/

























































































































































































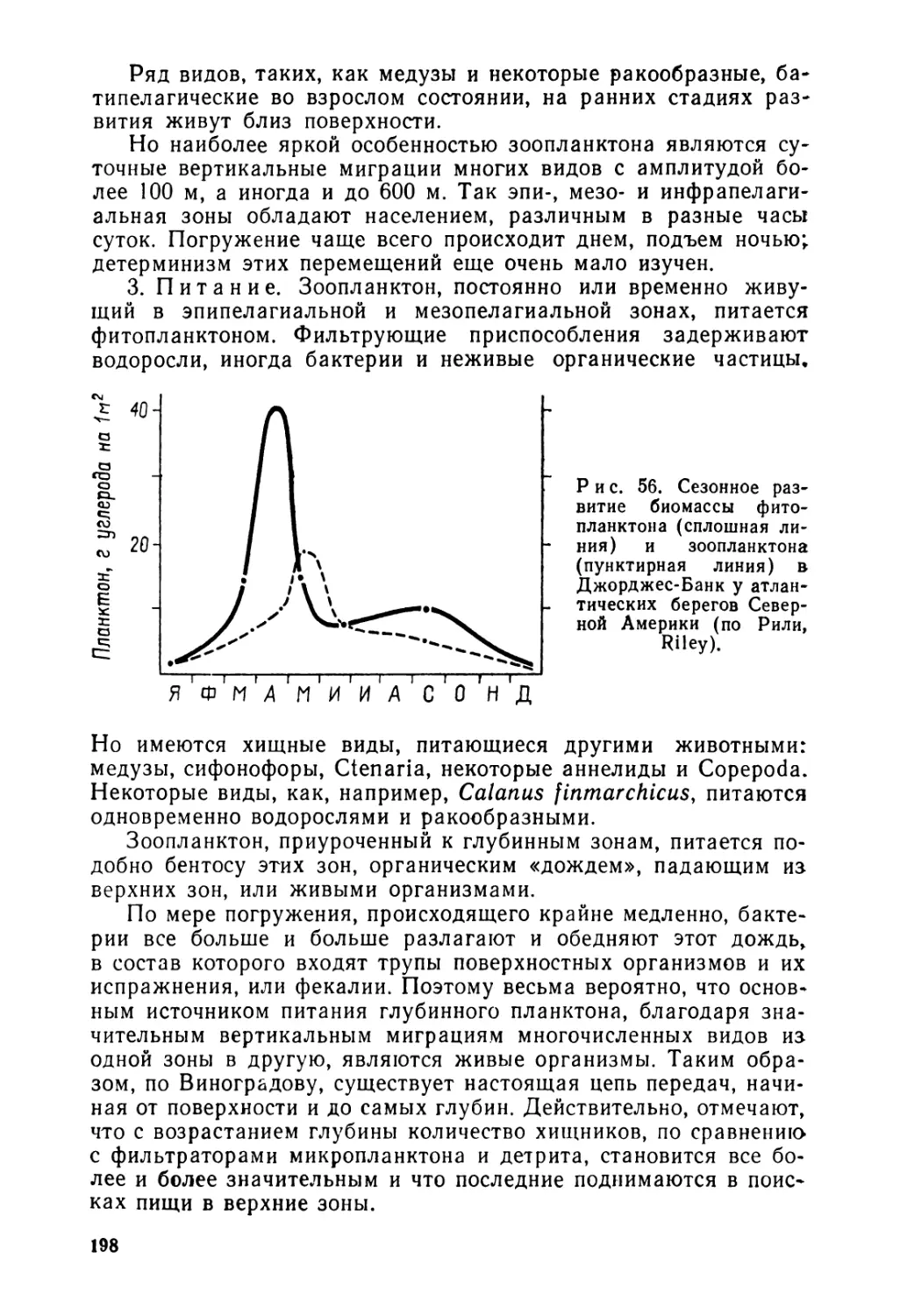

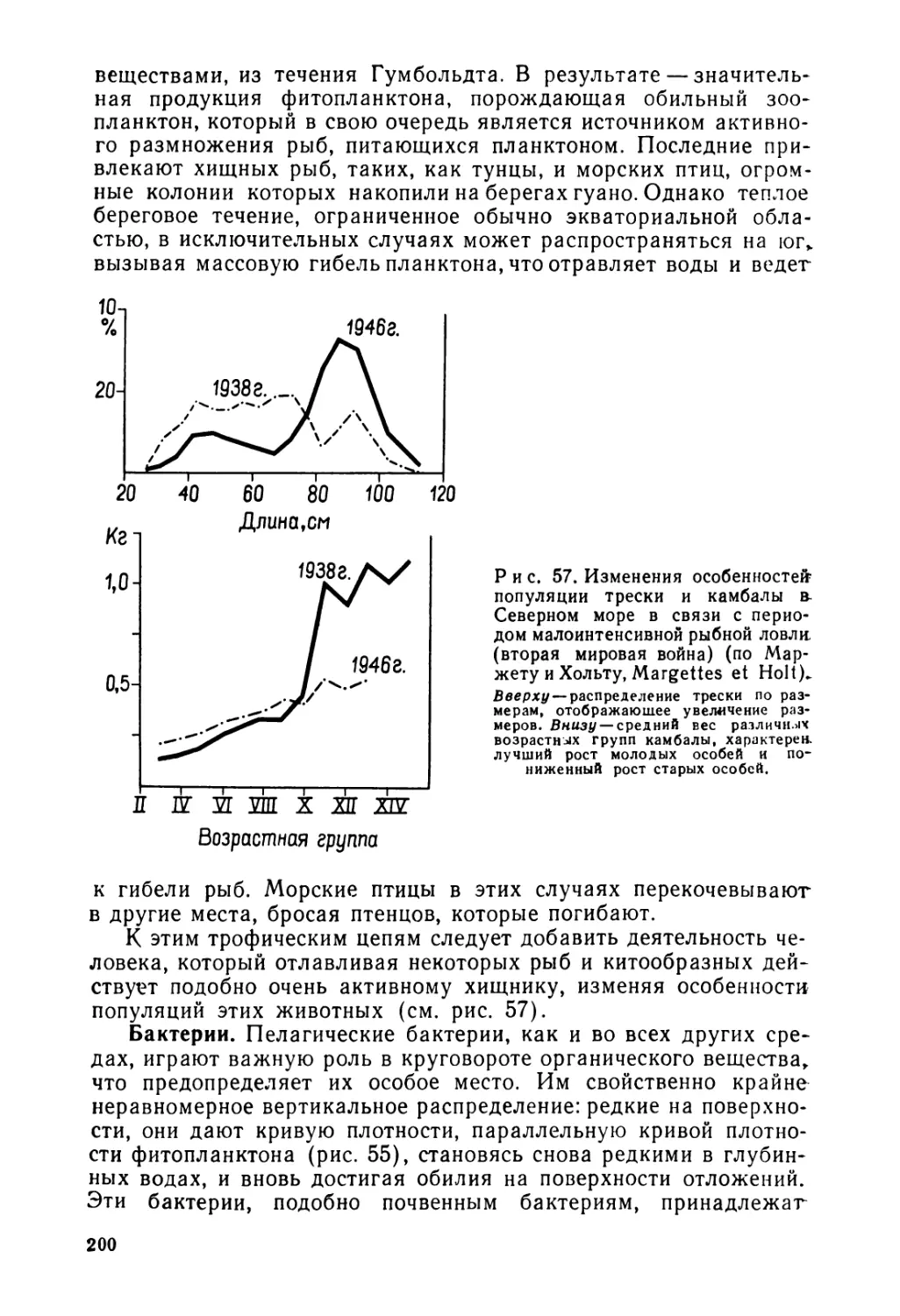





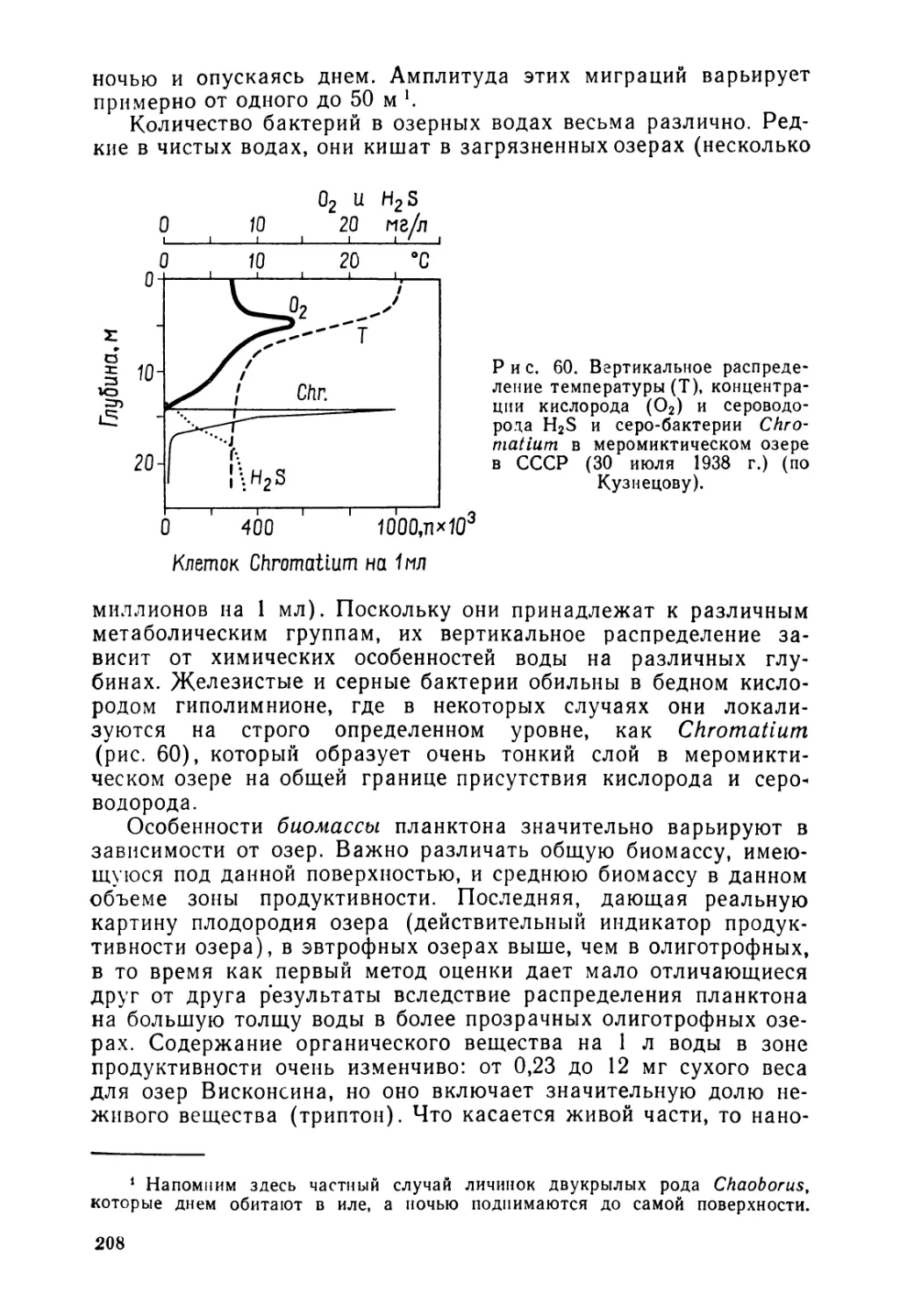






































































































Текст
...
ж
ОСНОВЫ
»
' "'’i
Ж
Ж
4;
ip.
> '. > illJJ'.;. ; .. jIH'. ' '.’ 'f! • ;1 !' I *. / .1' ?('
БИО
| I 1-н*гН<м»ма|й*а<мминин«шн«1И«тгн«>гм<1кки«(«шнн«1»ммн»(«<*жнм<1Ш<м«<ъапа»1«>ч«««<нн»
ГЕО
>ж*>минМмтнм<*нм«<ммм«и»«(Нмгн«^нж*<ч^«мпмйм»««*1«к««|Пнмн«ммкпп^и«ытчм№
ГРАФИН
;Ж'
til
Si;
Ssl
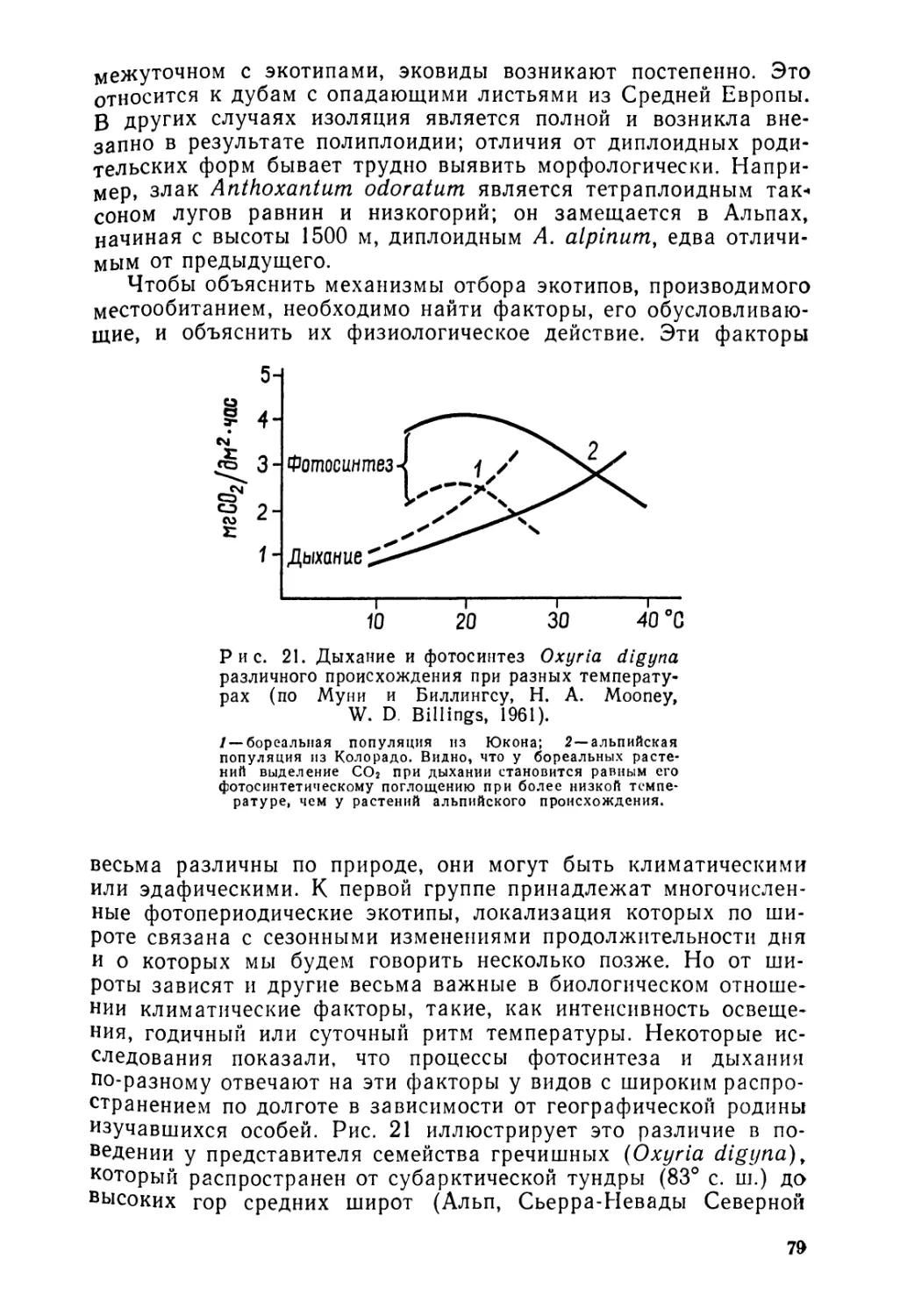
G. Lemee
Precis de biogeographie
Paris 1967
Ж. Леме
Основы биогеографии
Сокращенный перевод с французского Л, Г. Вольфензон, В. А. Потоцкой, Е. Г Мяло и Г А. Михайловой Редакция и послесловие профессора А. Г Воронова
Издательство „Прогресс*4, Москва 1976
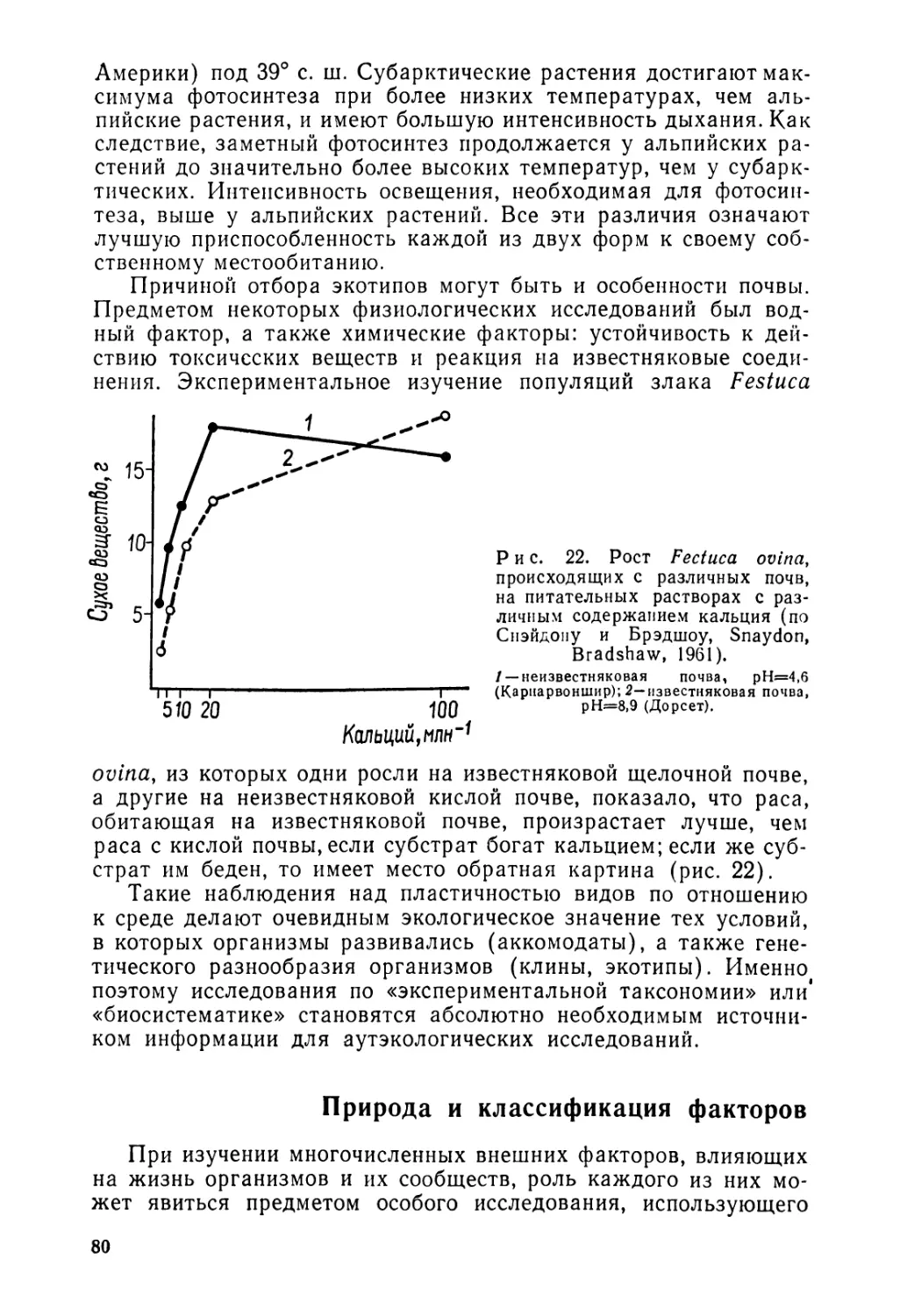
Рациональное использование территории нашей планеты, ее растительного и животного мира, а также остро стоящая проблема охраны природы обусловили быстрое развитие сравнительно молодой науки — биогеографии. Ее предмет — законы распределения, сочетания и внутренней структуры сообществ растений, животных и микроорганизмов.
Книга французского эколога профессора Ж. Леме представляет собой серьезное систематизированное изложение основных проблем биогеографии и является учебным пособием. В ней дана общая картина географического распределения живых организмов и показаны его причины; выявлена роль различных факторов среды в жизни организмов; раскрыты процессы формирования локализованных сообществ растений, животных и микроорганизмов; обрисованы крупные биогеографические формации земного шара.
Книга содержит богатый фактический и иллюстративный материал и может быть использована, при критической ее оценке, в качестве пособия студентами, преподавателями и научными работниками.
Редакция литературы по географии
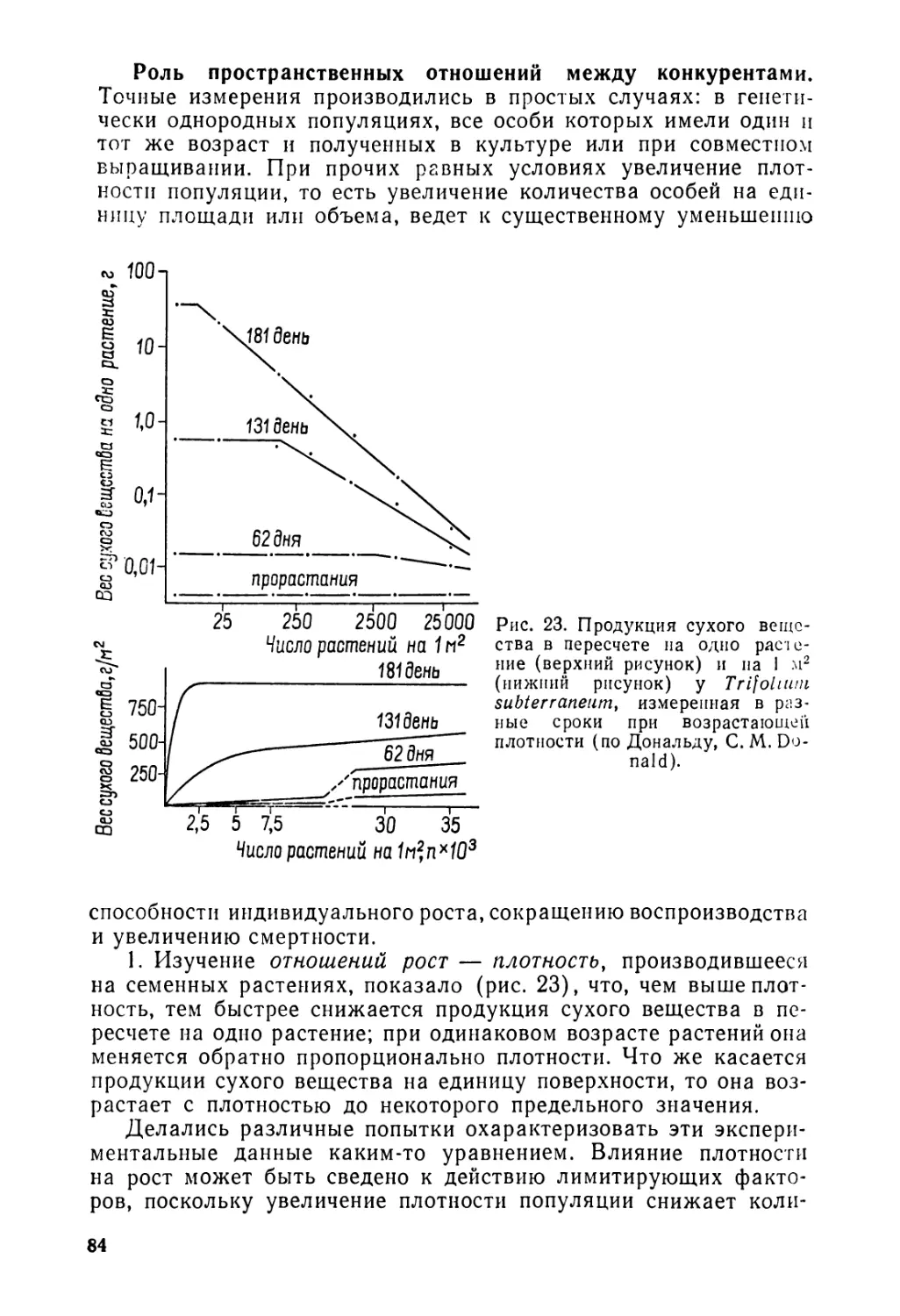
© Перевод на русский язык «Прогресс», 1976
п 20901—274
Л 006 (01)—76
45—75
Предисловие автора
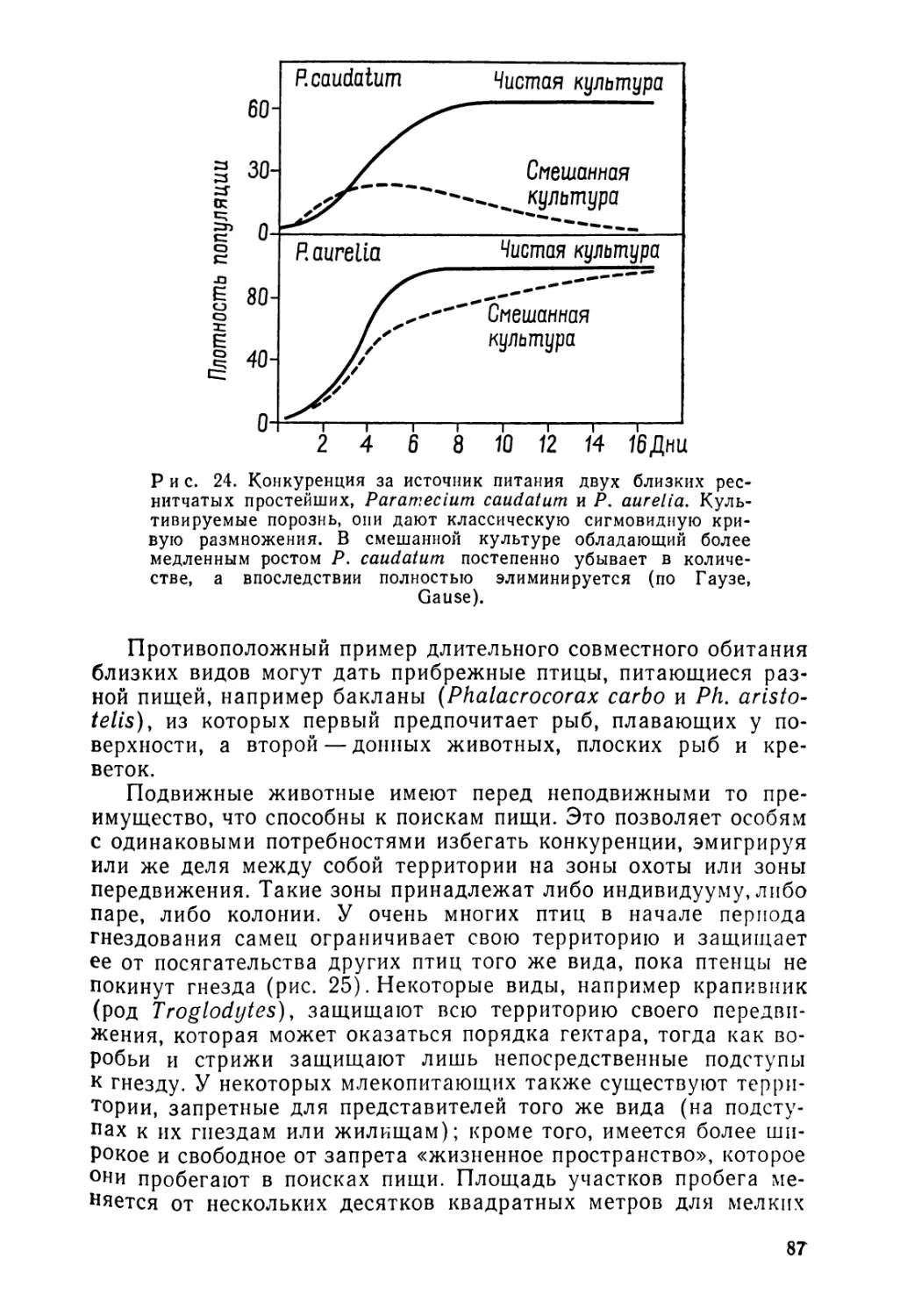
Публикация основ биогеографии вслед за геоморфологией, •климатологией, почвоведением и гидрологией в одной серии вполне логична. Будучи наукой синтетической, биогеография вынуждена обращаться к этим разнообразным дисциплинам, к их методике и достижениям, представляющим биологический интерес. Такая последовательность изданий позволяет избежать слишком подробной характеристики особенностей окружающей среды, отсылая читателя за необходимой информацией к соответствующим трудам. Именно благодаря им мы сумели ограничить обширный материал биогеографии размерами данного издания.
Как и предыдущие публикации, книга адресована студентам, специализирующимся в географических и естественных науках, а также тем исследователям, которые нуждаются в информации в области биогеографии. В результате мы вынуждены были опустить технические детали и пространные описания, но зато выбрать наиболее выразительные примеры и иллюстрации, а также снабдить книгу перечнем трудов общего характера и новых обзорных работ, в которых широко цитируются публикации исследователей. В столь сжатом объеме невозможно конечно представить всю совокупность поставленных задач и тем более осветить всю массу фактов, добытых в области распределения и экологии всех групп животных и растений — настолько велико их разнообразие. Нашей целью было в большей степени объяснить, чем научить. Поэтому, к сожалению, мы были вынужденв! сокра^ тить изложение некоторых аспектов биогеографии. Но, памятуя о том, что имеем дело с учебным пособием, мы постарались быть максимально объективными в области, где сталкиваются различные системы и концепции.
Следуя методу, использованному в некоторых трудах по биогеографии, выходящих за рубежом, мы рассматривали географию животных и растений в совокупности. Совместно обитающие организмы находятся в одинаковых внешних условиях и тесно взаимосвязаны. Многие особенности их географического и стадиального распределения вызваны общими причинами, в связи с этим для иллюстрации какого-либо факта мы имели возможность приводить рядом примеры из животного и растительного мира, чтобы подчеркнуть таким образом единство, лежащее в основе органического мира.
Введение
Биогеография — наука о географическом распределении живых организмов и причинах его изменения — словом, обо всех географических аспектах биологии.
Это биологическая наука, так как именно живые организмы составляют предмет ее изучения. Поэтому биогеографу необходимо иметь биологическое образование, в частности, он должен разбираться в физиологических аспектах поведения, законах развития, механизмах воспроизведения и наследственности живых организмов.
Но одновременно биогеография и географическая наука, так как она стремится установить связи между растительным и животным миром, с одной стороны, и географическими факторами— климатом, геоморфологией, почвами, деятельностью человека— с другой, чтобы получить синтетическую картину, характеризующую особенности земной поверхности. Для географа познание живой части ландшафта занимает первое по важности место в этом комплексе, так как, будучи связанной с другими элементами тесными взаимными отношениями, она выступает в качестве чувствительного индикатора особенностей географической среды.
Таким образом, биогеография занимается исследованием сложной области, в которой различают взаимозависимые дополняющие друг друга направления:
1) изучение географических ареалов таксономических единиц— видов, родов, семейств и т. д., их происхождения и изменения, а также флористических и фаунистических территорий, которые ими определяются, — составляет предмет хорологии;
2) изучение сообществ организмов, которые рассматриваются с точки зрения их организации, таксономического состава, динамики, географического распределения, — предмет биоценологии;
3) изучение связей организмов и их сообществ с окружающей средой — предмет экологии.
Происхождение этих трех главных направлений биогеографии различно. Хорология появилась тогда, когда натуралисты впервые занялись инвентаризацией живых форм, населяющих различные районы земной поверхности, стремясь одновременно уточнить границы их распространения. Предшественниками био-Ценологов были натуралисты-путешественники, которые пыта
7
лись определять и классифицировать важнейшие особенности растительности применительно к климату. Первыми экологами были великие агрономы XIX в., которые в поисках способов увеличения продуктивности культур пытались определить влияние, оказываемое на нее составом почвы и особенностями климата.
Но, несмотря на свое давнее происхождение, биоценология и экология развились лишь в последнее время. В значительной мере это объясняется экономической необходимостью: рациональное использование территории, улучшение качества сельскохозяйственной продукции, защита живых существ от истребления, оздоровление среды в связи с урбанизацией и индустриализацией— все это в значительной степени связано с экологией и биоценологией. Эти науки обладают теперь собственной методикой, что позволяет открывать законы, регулирующие отношения живых существ между собой и физической средой. Они завоевывают права гражданства в университетском образовании, у них есть свои периодические издания, ассоциации, проводятся международные конференции.
Часть первая Географическое распределение живых организмов и его причины: хорология
Каждый вид занимает сейчас па земной поверхности область, границы который определены противодействием, оказываемым каким-либо неблагоприятным фактором среды способности этого вила к распространению. Способность к распространению и лимитирующие условия не одинаковы для разных видов, вследствие чего их ареалы различаются по облику и протяженности. По тем же причинам генетически различные формы, из которых складывается вид, такие, как подвиды или разновидности, занимают несходные участки. Установление географических ареалов этих систематических единиц (таксонов) играет важнейшую роль в познании их происхождения, истории, экологии, равно как и эволюции палеогеографических условий. То же относится к таксономическим единицам более высокого ранга — таким, как роды или семейства, состоящие из более или менее близкородственных видов; ценные сведения об их эволюции можно получить, изучая их современные ареалы.
Мы займемся прежде всего средствами расселения организмов, точнее, их потенциальными возможностями к распространению. Далее будут описаны современные типы географических ареалов различных таксономических единиц. Затем специальному исследованию подвергнутся палеогеографические факторы, влияющие на размещение организмов в настоящее время. Экологические факторы будут рассмотрены во второй части книги. Наконец, мы дадим краткое описание больших биогеографических территорий, определяемых совпадением ареалов многочисленных таксонов.
Глава 1
Средства распространения организмов
Способность вида к распространению зависит от двух факторов: от быстроты размножения и способов распространения.
Способность к размножению
Различия в плодовитости разных видов колоссальны. У одноклеточных организмов скорость деления клеток велика. Промежуток между последовательными делениями может сокращаться до нескольких минут. Они представляют пример экспоненциально ускоряющегося (пока не вмешается внешний неблагоприятный фактор) размножения.
Грибы размножаются в основном спорами, порождаемыми обычно в избыточном количестве. После прорастания споры образуют мицелий, способный в свою очередь за несколько дней произвести новую массу спор. Подсчитано, что гриб Sclerospora, паразитирующий на кукурузе, может в течение ряда месяцев порождать около 6 млрд, спор на одно растение в день.
Водоросли также обладают замечательными способностями к размножению. Например, некоторые сине-зеленые свободно плавающие водоросли, которые летом вегетативно размножаются на поверхности озер, придают воде вид сине-зеленого пюре; это явление известно под названием «цветения воды».
Мохообразные и сосудистые споровые (Cryptogamae) могут производить большое количество спор; например, для плауна булавовидного (Lycopodium clavatum) и пантропического папоротника (Trichomanes radicans) установлено, что каждый их стебель может производить почти 30 млн. спор в год.
У семенных (Phanerogamae) органы распространения имеют крайне разнообразную природу: семена, плоды, соплодия, вегетативные отростки или почки и даже целые растения. Сернандер (R. Sernander) дал им общее название диаспор (от греческого 6taoneip(o — рассеивать).
Количество диаспор, образуемых на одном стебле в год, сильно варьирует в зависимости от вида. Оно мало у одноцветковых растений с крупными семенами, например у безвременника, лесной анемоны, и огромно у видов с мелкими семенами, например у орхидей, отдельные виды которых образуют многие десятки миллионов семян. Но для точной оценки способности данного вида к распространению необходимо учитывать, какое число генераций семян растение способно продуцировать. Монокарпические виды, преимущественно однолетние или двухлетние, производят лишь одну генерацию диаспор и гибнут, тогда
И
как отличающиеся долголетием деревья могут производить диаспоры почти каждый год в течение более века.
Среди яйцекладущих животных количество отложенных яиц сильно варьирует в группе в зависимости от вида. Например, среди ракообразных речной рак откладывает обычно 250 яиц, а травяной краб (Carcinus moenas) —свыше 100 тыс. Среди насекомых все рекорды побивает матка термитов — она кладет по одному яйцу в секунду в течение всей жизни. У рыб треска откладывает более 4 млн. икринок в год, сельдь — от 8 до 75 млрд., в то время как пресноводные рыбы гораздо менее продуктивны. Самые малые кладки у колюшки — 60—70 икринок. Дикие птицы отличаются очень малыми кладками: пара диких уток кладет в год 10—16 яиц, а некоторые виды — лишь одно» яйцо.
У млекопитающих численность детенышей в помете варьирует от единицы (приматы, китообразные, летучие мыши) до 20 (серая крыса), однако это количество может возрастать. К тому же мелкие млекопитающие достигают половой зрелости уже в возрасте нескольких месяцев. Но даже у слона, который является примером неблагополучия (лишь к 16 годам он достигает половой зрелости и слабо плодовит), потомство пары, огражденное от влияния неблагоприятных факторов, достигло бы, по подсчетам Дарвина, через 500 лет 15 млн.
Биологическое значение степени плодовитости. Формы, образующие громадное количество семян, изначально обладают несомненным преимуществом по сравнению со слабовоспроизво-дящими формами, так что виды, широко распространенные на Земле, обычно очень плодовиты.
Однако высокая плодовитость видов далеко не всегда влечет за собой их широкое распространение, так как они часто гибнут как на стадии зародыша, так и в процессе развития. Например, семена орхидей могут прорастать, лишь вступив в симбиоз с грибом определенного вида, который для этого должен присутствовать в почве по соседству; поэтому в естественных условиях прорастание орхидей происходит очень редко. Из нескольких сотен миллионов яиц, откладываемых маткой термитов, многие не достигают стадии имаго, а последние почти целиком состоят из неплодовитых рабочих и солдат.
Необыкновенная плодовитость многих видов почти полностью компенсируется эквивалентной гибелью под действием неблагоприятных факторов физической среды, отклонений в развитии,, паразитов, хищников, и это обеспечивает относительную стабильность их численности. Хвойные, ореховые деревья, дубы и буки наших лесов дают жизнеспособные семена в достаточном количестве лишь в некоторые годы, когда их продуктивность так велика, что семена не могут быть уничтожены птицами и грызунами. Точно так же лишь из одной икринки сельди на тысячу, и из четырех икринок макрели на миллион развиваются
12
мальки, не говоря уже о том, что не все мальки достигают зрелости. „
Это со всей очевидностью подтверждает высказывание Ире-нана (М. Prenant) о «...громадном значении смерти в естественных условиях... Это соображение, несмотря на его очевидность, зачастую оказывается незамеченным — столь велика поддерживаемая традиционным образованием привычка считать, что развитие организма протекает нормально от рождения до старости, но ведь именно это — исключительный случай».
Распространение организмов
Распространение — это перемещение индивидуумов, их диаспор или личинок. Оно может быть пассивным, если обеспечено внешним агентом, и активным, когда организм обеспечивает свое распространение какими-либо способами передвижения.
Пассивное распространение
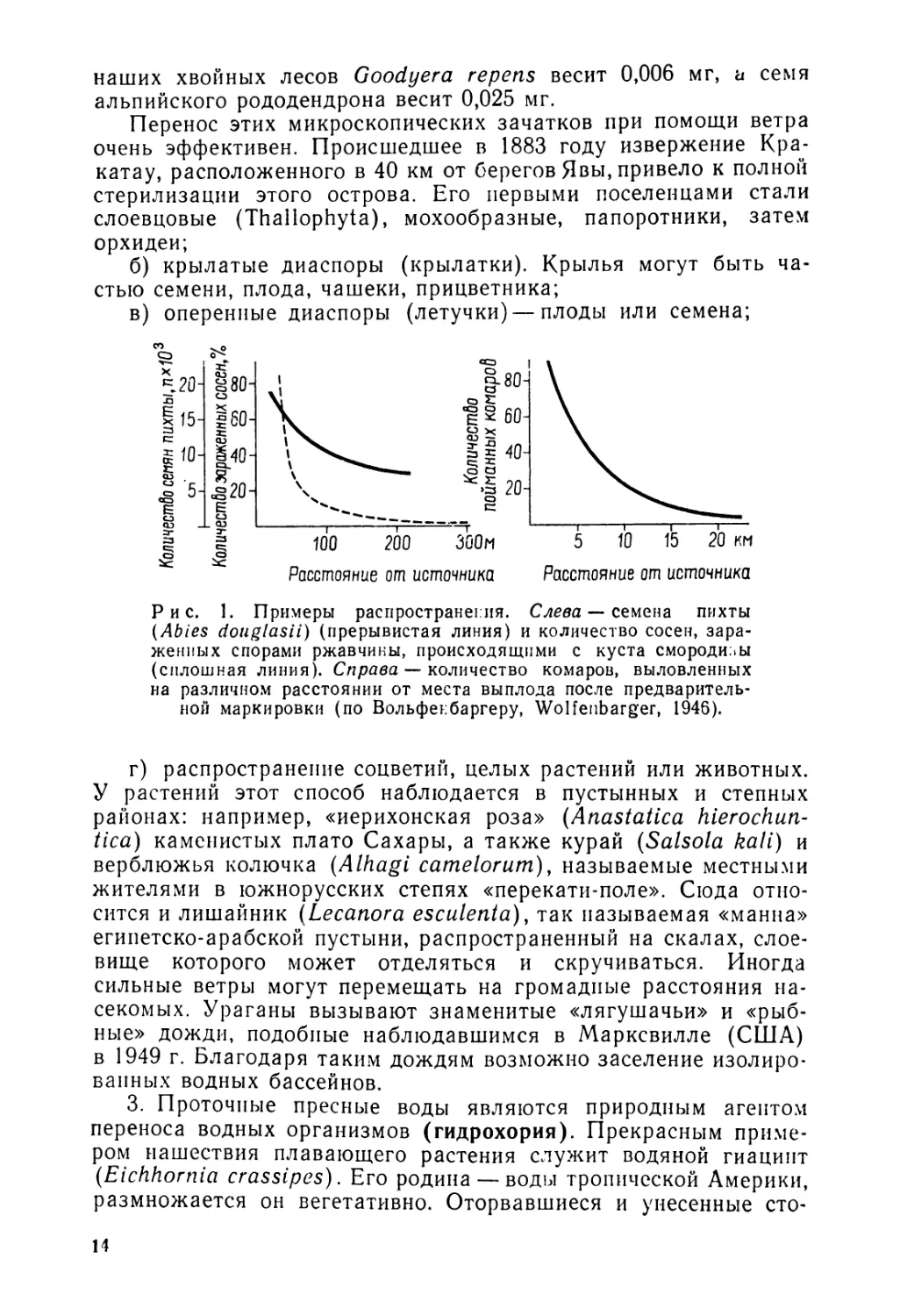
Согласно Вольфенбаргеру (Wolfenbarger), при пассивном распространении плотность распространения семян или организмов постепенно уменьшается пропорционально логарифму расстояния от места их происхождения. Таким образом, отношение между плотностью и расстоянием выражается на графике вогнутой кривой (см. рис. 1).
В то же время эффективность пассивного распространения сильно различается в зависимости от особенностей агентов переноса и от степени адаптации распространяющихся организмов.
1. Некоторые формы, как правило, не используют внешние источники энергии для распространения. Таковы тяжелые семена (барохоры), которые либо разбрасываются на небольшое расстояние при разрыве плода (дрок, бальзаминовые), либо погружаются в почву при помощи плодоножек (арахис, Linaria cymbalaria) (автохоры). В некоторых случаях плоды способны слегка перемещаться благодаря гигроскопическим деформациям (Erodium). Сюда же относится вегетативное размножение при помощи корневищ, столонов или луковиц. В этих случаях формы распространяются лишь на небольшие расстояния, но зато население оказывается относительно плотным и, таким образом, хорошо защищенным от конкуренции других видов.
2. Распространение при помощи ветра, или анемохория, обес* печивается разнообразными морфологическими особенностями. К ним относятся:
а) легкость зачатков: таковы цисты простейших и коловраток, споры споровых растений, пылевидные семена орхидей. Одно семя орхидеи-эпифита Dendrobium attenuatum или орхидеи из
13
наших хвойных лесов Goodyera repens весит 0,006 мг, а семя альпийского рододендрона весит 0,025 мг.
Перенос этих микроскопических зачатков при помощи ветра очень эффективен. Происшедшее в 1883 году извержение Кракатау, расположенного в 40 км от берегов Явы, привело к полной стерилизации этого острова. Его первыми поселенцами стали слоевцовые (Thallophyta), мохообразные, папоротники, затем орхидеи;
б) крылатые диаспоры (крылатки). Крылья могут быть частью семени, плода, чашеки, прицветника;
в) оперенные диаспоры (летучки) — плоды или семена;
Рис. 1. Примеры распространения. Слева — семена пихты (Abies douglasii) (прерывистая линия) и количество сосен, зараженных спорами ржавчины, происходящими с куста смородины (сплошная линия). Справа — количество комаров, выловленных на различном расстоянии от места выплода после предварительной маркировки (по Вольфеибаргеру, Wolfenbarger, 1946).
г) распространение соцветий, целых растений или животных. У растений этот способ наблюдается в пустынных и степных районах: например, «иерихонская роза» (Anastatica hierochun-tica) каменистых плато Сахары, а также курай (Salsola kali) и верблюжья колючка (Alhagi camelorum), называемые местными жителями в южнорусских степях «перекати-поле». Сюда относится и лишайник (Lecanora esculenta), так называемая «манна» египетско-арабской пустыни, распространенный на скалах, слоевище которого может отделяться и скручиваться. Иногда сильные ветры могут перемещать на громадные расстояния насекомых. Ураганы вызывают знаменитые «лягушачьи» и «рыбные» дожди, подобные наблюдавшимся в Марксвилле (США) в 1949 г. Благодаря таким дождям возможно заселение изолированных водных бассейнов.
3. Проточные пресные воды являются природным агентом переноса водных организмов (гидрохория). Прекрасным примером нашествия плавающего растения служит водяной гиацинт (Eichhornia crassipes). Его родина — воды тропической Америки, размножается он вегетативно. Оторвавшиеся и унесенные сто
14
лоны, ветви и листья могут дать начало новым поселениям. Завезенный в качестве декоративного растения в 1888 г. во Флориду, он распространился до Виргинии и в 1920 г. достиг Калифорнии. В 1890 г. он был привезен на Яву в ботанический сад Бюйтензорга \ откуда распространился по Индонезии, а затем проник в Австралию, на Филиппины, в Японию и на Гавайские острова. После его появления в 1902 г. в Ханое он колонизирует индокитайские воды, а в 1905 г. захватывает весь остров Цейлон. Неизвестно, когда это растение было привезено в Африку, но оно распространилось подобно чуме от Родезии до Камеруна. Таким образом, его ареал, будучи исходно ограниченным, становится пантропическим благодаря способности к расселению (правда, при первоначальном участии человека).
Представитель однодольных —элодея (Elodea canadensis), которая теперь образует настоящие луга на дне рек и каналов Европы, происходит из Северной Америки. Впервые она была замечена в 1836 г. в Ирландии, затем в 1842 г. ввозится в Англию, в 1845 г. она привлекла внимание во Франции в канале, соединяющем с морем город Кан, и в 1899 г. она достигает Прованса. Появившись в Бельгии и в окрестностях Берлина в 1860 г., элодея распространяется по Центральной Европе, и ее наблюдают в бассейне Дуная в 1865 г., в бассейне Вислы в 1867 г., в бассейне По в 1892 г.
При помощи плавучих диаспор растения распространяются тем дальше, чем дольше они могут находиться на плаву. Ридли (Ridley) составил длинный список рассеивающихся таким образом растений, отметив среди них большое количество представителей осоковых (Сурегасеае).
4. Морские течения переносят не только водоросли и планктон, но и диаспоры наземных растений. Но для того чтобы подобный способ распространения привел к расширению ареала данного вида, необходимо соблюдение ряда условий:
а) диаспоры должны находиться на плаву достаточное время, чтобы пройти расстояние между двумя берегами;
б) способность к прорастанию не должна нарушаться действием морской воды;
в) вид должен проявить способность к прорастанию и внедрению на новом берегу. Это означает, что такой способ миграции пригоден лишь для литоральных растений. Огромная масса зачатков, выбрасываемых реками в море, не имеет будущего.
Напротив, мангровые деревья, произрастающие в илистых тропических эстуариях и лагунах, подверженных ритмической смене приливов и отливов, распространяются по береговой линии. Некоторые из них распространены очень широко — Rhizop-hora mucronata, например, встречается от сенегальских берегов Африки до Адена, на Мадагаскаре, на Сейшельских и Комор-
1 Ныне Богор. — Прим. ред.
15
ских островах, па Цейлоне и па полуострове Индостан, па островах Малайского архипелага, в Австралии, на Тайване и большинстве полинезийских островов. Этот вид отсутствует лишь в мангровых лесах Америки, где он заменен Rhizophora mangle.
Способность некоторых литоральных растений распространяться с помощью морских течений была доказана прямыми опытами. Это относится к плодам кокосовых пальм, которые оказались способны развиваться на различных побережьях и, несомненно, колонизовали таким образом без помощи человека часть бесчисленных тропических островов Тихого и Индийского океанов — таких, как необитаемый остров Нассау, на котором в 1862 г. имелась одна кокосовая пальма, и остров Кракатау примерно через 20 лет после извержения. Человек также внес значительный вклад в распространение этого вида пальмы, происходящего с Тихоокеанского побережья Центральной Америки; в частности, начиная с XVI в. его распространяли испанцы.
Другая пальма, Nipa fruticans, растущая на илистых берегах эстуариев, рассеивает в воде свои плоды, которые прорастают и выбрасываются в большом количестве на малайские берега, где они часто образуют новые поселения. Напротив, огромный плод Lodoicea, распространенной на Сейшельских островах, может иногда достигать Мальдивских островов, но никогда сюда не внедряется, поскольку этой пальме не подходят условия местной литорали.
5. Распространение при помощи животных, или зоохория, встречается очень часто. С помощью подвижных животных могут распространяться бактерии, споры, цисты. Было показано, например, что перемещение мухи по стерильной питательной среде сопровождается появлением колоний бактерий. Что же касается яиц, мелких животных, диаспор высших растений, то они могут быть перенесены на самые различные расстояния в зависимости от того, какое животное служит переносчиком.
Особенно велика роль перелетных птиц. Благодаря голенастым и водоплавающим перелетным птицам широко распространились многие болотные и водные виды. Семена путешествуют либо в их пищеварительном тракте, либо погруженные в приставший к лапкам ил. Распространение в фекалиях может быть эффективным толькс! ‘в том случае, когда семена не теряют способности к прорастанию под действием пищеварительных соков. Особенно большое биологическое значение такой тип распространения имеет при заражении деревьев паразитирующими на ветках семенными (как омела и Loranthus). Такие паразитические растения встречаются в большом количестве в тропиках на самых разнообразных культивируемых видах — какао, манго и др. В Индии они распространяются при помощи Dicaeum eryth-rorhynchum— маленькой птички, которая извергает семена примерно через десять минут после заглатывания и обеспечивает, таким образом, лишь локальное распространение.
16
С помощью животных, имеющих шерстный покров, распространяются диаспоры, снабженные такими приспособлениями, как шипы, прицепки, липкий секрет и т. п. Особенно очевидна роль овец. Перегоняемые овцы оставляют на дорогах растения, происходящие из мест, по которым они только что проходили. Например, севеннские и косские скотогонные тропы покрыты видами средиземноморских равнин (такими, как Aegylops ovata и Aegylops triuncialis, Medicago hispida и Medicago arabica, Cynoglossum cheirifolium и Cynoglossum creticum и т. д.).
В места, где развита шерстяная промышленность, с помощью овец были завезены многие экзотические виды. Необычны богатство и история флоры, занесенной из Пор-Ювеналя в Монпелье. В течение XIX в. там было отмечено 527 иноземных видов. До 1830 года они происходили из средиземноморского бассейна. Затем была ввезена группа южноамериканских видов, потом — австралийских, новозеландских и южноафриканских. Надо признать, однако, что из всего этого набора лишь пять видов сумели натурализоваться.
Среди покрытых шерстью животных, распространяющих диаспоры лишь в ограниченном районе, можно отметить кролика, места пробега которого покрываются чернокорнем (Cynoglossum) и незабудками (Myosotis)—растениями с диаспорами, снабженными прицепками.
Среди беспозвоночных известны в качестве активных переносчиков многие виды муравьев (мирмекохория). И хотя пройденные муравьями расстояния ничтожны, все же в результате вокруг муравейников образуются особые небольшие цветники.
6. Наконец, человек начиная с доисторических времен становится все более и более эффективным агентом распространения.
а) в качестве сознательного агента распространения он расширил ареал, занимаемый культурными растениями и домашними животными. Благодаря отбору рас, адаптированных к новым климатическим условиям, границы этого распространения в последнее время раздвинулись. Бывает, что завезенные таким образом виды приживаются в новых районах вне тех участков, которые были предназначены для их культуры; это и называется натурализацией видов. Таковы во Франции растения-красители, например марена (Rubia tinctorum) и вайда (Isatis), которые более не культивируются. Lantana сатага, ввезенная из Тропической Америки в качестве декоративного растения, была распространена полюбившими плоды этого дерева птицами во всех районах тропиков.
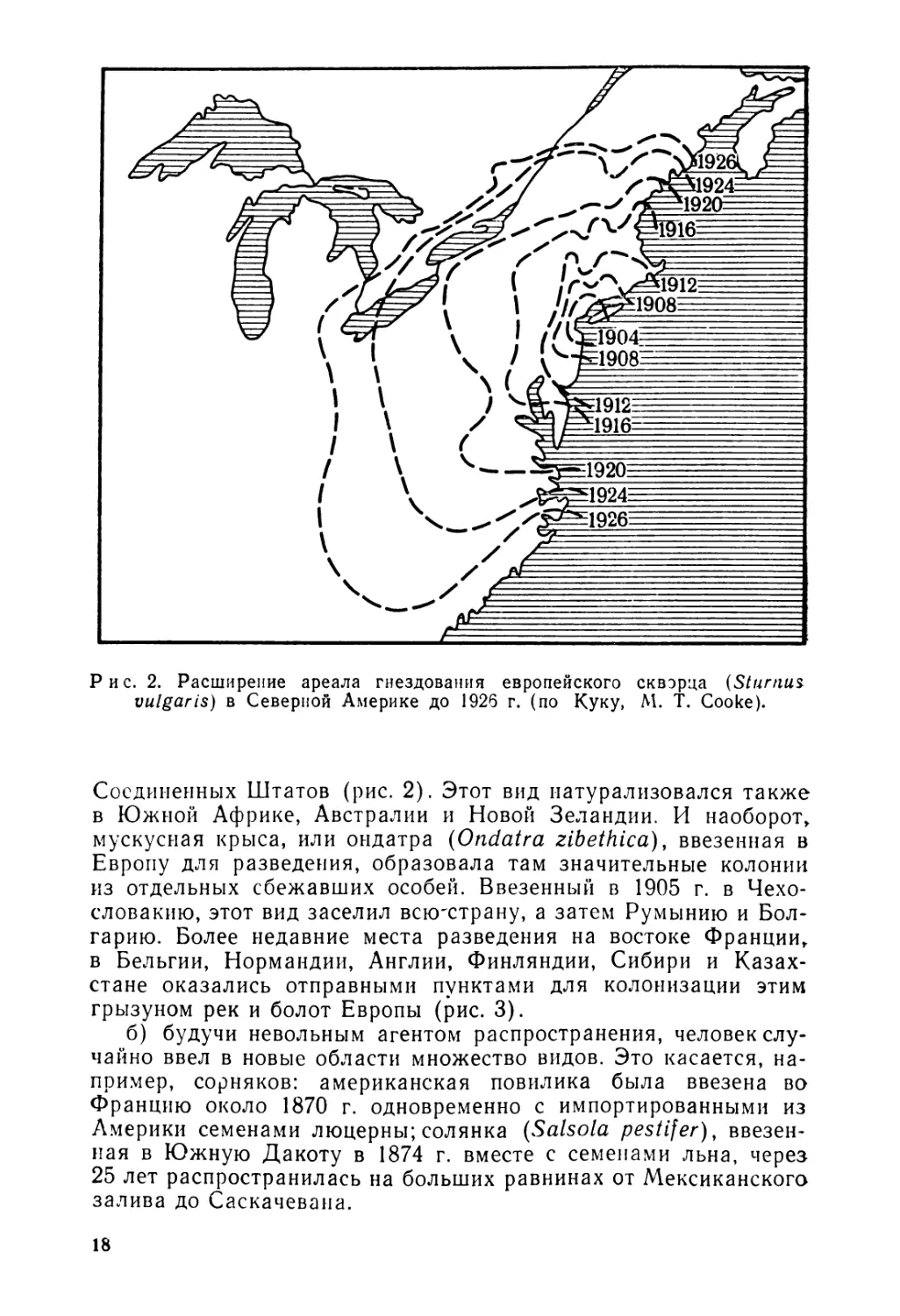
Таким же образом животные могут покидать места разведения и процветать в естественных условиях. Так, европейский обыкновенный скворец (Sturnus vulgaris), ввезенный в Центральный парк Нью-Йорка, начал там гнездиться в 1891 г. и оттуда распространился концентрически почти по йсему Востоку
17
Рис. 2. Расширение ареала гнездования европейского скворца (Sturnus. vulgaris) в Северной Америке до 1926 г. (по Куку, М. Т. Cooke).
Соединенных Штатов (рис. 2). Этот вид натурализовался также в Южной Африке, Австралии и Новой Зеландии. И наоборот, мускусная крыса, или ондатра (Ondatra zibethica), ввезенная в Европу для разведения, образовала там значительные колонии из отдельных сбежавших особей. Ввезенный в 1905 г. в Чехословакию, этот вид заселил вскгстрану, а затем Румынию и Болгарию. Более недавние места разведения на востоке Франции, в Бельгии, Нормандии, Англии, Финляндии, Сибири и Казахстане оказались отправными пунктами для колонизации этим грызуном рек и болот Европы (рис. 3).
б) будучи невольным агентом распространения, человек случайно ввел в новые области множество видов. Это касается, например, сорняков: американская повилика была ввезена во Францию около 1870 г. одновременно с импортированными из Америки семенами люцерны; солянка (Salsola pestifer), ввезенная в Южную Дакоту в 1874 г. вместе с семенами льна, через 25 лет распространилась на больших равнинах от Мексиканского залива до Саскачевана.
18
Растения и животные случайно расселялись также при помощи транспорта; так, железные дороги создали настоящую (Ьлору «балласта», а корабли переправили в Новый Свет черную и серую крысу. Вместе с культурными растениями ввозились их паразиты — такие, как червец (Icerya purchasii), который, выйдя из Австралии, занял весь мировой ареал цитрусовых.
Расселение вида может оказаться обеспеченным самыми различными способами в зависимости от сезона и от стации. В качестве примера Мюллер-Шнейдер (Р. Milller-Schneider) приводит облепиху (Hippophae rhamnoides), кустарник, колонизирующий аллювий альпийских потоков. Его ягоды в свежем виде рассеиваются птицами, в сухом — ветром или водой в период паводков.
В растительных сообществах, приуроченных к разным местообитаниям, преобладают различные способы расселения, что зависит от локальной эффективности разных агентов расселения. Так, Молинье и Мюллер (R. Molinier, Р. Miiller) показали эту характерную особенность биологии растительных сообществ1 в Провансе: на лужайках, поросших коротконожкой
Рнс- 3. Распространение мускусной крысы ондатры в Центральной Европе до 1У56 г. (по данным О. Е. Р. Р. — Европейской организации защиты растений).
____ 2-в 1932 г.; 2 —в 1951 г.; 3 — в 1954 г.; 4— освобожденные зоны.
м 1 ^P- groupement — группировка употребляется автором как синоним тер-Наца <<со°бщество». В русской литературе принято называть группировками альные стадии развития сообщества. — Прим. ред.
19
фактором расселения является ветер; роль животных здесь, напротив, очень мала — агентами расселения служат лишь муравьи, в то время как в дубравах расселение при помощи животных, в особенности птиц, становится преобладающим.
Активное распространение
Активное распространение сыграло, безусловно, большую роль в расселении видов, способных быстро и длительно передвигаться,— например, таких водных видов, как китообразные, таких наземных, как волк, таких крылатых, как странствующая саранча, которая была найдена за 600 км от берега.
Активные перемещения могут принимать характер массовых миграций (перелеты саранчи и бабочек, перемещения больших стад мелких грызунов — леммингов и серых крыс). В некоторых случаях эпизодические массовые миграции (которые следует отличать от сезонных) могут привести к внедрению вида на новую территорию. Так, по данным Жаннеля (R. Jeannel), бабочка (Danaus archippus) с Американского континента смогла в результате массовых миграций, насчитывавших миллионы особей, пересечь океан. На западе эта бабочка оказалась способной достичь в 1840 г. Новой Зеландии, в 1850 г. Гавайских островов и в 1870 г. Австралии; в восточном направлении она сумела пересечь Атлантику до Канарских островов (в 1880 г.), затем достигла Северной Африки, а недавно — Западной Европы.
Глава 2
Области географического распределения
Географический ареал таксономической единицы (вида, рода,, семейства и т. п.)—это часть земной поверхности, на которой данная единица представлена. Такой ареал состоит из элементарных, более или менее многочисленных и территориально близких участков, на которых действительно присутствует рассматриваемая таксономическая единица. Чтобы установить ареал в целом, необходимо предварительно произвести описание этих участков.
Картографическое воспроизведение
Самый точный метод состоит в изображении каждого местонахождения точкой на карте. Такая значковая карта 1 может быть создана лишь в очень крупном масштабе и годится только либо для очень редких форм чс узким географическим распространением, либо для небольшого участка обширного ареала.
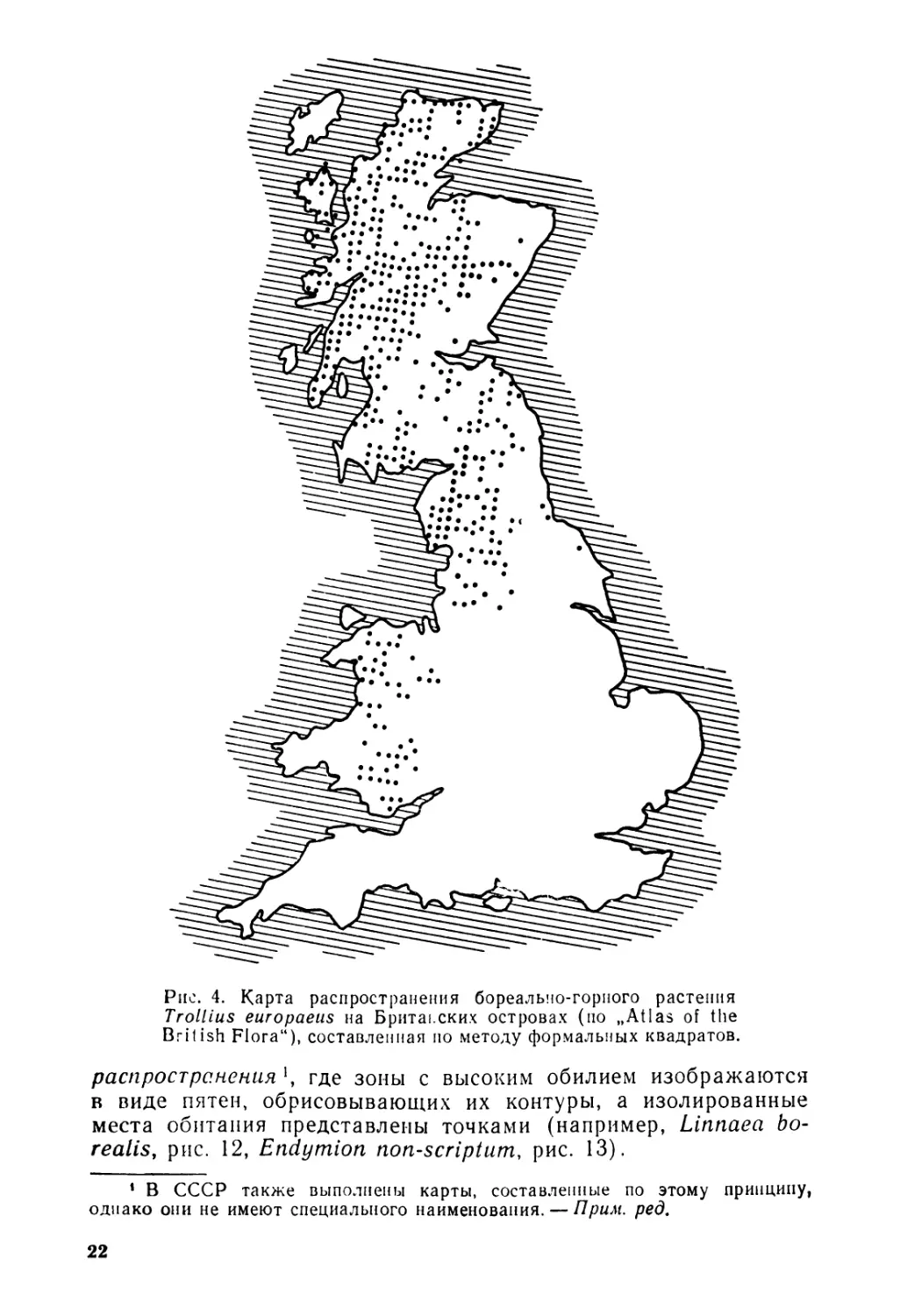
Однако в таком подробном представлении может не быть надобности или же оно может быть неосуществимым (если изу^ чаемая форма обычна или если размер территории требует более мелкого масштаба для своего изображения). Тогда можно разбить территорию на клетки, обозначив значком те из них, где данная форма встречается. Подобные карты, построенные методом формальных квадратов2, позволяют легко сравнивать распределение видов внутри какой-либо страны. Этот метод был использован в скандинавских странах, Нидерландах и Англии (рис. 4) для сосудистых растений.
Если мы хотим изобразить ареал, включающий в себя как зоны, где изучаемый таксон встречается очень часто, так и зоны, где он отсутствует или встречается лишь в немногих изолированных участках, то мы составляем карту площадей
1 Автор называет такую карту точечной. В русской литературе, однако, точечными называют карты, основанные на количественных данных, отнесен-Нь1* к площади. При этом каждая точка имеет определенный вес. — Прим.
2 Автор называет их «квадратизованными картами». Часто для построе-квадратов используется градусная сеть в меркаторской проекции. — Прим.
Ния ред.
21
Рис. 4. Карта распространения бореально-горного растения Trollius europaeus на Британских островах (по „Atlas of the British Flora"), составленная по методу формальных квадратов.
распространения где зоны с высоким обилием изображаются в виде пятен, обрисовывающих их контуры, а изолированные места обитания представлены точками (например, Linnaea borealis, рис. 12, Endymion non-scriptum, рис. 13).
1 В СССР также выполнены карты, составленные по этому принципу, однако они не имеют специального наименования. — Прим. ред.
22
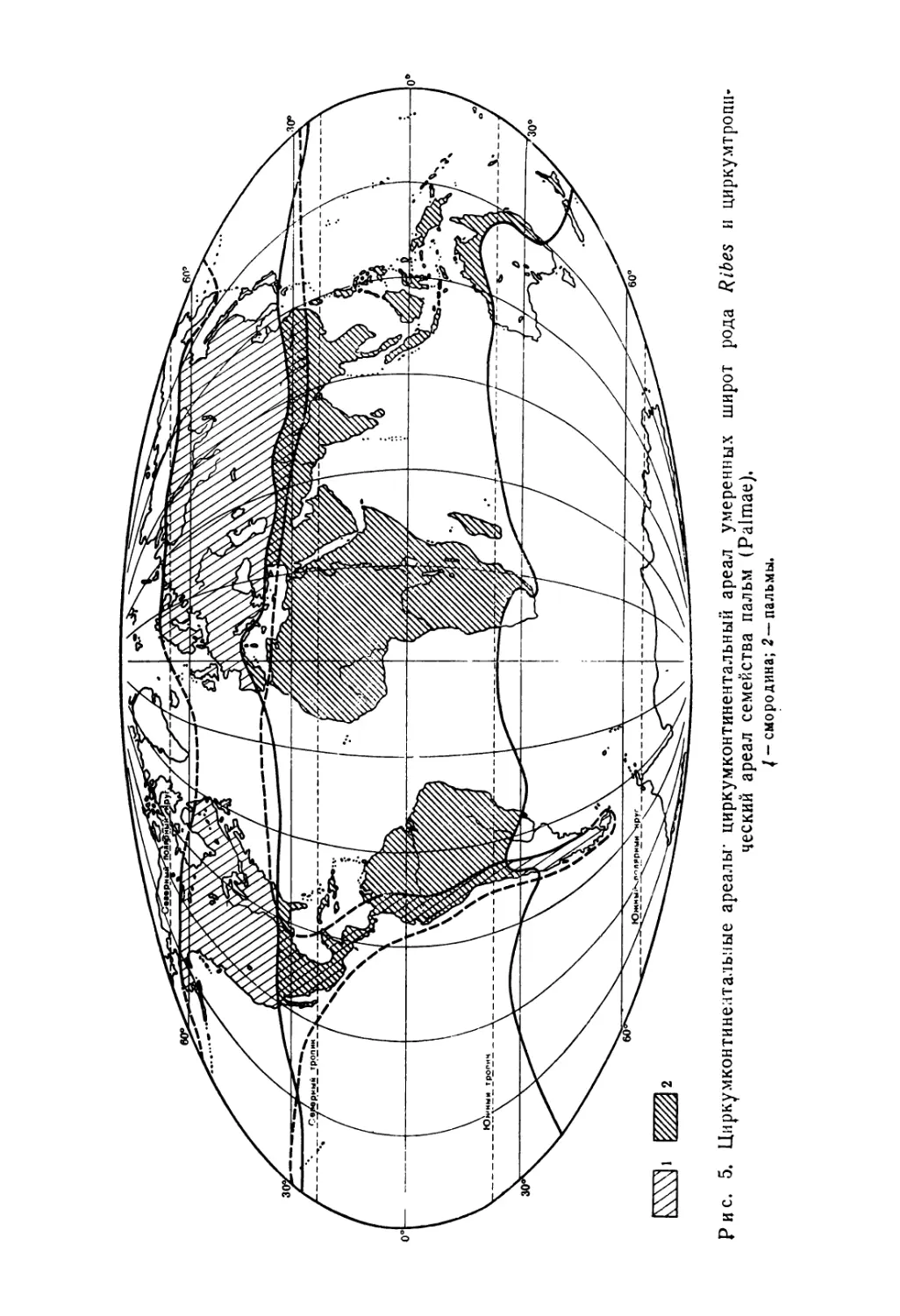
Рис. 5. Циркумконтинелтальные ареалы циркумконтинентальный ареал умеренных широт рода Rjbes и циркумтропи-ческий ареал семейства пальм (Paimae).
/ - смородина; 2 — пальмы.
Если площадь ареала велика или масштаб мал, то составляют карты абсолютных границ распространения, охватывающие всю совокупность местонахождений (например, род Ribes— смородина или семейство пальм, рис. 5). При таком синтетическом подходе не учитываются детали распространения, и поэтому только он пригоден в случае, когда ареал не полностью изучен (например род Verbascum, рис. 9).
Структура ареалов
^Многообразие мест происхождения, возраста и факторов, лимитирующих существование таксономических единиц, является причиной необычайного разнообразия протяженности и конфигурации их ареалов. Можно, однако, выделить несколько типов ареалов, различающихся по своим характеристикам.
Космополитные ареалы
Космополитными называют те ареалы, которые покрывают большую часть обитаемых зон земного шара. Частота распространения этого типа возрастает с повышением ранга таксономических единиц. Космополитны многие отряды и семейства: таковы коловратки среди животных, злаки и сложноцветные среди семенных растений. Напротив, роды и виды лишь очень редко бывают относительно космополитными. Для них более подходит предложенное Молишем (Molisch) наименование «семикосмополиты» (или полукосмополиты). Можно насчитать, например, около 20 видов семенных, ареал которых занимает более половины поверхности суши. В первую очередь к ним относятся вод-м ные или болотные растения, часто рассеиваемые перелетными птицами — например, ряска (Lemna), тростник {Phragmites сот-munis), рогозы, широколистный и узколистный (Typha latifolia и Т angustifolia). Вторую группу составляют распространяемые человеком сорные травы. К ним принадлежат однолетний мятлик, пастушья сумка, белая лебеда, одуванчик, большой подорожник. к животным космополитам относят тех, которые соседствуют с человеком, например комнатную муху или серую крысу. Сам человек, без сомнения, является видом, наиболее приближающимся к полному космополитизму.
Циркумконтинентальные и циркумокеанические ареалы
Так называют ареалы, занимающие совокупность земель или океанов, ограниченную какими-то широтными пределами. Двигаясь с севера на юг, можно выделить:
24
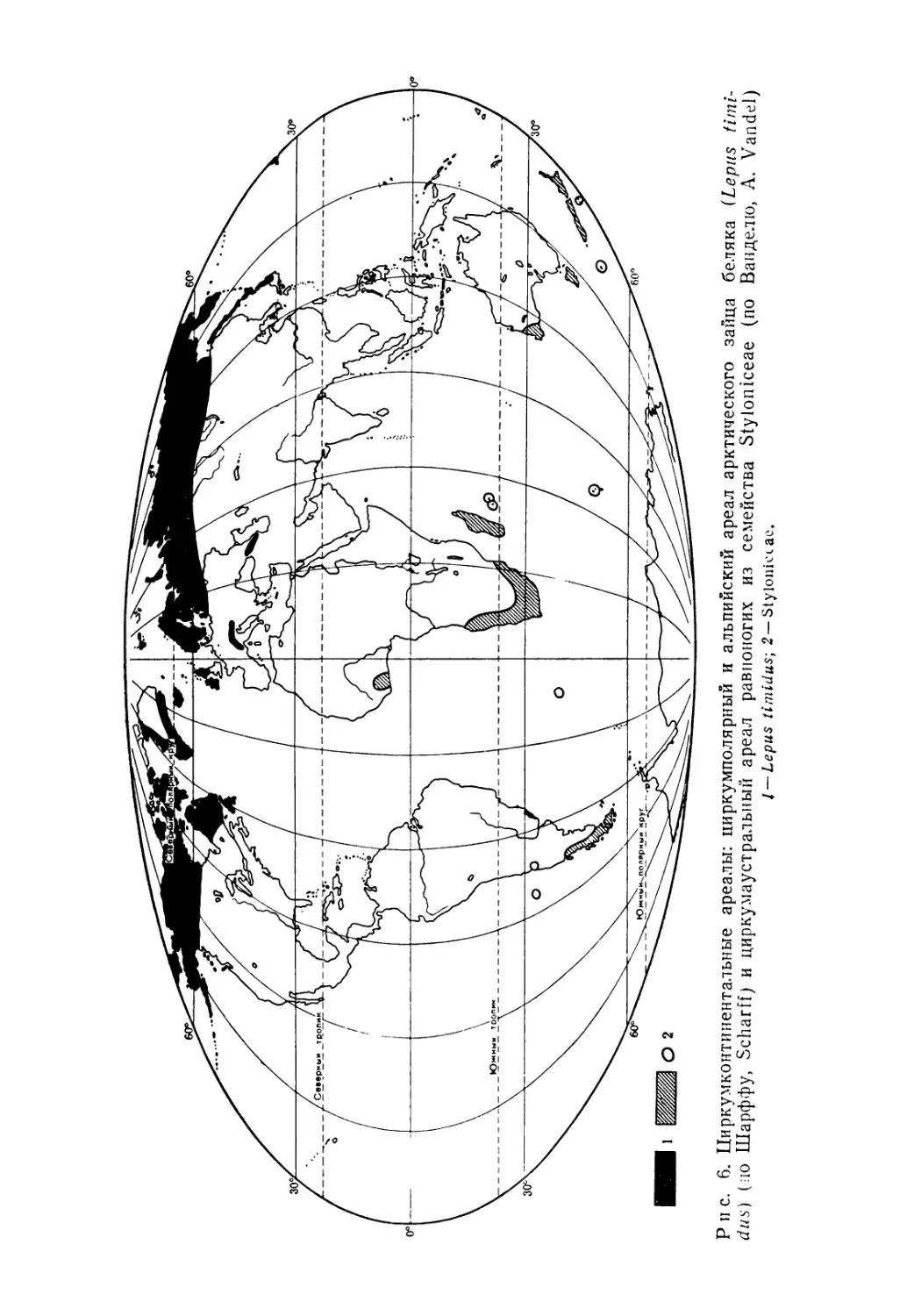
Рис. 6. Циркумконтинентальные ареалы: циркумполярный и альпийский ареал арктического зайца беляка (Lepus timi-dus} (ио Шарффу, Scharff) и циркумаустральный ареал равноногих из семейства Styloniceae (по Ванделю, A. Vandel) I — Lepus timidus-, 2— Styloniccас.
1) циркумполярные бореальные ареалы !, занимающие земли, моря или побережья вблизи Северного полярного круга, например ареалы некоторых тюленей (рис. 11), пингвинов, а среди цветковых растений — Carex lapponica или Rubus chamaemorus (рис. 11);
2) циркумконтинентальные ареалы умеренных широт северного полушария, например ареалы смородины (рис. 5) или Calt-ha palustris\
3) пантропические ареалы, часто встречающиеся среди семейств и родов семенных, например ареалы семейств акантовых и пальмовых (рис. 5) и ареалы родов Strychnos, Bauhinia, Hibiscus. Большая часть пантропических видов была распространена человеком, например Mimosa pudica, Coix lacryma jobi\
4) циркумконтинентальные ареалы1 2, расположенные южнее Тропика Козерога, немногочисленны и, естественно, весьма прерывисты. Ими обладают некоторые архаические группы — первичнотрахейные или онихофоры, а также равноногие ракообразные из семейства Slyloniceae (рис. 6).
Эндемичные ареалы
В противоположность космополитным эндемичные ареалы ограничены по территории. Площадь таких ареалов может быть очень разнообразной. Для рода, семейства или отряда она может быть относительно велика; например, Австралия, Тасмания и Новая Гвинея в совокупности образуют территорию, для которой эндемичен отряд однопроходных.3 Для вида или разновидности эндемизм может быть ограничен всего несколькими квадратными километрами или даже метрами: пещерные пауки были обнаружены лишь в одной пещере; семенное Lisimachia minoricensis занимает всего лишь несколько квадратных метров на острове Менорка.
С эволюционной точки зрения эндемизм может быть результатом двух совершенно разных процессов, которые необходимо различать:
1. Палеоэндемики (называемые также реликтовыми или консервативными эндемиками) представляют собой древние формы, ареал которых, первоначально весьма обширный, постепенно сократился под влиянием ухудшавшихся климатических условий или из-за конкуренции с более приспособленными видами. Такие виды обладают архаическими чертами, чаще всего занимают обособленное положение в классификации организмов и вытеснены в среду, где конкуренция слабая. Условия выживания палеоэндемиков очень ненадежны, и многие из них
1 Обычно различают циркумполярные ареалы, охватывающие зоны арктических пустынь, тундр и лесотундр, от бореальных, занимающих подзону тайги. — Прим. ред.
2 Иначе — южные циркумконтинентальные ареалы. — Прим. ред.
3 Правильнее считать эту группу подклассом. — Прим. ред.
26
ли уничтожены или находятся на пути к исчезновению из-за тродукиии иноземных видов или вмешательства человека.
ИН Материковые острова, длительное время находящиеся в изо-яции, образуют большей частью территории развития консервативного эндемизма. Как будет показано далее, хорошим примером этому служат Австралия, Новая Зеландия, Гавайские острова, Мадагаскар, Канарские острова. Напротив, для океанических островов, которые недавно образовались и никогда не были связаны с материком, эндемизм не характерен. Примером могут служить Полинезийские и Мальдивские острова. Что же касается островов, подобных Британским, лишь недавно отделившимся от материка, то на них могут существовать некоторые неоэндемики, речь о которых пойдет ниже.
Реликтовому эндемизму благоприятствуют также изолированные горные цепи. Гинкго, покрывавшее в третичном периоде все континенты, сохранилось сейчас в горах внутреннего Китая. Секвойи, некогда произраставшие в Европе, локализованы в настоящее время в прибрежной калифорнийской горной цепи. Небольшое семейство тропических растений геснериевых имеет изолированного представителя в Пиренеях — Ramondia pyrenaica. В верхнем поясе сахарских массивов Аххагар и Тассилин-Адджер изолированность которых усиливается аридностью области, эндемичны соответственно 40 и 60% растений.
2. Неоэндемики, которые иначе называют молодыми или прогрессивными эндемиками, — недавнего и локального происхождения и отличаются от родительских форм лишь некоторыми второстепенными чертами, что позволяет возвести их в ранг подвида (отсюда термин «микроэндемики», иногда используемый для их обозначения). Они принадлежат к эволюционирующим родам, сохранению новых форм внутри которых благоприятствует генетический или географический барьер. Например, сиги, обитающие в озерах Британии и Центральной Европы, представлены в каждом озере различными формами. На недавно изолированных Британских островах велик контингент неоэндемиков среди наземной фауны. Среди млекопитающих это белка (Sciurus vulgaris leucourus), которая отличается от континентальной формы несколько более светлым мехом, и землеройка (Sorex araneus castaneus), также несколько отличная от континентальных форм. Континентальные типы птиц также представлены на островах особыми расами — например, ирландская синица (Parus ater hibernicus) и шотландский клест (Loxia сиг-virostra scotica).
Викарирующие ареалы
Викарированием называют замещение таксономической единицы близкой единицей, происходящее либо в одинаковых Местообитаниях двух соседних регионов, либо в различных
27
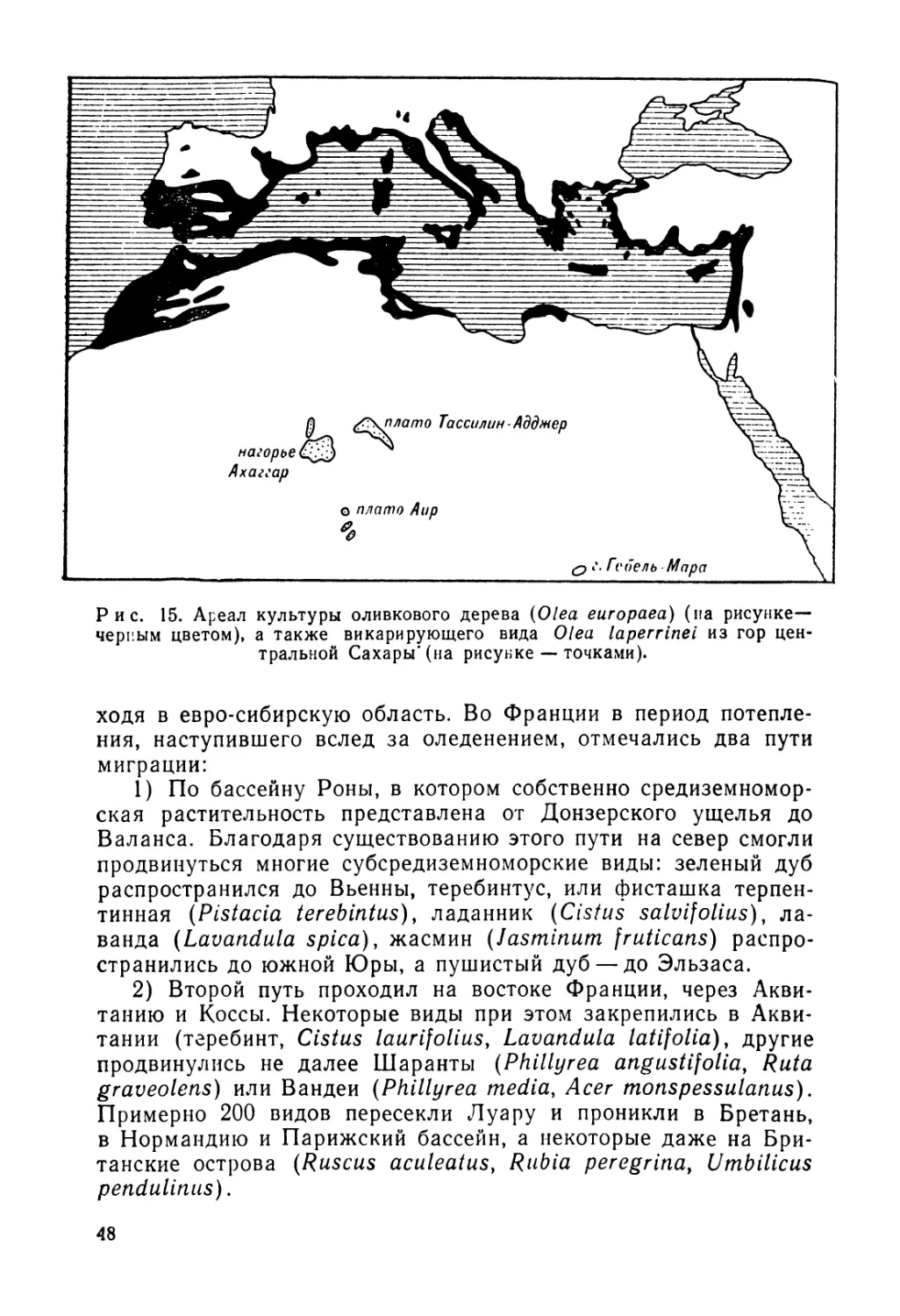
местообитаниях, расположенных на одной и той же территории В первом случае мы имеем дело с хорологическим, илигеографи ческим, викарированием, во втором — с экологическим викариро ванием. Примером первого случая может служить оливковоеде рево (Olea еигораеа) из средиземноморского бассейна, дика5 форма которого замещена в горных массивах Сахары очеш близким видом Olea laperrinei (рис. 15). Второй случай проил люстрируем на примере альпийских рододендронов (Rhododend гоп hirsutum и R. ferrugineum), из которых первый приурочеь к известняковому субстрату, а второй на нем не встречается Сейчас нас интересует первый случай викарирования.
Ареалы викарирующих видов могут быть полностью разоб щены, как показывает приведенный выше пример с оливковым! деревьями. Они могут также быть более или менее трансгрессив ными и дают тогда гибридные формы, часто в зоне совместногс распространения: например, осина (Populus tremula) из Европы и Западной Сибири и Populus tremuloides из Восточной Сибири \ вороны серая и черная из Северной и Южной Европы.
Викарирование может затрагивать таксономические единицы самых различных уровней. Приведенные выше примеры касаются близких видов, которые Майр и Шильдер (Мауг, Schilder) предлагают объединить в один таксон, называя его надвидом (super-espece). Классические примеры замещаемости видов представляют изолированные друг от друга горные массивы, например высокие горы Восточной Африки. Характерные для этих мест растения гигантские лобелии представлены там близкими формами, принадлежащими к одному эндемическому ряду. Многие млекопитающие — обезьяны, лемуры, антилопы, белки — представлены на горах Кения и Килиманджаро подвидами или вика-рирующими видами. Викарирование может проявляться и в водных средах, если они географически изолированы, как, например, озера: лососевые и сиговые рыбы ледниковых озер, Leu-cichthys из североамериканских озер представлены в каждом из них особой формой.
Замечательным примером океанического викарирования является фауна западных и восточных побережий Панамского перешейка, изолированных друг от друга в плиоцене. Многочисленные рыбы и моллюски представлены парами близких форм, каждая из которых встречается либо в атлантических, либо в тихоокеанских прибрежных водах.
Напротив, викарирование родов и семейств имеет реликтовый характер. Таковы саговники, роды которых разъединены
Populus tremuloides замещает нашу осину в Северной Америке и в Во--сточной Сибири не встречается; пример трансгрессивных викарирующих ареалов дают ель европейская (Picea abies) и ель сибирская (Picea obovata), область совместного обитания которых охватывает восток Европейской части Союза. — Прим. ред.
28
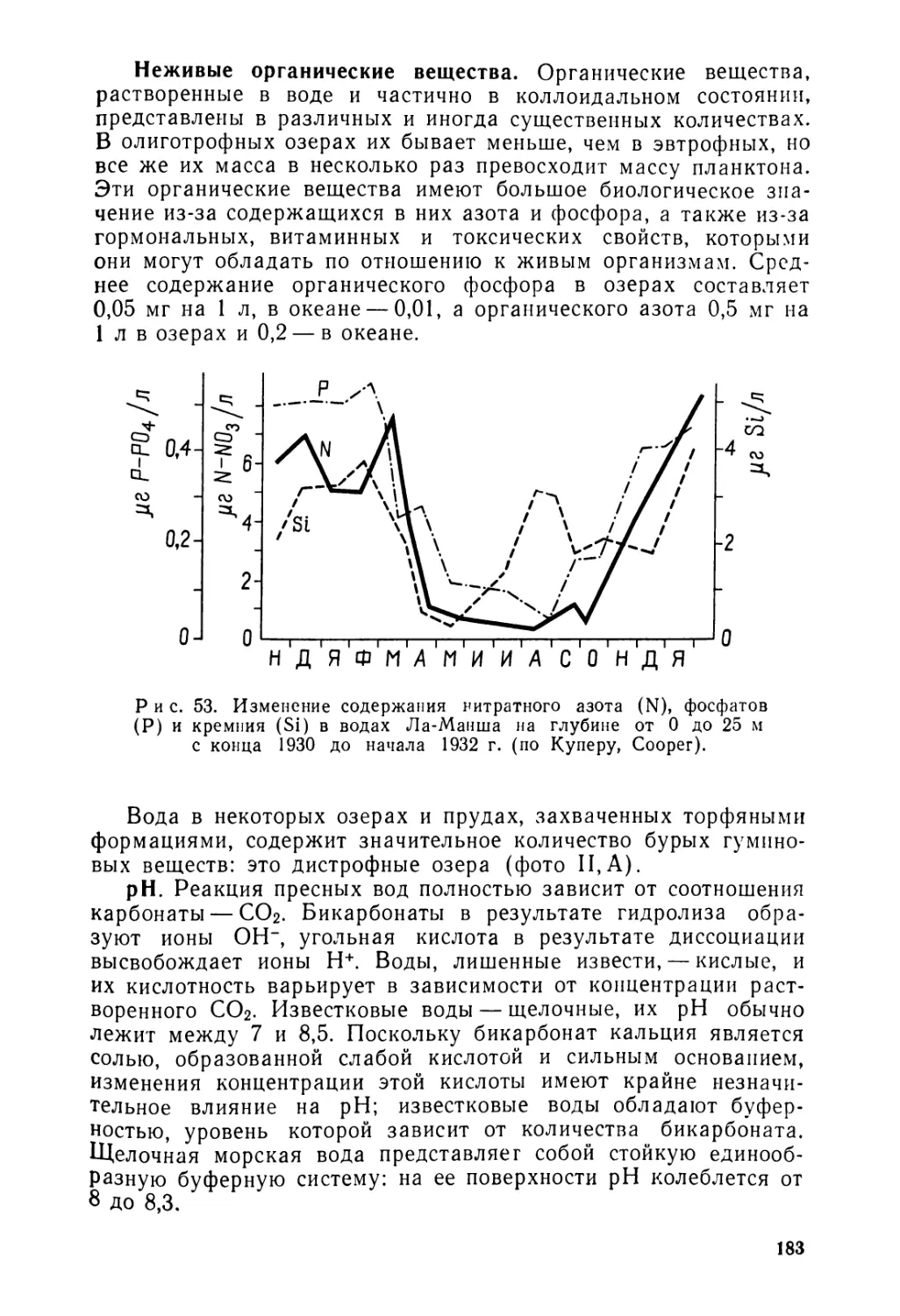
vr от друга в различных тропических областях; акантовые, ^оставляющие тропическое семейство, викарирующее с норичниковыми умеренных широт.
Случается также, что на различных территориях замещают уГ друга не родственные, но близкие по экологии и морфологии формы. Таковы американские кактусы и африканские сук-кулентные молочаи. В данном случае речь идет о ложном вибрировании (Rothmaler, Love), представляющем собой конвергенцию форм, созданных в одинаковых условиях среды.
Дизъюнктивные (разорванные) ареалы
При определении границ ареала на карте достаточно крупного масштаба для выявления деталей очертания редко остаются непрерывными. Чаще всего на периферии имеются островки меньшего размера. В этом случае мы говорим о расчлененных (фрагментированных) ареалах. Если же элементы ареала очень сильно удалены друг от друга, то ареал квалифицируется как разорванный, или разъединенный (дизъюнктивный).
Аналогично викарированию разъединение может затрагивать любые таксономические единицы. Так, класс первичнотрахейных представлен в Южной Африке, Конго, Гималаях, Малайе, Новой Гвинее, Австралии, Новой Зеландии, Центральной и Южной Америке. Семейство верблюдов (Camelideae) представлено верблюдами в Старом Свете, ламами — в Южной Америке. Род тапиров в настоящее время можно встретить в Южной Америке и в Малайе. Многие виды имеют аркто-альпийский ареал, ведущий начало с четвертичных оледенений, речь о которых пойдет ниже.
О причинах разъединения было много споров. В 1854 г. зоолог Дана (Dana) попытался объяснить присутствие некоторых видов в холодных морях обоих полушарий при их отсутствии в широкой полосе тропических вод, предположив, что эти виды образуются независимо по обе стороны экватора. Позднее ботаник Брикет (John Briquet) на вершинах гор Корсики обнаружил альпийские виды растений, не обладающие средствами переноса семян на большие расстояния. Не имея возможности объяснить их присутствие переносом с помощью наземных коммуникаций с Альп, Брикет предположил, что эти виды могли возникнуть сразу в нескольких точках своего ареала. Эта гипотеза политопического происхождения была подхвачена и стала известна под названием ологенеза\ однако она не получила широкого распространения и кажется нам маловероятной.
Другой причиной разъединения может оказаться миграция скачками. Напоминаем, какую роль в распространении растений играют перелетные птицы, морские течения, сильные ветры и человек. Но чаще всего разъединение наступает из-за фрагментации первоначально сплошного ареала, которая возникает °Д влиянием климатических изменений, или изменений конфи
29
гурации моря и суши, или конкуренции со стороны новых форм, имеющих большие способности к распространению, чем древние.
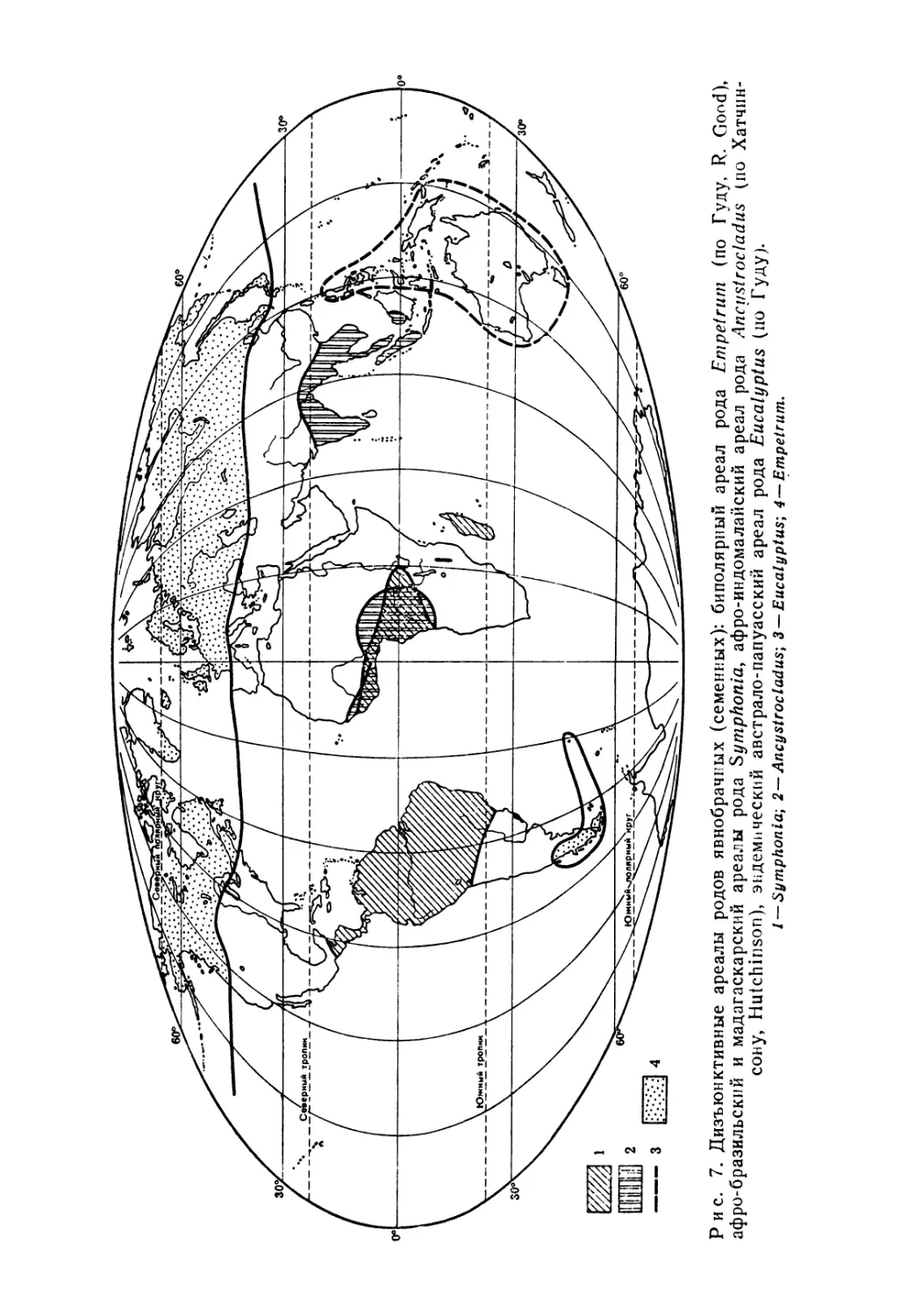
Особенно разительны биполярные разъединения, которые с большим трудом поддаются объяснению. Эти ареалы распределяются между северными и южными областями без промежуточных местонахождений. Например, такие растения, как Primula farinosa, Triglochin palustre, Gentiana prostrata, род Em-petrum (рис. 7), имеют преимущественно северный циркумбо-реальный ареал, но снова встречаются на южной оконечности Южной Америки. Среди попыток объяснить этот факт часто выдвигается предположение о переносе их на далекие расстояния перелетными птицами (известно, например, что ржанка бурокрылая мигрирует от Аляски до Магелланова пролива). Европейским и североамериканским букам {Fagus) соответствует близкий им род южных буков (Nothofagus); общие предки этих родов имели широкое географическое распространение в третичном периоде.
Среди морских животных также известны случаи параллельного распространения в умеренных и полярных областях обоих полушарий. Например, сардины, локализованные между летними изотермами 12 и 20° С, представлены двумя парами видов: одна пара в Северной и Южной Атлантике, другая — в северной и южной частях Тихого океана. Другой пример — арктические {Psolus squamatus) и антарктические {Psegregatus) голотурии. Для объяснения происхождения дизъюнктивных ареалов выдвигаются различные гипотезы: политопическое происхождение, древнее космополитное распространение, существование в глубине тропических вод {Sagitta hamata и некоторые рыбы), а также охлаждение в периоды оледенений L
Причины ограничения ареалов
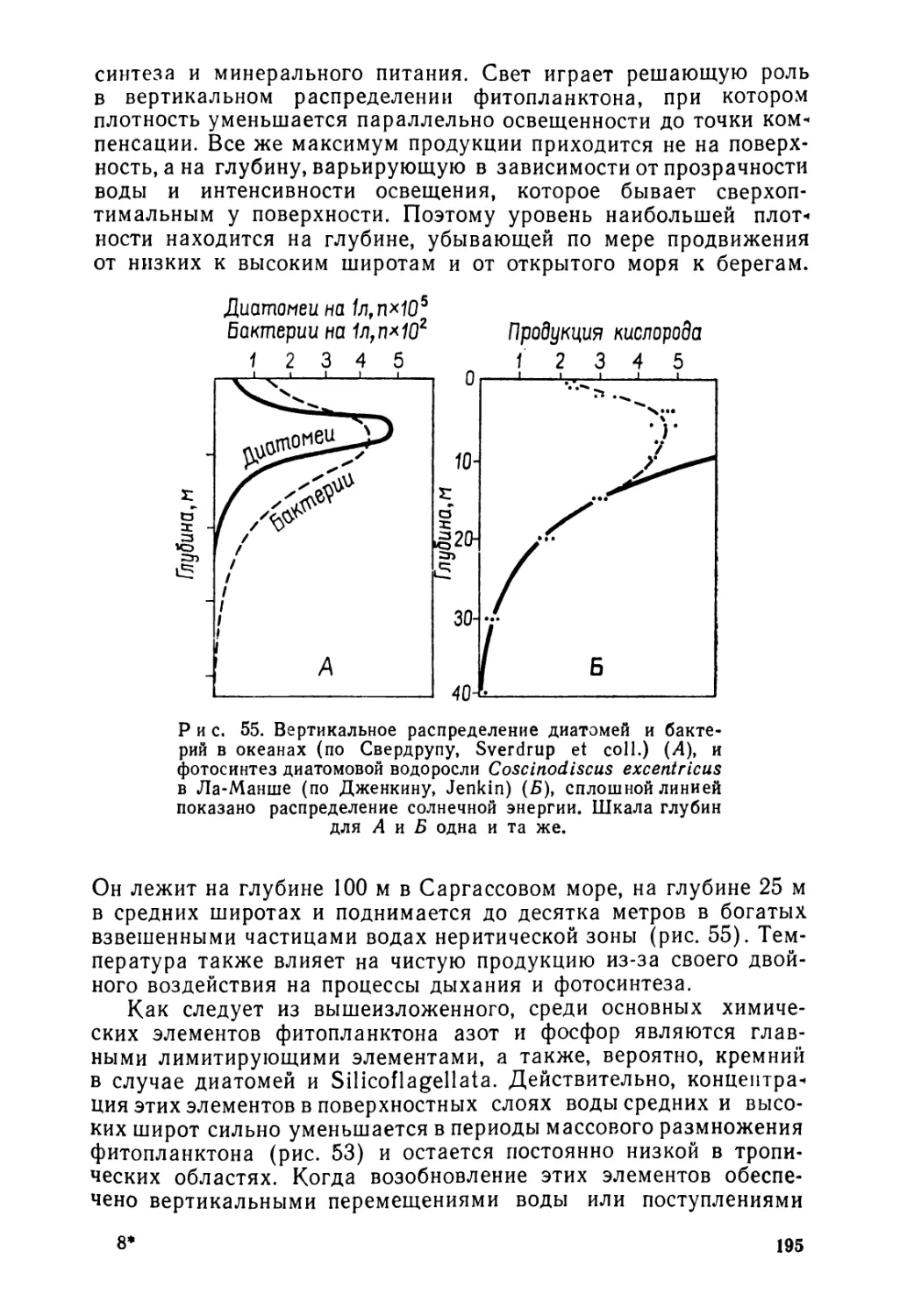
Такие причины имеют разнообразную природу.
1. Наиболее эффективной причиной является географический барьер. Эффективность морских барьеров зависит от способов перемещения видов. В этом смысле так называемые океанические острова, которые никогда не были связаны с материком,
1 Наиболее вероятно объяснение, данное известным русским зоологом и географом Л. С. Бергом, согласно которому в период четвертичного оледенения охлаждение тропических и экваториальных вод было достаточным, чтобы формы, обитавшие в бореальных водах северного полушария или потальных (аитибореальных) водах южного, могли преодолеть эту область; однако это похолодание было недостаточным, чтобы настоящие арктические и антарктические полярные формы расширили свои ареалы и приобрели биполярное распространение. Последующее потепление климата вызвало разрыв ареала. Считают, что бореальные формы были более активными в преодолении экваториального барьера, чем потальные. — Прим. ред.
30
Рис. 7. Дизъюнктивные ареалы родов явнобрачных (семенных): биполярный ареал рода Empetrum (по Гуду, R. Good), афро-бразильский и мадагаскарский ареалы рода Symphonia, афро-индомалайский ареал рода Ancustrocladus (по Хатчинсону, Hutchinson), эндемический австрало-папуасский ареал рода Eucalyptus (по Гуду).
1 — Symphonia; 2 — Ancystrocladus; 3 — Eucalyptus; 4 — Empetrum.
служат прекрасным примером отбора населения благодаря изоляции. Таков случай Галапагосских островов.
Наоборот, распространение многих морских видов лимитировано перешейком. Средиземное и Красное моря, разделенные Суэцким перешейком, имеют весьма различную фауну. Однако прорытие канала позволило некоторым видам Красного моря, например жемчужнице, перейти в восточную часть Средиземного моря.
Другим типом барьера для обитателей суши служит рельеф. Непрерывная горная цепь (например, Гималаи)— непроходимая стена для видов, обитающих на небольших высотах. И наоборот, горный массив изолирует сугубо горные формы, способствуя таким образом созданию эндемических островков (выше говорилось о восточноафриканских Lobelia).
Такие естественные препятствия, как озеро или река, могут быть непроходимыми барьерами для обитателей суши. Так, реки Лимпопо и Оранжевая служат границей распространения различных видов антилоп. К востоку и западу от озера Чад обитают различные расы антилопы (Bubalis major) и буйвола (Bubalus caff er).
Распространение некоторых животных может быть ограничено определенной растительной формацией, которая занимает обширное пространство и несовместима с образом жизни этих животных. Таковы, например, саванны для большинства лесных тропических видов и, наоборот, тропические леса по отношению к обитателям саванн (см. рис. 18).
2. Границы ареала в большинстве случаев бывают обусловлены климатическими условиями. Годовой цикл продолжительности дня, хода температур и влажности воздуха — основные лимитирующие факторы распространения видов. Ограничительное воздействие этих факторов проиллюстрировано на примере распространения сахарного клена (см. рис. 8).
3. В некоторых случаях конфигурация ареала определяется неблагоприятным субстратом. Много примеров, иллюстрирующих существование такого типа эдафических границ, доставляют высшие растения. Распространение видов, обитающих на известняковых почвах, останавливается в зоне контакта с почвами, богатыми кремнием, и наоборот. Примером этого служат места соприкосновения Армориканского массива с юрскими ореолами Парижского бассейна.
Если формы полностью адаптированы к субстрату, имеющему специфические химические свойства, контур ареала строго эдафичен. Примером служат виды, обитающие на соленых почвах, которые во Франции расположены за пределами литоралей по соседству с солеными источниками Лотарингии и Эльзаса.
4. В локализации видовых ареалов существенную роль играют также взаимовлияния организмов самой различной природы: па-
32
Рис. 8. Биоклиматические границы ареала сахарного клена (Acer saccha-гит) (сплошная линия) (по Дансеро, Dansereau).
/ — 100 см годовых осадков; 2 — средний абсолютный минимум —40°; 3 — средний абсолют* ный минимум —10°.
разитизм, комменсализм, конкуренция, которые можно объединить под общим названием биотических факторов. Любопытным примером этому служит совпадение ареалов аконитов и шмелей (Bombus)\ последние обеспечивают оплодотворение аконитов.
Если вид локализован в определенной формации, то судьба его ареала тесно связана с судьбой этой формации. Вырубка леса благоприятствует видам, обитающим на открытых пространствах за счет лесных видов. В Советском Союзе южная граница обитания таких лесных видов, как бурый медведь, росомаха, белая куропатка !, отодвинулась к северу; в то же время грызуны
1 Этот вид обитает в тундре, в лесной зоне живет на болотах, в лесостепи — в березовых колках и ольшаниках; правильнее было бы привести в ачестве примера лесных птиц глухаря и рябчика. — Прим, ред,
2 Ж. Леме
за
из травянистых формаций — суслик, заяц1 — распространились в северном направлении. Это было связано с вырубкой соседствовавших со степью лесов. Распространение пастбищ в северных степях способствовало тому, что к югу отодвинулись ареалы дрофы (Otis tarda), степного орла (Aquila тарах), байбака (Mar-mota bobac)—видов, связанных с покрытой кустарниками первобытной степью.
5. Анализ одних только современных географических и экологических условий не позволяет объяснить распределение живых существ. Это распределение возникло в результате длительного процесса эволюции живых существ при одновременно изменяющихся географических и экологических условиях. Для объяснения образования эндемических, викарирующих, разъединенных ареалов необходимо учитывать исторические факторы.
Палеогеографические факторы современного распространения живых существ
Исследование эволюции ареалов, или историческая биогеография, или эпионтология, — очень непростое дело. Необходимо сопоставить знания, приобретенные в самых различных областях, чтобы собрать достаточно аргументов в пользу наиболее правдоподобных гипотез.
Методы исследования
Методы исследования могут, во-первых, заключаться в изучении данных о геологическом прошлом Земли и, во-вторых, в анализе современного распространения таксономических единиц.
1. Лишь палеонтология и палеоботаника могут найти прямые свидетельства возраста видов и их распространения в прошлом. Но, к сожалению, имеются большие пробелы в возможностях этого метода. Такие группы видов, как черви, не поддаются фоссилизации. Другое обстоятельство заключается в том, что мы судим об ископаемых формах только по морфологическим чертам тех частей организма, которые могут быть законсервированы нами; поэтому палеонтологическое понятие вида не может не отличаться от биологического, применяемого к современным формам.
Наряду с изучением макроскопических остатков сравнительно недавно стало развиваться исследование микрофоссилий: игл губок, раковин фораминифер, водорослей, спор, пыльцы растений.
2. Палинология, наука о пыльце и ее использовании, в большей степени содействовала палеоботаническому изучению верхпе-третичног и четвертичного периодов.
1 Это относится преимущественно к зайцу-русаку. Зайцы ныне относятся к особому отряду. — Прим. ред.
34
Переносящаяся в атмосфере пыльца может оседать на по-еохностях, благоприятных для ее фоссилизации при быстром погребении в условиях, способствующих ее сохранению. Подобными свойствами обладают торфяники, днища озер и некоторые виды почв. Определение и подсчет зерен пыльцы в образце, взятом в определенном месте, позволяет воссоздать средний состав пыльцевого дождя в ту эпоху, когда исследуемый уровень выходил на поверхность, и представить себе окружающий растительный пейзаж. Если взять некоторое количество таких «пыльцевых спектров» на различных уровнях отложений, то можно воссоздать эволюцию растительности за время образования этих отложений.
Было разработано несколько методов изображения результатов этих определений.
Особое значение придают выявлению деревьев. Это связано с их большой палеоклиматической ролью, с тем, что они являлись определяющей частью растительного пейзажа, и с тем, что их пыльцу сравнительно легко определить с точностью до рода. В настоящее время прилагают большие усилия к тому, чтобы четко определять пыльцу как видов деревьев, так и кустарниковых и травянистых форм, так как это позволило бы более глубоко изучить экологические и климатические условия в прошлом.
3. Палеогеография призвана воссоздать характер поверхности земного шара в далеком прошлом, распределение суши и моря, рельеф. С помощью этой науки мы можем узнать, каковы были возможности миграции морских и наземных видов в различные геологические эпохи. К сожалению, данные ее остаются очень фрагментарными, а часть аргументов базируется именно на знании распространения систематических групп растений и животных в прошлом и настоящем.
Что же касается способов, которыми осуществлялись временные коммуникации, то это — геологическая проблема, и мы не будем здесь подробно останавливаться на предлагавшихся гипотезах. Напомним лишь основные объяснения этих связей, имеющиеся в настоящее время.
а) Теория межконтинентальных мостов, которые временно связывали континентальные щиты. Примером современных континентальных мостов, широко используемых живыми существами, служат Панамский и Суэцкий перешейки, Берингов пролив, Па-де-Кале, проливы между полуостровом Малакка, Явой, Суматрой и Калимантаном поднимались над водой в плиоцене, и достаточно было бы небольшого понижения уровня моря, чтобы вернуть им роль мостов для наземных видов. Однако биогеогра-фь! слишком увлеклись гипотезами, объяснявшими современное Разобщение прямыми временными связями в прошлом. Часто такие гипотезы выдвигались без достаточного географического или геологического обоснования, и последующие палеонтологические открытия полностью их обесценивали. Например,
2*
35
современное распределение лемуров в Африке, на Мадагаскаре, в Индии и Индонезии объяснялось существованием гипотетического древнего континента — Лемурии, который якобы занимал часть Индийского океана и связывал Индию с Африкой. Открытие ископаемых лемуров в Европе и Северной Америке в эоцене доказало их широкое распространение в древности и сделало эту гипотезу ненужной.
б) Теория «дрейфа континентов» Вегенера (A. Wegener). Она предполагала, что соединенные друг с другом материки представляли собой обширный блок, который сначала подвергался перемещениям относительно экватора, а потом претерпел фрагментацию, после чего материки удалились друг от друга. Эта теория, которая в свое время была принята с энтузиазмом, сегодня забыта !.
4. Палеоклиматология, или изучение древнего климата, также необходима для понимания современного распространения организмов. Как и в случае палеогеографии, климатические реконструкции в большей своей части основываются на изучении распространения флоры и фауны в древности, что делает выводы ненадежными. Для эпох, предшествующих четвертичному периоду, речь идет почти исключительно о вымерших формах, но их потребности могли в значительной степени отличаться от потребностей нынешних, даже наиболее близких им видов. Кроме того, нужно учесть, что вследствие изменения высоты над уровнем моря могли меняться местные климатические условия.
Однако для восстановления эволюции климата имеются данные другого порядка: это древние формы рельефа, характер отложений и их чередование, ископаемые почвы и даже химические исследования1 2.
5. Методы, которые позволяют восстановить историю эволюции таксономической группы или вида — их филогению, — могут косвенно содействовать восстановлению их прошлого распространения. Они основаны на сравнительном изучении морфологии, кариологии и ареалов родственных форм.
а) Среди данных, способных указать на возраст и преемственность форм, особое значение имеет хромосомная характеристика. Формы с большим числом хромосом обычно происходят от предков с низким числом хромосом либо при помощи простого удвоения этого числа (автополиплоидия), либо, чаще всего, при помощи скрещивания (аллополиплоидия). Во многих родах и видах можно поэтому видеть серии форм, где наряду с диплоидными встречаются формы с высокой степенью полиплоидии. Это позволяет восстановить область возникновения и пути
1 В настоящее время наблюдается повое оживление интереса к этой гипотезе. сильно модифицированной геологами. — Прим. ред.
2 Речь идет о содержании радиоактивных изотопов углерода С13 и кислорода О18 в известняках скал и окаменелостей, которое зависит от температуры в период отложения.
.36
расселения этих форм. Оказывается также, что полиплоидные Формы имеют обычно большую экологическую амплитуду, что позволяет им распространяться в областях, не подходящих по климату их диплоидным предкам. Случается даже, что эти формы выживают после гибели диплоидных родителей в климатически неблагоприятные периоды.
Приведем примеры, иллюстрирующие эти факты. Однолетний мятлик (Роа annua) представлен двумя однолетними подвидами: диплоидным подвидом exilis (2м = 14), обитающим в
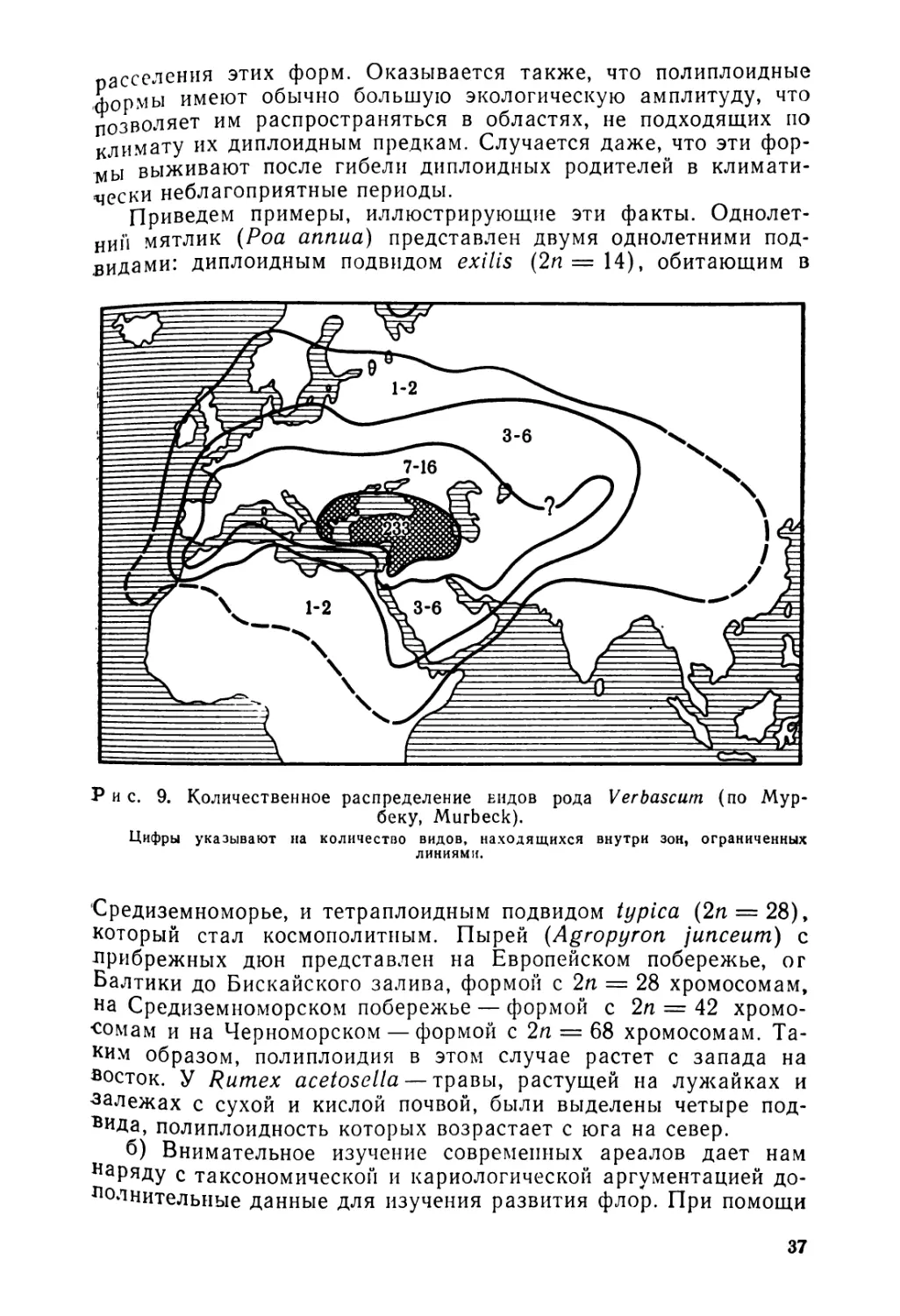
Рис. 9. Количественное распределение ендов рода Verbascum (по Мур-беку, Murbeck).
Цифры указывают на количество видов, находящихся внутри зон, ограниченных линиями.
Средиземноморье, и тетраплоидным подвидом typica (2м = 28), который стал космополитным. Пырей (Agropyron junceum) с прибрежных дюн представлен на Европейском побережье, ог Балтики до Бискайского залива, формой с 2п = 28 хромосомам, на Средиземноморском побережье — формой с 2п = 42 хромосомам и на Черноморском — формой с 2м = 68 хромосомам. Таким образом, полиплоидия в этом случае растет с запада на восток. У Ритех acetosella — травы, растущей на лужайках и залежах с сухой и кислой почвой, были выделены четыре под-Бида, полиплоидность которых возрастает с юга на север.
б) Внимательное изучение современных ареалов дает нам наряду с таксономической и кариологической аргументацией дополнительные данные для изучения развития флор. При помощи
37
хорологических методов стало возможным исследование центров расселения.
Богатство какой-либо таксономической единицы в пределах ее ареала неодинаково. Область наибольшего разнообразия, или центр максимального разнообразия, является центром расселения. Не надо, однако, думать, что такой центр обязательно должен быть областью возникновения данной таксономической единицы. Последняя может оказаться значительно более древней, и тогда современный центр расселения является вторичным. На карте (рис. 9) показан центр расселения рода Verbascum, представленного наибольшим числом видов в Юго-Восточной Европе и на Ближнем Востоке.
Выводы из наблюдений об относительном таксономическом богатстве в различных участках родового ареала часто подтвер-» ждаются изучением особенностей хромосом.
Род Iris, который включает около 250 видов, занимает непрерывный ареал в умеренных областях северного полушария. Однако наибольшее число видов и наибольшее разнообразие морфологических признаков наблюдается в восточной части средиземноморского бассейна. Большей частью у видов из этой области число хромосом невелико (от 2п = 16 до 2п = 24). Эта область была «колыбелью» рода Iris. Отсюда пошли все более полиплоидные формы, которые распространились в Европе и Восточной Азии вплоть до Северной Америки, где существует примерно 30 эндемичных форм, имеющих более 40 хромосом.
Глава 3
Флористические и фаунистические территории
Сравнение многочисленных ареалов позволяет выявить зоны, где накладывается друг на друга особенно большое число их границ. Эти пограничные зоны, появление которых вызвано некоторой, существовавшей в прошлом или в настоящее время, причиной географического или климатического характера, определяют контуры территорий, внутри которых состав флоры и фауны обладает некоторой однородностью. Эти территории определяются таксономическими единицами, которые только им свойственны или находят в их пределах наибольшее развитие. Совокупность таксономических единиц, характеризующих территорию, часто называют «элементом».
Таксономические единицы образуют иерархию. Точно так же территории, обнаруживающие между собой наибольшее сходство, объединяются в хорологические единицы более высокого ранга. Таким образом, существует иерархия территорий. Ступени этой иерархии получали различные наименования. Более всего приняты следующие: царства, области (или регионы), подобласти, секторы, участки !. Царства или области характеризуются крупными таксономическими единицами — например, целыми семействами, для подобласти скорее характерен род, а для сектора — вид.
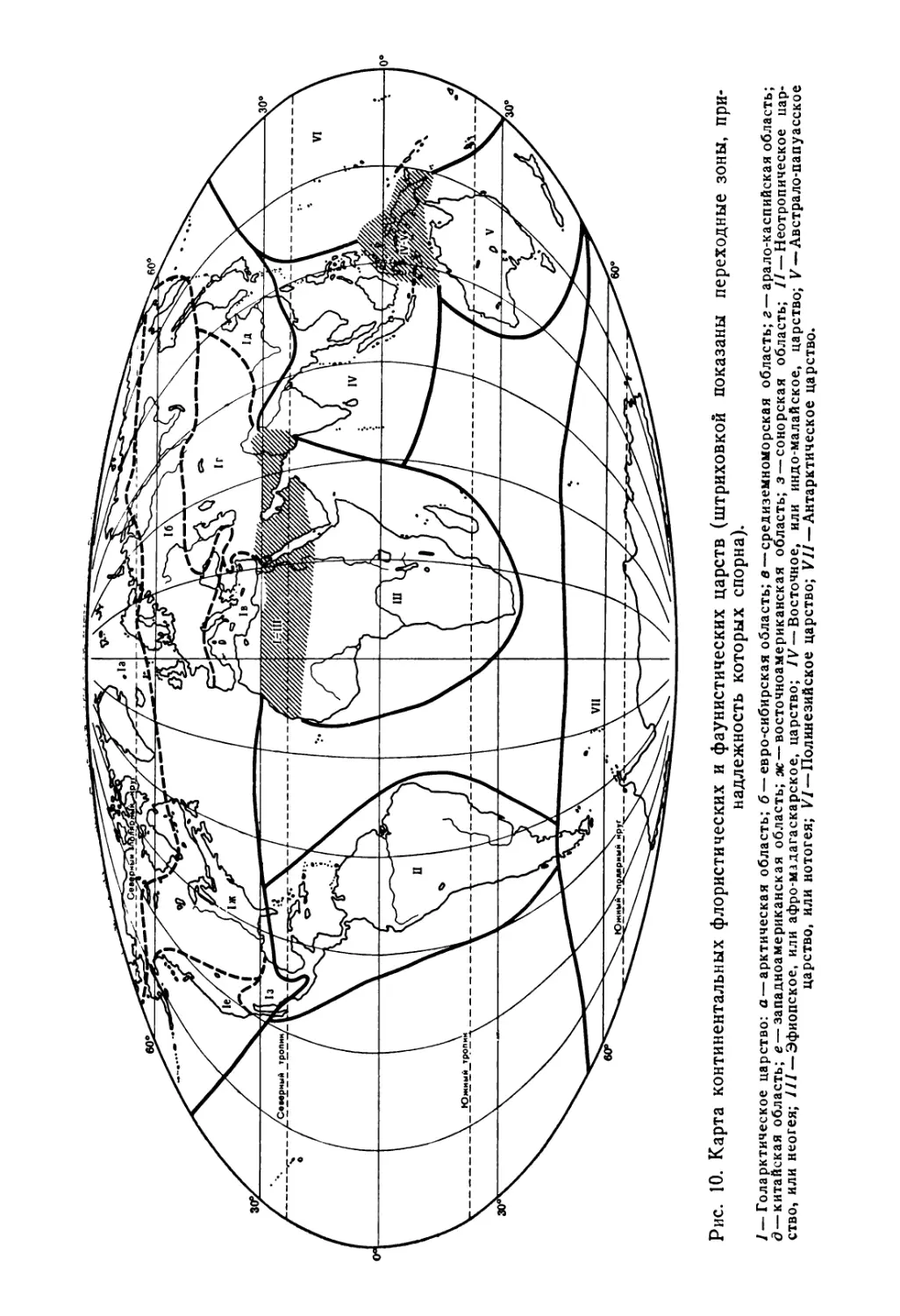
Естественно, что в зависимости от того, с какой систематической группой приходится иметь дело, границы и мера сходства биогеографических территорий могут различаться сообразно с экологией и историей данной группы. Однако в общем плане континентальные территории можно разделить на шесть крупных царств: голарктическое, неотропическое, эфиопское (афро-мадагаскарское), восточное (индо-малайское), австрало-папуасское и антарктическое; иногда выделяют также спорное полинезийское царство (рис. 10).
1 В качестве синонима «царства» автор предлагает регион; в качестве синонима региона (области)—провинцию. В русской литературе провинция — ДинИца> следующая за подобластью. Под подобластью автор понимает do-1\^1Пе (°бласть): в русской литературе это крупная категория, охватывающая ЧацКОлько областей. Наконец, вместо термина «сектор» в русской литературе Упо применяется «округ». Таким образом, в русской литературе наиболее Ласт^е^ИТельпа слеДУюЩая иерархическая система территорий: царство, об-ь» подобласть, провинция, округ, участок. — Прим. ред.
39
Голарктическое, или бореальное, царства
В целом к этому царству относят области, лежащие к северу от тропика Рака. Но лишь в Центральной Азии, где проходит гималайский барьер, границы этого царства с тропическим миром можно считать четкими. В Северной Америке, Сахаре и Китае имеются большие переходные зоны.
На этой громадной территории сосредоточены центры ареалов или же подавляющее большинство представителей многих групп животных и растений. Среди животных характерны семейства собачьих, медведей, бобров, кротов, а также хвостатые амфибии, лососевые и осетровые рыбы, жужелицы; из растений — большие семейства лютиковых, розоцветных, крестоцветных, зонтичных, ивовых.
Однако голарктические фауна и флора значительно беднее по видовому составу, чем тропические, которым они постепенно пришли на смену в результате изменения (похолодания) климата в третичном периоде. Их происхождение связано либо с адаптацией древних форм, либо с возрастанием разнообразия современных групп.
Биологическое население голарктического царства, несмотря на многочисленные общие черты, сохраняет большое разнообразие. Причины этого кроются прежде всего в разнообразии современного климата, а также в четвертичных оледенениях и непостоянстве связей между Евразией и Северной Америкой. Различают несколько областей на каждом из этих двух континентов.
Арктическая область
Это область бореальной тундры. Немногочисленные ее виды-были отобраны из наиболее холодоустойчивых форм аркто-тре-тичной флоры и фауны, в особенности из болотных, горных и прибрежных форм. Во время оледенений они либо отступили перед ледниками, либо сохранились в не затронутых ледниками убежищах — например, на Аляске, которая, будучи в то время отрезанной от других частей Северной Америки, была связана с северо-восточной азиатской тундрой через поднявшийся Берингов перешеек. Особенностями связей и разобщений того времени по сравнению с имеющими место сегодня и объясняется происхождение современных ареалов. Многие виды имеют циркумполярный ареал; они представлены иногда различными местными расами, как видно на примере тюленя (Phoca hispida) (рис. 11). Одни и те же роды могут быть представлены на севере Евразии, с одной стороны, и в Северной Америке и Гренландии— с другой, викарирующими видами. Таковы северный олень и карибу, которые являются двумя расами одного и того же вида, заяц-беляк и лемминги. Некоторые виды, которые были когда-то циркумбореальными, локализованы в настоящее время
40
Рис. 10. Карта континентальных флористических и фаунистических царств (штриховкой показаны переходные зоны, принадлежность которых спорна).
/ — Голарктическое царство: а — арктическая область; б — евро-сибирская область; в — средиземноморская область; г —арало-каспийская область; д — китайская область; е — западноамериканская область; ж — восточноамериканская область; з — сонорская область; //— Неотропическое царство, или неогея; /// — Эфиопское, или афро-мадагаскарское, царство; IV — Восточное, или индо-малайское, царство; V — Австрало-папуасское царство, или нотогея; VI — Полинезийское царство; VJJ — Антарктическое царство.
на более или менее ограниченных территориях. Например, мускусный овцебык, еще недавно обитавший на Аляске, а во время последнего оледенения — и в Центральной Европе, встречается в настоящее время лишь в Гренландии и на северо-востоке Канады.
И наоборот, многие виды распространены также и в высокогорных областях средних широт, так что невозможно установить,
Rubus chamaemorus
Рис. 11. Ареалы двух арктических циркумполярных видов (по Вальтеру, Н. Walter).
Слева — тюлень (Phoca hispida), представленный множеством викарирующих подвидов (по Дэвису, J. L. Davies); справа —морошка (Rubus chamaemorus).
Слева: / — тип; 2 — ssp. beaufortiana; 3 —ssp. krascfieninikovi', 4 —ssp. ос hot e fl-sis; 5 —ssp. birulai-, 5 —ssp. botnica‘, 7 —ssp. pomor or um.
происходят ли эти аркто-альпийские виды от северных или горных видов. И наконец, многие виды являются холодоустойчивыми формами евро-сибирской и борео-американской областей.
Оседлые позвоночные, как и моллюски, крайне малочисленны; из насекомых относительно хорошо представлены двукрылые и ногохвостки (Collembola) в отличие от жесткокрылых и чешуекрылых. Среди семенных растений циркумполярных видов очень мало (например, Rubus chamaemorus, см. рис. 11). В отличие от них хорошо представлены мохообразные и лишайники.
Евро-сибирская область
Эта обширная область представлена в Европе несколькими четко очерченными подобластями.
1. Для циркумбореальной подобласти, лежащей на севере, характерны несколько видов хвойных, которые создают основу
42
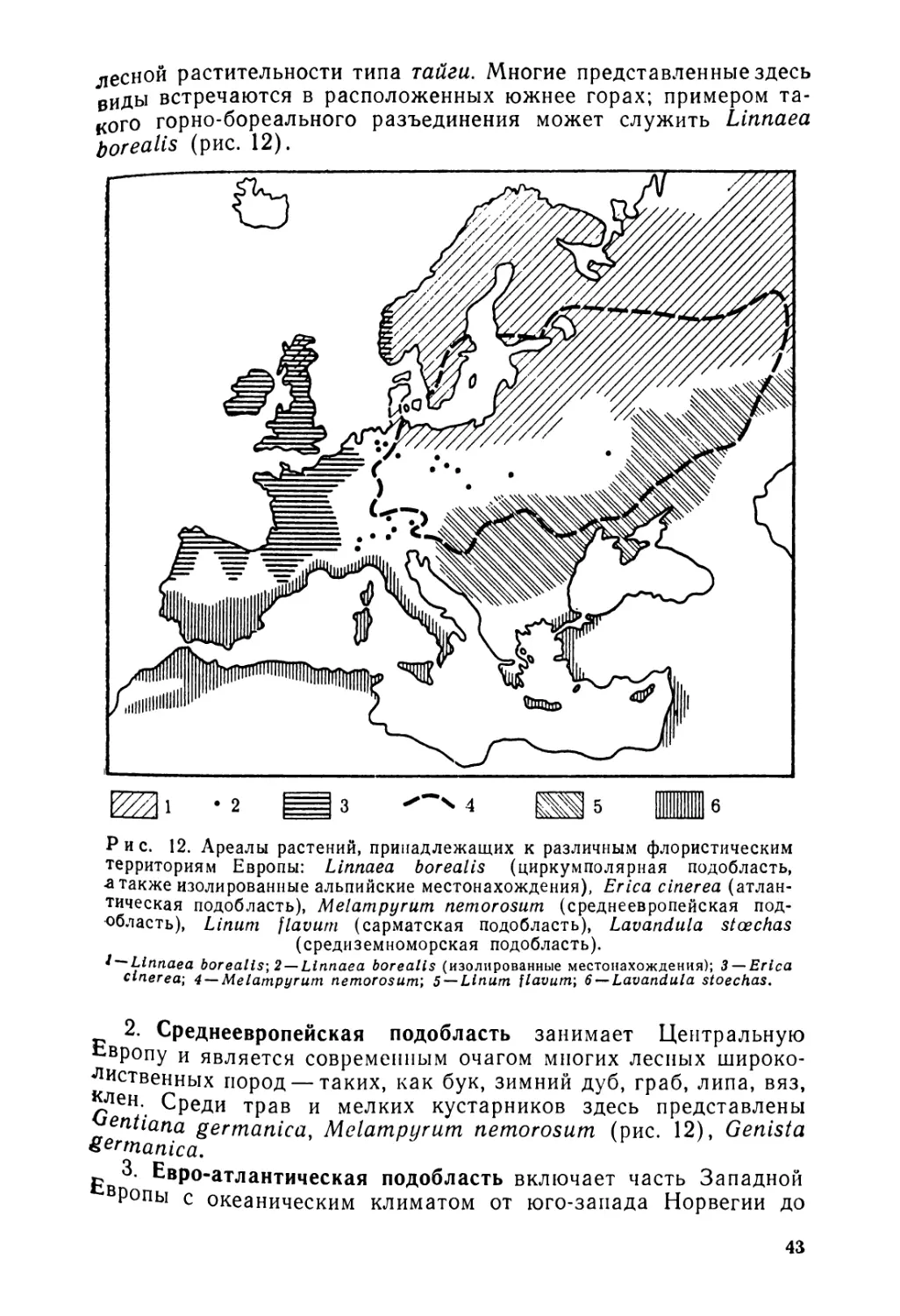
десной растительности типа тайги. Многие представленные здесь виды встречаются в расположенных южнее горах; примером такого горно-бореального разъединения может служить Linnaea borealis (рис. 12).
Рис. 12. Ареалы растений, принадлежащих к различным флористическим территориям Европы: Linnaea borealis (циркумполярная подобласть, -а также изолированные альпийские местонахождения), Erica cinerea (атлантическая подобласть), Melampyrutn nemorosum (среднеевропейская подобласть), Linum flavum (сарматская подобласть), Lavandula stcechas (средиземноморская подобласть).
i — Linnaea borealis-, 2 —Linnaea borealis (изолированные местонахождения); 3 —Erica cinerea; 4 —Melampyrutn nemorosum; 5 —Linum flavum; 6 —Lavandula stoechas.
2. Среднеевропейская подобласть занимает Центральную Европу и является современным очагом многих лесных широколиственных пород — таких, как бук, зимний дуб, граб, липа, вяз, ^лен. Среди трав и мелких кустарников здесь представлены entiana germanica, Melampyrutn nemorosum (рис. 12), Genista %ermanica.
Ев
3. Евро-атлантическая подобласть включает часть Западной Р°пы с океаническим климатом от юго-запада Норвегии до
43
северо-запада Пиренейского полуострова. Во Франции ее граница проходит через восточную часть Парижского бассейна, Морван, восточную окраину Центрального массива и Центральные Пиренеи. Этим районом ограничен ареал многих растений. Среди них собственно атлантические растения, например утесник карликовый (Ulex nanus), Erica ciliaris, vagans, tetralix, Endymion non-scriptum (рис. 13). Другие растения, однако, распространены дальше на восток, к Балтике, Центральной и Южной Европе и даже к Северной Африке: эта такие субатлантические виды, как пурпурная наперстянка, Sarothamnus, Genista anglica (рис. 13). Ланды, поросшие утесником, дроком и вереском, наиболее характерны для пейзажа этой области. На этой территории наблюдается загадочный тип разъединения: относящиеся сюда виды встречаются одновременно в Ирландии и северо-западной части Пиренейского полуострова, а иногда и на юго-западе Франции. Среди семенных — Daboecia polifolia и Saxifraga geum, среди моллюсков — Geomalacus maculosus, среди многоножек — Polydesmus gracilis.
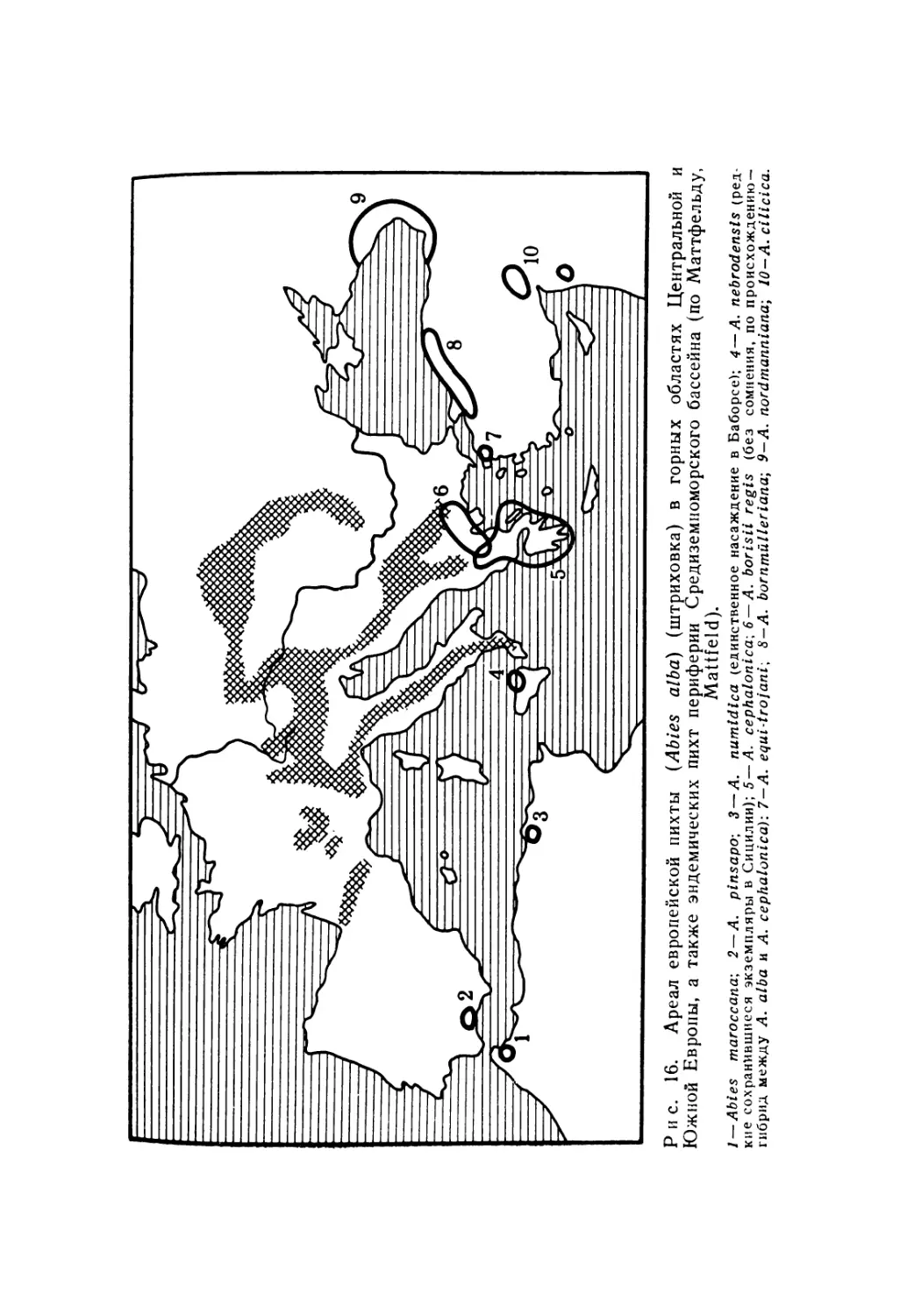
4. В особые подобласти выделяют верхние пояса высокогорья. В Европе к ним относят альпийскую подобласть (европейское высокогорье), а на юго-востоке — кавказскую подобласть. Дей* ствительно, здесь почти не встречаются равнинные виды. Напротив, очень многие роды представлены здесь большей частью своих видов; например, средн семенных таковы горечавки (Gen-Напа), проломник (Androsace), камнеломки (Saxifraga), соль-данеллы (Soldanella) (рис. 14). Альпийские виды, относящиеся к родам, хорошо представленным и на равнине, имеют, однако, лишь отдаленное родство с равнинными формами. Большая часть населения европейского высокогорья относится к аркто-альпий-скому элементу, современное распределение которого является следствием четвертичных оледенений. К нему относится почти 2/з семенных альпийского пояса в Центральных Альпах. Примеры такого распространения приведены на рис. 6.
Что же касается эндемиков, которые отсутствуют в северных областях, то они, по-видимому, третичного происхождения. Г и М. Брокман-Ерош (Н. et М. Brockmann-Jerosch) различают в альпийской флоре бореальную и южную группы. Представители первой группы, примером которых могут служить горечавки, покрывали, по-видимому, средние широты Евразии, и, таким образом, изолированные оледенениями родственные виды встречаются в Гималаях, горах Центральной Азии и даже на равнинах Дальнего Востока. Ко второй группе относят роды средиземноморского происхождения — такие, как льнянка (Li-naria), молодило (Sempervivum), Globularia. Изоляция фауны и флоры в различных горных цепях в межледниковые и постледниковые периоды привела к неоэндемизму. Например, заяц-беляк (Lepus timidus) представлен в Альпах и на Кавказе различными подвидами. Альпийская серна (Rupicarpa rupicarpa).
44
tu1 C*( Ареал АВУХ евро-атлантических растений: Endymion поп-scrip-tn (собственно атлантическое) и Genista anglica (субатлантическое) (ria Дюпону, Р. Dupont).
naymion non-scri ptum (точкой показано изолированно? местоположение)^ Genista anglica (крестиком показано изолированное местоположение).
Р II с. 14. Ареал альпийского рода Soldanella: показан вертикальной штриховкой, а для изолированных местонахождений — крестиками вид Soldanella alpina', точками обозначены другие виды этого рода (по Фирхап-перу, Vierhapper, упрощенно).
почти не отли.чается от пиренейской серны. Козлы, обитающие в Альпах (Capra ibex), Пиренеях и Иберийских горах (С. руге-naica), на Крите и Кавказе (С. hircus), находятся в близком родстве между собой.
Западно- и центральноазиатская, или арало-каспийская, область
Эта область включает степные и аридные территории Украины и Малой Азии вплоть до пустыни Гоби. Характер ее населения определяется резко выраженным континентальным климатом. Здесь представлено много различных форм: роды злаковых (Stipa, Aristida, Agropyron), примерно 15 эндемических родов маревых, бобовые (1600 видов Astragalus, а также Caragana, Oxytropis, Alhagi), гречишные (19 видов Calligonum из двадцати эндемичны), полыни и тамариски. На транскаспийской территории примерно 74 семенных видов эндемична.
Животный мир здесь также своеобразен и включает таких представителей, как кулан (Equus hemiones), антилопа сайга, верблюд дромадер, разнообразные грызуны (сурки, суслики, хомяки), дрофы, рептилии, многочисленные насекомые, например саранчовые.
Европейская часть этого биогеографического района составляет понтийскую, или сарматскую, подобласть, в состав которой
46
входят степи южнорусские и Дунайского бассейна до Венгрии. Примером сарматского ареала может служить ареал Linum flavum (рис. 12). Западнее этих мест можно обнаружить изолированные колонии сарматских видов, занимающие небольшие, благоприятные для степной флоры территории. Можно назвать, например, долины Центральных Альп, где имеются лужайки, поросшие Stipa, Astragalus, Oxytropis pilosa и т. п., и — в меньшей степени — южное известняковое плато Центрального массива Франции (Коссы), где встречаются Stipa, Scorzonera purpurea, Adonis vernalis. В окрестностях Майнца на сыпучих песках сохранилась большая колония степной флоры. Выше мы указали на палеоклиматическое происхождение этих разъединений.
Средиземноморская область
Эта область, определяемая средиземноморским климатом, очень четко ограничена с севера, так как горные цепи проходят вдоль берега. Но на равнинах и широких долинах переход к евроазиатской области носит значительно более плавный характер. Африканский и восточный участки этой области непосредственно переходят в засушливые пространства Сахары и арало-каспийской области.
От третичного периода здесь сохранилось лишь несколько реликтов, принадлежащих к тропическим семействам: мартышкообразные (макака Гибралтара и Северной Африки), пальмы (Chamaerops), из лавровых — лавр благородный (Laurus nobi-lis), марокканское железное дерево (Argania) из семейства Sapotaceae, из цезальпиниевых — рожковое и иудино дерево. Большая часть этих реликтов принадлежит к африканской части области. Оледенения четвертичного периода привели к значительному похолоданию в Северной Африке, что сопровождалось эмиграцией в этот район среднеевропейских и северных видов. Часть этих видов выжила (как, например, Cardamine pratensis, Ribes alpinum и т. п.), другие исчезли (например, граб, белая ива, ломкая крушина, остатки которой были найдены в окрестностях Алжира).
Сильно выражен эндемизм — около тысячи цветковых растений. Многие роды имеют здесь центр расселения. Таковы Muscat, Ophrys, Brassica и Sinapis, Coronilla и Ononis и т. д. Считается, что границы ареала культуры оливкового дерева точнее всего совпадают с границами средиземноморской флористической области (рис. 15; см. также ареал лаванды — Рис. 12).
Помимо собственно средиземноморских видов, ареал которых целиком находится внутри этой области, очень важный контингент простирается на значительное расстояние к северу, за-
47
Рис. 15. Ареал культуры оливкового дерева (Olea еигораеа) (па рисунке— черным цветом), а также викарирующего вида Olea laperrinel из гор центральной Сахары'(на рисунке — точками).
ходя в евро-сибирскую область. Во Франции в период потепления, наступившего вслед за оледенением, отмечались два пути миграции:
1) По бассейну Роны, в котором собственно средиземноморская растительность представлена от Донзерского ущелья до Баланса. Благодаря существованию этого пути на север смогли продвинуться многие субсредиземноморские виды: зеленый дуб распространился до Вьенны, теребинтус, или фисташка терпентинная (Pistacia terebintus), ладанник (Cistus salvifolius), лаванда (Lavandula spica), жасмин (Jasminum fruticans) распространились до южной Юры, а пушистый дуб —до Эльзаса.
2) Второй путь проходил на востоке Франции, через Аквитанию и Коссы. Некоторые виды при этом закрепились в Аквитании (теребинт, Cistus laurifolius, Lavandula latifolia), другие продвинулись не далее Шаранты (Phillyrea angustifolia, Ruta graveolens) или Вандеи (Phillyrea media, Acer monspessulanus). Примерно 200 видов пересекли Луару и проникли в Бретань, в Нормандию и Парижский бассейн, а некоторые даже на Британские острова (Ruscus aculeatus, Rubia peregrina, Umbilicus pendulinus).
48
Из-за альпийского орогенеза в третичном периоде географический облик средиземноморского бассейна подвергся глубоким изменениям; в четвертичном периоде последовали климатические изменения; эту область рано заселил человек. Весь этот комплекс факторов привел к многочисленным случаям эндемизма и дробления ареалов внутри средиземноморской области. Особенно большое влияние оказали эти события на население горных цепей и островов. Многочисленные виды, свойственные горам Средиземноморья, представлены викарирующими формами, которые изолированы на различных горных массивах. На карте (рис. 16) представлен классический пример распространения средиземноморских пихт.
Хорошим примером островного эндемизма является население Корсики и Сардинии. Эти острова отделены от Италии водным пространством с небольшими, не превышающими 105 м, глубинами, из которых к тому же выступает Тосканский архипелаг, и вполне возможно, что они являются свидетелями существования той суши, которая была связана с континентом до плейстоцена и получила название Тирренис (Tyrrhenis). О наличии таких, хотя бы временных, связей свидетельствует видовой состав млекопитающих в плейстоцене и в настоящее время. В период плейстоцена на этих островах обитали макаки и антилопы, карликовая раса слона (Elephas antiquus) и кистеухие или речные свиньи, Potamochoerus,— все формы, характеризующиеся тропическими родственными связями. Напротив, недавно исчезнувший бурый медведь и мелкое зайцеобразное Prolagus, которое должно было играть важную роль в питании неолитического человека, имели европейское происхождение. Из современных млекопитающих, которые насчитывают лишь около 30 видов, преобладает европейский элемент, представленный иногда местными расами, как у кабана; африканские черты присущи степной (пятнистой) кошке и средиземноморскому зайцу-русаку с Сардинии; благородный олень и корсиканский муфлон (эндемичный вид) имеют восточное происхождение. Что касается птиц, то хотя на Корсике и Сардинии нет эндемичных рас, здесь имеются как многочисленные оседлые расы перелетных видов, например обыкновенная ворона и соловей, так и оседлые виды, например сарыч и серая куропатка.
В сравнении с позвоночными животными эндемизм флоры выражен ярче. Имеется около сорока видов семенных, а также многочисленные подвиды и разновидности. В их состав входит несколько палеоэндемиков, среди которых одни принадлежат к родам, имеющим североафриканские родственные связи {Morisia, Nananthea), а другие таксономически изолированы внутри рода. Встречаются, однако, и неоэндемики, представленные на континенте такими викарирующими формами, как Ranun* cuius revelierii и Lamium corsicum.
49
Североамериканские области
Животное и растительное население Северной Америки по сравнению с населением Европы и внетропической Азии характеризуется большим сходством семейств и родов; однако видов, общих для этих территорий, мало. Это хорошо видно на примере лесных пород, представленных различными видами одних и тех же родов — буков, дубов, кленов, вязов, лип, сосен, пихт, елей. То же относится к млекопитающим — волкам, медведям, куницам, оленям, зайцам и т. п. европейский бизон отличается от американского.
Если один и тот же вид встречается на двух континентах, то, как правило, он представлен двумя викарирующими расами, например, лоси хвойных лесов Северной Америки несколько отличаются от своих сородичей из евразиатских лесов.
Эти факты указывают на то, что вплоть до относительно недавнего времени между Евразией и Северной Америкой существовали связи, после разрыва которых потомки форм, населявших эти два континента, эволюционировали различно. По-видимому, эти связи осуществлялись главным образом через Берингов пролив, малая глубина которого в настоящее время позволяет предположить, что в некоторые эпохи он играл роль межконтинентального моста. Несмотря на весьма северное расположение этого моста, в периоды межледниковых потеплений через него смогли проникнуть формы умеренного климата. Роль Берингова моста в миграции млекопитающих третичного периода общеизвестна.
Помимо различий, вызванных дивергентной эволюцией населения обоих континентов, наблюдаются различия, связанные с вымиранием некоторых форм на одном из континентов после их разделения. Население Европы было обеднено в результате оледенений, при которых исчезли многочисленные таксоны тропического происхождения, сохранившиеся в Северной Америке: Sequoia, Taxodium, Magnolia и т. п. Но, согласно современным палеонтологическим данным, имели место случаи вымирания и в Северной Америке: так было с верблюдами и ламами, с антилопой сайгой, которая сейчас обитает в Центральной Азии, — один вид этих антилоп населял в плейстоцене Аляску.
Другая особенность североамериканских биологических районов, и особенно сухих областей на юго-западе (Сонорская область), состоит в том, что они не так давно (по-видимому, в плиоцене) обогатились южноамериканскими тропическими формами: броненосцами (Dasypus novemcinctus), дикобразами (Erethizon dorsatum), сумчатыми (опоссумы), птицами (колибри, танагры), игуанами (Phrynosoma), ящерицами (Teiideae), кактусами и т. п.
50
Рис. 16. Ареал европейской пихты (Abies alba) (штриховка) в горных областях Центральной и Южной Европы, а также эндемических пихт периферии Средиземноморского бассейна (по Маттфельду, Mattfeld).
1 — Abies maroccana; 2— A. pinsapo; 3 — A. numidica (единственное насаждение в Баборсе); 4 — A. nebrodensis (редкие сохранившиеся экземпляры в Сицилии); 5 — A. cephalonica-, 6 —A. borisii regts (без сомнения, по происхождению — гибрид между A. alba и A. cephalonica): 7 —A. equi-trojani, S — A. bornmtilleriana; 9—A. nordmanniana; 10 —A. cilicica.
Необычайное разнообразие биологического населения области Северной Америки, расположенной между арктическим царством и тропической Мексикой, привело к разделению ее на несколько подобластей. Причины такого разнообразия — отчасти исторические (распространение канадского оледенения на Великие равнины), отчасти географические (горный барьер системы Кордильер на западе, к которому примыкает аридная зона), отчасти климатические (разнообразие современных климатических условий).
Неотропическое царство, или неогея
Эта биогеографическая территория простирается от Центральной Америки и Антильских островов до Патагонии и включает крайне разнообразные климатические зоны. Ее объединение в оригинальное единое целое является результатом истории. В меловом и в начале третичного периода она была связана с Северной Америкой и, возможно, с Антарктидой и Африкой, но затем оказалась изолированной вплоть до плиоцена. В течение длительной изоляции имевшиеся на ней формы эволюционировали самостоятельно, и некоторые из них развились в своеобразные филумы, изменчивость которых позволила им заселить разнообразные среды обитания.
У некоторых групп мы обнаруживаем ископаемые формы лишь на территории Южной Америки, где эти группы обитают и поныне. По-видимому, эта территория и была местом их возникновения. Таков среди млекопитающих отряд неполнозубых (Edentata или Xenarthra). Две ветви этого филума прекратили существование лишь в конце плейстоцена: это глиптодонты (Glyptodontideae) и близкие к ленивцам Gravigrada. В настоящее время остались лишь броненосцы, муравьеды и ленивцы. Таковы и широконосые обезьяны (Plathyrrhini), происхождение которых неизвестно.
Другие группы, распространенные первоначально в северном полушарии, впоследствии там исчезли: таковы сумчатые, представленные в настоящее время в Южной Америке семейством Didelphyidae, ископаемые формы этих животных существовали с мелового периода в Северной Америке, а в третичном периоде— в Европе L В третичном периоде в Южной Америке развились особые филумы копытных: это южноамериканские No-tungulata, почти неизвестные в северном полушарии, и отряд Litopterna. Они претерпели эволюцию, аналогичную эволюции североамериканских лошадей. Достигнув замечательного разнообразия, эти группы исчезли в плейстоцене.
Подобные случаи палеоэндемизма можно найти для всех
1 Кроме того, в Южной Америке имеется эндемическое семейство сумчатых — ценолестовые. — Прим. ред.
62
групп животных: из птиц можно назвать примитивное семейство тИнаму (Tinamiformae), туканов, нанду; среди рептилий —семейство ящериц Teiideae и кайманов; среди амфибий — несколько родов безногих (Apoda) и Pipa\ среди рыб — двоякодышащего Lepidosiren, электрического угря и т. д.
Высок уровень эндемизма и среди сосудистых растений. Только среди тропических покрытосеменных Гуд (R. Good) насчитал 25 семейств1 и около 3500 эндемических родов, таких, как канна, настурция, какао, хевея, хинное дерево, Cattleya.
В некоторых случаях южноамериканское третичное население имеет современных потомков и в других частях света. Это заставило биогеографов предположить, что в какие-то времена существовали прямые межконтинентальные мосты между этими частями света и Южной Америкой. Изучая ископаемые остатки, можно прийти к выводу, что такое распространение представляет собой остаточный ареал форм, имевших некогда широкое распространение. Например, курьезное современное распространение черепах Podocnemus из семейства Pelomedusideae в Южной Америке и на Мадагаскаре — реликт некогда существовавшего межконтинентального распределения. Это семейство было обнаружено в меловых отложениях в Северной Америке, в эоцене и в миоцене — в Африке и Европе и в эоцене — в Индии. Современное распределение грызунов Histricoidea, которые внезапно появились в миоцене в Патагонии и широко распространены в настоящее время в Южной Америке (агути, шиншилла, морская свинка, дикобраз), представляется несколько загадочным. Известны представители этой группы в Африке, но ископаемых остатков нигде найдено не было. Это послужило аргументом для палеогеографов в пользу существования в олигоцене африканобразильского моста 2.
И действительно, примеры подобного распределения семейств, родов и даже видов можно найти в самых разнообразных группах. Оно характерно для семейства черепах (Chelidideae), рыб — цихлиды (Cichlideae) и харациды (Characideae), пластинчатожаберных моллюсков (Mutelidae), птиц, которые представлены по обе стороны Атлантики идентичными или викарирующими видами (в особенности среди гусиных), и наконец, для родов семенных (Symphonic^ см. рис. 7).
1 Три больших семейства, рассматриваемые как эндемические, не являются, однако, вполне таковыми: это Bromeliaceae, представитель которых был недавно найден на массиве Фута-Джаллон (Африка), кактусовые, среди которых один вид, эпифит Rhipsalis baccifera, имеет пантропическое распределение, и Vochisiaceae, представленные локализованным в Конго моноспеци-Фическим родом.
По Вуду (A. Wood), две эти ветви Histricoidea — африканская и гожно-ернканская — произошли в действительности в результате параллельной и ависимой эволюции группы Paramyida, широко распространенной в эоцене "а всем северном полушарии.
53.
В плиоцене это своеобразное архаичное население Южной Америки претерпело изменения из-за образования нового моста между Северной и Южной Америкой (об этом свидетельствует теперь Панамский перешеек); это событие было вызвано крупными эпейрогеническими движениями земной коры. Обмен формами происходил в обоих направлениях. Выше приводились примеры проникновения из Южной Америки в Северную, но и
Рис. 17. Высотная локализация американских чешуекрылых в зависимости от широты. Показана роль ' экваториального моста, которую играют высокие горные цепи (по Хованицу, Hovanitz).
Южная Америка в свою очередь наводнена голарктической фауной. Там встречаются такие животные, как ягуары и пумы, тапиры, ламы (гуанако и вигони), пекари, олени (Cervidae), медведи (Tremarctos), носухи (Procyonidae), черепахи (Chelydra, Chrysemus) и с недавнего времени лягушки (Rana). Среди растений назовем смородины и горечавки (Gentiana), которые представлены в Аидах вплоть до Огненной Земли. Кордильеры облегчили переход через высокогорные области из одного полушария в другое видов, свойственных умеренным широтам (рис. 17).
Влияние этого двойного потока на смешение северо- и южноамериканской фауны было количественно оценено Симпсоном (G. Simpson) для млекопитающих: до плиоцена в Северной Америке было 27 семейств, а в Южной — 29, при этом ни одно из них пе было общим. В плейстоцене 22 семейства оказались общими, из них 14 происходили из Северной Америки, 7 — из Южной, не ясно происхождение лишь одного семейства. Другим примечательным фактом является исчезновение в Северной Америке некоторых видов после их проникновения в Южную, для которой они в настоящее время эндемичны. Таковы тапиры,
£4
ламы, пекари, очковый медведь (Tremarcios ornatus), единственный близкий ему вид — Т tibetanus — обитает в Центральной Азии, а также черепахи из рода Geomyda, которые в настоящее время встречаются еще лишь в индо-малайской области.
Вслед за этим обогащением Южной Америки североамериканской фауной последовало резкое обеднение автохтонной южноамериканской фауны; причины его, без сомнения, разнообразны. Согласно Фурону (Furon), оно отчасти было связано с «нарушением биологического равновесия и жестокой конкуренцией между группами с одинаковым образом жизни». Возможно, наконец, что косвенной причиной этого кризиса в конце плейстоцена послужили в большей или меньшей степени и оледенения; этот кризис продолжался, по существу, не более десятка миллионов лет и ознаменовался массовым исчезновением автохтонных третичных групп (Gravigradae \ Glyp-todontidae, Notungulata, Litopterna) и групп, эмигрировавших в плиоцене (лошади, мастодонты и крупный хищник Smilodon).
С неогеей связывают три группы островов: Антильские, Галапагосские и острова Хуан-Фернандес, современное население которых характеризуется ярко выраженным эндемизмом. Антильская фауна сохранила примитивных насекомоядных, Sole-nodon (один вид на Кубе, другой на Гаити), несколько крупных летучих мышей, наземных крабов и т. д. Население Галапаго-сов, изолированных вулканических островов, удаленных от Американского побережья на 700 км, поразило Дарвина в 1835 году удивительным сочетанием бедности и оригинальности населения. Туземная фауна позвоночных включает гигантскую сухопутную черепаху (по которой и названы острова), два эндемичных рода игуан, одну змею, одну ящерицу, южноамериканского геккона, эндемичный род крыс, а также птиц. Последние представлены южноамериканскими видами, пингвинами (единственный тропический вид этой группы) и птицами неизвестного происхождения— семейство Geospizidae (галапагосские вьюрки), которые адаптировались к различным местообитаниям этих островов. Трудно представить, что черепаха, крыса и игуаны пересекли морской пролив шириной 700 км. Это заставило некоторых биогеографов предположить, что некогда существовала связь этих островов с континентом, но сторонники теории исключительно океанического заселения отмечают, что морские течения могут Доставить плавающие предметы от американских берегов до островов за несколько дней.
1 Один представитель этой группы — Neomylodon — стал знаменит, когда скелет с остатками шерсти, лежащий на охапке ископаемого сена, был ^наружен в патагонской пещере. Это позволяет предположить, что животные Ыли одомашнены еще доисторическими людьми.
55
Эфиопское, или афро-мадагаскарское, царство
На фоне слабого сходства фауны и флоры тропической Африки с южноамериканскими настолько отчетливо выступает их сходство с населением тропической Азии, что фитогеографы объединили тропическую Африку и тропическую Азию в одно обширное палеотропическое царство. Напротив, население Южной Африки и Мадагаскара весьма самобытно, и поэтому эти территории часто выделяют из палеотропического царства; они будут рассмотрены нами раздельно. Мы также выделим особо Сахару, которая представляет собой зону перехода к па-леоарктическому царству.
Сходство черт населения тропической Африки и тропической Азии имеет в своей основе постоянство их наземных связей, свидетельством которых служит в настоящее время Суэцкий перешеек.
Как и на других континентах, лесистые области и области саванн или степей представляют собой совершенно различные биогеографические территории. Густые леса в Африке локализованы в настоящее время вблизи экватора и четко отделены от азиатских лесных тропических областей. Однако так было не всегда. В третичном периоде, быть может вплоть до плиоцена, преобладал влажный климат, и тогда леса тропического типа достигали Ближнего Востока, но этот лесной мост давно исчез. Поэтому среди лесного населения Африки и Восточной Азии есть общие семейства, но очень мало общих родов и почти нет общих видов (если не считать ввезенных человеком). Среди животных примером могут служить человекообразные обезьяны ], семейство Lorisidae1 2, панголины, оленьки, слоны; среди растений— род деревьев Ancystrocladus (рис. 7).
Равным образом продолжительная изоляция этих областей друг от друга сказывается на отсутствии в них некоторых групп. В лесной тропической Африке встречаются только свойственные ей семейства: чешуехвостые белки (Anomalurideae) и птицы турако. Это область излюбленного обитания некоторых групп, из птиц, например, ткачиков (Ploceidae) и мышиных птиц (Coli-idae). Формы, характерные для тропической Азии, будут упомянуты ниже.
С возрастанием сухости климата лесная зона Африки оказалась сначала изолированной, а потом—фрагментированной. С запада на восток прослеживаются массивы Либерии и западной части Берега Слоновой Кости, затем массив восточной
1 Горилла и шимпанзе — африканские животные, орангутан — восточное.
2 Семейство лемуров, представленное в Западной Африке потто, а в Восточной Азии — лори.
56
Сти Берега Слоновой Кости и Ганы1, и наконец централ ыю-(Ьриканскнй массив, который простирается от низовий Нигера яо бассейна Конго. Некоторые виды локализованы в одном из Перечисленных массивов или же представлены в них викарирую-птими формами. Среди животных, например, это один вид обезьян — Cercopithecus albogularis.
В противоположность лесным зонам, громадные пространства африканских степей и саванн были связаны с азиатскими вплоть до позднего плейстоцена, и эта связь была прервана лишь послеледниковой аридизацией Сахары и Ближнего Востока. Однако сходства населения от Сенегала до Индостанского полуострова вполне достаточно для установления единой суданодеканской области. Здесь находят наилучшие условия для развития сорговые (Andropogoneae) и осоковые (Сурегасеае). Из фауны африканских и азиатских саванн упомянем антилоп, носорогов (которые были широко распространены в северном полушарии в третичном периоде), слонов и гепардов (Aciotnyx jubatus), которые встречаются от Восточной Африки до Индии.
Однако фауна африканских саванн богата эндемичными группами. Это — бабуины, жирафы, окапи, зебры, гиппопотамы, трубкозубы, выдровые землеройки (Potanwgale) и т. п. В прошлом распределение некоторых из них было значительно шире, так что их эндемизм носит реликтовый характер. Так, жирафы обитали в нижнем плиоцене в Китае, Индии, Иране и Греции. Древнейшие остатки гиппопотамов в Сиваликских горах в Индии также датируются нижним плиоценом. В этом смысле Африка сыграла роль, близкую Южной Америке, но в отношении более поздней фауны.
Часть населения саванн, возможно, имеет лесное происхождение. Об этом свидетельствует наличие в них видов, родственных группам, распространенным в африканских лесах: это мартышки (Cercopithecus) среди обезьян, Xerus среди белок. Многие деревья саванн принадлежат к лесным родам. Так, видам суданской саванны Lophira alata, Khaya senegalensis, Parkia biglobosa, Erythrophleum guineense соответствуют L. procera, K- ivorensis, P. bicolor, E ivorense лесов Берега Слоновой Кости.
Изменения климата в четвертичном периоде затронули как фауну саванн, так и фауну лесов; это привело к появлению географических рас, изолированных в результате расширения площади лесов в плювиальные эпохи. Этот феномен очень ярко ил* люстрируют примеры некоторых птиц (рис. 18).
Что касается водной фауны, то здесь наблюдается несколько замечательных случаев палеоэндемизма; это рыбы Polypterus и Calamoichtys, последние потомки Paleoniscidae из пермо-триаса;
б Эти два массива, объединенные ныне в Низкий Берег Слоновой Кости» Ь1ли, видимо, еще недавно разделены саваннами, которые простирались да ам°го побережья.
57
семейство примитивных костистых рыб Mormyridae; подотряд Siluroidea (сомы), все четыре десятка родов которых, за исключением одного, эндемичны.
Еще более, чем в других частях света, для гор тропической Африки характерны эндемизм и замечательные разъединения; без сомнения, это связано с тем, что они представляют собой изолированные массивы, тогда как индо-малайские горы и американские Кордильеры непрерывно простираются вплоть до голарктических областей. Высокие горы Восточной Африки, Абиссинии, Кении, Килиманджаро и Рувензори объединены тем, что заселены древовидными лобелиями и крестовниками. Эти роды,
Р и с. 18. Распределение воробьиных (Latnprocolius chloropterus), обитающих в саваннах. Современное разъединение связано с расширением территорий, занятых лесами, в дождливые периоды четвертичной эры. Каждый из трех ареалов занят одним викарирующим подвидом (по Штрезе-ману, Е. Stresemann, и Гроте, Н. Grote).
/ — подвид cyanogenis’, 2-подвид аси-ticaulis; 5 —подвид elisabethae.
имеющие широкое географическое распространение, представлены здесь двадцатью гигантскими видами, локализованными на разных вершинах. Здесь также обильны верески и виды Swer-tia — рода, близкого нашим генцианам.
В Западной Африке такие вершины, как Камерун, пик Кларенс в Ферпандо-По и пик Сан-Томе, превышают 2000 м. В состав их флоры, помимо таких эндемиков, как Podocarpus man-nii (единственное хвойное растение Западной Африки), входят виды с сильно разъединенным ареалом. У одних наблюдается разъединение с Южной Африкой (это Helichrysum), у других — с горами Восточной Африки, у третьих — с Северной Африкой и Европой (Sanicula europaea, Succisa pratensis и т. д.), и даже с более удаленными районами (Hydrocotyle moschata, известный в Новой Зеландии и на Реюньоне). Чтобы объяснить присутствие европейских видов, Шевалье (A. Chevalier) выдвинул гипотезу скачкообразного распространения к югу, с массива па массив, с помощью перелетных птиц в холодное время года.
На невысоких горах Западной Африки представлена замечательная горная фауна и флора: например, гималайский предста-
58
ритель семейства ароидных — на Фута-Джаллоне, а на массиве Лимба — эндемичные жаба и сверчок, родственные видам из Восточной Африки, и муравей, основной ареал которого находится в Натале.
Мадагаскар
Этот большой остров дает возможность увидеть замечательный пример влияния, оказываемого древней географической изоляцией на состав современного населения. Считают, что остров отделился от Африки уже в начале третичного периода. Гипотезу о существовании древнего континента — Лемурии, — соединявшего его с Индией, мы обсуждали выше.
Для Мадагаскара, давно обособившейся территории, характерно население, в котором отсутствуют недавно возникшие и сохранились архаические группы.
Среди млекопитающих (расцвет которых падает на третичный период) отсутствуют обезьяны, крупные хищники, жвачные, копытные (за исключением Potamochoerus}, грызуны (за исключением крыс Rhizomys), но зато здесь весьма велико разнообразие лемуров и насекомоядных тенреков, а также имеется маленький примитивный хищник Cryptoprocta. Однако современное наличие Potamochoerus, а в плио-плейстоцене карликового гиппопотама, то есть двух форм Африканского континента, дает основание предположить, что в недавнем прошлом существовала «цепь островов и болот, представлявшая собой труднопроходимую и наполовину водную связь» с Африкой.
Изучение других групп приводит к аналогичным выводам. Среди наиболее известных эндемиков можно назвать недавно исчезнувших бегающих птиц Aepyornis и Mullerornis, а также род гигантской наземной черепахи Pyxis, почти исчезнувшей в настоящее время. Однако на острове отсутствуют ядовитые змеи, очень мало амфибий (нет квакш, лягушек, жаб и безногих амфибий), мало рыб.
Из растений имеется шесть семейств семенных, среди них около тридцати родов Chlaenaceae, а также интересное семейство Didiereaceae. Можно также отметить «дерево путешественников» Ravenala — мадагаскарский представитель банановых, введенный повсеместно в Африке.
Капская область
Обладая климатом средиземноморского типа, эта территория столь же отличается по флоре от основного африканского мае* сива, сколь и северная оконечность Африки. Здесь мало эндемичных семейств, но очень много эндемичных родов и видов. Широко представлены мезембриантемум и несколько родов вересковых и рестионовых. Эта небольшая территория, обладающая богатым видовым составом, служит источником многих садовых Растений (Asparagus plumosus, Freesia, разнообразные гладио
69
лусы, Lobelia erinus и т. д.) Выявить происхождение этой разнообразной по своим связям флоры — нелегкая задача. Расположение этой области на крайнем юге Африки наводит на мысль, что в этом «тупике» собраны виды, оттесненные с территорий^ лежащих к северу.
Сахаро-синдская область
Так называют широкую пустынную зону, в которую входят Сахара, Ливийская и Аравийская пустыни, южный Иран, Белуджистан и южный Пакистан. Проблема автономии этой территории как биогеографической области или ее принадлежности к голарктическому или афро-мадагаскарскому царствам, а также разделение между ними в зависимости от степени их относительного влияния — все это вопросы, на которые получены различные ответы, в зависимости от того, какие группы организмов брались за основу при исследовании.
Эта обширная область отличается специфическими условиями среды, но все же ее биологическое население слабо индивидуализировано; эндемизм здесь наблюдается на уровне видов, но мало заметен на уровне родов, почти полностью представленных в периферических областях. Состав современного населения этой области может быть объяснен лишь климатическими изменениями в позднем четвертичном периоде, о чем свидетельствуют археологические, геоморфологические, палеонтологические и, с недавнего времени, палинологические изыскания, результаты которых хорошо согласуются между собой. Уже в озерных виллафранкских отложениях Ахаггара была обнаружена исключительная смесь флор, содержавшая пыльцу тропических форм из восточного и западного Средиземноморья. В начале послеледникового периода в горах центральной Сахары господствовал влажный климат, при котором могла существовать мезофильная средиземноморская растительность — с дубами, кленами, липами, ольхами и кедрами. Усиление аридности климата в неолитическую эпоху проявилось в преобладании ксе-рофильных форм — каменного дуба, оливы, кипариса, можжевельника, алеппской сосны, каркаса (Celtis), фисташки, ююбы. В южном Тибести эта средиземноморская флора встретилась с сахельскими тропическими формами — акациями, фикусами; таким образом, собственно пустынной зоны в то время не существовало. В третьем тысячелетии до нашей эры аридное! ь климата усилилась, и наиболее ксерофильные таксоны средиземноморского и сахельского происхождения распространились на пространствах, покинутых мезофильными формами, небольшое число которых сохранилось на высоких горах. Аххагар и Тассилин сохранили эндемичные виды со средиземноморскими чертами — Olea laperrinei (вид, близкий к О. europaea), Cupres-sus dupreziana (близкий к С. sempervirens), Myrtus nivell<?a (близкий к М. communis). Южнее на массиве Эннеди сохрани'
60
дись те же тропические виды. Но, согласно Эйгу (Eig), «большая часть родов сахаро-синдского элемента является голарктической».
Напротив, среди фауны голарктические или африканские связи доминируют в зависимости от группы. Согласно де Бальзаку (Heim de Balzac), «млекопитающие Сахары имеют, по-видимому, тропическое, по большей части эфиопское, происхождение». То же относится к птицам. Что же касается жесткокрылых насекомых, то Пейеримгоф (Р. de Peyerimhoff) констатирует, что «средиземноморский элемент доминирует над тропическим в центральной Сахаре, что позволяет, без сомнения, присоединить этот ансамбль к палеарктической области». Шопар (Chopard) придерживается того же мнения в отношении прямокрылых насекомых. Жермен (L. Germain) считает, что если ограничиться моллюсками, то всю Сахару можно отнести к голарктической области.
На самом деле речь идет о промежуточной области, ее автономность определяется климатическими, а не историческими причинами, и протяженность изменялась с эволюцией климата.
Восточное, или индо-малайское, царство
В то время как северный и южный края этой территории четко обозначены гималайской цепью и океаном, ее восточная и западная границы — расплывшиеся и спорные. Восточные элементы смешиваются с эфиопскими в засушливых областях Инда и Раджпутана и с голарктическим — в Южном Китае. Кроме того, разделение индонезийских островов между восточным и австрало-папуасским царствами весьма спорно. Население позвоночных больших островов — Суматры, Явы, Калимантана, безусловно, имеет континентальное азиатское происхождение. От-
метим присутствие тигров, слонов, носорогов, крокодилов, питонов. Эти острова покоятся на континентальной платформе, нижний край которой в настоящее время уходит на глубину 100—200 м, но в плиоцене и плейстоцене этот край с большей или меньшей полнотой выступал над водой (рис. 19). Именно тогда недавняя континентальная фауна перешла на Зондские острова, о чем свидетельствуют их окаменевшие останки. К востоку платформа Зондского архипелага заканчивается глубоководьем, которое отделяет архипелаг от Филиппинских островов, Сулавеси (пролив Макассар) и от острова Ломбок (пролив Ломбок). Эта биогеографическая граница была выявлена в конце прошлого века английским биологом Уоллесом (Wallace) и поручила название «линии Уоллеса». Она играет роль строгой гра-ицы для млекопитающих и плохо летающих оседлых птиц (на-Ример, для куриных). Однако ареал тех групп животных и
61
Р и с. 19. Индонезийские острова и подводный рельеф, а также биогеографические границы между малайским и австрало-папуасским царствами (по Хосокава, Hosokawa, и Фосбергу, Fosberg).
У/, У2 — линия Уоллеса; В1, В2 — линия Вебера.
растений, которые издревле были дифференцированы или которые обладают средствами расселения на большие расстояния, простирается далее к востоку вплоть до Новой Гвинеи и даже до островов Полинезии. Таким образом, восточная граница индомалайского царства меняется в зависимости от изучаемой группы. Фитогеографы включают туда Новую Гвинею. Мы вернемся к этой переходной зоне при изучении австралийской и тихоокеанской областей.
Хотя для индо-малайской территории характерны многочисленные таксономические группы, встречающиеся и в Африке, у нее есть также собственный значительный палеоэндемический контингент, филумы которого развились в голарктических областях в третичном периоде, а затем были оттеснены на юго-восток Азии из-за похолодания климата. Это гиббоны, жившие в Европе в плиоцене, долгопяты (Tarsiidae), обнаруживаемые в Европе и Северной Америке, начиная с эоцена, и шерстокрыл (Galeopit-hecus)—летающее насекомоядное1. Среди растений это саговник (Cycas), пальмы Nipa и Kentia. Другое резкое различие между африканским и восточным царствами — это присутствие
1 В настоящее время шерстокрылов выделяют в особый отряд. — Прим-ред.
62
оследнем — благодаря связям с бореальными подобластями — В Плов, общих или родственных с голарктическими. В тропиче-Р их областях Востока встречаются медведи, кабаны, буйволы, сКеНи а среди растений — дубы и сосны, то есть роды в основам голарктические.
В силу этих разнообразных причин индо-малаиские фауна и жлора чрезвычайно богаты. Особенно много змей, широко распространены некоторые семейства птиц, например фазановые ^фазаны, павлины, куры), семенные растения, среди которых имеются сахарный тростник, цитрусовые, манговое дерево, пряности— гвоздика, имбирь, перец и ряд других культурных растений.
Австрало-папуасское царство, или нотогея
Под этим названием мы объединяем Австралию, Тасманию, Молуккские острова, Сулавеси, Новую Гвинею и Новую Зеландию. Единство этой биогеографической области основано прежде всего на фауне позвоночных и значительно слабее выражено у беспозвоночных и растений.
Хотя климатические условия на этих островах очень разнообразны, все же общим фактором является их очень древняя географическая изоляция, восходящая к началу третичного периода. Между собой Австралия и Новая Гвинея соединены не очень глубоким континентальным плато, носящим название Саульской платформы, которое поднималось в плиоцене и плейстоцене. На этой же платформе расположены Молуккские острова и остров Сулавеси. Это объясняет присутствие на них элементов австралийской фауны. В отличие от них Новая Зеландия была в течение всего времени изолирована.
Австралия и Новая Гвинея
Автохтонная фауна млекопитающих сводится здесь к самым примитивным формам, лишенным плаценты (Aplacentalia). Это сумчатые и однопроходные. Сумчатые адаптированы к самым Разнообразным способам существования, к которым в других местах адаптируются самые различные семейства млекопитающих. Отсюда и их большое разнообразие. У птиц 7з родов здесь эндемична: особенно интересны австралийские эму, лирохвост, Райские птицы и многочисленные голуби Новой Гвинеи. Из рыб в настоящее время в двух реках Квинсленда сохранился Epice-r^todus — потомок триасового Ceratodus. Другое доказательство Удивительного выживания местной фауны в течение миллионов ет насекомые и равноногие ракообразные, очень близкие ныне живущим в Австралии видам, обнаруженные в залежах Риасового угля Квинсленда.
63
Древность австрало-папуасского населения подтверждаемся и наличием необычайных разъединений. Напомним о существо, вании сумчатых в Америке. Такое же распространение имеюг два семейства бесхвостых амфибий. Два австралийских ро-щ миног встречаются и в Чили; точно так же в Южной Америке встречаются водяные черепахи из семейства змеешеие (Cheli-deae). Напомним также о группах, которые имеют панаустраль-ное распределение, таких, как род перипатус (Peripatus), рав-ноногие раки из семейства Styloniceae, рыбы из семейства Gala-xiidae. Последние встречаются в Австралии, Тасмании и Новой Зеландии, в Новой Каледонии, Патагонии, на Фолклендских островах и в Южной Африке. В противоположность этому отсутствие многочисленных широкораспространенных групп подчеркивает характерную для этих областей изоляцию.
На это очень древнее население накладывается контингент животных сравнительно недавнего малайского происхождения: водяная черепаха из семейства кожистых черепах (Trionychi-dae), которая оказалась способной пересечь морской пролив и достичь Новой Гвинеи; различные Rana, распространенные по всему свету, проникли в Новую Гвинею и достигли северной части Австралии, тогда как Bufo, которые также являются полукосмополитами, отсутствуют в австрало-папуасском населении. И наконец, человек осознанно или неосознанно ввел недавно значительный контингент животных, нарушивший состав коренного населения. Предполагают, что дикая собака динго, распространенная сейчас в Австралии, была туда интродуцирована доисторическим человеком. Что же касается пресноводных рыб, то почти все они принадлежат к семействам морского происхождения.
Единство и архаический характер современной австрало-папуасской флоры значительно менее ярко выражены. Зона тропического леса носит явный индо-малайский характер. Однако несколько родов и семейств семенных имеют центром расселения Австралию или локализованы в ней. Это эвкалипты (рис. 7), естественная западная граница которых совпадает с «линией Уоллеса»; в Австралии насчитывается 350 их видов. Архаический род Casuarina населяет Малайский архипелаг. Banksia и Gre-villea населяют Австралию и Новую Гвинею, Hakea — только Австралию. Семейство эпакридовых — в основном лишь австралийское.
Молуккские острова и Сулавеси
Расположенная между Новой Гвинеей и Калимантаном, эта группа островов имеет смешанное индо-малайское и австралопапуасское население. Молуккские острова, расположенные восточнее, имеют более богатый австрало-папуасский контингент; например, на Сераме, самом крупном из них, обитают 22 рода млекопитающих этого происхождения и только 9 — индо-малай-
64
кого. На Сулавеси индо-малайские черты населения более за-метньг, например, из млекопитающих там обитают дикобраз, олень/буйвол, кабан, долгопят, отсутствующие на Молукках, как обитавший здесь в плейстоцене слон.
Эти острова, расположенные между Зондской и Сахельской платформами, — сравнительно недавнего происхождения и имели, видимо, лишь непродолжительные связи с Филиппинами, Зондскими островами и Новой Гвинеей. Вебер (Max Weber), пораженный присутствием малайских элементов, счел спорной «линию Уоллеса» на западе и предложил перенести ее к востоку между Сулавеси и Молукками или между Молукками и Новой Гвинеей; эта новая граница получила название «линии Вебера» (см. рис. 19).
Новая Зеландия
Из-за бедности видового состава и резко выраженного палео-эндемического характера автохтонного населения оба больших новозеландских острова, как и окружающие архипелаги Окленд, Чатем и другие, демонстрируют яркий пример того, какое влияние на состав населения оказывает длительная географическая изоляция. Здесь можно также видеть, как может нарушиться непрочное равновесие архаической фауны при введении человеком видов сравнительно недавнего происхождения.
Если не считать интродуцированных видов, не существует ни одного собственно новозеландского млекопитающего. Рептилии
представлены лишь несколькими ящерицами и последним выжившим представителем распространенной в триасе архаической группы хоботноголовых (Rhynchocephalia)— гаттерией (Sphenodon), обитающей в настоящее время лишь на двух маленьких островах. Единственные амфибии — это два вида ля-
гушек, принадлежащие к эндемичному роду.
Очень интересны птицы — из-за разнообразия их биогеографических связей и преобладания примитивных эндемичных форм. К таким формам относится семейство бегающих птиц Dinornit-hidae, которые достигли на этих островах большого разнообразия; из них наиболее известны моа (Dinor nis) — вымершая лишь несколько сот лет назад гигантская форма; Apteryx, или киви, Родственный моа, но маленького размера, условия существования которого все более и более неблагоприятны. Очень близкая форма, как было обнаружено, обитала в плейстоцене в Квинс* Ленде. Это дало основание для гипотезы об очень древних cb^j Зях с Австралией. Гомогенная в других местах группа — попугаи— здесь представлена двумя особыми новозеландскими ро-Дами Nestor и Strigops\ у последних отсутствует грудной киль, почти атрофированы крылья, и они ведут ночной наземный образ Жизни.
Среди иноземных по происхождению групп встречаются осед-Ь1е Виды, родственные перелетным, имеющим более или менее
3 Ж. Леме
65
удаленный ареал. Это представитель фазановых Coturnix novae-zelandicae, исчезнувший в прошлом столетии; группа, к которой он относится, представлена в палеоарктических областях видами, совершавшими большие перелеты. То же относится кутке (Mergus australis) с Оклендских островов, близкими родственниками которой являются мигрирующие виды северного полушария. Без сомнения, здесь дело в древних миграциях. Но известен и недавний случай: это австралийский вид Zosterops lateralis, который обосновался в Новой Зеландии с 1862 г. и достиг островов Чатем и Окленд в 1865 г.
Третья интересная категория птиц представлена литоральными птицами, принадлежащими к южным группам: это пингвины и бакланы. Напомним в связи с этим сходство новозеландских островов с другими островами южной части Тихого океана и с крайним югом Южной Америки.
Флора дает несколько иную картину, чем фауна. Видовой эндемизм очень велик (примерно 80% автохтонных семенных), однако родовой эндемизм выражен слабо (имеется примерно 55 родов), а эндемичных семейств нет совсем. Это указывает на слишком большую древность населения. Большинство неэндемичных видов представлены в Австралии. Однако с появлением белого человека здесь натурализовались многочисленные пришлые растения. В 1922 г., по данным Томсона (G. М. Thomson), их насчитывалось более 600 видов — почти все сорные травы и доминирующие виды Европы.
Полинезийское царство
Население океанических островов неоднородно, в связи с чем индивидуальность полинезийского царства весьма спорна. Если не считать весьма своеобразных Гавайских островов и родственных по населению Южной Америке Галапагосов и островов Хуаи-Фернандес, то население этой области включает элементы индо-малайского и австрало-папуасского происхождения, постепенно обедняясь к востоку. Новой Каледонии, самому большому и наиболее выдвинутому на запад острову из этой группы, присущ значительный родовой и видовой эндемизм, по с отчетливо выраженными западными чертами (исключая Placostylus — представителя брюхоногих моллюсков Gastropoda, принадлежащего к южноамериканскому семейству).
Дальше на восток, на островах Фиджи, население все еще разнообразно, однако Самоа — значительно беднее. Это резкое обеднение к северу и востоку от островов Бисмарка, Соломоновых п Фиджи совпадает с исчезновением крупных островов и с границей между извержениями андезитов на западе и базальтов— на востоке; под ней многие геологи подразумеваю! границу древнего «меланезийского континента». Присутствие в Меланезии наземных брюхоногих, амфибий, одной автохтон-
66
«ой игуаны свидетельствует о существовании в прошлом связей Н континентом. Топография тихоокеанского дна (на глубине Ю00___1500 м прослеживаются многочисленные плато) в соче-
тании с его очевидным и значительным опусканием подтверждает эту гипотезу. Однако для таких биогеографов, как Майр (Е. Мауг) и Фосберг (F. R. Fosberg), «нет абсолютной необходимости предполагать реальные наземные связи». Аргументом в пользу океанического происхождения островов является отсутствие строго пресноводных групп рыб.
Та же проблема связей встает при рассмотрении Гавайских островов — обособленного вулканического архипелага, удаленного на 4000 км от американского побережья и на 1000 км от ближайшего атолла. Их фауна существенно отличается от фауны остальных полинезийских островов, но также крайне бедна, если не считать насекомых. Здесь также сильно выражен эндемизм: в частности, можно привести пример особого семейства воробьиных — гавайских цветочниц (Drepanididae), в котором выделились приспособленные к разнообразным условиям жизни виды; это указывает на древность этого семейства. Другим примером служит семейство брюхоногих моллюсков Achatinellidae. Флора, подобно фауне, характеризуется столь же высоким эндемизмом (имеется более 700 семенных среди 860 автохтонных видов); для нее характерны черты сходства с американской флорой.
Антарктическое царство
Южные океанические острова (Кергелен, Амстердам, Тристан-да-Кунья, Южная Георгия, Фолклендские и т. п.), юг Анд и Патагонии, а также Антарктический континент обладают в совокупности таксономически изолированными эндемическими родами и видами, что побудило биогеографов выделить автономную антарктическую область.
Из флоры наиболее примечательна «кергеленская капуста» (Pringlea antiscorbutica, Cruciferae), Асаепа (Rosaceae), Azo-rella (Umbelliferae), Jovellana (Scrophulariaceae) и в меньшей степени роды Hebe (Scrophulariaceae) и Colobanthus (Caryophy-Пасеае), которые распространены вплоть до Новой Зеландии и юго-востока Австралии. Это позволяет предположить о существовании в прошлом связей между всеми этими южными территориями. Эндемизм усугубляется исключительной по сравнению с арктическими территориями бедностью: около 30 местных семенных — на Каргелене, и всего два — на Антарктическом континенте, почти полностью покрытом льдами: Colobanthus cras-Slfolius и Deschampsia antarctica.
3»
Часть вторая
Факторы среды и их роль в распространении и существовании организмов: экология
Способность к размножению и расселению, которой обладают виды, позволила бы им равномерно распространиться по всей земной поверхности, если бы на их пути не существовали барьеры, воздвигнутые условиями среды и совместного обитания с другими видами. Границы ареала вида (там, где они стабильны), а также границы территорий, на которых вид локализован внутри ареала, определяются тем, с какими физико-химическими или биологическими условиями существование вида несовместимо.
Изучение отношений между организмами и окружающей их внешней средой составляет предмет экологии (термин, введенный Эрнстом Геккелем в 1869 г., от греческого oixo£ — дом, место обитания).
Глава 4
Общие черты отношений между экологическими факторами и организмами
Исследование связей между организмами и средой имеет два .•аспекта: 1) изучение характера среды, в контакте с которой живут организмы, мезологическая экология, или мезология\ 2) изучение поведения и реакций организмов в этой среде — этологическая экология, или этология (термин, введенный в 1859 г. •Сент-Илэром, Jeoffroy Saint-Hilaire), от греческого слова eftog — нрав, поведение). Каждый из этих двух аспектов исследования ставит общие проблемы, к изучению которых мы и перейдем.
Мезологическая экология
Совокупность энергетических, физических, химических и биологических условий, которые господствуют в непосредственном соседстве с организмом, составляет его окружение (environment англосаксонских экологов). Однородный по своим свойствам участок окружающей среды образует местообитание, или стацию \ или, согласно излюбленной зоологами терминологии, биотоп1 2.
Термин среда более точно характеризует природу вещества, образующего местообитание: например, водная, морская, подземная, воздушная среда.
Местообитание характеризуется множеством черт или факторов различной природы. Некоторые из этих факторов являются объектом изучения специальных дисциплин: климатологии и метеорологии, почвоведения и химии почв, гидрологии, физической океанографии. Но эти специальные дисциплины имеют <вою область исследования и изучают среду каждая в своем плане, тогда как эколог ставит в центр исследования живые организмы, а от небиологических дисциплин он заимствует лишь
1 Определенная так стация четко отличается от местонахождения, которое содержит лишь географическое обозначение места.
2 Термин «местообитание», или «стация», правильнее относить к определенному виду животных или растений, понимая под ним совокупность усло-Ии» в которых вид существует; под биотопом же понимают совокупность дТОРов неживой природы, с которой связан определенный биоценоз.— Прим.
71
методы изучения окружающей их среды и те их наблюдения, которые важны для понимания условий существования организмов.
В целом можно сказать, что мезология стремится определить, какое количество энергии и первичной материи из окружающей среды оказывается ежесекундно (или в течение данного периода) в контакте с организмом, и следовательно, в его распоряжении.
Этологическая экология
Под этим термином мы понимаем раздел экологии, изучающий поведение организмов в связи с условиями местообитания. Лишь наблюдение in situ процессов поведения, метаболизма, роста, воспроизведения, активности, перемещения и т. п. позволяет дать им точную количественную оценку и показать их эволюцию в естественных условиях. Такое наблюдение при использовании корреляционных методов позволяет выявить, какие из этих условий имеют решающее или лимитирующее значение. Но разнообразие этих факторов и их изменчивость во времени и пространстве приводят к необходимости использования экспериментальной методики для точного выяснения роли каждого из них. Такого рода эксперименты могут осуществляться с помощью физиологических методов, суть которых состоит в том, что все факторы, за исключением того, чье действие подлежит выяснению, поддерживаются на постоянном благоприятном уровне, тогда как этому единственному фактору придаются различные значения. Другой, значительно более близкий к естественным условиям способ заключается в том, чтобы контролировать фактор или группу факторов, придавая им различные значения в каждой выборке, тогда как остальные факторы флуктуируют естественным образом, по одинаково для всех выборок. За последние годы благодаря техническому прогрессу удалось частично преодолеть неполноценность описанных методов при помощи особых устройств с заданной программой, своего рода оранжерей, в которых факторы окружающей среды находятся под контролем так, чтобы схематически воспроизводить различные климатические ритмы. Наиболее совершенный пример такого рода устройств представляют собой фитотроны.
Различные наблюдения и опыты привели к установлению законов и принципов, определяющих взаимоотношения между организмами и средой.
Влияние изменения одного фактора. Прямые наблюдения (а с еще большей точностью экспериментальные исследования) показывают, что биологический процесс может осуществляться не при любых изменениях какого-либо фактора окружающей среды, но лишь между двумя его предельными значениями, минимумом и максимумом, которые и представляют собой границы толерантности данного процесса относительно данного фактор^
72
Внутри этих границ значения фактора не одинаково благоприятны: существуют значения или области значений, оптимальные для данного процесса. Эти три величины (минимум, оптимум и максимум значений изучаемого фактора) составляют три «кардинальные точки» Сакса (Sachs). Они не совпадают для различных процессов жизнедеятельности одного и того же организма. Особенно ясно это можно видеть на примере температурного фактора. Двигательная активность комнатной мухи возрастает с увеличением температуры почти до 40°, но продолжительность ее жизни максимальна между 15 и 28°; икра макрели выживает долее всего при 15°, но развивается после оплодотворения быстрее всего при 21°; у зеленых растений фотосинтетическая ассимиляция углекислого газа протекает оптимально между 20 и 30°, тогда как интенсивность процессов дыхательного окисления возрастает вплоть до температур, близких к летальным, то есть 40-45°.
При изменении какого-либо фактора поведение у разных видов может быть весьма различным. Чтобы выразить большую амплитуду толерантности, употребляют приставку эври\ чтобы отразить большую близость минимального и максимального пределов, употребляют приставку стено. Например, развитие икры лягушки (Rana pipiens), которое может протекать в интервале от 0 до 30°, является эвритермным, тогда как развитие икры лосося, ограниченное 0 и 12°, рассматривается как стенотермное.
У одного и того же вида ограничивающие и оптимальные
условия могут меняться на последовательных стадиях жизненного цикла; так, икра форели может развиваться, начиная с температур, близких к 4°, с оптимумом развития при 8°, тогда как взрослые особи при температурах ниже 10° не могут питаться достаточно интенсивно, чтобы расти, а оптимум роста для них лежит в области 13—16° Вообще говоря, границы толерантности
у молодых особей менее жестки, чем у взрослых.
Таким образом, полная реализация жизненного цикла организма требует определенной согласованности с сезонными ритмами факторов местообитания, или, когда речь идет о подвижных видах, их перемещения из одного местообитания в другое.
С другой стороны, влияние некоторого фактора в данный момент зависит от его предшествующего изменения. Например, сопротивляемость растения холоду зависит от скорости, с которой падает температура, и ткани способны выдержать более низ-кУ*о температуру, если охлаждение происходит постепенно.
Наконец, действие фактора может при одном и том же его значении зависеть от значений других факторов в тот же момент, ^тот аспект мы рассмотрим в следующем параграфе.
Взаимодействие нескольких факторов. Воздействие данного Фактора на организм может сильнейшим образом варьировать зависимости от состояния других факторов окружения. Как предельные, так и оптимальные точки на кривой, выражающей
73
воздействие фактора, могут менять свое положение. Так, возвращаясь к случаю устойчивости при низких температурах, укажем, что на примере многих культурных растений было замечено, что богатство почвы минеральным азотом повышает летальную температуру, тогда как большое содержание калия или фосфора ее снижает.
В 1840 г. немецкий агроном Либих (Justus von Liebig), изучавший относительное влияние различных питательных веществ на продуктивность возделываемых растений, отметил, что в основном на нее влияют количественные изменения не тех веществ, содержание которых и так велико, а веществ, содержащихся в дефиците. Ему мы обязаны первым выражением этого правила, известного как «закон минимума», который мы можем сформулировать следующим образом: «продуктивность культуры зависит только от того питательного элемента, который имеется в среде в наименьшем количестве» Ч
Например, продуктивность почвы, в которой отмечено недостаточное количество усвояемого азота, но прочие питательные вещества содержатся в достаточном количестве, не может быть повышена внесением этих последних, но лишь добавлением азотистых удобрений.
В 1905 г. Блэкман (F. F. Blackman) обобщил этот закон для совокупности факторов, влияющих на рост растений, назвав его «законом лимитирующих факторов»: «Проявление биологического процесса, на скорость течения которого оказывают влияние многие факторы, ограничено тем из них, который наиболее слабо представлен в окружающей среде» (по отношению к его оптимальному значению). Блэкман установил этот закон, изучая влияние интенсивности освещения, температуры и содержания СО2 в воздухе на интенсивность фотосинтеза. Последняя оказывается пропорциональной значению лишь одного из этих факторов, именуемого «лимитирующим», пока его место не занимает другой фактор, становясь в свою очередь лимитирующим (рис. 20).
Эти законы, установленные в экспериментальных условиях, естественно, проявляются в географическом распространении и распределении по стациям живых существ. Так, в Северной Африке, где сухость климата представляет ограничивающий фактор, различие в уровне осадков в двух точках па 0'1 см существенно влияет на живое население, тогда как на берегах Гвинейского залива, где осадки значительны, такая разница остается без последствий.
Однако в некоторых случаях действие факторов может зависеть от их относительного, а не абсолютного значения. Это
1 Выражение «в наименьшем количестве» следует понимать не буквально, а по отношению к оптимальному значению, иначе те элементы, действие которых наиболее благоприятно при очень малых дозах (микроэлементы), например бор и марганец, всегда будут казаться определяющими.
74
относится к содержанию в окружающей среде некоторых органических веществ: увеличение содержания одного из них может подавлять рост, уменьшая степень усвоения других элементов; -такое явление называют ионным антагонизмом. Кроме того, при одной и той же концентрации элементы могут оказывать либо -токсическое действие, либ оставаться нейтральными в зависимости от концентрации других элементов.,
Взаимодействие живых организмов. Экспериментальное изучение реакции организмов на различные факторы внешней среды должно производиться на изолированных особях или в достаточно разреженных популяциях, чтобы исключить эффекты взаимодействия— такие, как затенение или борьба за пищу; кроме
Рис. 20. Лимитирующее воздействие интенсивности освещения и температуры на фотосинтез культур пресноводной водоросли хлореллы. Выделение кислорода пропорционально интенсивности •освещения при малых ее значениях и достигает более высокого уровня с повышением температуры (данные Вас-синка, Wassink).
Световая энергия, эрг/сн2-сек
того, условия должны быть таковы, чтобы исключать нападение «паразитов и хищников.
Но в естественных условиях организмы живут в более или менее плотных и разнообразных в таксономическом отношении популяциях, и это сосуществование изменяет их экологическое поведение в благоприятную или неблагоприятную, в зависимости от обстоятельств, сторону. Жизнь в сообществе приводит, как правило, к сужению экологической амплитуды реакций организма на факторы среды, а иногда — к смещению оптимальных условий. К абиотическим факторам окружающей среды, связанным со свойствами неживой материи, прибавляются биотические фак* торы — прямое влияние организмов друг на друга, и в этом заключается еще одна причина того, что выводы, полученные в экологических экспериментах на отдельных организмах, нельзя 'бесконтрольно распространять на естественные условия. Биотические факторы составляют особый предмет изучения, равно Как и факторы неживой среды.
Более того, физико-химические свойства среды могут измениться просто из-за близости организмов друг к другу. Особенно убедителен в качестве иллюстрации этого «группового
75
эффекта» пример деревьев: отдельные деревья лишь слабо изменяют микроклиматические особенности температуры, влажности, ветра и даже освещенности, но в пространстве, занятом лесом, создается сильно измененный микроклимат. Таким образом, лишь вследствие того факта, что они живут в сообществе, организмы находятся в особых экологических условиях, созданных самим сообществом и отличных от условий подобной, но необитаемой среды.
Это взаимное влияние живых организмов, прямое и косвенное, приводит к тому, что экологи различают в своих работах два взаимодополняющих аспекта: аутэкологию, которая стремится определить поведение изолированного организма по отношению к факторам среды, и синэкологию, которая изучает поведение организмов в сообществе или поведение сообществ в их совокупности.
Экологическая пластичность видов
Большая часть видов обнаруживает в своих морфологических и физиологических свойствах изменения, зависящие от занимаемой ими части ареала или от локальных условий местообитания. Эта связь между внутривидовыми изменениями и экологическими условиями может быть обусловлена двумя причинами: а) реакцией организма в пределах его фенотипической пластичности; это не наследуемая реакция, которая лежит в основе приспособляемости, аккомодации, к особенностям стации; б) производимым средой обитания отбором вариаций генотипа 1 вида, приспособленных к данной среде обитания. В первом случае речь идет об аккомодатах, во втором — об экотипах или экоклинах.
Аккомодаты. Они являются результатом воздействия внешних факторов на рост организмов. Эти факторы вызывают лишь количественные изменения, но они могут быть весьма заметными и играть большую роль в судьбе данной популяции. Интенсивность освещения, влажность и водная среда — основные факторы, вызывающие появление аккомодатов. Различие во внешнем виде и анатомии листьев растений, выросших в тени и при полном освещении, носит классический характер. Иногда копт-* расты еще более очевидны у растений-амфибий, относящихся к одному и тому же генотипу, растущих в подводных и надводных условиях. Если части растения находятся в условиях, различающихся в отношении этих морфогенетически важных фак
1 Генотип организма — совокупность переданных родителями наследственных особенностей. Он образован двумя родительскими геномами (совокупностями n-хромосом, содержащихся в каждой гамете). Фенотип — совокупность проявляющихся черт генотипа, которые в отличие от геномов могут изменяться под воздействием внешних факторов.
76
торов, то они оказываются морфологически различными: на одном и том же буке имеются «солнечные» листья во внешней части кроны и «теневые» — во внутренней; листья одного и того» же стрелолиста имеют лентовидную форму под водой, округлую форму, когда они плавают на поверхности, и стреловидную, когда находятся в воздухе. Эта морфологическая пластичность сопровождается изменениями функциональных особенностей и реакций на экологические условия. Знание таких изменений необходимо для понимания амплитуды выносливости видов по отношению к среде.
Экотипы, или экологические расы, и экоклины. В отличие от аккомодатов являются наследственными, генотипическими проявлениями пластичности видов.
1. Клин (термин, введенный Хаксли, J. S. Huxley, в 1938 г.) характеризует постепенные морфологические изменения соответственно некоторому градиенту; когда этот градиент связан с экологическим градиентом, говорят об экоклине. Так, Грегор (J. W Gregor) установил у Plantago maritima существование экоклинов, связанных с климатическими изменениями (которые он называет топоклинами), а также с локальными градиентами солености. Постепенные морфологические и физиологические изменения, следующие за изменениями климатических факторов, были выявлены у древесных видов, таких, как сосна обыкновенная (Pinus Silvestris), у популяций которой различного происхождения, культивируемых в Стокгольме, обнаружилась весьма четкая зависимость содержания сухого вещества в иглах от продолжительности сезонного периода роста на их родине. Эта продолжительность определялась как число дней со средней температурой выше 6° (Лангле, Langlet).
Особая форма клинальных изменений представлена у полиморфных видов, в популяциях которых численные соотношения между различными формами изменяются в зависимости от географического градиента. Классическим примером может служить клещевина, акклиматизированная в Перу, которая представлена смесью сизой формы с восковидной кутикулой и формы несизой, причем их соотношение меняется с высотой в пользу первой от 0,15 до 100% (Харланд, Harland).
2. Экотипы, по определению Турессона (Turesson), автора этого термина, — это более или менее морфологически различные формы, разделенные одна от другой прерывистостью вариации и приспособленные к различным местообитаниям, хотя и способные давать гибридное потомство. Классические работы Клаузена (Clausen) о Potentilla glandulosa и Achillea millefolium в Калифорнии иллюстрируют эту концепцию: в калифорнийской Сьерра-Неваде встречаются 11 климатических экотипов Achillea millefolium, из которых четыре принадлежат к гекса-плоидному подвиду A. borealis и семь к тетраплоидному подвиду д, lanulosa. В пределах каждой формы также существует
77
некоторая изменчивость, но одна форма отделена от другой как в смысле локализации по высоте, так и морфологически: равнинные формы больше по размерам, высота ветвей у них порядка метра; с высотой средний рост уменьшается, и на высоте 3000 м над уровнем моря представлены карликовые популяции растений в форме распластанных розеток; непрерывная активность роста прибрежных экотипов уступает место состоянию зимнего покоя у горных, и это сочетается с различиями в периодах цветения. Высаженные по соседству в идентичных условиях местообитания, эти экотипы сохраняют свои характерные черты, хотя и отличаются от популяций in situ, так как приспосабливаются к местным условиям.
Различие между клипами и экотипами не всегда столь просто: описаны случаи сосуществования дискретных и непрерывных изменений при анализе в масштабе популяций, объединенных ограниченными периметрами. Так, Брэдшоу (A. D. Bradshaw), изучив около тридцати колоний злака Agrostis tenuis в Уэльсе, обнаружил у него морфологические и физиологические различия генотипической природы, причем формы постепенно переходили друг в друга или были разделены, смотря по тому, были ли изменения экологических условий резкими или постепенными.
Дискретный или непрерывный характер генотипических вариаций в зависимости от географического положения или стации имеет не только экологические, но и биологические причины. Аллогамия, вызывающая рекомбинацию важных генов, способствует постепенным изменениям в пространстве, тогда как аутогамия, апомиксис, вегетативное размножение, которые ограничивают эту рекомбинацию или препятствуют ей, способствуют генетической дискретности. Но и здесь могут вмешиваться экологические факторы, оказываясь благоприятными для аутогамных или аллогамных форм. Например, сообщества, существующие в стабильных экологических условиях, благоприятствующих видам со значительной продолжительностью жизни, обеспечивая одновременность цветения, лучше способствуют установлению непрерывных клинальных изменений, чем местообитания, быстро развивающиеся или с сильно флуктуирующими особенностями. Кстати, один и тот же вид может, в зависимости от условий среды обитания, быть хазмогамным, клейстогамным и даже апо- • миктическим; такие случаи отмечены у злаков.
Группы фертильных по отношению друг к другу популяций, приуроченные к определенной географической области или характерной стации, которые не могут скрещиваться с таксономически очень близкими им, но локализованными в других районах или стациях группами, составляют, по выражению Турессона, эковиды (ecoespeces). Их часто относят к разным видам. В некоторых случаях, когда внутренние изолирующие барьеры не столь непроницаемы, гибридизация еще возможна; в этом случае, про
78
межуточном с экотипами, эковиды возникают постепенно. Это относится к дубам с опадающими листьями из Средней Европы. В других случаях изоляция является полной и возникла внезапно в результате полиплоидии; отличия от диплоидных родительских форм бывает трудно выявить морфологически. Например, злак Anthoxantum odoratum является тетраплоидным так-соном лугов равнин и низкогорий; он замещается в Альпах, начиная с высоты 1500 м, диплоидным A. alpinum, едва отличимым от предыдущего.
Чтобы объяснить механизмы отбора экотипов, производимого местообитанием, необходимо найти факторы, его обусловливающие, и объяснить их физиологическое действие. Эти факторы
Рис. 21. Дыхание и фотосинтез Oxyria digyna различного происхождения при разных температурах (по Муни и Биллингсу, Н. A. Mooney, W. D Billings, 1961).
/ — бореальная популяция из Юкона; 2—альпийская популяция из Колорадо. Видно, что у бореальных растений выделение СО2 при дыхании становится равным его фотосинтетическому поглощению при более низкой температуре, чем у растений альпийского происхождения.
весьма различны по природе, они могут быть климатическими или эдафическими. К первой группе принадлежат многочисленные фотопериодические экотипы, локализация которых по широте связана с сезонными изменениями продолжительности дня и о которых мы будем говорить несколько позже. Но от широты зависят и другие весьма важные в биологическом отношении климатические факторы, такие, как интенсивность освещения, годичный или суточный ритм температуры. Некоторые исследования показали, что процессы фотосинтеза и дыхания по-разному отвечают на эти факторы у видов с широким распро-странениехМ по долготе в зависимости от географической родины изучавшихся особей. Рис. 21 иллюстрирует это различие в поведении у представителя семейства гречишных (Oxyria digyna). который распространен от субарктической тундры (83° с. ш.) до высоких гор средних широт (Альп, Сьерра-Невады Северной
79
Америки) под 39° с. ш. Субарктические растения достигают максимума фотосинтеза при более низких температурах, чем альпийские растения, и имеют большую интенсивность дыхания. Как следствие, заметный фотосинтез продолжается у альпийских растений до значительно более высоких температур, чем у субарктических. Интенсивность освещения, необходимая для фотосинтеза, выше у альпийских растений. Все эти различия означают лучшую приспособленность каждой из двух форм к своему собственному местообитанию.
Причиной отбора экотипов могут быть и особенности почвы. Предметом некоторых физиологических исследований был водный фактор, а также химические факторы: устойчивость к действию токсических веществ и реакция на известняковые соединения. Экспериментальное изучение популяций злака Festuca
Рис. 22. Рост Fectuca ovina, происходящих с различных почв, на питательных растворах с различным содержанием кальция (по Снэйдону и Брэдшоу, Snaydon, Bradshaw, 1961).
/ — неизвестняковая почва, рН=4,6 (Карнарвоншир); 2— известняковая почва, рН=8,9 (Дорсет).
ovirta, из которых одни росли на известняковой щелочной почве, а другие на неизвестняковой кислой почве, показало, что раса, обитающая на известняковой почве, произрастает лучше, чем раса с кислой почвы, если субстрат богат кальцием; если же субстрат им беден, то имеет место обратная картина (рис. 22).
Такие наблюдения над пластичностью видов по отношению к среде делают очевидным экологическое значение тех условий, в которых организмы развивались (аккомодаты), а также генетического разнообразия организмов (клины, экотипы). Именно поэтому исследования по «экспериментальной таксономии» или' «биосистематике» становятся абсолютно необходимым источником информации для аутэкологических исследований.
Природа и классификация факторов
При изучении многочисленных внешних факторов, влияющих на жизнь организмов и их сообществ, роль каждого из них может явиться предметом особого исследования, использующего
80
соответствующую данному случаю технику. Такое исследование необходимо для полного понимания причин локализации и поредения организмов в естественных условиях. Однако географическое распространение организмов и их состояние в каждый данный момент является результатом одновременного действия множества факторов, и это обстоятельство необходимо иметь в виду при дальнейшем исследовании отдельно взятых экологических факторов.
При классификации многочисленных экологических факторов можно следовать приводимым ниже двум концепциям:
1. Классификация факторов в зависимости от среды, в которой они действуют, приводящая к основному делению на наземную и водную среду. К ним относят:
а) климатические и атмосферные факторы: свет, температура воздуха, атмосферная влажность, осадки, ветер, газовый состав воздуха;
б) эдафические факторы, характеризующие субстрат: температура почвы, влага и воздух в почве, ее физические и химические свойства;
в) топографические факторы, связанные с орографией земной поверхности: их влияние сказывается лишь при посредстве климатических и эдафических изменений;
г) биотические факторы, к которым относят взаимодействие организмов между собой, и в частности воздействие человека.
2. Классификация, основанная на природе и характере физиологического воздействия факторов независимо от их локализации. Здесь различают:
а) энергетические факторы: излучение — в частности, свет и тепло;
б) водные факторы, атмосферная и почвенная вода;
в) химические факторы: газовый состав атмосферы и почвы, газы и другие вещества, растворенные в воде, минеральные и органические вещества почвы;
г) механические факторы: ветер, эрозия, снег, огонь;
д) биотические факторы: взаимодействие организмов.
Мы будем следовать последней классификации.
Глава 5
Биотические факторы
Под биотическими факторами мы понимаем всю совокупность прямых воздействий организмов друг на друга. Мы исключаем лишь непрямые влияния, возникающие вследствие изменения физических и химических свойств окружающей среды.
Эти взаимовлияния крайне разнообразны как по природе, так и силе воздействия. Между полной независимостью и обязательной и непрерывной взаимозависимостью имеются все градации, которые могут характеризоваться факультативной или временной зависимостью. С экологической точки зрения необходимо рассмотреть, к каким последствиям приводят эти взаимодействия, или коакции, по Клементсу и Шелфорду (Clements, Shelford), для выживания и процветания вовлеченных в них организмов. Приводимая ниже таблица характеризует различные возможности таких последствий.
Таблица 1
Изолированные организмы Сосуществующие организмы
А Б А Б
Независимость, нейтрализм 0 0 0 0
Антагонизм, аменсализм, антибиоз: А элиминируется из-за воздействия Б 0 0 0
Конкуренция: А и Б взаимодействуют 0 0 — —
Эксплуатация: А — паразит или хищник Б — 0 + —
Кооперация: А и Б взаимно выигрывают от сближения 0 0 + +
Комменсализм: благоприятная или облигатная для А зависимость от его хозяина Б 0 + 0
Мутуализм и симбиоз: облигатное или взаимовыгодное объединение А и Б — + +
Примечание: 0 означает отсутствие влияния на развитие организмов А или 5; Ч—развитие улучшается или делается возможным благодаря взаимодействию; ----------------развитие ухудшается или становится невозможным из-за
взаимодействия.
82
Кажущаяся простота этой классификации, основанной на результатах отношений, не должна заслонять факт существования многих промежуточных или сложных случаев, когда принадлежность к той или иной из вышеприведенных категорий не ясна. Поэтому мы предпочитаем рассматривать взаимодействия между организмами, исходя из природы этих взаимодействий, а не их последствий. Одни из этих взаимодействий основаны просто на близком соседстве независимых организмов, которое может оказаться благоприятным для них (кооперация) или неблагоприятным (конкуренция). В основе других — выделение в среду веществ, влияющих на соседние организмы. В зависимости от того, благоприятно или неблагоприятно это влияние, мы имеем случаи либо комменсализма, либо антагонизма. Некоторые отношения сводятся к простому контакту, который либо безразличен для организма, способного выдержать этот контакт (комменсализм), или же выгоден ему (мутуализм). И наконец, могут быть взаимоотношения, при которых проявляется алиментарная эксплуатация одного организма другим: это хищничество и паразитизм 1»2.
Взаимодействия, зависящие от плотности организмов
Количество особей, находящихся на данной поверхности или в данном объеме, оказывает различное воздействие: от него могут зависеть количественные характеристики внешних факторов, оно влияет на конкуренцию из-за источников энергии и вещества, и у животных оно вл;пет на психофизиологическое и социальное поведение.
Конкуренция
Это выражение используется биологами в довольно широком смысле. Мы будем применять этот термин в его этимологическом значении, понимая под ним конкуренцию нескольких организмов за источник энергии и вещества, если потребность в них больше, чем имеется в наличии.
Факторы, лежащие в основе конкуренции, могут быть трех типов: пространственные взаимоотношения особей в сообществе; специфические факторы, связанные с морфологией и собственно биологией таксономических форм, и наконец, энергетические и физико-химические условия внешней среды.
1 Мы не выделяем в особую категорию тип симбиоза, который в зависимости от случая можно рассматривать либо как идеальный мутуализм, либо как паразитизм, ставший необходимым хозяину.
2 Чаще симбиоз понимают как отношения, благоприятные для обоих организмов или благоприятные для одного из них, но не ухудшающие развитие второго, то есть как понятие, объединяющее отношения комменсализма и мутуализма.— Прим, ред.
83
Роль пространственных отношений между конкурентами. Точные измерения производились в простых случаях: в генетически однородных популяциях, все особи которых имели один и тот же возраст и полученных в культуре или при совместном выращивании. При прочих равных условиях увеличение плотности популяции, то есть увеличение количества особей на единицу площади или объема, ведет к существенному уменьшению
си 100-1
25 250 2 5 0 0 25 000
Число растений на 1 н2
Нисло растений на 1гг*п*103
Рис. 23. Продукция сухого вещества в пересчете на одно растение (верхний рисунок) и на 1 м2 (нижний рисунок) у Trifoliuni subterraneiim, измеренная в разные сроки при возрастающей плотности (по Дональду, С. М. Donald).
способности индивидуального роста, сокращению воспроизводства и увеличению смертности.
1. Изучение отношений рост — плотность, производившееся на семенных растениях, показало (рис. 23), что, чем выше плотность, тем быстрее снижается продукция сухого вещества в пересчете на одно растение; при одинаковом возрасте растений она меняется обратно пропорционально плотности. Что же касается продукции сухого вещества на единицу поверхности, то она возрастает с плотностью до некоторого предельного значения.
Делались различные попытки охарактеризовать эти экспериментальные данные каким-то уравнением. Влияние плотности на рост может быть сведено к действию лимитирующих факторов, поскольку увеличение плотности популяции снижает коли-
84
чество энергии и вещества, имеющихся в распоряжении каждой особи. К этому процессу было применено классическое уравнение Митчерлиха (Mitscherlich):
где У — мера роста, \у — увеличение роста, вызванное увеличением Ах меняющегося факторах (здесь — величина пространства, приходящегося на одно растение, то есть величина, обратно пропорциональная плотности), А — максимальная продукция при отсутствии конкуренции, с — постоянная, характеризующая действие фактора х. Интегрирование дает следующую формулу:
у = А(\ — е~сх).
Продукция на единицу поверхности У может быть выражена при помощи следующего видоизменения этой формулы:
Y = Ап(1 — е~с'п),
где п— число растений на единицу поверхности.
Поскольку в некоторых случаях эта формула не вполне точно отражает экспериментальные факты, были предложены другие формулы, в частности, японским исследователем Кира (Kira) и его сотрудниками.
2. Конкуренция влияет и на плодовитость. Продукция цветков и семян на одно растение тем меньше, чем больше плотность. У животных при очень большой плотности населения рождаемость часто снижается.
3. Рост плотности популяции может повышать уровень смертности за счет вызванного им увеличения конкуренции. Это обстоятельство играет важную роль в лесных насаждениях, где менее крепкие деревья постепенно отмирают при старении этих насаждений; так, в неэксплуатируемых буковых лесах было обнаружено, что через 120 лет оставался лишь один бук из 2000 деревьев плотной молодой популяции 10-летнего возраста.
У некоторых животных, например у Triboliuni — насекомых, живущих в муке, важный фактор в увеличении смертности при увеличении плотности — каннибализм.
Таким образом, влияя одновременно на рождаемость и смертность, конкуренция является важнейшим фактором регулирования плотности вида.
Значение для конкуренции морфологических и собственно биологических свойств организмов. 1. Сосудистые растения. В том исключительном случае, когда все индивидуумы в Данной среде абсолютно идентичны, их взаимная конкуренция приводит к одинаковому замедлению роста и уменьшению плодовитости. Но чаще даже в пределах генетически однородной популяции одного вида различия в возрасте и положении
85
приводят к тому, что индивидуумы по-разному реагируют на конкуренцию. Однако наибольшее разнообразие воздействии возникает в случае межвидовой конкуренции.
Следует отметить наиболее важные при конкуренции морфологические и биологические характеристики:
скорость роста: виды с большей скоростью роста имеют преимущество по сравнению с более медленно растущими видами;
способ использования пространства: виды, колонизирующие одинаковые уровни в атмосфере или почве, конкурируют значительно активнее, чем занимающие разные уровни. Виды, способные к быстрому распространению вширь посредством столонов или ризомов, имеют преимущество при захвате свободного пространства;
сезонная периодичность: несовпадение периодов активности видов, так же как несовпадение занимаемого ими пространства, ослабляет конкуренцию. Например, весенние лесные травы — пролеска (Scilla), чистяк (Ficaria), ветреница (Anemone) — впадают в состояние покоя в тот период, когда осенние, например злаки и губоцветные, находятся в состоянии активного роста. Поэтому между ними не возникает конкуренции;
продолжительность жизни: это важный фактор для конкуренции, поскольку виды с короткой продолжительностью жизни, например однолетние растения, должны, прорастая, очень быстро укореняться;
количество семян, производимое растением;
способность к регенерации после срезания, потравы или пожара. В наиболее благоприятном положении оказываются виды, у которых быстрее возобновляется рост.
2. Животные. Так же, как и растения, сосуществующие в одном сообществе, животные способны вступать в конкурентные взаимоотношения в самой различной степени. Если два вида нуждаются в разной пище или же их активность падает на разные часы или на разные сезоны, то конкуренции между ними не возникает; но конкуренция тем больше, чем более совпадают характер питания и периоды активности.
Классическим примером такого совпадения служат опыты Гаузе (Gause) (рис. 24) на двух видах парамеции (род простейших), регулярно получавших в качестве пищи бактерии. При культивировании видов порознь у них наблюдались сходные кривые роста популяций: разница была лишь в несколько большей скорости роста у одного из них. Когда же оба вида смешивались в равных количествах, то у того из них, скорость размножения которого исходно была больше, наблюдалось ее замедление, однако оно было гораздо менее заметно, чем у второго, количество которого через 5—6 дней уменьшалось и через 16 дней становилось равным 0. Здесь мы имеем бесспорный случай конкуренции, поскольку оба вида не нападали друг на друга и не вырабатывали токсических веществ.
86
60
§ 30
S’
E
f 80 О X E g 40 сг
0
2 4 6 8 10 12 14 16Дни
Рис. 24. Конкуренция за источник питания двух близких реснитчатых простейших, Paramecium caudatum и Р. aurelia. Культивируемые порознь, они дают классическую сигмовидную кривую размножения. В смешанной культуре обладающий более медленным ростом Р. caudatum постепенно убывает в количестве, а впоследствии полностью элиминируется (по Гаузе, Gause).
Противоположный пример длительного совместного обитания близких видов могут дать прибрежные птицы, питающиеся разной пищей, например бакланы (Phalacrocorax carbo и Ph. aristo-telis), из которых первый предпочитает рыб, плавающих у поверхности, а второй — донных животных, плоских рыб и креветок.
Подвижные животные имеют перед неподвижными то преимущество, что способны к поискам пищи. Это позволяет особям с одинаковыми потребностями избегать конкуренции, эмигрируя или же деля между собой территории на зоны охоты или зоны передвижения. Такие зоны принадлежат либо индивидууму, либо паре, либо колонии. У очень многих птиц в начале периода гнездования самец ограничивает свою территорию и защищает ее от посягательства других птиц того же вида, пока птенцы не покинут гнезда (рис. 25). Некоторые виды, например крапивник (род Troglodytes), защищают всю территорию своего передвижения, которая может оказаться порядка гектара, тогда как воробьи и стрижи защищают лишь непосредственные подступы к гнезду. У некоторых млекопитающих также существуют территории, запретные для представителей того же вида (на подступах к их гнездам или жилищам); кроме того, имеется более широкое и свободное от запрета «жизненное пространство», которое они пробегают в поисках пищи. Площадь участков пробега меняется от нескольких десятков квадратных метров для мелких
87
позвоночных и насекомых до нескольких квадратных километров для крупных млекопитающих.
Массовые миграции, по-видимому, вызываются у некоторых видов психическим состоянием, связанным с увеличением плотности или с нарушениями питания. Примером могут служить лемминги, мелкие грызуны тундры и бореальных лесов, некоторые виды саранчовых из аридных зон, например перелетная саранча. Если особи последнего вида живут разбросанно, то они
О 100м
।------1------J
Рис. 25. Карта районов охоты пар трех видов воробьиных из семейства Fringillidae, обитающих в разреженном лесу Онтарио в июне. Частичное наложение районов наблюдается лишь у разных видов (по Кендею, Kendeigh).
Zonotrichia albicollls (показана точками); Ме-lospiza melodia (показана косой штриховкой); Melosplza lincolnli (вертикальной штриховкой).
ведут оседлый образ жизни, пока не наступают условия, благоприятные для массового размножения. У взрослых особей при этом меняются морфология и окраска (по сравнению с одиночными особями), а также развиваются стадные и миграционные инстинкты. Перелеты саранчи охватывают обычно области, мало благоприятные для ее питания, поэтому она более или менее быстро погибает. Тогда снова одиночная фаза в другом месте дает начало стадной. Среди видов, для которых характерны такие непериодические миграции, находящиеся в зависимости от скорости их размножения, можно отметить спрингбока — южноафриканскую газель; исчезнувшего ныне североамериканского странствующего голубя; таких бабочек, как Vanessa cardui, которая при некоторых перелетах пересекает Средиземное море из Африки в Европу, капустниц; стрекоз, вызывающих «стрекозиные бури» в пампасах Патагонии; личинок двукрылых Sciara militarise которые мигрируют тесными рядами,
68
образующими длинную ленту. С повышением плотности связано (у насекомых) и другое психическое явление: каннибализм, являющийся одним из элементов, регулирующих плотность.
Экологические факторы, лежащие в основе конкуренции.
1. Зеленые растения. Свет — обычная причина конкуренции, поскольку растения затеняют друг друга; это затенение тем больше, чем больше плотность и чем продолжительнее стадия роста. В этой конкуренции за свет скорость роста в высоту приобретает решающее значение.
Конкуренция за воду возникает в тех случаях, когда запас почвенной влаги недостаточен для удовлетворения нужд всего растительного покрова. Способность растений к такой конкуренции зависит от многочисленных морфологических и физиологических особенностей, которые регулируют их водный баланс.
Еще одним объектом конкуренции между растениями является запас ассимилируемых минеральных веществ в почве. В зависимости от содержания в почве тех или иных элементов результаты такой конкуренции могут быть прямо противоположными. Это ясно показывают экспериментальные данные Штерна и Дональда (Stern, Donald): если высадить смесь одинаковых количеств Lolium rigidum и Trifolium subterraneum па почву, бедную азотом, то ее урожай в 10 раз превосходит урожай Lolium\ прибавление значительных количеств нитратного азота обращает эту пропорцию. Основной причиной такой инверсии является то, что злак плевел больше выигрывает от прибавления азота: увеличивается его рост и поверхность листьев; таким образом, он лишает клевер большей части световой энергии. Этот пример показывает, кроме того, как экологический фактор, влияющий на конкуренцию, может заместить другой фактор под влиянием изменения последнего.
2. Микроорганизмы. Пищевая конкуренция между микроорганизмами — общее явление, особенно конкуренция за источники углерода и азота. Например, одновременное высаживание Agrobacterium radiobacter и гриба Fusarium oxysporum в стерилизованную почву, бедную минеральным азотом, приводит к исчезновению гриба, тогда как прибавление нитрата вызывает обратный эффект.
3. Животные. Обилие и характер продуктов питания являются важнейшим внешним фактором конкуренции животных. Замена одного источника питания другим может привести к изменению результатов; если два вида ветвистоусых раков (С1а-docera)— Daphnia pulicaris и D. magna — воспитываются вместе, то первый элиминирует второй; однако эта элиминация сильно замедляется, если их кормят дрожжами, а не водорослями.
У некоторых сидячих колониальных животных, таких, как морской желудь (Balanus), мидии, устрицы, место, необходимое Для фиксации колонии на поверхности опоры, представляет другой объект конкуренции.
89
Биологические и биогеографические последствия конкуренции. Конкуренция — важный фактор, влияющий на ареал, на локализацию по местообитаниям внутри ареала, на состав и эволюцию сообществ организмов.
1. Влияние на ареал. По мере того как какой-либо вид удаляется от своего климатического оптимума, другие виды одновременно к этому оптимуму приближаются, этот вид из-за снижения вегетативной мощности и способности к воспроизведению элиминируется из все большего количества местообитаний в результате возрастающей конкуренции с другими видами. В отсутствие такой конкуренции обусловленный климатом ареал видов был бы более обширным, на что указывает выживание интродуцированных растений на защищенных от вторжения местной флоры участках. Среди наземных растений весьма обширный ареал имеют сорняки культурных площадей и пустырей, на которых конкуренция очень ограничена из-за прерывистого характера растительного покрова.
Другим доказательством влияния конкуренции на ограничение географических ареалов служит современная регрессия некоторых видов под воздействием этого фактора. Так, злак Spar-tina stricta с засоленных илов Западной Европы был вытеснен из большей части своего ареала новейшим пришельцем, более мощным злаком Spartina townsendii.
На рис. 26 показано прежнее и современное распространение в США мушки-паразита пшеницы (Cephas tabidus), ввезенной из Европы; она отступила на север своего ареала под воздействием близкого вида того же происхождения Cephas pyg* maeas, который откладывает яички на пшенице неделей раньше.
2. Влияние на локализацию по стациям. Если у двух видов одновременно проявляются одинаковые требования к среде и если у них к тому же одинаковые способы удовлетворения этих требований, то один из них элиминирует другой. Этот процесс, на который было обращено внимание уже Дарвином и который экспериментально изучался Гаузе, известен как «правило Гаузе». Например, берега прудов и проточных вод колонизуются большими видами-амфибиями — тростником, озерным камышом, рогозом (Typha), ежеголовником (Sparganiam), ирисом; эти растения образуют по берегам плотные заселения, которые часто соприкасаются, но никогда не смешиваются. Роль, которую играют экологические факторы в определении исхода соревнования очень близких форм, иллюстрируется опытами Харлама и Мартини (Harlam, Martini). Они культивировали 11 разновидностей ячменя в десяти местообитаниях, рассеянных в различных районах США: в первый год в каждую парцеллу была посеяна смесь этих разновидностей в равной пропорции, и каждый следующий год высевалась смесь, полученная от предыдущего урожая. Через несколько лет соотношения между
90
Рис. 26. Распределение двух взаимоисключающих видов насекомых, вывезенных в США в конце прошлого века и паразитирующих в стеблях ржи. Cephas tabidus был вытеснен из северной части своего ареала (пунктир) видом С. pygmaeus, личинки которого вследствие более ранней кладки уничтожают яйца первого (по Удину, Е. J. Udine).
разновидностями резко отличались от начальных, но по-разному в различных местообитаниях.
Сужение амплитуды естественного распределения вида по сравнению с амплитудой, определяемой его физиологическими возможностями, которое происходит под влиянием внешних факторов, может сопровождаться также смещением оптимальных условий; это показали де Фриз и Элленберг (О. de Vries, Н. ЕЬ lenberg). Рис. 27 иллюстрирует опыт по выращиванию двух однолетних сорняков при различных pH; при культивировании порознь наблюдается широкая зона оптимума между pH = 5 и 7, тогда как в смеси дикий цедис имеет четкий оптимум при pH = 5, а горчица —при pH = 7 и 8. В естественных условиях редис ацидофилен, а горчица нейтрофильпа.
В животном мире имеется множество подобных примеров сужения экологической амплитуды из-за конкуренции. Например, Одум (Odum) отмечает, что представитель воробьиных, синица
9L
(Partis atricapillus), обитающая в Северной Америке в лесах различного типа, что связано с отсутствием там близких видов, вынуждена в Европе во время сезона гнездования довольствоваться влажным лесом, поскольку в остальных лесах многочисленные близкие ей виды ведут сходный образ жизни.
2. Влияние на эволюцию сообществ. Конкуренция — важнейший двигатель эволюции сообществ. Замещение сообществ, богатых однолетними растениями, сообществами многолетних трав, а последних — сообществами кустарников и деревьев в большой мере связано с конкуренцией многолетних и
Р и с. 27. Рост дикой редьки (Raphanus raphanistrum) и горчицы полевой (Sinapis arvensis) на почвах с различными значениями pH. В чистой культуре у первой (слева) оптимум при pH = 5, у второй (в центре) при pH = 7; в культуре при смешении в равных долях (справа) оптимумы сдвинуты в сторону большей кислотности для редьки и большей щелочности для горчицы (по Элленбергу, El lenberg).
однолетних растений, а затем древесных пород — с травами. Вид, вытеснивший другие виды, может оказаться вытесненным в свою очередь, и так до тех пор, пока не установится ансамбль видов, гармонично поделивших пространство, источники энергии, воду и питательные вещества. В этих заключительных сообществах, возникающих в результате прогрессивной эволюции, межвидовая конкуренция достигает состояния равновесия.
Кооперация
Если конкуренция при повышении плотности индивидуумов всегда приводит к неблагоприятному результату, то под влиянием других разнообразных факторов их сближение может оказаться полезным или даже необходимым.
Высокая плотность населения может изменить условия среды в сторону, благоприятную для данного вида. Так, деревья в мае--сивах могут сохраняться и возобновляться в климатических условиях, неблагоприятных для отдельных деревьев того же вида.
92
Точно так же мелкие животные, например мыши или пчелы, лучше выживают при низких температурах, если образуют тесные группы, внутри которых поддерживается необходимая температура. Это явление называют «эффектом массы».
Как борьба при межвидовой конкуренции, так и защита от врагов могут лучше осуществляться в условиях сообщества. Также живущие стаями хищники — волки, койоты — могут убивать таких крупных животных, с которыми каждый из них в одиночку не справился бы.
В некоторых условиях образование видами значительных групп может также играть благоприятную роль для воспроизведения, что связано с сенсорными стимулами («групповой эффект»). Опыты, производившиеся на популяциях плодовой мушки дрозофилы различной плотности, снабженных обильной пищей с целью устранения конкуренции, дали следующие результаты:
Число пар мушек на сосуд 1 2 4 8 16 64
Число яиц, отложенных одной 102 121 148 204 158 100 самкой за первые десять дней
Видно, что существует оптимальная плотность для откладки яиц.
Необходимость плотности популяции, достаточно высокой для ее выживания, является фактом, который можно наблюдать в различных группах животных. Следует назвать колонии литоральных птиц, которые гнездятся и откладывают яйца, лишь собираясь в большие группы, что связано с взаимным сенсорным возбуждением. По Хатчинсону (Hutchinson), например, для сохранения колоний бакланов на островах вдоль побережья Перу, образованных гуано, они должны были включать не менее 10 тыс. особей при плотности порядка трех гнезд на 1 м2.
В целом популяции животных не могут возрастать и даже сохранять свою численность, если их размер падает ниже некоторого критического значения: это «принцип минимальной популяции». Особенно хорошо видно проявление этого принципа при попытках защиты видов, находящихся на грани исчезновения. Так обстоит дело с североамериканским белым журавлем, который, хотя и находится под охраной в канадском заповеднике, в течение многих лет его представляют всего 30—40 особей. Еще драматичнее пример других североамериканских птиц: в 1920 г. оставалось около 50 пар глухарей, в 1927 г. их количество уменьшилось до 20 экземпляров, а последнего из них видели в 1932 г. — и это несмотря на защиту их от истребления. К факторам, лежащим в основе этой регрессии, нужно отнести Не только уменьшение рождаемости, но и увеличение смертности вследствие ухудшения защиты от вредных влияний. Странствующий голубь (Ectopistes migratorius) был чрезвычайно широко Распространен на востоке США вплоть до середины XIX в.;
93
затем началось уничтожение его человеком и, по-видимому, какими-то эпизоотиями. Последние экземпляры этого вида, сохранившиеся в зоологическом саду, оказались неспособными предохранить вид от полного вымирания, наступившего в 1914 г.
Наибольшего значения достигает кооперация в сообществах наиболее специализированных насекомых, например пчел, ос, муравьев и термитов, у которых социальные взаимоотношения достигают крайне высокого уровня сложности. Он необходим для поддержания их колоний, включая коллективную постройку жилища, перенос материалов, удаление отходов, вентиляцию, уход за потомством. Эти коллективные действия координированы и обычно сочетаются с полиморфизмом.
Приведенные выше случаи относятся к кооперации, связанной со сближением особей одного и того же вида. Существует также межвидовая кооперация, которая, если становится необходимой, постепенно переходит в симбиоз. Множество подобных случаев наблюдается у низших организмов, например у микроорганизмов с комплементарными потребностями, которые могут образовывать настоящие ассоциации. Так, азотфиксирующие Azotobacter и Clostridium размножаются в присутствии целлюлозы, если она разлагается под действием целлюлолитических бактерий и грибов. Последние, таким образом, снабжают азотфиксирующие бактерии необходимыми им углеводами, а взамен получают азот в непосредственно усвояемой форме.
Взаимодействия, вызванные веществами, выделяемыми организмами
Мысль о влиянии выделяемых организмом химических соединений на соседние организмы была высказана в 1832 г. де Кандоллем (Auguste Pyrame de Candolle). Однако в XIX в. все исследования были направлены на изучение действия питательных веществ, необходимых для роста растений, и только к 1930 г. вновь обратили внимание на действие продуктов метаболизма, выделяемых организмами во внешнюю среду.
Эти выделения могут оказывать то благоприятное, то вредное воздействие на организмы-рецепторы. В последнем случае такие воздействия называют аллелопатией, если речь идет о влиянии одного высшего растения на другое, и антибиозом, если вещества выделяются микроорганизмами, а в общем случае вредные воздействия называют телетоксией.
Многочисленные эксперименты in vitro выявили полезные или вредные влияния экскретов на рост организмов. Однако их экологическое значение, то есть существование этих явлений в естественных условиях, не может быть доказано таким образом. И действительно, как в почве, так и в водной среде множество конкурирующих агентов препятствует этим воздействиям: это
94
разжижение, унос водой, фиксация на глинистых и органических коллоидных частичках, модификация или метаболизация активных молекул микроорганизмами, антагонизм или синергизм веществ в смеси и т. д.
Все разнообразие таких взаимодействий мы покажем на примерах различных групп организмов.
Химическое взаимодействие между сосудистыми растениями
Выделение активных продуктов может производиться либо живыми подземными или воздушными органами растения, либо свежими остатками, мертвыми корнями, опавшими плодами, мертвыми листьями или корой.
Хорошо изучен пример ореховых деревьев. Давно замечено, что иод этими деревьями хиреют томаты, картофель, люцерна. Естественные насаждения Juglans regia в Азии и Juglans nigra в Северной Америке обладают очень бедным травяным покровом. Дэвис (Davis) высказал гипотезу о том, что причиной этого явления служит присутствие во всем растении хининового соединения юглона. Орошение томатов и люцерны раствором юглона оказывает на них токсическое действие. Однако корни ореховых деревьев не выделяют этого вещества; лишь дождевая вода, стекая по листьям и стволам деревьев, обогащается юглоном и действует токсически на растения.
Возможно, что в аридных областях аллелопатические воздействия более распространены, чем во влажных, где почва подвержена выщелачиванию и где проявляется значительная микробиологическая активность. Корни гваюлы (Parthenium ar gent а,-turn) и листва Encelia farinosa, кустарников североамериканских полупустынь, оказывают, по-видимому, фитотоксическое действие, которое, однако, не вполне доказано. Во Франции в засушливых местообитаниях ястребинка и розмарин таким же образом элиминируют однолетние виды.
Примером вызванной корневыми выделениями стимуляции роста служат злаки, азотное питание которых в смеси с бобовыми улучшается из-за выделения последними аминокислот. Другой случай — это прорастание паразитических семенных растений: так, Striga lutea, паразит хлебных культур Южной Африки, прорастает лишь благодаря присутствию в воде, омывающей корни его хозяина, сахара из группы пентоз — ксилозы. Напротив, другие культурные растения — подсолнечник, турецкий горох (нут), арахис — экскретируют вещества, тормозящие прорастание Striga,
Химическое взаимодействие между микроорганизмами
Микроорганизмы в ходе высокоактивного метаболизма вырабатывают и выделяют во внешнюю среду многочисленные молекулы, способные благоприятно или неблагоприятно влиять на
95
другие виды: пищевые вещества, факторы роста, антибиотические вещества.
Образование трофических веществ. Иллюстрацией таких процессов служат биогеохимические циклы минеральных элементов в почве, в результате которых эти элементы переходят от одной группы микроорганизмов к другой, но в иной химической комбинации.
Если речь идет, например, об азоте, то протеолитические бактерии высвобождают аммиак, давая энергию микроорганизмам, окисляющим его в нитрит; последний процесс в свою очередь служит источником энергии для других нитрифицирующих микроорганизмов, переводящих его в нитрат. Таким образом, существует тесная зависимость между различными по характеру метаболизма группами почвенных бактерий.
Выделение факторов роста. Было обнаружено, что многие микроорганизмы могут развиваться лишь в присутствии других микроорганизмов, вырабатывающих для них факторы роста; эти вещества (состав которых не во всех случаях изучен) они не могут синтезировать сами. В отношении бактерий чаще всего речь идет о тиамине, биотине и витамине Bj2. В некоторых случаях химизм двух видов является комплементарным, так что они могут существовать лишь совместно, образуя подлинный трофический (пищевой) симбиоз.
Например, Шопфером (Schopfer) было показано, что два гриба (Mucor ramannianus и Rhodotorula rubra), из которых первый способен синтезировать лишь пиримидиновую фракцию молекулы витамина Bj2, а второй — лишь его тиазоловую фракцию, не могут расти изолированно в лишенных этого витамина средах. Вместе же они превосходно развиваются, обмениваясь этими фракциями. Аналогично этому, бактерии Streptococcus faecalis и Lactobacillus arabinosus не могут расти порознь в среде с недостаточным содержанием аминокислоты фенилаланина и витамина — фолиевой кислоты, поскольку первая нуждается в витамине, а вторая — в аминокислоте, то есть в веществах, которые они сами не синтезируют. Однако эти бактерии могут жить в смешанных культурах в той же среде, обмениваясь этими двумя веществами (Нурмикко, Nurmikko).
Выделение антибиотических веществ. Эти вещества многочисленны и вырабатываются многими грибами, актиномицетами и почвенными бактериями в чистой культуре. Например, гриб Trichoderma viride вырабатывает виридин (антифунгическое вещество) и глиотоксин (антифунгическое и антибактериальное вещество). Актиномицет Streptomyces griseus вырабатывает антибактериальные стрептомицин и гризеин, а также антифунгическое вещество актидион. Bacillus subtilis выделяет бацитрацин и бациллин (антибактериальные), бацилломицин (антифунгическое) и, наконец, субтилин, обладающий антибактериальным, антифунгическим и антипротозойным действием.
96
Однако, хотя выход антибиотиков в изолированных штаммах, культивируемых в лабораториях в оптимальных условиях на богатых средах, достаточно велик, иначе дело обстоит в почве. Уже само отсутствие преобладания вырабатывающих антибиотики микроорганизмов — аргумент против значительности антибиотического воздействия. Антибиотики, вырабатываемые в меньшем количестве, могут быть инактивированы другими микроорганизмами, или абсорбированы коллоидами, или унесены водой.
Микроскопические водоросли и пресноводные простейшие могут наряду со стимулирующими веществами вырабатывать токсины. Так, среда, содержащая культуру хлореллы, тормозит рост диатомовой водоросли Nitzschia frustulum и обратно. Было показано, что эти токсические воздействия могут быть одной из причин, определяющих состав и эволюцию популяций водорослей в небольших водоемах.
Химическое взаимодействие между высшими растениями и свободными микроорганизмами
Корни сосудистых растений окружены популяциями микроорганизмов, с которыми устанавливается взаимодействие.
Влияние корней на микропопуляции почвы. По соседству с корнями популяции микроорганизмов, находящихся в активном состоянии, обычно более многочисленны и имеют иной состав. Зону действия корней Гильтнер (Hiltner) в 1904 г. назвал ризосферой. Это воздействие связано с разнообразными изменениями эдафической среды, вызванными корнями, — изменением влажности, структуры, pH, выделением СОг и различных органических молекул, играющих трофическую роль, стимуляторов или ингибиторов роста, в частности аминокислот, сахаров, гетерози-дов, органических кислот, энзимов, витаминов.
Количество микроорганизмов прогрессивно возрастает по мере приближения к корням, но неодинаково для разных групп. Среди бактерий более всех стимулируются те формы, которым необходимы аминокислоты, а также спорообразующие формы. Под влиянием выделений корней некоторых видов растений интенсифицируется прорастание спор или склероциев патогенных форм грибов.
Влияние почвенных бактерий на сосудистые растения. Под влиянием стимулируемых корнями популяций почвенных микроорганизмов в свою очередь изменяются условия среды, в которой находятся корни, что влияет на жизнь растения в целом. Это влияние может быть различным — благоприятным или неблагоприятным. Особенно велика роль микроорганизмов в обеспечении минерального питания растений, и прежде всего — азотного питания. Однако сейчас известно, что молекулы разнообразных органических веществ, возникающих в процессе метаболизма микрофлоры, способны проникать в растения. Таковы, например»
4 Ж. Леме
97
естественный ауксин (уксусноиндоловая кислота), антибиотики.
Напомним здесь случаи, когда между микроорганизмами и корнями высших растений устанавливаются симбиотические отношения: бактериальные клубеньки на корнях бобовых, микориза многих видов — орхидей, вересковых, лесных деревьев и др. Велико значение ризосферы при заражении корней патогенными зародышами. Развитие паразитических микроорганизмов можег быть приостановлено не только действием корневых выделений (о чем шла речь выше), но и сапрофитными микроорганизмами ризосферы, защищающими, таким образом, высшие растения от инфекции. Например, в посевах клевера, растущего на подзолистых почвах СССР, обильны миколитические бактерии, разрушающие мицелий различных грибов, паразитирующих на сосудистых растениях; однако они отсутствуют в культурах пшеницы и льна.
Химическое взаимодействие между растениями и животными
Распространение во внешней среде многих продуктов метаболизма растений оказывает влияние на животных. Было обнаружено в лабораторных условиях, что старые культуры зеленых или диатомовых водорослей оказывают токсическое воздействие на дафний. Быстрое внезапное размножение летом сине-зеленых планктонных водорослей в озерах или прудах, называемое цветением воды, приводит вследствие интоксикации к гибели планктонных видов животных, рыб, а иногда даже пьющего эту воду скота. Это связано с выделением таких токсических веществ, как гидроксиламин, а также продуктов распада этой массы водорослей.
Взаимодействия, основанные на контакте или на предоставлении убежища: комменсализм и мутуализм
Существует два типа таких объединений организмов:
1) комменсализм, когда в преимущественном положении находится лишь тот из участников, который поселяется на другом;
2) мутуализм, когда от сближения выигрывают оба партнера, в этом случае можно говорить о симбиозе в широком смысле слова.
В действительности часто бывает трудно решить, какому из типов отдать предпочтение. Существуют, кроме того, переходные ступени от комменсализма к паразитизму, в то время как мутуализм представляет собой разновидность симбиоза.
Примером постоянного комменсализма в растительном царстве служат растения-эпифиты. Объединение эпифита с его живой опорой не случайно; количество и видовой состав эпифитов
98
на деревьях меняются в зависимости от породы и окружающих условий. Сосны и платаны, кора которых ежегодно полностью обновляется, почти не имеют эпифитов, тогда как дубы и пихты, для которых характерна шероховатая и постоянная кора, покрыты ими. Эпифиты, кроме того, очень чувствительны к атмосферной влаге, так что одна и та же порода деревьев может нести различную эпифитную флору в зависимости от микроклиматических условий.
Форезией называется использование одним животным другого просто в качестве средства передвижения. Навозные жуки (Geo-trupes) часто перевозят таким образом клещей из одной навозной кучи в другую.
Комменсалами являются также животные, которые живут в жилищах других животных, терпящих их у себя, и которые находят там убежище, подходящий микроклимат и обеспеченное питание. Фауны насекомых из нор млекопитающих, многочисленные животные, живущие в домах, — домовый паук, домовый сверчок, мышь, комнатная муха, — тоже комменсалы.
Очень трудно классифицировать организмы, живущие во внутренних полостях животных. Маленький краб (Pinnotheres pi-sum), живущий внутри раковин живых мидий, которая защищает его от хищников, является, по-видимому, просто комменсалом. Значительно более спорными являются взаимоотношения между кишечными популяциями микробов и хозяином: бактерия Escherichia coli и многие простейшие считаются простыми комменсалами (за исключением частных случаев, когда они могут вести себя как паразиты), однако целлюлолитические бактерии могут служить примерохм мутуализма, поскольку они вызывают разложение целлюлозы на ассимилируемые организмом сахара.
То же касается своеобразных объединений морских организмов, у которых особенности взаимной выгоды не всегда достаточно выяснены. Например, какую пользу приносят некоторым крабам закрепившиеся на них водоросли, губки, актинии? Камуфляж это или приманка для добычи? Как известно, рак-отшельник сам помещает актинию на раковину, в которой поселяется, и переносит ее на другую раковину при перемене жилища. Между прочим, актиния (Adamsia palliata) объединяется исключительно с Pagurus prideauxii, причем по мере того, как последний растет, актиния может защитить его при помощи выделения мембраны, удлиняющей отверстие раковины. Здесь, таким образом, наблюдается не просто сближение, но настоящее объединение двух организмов.
Взаимная выгода особенно отчетливо заметна при некоторых сближениях животных, например птиц и крупных травоядных: первые питаются эктопаразитическими насекомыми вторых. Тоже касается настоящих грибных плантаций, созданных некоторыми насекомыми, общественными или обитающими в древесине.
4*
99
Так, муравьи американского рода Atta срезают листья и переносят их куски как зонтики до гнезда. Там они растирают листья в кашицу, которую высевают вместе с грибами. Высеваются такие грибы, как, например, Rozites gongylophora, мицелий которых может служить пищей муравьям; в то же время зародыши непригодных в пищу грибов уничтожаются. Жуки-заболонники (Scolytidae) прогрызают в древесине галереи, где опилки служат средой, в которой развиваются различные группы грибов. Еще до определения их таксономической принадлежности все эти грибы объединили под названием «амброзия». Грибы развиваются на экскрементах личинок, питающихся мицелием грибов; грибы обеспечивают личинкам азотное питание, а «амброзия» находит в этих галереях благоприятный ей субстрат и влажность.
Опыление цветков насекомыми тоже может служить примером мутуализма другого типа. Насекомые, питающиеся на цветах, собирают с них нектар и пыльцу, одновременно обеспечивая опыление. Многие антофильные насекомые используют лишь один вид растений, что ведет к взаимной выгоде: посещение цветков одним и тем же видом насекомых — благоприятное условие для перекрестного опыления, так как насекомое, будучи хорошо знакомым с топографией цветка, берет взяток в минимальные сроки, что обеспечивает высокий сбор. Такое сочетание возникает случайно в результате разнообразных встреч насекомого с цветками; оно продолжается в период цветения, а затем возникает второе сочетание с более поздно цветущим цветком. Особенно характерно такое постоянство для пчел. Многие цветки посещает весьма ограниченное число видов. Примером могут служить акониты, которые называют «шмелиными цветами»: ареалы обоих совпадают. Желтый ирис (Iris pseuda-corus) имеет два сорта цветков; первый оплодотворяется шмелями, а второй — сирфидами (Syrphidае) из отряда двукрылых.
Ваниль у себя па родине оплодотворяется особым перепончатокрылым. Поскольку интродукция этого растения в другие области не сопровождалась введением туда соответствующего насекомого-опылителя, то приходится прибегать к искусственному опылению.
Напротив, соцветия сложноцветных и зонтичных посещаются насекомыми самых разнообразных видов.
Пищевое использование одних организмов другими: хищничество и паразитизм
Использование живых организмов для питания — банальный случай биотических воздействий, которые могут проявляться по-разному. Хищники — это свободные организмы, занимающиеся поисками живой животной или растительной пищи. Паразиты
100
живут всегда или почти всегда на поверхности (эктопаразиты) или внутри (эндопаразиты) живых организмов. Они потребляют живое вещество или продукты его метаболизма, не убивая (во всяком случае, предварительно) своего хозяина. Именно поэтому можно было сказать, что «хищник живет за счет капитала, а паразит — за счет прибыли». С экологической точки зрения эти различия между хищниками и паразитами менее важны, чем причины и следствия отношений, возникающих между эксплуататором и эксплуатируемым, которые можно выразить в одинаковых терминах как для хищников, так и для паразитов.
Виды зависимости хищника или паразита от жертвы или хозяина
Очевидно, что присутствие хищника или паразита в каком-.либо местообитании возможно только при наличии там используемых им организмов. Степень специфичности жертвы или хозяина различна у разных видов.
1. Полифагия. Распространена среди хищников, которые могут быть всеядными, как, например, бурый медведь; в зависимости от сезона он питается мелкими млекопитающими, медом, ягодами, даже рыбой. Чаще всего характер питания хищника в большой степени зависит от относительного обилия особей потребляемых им видов. Например, частичное вымирание кроликов из-за миксоматоза заставило лис питаться главным образом мелкими грызунами, птицами и даже рыбами. Мангусты, ввезенные на Ямайку для уничтожения крыс, стали после их исчезновения питаться домашними животными, птицами, рептилиями, амфибиями, наземными крабами.
Среди паразитических растений виды-полифаги часто состоят из физиологических рас, адаптированных к узкому кругу видов хозяев. Например, омела, паразитирующая более чем на 50 видах деревьев и кустарников, на самом деле состоит из различных рас, каждая из которых может паразитировать лишь на одном или нескольких видах. Линейная или стеблевая ржавчина пшеницы (Puccinia graminis tritici) состоит из многочисленных физиологических форм, каждая из которых способна инфицировать определенную разновидность пшеницы. После того как формы пшеницы, инфицированные ржавчиной, были последовательно заменены адаптированными к ней сортами, в конце концов появились формы ржавчины, адаптированные к новым сортам пшеницы.
2. Олигофаги. Живут за счет нескольких, часто родственных видов. Среди семенных паразитов корней — заразих — многие являются монофагами, однако заразиха (Orobanche minor) селится на многих бобовых. Дубы и ивы покрыты многочисленными галлами, образованными насекомыми из отряда перепончатокрылых— орехотворками (Cynipidae), которые нападают на
101
различные виды одного из этих родов. Личинка Taenia solium является паразитом свиньи, а личинка Taenia saginata — быка, тогда как личинка Taenia echinococcus * 1 может паразитировать на человеке, свинье, хищных и грызунах.
3. Монофаги. Ограничены одним хозяином. Монофагами являются многие насекомые. Среди бактерий дифтерийная палочка и пневмококк паразитируют лишь на человеке. Из вирусных заболеваний корь специфична для человека, а миксоматоздля кролика. В таких случаях ареал паразита или хищника целиком в пределах ареала используемого вида.
Иногда виды приспосабливаются к новому хозяину. Виды-фитофаги, нападающие на культурные растения, жили когда-то на диких растениях. Насекомые, уничтожающие культурные злаки, в большинстве своем обитали ранее в травянистых формациях тех же районов. «Подсолнечник, происходящий из Америки и ввезенный в Россию в XVIII в., стал добычей более чем пятидесяти вредных насекомых, принадлежащих к семи различным отрядам и живущих обычно на лесных диких сложноцветных» (Uvarov). Блохи, паразитировавшие на одном сумчатом животном, стали паразитами кроликов после их интродукции в Австралию, в то время как блохи, паразитировавшие на кроликах в Европе, не сумели адаптироваться в Австралии.
Во всех случаях необходимо, чтобы хищники в сезон своей активной жизни располагали организмами, поставляющими им пищу. Такое приспособление осуществляется в различные периоды, в зависимости от климата. Например, у луковой мухи {Hylemyia antiqua^ живущей на луке и диком чесноке в Европе и Северной Америке, происходит окукливание и начинается диапауза в конце лета, а насекомое вылупляется следующей весной. В то же время на Ближнем Востоке это насекомое вылупляется и откладывает яйца на молодых побегах чеснока, начиная с ноября — декабря, а последующее поколение окукливается в конце зимы, когда чеснок желтеет. Муха, паразитирующая на тлях {Epistrophe balteatd), проходит стадию диапаузы в Центральной Европе зимой, а в Израиле — летом: это именно те сезоны, когда в соответствующих районах исчезают тли.
В случае, когда паразит сменяет при прохождении своего жизненного цикла нескольких хозяев, необходимо, чтобы все они в нужные периоды находились недалеко друг от друга. Классическим примером этому в растительном царстве служат многочисленные ржавчинные грибы, а в животном — ленточные черви (Cestodes) и сосальщики (Trematoda).
Большинство паразитов является облигатными, однако некоторые из них факультативны, в частности грибы и бактерии,
1 Этого паразита относят сейчас к особому роду Echinococcus и называют
Е. granulosus. — Прим. ред.
102
которые могут постоянно существовать в качестве сапрофитов в почве. Таковы столбнячная палочка, симбиотические бактерии бобовых, микоризные грибы, гриб Fusarium oxysporum 1. В животном царстве примером факультативного паразитизма являются виды комаров, способные жить исключительно за счет растительных соков, но в некоторых случаях питающиеся кровью млекопитающих 2.
Последствия паразитизма для хозяина, а хищничества — для жертвы
Паразиты и хищники могут вызвать уничтожение используемого вида. Напомним здесь, что разрушительная деятельность человека привела к полному исчезновению многочисленных видов или к их истреблению на большей части их исходного ареала.
Но чаще активность паразитов и хищников вызывает лишь более или менее продолжительное уменьшение численности их жертв, поскольку последние оказываются при этом защищены от полного истребления самим фактом уменьшения их численности или же в силу того, что оставшиеся особи становятся резистентными благодаря иммунологическим процессам. В странах, где развился миксоматоз, это заболевание уничтожило 90% кроликов, однако выжившие особи оказались резистентными и дали жизнь новым поколениям. Другое возможное последствие паразитизма, существенное для выживания вида, это «паразитическая кастрация», то есть остановка развития гонад, приводящая к снижению его воспроизводящей способности.
Один и тот же паразит может оказывать различное влияние на выживание своих хозяев. Например, ареалы трипаносом — возбудителей сонной болезни — являются запретными районами для рогатого скота и лошадей, которые от этой болезни гибнут, но не опасны для обитания антилоп3.
О том, насколько паразитизм и хищничество лимитируют развитие вида, могут свидетельствовать результаты искусственной защиты этого вида от врагов, а также результаты введения этого вида на территорию, где такие враги отсутствуют. Банальным примером первого случая служит защита растений от травоядных с помощью изгородей. Истребляемые травоядными виды растений становятся более обильными и постепенно устанав
1 Этот гриб имеет, однако, наряду с чисто сапрофитными расами расы, способные паразитировать на корнях.
2 У комаров кровососущими являются самки, для которых необходимо это питание: без него невозможно развитие и откладка яиц; растительными соками питаются, как правило, самцы, не способные к кровососанию. — Прим, ред.
8 Болезнь крупного рогатого скота, близкая к сонной болезни человека, называется наганой,— Прим. ред.
103
ливается новое флористическое равновесие. Введение какого-либо вида без его врагов на новую территорию приводит к значительно большему его распространению, чем на родине, где его-жизнеспособность лимитирована паразитами и хищниками. Австралийский червец (Icerya purchasii), ввезенный случайна
Рис. 28. Взаимодействие популяций Paramecium caudatum и ее-хищника Didinium nasutum в различных экспериментальных условиях.
/ — среда лишена укрытия для парамеций; сначала гибнут парамеции, потом; хищник; II — в среде имеется укрытие для парамеций; погибает только хищник; /// — укрытия нет; периодически в среду вводятся парамеции (отмечено-стрелками). Наблюдается чередование-максимумов численности жертвы и хищника (по Гаузе).
в Южную Африку, средиземноморскую область и Америку,, истребил там плантации цитрусовых, тогда как на своей родине он оказался обезвреженным хищником — божьей коровкой (Novius cardinalis). Введение божьей коровки на плантации цитрусовых приостановило интенсивное размножение червеца. Такой контроль над распространением видов с помощью их естественных врагов стал средством биологической борьбы с нежелательными видами.
Количественные аспекты взаимозависимости хозяина и паразита, жертвы и хищника
Томпсон, Вольтерра и Лотка (Thompson, Volterra, Lotka) провели математическое исследование совпадающих флуктуаций численности вида и его хищника, Вольтерра сформулировал следующие три закона:
104
1) закон периодических циклов', флуктуации численности «обоих видов периодичны;
2) закон сохранения средних величин: при неизменности среды и поведения сосуществующих видов их средняя численность остается постоянной;
3) закон смещения средних величин: уничтожение части особей обоих видов, пропорциональной количеству каждого из них, .приводит к возрастанию средней численности используемого вида и уменьшению средней численности хищников.
Экспериментальная проверка этих постулатов, произведенная Гаузе, подтвердила их удовлетворительность. Две реснитчатых
•Р и с. 29. Зависимость между плотностью куколок Choristoneura fu-miferana (Microlepidoptera), питающихся почками ели, и процентом куколок, на которых паразитируют перепончатокрылые. Из рисунка видно, что этот процент уменьшается даже при очень небольшой плотности (по Миллеру, С. A. Miller).
Число куколок на 1м2лист8ы
.инфузории Paramecium caudatum и ее хищника (Didinium nasu-tum) объединяли при различных условиях. Парамеция получала неограниченное количество бактериальной пищи (рис. 28). Если у парамеции не оказывалось никакого укрытия, то введение в культуру Didinium приводило к полному ее истреблению, за которым следовало и исчезновение хищника из-за отсутствия пищи. Когда же часть популяции парамеции защищена от хищника в каком-то участке сосуда, то хищник размножается до тех пор, пока остаются незащищенные парамеции, а потом исчезает, тогда под защитой укрытия особи парамеции вновь занимают все заданное пространство. В третьей серии опытов периодически добавляли в культуру лишенных укрытия парамеций. Наблюдались максимумы и минимумы численности хищника, чередующиеся с максимумами и минимумами численности парамеции.
Сходные флуктуации имеют место и в естественных условиях. -Например, мидии на отмелях интенсивно истребляются морскими звездами, число которых резко увеличивается. После истребления мидий количество звезд снижается, после чего восстанавливается популяция мидий. Затем цикл возобновляется.
Изучение количественных соотношений между численностью хозяина и паразита показало, что относительное количество подвергшихся нападению паразитов особей возрастает
105
с увеличением их плотности, если эта плотность ниже некоторого предела, после преодоления которого наблюдается обратная зависимость (рис. 29).
Трофические и энергетические циклы внутри сообществ организмов: понятие экосистемы
Круговороты веществ между организмами и внутренней средой какого-либо сообщества настолько множественны и разнообразны, что возникает необходимость общей определенной трофической и энергетической организации. Рассматриваемое именно с точки зрения распределения, обмена материи и энергии сообщество представляет собой экологическую систему, или, по выражению Тэнсли (Tansley, 1935), экосистему.
Цепи питания в экосистеме
Между различными элементами экосистемы устанавливаются настоящие цепи переноса молекул — носителей энергии. Происходящие при этом значительные потери энергии компенсируются притоком ее извне за счет солнечного излучения (рис. 30). Привнесение энергии в систему обеспечивается зелеными растениями, которые синтезируют органические молекулы из минеральных соединений, находящихся в среде. Зеленые растения, таким образом, являются в экосистеме производителями, или продуцентами.
Накопленные органические молекулы служат прямым источником энергии для травоядных животных, а также для паразитов и сапрофитов и косвенным — для плотоядных. Совокупность всех этих гетеротрофных организмов образует группу потребителей. или консументов, из которых травоядные являются консументами первого порядка, а плотоядные — консументами второго и третьего порядка.
Большая часть не подвергшихся истреблению хищниками организмов каждого звена этой цепи, консументы, находящиеся на самом конце цепи, а также отходы животных — все это в совокупности источник энергии для огромного количества организмов-сапрофагов, которыми в основном являются бактерии; они разлагают все вышеперечисленное на различные органические и минеральные продукты, часть которых вновь поглощается живыми организмами. Эти сапрофаги называются разрушителями, деструкторами, или редуцентами.
Таким образом, цепи питания экосистемы состоят из четырех звеньев: 1) веществ, используемых организмом; 2) продуцентов; 3) консументов; 4) редуцентов. Небольшая поверхность стоячей воды, например пруд, являет собой пример хорошо очерченной экосистемы: используемыми молекулами служат вода, газообразные вещества, соли и органические вещества, раство-
106
репные в ней, а также донные осадки. Продуценты — это планктонные и донные водоросли, а также водные сосудистые растения. Консументами первого порядка являются мелкие животные, в особенности ракообразные, моллюски, черви. Консументами второго порядка служат рыбы, хищные личинки насекомых. Возможно и существование консументов третьего порядка: в виде рыб, поедающих других рыб. Редуцентами служат водные простейшие, бактерии и грибы, которые в громадном количестве
Солнечная энергия
Растворимые
Плотоядные-(консументы 3-гопоряЗка)
Плотоядные (консументы 2-го порядка)
S-I ческие ^шлатки^
Рис. 30. Схема циркуляции энергии внутри экосистемы.
Травоядные (консументы 1-го поряока)
Зеленые растения (продуценты)
размножаются на поверхности органического донного ила, образованного остатками свободно перемещающихся или сидячих организмов, а также самих редуцентов. В действительности же многие животные всеядны, и для них характерно по преимуществу животное, растительное питание или питание детритом в зависимости от имеющегося источника питания, а также от их возраста, что мы увидим на примерах в четвертой части.
Использование энергии в экосистемах
Часть энергии, фиксированной для производства органических молекул, или общей (полной) продукции (брутто-продук-Ции), рассеивается в процессе дыхания. Остающаяся чистая продукция, которую могут использовать консументы и редуценты, составляет примерно половину общей продукции (см. схему на Рис. 31). Отношение этой продукции к глобальному излучению {при выражении того и другого в калориях) носит название
107
эффективности фотосинтеза, или первичной эффективности. Ее относят либо ко всему спектру солнечного излучения, либо к энергии только видимого спектра, которая составляет примерно половину общей энергии. Продуценты улавливают лишь очень малую часть солнечной энергии. В среднем наземная растительность использует 1 % падающей на нее энергии (хотя при этом для некоторых культур этот процент оказывается значительно выше), в океане используется 0,2%, а в пресных водах — от 0,1 до 1% энергии. Таким образом, эффективность фотосинтеза очень мала по сравнению с эффективностью (КПД) моторов, но-
Дыхание
Тепло Дыхание
Рис. 31. Использование солнечной энергии зелеными растениями (продуцентами) и передача ее первичным консументам (по Одуму, Odum).
необходимо подчеркнуть, что для фотосинтеза используется внеземной источник энергии, что фиксированная энергия является для Земли чистой прибылью и что «растительная фабрика» покрывает громадные пространства и функционирует многие дни в году или даже весь год.
Передача потенциальной энергии от продуцентов к консументам первого порядка или между консументами различных трофических уровней имеет большую эффективность. Например, продукция крупного рогатого скота на пастбищах соответствует примерно 8% выработки чистой энергии, а продукция рыбоводческих хозяйств—15%. При дыхании часть исходной энергии расходуется на каждом уровне, так что при переходе в цепи питания с одного уровня на другой общая продукция живой материи уменьшается.
Редуценты в противоположность травоядным и плотоядным консументам обладают очень высокой метаболической актив* ностью, расходуя значительное количество энергии.
108
В конечном итоге остается некоторое количество неизрасходованных органических веществ, представляющее собой часть неиспользованной потенциальной энергии. Это торф, органический озерный ил, а также образовавшиеся в различные геологические эпохи угольные залежи.
Существование внутренних циклов в экосистемах не превращает их в системы, функционирующие по замкнутому циклу. Кроме солнечной энергии, в экосистему могут быть введены минеральные элементы, как растворенные в проточной или дождевой воде, так и в виде удобрений. Потери энергии могут быть
Рис. 32. Пирамиды биомасс (сверху) и энергии (снизу) в водной среде «Серебряных источников» во Флориде. Биомассы приведены в граммах сухого вещества на 1 м2. Используемая на каждом уровне энергия выражена в килокалориях на 1 м2 в год.
П — продуценты; Г —травоядные (консументы 1-го порядка); МП — мелкие плотоядные (консументы 2-го порядка); КП — крупные плотоядные
(консументы 3-го порядка); Р — редуценты (по Одуму, Н. Т. Odum).
КП = 1,5
МП=11
Т=37
] П=809
I КП =21
Р=5060 ПМЛ=383
|Т=3368
П=20810
следствием пожаров, потери минеральных веществ — следствием уноса просачивающимися водами; земледельческие культуры, эксплуатация лесов, скотоводство, охота и рыбная ловля выводят из системы энергию, аккумулированную на уровне продуцентов или консументов. Наконец, происходит обмен между сообщающимися друг с другом экосистемами.
Количественные особенности экосистем
Экосистему можно охарактеризовать количественньпм распределением энергии и элементов на ее последовательных уровнях, а также потоками этих величин, переносимых с одного уровня на другой; эти характеристики отличают одну экосистему от Другой.
1. Прежде всего необходимо определить массу живого вещества, приходящуюся на единицу площади, или биомассу, как Для всей системы в совокупности, так и для каждого трофического уровня. Результат можно графически выразить в виде пирамиды биомасс, ступени которой расположены снизу вверх в следующем порядке: продуценты, консументы первого, второго и третьего порядков. Ширина каждой ступени при этом пропорциональна соответствующей биомассе. На рис. 32 воспроизведен типичный пример такой пирамиды, на котором видно значительное
109
уменьшение биомассы снизу вверх. Изображенная сбоку масса редуцентов составляет лишь ничтожную часть общей массы. В нижеприводимой таблице даны некоторые примеры биомасс растительности и животного населения (см. также фотоУ).
Таблица 2
Сухое вещество, т на 1 га
Растительный покров (воздушная часть) североамериканская высокотравная
«прерия» 6,550
бородачевая (Andropogoneae) саванна в Гвинее 5-10
посадки сосны обыкновенной 35-летнего возраста, 163
строевой еловый и буковый лес 120-летнего воз- 275
раста, Бельгия вторичный 40—50-летний тропический лес, Гана 360
первичный тропический лес, Конго 1000
Беспозвоночные (напочвенные) кг на 1 га
лес из сосны обыкновенной, Англия 0,1-5
(сухое вещество)
бородачевая саванна, Гвинея 250
(сырое вещество)
Млекопитающие кг сырого вещества
на 1 га
аридная степь в СССР с антилопами сайга 0,35
сосняк, Шотландия 1
дубово-буковый лес, Бельгия 7
кустарниковая саванна, Уганда Птицы 195
леса, Чехословакия 0,5-1,2
дубово-буковый лес, Бельгия Рыбы 1,3
форельные озера, США 62,5
Ла-Манш на широте Плимута 162
коралловые рифы, Бермуды 490
старицы рек, США 625
Почвенные бактерии и грибы кг сухого вещества
на 1 га
культуры из Ротамстеда, Англ1я (примерный 700-1120
расчет)
Почвенная фауна
дождевые черви (Lumbricidae) из 120-летнего ду- 600
бово-буковою леса, Бельгия
Объем биомассы зависит не столько от фиксируемого ежегодно количества энергии, сколько от продолжительности жизни организмов. При годовой продукции одного и того же порядка де
по
ревья в лесных формациях аккумулируют значительно большую массу, чем травы. Вес морского или озерного зоопланктона (консументы) может оказаться большим, чем вес фитопланктона (продуценты), что связано с большей продолжительностью жизни ракообразных по сравнению с водорослями.
2. Нельзя сделать выводы об энергетических особенностях экосистемы на основании пирамиды биомасс, так как вещества, составляющие организмы, обладают неодинаковым энергетическим потенциалом. При равном весе липоиды содержат больше энергии, чем углеводы и белки. Определяя количество энергии, фиксированной в данный промежуток времени (общую энергию, или брутто-энергию), а также количество энергии, остающееся после затраты части ее на функциональные нужды (чистую энергию, или нетто-энергию), можно построить энергетическую пирамиду. Реальное построение такой пирамиды, однако, сопряжено с большими трудностями и возможно лишь с некоторым приближением. На рис. 32 параллельно пирамиде биомасс дана относящаяся к той же экосистеме энергетическая пирамида. Здесь также видно быстрое уменьшение количества энергии, начиная от продуцентов и кончая последним звеном пирамиды — плотоядными; однако отношения между звеньями здесь иные, чем в пирамиде биомасс, и изменяются в пользу травоядных. Можно также видеть, сколь значительна по сравнению с массой та часть энергии, которая используется редуцентами. И наконец, можно отметить, что редуценты используют для производства собственной биомассы лишь небольшую часть энергии, тогда как организмы других уровней затрачивают для этих целей примерно половину энергии.
3. Имеется очень мало точных энергетических данных об экосистемах в их совокупности. Однако мы имеем точные данные о чистой продукции1 зеленых растений, выраженной в количестве сухого вещества, а в случае культурных растений — о первичной чистой продукции2. Продукцией называют количество живого вещества, вырабатываемого в определенный отрезок времени, независимо от исходной и конечной биомассы. Сообщество, обладающее меньшей биомассой, чем другое, не обязательно оказывается менее продуктивным. Для этого достаточно, чтобы продуцирующие организмы обладали коротким сроком жизни (планктон) или частично употреблялись в пищу (травы пастбищ);
1 Не следует путать продукцию и урожайность, или урожай. Последние являются сельскохозяйственными терминами. Урожайность — это продукция веществ, потребляемых человеком.
2 Автор применяет в качестве синонимов термины «продуктивность» и «продукция». Одпако, по терминологии, принятой Международной биологической программой (МБП), первый термин следует применять лишь в общем смысле, понимая под ним способность организмов и сообществ давать продукцию; для обозначения конкретных величин производимого организмами и сообществами вещества рекомендуется применять термин «продукция».— Прим. ред.
1И
тогда переходы от одного уровня к другому происходят быстрее. Табл. 3 дает значения чистой продукции, выраженной в количестве сухого вещества зеленых растений из естественных сообществ и культур в различном климате.
Таблица 3
Чистая первичная продукция, кг/га
средняя за день в период роста
Культурные растения
пшеница, мировое среднее 3 440 23
пшеница, самый высокий валовый сбор 12 500 83
сахарная свекла, мировое среднее 7 650 43
сахарная свекла, самый высокий валовый сбор 16 000 89
сахарный тростник, мировое среднее 17 250 47
сахарный тростник, максимум в интенсивной культуре, Гавайи 67 000 184
люцерна, экспериментальная культура Травянистые формации 33 660 187
альпийские луга 200-1100 30
высокотравная «прерия», Северная Америка 4460
болотистые осоковые луга, Европа 4000—6300
удобренные злаковые луга, Германия 5590—15 500
саванны, поросшие злаком (Arundo donax), Япония Пустыни 52 000
юго-запад США (12 см осадков) Леса 400 2
сосновые леса с Pinus Silvestris, Европа 1750-7000
дубовые и буковые леса, Центральная Европа 4000—14 000
посадки эвкалиптов, Марокко 2500-5000
вечнозеленые тропические и экваториальные леса Водные сообщества 8520—25 0С0
Саргассово море 959 2,6
Различные озера (общая продукция) «Серебряные источники» 1100-18000 3-50
(Silver Springs) 26 930 74
Изучение этой таблицы позволяет сформулировать ряд важных положений. Существуют громадные различия в первичной продукции, связанные не с природой среды — наземной, морской или пресноводной, — а с богатством этой среды — энергией, водой и питательными веществами. Наименее продуктивны большие массы воды и пустыни; бореальные формации менее про-
112
дуктивпы, чем формации влажного тропического климата. Видно также, как огромна теоретическая возможность повышения продуктивности культур 1.
Что касается водных сообществ, маленькие пруды и коралловые рифы достигают размеров первичной продукции, равной продукции тропических лесов.
Данные о вторичной продукции показывают, что и она подвержена большим различиям. Это хорошо иллюстрируется на примере скотоводства. Если при экстенсивном методе ведения хозяйства для прокорма одной коровы требуется более 10 га, луг хорошего качества (1 га) может прокормить по крайней мере двух коров. Такие же контрасты наблюдаются при изучении продукции рыб. Интенсивный лов вдоль некоторых американских побережий позволяет установить, что на этом уровне энергетической пирамиды эффективность равна лишь 0,00015%, что связано как с тем, что водоросли задерживают лишь малую часть энергии излучения, так и с потерями в длинной цепи питания. Напротив, в рыбоводческих хозяйствах эффективность весьма велика. Что же касается общей продукции, то, насколько известно, она выше всего на коралловых рифах и составляет 81 т на 1 га в год.
4. Различная степень подвижности молекул живой материи и составляющих ее элементов может быть измерена отношением количества полученных или потребленных ею веществ в единицу времени на разных трофических уровнях: это так называемая скорость оборота {turnover rate). Время, необходимое для возобновления количества вещества, равного биомассе, называют временем оборота (turnover time). Например, в густом тропическом лесу, в котором установились постоянные значения биомассы и количества органического вещества в почве, при биомассе 250 т на 1 га и годовой продукции различных остатков, мертвых листьев, мертвой древесины и т. п. — Ют на 1 га, годовая скорость оборота сухого вещества составляет4%, а время оборота — 25 лет.
Циркуляция элементов в экосистеме также может быть объектом точного количественного изучения. Для различных лесных насаждений экологи определили количество ежегодно поглощаемых, а также удерживающихся или возвращающихся в почву элементов: количество вещества на 1 га в год увеличивается до определенного возраста, а затем снижается. При старении уменьшается и удержанная часть вещества. Например, в березовых насаждениях 30-летнего возраста деревья задерживают 30— 50% поглощенных элементов, тогда как в 67-летних березняках они задерживают лишь 10—20%.
1 На основании произведенных в Голландии оценок, основанных на фотосинтетической активности культурных растений и измерении потока солнечной энергии, возможный максимум первичной продукции за год при полном покрытии почвы растениями составил 36 т на 1 га.
113
Количество элементов, ежегодно возвращающееся в почву через лесную подстилку, сильно варьирует в зависимости от типа леса.
Таблица 4
Элементы, кг па 1 га/год
N р К Са Mg
Лиственные породы Северной Америки 19 4 15 73 10 1 ! Лютц и
Хвойные породы Северной Аме- 26 2 7 30 5 [ Шандлер
рики
Т ропический полулистопадный 200 7 68 206 45 Най
лес, Гана
Вторичный «зонтичный» лес из Musanga smithii, Конго 140 4 104 127 43 Лоделу и Мейер
Видно, что тропические леса возвращают почве больше элементов, чем леса умеренного климата; это особенно ярко выражено в отношении азота, исключением является фосфор.
Таблица 5
Элементы кг/га
N Р К Са Mg
Pinas sylvestris (47 лет, Великобритания)
живые растения 364 34 304 204 43
подстилка и гумус 192 9 И 25 9
минеральная почва Европейская лиственница (46 лет, Великобритания) 7300 107 250 437 300
живые растения 525 54 2:6 198 43
подстилка и гумус 594 25 23 104 27
минеральная почва Дуб (Quercus robur) (47 лет, Великобритания) 6125 463 223 404 254
живые растения 493 35 246 257 45
подстилка и гумус 71 11 8 35 5
минеральная почва Тропический дождевой лес (50 лет, Гана) 7476 42 329 424 300
живые растения 2000 135 875 2622 384
подстилка п гумус 35 1 10 45 6
минеральная почва 4590 13 650 2573 370
114
Возвращение элементов в почву происходит не только за счет мертвых частей, но также при стекании с растений дождевой воды; особенно это касается калия, который поступает в почву при дожде в количестве, равном примерно 20 кг в год на 1 га в умеренных лесах, и более 200 кг — в лесах Ганы.
В некоторых случаях было определено распределение минеральных элементов в различных фазах экосистемы. Выше (см. табл. 5) приведены примеры для лесных насаждений.
Из таблицы видно, что основные элементы встречаются в разных количествах и пропорциях даже в трех английских насаждениях, находящихся в одинаковых условиях и имеющих одинаковый возраст. Что касается растительных остатков, то накопление в них минеральных элементов зависит не столько от ежегодного поступления, сколько от степени их разложения. Например, в лесу Ганы, где в почву возвращается гораздо большее количество элементов, количество их, сохраненное в виде органических веществ, весьма невелико.
Часть третья
Сообщества организмов: биоценология
В естественных условиях организмы образуют более или менее сложные комплексы, в которых сосуществуют животные, растения и микроорганизмы. Биосфера включает множество подобных сообществ и их изучение — один из наиболее сложных аспектов и одна из основных целей биогеографии. Подобно тому как организм — не простое соединение тканей и органов, но единое целое, обладающее свойствами, которые не присущи отдельным его составным частям, так же и сообщество организмов является качественно новым единством, а не случайным сочетанием индивидуумов и обладает только ему присущими структурой, функциями и эволюцией.
Познание этого уровня организации представляет большие трудности, обусловленные различными причинами. Так, большая таксономическая неоднородность популяции сопряжена с необходимостью учета всех ее компонентов и сотрудничества многих специалистов. Вариации в составе популяций в пространстве и во времени ставят сложную проблему разграничения и классификации, а многообразные взаимодействия между организмами и средой затемняют детерминизм локализации и эволюции сообществ. Поэтому не удивительно, что этот объект исследования вызвал жесточайшие споры и до сих пор ученые не пришли к единому решению проблемы.
Изучение сообществ предполагает три последовательных этапа — разграничение, описание и классификацию, что позволяет перейти к исследованию их модификаций и картографическому изображению. Мы последовательно рассмотрим эти раз-.личные аспекты.
Глава &
Разграничение и организация сообществ
Разграничение сообществ
При разграничении сообществ сразу же возникает проблема выбора характерных признаков, которые подразделяют на физиономические, таксономические и экологические.
На основании физиономических признаков можно выделить площади с растительностью, сходной по своему облику, поскольку здесь сгруппированы в одинаковых пропорциях растения идентичных морфологических типов и сходного сезонного развития. Эта концепция ведет нас к понятию формации.
Таксономические критерии основываются на преобладающих единственных видах или на некоторой совокупности видов, на более или менее полном наборе имеющихся видов растений, животных или обоих этих царств. Такие группировки чаще всего обозначают термином сообщество.
Наконец, можно осуществить разграничение поверхностей, имеющих одни и те же биологически важные физико-химические признаки, хотя подобный способ очень труден, так как для его осуществления необходимо огромное количество измерений.
Однако, какой бы метод ни применялся, гомогенность — понятие весьма относительное. Абсолютного единообразия в сообществах не существует, но можно выделить различные уровни относительного единообразия в зависимости от размеров ассоциированных организмов. Так, почвенные организмы изменяют состав популяции в очень незначительных объемах по отношению к различиям в их размерах в зависимости от условий окружающей среды; мхи и лишайники, растущие на коре в одном лесу, могут быть сгруппированы на стволе различным образом в зависимости от вида несущего их дерева, его высоты и экспозиции, в то время как деревья этого же леса могут образовывать гомогенные популяции на протяжении гектаров. Насекомые образуют иногда однородные сообщества только на небольших поверхностях, в то время как птицы или крупные млекопитающие могут создавать одинаковые сочетания на обширных площадях.
Выработаны различные статистические тесты относительной гомогенности сообществ.
Другой источник трудностей — определение границ. Во многих случаях, особенно если они связаны с вмешательством человека, переходы от одного сообщества к другому бывают
11»
резкими; таковы границы между лесом и посевами, между выжженными и неповрежденными участками леса. Однако в других случаях возрастающие различия по градиенту одного важного экологического фактора, такого, как изменение климата с высотой или изменение влажности на пологом склоне, может вызвать более или менее непрерывное изменение растительного покрова, в котором некоторые экологи, как Уиттэкер и Куртис (Whittaker, Curtis), видят не соседствующие сообщества, а непрерывно изменяющийся растительный покров, который они называют континуумом. В действительности, как нам кажется, анализ совокупности флоры на площадях, достаточно ограниченных для того, чтобы единообразие было реальным, выявляет по крайней мере относительную прерывистость.
Организация популяций одного вида
При изучении организации сообщества различают два уровня: это прежде всего особи, относящиеся к одной таксономической форме, между которыми существуют родственные связи и которые сходным образом реагируют на внешнюю среду, и вся совокупность организмов, живущих в сообществе или по крайней мере значительная часть ее, такая, как сосудистые растения, насекомые, почвенная микрофлора.
Популяция одного вида в данном месте может служить объектом количественных уточнений, когда это касается числа особей и их флуктуаций, или пространства, занимаемого ею, распределения особей по отношению друг к другу.
Численность особей в популяции одного вида
Ее величина. Она может быть установлена с большей или меньшей степенью точности.
1) Относительное обилие. Это оценка количества экземпляров вида, имеющихся на исследуемой поверхности. Геоботаники используют обычно пятибалльную шкалу: 1—очень редко; 2 — редко; 3 — немного Н 4 — обильно; 5 — очень обильно1 2.
Обилие животных оценить труднее; для мелких видов его определяют при помощи взятия проб, в которых устанавливается пропорция индивидуумов для каждого вида. Крогерус (Kroge-rus) предложил три категории обилия:
доминанты — составляющие более 5% общего количества индивидуумов;
инфлуенты — составляющие от 2 до 5%;
рецеденты — менее 2%.
1 В нашей литературе — довольно обильно. — Прим. ред.
2 Это — шкала абсолютного балльного обилия, а не относительного обилия, как полагает Леме. — Прим. ред.
120
В наземных сообществах животные-фитофаги всегда преобладают над хищниками, и среди них виды-полифаги преобладают над видами со специализированным питанием.
Независимо от того, идет ли речь о животных или о растениях, обилие особей одного вида бывает обычно выше в местообитаниях, характеризующихся наличием неблагоприятных факторов, таких, как водоемы с повышенной соленостью или дефицитом кислорода, в связи с ограниченным числом видов в этих условиях.
2) Встречаемость. Предыдущий критерий, субъективный и неточный, можно заменить встречаемостью, статистическим понятием, которое основано на наличии или отсутствии особей одного вида на пробных площадках, взятых наугад внутри исследуемой площади:
р__ количество пробных площадок, на которых имеется вид \z । qq
общее количество пробных площадок
3) Плотность. Это количество особей на единицу площади. Для ее определения существует множество методов. Из плотности можно вывести среднюю площадь, приходящуюся на одну особь, величину, обратную плотности, и среднее расстояние между особями.
Вариации популяции. Численность особей одного вида на данной площади не является постоянной величиной; она — производное от рождаемости и смертности, от эмиграций и иммиграций. Эти процессы сильно варьируют в зависимости от условий существования и плотности популяции.
Допустив, что миграции отсутствуют, можно представить себе оптимальные условия, обеспечивающие популяции максимум рождаемости (или прорастания) и минимум смертности. Этот теоретический процент прироста был назван Чэпменом (Chapman) «биотическим потенциалом», другие авторы именуют его «потенциальным приростом». Но неблагоприятные условия, обусловленные различными лимитирующими факторами, снижают процент прироста, сводят его на нет или даже вызывают численное уменьшение популяции, поскольку реальная рождаемость бывает ниже потенциальной, а реальная смертность выше потенциальной смертности. Разница между процентом потенциального прироста и процентом истинного изменения, которая является оценкой значения неблагоприятных факторов, составляет то, что Чэпмен назвал «сопротивлением среды». Последнее бывает обусловлено как факторами, не зависящими от плотности, такими как холод, так и факторами, которые зависят от нее: ограничение пищевых ресурсов, насыщение воды кислородом, Действие паразитов и хищников и т. п. В этих условиях, когда вид занимает новую площадь, его плотность после быстрого или Постепенного повышения возрастает затем более медленно, пока,
121
наконец, не достигает потолка; рост популяции идет тогда по S-образной или сигмовидной кривой.
Другим условием, важным для будущего популяции, является эволюция возрастного состава составляющих ее особей. Действительно, размножение происходит обычно в определенных возрастных границах, и процент смертности данного вида варьирует в зависимости от возраста. Форма пирамиды возрастов зависит от соотношения между рождаемостью и смертностью. Старые древесные насаждения, в которых возобновление отсутствует, такие, как некоторые кедровники и можжевельники Северной Африки, обречены на вымирание, если не появится какой-либо благоприятный фактор. При разведении пашенной полевки (Micro-tus agrestis) количество особей от 0 до 2 лет составляло 60% в быстрорастущей популяции и всего 12% в стабилизированной популяции.
Размеры популяции зависят также от сезонных миграций, которые являются составной частью характерных годовых ритмов сообществ; существует рассеяние диаспор и расселение молодых животных за пределами сообщества; некоторые животные осуществляют частные и нерегулярные перемещения, рассматриваемые как номадизм; наконец, некоторые виды, такие, как скандинавские и канадские лемминги, саранча аридных зон, совершают особенно эффектные массовые миграции.
Пространственное и весовое заполнение сообщества совокупностью особей одного вида
Для характеристики облика и экологических особенностей сообщества гораздо более важное значение приобретает место, занимаемое особями, чем количество индивидуумов, особенно когда размеры последних сильно варьируют, как это наблюдается у древовидных растений и рыб. Для растений наиболее распространенной методикой является установление их степени покрытия, площади, занимаемой горизонтальной проекцией наземных частей совокупности растений, относящихся к одному виду, по отношению к общей площади исследуемого участка. Были предложены многочисленные пятибалльные шкалы. Согласно приводимой ниже, которой обычно пользуются в Центральной и Западной Европе, покрытие составляет (в баллах):
до 1/20 площати — 1, от более 1/20 до 1/4 — 2, от более 1/4 до 1/2 — 3, от более 1/2 до 3/4 — 4, болзе 3/4 площади — 5.
Виды, для которых характерен наиболее высокий процент покрытия, называют доминантами. Коэффициент смешанного
122
характера, так называемый коэффициент обилия — доминантно-сти, был введен Браун-Бланке и принят геоботаниками, которые придерживаются его методов. Механизм вычисления коэффициента базируется на том факте, что покрытие играет более существенную роль, чем обилие, в тех случаях, когда степень его высока, в то время как обилие становится определяющим в противоположном случае:
Вид покрывает максимум 1/20 площади особи редки: + особи мало обильны: 1 особи обильны: |
Вид покрывает более 1/20: } 2
от более 1/20 до 1/4: )
от более 1/4 до 1/2: 3
от более 1/2 до 3/4: 4
более 3/4 5.
Оценка степени покрытия площади может дать неправильное представление о реально занятом видом пространстве, которое можно тогда уточнить изменениями объема и веса сырого или сухого вещества, или какого-либо основного элемента живого вещества — углерода или азота. Весовые величины являются не чем иным, как биомассами (см. параграф, посвященный экосистемам), примененными к отдельным видам, чаще всего культурных или луговых растений. Измерение объема древесных стволов также широко используется в практике лесоводства.
Распределение особей на площади, занимаемой сообществом
Распределение особей одного вида внутри сообщества может носить случайный характер, а может иметь тенденцию либо к равномерному распределению, либо к соединению в группы. Случаи равномерного распределения в естественных условиях редки, ибо они являются следствием жестокой конкуренции и, напротив, во многих культурах такое распределение часто создается искусственно.
Случайное распределение встречается сравнительно редко. Тенденция к образованию групп, напротив, частое явление. Такая тенденция является следствием трех причин:
1) условия местообитания неоднородны и вид попадает в сре-ДУ, неодинаково благоприятную;
2) соседство неподвижных индивидуумов и их потомков, возникших в результате размножения: вегетативного или диаспорами, не способными к рассеиванию;
3) стадность связана с образом жизни: сближение пар, семейный образ жизни, более или менее организованные
123-
сообщества животных, возникающие в результате взаимного притяжения.
Характер распределения организмов имеет большое экологическое значение: жизнь более или менее плотной колонией увеличивает способность вида к защите и экспансии, благодаря чему индивидуумы менее уязвимы для хищников и конкуренции с другими видами. Помимо этого, характер распределения организмов представляет собой причину относительной гетерогенности внутри сообщества, которое предстает тогда в виде мозаики элементарных сообществ, отличающихся друг от друга обилием— доминантностью некоторых видов: великолепным примером могут служить пятна Hieracium pilosella на сухих известняковых луговинах Парижского бассейна.
Для определения степени агрегации или дисперсии особей прибегают к различным статистическим методам. Европейские фитоценологи 1 вслед за Браун-Бланке пользуются шкалой так называемой общественности (характера размещения) от балла 1, который применяется к видам, особи которых изолированы, до балла 5, когда индивидуумы образуют плотные и чистые группировки. В таблицах ассоциаций, составленных по методу Браун-Бланке, этот балл в каждом флористическом описании ставится после коэффициента обилия — доминантности вида (см. табл. 6).
Структура совокупности компонентов сообщества
Морфологическая структура сообществ определяется пространственным и временным распределением совокупности составляющих их организмов.
Способ группировки видов
В сообществе особи различных видов могут быть совершенно независимыми в своем распределении, но могут также обнаруживать более или менее сильную тенденцию к объединению или взаимному исключению. Помимо облигатных взаимосвязей между хозяевами и паразитами, сближение может иметь место, когда один вид создает локальную среду, благоприятную для другого вида. Возможно также, что небольшие отличия в физико-химических факторах локализуют на одних и тех же площадях виды, одинаково реагирующие на эти факторы. Наобо
1 Автор употребляет термины «фитосоциолог», «фитосоциология», принятые в зарубежной литературе, которые неудачны, так как дают основание проводить ненаучные параллели между наукой о человеческом обществе — социологией и наукой о растительных сообществах — фитосоциологией. В советской литературе применяют соответственно наименования фитоценолог, фитоценология. — Прим. ред.
124
рот, при наличии причин противоположного характера возможно более или менее полное исключение видов.
Простой способ для выявления таких пространственных взаимосвязей между двумя видами состоит в установлении среди многочисленных пробных площадок достаточного количества тех площадок, где имеются оба вида, и количества площадок, где присутствует только один вид, и в их сопоставлении с теоретиче-
р и с. 33. Примеры связей между величиной площадей и количеством видов.
Вверху, виды птиц в смешанных лесах Финляндии (по Палмгрену, Palm-gren). Внизу, кривая 1—ксерофиль-ный известняковый луг Парижского бассейна; кривая 2 —открытое сообщество щебнистых склонов северной Сахары. Видно, что при мало-различающемся общем количестве видов „ареала-минимума** луга равен примерно 2 м2, а для сообщества на щебне превышает 20 м2.
скими величинами, соответствующими гипотезе независимого распределения. В последнем случае следующая связь:
где N— общее количество обследованных площадок; Na — число площадок, на которых отмечен вид a, Nb — вид b\ Nab — число площадок, на которых отмечены одновременно виды а и Ь. Если разница между двумя величинами статистически значима, то виды имеют тенденцию к взаимному сближению или исключению в зависимости от того, является ли наблюдаемая величина большей или меньшей, чем величина вычисленная.
Взаимосвязь между площадью и количеством видов
Эта связь имеет логарифмический характер, поскольку количество новых таксонов при равном увеличении площади бывает все более и более незначительным. Ареалом-минимумом (минимальным ареалом) сообщества называют самую маленькую площадь, необходимую для того, чтобы на ней было представлено
125
большинство видов1. Были предложены различные математические критерии, для того чтобы внести большую объективность в это понятие. Ареал-минимум зависит от количества присутствующих видов, но также и от характера распределения и от степени заселенности территории. Рис. 33 дает примеры этой взаимосвязи.
Взаимосвязь между количеством видов и количеством особей
Соотношение между числом видов и числом особей на определенной площади может варьировать в различных сообществах. Вильямс (С. В. Williams) назвал индексом разнообразия частное от деления количества видов на логарифм количества наличных особей. Низкий индекс наблюдается в условиях среды, неблагоприятной для жизни, высокий индекс — в местах, лишенных значительно лимитирующих факторов, таких, как влажные тропические и экваториальные леса.
Ярусность
Этот термин обозначает вертикальное распределение организмов по различным уровням или ярусам. Надземная ярусность растений часто хорошо заметна. В ней различают:
криптогамный ярус, самый нижний, состоящий преимущественно из мохообразных и лишайников;
травяной ярус различной высоты, в котором иногда можно выделить несколько уровней2, как, например, на выкашиваемых лугах, где нижний уровень образован в основном мелкими бобовыми, средний — мелкими злаками, такими, как райграсе или душистый колосок, а верхний — высокими злаками, такими, как ежа, высокий райграсе или луговая овсяница;
кустарниковый ярус, включающий древовидные растения высотой не более нескольких метров, который может быть в некоторых случаях подразделен на два уровня, как в вересковых лайдах, заселенных молодыми березками;
древесный ярус, однородный в посадках и некоторых лесах, где ведутся рубки, но часто состоящий из нескольких хорошо выраженных уровней, имеющих значительную разницу в высоте как в лесах среднествольного хозяйства, или характеризующийся постепенным изменением высот, при котором в тропических лесах были произвольно выделены три уровня.
Растительное сообщество может состоять из одного или нескольких ярусов, и один и тот же вид может входить в несколько ярусов.
1 Правильнее эту площадь называть площадью выявления видового состава. Ареалом-минимумом обычно называют площадь, достаточную для выявления константных видов (понятие, введенное скандинавской школой фито-ценологов). — Прим. ред.
2 Можно говорить о подъярусах или даже об отдельных ярусах в пределах травостоя, древостоя и т. д. — Прим. ред.
126
Водная растительность с ее поднимающимися над поверхностью, плавающими и погруженными видами обладает не менее четко выраженной ярусностью.
Это вертикальное распределение имеет большое биологическое значение для видов, которые в зависимости от занимаемого яруса обитают в различных микроклиматических условиях.
Ярусность подземных органов также очень важна вследствие вертикальных вариаций физико-химических особенностей почв и конкуренции, которая может иметь место на уровнях корневых систем за воду, кислород и минеральные элементы (см. рис. 34),
Р и с. 34. Подземная ярусность растений травяного покрова дубово грабового леса в Парижском бассейне (по Жове, Р. Jovet).
Mp — Mercurialis perennis; Vm—Vinca minor; S—Scilla bi folia; Cm-Con-vallaria maialis.
Внутри этого растительного сообщества животные живут преимущественно или исключительно в одном ярусе, даже когда речь идет об очень мобильных видах. Так, птицы вьют гнезда и летают на определенных уровнях, различающихся в зависимости oi вида: одни предпочитают вершины больших деревьев, другие защиту подлеска. Из 240 видов насекомых, паукообразных и многоножек, зарегистрированных в дубовом лесу в США, 181 вид был собран в одном ярусе и только 3—5 видов обитали на всех Уровнях. Даже в травянистых формациях часть животных живет преимущественно на вершинах растений, а часть — у их основания. Водные животные также обитают в пределах определенных вертикальных границ. Но какой бы ни была среда обитания, у Некоторого количества видов наблюдаются вертикальные перемещения, подчиняющиеся суточному или сезонному ритму, как это будет видно из последующего изложения.
127
Степень развития растительного покрова
Растительный покров может быть сплошным или прерывистым или, как иначе говорят, сомкнутым или открытым. Это имеет большое значение для таких экологических особенностей местообитания, как термический микроклимат, влажность почвы, возможность эрозии, конкуренция между растениями и т. д. Степень покрытия в каждом ярусе является важнейшим элементом структуры сообщества.
Периодичность
Периодические изменения состояния и активности организмов вызывают более или менее явственные изменения аспекта сооб-
ществ, их экологических условий и взаимоотношений между ор-ганизмами. Эти изменения могут иметь । । 1 II I. 1 । Il । суточный, сезонный ритм или соответ
Р и с. 35. Сезонные вариации обилия нескольких видов комаров (пойманных днем на человеке за 1 час) в восточной Колумбии. Флуктуации связаны с сезонной влажностью, за исключением одного вида (по Бейтсу, Bates).
I — Haemagogus capricorni; Il — Aedes serratus; IH-Pso-rophora ferox-, IV-Sabethoi-des imperfectus (no Bates).
ствовать лунному календарю, как в морской воде, и даже образовывать многолетние циклы с менее регулярной периодичностью.
Суточные ритмы. У растений они очень важны в физиологическом отношении (фотосинтез, транспирация), но внешне не заметны, за исключением сумеречных движений раскрытия и закрытия цветков. У животных часы бодрствования и сна различны у разных видов; перемещения животных могут быть подчи
нены суточному ритму: так, у многих водных организмов наблюдаются дневные вертикальные перемещения, которые могут играть важную роль, как, например, перемещения рыбы Chauliodus sloanei, которая в Тихом океане опускается днем до 2500—3000 метров и поднимается ночью до глубины менее 500 метров.
Сезонные ритмы. Они более заметны в связи с чередованием теплых и холодных или влажных и сухих сезонов. Развитие листьев и листопад, которые связаны с ними, составляют характер
нейшую особенность лесных сообществ, состоящих из деревьев с опадающей листвой. Последовательные волны цветения лугов придают им ярко выраженный сезонный аспект. У оседлых животных значение и активность популяций одного вида
варьирует по сезонам различным образом в зависимости от видов; это наглядно проиллюстрировано на рис. 35. Мигранты являются для сообщества сезонными гостями. На обрабатываемых
128
площадях сезонные вариации фауны тесно связаны с календарем сельскохозяйственных работ: так, на посевах люцерны второй покос производится в начале августа, в то время, когда фауна очень богата; он приводит к исчезновению трипсов и цикадок. Первые исчезают до конца года, а вторые временно. Количество Microlepidoptera сначала уменьшается, затем снова увеличивается. Наблюдается массовое размножение тлей, в то время как в стогах образуется своеобразная фауна, богатая как по числу видов, так и по числу особей (Р. Шовен, R. Shauvin).
Жизненные формы
Если среда может лишь в слабой степени изменять формы растений, она в противовес этому играет огромную роль в распределении этих форм, благоприятствуя или исключая в различных случаях те из них, которые обладают существенными экологическими особенностями. Например, древовидные формы отсутствуют в высокогорье и аридных зонах, а однолетники, обычно не встречающиеся в высокогорье, широко представлены в аридных зонах. Совокупность морфологических особенностей, которые играют важную роль в локализации видов, образует их жизненную форму (life form или growth form английских авторов). Следует отметить, что общепринятой квалификации жизненных форм не существует, поскольку лимитирующие факторы различны для разных климатических условий и местообитаний.
Первая попытка классификации принадлежит Александру Гумбольдту, который в 1805 г. выделил 15 биологических типов. Из многочисленных классификаций, которые были с тех пор предложены, чаще всего применяется классификация датского ученого Раункиэра из-за ее экологических достоинств для климатов с неблагоприятными сезонами (сухими и холодными). Действительно, она основана на степени защищенности почек возобновления, которые дают начало новым ветвям после неблагоприятного периода. В зависимости от степени защищенности было выделено пять групп.
1. Фанерофиты. Растения, почки возобновления которых находятся на некотором расстоянии от поверхности земли; это деревья и кустарники, которые делятся на несколько групп в зависимости от их высоты. Самые низкие, высотой от 25 см до 2 м называют нанофанерофитами. Подразделения этой группы были сделаны на основании наличия или отсутствия почечных чешуй, типа листвы — опадающей или остающейся, размера и формы листьев.
2. Хамефиты обладают почками возобновления, расположенными близко над поверхностью земли (меньше 25 см), таким образом, они защищены окружающей растительностью, растительными остатками или снегом. Среди подразделений, которые были выделены, подушечные формы особенно замечательны
5 Ж. Леме
129
многочисленными прижатыми друг к другу деревянистыми веточками одинаковой высоты. Подобное строение обеспечивает сопротивление ветру, высыханию, абразии, тяжести снега и наблюдается чаще всего в высокогорьях в самых различных родах (Silene, Saxifraga, Draba, Androsace).
3. Гемикриптофиты обладают почками возобновления, расположенными на уровне поверхности земли, где они защищены растительными остатками, часто их собственными мертвыми листьями или снегом. Многие из них сохраняют зимующую розетку зеленых листьев.
4. Криптофиты обладают почками возобновления, погруженными в почву или в воду, отсюда две следующих подгруппы:
Геофиты являются луковичными, корневищными и клубневыми растениями. *
Гидрофиты — укореняющиеся на дне водоема или свободно плавающие растения, однако у последних почки или диаспоры в неблагоприятное время года расположены в глубине водоема.
5. Терофиты — однолетние растения, переживающие неблагоприятный период в состоянии диаспор.
Процент видов каждой категории на данной территории образует ее биологический спектр. Для семенных растений земного шара Раункиэр дает следующие процентные соотношения:
Фанерофиты (Ф) — 46
Хамефиты (X)— 9
Гемикриптофиты (Г) —26
Геофиты (Ге) — 4
Гидрофиты (Ги)— 2
Терофиты (Т) — 13
Таблица 7
Количество видов ф X г Ге Ги т
Климат фанерофитов Сейшельские острова 258 1 61 6 12 4 1 16
Климат терофитов Французская средиземномор- __ 7 13 29 8 0 42
ская область Гардая (северная Сахара) 300 6 16 20 3 0 58
Климат гемикриптофитов Французский Вексен (Париж- 8 6,5 51,5 25 9
ский бассейн) Степи нижнего Дуная 7 5 55 10 0 23
Климат хамефитов Юг Гренландии 169 1 21 60 7 7 4
Альпы (2400 до 2550 м) 323 1 24 67 4 0 4
В различных климатических зонах эти пропорции сильно меняются, что позволило этому автору выделить четыре основных
130
«фитоклимата»; климат фанерофитов — влажных тропических областей, климат гемикриптофитов — умеренных, неаридных областей, климат терофитов — жарких и умеренно теплых областей с более или менее длительным сухим периодом, климат хамефитов — высоких широт и высокогорий. Это наглядно иллюстрирует табл. 7.
Установлен также биологический спектр растительных сообществ, исходя из количества видов каждой группы, но этот метод не дает никакого представления о реальной физиономической характеристике. Например, в лесах умеренного пояса, бедных древесными видами, пропорция фанерофитов в биологическом спектре будет очень незначительной.
Глава 7
Критерии и классификации сообществ
По растительным сообществам имеется гораздо больше работ, чем по смешанным сообществам или сообществам животных. Это объясняется стабильностью растительных сообществ и большей легкостью их систематической инвентаризации. Количество таких исследований и разнообразие развиваемых в них концепций делает невозможным их исчерпывающее изложение в рамках данной работы, и мы выделяем здесь лишь основные направления подобных концепций.
Растительные сообщества
Системы, используемые для различения и классификации растительных сообществ, могут быть физиономического, флористического, экологического или динамического характера, или комбинациями этих аспектов.
Физиономические методы
Основа заложена Гумбольдтом, который в 1806 г. ввел термин формация для совокупности сообществ, имеющей четко выраженную физиономичность. В этот термин вкладывали различные значения. В настоящее время его снова применяют с большей точностью в его первоначальном смысле: это совокупность сообществ, то есть группировка \ определяемая доминированием одинаковых жизненных форм, сходной ярусностью и периодичностью. Вследствие того что этому термину придавались различные значения, многие геоботаники предпочитают выражение тип растительности1 2 * * * * * В.
1 В отличие от сообщества, предмета конкретного, мы обозначаем термином «группировка» совокупность сообществ, сходных по определенным признакам, выбранным для сравнения, какими бы пи были эти признаки. Следовательно, группировка является абстракцией аналогично виду по отношению к отдельным индивидуумам.
2 В советской литературе термин «группировка» применяется к начальным
стадиям развития еще не сформировавшегося растительного покрова.
«Сообщество» — термин, не имеющий определенного таксономического
значения. Сообществом является и лес вообще, и еловый лес, например, и еловый лес-зеленомошник.
«Фитоценоз» — конкретная, небольшая по размерам, наиболее однородная единица растительного покрова, занимающая на поверхности планеты площадь одного биогеоценоза. В качестве типологической единицы, аналогичной виду,
применяется термин «ассоциация». Ассоциация есть тип фитоценоза.
В дальнейшем сообразно этой концепции мы будем в качестве неопределенной по объему категории применять термин «сообщество». — Прим. ред.
132
В методике изучения приходится сталкиваться с трудностями двоякого рода — по существу и форме. Глобальный физиономический характер формации плохо поддается сжатому описанию и точному определению, поскольку он бывает неизбежно основан ла многочисленных аналитических признаках1. Проблема названия решалась различными способами: географы и некоторые ботаники предпочитают употреблять обычные термины, если последние обладают достаточно точным смыслом или дополняют ях различными определительными уточнениями, такими, как «листопадный лес с периодом покоя в холодное время года» или «степь колючекустарниковая». Часто возникает большое искушение использовать для описания каждой области местные названия, это оправдывает себя в случае, если эти названия соответствуют хорошо определимым частным особенностям. К сожалению, одни и те же названия могут соответствовать у разных авторов различным формациям. Не следует также смешивать формации, которые являются гомогенными образованиями, и ландшафты2, которые могут быть единообразной мозаикой формаций на больших площадях и которые представляют собой региональные географические единицы. Так, бразильская каатинга является обширной по площади совокупностью формаций мелколесья, древесных саванн, чащ и колючекустарниковых степей; галерейный дес — это лес, локализованный по днищахм долин в саванном ландшафте, но в его состав могут входить различные формации — затопляемые, болотистые и сухие.
Для того чтобы исключить разночтения и путаницу, которые могут быть связаны со словарями различных языков, некоторые геоботаники предложили оригинальную терминологию, состоящую из сложных словосочетаний. Так, для швейцарцев Брокман-Ероша (Brockmann-Jerosch) и Рюбеля (Riibel) летние листопадные леса, например, составляют группу формаций Aestisilvae, вечнозеленые луга — Sempervirentiherbosae и т. д. Другие, как Дансеро (Dansereau), пытаются создать универсальную систему при помощи условных формулировок и графиков.
Формации обычно объединяют в группы или типы формаций, выделяемые по характеру доминирующего яруса:
1. Сомкнутые леса. Древесный ярус сплошной, от 7 до 10 м в высоту, согласно данным различных авторов. Включает многочисленные формации, одни связанные с климатическими условиями, ареал которых имеет зональное распределение по поверх
1 В отличие от автора, который принимает термин «формация» в довольно неопределенном значении, в нашей литературе этому термину придается совершенно определенный смысл. Под формацией понимают совокупность сообществ, в которых эдификатором (средообразователем) является один вид растения. Например, формация ели сибирской, формация бука лесного, формация ковыля красноватого. — Прим. ред.
2 У автора «растительный пейзаж» — термин, не имеющий широкого распространения в литературе. — Прим. ред.
133
ности земного шара, другие значительно более локальные, свя* занные с местными эдафическими условиями. К первым относят: тропические дождевые леса, вечнозеленые и полулистопад-
ные (фото X),
тропические сухие листопадные леса с периодом покоя в сухое время года,
вечнозеленые склерофильные средиземноморские леса (фото IX, Б),
листопадные леса умеренного пояса с периодом покоя в холодное время года,
хвойные вечнозеленые бореальные леса (фото XI, А).
Из второй группы отметим тропические заболоченные леса и мангры.
2. Редколесья. Древесный ярус прерывистый, покрывает от 74 до 2/3 поверхности, имеется подстилающий злаковый ярус (фото VI, Б).
3. Чащи (синонимы: скраб, кустарниковая формация). «Тип древовидной растительности, сомкнутой, вечнозеленой или листопадной, обычно труднопроходимой, часто раздробленной на части, с отсутствующим или прерывистым злаковым покровом». Многочисленные формации, носящие местные названия, как южноафриканский thornbush (колючий кустарник), sagebrush (полынник) юго-востока Соединенных Штатов, калифорнийский чапараль, средиземноморский маквис, испанский маттораль^ ланды Западной Европы и т. д.
При деградации в результате выпаса или пожара этот тип растительности дает начало низким прерывистым кустарничковым группировкам с обильными травянистыми растениями, таким, как средиземноморские гариги на известняках (фото VIII, А).
4. Саванны. «Травянистая формация с верхним сплошным травяным ярусом по крайней мере 80 см высотой, который оказывает влияние на нижний ярус». Низкие деревья и кустар* ники расположены обычно изолированно или небольшими группами. Речь идет о тропических формациях. Американские ученые дали иное толкование этому термину: саванна для них характеризуется наличием низких деревьев, изолированных или образующих маленькие группы с пространством между ними, занятым высокими травами — сезонными (тропики) или постоянными (умеренные зоны) или кустарниками и густым моховым ярусом (холодные зоны); в последних случаях для европейских ученых речь идет о лугах, ландах или кустарниковой тундре. В тропических саваннах имеются все градации, начиная от редколесья до отсутствия древесной растительности: саванны редколесные,, с единичными деревьями, кустарниковые (фото V, Б, VI и VII).
5. Степи. Это слово русского происхождения обозначает открытые травянистые формации, в которых доминируют ксеро-фильные злаки с зимним периодом покоя, высыхающие в течение
134
лета. Между ними вкраплены другие жизненные формы: однолетники, высокие многолетние травы, низкие кустарники.
Североамериканская прерия (фото V, А), аргентинская пампа, венгерская пушта и даже североафриканские сообщества альфы очень близки степям. Но это название было распространено на совокупность открытых формаций аридных зон, где ксерофильные злаки могут быть крайне редки, если не отсутствуют полностью, и могут быть заменены такими деревянистыми растениями, как полыни (фото VIII, Б), или такими суккулентами, как молочай на юге Марокко.
Со степями можно также сопоставлять низкие и прерывистые (за исключением момента развития однолетников во влажный период) луговины, образующиеся в результате деградации деревянистых растений, известные в Средиземноморье под названием егте.
6. Луга и луговины. Травянистая формация гемикриптофитов, полностью покрывающая почву в течение всего года, состоящая главным образом из злаков, иногда из осоковых (Суре-гасеае) или из ситника и достигающая, по крайней мере в период активной вегетации, высоты порядка одного метра (иногда более). Луга бывают естественными эдафическими формациями, такими, как осоковые и тростниковые болота влажных или затопляемых местообитаний или искусственными формациями, созданными человеком, как, например пригодные для покоса луга.
Луговины — это низкие луга с низкой продуктивностью, связанной с эдафической сухостью, как ксерофильные луговины французских долин, или с коротким сезоном активности, как летние горные пастбища. В последнем случае они могут быть естественными, но часто являются следствием деградации естественных формаций.
7. Пустынные формации. Характерным для них является исключительно слабо выраженный постоянный покров, варьирующий в зависимости от локальных условий: злаки, кустарнички, суккуленты. Они представляют собой крайнюю степень разреженности индивидуумов по сравнению со степями, чащами, саваннами, возникающую под воздействием сухости (фото IV).
8. Криптогамные формации. Криптогамный ярус является •единственным или резкопреобладающим. Сюда относятся лишайниковая тундра и сфагновые болота.
Эта классификация, как большинство биологических классификаций, содержит переходные термины, которые делают необходимым использование произвольных критериев высоты, покрытия и т. д. Так, от центральной Сахары до Гвинейского залива часто можно наблюдать постепенные переходы от пустыни к степи, затем от степи к саванне, затем к редколесью, к полу-листопадному лесу и, наконец, к вечнозеленому лесу.
Несмотря на различия во флористическом материале между Двумя фитогеографическими территориями, идентичность аспекта
135
формаций, наблюдающаяся в сходных условиях среды, часто бывает ярко выражена. Так, Манжено (G. Mangenot) подчеркнул полную гомологию между вечнозелеными дождевыми лесами Амазонки — гилеей и такими же лесами Западной Африки.
Такие формации, физиономически сходные, несмотря на разницу во флористическом составе, благодаря воздействию аналогичных экологических условий Амберже (Emberger) назвал ho-moecies.
Родственные формации могут быть объединены в более крупные единицы, такие, как группы, о которых будет сказано ниже и для которых предложен термин типы формаций.
Физиономические группировки можно упрекнуть в известной расплывчатости и независимости от истории локальных флор» но, с другой стороны, эта независимость позволяет сближать территории, основные экологические признаки которых сходны благодаря отбору этими сходными средами одинаковых жизненных форм, возникших на основе различного флористического материала. Возможность быстрого распределения формаций на обширной территории позволяет расценивать этот метод как очень ценный при первом обследовании растительного покрова. Поэтому эта методика часто применяется в тропических областях.
Флористические методы
Мы имеем в виду многочисленные методы, которые опираются главным образом на знание видового состава с привлечением физиономических и экологических признаков, а также данных по динамике сообществ. Группировки сообществ, основанные на этих критериях, получили название ассоциаций, хотя концепции, которыми руководствуются при их выделении, весьма различны. Мы классифицировали эти методы по главным критериям, на которых они основаны.
Методы, основанные на доминантных видах. Они особенно часто используются авторами, которые пишут на английском языке, и скандинавами.
1. Английские фитогеографы считают высшими единицами формации, которые состоят из некоторого количества флористических единиц, объединяющих сообщества, имеющие одни и те же доминантные виды в основном ярусе. Эти единицы обычно называют ассоциациями (или консоциациями в случае многих доминантов). Чтобы избежать путаницы с другими концепциями, Мак-Лин (Me Lean) предложил термины доминион и кондо-минион. Этот метод оправдан особой значимостью видов, господствующих в верхних ярусах, в особых условиях местообитания и в динамике сообщества. Однако подобный подход может привести к объединению популяций весьма различных видов, чье объединение в одно сообщество более чем сомнительно. Так, пихтарники в горах Франции обладают разнообразным подле
136
ском, состав которого тесно связан с особенностями почвы (рис. 37); в других местах тот же флористический состав, несмотря на разницу в древесных породах, может наблюдаться в буковых лесах, особенности почв которых аналогичны таковым в пихтарниках с тем же подлеском. Таким образом, доминирующие виды часто играют весьма незначительную роль в качестве индикаторов условий местообитания вследствие их большой экологической амплитуды.
2. Американские экологи, среди которых Клементс (F. Е. Clements) занял ведущее положение, разработали своеобразную систему, обратив внимание при обследовании больших однородных площадей на общий облик и эволюционные тенденции растительного покрова. Считая, что единственными стабильными сообществами являются заключительные сообщества, находящиеся в равновесии с климатом, которые он назвал климаксами, Клементс называет их формациями, дает им физиономическую характеристику и подразделяет на ассоциации, выделяемые на основе совокупности доминирующих видов или родов. Эти доминирующие таксоны могут сменять друг друга или сочетаться различным образом на территории, занятой ассоциацией. Данное сочетание образует фациацию (от слова «facies» — фация. Суффикс— ation указывает на то, что сообщество является климатическим). Сама фациация может быть представлена вариантами меньшего масштаба, которые составляют лоциацию (от locus — место). Сходные сообщества, которые представляют собой ступени эволюции к климаксу или деградации последнего, получили соответственно названия ассоций, фаций и лоций. Это теоретическое положение о единственном климаксе (моноклимаксе), неприменимое, в частности, для аридных зон и высокогорья, было отвергнуто многими американскими фитогеографами, которые распространяют понятие формации и ассоциации на все совокупности стабильных сообществ и чья точка зрения приближается, таким образом, к концепции англичан.
3. В Скандинавии постепенно усилиями Хульта, Серпан-дера, а затем Дю Рие, Освальда и Нордхагена (Hult, Sernander, Du Rietz, Oswald, Nordhagen) наряду с другими был разработан оригинальный метод анализа растительности, основанный на доминировании в каждом отдельно взятом ярусе. Самая маленькая единица при этом — социон (соция)—площадь, занятая одним и тем же видом — доминантом в одном ярусе независимо от других ярусов. В многоярусном сообществе имеется столько же расположенных друг над другом соций, сколько и ярусов. Со-циация — это сообщество, характерной чертой которого является одинаковое расположение (суперпозиция) соций, она представляет собой основную единицу системы и вначале называлась ассоциацией. Когда единственная соция в одном ярусе накладывается на несколько соций в другом ярусе, совокупность последних образует консоцию. Консоциация — это сообщество,
137
в котором по крайней мере один ярус (обычно верхний) обладает гомогенностью соции, а другие представляют собой консоции.
Этот метод применяется в скандинавских странах, где довольно однообразная и бедная видами растительность не позволяет выделить четко ограниченные сообщества при помощи концепции верности, применяемой в Средней и Южной Европе. На флористически богатых территориях скандинавский метод неприменим из-за огромного количества комбинаций соций, которые он выявляет. Даже для шведских торфяников Освальд выделил более 150 социаций.
Методы, основанные на полном флористическом составе. Эти методы получили развитие в Средней присредиземноморской Европе. Первые указания на них можно найти в определении, данном на ботаническом конгрессе в 1910 г. французом Флао и швейцарцем Шрётером (Flahault, Schroeter): «ассоциация является растительным сообществом, с определенным и относительно постоянным в пределах данной площади флористическим составом, обладающим единообразной физиономичностыо и развивающимся в одинаковых условиях местообитания». Мы видим, что в этом определении фигурируют физиономические и экологические критерии. Швейцарский геоботаник Браун-Бланке 1 поставил во главу угла анализ полного флористического состава, что позволило ему выделить понятие характерных видов в качестве основного критерия ассоциации. По методу, предложенному этим автором, было осуществлено большое количество работ. Их часто называют работами школы Цюриха — Монпелье, по имени двух центров, где в основном работал автор. В последние годы усилия многих авторов были направлены на то, чтобы разграничивать флористические сообщества с максимальной объективностью.
1. Отбор проб. Первая задача состоит в установлении списка видов индивидуумов ассоциации, участков с единообразным флористическим составом при помощи пробных площадок, выделяемых статистическими методами, такими, как многочисленные мелкие площадки, отобранные случайно, или метод фито-цепологического описания, который является количественным флористическим анализом площади, большей по размеру, чем минимальный ареал, причем эта площадь должна быть как можно более гомогенной и репрезентативной.
2. Флористическое сравнение индивидуумов ассоциации. Эта вторая фаза состоит в установлении степени флористического сродства сообществ с тем, чтобы классифицировать их по группам. Для этого были разработаны различные методы.
а) Методы статистического исследования межвидовых связей состоят в том, что в различных индивидуумах ассоциации уста
1 Браун-Бланке основал фитоценологическую школу в городе Монпелье, во Франции. — Прим, ред,
138
навливается степень зависимости видов, взятых попарно, по методу, который может быть таким же, как для изучения характера сочетания видов внутри данного сообщества (см. выше). Таким образом, в полном списке встречающихся видов можно выявить виды, которые имеют тенденцию к объединению в одних и тех же сообществах, и те, которые проявляют тенденцию к раздельному существованию в разных сообществах.
б) Методы исследования флористического сходства между фитоценологическими описаниями состоят в установлении пропорции общих видов в попарно взятых описаниях. Сюда относят коэффициент общности Жаккара (Jaccard) — отношение количества видов, общих для обеих площадей, к общему количеству видов на обеих площадях:
о __ Nxy ху + ’
где Nx — количество видов, имеющихся на пробной площади х, Ny — количество видов, имеющихся на пробной площади у, и Nxy — количество видов, общее для х и у. Эта величина варьирует от 0, если Nxy равно 0, до 1, если все виды являются общими.
Другие коэффициенты, более точные, но с более сложной •формулой, учитывают обилие видов на сравниваемых площадях.
Польские геоботаники Кульчинский, Матушкевич, Чеканов-ский (Kulczinski, Matuszkiewicz, Czekanowski) использовали установленные таким образом коэффициенты для сравнения многочисленных пробных площадей при помощи таблицы совпадений. В этой таблице каждой из пробных площадей, расположенных в одинаковом порядке, соответствует строчка и столбец. В местах пересечения помечены соответствующие коэффициенты общности. Меняя порядок пробных площадей в строчках и колонках, стараются перегруппировать те из них, которые обладают наиболее высоким коэффициентом общности и поместить их вблизи диагонали. Каждая из таких групп, расположенных вдоль диагонали, представляет собой фитосоциологическую единицу (см. рис. 36).
3. Синтетические признаки сообществ.
а) Присутствие. Описания, относящиеся к одному и тому же флористическому сообществу, можно изобразить в виде таблицы, где виды занимают строчки, а пробные площади — столбцы. Тогда можно видеть, что виды встречаются в неодинаковом количестве пробных площадей. Константностью, или присутствием вида, называется процент описаний, в которых он встречается. Браун-Бланке установил пять классов константности, в которых отношение числа описаний с наличием вида к общему их числу увеличивается от класса к классу. Виды V класса называют константами.
139
1 8 2 9 1011 3 4 7 291218202217212827262516192423153031321413
Рис. 36. Пример таблицы дифференциального анализа по методу Чеканов-ского (Czekanowski) для 32 пробных площадей (числа по краям указывают их номера) растительных сообществ гариги в горах Тель-Атласа. Видны две группы ассоциаций (по Гиноше, Guinochet).
Было доказано, что кривая распределения по различным классам относится к типу I обобщенного уравнения вероятностей Пирсона (К. Pearson), если данные, которые послужили для ее составления, были гомогенными. Эта кривая может быть U-образной или последовательно понижающейся в зависимости от того, многочисленны или нет константные виды.
140
б) Верность. Посредством сравнения флористического состава различных сообществ данной территории выявляют локализацию или просто предпочтение некоторыми видами определенных сообществ. Предпочтение проявляется в большем обилии или в большей жизненности вида в этих сообществах по сравнению с другими, где существует данный вид. Браун-Бланке назвал виды, верные сообществу, характерными. Нехарактерные для сообщества (но могущие быть характерными для другого сообщества) виды называют сопутствующими видами. Среди последних некоторые могут объединяться только на части пробных площадок, в то время как другая часть пробных площадок может иметь другие сочетания сопутствующих видов. Эти виды Браун-Бланке назвал дифференциальными. Они также, как и характерные виды, являются индикаторами экологических различий между группами пробных площадок, но отличаются от характерных видов тем, что широко представлены в других сообществах.
4. Номенклатура сообществ. Ассоциация является основной единицей, которую вместе с Гиноше (М. Guinochet) можно определить как «статистически однородную флористическую группу... пробных площадок, взятых на флористически гомогенных площадях, обладающих по меньшей мере одним характерным видом».
Ассоциация обозначается одним или двумя видами, взятыми среди наиболее репрезентативных, характерных, более константных видов. К названию рода прибавляют суффикс etum, видовое название ставится в родительном падеже. Так, обычная ассоциация мезофильных удобренных сен жосных луков с Arrhena-therum elatius обозначается Arrhenatheretum elatioris, а ассоциация гариги с розмарином и Lithospermum fruticosum — как Rosmarineto-Lithospermetum fruticosae.
Сравнение описаний данной ассоциации часто позволяет выявить группы дифференциальных видов, индикаторов слегка различных экологических условий; единицы, определяемые этими видами, являются субассоциациями. Для того чтобы обозначить субассоциацию, после названия ассоциации ставят название дифференциального вида с окончанием — etosum. Так описана Arrhenatheretum elatioris, Primuletosum verts, богатая Primula verts и Rosmarineto-Lithospermetum, Quercetosum cocciferae, в которой хорошо представлен кермесовый дуб.
Что касается доминирующих видов, которые могут быть различными на разных пробных площадях одной и той же ассоциации, то они образуют простые фации.
Между ассоциациями существует большее или меньшее флористическое сходство, которое можно выявить при помощи тех же тестов, что и при сравнении описаний. Близкие ассоциации объединяют в союз. Сравнение союзов выявляет существование видов, которые в них локализованы или лучше представлены,
141
это характерные виды союзов. Союзы обозначают при помощи одного или двух видов с суффиксом ion в названии рода. Так Rosmarinion объединяет несколько ассоциаций средиземноморских гариг, в которых розмарин особенно обилен.
Флористически сходные союзы в свою очередь объединяют в порядки со своими характерными видами. Их обозначают по одному из них при помощи суффикса etalia.
Наконец, родственные порядки объединяют в классы. Один или два характерных вида с суффиксом etea служат для их обозначения.
На практике флористические описания, относящиеся к одной фитоценологической единице, объединяют в столбцах в таблицах, где виды классифицируются по их верности, как характерные для ассоциации, союза, порядка, как дифференциальные для субассоциаций и вариантов и как сопутствующие (см. табл. 6).
Таблица 8
Комбинации экологических групп в ассоциациях
Ассоциации i 2 3 4 5
a 6 a 6 a 6 a 6 a 6
Экологические группы Строгие кальциефилы: группа Bupleurum » Conringia » Falcaria » Delphinium » Sherar dia Предпочтительно кальциефилы: группа Sinapis » Sonchus arvensis Предпочтительно ацидофилы: группа Matricaria chamomilla группа Raphanus Облигатные ацидофилы: группа Scleranthus Гигрофилы: группа Gnaphalium uliginosum группа Juncus bufonius » Ric&ia HIM III II II 1 1 III III III III Hill hi ilium mi Hill III III III Hill i iiiiiiii и II IIIIIIII 1 Hill 1 II III III II 1 II III IIHIII Hill II II 1 II III Hill 1 II Hill Hill III II II
= доминирующая группа; = широко представленная группа; — слабо представлен 1 руппа; — спорадически представленная группа.
142
Флористическо-экологические методы. Изучение собственно экологических особенностей видов, сосуществующих внутри одного сообщества, показывает, что сообщество, хотя оно и существует в единообразных условиях местообитания, состоит из видов с различными экологическими требованиями при условии, что физические и химические условия этого местообитания находятся в пределах толерантности данных видов. Так, сорняки культуры, растущей на кремнисто-песчаной хорошо дренированной почве, состоят из видов — кальциефобов и индифферентных к кальцию, ацидофильных и толерантных к pH, псаммо-фильных и индифферентных к механическому составу почвы, ксерофильных и малочувствительных к влажности.
Составляющие ассоциацию виды можно объединить в группы с более или менее сходными экологическими потребностями. Это экологические группы — термин, введенный в геоботанику Дювиньо (Р. Duvigneaud). Сообщества характеризуются определенным составом, или «спектром», экологических групп, которые одновременно выявляют свое экологическое сходство и позволяют установить классификацию по условиям местообитания, показателями которых являются растения.
Элленберг (Ellenberg) предлагает именовать выделенные таким образом сообщества по названию растений, которые были избраны для обозначения наиболее широко представленных экологических групп.
Вот выдержка из таблицы комбинаций экологических групп в ассоциациях. Группы определены по методу вышеупомянутого автора при скашивании луга в Вюртемберге (см. табл. 8).
Сообщества животных и их связи с растительными сообществами
Первые исследования сообществ животных были осуществлены на примере донных морских сообществ, в населении которых часто преобладают сидячие или прикрепленные формы животных. В 1877 г. Мёбиус (К. Mobius) при исследовании биологии устричных банок ввел выражение биоценоз', «сообщество живых существ, соответствующее по составу и количеству видов и особей определенным средним условиям среды; сообщество связанных взаимной зависимостью организмов, которое сохраняется благодаря постоянному воспроизведению». Биоценозы животных, или зооценозы, отличаются от временных или случайных скоплений, связанных с сенсорным или пищевым привлечением, таких, как скопления ночных насекомых вокруг источника света, которые именуются скопищами (foules) Их следует отличать об обществ (societes), которые являются длительно существующими скоплениями и обязаны своим возникновением взаимному привлечению, а не случайной особенности
143
физической среды. Уровень сложности этих обществ различен, начиная от неорганизованных групп временного или факультативного характера, таких, как скопления тараканов, и кончая строго организованными обществами, как муравейники, ульи и термитники. Имеются и промежуточные формы, такие, как стада или стаи мигрирующих животных, где взаимное привлечение твердо установлено, а также общества низшего порядка, в которых имеет место постройка общего гнезда, но нет разделения труда, как у гусениц. Биоценоз, каким бы сложным он ни был, не обладает этим признаком, ибо общественные отношения у большинства его компонентов отсутствуют. Все же «общественная среда вторично приводит к формированию биоценоза, потому что многочисленные виды используют среду, не приобретая прямых связей ни с членами общества, ни даже с сопутствующими им организмами» (Грасс, Р. Grasse). Так, в муравейниках обитают мирмекофилы — виды, которые находят там благоприятные микроклиматические условия и пищу.
Популяции животных, как мы это видели выше, поддаются количественному и качественному описанию, как популяции растений.
Однако их исключительное фаунистическое разнообразие, подвижность, перемена местообитания в зависимости от стадии развития, сезона и даже времени суток усложняют их изучение.
Для разграничения и характеристики биоценозов животных были приняты методы и критерии, подобные тем, которые применялись для растительных сообществ: коэффициент фаунистической общности, доминирование, константность, верность видов. Так, Петерсон (Peterson), опираясь на многочисленные учеты, проведенные на протяжении многих лет, во все времена года на участках площадью 0,25 м2, выделил 9 типов сообществ на сублиторали возле датских берегов. Эти типы сообществ основаны на комбинации нескольких константных и характерных видов, в основном моллюсков и иглокожих. Гислер (Т. Gis-ler) подразделил сообщества североамериканской тихоокеанской литорали на ассоциации, характеризующиеся доминирующими видами, и на формации, характеризующиеся жизненными формами. Невелл (N. D. Newell) применяет к бентосу Багамских островов таксономические критерии, основанные, как у Петерсона, на комбинациях видов, и экологические критерии— на признаках местообитания и сравнивает установленные таким способом «сообщества организмов» с «сообществами местообитания».
Для наземных условий прямые и косвенные связи животного мира с растениями очевидны. Попытки описания сообществ животных и растений как единого целого были сделаны в соответствии с различными концепциями, разработанными местными фитоценологическими школами. Американские экологи, последователи Клементса, взяли за основу растительную
144
формацию, в которую включили позвоночных. Эти «растительно-животные формации» (plant-animal formations) получили от Клементса и Шелфорда (Clements, Shelford) название биомов — единиц, характеризующихся жизненными формами и доминирующими видами. Совокупности физиономически сходные, но таксономически различные, вследствие их локализации на фитогеографически различных территориях, объединены в типы биомов, которые соответствуют типам растительных формаций.
Попытки установить соответствие животных сообществ растительным ассоциациям, определяющимся доминирующими или характерными видами, дают противоречивые результаты. Очень подвижные животные—млекопитающие, птицы, крылатые насекомые— больше связаны с формациями, чем с ассоциациями. Многие даже предпочитают опушки (экотоны зоологов) в соответствии с их образом жизни, при котором их нора находится в одном сообществе, а охотничья территория — в другом. В этом случае они являются членами двух или нескольких сообществ, сменяя их в течение суток. Мелкие малоподвижные животные, напротив, образовывали бы чаще всего сообщества, которые можно наложить на растительные ассоциации, в понимании школы Цюрих — Монпелье и различимые, как и растительные ассоциации, по характерным и дифференциальным видам или при наложении на социации скандинавов, различаемые по константности и доминантности.
Эти смешанные сообщества получили в Европе, особенно после работ Мёбиуса (Mobius), название биоценозов. Советские биогеографы, в частности Сукачев, приняли это общее понятие и распространили его на всю совокупность живых организмов, включая почвенную микрофлору, тесно связав его с факторами местообитания (биогеоценоз).
Упомянем здесь также термин экосистема, очень близкий по общему характеру к биогеоценозу, который, однако, рассматривает сообщества с точки зрения обмена веществ и энергии L
1 Биогеоценоз — наименьшее подразделение биосферы, совокупность живых и неживых компонентов наиболее однородного участка. Экосистема — совокупность живых и неживых компонентов различного объема — «от капли до океана». — Прим. ред.
Глава 8
Экология и эволюция сообществ
Отношения сообществ со средой
Границы сообщества, находящегося в состоянии равновесия с соседними сообществами, обозначены модификацией некоторых факторов среды, которые играют решающую роль в пределах этих границ. На площади, занятой таким сообществом, особенности местообитания, напротив, претерпевают изменения, совместимые с существованием и сохранением сообщества. В каждой точке, где сообщество существует, определяющие факторы сходны, и обратно.
Таким образом, группировка, то есть совокупность сообществ, сходных по выбранным критериям, характеризуется аналогичными экологическими условиями. Степень сходства варьирует сообразно этим критериям.
Для сообществ, выделенных по жизненным формам, формациям и типам растительности, характерен отчетливо выраженный параллелизм в распространении с крупными климатическими и почвенными зонами земного шара независимо от их флористического состава. Степи умеренных областей соответствуют резко континентальному климату, сходному в центральной Евразии и на Великих равнинах Северной Америки, и расположены на аналогичных почвах, определяемых как черноземы. По мере приближения к более влажным областям эти степи переходят в лесные формации на бурых лесных почвах или на деградированных черноземах. В более сухих областях они становятся более низкими и более открытыми и связаны с каштановыми почвами. Хвойные леса бореального типа Канады и Северной Европы развиваются в условиях холодного климата и на сходных оподзоленных почвах.
Сообщества, основанные на преобладающих видах, связаны с более или менее разнообразными экологическими условиями. В сообществах с одним и тем же доминирующим видом эдафи-ческие условия часто бывают весьма различными. Примеры э^ого мы видим на рис. 37 и 38. Первый из них относится к пихтарникам, сообщества которых указывают только на условия влажного горного климата, а второй — к лугам с молинией, которые повсеместно являются индикаторами некоторой влажности почвы и отсутствия удобрений, но встречаются при самых различных водных режимах и значениях pH. Несомненно, однако, что доминирующий вид сообщает местообитанию некоторые
146
особые признаки, такие, как условия затененности, качество гумуса, характер конкуренции.
Сообщества, выделенные по совокупности флоры или фауны и определенные статистически, точнее всего характеризуют экологические условия, так как здесь принимаются во внимание виды с небольшой экологической амплитудой, так называемые характерные виды. Как бы то ни было, амплитуда толерантности видов, объединенных в ассоциации, обычно больше, чем амплитуда толерантности самой ассоциации. Пальман (J. Ра1-mann) приводит пример вереска (Erica сагпеа) субальпийского пояса Центральных Альп с очень большой амплитудой по отношению к pH, поскольку она находится в границах между 4,5
Рис. 37. Связи между фациями во-гезского пихтарника и физико-химическими свойствами гумуса (по Дю-шофуру, Р. Duchaufour).
Реакция, отношение C/N, процент катионной насыщенности коллоидного комплекса в пихтарниках с: 1—Mercurialis perennis; 2—Festuca silvatica; 3 — Luzula albida', 4—Vaccinium myrtillus.
и более 8, но который встречается в субальпийском поясе в различных ассоциациях и субассоциациях, локализованных в гораздо более узких зонах реакции. Рис. 37 и 38 — наглядное свидетельство этого факта.
Таким образом, всякое статистически и таксономически определенное сообщество, растительная ассоциация (в понимании школы Цюрих — Монпелье), сообщество животных или биоценоз является гораздо более чувствительным и надежным показателем условий местообитания, чем виды индикаторы, экологическая локализация которых зависит от многочисленных факторов различного характера — генотипических различий, взаимодействия и эволюции внешних факторов, а также исторических факторов.
Эта связь между сообществами организмов и особенностями их местообитания является результирующей многочисленных причинно-следственных связей, при которых участники отобраны согласно этим особенностям, но одновременно в большей
147
или меньшей степени обусловливают их в случае наземных многоярусных растительных сообществ или организованных сообществ животных. Так, каждое сообщество существует в определенных микроклиматических условиях.
Многочисленные наблюдения над режимом температуры или влажности воздуха иллюстрируют это положение. Изучая термический микроклимат в ассоциациях ланд и горных сосен субальпийского пояса швейцарских Альп на высоте около 2000 м»
Рис. 38. Зональная связь между растительными сообществами и почвенными профилями в болотистой впадине равнины Эльзаса (по Шеферу, Schaefer).
Растительные сообщества: / — сообщество Filipendula ulmaria; 11 — Molinietum, субассоциация с Carex hostiana-, 111 — Moll-nietum, субассоциация c Carex tomentosa; IV — Mesobrometwn, субассоциация c Molinia; F — типичный Mesobrometum. Горизонты почвенного профиля НС —кальциевый влажный мягкий гумус; МС —кальциевый мягкий гумус; А—анмоор (торф); G f —восстановленный глей; Go —окисленный глей; Fe —железистые конкреции; Са —известняковые конкреции; N —средний уровень грунтовых вод. Стрелка указывает направление современной эволюции в результате осушения.
Пальман (Palmann) смог их классифицировать по убывающей термофилии, связанной с экспозицией и рельефом (рис. 39). Подобные же связи между микроклиматическими условиями и локализацией сообществ организмов встречаются на уровне сообществ споровых растений и мелких животных. Например, на стволах изолированных деревьев поверхности северной и южной экспозиции часто несут различную флору мхов, лишайников и водорослей!. Животные, населяющие мертвые стволы де-
1 В советской литературе совокупность растений на стволе дерева или насекомых, живущих в стволе, рассматривается не как самостоятельное сообщество, а как часть такого сообщества. — Прим, ред, 148
ревьев, также испытывают значительное влияние микроклимата, связанного с экспозицией. Так, колонии насекомого-ксилофага (Ips typographus) вследствие колебаний температуры и влажности, которые наблюдаются в различных участках поверхности ствола, развиваются в четко локализованных секторах лежащих на земле стволов (рис. 40).
Рис. 39. Средние температуры в субальпийских лайдах и сосняках Швейцарских Альп, взятые на высоте 1,5 м над землей (I), на почве (II) и на глубине 10 см от поверхности (III).
1 — Pineto-Caricetum humllis; 2а — Muglieto-Ericetutn Carl-cetosum humilis, восточная экспозиция; 26 — Mugheto-Eri-ceturn Caricetosum hutnilis, южная экспозиция; 3 — Mugheto' Ericetum Hylocomietosunv, 4-Mugheto-Rhodoreturn hlrsutr, 5 — Rhodoreto-Vaccinietum.
Почвы, как и микроклимат, столь тесно связаны с растительными сообществами, что Браун-Бланке в результате различных фитоценологических и почвенных исследовании, проведенных одновременно, выдвинул предположение, что «каждой ассоциации соответствует почвенный профиль со своими физико-химическими и биологическими особенностями». Примеры этому мы
Рис. 40. Зоны различного развития жука Ips typographus в лежащих стволах в зависимости от микроклиматических условий.
/ — наиболее освещаемая солнцем зона (температура в коре достигает 50°): кладки отсутствуют; 2 — яйца отложены, но убиты; 3 — личинки развиваются, но погибают; 4 — полный цикл развития; 5 —повышенная смертность вследствие избыточной влажности.
видим на рис. 37 и 38: буково-пихтовый лес Вогезских гор на кислых материнских породах состоит из ассоциаций и субассо-Циаций, которые можно наложить на серию различных почв — бурых олиготрофных, наименее кислых, охристых подзолистых почв, подзолистых почв и, наконец, наиболее кислых подзолов. На пологих склонах заболоченных впадин равнины Эльзаса также наблюдается параллелизм зональности растительных
14»
сообществ и все менее гигрофильных почв. Но это правило не является абсолютным, так как многие виды заселяют только верхние горизонты, где влияние климатических факторов и организмов в развитых почвах превалирует над свойствами материнской породы до такой степени, что одно и то же сообщество может располагаться на профилях с различным нижним горизонтом и являющихся следствием различного педогенеза («аналогичные» почвы Пальмана). Таковы субальпийские ланды с Vacciniutn и альпийским рододендроном, или Rhodoreto-Vaccinietum, которые встречаются на «сильно деградированных гумусово-карбонатных почвах», на подзоле или на псев-доальпийском ранкере1 в зависимости от того, состоит ли подпочва из рыхлого известняка, силикатно-алюминиевых горных пород или скал (см. также Ph. Duchaufour, Precis de Pedologie)2.
Методы изучения эволюции сообществ
Стабильность очень многих сообществ весьма относительна. Со временем сообщества могут претерпевать глубокие изменения, детерминизм и последовательность которых составляет важный аспект биоценологии. В некоторых случаях изменения протекают достаточно быстро для того, чтобы на протяжении нескольких лет были возможны прямые наблюдения. Наиболее точные данные о растениях получают при помощи маленьких постоянных площадок, на которых периодически описывается флористический состав (рис. 41). Так, в открытых быстро эволюционирующих сообществах можно довольно скоро уловить тенденции различных видов к распространению или регрессии. На уровне формаций или для популяций крупных травоядных ценным документом является периодическая аэрофотосъемка. Иногда можно с успехом использовать исторические документы или научные работы, выполненные с достаточной точностью.
В других случаях известен возраст первоначально оголенных территорий: таковы участки, отвоеванные у моря, — голландские польдеры, заброшенные культуры, почвы, оголенные эрозией, моренные отложения, обнажившиеся в результате недавнего отступания ледника, такие катаклизмы, как грязевые потоки (на грязевых потоках Шасты в Калифорнии исследовали эволюцию почвы и растительности) или извержения вулкана (извержение Кракатау в 1883 г. дало возможность провести многочисленные наблюдения над восстановлением растительности и фауны).
1 Термин «ранкер» применяют к некоторым почвам высокогорных лугов или тундр. — Прим. ред.
2 Имеется русский перевод: Ф. Д ю ш о ф у р, Основы почвоведения, М.» 1970. — Прим, ред,
150
▲ Sarothamnus scoparius
+ Polytrichum piliferum To же, fl сообществах
Рис. 41. Эволюция растительных сообществ га постоянных площадках, взятых на континентальной дюне в лесу Фонтенбло. / — зона эрозии; молодые растения исчезают вследствие обнажения; // — зона в процессе закрепления; большинство молодых Corynephorus закрепилось, и разрастается кукушкин лен, лишенный каких бы то ни было адаптаций к подвижным субстратам. Размер площадок 0,25 м2.
° Corynephorus canescens,прорастание О To же, старые стерильные растения <5 То же, плодовитые растения
Болота являются прекрасным объектом для реконструкции их эволюции при помощи сохранившихся растительных остатков, пыльцевого анализа и определения процента С14. Но чаще всего приходится восстанавливать сукцессии при помощи современных данных: скрупулезное изучение переходов м жду соседними сообществами, поиски факторов, определяющих преобразование, позволяют получить указания относительно тенденций развития сообществ.
Факторы и механизмы преобразования сообществ
Такое стабильное сообщество, как биоценоз, находится в состоянии некоторого равновесия с факторами местообитания. Изменение одного из этих факторов благоприятствует одним
151.
видам, которые становятся более обильными или проникают в сообщество, и неблагоприятно для других видов, которые при этом регрессируют и могут быть даже полностью вытеснены. Эти изменения состава сообщества в свою очередь изменяют некоторые факторы, что вызывает очередные перестройки до тех пор, пока не будет достигнуто новое равновесие. Таким образом, следствия становятся в свою очередь причинами; этот непрерывный ряд противодействий является результатом первоначального нарушения.
Эти изменения могут быть слишком незначительными и кратковременными, чтобы достичь размеров, за пределами которых равновесие было бы окончательно нарушено. Во всех сообществах наблюдается более или менее явное колебание обилия видов, и их равновесие является в действительности флуктуирующим около некоторого среднего значения. В других случаях изменения затрагивают достаточную часть компонентов сообщества, чтобы создалось новое сообщество, неспособное самостоятельно вернуться к предшествующему состоянию. Мы последовательно рассмотрим оба эти случая.
Временные изменения внутри сообществ
Причины их различны. Исключительные метеорологические условия, такие, как суровая зима или затянувшийся засушливый сезон, могут на некоторое время изменить состав биоценоза. Так, необыкновенные холода в феврале и марте 1956 г. убили на границе их ареала, определяемой низкими температурами, много экземпляров многолетних видов, таких, как падуб (Ilex) и Sarothamnus в Центральной Европе, оливы и многие кустарники на юге Франции; была также отмечена повышенная смертность птиц. Необычные морозы на побережье Ла-Манша в начале 1963 г. уничтожили многих животных. Такое истребление может привести к увеличению численности видов, которыми питались эти животные: так, вслед за уменьшением количества осьминогов после холодной зимы последовало увеличение количества крабов и других ракообразных, которые составляют обычную пищу осьминогов. В аридных зонах обилие однолетних растений связано с количеством осадков в благоприятном для их развития сезоне.
Другой причиной нарушений в биоценозах являются эпидемии и инвазии среди животных. Так, быстрое распространение вируса миксоматоза, занесенного во Францию в 1952 г., вызвало массовую гибель кроликов; следствием этого для растений явилось распространение ранее поедавшихся видов за счет остальных, а для животных — модификация цепей питания в биоценозах, в которых кролики составляли существенный компонент питания хищных позвоночных. Так, лисицы в Камарге стали питаться птицами и рыбой, а в Англии и в Парижском бассей-
152
Рис. 42. Циклические вариации обилия американского зайца {Lepus americanus) и его основного хищника канадской рыси (Felis canadensis) по количеству шкур, сданных охотниками за год „Компании Гудзонова залива“ (по Мак-Луличу, Mac Lulich, 1937).
не — полевками. Сарыч, который также в основном поедает кроликов, в зонах распространения миксоматоза лишился важного источника питания, что привело к падению плодовитости. Антипаразитные обработки могут привести к таким же цепям последствий. В отдельных случаях, когда вредитель менее чувствителен к обработке, чем поедающий его хищник, результат таких обработок может оказаться прямо противоположным желаемому. Так, в Конго наблюдалось резкое увеличение численности клопа — вредителя кофейных деревьев после их обработок ДДТ, в результате которых гораздо больше пострадал другой клоп — хищник вредителя.
Некоторые изменения могут носить периодический характер. При этом численность видов колеблется с более или менее постоянными интервалами между максимумом и минимумом. Выдвигались различные теории для объяснения этих циклов, которые чаще всего бывают трех-четырех или девяти-десятилет-ними. Были установлены климатические колебания, состоящие из последовательно чередующихся сухих и влажных лет; их связывают с примерно десятилетним циклом солнечной активности. Эти чередования дают особенно наглядный эффект в биоценозах сухих зон.
В центральных североамериканских равнинах засухи 1860— 1862, 1870—1873, 1893—1895, 1917—1918, 1933—1935 годов сильно повлияли на экономику и эмиграцию в некоторых штатах, например в Канзасе. Пагубное влияние этих засух сказывается не только на сельскохозяйственных культурах, но и на естественной растительности: травяной покров резко сокращается и достигает своего первоначального уровня только по прошествии двух-трех влажных лет, а его флористический состав возобновляется еще медленнее. В прерии и по ее краям среди
153
изолированных деревьев наблюдается более высокая смертность, в результате которой деревья теряют территорию, завоеванную в предшествующие влажные годы. Такие циклы проявляются также весьма явственно в периферических районах Сахары, где несколько следующих друг за другом влажных лет способствуют значительному росту многолетних растений и достижению высокого уровня обилия однолетних. Животноводы пользуются этим, чтобы увеличить по возможности поголовье скота; наступают засушливые годы, и пастбища уже не могут прокормить увеличившиеся стада, которые приходится тогда значительно сокращать.
Цикличность в обилии некоторых животных может быть связана с их собственным циклом развития. Таковы, например, «годы майского жука», которые повторяются каждые три года и объясняются двумя причинами: с одной стороны, тем, что личиночная стадия этого жука длится три года, а с другой — тем, что лучшие климатические условия благоприятствовали одной генерации по сравнению с другими. Если среди потомков этой генерации будет значительная смертность вследствие неблагоприятных условий на стадии яйца, личинок или куколок, то другая генерация, попавшая в относительно благоприятные условия, сменит на некоторое время предыдущую в трехгодичных циклах.
Другая причина периодических колебаний может связывать хищников и их жертвы. Мы описали выше осуществленные в контролируемых условиях эксперименты, где было показано чередование в обилии жертвы и ее хищника. К уже приведенным примерам подобных флуктуаций в естественных условиях добавим пример американского арктического зайца-беляка (Lepus americanus) и рыси, обилие которых на севере Канады оценивалось в течение ряда лет по количеству шкур этих двух видов, полученных крупным торговым обществом — «Компанией Гудзонова залива» (рис. 42). Максимальная численность зайца ярко выражена каждые девять-десять лет, а максимумы численности его хищника приходятся на тот же год или с запозданием на год. Песец (Alopex lagopus) имеет ярко выраженный трех-четырехгодичный цикл в тех областях Гренландии, где он питается леммингами, грызунами, обилие которых подчинено периодичности именно этой длительности, однако в прибрежных областях, где он питается другими животными, у песца не наблюдается этих циклических вариаций.
Многочисленные более или менее периодические колебания еще не получили объяснения. Для таких питающихся травой животных, как арктический заяц и лемминг, была установлена связь с продуктивностью растительного покрова, которая сильно варьирует в циркумполярных областях. В других же случаях предполагают уничтожение эпизоотиями или даже говорят об изменениях эндокринной природы.
154
В лесных формациях периодические рубки или поляны, образовавшиеся в результате отмирания деревьев, мгновенно изменяют биоценозы, создавая благоприятные условия для светолюбивых и нуждающихся в обильном азотистом питании растений в ущерб ранее существовавшей растительности. Фауна при этом также меняется; было установлено, что в результате интенсивной рубки дугласии, дугласовой пихты (Pseudotsuga douglassii), в Скалистых горах значительно возросло количество медведей вследствие увеличения ягодных растений и пчелиных гнезд.
Изменения, ведущие к формированию новых длительно существующих сообществ
Преобразования внутри сообщества могут быть достаточно глубокими для того, чтобы привести к образованию нового сообщества.
Причиной подобной эволюции могут быть различные факторы.
Климатические изменения. Климатические изменения в четвертичном периоде сыграли существенную роль в эволюции флор и фаун. Так, в течение последних 10 тыс. лет подножия гор Центральной и Западной Европы последовательно были покрыты следующими климатическими формациями: тундрами, затем светлыми березово-сосновыми лесами, затем орешниками, дубовыми лесами, смешанными с различными лиственными породами, буковыми и пихтовыми лесами, наконец, для некоторых горных цепей — еловыми лесами.
В отложениях торфяников сохранились поддающиеся датировке следы остановок в развитии вследствие увеличения сухости климата, за которыми при новом возрастании влажности следует возобновление развития. Площади, подвергшиеся осушению, покрыты сильно гумифицированным торфом, на котором существовали более или менее облесенные ланды, в то время как увеличение влажности способствовало их замене сообществами сфагновых торфяников.
Эдафические изменения. В результате совместного воздействия климата и организмов постепенно меняются особенности почв. Так при влажном климате известняковые почвы все больше декальцинируются. Это приводит к увеличению кислотности, к изменению физических свойств и уменьшению плодородия. Параллельно с эволюцией почв идет эволюция растительности. На декальцинирующихся почвах сообщества, сначала кальциефильные и базифильные, уступают место все более и более кальциефобным и ацидофильным. С подобным явлением мы встречались на альпийских луговинах в Центральных известняковых Альпах (рис. 43). То же происходит в неподвижных дюнах, где вследствие их высокой проницаемости декальцинация при влажном климате протекает быстро: Солсбери (Salisbury)
155>
стадии
Рис. 43. Параллельная эволюция растительности (вверху) и почвы (внизу) в альпийском высотном поясе известняковых массивов в Центральных Альпах (по Браун-Бланке и Дженни, J. Braun-Blan-quet et Н. Jenny).
—>- Гумусойьш --------► Альпийская
поЗзол гумусовая
почва (климакс)
установил, что за три века в условиях английского климата происходит полная декальцинация и pH изменяется от 8 до 5,5 (рис. 44).
На новом скалистом или каменистом субстрате атмосферные факторы в зависимости от климата оказывают преимущественно механическое или химическое воздействие, которое ведет к образованию почв, сначала молодых, очень тонких, а затем все более и более мощных, на которых может закрепиться уже более густая растительность. Так, на морене, освободившейся в результате отступания тирольского ледника, процентное содержание глины и суглинка, первоначально равное нулю,
со g ci
Рис. 44. Декальцинация и оки- г° слепне грунтов дюн по мере увеличения возраста последних на берегах Англии (по- Солсбери, Е. J. Salisbury)
составляло через 60 лет соответственно 20 и 4% от мелкозема (<2 мм), и там возник еловый лес.
Изменения биологического происхождения. Они наиболее часты и быстры. Прямое воздействие оказывает появление или исчезновение какого-нибудь вида в сообществе. Оно может привести к значительному и длительное время существующему изменению последнего. Разница во флористическом составе пастбищ и прилегающих невыпасаемых территорий — яркий тому пример. Выпас в лесу может привести к его уничтожению вследствие отсутствия возобновления. Специальная интрод кция животного, уничтожающего вредный вид, — средство биологического метода борьбы с этим вредителем. Бурно распространяющиеся заросли травы Clidemia hirta, завезенной на острова Фиджи, были уничтожены путем интродукции маленького насекомого Thysanoptera, которое питается молодыми побегами растения и препятствует таким образом его росту и размножению, способствуя восстановлению автохтонной растительности.
Исключительно разнообразны косвенные воздействия организмов, осуществляющиеся путем изменения среды. Вторжение деревьев и кустарников в травянистую формацию создает для последней условия затенения, которые элиминируют
157
большинство слаботолерантных солнцелюбивых видов и содействуют распространению сциофильных (тенелюбивых) видов.
Органическое вещество, продуцируемое деревьями, отличается по своему происхождению, обилию, скорости разложения и химическим свойствам от органического вещества травяного покрова и содействует эволюции биоценоза, изменяя характер почвы. Так, лесные полосы, созданные в степной зоне СССР, изменяют черноземы, приближая их к бурым лесным почвам. Воздействие разных деревьев неодинаково вследствие различий в характере создаваемого ими затенения и подстилки. Так, во многих высокоствольных лесах французских равнин, если не проводятся осветляющие рубки ухода, бук имеет тенденцию вытеснять дуб. Молодые буки прекрасно развиваются в тени дубов, в то время как молодые дубы погибают в более густой тени буков.
Неоднократно отмечалось окисление и обеднение лесных почв при посадках хвойных пород после лиственных, при этом возникало или усиливалось оподзоливание. Напротив, лиственные породы, такие, как граб, липа, ольха, благодаря своей подстилке, относительно богатой азотом и минеральными веществами, улучшают почву.
Аккумуляция органических веществ некоторыми растительными сообществами является основной причиной их изменения: эволюция от луговин на известняках альпийского высотного пояса к ацидофильным сообществам происходит не только вследствие поверхностной декальцинации, но в основном из-за накопления все более мощного слоя кислых органических веществ (рис. 43). Эволюция торфяников вследствие одного только накопления торфа приводит к их осушению: первоначальные более или менее затопленные сообщества уступают место все менее и менее гигрофильным ассоциациям, до мезофильных сообществ, которые в некоторых случаях могут быть идентичны сообществам окружающих, незаторфованных территорий (рис. 45 и фото II).
Вспомним также о роли, которую играют некоторые растения в закреплении подвижных субстратов, например о роли во-лоснеца (Elymus arenanus) для литоральных дюн Франции, Spartina townsendii для илов эстуариев (фото 1,А), куропаточьей травы (Dryas) для известняковых осыпей Альп и т. д. Вследствие этого закрепления рассеянная флора незакрепленных субстратов исчезает и образуется новое сообщество. Животный мир дает подобные примеры фиксации подвижных субстратов литорали организмами, вырабатывающими известь, которые заменяют биоценоз подвижного дна биоценозом закрепленного субстрата.
Все эти виды, играющие важную роль в динамике, являются деструкторами по отношению к сообществам, в которые они внедряются, и эдификаторами новых сообществ.
158
MN MO Cl Cf
Свободная бода
Озерный ил
Осоковый торфяник
Сфагновый торсряник
Р и с. 45. Последовательное развитие озерных торфяников Центрального массива.
AI/V — ассоциация свободных вод с Nuphar и Myrlophyllum-, МС — инициальное сообщество с Menyanthes и Comarum', С/ —сплавина (плавающий торфяник) с Carlcetum lasiocarpae, подвергшаяся вторжению сфагнума; Sm — Sphagnetum medii, инициальная стадия сфагновых торфяников; Sf —Sphagnetum ZuscZ— поднимающегося сфагнового торфяника; Sec— Sclrpetum caespitosi высыхающих поверхностей; Cf-Ca-rlcetum fusel-ассоциация озерной кромки.
В сукцессии сообществ определяющие факторы преобразований могут быть различными по своей природе на разных этапах, как это показывает пример развития растительности литоральных дюн, которое начинается с фиксации, затем продолжается в виде декальцинации и окисления, для того чтобы завершиться развитием древесной растительности — деструктора лугов.
Биоценотические сукцессии
Их этапы
Когда оголенная поверхность избавлена от разрушительных воздействий, на ней более или менее быстро поселяется некоторое количество видов, образуя инициальную фазу развития живого населения, в которой большую роль играют случайности рассеивания зачатков и миграции животных.
Затем в результате воздействия различных факторов, рассмотренных в предыдущем параграфе, более или менее стабильные и длительно существующие сообщества, обычно все большей и большей структурной сложности, сменяют друг друга до тех пор, пока не будет достигнуто равновесие между климатом, почвой и сообществом. Это терминальная стадия эволюции, или климакс, которому соответствует почва определенного типа — педоклимакс.
Сукцессии, ведущие к терминальной стадии, называются прогрессивными, те, которые удаляются от нее, называются регрессивными. Прогрессивные сукцессии считаются первичными, если им не предшествовала регрессия, и вторичными — в противоположном случае. Прогрессивные сукцессии на дюнах, литоральных илах (фото I) и торфяниках (рис. 45 и фото II) часто бывают первичными, прогрессивные сукцессии в сельскохозяйственных районах — вторичные.
Таблица 9
Прогрессивные и регрессивные сукцессии французских равнин
Трминальная стадия-. Дубняк ацидофильный
Лес дубово-грабовый
Древесная стадия: Березняк • (эксплуатация)
А I
I i
Кустарниковая стадия: Сообщество Sarothamnus А : (пожар)
I Луговина с Anthoxantum и Festuca rubra
Дубняк из дуба пушистого А ?
Травянистая стадия
Чаща с Prunus spinosa | (эксплуа-
или Crataegus : тация)
(пожар) 4
I i
Луговина с Br achy podium :
pinnatum или Bromus erectus
Инициальная стадия:
Залежь с Rumex acetosella Залежь с кальциефильным и ацидофильными однолет- Galeopsis ladanum никами и т. д, А
А
Культуры (на кремнистой почве)
Культуры (на сухой известняковой почве)
160
Фото I. Заселение растительностью литоральных илов (фото автора).
А. Заселение двумя злаками Atropis mar it i та и Spartina townsendii (последний — на заднем плане слева) ила эстуария в верхней части зоны, заливаемой при каждом приливе. Дерновины, вначале изолированные, сливаются при разрастании и их центральные части при поднятии в результате заилевания заселяются другими видами. Таким образом, эти два вида обеспечивают эволюцию обнаженных илов приливной полосы «сликке» в галофильные луга «шорры» путем закрепления и аккумуляции этих илов (фотография сделана в момент отлива в устье Орпа).
Б. Мангры из Rhizophora, видны многочисленные ходульные корни (устье реки Казамансе, Сенегал).
Фото II. Стадии развития озерных торфяников (фото автора).
А. Начальные стадии образования торфяника из дистрофного озера: кайма Мепу-anthes trifoliata и Comarum palustre, постепенно захватывающая чистую воду; позади сменяющие ее заросли Carex lasiocarpa образуют плавающий торф, в котором укореняются несколько кустов ивы. Последовательно уплотняющийся торф заполнит озеро, затем его поверхность поднимется над уровнем озера, что приведет к образованию выпуклого болота (Мон-Дор, Овернь, около 1200 м).
Б. Заключительная стадия развития озерного торфяника: выпуклое болото с засыхающим сфагнумом, па переднем плане дерновины Eriophorum vaginatum, позади Cal-luna и Vaccinium, в глубине естественные заросли горной крючковатой сосны Pin us montana ssp. uncinata (болото в Барте, Мои-Дор, Овернь, около 1200 м).
Фото III. Пояса растительности скалистой литорали (фото автора).
А. Пояс супралиторали на скалистом берегу Котантена. Глыбы гранита, до которых долетают морские брызги, и в исключительных случаях заливаемые сильными приливами, покрыты лишайниками Ramalina, Xantho-па parietina и т. д. Внизу верхняя часть зоны с Caloplaca marina, корковым лишайником, маркирующим верхнюю часть пояса средней литорали. Основание рыхлого обрывистого берега с луговиной, где доминируют злаки и зонтичные, обдаваемой брызгами морской воды.
Б. Пояс средней литорали, расположенный на несколько метров ниже предыдущего (фото Л). Зона со студенистым лишайником Lichina pyginaea и бурой водорослью Pelvetia canaliculata, образующей более светлые пучки среди лишайника. Обнаженная поверхность скалы заселена прикрепленными к ней Chthamalus (Cirripedia) и морскими блюдцами, литоринами, брюхоногими моллюсками, поедающими водоросли и лишайники. Внизу видны три морских блюдца в их кормовой стации.
Б
। Фото IV. Пустынные формации Мавритании (фото Нежеле, A. Naegele).
^крепление эолового песка в виде холмиков злаком Aristida pungens. В глубине
>. Откп аллювиальная впадина с ксерофнльнымн кустарничками.
Рьиая степь на опесчанениом реге. Песок аккумулируется полосами со стороны, защищенной растениями от ветра.
A
Фото V Биоценозы травянистых формаций (фото Бурлиера, F Bourliere).
А. Североамериканская «прерия» (Национальный заповедник Бизон-Рейндж, Мои тана). Травянистое сообщество с преобладанием овсяницы (Festuca), где практикуется небольшой выпас, усеяно группами низких кустарников. Пастбищная нагрузка: бизоны и особенно вилорогая антилопа — примерно 35 кг на 1 га при биомассе злаков около 5 тыс. кг.
Б. Низкотравная разреженная саванна (регрессивная стадия с Thetneda и Hetero-pogon), равнины Руиндн— Ручуру в Национальном парке Киву [бывший Альберта, Конго (Киншаса)]. Пастбищная нагрузка: 11 видов копытных — около 240 кг на 1 га при биомассе злаков 6 тыс. кг Здесь заметен перевынас и нарастающее ухудшение ра~ стнтелыюстн. На фотографии представлены буйвол саванн и антилопа топи.
A
Фото VI. Саванны Сенегала (фото Трошена, J. Trochain).
А. Саванна с колючими («вооруженными») кустами Acacia seyal с открыт травяным ярусом в центральном Сенегале. Аспект сухого сезона после прохождсш огня: деревья лишены листвы, почва обнажена. Ежегодная сумма осадков от 600 ;к 700 мм.
Б. Саванна с густым древостоем с доминированием Combre-tum glutinosum в южном Сенегале. Аспект сезона дождей: заметен довольно редкий травяной ярус и многочисленные солнечные пятна на поверхности почвы. Такую формацию можно также квалифицировать как светлый листопадный лес. Ежегодная сумма осадков около 800 мм.
Фото VII. Саванна Берега Слоновой Кости.
А. Аэрофотоснимок Берега Слоновой Кости (север). Кустарниковые саванны суданского типа с изолированными лесными островками на повышенных участках, пересеченные глубокими долинами с узкими и прерывистыми галерейными лесами на их днищах. Видны следы прохождения стад.
Б. Саванна гвинейского типа в окрестностях Буаке, Берег Слоновой Кости (центральная часть). На первом плане участок, выгоревший в конце сухого сезона; вдали участок, выгоревший в начале сухого сезона; древесная растительность здесь густая в связи с менее губительным действием огня. Аспект влажного сезона. Ежегодная сумма осадков 1200—1300 мм (фото автора).
Фото VIII. Деградирующие растительные формации Средиземноморской области.
А. Гарига, или «бата», на известняках горы Монт-Кармель в Израиле. Одиночные вечнозеленые кусты — дубы (Quercus calliprinos) высотой в несколько метров. Межд' ними заметен эродированный каменистый субстрат с открытым разреженным покровов из низких кустарников, ладанников, фисташек, черноголовника (Poteriuin spinosum) и т. д. Климатически обусловленная формация — лес из Quercus calliprinos, высота 450 м, климат средиземноморский, субгумидный, ежегодная сумма осадков окол 800 мм (фото автора).
Б. Полынная (с Artemisia herba-alba) степь, развившаяся на скелетных почвах в связи с многовековым выпасом. Видны руины маслобойки римской эпохи; в настоящее время район совершенно лишен маслины (центральный Тунис: средиземноморский семиаридный климат, осадков примерно 300 мм в год) (фото Бэлди, Ch. Baldy).
A
Фото IX. Деградирующие лесные формации Средиземноморской области (фото автора).
А. Маквис на кремнистой толще: в центре пригодный к эксплуатации пробковый дуб, свидетель исчезнувшего естественного леса. На переднем плане Phillyrea, вереск, Calycotome\ немного дальше за ними молодые сосны.
Б. Остатки климатически обусловленного вечнозеленого жестколистного дубового леса на вершине Мопт-Кармель, высота около 500 м. Между этими многовековыми дубами (Quercus calliprinos) луговые поляны и маквис, куда внедряются алеппская сосна и дуб.
Фото X. Тропические тенелюбивые леса (фото Бурлиера, F. Boiirliere).
А. Горный пальмовый лес в Пуэрто-Рико. Стволы увешаны множеством эпифитов, бромелиевыми, орхидеями, папоротниками; почва покрыта папоротниками, которые указывают на большую атмосферную влажность.
Б
Б. Прогалина в экваториальном лесу Берега Слоновой Кости (заповедник Банко). Видны деревья, расположенные различными по высоте ярусами. В обрамлении прогалины можно узнать музангу, солнцелюбивый вид с большими пальчатовидными листьями. Обращает внимание густая тень под пологом леса.
Фото XI. Хвойные леса.
1. Бореальные хвойные леса Скалистых гор в Национальном нарке Глейше на границе Канады н Соединенных Штатов. Обращает внимание колоннообразная форма деревьев, приспособленных к обильным снежным заносам, и густота леса (фото Бурлнера, F. Bourlicre).
Б. Открытые насаждения алеппской сосны (Pinus halepensis) среди гариги с розмарином на известняковой почве в центральном Тунисе (субаридный средиземноморский климат: около 300 мм осадков в год). Эти насаждения не образуют флористически своеобразных сообществ, они, если не станут добычей огня, благоприятствуют восстановлению климатически обусловленного леса. В глубине равнина, поросшая алжирским ковылем (альфой) (фото Бэлди, Baldy).
Фото XII. Хвойный лес дождливого климата умеренных широт в Национальном парке Олимпик, штат Вашингтон, на северо-западе Соединенных Штатов. Огром ные деревья, Tsuga heterophylla, достигающие 60 м в высоту и 4,5 м в диаметре у основания ствола. Заметно обилие мхов и папоротников на почве. На заднем плане — прогалина с лиственными породами. В этом районе выпадает от 1250 до 2500 мм осадков в год в виде дождей.
л
Стадии сукцессии Залежь Кустарниковая стадия Сосняк 'Лиственный лес
Возраст, в годах 1 3 15 20 25 35 60 100 >150
Ammodramus savannarum Sturnella tnagna Geothlypis trichas Splzella pusilia Pipilo erythrophthalmus Richmondena cardinalis Dendroica pinus Piranga rubra Vireo olivaceus Hylocichla mustelina Dendroica virens
Количество видов Количество гнездящихся пар на 10 га 2 4 2 10 8 26 13 32 10 21 12 24 24 41 25 59 23 56
Рис. 46. Распределение количества видов и плотности гнездящихся птичьих пар на различных стадиях вторичной сукцессии в Джорджии от травяной залежи до клима-ксового леса из дуба и орешника (по Джонстону и Одуму, D. W. Johnston, Е. Р. Odum).
Табл. 9 дает пример прогрессивных и регрессивных сукцессий, часто наблюдаемых на французских равнинах вне средиземноморской области.
Таксономический состав и обилие в популяциях животных изменяются параллельно эволюции растительности и в значительной мере являются следствием этой эволюции. Эволюция может быть различной в разных группах: с возрастанием плотности растений и увеличивающегося разнообразия микроместообитаний количество видов обычно увеличивается на первых стадиях, а затем уменьшается, так что наиболее богатые видами сообщества относятся к промежуточным стадиям, а не к климаксу. Это мы видим на примере муравьев в растительной сукцессии на дюнах озера Мичиган и на комплексе насекомых в сукцессии на покинутых полях прерий с травой Грама (Bouteloua) в Оклахоме. В противовес этому в последней сук-Цессии обилие мелких млекопитающих уменьшается на промежуточных стадиях и снова возрастает на терминальных
7 Ж. Леме
161
стадиях. В другой сукцессии, идущей в Джорджии от сельскохозяйственных культур к лесу из дуба и Сагуа, количество птиц, судя по гнездам, последовательно возрастает, но максимальное количество их видов устанавливается до достижения климакса (рис. 46).
Внутри сложных систем, образующих крупные биоценозы, эволюция которых приводит к климаксным сообществам, устанавливается на уровне организмов особый тип сукцессий, которые можно назвать деструктивными и которые в экосистеме образуют пищевые цепи организмов-редуцентов. Описаны стадии разложения таких различных организмов, как труп млекопитающего, ствол дерева, желудь дуба, пресноводные и морские водоросли, выброшенные на берег. В случае трупов млекопитающих, например, первыми поселенцами являются мухи Musca и Calliphora, которые откладывают яйца на кожу, личинки превращаются в куколок примерно через неделю. Затем появляются привлеченные запахом мухи Lucilia и Sarcophaga-, запах прогорклого жира привлекает впоследствии кожеедов (жуки) и Aglossa (чешуекрылые), личинки которых питаются жиром. Затем разложение протеинов привлекает Necrobius (жуки) и Piophila (двукрылые); позднее аммиачные выделения привлекают многочисленных мух и жуков — Phora, Silpha, Necrophorus, Hister. Когда мумификация уже полностью закончена, появляются клещи, бабочки и новые жуки (Tyroglyphus, Tinea, Anthrenus); наконец, покрывающие кости остатки становятся добычей некоторых жуков — Ptinus и Tenebrio.
Понятие климакса
Это выражение обозначает терминальные сообщества растительных сукцессий, неспособные к дальнейшему развитию при отсутствии каких-либо воздействий. Такие сообщества могут быть выделены на основании различных концепций, как мы это видели выше, и их стабильность может быть следствием различных причин, результатом этого явились некоторые расхождения во взглядах на понятие «климакс».
Для Клементса (Clements) климакс — краеугольный камень его системы — формация, находящаяся в состоянии равновесия с данным климатом и зависящая только от него, откуда и пошел иногда употребляемый синоним «климатическая формация». Клементс признает, однако, что под воздействием некоторых локальных неклиматических факторов могут образовываться постоянные формации и ассоциации, он дает им специальные названия: дисклимакс в случае сообществ, нарушенных человеком и животными (фото VIII, А и Б); субклимакс, когда удерживается, чаще всего в результате пожаров, стадия, близкая к климаксу; преклимакс, когда повышенная местная влажность способствует образованию более гигрофильных сообществ,
162
чем это возможно в условиях данного климата; постклимакс в противоположном случае — на сухом местообитании.
В противовес этой теории моноклимакса европейские экологи придерживаются теории поликлимакса, понимая под климаксом все сообщества, по-видимому, не способные к дальнейшему развитию, которые остаются отчетливо различимыми в пределах территории климатически однородной по эдафиче-ским, топографическим и антропическнм условиям. Их совокупность образует комплекс климаксов. Так, в высокогорном поясе Альп этот комплекс включает не только Caricetum curvulae, терминальную стадию эволюции мезофильных лугов со снежным покровом средней продолжительности, но и Festucetum halleri слабо заснеженных лугов, бесснежные гребни с Loiseleu-rietum procumbens, сообщества снеговых долинок, Trichophore-turn торфяников, заканчивающих свое развитие (см. также фото II, Б). Стадии, удаленные от климакса, стабильность которых, однако, является практически неопределенной, такие, как сообщества озер, расщелин в скалах, выкашиваемых лугов, можно называть перманентными сообществами.
Во всех случаях понятие климакса является относительным. Сообщества, постоянные в масштабе человеческой жизни, очевидно, в масштабе истории не являются таковыми из-за климатических изменений или очень медленного развития некоторых эдафических факторов. Кроме того, некоторые климаксы образуют циклические фазы, которые в действительности представляют собой мозаику мелких, быстро развивающихся сообществ; их равновесие в высшей степени динамично, и их постоянство — в неопределенно продолжительном возобновлении этих циклов. Таковы лесные климаксы, в которых деревья, выпадающие из древостоя от старости или ввиду неблагоприятных условий, открывают поляны, на которых устанавливается особая фауна и флора до тех пор, пока молодые деревья не обеспечат зарастания (см. фото XII). Была также описана циклическая эволюция на известняковых склонах в Лапландии, которая заключалась в накоплении торфяного гумуса на горной породе лан-дами с Vaccinium, Dryas и Empetrum*, эта фаза завершалась растрескиванием вследствие высыхания, которое вызывало эрозию торфа и обнажение горной породы, на которой сызнова начинался новый цикл.
Наконец, терминальные стадии вторичных прогрессивных серий, весьма возможно, будут отличаться от терминальных стадий первичных серий вследствие необратимых изменений почвы, вызванных деятельностью человека. Так, хищническая эксплуатация лесов на выщелоченных слабокислых почвах с незначительными запасами питательных веществ и обменных оснований вызывает их значительное уменьшение, сопровождаемое окислением и оподзоливанием, которые допускают только восстановление лесных сообществ, отличных от предшествующих.
7*
163
Следует, таким образом, отличать «современную потенциальную растительность», завершающую прогрессивное развитие, от первичной естественной растительности, от которой она отличается более или менее глубоко в зависимости от величины необратимых изменений местообитания и полноты преобразований флоры и фауны, происшедших в это же время в данной области. В тропических областях, и в особенности в областях древней цивилизации, процессы деструкции и последующего возобновления естественной растительности могут повторяться по многу раз, и новая терминальная ассоциация, или климакс, будет все больше и больше отличаться от естественного климакса.
Глава 9
Картографирование растительности
Картографическое воспроизведение растительного покрова представляет собой рабочие документы, необходимые для многих исследований. Потребность в подобных документах особенно ощущается при экономической оценке территории, и это не наименее важная причина современного подъема картографирования растительности во многих странах. Впрочем, идея такого картографирования родилась давно: в начале XIX в. датчанин Скоу (Schouw) и француз де Кандоль (Aug. Р. de Candolle) предвидели пользу подобных работ. Но только в 1894 г. другой французский ботаник Флао (Ch. Flahaut) учредил Проект ботанических, сельскохозяйственных и лесных карт Франции, за которыми последовало опубликование листа карты Перпиньяна (Perpignan) в масштабе 1:200 000 — первой современной фитогеографической карты. Впрочем, подобная работа не могла быть предпринята раньше, так как прежде всего было необходимо, чтобы, с одной стороны, были ясно определены различные понятия, относящиеся к единицам растительности и их классификации, а с другой стороны, были созданы для исследуемых районов точные топографические карты нужных масштабов.
В последнее время аэрофотосъемка предоставила фитогеографам новый, особенно ценный источник документации. Точная интерпретация аэрофотосъемки делает необходимой применение разработанной техники фотограмметрии, которая позволяет с определенной точки зрения идентифицировать природу единиц растительности и установить их границы с большой точностью и значительной экономией времени. Все же аэрофотосъемка не сможет полностью заменить наземные исследования (фото VII, А).
При любом картографическом воспроизведении растительности последовательно возникают два вопроса. Что хотят изобразить и как это выполнить? Первый вопрос ставит проблему отображаемых фактов: флористических единиц, таких, как ассоциации, союзы, фации... физиономических единиц, таких, как формации, зоны растительности... конкретного распределения этих единиц или синтетического отражения доминантных единиц территории или естественной растительности, которую сменили или к которой вернутся культурные или деградированные сообщества.
165
Выбор масштаба карты находится в прямой зависимости от отображаемых объектов: так, для воспроизведения флористических сообществ и в меньшей степени формаций необходимы крупные масштабы (по меньшей мере 1 : 50000), синтетических представлений — средние (1 100000 до 1:250000) и мелкие масштабы (1:500 000 и мельче)
С другой стороны, степень разнообразия растительного покрова предопределяет использование более крупных масштабов в тех случаях, когда разнообразие велико, как в областях с сильно пересеченным рельефом, или, наоборот, более мелких масштабов, когда отмечается определенное единообразие растительного покрова, как на равнинах — лесистых, степных или пустынных.
Выбор цвета и значков для различных принятых единиц должен подчиняться определенной логике, следуя которой можно было бы рельефно выделить родственные связи и контрасты и сделать карту более выразительной.
Карты флористических сообществ
1. Метод флористического анализа сообществ, разработанный школой Цюрих — Монпелье, привел к созданию многочисленных карт, особенно в Центральной и Западной Европе, а также в Северной и Экваториальной Африке. Изображение in ех-tenso1 ассоциаций и их вариантов требует крупного масштаба. Во Франции первые карты были выполнены Люке (Luquet) (Мон-Дор) и Молинье (Molinier) (Сент-Бом) в масштабе 1:50 000, но с увеличением точности флористического анализа растительных сообществ службой Национального научно-исследовательского центра (CNRS) в Монпелье был принят масштаб 1 :20 000. В этом масштабе ею были изданы карты окрестностей Моппелье, Экса, Понтарлье, Клермон-Феррана, Альп в районе Лотаре, леса в окрестностях Страсбурга, долины Ба-Шелифа2. Картографирование в более крупном масштабе оказалось необходимым во многих случаях:
— мозаики или зонального распределения сообществ, сконцентрированных на небольших площадях, как, например, торфяники или берега озер или литораль (например, карта острова Гран-Рибо в масштабе 1 : 5000);
— сообщества с разнообразными фациями в процессе эволюции, как государственный лес в Сент-Бом, изображенный в масштабе 1 :2000;
— зоны сельскохозяйственных культур, детальное карти-
1 Полностью, во всех деталях. — Прим. ред.
2 Значительная часть Прованса была картографирована в этом масштабе Репе Молинье, по карта не опубликована.
166
ровапие которых имеет особое практическое значение. Центральная станция картографирования растительности в Штольценау и картографическая служба Немецкого географического института в Лейпциге часто проводили картирование в масштабе 1:5000, а карта холма эльзасского виноградника выполнена в масштабе 1 10 000.
Классификация ассоциаций по их флористическому сродству диктует выбор цветов для их изображения. Ассоциации одного союза изображаются одним цветом, но в зависимости от ассоциации более или менее густым или же различным образом заполненным черными значками. Союзы одного порядка изображают близкими оттенками. Обычно применяют желтые и красные дона — для ксерофильных сообществ, голубые и лиловые — для гигрофильных и водных сообществ, зеленые — для лесов и серые— для культивируемых территорий. Случаи деградации, переходы, мозаики отображают особым образом.
2. Исследование экологических особенностей сообществ путем использования их видов в качестве индикаторов и их объединение в «экологические группы» привели приверженцев этого метода к картографической классификации сообществ по экологическому сродству. Сообщества, в которых преобладают одни и те же экологические группы, изображаются одним цветом.
Так, на карте лесов бельгийской Лотарингии, выполненной в масштабе 1 :50 000, Дювиньо распределил цвета согласно факторам влажности и особенностей гумуса сообществ. Новый научный центр фитосоциологических и экологических исследований в Монпелье избрал тот же путь, намереваясь сделать возможным «практическое использование фитосоциологических карт для эксплуатации и оценки земель». На «экологических картах», понимаемых подобным образом, объединяются одним цветом территории, у которых сходны основные экологические признаки. Территории, объединенные таким образом, выделяются на карте различными оттенками или значками. Этот основной документ служит матрицей для различных калек, таких, как современное использование земель, сельскохозяйственное, лесохозяйственное и пастбищное использование и т. д.
3. Динамические отношения между флористическими сообществами можно изобразить картографически. Первая попытка такого рода была сделана Люди (W. Liidi), который опубликовал в 1921 г. карту сукцессий растительных сообществ в Лау-тербруннентале, в Швейцарии, в масштабе 1:50 000, где один Цвет соответствовал всем инициальным сообществам, другой — промежуточным сообществам и особый цвет — каждому из климаксов.
Айхингер (A. Aichinger) предложил в 1954 г. иной метод, изображая каждую динамическую серию определенным цветом, интенсивность которого тем ярче, чем ближе к климаксу изображаемое сообщество.
167
Именно эта задача вдохновляла Кунхольц-Лордата (Kuhn-holtz-Lordat) в его попытках создания «парцеллярной (дробной) картографии» с картами крупного масштаба, где на площадях с быстрой эволюцией растительного покрова, таких, как заброшенные сельскохозяйственные культуры, сукцессии общественных видов1 изображаются с такой точностью, что эти карты представляют собой настоящий архивный документ по динамике подобной растительности (рис. 47).
Рис. 47. Эволюция заброшенных сельскохозяйственных культур на грядках, обращенных к югу в Гар, Лангедок (масштаб: 1 : 1950, по Кунхольц-Лордату, Kuhnholtz-Lordat).
Верхняя, недавно заброшенная часть была захвачена свинороем (белый цвет), постепенно вытесненным Brachypodium phoenicoides и ладанником. В нижней части это луг с Brachypodium ramosum, который регрессирует ранее Br. phoenicoides и ладанника. Однако территория в целом постепенно захвытывается кермесовым дубом. Несколько каменных дубов предвосхищают климаксовый лес.
4. Картографирование сообществ, основанное на флористическом составе, может дать синтез для обширных территорий, что достигается использованием более мелкого масштаба. Возможны два решения задачи: изображение высших фитоценоло-гических единиц — союзов и порядков — или изображение на данной площади доминирующей ассоциации или двух-трех доминирующих ассоциаций.
Именно это последнее решение было использовано Лонгом (G. Long) для схематической карты растительных сообществ центрального Туниса в масштабе 1:200 000, где выбор цветов был продиктован доминирующими экологическими особенностями, вертикальные полосы двух чередующихся цветов указывали на наличие мозаик двух преобладающих сообществ.
1 Этот термин применяют к видам, образующим густые насаждения. Прим, ред.
168
Карты физиономических сообществ: формаций и доминирующих видов
Как и предыдущие, они дают изображение, детализированное— в крупном масштабе или обобщенное — в среднем и мелком масштабе.
Равно, как и предыдущие, они могут быть основаны па физиономическом сродстве (статичные) или на изменениях, вызванных в естественных сообществах деятельностью человека (динамичные) .
Карты, основанные на физиономическом сродстве и доминирующих видах. Многочисленные крупномасштабные детальные карты, составленные на основании этих критериев, были созданы в начале XX в., например, Друде (Drude) в Германии (в масштабе 1:20000), Бланком и Харди (Blanc, Hardy) в окрестностях Монпелье (в масштабе 1 : 50000), Рюбелем (Riibel) в Швейцарии, Гамсом (Gams) в Австрии, Адамовичем (Adamo-vitch) на Балканах, Кватреказасом (Quatrecasas) в Испании и т. д. Среди новейших карт отметим карту растительности острова Гуам Фосберга (Fosberg) в масштабе 1:50000 (1959 г.). Уже в масштабе 1:100 000 можно осуществить точное изображение только в случае довольно однородных территорий. Таковы выполненные в масштабе 1:93 750 карты земельного планирования («Land Planning») департамента внутренних дел Соединенных Штатов.
Зато масштаб 1 :200 000 хорош для составления синтетических карт и был использован для этого во многих странах. Такова карта Перпиньяна Флао (1897 г.), карта растительности Швейцарии Шмида (Schmid) (1950 г.), карта Марокко Амберже.
Были созданы карты более мелкого масштаба, дающие, естественно, более схематический обзор растительности.
Общая карта растительности Португалии (в масштабе 1:500000); карта растительного покрова Атласа Госсена (Н. Gaussen) в масштабе 1 1000 000; международная карта Всемирной службы использования земель («World Land Use Survey») в масштабе 1:1000 000; карты Северной Африки Амберже И Мэра (Embergcr, Maire) в масштабе 1 : 1500 000; карта зон растительности Нигерии Кейя (Кеау) (1959 г.) в масштабе 1:3 000 000; карта растительности Малайского архипелага ван Стеениса (С. van Steenis) (1958 г.) в масштабе 1:5 000 000.
Карты, основанные на динамике растительности. На этой концепции было основано средне- и мелкомасштабное картирование, осуществленное в Тулузе под руководством Госсена (Н. Gaussen). Основной единицей является серия растительности или растительный пояс в горах. Эта единица охватывает совокупность площадей, естественная растительность которых представлена одной и той же формацией, в которой преобладает один и тот же вид: так, в равнинах Центральной Франции это
169
серия дуба черешчатого, в горах — пояс бука. В результате использования этой растительности человеком были созданы, исходя из этих климаксов, многочисленные сообщества, которые можно объединить в типы растительности: культурная растительность, луга, фруктовые сады, ланды, чащи и т. д.
Каждой серии соответствует цвет, выбор которого определяется соответствующими климатическими условиями: синий обозначает влажный климат, черный — холодный, желтый — относительную сухость, оранжевый — сильную сухость, красный — жару. Так, для пояса бука, локализация которого определяется влажностью климата, был использован синий цвет, серии листопадных дубов при климате, переходном от влажного к сухому, обозначаются смесью синего и желтого, то есть зеленым, в котором желтый все больше и больше проступает в порядке: дуб че-решчатый, дуб сидячецветный, дуб пушистый. Жаркий и постоянно влажный экваториальный лес изображается лиловым цветом, который получается в результате смешения красного и синего.
Что касается типов растительности, которые признают идентичными в большей части серий, их выделяют благодаря разному способу использования цветов: культурная растительность — белый, участки лесовосстановления — горизонтальной штриховкой по цветовому фону серии, к которой относится порода дерева.
Карты-врезки дают дополнительные сведения о климате, почвах и их использовании, кантональной сельскохозяйственной статистике и т. д.
Эти карты составлены в двух разных масштабах:
а) Карта растительности, составленная службой центра CNRS в Тулузе в масштабе 1 :200 000;
б) Международная карта растительного покрова, составленная институтом Тулузского университета в масштабе 1 1 000 000; на этой карте, естественно, менее подробно, изображены территории со средиземноморским климатом.
Согласно вышеизложенным принципам, были также составлены карты растительного покрова бывшего Бельгийского Конго. Их масштаб в зависимости от степени сложности колебался между 1 100 000 и 1 1 000 000.
Карты зон растительности очень мелкого масштаба. Это карты, охватывающие часть земного шара, или даже карты мира, которые отображают зональность крупных типов климатических формаций: вечнозеленые экваториальные леса, тропические леса с периодом покоя в сухое время года, саванны и тропические степи, пустыни, склерофильные и хвойные средиземноморские леса и 1. д. (см. рис. 61).
Составленные на основании различных критериев географического или физиономического характера, с обозначениями в соответствии с разнообразной терминологией, эти карты отобра-
но
экают великий закон зональности климатов, растительности и почв, иллюстрируя удивительное соответствие между климатическими зонами, типами естественной растительности и климатическими почвами.
Из карт мира мы можем указать среди наиболее известных на карту Брокман — Ероша, опубликованную в Физическом настенном атласе (Physikalischer Wandatlas) в масштабе 1 25 000 000, а среди новейших — карту Кюхлера (Kiichler), опубликованную в Мировой географической серии эпеншида (Espenshade’s World Geography Series) в масштабе 1 : 22 800 000. Из карт различных частей света можно отметить карту растительности СССР в масштабе 1 10 000 000 и Азии в масштабе 1 17 000 000, карту Африки к югу от тропика Рака в масштабе 1 10 000 000 Кейя (R. Кеау, 1959), лесную карту Канады в масштабе 1:6336000 Холидея (Halliday, 1937) и др. h
1 Издана также карта растительности СССР под редакцией Е. М. Лав-.ренко и В. Б. Сочавы в масштабе 1 : 4 000 000.
Отметим здесь, что, к сожалению, и в других местах своей книги Лем$ (недостаточно использует советскую литературу. Между тем советская биогеографическая школа имеет большие научные достижения, обладает хорошими 'Традициями и в силу социально-экономических преимуществ нашего строя более тесно связана с народнохозяйственной практикой. Научные расхождения -с франко-швейцарской школой, представителем которой является Леме, связаны с такими понятиями биогеографии, как объем предмета исследований, •объем основных единиц растительного покрова — формаций, ассоциаций, типов растительности, а также соотношение растительных сообществ с сообществами животных, сущность антропогенного воздействия на среду, и неко--торыми другими.
Более подробно с точкой зрения советских биогеографов читатель может ознакомиться-, в частности, в книге А. Г. Воронова «Биогеография с основами ’биологии», М., 1963 (2 изд.— 1974 г.) и в труде П. П. Второва и Н. Н. Дроздова «Биогеография материков», М., 1974. — Прим, ред.
Часть четвертая.
Крупные биологические формации земного шара
Предыдущие части работы были посвящены рассмотрению факторов размещения живых организмов — географического, стадиального и по сообществам. Теперь необходимо показать, как сообщества в качестве биологических сущностей распределены в биосфере.
Мы остановимся только па сообществах, основанных на физиономических и структурных критериях, — формациях и типах растительности фитогеографов, биомах американских биогеографов. К тому же из этих сообществ мы сможем описать только основные совокупности, находящиеся в равновесии с климатическими условиями, к которым ведет естественная эволюция сообществ организмов и которые являются наиболее заметным выражением совокупности региональных географических факторов. Мы ограничимся тем, что определим их основные структурные, экологические и трофические свойства с единственной целью показать деятельность биогеографов в ее наиболее обобщенном виде. Мы последовательно рассмотрим формации морских и пресных вод, а также наземных сред.
Глава 10
Водные среды и сообщества
Общие свойства водных сред
Огромные различия в физико-химических свойствах, которые существуют между наземными и водными средами, создают в них совершенно разные условия жизни. Из основных свойств водной среды мы остановимся на тех, которые имеют особое экологическое значение.
Плотность, вязкость, давление. Плотность чистой воды при 0° и давлении 760 мм рт. ст. в 775 раз больше, чем плотность воздуха при тех же условиях, что значительно снижает расход энергии, затрачиваемой организмом, на создание опорных тканей и на то, чтобы выдержать собственный вес. Плотность живой материи составляет в среднем 1,03, то есть примерно равна плотности морской воды близ поверхности.
Разница в плотности воды и воздуха обусловливает большую разницу между вариациями давления по вертикали в обеих средах. Давление, испытываемое водными организмами, возрастает на 1 атм на каждые 10 м глубины.
Вязкость воды примерно в 60 раз выше вязкости воздуха при той же температуре. Поэтому вода оказывает гораздо большее сопротивление передвижению животных.
Проникновение солнечной радиации. Судьба солнечной радиации, падающей на поверхность воды, различна. Часть ее, становящаяся значительной при больших углах падения, отражается. Та часть, которая проникает в воду, частично поглощается и рассеивается. Степень проникновения солнечной радиации — выражение прозрачности — варьирует в зависимости от веществ, растворенных и взвешенных в воде. Она уменьшается по мере увеличения глубины согласно логарифмической функции I = IQe-h\ где / — интенсивность радиации на глубине /г, /о — интенсивность на поверхности, в — основание натуральных логарифмов (2,7), a k — коэффициент ослабления света.
Этот коэффициент варьирует в зависимости от длины волны. Инфракрасное излучение полностью поглощается на глубине 1 м. Максимум проникновения меняется в зависимости от водоемов: смещенный в сторону коротких волн в чистых водоемах, он отклоняется в сторону длинных волн в водоемах, богатых органическими веществами. Эти различия находят свое выражение в большом разнообразии цвета водоемов от голубого до темно-коричневого. Интенсивность освещения и спектральный состав
175-
света, проникающего на различные глубины, имеют огромное биологическое значение.
Распределение тепла. Источником тепла в скоплениях воды служит энергия солнечной радиации. Причиной охлаждения яв-ляется собственное излучение этих скоплений и испарение с их поверхности. Эти вариации затрагивали бы только незначителЬ' ный слой вследствие слабого проникновения радиации и большой удельной теплоты воды, если бы не было перемешивания, осуществляемого под воздействием вертикальных различий плотности течениями, ветром и конвекционными токами. В глубоких стоячих водоемах устанавливается термическая стратификация,
Рис. 48. Изменения по вертикали температуры и содержания кислорода в олиготрофном озере (оз. Констанс, по Ауербаху и др. Auerbach et al.) и эвтрофном озере (оз. Дженива, Висконсин, по Ьерджу и Джудей, Burge et Juday).
которая меняется по сезонам и варьирует в зависимости от климатов. Так, глубокие озера областей умеренного климата обладают годичным температурным циклом. Летом различают (рис. 48):
1) верхнюю зону различной толщины с температурой постояН’ ной и высокой вследствие перемешивания воды. Это слой перемешивания, или эпилимнион;
2) зону, где температура быстро понижается, называемую слоем скачка, термоклином, или металимнионом;
3) глубинную зону с постоянной температурой 4 °C, соответствующей максимальной плотности пресной воды; это глубинный слой, или гиполимнион.
Осенью похолодание вызывает выравнивание температур около 4 °C, сопровождаемое общим перемешиванием, продолжительность которого зависит от метеорологических условий и массы воды. Устанавливается зимняя ярусность, которая в случае замерзания поверхности вызывает стабилизацию вертикального градиента температуры: зимняя стагнация (застой воды). Вес
176
ной при нагревании поверхности происходит новое перемешивание. Таким образом, имеется двукратное чередование стагнации и обновления. В тропических районах разница температуры между эпилимнионом и гиполимнионом составляет всего несколько градусов, и перемешивание бывает частым или нерегулярным. В арктических и альпийских областях, где температура едва превышает 4°, имеется только один летний период циркуляции. В глубине некоторых озер вода бывает более богатой растворенными солями, и большая плотность, возникающая при этом,
Р и с. 49. Вертикальное распределение температур в центре Северной Атлантики (по Кларку, Clarke).
Сезонные изменения близ поверхности указаны для следующих месяцев: Л —апрель, Б —август, В —декабрь,
Г —февраль.
делает невозможным перемешивание. Эти озера были названы меромиктическими в противоположность озерам с полным сезонным перемешиванием, называемым голомиктическими.
Океаны также обладают термической стратификацией (рис. 49). В них различают:
1) верхний слой, подверженный перемешиванию благодаря вертикальным конвекционным токам и течениям, обусловленным Доминирующими ветрами, — это термосфера, или тропосфера моря. Температура в этом слое подвержена более или менее значительным сезонным флуктуациям в зависимости от климатических условий. В низких широтах и в теплое время года в средних широтах существует поверхностный теплый слой с довольно
177
единообразной температурой. Толщина этого слоя варьирует в зависимости от подвижности воды и обычно составляет 10— 30 м, но в тропических областях может превышать 100 м. Переход к холодным водам с постоянной температурой устанавливается под этим слоем на глубине порядка 300—500 м — это слой скачка, или термоклин. В морях умеренного климата зимнее охлаждение уничтожает этот термоклин. В полярных водах нет отчетливой термосферы — температура на всех глубинах остается близкой к 0°;
2) переходный слой, в котором температура, не испытывая сезонных изменений, равномерно и медленно понижается до глубины порядка 1500 м; это слой постоянного термоклина;
3) глубинный слой с относительно постоянной температурой от 3 до 4° в низких и средних широтах и несколько ниже 0° в полярных областях. На больших глубинах наблюдается небольшое повышение температуры адиабатического характера, вызванное сжатием воды.
На небольших расстояниях по горизонтали наблюдаются значительные различия в температуре морской воды, причиной этого могут быть следующие обстоятельства: теплые или холодные течения приводят в соприкосновение массы воды, которые не смешиваются вследствие своей температуры и различной солености; на континентальной платформе температура прибрежных вод океанов и эпиконтинентальных морей подвержена вариациям из-за незначительной и изменчивой глубины, прибрежных течений, приливов и отливов, впадения речных вод; подводный рельеф может разделять воды различной температуры, например, по обе стороны отмелей или порогов; таков порог Вивли-Томпсона в Северной Атлантике, к югу от которого простираются воды умеренной температуры (7° на глубине 1000 м), а к северу — холодные воды (менее 0° на глубине 1000 м). Эти резкие изменения температур образуют термические барьеры распределения организмов.
Растворенные газы. Процентное содержание в воде углекислого газа и кислорода меняется в противоположном направлении под воздействием процессов фотосинтеза и дыхания. В озерах воды эпилимниона, в которые проникает свет, насыщены (а иногда даже перенасыщены) кислородом в результате ассимиляции СО2. В гиполимнионе, где осуществляется только дыхательное окисление, концентрация СО2 держится на максимально высоком уровне, возможном при равновесии с содержанием бикарбоната, и в придонных водах, бедных известью, бывает даже в избытке (рис. 50). Концентрация кислорода, растворенного в гиполимнионе, зависит от того, насколько последний богат организмами (рис. 50): эта концентрация остается одинаковой на всех уровнях в бедных, олиготрофных озерах, но становится очень низкой — ниже термоклина — в высокопродуктивных, эвтрофных озерах. Сезонные изменения перемешивания воды влияют
178
на ее состав: весеннее и осеннее обновление заставляют воду, богатою кислородом, проникать в глубинные слои (до самого дна в голомиктических озерах). Температура воды в гиполимнионе также влияет на ее насыщенность кислородом, особенно посредством влияния на дыхание; так, в глубинах озер жарких стран кислорода почти нет.
Подобные же явления имеют место в океанах, где кислород, выделяемый водорослями, насыщает воду до глубины, зависящей от прозрачности воды. Поверхностное волнение и перемешивание обогащают поверхностные воды также и атмосферным кис-
Р и с. 50. Химическая стратификация вод эвтрофного меромиктического озера (Кроттен, Австрия) и олиготрофного голомик-тического озера (Грундль, Австрия) (по Кольраушу, Kohlrausch).
лородом. Его содержание уменьшается до постоянного термоклина, где оно достигает минимальной величины, различной в разных широтах: в тропической зоне оно часто бывает меньше 3 мл на 1 л, а иногда становится равным нулю, как в открытом море у берегов Аравии и Нижней Калифорнии. Содержание кислорода увеличивается по мере приближения к полюсам, где воды становятся все более и более холодными. В глубинных водах, глубже 1000 и до 1500 м, концентрация кислорода повышается До новой постоянной величины. Это повышение является, по-ви-Димому, следствием медленных холодных течений с океанической поверхности высоких широт (рис. 51). Что касается переходной зоны с минимальной концентрацией кислорода, то она, по-видимому, обязана своим происхождением разложению мертвых планктонных организмов, опускающихся с поверхности и скапливающихся на тех глубинах, где плотность воды становится
179
равной их плотности. Причиной полного отсутствия кислорода могут быть особые обстоятельства. Таков классический пример-Черного моря. Начиная с глубины 150 м и до самого дна (до-2200 м) вертикальному перемешиванию препятствуют значительные поступления на поверхность речных пресных вод, а обмен с глубинными водами Средиземного моря осуществляется только* через Босфорский порог, так что органические вещества скапливаются в глубине, где происходит их анаэробное разложение с выделением H2S.
Тотальная система СО2 океанических вод состоит главным образом из ионов СО3Н“ и СО3“.Этот тотальный СО2, как и
Рис. 51. Разрез Атлантического океана с севера на юг, показывающий распределение кислорода (в мл на 1 л) в воде и его связь с глубинными течениями, обозначенными стрелками (по Свердрупу, Sverdrup).
в озерах, содержится в незначительном, но колеблющемся количестве в водах эуфотической зоны и в больших количествах в афотической зоне.
Растворенные соли. Это признак, по которому пресные воды противопоставляются морским. Первые содержат очень мало минеральных веществ переменного, варьирующего состава, а вто^ рые характеризуются их высоким содержанием и постоянным составом.
1) Пресные воды содержат разное количество растворенных солей. Так, озера известняковых областей могут содержать до 800 мг сухого осадка на 1 л, а озера областей, богатых кремнием, менее 50 мг. Преобладающими анионами являются СО3Н_, затем SO4“, С1~; нитратов и фосфатов очень мало. В пресной воде присутствуют катионы Са++, Mg++, К+, Na+, Fe++ и следы других элементов. В тропических водах довольно много кремния. В водах, богатых кальцием, зеленые растения, поглощая при фотосинтезе углекислый газ, вызывают отложение извести. Таково происхождение туфа ручьев, кальциевых озерных илов и кальциевых включений живых растений Chara, Cyanophyceae, Cladophora.
180
В водах, содержащих кислород, имеются нитраты различного происхождения. Их количество частично зависит от активности автотрофных растений, поэтому в эвтрофных озерах нитраты могут исчезать летом из верхней части эпилимниона. В этих же озерах лишенный летом кислорода гиполимнион постепенно теряет нитраты и в то же время накапливает аммиак, образующийся в результате разложения органических остатков.
Неорганический фосфор содержится в этих водах в ничтожном количестве: часто менее 1 мг на 1 л. Это объясняется, как и в случае нитратов, быстрым поглощением неорганического фосфора растениями.
2) Морские воды. В открытом океане средняя концентрация растворенных солей равна 3,5%, но она бывает выше в тропических водах, подверженных сильному испарению, и ниже в полярных водах, обогащенных летом водой тающих льдов. Что касается эпиконтинентальных морей, то концентрация солей в них может быть весьма различной — более высокой в морях с интенсивным испарением, таких, как Средиземное и Красное моря> и более низкой в морях с низким уровнем испарения и значительным поступлением пресных вод, как Балтийское море. Соленость также бывает пониженной близ устьев крупных рек (см. рис. 52).
Средний состав морской воды:
СГ 19°/оо Na+ Ю,6<7оо
SOJ- 2,7 Mg** 1,3
СО3Н” 0,14 Са** 0,4
Вг- 0,064 К* 0,38
Количество малоподвижных элементов — железа, меди, марганца, бора и т. д. — довольно постоянно. Содержание кремния, необходимого диатомеям и многочисленным простейшим, различно: его бывает больше на литорали и особенно в глубинных водах (до 7 г на 1 м3), чем на поверхности, где диатомеи фиксируют его в значительных количествах и вызывают циклические изменения его концентрации, связанные с обилием диатомей (рис. 53).
Минеральные азот и фосфор, имеющие огромное значение для организмов, подвержены значительным пространственным и сезонным вариациям. Наиболее низкая и наиболее изменчивая концентрация нитратного азота и фосфатов наблюдается в верхнем слое, где значительное количество этих веществ метаболизируется в период массового размножения планктона (рис. 53). Концентрация нитратного азота и фосфатов, напротив, остается высокой в бедных организмами глубинных водах, где мертвые организмы планктона разлагаются, высвобождая азот и фосфор в минеральной форме.
181
Р и с. 52. Распределение солености поверхностных морских вод в Атлантическом океане и близ берегов Африки и Южной Америки (по Кларку, G. L. Clarke).
Неживые органические вещества. Органические вещества, растворенные в воде и частично в коллоидальном состоянии, представлены в различных и иногда существенных количествах. В олиготрофных озерах их бывает меньше, чем в эвтрофных, но все же их масса в несколько раз превосходит массу планктона. Эти органические вещества имеют большое биологическое значение из-за содержащихся в них азота и фосфора, а также из-за гормональных, витаминных и токсических свойств, которыми они могут обладать по отношению к живым организмам. Среднее содержание органического фосфора в озерах составляет 0,05 мг на 1 л, в океане — 0,01, а органического азота 0,5 мг на 1 л в озерах и 0,2 — в океане.
Рис. 53. Изменение содержания нитратного азота (N), фосфатов (Р) и кремния (Si) в водах Ла-Манша на глубине от 0 до 25 м с конца 1930 до начала 1932 г. (по Куперу, Cooper).
Вода в некоторых озерах и прудах, захваченных торфяными формациями, содержит значительное количество бурых гуминовых веществ: это дистрофные озера (фото II,А).
pH. Реакция пресных вод полностью зависит от соотношения карбонаты — СО2. Бикарбонаты в результате гидролиза образуют ионы ОН-, угольная кислота в результате диссоциации высвобождает ионы Н+. Воды, лишенные извести, — кислые, и их кислотность варьирует в зависимости от концентрации растворенного СО2. Известковые воды — щелочные, их pH обычно лежит между 7 и 8,5. Поскольку бикарбонат кальция является солью, образованной слабой кислотой и сильным основанием, изменения концентрации этой кислоты имеют крайне незначительное влияние на pH; известковые воды обладают буферностью, уровень которой зависит от количества бикарбоната. Щелочная морская вода представляет собой стойкую единообразную буферную систему: на ее поверхности pH колеблется от 8 до 8,3.
183
Помимо химического состава воды, ее температуры и существующего на больших глубинах давления, на pH влияет также биологическая деятельность организмов, изменяющая содержание СО2. В эуфотической зоне pH повышается днем в результате поглощения СО2 в процессе фотосинтеза и снижается ночью в процессе дыхания. Суточные изменения, однако, более значительны в слабо минерализованных, чем в соленых или известковых водах, образующих буферную систему. В озерах содержание СО2 в металимнионе повышается одновременно с понижением pH. В гиполимнионе содержание СО2 и pH обычно постоянно. В океанах обогащение углекислым газом промежуточных слоев ведет к понижению pH, которое достигает таким образом в Тихом океане 7,5 на глубине 800 м. На больших глубинах меньшая концентрация СО2 вызывает возрастание pH примерно до 8.
Крупные экологические группы водных организмов. Как и наземные, водные среды дают большое разнообразие условий местообитания, которое позволяет существовать разнообразным жизненным формам, образ жизни которых различен. Была разработана классификация жизненных форм, применимая ко всем достаточно крупным скоплениям воды.
а) Бентос включает организмы, локализованные на дне — сидячие, передвигающиеся по донной поверхности или зарывающиеся в субстрат. Следовательно, бентос сам является сложным комплексом, состоящим из различных биотопов1:
1) укорененные растительные организмы, которые одновременно проникают в свободную воду и в субстрат; в цепях энергии и вещества экосистемы являются продуцентами и локализуются в эуфотических зонах;
2) организмы, именуемые сидячими, часто микроскопические, прикрепленные к субстрату, но не проникающие в него. В немецком языке для них существует трудно переводимое наименование Aufwuchs. Растения и животные различного систематического положения, составляющие эту группу, прикрепляются либо к неживой опоре, органической или неорганической, либо к живЫхМ растениям. Последним авторы, пользующиеся английским языком, присвоили наименование перифитон (periphyton);
3) свободно живущие организмы, ползающие по дну или плавающие у дна. Одни из них очень малоподвижны — оседлые, се-дентарные, другие — вагильные, способные к значительным перемещениям, как ракообразные и рыбы;
4) организмы, живущие в донных отложениях. Эти отложения, обозначаемые общим названием 003 (ooze), имеют самый различный состав и происхождение и дают приют многочислен-
1 Вряд ли бептос, как и входящие в него группы организмов, можно назвать биотопом. Под последним понимают совокупность неживых компонентов биогеоценоза — почвы, рельефа, микроклимата и прочее. Совокупность биотопов морского дна носит название бенталь.— Прим. ред.
184
Рис. 54. Некоторые виды озерного и морского планктона.
a — Asterionella formosa (озерная диатомея); б —Planktonic На sol и в — Corethron hystrix (морские диатомеи); г — Ceratium hirundlnella (озерная Dinofiagellata); д — Ornithocerus splen-didus (морская динофлагеллята); е -— Thalaisicola pelagica (радиолярия); ж — Janthlna gracilis (морское переднежаберное Prosobranchia) со слизистым выростом, несущим яйца; з — То-mopteris oniscoformis (морская полихета); и —личинка „плу-теус“ морского ежа; к — Pleurobrachla (ктенофора).
ним микроорганизмам — редуцентам и хемоавтотрофам, а также, когда насыщение кислородом бывает достаточным, роющим животным.
б) Планктон состоит из живых организмов, свободно живущих в воде и пассивно перемещающихся в ней. В действительности, планктонные животные совершают плавательные движения, но эти движения обеспечивают им лишь незначительные по амплитуде, чаще всего вертикальные, перемещения и не позволяют противостоять течениям. Поскольку в состав планктона входят многочисленные зоологические группы (зоопланктон) и некоторые растительные группы, в частности водоросли
185
(фитопланктон), планктон включает особи самых разных размеров. Часть его, весьма значительную в весовом отношении, составляют микроорганизмы, не улавливаемые самыми тонкими ситами и сетями: это нанопланктон, состоящий из бактерий, простейших и очень мелких диатомей. Глубина погружения планктонных видов зависит от различных факторов. Освальд (W. Ostwald) в 1902 г. предложил следующую формулу для этого явления:
излишек веса
Глубина погружения =------------------------.
J сопротивление поверхности X вязкость воды
Числитель представляет собой разницу между плотностью плавающего тела и плотностью окружающей среды, умноженную на объем этого тела. У некоторых форм вакуолизация, наличие полостей, заполненных газом, студневидная оболочка или развитие студенистых тканей ведут к снижению излишка веса; известковый панцирь ракообразных, раковина Gastropoda очень тонкие или отсутствуют. Сопротивление поверхностей определяется отношением общей поверхности к объему и площадью ортогональной проекции тела на плоскость, перпендикулярную его траектории. Это сопротивление увеличивается дисковидно уплощенной и палочкообразно удлиненной формой, наличием отростков, крыловидных выростов и шипов. Подобные приспособления иногда осуществляются при образовании колоний с большой относительной поверхностью. На рис. 54 приведены примеры различных планктонных форм.
в) Нектон объединяет виды, обладающие достаточной способностью к перемещению для того, чтобы организмы не зависели от течений. Он объединяет главным образом рыб, но также и млекопитающих (китов, кашалотов, дельфинов и т. д.), черепах, ракообразных. Совокупность планктона и нектона противопоставляют организмам бентоса и называют пелагическими организмами.
г) Нейстон объединяет организмы, плавающие или перемещающиеся на контакте воздушной среды с водной поверхностью. Он состоит главным образом из насекомых и одноклеточных организмов и встречается в основном в пресных водах.
Население морей и океанов
Зональность биоценозов
Биологическое население морской среды распределено по зонам в зависимости от глубины вод и от дна. Неглубокие районы образуют неритическую провинцию, или зону. Они расположены на континентальной платформе, край которой, отмеченный, как правило, обрывом склона, находится обычно на глубине 200 м. Особенности воды здесь объясняются притоком вод с материка,
186
волнениями и незначительной глубиной. Характер дна различи пый, а население обильно и разнообразно. Области открытого моря образуют океаническую провинцию, или зону. Воды этой зоны довольно постоянны по своему физико-химическому составу, бедны живыми организмами и прозрачны.
Каждая из этих провинций состоит из бентической и пелагической областей.
Вертикальная зональность бентической области. Существует множество классификаций этой зональности. Классификация Пере и Пикара (Peres et Picard), базирующаяся на основных экологических признаках, а не на батиметрических критериях, имеет определенное преимущество. Следуя ей, выделяют основные биологические совокупности, свойственные всем морям земного шара. Эти авторы различают ряд зон.
1. Супралиторалъ. Зона заливается только в исключительных случаях, но часто увлажняется соленой водой (фото III,А).
2. Медиолитораль. Организмы этой зоны более или менее часто и довольно длительное время оказываются в надводном положении. В основном это приливо-отливная зона (фото III, Б) или «zone intertidale» (заливаемая приливом береговая полоса; английский термин, переделанный на французский лад). Ниже лежащие слои, обнажающиеся лишь в исключительных случаях, имеют тот же состав флоры и фауны, что и более глубокие, никогда не выступающие из-под воды уровни, примыкающие к ним !.
3. Инфралитораль. Зона никогда не выступает из-под воды или выступает лишь в исключительных случаях, но освещенность бывает достаточной для того, чтобы обеспечить существование солнцелюбивых водорослей и семенных семейства Zosteraceae. Нижняя граница этой зоны колеблется от 15 до 80 м в зависимости от прозрачности воды.
4. Окололиторалъная зона. Охватывает зону слабой освещенности, совместимой с существованием водорослей, наиболее толерантных к этому фактору. Эта зона простирается до края континентальной платформы (континентального шельфа) и вполне соответствует верхней неритической зоне некоторых океанологов.
5. Батиальная зона занимает континентальный склон и отлогие склоны, которые у его основания могут лежать выше 2000— 3000 м в зависимости от исследуемых районов и применяемых критериев. Для Бруна (A. F. Bruun) это изотерма 4 °C, Пере предпочитает считать границей резкую смену фауны, которая встречается на этих глубинах.
1 В других системах, где определяющим фактором является амплитуда наиболее мощных приливов и отливов, дело обстоит иначе. Так, Стефенсон (Т. A. Stephenson) включает в состав медиолиторальной зоны под названием инфралиторальной полосы участки, выступающие из-под воды в исключительных случаях.
187
6. Абиссальная зона включает население глубинных равнин, К}да не проникают виды, свойственные континентальной платформе.
7. Ультраабиссальная, или хабальная, зона. Очень бедная зона, включающая подводные впадины, расположенная на глубине 6000—7000 м.
Пере объединяет первые четыре зоны в литоральный, или фатальный, комплекс (по наличию в нем хлорофиллоносных растений), а три последние — в глубинный, или афитальный, комплекс. Эти два комплекса соответствуют эуфотической и афотической зонам.
Пелагическая область. Обладает вертикальной ярус-ностыо биоценозов, параллельной ярусности населения бентоса.
1. Эпипелагиаль, зона, нижняя граница которой соответствует минимальной освещенности, обеспечивающей фотосинтез, и представляет собой эуфотическую зону.
2. Мезопелагиаль, зона, в которую не проникает свет, но которая подвержена перемешиванию и сезонным изменениям температуры. Ее нижняя граница проходит в среднем на уровне около 200 м.
3. Инфрапелагиаль, зона от 200 до 500—600 м, затрагиваемая суточными вертикальными миграциями поверхностного планктона.
4. Батипелагиаль, зона от 500—600 м до 2000—2500 м, нижняя граница которой в средних широтах соответствует изотерме 4 °C.
5. Абиссо-пелагиаль, зона от 2000—2500 м до 6000—7000 м.
6. Хадопелагиаль, зона больших глубин.
Жизнь бентоса
Общая характеристика. Таксономическое разнообразие и плотность индивидуумов, очень высокие в медиолиторальной зоне, быстро снижаются с глубиной. Сначала в эуфотической зоне убывает количество и разнообразие водорослей, а затем — на больших глубинах — количество и разнообразие животных. Биомасса на скалистом берегу Калифорнии на глубине 1,5 м со« ставляет 4800 г/м2 (из них 4670 приходится на водоросли) и всего 980 г/м2 (из них 600 приходится на водоросли) на глубине 22 м. Петерсен (Petersen) наблюдал в норвежском фьорде на глубине 2 м сообщество, богатое Mytilus edulis, с биомассой около 19 кг/м2 (сырого веса), в то время как на глубине 320 м на илистом дне биомасса падает до 20 г. Уменьшение биомассы продолжается в батиальной и абиссальной зоне, для последней она составляет в среднем 1 г/м2 в Тихом и 0,7 г/м2 в Атлантическом океане. В тропических областях абиссальные зоны еще беднее, а большие впадины Средиземного моря почти необитаемы.
188
Эта высокая в верхних слоях плотность организмов выдвигает для прикрепленных организмов проблему свободного места на скалистых субстратах. Если эти субстраты оголены, на них возникает сукцессия смешанных популяций водорослей и животных, которая быстро обусловливает полное заселение.
Прикрепленные и неподвижные виды иногда сами несут на себе другие виды — водоросли, серпулы (Serpula), альционарии (Alcionaria) и т. д.
Ярко выраженная борьба за сбор пищи также является одним из следствий высокой плотности литорального бентоса. Она обусловливает, в частности, локализацию и обилие сидячих и некоторых роющих форм, которые подстерегают добычу или привлекают ее к себе. Большинство таких форм имеет приспособления для захвата и сбора: щупальца актиний, нематоциты Cnida-ria, липкую слизь, мерцательные реснички. Мшанки и неподвижные аннелиды, Serpula и Sabella, асцидии, пластинчатожаберные моллюски собирают пищу благодаря движению ресничек при помощи фильтрующей системы, которая позволяет извлечь взвешенные частицы и направить их в пищеварительную полость. По Дейлу (Dales), мидия, устрица, асцидия (Ciona intestinalis) могут профильтровать за 1 час количество воды в 50—100 раз большее, чем их собственный объем. Сердцевидки (Cardiuni edule), которые живут в песке, но сообщаются с поверхностью при помощи сифона, аккумулируют большое количество взвешенных частиц при помощи собирающего сифона и вызывают, таким образом, благодаря своим отбросам в случае высокой плотности этих организмов значительное повышение дна. Вследствие таксономического и этологического разнообразия населения бентосные сообщества образуют сложные экосистемы. Продуцентами главным образом являются водоросли (Chlorophyceae, Pheophyceae, Rhodophyceae, Diatomeae) и в значительно меньшей степени Cyanophyceae и покрытосеменные семейства Zoste-гасеае. В противоположность распространенному мнению, не существует четкой вертикальной зональности морских водорослей в зависимости от их добавочных пигментов, хотя эти пигменты в значительной степени модифицируют спектр поглощения солнечной энергии. Напротив, границы их толерантности по отношению к свету сильно различаются, и можно выделить фото-фильные и сциофильные водоросли с сильно варьирующей амплитудой толерантности. Наконец, сильная освещенность плохо переносится большинством водорослей, которые, впрочем, более или менее защищены от нее наличием обильной флоры эпифитов на их талломах 1. На скалистых субстратах в приливо-отливной зоне растительность, по данным Одума (Е. Р. Odum), вырабатывает от 0,5 до 1,5 г хлорофилла на 1 м2, то есть примерно столько,
Если бентосные морские бактерии обильны, то автотрофные формы их, по-видимому, играют несущественную роль, за исключением Rhodothiobacterium.
189
сколько дают луга и леса областей умеренного климата. То же относится и к чистой первичной продукции, которая колеблется (исключая коралловые рифы) между 0,5 и 3 г на 1 м2 в день.
Консументы бентоса имеют различные источники питания. Фитофаги, локализованные в неритических зонах, главным образом представлены брюхоногими моллюсками и морскими ежами. Одни, например литорины и Echinus, ощипывают крупные растения, другие — морские блюдца, хитоны, различные морские ежи (фото III, Б) — поедают микроскопические водоросли на поверхности субстрата и растений.
Хищники представлены в основном морскими звездами, которые поедают главным образом брюхоногих моллюсков, таких, как Мигех и Ригрига\ последние питаются другими брюхоногими, просверливая их раковины, ракообразными, Pycnogonidae, донными рыбами (скатами, камбаловыми и т. д.).
Редуценты представлены многочисленными бактериями в свободных водах, но больше всего их в верхнем (несколько сантиметров) слое морских отложений. Плотность бактерий, очень высокая на поверхности отложений, быстро уменьшается в глубине их, в то время как количество анаэробных бактерий возрастает (Zo Bell). Они разлагают органические вещества мертвых бентических или планктонных организмов.
Наконец, многие организмы питаются живыми и неживыми органическими частицами, взвешенными в воде, образующими пленку на поверхности дна или находящимися в толще отложений. Мы приводили выше пример сидячих и роющих форм, которые фильтруют морскую воду, для того чтобы задержать взвешенные частицы и микроорганизмы. Моллюски Scrobicularia и Turritella, полихеты из аннелид, такие Echinidae, как Echinocar-dium, офиуры, такие ракообразные, как Corophium, являются роющими формами, которые собирают детрит и микроорганизмы с донной поверхностной пленки. Многие роющие виды, так называемые илоядные формы (limivora), поглощают отложения, переваривая содержащиеся в них организмы и органические остатки. К этой группе относятся аннелиды, сипункулиды, голотурии, иглокожие.
Глубинный бентос, в котором отсутствуют организмы, способные к фотосинтезу, получает органические молекулы из двух источников:
1) медленно оседающие трупы пелагического планктона и нектона, большая часть которых используется бактериями частично во время оседания, а частично на дне;
2) автотрофные, способные к хемосинтезу бактерии, нитрифицирующие бактерии, или сульфобактерии. Бактерии больших глубин, приспособленные к царящим на данных глубинах большим давлениям, активны только при этих давлениях. Они составляют значительную долю в питании сидячих и роющих организмов глубинного бентоса, плотность которого, по-видимому,
190
связана с обилием «дождя» органических остатков и бактерий.
Разнообразие бентических биоценозов1. Существенные изменения особенностей морской среды по вертикали обусловливают зональность биоценозов, рамки которой были уточнены выше. Но для одного и того же уровня особенности субстрата — рыхлого или скалистого — обусловливают образование весьма различных сообществ. В меньшей степени модифицируют состав живого населения морской среды волнение воды, поступление материковых вод и терригенных осадков. Несмотря на таксономическое разнообразие, обусловленное древней и современной изоляцией океанов и наличием больших климатических зон земного шара, биоценозы на одной и той же глубине и одинаковом субстрате обладают весьма сходной структурой и организацией.
1. С у п р а л и т о р а л ь н а я зона. На скалистых берегах она заселена Cyanophyceac, лишайниками, морскими гастроподами, устойчивыми к длительному пребыванию вне воды, равноногими ракообразными (Ligia) и десятиногими ракообразными (крабы тропических областей) (фото III, А).
На песчаных берегах скопление растительных остатков на этом уровне обеспечивает питание обильной фауны, связанной с детритом. Это ракообразные, приспособленные к жизни в воздушной среде: Amphipoda (Talitrus, Orchcstia и др.), Isopoda, крабы (Ocypode, Ucd). Здесь же встречаются многочисленные детритофильные и хищные жуки.
Илы эстуариев и низких защищенных берегов заселены ассоциациями галофильных покрытосеменных.
2. Me д иол и тор ал ьн а я зона. Включает приливо-отливную зону (за исключением нижней части, выступающей из-под воды на очень короткое время). Представленные здесь виды распространены в зависимости от их способности выдерживать отсутствие воды или избегать его.
На скалистых субстратах вертикальное разнообразие горизонтов сильнее всего выражено на берегах с большой амплитудой приливов и отливов, но можно различить верхнюю подзону, часто выступающую из-под воды, для которой характерны термические вариации, прямое воздействие солнечных лучей, высушивание, вариации солености. Продуценты здесь представлены Cyanophyceac, Chlorophyceae, несколькими Pheophyceae, такими, как Pelvetia canaliculate. Наиболее характерными сидячими животными являются Cirripe-dia рода Chtamalus, подвижные животные представлены в основном Gastropoda: Patella, Littorina, Trochus (фото 1ПБ). Нижняя подзона в морях с небольшими приливами и отливами характеризуется преобладанием бурых водорослей, Melobesia, которые или просто покрывают скалы, или образуют настоящее сооружение, как карниз или тротуар, из Lithophyllutn побережий Средиземного моря. На берегах с сильными приливами и отливами различают пояса водорослей, в которых обычно первенствуют Pheophyceae; так, на скалистых берегах Западной Европы в медиолиторальной зоне сверху вниз представлены
1 В этом обзоре мы в значительной мере использовали работу Пере (J.-M. Peres), Oceanographie biologique et biologie marine, t I: La vie ben-thique.
191
Pelvetia canaliculata, Fucus spiralis, Ascophyllum nodosum (отсутствует на берегах с сильным волнением), Fucus vesiculosus, Fucus serratus, последний образует верхнюю часть инфралиторальной зоны. Именно у основания меднолиторальной зоны располагаются банки устриц и мидий.
Рыхлые субстраты, благодаря их свойству сохранять воду, позволяют животным, постоянно живущим или зарывающимся в их толщу, избежать выхода из воды и воздействия солнечных лучей. Помимо крупных видов, которые лишь временно зарываются в песок, таких, как иглокожие, ракообразные, рыбы, и видов, живущих в нем постоянно, таких, как аннелиды — полихеты, моллюски Gastropoda и Lamellibranchia, существует псаммофиль-ная фауна, состоящая из очень мелких видов, живущих исключительно в порах отложений и образующих оригинальные с таксономической точки зрения объединения: реснитчатые простейшие, коловратки, Gastrotricha, Archiannelides, нематоды, мелкие полихеты, веслоногие рачки и т. д. Размеры их меньше 1 мм, форма тела нитевидная. Их привлекает субстрат, и многим из них, как и их яйцам, позволяет прикрепляться к нему выделение слизи. Это формы большей частью слепые, однако они подвижны и питаются диатомеями, бактериями и органическим детритом. На большой глубине песок пропитан постоянной грунтовой водой различной солености и температуры и заселен видами, сходными с предыдущими по морфологическим и этологическим признакам.
Население илов, недостаточно насыщенных кислородом и состоящих из мельчайших частиц, которые закупоривают дыхательные пути и фильтрующие аппараты многочисленных роющих животных, отличается от населения чистых песков. Оно включает немногочисленные, но иногда довольно обильные виды. Для них характерны: аннелида (Nereis diversicolor), двустворчатые моллюски Муа arenaria и Scrobicularia (которая достигает плотности 500 на 1 м1 2), брюхоногий моллюск Hydrobia ulvae (до 6000 на 1 м2). Семенные укореняются в илах: травянистые растения сликке1 и шора2 в умеренных областях, деревья мангров в тропических областях (см. фото I).
Инфралитораль ная зона. Это самая верхняя из неосушаемых зон. Вследствие значительной освещенности она обладает высокой плотностью водорослей.
Наиболее характерными биоценозами скалистого дна этой зоны являются популяции светолюбивых бурых водорослей и известняковые образования, созданные колониальными животными. В состав первых входят такие крупные водоросли, как ламинарии, Lessonia, Macrocystis, талломы которых могут достигать более 10 м, Cystoseira, виды Fucus и Himanthalia в верхней части.
Формации фиксаторов извести образованы червями (Serpula), моллюсками (Vermetus) и в особенности мадрепоровыми кораллами, которые образуют коралловые рифы. Последние являются тропическими формациями, характерными для постоянно теплых, чистых, достаточно перемешиваемых вод с высокой соленостью. Участки, наиболее подверженные действию волн —
1 Сликке — приливная полоса, сильно увлажненная, часто с солеросом. — Прим. ред.
2 Шорре, шор — прибрежная полоса с галофильчой растительностью, заливаемая только при высоких приливах. — Прим. ред.
192
вершина склона рифа, «наветренная» поверхность рифа — состоят из массивных мадрепоровых кораллов, в то время как более глубокие участки внешнего склона, «подветренные» поверхности, склоны лагун слагаются из ветвистых форм. Эти кораллы служат приютом разнообразному и обильному населению: это свободноживущие водоросли, прикрепленные к мертвым частям постройки; зеленые водоросли, погруженные в известковый скелет полипов, симбиотические водорсли в тканях последних; Foraminifera, Alcyonaria, Bryozoa, Serpulidae, моллюски, которые способствуют укреплению рифа образованием извести; наконец, многочисленные животные, поселяющиеся в углублениях,— морские ежи, офиуры, аннелиды, ракообразные, рыбы. Этот весьма богатый биоценоз обладает высокой биомассой. По наблюдениям X. Т. Одума и Е. П. Одума (Н. Т. и Е. Р. Odum), на рифах Тихого океана она составляет примерно 700 г/м2 (сухого веса) для водорослей против 130 для растительноядных (включая полипы) и 10 для хищников. Очень высокая продукция составляет в год величину, в 12 раз превышающую биомассу. Она связана с повышенной общей фотосинтетической продукцией: 18 г на 1 м2 в день.
Рыхлые субстраты инфралиторальной зоны сильно различаются по богатству растительности. Встречаются густые «луга» из Zosteraceae (Zostera, Ро-sidonia, Cymodocea). Макроскопические водоросли, как правило, редки или отсутствуют, животные являются консументами и поглощают бактерий, диатомей, детрит, а также некоторые виды хищников. Петерсен (Petersen) описал многочисленные биоценозы днищ балтийских проливов.
Околол итора льн а я зона. Слабая освещенность благоприятствует здесь лишь сциофильным водорослям, часто встречаются Melobesia, известковые красные водоросли различного облика: на скалистом субстрате и рыхлом дне, укрепленном конкрециями, устанавливается так называемая «кораллогенная», образующая инкрустации, формация. На гравии и крупном песке, богатом детритом, встречаются густые популяции стоячих ветвистых Melobesia, известных в Ла-Манше под названием maerl1. Рыхлое дно занято различными, в зависимости от характера осадка, биоценозами.
Глубинные, или афитальные, зоны. Эти зоны, еще очень мало изученные, обедняются с глубиной.
В батиальной зоне Северной Атлантики твердые субстраты заняты биоценозом с крупными колониями мадрепоровых кораллов, с которыми связаны многочисленные прикрепленные и подвижные виды самых различных групп: губки, гидроиды, Alcyonaria, Crinoidea, Polychaeta, Bryozoa, моллюски, ракообразные, рыбы и т. д. Население илов состоит, как всегда, из роющих прикрепленных и подвижных форм, плотность которых, по Пере, непосредственно связана с количеством органических веществ, попадающих на дно.
Переход от батиальной к абиссальной зоне характеризуется изменением в видовом составе населения илов (твердые субстраты встречаются лишь в виде исключения), в котором представлены все крупные группы животных, свойственных рыхлому дну батиальной зоны, но биомасса становится очень низкой. По данным Филатовой, в северной части Тихого океана она состав-ляет от 0,01 до 1 г/м2.
1 Маёг!—«нуллипоровый песок». — Прим. ред.
8 Ж. Леме 193
Известно, что в хадальной зоне имеются голотурии, актинии, Echiuridae, полихеты, двустворчатые моллюски, Isopoda и Amphypoda из ракообразных, к ним надо прибавить популяцию барофильных бактерий, которые, несмотря на незначительную биомассу, равную, по Беллу, примерно 2 мг углерода на 1 м1 2, имеют огромное значение в цепях питания. Хищники на этих больших глубинах, по-видимому, отсутствуют.
В морях, сообщающихся с океанами через неглубокие пороги, афиталь-ный комплекс обеднен вследствие механического препятствия на пути обмена фауны и особого температурного режима вод. В Средиземном море, где температура глубинных вод близка к 13°, афитальный бентос беден, глубины Красного моря почти пустынны.
Фауна абиссалей, почти полностью эндемичная, обладает биологическими особенностями: это крупные размеры видов — часто они бывают гигантами для своей группы, — размеры, которые связаны, очевидно, с замедленным метаболизмом и с поздним половым созреванием; это развитие тактильных органов; это незначительное количество извести (хрупкие скелеты, тонкие панцири и раковины), а также уплощенная форма тела у подвижных форм.
Константность признаков глубоководной океанической среды и ее постоянство на протяжении геологических эпох обеспечили выживание довольно большого количества архаических форм: рыба целакант, недавно обнаруженные моллюски рода Neopilina, различные семейства десятиногих раков, известные из юрского периода, стебельчатые морские лилии, шестилучевые губки Hexactinellide 1.
Жизнь пелагиали
В ней можно различить четыре взаимосвязанных группы: фитопланктон, зоопланктон, бактерии и нектон.
Фитопланктон. 1. Состав. В состав фитопланктона входят Diatomeae, Dinoflagell ata, Silicof ligellata, Coccolithophoridae, некоторые Xanthophyceae и Cyanophyceae, а также бактерии и некоторые грибы, особое место в трофических цепях которых заставляет рассматривать их отдельно.
2. Распределение. Его вертикальное распределение, географическое распространение и сезонные изменения являются результатом многочисленных экологических факторов, которые влияют на размножение фитопланктона, поддержание его во взвешенном состоянии в эпипелагиальной зоне, его горизонтальные перемещения течениями и поедаемость животными2.
Размножение фитопланктона со скоростью деления клеток один раз в день зависит от лимитирующих факторов, его фото
1 Из этого не следует, что архаические формы отсутствуют в неритическом бентосе, где до сих пор, например, встречаются плеченогие Lingula и мечехвосты Limulus.
2 Были выведены формулы для расчета плотности фитопланктона по экологическим факторам. Наиболее полные формулы Рилея (Riley) учитывают комплекс вышеперечисленных факторов. Формулы Фрака (Fraca) применяются только к продукции с учетом световой энергии, температуры и отношения поверхности к объему рассматриваемых организмов.
194
синтеза и минерального питания. Свет играет решающую роль в вертикальном распределении фитопланктона, при котором плотность уменьшается параллельно освещенности до точки компенсации. Все же максимум продукции приходится не на поверхность, а на глубину, варьирующую в зависимости от прозрачности воды и интенсивности освещения, которое бывает сверхоптимальным у поверхности. Поэтому уровень наибольшей плотности находится на глубине, убывающей по мере продвижения от низких к высоким широтам и от открытого моря к берегам.
Рис. 55. Вертикальное распределение диатомей и бактерий в океанах (по Свердрупу, Sverdrup et coll.) (Л), и фотосинтез диатомовой водоросли Coscinodiscus excentricus в Ла-Манше (по Дженкину, Jenkin) (Б), сплошной линией показано распределение солнечной энергии. Шкала глубин для А и Б одна и та же.
Он лежит на глубине 100 м в Саргассовом море, на глубине 25 м в средних широтах и поднимается до десятка метров в богатых взвешенными частицами водах неритической зоны (рис. 55). Температура также влияет на чистую продукцию из-за своего двойного воздействия на процессы дыхания и фотосинтеза.
Как следует из вышеизложенного, среди основных химических элементов фитопланктона азот и фосфор являются главными лимитирующими элементами, а также, вероятно, кремний в случае диатомей и Silicoflagellata. Действительно, концентрация этих элементов в поверхностных слоях воды средних и высоких широт сильно уменьшается в периоды массового размножения фитопланктона (рис. 53) и остается постоянно низкой в тропических областях. Когда возобновление этих элементов обеспечено вертикальными перемещениями воды или поступлениями
8* 195
с континента, фитопланктон бывает особенно обильным. Подъемами глубинных вод, богатых минеральными элементами и связанных с течениями, объясняется обилие планктона в открытом море близ берегов Чили и Перу (течение Гумбольдта), у юго-западных берегов Африки (Бенгельское течение) и у берегов Калифорнии. Конвекционные токи, возникающие в результате охлаждения поверхности в областях с холодным сезоном, дают тот же эффект. Что касается поступления минеральных и органических веществ из рек, то возле устья эти поступления вызывают массовое размножение планктонных водорослей, такое, как, например, размножение диатомей летом в восточной части Средиземного моря в период паводка Нила или у берегов Норвегии во время таяния снегов.
Факторами, влияющими на уменьшение плотности фитопланктона, являются, помимо условий, неблагоприятных для его размножения, погружение фитопланктона в афотическую зону и поедание его животными. Несмотря на наличие приспособлений для парения, планктон имеет тенденцию к медленному погружению. Конечно, вертикальные подъемы воды, упоминавшиеся выше, снова выносят его на поверхность, но эти подъемы строго компенсируются эквивалентными погружениями. Что касается поедания фитопланктона хищниками, то этот вопрос будет рассмотрен в дальнейшем.
Складывающиеся в различных местах вариации факторов, воздействующих на обилие фитопланктона, являются причиной различий в его обилии. Сюда же следует добавить воздействие доминирующих ветров, которые перемещают к подветренным берегам поверхностные воды вместе с содержащимся в них планктоном.
3. Продукция. Количественная оценка продукции морского фитопланктона имеет большое экологическое и экономическое значение, принимая во внимание огромные площади, занятые экосистемами, первичную продуктивность которой он обеспечивает. Его биомасса очень изменчива, но всегда невелика. Она составляет 4 г сухого вещества на 1 м2 в Ла-Манше (Harvey), 16— в проливе Лонг-Айленд (Riley), но в пелагической области она гораздо ниже.
Эта плотность гораздо ниже плотности растений наземных экосистем. Что касается первичной продукции, то она сильно варьирует. Измерения, выполненные различными методами, дали крайне незначительные величины для тропической океанической провинции, воды которой очень бедны минеральными элементами (от 0,1 до 0,2 г С на 1 м2/день и менее 0,06 — в Атлантике; 0,5 г общей продукции сухого вещества на 1 м2/день в Саргассовом море) !. Сопоставление с высокой продуктивностью атоллов
1 Продуктивность по углероду была вычислена при помощи С14 Сорокиным и Кляшториным; общая первичная продукция оценивалась по выделению кислорода.
196
заставило Одума сравнить последние с «оазисами в пустыне». Наибольшее обилие питательных веществ в перемешиваемых течениями океанических водах сопровождается более высокой продукцией (от 0,25 до 0,5 г С на м1 2/день в Северной Атлантике и в зонах течений тропической Атлантики). Неритическая область с водами, богатыми питательными веществами, дает наиболее высокую и одновременно наиболее изменчивую первичную продукцию (от 0,8 до 3,7 г С на м2/день близ берегов Северной Атлантики; 3,2 г сухого веса общей продукции — в проливе Лонг-Айленд, 4,4 — в устье техасских эстуариев с зарегистрированным -максимумом, равным 23). Последние — величины того же порядка, что и в наземных экосистемах культур или естественной растительности в умеренных неаридных областях.
Зоопланктон. 1. Состав. Важно различать формы перманентно планктонные (голопланктон) и формы, которые входят в состав планктона только в определенный период своего развития (меропланктон); к первым относятся простейшие (форами-ниферы, радиолярии ресничные), кишечнополостные, Ctenaria, коловратки, Chaetognatha, Crustacea (особенно Copepoda), Tu-nicata, затем некоторые немертины, аннелиды и голотурии. В состав меропланктона входят личиночные стадии многих форм бентоса: кишечнополостных, немертин, мшанок, аннелид полихет, иглокожих, ракообразных, моллюсков, Enteropneusta.
Зоопланктон обладает различными приспособлениями к пелагическому образу жизни, среди которых наиболее оригинальны наполненные газом пузыри (Siphonophora) или слизистые выделения в форме плота (Janthina, рис. 54). Удельный вес уменьшается благодаря исключительно тонким раковинам и панцирям, •студенистому характеру тканей, которые у медуз содержат до ‘95% воды. Эпипелагические формы в большинстве случаев бывают прозрачными — невидимыми для хищников, в то время как формы афотической зоны более или менее ярко окрашены.
2. Вертикальное распределение. Распределение зоопланктона так же, как и распределение фитопланктона, характеризуется вертикальной стратификацией, а также горизонтальными и сезонными вариациями. В отличие от фитопланктона, зоопланктон доходит по вертикали до абиссали, однако плотность его гораздо выше в эпипелагиальной зоне, где локализованы живые водоросли, за счет которых он прямо или косвенно получает питательные вещества. Видовой состав зоопланктона меняется в зависимости от глубины; большинство видов локализовано в определенной зоне по вертикали в соответствии С их потребностями.
1 Известковые или кремневые скелеты этих двух групп накапливаются
на больших глубинах, где они образуют глобигериновые и радиоляриевые •Илы.
197
Ряд видов, таких, как медузы и некоторые ракообразные, ба* типелагические во взрослом состоянии, на ранних стадиях раз* вития живут близ поверхности.
Но наиболее яркой особенностью зоопланктона являются су* точные вертикальные миграции многих видов с амплитудой бо* лее 100 м, а иногда и до 600 м. Так эпи-, мезо- и инфрапелаги-альная зоны обладают населением, различным в разные часы суток. Погружение чаще всего происходит днем, подъем ночью; детерминизм этих перемещений еще очень мало изучен.
3. Питание. Зоопланктон, постоянно или временно живущий в эпипелагиальной и мезопелагиальной зонах, питается фитопланктоном. Фильтрующие приспособления задерживают водоросли, иногда бактерии и неживые органические частицы.
I I г I I I I I I Г I
ЯФМОИИАСОНД
Рис. 56. Сезонное развитие биомассы фитопланктона (сплошная линия) и зоопланктона (пунктирная линия) в Джорджес-Банк у атлантических берегов Северной Америки (по Рили, Riley).
Но имеются хищные виды, питающиеся другими животными: медузы, сифонофоры, Ctenaria, некоторые аннелиды и Copepoda. Некоторые виды, как, например, Calatius finmarchicus, питаются одновременно водорослями и ракообразными.
Зоопланктон, приуроченный к глубинным зонам, питается подобно бентосу этих зон, органическим «дождем», падающим из верхних зон, или живыми организмами.
По мере погружения, происходящего крайне медленно, бактерии все больше и больше разлагают и обедняют этот дождь» в состав которого входят трупы поверхностных организмов и их испражнения, или фекалии. Поэтому весьма вероятно, что основным источником питания глубинного планктона, благодаря значительным вертикальным миграциям многочисленных видов из одной зоны в другую, являются живые организмы. Таким образом, по Виноградову, существует настоящая цепь передач, начиная от поверхности и до самых глубин. Действительно, отмечают, что с возрастанием глубины количество хищников, по сравнению с фильтраторами микропланктона и детрита, становится все более и более значительным и что последние поднимаются в поисках пищи в верхние зоны.
198
4. Сезонные вариации и отношения с фитопланктоном. Несмотря на менее быстрый, чем у фитопланктонных одноклеточных, цикл развития видам зоопланктона обычно присущи значительные количественные флуктуации, связь которых с количественными флуктуациями фитопланктона бывает либо прямой, либо более или менее обратной. Подавляющее большинство зоопланктона живет за счет объедания водорослей, его количество пропорционально количеству водорослей. Однако если спрос превышает продукцию (расход водорослей в день может быть чуть меньше половины веса объедающих их животных), сокращается количество водорослей, а затем и число животных, лишенных необходимого количества пищи. Так, отношением хищник — жертва может быть объяснено чередование максимумов планктонных продуцентов и консументов, пример которого дает рис. 56. Это смещение можно также объяснить гораздо менее быстрым циклом развития животных, биомасса которых достигает своего максимума спустя много недель после массового размножения фитопланктона. Наконец, говорили о возможном вытеснении зоопланктона в плотных скоплениях фитопланктона вследствие выделения последним токсических веществ.
Нектон. Морской нектон состоит в основном из головоногих моллюсков, рыб и китообразных, к которым на литоралях можно добавить тюленей, моржей и ушастых тюленей, чистиков и пингвинов. Головоногие моллюски, молодь многих рыб и мелкие рыбы (сардины, анчоусы, сельди, макрели) а также усатые киты питаются планктоном. Киты Balaena питаются главным образом веслоногими рачками, эуфаузидами, мизидами. Особи средних промысловых размеров поедают от 1 до 1,5 т этих организмов в день, что возможно только в зонах высокой плотности зоопланктона, превышающей 1,5—2 г/м3. Пелагические рыбы, питающиеся планктоном, также разыскивают зоны, в которых он содержится в изобилии. Это, как мы видели, континентальные платформы и зоны подъема глубинных вод; здесь рыбы собирают планктон, фильтруя воду жабрами, которые играют роль сита.
Хищные пелагические рыбы, которые активно охотятся за такими животными, как мелкие рыбы, весьма многочисленны; особенно активны тунцы и акулы. К ним следует добавить кашалотов, китообразных, обладающих настоящими зубами, ластоногих, обитающих у берегов холодных морей, многочисленных морских птиц. На больших глубинах хищные рыбы компенсируют относительную редкость добычи способностью пожирать животных, почти равных им по величине. Таким образом, в океане устанавливается пищевая цепь, звенья которой взаимосвязаны. Берега Чили и Перу являются тому прекрасным примером; литоральные воды, сгоняемые в открытое море ветрами, дующими с материка, вызывают подъем холодных вод, богатых минеральными
199
веществами, из течения Гумбольдта. В результате — значительная продукция фитопланктона, порождающая обильный зоопланктон, который в свою очередь является источником активного размножения рыб, питающихся планктоном. Последние привлекают хищных рыб, таких, как тунцы, и морских птиц, огромные колонии которых накопили на берегах гуано. Однако теплое береговое течение, ограниченное обычно экваториальной областью, в исключительных случаях может распространяться на юг, вызывая массовую гибель планктона, что отравляет воды и ведет
Рис. 57. Изменения особенностей^ популяции трески и камбалы в Северном море в связи с периодом малоинтенсивной рыбной ловли (вторая мировая война) (по Мар-жету и Хольту, Margettes et Holt)^ Вверху — распределение трески по размерам, отображающее увеличение размеров. Внизу — средний вес различных возрастных групп камбалы, характерен, лучший рост молодых особей и пониженный рост старых особей.
к гибели рыб. Морские птицы в этих случаях перекочевывают в другие места, бросая птенцов, которые погибают.
К этим трофическим цепям следует добавить деятельность человека, который отлавливая некоторых рыб и китообразных действует подобно очень активному хищнику, изменяя особенности популяций этих животных (см. рис. 57).
Бактерии. Пелагические бактерии, как и во всех других средах, играют важную роль в круговороте органического вещества, что предопределяет их особое место. Им свойственно крайне неравномерное вертикальное распределение: редкие на поверхности, они дают кривую плотности, параллельную кривой плотности фитопланктона (рис. 55), становясь снова редкими в глубинных водах, и вновь достигая обилия на поверхности отложений. Эти бактерии, подобно почвенным бактериям, принадлежат
200
.к различным физиологическим группам: среди них выделяют фиксаторов молекулярного азота, автотрофных нитрификаторов и редуцентов различных органических веществ.
Континентальные стоячие водоемы: озера, пруды, лужи
Гидрологические свойства
Население стоячих вод зависит от их глубины, региональных условий климата и химических особенностей воды. Эти определяющие факторы послужили основой для их классификации.
Влияние глубины. В очень глубоких озерах перемешивание воды не достигает дна и существует постоянная застойная, лишенная кислорода зона с постоянными химическими особенностями и температурой. Такие озера называют меромиктическими, или озерами первого порядка. В менее глубоких озерах циркуляция один или несколько раз в году охватывает всю массу воды, но в последней устанавливается стратификация под воздействием временного застоя — это голомиктические озера, или озера второго порядка. Что касается скоплений воды, глубина которых не превышает нескольких метров, то в них перемешивание носит почти постоянный характер, и не возникает никакой выраженной термической и химической стратификации; это озера третьего порядка. Если поверхность невелика, их называют прудами или просто лужами.
Для этих различных категорий нельзя точно указать границы глубины, так как поверхность и ее форма, климат и иногда минерализация воды влияют на степень перемешивания.
Связь с климатом. Циркуляция озерных вод зависит так же ют термических особенностей климата. Хатчинсон и Лёффлер (Hutchinson, Loffler) разработали на этой основе следующую классификацию:
— мономиктические холодные озера арктических и альпийских областей с одним летним периодом циркуляции;
— мономиктические теплые озера субтропических областей, в которых перемешивание происходит во время холодного сезона;
— димиктические озера умеренных областей с двумя периодами обновления вод — весной и осенью;
— полимиктические озера области тропического климата с многочисленными периодами циркуляции;
— олигомиктическис озера, также тропические, с редкими и нерегулярными периодами циркуляции.
Химические особенности водоемов. Они обусловливают степень плодородия озер, и, следовательно, богатство организмами,
201
а также характер видов, населяющих эти водоемы. Тинеман и Науман (Thienemann, Naumann) разработали классификацию, основанную на этих свойствах.
1. Олиготрофные озера. Их воды бедны минеральным азотом и фосфором, первый встречается в нитратной форме на любой глубине, второй в виде следов (часто менее 1 рг/л) (рис. 50). Низкая плотность организмов обусловливает даже в гиполимнионе наличие растворенного кислорода и невысокое содержание СО2 (рис. 48), следствием чего в свою очередь является нерастворимость железа. Вода прозрачна, синего или зеленого цвета. К этому типу принадлежат глубокие озера с крутыми берегами (например, озеро Бурже, озеро Павен в Оверни, озеро Ретурне-мер в Вогезах).
2. Эвтрофные озера богаты азотом и фосфором. Обилие организмов приводит к такому истощению кислорода в глубинных водах во время фаз застоя, что азот встречается там в аммиачной форме, а железо — в виде углекислой закиси железа. Вода малопрозрачна, варьирует от зеленовато-коричневого до коричневого цвета. Небольшие глубины, обусловливая образование широких поясов растительности, благоприятствуют подобному типу озер, примером которых во Франции могут служить озера Нанюа (Юра) и Шамбон (Овернь).
3. Дистрофные озера. Это неглубокие озера с берегами, поросшими торфообразующей растительностью (рис. 45 и фото II, А). Воды окрашены в бурый цвет взвесью гуминовых веществ. Кислород в дефиците, реакция более или менее кислая. К этой группе принадлежат озера Лиспаш (Вогезы), Бурдуз и Шамбе-даз (Овернь).
Между этими типами можно найти все переходные стадии, и одно и то же озеро или пруд может переходить из одного типа в другой. Переход от олиготрофии к эвтрофии (мезотрофные озера)—довольно частое последствие расширения поясов литоральной растительности1, а в настоящее время — также следствие поступления сточных вод населенных пунктов и заводов. Так, озеро Леман еще ЗОлет назад было типично олиготрофным, поскольку его воды до самого дна были богаты кислородом, теперь же, на глубине, воды стали гораздо беднее им (Vivier). То же, поданным Миндера (Minder), произошло с Цюрихским озером. Неглубокие олиготрофные озера третьего порядка могут превращаться в дистрофные вследствие развития на их литорали торфянистых формаций, в этих случаях площадь свободной воды сокращается, причем иногда вплоть до полного исчезновения. Равным образом, неглубокие эвтрофные озера могут вследствие поступления аллювия сокращать свое водное зеркало, запол-
Изучение остатков животных и растений, сохранившихся в отложениях озер, в частности в Швеции, Австрии и Соединенных Штатах, выявило эту естественную эволюцию от олиготрофии к эвтрофии,
202
пяться все больше и больше и постепенно превращаться в болотные формации.
Деление на зоны. На дне озер наблюдается последовательная смена концентрических зон. Вследствие эрозивного воздействия волн берег представляет собой обрыв, у подножия которого формируется эрозионная терраса. Унесенные таким образом отложения переотлагаются на внутренний край этой поверхности
Рис. 58. Пояса растительности Фрас-несского пруда в 1944—1947 гг. (по карте растительных сообществ, масштаб
1 :20 000, Понтарлье, по Гиноше,
М. Guinochet).
От центра к берегам: / — погруженная и свободноплавающая ассоциация Myriophyletto* Nupharetum; 2—мозаика предыдущей и последующей ассоциации; 3 — полупогруженная ассоциация со Scirpus lacustris и тростником IScirpeto-Phragmitetum); ^-ассоциация с крупными осоками (Magnocaricion); 5 —внешний пояс ивовых зарослей (Salicetum cinereo-pentandraey, 5—пояс гигрофильного луга (Ange-liceto-Aconitetum napelli).
и увеличивают ее, образовывая аккумулятивную террасу. Верхняя часть, подверженная временным осушениям, образует эвли-торальную зону. Внутренняя часть и аллювиальный склон, который следует за ней, доступный для проникновения света, образуют сублиторальную зону. Ниже простирается глубинная зона.
Масса воды в зависимости от глубины и расстояния от берега обладает различными свойствами, и в ней различают литоральную и пелагическую зоны.
Литоральная растительность и население бентоса
Литоральная зона. Разделение литоральной растительности на зоны определяется длительностью осушения и глубиной, но сообщества различаются в зависимости от физических свойств субстрата и химического состава воды.
203
Полоса, прилегающая к воде, занята гигрофильной травянистой или древесной формацией. Литоральная зона эвтрофных озер и постоянных луж занята популяциями амфибиальных трав, осок (виды Carex), тростника, камыша (Scirpus lacustris) и т. д., которые на глубине порядка 1 м уступают место полностью погруженным или плавающим гидрофитам, таким, как Nympheaceae, рдесты, водные лютики (рис. 58). На глубине, превышающей 2—3 м, встречаются только харовые водоросли.
Часто литоральная растительность олиготрофных озер с песчаными или галечниковыми берегами бывает редкой и низкой и состоит из Litorella, Isoetes, харовых (особенно Nitella), Вгуор-hita (Fontinalis). Несмотря на прозрачность воды, сосудистые-растения не доходят до границ освещенности, допускающей успешную ассимиляцию; напротив, харовых и Fontinalis находили на глубине свыше 10 м.
Флора водорослей состоит из зеленых, диатомовых, сине-зеленых водорослей, прикрепляющихся к минеральному субстрату и растениям. В альпийских олиготрофных озерах была описана вертикальная зональность, характеризующаяся различными Суа-nophyceae, в то время как в эвтрофных озерах севера Германии наблюдается мощное развитие Cladophora. Близ нижней границы сублиторальной зоны, где освещенность крайне невелика,, бентосная флора водорослей приобретает оригинальный состав-и аспект: зеленые водоросли там почти отсутствуют, в то время как диатомовые и сине-зеленые водоросли образуют, вследствие обилия у них пигмента, ярко окрашенную мозаику. В частности, сине-зеленые водоросли богаты там красными пигментами, что заставило выдвинуть гипотезу хроматической адаптации.
Наиболее богата и разнообразна фауна в постоянных лужах и на богатых растительностью илах эвтрофных озер. В Мичигане, одном из озер, принадлежащих к этой группе, в сублиторальной зоне было найдено до 4400 макроскопических бентосных животных на 1 м2. Разнообразие местообитаний и питания благоприятствует этим высоким плотностям: виды, перемещающиеся по поверхности растений и дну (моллюски, личинки насекомых),, сидячие виды, гораздо менее многочисленные, чем в морском бентосе (сувойки, губки, мшанки), фауна, зарывающаяся в толщу отложений (личинки и куколки стрекоз, поденок, хиро-номид; кольчатые черви, нематоды). В подверженных осушению литоральных песках озер обитает, так же как и в литоральных песках морей, интерстициальная фауна, обладающая теми же приспособлениями и включающая простейших, коловраток, тихоходок, нематод и веслоногих рачков (Pennak).
Глубинная зона. Ее субстрат образован осаждающимися здесь отложениями. Они бывают озерного (автохтонные) или внешнего по отношению к озеру (аллохтонные) происхождения, минеральными или органическими. Минеральные отложения озерного происхождения — это в основном кальцит, образую
204
щийся из бикарбоната кальция или происходящий из раковин моллюсков, железо в виде гидрата окиси железа в аэрируемой воде или сернистого в анаэробных условиях, кремний биологического происхождения (раковинки диатомовых водорослей, скорлупки хризомонад, спикулы губок). Процентный состав этих компонентов минерального состава сильно варьирует в зависимости от характера озер.
Так же обстоит дело и с органическим веществом, которое играет существенную роль в биологических процессах озерных глубин. В олиготрофных озерах поступление органического вещества невелико и связано с планктоном. Оно попадает на глубины уже частично разложившимся, и его минерализация продолжается там в условиях периодически повторяющегося аэробиоза; ооз в этом случае, главным образом минеральный, обозначается в скандинавской терминологии как гиттия. В сильно эвтрофных и меромиктических озерах отсутствие окисления на глубине и среда, обладающая сильными восстанавливающими свойствами, допускают лишь неполное разложение органического вещества, которое образует сапропель. В озерах, в водах которых скапливаются гуминовые вещества в значительных количествах (особенно в дистрофных озерах, захваченных торфянистыми формациями), эти коллоиды осаждаются в виде хлопьев растворенными солями и оседают на дно в виде бурого ила, в котором биологическая деятельность выражена очень слабо; это дай скандинавских лимнологов.
В биологическом населении оозов бактерии играют важную роль, обеспечивая разложение и минерализацию органических поступлений сообразно с процессами, тесно связанными со степенью насыщения кислородом. Даже в глубинах, где имеется кислород, последний отсутствует в отложениях на глубине нескольких сантиметров, результатом чего является очень тонкая стратификация бактериальных форм.
Животный бентос глубинной зоны включает простейших, нематод, кольчатых червей, личинок насекомых, в частности Chi-ronomidae, которые живут в трубочках из ила, некоторых моллюсков и ракообразных.
Процент насыщения кислородом па поверхности отложений и его сезонные вариации также определяют состав бентических организмов. В богатых кислородом олиготрофных озерах имеются многочисленные бентические виды, среди которых наиболее характерными являются личинки Chironomidae рода Tanytarsus. В эвтрофных озерах обитают виды, приспособленные к низкому содержанию кислорода, например личинки Chironomus и Tendi* pes. Некоторые виды в период хорошей аэрации могут достигать высокой плотности. Так, на 1 м2 насчитывалось до 26 тыс. экземпляров Tendipes. Приспособление к полуанаэробному существованию в этих озерах реализуется различным образом: в крови личинок Chironomidae и кольчатых червей содержится гемоглобин
205
(пигмент, фиксирующий кислород, который содержится в незначительных количествах). Tubifex, сидячие кольчатые черви, строящие трубочки в иле, по мере уменьшения кислорода все более усиливают колебательные движения выступающей из трубки части тела. Личинки покидают глубинную зону на время летнего застоя; некоторые ночью поднимаются в эпилимнион (Chaobo-rus)\ веслоногие рачки и личинки Chironomidae летом впадают в спячку.
В сапропелях, где кислород почти полностью отсутствует, могут существовать только анаэробные простейшие. Постоянно низкая температура гиполимниона бореальных и альпийских озер обусловили выживание в них таких арктических форм, как ракообразные Mysis relicta, Pontoporeia affinis, и различные Ostracoda, которые выглядят как ледниковые реликты.
Озерный планктон
Лимнопланктон гораздо беднее видами, чем морской. Фитопланктон представлен водорослями из различных групп (сине-зелеными, диатомовыми, хлорококковыми, одноклеточными жгутиконосными водорослями, такими, как вольвоксовые, эвглено-вые, Peridinea и т. д.) (рис. 54), а также бактериями. Зоопланктон включает простейших, коловраток, веслоногих и ветвистоусых рачков, остракод; здесь встречаются также личинки различных видов, церкарии сосальщиков, личинки пластинчатожаберных моллюсков, личинки насекомых рода Chaoborus. В то время как кишечнополостные широко представлены в океанах, в пресных водах медузы встречаются очень редко (Limnocnida озер Восточной Африки, Craspedacusta Евразии и Северной Америки). Количество чисто планктонных видов в озере обычно не достигает и сотни, и лишь несколько видов, принадлежащих к различным группам, являются явственно преобладающими в любое время года.
Состав и обилие планктона подвержены сезонным изменениям, которые связаны с условиями среды и биологическими взаимодействиями различной природы. Во многих озерах умеренного климата весенний и осенний максимум чередуются с летним и зимним минимумом, но состав зависит от времени года. Каждый вид обладает пульсациями обилия, благодаря которым он может достигать на несколько дней или недель необыкновенно высокой численности. Так, в теплый летний период некоторые виды сине-зеленых водорослей могут достигать такой степения обилия, что даже меняется цвет воды («цветение воды»), и их разложение, которое лишает среду кислорода и отравляет ее ядовитыми веществами, может привести к гибели многих животных.
Кроме того, вертикальное распределение планктона в любой момент неодинаково. Некоторое количество организмов лока
206
лизовано на нижней стороне поверхностной пленки воды и держится на ней благодаря силе поверхностного натяжения: это нейстон, в котором Гейтлер (Geitler) различает воздушные формы, обитающие на поверхности (эпинейстон), и водные формы, обитающие под пленкой (гипонейстон); речь идет о водорослях, простейших, ракообразных, насекомых, личинках комаров. Погруженный планктон непосредственно зависит от освещенности
Рис. 59. Вертикальное распределение некоторых видов озерного планктона (по Финденегу, Findenegg).
Вверху — распределение планктона: Af — Asterionella formosa; Pm —Poly art hr а major, Ap — Asplanchna priodon; Dl — Daphnia longispina в озере Лунц, в Австрии в конце летней и во время зимней стагнации (по Рутнеру, Ruttner). Внизу', изменение распределения Oscillatoria rubescens и уровня изотерм 6 и 8° в озере Ворт в Каринтии.
в случае водорослей и лишь косвенно — в случае животных, которые питаются водорослями. В противоположность океанам, глубокие озера почти лишены планктона, за исключением бактериального в афотической зоне. Наибольшая плотность его наблюдается на небольшой глубине, но видовой состав в разное время меняется вместе с ней. Рис. 59 дает пример вертикального распределения нескольких видов в период сезонного застоя. Максимум обилия видов на данном уровне соответствует оптимальным значениям, которые имеют на этом уровне градиенты освещенности, температуры, насыщения кислородом и химического состава воды. Этот же рисунок дает пример сезонного развития вертикального распределения стенотермной холодноводной сине-зеленой водоросли под воздействием изменений температуры воды. Эта водоросль погружается летом вплоть до гиполимниона, где ее богатство фитоэритрином позволяет ассимилировать углекислый газ. Как и в океане, животные лимнопланктона осуществляют ежедневные вертикальные миграции, поднимаясь
207
ночью и опускаясь днем. Амплитуда этих миграций варьирует примерно от одного до 50 м 1.
Количество бактерий в озерных водах весьма различно. Редкие в чистых водах, они кишат в загрязненных озерах (несколько
Клеток Chromatlum на 1мл
Рис. 60. Вертикальное распределение температуры (Т), концентрации кислорода (О2) и сероводорода H2S и серо-бактерии Chro-matium в меромиктическом озере в СССР (30 июля 1938 г.) (по Кузнецову).
миллионов на 1 мл). Поскольку они принадлежат к различным метаболическим группам, их вертикальное распределение зависит от химических особенностей воды на различных глубинах. Железистые и серные бактерии обильны в бедном кислородом гиполимнионе, где в некоторых случаях они локализуются на строго определенном уровне, как Chromatium (рис. 60), который образует очень тонкий слой в меромиктическом озере на общей границе присутствия кислорода и сероводорода.
Особенности биомассы планктона значительно варьируют в зависимости от озер. Важно различать общую биомассу, имеющуюся под данной поверхностью, и среднюю биомассу в данном объеме зоны продуктивности. Последняя, дающая реальную картину плодородия озера (действительный индикатор продуктивности озера), в эвтрофных озерах выше, чем в олиготрофных, в то время как первый метод оценки дает мало отличающиеся друг от друга результаты вследствие распределения планктона на большую толщу воды в более прозрачных олиготрофных озерах. Содержание органического вещества на 1 л воды в зоне продуктивности очень изменчиво: от 0,23 до 12 мг сухого веса для озер Висконсина, но оно включает значительную долю неживого вещества (триптон). Что касается живой части, то нано-
1 Напомним здесь частный случай личинок двукрылых рода Chaoborus, которые днем обитают в иле, а ночью поднимаются до самой поверхности. 208
планктон может давать большую массу, чем масса собираемого ситами планктона более крупного размера. Биомасса планктона обычно бывает больше биомассы бентоса, но биомасса планктона не дает никакого представления о продуктивности озера вследствие короткой продолжительности жизни планктонных видов и высокого «коэффициента возобновления». Измерения, проведенные в озере Мендота (Висконсин), показали, что годовая его продукция составила 624 г сухого веса на 1 м2, из которых 585 г приходилось на фитопланктон, — величина в 140 раз большая, чем продукция бентоса того же озера.
Озерный нектон
Нектон озер и луж состоит в основном из рыб, а также в литоральной зоне из амфибий, черепах, змей и насекомых. Фауна различается в зависимости от типов озер и виды распространены в различных зонах. Олиготрофные озера благоприятны для лососевых, форелей, сигов, гольцов — два последних являются обитателями гиполимниона. В литоральной зоне живут окуни, щуки, плотва, уклейки, гольяны. Эвтрофные озера и пруды богаты карпами, рыбой, которая переносит дефицит кислорода, ее раз-ведение имеет коммерческое значение. Здесь встречаются также плотва, лещ, линь.
Пищевой режим озерных и прудовых рыб различен. Некоторые травоядны и ощипывают водные растения и их эпифиты (плотва); другие, питающиеся планктоном, снабжены тонкими жаберными ситами (сиги); многие питаются одновременно растениями и животными бентоса (осетр, карп, сом и т. д.). Рыбы, являющиеся в основном хищниками (форель, окунь, щука), поедают других рыб, когда они достигают достаточной величины. Десятисантиметровый окунь потребляет 0,15 г пищи (в сухом весе) в день, что эквивалентно 37 500 циклопам (36 в минуту).
Продуктивность промысловых рыб ввиду их экономического значения явилась объектом количественных исследований. Пруды й озера, не обогащенные удобрениями, дают от 5 до 12 кг сырого веса на 1 га в год в олиготрофных средах, до 390 —в эвтрофных водах с карпом. В рыбоводстве широко практикуется внесение фосфатных минеральных удобрений. Стимуляция фитопланктона, как результат такого мероприятия, увеличивает обилие зоопланктона и животных бентоса, а затем и рыб. Была таким образом зарегистрирована продукция от 500 до 1500 кг на 1 га за лето. Продолжительное обогащение вод органическими веществами может привести к огромному росту продукции: более Ю т с 1 га/год в Китае. Было установлено, что продукция травоядных рыб соответствует трофической эффективности порядка 20% чистой первичной продукции.
209
Проточные воды
Образуемая проточными водами среда значительно отличается от среды стоячих вод постоянным обновлением и более значительными контактами с наземными местообитаниями.
Экологические свойства проточных вод
Скорость течения и ее местные вариации являются фактором, определяющим распространение и условия жизни организмов, вследствие как механического влияния, так и косвенных воздействий на физические и химические свойства воды. И все же между стоячими и проточными водами существуют все переходные стадии: некоторые участки рек обладают очень медленным течением, в то время как некоторые озера пересекаются потоками воды со значительным дебитом.
1. Скорость течения увеличивается с уклоном поверхности, шириной и глубиной водного потока и зависит от неровностей и шероховатости дна. Скорость очень различна; она может быть медленной или составлять в потоках и на быстринах до 3 м/сек. Она уменьшается вблизи берегов и у дна, в непосредственной близости от которого скорость потока резко замедляется и почти равна нулю на поверхности. Эффект торможения особенно ярко выражен при слабых течениях и в холодных водах, обладающих повышенной вязкостью. Течение ламинарное вдали от дна и берегов, вблизи них становится турбулентным. В случае каменистого ложа пространства между камнями и поверхности глыб, обращенные к низовью, омываются водой, слабо обновляемой течением, но перемешиваемой турбулентными перемещениями.
2. Характер дна, имеющий большое биологическое значение, зависит от скорости течения. В быстрых потоках течение не сносит только камни. По мере замедления течения дно становится песчаным, затем илистым. При скорости меньше 20 см/сек оседают органические остатки, и органический ил, образовавшийся таким образом, образует биологически богатую среду.
3. Температура проточных вод обычно одинакова для всей толщи, но термический режим различается в зависимости от расстояния до истока. Температура у истоков постоянна и обычно близка к средней годовой температуре данного места. На неко« тором удалении от истока устанавливается суточная и сезонная флуктуация температуры.
4. Насыщенность кислородом проточных вод, даже при отсутствии зеленых растений, бывает хорошей вследствие перемешивания и значительной протяженности поверхности потока по отношению к объему. Постоянное обновление воды еще больше, чем богатство ее кислородом, обеспечивает организмам лучшие
210
условия дыхания, чем в стоячих водах, насыщенность которых кислородом может быть столь же высокой. Поступление органических веществ со сточными водами или с водами, которые сбрасывают фабрики и некоторые предприятия, вызывает уменьшение количества кислорода из-за массового размножения бактерий, несмотря на поступления кислорода из воздуха или за счет фотосинтеза L
5. Химический состав проточных вод разнообразен. Вблизи истоков он зависит от природы скальных пород, через которые проходят подземные воды; по мере удаления от истоков проточные воды пополняются минеральными и органическими веществами в растворенном и взвешенном состоянии, приносимыми ручьями. В промышленных районах наблюдается все больше и больше сбросов ядовитых веществ, кислот, хлористого натрия и т. д.
В общем, вместо концентрической зональности особенностей среды в замкнутых массах воды в реках происходит изменение вдоль их продольного профиля. Распределение биоценозов в этих двух местообитаниях отражает эти различия.
Биологическое население проточных вод
Мы последовательно рассмотрим различные секторы рек от источника до устья.
Население источников. Источник представляет для организмов крайне малоизменчивую среду, что является благоприятным условием для равновесия его населения и для выживания реликтовых видов. Однако существуют значительные различия в температуре и химических условиях источников: их температура варьирует почти от 0 до примерно 100° Их минерализация очень невелика в областях, где преобладают кремнистые и гранитные породы; в областях, богатых известью, в водах источников много бикарбоната кальция (который на поверхности образует отложения известкового туфа). Горячие источники богаты солями и насыщены газами. Растворенный кислород, как правило, в дефиците или отсутствует полностью.
В холодных источниках живут такие водоросли и стенотермные животные холодных вод, как Hydrurus foetidus семейства Chrysophyceae и планария (Planaria alpina). Горячие источники дают приют фауне, видовой состав которой по мере повышения температуры все более обедняется: зеленые водоросли исчезают
1 Падение содержания кислорода вследствие поступления органических веществ может ощущаться значительно ниже по течению. Так, вследствие периодического слива из отстойных бассейнов Вербуа, в которые поступают сточные воды Женевы, ухудшение качества воды в Роне дает о себе знать вплоть до Кюлоца, в 60 километрах от плотины (Р. Vivier).
211
примерно при 38°, диатомовые при 45°, сине-зеленые существуют при температуре свыше 60° (Synechococcus elongatus встречалась при температуре 66—69°), бактерии — до 77°; почти все эти виды эвритермны. Животные представлены простейшими, коловратками, нематодами, ракообразными, насекомыми и моллюсками; большинство их эвритермны и не встречаются при температурах выше 40° Однако личинка насекомого сетчатокрылки (Dasyhelea tersa) живет в термальных водах Зондских островов при температуре до 50° С, а ракообразное Thermasbaena mirabi-lis обнаружено только в термальном источнике с температурой 45° в Тунисе. В термальных, исключительно кислых источниках сольфатар, все же живут некоторые водоросли (красная водоросль, Cyanidium caldariorum, до 45°) и личинка Dasy-helea.
Население потоков. Планктон в них отсутствует, нектон не очень разнообразен, однако флора и фауна бентоса обильны.
Формы, прикрепленные к камням и растениям, или обрастания (Aufwuchs) представлены зелеными водорослями (Cladop-hora), красными водорослями (Lemanea), диатомовыми, сине-зелеными, водными мхами, губками, мшанками (Plumatelia). Подвижные формы обладают приспособлениями для сопротивления силе течения: сплющенная форма, обеспечивающая наименьшее сопротивление течению, мощные средства для закрепления, положительный тигмотаксис (склонность к плотным субстратам). Самые мелкие формы — простейшие, коловратки, нематоды — не обладают специальными морфологическими особенностями, но их мелкие размеры позволяют им целиком держаться в тонком слое неподвижной воды на поверхности опоры или в небольших неровностях. Полости между камнями, их нижняя поверхность, пучки нитчатых водорослей и мхов дают надежную защиту и обильно заселены видами, лишенными способности противостоять течению (раки, личинки слепней Tabani-dae и поденок Ephemeroptera, тихоходки, водные клещи Hydracarinae) .
Рыбы быстротекущих вод являются хорошими пловцами, как форель или лосось, или донными рыбами, как Cottus gobio и Etheostomidae — североамериканское семейство, у видов которого атрофированный плавательный пузырь увеличивает удельный вес, а широкие и горизонтально вытянутые у основания тела грудные плавники обеспечивают соприкосновение с дном. У животных, обитающих в стремительных потоках, жабры и дыхательные придатки развиты слабо, при перенесении в стоячие воды у многих из них быстро появляются признаки удушья.
Население речек и рек. Если верхняя часть водных потоков лишена настоящего планктона и несет только организмы, оторвавшиеся ото дна, то в реках большой протяженности с медленным течением, таких, как Волга, в нижнем течении имеется на
212
стоящий планктон или потамопланктон Что касается озерного планктона, захваченного водоотводными каналами, то он исчезает очень быстро.
Донное население медленно текущих рек резко отличается от донного населения быстрых потоков. Семенные (Phanerogamae) многочисленны и богаты эпифитами. Донные животные лишены приспособлений для закрепления. Пески, которые представляют ненадежную опору для фиксации и для перемещения по их поверхности, мало заселены. В промежутках между песчинками, погруженными в воду или просто увлажненными вследствие капиллярности, живет, как в аналогичных местообитаниях морских и озерных литоралей, сообщество мелких организмов, известное под названием псаммон. Илы заселены роющими видами: кольчатыми червями, пластинчатожаберными моллюсками (Sphae-rium и Pisidium, Unio и Anodonta), наядами поденок (Hexage-nia), личинками Chironomus. Только к раковинам прикрепляются губки (Ephydatia fluviatilis) и мшанки (Plumatella').
С менее хорошей аэрацией связаны действенные респираторные приспособления. Брюшные жабры наяд поденок крупнее и более многочисленны у видов, приуроченных к стоячим водам, чем у видов проточных вод; эти жабры подвижны у первых и неподвижны у вторых.
Рыбы от истока до устья занимают в реках различные секторы. В Европе от верховьев к низовьям различают зоны: форелей в прохладных и быстрых водах; хариуса; усача с пескарем, окунем, ельцом, голавлем, где течение еще достаточно быстрое, с хорошей аэрацией и песчаным или гравийным дном. Рона выше Лиона, значительная часть Сены и Рейна принадлежат к этой зоне. Зона леща и карпа, предпочитаемая линями, плотвой, уклейками, щуками, соответствует рекам с медленным течением. Дно илистое, покрытое тиной или мелким песком. К этому типу относятся Сона и нижнее течение Эльбы. Тропические реки, кроме того, кормят черепах, ящериц1 2 и млекопитающих (сирен, дельфинов).
В противоположность озерам реки образуют экосистемы, сообщающиеся с морем, и между ними и морем осуществляется обмен мигрирующими рыбами. Некоторые рыбы отправляются на нерест в океан и в реку возвращаются молодые рыбы: таковы стекловидные (стеклянные) угри или молодые угри, которые
1 В реках с быстрым течением, как средний Рейн, поверхностные воды которого пробегают расстояние между Балем и Мангеймом за 1 день с четвертью, планктонные организмы практически не могут размножаться. В медленно текущей реке, как Москва-река, которая проходит в день от 14 до 30 км, напротив, у быстро размножающихся видов достаточно времени, чтобы увеличить свою численность прежде, чем они достигнут устья.
2 Полуводный образ жизни ведут немногие представители семейств агам, игуан, тейид (например, каймановые ящерицы), варанов. К перечню, данному Ж. Леме, следует присоединить и крокодилов. — Прим. ред.
213
весной поднимаются вверх по течению. Другие, родившиеся в пресных водах, возвращаются туда для нереста: это осетры, миноги, сельди (A/osa), лососевые; последние в огромных количествах поднимаются вверх по рекам Тихоокеанского побережья Северной Америки и Сибири, причем многие при этом погибают. Кроме того, в нижнее течение впадающих в море и прибрежных рек заходят морские рыбы, способные в течение некоторого времени существовать в пресной воде, но не размножающиеся в ней, — это кефали (Mugil) и речная камбала1 (Platichtys) в средиземноморских реках, а также акулы тропических рек.
В противоположность озерам у рек соотношение размеров береговой полосы и водной массы благоприятствует образованию фауны береговых откосов — амфибий, птиц и особенно млекопитающих; среди последних отметим бобра, выдру, водяную крысу и водяную бурозубку (Sorex palustris).
Это естественное распределение биоценозов в речных водах все больше и больше нарушается деятельностью человека; прибрежные поселения и фабрики загрязняют воду. Если воды достаточно аэрированы, то бактерии и грибы2 производят естественную очистку от органических веществ, но, если кислород отсутствует, вредные продукты анаэробного брожения могут уничтожить все живое, за исключением бактерий. Таким образом были стерилизованы некоторые реки промышленных районов Франции.
Эстуарии. Нижние части течения рек, куда проникают морские воды, обладают особыми свойствами.
1. Изменения уровня, связанные с приливами и отливами и с режимом реки. Следствием этого является вертикальная зональность, непосредственно связанная с вертикальной зональностью океанов, от глубинной части фарватера и до наиболее высоких мест, которых достигают самые высокие приливы.
2. Сложный режим течений, направление и интенсивность которых постоянно меняются.
3. Накопление отложений тонких и нестабильных; чаще всего речь идет о богатых коллоидальной глиной плотных и вязких илах материкового происхождения.
4. Изменчивость солености во времени и пространстве. Более плотная морская вода проникает под пресные воды относительно далеко и достаточно толстым слоем в зависимости от приливов и отливов; на их контакте образуется слой турбулентного перемешивания; результатом является вертикальная стратификация солености и уменьшение последней по направлению к верховью. В каждом участке она варьирует в зависимости
1 Речные камбалы распространены в северной части Тихого океана и у берегов Европы в Атлантическом океане. — Прим. ред.
2 Грибы Sphaerotilus, белые нитевидные колонии которых иногда покрывают дно, могут в течение часа удалить до 3 кг органических веществ на площади 1 га.
214
от приливо-отливного ритма, его амплитуды, а также от дебита реки: так, в период низкой воды и в засушливые годы средняя соленость бывает выше.
5. Богатство вод пищей, обусловленное накоплением, вследствие столкновения противоположных течений, взвешенных частиц, принесенных пресными и морскими водами. Виды, живущие в эстуариях, — это преимущественно морские формы, некоторые пресноводные формы и виды, локализованные в солоноватых водах. Для всех этих видов характерна определенная толерантность к изменениям солености, которая является главным фактором в их локализации наряду с периодичностью осушения (для прикрепленного бентоса).
а) Бентос охватывает литоральные виды, которые простираются более или менее далеко в зависимости от их толерантности к слабой солености. Так, в устье Раиса Фишер-Пиет (Fi-scher-Piette) смог установить постепенность проникновения различных водорослей и животных. Илы заняты лужками из гало-фильных семенных растений различного состава в зависимости от уровня (см. фото I). В них обильны роющие формы, морские, как моллюски Муа arenaria и Cardium edule, краб (Carcinus maenas), кольчатые черви Nereis cultrifera и Arenicola marina; формы, приуроченные к солоноватым водам, как скробикулярии (моллюски), Corophium volutator (ракообразные, бокоплавы), кольчатый червь (Nereis diversicolor). К твердому субстрату прикрепляются усоногие раки Chthamalus и Balanus, моллюски, устрицы, мидии и морские блюдца. На водорослях живут брюхоногие моллюски, морские (Littorina) и солоноватых вод (Hyd-robia ulvae).
б) Планктон эстуариев обилен вследствие накопления потамопланктона и приноса морских форм. Так, в Эльбе, вверх по течению от Гамбурга, в 1 м3 воды содержится несколько тысяч ветвистоусых рачков (Cladocera), а в нижнем течении они составляют несколько миллионов, и к ним добавляются морские эвригалинные диатомовые.
в) Нектон состоит из рыб, которые в отличие от предыдущих групп благодаря своей подвижности обладают способностью-держаться в условиях, которые им соответствуют. Пресноводные виды обитают больше в континентальных поверхностных водах, в то время как морские виды предпочитают глубинные соленые воды. Но многие рыбы, такие, как султанка (Mullus) и морские камбалы (Pleuronectes), широко эвригалинны.
Пищевые цепи и продуктивность
Пищевые цепи в проточных водах отличаются от пищевых Цепей в стоячих водах обеднением базы. Хотя бентосные растения имеются в обеих средах, в проточных водах обилие фитопланктона ничтожно; в противовес этому проточные воды несут
21&
многочисленные органические остатки, количество которых увеличивается от верховьев к низовьям и которые являются пищей для многих видов. Организмы, которые поглощают взвешенные частицы, обладают различными органами для их сбора: мерцательные реснички губок, щупальца гидр, сифоны пластинчатожаберных моллюсков, придатки личинок мошек, снабженные веерами щетинок и т. д. Личинки ручейков (Trichoptera) сооружают шелковые ловчие сети, которые собирают пищевые частицы, переносимые течением.
Формы, объедающие микрофлору бентоса, представлены многочисленными личинками насекомых, брюхоногими моллюсками, планариями. Брюхоногие моллюски и рыбы поедают водные растения.
Многие виды являются всеядными, поедая в зависимости от места или времени года растения, животных или детрит. Таковы карпы, султанки, сомы, раки, личинки насекомых; среди последних наяды веснянок (Plecoptera), травоядные зимой, бывают всеядными или хищными летом.
Хищные виды разнообразны: насекомые и их личинки, такие рыбы, как форели, окуни, камбала 1 (Pleuronectes)\ водными животными питаются также хищные млекопитающие, такие, как выдра, и птицы, питающиеся рыбой, как зимородок и чайки.
Биомасса и продуктивность рек сильно варьируют в зависимости от их физических и химических особенностей. Форельные реки, холодные, олиготрофные, малопродуктивны, в то время как реки, которые пересекают возделанные плодородные области, обогащены минеральными и органическими веществами и сами плодородны. Неглубокие эстуарии с топкими отложениями имеют повышенную потенциальную продуктивность; благодаря богатству органическими веществами и планктоном они оказались особенно хорошим местом для разведения устриц, поскольку устрицы питаются мелкими органическими частицами и планктоном.
Первичная продукция проточных вод может быть равной продукции интенсивных культур. В «Серебряных источниках» Флориды на илистом дне чистая продукция в среднем составляет 7,5 г на 1 м2 в день.
Пищевой режим этих рыб меняется в зависимости от возраста; молодые форели и окуни питаются личинками насекомых и мелкими рыбами в более позднем возрасте. Молодые камбалы, ершоватки, лиманды (Limanda) питаются только ракообразными, потом во все возрастающей пропорции многощетинковыми кольчатыми червями.
Глава If
Наземная среда и наземные сообщества
Тундры
Под наименованием тундра объединяется зона растительности от естественной границы древесной растительности по направлению к полюсам (рис. 61). Помимо этих арктических и антарктических тундр, иногда выделяют, ввиду их очевидного сходства, альпийскую тундру, которая расположена выше границы древесной растительности в высокогорьях средних широт.
Тундра высоких широт
Климатические и экологические факторы. По направлению к полюсам безморозный период составляет меньше трех месяцев в году, а средняя температура наиболее теплого месяца опускается ниже 10 °C, что приводит к исчезновению деревьев. В течение этого короткого лета почва оттаивает не полностью, и ее вечно мерзлые слои (tjale или permafrost) препятствуют дренажу талых вод, благоприятствуя явлениям солифлюкции и вспучивания.
В Антарктике с более островным, чем в Арктике, климатом, зимы менее холодные и лето менее теплое, со средней более ровной температурой, тогда как в Арктике ее амплитуда значительна. Осадки существенно варьируют, в Арктике они иногда очень незначительны, однако, большей частью в виде снега. Сильные ветры как правило господствуют в Антарктиде. Во время наиболее теплого периода, когда воцаряется длинный полярный день, солнце, однако, остается низко над горизонтом. Эти климатические особенности благодаря незначительным неровностям поверхности способствуют разнообразию микроусловий.
Типы растительности. Тундра в широком смысле представляет мозаику сообществ, локализация которых в зна-i.. степени определяется продолжительностью периода вегетации и водным режимом почв. Результатом этого являли ня значительные физиономические различия арктической тундры: густые кустарниковые формации ив и берез в зоне, соседствующей с границей древесной растительности, формации низких пустошей с Ericaceae и мелкими ивами, особенно в менее холодных
217
частях, луговины с осоковыми и Juncaceae, открытые степи !, более или менее богатые споровыми, прерывистые формации мхов и лишайников, наиболее развитые в особенно холодных территориях, вблизи постоянных снежников.
Сукцессии также различны:
1. Гигрофильные серии развиваются на малодренированных равнинах и в понижениях, где в общем почвы оттаивают на меньшую глубину или водонепроницаемы. Они включают понижения, более или менее затопленные, с пушицами и осоками, сфагновые торфяники1 2, сообщества кустарничков из карликовых берез и мелких ив.
2. Тундра мезофильная распространяется на проницаемых, хорошо дренированных почвах, которые могут оттаивать на глубину около метра. Она включает луга, растения достигают высоты 15—30 см; здесь господствуют ссоковые и злаки, часто сочетающиеся с кустарничками, мхами и лишайниками. На Крайнем Севере эти сообщества исчезают и всецело господствуют лишайники.
3. Ксерофильные серии развиваются на возвышенных участках, сложенных грубым материалом; это степи, богатые лишайниками, с дриадами, ивами, осокой и ожикой.
4. В местах, где зимой скапливается снег, растительность располагается полосами в зависимости от средней продолжительности периода активной вегетации.
5. Нестабильные субстраты, подверженные действию солифлюкции и вспучивания, остаются частично обнаженными; растительность приурочена • к участкам, наименее подвижным, к бордюрам полигонов полигональных почв, каменным кольцам и линиям, бордюрам бугорков солифлюкции.
6. Каменистые, более или менее ровные поверхности, покрывающие обширные пространства в наиболее холодных частях, характеризуются весьма открытой растительностью с доминированием лишайников.
Животное население. Защита от холода и достаточное обеспечение питанием — наиболее самобытные черты приспособления животных к арктическим условиям. Гомотермы лучше защищены от холода, чем в более низких широтах: длинный шерстный покров, толстый подкожный жировой слой, невысокое отношение поверхности к объему тела. Близость вечной мерзлоты и обычно недостаточный дренаж исключает укрытие в глубоких норах. Мелкие животные, грызуны, горностаи, укрываются в углублениях под снегом; только суслик зимует в норах, вырытых на участках с песчаной почвой и глубокозалегающей вечной мерзлотой. Но многие гомотермные виды мигрируют — северные олени и карибу перекочевывают стадами в более южные части
1 В зоне тундр степей пет; имеются лишь отдельные представители степной флоры и немноговидовые степные комплексы, особенно богато представленные в тундрах Восточной Сибири. — Прим. ред.
2 Сфагновые болота особенно характерны для северной части лесной зоны; они развиты в лесотундре и южной тундре, а в северной постепенно уступают место гипновым болотам. — Прим. ред.
218
Рис. 61. Карта типов растительности земного шара (частично по Шимперу и Фаберу, Schimper et von Faber, частично по Кюхлеру, A. W. Kuchler).
/ — тундра; 2 — хвойные бореальные леса; 3 — смешанные леса из лиственных и хвойных пород; 4 — листопадные леса умеренного пояса; 5—средиземноморские вечнозеленые леса; 6— континентальные и средиземноморские степи; 7 —пустыни; 5 —степи с листопадными и колючими кустарниками; 9 — травяные степи с акациями в Африке; 10 — саванны и светлые леса; 11— ксерофильные леса; 12 — тропические листопадные леса; /.? —странны и полулиртопадцые леса; /^—тропические вечнозеленые леса; /5—субтропические речнозеленые леса; 16 - вечнозеленые леса умр-* ' рещкяо П9«с% > “ ‘ 1 *
тундры й даже в хвойные леса; большинство птиц — мигранты, за исключением белой куропатки, снежной совы (Nyctea scan-dica), ворона, морских птиц, там, где море не замерзает.
Эти животные, продолжающие активную жизнь на месте, должны прокормиться. Травоядные ищут лишайники и зеленые части растений в наименее заснеженных участках тундры. Многие хищники всеядны: полярный медведь заменяет тюленей рыбой, птичьими яйцами и даже растениями; лисица 1 питается моллюсками и другими морскими животными и даже остатками тюленей— добычи медведей; кормом белой куропатки являются ягоды и листва Vaccinium.
Среди пойкилотермных животных рептилии и амфибии представлены очень малым числом видов; насекомые, напротив, многочисленны. В холодный сезон эти животные зимуют и остаются невредимыми при температуре ниже нуля в течение долгих месяцев. С оттаиванием они становятся активными: насекомые вылупляются из яиц и куколок, и в тундре летом наблюдается необычайное размножение комаров и мух.
Однако краткость сезона активной жизни препятствует завершению полного цикла, и многочисленным насекомым необходимы два или три последовательных лета.
Многие гусеницы — полифаги. Шмели (Bombus), обильные в тундре, собирают нектар в течение всего продолжительного дня и могут накопить достаточные запасы для развития личинок в течение года.
Цепи питания. Продуктивность растительного покрова низка. Поэтому крупные травоядные, северные олени и карибу, чтобы обеспечить себе пропитание, нуждаются в значительной площади (8 км2 на особь), и их плотность низка. Напротив, мелкие грызуны образуют обильные популяции на лугах и мезофильных пустошах. За первыми охотится волк и человек, за вторыми: лисица2, горностай, хищные птицы. Летом с появлением мигрирующих птиц и насекомых устанавливаются более сложные цепи питания. Однако ввиду малого числа трофических уровней и незначительного разнообразия добычи, доступной каждому виду, любое значительное колебание обилия данного вида оказывает воздействие на обилие связанных с ним хищников, и это, без сомнения, — основная причина больших периодических флуктуаций, наблюдаемых у видов арктической фауны. Выше была описана связь лемминга и арктической лисицы.
В антарктических тундрах, где местные млекопитающие отсутствуют, наземные трофические связи еще проще.
Но наряду с этими исключительно наземными экосистемами в арктических и антарктических регионах существуют литораль
1 Вероятно, здесь и далее речь идет о полярной лисице, песце, а не о лисице обыкновенной, хотя последняя также населяет значительные территории тундры и отсутствует лишь в ее северных частях. — Прим. ред.
2 См. примечание выше. — Прим. ред.
220
ные экосистемы, для которых характерно обилие плотоядных, птиц, тюленей, медведей, лисиц, использующих пищевые ресурсы моря. Это изобилие животных само по себе — причина, определяющая примерную локализацию поселений человека на побережьях.
Горная тундра: альпийский пояс
С возрастанием высоты и изменением условий меняется состав горной флоры и фауны. На некоторых уровнях эти изменения выражены наиболее отчетливо, что позволило выделить последовательные пояса растительности — предгорный (или холмистый) горный, субальпийский, альпийский и нивальный. Эти понятия включают в действительности климатические и флористические признаки, весьма разнящиеся от массива к массиву В горах средних широт предгорный и горный пояса характеризуются лесами из лиственных пород, субальпийский пояс — хвойными лесами, альпийский пояс проходит от верхней! границы древесной растительности до постоянных снежников. Высотные границы и господствующие виды поясов различны от массива к массиву. Рассмотрим последовательную смену поясов на примере Альп и Пиренеев:
Таблица 10
Верхняя граница, м Климаксные сообщес
Центральные Альпы
Альпийский пояс Субальпийский пояс Горный пояс Предгорный пояс 2500-3200 1700-2300 1300-1700 600-800 Луга Г Лес из листвеш.иц, кедровой { и горных сосен [ Лес из елей Пихтово-буковый лес Дубовые и каштановые рощи
Восточные Пиренеи
Альпийский пояс
Субальпийский пояс
Горный пояс
Пояс равнинный и холмистый
3000
2400
1600
1000
Луга
Лес из горных сосен
Пихтово-буковый лес
(Лес из дубов листопадных, лес из каменного дуба, [ лес из пробкового дуба
Были попытки дать этой терминологии более широкое толкование, распространив ее на горы более низких широт на основании физиономических, чисто поверхностных характеристик.
Что касается горного климата, то он представляет видоизменение регионального климата с высотой, и Амберже предложил соответствующую климатическую терминологию. Например, в
221
средиземноморский высокогорный пояс он включает части марокканского Атласа, расположенные выше 2000 м, с семиаридным климатом и летней засухой, с флорой и фауной, отличной от аналогичной для тех же высот в Альпах. Точно так же высокогорья экваториальных областей характеризуются ровной температурой и повышенной влажностью, типичной для этих областей.
Границы поясов не проходят по горизонталям, их высота варьирует под влиянием различных климатических условий. В Альпах границы поднимаются от периферических массивов к центральным, где климат характеризуется меньшей влажностью, высокой инсоляцией и более высокими летними температурами. В условиях этого континентального климата наименее
Северные Южные
Предельны Центральные Альпы Предельны
2400
2200
2000
1800
1600
Рис. 62. Соотношение высоты верхней границы древесной растительности со средней температурой июля 10°, с севера на юг в Центральных Альпах, (по Брокман-Ерошу, Brockman-Jerosch),
требовательным деревьям обеспечен необходимый минимум — три месяца активной вегетации на наибольших высотах благодаря более высоким дневным температурам и наибольшей продолжительности солнечного сияния. Изотерма 10° наиболее теплого месяца — июля, рассматриваемая как показатель необходимого минимума для роста деревьев, в действительности ниже их высотной границы в Центральных Альпах и выше этой границы в периферических цепях (рис. 62). В средних широтах, например в Тибете, с резко континентальным климатом, деревья достигают наибольшей высоты 4500 м. По направлению к полюсам граница леса прогрессивно снижается от 1500 м в Юре до 250 м в северной Норвегии, и затем теряется в субарктической тундре.
Переход хвойного бореального леса в субарктическую тундру, так же. как и переход субальпийского леса в альпийскую тундру, прогрессивен. В обоих случаях прослеживается переход
222
ная зона типа пустоши, представленная низкорослыми кустарниками. В Альпах это преимущественно рододендроны, верески, Vaccinium, в сочетании с низкорослыми или увечными деревьями; этот верхний горизонт альпийского пояса —«зона сражений» (Kampfgegend) последних деревьев.
В небольших по высоте массивах Западной Европы, Центральном массиве и Вогезах граница древесной растительности образована горными буковыми лесами, на высотах 1500 и 1300 м соответственно. Пустоши и луга, следующие за ними, физионо-мически аналогичны пустошам и лугам альпийского пояса, но в действительности занимают место субальпийского пояса хвойных лесов, здесь отсутствующего. Поэтому Флао считал эти формации «псевдоальпийскими».
Условия существования. По разным причинам условия существования для организмов суровы.
1. Уменьшенное давление кислорода оказывает на больших высотах сильное влияние на гомотермных животных. Млекопитающие, способные обитать на больших высотах, немногочисленны; однако в Тибете тем не менее як и горный козел (Ibex)1 достигают высоты 5800 м. В Тибете же человек вел оседлый образ жизни выше всего (4860 м) и гонял стада еще выше (5500 м). Все-таки речь идет о популяциях, адаптированных к разреженному воздуху: Гумбольдт в начале прошлого века констатировал у индейцев высоких Анд большую емкость легких, чем у индейцев низких высот.
2. Прямое солнечное излучение, в связи с разреженностью воздуха, интенсивное; в то же время рассеянное излучение уменьшено.
3. Средняя температура воздуха понижается с высотой на 1° на каждые 140 м в Центральных Альпах, на 165 м — на Кавказе, на 200 м — в экваториальных Андах. В различных массивах на нижней границе альпийского пояса средняя годовая температура различна. В Центральных Альпах она составляет от —2° в центре до +3° в периферических частях. Причина этого явления была вскрыта выше.
Низкие зимние температуры оказывают неблагоприятные физиологические воздействия только на участках, где снег не задерживается; к тому же организмы зимуют при температурах немного ниже 0° На ход температуры в бесснежный период вследствие значительной роли прямого излучения сильное влияние оказывает экспозиция.
4. Влажность воздуха подвержена сильным колебаниям. Изолированные вершины, горные цепи, экспонированные прямо к океаническим ветрам, окутаны облаками и получают осадки, увеличивающиеся по мере подъема влажного воздуха вверх по склону.
Ibex — подрод рода козлов Capra. — Прим. ред.
223
Напротив, такие обширные приподнятые плато, как Тибет, высокие Центральные Анды, Большой Бассейн в США аридны.
5. Снежный покров более или менее обилен в зависимости ог зимней влажности. Распределяется он крайне неравномерно в связи с неровностями рельефа, так гребни остаются голыми, а снег скапливается позади них — в укрытых от ветров расщелинах и ложбинах, где образуются снежники, сохраняющиеся до лета и резко сокращающие период активной вегетации.
6. Наконец, ветер, часто очень сильный, является серьезным препятствием для птиц и насекомых, которые способны летать только в защищенных пространствах или в безветренную погоду.
Для млекопитающих к трудностям, связанным с климатом, добавляются естественные препятствия, так как здесь изобилуют крутые склоны, обрывистые стены, неустойчивые осыпи. Серны, ибексы и козлы, обитающие в высокогорьях, приспособились к этим опасным местам.
В почвах альпийского пояса, как и арктических тундр, проявляются процессы вспучивания и широко распространенные склоны превращают их в объекты эрозии. Устойчивые почвы развиваются путем накопления органического вещества при кратком периоде оттаивания, препятствующем полному разложению, и при окислении под воздействием значительных осадков (рис. 43). Конечное звено — альпийская гумусовая почва, или альпийский ранкер1. Таким образом, альпийская среда обнаруживает черты большого сходства, и притом существенно важные для жизни, с тундрой высоких широт: микротермный климат, продолжительный снежный покров, почвообразование, регулируемое физическими процессами и органическим веществом. Но альпийская среда характеризуется низким атмосферным давлением, особенностями годичного цикла продолжительности дня, интенсивностью излучения, более высокими максимумами летней температуры, пересеченным рельефом.
Растительность. В ней представлена значительная часть видов, общих с циркумполярной тундрой, что является следствием существования ледниковых периодов, но также имеются многочисленные собственные виды; эти флористические особенности и их причины рассмотрены в первой части.
Значительное число форм, представленных в альпийском поясе, перебралось сюда из нижних поясов; некоторые встречаются даже на равнинах: однако в альпийской среде они характеризуются обычно иной морфологией, для которой среди прэ-
1 Ранкер, согласно определению О. С. Гребенщикова, — почва, состоящая из одного гумусового горизонта (грубого гумуса), лежащего непосредственно на кремнистой материнской породе (см.: О. С. Гребенщиков, Геоботани-. ческий словарь, М., 1965). — Прим. ред.
224
чего типичны карликовые формы. Боннье (G. Bonnier) проанализировал в классических экспериментах морфологические модификации, последствия измененного метаболизма, вызванные альпийским климатом. Однако в естественной флоре высогорий формы какого-либо вида не являются простыми аккомодациями, а имеют особую генотипическую конституцию.
Альпийская среда, как и любая другая неблагоприятная для жизни среда, допускает только виды, относящиеся к жизненным формам, хорошо к ней адаптированным. Биологический спектр альпийского пояса Альп, так же как спектр гренландской тундры, не включает ни одного вида дерева, низкие деревца, образующие основу растительности верхнего субальпийского пояса, здесь крайне редки. На камнях и скалах широко представлены плотные подушки хамефитов, принадлежащие к разным семействам, устойчивые против снега, ветра, обнажения корневых систем. У гемикриптофитов почки защищены чехлом сохраняющихся прошлогодних листьев. Однолетники чрезвычайно редки в силу непродолжительности периода роста. Эта краткость благоприятна для растений с мощными способностями к вегетативному размножению, этой способности лишены некоторые виды, сохраняющие листву, как Ericaceae, или горечавки (Gen-tiana), образующие уже осенью бутоны цветков, развивающиеся вслед за исчезновением снега следующей весной (С. Fabar-ger).
Физиономически монотонные растительные сообщества флористически различны. Размещение их строго подчинено рельефу, воздействующему на микроклимат, продолжительность снежного покрова, ветер, дренаж, физические и химические свойства субстрата. Таким образом, если здесь можно распознать прогрессивно развивающиеся серии, такие, как серия, ведущая от известняковых лугов к кислым, в местах, где склоны достаточно пологи для формирования мощной почвы (рис. 43), в действительности существует и мозаика многочисленных постоянных сообществ: сообщества известняковых и кремнистых скал; сообщества осыпей, различающиеся в зависимости от гранулометрического и химического состава; луговые сообщества, различающиеся в соответствии с субстратом, микроклиматом и продолжительностью залегания снежного покрова; концентрические пояса сообществ вокруг ложбин со снегом, озер, торфяников и по берегам потоков.
Жизнь животных. В отличие от растений среди животных альпийского пояса мало видов, общих с видами циркумполярных тундр. Большинство видов к тому же распространяется в нижние пояса. Возможности их жизни в альпийском климате реализуются различным образом в разных группах.
Бескрылых насекомых в связи с сильными ветрами пропорционально больше. Поскольку их активность лимитируется
9 Ж. Леме
225
температурами выше О01, ночные виды отсутствуют или же ночная активность заменена дневной.
Краткость периода активности сокращает число поколений. У таких бабочек, как капустница и махаон, дающих на равнине два поколения в год, в альпийском поясе остается одно; другие, как Pieris callidia, завершающие на небольших высотах развитие за год, в высоких Альпах осуществляют его за два года.
Высота, которой достигают амфибии в Альпах, определяется равенством периода существования незамерзших вод и продолжительности их личиночной стадии.
Лягушка (Rana temporaria), у которой продолжительность личиночной жизни составляет 85—95 дней, поднимается до 2500 м, в то время как жаба (Bufo vulgaris), с периодом личиночной стадии 110—120 дней, поднимается не выше 2000 м.
Редкие рептилии альпийского пояса представлены здесь живородящими формами; таковы три альпийских вида: гадюка, медяница и живородящая ящерица; в Центральной Азии и Центральной Америке роды ящериц представлены яйцекладущими или яйцеживородящими видами на равнине и живородящими в высокогорьях. Из яиц, отложенных здесь в определенном месте и не получающих достаточного тепла, детеныши не вылупляются, хотя самки подыскивают наиболее благоприятные условия микроклимата.
Гомотермные животные более независимы от температуры, они либо проводят зиму в норах под снегом, где они впадают в спячку, как сурок, либо мигрируют в лесную зону, как серна и лань.
Почти все млекопитающие альпийского пояса травоядны; снежный барс Центральной Азии и росомаха Скалистых гор — исключение.
Аридные зоны
Аридные зоны — это территории с ничтожной или очень малой плотностью активной жизни. Если довольно легко представить себе пустынный пейзаж, с его своеобразной морфологией, разреженной растительностью, бедностью животного мира и очень своеобразными формами деятельности человека, то точное определение, несомненно, иллюзорно в силу различий используемых критериев и непрерывности градации последних по степени аридности. На этом основании мы не отождествляем формации, рассматриваемые в этой главе, с пустынными, понимание которых разными авторами более или менее широко.
1 Ногохвостки (Collembola) составляют исключение; они очень многочисленны в альпийских формациях, до уровня вечных снегов, как, например, Jsotoma saltans, которую находят в Альпах на высоте 3800 м, где она живет за счет пыльцы, приносимой ветром.
226
Аридная среда
Аридные климаты. Существуют климатические факторы, определяющие аридность; пытались так же объективно охарактеризовать ее с помощью климатических данных. Нам известно, что эффективная влажность климата зависит одновременно от осадков и испаряющей способности воздуха, наилучшее представление о которой дает потенциальная эвапотранспирация.
Одним из индексов, наиболее широко используемых для аридных регионов, является индекс влажности Торнтвейта (Thornthwaite, 1948), введенный для обоснования классификации климатов:
f ___ ЮОе - 60d lfl~ ЕТр
где ЕТР — потенциальная эвапотранспирация, е — избыточная влага осадков во влажные периоды (удерживающая способность почвы произвольно принята за 100 мм), d — дефицит влаги в сухой период. Индексы в значении —20 и —40 характеризуют семиаридные климаты, индексы ниже —40 климаты аридные. Мейгс в 1952 г. использовал этот критерий при составлении карты распространения аридных климатов земного шара (рис. 63), различая среди прочих зоны экстремально аридные или гипераридные, где осадки не имеют сезонного ритма и где на протяжении всего года не выпадает дождей; для этих зон индекс ниже —57.
Согласно различным авторам, пустыни соответствуют совокупности аридных и гипераридных зон, определенных таким образом (Meigs), или только зонам гипераридным (настоящие пу« стыни Амберже).
Эти годичные характеристики не дают точного представления о степени биологической сухости, которая зависит от сезонного соотношения осадков и нерегулярности их выпадения по годам. Для аридных зон характерны различные режимы выпадения осадков, влияние которых на условия жизни велико:
1) пустыни с осадками в холодный период (средиземноморский режим): северная Сахара (например, Гардая, рис. 64), север Аравии, Иран, Каракумы, Карру, запад большой пустыни Виктория в Австралии, чилийское побережье у 30° южной широты, пустыни Сонорская и Мохаве между США и Мексикой;
2) пустыни с осадками в теплый период (тропический режим): южная Сахара (например, Тесалит, рис. 64), Сомали, пустыня Тар в Пакистане, побережье Анголы и Калахари, пустыня на севере Австралии, пустыни Чиуауа и Тамолипан на севере Мексики;
3) пустыни с дождями во все сезоны: Арало-Каспийская низменность (например, Астрахань), пустыни Гоби, Намиб, запад пустыни Виктория, Патагония, андийское плато (пуна) Перу, севера Чили и Боливии;
4) пустыни с нерегулярными осадками, или экстрарридные, по Мейгсу: Центральная Сахара (например, Ин-Салах, рис. 64), юг Аравии, Такламакан в Центральной Азии, побережье Перу и севера Чили.
9*
227
Рис. 63. Карта мирового распределения аридных зон земного шара (по Мейгсу, Р. Meigs).
1 — семиаридные зоны; 2 — аридные зоны; 3 — гипераридные зоны.
Термический режим аридных зон очень различен в силу их значительного широтного простирания: Арало-Каспийская и Центральноазиатская впадины, Большой Бассейн в Кордильерах характеризуются резко континентальным климатом с суровой зимой, биологическое значение которой тем более велико, что снежный покров отсутствует или ничтожен.
Гордая, 526 м 21,4°
32°с.ш. 68 мм
Рис. 64. Омбро-термические диаграммы трех сахарских станций, расположенных с севера на юг (по Вальтеру и Лигу, Н. Walter, Lieth).
Зима средиземноморских аридных зон прохладна. Вблизи эк а юра постоянно тепло, и сезонная периодичность зависит от периодичности дождей. Равным образом, большое биологическое значение приобретает дневной ход температуры, большая амплитуда которого определяется интенсивным излучением, малой облачностью и сухостью почвы.
Атмосферная влажность в аридных зонах вообще очень низка на удаленных от моря территориях, но увеличивается по соседству с ним, что выражается в росах, совсем немалых, как
229
в мавританской Сахаре и на побережье Намиб, или даже в образовании настоящих туманов. Эти зимние туманы оказывают существенную поддержку растительности, которая в результате оказывается здесь сильно измененной: таковы «оазис туманов» Эркови в центре пустыни Нубии, где конденсируется влага иэ воздуха, приносимого с Красного моря, и пустыня Тихоокеанского экваториального побережья Южной Америки, увлажняемая за счет конденсации влаги, обусловленной холодным прибрежным течением, благодаря чему ксерофильные кустарники» суккулентные или колючие, покрыты эпифитами. Некоторые части этого последнего района, лишенные настоящих осадков, целиком зависят от атмосферной влажности и при отсутствии деревянистых растений бромелиевые, особенно эпифиты рода Til-landsia, произрастают прямо на почве с повернутой к морскому воздуху розеткой листьев.
Наконец, ветер является немаловажным элементом пустынных климатов, что связано не столько с его силой, сколько с постоянством на уровне почвы, связанным с недостатком растительных преград. Некоторые ветры, теплые и сухие, как сахарский сирокко или египетский хамсин, оказывают определенное физиологическое воздействие.
Различие субстратов. В аридном климате особенности топографии и свойства субстрата оказывают особое влияние на плотность и локализацию организмов.
В менее засушливых участках процессы почвообразования приводят к формированию почв, известных под названием серых, красных полупустынных и сероземов. Эти почвы очень бедны органическим веществом и представлены горизонтами, уплотненными аккумуляцией глины, извести или гипса. Но в результате развития эрозии на громадных пространствах обнажаются материнские породы или древние известняковые и гипсовые «коры»» сформировавшиеся в условиях более влажного климата; в дру-гих местах накапливаются отложения, образующие неподвижный субстрат.
В гипераридных зонах эоловые процессы или деятельность локальных водотоков лежат в основе всех поверхностных образований. Зоны абляции представлены каменистыми и скалистыми плато, называемыми в Сахаре гамада, и обширными аллювиальными пространствами, скатами, областями разлива вод, мелкий материал которых вынесен на поверхность ветром и известных в Сахаре под названием рег (фото IV, Б).
Отложения регов вблизи поверхности часто сцементированы известковыми и гипсовыми конкрециями.
Зоны эоловой аккумуляции занесены песком более или менее значительно: простые тонкие покровы, изолированные микродюны, нарастающие вокруг деревцев (небкха в Сахаре, фото IV. А), дюны, имеющие форму полумесяца, или барханы, обширные пространства вытянутых в длину дюн, образующих эрги.
230
В замкнутых депрессиях, области разлива вод, накапливаются отложения водотоков, вызываемых редкими ливнями, илы и глина, незасоленные в дайях, обогащенные растворимыми солями в себкхах, где развиваются засоленные почвы, описанные в предыдущей главе. Водный режим этих различных субстратов так же весьма различен. Крупные несцементированные отложения гамад, осыпей, эргов полностью поглощают влагу осадков, скапливающуюся на глубине, тогда как сцементированные реги, илистые дайи водонепроницаемы. Топография играет роль в распределении влаги дождей путем концентрации вод подземного стока и потоков в депрессиях и тальвегах. Водотоки вади временны, хотя иногда ежегодны, и питают грунтовые воды аллювиальных отложений. Сухость, и более ограниченно, тенденция к засолению и эоловой абразии — главные эдафические факторы, неблагоприятные для жизни в аридных регионах.
Флора
Флора аридных регионов родственна флоре соседних территорий. При высоком видовом эндемизме здесь, напротив, мало самобытных родов. Различия в климатических режимах и ме-
Р и с. 65. Сезонное развитие массы транспирирующих органов некоторых многолетних видов палестинской пустыни (по Оршану, G. Orshan).
Гистограмма отображает осадки за месяц.
Aha — Artemisia herba alba; На— Haloxylon articulatum; Nm — Noea mucronata; Zd— Zygophyl-lum dumosum.
стных условиях топографии и субстрата в этих регионах способствовали формированию здесь относительно богатой флоры, относящейся к различным жизненным формам и образующей различные типы растительности. Жизненные формы растений пустыни относят к двум противоположным группам. Одни, избегая засухи, проходят целиком цикл развития во влажные сезоны; Другие, хотя бы частично, переносят длинный период засухи в состоянии метаболической активности. К первой группе относят однолетние и многолетние виды с постоянными подземными органами.
231
Виды однолетние. Пустыня покрывается их цветками после достаточно интенсивных дождей. Это ашеб Сахары. Большинство этих терофитов распускает цветки, как только сформируется несколько листьев, семена следующей генерации созревают очень быстро: Bouteloua aristidoides, летний однолетний злак Калифорнии рассеивает семена четыре недели спустя; Воег-havia repens, зимний однолетник Сахары, может образовывать первые семена спустя десять дней после прорастания.
Эфемероиды. Они немногочисленны, однако исключительно приспособлены к аридному климату; их подземные органы развивают листья и цветки в течение нескольких недель влажного сезона; таковы корневища Carex physodes и С. desertorum в Туркестане, луковицы Dipcadi в Сахаре.
Многолетние виды с надземными постоянными органами. Их относят к различным жизненным формам, среди которых различают следующие:
1) деревья и высокие кустарники, корпи которых проникают на большую глубину до постоянного горизонта грунтовых вод, в то время как листья редуцированы и имеют приспособления против транспирации: это акации Сахары и Австралии;
2) низкие кустарники, панофанерофиты и хамефиты, период активного роста которых приходится на прохладный и влажный сезон, с длинными ветвями, снабженными большими листьями; в начале сухого сезона их рост замедляется, развиваются короткие ветви, несущие мелкие ксероморфные листья, и наконец, к середине или к концу засушливого периода листья отмирают в акропетальной последовательности \ так же, как иногда и часть ветвей (рис. 65). Следует отметить очень многочисленные виды этой группы: полыни, смирновию (Smirnovia turkestanica), Larrea divaricata (креозотовый куст североамериканских полупустынь);
3) безлистные или с редуцированными чешуйчатыми листьями кустарники, такие, как Ephedra, Calligonum, Arthrophytum («саксаулы» Туркестана), Anabasis articulata, Haloxylon articulatum, которые сокращают транспири-рующую поверхность в сухой сезон путем опадания молодых длинных ветвей или высыхания корковых тканей (рис. 53). Промежуточная с предыдущей группа образована видами, распускающими очень мелкие рано опадающие листья во влажный период, как ретам (Retama raetarn), или только в молодом возрасте, как колючее крестоцветное Zilla macrocarpa\ эти растения имеют зеленые ассимилирующие стебли;
4) злаки с листьями, свернутыми в трубку, как Aristida pungens (дрин сахарских дюн) (фото IV, А) и бородачи (Andropogon), мощные корни которых, окруженные чехлом из песчинок, проникают в глубь песка;
5) растения суккуленты, кактусы Нового Света, африканские молочаи с резервуарами для накопления воды, с крайне слабой транспирацией, поверхностные корни которых охватывают большую площадь; эта жизненная форма, которая нуждается в снабжении водой через достаточно короткие интервалы, выпадает в гипераридных зонах.
1 От верхушки побега к его основанию. — Прим. ред.
232
Фауна
Жизнь животных в аридных зонах сопряжена с теми же трудностями, что и жизнь растений, это проявляется в различных формах их поведения.
Снабжение водой. Различные потребности в водоснабжении— главный фактор в распространении наземных животных аридных зон. Животные, которые испытывают регулярную потребность в питьевой воде, в пустынях отсутствуют или поселяются вблизи постоянных источников воды. Таковы, во-первых, многочисленные млекопитающие, как слон, носорог, лев, леопард, которые обитали в степной части Сахары в плювиальные эпохи, а ныне оттеснены к ее периферии; во-вторых, рукокрьь лые, многие птицы, которые не удаляются от оазисов.
Животные, пьющие нечасто, — обычные хозяева аридных регионов. Среди них хорошо изучен верблюд; антилопам и газелям свойствен тот же тип поведения. Многие животные потребляют только воду других организмов. Среди травоядных отметим двух грызунов: песчанка (Psammomys obesus), роющая норы на юге Орана среди многочисленных суккулептпых растений, которые она поедает; неотома («древесная крыса») (Neotoma albi-gula), не имеющая никаких приспособлений к экономному расходованию воды, но добывающая ее в значительных количествах из кактусов, которыми она кормится. Некоторые из этих травоядных могут довольствоваться сухой пищей, по крайней мере временно, как антилопа аддакс, вес которой может достигать 100 кг, или постоянно, как песчанки (Meriones) — грызуны, которых можно кормить исключительно зерном без потери веса.
Для животных, способных впитывать влагу всей поверхностью тела, атмосферная влажность является фактором, ограничивающим их распространение. Так, моллюски отсутствуют в пустынях, где воздух почти постоянно сухой (вне оазисов), но могут проникать в зоны, где господствуют туманы и росы, как Helix desertorum на атлантических побережьях Сахары.
Защита от перегрева. В жаркие часы дня активность на поверхности почвы, температура которой может достигать 65°, приостанавливается. Поскольку летальная температура в общем колеблется между 40 и 50°, вынужденное извлечение на поверхность почвы приводит к гибели. Продолжительность периода высоких температур, который способно перенести животное, зависит от его цвета и роста; черный цвет, распространенный У насекомых Сахары, не является, однако, пагубным, так как их пигменты перехватывают большую часть инфракрасных лучей. Животные избегают перегретой поверхности почвы временным прекращением активности и защитой от инсоляции. Именно поэтому многим видам: грызунам, рептилиям, многочисленным насекомым, присуща ночная активность, многие из них, как
233
пустынный еж, тушканчики и песчанки, в жаркий период впадают в спячку, известную под названием летней спячки L
В качестве защиты от дневной жары достаточно укрыться в норах, под камнями, в трещинах скал или просто в тени растений; птицы, некоторые ящерицы и кузнечики удаляются от поверхности почвы, взбираясь на растения, ящерицы и прямокрылые, обитающие днем на поверхности почвы, обладают длинными лапами, отделяющими их тела от перегретой поверхности. Птицы особенно активны утром и вечером, когда ощущается прохлада; кладка приходится на прохладный сезон во избежание крайнего перегрева яиц, которые родители постоянно накрывают.
Пищевые проблемы. Животные пустынь больше, чем где бы то ни было, зависят от растительности. Развитие растительности после дождливых периодов дает толчок быстрому размножению животных, связанных с этим изобилием, вначале насекомых фитофагов и живущих на цветах, проводящих осталь-* ную часть года в диапаузе, потом насекомоядных и плотоядных.
Число же животных, потребляющих многолетние растения, относительно постоянно, но подвержено, однако, изменениям в более засушливые или влажные годы.
Это очень характерно для стад домашних животных — верблюдов, лошадей, овец, существующих в избыточном количестве за счет зеленых растений, их поголовье падает в очень засушливые годы.
Пищевой режим фауны различен. Некоторые хищные в действительности полифаги в силу необходимости, как фенек и шакал, которые потребляют грызунов, насекомых, растения. Напротив, отмечены газели и песчанки, питающиеся кузнечиками.
Основные биоценозы
Пустыни — крайнее выражение тенденции к засушливости, свойственной климатам прилегающих к пустыням районов. Равным образом и типы растительности, здесь развивающиеся, произошли от типов растительности более влажных районов — степей, саванн, сообществ кустарников, светлых лесов. С переходом к пустыне растительность становится все более редкой, что ведет к очень разреженным сообществам и, в конечном итоге, едва ли не к отсутствию жизни, как в сахарских танезруфтах.
В неблагоприятных климатах местные различия в термических условиях и условиях увлажнения приобретают решающее значение в распространении живых существ, образ жизни и плотность которых существенно изменяются от одного местообитания к другому. Несмотря на различие биогеографических территорий
1 Автор ошибается, включая песчанок в перечень впадающих в спячку животных. Из тушканчиков большая часть видов впадает только в зиМнюк> спячку; некоторые имеют и летнюю и зимнюю спячки. — Прим. ред.
234
и климатических режимов, представленных в аридных регионах, местообитания, сходные по топографическому положению и свойствам субстрата, характеризуются биоценозами, гомологичными по их облику и приспособлениям их компонентов. Охарактеризуем основные биоценозы аридных зон северного полушария.
Сахара, Ближний Восток и Центральная Азия. Сахара характеризуется относительно богатой фауной и флорой, преимущественно средиземноморского происхождения на севере и тропического на юге. Здесь представлено примерно 20 видов ящериц, 70 видов оседлых птиц, 65 видов млекопитающих и т. д. Континентальные пустыни Азии, характеризующиеся большими амплитудами температур в течение года, длинной и суровой зимой, жарким и сухим летом, напротив, гарантируют только короткий период, благоприятный для активной жизни, заключенный между двумя неблагоприятными. Результатом этого является суровый отбор, заметно сказывающийся как на степных систематических группах — грызунах и лошадиных, так и субтропиче-<ких — газелях и антилопах.
Всем пустынным зонам присуще, однако, аналогичное разнообразие топографических форм: песчаные пространства, каменистые плато, каменистые равнины, илистые незасоленные водоемы, .засоленные понижения, аллювиальные долины и равнины.
Дюнные поля. Они осваиваются растительностью, устойчивой к дефляции и засыпанию песком; в сахарских эргах особенно распространены злак дрин (Aristida pungens) и ретам из бобовых (Retama raetam). С этими растениями связана своеобразная фауна. На дрине вьет свои гнезда карликовая малиновка; саранчовое Platypterna живет в дернинах дрина, первичнобескрылые насекомые Thysanura обильны в песке близ основания стеблей. Небкха — это всхолмленные пространства эоловых песков на аллювиальных отложениях, закрепленные Limoniastrum, Salsola, Nitraria, Panicum и т. д.; на юге Алжира эти холмы — местообитания разнообразной фауны: различных насекомых, одного вида ящериц, одного ночного геккона, двух змей, птиц, одного грызуна, роющего здесь норы.
Подвижный характер субстрата способствует выработке у животных своеобразных приспособлений для передвижения. Многочисленные в песках рептилии либо бегают, либо ползают. Первые — ящерицы — исключительно быстрые, их пальцы обрамлены чешуйками. Вторые — змеи и ящерицы — передвигаются как бы плавая с помощью боковых и вертикальных движений туловища, с головой, частично погруженной или поднятой над песком; глаза, ноздри, рот у этих видов защищены покровами от проникновения в них песка; лапы их редуцированы или отсутствуют вовсе, как у Typhlosaurus из Негева. Многие членистоногие обладают волосистыми плюснами или очень развитыми чешуями, облегчающими передвижение по песку !; жесткокрылые насекомые Tenebrionidae сильно сплюснутой формы, с короткими членами
1 В настоящее время наличие на лапках каймы из чешуек, волосяных Щеточек рассматривается нередко как приспособление не к передвижению но песку, а к зарыванию в рыхлый субстрат, — Прим. ред.
235
способны необычно быстро прикрываться тонким слоем песка; скорпионы, пауки роют в подвижном песке углубления, в закрепленном песке — подземные ходы.
Скалистые плато и каменистые равнины. Это соответственно гамады и реги Сахары. Формации очень бедные; это крайне открытые степи1 со злаками (видами Aristida), полукустарниками с опадающей листвой, безлистными маревыми Haloxylon salicornicum (фото IV, Б). В южном Оране встречается любопытный Anabasis aretioides из маревых, с глубоким стержневым корнем и листьями в виде сжатых чешуй, собранных в плотную подушечку.
Эти территории пересечены более или менее опесчаненными ложбинами, где растительность становится более густой, с крупными вкраплениями злаков, полыней, ретама и др. и особенно с акациями в южной Сахаре.
Плотность почвенного субстрата регов препятствует поселению здесь роющих животных.
Глинистые равнины. В силу непроницаемости почв растительность здесь чрезвычайно разрежена.
Засоленные понижения. Как и в других регионах земного шара, галофильная пустынная растительность обнаруживает замечательные черты, к которым присоединяются признаки ксерофитов: безлистность некоторых из Limonium, удаляющих соль с помощью выделительных желез; безлистность саксаулов, любопытных деревцев из маревых в пустынях Туркестана, у которых развиваются весной ассимилирующие травянистые побеги; микрофилия тамарисков (Tamarix), корни которых извлекают влагу из глубинных слоев почвы, а надземные части удаляют соль; микрофилия франкений (Frankenia) с эрикоидными листьями; нередко усиливающееся в течение засушливого периода опадение листьев у Suaeda\ листовой диморфизм с листьями широкими и опадающими во влажный сезон и мелкими и сохраняющимися в сухой сезон, как у Alhagi maurorum, к тому же очень глубоко укореняющейся.
Рыхлые отложения долин и незасоленных пониже-п и й. Долины, в которых иногда скапливаются проточные воды (уэды, вади, арройо), и замкнутые собирающие воды аллювиальные понижения (дайя), обладают в пустынях наибольшей плотностью флоры и фауны. Наиболее богато представлены сообщества злаков, Panicuni, Pennnisetum и т. д. с разреженными акациями и бетоумом (Pistacia atlanticd) в северо-западной Сахаре; в других местах развиваются сообщества колючих кустарников: Zi-ziphus lotus, Zilla macroptera, Lycium aegyptiacum, и при небольшом засолении появляются тамариски (Tamarix) и селитрянка (Nitraria). По мнению Кецеля (Quezel), эта растительность представляет обедненный реликт более влажного климата. К этим местообитаниям приурочена фауна, нуждающаяся в растительной пище, богатой водой, таков Psammomys obesus северной Сахары, роющий норы в дайях и днищах уэдов.
Постоянно влажные понижения. По существу, оазисы не принадлежат к аридной среде, но их сообщества тем не менее своеобразны: рощи
1 Автор понимает термин «степи» весьма широко. С точки зрения большинства авторов, степь — тип растительности с доминированием ксерофиль-ных злаков, не сомкнутой с летним перерывом вегетации. — Прим. ред.
236
финиковых пальм Сахары, тополь евфратский Центральной Азии, вязы восточной Гоби.
Северная и Центральная Америка. К «пустыням» в Соединенных Штатах часто относят внутренние долины Большого Бассейна, получающие от 12 до 25 см осадков в год. Они характеризуются довольно густой растительностью с господством полыни Artemisia tridentata и могут возделываться без орошения. По существу это полынные степи, которые будут рассмотрены дальше. Южнее расположены другие бассейны, более засушливые и жаркие; пустыня Мохаве, со знаменитой Долиной Смерти, южнее пустыни Сонора, Чиуауа, Тамолипан — по большей части мексиканские. Сумма годовых осадков колеблется от 7 до-15 см, но сезонное их распределение различно: они выпадают либо зимой или ранней весной, как в пустынях Мохаве и Соноре,, либо в конце лета и осенью, как в двух последних; иногда, особенно в пограничной полосе, осадки выпадают в два периода.
Среди растений господствует «креозотовый куст» (Larrea divaricata), высокий кустарник от одного до полутора метров, покрытый мелкими листьями, частично не опадающими в сухой сезон, с глубоко проникающей корневой системой; кусты эти отстоят друг от друга па расстоянии от 5 до 15 м, но корни их, как правило, охватывают большую часть этих мнимых пустот. Часто совместно с этим видом встречается другой, более мелкий кустарник Franseria dumosa, так же с неопадающей листвой. Этим видам сопутствуют и другие жизненные формы: однолетние эфемеры влажного сезона, суккуленты из семейства кактусовых, древовидные юкки.
Подножия склонов и днища долин с почвами более влажными и плодородными несут иную растительность, среди которой на юге господствует «мескит» (Prosopis), древовидное растение из бобовых, достигающее 12 м в высоту, в сочетании с акациями, Parkinsonia, Celtis. Крутые склоны и каменистые холмы одеты разнообразной суккулентной растительностью: в северных частях особенно характерны агавы, Dasylirion, юкки и опунции, в-более южных частях — кактусы, среди которых примечателен гигантский кактус Carnegia gigantea, достигающий 15 м, удивительная Fouquieria splen-dens, пучкообразпые стебли которой, сукулентные и колючие, несут листья только во влажный сезон.
Фауна, отличающаяся в видовом отношении от фауны Старого Света,, имеет те же этологические и адаптивные свойства. Во влажный период обильны насекомые. Грызуны многочисленны и размещаются в зависимости от своих пищевых потребностей: одна из «древесных крыс» (Neototna albigula)* не способная экономить воду и сопротивляться обезвоживанию, питается кактусами; грызун перогнат (Perognathus penicillatus), испаряющий в тех же-условиях в 3 раза меньше воды, чем белоногий хомячок (Peromyscus tornil~ lo), обитает в наиболее сухих местообитаниях, в то время как второй размещается в травянистых, более прохладных понижениях. Койоты способны оставаться некоторое время без воды, питаясь зайцами и яйцами. Хотя птицы в общем нуждаются в регулярном утолении жажды, один вид, Carpodactus-mexicanus, способен существовать только за счет воды суккулентных растений»
237
которыми он питается. Тесная привязанность животных к некоторым пустынным растениям может быть показана на примере гигантского кактуса Carnegia gigantea, плоды которого потребляют примерно 50 видов птиц и который дает приют двум видам дятлов, устраивающих в его дуплах гнезда; когда же дупла пустеют, в них располагаются другие виды, в особенности ночные и дневные хищники.
Роль человека
При низкой плотности населения в пустынях присутствие человека, однако, проявляется в нарушениях, вызванных экстенсивным выпасом скота и использованием естественных растительных ресурсов. Пастух в аридных зонах — неизбежно кочевник, постоянно перемещающийся с места на место в поисках пастбищ, где выпали сезонные или случайные дожди. Вследствие незначительной густоты растений и краткости периода их роста выпас скота, даже временный, ощутим для видов, предпочитаемых4 животными; в результате эти виды порой сведены до минимума. Эоловая эрозия, один из бичей пустыни, усиливается, укрытие для молодых проростков уменьшается. В Сахаре насаждения из акаций, бетоума (Pistacia atlantica), где не встречаются молодые особи, в результате заготовки корма и дров обречены на исчезновение. Acacia апеига, или мелга (mulga) австралийской пустыни, так же исчезла в некоторых местах.
Оставшиеся виды, напротив, могут распространиться в результате уменьшения конкуренции. Так, американские экологи полагают, что кустарниковые формации сонорских пустынь заменили в результате выпаса и огня злаковые прерии (Bouteloua, Aristida, Hilaria).
Кое-где человек может уменьшить интенсивность распространения некоторых видов, собирая их плоды и семена в качестве добавочного корма или запаса на более сухие годы; таковы в Сахаре урожаи колосьев проса (Panicum), дрина (Aristida pun-gens), креба (Urochloa helopus), семян тауи (Aizoon canariense), плодов ююбы (Zizyphus), бутонов каперсов и т. д.
Степи
Можно определить степь как тип растительности, характеризующийся преобладанием ксероморфных, с интенсивным укоренением злаков, не покрывающих поверхность почвы сплошь, в промежутках могут развиваться различные жизненные формы: однолетние, луковичные геофиты, многолетние травянистые двудольные, иногда полукустарники.
Определенные таким образом степи существуют в двух весьма различных типах климата, для которых характерны периоды продолжительной засухи. Это континентальные климаты умеренных широт с суровой зимой и засушливым концом лета,
238
а также средиземноморские субаридные климаты, с зимой мягкой или холодной, но с продолжительным сухим периодом.
Термин «степь» распространяют на различные формации субаридных тропических климатов, представляющие климатические переходы к другим типам растительности или стадии их деградации. Таковы сахельские «степи» по южным границам Сахары, субаридные регионы юго-западной Африки и Австралии, представляющие крайне разреженные сообщества кустарников, часто колючих, промежутки между которыми заняты однолетними или многолетними низкорослыми злаками, что приближает их в этом случае к саваннам. Трошен (Trochain) предложил для этих формаций термин псевдостепи (фото VI, А).
Географическая и климатическая приуроченность
В Евразии степи образуют полосу, протянувшуюся от Молдавии и Украины к восточной Монголии, между листопадными, иногда хвойными лесами на севере и аридными и пустынными зонами на юге. Венгерские пушты представляют изолированный энклав (остров) степей в Центральной Европе. Для них характерен резкоконтинентальный климат с суровой зимой, с различной, иногда достигающей ста дней в году, продолжительностью снежного периода, теплое лето. В этих условиях среднегодовая температура имеет небольшое значение, как показывает ее колебание: от 0,5° в Сибири до 9° на Украине и даже до 11° в Венгрии. Осадки невелики: от 25 до 50 см; распределяются в течение всего года, летом, однако, их количество меньше величины потенциальной эвапотранспирации. Недостаточное количество осадков усугубляется низкой относительной влажностью воздуха (менее 50% в августе) и постоянными ветрами.
В Северной Америке степные формации занимают широкую срединную полосу от Манитобы и Саскачевана на севере до Мексиканского залива на юге и от Мичигана на востоке до Скалистых гор на западе. Эти формации в их восточной части называют «прериями», в западной — «Великими равнинами» (фото V,А). Изолированные острова степей прослеживаются и западнее; «тихоокеанская прерия» Калифорнии, «прерии Па-лузы» на северо-западе США. В степях СССР градиент температуры и сухости возрастает с севера на юг, тогда как термический градиент и градиент увлажнения в степях Северной Америки ориентированы различно: первый с севера на юг, второй с востока на запад, в связи с существованием барьера тихоокеанских горных цепей, задерживающих влажные ветры. На востоке граница между прерией и лесом приходится в северной части на районы с годовым количеством осадков от 60 до 75 см и от 80 до 100 см в южной части; переход к аридным формациям Запада происходит при 20—25 см осадков. Их сезонное распределение также весьма различно: на севере 3/4 осадков выпадают
239
в вегетационный период, с апреля по сентябрь, в Аризоне только половина, в Калифорнии от 10 до 20%, дожди здесь зимние.
В Канаде зимы длинные и суровые, средние температуры достигают 15°, период активной вегетации меньше 4-х месяцев, тогда как в Техасе и Калифорнии, где средняя годовая температура составляет 18°, отмечаются только редкие заморозки.
Итак, североамериканская прерия охватывает территории, характеризующиеся большим различием климатических условий, соответствующих, несомненно, лесному типу растительности в наиболее влажной зоне. Таковы канадские острова прерий в зоне хвойных лесов и полуостров прерий в Иллинойсе среди листопадных лесов. Речь идет об остатках степи, занимавшей обширные территории в «ксеротермическую» эпоху древнего по-слеледниковья и уцелевшей от поглощения лесом благодаря применению индейцами палов и последовавшему затем осушению почвы в связи с сельскохозяйственным использованием этих регионов.
В Южной Америке пампасы Аргентины, степи, подвергающиеся интенсивной эксплуатации и получающие более 60 см осадков при среднегодовой температуре 14—17°, также рассматривают как реликт, не соответствующий условиям современного климата. Южнее, в Патагонии, подножье андийских Кордильер и побережье залива Сан-Хорхе в условиях климата с холодной зимой и распределенными в течение всего года осадками покрыты степями *.
Почвы степей
Их свойства весьма тесно связаны со степными биоценозами. Разветвленные и глубоко проникающие в почву корни злаков ежегодно частично отмирают, обогащая почву органическими веществами, содержащими азот и минеральные элементы. Они образуют насыщенный гумус, который с частицами глины дает мелкохлопьевидное соединение; отсюда комковатая структура, создающая великолепную проницаемость и аэрацию почвы. Соединение с глиной в то же время «защищает» гумус от минерализации бактериями, это объясняет его плодородие. Для роющей фауны степей характерна большая активность, что приводит к сглаживанию различий в распределении гумуса по профилю. Отсюда термин изогумусные, данный степным почвам.
Тесная зависимость почвы и степной растительности подчеркнута, на границе степи с лесом, быстрой эволюцией почвенных профилей в бурые лесные почвы при лесоразведении и обратным ходом процесса, если восстанавливается травянистая формация.
Климатические условия воздействуют на свойства почвы частично непосредственно, через движение почвенных растворов,
1 По современным представлениям большая часть Патагонии покрыта полупустынями. — Прим, ред,
240
особенно извести, по профилю, частично воздействуя на густоту растительного покрова.
Таким образом, различают:
1) черноземы, или «черные земли», в срединной зоне континентальных степей с холодным сезоном, достигающие при оптимальном развитии мощности порядка 1,5 м, с темным бескарбонатным горизонтом от 60 до 80 см, в основании которого появляются известняковые конкреции; в направлении к климатам более влажным или более сухим они постепенно сменяются следующими почвами;
2) в более влажной переходной к лесу зоне в черноземах обнаруживается менее мощный гумусовый горизонт и начальные стадии миграции глины и железа под действием ненасыщенных гуминовых кислот, имеющих тенденцию к рассеиванию; это выщелоченные, или северные черноземы русского почвоведа Тулина;
3) в США почвы восточных прерий при более влажном климате еще менее насыщены, с нижним бескарбонатным горизонтом, уплотненным благодаря обогащению глиной; это «почвы прерий» Марбута (Marbut), или брюнизем (черноземовидные почвы прерий);
4) вблизи аридных зон растительность более разрежена, почвенные профили менее мощные, менее гумусированные, повышается содержание карбонатов в почве; это вначале южные черноземы, затем каштановые почвы;
5) в средиземноморском климате степные почвы отражают существенные климатические изменения недавнего четвертичного периода (полициклические почвы); мощный гумусовый горизонт одновременно красноватый, залегающий на горизонте накопления извести; для этих почв, близких к каштановым, И. П. Герасимов предлагает термин «почвы маррон», коричневые почвы сухих лесов и кустарников, во временно гидроморфных условиях понижений они более темные и переходят в тирс — черные слитые почвы.
Растительность
В ней представлены особые биологические типы, объединенные в формации, систематический состав и физиономический облик которых изменяется со свойствами климата и биотическими влияниями. Господствующей жизненной формой являются многолетние ксерофильные злаки, особенно относящиеся к родам ковыль (Stipa), пырей (Agropyron), тонконог (Koeleria), овсяница (Festuca), бородач (Andropogon), волоснец (Elymus). Их глубокие и сильно разветвленные корни очень эффективно впитывают влагу из почвы и достигают горизонтов, в которые проникают атмосферные осадки. Поэтому глубина проникновения корней уменьшается от регионов более влажных (около 2 м) к более сухим (около 0,5 м). Надземные органы развиваются аналогично подземным: в более влажных зонах господствует высокотравная, свыше 2 м, прерия, на востоке США с преобладанием бородачей (Andropogon), уменьшающиеся с возрастанием засушливости высота и биомасса приводят к смешанной, затем
241
пизкотравной прерии, где господствуют Buchloe dactyloides («трава бизонов») и Bouteloua gracilis. Степные злаки обладают склерофильными, сильно кутинизированными листьями, скрученными в трубку или сворачивающимися под действием сухости; в течение лета они более или менее быстро высыхают.
Прочие многолетние растения, травянистые или деревянистые у основания, снабжены мощными корнями, достигающими в более влажных зонах глубины порядка 3—4 м; бобовые и сложноцветные здесь особенно обильны.
Однолетние растения, малочисленные в менее аридных зонах, в более сухих регионах могут получить преобладание во влажный сезон, когда в результате выпаса скота наблюдается изре-живание многолетних видов. В Техасе и Аризоне сообщества летних однолетних злаков, относящихся преимущественно к родам Aristida и Bouteloua, покрывают обширные площади.
В идентичных климатических условиях при большей или меньшей проницаемости почвы изменяется глубина проникновения корней и флористический состав. Так на плотных илах развивается низкотравная степь, на песках — более пышная.
Таблица 11
Чистая продукция
г на 1 м2 за год г на 1 м2 за день вегетации
Высокотравные прерии, Оклахома и Небраска 450 3
Низкотравные прерии, Вайоминг (32 см осадков) 70 0,5
Кустарниковая полупустыня, Невада (12 см осадков) 40 0,2
Кроме распределения корней по разным почвенным горизонтам, различие в периодичности сезонного роста сводит к минимуму конкуренцию между видами одного сообщества. У родов северного происхождения Stipa, Роа, Agropyron развитие раннее, прерываемое наступлением лета, тогда как южные роды, Andropogon, Bouteloua, трогаются в рост позже и достигают максимальной продуктивности в конце лета или осенью. Полное соответствие сообществ местным условиям увлажнения делает их весьма чувствительными к циклическим колебаниям климата; засушливые годы приводят к отступанию более требовательных видов, что благоприятно сказывается на видах менее требовательных, тогда как случаи возврата относительно влажного климата в течение ряда лет благоприятствуют первым, обладающим большей способностью к распространению.
242
Чистая продукция степей сильно варьирует в зависимости от степени аридности климата; в США получены следующие показатели (см. табл. 11).
Жизнь животных
Фауна степей сталкивается с теми же, что и в пустыне, трудностями, хотя и в меньшей степени: сезонные жара и сухость, дефицит поверхностных вод, сильные ветры, периодически недостаточные пищевые ресурсы. Население различных местообитаний, сезонная периодичность, цепи питания замечательно приспособлены к степной среде.
Население различных местообитаний. Условия жизни существенно различаются в зависимости от того, остаются ли виды на поверхности или хотя бы временно ведут подземный образ жизни.
1. Животные, обитающие на поверхности. К этой группе относятся крупные травоядные и хищники — очень подвижные, быстрые, выносливые животные. Травоядные живут более или менее многочисленными стадами, ежедневно перекочевывая с места на место в поисках воды, и сезонно, чтобы избежать холода или засухи.
Таковы различные газели, антилопа сайгак, кулан, дикие лошади— в евразиатских степях; бизон, американские олени и вапити, вилорогая антилопа (Antilocarpa americana) —в североамериканских прериях; гуанако и олень пампы (Odocoileus be-zoarticus)—в степях Аргентины. Первоначальная численность бизонов до их истребления составляла приблизительно 50— 60 млн. голов; перемещались они стадами, насчитывающими более 2000 голов. Вилорогая антилопа, обитающая в более сухих степях, зимой укрывается в близлежащих кустарниковых и лесных формациях.
Степные птицы вынуждены гнездиться на почве и зачастую приспособились к ходьбе, как куриные или дрофа в Азии. Большинство, например журавли, мигрируют на холодный или сухой период в более умеренные или более влажные регионы.
Весьма многочисленные в степях насекомые, особенно саранчовые, после вылупления из яиц живут вне почвы. К ним относятся многочисленные мигрирующие виды, которые уничтожают растительность по пути своего следования, там, где они приземляются; это Locusta migratoria в Центральной Азии, Stauronotus maroccanus в Северной Африке, различные Melano-pus Северной Америки, Schistocerca paranensis в пампасах. Их яйца, отложенные в почву, могут сохраняться в течение нескольких лет, пока не установится благоприятная для вылупления влажность; молодые личинки, не защищенные от обезвоживания, так же нуждаются во влажных условиях; затем совершенно необходимой для них становится сухость воздуха.
243
2. Роющие животные. Обилие их — наиболее яркая черта фауны степных биоценозов. Принадлежат они к весьма различным группам.
Особенно обильны грызуны; в жаркий период они ведут по большей части ночной образ жизни, закрывая иногда отверстия нор па день; в зимний период они впадают в спячку или используют пищу, запасенную в подземных ходах. На разных континентах их виды различны; в евразиатской степи особенно характерны суслик, хомяк, тушканчик, сурок байбак; в американской прерии — луговые собачки (Cynomys), колонии которых могут занимать много квадратных километров, гоферы (Geomys), кролики и т. д.; в Патагонии встречаются туко-туко (Ctenomys).
Хищники, такие, как лисица, койот, также обитают в норах, ведя ночной образ жизни, а земляные белки (Citellus)—дневной.
Даже птицы представлены в степях несколькими видами,, устраивающими гнезда во входах необитаемых нор. Ящерицы и змеи на холодный сезон прячутся под землей.
Из насекомых в степях весьма многочисленны осы, пчелы и муравьи. Многие пчелы и осы живут вне почвы, однако встречаются и роющие. Муравьи строят подземные гнезда, иногда более метра глубиной, и играют весьма активную роль в перемешивании и повышении проницаемости почвы.
Во влажных зонах эту роль выполняют дождевые черви (Lumbricidae).
Цепи питания. Обилие животных находится в тесной связи с густотой растительного покрова, за счет которого существует обильная фауна травоядных, грызунов, насекомых-фитофагов и живущих на цветках. Так, например, степи Аризоны населены колониями луговых собачек, которые потребляют до 87% продукции ее растительного покрова. Бизон съедает почти столько же травы, сколько корова, и на половину больше, чем вапити. В прерии Аризоны на 1 км2 может прокормиться от 6 до 7 бизонов, что равно 3 т биомассы. В действительности у травоядных и грызунов, предпочитаемые корма которых не полностью совпадают, конкуренция оказывается ослабленной: так, если крупный рогатый скот поедает злаки, американские олени и вилорогие антилопы предпочитают двудольные травы и кустарники.
Если крупным травоядным почти не приходится опасаться в степях хищников, кроме волка в Старом Свете, то у грызунов есть многочисленные враги: койоты, лисицы, куньи, ночные хищные птицы, змеи. Некоторые из этих хищников, как и в пустынях, всеядны и могут, как койот и лисица, потреблять растительную пищу.
Травоядные насекомые, как, например, муравьи, — пищевой источник для целой серии вторичных консументов, хищных и паразитических насекомых, ящериц, птиц.
244
Воздействие человека
Интенсивное использование степей человеком началось сравнительно недавно. Здесь вначале получило развитие скотоводство, затем обработка земли. Плодородные почвы обеспечивают пшенице и маису наилучшие условия. Экстенсивное скотоводство ведет к деградации растительности; мелкотравье приходит на смену средним и высоким злакам, распространяются виды пустынных зон; грызуны становятся обильнее, используя больше семян однолетних растений и клубневидных корней; возрастает численность насекомых, особенно кузнечиков и саранчи. Уваров установил, что бедствия, связанные с перелетами саранчи, по большей части порождены перевыпасом, и некоторые виды, до недавнего времени не встречавшиеся в степях, теперь наводнили распаханные территории; наконец, перевыпас ведет к эрозии почвы.
Распашка степи полностью меняет экологические условия. Исчезают многие насекомые, но некоторые приспосабливаются к обработанным полям и становятся их бичом, как земляные блошки, пшеничные трипсы, жуки-щелкуны (Elateridae) юга России. Залежи, следующие за прекращением распашки, представляют очаги массовых размножений саранчовых.
Степные пожары в зависимости от климатических условий оказывают различное воздействие: в пограничных с лесом зонах они, по-видимому, препятствуют распространению последнего, тогда как в более засушливых зонах покров травянистых многолетников уступает место пустынным кустарникам: полагают, что формация трехзубчатой полыни (Artemisia tridentata) и более южная формация с крезотовым кустом (Larrea) распространились за счет прежних «прерий».
Тропические саванны
Ранее (см. гл. 7) при определении саванн как типа растительности мы отличали первоначальное их понимание от расширенного, принятого некоторыми фитогеографами.
Мы сохраняем здесь за ними первоначальное значение тропической формации с развитым травяным ярусом, представленным преимущественно многолетними злаками, поднимающимися на несколько дециметров над поверхностью почвы, с периодом покоя, приходящимся на сухой сезон, с различной примесью низкорослых трав и небольшими деревьями или кустарниками, растущими поодиночке или группами.
Саванны представлены во всех тропических климатах, от самых влажных до самых сухих; они граничат с такими различными формациями, как вечнозеленые, полувечнозеленые или листопадные леса, светлые леса или ксерофильные чащи. Самые
2Ш
большие площади они занимают в Африке, по имеются также в муссонной Азии, северной Австралии и тропической Америке. В горах тропиков саванны могут достигать верхних поясов растительности; так, на горе Камерун между 2000 и 3000 м располагается пояс формаций высоких многолетних злаков, лишенный деревьев: на границе с андийским лесом, к северу от экватора между 3000 и 4000 м во влажном и постоянно холодном климате, располагается парамос, травянистые пространства с ксероморфными злаками, Stipa, Festuca, Calamagrosiis.
Почвы саванн так же различны, как и климат: железистые тропические и ферралитные почвы, с панцирем или без него; латеритные гравий и панцирь, почвы постоянно или временно гидроморфные. Эти экологические различия связаны с различием происхождения, хотя оно и остается в большинстве случаев гипотетическим.
Растительность
В травяном ярусе заметно преобладают один или два фоновых вида злаков, относящихся чаще всего к родам Andropogon, Loudetia, Hyparrhenia, Pennisetum, Imperata, Aristida и в Австралии Themeda, Astrebla, Triodia. Размеры этих растений варьируют: в Африке «слоновая трава» (Pennisetum purpureum) может достигать в высоту нескольких метров, но ее обычные размеры— порядка метра; в зонах с весьма продолжительным сухим сезоном травы ниже и менее густые, распространены однолетники, придающие ландшафту облик степи, и некоторые исследователи считают их тропическими степями, или степными саваннами, или лесостепными формациями, если обильно представлены древесные породы (A. Aubreville) (фото VI,А). Для этих переходных формаций был также предложен термин псевдостепи. Они занимают обширные площади между собственно саваннами и пустынями; к ним относят сахельскую степь вдоль южной границы Сахары, замбезийскую степь по периферии Калахари; в засушливых Андах, к югу от экватора, на высотах около 4000 м парамос влажных более северных Анд сменяется пуной, мозаикой кустарниковых, кактусовых формаций и злаковых степей.
Для злаков саванн характерен более или менее выраженный ксероморфизм, вегетативные части представлены плотными дер-н^винами или длинными корневищами, укоренение плотное, семена обильные.
Кустарниковый и древесный ярусы развиты весьма различно и положены в основу физиономической классификации саванн:
1) травянистые саванны без деревянистой растительности: прерии, кампос-лимпос и кампинас (небольшие кампос) Амазонии;
2) саванны с рассеянными кустарниками и деревьями: саванны кустарниковые и древесные (фото VI,А), бразильские кампос-серрадос\
246
3) саванны с обильными деревьями; облесенные саванны, бразильские серрадос. Если кроны деревьев почти смыкаются, а травяной ярус сохраняется, в этом случае формацию относят к светлому лесу (фото VI, Б). В действительности, в некоторых регионах, например на юго-востоке Азии, светлые леса из двукрылоплодниковых образуют с более или менее открытыми саваннами мозаику со всеми переходами. Так же и серрадос Бра зилии переходят в чащи и более или менее густые низкорослые леса, называемые серрадао. Льяносы Венесуэлы — это мозаика тех же формаций, что и субтропический вельд Южной Африки.
Флора деревянистых растений саванн включает множество видов, большая часть которых имеет общий внешний облик и своеобразные морфологические признаки, резко отличающиеся от видов густых лесов: мощная корневая система может достигать значительной глубины даже у низких кустарников, как у Andira humilis, бобового растения кампос-серрадос\ стволы низкие и извилистые, кроны раскидистые; толстые деревянистые корки защищают камбий от пожаров; листья в общем довольно крупные, но плотные, кожистые, защищенные от чрезмерной транспирации. Это деревья обычно листопадные, опадение листвы происходит в сухой сезон; замечательное исключение — акация суданских саванн Acacia (Faidherbia) albida, сохраняющая листья в сухой сезон.
Так же, как травянистая флора, эти ксероморфные деревья не имеют близкого систематического родства с лесной флорой. В Африке обильные бобовые: акации (особенно в более засушливых зонах), Albizzia, Prosopis, Afzelia, Celtis и т. д. Для суданских саванн характерны виды Combretum, масляное дерево (Ви-tyrospermum parkii), баобаб (Adansonia digitata), Balanites aegyptiaca. В саваннах существует несколько пальм, как, например, борассовая пальма (Borassus flabellifer) в гвинейских саваннах, виды Hyphaene в Судане и на западе Мадагаскара, виды Mauritia в Гайане и Бразилии. В саваннах также существуют мясистые кустарники, кактусы — в Южной Америке, канделяброобразные молочаи — в Восточной Африке. Протейные распространены в саваннах южного полушария, листопадные эвкалипты— в саваннах Австралии. Для саванн Юго-Восточной Азии характерны двукрылоплодниковые, в Новой Каледонии —ниаули (Melaleuca leucodendron).
Наряду с этой оригинальной флорой существуют виды, викарные лесным, иногда трудно отличимые от них морфологически; примерами являются Khaya, Lophira, Terminalia, Bombax, Par-kia. Прерывистое распространение древесных растений саванн не связано с недостаточностью способов рассеивания семян. Изученные виды показывают значительную семенную продуктивность (Acacia karroo может дать за сезон 20 тыс. семян, 90% которых способны к прорастанию), причем семена характеризуются временными периодами покоя и значительной долговечностью.
247
Животные повреждают многие из них, а прорастание весьма затруднено вследствие интенсивной конкуренции травянистых растений и опустошений от пожаров.
Фауна саванн
Фауна саванн, как и флора, резко отличается от фауны прилегающих лесных формаций и различна в разных биогеографических регионах и климатических зонах. Однако она обладает общими этологическими признаками.
Изобилие травянистой растительной массы во влажный сезон определяет высокую плотность крупных травоядных, тогда как отсутствие холодного периода, в отличие от степей, повышает их значение по сравнению с подземными популяциями грызунов. Иногда обширные, иногда смешанные стада крупных травоядных представляют средство защиты от их врагов — хищников. В Восточной Африке крупная фауна, сохраненная благодаря созданию национальных парков, особенно обильна и разнообразна: много видов антилоп и газелей, гну, импала, зебры, буйволы, слоны, носороги, бородавочники (фото V, Б).
Это разнообразие травоядных определяет и разнообразие хищников: в Восточной Африке насчитывается от 20 до 25 видов, нападающих в соответствии с их размерами на различные жертвы. Львы предпочитают среднего размера копытных — гну, зебр, газель Томсона и молодняк более крупных видов — слонов, жирафов, буйволов; леопарды и гепарды охотятся за более мелкими жертвами: импалой, антилопой-канной, зайцами, так же, как гиеновые, или «дикие» собаки, которые поедают газелей и антилоп; шакал охотится за зайцами и куриными; сервал — за грызунами и цесарками; циветта — за мышами, птицами, мелкими рептилиями, насекомыми; гиены и отчасти шакал поедают падаль. Некоторые травоядные вынуждены терпеть высокое «давление хищника»; так, за газелью Томсона в некоторых частях ее ареала охотится до десятка плотоядных.
Саванны — область распространения бегающих птиц: страус— в Африке, нанду — в Америке, эму — в Австралии, казуар — в Новой Гвинее. Некоторые зерноядные виды демонстрируют замечательную способность к образованию стай, как, например, ткачики и просоеды (Quelea) в Африке.
Деревья представляют своеобразные местообитания для птиц, среди млекопитающих здесь представлены обезьяны (бабуины в Африке, Hapale penicillata в кампос-серрадос).
Существенной проблемой для позвоночных саванн является водоснабжение. Некоторые виды поедают сочные части растений; так, антилопы Южной Африки в начале сухого сезона поглощают плоды Citrullus coffer из семейства Cucurbitaceae, позже выкапывают клубни, богатые водой. Наиболее требовательные виды, как, например, носорог, водяной козел (Cobus cobus) или в Австралии птица Taeniopygia castanotis обитают вблизи воды.
1248
На водопой млекопитающие ходят стадами ночью, птицы летают стаями днем.
В сухой сезон птицы и многие млекопитающие мигрируют к более влажным территориям, как, например, слоны, зебры, антилопы. Эти перемещения, особенно в засушливые годы, сопряжены с риском; так, известен, например, случай гибели от голода и жажды в 1863 г. тысяч антилоп вида Cephalolophus mergens в Южной Африке.
Насекомые в саваннах обильны, особенно термиты, саранчовые, муравьи. Первые сооружают термитники различной формы, в зависимости от видов термитов, иногда приподнятые на несколько метров за счет перемещений из глубины почвы большого количества земли!, выносимой на поверхность после ее измельчения; эти почвы термитников более плодородны, чем окружающие участки, и часто заселяются видами деревянистых растений. Термиты — основная пища многочисленных хищников: насекомых, амфибий, ящериц, змей, трубкозуба.
Саранчовые тропических саванн многочисленны; некоторые, как пустынная саранча (Schistocerca gregaria) и перелетная саранча (Locusta migratorioides) в Африке, образуют странствующие стаи. Они составляют основу питания множества видов рептилий и птиц; некоторые птицы следуют за кочующими стаями саранчи, и уменьшение последней — одна из главных причин миграций птиц в Африке.
Происхождение и эволюция саванн
Происхождение саванн ставит весьма интересную, сложную и живо дискутируемую проблему. Конечно, происхождение их различно, так же как различны современные экологические условия их устойчивости или развития. Несколько схематичным образом можно, вслед за Робинсом (Robyns), назвать три возможные причины возникновения саванн: климатические, эдафические и антропогенные.
Саванны климатические. Фитогеографы XIX в., такие, как Энглер (Engler) и Шимпер (Schimper), определяли саванны как естественные (первичные) формации регионов с климатом слишком сухим для развития густых тропических лесов. Эта концепция не может относиться к саваннам, образующим прогалины в глубине густых лесов экваториального, или влажного тропического, климата, таких, как кампос амазонского леса или прибрежные саванны Берега Слоновой Кости.
В зонах с продолжительным сухим сезоном распространение саванн вслед за распашкой и их устойчивость к пожарам заставляет рассматривать их как вторичные формации, пришедшие
1 Согласно Грассе и Нуаро, объем гигантских термитников Bellicositertnes rex в Убанги-Чари колеблется от 100 до 1200 м3.
249
на смену в разных случаях густым полулистопадным, листопадным или светлым лесам. Однако, пораженные самобытностью ксерической флоры саванн этих регионов и их устойчивостью, некоторые фитогеографы, как Трошен для суданских саванн и Тидмарск (Tidmarsk) и Дайер (Dyer) для травянистого южноафриканского вельда, признают за ними характерные черты современных климатических формаций.
Эдафические саванны. В зонах густых лесов саванны по большей части приурочены к субстратам, неблагоприятным для развития леса:
1. Скалистые вершины, или ожелезнепные освобожденные (рис. 66).
островные горы («inselbergs»), и древние
эрозией коры (бове Западной Африки)
Низкий.
лес
Густой | Саванна
Рис. 66. Профиль через гору Оромбо-Бока, Берег Слоновой Кости, отображает у подножия вторичную саванну, следующую за распашками и пожарами, на склонах — реликтовый мезофильный лес и на вершине на древней денудированной коре — эдафическую саванну и низкий лес (по Шнеллю, Schnell).
2. Почвы с залегающим близко к поверхности непроницаемым уплотненным глинистым горизонтом, испытывающие сезонное чередование насыщения влагой и иссушения, неблагоприятные для лесных древесных пород; саванны такого рода отмечены на острове Тринидад, в Суринаме, Северной Родезии.
3. Бесплодные фильтрующие пески аллювиальных пластов: в этих местообитаниях выделяют малораспространенные первичные саванны, отмеченные в Суринаме и Гайане, Амазонии и Конго («esobes»).
4. Постоянно влажные гидроморфные, периодически затопляемые, сильно гумусированные почвы с весьма своеобразной растительностью из Сурегасеае, Xyris, Burmannia, Eriocaulon, Utricularia, Lycopodium, сфагновых мхов; эти болотистые саванны с весьма сходной флорой существуют во многих тропических регионах: побережья Суринама и Гайаны, «кампос де варзеа» в Бразилии, прибрежная полоса Берега Слоновой Кости, влажные «esobes» Конго.
К тому же некоторые из этих саванн, за исключением относящихся к последней группе, могут возникнуть в результате деградации светлых или низкорослых лесов с травяным нижним ярусом, в которые они иногда переходят.
250
Высокая степень родства флористического состава многочисленных эдафических саванн и сухих низкорослых лесов, включенных в массивы густого леса, и обширных саванн в более сухом климате, как между серрадос амазонского леса и центральной Бразилии, позволяет рассматривать их как свидетелей более сухих климатических периодов. Становление этой флоры, многочисленные роды которой имеют пантропическое распространение (Acacia, Cochlospermum, Prosopis, Jatropha и т. д.), относят к началу третичного периода, и имеется множество доказательств различного рода последовательных смен сухих и влажных периодов до недавнего времени; в четвертичном периоде также наблюдалось чередование нескольких «плювиальных» эпох с сухими. Эти пятна саванн во влажных тропических зонах, по-видимому, свидетели сухих периодов, сохранившиеся благодаря неблагоприятным для развития густого леса эдафическим условиям: постклимаксы, согласно терминологии американцев. В некоторых случаях они находятся в устойчивом состоянии, как показывает опыт охраны одного из участков сер-радо в штате Сан-Пауло; в других эдафических саваннах отмечают очень медленное внедрение леса. В частном случае прибрежных саванн Берега Слоновой Кости на песке Миеж (Miege) отметил продвижение леса (до 1 м в год), но это никак не свидетельствует о естественном, первичном характере этих саванн.
Шнель (Schnell) весьма справедливо привлек внимание к возможному влиянию древнего сведения лесов, даже в регионах ныне весьма малонаселенных; в неблагоприятных эдафических условиях распашка почв может способствовать их деградации, под воздействием эрозии и истощения, что препятствует восстановлению густого леса. В этом случае речь идет о вторичных саваннах эдафо-антропогенного происхождения.
Вторичные саванны. Сведение леса приводит к тому, что его восстановление после прекращения распашки более затруднено, чем в менее благоприятных климатических условиях. Во влажных зонах сокращение продолжительности существования залежей между обработкой благоприятствует становлению переходной травянистой растительности типа саванн: «соломенная трава» императа (Imperata cylindrical, быстро захватывающая территорию, поселяется на залежах всех тропических регионов. Во влажной Африке наиболее часто развивается высокая мезофильная саванна со «слоновой травой» (Pennisetum purpureum)\ ревянистая растительность здесь успешно восстанавливается. Но если эти послепахотные саванны регулярно подвергаются пожарам, они переходят в саванны постоянные (рис. 66).
В наиболее сухих зонах или на неблагоприятных субстратах, где развивается только низкорослый, особенно легко подверженный пожарам лес, последние могут непосредственно привести к возникновению саванн.
251
Некоторые фитогеографы, как Обревиль, Шнель, полагают, что многие саванны имеют именно такое антропогенное происхождение. Обревиль так описывал «кампины» амазонского леса: «Чащи, возникшие на этих проницаемых, бедных и очень сухих почвах, безусловно, очень уязвимы для пожаров. Так объясняется наличие вырожденных лесных плохорастущих формаций при осадках от 2 до 3 м и их превращение в травянистые, иногда очень обширные саванны». Аналогичной точки зрения придерживается Холмс (Holmes) на «патана» — горные саванны Цейлона, Морелло (Morello) —на «кампосы» возвышенных участков Чако и т. д.
Что саванны, расположенные в зонах климата густого тропического леса, представляют, по-видимому, пережитки предыдущего сухого периода, существенно подтверждается их устойчивостью к пожарам (фото VII, Б). Наблюдения и точные опыты, проводившиеся на Береге Слоновой Кости, демонстрируют быстрое становление леса при отсутствии пожаров в климатической зоне густого леса.
Формации деревянистых, но не древесных растений
Формации, объединенные под этим названием, составляют разнородный набор и обладают лишь одним общим признаком— их доминирующий ярус образован в основном деревянистыми низкорослыми либо невысокими растениями. Различают все переходы к другим сообществам:
1) от мелколесья их отделяют по произвольному рубежу высоты господствующего яруса, располагающемуся в основном между 6 и 8 м или же по характерному обильному ветвлению этих деревянистых растений от самого основания, что и определяет труднопроходимость этих формаций;
2) от травянистых формаций — степей и саванн — они отделяются по возрастающей роли деревянистых растений в их формировании.
Разнообразие этих формаций порождается как различием в высоте основного яруса (от нескольких дециметров до нескольких метров), его ассоциированием с другими ярусами — ярусом единичных деревьев или травяным ярусом, так и морфологическими особенностями деревцев, такими как вечнозеленая или опадающая листва, колючие ветви, суккулентность. Богатая терминология, часто регионального характера, отражает это разнообразие: чащи, кустарники, ланды, маквис, гарига — таковы лишь французские названия. Общий термин для них — кустарники, наименование формации полукустарников оставлено за сообществами низкорослых деревянистых растений.
Эти формации, естественные и вторичные, различие между которыми в некоторых случаях очень трудно установить, ветре-
252
чаются почти во всех климатических поясах. Нижеследующие описания формаций расположены в соответствии с их происхождением и климатической приуроченностью.
Естественные кустарниковые формации
Их существование определяют климатические или эдафиче-ские факторы.
Формации климатические располагаются между собственно лесными зонами и зонами, где исключено само существование фанерофитов:
1. Между бореальными лесами и тундрой распространены сообщества листопадных кустарников — берез, ив, ольхи, обычная высота которых колеблется от 50 см до 1 м, но может достигать и 2 м.
2. Выше естественной древесной границы в горах часто существуют кустарниковые полосы: чащи зеленой ольхи в Альпах, рябин и берез в Оверни, ланды из карликового можжевельника, горной сосны (Pinas mughus) в восточных Альпах. Если же к холоду добавляется сухость, доминирующей жизненной формой становится густой полукустарник подушковидной формы, иногда колючий, сообщества которого часто называют степями: таковы «степи» из Alyssum spinosum в марокканском Атласе и большая часть пуны Анд к югу от экватора на высоте 4000 м с господством Adesmia, Azorella, Opuntia и др.
3. Между лесами умеренного климата и континентальными степями может простираться полоса низких, более или менее густых сообществ полукустарников, называемых полынниками. Классическим примером таких сообществ служат полынники из Artemisia tridentata Большого Бассейна в Кордильерах Северной Америки. В обширных субаридных районах Патагонии распространены «степи» из колючих или густоветвящихся кустарников Herberts, Lycium, Adesmia и других с ксерофильпыми злаками, такими, как Stipa и Festuca.
4. В сухих областях регионов со средиземноморским климатом жестколистные леса могут уступать место естественным чащам вечнозеленых кустарников. Таковы чапараль в Калифорнии, «деревянистые степи» из ююбы (Zizyphus) или полыни по северной границе Сахары, чащи колючих и суккулентных растений Наталя, мульга-скраб с господством видов акаций немала с господством кустарниковых видов эвкалиптов в Южной и Юго-Западной Австралии.
5. В тропических районах естественные кустарниковые и полукустарниковые формации встречаются на территориях с субаридными климатическими условиями: колючие чащи юго-востока Мадагаскара, скраб, образованный видами эвкалиптов и акаций Квинсленда, каатинга на северо-востоке Бразилии.
253
Естественные кустарниковые формации в собственно лесных климатических условиях встречаются на субстратах, по разным причинам неблагоприятных для произрастания древесных пород:
1) вследствие их особых химических свойств, как на серпентиновых холмах в Новой Каледонии или во влажных солончаковых понижениях, где формируются чащи тамариска;
2) вследствие маломощности или скелетности почв таких, как, например, почвы на тропических железистых панцирях (рис. 66);
3) в местообитаниях, подверженных сильным ветрам: в горах, например на гребнях Вогезских гор, в поясе бучин, или на побережьях, как на датских берегах в области распространения буковых лесов, на тосканском побережье, где развиты леса из каменного дуба, на берегах Ла-Манша, где встречаются, например, заросли бирючины, облепихи и других кустарников.
Вторичные кустарниковые формации
Они сформировались на месте различных климатических сообществ.
1. Стадии деградации лесов или стадии их восстановления обычно включают фазу кустарников. Таковы в Средней Европе на карбонатной почве чащи из Prunus spinosa, самшита, можжевельника, на кислых почвах — ланды из вересковых и бобовых полукустарников. В средиземноморской Европе на известняковых почвах — гарига из кермесового дуба, розмарина и других видов, на кремнистых почвах—маквис (фото VIII и IX), в Калифорнии — чапараль. В тропических зонах — залежи с кустарниками, приходящие на смену сельскохозяйственным культурам, чащи, заменяющие мелколесья, такие, как сухие леса бразильских серрадао, леса из квебрахо и палосанто в Чако, либо, наоборот, как сеха Анд, очень влажный лес, также деградировавший в чащу, — маторраль.
2. Степи, где практикуется чрезмерный выпас, в результате ослабления конкурентной способности трав могут быть замещены более или менее густыми сообществами формаций полукустарников. Так, по-видимому, формация полынников Запада Соединенных Штатов заменила древние формации прерий. Под воздействием постоянного, со времен появления европейских колоний, выпаса овец кустарниковые вельды Южной Африки заняли место травянистых вельдов путем расселения необъедае-мого деревца Elythropappus rhinocerotis. Согласно Пури и Вальтеру (Puri, Н. Walter), колючие кустарники сухих областей Декана и Пенджаба вследствие интенсивного выпаса сменяют саванны. В Чако (Аргентина) первичные степи из Pennisetum, Trichloris, Setaria в начале XX в. в результате выпаса крупного рогатого скота частично были превращены в колючие чащи из Acacia, Mimosa, Cellis (Morello). Таким образом, выпас оказывает действие, противоположное действию пожаров, которые
254
благоприятствуют распространению травянистых формации в ущерб деревянистым.
Мы остановимся на некоторых наиболее важных из перечисленных формаций.
Некоторые кустарниковые формации
Ланды Западной и Центральной Европы. Ланды образуют тип растительности, наиболее характерный для атлантической подобласти евро-сибирской области. В условиях океанического климата они могут покрывать обширные пространства на территориях с бедными кислыми почвами — от Скандинавии и Шотландии до северо-запада Пиренейского полуострова,— внедряясь в тундры на севере и в маквис на юге. Флористически обедняясь, они распространяются в Центральную Европу.
Физиономические и флористические особенности. Это формации низких кустарников и полукустарников, относящихся к семействам вересковых и бобовых: первые представлены вересками и брусничными, вторые — утесниками (Ulex), метельчатым ракитником (Sarothamnus) и дроками (Genista). Эти виды, обладающие по происхождению ярко выраженной ксероморфией, произрастая во влажном климате и на почвах часто со значительными запасами воды, не являются настоящими ксерофитами. Некоторые из этих ксероморфов, как Erica tetralix и Genista anglica, приурочены к влажным и торфянистым ландам. Бедность питательными веществами и кислотность почвы определяют жизненную основу этих видов, их медленный рост, несмотря на постоянное наличие зеленых органов, позволяющих ассимилировать углерод большую часть года, и облигатный микоризный симбиоз вересковых.
Почва между группами деревянистых растений обычно не обнажена, а покрыта травами или чаще мхами и лишайниками, в особенности гипновыми мхами (Нурпасеае) и видами рода Cladonia.
Березы, сосны, рябины и другие деревья, возвышающиеся местами в лайдах, являются либо остатками уничтоженных лесных формаций, либо возможными их пионерами.
Экологические особенности. Ланды связаны с формированием поверхностного горизонта чистого органического вещества, слабо разложившегося, очень кислого, который квалифицируют как грубый гумус, или мор. На скалистых или каменистых субстратах этот органогенный горизонт образует основу почвенного профиля, почвы в этом случае относят к типу ранкер с грубым гумусом. На рыхлых мощных почвах кислый гумус, перемещаясь по профилю, вызывает процессы подзолообразования, которые завершаются формированием железистогумусовых подзолов.
255
Происхождение и эволюция. Первичные естественные ланды встречаются лишь на побережьях, где сила ветра препятствует поселению деревьев. В других местах ланды появляются вслед за повторными вырубками леса на выщелоченных и подзолистых почвах. Их естественная эволюция приводит к более или менее быстрому восстановлению леса. Человек может реализовать эту возможность путем насаждения хвойпых пород, но он также может и бесконечно препятствовать этому процессу (выпас скота или пожары). Ланды — формация, весьма подверженная пожарам. Огонь уничтожает в лапдах полукустарники, способствуя расселению геофитов, таких, как папоротник-орляк, и гемикриптофитов, подобных молинии. Однако растущие группами полукустарники быстро внедряются вновь.
Средиземноморские чащи. Большинство их — вторичные формации, значительно отличающиеся друг от друга по внешнему облику. Соваж (Ch. Sauvage), однако, для всей их совокупности предложил общее наименование — маторралъ, испанское название южноамериканского происхождения. Различие в облике этих формаций выражается: 1) в различной высоте яруса деревянистых растений (от менее метра до нескольких метров при переходе в настоящий лес); 2) в различной густоте этого яруса (от максимальной сомкнутости до той степени разреженности, при которой сообщества представляют собой уже луговины, залежи или степи с отдельными деревцами); 3) в самой возможности существования одиночных деревьев; 4) в морфологических особенностях доминирующих деревцев. Эти деревца, в общем, характеризуются густым ветвлением, неопадающей листвой и различными ксероморфными признаками, среди которых наиболее широкое распространение получили незначительные размеры и жесткость листьев. Их листья иногда колючие, как у кермесового дуба, иногда снабжены волосяным покровом различных типов, как у маслины, лаванды, а часто гладкие и с толстой кутикулой, как у фисташки-лешискус и у Phyllirea. Листья чаще всего так расположены по отношению к прямым солнечным лучам, что это позволяет им избегать чрезмерной транспирации или нагревания в полдень. Растения с очень маленькими листочками, завернутыми на нижнюю сторону (розмарин, верески) здесь так же обычны, как и виды с колючими листьями, например утесник (Ulex), Calycoioma. Голосеменные в этих формациях представлены видами можжевельника с игловидными или чешуйчатыми, расположенными наподобие черепицы, листьями.
Глубоко проникающая корневая система растений позволяет полностью использовать все резервы влаги, накопленные за зиму.
Физиологическая активность растений этих формаций отличается ярко выраженной периодичностью с двумя сезонами относительного покоя — прохладной зимой и сухим летом.
256
Среди основных формаций чащ средиземноморского климата отметим следующие:
— гариги, пространства низких кустарников на известняковых и мергелистых почвах, где доминирующими видами являются кермесовый дуб, розмарин, ладанники (Cistus) (фото VIII, А);
— маквис, высокие, труднопроходимые чащи па кремнистых почвах, образованные земляничным деревом, фисташкой-лентпскусом, миртом, видами рода Phyllirea, Calycotome, древовидным вереском (Erica arborea), над которыми часто возвышаются древесные породы — пробковый дуб или сосны (фото IX, А) ;
— чапараль, маквис сухих склонов Калифорнии, где осадки, особенно зимние, не превышают 300—500 мм, чаща высотой от 1 до 3 м, обычно образованная Adenostoma fasciculatum, видами родов Ceanothus, Arctostaphylos, маленьким вечнозеленым дубом Quercus dumosa',
— мали Южной Австралии существует при ежегодных, обычно зимних, осадках менее 400 мм на песках с известняковым слоем в глубине; в тех случаях, когда формация не деградирует, опа образована высокими, от 1 до 3 м, растениями: ветвящимися от основания эвкалиптами, видами Melaleuca, Acacia, Casuarina, протейными, миртовыми.
Животные в этих формациях особенно активны весной. Здесь множество грызунов, птиц, ящериц, змей и насекомых.
За исключением мали и, вероятно, некоторой части чапараля, эти сообщества образуются в результате истребления лесов из склерофильных дубов или хвойных; деревья эти могут постоянно присутствовать в виде открытых популяций, придавая своеобразие их внешнему облику. Американцы называют эти ландшафты «лесистой местностью» (woodlands), а Соваж предлагает для них термин — маторраль с деревьями (фото IX, Б).
Эти обезвоженные летом чащи богаты чувствительными к огню растениями, часто их уничтожающему. Наземные части в этом случае погибают, но пни и часто корни дают новые побеги. Из-за этих-то молодых побегов, полюбившихся овцам, пастухи устраивают пожары. Кроме того, палы способствуют прорастанию и восстановлению некоторых пород, таких, как Rhus, кустарник чапараля.
Тропические мелколесья и колючие чащи. Сообщества этих формаций занимают обширные пространства в сухих тропических и субтропических областях, где среднегодовые осадки колеблются от 400 до 800 мм. Это отнюдь не малые величины, но осадки выпадают в течение короткого периода, и их количество из года в год значительно меняется. Эти сообщества, образующие основной элемент растительного покрова в «полигоне сухости» в северо-восточной Бразилии, известные под названием каатинга, представлены в Чако и в Монте — в Аргентине, в Мексике, на юго-западе Мадагаскара и в Южной Африке, на полуострове Декан и в Пенджабе, а также в Австралии.
7210 Ж. Леме
257
К формациям этого типа применяют английские наименования «скраб» и «колючие чащи» (thornbrush).
Высота их варьирует и может превышать 10 м, тогда эти сообщества именуют сухими колючими лесами. Это закрытые формации; при деградации они переходят в открытые сообщества, превращаясь в колючекустарниковые степи, и возможно, что эти последние, покрывающие большие пространства в областях с сухим африканским климатом, возникли из лесов. Различные жизненные формы отличаются наибольшей ксероморф-ностью: низкие деревья и деревца, безлистные в сухой сезон, с колючими ветвями, в особенности бобовые растения; растения с колючими суккулентными побегами — кактусы всевозможной высоты, мясистые африканские молочаи, Didieraceae Мадагаскара; «деревья-бутыли» с непропорционально вздутыми стволами, сужающимися к основанию и вершине (Cavanillesia arbo-геа в Бразилии, Adansonia на Мадагаскаре, Brachychiton ги-pestris в Австралии); в каатинге бромелиевые с розетками огромных листьев, окаймленных шипами.
Лианы и эпифиты, за исключением Tillandsia, в некоторых сообществах каатинги отсутствуют. Деревянистые виды имеют глубокопроникающую корневую систему, благодаря которой они добираются до запасов влаги легко водопроницаемой в этих формациях почвы. Во время сухого сезона растительность здесь находится в состоянии покоя, который прерывается с первыми дождями буйным, в несколько дней, распусканием листвы.
Леса
Лесные формации представляют собой древесные насаждения определенной густоты, благодаря которой под пологом создаются условия, определяющие существование всех растений подчиненных ярусов. Кроны в этом случае должны соприкасаться полностью или оставлять лишь небольшие незаполненные пространства, либо временные окна, образовавшиеся вследствие гибели одного из деревьев. Если между деревьями существуют значительные незатененные поверхности, то это не лесные сообщества, несмотря на внешний лесной облик. Таковы ланды с березами в Западной Европе, гарига с соснами и маквис с пробковым дубом в средиземноморской области, «лесистые местности» на юго-западе Соединенных Штатов, древесная саванна и т. д. Эти формации с открытым древесным ярусом представляют собой либо предлесные стадии, либо стадии деградации леса. По высоте деревьев и по способу их ветвления леса также образуют ряд переходов к кустарниковым формациям. И если невозможно установить между этими формациями минимально точную и единообразную по высоте границу, то можно, и наоборот, рассматривать лес как формацию с деревянистыми
258
растениями в верхнем ярусе, которые обладают свободными от боковых ветвей стволами.
На земном шаре, еще не обжитом человеком, земли были покрыты лесами, за исключением областей, где климат или местные особенности не позволяли существовать деревьям: в очень холодных или аридных областях, в местах, подверженных частым сильным ветрам, или с неустойчивыми субстратами, на засоленных почвах и т. д. На территориях, благоприятных для развития леса, его локализация на ограниченных участках или даже полное отсутствие — следствие обезлесения, часто очень древнего. Отказ от использования человеком территорий, приемлемых для произрастания леса, ведет к восстановлению последнего с самой различной скоростью, зависящей от местных условий.
Леса представляют собой совокупность различных типов растительных сообществ, географическое размещение которых, за исключением местообитаний с очень своеобразными субстратами, непосредственно подчинено климатическим условиям. Зоны растительности совпадают с климатическими зонами, что отражено на рис. 61. Именно в такой последовательности мы опишем различные типы тропических лесов: средиземноморских жестколистных лесов, вечнозеленых и листопадных лесов умеренной зоны, хвойных лесов. Все эти формации обладают общими чертами структуры и местообитаний, которые и будут рассмотрены в первую очередь.
Общие особенности экологии и строения лесных сообществ
Древесный ярус определяет экологические условия организмов, обитающих в его тени. Особенности микроклимата и атмосферы по вертикальному градиенту сверху вниз испытывают возрастающие изменения, связанные с густотой древостоя и с характером размещения листового экрана:
1) прогрессивное уменьшение видимого солнечного излучения и изменение его спектрального состава;
2) прогрессивное выравнивание температуры, в особенности благодаря снижению максимумов;
3) прогрессивное повышение и выравнивание атмосферной влажности;
4) уменьшение движений воздуха;
5) уменьшение осадков па почве и их механического воздействия.
На рис. 67 приведены примеры этих изменений.
Лесная почва сразу или в течение всего года обогащается за счет опада листвы и мертвых остатков, которые, не будучи развеяны ветром и разлагаясь на месте, превращаются в гумус, соответствующий данным древесным породам.
26»
7аЮ*
Средние максимальные темпе-
22 24 26 28 °C ратуры воздуха,П-Ц 1959г.(Щ
2 4 6 8 мм Средний дефицит насыщения,
—I---1----1---1----1 717-Л7Г 1 ПК По /ТТЛ
10- / -8
20 40 60 80 100 1 2 3 4 5
Относительная Скорость ветра, м/сек освещенность
Рис. 67 Изменения по вертикали средних значений некоторых микроклиматических показателей в различных типах леса. Л —экваториальный вечнозеленый лес Берега Слоновой Кости (по Ка-шану, Cachan); Б —высокоствольный буковый лес в возрасте 120—150 лет (по Траппу, Е. Trapp); В—старые дубы в нижнем ярусе букового леса перед (пунктир) и во время (сплошная линия) периода распускания листвы (по Гейгеру и Аманну, Geiger et Amann).
Лесная растительность накапливает самую большую биомассу на единицу поверхности, не располагая непременно большей ежегодной продукцией, чем у травянистых формаций. Растения нижнего яруса тенелюбивы или обладают более или менее ярко выраженной теневыносливостью, за исключением части растений листопадных лесов, которые совершают цикл своего развития в период, когда деревья не одеты листвой. Цветение и созревание семян у растений нижних ярусов плохо обеспе-
260
чены, поэтому виды с активным вегетативным размножением сами себе обеспечивают благоприятные условия распространения. Что касается способа распространения семян, то в нижних ярусах анемохоры отсутствуют, в то время как зоохоры и автохоры широко представлены. Растения-сапрофиты, сосудистые бесхлорофилловые растения и грибы часто встречаются на перегное и разлагающейся древесине. В лесах, в частности, процветают два своеобразных биологических типа, зависящих от опоры и благоприятных микроклиматических условий, предлагаемых деревьями — эпифиты и лианы. Разнообразие микроместообитаний, создаваемое лесной растительностью, создает
Рис. 68. Сравнение морфологических особенностей трех лесных видов ногохвосток (Collembola), обитающих в разных природных условиях (по Дебуттевилю, Deboutteville).
I — Campy lot hor ах, longicornis, вид, живущий вне почвы; 11 — Uzelia setifera — геми-эдафический вид; 111 — Parajapyx gestria-nus — эуэдафический вид.
разнообразие фауны, которая там обитает и распределяется по вертикали от почвенной среды до верхушек больших деревьев:
1. В лесных почвах, населенных грызунами и представителями мелкой фауны — червями, членистоногими, простейшими, образующими эдафон, активность животных может быть большой. Согласно Гисину (Gisin), необходимо различать эуэдафон, который постоянно обитает в почве, в отличие от гемиэдафона, который сосредоточен на ее поверхности или лишь временно живет на ее глубине. Виды первой группы были названы также геобионтами, виды второй — геофилами. Членистоногие эуэда-фона (рис. 68), живущие в порах почвы и перегноя, представлены мелкими видами, узкими и продолговатыми, придатки которых (усики, лапки и щетинки) менее развиты, чем у форм, обитающих на поверхности; их глаза редуцированы или атрофированы и лишены пигмента. Чувствительные к сухости, эти виды держатся в пленке воды влажных горизонтов или обнаруживают, как нематоды и тихоходки, способность выживать В обезвоженном состоянии. Дыхательные органы редуцированы
10 Ж. Леме
261
или отсутствуют, газообмен осуществляется всей поверхностью тела.
Эдафон образует внутри экосистемы леса комплексное звено: основой его энергии являются остатки зеленых растений, хотя эдафон и включает водоросли и некоторые автотрофные бактерии. Начиная с этой получаемой доли органических молекул цепи питания различны: некоторые виды специализируются на определенной пище, другие, как дождевые черви, поглощают почву вместе с ее органической частью и населением микроорганизмов; большая часть зоологических групп, как нематоды, клещи, моллюски, коловратки, потребляют в зависимости от вида органические остатки, микроорганизмы или других животных. Велика роль фауны лесных почв в разложении поверхностных растительных остатков и во включении органического вещества в минеральную часть почвы. Деятельность дождевых червей, а в тропических областях термитов и муравьев, служит основой для формирования свойств лесных почв и их эволюции.
2. Мертвые стволы деревьев, разлагающиеся пни, неровности коры и дупла, вызванные шелушением ее верхних слоев, гумусовые отложения на ветвях деревьев в тропическом лесу («висячие почвы») образуют множество лесных микроместообитаний, населенных постоянно или временно особыми видами. Стволы погибших деревьев заселяются многочисленными насекомыми, особенно жуками, с различными режимами питания: флоэмой и камбием, древесиной, соком, грибами; часть из них — хищники или паразитируют на других животных.
3. Животное население, локализованное на поверхности, если говорить о млекопитающих, имеет небольшую плотность и образовано особями, живущими поодиночке или очень маленькими группами. Чем меньше плотность, тем большими размерами обладают виды животных. На 100 га лиственных лесов французских равнин могут прокормиться по крайней мере 7 косуль, но только 2 оленя. Впрочем, животные крупных размеров — явление исключительное; таковы слон и буйвол тропических областей, лось в северных лесах, тогда как многие группы млекопитающих представлены видами меньших размеров, такими, как карликовый слон Конго, оленьки и карликовые олени тропической Азии (Tragulus), Африки (Hyaemoschus), Чили (Pudu, humilis). Вообще рога оленей и других животных уменьшаются. Лесные животные имеют слаборазвитое зрение и обоняние, но очень тонкий слух.
4. Древесные местообитания заселены множеством насекомых и птиц, но в тропических лесах там обильны и млекопитающие, рептилии и амфибии. Позвоночные обладают различными приспособлениями к древесной жизни: сильные когти, противопоставляющиеся друг другу пальцы у обезьян, сумчатых, лазающих птиц, хамелеонов, ящериц и тропических лягушек; присасывающиеся подушечки пальцев у гекконов и тропических
262
350
Рис. 69. Соотношение типов растительности с годовым количеством осадков и продолжительностью сухого сезона в Индии (по Вальтеру, Н. Walter).
./ — вечнозеленый тропический лес; // —полулистопадный
тропический лес; ///—муссонный лес (Д —влажный, Б —сухой); IV — саванна; V — пустыня.
рукокрылых и подошвы лап у долгопятов, дикобразов Erethizon, .древесных даманов Dendrohyrax, обезьян Inuus\ цепкие хвосты хамелеонов, ящериц и древесных змей, обезьян Америки, грызунов и сумчатых; разрастание кожи между конечностями, играющее роль парашютов, у весьма удаленных друг от друга в биологическом отношении групп: шерстокрылов белок, сумчатых, ящериц {Draco). Среди насекомых общественные виды — термиты, муравьи, пчелы, осы — строят свои гнезда па деревьях, либо в дуплах стволов мертвых или чахнущих деревьев.
5. Воздушные местообитания внутри лесов, где не бывает сильных ветров, освоены бабочками со слабыми способностями к полету, и птицами, добывающими себе пищу на разных
1 Автор относит этот род к лемурам. В настоящее время он выделяется ® особый отряд. — Прим. ред.
10*
263
уровнях в зависимости от видов, — от поверхности земли и до' свободных воздушных пространств над деревьями.
Воздушное население, кроме гнездящихся пар, включает подвижных, не участвующих в размножении птиц обоих полов, поскольку всегда наблюдается дефицит территории вследствие деления ее согласно занятым гнездам. Эти лишние особи образуют резерв для обеспечения максимально возможной плотности, которая составляет в зависимости от особенности леса от 2 до 10 на 1 га.
Плотность и разнообразие животного населения внутри лесных массивов меньше, чем на их опушках. Население лесов обогащается за счет тех видов, которые находят там приют для ежедневного отдыха или в плохое время года, избегая их в остальное время.
Тропические леса
В тропических странах с небольшими абсолютными высотами температура воздуха держится на уровне, благоприятном для активной жизнедеятельности растений, однако режимы выпадения дождей здесь очень разнообразны: начиная с зон чисто экваториального режима с двумя короткими, относительно сухими сезонами до зон, где режим характеризуется одним коротким дождливым сезоном (краткий тропический режим, по Пегюи), включая зоны переходного экваториального режима, когда один из сухих сезонов усилен, а другой сглажен, и зоны длительного тропического режима, при котором единственный дождливый сезон значительно продолжительнее сухого сезона.
Структура и флористический состав внутритропических лесных формаций на почвах с хорошим дренажем очень чувствительны к режиму дождевых осадков и к их годовому количеству, но в различной степени: продолжительность сухого сезона является определяющим фактором в гумидных зонах, количество осадков имеет большее значение в наиболее засушливых зонах, по Вальтеру. На рис. 69, где показано соотношение этих двух климатических факторов с типами индийской растительности, со всей очевидностью вскрыта эта связь.
Попытки биологической характеристики тропических климатов с помощью климатических индексов основывались на различных способах выявления этих двух элементов. Сошлемся на классификацию Кеппена (Корреи), который в группе А, влажных тропических климатов (среднемесячная температура самого теплого месяца 18°, наивысшее количество годовых осадков 2/4- 14), различает три подгруппы:
Af — среднемесячное количество дождевых осадков самого сухого месяца более 60 мм;
Ат — дождевые осадки составляют менее 60 мм, но в целом количество* осадков достаточно, чтобы компенсировать сезонную сухость;
Aw — общее количество осадков не компенсирует сезонную сухость.
264
Система Торптвейта (Thorntwaite), основаннная на эвапотранспирации, с учетом количества осадков и их изменений в течение года, приводит к более сложной классификации. Сошлемся еще на индекс, установленный Манжено (G. Mangenot) для тропических районов:
. _ Р1\№ + Ms + t/x/5
1 ns + 500/t/n
где P— годовое количество осадков, в мм, Ms — среднее количество дождевых осадков сухих месяцев (с осадками до 50 мм), п3— число сухих месяцев, Ux — максимальная среднегодовая относительная влажность, Un^ минимальная среднегодовая относительная влажность. Введение значений атмосферной влажности придает этой формуле элемент большого биологического значения.
Лесные тропические формации, приуроченные к соответствующим климатическим условиям, в литературе известны под различными названиями. В нижеприводимой табл. 12 отображены наиболее употребительные термины и их эквиваленты.
В зонах перехода одного типа формаций в другой водные свойства почв могут способствовать размещению каждого типа соответственно их климатическим потребностям: наиболее гигрофильные занимают почвы, богатые тонкими минеральными частицами, а наименее гигрофильные — субстраты более грубого механического состава. Рельеф оказывает подобное воздействие: к тальвегам и основаниям склонов приурочены наиболее требовательные к влаге сообщества.
Леса гигрофильные (или вечнозеленые) и мезофильные (или полулистопадные). 1. Растительность. Густота сообществ этих формаций, разнообразие жизненных форм и флористическое богатство создают сначала впечатление путаницы (фото X, Б).
а) Деревья характеризуются чрезвычайно разнообразным видовым составом, но из множества видов ни один заметно не доминирует1. Деревья распределены по всем уровням, но все же можно произвольно выделить три яруса. Самый верхний, изолированно выступающий над всеми ярусами, достигает в высоту 40—50 м (рис. 70). Высокие стволы иногда бывают снабжены множеством конических выпуклостей. Основания большинства деревьев подкреплены мощными узкими контрфорсами, которые иногда переходят в досковидные корни, вертикальные плоскости, извивающиеся по поверхности почвы. Другие виды несут придаточные корни, наискось отходящие от основания
1 В лесах Малайского архипелага описано около 2000 видов деревьев, на Берегу Слоновой Кости — 600. В Амазонии на 1 га обнаружено 423 дерева, относящихся к 87 видам. Их обычное дисперсное расположение отдельными экземплярами затрудняет эксплуатацию определенных, пород. Так, в среднем -встречается одно пригодное к эксплуатации дерево кайя (Khaya ivorensis) на Ю га; сборщикам каучука («серингерос»), которые собирали латекс гевеи в Амазонии, приходилось пробегать большие расстояния, чтобы посетить все свои -Деревья.
265
266
Таблица 12
Климатические особенности
Физиономические признаки
Английские термины
Климаты
Индекс
по Кеппену по Манжено
Леса:
гигрофильные вечнозеленые I1 влажные дождевые леса (rain forest) Af >7
субгигрофильные вечнозелееые сезонные влажные вечнозеленые леса (wet evergreen) (смешанные леса) (mixed forest) Am 1 i 4-7
мезофильные сухие (муссонные) полулистопадные . П2 листопадные сухие вечнозеленые леса (dry evergreen) влажные листопадные леса (wet deciduous) сухие листопадные леса (dry deciduous) Aw J 1-4
ксерофильные колючие
колючие леса (thornforest)
1 I — омброфильные леса.
2 Ц —тропофильные лесд.
ствола (корни-ходули) или спускающиеся от ветвей. Однако их корневые системы проникают в почву неглубоко, и тропические вихри легко валят эти деревья. Листья (или листочки сложных листьев) почти всегда лаврового типа: жесткие, крупные, зубчатые и длиннозаостренные. Их сезонный ритм (от опадения до нового распускания) зависит от климатических условий:
6 5 10 м
Рис. 70. Диаграмма-профиль, демонстрирующая распределение по высоте видов деревьев первичного вечнозеленого леса в Гайане (по Дэвису и Ричардсу, Davis, Richards).
в гигрофильных вечнозеленых лесах все деревья имеют постоянно облиствленные кроны; в полулистопадных лесах заметная часть деревьев в сухой сезон лишена листьев, но деревья теряют листву не одновременно и на короткое время, а маленькие деревья и деревца сохраняют листву; в вечнозеленых сезонных лесах, занимающих промежуточное положение, деревья, сбрасывающие листву, встречаются более или менее изолированно. Появление цветков прямо на стволах деревьев, или каулифлория, редчайшее явление в других сообществах, здесь встречается часто у представителей самых разных семейств
267
(у деревьев какао, хлебных деревьев, различных Cola, Diospy-ros, Ficus и других).
б) Деревца формируют обычно густой ярус из подроста или видов кустарников, причем некоторые из них, как Dracaena, имеют неветвистый ствол. Их листва обладает теми же особенностями, что и листва деревьев.
в) Травяной ярус редкий, образован автотрофными очень тенелюбивыми видами — папоротниками, широколистными зла-ками, бегониями — и гетеротрофными видами — сапрофитами или паразитами (Rafflesia, виды из семейства баланофоровых). Папоротники, более чувствительные к атмосферной сухости, в полулистопадных лесах менее обильны, чем в вечнозеленых лесах.
г) Лианы, большей частью деревянистые, получают здесь значительное развитие; они принадлежат к весьма разнообразным семействам и обладают всевозможными способами прикрепления к деревьям-опорам: крючками, усиками, вьющимися стеблями, опорными корнями. Некоторые лианы, поднимающиеся до крон высоких деревьев, достигают в длину более 100 м, связывая деревья друг с другом.
д) Эпифиты особенно обильны в вечнозеленых лесах, постоянно влажная атмосфера которых благоприятствует их развитию. Некоторые деревья буквально покрыты ими. Эти растения относятся ко многим семействам, но преимущественно к папоротникам, орхидным, ароидным, а в Америке — к бро-мелиевым. Виды, обитающие на нижних частях стволов, гигрофильные и тенелюбивые, отличаются от солнцелюбивых видов, поселяющихся на вершинах высоких деревьев. Среди первых преобладают папоротники, мхи, среди вторых — бромелиевые, кактусовые из рода Rhipsalis, Эпифиты обладают различными приспособлениями, обеспечивающими их водное питание. Некоторые, как платицериумы (Platycerium), создают настоящие органические висячие почвы; у других, как у бромелиевых и Asplenium nidus, широкие основания листовых розеток образуют своеобразные «цистерны». Среди эпифитов особенно примечательны фикусы-душители, первоначально развивающиеся как скромные эпифиты. Они образуют на поверхности ствола-опоры корни, которые, достигнув почвы, увеличиваются в объеме, анастомозируют до образования полого цилиндра и душат таким образом приютившего их хозяина. Сами листья в насыщенной влагой атмосфере нижних ярусов могут давать приют водорослям, лишайникам, печеночникам.
После сведения гигрофильных лесов и на заброшенных сельскохозяйственных землях деревянистая растительность очень скоро восстанавливается солнцелюбивыми быстрорастущими видами, такими, как Macaranga, Trema, Musanga или африканские зонтичные деревья Cecropia, южноамериканский вариант предыдущего, и другие. Под покровом этих сообществ
268
начальных стадий восстановления вновь появляются деревья густого леса и мало-помалу восстанавливается вторичный лес физиономически, но не по флористическому составу, подобный первичному или очень давно восстановившемуся лесу. Уничтожение мезофильных лесов на территориях, где климат отличается более ярко выраженным сухим сезоном, приводит к более или менее быстрому внедрению саванн типа «гвинейских».
2. Фауна. Фауна густых тропических лесов характеризуется оригинальными чертами: она обильна и особенно разнообразна, следуя разнообразию микроместообитаний и ресурсов питания, которые они предлагают, так же, как и флора, в связи с большой древностью этих биоценозов, развивавшихся на месте без значительных изменений в течение всего третичного периода. Поэтому здесь сохранились такие примитивные группы, как первичнотрахейные (Peripatus). Атмосферная влажность в этих ле-сах благоприятствует существованию червей, в других местообитаниях представленных водяными формами: планарий, пиявок, немертин и даже на Яве — кольчатому многощетинковому червю, способному выходить из текучих вод и перемещаться среди разлагающихся растительных остатков.
Многочисленные группы животных населяют различные ярусы растительности или попеременно перемещаются из одного яруса в другой. Термиты в зависимости от вида строят свои термитники начиная от глубоких слоев почвы и кончая ветвями крупных деревьев. В этих лесах также множество муравьев. Одни из них живут на поверхности почвы, как бродячие муравьи (африканские Апотта, Eciton Америки), перемещаясь с места на место неисчислимыми колоннами, или как муравьи-листорезы (Atta). Муравьи-портные (Oecophylla), кампо-нотусы (Camponotus) мастерят свои муравейники в разветвленных стволах, другие устраивают их на ветвях и населяют эпифитами («муравьиные сады»). Некоторые живут в полостях живых органов, в полых стеблях Cecropia, в полостях корневищ эпифитов, таких, как Myrmecodia, в домациях, или «мешках для муравьев», листьев меластомовых и т. д. Обильные и разнообразные комары (на 10 км2 в Колумбии приходится 150 видов) распространены на разных высотах: Anopheles gambiae держится на высотах менее 7 м, а близкий вид — A. africanus — наиболее обилен на высоте от 13 до 25 м. Множество земноводных являются древесными видами и откладывают икру в ли* стве, некоторые помещают икру в резервуары для воды эпифитов из семейства бромелиевых. Среди животных, ведущих древесный образ жизни, следует назвать обезьян и других млекопитающих; на деревьях также обитают грызуны, хищные, сумчатые.
Чрезвычайно интересно распределение активности животных как во времени, так и в пространстве. Жизнь ежедневно развертывается по явно выраженным циклам. В то время как днем
269
относительно тихо, сумерки и ночь наполняются треском цикад, голосами земноводных, ночных птиц, обезьян и т. д. Активность животных наблюдается в течение всего года без определенных периодов покоя или миграции. Если в областях умеренного климата у насекомых наблюдается лишь один или два периода размножения в год, то во влажных тропических зонах у них несколько таких периодов, не разделенных перерывами.
Однако распределение дождей в течение года может управлять периодами размножения: в тех случаях, когда выражен сухой сезон, размножение животных чаще всего приурочено к началу влажного сезона, но в чрезмерно дождливых лесах Калимантана спаривание у млекопитающих приурочено преимущественно к периоду, характеризующемуся минимумом осадков.
В тропических лесах наблюдается некоторый номадизм, связанный с поисками пищи, в частности у обезьян и плодоядных птиц, которые стаями переходят с дерева на дерево в поисках фруктов.
3. Почвы. Контраст между богатством леса и бедностью почв бросается в глаза. Это ферралитные или глинистые латеритные почвы, где аккумулируются полуторные окисли в результате разрушения и выщелачивания соединений кремнезема и оснований. Эти почвы отличаются низким плодородием. Измерения распределения питательных элементов в экосистемах тропических лесов показали, что ежегодный возврат органических веществ в почву, сопровождающийся очень быстрым разложением, составляет 10—15 т на 1 га: 40—60% в год в колумбийских лесах, по Дженни (Н. Jenny), 63—70%—в Конго, по Ло-делу и Мейеру (Laudelout, Meyer). В результате быстрого раз* ложения высвобождаются минеральные элементы, которые тут же вновь поглощаются густой сетью поверхностных корневых систем деревьев. Таким образом, преобладающая часть запасов минеральных элементов включена в живую биомассу и нс накапливается в почве, как в лесах умеренных климатических областей. Но если тропический лес сведен для возделывания сельскохозяйственных культур, то органический запас в почве используется этими культурами лишь частично в связи с иммобилизацией этих элементов микроорганизмами — редуцентами (в частности, N и Р) —и вовлечением их в процесс выщелачивания (особенно К, Са и Mg). Если вырубка леса сопровождается выжиганием, то это приводит к большой потере органического азота и минеральных элементов путем рассеивания и частичного растворения золы. Вследствие этого урожайность сельскохозяйственных культур быстро падает. В зонах со значительным сухим сезоном длительное отсутствие лесного покрова вызывает уплотнение некоторых почвенных горизонтов в результате аккумуляции полуторных окислов железа, их дегидратации и кристаллизации. Этот процесс образования латеритной коры является как
270
бы «одеванием почв в панцирь». Саванна в этом случае заменяет уничтоженный лес.
4. Эдафические и топографические варианты. Внутри одной и той же климатической зоны, как обычно, рельеф и естественные особенности почвы вносят изменения в облик и флористический состав влажных тропических лесов. На кремнистых бесплодных, оподзоленных песках в разных областях опи-* саны невысокие, бедные леса, такие, как лес Уаллаба в Гайане, низкие амазонские леса на белых песках, иногда именуемые «каатингами», несмотря на их глубокое различие с каатингами бразильского северо-востока, «пустошный лес» с видами из семейства диптерокарповых в Индонезии и на Калимантане. Последние при деградации переходят в устойчивые полукустарниковые группировки.
Рельеф, способствующий формированию почвенных рядов на склонах и концентрации вод в долинах, разнообразит лесную растительность. Ее состав очень чувствителен к продолжительности и вертикальной амплитуде насыщения почв водой. Наблюдаются заметные физиономические и флористические различия между постоянно и временно затопляемыми лесами; различия конкретизированы в соответствующих (бразильских) терминах — игапо и варзеа.
Эти болотистые леса характеризуются небогатым видовым составом, обилием деревьев с подпорками и существованием воздушных дыхательных корней-пневматофоров.
5. Высотные и широтные климатические варианты. С увеличением абсолютных высот местности температура понижается и влажность возрастает, что приводит к образованию поясов растительного покрова. Горные тропические леса более низкорослы, характеризуются однообразием верхнего яруса и более искривленными стволами деревьев. Здесь наблюдается, однако, обилие эпифитов, которые иногда образуют вокруг стволов плотную муфту, в которой преобладают мхи и лишайники. Древовидные папоротники и бамбук там обычно обильны. Верхняя граница этих лесов варьирует от 2000 м в изолированных горах до 4000 м во влажных Андах, где ее образует очень низкий лес — сеха.
На небольших высотах мезофильные леса вблизи границы своего распространения, определяемой климатической сухостью, так же как сухие леса, часто погибают и замещаются саваннами, за исключением днищ долин, которые благодаря постоянной эдафической влажности остаются облесенными. Среди этих галерейных лесов, исключительно четко оконтуривающих гидрографическую сеть на аэроснимках (фото VII,А), различают вечнозеленые, полулистопадные, болотистые, приречные леса.
Гигрофильный вечнозеленый лес может выходить за пределы тропиков в тех случаях, когда осадки остаются обильными и выпадают в течение большей части года. Это субтропический
271
вечнозеленый лес (см. рис. 61). Эти леса формируются на восточных фасадах континентов: во Флориде, на юге Бразилии, юго-востоке Африки и Австралии, в Южном Китае, Тайване, Гаванских островах. Деревья принадлежат к тропическим группам растений — сапотовым, Ficus, Cinnamomum, пальмам, древовидным папоротникам и другим. Лианы немногочисленны, но эпифиты обильны, особенно из бромелиевых и орхидейных среди цветковых; известен даже фикус-душитель во Флориде. Некоторые из этих лесов образованы голосеменными: леса из Araucaria angustifolia на юге Бразилии, из высокогорных сосен на Паране. Встречаются насаждения голосоменных, являющиеся стадиями восстановления леса, таковы леса из Pinus carybaea во Флориде или из других сосен на Дальнем Востоке; возможно, что к таким насаждениям, согласно Гуэку (Hueck), следует отнести даже часть лесов из Araucaria в Бразилии.
Сухие листопадные леса. В отличие от предыдущих лесов их деревья теряют свою листву полностью в период более длительного и резко выраженного сухого сезона. К этим группам формаций относятся муссонные леса, светлые тропические леса и колючие леса. Описание последних, очень часто переходящих в формации деревцев было приведено ранее при характеристике этих формаций.
Зона муссонных лесов занимает значительную часть Индии, с ежегодным количеством осадков от 600 до 1700 мм и с продолжительностью засушливого сезона от 4 до 8 месяцев. По степени климатической влажности различают два типа таких лесов:
Влажные муссонные леса, в которых листва опадает полностью и притом на короткий срок лишь в верхнем древесном ярусе, достигающем высоты 30—35 м. Наиболее характерными видами являются сал (Shorea robusta), тиковое дерево (Tectona grandis), виды Termirtalia. Нижние ярусы сохраняют свою листву. Лианы и эпифиты малообильны, бамбуки встречаются часто.
Сухие муссонные леса, высотой всего 15—20 м, их деревья сбрасывают листву полностью, а деревца лишь частично. Деревья в основном принадлежат к тем же видам, что и во влажном типе. Бамбуки редки. Травяной ярус, также характеризующийся сезонным развитием, приобретает значение лишь во влажном сезоне.
Светлые леса характеризуются несплошным невысоким древесным сводом, по высоте довольно единообразным, с почти полностью листопадными видами. Травяной ярус сплошной, с господством злаковых, что позволяет относить эти леса к неогнестойким. Кажется, что в большинстве областей эти светлые леса заместили сухие тропические леса с нижними ярусами из деревцев и редким травяным покровом: на азиатском юго-востоке они в близких экологических условиях часто соседствуют с тропическими лесами, где выпадает более 1500 мм дождевых
272
осадков за год; в Катанге участки тропического леса существуют среди светлых лесов с Brachystegia, которые, если они защищены против огня, эволюционируют в сторону тропического леса. Однако, по Шмиду (Schmid), часть светлых лесов с дип-терокарповыми во Вьетнаме, возможно, является эдафическим климаксом на скелетных почвах. Светлые леса с Isoberlinia doka в суданской зоне, леса африканского востока в Замбии, в более ярко выраженных засушливых условиях, возможно климатического происхождения.
Мангры. Мангры — тропическая формация1 с господством деревьев или деревцев, уникальная по своей экологической приуроченности к илам эстуариев и болотистых лагун в пределах приливо-отливной зоны морей (фото I, Б).
Немногочисленные виды растений подразделяются на четко выраженные зоны. Внешняя зона, вдоль побережий, наиболее подверженная приливам, часто заселена ризофорами (Rhizophora), высота которых может превышать 15 м. Они снабжены многочисленными изогнутыми ходульными и придаточными корнями, несущими ветви и укореняющимися в иле. Семена их созревают па дереве, образуя длинный корень, вздутый как булава, который, когда проросток отделится от дерева, помогает ему во время отлива внедриться в ил, а при приливе — плавать и способствует таким образом распространению вида.
Обычно в средней и верхней частях приливо-отливной зоны поселяются Avicennia (из вербеновых), Sonneratia (из дербенниковых), Bruguiera (из ризофоровых), подземные стелющиеся корни которых образуют множество вертикальных корней, выступающих из ила и обеспечивающих аэрацию подземных частей (пневматофоры). Avicennia, кроме того, выделяют соли на поверхности листьев.
Между границей средних по высоте приливов и границей исключительно высоких приливов находится зона с переменной соленостью, обычно более слабой по сравнению с соленостью зон, расположенных ближе к внешнему краю и омываемых водой, приближающейся по своему составу к морской Там представлены своеобразные деревянистые виды: Laguncularia и Сопо-carpus во Флориде, пальма (Nipa fruticans) и Sonneratia acida на Малайском архипелаге и другие. Часто там встречается папоротник (Acrostichum аигеит). За пределами границ, которых достигают соленые воды, начинается континентальный лес.
Растительность мангров играет основную динамическую роль в аккумуляции и закреплении прибрежных илов. Она, следовательно, создает особую среду, с которой связаны виды животных, обладающие приспособлениями, обычными для приливо-отливной зоны морей, но являющиеся оригинальными в отношении использования деревьев в качестве опоры. Действительно, на ходульных корнях мангровых обитают моллюски, устрицы, литорины.
1 Мангры, обедняясь, могут распространяться за пределы тропиков по берегам, омываемым достаточно теплыми водами: во Флориде и на Бермудских островах, на юге Японии, севере Новой Зеландии, на побережье Наталя, в заливе Акаба в Красном море.
273
Крабы циркулируют в воде и по деревьям. Любопытная рыба, прыгун (Ре-riophthalmus), может благодаря своим измененным грудным плавникам перемещаться вне воды, а ее глаза, помещенные на верхней части черепа, расположенные на выростах, способных сокращаться, видят одинаково хорошо как в воздушной среде, так и в воде.
Средиземноморские леса
Леса средиземноморского климата образованы деревьями с вечнозеленой листвой, из двудольных или голосеменных. Первые представлены низкими деревьями, редко превышающими 20 м, с маленькими кожистыми листочками (склерофильные
Рис. 71. Лес из каменного дуба на плотноизвестняковом субстрате и последовательные стадии его деградации в Лангедоке (по Браун-Бланке, J. Braun-Blanquet).
1 — Quercetum ilicis; 2 —гарига из кермесового дуба (Quercetum cocci-ferae); J —низкотравные выпасаемые луговины (Brachypodietum ramosi);
4 — петрофитные открытые сообщества из молочаев (Euphorbia) и коровяка (Verbascum).
растения). В зонах с достаточным количеством осадков в западной части Средиземноморского бассейна обычной древесной породой является каменный дуб (Quercus ilex). Его насаждения богаты деревцами, так же вечнозелеными, как и он сам, таковы Phillyrea, земляничное дерево, фисташка-лентискус, Viburnum tinus и другие. Травяной и моховой ярусы разрежены. Каменный дуб может быть замещен другими видами дуба также с колючими вечнозелеными листьями, как, например, пробковый дуб на кремнеземистых грунтах, Quercus mirbeckii в Марокко, Quer-cus calliprinos в Палестине (фото IX, Б). В более теплых частях Средиземноморья дубовые насаждения сменяются лесами из дикой маслины (дикого оливкового дерева), фисташки-лен-тискуса и цератонии, лесами из аргании на юго-западе Марокко.
Хвойные климатические леса характерны для горных поясов: кедровые леса марокканского Атласа и Ливана, пихтовые среди
274
земноморские леса (см. рис. 16), леса из географических рас черной сосны или Pinus laricio или из Callitris Северной Африки. На равнине различные сосны (Pinus halepensis, Р. pinea, Р. pinaster), дикорастущие или интродуцированные, образуют насаждения, которые могут замещаться лиственными лесами или снова восстанавливаться (фото XI, Б).
Бассейн Средиземноморья, область древней цивилизации, — в основном пасторальной !, обладает лишь лесами, нарушенными пожарами, выпасом, эксплуатацией, вследствие чего эрозия уничтожила почвы более или менее полно (фото VIII, А). В климатической области распространения каменного дуба на мергелях и известняках первая стадия деградации — это низкая чаща кермесового дуба (Quercus coccifera), сообщества которого были описаны в главе о формациях деревцев. Измененные вследствие выпаса овец и палов, эти насаждения деградируют в луговины, богатые однолетниками, или в ланды с розмарином, лавандой, вереском и другими видами. Последняя стадия деградации представлена сообществами видов, не поедаемых скотом, которые встречаются среди камней, таких, как асфодели, молочаи, колючие чертополохи (рис. 71). Сообщества с кермесовым дубом и розмарином способствуют естественному переходу к Quercetum ilicis, а восстановление сообществ каменного дуба благоприятствует и ускоряет рост алепской сосны на плантациях, создаваемых под его покровом.
Известно, что могут существовать промежуточные сообщества между закрытыми лесами и формациями деревцев с рассеянными, отдельно стоящими деревьями. В Средиземноморье они покрывают особенно значительные пространства. Большая часть лесов там действительно представлена светлыми лесами.
Лесные формации и стадии их деградации в калифорнийской провинции со средиземноморским климатом физиономически очень схожи с соответствующими растительными сообществами Средиземноморского бассейна. Эти леса образованы дубами с колючими вечнозелеными листьями, высотой от 10 до 20 м, с примесью некоторых листопадных дубов и растений, относящихся к родам Castanopsis и Arbutus, Эти склерофильные леса почти всегда представляют собой открытые сообщества (woodlands и саванны, согласно терминологии американских авторов) и переходят при деградации в чащи, подобные маквису, известные в Калифорнии под названием чапараль.
Приречные или болотистые леса в условиях средиземноморского климата — это гигрофильные листопадные формации, хорошо отличающиеся от предыдущих. Образуют их тополя (Populus alba в Лангедоке), ясени, ивы.
В настоящее время, например, поголовье овец в Ливане насчитывает около 75 тыс., коз — до 500 тыс
275
Дождевые леса умеренного климата
Благодаря локальному распространению до умеренных широт постоянно влажного климата без резко выраженного холод-hi го сезона субтропические вечнозеленые леса переходят в растительные формации, имеющие с ними много общих признаков. Они распространены на юге Японии, Кореи и Китая, в «провинции береговых равнин» от Флориды до Северной Каролины, на юге Чили, в Новой Зеландии. Осадки там обычно превышают 1500 мм, и морозы практически неизвестны.
Деревья этих лесов в большинстве случаев с вечнозелеными кожистыми листьями, но более мелкими, чем в дождевых лесах низких широт. Лес там более низкий и менее густой. Присутствие листопадных видов вносит отчетливый сезонный аспект. Нижние ярусы густые, с деревцами, травянистыми лианами, травами и мхами. На деревьях цветковые и многочисленные споровые эпифиты.
Флористический состав примечателен наличием форм тропического происхождения: древовидных папоротников в Новой Зеландии, американской капустной пальмы (Sabal) и Tillandsia usneoides (эпифит из бромелиевых) на юго-востоке Соединенных Штатов, бамбука из рода Chusquea в Чили. Из деревьев характерны дубы, магнолии в северном полушарии, южный бук (Not* hofagus) и голосеменные (Agathis, Podocarpus) в южном полушарии, все породы с вечнозеленой листвой.
На юго-востоке Соединенных Штатов деградация этих климатических лесов благоприятствует распространению сосновых лесов, образующих теперь растительный покров на больших пространствах. Сосновые леса возникают и сохраняются благодаря пожарам.
Листопадные леса умеренного климата
В условиях умеренного климата с периодом покоя, приходящимся на холодный сезон, леса с опадающей листвой образуют хорошо развитую зону в Западной и Центральной Европе между хвойными бореальными лесами и Средиземноморской областью. Б Восточной Европе она сужается по широте, будучи ограниченной на севере тайгой, а на юге — степью. Она почти отсутствует в Сибири и снова появляется в Восточной Азии. В Северной Америке климаксовые сообщества листопадного леса занимают весь восток Соединенных Штатов, переходя в зону хвойных бореальных лесов на широте Великих озер, на западе — в прерию, на юге и юго-востоке — в вечнозеленые леса. В южном полушарии они представлены лишь на юге Нового Света.
Климатические условия. Биоклиматические границы этого типа растительности определяются комплексом природных факторов. Термические факторы: удлиненный холодный период или недостаток летнего тепла — регулируют переход к хвойным бо
276
реальным лесам. Контакт с травянистыми степными формациями и средиземноморскими сообществами связан с большой летней сухостью. Границы с зоной дождевых лесов менее ясны, но можно сослаться на физиологические потребности соответствующих древесных пород, которые не удовлетворяются в холодный период (рис. 61).
Можно сказать, что во всех областях распространения этих лесов наблюдаются морозы и осадки в виде снега. Общее количество осадков превышает 500 мм; они довольно равномерно распределяются в течение года. Тем не менее различия в климатических условиях довольно значительны, что и обусловливает зональность в распространении доминирующих древесных пород.
Флора и растительность. Листопадные леса умеренного климата почти повсюду изменены эксплуатацией, которая благоприятствует одной породе и уравнивает возраст деревьев. В неэкс-плуатируемых лесах древесный ярус довольно разнороден и по высоте может превышать 40 м. Деревья лишены выступов-подпорок, у них опадающие листья, — мелкие, тонкие, их почки защищены чешуйками. Эпифиты представлены лишь споровыми растениями, они более или менее полностью покрывают основания стволов. В старых насаждениях ярус деревцев часто разрежен и образован преимущественно видами с опадающей листвой. Однако некоторые виды имеют вечнозеленые листья, как, например, падуб в Европе, Drymis winter'd в Южной Америке. Лианы редки, они деревянистые, как обыкновенный плющ, ломонос, или травянистые, как жимолость, хмель. Травяной ярус так же, как и моховой, варьирует по густоте и составу. Травянистые растения и мхи в чистых и смешанных насаждениях образуют видовую мозаику — маленькие, хорошо разграниченные группы, сформировавшиеся, возможно, в соответствии с различной степенью межвидовой конкуренции.
Внутри зоны листопадных лесов умеренного климата климатические и эдафические различия определяют локализацию древесных пород и их ассоциаций. В Центральной и Западной Европе в срединных равнинных областях господствуют дубы, сидячецветный и черешчатый, граб, ясень, липа, ильм. В областях контакта со Средиземноморьем пушистый дуб замещает перечисленные древесные породы и смешиваетс/i часто с каменным дубом. В южной части атлантической подобласти встречается дуб пиренейский, или тоза (Quercus pyrenaicus). В горном лесном поясе царствует бук, образуя чистые или смешанные насаждения с пихтой. Севернее, от Нормадии до южной Скандинавии, буковые леса спускаются на равнины до уровня моря. Далее на север, как и выше в горах, они переходят в хвойные леса.
Флористический состав этих лесов значительно варьирует в зависимости от свойств почвы. Так, буковые леса на карбонатных почвах (рендзинах), на некарбонатных, некислых или
277
слабокислых почвах (бурых лесных почвах) и на кислых подзолистых почвах распределяются, согласно номенклатуре школы Цюрих — Монпелье, по трем различным союзам, соответственно — Cephalanthero — Fagion, Asperulo — Fagion и Luzulo — Fagion.
В Северной Америке отмечается значительно большее разнообразие древесных видов. Там описаны три больших климатических группы лесов:
1) на севере леса с буком (Fagus grandifolia) и кленом (Acer saccha-rum), к которым часто примешивается хвойная древесная порода, свойственная соседним бореальным лесам (Tsuga canadensis);
2) на юго-востоке леса с доминированием дубов (Quercus moniana, Q. coccinea, Q. alba и других) и каштана (Castanea dentata);
3) на юго-западе, по соседству с прерией, леса из дубов (Quercus borealis, Q. velutina, Q. alba, Q. macrocarpa и других) и ореха (Carya).
Все эти виды проникают далеко за пределы климатической области, где они господствуют. Ряд североамериканских родов отсутствует в Европе, но встречается частично на Дальнем Востоке; таковы Carya, Liquidambar, Li-riodendron.
Фауна. Фауна лесов с опадающей листвой умеренного пояса занимает в них различные ярусы и многочисленные микроместо-обитания. Зоологические группы представлены множеством видов, свойственных этим лесам, которые заменяют друг друга в одних и тех же экологических нишах разных биогеоценозов, демонстрируя замечательные примеры экологического викари-рования.
Оригинальные особенности этих лесов по сравнению с лесами более теплых климатов проявляются в очень резко выраженной сезонной периодичности. Зима — с низкими температурами, короткими днями, снежными заносами, с ухудшившимися возможностями пропитания — создает неблагоприятные условия для существования животных. Большую часть животного населения составляют мигрирующие или впадающие в зимнюю спячку виды. К первым относятся птицы, ко вторым — мелкие млекопитающие, рептилии, земноводные, моллюски, членистоногие. Что касается млекопитающих и птиц, которые в зимнее время продолжают вести активный образ жизни, то в суровые зимы значительная их часть погибает.
Плотность и локализация животного населения лесов в зимнее время отличается от его плотности и локализации в благоприятный сезон. Зимой надземные ярусы пустеют, их обитатели укрываются в почве, которая создает благоприятные условия животным, впадающим в спячку. Опушки в короткое время обогащаются видами соседних, не покрытых лесом территорий. Эти животные находят здесь лучшие условия, чтобы пережить суровое время года.
Каждая большая группа животных занимает различные ярусы. Среди млекопитающих встречается множество землероев-
278
грызунов (полевки, мыши) и насекомоядных (землеройки, кроты), а также наземные травоядные виды (олени, кабаны), хищные (лисицы, хорьки, барсуки и другие), всеядные (медведи), древесные виды — грызуны (белки) и хищные (дикие кошки, рысь).
Множество грызунов зимует в почве или в дуплах деревьев. Их насчитывается в разных лесах и в разные сезоны от 25 до 150 экземпляров на 1 га, а в виде исключения до 500. Их биомасса, порядка 4—25 кг на 1 га, превышает биомассу крупных млекопитающих. Оптимальная плотность последних в Европе —
С1
СЭ
X
НА М ИИАСОНДЯФ
Рис. 72. Сезонные изменения населения птиц кленово-букового леса в Огайо (по Вильямсу, А. В. Williams).
до двух пар оленей на 100 га и от 7 до 15 косуль в зависимости от достоинств лесов L
Птицы также распределяются по различным уровням. Тур-чек (Тигсёк) в грабово-дубовом лесу в Словакии насчитал 15% видов, гнездящихся на земле, 25 — в траве и кустарниках, 31 — на стволах или в стволах деревьев и 29%—в кроне деревьев. Однако больше половины этих видов добывают себе пищу на поверхности почвы. Плотность птичьего населения сильно колеблется вследствие временного присоединения к оседлым видам перелетных, пролетных и кочующих видов. Перелетные виды являются летними или зимними жителями, первые мигрируют
1 Необходимо отличать эти оптимальные плотности или «биологически терпимые», которые позволяют развиться максимальному количеству индивидуумов, от «плотности экономически .сносной», более слабой, причиняющей лишь малозначительный экономический убыток, который может бы!ь устранен без затруднений.
279
из самых низких широт, вторые — из более высоких широт. Пролетные — это виды птиц, которые весной транзитом летят к своим летним местообитаниям, а осенью — к зимним. Некоторые виды перемещаются на небольшие расстояния в течение года, за исключением сезона гнездования. Существуют виды частично оседлые, частично мигрирующие. На рис. 72 приведен пример этих сезонных изменений авифауны кленово-букового леса в Огайо. Биомасса птичьего населения меньше биомассы млекопитающих. В одном лесу Турчек (Turcek) оценил ее в 1,2 кг 16 экземпляров на 1 га, против 6,6 кг на 1 га млекопитающих.
Насекомые многочисленны, особенно на поверхности почвы в сезон активности. Зимой они углубляются в почву, в пни, под кору. Кроме сезонных изменений, связанных с жизненными циклами каждого вида, наблюдаются значительные качественные изменения от года к году в зависимости от метеорологических условий и взаимоотношений конкуренции и хищничества. Насекомые, живущие в почве, менее обильны, чем в тропических лесах, и их группы представлены различно: ногохвостки и трипсы наиболее многочисленны, муравьев значительно меньше, термиты редки или отсутствуют вовсе.
Кольчатые черви принадлежат к двум группам: большие земляные черви из семейств настоящие дождевые черви (Lumb-ricidae), и гигантские дождевые черви (Megascolecidae), и мелкие черви из группы энхитреид (Enchytraeidae). Первые могут достигать высокой плотности — до 100 на 1 м2 с биомассой более 50 г. Вторые могут быть гораздо более многочисленны и достигать летом биомассы порядка 13 г на 1 м2.
Большие различия в обилии и составе лесного эдафона существуют в зависимости от растительных сообществ и типов почв. Некислый гумус при высоком насыщении основаниями способствует развитию обильного населения, богатого бактериями, простейшими, дождевыми червями (Lumbricidae), моллюсками, обеспечивающими активную гумификацию и минерализацию органических веществ с одновременным эффективным перемешиванием с минеральными частицами почвы. Гумусовый горизонт носит характер мюлля. Гумус кислый, слабо насыщенный основаниями, заселяется преимущественно грибами; он богат мелкими животными, ногохвостками, клещами, кольчатые черви отсутствуют, и органические вещества, которые разлагаются медленно, накапливаются на поверхности и приобретают характер мора, или грубого гумуса.
Эксплуатация и эволюция. Листопадные леса умеренного климата испытали в течение четвертичного периода множество перемен. После последнего оледенения вмешательство человека становилось все более и более значительным. Рубки, с целью получения древесины или устройства пашен, пожары, охота, сбор ягод, выпас скота в лесу — такова деятельность человека, восходящая к предыстории. Однако есть некоторые заповедники,
280
с давнего времени неэксплуатируемые, которые, несомненно, близки, если не идентичны, настоящим первичным лесам.
В умеренных зонах все территории, которые освобождаются от вмешательства человека, более или менее быстро превращаются в лес. Таким образом, существует множество сукцессионных рядов растительных сообществ, направленных к лесным климаксам. Без сомнения, множество современных облесенных территорий претерпело несколько циклов распашки и оставлено под залежь.
Листопадный лес умеренного климата играет существенную роль в экономике этих областей, плотно населенных и весьма индустриализированных. Он является, таким образом, объектом интенсивной эксплуатации. Эксплуатация ведется по различным «режимам» — низкоствольный, высокоствольный и средний лес вырубаются различными способами и с различными интервалами, в зависимости от потребностей эта эксплуатация изменила более или менее значительно естественные биогеоценозы. Рубки леса, когда они сплошные и частые, изменяют микроклимат почвы, что вызывает нарушение биологического равновесия, увеличение выщелачивания, местами — эрозию, приводит в случае кислых малоплодородных почв к деградации прироста. Эксплуатация вызывает, кроме того, развитие светолюбивой и более или менее нитрофильной растительности, образующей переходные ассоциации, которые характеризуются такими растениями, как крестовник, красная наперстянка, кипрей (Иван-чай), белладонна, малина, ежевика, крапива. Там могут укорениться обычно вытесняемые из нижних ярусов такие светолюбивые деревья, как березы, сосны, ясень. Проводимые надлежащим образом вырубки позволяют сохранять или распространять определенные древесные породы, поддерживая их в конкурентной борьбе с другими видами. Именно таким образом на большей части Франции достигнута поддержка дуба против вытеснения его буком, более толерантным к затенению.
Вмешательство человека в эволюцию листопадных лесов проявляется и в других родах деятельности, имеющих тенденцию ко все большему и большему окультуриванию лесов. Это расчистка нижних ярусов, внесение удобрений, интродукция или замена естественных древесных пород селекционными вариациями и иноземными видами. Весьма распространившаяся практика— введение хвойных пород, чаще всего сосен, либо по причине их рыночной ценности, либо для восстановления сильно деградировавших лесных массивов. Ландовый сосняк, плантации сосен в бедной Шампани, Солони и других местах являют примеры этому. Плантации хвойных могут ускоренно расти благодаря благоприятному микроклимату и естественному восстановлению лиственного леса. Но эти плантации легко подвержены пожарам и, кроме того, за счет привноса ими кислого оргайиче-ского вещества обедняют почву.
Ц Ж. Леме
281
Бореальные и субальпийские хвойные леса
Хвойные бореальные леса, или тайга (слово русского происхождения), образуют непрерывный пояс на равнинах двух континентов между тундрой на севере, листопадными лесами или континентальными степями на юге. В самые низкие широты они проникают благодаря горным цепям (фото XI, А), где и образуют высотный пояс растительности — субальпийский пояс, располагающийся между альпийской тундрой и лиственными или смешанными горными лесами. Эта поясность точно воспроизводит широтную зональность.
Растительность и экологические условия. Хвойные породы этих лесов представлены разными видами сосен, пихт, елей и лиственниц. Все это деревья с иголками, вечнозелеными у трех первых и опадающими у лиственниц. Благодаря слабой транспирации вечнозеленой хвои и осеннему опаду хвои у лиственниц эти деревья могут выдерживать зимние условия без чрезмерного обезвоживания, хотя промерзшая почва не снабжает их водой. В этих лесах могут расти некоторые двудольные деревья: березы, виды рода Populus, ольхи.
Нижние ярусы обычно менее богаты и менее густы, чем в листопадных лесах вследствие более сурового климата и особенно в результате постоянного затенения, создаваемого деревьями (за исключением лиственниц), и накопления толстого слоя не-разложившейся подстилки. Нижние ярусы состоят из кустов ив, дёрена, смородин, кустарничков (Vaccinium, Ledum, Empetrum и др.), мохового или лишайникового ковра; травы (грушанки, папоротники и др.) редки.
Период активной жизни может быть сокращен до 4-х месяцев вследствие выпадения снега и глубокого промерзания почвы. В наиболее холодных районах почва бореального хвойного леса остается мерзлой на глубине в течение всего года. На Аляске долинные ледники спускаются в глубь этого леса. Однако он требует не менее одного месяца со средней температурой выше 10° и двух безморозных месяцев. Количество осадков сильно варьирует. Очень мало их на континентальных равнинах, порядка 250 мм, в горах выпадает до 1000 мм, где значительные снегопады позволяют существовать лишь расам с узкой кроной и колоннообразными стволами, удерживающими мало снега на своих ветвях.
Почвы, сформированные чаще всего на отложениях ледникового происхождения, характеризуются малой мощностью и сильной оподзоленностью. Бедность растительных остатков азотом и минеральными элементами в сочетании с суровостью климата способствует крайне медленному разложению органического вещества, которое накапливается в почве в виде грубого гумуса или очень кислого мора. Здесь поселяются главным образом
282
мицелии грибов; некоторые из грибов связаны с хвойными симбиотическим микоризным содружеством.
Показатели биомассы и первичной годовой продукции этих лесов ниже соответствующих показателей для более южных лиственных лесов; к северу и биомасса и продукция значительно уменьшаются. Примером, согласно Родину, служат следующие их значения, полученные в СССР:
Биомасса, т на 1 га Первичная чистая продукция, т на 1 га
Сосняки северотаежные 80 —
Пихтарники северотаежные 100 4,5
Сосняки южнотаежные 280 6,1
Пихтарники южнотаежные 330 8,5
Дубняки 400 9
Состав древесного яруса различается в зависимости от палеогеографических условий и современного климата. В Евразии господствующими породами с запада на восток являются: Pinus Silvestris, затем Picea excelsa, затем Picea obovata — Abies sibirica — Larix sibirica и, наконец, Larix dahurica — Pinus pumila. В Северной Америке описаны три типа: на Аляске и в Британской Колумбии климаксовые сообщества образуют сосны и ели, от Скалистых гор до Лабрадора — ели и лиственница, на обширной территории вокруг Великих озер — сосны и тсуга, «хемлок» (Tsuga canadensis).
На своей северной границе хвойный лес становится прерывистым, образуя в глубине тундры острова и галереи в долинах. Южнее своих равнинных границ хвойный лес формирует высотные формации. Именно поэтому Гималаи, Кавказ, Карпаты, Альпы, Пиренеи, Скалистые и Каскадные горы имеют верхний лесной пояс, образованный теми же родами — и особенно теми же видами, — что и бореальные хвойные леса. Таким образом, в Альпах этот субальпийский пояс представлен насаждениями из Picea excelsa, Larix europaea, Pinus montana и P. cembra. Ниже горные пояса часто образованы смешанными хвойно-лиственными лесами, подобно буково-пихтовым лесам в Западной и Центральной Европе.
Фауна. Распространение и цепи питания фауны хвойных лесов аналогичны распространению и цепям питания фауны листопадных лесов, однако представлены в большинстве случаев разными видами, к тому же сезонная периодичность выражена в еще большей степени вследствие того, что холодный сезон более продолжителен и суров.
Фауна в большинстве своем представлена беспозвоночными животными, грызунами и мелкими хищными, которые зимуют в одетой снегом почве. Большинство птиц на зиму переселяются в более низкие широты. Крупные травоядные — лоси, северные
11* 283
олени и другие представители семейства оленей (Cervidae) кормятся почками, корой, растущими на коре лишайниками. Оседлые птицы отыскивают насекомых под корой или извлекают семена из шишек; если случается, что эти последние отсутствуют, птицы во множестве перемещаются к югу.
Весной жизнь пробуждается внезапно. В течение всего благоприятного сезона насекомые быстро размножаются, особенно мухи и комары, которые часто вынуждают лосей держаться в воде. При этом лоси кормятся по соседству болотными травами, ветками ив, ольхи, берез, кувшинками. Северные олени поедают преимущественно мхи и лишайники, благородные олени— листву деревянистых растений и травы. Оптимальная плотность этих крупных травоядных более низкая, чем в лиственных лесах: в Швеции для лосей она составляет 0,5 на 100 га.
Хищные млекопитающие представлены различными видами собачьих (Canidae)—волк, красный волк (Cuon)1, лисица, а также росомахой, рысью, американской норкой, куницами, ласками и другими, которые охотятся главным образом на грызунов разного размера. Волки при случае нападают на крупных травоядных, а другие хищные уничтожают зайцев и кроликов. Медведи всеядны, они потребляют мелких млекопитающих, рыбу, насекомых, корни, плоды. Обилие каждого из этих млекопитающих испытывает более или менее периодические колебания, определяемые взаимоотношениями добычи и хищника (см. рис. 42, пример взаимозависимости между рысью и зайцем) и конкуренцией между видами с одним и тем же режимом питания. Человек нарушает эту взаимозависимость, вытесняя многие виды (охота) и эксплуатируя лес. В результате эксплуатации леса уменьшается количество северных оленей, куниц, рысей, которые держатся в наиболее густых лесных массивах, тогда как число лосей, предпочитающих прогалины в лесу и открытые лесные участки, увеличивается. Охота приводит к сокращению количества животных с дорогим мехом, которыми изобилует бореальный лес.
Среди насекомых некоторые группы или виды особенно вредоносны из-за их постоянного или эпизодического изобилия. Насекомые-ксилофаги (поедающие древесину), рогачи, или жуки-олени (Lucanidae), короеды (Scolytidae), златки (Вир-restidae) принадлежат к первым; среди вторых отметим лиственничных пилильщиков, которые уничтожают в отдельные годы молодые листья этих деревьев на больших площадях, гусеницу Choristoneura fumiferana, которая, поедая почки ели и бальзамической пихты (Abies balsamea), в Северной Америке погубила их на обширных пространствах в несколько приемов с начала XX в., гусениц походного шелкопряда (Cnet*
1 У Ж. Леме ошибочно указана африканская гиеновая собака (Ly-саоп). — Прим. ред.
284
hocampa), нападающих на сосны, гусениц Bupalus, количество которых в сосняках Германии колеблется по годам от 1 до 10 тыс. на 1000 м2.
Дождевой хвойный лес
Вдоль Тихоокеанского побережья Северной Америки, от Британской Колумбии до севера Калифорнии, склоны Берегового хребта и Каскадных гор от уровня моря до высоты 1000 м покрыты хвойным особенно пышноразвитым лесом, называемым здесь «береговым» (фото XII).
Климатические условия там существенно отличаются от климатических условий бореального леса. Климат в этих местах мягкий, температуры незначительно изменяются в течение года. Влажность воздуха высокая: годовые осадки, преимущественно зимние, составляют 1000—3000 мм, и лес купается в частых туманах. Самые высокие деревья.на склонах, обращенных влажным ветрам, перехватывают в год более 1000 мм влаги, образуемой этими туманами.
Наиболее характерны — тсуга, или «западный хемлок» (Тsaga heterophylla), туя, или «красный кедр» (Thuja plicata), дугла-сова пихта (Pseudotsuga taxifolia), красное дерево (Sequoia sempervirens), ситхинская ель (Picea sitchensis), а также Cha-maecyparis lawsoniana.
Некоторые из этих деревьев достигают в высоту почти 100 м и 6 м в диаметре. Три последних из указанных деревьев наиболее гигрофильны; дугласова пихта наиболее ксерофильна и входит в сочетании с соснами в состав горного леса, который располагается выше берегового леса.
Густой кустарниковый ярус образован множеством двудольных видов. Среди травянистых растений обильны папоротники. Моховой ковер часто плотный.
Подобно флоре, фауна этих лесов значительно отличается ют фауны бореальных хвойных лесов.
Послесловие
Книга профессора Парижского университета Орсэ Ж. Леме «Основы биогеографии» посвящена проблемам науки о закономерностях распространения и сочетания сообществ и их компонентов— животных, растений и микроорганизмов.
В настоящее время роль этой науки все более возрастает. Рост населения нашей планеты, развитие промышленности непрерывно увеличивают потребность в природных ресурсах и усиливают всестороннее воздействие человека на географическую среду. Охрана окружающей среды, восстановление нарушенных ландшафтов становятся важной предпосылкой существования и развития общества.
Сложные проблемы перестройки и сохранения географической среды порождены современными процессами урбанизации, которые все более ограничивают площади, занятые естественной, а нередко и культурной растительностью. Существенно влияет на эту среду также индустриализация (особенно механизация и химизация) сельскохозяйственного производства.
Трудно переоценить все усиливающееся значение природной среды как противовеса возрастающим психическим перегрузкам, свойственным напряженному современному ритму жизни. Активный отдых на лоне природы стал массовой, постоянной потребностью, возникла целая «индустрия туризма», а вместе с ней — новая отрасль науки — «рекреационная география».
В целом, усложняя задачу охраны и восстановления географической среды, научно-технический прогресс вместе с тем дает в руки человека все более мощные средства для решения этой проблемы, которые широко используются в условиях социалистического общества.
Биогеография является необходимым звеном в системе научных знаний, без которых невозможно вести целенаправленную работу по рациональному использованию естественных ресурсов, по охране и восстановлению природной среды.
Остановимся подробнее на важнейших проблемах биогеографии. Всевозрастающая потребность в растительном и животном сырье не может быть покрыта за счет выпускаемых промышленностью синтетических материалов и продуктов. Древесина, к примеру, еще долгое время будет источником сырья для различных
287
отраслей промышленности; спрос на пушнину сохранится, несмотря на производство искусственного меха, а естественным продуктам люди будут отдавать предпочтение перед синтезированными. В силу этого интерес к возобновимым ресурсам растительного и животного сырья вполне оправдан и будет непрерыв* но расти. Однако их восстановление возможно лишь в случае,, если изъятие не превышает прироста. Это относится к лесам, промысловым животным — наземным и водным — и полезным дикорастущим растениям.
Между тем темпы возобновления природных ресурсов растительного и животного сырья и их запасы в разных географических регионах неодинаковы. Вот почему исследования, прово-дившиеся в течение ряда лет по Международной биологическо/г программе (МБП) и ныне продолжающиеся в рамках международной программы «Человек и биосфера» (ЧИБ), осуществляемой под эгидой ООН, предусматривают прежде всего определение биомассы (запаса живого органического вещества в момент исследования) и продукции (прироста органического вещества за единицу времени с единицы площади) в различных пунктах земного шара. Определения биомассы и продукции проводятся в разных странах по единой методике. Такие работы должны быть положены в основу при составлении годичных планов лова-рыбы, норм отстрела пушных зверей и дичи, допустимой нагрузки на пастбища, норм рубки леса, сроков охоты, рыбной ловли, выпаса скота, с учетом региональных особенностей, что позволит избежать недоиспользования, равно как и чрезмерного использования, которое ведет к сокращению запаса того или иного ресурса, а в конечном итоге к его исчезновению с лица Земли. Так, известны многочисленные примеры вымирания ценных видов животных и растений. В то же время далеко не все ресурсы освоены человеком. На наших глазах человек открывает новые лекарственные растения, новые источники пищи и кормов домашних животных, производит ранее неизвестные антибиотики из бактерий и грибов.
В СССР при заметном участии биогеографов определяются запасы новых источников ресурсов, выясняются условия их существования и области распространения. Без этого невозможно их рациональное использование, а попытки введения в культуру, скажем, полезных растений кончаются неудачей.
Рациональное использование природных ресурсов немыслимо-без их охраны. Использование и охрана — две стороны одного-процесса. При этом охрана природы должна осуществляться повсеместно— и на участках, постоянно или временно изъятых и* хозяйственного пользования, в заповедниках и заказниках, и на территории, подвергающейся интенсивному хозяйственному воздействию. Заповедники должны служить эталонами нетронутой природы, что приобретает особенно большое значение в эпоху научно-технической революции. Они должны с возможной пол
288
нотой отражать природные условия различных географических зон и провинций. В создании сети заповедников видное место наряду с учеными других специальностей занимают биогеографы. В Советском Союзе с его разнообразием природных условий задачи охраны природы этим не исчерпываются, поскольку заповедники даже в передовых в этом отношении странах занимают 2—4% общей площади страны, остальная же находится в хозяйственном пользовании. Деятельность человека в настоящее время привела к столь существенным изменениям в составе природных сообществ, что практически невозможно найти участок территории, сохранивший свой первоначальный облик. Стало настоятельно необходимым восстановление численности некогда широко представленных, а ныне находящихся на грани исчезновения растений и животных, а также обогащение фауны и флоры различных областей путем восстановления ранее существовавших в этих областях животных и растений или путем завоза новых форм из других регионов. При этом биогеограф принимает участие в составлении так называемой «Красной книги», издаваемой Международным союзом охраны природы (МСОП), которая представляет собой перечень видов, находящихся под угрозой полного исчезновения и требующих повсеместной охраны. При этом именно биогеограф, а никто другой, должен определить, в какие отношения животное или растение, восстанавливаемое на данной территории, вступит с другими обитающими здесь видами, как это отразится на существовании сообщества в целом и не приведет ли акклиматизация или реакклиматизация к вредным для человека последствиям. Следует помнить, что охрана одного вида может послужить причиной снижения численности других видов.
Таким образом, знание биогеографических закономерностей должно лежать в основе всех народнохозяйственных планов и мероприятий, связанных с использованием растительных и животных ресурсов и их охраной.
На современном этапе перед человечеством встает задача не только сохранения и оптимизации природных условий, но и их рекультивации. Под рекультивацией понимают восстановление территорий, нарушенных деятельностью человека в такой степени, что их дальнейшее использование становится невозможным. Имеются в виду отвалы, терриконы, участки, подвергшиеся действию ветровой, водной эрозии или селевых потоков. Для техногенных ландшафтов нередко характерно высокое содержание в поверхностных слоях грунтов токсических веществ, препятствующих зарастанию этих участков. При восстановлении плодородия таких земель, их рекультивации, помимо механических и химических способов, существенное значение приобретают биологические методы, в том числе рациональный подбор растений. Знание экологических особенностей таких растений
289
и их географического распространения позволит биогеографу дать обоснованные рекомендации.
Все возрастает роль биогеографии в повышении продуктивности сообществ культурных растений и стад домашних животных. Поиски и изучение центров происхождения культурных растений и домашних животных ведут к увеличению разнообразия сортов культивируемых растений и пород скота, а также способствуют успешному географическому их районированию. Однако этим участие биогеографов в создании культурных сообществ растений и животных не исчерпывается. Дело в том,, что использование природных ресурсов особями одного вида ограничено. Так, в одновидовых посевах пшеницы или ржи все растения потребляют одни и те же питательные вещества. Стада крупного рогатого скота на пастбище поедают одни и те же виды растений. Гораздо эффективнее используются питательные вещества почвы травосмесями; поэтому создание многовидовых сообществ культурных растений и стад домашних животных — одна из важнейших задач сельскохозяйственной науки и биогеографии. В этом направлении сделаны пока первые шаги. Примерами могут служить посевы злаково-бобовых травосмесей^ сочетание ягодных кустарников и огородов с плодовыми садами, и прочее.
Хотя создание многовидовых сообществ сопряжено со значительными трудностями и требует участия многих специалистов,, оно обещает большие успехи в сельском хозяйстве.
Не следует умалять роль биогеографии в борьбе с вредными для человека и его хозяйства животными, сорными и паразитическими растениями. В отличие от широко распространенных я настоящее время химических способов борьбы, применение ко* торых во многих случаях сопряжено с загрязнением среды и с появлением вредителей, устойчивых к тем или иным химическим веществам, биологические меры борьбы не приводят к вредным последствиям. Применение биологических способов подавления численности вредных животных нередко связано с завозом их врагов — хищников и паразитов — из далеких стран. Поэтому необходимо знать их современное распространение и возможности существования в разных географических условиях. Решение этих вопросов требует участия не только биолога и специалиста по вопросам сельского хозяйства, но и биогеографа.
Изменение природных условий человеком, проникновение его в ранее не обжитые районы порождает круг проблем, связанных с появлением новых, ранее не известных болезней и распространением некоторых уже известных.
В исследовании природных и физиоантропных очагов видное место занимает характеристика биотических условий, к которым приурочены очаги, а также выяснение экологических особенностей и распространения носителей и переносчиков — все это за* дачи биогеографические.
290
Наконец, следует остановиться на индикационных исследованиях, при которых плохо различимые или неразличимые (так называемые деципиентные) компоненты ландшафта выявляются на основе изучения хорошо заметных физиономичных компонентов. К последним относят прежде всего растительные сообщества, а иногда и отдельные виды растений. Они могут служить индикаторами особенностей геологического строения, уровня залегания, степени минерализации грунтовых вод, характера почвенного покрова, присутствия тех или иных полезных ископаемых и т. д. Применение этих методов исследования ускоряет и удешевляет геологическую разведку и позволяет составить обоснованные планы использования сельскохозяйственных земель с учетом особенностей почв.
Таковы некоторые основные проблемы биогеографии, которые свидетельствуют о все возрастающей роли этой науки в жизни современного человечества. Ее изучают в университетах и педагогических институтах, ею интересуются люди различных профессий, имеющие дело с природными и культурными ландшафтами.
Книга Ж. Леме удовлетворяет интерес к проблемам биогеографии и может, при критической ее оценке, одновременно служить учебным пособием.
В структурном плане книга распадается на четыре части. Первая часть посвящена географическому распределению организмов и его причинам (хорология). Здесь рассматриваются типы ареалов, причины, обусловливающие их границы, значение палеогеографических факторов для современного расселения организмов, фаунистические и флористические царства и их подразделения.
Во второй части рассмотрены взаимоотношения организмов со средой (экология).
В третьей части рассмотрены проблемы науки о сообществах растений и животных (биоценология). Автор приводит определение популяции, методы описания сообществ и критерии их классификации. Специальные разделы посвящены сукцессиям — сменам сообществ и картографированию растительности — той области биогеографии, в которой особенно велики заслуги французских ученых.
Наконец, в четвертой части дана характеристика сообществ организмов, или, согласно классификации Леме, крупных биологических формаций Земли, которые соответствуют основным географическим зонам. В этом разделе при переводе опущена характеристика водных сообществ, которая менее органично связана с остальным текстом; кроме того, на русском языке имеются специальные работы, посвященные этим вопросам.
Среди сводок по биогеографии на русском языке следует назвать вышедшую в 1963 г. книгу «Биогеография с основами биологии» и опубликованный в 1974 г. труд «Биогеография мате
291
риков», посвященный характеристике главнейших сообществ-суши.
Сводку Ж. Леме характеризует полнота охвата основных проблем биогеографии, четкость изложения, яркость характеристик сообществ, оригинальность приводимых примеров. Она дает полное представление об основных закономерностях биогеографии и ключевых понятиях этой науки.
Несмотря на некоторые расхождения во взглядах с советской школой биогеографов, которые будут рассмотрены ниже, значительная часть положений, развиваемых Леме, может быть принята советскими учеными. Читатель найдет в этой книге современные представления об ареале, о причинах распространения организмов, о зональных сообществах суши земного шара. Кроме-того, здесь ярко охарактеризованы взаимоотношения оргаши-мов, в частности химическое воздействие одних организмов на другие, которому обычно уделяют мало внимания.
Основным достоинством книги является то, что она представляет руководство по биогеографии в противоположность ряду сводок, в которых проблемы ботанической географии и зоогеографии рассматриваются порознь. Какого бы вопроса ни касался Леме, он рассматривает его комплексно, что нередко бывает трудно сделать. Особенно это касается флористико-фаунистических характеристик различных регионов. Леме принадлежит одна из первых попыток таких комплексных характеристик. Книга содержит богатый справочный материал по растительному покрову и животному населению земного шара. Наконец, моно* графия включает обширную библиографию, что, несомненно^ повышает ее ценность.
При подготовке русского варианта книга была дополнена ком* ментариями, которые, как нам представляется, объясняют неизбежные различия в понимании тех или иных вопросов биогео* графии Леме и другими учеными. Основа этих различий кроется в принадлежности ученых к различным биогеографическим школам и, следовательно, в неодинаковом понимании объема науки.
Остановимся вкратце на основных различиях.
Леме, подобно основоположнику франко-швейцарской школы Рюбелю, понимает биогеографию как совокупность трех разделов: хорологии, биоценологии и экологии. Такое расширенное-толкование объема биогеографии нам представляется неправильным. Экологию принято выделять в самостоятельную науку. Биогеография использует те положения экологии, которые посвящены изучению влияния среды на распространение организмов и их сообществ. Это направление биогеографии именуют-экологической биогеографией.
В отличие от экологии биоценологию относят к числу пограничных наук — это биологическая и географическая наука одно* временно. В состав биоценологии входят: морфология биоценозов, их динамика, экология, история и географическое распро*
292
странение. Экология биоценозов составляет предмет как биоценологии, так и экологии, а географию биоценозов изучают одновременно биоценология и биогеография. Таким образом, предмет биоценологии и биогеографии частично совпадает, однако биоценология не является частью биогеографии, поскольку это различные науки.
Биогеография, подобно биоценологии, — пограничная между биологией и географией наука, она не имеет чисто биологических или чисто географических разделов. Ее задачи: изучение влияния среды на распространение сообществ и их компонентов на земной поверхности — экологическая биогеография; изучение влияния исторических причин на распространение сообществ и их компонентов — историческая биогеография; изучение фауны и флоры различных регионов — флористико-фаунистическая биогеография; выделение регионов различного ранга, отличающихся друг от друга по характеру фауны и флоры или по набору и строению сообществ — региональная биогеография. Иногда в качестве особого отдела выделяют количественную биогеографию, однако количественные методы исследования, развившиеся особенно в последние десятилетия в связи с запросами практики, пронизывают все разделы биогеографии.
Уже сам перечень основных направлений биогеографической науки показывает, что имеются два подхода. С одной стороны, могут изучаться фитоценозы, зооценозы, микробоценозы и биоценозы в целом, а также фауна и флора определенных территорий и акваторий; с другой стороны, объектом исследования становится географическое распространение сообществ, а также видов, родов и других таксономических единиц.
Таким образом, биогеография имеет дело с распространением сообществ организмов (биомов) и совокупностей флоры и фауны (биот) определенных территорий и акваторий.
Из этого следует, что биогеография представляет собой не простую совокупность ботанической географии и зоогеографии, как считают одни ученые, и не надстройку над двумя вышеупомянутыми науками, изучающую только те закономерности, которые находятся вне поля зрения ботанической географии и зоогеографии, как полагают другие ученые. Вся совокупность законов, определяющих географическое распространение организмов и их сообществ, составляет предмет биогеографии.
Хотя Леме и сообщает сведения, необходимые для уяснения основных проблем экологии и биоценологии, предметом его исследования является биогеография в точном понимании этого слова: влияние среды на распространение организмов, зональное распределение основных сообществ, проблема ареалов и флористико-фаунистических областей.
Рассматривая причины ограничения ареалов различных видов, Леме справедливо выделяет географические барьеры, отсутствие подходящего субстрата, биотические, климатические и
293
исторические факторы. К этому, однако, необходимо добавить, что как животные, так особенно растения во многих случаях в своем распространении не доходят до своих климатических пределов, поскольку неблагоприятные условия близ границ ареалов ослабляют организм, снижают его жизнедеятельность и делают менее устойчивым в конкурентной борьбе с организмами других видов.
Специальный раздел Леме посвящает фаунистическим и флористическим территориям.
Система принятых им подразделений при характеристике последних не вполне соответствует тем, которые распространены в работах советских исследователей. Леме выделяет царства, регионы, или области (regions); регионы, или провинции; доминионы (domaines), сектора, округа. Однако термин «регион» применяют в нашей литературе для обозначения единицы неопределенного объема. Регионом может быть и царство, и провинция, и округ. Термин «доминион» в советской литературе применяют редко. И если в «Физико-географическом атласе мира» Е. М. Лавренко доминион предстает в качестве синонима царства, то есть самой крупной единицы, то Леме относит его к третьей по объему категории. А. Н. Толмачев 1 предлагает выделять флористические царства, области, провинции, округа и районы. Эта система единиц принята большинством флористов и фаунистов советской школы; лишь наиболее мелкую единицу зачастую называют участком, а не районом.
Леме справедливо указывает, что, чем обширнее выделяемая территория, тем более крупными систематическими категориями она характеризуется: царства и области — семействами, доминионы — родами, сектора — видами. Этой же точки зрения придерживаются и другие биогеографы. Однако в последнее время наметилась тенденция характеризовать те или иные регионы не только эндемичными, лишь им свойственными формами и группами, но и характерными таксонами, то есть встречающимися за пределами региона, но на его территории достигающими высокой численности. В связи с этим все чаще прибегают к количественным оценкам численности видов, характерных для того или иного региона.
Растения и животные в той или иной местности находятся в одинаковых условиях среды, они входят в состав одних и тех же биоценозов. Однако различие в средствах расселения, подвижность животных и неподвижность растений, существование внутренней среды у высших гомотермных позвоночных животных— вот причины, определяющие различие границ ареалов растений, с одной стороны, животных — с другой. Более того, границы ареалов представителей различных типов и классов животных могут различаться весьма резко.
1 См. «Введение в географию растений», Л., 1974.
294
В ряде случаев принципиально невозможно проведение общих флористико-фаунистических границ, основанных на распространении цветковых и папоротникообразных растений и высших позвоночных животных, то есть представителей систематических групп, по которььм обычно строятся системы фаунистических и флористических регионов, но границы распространения растительноядных насекомых совпадают с пределами ареалов растений.
Попытка установления единых флористико-фаунистических областей, предпринятая Леме, весьма интересна, хотя и спорна. Неизбежная в работе такого объема краткость изложения принципов выделения регионов различного ранга не позволяет судить о степени обоснованности этого выделения. В некоторых случаях, видимо, идет речь о том, что выделенные, скажем, на основании флористических особенностей территории затем получают фаунистическую характеристику, или наоборот. Так, на приводимой Леме карте нанесены только границы царств, а границы регионов даны лишь в пределах голарктического царства. В одних случаях Леме отдает примат фаунистическому подразделению суши, например при выделении трех самостоятельных царств — эфиопского, или афро-мадагаскарского, индо-малайского и полинезийского, там, где флористы выделяют одно палеотропическое царство. В других случаях он следует за флористами, например при выделении границы между неотропическим царством (неогеей) и антарктическим. Подобный принцип заслуживает внимания и с дидактических позиций как способ подачи материала учащимся.
Раздел, посвященный сообществам организмов, Леме начинает с описания основных свойств растительных сообществ и методов их характеристик. И это обоснованно, поскольку растительное сообщество представляет структурную основу любого наземного сообщества. Различное толкование термина «растительное сообщество» свидетельствует о разнообразии взглядовэ свойственных различным фитоценологическим школам.
Автор приводит краткую характеристику взглядов британских и американских фитоценологов, которых, несмотря на незначительные расхождения, можно было бы объединить в одну англо-американскую школу, анализирует взгляды представителей скандинавской школы и флористико-экологической школы Дювиньо — Элленберга; детально рассматривает методы флористической школы Браун-Бланке. К сожалению, автор не касается методов, применяемых советской школой исследователей. Поэтому целесообразно провести сравнительный анализ концепций различных фитоценологических школ, учитывая, что в каждой из них можно различать разнообразные взгляды, а иногда и значительные расхождения.
Флористическая школа, к которой принадлежит Леме, ведет свое происхождение от франко-швейцарской фитоценологиче-
295
ской школы. Для нее характерно выделение основной единицы — ассоциации по флористическому признаку. Каждой ассоциации присущи характерные (свойственные только ей) виды и дифференциальные виды, которые встречаются лишь в некоторых сообществах данной ассоциации, что позволяет на их основании выделять субассоциации и варианты. Ассоциации по флористическому признаку объединяют в более крупные категории: союзы, порядки, классы, округа. Эти категории отсутствуют у авторов, принадлежащих к другим школам. Характерные виды могуг иметь невысокое обилие, более того, они редко попадают в число доминирующих видов. Поэтому к одной ассоциации представители школы Браун-Бланке могут отнести сообщества с господством бука или европейской ели, если в составе их травяного яруса будут присутствовать общие характерные виды. Подобные виды в отличие от доминантов, обладающих обычно широкой экологической амплитудой, индуцируют определенные экологические условия.
Ассоциация представляет абстрактную категорию, поскольку не учитывается обилие характерных видов и к одной ассоциации относят как сообщества, в которых данный вид единичен, так и те, в которых он характеризуется высоким обилием. Объем ассоциации оказывается весьма значительным.
Конкретные описания сообществ, проводимые по методике школы Браун-Бланке, характеризуются высокой степенью детализации, применяются простые методические приемы — шкала для одновременной оценки обилия и покрытия, полные флористические списки, методы сравнения сообществ при помощи коэффициентов общности и дифференциации и прочее. Большое внимание уделяют динамике растительного покрова.
В последние годы эта школа расширяет свое влияние во многих странах мира; работы с применением ее методики проводи’* лись, в частности, и у нас, в Советском Союзе.
Фитоценологи Швейцарии по сравнению с их французскими коллегами придерживаются несколько иных взглядов, хотя в 20-х годах текущего столетия они входили в одну школу, получившую название франко-швейцарской, или школы Цюрих — Монпелье. В отличие от сторонников Браун-Бланке представители этой школы при характеристике сообществ опираются не только на флористический состав, но и на количественные соотношения между видами, на принадлежность их к тем или иным жизненным формам, а также на место каждого сообщества в ряду смен в пространстве и во времени.
Эту так называемую физиономико-экологическую школу, взгляды которой во многом разделяют и советские ученые, представляют Гаме, Мейзель, Гиноше, Люди, Шмид и др.
Если в Западной Европе к тому времени, когда получили развитие фитоценологические исследования, естественная растительность на значительных территориях уже сменилась куль
296
турной, то в Северной Америке, как и в России, сохранились еще обширные, не освоенные человеком пространства. Отсюда, видимо, и те широкие обобщения и динамический подход к растительному покрову, которые характерны как для англо-американской, так и для русской (позже советской) фитоценологиче-ской школы.
Для представителей англо-американской школы: Клементса, Уивера, Шанца и др. — в США, Тенсли, Ричардса — в Великобритании, определяющим свойством растительного покрова оказывается его динамичность. Сторонникам этого направления принадлежит детальная разработка учения о сменах-сукцессиях, установление различных типов (рядов) сукцессий. Следует отметить, что проблеме сукцессий посвящены серьезные исследования русских специалистов. Ученые англо-американской школы при этих исследованиях широко применяют стационарные методы. При этом все сообщества делят на две категории: постоянно сменяющиеся стадии сукцессионных серий и неизменные в течение длительного периода, находящиеся в равновесии с климатическими условиями сообщества — климаксы. Первоначально считали, что растительный покров всей зоны или подзоны в целом стремится к одному и тому же климаксу, впоследствии пришли к заключению, что каждый ряд сукцессий заканчивается своим климаксовым сообществом. На этой последней точке зрения стоят и представители школы Браун-Бланке; она близка к взглядам советских ученых.
Для представителей англо-американской школы характерно необычайно широкое понимание основных единиц растительного покрова — формаций и ассоциаций. Формации соответствуют климаксовой растительности каких-либо зон, а ассоциации — растительности подзон. Такое же широкое понимание формаций свойственно и Леме, согласно которому формацию в отличие от ассоциации, выделяемой по флористическим особенностям сообщества, устанавливают в соответствии с физиономическим принципом. В этом Леме отдает дань англо-американским фито-ценологическим исследованиям — французская школа формаций не признает.
Как мы увидим далее, позиции ученых советской фитоцено-логической школы в отношении объема упомянутых двух единиц и принципов их выделения существенно отличаются от подобного толкования.
Представители шведской (финно-скандинавской, упсальской) школы, которую представляют Каяндер, Хульт, Килин, Дюръе, при характеристике растительных сообществ огромное значение придают константам — видам, присутствующим во всех фитоценозах, относящихся к одной типологической категории. Константные виды, как и характерные виды французской школы, выделяют без учета их обилия и других количественных показателей. В связи с этим большое внимание уделяют определению
297
величины ареал-минимума, то есть площади, на которой выявляются все константы данной ассоциации. Наименьшей классификационной единицей шведской школы являются социации, выделяемые по доминирующим видам различных ярусов. Социа-ция — более мелкая единица по сравнению с ассоциацией, принятой советскими учеными.
Советская фитоценологическая школа в качестве основной единицы принимает ассоциацию. Однако это понятие не идентично аналогичному термину французской школы, которую представляет Леме. Ассоциация — это тип фитоценоза. Если фитоценоз конкретен, то ассоциация представляет собой некую абстракцию как тип фитоценоза и в то же время определенную конкретность как совокупность фитоценозов. При этом каждый из фитоценозов, относящихся к определенной ассоциации, обладает общими признаками, которые свойственны всем однородным фитоценозам, и случайным признаком, свойственным только ему. При характеристике ассоциации индивидуальные признаки, принадлежащие отдельным фитоценозам, отбрасывают. В одну ассоциацию объединяют фитоценозы, характеризующиеся однородным видовым составом, определяющимся доминирующими растениями каждого яруса, однородной структурой и динамикой, а также характеризующиеся одинаковыми взаимоотношениями между растениями, образующими фитоценоз, и между растениями и окружающей их средой. Таким образом, для советской фитоценологической школы характерен комплексный подход к определению этого понятия. При этом ведущая роль принадлежит доминантам — господствующим растениям и особенно эдификаторам.
Вся классификационная система строится по принципу фито-ценотического сходства, которое выражается в общности доми-нантов, эдификаторов и жизненных форм, а для единиц более высокого ранга — в экологической, морфологической и физиологической близости эдификаторов.
Вторая основная категория — формация. В отличие от широкого понимания формации представителями англо-американской школы и неопределенного объема формации у Леме советские ученые относят к одной формации все сообщества, характеризующиеся одним и тем же эдификатором, например все леса из обыкновенного дуба, все степи с господством ковыля красноватого и т. д.
Наконец, наиболее крупной таксономической категорией является тип растительности. Типы выделяют на основе экологоморфологической близости компонентов растительного покрова. Примерами типов, которые, в общем, довольно близки к «крупным формациям» Леме, хотя и далеко не всегда совпадают с ними, включая каждый по нескольку «крупных формаций», могут служить степи, луга, болота, леса и прочее.
298
Таким образом, основные классификационные единицы рас-тигельного покрова советских ученых резко отличаются от основанных на флористических признаках ассоциаций Леме.
Советской школе свойственно динамическое понимание растительного покрова, роднящее ее с англо-американской и французской школами, представление о сочетании зональных, интра-зональных и экстразональных сообществ, успешное развитие картографирования растительности.
Применяя термин «сообщество» в конкретном смысле, Леме наименование «группировка» резервирует для совокупности идентичных сообществ, считая группировку столь же абстрактным понятием, как вид по сравнению с индивидуумами, его составляющими. Советская фитоцепологическая школа под группировкой понимает определенную стадию развития растительного покрова на лишенном растительности участке. При этом различают: пионерную группировку, соответствующую первой -стадии зарастания, характеризующуюся несомкнутым растительным покровом, простую группировку — следующую стадию развития с сомкнутым, но не диффузным распределением растений, относящихся к одной жизненной форме, и, наконец, сложную группировку, с сомкнутым, но не замкнутым растительным покровом, образованным растениями, относящимися к различным жизненным формам. Поэтому термин «группировка» в тексте книги был заменен термином «сообщество».
Как Леме, так и все перечисленные представители различных фитоценологических школ стоят на точке зрения дискретности, прерывистости растительного покрова, который распадается на сообщества различного ранга. Это позволяет на любой территории выделить наименьшую конкретную единицу — фитоценоз. Однако ряд ученых — Л. Г. Раменский и его последователи в СССР, Куртис, Уиттэкер и др. в США — полагают, что растительный покров непрерывно и постепенно меняется от места к месту и поэтому любое выделение фитоценозов носит субъективный характер. В действительности, как нам представляется, растительный покров представляет единство непрерывности и прерывистости, в нем выделяются участки, однородные по составу и динамике. Переходные полосы между такими участками, как правило^ много уже самих участков. Таким образом, фитоценозы— реальность, они существуют в природе, а не представляют собой результат произвольных абстракций исследователя. Поэтому совершенно закономерна точка зрения Леме, выделяющего сообщества, реально существующие в природе.
Некоторую сложность представляет проблема соотношения растительных сообществ с сообществами животных. Леме полагает, что отдельные группы животных могут быть связаны ассоциациями, а могут (для таких мобильных животных, как млекопитающие, птицы, летающие насекомые) быть связаны с крупными сообществами, например с формациями. Кроме того.
299
ряд видов предпочитает переходные полосы (экотоны), то есть принадлежит к нескольким ассоциациям (например, птицы, питающиеся в одном сообществе и гнездящиеся в другом). Такая точка зрения, развиваемая и некоторыми другими биогеографами, представляется неправильной. В этом случае пришлось бы выделять зооценозы, различные для разных групп животных: одни — для малоподвижных насекомых, например тлей и червецов, другие — для мелких грызунов, третьи — для копытных, четвертые — для перелетных птиц и т. д. Нам представляется, что, поскольку ценоз, как фитоценоз, так и зооценоз,— пространственная категория и поскольку растения, будучи неподвижными, определяют структурные особенности наземного биоценоза в целом, следует говорить не о различных зооценозах, образуемых разными видами и группами животных, а о смене биоценозов в процессе жизни животных. Эта смена может осуществляться в течение цикла развития, по сезонам года и в течение суток.
В соответствии с этим и категории классификации биоценозов должны соответствовать категориям классификации растительного покрова. Так, можно говорить о биоценотических ассоциациях, формациях и типах.
Поэтому термину «биом», который, как полагает Леме соответствует по объему формации, правильнее придавать общее значение, считая его объем синонимом понятия «сообщество».
Анализ основных различий во взглядах Леме и советских ученых имеет целью облегчить читателю понимание наиболее широко распространенных в нашей стране представлений в области биогеографии и смежных с нею наук. Это позволит критически оценить идеи, развиваемые Леме и в целом соответствующие воззрениям представителей французской школы Монпелье, школы Браун-Бланке.
При этом детальное рассмотрение различий не должно заслонить достоинств книги Леме. Более того, мы должны иметь в виду, что факты и закономерности, излагаемые в книге, независимо от взглядов автора существуют в природе и поэтому основные данные, характеризующие географическое распространение сообществ, экологические и исторические причины их распределения, представляют большой интерес. Однако даже не в этом ее основное достоинство. Написанная на современном уровне науки, она будит мысль. Несколько перефразируя известный афоризм, можно сказать, что человек, приступающий к изучению какой-либо науки, — не сосуд, который следует наполнить,, а светильник, который надо зажечь. И книга Леме выполняет эту задачу, заставляя читателей думать, сравнивать, обобщать..
Профессор А. Г. Воронов
Литература
Биогеография. Работы общего характера
Chevalier A., Cuenot L., Biogeographie, dans De Martonne. Traite de Geographic physique, Paris, 1925.
Gaussen H., Geographie des plantes, Paris, 2 ed., 1954.
Guinochet M., Logique et Dynamique du peuplement vegetal, Paris, 1955.
О z e n d a P., Biogeographie vegetale, Paris, 1964.
Polunin N., Introduction to plant geography, London, 1960.
P renant M., Geographie des animaux, Paris, 1933.
Schimper A., von Faber F., Pflanzengeographie auf physiologische Grundlage, lena, 2 Bds. 3 Aufl. 1935.
Walter H., Grundlagen der Pflanzenverbreitung, Stuttgart, 3 Bds, 1951, 1954, 1960.
Часть первая
Хорология
de Beaufort L. F., Zoogeography of the land and inland waters, London, 1951.
C a i n S. A., Foundations of plant geographv, New York, 1944.
Colloque sur la Caryosystematique et la Taxonomie experimentale, «Rev. de Cytol. et Biol, veget.» Paris, 25, 1962.
E1 h a i H., Biogeographie: les paysages vegetaux au Quaternaire en Europe occidentale, «С. D. U.», Cours de Sorbonne.
Fur on R., Causes de la repartition des etres vivants, Paris, 1958.
Good R., The Geography of the flowering plants, London, Green and Co., 2 ed., 1953.
Rothmaler W., Allgemeine Taxonomie und Chorologie der Pflanzcn, lena, 1950.
Walter H., Einfiihrung in die Phytologie, III, Grundlagen der Pflanzenverbreitung, II Teil: Arealkunde, Stuttgart, 1954.
Wulff E. V., An Introduction to historical Plant Geography, U. S. A., Chronica Botanica, 1950.
Часть вторая
Экология
Работы общего характера
Allee W. С., Emerson А. Е., Park О., Park Т., Schmidt К. Р.» Principles of Animal Ecology, Philadelphia, 1961.
Bodenheimers F. S., Precis d’Ecologie animale, Paris, 1955.
Clarke G. L., Elements of Ecology, New York, London, 2 ed., 1957.
Combes R., La forme des vegetaux et le milieu, Paris, 1946.
Daubenmire R. F., Plants and Environment. An textbook of plant autecology, New York, London, 2 ed., 1959.
301
Heslop-Harrison J., Fourty years of Genecology, Advances in Ecological Research, vol. 2, 1964, p. 159—247.
Hesse R., Allee W. C., Schmidt К. P., Ecological animal Geography, New York, London, 4 ed., 1949.
К e n d e i g h S. C., Animal Ecology, U. S. A. 1961.
Macfadyen A., Animal Ecology, Aims and Methods. London, 1963.
-Odum E. P., О d u m H. T., Fundamentals of Ecology, Phila del phie, 2 ed.,
1959.
Weaver J. E., Clements F. E., Plant Ecology, New York, 2 ed., 1938.
Факторы климатические и гидрологические
Bagnouls F., Gaussen H., Les climats biologiques et leur classification, «Annales de Geographie», Paris, 66, 1957, p. 193—220.
Ember ger L., Une classification biogeographique des climats, «Recueil des Trav. des lab. de Botan.“, Geol. et Zool. Faculte des Sci. Montpellier. Serie Botanique, fasc. 7, 1955, p. 3—44.
e i g e r R., The climate near the ground, Cambridge, Massachusetts, 1959. [Имеется русский перевод: Гейгер Р. Климат приземного слоя воздуха, М.. 1962.]
-«Handbuch der Pflanzenphysiologie, Bd. Ill: Pflanze und Wasser, Berlin, 1956.
«Recherches sur la zone aride», XV: Echanges hydriques des plantes en milieu aride ou semi-aride, UNESCO, Colloque de Madrid, 1961.
Russell M. B., Water and ils relation to soils and crops. New York, London, Academic Press. 1959.
SI at yer R. O., Me 11 г о у I. C., Practical Microclimatology. UNESCO, 1961.
T r a n q u i 11 i n i W., Das Lichtklima wichtiger Pflanzengesellschaften. «Hdb. der Pflanzenphysiologie», Bd. V/2: Die CO2 Assimilation, Berlin, 1960, S. 301—338.
Эдафические факторы
Bernstein L., Les sols halomorphes et leur vegetation, cm.: Les Problemes de la zone aride, UNESCO, Actes du Colloque de Paris, 1962, p. 151—190.
В 1 a c k C. A., Soil-Plant Relationships, New York, 1957.
Demol on A., Principes d’Agronomie, 1. La dynamique de sol, Paris, 1952.
D e m 0 1 0 n A., Principes d’Agronomie, II, Croissance des vegetaux cultive.
Paris, 5 ed., 1956. I
Duchaufour P., Precis de Pedologie, Paris, Masson, 2 ed., 1965. [Имеется русский перевод: Дюшофур Ф., Основы почвоведения, М., 1970].
D и v i g n е a u d Р., Den a ever • de Smet S., Cuivre et vegetation au Katanga, «Bull. Soc. roy Botan. de Belgique», 96, 1963, p. 93—231.
Hayward H. E., Bernstein L., Plant-growth relationships on salt-affected soils, «Botanical Review», t. 24, 1958, p. 584—635.
Russell E. W., Soil conditions and plant growth, London, 9, ed., 1961.
Soil, The Yearbook of Agriculture, Washington, 1957.
Механические факторы
Ahlgren I. F., Ahlgren С. E., Ecological effects of forest fires, «Botanical Review», t. 26, 1960, p. 483—533.
< a i 1 1 e u x A., T r i с a г t J., Cours de Geomorphologie, C. D. U., Cours de Sorbonne.
Fur on R., L’erosion du sol, Paris, 1947.
К u h n h 0 11 z - L о r d a t G., La terre incendiee, Essai d’Agronomie comparee, Nimes, Ed. Maison-Carree, 1938.
T г i с a r t J., Climat, vegetation, sol et morphologie, Vol. jubil. 50 anniv. labor, de Geogr. de Rennes, 1952, p. 229—239.
-302
Биотические факторы
Borner Н., Liberation of organic substances from higher plants and their role in the soil sickness problem, «Botanical Review», 26, 1960, p. 393—424. Dommergues Y., Les cycles biogeochimiques des elements mineraux dans les formations tropicalcs, «Bois et Forets des Tropiqus», 87, 1963, p. 10—24.
Duvigneaud P. et Tanghe M., L’evolution, Science moderne de syn-these, vol. 2: Ecosystemes et Biosphere, Bruxelles, Document Ministree de 1’Education Nationale et de la Culture 23, 1962. [Имеется русский перевод:
Дювиньо Р. и Танг М., Биосфера и место в ней человека, М., 1968 и 1973 г. (2 изд.).]
Elton Ch. S., The Ecology of invasions by animals and plants, London, 1958.
Jaeger P., Les aspects actuels du probleme de 1’entomogamie, «Bull. Soc. potan. de France», t. 104, 1957, p. 179 et 352.
Красильников H. А.. Почвенные микроорганизмы и высшие растения, изд-во АН СССР, М., 1958.
Lie th М., Die Stof f produktion der Pflanzendecke, Stuttgart, 1962.
Ovington J. D., Quantitative Ecology and the woodland ecosystem concept, «Adv. in Ecology», t. 1, 1963, p. 103—192.
Pochon J., de Barjac H., Traite de Microbiologie des sols, Paris, 1958.
Structure et physiologie des societes animales, Colloques internat. du C. N. R. S., Paris, XXXIV, 1952.
Часть третья
Сообщество организмов: биоценология
Becking R. W., The Zurich — Montpellier School of Phytosociology, «Botan. Review», 23, 1957, p. 411—488.
Braun-Blanquet J., Fuller G. D., Conard H. S., Plant Sociology, The study of plant Communities, New York, 1932
Braun-Blanquet J., Pflanzensoziologie, Vienne, 3 Aufl., 1964.
Cain S. A., Life-forms and phytoclimate, «Botan. Review», 16, 1950, p. 1—32.
Clements F. E., Dynamics of Vegetation, New York, 1949.
Dagnelie P., Contribution a 1’etude des communautes vegetates par Гапа-lyse factorielle, «Bull. Service de la Carte phylogeographique», Serie B, t. 5, 1960, p. 7—71, 93—195.
Dansereau P., Description and recording of vegetation upon a structural basis, «Ecology», t. 32, 1951, p. 172—229; «Bull. Serev. de Biogeogr, Montreal», № 8, 1953.
Dansereau P., A universal system for recording vegetation, «Contrib. Inst., botan. Univ, de Montreal», № 72, 1958.
G о u n о t M., Les methodes d’inventaires de la vegetation, «Bull. Service de la Carte phytogeogr.», Serie B, t. 6, 1961, p. 7—73.
Greig-Smith P., Quantitative Plant Ecology, London, 2 ed., 1964.
Kuchler A. W., A physiognomic classification of vegetation, «Assoc, amer. Geogr., Ann», t. 39, 1949, p. 201—210.
Poore M. E., The method of successive approximation in descriptive Ecology, «Adv. in Ecol. Research», t. 1, 1963, p. 35—68.
Rubel E., Plant Communities of the World, Essays in Geobotany in honour of W. Setchell, Univ. Calif. Press, 1936, p. 263—290.
Tischler W., Synokologie der Landtiere, Stuttgart, 1955.
Trochain J., Nomenclature et Classification des milieux vegetaux en Afri-que noire fran^aise, dans: Les Divisions ecologiques du Monde, Colloque de C. N. R. S., Paris, 1954, p. 73—93, Annee Biologique, t. 31, 1955, p. 317—334.
Wittaker R. H., Clasification of natural communities, «Botan. Review», t. 28, 1962, p. 1—239.
303
Часть четвертая
Крупные биологические формации земного шара
Морские воды
Moore Н. В., Marine Ecology, New York, London, 1958.
Peres J. M., Oceanographie biologique et biologie marine, Presses univ. de France, Paris. 2 vol., 1961.
Ra у mont J. E., Plankton and productivity in the oceans, London, 1963. Sverdrup H. U., Johnson M. W., Fleming R., The oceans. Their physics, chemistry and general biology, New York, 3 ed., 1949.
Континентальные воды
Barnach J., Downstream, A natural history of the river, New York, London, 1964.
D u s s a r t B., Limnologie, Paris, 1966.
Kuznetsov S. J., Die Rolle der Mikroorganismen im Stoffkreislauf der Seen (traduit de russe par A. Pochmann), VEB Deutsch, Verlag d. Wis-sensch., Berlin, 1959.
Reid G. К.» Ecology of inland waters and estuaries, New York, Reinhold, 1961.
R u 11 n e r F., Fundamentals of limnology, 3 ed., trad. G. D. Frey et F. E. Fry, Univ. Toronto Press, 1963.
Thienemann A., Die Binnengewasser, Stuttgart, 1925.
V i v i e r P., La vie dans les eaux douces. Que sais-je? Paris, 1961.
Тундра и альпийский пояс
Fa v ar ger C., Flore et Vegetation des Alpes. I. Etage alpin. II. Etage subalpin. Neuchatel, Deulachaux et Niestle, 2, 1956.
Polunin N., Botany of the Canadian eastern Arctic. Part III: Vegetation and Ecology, Ottawa, Nat. Museum Bull., № 104, 1948.
Аридные зоны
Biology of Deserts, London, Inst. Biol., 1954.
С 1 о u d s ley - Th о m p so n J. L., Chadwick M. J., Life in Deserts, London, 1964.
D e k e у s e r P. L., D e r i v о t J., La vie animale au Sahara, Paris, 1959.
Desert Research, Internat. Symposium Jerusalem. Research Council Israel, Spec. Public, № 2, 1953.
Kachkarov D. N., Кого vine E. P., La vie dans les deserts. Ed. fran-^aise par Th. Monod, Paris, 1942.
Killian Ch., Lemee G., Les xerophytes: leur economic d’eau, «Handb. d. Pflanzenphysiologie», III, 1956, p. 788—824.
«Recherches sur la zone aride», Actes des Colloques et Comptes-rendus de recherches, 21 vol., publies par 1’UNESCO. En particulier: VIII: Ecologie humaine et animale. Compte rendu de recherches. X: Climatologie. Compte rendu de recherches. XI: Climatologie et microclimatologie. Actes du Collo-que de Canberra. XV: Echanges hydriques des plantes en milieu aride et semi-aride. Compte rendu de recherches. XVI: Echanges hydriques des plantes en milieu aride et sumi-aride. Actes du Colloque de Madrid. XVIII: Les problemes de la zone aride. Actes du Colloque de Paris, 1962.
Troll C., Das Wasser als pflanzengeographischer Faktor, «Handb. d. Pflanzenphysiologie»; Bd. Ill, 1956, S. 750—786.
304
Степи
Carpenter J. R., The grassland biome, «Ecol. Monogr.», 10, 1940, p. 617— 684.
Haviland M. D., Forest, steppe and toundra, Cambridge, Univ. Press, 1926.
Weaver J. E., North American Prairie, Lincoln, Nebr., 1954.
Weaver J. E., Albertson F. W., Grasslands of the Great Plains, Their nature and use. Lincoln, Nebr., 1956.
Тропические формации
Aubert de la RiieE., BourliereF., HarroyJ. P., Tropiques (La nature tropicale). Horizons de France, Paris, 1954.
Aubreville A., Climats, forets et desertification de 1’Afrique tropicale, Paris, Soc. ed. marit, et colon., 1949.
Aubreville A., Les forets du BresiL Etude phytogeographique et forestiere, «Bois et Forets des Tropiques», № 59, 60, 1958.
Bunning E., Der tropische Regenwald, Berlin — Gottingen — Heidelberg, 1956.
Mangenot G., Etude sur les forets des plaines et plateaux de la Cote-d’lvoire, «Etudes eburneennes», IFAN Abidjan, IV, 1955, p. 5—61.
Mertens R., Die Tierwelt des tropischen Regenwaldes, Frankfurt, Kramer, 1948.
P a u 1 i a n R., Un naturaliste en Cote-d’Ivoire, Stock, Paris, 1948.
«Recherches sur la zone tropicale humide», «L’etude de la vegetation tropicale. Actes du Colloque de Kandy, 1956, UNESCO, 1958.
«Recnerches sur la zone tropicale humide», Sols et vegetation des regions tropicales. Travaux du Colloque d’Abidjan, 1959, UNESKO, 1961.
Richards P. W., The tropical rain forest. An ecological study, Cambridge Univ. Press, 1957. [Имеется русский перевод: Ричардс П. В., Тропический дождевой лес. М., 1961].
Schnell R., La foret dense. Manuels Ouest-Africains, Paris, 1950.
Schnell R., Le probleme des homologies phytogeographiques entre 1’Afri-que et I’Amerique tropicales, «Memoires Museum nat. d’Hist, nat.», Serie Botanique, t. XI, fasc. 2, 1961.
Simposio sobre о Cerrado. Editora da Universidade de Sao Paulo, 1963.
Troll C., Das Pflanzenkleid der Tropen in seiner Abhangigkeit von Klima, Boden und Mensch, «Deutscher Geographen Tag», Frankfurt, 28, 1951, S. 67.
Walter H., Die Vegetation der Erde. Bd. 1: Die tropischen und subtropischen Zonen, Stuttgart, Gustav Fischer, 1964. [Имеется русский перевод: Вальтер X., Растительность Земного шара. т. I: Тропическая и субтропическая зоны, М., 1969.]
Леса средних и высоких широт
Berg L. S., Natural Regions of the U. R. S. S., New York, 1950.
Braun E. L., Deciduous forests of eastern North America, Philadelphia, 1950.
Braun-Blanquet J., La chenaie d’yeuse mediterraneenne, «Mem. Soc. d’Etude des Sc. nat. de Nimes», № 5, «S. I. G. M. A.», communic. № 45, 1936.
Cochet P., Hu ch on H., T u r p i n P., V e 1 a у L., La foret, Paris, 1963.
FAO. influences exercees par la foret sur son milieu, Rome, FAO, 1962.
Lutz H. J., Chandler R. F., Forest soils, New York, 8 ed., 1959.
P a r d e L. M. et J., Arbres et forets, Paris, 2 ed., 1965.
Shelford V. E„ Sere, climax and influent animals wich special references to the transcontinental coniferous forest of North America, «Ecology», 16, 1935, p. 375—402.
305
Основные периодические издания по биогеографии
«Advances in Ecological Research», London and New York, Acad. Press.
«Compte rendus des Seances de la Societe de Biogeographie, Paris.
«Ecology», Publiee par Ecological Soc. of America et Duke Univ. Press.
«Ecological Monographs», Duke Univ. Press, Durham, U. S. A.
«Journal of Ecology», London.
«La Terre et la Vie», Revue d’Ecologie appliquee, Paris.
«CEcologia plantarum», Paris.
«Oikos», Acta oecologica scandinavica. Copenhague, Mundsgaard.
«Revue d’Ecologie et de Biologie du Sol», Paris.
«Vegetation», La Haye.
Оглавление
Предисловие автора 5
Введение 7
Часть первая
Географическое распределение живых организмов и его причины: хорология (перевод Л. Г. Вольфензон)
Глава 1. Средства распространения организмов 11
Способность к размножению 11
Распространение организмов 13
Глава 2. Области географического распределения 21
Картографическое воспроизведение 21
Структура ареалов 24
Причины ограничения ареалов 30
Палеогеографические факторы современного распространения живых существ 34
Глава 3. Флористические и фаунистические территории 39
Голарктическое, или бореальное, царство 40
Неотропическое царство, или неогея 52
Эфиопское, или афро-мадагаскарское, царство 55
Восточное, или индо-малайское, царство 61
Австрало-папуасское царство, или нотогея 63
Полинезийское царство 60
Антарктическое царство 67
Часть вторая
Факторы среды и их роль в распространении и существовании организмов: экология (перевод Л\ Г Вольфензон)
Глава 4. Общие черты отношений между экологическими факторами и организмами 71
Мезологическая экология 71
Этологическая экология 72
Экологическая пластичность видов 76
Природа и классификация факторов 80
Глава 5. Биотические факторы 82
Взаимодействия, зависящие от плотности организмов 83
Взаимодействия, вызванные веществами, выделяемыми организмами 94
Взаимодействия, основанные на контакте или на предоставлении убежища: комменсализм и мутуализм 98
(перевод В. А. Потоцкой)
Пищевое использование одних организмов другими: хищничество и паразитизм 100
Трофические и энергетические циклы внутри сообществ организмов: понятие экосистемы 106
Часть третья Сообщества организмов: биоценология
(перевод В А. Потоцкой)
Глава 6. Разграничение и организация сообществ 119
Разграничение сообществ 119
Организация популяций одного вида . „ 120
Пространственное и весовое заполнение сообщества совокупностью особей одного вида 122
Структура совокупности компонентов сообщества 124
Глава 7. Критерии и классификации сообществ 132
Растительные сообщества 132
Сообщества животных и их связи с растительными сообществами 143
Глава 8. Экология и эволюция сообществ . 146
Отношения сообществ со средой 146
Методы изучения эволюции сообществ . 160
Факторы и механизмы преобразования сообществ 151
Биоценотические сукцессии 160
Глава 9. Картографирование растительности 165
Карты флористических сообществ 166
Карты физиономических сообществ: формаций и доминирующих видов 169
Часть четвертая
Крупные биологические формации земного шара
Глава 10. Водные среды и сообщества (перевод В. А. Потоцкой) 175
Общие свойства водных сред 175
Население морей и океанов 186
Континентальные стоячие водоемы: озера, пруды, лужи . .201
Проточные воды 210
Глава 11. Наземная среда и наземные сообщества (перевод Е. Г. Мяло) 217
Тундры . 217
Аридные зоны 226
308
252
258
287
301
Степи 238
Тропические саванны . . . 245
Формации деревянистых, но не древесных растений (перевод Г. А. Михайловой)
Леса Послесловие . Литература
ж. Леме
ОСНОВЫ БИОГЕОГРАФИИ
Редактор Т А. Ольсевич
Мл. редактор Э. В. Рас ши в алое а Художник Э. И. Марков Художественный редактор В. А. Пузанков Технический редактор А. М. Токер Корректор Р. X. Пунга
Сдано в производство 10.3.1975 г.
Подписано к печати 7.1. 1976 г.
Бумага тип. № 1 60x9Jl/ie. Бум. л. 9,75.
Печ. л. 19,5+1,6 п. л. вклеек. Уч.-изд. л. 21,31.
Изд. № 17765. Цена 1 р. 77 к. Заказ № 626.
Тираж 14 000 экз.
Издательство «Прогресо
Государственного комитета Совета Министров СССР по делам издательств, полиграфии и книжной торговли.
Москва, Г-21, Зубовский бульвар, 21
Ордена Трудового Красного Знамени
Ленинградская типография № 2 имени Евгении Соколовой Союзполиграфпрома при Государственном комитете
Совета Министров 'СССР по делам издательств, полиграфии и книжной торговли.
198052, Ленинград, Л-52, Измайловский проспект, 29,
Флористический состав союза Salicion herbaceae снежных долинок восточных Пиренеев
Таблица в
Polltrichetum Saliceto — Anfhellelum Gnaphalieto — Sedetum candollei
1 Нпмрпя пооб 12 3 4 5 6 7 8 У 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29
Высота, м над уровнем 2240 2310 2190 2240 2360 2500 2400 2460 2280 2300 2400 2400 2510 2300 2430 2180 2300 2230 2500 2450 2470 2160 2480 2300 2240 2200 2480 2530 2420
моря Экспозиция, уклон 3 10° ЮВ2°Лож- Рав- В 10° Рав- СЗ СВ 20° С 10° С 5° С10° Рав- 3 5° 315° Ю 20° В 5° СЗ 5° 3 2° В 0° СВ 2° СВ 5° СВ 10° ЮВ 5° В 2° С 10° СВ 2° С 12° СВ 30° С 30е
бина нина ннна нина
Покрытие, % 100 95 90 60 70 95 85 90 80 95 90 100 70 40 60 40 75 80 80 65 60 50 65 60 60 60 50 50—7<
Площадь проб, кв. м 50 50 50 50 444 441 444 4 4 4 4 4 4 4 4 4 4 4 4 4 4 4
Характерные виды ассо-
циаций
Polytrichum sexangulare 5-5 5-5 4-4 2-3 • ••••• ••• • • • 1 ' • ••»•••••••
Dicranum falcatum 2-3 1-2 3-3 + • 2 4" • • “Ь “F • - • • -1- » • • “jH • • • • "4” • *4Н
Anthelia juratzkana 2*2-3 . 5-5 3-3 3*2 2-3 2-2 (+) +*2 1-2
Salix herbacea 4*3 3*4 5-4 4-4 4*4 5-5 5-5 3-2 3-3 3-3 • • а • • а , • а а а • « •
Sedum candollei + •1 . 2*2 1,-11 3-1 2-2 4-3 3-2 4-3 3*3 3-2 3-2 2-3 2-2 2-3 + 2-2-3 1-1 2-1-2
Cardamine alpina р • 4- 1*1 . . - 1*1 141 4- 1.1 2-1 1-1 1-1 1-1 2-2 1*1 1-1 1*1 2-2 1-1 -Ь 2-1-3
Arenaria biflora • • • • а а а • • а а а а а 3*2 2*2
Phleum ger ar di • • • • • "4" • • • »•••• + р . . + . . . . 1*1 2*2 2-3 2*2
Carex macrostyla • • • • . . . . . . . . • . + . . . 4- • 2
Дифференциальные вн-
ды ассоциаций
Sapina saainoides . 1+1+ 1+1. 1 + 1 1-1 1 + 1 + 1 . . а а а а • •• • • • 1 • • • •
Polytrichum piliferum • • • • 1-2 4- ' . 2-2 4- - 1'2. 1'2
Trifolium alpinum • • • • . 4“ 4“ 4“ • 4“ 1 “Г) п • п • 1 2 1-Г) 4- 4- 4-2-21-22-2
Plantago alpina р . +
Характерные виды союза
(Salicion herbaceae)
Gnaphalium supinum р сч сч СЧ 1 сч сч сч сч сч 4-+ сч • сч с « сч . сч со СЧ ( сч • сч сч сч сч СЧ ( + • СО СО < о* . сч < • го « со . ** . ( • -“ч сч сч •
Carex pirenaica р . + 4- + (+)2-2 . . . 2*31*2 4- 4- . 4- 1*2 (4-) 1*2 4-*2 . 4- • 2 . . 1*2 4--2
Pohlia commutata 2*2 . . + • 2 + 1*2 2*2 . 4--1 .... + 1*2 2*3 (4-) . 1*2 4- 1-2 . 4- •
Cerastium cerastoides 4- 4- 4- • . . 4- 1*1 4- 1*1 1*12'1 . 1*1 . . 4- . 4- 4-
Sibbaldia procumbens 4-4- +1*2 1-2 . . 2*2 1*2 (4-) . . • 1*2 . 2-2 . (4-)
Sedum alpestre 4- 4- . 4- 1-1 4- . (4-) 4- • 2-1
Solorina crocea • Ф • 4"1 • • • • • • ••••••«а». ”4”” • *
Сопутствующие виды
Роа alpina 4- 1-2 4- 4- . 4- 4- 4- 1-2 4- . 4* • . . 4- (4-) 4- 4" • 1-11-1 4- 4-
Agrostis rupestris • • • 4“ 4- 4- 4- 4--2 1*1 (4-) 2-21*2 4- . 4- • + • 2 *2 . 2-2 . . 1-2 4-
Polytrichum juniperinum 4- 4- Ы 1*1 1*1 4- 1 • 1 . •2 . . 4- 1-11-1
Veronica alpina р (4-) 4- 1*1 . . - • Ы 4- . . • . . . (4~) . 4" • 1 • 1 . . 4-4-
Sesleria dis tic ha • • • . 4-*. ..4-4-.... .4-4-
Bryum sp. 4- 4- 3*2 . + 4- • ......
Sisymbrinum pinnatifi- • • • ••••••• -J- “4“ 4" 4~ • • • • •
dum
Chrysanthemum alpinum • • ••••»•*• 4-. . .4-. •' • • • .4-4-.
Leontodon pyrenaicus 4" • • 4- • • (4-) 4-
Taraxacum pyrenaicum 4- . . (4-) 4- • 4-
Lotus corniculatus 1 а • • • 4- . 4- 1*1 • 4- «•••••••• • • • а •
Примечание: •—отсутствует, +—редко, Р —крайне редко.