом
алкалоидов, главный из которых — хинин, важнейшим видом по
содержанию алкалоидов (до 15%) является С. Ledgeriana (интересно,
что оно было найдено лишь один раз в 1865 г., тогда же с него были
собраны семена).
Культуры хинного дерева в настоящее время развиты главным
образом на Яве (90% всей хинной коры, поступающей на рынок).,
также на Цейлоне, в Гималаях, именно в тех местностях, которое
39S
по климатическим условиям близки к условиям восточных склонов
Анд (очень большая влажность, период дождей до девяти месяцев?
средняя годовая температура 12—13° и др.). Культивируют как почти
не изменённые дикие виды, так и ряд разновидностей, полученных
скрещиванием, закреплённых черенкованием и др. Сначала исполь-
зовали кору лишь ствола и ветвей хинного дерева, в дальнейшем—
и кору корней. Очень интересна С. robusta (до 26% алкалоидов):
она в дикой природе не существует и является гибридом между
С. Ledgeriana и С. succirubra. Здесь имеет место процесс п р е-
в р а щен и я дикого растения в культурное.
В России пытались ввести хинное дерево в 1888 г, в Батуми^
но неудачно. В настоящее время удачные опыты по культуре ве-
дутся в Аджарии и в Абхазки; они идут в направлении подбора
более холодостойких сортов, а также в направлении выработки
однолетних и двулетних культур. Используется не только корь,
но также древесина, листья и др.
Таковы данные по некоторым культурным растениям. Мы видим,
что в одних случаях культурное растение отличается ещё очень мало
от своего дикорастущего предка (хинное дерево, чайное дерево),
в других— это отличие значительно и иногда связь с дикорастущими
видами не столь ясна (картофель, подсолнечник, и др.), в третьих—
эта связь совершенно утеряна (некоторые пшеницы, просо). Вообще
же культурные растения в дикой природе никогда не существовали
и являются д е л'о м рук человеческих. Здесь человек
не только переделывает природу, но делает, творит её,
В заключение приводим несколько данных по площадям и вало-
вой продукции основных культур и их групп.
Таблица i
Посевные площади культур в СССР по основным группам (в или. га)1.
Культуры 1913 г. 1938 г. 1938 г. в % к 1913 г,
Вся посевная площадь 104,8 136,9 130,4
Зерновые - 94,4 102,4 108,5
В том числе:
Пшеница 1 31,6 41,5 131,3
Технические ...... 4,5 11,0 244,4
Огородно-бахчевые и картофель . 3,6 9,4 247,4
Кормовые 2,1 14,1 671,4
1 XXII годовщина Великой Октябрьской социалистической революции
(Материалы для агитаторов и пропагандистов, 1939).
399
Таблица г
Валовая продукция зерновых н технических культур (в млн. центнеров)1
Культуры 1913 г. 1929 г, 1933 г. 1937 г. 1937 г. В % К
1913 г. 1933 г.
Все зерновые культуры .... 801,0 717,4 898,0 1202,9 150 134
В том числе: Пшеница озимая и яровая . . . 262,0 188,8 277,3 ' 463,6 179 169
Технические культуры: Хлопок-сырец ......... 7,4 8,6 13,2 ‘ 25.8 347 196
Сахарная свёкла (фабричная) - 109,0 62,5 89,9 218,6 201 243
Картофель 233,1 456,3 492,5 656,3 282 133
Таблица з
Удельный вес СССР в мировом сельскохозяйственном гроизводстве
в процентах 1
Культуры Посевная площадь Валовая продукция
1929 г. 1933 г. 1937 г. 1929 г. 1933 г. 1937 г.
Пшеница 22,6 24,6 27,9 16,1 21,1 31,1
Ячмень 20,5 20,8 , 24,3 16,1 19,7 24,7
Овёс за, 1 29,6 31,5 22,9 25,3 31,5
Хлопок ............ 3,0 6,7 5,5 4,6 7,1 10,3
Сахарная свёкла ... 25,, 7 37,2 35,4 10,6 14,8 26,5
1 Социалистическое строительство Союза ССР (1933—1938 гг,), Госплан-
издат, М.—Л., 1939 г.
ПРИЛОЖЕНИЕ 2
СИСТЕМАТИЧЕСКИЙ СПРАВОЧНИК
(Краткие характеристики семейств цветковых растений и папоротников,
упомянутых в книге)
I. Асегасеае (кленовые). Семейство раздельнолепестковых двудоль-
ных из порядка фиста шинковых (Terebinthales). Сюда же относятся семейства
Rutaceae (рутовые), Sapindaceae (сапиндовые) и др, Некоторые авторы клено-
вые помещают в порядок сапиидоцветных (Sapindales). Порядок фисташнико-
вых очень близок к геранецветным, но отличается присутствием нектароносно-
го диска в цветке. У кленовых листья супротивные, цветки иногда однополые,
пятерного типа, тычинок вдвое больше, чем лепестков; плод двойной, крылатка.
Деревья в северном полушарии, несколько видов в тропической Азии. Род
Acer — клен (120 видов), у нас ряд видов.
2. Actinidiaceae (актинидиевые). Небольшое семейство раздель-
нолепестковых двудольных из порядка камеденосных (Guttiferales). Тропики
н субтропики. Стелющиеся или лазящие деревянистые виды. Род Actinidia —
в Восточной Азии.
3. Ai/.oaceae. Семейство раздельнолепестных двудольных из порядка
центральносеменных (Centrospermae). К этому же порядку относятся гвоздич-
ные, маревые, кермесовые и пр. Семейство по своим признакам свя-
зывает данный порядок с порядком Opuntiales (сем, кактусовых), Травы или
полукустарники, листья нередко суккулентны. Наиболее обширный род Ме-
sembrianthemum — рис, 173 и 174 (солнечник) имеет до 400 видов — цветки с
многочисленными лепестка,ми и тычинками, многочисленными гнездами завязи
(нижней). Всего около 600 видов в сухих или пустынных областях тропического
и субтропического поясов; большинство видов солнечника в Южной Африке.
4. Alisma'taceae (члстуховые). Небольшое (около SO видов) семей-
ство однодольных из порядка водных (Helobiae). Травянистые растения
сырых мест или растущие в воде. Похожи на лилейные, однако листья с сет-
чатым жилкованием. Цветки правильные, околоцветник расчленяется на ча-
шечку и венчик. Роды: Alisma (частуха), Sagittaria (стрелолист). Некоторые
виды распространены по всей земле.
5, Amaryllidaceae (амариллисовые). Семейство однодольных из
порядка лилиецветных. По своим признакам очень близко к сем. лилейных, от-
личаясь нижней завязью; тычинки иногда i-мэют придатки, которые
срастаясь, образуют «придаточный венчик^. По внешности похожи на лилей-
ные, у большинства — розетки прикорневых листьев, у многих луковицы.
Почти исключительно многолетние травы, Около 1 000 видов, особенно в Юж-
ной Африке и в Мексике. Роды- агава (Agave) — листовой суккулент, нарцисс
(Narcissus), подснежник (Galanthus — рис. 109) и др. Ряд декоративных рас-
тений (садовая и комнатная культура).
6. А п а с a rd i а с е а е. Семейство раздельнолепестных двудольных из
порядка фисташнпковых (Terebinthales). Близко к сем. ~ Meliaceae. Глав-
ным образом деревья теплых, реже умеренных стран, около 500 видов. Листья
простые или сложные, Цветки пятерные, иногда однополые. Роды: Pistacia
(Р. vera — фисташник), Rhus (сумах), Cotinns и др. В семействе очень много
полезных и ядовитых растений.
26 География растений
401
7i Annnacear <аноновые). Семейство раздельнолепестных двудольных
из порядка многоплодниковых (Polycarpicде). Близко к сем. магнолиевых;
деревянистые растения с листьями, похожими нт листья магнолиевых, но без
прилистников, Цветки обычно крупные с трехчтенными кругами. Главным
образом тропические, реже субтропические страны (около 80о видов),
8. Ар осу па сеа е (кутровые). Семейство спайнолепестных двудо 1ь-
ных из порядка скрученных (cont)rt.e). Много тетние травы, t а паны, кустарник !,
деревья, обычно вечнозеленые. Около 1 000 вщоз, главным образом в жарких
областях. Растения с млечным соком, листья супротивные, цельнокрайние.
Цветки правильные, крупаые. воропковпдные илн колесовидные, венчик в бу-
тоне скрученный, цветки четырех- или пятичленные, Плодопистжи сраста-
ются часто лишь в верхних частях Роды Vjпса (барвинок), Nenum (N, olean-
der— оаеаидр) и др.
0. А р о цо ge tonaceae (апоногетовыс), Очень небольшое (около 20
видов) семейство однодольных нз порядка водных (Не I obi ле). Сюда же относят-
ся сем. Pot ’mogetoirceae, Hydioch'”! t се ш и др, J-ick чочитепьно водные
растения (в пресных водах), с клубневидным корневищем п пучком n.i-
вдоших илн погруженных листьев. СопЕеиая выдаются изводы. У рола Аршао-
g Б п некоторые виды имеют листья сетчато-продырявленные (рис. 39), Тро-
пические и субтропические виды: Африка до Австралии.
1J. Aauifoliaceae (падубовые). Семейство раздельнолепестных из
порядка крушпноцветпых (Rbninn дез). Положение семеиствд не совсем ясно, и
некоторые систематики отнтсят ею к сап индо постным (Sipind ties). Деревя-
нистые растения, обычно с кожистыми листьями, Почти все виды дв\домны.
Цветки правиж ные, четверного (реже пятерного) типа, Г кавным обртзомтро-
шшсские и субтропические виды (всего око ю 170 видов!, Род [lex — падуб,
мноне виды падуба декоративные растения. Ilex aquifoliiim— в Западной
Еьропе.
11. А г а с е а е (ароидные). Важное тропическое семейство из
однодольных. Порядок аронннковых. Характерны сопветия-почятки с ме ткиыи
цветками на уто .шенной оси; при основании соцветия — крупный прицветный
лист, нередко ярьо окрашенный все соцветие можно принять за цветок-Листья
со средней жилкой и перистым жилкованием, как у двудольных. Цветки сходны с
лилейными, но иногда редуцированы до одной тычниГки или до одного пестика;
вообще, сравнительно с литейными, семейство представляет очень уклонившую-
ся ветвь. Некоторые авторы выводят из перцецветных (Pipenles) и отрицают
связь с лилчегвегными, Главным образом в тропических странах (до 1000 видов);
травы с клубневидным корневищем, деревянистые растения, часто лианы,
стоаь характерные для тропических лесов. В СССР несколько 'родоз (СгПя,
Arum, .Acorns). Тропические лианы- Philodendron, Monstera (V.. deliciosa —
комнатный филодендрон) и ip.
12. А га I i а сеа е (аралиевые). Семейство раздельнолепестных двудоль-
ных из порядка зонтпкоцветных (Unabelliflorae). К этому же порядку относит-
ся сем. Unabelbferae (зонтичные), Главным образом деревья, реже травы, ли-
стья простые или пальчатосложные, Цветки правильные, пятичленные, чашеч-
ка редуцирована. Семяпочки с одним покровом, завязь нижняя (очень редко
верхняя). Соцветия зонтиковидные или головчатые. Около 700 видов, в теплых
странах. Роды: Aralia (аралия), Hedera (плющ). Рапэх (жень-шень — рис.
186). В СССР отдельные представители в Крыму, на Кавказе, на Дальнем Во-
стоке.
13. Aristolochiaceae (к-tn сазтяззыз). Семейство двудо дьных
систематически довольно неопределенного ппложения, но в тследчдз время
многие систематики включают его в порядок мчэгопледниковых (Ро I year pic ле);
травы иди деревянистые лианы с почковидными или сердцевидны я я листьями,
Цветки правильные или неправильные с венчиковидным спайная ютныя око-
лоцветником. Тыш-нок б или 12. Около 200 видов, главным образам тропи-
ческих. Роды: Aristolochia (кирказон). Asarum (копытень).
13. Asclepiadaceae (дастачч а сазыз). Сзи?1?тэо дч/дальчых,
подкласс спайнолепестных. Порядок скрученных (Contirtae) — венч i г в бу-
тоне скрученный. Почти исключительно в тропиках и субтропиках (до 1 700
402
видов, особенно многочисленных в Африке. Травы, кустарники, деревья. Цветки
имеют с. о кное строение, так как лепесаки и тычинки иногда дают придатки,
образующие придаточные венчики. Пылинки в пыльцевых мешках нередко
слипаются и образуют п о л л и н и и. Эти и другие особенности в цветках
представляют сложные приспособ юния для опыления насекомыми К этому
семейству относится днсхидия (Dischiclia — рис. ^5), а также многие африкан-
ское кактусовидные виды. Одно jts наиболее специализированных семейств
сп ч инолепестных. В С СУР очень немного видов.
15. В a I s a m i п а с е а е (бальзаминовые), Семейство раздельнолепест-
ных двудольных из порядка геранецветных (G.ramales), К этому же порядку
относятся гераниевые, настурциевые (Tropaeol iceae — рис. 20 ?), леновые(Ь1-
п'сеае) и др. Травы голые и обычно сочные, с простыми листьями, цветки не-
правильные; чашечка нередко венчиковидная из 3—5 листочков, один листо-
чек в виде шпорца. Тычинок 5 с пыльниками, плотно прикладывающимися к
3‘илзн. Плод — коробочка, раскрывающаяся эластически. Род Inip°tiens (не-
дотрога) — главным образом тропические виды (всего около 403). Impitiens
trlsamina (Ост-Индия) — родоначальник наших садовых бальзаминов. Неко-
торые систематики относят семейство к S’pir.d 1 les.
16. Begoniaceae (бегонневые). Семсйе-вэ раздельнолепестных дву-
до |ьных, из порядка постенносеменных (Paricties— семяпочки расположе-
ны по стенкам завязи, а'не по краям плодолистиков). Главным 'образом
тр1вы (тропическая Амер| ка, Африка, Китай, Япония). Около 4)0 ви-
дов Цветьи однополые, однодомные, обычное яркоокраш^нным околоцветни-
кам. . ’вязь нижняя, с тремя крыльями. Легко дают помеси Везде культиви-
руются из-за ярких цветков и пестрых листьев. Род Begonia (бегония)
Г/, Berberidaceae (барбарисовые). Семейство раздельнолепестных
двудг |,ьных, из пор* д ча многопледниковых (Polycarpicae). К этому же порядку
относятся; магнолиевые, лютиковые, нимфейные, лунносеменннковые и др.
Семейство представляет переход от семейств многоплодниковых с древесными
представителями к семействам с травянистыми видами. Травы или кустарни-
ки. Некоторые травянистые виды имеют значительное число тычинок и пести-
ков, почему с трудом отличил ы от лютиковых. Цветки трехчленные, обое-
полые, пьльники раскрыв югся створками. Около 250 видов в умеренных стра-
нах. Роды Berberis, Lecnttee, A'^hmia.
IS. В e txu I а с e a e (березовые). Семене во двудольных, раздельноле-
пестных, из по ряде а букоцьетных (Fag-les), куда входит tw семейство
буковых. Деревья и кустарники северного полушария (около 100 видов),
под тропики почти не заходят. Цветки однополые, околоцветника нет или
он сильно редуцирован. Мужские цветки собраны в сережки. Опыаенче вет-
ром. Престо устроенные цветки, по всей вероятности, нужно рассматривать
как результат редукции. Роды; Betula (береза), Alnus (ольха), Corylus (ле-
щина), Carpinus (гр-б), Ostrya и др. г
19, Bignoniaceae (бнгнониевые). Семейство спайнолепестных дву-
дольных, j з порядка маскоцветных (Personatae). Близко родственно сем. норич-
никовых, относящемуся к тому же порядку, однако здесь главным образом де-
ревья, также лианы с различными прицепками, редко травы. Почти исключи-
тельно под тропиками (до 500 видов). Цветки неправильные, крупные. Семена
с крыловидными придатками. У нас на юге культивируются; Catalpa (дерево),
Campsis (прикрепляется корнями к стенам) и др.
20. Bombacaceae (баобабовые), Семейство раздельнолепестных
двудольных, из порядка мальвоцветных (Мл 1 vales); к последнему относятся так-
же семейства мальвовых и липовых. Тропические деревья. Очень близко к сем.
мальвовых; цветки пряш^льные, пятерные; тычинки многочисленные, спаянные
в трубку, но пыльники обычно четырех- или многокамерные (v мальвовых 2
гнезда). Роды: Bombax Adansonia digitata—баобаб (рис, 138), Durio —дуриан
(Малайский архипелаг), Cavanillesia— рис, 97дСе1Ь1 (С. pentandn дает ка-
пок) и др.
21. Borraginaceae (бурачниковые). Семейство спайнолепестных дву-
дольных цз порядка бурачниковидных (Borraginales), состоящего всего из двух
семейств. Ряд авторов относит бурачниковые к большому порядку трубкопвет-
ных(Tubiflorae). Почти исключительно травы. Растения семейства легко узна-
ются по ряду признаков: более или менее сочные стебли, жесткие волоски
по всему растению, цельнокрайние, обычно спиральные, листья, соцветия —-
Завитки, цветки чаще всего голубые, правильные, пятерные. Очень характер-
на четырёхлопастная завязь, после созревания распадающаяся на четыре ореш-
ка. Около 1500 видов, главным образом в умеренных странах северного полу-
шария, много видов в Средиземноморье, реже в тропиках. У нас много родов:
Anchusa, Cynogiossum (чернокорень), Echium (румянка), Echinospermum (ли-
пучка), Myosotis (незабудка), Eritnchium, Pulmonaria (медуница) и др.
22. В го me i i а с е а е (бромелиевые, ананасовые). Семейство однодоль-
ных, из порядка лилиецветных (Liliiflorae). Чаще всего эпнфитные травы, е
жесткими, мечевидными листьями, собранными в розетку; цветки типа лилей-
ных, собраны в соцветия, выходящие из пазух розеточных листьев. При цветках
нередко окрашенные прицветники. Исключительно в тропической Америке,
около 1000 видов. Роды: ананас, Tjllandsia— рис, 132, Njdularium— рис. 88 и др'
23. But о m а с е а е (сусаковые). Небольшое (7 видов) семейство однодоль-
ных, часто соединяемое с сем. Пастуховых ^Aiismataceae). Водно-болотные
растения. Род Butomus (сусак), Растения умеренных п тропических стран.
24. Вихасеае (самшитовые). Семейство раздельнолепестных двудоль-
ных. Систематическое положение недостаточно определённо: одни авторы по-
мешают в порядок Тпсоссае, другие — в Sapindales, третьи — в Rhamnales.
Всего около 30 деревянистых видов в тропиках и субтропиках, Листья вечно-
зелёные, цельнокрайние. Цветки редуцированные, без околоцветника. Род
Buxus (В. sempervirens — Средиземноморье, Кавказ — самшит, кавказская
«пальма»; прекрасная древесина; рис. 18),
25. Cactaceae (кактусовые). Семейство раздельнолепестных, двудоль-
ных, из порядка Opuntiales (опунциевые), Другие относят к порядку централь-
носеменных (Centrospermae). Между этими порядками имеется тесная
связь. Во всяком случае, кактусовые — одна из конечных ветвей раздельно-
лепестных. В тёплых областях Америки, редко в Африке (всего около 1 500
видов), Характерная форма суккулентов, обычно листья превращены в
колючки, Цветки по большей части крупные, с многочисленными листочками
околоцветника и многими тычинками. Роды: Cereus, Echinocactus, Mamillaria,
Opuntia, Rhipsalis п др.
26. C a ly ca n thaceae (каликантовые). Семейство раздельноле-
пестных двудольных, из порядка многоплодниковых (Polycarpicae). Кустар-
ники. Умеренная Восточная Азия, Северная Америка. Очень небольшое семей-
ство (всего 5 видов). Близко к магнолиевым — много листочков околоцветни-
ка (венчиковидно окрашенных), много тычинок и пестиков.
27. Campanulaceae (колокочьчиковые). Семейство спайнолепе-
стных двудольных, из порядка сшйнопыльннко ых (Synantheraies, Synandrae).
К этому же порядку относятся сложноцветные, которые можно выводить из
колокольчнковых. Травы, полукустарники, реже деревянистые растения, около
1 200 видов, в умеренных и субтропических странах. Цветки обычно правиль-
ные, пятичленные, пыльники у распускающихся цветков обычно склеены (ср.
сложноцветные), завязь нижняя. Растения с млечным соком. Большой род ко-
локольчик (Campanula), у нас много видов. У рода букашника (Jasione), цветки
собраны, как v сложноцветных, в корзинки.
28. Cannabinaceae (коноплевые). Семейство очень близко к туто-
вым (Могасеае). отличаясь отсутствием млечного сока, сухими орешковидными
плодами и др. Всего два рода: конопля (Cannabis), хмель (Hummus),
29. С а и пасе а е (канновые). Семейство однодольных, из порядка
Scitamineae (к последнему относятся также семейства банановые, имбирные
и Др.). Исключительно в тропической Америке; многолетние травы (60 ви-
дов) с крупными перпстонервными листьями, Цветки трехчленные, представ-
ляют очень сильно измененный тип лилейных, асимметричны; тычинки все
кроме одной лепестковидны, но и единственная плодущая плодуща лишь
наполовину (одкогнёздный пыльник), другам половина лепестковидна; завязь
нижняя. Одна из конечных ветвей однодольных.
30. Caprifoliaceae (жимолостные). Семейство спайнолепестных
двудольных, из порядка мареноцветных (Rubiales). Очень близко к мареновым,
резких различий между этими семействами не существует. Деревья или кустар-
ники с супротивными, простыми или перистое ложными листьями, прилистни-
ков обычно нет (отличие от мареновых). Цветки правильные или неправильные,
пятичленные; завязь нижняя. Около 350 видов; главным образом во внетропи-
404
ческих частях северного полушария. Роды: Lonicera (жимолость), Sambucus
(бузина), Linnaea (лнннея) и др.
31, Caricaceae (nai а-вые), Семейство раздельнолепестных двудоль-
ных, из порядка постенносеменных (Parietales). Небольшие деревья, мало ветви-
стые, на концах стеблей пучки листьев, обычно пальчаторассечённых. Цветки
чаше однополые. Плоды похожи на дыню. Во всех частях растений (и в частно-
сти в плодах) млечный сок. Всего около 30 видов, главным образом в тропи-
ческой Америке. Род Carica (в Америке) — С. papaya (отсюда название семей-
ства) — дынное дерево, из-за своих плодов столь распространённое под тропи-
ками (рис, 201).
32. Caryophyl laceae (гвоздичные). Семейство раздельнолепест-
ных, двудольных, из порядка центральносеменных (Centrosper'mae), так как
семяпочки (нередко многочисленные) сидят иа центральном семяносце. Глав-
ным образом в умеренном климате, в общем космополиты (до 1 500 видов). Тра-
вы пли полукустарники. Передков цветке пять пятичленных кругов, в других
случаях— редукция тех или других ч стей. Род : песчанка (Arenaria), гвозд i-
Ко (Dianthus). смолевка (Silene). Gypsoph /1а, звездчатка (Stell irга). Lyailia г др.
В СССР сем, очень распространено. Порядок Centrospermie легко выводился из
безлепестных примитивных раздельнолепестных, представляя особую конеч-
ную ветвь с полно развитым цветком.
33. Casuarinaceae (казуариновые). Семейство раздельнолепест-
ных двудольных, из порядка Verticillatae (мутовчатоцветные). Этот порядок (в
нём лишь одно сем. казуариновых) стоит особняком--от других й некоторыми
авторами считается наиболее примитивным среди двудольных растении и свя-
зывающим последние с голосеменными. С этой точки зрения семейство очень
важно в филогенетическом отношении, Исключительно деревья с А1утовчато-
расположеннымн редуцированными (до чешуек) листьями; прутьевидные стеб-
ли очень напоминают (по внешности) хвощи (рис. 213). Цветки однополые н
очень просто устроенные; мужские имеют одну тычинку и собраны в серёжко-
видные соцветия, женские имеют один пестик и собраны небольшими голов-
ками. Помимо очень просто устроенных однополых цветков казуарино ые с голо-
семенными сближаются также рядом анатомических признаков. Однако другие
авторы считают казуарпновые не за примитивные, но за редуцированные фор-
мы, что, повидимо.му, более правильно. Род Casuarina имеет около 25 видов,
произрастающих в Австралии, на Зондских, Маскаренских и некоторых дру-
гих островах, на Малайском полуострове.
34. Cephalotaceae (цефалотовые). Семейство раздельнолепест-
ных двудольных, на порядке- многоплод иковых,Имеет определённые связи с та-
кими семействами насекомоядных растений, как непецтесовые и саррацение-
вые, вследствие чего эти три семейства иногда объединяют в особый поря-
док непентесоцветных (Nepentliales), примыкающий к многоплодниковым. Но
положение семейства в филогенетической системе не вполне ясно, иногда его
помещают в порядке розоцветных. Семейство представлено всего одним ро-
дом и видом: Ceplnlotus follicularis, растущим на болотах Западной Австралии
(австралийский эндемик — рис. 211). Небольшое растение с розеткой прикор-
невых листьев, часть которых превращена в кувшинчики, в которые попадают
насекомые. Цветки имеют шестилистный околоцветник, 12 тычинок, б свобод-
ных плодолистиков.
35 Сега top h у I laceae (роголистниковые). Семейство раздельно-
лепестных двудольных, из порядка многоп.лоднпковых (Polycarpicae). Водные
погружённые растения, совершенно лишённые корней. Листья по четыре в му-
товке, очень сильно рассеченные (рис. 26). Цветки раздельнопоаые, редуциро-
ванные вследствие обитания в водной среде. Цветки цветут подводой. Несколь-
ко видов рода Ceratophyllum. три из них широко распространены по всей земле,
Зб, Chenopodiaceae (маревые, лебедовые),. Семейство раздель-
нолепестных двудольных, гз порядка центральносеменных (Centrospermae).
К данному порядку относятся также т-жие семейства: гвоздичные, порту-
лаковые, кермесовые и др. Для всех них характерен центральный семя-
носец (в завязи), на котором располагаются семяпочки, Сем. маревых име-
ет 1 400 видов (в СССР до 350 видов), распространённых во всех частях света,
главным образом в степях, полупустынях ц пустынях, особенно на засолённых
почвах (галофиты). Травы, а также кустарники и даже деревья (саксаул в Сред-
405
ней Азин). Цветки мелки и невзрачны с простым чашечковидиым околоцветни-
ком, тычинки против листочков околоцветника, завязь одно’пездная с одним
семенем. Маревые представляют переходное семейство от без |елесгных двудоль-
ных (порядок крапивоцветных) к семействам с хорошо р1звптьм! цветками
(с чашечкой и венчиком), мепно к сем. гвоздичных. Роды: Ал ab ists, Atriplex,
Chenopodium (марь), Eur.atia, Kochii, Saticoinia (солерос — рис. эр, Hiloxy-
lun (саксаул), Hilocnemum, Haiostachys, Salsola, Suaeda и др. Много сорня-
ков, почти космополитных.
37 Cistaceae (ладанниковые). Семейство раздельнолепестных дву-
дольных. из порядка постенносеменных (Panetales). Небольшие деревянистые
растения, реже — травы, около I6J видов в умеренных странах, особенно в
Средиземноморье. Листья простые, цветки правильные, пятерного или трапного
типа. Тычинки многочисленные. Семяпочки постенные. Часто же 1ез !стые
волоски с эф 1рными м юлами. Роды: Helianthemum (солнечник), Cistus ( ндш-
НИК) п пр.
33. С I u s I а с е а е. Семейство раздельнолепестных двудо шных, из пэпяд-
ка Guttifcrales. Чаще это семейство трактуют как подсемейство сеч Guthfe-
г.ае Деревья со смоляными или слизесмоляными ходами в троп;нюе,.их
лесах.
39. С о m b г е t а с е а е. Семейство раздельнолепестных двудольных, из
порядка мнртоцветных (MyrtTLes), сюда же относятся сем M/rticeae, L thra-
ceae, Onagiiceie, Punicaceae и др. Исключительно тропические деревянистые
растения, чщто шалы. У растений побережий плоды распространяются водой
(воздухосодержащая ткань), например, род Lagunciilam. Всего до 5i> видов.
4П. С о гл р о s 1 t а е (сложноцветные). Семейство спайнолепестных дву-
доаmbх, । з порядкаспаннопыльниковых (Synantheraks, S?nandsoe) Ь >тому
же порядку относятся семейства корт ольчпковых, лсбелиевых и др, Огром-
нее гА 11 слу видов семейство, до 15 000 видов, в последнее время считают даже
до 25 000 видов, распространено во всех частях света (очень обильно представ-
лено в СССР). Травы, кустарники, лианы, небольшие деревья.
Несмотря на разнообразие жизненных форм, растения, относящиеся к дан-
ному семейству, очень легко узнаются по характерному строению цветков и
соцветии. Посреднне по своему внешнему виду очень похожи на цветки и в об-
щежитии и называются цветками «цветок» ром пики, «цветок» вчичны и др.
Однако этот «цветок» представляет соцветие, так назыв "еиую корз тку, и со-
стоит из очень многочисленных (в большинстве случ >ев), небольших цветков,
тесно ci ученных на расширенном конце осп (цветоложе). Отсюда название се-
мепствэ. Цветки в типе состоят из пяти сросшихся в длинную трубку лепестков,
из пяти тьчинок, спаянных своими пыльниками (отсюда название всего по-
рядка), завязь нижняя, одногнездная, с одним семенем. Плод — семянка (круп-
ная семянка, например, у подсолнечника) Чашечка нередко отсутсавует п вме-
сто нее развиваются волоски или щетинки, совокупность которых представ аяет
так п ‘Зываеиую летучку или хохолок (например, аетучка v од 'влнчнка) — при-
способление для распространения плодов ветром. Цветки в т ine право жные,
но наверху трубки венчика лепестки могут в виде язычка отгибаться в сторону,
и тогда цветок будет не «трубчатый», а «язычковый(например цветки в кор-
31 нт е у одуванчика) Нередко в корзинке внутренние цветки трубчатые, а
краевые язычковые (например у ромашки, у мфгарпткн и многих других).
Встречаются и другие видоизменения цветков и нх комбинац ий в корзинках.
Все систематики согласны в том, что сложноцветные представляют одну из
высших конечных ветвен спайнолепестных. \ нрс встречается очень много
родов ромашка, в^силж, подсолнечник, одч, венчик, рстиа, ичкорчй, осот,
полынь и др., о^Снь много полезных растений земляная гр чца, салат, арти-
шок, ктрдон, агн та и т. д. Много декоративны растений георгины, криз >н-
темы, цинерарии. астры и др.
В книге были приведены а''кие роды: Antennarra (кошачья лапка), Art“misia
(полынь), Aster (астра), Bellis (маргаритка), Centmrea (василек), С1т»п killa
(хондрилл), ChrysHitliemum (п-'по зн in), Cirsium (б/дя i), Coreopsis, Cousinia
(куэнння), Суплгл, Errgjron (мел.чО лепестнчк), Eupatarium (посконник), Gni-
phMium (счшеннца), Helianthus ‘ (подсолнечник), HHrchr/sum (цмип), I -nla
(девяси ), Junnea (юрпнея), L юtnca (латук), L^ont don (i у аьб 'ба), L'nc oath
mum (hiikhihk), Linosyris (грудница), Matrcain (ромашка), Octant spermnm,
406
Onopordon, Petasites (подбел), Pyrethrum (пиретрум), Rudbeckia (рудбекия),
Scorzonera (козелец), Senecio (крестовник), Serratula (серпуха), Silphium,
Sohdago (золотая розга), Sonchus (осот), Taraxacum (одуванчик), Telekia (те-
лекич), Tragopogr n (козлобородник), Vernonia.
41—42 Coniferales (хвойные) Класс голосеменных растений (другие
классы Cycadales, Ginkgoales, Gnetales). Наиболее обширный класс из ныне
живущих голосеменных — около 370 видов в субтропических и главным образом
умеренных странах северного полушария. Исключительно деревья, реже кустар-
ники. Ствол сильно разветвленный, листья мелкие игольчатые (сосна) или
чешуйчатые (кипарис), реже (у ряда видов в южном полушарии) ш ipoKiie,
очень схожие с обыкновенными листьями двудольных. Сосуды в древесине
отсутствуют. Цветки однополье. однодомные, редко двудомные, мужские цветки
сережковидны, женские нередко в виде шишек. Проростки обычно со многими
семядолями. Чаще всего хвойные делят на два семейства тиссовые (Тахасеае)
и сосновые (Ршгсеае). Женские цветки первого шишек не образуют, у второго—
шишки. К первому семейству огносяг такие роды. Cephalotaxus, Dacrydium,
Phyl locladus, Podocarpus, S^xeg^thaea, Taxus, Torregi; ко второму. Ag this,
Araucaria, Cedius (кедр), Chamaecyparis, Cryptamena, mnninghimia, Cupressus
(кипарис), Jumptrus (можжевельник), Larix (лиственница), Libocedrus, Micro-
be ta, Picea (ель), Puius (сосна), Pseudotsuga, Sequoia (секвойя, Мамонтове
дерево), Taxodnim, Thuj i, Tsugvn др.
43. Convolvul iceae (вьюнковые); Семейство спайнолепестных дву-
дольных, из порядка Convolvub les (вьюнковидные). Ряд авторов относит семей-
ство к большому порядку Tubiflorae (трубкоцветные), куда включаются все
семейства вьюнковидных. В большинстве случаев вьющиеся травы, реже кустар-
ники и деревья с млечным соком. Листья часто сердцевидные, венчик обычно
крупный, воронковидный со сросшимися лепестками, в бутоне складчатый вдоль
и обычно скрученный вправо. Цветки правильные, пятерного типа, завязь дву-
гнездная, верхняя. Около 1 100 видов, преимущественно в теплых странах.
Роды Convolvulus (вьюнок), Calystggia и др. У нас несколько вадов.
44. Согпасеае (деретые) Огюсится к порядку Un be! 1 if lurae, иногда
выделяется в особый п рядо < Cornales Доенья, р$же куст 1рник i с прш сами
счппотивными ш стьями Цветки чаще все о четыпехчленные, со ве ня ино' да
зонтш ои дны Ок< ло 120 видов. Роды:Согпиз, Aucuba и др. Главным бра-
зом и сев. полушарии.
45. С га ss u I а с е а е (толстянковые). Семейство раздельнолепестных
двудольных, из порядка Rosales (розоцветные); к этому же порядку оросятся
сем. розанных, камнеломковых и др. Иногда семейство помещают в порядок
камнеломковидных(Saxifraginae), близкий к розоцветным. Толстянковые связы-
вают розоцветные с многоплодниковыми (Polycarpicae). Мясистые травы или по-
лукустарники (рис. 173). Около 1500 видов в умеренных теплых странах, растут
на сухих солнечных местах, на скалах, крышах и т. д.; много видов в горах.
Цветки правильные с чашечкой и венчиком, иногда с больш im числом частей в
кругах (особенно многочисленны тычинки), плодолистики чаще свободные, цвет-
ки близки по признакам к розанным. Роды Sedum (очиток), S=mpervivum (моло-
дило), Crassula (толстянка) и др. У нас много видов, главным образом в горах.
46 Cruciferae (крестоцветный Семейство раздедьнэтепгетных дву-
дольных, из порядка макоцветных (Rhaeadiles). К этому же порядку отно-
сятся семейства маковых, резедовых и др. Порядок маконветных выводится из
порядка многоплодниковых при посредстве семейства маковых. Сем крестоцвет-
ных представляет одну из конечных ветвей раздельноаепестных. Распростра-
нено главным образом в умеренных и холодных об остях северного полушария,
содержит около 3 б00 видов. Почти исключительно травы (редко поаукустарни-
кп). В СССР очень большое число родов и видов, представ аенных нередко огром-
ныл| числом особей. Растения семейства Очень легко узнаются по их цветкам, а в
отцветшем состоянии по плодам. Цветки правильные, обычно в соцветиях (кис-
ти), чашелистников и лепестков венчика по четыре, расположенных крест-на-
крест (отсюда название семейства), тычинок шесть (две более коротких), пестик
один (завязь верхняя). Плотнаще всего стручок (длина много раз превышает ши-
рину) или стручочек, раскрывающийся двумя створками, семена остаются
на перегородке. Среди крестоцветных очень много полезны': растений: капуста,
репа, редька, горчица, рыжик и др., много декоративных: левкой, желтофиоль и
др.; немало сорняков- сурепка, пастушья сумка, икотник, клоповник и т. д.
407
В книге приведены роды: Alyssum (бурачок), Anastatica, Вгауа ,Саг-
damine (сердечник), Cochlearia (хрен), Dentaria (зубянка), Draba (крупка),
Pringlea.
Al. Cucurbit а сеае (тыквенные). Различными систематиками эт»
семейство ставится в разных отделах филогенетической системы, одни отно-
сят его к спайнолепестным двудольным (особый порядок тыквенноцветных—
Cucurbitales),другие помещают его среди порядка постенносеменных (Parietales),
включающего главным образом семейства с раздельнолепестным венчиком
(раздельнолепестные двудольные). Правильнее всего смотреть на тыквенные
как на особую ветвь постенносеменных, эволюционировавшую в направлении
спайнолепестности венчика. С другими семействами спайнолепестных вряд ли
имеется какое-либо родство, хотя некоторые авторы помещают тыквенные вблизи
от колокольчиковых (порядок Synandrae) по ряду признаков: спайка тычи-'
нок и др. Семейство тыквенных распространено главным образом в тропических
странах (700—f Обсидев) преимущественно Старого Света. В СССР лишь несколь-
ко дикорастущих видов. По большей части травы лиановпдные с усиками,
реже кустарники или небольшие деревья. Цветки правильные, чаще однополые,
пятерного типа, венчик обычно спайнолепестный. Тычинок пять или свободных,
или сросшихся, или же сросшихся попарно. Очень характерны одногнёздные
пыльники (не двугнёздные, как обычно). Семяпочки постенные. Плод ягодо-
образный. К сем. тыквенных, кроме тыквы, относятся: арбуз, дыня, огурцы,
кустарник Acanthosicyos horrida с корнями до 15 м длиной (пустыня Намиб).
48. Cyatheaceae. Семейство папоротников. В большинстве случаев
деревья. Около 400 видов в тропических и субтропических странах, редко в
умеренных. Роды: AIsophila, Cyathea, Dicksonia и др.Спорангии с полным косым
кольцом и с поперечной трещиной. Характерные растения тропических и суб-
тропических лесов (рис. 87).
49. С у с a d асе ае (саговники). Семейство голосеменных, у ко-
торых семяпочки не заключены внутри завязи (кдк у покрытосеменных), а рас-
положены открыто на плодолистиках. К типу голосеменных относятся также
хвойные, гннкговые и др. Саговники представлены лишь одним родом Сусаа
(15 видов) в Восточней Азии, Восточной Африке и в Австралии.Это вымирающие
растения, особенно обильное распространение имевшие в мезозое. Иногда на-
зываются «саговыми пальмами,) из-за некоторого внешнего сходства с паль-
мами (но последние—покрытосеменные растения, относящиеся к классу одно-
дольных). Они имеют довольно толстый, не ветвящийся ствол (у пальм
он обычно тонкий и иногда слабо ветвится), несущий наверху пучок больших
перистых листьев (до 2 м). Особи раздельнополы, одни мужские, другие жен-
ские. Мужские цветки имеют вид шишки, состоящей из споролистиков с сидя-
щими на них микрослорангиями, женские — тоже состоят из споролистиков,
но с макроспорангнями (семяпочками), однако шишки не образуют и точка
роста стебля прорастает сквозь женский цветок, давая затем обыкновенные
зелёные листья. Этим сем. саговников отличается от сем. замиевых, у
которого женские шишки являются конечными, как и мужские. Очень инте-
ресно оплодотворение при посредстве движущихся сперматозоидов. В СССР
саговники совершенно отсутствуют, но культивируются в оранжереях, а на
открытом воздухе на южном берегу Крыма и на Черноморском побережье. Род
Cycas.
С у с a da les (цикадовые, саговники). Класс голосеменных растений.
В настоящее время всего 9 родов и около 85 видов, почти исключительно в тро-
пических и субтропических странах. Класс подразделяется на два семейства —
Cycadaceae и Zamiaceae.
50. Cyclanthaceae (цпклантовые). Семейство однодольных,из порядка
пальмовых (Palmales). Всего около 40 видов в тропической Америке По своим
признакам семейство совмещает признаки пальм (веерные листья) и ароидных
(соцветие — початок). По внешнему виду похожи на небольшие веерные пальмы.
Род Carludovica (листья некоторых видов дают материал для плетения).
51. Су регасеае (осоковые). Семейство однодольных, из порядка
осокоцветных (Cyperales), который содержит только это одно семейство. Многие
авторы сближаю г сем. осоковых с сем. злаков и соединяют в особый порядок
чешуецветных (Glumiflorae), однако осоковые и злаки имеют целый ряд с.чпе-
408
ггвенных отличий, что оправдывает их разъединение и отнесение к различным
«орядьам.
По своемувнешнему виду осоки всё же довольно схожи со злаками, относясь,
к одной и той же жизненной форме злаковидных растений: узкие длинные ли-
шенные листья, невзрачные соцветия и цветки, опыляемые ветром, и др. Однак*
стебель всегда трёхгранный, плотный, с листьями, расположенными на стебле
в три продольных ряда (у злаков стебель округлый, полый, листья сидят в два
ряда). Цветки в колосовидных соцветиях, обоеполые или однополые, с редуци-
рованным околоцветником или совсем голые. Тычинок три. Плод — орешек, у
рода осоки (Сагех) окруженный особым мешочком. Цветок во всех частях обнару-
живает явную редукцию (в результате перехода к ветровому опылению) и доволь-
но легко может быть выведен из цветка лилейных. В общем осоковые представля-
ют особую конечную ветвь однодольных, перешедших к ветровому способу опы-
ления. Всего более 3 600 видов, из которых род осока (Сагех) имеет до 2 000
видов (в СССР около 400 видов). Распространено семейство главным образом
в умеренных широтах, однако также имеются представители в тропических и
холодных областях. Некоторые виды осоковых развиваются в большом коли-
честве особей,.сплошными зарослями, определяя ландшафт, например, заросли
осок на болотах, заросли камышей (Scirpus) по берегам озёр, заросли пушицы
(Eriop horum) на болотах, заросли папируса (Cyperus papyrus) на реках и озё-
рах Центральной Африки и др. Осоковые обычно связаны с влажными место-
обитаниями, хотя отдельные виды встречаются в степях (Сагех humilis) и пусты-
нях (С. physocles и др). Другие роды: Cobresia, Elyna, Tricliophonim и др.
52. Dioscoreaceae (бататовые). Семейство однодольных, из порядка
лилиецветных (Liliiflorae). По ряду признаков отличается от других семейств
порядка: это травянистые растения с вьющимися или ползучими стеблями,
часто с мощными клубнеобразными корневищами, листья с сетчатым жилко-
ванием (у однодольных обычно жилкование параллельное или дугонервное),
нередко с стреловидным основанием. Цветки невзрачные, однополые; 220
видов, в большинстве случаев тропических. Dioscorea (мощные клубни, иногда
до 2 м, съедобные), Tamus (у нас — Крым, Кавказ, высокий, вьющийся стебель).
53. Dipsacaceae (ворсянковые). Семейство спайнолепестных двудольных,
из порядка Rubiales (мареноцветные). По своим корзинковидным соцветиям
очень сходно с сем. сложноцветных, почему некоторые авторы сближают эти
два семейства. Однако правильнее смотреть на это семейство как на одну из
конечных ветвей мареноцветных. Травы умеренных широт, особенно мног»
видов в Средиземноморье. Листья супротивные без прилистников. Цветки в
корзинковидных соцветиях, окружённых обвёрткой верхушечных листьев.
Цветки неправильные, четырёх -, пятнчленные. Из сросшихся прицветников
образована ложная чашечка. Плод с одним семенем (семянка). Около 160 видов.
Роды: Seabiosa (скабиоза), Succisa (сивец) и др.
54. Dipterocarpaceae (двукрылоплодные). Семейство раздель-
нолепестных двудольных, из порядка камеденосных (Guttiferales); однако ввиду
близости этого порядка к порядку постенносеменных некоторые систематики
объединяют оба порядка вместе в порядок постенносеменных (Parietales).
Троп п ческое семейство, главным образом в Южной Азии и на прилегаю-
щих островах (Борнео, Суматра, Филиппины и Др.), отчасти в тропической
Африке. Деревья с вечнозелёными листьями (с прилистниками). Многие богаты
смолой (отсюда название порядка).Очень характерна чашечка, которая остается
при плоде, причем два чашелистика (а иногда и все) сильно разрастаются в
крылья (отсюда название семейства). Цветки обоеполые, пятичленные, тычи-
нок иногда много. Плод — орех. Всего около 320 видов, некоторые развиваются
в большом количестве особей и образуют леса (например на Филиппинах).
Род Dipterocarpus и др.
55. D г о з е г а с е а е (росянковые). Семейство раздельнолепестных дву-
дольных, из порядка постенносеменных (Parietales). Нередко относят семейств»
кдр>г,|М порядкам, в частности к Sarraceniales. сближая с такими семействами,
как Sarraceniaceae. Поводом для подобного сближения служит то, что пред-
ставители этих семейств являются насекомоядными растениями. Все виды
росянковых насекомоядны. Травы или полукустарники, растут на болотах
или в воде. Листья покрыты волосками и выростами, выделяющими клейкий
сок, способный растворять бечковые вещества животных. Цветки правильные,
46*
четырех-,пятичленные. До 100. видов тропических и умеренных стран. Роды:
Drosera (росянка —у н дс три вида, особенно много видов в Австралии и Новой
Зел шдин), Dion.iea—мухоловка (один вид в> Северной Америке—рис. 162), Aldro-
vandi (один вид с разорванным ареалом — ряс. 2).
L6 Е be п асе а е (эбеновые). Семейство спайнолепестных двудольных,
из порядка эбеноцветных (Ebenales). Систематическое положение порядка не
вполне ясно Исключительно тропические и субтропические деревья и кустар-
ники. Сем. эбеновых содержит до 320 видов — вечнозеленые деревья. Цветки
обычно однополые, трех-семичленные. Многие виды дают ценную древесину
(виды Diospyros), другие виды ценятся, как плодовые деревья: хурма (Diospyros
lotus) и др.
57. Empetraceae (ворсинковые). Семейство раздельнолепестных
двудольньх, ш порядка крушиноцветных (Rhamnales). Иногда относится к
порядку молочайных на основании некоторых признаков цветка. Очень не-
большое семейство, всего несколько видов, в умеренных и холодных странах
Европы, Азии и Америки. Небольшие кустарнички, с узкими листьями с за-
вернутыми книзу краями, очень напгм шлющие растения сем. вересковых.
Цветки очень мелкие, трехчленные, чаще однополые, тогда растения двудомные.
Плод—ягода. В СССР вороника (Empetrnni nigrum), часто на болотах севера.
Epacridaceae (эпакридовые). Сечейсто спайнолепестных дву-
дольных, и порядка верес ко цветных (Eric lies). К этому порядку относя 1ся
также семейства: вересковые, грушанковые и др. Мелкие кустарники или
полукустарники, распространены почти исключительно в Австралии (включая
Тасманию и Новозеландию), немного видов на южных островах до Южной
Америки. По внешнему виду очень похожи на растения сем. вересковых (особен-
но на род Erica), имея мелкие, узкие листья. Цветки от вересковых нескольк®
отличаются: они более крупные, шчинок 5 в одном кругу (у вересковых—10
в двух кругах). Всего около 350 видов. Некоторые виды разводятся для деко-
р ставных целей.
t г i с а сеае (вересковые). Семейство спайнолепестных двудольных,
нз порядка верескоцветных (Ericales). Основное семейство порядка (другие
семейств!: грушанковые, эпакридэвые); с раздельнолепестными имеется связь
при посредстве порядка камеденосных (Guttiferales). Семейство содержит около
1 ЗоО вадов, распространенных почти по всей земле. Немало видов имеется и в
СССР. Kyci 1рникн и полукустарники, редко деревья, в больши 1Стве случаев
с вечнозелеными листьями. Листья иногда крупные (рододендроны;, но чаще
мелкие и узкие вересковидного, «эрикон дного<> типа. Цветки правильные, че-
тырех-, пятичленные. Тычинок 10в двух кругах; они раскрываются порами. Не-
которые виды развиваются в большом количестве особей, давая определенные
ландшафты; у нас в Европейской части (особенно на северо-западе) сплошные
заросли дает обыкновенный вереск (Callum vnlg-ris — рис. 133), в Западной
Европе вереццтники образованы так не видами рода* Eric'1. На горах большие
заросли д^т рододендрон (Rhododendron) в азалия (Az 1с ) На болотах мас-
сами встречаются: клюква (Oxycoccos), голубика (Vaccinium uliginosun), андро-
медэ и др.,'в лесах — брусника, черника (Vaccinium vttJS idiea и V. myrtil-
lus). Многие виды дают съедобные плоды, другие — декоративны.
Другие роды, приведенные в книге: Arbutus (землян тчное дерево), Arctosta-
phy 1оз(толокняка), Arctons, Cassandra (Кассандра), Cassiope, Ledum (багульник),
Loiseleuria, Phyllodoce.
би. E r i о c.a u 1 а с e a e. Семейство однодольных из порядка Enantio-
Ы ’st?e (сюда же относятся сем. Rcstionaceae, Xyridaceae и др-). Около
6i‘(j видов в субтропических п тропических странах, особенно в Южной Америке.
По большей части (многолетние травы с узкими листьями, часто прикорневылш,
соцветие очень похоже на соцветие сложноцветных, на длинной обычно стрелке,
цветки очень меткие, однополые, редуцированные. Род Eriocaulon (в Европе
только одни вид),
61. Erythroxylaceae (кокаиновые). Семейство раздельнолепест-
ных двудольных, из порядка геранецветных (Geraniales). Очень близко к сем.
леновых, но все виды являются деревянистыми. Растения теплых стран, глав-
ным образом в Америке. Около -UU видов. Род Ery throxy Ion (Е. coca — кока —
в листьях алкалоид кокаин). Некоторые систематики сближают семейство с
кленовыми.
410
62. Euphorbiaceae (молочайные). Семейство относится к раздельно-
лепестным двудольным, к порядку трехорешковых (Trieотеле), однако система-
тическое положение недостаточно о пре цел нно, и различные систематики поме-
шают семейство в разные части системы (например в порядок геранецветных
и'др.)-Семейство имеет до 4 500 видов, главным образом в тропических, также
в умеренных странах. По своему внешнему виду растения семейства крайне
разнообразны, относясь к различным жизненным формам: травы, кустарники,
деревья, лианы, кактусовидные мясистые растения (суккуленты). Последние
с трудом отличимы от настоящих кактусов, но при поранении дают млечный
сок, характерный вообще для молочайных. Листья у подобных суккулентов
или редуцированы или роль листьев переходит к стеблю — развиваются кладо-
дии (утешенный листовидный стебель). Кактусовндные молочаи предел ж аяют
прекрасный пример того, что в различных семействах, далеко расположенных
в филогенетической системе, могут вырабатываться в однородных внешних
условиях и однородные жизненные формы (кактусовидные формы имеются также
в сем. лэсточннковых — Asclepiaclaceae, сложноцветных). Цветок молочайных
иногда очень сильно редуцирован, и, например, у рода мо ючая (Euphorbia)
каждый мужсюй цветок состоит лишь из одной тычинки, женский — из одного
пестика; однако эти редуцированные цветки располагаются группами, окружен-
ными особым покрывалом, напоминающим как бы венчик (это «ложный цветок*,
соцветие). У других молочайных цветки менее ред- ц роваиы. Для всех них
характерна трехгнездная коробе чка, разрыв >ющаясл (иногда с очень большой
• (лой) при созревании. Имеется целый ряд полезных (астений каучуконосы
। находится в млечном соке)— Hevea br asi liens,s, M inih G1 izi.wii
(оба в Ьразилии); клещевина, Аает касторовое дерево (Ricinus 'communis). У
нас в СССР много видов молочая (Euphorbia), также прыеск! (Mercuri ilis).
63. Fagace а е (буковые). Семейство разде ibhj идесгных дв/дош-
ных. из порядка букощктных (Fagales). К этому же поря ку опюсигся сем.
березовых.Распространено в умеренных и тропических стр шах (кроме Центр t 1ь-
ной п Южной Африки), около 350видов, исключительно деревья, редко кустар-
ники. Цветки невзрачные, очень просто устроенные, обычно однополые, около-
цветник простои, мало заметный. Невзрачность цветков связана с опылением
при помощи ветра. Мужские цветки собраны в сережковидпые соцветия. Очень
характерной является чашевидная плюска, окружнощ in женские цветки
и состоящая из сросшихся кроющих листьев; тюск i ршрютается при плодах
(например плюска желудей у дубов). Роды: дуб (Qaercus) с многочлсаенныяи
видами до 200(в СССР 19 видов), 6yK(Figus — в ССсР трл вид1), каштан (Cas-
tanea). Все это основные породы широколиственных лес ж. Для южного полу-
шария характерен южный бук — NHhofigus. По мнению одних системпмков,
буковые являются примитивными двудольными (простые цветки —это лри.и-
тивные цветки) и располагаются в начале системы двудольных, другие ечн г нот
их редуцированными из более высоко организованных в результате переход
к ветровому опылению (простой цветок —-результат упрощения). Вторая точка
зрения более правильна.
64. Frankeniaceae (франкенпевые). Семейство раздельнолепестных
двудольных, из порядка постенносеменных (Parieles). К этому же порядку
етносктся сем. тамарисковых, с которыми франкенпевые близко родственны.
Очень неболниое семейство (около 60 видов), главным образом ксерофитов и
галсфитсв (Средиземноморье, Азия, Австралия, Америка). Травы и полукустар-
ники с л едкими листьями, лишенными суккулентностп — не мясистые «со ih;i-
ки.>. На вегетативных органах имеются железки, выделяющие капельки горько-
соленого раствора (род Frankonia— в СССР ряд ендов на юго-востоке Евро-
пейской части, в Средней Азии).
65. Fumariaceae (дымянковые). Семейство раздельнолепестных дву-
НОЛ1НЫХ, из порядка мэкопветных — Par.*ver'’ks Очень часто это семе ют в*
рассматривают как подсемейство сем. маковых — это показывает очень е.п ie
связь этих семейств. Цветки неправильные (у маковых — пртвильные), два
или один лепесток со шпорцами, шесть тычинок в двух пучках. Около 1д0
видов в умеренных странах северного полушария. Роды: Coryd ilis — хохлатка,
Fumaria — дымянка.
66. Gen tia пасеа е (горечавковые). Семейство спайнолепестных дву-
дольных, из порядка скрученных (Contortie). К этому'же порядку относятся
411
сем. маслиновых, кутровых, ласточниковых и др. Травы (однолетние и много-
летние), реже кустарники, Около 800 видов, главным образом в умеренных
странах. Растения голые, обычно с супротивными цельнокрайними листьями.
Цветки нередко крупные и ярко окрашены. Венчик скрученный в бутоне. Цвет-
ки правильные, четверного или пятерного типа. Завязь верхняя, одногнёздная.
Роды: Gentiana (горечавка — особенно на альпийских лугах—рис. 153), Chlora
и др. У нас много видов горечавки, также и другие роды. Многие виды содержат
горькие вещества.
67. Oeraniaceae (гераневые). Семейство раздельнолепестных двудоль-
ных, из порядка геранецветных (Geraniaies). В умеренных и субтропических
странах, около 630 видов, главным образом травы. Цветки правильные, пяти-
членные, тычинок Ю и 15, плод из пяти сросшихся плодолистиков с сильно
вытянутым столбиком; при созревании он расщепляется на пять продольных
полосок (центральная колонка остаётся), которые иногда с силой разбрасы-
ваются (распространение семян). Роды: герань (Geranium) цветки правильные,
пеларгонпум (Pelargonium) — цветки неправильные (особенно много видов в
Южной Африке), некоторые виды дают гераневое эфирное масло.
68. Gesneriaceae (геснериен^е). Семейство спайнолепестных дву-
дольных, из порядка маскоцветных (Personatae). К этому порядку относятся
также семейства: норичниковые, паслёновые, бигнониевые. Исключительно
тропическое семейство, лишь два рода в Южной Европе. Всего около 1 100
видов, почти исключительно травянистых растений, с обычно супротивными
листьями и яркими цветками. Очень много декоративных видов, других полез-
ных нет. Цветки неправильные. Очень близко к норичниковым, от которых
особенна отличается одногнёздной завязью (у норичниковых два гнезда).
69. Ginkgoales (гинкговые). Класс голосеменных (подобно саговникам
и хвойным). Имеет в настоящее время одно семейство (Ginkgoaceae) с одним
родом и одним Видом — гинкго двулопастное (Ginkgo bilob.i). Это стройное
дерево сохранилось в культурном состоянии в Китае и Японии, а затем стал»
разводиться и в других тёплых странах (у нас в Крыму и на Кавказе); в диком
состоянии, повидимому, встречается в западном Китае. Имеет крайне
оригинальные листья, не похожие на листья других семенных растений; они
веерообразны, часто двулопастные (откуда название вида) и имеет д и х о-
т о м и чески ветвящиеся жилки (рис. 187). На зиму листья опадают. Муж-
ские и женские споролистики находятся на различных растениях (двудомность):
первые собраны в небольшие серёжки, вторые — развиваются на укороченных
веточках. Оплодотворение при помощи подвижных сперматозоидов (как у сагов-
никовых). Гинкго является последним представителем класса, некогда (в мезо-
зое) широко распространенного; это как бы «живое ископаемое^.
7и. Gleicheniaceae. Одно из семейств папоротников. Распростра-
нено в тропиках и субтропиках, главным образом в южном полушарии. Листья
многократно дихотомически ветвятся, растут своей верхушкой, вследствие
чего достигают очень больших размеров, до 10— 12 м длины. Спорангии собраны
в кучки и имеют почти поперечное неполное кольцо. Представители семейства
известны с юры. Современные виды (до 30) — это остаток прежде сильно-
распространённого семейства.
71— 73. Gneta les (гнетовые). Класс голосеменных растений. В отличие
от других голосеменных цветки (мужские и женские) имеют особый покров,
похожий на околоцветник покрытосеменных, что вместе с другими признаками
даёт основание некоторым ботаникам выводить покрытосеменные из растений
этого класса. Другие считают гнетовые, особой боковой конечной ветвью голо-
семенных. Когда-то гнетовые были несомненно широко распространены, ттк как
их современные представители (всего около 50 видов^разбросаны по всем частям
света. Всего три семейства и три рода: Ephedra (сем. хвойннковые), Welwi-
tschia или Tumboa (сем. вельвичиевые) и Gjaetum (сем. гнетовые). Первый род
представлен кустарниками с прутьевидными стеблями и чешуйчатыми редуци-
рованными листьями, второй — имеет лишь один вид совершенно своеобраз-
ного внешнего вида (рис. 199), представители третьего рода (деревца или
лианы) очень похожи по внешности на покрытосеменные (широкие листья с
перистым жилкованием и др.).
74. Goo de ni а се а е (гоодениевые). Семейство спайнолепестных дву-
дольных, из порядка спайнопыльниковых (Synandrae). Это сравнитевыю небшаь-
412
шое семейство, около 300 видов, распространённых главным образом в Австра-
лии, но также в мангровых зарослях по побережьям Тихого и Атлантического
океанов. Травы, кустарники и полукустарники. По своим признакам очень
схоже с семейством колокольчиковых, из которого обычно выводится.
Отличия от колокольчиковых: ясно неправильные иветки, отсутствие млечног»
«ока, очень характерно чашевидное расширение под рыльцем, где собирается
пыльца.
75. Q г a m i п е а е (злаки). Семейство однодольных растений, из порядка
злакоцветных (Graminales). Ряд*-авторов относит это семейство к порядку че-
юуецветных (Glumiflorae), к которому относят также и сем. осоковых. Од-
нако более правильно, повиднмому, разъединять эти два семейства, так как они
имеют ряд существенных различий. Злаки представляют большое (до 7000
видов) семейство, крайне важное во многих отношениях. Распространены п«
всей земле, но особенно подтропиками. По внешности злаки относятся к «зла-
ковидной у жизненной форме (тонкий стебель, узкие длинные листья) и нескольк*
похожи на растения Семейства осоковых, но отличаются округлым, полым (с
перегородками на узлах) стеблем (соломина) и листьями, расположенными на
стебле в два продольных ряда. Нижняя часть листа охватывает (нередко на
большом протяжении) стебель, образуя так называемое влага пище; в месте
перехода влагалища в верхнюю плоскую ласть листа находится особый вы-
рост — язычок. Травы однолетние пли многолетние, иногда со стеблем, не от-
мирающим ежегодно (бамбук — рис. 205). Цветки очень упрощены и невзрачны,
так как опыляются.ветром; собраны сначала в небольшие колоски, а затем
в соцветия разных типов (сложный колос у ржи, пшеницы, метёлка у овса,
проса и др.)- Плод — зерновка. Цветки злаков по их строению могут быть
выведены из типичного цветка однодольных (из цветка лилейных) путём редук-
ции ряда частей, и потому сем. злаковых можно считать очень редуцированной
(в строении цветка), приспособившейся к ветровому опылению ветвью одно-
дольных. Многочисленные виды злаков важны в практическом отношении —
хлебные злаки, кормовые растения. В СССР злаки очень широко распространены,
нередко своим массовым развитием определяя ландшафт на степях, лугах,
болотах. Род мятлик (Роа) имеет в СССР более 100 видов, роды ковыль (Stipa),
вейник (Calamagrostis) — до 60, роды костер (Bromus), пырей (Agropyrum)—
до 50—ь0 видов и т. д.
Другие роды, приведенные в книге: Agrostis (полевица), Alopecurus (лисо-
хвост), Andropogon (бородач), Anthoxanthum (пахучий колосок), Aristicla,
Avena (овёс), Bambusa (бамбук), Beckmannia (бекмания), Bouteloua, Brachy-
podium (коротконожка), Bri/a (трясунка), Buchloe, Cymbopogon, Deschampsia
(шучка), FestLica (овсяница), Gynerium, Hierochloe (зубровка), Hordeum (яч-
мень), Hymmanche, Ischaemum, Koeleria (тонконог, келерия), Molinia, Nar-
dus (белоус), Panicum (просо), Paspalum, Pennisetum, Phalaris (канареечник),
Phragmites (тростник), Sesleria, Spinifex, Sporolobus, Triodia.
76. Guttiferae (камеденосные)- Семейство раздельнолепестных дву-
дольных, из порядка Guttiferales. К этому же порядку относятся семейства
двукрылоплодных, маркгравиевых и др. Главным образом тропическое семей-
ство — деревья, редко травы (более 80U видов), у нас встречается лишь травя-
нистый род зверобой (Hypericum). Для семейства (и для всего порядка) харак-
терно присутствие особых вместилищ во всех органах, где находятся смолы,
масла и др. Много полезных растений, содержащих камеди, жиры, у других
венная древесина, прекрасные плоды (например мангустан — Garcinia Mango-
strna). Цветки имеют сходство с цветками порядка постенносеменных, из
которых это семейство обычно выводится.
” 77. Halorrhagidaceae. Семейство раздельнолепестных двудоль-
ных, из порядка миртоцветных (Myrtales). К этому порядку относятся также
такие семейства: миртовые, дербенниковые, онагрнковые, Sonnentiaceae,
Rhizophora се? е и др. Как и весь порядок миртоцветных, данное семейство ха-
рактеризуется четырёхчленными частями цветка (реже двух- и трёхчленными),
но обнаруживает целый ряд упрощений в связи с переходом к водному образу
жизни и к опылению при помощи ветра (околоцветник иногда отсутствует,
етк ев зрачны и др.). Небольшое семейство (около 150видов), в субтропиче-
ских и умеренных странах, главным образом в южном полушарии. Травы очень
различного внешнего рг'1'1. Род Gunnera (Австралия) с крупными листьями
413
(рис. 216 и 217), род MyriophyllLim (уруть) — у нас везде в реках, с листьями,
рассеченными на узкие дольки (рис. 26).
i8. Н a ma m е 1 i d а сеа е. Небольшое семейство раздельнолепестных
двудольных (около 50 видов). Исключительно деревянистые растения (деревья
и кустарники) в тропических, субтропических,отчасти умеренных стран ix.Систе-
матическое положение не вполне ясно; одни авторы относят семейство к Ro-
8 des, другие — сближают с Pol^carpicae, третьи выделяют в самостоятельный
порядок На ma me lidales. Очевидно, семейство является связующим звеном
ряда порядков. Род Parrotia (Персия, Талыш) и др.
7у. Н е I о b i а е (водные). Порядок однодольных, куда относится ряд
семейств (частуховые — Alism itaceae, сусаковые — Butomaceae, рдестовые—
Pot'mugctcnaceae и др.) с представителями, растущими на болотах, в реках,
нередко совершенно погруженными в воду. Порядок имеет большое филогене-
тическое значение, так как многие систематики ставят его в 'основание всех
однодольных и ьыводяжз него все остальные порядки однодольных, кроме
того, порядок хорошо выводится из двудольных, именно из порядка много-
п юдниковых (Polycarpicae). Таким образом. порядок водных представляет-
как бы мост между однодольными и двудольными (первые произойти от вторых).
Водные имеют целый ряд признаков двудольных (многоплодниковых): листья
с перистым жилковшием, цветок с чашечкой и венчиком, много тычинок,
много пестиков и др.
8'J. Hydrocharitaceae (водокрасовые). Небольшое семейство (до
60 видов), плавающих «jin not ружейных в воду растений. Семейство однодоль-
ных из порядка водных (Helobiae). Цветки плавающих видов очень сходны с
частуховыми и сусаковыми, имея чашечку и венчик. Цветки погруженных
видов сильно редуцированы. Роды: Hydrochnris (водокрас), Villisneria (вал-
лиснерия — рис. 40), EUodea (водная чума). Тропические и умеренные страны.
8>. Hymenophyllaceae (тонколистные). Семейство папоротни-
ков. Это по большей части небольшие, очень нежные и изящные растения, с
тонкими листьями с ОДНИЛ1 слоен клеток мезофилла. Распространены главным
образом в тропиках на сильно влажных местах и в качестве эпифттов. До 2йб
видов. Однако некоторые виды имеются и в Европе, вСССРдва вида на Дальнем
Востоке. Спорангии с поперечным или косым полным кольцом.
12. Iridaccae (ирисовые) Семейство однодольных из порядка лнлие-
цвегных, куд» относятся также семейства лилейных, амариллисовых, анана-
совых и др- Очень близко к семейству амариллисовых, но в цветках имеется
л 1шь три тычинки (три тычинки внутреннего круга выпадают). Завязь нижняя.
Листья обычно двурядные, между ними появляется соцветие. Цветки пр шн яв-
ные пли неправильные, нередко яркой окраски. Многолетние травы или полу-
кустарники. Распространены особенно в Южной Африке, в Средиземномор-
ской области, также в Америке. Число видов до 900. Роды: Iris (ирис, касатик,
петушке) — обычно культивируется в садах, Crocus (шафран — рис. 152 и 154),
Sisynnchiuin, Alophia, Cypella и др.
83 Juglandaceae (ореховые). Семейство раздельнолепестных дву-
дол них, из порядка орехоцветных (Jughndales). К этому порядку относится
также сем. юлианпевых (Julianiaceae). Порядок по своим признакам очень
близок к порядку букоцветных, с которым он иногда соединяется, но
отличается перистыми листьями без прилистников и рядом признаков в цветке.
Семейство ореховых распространено в умеренных областях сев. потуиирия
(отчасти в тропической Восточной Азии), имеет около 35 видов. Исключительно
деревья с перистыми листьями, мелкими цветками, собранными
в сережки Околоцветник или ч.ашечковидный или отсутствует. При основании
женских цветков образование вроде плюски из сросшихся кроющих листоч-
ков Большое значение имеет род Jifglans—известный вид- грецкий орех (J.
regii). Американское дерево гикори (Caiya). Род Pterocarya (один вид в Закав-
казье). Простое устройство цветков правильнее считать за результат редукции,
как и у букоцветных.
84 . Julianiaceae (юлианиевые). Семейство раздельнолепестных дву-
дольных, из порядка орехоцветных (Jughndales). Небольшое семейство,
исключительно американское (всего пять видов в Мексике и Перу); по при-
знакам близко к ореховым. Деревья с перистыми листьями,
414
имеют смоляные ходы; последнее обстоятельство (и другие признаки) отличает
их от сем. ореховых.
85 Ju пс л селе (ситниковые). Семейство однодольных, из порядка лилие-
цветных (Liliif 1 аг'е) Цьеток устроен по типу лилейных, по, в связи с
ветровым опылением, околоцветник состоит из чешуевидных не окрашенных
листочков. По своим вегетативным органам схожи с осоками и отчасти со зла-
ками, почему некоторые систематики счигцог семейство сити жовых переход-
ным от йнанецветных к осокоцне’тн ям. Гл'BhLim образом в северном полушд-
рии Число видов около 300 Роды Juncus (ситник), Luziih (ожика) и др.
f-6. Labiitae (губоцветные) семейство спайнолепестных двудольных.
Различными авторами относится к разным порядкал) (в зависимости от более
узкого и ан более широкого понимания порядков), то к порядку вербеноцвет-
ных (Verbenales) — в нем удва семейства, то к трубкоцветным (Tubifl >гае),
куда иногда относят до 25 семейств Распространено главным образом в сухих
и теплых умеренных об частях (особенно в Средиземноморье), но очень обильно
представлено и в СССР (всего до 3 000 видов). Травы пли кустзрнчки. Для
Семенова характерно присутствие эфирных м^сел, от чего часто растения
являются сильно пахучими; стебли всегда четырехгранные с супротивными
листьями. Цветки всегда неправильные, обычно с двугубым вепч жом (откуда
название семейства), тычинок четыре (реже две), п аод из четырех распадающ жся
орешков. Цветки 'собраны в так называемые мутовки (в п^з/хах двух
с} противных листьев сидят пучки цветков, которые соприкасаются)? Форма
цветка определенно j кззыь^еа на опыление насекомыми (пчеаы. шмеаи и др-).
Много лекарственных растений (шалфей — Sdvia, мята — Menth '), хорош ix
эфироносов (розмарин, л'ванд% базилик), декоративных (кр юнэцветныи
шалфей). Ближе всего стоит к сем. вербеновых.
Другие роды, приведенные в книге: Aj и*! (Ж]вучка), Brundh (черн тон-
ка), GUeobdolon (зеленчук), Glech'ina (u.-uoai, LwinJuIi ( 11 в > и д j a i), N'peta.
(котов.н<), Phlomis (зопник), R)smirinus (розмдрш), StaJi/s (чюец), Тейс-
rinrn, Th/mus (тияьян).
87 Lauraceae (лавровы'е). Семейство раздельнолепестных двудольных, из
порядк 1 -п-кголледниковых Сюда же относятся семейств! м чноч цвые, шоко-
вые, мускатные, лютвьовыс и др., которые все вместе поедет шаяют о i ,н б шзко
родственный пика. К сем? лавровых относятся почти кс^аюч [те mi а деревья
(всего окоао 1 000 в-гдов\ растущие в тропических и субгропдчесц ix об пстях
всего света. Нередко в этих лесах они играют очень большею pni, Д-ревья
имеют эл шптическпе, вечнозеленые, кожистые листья, содсржчцш оф |риые
мае-1. Цвети) небольшие, чине всего из грехчаенных кругов, стбршных в
готовки или метелки. Роды: лавр (в Европе один вид — Laurus г. «bills), Cin-
nam mum — дает камфору, корицу и др.
18. Lecythidaceae. Семейство раздельнолепестных двуаэ шных, из
порядка миртоцвегных (Myrt des) Около IJ0 вчдэв тропических деревянистых
растений. Роды Lecythis, LSertholfetia (два южлоамерикшекях ь иа — дэюг
съедобные «бразильские орехи»).
<9— I Leguminosae (бобовые). Семейство р идет? то i и ::ч я <
двудольны^. Одни систематики считают бобовые семекгвэм, огн >гащ imch к
порядку розоцветных (Ff ’sMes), другие считают самостоятельны.1 порядком,
тесно связпнным с розоцветными. Это большое семейство (или парадах) одер-
жит до 15 000 видов, распространенных по всей земле и относящихся к самым
разнообразным жизненным формам: травы, полукустарники, кустарники,
лианы, деревья. Очень характерны для всех листья — в большинстве случаев
перистые с прнлис'никами. Но наиболее характерец плод — боб (отсюда на-
звание семейства). Цветок правильный иан неправильный, четырех-пятичлен-
ный, тычинок обычно 10, нередко сросшихся нитями. Среди бобовых нужно
различать три подсемейства (пли семейства): а) мимозовые, б) цезыьпиниевые,
в) мотыльковые, близко родственные друг другу
а) Мимозовые (Mimosaселе). Они ближе всего стоят к розоцветным. Цветки
правц аьные, тычинки, не сросшиеся нитями, их 10 или много (реже мало), они
ярко окрашены и делают цветок очень заметным Листья сложнолерпстые, при-
листники часто превращены в колючки Наиболее вашей род акация (Acacia) —
около 500 видов, из которых больше половины в Австралии. Акации играют
важную роль в саваннах (рис. 137), в скрэбе. В общежитии цветки акаций
415
еправильно называются «мимозой?). Род Albizzia — декоративные деревья,
культивируемые в Крыму и на Черноморском побережье, тоже называются
«мимозами »>. Настоящая мимоза (Mimosa) имеет всего четыре или восемь тычи-
нок. Около 350 видов. Другие роды: Parkia, Prosopis.
6) Цезальпиниевые (Caesalpiniaceae). Цветки неправильные, верхний лепе-
сток венчика в почке прикрывается всеми остальными. Деревья, главным об-
разом тропические (всего около 1 000 видов). В Крыму и.на Кавказе культи-
вируются: иудино дерево (Сек is siliquastrum) и цареградские рожки, или рож-
ковое дерево (Ceratonia siliqua).
Другие роды: Amherstia, Bauhinia, Berlinia, Caesalpinia, Cassia, GlQditschia,
Parkinsonia.
в) Мотыльковые (Papilionaceae). Очень многочисленное семейство, особення
распространённое в умеренных странах, а также под тропиками. Травы, под
тропиками обычно деревянистые виды. Очень характерно устройство цветка,
называемого мотыльковым (отсюда название семейства); он неправильный,
верхний лепесток венчика самый большой (парус), два боковых (крылья), два
нижних отчасти сросшихся лодочковидно (лодочка), причём верхний лепесток
охватывает (в почке) все остальные (обратно тому, что имеет место у цезаль-
пиниевых). Тычинок обычно десять, из них чаще всего девять срослись нитями,
одна (верхняя) свободная. В СССР семейство широко распространено. Из пище-
вых растений: горох, бобы, фасоль. Ряд декоративных растений. ЛАотыльковые
представляют одну из конечных ветвей раздельнолепестных с неправильными
цветками.
Роды, приведённые в книге: Ammodendron, Amorpha, Astragalus (астрагал),
Вгуа, Butea, Caragana, Chorizema, Cytisus (ракитник), D diibergia, Dipterix,
Erythrina, Genista (дрок), Giycyrrhiza (солодка), Halimodendron, J cksonia,
Lupinus (люпин), Maackia, Machaeriun, Onobrychis (эспарцет), Orobus (сочевич-
ник), Oxytropis (острокильник), Psoralea, Robinia. Sophora, Soji (соя), Tri-
folium (клевер), Ulex, Vicia (горошек)
92. Lemnaceae (рясковые). Небольшое (до 25 видов) семейство одно-
дольных, из порядка аронниковых (Arales). Сюда же относится сем Агасеае.
Очень сильно редуцированные водные растения, плавающие на воде или
совсем погруженные в воду. У рода Wolffia всё растение состоит из маленькой
(о.коло I мм) чечевички — нет ни листьев, ни корней; цветки крайне редуциро-
ваны, лишены околоцветника; мужской цветок имеет одну тычинку, женский—
один пестик. Род Lemna (ряска) — местами сплошь покрывает водоемы. Неко-
торые виды космополиты.
'93. Liiiaceae (лилейные). Семейство однодольных, из порядка лилие-
цветных (Liliiflorae). К этому же порядку относятся семейства: амариллисовые,
ананасовые, ирисовые и др. Как порядок, так и семейство являются
наиболее «типичными») среди всех однодольных; из них хорошо выво-
дятся все остальные однодольные, а с другой стороны, лилиецветные легко
связываются с многоплодниковыми, т. е. с двудольными. Другие ботаники
связывают однодольные и двудольные при посредстве порядка водных (Не(о-
biae). Главным образом многолетние травы с сильным развитием подземных орга-
нов (корневища, луковицы, клубни), редко деревянистые растения (древовидные
или вьющиеся). Всего до 3 000 видов, распространенных оттропнческихдо хо-
лодных областей, очень разнообразных по вегетативным органам, но крайне
однотипных по строению цветка; цветок правильный, из пяти трехчленных,
взаимно чередующихся кругов (околоцветник из двух кругов — деления на
чашечку и венчик нет, два круга тычинок, завязь верхняя).
Роды, приведённые в книге: Allium (лук), Aloe (алоэ), Colchicum, Conval-
laria (ландыш рис. ИЗ), Dame, Dasylirion, Dracaena, Gigea (гусиный лук),
Hyacinthel 11 (гиацинт), Lilium (лилия), Narthecium, Ornithogalum (птицемлеч-
ник), Par.is (вороний глаз), Phormium (P. tenax — новозеландский лён,
рис. Polygomtum (купен11, рис. 114), Ruscus (иглица, ряс. 191), Smihx,
Scilh, Tulipa (тюльпан), Veratrum (чемерица), Xanthorrhoea (травяное дерево—
в Австралии, рис. 140), Yucca (юкка, рис. 209).
94. L1 пасеае (леновые). Семейство раздельнолепестных двудольных,
из порядка геранецветных (Geraиiales), к этому же порядку относятся сем.:
гераневые, кисличные, настурцневые, бальзаминовые и др. Травы или кустар-
ники с узкими цельнокрайними листьями. Цветки правильные, пятерные или
416
четверные. Тычинки при основании срослись. Гнёзда завязи в ложной перего-
родкой. Около 150 видов, в умеренных и тропических странах. Наиболее богатый
видами род Linum (лён), у нас ряд видов.
95. Lobeliaceae (лобелпевые). Семейство спайнолепестных двудоль-
ных, из порядка спайнопыльниковых (SynantheraJes). К этому же порядку
относятся также сем. сложноцветных и колокольчиковых. Лобелпевые очень
тесно связаны с колокольчнковыми и отличаются от последних главным об-
разом только неправильным венчиком н более плотцо спаянными пыльниками.
Около 450 видов, главным образом в тропиках и субтропиках. Травы, реже
древесные формы. Род Lobelia — многие виды декоративны.
9G. Lycopodiaceae (плауновые). Семейство сосудистых споровых
растений. Травянистые растения с мелкими, спирально расположенными листья-
ми. Спорангии расположены по одиночке в пазухах илн в нижней части ли-
стьев. Нередко спорангии собраны в колоски. Около 200 видов, в тропических
и умеренных странах. Род Lycopodium (плаун) — у нас ряд видов.
97. Lythraceae (дербенниковые). Семейство раздельнолепестных дву-
дольных, из порядка миртоцветных (Myrtales). К этому же порядку относят-
ся миртовые, Sonneratiaceae и др. Весь порядок легко связывается с розоцвет-
ными, но отличается некоторыми признаками, в частности, четырёхчлен-
ными цветками. Сем. распространено в умеренных и тропических странах, очень
многочисленно в Америке; около 450 видов деревьев и трав. В СССР ряд тра-
вянистых видов. Цветки правильные, четырёхчленные (реже шестичленные),
между листочками чашечки особые выросты. Род Lythrum и др.
98. Magnoliaceae (магнолиевые). Семейство раздельнолепестных
двудольных, из порядка многоплодииковых (Polycarpicae). К этому же порядку
относятся семейства: аноновые, лавровые, мускатные, лютиковые и
др. Магнолиевые — это наиболее типичное семейство порядка и в то же вре-
мя крайне важное в филогенетическом отношении; многие систематики
ставят его в основании всей системы покрытосеменных растений, связы-
вая с голосеменными. Строение цветка обнаруживает ряд примитивных
признаков: части цветка обычно располагаются спирально, тычинок и пестиков
много, часто сильно выпуклое цветоложе н др. Цветки обычно с чашечкой и вен-
чиком, иногда очень крупные. В тропических и субтропических странах (кроме
Африки); всегда деревянистые растения с крупными простыми листьями. Около
100 видов. Роды: магнолия (Magnolia), тюльпанное дерево (Liriodendron)—
оригинальные листья (см. рнс. 117); прекрасные декоративные деревья
Drimys(CM. рис. 11), Schizandra и др.
Последние два рода нередко выделяют из семейства магнолиевых, относя
род DrimysKCeM. Winteraceae, род Schizandra—к сем. Schizandracae (см. ниже).
99. Malvaceae (мальвовые). Семейство раздельнолепестных двудоль-
ных, из порядка мальвоцветных (Maivales). Сюда же относятся сем. баоба-
бовых, липовых и др. Распространено в тропических и умеренных обла-
стях; около 900 видов. Травы иди деревянистые растения. Листья простые иди
лопастные с прилистниками. Цветки обычно крупные, правильные, пятерные,
часто внизу чашечки имеется ещё внешняя чашечка, тычинок нередко много,
срастающихся нитями в цилиндрическую полую трубку, внутри которой
проходят тоже многочисленные столбики. Пыльники тычинок имеют не четыре,
как обычно, а два гнезда. Наиболее важный род хлопчатник (Gossypium), у
нас встречаются виды мальвы (Maiva), алтей (Althaea).
100. Marattiaceae (мараттневые). Семейство папоротников. Исключи-
тельно под тропиками (всего 60—70 видов). Судя по многочисленным ископа-
емым остаткам, представляет вымирающую (реликтовую) группу. Нередкя
листья имеют до 2—3 м и обычно сложноперистые. Очень характерны при ос-
новании листьев прилистники. Спорангии обычно срастаются в группы.
101. Markgraviaceae (маркгравиевые). Семейство раздельнолепест-
ных двудольных, из порядка камедеибсных (Guttiferales). Иногда его относят
к порядку постениосеменных. Небольшое, чисто американское, тропическое
семейство (около 50 видов). Деревья, кустарники, часто лианоподобные растения
илн эпифиты. В соцветиях часть цветков превращается в оригинальные меш-
ковидные или шпорцевидные образования, содержащие нектар. Опыление
нередко птицами.
102. Meliaceae. Семейство раздельнолепестных двудольных из порядке
27 Гсогпп<1>ип_
фистаигннковых (Terebinthsles). Близко no своим признакам и сем. рутовых
того же порядка. Деревья, реже кустарники тропических и теплых стран.
Листья обычно перистые. Как и у рутовых, железки с маслом (листья имеют
просвечивающие точки). Мелкие цветки в соцветиях. Тычинки срощены нитями
в трубочку. Роды: Melia, Xylocarpus и др. Всего около 700 видов, многие дают
ценную древесину.
103. Me n ispermac еае (лунносеменпые). Семейство раздельнолепестных
двудольных, из порядка многоплодниксвых. Это одно из связующих семейств
между древесными и травянистыми семействами этого порядка. Чаще всего
вьющиеся деревянистые растения (реже деревья), главным образом в тропи-
ческих странах. Цветки с трехчленными кругами, правильные (двудомные).
В семени зародыш часто согнутый (отсюда название семейства). Около 300 видов.
104. Могасеае (тутовые). Семейство раздельнолепестных двудольных,
из порядка крапивоцветных. Это одно из важнейших тропических семейств, в
умеренном климате — лишь немного видов. Деревянистые растения, редко
травы. Важный признак — млечный сок. Цветки однополые, очень невзрачные,
мелкне, собранные в крайне оригинальные соцветия с сильно утолщенной осью,
принимающей форму блюдца, шара и др. Так, например, винная ягода — это
грушевидное соцветие с мелкими цветками внутри нее. Околоцветник чаще
всего из четырех листочков, тычинок четыре, пестик нз двух плодолистиков.
Наиболее обширным! (до 600 видов) и важным является род фикус (Ficus), даю-
t щий каучук, винные ягоды (инжир), шеллак и др ; род Morus (шелковица),
листья которого служат кормом для шелковичного червя, хлебное дерево (Лг-
tocarpus), молочное дерево (Galactodendron), мирмекофпльные виды рода Сссго-
pia, каучуконос Castilloa elastica н др.
105. Musaceae (банановые). Семейство однодольных, нз порядка Sciti-
mineae. К этому же порядку относятся имбирные, канновые и др. Исключитель-
но в тропиках. Крупные травы или деревья с очень большими листьями (всего
около 80 видов). Цветки обоеполые, реже однополые, неправильные, легко вы-
водятся из лилейного типа. Иногда околоцветник спайнотистный. Из шести
тычинок одна может быть бесплодной. Обычно цветки в сложных соцветиях.
Наиболее богатый видами род Musa (банан), дает съедобные плоды, также пень-
ку. Дерево путешественников (Ravenala — см. рис. 200) — один вид на Мада-
гаскаре, другой — в Гвиане и Бразилии.
106. Му о р о г а се а е (миопоровые). Семейство спайнолепестных дву-
дольных, из порядка маскоцветных (Personatae). Сюда же относятся норични-
ковые, пасленовые, бигнониевые, геснериевые и др. Небольшое семейство,
около 90 видов в Австралии, на океанических островах и в Восточной Азии.
Близко родственно норичниковым, от которых отличается двух- или много-
1 нездной завязью и маслянистыми железками в тканях. Цветки правильные или
неправильные, пятичленные. Род Myoporum. Ряд декоративных растений.
107. М у г 1 с а се а е (восковниковые). Небольшое семейство раздельно-
лепестных двудольных, большинством авторов выделяемое в особый порядок
kyricales. Последний обнаруживает близость к порядку орехоцветных (Juglan-
cLks), к которому иногда и относят сем. восковниковых. Деревья и кустарники,
чаще ьссго субтропических стран, с очень редуцированными цветками, собран-
ныди в колосья. Околоцветника нет, цветки однополые, одно-или двудомные.
Некоторые авторы простоту устройства цветков считают примитивным явле-
нием, однако правильнее считать, чю простота строения — эго результат ре-
дукции. Один род Myrica (40 видов); у нас лишь М. gale — в прибалтийских
странах (восковник, болотная мирта).
1G8. Myristicaceae (мускатные). Семейство раздельнолепестных
двудольных, из порядка многопчодниковых. Очень близко к сем. аноновых.
Вечнозеленые деревья тропических стран, главным образом в тропической
Азии (около 255 видов). Цветки правильные, однополые, обычно трехчленные.
Оиень характерны семена с лопастным эндоспермом; маслянистые железки в
тканях. Среди полезных растений — мускатный орех (Myristica fragrans), упо-
требляется в медицине и как пряность, родина Молуккские острова.
109. Myrtaceae (миртовые). Семейство раздельнолепестных двудоль-
ных, из порядка мнртоцветных (МугЫез). К этому же порядке относятся дер-
бенниковые. гранатовые и др. Большое тропическое семейство; деревья с ьечно-
о ААЛ —LJ^n'i Ч птлрТ1.д тлмРТАТ тАиПНЫе ЖЕЛвЗКП»
вследствие чего обладают сильным ароматным запахом. Цветки четырехчлен-
ные, тычинок много. Ряд важных родов: мирга (Myrtus communis) — в Ев-
ропе, эвкалипт (Eucalyptus—Австралия и отчасти Новая Гвинея) — до 4J0
видов, образует леса в саванновые леса (рис. 103), большую роль играет в скрэбе;
Melaleuca (до 100 видов), один вид дает каепуговое масло, гвоздичное дерево
(Caryophy Низ arcmat’cus) цветки дают «гвоздику(пряность) и гвоздичное масло.
НО. Nepenthaceae (непентесовые). Семейство раздельнолепестных
двудольных, из порядка многоплодниковых (Polycarpicae). Связь с последи im
порядком не является вполне достоверной, и потому иногда семейство ставят
в другие части системы. Исключительно под тропиками Старого Света (см.
рис. 203), от Австралии до Мадагаскара (около 60 видов). Лазящие кустарники,
цепляющиеся черешками листьев. Единственный род Nepenthes со своеобразны-
ми листьями-кувшинами. Цветки однополые, двудомные, четырехчленные.
Насекомоядные растения (рис. 202).
111. Nyctaginaceae. Семейство раздельнолепестных двудольный, из
порядка центральносеменных (Ceiitrospermae). К этому порядку относятся
семейства: гвоздичные, маревые, кермесовые и др. Тропическое семейство,
главным образом в Америке, до 160 видов. Травы и деревья с обычно супротив-
ными листьями. Околоцветник венчиковидный или довольно невзрачный, но
окруженный яркоокрашенными прицветными листьями.Декоративные растения:
ночная красав! ца (Mirabilis), буженвиллея (Bougainvillea spectabihs) с пре-
красно окрашенными прицветными листьями.
112. Nynaphaeaceae (кувшинковые, нимфейные). Семейство раз-
дельнолепестных двудольных, из порядка многоплодниковых. Это небольшое
семейство (около 1G0видов), распространенное главным образом подтропиками,
но вообще встречающееся везде. Исключительно водные растения с очень харак-
терной жизненной формой (плавающие по большей части листья, крупные
одиночные цветки). Цветки (до 35 с.и в диаметре) с большим количеством тычи-
нок и плодолистиков, сросшихся в общий пестик. В СССР белая водяная лилия
(Nymphaea alba н N. Candida — рис. 158), желтая кувшинка (Nuphar luteum)
и др- Другие виды: виктория-регия (Victoria regia — рис. 89) на р. Амазонке,
египетский лотос (Nymphaea lotus), индийский лотос (Nelumbium nuciferum—
рис. 194), американский род Cabomba (рис. 21) и др. Водный образ жизни ним-
фей. 1ь,х—это вторичное явление, так как почти все многоплодниковые наземные
ВИДЫ.
113. N у s s а с е а е. Очень небольшое (всего 8 видов) семейство раздельно-
лепестных двудольных, из порядка миртоцветных (Myrta les). Тропические и суб-
тропические кустарники с очередными листьями и мелкими цветками. Род
Nyssa (6 видов)—Ост-Индия, Восточная Азия, Северная Америка (восточные
части).
114. О 1 е а с е а е (масличные). Семейство спайнолепестных двудольных, из
порядка скрученных (Contartae). Сюда же относятся ласточниковые и др.
Иногда семейство выделяют'э особый порядок Li gustrales. В тропических и уме-
ренных странах; около 4000 видов. Деревянистые растения, редко травы. Харак-
терны четырехчленные (по большей части) цветки с двумя тычинками и двух-
гнездной завязью. Роды; ясень (Fraxinus), маслина (Olea), сирень (Syringa),
жасмин настоящий (Jas mi num), бирючина (Li gust rum), Phyllirea и др.
115. Onagraceae (онагриковые). Семейство раздельнолепестных дву-
дольных, нз цсрядка миртоцветных (Myrtales). Сюда же относятся сем.: мир-
товые, дербенниковые, гранатовые и др. Травы или реже кустарники. Цветки
правильные, обычно четверного типа (во всех кругах). Завязь нижняя с цветоло-
жем, нередко вытянутым в длинную трубку. Около 500 видов, главным обра-
зом в северном полушарии, в умеренных и субтропических странах, реже —
под тропиками. Роды Oenothera (ослинник) — Северная Америка, Тгара
(водяной орех), Epilobium (кипрей) — у нас много видов.
116. Orchidaceae (орхидные). Семейство однодольных, из порядка
Gynandrae (женомужние), имеющего только одно это семейство. Особенно бо-
гато представлено в тропиках, но немало видов и в умеренных областях (в част-
ности в СССР). Исключительно травы, под тропиками много эпифитов, украшаю-
щих влажнотропические леса. Одно из крупнейших семейств — до 15 000 ви-
дов. Орхидные представляют одну из наиболее специализированных групп
однодольные в паправ ’Н’п > приспособ^е-тя к опытыгно при помощи нагеко-
27* 419
мых; отсюда — неправильные крупные цветки с самой разнообразной раскрас-
кой (мелкие цветки всегда в соцветиях), один листок околоцветника в виде
губы, куда садятся насекомые, обычно сильный запах, нектар в особых, иногда
очень длинных шпорцах (До 50 см), пыльца склеена в одну массу (поллинии)
и др. Цветок легко выводится из лилейных (околоцветник из двух трёхчленных
кругов), ио тычинок одна или две (вместо шести), завязь нижняя. Очень свое-
образен орган, образованный из срастания тычинок н пестика, для обеспечения
перекрёстного опыления. Многочисленные орхидные культивируются в оран-
жереях из-за оригинальных, красивых цветков. Наши дикие орхидные довольно
невзрачны за исключением некоторых (например, венерин башмачок — Сур-
ripedium и др-).
В книге приведены такие роды: Calypso, Cypripedium, Epipogon, Goodyera,
Gymnadenia, Jonopsis, Orchis (ятрышник), Stanhopea, Vanilla (ваниль) и др.
117. Os mu n d aceae. Семейство папоротников. Около 15 видов в тропи-
ческих и умеренных странах. Спорангии с группой утолщенных клеток близ
вершины (кольца нет). Роды: Osmunda, Todea и др.
118. Oxalidaceae (кислицевые). Семейство раздельнолепестных дву-
дольных, из порядка геранецветных (Geraniales). К этому же порядку отно-
сятся семейства: гераневые, леновые, настурциевые, парнолистные и др. В
основном — у всех цветки пятикруговые, пяти членные, в большинстве случаев
правильные. К семейству относится около 340 видов тропических и умерен-
ных стран, преимущественно в Южной Африке и тропической Америке. Травы,
редко деревья. Очень обилен видами род кислица (Оха lis), разнообразный по
жизненным формам; у нас в хвойных лесах Oxalis acetosella.
119. Ра I mae (пальмовые). Семейство однодольных, из порядка початкоцвет-
ных (SpadicifЮгае). Сюда относят также сем. аронниковых. Другие системати-
ки последнее семейство (вместе с некоторыми другими)выделяют в особый по-
рядок Arales. Для початкоиветных характерны соцветия в виде початков и
верхушечные листья в виде особого покрывала около соцветия, или защищаю-
щего молодое соцветие или своей окраской привлекающего насекомых для опы-
ления. Сем пальмовых исключительно распространено под тропиками, в Ев
репе лишь один вид (Chamaerops humilis). В большинстве случаев стройные
деревья с неветвящимся или слабо ветвящимся стволом, реже кустарники или
лианы; в тропических лесах играют иногда очень большую роль. Листья у палым
в виде пучка на терч-ине ствола, веерные или перистые, иногда до К м длины,
Цестки мелкие, типа лилейных, собранные в крупные соцветия, окружённые
покрывалом. Опыление ветром или насекомыми. Пальмы играют очень боль-
шую роль в жизни населения тропических стран (стволы, как строительный
материал, листья, как волокнистые вещества, плоды употребляются в пищу,
дают масло н т. д.). Пальм около 1 500 видов.
В тексте книги приведены такие роды: Attalea, Areca, Astrocaryum, Boras
sus, Calamus (роганг), Chamaerops, Cocos (C, nucifera — кокосовая пальма -
рис. 204), Copernicia, Corypha, Euterpe (рис. 79), Elaeis (E. guinensis—масля-
ная пальма), НурЮепе, Klopstcckia^ Levistona, Manicaria, Nipa (рис. 93).
Phoenix (P. dactylifera, финиковая пальма — рис. 170, P.Jubal), Phytele
phas, Sabal, Trachycarpus.
120. Pandanaceae (панданусовые). Семейство однодольных. Одни
авторы относят семейство к порядку початкоцветных, другие выделяют £
особый порядок панданусоцветных (Pandanales), однако считают, что между
этими порядками имеется тесная связь и выводят последние из первых. Панда-
нусовые отличаются от пальмовых длинными линейными листьями и ещё бо-
лее редуцированными цветками. Панданусовые исключительно тропические
растения — от Африки до Полинезии, в Америке отсутствуют. Около 22(
видов деревьев, кустарников и лиан. Цветки однополые, обычно без околоцвет-
ника, мелкие и невзрачные, собраны в колбовидные соцветия, одетые окра-
шенными верхушечными листьями. Род панданус (Pandanus) с многочисленны-
ми видами: листья дают волокна,плоды некоторых съедобны. У нас культиви-
руется в оранжереях.
Papilionaceae (мотыльковые) см. Leguminosae.
121. Papaveraceae (маковые). Семейство раздельнолепестных двудоль-
ных, из порядка макоцветных (Papaverales). К этому же порядку отно-
сятсяСгисКегае — крестоцветные, Fumariaceae — дымянковые н др. Ма-
420
ковые хорошо связывают данный порядок с порядком многоплодниковых.
Травы, реже кустарники, очень редко деревья; обычно с млечным соком.
Цветки правильные, дв? чашелистика, четыре лепестка, тычинок много, завязь
из двух или многих плодолистиков. Около 450 видов, почти исключительно
во впетропическпх странах северного полушария. Роды; Papaver (мак), Cheli-
donium (чистотел).
122. Passi floraceae (страстноцветные). Семейство раздельнолепест-
ных двудольных, из порядка постенносеменных (Parietales); к этому же поряд-
ку относятся семейства: тыквенные, тамарисковые, бегоняевые и др. Особен-
ная близость к сем. тыквенных. Распространено под тропиками, особенно в
Америке; до 400 видов, главным образом деревянистые и травянистые лияиы,
цепляющиеся усикамн. Очень красивые, большие црегки пятерного типа.
В центре цветка находится особая колонка, на которой развивается придаточ-
ный венчик из многочисленных лепестковидных образований. Тычинки чаете
срастаются с колонкой. Многие виды Passiflora очень декоративны, плоды не-
которых съедобны.
123. Phyto laccaceae (кермесовые). Семейство раздельнолепестных
двудольных, из порядка центральносеменных (Centrospermae). Около 116 ви-
дов, распространенных в тропических странах, особенно в Америке. Травы
или деревянистые растения. Семейство интересно тем, что является переход-
ном между более простыми и более совершенными семействами порядка цен-
тральносеменных. Цветок правильный, с простым околоцветником, число ты-
чинок сильно варьирует, пестики свободные или срастаются. Phytolacca dode-
ca ndra культивируется из-за ягод, дающих тёмнокрасный сок (подкраска вина).
124. Р iperaceae (перцовые). Семейство раздельнолепестных двудольных.
Положение в системе довольно различно у разных авторов. Одни ставят их ря-.
дом с гречишными, другие сближают с многоплодииковымн, третьи ставят вбли-
зи бу ко цветных, ещё иные считают перцевые за одну из исходных групп покры-
тосеменных. Растения главным образом тропические. Травы, кустарники, не-
редко лианы и эпифиты; образуют довольно важный элемент тропических ле-
сов Более 1 000 видов, особенно в Ю «ной Америкен Индии. Цветки невзрач-
ные, без околоцветника, тройного типа, на сильно редуцированные, в густых
колосьях. Два больших рода: Piper (перец) и Peperomia.
125. Р irolaceae (грушанковые). Семейство спайнолепестных двудоль-
ных,из порядка вересковидных (Ericales). Сюда же относятся сем. Ericaceae,
Epacridaceae н др. Травы с вечнозелёными кожпетымн листьями или сапрофи-
ты, лишённые хлорофилла. Несколько десятков видов (около 35) в умеренных
странах северного полушария. Цветки правильные, венчик раздельнолепест-
ный или спайнолепестный. Круги в цветках пяти- или чегырёхчленные, тычи-
нок 10, в два круга. Тычинки открываются порами. Роды: Pirola (грушанка,
у нас ряд видов), Monotropa (подъельник) и др. Грушанкн — характерные ра-
стения наших еловых н сосновых лесов.
126. Platanaceae (платановые). Небольшое семейство раздельноле-
пестных двудольных; всего один род Platanus (платан, чинар), насчитывающий
лишь 6 видов. Родственно семейству Hamamelidaceae, относясь к порядку
Hamamelidaies (однако систематическое положение не вполне определённо).
Деревья, листья напоминают листья клёна. У листьев два прилистника, сра-
стающихся в колпачок, закрывающий почку. Цветки мелкие, однополые, в
плотных шаровидных соцветиях на длинных ножках. Виды платана распро-
странены от восточного Средиземноморья до Гималаев и в Северной Америке.
127. Plumbaginaceae (свинчатковые). Семейство спайнолепест-
ных двудольных. Положение в системе не вполне определенно; одни авторы по-
мещают в порядок первоцветных (Primulales), другие — в особый порядок
свинчаткоцветных (Plumbaginafes), сближая последний с центральносемен-
ными (раздельнолепестными). Свинчатковые — это как бы спайнолепестные
центральносеменные. Цветки правильные, пятичленные, тычинки против ле-
пестков, завязь одногнёздная с одной центральной семяпочкой. Семейств*
распространено в степях, полупустынях, на засолённых территориях, также в'
Средиземноморье. Травы многолетние, полукустарники, кустарники, до 300
видов. Роды; кермек (Statice — рис. 144), свинчатка (Plumbago), Acantholi-
mon — образующий огромные подушки.
128. Podostemonaceae. Семейство раздельнолепестных двудольных,
из порядка розоцветных (Rosales). Небольшое тропическое семейство (около
150 видов) травянистых растений, растущих в текущей воде (водопады, быст-
рые потоки), крайне оригинального внешнего вида (см. рис. 164); многие из
них похожи на водоросли или мхи. Плоские или нитевидные стебли прикреп-
ляются к подводным скалам при помощи расширенных плоских зелёных (!)
корней. Весь облик вызван приспособлениями к условиям существования.
Цветки тоже крайне редуцированы, часто без околоцветника, с одной тычин-
кой и пр. Главным образом в Америке н Азии. Роды: Основе, Podostemon,
Tristicha.
1^9. Polemoniaceae (спнюховые). Семейство спайнолепестных дву-
‘дольных, нзпорядка Convolvulales (вьюнковидные). По своим признакам близ-
ко к сем. Convolvulaceae (вьюнковые), отличаясь отсутствием млечного
сока, супротивными листьями, невьющчмися стеблями и обычно трубчатым
венчиком с отгибом. Некоторые авторы сближают с бурачниковыми (Borngina-
les). Главным образом травы, реже деревянистые растения. Около 270 видов,
преимущественно американских. Тропические н умеренные страны. Роды: Ро-
lemonium, у нас Р. coeruleum (синюха), Plilox (блоке) и др. Много декоративных
видов.
130. Ро lygonaceae (гречишные). Семейство раздельнолепестных
двудольных, из порядка гречихоцветных (Polygamies). В порядке лишь одно
семейство. Порядок сближается с Urticales (крапивоцветные)— сросшиеся
прилистники, и с Poiycarpicae (многоплодниковые) — трёхчленные цветки.
Травы, главным образом в севеоных умеренных странах, под тропиками ку-
старники, даже деревья. Около 750 видов. Характерны при основании листьев
срастающиеся прилистники, образующие трубку (раструб). Цветки мелкие,
правильные, с простым околоцветником, в сложных соцветиях. По своему
устройству цветки схожи с однодольными (тройной тип). Плод — обычно трёх-
гранный орешек. Роды: Polvgonum (гречиха), Rumex (щавель), Calligonum
(в Соедней Азии), Triplaris (Южная Америка — деревья) и др.
131. Р о 1 у р о d i а с е а е. Семейство папоротников. Спорангии с верти-
кальным неполным кольцом. Самое крупное семейство папоротников — до
3 000 видов; в тропических, субтропических и умеренных странах. Почти все
наши папоротники относятся к это)лу семейству. Роды: Cystopteris, Davallia,
Dryopteris, Polypodium, P-teridium (орляк), Struthiopteris н др.
132. Portu lacaceae (портулаковые). Семейство раздельнолепест-
ных двудольных, из порядка центральносеменных (Centrospermae). Небольшое
семейство (более 200 видов), являющееся переходным между кермесовымя
н гвоздичными, т. е. от более простых семейств к более совершенным (внутри
порядка центральносеменных). Травы или кустарники с мясистыми листьям я
и прилистниками. Цветки правильные/ с венчик; зидным околоцветником (4-
5 листочков). При цветке два чашечкокидных прицветника. Семена на централь-
ном семяносце. Распространение главным образом в Америке, но некоторые ви-
ды космополиты (портулак огородный). Ряд аративных видов (виды порту-
лака).
133. Р о t a mogetonaceae (рдесте . Семейство однодольных, из
порядка водных (Helobiae). Около 100 с лиц видов, погружённых в воду,
выставляющих из воды лишь соцветия во врег , плетения, или же погружённых
и во время цветения. Листья узкие, длинные, злаковядные. Цветки невзрачные,
опыляемые ветром (реже водой), чаще четверного типа с четырьмя членами г
каждом кругу. Роды: Potamogeton (рдест-ж. 26)—наиболее распро-
странён; Zestera (морская трава) н PosidoniJ — растут в морской вод1
(рнс. 163). Некоторые виды космополиты.
134. Primulaceae (первоцветные). Семейство спайнолепестных дву-
дольных из порядка первоцветных (Primulales). Иногда этот порядок связыва
ют с верескоцветными из спайнолепестных, но, невидимому, больше основание
выводить, его из центральносеменных, как особую ветвь последних, давших
спайнолепестный венчик. Семейство имеет более 500 видов, распространённых
по всей земле, но главным образом в северном полушарии, во внетропических
областях. Цветки правильные, хорошо выраженного пятерного типа, семей-
ство легко узнаётся по противовенчиковым тычинкам и одногнёздной завязи с
центральным семяносцем. В СССР целый ряд родов. Декоративны; примулг
422
(Primuij), цикламен (Lyciamen) и др. Другие роды: проломник (Androsace)—
рис. 176, Soldanella — рис. 42, седмичник (Trientalis).
135. Proteaceae (протейные). Семейство раздельнолепестных, из по-
рядка протейноцветных (Proteales). Порядок имеет лишь .одно семейство про-
течных. Положение порядка в филогенетической системе довольно неопреде-
лённо; одни считают его примитивным,.но почти не связанным сдругими по-
рядками, другие — упрощенным. Это — деревья с обоеполыми или однопо-
лыми цветками, собранными в соцветия (кисти, колосья и др.). Околоцветник
венчиковидный, четырехчленнып. Главным образом в южном полушарии (все-
го до 1 100 видов), до 3/3 видов в Австралии (роды Grevillea, Наkea, Banksia,
Dry an dra), более 1/i в Южной Африке (Leucadendron, Pro tea). В Австралии
играют значительную роль в растительных ценозах.
136. Punicaceae (гранатовые). Семейство раздельнолепестных дву-
дольных, из порядка мнртоцветных (Myrtales). Близко <тоит к сем. миртовых,
отличаясь отсутствием железок с эфирными маслами. Деревянистые растения
с правильными цветками, чашелистиков и лепестков 5—7, тычинок много, пло-
долистиков много, расположенных в два этажа, завязь нижняя с мясистым
красным цветоложем. Лепестки красные. Единственный род Punica (гранат),
лишь с двумя видами; Р. grana turn — гранат (от Средиземноморья до Гималаев),
везде культивируется в тропиках и субтропиках.
137. Ranunculaceae (лютиковые). Семейство раздельнолепестных
двудольных, из порядка многоплодниковых. Наиболее богатое семейство этог®
порядка (до 1 500 видов), распространено в умеренных и холодных странах,
в том числе и в СССР, где представлено очень большим числом родов и видов.
Подобно тому, как сем. магнолиевые является основным среди древесных се-
мейств мпогоплодниковых, так сем. лютиковых наиболее важным среди травя-
нистых семейств этого порядка. Краткая характеристика лютиковых очень
трудна. Это — травы с правильными или неправильными цветками и чаще
всего со спиральным расположением частей цветка. Чашечка и венчик обычно
пягпчленны, но тычинок и пестиков чаще всего много (отсюда название поряд-
ку. В цветках нередки нектарники, "в которые превращаются наружные ты-
чинки. У нас такие роды: лютик (Ranunculus — рис. 156), ветреница (Anemone
—рис. 70), борец (Aconitum), водосбор (Aquilegja), пеон (Paeonia), адонис (Ado-
nis), живокость (Delphinium), перелеска (Hepatica), чистяк (Ficaria), прострел
(PuIsatilla) и др.
138. Restionaceae. Семейство однодольных, из порядка Enantiob-
lastae (для порядка характерно положение зародыша, лишь прилегающее®
к эндосперму, причём последний является мучнистым). По своему внешнему
виду растения семейства очень напоминают злаки: стебли со вздутыми узлами,
и’Стья с влагалищами и язычками. Соцветия— колоскн, собранные в различ-
ные соцветия. Цветок —сильно упрощенный цветок лилейных. Основное рас-
пространение в Австралии и Южной Африке (вообщ^в южном полушарии) в
субтропических и умеренных областях. Около 250 видов. По большей части
многолетние травы на болотах и сухих местах.
139. Rhamnaceae (крушиновые). Семейство раздельнолепестных двудоль-
ных, из порядка крушин оцветных (Rhamnales). Сюда же относятся: Aquifolia-
ceae, Empctraceae, Ampelidaceae и др. Деревянистые растения,
кустарники, лианы, реже травянистые растения. Листья простые с п рил истин-
ками. Цветки правильные, четырех-,пятичленные с плоским или вогнутым цве-
толожем. Лепестки небольшие, иногда совсем отсутствуют. Тычинки невеж-
ду, а против лепестков. Около500 гидов, в умеренных и тропических странах.
Роды: Rhamnus (крушина), Paliurus (де ржи-де рев о), Ceanothus и др.
140. Rhizophoraceae (ризофоровые). Семейство раздельнолепест-
ных двудольных, из порядка мнртоцветных (Myrtales). Сюда же относятся се-
мейства: миртовые, дербенниковые и др. Тропическое семейство, около60 ви-
дов; деревья. Большое сходство с дербенниковыми, но листья имеют прилист-
ники. Целый ряд видов образуют мангровые заросли на морских побережьях
и имеют своеобразные приспособления (рис. 52, 90—92). Роды — Rhizoplie-
ra, Anisophylh, Brugiiiera, Ceriops п др.
141. Rosaceae (розянные). Семейство раздельнолепестных двудольных, из
порядка розоцветных (Rosales). Различными систематиками порядок розо-
цветных понимается то более узко, то более широко; в последнем случае к нему
423
присоединяют все семейства бобовых. Во всяком случае между розоцветными
и бобовыми имеется очень тесная связь. Розоцветные хорошо выводятся из мно-
гой ледниковых, дальнейшая ветвь их эволюции — миртоцветные. Розанные —
обширное семейство — более 2 000 видов трав, кустарников, деревьев; главным
образом в северном полушарии. Характеристика семейства очень трудна, так
как почти ^каждый признак сильно варьирует, но в общем цветки правильные^
€ чашечкой и венчиком пятичленного типа, тычинки по числу лепестков или в
кратном ч еле (5, 10,15, 20 и больше), пестиков чаще много. Части цветка распо-
лагаются кругами, не спирально, каку многоплодниковых. Цветоложе то плос-
кое, то выпуклое, то вогнутое, как бокал. Семейство обычно подразделяется на
ряд подсемейств, трактуемых иногда как семейства. В СССР многочисленные
представители: шиповник, малина, земляника, плодовые деревья —яблоня,
груша, вишня, слива, абрикос н др. Многочисленные культурные сорта.
В книге приведены такие роды: Асаепа, Adenostoma, Agrimonia (репейник),
AlchimiIla (манжетка), Amygdaius (миндаль), Dryas (дриада — рис. 167), Fi-
lipendula (таволж^нка), Geum (гравилат), Potantilla (лапчатка), Poterium
(черноголовник), Rubus (ежевика, малина), Sibbaldia (зиббальдия), Spiraea
(спирея), Qnilhja, Primus (слива, вишня).
142. Rubiaceae (мареновые). Семейство спайнолепестных двудоль-
ных, из порядка мареноцветных (Rubtales). К этому же порядку относятся
семейства: Caprifoliaceae — жимолостные, Valerianaceae — валериановые, Dip-
sacaceae — ворсянковые и др. Очень большое семейство, более 4500 ви-
дов. Травы и древесные растения, лианы и эпифиты, главным образом в тропи-
ческих странах. Листья супротивные, цельнокрайние; нередко прилистники
не отличимы от листьев, и тогда имеется мутовка из «листьев^. Цветки правиль-
ные, четырёх- или пятичленные. Чашечка редуцирована, завязь нижняя, двух-,
трёхгнёздная. В тропиках растения данного семейства играют большую роль,
роды: Cinchona (хинное дерево, в Андах Южной Америки — рис. 210), Coffea
р ево — рис. 193), Exostemma, Oxyanthus, Hydnophytum, Myrme-
codia и др. У нас виды родов Asperula (ясменник), Galium (подмаренник). Мин-
го полезных (декоративных, медицинских н др.) растений.
143. Rutaceae (рутовые). Семейство раздельнолепестных двудольных,
из порядка фисташниковых (Terebinthales). К этому же порядку относятся;
Асегасеае (кленовые), Sapindaceae (сапиндовые) и др. Порядок тесно связывает-
ся с геранецветными. Рутовые главным образом деревянистые растения. Осо-
бенно характерно присутствие (преимущественно в листьях) желёзок с эфирны-
ми маслами и смолами. Цветки правильные, пятичленные, обычно сильно пах-
нут. Диск в цветке (что характерно для фисташниковых) выделяет много нек-
гара, привлекающего, помимо насекомых, также колибри в Америке. Около
•00 видов, главным образом в тропических и субтропических странах. Роды:
Citrus (лимоны, апельсины и др.), Phellodendron (Р. atnurense — бархатное
дерево), Agathosma, Coleonema и др.
144. Salicaceae (ивовые). Семейство раздельнолепестных (первично-
покровных) двудольных, из порядка ивоцветных (Salicales). В этом порядке
находится лишь одно данное семейство. В системе оно стоит довольно изолиро-
ванно, но имеет связи с такими примитивными (или упрощенными) порядками,
как орехоцветные, букоцветные н др. Некоторые авторы ставят семейство со-
вершенно особняком от всех покрытосеменных. Это — деревья или кустарники
с простыми листьями с прилистниками. Цветки однополы^, двудомные, очень
редуцированные, собранные в кисти н серёжки. Околоцветника обычно нет.
Главным образом в умеренных областях северного полушария. Около 400 вид-
дов. Роды: тополь (Populus), ива (Salix — род, богатый видами, в СССР около
170 видов). Ивы и тополя играют роль в ландшафте, встречаясь обычна
сплошными насаждениями.
145. Sa 1 so laceae (солянковые). Чаще трактуется не как отдельное-
«емейство, а как одно нз подсемейств маревых — Chenopodiaceae (см. выше),
©бширный род Sa Isola — в СССР более 70 видов, A - a basis и др. Характерные
растения для полупустынь, пустынь, для засолённых почв.
146. Sapindaceae (сапиндовые). Семейство раздельнолепестных
двудольных, из порядка фисташниковых (Terebinthales). Некоюрые авторы от-
носят к порядку Sapindales. Исключительно тропические деревянистые расте-
ния; до 1 000 видов, среди которых нередки лианы. Порядок имеет определён-
424
мые связи с порядками геранецветных и мальвоцветных. Характерным для
порядка является присутствие в цветках нектарников в виде диска. Цветки
у сапиндовых часто неправильные. В вегетативных органах особые клетки с*
смолистым или млечным содержимым. Плоды некоторых видов превосходны.
147. Sarraceniaceae (саррацениевые). Семейство раздельноле-
пестных двудольных, из порядка многоплодниковых. Иногда выделяют особый
порядок Scrraceniales. Семейство насекомоядных растений, исключительн»
американское, всего 3 рода (8 видов); распространение см. рис. 197; наиболее
распространён род Sarracenia. Небольшие травянистые растения с листьями
в розетке. Трубчатые листья служат для ловли насекомых. Семейство сбли-
жается с другими семействами насекомоядных растений — цефалотовыми ж
непен тесовыми.
148. Saxifragaceae (камнеломковые). Семейство раздельнолепест-
ных двудольных, из порядка розоцветных (Rosales). Очень близко к семейству
роза иных. Травы или деревянистые растения. Цветки обычно правильные, а»
большей части пятичленные (реже 4—12-членные), но плодолистиков чаше всегв
два. Завязь верхняя или нижняя. Распространено почти по всей земле: околв
650 видов. По своим жизненным формам очень разнообразно; обычно делится
на несколько подсемейств. Роды: камнеломка (Saxifraga), садовый жасмин
(Philadelphus), смородина, крыжовник (Ribes), дейция (Deutzia), гортензия
(Hydrangea) и др.
149. Scrophulariaceae (норичниковые). Семейство спайноле-
пестных двудольных,из порядка маскоцветных (Personatae). Сюда же относят-
ся семейства: паслёновые, бнгнониевые, геснериевые; и др. Очень боль-
шое семейство — до 2 600 видов, распространено по всей земле, очень ши-
роко представлено в СССР. Травы, кустарники; деревья и лианы — редк».
Целый ряд полупаразитов и паразитов. Цветки всегда более или менее непра-
вильные, в типе пятичленные (венчик нередко двугубый), но тычинок четыре
или две, завязь двугнёздная с многочисленными семяпочками (у губоцветных,
с которыми имеется некоторое внешнее сходство, завязь четырёхлопастная).
Немало декоративных растений: Anthirrhinum (львиный зев), Calceolaria (каль-
цеолария), Mimulus (мимулус), дерево павловния (Paulownia) и др.
У нас много родов: Alectorolophus (погремок), Castilleja, Cymbaria, Euphra-
sia (очанка), Lathraea (петров крест), Limosella (лужайник), Linaria (льнянка),
Melairpyrum (марьянник), Odontites (зубчатка). Pedicularis (мытник), Scrophu-
laria (норичник), Verbascum (коровяк), Veronica (вероника).
150. Si mar u bacea e. Семейство раздельнолепестных двудельных,
из порядка фисташниковых (Terebinthales). Очень близко к сем. рутовых тог»
же порядка. Отличия от рутовых: отсутствуют желёзки с эфирными маг-
лами, цветки однополы. Деревья тёплых стран обычно с перистыми листьями;
еколо 130 видов. Род: Ailanthus (A. glandulosa — разводится у нас на юге, в
Крыму, на Кавказе) н др. Во всех вегетативных частях горькие вещества.
151. Solanaceae (паслёновые). Семейство спайнолепестных двудоль-
ных из порядка маскоцветных (Personatae). Стоит близко к норичниковым,
но цветок почти вполне правильный, пяти членный, лишь завязь двугнёздная
с косой (по отношению к срединной плоскости цветка) перегородкой. Травы или
кустарники. Около 1 700 видов; в тропических и умеренных странах. Присут-
ствие алкалоидов. Роды: табак (Nicotiana), паслён (Solanum), дурман (Daturai
и др. Ряд полезных растений: картофель, томаты, баклажаны и др. Ядовитые:
белена, беладонна и др.
152. So и п е га ti а сеа е. Семейство раздельнолепестных двудольных,
нз порядка миртоцветных (Му г t a les). Очень небольшое семейство тропических
деревьев с супротивными цельнокрайними листьями. Всего 12 видов в тропи-
ческой Африке и Азии. Цветки правильные, иногда однополые, четырёхчлен-
иые (или даже восьмичленные) Виды рода Sonneratia играют большую роль в
мангровых зарослях, корни их (дыхательные) растут из ила вертикально вверх
выше 1 м (рис. 92).
153. Та mar icace ае (тамарисковые). Семейство раздельно лепест-
ых двудольных, из порядка постенносеменных (Parietales); к этому же порядку
•тносятся франкениевые, фиалковые, страстноцветные и др. Распространен»
главным образом в Средиземноморье, в Центральной н Средней Азии, околв
100 видов деревьев и кустарников с мелкими чешуйчатыми или шиловидными
4Ж
листья.ми, густо покрывающими ветви, что придает нм своеобразную внешность.
Цветки довольно мелкие, но в густых соцветиях, четверного типа (иногда пя-
терного). Семена с волосками, похожи на семена ив. Растения преимуществен-
но засоленных мест, немясистые «солянки». Род Tamarix. Некоторые виды вы-
деляют на поверхности поваренную соль (Raaiinntria).
154. Т i 1 i а с е а е (липовые). Семейство раздельнолепестных двудольных,
из порядка мальвоцветных (Ma Iva les). Близко к сем. мальвовых и баоба-
бовых, отличаясь свободными тычинками, не сросшимися в одну колонку.
Тычинки иногда срастаются пучками или совершенно свободны. Пыльники все-
гда четырех гнездные. Деревья, реже травы, в теплых, реже умеренных стра-
нах; около 350 видов Род Tilia (липа) — у нас ряд видов.
155. Theaceae (чайные). Семейство раздельнолепестных двудольных,
из порядка Guttiferales (сюда же относятся сем. Guttiferae, Dipterocarpa-
сеае и др.). Деревья н кустарники, листья обычно вечнозеленые, очередные,
простые. В листьях, как и у всего порядка, имеются особые вместилищ! выде-
лений, благодаря чему листья ароматичные Цветки правильные, чашечка име-
ет 5-7 листочков, венчик — 5-9 лепестков, тычинки обычно многочисленные.
Опыление насекомыми. Около 209 видов (тропики и субтропики). Род Thea (чай-
ный куст), некоторые систематики называют этот род Camelia — камелия.
156. Thymelaeaceae. Семейство раздельнолепестных двудольных,
из порядка м фгоцзетных (Myrtales). Однакэ систематическое положение не
сопсем ясно, и некоторые авторы относят его к особому порядку Thymelaeales
и указывают на связь с круш шоцветныил (Rhamnales). Деревья и кустарники.
Цветки четверного типа, лепестки чаще всего совеем отсутствуют. Чашелисти-
ки нередко ярко окрашены. Цветоложе сильно вогнуто н нередко окрашено.
До 400 видов в Европе, Азан, Абраке, Австралии, также в Вест-Индии. Роды
Daphne — у нас несколько видов, Gaidia и др.
157. Tropaeo laceae (настурцлевые, капуцчнэвые). Семейство раз-
дельнолепестных двудольных, из порядка геранецветных. Сюда же относятся
семейства: кис лицевые, гераневые, парно тлстяые и др. Небольшое (до 50
видов) американское (главным образом южноамериканское, см. ряс. 293) се-
мейство. Травы, выощаеся или цепляющееся. Цветки пятерные, но неправиль-
ные (чю у геранецветных редко) со щпорцем, 8 тычинок (не 10), плодолисти-
ков 3 (не 5, как гераневых). У пае обычна в садах настурция (Tropaeolum).
158 Typhaceae (рогозовые). Семейство однодольных, из порядка
пачданусоцветиых (Pandanales). Лишь один род Typha (рогоз), распростра-
ненный почти по всей земле; всего 9 видов. Травы с корневищами, растут на
сырых и болотистых местах, с узкими двурядными листьями. Цветки одиопо-
<е, очень сильно редуцированные (вместо околоцветника — волоски), в длин-
ах цилиндрических соцветиях, расположенных одно над другим (нижиее —
некое, верхнее—мужское). Опыление ветровое.
159. Umbel liferae (зонтичные). Семейство раздельнолепестных
> дольных, из порядка зонтикоцветных (Umhelliflorae). К этому же порядку
’осятся семейства аралиевые и деренные. Очень большое семейство (более
-90 видов); имеет для нас особое значение, так как хотя распространено по
-му земному шару, но главным образом в умеренных областях северного по-
шария Травы, одно-, двух- и многолетние. Стебель полый, листья чаще все-
10 пальчатосложные, плюют вздутые влагалища. Мелкие цветки собраны в
t южный зонтик (отсюда название семейства), пятпчлепные, чашечка 'часто
не развивается, молодые тычинки загнуты внутрь цветка, завязь нижняя, из
двух плодолистиков, при созревании распадающихся; плод — двусемянка.
Цветки очень однообразны во всём семействе (обычно белые ила желтые) и для
определения до вида необходимо знание устройства плодов, которые сильно
варьируют. В стенках плодов развиваются особые ходы с эфирными маслами,
от чего плоды зонтичных сильно пахнут (анис, тмин, кориандр и др). Среди
зонтичных много полезных растений: морковь, петруш <а, укроп, сельдерей,
и др., но имеются крайне ядовитые — омег (Conium macuhtum), цикута (Ci-
cuta virosa) и др. Другие роды, приведенные в книге: Azorella, Bolax, Негас-
leum (борщгв ik), Seseli, Trinia.
160. U 1 m а с е а е (вязовые). Семейство раздельнолепестных двудоль-
ных, из порядка крапивоцветных (Urticales). К этому же порядку отно-
сится Moraceae, Urticaceae и др. Исключительно деревья, без млечного сока.
426
Семейство близко примыкает к сем. крапивных, отличаясь ебоеполыми цвет
ками. Цветки мелкие, опыляются ветром, цветут до появления листьев. Роды
Ulmus — вяз (около 20 видов умеренных стран северного полушария и в горах
тропической Азии), Celtis (60 видов — умеренные и тропические страны), Zel-
kowa (4 азиатских вида).
161. Urticaceae (крапивные). Семейство раздельнолепестных дву-
дольных, из порядка крапиврцветных (Urticales). Положение порядка не вполне
•пределенно, его часто сближают с букоцветными (Fagales), но некоторые свя-
зи имеются с порядками розоцветных, центральносеменных и др. В большинстве
случаев травы, реже деревья в тропических и умеренных странах (всего до 350
видов). Цветки однополые, однодомные, невзрачные, с чашечковидным около-
цветником; круги четырехчленные. Характерны особые жгучие волоски, при-
чиняющие иногда даже смерть (тропическая Laportea). У нас обычный род кра-
пива (Urtica).
162. Ur tic u I a ri ас е а е (пузырчатковые). Семейство спайнолепест-
ных двудольных, нз порядка маскоцветных (Perso па tae). К этому же порядку
относятся сем. Scrophulariaceae — норичниковые, Solanaceae — паслено-
вые и др- Очень близко к норичниковым, отличаясь одногнездной (ча-
ще всего завязью) с центральным семяносцем. Цветки неправильные, венчик
ебычно двугубый. Все виды насекомоядны, водные пли болотные растения,
исключительно травы. Около 250 видов, в теплых и умеренных странах. Ро-
ды: Urticularia (пузырчатка — у нас ряд видов), Pinguicula (жирянка).
163. Vaierianaceae (валериановые). Семейство спайнолепестных
двудольных, из порядка мареноцветных (Rubiales). К этому же порядку отно
сятся мареновые, жимолостные, ворсянковые и др. Валериановые тесно при-
мыкают к жимолостным, но сравнительно с ними обнаруживают ряд редукций
вместо пяти тычинок имеется три илн одна, из трех гнезд в завязи лишь в одном
развивается семя. Цветки асимметрические, чашечка, как и у всего порядка,
почти не развита или вместо нее перистый хохолок. Травы или полукустарни-
ки с супротивными листьями без прилистников. До 350 видов в северном по-
лушарии, но по Андам заходят и в южное полушарие. Род Valeriana (валериа-
на — у нас ряд видов) н др.
164- Verbenaceae (вербеновые). Семейство спайнолепестных дву-
дольных, которое вместе с сем. губоцветных иногда относят к порядку Verbe-
nales (вербеноцветных), но чаще, присоединяя н ряд других семейств (до 25)
•тносят к порядку трубкоцветных (Tubiflorae). Близко стоит к губоцветным
Особенно в тропических и умеренных странах южного полушария: всего до 800
видов. Преимущественно кустарники, реже деревья и травы. Листья супротив-
ные или мутовчатые. Цветки четырех-или пятичлеиные в более или менее слож-
ных соцветиях, обычно неправильные. Венчик нередко двугубый с длинной
трубкой, тычинок четыре или две, плод или костянка или распадается на че-
тыре плодика. К вербеновым относятся- тиковое дерево (Tec ton a gran dis), ман-
гровые деревья Avicennia (рис. 92), Verbena — декоративные виды, Vitex—
декоративные кустарники.
165. Vi о 1 ас еа е (фиалковые). Семейство раздельнолепестных двудоль-
ных, из порядка постенносеменных (Parietales). К этому же порядку относя-
тся семейства тамарисковые, страсти о цветные, бегониевые и др. Травы, кус-
тарники, редко деревья, часто с большими прилистниками. Цветки пятер-
ного типа, чаще неправильные, при этом один лепесток вытянут в шпо-
рец. Семяносцы с семенами постенные. Около 500 влдов в тропиках, субтро-
пиках н в умеренных странах. Наиболее богат видами род Viola (фиалка) -
у нас много видов.
166. V i t а с е а е (виноградные). Семейство раздельнолепестных двудоль-
ных, из порядка крушиноцветных (Rhamnales). Иногда семейство называется
Ampelidaceae. По большей части деревянистые лианы с лопастными или паль-
чатосложнымп листьями. Характерны усики, расположенные супротивно ли-
стьям. По признакам сходно с крушиновыми, но лепестки на вершине сра-
стаются и спадают в виде колпачка. Плод—ягода (у крушиновых — костянки
или коробочки). Около 500 видов,э тропических, субтропических, реже умерен-
ных. странах Роды: Vitis (виноград), Parthenocissus (американский виноград)
и Др.
427
167. Xyridaceae. Семейство однодольных, из порядка EnantioMastae.
€юда же относится сем. Ristionaceae. Небольшое семейство (около 50 ви
дов), преимущественно в тропической и субтропической Америке. Травы (мно-
голетние) с узкими прикорневыми листьями. Цветки правильные или непра-
вильные, типа лилейных,с рядом редукций. Соцветия в виде головки или колоса.
168. Winteraceae. Семейство раздельнолепестных двудольных, и®
ворядка Polycarpicae. Очень близко к Magnoliaceae, причем многие авто-
ры присоединяют его к магнолиевым (в широком смысле слова). Семейств»,
несомненно, является дальнейшей стадией эволЛции магнолиевых (прилистни-
ки отсутствуют, пестики в один круг). Тропики и субтропики, особенно южног»
полушария (в Африке отсутствует). Около 20 видов. Род Drimys и др.
169. Zamiaceae (замневые, саговники). Семейство голосеменных
растений, класс Cycadales. К этому же классу относится и семейство Су-
eadaceae, от которого сем. заммевых отличается тем, что на плодолистиках
видят всегда две семяпочки, причем женские шишки, так же как и мужские,
являются конечными (не прорастают). Несколько родов в тропической и суб-
тропической Африке, Америке, Австралии. Внешний вид, как у Cycadaceae.
Замневые представляют вымирающие растения, в настоящее время из-
вестно около 70 видов (8 родов — Ceratozamia, Macrozamia, Dioon и др.).
170. Zingiberaceae (имбирные). Семейство однодольных-, из поряд-
ка сцитамнновых (Scitamineae). К этому же порядку относятся сем. каиновых,
банановых и др. Исключительно тропические растения, главным обра-
зом Старого Света, играют большую роль в тропическом лесу (около 350 ви-
дов). Многолетние травы с корневищами, с большими листьями.(с влагалища-
ми и язычками), двурядиымн или спирально расположенными. Цветки типа
лилейных, ио из шести тычинок лишь одна плодуща, одна недоразвивается,
•стальные — лепестковидные, окрашенные. Две из последних образуют губу.
Завязь, как и у всего порядка, нижняя. Цветки яркие, собранные в соцветия.
Корневища ряда видов съедобны нлн содержат пряные вещества — нмбнрь
Zingiber officinale), кардамон (Elettaria). Некоторые виды декоративны.
171. Zygophyllaceae (парнолистные). Семейство раздельноле-
естных двудольных, нз порядка геранецветных (Geraniales). Главным образом
в теплых странах (около 200 видов); деревянистые растения: кустарники, де-
ревья, реже —травы. Характерны парноперистые супротивные листья. Цвет-
ки пятерного типа, правильные, инти тычинок внизу нередко расширены ли-
гговидыо. Представители семейства часто являются ландшафтными расте-
ниями в пустынях и на засоленных местах (Nitraria). Некоторые виды дают
денную древесину. В СССР несколько травянистых и кустарниковых видов.
ОСНОВНАЯ ЛИТЕРАТУРА
Отдел I. Учение об ареале
1. De Candolle A., Geographic botanique raisonn^e, I—H, 1^55.
2. Г ре б ие p П., География растений (перев. с немецкого), М., 1914.
S. Ди льс Л., Ботаническая география (перев. с немецкого), Птр., 1916-
4. Вульф Е., Введение в историческую географию растений, изд. 2,
И. -Л., 1933.
5. В у л ь ф Е., География растений, М. - Л., 1936.
6. В а л ь т е р Г., — Алехин В. В., Основы ботанической географии,
Биомедгиз, М. - Л., 1936.
7. Hanni g Е. и Winkler Н., Die Pflanzenareale (Собрание карт
ареалов). Выходит выпусками с 1926 г.
8. S с й г о t е г С., Genetisclie Pflanzengeographie, Handworterbuch d. Na-
turwiesenschaften, 1934.
Отдел II. Учение о взаимосвязях между растениями и средой
1. Бармин г Е., Ойкологическая география растений, иерэв. Годен
кина М. И. и Арнольди В. М., М., 1900. Та же книга, но в переводе Генкеля
А. Г., носит другое название — «Распределение растений в зависимости от
внешних условий», СПБ, 1902 (с дополнительным разделом Танфильева Г. И.,
ем. отд. .IV).
2. Neger F. W., Biologie der Pflanzen aut experimentaller Grundlage,
1913.
3. Мак-Дуго л л В. Б., Экология растений (перев. с английского),
М., 1935.
4. Кашкаров Д. Н., Среда и сообщество, М., 1933.
5. А ц ц и Д., Сельскохозяйственная экология (перев. с итальянского),
М. - Л., 1932.
б. Малеев В. П., Теоретические основы акклиматизации, 1933.
7. Максимов Н. А., Физиологические основы засухоустойчивости
растений, 1926.
8. Лысенко Т. Д., Теоретические основы яровизации, М., 1936.
9. Л у н д е г о р т Г., Влияние климата и почвы иа жизнь растений
(перев. с немецкого), М., 1936.
10. Поплавская Г. И., Краткий курс экологии растений, Л., 1937,
11. Г е й г е р Р., Климат приземного слоя воздуха (перев. с немецкого),
М. ~ Л., 1931.
12, We a ve г J. Е., С I е m е п t s F. Е., Plant ecology, 1938.
13. Гумбольдт А., География растений, М. - Л., 1936 (перев. с пе-
мецкого и латинского); в оригинале — в 1806—1845 гг.
Отдел III. Учение о фитоценозах
1. Пачоский И. К., Основы фитосоциологии, Херсон, 1921.
Я. Алехин В. В. и др., Методика геоботанических исследований, М
1025. ' ’
3. D u R i е tz G. Е., Zur methodologischen Grundlage der modernen
Fflanzetuoziologie, Upsala, 1921.
42f
4. Braun-Bia n que t I
5. С у к а ч ' - - ” Л
0. Rubel
d. Naturwiss.,
-ч « х а. j., Pflanzensozioiogie, 1926.
ёв В. Н., Растительные сообщества, изд. 4-е, Л., 192?.
IM ’ ^eo^raP^ie der Pflanzen. 3. Soziologie, Handworte buck
Отдел IV. Типы растительного покрове
а) Растительность земного шара
1. Гризебах А., Растительность земного шара, СПБ., 1874—1877 (шгев.
с немецкого оригин. 1872 г.).
2. Бекетов А. Н., География растений, СПБ., 1896.
Красное А. Н., География растений. Основы землеведения, в7 п.
IV, я. I, Харьков, 1899.
« Р b е 1 1 D. Н., An outline of plant geography, New-York, 19 6.
a. Rubel E., Pflanzengesellschaften der Erde, 1930.
6. F a b e г F. C., S c h i m p e r A. F. W., Pfianzengeographie auf physiology
scher Grund la ge, 3-е изд., 1935.
c I903^arSten Schenck H., Vegetationsbilder. Выходит.выпусками
8. Engler A., Drude O., Die Vegetation der Ercle. Монографии, вы-
ходят с 1896 г.
9. Мартоин Э., Основы физической географии, т. III, Биогеогра-
фия, М., 1940 (перев. с французского).
10. И л ь и и с к и й А. П., Растительность земного шара, Л., 1937.
б) Растительность СССР
1. Т а и ф и л ь е в Г. И., Главнейшие черты растительности России, 1902,
Приложение к книге Варминга Е. (см. выше, отд. II).
2. Коркинский С. И., Растительность России. Энцнкл. словарь Брок-
гауза - Ефрона. Том «Россия», XXVII, 1899.
3. Б у ш Н. А., Ботанико-географический очерк Европейской части СССР
п Кавказа, М. - Л., 1936.
4. Алехин В. В., Растительность СССР в основных зонах, 1936 (см.
зыше Вальтер-Алехин, отд. I).
5. С у к а ч ё в В. Н., Дендрология с основами лесной геоботаники, изд.
2-е, Л., 1938.
6. К е л л е р Б. А., Растительный мир русских степей, полупустынь и
пустынь, И., 1923.
7. Коровин Е. П., Растительность Средней Азнн, М. - Ташкент,
1934.
8. Растительность СССР: г. I (история растительности и флоры,
тундры, болота, луга), 1938# т. II (степи, полупустыни и пустыни, сорная ра-
стительность), 1940; т. III (леса, растительность Кавказа, Крыма, Средней
Азин, Дальнего Востока, — еще не вышел). Изд. Акад, наук СССР.
Отдел V. Флористические области
1. Кузнецов Н. И., Курс географии растений, Симферополь, 1920.
2. Н а у е k A., Allgemeine Pfianzengeographie, 1926.
3. К о м а р о в В. Л. Происхождение культурных растений, J938.
4. Т а н ф и л ь е в Г. И., Очерк географии и истории главнейших куль-
турных растений, Одесса, 1923.
Примечание. Приведенная литература во многих случаях «т-
носится не к одному какому-либо отделу, но также и к другим.
указатель русских терминов
А
Агенты распространения растений
75, 01, 121.
Акклиматизация, 105.
Аллювиальные почвы 370.
Альпийские кс.вры 278; луга 274;
лужайки 278, 279.
Аналитические признаки фитоцено-
зов 153.
Анаэробиоз 115.
Аяемоморфные листья 246.
Апсфнты 128.
Ареал 8. 20; его устойчивость 29,
его формирование 29; дизъюнк-
Ti вный 24; лабильный 29; ленточ-
ный 25; несплошной 24; преры-
вистый 25; разъединенный 24;
реликтовый 37; связанный 26;
сплошной 24; стабильный 29; че-
респолосный 31.
Ареалологическая география 8.
Арктотретпчная флора 327.
Архесфиты 128
Аспект 145, 258; непостоянный 157;
постоянный (константный) 157.
Ассоциация И, 17, 161; ее название
165.
Аутэкология 9.
Афилльные растения 308.
Ацидофильные растения ПО.
Аэрация 115.
Аэренхима 70.
Б
Еаэпфильные растения ПО.
Безлесье степей 265; тундр 304.
Белтовскне тельца 124.
Бентос 293.
Березовые колки 227.
Биотип 64.
Болота 285; их образование 289;
березовые 228; верховые 287;
гипновые 286; лесные 285; ман-
гровые 285; моховые 285; олиго-
трофные 286. 289; с соковые 286,
ольховые 228; переходные 287;
сосновые 243; сфагновые 289;
травяные 285; эвтрофные 228.
2(6; низинные 228. 285.
Болотные растения 60.
Бор-беломошник 236; брусничник
165, 226; долгомошни* Z36; зеле-
номошннк 162, 236; кисличник
165; черничник 165, 236.
В
Ведьмины кольца 322.
Вечнозеленость 181.
Вересковые пустоши 247; фитоцено-
зы 245.
Верещатники 245.
Верные виды 236.
Ветровал 74.
Ветроопыляемые растения 74.
Виды — см. растения.
Викарирующ”е виды 40, 41.
Влагоемкость 114; относительная! 14
полная 114.
Влажнотропические леса 171,184
Водные растения 60; плавающее
G0; погруженные 60.
Вредные газы 72.
Высокотравие 272.
Вымерзание растений 95.
Г
Газоустойчивость 72.
Галлерейные леса 227, 269.
Гапсфнт 111, облигатный ИЗ, 191
факультативный 113.
Гарига 206.
ГелиосЬчт 79.
Гелофит 100.
Гемикриптофит 100.
Геоботаника 13. 140.
География растений 7; фитоценозов
8,139; ареалогическая 8; генетн-
чеекая 8; историческая 8; флори-
стическая 8; экологическая 8,
Геофит 190,
Гетерофилдьные растения ЗОЭ.
Гибридизация 384; межвидовая 384;
межродовая 384,
Гибриды 384.
Гигрофиты 84, 89.
Гидрофильные коллоиды 87,
Гидрофиты 84, 90, 100.
Гида годы'90.
Гнлея 185, ЗС8.
Группировка растений 167, 192.
Д
Двулетник 60.
Деревья 57; их генезис 62; летне-
зеленые 57, 213; лавровые 193;
вечнозеленые 57, 181; зимнезеле-
ные 197.
Дерезняки 229.
Диаспора 29.
Дизъюнкция ареааов 24, 32; евра-
зипско-североамерикэяск''я 33;
австралийская 37;антаркгическая
35; ь рымско-малоазкйская 37;
неотропическая 34; пантропиче-
вкая Зл; палеотропическая 34;
северо-атлантическая 33; северо-
тнхоокеанская 33, средиземно-
морская 36; южно-аглачгическая
34: южно-тихоокеанская 34.
Диморфизм 270.
Джунгли 360.
Дождевые леса 171.
Доминант 151; временный 151; по-
стоянный 151.
До ковидные корни 175.
Дубовые леса 220.
Дубравы 220; зеленчуковые 223;
снытевые 223; осоковые 223.
Дыхательные корми 71,190.
Е
Ельник болотиотравяной 234; долго-
мошник 234; дубравный 234; брус-
ничник 236; зеле^омошник 234;
кисличник 236; ключевой 243;
сложный 234; сфагновый 234;
черничник 236.
Ж
Жестколистные леса 202.
Живое ископаемое 337, 359.
Живородящее растение 190,
Жизненная форма 55; их система
57; их генезис 62; альпийских
лугов 275; Раункиэра 98; тундр
432
299; степей 25б; лесов 296; пус-
тынь 307.
Жизненность 153.
3
Закаливание растений 96.
Заключительная стадия 144; фито-
ценозы 158.
Закон Заленского 87; .минимума Ли-
биха 134; относитечьности 134.
Заливные луга 269.
Замещающие виды 41.
Замещение факторов 134, 135.
Заросли чистые 149.
Засухоустойчивость 89.
Зимнезеленые леса 197; растения
236,278,281.
Злаковицные формы 57; травы 57, 59.
Зона растительности 168; снеговая
301.
Зональная растительность 227.
Зоохория 121.
И
Игольчато-хвойные леса 239.
Изошты 102.
Изолатеральные листья 209.
Изофены 102.
Изоценоз 166.
Индикаторы почв 14, ЮЗ.
Индиферентные растения ЮЗ.
Ингрззоиальная растительность 227.
235.
Интродукция 106.
К
Каатинга 201.
Кактусовидиая форма 56.
Кальцифильные растения 108
Кальцнфобные растения 108.
Кампосы 254.
Капельные острия 90.
Капил аярность 114.
Кардинальные точки температуры94.
Каулифлория 182.
Квадраты обилия 152.
Кислые растения ПО.
Классификация фитоценозов 163.
Климакс 159.
Клима тип 64.
Климаты — их классификация 92.
Ковыльник бескрасочный 262; кра-
сочный 262
Колония растений 142.
Колючие ксерофпльные фнтоценозы
199.
Компасные растения 82.
Компенсационная точка 79,
Конкуренция растений 12, 136; вну-
тривидовая 136; межвидовая 136.
Коренные фитоценозы 159.
Корпи втягивающие 107; досковид-
ные 175; дыхательные 71; пустын-
ных растений 308; ходульные
189, 228.
Космополитные виды 20; фитоценозы
21.
Коэфициент родовой 52.
Криволесье 303.
Криптофит 100.
Ксероморфная структура 87, 300;
болотных растений 290.
Ксерофильные леса 199.
Ксерофиты 81; их признаки 85—88.
Культивируемые растения 379.
Культурные растения 379 , 399.
Культуры вторичные 387, 390, 392;
первичные 387.
Кустарники 58; вечнозеленые 58;
летнезеленые 58; вересковые 245;
жестколистные 206, 209; мангро-
вые 188.
Кустарнички 58; вечнозеленые 58;
летнезеленые 58; эрякоидные 58.
Л
Лавровые леса 193; двудольные 193,
195, хвойные 195.
Леса байрамные 223; березовые 226;
болотистые 227, 242; буковые216;
влажнотропические 171; вторич-
ные 226; дубовые 220: еловые 233;
зимнезеленые 197; игольчато-
хвойные 230; ксерофильные 199;
лавровые 193; лиственничные 237;
летнезеленые 212; мелколиствен-
ные 226; муссоновые 198; осино-
вые 226; первичные 226; пихто-
вые 238; пойменные 227, 242;
саванновые 198; светаохвопные
231; субтропические 186; темно-
хвойные 291; умеренно-субтропи-
ческие 186; хвойные 230; широко-
лиственные 213.
Лесостепь 223, 265.
Лесотундра 298, 303.
Лианы 59, 179.
-Литоральный пояс 293.
Литофит 116.
Луга 268; альпийские 274; заливные
269; кобрезиевые 278; пустошные
284; субальпийские 271; сухо-
дольные 270; тундровые 282.
Льяносы 254,
М
Маквис 206, 207.
Мангровая растительность 188.
Матты 274, 277.
Мегафанерофит 100.
Мезофанерофит 100.
28 География растений
Мезофит 84.
Мелкозем 114.
Мелколиственные леса 213, 226, 23 X
Мерзлота вечная 299.
Мертвая лесная тень 79.
Местонахождение 67.
Местообитание 68.
Местопроизрастание 68.
Метод ментора 386. подбора пар
растений-производителей 384; по-
средника 384; предварительного
вегетативного сближения 384;
пробных площадок 165; экологи-
ческих рядов 119.
Миграция видов 29.
Микрофанерофит 100.
Микрофиллия 300.
Мирмекофильные растения 123.
Ммрмекохория 122.
Многолетник 59.
Модификация 65, 66.
Мозаика листовая 81.
Монокарпические растения 59.
Монотипные роды 52.
Морозостойкость 95.
Морфология фнтоценозов 145.
Мочажины 292.
Муравьи-листорезы 123.
Муссоновые леса 198.
Мутация 386.
Мюллеровские тельца 124.
Н
Нэнофаиерофпт 100.
Насекомоядные растения 122.
Насыщенность видовая экземпляр-
ная 136; ярусная 150.
Натурализация 105.
Нейтрофильные растения ПО.
Неофиты 128.
Нитратные растения 108.
Нунатаки 324
О
Оазисы 311.
Обилие 151; его учет 152; шкала
обилия 152.
Облигатные галофиты 113, 191
Обнаженное место 140; первичное
140; вторичное 141.
Обобщенная схема лесов 238.
Однолетник 60.
Оксифильные растения И0.
Октоплоиды 387.
Олиготрофные растения 108; болота
286, 289.
Ольшаиники 228.
Осинники 230.
Осиновые кусты 226.
Организационные признаки 54.
Осмотическое давление 86, 88.
433
Основные формы растений 58.
Островная флора 39.
Отбор бюсознателыгый 127, 383*
искусственный 38з; сознательный
130, 384.
Оттаивание 95.
Очаг происхождения культурных
растений 381.
П
Пампасы 255.
Паразиты 125.
Пастбища 273, летние 284, 305; зим-
ние 305.
Перекати-поле 260.
Периодичность роста 181.
Плавательные пузыри 91; ткаин 91.
Плакорные пространства 119.
Планктон 295.
Плейстон 294.
Пневматофоры 190.
Подводные растения 70.
Подвой 386.
Подзоны 301.
Подснежное развитие 98, 222.
Подушки-растения 58, 275, 277.
Покрытие 153.
Поликарпическне растения 59.
Полиплоиды 386.
Полукультурные формации'273.
Полукустарники 58
Полусгланик 303.
Потребность в свете 79.
Почва 107; ее вла гоемкость 114;
капиллярность 114; проницае-
мость 114; структура 116; физи-
ческие свойства 107; химические
107.
Почвенное дыхание 71.
Почвопостоянные растения 108,
Почвопредпочитающие 108.
Пояс растительности 118, 293.
Правило предварения 120.
Прерии 262; высокозлаковые 264;
иизкозлаковые 264.
Привой 386.
Признаки растений: приспособитель-
ные 54; организационные 54.
Пробные площадки 165.
Проницаемость почвы 114.
Пропорции 51.
Псаммофиты 115.
Психрофиты 274.
Пуны 280, 368.
Пустоши вересковые 247.
Пустыни ЗОо; арктические 323; био-
тические ЗОб; высокогорные 320;
глинистые 313; каменистые 315;
кустарппковые315; песчаные 315;
подвил;ные 306; полынные 313;
солончаковые 314; солянковые
314; сухие 306; холодные 320;
эфемерные 313.
Пушты венгерские, 262.
Р
Растения ацидофильные ПО; бази-
фильные ПО; водные 60; галофиты
111; длинного дня 83; индиферент-
ные 108; кальцифильпые 108; ко-
роткого дня 83; мирмекофильные
123; насекомоядные 122; нейтро-
фильное НО; нитратные 106; ок-
сифильные ПО, 390; олиготроф-
ные 108; почвопостоянные 108;
почво-предпоч^тающие 108; при-
шлые 128; сегетальные 129; си-
нантропнь е 128; солнечные 79;
солончаковые 111; сорные 128;
рудеральные 129; теневые 79;
эвтрофные 108.
Растительные формы 58.
Реакция среды ПО.
Редколесье 303; колючее 197, 199.
Режим почв 115; водный 115; воз-
душный 115.
Реликтовые ареалы 37.
Реликты 37, 38; ксеротермические
38; ледниковые 38; третичные 38.
Рельеф 118.
Ритмы развития 133.
Родовой коэфицнент 52.
С
Саванновые леса 198.
Саванны 249.
Сапрофиты 236.
Свет верхний 77; нижний 77.
Седиментация 270.
Серия 158; водная 158; глинистая
158; каменистая 158; песчаная
158.
Сероольшаниики 244.
Симбиоз 123.
Синантропные растения 128.
Сингенетическая сукцессия 144.
Син зоохория 122.
Синузия 126.
Синэкология 9.
Систематика фитоцеиозов 161.
Ситниковидные растения 60.
Скрещивание 384, 385.
Скрзб 209.
Склерофиллы 85.
Смены фитоцеиозов 157, 160; мете-
орологические 160; фенологиче-
ские 153, 160.
Смешанные леса 333.
Снеговые ложбинки 280.
Солнечные блики 79; растения 79.
Солончаки 112.
Солончаковые растения 111.
434
олонцы 112.
оснсгые леса 237.
оциация 162.
Сочетание растений 167.
Спектр биологический 101; флори-
стический 51.
Среда растений 67.
Стадия в развитии растений — свето-
вая 83; яровизации 104, 131.
Стадия сукцессии 143; бурьянная
143; корневищная 144; заключи-
тельная 144, 158.
Стенотопные виды 23.
Стенохорные 21.
Степи 255, 257.
Стланики 303.
Субдоминант 151.
Сублиторальный пояс 294.
Субстрат 107.
Субтропические леса 186.
Суккулент 59, 86, 88; листовой 59,
86; стеблевой 59, 86.
Сукцессия 142, 158, 287; ее стадии
143, 158; антропогенная 159; ве-
ковая 160*вторичная 158; зооген-
ная 159; климатогенная 159; пер-
вичная 158; прогрессивная 160;
регрессивная 160; сингенетиче.
ская 144; филоцеиогеиетиче-
ская 160; фитсгенная 159; ценц-
генетическая 160; эдафогенная
159; эндодииамическая 143, 160;
экзодинамическая 143, 160.
Сухость физическая 84; физиологи-
ческая 84.
Сфагновые болота 289.
Сциофиты 79.
Т
Танга 236.
Теневые растения 79.
Теневыносливость 80; методы учета
80.
Теория Вегенера 13, 30, 327, 356;
мостов 29; формирования ареалов
31.
Терофит ЮО.
Тетраплоиды 387.
Тип растительности 162.
Томи^ляры 207.
Травы 59; их генезис 62; вечнозеле-
ные 59; высокие59; зимнезеленые
59; злаковидные 59; папоротни-
ковидные 60; розеточные 60; сит_
никовидные 60; типа перека-
ти-поле 60.
Третичные растения 345.
Триплоиды 387.
Тропические леса 171, 184.
Тугайные леса 227.
Тундра 298; ее подзоны 207; алекто-
риевая 297; арктическая 301;
бугристая 302; вторичная 305;
дериовкная 283; дриадовая 302;
кладонневая 297; кочкарная 296;
кустарниковая 301; лишайниковая
297; луговинная280;моховая 29G;
осоковая 302;первичная 305; по-
лигональная 302; пятнистая 302;
цетрарневая 305; ягельная 297,
303.
Туника 97.
У
Умеренно-субтропические леса 186.
Условия существования 67.
Устойчивость фитоценозов 160.
Устьица ксерофитов 86.
Участок ассоциации 162, 165.
Ф
Факторы 68; биотические 68, 121;
биологнчески-измененные 69, вто-
ричные 69; исторические 68, 132;
климатические 68, 70; косвенно-
действующие 69; ограничиваю-
щие 134; орографические 68; пер-
вичные 69, прямодействующие 6'9;
эдафическве 68, 106; энтопиче-
ские 69; экологические 70; эколо-
гически действующие 134; их
совместное действие 135; их заме-
щение 135; экзогенные 69; эндо-
генные 69.
Фанерофит 98.
Фенологические явления 101; ка-
лендарь 103; смены 155.
Фенотип 66.
Физиономические жизненные формы
62.
Физиономия фитоценозов 138.
Филлодии 209.
Фитаэрон 164.
Фитобеитос 293.
Фитопланктон 164, 293.
Фитосоциология 18.
Фитоценоз 7, 139, 162, 167; заключи,
тельный 158; закрытый 150; ко-
ренной 158; многоярусный 150;
открытый 150; одноярусный 150-
простой 150; сложный 150; ярус-
ионасыщенный 150; его формиро-
вание 140,144; его ярусность 147.
Фитоценология 7, 9.
Фнтоэдафон 164.
Флагообразные формы 73.
Флористические карты 171, 325; под-
область 327; провинция 327; со-
став 145; спектр 51, область 327;
царство 327.
Флорологпя 17.
435
28*
формация 15, 16,17; класс формаций
163, 165; формация «типа» 193.
Фотопериодизм 83.
Фригана 207.
X
Хамефит 100.
Хасмофит 116.
Хвойные леса 230,
Ходульные корни 189.
Холодные пустыни 320.
ц
Цветочные часы 81.
Ценотип 64.
Центр ареала 32; возникновения 32;
геометрический 32; обилия 32.
Центр происхождения культурных
растений 381.
Ч
Чапараль 207.
Чистые заросли 149.
Ш
Шибляк 207, 220.
Широколиственные леса 213; поро-
ды 334.
Шнрокотравие 221.
Шкала обилия 152; теневыносливо-
сти 80.
Шохра 243.
Шпалерные формы 300.
щ
Щелочные растения ПО.
Э
Эвкалиптовые леса 208.
Эвритопиые виды 23.
Эврихорные виды 21.
Эвтрофные болота 223, 286; растения
108.
Эдафотпп 64.
Эдпфнкатор 152.
Экзодинамическая сукцессия 143.
Экологически равноценные виды14б;
разноценные 146.
Экологический ряд 118.
Экология 6, 13, 14, 52; фитоценозов
139.
Экотип 64.
Экспозиция 119.
Элемент флоры 41; арктический 42;
атлантический 44; генетический
42; географический 42; парнас-
ский 47; иранский 46; истори-
ческий 42; кавказский 46; кол-
хидский 46, 47; маньчжурский
46; переднеазиатский 45; понти-
ческий 47; сарматский 45; се-
верный (бореальный) 42; среди-
земноморский 45; среднеевропей-
ский 43; туранскнй 46; цен-
тральноазиатский 46.
Элиторальный пояс 294.
Эндемизм 38; консервативный 39;
прогрессивный 39, 40; реликто-
вый 39.
Эндемические виды (эндемики) 21,
38; горные 39; роды 64; островные
39; фнтоценозы 22.
Эндодннамическая сукцессия 143.
Эидозоохория 122.
Эндотрофная микориза 126.
Энтопнческие факторы 69.
Эпизоохория 121.
Эпифиллы 60, 125, 180.
Эпифиты 60, 61, 125, 180; цистер-
новые 185.
Эрикондные растения 58, 245.
Этиолированные растения 77.
Эфемеровая пустыня 310.
Эфемеры 89; пустынные 309.
Я
Яровизация 104, 131.
Ярусность фито ценозов 16, 147; во
времени 157; подземная 148; тро-
пических лесов 175.
Указатель латинских Названий
А
Abiegneta 238.
Abies alba 218, 238, 239, 245, 332.
— balsamea 241,.351.
- Borissii regis 239.
— Bornmulleriana 239.
— cephalonica'239, 340.
— cilicica 239, 343.
— gracilis 21.
— nebrodensis 239.
— Nordmanniana 233, 239, 332,
333.
— numidica 343.
— Pinsapo 239.
— Semenovi 332.
— sibirica 233, 236.
Acacia 124. 199, 311, 358, 371, 415.
— arabica 347.
— catechu 360.
— floribunda 210.
— spirocarpa 251.
Acaena 299, 369, 376, 424.
Acantliolimon 322, 421.
— diapensioides 340.
Acanthosicyos 408.
*=• horrida 317, 359.
Acer 401.
— campestre 334.
— ginnala 334.
— hircanum 334.
— ibericuni 334.
— negundo 353.
— manshuricum 334.
— pensilvanicum 352.
— platanoides 333.
— pseucloplqtanus 334.
— saccharum 219, 352.
— tataricum 334.
— tegmentosum 334.
— Trautvetteri 334.
— ukurunduense 334.
(видов, родов, семейств1, фитоцеиозов и др.
Aceraceae 401.
Achillea millefolium 263.
Aciculifruticeta 163, 301.
Aciculilignosa 163, 165, 170, 230.
Aciculisilvae 163, 165, 230.
Aconitum 423.
— nasutum 272.
Acorus calamus 130.
Actinidia arguta 338.
— kolomicta 338.
Actinidiaceae 401.
Adansonia 403.
— digitata 252, 253. 358.
Adenostoma fasciculatum 207, 208.
Adonis 339, 423.
— vernalis 32, 44, 154, 156.
Aegilops 388.
Aerrantia 164.
Aestiduriherbosa 255, 268.
A.estifruticeta 163, 228, 283,
Aestilignosa 163, 170, 212.
Aestisilvae 163, 213.
Agathis 231, 362, 363, 407.
- -- loranthifolia 374.
Agathosma capita turn 211.
Agave 366, 401.
— americana 33, 366.
Agrimonia 122, 424.
Agropyrum 262, 263, 264, 388, 413.
— repens 23, 142, 144.
Agrostemma githago 128.
Agrostis 277, 328, 413.
— alba 269.
planifolia 27L
— Syreistschikowi 137, 261.
— vulgaris 270.
Ailanthus glandulosa 225.
Aira antarctica 376.
Aizoaceae 401.
Ajuga reptans 215.
Aibizzia 377,416.
— julibrissin 47.
1 Последняя цифра у рода указывает страницу, где дана краткая ха-
рактеристика ссмейстЕа, куда этот род относится.
Alcllimilla 273. 369, 424.
Aidrovanda vesiculosa 23, 25.
Alectoria 305.
— nigricans 279.
— ochroleuca 297, 303, 329.
Alectorolophus 270, 425.
— majar var. apterus 130.
Alisma 288, 401.
— plantago aquatica 20.
Alismataceae 401.
Allium 339, 343, 416.
— ursinum 218.
Alnetum viridis 229.
Alnus fruticosa 301, 303.
l— incana 244 .
- glutinosa 228.
Aloe 359, 416.
Aiopecurus 413.
— alpinus 302.
mucronatus 340.
— pratensis 269. 283.
— ventricosus 269.
Alopliia 267, 414.
Alsophila 368, 372, 408.
— glauca 184.
Altherbosa 163, 268.
Alyssum 89, 407.
Amaryliidaceae 374, 401.
Amentiflorae 112.
Amherstia nobilis 86.
Ammodendron 315. 340, 416.
— conollyi 314.
Amorpha 353, 416.
Ampelidaceae 427.
Amygdalus nana 229.
Anabaena Nordenskioeldi 324.
Anabasis 340, 406.
— salsa 312, 313.
Anacardiaceae 401.
Anastatica 313, 407.
Anchusa ochroleuca 31.
— officinalis 31.
Andromeda 37.
Andropogon 253, 254, 263, 266, 353,
358, 413.
— furcatus 263.
— nutans 263.
— saccharoides 263.
— scoparius 262, 263.
Androsace alpina 277, 321.
— Helvetica 277.
Anemone 98, 423.
— narcissiflora 271.
- nemorosa 151, 152,215,217.
— ranunculoides 222.
- silvestris 156, 261.
Anisophylla 34, 35, 423.
Anonaceae 328. 356. 357, 402.
Antennaria campestns 263.
Anthoxanthum odoratum 271, 283.
Anthurium 177
Apocynaceae 402.
Aponogeton 71. 402.
— crrstralis 90.
Aponogetonace ie 402.
Aposeris foetida 218.
Aquerrantia 164, 293, 295.
Aquifoliaceae 402.
Aquiherbosa 163. 164.
Aquilegia 336, 423.
Arabis petraea 40.
Araceae 186, 402.
Aralia 226. 402.
— manshurica 46, 337.
Araliaceae 402.
Araucaria 407.
— imbricata 376, 378.
Arbutus 195. 207.
— andrachne 37. 45, 344.
— Menziesii 207.
— unedo 205, 206, 220, 342.
Arctagrostis latifolia 297.
Arctium lappa 128. 129.
Arctos taphylos 208, 246, 249, 410.
Arctous alpina 25, 297.
Areca catechu 361.
Arenaria 266, 405.
— musciformis 321.
Arisfida 254,266,313. 413.
— brevifolia 359.
— Karelini 308.
— purigens 311.
Aristolochia 402.
Aristolochiaceae 402.
Artemisia 58. 340, 353, 406.
— arenaria 314.
- - austriaca 128. 259.
— campestris 128.
— granatensis 276.
— herbaalba 315.
— hololeuca 21.
— Scorniakovii 340.
— terrae albae 46. 312, 313,315.
— tridentata 317, 354.
Artocarpus 182, 418.
Arum 402.
— maculitum 215.
Asarum 402.
— europaeum 95, 221.
Asclepiadaceae 356. 402.
Asparagus tenuifolius 27.
Asperula cynanchica 262.
— glauca 86, 88.
— odorata 88, 218.
— tinctoria 88.
Aster 353, 406.
— amellus 66.
— tripolium 112, 113.
Astragalus 339. 369, 416.
— • alpinus 283.
— arenarius 45. 116.
— austriacus 108.
— Musch ketovi 322.
Astrocaryuni 186, 420.
Atripkx 340, 406.
— сапа 112. 309, 313.
— hastaM 128.
— verrucifera 112
A+tilea 186, 420.
Aucubi 337, 407.
Aulacomnium 301.
— palustre 329.
— turgidum 296, 329.
Avena abyssinica 392.
— fatua 392.
— nuda 392.
— sativa 391.
A venastrum desertorum 340.
— pubescens 259.
— Schelliana 261.
versicolor 278.
Avicennia 189, 191, 193, 427.
— marina 191.
Azalea 37, 410.
Azorelia 299, 369, 376, 426.
В
Balsaminaceae 403.
Bambusa 413.
Banksia 371, 423.
' — serrata 210.
Barbella javanica 179.
Beckmannia eriiciformis 269.
Begonia imperialis 90.
Begoniaceae 356, 403.
Bellis perennis 94.
Berberidaceae 403.
Berberidetum vulgaris 229.
Berlinia 199, 416.
Berholletia excelsa 186.
Betornca grandiflora 273.
Betula exilis 230. 297, 301, 328.
— Ermanii 226.
— Medwedewi i 47.
— Middendorfii 303.
— лапа 38, 230, 248, 249, 290,
297, 301, 328.
— papyrifera 351.
— tortuosa 226, 304, 330.
Betulaceae 327, 403.
Bidens 142.
Bignonmceae 328, 356, 403.
Bolax 369, 376. 426.
Bombaeaceae 357, 403.
Boraginaceae 403.
Borassus 3t2, 347. 358, 420.
Boutgloua 262, 263, 254, 413.
— gracilis 264, 265.
— hirsuta 263.
— racemosa 263, 264, 353.
Bowenia 373.
Brachypodium 413.
— pinnatum 271.
— silvaticum 222.
Braya 328, 407.
Briza 266. 413.
Bromus 266. 339, 413.
- - Benekeni 222.
• - erectus 136. 137, 258, 259.
— inermis 23, 142.
— secalinus 128.
— tectorum 262.
— variegatus 271.
Bromeliaceae 356. 364, 404.
Bruguitra 190, 423.
- . gvmnorh’za 10$.
Brunella grandiflora 109.
Brya ebenus 370.
Bryopogon divergens 303.
Buchloe 264. 413.
— dactyloides 264, 265, 353.
Butea monosperma 198.
Butomaceae 404.
Butomus 288, 404.
Buxaceae 404.
Buxus sempervirens 45, 345, 404.
C
Cabomba 54, 419.
Cactaceae 328. 404. 536, 564.
Caesalpinia echinata 370.
Caesalpiniaceae 416.
Calamus 180. 361, 420.
Caliiergon 286.
Calligonum 308, 314, 339, 422.
Calluna 33, 246.
— vulgaris 108. 245, 247, 410.
Calycanthaceae 34, 404.
Calypso 236, 420
— bulbosa 24.
Calystegia 59, 407.
Camelia 337, 426.
— japonica 225.
— theifera 397.
Camelina glabrata 83.
— linicola 130.
Campanula 279, 404.
— lactiflora 272.
Campanulaceae 404.
Camphora 226.
Camp to thee turn trichoides 296, 329.
Cannabinaceae 404.
Cannabis 404.
Cannaceae 328, 356, 365, 404.
Caprifoliaceae 404.
Caragana frutex 229.
Cardamine alpina 281.
Carduus hamulosus 128.
Carex 328, 335, 340, 409.
-- aquatilis 269.
— caespitosa 228.
— curvula 277, 279.
— Goodenoughii 269.
— graciiis 288.
— firma 269, 277.
- flava 33.
439
Carex Huetiana 279.
- humilis 156, 409.
— limosa 290.
— oreophila 279.
— pachystylis 313.
— paradoxa 228.
— physodes 409.
— pilosa 222.
— rigida 296, 297, 302.
— rostrata 288.
- sempervirens 277.
— Soczavaeaiia 296.
— tristis 279.
— vesicaria 269.
Caricaceae 405.
Caricetum curvulae 278.
— firmae 278.
Carica papaya 360, 405.
Carludovica 370, 408.
Carpine turn duinensis 229.
Carpinus betulus 215, 218, 384.
— cordata 225, 334.
Carya 223, 414.
— cordiformis 352.
— ovata 352.
Caryocar vi-llostim 186.
Caryophyllaceae 51, 327, 340, 405.
Caryophyllus 362, 419.
Caryota urens 360.
Cassandra 37, 410.
Cassia 201, 416.
Cassiope 249, 328, 410.
— tetragona 248, 303.
Castanea vesca 342. 345.
Castilleja pallida 240.
Castilloa elastica 370, 418.
Casuarina 309, 373, 405.
— equisetifolia 373.
Casuarinaceae 405.
Catalpa 33, 403.
Cavanillesia arborea 201, 202.
Ceanothus 208, 423.
— cuneatus 208.
— papillosus 208.
Cecropia 186, 418.
— adenopus 124.
— cinerea 124.
Cedrus 242, 407.
— atlantica 343.
— deodara 337.
— Libani 343.
Celtis 224, 427.
Cembreta 238.
Centaurea 339, 406.
— cyanus 128.
— diffusa 128.
— marschalliana 26.
— margaritacea 46.
— orientalis 258.
Centaurea ruthenica 86, 108.
— trinervia 82.
Cephalaria tatarica 272.
Cephalotaceae 371, 405.
Cephalothaxus 194, 407.
— drupacea 225.
Ceratonia siliqua 346, 382, 416.
Ceratophyliaceae 405.
CeratophyHum 61, 70, 287, 405,
Ceratozamia 366, 428.
Cereus 201, 354, 366, 404.
— giganteus 202, 309,
Ceriops 191. „
Cetraria cucullata 297, 301,
— islandica 329.
— nivalis 297.
Chaerophyllum confusum 272.
Chamaecyparis 350. 407.
— nutkatensis 195,241,354.
— thyoides 243.
Chamaerops humilis 206, 342, 420,
Chenopodiaceae 112, 142, 327, 339,
405.
- album 128.
Chenopodium maritimum 112.
Chimaphila umbellata 59.
Chlora 253, 412.
Chondrilla jimcea 82.
Chorizema triangularis 210.
Chrysanthemum alpinum 278, 282.
Cicuta 288, 426.
Cinehona 370. 398, 424.
— Ledgeriana 398.
— officinalis 369.
— robusta 399.
— succirubra 399.
Cinnamomum 362. 415.
Cirsium 59, 284, 406.
— arvense 143.
Cistaceae 406.
Cistus 205, 206, 343, 344, 349, 406.
— tauricus 344.
Citrus trifoliatus 424.
Cladium effusum 244.
Cladonia 61, 305.
— alpestrina 297, 303.
— gracilis 301.
— rangiferina 297, 329.
— silvatica 297, 301,
— uncialis 297.
Clusiaceae 186, 406.
Cobresia 340. 409,
— Bellardii 60, 278, 279.
— schonnoides 278, 279.
Cochlearia 328, 408.
— arctica 94, 300.
Cocos nucifera 362. 420.
Coffea arabica 347, 360, 382, 397.
— Liberica 397.
Colchicum 139,386, 416.
Coleonema album 211.
Combretaceae 406.
440
Compositae 51, 328, 406.
Coniferalel 407.
Convailaria majalis 222.
Convolvulaceae 407.
Convolvulus lineatus 86.
Copernicia tectorum 254.
Coreopsis 353, 355, 406.
Corydalis 98, 411.
— cava 218.
- solida 222.
Cornaceae 407.
Cornus suecica 283.
Corylus avellana 215.
— heterophylla, 46.
- manshurica 47, 337.
Corypha gebanga 361.
— umbraculifera 360.
Cotinetum coggygriae 229.
Cotinus coggygria 344.
Cousinia 339, 406.
Crassula columnaris 316.
— pyramidalis 316.
Crassulaceae 407.
Crocus 98, 133, 342, 343, 414.
— albiflorus 278.
— Scharojani 280.
Cruciferae 51, 327, 407.
Cryptomeria japonica 225.
Cucurbitaceae 356, 408.
Cunninghamia 226, 407.
Cupressus 343 , 407.
— sempervirens 242, 346.
Cyathea 184, 363, 368, 372, 408.
Cyatheaceae 328, 408.
Cycadales 37, 408.
Cycadaceae 328, 408.
Cycas 373, 408.
— revoluta 226.
Cyclanthaceae 408.
Cymbaria borysthenica 37,
Cymbopogon 358. 413.
Cynara cardunculus 267.
Cynoglossum 122.
Cyperaceae 51, 328, 408.
Cyperus 286, 409.
Cypella 267, 414.
Cypripedium calceolus 109.
Cystopteris sudetica 25.
Cytisus nubigens 349.
— ruthenicus 44.
D
Dacrydium 363, 373, 407.
Dalbergia 370, 416.
Danae 47, 345, 416,
Daphne 349, 426.
— altaica 37.
— cneorum 22.
— gnidium 206.
~ Sophia 37.
Dasylirion 366, 416.
Datura stramonium 425.
Davallia canariensis 195.
Delphinium 272, 336, 423.
— cuneatum 157.
Dentaria 217, 222, 408.
— bulbifera 218.
- quinquefolia 218.
Deserta 163, 306.
Deschampsia arctica 302.
— alpina 283.
— caespitosa 110, 269,. 27L
— flexuosa 283, 329,
Desmodium 266, 353.
Deutzia 225, 327, 425.
Dianthus arenarius 116.
— brachyanthus 276.
Dicksonia 184, 363, 372, 408.
Dicranum 301.
— elongatum 296, 329,
Digitalis purpurea 218.
Dionaea 123, 292, 410.*
Dioon 366, 428.
Dioscoreaceae 409.
Diospyros 182, 410.
Dipsacaceae 409.
Dipterocarpaceae 185, 361, 409.
Dipterix odorata 186.
Dischidia 181, 403.
— rafflesiana 180.
Draba 328, 408.
— alpina 300.
— bryoides 275.
— glacialis 42, 329.
— hirta 329.
— verna 89, 262.
Dracaena draco 349, 350.
Drepanocladus 286.
— uncinatus 239.
Drimys 34, 35, 376, 417, 428.
Drosera 108, 123, 290, 292, 374, 410.
Droseraceae 409.
Dryandra mucronulata 210.
Dryas 248, 328, 424. ,
- octopetala 283, 302.
— punctata 296.
Dryopteris spinulosa 228.
— Thetypteris 228,
Dulrchium 286.
Duriherbosa 163, 170.
Durifruticeta 163, 202.
Durilignosa 163, 170, 202.
Dur io 403.
Durisilvae 163, 202.
Durvillea 378.
E
Ebenaceae 356, 410.
Echidnopsis cereiformis 56.
Echinocactus 366. 404.
Echium 349, 404.
— rubrum 339, 340.
- vulgare 106.
441
Ecliinocereus Blankii 56.
Echinospermum 122. 404.
Elaeis guineensis 357, 420.
Elodea canadensis 128, 296.
Elymus 391.
— giganteus 319.
Elyna 277, 409.
Eiynetum myosuroides 278.
Emersiherbosa 163, 285, 287,
Empetraceae 246, 410.
Emoetrum 246.
— nigrum 38, 248, 249, 290,
292, 335, 410.
— rubrum 249.
ncridaceae 246, 371, 374, 410.
ihedra 412.
jipogon 236, 420.
— aphyllus 24.
ca 58, 208, 245, 247, 343, 359,
374, 410.
— arboria 205.
— cinerea 335.
- tetralix 245.247, 248.
icaceae 51, 328, 410.
icifruticeta 163, 245.
' cilignosa 163, 245.
.'igeron canadensis 33, 128.
— ramosus 263.
'iocaulaceae 410.
'iocauron 286, 410.
“iopholum vagina turn 290, 296.
ritrich um nanum 326.
— villosum 300.
rrantia 163, 1-64.
Erythrina 199, 416.
Erythroxylaceae 410.
Erythroxylon coca 369, 370.
Eubrvosa 283. 296.
Eucalyptus 362, 371, 372, 419.
— diversicolor 208.
— marginata 208.
reclunca 208.
Euchlaena mexicana 394.
Eupatorium 254, 406.
Euterpe 186, 420.
— edulis 174.
Euphorbia 349, 411.
— acanthothamnos 207.
beau mi erar 56.
— t’.rukalli 86.
Euphorbiaceae 365, 411.
Euphrasia 270, 425.
Eurotia ceratoides 322. 340.
Exostema floribundum 120.
F
Fagaceae 327, 411.
Fag.turn alliostim ursini 217.
— as pern'osum odoratae 217.
— athyriosum filix fem'nae 217.
— bromosum nmosi 217.
44fi
Fagetumcaricosum albae 217.
— filicosum 219.
- - laurocerasosum 219.
— subalpinum 218, 219.
— trachystemonosum 219.
Fagopyrum esculentum 392.
— tataricum 393.
Fagus americana 219.
— orientalis 216, 217, 334.
— silvatica 216. 217. 334.
— taurica 216, 217, 334.
Falcaria vulgaris 128.
Festuca 266, 277, 339, 340.
— brevifolia 329.
— duriuscula 40.
— pratens's 269.
— rubra 119.
— sulcata 24, 40, 128,144,258,339.
— supina 283, 302,
Ficaria 98. 423.
— ranunculoides 98, 222.
- verna 215.
Ficus bengalensis 106.
— cordifolia 175.
- inaequalis 125.
— religiosa 90. 360.
— variegata 175.
Filipendula hexapetala 32, 136, 137,
157, 261. 339.
Fitzroya 376.
Fluviislvae 227.
Frankenia 113, 315.
Frankeniaceae 112, 411.
Fraxinus excelsior 219. 333.
— manshurica 225, 334.
— oxycarpa 334.
— rynchophylla 225, 334.
Frigidohumiliherbosa 274.
Frigorideserta 164, 170, 274, 302,
306. 320.
Fucus 378.
Fumaria officinalis 128.
Fumariaceae 411.
G
Gagea 282, 339, 416.
— erubescens 122.
— pusilia 262.
Galactodendron 370, 418.
Galanthus 133. 342, 401.
— nivalis 217.
Galeoldolon luteum 59, 95, 215.
Galium 366, 424.
- - verum 157, 262.
Genista 246, 247, 416.
— anglica 335.
— acanthoclada 207.
• - scoparia 203.
Gentiana 280, 336, 369, 412.
~ienfiana acaulis 279.
— bavarica subacaulis 321.
— cruciata 109.,
— tenella 283.
— vcrna 283.
Gentianaceae 411.
Geraniaceae 412.
G;r'(ilium collinum 271.
Gasneriaceae 328, 412.
Geum 122, 424.
jinkgo biloba 37, 337, 338, 458.
linkgoales 37, 337, 412.
llechoma hederacea 215.
jieditscliia caspia 47-
— sinensis 225.
— triacanthos 358.
Gleicheniaceae 328. 412.
Glycyrrhiza 339. 416.
Glyptostrobus 226.
Gnanhalium pusilium 281.
Gnetales 359, 412.
Gnidia pinifolia 211.
Ganocaryon pyriforme 90.
Goodeniaceae 371, 412.
Goodyera 236 , 420,
— repens 75.
Gossypium arboreum 396.
— barbadense 395.
— herbaceum 395.
— • hirsutum 83. 395.
— peruvianum 395.
Gramineae 5!, 328, 413.
Grevillea 371, 423.
Gunnera 376, 413.
— scabra 376.
Guttiferae 361, 413.
Gymnadenia conopea 75.
Gynerium 413.
Gypsophila altissima 108.
— paniculata 26, 32, 258.
H
Hakea 371, 423.
— saligna 210.
— suaveolens 86.
Halimodendron 315, 416.
Halocnemum strobilaceum 315.
Halorrhagidaceae 376, 413.
Halostachys caspia 315.
Haloxylon aphyllum 313, 340.
— persicum 315, 340.
Hamamelidaceae 414.
Hedera colchica 345.
— helix 344.
Helianthemum 343. 406.
Helianthus 263, 406.
— annuus 394.
Helichrysum 374, 406.
Helle jofus purpurascens 218.
del )biae 414.
depatica triloba 217.
Heracleum 426.
- - ponticum 272.
Herbosa 163.
Hevea brasiliensis 186, 370, 411.
Henrnia Penzigii 56.
Hiemiduriherbosa 249.
Hiemifruticeta 163, 197.
Hiemiligrtosa 168, 169, 197.
Hiemisilvae 163. 197, 357.
Hierochloa odorati 144,
— alpina 297.
HipDomarathrum crispum 261.
Hordeum distichum 391.
— polvstichum 391.
— sativum 391.
— secalium 340.
— spon taneum 391.
Humboldia 'Jaurifolia 125.
Humiliherbosa 283.
Hyacinthe!Io leucophaea 156, 340.
Hydnophvtum 125. 424,
Hydrangea 225.425
Hydrocharis morsus ranae 50.
Hvdrocharitaceae 414.
Hylocomium 301.
proliferum 162, 165, 296,
329.
Hymenophyllaceae 195, 414.
Hymenophyllum 220.
Hymmachne 254, 413.
Hypericum 413.
Hyphaene 253, 420.
thebaica311.
I
Ilex aquifolium 217, 220, 345, 402.
Impatiens parviflora 128, 129.
Imperata arundinacea 188.
Inula ensifolia 66. 82.
— germa nica 27.
— magnifica 272.
Iridaceae 414.
Iris 339, 414.
— aphylla 156,
— pumila31.
— pseudacorus 288.
Ischaemum 266, 413.
J
Jacksonia 36, 416.
Jasione 404.
Jasminum fruticans 45, 344.
Jonopis paniculata 125.
Juglandaceae 350. 414.
Jugians 213, 223. 224, 327, 352, 414.
— cinerea 353.
— manshurica 46, 225, 233.
— regia 233.
Julianiaceae 365, 414.
Juncaceae 415.
Juncus 142, 415.
443
Jun5penis 33 3, 407.
— barbadensis 370.
— depressa 242.
— excelsa 73, 74, 242, 341, 343.
— occidentals 352.
—- virginiana 370.
Jurinea arachnoidea 116, 258.
— cyanoides 116.
К
Klopstockia 86, 420.
lochia arenaria 113, 116.
Koeleria 262, 263. 266, 339, 353, 413.
— Delavignei 119.
— glauca 40. 86.
— gracilis 40, 256, 258, 261,
262, 339.
L
Lactuca scariol? 82.
Laguncularia racemosa 193.
Laminaria 378.
Laricetum dahuricae 238.
— pratosum 238.
— sibirtcae 238.
Larix americana 241, 351.
~ dahurica 233, 236, 333.
— europaea 333.
— laricina 292.
— sibirica 233, 236, 333.
Lathraea squamaria 44.
Lathyrus tuberosus 128.
Lauraceae 361, 415.
Laurifruticeta 163, 195.
Laurilignosa 163, 169, 193.
Laurisilvae 163, 193.
Lauras canariensis 195, 197, 349.
— nobilis 196, 415.
Lavandula 207, 415.
Lecanora esculenta 313.
Lecythidaceae 415.
Lecythis 370, 415.
Ledum 37, 38, 410.
— decumbens 297.
— palustre 165, 297.
Leguminosae 415.
Lemna minor 90.
Lemnaceae 416.
Leontice altaica 37.
Leontopodium alpinum 281.
Leontodon autumnalis 66.
Lessonia 378.
Leucadendron argenteum 374.
— Levisancus 211.
Leucanthemum vulgare 157.
Liatris 263.
Libocedrus 363, 376, 407.
Lichenosa 283, 296.
Ligustrum 225, 419.
Lignosa 163.
444
Liliaceae 416.
Lilium Szowitsianum 372.
Limosella aquatica 20.
Linaceae 416.
Linaria 343, 425.
— cretacea 21, 109.
— nevadensis 276.
Linosyris villosa 259.
Linum perenne 82.
Liriodendron 352, 417.
— tulipifera 34, 224.
-- cliinense 34, 225.
Litorideserta 164, 306.
Livistona chinensis 226.
Lobelia dortmanna 33, 44.
Lobeliaceae 417.
Loiseleuria procumbens 249, 410.
Lolium linicola 130.
Lonicera 225, 405.
— nigra 218.
Lupi nus 266, 416.
Luzula confusa 297, 303.
Lyallia 299, 405.
Lycopodiaceae 417.
Lycopodium inundatum'33,
Lysimachia nemorum 218.
Lythraceae 267. 417.
Lythrum 60, 417.
— salicaria 66, 288,
M
Maackia amurensis 337.
Macherium 370, 416.
Macrocystis 378.
Macrozamia 373, 428.
Magnolia kobus 225.
-- grandiflora 243.
Magnoliaceae 417.
Mahonia aquifolia 355.
- - nervosa 195.
Malva rotundifolia 81.
Malvaceae 417.
Mammilaria 366, 404.
Manicaria 186, 420.
Manihot dicliotoma 201.
Marattiaceae 328, 417.
Marcgraviaceae 356, 417.
Matricaria 406.
Medicago falcp ti 128.
Melaleuca 210, 362, 419.
Melampvrum pratense 290.
Melia 226, 418.
Meliaceae 417.
Meliea 266.
Menispermaceae 356, 418.
Mentha 415.
Mercurialis 411.
perennis 98.
Mesembrianthemum 316, 318, .359,
374, 401.
Bolusii 316.
Mesernhriantliemiim calcareum 316,
Microbiota 331, 407.
— decussa ta 338.
Mimosaceae 415.
Mimulus luteus 128.
Atobilideserta 164, 306.
Molinia coerulea 110.
Moraceae 357. 418.
Musa 362, 418.
Musaceae 418
Myoporaceae 371, 418.
Myoporum tuberculatum 210.
Myosotis 280, 336, 404.
— alpestris 283, 329.
— silvatica 156.
Myricn 335, 418.
— gale 44, 418.
Myricaceae 418.
Myristica 362.
Myristicaceae 328, 418.
.Myriophyllum 61, 70, 288, 414.
Myrmecodia 124, 125, 424.
Myrtaceae 246, 267. 356, 365, 418.
Myrtus communis 205, 206, 342,419.
N
Narcissus 401.
Nardus stricta 108, 270, 273, 329.
Narthecium 342, 416.
Nelumbium nuciferum 347, 348,419.
Nepentliaceae 328, 356, 361, 419.
Nepenthes 34, 362, 419.
— rafflesiana 361.
Nepeta ucrainica 258.
Nerium oleander 205, 342, 402.
Nicotiana 395.
— rustica 395.
— tabactim 395.
Nidularium Innocenti! 185.
Nipa 361, 420.
— fruticans 86, 192, 193.
Nothofagus 35, 226, 363,373, 376, 411.
— betuloides 195.
Nuphar 288, 419.
Nyctaginaceae 365, 419.
Nymphaea 288.
— alba 2S8, 419.
— lotos 347, 419.
Nymphaeaceae 419.
Nyssa biflora 244.
Nyssaceae 419.
О
Odontites lutea 259.
Odontospermum 313, 406.
Oenone flexuosa 295.
Oenothera biennis 128.
Olea europaea 203, 342.
Oleaceae 419.
Onagraceae 419.
Onobrychis arenaria 157, 262.
Onopordon 59. 407.
Opuntia 366, 404.
Orchidaceae 112, 419.
Orchis militaris 109.
Ornithogilum 399, 416.
Orobus albus 156.
Oryza sativa 396.
Osmuiida 220, 420.
— regulis 345.
Osmundaceae 420,
Ostrya 224, 403.
Oxalis acetosella 165, 235, 420.
Oxalidaceae 374, 420.
Oxyanthus hirsutus 120.
Oxycoccus palustris 38. 410.
Oxytropis 322, 340, 416.
P
Paliureturn australis 229.
Palmae 328, 356, 361, 420.
Paludisi Ivae 227.
Panax ginseng 337.
Pandanaceae 356. 361, 420.
Panicum 253, 254, 266. 413.
— miliaceum 392.
Papaver radicatum 323, 324.
— somniferum 83.
Papaveraceae 420.
Papilionaceae 416.
Paris quadrifolia 235.
Parkia africana 175.
Parkinsonia 199, 416.
Parrotia persica 47, 345.
Parthenocissus quinquefolia 220.
Paspalum 254, 266, 413.
Passifloraceae 365, 421.
Paxvlownia imoerialis 225,
Pedicularis 280, 425.
— comosa 339.
— hirsuta 297.
— sudetica 233.
— verticillata 283.
Pe’argonium 359, 374, 412.
Peltigera 301.
— aphtnsa 329.
Pennisetum 358, 359, 413.
Permentiera cereifera 183.
Petasites 142, 407.
— spurius 23, 25.
Phalaris arundinacea 269
Phellodendron amurense 46, 225, 226,
233, 424.
Phillyrea 206, 419.
Philodendron 180, Л02.
Phlomis 343, 415.
— pungens 258.
Ph or mi um tenax 363, 365, 416.
Phoenix d.ictylifera 311, 420.
• - Jubae 349, 420.
— silvestris 360.
Phragmites 286, 335, 413.
— communis 20, 288.
445
Phyllocladus 373, 407.
— rhomboidalis 374.
Pliyllodoce 328, 410.
— coerulea 248.
Phytelephas 186, 420.
— macrocarpa 370.
Phytolaccaceae 365, 421.
Picea 407.
— alba 351.
— canadensis 74.
— excelsa 31, 98, 218, 233, 236,
331, 351.
• - fennica 236, 331.
— Glehni 332.
— jezoensis 331.
— kamtchatkensis 332.
— koraiensis 332.
— nigra 351.
— obovata 31, 233, 236, 331, 351.
— orientalis 233, 311.
— Schrenkiana 233, 332.
— sitchensis 244, 354.
— tianschanica 332.
Piceeta 234.
— composita 234.
— hylocomiosa 234.
— nemorosa 234.
— polytricliosa 234.
— sphagnosa 234.
— uliginoso-herbosa 234.
Piceetum excelsae 165.
— fontinale 243.
— rivulare 243.
Pineta 236, 238.
— cladinosa 165, 236.
— nemorosa 165.
— silvestris hylocomiosa 165.
Pinetum myrtillosum 165.
— silvestris 165.
— vacciniosum 165.
Pinguicula 123, 292, 427.
Pinus 166, 407.
— Banksiana 241,351.
— canariensis 349.
— cembra ssp. sibirica 332.
— densiflora 76.
— eldarica 21, 332.
— halepensis 241, 341.
— Montesumae 534, 566.
— mughus 332.
— murreyana 241, 351.
— nigra 240.
— pallasiana 37, 241, 343.
— palustris 352.
— рейсе 343.
— pinaster 341, 342.
— pinea 341, 342.
— pithyusa 332.
— ponderosa 240.
— pumila 233. 242, 303, 331, 332.
— resinosa 241.
- regida 241, 352.
Pinus sibirica 233, 236.
— silvestris 166, 236, 35 L
— — f. pumila 291.
— Stankeviczi 21, 344.
— strobus 241, 352, 353.
— taeda 241, 352.
— uncinata 74, 240.
Piper 180, 226, 362, 421.
Piperaceae 361, 421.
Piroia 366, 421.
— rotundifolia 59.
Pirolaceae 421.
Pistacia lentiscus 206.-
— mutica 341, 343.
Platanaceae 421.
Platanus orientalis 342.
Pleurozium 162.
— Schreberi 162, 1C6.
Plumbaginaceae 112, 340, 42L
Pluv iifruticeta 163, 188.
Pluviilignosa 163, 169, 171.
Piuviisilvae 163, 171.
Poa 277, 328, 413.
— alpigena 283, 329.
— alpina 278, 283, 329.
— angustifolia 136, 137.
— annua 93.
• - arctica 283, 296, 329.
— attenuata 340.
— bulbosa 313.
— palustris 269.
— pratensis 23, 269.
Podocarpus 226, 231, 362, 363,
407.
— purdicana 374.
Podostemon olivaceus 295.
Podostenionaceae 294, 421.
Polemoniaceae 422.
Polemonium liumile 323.
Pollinia gryllus 262.
Polygonaceae 422.
Poly gona turn multiflorum 222
Polygonum aviculare 129.
— linicola 130.
— viviparum_278, 296.
Polypodiaceae 422.
Polypodium lineare 338.
— vulgare 21, 345.
Polytrichum 61, 234, 301.
— strictum 296, 329.
Populus balsamifera 351, 353.
— canadensis 353.
— tremula 226.
— tremuloides 351.
Portulacaceae 266, 422.
Posidonia ocaanica 294.
Potamogeton 20, 61, 422.
— lucens 288.
~ natans 288.
— perfoliatus 288.
Pot.imogetonaceae 422.
Potentiila 328, 424.
446
Potentilla alpestris 283.
— pratensis 262.
Poterium spinosum 207.
Pratoherbosa 268.
Primula 280, 282, 336, 423.
— acaulis 215.
— officinalis 156.
Primulaceae 328, 422.
Pringlea 299, 408.
Prosopis 199, 416.
— glandulosa 202.
Proteaceae 209, 359, 37], 372, 374,
376, 423.
Prunus fruticosa 41, 229.
— laurocerasus 196.
— spinosa 229.
Pseudotsuga 195, 240, 407.
— Douglasii 241, 354.
Psoralea floribunda 263.
Pteridium aquilinum 21, 363.
— esculentum 363.
Pterocarya flaxinifolia 345.
Ptilidium ciliare 296.
Pulmonaria officinalis 43, 44.
Pulsatilla 297, 423.
— patens 156, 158.
— pratensis 262.
Punica grana turn 345, 423.
Punicaceae 423.
Pyrethrum 407.
— Buschianum 272.
Q
Quercetum aegopodiosum 223.
- й caricosum piiosae 223.
— ‘ galeobdolosum 223.
— pubescentis 229.
— tilicsum 166.
fiuerctis 216, 411.
agrifolia 207.
— coccifera 206, 341.
_ densiflora 207.
— dentata 225.
— dumosa 208.
— Hartwisiana 334.
— iberica 334.
— ilex 205, 341, 342.
— macranthera 334.
— mongolica 225, 233.
petraea 44, 220, 245, 334.
— pontica 47 , 334.
— pubescens 220, 334.
— robur 215, 219.
— suber 205.
Quillaja saponaria 369
R
Ramischia secunda 59.
Ranunculaceae 51, 327, 423.
Ranunculus 70, 423.
— acetosellaefcrmis 276.
Ranunculus alpestris 281, 282.
— auricomus 215.
-7 borealis 283; 329.
— glac<a 1 is 328.
— nivalis 283, 302.
— pedatus 262.
— pigmaeus 283.
Rauoulia eximia 363, 364.
Ravenala 34, 360, 418.
— madagascariensis 57, 35?.
Reaumuria 315, 426.
Restionaceae 371, 374, 423.
Rhamnaceae 423.
Rhipsalis 186, 404.
Rhizophora 189, 191, 423.
— mangle 193.
— mucronata 180, 191.
- stylosa 190.
Rhizophoraceae 35, 189.
Rhododendron 225, 410.
— caucasicum 196.
— ferrugineum 197.
— hlrsututn 197.
— ponticum 197, 345.
— Smirnowii 47.
Rhuretum coriariae 229.
Rhus coriaria 45, 344.
Ribes 33, 34, 425.
Ricinus communis 83, 411.
Robinia 416.
—’ pseudacacia 353.
Rosaceae 327, 423.
Rosmarinus 206, 207, 415.
Rozites gangylophora 123.
Rubiaceae 368, 424.
Rubiis arcticus 335.
— chamaemorus 335.
Rudbeckia 353, 407.
Rumex 273. 422.
— acetosella 144.
Rupideserta 164, 306.
Ruscus 342, 345, 416.
— ponticus 207, 344.
Rutaceae 424.
S
Sabal 352, 420.
— Adansonii 244.
Saccharum spontaneum 188.
Sagittaria 401.
Salicaceae 33, 327, 424.
Salicetum 228.
— herbaceae 281.
Salicornia 406.
— herbacea 59, 112, 113, 369,
315-
Salix 328, 424.
— baicalensis 301, 328.
— giauca 297, 301, 328.
447
Salix herbacea 281, 300.
— lanata 301, 328.
— lapponum 301, 328.
— phylicifolia, 301.
— polaris 296, 300, 328.
— pulchra 297, 301, 328.
— reptans 328.
— reticulata 296. 299, 300, 328.
— triandra 142.
Salsola arbuscula 315.
— kali 113.
— laricifolia 315.
S.alsolaceae 340. 424.
Salvia 339, 415.
aethiopis 86.
— nemorosa 128.
— nutans 119, 258, 340.
— platensis 137, 155, 156, 261
339.
Sapindaceae 356, 424.
Sargassum 294. 378.
Sarracenia 291, 425.
Sarraceniaceae 425.
Satissurea 322.
Saxegothea 376, 407.
Saxideserta 164, 306.
Saxifraga 25. 277. 369, 425.
— tridaetylis 262.
Saxifragaceae 51, 425.
Scabiosa 409.
— caticasica 336.
— ochroleuca 44.
Schizandra chinensis 338.
Schizandraceae 417.
Scilla 416.
bifolia 217.
- - cerntia 151, 222.
- - nutans 215.
Scirpus 60, 409.
— lacustris 288.
— maritimus 113.
Scorzonera tau-saghiz 21.
Serophularia cretacea 109.
Scrophtilariaceae 328, 425.
Secaie 388.
- - cereale 390.
- - fragilis 390.
Sedum 86, 407.
Selaginella 177.
Sempervivum 59. 86, 349, 407.
Sempervirentiherbosa 163, 170, 268,
274, 282.
Senecio 266, 336, 366. 407.
— campester 156.
- - netrurensis 272.
— silvaticus 111.
— vulgaris 21.
Sequoia 37, 240, 407.
— gigantea 241, 354.
— sempervirens 194, 195, 231,
354.
Serra tula 339, 407.
— centauroides 82-
Seseli hyppomara thrum 340.
Sesleria 277, 413
— disticha 278.
Sibbaldia procumbens 281, 283.
Siccideserta 163, 170, 306.’
Silene 266, 339, 405.
— acaulis 277.
— cretacea 21, 109.
— densiflora 41, 116.
— parviflora 25, 41, 116.
Silphium 82. 353, 407.
Simarubaceae 425.
Sisymbrium Loesellii 129.
Sisyrinchium angustifolium 129.
Smilax 205, 416.
Solanaceae 267, 365, 425.
Solanum tuberosum 393.
Soldanella 98, 282. 423.
— alpina 97.
— pusilia 281.
Solerrantia 164.
Solidago 247, 353, 407.
— virga aurea 66.
Sonchus 349f 407.
— oleraceus 21.
Sonneratia 189, 191,“ 425.
— alba 115, 191.
Sonneratiaceae 189, 425.
Sophora japonica 225.
Soya hispida 83.
Spartium junceum 85.
Sphagniherbosa 163, 287, 289.
Sphagnum 24, 61. 165. 335.
- - fusctim 289.
— . Girgensohnii 329.
— medium 288.
— squarrosum 329.
Spinifex 318, 413.
Spiraea crenifolia 229.
— hipericifolia 229.
Sporobolus 358, 413.
Stachys recta 44, 340-
Stanhopea oculata 75.
Statice 113. 309, 421.
— Gmelini 113.
— tatarica 258, 260.
Stellaria 405.
— media 21, 93. 94, 95.
Stereocaulon alpinum 329.
Stipa 256, 263, 266, 413.
— capillata 119, 150, 258, 262.
339.
— ichu369.
— Joannis 256, 259, 261, 262, 339.
— Lessingiana 32, 258, 259, 261,
339.
— orientalis 340.
— rubens 257, 261, 339, 340.
— spartea 262.
448
Stipa stenophylla 32, 256 259, 261
339.
- ucrainica 259, 261, 339.
Struthiopteris 60, 422.
Suaeda maritima 112, 113.
— splendens 138.
Submersiherbosa 163, 287, 293.
Succisa praemorsa 66.
Symphytum tauricum 36.
Syringa 225, 419.
— amtirensis 337.
Syringetum vulgaris 229.
T
Tamaricaceae 339, 340, 425.
Tamarix 113, 309, 315, 426.
Tamus 409.
— communis 205.
Taraxacum 407.
— serotinum 259.
— vulgare 21.
Taxodium 37. 350. 407.
— distichum 194, 243, 327,
352, 368.
— mucronatum 366.
Taxus 194, 407.
— baccata 217, 218, 245.
Tectona grandis 198, 361, 362, 427.
Telekia speciosa 272, 336.
Teucrium 343, 415.
Terriherbosa 163, 165.
Theaassamica 397.
— sinensis 397.
Theaceae 426.
Thiaspi arvense 128.
Theobroma cacao 298.
Thuja 240, 407.
— gigautea 195, 354.
— heterophylla 194.
— occidentalis 194, 241.
Thyme laeaceae 426.
Thymus 206, 207, 415.
— Marschallianus 40.
( — odoratissimus 40.
Thyphaceae 426.
Tilia amurensis 225, 334.
— argentea 334.
— cordata 233, 334.
— grandiflora'334.
- manshurica 225, 334.
Tiliaceae 426.
Tillandsia 214, 243, 404.
— usneoides 201, 352.
Torreya 33, 407.
Trachycarpus excelsa 226.
Trachystemon orientalis. 219.
Tragopogon orientalis 156.
Tribulus terrestris 128.
Trichophorum alpinum 38.
Trientalis europaea 235.
Trifolium ambigutim 271.
canescens 271.
- - montanum 137, 157, 340.
- fragiferum 91.
Trinia Henningii 258.
Triodia 318, 413.
Triplaris 186 , 422.
Trisetum 277.
— flavescens 271, 279.
Tristicha hypnoides 295.
Triticum 387.
— aegilopoides 388, 389.
compactum 387.
dicoccoides 383, 389.
dicoccum 388.
durum 388.
monococcum 388.
— spontaneum 388.
— turgidum 388.
— vulgare 388.
Trollius europaeus 283.
Tropaeolaceae 365, 426.
Tsuga 240, 350, 407.
canadensis 352.
heterophylla 195, 241.
— mertensiana 241, 354.
— occidentalis 241.
Tulipa 339, 416.
— Sehrenkii 31.
Typha 426.
— angustifolia 286.
U
Ulex 335, 416.
— Gallii 248.
Ulmaceae 426.
Ulmus 334, 427.
— foliacea 334.
— laevis 334.
scabra 333, 334.
— suberosa 334.
Umbelliferae 327, 426.
Urtica cannabina 129.
— dioica 21.
- urens 21.
Urticaceae 112. 427.
Utricularia 123, 427.
Utriculariaceae 427.
V
Vaccaria parviflora 129.
Vaccinium 249, 328. 410.
— myrtillus 166, 410.
— uliginosum 296, 297.
vitis idaea 38, 296.
Valeriana 336, 366 . 427.
— alliariaefolia 272.
— capitata 329.
Valerianaceae 427.
Vallisneria sniralis 91.
449
Vanilla 180, 201, 420.
— planifolia 367.
Veratnim lobelianum 273, 283.
— nigrum 157.
Verbascum phoeniceum 44.
Verbena chamaedryfolia 266.
Verbenaceae 189, 427.
Veronia 254.
Veronica 339, 425.
— alpina 281.
— incana 136.
— montana 218.
verna 262.
Vinca herbacea 262.
Vicia 266, 416.
— pisiformis 45.
Victoria regia 186, 419.
Viola 336, 3G9, 427.
— biflora 283, 329.
collina 45.
— nevadensis 276.
rupestris 136, 137.
— tricolor 93, 94.
Violactae 427.
Vitaceae 427.
Vitex agnus castus 342.
Vitis 81, 220, 427.
— amurensis 338.
W
Welwitschia 412.
- mirabilis 317, 358, 359,
Winteraceae 428.
X
Xanthorrhoea 254, 255, 318, 373, 416.
Xylocarpus 190, 418.
— granatum 190.
Xyridaceae 365, 428.
Y
Yucca 354, 416.
— Whippei 367.
Z
Zamia floridana 368.
Zamiaceae 34, 328, 428.
Zea mays 393.
Zelkova crenata 345.
Zingiberaceae 176, 361, 428.
Zostera 293, 422.
— marina 294.
Zygophyllaceae 339, 428.
СОДЕРЖАНИЕ
Стр.
Предисловие к первому изданию...................................... 3
Предисловие ко второму изданию..................................... 5
Введение .......................................................... 6
Краткий очерк истории фитогеографии, экологии
и фитоценологи и .........................................• . . . 10
ОТДЕЛ I
Учение об ареале
Ареал .............................................................20
Размеры ареала. Растения космополиты. Размещение индивидуумов
по площади ареала. Методические указания.
Типы ареалов ......................................................24
Основные типы ареалов. Причины различных конфигураций и границ
ареалов. Устойчивость ареалов.
Формирование ареалов............................................. 29
Миграции. Центры ареалов.
Разъединение (дизъюнкция) ареалов..................................32
Причины разьединении. Примеры разъединений. Реликтовые ареалы
и реликты. Реликты и их возраст.
Явление эндемизма................................................; 38
Эндемизм и эндемические виды (эндемики). Островные флоры. Гор-
ные страны и эндемизм. Эндемизм прогрессивный и викарирующие
виды.
Элементы флоры. ..............................~....................41
Главнейшие элементы флоры СССР. Элементы флоры Кавказа.
Число видов в различных частях земной поверхности .................47
Закономерности в распределении видов. Соотношения систе.матических .
единиц, флористические спектры и пропорции. Родовой коэфициент.
ОТДЕЛ II
Учение о взаймосвязях между растениями н средой (экология)
Изменчивость растений .............................................53
Жизненные форды ...................................................55
Системы жизненных форм. Вопросы генезиса жизненных форм.
Биотипы и экотипы..................................................64
Среда растений. ..................'................................67
Основные факторы среды.
Климатические факторы..............................................70
1. В о з д у х. .................................................. —
Кислород. Углекислый газ. Примеси воздуха. Движение воздуха.
Положительное действие ветра.
2. С в е т. .......................................................7о
Солнечные и теневые растения. Потребность в свете у различных расте-
ний (Liclitgenuss). Направляющее действие света. Свет как ботанико-геогра-
фический фактор.
3. В о д а........................................................83
Ксерофиты и их признаки. Гигрофиты и их признаки. Гидрофиты и
их признаки. Вода к распространение растений.
29* 451
Ci p.
4. Ten ло. . .......................................................gz
Классификация климатов. Три кардинальные точки. Морозостойкость.
Система жизненных форм Раункиэра. Фенологические явления. Яро-
визация. Акклиматизация и натурализация.
Эдафические факторы. ..................................................
Растения-индикаторы. Кальцифильные растения. Реакция среды.
Солончаковые растения - галофиты. Физические свойства почв.
Псаммофиты. Литофиты. Хасмофиты. Структура почв.
Орографические факторы..............................................П7
Влияние рельефа. Экспозиция склонов.
Биотические фактсры. ...............................................121
Растения и животный мир. Влияние растений друг на друга. Синузии.
Влияние человека. ..................................................127
Синантропные растения. Сознательное обогащение флоры человеком.
Человек и создание новых растений.
Исторические факторы................................................132
Совокупное влияние факторов . . ..............................134
Замещение факторов.
ОТДЕЛ Ш
Учение о фитоценозах
Конкуренция между растениями .......................................13®
Закономерные сложившиеся сочетания растений— фитоценозы.............138
Основные проблемы при изучении фитоценозов.
Формирование фитоценозов............................................140
Сукцессии.
Морфология фитоцеиозов..............................................145
Флористический состав. Ярусность. Обилие. Методические указания
к учету обилия. Покрытие. Жизненность.
Подвижность фитоценозов.............................................155
Фенологические смены. Смены в связи с метеорологическими условия-
ми. Сукцессии, их различные типы.
Устойчивость фитоценозов............................................16®
Систематика фитоценозов. ...........................................161
Ассоциации. Основные таксономические единицы. Классификация
Брокман-Ероша и Рюбеля. Пример классификации для лесов. Соста-
вление названий социаций и ассоциаций.
Фнтоцеиозы н история страны.........................................166
ОТДЕЛ IV
Основные типы растительного покрова
Схема идеального континента. .......................................168
I. Класс формаций — влажнотропические древесные н кустарниковые фи-
тоценсзы (Рluviilignosa).......................................171
i. Группа формаций — влажнотропические леса (Pluviisilvae). . . —
Общая характеристика. Фито це но логи чес кая структура. Лианы. Эпи-
фиты. Вечиозелёность тропического леса и периодичность роста. Срав-
нительная бедность цветами тропических лесов. Каулифлория. Крат-
кий обзор тропических лесов в разных странах. Субтропические и уме-
ренно-субтропические леса. Влажнотропический лес и человек.
2. Группа формаций —влажнотропические кустарниковые фитоце-
нозы (Pinviifruticeta) или мангровая растительность. ..........18*
Общая характеристика. Ходульные корни. Дыхательные корни. Ман-
гровые растения —галофиты.
II. Класс формаций — древесные и кустарниковые фитоцеиоэы лаврового
типа (Laurilignosa). ...............................................
1. Группа формаций — лавровые леса (Laurisii/ае).
452
Стр.
Общая характеристика. Двудольные «лавровые* леса. «Лавровые*
леса из хвойных деревьев.
2. Группа формаций— кустарниковые фитоценозы лаврового типа
(Laurjfruticeta). ..............................................'. . . 195
Отдельные примеры «лавровых* кустарниковых фитоценозов.
III. Класс формаций — знмнезелсные древесные и кустарниковые фнто-
ценозы (Hjemilignosa)..................................................... ЮТ
Муссоновые леса. Саванновые леса. Колючие ксерофильные леса и ко-
лючие кустарниковые фитоценозы. Краткий обзор ксерофильных ле-
сов в разных странах.
IV. Класс формаций —жестколистные древесные и кустарниковые фито-
ценозы (Durilignosa)..............................................202
Обща^ характеристика. Жестколистные формации в Средиземноморье.
Обзор жестколистных формаций в других Лранах. Жестколистные
леса и человек.
V. Класс формаций — летнезеленые древесные и кустарниковые фитоце-
нозы (Aes til ignosa).........................................212
1. Группа формаций—летнезелёные леса (Aestisil vae)...........213
Общая характеристика. Буковые леса. Дубовые леса. Широко-
лиственные леса востока Азии. Широколиственные леса южного
полушария. Мелколиственные леса (берёзовые и осиновые).
Леса пойменные и болотистые.
2. Группа формаций—летнезелёные кустарниковые фитоценозы (Aes-
tifrnticeta)....................................................228
Шибляк. Дерезняки. Летнезелёные кустарниковые фитоценозы
тундровой зоны. Летнезелёные леса и человек.
VI. Класс формаций — игольчатохвойиые, древесные и кустарниковые
фитоценозы (Aciculilignosa). ........................................230
1. Группа формаций — нгольчатохвойные леса — Acictilisilvae.. . . —
Общая характеристика. Леса еловые. Леса сосновые. Еловые и
сосновые леса и их взаимоотношения. Хвойные леса Центральной
и Восточной Сибири. Обобщенная схема типов хвойных лесов
в СССР. Хвойные леса Западной Европы. Хвойные леса Север-
ной Америки. Средиземноморские хвойные леса.
2. Группа формаций — хвойные кустарниковые фитоценозы (Aciculi-
fruticeta)......................................................242
Леса пойменные и болотистые (хвойные). Хвойные леса и человек.
VII. Класс формаций — деревянистые фнтоценозы с листьями верескового
типа (Ericilignosa)............................................... 245
Общая характеристика. Приатлантнческие верещатники. Горно-аль-
пийские и арктические верещатники. Верещатники внеевропей-
ских стран.
VIII. Класс формаций — травянистые зимнезелёиые ксерофильные фито-
ценозы (Hiemiduriherbosa) Саванны...................................249
Общая характеристика. Африканские саванны. Южноамериканские
Саванны. Австралийские саванны. Саванны (также саванновые
леса) и человек.
IX. Класс формаций — травянистые летиезелёные ксерофильные фитоце-
иозы — степи........................................................255
(Aestiduriherbosa).
Общая характеристика. Степи СССР. Венгерские пушты. Северо-аме-
риканские прерии. Взаимоотношения степи и леса. Пампасы Южной
Америки. Южноафриканские степи. Степи и человек.
X. Класс формаций — травянистые мезофильные фитоценозы с более или
менее высоким травостоем — луга (Pratoherbosa)......................268
452
Луга заливные. Луга суходольные или материковые. Луга субальпий-
ские. Высокотравие. Луга и человек.
XI. Класс формаций — травянистые альпийско-тундровые фитоценозы с
низким травостоем (Frigidohumiliherbosa)..............................271
Альпийские луга. Альпийские лужайки. Снеговые ложбинки. Тун-
дровые луга (луговинные тундры). Дерновииные тундры. Альпийские
и тундровые луга и человек.
XII. Класс формаций — травянистые гигрофильные фнтоценозы — травя-
ные болота (Emersiherbosa). ..........................................285
Общая характеристика. Низинные травяные болота. Сукцессии на боло-
тах.
XIII. Класс формаций — сфагновые олиготрофные фнтоценозы — сфаг-
к новые (верховые) болота (Sphagtiiherbosa)......................... 289
Общая характеристика. Основные типы сфагновых болот. Травяные
и сфагновые болота и человек.
XIV н XV. Классы формаций — фнтоценозы водных растений, погружён-
ных в воду, плавающих на поверхности воды или взвешенных в воде
(Submersiherbosa и Aquerrantia)...................................299
Бентос. Плейстон. Планктон. Водная растительность и человек.
XVI н XVII. Классы формаций — моховые и лишайниковые тундровые
фнтоценозы с господством зелёных мхов н лишайников (Eubryosa
и Lichenosa).................................................... 296
Моховые тундры, Моховые тундры с кустарничковым ярусом. Лишай-
никовые тундры. Краткая характеристика тундров'й золы. Жизнен-
ные формы тундр. Растительность тундр СССР. Взаимоотношения
тундры и леса. Тундры и человек.
Пустыни (Deserta)................/....................................306
XVIII. Класс формаций — фнтоценозы с разреженной раститель-
ностью, обусловленной сухим и тёплым климатом — сухие пустыни
(Siccideserta).........................................___...........
Географическое распределение сухих пустынь. Жизненные формы
пустынь. Различные типы пустынь. Сахара. Аравийская пустыня. Сред-
неазиатские пустыни. - Центральноазиатские пустыни. Южноафрикан-
ские и американские пустыни. Австралийские пустыни. Пустыни
и человек.
XiX- Класс формаций — фнтоценозы с разреженной растительностью, об-
условленной холодным климатом - холодные пустыни (Frigorideserta). 320
Высокогорные холодные пустыни. Арктические холодные пустыни.
Хслсдиые пустыни и человек.
ОТДЕЛ V
Флористические области и подобласти
I. Голарктическая область..........-..................................327
Общая характеристика. Арктическая подобласть. Ев рос и бирс кая под-
область. Китаиско-япоиская подобласть. П о нти чес ко-цен траль но-
азиатская подобласть. Средиземноморская подобласть. Североафрикан-
ско-индпнекая пустынная подобласть. Макэронезпйская переходная
подобласть. Североамериканская атлантическая подобласть. Под-
область североамериканских степей (прерий). Североамериканская па-
цифическая подобласть.
JI. Палеотпопнческая область..........................................355
Общая характеристика. Индоафриканская подобласть. Малезийская
подобласть. Новозеландская подобласть. Подобласть Сандвичевы
(Гавайских) островов. Подобласть африканских островов.
Л. Неотропнческая область..........................................
Общая характеристика, Центральноамериканская подобласть. Тропи-
ческая подобласть. По ' асть Анд.
IV. Австралийская область
45
Стр.
V. капская ооласть.................................................. 374
уЪ Антарктическая область............................................375
VII- Океанические области............................................378
Приложение т. Культурные растения и их происхождение.................379
Культурные и культивируемые растения. Возраст культурных расте-
ний. Основные очаги происхождения культурных растений. Проис-
хождение культурных растений.
I. Некоторые культурные растения умеренной полосы....................387
Пшеница. Рожь. Ячмень. Овес. Просо обыкновенное. Гречихл. Кар-
тофель.
II. Некоторые более южные культуры...................................393
Кукуруза (маис). Подсолнечник. Табак.
III. Некоторые тропические н субтропические культуры.................395
Хлопчатник- Рис. Чайный куст. Кофейное дерево. Шоколадное дерево.
Хинное дерево.
Приложение 2. Систематический справочник (Краткие характеристики
семейств цветковых растений, упомянутых в книге)...................401
Основная литература. ............................................429
Указатель русских терминов ..................................... 431
Указатель латинских названий и видов, семейств и флтоценозов) 437
Карта растительности СССР.
Карта растительности земного шара (по Брокман-Ерошу)
Карта растительности зелшого шара (по Б. Сов. атласу).