/









































































































































































































































































































































































































































































Текст
АКАДЕМИЯ НАУК СССР
ИНСТИТУТ ФИЗИОЛОГИИ РАСТЕНИЙ им. К. А. ТИМИРЯЗЕВА
А. Ф. КЛЕШНИН
РАСТЕНИЕ И СВЕТ
ТЕОРИЯ И ПРАКТИКА
СВЕТОКУЛЬТУРЫ
РАСТЕНИЙ
http://biotex.ibss.org.ua
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
Москва — 1954
ОТВЕТСТВЕННЫЕ РЕДАКТОРЫ:
академик А. Л. К У Р С А Н О В,
проф. А. А. Н И Ч И II О Р О В И Ч
Светлой памяти
Николая Александровича Максимова
посвящает этот труд автор
ОТ АВТОРА
Настоящая работа была задумана первоначально как заключитель-
ная часть большой монографии «Растение и свет». Однако актуальность
проблемы светокультуры растений и перспективы применения электри-
чества в сельском хозяйстве СССР побудили нас заняться прежде всего
вопросами светокультуры растений. Освещение общих вопросов свето-
физиологии растений — оптических свойств растительных тканей и пиг-
ментов, обмена веществ, роста и развития и пр. — было перенесено по-
этому в последующие части монографии, а в настоящей работе они рас-
сматриваются попутно и лишь в той мере, в какой это необходимо для
понимания вопросов светокультуры растений.
В настоящее время отсутствуют работы, обобщающие и сводящие
воедино всю совокупность обширной и часто противоречивой литературы
по интересующей нас проблеме. Поэтому мы стремились к наибольшей
библиографической полноте, старались охватить возможно полнее вею
существующую литературу. Это поможет работникам советской науки
избежать повторений и ошибок и решать вопросы светокультуры, опи-
раясь на опыт предыдущих исследователей.
В основу работы положен единый принцип — понятие о физиологи-
ческой радиации, выдвинутое и разрабатываемое советскими учеными.
Это понятие, в свою очередь, основано на одном из главных положений
К. А. Тимирязева: зеленое растение использует лишь ту часть лучистой
энергии, которая поглощается пигментами листа и, в особенности, хлоро-
филлом. Понятие о физиологической радиации проходит красной нитью
через все разделы работы. В свете этого понятия рассматриваются источ-
ники освещения, солнечная радиация, методы измерения лучистой энер-
гии, общие и частные вопросы светокультуры растений. Всюду, где это
было возможно, наряду с единицами освещенности, совершенно непри-
емлемыми для целей физиологии растений, но обычно применяемыми
большинством авторов, в скобках приводится интенсивность физиологи-
ческой радиации, выраженная в объективных единицах (эрг/см2-сек).
В связи с тем, что светокультура растений находится на границе двух
наук — физиологии растений и физики — в первой части нашей работы
рассматривается учение о лучистой энергии и дается возможно полная
характеристика искусственных источников радиации. Здесь же характе-
ризуется естественная солнечная радиация и рассматриваются методы
измерения лучистой энергии. Это тем более необходимо, что подобная
характеристика в свете понятия о физиологической радиации дается
впервые.
1*
4
От автора
Вторая часть работы посвящена общим вопросам светокультуры —
выращиванию растений исключительно при искусственном освещении
и их культуре при дополнительном освещении. Основное внимание в этой
части сосредоточено на вопросах значения спектрального состава и ин-
тенсивности радиации и длины дня. Здесь же рассмотрена также история
вопроса.
В третьей части монографии освещены вопросы частной светокуль-
туры растений и ее практического применения и дана, по возможности,
исчерпывающая библиография.
Мы отдаем себе отчет в том, что предлагаемая работа не лишена оши-
бок, тем более что подобное обобщение в виде отдельной монографии
делается впервые. Все замечания и указания на недочеты работы будут
приняты с благодарностью.
В заключение автор считает своим долгом выразить искреннюю благо-
дарность и признательность академику А. Л. Курсанову, заслужен-
ному деятелю науки проф. С. О. Майзелю, проф. П. А. Генкелю,
проф. А. А. Ничипоровичу, канд. биол. наук В. М. Леману, сотруднику
Главного ботанического сада Академии Наук СССР В. Н. Ворошилову
и Н. Н. Протасовой за внимательный просмотр рукописи настоящей
работы и ценные советы и указания.
А. Клешнин
17 октября 1952 г.
Институт физиологии растений
им. К. А. Тимирязева
Академии Наук СССР
Часть I
ФИЗИЧЕСКИЕ
И ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ
СВЕТОКУЛЬТУРЫ РАСТЕНИЙ
Запросы жизни всегда являлись первыми стиму-
лами, побуждавшими искать знания, и, в свою оче-
редь, степень их удовлетворения служила самым до-
ступным, самым наглядным знамением его успехов.
К. А. Тимирязев. Столетние итоги физиологии расте-
ний, 1901 (Соч., т. V, 1938, стр. 423)
ВВЕДЕНИЕ
Неузнаваемо изменилась за последние годы экономическая география
нашей страны. Там, где шумела тайга, создаются промышленные центры,
на месте глухих лесов возникают города, закладываются шахты и руд-
ники. Все более и более актуальной становится проблема бесперебойного
круглогодичного снабжения свежими фруктами и овощами беспрерывно
возрастающего населения наших северных городов и новых промышлен-
ных центров. Перед работниками сельского хозяйства поставлена задача:
«Увеличить производство овощей, картофеля и продуктов животновод-
ства в пригородных зонах Москвы, Ленинграда, городов Урала, Донбасса,
Кузбасса и других промышленных центров и крупных городов; создать
картофельно-овощные и животноводческие базы в новых промышленных
районах»1.
К. А. Тимирязев, определяя экономическую сущность земледелия,
писал: «Земледелец из дарового сырого материала — воздуха и даровой
силы — солнечного света изготовляет ценности; в этом главная тайна
производительности его труда» (Соч., т. III, 1937, стр. 86). Однако, хотя
содержание углекислого газа редко снижается до предела, делающего
невозможным культуру растений, количество лучистой энергии солнца
часто бывает настолько мало, что совершенно исключает нормальное
выращивание растений.
Культура растений в закрытом грунте в зимние месяцы в большей
части районов СССР возможна лишь при дополнительном искусственном
освещении. Следовательно, проблема светокультуры, или выращивания
растений при помощи искусственных источников радиации, представляет
собою не только теоретический, но и практический интерес. Большие
возможности для решения этой проблемы имеются в новых промышлен-
ных районах, богатых источниками энергии и расположенных близ круп-
ных электростанций (Волхов, Днепропетровск, Куйбышев, Сталинград),
около больших промышленных городов (Москва, Ленинград) и т. д.
С построением мощных волжских гидроэлектростанций в Куйбышеве и
Сталинграде, которые будут давать стране 20 млрд, квт-час. электро-
энергии в год [241—243], светокультура растений из стен опытных
и научных учреждений должна переместиться в производственные теп-
лицы и стать достоянием широкой сельскохозяйственной практики.
Уже сейчас в передовых колхозах и совхозах строятся большие электри-
1 Директивы XIX съезда партии по пятому пятилетнему плану развития СССР
на 1951—1955 годы. Госполитиздат, 1952, стр. 15—16.
8
Физические и физиологические основы светокультуры растений
фицированные теплицы, и освещение растений люминесцентными лам-
пами входит в колхозную практику [143].
В этой связи следует подчеркнуть, что светокультура растений как
самостоятельная проблема физиологии растений, имеющая большое
теоретическое и практическое значение, возникла в СССР. Приоритет
советских ученых в данной области не подлежит сомнению. Особенно
много сделали академик Н. А. Максимов, проф. Н. А. Артемьев,
В. П. Мальчевский и др., положившие начало изучению этой проблемы
и разрешившие многие вопросы светокультуры растений.
Проблема светокультуры растений состоит из нескольких более или
менее самостоятельных вопросов. Прежде всего укажем на выращивание
растений целиком на искусственном свете. Этот вопрос, помимо чисто
теоретического значения, может представлять и большой практический
интерес (культура растений на Крайнем Севере в условиях полярной
зимы, озеленение станций метрополитена и т. п.).
Другим, более важным по практическому значению вопросом яв-
ляется выращивание растений при естественном солнечном свете с д о-
полнительным освещением в ночные часы или в пасмур-
ные и зимние дни, когда интенсивность солнечной радиации недостаточна.
Это имеет большое значение в умеренной полосе СССР для культуры ра-
стений в закрытом грунте, ранней выгонки рассады, быстрого получения
сеянцев древесных пород, декоративного и комнатного цветоводства,
селекции и семеноводства и т. д.
Наконец, может возникнуть специфическая задача удлинения
короткого дня, недостаточного для нормального развития длинноднев-
ных растений. Эта потребность имеется в южных районах страны, где,
несмотря на сравнительно высокую интенсивность радиации, культура
многих растений, особенно в зимнее время, не удается из-за слишком
короткого дня.
Таким образом, светокультура растений в наши дни приобретает
все большее и большее значение. Между тем, литература, обобщающая
опыт выращивания растений при помощи искусственных источников
радиации, сравнительно невелика и в значительной части устарела [81,
363, 646, 745]. Настоящая работа представляет собою попытку восполнить
этот пробел.
Глава I
ЛУЧИСТАЯ ЭНЕРГИЯ
И ОПТИЧЕСКИЕ СВОЙСТВА РАСТЕНИЙ
1. ЛУЧИСТАЯ ЭНЕРГИЯ
Лучистая энергия есть энергия электромагнитных колебаний, воз-
никающих в результате колебательных движений электронов возбужден-
ных атомов и молекул. Как и всякие другие колебания, электромагнит-
ные колебания характеризуются амплитудой (А), длиной волны (X),
скоростью распространения (с), периодом (Т) и частотой (v). Соотношение
между скоростью распространения, длиной волны и частотой выражается
следующим уравнением:
r = vX, (1)
причем скорость есть величина постоянная:
с = 2,998 • 1010 см/сек, .
а длина волны л и частота у лежат в широком значении величин — от
О до оо . Единицами измерения первой служат ангстрем (А), миллимикрон
(пш) или микрон (у), а единицей частоты — герц (гц), отвечающий одному
колебанию в секунду; между единицами длины волны существуют сле-
дующие соотношения:
1А = 10—1 ту = 10~4 у = 10—8 см.
В настоящей работе мы всюду, за небольшим исключением, будем
пользоваться в качестве единицы длины волны миллимикроном и лишь
в специальных случаях будем прибегать к помощи ангстремов или микро-
нов.
Собственно свет, или область электромагнитных колебаний, восприни-
маемая средним человеческим глазом, лежит в пределах длин волн
от 400 до 800 ту с частотою от 3,74-1014до 7,49*1014 гц. Более высокая
частота характеризует ультрафиолетовые, а более низкая — инфракрас-
ные лучи.
Согласно современным представлениям, лучистая энергия излучается
и распространяется не беспрерывно, а в виде отдельных неделимых пор-
ций — квантов, или фотонов, величина которых определяется длиною
волны или частотой:
E = hv = %-, (2)
А
I. Лучистая энергия и оптические свойства растений
11
Таблица 1
Xарактеристика электромагнитных колебаний
Излучение Длина волны Частота коле- Энергия кванта (в электрон- вольтах) Величина Эйнштейна (в г-кал) 101 Число кван- тов в 1 эрге 1012
( в m й : оании (в гц) 1 1014
Ультрафиолетовые лучи
короткие <300 9,99 >4,11 >9,50 <0,15
длинные 300—400 9,99—7,49 4,11—3,08 9,50—7,12 0,15—0,20
Видимые лучи (свет)
фиолетовые .... 400—440 7,49—6,80 3,08—2,80 7,12—6,46 0,20—0,23
синие 440—490 6,80—6,12 2,80—2,52 6,46—5,82 0,23—0,25
зеленые 490—565 6,12—5,30 2,52—2,18 5,82—5,02 0,25—0,29
желтые 565—595 5,30—5,04 2,18—2,08 5,02—4,80 0,29—0,30
оранжевые .... 595—620 5,04—4,84 2,08—2,02 4,80—4,69 0,30—0,32
красные 620—800 4,84—3,74 2,02—1,54 4,69—3,51 0,32—0,41
Инфракрасные лучи . >800 <3,74 <1,54 <3,51 >0,41
2. ОСНОВНЫЕ ЗАКОНЫ ФОТОХИМИИ
Согласно основному закону фотохимии — закону Гроттгуса, всякий
фотохимический процесс, будет ли это простая химическая реакция ил£
сложное физиологическое явление, совершается лишь под действием
тех лучей, которые поглощаются реагирующей системой. Конкретный
физический смысл этой общей закономерности придает закон Эйнштейна,
согласно которому для каждого элементарного фотохимического акта
необходимо поглощение каждой реагирующей молекулой одного кванта.
Это означает, что для фотохимического превращения одного грамм-
моля вещества необходимо количество энергии, определяемое уравнением:
En = Nhi=^, (4)
где N — число Авогадро, равное
N = 6,061 • Ю23 моль-1,
h — постоянная Планка, у — частота, с — скорость света и X — длина
волны. Иными словами, энергия грамм-молярного числа квантов, полу-
чившая наименование эйнштейна, определяется частотой колебаний или
длиной волны:
= 3,969. 10-^ч=1,191 • 108 у. (4а)
Следует подчеркнуть, что фотохимический акт происходит лишь в том
случае, если энергия поглощаемого кванта достаточно велика, т. е. до-
стигает определенного значения q, характеризующего данный фотохими-
ческий процесс:
hv q. (5)
Если энергия фотона выше величины q, ее избыток деградирует в виде
тепла. Из сказанного следует, что наиболее эффективно для фотохими-
ческих реакций излучение, кванты которого бесконечно приближаются
к величине q. В этом случае деградирует в виде тепла наименьшее коли-
чество поглощенной энергии. При прочих равных условиях в таком
12
Физические и физиологические основы светокультуры растений
случае следует ожидать максимальной скорости фотохимической реак-
ции или физиологического процесса, в основе которого лежит соответ-
ствующая фотохимическая реакция.
3. ОПТИЧЕСКИЕ СВОЙСТВА РАСТЕНИЙ
Подобно тому как корни являются органом, посредством которого
растения получают из почвы воду и минеральные вещества, органом вос-
приятия лучистой энергии, необходимой для нормального хода физио-
логических процессов, служит лист. Именно в листе совершаются все
Фиг. 2. Поглощение лучистой энергии листьями растений при одинаковой спек-
тральной интенсивности
1 — недотроги (Im-patiens noli tangere); 2 — кувшинки (Nimphara alba); 3 — периллы (Perilla ocy-
moides); 4 — пеларгонии (Pelargonium zonal?); 5 — тополя (Povutus saiveolens); 6 — салата (Laciuca
sativa); 7 — фикуса (Ficus sp.); 8 — теоретическая кривая поглощения энергии "средним» листом
важнейшие фотохимические акты и процессы, зависящие от них. Именно
здесь совершается фотосинтез, образуются всевозможные физиологи-
чески активные вещества; здесь возникает фотопериодический импульс,
вызывающий переход растений к цветению, и т. д. Зеленый лист, содер-
жащий хлорофилл, обеспечивает энергией весь растительный организм.
Процесс поглощения лучистой энергии листьями высших растений
в настоящее время изучен сравнительно хорошо [85, 349, 693, 749]. Как
непосредственные экспериментальные данные (фиг. 2, 3—7), так и теоре-
тические расчеты (фиг. 2, 7—2) показывают, что лист поглощает в основ-
ном видимые и ультрафиолетовые лучи (300—750 ту), причем максимумы
поглощения приходятся на оранжево-красные (600—680 ту) и сине-
фиолетовые и ультрафиолетовые лучи (300—500 ту), а минимум — на
зеленые лучи (500—600 ту). В области 750 ту находится граница, за пре-
делами которой поглощение лучистой энергии практически равняется
нулю (фиг. 2). Поглощение возникает лишь в далеких инфракрасных
лучах; в основном оно совершается водою, содержащейся в тканях листа
(фиг. 3, 3).
В зоне 300—750 ту лист поглощает от 60 до 80% энергии падающих
лучей; наименьшим поглощением обладают теневые, или тенелюбивые,
а наибольшим — световые, или светолюбивые, растения. Растения, вы-
I. Лучистая энергия и оптические свойства растений
13
ращиваемые в условиях светокультуры, следует отнести к первой группе.
Лист, как и всякий другой орган растения, состоит в основном из
углеводов, белков и жиров или их производных. Перечисленные веще-
ства составляют основу как протоплазмы, так и клеточных оболочек
растений. Все это — вещества, которые поглощают лишь «короткие»
ультрафиолетовые « 300 ту) и далекие инфракрасные лучи (порядка
-5—6 у). Поглощение в области 300—750 ту обусловлено исключительно
<Лиг. 3. Поглощение лучистой энергии пигментами, протоплазмой и водою листа
при одинаковой спектральной интенсивности
1 — хлорофилл и каротиноиды; 2 — хлорофилл; 3 — каротиноиды; 4 — протоплазма в отсутствие
пигментов (масштаб увеличен); 5 — вода
I — зона физиологической радиации, поглощаемой хлорофиллом и каротиноидами; II — зона
«физиологической радиации, поглощаемой хлорофиллом; III — эона инфракрасной радиации,
не поглощаемой листом; IV — зона инфракрасной радиации, поглощаемой листом.
аб — граница зоп I и II; вг — граница зон II и III
пигментами пластид — хлорофиллом и каротиноидами, а в некоторых
случаях также и пигментами клеточных оболочек и вакуолей — флаво-
ноидами и антоцианами.
Таблица 2
Содержание пигментов в пластидах листа
по Зейбольду и Эгле [750]
Содержание пигментов в пластидах растений
световых теневых
г'смг г-мол/см2 г/см2 г-мол'см'2
10-5 10-8 1 о— IO-»
Хлорофилл а ... . 5,2 5,8 3,7 4,8
Хлорофилл b 1,2 I,3 1,3 1,4 I
Хлорофилл а - г Ь . . 6,4 7,1 М 6,2
Ксантофилл 1,2 2,1 0,94 1,7
Каротин 0,24 0,45 0,19 0,35
1 т, 1 Каротиноиды .... 1,44 2,55 1,13 2,05
Все пигменты .... 7,84 6,13
14
Физические и физиологические основы светокультуры растений
Как показывает табл. 2, содержание пигментов в пластидах листьев,,
отнесенное к единице поверхности (поверхностная концентрация), не-
сколько выше у растений, развивающихся в условиях полного солнечного-
освещения, чем у растений, произрастающих в тени или выращиваемых
в условиях светокультуры.
Если принять, что хлорофилл и каротиноиды распределяются равно-
мерно относительно друг друга, то поглощение энергии каждым из этих
пигментов можно выразить уравнением:
< = (б>
а общее поглощение обоими пигментами будет равно:
/.=/.(1-10-^, ' (7),
где 10 — интенсивность падающей радиации; 1а — энергия, поглощаемая,
всеми пигментами; 1'а — энергия, приходящаяся на данный пигмент,.
Е — суммарная оптическая плотность; Е' — оптическая плотность дан-
ного пигмента.
В свою очередь, оптическая плотность Е' пигмента, входящего в общую-
систему пигментов, определяется уравнением
Е' ~ кс, (8);
а суммарная оптическая плотность Е представляет собой сумму всех
оптических плотностей:
Е = -ф- Л2с2-|- к3с3 -ф- ... кпс„, (9)i
где с есть поверхностная концентрация соответствующего пигмента,
(в г/см2 или г-мол/см2), а к — коэффициент поглощения, отвечающий,
единице поверхностной концентрации пигмента, зависящий от длины
волны и определяемый из кривой поглощения лучистой энергии пигмен-
том.
Подобным методом при помощи табл. 2 получены кривые 1, 2 и 8 на.
фиг. 2 и кривые 1, 2, 3 и 5 на фиг. 3. В последнем случае кривая 1 выра-
жает поглощение энергии всеми пигментами пластид, кривая 2 —• погло-
щение хлорофиллом, кривая 3 — поглощение каротиноидами, а кривая
5 — поглощение лучистой энергии водою, содержащейся в тканях «сред-
него» листа.
Энергию, падающую на лист, графически можно разделить на четыре,
зоны, имеющие различное физиологическое значение (фиг. 3). К первой
зоне (Z) относится область, лежащая в пределах 300—520 шр и поглощае-
мая целой системой веществ: хлорофиллом, каротиноидами, протоплаз-
мой, ферментами, флавоноидами и др. Большая часть поглощаемой энер-
гии приходится здесь на хлорофилл и каротиноиды. Именно этим следует
объяснить разнообразие физиологических реакций, которыми отвечает
растение на действие ультрафиолетовых и сине-фиолетовых лучей.
Вторая физиологически важная область излучения (ZZ) расположена:
в пределах 520—700 шр-. Поглощение здесь связано исключительно с хло-
рофиллом, и лишь ничтожная доля приходится на некоторые ферменты.
Это те самые оранжево-красные лучи, значение которых для фотосинтеза:
выяснено блестящими работами К. А. Тимирязева. Это радиация, имеющаяз
I. Лучистая энергия и оптические свойства растений
15-
едва ли не решающее значение буквально во всех физиологических про-
цессах: в фотосинтезе, развитии, формообразовании, синтезе пигментов и др._
Инфракрасные лучи с длиною волны в пределах 750—1050 шц (III)
можно назвать абиотической радиацией. В жизни растений они, повиди-
мому, играют ничтожную роль вследствие незначительного поглощения
тканями листа.
Инфракрасная радиация, лежащая за пределами 1050 шц (27), яв-
ляется мощным фактором теплового режима растений. Поглощаясь водою,
протоплазмой и другими составными частями клетки (особенно в области
5—6 р), эта часть радиации оказывает сильнейшее воздействие на многие
стороны жизнедеятельности растений, хотя, по всей вероятности, не-
играет самостоятельной роли в фотохимических процессах.
Фиг. 4. Относительное поглощение квантов пигментами «среднего» листа
1 — все пигменты; 2 — хлорофилл; 3 — каротиноиды
Согласно закону Эйнштейна, интенсивность фотохимических процес-
сов зависит не от абсолютного поглощения энергии безотносительно
к ее квантовой природе, а от числа поглощенных квантов определенной
величины. При одном и том же поглощении энергии количество погло-
щенных квантов, как известно, возрастает вместе с длиною волны (фиг. 1).
На фиг. 4 показано относительное поглощение квантов разной величины
различными пигментами пластид при одинаковой интенсивности падаю-
щей радиации. Кривые получены в результате перемножения ординат
кривой 2 (фиг. 1) на ординаты соответствующих кривых фиг. 3. Ниже
будет показано, что кривые относительного поглощения квантов пиг-
ментами пластид лежат в основе спектрограмм многих физиологических
процессов (фиг. 5, 6, 9, 10, 13, 14 и 18).
4. ВЫВОДЫ
1. Лучистая энергия излучается в виде дискретных порций — квантов-
или фотонов, величина которых прямо пропорциональна частоте и обратно-
пропорциональна длине волны.
16 Физические и физиологические основы светокультуры растений
2. Для всякого элементарного фотохимического акта требуется опре-
деленная величина кванта; общая скорость реакции пропорциональна
количеству квантов, поглощаемых в единицу времени.
3. Листья высших растений поглощают ультрафиолетовые (300—
400 ту), видимые (400—750 ту) и далекие инфракрасные лучи (> 1050 ту).
В области видимых и ультрафиолетовых лучей величина кванта нахо-
дится в пределах 1,65—4,11 eV, а в инфракрасных лучах она снижается
за пределы 1,2 eV.
4. При прочих равных условиях, когда поглощение лучистой энергии
одинаково, а величина квантов достаточна для определенного фотохими-
ческого или физиологического процесса, длинноволновые лучи, несущие
большее количество квантов, более полно используются растением в фо-
тохимическом процессе.
Глава II
ФИЗИОЛОГИЧЕСКАЯ РАДИАЦИЯ
Зеленые растения, содержащие хлорофилл, к которым относятся и
высшие растения, являются единственными организмами, способными
создавать органические вещества при помощи лучистой энергии Солнца.
Органом, где совершается этот сложный процесс, у высших растений
является лист, а акцептором лучистой энергии, посредством которого
она поглощается и превращается в химическую энергию органических
веществ, служит хлорофилл — пигмент, содержащийся в пластидах листа.
Наряду с хлорофиллом, в качестве дополнительных акцепторов лу-
чистой энергии выступают и каротиноиды — ксантофилл, каротин и дру-
гие, которые вместе с хлорофиллом содержатся в хлоропластах и опре-
деляют всю совокупность фотохимических процессов, совершающихся
в растении.
Лучистая энергия, поглощаемая пигментами пластид (300—700 иг,.),
есть физиологическая радиация1. Она имеет решающее
значение во всех жизненных отправлениях растений. Впервые это понятие
ввел Л. А. Иванов [81], и в настоящее время целесообразность подобного
понятия обосновывается большим экспериментальным материалом [ИЗ,
123].
1. ФОРМИРОВАНИЕ И ДВИЖЕНИЕ ЛИСТЬЕВ II ПЛАСТИД
Для организма, жизнь которого невозможна без притока лучистой
энергии, первой ответной реакцией на ее действие является создание
соответствующего оптического аппарата, позволяющего наиболее целе-
сообразно и рационально использовать энергию падающих на растение
лучей.
Как показывают экспериментальные данные [660, 842], формирование
листьев есть процесс, зависящий в значительной степени от лучистой
энергии, в особенности от оранжево-красных лучей (фиг. 5, 6 и 10 и табл. 3).
В условиях одинаковой интенсивности различных областей спектра
физиологической радиации средние размеры листа, его вес и общая листо-
вая поверхность растения выражаются спектральной кривой, в общем
виде отвечающей кривой поглощения квантов хлорофиллом (фиг. 4). 1 2
1 Понятие физиологической радиации, данное в настоящей работе, отличается
от принятого в общей биологии. В последней под физиологической радиацией чаще
всего принимают излучение самих организмов (свечение микроорганизмов, митогене-
тические лучи и т. д.).
2 А. Ф. Клешнин
18
Физические и физиологические основы светокультуры растений
Для получения одинаковой листовой поверхности интенсивность сине-
фиолетовых лучей должна быть примерно в два раза выше интенсивности
оранжево-красных лучей (фиг. 6, 4, 3).
Фиг. 5. Репа (Brassica гара) Петровская, выращенная при одинаковой
интенсивности физиологической радиации под люминесцентными лам-
пами разного света
1 — синего; 2 — зеленого; 3 — красного
Давно известно, что синтез хлорофилла у высших растений представ-
ляет собою в подавляющем большинстве случаев фотохимический про-
цесс, хотя непосредственный предшественник этого пигмента и образуется
Фиг. 6. Огурцы (Cucumis sativus), выращенные при одинаковой
интенсивности физиологической радиации под люминесцентными
лампами разного света
1 — синего; 2— зеленого; 3— красного; 4— синего при интенсивности
физиологической радиации в два раза .выше, чем в других вариантах.
Размеры листа под синими и красными лампами одинаковы, если интенсив-
ность синих лучей в два раза выше, чем интенсивность красных
в темноте. Как показывают исследования Франк 1494], наиболее эффек-
тивны в этом процессе оранжево-красные и сине-фиолетовые лучи (фиг 7).
Невидимому, акцептором лучистой энергии для названного процесса
являются хлорофиллоген или протохлорофилл и пигменты, близкие к хло-
рофиллу и образующиеся в темноте. В темноте же образуются и кароти-
ноиды, но в меньшем количестве, чем на свету. Есть основания полагать,
что наиболее эффективны в последнем случае сине-фиолетовые лучи.
Оптическая система растений представляет собою чрезвычайно по-
движный аппарат, зависящий как от интенсивности, так и от спект-
рального состава радиации. Помимо размеров листьев и количества
пигментов, под влиянием лучистой энергии изменяются также размеры,
II. Физиологическая радиация
19?
Таблица
Формирование листьев в зависимости от спектрального состава физиологическое;
радиации при выращивании растений под люминесцентными лампами с разным
спектром излучения '
Воз- Сухой вес одного листа (в мг) Площадь одного листа (в см2) Общая листовая по- верхность растения (в см2)
Растение раст (в Свет
днях) СИНИЙ зеле- крас- СИНИЙ зеле- крас- СИНИЙ зеле- крае -
ный ный НЫЙ ный ный ный
Салат (Lactuca sa- tiva) Московский Кольраби (Brassica * 0,57 0,45 0,94 — - — — —
caulorapa) Вен-
скан 51 625 430 509 — — — — —
Репа (Brassica та- ра) Петровская. Редис (Raphanus 45 100 67 350 — — — — — —
sativus f. radi- cula) розово- красный .... Огурцы (Cucumis 17 — — — 41,4 29,6 75,0 104,0 248,1 148,1 448,2
sativus) .... 1 — — — 94,7 37,9 568,6 227,6 6224
* В возрасте 4 листочков.
число, химическое состояние и положение пластид. Зависимость диаметра*
и числа хлоропластов от спектрального
состава излучения выражается кривой,
отвечающей поглощению квантов хлоро-
филлом (фиг. 4), а положение пластид
определяется главным образом ультра-
фиолетовыми и сине-фиолетовыми лу-
чами— в соответствии с фиг. 4, 3 [828].
400 500 500 100
Длина вилнО/ ( 0 тр/
Фиг. 7. Синтез хлорофилла в зависимости
от спектрального состава физиологическом
радиации [494]
Фиг. 8. Положение хлоропла-
стов в зависимости от освеще-
ния листьев
А — в темноте; Б — при слабой
интенсивности радиации; В — при
высокой интенсивности радиации
2'
20
Физические и физиологические основы светокультуры растений
Пластиды, представляющие собою диски диаметром в 4—6 в условиях
низкой и оптимальной интенсивности радиации занимают положение,
перпендикулярное направлению лучей. При чрезмерно высокой интенсив-
ности пластиды перемещаются на боковые стенки клеток и распола-
гаются параллельно направлению лучей (фиг. 8). Это обстоятельство
позволяет растениям регулировать поглощение лучистой энергии, под-
держивая его на определенном более или менее постоянном уровне.
Известно, что листья растений обладают диафототропической способ-
ностью, т. е. способны занимать положение, перпендикулярное направ-
лению лучей. Как показали исследования Бюнинга [432], фототропизм
«гак высших, так и низших растений связан с поглощением лучистой энер-
гии каротиноидами 1 (фиг. 4, 3). Иными словами, положение листовой
пластинки определяется главным образом сине-фиолетовыми и ультра-
фиолетовыми лучами.
Таким образом, как формирование, так и движение пластид и листьев
растений связаны главным образом с физиологической радиацией, лежа-
щей в пределах 300—700 шт, причем максимальной эффективностью об-
ладают оранжево-красные и сине-фиолетовые лучи. Главная роль в фор-
мировании оптического аппарата принадлежит оранжево-красным, а
в движении листьев и пластид — сине-фиолетовым лучам.
2. ФОТОСИНТЕЗ
Основным физиологическим процессом, который в конечном итоге
определяет всю совокупность жизнедеятельности растений и их урожай,
Фиг. 9. Относительная интенсивность фотосинтеза у пшеницы
Triticum sp. в зависимости от спектрального состава
физиологической радиации [553]
1—при одинаковом количестве квантов; 2 — при одинаковой пптеп-
сивности радиации
1 В последнее время появились исследования, которые показывают, что фото-
тропизм связан с другими желтыми пигментами (рибофлавином).
II. Физиологическая радиация
21
является фотосинтез — фотохимический процесс образования разнообраз-
ных органических вегцеств из углекислого газа и воды за счет энергии,
поглощаемой хлорофиллом, а по некоторым новейшим данным — и каро-
тиноидами. В настоящее время не подлежит сомнению, что в процессе
фотосинтеза образуются не только углеводы, но и самые различные веще-
ства, определяющие процессы роста, развития и формообразования ра-
стений [37, 38, 236, 237].
Со времени классических исследований К. А. Тимирязева проведено
немало работ, посвященных изучению сравнительной эффективности раз-
личных лучей в процессе ассимиляции растением углекислого газа. Как
показывает фиг. 9, фотосинтез совершается исключительно в области фи-
зиологической радиации (400—700 ту), причем максимальной эффектив-
ностью обладают оранжево-красные и сине-фиолетовые, а минимальной —
зеленые лучи [553].
3. РАЗВИТИЕ РАСТЕНИЙ
Согласно теории стадийного развития растений, созданной академиком
Т. Д. Лысенко [175], развитие растений — процесс, совершающийся
в точках роста и вызывающий переход растений к заложению цветков,
цветению и плодоношению, — про-
исходит стадийно, в виде определен-
ных, выработанных в процессе эволю-
ции, качественных изменений. Для
прохождения растением каждой ста-
дии — стадии яровизации и световой
стадии — необходима определенная
совокупность внешних условий; все
стадии развития растений соверша-
ются строго последовательно и не-
обратимо.
Отличительной особенностью пер-
вой стадии — стадии яровизации —
является то, что ее нормальное про-
хождение находится в сильнейшей
Фиг. 10. Скорость фотопериодиче-
ского процесса и площадь листьев
в зависимости от спектрального
состава физиологической радиации
зависимости от температуры. У ози-
мых, двулетних и большинства многолетних растений процесс яровизации
совершается в условиях низких температур (3—5°). В дальнейшем,
при изложении вопросов частной светокультуры растений, будут рас-
смотрены примеры, иллюстрирующие значение низких температур для
нормального развития растений, т. е. для прохождения стадии яро-
визации.
Светокультура растений имеет дело главным образом со второй ста-
дией — световой, которая начинается лишь после завершения стадии
яровизации. Для прохождения этой стадии необходим свет. Иными сло-
вами, ее прохождение определяется интенсивностью излучения, спект-
ральным составом его и длиной дня.
В настоящее время все высшие растения принято разделять на три
группы: растения длинного дня, цветущие значительно раньше или исклю-
чительно в условиях длинного дня (более 12 часов); растения короткого
дня, цветущие значительно раньше или исключительно в условиях
короткого дня (менее 12 часов), и нейтральные растения, цветущие незави-
симо от длины дня. Подавляющее большинство наших сельскохозяйствен-
ных и декоративных растений относятся к первой группе, причем в пре-
делах почти каждого вида встречаются сорта и разновидности, дающие
1К! физические и физиологические основы светокультуры растений
самую различную степень реакции — от полного безразличия к длине
дня (нейтральность) до цветения исключительно на длинном дне. Эти
растения лучше всего развиваются в условиях 17—18-часового дня или
беспрерывного освещения. Все они в зимнее время в условиях закрытого
грунта даже на юге нуждаются в дополнительном освещении ночью.
Значительно меньшее количество видов входит в группу растений
короткого дня. Здесь, как и в группе длиннодневных растений, в пределах
большинства видов встречаются как нейтральные формы, так и формы,
цветущие лишь в усло-
виях короткого дня.
У короткодневных ра-
стений дополнительное
освещение в ночные часы
вызывает или задержку в
наступлении цветения, или
полное отсутствие цвете-
ния; причем это явление
сопровождается интенсив-
ным вегетативным ростом.
Собственно нейтраль-
ных растений, не реаги-
рующих на дополнитель-
ное освещение в ночные
часы ускорением или за-
медлением в наступлении
цветения, сравнительно
немного. К ним отно-
сятся, например, аспара-
гус (Asparagus plumosus,
A. Sprengeri), долихос (Do-
lichos lablab}, комнатный
клен (Abutilon hybridum
maximum), малина (Rubus
idaeus), маргаритки (Bellis
perennis), мимоза (Mimosa
pudica), ночная красавица
Фиг. 11. Салат (Lactuca sativa), выращенный (Mirabilis jalapa), напер-
под люминесцентными лампами разного света стянка (Digitalis ригри-
1 — синего; 2 — зеленого; 3 — красного геа), ОрлИКИ (Aquilegitt са-
nadensis, A. hybrida), ро-
додендрон (Rhododendron roseus elegans), ciLa^7Ka.(Asparagus officinalis) и др.
Клубнеобразование у всех без исключения растений — георгин (Dahlia
variabilis), топинамбура (Helianthus tuberosus), земляного миндаля (Су-
perus esculentus), картофеля (Solanum tuberosum) и др. — совершается
или исключительно, или значительно быстрее и обильнее в условиях
короткого дня. Короткий день необходим также для образования корне-
плодов у редиса и формирования кочана или листовой розетки у салата.
Луковицы, напротив, образуются исключительно в условиях длинного дня.
Более подробные сведения по влиянию длины дня на растения можно
найти в сводке Ю. А. Самыгина [285], в которой рассмотрено около
600 видов растений и приводятся многочисленные данные по сортам
важнейших сельскохозяйственных и декоративных растений. К этому
вопросу мы вернемся при рассмотрении частной светокультуры расте-
ний (часть третья).
II. Физиологическая радиация
23
Для удлинения короткого естественного1 дня’ с целью ускорения
или замедления цветения нужны исключительно низкие интенсивности
дополнительного освещения. Часто бывает достаточна интенсивность
порядка 1,5—3 эрг/см2-сек (0,25—0,5 лк), чтобы значительно ускорить
цветение длиннодневных растений, например куколя (Agrostemma Githa-
gO’, ячменя (Hordeum vulgare), стенника (Iberis amara), или задержать
этот процесс у растений короткого дня—проса (Panicum mileaceum), сои
(Soja hispida). Обычно для получения полного эффекта пользуются интен-
сивностью 500—1500 эрг/см2 • сек (100—300 лк). Наиболее эффективны
в обоих случаях — как для длиннодневных, так и для короткодневных
растений — оранжево-красные и фиолетовые лучи (фиг. 10; 364, 365, 580).
Фиг. 12. Уменьшение длины меж-
доузлий овса (Avena saliva) в за-
висимости от спектрального со-
става физиологической радиации
; В условиях искусствен-
ного освещения при благо-
приятной длине дня разви-
тие растений совершается
наиболее интенсивно в оран-
жево-красных и наименее ин-
тенсивно в зеленых лучах.
Длиннодневные растения при
беспрерывном освещении или
длинном дне, как и коротко-
дневные растения в условиях
короткого дня, зацветают
раньше всего при выращива-
нии в красных, затем в синих
и в последнюю очередь в зеле-
ных лучах (фиг. 11). Растения
короткого дня в условиях бес-
прерывного освещения или
длинного дня зацветают вна-
чале при выращивании в си-
Фиг. 13. Формирование стеблевых утолщений
кольраби (Brassica caulorapa), выращенной
под лампами разного света
1 — синего; 2 — зеленого; 3 — красного
них, а затем в оранжево-
красных лучах. Последнее находится в связи с большей фотопериоди-
ческой активностью оранжево-красных лучей (фиг. 10).
4. ФОРМООБРАЗОВАНИЕ РАСТЕНИИ
Лучистая энергия является мощным фактором формообразования,
в значительной степени определяющим общий вид растений. Мы уже
24
Физические и физиологические основы светокультуры растений
Накопление сухой массы растений при выращивании под
Растение Возраст (в ДНЯХ) Листья свет Стебли свет
синий зеле- ный крас- ный синий зеле- ный крас- ный
Редис розово-красный (Hapharius sativus f. radicula) Репа Петровская (Brassica гара) . Капуста белокочанная № 1 (Bras- sica oleifera f. capitala) Кольраби венская (Brassica caulo- rapa) Патисоны (Cucurbita pepo) .... Салат московский (Lactuca sativa) To же 45 72 60 45 34 51 в возрасте 4 листьев 1,02 3,55 2,30 1.00 2,24 1,69 0,314 0,72 2,26 2,39 0,74 1,72 0,63 0,241 1.61 7,10 5,88 2,79 2,05 0,82 0,431 0,52 0,70 0,10 0,54 1,04 0,064 0,46 1,00 0,07 0,40 0,28 0,072 1,08 2,30 0,52 0,46 1,00 0,125
отмечали (табл. 3, фиг. 5 и 6) значение различных областей спектра для
формирования листьев, которые вообще образуются лишь на свету,
оставаясь в зачаточном со-
стоянии в темноте. Свет, пред-
отвращая чрезмерное вытя-
гивание междоузлий, влияет
на размеры стебля. В лите-
ратуре обычно указывается,
что фактором, устраняющим
этиоляцию растений, явля-
ются сине-фиолетовые лучи.
Между тем, новейшие дан-
ные не оставляют сомнений
в том, что наиболее мощ-
ные и приземистые расте-
ния образуются в оранжево-
красных лучах [111, 363,
515, 516, 842]. Сказанное
иллюстрируется фиг. 12, где
показана эффективность раз-
Фиг. 14. Формирование корнеплодов репы
(Brassica гара)
1 — в синем; 2 — в зеленом; 3 — в красном свете
ных лучей спектра в предотвращении вытягивания стеблей [515, 516].
Невидимому, преувеличение значения сине-фиолетовых лучей в ростовых
процессах, характерное для старой физиологической литературы, осно-
вано на методической ошибке — недостаточной монохроматизации на-
званных лучей. Эффект, приписываемый сине-фиолетовым лучам, в дей-
ствительности вызывался ультрафиолетовой радиацией.
Различные образования, связанные с утолщением стебля, также нахо-
дятся в связи с действием лучистой энергии. Так, формирование стеб-
левого утолщения кольраби (Brassica caulorapa) совершается раньше и
наиболее интенсивно в оранжево-красных лучах (фиг. 13). То же на-
блюдается и при образовании корнеплодов репы (Brassica гара) (фиг. 14),
свеклы (Beta vulgaris), брюквы (Brassica napus), моркови (l)aucus carota),
редиса (Raphanus sativus f. radicula) и др. Последнее не находится в про-
II. Физиологическая радиация
25
лучами разных областей спектра (вес одного растения в г)
Таблица 4
Корнеплоды Корни Все растение
свет свет свет
синий зеленый красный синий зеленый | красный синий зеленый красный
0,33 0,66 0,00 0.20 0,76 2,55 0,47 1,16 0,15 0,75 0,66 2,52 2,34 5,37 1,33 3,21 4,1! 12,17
— — — 0,90 1,02 3,32 3,90 4,41 11,50
— — — 0,50 0,2!) 2,51 1.60 1,10 5,82
— — — 1,17 0,64 0,75 3,95 2,76 3,26
— — . — 0,70 0,15 0,36 3,43 1,06 2,18
— — — 0,043 0,031 0,108 0,421 0,344 0,664 •
тиворечии с общеизвестным фактом: в условиях длинного дня корнеплод
редиса никогда не образуется под лампами накаливания (фиг. 15),
излучение которых, как изве-
стно, сосредоточено главным
образом в оранжево-красных лу-
чах. Дело в том, что в этом слу-
чае процессы развития совер-
шаются настолько интенсивно,
что растения образуют стрелки
и зацветают, не успев образо-
вать корнеплодов (фиг. 16).
Однако достаточно к оранжево-
красным лучам добавить опре-
деленное количество сине-фио-
летовых лучей, задерживаю-
щих процессы развития, сохра-
нив без изменения общую ин-
тенсивность радиации, чтобы
началось образование корнепло-
дов, причем значительно рань-
ше, чем при выращивании ре-
диса в синих лучах.
Луковицы лука (Allium сери)
образуются исключительно под
влиянием оранжево - красных
лучей — под лампами накали-
вания (фиг. 17), под люминес-
центными лампами красного
света (фиг. 18) и другими, и
совершенно не образуются при
Фиг. 15. Образование корнеплодов редиса
(Raphanus salivus f. radiculd)
1 — под люминесцентными лампами белого света
(корнеплод); 3 — под лампами накаливания
(пет корнеплода)
выращивании лука в синих или
зеленых лучах. То же самое следует сказать относительно формирова-
ния кочана у капусты (Brassica oleracea f. capitula} пли кочанного
26
Физические и физиологические основы светокультуры растений
салата (Lactuca sativa) (фиг. 19). Однако листовая розетка обычного
салата (Lactuca sativa) формируется исключительно при выращивании
под лампами белого света, богатыми синими лучами (фиг. 20), или под
лампами синего света. Здесь, так же как и у редиса, быстрый переход
к цветению препятствует образованию розетки в оранжево-красных лучах.
Из сказанного следует, что оранжево-красные лучи, лежащие в
области максимального поглощения лучистой энергии хлорофиллом
(фиг. 3 и 4), являются физиологически наиболее эффективными в самых
разнообразных процес-
сах. Достаточно про-
стого перечисления раз-
личных физиологичес-
ких процессов — фото-
синтез, переход к цвете-
нию, синтез хлорофил-
ла, формирование ли
стьев, развитие корней,
образование луковиц,
кочанов, корнеплодов и
т. д., — где решающая
роль принадлежит оран-
жево-красным лучам,
чтобы убедиться в их ис-
ключительном значении
Как правило, при
одной и той же интен-
сивности излучения ма-
ксимальное накопление
сухого вещества, неза-
висимо от того, прихо-
дится ли оно на листо-
вую массу, корнеплоды,
плоды или другие орга-
ны, получается в оран-
жево-красных лучах
(табл. 4); исключение
составляют лишь расте-
Фиг. 16. Редис (Raphanus sativus f. radicula), выра-
щенный под люминесцентными лампами белого света
и под лампами накаливания
ния, которые под влиянием этой радиации чрезмерно быстро завершают
свой жизненный цикл, рано зацветая и образуя плоды (салат, редис и др.).
Однако и здесь на ранних фазах развития накопление сухой массы про-
исходит наиболее интенсивно в оранжево-красных лучах, и лишь позднее,
по мере прохождения стадий развития и старения растений, этот процесс
становится более интенсивным в сине-фиолетовых лучах, под влиянием
которых растения продолжают вегетативный рост.
Повидимому, сине-фиолетовые лучи следует рассматривать как фактор
своеобразного регулирования, под влиянием которого устанавливаются
нормальные соотношения между разными процессами без чрезмерного
преобладания какого-либо одного из них. По всей вероятности, это на-
ходится в связи с тем, что названные лучи поглощаются не одним хлоро-
филлом, а целым комплексом веществ (фиг. 3). Что касается зеленых лу-
чей, наименее поглощаемых пигментами пластид, то им в жизни растений
принадлежит незначительная роль. Как правило, источники радиации,
излучающие зеленые лучи (например, люминесцентные лампы зеленого
х'вета), мало пригодны для культуры растений.
II. Физиологическая радиация
27
В последнее время Б. С. Мошков [213—224] выступил с серией работ,
в которых он пытается доказать исключительно большое значение зеленых
лучей. Этот автор нашел, что максимальный
вес растений земляники {Fragaria sp.), живо-
родки {Bryophyllum sp.) и периллы наблюдает-
ся при культивировании их в зеленых лучах.
К сожалению, в своих первых сообщениях
Б. С. Мошков не приводил методики исследо-
вания, что значительно обесценивало результа-
ты его работы и внушало серьезные сомнения
в достоверности экспериментальных данных.
Фиг. 18. Образова-
ние луковиц у лука
(Allium сера), выра-
щенного под люми-
несцентными лампа-
ми разного света
1 — синего; 2 — зелено-
го; 3 — красного
Фиг. 17. Образование луковиц-у лука
(Allium сера) Каба болгарский ।
1 — под люминесцентными лампами белого света
(нет луковицы); 2 — под лампами накаливания
(луковица)
Его последняя работа [224], где приведена,
наконец, методика исследований, показала, что
указанные выше сомнения имели основания. Пользуясь для получения
«оранжево-красных» лучей лампой накаливания, водным экраном и све-
тофильтром «КС-5», пропускающим лучистую энергию начиная с 590 Шд,
Фиг. 19. Образование кочана у салата Ромэн (Lactuca sativa),
выращенного под люминесцентными лампами разного света
1 — синего; 2 — красного
Фиг. 20. Развитие салата (Lactuca sativa) Московский
1 — под люминесцентными лампами белого света (розетка);.
2 — под лампами накаливания (стрелка)
II. Физиологическая радиация
29
Мошков в действительности получал не оранжево-красные лучи, а излу-
чение, лежащее в пределах 590—1100 ту. Расчеты показывают, что фи-
зиологически активные оранжево-красные лучи (590—700 птл) составляли
при этом лишь 20%, а остальные 80% излучения приходились на физио-
логически неактивные инфракрасные лучи (700—1100 ту), даже не по-
глощаемые тканями листа (фиг. 3). Не удивительно, что результаты,
полученные Б. С. Мошковым, находятся в явном противоречии как с тео-
ретическими положениями, так и с экспериментальными данными
других авторов.
5. ЗНАЧЕНИЕ УЛЬТРАФИОЛЕТОВОЙ РАДИАЦИИ
Значительно менее определенны наши знания относительно роли ультра-
фиолетовых лучей. Вне всяких сомнений стоит лишь тот факт, что «корот-
кие» ультрафиолетовые лучи с длиною волны менее 290 ту. вызывают
гибель всякого растительного организма, будь то гриб или высшее расте-
ние. Речь может итти лишь о большей или меньшей устойчивости тех или
иных видов растений. Большинство растений под влиянием этой радиации
погибает почти мгновенно [513], и лишь незначительное меньшинство,
например кукуруза (Zea mays), пшеница {Triticum vulgare), некоторые
альпийские формы — кипрейник (Epilobium alpinum), смолевка (Silene
inf lata, S. maritima) и др., отличаются относительной устойчивостью,
развиваясь более или менее нормально при известной интенсивности этих
лучей [670,671]. Несомненно, что данное явление находится в связи
с поглощением коротких ультрафиолетовых лучей белками протоплазмы
и денатурацией белков под влиянием квантов, превышающих 4 eV. Мы
предлагаем назвать эти лучи витацидной радиацией, т. е. излучением,
несущим смерть всякому живому организму.
Большинство авторов [670, 671] считает, что «длинные» ультра-
фиолетовые лучи, лежащие в пределах 300—400 ту, играют незначитель-
ную роль в жизни растений и не влияют на их развитие и урожай. Имеются,
однако, указания, что ультрафиолетовая радиация, сосредоточенная
в узкой области между 290—315 Шу, имеет исключительно важное зна-
чение для формирования нормальных растений [513]. Во всяком случае,
следует заметить, что априори, принимая во внимание многообразие
веществ, поглощающих «длинные» ультрафиолетовые лучи (протоплазма,
ферменты, хлорофилл, каротиноиды и др.), допущение о незначитель-
ной роли названной радиации, особенно в отношении химического состава
растений, мало вероятно. Необходимо дальнейшее тщательное исследо-
вание этого вопроса.
6. ЗНАЧЕНИЕ ИНФРАКРАСНОЙ РАДИАЦИИ
Точно так же неопределенны наши знания о значении инфракрасных
лучей. Несомненно, что ближняя инфракрасная радиация (750—1000 пл),
мало поглощаемая тканями листа (фиг. 2 и 3), не может оказывать суще-
ственное влияние на процессы, протекающие в растении. Речь может
итти лишь о более далеких инфракрасных лучах (более 1000 пн.), погло-
щаемых водою, которая содержится в тканях листа, пигментами пластид
и протоплазмой. Положительное значение этой радиации отмечалось
неоднократно [213, 215, 216, 218, 219, 223, 224, 670, 820]. Так, в опытах
Пиршле и Веттштейна [670] по выращиванию растений целиком при
искусственном свете ламп накаливания устранение инфракрасной радиации
из спектра этих ламп посредством водного экрана в большинстве случаев
30
Физические и физиологические основы светокультуры растений
приводило к значительному снижению сухой массы растений. Особенно,
велико это снижение было у томатов (Solanum lycopersicum) (37%), куку-
рузы (Zea mays') (45%), льна (Linum usitatissimum) (71%) и др. По дан-
ным Б. С. Мошкова [212, 213, 215, 218, 219, 223, 2211, положительный
эффект от добавления «далеких» инфракрасных лучей получается тем
выше, чем ниже температура окружающей среды. Несомненно, что этот
эффект находится в связи с тепловым режимом листьев и других органов
растений.
7. ТЕМПЕРАТУРА ЛИСТЬЕВ
До последнего времени не было систематических исследований, посвя-
щенных температурному режиму листьев под влиянием тех или иных
ламп. Имелись лишь отдельные случайные наблюдения. Так, Ульрих
[820] заметил, что лампы солнечного света повышают температуру листьев
по сравнению с температурой воздуха на 5—6°. По данным Уитроу [854],
это повышение для ламп накаливания равняется 9—11°, а для люминес-
центных ламп составляет 4—5°. Вольтова дуга с углями «Солнечный
свет» при интенсивности физиологической радиации в 155 тыс. эрг/см2-сек:
повышает температуру листьев на 3,5° [661].
Первое систематическое исследование в этом направлении принадле-
жит В. Г. Карманову [97], который изучал температуру листьев томатов.
(Solanum lycopersicum) и лимонов (Citrus limonum) в зависимости от инте-
гральной интенсивности падающей радиации, полученной посредством,
ламп накаливания в 750 вт, и температуры окружающей среды. При изме-
нении интегральной интенсивности в пределах 0,28—1,43 г-кал/см'-мин.
или 200—1000 тыс. эрг/см^сек1, разница между температурой воздуха и
температурой листа возрастает прямо пропорционально интенсивности
радиации, достигая при 1000 тыс. эрг/см'-сек 18—22°. Для повышения
температуры на 1° на лист должно падать около 40—45 тыс. эрг/см2-секу
или 0,057—0,064 г-кал/см2-мин 2. Опыт показывает, что в пределах тем-
ператур от —6° до +18° повышение температуры листьев по сравне-
нию с температурой воздуха практически не зависит от температуры
последнего.
В наших опытах [119] в качестве объектов служили филлокактус
(Phyllocactus hybrida), амариллис (Amarillis hybrid а), огурцы (Сиси-
mis sativus) и пеларгония (Pelargonium zonule). Измерение темпера-
туры термоиглами В. П. Кислова [104, 105] производилось под лампами
накаливания в 500 и 1000 вт, под ртутной лампой Игар-2 (500 вт, 220 в,.
4—5 а), под неоновой лампой НД-1 (475 вт, 220 в, 6,5 а) и под люминес-
центными лампами белого, синего, зеленого и красного света (15 вт).
Каждое измерение производилось в течение 10—25 минут двумя термо-
иглами: одну иглу вводили в лист, а другую помещали под листом на
расстоянии 1 см от него. Температура воздуха равнялась 19—21° и воз-
растала во время измерения на 1—6°. Разницу между температурой листа
и температурой воздуха (Д1°) определяли как разницу в показаниях
между двумя названными иглами. Интегральную интенсивность радиа-
ции измеряли микротермостолбиком (табл. 5—8 и фиг. 21—25).
Как видно из кривых фиг. 21, показывающих характер изменения
температуры листьев после включения ламп, разница между темпера-
турой листьев и температурой воздуха быстро возрастает и достигает
максимума через 5 минут. При низких интенсивностях радиации и низких 1 2
1 Это соответствует освещенности порядка 5—25 тыс. лк.
2 Около тысячи люксов.
II. Физиологическая радиация
31
значениях температуры листа последняя остается неизменной независимо
от времени горения ламп (фиг. 21, 1—5). При высоких значениях назван-
ных величин температура листьев начинает постепенно падать (фиг. 21,6),
приближаясь к некоторой величине, характеризующей состояние равно-
весия. Названное явление находится в связи с появлением водного дефи-
цита в листе и уменьшением слоя воды, содержащейся в нем, и наблюдается,
как при выращивании под лампами накаливания, так и при культивиро-
вании растений под неоновыми и ртутными лампами.
Фиг. 21. Изменение температуры листьев огурцов (Cucumis satirus)
при различной интенсивности радиации в зависимости от продолжитель-
ности освещения лампой накаливания в 1 квт
1 — при 65 тыс. эрг/см! сек (100 см); 2— при 100 тыс. эрг/см2 сек (75 см); 3 — при
200 тыс. эрг/см’-сек (50 см); 4— при 275 тыс. эрг/см сек (40 см); 5 — при 425
тыс. эрг,см - сек (30 см); в — при 700 тыс. эрг'см - сек (20 см)
В скобках указано расстояние листа от лампы
В наших опытах подобное падение температуры листа наблюдалось
лишь у двудольных с обычными мезофильными листьями (огурцы, пелар-
гония). У слабо транспирирующих суккулентов (филлокактус) или одно-
дольных с мощной проводящей системой (амариллис) температура листьев
поддерживается на постоянном уровне, независимо от интенсивности излу-
чения и экспозиции (фиг. 22). У суккулентов с толстыми, мясистыми листь-
ями наблюдается значительно более медленное нагревание листа, чем у
других растений; равновесие достигается не через 5 минут, как, например,
у огурцов, амариллиса или пеларгонии, а через 25—30 минут (фиг. 22).
В свете приведенных данных следует различать максимальную и ста-
ционарную температуру листа. Первая наблюдается через 5 минут после
включения ламп и характеризует неустойчивое и кратковременное состоя-
ние, когда чрезмерное нагревание листа вызывает усиление транспирации
и появление водного дефицита. Вторая наступает через 20—25 минут и,
вероятно, отвечает установлению подвижного равновесия, когда транс-
32
Физические и физиологические основы светокультуры растений
пирация приходит в соответствие с температурой листа и устанавливается
определенная степень насыщенности тканей листа водою. На фиг. 23
показана зависимость максимальной и стационарной температуры листа
от интенсивности интегрального излучения. Первая выражается прямо-
линейной зависимостью, а вторая — кривой, стремящейся к пределу.
Фиг. 22. Изменение температуры листьев при интенсивности радиации
в 400 эрг'см2-сек, получаемой посредством ламп накаливания в 500 вт.
Расстояние листа от лампы — 20 см
1 — филлокактус (Phyllocactus hybrida); S — амариллис (Amarillis hybrida);
3 — пеларгония (Pelargonium zonal?) — зеленый лист; 4 — то же, белый лист
Температура срезанных листьев несколько выше, чем температура
.листьев, находящихся на растении (табл. 8). В опытах с пеларгонией
при интенсивности радиации в 400 тыс. эрг/см’-сек температура зеле-
ного листа в первом случае превышала температуру воздуха на 11 °, а во
втором на 8°.
Таким образом, температура листьев в значительной степени опреде-
ляется водным режимом растений. Она пропорциональна содержанию
воды и обратно пропорциональна интенсивности транспирации. В усло-
виях светокультуры, как и в естественных условиях, вода регулирует
температурный режим листьев. Завядание листьев вызывает снижение
транспирации не только вследствие уменьшения количества свободной
воды в протоплазме, но и благодаря снижению температуры листьев.
В этом — физиологическое и приспособительное значение водного дефи-
цита и завядания растений.
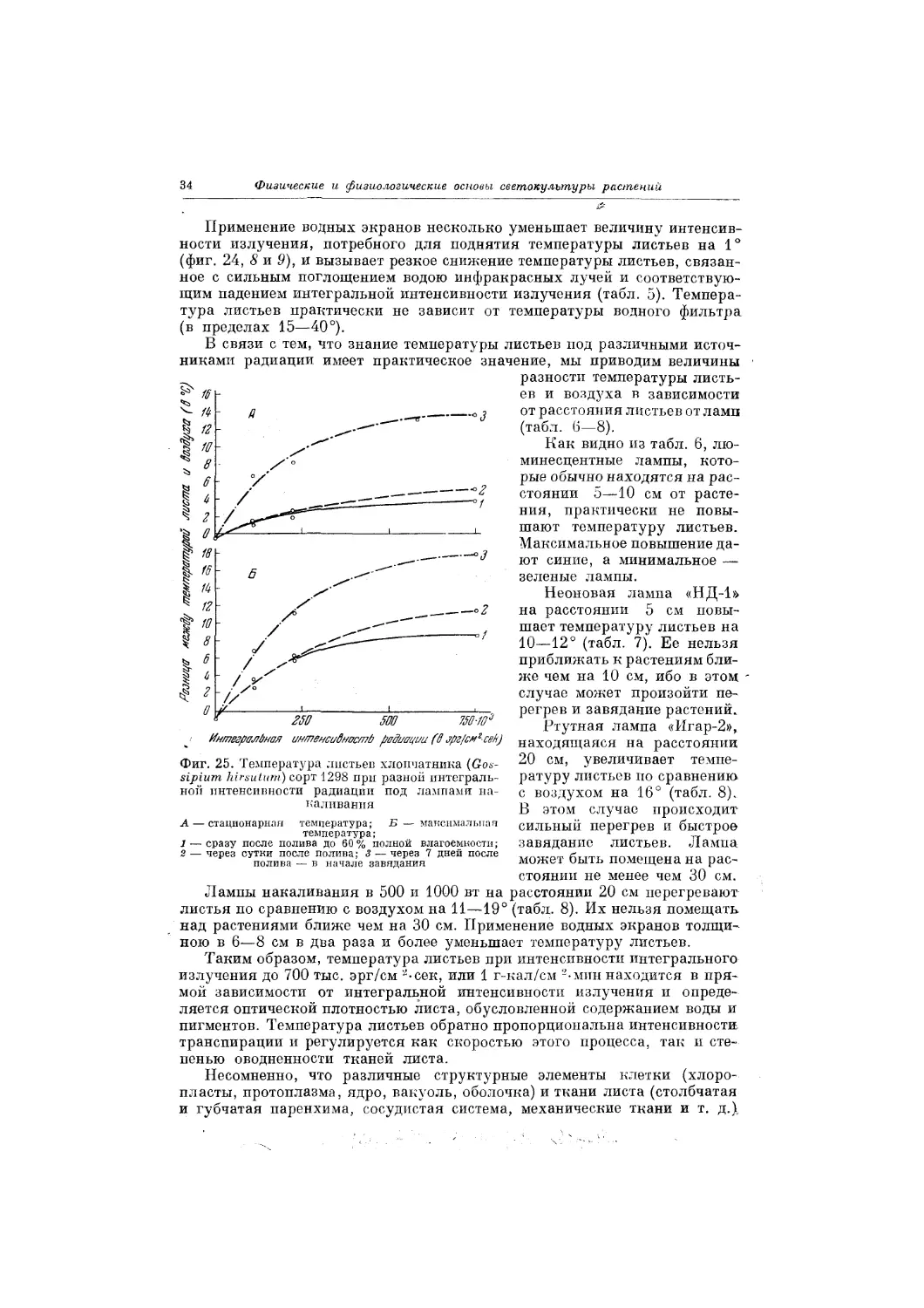
Максимальная температура листьев прямо пропорциональна интенсив-
ности интегрального излучения (фиг. 24). Величины интенсивности ра-
диация от разных ламп, потребные для поднятия температуры листьев
II. физиологическая радиация
33
%
разные растений на 1°, показаны в табл. 5. У сильно транспирирую-
щих растений, например, у хлопчатника, пропорциональность не на-
блюдается (фиг. 25).
Сопоставление температуры зеленых и белых листьев пеларгонии
(табл. 5, 7 и 8; фиг. 24, 7, 10, 12 и 13) не оставляет сомнений в том, что
г-Лал'смгли/и
I____ t________I I 1 , t > t
О w ZOO ООО Ш SOO 600 700 ю1
spj/c**. сек
Фиг. 24. Зависимость максимальной
температуры листьев от интенсивности
интегрального излучения
1 — амариллис (Amarillis hybrida), люминес-
центные лампы; 2 — филлокактус (Phyllocactus
hybrida), неоновая лампа НД-1; 3 — огурцы
(Cucumis sativus), неоновая лампа НД-1;
4 — черная бумага, лампа накаливания; 5 —
филлокактус (Phyllocactus hybrida), лампа
накаливания; 6 — огурцы (Cucumis sativus),
ртутная лампа Игар-2; 7 — пеларгония (Pelar-
gonium zonale, зеленый лист), неоновая лампа
НД-1; S—амариллис (Amarillis hybrida), пам-
па накаливания с водным фильтром (6—8 см);
.9 — амариллис (Amarillis hybrida) и огурцы
(Cucumis sativus), лампа накаливания; 10 —
пеларгония (Pelargonium zonale, белый лист),
неоновая лампа НД-1; 11—белая бумага,
лампа накаливания; 12 — пеларгония (Pelar-
gonium zonale, зеленый лист), лампа накали-
вания; 13 — пеларгония (Pelargonium zonale,
белый лист), лампа накаливания
'Фиг. 23. Зависимость температуры
листьев огурцов (Cucumis sativus) от
интенсивности интегрального излуче-
ния, получаемого посредством лампы
накаливания в 1000 вт
г — максимальная температура; 2 — стацио-
нарная температура
наряду с водой большую роль играют
и пигменты листа. Чем больше их со-
держание, т. е. чем больше поглоще-
ние лучистой энергии, тем выше тем-
пература листа, тем круче подъем
кривых и тем меньше интенсивность
излучения, необходимая для повы-
шения температуры листьев на 1°.
Огромное значение имеет толщина листа. Достаточно сопоставить ли-
стообразные стебли филлокактуса с листьями амариллиса или огурцов
(табл. 5, 7, 8; фиг. 24, 2, 3 и 5, 9), чтобы убедиться, что вместе с увеличе-
нием толщины листьев возрастает их температура и снижается интенсив-
ность радиации, потребная для поднятия температуры листьев на 1°.
Соответственно поглощению листьями лучистой энергии, излучаемой
разными лампами, наибольшее повышение температуры листьев наблю-
дается под люминесцентными и неоновыми лампами, наименьшее —
под лампами накаливания, а ртутная лампа занимает промежуточное
положение (табл. 5; фиг. 24, 1 и 9, 2 и 5, 3, 6 и 9, 7 п 12, 10 и 13).
3 А. Ф. Клешнин
34
Физические и физиологические основы светокультуры растений
Применение водных экранов несколько уменьшает величину интенсив-
ности излучения, потребного для поднятия температуры листьев на 1°
(фиг. 24, 8 и 9), и вызывает резкое снижение температуры листьев, связан-
ное с сильным поглощением водою инфракрасных лучей и соответствую-
щим падением интегральной интенсивности излучения (табл. 5). Темпера-
тура листьев практически не зависит от температуры водного фильтра
(в пределах 15—40°).
В связи с тем, что знание температуры листьев под различными источ-
никами радиации имеет практическое значение, мы приводим величины
Интегральная интенсивность радиации (в арг/смгсеЬ)
Фиг. 25. Температура листьев хлопчатника (Goi-
sipium hirsulum) сорт 1298 при разной интеграль-
ной интенсивности радиации под лампами на-
каливания
А — стационарная температура; Б — максимальная
температура;
1 — сразу после полива до 60 % полной влагоемности;
2 — через сутки после полива; 3 — через 7 дней после
полива — в начале завядания
разности температуры листь-
ев и воздуха в зависимости
от расстояния листьев от ламп
(табл. 6—8).
Как видно из табл. 6, лю-
минесцентные лампы, кото-
рые обычно находятся на рас-
стоянии 5—10 см от расте-
ния, практически не повы-
шают температуру листьев.
Максимальное повышение да-
ют синие, а минимальное —
зеленые лампы.
Неоновая лампа «НД-1»
на расстоянии 5 см повы-
шает температуру листьев на
10—12° (табл. 7). Ее нельзя
приближать к растениям бли-
же чем на 10 см, ибо в этом -
случае может произойти пе-
регрев и завядание растений.
Ртутная лампа «Игар-2»,
находящаяся на расстоянии
20 см, увеличивает темпе-
ратуру листьев по сравнению
с воздухом на 16° (табл. 8).
В этом случае происходит
сильный перегрев и быстрое
завядание листьев. Лампа
может быть помещена на рас-
стоянии не менее чем 30 см.
Лампы накаливания в 500 и 1000 вт на расстоянии 20 см перегревают
листья по сравнению с воздухом на 11—19° (табл. 8). Их нельзя помещать
над растениями ближе чем на 30 см. Применение водных экранов толщи-
ною в 6—8 см в два раза и более уменьшает температуру листьев.
Таким образом, температура листьев при интенсивности интегрального
излучения до 700 тыс. эрг/см 2-сек, или 1 г-кал/см 2-мин находится в пря-
мой зависимости от интегральной интенсивности излучения и опреде-
ляется оптической плотностью листа, обусловленной содержанием воды и
пигментов. Температура листьев обратно пропорциональна интенсивности
транспирации и регулируется как скоростью этого процесса, так и сте-
пенью оводненности тканей листа.
Несомненно, что различные структурные элементы клетки (хлоро-
пласты, протоплазма, ядро, вакуоль, оболочка) и ткани листа (столбчатая
и губчатая паренхима, сосудистая система, механические ткани и т. д.).
II. Физиологическая радиация
35
Таблица 5
Интенсивность радиации, необходимая для поднятия температуры листа на 1°
(в тыс. эрг/см2 • сек) *
Объект исследования Источники радиации
люминес- центные лампы неоновая лампа «НД-1» 475 вт ртутная лампа «Игар-2» 500 вт Лампы накаливания 500—1000 вт
водный фильтр без водно- го фильтра
Черная бумага — — — — 19
Филлокактус (Phyllocacius hybrida) 13 — — 23 (575)
Амариллис (Amarillis hybrida) . . . 10 — — 28 (2300) 34 . (850) -
Огурцы (Cucumis sativus) — 16 24 — 34 (850)
Хлопчатник (Gossipium hirsutum) . Пеларгония (Pelargonium zonale) — — — 37 (930)
зеленый лист — 26 —. — 50 > (1250)
белый лист . ' — 40 — — 100 J (2500) ..
Белая бумага — — — — 42
* Цифры в скобках показывают освещенность (в лк).
Таблица 6
Разница между температурой листьев амарил-
лиса (Amarillis hybrida) и воздуха при
освещении люминесцентными лампами (15 вт)
белого, синего, зеленого и красного света
Расстояние от ламп (в см)
Тип ламп 1 2,5 5
«0С-15» ...... 3,2 — 0,6
«СС-15» 3.6 I,6 0,6
«ЗС-15» 2,6 1,0 О/
«КС-15» 3.2 1,2 0,6
имеют совершенно различный температурный режим, зависящий от ха-
рактера поглощения лучистой энергии этими элементами структуры и
тканями. Не исключена возможность, что именно эта микротемпература
И наличие градиентов температур в пределах клетки, особенно в хлоро-
пластах, а не суммарная температура листа имеют решающее значение
для определенного направления физиологических процессов и соответ-
ствующего развития растений.
з*
36
Физические и физиологические основы светокультуры растений
Таблица 7
Разница между температурой листьев и воздуха при освещении неоновой лампой
НД-1 (500 ит, 220 в, 6,5 а)
Расстояние от лампы (в см)
5 10 15 20 30 40 50
Филлокактус (Phyllocactus hybrida) . 17,5 — 6,5 — 4,4 — —.
Огурцы (Cucumis sativus) Пеларгония (Pelargonium zonale) 12,0 9,0 — 5,0 3,5 2,2 1,0
зеленый лист 9,6 4,5 — 3,0 — — —
белый лист 7,5 3,5 — 1,5 т а б л и ц а 8
Разница между температу рой листьев и воздуха при освещении лампами накаливания
(500—1000 вт, 220 в) и ртутной лампой «Игар-2» (500 вт, 220 в, 4—5 а)
Источник радиации Растение / С Расстояние от ламп (в см)
20 30 | 40 50 70 100
Ртутная лампа Огурцы (Cucumis sativus) 16,0 8,0 6,0 4,0 2,0 0,6
«Игар-2» Лампа накалива- Филлокактус (Phyllocactus hybrida) 20,0 8,0 — 3,4 1 —
ния 500 вт: без водного экра-
на Амариллис (Amarillis hybrida) . . 11,5 6,0 4,0 — — ! —
Огурцы (C. sativus) Пеларгония (Pelargonium zonale): 12,0 8,0 4,5 3,0 2,0 . 1,0
зеленый лист 11,6 3,5 — 1,6 1 — —
белый лист 6,6 2,5 — 0,5 — —
с водным экра- ном в 6—7 см Амариллис (A. hybrida) 3,0 1,2 0 — — —
Лампа накалива-
ния 1000 вт:
без водного экра- Амариллис (A. hybrida) 19,5 11.0 7,5 — —
на Огурцы (С. sativus) . Пеларгония (Р. zonale) 19,0 12,0 8,0 5,0 — 2,0
зеленый лист 19,5 8,0 — — — —
белый лист 7,0 — — — — —
зеленый лист (срезанный) .... 17,5 11,0 — — — —
белый лист (срезанный) Бумага: 17,0 — — — — —
черная 41,0 18,5 11.0 —. — —
белая 16,5 7,5 40 — — —
с водным экра- Амариллис (A. hybrida) 9,0 2,5 1,0 — — —
ном в t;—7 см Пеларгония (Р. zonale):
зеленый лист 5,5 1,5 — — — —
белый лист 1 2,0 i i — — i —,
II. Физиологическая радиация
37
8. ЗНАЧЕНИЕ ТЕМПЕРАТУРЫ СРЕДЫ
ДЛЯ ФИЗИОЛОГИЧЕСКИХ ПРОЦЕССОВ РАСТЕНИЯ
Указывая на значение температуры для светокультуры растений,,
следует иметь в виду главным образом ее влияние на рост растения, его
дыхание и ассимиляцию углекислоты. Именно соотношением этих про-
цессов определяется в конечном итоге урожай. Как показывают опыты,
скорость роста растений определяется правилом Вант-Гоффа в пределах
температур от 0 до 35°. В этом случае скорость роста удваивается при
повышении температуры на каждые 10°, а максимальная скорость наблю-
дается при 25—40°. Оптимальной температурой, при которой бывает
максимальный рост, для культурных растений умеренных широт: ячмень
{Hordeum vulgare), рожь {Secale cereale), овес {Avena sativa), пшеница
{Triticum vulgare), гречиха (F ago руги m esculentum), тимофеевка {Phleum ar-
vense) и др., является температура воздуха порядка 25—30°. Для тепло-
любивых растений: подсолнечник {Helianthusannuus), кукуруза {Zea mays),
тыква {Cucurbita pepo), дыня {C. melo), огурцы {Cucumis sativus), горчица
{Sinapis alba) и др., температурный оптимум лежит в пределах 30—40°.
Дыхание растений, подобно ростовым процессам, определяется также
правилом Вант-Гоффа и достигает максимума при температуре воздуха
30—40°. В этом случае наблюдается максимальная потеря органиче-
ского вещества: у листьев 1—3, у почек и корней 2—6, а у растущих ко-
решков 4—8 мг на 1 г сухого вещества в час. Если принять во внимание,
что максимальная интенсивность фотосинтеза колеблется в пределах 40—
80 мг углеводов на 1 г сухого веса листьев в час, то потеря в листьях,
связанная с дыханием, составляет 2—4 % образующихся в процессе фото-
синтеза веществ. У травянистых видов, где листья составляют около
50% сухого вещества растения, потеря на дыхание для всего растения
будет равняться примерно 5—10%.
Значительно более сложна зависимость интенсивности фотосинтеза
от температуры. При естественном освещении (400 тыс. эрг/см2-сек) и
обычном содержании углекислого газа в воздухе окружающей среды
(0,03%) максимальная интенсивность фотосинтеза у таких растений, как
картофель {Solanum tuberosum), томаты (5. lycopersicum), сахарная
свекла {Beta vulgaris), конские бобы {Vida faba) и др., наблюдается
при температуре воздуха 18—20°. При снижении интенсивности освеще-
ния до 4% полуденной летней радиации (16 тыс. эрг/см-сек) температур-
ный оптимум фотосинтеза снижается до 10—17°. При повышении содер-
жания углекислого газа до 1 % температурный оптимум перемещается
в область 30—36°.
Таким образом, исходя из данных о зависимости роста, дыхания и
фотосинтеза от температуры окружающей среды, следует принять, что при
обычном содержании углекислоты оптимальной температурой для накоп-
ления органического вещества и, следовательно, для урожая растений
умеренных широт будет температура воздуха порядка 15—25°. При обо-
гащении атмосферы углекислотой необходима температура порядка 30°.
В случае применения искусственных источников радиации в период их
действия температура воздуха должна быть снижена в соответствии с спо-
собностью этих источников повышать температуру листьев.
9. ЗНАЧЕНИЕ ИНТЕНСИВНОСТИ ФИЗИОЛОГИЧЕСКОЙ РАДИАЦИИ
Для прохождения большинства физиологических процессов нужна
сравнительно низкая интенсивность радиации. Исключением являются
лишь развитие и фотосинтез растений. Именно эти процессы и определяют,
38
Физические и физиологические основы светокультуры растений
какая интенсивность физиологической радиации необходима для культуры
растений. Опыты В. О. Казаряна [89] с красной периллой (Perilla nanki-
nensis) и сафлором (Carthamus tinctorius) показывают, что в условиях
естественного освещения максимальная скорость развития наблюдается
в первом случае — при освещенности в 10 тыс. лк, а во втором — при
25 тыс. лк. Это соответствует интенсивности физиологической радиации
порядка 50—125 тыс. эрг/см 2-сек (фиг. 26). В опытах Бенедикта [406]
максимальное количество цветущих растений пырея (Agropyrum cristatum
и A. smithii) наблюдалось при освещенности в 60%, а у бутелуи (Boute-
loua gracilis) — лишь при полной солнечной радиации.
Фиг. 26. Значение интенсивности солнечной радиации для развития
и урожая растений [89]
/ — спорость развития красной периллы (Perilla nankinensis); 2 — то же сафлора
(Carthamus tinctorius); 3 — величина урожая семян красной периллы; 4 — то же
сафлора
Таким образом, из приведенных данных следует, что интенсивность
физиологической радиации, потребная для наиболее быстрого перехода
растений к цветению, в зависимости от вида, лежит в широком диапа-
зоне — от 50 до 400 тыс. эрг/см 2-сек.
Следует, однако, иметь в виду, что при употреблении источников ра-
диации, богатых оранжево-красными лучами, интенсивность освещения
может быть значительно ниже. Это относится, в частности, к лампам
накаливания, неоновым лампам, люминесцентным лампам тушеного света
и др.
Интенсивность физиологической радиации, необходимую для фотосин-
теза, можно рассчитать, исходя из абсолютной величины ассимиляции
углекислоты. Известно, что в естественных условиях фотосинтез у боль-
шинства культурных растений — фасоли (Phaseolus vulgaris), конских
бобов (! icia faba), шпината (5pinacea oleracea), сахарной свеклы (Beta
vulgaris), томатов (Solanum lycopersicum), картофеля (5. tuberosum), Лквы
(Cucurbita pepo), подсолнечника (Helianthus annuus) и др. — равняется
1—2 г сухого вещества на 1 м2 листовой поверхности в час. Теплотворная
способность сухого вещества растений лежит в пределах 4000—5000 кал/г;
II. Физиологическая радиация
39
следовательно, в процессе фотосинтеза каждый 1 см1 листа ежесекундно
аккумулирует 5—10 тыс. эргов энергии, падающих на лист в виде физио-
логической радиации. Поскольку при этом из 12 квантов поглощенной
энергии используется лишь 4 кванта, а остальные деградируют в виде
тепла, то поглощаемая энергия должна быть равна 15—30 тыс. эрг/см2-сек.
В случае гипотетического источника, излучающего одинаковое количе-
ство энергии во всех областях спектра физиологической радиации, поглоще-
ние лучистой энергии хлорофиллом составит примерно 50% всей энергии,
падающей на лист в виде физиологической радиации. Это означает, что
Физиологическая радиация (варг/смг се!)
Интегральная рааиация ( г-нал/аи мин;
О ~1д ^7 ~дд Тд Д7 Ж М № _ Sd >7
Солнечная радиация (в °/0 h максимальной)
Фиг. 27. Интенсивность фотосинтеза в зависимости от интенсивности
солнечной радиации
1 — шпинат (S-pinacea о Oracea); 2 — картофель (Solanum tuberosum); 3—жерушник
(Nasturtium patustrn); 4 — сосна (Pious sp.); 5 — папоротник (Druopteris austriaca )
теоретически интенсивность физиологической радиации на уровне расте-
ний должна равняться 30—60 тыс. эрг/см2 • сек. Практически она должна
быть несколько выше, так как часть энергии не попадает на растения
и поглощается почвой, поглощение энергии в листе часто падает в связи
с перемещением хлоропластов на боковые стенки клеток, различные ярусы
листьев находятся в неодинаковых условиях освещения и т. д.
Величина интенсивности физиологической радиации, при которой
наблюдается максимальная интенсивность фотосинтеза, повышается
в случае применения источников радиации, богатых сине-фиолетовыми
лучами, и снижается, если в спектре излучения источника содержатся
в основном оранжево-красные лучи. Более подробно этот вопрос будет
рассмотрен ниже.
Приведенные выше теоретические расчеты показывают, что в процессе
фотосинтеза усваивается около 17% лучистой энергии, падающей на рас-
тение в виде физиологической радиации. Для солнечного излучения,
40
Физические и физиологические основы светокультуры растений
содержащего примерно 50% инфракрасных лучей, теоретический коэф-
фициент использования составит, следовательно, 8—9% общего коли-
чества энергии. Однако реально, как показывают опыты, этот коэффи-
циент не превышает величины порядка 5%. Это означает, что в рассмот-
ренном нами гипотетическом случае интенсивность физиологической
радиации на уровне растений в условиях светокультуры должна быть
равна 60—120 тыс. эрг/см2-сек. Интенсивность фотосинтеза у большин-
ства высших растений, исключая светолюбивые формы, в этом случае
достигает максимума, и дальнейшее увеличение интенсивности радиации
уже не оказывает большого влияния на накопление органического веще-
ства (фиг. 27).
Следовательно, исходя из данных о зависимости процессов развития
и фотосинтеза от интенсивности радиации, следует принять, что интенсив-
ность физиологической радиации, оптимальная для нормального роста и
развития большинства растений, лежит в пределах50—120 тыс. эрг/см 2-сек,.
или 0,07—0,17 г-кал/см2-мин. Изменение интенсивности в сторону
меньших величин влечет снижение урожая растений, а повышение интен-
сивности или не влияет на урожай, или дает незначительный эффект, не
оправдывающий экономических затрат на энергию. Лишь в некоторых спе-
цифических случаях, когда культивируют тенелюбивые растения, возможно-
применение более низкой интенсивности, а при культуре крайне светолю-
бивых растений, например хлопчатника, подсолнечника или кукурузы,,
следует рекомендовать интенсивность более 120 тыс. эрг/см 2-сек.
.•> 10. ВЫВОДЫ
1. Для нормального роста и развития растений реальное значение-
имеет главным образом излучение, лежащее в пределах 300—750 ту и
поглощаемое пигментами пластид. Это — физиологическая радиация.
2. Максимальной активностью отличаются оранжево-красные лучи
(600—700 ту), поглощаемые исключительно хлорофиллом. В этих лучах
с максимальной скоростью совершаются фотосинтез, образование листьев,
формирование корнеплодов, луковиц, стеблевых утолщений, переход
растений к цветению, фотопериодический процесс и т. д. Здесь, как пра-
вило, наблюдается максимальный урожай.
3. Минимальной активностью обладают зеленые лучи (500—600 ту),
где все перечисленные процессы идут наименее интенсивно и выращива-
ние растений нерационально из-за большого расхода лучистой энергии.
4. Специфическую роль выполняют сине-фиолетовые лучи (400—500ту),
поглощаемые хлорофиллом, каротиноидами, ферментами и другими ком-
понентами клетки. Эффективность этой радиации в два раза меньше,
чем оранжево-красных лучей. В большинстве случаев они задерживают
переход растений к цветению, благоприятствуют синтезу белков и, по-
видимому, вообще сильно влияют на химический состав растений.
5. Реальное значение для синтеза органического вещества и создания
урожая растений имеет лишь область, лежащая в пределах 400—700 ту,
где происходит ассимиляция углекислоты (фиг. 10).
6. Ультрафиолетовые лучи с длиною волны 300—400 ту, невиди-
мому, выполняют лишь формативную роль, предотвращая чрезмерное
вытягивание растений. Особенной эффективностью в этом отношении
отличаются лучи с длиною волны, равной 290—315 ту, поглощаемые
обычным стеклом. Не влияя значительно на урожай, эта часть радиации,
тем не менее, несомненно, оказывает влияние на химический состав расте-
ний, повышая содержание белков и витаминов в тканях растений.
II. Физиологическая радиация
41
7. Короткие ультрафиолетовые лучи, лежащие за пределами 300 шр,
несут гибель всякому как животному, так и растительному организму.
Это — витацидная радиация, вредная для растений.
8. Инфракрасные лучи с длиною волны 750—1000 тр., не поглощае-
мые или мало поглощаемые пигментами и тканями растений, практически
не имеют значения для физиологических процессов. Их следует назвать
абиотической радиацией.
9. Инфракрасные лучи, начиная с 1000 тр, в основном поглощаемые
водою тканей листа, имеют значение температурного фактора, определяю-
щего тепловой режим тканей растении. Они, несомненно, влияют на ско-
рость и направление физиологических процессов. Роль этих лучей поло-
жительна в условиях низких температур воздуха (менее 20°) и отрица-
тельна при высоких температурах (более 30е).
10. «Далекие» инфракрасные лучи с длиною волны порядка 4—6 м,
поглощаемые хлорофиллом и другими составными частями протоплазмы,
невидимому, могут иметь и самостоятельное значение, активируя фото-
химические процессы, которые могут совершаться при действии квантов
незначительной величины (0,2—0,3 eV).
11. Оптимальная интенсивность физиологической радиации, потреб-
ная для нормального развития и роста растений в условиях светокуль-
туры, лежит в пределах 50—100 тыс. эрг/см2-сек.
12. Интенсивность физиологической радиации, необходимая для до-
полнительного освещения растений в ночные часы в целях удлинения
дня и ускорения развития длиннодневных или замедления развития ко-
роткодневных растений, равняется 500—1500 эрг/см 2-сек.
Глава Ш
ИСТОЧНИКИ РАДИАЦИИ
Всякий источник лучистой энергии, применяемый для культуры рас-
тений, должен прежде всего обладать спектральным составом и мощностью,
отвечающими требованиям растений вообще и данной культуры в особен-
ности. Кроме того, этот источник должен быть экономичен, достаточно
долговечен и прост в эксплуатации. Этим требованиям в большей или
меньшей степени удовлетворяют лишь электрические источники радиации:
люминесцентные, газосветные и дуговые лампы и лампы накаливания.
Лучистая энергия, представляя собою энергию электромагнитных
колебаний, вызывается колебательными движениями электрических за-
рядов, т. е. электронов и других заряженных частиц, входящих в состав
твердых, жидких или газообразных веществ. У твердых и жидких тел,
где движения зарядов, влияя один на другой, носят самый разнообразный
характер, излучение имеет сплошной спектр, часто захватывающий об-
ласть ультрафиолетовых, видимых и инфракрасных лучей. У газов, где
молекулы относительно свободны и движение электронов определено лишь
внутримолекулярными или внутриатомными силами, спектр носит опреде-
ленный линейчатый или линейчато-полосатый характер, специфический для
каждого отдельного вещества. Типичными источниками сплошного спектра
являются лампы накаливания. У газосветных ламп, как правило, на-
блюдается линейчатый спектр. Люминесцентные лампы, дуговые лампы
и некоторые другие источники имеют спектр смешанного характера,
где на основной сплошной фон накладываются отдельные линии излу-
чения.
Для всякого излучения, представляющего собою беспрерывное рас-
ходование энергии, необходим систематический приток энергии извне.
Этот процесс осуществляется или посредством нагревания тел (темпера-
турное или тепловое излучение), или их облучением (фотолюминесценция),
или в результате разгона и столкновения заряженных частиц в электри-
ческом поле (электролюминесценция газовых разрядов). В зависимости
от природы физического явления, лежащего в основе излучения, источ-
ники радиации принято разделять на температурные, люминесцентные,
газосветные и др. К источникам температурного, или теплового, излуче-
ния относятся лампы накаливания, к газосветным источникам — неоно-
вые, ртутные, натриевые и другие лампы. Люминесцентные и дуговые
лампы представляют собою источники смешанного типа: у первых комби-
нируется излучение газового разряда с фотолюминесценцией, а у вторых —
температурное излучение с излучением газового разряда. Ниже будут
III. Источники радиации
43
указаны некоторые другие источники, излучение которых носит также
• смешанный характер.
Источники радиации изготовляются или в виде стеклянной колбы,
имеющей шарообразную или другую форму, или в виде длинной стеклян-
ной трубки. Первые мы будем называть шарообразными, а вторые — труб-
чатыми источниками излучения.
1. ЭНЕРГЕТИЧЕСКИЕ И СВЕТОТЕХНИЧЕСКИЕ ХАРАКТЕРИСТИКИ
ИСТОЧНИКОВ РАДИАЦИИ
Основными энергетическими характеристиками всякого источника
радиации являются мощность и лучистый поток. Номинальная мощность
источника (IF), выражаемая обычно в ваттах, характеризует расходование
энергии источником, но отнюдь не энергию излучения. Последняя харак-
теризуется лучистым потоком, или мощностью излучения (Р).
Лучистый поток (Р) есть количество энергии, излучаемое источником
радиации в единицу времени (Z) во всех направлениях. Величина потока
обычно выражается в ваттах, милливаттах, микроваттах, эрг/сек или
кал/мии.
1 вт—103 мвт=106 мквт=107 эрг/сек = 14,34 кал/мин.
Если речь идет о суммарном излучении всех длин волн, то говорят:
интегральный лучистый поток; если дело касается
определенного вида излучения, лежащего в пределах некоторых длин
волн, то употребляют выражения: поток инфракрасной
радиации (более 750 тр), поток витацидной радиа-
ции (менее 300 шц) и т. д. В нашем случае речь всюду будет итти, глав-
ным образом, о потоке фитофизиологической радиации, лежащей в пре-
делах 300—750 гпц.
В литературе, справочниках, каталогах и тому подобных изданиях
приводится обычно не лучистый поток источников, а так называемый
световой поток (Ф), т. е. поток лучистой энергии, оцениваемый по свето-
вому ощущению средним человеческим глазом. Единицей светового по-
тока является люмен (лм)1 — световой поток, испускаемый полным из-
лучателем (абсолютно черным телом) при температуре затвердевания
платины (2042° К) с площади 0,5305 мм’. Световой поток есть фотометри-
ческий аналог лучистого потока. Являясь субъективной оценкой лучи-
стого потока, он определяется, следовательно, спектральной чувствитель-
ностью глаза или видностыо (И).
Абсолютная видность, или световая отдача лучистого потока, есть
отношение светового потока, выраженного в люменах, к лучистому
потоку, выраженному в ваттах:
Максимальное значение видности (Гмак0), принимаемое за единицу,
наблюдается при длине волны в 555 шр. В этом случае лучистый поток в 1 вт
соответствует световому потоку в 621 лм. Иными словами
I ма1.с = 62'1 лм/вт.
Отсюда следует, что световой поток в 1 лм, излучаемый в виде линии
555 m/i, равняется 1,61 ИО-3 вт. Эта величина получила наименование
механического эквивалента света:
М = 0,00161 вт/лм.
1 В немецкой литературе употребляется обычно люмен Гефнера (Him), равный
0,88 международного люмена.
44
Физические и физиологические основы светокультуры растений
Обычно спектральная чувствительность глаза выражается в виде
относительной видности (К.,). Относительная видность есть отношение
видности любой длины волны (Кд) к максимальной видности (I макс)'
7 ~ К>. = ^. (10а)
* макс
Величины относительной видности даны в табл. 9, а кривая относи-
тельной видности — на фиг. 28.
Фиг. 28. Спектральная чувствительность среднего
человеческого глаза
Т а б л и ц а 9>
Коэффициенты относительной видности (К ) среднего человеческого глаза
Длина волны (В Ши.) Длина волны (в ти) К/. Длина волны (в mu.) А’/.
400 0,0004 520 0,710 (0,80) 640 0,175 (0,12)
410 0,0012 530 0,862 650 0,107
420 0,0040 540 0,954 (0,98) 660 0,061 (0,085)
430 0,0016 550 0,995 670 0,032
440 0,023 560 0,995 (1,00) 680 0,017 (0,024)
450 0,038 570 0,952 690 0,0082
460 0,060 (0,022) 580 0,870 (0,82) 700 0,0041 (0,002),
470 0,091 590 0,757 710 0,0021
480 0,139 (0,165) 600 0.631 (0,566) 720 0,00105 ’
490 0,208 610 0.503 730 0,00052
500 0,323 (0,490) 620 0,381 (0,333) 740 0,00025
510 0,503 630 0,265 750 0,00012
760 0,00006
В связи с тем, что для светофизиологии и светокультуры растений
представляет интерес главным образом поток физиологической радиации,.
III. Источники радиации
45
а в справочниках обычно дается световой поток, возникает необходимость
перевода величин светового потока в величины лучистого потока.
Величина светового потока источника есть результат перемножения
ординат кривой относительной видности на соответствующие ординаты
спектральной кривой излучения источника. Кривая, полученная в резуль-
тате подобного перемножения, показывает чувствительность глаза к из-
лучению данного источника, а площадь, лежащая между этой кривой и
осью абсцисс, соответствует величине светового потока источника. На
фиг. 29 в качестве примера приводятся соответствующие кривые для лампы
Фиг. 29. соотношение лучистого и светового потока для ламп г, г
накаливания в 300 вт
1 — распределение энергии в области физиологической радиации спектра лам-
пы накаливания; 2 — относительная видность спектра лампы накаливания
аб — граница между ультрафиолетовыми и видимыми лучами; eg — граница
между видимыми и инфракрасными лучами
накаливания мощностью в 300 вт: кривая 1 показывает распределение
энергии в спектре излучения, а кривая 2 — результат перемножения
ординат цервой кривой на ординаты кривой относительной видности
глаза. Здесь площадь (Sp), ограниченная кривой 1, осью абсцисс и линиями
об и вг, соответствует величине лучистого потока в видимой части излу-
чения лампы накаливания (Pv) (400—760 my), а площадь между
кривой 2 и осью абсцисс — величине светового потока Ф.
Для перехода от светового потока, выраженного в люменах, к лучи-
стому потоку видимого излучения, выраженному в ваттах, необходимо
знать отношение
4'- = К. (И)
Оно легко определяется, если известно распределение энергии в спек-
тре источника, методом построения кривых и определения площадей
(фиг. 29).
Для любого источника радиации справедливо уравнение
1,61 • 10~3Л'Ф, (12)
46
Физические и физиологические основы светокультуры растений
Таблица 10
Значения 1 лм (в вт) и переходных коэффициентов К для разных источников
искусственной радиации
Лампы Мощность ламп (в вт) Поток физио- логической радиации, эквивалент- ный 1 лм (в вт) Переходный коэффициент К
для потока физиологиче- ской радиа- ции для интег- рального лу~ чистого потока
А. Лампы накаливания
10 8,40 • 10-3 5,21 —
25 8,05 • 10-3 5,00 —
1. Пустотные 40 7,20 • 10-3 4,41 50,7
60 6,68 • 10-з 4,14 —
50 6,61 • 10—3 4,11 —
75 6,30 • ю-з 3,91 34,3
100 6,15 • 10-з 3,82 32,1
200 5,97 • 10-3 3,71 29,0
2. Газополные 300 5,86 • Ю-з 3,64 27,6
500 5,67 • Ю-з 3,52 24,6
1000 5,54 • 10-3 3,44 22,6
2000 5,46 • 10-3 3,39 22,0
3. Проекционные 1000—3000 4,82 • 10-3 2,99 15,5
Б. Люминесцентные —
лампы -
1. Низковольтные . '
дневного света 15—100 4,03 • Ю-з 2,50 —
мягкобелого света 15—40 4,25 • Ю-з 2,64 —
белого света 15—100 • 3,55 10 3 2,20 —
синие * (CaWOj 15—30 5,15 • 10-3 3,20 3,20
» (MgWO4) 15—30 4,06 • 10-3 2,54 2,54
зеленые (ZnSiO3) 15—30 2,22 • 10-3 1,38 1,38
красные (CdB2O,5) 15—30 4,51 • Ю-З 2,80 —
золотистые 15—30 3,42 • 10-3 2,12 —
2. Высоковольтные:
дневного света 50 4,60 • 10-3 2,85 —
мягкобелого света 100 4,30 • 10-3 2,67 —
белого света 100—120 3,85 • 10-3 2,39 —
3. Высокого давления
(«HgL») 75—120 2,24 • 10-3 1,56 —
В. Лампы смешанного
т it п а
«S-1» 315 6,45 • 10-3 4,00 24,8
«HWA-500» 255 4,72 • 10-3 2,93 17,5
* В скобках поставлены люминофоры.
III. Источники радиации
47
Таблица 10 (продолжение)
Лампы Мощность ламп (в вт) Поток физио- логической радиации, эквивалент- ный 1 лм (в вт) Переходный коэффициент К
для потока физиологиче- ской радиа- ции Для интег- рального лу- чистого потока
Г. Газосветные лампы
1. Ртутные лампы ;
«СН» 270 4,38 • 10-3 2,78 3,25
«Игар-2» 500 3,98 • Ю-з 2,47 —
«Н-2» 250 3,09 • 10-3 1,92 —
«Н-1» 400 2,96 • 10-3 1,84 —
«ПРК-4» 220 4,26 • 10-з 2,65 3,61
«ПРК-2» 375 4,14 • 10-3 2,57 3,45
«ПРК-7» 1000 4,34 • 10-3 2,70 3,76
«Н-5» 250 3,32 • 10-3 2,06 —
«СВД-125» 125 3,71 • 10-з 2,30 —
«СВДШ-250» 250 3,70 • 10-3 2,30 3,32
«н-з» 85 3,83 • Ю-з 2,38 —
«Н-4» 100 3,88 • Ю-з 2,41 —
«HgH-ЮОО» 265 2,43 • Ю-з 1,51 1,65
«HgQS-500» (матовое стекло) . 120 4,21 • 10-з 2,62 2,85
«HgHS-8000» (увиолевое стекло) 1500 3,62 • Ю-з 2,25 2,45
«HgQS-ЗОО» (увиолевое стекло) 75 5,00 • 10-з 3,10 3,49
«HgQS-500» Bl. (NiO стекло) . 120 1,69 • 103 1,05 • 105 1,08 • 105-
«HgQS-500» (NiO -J-Сг2О-стекло) 120 1,93 • 1©2 1,20 • 105 1,34 • 105
2. Натриевые лампы 45—140 1,88 • Ю-з 1,17 1,50
3. Цинковые лампы низкого дав-
ления — 2,33 • 10-з 14,45 15,50
4. Кадмиевые лампы
«КК-1» 460 1,06 • 10-2 6,60 13,20
низкого давления — 2,26 • 10-2 14,10 15,50
5. Цезиевые лампы 500 3,74 • 10-2 21,50 181,00
6. Неоновые лампы 28—485 7,35 • Ю-з 4,62 4,70
7. Гелиевые лампы — 5,77 10-3 3,58 —
8. Криптоновые лампы — 3,56 • 10-3 2,21 —
9. Ксеноновые лампы 150—700 1,48 • 10-2 9,20 —
Д. Вольтовы дуги
1. Открытые с чистыми углями . — 7,23 • 10-3 4,49 —
2. Открытые с пламенными углями — 7,01 • 10-3 4,36 — , ,
3. Открытые с углями «Солнеч-
ный свет» -— 7,25 • 10-3 4,51 —
где К — константа, определяемая формулой (11) и специфическая для
каждого источника. В случае светового потока, выраженного в люменах
Гефнера, вместо множителя 1,61 в формулу входит другая величина,
равная 1,44.
Если линии аб и вг переместить в другие точки оси абсцисс, то соот-
ветственно изменятся значение К и спектральные границы измеряемого
лучистого потока, который можно обозначить как /\2_Х1. Пользуясь
подобным методом, можно легко перевести величины светового потока
в величины лучистого потока, лежащего в любой части спектра, — от
48
Физические и физиологические основы светокультуры растений
узких полос до всего спектра. Заметим, что аналогичный метод суще-
ствует для пересчета единиц освещенности в объективные единицы лучистой
энергии [см. главу VI, фиг. 94 и 95, табл. 59, формулы (39) и (40)].
В табл. 10 приводятся значения потока фитофизиологической радиации
от разных источников излучения, эквивалентные 1 лм. Там же приводятся
и величины коэффициентов К, позволяющие при помощи формулы (12)
'Фиг. 30. Распределение силы света лампы на-
каливания со световым потоком в 1000 лм
I — в горизонтальной плоскости; 2 — в вертикальной
плоскости
сделать перевод светового по-
тока в величины интеграль-
ного лучистого потока и по-
тока физиологической радиа-
ции. Эта таблица составлена
по методу, который иллю-
стрируется фиг. 29.
Очень важной характери-
стикой источников радиации
является угловая плотность
лучистого потока (F) — коли-
чество лучистой энергии, из-
лучаемое в единицу времени
внутрь единицы телесного
угла(ы) — стерадиана (стр.)1.
Угловая плотность характе-
ризует величину лучистого
потока в данном направле-
нии. Единицами угловой
плотности потока являются
вт/стр, эрг/сек-стр. и т. д.
Фотометрический аналог
угловой плотности — сила
света (/) — световой поток,
излучаемый внутрь единицы
телесного угла—стерадиана.
Сила света измеряется в све-
чах (св). Международная
свеча, принятая во всех стра-
нах за исключением Герма-
нии, есть сила света точеч-
ного источника в тех направ- (
лениях, в которых он испу-^г
скает световой поток в 1 лм,
равномерно распределенный
внутри телесного угла в 1 стерадиан. Международная свеча равняется
1 лм/стр. В немецкой литературе обычно употребляется свеча Гефнера
(НК), т. е. сила света пламени амилацетата шириною в 8 мм и высотою
в 40 мм. Свеча Гефнера равняется 0,88 лм/стр (международной свечи)
и обозначается 1 Hlm/Ster.
Шарообразные источники радиации — лампы накаливания, дуговые
лампы, ртутные лампы сверхвысокого давления и другие — обычно ха-
рактеризуются средней сферической угловой плотностью лучистого
потока (Fo) или средней сферической силой света 1 2 (Zn):
1 Стерадиан есть телесный угол, вырезающий на сфере с радиусом R поверхность
с площадью R-.
2 Средняя сферическая сила света соответствует освещенности (в лк) на рас-
стоянии 1 м от источника.
III. Источники радиации
49
А=-^=0,08Р (13)
/„ = ^- = 0,08Ф, (13а)
где Р — лучистый поток, а Ф — световой поток источника.
Характеристикой трубчатых источников радиации — газосветных
трубок, люминесцентных ламп, софитных ламп накаливания и других —
Фиг. 31. Распределение силы света прожекторной лампы со световым
потоком в 1000 лм. На осях аб и вг указана сила света (в св)
1 — в горизонтальной плоскости; 2 — в вертикальной плоскости
является максимальная горизонтальная угловая плотность лучистого
потока (/’w) или максимальная горизонтальная сила света (ZTO) — плот-
ность лучистого потока или сила света в направлении, перпендикулярном
к оси трубки:
^,„=^г = 0,1Р; (14)
/я> = ^=0,1Ф.
(14а)
Распределение лучистого или светового потока в пространстве обычно
изображается графически. Берут условный источник радиации, дающий
световой поток в 1000 лм (или лучистый поток в 1000 вт), и в различных
направлениях от излучающего центра откладывают величины, пропор-
циональные силе света (пли угловой плотности лучистого потока) в дан-
ном направлении. Направление определяется углом а относительно верти-
кальной или горизонтальной оси источника.
4 А. Ф. Клешнин
50
Физические и физиологические основы светокультуры растении
Фиг. 32. Распределение силы света газосв
ной трубки со световым потоком в 1000
1 — в горизонтальной плоскости; 2 — в вер
калькой плоскости (вдоль трубки)
На фиг. 30—33 приводятся наиболее типичные случаи распределения
силы света в горизонтальной и вертикальной плоскости для источников
радиации, употребляемых в светокультуре растений. Фиг. 30 характе-
ризует обычные лампы накаливания с кольцеобразной нитью накала,
расположенной в горизонтальной плоскости; фиг. 31 — проекционные
лампы накаливания, нити которых расположены в виде решетки в верти-
кальной плоскости; фиг. 32 — трубчатые лампы с равномерно светящейся
поверхностью, а фиг. 33 — точеч-
ные дуговые лампы с электродами
Ю0 в виде шариков, расположенных в;
плоскости, перпендикулярной от-
носительно оси лампы.
Источники радиации, обладаю-
120 щие сравнительно большой излу-
чающей поверхностью (5,),—газо-
светные трубки, люминесцентные
//у лампы и другие — характеризуются
также поверхностной плотностью
лучистого потока (Н} — величиной
100 излучения, испускаемого во всех
направлениях единицей площади
' "светящейся поверхности лампы.
Единицами поверхностной плот-
ности служат вт/см'2, эрг/см'2-сек
ди и др. Аналогичным фотометриче-
ским понятием является свети-
мость (А) — световой поток, пспус-
70 каемый единицей поверхности. Све-
тимость измеряется в лм/см'2.
Распределение энергии в спск-
дд тре излучения источника харак-
теризует спектральная интенсив-
ность (2?х)—величина поверхност-
ной плотности лучистого потока,
50 отнесенная к единице длины волны.
Обычно спектральная интенсив,-
ность выражается или в вт/см'2-А
т_ или в процентах к максимальному
[м излучению (£>п).
п Для наших целей важным поня-
тием является яркость (/?) — лучи-
стый поток, испускаемый единицей
поверхности внутрь единицы телесного угла — стерадиана. Энергетиче-
ская яркость (7V) измеряется в вт/см2-стр, а световая яркость—в стиль-
бах (сб). Стильб равняется 1 св/см2.
Согласно закону Ламберта, яркость поверхности одинакова во всех
направлениях. Это означает, что сила света элемента поверхности (Д) из-
меняется пропорционально косинусу угла между нормалью к поверхности
и данным направлением:
Z —I cos р.
(15).
Яркость трубчатых источников в направлении, перпендикулярном
к оси трубки, определяется уравнением
III. Источники радиации
51
p
dl ’
2V = ^”’=(),1
п I
ИЛИ
Im ___ ,< Ф
dl — ~dl
(16)
(16a)
где N и В — энергетическая и световая яркость, Р и ф — лучистый и
световой поток, Fm — максимальная горизонтальная угловая плотность
лучистого потока, 1т — максимальная горизонтальная сила света,
a d и I — диаметр и длина трубки (в см).
Фиг. 33. Распределение силы света точечной
дуговой лампы со световым потоком в 1000 лм
1 — в горизонтальной плоскости; 2 — в вертикальной
ПЛОСКОСТ.
Экономичность источников света характеризуется лучистой или све-
товой отдачей (/)) — отношением лучистого или светового потока к номи-
нальной мощности источника. Лучистая отдача выражается в процентах,
а световая отдача — в лм/вт. Чем выше эти показатели, тем более эконо-
мичен источник. В табл. 11 сопоставляются основные энергетические
(радиометрические) и фотометрические величины и понятия.
4*
Физические и физиологические основы светокультуры растений
Таблица 11
Энергетические (радиометрические) и фотометрические величины
Энергетические понятия Символы | Единицы измерения Фотометрические понятия Символы Единицы измерения
Лучистая энергия и Джоуль 1 Световая энергия Q лм сек
Лучистый поток . . Угловая плотность Р = ~~ t р ; Ватт i ! Световой поток лм
лучистого потока . Поверхностная плот- ность лучистого F=~~ со вт/стр Сила света 7 = -ф- G) св
потока Спектральная интон- dH вт/см- Светимость А^ ф Si лм/см2
сивность Энергетическая яр- h'' = djk p вт/см2 • А — Р, = -<L_ о / • (о —
кость N = -. 0 i • (♦) вт/см2 • стр Яркость со
Лучистая отдача . . Интенсивность ра- p r‘~'w p 0/ /0 Световая отдача W лм/вт
диацип * . ... h вт/см- i Освещенность * L — - S лк
Об интенсивности радиации и освещенности речь будет итти в главе V.
2. ОБЩИЕ ЗАКОНЫ ТЕПЛОВОГО ИЗЛУЧЕНИЯ
Рассмотрение различных источников радиации мы начнем с темпера-
турных, или тепловых, источников излучения, к которым относятся лампы
накаливания и дуговые лампы. В основе излучения этих источников лежат
законы так называемого черного излучения [14, 48, 78, 152, 240, 261].
Основным законом всякого температурного излучения является закон
Кирхгофа, который гласит, что отношение излучательной (елГ) и поглоща-
тельной а,Т способностей, взятых для одной и той же точки тела, одного
направления излучения, одной длины волны и одной плоскости поляри-
зации. есть величина постоянная для всех тел, равная излучательной
способности абсолютно черного тела при той же температуре (Д.т):
Для интегрального излучения тела, относящегося ко всем длинам
волн, уравнение Кирхгофа имеет вид:
~ = " (17а)
Абсолютно черное тело есть такое тело, которое целиком поглощает
все падающие на пего лучи независимо от длины волны. В качестве есте-
ственных тел, приближающихся к абсолютно черному телу, можно назвать
III. Источники радиации
53
лишь немногие тела, — сажу, платиновую или висмутовую чернь, коэф-
фициенты поглощения которых равны 0,96—0,99. Поглощательная спо-
собность других тел значительно ниже. Для вольфрама она определяется
следующей эмпирической формулой:
аХ7.=х1,79-\“о’2~2,5.1О-5Т, Д8)
где Т как и всюду, есть абсолютная температура тела (в °К).
Универсальным уравнением, выражающим излучение абсолютно
черного тела, является уравнение Планка:
<1!))
ЮЛ'
в — 1
где с — скорость света, h — постоянная Планка, а— длина волны (в см),
Т—абсолютная температура; с — основание натуральных логарифмов,
равное 2,718; К — постоянная Больцмана.
К = 1,371 • эрг/град.
При введении обозначений
= 2№/г = 3,707 • 105эрг/см’2 • град
и
г/г . .....
1,433 см • град
уравнение Планка принимает более простую форму:
(19а)
Получаемое по формуле Планка излучение выражается в эрг/см3 • сек.
Для практических целей пользуются, однако, не уравнением Планка,
а его водоизменением:
/ 4,9651 \ —1
//= 142,32, (20)
где величины х и у равны:
*=7-, (21)
а
Переменные х и у в этом случае изменяются ио тем же законам, что и
переменные Е\т и л в уравнении Планка. Иными словами, кривые обоих
уравнений совершенно идентичны. Величины а,„ и Еш в каждом отдельном
случае определяются по формулам Вина.
Согласно первой формуле Вина, выражающей закон смещения мак-
симума, произведение длины волны максимума (лт) и абсолютной
температуры (71) есть величина постоянная:
(23)
где Л равно
J = 0,2884 см • град.
54
Физические и физиологические основы светокультуры растений
Из уравнения следует, что с увеличением температуры тела максимум
излучения смещается в сторону более коротких длин волн:
>„ = 0,2884-1-. (23а)
Второй закон Вина позволяет определять максимальное значение из-
лучения (Ет) абсолютно черного тела, отнесенное к единице длины волны:
Е,п = НТ\ ' (24)
где В — постоянная величина, равная
В — 1,315 • 10~4 эрг/см" • сек • град.
Фиг. 34. Распределение энергии излучения в спектре абсолютно черного тела
Графически уравнение Планка выражается изотермической кривой,
представленной на фиг. 34. Площадь, ограниченная этой кривой и осью
абсцисс, есть интегральное излучение абсолютно черного тела. Согласно
закону Стефана-Больцмана, оно пропорционально четвертой степени
абсолютной температуры
Е = .Т\ . (25)
здесь <7 — постоянная, равная
5,74 • 10-5 эрг/см2 • сек • град.
Обычно на практике пользуются не уравнением (20), а специальной
таблицей, где приводятся все практически возможные значения пере-
менных х и у (табл. 12). Подобная таблица, кривая на фиг. 34 и уравнения
Вина (23 и 24) позволяют получить полную характеристику излучения
абсолютно черного тела при любой температуре. Подобная характеристика
для некоторых температур приводится в табл. 13.
Строго говоря, перечисленные выше законы не приложимы к реальным
так называемым серым телам с поглощательной способностью менее
единицы (а,<И), хотя, несомненно, в основе их излучения лежат те же
закономерности. Па практике обычно в формулы черного излучения вно-
III. Источники радиации
55
Т а б л и ц а 12
Значения величин х и у в уравнении Планка (20)
X V X V X
0,10 4,70 • 10-15 i 1,35 0,8230 3,10 0,1257
0,15 7,91 • 10-9 1 40 0,7853 3,20 0,1141
0,20 7,37 • 10-6 1,45 0,7476 3,30 0,1038
0,25 ' 3.45 • 10-1 1.50 0,7104 3,40 0,0947
0,30 0,38 > 10-2 1,55 0,6738 3,50 0,0866
0,35 1,87 ’ 10—2 1,60 0,6380 3,60 0,0792
0,40 5,65 - IO-2 1,65 0,6038 3,70 0,0726
0,45 0,1245 1,70 0,5711 3,80 0,0667
0,50 0,2217 1 I,75 0,5398 3,90 0,0614
0,55 0,3395 ! 1,80 0,5099 4,00 0,0565
0,60 0,4663 ' 1,85 0,4820 4,50 3,83 • Ю-з
0,65 0,5908 1,90 0,4546 5,00 2,68 • Ю-з
0,70 0,7042 1,95 0,4297 6,00 1,42 • Ю-з
«0,75 0,8005 ; 2,00 0,4054 7,00 0,82 • Ю-з
о,so 0,8774 2,10 0,3614 8,00 0,50 • Ю-з
«0,85 0,9345 2,20 0,3228 9,00 0,33 • Ю-з
0,90 0,9724 2,30 0,2887 i 10,00 0,22 • Ю-з
«0,95 0,9934 2,40 0,2585 15,00 4,78 • 10—1
1,00 1,0000 2,50 0,2318 i 20,00 1,58 • 10-1
1,05 0,9944 2,60 0,2083 i 30,00 3,25 • 10-5
1,10 0,9788 2,70 0,1875 40,00 1,05 • IO—5
1,15 0,9563 2,80 0,1688 50,00 4,36 • IO-8
1,20 0,9277 2,90 0,1528
1,25 0,8951 3,00 0,1383 ।
1,30 0,8598 (
'сятся некоторые поправки, а вместо истинной температуры подставляется
так называемая цветовая температура (Тс).
Цветовая температура есть температура абсолютно черного тела, при
которой последнее имеет тот же спектр излучения, что и реальное тело
при данной температуре.
Для вольфрама, который в качестве тела накала употребляется в со-
временных лампах накаливания, формула Планка имеет вид:
Лб(е^с_ 1 )
где у, в свою очередь, равно
у =0,423 — 2,7 • 10-5Г. (27)
Оба уравнения справедливы для температур, лежащих в пределах
1000—3500° К.
Уравнение Стефана—Больцмана также видоизменяется в эмпири-
ческую формулу Гельфготта:
= —с-«7); (28)
входящая в это уравнение постоянная у. для вольфрама равняется
а=1,47-10<
56
Физические и физиологические основы светокультуры растений
Т а б л и к а 13
Характеристика излучения абсолютно черного тела ,
Абсолютная температура (в »К) Интегральное излучение (в эрг/см- • сек) Максимальное излучение (в эрг/см- • сек) Длина волны максимума (В ?.) Границы излучения *
начало излу- чения (в mu.) конец излуче- ния (в у)
273(0° С) 3,19 • 105 1,96 10.55 2430 260
300 (27° С) 4,65 • 105 3,07 9,60 2205 236
310 (37° С) 5,31 • Ю5 3,76 9,30 2140 228
320(47° С) 6,01 • 105 4,42 9,00 2007 221
373 (100° С) 1,28 • 10» 9,54 7,73 1775 190
400 1.47 • 106 13,54 7,20 1655 177
500 3,59 • 106 41,1 5,76 1360 141
600 7,46 • 106 95,6 4,80 1120 118
700 1.38 • 107 221 4,12 94 7.5 101
800 2.36 • 107 432 3.60 827.5 88,5
900 3,77 • 107 776 3,21 736 79,0
1000 5,74 • 107 1 315 2,88 692,5 70,8
1100 8,44 • 107 2 120 • 2,62 602,5 64,4
1200 1.18 • 10* 3 280 2,40 552.5 59.0
1300 1,64 10* 4 900 2 22 510 52,1
1400 2.19 • 108 7 060 2,06 4 73 50,6
1500 2,92 • 108 10 000 1,92 441 47,2
1600 3.74 • 108 13 810 1,80 ' 414 43,2
1700 4.77 • 10* 18 700 1,70 390 41.8
1800 5,99 • 10* 24 800 1,60 368 39,3
1900 7,44 • 108 32 500 1,52 348,5 37,4
2000 9,13 • Ю8 42 100 1,47 331 35,4
2100 1,11 • 106 53 700 1,37 315 33,6
2200 1,34- 109 67 750 1,31 301 32,2
2300 1,60 • 109 85 000 1,25 288 30,7
2400 1,89 • 109 104 400 1,20 276 29,5
2500 2 23— -109 1 28 200 1,15 267 28,2
2600 2,61 • 109 169 900 1,11 254,3 27,2
2700 3,03 • 109 188 100 1,07 245,5 26,3
2800 3,51 • 109 226 000 1,03 236,3 25,3
2900 4,04 • 109 270 000 0,994 228,5 24,4
3000 4,62 • 109 320 000 0,960 220,5 23,6
3100 5,31 • К)9 376 500 0,930 214 22,9
3200 6,01 • 109 4 42 000 0,900 207 22?2
3300 6,84 • 109 514 000 0,874 201 21,5
3400 7,58 • 109 597 000 0,857 197 21,1
3500 8,57 • 109 691 000 0,824 189,5 20,3
3600 9.77 • 109 792 000 0,800 184 19,7
3700 1,08 • 10™ 910 000 0,779 179,2 19,2
* За начальное и конечное излучение принято излучение в 0,01% /%.
соответствует 0,2.3, а л2 — 24,6 л/%.
III. Источники радиации
57
Характеристика излучения вольфрама приводится в табл. 14.
Таблица 14
Xарактеристика излучения вольфрама
Истинная темпе- ратура (в °К) Цветовая темпе- ратура (в °К) Интегральное излучение (в эрг/см= • сек) Длина волны максимума (в У) Световая отдача (в лм/вт)
300 — 1,5 • ИЙ ' — —
400 — 6,0 • 101 — —
500 — 1.9 • Ю3 * 5 * — —
600 — 4,8 • ]05 — —
700 — 1,05 • 10" — —
800 — 2,06 • 10» — —
900 — 3,79 • 106 — —
1000 1006 6.54 IO" 2,72 —
1100 1108 10,72 • И)'1 2,60
1200 1210 16,91 • 106 2,38 —
1300 1312 25,76 • И)" 2,20 —
1400 1414 3.82 • 107 2,04 0,09
1500 1517 5,55 • 107 1,90 0,20
1600 1619 7,77 • 107 1,78 0,40
1700 1722 10,59 • 107 1,67 0.71
1800 1825 14,22 • 107 1,58 1,16
1900 1929 18,55 • Н)7 1,49 1,88
2000 2033 23,72 • 107 1,42 2,78
2100 2137 29.82 • И)7 1,35 3,95
2200 2242 37.18 • 107 1,28 5,47
2300 2347 4.59 • 106 1,23 7,25
2400 2452 5.58 • 106 1,18 9,37
2500 2557 6,76 • 108 1,13 11,67
2600 2663 8,08 • 108 1,08 14,28
2700 2770 9,62 • 108 1,04 17,26
2800 2878 11,29—108 1,00 20,43
2900 2986 13,21—108 0,965 23,80
3000 3094 15,39—108 0,930 27,10
3100 3202 17.75—108 0,900 31,0
3200 3311 2,03—109 0,872 34,6
3300 3422 2,32—10® 0,842 38,5
3400 3533 2,64—109 0,816 42,6
3500 3646 3,00—109 0,781 45,9
3655 3817 3,60—10» 0,756 53,1
Табл. 14 показывает, что как и у абсолютно черного тела, вместе’
с увеличением истинной температуры вольфрама возрастает как интеграль-
ное излучение, так и световая отдача. Одновременно перемещается макси-
мум излучения в сторону более коротких длин волн и увеличивается
разрыв между истинной и цветовой температурой.
3. ЛАМПЫ НАКАЛИВАНИЯ
Наиболее простыми и распространенными источниками радиации
являются лампы накаливания [9, 28, 48, 54, 69, 72, 78, 98, 203, 298, 321].
К этим лампам как температурным источникам излучения приложимы
58
Физические и физиологические основы светокультуры растений
законы черного излучения с теми оговорками, которые отмечались выше.
В качестве излучающего тела у современных ламп накаливания употреб-
ляется вольфрам, раскаляемый в результате прохождения электрического
тока до температуры 2355—3350° К. Более высокая температура воль-
'фрамовой нити невозможна из-за распыления и размягчения металла,
температура плавления которого лежит при 3655° К.
Колбы обычных ламп накаливания изготовляются из стекла «Флинт-
глясс», которое прп толщине в 0,8 мм пропускает лучистую энергию, как
это показано на фиг. 35, 4. Там же приводятся кривые пропускания увиоле-
<вого стекла, стекла «витаглясс» и кварца, применяемых для изготовления
Фиг. 35. Пропускание лучистой энергии разными стеклами, водой и кварцем
1 — стеклом «новиоль-О»; 2 — оранжерейным стеклом; в — оконным стеклом;
4 — ламповым стеклом («флпптглясс») 0,8 мм; 5 — стеклом «витаглясс»; 6 — увпо-
левым стеклом; 7 — кварцем; 8 — водой
некоторых газосветных ламп и ламп накаливания. Как видно из приведен-
ных кривых, лампы накаливания в лучшем случае, прп увиолевой колбе,
имеют границу излучения около 280 ту. В лампах из обыкновенного
стекла эта граница находится около 300—310 ту.
Как показывает фиг. 36, лампы накаливания являются источником
главным образом инфракрасных лучей с максимумом излучения в зави-
симости от мощности при 875—1210 ту. По мере увеличения мощности
ламп, максимум смещается в сторону более коротких волн. Сопоставляя
кривые фиг. 36 и фиг. 3, легко убедиться, что максимум излучения прихо-
дится на область, наименее поглощаемую тканями листа. Лишь у ламп
с небольшой номинальной мощностью этот максимум смещается в сторону
больших длин волн, поглощаемых водою, содержащейся в тканях листа.
Инфракрасная радиация в спектре ламп накаливания составляет 80—
90% всего излучения, достигая наибольшего значения (91%) у мало-
мощных пустотных ламп и снижаясь до 81 % у проекционных. У обычных
осветительных ламп мощностью в 100—1000 вт она колеблется в преде-
лах 85—88%.
Физиологическая радиация в спектре ламп накаливания составляет
10—20% общего излучения и представлена в основном оранжево-крас-
ными лучами. Относительно возрастая , при увеличении мощности ламп,
III. Источники радиации
59
она достигает у осветительных ламп в 100—1000 вт 11,5—15,5% общего
лучистого потока. Лампы накаливания бедны спне-фполетовыми лучами
Фиг. 36. Распределение энергии в спектре излучения ламп накаливания
1 — вакуумная лампа 10 вт; 2 — то же 60 вт; з — газополная лампа 50 вт;
/ — то же 2 квт; 5 — кинопроекционная лампа 10 квт
и почти не содержат ультрафиолетовых лучей (фиг. 37, табл. 15). Послед-
ние в лучшем случае, у проекционных ламп, достигают 3% физиологи-
1 — пустотная лампа 10 вт; .? — то же 60 вт; 3 — газополная лампа 50 вт; 4 — то же,
100 вт; 5 — то же, 300 ит; в — то же 2 квт; 7 — кинопроекционная лампа 10 квт
ческой радиации, или 0,5% всего излучения, а у маломощных пустотных
ламп снижаются до 0,5% физиологической радиации, или 0,1 % общего
излучения. Обычные лампы в 100—1000 вт содержат ультрафиолетовых
лучей от 0,6 до 1,5% физиологического излучения.
60
Физические и физиологические основы светокультуры растений
Таблица 15
Спектральный состав физиологической радиации ламп накаливания в 300 вт
Лучи
ультра- фиолето- вые фиолето- вые синие зеленые желтые оранжевые красные
Длина волны В ПЦХ 300—400 400—4 40 440—490 490—565 565—595 595—620 620—800
Доля излучения (в % ко всей физиологиче- ской радиации) 1Д 1,8 4,6 14,0 7,9 8,0 62,6
Мощность ламп мало влияет на распределение энергии в спектре
физиологической радиации. По мере увеличения мощности растет лишь абсо-
лютная и относительная величина физиологической радиации, а граница
излучения несколько смещается в сторону более коротких ультрафиоле-
товых лучей.
Физическая характеристика обычных осветительных ламп приводится
в табл. 16, которая показывает, что с увеличением номинальной мощности
ламп растет как истинная, так и цветовая температура нити. Последнее,
в свою очередь, налагает отпечаток на все остальные характеристики ламп,
изменение которых было отмечено выше.
Листья растений, находящихся под лампами накаливания, поглощают
60—65% падающей на них физиологической радиации (табл. 48). В основ-
ном это оранжево-красные лучи. Основная часть поглощения приходится
Физическая характера
(Оден
Лампы Мощность (В ВТ) Температура нити (в СК) Излуче
истинная цветовая общее (в вт/см2)
10 — 2390 49,9
25 — 2493 60,4
Пустотные ' 40 2445 2500 62,0
60 — 2509 64,0
50 — 2670 81,9
75 2640 2705 89,2
100 2670 2740 94,6
200 2740 2810 106,0
Газополные 300 2765 2810 111,5
500 2840 2920 126,6
1000 2895 2980 138,9
2000 2915 3000 143,4
10000 3190 3300 221,6
Проекционные I 30000 3190 3300 221,6
Ill. Источники радиации
61
на хлорофилл, который поглощает 54—57% падающей на лист физиоло-
гической радиации. Характеристика физиологической радиации, по-
глощаемой пигментами листа, показана на фиг. 38.
Фиг. 38. Поглощение лучистой энергии пигментами листа
под лампами накаливания
7— всеми пигментами (хлорофплл-F каротиноиды); 2 — хлорофиллом; .?— 1;о-
лпчество квантов, поглощаемое хлорофиллом
Лампы накаливания, выпускаемые в СССР, отличаются большим
многообразием. Для культуры растений следует рекомендовать прежде
стика ламп накаливания
[646])
Таблица 16
ние нити 1 Максимум излучения Лучистый поток Используемая энергия (в % к по- требляемой)
видимое Длина волны максимума (в и) максимальная!
В ВТ; см2 % I общему спектральная общий интенсивность' (и вт) (в вт/см2-А) | 1 видимый (В ВТ) общая ! | видимая
— .— 1,210 1,03 • 10-2 — 0,65 — i
— — 1,160 1,26 • IO-2 — 1,97 — i -
о.З о 8,7 1,155 1,28 • 10-2 ! 38,35 3,32 95,9 ! 8,3
— — 1,150 1,31 • 10-2 I 4,01 — 1
— — 1,108 1,74 • 10-2 3,30 — —
10,15 1 1,4 1,107 1,93 • 10-2 57,00 6,50 76,0 8,7
11,20 11,9 1,055 2,00 • К)-2 76,70 9,05 76,7 9,1
13,65 12,8 1,030 2,30 • 10-2 164,50 21,15 82,3 10,6
14,75 13,2 1,020 2,42 • IO-2 253,00 33,41 84,4 11,1
18,15 14,3 0,990 2,80 • 10-- 416,50 59,60 83,4 11,9
21,10 15,2 0,970 3,06 • 10~2 820,40 129,00 82,4 12.9
22,15 15,4 0,964 3,20 • IO-2 1750,00 270,00 87,5 13,5
42,65 19,3 0,875 5,14 • IO-2 9061,00 1740,00 90,6 17,5
42,65 1 19,3 0,875 5,14 • 10-2 27183,00 5220,00 90,6 17,5
62
Физические и физиологические основы светокультуры растений
всего обычные стандартные осветительные лампы, со сроком службы
в 1000 часов, рассчитанные на напряжение в 120 и 220 в (табл. 17).
Т аблпца 17
Светотехническая характеристика нормальных осветительных ламп
(ОСТ 5154)
Лампы Номинальная МОЩНОСТЬ (в ВТ) Световой поток (в лм) Световая отдача (в лм/вт) Средняя сфери- ческая сила света (в св) Удельная мощ- ность (в вт св) Итенсивность физиологиче- ской радиации в эрг,см сек на расстоянии (в см)
100 50 25
10 66 6,60 5,2 1,93 44 176 704
15 124 8,25 9,9 1,52 82 328 1312
25 225 9,00 17,9 1,40 144 576 2304
40 380 9,50 30,3 1,32 218 872 3488
60 645 10,93 51,4 1,17 336 1344 5376
Стандартные 75 881 12,07 70 1,07 441 1764 7056
лампы 110, 120 100 1275 13,01 101 0,99 620 2480 9’220
и 127 в 150 2175 14,80 173 0,87 1050 4200 16800
200 3050 15,64 243 0,82 1450 5800 23200
300 4875 16,58 388 0,77 2270 9080 36320
500 8725 17,81 695 0,72 3940 13760 55040
750 13690 18,50 1090 0,69 6100 24400 97600
1000 19000 19,19 1513 0,66 8400 33600 134400
15 95 6,33 7,5 2,00 62 248 992
25 191 7,65 15,2 1,64 122 188 1952
40 336 8,40 26,8 1,49 185 740 2960
60 540 9,00 43,0 1,40 275 1100 4400
75 671 9,19 53,5 1,40 3 3 / 1348 5792
Стандартные 100 1000 10,42 79,6 1,26 495 1980 7220
лампы 220 в 150 1710 13,87 136 1,10 725 2900 11600
200 2510 13,07 200 1,00 1200 4800 19200
300 4100 14,24 326 0,92 1900 7600 .30400
500 7560 15,59 602 0,83 3510 14040 56160
750 12230 16,80 974 0,77 5450 21800 87200
1000 17200 17,73 1370 0,73 7500 30000 120000
55 650 11,80 51,6 1,06 324 1296 5184
Бисниральныс 71 900 12,7 71,6 0,99 440 1760 7040
лампы 127 в 96 1300 13,5 103,0 0,93 630 2520 10080
Биспиральные J 82 82 11,0 71,6 1,15 440 1760 7040
лампы 220 в | 109 109 11,9 103,0 1,07 630 2520 10080
Маломощные лампы в 10—40 вт относятся к пустотным, или вакуум-
ным, а остальные (60—1000 вт) — к газополным лампам, наполняемым
смесью аргона (86%) и азота (14%). Газонаполненность или пустотность
ламп не влияет на их спектральный состав излучения.
Для выращивания растений пригодны и другие типы ламп со сроком
службы в 1000 часов — судовые, трамвайные и цилиндрические (табл.18).
III. Источники радиации
63
Табл it ц а 18
Светотехническая характеристика ламп накаливания специального назначения
(Б. JJ. Балашпнскип и др. [9])
Лампы Номинальная мощность (в ВТ) 1 Световой поток (в ЛМ) Световая отдача (в лм,вт) Средняя сфери- ческая сила света (в св) Удельная мощ- ность (в вт.'св) Интенсивность физиологиче- ской радиации в эрг/см • сек на расстоянии (в см)
100 50 25
Судовые 110 в 25 180 7,2 14,3 1,75 115 460 1840
40 304 7,6 24,2 1,65 174 696 2784
60 516 8.6 41,0 1,46 274 1096 4384"
Судовые 220 в 25 153 6.1 12,2 2,04 97 388 1552
40 263 6.7 21,0 1,90 151 604 2416
60 432 7.2 35,2 1,71 267 1068 4272’
Трамвайные 120 в 60 390 (>.5 — — .. —
Цилиндрические 15 105 7.0 — —
110 п 127 в 25 190 7.6 — — — — —-
То же 220 в 25 157 6,3 — — —
Т а б л и и а 19
Светотехническая характеристика софитных ламп 12ч, 127
и 220 в.
(Б. Л. Балашпнскпй и др. [9])
Мощность ламп (в вт) Световой поток (в лм) Световая отдача (в лм/вт) Горизонталь- ная сила света (в св)
15 109 7,3 10,9
25 187 / .') 18,7
40 312 7,8 31,2
Т а б л и ц а 20*
Светотехническая характеристика зеркальных ламп
(Б. Л. Балашпнскпй и др. [9])
Заводской шифр ламп Номинальное напряжение (в в) Номинальная мощность (в ВТ) Световой поток (в лм) Световая отдача (в лм/вт)
«ЗН-1» 127 200 2700 13,5 •
«ЗН-2» 127 300 4300 14,3
«зп-з» 220 200 2100 11,0
«ЗН-4» 220 300 3600 12,0
Отличаясь малой мощностью (15—50 вт) и сравнительно небольшой
световой отдачей (6—8,5 лм/вт), они менее экономичны, чем обычные
осветительные лампы большой мощности (100—1000 вт), световая отдача
64
Физические и физио логические основы светокультуры растений
Светотехническая характера
(Н. 11. Ермо
Лампы Мощность (в ВТ) Световой поток (в лм) при напряже- нии (в вольтах) Световая отдача (в лм/вт) при напря- жении (в вольтах)
ПО—127 220 110 — 127 220
Концентрированного свегораспрсделе-
ния 300 500 4000 7800 3700 6800 14,6 15,6 12,4 13,6
Преимущественно направленного сне-
тораспределения 200 2700 2200 13,5 11,0
300 4300 3600 14,0 12,0
Широкого светораснределения .... «Кососвет» концентрированного свето- 300 4400 3700 ! 14,6 12,4
распределения 300 1 4400 3700 14,6 12,4
j 500 ( 7800 6800 1 i i 15,6 i 13,6
которых колеблется в пределах 10—19лм/вт. Судовые, трамвайные и цилин-
дрические лампы можно поэтому рекомендовать лишь для небольших
опытов в лабораторных условиях или в тех случаях, когда требуется
сравнительно небольшая интенсивность радиации, например при удлине-
нии естественного дня с целью вызвать фотопериодический эффект.
Сказанное относится также к софитным лампам с нитью накала, рас-
положенной вдоль цилиндрической трубки лампы. Эти лампы изготовля-
ются в виде стеклянной трубки длиною в 25 см и с диаметром в 3 см: длина
инти — 16,5 см. Номинальный срок службы ламп — 1000 часов (табл. 19).
Исключительного внимания заслуживают зеркальные лампы, колбы
которых в верхней части покрыты изнутри зеркальным слоем (табл. 20
и 21). Поэтому лучистый поток ламп целиком направляется в ниж-
нюю полусферу (фиг. 73). Эти лампы не нуждаются в дополнительной
осветительной арматуре, обычно сильно затеняющей растения в тепли-
цах в дневные часы.
Б. С. Мошков [216] рекомендует для выращивания растений мало-
мощные низковольтные лампы с последовательным включением в обыч-
ную осветительную сеть с напряжением в 120 или 220 в. По его данным,
обычные автомобильные лампы, работающие на небольшом недокале*,
дают очень хороший эффект. Характеристика разных низковольтных ламп
приведена в табл. 22 и 23.
Невидимому, предпочтение следует отдать судовым, железнодорожным
и другим низковольтным лампам, имеющим большой срок службы и об-
ладающим большей световой отдачей, чем автомобильные стоп-лампы.
По спектральному составу физиологической радиации низковольтные
лампы не имеют преимуществ перед обычными осветительными лампами
накаливания.
В кратковременных лабораторных опытах, особенно в случае необ-
ходимости получения мощного пучка лучистой энергии, следует обра-
щаться к проекционным и прожекторным лампам, отличающимся большой
* В этом случае лампы могут служить около 500 часов.
III. Источники радиации
65
стика зеркальных ламп ВЭИ
линский [72])
Таблица 21
Коэффициент полезного действия Коэффициент усиления Угол макси- мальной силы света (в °) Защитный угол (в °) Размеры колбы (в см) Срок службы (в час.)
диаметр длина
90 9,5 15 30 17,8 26,5—27,0 800
90 9,5 15 30 17,8 26,5—27,0 800
89 4,5 25 30 12,0 23,0—25,5 1000
89 4,5 25 30 12,0 23,0—25,5 1000
91 4,0 70 12 13,2 25 1000
90 10 10 0—40 17,5 26,5—27,0 800
90 10 10 0—40 17,5 26,5—27,5 800
Таблица 22
Светотехническая характеристика автомобильных ламп *
(А. С. Соколов [298])
Заводской шифр ламп Номинальное напряжение (в вольтах) Расчетное напряжение (в вольтах) Мощность (в ВТ) Световой поток (в лм) Световая отдача (в лм/вт) Средняя сфе- рическая сила света (в св) Освещен- ность на рас- стоянии 10 см (в лк) Срок службы (в час.)
«А-16» 6 7,5 1,88 12,6 6,70 1 100 500
«А-32» 6 6,8 3,00 25,5 8,50 2 200 75
«А-15» 6 6,8 3,00 25,0 8,30 2 200 100
«А-19» 6 7,0 3,50 25,1 7,15 2 200 300
«А-1» 6 6,8 4,50 38,0 8,40 3 300 100
«А-17» 6 7,0 4,83 37,7 7,80 3 300 300
«А-18» 6 6,5 7,53 75,4 9,75 6 600 300
«А-5» 6 6,8 11,50 125 10,50 10 1000 100
«А-3» 6 6,8 14,30 190 13,30 15 1500 125
«А-4» 6 6,5 18,90 265 14,00 21 2100 105
«А-20» 6 6,4 21,30 264 12,40 21 2100 150
«А-22» 12 14,4 2,09 12,6 6,00 1 100 500
«А-23» 12 14,0 3,18 18,8 5,92 1.5 150 500
«А-9» 12 13,5 4,50 38,0 8,40 3 300 100
«А-24» 12 13,0 5,90 37,7 6,40 3 300 500
«А-25» 12 13,0 8,25 75,4 9,13 6 600 500
«А-10» 12 13,5 14,5 190 13,3 15 1500 125
«А-20» 12 12,8 18,6 264 14,2 21 2100 300
« А-11» 12 12,5 18,9 265 14,0 21 2100 125
поток, сила света,
срок службы и
* Все параметры — мощность, световой
другие даются для расчетного напряжения.
5 А. Ф. Клешнин
66
Физические и физиологические основы светокультуры растений
Таблица 23
Светотехническая характеристика низковолътовых ламп, накаливания
(Б. Л. Балашинский и др. {9])
Лампы Заводской шифр ламп Номинальное напряжение (в вольтах) Номиналь- ная мощ- ность (в ВТ) Световой поток (в лм) Световая отдача (в лм/вт) ОЗ —( X Н К е со н С_) и s W &G3 СО £ « ч ° S 1 Освещен- ность на рас- стоянии 10 см (в лк) Срок служ- бы (в час)
«МО-9» 12 11 100 9,1 7,95 795
«МО-10» 12 20 200 10,0 15,90 1590
Местного «МО-11» 12 40 500 12,5 39,80 3980
освещения «МО-12» 36 14 100 7,1 7,95 795 1000
«МО-13» 36 25 200 8,0 15,90 1590
«МО-14» 36 50 500 10,0 39,80 3980
«Ж-10» 50 10 70 7,0 7,56 756
«Ж-5» 50 15 120 8,2 9,80 980
Железно- «Ж-7» 50 25 220 8,8 17,50 1750 1000
дорожные «Ж-8» 50 50 550 11,0 43,70 4370
«Ж-1» 24 10 70 7,0 7,56 756
«ж-з» 24 25 250 10,0 19,90 1960 800
«С-12» 24 25 237 9,5 18 85 1885
«С-13» 24 40 440 11,0 35,00 3500
«С-14» 24 60 720 12,0 57,30 5730
«С-15» 24 100 1400 14,0 111,20 11120
Судовые «С-16» 24 150 2200 16,0 175,00 17500 1000
«С-1» 55 25 198 7,9 15,75 1575
«С-2» 55 40 332 8,3 26,40 2640
«С-3» 55 60 564 9,4 44,85 4485
«СМ-11» 13 5 40 8,0 3,18 318 j
«СМ-12» 13 10 105 10,5 8,35 835
«СМ-13» 13 15 180 12,0 14,40 1440 125
«СМ-17» 13 25 362 14,5 28,80 2880
Самолетные «СМ-14» 26 5 40 8,0 3,18 318
«СМ-15» 26 10 80 8,0 6,36 636 )
«СМ-10» 26 15 142 9,5 11,30 1130 «. 100 J
«СМ-18» 26 25 362 14,5 28,80 2880 125
( «К-3» 5 35 77 2,2 5,72 572 500
Кинопроек- I «К-7» 12 30 528 17,6 42,00 4209 50
ционные । 1 «К-9» 12 50 1С0Э 20,0 79,60 7960 50
( «ПЖ-1» 12 50 1000 20.0 79,60 7960 50
Прожектор- 1 «ПЖ-8» 23 100 2050 20,5 163,00 16300 30
ные । 1 «ПЖ-10» 50 25 250 10,0 19,90 1990 300
световой отдачей и содержащим относительно больше сине-фиолетовых
и ультрафиолетовых лучей. Эти лампы могут работать лишь в строго
фиксированном положении — вертикальном или горизонтальном, откло-
нение от которого допускается не более, чем на 15° (табл. 24).
В целях обогащения ламп накаливания ультрафиолетовыми лучамж
колбы ламп изготовляют из увиолевого стекла, пропускающего более
III. Источники, радиации
67
Таблица 24
Светотехническая характеристика проекционных и прожекторных ламп накаливания
(Б. Л. Балашинский и др. [9])
Тип ламп Заводской шифр ламп Номи- нальное напряже- ние (в вольтах) Номи- нальная мощность (в ВТ) Световой поток (в лм) Световая отдача (в лм/вт) Срок службы (в час)
«К-12» 110 300 6450 21,5 50
Кинопроек- «К-18» 127 300 6450 21,5 50
ционные «К-14» * 110 500 11000 22,0 50
«К-15» * 110 750 17250 23,0 50
«ПЖ-13» 110 500 10500 21,0 100
«ПЖ-14» 110 1С0Э 22200 22,2 100
«ПЖ-20» 220 500 9800 19,6 100
Прожекторные «ПЖ-21» 220 1000 21000 21,0 100
«ПЖ-11»** 50 4000 4000 16,0 500 .
«ПЖ-23» 50 11000 11000 22,0 100
«ПЖ-15» 110 1500 34500 23,0 100
«ПЖ-16» 110 2000 47400 29,7 100
Мощные про- «ПЖ-17» 110 3000 72300 24,1 100
жекторные «ПЖ-20» 110 3500 94500 27,0 100
«ПЖ-18» 110 5000 127500 25,5 100
«ПЖ-19» 110 10000 280000 28,0 100
* Лампы эксплуатируются в вертикальном положении цоколем вверх.
* * Лампа эксплуатируется в горизонтальном положении.
Остальные — только в вертикальном положении цоколем вниз.
короткие ультрафиолетовые лучи, чем лампы из обычного стекла (фиг. 35
и табл. 25).
Таблица 25
Светотехническая характеристика ламп накаливания
в увиолевой колбе
(А. П. Иванов [78])
(Напряжение 28—32 и 110—120 в; срок службы — 500 часов)
Мощность ламп (в вт) Световой поток (в лм) Световая отдача (в лм/вт) Сила тока (в а) при напряжении в 110—120 в
60 785 13,1 0,5
300 6000 20,0 2,6
500 10750 21,5 4,3
Специального интереса заслуживают лампы-термоизлучатели («ЗС-2»,
«ЗС-1» и «ЗС-З»), излучение которых целиком сосредоточено в инфра-
красных лучах с максимумом в пределах 0,75—3 у. Лампы изготовляются
мощностью в 250 и 500 вт и рассчитаны на напряжение в 127 и 220 в.
Продолжительность службы —• 2000 час. Колбы ламп имеют параболи-
ческую форму и в верхней части содержат зеркальное покрытие, позво-
ляющее обходиться без осветителей.
68 Физические и физиологические основы светокультуры растений
4. ЭЛЕКТРОДОСВЕТНЫЕ ЛАМПЫ
К лампам накаливания примыкает другая обширная группа источ-
ников радиации, которые также относятся в основном к температурным
источникам излучения. Мы имеем в виду электродосветные или дуго-
вые лампы [48, 79, 169]. Ограничимся рассмотрением некоторых дуговых
ламп с разрядом, совершающимся в атмосфере инертных газов — азота,
аргона или неона и паров ртути, а также некоторых дуговых ламп с раз-
рядом в атмосферном воздухе.
В современных электродосветных лампах применяются вольфрамовые
электроды, изготовляемые в виде двух небольших шариков. В зависи-
мости от типа ламп рабочая температура электродов колеблется в пре-
делах 3100—3400° К, что соответствует цветовой температуре порядка
3200—3500° К. Высокая температура электродов определяет положение
максимума излучения, который по сравнению с лампами накаливания
смещается влево—в область 800—900 ту. Это означает обогащение спектра
ультрафиолетовыми лучами. В остальном по характеру физиологической
радиации дуговые лампы не отличаются от ламп накаливания. Физи-
ческие и светотехнические показатели некоторых дуговых ламп перемен-
ного тока приведены в табл. 26.
Таблица 26
С термоэлектронным
ионизатором * . .
— 500—1OW
1030 300—400
1280 300—400
1530 300—400
1750 300—400
1930 300—400
_ । —
i
С ионизатором тлею-
щего разряда фир-
мы Филипс
С контактным зажи-
ганием
120 120—220 55,0 1 2,2 >1880 150 0,800
51,5 120—220 34,0 1,5 — — 59 0,870
57,0 120—220 28,5 2,0 — — 73 0,780
62,5 120—220 25,0 2,5 — — 87 0,720
67,5 120—220 22,5 3,0 — — 99 0,680
73,0 120—220 20,5 3,5 — — 110 0,665
— 120—220 50—60 1,3 400 • 40 —
120—220 50—60 2,5 750 — 75 —
горизонтальное распределение силы света для этой лампы
* Вертикальное и
приведено на фиг. 33.
В отличие от ламп накаливания, дуговые лампы имеют отрицатель-
ную вольтамперную характеристику; при включении их в сеть необходимо
балластное сопротивление, которое в виде обычного сопротивления или
дросселя включается в сеть последовательно.
Лампы накаливания и дуговые лампы, даже те их типы, которые до-
стигают максимально возможной цветовой температуры, все же бедны
коротковолновыми ультрафиолетовыми и сине-фиолетовыми лучами. Для
устранения этого недостатка оба типа ламп комбинируются с ртутными
лампами [79]. Такими источниками смешанного излучения являются
III. Источники радиации
69
комбинированные лампы («S-l», «S-2» и «HWA-500»).
Лампы «S-1» и «S-2», или «солнечные лампы», представляют собой
комбинацию дуговых ламп с вольфрамовыми электродами с ртутными
лампами высокого давления и лампами накаливания. Поток физиоло-
гической радиации, испускаемый лампой «S-1», в основном излучается
электродами (84%) и парами ртути (13,5%); вольфрамовая нить излу-
чает лишь 2,5%. Колбы ламп изготовляют из тугоплавкого стекла, про-
пускающего ультрафиолетовые лучи; эти лампы применяют в физио-
терапии (табл. 27).
Таблица 27
Светотехническая характеристика дуговых вольфрамовых ламп с электродами
в атмосфере ртутных паров
(А. П. Иванов [79])
Лампы
«S-2»
«S-1»
Лампы
«S-2» .
«S-Ь .
Мощность
(в вт)
и
si
Сила тока
(в а)
S S
О. и
в и
я
о
Я
ЕС
л
И
Световой поток (в лм)
Световая от-
fl ача (в лм/вт)
I I
130 175 — — | 15 I 3
315 450 0,21 0,54,10—11' 10
I I
Лучистый поток
8,5
30,0 4900
1000
100
1600
1000
12,5
18,3
и
й Qi
физиологиче-
ской радиации
витапидной
радиации
Н
£ ss'g
о як
Н
инфракрасной
радиации
Температура (в ° К)
3260 2330 285
38,79 16,20
0,08 I 0.03
’ I
201,131 83,77
I
240 : 8,65
i
300
s
й
Солнечные лампы типа «МЭЛЗ» или «HWA-500» состоят из лампы
накаливания и ртутной лампы сверхвысокого давления, заключенных
в колбу из матированного стекла. Спектр ламп показан на фиг. 39, а ха-
рактеристика — в табл. 28.
Из многочисленных типов вольтовых дуг для культуры растений при-
годны лишь дуги закрытого и открытого горения с чистыми и пламен-
ными углями, работающие на переменном токе. Электроды первых по-
мещают в стеклянные колбы, электроды вторых остаются открытыми.
Пламенными углями называются угли, содержащие различные химиче-
ские примеси. Для получения белого света примешивают CaF3 и CaF4,
для желтого света — CaF., для красного — SrF3 и т. д. Добавление же-
леза обогащает спектр лучами, лежащими в пределах 230—320 ту, до-
бавление Fe, Ni и Si — лучами с длиною волны 270—320 ту, а добав-
ление Со — лучами от 310 до 350 ту. Дуги интенсивного горения, так же
как и дуги постоянного тока, непригодны для светокультуры растений
из-за технических трудностей эксплуатации. Характеристика дуговых
,70 Физические и физиологические основы светокультуры растений
ламп приводится в табл. 29, а распределение энергии в спектре — на
фиг. 40. Спектр дуги высокоинтенсивного горения представлен на фиг. 94.
Для вольтовых дуг, независимо от их типа, необходимо балластное
сопротивление, которое потребляет 15—30% мощности, и специальный
регулятор, посредством которого поддерживается постоянное расстояние
между углями.
Фиг. 39. Распределение энергии в спектре излучения солнечной лампы «HWA-500»
Вольтова дуга представляет собою источник температурного типа, ра-
бочая температура которого при давлении паров в 1 атм колеблется
Таблица 28
Светотехническая характеристика солнечных ламп
Лампы Мощность (в ВТ) Напряжение (в вольтах) Сила тока (в а) Время раз- горания (в- мин.)
спирали накала разряд- ной трубки лампы в целом на спи- рали накала на раз- рядной трубке на лампе при за- жигании стацио- нарная при за- жигании
«мэлз» . . . — — 300 — -— 220 180-220 1,35—1,40 1,70 4,0
«HWA-500» . . 185 70 255 156 78 220 170 1,19 1,40 4,5
Лампы Световой поток (в лм) Световая отдача (в лм, вт) Лучистый поток Физиологическая радиация (в % к подводимой мощности)
спирали накала разрядной трубки общий физиологиче- ской радиации инфракрасной радиации общий (в вт)
В ВТ в % к общему В ВТ в % к общему
«МЭЛЗ» . . . — — 5500 18,5 — — — — — —-
«HWA-500» . 2700 2800 5500 21,6 26,0 16,8 129,0 83,2 155,0 10,2
Фиг. 40. Распределение энергии в спектре излучения вольтовых дуг
Фиг. 41. Характеристика открытой вольтовой дуги с чистыми углями
1 — распределение квантов в спектре; 2 — поглощение квантов «пигментами
пластид; 3 — поглощение квантов хлорофиллом
Длина Пали/;/ (в т/а)
Фиг. 42. Характеристика открытой вольтовой дуги с пламенными
углями желтого света
1 — распределение квантов в спектре; 2 — поглощение квантов пигментами
пластид; 3— поглощение квантов хлорофиллом
Таблица 29
Светотехническая характеристика дуговых ламп с разрядом в атмосферном воздухе, работающих на переменном токе
Дуги Мощность вместе с бал- ластным сопротивле- нием (в вт) Напря- жение на дуге (в воль- тах) Сила тока (в а) Световой поток (в лм) Световая отдача (в лм/вт) Полусфериче- ская сила света (в св) Удельная мощность (в вт/св) Продолжи тельность горения углей (в час)
С чистыми углями
Открытые
с вертикальными углями 200—1200 27—33 6—25 680— 8300 3,4—7,0 90—1100 2,2 —1,1 10—18
с наклонными углями . . , . . . . 300— 580 40—50 8—12 1325— 2650 4,0—4,6 200— 400 1,65—1,45 8-20
Закрытые
с вертикальными углями 1400—2650 40—50 6—-15 3700— 9700 2,65—3,65 450—1200 3,1 —2,2 100—200
с наклонными углями 1700—2200 40—50 8—12 3700— 6650 2,15—2,2 550—1000 3,1 —2,2 20— 35
экономичные 485—1900 40—50 3— 8 1800— 9600 3,7 —5,0 270—1450 1,8 —1,3 10— 22
С ила ме н н ы ми угля м и
Открытые
с вертикальными углями 270— 750 28—24 8—15 5250—20400 19,5—27,2 700—2700 0,39—0,28 10— 22
с наклонными углями 270— 750 — — 4650—18000 17,2—24,0 700—2700 0,39—0,28 —
Закрытые
с вертикальными углями 310— 840 40—60 8—15 6500—24500 21,0—29,2 3000 0,39—0,28 80—120
с наклонными углями 310— 840 40—60 8—15 5300—20000 17,0—24,0 •— 0,39—0,28 80—120
Титанкарбидная 245—350 60—70 3—4,5 5100— 9000 20,8—25,8 625—1100 0,39—0,32 50— 75
Физические и физиологические основы светокультуры растений
Распределение энергии в
(А. П. Ив
Лампы Мощность (в ВТ) Давление паров ртути (в атм) Единицы измерения интенсив- ности линий
витацидной радиации
237,8 240,0 248,3 253,7 265,2 269,9 275,3 280,4 289,4 29
Низкого давления «СН» 270 0,0003 Эрг./см2 сек.* — .
Высокого давления «11-5» 250 О,4 То же > — — — — — — — —. —
«Н-2» 250 0,5 » — — — — — — — — —
«11-1» 400 1,0 » » — — — — — -— — — —
«11РК-4» 70 — » » 9,3 9,3 32,8 91 105,6 22,3 20,4 52,3 30,0
«ПРК-2» 375 » » 11,0 11,0 51,0 180 185,0 36,6 31,1 95,2 63,5
«ПРК-7» 1000 — » » 117,1 117,1 370,0 G72 1073,0 320,0 210,0 655,0 281,5
Сверхвысокого давления «СВД-125» 125 25 » » — — — — —
«СВДШ-250» 250 30-35 » » — — — — — — — — —
«н-з» 85 30 » » — — — — — — — — —
«Н-4» 100 8 » » — — — — — — — — —
«HgH-ЮОО» 265 — % * * — — — — — — — — —
«HgQS-500» 220 — То же — — — — — — — — —
«HgHS-8000» 1500 — » » — — — — — — .— — —
«HgQS-ЗОО» 75 — » » — — — — 4,0 — 1,5 3,0 3,0
«HgQ-ЗОО» 75 — » » — — — —. — — — — —
«HgQS-500 Bl»' 120 — » * «и и» — — —. — — — — —
«HgQS-500» 120 — » si» — — — — — — — — 8,0
* На расстоянии 1 м.
** В % к интенсивности линии 546,1 тр.
*** В % к интенсивности линии 365,0 тр.
**** В % к интенсивности линии 312,6 тр.
’ анергии в спектрах ртутных ламп
(А. П, Иванова [79])
Таблица 30
Длина волны линий (в т;1)
физиологической радиации инфракрасной радиации
289,4 292,5 296,7 302,2 312,6 334,1 365,0 390,6 398,4 404,7 407,8 435,6 491,6 546,1 577,0 690,7 1014,0 1128,7 1367,3 1529,5 1711,0
— — — — — — 300 — — 590 — 820 — 810 263 — — — — — —
— — —- -— — — 700 -— -— 382 — 720 43 905 16'40 — — — —. —
— — — — •— —. 225 — — 236 —. 445 15 575 550 — — — — — —
— — — — — — 410 —, — 445 — 920 30 1310 1270 — •— — — — —
30,0 — 73,6 137 329 40,6 516 8,7 11,8 175,5 — 292 ' 9,6 310 348 5,0 124,0 73,4 75,9 28,8 43,4
2 63,5 — 136,0 269 604 92,5 1040 29,0 34,5 382,0 — 606 17,9 690 800 8,9 277,5 158,6 164,2 62,0 109,0
3 281,5 — 782,5 1725 3270 542,5 5740 93,0 138,4 1749,0 — 2800 162,0 3000 4470 93,0 1780,0 1005,0 1023,0 540,0 669,0
. — — — 278 — — 135 263 11,5 370 225 28,9 145,0 104,0 129,5 100,0 102,0
— -— —- — .— —. 695 — — 457 657,0 102,0 850 838 174,2 645,0 445,0 417,0 — —-
— — —• — — — 257 — — 145,0 — 271,0 — 335 192 — — — — — —
— —- — — — — 245 — — 142,0 — 269,0 16,0 336 305 — — — — — —
— .— — — — — 3 — — 18,0 3,0 55,0 °,4 100 87 1,7 24,0 — — — —
— — — — 2 5,0 112 — — 46,0 8,0 91,0 1,5 100 79 1,5 39,0 — — — —
— — 1,5 3 14 5,0 106 — — 37,0 8,0 72,0 °,4 100 123 2,0 42,0 — — — —
3,0 1,0 14,0 21 53 9,0 109 — — 43,0 7,0 86,0 I,4 100 68 I,3 35,0 — — — —.
— — — — 3 5,0 — — — — — — -— — — — — — — — —
— — 0,3 2 17 9,0 100 — — 2,0 ; — — —* —— — —, — — — —
8,0 0,3 9,0 26 100 9,0 12 — — 2,0 — — — — —. — — — — — —
III. Источники радиации
73
в пределах 3900—4200° К. Спектральный состав излучения зависит от ре-
жима горения и типа применяемых углей.
Как видно из фиг. 40, для культуры растений наиболее пригодными
следует признать открытые дуги с желтыми пламенными углями. Спектр
этих ламп, богатый оранжево-красными и фиолетовыми лучами, весьма
близок спектру поглощения лучистой энергии хлорофиллом листа. Рас-
пределение квантов в спектре названных ламп и их поглощение пигмен-
тами, содержащимися в пластидах листа, показано на фиг. 41 и 42. Пиг-
менты поглощают в обоих случаях около 70%, а хлорофилл—60%
всех квантов физиологической радиации, падающей на лист (табл. 48).
В виде лучистой энергии в лампах с чистыми углями расходуется
около 20%, а в лампах с пламенными углями до 30% всей подводимой
мощности. На физиологическую радиацию приходится около 10% всего
излучения в первом случае и примерно 30% — во втором. Инфракрасная
радиация (длина волны — до 1200 ту.) составляет 90% у дуг с чистыми
углями и 70% у дуг с пламенными углями, излучающими желтый свет.
Всякая открытая дуга, независимо от типа углей, излучает некоторое
количество витацидных лучей. У ламп с чистыми углями энергия этих
лучей составляет около 0,9%, а у дуг с пламенными углями — до 2,6%
всего излучения. По этой причине при культуре растений открытую дугу
надлежит помещать за специальным экраном, поглощающим витацидные
лучи. Обычно для этой цели применяется обыкновенное оконное стекло
(фиг. 35), из которого и изготовляется фонарь, содержащий дугу.
Кроме того, при сгорании электродов образуются газообразные ве-
щества, вредно действующие на растения. Поэтому непременным усло-
вием эксплуатации дуги является обязательная вентиляция осветитель-
ного фонаря, устраняющая вредные газы.
5. РТУТНЫЕ ЛАМПЫ
Третьей многочисленной и разнообразной группой источников ради-
ации являются газосветные лампы, в основе излучения которых
лежит газовый разряд — прохождение электрического тока через газовую
среду [79, 84, 151, 245]. Излучение возникает в результате возбуждения
атомов или молекул при столкновении с электронами или друг с другом.
Характерной особенностью газосветных ламп является линейчатый
спектр, представленный или отдельными изолированными линиями, или
определенными группами линий. Относительная интенсивность линий
определяется давлением газа и типами стекла, обладающими различной
степенью поглощения. Мы рассмотрим ртутные, натриевые, неоновые и
некоторые другие лампы.
Современные ртутные лампы — источники радиации с разрядом, про-
текающим в атмосфере ртутных паров, разделяются на лампы низкого,
высокого и сверхвысокого давления [79, 84, 86, 87, 151]. К первым от-
носятся лампы с давлением ртутных паров до 0,3 мм рт.ст. (0,004 атм),
вторые обладают давлением порядка 0,5—1,5 атм, а третьи — бо-
лее 1,5 атм.
Лампы имеют оболочку из стекла (ртутно-стеклянные лампы) или кварца
(ртутно-кварцевые лампы). Рубашки ламп низкого давления обычно из-
готовляют из обыкновенного стекла, ламп высокого давления — из стекла
или кварца, а ламп сверхвысокого давления — из разнообразных специ-
альных стекол, имеющих различные коэффициенты поглощения.
Спектр излучения разных ламп показан в табл. 30 и на фиг. 43—
46, а характеристика ламп — в табл. 31—34.
74
Физические и физиологические основы светокультуры растений
Светотехническая характеристика
Мощность (в ВТ) Напряже- ние на лампе (в воль- тах) Сила тока (в а) Плотность тока (в а/см2) Световой поток (в лм) Световая отдача (в лм/вт) Горизон- тальная сила света (в св) Яркость (в сб)
197 — 3,5 0,660 4700 23,8 462 1,90
202 — 3,5 0,715 4780 23,7 470 2,01
207 — 3,5 0,280 4850 23,4 477 2,13
212 — 3,5 0,740 4900 23,1 482 2,24
217 — 3,5 0,920 4930 22,7 485 2,35
270 * 74 3,65 6800 25,0 680 2,10
* Лампа «СН».
Светотехническая характеристика ртутных
Лампы Мощность (в ВТ) Напряжение (в в) Сила тока (в а) Световой поток (в лм)
лампы отдельно вместе с дросселем на лампе | зажигания стационар- ная зажигания
мэлз «Игар-4» 250 281 125 180 2,20 4,0—4,5 7000
«Игар-3» 400 438 125 180 3,52 5,6—6,2 14000
«Игар-2» 500 540 125 180 4,40 7,5—8,2 16000
Фирм «Осрам» («HgH») и «Филора» («НО») «HgH-ЗОО» («НО-ЗОО») . . . 90 100 110 180 0,93 1,58 3000
«HgH-500» («НО-500») . . . 140 150 130 180 1,20 1,90 5000
«HgH-ЮОО» («НО-ЮОО») . . 265 280 130 180 2,20 3,70 10000
«HgH-2000» («НО-2000») . . 450 475 130 180 3,70 6,30 20000
«HgH-5000» («НО-5000») . . 1000 1060 — — — — 50000
Фирмы «О сир а» * «МА-400» 400 425 130 180 3,50 5,50 18000
«МА-650» 650 685 130 180 6,00 9,40 31000
Фирмы «Вестингхауз» «Н-5» 250 135 2,06 — 10000
«Н-2» 250 — 70 — 3,90 — 7500
«Н-1» 400 -— 143 —. 3,00 — 16000
Лампы «МА-150» и «МА-250» имеют примерно те же характеристики, что
и лампы «HgH-500» и «HgH-ЮОО».
£ь со rfx сл >и со со со со со ю о О О *4 СЛ О sU *4 СЛ СО ЬЭ СЛ СО 'о 'о О 'kj 'ф 'о 'Vi **4 W ООО лампы отдельно Светова! (в л;
25,0 32,0 29,6 30,0 33,3 35,7 42,1 47,1 42,3 45,5 вместе с дросселем я отдача м/вт)
о © 8 II Mill III Яркость (в сб)
63,7 33,2 23,2 47,4 лучистый поток (в ВТ) Физиоло ради
11,8 11,0 8,4 10,8 В % К ПОД- ВОДИМОЙ мощности гическая ация
а
05
й
й
$а
II 1 1 I 1 1 ill Давление паров ртути (в атм)
3—8 3—8 3—8 4,0 4,5 5,0 4,0 4,0 3,5 Время зажигания (в мин)
1 10,0 13,5 18,5 4,0 7,0 11,0 14,4 15,0 21,5 4,3 10,0 15,25 1 Расстояние между электродами (в см)
1 2,7 2,9 3,1 2,0 2,3 3,0 3,3 3,0 3,3 1,3 2,85 3,5 1 7 Диаметр разрядной трубки (в см)
23,2 28,0 33,0 16,0 21,2 25,0 31,0 зо, 43,0 1 длина Размерь (в
3,7 3,9 4,1 4,0 4,0 4,6 5,0 4,6 5,5 диаметр ,т лампы см)
«11111 00 В ВТ физиологиче- ской радиации | Лучистый поток
1 85,6 1 в % к общ.
5,0 В ВТ инфракрасной радиации
14,4 я о» О X® м
..“Mill 00 общий (в ВТ)
1 I S‘8 1 в % к под- водимой мощности Физиологическая радиация _ I
Mill! со 1 интенсив- ность на расстоя- 1 НИИ 1 м (в эрг/см2 • , 1 -сек) !
f 1 1 1 1 I co Давление паров ртути (в атм) | 1
__ 93,5 127 1 1 длина Размеры раз- рядной трубки (в см)
2,9 2,8 2,7 2,6 2,5 2,5 диа- । метр 1
Ci C5 Ф C5 05 05 © о о О О о © Ф о о о о Срок службы (в час)
76
Физические и физиологические основы светокультуры растений
Таблица 33'
Светотехническая характеристика ртутных ламп высокого давления в кварцевой
оболочке МЭЛЗ
Лампы Мощность (в ВТ) Напряжение (в вольтах) Сила тока (в а) Время зажигания (в мин) Длина разрядного столба (в см) Размеры труб- ки (в см) Срок службы (в час.) Световой поток (в лм) Световая отдача (в ЛМ,ВТ)
на лампе яв сети стационар- ная ! зажигания ! длина диаметр
«ПРК-4» «ПРК-5» «ПРК-2» «ПРК-7» 220 240 375 1000 70 120 120 135 120 220 220 220 3,75 2,30 3,75 8,05 5,0 4,2 6,0 14,0 10 15 15 10 6,0 10,5 12,1 17,5 17,3 22,7 25,7 31,8 1,8 1,8 2,0 3,0 800 800 800 800 4900 11000 51250* 25 28 45
Л ампы Лучистый поток - Физиологическая радиация (% к под- водимой мощности) Интенсивность радиации на расстоянии 1 м (в эрг/см2 • сек)
физиологи- ческой витацидной падиации инфракрас- ной радиации & - Л а ях
В ВТ s £ в % 4 к общему Е В ВТ | в % к общему ! —— общий (в ВТ) физиологиче- ской витацидной 1 инфракрас- ной общей
«ПРК-4» 20,8 «ПРК-5» — « ПРК-2»| 45,5 «nPK-7»j222,O 73,5 74,6 71,8 4,2 7,9 44,5 15,0 13,0 14,4 3,2 7,5 42,5 11,5 12,4 13,8 28,2 60,9 309,0 8,7 11,0 20,0 2183,8 4563,8 33782,0* 447,6 800,4 4592,2* 345,5 771,3 3415,0* 2976,3 6135,5 40789,2*
* Данные относятся к лампам, горевшим 1000 часов.
Ртутные лампы — источники по преимуществу коротковолновых ультра-
фиолетовых и сине-фиолетовых лучей. Поток инфракрасной радиации,
лежащей в области 1,014—1,711 р, в лучшем случае, у ламп сверхвысо-
кого давления, составляет 30% (фиг. 46), обычно же, у ламп низкого
и высокого давления, колеблется в пределах И—15% (фиг. 43 и 44),
а у ламп из специального стекла (табл. 34) может снижаться до нуля,
если пренебречь излучением самого стекла.
Физиологическая радиация представлена желтой (577 тр), зеле-
ной (546 тр), тремя фиолетовыми (404,7; 407,8 и 435,6 тр) и несколькими
ультрафиолетовыми (302,2; 312,6; 334,1; 365,0; 390,6 и 398,4 тр) лини-
ями. Интенсивность линий разных типов ламп показана в табл. 30. Лампы
с давлением ртутных паров менее 1 атм совершенно не содержат оран-
жево-красных лучей. Последние появляются в виде сплошного фона лишь
при более высоком давлении.
Витацидная радиация, испускаемая лишь ртутно-кварцевыми лампами
или лампами из специального стекла, представлена большим числом
линий, которые простираются до 237,8 тр (табл. 30). У ртутно-кварце-
вых ламп она составляет 13—15% всего излучения.
Лампы низкого давления изготовляются в виде длинных стеклянных
трубок (порядка 1 м) с диаметром 2,5— 2,9 см и мощностью 200—300 вт.
III. Источники радиации
77
Номинальный срок службы — 600 часов. Лампы переменного тока рас-
ходуют 30—40% энергии сверх номинальной мощности в пусковой
Фиг. 43. Распределение энергии в спектре излучения ртутной лампы
низкого давления
Р=0,01 мм рт. ст.; I = 1,0 а/см-.
Общая высота линий соответствует их относительной интенсивности, сплошные ли-
лии — поглощению, а пунктир — пропусканию и отражению лучистой энергии хлорофиллом*.
арматуре. Характеристика некоторых ламп низкого давления, изготовлен-
ных Московским электроламповым заводом (МЭЛЗ) и фирмой «Джене-
Фиг. 44. Распределение энергии в спектре излучения ртутной лампы высокого
давления «Игар-2»
P=t атм; 1=4,4 a; v=6,8 в/см; W</=27 вт/см.
рал электикэл компани» (лампа «СН»), показана в табл. 31, а спектраль-
ная характеристика—в табл. 30 и на фиг. 43.
* Этот прием изображения поглощения лучистой энергии хлорофиллом при-
меняется и дальше на фиг. 45, 47—50 и 52—54.
78
Физические и физиологические основы светокультуры растений
Как видно из табл. 31 и фиг. 43, излучение названных ламп представ-
лено в основном физиологической радиацией (85%), а излучение инфра-
красных лучей составляет лишь 15%. Физиологическая радиация со-
стоит главным образом из видимых (85—90%) и частично из ультра-
фиолетовых лучей, представленных единственной линией в 365 ту
(10—15%). Фиолетовые лучи составляют 50%, а желто-зеленые — 35—
40% всего физиологического излучения.
В настоящее время наиболее распространенным типом ртутных ламп,
вытесняющим лампы низкого давления (с жидким патодом), являются
лампы высокого давления с оксидным катодом и давлением ртутных па-
ров порядка 0,5—1,5 атм. Лампы мощностью в 90—1000 вт изготовляют
Фиг. 45. Распределение энергии в спектре излучения ртутно-кварцевой
лампы 11РК-7
в стеклянной или кварцевой оболочке; они рассчитаны на напряжение
в 120—220 в. Полезный срок службы ламп — 1000—2000 часов. В пуско-
вой арматуре лампы этого типа потребляют 8—12% энергии сверх своей
номинальной мощности. Лампы представляют собою короткие стеклян-
ные или кварцевые трубки длиною 15—40 см с диаметром 1,8—5,5 см.
Распределение энергии в спектре некоторых отечественных и загра-
ничных ламп показано в табл. 30 и на фиг. 44 и 45, а характеристика
ламп — в табл. 32 и 33.
Отличительной особенностью спектра ртутных ламп высокого дав-
ления является расширение линий и появление сплошного фона, дости-
гающего 5—12% всего излучения (фиг. 44). Ртутно-стеклянные лампы,
как и лампы низкого давления, служат в основном источником физиоло-
гической радиации, состоящей из видимых (80—90%) и ультрафиолетовых
лучей (линия 365 вщ — 10—20%). Сине-фиолетовые лучи составляют
30—35%, а желто-зеленые — 50—60% всей излучаемой энергии. Кроме
того, в спектре этих ламп в виде сплошного фона появляются оранжево-
красные лучи.
Ртутно-кварцевые лампы, в отличие от ртутно-стеклянных, имеют
спектр, линии которого простираются далеко в область коротких ультра-
фиолетовых лучей (237,8—296,7 пщ), вызывающих гибель растений (фиг.45
и табл. 30). Эта витацидная радиация составляет 13—15%, физиологиче-
Светотехническая характе ристика
Лампы Мощность (в ВТ) Напряжение (в вольтах) Сила тока (в а) Световой поток (в лм) Световая отдача (в лм/вт) Горизон- тальная сила света (в св) Яркость (в сб)
лампы отдельно вместе с дроссе- лем стацио- нарное зажи- гания стацио- нарное зажи- гания лампы отдельно вместе с дроссе- лем
«мэлз» «СВД-125» 125 190 0,7 4500 36,0
«СВДШ-250» 250 — 65—78 120—150 3,7—4,4 7,3 11500 46,0 — 1250 25000
Фирм «Вестингхауз» и «Дженерал электрик ком- пани» «Н-3» 85 250 о,4 3000 35,0 300
«11-4» 100 — 130 — 0,86 — 3500 35,0 — — 400
Фирмы «Филипс» «НР-300-1» 75 90 230 410 0,4 — 3000 40,0 33,4 300 1000
«НР-300-П» 75 83 115 180 0,75 1,25 3000 40,0 36,0 300 1000
«Н Р-500» 120 130 115 180 1,15 1,9 5000 41,7 36,5 — —
«ПР-400» 950 1050 1800 — 0,56 —. 40000 42,0 — —
Фирмы «Осрам» «HgQ-ЗОО» (матовое стекло) 75 — 0,7 . . .
«HgQS-ЗОО» (увиолевое стекло) 75 — — — ОД — 3000 40,0 -— —. —
«HgQS-500» (матовое стекло) 120 — — 1,1 — 5000 41,6 «— — —
«HgQS-500 В1» 120 — — — 1,1 — — — — — —
«HgQS-500» (МО и Сг2О— стекло) 120 — — 1,1 — — .— —. —. —
«HgH-ЮОО» 265 — — — 2,2 —. 10000 43,2 — — —
«HgHS-8000» (увиолевое стекло) 1500 — — —. 8,0 — 80000 53,3 — — —
« Н gB-500» 500 — 70—90 220 6,0 7,0 22500 45,5 — 2250 32000
Фирмы «Сирэй» 500 — — — — — 25000 45,0 — — 20000
Табл п ц а 34
(актериетика ртуптых ламп сеерхаысокогп давления
дзон- ьпая света св) Яркость (в сб) Лучистый поток Физиоло- гическая радиация (в % К ПОДВО- ДИМОЙ мощности) Интенсивность радиации на расстоя- нии 1 м (в эрг'см2-сек) Давление паров ртути (в атм) Время зажига- ния (в мин) Расстоя- ние между электро- дами (в см) Диаметр разрядной трубки (в см)
физиологиче- ской радиации витацидной радиации инфракрасной радиации общий (В ВТ)
В ВТ В % к об- щему В ВТ В % к об- щему В ВТ п о/ в /о к об- щему физиоло- гической витацид- ной инфра- красной общей
16.7 69.4 7,3 30,6 24,0 12,2 1311 580 1892 25
50 25000 42,5 71,5 — -— 16,9 28,5 59,4 15,5 3773 — 1507 5281 30—35 5—8 0,4—0,6 2,0—2,3
300 11,5 .— . - _ . 12,3 1200 — . 30 1,8 0,4
- 400 13,6 — — — — — — 12,4 1313 — — — 8 — 2,4 0,75
00 1000 — — —- —- — — — — — — — —. — — 1,8 0,45
00 1000 — — — — — — — — — — — — — 4,5 2,2 0,65
- — — — — — — — — — •— — — — 3,0 2,8 0,75
— — —. — - — —. • — — — — — — — — — 25,0 —
- — — 100 — — — — — — — — — — — — — —
- — 15,0 89,0 0,8 4,8 1,05 6,2 16,85 18,2 — — — — — — — —
- — 21,0 92,0 0,0 0,0 1,8 8,8 22,8 15,9 — — — —. — — — —
- — — 97,7 —• 2,3 —- 0,0 — — — — — — — — — —
- — —- 89,7 — 10,3 — 0,0 —- — — — — — — — —
- — 24,3 91,7 0,0 0,0 2,2 8,3 26,5 8,4 •— — — — — — — —
- — 289,6 91,6 0,6 0,2 25,9 8,2 316,1 17,5 — — — —. — — -— —
50 32000 — — —• — — — — — — — — — 50 10 0,5 3,0
- 20000 — — — — — — — — — — — — — — 0,4—0,6 —
III. Источники радиации
ская радиация— 70—75%, а инфракрасные лучи— 11—14% всего из-
лучения. Физиологическая радиация содержит главным образом ультра-
фиолетовые лучи в виде линий 302,2; 312,6; 334,1; 365,0; 390,6 и
398,4 шц (45—50%). Видимая часть представлена желто-зелеными
(30%) и сине-фиолетовыми (20%) лучами.
Культура растений при помощи ртутно-кварцевых ламп возможна
лишь при наличии экрана из обычного или увиолевого стекла, устраняю-
щего витацидные лучи. Позволяя несколько увеличить ультрафиолетовое
излучение, эти лампы имеют преимущество перед ртутно-стеклянными
лампами только в том случае, если защитный экран состоит из увиоле-
вого стекла или стекла «витаглясс». В дру1их случаях их действие не
отличается от действия ртутно-стеклянных ламп.
Фиг. 46. Распределение энергии в спектре излучения ртутной лампы сверхвысокого
давления СВДШ-250
Лампы сверхвысокого давления (4—50 атм) с воздушным охлажде-
нием, мощностью в 75—5000 вт, изготовляются из различных стекол.
При мощности до 125 вт лампы имеют давление ртутных паров 10—15 атм,
световую отдачу 40—42 лм/вт и яркость 1000—1300 сб. Срок службы —
около 2000 часов. Более мощные лампы (250—500 вт) с давлением паров
30—50 атм имеют световую отдачу 40—45 лм/вт и яркость 20000 сб. Срок
службы этих ламп — 500 часов. Дальнейшее увеличение мощности тре-
бует высокого напряжения (600—900 вольт) и связано с еще большим
сокращением срока службы (25—100 часов). Такие лампы для культуры
растений мало пригодны.
Характеристика ламп дана в табл. 34, а распределение энергии
в спектре — в табл. 30 и на фиг. 46. В лампах из обычного стекла (табл.
30) физиологическая радиация представлена видимыми и ультрафиолето-
выми лучами, а инфракрасные лучи составляют 30% всего излуче-
ния. Ультрафиолетовые лучи (линия 365 шр) составляют 20%,
сине-фиолетовые — 30—35%, желто-зеленые — 45—50%, а оранжево-
красные лучи — в виде сплошного фона и линии 690,7 шр — не превы-
шают 5—10% всего физиологического излучения. У некоторых ламп,
изготовленных из специального стекла (например «HgH-ЮОО», табл. 30),
физиологическое излучение практически состоит из одних видимых
80
Физические и физиологические основы светокультуры растений
лучей, причем желто-зеленые лучи (70%) преобладают над сине-фиоле-
товыми (28%), у других же ламп — из увиолевого стекла («HgQS-ЗОО»,
табл. 30) — линии ультрафиолетовых лучей простираются до 253,7 тр
и в спектре содержится некоторое количество витацидных лучей (5%).
Изготовляются также лампы, излучающие лишь одни ультрафиолетовые
лучи, относящиеся к физиологической радиации («HgQ-ЗОО», «HgQS-500 В1»
и «HgQS-500», табл. 30).
Растения, выращиваемые под ртутными лампами, получают исклю-
чительно ультрафиолетовые, сине-фиолетовые и желто-зеленые лучи,
причем поглощение энергии всеми пигментами листа, в зависимости от
спектра излучения ламп, составляет 70—95%, а хлорофиллом — 60—90%
всего физиологического излучения, падающего на лист (табл. 48). Каро-
тиноиды получают всего лишь 5—10%.
Инфракрасные лучи, содержащиеся в спектре ртутных ламп, прихо-
дятся на область (1—1,7 р), которая поглощается тканями листа.
Из всего многообразия ртутных ламп для культуры растений следует
рекомендовать лампы высокого и сверхвысокого давления, которые, как
это видно из табл. 32—34, превращают в физиологическую радиацию
относительно большее количество подводимой мощности. Предпочтение
следует отдать лампам из увиолевого стекла или ртутно-кварцевым лам-
пам, применяя в последнем случае увиолевый экран. Эти типы ламп
излучают линии с длиною волны порядка 290—320 тр, которые, как уви-
дим позднее, вызывают у растений благоприятный эффект.
6. НАТРИЕВЫЕ ЛАМПЫ
Из других газосветных ламп заслуживают внимания натриевые и
неоновые лампы, дающие излучение в области длинноволновых желтых
и оранжево-красных лучей.
Натриевые лампы являются источниками лучистой энергии, которая
почти целиком представлена в виде одной дублетной линии в 589—
Светотехническая характе
Лампы Мощность (в ВТ) Напряжение (в вольтах) Сила тока (в а) Свето- вой поток (в лм) Световая отдача (в лм/вт)
лампы отдель- но вместе с транс- форма- тором ста- цио- нарное зажи- гания В лампе в сети лампы отдельно вместе с транс- формато- ром
«На-2» ....... 75 130 420 0,55 — 3200 — 42,0
«На-1» — 110 140 420 0,65 — 4400 — 40,0
«SO(Na)-250» .... 45 65 80 340 0,6 1,4 2500 55,5 38,5
«SO(Na)-300» .... 53 63 65 180 0,9 3000 56,5 47,5
«SO(Na)-400» .... 60 80 110 340 0,6 1,4 4000 66,5 50,0
«SO(Na)-500» .... 80 94 75 190 1,2 —• 5000 62,5 53,0
«SO(Na)-650» .... 85 105 165 400 0,6 1,4 6500 76,5 62,0
«SO(Na)-1000» . . . 140 165 165 400 0,9 2,2 10000 71,5 61,0
« Na-300» 56 68 50 <220 1,15 — 2700 48,0 40,0
«Na-600» 100 118 60 <220 1,8 — 5400 54,0 46,0
«DA-90» 90 105 76 <220 1,02 —— 4200 47,0 40,0
III. Источники радиации
81
589,6 тр, содержащей 76% всего излучения, или 98% физиологических
лучей.
Инфракрасная радиация в спектре натриевых ламп представлена
линиями 819,5 и 1140,4 тр и составляет 22% всего изучения. Распределе-
ние энергии в спектре показано в табл. 35, а характеристика отечествен-
ных и некоторых заграничных ламп в табл. 36.
Т а б л и ц а 35
Распределение эн°ргии в спектре натриевых ламп
Длина волны (в m,j.) Относительная интенсив- ность линий (в % к интен- сивности линии 589, 6 т;х) Длина волны (в т>.) От осительная интенсив- ность линий (в % к интен- сивности линии 589, 6 ту)
498,3 0,2 616,1 0,3
515,4 . 0,1 819,5 19,0
568,8 1,2 1140,4 10,0
589,6 100,0 -
Лампы изготовляются мощностью в 45—140 вт; для них необходим
пусковой трансформатор, который потребляет от 17 до 44% энергии сверх
номинальной мощности ламп. Срок службы ламп — 2000—3000 часов.
7. НЕОНОВЫЕ ЛАМПЫ
Из современных неоновых ламп рассмотрим лишь высоковольтные
трубки положительного свечения (с холодным катодом) и дуговые лампы
интенсивного горения. Первые употребляются для рекламы, а вторые —
в качестве фонарей авиационных маяков.
ристика натриевых ламп
Табл и ц а 36
Лучистый поток Физиоло- гическая радиация (в % к под- водимой мощности) Расстоя- ние между- электро- дами (в см) Диаметр разрядной трубки (в см) Размеры вакуумной рубашки (в см)
физиологической радиации инфракрасной радиации общий (в ВТ)
в ВТ в % к общему в вт в % к общему длина диаметр
6,0 78,0 1,70 22,0 7,70 7,9 30,0 1,6 20,0 6,3
8,3 78,0 2,34 22,0 10,64 7,5 60,0 1,6 35,0 6,2
4,7 78,0 1,32 22,0 6,02 7,1 26,0 1,4 24,5 5,0
5,75 78,0 1,62 22,0 7,37 8,8 24,2 2,1 27,0 6,0
7,5 78,0 2,12 22,0 9,62 9,4 37,5 1,4 30,5 5,0
9,4 78,0 2,65 22,0 12,05 10,0 35,0 2,4 32,0 7,0
12,2 78,0 3,40 22,0 15,62 11,6 61,0 I,4 42,0 5,0
18,8 78,0 5,03 22,0 23,83 11,5 81,0 1,7 52,0 6,5
5,1 78,0 1,44 22,0 6,54 7,5 20,0 2,0 33,5 3,8
10,1 78,0 2,82 22,0 12,92 8,7 30,0 2,5 44,5 4,5
7,9 78,0 2,23 22,0 10,13 7,5 45,0 2,4 17,5 8,0
6 А. Ф. Клешнин
82
Физические и физиологические основы светокультуры растений
Излучение неоновых ламп сосредоточено целиком в области физио-
логически наиболее ценных оранжево-красных лучей (фиг. 47, табл. 37)
и поглощается исключительно хлорофиллом листа. Поглощение состав-
ляет 75% падающих лучей (табл. 48).
Характеристика некоторых неоновых ламп приведена в табл. 38.
Для культуры растений пригодны лишь дуговые и некоторые реклам-
750
ООО
Фиг. 47. Распределение энергии в спектре излучения неоновой лампы
(Р=2 мм рт. ст.;Т=0,75 а; V—1,59 в/см; Ш<г=1,19 вт/см)
Светотехническая характери
Лампы Мощность • (в ВТ) Напряже- ние - (в воль тах) Сила тока (в а) Плотность тока (в а/см3)
всей трубки 1 см раз- рядного столба
Высокого напряжения с холодным катодом «Ne-2250/2,0» 28 0,115 810 0,05 0,018
«Ne-2250/2,5» 34 0,115 990 0,05 0,018
«Ne-22500/2,0» . 163 0,68 455 0,50 0,018
«Ne-22500/2,5» 197 0,68 555 0,50 0,018
Дуговые, интенсивного горения (фир- мы «Филипс») «4309» 485 — 220 6,40 —
ные лампы положительного свечения. Последние могут употребляться
лишь в тех случаях, когда искусственное освещение применяется допол-
нительно к естественному дневному свету.
8. ГЕЛИЕВЫЕ ТРУБКИ
В заключение характеристики газосветных ламп необходимо сказать
несколько слов об источниках, имеющих не столько практическое, сколько
III. Источники радиации
83>
принципиальное теоретическое значение. Прежде всего следует отметить
гелиевые трубки [79] положительного свечения, излучение которых'
почти целиком представлено оранжево-красными (45%) и фиолетовыми'
лучами (50%). Обе половины спектра (фиг. 48) как раз приходятся на
максимумы поглощения лучистой энергии хлорофиллом. Пигменты листа
поглощают 90% энергии, излучаемой лампой, причем 80% приходится
на долю хлорофилла (табл. 48). Это, несомненно, чрезвычайно благопри-
Таблица 37
Распределение энергии в спектре неоновых ламп
(А. П. Иванов [79])
Длина волны (в тр.) Относительная интенсивность линий Длина волны (в тр.) Относительная интенсивность линий Длина волны (в тр) Относительная интенсивность линий
585,2 11,0 621,7 6,3 667,8 29,0
588,2 8,2 626,6 18,3 ;. 671,7 17,8
594,4 13,8 630,5 6,9 692,9 27,5
597,5 3,2 633,4 29,2 702,4 0,5
603,0 3,7 638,3 23,0 703,2 47,0
607,4 13,6 640,2 100 717,4 6,8
.609,6 21,0 650,4 43,2 724,5 18,8
612,8 0,5 653,3 11,0 743,9 4,7
614,3 35,8 659,2 16,0 808,2 8,5
616,4 9,0 665,2 0,5
' Т а б л и ц а 38
стика неоновых ламп
Световой поток (в лм) Световая отдача (в лм/вт) Яркость (в сб) Лучистый поток (в вт) Физиоло- гическая радиация (в % к ПОД- ВОДИМОЙ мощности) Срои службы (в час.) Размеры труб- ки (в см)
всей трубки 1 см раз- рядного столба всей трубки i см раз- рядного столба длина диа- метр
100 0,50 3,78 0,027 0,73 0,0037 2,64 2000 2,0 2,2
130 0,52 3,87 0,027 0,96 0,0037 2,80 2000 2,5 2,2
2550 12,75 15,7 0,62 18,70 0,093 11,50 2000 2,0 2,2
3200 12,80 16,2 0,63 23,50 0,093 11,90 2000 2,5 2,2
6500 -— 13,4 2,0 47,6 — 9,85 — 1,25 3,5
ятно для нормального прохождения физиологических процессов в ра-
стении.
9. КРИПТОНОВЫЕ И КСЕНОНОВЫЕ ЛАМПЫ
Аналогичным спектром обладают также криптоновые и ксеноновые
трубки (фиг. 49 и 50). В спектре видимого излучения у первой преобладают
оранжево-красные (55%), а у второй — синие лучи (80%).
6*
84
Физические и физиологические основы светокультуры растений
Сине-фиолетовая радиация криптоновой лампы поглощается в основ-
ном хлорофиллом, а. ксеноновой лампы — почти целиком каротино-
идами. В обоих случаях пигменты листа поглощают 80—90% энергии,
причем на долю хлорофилла приходится у криптоновых ламп 70%,
Фиг. 48. Распределение энергии в спектре излучения гелиевой лампы
а у ксеноновых ламп — 50% всех падающих физиологических лучей
(табл. 48).
Фиг. 49. Распределение энергии в спектре излучения криптоновой лампы
К сожалению, существующие в настоящее время типы перечислен-
ных ламп из-за малой мощности и незначительной поверхностной плот-
ности лучистого потока непригодны для культуры растений.
В последнее время появились компактные ксеноновые лампы, мощ-
ностью в 150, 400 и 700 вт, с дуговым разрядом длиною в 3,5; 6,5 и 6,5 мм
при давлении ксенона в 20, 30 и 40 атм (табл. 39).
III. Источники радиации
85
Эти лампы имеют сплошной спектр (фиг. 51), содержащий 8—15%
физиологической радиации. Поглощение этой радиации листом состав-
ляет 82%, а хлорофиллом — 64,5%.
Фиг. 50. Распределение энергии в спектре излучения ксеноновой
лампы с холодным катодом
Фиг. 51. Характеристика дуговой ксеноновой лампы
I — распределение энергии в спектре; 2 — распределение квантов в спектре; 3— поглощение
квантов лучистой энергии листом; 4 — поглощение квантов лучистой энергии хлорофиллом'
Инфракрасцая радиация обусловлена нагревом электродов (2580° К)
и оболочки (1100° К) и Теилцвыми явлениями, сопровождающими газовый
разряд.
86
Физические и физиологические основы светокультуры растений
Таблица 39
Светотехническая характеристика дуговых ксеноновых ла&ьп
Мощность (В ВТ) Световой поток (в ВТ) Световая отдача (в лм/вт) Сила света (в св) Лучистый поток Давление ксенона (в атм) Размеры лампы (в см)
Физиологиче- ской радиации витацидной радиации инфракрасной радиации Общий (в вт)
В ВТ В % к общему В ВТ । в % | к общему В ВТ в % к общему Длина диаметр
150 2800 18 220 6,85 7,85 1,25 - 3,85 60,7 88,'3 68,80 20 0,35 0,10
400 11030 27 870 25,60 13,50 4,30 6,65 117,0 79,8 146,90 30 0,65 0,18
700 15000 35 2000 59,15 14,85 9,85 7,35 240,0 77,7 309,00 44 0,65 0,20
10. ЦИНКОВЫЕ И КАДМИЕВЫЕ ЛАМПЫ
Большой интерес представляют также цинковые и кадмиевые лампы
(79] низкого и высокого давления. Отличительной особенностью спектра
Фиг. 52. Распределение энергии в спектре излучения кадмиевой лампы высокого
давления
(Р=200—800 мм рт. ст.)
этих ламп является наличие трех групп линии — в красных, зеленых
и ультрафиолетовых лучах (фиг. 52 и 53, табл. 40).
Первые совпадают с максимумом, а вторые — с минимумом поглощения
лучистой энергии хлорофиллом. Кадмиевая лампа высокого давления
(«КК-1»), изготовляемая Московским электроламповым заводом в квар-
цевой оболочке, содержит в спектре излучения 20% витацидных,
50% физиологических и 30% инфракрасных лучей. Физиологическая
радиация представлена ультрафиолетовыми (35%), сине-зелеными (40%) и
красными (25%) лучами. Содержание этих лучей в спектре ламп низ-
кого давления равно соответственно 70, 25 и 5%. Примерно такое же
III. Источники радиаЦии
87
распределение энергии наблюдается в спектре цинковых ламп низкого
давления: 45, 50 и 5%.
Поглощение лучистой энергии названных ламп пигментами листа
равняется 80—95%, причем хлорофилл поглощает 60—70% падающих
физиологических лучей (табл. 48).
W 500 000
Длина Оолнб/ (0 та)
~700
Фиг. 53. Распределение энергии в спектре излучения цинко-
вой лампы низкого давления
(Р = 0.01 мм рт. ст.)
Характеристика кадмиевой лампы высокого давления «КК-1» при-
водится в табл. 41.
Распределение энергии в спектре кадмиевых ламп
(А. П. Иванов [79])
Таблица 40
Длина » волны' (в.тр.) Относительная интен- сивность линий Длина волны (в mix) Относительная интен- сивность линий Длина волны (в mix) Относительная интен- сивность линий
Лампа низкого Давления (0,00001 атм) «КК-1» (давление 0,22—1,0 атм) Лампа низкого Давления (0,00001 атм) «КК-1» (давление 0,22—1,0 атм) Лампа низкого давления (0,00001 атм) «КК-1» (давление 0,22—1,0 атм)
228,8 18,5 308 — 2,9 508,6 100,0 85,0
232,9 — 18,5 326,1 610,0 12,0 643,8 35,0 100,0
267,4 — 46,4 340,3 25,0 38,9 739,9 — 4,15
276,3 — 5,6 346,6 57,0 39,8 1039,5 46,0 58,6
283,6 — 37,0 361,0 58,0 67,5 1432,7 — 44,0
288 — 64,7 467,8 56,0 42,6 1515,4 — 31,2
298 —. 9,2 480,0 104,0 62,0 1643,2 — 22,1
88 Физические и физиологические основы светокультуры растений
Таблица 41
Светотехническая характеристика кадмиевой лампы высокого давления «КК-1»
Мощность (в вт)...................................................... 460
Напряжение на лампе (в вольтах)
стационарное ...................................................... 120
при зажигании................................................... 600—1200
Сила тока (в а)
стационарная..................................................... 4,55
при зажигании..................................................... 8,75
Световой поток (в лм)............................................. 6000
Световая отдача (в лм/вт)......................................... 13,0
Лучистый поток
физиологической радиации (в вт) ................................ 63,6
то же (в % к общему)............................................ 49,8
витацидной радиации (в вт)........................................ 27,8
то же (в % к общему)............................................ 21,8
инфракрасной радиации (в вт).................................... 36,2
то же (в % к общему)............................................ 28,4
общий (в вт)...................................................... 127,6
Физиологическая радиации (в % к подводимой мощности).............. 13,8
Время разгорания (в мин.).......................:................. 20,0
Давление паров (в атм) ...........................................о,22—1,0
Диаметр трубки (в см).......................,..................* • 1,0
И. ЦЕЗИЕВЫЕ ЛАМПЫ
Цезиевые лампы представляют собой источник красных и инфракрас-
ных лучей, мало или совсем не поглощаемых пигментами листа (фиг. 54).
Физиологическая радиация здесь составляет всего лишь 12%,’ остальное
приходится на инфракрасные лучи, большая часть которых (74%) не погло-
щается даже водою, содержащейся в тканях листа. Лампа мощностью
III. Источники радиации
89
в 500 вт представляет собою стеклянную трубку длиною 40 см, диамет-
ром 5 см. Лампа имеет стационарное напряжение 50 в и потенциал за-
жигания 300 в. Срок службы — 100—200 часов.
12. ЛЮМИНЕСЦЕНТНЫЕ ЛАМПЫ
Обширной группой источников радиации являются люминесцентные
лампы [28, 39, 79, 166, 167, 177, 178, 235, 245, 252, 274, 292, 317, 318].
Это лампы газового разряда, у которых внутренние стенки трубок по-
крыты люминофором, светящимся под влиянием ультрафиолетовых лучей,
возникающих в результате разряда. В качестве люминофоров обычно
Фиг. 55. Характеристика люминесцентных ламп дневнрго света
1 — распределение энергии в спектре; 2 — распределение квантов в спектре;
3 — поглощение квантов лучистой энергии пигментами пластид; 4 — поглоще-
ние квантов хлорофиллом
используются следующие соединения: CaWO4, MgWO4, ZnSiO3r
ZnBeSiO3, CdSiO3, CdB,O5, ZnSiO4 и др.
Согласно закону Стокса, излучение, возникающее в результате люми-
несценции, имеет большую длину волны, чем излучение, поглощаемое
люминофором. Иными словами, в процессе люминесценции поглощае-
мая люминофором лучистая энергия трансформируется в излучение
с меньшей частотой и, следовательно, с меньшей величиной квантов.
Все перечисленные выше люминофоры имеют область поглощения в пре-
делах 220—300 шр. с максимумом около 240—285 шр. Их излучение,
в зависимости от вида люминофора, приходится на синие, зеленые или
красные лучи.
В качестве разрядных трубок применяются неоновые и ртутные лампы.
В ртутных лампах низкого давления используется в основном линия
253,7 шр, в лампах высокого давления — линия 365 шр-, а в неоновых
лампах — линии 250 и 380 шр-, которые и вызывают свечение соответ-
ствующих люминофоров. Рассмотрим лишь высоковольтные и низко-
вольтные фотолюминесцирующие ртутные лампы низкого давления и
ртутные лампы высокого давления.
Высоковольтные люминесцентные лампы, представляющие собою
стеклянные трубки, характеристика которых приведена в табл. 42, могут
«О
Физические и физиологические основы светокультуры растений
иметь самый различный спектр излучения. Наиболее распространены
лампы дневного света и розовато-белые лампы, спектр излучения которых
Фиг. 56. Характеристика люминесцентных ламп мягкобелого света
1 — распределение энергии в спектре; 2 — распределение квантов в спектре;
3— поглощение квантов лучистой энергии пигментами пластид; 4— поглоще-
ние квантов хлорофиллом
близок к спектру низковольтных люминесцентных ламп дневного, мягко-
белого и белого света (фиг. 55—57).
Фиг. 57. Характеристика люминесцентных ламп белого света
1 — распределение энергии в спектре; 2 — распределение квантов в спектре;
3 — поглощение квантов лучистой энергии пигментами пластид; 4 — погло-
щение квантов хлорофиллом
Низковольтные лампы, также представлящие собою стеклянные
трубки, рассчитаны на работу от сети с напряжением в 110 и 220 в. Срок
службы ламп — 1500—2500 часов. Размеры и физическая характеристика
III. Источники радиации
91
трубок приведены в табл. 43, светотехнические показатели ламп —
в табл. 44, а распределение энергии в спектре — на фиг. 55—61 и 95.
Фиг. 58. Характеристика* люминесцентных ламп синего света
1 — распределение энергии в спектре; 2 — распределение квантов в спе-
ктре; 3 — поглощение квантов лучистой энергии пигментами пластид;
4 — поглощение квантов хлорофиллом
Там же показано поглощение лучистой энергии пигментами листа. Лучше
всего лампы работают при температуре воздуха 23° [292].
Фиг. 59. Характеристика люминесцентных ламп зеленого света
1 — распределение энергии в спектре; 2 — распределение квантов в спе-
ктре; 3 — поглощение квантов лучистой энергии пигментами пластид;
4 — поглощение квантов хлорофиллом
Наиболее распространены лампы белого, мягкобелого и дневного
света. Из других ламп следует назвать синие *, зеленые и красные лампы,
1 У отечественных синих ламп в качестве люминофора применяется MgWO4.
Спектр этих ламп приведен на фиг. 95.
92
Физические и физиологические основы светокультуры растений
в которых в качестве люминофоров используются CaWO4, ZnSiO, и
CdB2O5.
Фиг. 60. Характеристика люминесцентных ламп красного света
1 — распределение энергии в спектре; 2—распределение квантов в спектре;
3 — поглощение квантов лучистой энергии пигментами пластид
Как видно из упомянутых выше рисунков, спектр люминесцентных
ламп слагается из основного сплошного фона (94%) и отдельных линий %
Фиг. (51. Характеристика золотистых люминес-
центных ламп
1 — распределение энергии в спектре; 2 — распределение
квантов в спектре; 3 — поглощение квантов лучистой энер-
гии пигментами пластид
возникающих в результате
разряда в атмосфере ртутных
паров (6%). Лишь у золотис-
тых ламп ртутные линии по-
глощаются специально подо-
бранным стеклом. Излучение
люминесцентных ламп со-
ставляет 26,5% подводимой
мощности, 53% теряется на
теплопроводность и конвек-
цию, а 20% приходится на
балластное сопротивление
или дроссели [178].
В зависимости от типа
ламп листья растений погло-
щают от 65 до 90% излуче-
ния, причем хлорофилл по-
глощает 40—70% всех падаю-
щих на лист лучей (табл. 48).
Наибольшее поглощение лу-
чистой энергии листьями рас-
тений происходит в спектре
излучения белых, красных и
золотистых ламп, наимень-
шее — зеленых ламп.
1 На фиг. 55—61 и 95 отдельные линии представлены в виде столбцов, площади
которых соответствуют интенсивности линий.
III. Источники радиации
93
Т а б л и ц а 42
Светотехническая характеристика высоковольтных люминесцентных ламп
Лампы
Физиологиче-
ская радиция
Размеры
трубок
Белого света . . . .
Мягкобелого света
Дневного света . . .
121 307
100 252
50 230
50 j 230
0,5
0,177 3600
, । 29,7
0,177'2880- 28,8
0,25 0,031 2000 40,0
0,250,031.2000) 40,0
0,76
0,76
0,31
0,31
14,60
11,80
8,60
9,20
12,00 2000
11,68 2000
17,20 2000
18,40 '2000
2,5
2,0
2,0
2,0
2,2
2,2
3,5
3,5
Фиг. 62. Распределение энергии в спектре излучения люми-
несцентных ламп высокого давления («HgL=300»)
Табл и ц а 43
Техническая характеристика низковольтных люминесцентных ламп
Мощность (в ВТ) Напряжение (в вольтах) Сила тока (в а) Длина трубки (в см) Диаметр трубки (в см) Площадь светящейся поверхности (в см) Площадь светящейся проекции лампы (в см)
на лампе в сети
6 45 0,15 23,0 1,59 115,0 36,6
8 54 0,18 30,5 1,59 152,0 48,5
14 41 0,37 38,0 3,80 455,0 144,5
15 56 110—125 0,30 45,0 2,54 360,0 114,2
15 48 0,33 45,0 3,80 540,0 171,0
20 62 0,35 60,5 3,80 725,0 230,0
30 103 220—250 1 0,34 91,0 2,54 728,0 231,0
40 108 0,41 121,5 3,80 1450,5 460,0
65 50 110—125 1,35 91,5 5,40 1550,0 492,0
100 72 220—250 1,45 152,5 5,40 2600,0 825,0
94
Физические и физиологические основы светокультуры растений
Таблица 44
Светотехническая характеристика низковольтных люминесцентных ламп
Лампы Мощность (в ВТ) Световой поток (в лм) Световая отдача (в лм/вт) Яркость (в сб) Физиологическая радиация
лучистый поток (в вт) в % к подво- димой мощности
Дневного света
«ДС-15» ...... 15 495 33,0 0,60 1,99 13,30
«ДС-20» 20 760 38,0 0,48 3,06 15,30
«ДС-30» 30 1230 41,0 0,73 4,96 16,50
«ДС-40» ..... 40 1800 45,0 0,51 7,25 18,10
«ДС-65» 65 1800 27,0 0,36 7,25 11,10
«ДС-1ОО# 100 3350 33,5 0,40 13,50 13,50
Мягкобелого света
«МБС-15» 15 435 29,0 0,53 1,85 12,30
«МБС-20» > 20 640 32,0 0,41 2,72 13,60
«МБС-30» 30 1100 37,0 0,65 4,67 15,50
«МБС-50» 40 1500 37,0 0,43 6,38 16,00
Белого света
«БС-15» 15 585 39,0 0,71 2,08 13,85
«БС-20» 20 900 45,0 0,57 3,20 16,00
«БС-ЗО» 30 1440 46,0 0,85 5,10 17,00
«БС-40» 40 2120 53,0 0,60 7,50 18,80
«БС-65» ..... 65 . 2100 32,0 0,43 7,43 11,40
«БС-100» 100 4200 42,0 0,51 14,90 14,90
15 315 21,0 0,39 1,62 10,90
Синие 20 460 23,0 0,29 2,38 11,90
30 780' 26,0 0,47 4,01 13,40
15 900 60,0 1,10 2,00 13,30
Зеленые . . . . 20 1300 65,0 0,83 2,88 14,40
30 2250 75,0 1,35 5,00 16,70
15 300 20,0 0,36 1,35 9,00
Красные . ... • 20 440 22,0 0,28 1,98 9,90
30 750 25,0 0,45 3,38 11,30
- • 15 375 25,0 0,57 1,25 8,35
Золотистые . . . • 20 540 27,0 0,34 1,79 8,95
30 930 31,0 0,55 3,08 10,30
Таблица 45
Светотехническая характеристика люминесцентных ламп высокого давления
Лампы Мощность (в ВТ) Напряже- ние на лампе (В в) Сила тока (В а) Световой поток (в лм) Световая отдача (в лм/вт) Физиологическая радиация
лучистый поток (в ВТ) в % К подво- димой мощности
«HgL-ЗОО» 75 125 0,75 3300 44,1 74,0 9,9
«HgL-500» 120 125 1,20 5500 46,0 126,0 10,5
III. Источники радиации
95
Большой интерес представляют люминесцентные лампы высокого
давления [274], по форме и размерам (диаметр колбы 11—13 см) сходные
с обычными лампами накаливания. Люминесцентные лампы этого типа
представляют собой ртутные лампы высокого давления, разрядные трубки
которых, изготовляемые из кварца, помещают в стеклянную колбу,
покрытую изнутри люминофором (ZnCdSCu). Характеристика ламп пред-
ставлена в табл. 45, распределение энергии — на фиг. 62.
13. ВЛИЯНИЕ НАПРЯЖЕНИЯ НА СВЕТОТЕХНИЧЕСКИЕ ПОКАЗАТЕЛИ ЛАМП
Характеристики ламп, приведенные в соответствующих таблицах,
всюду соответствуют номинальному напряжению в сети. В действитель-
ности же как в производственных, так и в опытных условиях лампы
часто работают на другом напряжении. Эти отклонения от номинала
особенно велики в больших городах и индустриальных центрах в рабочие
и вечерние часы. Изменения напряжения, влияя на рабочую температуру
нити накала или газовый разряд, приводят к изменению и других пока-
зателей ламп: силы света, светового потока, мощности, интенсивности
излучения и т. д.
Приближенно, при небольших изменениях напряжения (±10%),
зависимость основных параметров ламп накаливания от напряжения
в сети выражается следующим уравнением:
Н____/ V \т
(29)
где т — константа, зависящая от рассматриваемой характеристики
ламп (табл. 46), 1'0 и V — номинальное и реальное напряжение, а
и Н — номинальная и реальная величина соответствующей характери-
стики.
Таблица 46
Величина, коэффициента т для различных характеристик ламп накаливания
с вольфрамовой нитью
(Иванов [78])
Лампы Сила тока (в а) Цветовая темпера- тура (в °К) Свето- вая отдача (в лм/вт) Коэффициент m для
мощ- ности " силы света и светового потока темпера- туры нити срока служ- бы СИЛЫ тона
Пустотные .— 2450 10 1,581 3,52 0,345 —13,5 0,585
Газополныо .... <2,5 2800 12 1,526 3,67 0,398 —14,8 0,526
.... >2,5 •2950 16 1,541 3,47 0,391 —13,9 0,541
В случае больших изменений напряжения для ламп накаливания
применимо другое уравнение:
1^ = А'(1«к)2 + z'k’ IV (30)
Входящие в это уравнение константы К и Р приведены в табл. 47.
Изменения мощности и светового потока в зависимости от напряжения
сети показаны для других ламп на фиг. 63 и 64.
В общем следует принять, что с увеличением напряжения происходит
увеличение мощности, светового потока и силы света и уменьшение срока
96
Физические и физиологические основы светокультуры растений
службы ламп. В производственных условиях или в опытах, где не
требуется большая точность, можно пренебречь изменениями перечислен-
ных величин, но в точных опытах колебания их следует учитывать.
Напряжение на лампе (8 °/а Н номиналЬнамрJ
Фиг. 63. Изменение мощности ламп
в зависимости от напряжения сети
1 — неоновая трубка с холодным катодом;
2—-ртутная трубка низкого давления; з— на-
триевая лампа «SO(Xa)-650»; 4 — лампа нака-
ливания; 5 — ртутная лампа высокого давле-
ния; 6 — солнечная лампа («HWA-500»);
7 — люминесцентная лампа высоковольт-
ная; 8 — низковольтная люминесцентная
|лампа
Напряжение на лампе ( 9 % /< наминалднамр)
Фиг. 64. Изменение светового потока
ламп в зависимости от напряжения
в сети
1 — неоновая трубка с холодным катодом;
2— ртутная трубка низкого давления; 3 — на-
триевая лампа «SO(Na)-650»; 4 — лампа нака-
ливания; .5 — ртутная лампа высокого давле-
ния; 6 — ртутная лампа сверхвысокого давле-
ния; 7—высоковольтная люминесцентная лам-
па; 8 — низковольтная люминесцентная лампа
1\а б лица 47
Величина коэффициентов К и Р для различных характеристик ламп накаливания
с вольфрамовой нитью
Лампы Мощность (в ВТ) Свето- вая отдача .(в лм/вт) Коэффициенты для
МОЩНОСТИ силы света и светового потока световой отдачи силы тона
Кр рр ф Р* Л; Kt Pi
Пустотные 15— 60 10,0 —0,028 1,580 —0,946 3,513 —0,918 1,932 —0,028 0,580
40— 50 12,5 0,057 1,523 —1,425 3,685 —1,482 2,162 0,057 0,523
Газопол- 60—150 12,5 0,057 1,523 —1,669 3,613 —1,726 2,090 0,057 0,523
200—500 16,0 0,083 1,543 —1,607 3,384 —1,690 1,841 0,083 0,543
14. СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА ФИЗИОЛОГИЧЕСКОЙ
ЭФФЕКТИВНОСТИ ЛАМП
Физиологическая эффективность всякого источника радиации опре-
деляется долей энергии, излучаемой в виде физиологических лучей,
степенью их поглощения пигментами пластид (особенно хлорофиллом)
и количеством квантов, содержащихся в единице энергии.
>
е
Клешнин
Таблица 48
Сравнительная характеристика физиологической эффективности источников
ч Лампы Мощность ИСТОЧНИК:; (в вт) Физиологическая радиация (в % от под- водимой мощности источника) Поглощение лучистой энергии пигментами пластид «среднего» листа (в % к падаю- щей) Теоретический коэф- фициент использова- ния подводимой энергии Количество квантов в единице энергии физиологической радиа- ции Теоретическая эффек- тивность источников радиации по отношению к лампе накаливания в 300 вт Теоретическая удельная мощность, потребная для куль- туры растений (в квт/м2)
всеми . , пигментами 1 хлорофиллом в % к мощ- ности источника в относитель- ных единицах но сравнению с лампой накаливания в 300 вт в 1 эрге й « 5s £«« Е- И S>S “ с. s s И О и g о «о о r «н о О С к ? go £**"S*S © kC Я ft R W И Д и о о s и
I. Лампы накаливания 1. Пустотные 2. Газополные 3. Проекционные II. Дуговые лампы 1. Открытые с чистыми углями 2. Открытые с пламенными углями (желтые) 3. Лампа «S-l>> 10 25 40 60 75 100 200 300 500 1000 2000 1000-3000 i 315 8,7 ЭД 10,6 ИД 11,6 12,5 13,5 17,5 2,5— 5,0 13,6-19,1 8,6 59,0 59,7 60,0 61,5 60,8 61,1 62,0 63,0 63,2 63,6- 64,5 68,3 67,6 72,1 68,9 53,7 54,3 54,5 55,6 54,0 54,1 54,6 55,0 55,1 55,5 56,2 57,4 57,8 59,0 61,5 4,7 4,9 5,8 6,1 6,5 7,1 7,6 10,1 1,4— 2,9 8,0—11,3 5,3 0,77 0,80 0,95 1,00 1,06 1,16 1,24 1,69 0,23—0,47 1,31—1,85 0,77 3,36 1011 3,32 • ЮН 3,28 • ЮН 3,22 • ЮН 3,13 • юн 3,13 • юн 3,11 • юн 3,10 • юн 3,09 • ЮН 3,08 • 10” 3,07 • 10” 3,06 • 10” 2,84 • 10” 2,84 • 10” 2,90 • 10” 1,08 1,07 1,06 1,04 1,02 1,00 1,00 1,00 1,00 1,00 0,99 0,99 0,92 0,92 0,93 0,79 0,80 0,95 1,00 1,06 1,16 1,23 1,62 0,21—0,43 1,21—1,70 0,72 0,63—1,52 0,62—1,50 0,52—1,26 0,50—1,20 0,47—1,13 0,43—1,03 0,40—0,98 0,31—0,74 2,40—2,80 0,41—0,70 0,70—1,67
S
о
£
£
Чз
&
си
£
&
4=
£
£
Таблица 48 (продолжение)
Й s S' о кая /о ОТ ПОД- юсти Поглощение лучистой энергии пигментами пластид «среднего» листа (в % к падаю- щей) Теоретический коэф- фициент использова- ния подводимой энергии Количество квантов в единице энергии физиологической радиа- ции 2 I S К и Я S ф ft Ф S eg = и fS ° S й j; Д и ". ? а щость, я куль- а. (в квт/м2)
Лампы a 8 зиологичес нация (в ® ИМОЙ М0Щ1 очника) 1тами © ч ч S мощ- ика штель- иницах знению' [ОЙ | 1ВЭНИЯ ВТ 1 о ч । Й h § S И ° й и X Ч ф О Д 03 2 « я к § “о ичеыш ть исто ши по ( хе пака; ВТ я 5 ₽» s £ 2 «я Ф Й Ей Д Я Я О ДЯДЯ
*Г> ф S s о а к а ? X н- Ф ** *-* О Ч Л t g®sg а © & Й Е ct о Ф Д ф Вч
5 я S ь 0х О Н oS Ф £ И я Ч со ° Ш М Чсо О ф ь А
ваши Д Й и и Й S 3 и _A Д Д R О й Я CQ Я И й о‘ о к ® 8 ЬйК я Н Р» к н
III. Ртутные лампы
1. Низкого давления «СИ» 2. Высокого давления ‘270 8,3 82,8 70,3 5,8 0,95 2,37 • 10й 0,76 0,73 0,68—1,64
«Пгар-2» 500 11,8 82,6 74,6 8,8 1,44 2,38 • ion 0,71 1,11 0,45—1,08
«Н-5» 250 11,0 77,0 68,5 7,5 1,23 2,48 • юн 0,80 0,98 0,51—1,22
«Н-2» 250 8,4 76,1 67,1 5,6 0,92 2,52 • юн 0,81 0,74 0,73—1,75
«Н-1» 400 10,8 74,8 66,4 7,2 1,18 2,54 • юн 0,82 0,97 0,56—1,33
«ПРК-4» 220 8,7 82,6 73,0 6,3 1,03 2,38 • IO” 0,77 0,79 0,63—1,52
«ПРК-2» 375 11,0 81,5 73,0 8,0 1,31 2,34 • 1011 0,75 0,98 0,51—1,23
«ПРК-7» . . . 3. Сверхвысокого давления 1000 20,2 82,7 73,1 14,7 2,41 2,33 • 1011 0,75 1,81 0,28—0,66
«СВД-125» 125 12,2 79,7 70,0 8,5 1,39 2,44 • 10'1 0,79 1,10 0,45—1,09
«СВДШ-250» 250 15,5 77,6 69,7 10,8 1,72 2,52 • 1011 0,81 1,39 0,36—0,87
«н-з» 85 12,3 75,3 70,6 8,7 1,42 2,39 1011 0,77 1,17 0,43—1,20
«Н-4» 100 12,4 78,9 68,9 8,5 1,39 2,53 • 1011 0,81 1,21 0,42—0,99
«HgQS-ЗОО» 75 18,2 84,1 73,6 13,6 2,23 2,23 • 1011 0,72 1,60 0,31—0,75
«HgQS-500» 120 15,9 82,2 72,2 11,5 1,89 2,38 • 1011 0,77 1,45 0,35-0,83
«HgH-ЮОО» 265 8,4 70,0 63,3 5,2 0,85 2,66 • 1011 0,86 0,73 0,68—1,64
«HgHS-SOOO» 1500 17,5 79,0 71,0 12,4 2,04 2,41 • 1011 0,79 1,61 0,31—0,75
«IIgQ-ЗОО» 75 — 95,0 85,0 — — 1,66 • 1011 0,54 — —
«HgQS-500Bl» 120 — 96,6 88,5 — — 1,81 • 1011 0,58 — —
«HgQS-500» * 120 — 95,6 81,6 — — 1,65 • 10П 0,53 — —
* Колба выполнена из стекла, содержащего NiO и Сг.,О.
Физические и физиологические основы светокультуры растений
Т а Г» л и ц а 48 (продоллсение)
сз X S X о 1 :ная | % от под- , дости Поглощение лучистой энергии пигментами пластид «среднего» листа (в % к падаю- щей) Теоретический коэф- фициент использова- ния подводимой энергии Количество квантов в единице энергии физиологической радиа- ции ч эффек- чников шюшению ливания Теоретическая удельная мощность, потребная Для куль- туры растений (в квт/м2)
Лампы Мощность ист (В ВТ) Физиологичес радиация (в 1 ВОДИМОЙ М0Щ1 источника) 1 1 всеми । пигментами j хлорофиллом! в % к мощ- ности 1 источника j в относитель- ных единицах по сравнению с лампой накаливания в 300 вт 1 в 1 Эрге в относитель-| ных едини- цах по сравнению с лампой на- 1 наливания | в 300 вт i Теоретическая тивность исто радиации по ( к лампе пака, в 300 вт
IV. Солнечная лампа«Н Л Л-500» V. Натриевые лампы «На-2» «На-1» «SO(Na)-250» «SO(Na)-300» «SO(Na)-400» «SO(Na)-500» «SO(Na)-650» «SO(Na)-1000» «Na-ЗОО» «Na-600» «Na-90» VI. Неоновые лампы 1. С холодным катодом . . . 2. Дуговые интенсивного го- рения VII, Ксеноновые дуговые лампы- А III. Гелиевые лампы IX. Криптоновые лампы ,• . . . 255 75 110 45 53 60 80 85 140 56 100 90 1 50 - 200 500 150 400 700 10,2 7,9 7,5 7,1 8,8 9,4 10,0 11,6 11,5 7,5 8,7 7,5 11,5 10,0 10,2 15,2 17,0 68,5 70,0 70,0 70,0 70,0 70,0 70,0 70,0 70,0 70,0 70,0 70,0 75,3 75,3 82,0 82,0 82,0 89,4 80,5 60,0 70,0 70,0 70,0 70,0 70,0 70,0 70,0 70,0 70,0 70,0 70,0 75,3 75,3 64,5 64,5 64,5 79,3 70,0 6,1 5,5 5,2 5,0 6,2 6,6 7,0 ’ 8,0 8,0 5,2 6Д 5,2 8,6 7,5 6,6 9,8 11,0 1,00 0,90 0,85 0,82 1,02 1,08 1,15 1,33 1,31 0,85 1,00 0,85 1,41 1,23 1,08 1,6 1,8 2,82 • 10’1 2,95 -10” 2,95 • 10’1 2,95 • 10” 2,95 • 10” 2,95 • 10” 2,95 • 10” 2,95 • 10” 2,95 • 10” 2,95 • 10’1 2,95 10” 2,95 • 10” 3,32 10” 3,32 • 10” 2,54 • 10” 2,54 • 10” 2,54 • 10” 2,52 • 10” 2,64 • 10” 0,90 0,95 0,95 0,95 0,95 0,95 0,95 0,95 0,95 0,95 0,95 0,95 1,07 1,07 0,83 0,83 0,83 0,81 0,85 0,91 0,85 0,81 0,78 0,97 1,03 1,09 1,26 1,24 0,81 0,95 0,81 1,51 1,32 0,90 1,33 1,49 0,55-1,32 0,59-1,41 0,62 -1,48 0,64—1,54 0,51—1,24 0,49—1,17 0,46—1,10 0,40—0,95 0,40—0,97 0,62—1,48 0,53—1,26 0,62—1,48 0,33-0,80 0,38—0,91 0,56—1,34 0,38—0,90 0,34—0,81
Таблица 48 (продолжение)
а JC Я 5 с Физиологическая радиация (в % от под- водимой мощности источника) Поглощение лучистой энергии пигментами пластид «среднего» листа (в % к падаю- щей) Теоретический коэф- фициент использо- вания подводимой энергии Количество квантов в единице энергии физиологической радиации а 1 ЕЙ м ЕС да Д Е e-g § ЕС Теоретическая удельная мощность, потребная для куль- туры растений (в квт/м2)
Лампы Мощность ист (В ВТ) всеми пигментами хлорофиллом В % К мощ- ности источника в относитель- ных единицах по сравнению с лампой накаливания в 300 вт в 1 эрге Л й § s S М « S S д о И g ЕС С Й ® Й X и, е * х Q х «5 е; ° Л a W Яй EJQOS ® Теоретическа; тивность истс радиации по с к лампе нака. в 300 вт
X. Кадмиевые лампы 1. Высокого давления «КК-1» 2. Низкого давления XI. Цинковые лампы низкого давления ХШ. Люминесцентные лампы 1. Высоковольтные а) дневного света б) мягкобелого света . . . в) белого света 2- Низковольтные «ДС-15» . . «ДС-20» . . а) дневного «ДС-30» . . света «ДС-40»* . «ДС-65» . . «ДС-100» . «МБС-15» . б) мягкобе- «МБС-20» . лого света ' «МБС-30» . «МБС-40» . 460 50 50 ; юо 15 20 30 40 65 100 15 20 30 40 13,8 18,4 17,2 12,0 13,3 15,3 16,5 18,1 11,1 13,5 12,3 13,6 15,5 16,0 77,3 96,5 95,4 49,2 78,0 76,0 75,5 80,0 80,0 80,0 80,0 80,0 80,0 84,1 84,1 84,1 84,1 62,3 72,1 62,4 49,2 58,5 61,5 68,0 59.4 59,4 59,4 59,4 59,4 59,4 61,9 61,9 61,9 61,9 . 8,6 10,7 10,6 8,2 7,9 9,1 9,8 10,7 6,6 8,0 7,6 8,4 9,6 9,9 1,41 1,76 1,74 1,34 1,30 1,49 1,61 1,75 1,08 1,31 1,25 1,38 1,57 1,62 2,42 10п 1,88 • 10п 2,12 • 1011 3,54 • 1011 2,76 • 1011 2,86 • 1011 2,90 • 1011 2,75 • 10” 2,75 • 1011 2,75 10й 2,75 • 10п 2,75 • 10й 2,75 • 10п 2,82 • 10п 2,82 • 10й 2,82 • 101] 2,82 • 1011 0,78 0,60 0,68 1,14 0,89 0,92 0,93 0,88 0,88 0,88 0,88 0,88 0,88 0,91 0,91 0,91 0,91 1,10 1,51 1,60 1,25 1,15 1,32 1,43 1,55 0,90 1,16 1,14 1,25 1,43 1,47 0,45—1,09 0,33—0,80 0,31—0,75 0,40—0,96 0,43—1,04 0,38—0,91 0,35—0,84 0,32—0,78 0,52—1,25 0,43—1,04 0,44—1,05 0,40—0,96 0,35—0,84 0,34—0,82
100 Физические и физиологические основы светокультуры растений
Таблица 48 {продолжение)
Лампы Мощность источника (В ВТ) Физиологическая радиация (в % от под- водимой мощности источника) Поглощение лучистой энергии пигментами пластид «среднего» листа (в % к падаю- щей) Теоретический коэф- фициент использо- вания подводимой энергии Количество квантов в единице энергии физиологической радиации ретическая эффек- ность источников нации по отношению ампе накаливания 00 вт Теоретическая удельная мощность, потребная для куль- туры растений (в квт/м2)
всеми пигментами хлорофиллом в % К мощ- ности источника в относитель- ных единицах по сравнению с лампой накаливания в 300 вт в 1 эрге в относитель- ных едини- цах по сравнению с лампой на- каливания в 300 вт
о о Ен о К Чсо s л
- «БС-15» . 15 13,8 73,5 67,5 9,3 1,52 2,98 • юн 0,99 1,51 0,33—0,80
«БС-20» • 20 16,0 73,5 67,5 10,8 1,77 2,98 • юн 0,99 1,76 0,28—0,68
в) белого «БС-30» • 30 17,0 73,5 67,5 11,4 1,87 2,98 • 1011 0,99 1,86 0,27—0,65
света «БС-40»< 40 18,8 73,5 67,5 12,6 2,06 2,98 • 1011 0,99 2,05 0,24—0,59
«БС-65» . 65 11,4 73,5 67,5 7,7 1,26 2,98 • 1011 0,99 1,25 0,40—0,96 § о
«БС-1С0» 100 14,9 73,5 67,5 10,1 1,65 2,98 • 1011 0,99 1,64 0,30—0,73 £
15 10,9 88,3 54,3 5,9 0,97 2,41 • ЮН 0,78 0,76 0,65—1,58 Я
г) синие (CaWO4) * . . . 20 11,9 88,3 54,3 6,5 1,06 2,41 • 1011 0,78 0,84 0,60—1,43
30 13,4 88,3 54,3 7,3 1,20 2,41 • 10” 0,78 0,91 0,55—1,32
15 13,3 66,2 43,0 5,7 0,93 2,57 • 10” 0,83 0,77 0,65—1,56 S а
д) зеленые (7nSiO3) . . . 20 14,4 66,2 43,0 6,2 1,02 2,57 • 10” 0,83 0,85 0,59—1,41 £ s:
30 16,7 66,2 43,0 7,2 1,18 2,57 • 10” 0,83 0,99 0,51—1,21
15 9,0 74,5 72,5 6,5 1,07 3,03 • 10” 0,98 1,05 0,47—1,14
е) красные (CdB2O5) . . . 20 9,9 74,5 72,5 7,2 1,18 3,03 • 10” 0,98 1,15 0,43—1,04
30 11,3 74,5 72,5 8,2 1,34 3,03 • 10” 0,98 1,31 0,38—0,92
15 8,3 70,5 69,4 5,7 0,93 3,04 • 10” 0,98 0,92 0,54—1,30
ж) золотистые . 20 8,0 70,5 69,4 6,2 1,02 3,04 • 10” 0,98 1,00 0,50—1,20
30 10,3 70,5 69,4 7,1 1,16 3,04 • 10” 0,98 1,14 0,44—1,05
3. Высокого давления
«HgL-ЗОО» . . 75 9,9 74,0 65,8 6,5 1,06 2,84 • 10” 0,91 0,97 0,51—1,22
«HgL-500» . . 120 10,5 74,0 65,8 6,9 1,13 2,84 • 10” 0,91 1,03 0,49—1,17
* В скобках поставлены люминофоры.
102
Физические и физиологические основы светокультуры растений
Как видно из табл. 48, физиологическая радиация у современных
ламп составляет 8—18% всей мощности, потребляемой лампами. Для
однотипных ламп ее доля, как правило, растет вместе с увеличением их
мощности. Для ламп накаливания одной и той же номинальной мощности
она примерно на 20% выше у ламп, рассчитанных на напряжение 120 в,
чем у ламп, рассчитанных на 220 в. Ртутные лампы высокого и сверх-
высокого давления более выгодны в отношении расхода электроэнергии,
чем лампы низкого давления.
Из всего многообразия ламп по степени использования подводимой
энергии предпочтение следует отдать люминесцентным лампам дневного,
мягкобелого и белого света мощностью в 30—40 вт, где физиологическая
радиация, почти целиком излучаемая в области видимых лучей, дости-
гает 15—19% всей подводимой мощности.
Поглощение хлорофиллом физиологической радиации различных ис-
точников колеблется в пределах 50—90%. Наибольшей величины это
поглощение достигает в спектре ртутных, неоновых, натриевых и некото-
рых люминесцентных ламп (белого и красного света) и наименьшей вели-
чины — в спектре ламп накаливания и люминесцентных ламп синего
и зеленого света.
Количество квантов в единице энергии физиологической радиации
для рассмотренных нами источников излучения равняется величине
порядка 1,65-1О31 — 3,54-Ю11 эрг-1. Больше всего квантов содержится
в излучении ламп накаливания, неоновых, натриевых и люминесцентных
ламп красного и белого света и меньше всего — в спектре ртутных и люми-
несцентных ламп синего и зеленого света. Заметим для сравнения, что
в 1 эрге физиологической радиации прямых солнечных лучей в летние
полуденные часы содержится 2,89-1011 квантов.
Произведение лучистой отдачи источника радиации на величину по-
глощения лучистой энергии хлорофиллом листа мы предлагаем назвать
теоретическим коэффициентом использования энергии. Этот коэффициент
отвечает использованию лучистой энергии растением, выраженному
в процентах от мощности источника радиации. Этот коэффициент рав-
няется 5—15%, достигая наибольшего значения у некоторых ртутных
и люминесцентных ламп и опускаясь до минимума у ламп накаливания
и натриевых ламп.
Для сопоставления эффективности разных источников радиации
в качестве эталона возьмем лампу накаливания в 300 вт, приравняв
единице характеризующие ее коэффициент использования и количество
квантов в единице энергии физиологической радиации. Произведение,
относительной величины коэффициента использования на относительное
количество квантов в единице энергии физиологической радиации позво-
ляет получить теоретическую эффективность ламп, выраженную в относи-
тельных единицах по отношению к лампе накаливания мощностью
в 300 вт.
Как видно из табл. 48, наиболее эффективными источниками радиации
следует признать ртутные лампы сверхвысокого давления, люминесцент-
ные лампы дневного, белого, мягкобелого и красного света (особенно
мощностью в 30 и 40 вт), неоновые лампы, некоторые натриевые лампы
и проекционные лампы накаливания.
Если исходить из интенсивности солнечной радиации, потребной
для нормального роста и развития тенелюбивых и светолюбивых растений,
в 50—120 тыс. эрг/см2-сек, что для прямых солнечных лучей в летние
полуденные часы соответствует падению 1,45-1016 — 3,47-Ю16 квантов
на 1 см2 в секунду, то следует признать, что растения для нормального
III. Источники радиации
103
развития и роста нуждаются в поглощении хлорофиллом листа
0.80-1015 16—1,90-10’’ квантов на 1 см2 в секунду. Как показывают расчеты,
чтобы обеспечить это условие посредством лампы накаливания в 300 вт
и идеального осветителя с коэффициентом полезного действия в 0,85,
направляющего весь лучистый поток в нижнюю полусферу и равномерно
распределяющего его на площади в 1 м2, необходима затрата удельной
мощности в 0,5—1,2 квт/м2. Теоретическая удельная мощность других
источников, необходимая для выращивания растений целиком на искус-
ственном свете, обратно пропорциональна эффективности ламп, выраженной
в относительных единицах по сравнению с лампой накаливания в 300 вт. В
табл. 48 показана удельная мощность для некоторых ламп, лучистый поток
которых посредством зеркального осветителя (коэффициент полезного
действия которого равен 0,85) целиком направляется в нижнюю полу-
сфер)- и равномерно распределяется на площади в 1 м2.
Приведенные в этой таблице удельныемощности сравнительно невелики.
Во всяком случае, они значительно меньше реальных мощностей, которые
употребляются в обычной практике и служат причиной ограниченного
распространения светокультуры растений. Мы приводим их для того,
чтобы, во-первых, сопоставить экономичность разных источников излу-
чения, а во-вторых, показать, что успех светокультуры растений в значи-
тельной степени определяется правильным подбором источника радиации
и отражателя, позволяющего с наименьшими потерями использовать
лучистый поток. Современные отражатели, разработанные для других
целей, мало пригодны для культуры растений. Задача заключается в том,
чтобы создать отражатель, удовлетворяющий специфическим запросам
культуры растений при помощи искусственных источников радиации.
15. РАЗМЕЩЕНИЕ ИСТОЧНИКОВ РАДИАЦИИ
При удельных мощностях разных источников света, приведенных
в табл.’48, нельзя ожидать одинакового накопления сухого вещества
растениями вследствие различий их формы (розеточная, стеблевая) и
разной реакции на спектральный состав излучения. Одинаковое накопле-
ние сухого вещества может происходить лишь у видов, имеющих розе-
точную форму, не прошедших стадии яровизации, или фотопериодически
нейтральных и, следовательно, не зависящих в своем развитии от спек-
трального состава радиации (земляника, репа и др.).
Благодаря расположению листьев на одном уровне и в одной пло-
скости розеточные растения могут быть равномерно освещены при помощи
источников, расположенных над растениями на некоторой высоте. Поло
жение меняется в случае культур типа томатов, огурцов и других ра-
стений, обладающих многоярусной системой побегов и листьев. Эти ра-
стения практически невозможно равномерно осветить при расположении
источников над растениями. В этом случае нормально функционируют
лишь верхние листья, получающие достаточное количество света; ниж-
ние же листья находятся в тени. Наиболее целесообразным в этом случае
следует признать расположение источников в вертикальной плоскости
в междурядиях растений. При подобном размещении источников радиации
удельная мощность, т. е. мощность, отнесенная к единице освещаемой пло-
щади (вт/м2), может быть в несколько раз больше, чем при расположении
источников над растениями. Конкретное решение проблемы в данном
случае определяется характером культур, высотой растений, их густо-
той, расположением и т. д.
104
Физические и физиологические основы светокультуры растений
Таким образом, имеются два принципиально различных приема раз-
мещения источников излучения: над растениями и в междурядиях ра-
стений. Оба приема имеют свои достоинства и недостатки, и выбор приема
зависит от высоты растений.
Сравнительно просто решается задача размещения трубчатых источ-
ников радиации, имеющих холодные стенки (люминесцентные лампы,
неоновые лампы с холодным катодом, ртутные лампы низкого давления
'с?' /сГ' ''о4 ''о
Фиг. 65. Размещение трубчатых источников радиации
для культуры растений
а — в горизонтальной плоскости над растениями; б — в верти-
кальной плоскости между растениями (лампы стоят вертикально);
в —-то же, лампы расположены горизонтально
и др.). Так как соприкосновение растений с этими источниками радиации
не вызывает ожогов, их можно размещать в непосредственной бли-
зости от растений (на расстоянии 5—10 см) как в горизонтальной
плоскости, над растениями, так и в вертикальной плоскости, в между-
рядиях растений (фиг. 65—69). Последнее следует особенно рекомендо-
вать для теплиц, так как в этом случае лампы не мешают освещению
естественным солнечным светом, не затеняют растений (фиг. 66).
Сами лампы могут быть расположены как вертикально, так и гори-
зонтально. Для коротких трубок типа люминесцентных ламп мощ-
ностью в 15 вт предпочтение следует отдать вертикальному положению
(фиг. 65, б), а длинные трубки (более 1 м) удобнее располагать горизон-
тально (фиг. 65, в). В этом случае, как показано на схеме (фиг.65), они
включаются по мере роста растений и выключаются снизу по мере
III. Источники радиации
105.
старения и отмирания нижних листьев. Рамы размещают на расстоянии
30—50 см одна от другой.
Фиг. 66. Теплица, освещаемая люминесцентными лампами, расположенными
в междурядиях растений
(Лаборатория искусственного климата Московской сельскохозяйственной академии
им. К. А. Тимирязева)
Фиг. 67. Рама для вертикального расположения люминесцентных .ламп
Переносная конструкция для вертикального размещения люминес-
центных ламп в вертикальной плоскости в междурядиях растений
106
Физические и физиологические основы светокультуры растений
показана на фиг. 67. Длина рамы равняется 82 см, а расстояние между
лампами — 7 см.
В подобной раме длинные люминесцентные лампы могут быть разме-
щены и горизонтально (фиг. 68). Раму прикрепляют к дросселям, кото-
рые одновременно служат подставкой, придающей раме устойчивое
Фиг. 68. Рамы для горизонтального расположения люми-
несцентных ламп
а — коробка, закрывающая дроссели
(Лаборатория искусственного климата Московской сельскохозяй-
ственной академии им. К. А. Тимирязева)
положение. На наших рисунках дроссели видны под рамой в нижних частях
рисунков. На фиг. 68 они не видны, так как закрыты крышкой (а). Такими
рамами, по нашему предложению, оборудованы теплицы Лаборатории
искусственного климата Московской сельскохозяйственной академии
им. К. А. Тимирязева.
В некоторых случаях дроссели можно заменить обычными лампами
накаливания. Последние, работая на недокале, одновременно служат
источником оранжево-красного излучения. Для люминесцентных ламп
мощностью 15 вт в качестве подобного балластного сопротивления могут
служить лампы накаливания с номинальной мощностью 75 вт (при
III. Источники радиации
107
напряжении в 127 в). Включенные последовательно с люминесцентными
лампами, они приобретают мощность порядка 30 вт. Рама, частично со-
держащая вместо дросселей лампы накаливания, показана на фиг. 69.
Одна половина люминесцентных ламп смонтирована с дросселями, рас-
положенными внизу рамы, а вторая половина имеет балластное сопротив-
ление в виде ламп накаливания (вверху рамы).
В настоящее время возникла настоятельная необходимость в кон-
структивной разработке универсальных осветительных рам для размеще-
Фиг. 69. Рама для вертикального расположения люминесцентных
ламп с частичной заменой дросселей лампами накаливания
ния люминесцентных ламп в зависимости от вида растении как в горизон-
тальной, так и в вертикальной плоскости.
Б. ('.. Мошков 1216] для создания равномерного пространственного
распределения лучистой энергии рекомендует употреблять низковольт-
ные маломощные автомобильные и’ автотракторные лампы накаливания,
равномерно распределяя их в пространстве, занимаемом растениями.
Более сложная и трудная задача — размещение мощных источников,
выделяющих значительное количество тепла (лампы накаливания, нео-
новые дуговые лампы, ртутные лампы высокого и сверхвысокого давле-
ния и др.). Высокая температура стенок ламп не позволяет размещать
эти источники в непосредственной близости от растений. Повидимому,
единственным решением является помещение ртих источников радиации
над растениями на определенной высоте. В связи с большей лучистой
отдачей более выгодно употребление мощных источников, позволяющих,
кроме того, освещать большую площадь наименьшим количеством
осветительных точек.
108 Физические и физиологические основы светокультуры растений
16. ОСВЕТИТЕЛИ (СВЕТИЛЬНИКИ)
Размещение источников радиации над растениями требует применения
осветителей, направляющих весь лучистый поток в нижнюю полусферу,
на площадь, занимаемую растениями [13, 69, 203, 3211. Осветитель, или
светильник, пригодный для культуры растений, состоят из источника
радиации и отражателя (рефлектора). Последний представляет собою
1 —лампа без отражателя; 2 —• железный эмалированный отражатель «Аль-
фа»; 3 — железный эмалированный глубокоизлучатель; 4 — зеркальный
глубокоизлучатель; 5 — зеркальная лампа ЗН-6
определенной формы колпак с белой или зеркальной внутренней поверх-
ностью, отражающей лучистый поток в нужном направлении. Всякий
осветитель характеризуется коэффициентом полезного действия, коэф-
фициентом усиления, кривой распределения лучистого потока в верти-
кальной (и горизонтальной) плоскости и величиной телесного угла,
внутрь которого излучается лучистый поток.
Коэффициент полезного действия осветителя есть
отношение лучистого потока осветителя (Р) к лучистому потоку лампы (PL)
1 = ^- (31)
III. Источники радиации
109
В лучшем случае (у зеркальных отражателей) коэффициент достигает
значения 0,85, у обычных же эмалированных отражателей колеблется
в пределах 0,55—0,60.
Коэффициент усиления есть отношение максимальной
силы света (/,„) осветителя к средней сферической силе света лампы (10):
3 = ^. (32)
-'о
У зеркальных светильников коэффициент усиления может достигать
5,5, а у эмалированных осветителей он равняется 2,6.
УглЬ/ (S’) Mej/cfy асб/о ламаЬ и направлением лучей
Фиг. 71. Изменение освещенности, создаваемой условной лампой
в 1000 лм, в зависимости от расстояния от вертикальной оси лампы.
Высота подвеса — 100 см
1 — лампа беа отражателя; 2 —. пампа с отражателем «Альфа»; 3 — лампа
с эмалированным глубокоиэлучателем; 4 — лампа с зеркальным глубоко-
излучателем
Распределение лучистой энергии в пространстве, получаемое в резуль-
тате действия осветителя, обычно характеризуется кривой распределения
силы света в вертикальной плоскости. Положение каждой точки, лежащей
на кривой, соответствует величине силы света в данном направлении.
Последнее, в свою очередь, определяется величиной угла а по отношению
к оси осветителя. Примеры подобных кривых можно видеть на фиг. 70.
В настоящее время отсутствуют отражатели, удовлетворяющие тре-
бованиям светокультуры растений. Обычно применяемые для ламп нака-
ливания отражатели типа глубокоизлучателей, распределение энергии
которых приведено на фиг. 70, а характеристика — в табл. 49, мало при-
годны из-за неравномерной освещенности, создаваемой светильниками,
и слишком большого телесного угла, в пределах которого излучается
энергия.
Как видно из фиг. 71, освещенность, создаваемая на горизонтальной
поверхности осветителями, отличается исключительной неравномер-
110
Физические и физиологические основы светокультуры растений
ностью, являясь максимальной непосредственно под лампами и сильно
снижаясь уже при незначительном удалении в сторону от нее. В связи
с большим телесным углом светильников большое количество энергии
излучается в стороны и пропадает для растений.
«/ (в °)
Фиг. 72. Распределение в вертикальной плоскости силы
света идеального светильника для культуры растений
Таблица 49
^Характеристика отражателей для ламп накаливания
Отражатели Мощность ламп (в вт) Защитный угол (в с) Телесный угол (в стр) Коэффи- циент полез- ного действия Коэффи- циент усиле- ния Сила света (в св) в разных направлениях (в ° угла)
0° 5° 15° ! I 25° | 35° 45° 55? 65°
Железный эма- лированный «Альфа» . . <60 31 2,93 0,57 2,6 212 212 210 200 193 170 102 34
Железный эма- лированный глубокоизлу- чатель . . . <1000 27 3.45 0,58 2,6 263 269 259 237 214 179 91 30
Зеркальный глубокоизлу- чатель . . . <500 35 2,80 0,85 5,5 392 436 378 361 382 265 90 0
Ill. Источники радиации
111
Идеальным осветителем для культуры растений следует признать
светильник с вертикальным распределением силы света, выражающимся
кривой следующего уравнения:
где Zo — сила света в направлении оси светильника (7.7=0), а I — сила
света в направлении под углом а.
Фиг. 73. Распределение в вертикальной плоскости силы света для
зеркальных ламп ВЭИ
1 — лампа широкого светораспределепия; 2 — лампа преимущественно направ-
ленного светораспределения; Я — лампа концентрированного светораспределе-
ния; 4—5 лампа «Кососвет» концентрированного светораспределения (4 — в
плоскости симметрии; .5— в плоскости, перпендикулярной плоскости симметрии)
В случае подобного распределения лучистой энергии (фиг. 72) ос
вещенность в пределах телесного угла будет равномерной. Из всех
существующих в настоящее время светильников наиболее простыми
следует признать зеркальные лампы, для которых не нужны отражатели.
Зеркальные лампы широкого светораспределения (фиг. 73,1) при низком
подвесе дают наиболее равномерное освещение (фиг. 107). Полной равно-
112 Физические и физиологические основы светокультуры растений
мерности освещения можно добиться лишь при помощи подвижной си-
стемы ламп или более частым размещением ламп при соответствующем
уменьшении их мощности и высоты подвеса.
Сказанное относится к осветителям с точечными источниками радиа-
ции (лампами накаливания, дуговыми и ртутными лампами сверхвысо-
кого давления и др.), размеры которых относительно невелики по сравне-
нию с расстоянием между источником и освещаемым объектом. Значи-
тельно хуже обстоит дело с отражателями к трубчатым источникам
излучения — неоновым, люминесцентным, некоторым ртутным лампам
и др. Перераспределения лучистого потока здесь удается добиться лишь
в направлении, перпендикулярном к оси лампы. Его распределение
в продольном направлении изменить посредством обычных корытообраз-
ных отражателей не удается.
Рациональным решением вопроса здесь может служить лишь под-
вижная система трубок, располагаемых параллельно одна другой вдоль
направления движения, и применение отражателей с распределением лу-
чистого потока в плоскости, перпендикулярной к оси трубки, выражаемым
уравнением (33). Подобным приемом можно добиться наиболее полного
использования лучистого потока и создания равномерной интенсивности
радиации в различных точках стеллажа.
17. ОХЛАЖДЕНИЕ ИСТОЧНИКОВ РАДИАЦИИ
Большинство источников радиации, пригодных для культуры растений,
одновременно являются источниками большого количества тепла, которое
выделяется или в виде инфракрасных лучей, или путем конвекции от
стенок ламп и приборов включения. Достаточно сказать, что темпера-
тура разряда натриевых ламп равняется 540° К; у ртутных ламп низкого
давления она колеблется в пределах 600—800° К, а у ламп высокого дав-
ления достигает 6400° К. Мы уже отмечали, что рабочая температура
ламп накаливания может достигать 3200° К, а у дуговых ламп 4000° К.
Следовательно, в связи с высокой температурой процессов лучеис-
пускания, всякий источник радиации в спектре излучения должен со-
держать и содержит определенное количество тепловых инфракрасных
лучей. Различие между источниками заключается лишь в том, что у одних
максимум инфракрасного излучения приходится на область 3—6 р.
(натриевые лампы и ртутные лампы низкого давления), у других же он
смещается в сторону более коротких волн — 0,8—1,5 р (лампы накали-
вания, дуговые лампы и др.). Даже у относительно холодных люминес-
центных ламп инфракрасная радиация составляет значительную часть
излучения, а у ламп накаливания она достигает 90% всех испускаемых
лучей. У ртутных ламп сверхвысокого давления и натриевых ламп со-
держание инфракрасных лучей снижается до 30%, а у ртутных ламп
высокого и низкого давления — до 10—15%.
Обычно стенки ламп, будь это лампы накаливания или ртутные трубки,
обладают сравнительно высокой температурой, вызывающей гибель и
отмирание тканей растений при их соприкосновении с лампами. Стенки
ламп накаливания мощностью в 50—1000 вт нагреваются до 125—188°.
То же самое следует сказать о дуговых неоновых лампах, ртутных лампах
высокого и сверхвысокого давления и других. Лишь стенки люминес-
центных ламп нагреваются незначительно по сравнению с температурой
окружающего воздуха, и соприкосновение этих ламп с органами растений
не приносит вреда последним.
III. Источники радиации
113
‘ Таким образом, охлаждение ламп или, по крайней мере, устранение
отрицательного аффекта инфракрасных лучей — чрезвычайно важная
проблема в светокультуре растений.
Одним из наиболее ранних и чаще всего употребляемых приемов
охлаждения как ламп накаливания, где в каждую минуту на каждые
1000 вт потребляемой мощности выделяется около 10 тыс. кал тепла, так
и других ламп является водный экран. Предложенный впервые в 1911 г.
[268], этот способ получил затем широкое распространение в лаборатор-
ных опытах, особенно по выращи-
ванию растений целиком на искус-
ственном свете [381, 449,548, 848].
Фиг. 74. Способы применения водных
экранов для охлаждения ламп накали-
Фиг. 75. Лампа, охлаждаемая водою
(общий вид)
вания
А — лампы над водным экраном; Б — лампы
погружены в воду
а — лампы; б — приток воды; в — слой про-
точной воды; г — отток воды
Метод водного экрана заключает-
ся в том, что лампы накаливания
помещают над потолком из тол-
стого стекла по которому беспре-
рывно течет слой холодной воды толщиной в 1—10 см.
В настоящее время лампы накаливания для охлаждения и устране-
ния инфракрасных лучей просто погружают в проточную воду вплоть до
наиболее горячей цилиндрической части колбы [109, 184, 216, 218, 219].
В этом случае значительно снижается также потеря лучистой энергии
в результате устранения отражения лучей на границе между воздухом и
водой, достигающего значительных размеров (фиг. 74).
Крупным недостатком описанного приема является невозможность
его применения в производственных условиях — в теплицах
Необходимость периодического очищения стеклянного потолка от
минеральных осадков и водорослей, интенсивно развивающихся в воде,
ограничивает применение этого метода и в лабораторных условиях.
Оригинальным решением проблемы является метод охлаждения каж-
дой лампы посредством помещения ее в специальную стеклянную колбу
с дестиллированной водой, которую охлаждают током холодной водопро-
8 А. Ф. Клешнин
114
Физические и физиологические основы светокультуры растений
водной воды, пропускаемой через трубку, спирально располагаемую
вокруг лампы 1517, 785]. Конструкция, подобного охлаждения для ламп
Фиг. 76. Схема лампы, охлаждаемой
водою
а — патрон; б — фарфоровый цоколь; е—стек-
лянная колба, наполненная дестиллированной
водой; г—медная спираль стоком водопро-
водной воды; д — кольцо; е — лампа накали-
вания; ж — предохранительный клапан; з —
наполнитель
Коэффициенты поглощения (ai
накаливания показана на фиг. /о и
76. Этот метод, лишенный недостат-
ков метода простого водного экрана,
заслуживает большого внимания, так
как его можно применять в теплицах.
В основе метода устранения ин-
фракрасных лучей посредством тока
холодной воды лежит свойство по-
следней поглощать инфракрасные лу-
чи без значительного ослабления
физиологической радиации.
Зная коэффициенты поглощения
(экстинкции) (К) воды (табл. 50), по-
средством формулы Буге-Ламберта
ЕТ=Ш-е~ке1 (34)
или
£у.= 100 • 10-АЧ (34а)
можно легко рассчитать пропускание
лучистой энергии (7:т) слоем воды
толщиною в I сантиметров, выражен-
ное в процентах к падающей энергии.
На фиг. 77 приведены кривые, по-
лученные подобным способом.
Как видно из фиг. 77, применяя
воду, можно устранить лишь инфра-
красные лучи с длиною .волны более
1 у. Полное устранение инфракрас-
ной радиации невозможно даже при
помощи слоя в несколько метров.
Фиг. 77, 7 дает представление о спек-
тре излучения лампы накаливания
Таблица 50
пинкции) дестиллированной воды
Длина волны (в та.) Для основания натуральных логарифмов Ке Для основания десятичных логарифмов Кю ' Длина волны (в та.) Для основания натуральных логарифмов К в Для основания десятичных логарифмов К1о
250 0,316 0,137 900 0,0186 0,00807
300 0,100 0,0434 1000 0,447 0,193
400 0,0263 0,0114 1050 0,186 0,0807
500 0,0100 0,00434 1100 1,21 0,525
600 0,000263 0,000114 1180 31,6 13,7
650 0,00011 $ 0,0000476 1200 5,89 8,65
700 0,00012 0,0000521 1300 129,0 56,0
800 0,00144 0,000608 1350 20,4 8,86
870 0,0263 0,0114 1400 100,0 42,4
III. Источники радиации
115-
в 2000 вт, получаемого в результате прохождения лучей через водный
экран толщиною в 10 см. Физиологическая радиация проходит почти без
изменения, если не считать некоторого поглощения ультрафиолетовых
и сине-фиолетовых лучей, а из инфракрасных лучей остаются лишь корот-
кие лучи, примыкающие к видимой части спектра и не поглощаемые
тканями листа (750—1000 т;л).
Спектр лампы накаливания после прохождения лучей через водный
экран в 10 см содержит 51 % физиологической радиации и 49% инфра-
красных лучей. Слой воды в 10 см устраняет инфракрасную радиацию
примерно на 85%. Это означает, что для охлаждения лампы в 1000 вт,
Фиг. 77. Пропускание лучистой энергии дестиллированной
водой при разной толщине ее слоя
1 — 1 см; 2 — 2 см; 3 — 5 см; 4 — 10 см; 5 — 2 м; 6 — спектр излучения
лампы накаливания в 2 квт; 7 — то же при наличии водного экрана
в 10 см
выделяющей ежеминутно 10000 г-кал тепла, требуется ток воды со ско-
ростью 8,5 л/мин.
Так как излучение лампы накаливания в результате прохождения
через водный экран содержит физиологические и инфракрасные (абиоти-
ческие) лучи в отношении 1:1, то многие исследователи, например
Б. С. Мошков [224], полагают, что при употреблении водного фильтра
спектр ламп накаливания становится идентичным спектру естественного
солнечного излучения в летние полуденные часы. Подобное утверждение,
с точек зрения физиолога и физика, не выдерживает никакой критики,
так как спектральный состав излучения в обоих случаях остается совер-
шенно различным. Достаточно сопоставить фиг. 77, 7 с фиг. 80, чтобы
убедиться в неверности этого утверждения.
Другой, практически более целесообразный прием устранения отри-
цательного действия инфракрасных лучей — метод движения ламп. Этот
метод, кстати сказать, является единственным для дуговых неоновых
ламп или ртутных ламп высокого и сверхвысокого давления. Впервые
он был применен в Лондонском ботаническом саду в 1911 г. для культуры
растений в оранжерее [268]. Мощный осветительный фонарь помещали
под крышей, и он медленно двигался взад и вперед вдоль теплицы. Позднее
8*
116
Физические и физиологические основы светокультуры растений
этот принцип использовали Трюфо и Тюрнессан [814, 815], в опытах
которых лампы накаливания находились во вращательном движении
в горизонтальной плоскости над растениями.
Инженер И. Н. Финкельштейн [324, 325, 564] разработал простую
и оригинальную конструкцию подвеса ламп, посредством которой им при-
дают возвратно-поступательное движение вдоль стеллажа. Эта система
в Советском Союзе находит все более и более широкое распространение
в теплицах научных учреждений и овощных хозяйств [18, 230, 351].
Помимо устранения перегрева растений, она обеспечивает и равномер-
ность их освещения.
Лампы накаливания, как и другие источники радиации, излучающие
большое количество тепловых лучей, позволяют выращивать растения
в условиях сравнительно невысоких температур при наличии специаль-
ного подогрева почвы. Возможно, что применение холодных оранжерей
с почвенным обогревом окажется в дальнейшем одним из радикальных
решений проблемы отрицательного влияния инфракрасной радиации
ламп. К сожалению, ни тепловой баланс листьев, ни температурный
режим «холодных» теплиц, освещаемых лампами накаливания или дру-
гими аналогичными источниками, до сих пор не привлекли должного
внимания исследователей.
18. ВЫВОДЫ
1. Для культуры растений пригодны любые современные источники
радиации, если в их спектре не содержится витацидной радиации и если
мощность источников достаточно велика.
2. Все существующие в настоящее время лампы недостаточно эконо-
мичны. Физиологическая радиация обычно колеблется в пределах 5—20%
подводимой мощности, а поглощение этой радиации хлорофиллом состав-
ляет 50—90%. Иными словами, в лучшем случае растения могут исполь-
зовать на создание органического вещества не более 2,5—18% энергии,
потребляемой лампами.
3. В настоящее время важнейшей проблемой светотехники, помимо
создания более экономичных источников излучения, является разработка
рациональных осветителей, дающих строго направленный пучок и со-
здающих равномерное освещение на определенной площади.
4. Другая важная проблема — размещение источников радиации и
устранение избытка инфракрасных лучей, вызывающих перегрев растений.
Глава IV
СОЛНЕЧНАЯ РАДИАЦИЯ
Солнечная радиация — ее интенсивность, спектральный состав и су-
точная продолжительность — представляет собою решающий экологиче-
ский фактор местообитания растений. Знание этого фактора необходимо
для разработки рационального реяшма искусственного освещения ра-
стений в теплицах, парниках и других сооружениях закрытого грунта.
1. ОБЩАЯ ХАРАКТЕРИСТИКА
Солнце есть тепловой источник радиации, на поверхности которого
температура равняется 5730° К. Солнечное излучение подчиняется зако-
нам черного излучения, которые были рассмотрены нами в предыдущей
Фиг. 78. Схема прохождения солнечных лучей через атмосферу
Линия аб — поверхность Земли; вд — граница атмосферы; ог — толщина атмосфе-
ры; h— высота солнцестояния; т;, — масса атмосферы при высоте солнцестояния h;
mz — масса атмосферы при положении Солнца в зените; т„ — масса атмосферы
при h = 0°
главе. Как интенсивность, так и спектральный состав солнечной радиа-
ции, достигающей поверхности Земли, определяется в основном высотой
солнцестояния. Метеорологические факторы — облачность, содержание
водных паров в атмосфере, мутность и прозрачность последней — играют
значительно меньшую и подчиненную роль.
Высота солнцестояния выражается углом/г между направлением лучей
и линией горизонта (фиг. 78). Эта величина изменяется на протяжении
суток, зависит от времени года и, кроме того, определяется географиче-
ской широтой. Полуденные значения h показаны в табл. 51.
Из табл. 51 видно, что в пределах СССР максимальная высота солнце-
стояния наблюдается 23 июня в районе ст. Кушка (Туркменская ССР),
где в это время она достигает 78°, а минимальная — 23 декабря на Земле
Франца-Иосифа, где в зимнее время солнце опускается на 8—10° ниже
линии горизонта. В средней полосе СССР, лежащей на линии 55-й парад-
118 Физические и физиологические основы светокультуры растений
Таблица 51
Высота солнцестояния в полуденные часы на различных широтах
в пределах СССР
(В. В. Шаронов [342])
Дата Широта (в °)
35 40 45 50 55 60 65 70
Январь 1 31 26 21 16 11 6 2 —3
11 33 28 23 18 13 8 3 —1
21 35 30 25 20 15 10 5 0
31 38 33 28 23 18 13 8 3
Февраль 10 40 36 31 26 21 16 11 6
21 44 39 34 29 24 19 14 9
Март 2 47 42 38 32 27 22 17 12
12 51 46 41 36 31 26 21 16
22 55 50 45 39 35 30 25 20
Апрель 1 60 55 50 44 40 35 30 25
11 64 58 54 48 44 38 34 29
21 67 62 57 51 47 42 37 32
Май 1 70 66 60 54 50 45 40 35
11 73 68 62 57 53 48 43 37
21 75 70 64 59 55 50 45 40
31 76 71 65 60 57 52 47 41
Июнь 10 77 72 67 62 57 53 48 42
20 78 73 68 63 58 53 48 43
30 77 73 67 63 58 53 48 43
Июль 10 76 72 66 62 57 52 47 42
20 75 70 65 60 59 50 45 40
30 73 68 63 58 53 49 43 39
Август 9 70 66 61 56 51 46 41 36
19 67 63 58 53 48 43 38 33
29 64 59 54 49 44 39 34 29
Сентябрь 8 61 56 51 46 41 35 31 26
18 57 52 47 42 36 32 27 22
28 54 48 43 38 33 28 23 18
Октябрь 8 49 44 39 34 29 24 19 14
18 45 40 35 30 26 20 15 11
28 41 36 31 26 22 16 11 6
Ноябрь 7 38 33 28 23 18 13 8 2
17 36 31 26 22 16 11 6 1
27 34 29 24 19 14 8 4 —1
Декабрь 7 32 27 22 17 12 7 2 —2
17 31 26 21 16 11 6 2 —3
27 31 26 21 16 11 6 2 —3
IV. Солнечная радиация
119
лели (Москва—Казань—Свердловск—Новосибирск—Николаевск-на-
Амуре) полуденная высота Солнца над горизонтом в летнее время
достигает 58°, а зимою снижается до 11°.
Как видно из фиг. 78, высота солнцестояния (/г) определяет массу
атмосферы (т) — путь, проходимый солнечными лучами через толщу атмо-
сферы Земли. Значение т для различных высот солнцестояния приведено
в табл. 52 и на фиг. 79. Именно от величины этого пути зависит интенсив-
ность и спектральный состав солнечных лучей, достигающих поверх-
ности Земли. Масса атмосферы достигает наименьшего значения при
Фиг. 79. Влияние высоты солнцестояния (Л) па массу атмо-
сферы (т) и интенсивность радиации (/. г-кал/см2-мин)
1 — масса атмосферы; г — интенсивность прямых солнечных лучей
на нормальной поверхности; 3 — то же на горизонтальной поверх-
ности; 4 — интенсивность диффузной радиации
положении Солнца в зените (znz) и увеличивается по мере снижения
высоты солнцестояния.
Т а б л и ц а 52
Зависимость массы атмосферы опг высоты солнцестояния
h (в ') 90 60 . 30 10 5 3 0,5
гп 1 1,2 2,0 5,6 10,4 15,4 35,4
Масса атмосферы влияет на солнечную радиацию двояким образом.
Прежде всего следует отметить рассеивание прямых солнечных лучей
молекулами газов, входящих в состав атмосферы. Этот процесс совершается
по закону Рэлея, который гласит, что интенсивность рассеянной радиа-
ции равняется величине, обратно пропорциональной четвертой степени
длины волны:
120
Физические и физиологические основы светокультуры растений
Это означает, что область излучения с длиною волны в 340 ту рассеи-
вается в 16 раз интенсивнее, чем область максимума поглощения лучи-
стой энергии хлорофиллом (680 ту). Из этого следует далее, что по мере
снижения высоты солнцестояния прямая солнечная радиация относи-
тельно обедняется коротковолновыми ультрафиолетовыми и сине-фиоле-
товыми лучами и обогащается длинноволновыми оранжево-красными
лучами. Одновременно увеличивается доля рассеянно!! радиации.
Наряду с рассеиванием,в атмосфере происходит поглощение лучей не-
которых областей солнечного спектра и, следовательно, изменение спек-
трального состава солнечной радиации. Озон, содержащийся в верхних
слоях атмосферы, целиком поглощает короткие ультрафиолетовые лучи.
Благодаря этому обстоятельству вредные витацидные лучи с длиною волны
короче 295 ту не достигают поверхности Земли.
Большое влияние на интенсивность и спектральный состав солнечной
радиации имеет содержание в атмосфере водных паров, поглощающих
главным образом инфракрасные лучи. Водные пары, количество которых
колеблется в пределах от 0,01 до 4%, поглощают до 20% всей энергии
падающих солнечных лучей. Это поглощение представлено на фиг. 80
в виде широких впадин у нижней кривой. Во избежание чрезмерного
усложнения диаграммы мы опускаем их у других кривых.
2. СПЕКТРАЛЬНЫЙ СОСТАВ СОЛНЕЧНОЙ РАДИАЦИИ
Спектральный состав солнечной радиации на границе земной атмо-
сферы показан на фиг. 79,7. Солнечная радиация практически пред-
Фиг. 80. Распределение энергии в солнечном спектре при разной
высоте солнцестояния
1 — на границе земной атмосферы; 2— 5—на поверхности Земли при высоте
солнцестояния: 2— 65'; 3— 30 ; 4— 19,3; ,5— 11,3 Впадины (вп) отвечают
поглощению лучистой энергии парами воды атмосферы. Вертикальные линии —
границы видимой радиации
ставлена здесь излучением, лежащим в пределах 150—3000 ту с мак-
симумом около 490 ту. В результате влияния земной атмосферы, по-
верхности Земли достигают лишь лучи с длиною волны более 295 ту.
IV. Солнечная радиация
121
Спектральная характеристика солнечной радиации на поверхности
Земли представлена на фиг. 80 и 81 и в табл. 54. Как видно из этих кривых,
спектральный состав, положение максимума и граница излучения
в области ультрафиолетовых лучей определяются высотой солнцестояния.
При уменьшении высоты солнцестояния солнечный спектр обогащается
оранжево-красными лучами, а максимум излучения и его ультрафиоле-
товая граница смещаются в сторону более длинноволновых лучей (фиг. 80 и
табл. 53). При положении Солнца в зените максимум приходится на
490 шщ, а при солнцестоянии в 11° он смещается в область 710 тц.
Фиг. 81. Коэффициент прозрачности сухого воздуха и распределение энергии
в спектре физиологической радиации пряных солнечных лучей и рассеян-
ного света при высоте солнцестояния:
2 — 65°; 2—30?; 3— 19\3; 4—11°,3; 5—-рассеянный свет голубого неба;
6 — рассеянный свет при сплошной низкой облачности; 7 — коэффициент прозрачности
сухого воздуха
Т а б л и ц а 53
Влияние высоты солнцестояния на границу излучения в области ультрафиолетовой
радиации
Сопоставление табл. 51 и 53 позволяет сделать заключение, что в сред-
ней полосе СССР в спектре солнечной радиации в зимние месяцы совершенно
отсутствуют ультрафиолетовые лучи короче 306 шц. Зато в полуденные
часы летних месяцев спектр ультрафиолетовой радиации простирается
вплоть до 295 тц. В этой связи следует подчеркнуть, что в условиях
теплиц и парников, застекленных обычным стеклом, к растениям никогда
не доходят ультрафиолетовые лучи с длиною волны менее 340 пщ
(фиг. 35).
Помимо прямых солнечных лучей, падающих на земную поверхность
под углом h, солнечная радиация содержит также определенное количе-
ство диффузной радиации, возникающей в результате рассеивания лучей
атмосферой; эти лучи падают под самыми различными углами. Спектраль-
ный состав этого вида солнечного излучения не отличается постоянством.
Он зависит от прозрачности воздуха, облачности и других метеорологиче-
122
Физические и физиологические основы светокультуры растений.
ских факторов. Спектр рассеянной радиации голубого неба и при мощ-
ной облачности показан на фиг. 81, <5, 6. Заметим кстати, что первую легко
Фиг. 82. Относительное изменение спектрального состава солнеч-
ной радиации в зависимости от высоты солнцестояния
1 — ультрафиолетовые лучи (<400 ту.); 2 — фиолетовые лучи (400—440 т?);
3 — синие лучи (440—490 т;,.); 4 — зеленые лучи (490—565 тн);
5 — желтые лучи (565—595 т;>.); 6 — красные лучи (595—760 тл); 7 — ин-
фракрасные лучи (>760 т;,.)
имитировать люминесцентными лампами синего света с люминофором из
MgW(b (фиг. 95).
Т а б л и ц а 54
Относительная интенсивность различных областей спектра солнечной радиации
при разной высоте солнцестояния
(в % к общему излучению)
Высота солнцестояния (в с)
Излучение 0,5 5 10 20 30 50 90
Ультрафиолетовое (295—400 ту.) . . . 0 °,4 1,0 2,0 2,7 3,2 4л
Видимое (400—760 ту.) 31,2 38,6 41,0 42,7 43,7 43,9 45,3
В том числе:
фиолетовое (400—440 ту.) 0 0,6 0,8 2,6 3,8 4,5 5,4
синее (440—490 ту) 0 2,1 4,6 7,1 7,8 8,2 9,0
зеленце (490—565 ту.) 1,7 2,7 5,9 8,3 8,8 9,2 9,2
желтое (565—595 ту) 4Д 8,0 10,0 10,2 9,8 9,7 10,1
красное (595—760 ту) 25,4 25,2 19,7 14,5 13,5 12,2 11,5
Инфракрасное (>760 ту) 68,8 61,0 58,0 55,3 54,6 52,9 50,0
IV. Солнечная радиация
123
Физиологическая радиация в солнечном излучении содержит ультра-
фиолетовые и видимые лучи. В зависимости от высоты солнцестояния
Длина валнЬ/ (в тр)
Фиг. 83. Поглощение солнечной энергии пигментами пластид при высоте солнце-
стояния 65°
1 — распределение энергии в солнечном спектре; 2 — распределение квантов в солнечном спектре;
3 — поглощение нваптов листом (хлорофилл+каротипопцы); 4— поглощение квантов хлорофиллом
количество первых в общем лучистом потоке колеблется от нуля до 5%,
а вторых — от 30 до 45%. Остальные 50—70% приходятся на инфракрас-
Фнг. 84. Поглощение солнечной энергии пигментами пластид
при высоте солнцестояния И°,3
1 — распределение энергии в солнечном спектре; 2 — распределение квантов в сол-
нечном спектре; 3—поглощение квантов листом (хлорофилл+каротиноиды);
4 — поглощение квантов хлорофиллом
ные лучи (табл. 54, фиг. 82). В естественных условиях по мере уменьше-
ния высоты солнцестояния, что бывает на протяжении суток, при пере-
124
Физические и физиологические основы светокультуры растений
ходе от лета к зиме и при перемещении от экватора к полюсам, происхо-
дит относительное увеличение содержания инфракрасных и красных
лучей и уменьшение содержания желтых, зеленых, сине-фиолетовых и
ультрафиолетовых лучей. Эти изменения в соотношении различных обла-
стей спектра совершаются настолько незначительно и постепенно, что
можно говорить о практической неизменности спектрального состава
солнечной радиации при изменении солнцестояния от 90 до 20°. Лишь
дальнейшее уменьшение солнцестояния, за пределы 20°, имеет реальное
значение для спектрального состава солнечной радиации. Сказанное сле-
дует иметь в виду, когда идет речь о сезонной, суточной и широтной
изменчивости спектрального состава солнечного излучения.
На фиг. 83 и 84 показано распределение квантов в спектре солнечной
радиации и их поглощение пигментами пластид при различной высоте
солнцестояния. Кривые показывают, что даже в полуденные часы летом, при
максимальной высоте солнцестояния в средних широтах, наибольшее коли-
чество квантов приходится на оранжево-красные лучи — максимум погло-
щения лучистой энергии хлорофиллом. В этом случае количество квантов
в 1 эрге физиологической радиации равно 2,89-1011. С уменьшением
высоты солнцестояния это количество увеличивается и при высоте в 11°
достигает величины 3,14-1011 на 1 эрг.
Поглощение физиологической радиации пигментами пластид рав-
няется 60—70% от падающей физиологической радиации. Процент по-
глощения минимален при максимальной высоте солнцестояния и растет
по мере уменьшения высоты солнцестояния, т. е. в утренние и вечерние
часы. Хлорофилл поглощает 50—55% всех падающих на лист физиоло-
гических лучей. Остальные 10—15% поглощаются каротиноидами. Сопо-
ставляя фиг. 83 и 84, нетрудно заметить, что утром и вечером растения
получают относительно больше оранжево-красных лучей, чем в полуден-
ные часы.
Следует признать, что при изменении высоты солнцестояния в пре-
делах 20—90° количество квантов в единице энергии физиологической
радиации и поглощение последней пигментами листа остается практи-
чески неизменным.
3. ИНТЕНСИВНОСТЬ СОЛНЕЧНОЙ РАДИАЦИИ
Интенсивность солнечной радиации на границе земной атмосферы харак-
теризуется солнечной постоянной, которая равняется 1,88 г-кал/см2-мин,
или 1,31-103 * * 6 эрг/см2-сек. Интенсивность радиации на земной поверх-
ности определяется высотой Солнца над горизонтом и, следовательно,
зависит от географической широты, времени года и времени суток. Кроме
того, на эту величину оказывают влияние облачность, содержание в атмо-
сфере водных паров, прозрачность воздуха и другие метеорологические
и климатологические факторы. В лучшем случае — летом, в ясные сол-
нечные дни, в полуденные часы — земной поверхности достигает
70—80% энергии, а 20—30% теряется при прохождении через атмосферу
(табл. 55).
Различают интенсивность радиации на нормальной и горизонтальной
поверхности. Первая характеризует количество лучистой энергии, падаю-
щее на поверхность, перпендикулярную направлению лучей, а вторая —
количество энергии, получаемой горизонтальной поверхностью, находя-
щейся под углом h к направлению лучей. Зависимость обеих величин
от высоты солнцестояния показана на фиг. 2 и 3.
73
IV. Солнечная радиация
125
Таблица 55
Максимальная интенсивность солнечной радиации в СССР
(Н. Н. Калитин [93])
Географические пункты Географиче- ская широта 9 Максимальная интенсив- ность радиации
в г-кал/см2- мин в % к солнеч- ной постоян- ной
Бухта Тихая 80,7 1,30 69
Баренцбург (о-в Шпицберген) . 78,1 1,36 72
Маточкии шар (о-в Новая Земля) 73,7 1,30 69
Уэллен (Чукотка) 65,2 1,39 74
Якутск 62,0 1,47 78
Павловск (Ленинград) 60,0 1,43 76
Москва 55,8 1,48 79
Курск 53,2 1,51 80
Нерчинск 52,0 1,45 77
Евпатория 45,2 1,45 77
Тбилиси . 41,7 1,51 80
Ташкент 41,3 1,50 80
Интенсивность солнечной радиации на нормальной поверхности вы-
ражается формулой Буге:
= (36)
где /0 — солнечная постоянная, 1п — интенсивность солнечной радиации
на нормальной поверхности, т — масса атмосферы, а Р — коэффициент
прозрачности, зависящий от длины волны (фиг. 81, 7).
Коэффициент прозрачности есть величина, выражающая пропускание
солнечной радиации при прохождении солнечными лучами массы атмо-
сферы, принятой за единицу. Суммарный коэффициент р, выражающий
пропускание для всего спектра, колеблется в пределах 0,75—0,85. Макси-
мального значения он достигает зимой, а летом становится минимальным.
Интенсивность солнечной радиации на горизонтальной поверхности
можно выразить уравнением
Л = Цр™ sin h, (37)
где 1/; — интенсивность солнечной радиации на горизонтальной поверх-
ности.
Для реальной атмосферы, всегда содержащей пыль и водные пары,
которые вызывают помутнение атмосферы, в формулу Буге вносится по-
правка, и формула приобретает следующий вид:
‘ (38)
где т — коэффициент мутности, показывающий условное удлинение пути
солнечных лучей через атмосферу, связанное с наличием мутности.
Этот коэффициент колеблется в пределах 2,1—3,5, достигая максимума
в июле и снижаясь до минимума в январе. Для периода летней вегета-
ции (апрель—сентябрь) он равняется в среднем 3,2.
Сезонная изменчивость интегральной интенсивности солнечной радиа-
ции приведена на фиг. 85 и 86. Как видно из кривых (фиг. 86), интенсив-
ность на нормальной поверхности достигает максимума в апреле—мае,
а минимума — в декабре. В летние месяцы она почти не зависит от геогра-
126
Физические и физиологические основы светокультуры растений
фической широты: при переходе от 35° к 70° с. ш. интенсивность умень-
шается лишь на 15%. Совершенно иная картина сильнейшей зависи-
мости интенсивности радиации от широты наблюдается зимою. Чем
больше широта, тем больше падение интенсивности зимою, и тем больше
различие между летней и зимней интенсивностью радиации.
В общем, если пренебречь местными климатическими факторами
(облачность, мутность атмосферы и др.), то следует принять, что
Фиг. 86. Сезонная изменчивость инте-
гральной интенсивности полуденной
солнечной радиации на горизонтальной
поверхности для различных географи-
ческих широт
Фиг. 85. Сезонная изменчивость инте-
гральной интенсивности полуденной сол-
нечной радиации на нормальной поверх-
ности для различных географических
широт
1—35°; 2—40°; 3—45°; 4—50 = ; 5—55°; 7 — 35°; 2—40°; 3—45°; 4—50°; 5—55°;
в — 60°; 7 — 65°; 3—70° 6 — 60°; 7—65°
в пределах СССР интенсивность' интегральной радиации, падающей
на нормальную поверхность в течение вегетационного периода (апрель—
сентябрь) в полуденные часы, практически остается неизменной, колеблясь
в пределах 1,20—1,44 г-кал/см2-мин, или 840—1000 тыс. эрг/см2-сек.
Это соответствует освещенности порядка 80—100 тыс. люксов. В декабре
в зависимости от широты интенсивность колеблется от 0 до 1,37кал/см2-мин,
или от нуля до 950 тыс. эрг/см2-сек. Несколько иначе обстоит дело
с сезонной изменчивостью интегральной интенсивности радиации, падаю-
щей на горизонтальную поверхность (фиг. 86). В этом случае зависимость
от географической широты наблюдается как зимой, так и летом, хотя
в первом случае она выражена сильнее. В июне при переходе от 35°
к 70° с. ш. полуденное значение интенсивности уменьшается от 1,37
до 0,85 г-кал/см2-мин, т. е. на 38%, а в декабре — от 0,7 до 0 г-кал/см2-мин.
1 V. Солнечная радиация
127
Суточная изменчивость интенсивности солнечной радиации приведена
на фиг. 87. Она достигает максимума в полуденные часы и снижается при
переходе к утренним и вечер-
ним часам, практически до-
стигая нуля с наступлением
навигационных сумерек, т. е.
при опускании Солнца ниже
горизонта на 12°.
Принято считать, что доля
диффузной радиации в общем
потоке солнечного излучения
сравнительно невелика, а
роль рассеянного света в
жизни растений незначитель-
на. В действительности же
наблюдается совершенно
иная картина. Как видно из
табл. 56, в которой дана ин-
тенсивность диффузной ра-
диации для различных высот
солнцестояния, процент этого
излучения в потоке физиоло-
гической радиации в ясный
солнечный день составляет
не менее 15. Роль рассеянной
радиаций увеличивается еще
более в туманные и облачные
дни, когда солнечная радиа-
ция может состоять целиком
Фиг. 87. Суточная изменчивость интенсивности
солнечной радиации
1—на нормальной поверхности; 2—на горизонтальной
поверхности; 3 — диффузная радиация; 4— при легкой
облачности; 5 — при густых дождевых облаках
из рассеянных лучей.
В естественных условиях интенсивность диффузной радиации больше
зависит от состояния атмосферы, ее мутности, наличия облачности
Табл и ц а 56
Зависимость интенсивности диффузной солнечной радиации от высоты
солнцестояния
Высота солнце- стояния (В °) Интенсивность диффузной солнечной радиации
в тыс. эрг/см2-сек в кал/см- • мин В тыс. лк в % к общему излучению в % к физио- логической радиации
5 15 0,022 3 73,4 87,5
10 20 0,030 4 42,9 65,2
15 31 0,044 6 36,6 57,1
20 36 0,052 7 29,0 47,6
25 41 0,059 8 23,6 40,0
30 46 0,067 9 21,0 36,0
35 51 0,074 10 18,3 32,7
40 62 0,089 12 18,0 31,8
45 67 0,096 13 16,8 30,1
50 72 0,100 14 15,4 27,0
55 77 0,110 15 15,0 26,2
128
Физические и физиологические основы светокультуры растений
или тумана, чем от географической широты, времени года и времени
дня. Как видно из фиг. 87, диффузная радиация после восхода Солнца
быстро достигает своего максимального значения и на этом уровне
держится почти до конца дня, начиная уменьшаться лишь незадолго
до захода Солнца.
Относительная величина диффузной радиации возрастает в зимние
месяцы и при увеличении географической широты, хотя при этом абсо-
лютное значение диффузной радиации в обоих случаях падает. Сказанное
становится ясным при сопоставлении табл. 51 и 56.
Огромное влияние на интенсивность солнечной радиации имеет облач-
ность. Всякие облака, покрывающие небосвод, но оставляющие Солнце
незакрытым, увеличивают освещенность на 5—25°/0. Облачность,
Фиг. 88. Влияние облачности на величину естественной солнечной осве-
щенности
Ci—перистые облака; С©—перисто-кучевые облака; Ас—высоко-кучевые облака;
Си—-кучевые облака; Cs—-слоисто-кучевые облака; СЬ — кучево-дождевые облака;
St — сплошные слоистые облака; SN — сплошные дождевые облака
А — Солнце не закрыто облаками; Б — Солнце за облаками
Дробные числа показывают степень покрытия неба облаками
закрывающая солнечный диск, вызывает, напротив, сильное уменьше-
ние освещенности, достигающее при сплошной дождевой облач-
ности 80%. Как уменьшение, так и увеличение интенсивности солнечной
радиации зависят от высоты солнцестояния, типа облачности и степени
покрытия небосвода облаками (фиг. 88). На этой фигуре пунктирная ли-
ния, параллельная оси абсцисс, соответствует освещенности в отсутствие
облаков (100%); верхние линии показывают увеличение освещенности при
наличии соответствующей облачности и свободном солнечном диске,
а нижние линии — уменьшение освещенности под влиянием облачности,
закрывающей солнечный диск.
До сих пор, говоря об интенсивности солнечной радиации, мы имели
в виду исключительно интегральный лучистый поток в условиях есте-
ственного местообитания растений. Между тем для светофизиологии и
светокультуры растений наибольший интерес представляет интенсивность
солнечного излучения не на открытом месте, а в условиях закрытого
грунта — в теплицах и парниках, и притом не всего лучистого потока,
а лишь физиологической радиации.
IV. Солнечная радиация
129
Общеизвестно, что в теплицах, в результате потери из-за отражения
и поглощения стеклом и каркасом теплицы, интенсивность радиации
равняется 40—60% интенсивности на открытом месте вне оранжереи
[646]. Следовательно, для того чтобы получить представление о количестве
лучистой энергии в теплицах, необходимо все величины, приведенные
на фиг. 85 — 88 и в табл. 55 и 56, уменьшить вдвое. Следует помнить
при этом, что физиологическая радиация составляет 40—50% интеграль-
ного солнечного излучения (фиг. 82). Величины полуденной интенсивности
физиологической радиации в теплицах для различных географических
широт и различного времени года показаны на фиг. 89. Там же приведена
интенсивность, потребная для развития теневых и светолюбивых растений
(линии аб и вг).
Фиг. <89. Интенсивность физиологической радиации в теплицах в полуденные часы
в разные месяцы на различной географической широте в пределах СССР. Римские
цифры означают месяцы
аб—интенсивность, потребная для развития теневых растений; се — интенсивность, потребная
для развития светолюбивых растений
Как видно из кривых, интенсивность физиологической радиации, по-
лучаемая растениями в теплицах в полуденные часы, настолько велика,
что на всей территории СССР позволяет выращивать светолюбивые растения
на протяжении пяти месяцев, с апреля по август, а теневые растения —•
с марта по сентябрь. В остальные месяцы (октябрь—февраль) культура
растений в теплицах возможна лишь при помощи дополнительного искус-
ственного освещения.
Последнее для светолюбивых растений необходимо в умеренной полосе,
лежащей севернее 50° с. ш., с октября по февраль, а для теневых растений
с ноября по январь. Интенсивность искусственного освещения зависит
от вида растений и географической широты. На юге СССР полуденная
интенсивность физиологической радиации в теплицах достаточна для
выращивания даже светолюбивых растенш! на протяжении круглого года;
ограничивающим фактором здесь является длина дня.
В связи с комнатной культурой растений специального внимания
заслуживает вопрос о спектральном составе и интенсивности радиации
на окнах и в комнатах жилых и служебных помещений. В тех случаях,
когда на окна падают прямые солнечные лучи (их спектральный
9 А. Ф. Клешнин
130 Физические и физиологические основы светокультуры растений
состав показан на фиг. 80 и 81), вопрос об интенсивности решается
так же, как и при культуре растений в теплице. Но в большинстве
случаев, особенно на северной и, частично, на западной и восточной сто-
ронах зданий, комнатные растения получают лишь диффузную радиацию,
величину которой показывает фиг. 90. На оси абсцисс здесь отложено
80'
70
0 --------------1--------------1-------------I-------------L.
0,5 1 1.5 Z
Расстояние от о/гна (в м)
Фиг. 90. Изменение интенсивности естественной радиации
в комнате в зависимости от расстояния от окна
расстояние от окна (в метрах), а на оси ординат — интенсивность радиа-
ции (в процентах к интенсивности диффузной радиации за окном). Фиг. 90
показывает, что при отсутствии попадания прямых солнечных лучей интен-
сивность радиации даже на окне не превышает половины интенсивности
естественной радиации, а в глубине комнаты, на расстоянии 2 м от окна,
снижается до 7,6%. Измерения показывают, что в лучшем случае, в полу-
денные солнечные часы, освещенность в глубине комнат не превышает
700 лк и обычно колеблется в пределах 30—300 лк [677, 678, 681].
4. ДЛИНА ДНЯ
Длина дня, как и интенсивность радиации, подвержена сезонной
изменчивости и зависит от географической широты. На юге СССР она
колеблется в пределах от 10 до 14 часов, а в полярных районах — от
нуля до 24 часов. Максимальная длина дня наблюдается летом (26 июня),
а минимальная — зимой (23 декабря). Независимо от географической
широты длина дня становится больше 12 часов с 18 марта и менее 12
часов — с 25 сентября. Изменения длины дня на различных широтах при-
ведены в табл. 57 и на фиг. 91.
Однако астрономическая длина дня, на протяжении которой интен-
сивность радиации изменяется от нуля до соответствующего максимума,
не дает полной характеристики условий развития растений. Для куль-
туры растений большее значение имеет длина дня, отвечающая интен-
сивности физиологической радиации, потребной для нормального роста
и развития растений (50 тыс. эрг/см3-сек, или 0,07 г-кал/см2-мин). Ха-
рактеристика подобной эффективной длины дня для теплиц приведена
на фиг. 92. На юге СССР она колеблется в пределах от 6 до 11 часов,
а в полярных областях — от нуля до 14 часов.
Фиг. 91. Сезонное изменение длины дня на различных широтах
в пределах СССР
Фиг. 92. Сезонное изменение длительности освещения на раз-
личных широтах, соответствующего в условиях теплиц 50 тыс.
эрг/см2-сек физиологической радиации
Таблица 57
Астрономическая длина дня в пределах СССР
(В. В. Шаронов [342])
Даты Географическая широта (в °)
35 | 45 50 55 60 65 70
Январь, 20 10 ч. 10 м. 9 ч. 46 м. 9 ч. 18 м. 8 ч. 45 м. 8ч. 00 м. 7 ч. 01 м. 5 ч. 32 м. 0 ч. 00 м
Февраль, 20 И ч. 06 и. 10 ч. 54 м. 10 ч. 40 м. 10 ч. 23 м. 10 ч. 03 м. 9 ч. 37 м. 9 ч. 03 м. 8 ч. 09 м.
Март, 17—18 12 ч. 00 м. 12 ч. 01 м. 12 ч. 01 м. 12 ч. 00 м. 12 ч. 00 м. 11 ч. 58 м. 12 ч. 01 м. 12 ч. 00 м.
Апрель, 20 13 ч. 13 м. 13 ч. 27 м. 13 ч. 43 м. 14 ч. 02 м. 14 ч. 27 м. 14 ч. 59 м. 15 ч. 45 м. 16 ч. 58 м.
Май, 20 14 ч. 07 м. 14 ч. 31 м. 15 ч. 01 м. 15 ч. 37 м. 16 ч. 23 м. 17 ч. 30 м. 19 ч. 22 м. 24 ч. 00 и.
Июнь, 19—22 14 ч. 31 м. 15 ч. 02 м. 15 ч. 38 м. 16 ч. 23 м. 17 ч. 23 м. 18 ч. 53 м. 21 ч. 49 м. 24 ч. 00 м.
Июль, 20 14 ч. 12 м. 14 ч. 38 м. 15 ч. 09 м. 15 ч. 47 м. 16 ч. 36 м. 17 ч. 47 м. 19 ч. 44 м. 24 ч. 00 м.
Август, 20 13 ч. 20 м. 13 ч. 36 м. 13 ч. 52 м. 14 ч. 14 м. 14 ч. 41 м. 15 ч. 16 м. 16 ч. 08 м. 17 ч. 28 м.
Сентябрь, 25—26 12 ч. 02 м. 12 ч. 01 м. 12 ч. 00 и. 12 ч. 00 м. И ч. 58 м. И ч. 58 м. 11 ч. 58 м. 12 ч. 04 м.
Октябрь, 20 11 ч. 10 м. 10 ч. 59 м. 10 ч. 47 м. 10 ч. 31 м. 10 ч. 12 м. 9 ч. 46 м. 9 ч. 17 м. 8 ч. 27 м.
Ноябрь, 20 10 ч. 13 м. 9 ч. 51 м. 9 ч. 23 м. 8 ч. 51 и. 8 ч. 09 м. 7 ч. 12 м. 5 ч. 47 и. 2 ч. 32 м.
Декабрь, 20—23 9 ч. 48 м. 9 ч. 19 м. 8 ч. 44 и. 8 ч. 06 м. 7 ч. 09 м. 5 ч. 51 м. 3 ч. 43 м. 0 ч. 00 и.
132 Физические и физиологические основы светокультуры растений
IV. Солнечная радиация
133
5. КОЛИЧЕСТВО ЛУЧИСТОЙ ЭНЕРГИИ
Рассмотрим вопрос о количестве энергии физиологической радиации,
падающей на горизонтальную поверхность в течение дня. Эта величина,
зависящая от интенсивности радиации и длины дня, служит важной
Фиг. 93. Количество физиологической радиации, падающее за день на
горизонтальную поверхность в теплицах в различные месяцы на разной
географической широте в пределах СССР. Римские цифры означают
месяцы
аб — количество энергии, необходимой для нормальной культуры растений
Таблица 58
Ежемесячное и годовое количество солнечной энергии, падающее на единицу площади
горизонтальной поверхности в пределах СССР
Месяц Ленинград Москва Киев
в кал/см'2 в КВТ-Ч/М2 в кал/см2 в КВТ-Ч/М2 в к 5 л/см2 в квт-ч/м-
Январь 0,1 1,16 °,2 2,3 0,8 9,3
Февраль .... 0,7 8,1 1,0 11,6 1,9 22,0
Март 2,6 30,2 3,1 36,0 3,0 34,8
Апрель 4,4 51,0 5,8 67,2 3,5 40,5
Май 7,5 87,0 9,7 112,5 9,9 115,0
Июнь 8,2 95,0 10,2 118,1 8,9 103,0
Июль 7,9 91,6 9,7 112,5 10,9 126,0
Август 4,4 51,0 7,1 82,4 9,4 6,8 4,0 109,0
Сентябрь .... Октябрь 2,7 1,0 31,3 11,6 4,0 1,7 46,4 19,7 79,0 46,4
Ноябрь 0,1 1,16 0,3 3,5 1,0 11,6
Декабрь .... 0,1 1,16 0,1 1,16 0,4 4,6
Январь—декабрь 39,8 459,0 52,9 615,0 60,8 705,0
134
Физические и физиологические основы светокультуры растений
характеристикой условий местообитания растений. Количество физиологи-
ческой радиации, падающей на горизонтальную поверхность в теплицах,
показано на фиг. 93.
Линия аб на фиг. 93 показывает количество энергии, получаемое
растениями в условиях светокультуры при интенсивности радиации
в 50 тыс. эрг/см2-сек и длине дня в 17 часов (70 г-кал/см2*день) и
достаточное для нормального развития растений. В пределах СССР
в условиях закрытого грунта достаточное количество энергии расте-
ния получают лишь с апреля по август. Весною (февраль—март)
и осенью (сентябрь—октябрь) это количество достигает необходимого
уровня лишь в южных районах страны, а зимою недостаточно для нор-
мального развития растений даже на юге СССР. В зимние месяцы
(а в умеренной полосе и полярных областях также и ранней весною и
поздней осенью) недостаток энергии должен быть восполнен искусствен-
ными источниками радиации.
В табл. 58 приведено количество лучистой энергии, получаемое еди-
ницей площади земной поверхности в виде солнечных лучей.
Солнечная энергия, падающая в течение года на всю террито-
рию СССР, равняется 12,72-1015 квт-час. Она в 3600 раз превышает миро-
вое производство электроэнергии и соответствует энергии, заключенной
в 2200 млрд, т каменного угля или 1100 млрд, т нефти*.
6. ВЫВОДЫ
1. Солнечная радиация представляет собою чрезвычайно изменчивый
фактор, зависящий от времени дня, времени года, географической ши-
роты, состояния атмосферы, облачности и других факторов.
2. Солнечная радиация есть излучение, лежащее практически в пре-
делах 295—3000 шд. с максимумом около 490 шц. Максимум смещается
в сторону красных лучей при уменьшении высоты солнцестояния, т. е.
с увеличением географической широты, при переходе от летних месяцев
к зимним и в утренние и вечерние часы.
3. Интенсивность солнечной радиации в летние полуденные часы
равняется 840—1000 тыс. эрг/см3-сек., что соответствует 80—100 тыс. лк.
Физиологическая радиация составляет 50% всего излучения (420—
500 тыс. эрг/см3-сек).
4. В условиях закрытого грунта, так же как и в естественной обста-
новке, поздней осенью, зимою и ранней весною ограничивающим факто-
ром являются интенсивность радиации и длина дня. В этот период для
нормального роста и развития растений необходимо дополнительное
искусственное освещение.
* В 1937 г. мировое производство электроэнергии равнялось 364 млрд, квт-час,
мировая добыча каменного угля — 1315,36 млн. т, а нефти — 281,3 млн. т.
Глава V
МЕТОДЫ ИЗМЕРЕНИЯ ЛУЧИСТОЙ ЭНЕРГИИ
Измерение лучистой энергии для целей светофизиологии. и свето-
культуры растений есть проблема измерения в основном физиологиче-
ской радиации и энергии некоторых других лучей, физиологическое
значение которых еще недостаточно изучено. Как видно из фиг. 2
и 3, измерению подлежит прежде всего область, лежащая в пределах
300—700 ту. и поглащаемая пигментами пластид. С физиологической
точки зрения целесообразно учитывать отдельно следующие области
спектра:
1. Область 500—700 ту., поглощаемую исключительно хлорофиллом.
2. Область 400—500 ту, поглощаемую хлорофиллом, каротиноидами,
протоплазмой и ферментативной системой листа.
3. Ультрафиолетовые лучи (295—400 ту.), поглощаемые пигментами
пластид, антоцианами и флавоноидами, ферментативной системой и дру-
гими компонентами протоплазмы.
4. Инфракрасные лучи, особенно начиная с длины волны в 1050 ту,
поглощаемые тканями листа (главным образом содержащейся в них
водою).
В настоящее время отсутствуют измерительные приборы, позволяю-
щие измерять физиологическую радиацию. Создание подобных прибо-
ров — дело будущего. В подавляющем большинстве случаев измеряется
или интегральный лучистый поток (например, в актинометрии), или ос-
вещенность (чаще всего в физиологических исследованиях). Однако до-
статочно бросить общий взгляд на кривые на фиг. 2, 3, 7, 9, 10 и 12, чтобы
убедиться в том, что ни интегральная радиация, ни, тем более, освещен-
ность не могут служить показателем количества лучистой энергии, ис-
пользуемой растением для физиологических процессов.
При измерении интегрального лучистого потока вместе с физиологи-
чески активными лучами учитываются и неактивные лучи. Это обстоятель-
ство может совершенно исказить действительную картину явления. При
измерении освещенности учитываются, собственно, не лучистая энергия,
а ее восприятие средним человеческим глазом в соответствии с его спек-
тральной чувствительностью (фиг. 28).
1. ИНТЕНСИВНОСТЬ РАДИАЦИИ
В области светофизиологии и светокультуры растений измерение лу-
чистой энергии сводится в основном к измерению интенсивности ради-
ации на уровне растений.
136
Физические и физиологические основы светокультуры растений
Интенсивность радиации (Е) есть количество лучистой энергии, выра-
женное в объективных единицах и падающее на единицу освещаемой по-
верхности в единицу времени (табл. 11). Обычно интенсивность радиации,
будет ли это физиологическая или интегральная радиация, выражается
в эрг/см’-сек, кал/смг-мин или вт/см2. Отношение между этими едини-
цами выражается следующими равенствами1: 1 эрг/см2-сек=1,43-10~6
кал/см2-мин = 1-1О-7 вт/см2; 1 кал/см2-мин = 6,96-105 эрг/см2-сек =
=6,96-10“-вт/см2; 1 вт/см! = 1-107 эрг/см2-сек = 14,3 кал/см-’-мин.
2. ОСВЕЩЕННОСТЬ
Так как в ботанической литературе часто вместо интенсивности ради-
ации указывается ее фотометрический аналог — освещенность (табл. 11),
то остановимся на ее определении, единицах измерения и их отношении
к единицам интенсивности радиации.
Освещенность (L) есть отношение светового потока к площади осве-
щаемой поверхности (S). Практической единицей освещенности является
люкс (лк)—освещенность, создаваемая источником радиации с силою света
в 1 св на расстоянии 1 м; 1 лк равняется 1 лм/м2. Лампа в п свечей на рас-
стоянии 1 м создает освещенность в п люксов. В немецкой литературе
вместо международного люкса употребляется люкс Гефнера (Н1х), который
> равняется 0,88 лк, а в американской литературе — большей частью футо-
свеча (fc), равная освещенности от источника в 1 св на расстоянии 1 фута
(30,48 см), т. е. 10,76 лк. В научной светотехнической литературе едини-
цей освещенности обычно является фот (ф) — освещенность, создаваемая
источником радиации в 1 св на расстоянии в 1 см. Иными словами, 1 фот
равняется 1 лм/см2, или 10 тыс. лк.
Освещенность есть интенсивность радиации, оцениваемая средним че-
ловеческим глазом. Иными словами, объективно величина освещенности,
получаемая при помощи того или иного источника радиации, есть ре-
зультат перемножения ординат кривой относительной видности (фиг. 28,
табл. 9) на ординаты кривой распределения энергии в спектре источника
(фиг. 94, 95 и 97). Ясно, что в зависимости от источника радиации со-
держание энергии в 1 лк будет различно. Это обстоятельство абсолютно
исключает возможность выражения интенсивности радиации в единицах
освещенности — люксах или футосвечах, что допустимо лишь для одно-
типных источников с одинаковым спектральным составом излучения или
в случае, когда известен переводный коэффициент от единиц освещенности
к объективным единицам физиологической радиации.
3. ПЕРЕХОД ОТ ОСВЕЩЕННОСТИ К ИНТЕНСИВНОСТИ РАДИАЦИИ
Рассмотрим несколько типичных случаев перевода единиц освещен-
ности в единицы интенсивности радиации. На фиг. 94,7 представлено
распределение энергии в спектре вольтовой дуги высокоинтенсивного го-
рения, а на фиг. 94,2 — произведение ординат кривой 7 на ординаты
кривой относительной видности. Линии аб, вг и де на фиг. 94 являются
границами витацидной, «длинной» ультрафиолетовой, видимой и инфра-
красной радиации, а площади S2, S;J, Sj и S4 (фиг. 94), ограниченные этими
1 Эти отношения непосредственно вытекают из отношений между единицами
энергии: 1 эрг—10_7 дж=2,39-10—8 кал=2,39-10~11 ккал; 1 дж=1Ь7 эрг—0,24
кал—2,4» 10—* ккал; 1 кал—4,18-107 эрг=4,18 дж=10—3 ккал; 1 ккал=4,18-10го
эрг=4,18*103 дж—103 кал— 1,1(М0—3 квт-ч; 1 квт-ч—862 ккал.
И. Методы измерения лучистой, энергии
137
линиями и соответствующими участками кривой 1 и оси абсцисс, соот-
ветствуют интенсивности радиации перечисленных областей спектра.
Площадь S„, лежащая в пределах кривой 2 и оси абсцисс, отвечает ос-
вещенности, создаваемой названной лампой. С помощью планиметра или
иным способом нетрудно установить отношения:
А-„=^ и * = -£-, - О»)
0?’ 0 0 ‘
где 6,=5J-j-3'24-iS'g4-54 — площадь, отвечающая интегральной интенсив-
ности излучения, a Sn — площадь, соответствующая интенсивности
любой произвольно взятой области спектра.
Длина ВолнЗ/ ( 3 тр)
Фиг. 94. Переход от освещенности к интенсивности радиации
1 — распределение энергии в спектре излучения вольтовой дуги высокопнтепспвпого
горения; 2 — относительная видность спектра вольтовой дуги; линии
аб, вг, де, — границы витацидпой, ультрафиолетовой, видимой и инфракрасной радиа-
ции; Si, S2, S., и Si — площади, соответствующие интенсивности видимой, вита-
цидпой, ультрафиолетовой п инфракрасной радиации; Sr — площадь, соответствующая
освещенности
Отношение между освещенностью и интенсивностью соответствующих
областей спектра выражается уравнением:
# = 1,61.#L, ' (40)
где Е — интенсивность радиации в эрг/см!-сек; /. — освещенность(в лк),
а К — коэффициент, определяемый по формуле (39). Если освещен-
ность выражена в люксах Гефнера, вместо величины 1,61 в формулу
вставляется множитель 1,44, если же в футосвечах, то — 17,32. Легко
убедиться, что отношение освещенности к интенсивности радиации вы-
ражается тем же уравнением, что и отношение светового потока к лу-
чистому потоку [глава IV, формулы (11) и (12), фиг. 29].
Для лампы, характеризуемой фиг. 94, значение коэффициентов равно:
#,=3,10, #,=0,0208, #3=0,42, V =4,83 и #=8,36; значение 1 лк
равно 4,96 — для видимой, 0,0333 — для витацидной, 0,67 — для уль-
трафиолетовой, 7,72 — для инфракрасной радиации и 13,38 эрг/см!-сек —
для интегрального излучения.
138
Физические и физиологические основы светокультуры растений
100
Длина волнО (в тр)
Фиг. 95. Переход от освещенности к интенсивности радиации
7 — распределение энергии в спектре люминесцентной лампы синего света
(MgW0,);2— относительная видпостьспектра люминесцентной лампы синего света;
S — площадь, соответствующая интенсивности излучения; Sv— площадь, соот-
ветствующая освещенности
Таблица 59
Определение коэффициента К для перевода единиц освещенности в единицы интен-
сивности радиации для ртутной лампы низкого давления (0,01 мм рт. ст.) в квар-
цевой оболочке
Длина волны (в т;л) Относительная интенсивность линий (в услов- ных единицах) Коэффициент относительной видности (в %) Относительная интенсивность света Переходный коэффициент Значение 1 лк (в эрг/ см2 • сек)
253,7 296,7 100 1,86 0 0 0 0
253,7—296,7 312,6—313,2 365 —366,3 101,86 (= S2) 8,06 7,56 0 0 0 0 0 5,80 (=S'2 Л 9,35
312,6—366,3 404,7 407,8 435,6 546,1 577—579 15,62 (=^3) 8,60 1,23 14,25 13,45 4,64 0,062 0,088 1,777 98,430 88,900 0 0,005 0,011 0,254 13,200 4,120 0,89 1,43
404,7—579,0 1014 42,17 ( = 8^ 8,06 0 17,575 (=SV) 0 МО \ 8Д 3,80
1064 8,06 (= 61!) — 0 0,46 0,74
253,7—1014 167.81 (= £) ... — 17,575 (=6\,) 9,55 15,38
К Методы измерения лучистой энергии
139
Таблица 60
Перевод единиц освещенности в единицы интенсивности физиологической радиации
для разных типов ламп
Типы ламп Мощность лампы | (в ВТ) 1 лк (в эрг/см- • сек) 1 эрг/см'2 сек (в лк)
А. Лампы накаливания
( ю 8,40 . 0,119
| 25 8,05 0,124
1. Пустотные j 40 7,20 0,139
( 60 6,68 0,152'
50 6,61 0,154
75 6,30 0,158
100 6,15 0,162
200 5,97 0,167
2. Газополные 300 5Ж 0,171
500 5,67 0,176
1000 v/ 5,54 0,180
2000 5,46 0,183
3. Проекционные 1000—3000 4,82 0,207
Б. Л то м и н е с ц е н т ц ы е лампы
1. Низковольтные: * -
дневного света 15—100 4,03 0,248
мягкобелые 15—40 4,25 0.235
белые 15—100 3,55 0,220?
синие (CaWO4) . 15—30 5,15 0,194
синие (Mg\VO4) 15—30 4,06 0.246
зеленые 15—30 2,22 0,450
красные 15—30 4,51 0,222
золотистые 15—30 3,42 0,292 '
2. Высоковольтные:
дневного света . . 50 4,60 0,218
мягкобелые 100 4,30 0,232
белые 100—120 3,85 0,260
3. Высокого давления («HgL») . . . 75—120 2,24 0,445
В. Солнечные лампы
«S-1» 315 6,45 0,155
«HWA-500» 255 4,72 0,212
Г. Газосветные лампы
1. Ртутные лампы:
«СН» 270 4,38 0,228
«Игар-2» 500 3,98 0,251
«Н-2? 250 3,09 0,324
«Н-1» 400 2,96 0,338
«ПРК-4» 220 4,26 0.234
«ПРК-2» 375 4,14 0,242
140
Физические и физиологические основы светокультуры растений
Таблица 60 (продолжение)
Типы ламп Мощность лампы (в ВТ) 1 лк (в эрг/см2 сек) 1 эрг/см2 • сек (в лк)
«Ш’К-7» 1000 4,34 0,220
«Н-5» 250 3,32 • 0,301
«СВД-125» 125 3,71 0,269
«СВДШ-250» 250 3,70 0,270
«Н-3» 85 3,83 0,261
«Н-4» 100 3,88 0,257
«HgH-ЮОО» 265 2,43 0,410
«HgQS-500» (матовое стекло) .... 120 4,21 0,238
«HgHS-8000» (увиолевое стекло) . . 1500 3,62 0,276
«HgQS-ЗОО» (увиолевое стекло) . . . 75 5,00 0,200
«HgQS-500Bl» (NiO—стекло) .... 120 169 000 0,590 • IO'5
«HgQS-500» (РИО и Сг2О стекло) . . 120 193 000 0,517 • 10~5
2. Натриевые лампы: 45-140 1,88 0,531
3. Цинковые лампы высокого давления 23,3 0,043
Кадмиевые лампы —
4. КК-1 460 10,6 0,094
низкого давления — 22,6 0,044
5. Цезиевые лампы 500 37,4 0,027
6. Неоновые лампы 28—485 7,35 0,136
7. Ксеноновые дуговые лампы .... 150—700 5,65 0,177
8. Гелиевые лампы — 5,77 0,173
9. Криптоновые лампы — 3,56 0,380
Д. Вольтовые дуги
1. Открытого горения
Чистые угли 7,23 0,138
Пламенные угли «солнечный свет» . — 7,25 0,138
Желтые пламенные угли — 7,01 0,142
2. Закрытого высокоиитенсивного го-
рения . . — 5.61 0,178
Т а б л и ц а 61
Значение 1 лк (в эрглм- • сек физиологической радиации) и 1 эрг^м’ - сек
физиологической радиации (в лк) для естественной солнечной радиации при
различной высоте солнцестояния f64_6J
Высота солнцестоя- ния (в с) 1 лк (в эрг/см2 • сек) 1 эрг/см2 сек (в лк)
прямой солнечный свет прямой + Диффузный солнечный свет равномерно облачное небо прямой солнечный свет прямой 4- диффузный солнечный свет равномерно облачное небо
65 6,20 6,15 0,161 0,163 —
47,2 6,05 5,94 — 0,165 0,168 —
30 5,94 5,69 — 0,168 0,175
22,4 5,89 5.55 — 0,170 0,180
19,3 5,80 5,51 6,05—6,57 0,172 0,192 0,153—0,163
16,4 5,75 5,47 — 0,174 0,183 —
14,3 5,69 5,47 — 0,176 0,183 —
12,6 5,65 5,47 — 0,177 0,183 —
11,3 5,60 5,47 — 0,178 0,183 —
V. Методы измерения лучистой энергии
141
Значительно проще обстоит дело для источников, излучение которых
сосредоточено целиком в области видимых или физиологических лучей.
Подобный случай представлен на фиг. 95. Для характеристики лампы
здесь достаточно знать лишь одно значение К, которое будет соответ-
ствовать и физиологической радиации, и интегральному излучению. Для
лампы, кривая которой приведена на фиг. 95, 76=2,54, а 1 лк—
4,06 эрг/см2-сек физиологической радиации или суммарного излучения.
Еще меньшей сложностью отличается метод перевода единиц освещен-
ности в единицы интенсивности радиации для газосветных ламп, имеющих
линейчатый спектр. В качестве примера рассмотрим ртутную лампу низкого
давления в кварцевой оболочке. Подобные расчеты приведены в табл. 59.
Этим способом можно рассчитать величину излучения в любой области
спектра, вплоть до отдельных линий, для любого источника, имеющего
линейчатый спектр.
Значения коэффициентов К для физиологической радиации и инте-
грального излучения приведены в табл. 10, а значения 1 лк в эрг/см2>сек
физиологической радиации для различных источников ради-
ации — в табл. 60. Значение люкса естественной освещенности зависит
от высоты Солнца над горизонтом, облачности и других факторов (табл.61).
4. ПРИМЕНЕНИЕ СВЕТОФИЛЬТРОВ
Наибольшую сложность представляют расчеты в случае применения
различных светофильтров. Последние, обладая избирательной поглоти-
тельной способностью, вносят сильнейшие изменения в спектральный со-
став излучения, проходящего через фильтр и достигающего растений.
В качестве светофильтров применяются или стеклянные фильтры, спек-
тральная характеристика которых приводится обычно в виде кривых
пропускания (фиг. 35 и 98), или желатиновые и жидкие фильтры, оп-
тические свойства которых определяются уравнением:
1, = 10к- Ю-^, (41)
где Ех — оптическая плотность красящего вещества; Znx — спектраль-
ная интенсивность радиации, падающей на светофильтр; —- спектраль-
ная интенсивность пропускаемой светофильтром радиации.
В свою очередь, оптическая плотность равняется произведению кон-
центрации вещества (с), толщины слоя (<7) и коэффициента поглощения
(экстинкции) — к, зависящего от длины волны:
Ex = kcd. (42)
Толщина слоя жидких светофильтров обычно показывается в см, кон-
центрация — в процентах, г/л или мол/л, а к изменяет свое значение
в зависимости от способа выражения концентрации вещества.
Как видно из формулы (41), для получения идентичных спектраль-
ных кривых пропускания нет необходимости придерживаться постоян-
ной концентрации вещества и постоянной толщины слоя раствора. Для
этой цели важно обеспечить лишь постоянство произведения названных
величин, т. е. постоянство поверхностной концентрации красящего ве-
щества. Ни толщина слоя, ни концентрация не влияют на спектральную
кривую пропускания, если остается постоянной поверхностная концен-
трация красителя. Спектр пропускания светофильтра определяется- ис-
ключительно коэффициентом экстинкции и поверхностной концентра-
цией вещества.
142
Физические и физиологические основы светокультуры растений
Следовательно, определив однажды при помощи спектрофотометра
кривую пропускания лучистой энергии красителем для любой произ-
вольно выбранной толщины слоя и концентрации, можно, пользуясь фор-
мулами (41) и (42), установить коэффициенты экстинкции, а при помощи
последних, опять-таки по тем же формулам, рассчитать и построить кри-
вые пропускания для любой поверхностной концентрации вещества.
Поверхностная концентрация вещества выражается обычно в гюб-
лях (гб). 1 гюбль = 1 г/м’, или 0,1 мг/см2. Если концентрация раствора
Фиг. 96. Распределение энергии в спектре лампы накаливания в 1 квт
1 — без экрана; 2 — с экраном из CuSO4, 4 000 гюблей (10%, 4 см); 3 — с экраном из К2Сг2О7
4 000 гюблей (10%, 4 см); спектры пропускания: 4 — раствора CuSO4, 4 000 гюблей; о—рас-
твора К2Сг2О7, 4 000 гюблей. Линия аб — граница видимой радиации
дана в процентах, то поверхностная концентрация светофильтра бу-
дет равна
cH=l00’~q = i00md, (43)
где т — содержание красителя (в%); q — количество раствора (в см3);
5—площадь светофильтра (в см2); d — толщина слоя (в см); с„—по-
верхностная концентрация (в гб). Если же содержание красителя дано
в г/л, то формула принимает вид
св= ™q = 10md, (43а)
где т — содержание красителя (в г/л).
После того как подобран соответствующий светофильтр с желаемой
кривой пропускания, производится определение спектрального состава
излучения, падающего через данный фильтр на растение от того или
иного источника радиации. Для этой цели ординаты спектральной кривой
излучения источника умножают на ординаты кривой пропускания свето-
фильтра. Кривая, полученная в результате подобного перемножения, ха-
рактеризует спектральный состав излучения, получаемого растениями
от данного источника через определенный светофильтр. Сказанное
V. Методы измерения лучистой энергии
143
иллюстрируется примером расчета для лампы накаливания и раство-
ров CuS04 и К2Сг2О7 (фиг. 96). Оба случая в лабораторной практике
встречаются наиболее часто. Кривые 4 и 5 показывают пропускание
лучистой энергии растворами CuSOi и К.,Сг2О7 при поверхностной кон-
центрации в 4000 гб, а кривые 2 и 3 (фиг. 96) — распределение энергии
в спектре от лампы накаливания в присутствии названных фильтров.
Площади, ограниченные осью абсцисс и кривыми 2 и 3, отвечают интен-
сивности радиации, получаемой при помощи соответствующих экранов,
Фиг. 97. Переход от освещенности к интенсивности физиологиче-
ской радиации.
Распределение энергии в спектре лампы накаливания в 1 квт с фильтрами
1 — из CuSO,. 4000 гюблей; 3 — из К2Сг2О7, 4000 гюблей;
относительная видность спектра после фильтров: 2 — из CuSO4; 4—из К2Сг2О7
S, и S2 — площади, соответствующие интенсивности излучения. Sv, и Sv2 —
площади соответствующие освещенности
причем CuSO4 выделяет исключительно физиологическую радиацию,
а К2Сг9О7 — физиологическую радиацию (32,3 %) и ближние инфракрас-
ные лучи (67,7%). Отношение энергии физиологической радиации за эк-
ранами из CuSO4 и К2Сг2О7 равняется 1:4,64, а интегральной радиа-
ции — 1:14,65. Заметим попутно, что в нашем случае раствор CuSO4 про-
пускает 15,6%, а К9Сг2О7 — 72,2% энергии видимых лучей лампы на-
каливания.
Для перевода единиц освещенности в единицы интенсивности физио-
логической радиации обычным способом, описанным на стр. 136—137,
устанавливают коэффициент (фиг. 97) и по формуле (40) определяют
значение 1 лк. В нашем случае с лампой накаливания величина коэф-
фициента для CuSO4 равняется 2,24, а для К2Сг2О7 — 5,70. Соответствую-
щие значения 1 лк равны 3,68 и 9,10 эрг/см2-сек. Подобным методом
можно пользоваться в случае любого источника радиации и любого све-
тофильтра — жидкого, стеклянного или в виде желатиновой или иной
пленки. При этом необходимо знать кривую пропускания фильтра и кри-
вую распределения энергии в спектре источника.
144
Физические и физиологические основы светокультуры растений
5. ИЗМЕРИТЕЛЬНЫЕ ПРИБОРЫ
Наиболее пригодным и удобным прибором для измерения освещен-
ности как в лабораторных, так и в производственных условиях является
люксметр, в основе которого лежит селеновый фотоэлемент с внутрен-
ним фотоэффектом и зеленый светофильтр «ЗС-1» или «Gg-1» (фиг. 98,3).
Спектральная чувствительность этого прибора (фиг. 98,2) сравнительно
неплохо воспроизводит чувствительность среднего человеческого глаза
(фиг. t28). В этом легко убедиться из табл. 9, где в скобках приведены
Фиг. 98. Спектральная чувствительность селенового фотоэлемента
с внутренним фотоэффектом
1—фотоэлемент без фильтра; 2 — фотоэлемент с зеленым фильтром «ЗС-1» или
«Gg-1»; з — пропускание светофильтра «ЗС-1» или «Gg-1»
цифры, показывающие чувствительность люксметра. Для целей светокуль-
туры не следует пользоваться люксметрами Агрофизического инсти-
тута, спектральные кривые которых вследствие отсутствия зеленого
фильтра совершенно не совпадают с кривой чувствительности человече-
ского глаза (фиг. 98,7). Эти приборы пригодны лишь для измерения
освещенности, получаемой от ламп накаливания, при помощи которых
они градуируются.
Мы остановились столь подробно на методах измерения освещенности
по двум причинам. Во-первых, чтобы показать, что измерение освещен-
ности для фитофизиологических исследований имеет значение лишь в тех
случаях, когда возможен перевод единиц освещенности в объективные
единицы интенсивности физиологической радиации. Во-вторых, чтобы об-
легчить перевод первых в последние, так как в большинстве исследова-
ний интенсивность радиации выражается в люксах.
Оставляя в стороне многочисленные методы измерения интенсивности
радиации в объективных единицах энергии, рассмотрим лишь некоторые
из них, наиболее пригодные для наших целей. Речь идет о термоэлектри-
ческом и фотоэлектрическом методах измерения. Приборы первого типа
обладают интегральной, а приборы второго — селективной чувствитель-
ностью. Это означает, что первые, будучи отградуированными однажды,
V. Методы измерения лучистой энергии
145
пригодны для любого источника радиации, независимо от спектрального
состава излучения, вторые же нуждаются в градуировке или установле-
нии специального коэффициента для каждого отдельного типа источника
радиации.
Термоэлектрический метод основан на явлении возникновения элек-
трического тока при неодинаковом нагревании двух спаев различных
металлов. Обычно один спай находится при температуре окружающего
воздуха, а другой нагревается под влиянием поглощаемой им лучистой
энергии. Возникающая при этом термоэлектродвижущая сила пропор-
циональна разности температур, т. е. количеству поглощаемой, а следо-
вательно, и падающей, радиации. Наиболее пригодны термопары, со-
стоящие из константана в качестве одного металла и меди, золота или
серебра — в качестве другого. Во всех трех случаях при разности тем-
ператур в 1° возникают электродвижущие силы порядка 41,5 —
41,7 микровольт. Измерительный прибор состоит из термостолбика, пред-
ставляющего собою батарею последовательно включенных термопар,
низкоомного гальванометра (10—15 ом) с чувствительностью порядка
10-6 в, или 10~" а, и набора стеклянных светофильтров, позволяющих
выделять различные области спектра. Благодаря зачернению наружных
спаев приемника прибор обладает неселективной чувствительностью в ши-
роком диапазоне длин волн и позволяет измерять интенсивность как
интегрального, так и монохроматического потока лучистой энергии.
Для ботанико-физиологических целей желательно иметь набор б диф-
ференциальных светофильтров, спектральные характеристики которых
представлены на фиг. 99. Фильтры «БС-4» или «БС-5» (шотовские
«Wg-5» или «Wg-б») совместно с «Кс-19» (шотовский «Rg-8») позволяют
измерять физиологическую радиацию, лежащую в пределах 300—700 ту,
фильтры «БС-8» или «ЖС-4» (шотовский «Gg-З») позволяют измерять
ультрафиолетовые лучи, а фильтры «ЖС-18» и «ОС-11» (шотов-
ские «Gg-И» и «0g-l») разделяют физиологическую радиацию на коротко-
волновые и длинноволновые лучи по линии аб (фиг. 3).
Метод измерения с помощью дифференциальных светофильтров заклю-
чается в том, что интенсивность соответствующей области излучения
измеряется путем определения разности между излучениями, измеряе-
мыми в результате применения соответствующих светофильтров Пусть
интенсивность радиации, измеряемая светофильтром «БС-4», равняется г,
«Кс-19» — к, «БС-8» — I, а при помощи «ОС-11» — т. Тогда интен-
сивность физиологической радиации будет равна п=i — к; интенсивность
ультрафиолетовых лучей: p = i — Z; интенсивность сине-фиолетовой
радиации: g = i — т; интенсивность оранжево-красной радиации:
г=т — к и т. д. Нетрудно убедиться, что количество инфракрасных
лучей выражается величиной к.
Прибор, основанный на термоэлектрическом эффекте,'сконструирован
в Институте физиологии растений им. К. А. Тимирязева Академии Наук
СССР. Он представляет собою комбинацию двух термостолбиков с одина-
ковым внутренним сопротивлением (~17—20 ом) и равными электродви-
жущими силами (~7 микровольт на 0,001 кал/см2-мин), направленными
одна против другой. В одном термостолбике в качестве светофильтра
применяется фильтр «Wg-5», в другом — «Rg-8». Такой компенсацион-
ный термостолбик позволяет неселективно измерять всю физиологиче-
скую радиацию в пределах 300—700 ту и в соединении с гальваномет-
ром, обладающим внутренним сопротивлением в 11,2 ома и ценой деле-
ния в 0,0235 pA/mm, имеет чувствительность порядка 200 эрг/см2-сек,
или 0,282-10~3 кал/см‘2-мин на 1 мм шкалы.
10 А. Ф. Клешнин
146
Физические и физиологические основы светокультуры растений
Недостатком этого метода является наличие ошибки, вносимой соб-
ственным излучением светофильтров, которое возникает в результате
нагревания последних. Единственным способом устранения этой ошибки
является кратковременность измерения и введение светофильтров в общий
корпус прибора, изготовляемый из материала с достаточно высокой тепло-
проводностью. Это позволяет свести до минимума разность между темпе-
ратурой светофильтров и температурой внутренних спаев термобатареи.
Фотоэлектрический метод основан на свойстве некоторых металлов
и их соединений освобождать электроны под влиянием трх или иных
Фиг. 99. Спектральная характеристика светофильтров
для измерения физиологической радиации и ее частей
7 — «БО5» («Wg-б»); 2— «БС-4» («W&-5»); 3— «КС-19» («Rg-8»);
7 — «ОС-11» («Og-1 ’>); 5 — «ЖС-1Я» («GiX-1 1 •>); 6 — «ЖС-Л» («Crg-З );
7 — ацетил-целлюлозная пленка, обработанная раствором КОН; 8 — хло-
рофилл в растворе; 2хлорофилл в листе
лучей. Приборы этого типа обладают селективной чувствительностью;
при пользовании ими следует прибегать к специальным коэффициентам
для каждого типа источников радиации. Наиболее простым и удобным
прибором является селеновый фотоэлемент с внутренним фотоэффектом;
спектральная характеристика чувствительности этого фотоэлемента пред-
ставлена на фиг. 98,7. Как видно из этой кривой, максимум чувствитель-
ности приходится на 590 ту, и чувствительность снижается в оба конца
спектра, медленно в сторону коротковолновых и быстро — в сторону
длинноволновых лучей.
Прибор состоит из приемника — фотоэлемента, заключенного в метал-
лический корпус с круглым окном из матового стекла, гальванометра
с чувствительностью порядка 10 '"а и внутренним сопротивлением в пре-
делах 10—100 ом и набора диафрагм, допускающих измерение лучистых
потоков в широком диапазоне интенсивностей — от 100 до 500 тыс.
эрг/см-’-сек. Прибор пригоден для непосредственного измерения лишь
V. Методы измерения лучистой энергии
147
физиологической радиации источников с известной спектральной кри-
вой излучения. Отдельные области излучения, так же как и интегральный
лучистый поток, определяются методом расчета.
Методы расчета и градуировка прибора аналогичны расчетам и гра-
дуировке люксметра, описанным выше (§ 3). Обозначим интенсивность фи-
зиологической радиации через 1, а показания гальванометра через т;
тогда интенсивность радиации будет равна:
I — у; кт. (44)
Здесь величина к есть отношение величины падающего лучистого по-
тока в пределах 300—700 ту. к величине лучистого потока, восприни-
маемого фотоэлементом, а ц — коэффициент, определяемый опытным
Фиг. 100. Система фотоэлементов для измерения лучистой
энергии, поглощаемой пигментами пластид листа
1 — поглощение лучистой энергии пигментами пластид; спектральная чув-
ствительность фотоэлементов: 2 — натриевого, 3 — окепдно-цезиевого,
4 — гидридно-калиевого, 5 — рубидиевого
путем в результате градуировки. Последний соответствует величине
1,61 в формуле (40). Практически коэффициент К, зависящий от источника
радиации, определяется описанным выше методом (фиг. 94, 95 и 97 и табл.
59). Различие заключается лишь в том, что в данном случае при состав-
лении кривой и определении площади Sp (см. стр. 45 и 136 — 137)
вместо коэффициентов относительной видности К применяются соответ-
ствующие значения ординат кривой 1 на фиг. 98. Это сказывается на
величине площади S„ и, следовательно, на величине
Коэффициент г, не зависит от источника радиации и определяется
с помощью эталонной лампы с известной кривой распределения энергии
в спектре, известной величиной I и, следовательно, известным значением К.
Практически в качестве эталонной лампы может служить прожектор-
ная лампа' накаливания мощностью в 500 вт, отградуированная на цве-
товую температуру и силу света. Градуировку можно произвести в Палате
мер и весов, в Государственном оптическом институте (Ленинград) или
во Всесоюзном электротехническом институте (Москва). Подобная лампа,
работающая при напряжении порядка 90 в, обладает цветовой темпера-
турой около 2850°К и силой света в 500—600 св. На расстоянии 1 м лампа
создает интенсивность физиологической радиации в 3630 эрг/см2-сек.
10*
148
Физические и физиологические основы светокультуры растений.
Большой интерес представляет попытка Л. А. Иванова [80] создать
измерительный прибор, кривая чувствительности которого воспроизводит
кривую поглощения лучистой энергии пигментами пластид. В своем
Фиг. 101. Система фотоэлементов для измерения лучистой
энергии, поглощаемой хлорофиллом листа
1 — поглощение лучистой энергии хлорофиллом листа; спектральная чув-
ствительность фотоэлементов: 2 — натриевого, 3 — оксидно-цезпевого,
4 — гидридно-калиевого
первоначальном варианте этот прибор представлял собою комбинацию
двух термометров; один из них наполняли толуолом, а другой — рас-
твором хлорофилла в этой жидкости. Будучи отградуирован, прибор по
Фиг. 102. Система фотоэлементов для измерения лучистой
энергии, поглощаемой каротиноидами листа
1 — поглощение лучистой энергии каротиноидами листа; спектраль-
ная чувствительность фотоэлементов: 2— рубидиевого, 3— медно-
закисного
разности температур показывал количество лучистой энергии, поглощае-
мое пигментами листа. В настоящее время в подобном фито актинометре
проф. Л. А. Иванов применяет термостолбики, экранированные раствором
хлорофилла и соответствующим растворителем.
Создание фитоактинометра, позволяющего измерять интенсивность
физиологической радиации, поглощаемой как всей системой пигментов
V. Методы измерения лучистой энергии
149
пластид, так и отдельными ее компонентами, несомненно, представляет
огромный интерес. Но, невидимому, здесь по ряду обстоятельств следует
отказаться от экранирования приемника растворами пигментов, а надо
пойти по пути комбинирования различных светофильтров и фотоэлемен-
тов с внешним фотоэффектом с целью воспроизведения кривой чувстви-
тельности прибора идентичной кривой поглощения лучистой энергии
пигментами листа. Подбором различных площадей фотоэлементов можно
довольно точно воспроизвести любую заданную кривую чувствительности.
Подобные системы фотоэлементов представлены нами на фиг. 100—102.
Система, показанная на фиг. 100, отвечает поглощению лучистой энергии
всеми пигментами пластид, на фиг. 101 — хлорофиллом, а на фиг. 102 —
каротиноидами «среднего» листа.
Необходимо конструирование соответствующих приборов, для чего
нужна совместная работа физиологов растений с физиками и техниками.
6. ОПРЕДЕЛЕНИЕ КОЛИЧЕСТВА КВАНТОВ
В современной физиологии растений возникает необходимость из-
мерения не только интенсивности радиации, но и количества квантов,
1 — распределение энергии; 2 — распределение квантов; St и S2 — площа-
ди, соответствующие интенсивности радиации и числу квантов;
аб — граница видимой радиации
получаемых растениями. В настоящее время приборов для определения
этой величины в области физиологической радиации не имеется, хотя
его конструирование не связано с большими техническими трудностями.
Для этой цели приборы, характеристики которых представлены на
фиг. 100—102, необходимо лишь снабдить дополнительным светофиль-
тром, кривая пропускания которого (фиг. 99,7) для физиологической
радиации соответствовала бы формуле (3) или кривой 2 на фиг. 1. После
соответствующей градуировки названные приборы позволят измерять, по-
мимо интенсивности физиологической радиации, и количество квантов.
150
Физические и физиологические основы свепгокулътуры растений
Количество квантов, падающих на растение и поглощаемое растением,
легко рассчитать, если известен спектральный состав излучения и его
интенсивность. Метод расчета приведен на фиг. 103. Кривая 1 здесь соот-
ветствует распределению энергии, а кривая 2 — распределению квантов
в спектре лампы накаливания. Последняя получается в результате пере-
множения ординат кривой 1 (фиг. 103) на ординаты кривой 2 (фиг. 1).
Пусть кривые фиг. 103 перекрещиваются в области 680 шц, где спектраль-
ную интенсивность условно примем равной 1 эрг/см2-сек/А. Площадь
iSj (фиг. 103), ограниченная кривой 1, осью абсцисс и линией аб, будет
соответствовать определенной интенсивности физиологической радиации,
а площадь S„, ограниченная кривой 2, осью абсцисс и линией аб, — коли-
честву квантов, соответствующему данной интенсивности. Если отноше-
ние площадей есть
Л- = ^, ( (45)
то количество квантов в единице энергии физиологической радиации
данного источника, согласно формуле За, будет равно
п? = 5,09 • 1015 0,68.10~4 -к = 3,46- Ю11^ ‘ (46)
Количество квантов в 1 эрге физиологической радиации для разных
источников излучения, рассчитанное указанным методом, представлено
в табл. 48. С помощью этой таблицы нетрудно также определить коли-
чество квантов, поглощаемое листом или отдельными компонентами пиг-
ментной системы пластид. ,
х 7. РАСЧЕТЫ ИНТЕНСИВНОСТИ РАДИАЦИИ И ОСВЕЩЕННОСТИ
В практике светофизиологии и светокультуры растений часто при-
ходится рассчитывать интенсивность радиации или освещенность в за-
висимости от расстояния источника и удельной мощности ламп. Зависи-
мость интенсивности от расстояния для точечных источников выражается
законом обратных квадратов и законом косинусов. Эти законы гласят,
что на плоскости, находящейся под определенным углом к направлению
лучей, интенсивность радиации (или освещенность), получаемая с по-
мощью точечного источника, равняется величине, обратно пропорцио-
нальной квадрату расстояния от источника и прямо пропорциональной
косинусу угла между направлением лучей и нормалью к освещаемой
поверхности:
где Zo — интенсивность радиации при г=1 и а = 0.
Практически всякий реальный источник можно считать точечным,
если линейные размеры его светящейся поверхности не превышают 0,1
расстояния от источника до освещаемого объекта. Ошибка в этом случае
составляет всего лишь 0,25%.
Из определения освещенности следует, что сила света источника
в определенном направлении, выраженная в свечах, отвечает освещенности
на расстоянии 1 м в том же направлении, выраженной в люксах. Следо-
вательно, зная силу света источника (табл. 17, 18, 22, 23 и 26) и ее
распределение в пространстве(фиг. 30—33, 70, 72, 73), можно без большого
труда рассчитать освещенность (а следовательно, и интенсивность радиа-
ции) на любом расстоянии и в любом направлении от источника.
V. Методы измерения лучистой энергии
151
Пусть, как это показано на фиг. 104, источник радиации находится
в положении I на высоте R от горизонтальной поверхности, совпадающей
с осью абсцисс. Освещенность /. в точке а, находящейся на расстоянии г
в направлении, определяемом углом а, как и в любой другой точке, будет
равна величине, определяемой формулой (47). Поскольку r=cos~> то
удобнее пользоваться уравнением
Фиг. 104. Интенсивность радиации в зависимости от высоты подвеса ламп
I—III точки подвеса ламп па высоте 1 м; IV—VI — то же на высоте 0,5 м
1—6 — освещенность, создаваемая этими лампами в отдельности
А и Б — суммарная освещенность, создаваемая лампами при подвесе па высоте 1 и 0,5 м
На фиг. 104 в качестве примера приведена освещенность в различных
точках стеллажа при освещении лампами накаливания в 500 вт (220 в),
находящихся в обычных эмалированных глубокоизлучателях (фиг. 70, 3)
на расстоянии 1 м одна от другой и при высоте подвеса в 1 м (положения
I, II, III), и в 0,5 м (положения IV, V, VI). Кривая А отвечает суммар-
ной освещенности при высоте подвеса ламп в 1 м, кривая Б —• при высоте
в 0,5 м. Кривые 1—6 соответствуют освещенности, создаваемой каждой
лампой в отдельности и рассчитанной по формуле (48). о
Как видно из кривых А и Б (фиг. 104), более равномерное освещение
создается при большей высоте подвеса ламп. В случае низкого положения
ламп равномерность освещения может быть обеспечена лишь путем умень-
шения расстояния между лампами.
Для трубчатых источников радиации — неоновых ламп, ртутных
ламп низкого давления, люминесцентных ламп и других — можно предло-
жить следующую формулу, выражающую зависимость освещенности
(или интенсивности радиации) от расстояния освещаемой точки от лампы
(7?), диаметра (</) и длины (Z) лампы:
/у — __£1 _1___-I2-.. (49)
152
Физические и физиологические основы светокультуры растений
где L — освещенность в фотах, 1
расстояние и линейные размеры
Расстояние по длине агампд/ (S см)
Фиг. 105. Освещенность, создаваемая неоно-
вой лампой в 500 вт
J — па расстоянии 1 м; 2 — на расстоянии 0,5 м.
Длина светового столба — 100 см, диаметр лампы —
3,5 см, яркость — 2 сб, световой поток — 6500 лм
— яркость в стильбах, a R, d и I —
тр и длина) лампы (в см). Зна-
чение 1г и Z, ясно из фиг. 105.
Как видно из фиг. 105, где
представлена освещенность в
различных точках, находящих-
ся непосредственно под неоно-
вой лампой, располагаемой на
расстоянии 50 и 100 см от осве-
щаемой поверхности, интенсив-
ность радиации снижается в
сторону обоих концов лампы,
причем это снижение тем боль-
ше, *чем меньше расстояние
между лампой и освещаемой
плоскостью.
Более сложную проблему
представляет собой расчет ин-
тенсивности радиации или осве-
щенности, создаваемой системой
трубчатых источников радиа-
ции, расположенных в виде ре-
шетки на некоторой высоте над
освещаемой поверхностью. Это
имеет место при освещении ра-
стений люминесцентными лам-
пами.
Обычно в этом случае освещенность или интенсивность радиации
определяется методом непосредственных измерений.
Фиг. 106. Распределение освещенности поперек стеллажа (1,4 м) при освещении зер-
кальными лампами ВЭИ (300 вт, 120 в) широкого светораспределения, располо-
женными на высоте в 25 см и на расстоянии в 75 см одна от другой (Li п £2)
1 и 2 — освещенность от каждой лампы (L) в отдельности; 3 — суммарная освещенность
На фиг. 106 и 107 показано распределение освещенности поперек
стеллажа, получаемой при помощи зеркальных ламп ВЭИ. Из сравнения
этих рисунков видно, что посредством названных ламп при соответствую-
V. Методы измерения лучистой энергии
153
щих расстояниях между лампами и высоте подвеса их можно получить
сравнительно равномерную освещенность, причем лампы концентрирован-
ного светораспределения более пригодны для узких стеллажей (1 м),
а лампы с широким светораспределением —- для широких стеллажей
(1,5 м).
Фиг. 107. Распределение освещенности поперек стеллажа
(1 м) при освещении зеркальными лампами ВЭИ (500 вт,
120 в) концентрированного светораспределения, расположен-
ными на высоте в 50 см и на расстоянии в 50 см одна
от другой (Ь1 и L2)
1 и 2 — освещенность от каждой лампы в отдельности; 3 — суммарная
освещенность от этих ламп; 4— суммарная освещенность от аналогич-
ных ламп в 300 вт, 120 в
8. ИЗМЕРЕНИЕ ТЕМПЕРАТУРЫ ЛИСТЬЕВ
При изучении вопросов светокультуры растений, кроме интенсивности
падающей и поглощенной радиации, полезно также знать и температуру
листьев. Для этой цели в настоящее время применяются специальные
термоиглы, вводимые в лист. В основе работы этих термоигл лежит термо-
электрический эффект. Подобные иглы сконструированы В. П. Кисловым
[104, 105], который в качестве одного спая брал константан, а в качестве
другого наносил амальгаму золота. Прибор имеет термоэлектродвижу-
щую силу в 41 p-v на 1°. Сотрудник Института физиологии растений
им. Тимирязева инж. Алексеенко предложил термоиглу, сконструирован-
ную из остро отточенной медицинской иглы из нержавеющей стали (от
шприца), внутрь которой введен константан. Термоэлектродвижущая
сила такой иглы равняется 52—56 p-v на 1°.
Недостаток описанного метода заключается в том, что при введении
иглы внутрь листа он повреждается. Поранение, несомненно, отражается
на транспирации, а следовательно, и на температуре листа.
В качестве другого прибора для измерения температуры листьев сле-
дует рекомендовать термистор — прибор, работа которого основана на
изменении сопротивления полупроводника при изменении температуры.
Термистор не надо вводить внутрь листа, а достаточно прикладывать
к его поверхности. Недостатком прибора является то, что он показывает
не внутреннюю температуру листа, а температуру, среднюю между
154 Физические и физиологические основы светокультуры растений
температурой поверхности листа и температурой воздуха. Оба типа при-
боров изготовляет Ленинградский агрофизический институт.
9. ВЫВОДЫ
1. Необходимо отказаться от измерения лучистой энергии в единицах
освещенности. Следует пользоваться объективными единицами интенсив-
ности физиологической радиации. Единицами освещенности можно поль-
зоваться лишь в тех случаях, когда возможен их перевод в единицы
интенсивности радиации.
2. В качестве наиболее простых методов измерения необходимо реко-
мендовать термоэлектрический и фотоэлектрический методы. Приборы
первого типа, снабженные соответствующими светофильтрами, позволяют
измерять радиацию любых областей спектра, а приборы второго типа
измеряют лучистую энергию, поглощаемую листом и компонентами его
пигментной системы.
Часть II
ОБЩАЯ СВЕТОКУЛЬТУРА
РАСТЕНИЙ
Глава VI
ВЫРАЩИВАНИЕ РАСТЕНИЙ ИСКЛЮЧИТЕЛЬНО
ПРИ ИСКУССТВЕННОЙ РАДИАЦИИ
Выращивание растений исключительно в условиях искусственного
освещения в настоящее время представляет собою главным образом тео-
ретическую проблему, имеющую ограниченное практическое значение.
Однако после успешного решения этот прием будет иметь исключительно
большое значение для северных районов СССР. Сейчас же речь может
итти лишь о создании лабораторий искусственного климата, озеленении
метрополитена, комнатном цветоводстве и т. п.
1. КУЛЬТУРА РАСТЕНИЙ ПРИ ПОМОЩИ ДУГОВЫХ ЛАМП
Культура растений исключительно при помощи искусственных ис-
точников радиации стала возможной лишь с появлением дуговых ламп,
хотя не было недостатка в попытках использовать для этой цели и дру-
гие источники излучения — магниевые фонари, керосиновые лампы,
газовые горелки и пр. [319, 812]. Исследования с перечисленными источ-
никами радиации в настоящее время представляют главным образом
исторический интерес, хотя керосиновые лампы и применяются в неко-
торых случаях [96, 127, 192].
Уже первые опыты по культуре растений при помощи вольтовой дуги
показали принципиальную возможность выращивания растений в усло-
виях исключительно искусственного освещения, если из спектра назван-
ных ламп посредством экрана из обычного стекла устранены вредные
ультрафиолетовые лучи « 300 шр) и если интенсивность радиации до-
статочно велика [466, 467, 612, 754, 755, 756]. В своей первой публикации
Сименс [754] торжественно писал, что «в будущем садовод сможет стать
независимым от солнечного света для производства плодов высокого ка-
чества во всякое время года». Выращивая при помощи угольной дуги
горчицу (5 inapis alba) и морковь {Caucus саго la) только на искусствен-
ном свете, он обнаружил отрицательное действие на растения открытой
вольтовой дуги без светофильтра, устраняющего вредные ультрафиоле-
товые лучи. В опытах Дегерена [466] тоже применялась угольная дуга
в 2000 св. При употреблении открытой дуги уже через несколько дней
обнаруживалось сильное угнетение растений канн {Саппа sp.), сирени
{Syringa sp.), хризантем {Chrysantemum sp.), ночной красавицы {Mira-
bilis jalapa), пеларгоний {Pelargonium sp.) и др. При наличии экрана из
158
Общая светокультура растений
обычного оконного стекла растения не повреждались, но и не росли.
У трав и гороха (Pisum sativum) наблюдалось вытягивание и полегание,
бобы (ticia faba) образовали лишь первые листочки, а у других растений
[лен (Linum usitatissimum), пеларгония (Pelargonium sp.), драцена (Dra-
caena sp.), камелия (Camelia faponica), фикус ( icus elastica), азалия
(Azalea indica), рододендрон (Rhododendron sp.)] наблюдался лишь очень
медленный рост. Ни одно растение, за исключением ячменя, не цвело.
Дегерен пришел к заключению, что отрицательный результат являлся
следствием малой мощности ламп, не обеспечивавших достаточной интен-
сивности фотосинтеза. Интенсивность радиации лишь поддерживала
нормальное состояние взрослых растений, но была недостаточна для
роста и развития молодых растений.
Исследования названных авторов, так же как и опыты Бэйли [398— .
400] и Боннье [416—418], о которых более подробно будет сказано в сле-
дующей главе, показали принципиальную возможность светокультуры
растений, но определили, однако, в общем отрицательное отношение
к вольтовой дуге, как источнику радиации для выращивания растений.
С появлением ламп накаливания названный источник излучения был
вообще забыт. Причиной этого было также и техническое несовершенство
дуговых ламп того времени.
Исследования [421, 422, 528, 626, 658, 661, 813], проведенные как с от-
крытыми дугами, так и с дугами, работающими в замкнутой атмосфере,
показали, однако, хорошие качества этого типа ламп. Так, в опытах Тоттин-
гэма [813] значительно больший сухой вес (21,6 мг) имели томаты (Sola-
num ly coper sicum), выращиваемые в течение 9 дней под лампами «солнеч-
ный свет», чем под лампами накаливания (17,4 мг). В обоих случаях расте-
ния выращивались в условиях 15-часового дня при температуре воздуха
22° и освещенности 21 500 лк. Подобная освещенность соответствует
интенсивности физиологической радиации порядка 110 тыс. эрг/см1 2-сек
под лампами накаливания и 140 тыс. эрг/см2-сск под «солнечной лампой».
Митчел [626] в опытах с томатами (Solanum lycopersicum) сопоставил
эффективность вольтовой дуги с углями «солнечный свет» мощностью
3000 вт (60а, 50в) с эффективностью лампы накаливания в 2000 вт. В одной
серии опытов растения выращивались при одинаковой освещенности 7500—
8600 лк,1 а в другой — при одинаковой интегральной интенсивности из-
лучения, равной 100 тыс. эрг/см2-сск2. В первом случае лучший эффект
давали лампы накаливания, а во втором — дуговые лампы (табл. 62).
Если рассчитать эффективность ламп (сухой вес одного растения) для
одинаковой интенсивности физиологической радиации, то нетрудно
установить, что лампы накаливания эффективнее вольтовых дуг. Сказан-
ное не нуждается в объяснении, если принять во внимание, что спектр
вольтовой дуги содержит главным образом сине-фиолетовые и ультра-
фиолетовые лучи (фиг. 40, 4), а в спектре ламп накаливания много оран-
жево-красных лучей (фиг. 37).
Паркер с сотрудниками [421, 422, 658, 661], пользуясь вольтовой
дугой открытого горения (75а, 55в) с различными углями и лампами на-
каливания в 200 вт, показали, что с помощью этих источников радиации
при 16 часах освещения в сутки можно выращивать сою (Sofa hispida),
ячмень (Hordeum vulgare), коксагыз (Taraxacum кок-saghys) и другие
1 Эта освещенность соответствует интенсивности физиологической радиации
в 5'1—58 тыс. эрг/см2-сек под дуговой лампой и 38—44 тыс. эрг/см2-сек под лам-
пой накаливания.
2 Это соответствует интенсивности физиологической радиации в 30—35 тыс.
эрг/см2-сек под дуговой лампой и 15—20 тыс. эрг/см"-сек— под лампой накаливания.
VI. Выращивание растений при искусственной радиации
159
Таблица 62
Сухой вес и содержание углеводов (в граммах) при 2-недельном выращивании
томатов под лампами накаливания и под дуговой лампой (Митчел [626])
Органы растения Сухой вес 100 растений Содержание углеводов в 100 растениях
при 100 тыс. эр г/см2 • сек при 7500 — 8600 лк при 100 тыс. эрг/см- сек при 7500-8600 лк
лампа накали- вания воль- това дуга лампа накали- вания воль- това дуга лампа накали- вания вольтова дуга лампа накали- вания вольтова дуга
Корпи . . 8,36 31,20 30,35 38,12 1,04 4,82 —
Стебли . . 23,97 30,80 66,25 49,31 3,69 6,69 — —
Листья . . 43,58 112,37 83,68 71,55 5,06 27,33 23,09 16,61
Все расте- ние . . . *ч - 75,91 174,37 180,28 158,98 9,79 38,84 42,43 31,45
растения не хуже, чем при естественном освещении в сентябре. В их опы-
тах применялись лампы с углями «солнечный свет» (фиг. 40, -5) и другими,
отличавшимися друг от друга различным содержанием в спектре излу-
чения оранжево-красных лучей. Освещенность на расстоянии 1 м, созда-
ваемая дуговой лампой, равнялась 21 500 лк (или 159 тыс. эрг/см2-сек
физиологической радиации), а при помощи ламп накаливания — 1700 лк
(10 тыс. эрг/см'2-сек физиологической радиации). В качестве фильтра
служило обычное стекло.
Авторы нашли, что независимо от типа углей добавление излучения
от ламп накаливания, несмотря на незначительное увеличение интенсив-
ности радиации, дает всегда положительный эффект, выражающийся
в большем сухом и сыром весе растений (на 30—40%) и лучшем накопле-
нии углеводов и органического азота. Растения сои в возрасте 21 дня
имели сухой вес 2.23 г в опытном варианте и 0,98 г при естественном осве-
щении в оранжерее в сентябре. Вольтовы дуги, имитирующие комбина-
цию дуги «солнечный свет» с лампами накаливания, дают меньший эффект,
чем подобная комбинация. Одной из причин названного явления, по
мнению этих авторов, является повышение температуры листьев. При
температуре воздуха 21 ° температура листьев равнялась 22°,8 под лампами
накаливания, 24°,4 — под вольтовой дугой и 26°,1 —под их комбинацией.
Таким образом, на основании новейших исследований следует придти
к заключению, что, вопреки общепринятому мнению, вольтовы дуги
открытого горения при наличии стеклянного экрана, устраняющего
вредные ультрафиолетовые лучи, являются хорошим источником радйа-
ции для культуры растений. Развивающиеся при этом растения по своему
внешнему облику и химическому составу не отличаются от растений,
произрастающих в естественных условиях. Благодаря своей исключитель-
ной мощности угольные дуговые лампы имеют много преимуществ перед
другими источниками света, позволяя выращивать нормальные растения
при небольшом числе ламп.
2. КУЛЬТУРА РАСТЕНИЙ ПРИ ПОМОЩИ ЛАМП НАКАЛИВАНИЯ
С появлением ламп накаливания большинство исследований прово-
дилось с этим типом ламп. Полученные при этом результаты представля-
ют большой теоретический и практический интерес. Первая попытка
158
Общая светокультура растений
обычного оконного стекла растения не повреждались, но и не росли.
У трав и гороха {Pisum sativum) наблюдалось вытягивание и полегание,
бобы (г icia faba) образовали лишь первые листочки, а у других растений
[лен {Linum usitatissimum), пеларгония {Pelargonium sp.), драцена {Dra-
caena sp.), камелия {Camelia faponica), фикус ( icus elastica), азалия
{Azalea indica), рододендрон {Rhododendron sp.)] наблюдался лишь очень
медленный рост. Ни одно растение, за исключением ячменя, не цвело.
Дегерен пришел к заключению, что отрицательный результат являлся
следствием малой мощности ламп, не обеспечивавших достаточной интен-
сивности фотосинтеза. Интенсивность радиации лишь поддерживала
нормальное состояние взрослых растений, но была недостаточна для
роста и развития молодых растений.
Исследования названных авторов, так же как и опыты Бэйли [398— .
400] и Боннье [416—418], о которых более подробно будет сказано в сле-
дующей главе, показали принципиальную возможность светокультуры
растений, но определили, однако, в общем отрицательное отношение
к вольтовой дуге, как источнику радиации для выращивания растений.
С появлением ламп накаливания названный источник излучения был
вообще забыт. Причиной этого было также и техническое несовершенство
дуговых ламп того времени.
Исследования [421, 422, 528, 626, 658, 661, 813], проведенные как с от-
крытыми дугами, так и с дугами, работающими в замкнутой атмосфере,
показали, однако, хорошие качества этого типа ламп. Так, в опытах Тоттин-
гэма [813] значительно больший сухой вес (21,6 мг) имели томаты {Sola-
num lycopersicum), выращиваемые в течение 9 дней под лампами «солнеч-
ный свет», чем под лампами накаливания (17,4 мг). В обоих случаях расте-
ния выращивались в условиях 15-часового дня при температуре воздуха
22° и освещенности 21 500 лк. Подобная освещенность соответствует
интенсивности физиологической радиации порядка 110 тыс. эрг/см‘1 2-сек
под лампами накаливания и 140 тыс. эрг/см‘2-сек под «солнечной лампой».
Митчел [626] в опытах с томатами {Solanum lycopersicum) сопоставил
эффективность вольтовой дуги с углями «солнечный свет» мощностью
30ОО вт (60а, 50в) с эффективностью лампы накаливания в 2000 вт. В одной
серии опытов растения выращивались при одинаковой освещенности 7500—
8600 лк,1 а в другой — при одинаковой интегральной интенсивности из-
лучения, равной 100 тыс. эрг/см'2-сек2. В первом случае лучший эффект
давали лампы накаливания, а во втором — дуговые лампы (табл. 62).
Если рассчитать эффективность ламп (сухой вес одного растения) для
одинаковой интенсивности физиологической радиации, то нетрудно
установить, что лампы накаливания эффективнее вольтовых дуг. Сказан-
ное не нуждается в объяснении, если принять во внимание, что спектр
вольтовой дуги содержит главным образом сине-фиолетовые и ультра-
фиолетовые лучи (фиг. 40, 4), а в спектре ламп накаливания много оран-
жево-красных лучей (фиг. 37).
Паркер с сотрудниками [421, 422, 658, 661], пользуясь вольтовой
дугой открытого горения (75а, 55в) с различными углями и лампами на-
каливания в 200 вт, показали, что с помощью этих источников радиации
при 16 часах освещения в сутки можно выращивать сою {Sofa hispida),
ячмень {Hordeum vulgare), коксагыз {Taraxacum кок-saghys) и другие
1 Эта освещенность соответствует интенсивности физиологической радиации
в 51—58 тыс. эрг/см‘--сск под дуговой лампой и 38—44 тыс. эрг/см2-сек — под лам-
пой накаливания.
2 Это соответствует интенсивности физиологической радиации в 30—35 тыс.
эрг/см2-сек под дуговой лампой и 15—20 тыс. эрг/см--сек — под лампой накаливания.
VI. Выращивание растений при искусственной радиации
159
'Г а б л и ц а 62
Сухой вес и содержание углеводов (в граммах) при 2-неделъном выращивании
томатов под лампами накаливания и под дуговой лампой (Мигчел [626])
Сухой вес 100 растений Содержание углеводов в 100 растениях
Органы растения прп 100 тыс. эрг/см2 • сек при 7500— 8600 лк при 100 тыс. эрг/см- • сек при 7500 — 8600 лк
лампа накали- вания воль- това дуга лампа накали- вания воль- това дуга лампа накали- вания вольтова Дуга лампа накали- вания вольтова дуга
Корни . . 8,36 31,20 30,35 38,12 1,04 4,82 —
Стебли . . 23,97 30,80 66,25 49,31 3,69 6,69 — —
Листья . . Все расте- 43,58 112,37 83,68 71,55 5,06 27,33 23,09 16,61
ние . . . - 75,91 174,37 180,28 158,98 9,79 38,84 42,43 31,45
растения не хуже, чем при естественном освещении в сентябре. В их опы-
тах применялись лампы с углями «солнечный свет» (фиг. 40, 5) и другими,
отличавшимися друг от друга различным содержанием в спектре излу-
чения оранжево-красных лучей. Освещенность на расстоянии 1 м, созда-
ваемая дуговой лампой, равнялась 21 500 лк (или 159 тыс. эрг/см‘2-сек
физиологической радиации), а при помощи ламп накаливания — 1700 лк
(10 тыс. эрг/см2-сек физиологической радиации). В качестве фильтра
служило обычное стекло.
Авторы нашли, что независимо от типа углей добавление излучения
от ламп накаливания, несмотря на незначительное увеличение интенсив-
ности радиации, дает всегда положительный эффект, выражающийся
в большем сухом и сыром весе растений (на 30—40%) и лучшем накопле-
нии углеводов и органического азота. Растения сои в возрасте 21 дня
имели сухой вес 2.23 г в опытном варианте и 0,98 г при естественном осве-
щении в оранжерее в сентябре. Вольтовы дуги, имитирующие комбина-
цию дуги «солнечный свет» с лампами накаливания, дают меньший эффект,
чем подобная комбинация. Одной из причин названного явления, по
мнению этих авторов, является повышение температуры листьев. При
температуре воздуха 21 ° температура листьев равнялась 22°,8 под лампами
накаливания, 24°,4 — под вольтовой дугой и 26°,1 — под их комбинацией.
Таким образом, на основании новейших исследований следует придти
к заключению, что, вопреки общепринятому мнению, вольтовы дуги
открытого горения при наличии стеклянного экрана, устраняющего
вредные ультрафиолетовые луйи, являются хорошим источником радйа-
ции для культуры растений. Развивающиеся при этом растения по своему
внешнему облику и химическому составу не отличаются от растений,
произрастающих в естественных условиях. Благодаря своей исключитель-
ной мощности угольные дуговые лампы имеют много преимуществ перед
другими источниками света, позволяя выращивать нормальные растения
при небольшом числе ламп.
2. КУЛЬТУРА РАСТЕНИЙ Ш’И ПОМОЩИ ЛАМИ НАКАЛИВАНИЯ
С появлением ламп накаливания большинство исследований прово-
дилось с этим типом ламп. Полученные при этом результаты представля-
ют большой теоретический и практический интерес. Первая попытка
160
Общая светокультура растений
выращивания растений с помощью ламп накаливания принадлежит Телену
[796], который для этой цели применял калильную лампу Лодыгина1.
Слабая интенсивность радиации не позволила этому автору получить
нормальные растения [овес (Avena saliva), гречиха ( agopyrum esculentum),
бобы (I icia faba), сахарная свекла (Beta vulgaris), горчица (Sinapis alba),
горох (Pisum sativum) и элодея (Elodea canadensis)] и довести их до цвете-
ния. У растений наблюдались явные признаки этиоляции (вытягивание
междоузлий и слабое развитие листовых пластинок).
Гарвей [540, 541], применяя лампы накаливания в 200 и 1000 вт при
беспрерывном освещении в условиях различной температуры (14 и 25—30°)
и освещенности (4100,4900, 7300 и45 000 лк1 2), довел до цветения, плодо-
ношения и созревания семян большое число самых разнообразных расте-
ний. Семена отличались хорошим качеством и всхожестью и содержали
большое количество крахмала. Капуста (Brassica oleifera var. capitata)
образовала нормальный кочан, а картофель (Solanum tuberosum) —
клубни. Некоторые растения, например томаты (Solanum lycopersicum),
лишь цвели, но не плодоносили, а капуста, нуждаясь в низких темпера-
турах для прохождения стадии яровизации, не цвела совсем.
В своей следующей работе [546] названный автор, расширив ассорти-
мент растений, показал, что для нормального развития и плодоношения
различные виды и сорта растений нуждаются в разной интенсивности
радиации, лежащей в очень широком диапазоне — от 2700 до 130 тыс. лк,
или 16—780 тыс эрг/см2-сек физиологической радиации (табл. 63). Неко-
торые растения, указанные в конце таблицы, лишь цвели, но не плодо-
носили.
Опыты Гарвея вскоре были повторены Мунерати [633] и Гиббеном
[548]. Опыты Гиббена интересны тем, что в некоторых вариантах для
устранения инфракрасных лучей был применен водный экран, а в дру-
гих — стеклянный экран, устраняющий ультрафиолетовые лучи с дли-
ною волны короче 350 шд. Кроме того, этот автор сравнил растения,
выращиваемые под лампами накаливания, с растениями, культивируе-
мыми при естественном освещении в теплице ( в феврале—марте), и по-
казал, что опытные растения — тюльпаны (Tulipa sp.), гиацинты (Нуа-
cynthus sp.) и др. — под лампами накаливания развиваются значительно
быстрее, чем в контроле. Наличие водного экрана или устранение ультра-
фиолетовой радиации не влияло на развитие растений, если не считать
несколько лучшего развития растений в отсутствии инфракрасных лучей,
повышающих температуру растений.
В Советском Союзе выращивание растений при помощи искусствен-
ной радиации связано с именем академика Н. А. Максимова [179—183,
188], опыты которого в этом направлении начались в 1923 г. В своих
первых опытах [179] при помощи ламп накаливания в 500 и 1000 вт (на
высоте 1 м) он выращивал в условиях беспрерывного освещения и
12-часового дня озимые и яровые формы пшеницы (Triticum vulgare), ячмень
(Hordeum vulgare), горох (Pisum sativum), сою (Sofa hispida), гречиху
(Fagopyrum esculentum), томаты (Solanum lycopersicum), огурцы (Cucumis
sativus), кабачки (Cucurbita pepo) и подсолнечник (Helianthussp.). В резуль-
тате длительных опытов было установлено, что растения, выращиваемые
под лампами накаливания, развиваются значительно скорее, чем растения,
1 В иностранной литературе называется лампой Нернста (1897); в действитель-
ности же эта лампа изобретена Лодыгиным (1872).
2 Соответствует 25, 29, 44 и 270 тыс. эрг/см2>сек физиологической радиации.
VI. Выращивание растений при искусственной радиации
161
Таблица 63
Интенсивность радиации, потребная для цветения и образования семян
у различных растений
(Геидриас и Гарвей [546])
Растение Темпера- тура воздуха (в CC) Освещенность (в тыс. лк) Интенсивность физиологической радиации (втыс. эрг/см2-сек*)
Шалфей (Salvia sp.) 20 3,2 19,0
Горох (Pisum sativum) 14 4,3 26,0
Гвоздика (Dianthus barbatus) 20 7,0 42,0
Паслен (Solanum nigrum) 20 8,0 48,0
Смолевка (Silene latifolia) 25 8,0 52,0
Молочай (Euphorbia splendens) 20 8,6 52,0
Фасоль (Phaseolus vulgaris) 25 9,1 54,0
Звездчатка (Slellaria media) 2 5 9,9 59,0
Овес (Avena sativa) 14 10,0 60,0
Болтония (Boltonia asteroides) 25 10,0 60,0
Мелколепестник (Erigeron canadensis) . . 25 11,5 69,0
Донник белый (Melilotus alba) 14 12,5 75,0
Оксидафус (Oxidaphus nyctagineus) .... 20 14,5 87,0
Редис (Raphanus sativus) 14 15,0 90,0
Клевер луговой (Tri folium pfatense) . . . 14 15,0 90,0
Донник аптечный (Melilotus officinalis) . 14 15,0 90,0
Лилия (Li Hum longi folium) 20 130,0 780,0 .
Настурция (Tropaeolum minus) 20 2,7 — 4,8 16,0 — 29,0
Тыква мускатная (Cucurbita moschata) . . 20 2,7 — 5,4 16,0 — 32,0
Гречиха (Fagopyrum escidentum) 20 3,2 - 4.3 19,0 — 25,7
Конопля (Cannabis sativa) 20 3.2 — 4.8 19,0 — 28,7
Фасоль (Phaseolus vulgaris) 20 3,2 — 5,9 19,0 — 35,3
Пеларгония (Pelargonium sp.) 20 3.7 — 5,9 22,5 — 35,3
Клевер шведский (Trifolium hybrid umf 20 4,3 — 6.4 25,7 — 38,3
Лен (Linum usitatissimum) 20 5,35— 11,0 32,0 — 66,0
Астра (Aster sp.) 25 5,35— 22,2 32,00—133,00
Кукуруза (Zea Mays) 25 5.35 - 32,0 32,00—190,00
Марь (Chenopodium album) 20 5,35—114,0 32,00—664,00
Щирица (Amaranthus retrojlexus) 20 5.35—114,0 32,00—664,00
Гибискус (Hibiscus trionum) 25 6,15 — 9.65 38,60— 58,00
Рожь (Secale cereale) 14 6.45 -13,00 38,00— 78,00
Ячмень (Hordeum vulgare) 14 7,00— 8,6 42,00— 51,50
Пшеница мягкая (Triticwn vulgare') . . . 11 7,00— 8,6 42,00— 51,50
Пшеница твердая (Triticum durum) . . . Г! 7,00— 8,6 42,00— 51,50
Табак (Nicotiana tabacum) 20—25 8,60—108,0 51,50— 64,80
Салат (Lacluca sativa) 25 9.65— 14,5 57,75— 87,00
Земляника (Fragaria sp.) 20 16.00— 22,2 96,00—133,20
Тыква мускатная (Cucurbita moschata) ** . 25 10,00 60,00
Анютины глазки (Viola tricolor) ** . . . . 25 10,75 64,50
Картофель (Solanum tuberosum) **.... 25 4,80— 5,40 28,70— 32,30
Тыква обыкновенная (Cucurbita pepo) ** . 25 5.40— 8,00 32,30— 48,00
* Рассчитано нами.
* * Цвели, по не плодоносили.
11 А. Ф Клешнин
162
Общая светокультура растений
произрастающие в естественных условиях. Это обстоятельство позво-
ляет выращивать 4—5 поколений растений в год. Несмотря на сравнительно
невысокую освещенность (порядка 8000 лк), вес зерен был почти равен или
равен их весу при произрастании растений в естественных условиях.
Так, у пшеницы вес 100 зерен опытных растений равнялся 3,05 г, а ис-
ходных семян — 4,41 г, у ячменя — соответственно 3,10 и 3,29 г. Одно-
временно автор отмечал пожелтение листьев в условиях беспрерыв-
ного освещения у томатов и бобов и ненормальное вытягивание листьев
злаков и стеблей у двудольных растений под влиянием ламп накаливания,
несмотря на нормальную зеленую окраску растений. Позднее Н. А.
Максимов [180], расширив ассортимент растений [огурцы (Cucumis
sativus), салат (Lactuca sativa), редис (Raphanus sativus var. radicula),
лен (Linum usitatissimum), лук (Allium сера), шпинат (Spinacea oleracea),
земляника (I- ragaria sp.) и др.], показал, что большинство растений пре-
красно развивается под лампами накаливания при 8000 лк (48 тыс.
эрг/см2-сек физиологической радиации), однако крестоцветные, салат
и шпинат отличаются при этом чрезмерным вытягиванием подсемядоль-
ного колена. Пшеница Новинка, созревающая в естественных условиях
через 80 дней, под лампами накаливания давала урожай через 45—48
дней, а лен вместо 90 дней — через 60—65 дней.
После исследований Н. А. Максимова и Гарвея появилось большое
количество работ по выращиванию растений при помощи ламп накали-
вания. Эти исследования продолжаются и в настоящее время. Рассмотрим
некоторые из них, имеющие принципиальное значение. Подавляющее
большинство исследователей вслед за Н. А. Максимовым [179] отмечает
значительное ускорение в развитии растений под влиянием ламп накали-
вания, выражающееся в быстром переходе растений к цветению, плодо-
ношению и созреванию семян. Непременным условием при этом является
устранение перегрева растений, обычного под лампами накаливания.
В опытах Артура и Гутри [376, 378] при беспрерывном электрическом
свете от ламп накаливания семена клевера (Ttifolium sp.) были получены
через 38 дней после посева, а зрелые колосья ячменя (Hordeum vulgare),
овса (Avena sativa) и яровой пшеницы (Triticum vulgare) — через 47 дней.
Трюфо и Тюрнессан [814, 815] посредством ламп накаливания, при
освещенности 37—40 тыс люксов (200—220 тыс. эрг/см2-сек физиологи-
ческой радиации) и температуре 22—28°, выращивали табак (Nicotiana
tabacum), овес (Avena sativa), землянику (Fragaria sp.) и фасоль (Pha-
seolus sp.). Зрелые ягоды земляники отличного качества были получены
через 40 дней вместо обычных для этой культуры 80 дней. Фасоль дала
нормальные зрелые семена.
В опытах, проведенных в целях селекции и семеноводства. Гиорт
1551] показал, что при помощи ламп накаливания при беспрерывном
освещении и удельной мощности 750 вт/м2 можно получить обильный
урожай семян значительно быстрее, чем в естественных условиях, у мно-
гих растений: тысячеголова (V accaria segetalis), куколя (Agroslemma
githago), ноланы (Nolana sp.), чернушки (Nigella sativa), смолевки (Silene
pendula), льна (Linum usitatissimum), ячменя (Hordeum vulgare), коллин-
зии (Collinsia bicolor), кларкии (ClarKia pulchella, Clarnia elegans), гилии
(Gilia achillifolia), львиного зева (Antirrhinum majus), бобов (I icia. faba),
фасоли (Phaseolus vulgaris) и гибискуса (Hibiscus trionum).
Созревание семян наступает значительно быстрее, чем в естественных
условиях: у тысячеголова — через 50 дней после посева, у ячменя —
через 47—57 дней, а у коллинзии — через 60 дней. В течение года автор
получал около четырех поколений растений.
VI. Выращивание растений при искусственной радиации 1б;>
В. П. Мальчевский [191, 192] исследовал около 165 видов и сортов
растений и нашел, что у 85% испытанных им растений при выращивании
под лампами накаливания вегетационный период сокращается на 25—50%.
У томатов {Solanum, lycopersicum) вегетационный период сокращается
с 115—160 до 92—100 дней, у лука {Allium сера) — с 50 до 35, у салата
{Lactuca sativa) — с 50—60 до 35—-40, у кабачков {Cucurbita pepo) —
с 95 до 66—75, у кольраби {Brassica caulorapa) — с 105—110 до 90—100,
у перца {Capsicum sp.) — с 160—175 до 115, у баклажан {Solanum melon-
gena) — с 160—175 до 110, а у пшеницы {Triticum vulgare) — с 90—112
до 45—68 дней. Число зерен в колосе пшеницы, выращенной под лампами
накаливания при освещенности 4—8 тыс. люксов (24—48 тыс. эрг/см2-сек
физиологической радиации), равнялось 15—20, а в контроле 12—17.
Вес 1000 зерен соответственно равнялся 29—35 и 29—34,4 г.
В опытах В. П. Мальчевского кукуруза {Zea mays) давала 2—3, то-
маты— 3, просо {Panicum miliaceum) — 4, лен {J.inum usitatissimum) —
4—5, яровая пшеница — 6—7, а гречиха {В agopyrum esculentum)— 7 уро-
жаев в год. Томаты «Дварф стоун», по данным В. II. Мальчевского [192],
при беспрерывном освещении зацветают на 49-й день, а зрелые плоды
получаются на 104—110-й день. В естественных условиях этот сорт
цветет через 80 дней, а зрелые плоды получают через 165 дней.
Исключительно интересные данные получены Е. Г. Победимовой
[248]. Выращивая растения при освещенности 2—3 тыс. люксов (12—18
тыс. эрг/см2-сек физиологической радиации), получаемой от ламп нака-
ливания, она нашла, что многолетник мерингия {Moehringia trinervia и
М. diversifolia) зацветает на первом году, многолетники дряква {Cycla-
men соит), ясколка {Cerastium dahuricum) и крашенинниковия {Кга-
scheninniitovia Borodinii) цветут дважды в году, а однолетний паслен
{Solanum nigrum) превращается в многолетник. Томаты при беспрерывном
освещении также становятся многолетниками [59]. В опытах В. П. Маль-
чевского [196] шиповник при беспрерывном освещении от ламп накали-
вания зацветал в первом году культуры и цвел дважды во втором.
Большой интерес представляет сообщение И. II. Никитина [232,
233], который выращивал сеянцы дуба {Quercus robur) при беспрерывном
освещении под лампами накаливания, температуре 25—30° и освещенности
3,5—4 тыс. лк (21—24 тыс. эрг/см2-сек физиологической радиации).
Через 5 месяцев, по достижении сеянцами высоты 2,1 м, последние были
высажены в грунт, где и развивались в естественных условиях в течение
нескольких лет. Дубки зацвели на седьмом году, а в следующем году
были получены первые зрелые жолуди. Обычно же дуб начинает плодо-
носить лишь через 50—80 лет.
Интересные результаты получены Б. С. Мошковым [213, 216, 218,
219, 221, 224], который при помощи ламп накаливания получает зрелые
плоды томатов {Solanum lycopersicum) через 45—50 дней после появления
всходов. Применяя лампы накаливания в 200—300 вт (удельная мощность
3,2—4,8 квт/м2) и водный экран, нагреваемый погруженными лампами
до 45°, этот автор установил, что томаты Пушкинские и Движение на
север, обычно дающие зрелые плоды через 110—120 дней, плодоносят
через 45—50 дней, формируя за 2 месяца 5—8 плодов с общим весом
250—300 г на растение (табл. 123).
В другом опыте с теми же лампами при удельной мощности 10,8 квт/м2
и интегральной интенсивности радиации 250—300 тыс. эрг/см2-сек, из
которой половина приходится на физиологическую радиацию, а другая
половина — на инфракрасные лучи, Б. С. Мошков культивировал ветви-
стую пшеницу {Triticum turgidum), сравнивая ее с той же пшеницей,
п*
164 Общая светокультура растений
одновременно произраставшей в оранжерее (в мае—августе). Под лампами
накаливания пшеница начала колоситься через 32, а в контроле через
53 дня. Через 70 дней под лампами был получен урожай семян по 3—4,5 г
с колоса; в теплице созревание семян наступило через 112 дней. При
помощи ламп накаливания Б. С.
Мошкову [224] удалось получить
в течение года пять поколений
X*. ветвистой пшеницы.
Фиг. 108. Горчица (Sinapis alba),
выращенная под разными лампами
1 — под люминесцентными лампами;
2 — под лампами накаливания
•Dili". 10!). Огурцы (Cucumis sativus),
выращенные под разными лампами
1 — под люминесцентными лампами;
2 — под лампами накаливания
Хлопчатник (Gossypium hirsutum) Одесский в опытах Б. С. Мошкова
[224] бутонизировал под лампами накаливания при удельной мощности
4,8 квт/м2 в условиях 18-ча-
Фиг. 110. Просо (Panicum miliaceum)
Арнольд 0853, выращенное под разными лампами
z — под люминесцентными лампами; 2 — под лампами
накаливания
сового дня через 21 день по-
сле всходов, цвел — через 44,
а зрелые коробочки дал через
85 дней. Б обычных условиях
он бутонизирует через 40,
цветет через 70, а созревает
через 115 дней.
Некоторые растения при
круглосуточном освещении
лампами накаливания цветут
почти беспрерывно. Сказан-
ное отмечается, например,
для примулы (Primula sp.),
орхидей и др. [8, 30].
Отличительной особен-
ностью растений, разви-
вающихся под лампами на-
каливания, является чрез-
мерное вытягивание стебля
(фиг. 108), в особенности под-
семядольного колена (фиг. 109), а у однодольных — вытягивание листьев
(фиг. 110). Обычно это явление сопровождается слабым развитием меха-
нических тканей, что часто приводит к полеганию растений (рис. 111).
Этот факт отмечается всеми без исключения исследователями, имеющими
дело со светокультурой растений [8, 30, 179, 180, 247, 248, 428, 540,
541, 5511
VI. Выращивание растений при искусственной радиации
165
Особенно плохо удается с помощью ламп накаливания культура
крестоцветных, подсемядольное колено которых чрезвычайно чувстви-
тельно к спектральному составу лучей и интенсивности излучения (фиг.
108 [180, 428]). Многие растения, имеющие в естественных условиях
розеточную форму, под лампами накаливания образуют стебель. Таковы,
Фиг. 111. Томаты (Solanum lycopersicum) Печорские,
выращенные под разными лампами
1 — под люминесцентными лампами; 2 — под лампами накаливания
например, салат (фиг. 20) или редис (фиг. 16), обычно под лампами нака-
ливания сразу же дающие цветочный стебель [184]. Росянка (Drosera
sp.) — типичное розеточное растение — также в результате вытягивания
междоузлий приобретает стеблевую форму [30].
Некоторые данные позволяют думать, что в результате длительной
культуры изменения, возникающие под лампами накаливания, стано-
вятся наследственными. Н. А. Артемьев (8) в результате многолетних
опытов по выращиванию растений при помощи ламп накаливания нашел,,
что огурцы (Cucumis sativus) Муромские от поколения к поколению из-
меняют форму и размеры плодов. Последние становятся мельче и приоб-
ретают грушевидную форму. В 4-м поколении они почти совсем не имеют
166
Общая светокультура растений
семян и по качеству становятся лучше тепличных. Растения, высаженные
из люминостата в грунт, обычно погибают.
Быстрый рост в высоту под влиянием ламп накаливания открывает
широкие перспективы для применения этого источника при выращивании
сеянцев древесных растений.
В опытах Ширли [753], который в условиях различной освещенности
(от 320 до 3500 лк) выращивал Мамонтове дерево (Sequoia sempervirens)
и сосну (Pinus Taeda), было установлено, что для нормального роста
Мамонтова дерева необходима освещенность порядка 3500 лк.
Одновременно Редингтоном [703, 704] были проведены опыты по вы-
ращиванию бука (' agus silvatica). Культивируя названное растение
в течение 134 дней под лампами накаливания в 200 вт, автор нашел, что
в условиях беспрерывного освещения бук отличается интенсивным и
беспрерывным ростом.
Позднее Е. Г. Победимова [248] в течение нескольких месяцев в усло-
виях беспрерывного освещения (около 3000 лк, или 18 тыс. эрг/см~-сек
физиологической радиации) выращивала калину (> iburnum Lantana),
ель (Picea alba) и лиственницу (Larix sibirica), получив хороший при-
рост в высоту.
Нормальное развитие под лампами накаливания акации, лимона и
розы отмечает также Н. А. Артемьев [8].
По данным И. Н. Никитина [232, 233], дуб (Quercus robur) под лампами
накаливания в течение З1 месяцев достигает высоты в 140—250 см и
дает прирост по 1—2,5 см в день.
Широкие исследования в области светокультуры древесных пород
провел В. П. Мальчевский [196]; применяя освещенность в 4—6 тыс. лк
(24—36 тыс. эрг/см2-сек физиологической радиации), он установил, что
при беспрерывном освещении лампами накаливания можно в течение
года получить такие саженцы, которые в естественных условиях получают
лишь через несколько лет (табл. 64).
Т а б л и 11, а 6 4
Высота сеянцев древесных в условиях светокультуры
(Мальчевский [196])
ч Растение т (в ДНЯХ) Высота растений
ль (есте- ое осве- см лампы накалива- лампы накалива- пин; беспрерыв- ное освещение лампы накалива- ния 20 часов и неоновые лампы— 4 часа
ния; 14-часовой день
Возрас контро ствснн щение) см % к кон- тролю см % к кон- тролю см % к кон- тролю
Ель (Picea sp.) .... 120 4,7 5,1 1 104 8,8 180 t"' / 118
Сосна (Pinus sp.) . . . 150 5,8 14,8 255 8,1. 140 10,0 172
Лиственница {Larix. sp.) 120 4,7 14,5 308 27,2 570 22,2 466
Береза (Betula sp.) . . 150 19.7 67,1 340 74,3 376 76,6 336
Для культуры ели достаточно 4 тыс. лк (24 тыс. эрг/см‘2-сек физио-
логической радиации), но непременным условием быстрого роста является
беспрерывное освещение. Для сосны нужна освещенность 6 тыс. лк
(36 тыс. эрг/см2-сек физиологической радиации) и 14-часовой день.
Исключительно светолюбивым деревом является лиственница, которая
отрицательно реагирует как на уменьшение освещенности ниже 6 тыс. лк,
VI. Выращивание растений при искусственной радиации 167
так и на сокращение периода освещения. Древесные растения, культиви-
руемые в условиях беспрерывного освещения под лампами накаливания,
характеризуются гипертрофированной толщиной годичных колец и круп-
ными размерами хвои.
В другом опыте В. П. Мальчевский [196] исследовал рост однолетних
сеянцев древесных пород, которые в течение 5 месяцев (20 апреля—20
сентября) выращивались в условиях 12-часового дня под лампами накали-
вания (2,2—3,0 тыс. лк, или 13—18 тыс. эрг/см2-сек физиологической
радиации), при температуре 23—28° (табл. 65).
Т а б л и ц а 65
З-.нлсячный прирост однолетних сеянцев древесных пород в условиях светокультуры
(Мальчевский [196])
; Прирост сеянцев
Растение - контроль (естественное освещение) лампы накали- вания. верхнее освещение лампы накали- вания, боновое освещение
в % к кон- ВСМ | тролю в см в % к кон- тролю
1 Лещина (Corylus arellana) 17,5 60.0 343 62,4 357
Липа (Tilia cordata') 10,2 70,8 G95 75,1 738
Желтая акация (Caragana arborescens) . i 22,6 77,5 344 73,5 326
Боярышник (Crataegus oxyacantha) . . | 13,8 48,0 348 40,2 292
Барбарис (Berbej-is vulgaris) 12,2 32,3 265 80,0 246
Ясень (Fraxinus exelsa) j 8,0 20,1 251 16,4 205
Характерной особенностью древесных пород при непрерывном осве-
щении лампами накаливания является длительный рост, равномерный
или прерываемый кратковременными периодами покоя. Впервые на этот
факт обратил внимание Клебс [578], установивший его для бука (Fagus
silvatica}, а позднее Редингтон [703, 704] и В. И. Мальчевский [196].
В опытах Н. А. Максимова и В. М. Лемана [183] дуб (Quercus pedunculate)
давал около 10 приростов в год и достигал высоты в 1,5 м, а яблоня (Ма-
lus sp.), вишня (Cerasus sp.), клен (Acer sp.), ясень ( raxinus sp.), липа
(Tilia sp.), ракитник (Cytisus sp.), пузырник (Colulea sp.) и бересклет
(Evonymus sp.) сбрасывали листья и прекращали рост на 1—3 месяца
позднее и трогались в рост на 1 — 14, месяца ранее, чем контрольные
растения. По данным В. М. Лемана [155], рост дуба в условиях беспрерыв-
ного освещения состоит из ритмично чередующихся периодов интенсив-
ного роста в высоту и периодов относительного покоя. Каждый цикл,
в зависимости от вида растения, равняется 17—50 дням, причем на период
роста приходится 10—40 дней, а на период покоя 7—10 дней. Сказанное
установлено как для Quercus pedunculate,, так и для других видов: Q. сег-
ris, Q. suber и О. ilex-, конечная высота растений определяется соотноше-
нием между периодами роста и периодами покоя (табл. 66).
Типичная картина поведения древесных пород в условиях беспре-
рывного освещения представлена на фиг. 112. Растения в среднем сосуде
выращивались под лампами накаливания и характеризуются признаками,
типичными для растений, культивируемых под названным источником
радиации: малым диаметром стебля, длинными междоузлиями, чрез-
мерным ростом в высоту и т. д.
Очень интересное исследование провел В. 3. Гулисашвили [53],
который беспрерывным освещением лампами накаливания в 1000 вт
168
Общая светокультура растений
нарушал состояние покоя у большого числа древесных пород раз-
личного географического происхождения. Контрольные растения разви-
вались в естественных условиях. Автор нашел, что нарушение покоя
совершается быстрее при более позднем перенесении растений под лампы
накаливания (9 января) и особенно после воздействия низкими темпера-
турами. У пород холодного климата период покоя нарушается труднее,
Фиг. 112. Сеянцы дуба (Qtiercus sp.), выращенные под разным!! лампами
1 — под люминесцентными лампами; 2 — под лампами накаливания; з — вазон первоначально
находился под люминесцентными лампами (первая розетка), а затем был перенесен под лампы
накаливания (вторая розетка)
чем у субтропических культур. Особенно упрямы: дуб (Quercus тасгап-
therd), граб (Carpinus betulus) и тюльпанное дерево (! iriodendron tulipifera).
Некоторые виды сосны (Pinus silvestris hamaid), эвкалиптов (Eucaliptus
sideroxilen) и кипариса (Cupressus cochmeriand) в условиях светокультуры
не удаются совсем.
Т а б л и ц а 66
ПродолжителъностЬ периодов роста и покоя у сеянцев дуба
при беспрерывном ;лектрическом освещении
(Леман [155])
Виды дуба Продолжительность (в днях) Высота (в см)
опыта роста ПОКОЯ
Quercus pedunculate . . 290 190 100 160
Q. cerris 360 130 230 39
Q. suber 360 180 180 87
Q. ilex 360 250 110 10Q
VI. Выращивание растений при искусственной радиации
169
В результате двухлетних опытов было установлено, что положитель-
ный эффект беспрерывного освещения от ламп накаливания в условиях
светокультуры отмечается у видов из умеренной полосы; виды холод-
ного климата реагируют слабо, у них лишь несколько увеличивается
период роста (облиствения), а некоторые субтропические вечнозеленые
культуры реагируют отрицательно.
По характеру реакции на беспрерывное освещение древесные породы
следует разделить на три группы (табл. 67).
К первой относятся растения, у которых в зависимости от длины дня
не изменяется число приростов, но меняется их величина. Это —
дуб (Quercus macr anther а), клен (Acer Trautvetteri, A. campestre}, граб
(Carpinus orientalis) и отчасти пробковое дерево (Phellodendron
amurense).
Вторую группу составляют растения, обладающие резко выраженной
периодичностью роста, у которых в условиях беспрерывного освещения
лампами накаливания увеличивается число приростов и растения стано-
вятся вечнозелеными. К этой группе относятся граб (Carpinus betulus),
бук (Fagus orientalist, дуб (Quercus ibericd) и др.
Т а б л и ц а 67
Рост древесных пород в условиях беспрерывного освещения лампами накаливания
(Гулисашвили [53])
Период облиствения (в днях) Число при- ростов в году Прирост
за год в см
Растение памп ания ! "к в ® в Я о мпами вания о? , и из S § s « 2 Я 05 Я К « е? 1 S & о
Я д Ч RfflR 2 g R д е;
оно О н О 0^0
0,0 ез * § « 0.0
сС 1 Г J сЗ
t£ X S Ь о !=[ К И К 2 l=£ X щ Е-< О
о « о й О 05 о о 3
G Я й И д
Дуб (Quercus macranthera) 265 184 1 1 9 6
Клен (Acer Trautvetteri) 193 184 1 1 2 16
Клен (Acer campestre) 231 245 1 1 17 5
Граб (Carpinus orientalis) 283 248 1 1 6 5
Пробковое дерево (Phellodendron amurense) 365 275 1 1 19 7
Бук (Fagus orientalis) 365 235 7 2 49 12
Граб (Carpinus betulus) 365 249 5 1 16 —
Дуб (Quercus iberica) 365 187 8 2 64 8
Береза (Betula verrucosa) . . .’ ~ 365 284 Р 1 53 10
Тисс (Taxus baccata) 365 365 Р Р 18 10
Катальпа (Catalpa speciosa) 296 241 Р 1 40 13
Криптомерия (Cryptomeria japonica) . . . 365 365 Р Р 11 10
Шелковица (Morus alba) 365 237 Р 1 11 8
Акация (Acacia melanoxylon) 365 365 Р Р 76 102
Акация (A. longifolia) 365 365 Р Р 36 108
Пинея (Pinus pinea) 365 365 Р 1 23 15-
Дуб (Quercus suber) 365 365 Р 1 28 14
Примечание. Р — равномерный постоянный или почти постоянный рост.
170
Общая светокультура растений
Третья группа состоит из вечнозеленых пород с круглогодичным
беспрерывно-равномерным или периодическим ростом; и те и другие
под лампами накаливания, в условиях непрерывного освещения, растут
беспрерывно, без периода покоя.
Подобной реакцией обладают дуб (Quercus- snber), пинея (Pinus pined},
акация (Acacia longifolia, Acacia melanoxylori}, шелковица (Morns alba},
криптомерия (Cryptomeria japonied), катальпа (Catalpa sp.), тисс (Taxus
baccata) и береза (Betula verrucosa).
Б. С. Мошков [216, 224] для выращивания лимона (Citrus limori) при-
меняет автотракторные лампы мощностью 20 вт. Лампы последовательно
включаются в сеть с напряжением 120 в, по 22—24 лампы из расчета
1200 вт/м’. Сеянцы лимона, выращиваемые в этих условиях, за 9 месяцев
(апрель—декабрь) достигают высоты 95 см и дают прирост в 50 ярусов;
те же лимоны, выращиваемые одновременно в оранжерее, дают прирост
лишь в 17 ярусов. Черенки черной смородины за 2 месяца достигали вы-
соты 50—60 см.
Выращенные в условиях светокультуры сеянцы древесных (дуб,
береза, ясень и др.) прекрасно приживаются при пересадке в грунт и
без существенных повреждений переносят наши суровые зимы [154—
157, 233]. По данным И. Н. Никитина [232], 3-летние сеянцы в этом случае
выглядят как 12-летние. В опытах В. М. Лемана [154, 156] 3-летние дубки
после пересадки в грунт достигали высоты 90—150 см (контроль — 26 см)
и имели внешний вид и размеры 8—10-летних деревьев. По более поздним
данным И. И. Никитина [233], 8-летние дубки (Quercus robur}, полученные
методом светокультуры, а затем пересаженные в открытый грунт, имеют
высоту 5,1 м, а контрольные растения — 1,35 м.
Лампы накаливания — неплохой источник радиации для культуры
мхов и других близких к ним растений. Мохообразные всегда появляются
в пещерах, которые время от времени освещаются лампами накаливания.
В подобных условиях замечены следующие растения: печеночница
Marchantia polymorpha, мхи Amblystegium juratzkanum, Amphidium Moug-
lotii, Brachyteciurn rutabulum, B. velutinum, Bryum caespiticum, B. capil-
lare, Eucalypta contorta, E. vulgaris, Fissidens bryoides, Funaria hyd-omet-
rica, Leptobryum piritarme, папоротники Asplenium trichomonas, Rhyncho-
stegia tenella, Scolopendrium officinale и др. [150, 464, 534, 631, 797,
798].
Таким образом, лампы накаливания были испытаны и оказались при-
годными для культуры большого числа самых разнообразных растений,
относящихся к различным группам растительного мира: водорослей,
мхов, хвойных, пальмовых, кактусов, злаков и других высших
растений.
Благодаря обилию оранжево-красных лучей, растения под лампами
накаливания отличаются быстрым развитием, ускоренным переходом
к цветению, плодоношению и созреванию семян. Это обстоятельство чрез-
вычайно благоприятствует применению названных ламп для культуры
растений, выращиваемых ради плодов или семян, а также для ускоренной
выгонки сеянцев древесных. По той же причине они мало пригодны или
совсем непригодны для выращивания растений, культивируемых с целью
получения вегетативных органов (листьев, корнеплодов и т. д.), как,
например, редис, салат и др.
Недостаток сине-фиолетовых и ультрафиолетовых лучей вызывает
чрезмерное вытягивание междоузлий и рост в высоту. По этой причине
лампы накаливания мало пригодны для культуры декоративных растений,
крестоцветных, для целей селекции и т. д. Обилие инфракрасной радиа-
VI. Выращивание растений при искусственной радиации 3 71
ции позволяет пользоваться лампами накаливания для нарушения зим-
него покоя древесных пород.
Так как лампы накаливания чрезвычайно богаты инфракрасными
лучами, которые в условиях светокультуры в закрытых помещениях
вызывают перегрев растений, при помощи этих ламп можно получить
лишь сравнительно невысокие интенсивности радиации. Повышение
интенсивности радиации возможно лишь при наличии водного экрана
из проточной воды (фиг. 74—76), поглощающего и устраняющего около
50% инфракрасного излучения ламп (фиг. 77), или в условиях значитель-
ного понижения температуры воздуха, устраняющего перегрев растений.
В последнем случае растения нуждаются в обогреве корневой системы.
3. ЗНАЧЕНИЕ ИНФРАКРАСНОЙ РАДИАЦИИ
В связи с применением водных экранов были проведены специальные
исследования, посвященные значению инфракрасных лучей. Пиршле
и Веттштейн [670] исследовали влияние на развитие растений устранения
тепловых лучей посредством водного экрана. В условиях 14-часового
дня при температуре 18—22° и освещенности 6,5—8 тыс. лк (33—48 тыс.
эрг/см‘~-сек физиологической радиации) они выращивали многие растения
и нашли, что в большинстве случаев применение водного экрана дает от-
рицательный эффект (табл. 68).
При наличии водного экрана растения, как правило, сильнее вытянуты
в высоту (томаты, табак, фасоль и др.) и имеют меньший свежий и сухой
вес. Присутствие инфракрасных лучей часто значительно увеличивает
сухой вес растений. Особенно велико это увеличение у льна — Linum
usitatissimum (350%), песчанки — Arenaria Marschlinsii (236%), A. serpyl-
Ufolia (208%), кппрейника — Epilobium alpinum (215?6), кукурузы —
7,еа mays (181 %), белены — Hyoscyamus niger (172%), львиного зева —
Antirrhinum majus (168%) и томатов — Solanum lycopersicum (163%).
Аналогичные результаты получены также Уитроу [854], которые
выращивали шпинат (Spinacea oleraced), сою (Soja hispidd) и томаты
{Solanum lycopersicum) под лампами накаливания в условиях 18-часового
дня при освещенности 5 и 7,5 тыс. лк и температуре 20°. Применение вод-
ного экрана в большинстве случаев вызывало вытягивание растений и
уменьшение свежего и сухого веса растений; последний под влиянием
инфракрасных лучей увеличивался: у томатов —• на 15%, у шпината —
на 40% и у сои — на 59 % (табл. 76).
В настоящее время трудно сказать, влияет ли прп устранении водного
экрана простое повышение температуры растений или же играет роль
какой-то специфический фотохимический процесс, вызываемый инфра-
красной радиацией. Есть основание полагать, что решающее значение
имеет дополнительный нагрев. Так, например, в опытах Б. С. Мошкова
[224] добавление инфракрасных лучей к спектру ламп накаливания,
охлаждаемых водою, или добавочное воздействие этими лучами в ночные
часы вызывало увеличение сухого веса лишь у теплолюбивых растений:
на 150% — у огурцов (Cucumis sativus') Клинские и на 70% — у живо-
родки (Bryophyllum sp.). У холодостойкой абиссинской капусты (Bras-
sica carinatd) в этих условиях вес снижался (на 25%).
Так как ближние инфракрасные лучи мало поглощаются или совсем
не поглощаются тканями растений, то несомненно, что положительный
эффект инфракрасных лучей связан с действием излучения, лежащего
далеко за пределами 1100 шц.
Таблица 68
Высота и вес растений, выращенных под лампами накаливания (6,5—8 тыс. лк, 14-часовой день, 18—22°)
(Пиршле и Веттштейн [670])
Растение Возраст (в Днях) Высота (в см) Сырой вес (в г) Сухой вес (в г) Сухое вещество (в %)
лампы без ВОДНОГО экрана лампы с водным экраном лампы без ВОДНОГО экрана лампы с водным экраном лампы без водного экрана лампы С ВОДНЫМ экраном
лампы без водного экрана лампы с ВОДНЫМ экраном
Бальзамин (Impaliens Balsamina) .... 45 48 — 51 40 - -43 23,48 24,25 1,022 1,065 4,36 4,39
Томаты (Solanum lycopersicum) 49 8,5- -11 9 - -12 3,95 2,88 0,360 0,221 9,12 7,68
Табак (Nicotiana tabacum) 44 12 - -14 14 - -16 2,53 3,13 0,172 0,162 6,81 5,18
Фасоль (Phaseolus multijlorus) 31 55 — -62 61 - -69 13,31 11,85 1,804 1,556 13,54 13,11
Кукуруза (Zea mays) 35 66 - 68 49 - -57 24,05 15,33 1,556 0,861 6,47 5,61
Ночная красавица (Mirabilis jalapa) . . . 42 32 - -36 43 - -45 4,56 5,21 0,399 0,395 8,75 7.58
Белена (Hyoscyamus niger) 42 14 - 17 11,5- -13 1,64 1,04 0,136 0,079 8,32 7,60
Львиный зев (Antirrhinum majus) .... 48 15 - -17 12,5- -14 0,77 0,47 0,077 0,046 9,98 9,78
Петуния (Petunia nyctaginiflora) 41 12,5— -14 14 -16 1,61 1,66 0,118 0,103 7,35 6,20
Лен (Linum usitatissimum) 31 35 - -39 19 - -22 0,41 0,15 0,049 0,014 11,88 9,36
Кипрейник (Epilobium alpinum) 46 2 — - о 2,5- - 3 0,193 0,093 0,021 0,0098 11,13 10,43
Смолевка (Silene inflate,) 41 10 - -13 15 - -17 0,701 0,494 0,052 0,043 7,38 8,62
Смолевка (б”, maritime) 41 9,5- -11 13 - -16 0,554 0,520 0,038 0,035 6,91 6,72
Песчанка (Arenaria serpyllifolia) 40 12 - -13 13 - -15 0,599 0,342 0,077 0,037 12,79 10,86
Песчанка (Л. Marschlinsii) 40 6 - - 7 8 - - 9 0,118 0,060 0,013 0,0055 10,95 9,21
172 Общая светокультура растений
VI. Выращивание растений при искусственной радиации
173
Артур [381] сопоставил влияние ламп накаливания в 500 вт с различ-
ной цветовой температурой, а следовательно, и различной световой от-
дачей (30 и 10 лм/вт). Выращивая гречиху (Лagopyrum esculentum) в тече-
ние 12 дней при освещенности 2 тыс. лк и температуре 25,5°, автор устано-
вил, что лучший эффект дают лампы с меньшей цветовой температурой.
В этом случае сухой вес растений на 11—13% выше, чем под лампами
с большей цветовой температурой. Это, несомненно, находится в связи
с относительно большим содержанием оранжево-красных и далеких инфра-
красных лучей при меньшей световой отдаче.
4. ЗНАЧЕНИЕ УЛЬТРАФИОЛЕТОВОЙ РАДИАЦИИ
Неоднократно делались попытки устранить основной недостаток
ламп накаливания — малое содержание сине-фиолетовых и ультрафио-
летовых лучей — путем комбинации их с другими источниками излуче-
ния. Артур, Гутри и Ньюэлл [379] для этой цели применяли вольтову
дугу и ртутные лампы. При освещенности в 12—14 тыс. лк они выращи-
вали ячмень {Hordeum vulgare), пшеницу {Triticum vulgare), редис {Ra-
phanus sativus var. radicula), капусту {Brassica oleracea f. capitata) и полу-
чали хороший результат. Ячмень и пшеница развивались значительно
лучше, чем в оранжерее весной (март—апрель). К сожалению, в их опытах
отсутствовал контроль, который дал бы возможность оценить положйтель-
ное значение добавления источников, богатых сине-фиолетовыми лучами,
к основному источнику — лампам накаливания.
Обстоятельные исследования по комбинированию ламп накаливания
с различными ртутными лампами провели Пиршле и Веттшейн [670,
671, 848]. В опытах этих авторов испытывались лампы накаливания
в 500 вт, которые создавали освещенность в 8—10 тыс. лк (48—60 тыс.
эрг/см2>сек физиологической радиации). В зависимости от варианта
(табл. 69) к лампам накаливания добавляли разные ртутные лампы сверх-
высокого давления: «HgQ-500», излучающие лишь «длинные» ультра-
фиолетовые лучи О 320 ту), и «HgQS-500», излучающие, кроме того,
«короткие» ультрафиолетовые лучи ( > 270 шц). На долю первых прихо-
дилось 1200—1400 лк, а на долю вторых — 650—800 лк.
Как видно из табл. 69, добавление ультрафиолетовых лучей с длиною
волны более 320 тр мало влияет на рост растений, лишь несколько по-
вышая или понижая вес и высоту растений. Добавление коротковолновой
радиации ()> 270 шр), напротив, вызывает резкий отрицательный эффект,
уменьшая высоту и вес растений. Такие растения, как томаты {Solanum
lycopersicum), горох {Pisum sativum), львиный зев {Antirrhinum majus),
петуния {Petunia hybrida), лен {Linum usitatissimum), песчанка {Arenaria
serpyllifolia, A. Marschlinsii), быстро при этом погибают; другие же —
бальзамин {Impatiens Balsamina), ночная красавица {Mirabilis jalapa),
колеус {Coleus sp.), белена {Hyoscyamus niger) и табак {Nicotiana tabacum)
обнаруживают явные признаки страдания. Лишь кукуруза {Zea mays),
пшеница {Triticum vulgare), дурнишник {Xanthium canadense) и альпий-
ские формы кипрейника {Epilobium alpinum) и смолевки {Silene inflata)
оказываются стойкими к этим лучам, выглядят нормальными, а смолевка
даже цветет и образует плоды.
Еще более подробный анализ значения ультрафиолетовых лучей для
роста растений был произведен Пиршле [671], который в условиях
14-часового дня при освещенности в 7—9 тыс. лк (42—54 тыс. эрг/см!-сек
физиологической радиации) выращивал растения под лампами накали-
вания с добавлением ртутных ламп сверхвысокого давления: «HgQ-500»,
174
Общая светокультура растений
Влияние ультрафиолетовых лучей на рост расте
(8—10 тыс. лк, 20°, относительная влажность 75—80%,
Растение
Высота растений (в см)
лампы нака- ливания лампы нака- ливания + + «HgQ- 500» (> 320 ту) лампы нака- ливания 4- + «HgQS- 500» (> 270 ту.)
Бальзамин (Impatiens Balsamina) .... 5!) 62—65 61—63 25—30
Томаты (Solanum lycopersicum) 68 29—33 35—42 15—20
Табак (Nicotiana tabacum) 72 41—48 47—56 11—12
Фасоль (Phaseolus multijlorus) 34 61—68 71—74 13—15
Горох (Pisum sativum) 27 82—86 80—85 6— 7
Кукуруза (Zea mays) 44 85—95 85—93 75—80
Пшеница (Triticum vulgare) 34 72—76 67—72 51—56
Ночная красавица (Mirabilis j ala pa) . . . 46—52 48—54 12—13
Белена (Hyoscyamus niger) 35—42 33—45 15—16
Львиный зев (Antirrhinum majus) .... 62 51—53 47—56 —
Петуния (Petunia hibrida) 46 25—37 24—37 —
Лен (Linum usitatissimum) 47 84—92 65—73 —
Дурнишник (Xanthium canadense) .... 41 42—50 30—38 26—28
Колеус (Coleus sp.) 24—30 30—35 8—15
Кипрейник (Epilobium alpinurn) 40 13—15 11—13 1—15
Смолевка (Silene injlata) 46 15—22 26—31 6—10
Смолевка (S. maritime,) 46 20—24 18—27 2— 4
Песчанка (Arenaria serpylli folia) 48 18—21 15—18 0,5
Песчанка (.4. Alarschlinsii) 48 9—И 9—10 2— 4
Влияние ультрафиолетовых лучей на роет росте
(7—9 тыс. дк, 18—19°, относительная влажность —
Растение Высота местообита- ния над уровнем моря (в м) Высота растений (в см) Сы
лампы накалива- ния (7,0—9,0тыс. лк) лампы накалива-| ния + «HgQ-500» (8,5 — 12,0 тыс. лк) лампы накалива- | ния + »IIgQS- | 500 В1» (7,5 — ; 8,0 тыс. лк) лампы накалива-: ния + «HgQS- 300» । лампы накалива- ния (7,0-9,0 тыс. ЛК)
Смолевка (Silene maritima) .... Звездчатка (Stellaria media) . . . . • Кипрейник (Epilobium collinum) . . Кипрейник (E. alpinurn) Песчанка (Arenaria serpylli folia) . . Смолевка (Silene inilata) Смолевка (.S’. alpina) * He растут. 0 35 2000-2100 ( 2200 j 2300 [ 35 1 100 j 270 | 900 { 400 ] 1200 [2400—2500 2850 18,0 18,5 8,14 8,33 9,61 24,2 17,9 14,3 17,7 19,2 17,5 9,3 25,8 13,8 16,4 6,45 6,03 6,74 15,1 16,0 9,3 15,7 17,4 14,9 7,3 20,1 14,0 12,7 6,44 6,20 7,64 15,0 14,7 8,7 13,7 16,5 15,0 8,2 19,6 - 5,04 4,16 6.42 х. .. 3,6 ! 4,1 1 1,653 2,014 0,576 1,375 0,847 1,793 1,235 1,052 1,038 1,695 1,614 1,552 1,338
VI. Выращивание растений при искусственной радиации
175
ний, выращиваемых под лампами накаливания длина дня —14 часов) (Пиршле и Веттштейн [670]) Табл и и а 69
Сырой вес (в г) Сухой вес (в г) Сухое вещество (в %)
лампы лампы лампы лампы лампы лампы
накалива- накалива- накалива- наналива- накалива накалива-
лампы иия _|_ ния _|_ лампы ния । ния । лампы ния । ния +
“киа™' +«HgQ- +«HgQS- накалива- 4. «HgO- 4-«HgQS- накалива- «HgQ- +«HgQS-
вания 500, 500;> иия 500» 500> 1 НИИ 500» 500»
(>320 ту) О 270 ту) (> 320 ту) (> 270 ту)] (> 320 ту) (>270 ту > II'
37,90 36,96 5,96 3,29 2,67 0,329 8,19 7,22 5,52
12,97 15,95 — 2,14 2,67 — 16,50 16,73 —
15,73 19,81 0,91 2,54 2,90 0,061 16,10 14,51 6,72
13,21 14,98 — 1,82 2,01 — 13,72 13,36 —
4,09 3,37 — 0,40 0,35 — 9,79 10,34 —
22,70 28,41 17,80 2,05 2,95 1,68 9,04 10,36 9,42
2,08 1,87 1,27 0,241 0,204 0,155 11,63 10,81 12,36
11,34 13,32 0,76 1,60 1,70 0,064 14,05 12,77 8,56
4,59 4,67 0,79 0,58 0,58 0,07 12,64 12,52 9,00
6,45 6,92 — 0,89 0,87 — 13,72 12,51 —
2,95 3,01 — 0,27 0,27 — 9,04 9,07 —
2,09 1,92 — 0,41 0,32 — 19,49 16,82 —
6,43 5,81 6,24 0,93 0,78 0,78 14,50 13,32 2,48'
41,39 40,80 6,03 3,49 3,22 0,43 8,4'4 7,87 7,04
0,66 0,60 0,025 0,073 0,068 0,0025 11,08 11,36 10,02
1,43 1,71 0,53 0,17 0,19 0,044 11,72 10,95 8,31
1,57 1,78 0,14 0,13 0,13 0,0097 8,02 < 7,52 6,92
1,05 0,84 — 0,154 0,102 — 1’4,70 12,44 —.
0,52 0,49 — 0,059 0,052 — 11,22 10,70 —
Т а б л и и а 70
нии, выращиваемых поо лампами накаливания
60—70%, длина дня — 14 часов) (Пиршле [671])
рой вес (в г) Сухой вес (в г) Сухое вещество (в %)
С5 | Я Iя га W га га га о i га га га
х ffl а и 3 х _ и а га 3 1 я X и
a s 1 д । s н । 1 ч<» чо та О н ; «О л га СУ га - — . SI и 1 Ч 1 сч Ч» 1 4“ яо« ко ' к «о I 56 s' 1 raO^ra'S' гаО
й ti> ) • 1 л CL-J е , on a х ьв I — -1) « к х txi X С5 ' X ЬС 1 X r-Г Я К Ы)
пы на 4- «Н - (8,5 тыс. пы на 4- «Н В1» (1 тыс. . пы на + «н, пы на (7,0- пы на + «II > (8,5- . ли) пы на 4- «И В1» (7 ТЫС. . пы на 4- «Н > пы на (7,0- пы на + «Н > (8,5 1 тыс. пы на + «н В!» ( тыс. ЯД
У5о°. Й®00 Я Ко - Ко о Я Коо я Ко s X л = 6 °, я ка
га S о см га 5 о » , я So j га S JG га й о х га х о - га й о га й X га Й о см га S о
[J X Ю |-а К lO QO 1 1-1 Д ОЭ Ч Д Ч Ч S ю Н Ч S л оо КИИ i-j X *4 6» н* 4 СЧ Й1ЛОО И Дега
1,761 1,478 182,8 215,6 155,5 * 11,7 12,2 10,6 *
2,147 1,598 * 191,6 205,7 151,9 * 9,6 9,6 9,3 V
0,577 0,493 0,136 83,4 85,6 64,0 16,4 14,5 14,8 161 . 12,1
1,241 1,215 0,115 141,2 143,7 139,0 11,8 10,7 10,7 П,4 10,2
0,731 0,756 0,346 92,4 101,9 81,9 34,0 10,9 13,9 11,1 9,9
1,065 1,294 * 327,8 207,9 234,9 * 18,3 19,5 18,2
1,365 1,098 141,8 148,2 125,6 * 11,5 11,3 И,4 *
0,850 0,625 132,1 98,3 72,6 * 11,8 11,5 11,6 *
1,048 0,778 * 152,0 161,0 108,0 * 14,6 14,5 14,0 V
1,687 1,499 0,119 260,3 272,6 200,3 15,4 15,4 16,2 13,4 13,0
1,324 1,509 0,099 254,2 256,7 217,9 12,4 16,5 19,5 14,2 12.4
1,073 0,964 0,363 226,5 167,6 148,3 38,7 14,8 15,6 14,5 10.6
1,469 1,265 0,065 155,8 195,3 139,0 8,0 1.1,6 13,3 11,1 12,2.
176
Общая светокультура растений
«HgQS-500 В1» и «HgQS-ЗОО». Первая излучает видимые и «длинные»
ультрафиолетовые (]> 320 mg), вторая— лишь «длинные» ультрафиолето-
вые (296,7—404,7 mg), а третья, кроме того, —и «короткие» ультрафиоле-
товые лучи (265,2—690,7 mg (табл. 70).
Добавление одних лишь «длинных» ультрафиолетовых лучей (]> 320mg),
в зависимости от вида растений, снижает накопление сухого веще-
ства от 10 до 45% и высоту растений от 12 до 40%. Присутствие видимых
лучей (лампа «HgQ-500»), повышая интенсивность радиации, смягчает
этот эффект, а в некоторых случаях даже увеличивает сухой вес растений;
высота растений при этом снижается примерно на ту же величину (на
10—35%). Добавление «коротких» ультрафиолетовых лучей (Д> 270 mg)
вызывает угнетение и гибель долинных растений — звездчатки (Stella-
ria media), песчанки (Arenaria serpyllifolia), смолевки (Silene maritima).
Высокогорные формы кипрейника (Epilobium collinum) и смолевки (Si-
lene inflata и S. alpina) обладают относительной устойчивостью против
витацидных лучей. Несмотря на резкое подавление ростовых процессов,
растения выглядят нормальными, а некоторые (Epilobium alpinum) даже
цветут.
Оберг [364] сравнил комбинацию ламп накаливания в 500 вт и ртутной
лампы «НО-1000» с неоновой лампой при интенсивности видимой радиа-
ции1 около 0,02 кал/см2-мин (14 тыс. эрг/см2-сек) и длине дня 16 часов.
Автор нашел, что при излучении ртутной лампы, равном 25% суммарного
излучения, названная комбинация для томатов (Solanum lycopersicum)
дает тот же эффект, что и неоновая лампа, если не считать несколько
лучшего развития листвы в условиях смешанного света и большей при-
земистости растений при неоновом освещении.
Таблица 71
Рост растений при различном сочетании ламп накаливания и ртутных ламп
.(8 тыс. лк, длина дня—15 часов; повторность — 21—45 растений). По Уитроу А. И.
и Уитроу Р. Б. [854]
Доля лучистого потока (в %) Соя (So) a max) Билокси; 25° Шпинат (Spinacea oleracea) Нобель; 20° Томаты (Solanum lycopersicum); 15°
СР CD и е --S О О
от ламп на- каливания L от ртутных ламп высота (в си о rt iacz о О ад К ►Й Д О О й сухой вес одного раса ния (в г) сырой вес одного paci ния (в г) сухой вес одного paci нпя (в г) число зре- лых листве высота (в ы сырой вес одного paci ния (в г) сухой вес одного paci НИЯ (В 1)
93—75 7—25 57 8,1 1,0 5,1 0,37 9,8 36,0 21,8 1,83
75—14 25—86 64 6,0 0,7 3,1 0,23 8.1 35,0 16,9 1,93
14— 5 86—95 64 5,2 0,6 1,4 0,10 6,3 22,0 10,5 0,57
Весьма обстоятельное исследование по комбинированию ламп накали-
вания (1 квт) и ртутных ламп высокого давления «Н-1» (400 вт) про-
вели Уитроу [854]. Эти авторы в условиях 15-часового дня при 8 тыс. лк
выращивали шпинат (Spinacea oleracea), сою (Soja max) и томаты (Solanum
lycopersicum), причем доля излучения ртутной лампы в общем лучистом
потоке варьировала от 7 до 95%. Как видно из табл. 71, увеличение доли
радиации ртутной лампы (содержащей лишь видимые и «длинные» ультра-
фиолетовые лучи) вызывает все большее и большее уменьшение высоты
1 Видимая радиация (400—700 mg) практически соответствует физиологической
радиации.
VI. Выращивание растений при искусственной радиации
177
и веса растений. Помимо непосредственного влияния излучения ртутной
лампы, здесь, конечно, влияло и уменьшение интенсивности физиологи-
ческой радиации. Это обстоятельство не нуждается в объяснении, если
принять во внимание, что 1 лк лампы «Н-1» содержит в два раза меньше
энергии физиологического излучения, чем 1 лк лампы накаливания
(табл. 60).
В. С. Мошков [224] комбинировал низковольтные лампы накаливания
(21 вт, 6 в) с люминесцентными лампами в 30 вт. При общей удельной
мощности 1,87 квт/м2 на долю ламп накаливания приходилось 1,5 квт/м2, а
на долю люминесцентных ламп — 0,3 квт/м2. Люминесцентные лампы мон-
тировались поочередно с рядами, состоящими из 12 последовательно вклю-
ченных низковольтных ламп накаливания, работающих при напряжении
5,3 в. Особенно хорошие результаты эта комбинация дала при выращи-
вании лимона {Citrus limori) и рассады томатов {Solanum lycopersicum).
Чтобы покончить с вопросом о комбинировании ламп накаливания
с газосветными трубками, следует назвать исследования С. И. Доброхото-
вой [59] и В. П. Мальчевского [196], которые к лампам накаливания
добавляли неоновые лампы. В опытах С. И. Доброхотовой в условиях
беспрерывного освещения ежесуточная ,4-часовая замена ламп накали-
вания неоновыми трубками вызывала ускорение развития томатов {So-
lanum lycopersicum) на 10—13 дней и увеличение сухого веса в 1'/2—2
раза. По данным В. П. Мальчевского, сеянцы ели {Picea sp.), сосны {Sinus
sp.) и березы {Betula sp.) также реагируют положительно на подобную
замену одного источника света другим. У березы и ели наступает при этом
более интенсивное ветвление и образуются более крупные листья.
Таким образом, посредством источников радиации, богатых коротко-
волновыми лучами (ртутные лампы, вольтовые дуги и др.), можно пол-
ностью устранить чрезмерное вытягивание растений, обычное под лампами
накаливания. Эффективными следует признать ультрафиолетовые лучи,
лежащие в пределах 290—400 шц. При этом вместе с уменьшением высоты
растений наблюдается некоторое снижение сухого и свежего веса расте-
ний. Для устранения «зеленой» этиоляции требуется сравнительно не-
большая мощность ртутных ламп — 40—60 вт на каждые 1000 вт, полу-
чаемых посредством ламп накаливания, или 75—120 bt/m>j. Что касается
комбинирования ламп накаливания с неоновыми лампами, то это соче-
тание следует признать малоудачным, так как оба источника имеют при-
мерно одинаковый спектральный состав (фиг. 37 и 47) и содержат по
преимуществу или исключительно оранжево-красные лучи.
/
5. КУЛЬТУРА РАСТЕНИЙ ПРИ ПОМОЩИ ГАЗОСВЕТНЫХ ЛАМП. ЗНАЧЕНИЕ
РАЗЛИЧНЫХ ОБЛАСТЕЙ СПЕКТРА ФИЗИОЛОГИЧЕСКОЙ РАДИАЦИИ
Первую попытку выращивания растений с помощью газосветных ламп
предпринял Телен [796], который для этой цели применял ртутную лампу
Шотта (За, 220 в) в оболочке из фосфорно-бариевого стекла, пропускаю-
щего «короткие» ультрафиолетовые лучи вплоть до 253 та. Вследствие
этого опыт дал отрицательные результаты. Первоначально применяв-
шиеся высоковольтные неоновые трубки (720) из-за невозможности по-
лучить радиацию высокой интенсивности также не могли служить источ-
ником излучения для выращивания растений на искусственном свету.
Лишь с появлением мощных высокоинтенсивных ламп стала возможной
культура растений при помощи этих источников радиации.
Первые опыты по сравнению растений, выращиваемых под различными
лампами, не могут претендовать на методическую безупречность в силу
12 А. Ф. Клешнин
178
Общая светокультура растений
невыравненности физиологической радиации в различных вариантах
опытов. Так, Артур и Стьюарт [382], выращивая гречиху (Fagopyrum
esculentum) в течение 8—11 дней в условиях беспрерывного освещения,
сопоставили эффективность лампы накаливания в 500 вт с эффективностью
ртутной, неоновой и натриевой ламп при интенсивности радиации, коле-
бавшейся в различных вариантах опыта в пределах 0,08—0,40 кал/см3-мин
(56—280 тыс. эрг/см3-сек). Экстраполируя, авторы нашли, что при одина-
ковой интенсивности радиации сухой вес растений, выраженный в отно-
сительных единицах, равняется: под лампами накаливания — 1,00, под
неоновыми — 1,41, под натриевыми — 1,20, а под ртутными лампами —
0,62. Метод экстраполяции в данном случае совершенно не приложим,
и полученные авторами данные вызывают серьезные сомнения.
То же самое следует сказать об исследованиях Восса [831] и Смита
[775]. Первый выращивал овес (Avena sativa) и горох (Pisum sativum)
под лампами накаливания (300 и 500 вт), натриевыми («Na-бОО», 118 вт),
неоновыми («Ne-бОО», 520 вт) и ртутными («HgHS-бОО», 150 вт) лампами,
причем энергия излучения, выраженная в единицах освещенности, колеба-
лась в пределах 880—1170 лк. Совершенно очевидно, что сравнивать натрие-
вую лампу с неоновой нет никакого смысла, если физиологическая радиа-
ция в первом случае равняется 1660, а во втором — 9200 эрг/см2-сек.
Некоторый смысл имеет лишь сравнение ртутных ламп «HgH-ЮОО»
(272 вт) и «HgHS-ЮОО» (272 вт) с лампами накаливания в 500 вт при
одинаковой освещенности (1930—2320 лк). В этом случае лучшее развитие
растений наблюдается под лампами накаливания, а худшее — под увио-
левой лампой «HgHS-ЮОО», излучение которой простирается в сторону
коротких ультрафиолетовых лучей несколько дальше, чем у лампы
«HgH-ЮОО».
Методически безупречными следует признать лишь некоторые опыты
Роденбурга [733], который выращивал огурцы (Cucumis sativus), томаты
(Solanum lycopersicum) и другие растения под лампами накаливания
(65 вт), натриевыми («Na-650», 100 вт), ртутными («НО-500», 150 вт) и
неоновыми (100 вт) лампами при одинаковой интенсивности видимой
радиации (4 тыс. эрг/см3-сек) и при одинаковой удельной мощности
(100 вт/м3). Автор нашел, что в обоих случаях наиболее мощные растения
получаются под неоновыми, а наихудшие — под ртутными лампами.
Промежуточное положение занимают лампы накаливания и натриевые
лампы, причем первые дают меньший эффект, чем вторые (табл.у72).
Другим методически безупречным исследованием следует признать
работу Оберга [364], который сопоставил ртутные, натриевые и неоно-
вые лампы при одинаковой интенсивности видимой радиации
(0,012 кал/см!-мин, или 8400 эрг/см3-сек) в условиях 16-часового дня
(табл. 73). Автор получил те же результаты, что и Роденбург. К сожалению,
в обоих случаях интенсивность радиации была сравнительно невелика.
Сравнительно немного имеется исследований по комбинированию раз-
личных газосветных ламп. М. С. Калинин [91] в опытах по выращиванию
шпината (Spinacea oleracea) соединял неоновые трубки с аргоновыми,
сопоставляя эту комбинацию с лампами накаливания. К сожалению,
отсутствие объективных показателей интенсивности лучистой энергии
совершенно обесценивает эту работу. Артур и Гарвилл [385], выращивая
при беспрерывном освещении под натриевыми лампами (20 тыс. лк) гиа-
цинты (Hyacynthus orientalis), гардению (Cardenia sp.), пеларгонию (Pe-
largonium zonale), хлопчатник (Gossypium hirsutum), гречиху (Fagopyrum
esculentum), бегонию (Begonia tuberosa) и другие растения, нашли, что
они в этом случае развиваются нормально лишь на начальных этапах,
VI. Выращивание растений при искусственной радиации
179
Табл и ц а 72
Развитие растений под равными источниками радиации при одинаковой
интенсивности видимой радиации
_(4 тыс. эрг/см'2 • сек) (Роденбург [733])
Варианты опыта Мощность ламп (в вт) Освещенность (в Н1х) Интенсивность ви- 1 димой радиации (в эрг/см2-сек.) Сырой вес одного растения (в г) Сухой вес одного растения (в г) Длина одного листа (в см) Длина одного междоузлия (в см) Высота растений (в см) Длина 3-го листа (в см) Длина гипокотиля (в см)
Томаты (Solanum lyco- persicum)
Неоновые лампы . . . 100 610 4100 5,34 0,21 3,8 2,3 — — —
Натриевые лампы . . . 100 2480 4500 4,43 0,16 3,4 1,8 — — —
Лампы накаливания . . 65 650 4100 3,79 0,14 2,9 1,7 — — —
Ртутные лампы .... Огурцы (Cucumis sa- tiva) 150 1600 4300 2,88 0,11 2,5 0,7 — — —
Неоновые лампы . . . 100 610 4100 — — — — 18,4 8,0 2,6
Натриевые лампы . . . 100 2480 4500 — — — — 9,6 4,9 3,0
Лампы накаливания . . Ртутные лампы «НО- 65 650 4100 — — — — 23,8 7,5 5,0
500» 150 1600 4300 — — — — 11,3 6,3 2,7
Таблица 73
Развитие растений под разными газосветными лампами при одинаковой интенсивности
видимой радиации
(около 8400 эрг/см'2* сек); повторность — 7—8 растений (Оберг [364])
/ Варианты опыта 1 Сырой вес одного расте- ния (в г) Сухой вес одного растения Д о S f Л О н S S F § Размеры 5-го листа (в см)
в г в % к сы- рому весу длина ширина
Томаты {Solanum lycopersicum)
Ртутные лампы • . . 4,69 0,249 5,3 8,4 1,77 1,12
Натриевые лампы 5,43 0,343 6,3 8,7 2,12 1,28
Неоновые лампы 5,88 0,405 6,9 8,9 2,10 1,30
Шпинат (Spinacea oleracea)
Ртутные лампы 3,97 0,219 5,5 9,9 — —
Натриевые лампы 5,74 0,317 5,7 10,3 — —
Неоновые лампы 5,84 0,352 6,0 10,7 — —
Салат {Lactuca sativa)
Ртутные лампы 2,17 0,084 3,8 8,0 1,38 0,49
Натриевые лампы 2,31 0,086 3,7 8,6 1,47 0,49
Неоновые лампы 4,06 0,155 3,8 9,5 1,56 0,59
12*
180
Общая светокультура растений
а в дальнейшем образуют исключительно белые и желтые листья, лишен-
ные хлорофилла. Для устранения этого явления авторы применяли ртут-
ную лампу «НЗ», включая ее на 2 часа ежесуточно. В этом случае они имели
совершенно нормальное развитие растений.
Пиршле и Веттштейн [670] также комбинировали натриевые лампы
«.Na-500», дававшие освещенность в 6,0—7,5 тыс. лк (11—14 тыс. эрг/см2-сек
физиологической радиации), и ртутную лампу сверхвысокого давле-
ния «Н-4», которая давала освещенность в 450—600 лк (1,7—2,3 тыс.
эрг/см2-сек). Они получили при этом нормальные растения без всяких
признаков этиоляции.
Оберг [364] сравнил комбинацию ртутной («HgH-500») и натриевой
(«Na-ЮОО») ламп с лампой накаливания и натриевой лампой («Na-650»)
при интенсивности видимой радиации 0,035 кал/см2-мин (24,5 тыс.
эрг/см2-сек) и нашел, что томаты (Solanum lycopersicum) развиваются
лучше при смешанном свете (приземистые мощные растения с интенсив-
ной зеленой окраской), чем под лампами накаливания (бледные, вытя-
нувшиеся растения) или под натриевыми лампами (вытянувшиеся ра-
стения с сильно закрученными листьями).
К сожалению, этим исчерпывается список работ по сочетанию раз-
личных газосветных ламп для выращивания растений. Повидимому,
имеет смысл комбинировать источники, излучающие различные области
физиологической радиации, как неоновые и ртутные лампы и т. п. Однако
экспериментальных данных настолько мало, что сделать какие-либо
определенные выводы пока нельзя.
6. КУЛЬТУРА. РАСТЕНИЙ ПРИ ПОМОЩИ ЛЮМИНЕСЦЕНТНЫХ ЛАМП.
ЗНАЧЕНИЕ РАЗЛИЧНЫХ ОБЛАСТЕЙ СПЕКТРА ФИЗИОЛОГИЧЕСКОЙ
РАДИАЦИИ
Люминесцентные лампы для культуры растений впервые применил
Нэйлор [635—637]. Этот автор в условиях 16-часового дня при освещен-
ности 6,5 тыс. лк (23—26 тыс. эрг/см2-сек физиологической радиации)
выращивал растения капусты (Brassica oleracea), кукурузы (Zea mays),
фасоли (Phaseolus sp.), сои (Soja hispida), табака (Nicotiana tabacum),
томатов (Solanum lycopersicum), укропа (Anethum graveolens), однолет-
ней свеклы (Beta vulgaris) и дурнишника (Xanthium sp.) под люмине-
сцентными лампами белого и дневного света мощностью 30 вт. Контроль-
ные растения росли в условиях естественного зимнего освещения в оран-
жерее. Опытные растения развивались значительно быстрее, имели более
короткие стебли с короткими междоузлиями и крупными листьями, обла-
дали большим сухим и сырым весом и более интенсивной зеленой окраской,
чем контрольные растения.
В СССР культура растений при помощи люминесцентных ламп нача-
лась почти одновременно в 1946 г. в Институте физиологии растений
им. К. А. Тимирязева Академии Наук СССР [112—115, 118—123, 184]
и в Агрофизическом институте ВАСХНИЛ [216, 224]. В настоящее время
люминесцентными лампами пользуется также лаборатория искусственного
климата Московской сельскохозяйственной академии им. К. А. Тимиря-
зева [160, 161].
Обширные исследования по сопоставлению люминесцентных ламп
с другими источниками радиации принадлежат Ульриху [820], Уитроу
[854] и А. Ф. Клешнину [ИЗ, 114, 118, 120, 121, 123, 184]. В своей первой
работе Ульрих [820] сравнивал два источника радиации: люминесцентные
лампы розовато-белого света «HNR-ЮО» мощностью в 100 вт и солнечные
VI. Выращивание растений при искусственной радиации 181
лампы «HWA-500», представляющие комбинацию лампы накаливания
(185 вт) с ртутной лампой сверхвысокого давления (70 вт). В первом случае
удельная мощность равнялась 900 вт/м2, а во втором — 765 вт/м2.
Растения — ячмень {Hordeum vulgare), соя {Soja hispida), люпин
Lupinus luteus, L. angustifolius), картофель {Solanum tuberosum), фасоль
{Phaseolus sp.), сосна {Pinussp.), львиный зев {Antirrhinum majus, A. loja),
кипрейник {Epylobium hirsutum) и печеночница {Sphaerocarpus Donellii)—
выращивались при освещенности в 2460 лк (10—11 тыс. эрг/см2-сек
физиологической радиации). В качестве контроля служили растения,
выращивавшиеся в оранжерее при естественном освещении в середине
зимы (декабрь—январь). Опыты показали, что интенсивность порядка
10 тыс. эрг/см2-сек недостаточна для культуры перечисленных растений,
за исключением львиного зева, который в обоих вариантах опыта нор-
мально развивался и цвел. При выращивании под люминесцентными
лампами во всех случаях наблюдались более приземистые растения с ин-
тенсивной темнозоленой окраской листьев. Цветение сои и фасоли насту-
пило раньше при выращивании под люминесцентными лампами, ячмень
не цвел вообще, а растения люпина из-за недостатка света погибли. У яч-
меня наблюдались явные признаки этиоляции, более сильные под сол-
нечными лампами. Сырой и сухой вес растений, выращенных под
люминесцентными лампами, был несколько меньше, чем у растений, куль-
тивировавшихся под лампами «HWA-500». Картофель развивался лучше
под люминесцентными лампами, давая в ряде случаев большее накопле-
ние сухой массы и образуя более приземистые растения с крупными
листьями.
Значительно больший интерес представляет другое исследование Уль-
риха [820], где производилось сравнение растений, выращиваемых под
люминесцентными лампами розовато-белого света («HNR», 0,75 а) при
освещенности в 7,5 тыс. лк (32 тыс. эрг/см2-сек физиологической радиации)
и в условиях естественного солнечного освещения летом (23 апреля-
31 июля). Длина дня в первом случае равнялась 15 часам, а во втором
постепенно возрастала от 14 до 17 часов. Опыт показал, что для нормаль-
ного развития люпина, сои и фасоли интенсивность физиологической
радиации порядка 30 тыс. эрг/см2-сек недостаточна. Растения желтого
и синего люпина заболели и погибли. У сои, выращиваемой под люми-
несцентными лампами, обнаружилось сильное уменьшение числа узлов,
чрезмерное вытягивание междоузлий и сильнейшее отставание в накопле-
нии сухой массы по сравнению с контролем. Фасоль зацвела на 7 дней
позднее и дала в два раза меньше цветков, чем в естественных условиях.
Люминесцентные лампы оказались прекрасным источником радиации
для культуры картофеля, который под этими лампами растет не хуже,
чем летом в оранжерее. Опытные растения образуют более крупные
листья с интенсивной зеленой окраской; при выращивании под лампами
наблюдается цветение, а клубнеобразование идет одинаково как при
искусственном освещении, так и в контроле.
То же самое следует сказать и о культуре ячменя, который, несмотря
на огромные различия в интенсивности радиации под лампами и в есте-
ственных условиях, в обоих случаях рос почти совершенно одинаково
(табл. 74).
Люминесцентные лампы оказались лучшим источником радиации для
культуры печеночницы {Sphaerocarpus Donellii), чем лампы накалива-
ния (500 вт).
Уитроу [854] сопоставили лампы накаливания мощностью 1000 вт,
ртутные лампы высокого давления «Н-1» (400 вт) и люминесцентные
182
Общая светокультура растений
Характеристика растений ячменя (Hordeum vulgare), вира
(У льрих
Варианты опыта Длина стебля (в см) Число междо- узлий Средняя длина междо- узлия (в см) Диаметр стебля (в мм) Длина колоса (в см) Число зерен в колосе
Абиссинский ячмень Люминесцентные лампы . . . 65 6 9,2 3,2 7,0 19
Естественное освещение (в оранжерее) 54 5 9,1 2,4 5,4 14
Ячмень Изарин Люминесцентные лампы . . . 60 6 10,2 2,1 8,5 13
Естественное освещение (в оранжерее) 42 5 8,4 2,2 6,3 18
лампы белого и дневного света (30 вт) при равной интенсивности види-
мой радиации, равной освещенности и равной удельной мощности.
В качестве объектов служили соя (Soja max), томаты (Solanum lycoper-
sicum), астры (Callistephus chinensis) и шпинат (Spinacea oleraced). Сравни-
вая источники радиации при затрате одинаковой удельной мощности,
авторы нашли, что эффективность ламп зависит от вида растений; у астр
лучший результат наблюдается под лампами накаливания, а у шпината
под люминесцентными лампами. Наименее экономичной в обоих случаях
оказалась ртутная лампа (табл. 75).
Таблица 75
Характеристика растений, выращенных .под разными лампами
при одинаковой удельной мощности
(50 дней; 15-часовой день; повторность — 15—20 растений) (Уитроу А. П.
и Уитроу Р. Б. [854])
Лампы Освещенность (в лк) Интенсивность физиологической радиации (в эрг/см2 • сек) Астры (Callistephus chinensis) Гарт оф Франс Шпинат (Spinacea oleracea) Нобель
высота (в см) сухой вес | (в г) число цве- точных стеб- лей высота (в см) сухой вес (в Г) число цве- точных стеб- лей
Лампы накаливания 1000 вт . 7600 42 000 46,9 8,1 9,5 5,2 1,4 0
Люминесцентные лампы бе- лого света 30 вт 13 000 35 000 21,4 4,8 3,0 52,3 1,8 0,74
Ртутные лампы «Н-1» 400 вт 13 000 38 000 18,6 3,8 1,0 14,7 0,6 0,18
Сопоставление люминесцентных ламп с лампами накаливания (табл. 76)
показало, что как при одинаковой освещенности (8 тыс. лк), так и оди-
наковой интенсивности видимой радиации (28—32 тыс. эрг/см2-сек) луч-
ший эффект по накоплению сухой массы дают лампы накаливания, осо-
бенно без водного экрана, устраняющего далекие инфракрасные лучи
(7 3> 1р). Однако растения, выращиваемые под люминесцентными лампами,
VI. Выращивание растений при искусственной радиации
183
* Таблица 74
щенны.х под люминесцентными лампами и в естественных условиях
[820])
Сухой вес одного растения (в г) Число листьев Размеры листьев (в см) Площадь листьев (в см-) Вес 100 зерен (в г)
зерен стеблей корней длина ширина средняя одного листа общая
0,67 4,75 1,05 9 25,0 0,99 23,0 207,0 3,53
0,65 5,97 0,90 8 ' 20,9 0,95 17,8 142,6 4,55
0,53 4,21 1,59 10 25,75 0,78 20,58 205,8 4,07
0,73 3,16 0,55 9 20,40 0,86 15,45 139,3 4,05
имеют нормальный вид без всяких признаков чрезмерного вытягивания
стебля, чего нельзя сказать о растениях, культивируемых под лампами
накаливания. У растений шпината, сои и томатов различий в сыром и
сухом весе при выращивании под люминесцентными лампами белого и
дневного света и одинаковой освещенности (8 тыс. лк) не оказалось.
Высота растений во всех случаях была несколько ниже при культивиро-
вании под лампами дневного света.
В опытах 1947 и 1948 гг. мы [ИЗ, 123, 184] произвели сравнение ламп
накаливания в 200 вт и люминесцентных ламп белого света в 15 и 30 вт.
12 ламп накаливания для охлаждения погружались в кювету с проточной
водопроводной водой (фиг. 74, Б), а люминесцентные лампы располагались
над растениями в виде горизонтальной решетки (фиг. 65, Л) из расчета
15 ламп на 1 м для ламп в 30 вт и 30 ламп на 1 м для ламп в 15 вт. Удель-
ная мощность равнялась 450 вт/м2 для люминесцентных ламп и 2400 вт/м2
для ламп накаливания. Эффективность ламп сравнивалась при одинаковой
освещенности в 8—10 тыс. лк, что соответствует интенсивности физиоло-
гической радиации в 28—35 тыс. эрг/см2-сек под люминесцентными лам-
пами и 48—60 тыс. эрг/см2-сек под лампами накаливания. Объектами опы-
тов служили: салат {Lactuca sativa), горчица {Sinapis alba), огурцы {Cu-
cumis sativus), томаты {Solanum lycopersicum), патиссоны {Cucurbita pepo),
шпинат {Spinacea oleracea), редис {Raphanus sativus var. radicula), дуб
{Quercus sp.), каланхое {Kalanchoe sp.), горох (Pisum. sativum), капуста
кочанная {Brassica oleraceavar. capitata), капуста цветная {Brassica oleracea
var. botrytis), кольраби {Brassica caulorapa), кок-сагыз {Taraxacum kok-
saghys), репа {Brassica rapa), брюква {Brassica napus), петрушка {Petro-
selinum sativum), свекла {Beta vulgaris), морковь (Daucuscarota), пук {Allium
сера), просо {Panicum miliaceum), пшеница {Triticum sp.) и другие растения.
Опыты с беспрерывным освещением показывают, что под люминесцент-
ными лампами развиваются совершенно нормальные приземистые и мощные
растения с крупными листьями и короткими междоузлиями, без всяких
признаков чрезмерного вытягивания стебля.
Растения, выращиваемые под лампами накаливания, характеризуются,
напротив, сильным ростом в высоту и вытягиванием междоузлий, в осо-
бенности подсемядольного колена. Сказанное иллюстрируется фиг. 108—
114. На фиг. ИЗ представлено дальнейшее развитие огурцов, показанных
184
Общая светокультура растений
Характеристика растений, выращенных под лампами накаливания и люминесц
физиологичес
(Уитроу А. II. и
Варианты опыта Длина дня (в час.) Длитель- ность опыта (в днях) Освещен- ность (в лк) Интенсивность видимой радиа- ции (в эрг, см2 • сен) Число растений
общее цветущих (в % к об- щему)
Шпинат (Spinacea oleracea) Нобель При равной освещенности лампы накаливания: с водным фильтром 18 55 8000 46 80
без водного фильтра 18 55 8000 — 56 85
люминесцентные лампы 18 55 8000 — 37 100
При равной интенсивности радиации лампы накаливания: с водным фильтром 24 42 5400 28000—32000 47 100
без водного фильтра 24 42 5400 28000—32000 45 89
люминесцентные лампы 24 42 9700 28000—32000 48 100
Соя (Soja max) При равной освещенности лампы накаливания: с водным фильтром 18 24 8000 48
без водного фильтра 18 24 8000 — 55 —
люминесцентные лампы 18 24 8000 — 51 —
При равной интенсивности радиа- ции лампы накаливания: с водным фильтром 18 31 5400 28000—32000 40
без водного фильтра 18 31 5400 28000—32000 42 —
люминесцентные лампы 18 31 9700 28000—32000 40 —
Томаты (Solanum. lycopersicum) Ин- диана Балтимор При равной интенсивности радиации лампы накаливания: с водным фильтром 15 31 5400 28000—32000 20
без водного фильтра 15 31 5400 28000—32000 20 —
люминесцентные лампы . ... . 15 31 9700 28000—32000 20 —
на фиг. 109. Бросается в глаза более интенсивная окраска листьев
и более раннее цветение растений, выращиваемых под люминесцентными
лампами, чем растений, культивируемых под лампами накаливания.
На фиг. 114 показаны растения двух видов периллы: Perilla ocymoides
(1, 2) и Р. nankinensis (3, 4), отличающихся один от другого окраской
листьев. Первый имеет зеленые листья, а второй отличается очень
темной красной окраской листьев, богатых антоцианами. Как видно
из рисунка, под лампами накаливания вытягивание стебля совершается
одинаково у обоих видов, а выращиваемая под люминесцент-
ными лампами красная перилла отличается значительно меньшей высо-
VI. Выращивание растений при искусственной радиации
185
Т а б лица 76
ентными лампами белого света при равной освещенности и равной интенсивности
кой радиации
Уитроу Р. Б. [854J)
Высота растений (в см) Сырой вес одного растения (в г) Сухой вес одного растения (в г) Сухое вещество (в % к сырому весу)
наземной массы корней общий наземной массы корней общий
9,0 40,8 2,3 43,10 3,00 0,19 3,19 7,4
3,0 64,5 3,2 67,70 4,20 0,28 4,48 6,6
24,4 44,3 1,3 45,60 2,50 0,09 2,59 5,6
68,0 32,5 1,1 33,6 2,30 0,11 2,41 7,1
26,8 28,1 1,2 29,3 2,30 0,10 2,40 8,0
31,3 24,8 0,8 25,6 1,90 0,07 1,97 7,6
ь
54,1 6,8 1,23 8,03 0,85 0,08 0,93 11,5
36,4 10,0 1,32 11,32 1,36 0,12 1,48 12,2
30,5 4,7 0,96 5,66 0,54 0,06 0,60 10,6
74,2 8,4 0,80 9,2 1,06 0,10 1,16 12,7
51,2 11,6 1,10 12,7 1,70 0,13 1,83 14,4
40,3 5,0 0,58 5,58 0,62 0,05 0,67 11,8
30,6 18,3 2,3 20,6 1,18 0,15 1,33 6,0
43,0 21,3 1,6 22,9 1,40 0,13 1,53 6,3
10,0 9,9 1,6 11,5 0,84 0,14 0,98 8,5
той, чем зеленая. Присутствие антоцианов в листьях Р. nankinensis ведет
к сильному уменьшению высоты стебля при освещении люминесцентными
лампами, значительно меньшему у Р. ocymoides. Повидимому, антоцианы,
поглощая сине-фиолетовые лучи, содержащиеся в спектре люминесцент-
ных ламп, не остаются в стороне от фотохимических процессов, оказы-
вающих влияние на рост растений.
По характеру развития при выращивании под люминесцентными
лампами и лампами накаливания растения можно разделить на две диа-
метрально противоположные группы, объединяемые рядом промежуточ-
ных форм. Одни растения — редис, салат, горчица, томаты и другие —
186
Общая светокультура растений
Таблица 77
Развитие растений под лампами накаливания и люминесцентными лампами
в условиях беспрерывного освещения при освегценности в 8—10 тыс. лк
' Растение Число дней до
стрелкования бутонизации цветения
под люминесцент- ными лампами под лампами нака- ливания под люминесцент- ; ными лампами ! под лампами нака- ливания под люминесцент- ными лампами под лампами нака- ливания
Редис (Raphanus sativus var. radicula) розовато-красный Редис (R. sativus var. radicula) Ледяная со- сулька Салат (Lactuca sativa) Московский Горчица белая (Sinapis alba) Горох (Pisum sativum) Урожайный Томаты (Solanum lycopersicum) Джон-Бэр . . Огурцы (Cucumis sativus) Вязниковские . . со 22 33 0 0 0 со 26 71 15 40 20 19 20 12 10 28 21 00 35 34 24 45 39 21 26 16 22 43 51
Таблица 78
Сухой вес растений под лампами накаливания и люминесцентными лампами
в условиях беспрерывного освещения при освещенности в 8—10 тыс. лк
Варианты опыта Возраст растений (в днях) Сухой вес одного растения (в г)
стеблей листьев корней общий
Брюква (Brassica napus)
Люминесцентные лампы 86 — 5,15 3,76 8,91
Лампы накаливания Томаты (Solanum lycopersicum) 86 — 6,78 13,76 20,51
Люминесцентные лампы 60 1,37 1,92 0,69 3,98
Лампы накаливания .... 60 3,56 3,07 1,54 4,28
Капуста кочанная (Brassica oleracea var. capitata)
Люминесцентные лампы 46 0,46 3,07 1,55 5,11
Лампы накаливания 46 0,82 3,74 1,58 6,14
Горчица (Si па pis alba)
Люминесцентные лампы 82 1,77 0,57 0,42 2,76
Лампы накаливания 82 1,07 0,12 0,07 1,27
Патиссоны (Cucurbita pepo)
Люминесцентные лампы 67 0,55 1,54 0,93 3,02
Лампы накаливания 67 0,40 1,03 0,42 1.85
VI. Выращивание растений при искусственной радиации.
187
Фиг. ИЗ. Огурцы {Cucumis saticus) Вязнцковские,
выращенные под разными лампами
7 — под люминесцентными лампами; 2 — под лампами накалива-
ния
Фиг. 114. Перилла, выращенная под разными лампами
1 — Perilla ocymoides, выращенная под люминесцентными лампами; 2 — то же под лампами
накаливания; 3 — Р. nankinensis, выращенная под люминесцентными лампами; 4 — то же
под лампами накаливания
188
Общая светокультура растений
зацветают и плодоносят раньше под лампами накаливания и позднее под
люминесцентными лампами (фиг. 16, 20 и 115). Различие в скорости раз-
вития, выражаемое разницей в длине вегетационного периода, здесь
может достигать величины от нескольких дней (горох) до нескольких
месяцев (салат). Это — растения длинного дня (табл. 77). Другие растения—
огурцы, соя (Sofa sp.), щирица (Amaranlus retroflexus) — в условиях
Фиг. 115. Горчица (Sbiapis alba), выращенная под люми-
несцентными лампами (7) и под лампами накаливания (2)
длинного дня зацветают раньше под люминесцентными лампами попозд-
нее— под лампами накаливания (фиг. ИЗ). Это — растения короткого дня
(табл. 77). На коротком дне, как увидим позднее, они зацветают, как и
длиннодневные растения, раньше под действием оранжево-красных лучей.
Накопление сухой массы растений при одинаковой освещенности,
как правило, совершается значительно интенсивнее при выращивании
под лампами накаливания, чем под люминесцентными лампами (табл. 78).
В этом нет ничего удивительного, так как при одной и той же освещен-
ности интенсивность физиологической радиации под люминесцентными
лампами значительно меньше, чем под лампами накаливания. Однако
конечный вес растений определяется темпами их развития. Растения,
быстро переходящие к цветению под лампами накаливания (салат, редис
горчица и др.), в этом случае обычно имеют значительно меньшие конечные
VI. Выращивание растений, при искусственной радиации
189
размеры, сухой и сырой вес (фиг. 115 и 116; табл. 78). Растения, находя-
щиеся почему-либо в вегетативном состоянии, обладают, напротив, боль-
шими конечными размерами при выращивании под лампами накаливания,
чем при культивировании под люминесцветными лампами (фиг. 114).
Луковицы лука образуются лишь при выращивании под лампами нака-
ливания и совсем не образуются
под люминесцентными лампами
белого и дневного света (фиг. 17).
Формирование корнеплодов зави-
сит от типа растений. У таких
растений, как редис, которые под
лампами накаливания быстро из-
растают и цветут, корнеплоды об-
разуются исключительно под лю-
минесцентными лампами (фиг. 15),
а у репы, не прошедшей стадии
яровизации, корнеплод лучше раз-
вивается при выращивании под
лампами накаливания и хуже —
под люминесцентными лампами
(фиг. 117). То же самое следует
сказать о формировании кочана
или листовой розетки у салата
(фиг. 19 и 20).
В 1949 г. опыты были продол-
жены с люминесцентными лампа-
ми красного, зеленого и синего
света мощностью в 15 вт, изготов-
ленными по специальному заказу
Московским электроламповым за-
водом. Спектральный состав излу-
чения ламп приведен на фиг. 59,
60 и 95. Как и в предыдущие годы,
лампы располагали в виде гори-
зонтальной решетки на высоте в
10—15 см над растениями (фиг.
65, А), исходя из удельной мощ-
ности в 450 вт/м!. Интенсивность
физиологической радиации во всех
вариантах была одинакова и ко-
лебалась на протяжении опыта в
пределах 25—40 тыс. эрг/см-'-сек.
Объектами служили те же расте-
ния, что и в 1947 и 1948 гг.
В результате этих опытов [115,
118, 123] было показано, что при
Фиг. 116. Салат (Lactuca sativa), выращен-
ный под люминесцентными лампами (/)
и под лампами накаливания (2)
одинаковой интенсивности физиологической радиации развитие растений
в условиях благоприятной длины дня наиболее быстро совершается в оран-
жево-красных и медленнее всего — в зеленых лучах. Это означает, что
длиннодневные растения в условиях длинного дня, а короткодневные
растения в условиях короткого дня раньше всего зацветают под люмине-
сцентными лампами красного света, затем — под синими лампами и в по-
следнюю очередь — в зеленых лучах (фиг. 11, табл. 79). Короткодневные
растения при беспрерывном освещении благодаря большой фотоперио-
190
Общая светокультура растений
дической активности оранжево-красных лучей (фиг. 10) зацветают вначале
под синими, а затем под оранжево-красными лучами. Это установлено для
тцирипы {Amaranthus retro-
jlexusp сои, огурцов и
других растений.
Подобная реакция ра-
стений длинного и корот-
кого дня на спектральный
состав излучения позволя-
ет сделать заключение, что
суточный ход процессов,
протекающих в листе на
световой стадии развития,
слагается из двух этапов.
Первый этап, одинаковый
для всех высших растений,
независимо от их принад-
лежности к той или иной
фотопериодической груп-
пе, соответствует основно-
му светлому периоду суток
(фиг. 118, /) и нуждает-
Фиг. 117. Корнеплоды репы {Brassica гара),
выращенные под разными лампами
1 — под люминесцентными лампами; 2 — под лампами
накаливания
ся в высокой интенсивно-
сти радиации.
Второй этап, требую-
щий различных условий у
растений длинного и ко-
роткого дня, обусловливает фотопериодический эффект, т. е. специфиче-
скую реакцию растений на длину дня. У растений длинного дня процес-
сы, характеризующие этот
этап, протекают на свету и
могут совершаться при не-
больших интенсивностях
радиации (фиг. 142 и 151
и табл. 102, 104 и 106)
[109, 488, 521, 596, 861];
у растений короткого дня—
требуют темноты и задер-
живаются уже при слабом
освещении (фиг. 118, II)
Эффективность различ-
ных областей спектра оди-
накова для процессов обо-
их этапов: наиболее эффек-
тивны оранжево-красные,
менее — сине-фиолетовые и
наименее эффективны зе-
леные лучи (фиг. 4, 2). Раз-
личие заключается лишь в
том, что на первом этапе
процессы, вызываемые те-
ми или иными лучами,
протекают в одном направ-
лении у всех фотопериоди-
в условиях длинного и короткого дня на скорость
развития растений длинного и короткого дня
I — период (12 час.), в который растения нуждаются в
высокой интенсивности радиации; II — период (12 час.),
в который растения нуждаются в слабой интенсивности
радиации (растения длинного дня) или в темноте (растения
короткого дня)
VI. Выращивание растений при искусственной радиации
191
ческих групп растений, а на втором—в диаметрально противоположных
направлениях (на фиг. 118 направление процессов обозначено тонкими
стрелками).
Скорость процессов развития, определяющая длину вегетационного
периода растений, представляет собою результат соотношения скоростей
процессов на обоих этапах. На фиг. 118 результирующая скорость разви-
тия обозначена жирными стрелками.
Схема на фиг. 118 дает теоретическое изображение скорости развития
в соответствии с экспериментальными данными, приведенными в табл. 77
и 79.
Таблица 79
Развитие растений под люминесцентными лампами красного, зеленого и синего света
Растение Длина дня (в час.) Число дней от всходов до бутонизации
красный свет зеленый свет синий свет
Салат (Lactuca satira) Московский . 24 33 54 45
Капуста цветная {Brassica oleracea. var. botrytis) 24 115 00 150
Щирица (Amaranthus retrojlexus) . . 8 47 69 51
To же 24 89 — 72
Накопление сухого вещества в большинстве случаев выражается
спектральной кривой, отвечающей кривой 2 на фиг. 4, т. е. происходит
интенсивнее всего в оранжево-красных и наименее интенсивно — в зе-
леных лучах (фиг. 13, 14 и 18 и табл. 80).
В некоторых случаях максимальное накопление сухого вещества
наблюдается в сине-фиолетовых лучах. Растения, развитие которых под
оранжево-красными лучами совершается быстрее, чем под сине-фиолето-
выми, успевают накопить за период вегетации значительно меньше
сухого вещества под оранжево-красными лучами, чем под сине-фиолето-
выми, где долгое время продолжается вегетативный рост (например,
салат).
Формообразовательные процессы — формирование листьев у большин-
ства растений, кочанов — у капусты и салата, стеблевого утолщения —-
у кольраби, корнеплодов — у репы, усов — у земляники и т. д., также
наиболее интенсивно протекают под влиянием оранжево-красных лучей,
если растения не переходят чрезмерно быстро к цветению, как, например,
салат (фиг. 5, 6, 13, 14, 18 и 19). Луковицы у лука образуются только
в оранжево-красных лучах (фиг. 18).
Последующие исследования, проведенные в институте физиологии
растений им. К. А. Тимирязева Академии Наук СССР в 1950—1951 гг.,
касаются не только значения спектрального состава физиологической
радиации для процессов роста, развития и формообразования растений,
но и значения интенсивности излучения для этих процессов. Опыты про-
водились с теми же люминесцентными лампами красного и синего света
мощностью в 15 вт, расположенными в виде горизонтальной решетки на
различной высоте над растениями (5—55 см). Интенсивность физиологи-
ческой радиации, измеряемая специально отградуированным селеновым
фотоэлементом, изменялась, в зависимости от расстояния, в пределах
5—30 тыс. эрг/см’-сек. Температура воздуха колебалась в пределах
21—27°, а температура листьев, в соответствии с фиг. 24, 1, превышала
температуру воздуха на 0°,5—3° (фиг. 119, В\
192
Общая светокультура растений
Таблица 80
Сухой вес растений под люминесцентными лампами красного, зеленого и синего света
при беспрерывном освещении
(25—40 тыс. эрг/см2 • сек)
Варианты опыта Возраст растений (в днях) Сухой вес одного растения (в г)
листьев стеблей корней корне- плодов общий
Редис {Raphanus sativus var. radicula) розово-красный
Лампы синего света 45 1,02 0,52 0,47 0,33 2,34
» зеленого » 45 0,72 0,46 0,15 0,00 1,33
» красного » 45 1,61 1,08 0,66 0,76 4,11
Репа {Brassica гара) Петров- ская *
Лампы синего света 72 3,55 — 1,16 0,66 5,37
» зеленого » 72 2,26 — 0,75 0,20 3,21
» красного » 72 7,10 — 2,52 2,55 12,17
Капуста кочанная {Brassica oleracea var. capitata) № 1
Лампы синего света 60 2,30 0,70 0,90 — 3,90
» зеленого » 60 2,39 1,00 1,02 — 4,41
» красного » 60 5,88 2,30 3,32 — 11,50
Кольраби {Brassica caulorapa) Венская
Лампы синего света 45 1,00 0,10 0,50 — 1,60
» зеленого » 45 0,74 0.07 0,29 — 1,10
» красного » 45 2,79 0,52 2,51 — 5,82
Патиссоны {Cucurbita pepo)
Лампы синего света 34 2,24 0,54 1,17 — 3,95
» зеленого » 34 1,72 0,40 0,64 — 2,74
» красного» » 34 2,05 0,46 0,75 — 3,26
Салат {Lactuca sativa) Москов- ский
Лампы синего света 51 1,69 1,04 0,70 — 3,43
» зеленого » 51 0,63 0,28 0,15 — 1,06
» красного » 51 0,82 1,00 0,36 — 2,18
В качестве объектов служили растения репы Петровская и салата
Берлинский и Майский. Репа в условиях нашего опыта, т. е. при высоких
температурах (20—30°), все время находилась в вегетативном состоянии,
так как не проходила стадии яровизации, и поэтому спектральный состав
излучения не влиял на ее развитие. Салат Майский, напротив, под влия-
нием красных лучей совершенно не образует розетки и, быстро пройдя
световую стадию, рано зацветает. Салат Берлинский — позднеспелый
сорт — занимает промежуточное положение. Посев репы производился
6 ноября, салата Берлинский — 24 ноября, а салата Майский — 9 марта.
Всходы появились соответственно 11 ноября, 27—29 ноября и 15 марта.
Опыт с репой длился 100 дней, с поздним салатом — 40 дней, а с ранним
салатом — 90 дней. Результаты опытов представлены на фиг. 119—133.
ИнтенсиЗнастб рималогичесЛий радиации
(в тбк арг/смг- сид)
Фиг. 121. ('.ухой вес растений репы (Bras-
sica гара} Петровская, выращенных под
люминесцентными лампами разного
света
1 — красного; 3 — синего
А — сухой вес листьев одного растения;
Б — сухой вес корнеплода одного растения;
В — сухой вес корней одного растения
Фиг, 119. Характеристика растений
репы (Brassica гара} Петровская, выра-
щенных под люминесцентными лампами
разного света
1 — красного; 2 — синего
А—число листьев на одном растении; Б —
листовая поверхность: а—-всего растения;
б — одного листа; И — температура воздуха
Интежс/днастб риаимагичесРой радиации
(ff тй/с. аре/аиг-аеР)
Фиг. 120. Вес растений репы (Brassica
гара} Петровская, выращенных под лю-
минесцентными лампами разного света
1 — красного; 2 — синего
А — сырой вес одного растения; Б—сухой
вес одного растения; В — содержание сухого
вещества
Фиг. 122. Характеристика растений са-
лата (Lactuca sativa) Берлинский, выра-
щенных под люминесцентными лампами
разного света
1 — красного; 2 — синего
А — сырой вес одного растения; Б — сухой
вес одного растения; В — число листьев
на одном растении
13 А. Ф. Клешнин
194
Общая светокультура растений
>00 -
ИнгпенсиОнастр (ршиа/гогичес/иай радиации
(6 то/с ирг/сиг-сек)
Как видно из сопоставления кривых на фиг. 119—122, рост и развитие
растений репы и салата Берлинский возможны лишь при интенсивности
физиологической радиации более 4—5 тыс. эрг/см!-сек. При увеличении
интенсивности радиации число и размеры листьев, общая листовая поверх-
ность, сырой и сухой вес растений возрастают пропорционально интен-
сивности радиации в пределах от 5 тыс. до 20—25 тыс. эрг/см2-сек. Отно-
шение ассимиляционного аппарата к неассимилирующим органам при
этом падает. В условиях нашего опыта увеличение интенсивности радиа-
ции за пределы 20—25 тыс. эрг/см2-сек часто вызывало уменьшение ве-
личин перечисленных показателей.
Повидимому, это находится в связи
с чрезмерным повышением темпера-
туры листьев при слишком большом
приближении растений к лампам (ме-
нее 5 см).
Сопоставление прямолинейных
участков кривых, выражающих из-
менение площади листьев, сырого и
сухого веса растений в зависимости
от интенсивности и спектра радиа-
ции, показывает, что у растений,
находящихся в вегетативном состоя-
нии (репа) или слабо реагирующих
па спектр излучения изменением ско-
рости развития (салат Берлинский),
наблюдается сильное влияние спек-
трального состава на накопление
органического вещества. Последнее
при выращивании растений под лам-
пами красного света, как правило, в
два раза выше, чем под синими лам-
пами. То же сайое относится и к
размерам листьев и общей листовой
поверхности растений.
Сказанное находится в полном
соответствии со спектральной кри-
вой фотосинтеза и отвечает погло-
щению квантов лучистой энергии
хлорофиллом листьев (фиг. 4,2у
Совершенно иные результаты на-
блюдаются у растений, развитие ко-
торых сильно зависит от спектра излучения. Например, опыты с салатом
Майский показывают, что растения, культивируемые под красными и
синими люминесцентными лампами, резко различаются длиной вегетацион-
ного периода и скоростью процессов развития (фиг. 123 и 124). Назван-
ный сорт салата при выращивании под красными лампами совершенно не
имеет розеточной формы и зацветает, в зависимости от интенсивности ра-
диации через 30—70 дней. Растения, культивируемые под синими лампами,
долгое время находятся в розеточном состоянии и зацветают лишь че-
рез 55—90 дней.
Это обстоятельство приводит к резкому изменению обмена и накопле-
ния органического вещества. Благодаря интенсивно протекающим про-
цессам развития, растения салата, культивируемые под красными лампами,
имеют ту же листовую поверхность и тот же сырой и сухой вес, что и
Фиг. 123. Характеристика развития
растений салата (Lactuca sativa) Май-
ский, выращенных под люминесцент-
ными лампами разного света
1 — красного; 2 — синего
А — число дней до бутонизации; Б — скорость
развития (величина, обратная длительности
вегетационного периода)
VI. Выращивание растений при искусственной радиации
195
под синими лампами (фиг. 125 и 126), а конечный вес растений значительно
выше в условиях освещения синим светом, чем красным (фиг. 126, 13).
Анализы, проведенные О. П. Осиповой и И. В. Тимофеевой, показы-
вают, что несмотря на большее количество пигментов при выращивании
Фиг. 124. Растения салата {Lactuca s.ativa) Майский, выращенные под люминесцент-
ными лампами разного света при различной интенсивности физиологической радиации
А — под лампами красного света: 1 — 5,5 тыс. эрг/см2• сек; 2—8 тыс. эрг/см2-сек; 3— 13 тыс.
эрг/см2 сек; 4—21 тыс. эрг/см2 сек; Б — под лампами синего света: 1 — \ тыс. эрг/см2-сек;
2— 9 тыс. эрг/см2• сек; 3— 16,5 тыс. эрг/см2-сек; 4 — 26 тыс. эрг/см2-сек; 5 — 32 тыс. эрг/см2• сек.
под красными лампами (фиг. 127, 128 и 129), содержание углеводов (рас-
творимые сахара и крахмал) и их отношение к белкам практически
одинаковы как под синими, так и под красными лампами (фиг. 130).
С увеличением интенсивности радиации количество углеводов уве-
личивается, а количество белков, большее под синими лампами, сни-
жается. Содержание общего и белкового азота также снижается (фиг. 131).
С увеличением интенсивности радиации значительно увеличивается
использование азота в синтезе белков, т. е. возрастает доля белкового
13*
Фиг. 127. Содержание хлорофилла в
листьях растений салата {Lactuca sativa)
Майский, выращенных под люминес-
центными лампами разного света
Фиг. 125. Характеристика растений са-
лата {Lactuca sativa) Майский, выращен-
ных под люминесцентными лампами раз-
ного света
1 — красного; 2 — синего
А—число листьев на одном растении; Б — сред-
няя площадь одного листа; В—-общая площадь
листьев одного растения
Интенсивность филиалогичесАой рсйиа^ии
(Й mb/с зрг/смгсей)
Фиг. 126. Вес растений салата {Lactuca
sativa) Майский, выращенных под лю-
минесцентными лампами разного света
7 — красного; 2 — синего
А — сырой вес одного растения; Б — отноше-
ние веса листьев к весу стебля; В — конечный
сухой вес одного растения
1 — красного; 2 — синего
Фиг. 128. Содержание хлорофилла
в листьях салата {Lactuca sativa) Берлин-
ский, выращенных под люминесцентны-
ми лампами разного света
7 — красного; 2 — синего
А — общее содержание хлорофилла; Б—со-
держание хлорофилла а; В — то же Ь;
Г — соотношение хлорофилла а и Ъ
VI. Выращивание растений при искусственной радиации 197
азота по отношению к общему (фиг. 131, В). У вегетирующих растений
содержание хлорофилла значительно меньше, чем у репродуцирующих;
разрыв в содержании хлорофилла у растений, культивируемых на синем
и красном свете, значительно больше у салата Майский, чем у салата
Берлинский (фиг. 127 и 128). Последнее также, несомненно, находится
в связи с различной скоростью развития названных сортов под красными
и синими лучами. :
Фиг. 130. Химический состав листьев
растений салата (Lactuca. sativa) Май-
ский, выращенных под люминесцентны-
ми лампами разного света
Фиг. 129. Содержание пигментов в .тисть-
ях растений салата (Lactuca saliva) Xtaii-
ский, выращенных под люминесцентными
лампами разного света
1 — красного; 2 — синего
А — содержание каротиноидов в единице площа-
ди листа; Б — содержание каротиноидов в еди-
нице сырого веса листа; В — молекулярное со-
отношение хлорофилла и каротиноидов
1 — красного; 2 — синего
А — содержание белков; Б — содержание
углеводов; В — соотношение белков и угле-
водов
Сопоставление отношения веса сухого вещества репы к интенсивности
физиологической радиации показывает, что коэффициент использования
лучистой энергии больше для красных ламп, чем для синих, и возра-
стает вместе с увеличением интенсивности до известного предела. Это
различие у салата Майский значительно меньше, чем у репы.
В дальнейшем опыты по аналогичной методике были продолжены
с астрами (Callistephus chinensis) (фиг. 132 и 133), бегонией {Begonia
semperflorens), колокольчиком (Campanula maji), комнатной розой (Rosasp.)
и двумя расами колеуса (красная и зеленая пестролистные формы Coleus
sp.). Во всех случаях накопление органического вещества шло значи-
тельно быстрее в красном свете, чем в синем. Астры реагировали так же,
как и салат Берлинский. Несмотря на более быстрое зацветание при вы-
ращивании под красными лампами, астры благодаря длительной розеточ-
ной фазе имели в этом случае больший сухой вес, чем при культивирова-
198
Общая светокультура растений
нии под лампами синего света. Присутствие антоцианов в листьях колеуса
вызывало уменьшение веса растений.
Рассмотрим еще несколько работ, имеющих более частный характер,
но показывающих широкие возможности применения люминесцентных
ламп. Гартман [537, 538] выращивал под люминесцентными лампами (30 в)
землянику (Fragaria sp.). В условиях длинного дня (16 часов) при ком-
бинации ламп белого и дневного света (5000 лк) земляника по мощности
ю го зо
Интенсиднастд фшиатэгииес/иш радиации
(6 mb/c эрг/смг сек)
Фиг. 131. Содержание азота в листьях
растений салата (Lactuca sativa) Май-
ский, выращенных под люминесцентны-
ми лампами разного света i
1 — красного; 2 — синего
А. — содержание общего азота; Б — содержа-
ние белкового азота; В — белковый азот в %
к общему
развития не уступала растениям, ра-
стущим летом в естественных усло-
виях под открытым небом. Гамнер
[529], применяя эти лампы, выращи-
вал томаты (Solanum lycopersicum),
сою (Soja max'), бобы (Vicia faba) и
другие растения. Он нашел, что луч-
ший эффект дает комбинация ламп
белого и дневного света при отноше-
нии мощностей 1:1. Томаты дают
хороший урожай плодов в течение
11 недель. Сухой вес растений лишь
на 30%, а урожай плодов на 50%
меньше, чем в условиях естественно-
го освещения летом. Ши-Вей лу [752]
использовал люминесцентные лампы
для культуры в стерильных условиях
бэрии (Baeria chrysostoma), а Снайдер
[781 ] —для выращивания подорожни-
ка (Plantago lanceolatus). Б. С. Мош-
ков [216] рекомендует люминесцент-
ные лампы для культуры теневынос-
ливых растений — земляники (Fraga-
ria sp.), гречихи (F agopyrum esculen-
tum), ивы (Salix sp.), винограда (i itis
vinifera), черной смородины (Ribes
nigrum) и лимона (Citrus limon).
При удельной мощности в 320 вт/м2
земляника Рощинская дает зрелые
плоды через 45 дней, гречиха Бога-
тырь и Тереховская бутонизируют
через 10 дней, а зрелые семена дают
через 40 дней; лимон за 6 месяцев достигает высоты в 51 см и дает
27 ярусов листьев.
Ван дер-Вен [827], пользуясь люминесцентными лампами белого света,
культивировал различные виды тополя (Populus alba, Р. robustrа, Р. та-
rylandica, Р. tremula, Р. trichocarpa, Р. serotina, Р. nigra и Р. lasiocarpa)
при интенсивности радиации порядка 10 мвт/см2 (100 тыс. эрг/см2- сек)
и длине дня 9, 12, 16 и 24 часа. В .одной серии опытов температура равня-
лась 30° днем и 20° ночью, а в другой —18° днем и 14° ночью.
Опыт показал, что, независимо от температуры, непрерывный рост на-
званных растений наблюдается лишь в условиях 16-часового дня и беспре-
рывного освещения. На коротком дне (9 и 12 часов) у растений прекра-
щается рост и они уходят в покой через 4—6 недель после начала роста.
При создании искусственной зимы (9-часовой день, 5°С) у растений, нахо-
дившихся в условиях длинного дня, лишь прекращался рост, но они
оставались зелеными, а растения с короткого дня переходили в состояние
VI. Выращивание растений при искусственной радиации
199
покоя и сбрасывали листья. При возвращении растений в первоначаль-
ные условия у длиннодневных вариантов рост начинался сразу, а у ко-
роткодневных — лишь через несколько недель.
Своеобразный метод эксплуатации так называемых «тонких люминес-
центных ламп» предложили Паркер и Бортвик [662], комбинируя назван-
ные лампы с лампами накаливания, выполняющими одновременно роль
Фиг. 132. Астры (Calllstephus chinensis), выращенные под люминесцентными
.дампами разного света при разной интенсивности физиологической радиации
7 — 8 тыс. эрг/см2 сек; 2 — 14 тыс. эрг/см'-сек; '3 — 21,5 тыс. эрг/см--сек; 4 — 16 тыс.
эрг/см2-сек; 5—20 тыс. эрг,см!-сек; 6—25 тыс. эрг/см--сек; 7— 38 тыс. эрг/см2-сек
А — под лампами красного света; Б — под лампами синего света
дросселей. Каждая лампа длиною в 2,44 м. нуждается в трех лампах нака-
ливания мощностью в 60 вт (115 в). При общей мощности такой системы
в 190 вт на долю люминесцентной лампы приходится 66 вт (35%), а на
долю ламп накаливания — 124 вт (65%). Осветительный щит размерами
в 1,22x2,67 м (3,26 м'-), оборудованный 18 люминесцентными лампами
и 54 лампами накаливания, при напряжении в 453 в и общей силе тока
в 8,3 а (общая мощность 3,420 квт; 1 квт/м-), на расстоянии 46 см создает
начальную освещенность в 13 тыс. лк. Через 300 часов освещенность сни-
жается до 11 тыс. лк и остается постоянной в течение 2000 часов.
200
Общая светокультура растений
ИтенсивностА физиологической радиации (0 тй/с орг фмг сек)
Авторы сравнили воздействие описанной системы освещения с освеще-
нием одними люминесцентными лампами и комбинированным освещением
от вольтовой дуги и ламп накаливания. В результате 3-недсльной культуры
сои (5'о/а max) Билокси эти авторы установили, что комбинация люми-
несцентных ламп с лампами накаливания дает значительно лучшие ре-
зультаты, чем одни люминесцентные лампы, и не уступает комбинации
вольтовой дуги с лампами накаливания, несмотря на то, что освещенность
в последнем случае была значительно выше (23 тыс. лк). Сырой и сухой
вес одного растения в первом случае равнялся соответственно 10,22 и
1,26 г, а во втором—8,77 и 1,03 г,
а под вольтовой дугой—11,87
и 1,66 г. Растения, выращенные
в оранжерее весною, имели сы-
рой вес 8,70 г и сухой вес —
1,16 г. Соответствующие цифры
для зимних оранжерейных ра-
стений равны 5,16 и 0,48 г.
Инженер Г. О. Алексеенко в
1951г. предложил аналогичный
метод для эксплуатации низко-
вольтных люминесцентных ламп
мощностью 15 вт, работающих
при напряжении в 120в, заменив
дроссели обычными лампами на-
каливания в 75 вт (127 в).
Посредством подобной комбина-
ции ламп (фиг. 69) в Институте
физиологии растений им. К. А.
Тимирязева Академии Наук
СССР в зимние месяцы культи-
вируются лимоны {Citru s limon).
Недавно М. X. Чайлахян и
Т. В. Некрасова [338] сравнили
воздействие люминесцентных
ламп «БС-15» и ламп накалива-
ния при выращивании сеянцев
лимонов {Citrus limon) и апель-
синов {С. sinensi). Выращивая
в 1200—1400 лк (7200—8400
под лампами накаливания и
4800—5600 эрг/см2-сек под люминесцентными лампами), эти авторы нашли,
что люминесцентные лампы дают значительно лучший эффект, чем лампы
накаливания. В этом случае образуются совершенно нормальные расте-
ния, имеющие значительно больше листьев и побегов и растущие более
интенсивно.
Б. С. Мошков [224], пользуясь осветительной рамой с люминесцент-
ными лампами (450 вт/м2), установил, что эти лампы особенно пригодны
для культуры салата {Lactuca sativa), шпината {Spinacea oleracea), укропа
(Anetam graveolens), земляники {Fragaria sp.) и других розеточных и низко-
рослых растений. В другом опыте, при помощи тех же ламп, Б. С. Мош-
ков сопоставил эффективность верхнего и бокового освещения при одина-
ковой интенсивности радиации. Он нашел, что у таких растений, как
лук (Allium сера) или злаки, лучший результат наблюдается при двусто-
роннем боковом освещении. Прп верхнем освещении сухой вес одного
Фиг. 133. Характеристика растений астр(С«7-
listephus chinensis), выращенных иод люминес-
центными лампами разного света
1 — красного; 2 — синего
А — скорость развития (величина, обратная длине
вегетационного периода); В — сухой вес одного ра-
стения
названные растения при освещенности
эог/см2-сек физиологической радиации
VI. Выращивание растений при искусственной радиации
201
растения ветвистой пшеницы равняется 0,84 г, при одностороннем боковом
освещении он увеличивается до 2,8 г, а при двустороннем боковом освеще-
нии достигает 4,02 г.
Таким образом, в настоящее время люминесцентные лампы нашли
широкое применение для культуры растений, особенно в научно-исследо-
вательских учреждениях. Благодаря применению этих ламп возможно
выращивать совершенно нормальные растения самых различных видов
в любое время года.
Сопоставление разных типов люминесцентных ламп показало, что
для культуры растений пригодны лампы белого, дневного, красного и
синего света и совершенно непригодны лампы зеленого света. Для целей
семеноводства, для ускоренного получения цветков, плодов и семян сле-
дует рекомендовать лампы красного и белого света, для селекционных
целей — лампы дневного света, а для культуры таких растений, как са-
лат или редис, лампы синего света.
7. ЗНАЧЕНИЕ ИНТЕНСИВНОСТИ ФИЗИОЛОГИЧЕСКОЙ РАДИАЦИИ
Специального внимания заслуживает вопрос об интенсивности радиа-
ции и длине дня в условиях светокультуры растений. Оден [646], пользуясь
лампами накаливания, пытался установить связь между интенсивностью
радиации и длиною дня. В его опытах интегральная интенсивность коле-
балась в пределах 0,05—0,5 кал/см1 2-мин \ а ее произведение на длину
дня равнялось 12—20 кал/см2. Большинство растений в этих усло-
виях погибало из-за недостатка света в следующие сроки: очиток {Sedum
lydium) — через 3 недели, салат {Lactuca sativa) — через 3—4, камне-
ломка {S axifraga decipiens) — через 4, бобы {Vicia faba) — через 4—5,
фасоль {Phaseolus vulgaris) — через 6 недель, а фиалка (Уiola faba) — после
цветения. Нормальное развитие душистого горошка {Lathyrus odoratus),
бобов {Vicia faba) и бальзамина {Impatiens balsamina) наблюдалось, не-
зависимо от длины дня, лишь при интенсивности радиации более 0,25
кал/см2-мин 2. Закон постоянства количества лучистой энергии (1. t =
= const) к светокультуре растений оказался нелриложимым.
Аналогичные данные несколько ранее были получены Дэвисом и
Хоэглендом [463], в опытах которых объектом исследования служили
проростки пшеницы {Triticum vulgare), а источником радиации — лампы
накаливания в 300 вт. Интенсивность радиации варьировала при этом
от 1200 до 3000 условных единиц, а длина дня — от 4 до 24 часов. Оказалось,
что растения для своего нормального развития нуждаются в определен-
ной интенсивности физиологической радиации и определенной длине дня,
причем одно не может компенсировать другое.
Интересное исследование с. синезелеными водорослями Phormidium
nucinatum autumnale и Ph. corium проведено Иггена [552]. Применяя
лампы накаливания (150—500 вт), охлаждаемые водою, и варьируя осве-
щенность в пределах 45—3000 лк, а длину дня от 1 до 24 часов, эта иссле-
довательница нашла, что при небольших количествах света (менее
900 лк X час, или 28 кал/см2) решающее значение имеет интенсивность радиа-
ции, а при большом количестве света (более 3000 лк X час, или 93 кал/см'2)—•
длина дня. В первом случае накопление сухого вещества возрастает при
увеличении освещенности, а во втором — с увеличением длины дня.
Из табл. 63 видно, что разные растения для нормального роста и раз-
вития нуждаются в совершенно различных интенсивностях радиации.
1 Около 3,5—35 тыс. эрг/см2-сек физиологической радиации.
2 Около 17,5 тыс. эрг/см2*сек физиологической радиации.
202
Общая светокультура растений
Для одних растений достаточно 19 тыс. эрг/см1 2 3 * 5-сек, для других необхо-
димо 650 тыс. эрг/см2-сек. Характер зависимости развития, урожая и
других показателей от интенсивности радиации выражается кривыми,
представленными на фиг. 119—135. Первоначально, при низких интен-
сивностях радиации, наблюдается прямая зависимость между развитием
или урожаем и интенсивностью или освещенностью, а затем увеличение
урожая, роста или другого показателя начинает отставать от увеличения
интенсивности. Это отставание тем
больше, чем выше освещенность. При
определенной величине интенсивно-
сти, различной для разных растений,
Интенсивность физиологической райит/ии, (В mine
орг/смгсеВ)
Интенсивность физиологичесЬсй риВиоции
( 6 тфе орг/см г сек)
Фиг. 134. Развитие растений в зависи-
мости от интенсивности физиологиче-
ской радиации
1 — число цветущих растений ириса (Iris sp.,
сорт Император); 2 — число бутонизпрующих
растений сои. (Saia max, сорт Билокси);
3 — число соцветий у сои; 4 — число разделив-
шихся чечевичек у ряски (Lemna minor);
5 — число дней до бутонизации у абиссинской
капусты (Brassica carinata)
Фиг. 135. ('.ухой вес одного растения
в зависимости от интенсивности’физиоло-
гической радиации
1 — огурцы (Cucumis sativus) Неросимые; 2 —
абиссинская капуста (Brassica carinata); 3 —
лук (Allium сера) Бессоновский (вес 100 ра-
стений); 4 — земляника (Fragaria sp.) Мысов-
с.а; 5 — томаты (Solanum Ipcoporsicwn) Пуш-
кинские; 6 — ряска — Lemrta minor (пес 1000
растений); 7 — кипрейник — Epilobium fiirsu-
turn (сырой вес 10 растений)
процессы развития или величина урожая перестают зависеть от интен-
сивности света; дальнейшее увеличение освещенности становится беспо-
лезным, а при некоторых условиях может дать даже отрицательный эф-
фект (фиг. 119—135).
Сказанное можно проиллюстрировать некоторыми опытами по выра-
щиванию растений в условиях различной освещенности. П&рвыо попытки
в этом направлении были предприняты Ширли [753], который в условиях
различной освещенности выращивал некоторые древесные породы, и
Роденбургом [720], выращивавшим при различной освещенности огурцы
(Cucumis sativus), томаты (Solanum lycopersicum), бегонию (Begonia sp.),
лобелию (Lobelia erinus), орхидею (Cattleya) и другие растения.
Эшби [392], культивируя ряску (Lemna minor) под лампами накали-
вания в условиях различной освещенности — от 800 до 16 000 лк
VI. Выращивание растений при искусственной радиации
203
(4,8—96 тыс. эрг/см1 2-сек физиологической радиации), нашел, что сухой
вес растений и скорость размножения возрастают лишь до 5000—8000 лк
(30—48 тыс. эрг/см2-сек физиологической радиации). Дальнейшее увели-
чение интенсивности уже не оказывает влияния на рост и размножение
ряски (фиг. 134, 4 и фиг. 135, 6‘).
Фюртауэр [498] установил, что у различных гибридов кипрейника
(Epilobium hirsutum, Е. luteum и Е. albiflorum) сухой вес растений возра-
стает вплоть до 13 000 лк (80 тыс. эрг/см2-сек физиологической радиации;
фиг. 135, 7), а высота достигает максимума при 5000—9000, лк (30—54
тыс. эрг./см2-сек физиологической радиации).
Гарнер [528] культивировал растения сои (Soja max, сорт Билокси)
под лампами накаливания в 1000 вт в условиях 10-часового дня при осве-
щенности 500, 1000, 1500, 2000, 4000 и 8000 лк’. Этот автор нашел, что
бутонизация сои начинается лишь при 1000 лк (5500 эрг/см2-сек физиоло-
гической радиации), но достигает 100% уже при освещенности в 1500 лк
(8250 эрг/см^сек физиологической радиации; фиг. 134, 2); количество
цветков продолжает возрастать вплоть до освещенности в 8000 лк (44 тыс.
эрг/см2-сек физиологической радиации; фиг. 134, 3).
Аналогичный опыт с длиннодневным растением укропом (Anetiirn
.graveolens} был поставлен Нейлором [637]. Этот автор, пользуясь лампами
накаливания, в течение 4 дней выдерживал укроп в условиях беспрерыв-
ного освещения при различной освещенности (до 10 000 лк, или 60 тыс.
эрг/см2-сек физиологической радиации). Он установил, что для стрел-
кования укропа необходима освещенность порядка 3000 лк^|8 тыс.
эрг/см2-сек физиологической радиации). Дальнейшее увеличение освещен-
ности не ускоряет этого процесса.
По данным Гардера и Витша [532], заложение цветков у каланхое
(Kalanchoe blossfeldiana} в условиях 9-часового дня происходит лишь при
освещенности 1000 лк (6 тыс. эрг/см2-сек физиологической радиации) и
совершается с наибольшей скоростью при 2000 лк (12 тыс. эрг/см2-сек
физиологической радиации).
Зивори и Вент [762], культивируя длиннодневное растение бэрию
(Baeria chry so stoma} под люминесцентными лампами в условиях беспре-
рывного освещения при освещенности 300, 740, 1180, 1620, 2060 и 2500 лк,
нашли, что названное растение может нормально расти и развиваться
лишь начиная с освещенности в 1600 лк.
По данным Б. С. Мошкова [216], перилла (Perilla ocymoides} в усло-
виях короткого дня под лампами накаливания нормально развивается
лишь при интегральной интенсивности радиации 100 тыс. эрг/см2-сек.
В другом опыте Б. С. Мошков [216, 224], применяя лампы накаливания
в 200, 300 и 500 вт и специальный экран из бумажной кальки 2, культи-
вировал огурцы (Cucumis sativus}, абиссинскую капусту (Brassica cari-
nata), лук (Allium сера}, землянику (Fragaria sp.) и томаты (Solanum
lycopersicum} в течение 15—25 дней в условиях 17—20-часового дня при
интегральной интенсивности радиации 50, 100, 200, 300, 400, 500, 600 и
800 тыс. эрг/см2-сек, т. е. при интенсивности физиологической радиации 3,
6, 13, 26, 39, 52, 65, 78 и 104 тыс. эрг/см2-сек. Оказалось, что наиболее те-
нелюбивы лук и земляника, у которых максимальный сухой вес наблю-
дается при интенсивности физиологической радиации в 52 тыс. эрг/см2-сек.
1 Соответствует интенсивности физиологической радиации в 2750, 5500, 8250,
11 000, 22 000 и 44 000 эрг/см2-сек.
2 Этот экран, помещаемый между лампами и верхушками растений и рассеиваю-
щий инфракрасные лучи, позволяет приблизить лампы почти вплотную к верхушкам
растений.
204
Общая светокультура растений
Дальнейшее увеличение интенсивности вызывает уменьшение сухого
веса растений, невидимому, из-за перегрева листьев (фиг. 135, 3 и 4).
У томатов наиболее быстрая бутонизация (через 10—12 дней после посева)
наблюдается уже при интенсивности физиологической радиации в 52 тыс.
эрг/см2-сек, а сухой вес растений продолжает возрастать вплоть до 78 тыс.
эрг/см2-сек (фиг. 135, 5). Очень светолюбивыми оказались огурцы и абис-
синская капуста, у которых сухой вес растений продолжает еще увеличи-
ваться пропорционально интенсивности радиации при 65 тыс. эрг/см2-сек
(фиг. 135,1 и 2). Растения абиссинской капусты цветут лишь при интенсив-
ности физиологической радиации 13 тыс. эрг/см2-сек. Число дней до цвете-
ния сокращается пропорционально увеличению интенсивности радиации до
39 тыс. эрг/см2-сек. Дальнейшее увеличение интенсивности уже не сказы-
вается на времени цветения (фиг. 134, 5).
Позднее Б. С. Мошков и В. Б. Карманов [222] в опытах с томатами
показали, что при пользовании лампами накаливания оптимум интенсив-
ности радиации определяется температурой воздуха. При 25° максималь-
ный сырой и сухой вес томатов был при удельной мощности в 1,2 квт/м2,
а при температуре воздуха 12—15° — при 4,8 квт/м2.
Гартзема и Лёйтен [536], пользуясь люминесцентными лампами,
нашли, что в условиях 16-часового дня при температуре 15—17° ирис
(Iris sp.) Император начинает цвести лишь при освещенности в 2500 лк
(10 тыс. эрг/см2-сек физиологической радиации). Увеличение освещен-
ности до 6200 лк (24,8 тыс. эрг/см2-сек физиологической радиации), прак-
тически не влияя на время зацветания, сильно повышает процент цвету-
щих растений (фиг. 134, 7). Эти исследовательницы нашли, что для ириса
оптимальная освещенность равна 4500—5000 лк (18—24 тыс. эрг/см2-сек
физиологической радиации). В этом случае развиваются нормальные расте-
ния, дающие 100% цветения через 133—143 дня.
В наших исследованиях, проведенных с салатом (Lactuca sativa), репой
(Brassica rapa), астрами (Callistephus chinensis) и другими растениями,
было установлено, что для этих растений оптимальная интенсивность
физиологической радиации равна 18—20 тыс. эрг/см2-сек (фиг. 119—133).
Положение оптимума зависит от спектрального состава излучения: в крас-
ных лучах он наступает при меньшей интенсивности, чем в синих.
К сожалению, этим исчерпывается список работ, посвященных значе-
нию интенсивности радиации для культуры растений целиком при искус-
ственном освещении.
8. ЗНАЧЕНИЕ ДЛИНЫ ДНЯ
Первая попытка изучения влияния длины дня на растения в условиях
искусственного освещения была предпринята еще Массаром [614], культи-
вировавшим под лампами накаливания в 400 вт мхи (Canocephalus canicus,
Lumularia cristata, Marchantia polymorpha, M. emarginata, Pellia epiphylla,
P. endiviaefolia), плауновые (Selaginella Helvetica) и цветковые: заячья
капуста (Sempervivum tectorum), сныть (Aegopodium podagraria), вербей-
ник (Lysimachia nummularia), барвинок (i inca minor), глехома (Glechoma
hederaceum), дубровка (Ajuga reptans) и колокольчик (Campanula persi-
folia), в условиях 6-, 12-, 18- и 24-часового дня.
Позднее Н. А. Максимов [179] установил, что для пшеницы (Triticum
vulgare), ячменя (Hordeum vulgare), гороха (Pisum sativum), бобов (\ icia
faba) и гречихи (F agopyrum esculentum) беспрерывное освещение является
более благоприятным, чем 12-часовой день. В первом случае сухой вес
растений в 1,6—5,8 раза больше, чем в условиях короткого дня.
VI. Выращивание растений при искусственной радиации
20
Пфейффер [667] дала анатомическую характеристику растений, вы-
ращиваемых при длине дня в 5, 7, 12, 19 и 24 часа под лампами накалива-
ния в 1500 вт при освещенности, падавшей по мере горения ламп от 4400
до 3500 лк (14,4—11,4 тыс. эрг/см-’-сек физиологической радиации). Этот
автор показал, что лучшее развитие стебля с наиболее развитой флоэмой и
ксилемой у гречихи (Fagopyrum esculentum)w. томатов (5"olanum lycopersicum)
и максимальная высота растений у ночной красавицы (Mirabilis jalapa)
наблюдаются при длине дня в 17 часов. Наибольший объем корневой
системы у последней наблюдался в условиях беспрерывного освещения.
Обстоятельное исследовачие этой проблемы провели Артуре сотрудни-
ками [376, 377, 379], которые выращивали растения под лампами накали-
вания, охлаждаемыми водным экраном, при различной интенсивности
радиации и длине дня (табл. 81).
Таблица 81
Сырой вес растений под лампами накаливания при различной длине дня
(Артур, Гутри и Ньюэлл [379])
Длина дня (в часах) Капуста (Brassica oleracea var. capitata) 1 (листья) Редис (Raphanus sativus var. rddicula) 8000 лк
1 4500 лк; 25° 9000 лк; 25°
сырой вес (в г) содержание воды (в %) сырой вес (в г) содержание сырой вес (в г) содержание воды (в %)
воды (в о/ \ /о)
5 94 90,4 362 92,5 — —
7 130 88,9 379 92,8 10 92,9
12 472 . 91,4 476 92,8 20 92,7
17 680 89,4 907 93,0 81 90,7
19 1 715 85,6 1032 92,0 30 90,7
24 | 490 86,2 — — 16 84,9
Оказалось, что наилучшее накопление сухого вещества наблюдается
при длине дня в 17—19 часов. Дальнейшее увеличение продолжитель-
ности освещения приводит к снижению сухого веса у растений (гречихи,
капусты, редиса, шалфея и др.).
Редингтон [703, 704], применяя лампы накаливания в 300 вт при тем-
пературе 22—28°и влажности воздуха 61—64%, выращивал большое число
растений и установил, что такие растения, как клейния (Kleinia arti-
culata), бёмерия (Boehmeria nivea), кукуруза (Zea mays), горох (Pisum
sativum), вика ('. icia sativa), качим (Gypsophila elegans), подмаренник
(Galium verum), конопля (Cannabis sativa), хмель (Humulus japonicus),
шалфей (Salvia splendens), маниго (Hibiscus manihot), тыква (Cucurbita
pepo), хлопчатник (Gossipium herbaceum), пеларгония (Pelargonium sp.),
маранта (Marantha arundinacea) и лен (Linum usitatissimum), растут лучше
при 16-часовом, чем при 8- и 24-часовом дне, хотя горох, вика, качим,
подмаренник, лен, тыква, хлопчатник, пеларгония и кукуруза цветут
раньше при 24-часовом дне, чем при 16-часовом.
Ши-Вей лу [752], культивируя длиннодневное растение берию (Baeria
chrysostoma) в условиях стерильной культуры при освещенности 4500 лк,
получаемой посредством люминесцентных ламп, нашел, что высота расте-
ний и число цветков увеличиваются вместе с увеличением длины дня
от 4 до 18 часов, а число листьев достигает максимума при 12—14-часовом
дне и затем, по мере дальнейшего увеличения длины дня, снова умень-
шается.
206
Общая светокультура растений.
равняется о—ти часам, для
рин — 9—14 часам. В этих
сои пеки]
условиях
узлов „ __ ____ .. . , ... , _ . . ,
цветения рудбекии (Rudbeckia bicnlor)
бутонизации __________ ____ ; *. "..
ело дней до бутонизации белены (Hyoscyamus niger)
7— число дней до цветения, высота растений и число
у сои (Sdja hispida) Мандарин; 4 — число дней до
7... ~ 5— число дней до
львиного зева (Antirrhinum majus); 6 — чи-
Стейнберг и Гарнер [786] выращивали при помощи ламп накаливания
в 1000 вт, охлаждаемых водою, при освещенности 13—28 тыс. лк (72—155
тыс. эрг/см2-сек физиологической радиации) растения сои (Sofa max),
свеклы (Beta vulgaris) и рудбекии (RudbecKia bicolor) в условиях различной
длины дня (от 4 до 24 часов) и различной температуры (от 15 до 32°). Эти
авторы нашли, что оптимальная длина дня для развития сои Билокси
~ '2 часам и для сои Манда-
я названных сортов зацве-
тают раньше всего, а удли-
нение пли укорачивание
дня вызывает запаздыва-
ние в цветении. Высота ра-
стений и число узлов воз-
растают вместе с длиною
дня и в большинстве слу-
чаев достигают максимума
при 15—17-часовом дне
(фиг. 136, 1—3). Сахарная
свекла цветет лишь при
длине дня более 15 часов,
причем при температуре
15° одновременно как в
условиях 18-часового дня,
так п при беспрерыв-
ном освещении, а при 18°
на 18 дней быстрее при
24-часовом дне. У рудбе-
кип переход к цветению
ускоряется по мере удли-
нения дня; раньше всего
она цветет при 18-часовом
дне. Дальнейшее увеличе-
ние длины дня уже не влия-
ет на скорость перехода
к цветению (фиг. 136, 4).
Бортвик с сотрудника-
ми [421], выращивая яч-
мень (Hordeum vulgare) под
вольтовой дугой в комби-
нации с лампами накаливания, при освещенности 23 тыс. лк в условиях
различной длины дня, нашли, что лучший эффект в отношении скорости
развития и урожая семян дает 16-часовой день. Беспрерывное освещение
и 18-часовой день, не ускоряя перехода к цветению, тем не менее вызы-
вают снижение урожая семян. В другой работе названные авторы [422J
показали, что при выращивании кок-сагыза посредством тех же источни-
ков радиации, при освещенности 13—19 тыс. лк, лучший рост и развитие
растений наблюдаются при тех же 16 часах ежедневного освещения.
Мельхерс и Ланг [589, 590] подробно исследовали значение длины дня
для развития белены (Hyoscyamus niger, Н. albas), львиного зева (Antirr-
hinum mafus) и дурнишника (Xanthium sp.), выращивая названные расте-
ния под лампами накаливания при различной длине дня. Как видно из
фиг. 136, о—6, для нормального развития растений длинного дня доста-
точно 17—20-часового, а для некоторых форм даже 14—16-часового
освещения в сутки. Дальнейшее увеличение длины дня или не дает эффекта,
VI. Выращивание растений при искусственной радиации
207
или последний настолько незначителен, что не имеет практического зна-
чения. Тот же результат получен в работе Клаэс и Ланга [445].
Б. С. Мошков [224] исследовал значение длины дня для земляники
{Fragaria sp.) Мысовка и томатов {Solanum lycopersicum) Пушкинские и
Грибовские. Землянику выращивали под лампами накаливания с водным
экраном при интегральной интенсивности радиации в 120—150 тыс.
эрг/см2-сек в условиях различной длины дня. Оказалось, что наиболее
быстрая бутонизация (через 12 дней) наблюдается при длине дня в 16 ча-
сов, наиболее раннее цветение (через 19 дней) — в условиях 16—18-ча-
сового дня, а наиболее быстрое созревание ягод (через 42 дня) — в усло-
виях 14—20-часового дня. Максимальный урожай ягод бывает при 14-ча-
совом дне. Томаты развивались под лампами накаливания при удельной
мощности 3,6 и 7,2 квт/м'2, при интегральной интенсивности радиации
в 100 и 200 тыс. эрг/см2-сек. Опыт показал, что при температуре 22—25°
максимальный сырой и сухой вес томатов получается при длине дня
в 20 часов.
Клаэс [444] показала, что для культуры белены {Hyoscyamus niger)
нет необходимости давать высокую интенсивность радиации в течение
всех 16—20 часов. Достаточно, если интенсивное освещение продолжается
9—10 часов, а в остальные часы интенсивность снижается до 20—30 лк
(120—180 эрг/см2-сек). В этом случае наблюдается нормальное стрелко-
вание и цветение растений.
Некоторые растения вообще не выносят беспрерывного освещения и
должны иметь ежесуточно 6—7 часов темноты. Таковы томаты {Solanum
lycopersicum), пеларгония {Pelargonium sp.), колеус {Coleus зр.),клейния
{Kleinia articulatd) и другие растения [59, 224, 376, 377, 379, 703, 704, 774].
Отрицательный эффект обычно выражается в медленном росте, разруше-
нии хлорофилла, скручивании листьев и появлении на последних желтых
пятен. У льна наблюдается лишь потемнение листьев, нс сопровождаю-
щееся угнетением растений [670].
Роденбурх’ [733] заметил, что угнетение томатов {Solanum lycopersicum)
в условиях беспрерывного освещения зависит от температуры. Более
подробно этот «фотопериодический хлороз» был недавно исследован Уитроу
[856]. Названные авторы нашли, что оптимальной длиной дня для томатов
является 18 часов. Дальнейшее удлинение дня даже при помощи слабых
источников радиации, дающих около 40 лк (240 эрг/см2-сек физиологи-
ческой радиации), приводит к угнетению растений и появлению пятнисто-
сти листьев. Обычно хлороз листьев начинается уже через 4 дня и стано-
вится ясно заметным через 7 дней. Угнетаются лишь молодые листья,
развивающиеся в условиях неблагоприятной длины дня. Старые сфор-
мировавшиеся листья угнетаются, как правило, мало. Непременным
условием «фотопериодического хлороза» томатов является высокая
температура воздуха. При температуре менее 12° этот хлороз мало
заметен.
С. И. Доброхотова. [59] показала, что страдание томатов {Solanum
lycopersicum) в условиях беспрерывного освещения наблюдается не у всех
сортов (табл. 124). Выращивая томаты при лампах накаливания в условиях
14-часового дня и при беспрерывном освещении при освещенности 7000—
8000 лк (42—48 тыс. эрг/см2-сек физиологической радиации), она нашла,
что особенно сильно страдает сорт Лукуллус, у которого цветение запазды-
вает на 48 дней и сильно снижается урожай плодов (2,6 г вместо 212 г
при 14-часовом дне). Менее отзывчив сорт Дварф стоун, у которого цвете-
ние задерживается на 27 дней и почти не снижается урожай плодов (130,4 г
против 145,2 г). Сорт Уинтер, зацветая в обоих случаях одновременно,
208 Общая светокультура растений
в условиях беспрерывного освещения дает несколько лучший урожай
(347,2 г), чем на 14-часовом дне (273,0 г).
По данным Б. С. Мошкова [224], сильная отрицательная реакция на
беспрерывное освещение наблюдается у томатов {Solanum lycopersicum)
сорта Грибовский; сорт Пушкинский испытывает лишь незначительное
угнетение.
Интересно поведение некоторых короткодпевных растений, дающих
максимальный сухой вес лишь в условиях короткого дня. А. И. Челядин-
цева [339], пользуясь лампами накаливания в 750 вт, висевшими на высоте
80 см от почвы, выращивала земляной орех {Arachis hypogaea) в условиях
6-, 10- и 14-часового дня. Она установила, что наиболее раннее цветение
и максимальное количество цветков у названного растения бывают при
10-часовом дне. Удлинение или укорочение дня дает одинаковый отри-
цательный результат. Максимальный сырой и сухой вес растений у одних
сортов наблюдается в условиях 10-часового, а у других — в условиях
14-часового дня.
В опытах Б. С. Мошкова [224] у лимона лучший прирост был в усло-
виях 12-часового дня, чем при 18- и 24-часовом дне; однако это происхо-
дило лишь при высоких интенсивностях радиации; при снижении интен-
сивности радиации лучший прирост у лимона, как и у других растений,
наблюдается в условиях 18-часового дня.
Нередко лучшие результаты наблюдаются при беспрерывном освеще-
нии.
Так, Смит [771], исследуя рост и развитие растений под лампами на-
каливания в условиях 6-, 12-, 18 и 24-часового дня, установил, что у яч-
меня {Hordeum vulgare), овса {Avena sativa), ржи {Secale cereale), пшеницы
{Triticum vulgare) и гороха {Pisum sativum) максимальный сухой вес и наи-
более раннее цветение наблюдаются в условиях беспрерывного освеще-
ния. Увеличение сухого веса по сравнению с таковым в условиях 18-
часового дня составляло 24—68%, а ускорение в наступлении цветения —
от 5 до 33% (на 5—27 дней). У овса и ячменя при беспрерывном освещении
наблюдалось также максимальное кущение растений.
У всех перечисленных растений положительный эффект беспрерыв-
ного освещения наблюдался лишь при их выращивании под лампами
накаливания при сравнительно невысокой интенсивности освещения. В по-
левых условиях беспрерывное освещение, создаваемое с помощью допол-
нительного освещения от ламп накаливания, давало, как правило, отри-
цательный результат. Максимальный вес корней и максимальное сахаро-
накопление у свеклы {Beta vulgaris) при освещенности в 9 тыс. лк, полу-
чаемой посредством люминесцентных ламп и ламп накаливания, наблю-
дается также при беспрерывном освещении [821].
По данным Б. С. Мошкова [224], формирование колоса у ветвистой
пшеницы {Triticum turgidum) лучше всего происходит в условиях беспре-
рывного освещения.
Аллард и Гарнер [374] показали, что различные растения даже в пре-
делах одной и той же длиннодневной группы по-разному относятся к длине
дня. У цинии {Zinnia angustifolia) максимальный вес растений наблюдает-
ся при беспрерывном освещении, а у укропа {Anetum graveolens) — в усло-
виях короткого 12-часового дня (табл. 82). Последнее, несомненно, на-
ходится в связи с быстрым стрелкованием укропа в условиях беспрерыв-
ного освещения.
Оберг [364], выращивавший растения в смешанном свете от лампы
накаливания (500 вт) и ртутной лампы «НО-ЮОО» при интенсивности ви-
димой радиации 0,025 кал/см2-мин (17,5 тыс. эрг/см2-сек; 75% от лампы
VI. Выращивание растений при искусственной радиации
209
Таблица 82
Влияние длины дня на рост и развитие растений
(Аллард и Гарнер [374])
Растение Дата посева Окончание опыта Длина дня (в час.) Дата Число дней до цветения Сырой вес (в г) Сухой вес (в г)
бутони- зации цветения наземной массы корней общий наземной массы корней общий
Циния {Zinnia ап- gustijolia) . . . 6/XII 9/Ш 6 2,0 0,5 9 5 -г-' 0,3 0,1 0,4
6/XII 9/Ш 12 24/11 7/III 101 233,0 7,0 240,0 22,0 1,5 23,5
6/XII 9/Ш 18 23/II 2/III 96 504,5 19,0 523,5 54,0 3,5 57,5
6/XII 9/Ш 24 23/Ц 1/Ш 95 681,0 20,0 701,0 77,0 4,5 81,5
Укроп {Anetum graveolens) . . . 5/1 19/Ш 12 4/Ш 18/Ш 71 810,0 35,0 845,0 89,0 5,0 94,0
5/1 19/Ш 24 1/II 8/II 34 285,0 7,0 292,0 84,0 4,0 88,0
накаливания и 25% от ртутной лампы), также установил различную реак-
цию растений на длину дня (табл. 83).
Таблица 83
Влияние длины дня на рост и развитие растений
(Оберг [364])
Растение Длитель- ность опыта (в днях) Длина дня (в час.) Сырой вес одного ра- стения (в г) Сухой вес одного растения Число листьев Высота ра- стений (в см) Длина гипо- котиля (в см)
в г R °/ /о к сырому весу
( 6 0,487 0,048 9,9 5,3 76
Томаты {Solanum lycopersicum) 21 12 18 0,552 0,725 0,072 0,105 13,0 14,5 4,9 5,1 80 71 —
24 0,579 0,084 14,5 4,7 71 —
6 0,463 0,040 8,6 5,5 — —
Шпинат {Spinacea oleracea) 20 ' 12 1 18 0,571 0,674 0,090 0,119 15,8 17,6 5,8 7,8 — _
24 0,739 0,138 18,6 7,5 — —
6 0,350 0,019 5,5 4,8 — —
Салат {Lactuca sativa) .... 21 .. 12 18 0,764 0,885 0,062 0,094 8,1 10,6 5,0 5,3 — —
24 0,950 0,118 12,4 5,5 — —
6 0,050 0,003 7,0 — 2,7
Кресс-салат {Lepidum sativum) 15 12 18 0,082 0,113 0,007 0,012 9,0 10,6 — Z 3,0 3,6
24 0,106 0,013 12,1 — 3,0
Для томатов оптимальным является 17-часовой день, а для салата,
шпината и кресс-салата (Lepidum sativum) — беспрерывное освещение.
Для древесных пород установлено, что беспрерывный рост происхо-
дит лишь при длине дня в 16—24 часов; при более коротком дне растения
уходят в покой после первого прироста [827].
14 А. Ф. Клешнин
210 Общая светокультура растений
Подводя итог, следует сказать, что для максимального накопления су-
хого вещества каждый вид растений нуждается в определенной длине дня.
В большинстве случаев сухой вес растений возрастает вместе с увеличением
длины дня и достигает максимальной величины при 16—19-часовом дне,
а у некоторых видов — даже при 10—12-часовом. Беспрерывное освещение,
повидимому, не является наиболее благоприятным условием для нормаль-
ного роста и развития растений. Во всяком случае, положительный эффект
от беспрерывного освещения, если он и имеется, может не оправдывать
затрат электроэнергии. Переход к цветению у короткодневных форм бы-
стрее всего совершается при длине дня в 10—14 часов, а у растений длин-
ного дня — при 16—20 часах. В последнем случае беспрерывное освещение
обычно дает незначительное дополнительное ускорение развития.
9. ЗНАЧЕНИЕ ПРЕРЫВИСТОГО ОСВЕЩЕНИЯ
Специального рассмотрения заслуживает вопрос о применении преры-
вистого освещения. Последнее может представлять и чисто практический
интерес в связи с применением подвижной системы инж. И. Н. Финкель-
штейна.
Гарнер и Аллард [504, 506], пользуясь лампами накаливания в 1000 вт,
охлаждаемыми током воды, выращивали растения при освещенности
20—40 тыс. лк (120—240 тыс. эрг/см‘2-сек физиологической радиации) при
различном соотношении света и темноты. В четырех сериях опытов, при-
меняя соотношение света и темноты 1:1, 2:1 и 1:2, эти авторы исследо-
вали влияние прерывистого освещения на рост и развитие растений и срав-
нивали его с влиянием короткого и длинного дня и беспрерывного
освещения.
В первой серии опыта были сопоставлены периоды освещения дли-
тельностью 15 секунд, 1 минута и 1 час, чередовавшиеся с равными
периодами темноты. Контрольные растения развивались при беспрерыв-
ном освещении и в условиях короткого дня (12 часов). Во второй серии
опыта периоды освещения и равные им периоды темноты равнялись 5
и 15 секундам, 1,5, 15 и 30 минутам, 1 и 12 часам (контроль). В третьей
серии опытов периоды освещения были следующие: 3,33, 10 и 40
секунд, 3,33, 10, 20 и 40 минут и 8 часов (контроль), а периоды темноты
были в два раза больше. В четвертой серии опыта периоды освещения
были, наоборот, в два раза больше периодов темноты, а контрольные
растения развивались в условиях длинного 16-часового дня.
В качестве объектов служили растения короткого дня — соя {Soja
max, сорта Билокси и Пекин), перилла {Perilla frutescens), колеус {Coleus
sp.) и космос {Cosmos sulphurous), растения длинного дня — рудбекия
{Rudbeckia bicolor}, шток-роза {Althaea rosea), шпорник {Delphinium ajacis)
и свекла {Beta vulgaris), нейтральные растения — соя (сорт Мандарин)
и гречиха {Fagopyrum vulgare).
Опыты длились, в зависимости от вида растения, от 20 до 98 дней.
Было установлено, что прерывистое освещение, состоящее из чередую-
щихся периодов света и темноты (от 3 секунд до 1 часа), действует на раз-
витие растений так же, как беспрерывное освещение или длинный день,
т. е. ускоряет или вызывает цветение длиннодневных растений и задержи-
вает или предотвращает цветение растений короткого дня. Сырой и сухой
вес растений, как видно из кривых на фиг. 137, независимо от фотоперио-
дической группы растений, имеет минимальное значение при чередовании
света и темноты продолжительностью от 15 секунд до 15 минут. Увеличение
или уменьшение продолжительности светлых и темных периодов за пре-
VI. Выращивание растений при искусственной радиации
21
делы названных величин, как правило, ведет к увеличению веса растений.
Чередование света и темноты в отношении 1:2 или 2:1 (третья и четвертая
серии опыта) также в большинстве случаев приводило к значительному
уменьшению веса растений по сравнению с таковым в условиях как корот-
кого, так и длинного дня.
Аналогичные данные для зеленых водорослей — гормидиум (Ног-
midium nitens), стихококкус (Stichococcus bacillaris), хламидомонас (Chia-
midomonas pseudococcum) и сценедесмус (Scenedesmus obliquus) получила
Иггена 1562], которая применя-
ла освещение от ламп накалива-
ния в 300 лк (1800 эрг/см'2-сек
физиологической радиации). Ва-
рианты прерывистого освещения
были те же, что и во второй се-
рии опытов Гарнера и Алларда.
Иггена нашла, что минимальное
накопление сухого вещества на-
блюдается при прерывистых
экспозициях длительностью от
15 секунд до 15 минут; накопле-
ние сухого вещества возрастает
как при увеличении, так и при
уменьшении частоты мигания.
Портсмут [682] сравнил эф-
фективность короткого дня,пре-
рывистого и беспрерывного ос-
вещения для растений огурцов
(Cucumis sativus) при освещенно-
сти 1000—3200 лк (5500—17 700
эрг/см2 - сек), полученной посред-
ством ламп накаливания. Этот
автор установил, что по высоте,
весу и листовой поверхности
растения, выращиваемые на пре-
рывистом освещении (1 минута
света — 1 минута темноты), зна-
чительно отстают от растений,
развивавшихся в условиях ко-
роткого дня (12 часов), а по-
Арадалжшлелбмюепй </ тетбех явриаШ
Фиг. 137. Влияние прерывистого освещения
па сухой пес растений
1 — рудбекил (Rudbeckia tricolor); 2 — гречихи (Fago-
pyrum esculentum); .3 — космоса (Cosmos sulphurous);
4— живокости (Delphinium ajacis); ,5—батата (Ipo-
ma a batalus); 6 — шток-розы (Althaea rosea)
следние почти не уступают растениям, культивируемым при беспрерыв-
ном освещении.
Прерывистое освещение, сокращая срок службы ламп, не дает, следо-
вательно, каких-либо преимуществ по сравнению с длинным днем.
10. ЗНАЧЕНИЕ ФОТОПЕРИОДИЧЕСКИХ ЦИКЛОВ
В естественных условиях растения обычно развиваются при регуляр-
ной смене света и темноты в пределах 24-часового цикла (естественный
фотопериодический цикл). Исключением являются лишь полярные об-
ласти, где летом в течение нескольких месяцев длится беспрерывный
день, а зимой — беспрерывная ночь.
Применение искусственных источников радиации позволяет выращивать
растения в условиях любого фотопериодического цикла — от нескольких
секунд (прерывистое освещение) до нескольких суток и даже недель (бес-
14* • -
212
Общая светокультура растений
прерывное освещение). Исследований в этом направлении очень немного,
но все они дают совершенно одинаковый результат.
Гарнер и Аллард [374, 504], пользуясь лампами накаливания в 1000 вт
при освещенности 13 тыс. лк (72 тыс. эрг/см'2-сек физиологической радиа-
ции), подробно исследовали значение циклов различной продолжитель-
ности (от 1 до 72 часов) для роста и развития растений длинного дня —
ук-ропа (Anetum graveolenS), пшеницы (Triticum aestivum), цинии(Zinnia
angustifolia) и рудбекии (Rudbeckia bicolor), растений короткого дня —
сои (Sofa max, сорта Билокси и Пекин) и нейтральных растений — сои
(Sofa max, сорт Мандарин). Их опыты показали, что у всех перечисленных
растений нормальный рост и нормальное развитие наблюдаются лишь
в условиях естественного 24-часового цикла.
Аналогичные данные получили Клаэс и Ланг [445], которые, пользуясь
лампами накаливания в 200 вт, сравнили циклы в 24, 48 и 72 часа для
белены (Hyoscyamus niger). В их опытах белена быстрее всего зацветала
в условиях 24-часового цикла при длине дня более 10 часов.
Снайдер [781 ] при помощи люминесцентных ламп (при освещенности
9000 лк) культивировал подорожник (Plantago lanceolatus) в условиях
10—46-часовых циклов. В этом опыте также было установлено, что подо-
рожник нормально развивается лишь в условиях длинного дня при 24-ча-
совом цикле.
Гамнер [528], пользуясь вольтовой дугой, культивировал сою (Sofa
max, сорт Билокси) при освещенности 13 тыс. лк в условиях различных
циклов — от 24 до 52 часов — и нашел, что соя быстрее всего зацветает
в условиях короткого дня при естественном 24-часовом цикле.
Аналогичные результаты для сои (Sofa max) Билокси и периллы (Pe-
rilla ocymoides) были получены Уорингом [836] при культуре названных
растений посредством люминесцентных ламп при 11 тыс. лк (47 тыс.
эрг/см2-сек физиологической радиации) в условиях 24—48-часовых циклов.
В опытах этого автора максимальное количество цветущих растений и
наибольшее количество цветков наблюдались лишь в условиях короткого
дня при 24-часовом цикле.
Таким образом, приведенные данные не оставляют сомнений в том,
что в условиях светокультуры наиболее благоприятным является есте-
ственный 24-часовой цикл при оптимальной длине дня, зависящей от фо-
топериодической группы растений. В этом случае наблюдается наиболее
быстрое зацветание и наибольшее количество цветков.
/ 11. РАЗЛИЧНЫЕ ТИПЫ ЛЮМИНОСТАТОВ
Выращивание растений целиком при искусственном освещении имеет
значение для культуры растений в условиях полярной зимы, озеленения
станций метрополитена и других закрытых помещений и для научно-ис-
следовательских работ. Более подробно эти вопросы мы рассмотрим в сле-
дующих главах. Здесь же остановимся лишь на проблеме освещения камер
искусственного климата (люминостатов) для выращивания растений
в строго определенных и строго постоянных условиях освещения, темпера-
туры, влажности, содержания углекислоты и т. д.
Первая попытка в этом направлении принадлежит А. С. Фамин-
цыну [319], который еще в 1868 г. сконструировал специальный фонарь, осве-
щаемый керосиновыми лампами в 12 св, для культуры низших растений.
Прототипом современных люминостатов является камера Клебса [578].
Этот автор построил специальные фанерные камеры высотою 82 см с пло-
щадью 0,7 м2, оборудованные лампами накаливания в 50 св, работавшими
VI. Выращивание растений при искусственной радиации
213
на постоянном токе с напряжением 65 в. Лампы помещались под потол-
ком, по 4—6 ламп на камеру. Температура колебалась в пределах 22,1 —
26,2°, а влажность воздуха — 55—70%. Судя по напряжению и силе
тока (6 а), удельная мощность, потребляемая в камерах, равнялась
550 вт/м2. При помощи этих камер Клебс проводил длительные опыты
по влиянию беспрерывного электрического света на рост и состояние
покоя у бука {Fagus silvatica).
Начиная с тридцатых годов нашего столетия люминостаты появились
во многих ботанических и сельскохозяйственных научных учреждениях
СССР, Германии, США и других стран. Обычно для этой цели исполь-
зуются подвальные помещения лабораторий или оранжерей, где обеспе-
чивается постоянство освещения, температуры, влажности, содержания
углекислоты и т. д. Первоначально в качестве источников радиации слу-
жили обычные лампы накаливания, подвешиваемые на определенной
высоте, причем избыток тепла устранялся вентиляцией. Первые люмино-
статы подобного типа были построены в 1926 г. в Тимирязевской сельско-
хозяйственной академии [8, 90, 128, 129]. Они представляли собою камеры
с площадью в 2 м2, освещаемые лампами накаливания в 500 вт, находив-
шимися на высоте 50—60 см. Постоянство условий было обеспечено двой-
ными стенками, герметичностью дверей и системой охлаждения. В этих
люминостатах Н. А. Артемьев, Е. Д. Корольков и М. С. Калинин произ-
вели большие исследования по электрокультуре и светокультуре растений,
вырастив несколько поколений растений огурцов {Cucumis sativus), то-
матов {Solanum lycopersicum), овса {Avena sativa), вики (i icia sativa) и др.
Вскоре большой люминостат был построен в подвале Стэнфордского
университета (США), в котором были произведены исследования по биоло-
гии пшеницы [737, 738]. Люминостат представлял собою комнату высотою
2,5 м, площадью 15,6 м2 (7,8x2 м), освещенную 15 лампами накаливания
в 300 вт, которые подвешивались на высоте 1,2 м от сосудов. На уровне
растений обеспечивалась освещенность в 6000 лк (35 тыс. эрг/см2-сек
физиологической радиации); температура воздуха поддерживалась с точ-
ностью до 0,°1, а относительная влажность — в пределах 57—58%.
Аналогичный люминостат, применяемый в лаборатории минерального
питания Калифорнийского университета, описали Дэвис и Хоэглэнд [462].
Он представляет собою стеклянную камеру объемом 1,4 м:’ с пло-
щадью 0,9 м2, освещаемую лампами накаливания в 300 вт, расположен-
ными вне камеры сверху и сбоку, со стороны боковых стенок. Через
камеру пропускается ток воздуха со скоростью 11,3 м"7мпн, причем
температура в камере равняется 20°, а освещенность — 26 тыс. лк
(156 тыс. эрг/см2-сек физиологической радиации).
Одновременно появились люминостаты, где для охлаждения ламп на-
каливания, помимо вентиляции, применяется водный экран, который
помещается непосредственно под лампами, на некоторой высоте над ра-
стениями. Этот прием при создании люминостатов был впервые применен
в 1925 г. в Институте Бойс-Томпсона (Нью-Йорк), где в подвале под оран-
жереей были построены большие камеры размером 3,35x3,35 м (11,6 м2).
Люминостаты освещаются 25 лампами накаливания по 1000 или 1500 вт,
находящимися над стеклянным потолком, по которому пропускается бес-
прерывный ток воды, поглощающий 50% инфракрасных лучей. Удельная
мощность, в зависимости от мощности ламп, равняется 2 или 3,2 квт/м2.
Освещенность на уровне растений при лампах в 1500 вт достигает 9000 лк
(54 тыс.эрг/см2-сек физиологической радиации). Температура обычно
поддерживается около 26°, а относительная влажность воздуха —- около
80%. В этих люминостатах, описанных Артуром и его сотрудниками
214
Общая светокультура растений
[144, 377, 379, 449, 667], было произведено большое число исследований
как по светокультуре растений, так и по другим вопросам физиологии
растений.
Последним наиболее совершенным люминостатом, основанным на
охлаждении ламп накаливания водным экраном, является люминостат,
построенный Биологическим институтом в Далеме (Берлин) и описанный
Веттштейном и Пиршле [670, 671, 848]. Он состоит из четырех камер,
каждая площадью в 1 м2. Камеры оборудованы четырьмя лампами накали-
вания по 500 вт (2 квт/м2), помещенными в специальных рефлекторах над
двойным стеклянным потолком, между стеклами которого, в 3 мм толщи-
ною, пропускается беспрерывный ток воды слоем в 10 см. Освещенность
достигает 10 тыс. лк (60 тыс. эрг/см2-сек физиологической радиации).
В зависимости от задач исследования, под стеклянным потолком непосред-
ственно над растениями можно помещать другие типы ламп (ртутные,
натриевые и пр.). Температура и влажность регулируются автоматически,
причем первая может быть установлена в пределах 0—45°, а вторая —
в пределах 20—80%.
Люминостаты подобного типа более простой конструкции в настоящее
время широко распространены и находят применение во многих физио-
логических лабораториях и ботанических учреждениях [364, 438,
545, 654].
Стейнберг [785] описал люминостаты, где лампы накаливания для
охлаждения помещались в стеклянные колпаки, наполненные дести лли-
рованной водой, которая охлаждалась беспрерывным током водопровод-
ной воды, текущей через спирали (фиг. 75 и 76). Этот автор применял
лампы мощностью 1000 вт, получая освещенность 10 тыс. лк (55 тыс.
эрг/см2-сек физиологической радиации) на расстоянии 1 м, 20 тыс. лк
(110 тыс. эрг/см2-сек физиологической радиации) — на расстоянии 25 см
и 40 тыс. лк (220 тыс. эрг/см2-сек физиологической радиации) — на рас-
стоянии 5 см от ламп. Этот способ охлаждения позволяет приближать
лампы к растениям на любое расстояние. В люминостатах с такими лам-
пами проведено много исследований по росту и развитию растений [374,
504, 786].
Совсем недавно Паркер [658, 661] описал люминостат, который осве-
щается вольтовой дугой, работающей при токе в 60 или 75 а и напряжении
50 в и создающей на расстоянии 1 м освещенность порядка 20 тыс. лк
(145 тыс. эрг/см2-сек физиологической радиации). Люминостат представ-
ляет собою комнату высотою 3 м и площадью 15 м2 (5,5x2,75 м). Темпе-
ратура и влажность воздуха в люминостате регулируются автоматически.
Вольтова дуга помещается посредине камеры, на высоте 1,8 м. Она обо-
рудована специальным параболическим рефлектором и помещена в фонаре
из обычного стекла, который устраняет вредные ультрафиолетовые лучи.
Вредные продукты горения устраняются из фонаря посредством особой
вентиляции. В качестве дополнительного источника оранжево-красных
лучей служат обычные лампы накаливания в 200 вт, которые, в количе-
стве 8 штук на камеру, располагают непосредственно над растениями и
на расстоянии 1 м создают освещенность 1600 лк (10 тыс. эрг/см2-сек
физиологической радиации). Растения помещают на полукруглых стел-
лажах, имеющих три различных уровня, уступами понижающихся
к середине камеры. Большинство работ этого автора и его сотрудников
проведено в описанном люминостате [421, 422, 426, 661, 663].
В настоящее время лампы накаливания в люминостатах постепенно
вытесняются люминесцентными лампами. В этом случае получаются
совершенно нормальные растения и, кроме того, значительно легче регу-
VI. Выращивание растений при искусственной радиации
215
лируется температура. Первое описание подобного люминостата, соору-
женного в Калифорнийском технологическом институте (США), сделано
Вентом [315, 843]. В его камерах в качестве источника радиации служили
люминесцентные лампы бело-
го или дневного света мощ-
ностью 40 и 80 вт, которые в
виде горизонтальной решетки
располагались над растения-
ми. При удельной мощности
240 вт/м2 на расстоянии 30 см
удавалось создать освещен-
ность 4—5 тыс. лк. В послед-
ние годы лаборатория зна-
чительно модернизирована
[844]. В этой усовершенство-
ванной лаборатории, полу-
чившей наименование фито-
трона, камеры оборудованы
люминесцентными лампами,
работающими на перекале.
При силе тока 500 та (вместо
нормальных 100—200 та)
лампы обеспечивают освещен-
ность 22 тыс. лк. Источником
оранжево-красных лучей слу-
жат лампы накаливания в
60 вт. Аналогичные люмино-
статы, оборудованные люми-
несцентными лампами, описа-
ны многими авторами [529,
537, 627, 826].
Следует отметить также
люминостаты, описанные
Ван-дер-Вон [826] и освещае-
мые люминесцентными лам-
пами мощностью 40 вт. В этом
случае 8 ламп (320 вт) рас-
полагаются под потолком, а
22 лампы —• на боковых стен-
ках, по И ламп (440 вт) на
каждой. При удельной мощ-
ности 800 вт/м2 автор получал
равномерную освещенность
10—12 тыс. лк. Это соответ-
ствует интегральной интен-
сивности радиации 60—70
вт/м2 (60—70 эрг/см2 • сек).
Температура регулируется в
пределах 17—35°. В назван-
108
Фиг. 138. Схема люминостата с комбинирован-
ным верхним и боковым освещением люминес-
центными лампами в 30 вт (размеры даиы в см)
а — лампы; б — держатели; в — защитные козырьки;
А — вертикальный разрез; Б — план
ных условиях нормально
развиваются, цветут и плодоносят огурцы (Cucumis sativus), томаты (Sola-
num lycopersicum), хризантемы (Chrysanthemum sp.), флоксы (Phlox sp.),
табак (Nicotiana tabacum), салат (Lactuca sativa), бальзамины (Impatiens
balsamina) и другие растения.
216
Общая светокультура растений
Митчелл [627] описывает люминостат в Институте ботаники Манче-
стерского университета, оборудованный 12 лампами накаливания по
150 вт и 12 люминесцентными лампами синего света по 30 вт. При удель-
ной мощности 4,7 квт/м2 (лампы накаливания — 3,9 квт/м2 и люминесцент-
ные лампы — 0,8 квт/м2) на расстоянии 46 см создается освещенность
в 22 тыс. лк. Несмотря на отсутствие водного экрана, в люминостате под-
держивается постоянная температура, регулируемая в пределах 4—20°.
Аналогичный принцип кладется в основу освещения в Лаборатории
искусственного климата Института физиологии растений им. К. А. Тимиря-
зева Академии Наук СССР [189, 190,322]. Здесь, кроме ламп накаливания,
которые размещаются в куполе камер над водным потолком, растения будут
освещаться люминесцентными лампами белого, красного и синего света.
Таким образом, в люминостатах в настоящее время в большинстве
случаев применяются лампы накаливания и люминесцентные лампы:
первые — при удельной мощности 2—3 квт/м2, а вторые — при 0,25—
0,5 квт/м2. Лампы накаливания для охлаждения помещают над водным
экраном или погружают в проточную воду, а люминесцентные лампы
охлаждают посредством вентиляции. При лампах накаливания обычно по-
лучают освещенность порядка 10 тыс. лк (около 60 тыс. эрг/см-2сек фи-
зиологической радиации), а при люминесцентных лампах — 5—7 тыс. лк
(20—30 тыс. эрг/см2-сек физиологической радиации). Для получения более
высокой интенсивности радиации люминесцентные лампы необходимо
располагать не горизонтально, как это обычно делается (фиг. 65, а),
так как в этом случае на 1 м2 можно затратить не более 500 вт, а верти-
кально, в междурядьях растений (фиг. 65, б и в), когда удельную мощность
можно довести до 1,5—3 квт/м2. Если люминесцентные лампы располагать
непосредственно над растениями, на расстоянии 10—12 см, то посредством
ламп в 30 вт можно получить освещенность 13 тыс. лк (52 тыс. эрг/см2-сек
физиологической радиации), а при помощи ламп в 40 вт — 21 тыс. лк
(84 тыс. эрг/см2-сек физиологической радиации) [529, 635, 636, 637].
Специальный интерес представляют собою люминостаты более упро-
щенной конструкции, употребляемь^ в средних школах для выращива-
ния растений в зимнее время в биологических и методических кабинетах
и для постановки простых опытов по биологии развития и физиологии
растений. Инициатива в этом направлении принадлежит Ф. Д. Сказкину
[295, 296], который построил подобный люминостат в Государственном
педагогическом институте им. А. И. Герцена (Ленинград). Люминостат
представляет собою фанерную камеру высотою 1 м и площадью 0,75x75 м
с конусовидной крышей, с отверстием для вентиляции. Внутренние стенки
для лучшего отражения лучистой энергии окрашивают в белой цвет.
Под потолком по углам люминостата находятся 4 лампы накаливания
общей мощностью 500 вт. В этих условиях автор производил опыты по
фотопериодизму растений, минеральному питанию, водным культурам и
удобрению углекислотой, выращивая ячмень {llordeum vulgare), пшеницу
{Triticum vulgare), овес {Avena sativa), кукурузу {Zea mays), редис {Rapha-
nus sativus var. radicula), фуксии {Fuchsia sp.) и другие растения.
Другой аналогичный люминостат, построенный в Ботаническом каби-
нете Ленинградской естественно-научной педагогической станции, описан
Н. М. Верзилиным [30, 31]. Наиболее удобны, по мнению автора, следую-
щие размеры камеры: высота—115 см, длина —110 см и ширина — 60 см.
На задней стенке камеры помещают комнатный термометр и психрометр
Августа, а в углу, для поддержания необходимой влажности воздуха,
банку с водою. Для вентиляции в полу камеры просверливают ряд отвер-
стий, а в потолке — одно отверстие диаметром 10 см. Все отверстия снаб-
VI. Выращивание растений при искусственной радиации
217
жают задвижками, посредством которых легко можно регулировать их
величину и скорость вентиляции. Внутренние стенки окрашивают в белый
цвет. Люминостат освещают обычными электрическими лампочками в
100—200 вт, исходя из удельной мощности 400—500 вт/м2. Для регулирова-
ния температуры, кроме того, служит обычная электрическая печка с рео-
статом, которую включают или выключают по мере надобности. Посредством
задвижек, регулирующих размеры отверстий, легко установить в камере
температуру 20—25°, если температура комнаты не превышает 15—16°.
При закрытых отверстиях вентиляции лампочка в 200 вт повышает темпе-
ратуру до 35—40°. Люминостатами этой конструкции с успехом пользу-
ются свыше 30 ленинградских школ.
Люминостаты позволяют организовать в школах самые разнообраз-
ные опыты по ботанике. Интересно в этом отношении сообщение И. И. За-
цепина [76], который, пользуясь люминостатом конструкции И. М. Вер-
зилина, освещенным лампочкой в 200 вт, организовал опыты по светокуль-
туре растений в 6-й образцовой школе г. Грозного.
Инициативу советских педагогов в этом направлении необходимо
поддержать, особенно в настоящее время, в связи с появлением люминес-
центных ламп, которые просты в эксплуатации и могут дать исключи-
тельно интересные результаты, в особенности если применять лампы
с различным спектральным составом — красные, синие и зеленые [121].
Схема простого люминостата с комбинированным верхним и боковым
освещением люминесцентными лампами дана на фиг. 138. При общей
удельной мощности в 1,32 квт/м2 на верхнее освещение приходится 510 вт
(17 ламп в 30 вт), а на боковые и заднюю стенки — по 270 вт (на каждую
стенку по 9 ламп в 30 вт); на боковых стенках лампы ставятся вертикаль-
но, а на задней — кладутся горизонтально.
В заключение следует упомянуть люминостаты для культуры мхов,
водорослей и других низших организмов. Первоначально для этих целей
применяли лампы накаливания в 100—500 вт, охлаждаемые током воды
[150, 327, 367, 415, 520, 539, 562, 570, 654, 690, 691, 793, 833]. Обычно
лампу накаливания помещают в стеклянный колпак из хорошего, про-
зрачного стекла, который в свою очередь находится в другом, аналогич-
ном колпаке большего размера. Между стенками этих колпаков пропу-
скают ток водопроводной воды слоем в 5—10 см. В некоторых случаях
лампу просто погружают в сосуд, наполняемый проточной водой [691].
Эту осветительную систему помещают в потолке термостата, в котором
размещают колбы, пробирки или другие сосуды с культурами водорослей.
Подобным способом в условиях беспрерывного освещения при освещенно-
сти 2—5 тыс. лк (12—30 тыс. эрг/см2-сек физиологической радиации)
успешно культивируются эвдорина (Eudorina elegans), пандорина (Рап-
dorina тогит), хлорелла (Chlorella sp.), улотрикс (Ulotrix oscillarind),
зигоклойиум {Sigoclonium subscinosum), эдогониум (Qedogonium capillare),
сценедесмус (Scenedesmus obliquus, S. quadricauddj, зигнема {Zygnema
sp.), формидиум (Phormidium corium, Ph. uncinatiim) и другие низшие
растения.
Иногда вместо ламп накаливания применяют неоновые лампы, оде-
ваемые в стеклянную рубашку. Лампу охлаждают проточной водой, про-
пускаемой между стенками рубашки и лампы. Осветительную систему
погружают в обычный аквариум, в котором и культивируются водоросли.
Температуру регулируют изменением скорости воды, протекающей через
рубашку. При подобном освещении нормально развивается, в частности,
нитчия (Nitzchia closterium) [572]. Высоковольтные неоновые лампы с хо-
лодными катодами применяются без водного охлаждения [42].
218
Общая светокультура растений
В последние годы перечисленные источники радиации в люминостатах
для культуры водорослей вытесняются люминесцентными лампами бе-
лого или дневного света, которые создают равномерное рассеянное осве-
щение и позволяют легко поддерживать требуемые температуры без вод-
ного охлаждения. При освещении люминесцентными лампами (2—5 тыс. лк)
удается хорошо культивировать эвглену (Euglena gracilis), хлореллу
(Chlorella pyrenoidosa), сценедесмус (Scenedesmus obliquus, S. acuminatus)
и другие, а также изолированные ткани высших растений [362, 368, 525,
788].
12. ВЫВОДЫ
Таким образом, подводя итог, следует сказать:
1. Для выращивания растений исключительно при помощи искусствен-
ных источников освещения пригодны любые источники радиации, если
в спектре излучения их содержатся лучи, лежащие в пределах 400—700шр.
(физиологическая радиация), если отсутствуют «короткие» ультрафиоле-
товые лучи с длиною волны менее 300 шр (витацидная радиация) и если
интенсивность физиологической радиации достаточно велика. Испытаны
и дали хорошие результаты лампы накаливания, дуговые лампы, ртутные
лампы (высокого и сверхвысокого давления), неоновые лампы (с холодными
электродами и дуговые интенсивного горения), люминесцентные лампы
дневного, белого, мягкобелого, красного и синего света, солнечные лампы
и др.
2. «Короткие» ультрафиолетовые лучи (<300 шр) — витацидная ра-
диация — вызывают гибель или значительное угнетение растений, а при
незначительной интенсивности или небольшой длительности экспозиции
приводят к снижению сухого веса и высоты растений. Поэтому вольтовые
дуги открытого горения или ртутно-кварцевые лампы надо снабжать
специальными экранами из обычного стекла, поглощающего названные
лучи.
3. Инфракрасные лучи, лежащие в пределах больших длин
волн (>1100 шр.), при интенсивности, не влекущей перегрева растений,
вызывают, как правило, увеличение сухого веса растений и некоторое
уменьшение их высоты. Применение водных экранов для ламп накали-
вания в большинстве случаев снижает урожай и приводит к некоторому
вытягиванию стеблей. Не исключена возможность, что последнее находится
в связи с устранением ультрафиолетовых лучей в результате их погло-
щения стеклянным экраном, неизбежным при применении водного
экрана.
4. Различные области физиологической радиации вызывают неодина-
ковый эффект. Лампы, богатые оранжево-красными лучами (лампы на-
каливания, неоновые, люминесцентные лампы красного света и др.), в усло-
виях благоприятной длины дня приводят, как правило, к значительному
ускорению развития растений, вызывая их более раннее цветение и пло-
доношение. Короткодневные растения при беспрерывном освещении этими
лучами зацветают позднее, чем при действии сине-фиолетовых лучей.
При отсутствии больших сдвигов в скорости развития при культивирова-
нии растений в оранжево-красных лучах наиболее интенсивно идет на-
копление сухой массы, формирование луковиц, корнеплодов, кочанов и
других органов растений.
Источники, богатые сине-фиолетовыми лучами (ртутные лампы, лю-
минесцентные лампы синего и дневного света и др.), вызывая задержку
в цветении растений, особенно благоприятны для культуры растений
<с целью получения вегетативных органов (салата, редиса и пр.). Однако
VI. Выращивание растений при искусственной радиации 219
для получения одной и той же зеленой массы мощность этих ламп должна
быть в 2 раза выше, чем мощность источников оранжево-красных лучей.
5. Длинные ультрафиолетовые лучи имеют значение для получения
нормальных растений. Для предотвращения вытягивания стеблей лампы
накаливания целесообразно комбинировать с ртутными лампами или
вольтовыми дугами, которые богаты этими лучами. Следует учесть, что
при этом одновременно с устранением чрезмерного вытягивания растений
наблюдается некоторое уменьшение сухого веса растений.
6. В условиях искусственной радиации возможна культура самых
различных растений — от низших водорослей до высших цветковых
растений.
7. Интенсивность физиологической радиации, необходимая для нор-
мального развития и роста растений, изменяется в очень широком диапа-
зоне величин и зависит от вида растений и типа ламп. Обычно она колеб-
лется в пределах 50—120 тыс. эрг/см2>сек.
8. Для накопления сухого вещества наиболее благоприятна длина дня
в 17—18 часов для всех фотопериодических групп растений, а для пере-
хода к цветению длиннодневных растений — 17—18 часов или беспрерыв-
ное освещение. Культура короткодневных растений с целью получения
цветков или семян возможна лишь при длине дня короче 12—14 часов.
Глава VII
ВЫРАЩИВАНИЕ РА СТЕНИЙ ПРИ ПОМОЩИ
ДОПОЛНИТЕЛЬНОГО ИСКУССТВЕННОГО ОСВЕЩЕНИЯ
Выращивание растений исключительно при искусственной радиации
представляет в настоящее время главным образом теоретический инте-
рес, культура же растений при помощи дополнительного к естествен-
ному свету искусственного освещения имеет большое практическое значе-
ние и распространена довольно широко, особенно в умеренной полосе.
Потребность растений в дополнительном искусственном освещении
в зимние месяцы определяется методом фитометра [566], заключающимся
в том, что растения высевают на протяжении года каждые 2 недели и вы-
ращивают в течение 25—30 дней. По весу растений (т. е. величине привеса),
сопоставляя его с весом растений в летние месяцы (июнь—июль), судят о по-
требности растений в дополнительном свете, аотносительную эффективность
ламп в разное время года определяют отношением отрезков вд и гд(фиг. 139).
Чем больше отношение вд/гд, тем выше эффективность дополнительного
освещения. Пользуясь этим методом, Н. А. Максимов [188] показал, что
в условиях закрытого грунта в Ленинграде фасоль {Phaseolus sp.) может
расти и накапливать органические вещества лишь в период с 12 февраля
по 20 октября, достигая максимального привеса сухого вещества (4,5 г)
и максимальной листовой поверхности (575 см2) во второй половине июня
(21 июня). В течение зимнего периода (20 октября — 12 февраля) вес
растений фасоли не превышает веса семян, так как растения существуют
за счет запасных веществ, отложенных в семенах (около 0,5 г).
Аналогичные данные для Москвы получила С. В. Тагеева [307], кото-
рая также нашла, что фасоль {Phaseolus sp.) в условиях Московской об-
ласти в зимние месяцы (до 8 февраля) может существовать лишь за счет
органических веществ, находящихся в семенах. Результаты, полученные
Тагеевой, представлены на фиг. 139.
Позднее мы будем встречаться с применением метода фитометра при
определении потребности в лучистой энергии для пшеницы {Triticum sp.),
огурцов {Cucumis sativus) и томатов {Solanum lycopersicum) (табл. 121 и
122 и фиг. 153) [130, 463, 775].
Как показывают фиг. 89 и 91—93, дополнительное искусственное осве-
щение необходимо как для усиления недостаточной интенсивности радиа-
ции, так и для удлинения коротких дней в осенне-зимние и весенние ме-
сяцы. Поэтому обычно этот способ освещения применяется в ночные часы,
а днем реже. В дальнейшем, говоря о дополнительном освещении, «мы
VII. Выращивание при дополнительном искусственном освещении
221
будем всюду иметь в виду дополнительное ночное освещение; дополни-
тельное дневное освещение будет всякий раз оговорено.
Нет необходимости доказывать, что дополнительное освещение в оран-
жереях значительно выгоднее беспрерывного освещения при культуре
растений целиком на искусственном свете. В этом случае основным источ-
ником радиации является солнце, лампы же служат лишь дополнитель-
ным источником излучения. В табл. 58 показано, что в пределах СССР
каждый квадратный метр площади ежегодно получает энергию, равную
500—700 квт-час. Ясно, что даже самое мощное искусственное освещение
не может заменить колоссальные количества энергии, которые получают
Фиг. 139. Сухой вес фасоли (Phaseolus sp.) в зависимости от времени
посева
1 — при дополнительном освещении лампой накаливания в 500 вт; 2— без до-
полнительного освещения
аб — исходный вес семян; гд — вес растения, выращенного без Дополнитель-
ного освещения; вд — вес растения, выращенного при Дополнительном освещении
растения в виде солнечных лучей. Дополнительное искусственное осве-
щение может иметь значение лишь в определенное время года и на опре-
деленных этапах развития растений, причем оно играет вспомогательную
роль.
Первая попытка дополнительного освещения растений была предпри-
нята Сименсом [754—756], который для культуры растений в зимнее
время на широте Лондона использовал угольную вольтову дугу в 4000 св,
находившуюся на расстоянии 2—3 м от растений и горевшую в ночные
часы, от захода до восхода солнца, и обеспечивавшую беспрерывное осве-
щение. Опыт производился в течение 6т/2 месяцев — с 23 октября 1880 г.
по 5 мая 1881 г. Этот автор нашел, что при употреблении открытых воль-
товых дуг необходим специальный экран из обычного стекла, устраняю-
щий вредные ультрафиолетовые лучи, и что растения могут нормально раз-
виваться совершенно без ночного «отдыха». Горох (Pisum sativum), по-
сеянный в конце октября, дал зрелые плоды 16 февраля. Виноград (Иг-
tis vinijera), малина (Rubus idaeus) и земляника {Fragaria sp.), пересажен-
ные в теплицу 16 декабря, нормально зацвели и дали зрелые плоды отлич-
ного качества: малина — 1 марта, виноград — 10 марта, а земляника —
14 марта. Банан (Musa paradisiaca) плодоносил в феврале. Дыня (Сиси-
222
Общая светокультура растений
mis melo) образовала плоды замечательных размеров и вкуса. Лишь злаки —
пшеница (Triticum vulgare), овес (Avena sativa) и ячмень (Hordeum vulgare),
несмотря на нормальный выход в трубку, не дали урожая и полегли.
Несколько худшие результаты получил Дегерен [466, 467], также
применявший угольную дугу и стеклянный экран и дававший дополни-
тельное ночное освещение. В его опытах растения из-за недостатка
света отличались чрезвычайно медленным развитием. Лишь взрослые,
крепкие растения чувствовали себя более или менее нормально.
Интересные результаты были получены Бэйли [398—400], который
при помощи вольтовой дуги в 2 тыс. св, находившейся на высоте 1—3 м,
удлинял естественный зимний день до 24 часов и наблюдал значительное
ускорение в развитии и созревании растений шпината (Spinacea oleracea),
цикория (Cichorium intybus), жерухи (Nasturtium officinale) и салата
(Lactuca sativa). Растения переходили к плодоношению настолько быстро,
что не успевали образовать достаточное количество съедобных листьев.
У контрольных растений образовались нормальные розетки листьев.
Бэйли, несомненно, является первым исследователем, установившим
экспериментально влияние длины дня на развитие растений.
Чтобы покончить с первым периодом в развитии светокультуры расте-
ний, который характеризуется применением вольтовой дуги, следует еще
назвать исследования Боннье [416—418], который, пользуясь этим источ-
ником радиации, выращивал зимой большое количество растений, главным
образом древесных, луковичных и корневищных, богатых запасами пи-
тательных веществ: морозник (Helleborus niger, Н. nutans), лютик (Ra-
nunculus bilbosus, R. asiaticus), кубышку (Nuphar luteum), кресс-салат
(Lepidum sativum), липу (Tilia silvestris), фиалку (i iola canina), пеларго-
нию (Pellargonium peltatum), явор (Acer pseudoplatanus), каштан (Aescu-
lus hippocastanum), настурцию (Tropaeolum majus), клокичку (Staphylea
pinnata), чину ( athyrus sativus), горох (Pisum sativum), вику (Vicia sativa),
миндаль (Amygdalus communis), землянику (Fragaria vesca), гравилат
(Geum urbanum), крестовник (Senecio . acobaea), золотарник (Solidago virga
aurea), пижму (Tanacetum vulgare'), горькушу (Centaurea facea), ястре-
бинку (Hieracium pilosella), первоцвет (Primula officinalis), ясень (Fra-
xinus excelsior), коровяк (> erbascum thapsus), норичник (Scrophularia
nodosa), веронику ( eronica officinalis), дубровник (Teucrium scorodonid),
чистец (Stachys tuberifera), подорожник (Plantago lanceolata), гречиху
(Polygonum fagopyrum), молочай (Euphorbia cyparissias), вяз (Ulmus cam-
pestris), бук (Fagus silvatica), граб (Carpinus betulus), тополь (Populus
alba), ландыш (Convallaria majalis), гиацинт (Hyacinthus orientalis), шаф-
ран (Crocus vernus), тюльпан (Tulipa gesneriana), элодею (Elodea cana-
densis), кукурузу (Zea myas), ель (Picea excelsa), сосну (Pinus austriaca),
папоротник (Pteris tremula) и.тисс (Taxus baccata). В большинстве слу-
чаев растения отличались несколько упрощенным анатомическим строе-
нием, чрезмерным вытягиванием стебля и другими признаками так назы-
ваемой «зеленой этиоляции», наблюдаемой обычно при недостатке света.
В общем, несмотря на отдельные неудачи, опыты перечисленных авто-
ров показали полную возможность выращивания растений в зимнее
время путем дополнительного искусственного освещения в ночные часы
и установили исключительно вредный эффект коротких ультрафиолето-
вых лучей.
В дальнейшем, после нескольких попыток использовать в качестве
источников радиации магниевые лампы [812] и газовые горелки Ауэра
[569, 699], дававшие некоторый положительный эффект, в светокультуре
растений прочно закрепились лампы накаливания. Так, например, Рэйн
VII. Выращивание при дополнительном искусственном, освещении
223
[698] для удлинения естественного дня пользовался семью угольными
лампами по 16 св. В результате увеличения длины дня наблюдалось зна-
чительное ускорение цветения шпината {Spinacea oleracea).
Несколько позднее с тем же результатом лампы накаливания были
применены Корбетом [452]. В его опытах под влиянием дополнительного
освещения наблюдался лучший рост у шпината {Spinacea oleracea), ка-
пусты {Brassica olerac a var. capitata), редьки {Raphanus sativus) и салата
{Lactuca sativa). В большинстве случаев наблюдалась тенденция к более
раннему цветению.
Телен [796] для удлинения естественного дня использовал лампы Ло-
дыгина. Начиная с середины августа, он в течение 6 недель выращивал
овес {Avena sativa), горох {Pisum sativum), гречиху {F agopyrum esculentum)
и горчицу {Sinapis alba) при помощи трех ламп по 250 св. За исключением
гречихи, растения чувствовали себя значительно лучше при дополнитель-
ном освещении, чем в контроле (северное окно). Увеличение веса по срав-
нению с контролем составляло: у овса — 11,4%, у гороха — 59% , а у гор-
чицы — 71,5%. Цветение растений наступило через 4% недели, а контроль-
ные растения не цвели совсем.
В наши дни лампы накаливания, после обширных исследований боль-
шого числа авторов [131—141, 191—193, 500, 556, 579, 646,811], прочно
вошли в практику овощеводства, цветоводства, селекции и семеноводства
и в другие отрасли культуры растений.
Почти одновременно с лампами накаливания в качестве дополнитель-
ных источников радиации стали применять ртутные и неоновые лампы.
В опытах Даджон [470, 692] для выращивания растений в оранжерее
в зимнее время была применена ртутная лампа низкого давления си-
стемы Купер—Юитт (на высоте 1,23 м). В качестве опытных объектов
служили соя {Soja max), морковь {Daucus carota), цветная капуста
{Brassica oleracea var. botrytis), кукуруза ( ea mays), салат {Lactuca
sativa), ячмень {Hordeum vulgare), пшеница (Triticum vulgare), различные
сорта гороха {Pisum sativum) и бобов ( icia (aba) и другие растения.
Этот автор отмечает сильный рост у бобов, кукурузы, цветной капусты,
пеларгонии, гвоздики, винной ягоды и томато;:. Земляника и роза
в опытном варианте цвели, а морковь дала урожай корнеплодов.
Неоновые лампы для дополнительного освещения растений ввел
Гёстерманн [206, 485, 554, 557], выращивая растения томатов {Solanum
lycopersicum) и огурцов {Cucumis sativus). Как видно из табл. 84, где
Т а б л и ц а 84
Влияние неоновых ламп на урожай томатов (Solanum lycopersicum)
и огурцов (Cucumis sativus)
(Гёстерман [554])
Растение Естественное освещение Естественное освещение 4- неоновые лампы
ЧИСЛО плодов вес плодов (в кг) число плодов вес плодов (в нг) увеличение веса пло- дов по сравнению с контролем (в %)
О и ы т 19 16 г.
Огурцы . . . 370 186,6 500 277,53 48,5
Томаты . . . 69,2 95,94 38,5
Опыт 19 17 г.
Огурцы . . . 370 163,4 485 230,22 85,0
Томаты . . . 70,0 — 101,00 44,0
224
Общая светокультура растений
приведены данные опытных и контрольных теплиц, применение неоновых
ламп вызывает увеличение урожайности на 44—50%.
Позднее большое число исследований с неоновыми лампами различных
типов провел Роденбург [717, 720, 722—725, 728, 733]. Кроме ламп с хо-
лодным катодом, которые употреблялись до него, он применил и нео-
новые лапы интенсивного горения [720].
Сравнительно недавно стали применяться натриевые [839] и люминес-
центные лампы [610], а также современные дуговые [649] и солнечные
лампы [603].
В Советском Союзе интерес к дополнительному освещению растений
возник в конце 20-х годов. Лампы накаливания впервые появились в ис-
следованиях И. Н. Коновалова [125], а неоновые лампы с холодным
катодом применил В. П. Мальчевский [192].
Современные неоновые лампы интенсивного горения, ртутные, люми-
несцентные и зеркальные лампы в качестве дополнительного источника
радиации впервые появились в работах А. Ф. Клешнина [123].
1. ЗНАЧЕНИЕ РАЗЛИЧНЫХ ОБЛАСТЕЙ СПЕКТРА ФИЗИОЛОГИЧЕСКОЙ
РАДИАЦИИ. СРАВНЕНИЕ РАЗЛИЧНЫХ ИСТОЧНИКОВ РАДИАЦИИ
Впервые сопоставление различных типов ламп при дополнительном
освещении растений произвел Роденбург [717], который в зимних усло-
виях Голландии испытал лампы накаливания, ртутные лампы из обыч-
ного стекла, ртутные лампы «Ультрасоль» с коротковолновой радиацией
(до 280 шд) и высоковольтные неоновые трубки, работающие при напря-
жении 5000 в. Автор сразу же отказался от ртутных ламп из-за слабого
эффекта (обычная лампа) или вредного действия на растения («Ультрасоль»)
и сосредоточил свое внимание на сравнении ламп накаливания и неоновых
ламп.
В опытах с земляникою (Fragaria sp.) Мадам Лефебр растения осве-
щались по 14 часов в сутки (с 17 до 7 час.) в течение 14 дней (28 января —
10 февраля), а затем по 9 часов (с 22 до 7 час.) в течение 35 дней (11 фев-
раля—18 марта), при температуре воздуха 20°. В качестве источников
радиации служили лампы накаливания по 100 вт и неоновые трубки
длиною 9 м, зигзагообразно изогнутые и работающие при напряжении
5000 в. Контрольные растения обогревались электронагревателем, под-
держивавшим в контроле ту же температуру (20 °)1, что и в опытных
вариантах. Освещенность под лампами накаливания равнялась 1000 лк
(6 тыс. эрг/см2-сек физиологической радиации), а под неоновыми
трубками — 1500 лк (11 тыс. эрг/см2-сек физиологической радиации).
Всего было использовано 152 куста земляники (25 кустов на 1 м2). Перед
началом опыта растения выдерживались при низкой температуре (15°).
Как видно из табл. 85, лучший эффект дают неоновые лампы, которые
увеличивают урожай ягод на 116%. Некоторое запаздывание в цветении
и плодоношении под неоновыми лампами автор объясняет более низкой
температурой, которая наблюдалась в этом случае (на 4° ниже, чем под
лампами накаливания).
В опыте с китайской капустой (Brassica pekinensis) автор применял
дополнительное освещение для культуры названного растения в зимние
1 Опыт Роденбурга с соответствующим подогревом контроля является единствен-
ным во всей мировой литературе по светокультуре растений. Это обстоятельство —
неравенство температур в опытных вариантах и контроле — другими авторами не учи-
тывалось.
VII. Выращивание при дополнительном искусственном освещении
225
Таблица 85
Влияние дополнительного освешения на культцри земляники (Fragaria sp.)
(Роденбург [717])
Варианты опыта Освещенность (в лк) Дата цветения Число расте- ний (в %) Число плодов на одном цвету- щем растении Число ягод со 100 растений в течение недель Уро;кай ягод к 14'IV
| ПЛОДОНО-: сящих 1 цветущих нецвету- щих 1-й 2-й 3-й ЧИСЛО В % к конт- ролю
Неоновые лампы . . 1500 20/11 75 75 25 6,7 88,0 258,0 168,0 514,0 216,0
Лампы накаливания 1000 18/11 58 58 42 139,0 115,0 31,8 285,8 120,0
Контроль — 25/П 39 55 45 4,1 22,7 122,5 83,4 238,6 100
месяцы. При 14-часовом дополнительном освещении (с 17 до 7 час.)в 5000 лк
(30тыс. эрг/см2-сек физиологической радиации под лампами накаливания
и Зэ тыс. эрг/см2. сек под неоновыми трубками) растения развивались
лучше под неоновыми лампами, чем под лампами накаливания. В по-
следнем случае растения сильно вытягивались. Роденбург на основании
своих многочисленных опытов приходит к заключению о том, что лампы
накаливания не выдерживают сравнения с неоновыми трубками; они
сильно перегревают растения, вызывают их чрезмерное вытягивание
и от них невозможно получить высокие интенсивности видимой радиа-
ции из-за обилия инфракрасных лучей. Этих недостатков нет у неоно-
вых ламп, которым и следует отдать предпочтение.
Позднее этот автор [733] провел большое число опытов по сопоставле-
нию разных источников радиации при дополнительном освещении
растений. В опытах с томатами (Solanum lycopersicum) Эльза Крэйг он
показал, что при выращивании рассады лучший эффект дают неоновые
лампы интенсивного горения, а разные типы ртутных ламп (низкого и
высокого давления) не отличаются один от другого. Во всех случаях
автор выравнивал или удельную мощность (вт/м2), или интенсивность
видимой радиации (эрг/см2-сек), освещая растения по 8 часов в сутки
(с 22 до 6 час.).
Из опыта по сравнению ламп накаливания (40 вт), ртутных ламп
сверхвысокого давления «НР-300» (75 вт) и натриевых ламп «SO-650»
(100 вт ) было установлено, что при интенсивности видимой радиа-
ции порядка 2160—2900 эрг/см2-сек наилучший эффект в отношении уси-
ления роста наблюдается под натриевыми лампами. Астры (Callistephus
chinensis) при посеве 8 декабря зацветали в результате дополнительного
освещения в конце февраля сначала под лампами накаливания и ртут-
ными лампами, а затем под натриевыми лампами. Контрольные растения
находились в стадии розетки.
В другом опыте были сопоставлены лампы накаливания и ртутные
лампы сверхвысокого давления «НР-300» при одинаковой удельной
мощности, равной 75 вт/м2. Лучшая рассада томатов была получена под
ртутными лампами; растения были более приземистыми, имели короткие
междоузлия, отличались крупными листьями, большим сырым и сухим
весом и содержали большее количество зольных элементов и органи-
ческого вещества.
В третьем опыте сравнивались неоновая лампа (100 вт), натриевая
лампа «SO-650» (100 вт) и ртутная лампа «НО-500» (150 вт). Интенсивность
* видимой радиации под лампами была соответствейно равна 3100, 2800 и
15 А. Ф. Клешнин
226
Общая светокультура растений
2600 эрг/см2-сек. Посев растений был произведен 9 октября, а освещение —
с 23 ноября по 6 января. Готовая рассада томатов имела следующую
высоту: контроль — 20 см, под неоновыми лампами — 32 см, под
натриевыми лампами — 33 см,, под ртутными лампами — 26 см. Даль-
нейшее развитие растений проходило без дополнительного освещения
в сосудах. Растения зацвели в начале марта. Полученный урожай был
сравнительно невелик (1,3 — 1,65 кг с куста), причем лучший эффект
дали неоновые лампы (на 27% больше, чем в контроле), а худший —
ртутные (на 12% больше контроля). Эффективность натриевых ламп была
несколько выше, чем ртутных (на 15% больше контроля).
Аналогичные результаты были получены в опыте при посеве 23 ноября
и освещении с 29 ноября по 15 февраля. В этом случае сравнивались
лишь неоновые и натриевые лампы при интенсивности видимой радиации
3200 эрг/см2-сек. Высота готовой рассады равнялась в контроле 27 см,
под натриевыми лампами 29 см, под неоновыми лампами 36 см. Цветение
сначала наступило у растений, выращиваемых под неоновыми лампами,
затем под натриевыми и в последнюю очередь — в контроле. Общий
урожай плодов на 21 июня во всех вариантах опыта был одинаков, но
плодоношение началось раньше у растений, культивируемых под неоно-
выми лампами.
В опытах с земляникой (сорт Дэйч Эверн) сравнивались лампы нака-
ливания, ртутные лампы низкого и высокого давления, натриевые и нео-
новые лампы при интенсивности видимой радиации порядка 150—170;
180—210 и 2600—3100 эрг/см2-сек. Для получения низких интенсивностей
применяли ртутные лампы низкого давления (60 вт), неоновые лампы
с дуговым разрядом (25 и 75 вт) и лампы накаливания мощностью 25 и
28 вт, помещаемые на высоте 60—100 см. Опыт показал, что при культуре
земляники зимою низкие интенсивности дополнительного освещения не-
достаточны для нормального произрастания растений. Растения цветут
и частично плодоносят во всех вариантах опыта, за исключением контроля,
но дают очень низкий урожай. Культура земляники возможна лишь
при интенсивности видимой радиации около 3000 эрг/см2-сек, получае-
мой при неоновых лампах интенсивного горения (100 вт), ртутных лампах
высокого давления (150 вт) и натриевых лампах (100 вт). Освещение
длилось с 5 октября по 9 февраля, и было установлено, что по интенсив-
ности цветения и количеству плодов лучший эффект дают натриевые,
а по качеству урожая — неоновые лампы. Плодоношение длилось с фев-
раля до середины марта, причем с одного растения был получен следующий
урожай: в контроле — 0,41 г, под неоновыми лампами — 20,9 г, под
натриевыми лампами 24,2 г, а под ртутными лампами — 18,2 г.
Рейнгольд 1707] в результате своих проведенных в Германии опытов
по выращиванию рассады огурцов (Cucumis sativus), томатов (Solanum
lycopersicum) и кольраби (Brassica caulorapa) и культуре редиса (Raphanus
sativa var. radicula) и салата (Lactuca sativa) при помощи ламп накали-
вания (700 вт) и неоновых ламп с дуговым разрядом (225 вт) пришел
к заключению, что наиболее пригодным источником радиации следует
признать неоновые лампы. В его опытах при высоте подвеса ламп 50—80 см
создавалась освещенность 500 лк (3000 эрг/см2-сек под лампами накали-
вания и 3670 эрг/см2-сек физиологической радиации под неоновыми
лампами). Лампы включались ночью на 6 часов (с 21 часа 30 мин. до
3 час. 30 мин.). В одном опыте томаты сорта Спот резистинг были высеяны
15 ноября и распикированы 20 ноября. Освещение производилось с 4 де-
кабря по 12 января. По окончании освещения растения были высажены
в грунтовую теплицу (табл. 86)
VII. ыращивание при дополнительном искусственном освещении
227
Таблица 86
Состояние рассады и урожай томатов при дополнительном освещении рассады
(Рейнгольд [707])
Варианты опыта Состояние рассады на 21/XII Урожай с одного куста Вес одного плода
высота (в см) ширина листа (в см) ЧИСЛО плодов вес плодов Г %
ЭК8. % кг %
Контроль 30 4 20,6 100 8,15 100 395 100
Неоновые лампы . . 60 7 24,0 116,5 9,70 119,3 405 102,3
Лампы накаливания 40 4 23,3 113,3 9,40 115,5 403 101,8
Аналогичные результаты были получены с сортом Туксвуд и с огур-
цами. Опыты с выгонкой рассады кольраби дали отрицательный результат.
В опытах с культурой редиса при посеве 13 января урожай корнеплодов
был получен 5 марта, причем применение ламп накаливания увеличило
урожай на 204,9%, а неоновых ламп — на 222,2%. Последние дали лучший
эффект и при культуре салата (13 января —• 28 февраля).
В опытах Смита [772, 773] с томатами {Solanum lycopersicum) и крас-
ным клевером {Trifolium rubens) лучший эффект в условиях Норвегии
также дали неоновые лампы. Если при одинаковой удельной мощности
относительную эффективность натриевых ламп считать за единицу, то
под лампами накаливания она будет равна 0,7, а под неоновыми лампами —
2,1. Величины того же порядка в опытах с крессом были получены
несколько ранее Рейнау [705].
Линденбейн [603] сопоставил лампы накаливания (250 и 500 вт), нео-
новые трубки (6 а, 380 в) и солнечные лампы при дополнительном осве-
щении томатов {Solanum lycopersicum) в осенний и весенний периоды
и бобов {Phaseolus sp.) — в зимние месяцы. Растения освещали по 6 часов
(с 3 до 9 час.) при температуре воздуха 20°. Опыт показал, что дополни-
тельное освещение ускорило цветение бобов, независимо от типа источ-
ника света, на 12—16 дней. Контрольные растения зацвели через 46,
а опытные — через 31—35 дней после начала освещения. Мощные и при-
земистые растения томатов получались под неоновыми и солнечными
лампами. Мощные, но чрезмерно вытянутые растения наблюдались и
под лампами накаливания в 500 вт, а под лампами в 250 вт растения были
слабые, с явными признаками этиоляции.
Лори и Пёш [596, 674] в опытах, проведенных в штате Огайо (США)
с цветочными культурами, сопоставили лампы накаливания, неоновые
и ртутные лампы мощностью в 450 вт, висевшие в центре стеллажа
на высоте 90 см и включаемые в вечерние часы, с 17 час. 30 мин. до
21 час. 30 мин. (на 4 часа). Эти авторы нашли, что для зимней культуры
цветочных растений (с 19 декабря по 24 апреля) наиболее пригодным
источником дополнительной радиации являются лампы накаливания,
при которых можно получить наиболее быстро максимальное количество
цветков. Ртутные лампы в опытах названных авторов давали наихудший
эффект (табл. 87).
В опытах Шаминад и Буше [430] при дополнительном освещении
в зимние месяцы (Франция) растений кальцеолярии {Calceolaria sp.) лампы
накаливания и неоновые лампы дали одинаковый эффект — ускорение
наступления бутонизации и цветения на 40 дней.
15*
228
Общая светокультура растений
Эффективность различных источников радиации,
(Лори и
Растения Начало цветения
К л. н. Ne Hg
Кальцеолярия {Calceolaria hybrida var. multi- jlora папа) 14/IV 7/Ш 25/Ш зо/ш
Анютины глазки {Viola tricolor) — — — —
Бархатцы {Tagetes patula) — — — —
Дидискус {Didiscus coerulea) — — — —
Лептозине (Leptosyne maritima) — — — —
Хризантемы {Chrysanthemum segetum) Ивнинг стар — — —- —
Морнинг стар — — — —
Норт стар — — — —
Хризантемы (С. Burridgeanum) — — — —
Эклипс — — — —
К — контроль, л. н.— лампы накаливания, Ne — неоновые лампы, Hg — ртутные
лампы.
Обширное и весьма интересное исследование провел Оберг [363],
который в условиях зимы в Швеции сопоставил эффективность дополни-
тельного освещения, получаемого посредством различных ламп. Этот
автор исследовал эффективность ламп накаливания в 500 вт, натриевых
ламп «SO-IOOO» (265 вт), неоновых ламп интенсивного горения (475 вт) и
ртутных ламп высокого давления «НО-ЮОО» (265 вт), включаемых на
16 часов (с 8 до 24 час.) и дававших интенсивность видимой радиации по-
рядка 11 тыс. эрг/см2-сек. При посеве томатов {Solanum lycopersicum)
26 сентября и дополнительном освещении с 31 октября контрольные расте-
ния вскоре погибли, а среди опытных лучшими оказались растения, куль-
тивируемые под натриевыми и неоновыми лампами, которые отличались
максимальной приземистостью и максимальным сырым весом. Состоя-
ние растений к концу опыта (25 января) характеризуется табл. 88.
Таблица 88
Характеристика растений томатов (Solanum lycopersicum) при выращивании
в зимний период в оранжерее посредством дополнительного освещения
(Оберг [363])
Варианты опыта Сырой вес (в г) Высота стебля (в см) Диаметр стебля (в мм) Междоузлия
корней стебля листьев общий ЧИСЛО длина (в см)
Лампы накаливания 1,1 37,0 35,5 73,5 141 5,2 22 6,4
Ртутные лампы . . 0,9 27,1 26,8 54,7 108 5,3 20 5,4
Натриевые лампы . 1,6 35,7 51,2 88,6 82 7Д 16 5,1
Неоновые лампы . . 1,8 34,9 46,2 83,0 80 7,1 17 4,7
Контроль Р а стен ИЯ по гиб л и
Другие опыты этого автора показали, что наиболее ранний и макси-
мальный урожай получают при освещении рассады неоновыми лампами.
VII. Выращивание при дополнительном искусственном освещении
229
Таблица 87
применяемых для дополнительного освещения
Пёш [596])
Первая срезка готовых цветков Число цветков Высота растения (в см)
К Л. я. Ne Hg К Л. я. Ne Hg К Л. я. Ne Hg
6/III 13/Ц 13/Ц 13/Ц 44 192 126 92 10,9 11,9 8,6 П,2
13/11 13/Ц 25/Ц 13/Ц 67 122 89 118 16,2 23,4 17,8 17,2
5/IV 6/Ш 18/Ш 18/Ш 78 164 147 116 29,6 24,4 24,4 20,3
28/II 19/11 19/Ц 19/Ц 71 86 41 22 32,4 24,4 19,8 21,4
1/IV 7/II 7/П 18/Ш 46 102 91 73 26,6 42,4 34,8 38,1
5/IV 7/П 9/11 18/Ш 51 115 122 57 27,2 34,0 32,5 33,0
11/IV зо/ш зо/ш 5/1V 36 71 58 31 51,0 57,2 48,0 50,3
11/IV ЗО/Ш зо/ш 5/IV 27 46 33 35 51,0 45,0 44,5 45,2
15/IV зо/ш зо/ш 5/IV 25 39 34 34 35,6 54,6 52,0 37,6
Так, при посеве томатов 14 октября и освещении в рассадный период
(31 октября — 27 января) по 14 часов в сутки (с 8 до 22 час.) плодоноше-
ние в контроле началось 6 июня, в варианте опыта с натриевыми лампами —
23 мая, а с неоновыми лампами — 4 мая, причем суммарный урожай
с куста (к 17 июля) соответственно равнялся 2,83; 4,23 и 6,35 кг.
Исследования Уитроу [855] в штате Индиана (США) показали, что
при дополнительном освещении в течение февраля—марта (с 18 час.
30 мин. до 0 час. 30 мин.) при интенсивности видимой радиации порядка
12 тыс. эрг/см2-сек эффективность различных ламп зависит от вида расте-
ний (табл. 89).
Таблиц а 89
Влияние дополнительного освещения ночью на рост растений
(Уитроу А. П. и Уитроу Р. Б. [855])
Варианты опыта
Томаты (Sola-
num lycoper-
sicum)
Индиана
Болтимор
(опыт — 30 дней)
Соя (Soja max)
Билокси
(опыт — 30 дней)
Шпинат (Spi-
nacea oleracea)
Нобель
(опыт — 70 дней)
Астры (Calli-
stephus chi-
nensis)
Гарт оф Франс
(опыт - 70 дней)
Лампы накаливания
500 вт................. 2260
Люминесцентные лампы
дневного света 480 вт
(16 ламп по 30 вт) .
Ртутные лампы высо-
кого давления «Н-1»
400 вт .............
Контроль (лампы нака-
ливания 10 вт) . . .
3890
3890
11
6,5 0,44 6,0 8,2 1,09 8,2 18,7 1,39 5,2 74,9 8,6 46,9
6,4 0,45 5,4 6,9 0,85 6,9 27,9 1,78 52,3 49,2 4,8 21,4
7,2 0,47 3,7 7,6 0,96 7,6 8,6 0,62 14,7 38,8 3,8 18,0
2,1 0,16 2,5 4,7 0,53 4,7 — — — — — —
230
Общая светокультура растений
Астры (Callistephus chinensis) и соя (Soja max) лучше всего разви-
ваются под лампами накаливания, шпинат (Spinacea oleracea) — под
люминесцентными, а томаты (Solanum lycopersicum) —• под ртутными
лампами.
Аналогичные выводы в свое время были получены Оденом [646,650],
который, пользуясь лампами накаливания, установил, что развитие го-
роха (Pisum sativum), малины (Rubus idaeus) и фасоли (Phaseolus sp.)
ускоряется длинноволновыми лучами (при недокале ламп), а редис, укроп
и салат предпочитают коротковолновые лучи (перекал ламп).
Гелин и Бюрстрём [509] исследовали реакцию растений овса (Avena
sativa), салата (I actuca sativa) и гороха (Pisum sativum) на дополнитель-
ное освещение (с 6 до 8 час. и с 16 до 22 час.), получаемое с помощью на-
триевых ламп «SO-IOOO» (140 вт), ртутных ламп «НО-250» (265 вт),
люминесцентных ламп белого света «ТБ» (40 вт) и солнечных ламп «МБ-500»
(250 вт). Каждая лампа освещала площадь в 1,44 м‘- и находилась на
высоте 50 см от почвы. Люминесцентные лампы были вмонтированы
в специальный рефлектор (по 5 штук). Посев был произведен 17 ноября,
освещение давалось с 20 ноября по 12 февраля.
Т аб лиц а 90
Влияние дополнительного освещения на рост, растений
(Гелин и Бюрстрём [509])
Варианты опыта Удельная мощность (вт/м2) Горох (Pisum sativum) Овес (Avena saliva) Салат (Lac- tuca saliva)
I сырой вес одного растения (в г) сухой вес одного растения (в мг) высота (в см) сырой вес одного растения (в г) сухой вес одного растения (в мг) высота (в см) сырой вес одного растения (в г) сухой вес одного j растения (в г)
ЛИСТЬЯ _ 1 стебель листья стебель
Контроль -— 1,77 71 79 55 0,65 36 18 30 1,76 84
Натриевые лампы .... 100 3,12 126 147 83 1,11 57 31 42 8,24 377
Ртутные лампы 185 2,74 102 130 80 1,02 51 29 40 7,51 325
Люминесцентные лампы . 140 3,56 182 179 86 1,26 68 40 44 12,64 537
Солнечные лампы .... 175 3,84 180 247 100 1,72 81 63 49 12,72 522
Как видно из табл. 90, лучший результат во всех случаях дают солнеч-
ные и люминесцентные лампы. Неплохой эффект получается и от натрие-
вых ламп, хотя удельная мощность в этом случае значительно ниже,
чем в других вариантах опыта. Различий во времени наступления цвете-
ния при дополнительном освещении, получаемом посредством перечислен-
ных ламп, не наблюдалось.
В 1948 г. нами совместно с В. В. Адоратским в оранжереях совхоза
«Марфино» (Москва) был поставлен опыт по выращиванию рассады тома-
тов (Solanum -lycopersicum.) Лучший из всех и огурцов (Cucumis sativus)
Клинские посредством дополнительного освещения люминесцентными
лампами и лампами накаливания [123]. Посев томатов производился
1 января, посев огурцов — 23 января. Растения днем находились на
естественном свете, который, в зависимости от погоды, колебался в по-
луденные часы в пределах 1000—9000 лк, а в ночное время в течение
VII. Выращивание при дополнительном искусственном освещении
231
9 часов (с 22 до 7 час.) освещались лампами нахваливания или люминес-
центными лампами «БС-15». Освещение продолжалось в течение 35 дней—
с 22 января по 26 февраля.
Опыт ставился по следующей схеме:
1) лампы накаливания 450 вт/м2;
2) лампы накаливания 150 вт/м2;
3) люминесцентные лампы белого света 450 вт/м2;
4) люминесцентные лампы белого света 150 вт/м2;
5) контроль (без дополнительного освещения).
Лампы накаливания мощностью в 300 вт помещались в софиты из белой
жести, служившие отражателями. Лампы двигались вдоль стеллажа по-
средством установки Финкельштейна в пределах 1 м. Люминесцентные
лампы (30 вт) располагались в виде горизонтальной решетки над расте-
ниями на высоте 10 см в количестве 15, или 5 трубок на 1 м. В варианте
с удельной мощностью 450 вт/м2 лампы были неподвижными, а в варианте
150 вт/м2 двигались вдоль стеллажа в пределах 12,5 см. Во избежание
затенения в дневные часы люминесцентные лампы снимали.
Таблица 91
Характеристика готовой рассады томатов (Solanum lycopersicum)
при дополнительном освещении
Варианты опыта
Контроль
Лампы
300 вт
Люминесцентные лампы
«БС-30».............
накаливания
Удельная мощность (в вт/м2) Освещенность (в лк) Интенсивность физиологической радиации (в эрг/см2-сек) Сырой вес одного растения (в г) Сухой вес одного растения (в г) Число листьев одного растения Площадь одного листа (в см2) Общая листовая поверхность (в см2) Высота растений (в см)
О (Х> &< О Е стебля корней общий средняя 1 макси- мальная
— 200* — 12,70 12,70 2,07 27,47 1,69 5,2 29,6 47,6 153,7 19,1
450 4600 28 000 17,91 16,90 2,71 37,51 2,65 6,3 32,7 49,8 200,6 28,4
150 1600 10 000 12,75 16,53 2,71 31,99 2,20 6,0 30,2 44,8 181,2 27,6
450 5000 18 000 40,25 22,78 6,49 69,52 5,17 6,7 56,4 91,1 377,4 25,1
150 2000 7 000 17,49)13,99 5,22^36,70 2,57 6,6 31,4'55,11207,1 1 1 17,3
засвечивания от соседних вариантов.
* Освещенность в результате
Табл и ц а 92
Характеристика готовой рассады огурцов (Cucumis sativus)
при дополнительном освещении
Варианты опыта Удельная мощность (в вт) Сухой вес одного растения (в г) Площадь листьев (в см2)
1-го 2-го общая
Контроль — 0,83 27,2 20,1 47,3
Лампы накаливания 300 вт . . 450 1,98 50,0 34,2 84,2
« « « 150 1,36 36,8 14,1 50,9
Люминесцентные лампы «БС-30» 450 2,60 57,4 42,4 99,8
« « « 150 1,66 56,3 35,3 91,6 -
232
Общая светокультура растений
В каждом варианте освещали площадь стеллажа в 2 м2, на которой
размещали 270 растений огурцов и 280 растений томатов. Каждый вариант
отделялся от другого защитным пространством длиною 2 м. Температура
была одинаковой во всех вариантах опыта и колебалась в пределах
20—28°.
По окончании дополнительного освещения растения были высажены
на постоянные места (27 февраля): огурцы — в ангарную, а томаты —
в грунтовую теплицы, где за каждым вариантом опыта было продолжено
наблюдение до конца плодоношения (2 августа). В каждом варианте было
Фиг. 140. Влияние дополнительного освещения на рассаду томатов
(Solanum lycopersicum') Лучший из всех
1—люминесцентными лампами «БС-15», 450 вт'м’; 2—люминесцентными лампами «БС-15», 15О ’вт/м:;
3 — лампами накаливания, 450 вт/м-; 4 — лампами накаливания, 150 вт/м2; 5 — контроль
по 140 растений томатов и 124 растения огурцов. В контроле количество
растений было соответственно равно 280 и 248.
Как видно из табл. 91 и 92 и фиг. 140, лучший результат дают люми-
несцентные лампы при удельной мощности 450 вт/м2. В этом случае
формируются наиболее мощные растения с хорошо развитой листовой
поверхностью и корневой системой. Дополнительное освещение не ска-
залось на сроке появления женских цветков у огурцов (24—26 марта),
но ускорило бутонизацию и цветение у томатов. Наибольшее ускорение
(на 10 дней) наблюдалось в варианте с люминесцентными лампами при
удельной мощности 450 вт/м2. В последнем случае действие дополнитель-
ного освещения, постепенно ослабевая, проявлялось вплоть до 8-й кисти
(табл. 93).
Дополнительное освещение не повлияло на время начала плодоноше-
ния (5 апреля) и урожай огурцов (32,2—35,5 плода на один куст за период
с 5 апреля по 31 июля) и вес плодов у томатов, который равнялся в сред-
нем 50 г, постепенно снижаясь, по мере плодоношения, от 66,6 до 18 г, но
ускорило плодоношение и увеличило урожай томатов (табл. 94). В среднем
за сезон (15 мая—2 августа) увеличение урожайности составило 33 % под
VII. Выращивание при дополнительном искусственном освещении
233-
Т а б л иц а 93
Динамика бутонизации и цветения томатов (Solanum lycopersicum)
при дополнительном освещении рассады
Варианты опыта Удельная мощность (В ВТ/М") Дата бутонизации кистей (снизу вверх)
1-й 2-Й Зй 4-й 5-й 6-Й 7-й 8-й
Контроль — 22/111 1/IV 8/IV 11/IV 18/IV 24/IV 29/IV 3/V
Лампы накаливания , 450 22/Ш 28/III 3/IV 11/IV 17/IV 22/IV 27/IV 3/V
150 23/1П 28/111 13/IV 15/IV 17/IV 23/IV 29/IV 5/V
Люминесцентные лам- ( пы «БС-30» ....-: 450 И/III 20/Ш 28/III 6/IV 13/IV 19/IV 23/IV 30/IV
150 19/UI 26/111 5/IV 13/IV 19/IV 21/IV 26/IV 30/IV
Дата цветения кистей (снизу вверх)
1-й 2-й 3-й 4-й 5-й 6-Й 7-Й 8-й
Контроль — 8/IV 14/IV 19/IV 21/IV 29/IV 5/V 5/V —
Лампы накаливания , 300 вт < 450 2/IV 11/IV 17/IV 22/IV 27/IV 2/V 2/V —
150 5/1V 11/IV 17/IV 22/IV 27/IV 2/V 2/V —
Люминесцентные лам- , пы «БС-30» .... 1 450 22/Ш 1/IV 11/IV 17/IV 23/IV 30/IV 5/V —
150 28/III 10/IV 16/IV 21/IV 26/IV 2/V 2/V —
люминесцентными лампами при 450 вт/м2 и 12—17% — в других вариан-
тах.
Таким образом, в нашем исследовании лучшим источником радиации
для дополнительного освещения рассады томатов и огурцов оказались
люминесцентные лампы как в отношении качества рассады, так и времени
плодоношения и урожая плодов.
Зимою 1950 г. в теплицах совхоза «Краснове» (Московской обл.) нами
было предпринято исследование по сравнению эффективности ламп на-
каливания в 300 вт, зеркальных ламп в 500 вт, неоновых ламп интенсив-
ного горения «НД-1» в 500 вт и ртутных ламп высокого давления
«Игар-2» в 500 вт [123]. Лампы помещали в корытообразные софиты
из белой жести, которые, будучи на высоте 30—40 см над растениями,
двигались вдоль стеллажа посредством системы И. Н. Финкельштейна
в пределах 2 м. Удельная мощность была одинаковой (265—290 вт/м2),
за исключением зеркальных ламп, где по техническим причинам она не
могла быть меньше 430 вт/м2. Лампы включались в ночные часы на 8 часов
(с 18 до 2 час.) с 30 января по 14 февраля.
Подопытными объектами служили огурцы {Cucumis sativus) Клинские
и Неросимые. Первые были посеяны 9 января, а вторые— 25 января.
По окончании освещения растения были высажены в грунт на постоян-
ные места в грунтовые теплицы, где за ними были обеспечены соответствую-
щий уход и наблюдения. Растения находились в рядках на расстоянии
35 см в рядке и 50 см между рядками. В каждом варианте опыта было по
три повторности, а в контроле — пять повторностей. В каждой повтор-
ности имелось шесть рядков по 10—11 растений (63 растения в по-
вторности).
Как видно из табл. 95 и фиг. 141, дополнительное освещение во всех
случаях дало значительный положительный эффект, увеличивая вес и
234
Общая светокультура растений
Таблица 94
Динамика плодоношения томатов (Solanum lycopersicum) при дополнительном
освещении рассады
Варианты опыта Удельная мощ- ность (в вт/м2) Число плодов с одного куста
15—31/V 1-15 VI 16-30/VI 1- 15/VII 16/VII—2/VIII Всего (15/V ЭКЗ. за сезон -2/VIII) % к конт- ролю
Контроль Лампы накаливания 300 вт { Люминесцентные лампы , «БС-ЗО» j 450 150 450 150 1 ,80 1,56 1,45 2,30 2,14 3,75 3,44 3,50 4,65 4,00 6,27 7,10 8,50 6,74 7,74 4,84 7,70 6,02 6,60 6,02 11,20 15,15 16,15 27,89 34,93 36,44 100 125 131
- Урожай плодов с одного куста (в кг)
15—31/V 1—15/VI 1 16-30/VI 1—15/VII IIIA/&-IIA/9I Всего (15, V КГ за сезон -2/VIII) в % к конт- ролю
Контроль Лампы накаливания 300 вт { Люминесцентные лампы Г «БС-ЗО» 1 450 150 450 150 0,122 0,103 0,096 0,153 0,143 0,230 0,222 0,218 0,281 0,243 0,350 0,405 0,420 0,370 0,447 0,262 0,316 0,286 0,301 0,301 0,199 0,300 0,229 0,235 0,230 1,161 1,346 1,249 1,540 1,364 100 116 112 133 117
Таблица 95
огурцов (Cucumis sativus) при дополнительном
освещении
Характеристика рассады
Варианты опыта Удельная мощ- ность (в вт/м2) Высота расте- ний (в см) Сухой вес одного растения (в мг) Число листьев Площадь листьев (в см2)
листьев стеблей и корней общий 1-я семя- доля 2-я семя- доля 1-й лист 2-й лист общая
Сорт Клинскис Контроль 8,9 33 11 44 2,0 6,1 6,6 14,7 2,5 29,9
Лампы накаливания . . . 265 10,5 75 37 112 2,0 6,3 7,1 26,5 11,6 50,5
Зеркальные лампы .... 430 10,8 75 25 100 2,0 5,7 7,6 24,0 10,9 48,2
Ртутные лампы «Игар-2» . 290 6,9 46 10 56 2,0 7,3 7,8 21,8 6,0 42,9
Неоновые лампы «НД-1» . 280 5,7 56 12 68 2,0 6,5 8,6 23,0 3,0 41,1
Сорт Неросимые Контроль — 11,0 24 16 40 1,0 4,8 5,8 6,7 0,0 17,3
Лампы накаливания . . . 265 11,2 40 18 58 1,7 5,4 6,0 10,5 0,7 22,6
Зеркальные лампы .... 430 9,2 42 18 60 2,0 4,2 5,8 13,1 1,2 24,3
Ртутные лампы «Игар-2» . 290 6,6 58 22 80 1,7 8,1 10,0 13,8 0,6 32,5
Неоновые лампы «НД-1» . 280 7,5 35 И 46 1,2 6,3 7,4 8,3 0,1 22,1
VII. Выращивание при дополнительном искусственном освещении
235
листовую поверхность растений. Наиболее эффективными для клинских
огурцов (по накоплению сухой массы) оказались лампы накаливания и
зеркальные лампы, а для огурцов Неросимые — ртутные лампы. Нормаль-
ная, без всяких следов вытягивания, рассада обоих сортов образуется
под ртутными лампами. Под лампами накаливания, как и под зеркаль-
ными лампами, рассада сильно вытягивается.
Фиг. 141. Влияние дополнительного освещения на рассаду огурцов
(Cucumis sativus) Клинские
1 — обычными лампами накаливания; 2 — зеркальными лампами накаливания;
3 — неоновыми лампами «НД-1»; 4 — ртутными лампами «Игар-2»; 5 — контроль
В табл. 96 показана динамика цветения растений.
Из табл. 96 следует, что дополнительное освещение во всех вариантах
опыта увеличивает количество как мужских, так и женских цветков, сме-
щая их соотношение в сторону последних. Максимальная эффективность
в смысле увеличения количества цветков принадлежит ртутным и зеркаль-
ным лампам. Появление завязей и женских цветков наступает раньше
всего также под ртутными лампами.
В табл. 97 приведена динамика плодоношения растений. Дополнитель-
ное освещение от ламп накаливания, зеркальных и ртутных ламп повы-
шает урожай на 40—50%. Лучший эффект дают ртутные лампы. Неоновые
лампы в наших опытах дали незначительный эффект в связи с ненормаль-
ным режимом горения: в результате падения напряжения лампы часто
выключались и вообще работали не на полную мощность. Различия между
вариантами, особенно сильные в первые две недели, постепенно сглажи-
ваются, однако для ламп накаливания, зеркальных и ртутных ламп эффект
сохраняется на протяжении всего сезона.
Зимою 1950/51 г. нами совместно с С. И. Китаевым была сделана
попытка зимней культуры томатов {Solanum lycopersicum) и огурцов
{Cucumis sativus) [123]. Опыт проводился в теплицах совхоза «Марфино»
при дополнительном освещении с 31 ноября по 25 января обычными
Та блица 96
Динамика цветения огурцов (Cucumis sativus) Неросимые при дополънителъном освещении рассады
Число структурных элементов на 100 растений на
Число Общее число' 1
Варианты опыта делянок растений 13/Ш 15/Ш 17/III 19/III 21411 23/III 25/Ш 27/111 29/111 31/Ш общее за сезон (в % к
контролю)
Мужские цветки
Контроль 5 336 8 32 43 118 148 188 130 126 140 159 100
Лампы накаливания 2 107 8 20 28 105 141 245 173 138 180 204 114,8
Зеркальные лампы 3 189 6 24 43 115 208 232 197 178 164 181 125,0
Ртутные лампы «Игар-2» .... 3 189 7 35 65 168 207 236 210 217 244 212 148,1
Неоновые лампы «НД-1» .... 3 ' 189 5 31 48 156 155 262 182 218 248 193 138,8
Завязи
Контроль 5 336 1,5 3,9 7,4 37 53 87 79 130 145 150 100
Лампы накаливания 2 107 3,8 6,5 — 35 45 95 92 100 144 191 102,3
Зеркальные лампы 3 189 4,2 6,3 10,0 81 106 137 107 141 234 146 140,0
Ртутные лампы «Игар-2» .... 3 189 7,4 11,6 10,5 68 93 146 136 185 227 238 161,5
Неоновые лампы «НД-1» .... 3 189 4,2 5,3 8,5 41 53 86 127 180 170 149 118,6
Женские цветки
Контроль 5 336 0,0 0,0 1,5 12 20 34 85 92 120 129 100
Лампы накаливания ...... 2 107 0,0 0,0 7,5 31 38 38 128 174 240 248 183,0
Зеркальные лампы 3 189 0,0 0,0 10,0 40 67 77 188 204 327 240 233,5
Ртутные лампы «Игар-2» .... D 189 0,5 1,0 10,6 32 65 114 115 144 223 241 191,5
Неоновые лампы «НД-1» .... 3 189 0,0 0,0 0,5 23 42 110 133 152 192 174 166,3
Отношение числа мужских и женских цветков
Контроль 5 336 СО оо 28,6 9,9 7,1 5,5 1,5 1,4 1,1 1,2 —
Лампы накаливания 2 107 со 00 3,7 3,4 3,7 6,5 1,3 0,8 о,7 0,8 —
Зеркальные лампы 3 189 со со 4,9 2,8 3,1 3,0 1,0 0,9 0,5 °,7 —
Ртутные лампы «Игар-2» .... 3 189 14,0 7,0 6,1 5,3 3,2 2,1 1,8 1,5 1,1 0,9 —
Неоновые лампы «НД-1» .... 3 189 00 ОО 96,0 6,8 3,7 2,4 1,3 1,4 1,3 1,1 —
236 Общая светокультура растений
Таблица 97
Динамика плодоношения огурцов (Cucumis Sativus) ПеросимЫе при дополнительном освещении рассады
Варианты опыта Число плодов с 10 растений за период
29—31/III 1-15/IV 16-30/IV 1-15,-V 16-31/V 1—15/VI 16-30/VI Сезон (29/III—30/VI)
экз. % К кон- тролю ЭКЗ. % к кон- тролю экз. % к кон- тролю экз. % К кон- тролю ЭКЗ. % К кон- тролю ЭКЗ. % К кон- тролю ЭКЗ. % К кон- тролю ЭКЗ. % к кон- тролю
Контроль 0,00 — 11,2 100,0 27,2 100,0 17,5 100,0 19,2 100,0 12,1 100,0 1,65 100,0 89,0 100,0
Лампы накаливания . . 0,20 — 15,9 142,0 36,8 135,0 22,9 130,5 26,7 139,1 19,5 161,0 2,23 135,0 124,3 140,0
Зеркальные лампы . . . 0,95 — 17,2 154,0 39,2 144,0 27,2 155,0 24,4 127,1 15,7 129,1 2,21 134,0 127,0 143,0
Ртутные лампы «Игар-2» 1,22 — 18,9 169,0 39,1 143,5 26,1 149,0 26,1 136,0 17,2 142,0 2,38 144,1 131,1 147,5
Неоновые лампы «НД-1» 0,42 — 9,4 84,0 29,2 107,0 18,7 106,5 24,3 126,5 16,4 135,0 2,27 137,5 100,8 113,0
Плодоношение огурцов (Cucumis sativus) Клинские при дополнительном освещении рассады
Таблица 98
Варианты опыта Удельная мощность (в квт/м) Пло- щадь (в м2) Число расте- ний Дата плодоношения Количество плодов с 10 растений Вес плодов с 10 растений Товарная продукция с 10 растений Вес одного плода (в г) Расход элек- троэнергии (в квт-час/кг)
ЭКЗ. % к кон- тролю нг % к кон- тролю КГ % к кон- тролю
Контроль — 3 27 9—23/XI 9,3 100,0 0,59 100,0 0,44 100,0 47,5 —
Лампы накаливания . . 0,9 4 33 9/XI—1/ХЦ 13,4 144,0 0,87 147,5 0,61 138,5 45,6 270
1,5 4 35 9/XI—13/1 15,6 168,0 0,89 151,0 0,74 168,0 47,4 365
Зеркальные лампы . . 1,0 2 15 9—23/XI 16,2 174,0 1,12 190,0 0,95 215,5 58,5 172
I,5 2 13 9/XI—13/1 24,6 264,1 1,31 222,0 1,26 286,0 51,4 203
Ртутные лампы «Игар-2» 1,0 2 17 9—23/XI 17,9 192,1 1,73 293,0 0,99 225,2 55,2 152
Неоновые лампы «НД-1» 1,0 1 10 9—23/XI 20,0 215,0 1,27 215,5 0,97 221,0 48,5 173
VII. Выращивание при дополнительном искусственном освещении 237
238
Общая светокультура растений
Н
Состояние рассады и урожай огурцов (Cucumis sativus) Клинские при дополнительном освещении в рассадный период
(Протасова [253, 254])
Число плодов за весь вегетационный период (с 15/III по 30/VIII) % к кон- тролю 100 97,0 111,0 120,5 140,9 124,0
с одного растения 28,0 27,1 31,0 36,0 39,3 34,6
с 1 м- 167,5 162,5 185,6 201,6 235,6 207,5 1
за первые полтора месяца (с 15/III по 30JV) . 2 с одного % к ион- с 1 м растения тролю 8,2 100 9,2 111,8 9,4 114,0 10,7 130,0 11,6 141,0 9,7 125,0
49,3 55,0 56,2 64,0 69,5 61,7
Состояние рассады на 12/11 1 вес одного растения сырой I сухой (в г) (в мг) 1 1 140 265 505 675 495 560
2,42 4,26 6,92 8,70 7,04 7,32
общая площадь листьев (в см ) 40,7 132,1 203,4 233,0 181,0 190,5
высота растений (в см) 12 20 22 27 14 15
(sraa • rno/jcie я) ЙиПеиивй Иояээыыоиоиеиф чюопаиэнэхни 17 000 41 000 52 000 40 000 51 000
(.w/xa я) чхэон -ШОК ВВНЧИЭ1ГЛ 225 450 750 500 475
Источник радиации Контроль ( Лампы накаливания .< 1 Ртутные лампы «Игар-2» Неоновые лампы «НД-1»
лампами накаливания в 300 и в
500 вт, зеркальными лампами на-
каливания в 500 вт, ртутными
лампами «Игар-2» (500 вт) и
неоновыми «НД-1» (500 вт). Кон-
трольные растения находились в:
условиях естественного освещения.
В качестве объектов служили огур-
цы Клинские и томаты Лучший
из всех. Опыт с томатами показал,
что культура этих растений зимою
возможна только при дополни-
тельном освещении, но одного лишь
ночного освещения для нормаль-
ного развития недостаточно. При
7-часовом дополнительном освеще-
нии (с 2 до 9 час.) и удельной
мощности в 1000—1500 вт/м2 уро-
жай плодов колебался в преде-
лах от 150 до 930 г с 10 кустов.
Опыт с огурцами показал,
что дополнительное освещение уве-
личивает урожай плодов на 40—
18 % , причем лучший эффект дают
ртутные, неоновые и зеркальные
лампы, повышающие урожай на
115—185% (табл. 98).
К сожалению, в наших опытах,
вследствие нарушений агротехни-
ки, у растений перестали завязы-
ваться плоды, и опыт не удалось
довести до конца.
В дальнейшем наши исследова-
ния по дополнительному освеще-
нию рассады томатов и огурцов
лампами разных типов посред-
ством подвижной установки
И. II. Финкельштейна были продол-
жены Н. Н. Протасовой [253, 254],
которая на протяжении 1950—
1952 гг. провела в этом направле-
нии серию опытов. В 1951 г. в сов-
хозе «Марфино» освещение расте-
ний производилось по 8 часов в
сутки (с 22 до6 час.). Огурцы(Cucu-
mis sativus) Клинские, посеянные
13 января, освещались с 16 января
по 13 февраля, а томаты (Solanum
lycopersicum) Маяк (посев 13 фев-
раля)— с 15 февраля по 15 марта
(табл. 99 и 100).
Опыты, поставленные в 1951—
1952 гг. в совхозе «Тепличный»,
были проведены по более широкой
VII. Выращивание при дополнительном искусственном освещении
239'
Таблица 100
Урожай томатов (Solanum lycopersicum) Маяк при дополнительном
освещении в рассадный период
(Протасова [253, 254])
Источники радиации
Контроль ..................
Лампы накаливания .........
Ртутные лампы «Игар-2» . . .
Неоновые лампы «НД-1» . . . .
Ртутные лампы «Игар-2» (500
вт/м!) неоновые лампы
«НД-1» (475 вт/м2).........
Ртутные лампы «Игар-2» (500
вт/м2)лампы накаливания
(300 вт/м2) ...............
Удельная мощ- ность (в вт/м2) Интенсивность физиологической радиации (в эрг/см2 сек) Урожай плодов
за первые полтора месяца (со 2/VI по 15/VII) за весь вегетационный период (со 2/VI по 14/IX)
с 1 м2 (в кг) с 1 ра- стения (в кг) В % К кон- тролю с 1 м! (в кг) с 1 ра- стения (в кг) В % к кон- тролю
— —. 5,70 0,95 100 13,50 2,25 100
450 41 000 6,06 1,01 106,0 14,30 2,38 106,0
500 40 000 6,72 1,12 117,8 15,00 2,50 111,0
475 51000 7,44 1,24 130,5 17,04 2,84 126,0
975 52 600 9,96 1,66 175,0 22,08 3,68 167,0
800 52 000 8,32 1,38 146,0 19,20 3,20 142,0
Таблица 101
Состояние рассады и урожай огурцов (Cucumis sativus) Влинские
при дополнительном освещении в рассадный период
(Протасова [253, 254])
Источники радиации Удельная мощность (в вт/м2) Интенсивность фи- зиологической ра- диации (в эрг/см2 • сек) Состояние рас- сады на 2/II Урожай плодов
высота ра- стений (в см) общая пло- щадь листьев (в см2) за первые 4о дпей (с 20/III по 1/V) за весь вегетационный период (с 20/III по 1/IX)
с 1 м2 (в кг) с 1 ра- стения (в кг) В % к кон- тролю с 1 м2 (в кг) с 1 ра- стения (в кг) в % к коп- 1 тролю
Контроль — — 14,0 44,0 3,0 0,50 100 9,1 1,52 100
Л ю ми п о сце HTIH.I е лампы синего све- та, 15 вт 200 65 000 9,0 107,0 4,98 0,83 166,0 14,46 2,40 159,0
Л юминесцептные лампы красного света, 15 вт . . . 200 54 100 10,5 104,5 4,56 0,76 152,0 13,56 2,26 149,0
Лампы накаливания 300 64 460 22,0 110,5 3,9 0,65 130,0 11,3 1,89 124,0
Ртутные лампы «Игар-2» 500 66 000 10,0 91,0 4,44 0,74 148,0 17,64 2,94 194,0
Неоновые лампы «НД-1» 475 66 150 11,0 95,0 5,64 0,94 188,0 14,22 2,37 156,5
Зеркальные ламп гл накаливания . . . 500 147 420 29,0 137,5 6,6 1,10 220,0 15,96 2,66 175,0
Ртутные лампы «Игар-2» (500 вт/м'2) + лампы накали- вания (200 вт/м2) . 700 67 000 18,0 126,0 5,82 0,97 194,0 17,82 2,97 195,8-
240
Общая светокультура растений
программе. Огурцы Клинские (посев 25/ХП) освещали в течение месяца
(с 1 января по 1 февраля) по 6 часов в сутки (с 22 до 4 час.). Схему и
результаты опытов мы приводим в табл. 101.
Как видно из табл. 99—101, при одинаковой интенсивности допол-
нительного освещения (порядка 40—70 тыс. эрг/см2. сек физиологической
радиации) лучший результат наблюдается у огурцов при освещении
ртутными, а у томатов — неоновыми лампами. Хороший результат дают
также люминесцентные и зеркальные лампы. Преимущество люминес-
центных ламп заключается в том, что они дают хороший результат уже
при небольшой удельной мощности. Зеркальные лампы позволяют полу-
чать высокие интенсивности дополнительного освещения.
Зимою 1951 г. (январь—март) Леман [160] сопоставил эффективность
люминесцентных ламп «ДС-30» с лампами накаливания при дополнитель-
ном освещении растений в теплицах. В обоих случаях лампы горели по
16 часов в сутки в ночные и дневные часы. В последнем случае они вклю-
чались при падении естественной освещенности ниже 5000 лк. Удельная
мощность равнялась 300—600 вт/м2 для люминесцентных ламп и 1000—
1500 вт/м2 для ламп накаливания. Лампы накаливания помещались
над растениями, а люминесцентные лампы—в виде вертикальных решеток
в междурядьях растений (при расстоянии между рамами 0,5 —1 м;
фиг. 66 и 68). Естественная освещенность колебалась в пределах 1000—
3000 лк, а люминесцентные лампы давали дополнительную освещен-
ность 5000—8000 лк (20 тыс.—32 тыс. эрг/см2-сек физиологической
радиации).
В качестве объектов служили пшеница (Triticum vulgare) Лютесценс 62
и ветвистая Кахетинская (Triticum turgidum), фасоль (Phaseolus sp.)
Латвия 800, земляника (Fragaria sp.) Ремонтантная и Дессертная и одно-
летние сеянцы инжира (Ficus carica), лимона (Citrus limori), хинного де-
рева (Cinchona sp.), дуба (Quercus sp.), березы (Betula sp.), лиственницы
(Larix sibirica), ели (Picea excelsd) и другие растения.
Автор нашел, что зимою в условиях теплиц в средней полосе СССР,
когда растения гибнут от недостатка естественного освещения, при до-
полнительном освещении люминесцентные лампы дают лучший эффект,
чем лампы накаливания. Под лампами накаливания растения значительно
быстрее зацветают и созревают, но отстают по другим показателям. Под
люминесцентными лампами растения более приземисты, отличаются ко-
роткими и толстыми междоузлиями, обладают нормальным циклом раз-
вития, имеют нормальную окраску, размеры и форму листьев, лучше
кустятся и ветвятся и дают лучший урожай. Урожай земляники был на
35% выше под люминесцентными лампами, чем под лампами накаливания.
Абсолютный вес семян пшеницы и фасоли на 20—30% больше, чем у расте-
ний под лампами накаливания, и на 5—35% больше, чем у исходных
семян.
Чтобы покончить с вопросом о применении различных источников
радиации, мы остановимся еще на комбинировании различных типов
ламп. Исследований в этом направлении очень немного.
Роденбург [732], культивируя землянику (Fragaria sp.) в зимние ме-
сяцы, создавал дополнительное освещение с помощью комбинации неоно-
вых ламп (450 лк, или 3000 эрг/см2-сек физиологической радиации) и
ламп накаливания (28 лк, или 175 эрг/см2-сек физиологической радиации).
Растения освещались с 7 октября по 8 часов в сутки. Автор нашел, что наз-
ванная комбинация дает худший результат, чем одна неоновая лампа. При
юборе ягод с 30 января по 13 марта с одного растения был получен в сред-
нем следующий урожай: в контроле — 1,25 г, при комбинации неоновой
VII. Выращивание при дополнительном искусственном освещении
241
лампы с лампами накаливания — 34,2 г, под неоновой лампой — 54,5 г.
Соответствующие цифры для числа ягод, полученных с одного растения,
равны: 0,20; 7,24 и 10,45.
Иные результаты получила Н. Н. Протасова (253, 254), комбинировав-
шая источники сине-фиолетовых и оранжево-красных лучей. Источни-
ками первых служили ртутные лампы «Игар-2», а источниками вторых —
неоновые лампы «НД-1» или лампы накаливания. Как видно из табл. 100
и 101, подобные комбинации дают значительно лучший эффект, чем те же
лампы в отдельности. Особенно хороший результат наблюдается при
комбинировании ртутных и неоновых ламп.
Шаминад и Буше [435] в опытах с дополнительным освещением иссле-
довали значение инфракрасных лучей для развития кальцеолярии (Cal-
ceolaria sp.). Пользуясь лампами накаливания (350 лк), авторы освещали
растения по 8 часов, причем в одном варианте лампы применялись без
водного экрана, а в другом — употреблялся водный фильтр. Устранение
инфракрасных лучей практически не имело значения для цветения расте-
ний, и в обоих случаях оно наступало одновременно — через 104 дня,
а бутонизация при наличии водного экрана запоздала на 7 дней по сравне-
нию с вариантом без этого экрана. Кроме того, при устранении инфракрас-
ных лучей продолжительность цветения увеличилась на 6 дней.
Таким образом, сопоставляя экспериментальные данные по дополни-
тельному освещению растений в ночные часы с целью увеличения длины
дня или увеличения интенсивности радиации, следует сделать заключе-
ние, что для этих целей пригоден любой источник радиации, если его излу-
чение сосредоточено в области физиологической радиации (400—700 ту)
и не содержит коротких ультрафиолетовых лучей (<С 300 тр). Положитель-
ный эффект установлен для обычных и зеркальных ламп накаливания,
солнечных, дуговых, люминесцентных, натриевых, неоновых и ртут-
ных ламп.
Однако, если положительное влияние дополнительного освещения не
подлежит сомнению, то вопрос о сравнительной эффективности различных
источников далеко еще не решен. Максимальной эффективностью обла-
дают то лампы накаливания [596, 855] или неоновые лампы [253, 254,
363, 707, 717, 733, 772], то люминесцентные [123, 160], солнечные
[509] или ртутные лампы [123, 253, 254]. Несомненно, что причиной по-
добной противоречивости экспериментальных данных, помимо методи-
ческих ошибок, является неодинаковая реакция различных видов растений.
Это положение можно проиллюстрировать прямыми опытными данными
ряда авторов [253, 254, 509, 855], в экспериментах которых разные расте-
ния вели себя различно (табл. 89, 90). В настоящее время ясно лишь
одно, что под влиянием ртутных ламп получаются приземистые растения,
а под лампами накаливания они отличаются чрезмерной вытянутостью
стебля. Следует также иметь в виду, что под ртутными лампами растения,
как правило, имеют наименьший вес, а лучший эффект по величине уро-
жая в большинстве случаев наблюдается под неоновыми лампами.
2. ЗНАЧЕНИЕ ИНТЕНСИВНОСТИ ДОПОЛНИТЕЛЬНОГО ОСВЕЩЕНИЯ
Другим принципиально важным вопросом культуры растений с по-
мощью искусственных источников радиации является интенсивность до-
полнительного освещения. Эта проблема, пожалуй, даже более сложна,
чем вопрос о выборе источника радиации. Дело в том, что в данном случае
имеет значение не только вид растения и конкретная цель выращивания,
но и естественная длина дня и интенсивность солнечной радиации.
16 Л. Ф. Клешнин
242
Общая светокультура растений
Грин и его сотрудники [521], пользуясь лампами накаливания мощ-
ностью 50, 100, 200 и 500 вт, изучали влияние дополнительного освещения
в 80, 180, 420 и 1250 лк (535, 1000, 2500 и 7000 эрг/см2-сек физиологи-
ческой радиации) на растения левкоя (Matthiola incana). ноготков (Calen-
dula officinalis), анютиных глазок (Viola tricolor), астр (Callistepkus chinen-
sis) и красной гвоздики (Dianthus sp.). Посев растений производился
осенью (октябрь—ноябрь). Развитие растений совершалось в условиях
Фиг. 142. Зависимость длины вегетационного периода
от интенсивности дополнительного ночного освещения
1 — ноготки (Calendula officinalis); 2 — астры (Callistephus chinensis);
3 — левкой (Matthiola incana); 4 — анютины глазки (Viola tricolor);
5 — могар (Setaria itaiica); 6 — ромашка (M atricaria capensis); 7 — ди-
дискус (Didiscus coerulea) и хризантемы (Chrysanthemum sgetum); 8-—
кларкия (Clarhia elegans); 9— куколь (Agrostemma Gitthago); 10 — ячмень
(Ilordeum vulgare); 11 — стениик (Iberis amara)
короткого зимнего дня при дополнительном освещении продолжитель-
ностью 5 часов (с 2 до 7 час.). Контрольные растения находились в усло-
виях естественного дня. Авторы установили положительное влияние на
все растения, за исключением ноготков; оно выражалось в сокращении
вегетационного периода на 10—20 дней и увеличении количества цветков
в несколько раз. Особенно эффективным дополнительное освещение ока-
залось для анютиных глазок и астр. Первые цвели на 10—14 дней раньше
контроля и увеличили количество цветков в 10—17 раз, а вторые цвели
только при дополнительном освещении (на 140-й—160-й день). Левкой
зацвел раньше на 18—23 дня, а количество цветков у гвоздики увеличи-
лось на 16—21%. Лишь ноготки не ускорили цветения (фиг. 142, 1) и не
увеличили количество цветков; они отличались лишь большей высотой
стебля (38—45 см вместо 27 см в контроле). У остальных растений раз-
личий по высоте или не наблюдалось, или они были незначительны. Астры
в контроле все время оставались в состоянии розетки.
Положительное влияние дополнительного освещения наблюдалось
уже при 80 лк (535 эрг/см2-сек физиологической радиации), а дальнейшее
VII. Выращивание при дополнительном искусственном освещении
243
увеличение освещенности уже не давало дополнительного эффекта
(фиг. 142?.
Уитроу [477, 857, 858], используя дополнительное освещение в 15,
40, 80, 160 и 1100 лк (90, 240, 480, 960, 1920 и 6100 эрг/см2-сек физиоло-
гической радиации), выращивал анютины глазки (Ftola tricolor), астры
(Callistephus chinensis) и левкой (Matthiola incana). Первые увеличивали
число цветков даже при посеве 29 июня (11—16 цветков на растении вместо
1,6 в контроле), вторые (посев 2 октября) ускоряли цветение на 39 дней,
а левкой (посев 28 декабря) — на 5—11 дней. Во всех случаях эффект
был одинаков как при 15 лк, так и при 1100 лк. В другом исследовании
этот автор [861], применяя различную освещенность (3, 10, 100 и 1000 лк,
или 18, 60, 600 и 6000 эрг/см2-сек физиологической радиации), увеличи-
вал продолжительность короткого осеннего и зимнего дня до 21 часа.
Астры и анютины глазки при всех интенсивностях дополнительного осве-
щения увеличили количество цветков (табл. 102) и ускорили наступление
цветения: первые — на 37—59 дней, а вторые — на 55—72 дня (фиг.
142, 2,4), левкой же вместе с сокращением вегетационного периода (на
5—45 дней; фиг. 142, 3) уменьшает количество цветков на растении в 2—5
раз (табл. 102). Как видно из соответствующих кривых на фиг. 142 и
из табл. 102, действие дополнительного освещения начинается ужо при
освещенности 3 лк (18 эрг/см2-сек физиологической радиации) и достигает
максимального значения при 100 лк (600 эрг/см2-сек физиологической
радиации).
Таблица 102
Xарактеристика растений в зависимости от интенсивности дополнительного
ночного осве.н ения
(Уитроу и Бенедикт [861])
Растения Освещенность (в лк) Посев Освещение Сырой вес на- земной массы одного растения (в г) Сухой вес (в г) Отношение веса наземной массы к весу корней Высота растений' (в см) [ 1 Число цветков на одном расте- нии
наземной 1 массы корней общий
Анютины глазки (Viola tricolor) Джиант суисс . 0 10/VIII 10/X—28/XI 3,95 0,52 0,25 0,77 2,0 10,6 4,9
3 10/VIJI 10/X—28/XI 5,3 0,63 0,26 0,89 2,4 11,7 4,7
10 10/VIII 10/X—28/XI 6,2 0,81 0,28 1,09 2,9 11,9 12,2
100 10/VUI 10/X—28/XI 7,9 0,80 0,23 1,03 3,э 11,6 27,0
1000 10/Vin 10/X—28/XI 8,0 0,76 0,15 0,91 5,2 13,4 32,3
Левкой (Matthio- la incana) Ли- лак Лавендер . 0 14/IX 10/X—21/XII 5,75 0,64 0,16 0,80 4,0 71,8 114,9
3 14/IX 10/X—21/XII 8,5 0,86 0,20 1,07 4,3 89,0 60,9
10 14/IX 10/X—21/XII 7,4 0,79 0,17 0,95 4,7 69,3 38,7
100 14/IX 10/X—21/XII 7,7 1,10 0,20 1,29 5,6 76,1 26,3
1000 14/IX 10/X—21/XII 6,4 0,86 0,14 1,01 6,1 74,7 20,3
Астры (Calliste- phus chinensis) Гарт оф Франс 0 6/IX 10/X—27/XII 3,3 0,41 0,23 0,65 1,8 17,5 2,7
3 6/IX 10/X—27/XII 8,3 0,99 0,44 1,44 2,2 12,7 10,6
10 6/IX 10/X—27/XII 9,6 1,21 0,61 1,81 2,0 15,7 8,7
100 6/IX 10/X—27/XII 8,3 1,06 0,41 1,47 2,6 17,1 9,2 8,8
1000 6/IX 10/X—27/XII 7Д 0,97 0,54 1,51 1,8 21,2
16*
244
Общая светокультура растений
Палилов [244] сопоставил эффективность дополнительного освещения
с помощью ламп накаливания в 200, 500 и 1000 вт, помещаемых на высоте
50 см, на расстоянии 1 м друг от друга. Судя по расчетам (табл. 17, фиг. 70),
освещенность в различных вариантах была равна 3, 9 и 20 тыс. лк (18,
54 и 120 тыс. эрг/см2-сек физиологической радиации). Подопытным объек-
том служили огурцы (Cucumis sativus) Клинские, которые высевались
26 января и после всходов освещались с 12 час. ночи до 9 час. утра. Ха-
рактеристика растений по состоянию на 21 марта представлена в табл. 103.
Автор нашел, что различия между вариантами 500 и 1000 вт/м2 настолько
незначительны, что наиболее выгодно и целесообразно пользоваться удель-
ной мощностью 500 вт/м2.
Т а б л и ц а 103
Характеристика рассады огурцов Клинские при дополнительном освещении
лампами накаливания
(Палилов [244])
Варианты опыта Удельная мощность (в вт,м) Длина стебля (в см) Коли- чество листьев па 1 ра- стение Общая площадь листьев (в см2) Ускорение в наступ- лении плодоно- шения (в днях)
Контроль — V 6 131 —
Лампы накаливания 200 13,6 7 365 14
» » 1000 30,0 12 641 30
Обстоятельное исследование по влиянию интенсивности дополнитель-
ного ночного освещения принадлежит Лори и Пёш [596], которые в зим-
ние месяцы (с 15 декабря) выращивали цветочные растения при освещении
с 17 до21 час., пользуясь лампами накаливания в 40, 75 и 150 вт. Резуль-
таты их опытов мы приводим на фиг. 142, 6—£и в табл. 104. Дополнитель-
ное освещение, не влияя на количество цветков, во всех случаях сокращало
вегетационный период на 20—60 дней. Эффективность освещения была
одинакова, независимо от его интенсивности. Аналогичные результаты
с теми же объектами были получены в другом опыте с применением ламп
накаливания в 25, 50 и 200 вт при освещенности 70, 130 и 200 лк (480,
780 и 1200 эрг/см2-сек физиологической радиации).
Остин [396] исследовал значение интенсивности дополнительного
освещения для перехода к цветению рудбекии (Rudbeckia bicolor) и кос-
моса (Cosmos sulphureus). Он нашел, что для предотвращения цветения
космоса и перехода к цветению рудбекии необходимо дополнительное
освещение, удлиняющее естественный зимний день, порядка 4—18 лк
(24—110 эрг/см2-сек физиологической радиации). Освещенность в
50—600 лк (300—3600 эрг/см2-сек физиологической радиации) вызывает
одинаковый эффект.
Большие исследования по выращиванию цветочных культур при раз-
личных освещенностях и удельных мощностях (300, 500 и 1000 вт/м2)
в зимнее время принадлежат Н. П. Красинскому и Е. И. Князевой [131,
134, 136], которые пользовались лампами накаливания в 300, 500 и 1000 вт
с рефлекторами — обычным «Универсала» и зеркальным «Электросила».
Лампы висели на высоте 50 см над растениями и горели в течение 16 ча-
сов (с 23 до 15 час.). Содержание СО2 доводилось до 0,2—0,3%, а темпе-
ратура во всех вариантах равнялась 15—16° С. Опыт показал (табл. 105),
VII. Выращивание при дополнительном искусственном освещении
245
Таблица 104
Характеристика растений при различной интенсивности дополнительного
ночного освещения
(Лори и Псш [596])
Растения Освещен- ность (в лк) Число цветков на одном растении Дата ; цветения Ускорение!
цветения по сравне- нию с контро- лем (в днях) \ Длина I стебля j (в см)
Хризантемы (Chrysanthemum sege- tum) Морнинг стар 0 42,0 1/1V — 40,6
130 41,5 20/II 39 43,2
220 38,0 20/11 39 48,3
430 44,0 18/11 41 46,2
Хризантемы (Chrysanthemum fru- tescens) Бостон Йеллоу 0 17,0 7/1II — 28,2
130 16,0 15/11 20 49,8
220 18,0 16/11 19 45,7
430 19,0 12/11 23 46,2
Дидискус (Didiscus coerulea) . . .. 0 34,0 1/V — 53,5
130 32,6 25/III 36 51,0
220 31,0 21/111 40 43,2
430 38,0 24/III 37 56,0
Кларкин (Clarkia elegans) 0 — 24/111 — —
130 — 24/II 28 —
220 — 20/11 32 —
430 — 20/II 32 —
Левкой (Matthiola incana) «Раби» . 0 1,0 13/Ш — —
130 1,0 19/11 22 102,8
220 1,0 14/11 27 96,8
430 1,0 18/11 23 99,5
Ромашка (Matricaria capensis) . . . 0 17,0 10/V — 24,1
130 16,2 1/Ш 61 36,0
220 17,0 1/Ш 61 30,3
430 19,0 1/Ш 61 36,0
что дополнительное освещение, действие которого особенно сильно ска-
зывается на левкое (Matthiola incana), ускоряет наступление цветения
практически одинаково при всех освещенностях, лежащих в пределах
5—25 тыс. лк (30—150 тыс. эрг/см2-сек физиологической радиации).
Данные по динамике цветения и фотографии левкоя, приводимые авторами,
говорят за то, что лучший результат имеет место при 12 000 лк, полу-
чаемых при помощи ламп накаливания в 500 вт с зеркальным рефлектором.
Применение ламп в 1000 вт дополнительного эффекта не давало, а значе-
ние рефлекторов сказывается лишь при удельной мощности в 300 вт.
Дополнительное освещение во всех случаях приводило к значительному
улучшению качества растений, которые в большинстве случаев не усту-
пали растениям, выращенным летом.
Т а б л и ц а 105
Влияние интенсивности дополнительного освещения на цветение декоративных растений
(Красинский [136])
Растения Тип отражателя Удельная мощность (в вт/м") Освещен- ность (в лк) Интенсив- ность физио- логической радиации (в эрг/см2-сек) Начало опыта Количе- ство растений Начало цветения Максимум цветения Число Дней до цветения Ускорение наступ- ления цветения (в днях) Площадь листьев (в см-)
Цинерария (Cineraria hybrida) 1-й опыт* 0 0 ю/хп 13—18 7/1 27/1 28
«Универсаль» 300 4 800 28 800 ю/хп 13—18 29/XII и/i 19 9 —
Зеркальный 300 7 200 43 200 Ю/ХП 13—18 29/XII 11/1 19 9 —
«Универсаль» 500 8 000 48 000 ю/хп 13—18 26/X 11 4/1 16 12 —
Зеркальный 500 12 000 72 000 ю/хп 13—18 26/XII 31/XII 16 12 —
«Универсаль» 1 000 16 000 96 000 ю/хп 13—18 26/XII 31/XII И 17 —
Зеркальный 1 000 24 000 144 000 10/XI 1 13—18 21/XII 31/XII 11 17 —
2-й опыт** — — 0 0 20/1 7—10 25/11 ll/III 36 —
«Универсаль» 300 4 800 28 800 20/1 7—10 13/11 25/11 24 12 —,
Зеркальный 300 7 200 43 200 20/1 7—10 13/11 25/11 24 12 —
«Универсаль» 500 8 000 48 000 20/1 7—10 13/11 22/11 24 12 —
Зеркальный 500 12 000 72 000 20/1 7-10 13/П 22/11 24 12 —
«Универсаль» 1 000 16 000 96 000 20/1 7—10 9/II 22/11 20 16 —
Зеркальный 1 000 24 000 144 000 20/1 7—10 9/II 19/11 20 16
Левкой (Matthiola incana) 1-й опыт — 0 0 ю/хп 12—21 9/111 25/111 85 —
«Универсаль» 300 4 800 28 800 ю/хп 12-21 16/1 31/1 33 52 —
Зеркальный 300 7 200 43 200 ю/хп 12—21 7/1 25/1 25 60 —
«Универсаль» 500 8 000 48 000 ю/хп 12—21 4/1 20/1 25 60 —
* Исходные растения бутопизировали.
* * Исходные растения в начале бутонизации.
246 Общая светокультура растений
Растения Тип отражателя Удельная мощность (в вт,м2) Освещен- ность (в лк) Интенсив- ность физио- логической радиации (в эрг/см2 • сек) Начало опыта
1-й опыт Зеркальный 500 12 000 72 000 ю/хп
«У ниверсаль» 1 000 16 000 96 000 Ю/ХП
Зеркальный 1 000 24 000 144 000 10/XII
- — 0 0 25/1
2-й опыт «У ниверсаль» 300 4 800 28 800 25/1
Зеркальный 300 7 200 43 200 25/1
«Уииверсаль» 500 8 000 48 000 25/1
Зеркальный 500 12 000 72 000 25/1
«У ниверсаль» 1 000 16 000 96 000 25/1
Зеркальный 1 000 24 000 144 000 25/1
.—' 0 0 з/ш
3-и опыт «У ниверсаль» 300 4 800 28 800 з/ш
Зеркальный 300 7 200 43 200 З/Ш
«Уииверсаль» 500 8 000 48 000 3/111
Зеркальный 500 12 000 72 000 3/111
«У ниверсаль» 1 000 16 000 96 000 3/111
Зеркальный 1 000 24 000 144 000 3/1 п
Лакфиоль (Chieran- thus chieri) — — 0 0 26/1
«Универсаль» 300 4 800 28 800 26/1
Зеркальный 300 7 200 43 200 26/1
«Уииверсаль» 500 8 000 48 000 26/1
Зеркальный 500 12 000 72 000 26/1
«Уииверсаль» 1 ООО 16 000 96 000 26/1
Зеркальный 1000 24 000 144 000 26/1
Т аб лица 105 (продолжение)
Ноличе- Начало Максимум Число Ускорение наступ- Площадь
ство дней до ления листьев
растений цветения цветения цветения цветения (в днях) (в см-) to
12—21 4/1 20/1 25 60 — © -£
12—21 4/1 20/1 25 60 — R сь
12—21 31/XII 16/1 21 64 — S
10 10/Ш 7/1V 43 — —
10 20/11 28/11 25 18 — S3 "13
10 17/11 25/11 23 20 — 53 Qj
10 17/11 23/11 23 20 — О a
10 17/11 25/11 23 20 — о
10 14/11 23/П 20 23 — £ 3
10 14/II 23/11 20 23 — §
10 16/1II — 13 0 — s о
10 16/1II — 13 0 —
10 16/1II — 13 0 — g
10 16/1II — 13 0 — g
10 16/111 — 13 0 — s
10 16/1II — 13 0 — 3
10 16/111 — 13 0 — HOM
4 7/П 22/] I 12 — — <3 Cb
4 4/11 13/11 9 3 — -s
4 4/11 16/ п 9 3 —
4 7/П 13/11 12 0 —
4 1/П 7/П 6 6 —
4 1/П 13/Ц 6 6 —
4 1/11 7/П 6 6 — 247
Растения Тип отражателя Удельная мощность (в вт/м2) Освещен- ность (в ли) Интенсив- ность физио- логической радиации (в эрг/см2 • сек) Начало опыта
Гортензия (Hydran- gea hortensis) 0 0 23/1
«Универсаль» 300 4 800 28 800 23/1
Зеркальный 300 7 200 43 200 23/J
«Универсаль» 500 8 000 48 000 23/1
Зеркальный 500 12 000 72 000 23/1
«Универсаль» 1000 16 000 96 000 23/1
Зеркальный 1000 24 000 144 000 23/1
Роза (Rosa indica) — — 0 0 23/1
«Универсаль» 300 4 800 28 800 23/1
Зеркальный 300 7 200 43 200 23/1
«Универсаль» 500 8 000 48 000 23/1
Зеркальный 500 12 000 72 000 23/1
«Универсаль» 1 000 16 000 96 000 23/1
Зеркальный 1000 24 000 144 000 23/1
Пеларгония (Pelargo- nium grandiflorum.) — — 0 0 26/1
«Универсаль» 500 7 200 43 200 26/1
Колокольчик (Campa- nula medium gran- dijlorum) — 0 0 26/1
«Универсаль» 500 7 200 43 200 26/1
Таблица 105 (продолжение)
Количе- ство растений Начало цветения Максимум цветения Число дней до цветения Ускорение наступ- ления цветения (в*днях) Площадь листьев (в;см-)
8 17/1II 84
8 22/Ш — 58 26 —
8 22/111 — 58 26 —
8 22/Ш — 58 26 —
8 15/1II — 51 33 —
8 15/Ш — 51 33 —
8 10/ III —- 46 38 —
10 22/Ш — 58 — —
10 12/Ш — 48 10 —
10 12/1II — 48 10 —
10 6/Ш — 42 16 —
10 12/Ш — 48 10 —
10 6/Ш — 42 16 —.
10 2/1II — 38 20 —
11 10/V —- 104 — 602
11 16/Ш -— 49 55 1 878
6 — — со —
6 13/1II — 46 СО —
248 Общая светокультура растений
VII. Выращивание при дополнительном искусственном освещении
249'
Фабиан [488,531 ] исследовала значение интенсивности дополнительного
освещения для развития длиннодневных растений — ячменя (Hordeum vul-
gare), стенника (Iberis атага), куколя (Agrostemma githago) и короткоднев-
ного растения — могара (Setaria italica), высеянных весною (апрель—май)
и выращивавшихся в условиях короткого дня (9 часов) с дополнительным
«ночным» освещением от ламп накаливания в 100 и 500 вт. Интенсивность,
освещения варьировала от 0,5 до 530 лк (3—3000эрг/см2-сек физиоло-
гической радиации). Опыт показал, что в этом случае освещенность до
1 лк (6 эрг/см2-сек физиологической радиации) воспринимается как тем-
нота, т. е. не вызывает цветения у длиннодневных и не задерживает его
у короткодневных растений. Дальнейшее увеличение освещенности вы-
зывает и постепенно усиливает фотопериодический эффект, который до-
стигает максимума при 100—400 лк (600—2400 эрг/см2-сек физиологи-
ческой радиации) (фиг. 142, 5, (й^11).
Бортвик и Паркер [419] установи-
ли, что для короткодневной сои (Soja
max) Билокси достаточно 10 лк (60
эрг/см2-сек физиологической радиации)
Таблица 106
Заложение цветков у сои (Sofa max)
Билокси в зависимости от интен-
сивности дополнительного освещения
дополнительного к короткому дню ос- вещения, чтобы полностью предотвра- тить заложение цветков. Количество последних уменьшается уже при интен- сивности дополнительного освещения порядка 0,01—0,2 лк, или 0,6—1,2 эрг/см2-сек физиологической радиации (Бортвик и Паркер [419])
Освещен- ность Число растений с цветочными зачатнами Число бутонов на одном растении
0 10 10,2
(табл. 106). 0,01—0,2 10 8,8
Позднее те же авторы [426] нашли, что кратковременное освещение (1 час) 0,5 10 * 8Д
1 8 5,8
в середине ночи вызывает цветение бе- 5 2 0,3
лены (Hyoscyamus niger) в условиях 10 0 0
короткого дня (12 часов) в том случае, 50 0 0
если интенсивность этого кратковремен- ного освещения достигает 30, а в неко-
торых случаях всего 4 лк (160 и 24 эрг/см2-сек физиологической радиа-
ции). Максимальная эффективность наблюдается при освещенности 250 лк
(1500 эрг/см2-сек физиологической радиации).
Н. Н. Константинов [126] исследовал значение интенсивности допол-
нительного освещения для короткодневного хлопчатника (Gossipium hir-
sutum). Пользуясь лампой накаливания в 300 вт, висевшей над растениями
в поле, автор нашел, что дополнительное освещение вызывает запаздыва-
ние бутонизации, цветения и созревания хлопчатника и увеличивает
высоту заложения первой плодовой ветви. Запаздывание бутонизации на-
ступает уже на расстоянии 3 м от лампы (около 50 лк, или 360 эрг/см2-сек
физиологической радиации), а полная задержка цветения — на рас-
стоянии 1 м (около 450 лк, или 2700 эрг/см2-сек физиологической
радиации).
К перечисленным выше исследованиям примыкает ряд работ по уста-
новлению величин удельной мощности, необходимых для дополнитель-
ного освещения растений.
Дэрроу и Уолдо [459], пользуясь лампами накаливания, культиви-
ровали более сотни сортов земляники (Fragaria sp.) при дополнительном
освещении в зимние месяцы. Лампы включались с наступлением темноты
и выключались в 22 час. В одном случае применялись лампы в 1000 вт
(700 вт/м2), которые помещались на высоте 90 см, а в другом варианте—•
250
Общая светокультура растений.
лампы мощностью 200 вт (260 вт/м2), находившиеся на высоте 70 см.
Авторы не приводят величин освещенности, однако, судя по приведен-
ным данным, она была равна, невидимому, 5000 и 1250 лк 00 и ^5 тыс.
эрг/см2-сек физиологической радиации). Контрольные растения раз-
вивались без дополнительного освещения. В одной серии опытов растения
подвергались дополнительному освещению сразу же с наступлением осен-
них холодов, до ухода в зимний покой, а в другой серии — лишь с начала
зимы, после кратковременного покоя.
В табл. 107 мы приводим в качестве примера экспериментальные дан-
ные для двух сортов. Из таблицы видно, что дополнительное освещение
действует эффективнее, если растения не успели уйти в покой. Цветение
наступает раньше при удельной мощности 700 вт/м2, причем в этом случае
на растении образуется больше цветков. Листовая поверхность в одних
случаях больше при 700 вт/м2, а в других — при 260 вт/м2. В другом
опыте авторы показали, что новообразование листьев в большинстве
случаев совершается интенсивнее при большой удельной мощности, чем
при малой или в контроле, а количество цветков пропорционально числу
листьев.
Таблица 107
Влияние дополнительного освещения на растения земляники (Fragaria sp.)
(Дэрроу и Уолдо [459])
1-я серия освещение 7/XI—10/111 2-я серия освещение 18/XII—8/III
Варианты опытов И й о К © tc те К те д §
о S К те к и m те д
S Я © s те «с и X Ч д те Ф о й те К 2 § ® И
3" Е- Ь й ф Т Хг- F ЙГсч L Лит лове (в СП Нач цвет Чис. цвет 27/1 Чис. цвет 8/II] S® ° чёг
Сорт «Блэйкмор»
Контроль 7/1 4,7 40,7 20/II 0 5,7 33,8
260 вт/м2 20/XII 11,0 65,0 8/II 0 9,2 68,8
700 вт/м2 7/XII 17,0 75,0 20/1 1,0 11,0 82,8
Сорт «Говард 17»
Контроль 21/III 0 18,8 20/11 0 2,2 15,8
260 вт/м2 7/XII 10,5 42,0 8/II 0 5,7 42,2
700 вт/м2 29/XI 12,0 14,7 3/II 0 9,3 84,2
В. В. Селихов [293], применяя лампы накаливания в 500, 1000 и 2000 вт
(по 1 лампе на площадь 10—15 м2), выращивал сахарную свеклу {Beta
vulgaris) в условиях беспрерывного освещения в зимнее время. Контроль-
ные растения, несмотря на нормальное прохождение стадии яровизации,
в условиях зимнего короткого дня не зацвели совсем, а опытные растения
перешли к стрелкованию, дали бутоны, зацвели и образовали зрелые
семена на первом году жизни. При посеве 31 октября стрелкование расте-
ний началось в феврале — через 96 дней при 200 вт/м2, через 98 дней при
100 вт/м2 и через 114 дней при 50 вт/м2, бутонизация в марте — соответ-
ственно через 125, 127 и 135 дней, цветение в апреле — через 150, 156 и
165 дней, а созревание семян в мае — через 182, 191 и 204 дня. Автор
рекомендует для зимнего семеноводства сахарной свеклы дополнительное
освещение при удельной мощности 100 вт/м2.
VII. Выращивание при дополнительном искусственном освещении
251
Таблица 108
Значение интенсивности дополнительного освещения для томатов
(Solanum lycopersicum) Бизон
(Курбатов и Васильева [147])
Варианты опытов
Урожай первого
месяца с куста
1-я серия*
•Освещение лампой на-
каливания 500 вт.. .
Контроль .. .. . .. , . _
2-я серия**
•Освещение лампами
накаливания .. . ..
Контроль . . . . . .. .
20/V 6,0 0,30 137
23/V 4,7 0,26 118
22/V 7,2 0,46 210
17/V 5,2 0,22 100
19/V
19/V
19/V
Общий урожай
с куста
(с 17/11 по 27/VII)
г
I ®
ЧИСЛО ПЛОДОВ (в кг) В % к тролю
17,4 0,83 148
17,8 0,88 157
17,7 0,95 170
11,9 0,56 100
12,8 0,64 168
13,2 0,60 158
8,0 0,38 100
* Освещенность согласно расчетам (табл. 17, фиг. 70) должна быть равна
соответственно 2100, 1300 и 500 лк.
* * Освещенность соответственно равна 2000 и 1250 лк.
И. М. Курбатов и Т. М. Васильева [147] исследовали значение интен-
сивности дополнительного освещения для томатов {Solanum lycopersicum)
сорта Бизон; растения ежедневно освещались по 6 часов (3 часа утром и
•3 часа вечером) с помощью ламп накаливания в 200, 300 и 500 вт, помещае-
мых на высоте 70 см. В первой серии опытов растения высевались 7 января
(всходы 16 января) и исследовалось влияние расстояния от лампы, а во
второй — при посеве 8 февраля (всходы 19 февраля) — исследованию
подвергалось значение удельной мощности. Результаты приведены в
табл. 108. По мнению авторов, для томатов существует определенный
оптимум удельной мощности (около 300 вт/м2), превышение которой может
иметь отрицательный эффект. Последнее, несомненно, находится в связи
•с перегревом растений под влиянием инфракрасных лучей. Устраняя
перегрев, например, с помощью подвижной системы ламп Финкельштейна,
можно, без сомнения, ожидать усиления положительного влияния допол-
нительного освещения при дальнейшем увеличении удельной мощности.
Исследования П. П. Протасовой [253, 254] в колхозе им. Сталина
полностью подтверждают это положение (табл. 109). В ее опытах с 6-ча-
совым дополнительным освещением с 10 января по 15 февраля стационар-
ная установка при 500 вт/м2 дала снижение урожая, а в подвижной
установке лучший результат наблюдался при удельной мощности 500 вт/м2.
М. А. Бассарская и П. В. Алехин [12] нашли, что колошение пшеницы
{Triticum гщ/^«ге)придополнительномосвещении от ламп накаливания в зим-
нее время наступает раньше контроля на 2—3 недели, но различий эффек-
тивности между удельными мощностями в 500,1000 и 2000 вт/м'2 практи-
чески нет (разница в 3— 4 дня).
252
Общая светокультура растений
Таблица 109‘
Состояние рассады и урожай огурцов (Cucumis sativus) Елинские
при дополнительном освещении рассады лампами накаливания
(Протасова [253, 254])
Варианты опытов Состояние рассады на 15/П Урожай плодов к 1/V (в % от контроля)
Высота растений (в см) ЧИСЛО листьев на рас- тении общая площадь листьев (в см-) сухой вес 1 растения (в мг).
Контроль 14,6 3 46,5 125 100
Подвижная установка Финкельштейна 300 вт/м2 21,2 5 147,3 420 120
500 вт/м2 16,0 5 234,0 690 172
1000 вт/м2 30,2 5 253,7 680 —
Стационарная установка 500 вт/м2 25,4 4 118,2 406 96
Таблица Г10'
Значение удельной мощности дополнительного освещения-
для культуры зимнего левкоя (Matthiola. incana)
(Стельмахович и Викулина [301])
Варианты опытов Удельная мощность (в вт/м ) Число дней до Ускорение цветения по сравнению 1 С контролем Число цветков Прирост побегов
бутонизации цветения в % к кон- тролю (в см) в % к кон- тролю
Освещение с 26/XI
Освещение лампами накаливания . . 100 21 42 62 18 150 20 250
200 21 31 73 24 200 24 300
300 21 31 73 24 200 26 325
500 21 31 73 25 208 28 350
Контроль — 42 104 — 12 100 8 1 100
О свещени е с 5/1
Освещение лампами накаливания .. . 100 31 59 24 63 252 12 133
200 31 51 32 62 248 26 289
300 31 46 37 65 260 27 300
500 31 41 42 68 272 32 356
Контроль — 54 83 — 25 100 9 100
О с в е щ е н и е с 4 /н
Освещение лампами накаливания . . 100 11 27 12. 23 192 11 138
200 11 26 13 24 200 12 150
300 11 25 14. 24 200 16 200
500 11 21 1.8 26 217 15 188
Контроль — 21 39 — 12 100 8 100>
VII. Выращивание при дополнительном искусственном освещении
253
М. Л. Стельмахович и Л. А. Викулина (301) сопоставили удельную
.мощность в 100, 200, 300 и 500 вт/м2 (высота подвеса 55—60 см) при до-
полнительном освещении (с 23 до 7 час.) в зимние месяцы левкоя зимнего.
Авторы установили, что дополнительное освещение с помощью ламп нака-
ливания значительно ускоряет наступление цветения, увеличивает коли-
чество цветков и усиливает прирост (табл. 110). Опыт показал, что для
нормального цветения растений достаточна удельная мощность 200 вт/м2,
а для вегетативного роста эффективность возрастает вплоть до 500 вт/м2.
С. Г. Сааков (280) изучал эффективность дополнительного освещения
в зимние месяцы в теплицах Ленинграда. В его опытах применялись
лампы накаливания в 300 и 500 вт (130 и 220 вт/м2), которые освещали
растения по 6 часов в сутки с 5 января. В качестве опытных объектов
служили цинерария (Cineraria hybrida}, гортензия (Hydrangea hortensis),
левкой (Matthiola incand), незабудка (Myosotis alpestris) и первоцвет
(Primula sp.). Цинерария подвергалась освещению в фазе бутонизации,
в фазе стрелкования и в розеточном состоянии, остальные растения —
Таблица 111
Значение удельной мощности дополнительного освещения для выращивания
цветочных растений
(Сааков [280])
Варианты Удельная мощность (в вт/м-) Число дней до Ускорение по сравнению с контролем (в днях)
бутониза- ции цветения
Цинерария (Cineraria hybrida) Опыт 1. Освещение в фазе бутонизации
Лампы накаливания 130 — 17 46
220 — 12 51
Контроль — — 63 —
Опыт 2. Освещение в фазе стрелкования Лампы накаливания 130 .— 30 39
220 — 20 49
Контроль — — 69 —
Опыт 3. Освещение в фазе розетки
Лампы накаливания 130 — 23
220 — 38 40
1{оптроль — — 78 —
Гортензия (Hydrangea hortensis)
Лампы накаливания 130 20 75 10
220 20 70 15
Контроль — 30 85 —
Левкой (Matthiola incana)
Лампы накаливания 130 20 57 44
220 20 45 56
Контроль — 69 101 —
Незабудки (Myosotis alpestris)
Лампы накаливания . . . 130 20 23 38
Контроль — 58 61 —
254
Общая светокультура растений
до перехода к бутонизации. Опыт показал, что для культуры названных
растений в зимние месяцы необходима удельная мощность порядка 200—
250 вт/м2 (табл. 111). В этом случае развиваются нормальные растения
с темнозелеными листьями. Благодаря дополнительному освещению
пропускная способность теплиц увеличивается в 2—3 раза.
Подводя итог сравнительно большого числа часто весьма обстоятель-
ных исследований по значению величины интенсивности дополнительного-
освещения для выращивания различных растений и потребной для этой
цели удельной мощности [21, 123, 126, 131, 134, 136, 147, 244, 280, 293,
301, 396, 419, 426, 459, 468, 488, 521, 531, 596, 607, 687, 717, 720, 732,
733, 775, 778, 787, 791, 846, 847, 857, 858, 861], следует признать, что
обе названные величины определяются целями выращивания растений,
видами и состоянием растений, местом и временем выращивания.
Для ускорения перехода растений длинного дня к цветению или
для задержания или предотвращения прохождения световой стадии у ко-
роткодневных растений интенсивность физиологической радиации при до-
полнительном освещении не должна превышать 1—2 тыс. эрг/см2-сек.
Для ламп накаливания это отвечает освещенности 200—300 лк и дости-
гается с помощью обычного широкоизлучателя при удельной мощности
в 50—75 вт/м2. Это установлено для цветочных растений, злаков, сои,
сахарной свеклы и других и, невидимому, может быть распространено
на остальные растения. В южных районах страны (Средняя Азия, Кав-
каз, Крым, Украина), где естественная интенсивность радиации в зимнее
время достаточна для вегетативного роста растений и задача дополни-
тельного освещения сводится лишь к простому удлинению дня с целью
получения фотопериодического эффекта, нет необходимости прибегать
к более высоким интенсивностям радиации.
В средней полосе СССР при сравнительно невысокой зимней интен-
сивности радиации цель дополнительного освещения сводится не только
к удлинению дня, но и к увеличению недостаточной естественной осве-
щенности. В этом случае для получения нормальных цветочных ра-
стений, для выращивания рассады томатов или огурцов и для культуры
других растений необходима несколько большая удельная мощ-
ность — порядка 500 вт/м2. Получаемая в этом случае освещенность от
ламп накаливания при равномерном распределении светового потока на
освещаемой площади равняется примерно 5000—8000 лк и соответствует
интенсивности физиологической радиации 30—48 тыс. эрг/см2-сек. Даль-
нейшее увеличение удельной мощности или интенсивности радиации
при употреблении ламп накаливания путем увеличения их мощ-
ности или уменьшения высоты подвеса обычно не вызывает усиления
положительного эффекта, а часто дает даже отрицательный эффект. По-
следнее, несомненно, находится в связи с перегревом растений под влия-
нием инфракрасных лучей.
3. ДОПОЛНИТЕЛЬНОЕ ДНЕВНОЕ ОСВЕЩЕНИЕ
В предыдущей главе мы отмечали, что интенсивность физиологиче-
ской радиации, потребная для нормальных роста и развития растений,
должна быть равна 50—120 тыс. эрг/см2-сек, что для прямой солнечной
радиации соответствует 10—25 тыс. лк. В тех случаях, когда естествен-
ная солнечная радиация ниже этих величин, возникает потребность в до-
полнительном освещении в дневные часы. Эта необходимость чаще всего
имеет место в условиях закрытого грунта в зимние и пасмурные дни
в умеренной и, особенно, в приполярной областях (фиг. 89). К сожа-
VII. Выращивание при дополнительном искусственном освещении 255
лснию, исследований в этом направлении почти совершенно нет. Нам
известны лишь две работы, посвященные значению интенсивности до-
полнительного дневного освещения, причем обе работы касаются пше-
ницы (Triticum vulgare) [12, 313]. М. А. Бассарская и П. В. Алехин [12],
пользуясь лампами накаливания при удельной мощности 500, 1000 и
2000 вт/м'2, в условиях Одессы в оранжерее выращивали пшеницу при
дополнительном освещении днем (с 8 до 16 час.), ночью (с 16 до 8 час.)
и беспрерывном. Посев пшеницы производился 26 ноября, а колошение
в большинстве случаев было во второй половине января. Оказалось,
что дополнительное освещение в дневные часы без удлинения короткого
зимнего дня практически не влияет на время колошения растений, уско-
ряя развитие по сравнению с контролем лишь на 3—6 дней; беспре-
рывное дополнительное освещение в ночные часы ускоряет развитие оди-
наково — на 15—18 дней, а образование нормальных семян имеет место
только при беспрерывном освещении или при дополнительном ночном
освещении при удельной мощности 1000 и 2000 вт/м2. При мощности
2000 вт/м'2 семена образуются в обоих цветках каждого колоска, при
1000 вт/м'2 — лишь в одном цветке колоска, а при 500 вт/м2 но завязы-
ваются вообще.
В опытах М. Я. Трегубенко [313] испытывались различные комби-
нации дневного и ночного освещения, осуществляемого с помощью ламп
накаливания. В дневные часы удельная мощность была равна 200, 300,
500 и 1000 вт/м2, причем лампы в большинстве случаев включались на
16 часов. Ночью (в остальные 8 часов) удельная мощность равнялась 40,
100, 200 и 500 вт/м2. В качестве рефлектора служил широкоизлучатель
«Универсаль», а высота подвеса ламп по мере уменьшения мощности
ламп снижалась от 100 до 35 см. Автор нашел, что на широте Харькова
пшеница, посеянная в конце октября, формирует нормальный колос
(12—15 зерен) и дает зрелые семена к середине февраля лишь при дополни-
тельном дневном освещении в течение 16 часов при удельной мощности
1000 вт/м2 и при дополнительном ночном освещении (остальные 8 часов)
при удельной мощности 200 вт/м2. В остальных вариантах, несмотря
на почти одновременное колошение и цветение, количество зерен в ко-
лосе не достигает нормальной величины и постепенно падает от 10 до 0
по мере уменьшения удельной мощности дневного освещения.
Если интенсивность дополнительного ночного освещения представляет
собою более или менее постоянную величину, определяемую лишь видом
растения, то величина дополнительного дневного освещения в сильней-
шей степени зависит от географической широты, времени года и погоды.
Необходимая интенсивность дневного освещения может быть определена
на основании кривых на фиг. 89. Для определенной широты, определяе-
мой осью абсцисс, и определенного времени года, определяемого кри-
выми I—XII, она равняется разности ординат линий аб или вг и одной
из кривых I—XII.
4. ЗНАЧЕНИЕ ДЛИТЕЛЬНОСТИ ДОПОЛНИТЕЛЬНОГО ОСВЕЩЕНИЯ
Вопрос о длительности дополнительного ночного освещения сводится,
в сущности говоря, к вопросу о длине дня и фотопериодической природе
растений. Нет необходимости говорить о том, что ночное освещение бу-
дет иметь положительное значение лишь для растений длинного дня и,
отчасти, для нейтральных растений, а для растений короткого дня имеет
смысл лишь в случае необходимости задержать цветение, т. о. в случае
культуры растений с целью получения вегетативной массы. Следовательно,
.256
Общая светокультура растений
длительность дополнительного ночного освещения определяется при-
надлежностью растений к определенной фотопериодической группе
и устанавливается в каждом конкретном случае в зависимости от вида
растения и задачи культуры.
Гаут [543], выращивая душистый горошек (Lathyrus odoratus) при
дополнительном освещении зимою с помощью ламп накаливания в 200 вт,
установил, что это растение лучше всего развивается в условиях 15-ча-
сового дня. В этом случае цветение наступает почти одновременно с цве-
тением на 21-часовом дне, а высота растений и количество цветков до-
стигают максимального значения. Несколько позднее Бомо и Уивер [402]
нашли, что для роста картофеля и максимального урожая клубней в усло-
виях дополнительного освещения необходим 15-часовой день. Умень-
шение, как и удлинение дня влечет снижение веса как всего растения,
так и клубней.
Обширное исследование значения длительности дополнительного осве-
щения для роста и развития растений было предпринято Смитом [771].
В условиях Норвегии этот автор исследовал влияние длины дня на ячмень
{Hordeum vulgare), овес (Avena sativa), рожь (Secale cereale), пшеницу
(Triticum vulgare), тимофеевку (Phleum arvense) и горох (Pisum sativum).
Естественный день в условиях опыта изменялся от 10 часов 37 минут
до 18 часов 44 минут, а недостающее до заданного количество светлых
часов добавлялось с помощью света от ламп накаливания в 200 вт, да-
вавших освещенность 600 лк (3600 эрг/см--сек физиологической радиации).
В результате исследования было установлено, что во всех случаях ма-
ксимальный сухой вес растений, максимальная высота, наиболее крупные
листья и наибольшее кущение наблюдались в условиях 16—18-часового
дня (фиг. 144, 8 и Р; фиг. 145, 1—3 и фиг. 146, 1—6). Беспрерывное осве-
щение давало худший эффект. Переход к цветению выражался кри-
выми 8 и 9 на фиг. 143 и быстрее всего совершался в условиях беспре-
рывного освещения. По сравнению с 18-часовым днем ускорение состав-
ляло для овса — 6 — 8, для ячменя — 5 — 7, а для ржи — 1 — 5 дней.
Тимофеевка и горох цвели одновременно как при беспрерывном осве-
щении, так и в условиях 18-часового дня. По сравнению с 12-часовым
днем ускорение равнялось для гороха —7—10, для тимофеевки —12—14,
для ячменя — 32—33, для ржи — 44 — 46 и для овса — 54—-64 дням.
Иными словами, для всех перечисленных растений оптимальным следует
признать 16—18-часовой день.
По данным М. X. Чайлахяна [335, 336] оптимальной длиной дня
для ячменя (Hordeum vulgare) следует признать также 17—18 часов, а не
беспрерывное освещение, так как в обоих случаях колошение наступает
одновременно и растения имеют одинаковый сухой вес.
Остин [395, 396], исследуя значение длины дня для роста и развития
бальзамина (Impatiens balsamina) при интенсивности дополнитель-
ного освещения 1000—1500 лк (6000—9000 эрг/см2-сек физиологической
радиации), получаемого от лампы накаливания в 200 вт, показал, что
бутонизация и цветение растений раньше всего наступают в условиях
короткого (8 часов) или естественного дня (12—15 часов), а максималь-
ные высоты, число узлов и сухой вес надземной массы и корней имеют
место при длине дня в 16 часов. В условиях 20-часового дня растения
по всем показателям несколько отстают или, во всяком случае, бывают
не лучше, чем при 16-часовом дне.
Синг и его сотрудники [758—761], давая дополнительное освещение
льну (Linum usitatissimum), пшенице (Triticum vulgare), овсу (Avena
sativa) и индийской конопле (Crotalaria juncea), изучили поведение
VII, Выращивание при дополнительном искусственном освещении 257
названных растений при длине дня в 1, 2, 4, 6, 8,12, 15, 18 и 24 часа. Они
нашли, что для льна и пшеницы оптимальным является 18-часовой, а для
индийской конопли — 12-часовой день (фиг. 143, 10—12; фиг. 144,
10—12', фиг. 145, 4—5, и фиг. 146, 7). У пшеницы при 18-часовом дне
наблюдаются минимальный вегетационный период, максимальный вес рас-
тений и максимальное кущение. Наибольший урожай семян получался
Фиг. 143. Число дней от всходов до цветения в условиях
различной длины дня при дополнительном освещении
1 — овес (Апппа saliva var. bysantina); 2 — ячмень (Hordcum vul-
garc); 3 — горох (Pisum sativum); 4 — фасоль (Phasmlus vulgaris);
5 — просо (Panicum miliaceum); 6 — конопля (Cannabis saliva);
7 — соя (Snja hisnida); 8 — рожь (S"cal" cr al"); 9 — тимофеевка
(PhPum arv -ns ); 10— кроталария (Crvta^aria junc a); n— леи
(Linum usitalissimum); 12—пшеница (Trilicum vulgare); 13— аф-
риканский кенаф (Hibiscus sabdariffa)
в условиях 15-часового дня. У льна максимальный вес растений имел
место при 24 часах, а минимальный вегетационный период и максималь-
ный урожай семян — при 18 часах. У индийской конопли минимальный
вегетационный период, максимальный вес семян и максимальное ветвление
наблюдаются при 12-часовом дне, а максимальный вес растений, макси-
мальное количество листьев, максимальная листовая поверхность и макси-
мальный урожай волокна — при беспрерывном освещении.
Ф. Э. Реймерс [272], применяя лампы накаливания (100 вт/м2) в ка-
честве источника дополнительного освещения, нашел, что у двух сортов
лука (Allium сера) — Каба болгарский и Даниловский — вес луковиц
17 А. Ф. Клешнин
258
Общая светокультура растений
и вес листьев достигают значительно большей величины в условиях
естественного летнего дня, чем при беспрерывном освещении. Одновре-
менно автор установил для салата Идеал, Эйер Гельбер, Троцкопф и
Гр. Блонд парессер, что стрелкование наступает почти одновременно
как при беспрерывном освещении, так и в условиях естественного летнего
дня, а в некоторых случаях — даже при 14-часовом дне.
Фиг. 144. Сухой вес одного растения в условиях различ-
ной длины дня при дополнительном освещении
1 —овес (Avena sativa var. bysantina); 2 — ячмень (Hordeum vulga-
re); 3 — горох (Pisum sativum); 4 — фасоль (Phas<>olus vulgaris);
5 — просо (Panicum miliaceum); 6 — конопля (Cannabis sativa);
7 — соя (Soja hispida); 8 — рожь (Secale cer al°); 3 — тимофеевка
(Phleum arvens-); 10 — пшеница (Triticum vulgare); ц — лен (Linwm
usitatissimum); 12 — кроталария (Crotacaria juncea)
Скинер [763] изучал прирост рододендрона (Rhododendron ponticum,
Rh. rodeum elegans) в условиях различной длины дня (12, 16, 18 и 24 ча-
сов) при дополнительном освещении с помощью ламп накаливания в 75 вт
(250—300 лк или 1500—1800 эрг/см'2-сек физиологической радиации).
Как видно из кривых 8 и 9 на фиг. 146, величина прироста возрастает
от короткого к длинному дню, достигая максимального значения при
беспрерывном освещении. Следует подчеркнуть, что дополнительный эф-
фект беспрерывного освещения по сравнению с 18-часовым днем в этом
случае невелик и далеко не соответствует затрате энергии при беспре-
рывном освещении.
VII. Выращивание при дополнительном искусственном освещении
259
В опытах Кларка и Ломбарда [446] по культуре картофеля (Solanum
tuberosum) в зимние месяцы было установлено, что для плодоношения
картофеля оптимальная длина дня равна 18 часам (фиг. 147). Авторы
применяли лампы накаливания в 60 вт, дававшие освещенность
200—300 лк (1200—1800 эрг/см2-сек физиологической радиации), и
нашли, что число цветков и плодов и вес семян на растении возрастают
вплоть до 18 часов. Дальнейшее увеличение дня дает или незначительный
положительный или отрицательный эффект.
А. И. Челядинцева [340] выращивала земляной орех (Arachis hypogaea)
в летние месяцы в условиях открытого грунта в районе г. Краснодара.
Длина дни (S vac.)
Фиг. 145. Кущение и ветвление растений в условиях
различной длины дня при дополнительном освещении
(число побегов на одном растении)
1 — ячмень (Hordeum uu/gare); 2 — овес (Arena safira); 3 — рожь
(Secal" cereale); 4— пшеница (Triticum vulgare); 5—нроталарня
(Crotalaria juncea, в % от числа побегов при 12-часовом дне)
Беспрерывное освещение создавалось с помощью ламп накаливания,
которые включались в ночные часы, обеспечивая освещенность 150 лк
(900 эрг/см2-сек физиологической радиации), а естественный день ко-
лебался в пределах 12—15\.? часов. Автор нашел, что длина дня почти
не влияет на время цветения растений, а число плодов, их вес и вес рас-
тений достигают максимума при естественной длине дня, снижаясь при
беспрерывном освещении и, отчасти, в условиях 10-часового дня.
Я. И. Потапенко и Е. И. Захарова [251] изучали реакцию различных
сортов винограда (. ills vinifera) на дополнительное ночное освещение
и нашли, что рост лозы, образование листьев и формирование корневой
системы лучше всего совершаются в условиях естественного летнего дня.
Беспрерывное освещение в большинстве случаев давало худший резуль-
тат и, задерживая созревание лозы, отрицательно сказывалось на бу-
дущей зимовке растений. Уоринг [835] показал, что у сосны (Pinus Sil-
vestris') максимальная длительность периода роста, максимальное число
листьев и максимальный прирост наблюдаются в условиях 20-часового
дня (фиг. 146, 10).
Как показывают опыты Паркера и его сотрудников [426], цветение
белены (Hyoscyamus niger) наступает лишь при длине дня более 12 часов,
260
Общая светокультура растений
причем максимальная скорость стрелкования имеет место в условиях
16-часового дня, а при длине 17 и 18 часов стрелкование совершается
значительно хуже.
По данным Талукдара [818], африканский кенаф {Hibiscus sabla-
riffa) быстрее всего зацветает в условиях 10-часового дня (фиг. 143, 13),
'Фиг. 146. Рост растений в условиях различной длины дня
при дополнительном освещении
1 — пшеница (Triticum vulgare); 2 — рожь (Secale cereale); .3 — овес
saliva); i — ячмень (Hordeum vulgare); 5—горох (Pisum sati-
vum); 6 — тимофеевка (Phleum arvens1); 7— лен (Linum usitatissimum);
8 — рододендрон (Rhododendron ponticum); 0— рододендрон (Rhodo-
dendron ros-um elegans); 10 — сосна (Pinus Silvestris); 1—7—высота
растений (в см); 8—10 — прирост растений (в см); 11 — высота расте-
ний (в % от высоты при 12-часовом дне)
максимальной высоты достигает при 14-часовом дне, а максимального
диаметра стебля — при 16 часах.
Некоторые растения, происходящие из тропических областей, для нор-
мального роста и развития нуждаются в коротком дне. В этом случае
они зацветают раньше, растут интенсивнее и дают максимальный урожай.
Таковы, например, тефрозия {Tephrosia Candida), цветущая лишь при
длине дня в 10—13% часов, или батат {Ipomoea batatas), дающий луч-
ший урожай клубней при той же длине дня [617, 618]. Рост хинного
дерева {Cinchona ledgeriana, С. succirubra) также интенсивнее всего со-
вершается при 10—12-часовом дне, а беспрерывное освещение снижает
вес этих растений в 5—6 раз [210]. Г. А. Евтушенко [68] показал, что
VII. Выращивание при дополнительном искусственном освещении
261
большинство сортов табака (Nicotiana tabacum) (Трапезонд, Тык-кулакт
Самсон, Виргиния, Гаванна, Мэрилэнд и др.) растет лучше, если длина
дня не превышает 10 часов. В опытах Сарторис [739] тростник {Saccharum
spontaneum) лучше всего рос при длине дня в 13 часов. Эта же длина
дня благоприятна для цветения названного растения. Скалли, Паркер
и Бортвик [746] нашли, что максимальный сухой вес лука {Allium сера)
получается в условиях 12-часового дня, а в ряде случаев в условиях
10-часового дня.
В некоторых случаях дополнительное ночное освещение, удлиняющее
день, вызывает уменьшение урожая
не в силу непосредственного влияния
на ростовые процессы, а в результа-
те резкого сокращения вегетацион-
ного периода, связанного с быстрым
переходом к цветению и с последую-
щей остановкой роста. Обычно это
наблюдается у однолетних длинно-
дневных растений, особенно при до-
полнительном освещении лампами
накаливания. Так, Гард-Каррер [560]
сопоставила развитие и урожай ра-
стений яровой и озимой пшеницы
{Triticum vulgare) в условиях корот-
кого (8 часов), естественного (9—-15
часов) и длинного (17 часов) дня
при различной температуре. Расте-
ния высевались 2 декабря и выращи-
вались при температуре 10—14° и
20—23°. В качестве источника радиа-
ции служили лампы накаливания в
100 вт, создававшие освещенность
400 лк (2400 эрг/см2-сек физиологиче-
ской радиации). Результаты опытов,
частично приведенные в табл. 112,
не оставляют сомнений в том, что
снижение урожая под влиянием до-
полнительного освещения (17-часо-
вой день) связано главным образом с
В другой работе [561], где в качестве
Фиг. 147. Характеристика растений
картофеля (Solanum tuberosum), выра-
щенных при различной длине дняj'
1 — число цветков на одном растении карто-
феля Себаго; 2— число цветков на одном
растении картофеля Эрлейн; 3—число плодов
на одном растении картофеля Себаго; 4— чи-
сло плодов на одном растении картофеля Эр-
лейн
чрезмерным ускорением развития,
источника радиации применялись
лампы накаливания в 300 вт, длина дня составляла 18 часов, а освещен-
ность равнялась 500 и 1000 лк (3000 и 6000 эрг/см2-сек физиологической
радиации), были получены в общем те же результаты. Дополнительное
освещение в зимнее время приводило к уменьшению кущения и снижению
урожая как общей массы, так и зерна, хотя качество последнего и не
ухудшалось. Аналогичные данные были получены Чайлахяном [335, 336]
для пшеницы {Triticum vulgare), рапса {Brassica napus var. oleifera) и
вики ( icia sativa), G. А. Алекперовым [3] для ржи {Sicale eereale), а Ден-
фером [468] — для горчицы {S inapis alba) и ячменя {Hordeum vulgare).
Мы лишены возможности рассмотреть другие работы [3, 25, 60, 260,
266, 267, 405, 419, 420, 422, 528, 571, 584, 591, 619, 653, 656, 666,
732, 746, 776, 838, 845, 847], в которых получены в общем те же ре-
зультаты.
В заключение исследований по вопросу о влиянии длительности допол-
нительного освещения мы рассмотрим работы В. М. Катунского [101]
262
Общая светокультура растений
Таблица 112
Влияние дополнительного освещения на развитие и структуру урожая озимой
и яровой пшеницы (Triticum vulgare)
(Гард-Каррер [560])
Яровая пшеница Озимая пшеница
21° С 12° С 12° С
О' О' 5*
ю ю ю
О' 1 ю О' о- 1 1Л у 1 ю О’
оо о? ** со О 00 о
Цветение 25/IV 18/Ш 13/1 9/V 3/IV 19/11 8/VI 16/V 13/IV
Число дней до цветения 144 106 42 158 123 79 188 165 131
Высота растений (в см) 82 84 48 90 98 66 110 115 101
Кущение (число побе- гов в кусте) 8,1 6,2 2,7 5,8 3,7 3,8 6,6 5,3 3,4
Число фертильных ко- лосьев ....... 2,9 4,1 1,6 3,9 3,5 1,3 2,0 4,3 2,9
Длина колоса (в см) . 8,5 7,8 5,4 9,5 8,6 5,4 13,7 10,0 8,4
Вес зерен в колосе (в г) 0,24 0,61 0,38 0,51 0,85 0,37 0,23 0,98 1,15
Вес 100 зерен (в г) . . 2,8 3,9 3,6 3,5 5,5 2,2 3,5 4,3
Урожай зерна на одно растение (в г). . . . 0,77 2,40 0,67 1,91 2,81 0,45 0,53 4,06 3,57
Сухой вес одного ра- стения (без зерна)(в г) 10,3 8,1 — 9,5 7,7 1,2 12,9 8,7 3,8
п Льюиса и Вента [599]. Первый, взяв в качестве исходного 10-часовой
день, удлинял его с помощью ламп накаливания до 16, 18 и 24 часов.
Интенсивность дополнительного освещения равнялась 65—85 лк (390—
500 эрг/см2-сек физиологической радиации). В качестве объектов служили
растения длинного и короткого дня. Результаты опытов мы представили
в виде кривых 7, 2, 3, 7, 7 на фиг. 143 и 144. Эти кривые ясно показывают,
что для быстрого перехода к цветению растения короткого дня нуждаются
в 10—12-часовом дне, а длиннодневные растения в 18—24-часовом дне.
Одни растения, как, например, фасоль {Phaseolus vulgaris), почти не
ускоряют наступления цветения при переходе от длинного дня к бес-
прерывному освещению, у других же — овса {Avena sativa) и ячменя
{Hordeum vulgare) — это может сопровождаться значительным сокраще-
нием вегетационного периода. Что касается веса растений, то в этом
отношении реакция растений на длину дня может быть различной.
У одних, например у гороха {Pisum sativum) и фасоли {Phaseolus vul-
garis, сорт «Осборн»), вес растений возрастает почти линейно вместе с уве-
личением длины дня; у других — сои {Soja hispida) и фасоли {Phaseo-
lus vulgaris, сорт «Золотая гора»)—он достигает максимума при 16—18 ча-
сах, а затем, при дальнейшем удлинении дня, начинает снижаться;
у третьих — конопли {Cannabis sativa) и проса {Panicum miliaceum) — макси-
мум наступает также при 16—18 часах, но беспрерывное освещение
не вызывает отрицательного эффекта. Существуют, наконец, и такие рас-
тения, как, например, овес {Avena sativa) или ячмень {Hordeum vulgare),
которые уменьшают свой вес по мере увеличения длины дня.
Льюис и Вент [599] при помощи люминесцентных и ртутных ламп,
дававших освещенность 4500 лк, удлиняли 7-часовой день до 8, 12, 14,18
VII. Выращивание при дополнительном искусственном освещении
263
и 24 часов. Авторы нашли, что для одних видов — бэрии {Baeria chrysostoma),
бэйлеи (Baileya multiradiata), кореопсиса {Coreopsis calif arnica), эшольции
(Escholtzia californica), годеции (Godetia quadrivulnera), линантуса (Linant-
hus montanus) и фацелии {Phacelia Parryi) — более благоприятно бес-
прерывное освещение, вызывающее наиболее раннее цветение, для дру-
гих — ментцелии {Mentzelia I indleyi) и монарделлы {Monardella lanceo-
lata) — 18-часовой день и беспрерывное освещение дают одинаковый
эффект, а для некоторых, например эшольции {Escholtzia lobbii), более
благоприятен 18-часовой день (табл. 113).
Таблица ИЗ
Число дней до цветения в зависимости от длины дня
(Льюис и Вент [599])
Растения Длина дня (в часах)
8 12 14 18 24
Бэрия {Baeria chrysostoma) .... 57 69 47 40 32
Бэйлея (Baileya multiradiata) . . . 00 00 00 00 103*
Кореопсис (Coreopsis californica) 103* 103* 77 54 43
Эшольция (Escholtzia californica) . 00 00 СО 98 56
Годеция (Godetia quadrivulnera) со 00 103* 97 85
Линантус (Linanthus montanus) . . со 00 00 74 56
Фацелия (Phacelia parryi) .... 00 103* 65 43 33
Ментцслия (Mentzelia Lindleyi) . . со 103* 103* 63 65
Монардолла (Monardella lanceolata) со СО СО 103* 103*
Эшольция (Escholtzia lobbii) . . . 103* 103* 99 59 72
Гилия (Gilia tricolor) 00 80 103* 97 85
Мадия (Madia elegans) 76 СО 103* 81 64
Энотера (Oenothera deltoides) ... . СО 00 00 СО 00
* Бутоны к концу опыта.
Судя по фотографиям, приведенным в работе, энотера {Oenothera del-
toides), мадия {Madia elegans var. densifolia) и фацелия {Phacelia Parryi)
имеют лучший рост при 18-часовом дне, а бэрия {Baeria chryso-
stoma) — в условиях 14-часового дня. Размеры листьев у эшольции
{Escholtzia lobbii) возрастают вместе с длиной дня и достигают макси-
мального значения при беспрерывном освещении. Максимальные конеч-
ные размеры фацелии {Phacelia Parryi) наблюдались при 10—12-часо-
вом дне.
Таким образом, приведенные данные не оставляют сомнений в том,
что для получения максимального урожая зеленой массы растений в боль-
шинстве случаев наиболее благоприятен 16—18-часовой день.
Лишь некоторые экваториальные растения для накопления наибольшей
массы нуждаются в коротком дне, а другие немногочисленные виды
предпочитают беспрерывное освещение. В последнем случае обычно беспре-
рывное освещение дает незначительный дополнительный эффект по сравне-
нию с длинным, 16—18-часовым днем. Как правило, наиболее быстрый пере-
ход к цветению у растений короткого дня наступает при 10—12-часовом
дне, а у растений длинного дня — при 17—18-часовом. Дальнейшее уве-
личение дня обычно дает незначительный эффект или совсем не влияет.
264
Общая светокультура растений
Для определения необходимой длительности ночного освещения сле-
дует руководствоваться кривыми на фиг. 91, добавляя недостающее число
часов освещения в зависимости от естественной длины дня, целей вы-
ращивания и вида растений.
5. ЗНАЧЕНИЕ ВРЕМЕНИ ДОПОЛНИТЕЛЬНОГО ОСВЕЩЕНИЯ
В большинстве случаев дополнительное ночное освещение дается в ве-
черние часы, хотя иногда и приходится пользоваться утренним осве-
щением [147, 262, 287]. Исследований, посвященных сравнительной
эффективности освещения в утренние и вечерние часы, сравнительно не-
много [468, 488, 521, 596], причем все авторы считают, что время допол-
нительного освещения практического значения не имеет. В опытах
Грина [521, 857] с астрами {Callistephus chinensis], анютиными глазками
(i iola tricolor) и ноготками {Calendula officinalis) был установлен не-
сколько лучший эффект дополнительного вечернего освещения (с 21 до
2 час.) по сравнению с утренним (с 2 до 7 час.). При освещенности 420 лк
(2500 эрг/см2-сек физиологической радиации), получаемой с помощью
ламп накаливания в 200 вт, в первом случае наблюдалось большее ко-
личество цветков (соответственно 42,5 и 36,6 у анютиных глазок, 4,2
и 1,7 у астр и 3,5 и 2,9 у ноготков), несколько большая высота растений
и более раннее наступление цветения (на 6 дней у анютиных глазок
и на 13 дней у астр), чем во втором.
У других авторов лучший эффект давало, напротив, освещение в ут-
ренние часы. Фабиан [4881, удлинявшая с помощью ламп накаливания
(10—400 лк, или 60—2400 эрг/см'2-сек физиологической радиации) ко-
роткий день (10 час.) в утренние (с 1 до 8 час.) или вечерние (с 18 до
1 час.) часы, нашла, что у ячменя {Hordeuт vulgare) и куколя {Agrostemma
Githago) колошение и цветение наступают на несколько дней раньше,
если дополнительное освещение приходится не на вечерние, а на утрен-
ние часы. У ячменя различие достигало 12 дней, а у куколя не превы-
шало 4 дней. У короткодневного растения могара {Setaria italica) раз-
личия в сроках цветения при утренн м и вечернем освещении достигали
20—25 дней, причем растения при утреннем освещении зацветали по-
зднее, чем при вечернем.
Более подробно этот вопрос исследовали Лори и Пёш [596], которые
установили, что для одних растений — левкоя {Matthiola sp.), скабиозы
{Scabiosa lovelines), венидиума {> enidium decurrens calendulaceum), хри-
зантемы {Chrysanthemum frutescens) и др. — время освещения не играет
никакой роли; для других растений — астры {Callistephus chinensis), хри-
зантемы {Chrysanthemum sp.) и др. — лучший результат наблюдается
при утреннем освещении, а в некоторых случаях, например у василька
{Centaurea imperialis alba'', — при освещении в вечерние часы.
Повидимому, вопрос о времени дополнительного освещения нуждается
в дополнительном изучении, хотя a priori следует предположить, что
определенных и больших различий между влиянием освещения в утрен-
ние и вечерние часы ожидать нельзя.
6. ЗНАЧЕНИЕ ПРЕРЫВИСТОГО И КРАТКОВРЕМЕННОГО НОЧНОГО
ДОПОЛНИТЕЛЬНОГО ОСВЕЩЕНИЯ
В предыдущей главе было отмечено, что для получения фотопериоди-
ческого эффекта совсем не обязательно давать дополнительное беспрерыв-
ное освещение, а можно ограничиться или прерывистым освещением или
VII. Выращивание при дополнительном искусственном освещении
265
кратковременным освещением в определенные часы ночи. В некоторых
случаях, особенно в садоводстве, это обстоятельство используют или
в целях ускорения цветения растений длинного дня, или для задержки
цветения короткодневных растений.
Серия исследований в этом направлении принадлежит Артуру и его
сотрудникам [144, 383, 387, 679, 680]. Эти авторы сконструировали
специальную теплицу — термостат. Эта теплица представляет собою
большой термостат (2,4 X 5,8 м) с глухими теплоизолирующими стенками
и односкатной стеклянной крышей, наклоненной на юг. Температура
в теплице поддерживается за счет солнечной радиации и 10 ламп накали-
вания в 500 вт (350 вт/м2), которые включаются или выключаются с по-
мощью реле по мере падения или повышения температуры. Опыты пока-
зали, что для поддержания температуры на уровне 16° достаточно,
чтобы в зимнее время лампы горели ежесуточно в среднем по 4 часа,
а количество расходуемой энергии равнялось 18 квт-час на теплицу
(14,1 м2) или 1,28 квт-час на 1 м2 в сутки. В этих условиях, при преры-
вистом освещении, нормально развиваются и цветут крестовник {Senecio
cruentus), душистый горошек {Lathyrus odoratus), львиный зев {Antirrhinum
majus), молочай {Euphorbia pulcherrima), паслен {Solanum pseudocapsicum),
башмачки {Calceolaria sp.), бегония {Begonia semper) lor ens), фуксия {Fuch-
sia hybrida), гардения {Gardenia jasminoides), гречиха {Fagopyrum
esculentum) и другие. Растения в теплице-термостате выглядят значи-
тельно лучше, чем в обычной теплице без освещения. Все растения, за
исключением короткодневных крестовника и молочая, цветут значительно
ранее, чем без дополнительного освещения [383].
В другом исследовании [387] авторы сравнили теплицу-термостат
с обычной неосвещаемой теплицей и теплицей, освещаемой также лампами
накаливания в 500 вт. В последнем случае лампы включались или выклю-
чались синхронно с лампами в теплице-термостате при помощи одного
и того же реле, регулирующего температуру в теплице-термостате. Послед-
няя в различные годы колебалась в пределах 13—20°. В среднем лампы
горели 3 часа 24 минуты в сутки в период с декабря по февраль и 1 час
36 минут — в ноябре или марте. Ежесуточный расход энергии на теплицу-
термостат, в зависимости от времени года, колебался от 13 до 18 квт-час.
В качестве объектов исследования служили те же растения, что и в преды-
дущем году. Кроме того, были испытаны бегония {Begonia rex), орхидея
{Cymbidium insigne), гвоздика {Dianthus caryophyllus), земляника (Fra-
garia chiloensis), гербера {Gerbera jamesoni), ’гладиолусы {Gladiolus
Childsii, G. gandavensis, G. primulinus), гиацинты {rlyacynthus orientalis),
лилии (Li Hum longiflorum var. giganteum, L.l. var. eximium, L. tennifoliam),
левкой {Matthiola annua), мак {Papaver orientale), пеларгония {Pelar-
gonium zonale), петуния (Petunia hybrida), первоцвет {Primula sp.), роза
{Rosa sp.). Как и в предыдущих опытах, все растения, за исключением
короткодневных молочая и крестовника, при прерывистом освещении
нормально развивались и цвели. Авторы нашли, что при дополнительном
прерывистом освещении вес растений был значительно выше, а цветение —
значительно раньше, чем в обычный неосвещаемой теплице (контроль).
Особенно большое различие наблюдалось в наиболее темные зимние ме-
сяцы при сильном падении наружной температуры. В этом случае, в связи
с частым включением ламп и незначительной естественной освещенностью,
растения в теплице-термостате развивались почти одинаково с расте-
ниями в обычной освещаемой теплице, несмотря на лучшее естественное
освещение в последней. В дальнейшем, с увеличением естественной осве-
щенности и уменьшением числа включений ламп, лучшие результаты
266
Общая светокультура растений
наблюдались в обычных освещаемых и даже неосвещаемых теплицах
(табл. 114).
Таблица 114
Сухой вес одного растения гречихи при дополнительном
прерывистом освещении в теплице-термостате и обычной
теплице
(Артур и Гарвил [387])
Сухой вес одного растения (в г)
Период выращивания Контроль (обычная теп- лица без осве- щения лампами) Обычная теп- лица, освещаемая лампами Теплица-термо- стат, освещаемая и отапливаемая лампами
7/Х1— 11/XII 0,22 0,27 0,24
11/XII —14/1 0,14 0,52 0,40
14/1— 4/1II 1,10 3,06 1,33
4/Ш—14/IV 1,32 1,92 0,56
В специальном опыте с гладиолусами эти авторы [388] нашли, что
с помощью прерывистого освещения в ночные часы (в сумме 3—5 часов)
можно заставить цвести в зимнее время многие сорта гладиолусов. При
посадке луковиц в середине или конце октября растения зацветают в ян-
варе—марте, т. е. через 80—160 дней, вместо обычных для летнего
времени 80—110 дней. Наперстянка (Digitalis purpurea), которая обычно
зимою не цветет, легко зацветает, если после воздействия низкими тем-
пературами получит прерывистое освещение [389].
Таким образом, в наиболее темные и холодные зимние месяцы
(ноябрь—январь) многие растения можно выращивать в теплицах-
термостатах, освещаемых и отапливаемых исключительно солнцем
и лампами накаливания, при прерывистом дополнительном освещении.
Юм [558] культивировал шпорник (Delphinium afacis), хризантемы
(Chrysanthemum segetum), левкой (Matthiola incana) и астры (Callistephus
chinensis) при дополнительном прерывистом освещении в зимние и
весенние месяцы. Он применял лампы накаливания в 25—75 вт (13,5—
55 вт/м2), создавая дополнительное освещение (8—500 лк, или 50—3000
эрг/см2>сек физиологической радиации), в сумме рагное 4—6 часам.
Опыты показали, что у астр и хризантем время цветения не зависит от
характера прерывистости, а у шпорника и левкоя цветение раньше всего
наступает при частой смене света и темноты (через 43—44 секунды). В этом
случае цветение наступает даже несколько раньше, чем при беспре-
рывном дополнительном освещении в течение 4 часов.
Стьюарт [481, 792] воспользовался кратковременным ночным освеще-
нием для задержки цветения короткодневных хризантем (Chrysanthemum
sp.). Автор нашел, что кратковременное освещение (30—60 минут) в сере-
дине ночи при интенсивности 160—1000 лк вызывает у многих сортов
хризантем задержку в наступлении цветения на 2—3 месяца, сдвигая
время цветения на январь—февраль.
В. М. Катунский [99], пользуясь лампами накаливания, выращивал
фасоль (Phaseolus vulgaris), просо (Panicum miliaceum) и горох (Pisum
sativum) при освещенности 5000 лк (20 тыс. эрг/см2-сек физиологической
радиации) в условиях длинного (18 часов) и короткого (12 часов) дня,
причем в обоих случаях в ночные часы темнота каждый час прерывалась
5-минутным освещением. Контрольные растения развивались на длинном
(18 часов 30 минут) и коротком (13 часов) дне и при беспрерывном освеще-
VII. Выращивание при дополнительном искусственном освещении
267
нии. Опыт показал, что при прерывистом ночном освещении растения
ведут себя, как при беспрерывном освещении или на длинном дне: раз-
витие растений длинного дня уск ряется, а цветение короткодневных
растений задерживается. В другом опыте с фасолью и горохом, когда
растения выращивались в условиях короткого дня (9 часов), а темный
(15 часов) период прерывался каждые 3, 5 и 7]/9 часа соответственно
3, 5 и 15 минутами света, В. М. Катунский нашел, что для ускорения
развития длиннодневных и задержки развития короткодневных растений
совсем не обязательно давать беспрерывное освещение, а достаточно
каждые 3 часа освещать растения
в течение 3 минут.
Более подробно этот вопрос иссле-
довали Клаэс и Ланг [445], которые
выращивали белену (Hyoscyamus ni-
ger) в условиях 9-часового дня с до-
полнительным «ночным» освещением,
даваемым в различные часы «ночи».
Как видно из кривых на фиг. 148,
наибольший эффект получается, если
дополнительное «ночное» освещение
давать через 8—10 часов после окон-
чания «дня», причем 2 часовое осве-
щение более эффективно, чем 1-часо-
вое. В первом случае стрелкование
наступает одновременно с контроль-
ными растениями, находящимися на
длинном дне, во втором случае —
позднее на 3 дня.
Аналогичные результаты для ко-
роткодневного растения каланхое
(Kalandiwi Blossfeldiana) получили
Гардер и Боде [533]. При экспози-
ции в 2—32 минуты и освещенно-
сти в 1000 лк наиболее сильный эф-
фект (задержка цветения) получался
при освещении через 6—9 часов по-
сле окончания «дня» (фиг. 149).
Эти авторы выращивали также каланхое в условиях 13-часового дня
о дополнительным освещением в середине ночи продолжительностью по
1 минуте или по 6 минут при освещенности 300, 700, 1500, 3000, 6000,
12 000, 25 000 и 50 000 лк. Авторы нашли, что полное устранение цветения
наблюдается лишь при количестве света, равном 9000 лк-мин (~ 3 000 000
эрг/см2). По мере уменьшения этой величины растет количество цветков
и длина соцветий (фиг. 150).
Специального интереса заслуживают так называемые «световые удары»
В. П. Мальчевского [193]. Сущность этого приема сводится к тому, что
растения, выращиваемые в обычных условиях, в оранжерее, подвергаются
ежедневным кратковременным и редким воздействиям лучистой энергии
с помощью источников радиации, богатых длинноволновыми лучами
(лампы накаливания, неоновые лампы). «Световые удары» производятся
в дневные часы каждые 2'/2—3 часа и длятся 5—20 минут. В. П. Маль-
чевский применял лампы накаливания в 1000 вт, создававшие освещен-
ность порядка 20 000 лк (120 000 эрг/см2-сек физиологической радиации),
и высоковольтные неоновые трубки, с помощью которых на уровне
I 10
О 1 г 3 4 5 6 7 8 S <0 IS 12 13 /4 15
Число часов ат начала „ ночного ’ периода
Фиг. 148. Число дней до стрелкования
белены (Hyoscyamus niger) в зависимо-
сти от дополнительного «ночного» осве-
щения, даваемого в различные часы
«ночного», периода
7 — длительность освещения 1 час;
2 — длительность освещения 2 часа
268
Общая светокультура растений
растений создавалась интенсивность физиологической радиации порядка
1000 эрг/см1 2-сек. На пути лучистого потока, получаемого с помощью ламп
накаливания, ставились водные экраны (0,5 см) с проточной водой,
Фиг. 149. Эффективность кратковременного освещения
каланхое (Kalanchoe Blossfeldiana) в различные часы ночи
1—число цветков на одном растении; 2— число дней до цветения
поглощающие инфракрасную радиацию. В опытах с фасолью (Phaseolus
vulgaris) «Золотая гора» и салатом {Lactuca sativa ) автор показал, что
Фиг. 150. Влияние количества света при кратковременном
«ночном» освещении (1 — 6 минут) на развитие каланхое
(Kalancho'e Blossfelcliana) в условиях 13-часового дня
1 — длина соцветия; 2 — число цветков
«световые удары» несколько ускоряют наступление цветения и увеличи-
вают сухой вес растений. При 20-минутном «ударе» с помощью ламп
накаливания бутонизация ускоряется на 7 дней, а сухой вес растения
увеличивается на 28%. Такой же «удар» светом неоновых ламп вызывает
ускорение бутонизации на 11 дней и увеличение сухого, веса на 94%.
VII. Выращивание при дополнительном искусственном освещении 269
К сожалению, исследования в этом направления не получили дальней-
шего развития, а в настоящее время, в связи со смертью В. П. Мальчев-
ского, полностью прекращены.
7. ЗНАЧЕНИЕ ВРЕМЕНИ ГОДА ДЛЯ ДОПОЛНИТЕЛЬНОГО ОСВЕЩЕНИЯ
Специального интереса заслуживает вопрос о значении и эффектив-
ности дополнительного освещения в зависимости от времени года. Ис-
следований в этом направлении очень немного. Помимо опытов Артура
и Гарвила [387], на которых мы уже останавливались (табл. 114), нам
следует рассмотреть еще несколько работ.
Освещенность (в лв)
Фиг. 151. Количество цветков у анютиных глазок {Viola tricolor)
в зависимости от интенсивности дополнительного освещения лампами
накаливания и времени года
1 — в декабре; 2 — в январе; 3 — в феврале; 4 — в марте
Грин и его сотрудники [521] при дополнительном освещении в разные
месяцы выращивали анютины глазки {Viola tricolor). Как видно из кри-
вых, приведенных на фиг. 151, оптимальная освещенность, полученная
с помощью ламп накаливания, равна 400—500 лк (2400—3000 эрг/см1 2-сек
физиологической радиации). Дальнейшее увеличение интенсивности
дополнительного освещения в любое время года или мало эффективно
(декабрь—январь) или дает даже отрицательный результат (февраль—
март). Количество цветков увеличивается по мере перехода от зимних
к весенним дням.
Аналогичные данные получил Ф. Э. Реймерс [271], который в различ-
ные сроки (2 ноября, 3 января и 10 марта) выращивал корнеплоды столовой
свеклы {Beta vulgaris) египетской, давая растениям дополнительное
освещение (330 вт/м2), Автор установил, что стрелкование как при освеще-
нии, так и в контроле наступает тем раньше, чем позднее высаживаются
корнеплоды. Несомненно, что здесь, помимо прохождения яровизации,
имела значение естественная освещенность в дневные часы. Специальные
опыты Фабиан [488] не оставляют сомнений в этом вопросе. Эта исследо-
вательница, затеняя растения в дневные часы и давая дополнительное
270 Общая светокультура растений
освещение ночью, нашла, что длина вегетационного периода как длин-
нодневных — ячменя (Яordeum vulgare), куколя (Agrostemma t.ithago), так
и короткодневного — могара (Setaria italica) растений тем короче,
чем выше интенсивность радиации в дневные часы (фиг. 152). Это обстоя-
тельство в условиях недостаточного дневного освещения ставит задачу
или увеличения интенсивности ночного освещения, или дополнительного
дневного освещения. В первом случае основное значение для растений
приобретает искусственное освещение, а естественный свет начинает
играть лишь второстепенную роль — как дополнительный свет, удли-
няющий искусственный день. Именно в этой связи следует отметить
Фиг. 152. Влияние интенсивности основного и дополнительного освещения
на колошение
А—ячмень (Hordeum sp.); Б —могар {Setaria italica)
1 — без затенения; 2 — при затенении на 50%; 3 — при затенении на 66% интенсив-
ности естественной летней радиации
указания ряда авторов, которые для северных районов (Голландия,
Скандинавия, приполярные районы СССР) рекомендуют сравнительно
высокие инсивности дополнительного ночного освещения.
С. В. Тагеева [307] высевала растения фасоли (Phaseolus sp.) каждые
12—30 дней, начиная с 30 ноября и кончая 6 апреля, и культивировала
их в течение 28—31 дня под лампами накаливания в 500 вт, горевшими
ночью, в зависимости от времени посева, по 16—9 часов. Контрольные
растения высевались в те же сроки и развивались без дополнительного
освещения. В результате этих опытов (фиг. 139) было найдено, что эффект
от дополнительного освещения, выражающийся в привесе сухой массы
растений по сравнению с весом семян, имеет место как в зимние, так и
в весенние месяцы (декабрь—апрель). При этом максимальный относитель-
ный эффект наблюдается зимой (декабрь—январь), а весною, по мерс
увеличения длины дня и интенсивности естественного освещения, сильно
уменьшается, хотя абсолютная величина эффекта, в соответствии с дан-
ными Грина и Реймерса, растет. В декабре—январе при дополнительном
освещении при помощи ламп накаливания в 500 вт растения развиваются
так же, как контрольные растения в конце*февраля или начале марта.
Применение ламп накаливания в 1000 вт в зимние месяцы (январь) зна-
VII. Выращивание при дополнительном искусственном освещении 271
чительно повышает эффективность дополнительного освещения по сравне-
нию с лампами в 500 вт; в этом случае опытные растения не уступают
контрольным при развитии последних в апреле.
8. ТЕМПЕРАТУРНЫЙ РЕЖИМ ПРИ ДОПОЛНИТЕЛЬНОМ ОСВЕЩЕНИИ
Исключительное значение в условиях светокультуры приобретает
вопрос о температурном режиме растений. Отправильного регулирования
температуры зависит как переход растений к цветению, так и величина
урожая.
Во-первых, многие растения нуждаются в определенных низких
температурах для прохождения стадии яровизации. Во-вторых, для
нормального роста одних растений необходимы высокие температуры
(теплолюбивые растения), другие же в этих условиях сильно страдают.
У многих растений прохождение световой стадии сильно зависит от тем-
пературы (табл. 115).
Таблица 115
Значение температуры воздуха для развития растений
(Робертс и Стракмэйер [716])
Температура воздуха
13° С
17—18° С
21—24° С
Растения
л
и
в
ч
и
«
Е
ч
к
ч
ч
Ч
к
к
к
Мятлик {Роа pratensis)......................
Гвоздика {Dianthus barbatus)................
Соя {Soja max Чиппева)......................
Люцерна {Medicago sativa)...................
Эпифиллум {Zygocactus truncalus)............
Бегония {Begonia semper jlorens) ...........
Шалфей {Salvia splendens).............. . .
Петуния {Petunia hybrida).............. . .
Шпинат {Spinacea oleracea)..................
Космос {Cosmos sulphureus)..................
Бувардия {Bouvardia humboldtii).............
Просвирник {Malva splendens)...............|
Дурнишник {Xanthium echinatum)..............
Хризантемы {Chrysanthemum morijoliuin) . .
Ипомей {Ipomaea purpurea)..................j
Молочай {Euphorbia pulcherrima)............i
Примечание. Знак означает цветение,
Перегрев растений, особенно при употреблении ламп, богатых инфра-
красными лучами, представляет собою явление, наиболее часто наблюда-
емое в условиях светокультуры. При этом имеет значение не столько тем-
пература среды, сколько температура тканей растений, особенно листьев.
Перегрев растений является тем фактором, который чаще всего огра-
ничивает возможности повышения удельной мощности за пределы 500 вт/м2
и вызывает страдание растений и низкий урожай. К сожалению, вопрос
272
Общая светокультура растений
о температурном режиме растений и температурном балансе листьев
в условиях светокультуры представляет наименее разработанную про-
блему. Несомненно лишь одно, что при употреблении ламп, богатых инфра-
красными лучами, растения нуждаются в несколько меньших темпера-
турах воздуха, чем в естественных условиях или при применении так
называемых «холодных» ламп, в спектре которых мало инфракрасных
лучей. При употреблении излучателя инфракрасной радиации (лампы
накаливания и другие) и наличии почвенного обогрева выращивание
растений возможно даже при температуре воздуха ниже нуля.
В этом отношении показательны опыты Бижо [300, 304, 413, 414,
440, 783], который, пользуясь специальной отопительной системой, в зим-
ние месяцы выращивал растения на открытом воздухе при температуре
—11° и при наличии снежного покрова в 10 см. Названная отопительная
система представляет систему стальных змеевиков, входящих в бетонные
блоки, располагаемые в междурядьях растений на расстоянии 0,4—1,3 м
друг от друга, и систему труб, проложенных в почве. Через змеевики
и трубы пропускается ток воды с температурой порядка 40°. Растения,
находящиеся между блоками, поддерживают свой температурный режим
за счет инфракрасной радиации бетонных стенок, нагреваемых водою.
Автор без всякого остекления добивался цветения и плодоношения многих
растений [азалии {Azalea indica}, цикламены {Cyclamen persicum), розы
{Rosa sp.), лилии {Lilium sp.), гортензии {Hydrangea hortensis}, камелии
{Camelia japonica) {Cerasus sp.), земляника {Fragaria sp.), цитрусовые
{Citrus sp. sp.), плодовые и овощные культуры] в середине зимы (февраль).
В. Г. Карманов [97, 217, 224] выращивал редис {Raphanus sativa var.
radicula} в зимние месяцы при температуре —5 —18°, пользуясь лампами
накаливания как источником инфракрасных лучей. В его опытах редис
в течении 20 суток образовывал корнеплод в 30 г. В последние годы лучи-
стый обогрев растений приобретает практическоЛзначение [6].
Говоря о температурном режиме растений в условиях светокультуры,
следует также иметь в виду, что растения, выращенные при дополнитель-
ном освещении, резко снижают свою морозостойкость. Это обстоятельство
следует учитывать при пересадке растений в грунт, особенно при неблаго-
приятных метерологических условиях ранней весной или поздней осенью.
Сказанное установлено для многих озимых растений: клевера {Tri-
folium sp.), люцерны {Medicago sp.), ржи (Secale sereale), пшеницы {Triti-
cum vulgare} и др. [309, 346, 347, 360, 819], а также для древесных: ивы
{Salix viminalis, S. philicifolia^, смородины {Ribus nigrum}, абрикоса
{Prunus armeniaca), абелии {Abelia grandiflora}, акации {Robinia pseudaca-
cia}, апельсина {Citrus sinensis}, лимона {C. li ron), мандарина {C. no-
bilis} и других [82,2 )7, 585]. Озимая рожь и пшеница, выращенные в обыч-
ных естественных условиях, вымерзают при —14 —16° на 20—50%,
а после пребывания под лампами накаливания в точение 15 дней погибают
почти целиком — на 93—100 % [347]. Величины того же порядка получены
для клевера и желтой люцерны [360].
Крамер [585], наблюдавший за ростом абелии {Abelia grandiflora}
и акации {Robinia pseudacacia}, находящихся близ уличных фонарей,
обнаружил, что наблюдается полнейший параллелизм между интенсив-
ностью освещения, ростом побегов и степенью их вымерзания зимою.
Из сказанного следует, что при высадке в грунт растений, находившихся
в условиях светокультуры, следует создавать для озимых условия за-
калки, а для древесных необходимо обеспечить условия для одревеснения
побегов (своевременная, достаточно ранняя высадка). В этих условиях
можно избежать вымерзания растений зимою.
VII. Выращивание при дополнительном искусственном освещении
273
Т а б л и ц а 116
Влияние дополнительного освещения и удобрения С02 на рост и развитие
растений
(Гардер, Кепплер и Ройс [530])
О И (-ко и ч: СЗ К Диаметр сопветий Длина побегов -это: Урожай ягод
1 с5 3 gs о •a '8 ° s ощад 11.1 ( И О со а (в см) 5 О на 27/XII (в см) плодов к 7/IV
Варианты опытов О X н макси- мальный сЗ
- Начало Число Д явления продукц Общая г листьев н Число ц 10 расте: средний Высота I (в см) средняя макси- мальная о ЧИСЛО 1 ягод к
Гортензия (Hydrangea
hortensis) Мадам Мульер
Контроль 25 XI 80 — 38 3,5 9,5 — — — — —
Удобрение СО2 .... 25/XI 80 — 44 4,2 11,5 — — — — —
Лампы накаливания . . Лампы накаливания + 25/XI 44 — 52 9,9 17,0 •— — — — — —
-г удобрение СО-2 • • 25/XI 44 — 46 11,4 16,0 — — — — — —
Шотар
Контроль 22/X CO 34,1 3 1,2 1,3 — — 16 17 . — -—
Удобрение СО2 .... 22/X oo 79,8 14 2,2 4,6 — 27 45 — — —
Лампы накаливания . . 22/X 67 143,8 40 6,9 12,0 — 75 150 — — —
Лампы накаливания Д
+ удобрение СО2 . . 22/X 57 1534,2 41 9,8 16,0 — 131 250 — — —
Маршал Фош
Контроль 22/X CO — 7 3,8 5,5 —— — — — —
Удобрение СО2 .... 22/X CO — 3 о,4 1,1 — — — —. — —
Лампы накаливания . . Лампы накаливания 22/X 73 — 28 8,1 14,0 — — — — — —
4- удобрение СО2 . . Цинерария (Cineraria 22/X 73 — 22 10,8 18,0 —- — — — — —
hybrida)
Контроль 3/1 54 — 10 — — — — — —- — —
Удобрение СО2 .... 3/1 33 — 42 — — — — — —. — —
Лампы накаливания . . Лампы накаливания 4- 3/1 33 — 135 — — — — — — — ——
4- удобрение СО2 . . ЗД 30 — 124 — —- —- — — — — —
Колокольчик (Campa-
nula media)
Контроль 2/1 CO — 0 — — 80 .— —. — — —
Удобрение СО2 .... 2/1 90 — 0 — — 147 — — —. — —.
Лампы накаливания . . Лампы накаливания 4- 2/1 55 — 10,4 — — 490 — — — — —
4 удобрение СО2 . . 2/1 48 — 13,8 — — 516 — — — — —
Касатик (Iris pumila) •
Контроль 2/1 20 — 3 — — — — — -
Удобрение СО2 .... 2/1 17 — 11 — — —- — — — — —
Лампы накаливания . . Лампы накаливания 3- 2/1 15 — 23 — — — — — — — —
4 удобрение СО2 . . 2/1 14 — 34 — — —• — — — — —'
18 А. Ф. Клешнин
274
Общая светокультура растений
Таблица 116 {продолжение)
Варианты опытов
Диаметр
соцветий
Длина
побегов
на 27/XII
(в см)
Урожай
ягод
к 7/IV
Земляника {Fragaria sp.)
Контроль ...........
Удобрение СО2 ....
Лампы накаливания . .
Лампы накаливания 4-
+ удобрение СО2 •
9. УДОБРЕНИЕ УГЛЕКИСЛОТОЙ
Удобрение растений углекислотой повышает эффективность свето-
культуры. Особенно остро необходимость в этом мероприятии возникает
в зимние месяцы в условиях защищенного грунта. Исследования в этом
направлении ведутся давно, и в настоящее время накоплен сравнительно
большой экспериментальный материал, позволяющий сделать определен-
ные заключения.
Наиболее обстоятельные исследования по совместному применению
С09 и дополнительного освещения произведены в цветоводстве. Артур
с сотрудниками [379] в опытах с настурцией (Tropaeolum majus), розами
(Rosa sp.), пеларгонией (Pelargonium sp.) и амброзией (Ambrosia arte-
misijolia) показали, что совместное действие освещения и СО2 дает лучший
эффект, чем каждый фактор в отдельности. Розы в их опытах цвели в апреле,
а вес растений амброзии под лампами накаливания при удобрении СО9
был значительно выше, чем под одними лампами или в контроле (табл. 119).
Раньше зацветают также настурции и пеларгония, причем настурция
страдает от дополнительного освещения, если одновременно атмосфера
не обогащена СО2.
Обширное исследование по этому вопросу принадлежит Гардеру с со-
трудниками [530]. Эти авторы с помощью ламп накаливания в 500 и 750 вт
выращивали в зимнее время львиный зев (Antirrhinum majus), бархатцы
(Tagetes patula папа), касатик (Iris pumila), колокольчик (Campanula
media), цинерарию (Cineraria hybrida) и гортензии (Hydrangea hortensis
сорта Шотар, Маршал Фощ и Мадам Мульер). Лампы висели над стел-
лажем на высоте 1 м и на расстоянии 1,2 м друг от друга (350 и 520 вт/м2).
Содержание СО2 колебалось от 0,15% ночью до 0,25% в пасмурные и
0,35% — в солнечные дни. Температура равнялась 20°. Схема и резуль-
таты опытов приведены в табл. 116. Обширные данные, полученные назван-
ными авторами, не оставляют сомнений в том, что лучший эффект в боль-
шинстве случаев дает совместное действие дополнительного освещения
и СО2, причем эффективность одного лишь освещения приближается
к эффективности совместного действия факторов, а эффективность одной
лишь СО2 настолько ничтожна, что растения по всем показателям почти
не отличаются от контроля. Аналогичные данные для зимней культуры
земляники (Fragaria sp.) получены голландскими авторами [518].
VII. Выращивание при дополнительном искусственном освещении
275
Таблица 117
Влияние дополнительного освещения и удобрения С02 на рост и развитие растений
(Красинский [131, 136])
Варианты опытов Число дней до цветения 1 Высота растений (в см) Площадь листьев (в см’) Длина по- бегов (в см) Число цветков Диаметр цветков (в см)
Цинерария (Cineraria hybrida) Опыт 1. Освещение в фазе бутонизации Контроль 60
Удобрение С0.2 45 — — — — —
Лампы накаливания ночью (23—7 час.) ф- ф- удобрение СО2 30 — — — — —
Опыт 2. Освещение в фазе стрелкования Контроль 75—90 32,1 955 — — —
Удобрение СО2 75—90 29,6 1028 — — —
Лампы накаливания днем (7—15 час.) . . — 37,2 1002 — — —
Лампы накаливания ночью (23—7 час.) . 60 44,5 912 — — —
Лампы накаливания днем (7—15 час.) ф- -|- удобрение СО2 — 40,7 1048 — — —
Лампы накаливания ночью (2.’>— 7 час.) -ф ф- удобрение СО2 45 45,0 991 — — —
Опыт 3. Освещение в фазе розетки Контроль 120 15,0 — — — —
Удобрение СО2 120 11,0 — — — —
Лампы накаливания днем (7—15 час.) . . — 26,7 — — — —
Лампы накаливания ночью (2-1—7 час.) . 90 22,0 — — —
Лампы накаливания днем (7—15 час.) -ф ф- удобрение СО2 — 26,1 — — — —
Лампы накаливания ночью (23 7 час.) ф- 4- удобрение СО2 60 41,0 — — — —
Гортензия (Hydrangea hortensis) Контроль 135—150 — 280 17,4 —.. -—
Удобрение СО2 135—150 — — 24,8 — —
Лампы накаливания днем (7—15 час.) . . 120 — 337 30,5 — -—
Лампы накаливания ночью (23—7 час.) . 105 — 359 29,5 — -
Лампы накаливания днем (7—15 час.) ф- -ф удобрение СО2 105 — 404 40,0 — •
Лампы накаливания ночью (23—7 час.) ф- ф- удобрение СО2 75 — 484 44,5 --
Роза (Bosa indica) Контроль 90 . --
Удобрение СО2 90 — — — — —
Лампы накаливания днем (7—15 час.) . . — — — — — —
Лампы накаливания ночью (23—7 час.) . 75 — — — —
Лампы накаливания днем (7 15 час.) ф- -ф удобрение СО2 60 — — — — —
276
Общая светокультура растении
Варианты опытов
Лампы накаливания ночью (23—7 час.)-|-
+ удобрение СО2 .....................
Первоцвет {Primula obconica)
Контроль ............................
Удобрение С02........................
Лампы накаливания днем (7—15 час.) . .
Лампы накаливания ночью (23—7 час.) .
Лампы накаливания днем (7—15 час.) -|-
+ удобрение С02 ...................
Лампы накаливания ночью (23—7 час.) -|-
-р удобрение С02.....................
Таблица '117 {продолжение)
Серия работ принадлежит Н. II. Красинскому [131—140J, который
в производственных условиях на массовом материале исследовал реакцию
большого числа растений — первоцвета {Primula obconica), цинерарии
{Cyneraria hybrida), розы {Rosa indica), левкоя {Matthiola incana, M. annua),
гортензии {Hydrangea hortensis), лилии {Lilium candidum), цикламена
{Cyclamen persicum) и др. — на дополнительное освещение и обогащение
атмосферы СО2. Автор применял лампы накаливания в 1000 вт, помещая
их на высоте 50—75 см от растений. Удельная мощность равнялась 450 вт/м2,
а содержание СО2 колебалось в пределах 0,2 — 0,3%. Опыт длился
с 26 ноября по 1 апреля. Лишь лилия и цикламен не дали положительной
реакции на дополнительное освещение и СО.,. Остальные же растения
под влиянием этих факторов значительно ускорили цветение и дали про-
дукцию лучшего качества (табл. 117). Роза дала крупные бутоны с пре-
красным ароматом, левкой — крупные, эффектные и сильно пахнущие
цветки, а растения примулы не уступали обычным весенним растениям.
Время освещения не играло никакой роли для левкоя, розы и примулы,
а цинерария и гортензия предпочитали ночное освещение. Дополнитель-
ное освещение без одновременного обогащения атмосферы СО2 действо-
вало всегда слабее, а одна углекислота эффекта, как правило, не давала.
Опыты Н. П. Красинского были продолжены М. Б. Равняем и
А. П. Сарычевой [262—264, 287]. Авторы применяли лампы накаливания
в 300 вт, которые находились на высоте 50—75 см от растений и на рас-
стоянии в 1 м друг от друга (300 вт/м2). Дополнительное освещение, начи-
навшееся во второй половине января, производилось дважды в сутки:
с 0 до 6 и с 12 до 14 час. Концентрация СО., равнялась 0,3 — 0,4%, а тем-
пература была одинаковой во всех вариантах и колебалась в пределах
13,2 — 23,0°. Как показывает табл. 118, дополнительное освещение и
СО2 действуют лучше совместно, чем в отдельности. Это выражается
в ускорении цветения, лучшем развитии растений, большем количестве
цветков и т. д. Аналогичные результаты были получены также в другом
опыте с лакфиолем {Cheiranthus cheiri), цинерарией {Cyneraria hybrida),
VII. Выращивание при дополнительном искусственном освещении
277
Таблица 118
Влияние дополнительного освещения и удобрения С02 на рост и развитие растений
(Сарычева [287])
Варианты опытов Высота рас- тений (в см) Площадь листьев (в с№)! Начало цветения Полное цветение Число дней до цветения Диаметр со- цветий (в см) Диаметр ро- зетки (в см) Число цветков Число соцветий
Гортензия (Hydrangea hortensis)
Контроль ........................
Удобрение СО2....................
Лампы накаливания ...............
Лампы накаливания С удобрение1 С О2
41,1
I 46,3
53,4
57,8
Цинерария (Cineraria hybrida)
Контроль .........................
Удобрение СО2 ....................
Лампы накаливания ................
Лампы накаливания + удобрение СО2
Глоксиния (Gloxinia sp.)
Контроль .........................
Удобрение С02 ....................
Лампы накаливания ................
Лампы накаливания + удобрение СО2
Левкой (Matthiola incana)
Контроль .........................
Удобрение СО2.....................
Лампы накаливания ................
Лампы накаливания удобрение СО2
Примула (Primula obconica)
Контроль .........................
Удобрение СО2 ....................
Лампы накаливания ................
Лампы накаливания-Г удобрение СО2
гортензией {Hydrangea hortensis), левкоем {Matthiola incana) и розой
{Rosa indica).
Значительно меньшее количество исследований проведено с овощами
и другими сельскохозяйственными растениями. Здесь можно назвать
лишь исследования Артура и его сотрудников [144, 379] и несколько
других работ. В опытах Артура применялись лампы накаливания в 1000 вт,
создававшие освещенность 7600 и 8190 лк (45 600 — 49 140 эрг/см2-сек
физиологической радиации). Лампы горели в течение 6 часов ежедневно,
а содержание СО2 повышалось до 0,3%. Опыты показали, что в зимнее
время наилучшие результаты в большинстве случаев имеют место при
дополнительном 6-часовом ночном освещении и обогащении атмосферы
углекислотой (табл. 119).
В опытах Гардера и его сотрудников [530] огурцы {Cucumis sativus),
посеянные 12 августа и с И ноября подвергавшиеся дополнительному
278
Общая светокультура растений
Таблица 119
Влияние дополнительного освещения и удобрения С02 на вес растений
(Артур, Гутри и Ньюэлл [379])
Растения Возраст (в днях) Вес одного растения (в г)
контроль дополни- тельное освещение со2 + до- полни- тельное освещение
Огурцы (Cucumis sativ us) — 134 244 315
Картофель (Solanum tuberosum) — 271 495 323
Пшеница (Triticum vulgare) 48 3,4 3,3 6,3
Овес (Avena sativa) 51 3,5 1Д 9,0
Ячмень (Hordeum, vulgare) ......... 30 К7 I,4 3,4
Соя (Soja max) Билокси . 40 14,2 52,8 50,8
» » » Токио 40 11,6 30,8 52,1
» » » Мандарин 40 7,8 35,8 32,1
Клевер красный (Trijolium pratense) .... 76 1,1 6,0 9,2
Табак (Nicotiana labacum) 38 866 1230 1295
Амброзия (Ambrosia artemisifolia) 60 260 333 598
освещению и воздействию С09, начали плодоносить на 18 дней раньше,
чем в контроле, повысив одновременно урожай на 11%. Мы видели
(табл. 116), что земляника (Fragaria sp.) также дает положительный
эффект.
Таблица 120
Плодоношение и урожай томатов Лучший из всех при дополнительном освещении
и удобрении СО2
(Джалилова [354])
Варианты опытов Начало плодоно- шения Ускорение плодоно- шения по срав- нению с контро- лем (в днях) Урожай плодов
в кг/м- R в /о к контро- лю
Опыт 1. Посев 1.1
Контроль 28/V — 8,62 100
Лампы накаливания — — 7,91 91,6
Лампы накаливания + удобрение СО2 . . . 5/V 23 8,99 104,5
О п ы т 2. Посев 20/1 /
Контроль 10/VI — 8,77 100
Лампы накаливания . . — — 9,41 107,4
Лампы накаливания -[ удобрение СО2 . . . 23/V 18 10,22 116,4
О п ы т 3. Посев 2/11
Контроль 15/VI — 7,72 100
Лампы накаливания — — 9,94 129,0
Лампы накаливания + удобрение СО2 . . . 8/ \ I 7 10,85 141,0
VII. Выращивание при дополнительном искусственном освещении
279
Р. А. Джалилова [354], пользуясь горшечными культурами томатов
(Solanum lycopersicum), с помощью дополнительного освещения и СО2
получала первые плоды 22 февраля (при посеве 1 ноября). При дополни-
тельном освещении длительностью 11 часов и при различных сроках
посева она получила результаты, приведенные в табл. 120.
С помощью дополнительного освещения и удобрения атмосферы СО.,
Джалиловой удалось в течение 9 месяцев провести две ротации и полу-
чить урожай томатов в 14 кг/м2.
Таким образом, опыты по совместному применению дополнительного
освещения и обогащения атмосферы углекислотой показывают, что оба
фактора действуют значительно лучше совместно, чем в отдельности.
В этом случае ускоряется переход к цветению и плодоношению, увели-
чивается продуктивность и урожай и улучшается качество продукции.
Обычно концентрация СО., доводится до величины порядка 0,2—0,3%.
Наиболее просто и удобно удобрение углекислотой производится обыч-
ным опрыскивателем типа «Автомакс». При давлении в 5—6 атмосфер
объема одного опрыскивателя (20 л) достаточно, чтобы довести содер-
жание С09 до 3% в теплице объемом 30—40 м3. Опрыскиватель в свою
очеред!» заполняется СО2 с помохцью редуктора из баллона, содержащего
жидкую углекислоту. Этот прием, предложенный А. В. Крыловым
в 1950 г., с успехом применяется в производственных условиях в теп-
лицах совхоза «Краснове» (Москва). М. Б. Равич [264] рекомендует
использование газовых отходов от печей беспламенного горения.
10. ВЫВОДЫ
Таким образом, экспериментальные данные по дополнительному
освещению растений позволяют сделать следующие общие выводы.
1. Необходимость в дополнительном освещении возникает всегда,
когда день становится слишком короток (менее 12 часов), а интенсивность
естественного освещения снижается до уровня, лежащего ниже мини-
мальных потребностей растений. Чаще всего эти обстоятельства возни-
кают в зимние месяцы в умеренной зоне и полярных областях. В зави-
симости от времени года и географической широты, растения могут нуж-
даться как в н о ч н о м, так п дневном дополнительном освещении
пли в том и другом одновременно.
2. Для дополнительного освещения растений пригодны любые источ-
ники радиации: лампы накаливания, люминесцентные, неоновые, ртут-
ные, натриевые и др. Невидимому, различные растения нуждаются
в определенных типах ламп.
3. При дополнительном ночном освещении, если интенсивность есте-
ственной дневной радиации достаточно велика и задачей дополнитель-
ного освещения является простое удлинение дня в целях получения
фотопериодического эффекта, интенсивность физиологической радиации
в большинстве случаев должна быть равна 1000—2000 эрг/см2-сек. Для
ламп накаливания это соответствует освещенности 200—400 лк и по-
лучается при удельной мощности 50—75 вт'м2.
4. В овощеводстве, прп получении рассады томатов пли огурцов, при
культуре земляники, где применяется не простое удлинение естественного
короткого дня, а одновременно и повышение недостаточной интенсив-
ности естественной радиации, рекомендуется для средней полосы интен-
сивность физиологической радиации 30—50 тыс. эрг/см2-сек, т. е. осве-
щенность 5—8 тыс. лк, и удельная мощность 500 вт/м2 (для ламп нака-
ливания). Дальнейшее увеличение интенсивности освещения дополнитель-
ного эффекта не дает.
280
Общая светокультура растений
5. Интенсивность дополнительного дневного или ночного освещения,
применяемого с целью повышения недостаточной интенсивности есте-
ственной радиации, определяется временем года, географической широтой
и видом растения. Она колеблется в пределах 6—100 тыс. эрг/см2-сек
(1—16 тыс. лк для ламп накаливания), что соответствует большой
удельной мощности, достигающей иногда 1000—2000 вт/м2.
6. При определении длительности ежесуточного освещения следует
исходить из естественной длины дня, задач выращивания и фотопериоди-
ческой группы растений. Максимальный вес у большинства растений
наблюдается при суммарной длине дня 16—18 часов. Наиболее быстрое
цветение и максимальный урожай у растений длинного дня обычно на-
блюдается также при длине дня в 16—18 часов. Растения короткого дня
наиболее быстро зацветают в условиях 10—12-часового дня.
7. В некоторых случаях, при достаточной естественной освещенности,
длительное ночное освещение можно заменить или прерывистым, или
кратковременным световым воздействием в середине ночи в течение
20—30 мин. Этот прием следует особенно рекомендовать в цветоводстве,
в особенности для предотвращения цветения растений короткого дня.
8. Дополнительное освещение особенно эффективно, если одновременно
псщышается содержание СО2 в атмосфере. Концентрацию последней сле-
дует повышать до 0,2—0,3%.
Часть III
ЧАСТНАЯ СВЕТОКУЛЬТУРА
РАСТЕНИЙ
Глава VIII
СВЕТОКУЛЬТУРА ОВОЩНЫХ И САДОВЫХ РАСТЕНИИ
Культура овощных и садовых растений, в отличие от культуры цве-
точных растений, имеет целью получение максимального урожая плодов,
луковиц, клубней, корнеплодов и других органов, т. е. максимальное
накопление органического вещества. Последнее, представляя собою раз-
ность между интенсивностью фотосинтеза и интенсивностью дыхания,
в конечном итоге определяется площадью листьев и интенсивностью фи-
зиологической радиации. Сказанное наглядно иллюстрируется наблюде-
ниями Девиса и Хоэглэнда [462], В. М. Маркова [130] и других.
Девис и Хоэглэнд [462] выращивали пшеницу (Triticum vulgare)
в различное время года в условиях различной естественной солнечной
радиации, но при строгом постоянстве остальных факторов (t=20,°o).
Авторы нашли, что величина сухого веса растений в сильнейшей степени
зависит от времени года (табл. 121) и, в конце концов, определяется
количеством естественной радиации.
Таблица 121
Влияние времени года на сухой вес пшеницы (Triticum rulgare)
(Девис и Хоэглэнд [462])
Период выращивания Сухой вес растений (в г) Период выращивания ; Сухой вес растений (в г)
22/XI—20/XJ | 25.7 25/Ш —22/IV 46,2
29/XII—27/1 15,2 2/V —30/V 97,5
18/11 —18/Ш 39,9 ' 17/VIII—13/IX 86,9
В. М. Марков [130] установил четкую зависимость между урожаем
огурцов (Cucumis sativus) и количеством лучистой энергии, полученной
растениями в клинских теплицах (табл. 122).
Из сказанного следует, что для получения большого урожая плодов
овощные и другие сельскохозяйственные растения нуждаются в высокой
интенсивности радиации. Чем выше интенсивность естественной пли
искусственной радиации, тем выше урожай.
В этой связи необходимо подчеркнуть, что светокультура в овоще-
водстве должна играть специфическую и подчиненную роль, а основным
источником лучистой энергии, создающим урожай, надо признать солнце.
284
Частная светокультура растений
Таблица 122
Зависимость урожая огурцов (Cucumis sativus) от количества лучистой
энергии
(Марков [130])
Период выращивания Количество лучистой энергии Урожай плодов
нал/м2 % 1ПТ/М2 О' о
15/Х — 1/II 26 885 18,4 40 20,0
J/Ш— i/vrn 148 785 100 200 100
«Земледелец из дарового сырого материала — воздуха и даровой силы —
солнечного света изготовляет ценности; в этом главная тайна производи-
тельности его труда» (Тимирязев, соч., т. III, 86).
Большое значение светокультура растений имеет, однако, в овоще-
водстве защищенного грунта. При получении ранней рассады весной,
в семеноводстве, при культуре растений в полярных областях и т. д.
можно и нужно прибегать к искусственному освещению.
В настоящее время еще рано говорить о культуре овощных растений
целиком при искусственном освещении. Исследования, проведенные
С. И. Доброхотовой [59] и Б. С. Мошковым [216, 218,219,224], показы-
вают, что в этом направлении предстоит еще очень большая работа. Не-
смотря на то, что Б. С. Мошкову с помощью большой интенсивности ради-
ации и обогащения лучистого потока инфракрасными лучами с большой
длиной волны1 удалось значительно ускорить плодоношение томатов
(Solanum lycopersicum), для получения 1 кг плодов трубуется, однако,
около 1000 квт-час (табл. 123). Интересно, что в опытах С. И. Доброхо-
товой при длине вегетационного периода 95—118 дней требовались зна-
чительно меньшие количества энергии (400—900 квт-час/кг, табл. 124).
Таблица 123
Характеристика по различным показателям томатов (Solanum lycopersi-
cum) сортов Движение на север и Пушкинский при культуре целиком
на искусственном освещении
Лампы накаливания 200—300 вт; удельная мощность 3,2—4,8 квт/м2;
длина дня 16—20 чае.; дневная температура 22—25°, ночная темпе-
ратура 15—18°
(Мошков [216])
1. Число дней от всходов до появления зрелых плодов .... 45—50
2. Урожай плодов за 60 дней культуры с одного растения
количество................,............................... 5—8
вес (в г)............................................ 250—300
3. Вес одного плода (в г).................................... 50—75
4. Урожай с 1 м2 (в кг)*.................................. 2,5—3,0
5. Количество энергии за вегетационный период (в квт-час/м2) 3500
6. Количество энергии на 1 кг плодов (в квт-час) **.......... 1000
* Рассчитано нами, исходя из 10 растений на 1 м2.
* * Рассчитано нами, исходя из 16—20-дневного рассадного периода
при 36 растениях на 1 м2 и 30—45-дневиой культуры на постоянном
место при 10 растениях на 1 м2.
1 Лампы накаливания погружаются в проточную воду, температура которой
повышается до 45°. Инфракрасная радиация с большой длиной волны излучается
водою за счет повышения температуры.
VIII. Светокультура овощных и садовых растений
285
Т а б л и ц а 124
Характеристика различных сортов томатов (Solanum lycopersicum) при культуре
( целиком на искусственном освещении
Лампы накаливания 300—500 вт, освещенность 7000—8000 лк
(42 000—48 000 эрг/см2 • сек), t = 22—25°
(Доброхотова [59])
Дварф стоун
14-час. день . . .
Беспрерывное ос-
вещение . . . .
Уинтер
14-час. день . . .
Беспрерывное ос-
вещение . . . .
Лукулл
14-час. день . . .
3/XJ 13/Х1 29/1 87 29/11 118 6 145,2 0,87 24,2 5,2 800 900
3/XI 13/Х] 2/1 60 15/11 104 4 130,4 0,78 32,6 5,0 1250 1600
3/XI 13/XI 26/XII 53 9/11 98 9 273,0 1,64 30,3 4,8 2700 420
3/Х1 13/XI 26/XII 53 9/11 98 10 374,2 2,24 34,8 4,5 1200 500
3/Х1 13/XI 21/XII 48 6/II 95 9 212,0 1,27 23,6 3,6 760 500
* Рассчитано нами, исходя из 6 растений на 1 м1 2.
* * Рассчитано нами, исходя из удельной мощности 500 вт/м2, потребной для
создания освещенности 8000 лк.
Несколько более выгодна культура огурцов (Cucumis sativus).
Е. Д. Корольков [128] показал, что клинские огурцы в условиях 16-ча-
сового дня при удельной мощности 250 вт/м2 в течение 70—75 дней дают
по 3—4 огурца, или по 270—360 г на растение. При густоте стояния
в 4 растения на 1 м2 это соответствует урожаю в 1,08—1,44 кг/м2. За-
трата электроэнергии составляет 18—25 квт-час на 1 огурец, или 194—
278 квт-час на 1 кг плодов.
Е. Д. Корольков [129] обращает внимание на то, что в описанных
условиях у огурцов наблюдается слабое развитие завязей (развивается
45—50%) и тенденция к преимущественному образованию мужских
цветков.
Как бы то ни было, приведенные выше величины расхода электро-
энергии, потребной для культуры томатов и огурцов, в настоящее время
сильно ограничивают возможность культуры названных растений це-
ликом на электрическом свете Ч
1 С приведенными выше величинами полезно сопоставить расход энергии, иду-
щей на отопление теплиц при обычной тепличной культуре растений. Дефицит тепла,
восполняемый за счет отопления, определяется уравнением:
Q=Kxts. (50)
Здесь т — перепад температуры между температурой теплицы и наружной, t — вре-
мя, 5 — площадь, а К — коэффициент пропорциональности, равный для одинарного
стеклянного перекрытия 5 кг-кал/м2-час. град. Для поддержания температуры в 20°
в умеренной полосе СССР ежемесячный расход энергии в зимние месяцы равняется
в среднем 105 000 кг-кал/м2, или 120 квт-час/м2. Для Верхоянска эта величина
возрастает до 260 000 кг-кал/м2, или 300 квт-час/м2.
286
Частная светокультура растений
Может быть, более перспективной, по крайней мере в ближайшее
время, окажется культура земляники (Fragraria sp.), которая прекрасно
растет и плодоносит под люминесцентными лампами при удельной мощ-
ности 450 вт/м1 2. То же самое следует сказать о культуре редиса (Ra-
phanus sativus var. radicula) и салата (Lactuca sativa), которые хорошо
растут под люминесцентными лампами синего, дневного или белого света.
Отсутствие обстоятельных производственных опытов в этом направлении
не позволяет ответить положительно о рентабельности названных куль-
тур в указанных условиях.
Иными словами, в настоящее время, говоря о культуре растений це-
ликом при искусственном освещении, можно иметь в виду лишь земля-
нику (Fragraria sp.), редис (Raphanus sativus var. radicula), салат (Lac-
tuca sativa), китайскую капусту (Brassica pekinensis), петрушку (Petrose-
linum sativum) (на зелень), щавель (Rumex acetosa), шпинат (Spinacea
oleracea), лук (Allium сера, A. fistulosum) (на перо), чеснок (Allium sa-
tivum), салатный цикорий (Cichorium endivia), сельдерей (Apium graveo-
lens) и некоторые другие. Имеющиеся в этом направлении исследования
С. В. Александрова [4, 5] и И. А. Веселовского 133] дают обнадеживаю-
щие результаты. Мы рассмотрим их в соответствующих разделах, по-
священных перечисленным растениям.
Успешное решение проблемы выращивания овощных растений в за-
щищенном грунте исключительно при искусственном освещении нуж-
дается еще в многочисленных исследованиях при совместном участии
агротехников, селекционеров, физиологов и светотехников. Задача
заключается в том, чтобы повысить урожайность, подобрать культуры
и сорта, разработать агротехнику и значительно снизить расходование
электроэнергии.
Поэтому в дальнейшем, говоря о светокультуре овощных и садовых
растений, мы будем иметь в виду главным образом культуру их при
дополнительном освещении в ночные часы или в пасмурные дни
в осенний, зимний и весенний периоды.
1. ОВОЩНЫЕ РАСТЕНИЯ
Томаты (Solanum lycopersicum или Lycopersicum esculentum)
Теплолюбивое и светолюбивое растение с длиною вегетационного
периода порядка 90—120 дней. Для нормального роста и развития
нуждается в температурах 20—25° днем и 15—20° ночью. При
повышении или понижении ночных температур урожай плодов резко
снижается [843]. В естественных условиях завершение световой стадии
происходит при получении растением 12 000 кг-кал/м2 лучистой энер-
гии [354]. По данным А. А. Авакяна [2], исследовавшего около 80
сортов, томаты относятся к группе коротко дневных пли нейтральных
растений с длиною световой стадии 25—30 дней и ускоряющих переход
к цветению на коротком дне максимум на 15 дней. По мнению Оберга
[363], томаты следует отнести к группе нейтральных растений, которые
в естественных условиях при нормальном освещении обнаруживают тен-
денцию к короткодневности, а в условиях недостаточного освещения по-
ложительно реагируют на увеличение длины дня. Оптимальная длина
дня для ростовых процессов равна 18—20 часам (табл. 83 [224, 364,
667]). При освещенности менее 4500 лк (18 тыс. эрг/см2-сек физиологи-
ческой радиации) томаты, несмотря на нормальный рост, образуют
очень мало цветков и не плодоносят [843]. Максимальный сухой вес
VIII. Светокультура овощных и садовых растений
287
растений наблюдается, в зависимости от сорта, при интенсивности
физиологической радиации порядка 50—70 тыс. эрг/см2-сек (фиг. 135, 5
[224]).
Томаты, к&к правило, отрицательно реагируют на беспрерывное осве-
щение, обнаруживая своеобразный «фотопериодический хлороз». Листья
при этом желтеют, растения прекращают рост, страдают и в конечном
итоге гибнут [59, 375, 379, 458, 670, 707, 717, 733, 772, 856]. Для куль-
туры томатов в условиях закрытого грунта при дополнительном осве-
щении рекомендуется 16—17-часовой день.
Первые попытки выращивания томатов с помощью искусственного
освещения предприняты давно [470, 511, 692, 754], и в настоящее время
литература по светокультуре томатов насчитывает сравнительно большое
число работ [2, 4, 5, 56, 57, 59, 83, 123, 130, 147, 191, 199, 205, 244, 273,
277, 283, 289, 290, 326, 350, 354, 363, 365, 376, 409, 447, 458, 465, 554—
557, 610, 613, 624, 707, 708, 717, ’720, 733, 770, 772—776, 782, 829, 843,
853, 860].
В большинстве случаев дополнительное освещение томатов приме-
няется в рассадный период в ночные часы, а дальнейшее развитие ра-
стений совершается исключительно при естественном освещении. Этот
прием позволяет получать готовую рассаду в любое время года, причем
значительно быстрее и лучшего качества (табл. 86, 88, 89 и 91 п фиг. 140).
При этом формируются более мощные и приземистые растения с хорошо
развитой листвой и корневой системой [83, 123, 244, 273, 283, 289, 363,
409, 603, 610, 613, 707, 717, 720, 733, 772, 773, 855].
Лучшее качество рассады, получаемое прп дополнительном освещении,
обычно приводит к более раннему плодоношению и повышению урожая.
По данным Лошаковой [605], ускорение плодоношения при освещении
в апреле (230 лк) составляет 19—21 день. В опытах Фогля [829] с тома-
тами Датский экспорт и Лучший боннский, посеянными 13 января, цве-
тение растений и созревание плодов в результате дополнительного осве-
щения наступили раньше по сравнению с контролем на 20—24 дня.
Соответствующий сдвиг наблюдается также и во времени наступления
максимума урожая. Н. Палилов [244] нашел, что дополнительное осве-
щение в течение 18 суток ускоряет цветение томатов на 12 дней.
В. М. Игнатьева [83] показала, что томаты Бизон в результате дополни-
тельного освещения в рассадный период ускоряют начало плодоношения
на 7 дней, а при освещении в течение всей вегетации — на 31—66 дней.
В последнем случае прп 13-часовом дополнительном освещении
(7—10 тыс. лк) с помощью ламп накаливания в 500 вт растения начали
плодоносить через 77 дней. Контрольные растения плодоносили через
143 дня. По данным В. П. Мальчевского [191], освещение рассады по
4 часа в сутки в течение 1 месяца вызывает сокращение вегетационного
периода на 10—15%. С. И. Доброхотова [59] нашла, что в условиях
открытого грунта рассада, полученная с помощью дополнительного осве-
щения (4 часа ежедневно, 4—5 тыс. лк), дает урожай плодов на
17—21 день раньше, чем обычная рассада. Томаты Эльза Грейг в опытах
Роденбурга [733] после освещения в рассадный период неоновыми лам-
пами начали плодоносить на 14 дней раньше, чем в контроле. Д. Л. Ро-
машенков [277] показал, что дополнительное освещение вызывает уско-
рение бутонизации на 4—8 дней. Оберг [363], выращивая рассаду томатов
при дополнительном 14-часовом освещении неоновыми лампами (475 вт)
в течение 3 месяцев (с 31 октября по 27 января), нашел, что в резуль-
тате дополнительного освещения плодоношение наступило раньше на
29 дней — 4 мая вместо 6 июня в контроле.
288
Частная светокультура растений
Лишь в небольшом числе работ отмечается отсутствие ускорения
плодоношения под влиянием дополнительного освещения в рассадный
период. Так, в наших опытах (табл. 93) ускорение бутонизации,
цветения и плодоношения наступало лишь при освещении рассады
люминесцентными лампами, а лампы накаливания эффекта не дали [123].
II. М. Курбатов и Т. М. Васильева [147] также отмечают отсутствие
ускорения в наступлении цветения и плодоношения под влиянием ламп
накаливания (табл. 108). В опытах Смита [772, 773] плодоношение на-
ступило практически одновременно как при дополнительном освещении
рассады различными лампами (лампы накаливания, неоновые и натрие-
вые трубки), так и в контроле.
Р. А. Джалилова [56] нашла, что ускорение в наступлении плодо-
ношения зависит от времени посева растений: чем раньше посев, тем
сильнее эффект (табл. 120). То же самое установил Оберг [363]; в его опытах
плодоношение томатов под влиянием неоновых ламп ускоряется па 11 дней
при посеве растений 6 декабря и на 33 дня — при посеве 14 октября.
Иными словами, эффект дополнительного освещения рассады томатов
в смысле ускорения цветения и плодоношения растений определяется
временем посева: при позднем посеве он может равняться нулю, а при
раннем посеве достигает 30 дней. При обычном посеве рассады в конце
декабря — начале января ускорение в наступлении урожая обыкновенно
достигает 10—14 дней.
Большинство авторов, за небольшим исключением [326, 605, 829],
отмечают то или иное повышение урожайности в результате дополни-
тельного освещения томатов. В опытах Рейнгольда [707] повышение
урожайности равнялось 10—15%, а урожай достигал 9,7 кг с куста
(табл. 86) при площади питания в 60X60 см. И. М. Курбатов и Т. М. Ва-
сильева [147] нашли, что дополнительное освещение рассады вызывает
увеличение урожая плодов на 48—70% (табл. 108). В опытах В. М. Иг-
натьевой [83] урожай томатов в результате дополнительного освещения
рассады достигал 10 кг/м2 и был выше урожая контрольных растений
на 28%; в первые две недели плодоношения различие достигало 88%.
По данным В. П. Мальчевского [191 ], повышение урожайности томатов
достигает 50—100% .С. И. Доброхотова [59], выращивавшая рассаду томатов
для посева в открытый грунт, нашла, что в этом случае дополнительное
освещение увеличивает урожай на 5—64%. В опытах Д. Л. Ромашен-
кова [277], выращивавшего рассаду томатов в теплицах и парниках и
затем высаживавшего растения в открытый грунт и в парники, урожай
томатов после дополнительного освещения колебался в пределах 0,45—
1,98 кг с куста, а прибавка от освещения варьировала от 9 до 37%. Ве-
личины того же порядка приводит Р. А. Джалилова ([56] табл. 120).
В опытах Оберга [363], который высевал семена 14 октября, а дополни-
тельное освещение давал с 31 октября по 27 января, урожай томатов
под влиянием неоновых ламп достигал 6,35 кг с куста, а увеличение
урожая в результате дополнительного освещения равнялось 50—124%.
Сравнительно высокие урожаи с куста получала также Уитроу [853]—
4,2—4,7 кг, хотя эффект дополнительного освещения был невелик
(3—10%). В наших опытах (табл. 94) увеличение урожайности под влия-
нием ламп накаливания было также сравнительно невелико (12—16%),
однако люминесцентные лампы повысили урожай на 17—33% [123].
В опытах Н. II. Протасовой [253, 254] 8-часовое дополнительное осве-
щение рассады в ночные часы давало у сорта Маяк повышение урожай-
ности, в зависимости от типа ламп, на 4—63%— с 13,5 кг/м2 в контроле
до 14,3—22.0 кг/м2 в опытных вариантах (табл. 100).
VIII. Светокультура овощных и садовых растений
289
Следует, таким образом, принять, что дополнительное освещение рас-
сады повышает урожайность томатов на 30—50%, а при правильной
агротехнике — даже на 70—125%.
В условиях закрытого грунта, при обычной для томатов площади
питания в 50x35 см (0,175 м2) и без дополнительного освещения рас-
сады, урожай плодов обычно колеблется в пределах 0,5—2,0 кг
с куста [147, 273, 326, 707, 829]. Это примерно та же урожайность,
что наблюдается и в условиях открытого грунта [2, 277, 354]. При
выращивании рассады с помощью дополнительного освещения урожай
томатов повышается в закрытом грунте до 0,8—3,0, а в условиях
хорошей агротехники даже до 6—8 кг с куста [123, 147, 253, 254,
273, 277, 289, 363, 853]. Иными словами, дополнительное освещение
рассады позволяет повысить обычную для томатов урожайность
с 3—12 кг/м2 до 4,8—18 кг/м2.
Для дополнительного освещения рассады пригодны любые источники
радиации, если они достаточно мощны и не содержат вредных витацид-
ных лучей. В настоящее время испытаны лампы накаливания, люминес-
центные лампы белого и дневного света, ртутные, натриевые и неоновые
лампы [123, 253, 254, 273, 283, 289, 290, 363, 603, 707, 717, 720, 733,
772, 773, 855]. Лучшие результаты получаются с богатыми оранжево-
красными лучами неоновыми лампами (табл. 86 и 88 [273, 289, 363, 603,
707, 772, 773]) и люминесцентными лампами (табл. 91 и 94 и фиг. 140
[123]). Особенно хорошие результаты получаются при комбинации ртут-
ных ламп с неоновыми лампами или лампами накаливания [253, 254].
Чаще всего применяются, однако, более простые для эксплуатации
лампы накаливания [56, 83, 130, 147, 244, 277, 326, 829, 854].
В настоящее время имеется очень немного исследований, которые
позволяют говорить об оптимальной для томатов интенсивности допол-
нительного освещения и величине потребной для этой цели удельной
мощности. И. М. Курбатов и Т. М. Васильева [147] нашли, что при упо-
треблении ламп накаливания и увеличении освещенности с 500 до
2 000 лк наблюдается соответствующее увеличение урожайности
(табл. 108). В опытах А. Ф. Клешнина [123] с лампами накаливания
и люминесцентными лампами было обнаружено, что качество рассады
и урожай продолжают возрастать до 5000 лк, или 30 тыс. эрг/см2-сек
физиологической радиации (табл. 91 и 94). Этой величине освещенности
соответствует удельная мощность 450 вт/м2. Смит [772] показал, что
дополнительное освещение томатов в условиях Норвегии должно созда-
вать освехценность порядка 4000 лк. Судя по кривой, которую приводит
автор, дальнейшее увеличение интенсивности дополнительного освеще-
ния не ускоряет получения рассады. В литературе освещенность обычно
колеблется от 500 до 10 000 лк (3—60 тыс эрг/см2-сек физиологиче-
ской радиации), а удельные мощности — от 300 до 750 вт/м2 [130, 147,
244, 273, 289, 290, 326, 409, 707, 733, 829, 853, 855]. Чаще всего
употребляются лампы мощностью 500 вт, висящие на расстоянии
1 м друг от друга и при высоте подвеса 50 см, создающие освещенность
порядка 8000 лк (40 тыс эрг/см2-сек физиологической радиации).
Принимая во внимание, что длина дня для томатов не должна пре-
вышать 18 часов, длительность дополнительного освещения, в зависи-
мости от длины естественного дня, должна лежать в пределах от 4 до
8 часов [123, 147, 326, 733, 829, 853, 854]. В зимние месяцы в средней
полосе СССР наиболее благоприятные результаты дает дополнительное
освещение продолжительностью 8 часов, создающее вместе с естествен-
ным освещением 15—18-часовой день.
19 А. Ф. Клешнин
290
Частотная светокультура растений
И. М. Курбатов и Т. М. Васильева [147] показали, что оптимальная
длительность периода освещения рассады равняется 30 дням. Дальней-
шее увеличение периода освещения или не усиливает положительное
влияние ci'o, или приводит даже к отрицательным результатам (табл. 125).
Т а б л и ц а 12э
Влияние длительности периода освещения на. уромсай томатов Вазон.
Лампы накаливания.
(Удельная мощность — 500 вт/см2). Осветитель «Универсаль». Высота подвеса —
70 см. Освещение—.‘/часа утром и 3 часа вечером. I = 18—20°
(Курбатов и Васильева [147])
Всходы тьность (а освещения х) ка в грунт й сбор зре- 1ОДОВ Урожай с 1 пуста
за первый месяц общий (с 10(V по 2/VIII)
§ t=t е Ё 1
о И ЧвЗ о 3 га Цве гаи “ 3 И R ШТ. кг % шт. i кг %
22/XII 2—4/1 0(К) 5/Ц 16/111 17/V 4,75 i 0,193 100 12,65 0,515 100
30 5/11 13/1II 13/V 8,11 0,357 185 19,00 0,780 151
40 15/11 7/Ш 16/V 6,86 0,283 147 18,60 0,784 152
50 25/II 20/Ш 16/V 4,69 0,204 106 17,61 0,748 145
60 7/Ш 10/III 16/V 4,62 0,175 90 19,90 0,776 150
71 15—17/1 0 22/II 25/III 17/V 5,19 0,225 100 11,95 0,555 100
/ 30 22/11 25/III 10/V 6,06 0,297 132 17,35 0,842 152
40 4/Ш 25/Ш 27 /V 4,75 0,227 101 14,45 0,657 119
50 14/Ш 18/1II 23/V 2,81 0,143 63 12,15 0,500 90
22/1 0 9/Ш 5/IV 4/VI 5,11 0,219 100 9,3 0,346 100
30 9/Ш 5/1V 3/VI 8,05 0,225 103 15,40 0,670 194
В производственных условиях большей частью придерживаются месяч-
ного срока. Этот же срок в большинстве случаев встречается it в литера-
туре [123, 192, 273, 707, 733, 853].
Таким образом, при ежедневном освещении в течение 8 часов, при
удельной мощности 500 вт/м2 и длительности периода освещения 30 дней
на каждый квадратный метр теплицы, содержащий 120—140 растений
томатов, затрачивается 120 квт-час энергии. Это означает, что на каждое
растение рассады потребляется около 1 квт-час.
Дополнительное освещение томатной рассады имеет смысл в зимние
месяцы и ранней весной (февраль). В марте оно практически уже не
эффективно. В производственных условиях выгонка рассады обычно про-
изводится в январе — феврале. При посеве семян в конце декабря или
начале января рассада при дополнительном освещении достигает готов-
ности в начале февраля, растения зацветают в середине марта, а первый
сбор зрелых плодов производится в мао (табл. 125).
Большое значение для культуры томатов имеет вопрос о времени вы-
ращивания рассады. Этот вопрос подробно исследован Смитом [7751.
Названный автор, высевая томаты в различные сроки (18 октября, 1, 4,
22, 27 декабря, 10, 31 января, 28 марта, 6, 12, 19 апреля и 13 мая), вы-
ращивал их в оранжерее при естественном освещении на широте г. Осло
VIII. Светокультура овощных и садовых растений
291
(11< рвегия, 60°с. ш.). Опыт показал (фиг. 153), что при посеве в октябре
томаты почти не растут и, в конце концов, начинают терять в весе: при посеве
в начале декабря они по-
настоящему трогаются в
рост лишь через 60 дней
после иосева и лишь при
посеве во второй половине
марта — начале апреля ра-
стения развиваются так
же, как и прп посеве в мае.
Как показывают кривые
на фиг. 154, нормальное
развитие томатов начи-
нается лишь при длине дня
в 12 часов и количестве
лучистой энергии 300
г-кал/см2 в день. Эти усло-
вия создаются только во
второй половине марта,
при высоте солнцестояния
около 30°.
Фиг. 153. Рост растений томатов (Solanum lycoper-
sicum) в зависимости от времени посева
1 — 18 октября; 2 — 4 декабря; 3 — 22 декабря;
7 — 19 января; 5 — 31 января; в — 19 апреля; 7 — 7 мая
Иными словами, томаты
нуждаются в дополнитель-
ном освещении лишь в
осенне-зимний и зимне-
весенний периоды до 2-й
половины марта. С середины марта рассада томатов дополнительного
освещения не требует.
Фиг. 15'5. Рост томатов (Solanum lycopersicum} в зависимости от количества лучистой
энергии
1 — число дней, необходимое для накопления 6 г зеленой массы; 2 — ежедневный прирост зеленой
массы (в г) (на основании кривой /); з — длина дня (в часах); 4— количество лучистой энергии
(в г-кал.'см2 в день); 5 — высота Солнца над горизонтом
19*
292
Частная светокультура растений
Б. С. Мошков и его сотрудники [50, 221, 224] разработали метод уско-
ренного выращивания рассады томатов, позволяющий выращивать рас-
саду в течение 20 дней в любом помещении и в любое время года. Метод
заключается в том, что растения выращиваются исключительно под лам-
пами накаливания в 200 или 300 вт, которые погружаются в слой про-
точной воды, нагреваемой лампами до 40—45°. или при комбинации низко-
вольтных ламп накаливания (1,5 квт/м2) с люминесцентными лампами
(0,3 квт/м2). Рассада томатов, культивируемая подобным способом и
высаженная на постоянное место в оранжерею в условия естественного
освещения весною, дает зрелые плоды через 60 дней и в течение 3 месяцев
дает урожай 750—1140 г с куста.
Применение подобного метода нуждается в затрате высоких удельных
мощностей (3,2—4,8 квт/м') и связано с большой затратой электро-
энергии: 32—34 квт-час на 1 растение рассады или 35—43 квт-час на 1 кг
плодов.
До сих пор, говоря о культуре томатов с помощью искусственного
освещения, мы имели в виду исключительно получение рассады. Что
касается культуры томатов в зимнее время с целью получения плодов,
то этот вопрос еще далек от разрешения. Исследований в этом направле-
нии очень немного.
Е. Д. Корольков [130], применяя дополнительное освещение с помощью
ламп накаливания по 13 часов в сутки (с 6 час. ЗОмин. до 19 час. 30 мин.)
при удельной мощности 750 вт/м2, получил зрелые плоды томатов в фев-
рале. В его опытах семена высевались 18 ноября, всходы появились
23 ноября, а рассада была получена к 15 декабря. С. И. Доброхотова
[59], применяя лампы накаливания в 500 вт (4000—5000 лк) при продол-
жительности освещения по 14 часов в сутки на протяжении всего вегета-
ционного периода, также получала зрелые плоды томатов (сорта Санрайз,
Дварф Стоун и Уинтер). При посеве семян 17 ноября контрольные расте-
ния в условиях ленинградской зимы погибли уже в конце декабря —
начале января, опытные зацвели в январе—феврале, а зрелые плоды
дали в марте—апреле: Уинтер — 11 марта, Санрайз — 21 марта, а Дварф
Стоун — 4 апреля. Р. И. Джалилова [354] с помощью ламп накаливания
и дополнительного удобрения СО2 выращивала томаты в условиях гор-
шечной культуры с 1 ноября по середину апреля, получив зрелые плоды
22 февраля. За две ротации — с 1 ноября по 15 апреля и с 1 февраля
по 1 июля — она собрала урожай томатов в 14 кг/м2.
К сожалению, ни один из названных авторов не приводит урожая
плодов для осенне-зимней ротации. В этом отношении выгодно отличаются
работы В. М. Маркова и Л. И. Марковой [199] и Оберга [363].
Первые применяли дополнительное освещение с помощью ламп на-
каливания (500 вт/м2) в период осенней ротации томатов в совхозе «Брат-
цево» под Москвой. Томаты Эльза Крэйг высевали 1 июня и до 22 августа
культивировали в естественных условиях в открытых парниках, затем
растения переносили в теплицу, где освещали в дневные (8 часов) или
в ночные часы (10 часов) с помощью ламп в 500 вт, находившихся на
высоте 40—60 см от верхушек растений, в течение 60 дней (4 октября —
4 декабря). Перед началом освещения верхушки растений прищипыва-
лись (13 сентября). При освещении в дневные часы растения получали
11—12-часовой день, а при освещении ночью находились в условиях
17—19-часового дня. Авторы нашли, что плодоношение томатов насту-
пает одновременно, независимо от времени освещения, через 120 дней
после посева и длится в течение 67 дней (28 сентября — 4 декабря);
урожай плодов равнялся 5,06 кг/м2 при дневном освещении, 4,67 кг/м2
VIII. Светокультура овощных и садовых растений
293
при ночном и 3,85 кг/м2 в контроле. Это соответствует 1,00, 0,94 и 0,78 кг
с куста. Опыт показывает, что при осенней ротации томатов на 1 кг плодов
затрачивается 47,5 квт-час при дневном освещении и 67,5 квт-час при
ночном.
Оберг в районе г. Упсала (Швеция), лежащего на широте Ленинграда,
произвел исследование по культуре томатов в зимний период. Посев
растений производился 6 июля, пересадка рассады на постоянное место —
15 августа, а дополнительное освещение — с 11 сентября, когда на ра-
стениях имелось уже по 3 кисти. Растения освещались по 18 часов в сутки
(с 6 до 24 час) с помощью натриевых ламп «So-ЮОО» мощностью 140 вт или
ртутными лампами «НО-ЮОО» по 265 вт, дававшими интенсивность
видимой радиации в 0,016 г-кал/см2-мин (11 000 эрг/см2-сек).
Плодоношение началось 23 декабря под ртутными лампами и 3 января —
под натриевыми лампами и в контроле. К 27 февраля с 4 кистей каждого
растения был получен урожай 0,68 кг под натриевыми, 0,58 кг — под
ртутными лампами и 0,30 кг — в контроле.
Опыты, проведенные нами совместно с С. И. Китаевым [123] в совхозе
«Марфино» по культуре томатов зимою (3 ноября — 9 февраля), пока-
зывают, что в этот период одного лишь ночного освещения (1000 вт/м2;
с 24 до 7 час.) недостаточно.
Таким образом, на основании исследований Маркова и Оберга сле-
дует сделать заключение, что в осенне-зимний период при дополнительном
освещении и густоте стояния 6 растений на 1 м2 можно получить урожай
томатов 0,6—1 кг с куста, или 4—5 кг/м2. При этом на протяжении веге-
тационного периода затрачивается энергия, равная 50—150 квт-час на
1 кг плодов.
Огурцы (Cucumis sativus)
Огурцы, как и томаты, относятся к группе светолюбивых и теплолюби-
вых растений. Урожай огурцов, особенно в тепличных условиях, нахо-
дится в теснейшей зависимости от количества лучистой энергии, полу-
чаемой растением (табл. 122). Сухой вес растений возрастает линейно
вместе с увеличением интенсивности физиологической радиации до вели-
чины более 70 тыс. эрг/см2-сек (фиг. 135, 7; [224]). При благоприятном
освещении растения огурцов нуждаются в температуре 25—30°. В этом
случае процессы роста и развития совершаются с максимальной ско-
ростью.
Подобно томатам огурцы следует отнести к группе нейтральных расте-
ний с тенденцией к короткодневности. Ускорение в наступлении цветения
в условиях короткого дня обычно равняется нескольким дням и не пре-
вышает 12 дней [697]. Исследования по светокультуре огурцов начались
давно [754], и в настоящее время дополнительное освещение огурцов
широко применяется в рассадный период для получения ранней и добро-
качественной рассады. Большинство исследований проведено именно
в этом направлении [18, 21, 46, 123, 130, 147, 149, 205, 230, 244, 273,
283, 289, 350, 351, 376, 379, 530, 555,556,579,587,621,706,707,717,719,
723, 729, 733, 782, 803, 804].
Несмотря на сравнительно большой список работ, литература по
светокультуре огурцов отличается меньшей определенностью выводов,
чем литература по томатам. Обстоятельных и хорошо поставленных и
подробно описанных опытов сравнительно немного. Однако все авторы
единогласно отмечают положительное влияние дополнительного осве-
щения на рост и развитие рассады. Обычно в этом случае наблюдается
быстрый рост рассады, большая мощность растений, большая площадь
294 Частная светокультура растений
листьев, более сильная корневая система и более интенсивная зеленая
окраска [46, 123, 149, 244, 253, 254, 289, 379, 556, 587, 706, 707, 717, 719,
723, 729, 733, 791]. Характеристика рассады в условиях различного
освещения приведена нами в табл. 92, 95, 99, 101, 103, 119 и на фиг. 141.
По данным Гильмера [550], при дополнительном освещении по 4 часа
в сутки готовая рассада огурцов может быть получена на 3 недели раньше,
чем без освещения. В. А. Брызгалов и М. И. Русин [21] нашли, что при
декабрьском посеве рассада становится готовой прп дополнительном
освещении через 30 дней, а в контроле — через 40—45 дней. В наших
опытах [123] при посеве семян в последних числах января готовая рассада
была получена 14 февраля (табл. 95; фиг. 141). Благодаря более сильной
корневой системе рассада, полученная с помощью дополнительного
освещения, лучше укореняется при посадке на постоянное место, а в ре-
зультате большей листовой поверхности быстрее развивается, раньше
начинает плодоносить и дает более высокий урожай.
Более раннее плодоношение огурцов при дополнительном освещении
рассады отмечают многие авторы. В опытах Роденбурга [720] под влия-
нием неоновых ламп ускорение плодоношения равнялось 7—14 дням.
По данным В. А. Брызгалова и М. И. Русина [21], плодоношение огурцов
при освещении рассады начинается раньше на 21—27 дней. Палилов
[244] в опытах с огурцами Клински? установил, что дополнительное
освещение в зависимости от мощности ламп дает ускорение плодоношения
на 14—30 дней (табл. 103). В другом опыте с различной длительностью
освещения (12- и 16-часовой день) он нашел, что при освещении огурцов
того же сорта с 19 января по 20 марта лампами накаливания в 500 вт
плодоношение опытных растений наступает на 66—67-й день (27 марта),
а плодоношение контрольных — на 94-й день (25 апреля). По сообщению
П. В. Шереметевского [343], дополнительное освещение рассады огурцов
ускоряет плодоношение на 30—36 дней. По данным Д. И. Нацентова
[228], ускорение плодоношения составляет 10—15 дней. То же самое
отмечает И. А. Шубов [351] на основании производственных опытов
в совхозе «Марфино». Более раннее плодоношение огурцов при освещении
неоновыми лампами отмечают также Рейнгольд [707] и Куфферат [587].
В наших опытах [123] с огурцами Елинские и Неросимые было найдено,
что первые не ускоряют плодоношения при освещении рассады, а вторые
начинают цвести и плодоносить раньше контрольных на несколько дней
(табл. 96 и 97).
Что касается урожайности огурцов при дополнительном освещении,
то последняя в большинстве случаев значительно повышается, если
освещение не было чрезмерным и рассада не оказалась изнеженной.
По данным Роденбурга [720], увеличение урожая в результате дополни-
тельного освещения рассады (с 12 ноября по 29 декабря) равняется 16—23 %.
Гардер, Кепплер и Ройсс [530] нашли, что при освещении рассады огурцов
урожай последних повышается на 11%. В опытах Строуда [791], при
дополнительном освещении в 1000—1700 лк по 12 часов в сутки (с 19
до 7 час.) с 4 января, количество огурцов при плодоношении в марте—
апреле увеличилось на 20%. В. А. Брызгалов и М. И. Русин [21] также
отмечают повышение урожайности при дополнительном освещении.
По данным П. Палилова 1244], в условиях дополнительного освещения
рассады наблюдается увеличение урожайности на 80—200%. В опытах
Рейнгольда [707] при посеве огурцов 7 декабря дополнительное освещение
рассады повысило урожай плодов с 8,4 кг до 12,8 кг на одно растение. Число
плодов на растении увеличилось при этом с 17 до 26 (табл. 126). Согласно
Куфферат [587], увеличение урожая огурцов при дополнительном осве-
VIII. Светокультура овощных и садовых растений
295
Таблица 126
Слияние дополнительного освещения неоновыми лампами на урожай огурцов
в зависимости от сроков посева
(Рейнгольд [707])
Варианты опыта Урожай плодов на одно растение
ШТ- 1 ; кг %
1. Контроль. Посев 7/Х11 17,1 8,4 100
2. Освещение неоновой лампой с 21 час. .30 мин. до 3 час. 30 мин. (6 часов; 500 лк) а) посев 7/XII 26,1 i |-'.7 151
б) посев 17/XI I 26,8 ' 12,8 152
в) посев 27/XTI J 8.9 ПО
щении рассады в среднем равняется 20 % , а по данным Д. И. I [ацентова[228]—
30%. Наши опыты [123] показали, что огурцы Перосимые, в зависимости
от источника радиации, повышают урожай в среднем за сезон на 13—47%
(табл. 97). В опытах Н. II. Протасовой [253, 254] дополнительное осве-
щение рассады огурцов Елинские увеличивало урожай плодов, в зависи-
мости от источника радиации, на 11—95% (табл. 99 и 101). Как правило,
особенно сильный эффект наблюдается в начале плодоношения: в даль-
нейшем он постепенно снижается (табл. 97 [123, 791]).
Таким образом, дополнительное освещение рассады огурцов вызывает
ускорение в наступлении плодоношения от нескольких дней до 1 месяца
и увеличивает урожай на 10—200%. Различия в эффективности освещения,
наблюдаемые у разных авторов, обусловлены неодинаковыми условиями
опытов, сортовыми особенностями и временем посева и освещения расте-
ний. Как видно из табл. 126, эффективность дополнительного освещения
тем выше, чем раньше произведен посев растений.
В некоторых случаях дополнительное освещение огурцов дает отри-
цательный результат, особенно при позднем и длительном освещении
(табл. 127). Сказанное может быть обусловлено целым рядом причин.
Во-первых, имеет место перегрев растений. Он особенно вероятен при
длительном освещении в весенние дни. Во-вторых, возможна изнежен-
ность растений, когда они вместо репродуктивного развития продолжают
интенсивный вегетативный рост. В-третьих, следует иметь в виду воз-
можность изменения соотношения между женскими и мужскими цвет-
ками. Последнее в условиях дополнительного освещения смещается
в сторону мужских цветков [128, 803].
При дополнительном освещении рассады огурцов испытаны лампы
накаливания (простые, зеркальные и из увиолевого стекла), неоновые,
ртутные и люминесцентные лампы [123, 253, 254, 273, 283, 289, 350, 587,
707, 717, 719, 723, 733, 782]. В большинстве случаев применяются, однако,
лампы накаливания. Отличительная особенность растений огурцов, выра-
щиваемых под лампами накаливания, состоит в чрезмерном вытягивании
подсемядольного колена (фиг. 141). Гильмер [550] рекомендует лампы
накаливания с колбой из увиолевого стекла, излучение которых содержит
больше ультрафиолетовых лучей, чем у обычных ламп. По его данным,
в этом случае развиваются приземистые интенсивно зеленые растения
огурцов. Ван дер Штадт [782] советует применять неоновые лампы. В его
опытах использовались неоновые трубки длиною 1,75 м и мощностью
296
Частная светокультура растений
Таблица 127
Влияние длительного освещения и сроков посева на урожай огурцов Куленкамп
(освещение лампами накаливания 500 вт, 3 часа утром и 3 часа вечером)
(Курбатов и Васильева [147])
Всходы Длитель- ность освещения (в днях) Высадка в грунт Цветение Начало плодоно- шения Урожай с одного куста к 19—23/VI
ШТ. КГ %
5/1 0 (К) 5/II 17/П 29/Ш 10,03 1,78 100
30 5/П 19/11 17/Ш 12,50 1,96 110
45 19/11 17/П 17/11 10,07 1,52 86
19/1 0 5/11 13/Ш 9/1V 13,00 2,58 100
15 5/П 17/Ш 4/1V 10,05 1,52 59
30 19/П 3/1II 26/1II 12,50 2,05 79
45 5/11Т 28/II 25/Ш 8,66 1,35 52
1/II 0 15/Ц 13/Ш 9/IV 14,20 2,44 100
15 15/11 16/11I 4/IV 10,00 1,65 68
30 2/1II 13/Ш 29/Ш 12,30 2,05 84
45 16/111 13/Ш 29/Ш 13,50 1,96 80
800 вт, ко торые нахе )дились на высоте 85 см и включа лист» на 8 часов
(с 23 до 7 час). В этом случае формируются крепкие, мощные и приземи-
стые растения огурцов. Родеибург [717, 722, 723, 733] и Куфферат [587]
также дают положительные отзывы неоновым лампам, рекомендуя их
для выращивания огурцов.
Рейнгольд [707] сопоставил лампы накаливания и неоновые трубки
и нашел, что лучший эффект в смысле ускорения плодоношения и увели-
чения урожая дают неоновые лампы. А. Ф. Клешнин [123] установил,
что для клинских огурцов значительно более эффективны люмине-
сцентные лампы белого света, чем лампы накаливания (табл. 92). В дру-
гом опыте А. Ф. Клешнин [123] показал, что для огурцов Неросимые
наиболее пригодны ртутные лампы «Игар-2», а лампы накаливания
значительно менее эффективны (табл. 95—97). В опытах И. II. Протасо-
вой [253, 254] .лучший урожай также наблюдался при освещении ртут-
ными лампами «Игар-2» (табл. 99 и 101).
Таким образом, следует признать, что для дополнительного освеще-
ния рассады огурцов пригодны все существующие типы ламп, если в их
спектре отсутствуют вредные ультрафиолетовые лучи. Наиболее эффек-
тивными следует признать источники радиации, богатые сине-фиолето-
выми и ультрафиолетовыми лучами — люминесцентные и ртутные лампы.
Исследований, посвященных интенсивности радиации и удельной
мощности, необходимых для культуры рассады огурцов, имеется очень
немного.
В опытах Строуда [791] освещенность в 1000 и 1700 лк одинаково
ускоряли плодоношение и увеличивали урожай. Растения были посеяны
29 декабря и начали освещаться с 4 января по 10 часов в сутки (с 21 до
7 час.). В результате значительно увеличилась листовая поверхность рас-
сады (в 5,2 раза при 993 лк и в 7 раз — при 1722 лк) и урожай плодов.
Если в контроле с одного растения было получено 0,1 огурца в марте и 16,1
VIII. Светокультура овощных и садовых растений
297
огурца в апреле, то для дополнительного освещения соответствующие
цифры были равны 4,6 и 19,3. Аналогичные данные при сравнении осве-
щенностей в 500 и 1000 лк, получаемых при помощи неоновой лампы,
получил Роденбург [720]. При освещении с 12 ноября по 29 декабря
(с 22 до 6 час.) плодоношение наступило 12 марта при освещенности в
1000 лк, 19 марта — при 550 лк и 26 марта в контроле. К концу марта
с каждого растения был получен следующий урожай плодов: при 1000 лк—
11,3 шт., при 500 лк — 10,5 шт., а в контроле — 5,4 шт. Общий урожай за
вегетационный период (март—июнь) соответственно равнялся 40,9, 43,3 и
35,2 шт. В. А. Брызгалов и Русин [21], применяя лампы накаливания
в 300 вт, висевшие на расстоянии 90 см друг от друга на высоте 30—40 см
от почвы, исследовали состояние рассады как непосредственно под лам-
пами (5200 лк), так и между ними (2000 лк). Авторы обнаружили при
этом, что в первом случае наблюдаются более приземистые растения с боль-
шей листовой поверхностью и большей толщиной стебля. В наших опытах
[123] с огурцами Клинские было найдено, что под лампами накалива-
ния при освещенности 4600 лк (450 вт/м2) рассада развивалась значи-
тельно лучше, чем при 1600 лк (150 вт/м2). Соответствующие различия
под люминесцентными лампами были значительно меньше (табл. 92).
Палилов [244] сопоставил влияние удельных мощностей в 200, 500 и
1000 вт/м2 при высоте ламп 50—100 см. Автор установил, что различия
рассады наблюдаются лишь между вариантами в 200 и 500 вт/м2. Раз-
личия между вариантами в 500 и 1000 вт/м2 были незначительны (табл.
103). По данным Н. Н. Протасовой [253, 254], испытавшей значение удель-
ных мощностей 300, 500 и 1000 вт/м2 при освещении рассады огурцов
Клинские подвижной установкой Финкельштейна, лучшая рассада и
максимальный урожай получаются при 500 вт/м2 (табл. 109). В другом
опыте И. II. Протасовой лучшая рассада и лучший урожай плодов на-
блюдались при удельной мощности 750 вт/м2, т. е. при интенсивности
физиологической радиации порядка 50 тыс. эрг/см2-сек (табл. 99). Таким
образом, для освещения рассады огурцов при помощи ламп накаливания
следует рекомендовать удельную мощность 300—500 вт/м2, т. е. осве-
щенность порядка 5000—8ОО0 лк или интенсивность физиологической
радиации порядка 30—50 тыс. эрг/см2-сек. Величины того же порядка,
повидимому, следует рекомендовать и для неоновых и ртутных ламп.
При употреблении люминесцентных ламп достаточна удельная мощность
порядка 200—400 вт/м2.
В большинстве случаев длительность ежедневного дополнительного
освещения колеблется в пределах 6—8 часов [123, 149, 273, 289, 290,
587, 707, 729, 733]. По данным Роденбурга [729], дававшего освещение
в течение 4 и 8 часов, размеры рассады были тем больше, чем больше
была длительность освещения. Палилов [244] нашел, что цветение и пло-
доношение огурцов наступают одновременно как при 12-часовом, так и
при 16-часовом дне, но урожай значительно выше во втором случае.
Если исходить из удельной мощности 500 вт/м2 и длительности осве-
щения 8 часов в сутки в течение 30 дней, то потребность в электроэнергии
будет равна 120 квт-час/м2. При густоте стояния рассады 135 растений
на 1 м2 на каждое растение затрачивается 0,9 квт-час. При густоте стоя-
ния на постоянном месте 4—6 растений на 1 м2 и выходе продукции
10—13 кг/м2 это соответствует затрате электроэнергии 0,27—0,54 квт-час
на 1 кг плодов.
В настоящее время совершенно отсутствуют опыты но культуре огур-
цов в зимние месяцы при помощи дополнительного освещения. Исследо-
вания, проведенные нами совместно с С. И. Китаевым [123] в совхозе
298
Частная светокультура растений
«Марфино» (Москва), показывают, что дополнительное освещение дает
положительный эффект, однако урожай плодов при ночном освещении
слишком незначителен, чтобы культура могла быть рентабельной (табл. 98).
Предстоит еще большая работа, особенно по вопросам селекции и
агротехники. Только при значительном повышении урожая можно будет
говорить о зимней культуре огурцов.
С. В. Александров [4, 5] сообщает, что огурцы хорошо растут в произ-
водственных условиях при освещении исключительно искусственным
светом (500 вт/м'2). Однако о величине урожая он ничего не пишет.
Автор получал зрелые огурцы с весом одного плода 80 г.
Баклажаны (Solanum melongena)
Но данным Артура [144, 376, 379], положительно реагируют на до-
полнительное ночное освещение и удобрение (X).,. При дополнительном
ночном освещении продолжительностью 6 часов, получаемом с помощью
ламп накаливания, и при освещенности 8200 лк растения цветут и плодо-
носят. Контрольные растения, находящиеся в условиях естественного
зимнего дня, не цветут.
Дыня (Cucumis melo)
13 опытах Сименса 1754, 755, 7561 при освещении дуговыми лампами
давала нормальные плоды в зимние месяцы при беспрерывном освещении.
Плоды отличались замечательным вкусом и ароматом. Те же результаты
получены Палиловым [244].
Тыква (Cucurbita pepo)
Длиннодневные или нейтральные растения. Оптимальная длина дня
для ростовых процессов равна 16 часам [703, 704]. Работ по дополнитель-
ному освещению названной культуры нет, если не считать исследования
Рэмэли [697], который выращивал тыкву в зимние месяцы при освещении
160—320 лк с 17 до 22 час. и нашел, что тыква зацветает одновременно,
как при освещении, так и в контроле, через 54 дня после начала опыта.
('. В. Александров [4, 5] пытался культивировать кабачки исключи-
тельно под лампами накаливания (500 вт/м2), но получил отрицательные
результаты.
Арбуз (Citrullus vulgaris)
Нейтральное растение. Зимою на дополнительное освещение (с 17 до
22 час., 160—320 лк) не реагирует, зацветая одновременно с контролем,
через 82 дня [697].
Перец (Capsicum)
Теплолюбивые нейтральные или короткодневные растения [320]. Для
роста и развития нуждаются в температуре 18—25°. На короткий день
реагируют лишь в возрасте от 20—30 до 55—75 дней [320].
При культуре в зимние месяцы мало отзываются на дополнительное
освещение в ночные часы. В опытах Октер и Гарли [394] перец зацветал
одновременно как на 12-часовом и 15-часовом дне, так и при беспрерывном
освещении. По данным Дите [465], дававшей дополнительное освещение
с 16 до 23 и с 514 до 914 час. от лампы накаливания в 100 вт, С. аппиит
зацветает почти одновременно как в контроле (при естественном дне),
VIII. Светокультура овощных и садовых растений
299
так г> при дополнительном освещении, но более обильно в контроле. Осве-
щаемые растения растут значительно интенсивнее. При посеве 16 сентября
опытные растения к 6 марта достигли высоты 29 см, а контрольные — 16 см.
В некоторых случаях дополнительное освещение в зимние месяцы ока-
зывает даже отрицательное влияние, вызывая опадение плодов и снижая,
следовательно, урожай [448].
В опытах Рэмэли [697] С. jrutescens при дополнительном освещении
160—320 лк с 17 до 22 час. зацветал через 105, а без освещения — через
125 дней.
Физалис (Physalis pubescens)
Длиннодневное светолюбивое растение.
При дополнительном освещении (с 17 до 22 час; 160—320 лк) в зимнее
время зацветает через 105 дней, на 20 дней раньше, чем без освещения
(697).
Салат (Lactuca sativa)
Типичное длиннодневное растение, цветущее исключительно на длин-
ном дне или значительно ускоряющее в этом случае наступление цветения
[269, 394, 557, 700, 715, 802]. Для получения листовой продукции салат
необходимо выращивать при сравнительно низких температурах (12—18°).
Салат может произрастать при сравнительно невысоких интенсив-
ностях радиации. Максимальный сухой вес растений наблюдается при
интенсивности физиологической радиации порядка 20—25 тыс эрг/см2-сек
(фиг. 126). Урожай салата, культивируемого в теплицах в качестве куль-
туры-уплотнителя, колеблется в пределах 0,6—1,2 кг/м2; при самостоя-
тельной культуре салата урожай повышается до 2—2,5 кг/м2.
В настоящее время литература по дополнительному освещению салата
представлена работами многих авторов [4, 5, 136, 225, 226, 244, 269, 273,
283, 376, 398, 399, 400, 428, 453, 470, 511, 512, 557, 579, 607, 646, 692,
700, 705, 707, 715, 717, 810]. Дополнительное освещение применяется как
для получения рассады [579], так и при культуре на постоянном месте
[136, 244, 273, 607, 646, 707].
Принимая во внимание, что при дополнительном ночном освещении
большинство сортов салата быстро выбрасывают стебель, часто не успев
сформировать листовой розетки или кочана [269, 394, 557, 700, 715, 802],
для культуры салата с помощью дополнительного освещения необходимо
отбирать сорта, обладающие большим вегетационным периодом, которые
успевают сформировать кочан задолго до перехода к стрелкованию. К та-
ковым относятся следующие сорта: Том Тумб, Рудольфе Либлинг, Уайт
Бостон, Берлинский, Гарткопф, Богемия, Гольдкопф, Ромэн и другие.
В случае раннеспелых сортов, таких, как Майский или Московский,
дополнительное освещение можно давать лишь в пределах короткого
12-часового дня.
Дополнительное освещение салата в зимние месяцы оказывает силь-
нейшее влияние на ростовые процессы. По данным Тестермана [557] при
6-часовом вечернем освещении в 300—900 лк рыночногодный кочанный
салат в зимние месяцы можно получить через 18 дней вместо 28—35 дней
в контроле. В опытах Клейна [579] салат, посеянный 17 января под лам-
пами накаливания, к 18 февраля достиг сухого веса 1,684 г. Контроль-
ные растения к этому времени достигли веса 0,200 г. По данным Криста
и Стаута [453], увеличение сырого веса в результате дополнительного
освещения достигает 10—65%, но происходит главным образом за счет
300
Частная светокультура растений
корней, а не листьев. Людвиг [607], освещая растения салата в течение
3 часов после захода солнца (220 лк), получил увеличение урожая на
20%. При посеве 2 марта готовые кочаны салата были получены 5 апреля.
Вес кочана при освещении равнялся 186 г, а вес контрольных кочанов —
154 г. Аналогичное увеличение урожая наблюдалссь также и при куль-
туре салата с 12 октября по 20 января в условиях 14-часового дня при
той же интенсивности дополнительного освещения. Тинкер [810] иссле-
довал несколько сортов салата, культивируя их зимою при дополнитель-
ном освещении. Во всех случаях освещение с помощью ламп накаливания
в 150 вт давало положительный результат. Палилов [244], культивируя
салат под лампами накаливания при удельной мощности 500 вт/м2 в усло-
виях 16-часового дня, нашел, что зимою (февраль—март) готовая продук-
ция получается через 31 день, а урожай равняется 2,2 кг/м2. Урожай
контрольных растений, развивавшихся без дополнительного освещения,
не превышал 0,3 кг/м2. Вкусовые качества салата оказываются значи-
тельно выше при дополнительном освещении, причем листья содержат
больше сахара. По данным Рейнгольда [707], при культуре салата Бот-
терс Трэйб с 13 января по 28 февраля получаются следующие результаты:
вес контрольных растений — 140 г (100%), под лампами накаливания —
366 г (261%), а под неоновыми лампами —417 г (298%). В опытах Кернике
[580] с кочанным салатом Майкониг средний вес месячного кочана
(29 марта—30 апреля) равнялся 270—274 г при освещении (71 вт/м2) и
171 г в контроле.
Для дополнительного освещения салата пригодны лампы накаливания,
неоновые, натриевые и ртутные лампы и люминесцентные лампы белого,
дневного и синего света. Лампы накаливания, неоновые лампы и другие
источники оранжево-красных лучей пригодны для дополнительного осве-
щения лишь для сортов с большим вегетационным периодом. Как видно
из приведенных выше данных Рейнгольда, лучше всего пользоваться нео-
новыми лампами, под которыми получается лучший урожай и лучшее
формирование кочана (фиг. 19). Рейнау [705] сопоставил неоновые и на-
триевые лампы и лампы накаливания из обычного и увиолевого стекла,
освещая салат в ночные часы по 5—6 часов. Было установлено, что при
одинаковой освещенности (300 лк) лучший результат по сравнению
с контролем дают неоновые и натриевые лампы, увеличившие урожай
листовой массы на 38 и 11%.
Что касается скороспелых длиннодневных сортов (Майский, Москов-
ский и др.), то здесь следует рекомендовать источники сине-фиолетовых
лучей (ртутные и люминесцентные лампы). Под влиянием источников
оранжево-красных лучей названные сорта быстро выбрасывают стебель,
не образуя листовой розетки (фиг. 11 и 20 [ИЗ, 118, 184, 646]).
Как показали опыты Гелина и Бюрстрема [509], накопление сухой
массы при дополнительном освещении лучше всего совершается под лю-
минесцентными и солнечными лампами, богатыми синими лучами (табл. 90).
Для дополнительного освещения салата обычно применяют небольшие
интенсивности освещения — порядка 200—500 лк [273, 607, 705, 707].
Это соответствует удельной мощности 25—50 вт/м2. Для умеренных ши-
рот СССР следует рекомендовать освещенность 5000—8000 лк (30—50 тыс.
эрг/см2*сек физиологической радиации), т. е. удельную мощность 500 вт/м2.
Что касается длины дня, то последняя чаще всего равняется 13—16 ча-
сам. В зависимости от естественной длины дня ежесуточное дополнитель-
ное освещение равно 6—8 часам [244, 273, 290, 509, 607, 646, 705, 707].
Если исходить из урожая салата 2,5 кг/м2, удельной мощности 100—
500 вт/м2 и длительности освещения 6 часов, то в течение вегетационного
VIII. Светокультура овощных и садовых растений
301
периода салата (30 дней) на 1 кг урожая затрачивается 7,2—36 квт-час
энергии.
В настоящее время можно рекомендовать выращивание салата пол-
ностью на искусственном свете. С. В. Александров [4, 5], пользуясь лам-
пами накаливания (500 вт/м2), горевшими в течение 11—12 часов, культи-
вировал салат в производственных условиях, получая урожай 2—2,3 кг/м2.
Для выращивания салата исключительно при искусственном освещении
особенно пригодны люминесцентные лампы [ИЗ, 118]. Для розеточных
форм (Майский, Московский) пригодны люминесцентные лампы белого,
дневного и синего света (фиг. И и 20), а для кочанных форм (Ромэн и др.)—
люминесцентные лампы красного света (фиг. 19). Для нормального
роста растений достаточно удельной мощности 450 вт/м2. Лампы разме-
щаются в виде горизонтальной решетки на высоте 20—25 см над расте-
ниями. В этом случае готовая продукция может быть получена примерно
через 30 дней. По данным Б. С. Мошкова [224], под люминесцентными
лампами салат особенно хорошо растет при беспрерывном освещении.
В его опытах свежий вес 20-дневного растения равнялся 6,2 г, а сухой —
270 мг; на 1 кг продукции было затрачено 300 квт-час.
Принимая во внимание, что салат не нуждается в высоких интенсив-
ностях освещения в зимние месяцы, можно без большого труда заниматься
селекцией и семеноводством салата. Под влиянием дополнительного осве-
щения с помощью источников радиации красного света салат быстро пере-
ходит к цветению и дает семена [802].
Салатный цикорий (Cichorium endivia)
И. А. Веселовский [33] рекомендует для культуры на зелень в условиях
беспрерывного освещения целиком под лампами накаливания в 200—300 вт
(80—120 вт/м)2.
Шпинат (Spinacea oleracea)
Типичное длиннодневное и светолюбивое растение, в большинстве слу-
чаев совершенно не цветущее на коротком дне. Культивируется ради
листьев. Техническая годность наступает после формирования трех пар
розеточных листьев (через 28 дней). Средний урожай через 60—70 дней
равняется 1—1,5 кг/м2. Исследований по светокультуре шпината немного
[4, 5, 136, 212, 244, 276, 289, 398, 428, 441, 500, 650, 698, 716, 717, 742, 855].
Обычно при дополнительном ночном освещении шпинат быстро, через
22 дня, выбрасывает стебель, а через 33 дня зацветает [276, 428, 441, 500].
По данным Б. С. Мошкова [212], шпинат зацветает лишь при длине дня
не менее 13 часов. Наиболее быстро цветение происходит при 17—24-часо-
вом дне — через 23 дня. Максимальный вес растений (14,38 г) наблю-
дается при длине дня в 14 часов, а беспрерывное освещение сильно снижает
накопление сухой массы растений, и вес растения снижается до 5,02 г.
Поэтому для культуры шпината следует рекомендовать короткий день
(12 часов), в пределах которого и следует давать дополнительное освеще-
ние. Длинный день (более 14 часов) выгодно применять лишь при семе-
новодстве шпината.
Дополнительное ночное освещение очень сильно влияет на рост шпи-
ната. В опытах Родникова [276] шпинат Голландский, посеянный 24 марта
и развивавшийся под лампой накаливания 1000 вт (445 вт/м2), достиг
к 21 апреля веса 4,7 г. Вес контрольных растений равнялся к этому вре-
мени 1,9 г. Как показывает табл. 89, для культуры шпината наиболее
Частная света пультура растений
ирш'идны люминесцентные лампы дневного света при освещенности по-
рядка 4000 лк.
С. В. Александров 14, 5] рекомендует шпинат для культуры исключи-
тельно при искусственном освещении. В его опытах под лампами нака-
ливания (500 вт/м2) был получен урожай 0,5—0,75 кг м2.
Французский шпинат (Alriplcx hortensis)
Нейтральное однолетнее растение. При дополнительном освещении
(с 17 до 22 час.; 160—320 лк) цветет одновременно с контролем— через
91 день 1697].
Укроп (Aiieihum grav colons)
Типичное длиннодневное растение, резко реагирующее на дополни-
тельное освещение. Бутонизирует при интенсивности физиологической
радиации порядка 8000 эрг/см'2-сек [637]. В качестве культуры уплотне-
ния дает урожай 0,4—0,5 кг/м'2, а при самостоятельной культуре — 1 кг/м‘2
[527, 637]. Для цветения нуждается лишь в 4-дневном дополнительном
ночном освещении и зацветает уже при 14-часовом дне [527, 637]. На ко-
ротком дне (менее И часов) долго находится в вегетативном состоянии.
Нормальный рост и развитие растений наблюдаются при интенсивности
дополнительного освещения 0,6 г-кал/см2-мин (42 тыс. эрг,см'2-сек [646]).
В этом случае при посеве 7 февраля готовая продукция получается через
6 недель. Для культуры укропа годятся лишь источники радиации, бо-
гатые сине-фиолетовыми лучами [646].
Для культуры укропа с целью получения листовой массы необходимо
рекомендовать короткий день (10—И часов), а для получения семян —
длинный день (14—15 часов). Невидимому, лучший эффект будут давать
ртутные лампы и люминесцентные лампы белого, дневного и синего света.
Александров (4, 5), выращивая укроп в производственных условиях
только при электрическом освещении (500 вт/м'2), получал урожай в 1 кг/м2.
Щавель (Rumex acetosa)
С. В. Александров [4, 5] культивировал щавель в производственных
условиях при 12-часовом освещении исключительно под лампами нака-
ливания (500 вт/м2), получая урожай 3,2—3,7 кг/м2.
Редис (Raphanus sativa var. radicula)
Длиннодневное растение, цветущее только на длинном дне. Корнеплоды
образуются исключительно на коротком дне. Максимальный вес растений
имеет место в условиях 17-часового дня (табл. 81 [379]). Температура до
образования корнеплодов должна равняться 12—15°, а после формирова-
ния корнеплодов 16—18°. Готовая продукция поступает через 40—45
дней после посева. Урожай равняется 2 кг/м'2.
Литература по дополнительному освещению редиса представлена не-
большим числом работ [3,4, 64, 136, 273, 283, 376, 394, 428, 441, 453, 500,
579, 646, 697, 700, 707, 717, 742, 822].
Как показывают опыты, дополнительное ночное освещение, удлиняю-
щее день, вызывает быстрое образование цветочного стебля и цветение
редиса и препятствует образованию корнеплода [64, 394, 500, 697, 700,
742, 822]. Поэтому дополнительное освещение редиса следует производить
или в пределах короткого дня пли прибегать к источникам радиации,
богатым сине-фиолетовыми лучами (ртутные лампы, люминесцентные лам-
VIII. Светокультура овощных и садовых растений 303
пы дневного, белого и синего цвета). Впрочем, некоторые сорта (например
Фрэнч Брэкфаст или Уайт Сайкл) даже при дополнительном освещении
с 17 до 22 час. (160—300 лк) зацветают лишь через 90—120 дней, а в усло-
виях естественного зимнего дня — через 139 дней или совсем не цветут.
Подобные сорта зимою, невидимому, можно культивировать и в условиях
дополнительного ночного освещения.
Клейн [579], применяя дополнительное освещение растений в ночные
часы (с 22 до 6 час.) в зимние месяцы (январь—март), выгонял рассаду
редиса с помощью ламп накаливания в 500 вт (100 вт/м2), висевших на
высоте 1,2 м. Расход электроэнергии в его опытах равнялся 0,75 квт-час/м2.
Редис при посеве 17 января достиг к 18 февраля веса 2,35 г. Контрольные
растения в это время весили 0,63 г.
Оден [646] и Рейнгольд [707] в зимнее время культивировали редис
на постоянном месте при дополнительном освещении до получения корне-
плодов. Оден нашел, что при источниках, богатых сине-фиолетовыми
лучами, и интенсивности освещения 0,5 г-кал/см2-мин готовые корнеплоды
редиса могут быть получены через 5 недель. Рейнгольд, применяя лампы
накаливания (200 вт) или неоновые трубки, получил значительное уве-
личение урожая редиса при освещении растений в ночные часы (с 21 час.
30 мин. до 3 час. 30 мин). При посеве 13 января рыночногодный редис был
получен 5 марта, причем в результате освещения наблюдалось увеличе-
ние урожая по количеству корнеплодов на 60—80%, а по весу корнепло-
дов — в 3 раза. Вес корнеплода в контроле равнялся 15,6 г, а под неоно-
выми лампами и лампами накаливания — 27,8—29,8 г.
В опытах Криста и Стаута [453] дополнительное освещение в ночные
часы удваивало вес растений.
При помощи люминесцентных ламп белого и синего света редис можно
культивировать целиком при искусственном освещении [ИЗ, 184, 224].
Для этих целей достаточна удельная мощность 450 вт/м2.
Петрушка (Petroselinum sativum)
По данным Веселовского [33], можно культивировать на зелень при
освещении только лампами накаливания в 200—300 вт (80—120 вт/м2).
При беспрерывном освещении срезка производится каждые 20 дней.
С. В. Александров [4, 5] культивировал петрушку на зелень в производ-
ственных условиях полностью при искусственном освещении на 12-ча-
совом дне при удельной мощности 500 вт/м2 и получал урожай 0,5—0,9 кг/м2.
Репа (Brassica гара)
Длиннодневное растение. Дополнительное ночное освещение в зимние
месяцы можно применять для целей селекции и семеноводства [136, 244,
651]. По данным Палилова [244], у репы Петровская зрелые семена в усло-
виях дополнительного освещения (500 вт/м2) при длине дня 16 часов
можно получить через 57 дней (27 апреля) — на 9 дней раньше, чем в конт-
роле (6 мая).
Как показал Оден [651], для получения семян репы зимою необходима
интенсивность дополнительного освещения 0,1 г-кал/см2-мин (70 тыс.
эрг/см2-сек), получаемая с помощью ламп накаливания в 500 или 750 вт.
В зимние месяцы (7 января—16 марта) для получения семян затрачивается
около 5,8 кг-кал/см2 (2,45 кг-кал/см2 за счет естественного света и
3,35 кг-кал/см2 — с помощью ламп). Урожай семян с 1 растения
в опытах Одена равнялся 6 г, а в полевых условиях — 12 г.
:04
Частная светокультура растений
Сельдерей (Apium graveolens)
В опытах И. А. Веселовского [33] выращивался при освещении только
лампами накаливания в 200—300 вт (80—120 вт/м2). Срезка зелени произ-
водилась каждые 20 дней.
Морковь (Daucus carota)
Несмотря на то что первые опыты по светокультуре моркови были
проведены давно [754], литература по дополнительному освещению ее
очень невелика [136, 244, 289, 290, 428, 470, 610, 613, 692, 736].
Магру, Маниго и Мариа [610, 613] указывают, что в зимние ме-
сяцы морковь можно выращивать с помощью люминесцентных ламп днев-
ного света, помещая их на высоте 10 см над растениями и увеличивая
естественную освещенность до 3000 лк. По их данным, таким способом
в январе можно выращивать растения, не уступающие растениям, которые
культивируются летом.
При дополнительном освещении зимою хорошо удается семеноводство
моркови [136, 244, 736]. Палилов 1244] под лампами накаливания (500 вт/м2)
получал столько же семян, сколько и в полевых условиях. Лучшие ре-
зультаты наблюдаются в условиях 16-часового дня. В этом случае зрелые
семена можно получить через 90 дней (5 апреля). Контрольные растения
дают семена на 15 дней позднее (20 апреля).
Кольраби (Brassica caulorapa)
Рейнгольд [707] в зимние месяцы с помощью дополнительного освеще-
ния (с 21 час. 30 мин. до 3 час. 30 мин.) (6 ноября—9 февраля) выращивал
рассаду кольраби. При посеве 6 ноября готовая рассада была получена
в январе. Положительный результат был получен лишь с помощью нео-
новых ламп (увеличение в весе растений на 113%). Лампы накаливания
положительного влияние не оказали. Высаженные на постоянное место
(9 февраля), растения во всех случаях дали ничтожный урожай (0,3кг/м2),
причем в результате дополнительного освещения в рассадный период
произошло снижение урожайности (на 10%).
Людвиг [607], применяя лампы накаливания (220 лк), пытался вы-
ращивать кольраби ранней весною (21 февраля—20 апреля). Несмотря
на положительное влияние освещения, урожай был получен плохой.
Повидимому, кольраби, являясь светолюбивым растением, нуждается
в интенсивном дополнительном освещении. Кернике и Линденбейн [580]
нашли, что кольраби при дополнительном освещении (4 апреля—3 мая)
слабее завязывает стеблевое утолщение, чем в контроле.
Капуста (Brassica oleracea)
| Оптимальная длина дня для ростовых [процессов равна 19 часам
(табл. 81 [379]).
Положительное влияние дополнительного освещения на цветную и
кочанную капусту отмечают Сименс [754], Артур [376] и Палилов [244].
Н. П. Красинский [136] в январе получил мощно развитые растения цвет-
ной капусты с помощью дополнительного освещения (с 6 до 9 и с 15 до
21 час.) от лампы накаливания в 1000 вт (500вт/м2). Диаметр головок
у опытных растений достигал 7—9 см. Контрольные растения имели
лишь зачаточные головки (2—3 см в диаметре).
VIII. Светокультура овощных и садовых растений
305
Палилов [244] и Красинский [136] рекомендуют дополнительное осве-
щение для семеноводства капусты в зимние месяцы. В опытах Палилова
семенники кочанной капусты № 1 и цветной капусты Снежный шар,
посаженные в начале января, под лампами накаливания (500 вт) дали
зрелые семена в конце марта — начале апреля. Лучший результат наблю-
дался в условиях 16-часового дня. В этом случае цветение кочанной ка-
пусты началось через 41 день (14 февраля), а зрелые семена были полу-
чены через 79 дней (24 марта); соответствующие цифры для контроля —
48 и 90 дней.
Оден [651] для целей семеноводства краснокочанной капусты реко-
мендует интенсивность радиации 0,1 г-кал/см2-мин (70 тыс. эрг/см2-сек),
получаемую с помощью ламп накаливания в 500 и 750 вт. По его данным,
для получения семян в зимние месяцы (7 января—16 марта) необходимо
затратить 5,8 кг-кал/см2, причем 2,45 кг-кал/см2 приходится на естествен-
ный свет, а 3,35 кг-кал/см2 — на дополнительное искусственное осве-
щение. В этом случае можно получить с одного растения примерно столько
же семян (9 г), сколько и в полевых условиях (11 г).
Китайская капуста (Brassica pekinensis)
Типичное длиннодневнос растение, цветущее через 36—48 дней в усло-
виях 18-часового дня и через 74 дня — при короткохм дне. Зимою при
дополнительном освещении с помощью ламп накаливания в 500 вт цве-
тет через 46—62, а в контроле — через 80—93 дня [431]. По данным Ро-
денбурга [717], зимою нуждается в дополнительном ночном освещении
в 5000 лк (30 тыс. эрг/см2-сек физиологической радиации).
Под лампами накаливания растения сильно вытягиваются [283, 717].
С. В. Александров [4, 5] культивировал это растение исключительно
при искусственном освещении с помощью ламп накаливания (500 вт/м2),
получая урожай 1,6—1,7 кг/м2.
Лук (Allium сера)
Лук представляет собою многолетнее растение, образующее луковицы
и зацветающее лишь в условиях длинного дня. Оптимальная интенсив-
ность физиологической радиации равна 50 тыс. эрг/см2-сек (фиг. 135,3
1224]). В зимнее время обычно выращивается в качестве выгоночной куль-
туры на перо. Без дополнительного освещения почти не дает увеличения
сухого веса, образуя зеленое перо исключительно за счет запасных ве-
ществ, отлагаемых в луковицах. Лишь начиная с февраля прирост сухого
вещества составляет 25—50%. Средний урожай лука-пера колеблется
в пределах 8—10 кг/м2.
Работ по светокультуре лука очень немного, причем большинство из
них находится в связи с изучением фотопериодической реакции [125,
205, 272, 428, 501, 611, 618, 646, 742, 746, 799, 851].
И. Н. Коновалов [125], применяя дополнительное освещение (14—
1 час.) от лампы накаливания в 300 вт, культивировал лук-севок
с ноября по январь. В результате освещения сухой вес подопытных расте-
ний был на 67% больше веса контрольных.
Бос [428], выращивая лук Зильберцвибель в условиях дополнитель-
ного ночного освещения с помощью ламп накаливания при различной
удельной мощности и длине дня, установил, что зимою (декабрь—январь)
нормальные растения лука и хорошее формирование луковиц наблюдаются
лишь при освещенности порядка 7000 лк и длине дня 17 часов. При осве-
щенности 3000 лк и длине дня 14—15 часов луковицы образуются
20 А. Ф. Клешнин
306
Частная светокультура растений
небольшие, и растения быстро уходят в покой, а при 1000 лк в условиях
12-часового дня растения хотя и растут, но очень медленно. Контрольные
растения этого сорта, выращиваемые без дополнительного освещения,
в зимнее время не растут и обычно погибают.
Магрэдер, Аллард и Макклеланд [611, 618], исследуя более 30 сор-
тов лука, нашли, что растения, полученные из семян, формируют нор-
мальные луковицы лишь в условиях длинного дня; оптимальной длиной
дня следует признать 16—18 часов. В условиях короткого дня (10—13 ча-
сов) растения образуют лишь зеленое перо. Аналогичные данные получены
Ф. Э. Реймерсом [272]. Этот автор показал, что беспрерывное освещение
дает худший эффект, чем 16—17-часовой день.
Таким образом, для культуры лука на перо растения нуждаются
в 10—13-часовом дне, а для формирования луковиц — в 16—18-часовом
дне. Для нормального роста в зимнее время лук, получаемый из семян
и из севка, нуждается в дополнительном освещении с интенсивностью
физиологической радиации порядка 40 тыс. эрг/см2-сек.
Источники радиации, богатые оранжево-красными лучами, благо-
приятствуют образованию луковиц, а источники, содержащие много
сине-фиолетовых лучей, задерживают и даже полностью приостанавли-
вают этот процесс (фиг. 17 и 18).
С. В. Александров 14, 5], культивируя лук только при искусственном
освещении лампами накаливания (500 вт/м2), получал урожай пера 13 кг/м2.
По данным И. А. Веселовского [33], для культуры репчатого лука на
перо исключительно с помощью ламп накаливания необходима удельная
мощность 80—120 вт/м2. В этом случае при беспрерывном освещении и
температуре 20—24° готовая продукция (перо длиною 30—40 см) может
быть получена через 14—15 дней. В течение 30 дней можно произвести
две срезки пера — по 7 и 3 кг/м2.
Лук-батун (A. fistulosum) в течение 45 дней дает три срезки пера —
по 8,5 и 2,5 кг/м2.
При культуре репчатого лука на перо на 1 кг продукции затрачивается
по 4,1—5,2 квт-часпри одной срезке и по 5,7—6,6 квт-час при двух срезках.
Соответствующие цифры для культуры лука-батуна равны: при одной
срезке — 3,6—5,4, при двух срезках — 4,4—5,1, а при трех срезках —
5,2—7,7 квт-час.
Дополнительное освещение в зимние месяцы можно применять и для
получения семян лука. Исследований в этом направлении нет, если не
считать отдельных работ по фотопериодизму названного растения [742,
746]. Лучший результат следует ожидать в условиях 18-часового дня.
Чеснок (Allium sativum)
С. В. Александров [4, 5] рекомендует для культуры на зелень при одном
искусственном освещении с помощью ламп накаливания (12 часов; 500
вт/м2). Он получал в этом случае урожай 6,0—7,3 кг/м2.
2. САДОВЫЕ РАСТЕНИЯ
Земляника (Fragraria)
48 Тенелюбивые растения; достигают максимального сухого веса
при интенсивности физиологической радиации 50 тыс. эрг/см2-сек
(фиг. 135, 4 [224]). Для закладки цветочных почек нуждаются в низких
температурах. В естественных условиях заложение бутонов имеет место,.
VIII. Светокультура овощных и садовых растений
307
когда минимальная (ночная) температура постепенно снижается от 16
до —4°, а максимальная (дневная) — от 32 до 4,5°. При ранней весенней
выгонке земляники в оранжереях рекомендуется температуру постепенно
повышать от 3—4° до 20°. Практически это означает еженедельное повы-
шение примерно на 2° вплоть до начала цветения.
Обычные сорта земляники относятся к группе растений короткого
дня. Они закладывают плодовые почки при длине дня 10—12 часов. Ре-
монтантные сорта являются нейтральными. Для выгоночной культуры
рекомендуются следующие сорта: Рощинская, Нобль, Саксонка, Коралка,
Иосиф Магомет, Ананасная белая, Мысовка и Миланская (клубника).
Выгонка производится в горшках диаметром 18—20 см.
Обычно при выгонке без дополнительного освещения получают урожай
100 г, а при хорошей агротехнике — 150—200 г с куста. При размеще-
нии 20 сосудов на 1 м2 это означает урожай 2—4 кг/м2. Выгонка начинается
в январе—феврале, цветение растений наступает через 30—40, а плодо-
ношение — через 60—90 дней.
Вопросом дополнительного освещения земляники в зимние месяцы
занимались многие авторы [32, 160, 283, 290, 386, 387, 457—461, 470,
530, 556, 692, 700, 717, 718, 726, 727, 729, 732, 733, 754—756].
В результате исследовании установлено, что при дополнительном
ночном освещении земляника лучше развивается, образует большую листо-
вую поверхность, раньше зацветает и плодоносит и дает более высокий
урожай. Роденбург [729] отмечает, что у земляники Дойтш Эверн в ре-
зультате дополнительного освещения с помощью ламп накаливания длина
листовой пластинки сравнительно с контролем возрастает на 46%, а ее
ширина — на 170%. В другой работе этот автор [732] показал, что увели-
чение размеров листовой пластинки наблюдается при дополнительном
освещении как неоновыми лампами, так и лампами накаливания и дости-
гает, в зависимости от интенсивности и длительности освещения, 25—250%.
Дэрроу и Уолдо [459], исследовавшие около 140 сортов земляники,
нашли, что дополнительное освещение от ламп накаливания в 200 и 1000 вт
в зимние месяцы увеличивает листовую поверхность по сравнению
с контролем в среднем на 120—160%. Аналогичные данные получены
в другой работе [461] и для ламп в 500 вт.
Обычно в результате дополнительного ночного освещения увеличи-
вается число цветоносов и цветков. В опытах Дэрроу и Уолдо [459], про-
веденных с 57 сортами земляники, было обнаружено, что количество
цветков увеличивается в среднем в 7 раз (табл. 107). Роденбург [727] от-
мечает, что при освещении земляники неоновой лампой количество цвето-
носов увеличивается на 70%. В других опытах [732] этот автор нашел,
что при освещении неоновыми лампами и лампами накаливания количество
цветков, в зависимости от условий освещения, увеличивается от 2 до 7 раз.
Увеличение размеров листовой поверхности и количества цветоносов
и цветков приводит к увеличению урожая ягод. Как видно из табл. 116,
это увеличение довольно значительно. Аналогичные данные приводит
Роденбург. Зимою оп получал урожай практически лишь при дополнитель-
ном освещении [717, 727, 733]. В опытах с земляникой Белль Альянс
этот автор нашел, что прп беспрерывном освещении (с 23 ноября) с помощью
ламп накаливания в 300 вт растения бутонизируют через 3 недели, а через
6 недель дают зрелые красные ягоды отличного аромата и качества. В дру-
гом опыте Роденбург установил, что земляника Мадам Лефэбр, освещае-
мая по 14 часов (с 17 до 7 час.) в сутки (28 января— 18 марта) с помощью
ламп накаливания или неоновых трубок, дает зрелые ягоды через 7 не-
дель, увеличивая урожай на 24% под лампами накаливания и на 120%
20*
308
Частная светокультура растений
под неоновыми трубками. В результате трехнедельного периода плодоно-
шения (к 14 апреля) было получено по 2,3 ягоды с куста в контроле, по
3 ягоды под лампами накаливания и по 5 ягод под неоновыми трубками.
При культуре растений с 5 октября по 9 февраля под различными лампами
был получен урожай около 200 г; с контрольных растений было полу-
чено 5 г [733].
При освещении (8 часов) со 2 октября с помощью неоновой лампы (475 вт)
или комбинации последней с лампами накаливания Роденбург получал
в течение 42 дней (с 30 января по 13 марта) урожай с каждого растения:
в контроле — 1,35 г, при комбинации ламп — 34,2 г, а под неоновой
лампой — 54,5 г. Соответствующие цифры для числа ягод были 0,2, 7,3 и
10,4, а для урожая — 20, 545 и 875 г/м2 [722]. В опыте с освещением
неоновыми лампами (8 часов) в 450 и 350 лк с 16 сентября и 1 октября
автор нашел, что в обоих случаях урожай ягод с одного растения равнял-
ся 38 г при освещении с 16 сентября и 30 г при освещении с 1 октября.
Урожай в контроле колебался в пределах 0,25—3,6 г. Количество ягод
при освещении равнялось 8,2—10,5, а в контроле 0,08—0,6 [722].
Подпочвенный обогрев в опытах этого автора давал отрицательный
результат. По данным голландских авторов [518], дополнительное освеще-
ние зимою увеличивает урожай земляники на 16%, а освещение вместе
с добавлением СО2 — на 40%.
В опытах В. М. Лемана [160] земляника Ремонтантная в зимние ме-
сяцы (январь—март) росла в теплице лишь при дополнительном освеще-
нии (16 часов), причем под люминесцентными лампами «ДС-30»
(300—600 вт/м2; 5000—8000 лк) давала урожай на 35% выше, чем под
лампами накаливания (1000—1500 вт/м2).
Дополнительное ночное освещение земляники в зимне-весенний пе-
риод значительно ускоряет наступление цветения и плодоношения расте-
ний. В опытах Ранке [700] и Гёстермана [556] земляника Дейтш Эверн,
освещаемая лампами накаливания с 10 ноября (600 лк, с 16 до 22 час.),
зацвела 1 января, а без освещения — 27 января. По данным Гардера
(табл. 116), ускорение наступления плодоношения под влиянием ламп
накаливания равняется 21—23 дням. Дэрроу и Уолдо [459], исследовав-
шие много сортов земляники, нашли, что, в зависимости от сорта, ускоре-
ние наступления цветения при культуре с 7 декабря по 10 марта колеб-
лется от 10 до 80 дней (табл. 107). Одни сорта, как, например, Миссионэри,
зацветают раньше на 10 дней, а другие (Эттерсбург 121, Fr. virginiana) —
на 10—35 дней. Ф. Д. Лихонос [164], ссылаясь на опыты Петергофского
плодо-овощного института и Ленинградского физико-агрономического
института, сообщает, что при дополнительном ночном освещении плодо-
ношение земляники наступает через I1/,, а без освещения — через 3 ме-
сяца после начала выгонки.
h . Таким образом, приведенные данные не оставляют сомнения в том, что
культура земляники, невозможная в зимние месяцы без дополнительного
освещения, становится возможной, если растения подвергаются освеще-
нию с помощью искусственных источников радиации. В качестве послед-
них могут применяться лампы накаливания, дуговые, неоновые, ртутные,
натриевые, люминесцентные и другие типы ламп. Обычно применяются
лампы накаливания [387, 458, 459, 530, 700]. Роденбург [717, 718,
727, 729, 733] усиленно рекомендует неоновые лампы, которые, по его
данным, дают ягоды лучшего качества, хотя по величине урожая лучший
результат наблюдается под натриевыми лампами (табл. 128). Повидимому,
и здесь лучшие результаты следует ожидать от люминесцентных ламп
[160].
VIII. Светокультура овощных и садовых растений
309
Таблица 128
Урожай земляники при дополнительном освещении 8 часов
(с 22 до 6 час. с 5 октября по 9 февраля) (г = 14—21°)
(Роденбург [733J)
Варианты опытов Мощность ламп (в ВТ) Интенсив- ность видимой радиации (в эрг/см- • сек) Урожай ягод за пе- риод с 1/III по 16/III
Г ШТ.
Неоновые лампы 100 3100 209 75
Натриевые лампы 100 2800 242 92
Ртутные лампы 150 2600 219 70
Контроль — — 5 1
Следует отметить, что в опытах по дополнительному освещению зем-
ляники в зимние месяцы в большинстве случаев наблюдается сравнительно
невысокий урожай. Это, несомненно, находится в связи с той сравнительно
невысокой интенсивностью радиации, которая обычно применяется при
культуре земляники. Ранке [700], применяя лампы накаливания в 200 вт,
получала освещенность 600 лк. В опытах Роденбурга [727, 732] с неоно-
выми лампами интенсивность радиации равнялась 480 лк. При сопоставле-
нии различных типов ламп этот автор [733] в лучшем случае пользовался
интенсивностью физиологической радиации порядка 3000 эрг/см2-сек
(табл. 128). В опытах Гардера [530] освещенность не превышала 2000 лк.
Между тем, как показывают опыты Дэрроу и Уолдо [459], частично пред-
ставленные в табл. 107, интенсивность радиации имеет большое значение
для культуры земляники. В опытах этих авторов лампы накаливания
в 1000 вт (700 вт/м2, около 4000 лк) давали значительно лучший эффект,
чем те же лампы в 200 вт (260 вт/м2, около 1200 лк). В первом случае
большей частью наблюдалось более раннее цветение (на 10—30 дней),
большее число листьев (на 40%) и цветков (на 60%) и большая площадь
листьев, чем во втором.
Повидимому, для культуры земляники зимою, как и для других расте-
ний, нужна освещенность порядка 8000 лк (48 тыс. эрг/см2-сек), т. е. при
высоте подвеса 50 см удельная мощность порядка 700 вт/м2.
Опыт показывает, что для земляники оптимальная длина дня равна
16—18 часам. Дэрроу и Уолдо [459] нашли, что при культуре земляники
с 8 января лучший эффект дает дополнительное освещение в течение 7 ча-
сов. Листовая поверхность в этом случае значительно больше (235,8 см2),
чем при дополнительном освещении в течение 4 часов (159,7 см2) или в те-
чение 14 часов (135,9 см2). В опытах Дэрроу [461] при освещении земля-
ники лампами накаливания в 500 вт (около 4500 лк) с 1 сентября, 15 сен-
тября и 1 января было найдено, что лучшая листовая поверхность (900—
1000 см2) наблюдалась в том случае, если длина дня равнялась 16 часам.
На практике обычно пользуются дополнительным освещением в течение
8 часов, т. е. длиною дня 16—18 часов. В опытах Роденбурга [732] в этом
случае был получен значительно лучший урожай, чем при освещении в
течение 4 часов (на 30—40%).
Артур и Гарвилл [387] при культуре земляники Дорсет применяли
прерывистое освещение в ночные часы (в сумме по 3 часа 24 минуты в сутки
в декабре—феврале и 1 час 36 минут в сутки — в ноябре и марте). При
310
Частная светокультура растений
освещении с 15 сентября и удельной мощности 350 вт/м2 авторы получали
зрелые ягоды к 15 декабря.
Роденбург [717] рассчитал, что при культуре земляники в зимние ме-
сяцы необходимо дополнительное освещение в течение 50 дней. При удель-
ной мощности 375 вт/м2 и длительности ежедневного освещения 9 часов
это означает расход электроэнергии 170 квт-час/м2. Если исходить из
густоты стояния 25 растений на 1 м2 и урожая по 5 ягод с куста, то на каж-
дую ягоду затрачивается около 1,35 квт-часа.
При надлежащей агротехнике и правильном подборе сортов земляника
может культивироваться полностью при искусственном освещении. Она
неплохо растет под люминесцентными лампами при удельной мощности
450 вт/м2.
Малина (Rubus idaeus)
Положительная реакция на дополнительное освещение отмечена еще
Сименсом [754—756]. Этот автор при освещении растений с 16 декабря
получал зрелые ягоды малины 1 марта. В опытах Одена [646] малина
Аскер после обработки ацетиленом и освещения с 22 октября обильно
зацвела в январе, начала плодоносить 17 февраля и плодоносила в тече-
ние февраля и марта. Средний вес ягод равнялся 2,5 г, а содержание
сахара 9,4% (вместо обычных 5—6%). Для выгонки малины необходимо
100 г-кал/см2 в день.
Банан (Musa paradisiaca)
В опытах Сименса [754—756] при дополнительном освещении с по-
мощью вольтовой дуги в зимние месяцы дал гроздь плодов весом 49 кг.
По данным автора, плоды отличались превосходным вкусом.
3. выводы
Подводя итог, следует отметить, что в настоящее время дополнительное
ночное освещение овощных и садовых растений применяется главным
образом для получения ранней рассады.
Этот прием позволяет получать рассаду многих овощных растений
в любое время года, причем получаются более мощные и крепкие растения,
значительно увеличивается урожай и ускоряется плодоношение.
Для получения рассады пригодны любые источники радиации —
лампы накаливания, ртутные, неоновые, натриевые и люминесцентные
лампы; лучший эффект наблюдается под люминесцентными лампами. Для
выращивания рассады необходима освещенность порядка 5000—8000 лк
(30—48 тыс. эрг/см2-сек физиологической радиации), для чего требуется
удельная мощность порядка 300—500 вт/м2. Лампы обычно помещаются
на расстоянии 1 м друг от друга и на высоте 50 см от почвы. В случае
применения подвижной системы Финкельштейна высота подвеса ламп
равняется 25—35 см. В качестве отражателей рекомендуются обычные
широкоизлучатели типа «Универсаль». Зеркальные лампы не нуждаются
в осветительной арматуре.
Оптимальной длиной дня для большинства растений следует признать
16—17 часов. Для этого в умеренной полосе в зимнее время необходимо
давать дополнительное ночное освещение в течение 7—8 часов.
При длительности освещения 30 дней и густоте стояния рассады
120 растений па 1 м2 на каждое растение рассады затрачивается 1 квт-час
энергии.
VIII. Светокультура овощных и садовых растений
311
Что касается культуры растений в зимние месяцы с целью получения
плодов, то этот вопрос еще далек от своего решения. В принципе можно
принять, что зимою, применяя дополнительное освещение, можно выра-
стить или выгнать любое овощное или садовое растение, если обеспечена
надлежащая интенсивность радиации и создан необходимый температур-
ный режим. Для этой цели пригодны те же источники радиации, что и
для получения рассады; лучший эффект получается под люминесцентными
лампами. Интенсивность радиации должна быть повышена до 10—15 тыс.
лк (60—90 тыс. эрг/см2-сек физиологической радиации). Практически
эти величины создаются с помощью ламп накаливания в 850—1000 вт,
помещаемых в рефлекторе «Универсаль» на расстоянии 1 м друг от друга
на высоте 50 см от растений.
В силу неэкономичности современных типов ламп, затраты большого
количества электроэнергии и дороговизны последней культура растений
в зимние месяцы, исключая получение рассады, в настоящее время не-
рентабельна. Этот прием станет рентабельным лишь при резком сокраще-
нии вегетационного периода растений, увеличении урожайности и созда-
нии экономичных типов ламп. Решение проблемы зависит, следовательно,
от совместных усилий селекционеров, агротехников и инженеров-свето-
техников.
Глава IX
СВЕТОКУЛЬТУРА ДЕКОРАТИВНЫХ РАСТЕНИЙ
Литература по светокультуре декоративных растений охватывает
большое число видов и сортов растений и представлена значительно боль-
шим числом работ, чем литература по культуре овощных растений.
В отличие от светокультуры овощных растений дополнительное осве-
щение цветочных и декоративных растений ставит своей целью не полу-
чение урожая плодов, ягод или других органов, а получение максималь-
ного количества цветков или растений, удовлетворяющих декоративным
требованиям и запросам эстетики. Иными словами, в данном случае имеет
значение не столько накопление органического вещества в процессе фото-
синтеза, сколько формообразовательный процесс и прохождение световой
стадии. Поэтому часто при освещении декоративных растений решающая
роль принадлежит не интенсивности радиации, а длительности суточного'
освещения и спектральному составу излучения.
Проблема светокультуры цветочных растений в настоящее время мо-
жет быть сведена к следующим трем вопросам: 1) выращиванию рассады
в зимний период, 2) зимней выгонке растений и 3) культуре растений
в комнатах и закрытых помещениях. Разбору этих вопросов и посвящена
настоящая глава.
1. ^ОДНОЛЕТНИКИ
Однолетники в большинстве своем исключительно светолюбивые?
растения. Поэтому их светокультура представляет наибольшие трудности.
С этих растений мы и начнем нашу главу.
Аброния (Abronia umbellata)
Оранжерейное многолетнее растение, культивируемое как однолетник,
относится к группе растений длинного дня. В зимние месяцы при допол-
нительном освещении (с 17 до 22 час.) в 150—300 лк зацветает через
120 дней от посева; цветение контрольных растений наступает через.
153 дня [697].
Горицвет («Уголек в огне») (Adonis aestivalis)
Длиннодневное светолюбивое, но теневыносливое растение. Культи-
вируется в открытом грунте. Рассада готовится в парниках. Цветет в мае—
IX. Светокультура декоративных растений,
313:
июне. В оранжерее зимою при дополнительном освещении в 150—300 лк
(с 17 до 22 час.) зацветает через 152 дня, без освещения— через 166 дней
[697].
Агератум (Долгоцветка) (Ageratum conyzoides)
Многолетнее длиннодневное растение, культивируемое как однолет-
ник. Теплолюб и светолюб. Рассада готовится в парниках. В открытом!
грунте цветет с июня до поздней осени. На дополнительное ночное освеще-
ние зимой реагирует мало. В опытах Рэмэли [697] при ночном освещении
в 150—300 лк (с 17 до 22 час.) зацветал одновременно с контролем (через,
98 дней).
Куколь (Agrostemma)
Длиннодневные и светолюбивые растения. В открытом грунте цветут
в середине и конце лета (июль—август). Исключительно сильно реаги-
руют на дополнительное освещение. Agrostemma githago, цветущая на
коротком 10-часовом дне через 85 дней, при дополнительном ночном осве-
щении в 16 лк зацветает через 48 дней (фиг. 142, 9 [488, 531]). Дальнейшее
увеличение интенсивности дополнительного освещения (до 500 лк) практи-
чески уже не влияет на время наступления цветения, ускоряя его всего^
лишь на 5 дней [488]. Agrostemma coelirosa при оранжерейной культуре
зимою в условиях беспрерывного освещения (интенсивность дополнитель-
ного освещения 100—200 лк) ускоряет цветение на 98 дней и цветет в на-
чале января [697].
Бурачок (Каменник) (Alyssum)
Светолюбивые, коротко дневные, нейтральные и длиннодневные расте-
ния. Высеваются в марте в парники или осенью в грунт. В первом случае
зацветают в первой половине лета (июнь—июль), во втором случае —
в июле—августе. A. maritima в одних случаях ведет себя как нейтральное
растение, зацветая одновременно (через 62 дня) при дополнительном
ночном освещении и в контроле, а в других случаях — как растение
короткого дня — зацветая через 137 дней при освещении (с 17 до 22час.;
320—620 лк) и через 60 дней — в контроле [697]. A. montanum ускоряет
цветение при дополнительном освещении 30—60 лк [496]. A. Benthaurii
в зимние месяцы при дополнительном ночном освещении (8 часов, 500 вт/м2)
цветет через 75, а в контроле — через 94 дня.
Амарантус (Amaranthus sp.)
Типичное короткодневное и светолюбивое растение. На дополнитель-
ное ночное освещение реагирует отрицательно, значительно запаздывая
с цветением. В зимнее время в оранжерее зацветает без освещения через
71 день после посева, а при освещении в 150—300 лк (с 17 до 22час.) —•
через 163 дня [697]. Дополнительное освещение можно рекомендовать
лишь в пределах короткого дня (т. е. днем), а в ночные часы — с целый
предотвращения цветения.
Очный цвет (Воробьиное перо) (Anagallis linifolia)
Многолетнее растение, культивируемое как однолетник в клумбах
и горшках. Типичное длиннодневное растение. Цветет с июля по сентябрь.
Зимою при дополнительном ночном освещении в 150—300 лк (с 17 до-
22 час.) зацветает через 108 дней, без освещения — через 207 дней [697 ].
'314
Частная светокультура растений
Львиный зев (Antirrhinum majus)
Типичное длиннодневное растение, не цветущее совсем или сильно
запаздывающее в условиях короткого дня. Оптимальная длина дня для
цветения равна 16 часам (фиг. 136, 5 [589, 590]). Нуждается в прямом
солнечном освещении, но может произрастать и в полутени. Наиболее
мощные и красивые растения образуются при температуре 7—13°. Будучи
многолетним растением, культивируется как однолетник. Высаживается
в грунт в виде рассады, которая готовится с конца февраля или начала
марта. При выгоночной культуре высевается в августе и в условиях
оранжереи зацветает в январе. Для выгонки нуждается в дополнительном
ночном освещении.
Литература по светокультуре львиного зева представлена фядом работ
[131—140, 383, 387, 530, 579, 594, 595, 598, 673, 678, 686, 688, 716, 857,
858]. Львиный зев сильно реагирует на дополнительное ночное освеще-
ние, увеличивая количество цветков и ускоряя наступление цветения.
По данным Гардера и других [530], A. majus ргаесох, посеянный 16 ав-
густа и освещаемый с 11 ноября, ускорил наступление цветения на
87 дней; при удобрении СО, и освещении он зацвел 20 декабря, а без осве-
щения — 7 марта. В опытах Лори и Пёш [595, 673] с сортом Шевиот
Мэйд при посеве 25 сентября и освещении с помощью ламп накаливания
в 150 вт цветение началось 6 марта — на 15 дней раньше контроля.
В опытах Уитроу [678, 857, 858] львиный зев сорта Сун Тан при
ежедневном освещении 10 часов (с 21 до 7 час.) с 7 октября по 24 февраля
•зацвел 30 ноября, а без освещения — 15 декабря. При этом наблюдалось
увеличение в количестве цветков на растении в 2,8 раза. Другой сорт
(Роз Куин), посеянный 23 августа, при освещении в 172 лк (с 27 октября
по 24 февраля) начал цвести 16 января, ускорив цветение на 19 дней и
увеличив количество цветков в 3,1 раза. По данным Поста [686, 688],
дополнительное освещение в 150 лк (с 17 до 21 час.), даваемое с 18 декабря,
ускоряет наступление цветения на 9—21 день. Контрольные растения
цветут 12 марта—4 апреля, а опытные — 3—21 марта. Эффективность
дополнительного освещения выражается сильнее при низких температу-
рах (ускорение на 17 дней при 10° и на 9 дней — при 15°).
В наших опытах A. majus папит (сорт Канариен фогель) и A. majus
maximum (сорт Шнеллыптурм) в зимние месяцы цвели лишь при допол-
нительном освещении (через 77'—129 дней) и совсем не цвели в контроле.
По данным Уитроу [678, 857], при зимней культуре львиного
зева затрачивается 46—50 квт-час на 100 цветков. Этот автор освещал
растения в течение 120—140 дней по 10 часов в сутки (с 21 до 7 час.), при-
меняя лампы накаливания, создававшие освещенность 172 лк. Лори и
Пёш [595] пользовались лампами накаливания в 150 вт, находившимися на
расстоянии 1,2 м друг от друга при высоте подвеса 38 см (100 вт/м2) и
освещали растения по 4 часа в сутки. По их данным, при культуре льви-
ного зева зимою на 1 цветок затрачивается 0,6 квт-час, а на 1 м2 —
22 квт-час.
Артур [383, 387] для зимней культуры львиного зева успешно приме-
нял прерывистое ночное освещение, в сумме составляющее 3 часа 24 ми-
нуты за ночь, используя лампы накаливания в 500 вт (350 вт/м2).
Клейн [579] с помощью дополнительного ночного освещения (с 22
до 8 час.) от ламп накаливания в 500 вт, находившихся на расстоянии
1,3 м на высоте 1,2 м (300 вт/м2), занимался выгонкой рассады львиного
зева. Выгонка производилась с декабря по февраль, причем ежедневно
на 1 м2 затрачивалось 2,4 квт-час электроэнергии.
IX. Светокультура декоративных растений
315
Портер и Ли [677, 678] рекомендуют львиный зев для комнатной куль-
туры вдали от окна. Растения находятся в хорошем состоянии, если по-
лучают дополнительное ночное освещение в 1000—5000 лк в течение
5—10 часов.
Однако следует иметь в виду, что культура львиного зева не всегда
удается при дополнительном освещении зимою. Так, в опытах Н. П. Кра-
синского [131—140] дополнительное освещение не дало никакого эффекта,
и культура львиного зева зимою не удалась. Пост [688] также не реко-
мендует прибегать к дополнительному освещению или высоким темпе-
ратурам (более 10°) при культуре львиного зева зимою. По его мнению,
в этих случаях формируются невысокие и слабые растения. Повидимому,
неудача в этих случаях связана с перегревом растений под лампами
накаливания. Кроме того, не все сорта пригодны для этой цели. Во всяком
случае, влияние освещения сказывается лишь на весенних и летних
сортах и не обнаруживается у зимних сортов [594].
Колючий мак (Argemone sp.)
Длиннодневное светолюбивое растение. При дополнительном ночном
освещении (с 17 до 22 час.; 160—320 лк) зимою цветет через 102 дня, без
освещения — через 150 дней [697].
Ясменник (Шерошница) (Asperula orientalis)
Длиннодневнэе тенелюбивое растение, нуждающееся в полутени.
При дополнительном освещении в зимние месяцы (с 17 до 22 час.; 150—
300 лк) цветет через 50 дней после посева, без освещения — через
132 дня [697].
Бэрия (Baeria chrysostoma)
Длиннодневное растение. Максимальная высота и наибольшее коли-
чество цветков наблюдаются в условиях 18-часового дня, а максималь-
ное количество листьев —• при 12—14-часовом дне [752]. В условиях
беспрерывного освещения с помощью люминесцентных ламп дневного
света или ртутных ламп Купер Хьюит (4500 лк) зацветает через 32 дня,
в условиях короткого дня (12 часов) — через 69 дней (табл. 113 [599]).
Череда (Biedens frondosa)
Короткодневное растение. Дополнительное ночное освещение зимою,
подавляя цветение, обусловливает мощный вегетативный рост; цветение
наступает через 30 дней после установления короткого дня [502]. При
переносе цветущего растения в условия дополнительного ночного осве-
щения снова начинается вегетативный рост [501].
Брахикома (Bra chi come iberidifolia)
Длиннодневное светолюбивое растение. Высевается в апреле (в пар-
ник), цветет во второй половине лета (июль—август). В зимнее время
в оранжерее при дополнительном ночном освещении (с 17 до 22 час.; 160—
300 лк) зацветает через 139 дней после посева, без освещения — через
153 дня [697].
316
Частная светокультура растений
Ноготки (Calendula officinalis)
Нейтральные растения с тенденцией как к длиннодневности, так и
короткодневности. Светолюбы, но выносят затенение. Для роста нуж-
даются в ночной температуре 7—10°. На дополнительное ночное освещение
в большинстве случаев реагируют слабо, не увеличивая количества цвет-
ков и не ускоряя цветения (фиг. 142; 1 [594, 59.5, 673, 678, 697, 857, 858]).
В опытах Рэмэли [697] один сорт оказался даже короткодневным и при
дополнительном освещении зимою зацвел через 76 дней вместо 61 дня
в контроле. Лишь в опытах Грина и его сотрудников [521] сорт Болле
Голд при ночном освещении (с 21 до 2 час.; 3000—5000 лк) в течение
89 дней зацвел раньше на 15 дней и увеличил количество цветков на41 %.
Сорт Оранж Кинг реагировал лишь увеличением числа цветков (на 32—
46%), фактически не ускорив наступления цветения. По мнению Поста
[688], дополнительное освещение вызывает удлинение стебля и измель-
ченность цветков.
По данным Грина [521], для ноготков достаточно 180 лк, причем лучший
эффект дает ночное освещение (с 21 до 2 час.), а не освещение в дневные-
и вечерние часы (с 16 до 21 час.). При ежедневном семичасовом освещении,
в течение 125 дней с помощью лампы накаливания в 100 вт (180 лк) на
каждый цветок затрачивается 0,2 квт-час.
Астры (Callistephus chinensis, С. hortensis, Aster chinensis,
Callistemmum chinensis)
Нормально растут и цветут при интенсивности 20—40 тыс. эрг/см2-сек
(фиг. 132 и 133). Для роста нуждаются в ночной температуре 10—15°,
а для заложения цветков — 24°. Большинство сортов относятся к длинно-
дневным растениям; некоторые сорта являются нейтральными и коротко-
дневными. По данным Поста [683], астры бывают длиннодневными лишь
на ранних фазах развития, а позднее они становятся коротко дневными.
Астры цветут во второй половине лета: с августа до наступления замороз-
ков. Посев производится в марте—апреле, а высадка в грунт — в конце
мая.
Литература по светокультуре астр представлена сравнительно боль-
шим числом работ и охватывает большое число сортов [123, 131—140,.
153, 290, 411, 412, 521, 552, 558, 594, 595, 596, 678, 688, 697, 733, 742,
854, 857—862]. Лишь в опытах Н. П. Красинского [131—140] куль-
тура астр не удалась при дополнительном освещении зимою. Коротко-
дневные формы (например Америкен Бьюти) дают, естественно, также
отрицательный результат при ночном освещении [496, 742]. В подав-
ляющем же большинстве опытов культура астр вполне удается при осве-
щении растений зимою. Одни сорта — Калифорния Жиант, Крего Дип
Роз, Крего Шэлл Пинк, Гарт оф Франс, Воган Соншайн, Рэйне дес Гал-
лес и другие — цветут только при дополнительном ночном освещении,
а в условиях короткого зимнего дня находятся в стадии розетки [290,
521, 595, 596, 678, 697, 733, 858]. Обычно при освещении эти астры, в за-
висимости от сорта и условий, зацветают через 100—165 дней от посева.
Другие сорта — Триумф Циннобер Кармин, Эрли Ройял Пэрпл, Куин
оф Маркет, Пурити, Дэйбрэк, Болле Уайт, Ройял Пэрпл, Лавендет Джем,
Америкен Брэнчинг, Комет и др. — при освещении зацветают на 5—49 дней
раньше, чем в контроле [678, 858—862]. У этих сортов при дополни-
тельном ночном освещении цветение в зимнее время наступает через
80—160 дней, а без освещения — через 90—180 дней. При посеве в октябре
астры, в зависимости от сорта, цветут в декабре—феврале. Сравнительно
IX. Светокультура декоративных растений
317
немногие сорта (Кинг и др.) относятся к нейтральным и в зимние месяцы
цветут одновременно — через 100—135 дней — как без дополнитель-
ного освещения, так и с освещением [595, 678].
При дополнительном освещении астр в зимнее месяцы даже при ни-
чтожно малой интенсивности освещения (10 лк) наблюдается значитель-
ное увеличение в весе наземной массы и листовой поверхности растений
(табл. 102 [412, 862]). В большинстве случаев бывает значительное уве-
личение диаметра цветков [596, 678, 858]. Обычно диаметр цветка увели-
чивается на 20—55%, с 5—6 до 7—9 см. При этом, как правило, наблю-
дается уменьшение количества цветков с 9—16 до 5—6 на 1 растение
[596, 678, 858].
Для дополнительного освещения астр пригодны лампы накаливания,
люминесцентные, ртутные и натриевые лампы. Наиболее высокие и мощ-
ные растения наблюдаются под лампами накаливания (табл. 89 [854]).
Как показали опыты Роденбурга [733], под натриевыми лампами многие
растения зацветают в розеточной форме, без вытягивания стебля.
Астры чрезвычайно отзывчивы на дополнительное освещение, реаги-
руя на ничтожные интенсивности радиации (фиг. 142, 2). Как показывают
опыты ряда авторов, хороший результат наблюдается уже при освещен-
ности порядка 100 лк [521, 858, 861]. Дальнейшее увеличениё освещен-
ности обычно не вызывает соответствующего ускорения в наступлении
цветения и увеличения размеров цветков. Для получения эффекта доста-
точно удельной мощности 50—100 вт/м2 при высоте подвеса 50—100 см.
Лучшие результаты наблюдаются при длительности дополнительного
освещения 8—10 часов, т. е. при длине дня 16—17 часов [521, 678, 733,
857, 858]. Некоторые авторы применяют 4—6-часовое дополнительное
освещение [594, 595, 596, 697, 854]. Юм [558] нашел, что прерывистое
освещение (350 лк) дает тот же результат, что и обычное дополнительное
освещение, ускоряя наступление цветения на 23—31 день.
Опыты показывают, что в дополнительном освещении астры нуждаются
на протяжении всего жизненного цикла — от появления первого листа
до бутонизации [552, 688, 858].
Грин и его сотрудники [521 ] нашли, что при зимней культуре астр
на 1 цветок затрачивается 0,2—0,8 квт-час, а по данным Портера
и Ли [678], на 1 растение расходуется от 4 до 8,5 квт-час.
Лори и Пёш [596] считают, что дополнительное освещение в утренние
часы значительно эффективнее освещения вечером. У других авторов
утреннее и вечернее освещение давало одинаковый эффект [857].
Дополнительное освещение в рассадный период в течение 7—26 дней
ускоряет последующее цветение растений при высадке их в грунт; для
сортов Эрли Ройял Пэрпл и Шэлл Пинк ускорение равняется 14 дням [552 ].
Для получения ранних летних цветков Пост [688] рекомендует
высевать астры в феврале—марте, давая им дополнительное освещение до
начала мая.
Кардиоспермум (Cardiospermum halicacabum)
Нейтральное растение. При дополнительном ночном освещении зимою
(с 17 до 22 час.; 160—300 лк) зацветает одновременно с контролем — через
127 дней [697].
Целозия (Celosia argentea)
Нейтральное теплолюбивое и светолюбивое растение. Высевается
в апреле в теплый парник; на постоянное место высаживается во второй
318
Частная светокультура растений
половине июля. Цветет во второй половине лета, с июля до осени. При
культуре зимою в оранжерее зацветает одновременно — через 137 дней —
как при ночном освещении, так и без него [697].
Василек (Centaurea)
Васильки [Centaurea cyanus, С. americana, С. moschata (С. saveolens)]
относятся к длиннодневным и светолюбивым растениям. Нуждаются
в ночной температуре 10—15° [688]. Высеваются в парники во второй
половине апреля, затем высаживаются на постоянное место. Цветут
с начала лета (июнь) до осени.
Литература по светокультуре васильков насчитывает несколько работ
[497, 595, 596, 598, 673, 678, 686, 688, 697, 716, 857].
В зимнее время при культуре в оранжерее васильки цветут или исклю-
чительно при дополнительном ночном освещении (С. americana) или
значительно ускоряют в этом случае наступление цветения [595, 596,
598, 673, 678, 686, 688, 697]. У С. cyanus ускорение составляет 15—74
дня, а у С. moschata var. rubra и С. moschata var. alba — 5—27 дней. При
дополнительном освещении васильки зацветают через 75—193 дня, а без
освещения — через 98—211 дней. Посеянные в конце сентября или начале
октября, они зацветают при освещении в декабре—марте, а без освеще-
ния — в январе—апреле. В результате освещения наблюдается увеличе-
ние количества цветков [594, 595, 678, 688].
Для дополнительного освещения применяются лампы накаливания
в 100—200 вт и создается освещенность от 150 до 400 лк. Длительность
дополнительного освещения у большинства авторов колеблется в пре-
делах 4—6 часов. Лори и Пёш [595, 596] показали, что у С. americana
(сорт Уайт) лучший эффект наблюдается при утреннем (с 2 до 7 час.),
а у С. moschata var. rubra (сорт Лавендер) и С. moschata var. alba — при
вечернем освещении (с 17 до 22 час).
По данным Поста [688], наиболее благоприятные результаты полу-
чаются при посеве растений в сентябре и дополнительном освещении
с января. В этом случае цветение растений начинается в марте.
В опытах Пёш [673] при культуре васильков зимою при освещении
лампами накаливания (4 часа; 100 вт/м2) с 4 декабря на 1 цветок затра-
чивалось для С. cyanus 0,003, а для С. moschata var. rubra {С. imperialis) —
0,047 квт-час. На 1 м2 в первом случае приходилось 24, а во втором —
29 квт-час.
Хризантемы летние (Chrysanthemum)
Обширная группа летних хризантем — Ch. Buridgeanum, Ch. coro-
narium, Ch. segetum и Ch. carinatum — относится к растениям длинного
дня. Для роста нуждаются в ночной температуре 10—12° [688]. Для куль-
туры в открытом грунте высеваются в парники в начале мая. Цветение —
с июля по сентябрь.
При дополнительном освещении культивируются зимой [594, 595,
596, 598, 673, 678, 686, 688, 697, 857, 858]. Ночное освещение вызывает
ускорение наступления цветения на 14—64 дня (табл. 87 и 104). Уско-
рение составляет у Ch. carinatum — 36, у Ch. coronarium — 20, у Ch.
Buridgeanum — 33—64, у Ch. segetum сорт Морнинг Стар — 39—57,
а у сортов Нортен Стар — 14—33, Султан — 37—45 и Доннети Добл
Скарлет — 34—48 дней. При посеве в октябре—ноябре растения зацве-
тают при освещении, в зависимости от сорта, в декабре—феврале (через
83—119 дней). Как правило, дополнительное освещение увеличивает
IX. Светокультура декоративных растений 31!й
количество цветков на растении в БД— 2 раза (табл. 87). По данным1
Поста [688], положительный эффект у Ch. coronarium наблюдается лишь,
при температуре 10—12° (ускорение цветения на 25—27 дней). При тем-
пературе 15° положительного влияния практически нет (ускорение на
3 дня). У Ch. segetum дополнительное освещение действует положительно
как при температуре 10°, так и при 15°. Ускорение цветения в обоих
случаях равняется 28—30 дням; увеличение количества цветков бывает,
однако, лишь при 10° (в 4 раза); при 12° этот эффект снижается до 57%,.
а при 15° дополнительное освещение вызывает уменьшение количества
цветков (на 43%). Без дополнительного освещения высокие температу-
ры (более 15°), не влияя на время цветения, увеличивают количество
цветков на 60—370%.
Для освещения применяются лампы накаливания в 100—200 вт, со-
здающие освещенность порядка 150—400 лк (табл. 87). Специальными
опытами Лори и Пёш [596] показали, что интенсивность радиации от
75 до 460 лк дает одинаковый эффект (табл. 104, фиг. 142, 7).
Длительность дополнительного освещения, в зависимости от есте-
ственной длины дня, колеблется от 4 до 10 часов. Юм [558] нашел, что
прерывистое освещение дает тот же эффект, что и обычное ночное осве-
щение; так, Ch. segetum (сорт Ивнинг Стар) при дополнительнохм осве-
щении — прерывистом и обычном — зацветает через 101 —112, а в конт-
роле — через 132 дня. По данным Лори и Пёш [596], лучший результат
дает утреннее дополнительное освещение (с 2 до 7 час).
По данным Уитроу [858] и Портер и Ли [678], при культуре хризан-
тем расходуется 0,19 квт-час на 1 цветок. В опытах Пёш [673] эта затрата
равнялась 0,1—0,21 на цветок, или 28—29 квт-час на 1 м2.
Кларкия (Clarkia elegans)
Длиннодневное светолюбивое растение. Для роста нуждается в ночной
температуре 15—21°, для заложения цветков— более 15°. При темпе-
ратуре менее 12° образует толстые суккулентныо стебли и крупные листья,
но не цветет. Высевается в апреле в холодные парники. Цветет с июля по
сентябрь. При дополнительном ночном освещении зимою ускоряет наступ-
ление цветения на 10—36 дней (табл. 104), зацветая через 95—109 дней
после посева [521, 595, 596, 598, 686, 697, 857]. При посеве в сентябре
цветет в декабре—январе; при освещении с января цветет в марте [686].
Ночное ’ освещение вызывает увеличение количества цветков при 10°
(на 67%) и уменьшение — при 15° (на 29%); при 12° количество цветков
не изменяется [686].
Лори и Пёш [596] нашли, что освещенности в 75 и 160 лк дают одина-
ковый результат (табл. Т04; фиг. 142, 8). Для дополнительного освещения
обычно применяют лампы накаливания в 100—200 вт, помещая их на
высоте 50—100 см. Грин с сотрудниками [521] пользовались лампами
в 500 и 1000 вт, создавая освещенность 3000—5000 лк. По их данным,
для культуры кларкии в зимнее время затрачивается 1,82—1,95 квт-час
на цветок.
Длительность дополнительного освещения обычно равняется 4—6 ча-
сам ежедневно.
Клеома (Cleome spinosa)
Длиннодневные, нейтральные и короткодневные растения. В опытах
Рэмэли [697] С. spinosa при дополнительном освещении зимою (с 17 до
22 час.; 160—300 лк) зацвела через 125, а без освещения — через 95 дней.
320
Частная светокультура растений
По данным Робертса и Стракмайера [716], зимою цветет лишь в условиях
длинного дня при^ дополнительном освещении 300—-800 лк и темпера-
туре 21 °.
Колломия (Collomia biflora)
Длиннодневное растение. При дополнительном ночном освещении
дс 17 до 22 час.; 160—320 лк) в зимние месяцы цветет через 90, а без осве-
щения — через 120 дней [697].
Вьюнок (Convolvulus tricolor)
Светолюбивое длиннодневное вьющееся растение. Цветет с июня
до осени. Зимою при дополнительном ночном освещении' с 17 до 22 час.;
160—320 лк) зацветает через 83 дня, без освещения — через 158
дней [697].
Ленок (Coreopsis или Calliopsis)
Длиннодневные светолюбивые растения. Высеваются в конце апреля
в парники. Цветут с начала лета до осени (июнь—сентябрь). С. tinctoria
при дополнительном освещении зимою зацветает через 139 дней, ускоряя
цветение на 52—76 дней [595, 598, 673, 678, 679]. Требует дополнитель-
ного освещения в 150—400 лк (100—200 вт/м3) при длительности осве-
щения 5—6 часов ежедневно. С. californica (С. Douglasii или Leptosyne
DouglasH) в условиях короткого дня (12 часов) бутонизирует лишь через
103 дня, а при беспрерывном освещении (4500 лк) с помощью ртутных
или люминесцентных ламп зацветает через 43 дня (табл. 113 [599]). По-
ложительный эффект ночного освещения для этого вида отмечают также
Лори и Киплинге р [598].
По данным Пёш [673], при культуре зимою необходима затрата
электроэнергии 0,002 квт-час на 1 цветок, или 29 квт-час на 1 м2.
Космеа (Красотка) (Cosmos)
С. bipinnatus и С. sulphureus—светолюбивые короткодневные расте-
ния. Для роста и заложения цветков нуждаются в ночной температуре
12—15° [688]. Высеваются в конце апреля в парники. Цветут с июля
до морозов.
Дополнительное ночное освещение, удлиняющее естественный зимний
день, или значительно задерживает наступление цветения или совсем
предотвращает цветение [396, 397, 412, 500—502, 609, 697, 715, 716,
742, 806, 857, 861, 862]. Поэтому дополнительное освещение космоса
можно допускать лишь в пределах короткого 10—12-часового дня. При
коротком дне растения зацветают через 50—100 дней, образуя макси-
мальное количество цветков. Опыт показывает, что количество цветков
на растении тем больше, а цветение тем раньше, чем больший период
растения получают короткий день. Длительность световой стадии рав-
няется 14 дням [609]. Высота растений возрастает вместе с увеличением
длины дня [806]. При дополнительном освещении увеличивается вес ра-
стений и листовая площадь [412].
?Для устранения цветения короткий зимний день обычно удлиняется
до 16—18 часов. По данным Остина [396], минимальная интенсивность
дополнительного освещения лежит в пределах 4—18 лк. При 4 лк
С. sulphureus цветет, при 18 лк находится в вегетативном состоянии.
Полный эффект имеет место уже при интенсивности дополнительного
IX. Светокультура декоративных растений
321
освещения в 10—50 лк [501, 862]. Обычно же применяются лампы нака-
ливания в 100—200 вт, создающие освещенность 150—300 лк.
Скерда (Крепис) (Crepis sp.)
Длиннодневное растение. Зимою цветет лишь при дополнительном
освещении в ночные часы при интенсивности порядка 1000—2000
эрг/см2-сек; цветение наступает примерно через 40 дней после начала
освещения [742].
Шпорник (Delphinium ajacis)
Светолюбивое и длиннодневное растение. Для роста необходима ноч-
ная температура 10—15° в первые 3 месяца и 15—21° в последующий
период; для заложения цветков необходима температ}ра более 15° [688].
Зацветает в середине лета (июнь). В зимние месяцы при температуре
менее 10° находится в розеточном состоянии. При дополнительном ноч-
ном освещении можно культивировать зимою [397, 595, 596, 598, 673,
686, 688, 697]. В этом случае при посеве в ноябре зацветает в феврале—
марте, через 90—135 дней, а при освещении с января цветет в марте—ап-
реле. Дополнительное освещение ускоряет наступление цветения (на 20—
50 дней) и увеличивает количество цветков (на 20—60%). По данным
Поста [686, 688], увеличение количества цветков наблюдается лишь при
температуре 10—12°. При 15° дополнительное освещение несколько
уменьшает количество цветков. Некоторые сорта на дополнительное ос-
вещение реагируют слабо [521].
Для дополнительного освещения необходима интенсивность 150—
400 лк. Лучший результат наблюдается при 20-часовом дне [397]. Пре-
рывистое освещение дает тот же эффект, что и обычное, ускоряя цве-
тение на 14—32 дня [558]. Как показывают опыты Лори и Пёш [584]
с сортами Кармин Кинг, Дэнтиппесс и Блу Спайр, утреннее освещение
(с 2 до 7 час.) действует лучше, чем вечернее (с 17 до 22 час.). По данным
Грина [521], при культуре шпорника зимою на 1 растение затрачивается
7,5—9 квт-час. В опытах Пёш [673] при освещении лампами накалива-
ния (100 вт/м2; 4 часа) затрата электроэнергии составляла 0,16 квт-час
на 1 цветок, или 29 квт-час на 1 м2. v •
Лобия (Dolichos lablab)
Нейтральное светолюбивое и теплолюбивое вьющееся растение.
Высевается в марте, на место высаживается в июле, а цветет во второй
половине лета (июль—август).
На дополнительное ночное освещение зимою не отзывается, зацветая
одновременно с контролем [697].
Эмилия (Emilia flammea) ।
Длиннодневное растение. Зимою при дополнительном ночном осве-
щении (с 17 до 22 час.; 160—320 лк) цветет через 99, а без освеще-
ния — через 110 дней [697].
Полынок (Eschscholtzia)
Длиннодневные и чрезвычайно светлолюбивые растения, раскрыва-
ющие цветы лишь в солнечные дни. Дополнительное ночное освещение
ТГ ттатттттита
322
Частная светокультура растений
ускоряет наступление цветения [496]. Е. calijornica при беспрерывном
освещении с помощью ртутных или люминесцентных ламп (4500 лк) за-
цветает через 56 дней, а при длине дня короче 14 часов не цветет совсем
(табл. ИЗ [5991).
Аналогичным образом ведет себя Е. lobbii, зацветая при беспрерыв-
ном освещении через 72 дня, тогда как на коротком дне бутонизирует
лишь через 103 дня (табл. ИЗ [599]).
Молочай (Euphorbia)
Е. marginata и Е. heterophylla — теплолюбивые, светолюбивые и ко-
роткодневные растения. Высеваются в парник в апреле.
Дополнительное ночное освещение (30—50 лк), удлиняющее короткий
зимний день, сильно задерживает или полностью устраняет цветение
[501, 697]. Е. marginata зимою без освещения зацветает через 155 дней,
при освещении — через 184 дня [697].
При культуре растений с целью получения цветков дополнительное
освещение можно рекомендовать лишь в пределах короткого 10—12-ча-
сового дня. Для получения растений, находящихся в вегетативном со-
стоянии, необходимо дополнительное ночное освещение в течение 7—8
часов (около 100—200 лк).
Фелиция (Felicia amelloides)
Нейтральное растение. При дополнительном ночном освещении
(400 лк; с 18 до 22 час.) цветет одновременно с контролем [595].
Гайлардия (Gailardia)
G. Lorenziana, G. aristala (G. grandijlora) и G. pulchella—светолюби-
вые и длиннодневные растения. Посев производится в апреле в парник.
Цветут на протяжении всего лета и осени.
При культуре зимою в оранжерее дополнительное ночное освещение
ускоряет наступление цветения на 11—75 дней и одновременно увеличи-
вает количество цветков в 1/4—6 раз [595, 598, 697, 858, 859]. При по-
севе в конце ноября растения цветут через 104—129 дней (в марте—
апреле).
Интенсивность дополнительного освещения 150—300 лк, длительность
ежедневного освещения 5—10 часов. По данным Лори и Пёш [595], луч-
ший эффект дает вечернее освещение (с 18 до 22 час.).
Для получения 100 цветков в зимнее время необходимо затратить
76 квт-час [859].
Гилия (Gilia)
Светолюбивые длиннодневные растения. Посев в марте—апреле. Вы-
садка в грунт в апреле—мае. Цветение с мая по август.
G. caespitosa в зимнее время в оранжерее при дополнительном ночном
освещении (с 17 до 22 час.; 150—300 лк) цветет через 77 дней; без освеще-
ния не цветет совсем [697]. G. tricolor в условиях 18—24-часового дня,
создаваемого с помощью ртутных или люминесцентных ламп (4500 лк),
цветет через 80 дней, а в условиях короткого дня (8 часов) не цветет со-
всем (табл. ИЗ [599]). G. grandijlora нуждается для роста в ночной темпе-
ратуре 12—18°. При дополнительном ночном освещении с 7 января (с 17
до 21 час.; 150 лк) ускоряет наступление цветения на 5—15 дней. Влия-
IX. Светокультура декоративных растений
323
ние заметно сильнее при 10° и слабее — при 15°. В первом случае при
дополнительном освещении наблюдается уменьшение количества цветков
(на 68%), а во втором — увеличение (на 50% [688]).
Годеция (Godetia)
Длиннодневные светолюбивые растения. Высеваются в апреле в пар-
ники. Цветут с июня по октябрь.
G. атоепа в зимнее время при дополнительном ночном освещении
(с 17 до 22час.; 150—300 лк) зацветает через 71 день, без освещения — че-
рез 85 дней [697]. G. quadrivulnera при беспрерывном освещении под
ртутными или люминесцентными лампами (4500 лк) цветет через 85 дней,
а на коротком дне (12 часов) остается в вегетативном состоянии
(табл. ИЗ [599]).
Качим (Gypsophila elegans)
Относится к группе растений длинного дня. Посев — рано весной или
в июне. Цветение — с августа. Зимою при дополнительном ночном ос-
вещении в оранжерее зацветает раньше, чем без освещения [598].
Подсолнечник (Helianthus debilis)
Среди многочисленных видов подсолнечника лишь очень немногие,
например Н. debilis (Н. cucumer if alias') относятся к растениям длин-
ного дня. Большинство видов — коротко дневные растения.
Подсолнечник — светолюбивое растение. Высевается в марте, цветет
с июля по сентябрь.
Дополнительное ночное освещение в зимнее время, удлиняющее есте-
ственный короткий день, применимо лишь к длиннодневным формам.
В опытах Уитроу и Бибел [860, 862] Н. cucumerifolius, посеянный 15 де-
кабря, при дополнительном освещении (с 14 января, 18-часовой
день, 10 лк) зацвел в оранжерее в апреле, через 126 дней. Без освещения
цветение наступило 26 дней спустя, через 153 дня.
Гелихризум (Helichrysum bracteatum)
Длинно дневное светолюбивое растение. Высевается в марте—апреле.
При дополнительном ночном освещении, удлиняющем короткий день,
зацветает раньше на 30 дней [496].
Гелиптерум (Helipterum Manglesii) >
Длиннодневное растение. Зимою при дополнительном освещении (с 17
до 22 час.; 160—300 лк) цветет через 83 дня, а без освещения — через
105 дней [697].
Стенник ’ (Iberis)
Iberis umbellate, и I. атага относятся к светолюбивым и длинноднев-
ным растениям. Высеваются осенью или весной. В первом случае цве-
тут в мае—июне, во втором — в июле—августе.
При дополнительном ночном освещении зимою зацветают через 60—
146 дней — на 26—60 дней раньше, чем в контроле [488, 496, 497, 531,
595, 673, 678, 697, 811].
21 *
324
Частная светокультура растений
Фабиан [488] нашла, что интенсивность дополнительного освещения
не должна быть ниже 80 лк (фиг. 142, 11). Обычно применяется интен-
сивность порядка 200—400 лк. Длительность дополнительного освеще-
ния — 5—6 часов.
При культуре зимою с помощью ламп накаливания (4 часа; 100 вт/м2)
на 1 цветок затрачивается 0,3 квт-час, или 29 квт-час на 1 м2 [673].
Бальзамин (Impaliens balsamina)
Светолюбивые короткодневные или нейтральные растения. Высеваются
в парниках в начале апреля. Цветение наступает в июне—июле.
В зимние месяцы в оранжерее дополнительное освещение в ночные
часы или не влияет на наступление цветения [397, 697] или вызывает
запаздывание в цветении на 14—44 дня [395, 496]. Зацветает через
50—90 дней.
Для вегетативного роста оптимальным является 16-часовой день.
В этом случае наблюдается максимальный сухой вес, максимальная вы-
сота и наибольшая листовая поверхность [395, 397].
Задержка цветения у короткодневных форм также наступает при
удлинении дня до 16 часов. Для задержки достаточно 17—320 лк [697,
395, 397].
Граммофончики (Ipomoea purpurea)
Короткодневное теплолюбивое и светолюбивое вьющееся растение.
Для вегетативного роста нуждается в ночной температуре 12—18°, для
заложения цветков —17—18° [688]. Высевается в апреле—мае. Цветет
с июля до осени.
По данным Рэмэли [697], при дополнительном ночном освещении зи-
мою зацветает через 300, а без освещения — через 61 день. По другим
данным, является короткодневным растением лишь при температуре
21—24°, при 17—18°,5 становится нейтральным, а при 13°— длинно-
дневным [715, 716]. Для задержки цветения применяется освещенность
160—320 лк. Дополнительное освещение можно применять для выгонки
рассады [579].
Душистый горошек (Lathyrus odoratus)
Светолюбивое длиннодневное растение. Для роста необходима ночная
температура 7—15° [688]. Высевается или в парники (в апреле) или
непосредственно в грунт (в мае).
Зимою культивируется при дополнительном ночном освещении [283,
376, 383, 387, 521, 543, 547, 556, 557, 594, 595, 646, 688, 697, 700,
717, 720, 725, 858]. В этом случае при освещении вегетационный период
сокращается на 15—22 дня, а количество цветков увеличивается на 15—
56%. Ускорение цветения, по данным Лори [594], равняется 14—
20 дням. По данным Роденбурга [283, 717, 720], дополнительное осве-
щение увеличивает количество цветков на 20—40%. В опытах Гаута [543]
душистый горошек, посеянный 15 ноября и освещавшийся с 1 ян-
варя по 21 февраля, зацвел 12—15 февраля, а в контроле 7—9 марта.
В первом случае на растении образовалось по 6—13 цветков, а во вто-
ром — 1—2 цветка. Йо данным Лори и Пёш [595, 596], у сортов Болле
оранж, Мистрис Гувер, Уайт Гармони и Жанн Мамитчпри освещении с 29
ноября готовая продукция была получена 27 января, а без освещения —
6 февраля. Различий в количестве цветков авторы не нашли.. Сорт Дварф
Купид зацветает при дополнительном освещении через 72, а без осве-
IX. Светокультура декоративных растений
325
щения — через 92 дня [697]. Артур и Гарвил [387], применявшие пре-
рывистое освещение, нашли, что душистый горошек зацветает 11 января,
а без освещения — 26 февраля. Количество цветков в первом случае
было больше на 54%.
Портер и Ли [678] приводят данные, из которых следует, что сорт
Пинки в зимнее время цветет лишь при дополнительном освещении,
а сорт Болле Роз зацветает одновременно как при освещении, так и
в контроле, но количество цветков в первом случае больше на 30%.
В опытах Грина и его сотрудников [521] с сортами Роз Куин и Блу бард
также было отмечено увеличение количества цветков на 15—38%. Ана-
логичные данные были получены Лори и Пёш [596] при дополнительном
освещении (550 лк) неоновыми лампами.
Таким образом, следует признать, что в зимние месяцы при дополни-
тельном освещении душистый горошек зацветает через 72—101 день.
При посеве в октябре растения цветут в декабре—январе, при освещении
с конца декабря зацветают в начале марта, а при освещении с середины
февраля —- в конце апреля.
По данным Лори и Пёш [596], испытавших лампы накаливания в 40,
75 и 150 вт, для культуры душистого горошка достаточно удельной мощ-
ности 40 вт/м2. Обычно применяют освещенность 160—320 лк. Роден-
бург [283, 717, 720] рекомендует неоновые лампы (500 лк).
Гаут [543] нашел, что оптимальным для душистого горошка является
16-часовой день. Оден [646] также установил, что лучший результат
наблюдается в условиях 15-часового дня при интенсивности ради-
ации 0,06 г-кал/см2-мин (42 тыс. эрг/см2-сек). В этом случае душистый
горошек, посеянный 20 сентября, зацветает 24 декабря, т. е. через
90 дней. При беспрерывном освещении растения выглядят нормально,
но бутоны опадают.
При зимней культуре при освещении в течение 163 дней с 21 до
7час. (160 лк) на 100 цветков затрачивается 22—47 квт-час [678,858, 859].
Клейн [579] в 'зимнее время пользовался лампами накаливания
(300 вт/м2) для выгонки рассады душистого горошка.
Садовая роза (Lavatera trimestris)
Исключительно светолюбивое, длиннодневное растение. Посев де-
лается в апреле в парники или позднее непосредственно в грунт. Цве-
тение — с июля по сентябрь.
Зимою в оранжерее при дополнительном ночном освещении (с 17 до
22 час.; 150—300 лк) цветет через 95 дней, без освещения — через
156 дней [697].
Лимнантёс (Limnanthes Douglassii)
Длиннодневное сильно реагирующее на дополнительное освещение ра-
стение. Зимой при дополнительном ночном освещении (с 17 до 22 час.;
160—300 лк) цветет через 60, а без освещения — через 114 дней [697].
Кермек (Limonium Suworowi или Statice Suworowi)
Нейтральное светолюбивое растение. Высевается в апреле, цветет
с июля по сентябрь.
На дополнительное ночное освещение не реагирует, зацветая одно-
временно как при освещении, так и в контроле (через 40 дней). В резуль-
тате освещения образуется лишь несколько больше цветков [595].
326
Частная светокультура растений
Линантус (Linanthus montanus или Gilia montana)
Длиннодневное растение, цветущее лишь в условиях длинного дня.
Наиболее быстро зацветает при беспрерывном освещении под ртутными
или люминесцентными лампами (4500 лк; табл. ИЗ [599]).
Льнянка (Linaria)
Светолюбивые и длиннодневные растения. Высеваются в парник во вто-
рой половине апреля. Цветут с июня по сентябрь.
L. тагоссапа в оранжерее при дополнительном ночном освещении
(с 17 до 22 час.; 150—300 лк) зимою зацветает через 84 дня, без освеще-
ния — через 109 дней [697].
L. bipartita и L. reticulata также хорошо реагируют на дополнительное
освещение зимою [811].
Лобелия (Lobelia erinus)
Длиннодневное растение. При удлинении короткого дня с помощью
ламп накаливания несколько ускоряет (на 10 дней) начало цветения 1496].
В наших опытах зимою Lobelia erinus compacta (сорт Кристаль Паласт)
цвела лишь при дополнительном ночном освещении (500 вт/м2). Цвете-
ние наступало через 66—129 дней.
Прекрасно растет при «нижнем» освещении с помощью люминесцент-
ных ламп белого света (600 вт/м2). В этих условиях она зацветает через
48 дней и цветет беспрерывно в течение нескольких месяцев [120]. Можно
рекомендовать для озеленения станций метрополитена, для создания там
газонов или клумб при помощи «нижнего» освещения.
Люпин (Lupinus Hartwegii)
Однолетнее нейтральное растение с тенденцией к длиннодневности.
Для роста нуждается в ночной температуре 10—15° [688]. По данным
Поста [688], на дополнительное ночное освещение зимою (с 7 января)
не реагирует, зацветая одновременно с контролем (8—20 апреля), вна-
чале при температуре 15° (5—8 апреля), а затем при 10° (12—20 апреля).
Горицвет (Lychnis ’coeli-rosa)
Длиннодневное светолюбивое растение. Высевается в конце апреля,
цветет с мая по сентябрь. При дополнительном ночном освещении за-
цветает через 70, а без освещения — черрз 160 дней [697].
Мадия (Madia elegans или М. corymbose)
Длиннодневные растения, не цветущие на коротком дне (12 часов) и
наиболее быстро (через 64 дня) зацветающие при беспрерывном освещении
ртутными или люминесцентными лампами (4500 лк; табл. ИЗ [599]).
Малькомия (Malcomia maritima)
Длиннодневное растение. Летом в условиях естественного дня или
при дополнительном ночном освещении цветет через 64, а на коротком
дне — через 72 дня [496]. Зимою при дополнительном ночном освещении
IX. Светокультура декоративных растений.
327
(с 17 до 22 час.; 160—300 лк) зацветает через 91 день, а в контроле —
через ИЗ дней [6971.
Ромашка (Matricaria)
Светолюбивые и длиннодневные растения. Высеваются в марте в пар-
ник; цветение — с июня по сентябрь.
В зимней культуре с помощью дополнительного ночного освещения
испытаны М. chamomilla и М. inodora [496, 595, 596, 598, 678]. При до-
полнительном освещении эти ромашки увеличивают количество цветков
(на 36%) и значительно ускоряют наступление цветения (на 57—70 дней;
табл. 104).
Лори и Пёш [596] нашли, что для дополнительного освещения до-
статочно 75 лк. Дальнейшее увеличение освещенности (до 460 лк) со-
ответствующего усиления эффекта не дает (фиг. 142; 6). Дли-
тельность ежедневного дополнительного освещения 5—6 часов.
Ментцелия (Mentzelia Lindleyi или Bartonia aurea)
Длиннодневное растение, сильно реагирующее на дополнительное осве-
щение. Зимою в опытах Рэмэли 1697] при освещении (с 17 до 22 час.;
160—320 лк) цвело через 63 дня, а без освещения — через 109 дней.
Аналогичные результаты получили Льюис и Вент [599], применявшие
ртутные или люминесцентные лампы (4500 лк; табл. ИЗ). Оптимальная
длина дня — 18 часов.
Монарделла (Monardella lanceolata)
Длиннодневное растение. В опытах Льюиса и Вента [599] бутонизи-
ровала (через 103 дня) лишь в условиях 18-часового дня и беспрерывного
освещения ртутными или люминесцентными лампами (4500 лк; табл. ИЗ).
Немезия (Nemesia versicolor)
Многолетнее растение, культивируемое как однолетник. Относится
к группе растений длинного дня. Светолюб. Для роста нуждается в ноч-
ной температуре 7—12° [688]. Высевается в апреле в парник.
Зимою при дополнительном ночном освещении (с 17 до 22 час.; 160—
320 лк) зацветает через 91 день, без освещения — через 131 день 1696].
Немофила (Nemophila Menziesii)
Длиннодневное растение. Высевается в апреле. Цветет с июня по июль.
Зимою при дополнительном ночном освещении в 160—320 лк (с 17 до
22 час.) зацветает через 52 дня, а без освещения — через 102 дня [696].
Чернушка («Девица в зелени») (Nigella damascene)
Растение длинного дня. В грунту зацветает через 2 месяца. Хорошо
реагирует на дополнительное освещение [806].
Зимою в оранжерее при дополнительном ночном освещении (с 17 до
22 час.; 160—320 лк) цветет через 79, а без освещения — через
119 дней [696].
Хорошо растет при «нижнем» освещении с помощью люминесцентных
ламп (600 вт/м2), зацветая'через 60 дней после всходов. Можно рекомен-
довать для создания клумб на станциях метрополитена.
328
Частная светокультура растений
Мак полевой (Papaver rhoeas)
Длиннодневное и светолюбивое растение. Высевается или осенью или
весной. В первом случае цветет с июня по август, а во втором — с июля
по сентябрь.
Без дополнительного ночного освещения зимою или не цветет совсем
[594, 678] или значительно запаздывает с переходом к цветению. При
дополнительном ночном освещении (с 17 до 22 час.; 160—320 лк) зимою
зацветает через 77 дней, без освещения — через 144 дня [696].
Петуния (Petunia hybrida)
Многолетнее растение, культивируемое как однолетник. Для роста
и заложения цветков нуждается в ночной температуре 10—24° [688].
Относится к группе растений длинного дня [716]. Светолюб, но может
развиваться и в полутени.
В опытах Бэйли [398] растения при освещении дуговыми лампами
росли мощнее и стройнее и цвели раньше и богаче, чем в контроле. Как
показывают современные данные, петуния при дополнительном освеще-
нии зимою (с 16 до 21 час.; 3000—5000 лк) ускоряет наступление цвете-
ния на 18 дней и увеличивает количество цветков на 50% [521].
Пост [688] рекомендует дополнительное освещение с октября по апрель
при температуре 17—18°.
Артур и Гарвилл [387] успешно культивировали петунию зимою при
прерывистом ночном освещении. .
Фацелия (Phacelia)
Ph. viscida и Ph. Whitlavia относятся к теневыносливым и длинно-
дневным растениям. Высеваются в мае прямо на месте. Цветут с июня
до осени.
При дополнительном ночном освещении (с 17 до 22 час.; 160—320 лк)
в зимние месяцы зацветают: Ph. Whitlavia — через 63, a Ph. viscida —че-
рез 92 дня; соответствующие сроки для контрольных растений — 125 и
150 дней [696].
Пламенник (Phlox Drummondii)
Длиннодневное растение. Для роста нуждается в ночной темпера-
туре 12—15° [688]. Высевается в марте в парник, цветет все лето (с июня)
до осени.
Зимою при дополнительном освещении (с 17 до 22 час.; 160—320 лк)
зацветает через 77—90 дней, без освещения — через 95—125 дней [697].
В опытах Поста [686, 688] при освещении с 7 января (с 17 до 21 час.;
150 лк) зацвел через 93—110 дней (10—27 апреля), а в контроле — че-
рез 118—129 дней (5—16 мая). Дополнительное освещение резко увели-
чивает количество цветков (на 35—270%). При температуре 15° зацве-
тает на 15—17 дней раньше, чем при 10—12°.
Квамоклит (Quamoclit coccinea или Ipomoea coccinea)
Длиннодневные и короткодневные растения. Первые на дополнитель-
ное ночное освещение зимою реагируют мало, зацветая при освещении
через 79, а без освещения — через 85 дней. Вторые в условиях
естественного зимнего дня цветут через 64 дня, а при дополнительном
освещении (с 17 до 22 час.; 160—320 лк) — через 135 дней [696].
IX. Светокультура декоративных растений
329’
Резеда (Reseda odorata)
Длиннодневное и светолюбивое растение. Для роста нуждается в ноч-
ной температуре 7—12° [688]. Высевается в парник в марте—апреле
или в мае непосредственно в грунт. Цветет с июня до осени.
Дополнительное ночное освещение зимою (длительность 5—6 часов
при освещенности 160—320 лк) несколько ускоряет наступление цветения
(на 10 дней) и вызывает лучший рост растений [521, 579, 696]. При осве-
щении зацветает через 124 дня. Однако, по данным Поста [688], до-
полнительное освещение, ускоряя рост, одновременно вызывает ослабле-
ние стебля. Этот автор не рекомендует дополнительное освещение для
культуры резеды.
Рудбекия (Rudbeckia bicolor)
Светолюбивое длиннодневное растение. Оптимальная длина дня для
цветения равна 18 часам (фиг. 136, 4 [786]). На коротком дне находится
в розеточном состоянии.
По данным Остина [397], при дополнительном освещении 1000—
1600 лк стрелкование наступает через 17 дней. Как показывают опыты
Функе [496], достаточна интенсивность дополнительного освещения
100—200 лк.
В опытах Остина [396] R. bicolor зацветала при длине дня 20 часов,
создаваемой с помощью дополнительного освещения не менее 15—20 лк.
Дополнительное освещение в 2,5—4 лк эффекта не давало.
Сальпиглоссис (Salpiglossis sinuata)
Длиннодневное и исключительно светолюбивое растение. Для роста
нуждается в ночной температуре 12—18° [688]. Высевается в начале
апреля, цветет с июня по сентябрь.
При дополнительном освещении зимою ускоряет цветение на 11 —
43 дня и увеличивает количество цветков [595, 598, 673, 678, 686, 688].
При посеве 23 ноября под лампами накаливания зацветает 23 марта—
25 апреля, через 120—132 дня.
При температуре 15° дополнительное освещение дает значительно
лучший эффект, чем при 10—12°. В первом случае цветение наступает
через 96 дней, а во втором — через 101—122 дня [686, 688].
Длительность освещения 5—6 часов, освещенность — 400 лк. Луч-
ший результат наблюдается при вечернем освещении (с 18 до 22 час.).
Зимою при культуре с помощью ламп накаливания (4 часа; 160 вт/м2)
на 1 цветок затрачивается 1,2 квт-час, а на 1 м2 — 29 квт-час [673].
Шалфей (Salvia)
Большинство видов относится к группе растений короткого дня.
Имеются длиннодневные и нейтральные формы. Светолюб, но может
развиваться и в полутени. Для роста нуждается в температуре 12—24°,
а для заложения цветков — 12—15° [688]. При посеве в конце февраля
зацветает в июле.
Наиболее распространенный вид S. splendens является растением
короткого дня и при дополнительном освещении в ночные часы не зацве-
тает [716, 742, 857, 860, 862]. Для устранения цветения достаточно осве-
щенности в 10 лк. При дополнительном освещении наблюдается усилен-
ный вегетативный рост [579].
330
Частная светокультура растений
S. runcinata относится к длиннодневным, а некоторые формы к ней-
тральным растениям [496, 697].
В условиях благоприятной длины дня в зимние месяцы шалфей зацве-
тает через 125—128 дней.
Можно культивировать в комнатах вдали от окна при ежедневном
дополнительном освещении (1000—5000 лк) в течение 5—10 часов [677,
678].
Мыльнянка (Saponaria calabrica или S. multiflora)
Длиннодневное светолюбивое растение. Высевается в апреле в пар-
ники.
Зимою при дополнительном ночном освещении (с 17 до 22 час.; 100—
320 лк) зацветает через 54 дня, без освещения — через 100 дней [696].
Скабиоза (Scabiosa atropurpurea)
Длиннодневное светолюбивое растение. Может культивироваться и
в полутени. Для роста нуждается в ночной температуре 10—15° [688].
Высевается в начале апреля, цветет с июля до осенних заморозков.
Зимою культивируется при дополнительном ночном освещении [595,
596, 598, 678, 688, 697, 857, 858, 862]. В этом случае растения зацветают
через 95—147 дней, на 20—78 дней раньше контроля. Некоторые сорта,
например Азур Фэйри, Аметист, без дополнительного освещения в зим-
ние месяцы не цветут совсем [687, 857, 862]. Освещение в ночные часы
увеличивает количество цветков в два раза [595].
Длительность дополнительного освещения у различных авторов
колеблется от 5 до 10 часов, освещенность — от 10 до 440 лк. Утреннее
и вечернее освещение дают одинаковый эффект [596].
По данным Портер и Ли [678], в зимние месяцы при ежедневном до-
полнительном освещении 10 часов (160 лк) в течение 71 дня на 1 цветок
затрачивается 0,29—0,31 квт-час.
Шизантус (Schizanthus)
Sch. pinnatus, Sch. retusus и Sch. wisetonensis — длиннодпевные свето-
любивые растения. Для роста нуждаются в ночной температуре 7—12°
[688]. Высеваются в апреле или мае. Цветут с июня по октябрь.
В зимние месяцы при дополнительном ночном освещении зацветают
значительно раньше, чем без освещения [521, 595, 673, 678, 697, 857].
При посеве 23 ноября Sch. pinnatus цветет 18 января при освещении и
4 марта — в контроле. Кроме того, под лампами накаливания наблюдается
увеличение количества цветков. Sch. wisetonensis при дополнительном
освещении (с 17 до 22 час.; 160—320 лк) зацветает через 66, а без освеще-
ния — через ИЗ дней [697].
По данным Поста [688], дополнительное освещение дает отрицатель-
ный эффект. Sch. wisetonensis сильно страдает; растения получаются сла-
быми и тонкими.
Дополнительное освещение необходимо ежедневно в течение 6—8 ча-
сов при освещенности порядка 440 лк.
На 1 растение при употреблении ламп накаливания в 500 и 1000 вт
в течение 42 дней затрачивается 1,34—1,42 квт-час [521]. По данным
Пёш [673], при удельной мощности 100 вт/м2 на 1 цветок затрачивается
0,08 квт-час или 17 квт-час/м2.
IX. Светокультура декоративных растений
331
Смолевка (Silene latifolia или S. vulgaris)
Длиннодневное растение. Высевается в марте, цветет с июня по сен-
тябрь. Зимою при дополнительном ночном освещении (с 17 до 22 час.;
160—320 лк) цветет через 120 дней, без освещения не цветет совсем [697].
Бархатцы (Tagetes)
Теневыносливые, длиннодневные, короткодневные или нейтральные
растения. Для роста нуждаются в ночной температуре 12—18° [688].
Фиг. 155. Бархатцы {Tagetes patula), выращенные
под люминесцентными лампами в камере-нише - . <
Для грунтовой культуры бархатцы высеваются в парникигво второй
половине апреля.
Длиннодневные формы (Т. erecta) зимою культивируются при допол-
нительном ночном освещении [123, 594, 598, 673, 678, 697, 716]. При
вечернем дополнительном освещении в течение 5—6 часов (160—320 лк)
цветение наступает через 72, а без освещения — через 98 дней [697].
В наших опытах Т. erecta сорта Гольдене Шёне в зимние месяцы цвел
лишь при дополнительном освещении в течение 8—12 часов (500 вт/м2).
В этом случае цветение наступало через 123—137 дней. Кроме того, до-
полнительное освещение вызывает увеличение количества цветков при-
мерно на 40% [595]. Пост [688] рекомендует давать дополнительное осве-
щение с октября по апрель.
Освещение короткодневных форм (Г. patula) возможно лишь в пре-
делах короткого дня (10—12 часов). Дополнительное освещение, удли-
332
Частная светокультура растений
няющее естественный зимний день, обычно вызывает запаздывание в цве-
тении на 21—22 дня [153, 697]. На коротком дне Т. patula зацветает через
49—54, а при освещении — через 70—72 дня. По данным Гардера и его
сотрудников [530], Т. patula папа при дополнительном освещении уве-
личивает свежий вес на 120, а количество цветков — на 60%.
Для задержки цветения необходима освещенность 160—320 лк при
длительности дополнительного освещения 8—10 часов.
По данным Пёш [673], при культуре зимою затрата электроэнергии
при освещении лампами накаливания (4 часа; 100 вт/м2) составляет
0,83 квт-час на 1 цветок, или 29 квт-час на 1 м2.
Т. patula папа (сорт Оранжефламме) хорошо растет в нишах и при
нижнем освещении с помощью люминесцентных ламп (600 вт/м2), при
интенсивности физиологической радиации 25 000 эргсм2-сек, форми-
руются мощные и красивые растения (фиг. 155). Можно рекомендовать
для создания клумб на станциях метрополитена [120].
Титония (Tithonia sp., Helianthus sp., Leighia sp.)
Короткодневное растение, зимою цветущее лишь в условиях есте-
ственного дня (через 116 дней). Дополнительное освещение в 10 лк, удли-
няющее день до 18 часов, полностью устраняет цветение [860, 862].
Дидискус (Trachymene coerulea, Didiscus coerulea)
Длиннодневное растение. Для роста нуждается в ночной температуре
10—15° в первые 2 месяца и 15—21° — 'в последующее время. Для зало-
жения цветков необходима температура более 15° [688]. Высевается
в апреле; на постоянное место высаживается в мае. С помощью дополни-
тельного ночного освещения может культивироваться зимою [521, 595, 596,
598, 673, 678, 686, 688, 697, 858]. При посеве в ноябре зацветает в фев-
рале—марте, через 102—126 дней; при посеве с января цветет в мае.
Дополнительное освещение сокращает вегетационный период на
16—45 дней и увеличивает количество цветков на 50—80% (табл. 87
и 104). В опытах Поста [688] при освещении наблюдалось уменьшение
количества цветков на 40—50%.
Лори и Пёш [596] показали, что в пределах освещенности от 75 до
460 лк дополнительное освещение дает одинаковый эффект (табл. 104;
фиг. 142, 7). Обычно применяются лампы накаливания в 100—200 вт,
дающие освещенность 150—400 лк.
Длительность дополнительного освещения колеблется в пределах 5—
10 часов. Лучший результат наблюдается при вечернем освещении [595].
По данным Грина [521], при культуре зимой необходима затрата эле-
ктроэнергии 9 квт-час на растение, а по данным Пёш [673], — 0,1 квт-час
на 1 цветок, или 29 квт-час на 1 м2. Уитроу [678, 858] нашел, что на 100
цветков необходимо 27 квт-час.
Настурция (Tropaeolum majus)
Исключительно светолюбивое, длиннодневное или нейтральное расте-
ние. Для роста нуждается в ночной температуре 10—21 °, для заложения
цветков — 10—18° [688]. Посев — во второй половине апреля.
Культура в зимнее время возможна лишь при дополнительном ноч-
ном освещении [376, 379, 598, 646, 678, 688, 715, 716, 857, 858]. Это ведет
к увеличению количества цветков [678, 858] или к ускорению начала
IX. Светокультура декоративных растений
333
цветения [376, 697, 716]. Цветение при освещении наступает через 60—73,
а без освещения — через 60—98 дней. В наших опытах Tr. majus
папитп сорта.Шарлах Кугель и Кайзерин фон Индия при дополнитель-
ном ночном освещении с 11 февраля по 3 июня (8—12 часов; 500 вт/м2)
цвели через 77—92 дня, но имели сильно этиолированный облик. Конт-
рольные растения не цвели совсем. Некоторые сорта, например Гольден
Глим, при дополнительном освещении не ускоряют цветения, но в два
раза увеличивают количество цветков [857, 858].
Пост [688] рекомендует культивировать настурцию в зимние месяцы
при температуре 17—18° при ночном освещении с октября по апрель.
По данным Артура и его сотрудников [379], дополнительное освещение
(около 8000 лк) особенно эффективно при обогащении атмосферы угле-
кислотой.
Портер и Ли [678] считают, что при зимней культуре настурций на
1 цветок затрачивается около 0,1 квт-час. Они рекомендуют дополнительное
освещение в течение 10 часов (с 21 до 7 час.) при освещенности 175 лк.
Цинния (Zinnia elegans)
Светолюбивое, коротко дневное или нейтральное растение. Для роста
нуждается в температуре более 15° [688]. Высевается в апреле—мае,
цветет с июля до морозов.
Дополнительное освещение, удлиняющее естественный зимний день,
может задержать или полностью устранить цветение растений [153, 697].
При ночном освещении наблюдается мощный вегетативный рост [688].
В условиях короткого зимнего дня цинния зацветает через 83—85
дней. Для задержки цветения достаточно 160—320 лк.
Дополнительное освещение следует рекомендовать лишь в пределах
короткого дня (10—12 часов).
2. ДВУЛЕТНИКИ
Двулетники представляют собою растения, зацветающие лишь на вто-
ром году. Цветение наступает обычно рано весной. Отличительной особен-
ностью двулетников является потребность в определенный период в низ-
ких температурах для прохождения стадии яровизации. В условиях вы-
соких температур процессы яровизации задерживаются, и растения могут
не зацвести.
Светокультура двулетников представлена сравнительно небольшим
числом работ, охватывающих немногие виды растений.
Колокольчик (Campanula medium)
Длиннодневное растение. Высевается в мае в парник, цветет в июне-
июле следующего года. В зимние месяцы в условиях светокультуры за-
цветает через 46—60 дней после начала освещения (табл. 105 и 116 [136,
530, 725]). Без освещения растения или не цветут или погибают.
При освещении растений с января цветение колокольчика начинается
в марте. Сильный эффект дает одновременное удобрение атмосферы угле-
кислотой. В этом случае цветение начинается несколько раньше, и уве-
личивается количество цветков (табл. 116 [530]).
Для освещения применяются лампы накаливания в 500 и 750 вт.
В опытах Н. П. Красинского [136] освещенность равнялась 12 000лк,
а освещение длилось в течение 16 часов (с 23 до 15 час.). Роденбург [725]
рекомендует неоновые лампы. , :
334
Частная светокультура растений
Энотера (Oenothera sp.)
Длиннодневные или нейтральные растения, предпочитающие полу-
тень. Положительно реагируют на дополнительное ночное освещение ,
ускоряя наступление цветения [716].
Анютины глазки (Viola tricolor)
Длиннодневное, светолюбивое растение. Для роста нуждается в ноч-
ной температуре 7—12° [688]. Многолетник, но культивируется как дву-
летник. Можно культивировать как однолетник, высевая в марте и вы-
саживая на постоянное место в мае. При посеве в июне цветет в следую-
щем году весной.
Сильно реагирует на дополнительное ночное освещение в зимние
месяцы [399, 521, 595, 596, 598, 673, 678, 697, 8^7, 859, 861], эффективность
которого зависит от интенсивности естественной радиации (фиг. 151).
При освещении значительно сокращается вегетационный период и уве-
личивается количество цветков (табл. 87). По данным Бэйли [399], уско-
рение цветения в результате ночного освещения составляет несколько
недель. В опытах Рэмэли [797] дополнительное освещение (с 17 до 22 час.;
160—320 лк) в зимние месяцы сократило вегетационный период на
128 дней — с 210 до 82 дней. Сорт Джиант СуиссБлу при дополнительном
освещении 80—1250 лк ускоряет цветение на 10—22 дня, зацветая через
98—107 дней вместо 117 дней [521]. Одновременно наблюдается увеличе-
ние количества цветков с 3 до 30—50. По данным Уитроу [857], дополни-
тельное освещение 10—15 лк с 21 до 7 час. увеличивает количество цвет-
ков на растении с 1,6 до 16,2. Лори и Пёш [595] нашли, что V. tricolor,
выкопанная из почвы и поставленная 1 января под лампы накаливания
(220 вт/м2), зацвела через 51 день (20 февраля), одновременно с контро-
лем, увеличив количество цветков почти в два раза (с 28,7 до 44,2).
Опыты показывают, что при пользовании лампами накаливания в 100—
200 вт оптимальная освещенность при дополнительном освещении равна
180—400 лк (фиг. 142, 4) (табл. [521, 857, 858, 862]).
Длительность дополнительного освещения 5—10 часов. Лучший эф-
фект наблюдается при освещении с 21 до 7 час.; освещение с 21 до 2 час.
дает несколько меньший эффект; при утреннем освещении (с 2 до 7 час.)
получаются наихудшие результаты [521, 858].
При освещении 195 или 450 лк, создаваемом в течение 10 или 5 часов,
и продолжительности дополнительного освещения 130 дней (с 13 но-
ября) на 1 цветок затрачивается 0,03 квт-час [521, 857].
По данным Пёш [673], при удельной мощности 100 вт/м2 и дополнитель-
ном освещении в течение 4 часов расходуется 0,05 квт-час на 1 цветок, или
23 квт-час на 1м2.
3. МНОГОЛЕТНИКИ
Многолетники — растения, цветущие длительное время, многие
с ранней весны и, часто, до поздней осени. Обладая органами запаса —
корневищами, клубнями, луковицами и другими, — они отличаются боль-
шей теневыносливостью и, следовательно, представляют собою объекты,
более пригодные для светокультуры в зимние месяцы.
Тысячелистник (Achillea millefolium roseum)
Длиннодневное светолюбивое растение. При освещении с 6 января
по 4 часа в сутки (с 18 до 22 час.; 380 лк) зацветает через 91 день
IX. Светокультура декоративных растений
335
(7 апреля); без освещения — через 120 дней (9 мая) [496, 595, 598, 673,
678].
При культуре зимою с помощью ламп накаливания (4 часа; 100 вт/м2}
затрачивается 0,15 квт-час на 1 цветок, или 23 квт-час на 1 м2 [673].
Анху за (Anchusa italica)
Длиннодневное растение. В естественных условиях цветет в июне—
июле. По данным Тинкера [807], при дополнительном освещении с начала
марта (5 часов; 20—40 лк) растения сорта Прайд оф Давер цветут одно-
временно как при освещении, так и в контроле, но при дополнительном
освещении растения выглядят несколько хуже, чем в контроле.
Ветреница (Anemone coronaria)
Нейтральное теневыносливое растение. Цветет с апреля по июнь.
В холодной оранжерее в горшках при посадке в октябре зацветает в ян-
варе—феврале.
По данным Одена [646], A. coronaria var. hollandica в зимнее время
в условиях 15—17-часового дня при дополнительном освещении 0,02—
0,03или0,07—0,16г-кал/см2-мин (14—21 тыс. или49—112 тыс. эрг/см2-сек}
не цветет.
Пупавка (Anthemis tinctoria)
Длиннодневное растение. Цветет с июня по октябрь. Ускоряет на-
ступление цветения в результате дополнительного освещения, удлиняю-
щего короткий день [496, 497, 715, 716].
Резуха (Arabis alpina)
В грунту цветет в апреле—мае. Зимою культивируется при дополни-
тельном освещении. В опытах Одена [646] при дополнительном освещении
интенсивностью 0,08 г-кал/см2-мин (56 тыс. эрг/см2-сек) с 26 ноября
зацвела через 14 дней (10 декабря) и цвела на протяжении всего января.
Астры (Aster) -
Исключительно светолюбивые короткодневные растения. В условиях
длинного дня задерживают цветение на 16—100 дней.
По данным Функе [496], A. linosyris цветет при дополнительном осве-
щении 30—60 лк (16-часовой день) через 107 дней, а без освещения
(8-часовой день) — через 91 день; соответствующие данные для A. mul-
ttflorus — 72 и 42 дня, а для A. ericoides — 122 и 80 дней.
Дополнительное освещение для астр можно рекомендовать лишь
в пределах короткого дня. Ночное освещение следует применять лишь
для задержки цветения или усиления вегетативного роста. Гарнер и Ал-
лард [501] применяли ночное освещение для нарушения покоя у A. U-
harifolius. При освещении с 27 ноября по 8 часов в сутки (с 16 час. 30 мин.
до 0 час. 30 мин.) с помощью лампы накаливания (100 вт на расстоянии
30 см) растения тронулись в рост на третьей неделе декабря; без освеще-
ния растения находились в покое.
Аквилегия (Aquilegia hybrida)
Нейтральное тенелюбивое растение.
В зимние месяцы на дополнительное освещение не реагирует, зацве-
тая одновременно с контролем [595]. д •
•336
Частная светокультура растений
Астильба (Astilbe japonica)
Светолюбивое растение, культивируемое на открытых местах. Цветет
в июле—августе. При внесении в оранжерею или в комнату в декабре—
январе зацветает в марте—апреле.
По данным Максимова [188], в оранжерее в зимние месяцы прекрасно
развивается под лампами накаливания 1000 вт.
Осреция (Aubrietia hybrida)
По данным Одена [646], в зимние месяцы нормально цветет при допол-
нительном освещении с 27 декабря по 31 января.
Маргаритка (Bellis perennis)
Многолетнее длиннодневное растение, культивируемое как двулетник.
Высевается в марте—апреле, пересаживается в августе и цветет ранней
весной (до наступления жарких дней).
Зимою при дополнительном освещении ускоряет цветение и увели-
чивает количество цветков [399, 595, 596, 857, 859]. Еще Бэйли [399]
отметил, что при освещении вольтовой дугой маргаритки зацветают на
несколько недель раньше контроля. В опытах Уитроу [678, 859] при еже-
дневном освещении (с 21 до 7 час.; 175 лк) в течение 135 дней (19 ноября—
3 апреля) маргаритка Шаста цвела раньше контроля на 22 дня и увели-
чила количество цветков на 154,6%. В этом случае на 1 цветок затрачи-
валось 0,57 квт-час.
Василек (Centaurea montana)
Длиннодневное светолюбивое растение. При дополнительном осве-
щении в 30—60 лк, удлиняющем день до 16 часов, ромажйГ зацветает на
14 дней раньше контроля [496].
Хризантемы (Chrysanthemum)
К группе многолетних хризантем относятся длиннодневные Ch. ma-
ximum, Ch. parthenium (Matricaria exima) и Ch. Parthenoides (Matricaria
capensis) и короткодневные Ch. leucanthemum и Ch. indicum. Последняя
в открытом грунте может культивироваться лишь на юге.
При дополнительном освещении (с 18 до 22 час.; 360 лк) зимою (с 6 ян-
варя) Ch. maximum зацветает через 69 дней (17 марта), а без освещения —
через 96 дней (13 апреля); одновременно наблюдается увеличение раз-
меров цветков (на 40%). В других опытах растения при дополнительном
освещении зацветали на 105-й день, причем ускорение составляло
56 дней, а количество цветков увеличивалось с 19,7 до 26,6 [595, 598,
673, 678]. Для получения названного эффекта необходима удельная мощ-
ность порядка 50—75 вт/м2 или освещенность в 360 лк.
По данным Пёш [6'3], для культуры Ch. maximum с помощью допол-
нительного освещения (4 часа; 100 вт/м2) необходима затрата электроэнер-
гии 0,17 квт-час на 1 цветок, или 18 квт-час на 1 м2.
Ch. Parthenium (Matricaria exima) зимою цветет лишь при дополни-
тельном освещении или ускоряет цветение на 48—57 дней. При освеще-
нии с 7 января (с 17 до 21 час.; 150 лк) цветение наступает 21—24 марта
при температуре 12—15° и 25 апреля при 10° [686, 688]. Этот вид реаги-
IX. Светокультура декоративных растений 337
рует одинаково как на вечернее, так и на утреннее освещение [596]. Ch.
Parthenoides {Matricaria capensis) также сильно реагирует на дополнитель-
ное освещение зимою (табл. 104), ускоряя цветение и увеличивая коли-
чество цветков [596]. Для получения эффекта в обоих случаях необходима
освещенность порядка 75 лк [596].
Дополнительное освещение короткодневных форм — Ch. leucanthemum
и Ch. indicum — можно рекомендовать лишь в пределах короткого дня
(12 часов), а в ночные часы — лишь с целью усиления вегетативного роста.
В последнем случае оба вида значительно — на 39—74 дня — задержи-
вают цветение [433, 496, 806]. По данным Мошкова [212], Ch. indicum
цветет лишь при длине дня короче 14 часов. В условиях 11-часового дня
бутонизация наступает через 23, а цветение — через 74 дня. Беспрерыв-
ное освещение полностью устраняет цветение. Ch. morifolium (Ch. hor-
torum), сорта Лилиан Дэти и Голден Скептр, тоже цветут лишь на корот-
ком дне [715, 716].
Дополнительным освещением в ночные часы пользуются для задержки
цветения с целью получения цветков в более поздние месяцы. Пёш [675],
культивируя хризантемы Валенсия, Кордова, Принстон, Робинда и Орчид
Бьюти, обычно цветущие лишь осенью, с уменьшением длины дня, давал
дополнительное освещение в течение 48 дней, начиная с 5, 12, 19, 26 ав-
густа и 2 сентября. Автор установил, что задержка составляла 16—
50 дней, и цветение наступало 29 ноября—15 января. Лишь при осве-
щении с 5 августа задержка в цветении была несколько меньшей (3—
40 дней). В аналогичном опыте Поста [688] с сортами Дороти Тэрнер,
Узания и Принстон при освещении с 20 августа по 20 сентября задержка
в цветении равнялась 14—20 дням. Контрольные растения цвели 25 ок-
тября—8 ноября, а при освещении — 16—22 ноября.
Пёш [673] показал, что наиболее сильный задерживающий эффект
наблюдается под лампами накаливания (50 вт; 40—140 лк); ртутные
лампы (450 вт; 30—400 лк) действуют значительно слабее. Сравнивая
влияние ламп накаливания мощностью от 15 до 150 вт (20—360 лк), автор
показал, что задержка цветения бывает лишь прц освещенности не менее
40 лк; освещенность 20 лк (15 вт) не влияет. В опытах Линка [604] хри-
зантемы при дополнительном освещении в 20—400 лк задерживали цве-
тение на 27—29 дней.
Для задержки цветения можно пользоваться и кратковременным ноч-
ным освещением. Стьюарт с сотрудниками [481, 792], применяя дополни-
тельное освещение в середине ночи (30—60 минут; 160—1000 лк), задер-
живали цветение у Мистрис Дэвид Пэй, Орчид Куин и других сортов на
60—90 дней. Вместо обычного цветения в ноябре хризантемы цвели в ян-
варе-феврале. У сортов Мистрис Г. Е. Киддер, Джозефин Бирнс, Кор-
нелия, Голд Куин и других при освещении в середине ночи с 6 сентября
(45 минут; 320—600 лк) цветение запаздывало на 29—32 дня.
Chr. coccineum па дополнительное ночное освещение не реагирует
[595].
Ландыш (Convaliaria majalis)
Тенелюбивое растение. Зацветает на третий год в мае—июне.
Широко применяется зимняя выгонка ландышей. Выгонка значительно
ускоряется с помощью дополнительного освещения [283, 289, 522—524,
547, 579, 587, 646, 650, 717, 718]. При зимней выгонке ландышей без осве-
щения обычно наблюдаются слабые и желтые растения. При освещении
образуются мощные растения с интенсивно зелеными листьями и круп-
ными цветками. В опытах Клейна [579] ландыши зацветали при допол-
22 А. Ф. Клешнин
338
Частная светокультура растений
нительном освещении через 8, а в контроле — через 14—18 дней. По дан-
ным Гербачека 1547], для выгонки ландышей при освещении с помощью
ламп накаливания в 500 вт достаточно 8 дней. Оден [646] нашел, что
лучший результат (самое большое количество цветков) имеет место при
12-часовом дополнительном освещении при интенсивности радиации
0,12 г-кал/см2 (84 тыс. эрг/см2-сек). В этом случае цветение наступает
через 13 дней при затрате лучистой энергии 1120 г-кал/см2 (86 г-кал/см2
в сутки). В другом месте этот автор рекомендует 10-часовое дополнитель-
ное освещение в 500 лк, указывая, что в этом случае при выгонке в де-
кабре—январе цветение наступает через 15 дней [650]. Беспрерывное
освещение дает худший эффект.
Для выгонки ландышей непригодны источники радиации, богатые сине-
фиолетовыми лучами (ртутные лампы, лампы накаливания при перекале
и т. д.). В этом случае наблюдается подавление образования цветочных
почек, и растения не цветут даже при переносе в нормальные условия [ 646].
Предпочтение следует отдать источникам оранжево-красных лучей (нео-
новые лампы, лампы накаливания и другие). Согласно Куфферат [587],
можно применять даже обычные неоновые рекламные трубки мощ-
ностью в 600 вт, создающие освещенность 500—1000 лк.
Ленок (Coreopsis)
К многолетним формам ленка относятся С. lanceolata и С. grandi-
jolia. Оба вида — растения длинного дня. Цветут с июня по сентябрь.
При дополнительном освещении (с 18 до 22 час.; 220—380 лк) в зимние
месяцы ленок зацветает через 105 дней, а без освещения — через 134 —
151 день [595, 673, 678]. При освещении с декабря—января цветение
наступает в марте—апреле.
При культуре зимою при дополнительном освещении лампами нака-
ливания (4 часа; 100 вт/м2) затрачивается 0,017 квт-час на 1 цветок, или
23 квт-час на 1'м2 [673].
Циноглоссум (Cynoglossum amabile)
Длиннодневное растение. При культуре растений зимою дополни-
тельное освещение (с 18до 22час.; 400 лк)ускоряет наступление цветения
на 26 дней [595, 673, 678].
При дополнительном освещении с помощью ламп накаливания (4 часа;
100 вт/м2) зимою затрачивается 0,002 квт-час на 1 цветок, или 29 квт-час
на 1 м2 [673].
Шпорник (Delphinium hybridum)
Светолюбивое длиннодневное растение. Цветет с июня по август.
Зимою при дополнительном освещении (с 18 до 22 час.; 380 лк) цветет
через 87 дней, ускоряя цветение на 9 дней и увеличивая количество цвет-
ков на 13% [595, 673, 678].
При освещении 4 часа и удельной мощности 100 вт/м2, на 1 цветок за-
трачивается 0,93, а на 1 м2 — 23 квт-час [673].
Гвоздика (Dianthus)
Многолетние светлолюбивые растения D. barbatus и D. caryophyllus
культивируются как двулетники. Первая для роста и заложения цвет-
ков нуждается в ночной температуре 12—18°, а второй для роста нужно
IX. Светокультура декоративных растений
339
7—12°, а для заложения цветков — более 7° [688]. Оба вида относятся
к группе растений длинного дня. Встречаются и нейтральные формы
(678, 716, 858]. Посев гвоздики производится в апреле—мае. D. barbatus
цветет в июне—июле, a D. caryophyllus — в июле—августе.
Зимою оба вида культивируются при дополнительном освещении
[387, 388, 521,594,673,695,697, 858, 859]. D. barbatus зимою при дополни-
тельном освещении (с 17 до 22 час.; 100—320 лк) зацветает через 80, а без
освещения — через 138 дней; ускорение равно 45—58 дням [695, 697].
По данным П. И. Лапина [153], дополнительное освещение имеет смысл
только по прохождении растениями стадии яровизации. В против-
ном случае растения зацветают лишь через год, одновременно с кон-
тролем.
В опытах некоторых авторов [521, 595, 673, 678, 858, 859]
D. caryophyllus на дополнительное освещение продолжительностью
5—10 часов при освещенности от 175 до 5000 лк реагировала лишь увели-
чением количества цветков на 16—25%; ускорения наступления цветения
не наблюдалось. В других случаях наблюдалось, однако, значительное
ускорение. Рэмэли [697] нашел, что при длительности дополнительного
освещения 5 часов (с 17 до 22 час.) и освещенности 160—320 лк гвоздика
зацветает через 162, а без освещения — через 182 дня. В опытах П. И.
Лапина [153] с сортом Гренадин при освещении растений (200 вт/м2)
после прохождения ими стадии яровизации цветение наступало через
194 дня, а в контроле, независимо от прохождения яровизации, — черев
414—417 дней. По данным Артура и Гарвилла [387], сорта Пурити,
Димити и Кримсон при дополнительном прерывистом освещении (в сумме
4—5 часов в сутки, около 1000 вт/м2) ускорили цветение на 14—54 дня.
Лучший результат наблюдался под натриевыми лампами, а лампы нака-
ливания и люминесцентные лампы были менее эффективны. В результате
освещения наблюдалось также увеличение количества цветков.
С. chinensis при дополнительном ночном освещении зимою (100—
200 лк) ускоряет наступление цветения на 34 дня [фЭТТ. £#>"
D. plumarius при дополнительном освещении (с 17 до 22 час.; 320 лк)
цветет через 128 дней, а без освещения — через 154 дня [695, 697].
Однолетняя форма D. caryophyllus (сорт Маргарита) также положи-
тельно реагирует на дополнительное освещение. В условиях 10-часового
дня она зацветает через 123 дня, а при дополнительном освещении
(200 вт/м2) — через 86 дней [153].
В большинстве случаев применяются лампы накаливания в 500 вт.
Рэмэли [697] пользовался освещенностью 160—320 лк, а Грин и его
сотрудники [521] — 3000—5000 лк. Длительность дополнительного осве-
щения 5—10 часов. В зимнее время на 1 растение затрачивается
27, 22 квт-час (0,3 квт-час на 1 цветок; [521]).
D. Heddewigii прекрасно растет при «нижнем» освещении с помощью
люминесцентных ламп (600 вт/м2), зацветая через 60 дней.
В группе с другими растениями можно рекомендовать для создания
клумб на станциях метрополитена.
Диклитра [Dicentra (Diclytra) spectabilis]
Грунтовое, оранжерейное и комнатное растение. В грунту цветет
в июне—июле, а в оранжерее при выгонке с середины декабря — в ян-
варе—феврале.
По данным Одена [646], для выгонки диклитры необходимо ежедневное
10-часовое дополнительное освещение интенсивностью 0,12 г-кал/см2-мин
22*
340
Частная светокультура растений
(84 тыс. эрг/см2-сек). В его опытах названное растение, посаженное
в начале октября и освещаемое с 7 января (с 23 до 7 час.) достигало пол-
ного цветения 5 марта.
Наперстянка (Digitalis purpurea)
Нейтральные или длинно дневные растения, мирящиеся с затенением.
Для роста нуждаются в ночной температуре 4—21°, для заложения цвет-
ков 5° [688]. Высеваются в мае в парник, зацветают через год в июне—
июле и цветут по август. Нейтральные формы на дополнительное осве-
щение зимою не реагируют [595], а длиннодневные цветут лишь при до-
полнительном освещении [389].
Дополнительное освещение рекомендуется давать после выдержива-
ния растений при 5° в течение 4 месяцев [688].
Шелковая трава [Digraphis (Phalaris) arundinacea]
Длиннодневное растение, при дополнительном ночном освещении
300—800 лк цветет, а в условиях естественного зимнего дня находится
в вегетативном состоянии [715]. По другим данным, не реагирует на до-
полнительное освещение (3000—4000 лк), зацветая одновременно с кон-
тролем [373].
Диморфотека (Dimorphotheca aurantiaca)
Теплолюбивое, светолюбивое и длинно дневное растение. Высевается
в ящики во второй половине апреля. Цветет с июня до осени. Зимою при
дополнительном ночном освещении зацветает через 84 дня после посева,
ускоряя наступление цветения на 45 дней [595, 697].
Дороникум (Doronicum)
Нейтральные растения с тенденцией к длиннодневности. В грунту
зацветают рано и цветут все лето.
D. caucasicum на дополнительное освещение (с 12 до 22 час.; 220 лк)
реагирует слабо, зацветая на 7 дней раньше, чем в контроле, и несколько
увеличивая количество цветков [595, 678]. Цветение наступает через 62 дня
при освещении и 69 дней — в контроле.
D. pardalianches совсем не реагирует на ночное освещение [496].
Эрлангея (Erlangea tomentosa)
Нейтральное растение, не реагирующее на дополнительное освещение.
При ночном освещении зимою (с 18 до 22 час.; 400 лк) цветет одновременно
с контролем [595].
Гайлардия (Gaillardia grandiflora)
Светолюбивое длиннодневное растение. Цветет с июня до осени.
Сильно реагирует на дополнительное освещение (с 18 до 22 час.;
380 лк) зимою, увеличивая количество цветков (в 4,9 раза) и ускоряя цве-
тение (на 39 дней) [595, 673, 678]. При освещении с 6 января гайлардия
зацветает через 85 дней (2 апреля), а без освещения — через 124 дня
(11 мая).
Зимою при дополнительном освещении в течение 4 часов и удельной
мощности 100 вт/м2 на 1 цветок затрачивается 0,053, а на 1 м2 23 квт-час
[673].
IX. Светокультура декоративных растений
341
Перекати-поле (Gypsophila paniculate)
Длиннодневное светолюбивое растение. Цветет с июня по август.
Зимою при дополнительном освещении (с 17 до 22 час.; 110—320 лк)
зацветает через 55, а без освещения — через 75 дней. Ускорение цветения
в результате освещения равно 20—36 дням [695, 697].
Подсолнечник (Helianthus)
Н. giganteus и Н. angustijolius— светолюбивые короткодневные формы.
При дополнительном ночном освещении в 30—50 лк или не цветут совсем
(Н. giganteus) или сильно задерживают наступление цветения (7/. angusti-
jolius) [501].
Ночная фиалка (Hesperis matronalis)
Тенелюбивое растение. Высевается в грунт в апреле или августе.
Цветет в июне—июле.
По данным Одена [646], при освещении 0,05 г-кал/см2-мин. (35 тыс.
эрг/см2-сек) со 2 ноября зацветает через месяц.
Гуннемания (Hunnemania fumariifolia)
Длиннодневное растение. Для роста нуждается в ночной температуре
15—27°, для заложения цветков — выше 15° [688]. Цветет поздним
летом и осенью.
При культуре зимою нуждается в дополнительном освещении. В этом
случае значительно ускоряет наступление цветения [688].
Гистрикс (Hystrix patula, Asprella hystrix)
Орнаментальное длиннодневное растение, положительно реагирующее
на дополнительное освещение [373].
Касатик (Iris)
Светолюбивые длиннодневные растения. Цветение начинается при
интенсивности физиологической радиации 6000 эрг/см2-сек и достигает
максимума (100% растений) при 23 тыс. эрг/см2 • сек. (фиг. 134, 1 [536]).
Для роста нуждаются в ночной температуре 7—12° [688], для заложения
цветков 23—31° [536]. Цветут с апреля по июль.
Дополнительное освещение зимою ускоряет цветение и увеличивает
количество цветков. В опытах Гарнера и Алларда [500] I. jlorentina,
посаженный 20 октября и освещаемый с 1 ноября по 8 часов в сутки
(с 16 час. 30 мин. до 0 час. 30 мин.), зацвел через 54—60 дней (26—30 де-
кабря); цветение в контроле наступило лишь 42 дня спустя (12 февраля).
I. pumila при дополнительном освещении зацветает через 14—15 дней,
а без освещения — через 17—20 дней (табл. 116); количество цветков
в первом случае равняется 24—34, а во втором 3—11 [530]. Особенно
сильное влияние дополнительного освещения наблюдается при одновре-
менном удобрении углекислотой (табл. 116). По данным «Лории Пёш [595],
длительность освещения для I. tingitana определяется состоянием
корневищ. Сорт Уэдж Вуд при посадке 29 ноября и освещении с 9 декабря
зацветает через 83 дня (3 марта), а при посадке 20 декабря и освещении
с 9 января — через 67 дней (17 марта). Соответствующие сроки для сорта
342
Частная светокультура растений
Император — 114 и 100 дней. В опытах Пёш [673] I. tingitana при осве-
щении с 13 декабря начал цвести через 75 дней (2 марта), одновременно
с контролем, но более обильно. Число цветущих растений в первом случае
равнялось 97 %, а в контроле — лишь 47 %. Аналогичные результаты
получили Гартзема и Лёйтен [536], освещавшие I. tingitana (сорт Импе-
ратор) люминесцентными и ртутными лампами в различные сроки. Осве-
щение производилось в вечерние часы, создавая суммарную длину дня
16 часов. Контрольные растения находились при естественной длине дня.
Как видно из табл. 129, дополнительное освещение, не влияя на время
зацветания, сильно увеличивает количество цветущих растений. Одно-
временно оказалось, что количество последних зависит от сроков осве-
щения: чем раньше начинается освещение, тем большее число растений
зацветает.
Таблица 129
Влияние дополнительного освещения на рост и развитие ириса
(Гартзема и Лёйтен [536])
Варианты Начало освещения Число дней от про- растания до начала освещения Средняя длина листьев к началу освещения (в см) Число дней от про- растания до цвете- ния Число цветущих растений (в % от общего) Средняя длина цветочного стебля (в см) Средняя длина листьев (в см)
Опыт первый
Люминесцентные лампы 12/XI 19 11,9 109 93 71,3 93,7
26/XI 47 36,6 106 61 73,4 92,0
ю/хп 61 55,1 108 64 67,9 91,1
7/1 75 73,5 116 6 64,3 89,2
Контроль — — — 110 3 38,5 87,1
Опыт второй
Люминесцентные лампы — 44 39,7 99 48 63,6 90,2
— 59 57,9 96 35 60,4 89,4
— 74 75,7 107 3 63,7 89,9
Ртутные лампы «Н0-2000» — 44 38,9 98 83 71,9 93,7
— 59 58,8 97 94 74,1 96,8
— 74 77,0 107 10 70,0 90,0
Контроль — — — 103 7 52,2 88,7
Сопоставление ртутных и люминесцентных ламп, произведенное при
одинаковой интегральной интенсивности радиации, показало, что луч-
ший результат дают ртутные лампы. Не исключена возможность, что этот
результат находится в связи с большим содержанием у ртутных ламп
инфракрасных лучей, доля которых составляет 60% и которые лежат
в далекой инфракрасной области. Вызывая дополнительный нагрев расте-
ний и почвы, инфракрасные лучи могли вызвать лучшее цветение ириса.
У люминесцентных ламп доля инфракрасных лучей составляет всего 10%
их радиации.
Лори и Пёш [595] считают, что для ирисов достаточно 80 вт/м2, однако
Гардер и его сотрудники [530] применяли лампы накаливания в 500 и
750 вт, исходя из расчета 350 и 520 вт/м2.
IX. Светокультура декоративных растений
343
Книпгофия (Kniphofia Pfitzerii)
Нейтральное растение. По данным Лори и Пёш [595], на дополнитель-
ное ночное освещение зимою не реагирует.
Лептосина (Leptosyne maritima)
Светолюбивое и длиннодневное растение.
В оранжерее при дополнительном ночном освещении зимою зацветает
на 23—43 дня раньше, чем без освещения [595, 596, 678]. При посеве
в середине сентября цветение наступает в ноябре, через 62 дня.
Для дополнительного освещения необходима освещенность порядка
400 лк и длительность освещения 5—6 часов. Утреннее освещение (с 2
до 7 час.) дает значительно лучший эффект, чем вечернее (с 17 до 22 час.;
[596]).
Кермек (Limonium sinuatum, Statice sinuata)
Нейтральное светолюбивое растение, цветет в июле. На дополнитель-
ное ночное освещение зимой не реагирует, зацветая одновременно с кон-
тролем через 100 дней после посева [697].
Люпин (Lupinus perennis)
Светолюбивое растение. Высевается в апреле—мае, зацветает на сле-
дующий год в июле—августе.
На дополнительное ночное освещение зимою не реагирует, зацветая
через 121 день как при освещении, так и без него [697].
Вербейник (Lysimachia ciliata)
Длиннодневное растение, ускоряющее наступление цветения при
дополнительном ночном освещении [497].
Губастик (Mimulus luteus)
Светолюбивое, но теневыносливое растение. Высевается в начале
апреля. Цветет с июня по октябрь. На коротком дне не цветет и поло-
жительно реагирует на дополнительное ночное освещение, удлиняющее
короткий день [496, 497].
Незабудки (Myosotis)
Тенелюбивые растения, предпочитающие полутень. Относятся к группе
длиннодневных растений. Высеваются в марте в парник.
Зимою при дополнительном освещении (с 17 до 22 час.; 160—320 лк)
М. scorpioid.es цветет через 43, а без освещения — через 95 дней [697]. В опы-
тах Саакова [279, 280] с 6- и 8-часовым дополнительным освещением с по-
мощью ламп накаливания (300—500 вт на высоте 1 м) ускорение цветения
М. alpestris равнялось 30 дням. При освещении с 10 января с помощью
ламп накаливания в 300 вт (190 вт/м2) растения цвели через 23 дня
(2 февраля), а в контроле — через 61 день (12 марта) (табл. 111).
344
Частная светокультура растений
Мак сибирский (Papaver nudicaule)
Нейтральное светолюбивое растение. Для роста нуждается в ночной
температуре 7—10°. При посеве весною цветет с июня по сентябрь, при
посеве осенью — с июня по август.
По данным Поста [686, 688|, на дополнительное освещение зимою
(с 7 января) реагирует отрицательно, уменьшая количество цветков в 2—
3 раза и зацветая одновременно с контролем (28 марта—4 апреля).
Мак восточный (Papaver orientale)
Светолюбивое длиннодневное растение. Высевается осенью, цветет
с мая по июль.
В зимнее время цветет лишь при дополнительном ночном освещении
[387].
Флокс (Phlox paniculata)
Светолюбивое нейтральное растение. При дополнительном освещении
зацветает одновременно с контролем [715], но растения развиваются более
мощными.
По данным Портер [677, 678], флоксы при ежедневном дополнительном!
освещении (1000—5000 лк) можно культивировать в комнатах вдали от
окна.
Первоцвет (Primula vulgaris, Р. acaulis)
Длиннодневное тенелюбивое растение. Цветет весной.
При выгонке зимою под лампами накаливания зацветает через 9,
а в контроле — через 14 дней; при освещении листья становятся крупнее
и обладают более зеленой окраской [811].
Роза (Rosa)
Для роста нуждается в ночной температуре 14—18° [688].
Несмотря на то что культурой роз при дополнительном освещении
зимою занимался еще Сименс [754—756], светокультура этого растения
находится в зачаточном состоянии и насчитывает небольшое число работ
[131 140, 263, 314, 376, 379, 387, 492, 523, 547, 579, 700, 859].
Дополнительное освещение применяется главным образом для зимней
выгонки растений. При этом в большинстве случаев наблюдается лучший
рост, большая облиственность, большее число цветков и более раннее
цветение. В опытах Ранке [700] при дополнительном освещении (с 16 до
22 час.; 600 лк) с 10 ноября цветение роз наблюдалось через 37 дней (17—
19 декабря), а без освещения — на 20—30 дней позднее (в начале января).
Клейн [579] нашел, что при дополнительном освещении с 17 января
(8 час.; 2000—3000 лк) цветение начинается через 36—43 дня (22—
29 февраля) — на 9—16 дней раньше контроля (9 марта). Артур и Гутри
[376] испытывали розы Премьер и Гузьер Бюттй и вьющиеся розы и
нашли, что при дополнительном освещении наблюдается более раннее
и обильное цветение по сравнению с контролем. По данным Гербачека
[547], розы зацветают под лампами накаливания в 500 вт через 5 недель.
Фишер [492] установил, что под лампами накаливания в 206 и 750 вт гор-
шечные розы полностью .распускают листья через 4 недели.
В опытах Н. П. Красинского [131, 136] с двумя сортами R. indica
(Ульрих Брюннер фис и Пернецианская) было установлено, что при
IX. Светокультура декоративных растений
345-
Фиг. 156. Комнатная роза (Нова sp.), выращенная
в камере-ннше при люминесцентном освещении
дополнительном освещении в течение 16 часов (с 23 до 15 час.) с по-
мощью ламп накаливания выгонка роз занимает 1/4 месяца; при этом
образуются крупные бутоны с прекрасным ароматом. При освещенности
24 000 лк цветение наступает через 38, а при 5000—46 000 лк — через
42—48, а в контроле — через 58 дней. При освещении со второй половины
октября розы цветут в конце декабря; при освещении с начала декабря
цветение наблюдается в начале февраля, а при освещении с конца ян-
варя — в первой половине марта. Особенно сильный эффект наблюдается
при одновременном удоб-
рении углекислотой (табл.
117). По данным Равича
и Сарычевой [263], допол-
нительное освещение с 24
до 7 час. ежедневно вызы-
вает более мощное разви-
тие роз и образование
большего количества буто-
нов (на 40%). В опытах
Грина и его сотрудников
[521] с розой Бриарклиф
также наблюдалось увели-
чение количества цветков
на 38%, но цветение про-
исходило одновременно с
контролем.
Артур и Гарвилл [387]
успешно культивировали
розы при прерывистом ос-
вещении с помощью ламп
накаливания в 500 вт.
Являясь светолюбивы-
ми растениями, розы ну-
ждаются в сравнительно
большой интенсивности
дополнительного освеще-
ния. Во всяком случае, в
опытах Н. П. Красипского
[136] лучший результат
наблюдался при освеще-
нии с помощью ламп нака-
ливания в 1000 вт и зер-
кального рефлектора, создававших освещенность 24 000 лк. Артур и его
сотрудники [379] также пользовались удельной мощностью 1000 вт/м2
(8200 лк). Однако другие авторы с успехом применяли лампы в 500 вт
и при освещенности 3000—5000 лк получали неплохие результаты [387,
521, 547, 579].
Длина дня для роз, невидимому, должна равняться 17—18 часам,
т. е. необходимо дополнительное освещение в течение 8—10 часов. По дан-
ным Грина и его сотрудников [521], при освещении роз в течение 119 дней
с помощью ламп накаливания в 500 и 1000 вт (3000—5000 лк) на 1 расте-
ние затрачивается 35,5 квт-час.
Комнатные розы хорошо растут в нишах при освещении с помощью
люминесцентных ламп порядка 25 тыс. эрг/см4-сек. В этом случае при
хорошем питании они цветут каждые 20—50 дней (фиг. 156 и 157). Их
546
Частная светокультура растений
можно рекомендовать для культуры на станциях метрополитена и в дру-
гих закрытых помещениях [120, 123].
Фиг. 157. Третье цветение комнатной розы {Rosa
sp.), выращенной при люминесцентном освещении
Рудбекия (Rudbeckia)
Типичные длиннодневные растения, цветущие лишь в условиях длин-
ного дня; на коротком дне находятся в розеточном состоянии. Светолюбы.
Высеваются в апреле, цветут с июля по октябрь.
Rudbeckia sp., посеянная 11 сентября, при дополнительном освещении,
зацветает 12 января, через 123 дня [496]. Аналогичная реакция наблю-
дается у R. lacinata и других видов [397, 502, 715, 716].
Саксифрага (камнеломка) (Saxifraga decipiens)
Длиннодневное светолюбивое растение, могущее развиваться и в тени.
Для культуры зимою S. decipiens var. grandiflora нуждается в допол-
нительном освещении 0,12 г-кал/см2-мин (84 тыс. эрг/см2-сек), удли-
няющем естественный день до 15—17 часов. В этом случае камнеломка
зацветает через 35 дней [646]. При освещении с 29 ноября цветение насту-
пает 2 января. При дополнительном освещении S. decipiens var. bathoni-
ensis образует цветки более крупные и в большем количестве [807].
Очиток (Sedum)
Длиннодневные светолюбивые растения. Положительно реагируют на
дополнительное освещение. S; aizoon, S. telephium, S. selskianum, S. spec-
tabile, S. lividum, S. kamtschaticum и другие виды цветут лишь в условиях
длинного дня, а на коротком дне находятся в розеточном состоянии [370,
497, 807]. A. coeruleum зимою при дополнительном освещении (с 17 до
IX. Светокультура декоративных растений 347
22 час.; 160—320 лк) цветет через 132, а без освещения — через 182 дня
[697]. По данным Алларда [370], оптимальная длина дня для S. tele-
phium равна 16 часам. В этом случае формируются наиболее мощные
растения.
Крестовник (Senecio cruentus)
Светолюбивое короткодневное растение. Высевается в конце апреля,
цветет с июля по октябрь.
При дополнительном ночном освещении, удлиняющем естественный
зимний день, задерживает наступление цветения [383]. Освещение можно
рекомендовать лишь в пределах короткого дня.
Золотень (Solidago)
Длиннодневные и короткодневные растения. S. virgaurea в условиях
короткого дня (8 часов) зацветает через 106 дней, а при дополнительном
освещении (16-часовой день) — через 128—133 дня [496]. A. juncea при
освещении с 15 ноября цветет через 120 дней (15 марта), а без освещения
остается в вегетативном розеточном состоянии [501].
Таволга (Spiraea vanhoutthei)
Цветет с весны до осени. Для перехода к цветению нуждается
в низких температурах (5°). Зимою при дополнительном освещении в
0,06 г-кал/см2-мин; (42 тыс. эрг/см2-сек) цветет в условиях 17-часового
дня через 21 день после начала освещения [646].
Купальница (Trollius hybridus)
Тенелюбивое растение. Цветет в мае. Зимою цветет при дополнитель-
ном освещении при интенсивности радиации 0,12 г-кал/см2-мин
(84 тыс. эрг/см2-сек) и длине дня 17 часов. При освещении с 21 ноября
зацветает 10 января, т. е. через 51 день [646].
Венидиум (Venidium calendulaceum)
Длиннодневное и светолюбивое растение. Высевается в парник в ап-
реле, цветет с июня до осени.
В зимнее время нуждается в дополнительном освещении в ночные
часы. При освещении зацветает на 47 дней раньше, чем в контроле [595].
Для освещения достаточно лампы накаливания в 100 вт. Наблюдается
одинаковый эффект как при утреннем (с 2 до 7 час.), так и вечернем осве-
щении .
Вербена (Verbena hybrida)
Светолюбивое длиннодневное растение. При культуре в зимние ме-
сяцы нуждается в дополнительном ночном освещении. Еще Бэйли [398]
наблюдал при освещении более раннее и богатое цветение. По современ-
ным данным, при освещении зацветает через 195 дней, на 105 дней раньше
контроля [646]. В наших опытах цвела лишь при дополнительном ночном
освещении с помощью ртутных ламп «Игар-2» (300 вт/м2). Цветение на-
ступило через 144 дня. Контрольные растения не цвели совсем. Необхо-
дима освещенность порядка 160—320 лк.
348
Частная светокультура растений
Барвинок (Vinca)
Тенелюбивые нейтральные растения. V. rosea зимою цветет одновре-
менно как при освещении, так и без него, через 195 дней [697].
Портер [677, 678] рекомендует V. elegantissima, V. minor и V. major
для культуры в глубине комнаты. Растения чувствуют себя хорошо,
если получают дополнительное освещение 1000—5000 лк в течение 5 ча-
сов (с 17 час. 30 мин. до 22 час. 30 мин.).
Фиалка (Viola)
Длиннодневные растения. V. cornuta зимою цветет лишь при допол-
нительном освещении (с 17 до 22 час.; 160—320 лк), зацветая через 145
дней [ 6971. V. odorata также положительно реагирует на дополнительное
освещение в ночные часы [598]. V. papilionacea (V. cucullata), находящаяся
в покое при переносе (3 сентября) в теплицу, под лампами накаливания
в условиях 18-часового дня трогается в рост и образует фертильные клей-
стогамные цветки; контрольные растения из периода покоя не выходят
[500]. Для роста нуждаются в ночной температуре 1,5—10° [688].
Смолка (Viscaria viscosa, Lychnis viscaria)
Длиннодневное растение. В зимние месяцы цветет лишь при допол-
нительном освещении (с 17 до 22 час.; 160—320 лк) —через 62 дня [697].
4. ДЕКОРАТИВНО-ЛИСТВЕННЫЕ РАСТЕНИЯ
В группу декоративно-лиственных растений мы выделяем растения,
культивируемые не ради цветков, а ради красивой окраски листьев.
Для выращивания названных растений имеет значение не столько длина
дня, сколько интенсивность и спектральный состав радиации, необхо-
димые для получения нормальных растений.
Ахирантес (Achyranthes brilliantissima)
Нейтральное растение. Зимою реагирует положительно на дополни-
тельное освещение, усиливая рост на 25—50% [579, 594]. Необходима
удельная мощность 100—300 вт/м2.
Колеус (Coleus)
Светолюбивые, но теневыносливые растения, нормально растущие
исключительно при искусственном освещении 20 тыс. эрг/см2-сек.
Хорошо развивается при дополнительном освещении [376, 379,
521, 579]. По данным Роденбурга [717], С. Hehnallianas цветет одновре-
менно как при дополнительном освещении, так и в контроле. Длина дня
не должна превышать 16—17 часов, так как растение сильно страдает при
беспрерывном освещении [290, 379]. Зимою нуждается в дополнительном
освещении в 8000 лк [376, 379].
Различные колеусы хорошо растут в нишах при освещении люминис-
центными лампами при интенсивности физиологической радиации порядка
25 тыс. эрг/см2-сек (фиг. 158). Можно культивировать на станциях
метро [120, 123].
Эхеверия (Echeveria retusa)
Светолюбивое растение. Обычно цветет в феврале. При дополнитель-
ном освещении интенсивностью 0,03 г-кал/см2-мин (21 тыс. эрг/см2-сек)
с декабря зацветает через 3 недели [646]. /
IX. Светокультура декоративных растений.
349
Иресина (Iresine Herbstii)
В опытах Лори и Пеш [595] при дополнительном освещении от лампы
накаливания в 150 вт, висящей на высоте 46 см, в течение 55 дней
(13 февраля — 9 апреля) наблюдалось лишь незначительное увеличение
в высоте растений.
Мята (Mentha)
Длинно дневные, нейтральные и короткодневные растения. М. rotun-
difolia и М. longifolia (М. silvestris) относятся к длиннодневным формам,
ускоряющим цветение при дополнительном освещении [496], М. citrato, —
к короткодневным формам, а М. аг-
vensis, М. piperita и М. spicata яв-
ляются нейтральными растениями [372].
Для задержки или ускорения цвете-
ния необходима освещенность порядка
3000—4000 лк (лампы накаливания
в 200 вт на высоте 30 см, на расстоянии
1 м друг от друга).
Хрустальник (Mesembryanthemum
pyropaeum var. album, M. Jineare)
Короткодневное светолюбивое ра-
стение. Зимою при дополнительном
освещении (с 17 до 22 час.; 160—320 лк)
цветет через 184 дня, а без освеще-
ния — через 119 дней [697].
Паникум (Panicum virgatum)
Орнаментальное короткодневное ра-
стение, цветущее лишь на коротком
дне. Дополнительное ночное освещение
(350 лк) полностью устраняет цветение,
сильно увеличивая вегетативный рост
[405].
Фиг. 158. Колеус (Coleus sp.), выра-
щенный в камере-нише при люминес-
центном освещении
Перилла (Perilla nankinensis)
Короткодневное светолюбивое ра-
стение, совершенно не цветущее при
длине дня более 13 часов. Заложение
цветков начинается при интенсивности
физиологической радиации 25 тыс.
эрг/см 2-сек, но максимальной скорости
достигает лишь при 500 тыс. эрг/см2-сек [89]. При дополнительном осве-
щении полностью устраняется цветение [212, 496].
Кипарисовая трава (Santolina sp.)
В опытах Лори [594] положительно реагировала на дополнительное
освещение (100 вт/м2), усиливая рост на 30%.
350
Частная светокультура растений
5. КЛУБНЕВЫЕ И ЛУКОВИЧНЫЕ РАСТЕНИЯ
Отличительной особенностью луковичных и клубневых растений
является наличие большого количества запасных питательных веществ,
отлагаемых в луковицах и клубнях. Это обусловливает большую тене-
выносливость названных растений и позволяет выгонять их в зимние
месяцы при сравнительно невысокой интенсивности дополнительного
освещения. Кроме того, большинство растений этой группы нуждается
в известный период в низкой температуре для прохождения стадии яро-
визации.
Шафран (Crocus vernus)
Светолюбивое длиннодневное растение. Для зимнего цветения луко-
вицы высаживаются в августе—сентябре. Шафран Уайт Лэди при осве-
щении с 11 февраля зацветает через 5 дней (16 февраля), а без освещения—
на 4 дня позднее (20 февраля); сорт Принцесс Луиз, при освещении с
13 января, цветет через 13 дней (26 января), а в контроле — через 22 дня
(4 февраля [811]).
Для дополнительного освещения необходимы лампы накаливания
в 200 свечей, которые подвешиваются на высоте 20—30 см и включаются
ежедневно на 8 часов (с 22 до 6 час.).
Георгины (Dahlia variabilis)
Светолюбивые растения. Среди многочисленных сортов представ-
лены длинно дневные, нейтральные и коротко дневные формы.
При дополнительном освещении зимою (с 21 до 7 час.; 175 лк) зацве-
тают через 130 дней, без освещения — через 160 дней; освещение увели-
чивает вес растений и количество цветков в несколько раз [598, 678, 806,
857, 858]. При освещении с 28 октября цветение георгин наступает 25 фев-
раля. Клубни лучше образуются в условиях короткого 12-часового дня
[806, 808].
На 1 цветок затрачивается за сезон 2,72 квт-час [858].
Подснежник (Galanthus sp.)
Неприхотливое растение, обычно цветущее ранней весной. При осве-
щении поздней осенью наблюдается более раннее цветение и образуются
более крупные цветки [673].
Шпажник (Gladiolus)
Светолюбивые длиннодневные растения. Для роста нуждаются
в ночной температуре 12—18° [688]. Летом, в зависимости от сорта,
зацветают через 80—111 дней.
Дополнительное освещение в зимние месяцы ускоряет наступление
цветения на 8—10 дней [594, 840, 849], увеличивает количество цветков.
[840] и число цветущих растений [568, 595, 673, 849].
Сказанное относится к сортам Виргиния, Галлей, Ф. Пендлтон,
Америка, Мистрис Ф. Кинг, Кольман, Санбин, Кримсон, Глоу и др.
Сорта Лос Анжелос и Миртл, высаженные в конце октября, цветут лишь
при дополнительном освещении.
Артур и Гарвилл [387, 388] культивировали гладиолусы зимою при
дополнительном прерывистом освещении (460—-510 вт/м2) и наблюдали
IX. Светокультура декоративных растений
351
цветение через 96—164 дня, т. е. на 20—63 дня позднее по сравнению
с растениями, развивающимися летом в грунту. Крупноцветные сорта
зимою цветут только при дополнительном
освещении. При высадке луковиц в сен-
тябре—октябре цветение наступает в ян-
варе—марте. Приведенные данные отно-
сятся к G. Childsii, G. gandavensis, G. le-
moni и G. primulinus, представленными
следующими сортами: А. Э. Амос, Аве
Мария, Бэтти Нутолл, Ч. Диккенс, Дэн-
тинесс, Дугласс, Эдит Робсон, Ф. Э. Бен-
нет, Флэммингсуорд, Джиант Нимф, Голд
Игл, Голден Дрим, Грилей, Галлей, Йергс
Уайт, Лос Анжелос, Менуэт, МаммотУайт,
Пендлтон, Пикарди, Ред Гонтлет, Сеньо-
рита, Уайт Баттерфлай и др.
Для выгонки гладиолусов обычно при-
меняются лампы накаливания в 100—150 вт
из расчета 400—500 вт/м2. Артур и Гар-
вилл [388] создавали освещенность по-
рядка 2000 лк. По данным Джонса [568],
лампы накаливания в 100—300 вт дают
одинаковый эффект. Сысина [305] приме-
няла для зимней выгонки гладиолусов
люминесцентные лампы (с 24 до 7 час.).
При посеве клубнелуковиц 30 сентября
растения цвели в январе—феврале.
Гладиолусы прекрасно растут в нишах
при освещении люминесцентными лампа-
ми (25 тыс. эрг/см2-сек), зацветая через 90
дней после посадки клубнелуковиц (фиг.
159). Их можно рекомендовать для озе-
ленения станций метрополитена.
Гиацинт (Hyacinthus orientalis)
Длиннодневные или нейтральные све-
толюбивые растения. Для заложения цвет-
ков нуждаются в температуре 25°; зало-
жение цветочных почек задерживается
при температуре менее 12 и более 29°
[688]. Цветут рано весной.
Зимою при дополнительном освещении
зацветают через 14—28 дней, а без осве-
щения на 5—И дней позднее [46, 547,
549, 550, 579, 594, 646, 650, 811]. В опы-
Фиг. 159.. Гладиолус (Gladiolus
sp.), выращенный под люминес-
центными лампами в камере-нише
тах Пёш [673] на дополнительное освещение зимою не реагировал.
Оден [646] исследовал влияние дополнительного освещения продол-
жительностью 4, 6, 9, 12 и 24 часа при соответствующем изменении
интенсивности его: 0,16, 0,11, 0,08, 0,05 и 0,02 г-кал/см2«мин; лучший
результат наблюдается при 6-часовом освещении, при интенсивности
0,11 г-кал/см 2-мин (77 тыс. эрг/см2-сек). В другом случае этот автор
[650] рекомендует 10-часовое дополнительное освещение в 250 лк. Бес-
прерывное освещение дает несколько худший эффект. Оден [646] устано-
352
Частная светокультура растений
вил, что при дополнительном освещении гиацинтов следует избегать источ-
ников коротковолновых лучей, подавляющих заложение бутонов.
Артур и Гарвилл 13871 для выгонки гиацинтов Энносанс, Бисмарк,
Гран Мэтр и Куин оф Блуз применяли прерывистое дополнительное осве-
щение (350 вт/м2) и получили хорошие результаты; цветение растений
в этом случае наступало через 14—28 дней.
По данным Портер [677, 678], при 5-часовом вечернем освещении
в 3000—5000 лк гиацинты можно культивировать в комнатах вдали от
окна.
Лилия (Lilium)
Длипнодневные или нейтральные тенелюбивые растения. Для роста
нуждаются в температуре 15—-21° [688].
По светокультуре различных видов лилий — L. candidum, L. tenuijo-
lium, L. longijlorum var. giganteum, L. longijlorum var. eximium (L. Har-
rissii) и L. longijlorum var. jormosum,—имеется ряд работ [131,225,226,
387, 521, 594, 595, 598, 646, 673, 678, 687]. В опытах Лори и Пёш [595,
678] с L. longijlorum var. giganteum было установлено, что при дополни-
тельном освещении (с 18 до 24 час.) с помощью ламп накаливания в 150 вт
цветение растений начинается через 35 дней (25 марта), а без освещения—
через 51 день (10 апреля). В других опытах с L. JIarrirsii, L. longijlorum,
сорт Эрабу и'Б. longijlorum var. giganteum цветение при освещении насту-
пало одновременно с контролем. В опытах Поста [688] с L. longijlorum
var. giganteum ускорение цветения при дополнительном освещении рав-
нялось 17—40 дням и наблюдалось только прп температуре менее 15е.
Оден [646] считает, что для зимней выгонки L. longijlorum var. jormosum
необходимо дополнительное освещение интенсивностью 100 г-кал/см2
в день; в этом случае цветение наступает через 30 дней. По данным Поста
[687], освещение при 1000 лк даст лучший результат, чем при 350 лк.
При прерывистом дополнительном освещении лампою в 500 вт L. Паг-
rissli зацветает через 82, L. longijlorum (Эрабу) — через 70, L. longi-
jlorum var. giganteum — через 86, a L. tenuijolium — через 60 дней после
прорастания; последняя в контроле цветет через 78 дней [387].
Интересно, что для лилий, невидимому, имеет значение не только свет,
но и повышение температуры. Во всяком случае, одно лишь повышение
последней с 12—15° до 22—23° вызывает тот же эффект, что и.освещение
[387, 595]. В этом случае, Л. longijlorum зацветает через 36 дней после
повышения температуры (26 марта), a L. //arrissii и L. longijlorum var.
giganteum образуют наиболее мощные и приземистые растения.
Таким образом, следует считать, что дополнительное освещение лилий
в зимние месяцы может вызвать ускорение цветения па 15—20 дней, при-
чем цветение, в зависимости от сорта, наступает через 30—80 дней. Для
дополнительного освещения рекомендуются лампы накаливания от 150
до 500 вт.
Мускари (Muscari botryoides)
Нейтральное растение. Прп дополнительном освещении с середины
декабря зацветает через 48 дней, одновременно с контролем (19 февраля;
[595]). При зимнем ночном освещении увеличивается количество стрел-
кующихся растений [673].
Нарцисс (Narcissus)
Длиннодневные или нейтральные светолюбивые растения. Зацве-
тают на 2—3-й год.
IX. Светокультура декоративных растений
353
При дополнительном освещении зимою в большинстве случаев уско-
ряют цветение на 6—7 дней, зацветая через 16—32 дня после начала
освещения [225, 226,443, 523, 549, 594, 595, 646,650, 673, 811]. Сказанное
относится к N. poeticus ornatus (Принцепс и др.) и N. pseudonarcissus
(Эмперор и др.). N. tazetta (Пэйпер Уайт и др.) при освещении зацветает
одновременно с контролем, через 30 дней. Желтый нарцисс зимою обычно
цветет через 14, а при дополнительном освещении — через 7 дней [549].
По данным Одена [646], для выгонки N. pseudonarcissus и N. poeticus
ornatus при освещении в январе необходимо 17 дней, причем лучший
эффект наблюдается при 9-часовом освещении и затрате 20 г-кал/см2
в день, т. е. при интенсивности радиации 0,05 г-кал/см 2-мин (35 тыс.
эрг/см2-сек). Беспрерывное освещение дает худший результат.
Орнитогаллум (Ornithogallum lacteum)
Нейтральное растение. На дополнительное ночное освещение не реаги-
рует, зацветая одновременно с контролем [595].
Тюльпаны (Tulipa Gesneriana)
Длиннодневные или нейтральные светолюбивые растения. В зимние
месяцы при дополнительном освещении наблюдается ускорение цветения
растений на 6—8 дней [46, 523, 549, 550, 579, 594, 646, 650, 770, 811].
В опытах Тьеббес и Упгоф [811] сорт Морилло при освещении с 13 января
зацвел через 23 дня (5 февраля), без освещения — через 29 дней (11 фев-
раля). По данным Гиббена [549], тюльпаны, обычно зацветающие через
21—24 дня, при освещении с помощью ламп накаливания (500 вт/м2)
цветут через 14—18 дней. Оден [646, 650] нашел, что Т. Gesneriana под
лампами накаливания цветет через 8—10 дней. Ускорение в 7 дней наблю-
далось в опытах Смита [770]. По данным Лори и Пёш [595], Т. Gesneriana
(сорта Уилльям Копланд и Клара Буфф) на дополнительное освещение не
реагирует, зацветая одновременно с контролем. То же самое установил
Роденбург [717] для сорта Энкель Вроэг и Тинкер [807] для сорта
Уилльям Питт.
По данным Одена [646, 650], выгонка тюльпанов не удается в декабре,
до прохождения периода покоя. В январе они зацветают лишь при допол-
нительном освещении в течение 6—12 часов, при затрате 40 г-кал/см2
в день (т. е. при интенсивности радиации 0,05—0,11 г-кал/см 2-мин или
при освещенности 250 лк). При беспрерывном освещении получается
худший результат, а при 4-часовом освещении растения не зацветают.
Общая затрата энергии за период выгонки равняется 320—400 г-кал/см2.
По данным Портер [677, 678], тюльпаны при дополнительном освеще-
нии (5 часов; 1000—5000 лк) пригодны для культуры в комнатах вдали
от окна.
6. ОРАНЖЕРЕЙНЫЕ И КОМНАТНЫЕ РАСТЕНИЯ
Группа оранжерейных и комнатных растений но однородна. В ней
представлены самые различные биологические формы — однолетники и
многолетники, кустарники и полукустарники, древесные, кактусы,
папоротники и другие. В большинстве своем это вечнозеленые тропиче-
ские и субтропические растения. Это наиболее теневыносливая группа деко-
ративных и цветочных растений. Многие являются нейтральными и обла-
дают способностью цвести зимою или ранней весной.
23 А. Ф. Клешнин
354
Частная светокультура растений
Комнатный клен (Abutilon hybridum)
Вечнозеленое древесное растение. Относится к группе светолюбивых
нейтральных растений. Цветет через 198 дней как при дополнительном
освещении, так и без него [697]. На дополнительное освещение зимою
реагирует усилением роста [579].
Этнонема (Aetliionema grandiflorum)
Длииноднсвпое многолетнее растение. При дополнительном освеще-
нии зацветает через 85, а па коротком дне — через 91 день [496].
Амариллис [Amaryllis (Ilippeastrum) sp.]
Луковичное многолетнее растение. Зимою при дополнительном осве-
щении с помощью ламп накаливания в 500 вт зацветает на 4—6 дней
раньше контроля [547, 579].
Антерикум (Anthericum sp.)
Многолетнее оранжерейное растение с клубнеобразным корневищем.
Рекомендуется для культуры в глубине комнат при дополнительном
освещении в течение 5—10 часов в 1000—5000 лк [577, 678].
Аспарагус (Asparagus)
Нейтральное многолетнее растение. A. Sprengeri в комнатах без допол-
нительного освещения культивируется около светлого окна. В глубине
комнаты можно культивировать лишь при дополнительном вечернем
освещении в 1000—5000 лк в течение 5 часов [595, 677, 678].
В оранжерее Л. Sprengeri, как и Л. plumosus, отвечает на дополни-
тельное освещение с помощью лампы накаливания в 150 вт (на высоте
45 см) усилением ростовых процессов на 12—38% [595].
Бэйлея (Baileya multiradiala)
Зимою цветет через 103 дня лишь в условиях беспрерывного допол-
нительного освещения с помощью люминесцентных и ртутных ламп
в 4э00 лк (табл. 113) [599].
Бегония (Begonia)
Многолетние травянистые растения и полукустарники. Относятся
к нейтральным и короткодневным формам. Тенелюбивы.
Из короткодновных форм следует назвать В. optima. Это растение
зимою при дополнительном освещении (с 17 до 1 часа) цветет менее обильно,
чем в контроле [733]. По данным Поста [688], дополнительное освещение
в 500 лк полностью задерживает заложение цветков у бегоний сорта
Люсилль.
Людвиг [607] давал дополнительное освещение (с 7 октября) растениям
В. hybrida (сорт Лоррэн). При 3-часовом освещении перед восходом солнпа
(120 и 835 лк) бегонии Конкурент и Ф. Паулииг достигали рыночной
годности через 69—71 день; у контрольных растений это состояние дости-
галось через 80—90 дней. В другом опыте этот автор нашел, что лучший
результат для названных сортов наблюдается при освещенности .120 лк
IX. Светокультура декоративных растений
355
и длине дня 12 часов; в этом случае рыночная годность наступает через
58—60 дней. Положительное влияние дополнйтельного освещения на
бегонию Лоррэн отмечает также Фишер [492]. По данным Роденбурга
[518, 722, 723] и Максимова [188] дополнительное освещение вызывает
у бегоний усиление роста.
По данным Одена [647], для выгонки бегоний зимою необходима
затрата энергии по 80 г-кал/см2 в день. Лучший эффект наблюдается
в условиях беспрерывного освещения.
Для дополнительного освещения бегоний достаточна интенсивность
в 160 —200 лк [575, 576, 607].
Артур и Гарвилл [387, 677] нашли, что В. гех и другие виды бегоний
хорошо развиваются при прерывистом дополнительном освещении с по-
мощью лампы накаливания в 500 вт; цветущие формы развиваются зна-
чительно мощнее, образуют более крупные цветы, но зацветают одновре-
менно с контролем.
В комнатах бегонии культивируются в местах, защищенных от пря-
мых солнечных лучей. При дополнительном вечернем освещении в 1000—
5000 лк В. гех и другие прекрасно развиваются в глубине комнат, вдали
от окна [595, 677, 678]. В. metallica, В. lawiana и В. Featsii хорошо растут
в нишах при освещении люминесцентными лампами (25 тыс. эрг/см2-сек
физиологической радиации [120, 123]).
Бегония месячная (Begonia semperflorens)
Длинно дневное и светолюбивое, но теневыносливое растение, нор-
мально растущее и цветущее при 20 тыс. эрг/см2-сек физиологической
радиации. Для роста нуждается в ночной температуре 12—18°, для буто-
низации — не более 21°. Культивируется в цветниках, комнатах и оран-
жереях. Для посадки в грунт (июнь) высевается в ноябре—декабре
в оранжерее.
Литература по светокультуре бегонии месячной насчитывает немного
работ [387, 547, 579, 646, 677, 678, 688, 716, 744]. Бегония хорошо отзы-
вается на дополнительное освещение, усиливая рост и ускоряя цветение
[716, 744]. При дополнительном освещении лампами накаливания расте-
ния обычно бывают в два раза мощнее и обладают более крупными цвет-
ками [579, 677].
Для выгонки бегонии в зимнее время необходимо дополнительное»
освещение в течение 8—15 часов [579, 716, 744]. По данным Одена [646],
лучший результат наблюдается при беспрерывном освещении. Артур
[383, 387] рекомендует прерывистое ночное освещение, в сумме равное
4 часам за ночь.
Что касается необходимой интенсивности радиации, то эта величина
у различных авторов сильно колеблется. Шнейдер и Фогль [744] приме-
няли освещенность в 120—220 лк. В опытах Робертса и Стракмэйера
[716] освещенность равнялась 320—860 лк, а в опытах Гербачека [547] —
8000 лк. Следует принять, что для выгонки бегоний, в зависимости от
времени года, погоды и географической широты, необходима удельная
мощность 300—500 вт/м2 при высоте подвеса ламп 50—100 см.
Но мнению Одена [646], для выгонки бегоний необходимо ежедневное
добавление лучистой энергии порядка 80 г-кал/см2. В опытах Клейна
[579] при получении рассады В. semperjlorens и В. elegans ежедневно
затрачивалось 2,4 квт-час/м2.
По данным Портера и Ли [677, 678], бегония пригодна для культуры
в комнатах вдали от окна. В этом случае растения ежедневно осве-
23*
356
Частная светокультура растений
щаются лампой накаливания (150 вт) в течение о—10 часов (1000—
5000 лк).
В. semperjlorens хорошо растет при нижнем освещении с помощью люми-
несцентных ламп (600 вт/м2) при интенсивности порядка 25 тыс. эрг/см2-сек.
Можно рекомендовать для культуры в клумбах и нишах на станциях
метрополитена.
Дерево жизни (Biota orientalis)
Хорошо растет в нишах, освещаемых люминесцентными лампами при
интенсивности физиологической радиации 25 тыс. эрг/см2-сек (фиг. 160).
Можно рекомендовать для культуры на станциях метрополитена.
Фиг. 160. Растения, находившиеся в камере-нише
при люминесцентном освещении в течение 4 месяцев
1 — биота (Biota orientalis); 2 — пеперомия (Peperomia argyraea)
Бувардия (Bouvardia Humboldtii)
Многолетнее, длиннодневное растение. Для роста и заложения цвет-
ков нуждается в ночной температуре 15—23° [6881. На дополнительное
освещение (300—800 лк) реагирует лишь при температурах порядка
17—18°; при более высокой температуре становится нейтральным [7161.
Броваллия (Browallia sp.)
Нейтральное, светолюбивое растение. Оптимальная температура 15—
21° [688]. Высевается в марте; на постоянное место высаживается в июне.
В зимние месяцы зацветает одновременно (на 174-й день) как при допол-
нительном освещении, так и без него [697]. Дополнительное освещение
имеет смысл лишь для усиления ростовых процессов.
Кактусы (Cactaceae)
Литература по дополнительному освещению и фотопериодизму как-
тусов почти отсутствует. Известно лишь, что Epiphyllum truncatum (Zygo-
caclus truncalus) при температуре 17—18° является короткодневным,
а при 13° становится нейтральным [716]. Это растение для роста нуждается
IX. Светокультура декоративных растений
357
в температуре 12—24°, а для заложения цветков — 12—18° [688]. По дан-
ным Гербачека [547], кактусы при дополнительном освещении лампами
накаливания в 500 и 750 вт прекрасно растут и цветут.
В комнатах кактусы можно культивировать вдали от окна, если
с наступлением сумерек освещать растения в течение 5—10 часов лампой
накаливания, создающей освещенность в 1000—5000 лк [677, 678]. Epi-
phyllum truncation (привитый на Peireskia), Opuntia и другие кактусы
хорошо растут при люминесцентном освещении в нишах при интенсив-
ности физиологической радиации порядка 25 тыс. эрг/см 2-сек.
Башмачки (Calceolaria)
Длинподневные и нейтральные растения. Тенелюбы, страдающие
под влиянием прямых солнечных лучей. Для роста и заложения цветков
нуждаются в ночной температуре 7—18°. С. pinnata — нейтральное рас-
тение [496], а С. hybrida и С. multijlora относятся к группе растений длин-
ного дня. Длиннодневные формы сильно реагируют на дополнительное
освещение зимою. С. hybrida в зимние месяцы находится в розеточном
состоянии, а при дополнительном освещении в 300—500 лк по 4—7 часов
зацветает через 43 дня после начала освещения [441,595, 598, 673, 677, 678].
С. multijlora при дополнительном освещении с 20 декабря по 5 часов
(с 17 до 22 час.) при помощи ламп накаливания в 25—150 вт зацветает
через 55 дней, а без освещения — через 101 день (табл. 87; [596]). В пер-
вом случае цветение наступает 13 февраля, а к 10 марта растения стано-
вятся совершенно готовыми; контрольные растения цветут лишь с 7 ап-
реля.
В опытах Шаминада и Буше [435] дополнительное освещение по
8 часов с 19 ноября лампами накаливания (150 и 350 вт) или неоновыми
лампами ускоряло цветение растений на 40—46 дней; контрольные рас-
тения цвели 17 апреля, а опытные 3—9 марта.
В опытах Артура [383, 387] при прерывистом дополнительном осве-
щении лампой накаливания в 500 вт башмачки зацветали в конце января,
а без освещения — на месяц позднее. При освещении растения были мощ-
нее и образовали значительно больше цветков. Для культуры назван-
ного растения авторы рекомендуют температуру 12—15°; при темпера-
туре 20° растения вытягиваются.
Портер [677, 678] рекомендует башмачки для культуры при допол-
нительном освещении в 10С0—50С0 лк в глубине комнаты вдали от окна.
Камелия (Camelia japonica)
Вечнозеленое дерево. Относится к группе длипподневиых, тенелю-
бивых растений. Несмотря на то что первые опыты по светокультуре каме-
лий были проведены еще Дегереном [467], литература по дополнительному
освещению этого растения почти совершенно отсутствует. В опытах
Функе [496] по дополнительному 8-часовому освещению (30—60 лк)
С. japonica var. Chandleri цветение последней при освещении с 13 октября
началось через 83 дня (4 января;, а без освещения — на 29 дней позднее
(2 февраля).
Колокольчик (Campanula isophylla)
Длиннодневное тенелюбивое растение. Цветет летом.
Сорт Стар еф Бетлеем зимою при дополнительном освещении неоновой
лампой зацветает раньше и образует больше цветков, чем в контроле,
358
Частная светокультура растений
на 61 % [596]. Более обильное цветение под влиянием неоновых ламп
отмечает также Роденбург [720, 725]. В его опытах колокольчик при осве-
Фпг. 161. Ко.юко.1ьч111' (Campa-
nula isophilla maji), выращенный
в камере-нише при люминесцент-
ном освещении
щенпи неоновой лампой со 2 декабря но
8 часов (600 лк) зацвел 1 марта, а в кон-
троле 15 марта. В первом случае на каж-
дом растении было по 25 цветков, а во
втором — по 15.
С. isophyIla maji хорошо растет и цве-
тет в нишах при люминесцентном освеще-
нии при интенсивности физиологической
радиации порядка 25 тыс. эрг/см2-сек
(фиг. 161). Можно рекомендовать для озе-
ленения станций метролитена.
Кассия (Cassia chamaeerista)
Длинно дневный многолетний кустар-
ник. Светолюб, цветет все лето. Зимою
при дополнительном освещении (с 17 до
22 час.; 160—320 лк) зацветает через 82,
а без освещения — через 103 дня [646].
Желтофиоль (Cheiranthus cheiri)
Длиннодневное многолетнее растение.
Зимою можно культивировать при допол-
нительном освещении [134—136, 263].
В этом случае цветение наступает на 7
дней раньше контроля (табл. 105) и зна-
чительно увеличивается количество цвет-
ков. При освещении цветение начинается, в зависимости от состояния
растений, через 5—19, а без осветцения — через 12—26 дней.
По данным II. П. Красинского [136], в умеренной полосе СССР необ-
ходима длительность дополнительного освещения 16 часов (с 23 до 15 час.)
и освещенность 5000 лк.
Хризантема (ромашка) (Chrysanthemum frutescens)
Длиннодневное грунтовое, оранжерейное и комнатное растение, цве-
тущее почти непрерывно. Для роста нуждается в ночной температуре
10—12° [688]. При дополнительном освещении в зимнее время зацветает
через 130 дней от посева, ускоряя цветение на 15—60 дней (табл. 104).
При посеве в середине сентября цветет в январе [595, 596]. При освеще-
нии с начала января цветет в конце марта [686, 688]. Дополнительное осве-
щение увеличивает количество цветков на 18—130% [686]. Лори и Пёш
[596] показали, что при изменении освещенности с 75 до 460 лк эффект
остается неизменным (табл. 104).
Рекомендуется длительность освещения в 5—6 часов, безразлично
в утренние пли вечерние часы.
По данным Поста [688], лучшие результаты получаются при освещении
с декабря по март при температуре 12°.
Хризантема (Chrysanthemum morifolium, Ch. chinensis)
По данным Лори и Киплингер [598], положительно реагирует на допол-
нительное ночное освещение (4—6 часов) от ламп накаливания в 40 вт при
высоте повеса 60 см (25 вт/м2).
/А. Светокультура декоративных растении,
359
Цинерария (Cineraria)
Длиннодневные и нейтральные растения. Для роста нуждаются в ноч-
ной температуре 7—15°, а для заложения цветков — нс более 18° [688].
Цветут зимой и ранней весной, через 7—8 месяцев после посева.
Зимою культивируются при дополнительном освещении [131—140,262,
263, 279, 280, 283, 287, 289, 314, 521,530,547,594,595,596,673,678,
717,720, 723, 725]. Наибольшее число исследований проведено с С. hybrida.
Ночное освещение растений вызывает раннее цветение и образование боль-
шего количества цветков. В опытах Гардера и его сотрудников [530] гото-
вая продукция цинерарий была получена при удобрении СО2 и дополни-
тельном освещении через 50 дней (12 февраля), при освещении без СО2 —
через 57 (19 февраля), а в контроле — через 64 дня (26 февраля). Коли-
чество цветков в первом случае было на 48% больше, чем в контроле
(табл. 116). Равич и Сарычева [262, 263, 287] нашли, что цинерария, осве-
щаемая с начала бутонизации (с 11 января), зацветает через 23 дня (3 фев-
раля), а в контроле — через 39 дней (19 февраля). Особенно сильный эффект
наблюдался при одновременном удобрении углекислотой; в этом случае
количество цветков по сравнению с контролем увеличилось на 45%.
По данным С. Г. Саакова [274, 289], дополнительное 6- и 8-часовое осве-
щение зимою с помощью ламп накаливания в 300—500 вт (при высоте
подвеса 1 м), удлиняющее естественный день до 12 и 14 часов, вызывает
ускорение цветения на 30—54 дня (табл. 111). По сообщению Куфферата
[587], освещение цинерарий с помощью неоновых ламп вызывает ускорение
цветения на 2—3 недели по сравнению с контролем, причем образуются
более крупные цветки.
Светокультура С. hybrida подробно изучена Н. П. Красинским [131—
140]. Этот автор с помощью ламп накаливания в 1000 вт (445 вт/м2) в тече-
ние 125 дней (26 ноября—1 апреля) освещал растения с 7 до 15 или
с 23 до 7 час., одновременно удобряя углекислотой. Лучший результат
наблюдался при освещении в ночные часы и удобрении СО2 (табл. 117).
При освещении растений1 с фазы розетки цветение наступало через 60,
а в контроле — через 120 дней. Освещение с фазы стрелкования вызвало
цветение через 45 дней вместо 75—90 дней в контроле. Соответствующие
сроки при освещении с момента бутонизации — 30 и 60 дней. Н.П. Кра-
синский считает, что при освещении цинерарий со второй половины ок-
тября цветущие растения можно иметь в конце ноября. В других опытах
Н. П. Красинский показал (табл. 105), что при освещении в течение
16 часов (с 23 до 15 час.) для культуры цинерарий необходима освещенность
12 000—16 000 лк, создаваемая при высоте подвеса 50 см с помощью ламп
накаливания в 1000 вт и осветителя «Уииверсаль» или при помощи ламп
накаливания в 500 вт и зеркального осветителя «Электросвет». Лампы
в 300 вт, создающие ^освещенность 5000—8000 лк, значительно менее
эффективны. При освещении растений с периода бутонизации, максимум
цветения под лампами в 500 и 1000 вт наступает одновременно — через
21—24 дня, под лампами в 300 вт — через 32 дня, а в контроле — через
48 дней. При освещении растений в более молодом возрасте (с фазы едва
намечающихся бутонов) максимум цветения соответственно наступает
через 33, 35 и 50 дней.
В общем для С. hybrida, в зависимости от времени года (середина или
конец зимы), необходимо дополнительное освещение 5000—8000 лк
(300 вт/м2, начиная с конца января) или 12 000—16 000 лк (500—1000 вт/м2,
в декабре). Длительность дополнительного освещения 8—10 часов.
1 При одновременном удобрении СО2.
360 Частная светокультура растений
Из других видов испытаны С. cruenta, С. multijlora и С. stellata [595,
678, 720]. Первая при освещении с 29 ноября лампой накаливания в 100 вт
(с 18 до 22 час.) зацветает через 58 дней (26 января), без освещения — через
83 дня (20'февраля), а последние ускоряют цветение на 8—20 дней [595,
673].
В опытах Роденбурга [720] С. multijlora зимою (декабрь—февраль)
цвела лишь при дополнительном освещении с 20 ноября и не цвела в конт-
роле. Этот автор нашел, что С. multijlora папа при дополнительном осве-
щении с 9 ноября неоновыми лампами при освещенности в 500 или 1000 лк
цвела одновременно 5—8 декабря, а в контроле — 30 декабря. Анало-
гичные результаты получены Лори и Пёш [596]. В их опытах цинерария
при освещенности в 550 и 1000 лк также зацвела одновременно, а кон-
трольные растения цвели позднее на 4 недели.
По данным Грина и его сотрудников [521], при освещении (3000—
5000 лк) в течение 60 дней с 21 до 2 или с 16 до 21 час. С. multijlora уско-
ряет цветение на 15 дней, причем на 1 растение затрачивается 4,28—
4,82 квт-час электроэнергии.
Кордилииа (Cordyline sp.)
Рингвальд [712] культивировал зимою при дополнительном освещении
с помощью ламп накаливания в 500 вт (125 вт/м2) по 4 часа в сутки (с 21
до 1 часа).
Кротон (Croton salicifolia)
Хорошо растет в нишах, освещаемых люминесцентными лампами при
интенсивности физиологической радиации в 25 тыс. эрг/см2-сек (фиг. 162).
Можно культивировать па станциях метрополитена.
Криптомерия (Cryptomeria japonica)
Кадочное деревцо. В условиях беспрерывного дополнительного осве-
щения в ночные часы лампами накаливания в 1000 вт растет почти бес-
прерывно, давая годовой прирост 11 см [53].
Цикламен (Cyclamen persicum)
Длинно дневное светолюбивое многолетнее растение. Для роста необ-
ходима ночная температура 12—18° в молодом возрасте и 10—16° во
взрослом состоянии 1688]. В комнатах и оранжереях цветет всю зиму,
с октября по март. Хорошо реагирует на дополнительное освещение, уско-
ряя цветение, увеличивая количество и размеры цветков и листьев и уси-
ливая рост [547, 575, 576, 579, 594, 595, 673, 678, 712].
Клейн [579] применял дополнительное освещение (300 вт/м2) для
выгонки рассады.
В опытах Н. П. Красинского [131] культура цикламена при дополни-
тельном освещении не удалась.
Циперус (Cyperus)
Комнатное и оранжерейное декоративное растение. По данным Лори
[594], не реагирует на 3—4-часовое дополнительное освещение с помощью
ламп накаливания в 150 вт (100—160 вт/м2). С. longus можно культиви-
ровать в глубине комнат, вдали от окна, с помощью дополнительного осве-
щения 1000—5000 лк в течение 5—10 часов [678].
IX. Светокультура декоративных растений
361
Фиг. 162. Растения, находившиеся в камерс-нншс
при люмплссцентлом освещении и течение 5 месяцев
1 — белокрыльник (Calla aci/iiopico.); 2 — кротон (Croton salicifolius)
Фиг. 163. Драцена (Dracaena deremensis), находив-
шаяся в камере-нише под люминесцентными "лампами
в течение 5 месяцев
362
Частная светокультура растений
Ракитник (Cytisus laburnum)
Светолюбивый кустарник. Цветет в конце зимы и ранней весной.
При дополнительном ночном освещении ускоряет цветение и зацветает
зимой [579].
Диффенбахия (Dieffenbachia seguine)
Декоративное теплолюбивое растение, погибающее при температуре
менее 10°.
Рекомендуется для культуры в глубине комнат при дополнительном
освещении 1000—5000 лк в течение 5—10 часов [677, 678].
Драконово дерево (Dracaena)
Тенелюбивое декоративное многолетнее растение. Первые опыты по
дополнительному освещению предприняты еще Дсгереном [467]. Порте})
[677, 678] рекомендует для культуры в глубине комнат при дополнитель-
ном освещении 1000—5000 лк в течение 5—10 часов. D. deremensis и дру-
гие хорошо растут в нишах при люминесцентном освещении при интен-
сивности физиологической радиации порядка 25 тыс. эрг/см2-сек
(фиг. 163). Можно рекомендовать для озеленения станций метрополитена
[120, 123].
Эхиум (Echiuni plantagineuin)
Длинно дневное растение, сильно реагирующее на дополнительное
освещение зимою. При освещении с 17 до 22 час. (160—320 лк) зацветает
через 66, а без освещения — через 112 дней [697].
Вереск (Erica meanthera)
Для роста нуждается в ночной температуре выше 4°. Обычно цветет
в ноябре—декабре. Дополнительное освещение рекомендуется для удли-
нения дня в зимние месяцы [688].
Посконник (Eupalorium coolest]um)
Короткодневное растение. Для роста нуждается в ночной температуре
15—21° [688]. Нормально цветет в начале октября. Дополнительное осве-
щение в ночные часы рекомендуется с 5 августа для передвижения цвете-
ния на более поздние сроки [688].
Молочай (Euphorbia fulgens)
Короткодневное растение. Для роста нуждается в ночной температуре
12—21°, для заложения цветков — от 14 до 18е [688]. Цветет с декабря
по апрель. Дополнительное ночное освещение вызывает усиленный веге-
тативный рост [688].
Пойнсеттия (Euphorbia pulcherrima, Poinsettia pulcherrima)
Короткодневное растение. Для роста нуждается в ночной температуре
12—18°, для заложения цветков — от 14 до 18° [688]. Цветет зимою. До-
полнительное освещение можно рекомендовать лишь в пределах короткого
IX. Светокультура декоративных растений
363
дня (10—12 часов). Ночное освещение в 300—800 лк задерживает наступле-
ние цветения [594, 716]. Такой же результат наблюдается при ночном
прерывистом освещении [383, 387].
Папоротники (Felices)
Тенелюбивые растения. Зимою хорошо развиваются при дополнитель-
ном освещении (например, Adiantum cuneatum, Nephrolepis sp., Pteris
•sp. и другие [523, 712]).
Nephrolepis exallata var. bpstoniensis и Polystichuni lanchytis рекомен-
дуются для культуры в комнатах вдали от окна при дополнительном
освещении с наступлением сумерек в течение 5—10 часов при освещен-
ности 1000—5000 лк [595, 677, 678].
Фикус (Ficus elastica)
Вечнозеленое тенелюбивое растение. В опытах Дегерена [466] на до-
полнительное освещение нс реагировал.
F. elastica и F. pumila (F. repens') можно культивировать в слабо осве-
щенных комнатах, вдали от окна, при дополнительном вечернем освещении
1000—5000 лк в течение 5—10 часов [677, 678].
Фризия (Freesia hybrida)
Луковичное растение. Цветет зимой через 6—7 месяцев после посева
семенами. При дополнительном освещении снижается количество цвет-
ков. Ускорения наступления цветения не наблюдается [595, 673].
Фуксия (Fuchsia hybrida)
Длиннодневное светолюбивое растение. Для роста нуждается в ноч-
ной температуре 15—21° [688]. Дополнительное освещение зимою уско-
ряет наступление цветения [387, 398, 496, 547, 579, 678, 715]. По данным
Клейна [547, 579], цветение фуксий зимою под лампами накаливания
в 500 вт начинается через 10—14 дней после начала освещения. В опытах
Артура [383, 387] фуксия при дополнительном прерывистом освещении
с помощью ламп накаливания в 500 вт развивалась значительно мощнее
и зацветала на 2 недели раньше, чем в контроле. По данным Макси-
мова [188], фуксия в зимние месяцы под лампами накаливания (1000 вт;
с 14 до 10 час.) быстро дает новые побеги, отличающиеся, однако, чрез-
мерной вытянутостью.
Фуксия хорошо растет в нишах при люминесцентном освещении при
интенсивности физиологической радиации порядка 25 тыс. эрг/см2-сек.
В этом случае она цветет беспрерывно в течение нескольких месяцев.
Можно рекомендовать для культуры на станциях метрополитена [120,
123].
Гардения (Gardenia)
Длинноднев-ные светолюбивые вечнозеленые растения. Цветут с мая
по октябрь.
По данным Артура [383, 387], G. jasminoides положительно реаги-
рует на прерывистое дополнительное освещение с помощью ламп накали-
вания в 500 вт, зацветая зимою. Для заложения бутонов нуждается в ноч-
ных температурах порядка 15—18°. При температурах выше 21° —
только вегетативный рост.
364
Частная светокультура растений
G. grandiflora и G. Veitchii также положительно реагируют на допол-
нительное ночное освещение [573, 598]. Дополнительное освещение в 400
и 1250 лк дает одинаковый эффект [573].
Дрок (Genista tinctoria)
Многолетнее полукустарниковое тенелюбивое растение. Реагирует на до-
полнительное освещение зимою усилением роста [594]. По данным Одена
[646], растения, ушедшие в зимний покой, при интенсивном дополнитель-
ном освещении распускают листья, но не цветут.
Портер [677, 678] рекомендует для культуры в глубине комнат при
дополнительном освещении 1000—5000 лк в точение 5—10 часов.
Гербера (Gerbera Jamesonii)
Многолетнее растение, цветущее зимою при дополнительном непре-
рывном освещении с помощью ламп накаливания в 500 вт [387].
Глоксиния (Gloxinia hybrida)
Тепличное клубневое растение. Тенелюб, не выносящий прямых сол-
нечных лучей.
Исследования по дополнительному освещению глоксиний [262, 263,
289, 333, 518, 574, 575, 587, 596, 622, 646, 678, 712, 720, 723, 826] дают
вполне определенные результаты. Глоксинию можно культивировать
зимою, причем дополнительное освещение значительно ускоряет наступле-
ние цветения, увеличивает количество цветков и улучшает качество
растений. Ван дер Веп [826] выгонял глоксинии с помощью люминесцент-
ных ламп. В опытах Рингвальда [712] при освещении лампами накали-
вания в 500 вт (125 вт/м2) по 4 часа в сутки (с 21 до 1 часа) глоксиния
зацвела на 14 дней раньше контроля. По данным Кинда [574], при 4-ча-
совом дополнительном освещении от ламп накаливания в 200 вт глокси-
нии цветут обильнее и на 4 недели раньше контроля. Роденбург [720] ис-
следовал влияние дополнительного освещения от неоновой лампы с 8 ян-
варя по 13 марта при освещенности 500 и 800 лк па развитие глоксинии
Кайзер Фридрих. Он установил, что освещение в 500 лк мало влияет
на растения, а освещение в 800 лк вызывает более обильное цветение
(по 9 цветков вместо 7 в контроле), наступающее значительно раньше,
чем в контроле. Максимум цветения в этом случае наступил через 96 дней
(11 апреля) и продолжался в течение 16 дней, а в контроле растения до-
стигли максимального цветения лишь через 110 дней (25 апреля) и нахо-
дились в этом состоянии 10 дней. Цветение при дополнительном освещении
началось 2 марта (в контроле 28 марта) и окончилось 25 мая, одновременно
с контролем.
Меурман [622] в условиях Финляндии при дополнительном освещении
с помощью неоновой лампы (475 вт; 900 лк; с 18 до 22 и с 6 до 10 час.) куль-
тивировал глоксинию с 4 декабря по 31 марта при температуре 22°. Этот
исследователь установил, что в зимние месяцы дополнительное освещение
увеличивает количество цветков па 26—68%, ускоряет наступление
цветения на 2—3 недели и значительно улучшает качество растений
(табл. 130). По данным этого автора, при посадке клубней в конце ноября
и освещении с начала декабря для глоксиний требуется суммарное допол-
нительное освещение в течение 400 часов. При мощности ламп в 475 вт
это соответствует затрате энергии в 2 квт-часа на 1 растение.
IX. Светокультура декоративных растений
365
Таблица 130
Слияние дополнительного освещения с помощью неоновых ламп при культуре
разных сортов глоксиний
(Меурман [622])
Варианты опыта
Кайзер Фридрих
Неоновая лампа .... 27/1
Контроль ...........
Кайзер Вильгельм
Неоновая лампа . . . .
Контроль ...........
Принц Альберт
Неоновая лампа . . . .
Контроль ...........
17/II
З/Ш
17/111
З/Ш
24/III
17/11 2/III
З/Ш 23/III
Ю/Ш 23/Ш
24/1II —
10/III 30/1П
54
75
89
103
7,2 5,7 112 1—8 30,0 0,0 46,3 30,4 23,7 69,6
11,1 8—16 100,0 0,0 0,0
8,0 1—12 42,9 38,1 19,0
16,6 12—20 46,2 50,0 3,8
9,9 1—12 0,0 40,0 60,0
Лори и Пёш [595] также показали, что при дополнительном освещении
870 лк неоновыми лампами глоксинии зацветают раньше на неделю и
увеличивают количество цветков на 30—37 % .
В опытах Куффората [287, 587] с дополнительным освещением глок-
синий неоновыми лампами (600 вт; 8 часов) в течение 40 ночей, растения
не реагировали на освещение в период опыта, но весной зацвели раньше
и дали больше цветков. М. Б. Равич и А. Н. Сарычева [262, 263] нашли,
что дополнительное освещение вызывает увеличение диаметра розетки
в 2,3, а площади листьев — в 4,3 раза по сравнению с контролем. При
одновременном удобрении СО2 наблюдается еще более сильный эффект;
диаметр розетки в этом случае увеличивается в 3,4 раза, а площадь листьев
в 6,8 раза (табл. 118).
По данным Одена [646], для зимней выгонки глоксиний необходима
ежедневная затрата лучистой энергии в 100—230 г-кал/см2, причем луч-
ший эффект наблюдается при беспрерывном освещении.
Таким образом, для зимней культуры глоксиний следует рекомендо-
вать неоновые лампы в 475 вт, которые следует помещать на высоте 1—
1,25 м. Длительность дополнительного освещения — 8 часов, а освещен-
ность — 800 лк. Клубни необходимо высаживать в ноябре, а освещение
производить сразу после появления побегов и до конца февраля. В этом
случае цветение наступает в феврале и на растение затрачивается около
2 квт-час.
Глоксинии рекомендуются для культуры в глубине комнат при еже-
вечернем дополнительном освещении в 1000—5000 лк в течение 5—10 ча-
сов [677, 678].
Гревиллея (Grevillea robusta)
Вечнозеленый светолюбивый кустарник.
Можно культивировать в глубине комнаты при дополнительном вечер-
нем освещении 1000—5000 лк в течение 5—10 часов [677, 678].
366
Частная светокультура растений
Плющ (Iledera)
Вьющееся вечнозеленое теневыносливое растение. JI. helix var. ати-
rensis и 11. colchica var. arborescens зацветают быстрее в условиях короткого
дня. Дополнительное освещение задерживает наступление цветения на
35—48 дней [497]. II. helix при дополнительном 4-часовом освещении
(110—160 вт/м') усиливает прирост на 50% [594].
Портер [677, 678] рекомендует II. helix для культуры в глубине ком-
нат при вечернем освещении 1000—5000 лк в течение 5—10 часов. Ана-
логичная рекомендация дается Лори
и Пёш [596].
Фиг. 1 (> ।. Китайский розан (// il>ivrus ro-
sa-siпенsis), выращенный в камере-нп-
nie при люминесцентном освещении
Гелиотроп (Helio tropium
peruvianum)
Длиннодневный светолюбивый по-
лукустарник. Сильно реагирует па
дополнительное ночное освещение
ускорением цветения и усилением
роста [496, 579, 594]. При 4-часовом
дополнительном освещении (110—160
вт/м2) рост усиливается на 25% [594],
а при 8-часовом освещении (с 22 до
6 час.: 300 вт/м2) — па 86—100% [579].
Гелксина (Helxine soleirolii)
Тенелюбивое растение. Рекомен-
дуется для культуры в глубине ком-
нат при дополнительном вечернем
освещении в течение 5—10 часов и
освещенности 1000—5000 лк [677.
678].
Розан китайский (Hibiscus
rosa-sinensis)
Древовидный кустарник. Обычно
цветет зимой.
Хорошо растет в нишах при осве-
щении с помощью люминесцентных
ламп (25 тыс. эрг/см2-сек физиологи-
ческой радиации) (фиг. 164). Можно
рекомендовать для культуры в нишах
станций метрополитена (120, 123].
Гортензия (Hydrangea horlensis)
Длиннодневный теневыносливый оранжерейный кустарник. Для роста
нуждается в ночной температуре 12—18°, для заложения цветков — менее
18° [6881. Является излюбленным объектом для зимней выгонки с помощью
дополнительного освещения [131—140, 249, 262, 263, 279, 280, 287. 314,
492, 530, 547, 575, 579, 595, 646, 744].
Гортензия хорошо реагирует на дополнительное освещение, увели-
чивая прирост побегов, количество и диаметр соцветий и ускоряя наступ-
ление цветения. Еще Клейн [547, 579] отметил более раннее цветение гортен-
зий при дополнительном освещении (на 10—14 дней). Затем Людвиг [607]
IX. Светокультура декоративных растений
3G7
показал, что при дополнительном утреннем освещении (с 24 декабря) в тече-
ние 3 часов (850 или 120 лк) готовая продукция у сортов Шотар и Вибраж
получается через 86—88 дней, а в контроле — через 94 дня. В другом
опыте с сортами Шотар и Эльж рыночно готовая продукция при длине
дня 12 и 15 часов и освещенности 120 или 220 лк была получена через 72—
77 дней, а в контроле — через 82—85 дней. По данным Фишера [492],
дополнительное освещение с помощью ламп накаливания в 200
пли 750 вт дает одинаковый эффект — ускорение цветения на 2—3 недели.
В опытах Гардера с сотрудниками [530] с сортами гортензий Мадам
Мульер, Маршал Фош и Шотар было найдено, что первый сорт при до-
полнительном освещении (с 22 октября или 25 ноября) зацветает через
37—44 дня против 72—80 дней в контроле, а два другие цветут лишь при
освещении: Шотар — через 57—67 дней, Маршал Фош — через 73 дня
(табл. 116). Н. П. Красинский [131—140] нашел, что в зимние месяцы
гортензия зацветает при дополнительном освещении и удобрении СО2—
через 75, при одном лишь освещении — через 105, а в контроле — через
135—150 дней (табл. 117). В опытах М. Б. Равича и А. П. Сарычевой [262,
287] при освещении с 13 января полное цветение растений насту-
пило через 47—52 дня (1—5 марта), а в контроле — через 76 дней (19 марта,
табл. 118). По данным С. Г. Саакова [279, 280], дополнительное освещение
от ламп накаливания в 200—500 вт ускоряет наступление цветения на
10—15 дней (табл. 111). Аналогичные результаты получили и другие ав-
торы [249, 595, 744].
В общем при дополнительном освещении гортензии зацветают зимою,
в зависимости от сорта, состояния растений и режима, через 40—100 дней,
ускоряя наступление цветения ио сравнению с контролем на 6—75 дней.
При освещении с начала октября растения зацветают в ноябре—январе.
Обычно в результате дополнительного освещения гортензии разви-
ваются значительно лучше [547,579], образуя значительно более крупные
листья и большее их число (табл. 116 и 118 [136,262, 287,530]). Это, уве-
личивая фотосинтез, приводит к значительному увеличению диаметра
соцветий и количества цветков (табл. 116 [262, 287, 530, 579]). Одновре-
менно наблюдается усиление прироста молодых побегов по длине в 5—8,
а по количеству — в 14—17 раз [530].
По данным Одена [646], для культуры гортензий необходим 17-часовой
день и интенсивность дополнительного освещения 0,12 г-кал/см2-мин
(84 тыс. эрг/см2-сек). В опытах Н. И. Красинского [136] лучший эффект
наблюдался под лампами накаливания в 1000 вт с зеркальной армату-
рой (24 000 лк). В этом случае наблюдалось наиболее раннее цветение
(через 46 дней, 10 марта) и наиболее мощные приземистые растения
(табл. 117). Хороший результат был получен и под лампами в 300 вт
(5000—7000 лк). Здесь при освещении с 23 января растения зацвели через
58 дней (22 марта); цветение контрольных растений было на84-й день
(17 апреля). В некоторых случаях бывает достаточно освещенности
в 120 лк, чтобы получить хороший результат [607].
Иными словами, для культуры гортензий в зимние месяцы необходимо
дополнительное освещение в течение 8—10 часов при удельной мощности
300—500 вт/м2 (5000—8000 лк). Лучший результат наблюдается при ноч-
ном освещении. Дневное освещение дает меньший эффект [131, 136].
Ионопспдиум (lonopsidium acaule)
Нейтральное растение. Зимою как при дополнительном освещении
(с 17 до 22 час.; 160—320 .лк), так и в условиях естественного короткого
дня зацветает одновременно — через 61 день [697].
368
Частная светокультура растений
Каланхое (Kalanchoe)
Светолюбивые короткодневные и нейтральные растения. Начинают
закладывать цветки прп интенсивности физиологической радиации 6 тыс.
эрг/см2-сек; максимальной скорости этот процесс достигает при 12 тыс.
эрг/см2-сек [532]. Для роста нуждаются в ночной температуре 10—21°
[688].
По данным Функе [496], К. laciniata и К. coccinea сильно задержи-
вают цветение при дополнительном освещении, удлиняющем короткий
8-часовой день. Задержка у К, coccinea составляет около 75 дней.
Для устранения цветения у К. Blbssfeldiana достаточно кратковремен-
ного освещения (1—32 минуты) в 1000 лк в середине ночи (через 5—9 ча-
сов после наступления темноты) (фиг. 149 [533]).
Каланхое рекомендуется для культуры в комнатах, вдали от окна,
при дополнительном вечернем освещении 1000—5000 лк в течение 5—
10 часов [596, 6781.
Маранта (Marantha sp.)
Оранжерейное растение. Оптимальная длина дня для ростовых про-
цессов равна 16 часам [703, 704]. Зимою под лампами накаливания
(1000 вт; с 14 до 10 час.) значительно ускоряет образование новых листьев
[188].
Левкой (Matthiola)
Светолюбивые и длиннодневные растения. Для роста нуждаются в ноч-
ной температуре 7—15°, а для заложения цветков — в температуре не более
18° [688].
Литература по зимней культуре левкоя с помощью дополнительного
освещения сравнительно большая [131—140, 153, 249, 262, 268, 279, 280,
287, 301, 387, 496, 521, 547, 558, 579, 595, 596, 598, 646, 686, 688, 697,
715, 716, 857, 858, 859]. Испытаны Cheira.nt.hus annuus (М atthiola annua)
var. anglica, M. bicornis и M. incana.
Дополнительное освещение в ночные часы увеличивает вес растений
на 10—70% (табл. 102 [596, 858, 859, 861]) и количество цветков [153]
и значительно сокращает длину вегетационного периода (табл. 104, 105,
111 и 118). В зависимости от условий и сорта левкоя, ускорение наступ-
ления цветения составляет 5—90 дней.
М. bicornis при дополнительном освещении зацветает через 104, а без
освещения — через 118 дней [697]. Н. П. Красинский [131—140] пока-
зал, что летние сорта при дополнительном освещении зимою удаются
плохо, а осенние сорта зацветают через 1 месяц, ускоряя наступление
цветения на 3 месяца против контроля. Зимние сорта М. incana при начале
освещения с 10 декабря зацветают в январе через 21—37 дней, в зависи-
мости от интенсивности освещения, ускоряя наступление цветения на
59—75 дней [136]. В опытах С. Г. Саакова [279, 280] с зимним левкоем
ускорение цветения при 6-часовом освещении составляло 41—54 дня.
При освещении с 7 января растения цвели через 45—57 дней (19 фев-
раля—6 марта), а в контроле — через 101 день (15 апреля).
По данным Грина и его сотрудников [521, 858], М. incana сорт Джиант
Бисмарк при посеве 17 ноября и начале освещения 18 февраля цветет
через 106—111 дней, а без освещения — через 129 дней; у сорта Бьюти
оф Пайс ускорение составляет 20, а у сорта Перпетуэл Брэнчинг и Лилак
Лавендер — 8—11 дней. При освещении с 15 декабря сорт Руби ускоряет
IX. Светокультура декоративных растений
369
наступление цветения на 22—27, а сорт Болле Уайт — на 18—22 дня [596];
У сорта Лилак Лавендер, посеянного 18 сентября, цветение насту-
пает в-январе—феврале: через 129 дней при освещении и через 158 дней —
в контроле [595]. В опытах Поста [686] ускорение начала цветения рав-
нялось, в зависимости от температуры, 0—14 дням: при температуре
10—12° ускорение было равно 14 дням, а при 15° ускорения не было.
В первом случае растения при освещении с 7 января цвели 28 февраля —
3 марта, а во втором — 28 марта. Контрольные растения зацветали
17—28 марта. По данным Полуяхтова [249], зимний левкой при освеще-
нии лампами накаливания в 300 вт (150 вт/м2) с 13 января по 8 часов
в сутки (с 23 до 7 час.) зацвел через 34 дня (15 февраля); контрольные
растения к этому времени находились в вегетативном состоянии.
Для зимней культуры левкоя применяются обычно лампы накалива-
ния в 100, 200, 300 и 500 вт. Некоторые авторы рекомендуют даже лампы
в 15—40 вт [596, 598, 859]. В свое время Оден [646] заметил, что лампы
накаливания, работающие при перекале, действуют на Cheiranthus annuus
лучше, чем при обычном режиме. Однако позднее было установлено, что
лампы из специального стекла, пропускающего ультрафиолетовые лучи,
не имеют преимуществ перед обычными лампами накаливания [596].
Устранение с помощью светофильтра инфракрасных лучей (Д> 1000 шр)
также не оказывает никакого влияния на культуру левкоя [521].
Как показывают опыты ряда авторов [521, 546, 858], исследовавших
влияние различных ламп накаливания (25—500 вт) на наступление цве-
тения левкоя, для дополнительного освещения достаточна освещенность
15 лк (фиг. 142, />)• Дальнейшее увеличение освещенности практически
не сказывается на времени цветения левкоя (табл. 104). II. П. Красин-
ский [136], изучавший действие различных ламп (300,500 и 1000 вт) и от-
ражателей (обычного «Уииверсаль» и зеркального «Электросвст»), устано-
вил, что для зимнего левкоя в средней полосе СССР в зимние месяцы
необходимо дополнительное освещение порядка 7000 лк (табл. 105). Это
создается с помощью лампы накаливания в 300 вт и зеркального рефлек-
тора «Электросвет» при высоте подвеса 50 см. Снижение освещенности
до 5000 лк приводит к запаздыванию цветения на 9—15 дней. Стельма-
хович и Викулина [301 ] нашли, что при освещении с 26 ноября и 5 января
необходима удельная мощность 300 вт/м2, а при освещении с 4 февраля —
100 вт/м2. При начале освещения в ноябре зимний левкой зацветает через
30 дней (27 декабря), при освещении с начала января — через 40—45 дней
(15 февраля), а при начале опыта в феврале — через 20—25 дней (25 фев-
раля— 1 марта; табл. 110). Аналогичные данные получены С. Г. Саако-
вым [280], в опытах которого М. incana при освещении с 6 января
(300 вт/м2) зацвел через 45 дней (19 февраля: табл. 111).
В зависимости от места 'и времени года длительность ежедневного
дополнительного освещения для левкоя колеблется в пределах 5—10 ча-
сов. Время освещения (утром или вечером) для названной культуры
значения не имеет [596].
По данным Н. II. Красинского [131—140], для выгонки левкоя доста-
точно 30—45 дней. Чем раньше начинается опыт, тем сильнее эффект;
при освещении с 10 декабря ускорение цветения составляет 59—75 дней,
при освещении с 25 января — 24—30 дней, а при освещении с 3 марта
эффекта нет (табл. 105).
При длительности освещения 60—70 дней и освещенности 3000—
5000 лк (лампы накаливания в 500—1000 вт) на 1 растение левкоя
затрачивается 1,86—2,12 квт-час [521]. В случае применения маломощных
ламп (25—150 вт) эти затраты снижаются до 0,12—0,72 квт-час [596].
370
Частная светокультура растений
Комнатный плющ (Mikania scandens)
Длиннодневное растение, цветущее лишь при длине дня 12—1(5 ча-
сов и не цветущее при более коротком или длинном дне [371, 501]. В усло-
виях 16-часового дня при дополнительном освещении лампой накалива-
ния в 200 вт зацветает через 67 дней после начала освещения (на 179-й
день после всходов).
Мимоза (Mimosa pudica)
Многолетнее оранжерейное растение, культивируемое как однолет-
ник. К длине дня относится безразлично. Зимою зацветает одновременно —
через 197—200 дней, как при дополнительном освещении (с 17 до 22 час.;
160—320 лк), так и в контроле [697]. По данным Максимова [188], в зим-
ние месяцы под лампами накаливания (1000 вт) трогается в рост и покры-
вается свежими листьями с нормальной чувствительностью.
Нэгелия (Naegelia cinnabarina)
Нейтральное или короткодневное растение. При дополнительном
освещении (300—800 лк) зацветает одновременно или позднее, чем в кон-
троле [716].
Орхидеи (Orchidaceae)
Тенелюбивые, многолетние травянистые растения. Для роста нуж-
даются в ночной температуре 12—21° [688]. Литература по дополнитель-
ному освещению орхидей очень бедна [547, 575, 579, 641, 678]. Хорошо
реагирует на дополнительное освещение Cattleya Trianae, цветущая
обычно в феврале—марте. При дополнительном освещении в течение
2—3 недель это растение зацветает в январе [579]. Cymbidium insigne
цветет зимою при дополнительном прерывистом освещении от лампы
накаливания в 500 вт [387]. Ускорение в цветении орхидей при дополни-
тельном освещении в 1450 лк отмечают также Портер и Ли [678]. По дан-
ным Кинда [575], орхидеи, культивируемые при дополнительном осве-
щении, зацветают на год раньше, чем в контроле.
По данным Ратцека [701], орхидеи (Laelia, Cattleya, Phalaenopsis),
освещаемые в ночные часы лампами накаливания в 40—75 вт (75—180 лк),
растут значительно быстрее, увеличивая вес на 30—40%.
Кислица (Oxalis sp.)
Короткодневное растение, нуждающееся в прохладных и светлых
помещениях. Не выносит, высоких температур. Зимою под лампами нака-
ливания (1000 вт, с 14 до 10 час.) сильно ускоряет новообразование листьев
1188].
Пальмы (Palmae)
Светолюбивые и тенелюбивые декоративные растения. Последние реко-
мендуются для культуры в глубине комнат (вдали от окна) при допол-
нительном освещении 1000—5000 лк в течение 5—10 часов [677, 678].
Винтовое дерево (Pandanus veitchii)
Декоративное вечнозеленое растение. Тенелюб. Можно рекомендо-
вать для культуры в глубине комнаты, вдали от окна, при дополнительном
освещении 1000—5000 лк в течение 5—-10 часов [596, 677, 678].
IX. Светокультура декоративных растений
Хорошо растет в нишах или прп нижнем освещении люминесцентными
лампами (600 вт/м2) при интенсивности физиологической радиации порядка
25 тыс. эрг/схг-сек (фиг. 165). Можно рекомендовать для культуры на
Фиг. 165. Винтовое дерево (Pandanus veitchii), выращенное
при нижнем освещении люминесцентными лампами (600 вт/м2)
станциях метрополитена в клумбах, нишах пли при декорировании кар-
низов [120, 123].
Пейреския (Pcireskia sp.)
Хорошо растет в нишах или прп нижнем освещении люминесцентными
лампами (600 вт/м2). Можно рекомендовать для культуры в нишах на
станциях метрополитена.
Пеларгония (Pelargonium)
Длиннодневные пли нейтральные многолетние растения. Оптимальная
длина дня для ростовых процессов равна 16 часам [703, 7041. Для роста
нуждаются в ночной температуре 10—15°, для заложения цветков •— более
15—18е [680]. В опытах по дополнительному освещению применялись
вольтова дуга [466, 467], ртутная лампа [470, 692] и лампы накаливания
[547, 579]. По данным Клейна [547, 579], зимою не цветет совсем, а
прп дополнительном освещений зацветает через 10—14 дней.
Современные исследования касаются Р. grandijlorum, Р. zonale и
Р. horlorum. Крупноцветная герань (Р. grandijlorum} — длиннодневное
растение, в зимнее время сильно реагирующее на дополнительное осве-
щение. В опытах Н. П. Красинского [136] при освещении в 8000 лк
с 26 января это растение зацветало через 49 дней (16 марта), а без осве-
щения — через 104 дня (10 мая; табл. 105). Количество побегов в период
цветения равнялось 7 при освещении и 4 — в контроле, а площадь листьев
20 марта была соответственно равна 1878 и 602 см'2. У других авторов
при освещении в течение 74 дней (с 16 до 21 час.; 3000—5000 лк) пеларго-
ния увеличивала количество цветков на 39% и ускоряла цветение на
16 дней [521]. Освещение с 21 до 2 час. давало меньший эффект. По данным
Артура и его сотрудников [379], цветение пеларгонии зимою наблюдается
24*
372
Частная светокультура растений
при дополнительном освещении в 8200 лк, причем растения страдают
в случае беспрерывного освещения. Для выгонки Р. gi'andijlorum, по дан-
ным Одэна [646], необходима
длина дня 17 часов и интенсив-
Фиг. 166. Пеларгония (Pelargonium zonale)
(сорт Метеор), выращенная под люминесцент-
ными лампами н камере-нише
ность дополнительного освеще-
ния 0,3 г-кал/см2-мин (21 тыс.
эрг/см2-сек).
Р. zonale относится к ней-
тральным растениям и при осве-
щении с 13 февраля во 9 апре-
ля увеличивает лишь размеры
цветков, не ускоряя цветения
1594, 595, 596, 720]. Р. hortorum
также не реагирует на дополни-
тельное освещение [673, 688].
Пеларгония рекомендуется
для культуры в глубине комнат
(вдали от окна) пртг вечернем
освещении 1000—5000 лк в те-
чение 5—10 часов [595,677, 678].
Пеларгония Метеор хорошо
растет и обильно цветет в ни-
шах и при нижнем освещении с
помощью люминесцентных ламп
(600 вт/м2) при интенсивности
физиологической радиации по-
рядка 25 тыс. эрг/см2-сек (фиг.
166). Можно культивировать на
станциях метрополитена [120,
123].
Пеперомия (Peperomia)
Комнатные декоративно-лиственные травянистые растения. Р. cor-
date, и Р. Sandersii можно культивировать в глубине комнат при дополни-
тельном освещении 1000—5000 лк в течение 5—10 часов [596, 677, 678].
Р. argyraea, хорошо растет в нишах, освещаемых люминесцентными лам-
пами, при интенсивности физиологической радиации 25 тыс. эрг/см'2-сек.
(фиг. 160).
Филодендрон [Philodendron (Monstera) sp.]
Декоративно-лиственное теневыносливое растение.
Портер [677, 6781 рекомендует филодендрон для культуры в глубине
комнат при дополнительном освещении 1000—5000 лк в течение 5—10 ча-
сов.
Пикерия (Piqueria trinervia)
Короткодневное растение. Для роста нуждается в ночной температуре
4—18° [688]. Дополнительное ночное освещение применяется для задержки
цветения. В этом случае дополнительное освещение следует давать
с 20 сентября [688].
IX. Светокультура декоративных растений
373
Плюмбаго (Plumbago capensis)
Прекрасно развивается и цветет при нижнем освещении люминес-
центными лампами (600 вт/м’2). В условиях 16-часового дня цветет каждые
30—40 дней. Каждое цветение продолжается неделю. Можно рекомен-
довать для декорирования ниш на станциях метрополитена 1120, 123].
Первоцвет (Primula)
Длиннодневные тенелюбивые растения, цветущие почти беспрерывно.
Зимою при дополнительном освещении цветут раньше и дружнее и
образуют больше цветков, отличающихся более крупными размерами
и более яркой окраской [131—140, 279, 283, 287, 547,579, 594, 595, 596,
678, 712, 717, 725].
Как показал Н. П. Красипский [131, 134—136] опытами с Р. obconica,
лучший результат наблюдается при дополнительном освещении с одно-
временным удобрением СО2 (табл. 117). В этом случае растения не усту-
пают весенним, обладая цветками с диаметром 4,5 см (в контроле 3,0—
3,3 см). По данным А. П. Сарычевой [287], при освещении с 27 января
цветение наступает через 23 дня (19 февраля), а к 29 мартана растениях
имеется следующее количество цветков: при освещении и удобрении СО2—
54, при освещении без дополнительного удобрения С09— 33,8, в кон-
троле— 23; соответствующее число кистей на 1 растение равно 4,7;
3,3 и 3,1 (табл. 118). В опытах С. Г. Саакова [279] при освещении в тече
ние 6—8 часов с помощью ламп накаливания в 300—500 вт ускорение
цветения составляло 25—30 дней.
Р. malacoides при дополнительном освещении неоновыми лампами
в 100 вт цветет значительно раньше, чем в контроле [596]. При освеще-
нии с конца ноября она зацветает в январе, причем цветет интенсивнее,
чем без освещения [720]. Однако, по данным Поста [684, 688], Р. mala-
coides относится к короткодневным растениям. При дополнительном осве-
щении при 10—15° она запаздывает с цветением на 10 дней, а при 15—21°
не цветет совсем.
Примулы можно культивировать при прерывистом освещении с по-
мощью ламп накаливания в 500 вт [387].
Для зимней культуры примул пригодны лампы накаливания, неоно-
вые лампы и другие при удельной мощности 450—500 вт/м2 и длительности
освещения 6—8 часов.
Вишня (Pi-unus)
Светолюбивое длиннодневное растение. Для перехода к цветению
нуждается в низких температурах. Р. triloba при дополнительном осве-
щении в условиях длинного дня становится вечнозеленым растением
и зацветает через 30 дней после начала освещения [646]. В опытах
Клейна [579] при освещении с 31 января зацвела через 9 дней (9 февраля),
а без освещения не цвела совсем. Другой влд — Р. japonica (Р. sinen-
sis) — при освещении со 2 февраля зацвел 11 февраля, а в контроле —
на 4 дня позднее (15 февраля).
По данным Одена [646], для выгонки Р. triloba необходима затрата
энергии 15 г-кал/см2 в день. Клейн [579] применял удельную мощность
300 вт/м'3, освещая растения но 8 часов в день (с 22 до 6 час.).
374
Частная светокультура растений
Рейнекня (Reincckia сагпеа)
Хорошо растет в нишах при осве:
при интенсивности физиологической
энип люминесцентными лампами
радиации в 25 тыс. эрг/см--сек
(фиг. 167). Можно культивиро-
вать па станциях метрополи-
тена.
Рододендрон (Азалия)
(Rododendrou, Azalea)
Тенелюбивые длпннодневные
вечнозеленые растения. Для ро-
ста нуждаются в ночной темпе-
ратуре 12—26°, для заложения
цветков 21—32°, а для их по-
следующего развития 4—12°
[688]. Дополнительное освеще-
ние обычно усиливает рост [646,
648]. Rh. carolinianum, Rh. ma-
ximum и Rh. macronulatum при
дополнительном освещении в
250—300 лк, увеличивающем
длину дня с 13 до 18 часов, уси-
ливают по сравнению с контро-
лем интенсивность роста на
6—39 %при температуре 10—12°
tr.7 ,, ... . х и на 40—8096 при 24—27°, но
Фиг. ]()/. 1 еииекия {Reineckia earned), выра- 1
щепная в камере-иише при люминесцентном 1 !1,ll,no задерживают цветение
освещении [763]. Rh. ponticum и Rh. roseum
elegans наиболее интенсивно ра-
стут в условиях беспрерывного освещения (фиг. 146, 8 и .9; [763]).
Дополнительное освещение зимою ускоряет цветение и увеличивает
количество и размеры цветков у Rh. hybridum, Rh. indicum, Rh. japoni-
cum и Rh. obtusum [466, 496, 523, 526, 579, 646, 648, 763, 770, 800]. Rh.
hybridum без освещения цветет в конце марта—начале апреля, а при
дополнительном освещении интенсивностью 0,03 г-кал/см2-мин.
(21 тыс. эрг/см2-сек) в условиях 15-часового дня зацветает в начале
февраля [646, 648]. 13 опытах Клейна [547, 579] цветение азалий при до-
полнительном освещении (с 22 до 6 час.; 1000—3000 лк) началось на 10 дней
ранее, чем в контроле. По данным Смита [770], дополнительное освещение
вызывает ускорение цветения на 14 дней. <Ьунке [496] нашел, что Rh.
indicum (сорт Мадам Петрик) при 8-часовом дополнительном освещении
в 30—60 лк зацветает через 55 дней, а без освещения — через 91 день;
в первом случае, при освещении с 13 октября, цветение начинается
7 декабря, а во втором — лишь 12 января, па 36 дней позднее.
В опытах Скиннера [763] с Rh. japonicum, Rh. obtusum растения росли
и цвели значительно интенсивнее в условиях 18-часового дня при допол-
нительном освещении в 250—300 лк, чем в контроле в условиях корот-
кого дня.
По данным Поста [688], дополнительное освещение не влияет на зало-
жение цветков, ио ускоряет их формирование.
При дополнительном освещении азалий необходимо тщательно избе-
гать перегрева растений.
IX. Светокультура декоративных растений
375
Узамбарская фиалка (Saintpaulia ionantha)
Комнатное и оранжерейное растение. Цветет через 6 месяцев после
посева. Можно культивировать в глубине комнат, вдали от окна, если
ежедневно освещать по 5—10 часов при интенсивности 1000—5000 лк-
1596, 677, 678].
Сансевьера (Sanseviera zeylanica)
Вечнозеленое комнатное растение. При
дополнительном освещении 1000—5000 лк
в течение 5—10 часов культивируется в
глубине комнат вдали от окна 1596].
Крестовник (Senccio articuiatus)
Тенелюбивое растение. Хорошо растет
в нишах при освещении люминесцентными
лампами при интенсивности физиологиче-
ской радиации порядка 25 тыс. эрг/см2-сек
(фиг. 168). Можно рекомендовать для куль-
туры на станциях метрополитена.
Паслен (Solanum)
Декоративно-ягодные растения. S.pseu-
docapsicum. зимою можно культивировать
при дополнительном прерывистом освеще-
нии лампами накаливания в 500 вт |383,
387].
Solanum Hendersonii хорошо разви-
вается и цветет при нижнем освещении лю-
минесцентными лампами (600 вт/м2; фиг.
169). Цветение наступает через 49 дней
и продолжается почти беспрерывно в те-
чение нескольких месяцев. Можно куль-
тивировать на клумбах на станциях мет-
рополитена.
Фиг. 1(>S. IxpocToBiHii; {Seneeio нг-
lit'u la tn s} r выращенный в камере-
нише при люминесцентном осве-
щении
Сирень (Syringa sp.)
Кустарник с опадающими листьями. Дополнительное освещение зимою
применяется для нарушения покоя. По данным Скока [764], при освеще-
нии в феврале лампами накаливания в 2000 вт при температуре 25° в усло-
виях 16-часового дня распускает листья через 8 дней. Как показал Клейн
[579], белая сирень при дополнительном освещении (300 вт/м2; с 22 до
5 час.) зацветает через 16 дней после тепловой ванны (2 февраля), а без
освещения— через 25 дней (11 февраля). В первом случае листья отли-
чаются более зеленой окраской, а раскрытие цветков более равномерно.
Сорт Юлиус Шпэйт при освещении цветет через 14 дней после ванны
(2 февраля), а в контроле — через 24 дня (12 февраля).
По данным Гооля и Нориндера [523], для выгонки сирени достаточно
освещенности 150—300 лк, получаемой от ламп накаливания в 100 вт.
Оден [646, 650] считает, что для цветения S. vulgaris в декабре—январе
376
Частная светокультура растений
Фиг. 169. .Декоративный соляиум (Solatium Hendersonii), выращенный
при нижнем освещении люминесцентными лампами (600 вт/м2)
необходима длительность дополнительного освещения 10 часов и осве-
щенность 500 лк. В этом случае цветение наступает через 20—22 дня после
тепловой ванны или воздействия ацетиленом и затрачивается
30 г-кал/см2 в день.
Тисс (Taxus baccata)
Вечнозеленый кустарник. По данным В. 3. Гулисашвили [33], в усло-
виях беспрерывного освещения при дополнительном освещении лампами
накаливания в 1000 вт растет медленнее, чем в контроле.
Тунбергия (Thunbergia alata)
Светолюбивое нейтральное вьющееся растение. Зимою цветет одно-
временно (через 83 дня) как при дополнительном освещении, так п без
пего [697].
Торения (Torenia Fournierii)
Однолетние длиннодневные или нейтральные растения. Первые зимою
при дополнительном освещении с 17 до 22 час. (160—320 лк) зацветают
через 164, а без освещения —через 184 дня; вторые в обоих случаях цветут
через 147 дней [697].
Традесканция (Tradescantia)
Короткодневные или нейтральные растения. Т. undata при дополни-
тельном освещении зацветает позднее, чем в контроле [496], а 7'. viridis
(Т. jluminensis') на дополнительное освещение не реагирует совсе.м [492].
Т. viridis рекомендуется для культуры в глубине комнат при дополнитель-
ном освещении 1000—5000 лк в течение 5—10 часов [677, 678]. Zebrina
pendula (Tradescantia zebrina), Tradescanlia virginica и Spironema (Tra-
descantia) jragrans прекрасно растут при нижнем освещении люминес-
центными лампами (600 вт/м2; фиг. 170 и 171). Можно рекомендовать
для культуры на станциях метрополитена: Tr. zebrina — в качестве
ампельного растения для декорирования карнизов, a Tr. virginica и
Spironema jragrans — для культуры на клумбах или в нишах.
IX. Светокультура декоративных растений
377
Фиг. 170. Традесканция {Tradescanlia lir^inica), выращенная при пни.нем
освещении люминесцентными лампами (600 вт/м2)
Фиг. 171. Традесканция {Tradescanlia zebrina),
выращенная при нижнем освещении .помииесщчп
ними лампами ((>00 вт;м2)
378
Частная светокультура растений
Калина (Viburnum opulus)
Светолюбивый кустарник с опадающими листьями. Дополнительное
освещение применяется для нарушения зимнего покоя. При темпера-
туре 21° в условиях дополнительного освещения, удлиняющего короткий
день до 16 часов, полное распускание листьев наступает через 10 дней [764].
По данным Одена [646], калина при 14-часовом дополнительном осве-
щении становится вечнозеленой и цветет зимою.
Белокрыльник (Zantcdeschia aethyopica, Galla aethyopica)
Светолюбивое нейтральное растение. Для роста нуждается в темпера-
туре 15—21° [688]. Зимою цветет одновременно и одинаково как при
освещении, так и без него [595, 673].
Хорошо растет в нишах при люминесцентном освещении при интен-
сивности физиологической радиации порядка 25 тыс. эрг/см2-сек (фиг. 162).
7. ДИКОРАСТУЩИЕ РАСТЕНИЯ
Хотя дикорастущая и лекарственная флора не имеет прямого отно-
шения к декоративному цветоводству, мы рассмотрим вопрос о ее свето-
культуре в настоящей главе. Целесообразность этого определяется, во-
первых, однородностью задач, а во-вторых, отсутствием четких границ
между дикорастущими растениями и декоративными цветочными куль-
турами.
Кроме того, следует иметь в виду, что литература по светокультуре
дикорастущих растений сравнительно невелика, и нет необходимости
рассмотрение этого вопроса выделять в виде специальной главы.
Щирица (Amaranthus retrollexus)
Короткодневпое растение. Зимою нормально растет при интенсивности
дополнительного освещения 300—800 лк и температуре 21°. В условиях
длинного дня вегетирует, па коротком дне цветет [715].
Амброзия (Ambrosia artemisil'olia)
Зимою хорошо культивируется при 6-часовом дополнительном осве-
щении в 8000 лк, особенно при удобрении атмосферы углекислотой [379].
Ветреница (Anemone)
Многолетнее растение. A. hepatica зимою цветет при дополнительном
освещении в 0,02—0,06 г-кал/см2-мип (14—42 тыс. эрг/см2-сек), a A. Sil-
vestris нормально развивается и бутонизирует при 0,08—0,18 г-кал/см2-мин
(56—126 тыс. эрг/см2-сек) [646].
Пупавка (Anthemis cotula)
Однолетнее длиннодневное растение. Зимою цветет лишь при допол-
нительном освещении в 300—800 лк [715, 716].
IX. Светокультура декоративных растении 379
Икотник (Berteroa incana)
Длиннодисвиое растение, в условиях дополнительного освещения
цветущее через 1'16 диен, а па коротком дне — через 131 день [496].
Марь (Blitum capitalism, Chenopodium capitatum)
Длинподневное растение. Зимою цветет при дополнительном осве-
щении (с 17 до 22 час.; 160—320 лк) через 93 дня, а без освещения —
через 171 день [697].
Черноголовка (Brunella vulgaris)
Многолетнее теневыносливое и длиниодневнос растение, зацветающее
раньше при дополнительном освещении [496].
Киирейпик (Epilobium adenocaulon)
Длппнодневное растение. Зимой цветет лишь при дополнительном
освещении (с 17 до 22 час.; 160—320 лк) через 150 дней [697].
Герань (Geranium silvaticum)
В условиях 17-часового дня при дополнительном освещении в
0,05 г-кал/см2-мип (35 тыс. эрг/см2-сек) зимою (январь) зацветает через
3 недели после начала освещения [646].
Цмип (Helichrysum rupestre, Gnaphalium lanceolatum)
Длинподневное растение. При дополнительном ночном освещении
ускоряет начало цветения [496].
Пазник (IJypochoeris radiculala)
*
Длппнодневное растение, ускоряющее наступление цветения при
дополнительном освещении [496].
Клоповник (Lepidium draba)
Длиннодневное растение, цветущее при дополнительном освещении и
нецветущее в условиях короткого дня [496].
Зюзник (Lycopus europaeus)
Многолетнее длиннодневное растение. При дополнительном освеще-
нии зацветает через 78 дней (2о июля), а в условиях короткого дня не
цветет совсем [496].
Просвирник (Malva verticil lata)
Однолетнее короткодпевное растение. При осеннем посеве цветет
в следующем году в июле—августе.
При дополнительном ночном освещении зимою не цветет [716]. Допол-
нительное освещение можно рекомендовать лишь в пределах короткого
дня. Задержка цветения наблюдается при освещенности 320—880 лк.
380
Частная светокультура растений
Муленбергия (Muhlenbergia)
М. mexicana относится к нейтральным растениям и цветет одновре-
менно как при дополнительном освещении, так и в контроле. М. Schre-
berii является длинно дневным- растением и в условиях 16 — 18-часового
дня при дополнительном освещении в 3000—4000 лк зацветает на
37—68 дней раньше, чем в контроле [373].
Гулявник (Nasturtium silvestre)
Длиннодневное растение. Цветет лишь при дополнительном освещении
в условиях длинного дня и но цветет на коротком дне [496].
Песлея (Neslea paniculata)
Длишюдновнос растение, раньше зацветающее при дополнительном
освещении, а затем в условиях короткого дня [496].
Фацелия (Phacelia Parryi)
Длиннодневное растение. При дополнительном освещении ртутными
или люминесцентными лампами (4800 лк) раньше зацветает при беспре-
рывном освещении, а в условиях короткого дня (12 часов) запаздывает
с цветением более чем на 70 дней (табл. 113 [599]).
Тростник (Saccharum spontaneum)
По данным Алларда [371], обильно цветет в условиях длинного дня
при освещении лампами накаливания в 200 вт при высоте подвеса 30 см.
В опытах Сарториуса [739] цветение наблюдалось лишь в условиях 13-
часового дня. В тех же условиях развивались наиболее мощные растения.
Норичник (Scrophularia nodosa)
Многолетнее тенелюбивое длиннодневное растение, ускоряющее цве-
тение при дополнительном освещении [497].
Паслен (Solanum’ rostralum)
Короткодневное растение, зимою в условиях естественного короткого
дня цветет через 106, а при дополнительном освещении (с 17 до 22 час.;
160—320 лк)—через 157 дней [697].
Ярутка (Thlaspi arvense)
Однолетнее нейтральное растение, не реагирующее на дополнительное
освещение (300—800 лк) и цветущее лишь при температуре не выше 13е
[716].
Вяжечка (Turritis glabra)
Растение длинного дня. При дополнительном освещении цветет через
54 дня, а в условиях короткого дня — более чем на Ю ;ineii позднее [496].
Дурнишник (Xantium)
Растения короткого дня, пе цветущие или сильно задерживающие
цветение при дополнительном освещении. X. pensylranicum, и X. echina-
IX. Светокультура декоративных растений
381
1ит зимою цветут лишь без дополнительного освещения [637, 716]. A. com-
mune в зимние месяцы при дополнительном освещении (с 17 до 22 час.;
160—320 лк) цветет через 152, а в контроле — через 53 дня [697].
8. ЛЕКАРСТВЕННЫЕ РАСТЕНИЯ
Горицвет (Черногорка) (Adonis vernalis)
Корневищное многолетнее растение, зацветающее в конце апреля и
цветущее по начало июня.
Для культуры зимою нуждается в длине дня 15—17 часов и интенсив-
ности радиации более 0,2 г-кал/см2-мин (14 тыс. эрг/см2-сек); при мень-
шей интенсивности растения хотя и бутонизируют, но не цветут и отли-
чаются чрезмерным вытягиванием стебля [646].
Чистотел (Chelidonium majus)
Тенелюбивое многолетнее растение. Зимою наблюдается мощное веге-
тативное развитие в условиях беспрерывного дня при дополнительном
освещении интенсивностью 0,005—0,012 г-кал/см2-мин (3500—
8400 эрг/см2-сек) [646]. По данным Н. А. Максимова [184], превосходно
развивается зимою под лампами накаливания (1000 вт, с 14 до 10 час.),
без всяких признаков вытягивания междоузлий.
Марь (Chenopodium anthelminticum)
Однолетнее растение, цветущее в июле—августе. Относится к коротко-
дневным растениям, не цветет или запаздывает с цветением в условиях
беспрерывного освещения (удельная мощность 500 вт/м2) и цветет через
53—70 дней в условиях 12-часового дня [330].
Дурман (Datura stramonium)
Однолетнее травянистое растение, короткодневное или нейтральное.
Зимою в условиях естественного короткого дня цветет и плодоносит,
а при дополнительном освещении (300—800 лк) при температуре 21°
дает сильный вегетативный рост [715, 716].
Желтушник (Erysimum cheiranthoides)
Нейтральное растение. На дополнительное освещение, удлиняющее
короткий день, ускорением цветения не реагирует [496].
Белена (Hyoseyamus niger)
Длиннодневное двухлетнее растение, цветущее лишь при длине дня
более 12 часов. Оптимальная длина дня 16 часов. Для перехода к цветению
в условиях короткого дня достаточно кратковременного ночного осве-
щения (1 час) при освещенности 10—30 лк; оптимум — при 500 лк [664].
Яснотка (глухая крапива) (Lamium album)
Многолетнее тенелюбивое растение. Зимою хорошо развивается в усло-
виях 17-часового дня при дополнительном освещении в 0,2 г-кал/см2-мин
(140 тыс. эрг/см2«сек); цветет через 79 дней после начала освещения [646],
382
Частная светокультура растений
Маршанция (Marchantia polyniorpha)
Тенелюбивое растение. Хорошо культивируется при дополнительном
освещении лампами накаливания в 200 вт. В условиях 18—20-часового
дня при дополнительном освещении в 1000 лк растения увеличивают
сырой и сухой вес и площадь таллома более чем в два раза по сравнению
с контролем [583]. Органы вегетативного размножения у маршанций
лучше развиваются па длинном дне, т. е. при дополнительном освещении
[583]. Архогонип образуются лишь на длинном дне и не образуются
в условиях короткого дня, а антеридии — как на длинном, так и на ко-
ротком дне, но более интенсивно в первом случае [742].
Для создания длинного дня в зимние месяцы необходима интенсивность
дополнительного освещения порядка 0,005 г-кал/см2-мин. (3500 эрг/см2- сек).
Базилик (Ocimum basilicum)
Многолетнее кустарниковое длппнодневное растение. В условиях
естественного летнего дня или при дополнительном освещении цветет
через 94 дня, на коротком дне — через 109 дней [496].
Подорожник (Plantago lanceolata)
Многолетнее длинподневное растение, цветущее лишь в условиях
длинного дня при дополнительном освещении, по крайней мерс, в течение
10 дней [781].
Дубровник (Tcucrium chamaedrys)
Многолетнее кустарниковое длиннодневное растение, ускоряющее
наступление цветения при дополнительном освещении [497].
Жгучая крапива (Urtica urens)
Многолетнее тенелюбивое растение. Прекрасно развивается в зимние
месяцы под лампами накаливания (1000 вт, с 14 до 10 час.), без всяких
признаков вытягивания междоузлий [188].
9. КОМНАТНОЕ ^ЦВЕТОВОДСТВО
Литература о дополнительном освещении в комнатном цветоводстве
очень бедна [436, 492, 596, 677, 678]. Известно, что нормальное естествен-
ное освещение растений в комнатах может быть обеспечено лишь на окнах,
расположенных на южной стороне. Па окнах, расположенных па восток
или запад, возможна культура лишь некоторых растении из числа тенелю-
бивых или теневыносливых, а на северных окнах и в глубине комнат
можно культивировать сравнительно немногие растения, отличающиеся
исключительной теневыносливостью. Г. Е. Киселев [102] приводит таб-
лицу изменения интенсивности радиации в глубине комнаты по мере уда-
ления от окна. Мы привели ее в виде кривой на фиг. 90. На осп абсцисс
здесь отложено расстояние от окна (в метрах), а на оси ординат — интенсив-
ность радиации, выраженная в процентах от интенсивности естественной
солнечной радиации за окном. Из графика видно, что при отсутствии
попадания прямых солнечных лучей интенсивность радиации даже на
окне не превышает половины естественной солнечной радиации, а в глубине
комнаты равняется лишь 7,6%. Измерения показывают, что в лучшем
случае, в полуденные часы в солнечные дни, освещенность в глубине
комнат не превышает 700 лк, в среднем колеблясь в пределах 30—300 лк
[677, 678, 681].
IX. Светокультура декоративных растений
383
Таблица 131
Растения, пригодные для комнатной культуры вдали от окна при дополнительном
осве/цении лампами накаливания в 100—200 вт
(1000—5000 лк) в течение 5—10 часов в сутки (с 17 час. 30 мин. до 22 час. 30 мин.)
(Портер и Ли [078])
А. Цветочные
Б. Д e к о p а т и в и о - л и с т в e и и ы e
1. Львиный зев (Antirrhinum sp.)
2. Бегония королевская (Begonia rex)
3. Бегония месячная (Begonia semper-
jlorens)
4. Кактусы (Cactacea)
о. Башмачки (Calceolaria sp.)
6. Ноготки (Calendula officinalis)
7. Цинерария (Cineraria sp.)
8. Цикламен (Cyclamen persicum)
9. Дрок (Genista sp.)
10. Глоксиния (Gloxinia sp.)
11. Гиацинт (Hyacynthus sp.)
12. Пеларгония (Pelargonium sp.)
13. Флокс (Phlox sp.)
14. Узамбарская фиалка (Saintpaulia
ionantha)
15. Шалфей (Sal via sp.)
16. Тюльпан (Tulipa sp.)
17. Барвинок (Vinca elegantissima)
18. Барвинок (Vinca ma for)
19. Барвинок (Vinca minor)
20. Белокрыльник (Zantedeschia aethio-
pica)
1. Антерпкум (Anthericum Ullage)
2. Аспарагус (Asparagus Sprengeri)
3. Аспидистра (Aspidistra lurida')
4. Толстянка (Crassula arborescens)
5. Циперус (Cyperus longus)
6. Диффенбахия (Dieffenbachia seguine)
7. Драцена (Dracaena sp.)
8. Эхеверия (Echeveria sp.)
9. Фикус (Ficus elastica)
10. Фикус (Ficus pumila)
11. Грэвпллея (Grecillea robusta)
12. Плющ (Hedera helix)
13. Гельксина (Tlelxine soleirolii)
14. Каланхое (Kalanchoii sp.)
15. Папоротник (Nephrolepis exaltata lo-
stoniensis)
16. Пальмы (Papnae)
17. Панданус (Pandanus Veitchii)
18. Пеперомия (Peperomia cordata)
19. Пеперомия (Peperomia Sandersii)
20. Филодендрон (Philodendron sp.)
21. Папоротник (Polystichum lonchllis)
22. Сансевьера (Sanseviera zeylanica)
23. Очиток (Sedum sp.)
24. Традесканция (Tradescantia viridis,
T. fluminensis)
13 свете вышеизложенного становится ясным, что культура растений
на северных окнах и в глубине комнат возможна лишь при дополнительном
освещении. Фишер [492] с помощью лампы накаливания в 200 вт пытался
культивировать гортензию (Hydrangea hortensis), бегонию (Begonia hyb-
rida) Глуар де Лоррэн, бальзамин (Impatiens sultani), настурцию (Тго-
paeolum sp.), циперус (Cyperus alternijolius), перец (Piper nigrum) и
годецию (Godetia hybrida) в глубине комнаты, вдали от окна, на расстоя-
нии 50 см от лампы. В его опытах растения страдали и быстро погибли.
Успешные результаты получили Портер [677, 678, 681] и Лори л Пёш
'[596]. Этим авторам удалось подобрать ряд цветочных и декоративных
растений, которые способны произрастать и цвести в глубине комнат,
вдали от окна, при искусственном освещении в течение 5—10 часов. Список
этих растений мы приводим в табл. 131. Большинство цветочных, приве-
денных в таблице, в названных условиях цвели. Лишь ноготки, цинера-
рия и цикламен оставались в вегетативном состоянии.
Не подложит сомнению, что дальнейшие исследования в этом направ-
лении позволят создать значительно лучшие условия и значительно
384
Частная светокультура растений
расширить список растений, которые можно культивировать как в глу-
бине комнат, так и на северных окнах. Широкие возможности открывают,
в частности, люминесцентные лампы. Однако и лампам накаливания здесь
принадлежит не последняя роль. Они незаменимы, в частности, для подо-
грева оранжереек, освещения и подогрева аквариумов и т. д.
10. ОЗЕЛЕНЕНИЕ СТАНЦИЙ МЕТРОПОЛИТЕНА
Специфическую задачу культуры цветочных и декоративных растений
с помощью искусственных источников освещения представляет собой
проблема озеленения станций метрополитена и других закрытых поме-
щений. Здесь, помимо общих трудностей выращивания растений в отсут-
ствие естественного света, приходится преодолевать ряд затруднений тех-
нического порядка, связанных с определенными архитектурными и эсте-
тическими требованиями. Выращиваемые растения должны быть нормаль-
ными, без всяких признаков этиоляции, вытягивания и пожелтения,
должны нормально цвести. Совершенно очевидно, что они должны пол-
ностью гармонировать со всем архитектурным ансамблем, составляя
с ним единое целое и выражая определенную архитектурную и художе-
ственную мысль. Следовательно, спектральный состав излучения должен
удовлетворять как физиологическим, так и эстетическим требованиям,
а источник света не должен бить в глаза и слепить публику своей чрез-
мерной яркостью. Отсюда необходимость в специальном расположении
и оформлении осветительной арматуры и ламп. Это особенно необходимо
для источников излучения с плохой цветностью, например для ртутных
ламп с их мертвенным светом или натриевых трубок, дающих неприятный
желтый цвет. Оба типа ламп вообще предпочтительно включать в такие
часы, когда в помещении отсутствует публика. Источники освещения
должны отличаться постоянным режимом горения и но должны требовать
беспрерывного наблюдения.
До последнего времени проблема озеленения метрополитена пред-
ставляла собою мечту, и лишь за последние несколько лет она встала на
вполне реальную физиологическую и техническую почву. Впервые мысль
об озеленении метрополитена была высказана акад. Максимовым в речи,
произнесенной по радио в 1945 г. В последнее время эта проблема посте-
пенно привлекает внимание общественности [17, 58, 122, 123, 234, 331,
332].
Проблема декоративного растениеводства с помощью исключительно
искусственных источников радиации ставится впервые, и сейчас трудно
предполагать конкретные пути, которыми опа будет решена. Можно лишь
предположительно наметить некоторые общие вопросы и указать возмож-
ные путл их решения.
В принципе, для озеленения метрополитена пригодны любые источники
радиации, однако по ряду причин от некоторых приходится отказаться.
Из-за непостоянства режима и необходимости беспрерывного наблюдения
совсем непригодны вольтовы дуги. Мало пригодны лампы накаливания.
Под ними, как было отмечено выше, происходит вытягивание растений,
что, конечно, не может являться положительным качеством растений,
выращиваемых для декоративных целей. Ртутные и неоновые лампы из-за
неблагоприятной цветности могут иметь лишь ограниченное применение.
Наиболее пригодными следует признать люминесцентные лампы. Именно
на них мы и сосредоточим свое внимание.
В условиях метрополитена исключительное значение имеет вопрос
о размещении источников света. Следует ограничить применение верхнего
IX. Светокультура декоративных растений
385
освещения, а остановиться на нижнем или боковом. Источники света
должны быть совершенно спрятаны от зрителей. Верхний свет, из-за боль-
шой яркости источников, потребной для выращивания растений, может
употребляться лишь в короткие ночные часы, когда отсутствует публика.
Озеленение метрополитена может быть произведено тремя различными
способами: 1) подбором теневыносливых растений, растущих без специаль-
ного дополнительного освещения; 2) при помощи дополнительного осве-
щения высокой интенсивности в ночной период (5—6 часов) и содержания
растений в дневные часы при обычном для станций освещении и 3) посред-
ством специального беспрерывного освещения.
В первом случае растения выращиваются в естественных условиях
или в оранжерее, а затем готовыми переносятся на станции метрополитена
и находятся там при обычном освещении до тех пор, пока не начнут жел-
теть и терять листья. Для восстановления нормального облика они снова
переносятся в оранжерею или в естественные условия. Совершенно оче-
видно, что этот прием может быть применим лишь к ограниченному числу
исключительно устойчивых к затенению растений. По данным Лори
и Пёш 1596], некоторые растения могут находиться в нормальном состоя-
нии от 3 до 8 месяцев совершенно без естественного освещения, исключи-
тельно под лампами накаливания в 40 или 150 вт, включенными на 17 ча-
сов в сутки (табл. 132). Несомненно, что с помощью специальных
Таблица 132
Результаты выращивания растений в условиях низкой освещенности
от ламп накаливания в 40 или 150 вт при 17-часовом дне
(Лори и Пёш [596])
Растения Длительность освещения (в месяцах) Качество растений
Аукуба {Aucuba japonica) 8 Отличное
Толстянка {Crassula arborescens) 8 »
Драцена {Dracaena iragrans massangeana) 8 »
Панданус {Pandanus Veilchii) 8 »
Сансевьера {Sanseviera zeylanica) 8 »
Портулакария {Porltdacaria aira) 6 »
Плющ {Iledera helix) 6 Хорошее
Алое {Aloe denliculala) 4 »
Толстянка {Crassula lycopoides) 4 »
Толстянка {Crassula pscudolycopoiodes) 4 »
Толстянка {Crassula tetragona) . . 4 »
Диффенбахия {Difjenbachia picla, D. brasillensis) . . 4 »
Фикус {Ficus elaslica) 4 »
Пеперомия {Peperomia cordala) 4 »
Папоротник {Nephrolepis exaltala bosloniensis) .... э Удовлетвори-
тельное
Антерикум {Anthericum liliastrum) 4 То же
Комнатный виноград {Cissus rhombi folia) 4 » »
Драцена {Dracaena tenninalis) 3 )> »
Аспарагус {Asparagus Sprengeri) 2 Плохое
Циртомпум {Cyrtomium falcatum) 1 »
Камнеломка {Saxifraga sarmentosa) 1 »
25 А. Ф. Клешнин
I
386
Частная светокультура растений
исследований список, приведенный этими авторами, можно значительно
увеличить, особенно за счет луковичных растений.
/7 В В
Фиг. 172. Светильник для горшечных культур на станциях метрополитена
А — дневное наложение (лампы выключены); К — положение ночью
(лампы включены); В — вид в плане
Значительно более перспективным следует признать второй способ —
метод дополнительного освещения. В этом случае растения также выра-
щиваются в оранжерее, а затем переносятся на станции метрополитена
и в ночные часы (5—6 часов) получают дополнительное освещение высо-
Фиг. 173. Схема верхнего н нижнего освещения клумб,
расположенных около стен па станциях метрополитена
А — ночное положение (все лампы включены); Б—положение днем
(включены лишь нижние лампы)
IX. Светокультура декоративных растений
387
кой интенсивности с помощью ртутных или люминесцентных ламп или
ламп накаливания. Несомненно, что в этом случае может быть подобран
широкий ассортимент растений и могут быть осуществлены самые разно-
образные приемы оформления. Способы применения люминесцентных
ламп в этом случае изображены на фиг. 172 и 173. Для горшечных культур
рекомендуется специальный светильник, представленный на фиг. 172.
Фиг. 174. Схема освещения люминесцентными лампами растений
в клумбах или газонах на станциях гметроиолптена
А — вид в плане; Б — вид сбоку
На день он опускается на вазон, а ночью включается и поднимается на
уровень растений. Для клумб, расположенных около стен, следует реко-
мендовать комбинированный нижний и верхний свет (фиг. 173). Нижние
лампы могут гореть беспрерывно, а верхние — только в ночные часы:
днем они поднимаются и прикрепляются к стене (на рисунке — справа).
Большой интерес представляет третий способ — выращивание пли
поддержание нормального состояния растений исключительно при помощи
искусственного освещения [120, 123]. В этом случае могут быть осуще-
ствлены самые разнообразные варианты, но исключительно с помощью
нижнего и бокового освещения, источники которого скрыты от зри-
телей с помощью специальных отражателей. На фиг. 174 представлена
схема клумбы или газона, освещаемого с помощью люминесцентных
25*
388
Частная светокультура растений
ламп. Лампы располагаются в междурядьях растений, почти непосред-
ственно на земле или даже при некотором углублении в почву и освещают
растения снизу. На фиг. 175 приведена схема озеленения пьедестала
с помощью газонных стелющихся или вьющихся растений, освещаемых
люминесцентными лампами, дающими нижний свет. От зрителей лампы
спрятаны специальными козырьками. Этот
устройства ложного окна, декорированного
Фиг.
же принцип применим для
растениями, которое приве-
дено на фиг. 176. На фиг.
177 мы показываем схему
выращивания растений в
нише, а на фиг. 178 — при-
мер декорирования зеле-
ными растениями колонн.
Число возможных схем
можно было бы еще увели-
чить, однако и приведен-
ных примеров достаточно,
чтобы показать полную
возможность озеленения
метрополитена, не приме-
няя дополнительного ко-
личества ламп, а лишь
соответственным образом
располагая те, которые
имеются на станциях. Спе-
цифическая трудность воз-
никает в четырех послед-
них случаях (фиг. 175—
178) в связи со стремлени-
ем растений располагать
свои листья в направлении
к источнику света, со-
вершенно оголяя ветви и
стебли со стороны публи-
ки. Для устранения это-
„ го недостатка необходимо
полагая
или дополнительно приме-
175. Схема декорирования пьедесталов пять
на станциях метрополитена
1 ники
чей (ртутные лампы), рас-
их в точках желаемого направления листьев и побегов, или
специальные источ-
сине-фиолетовых лу-
пользоваться в качестве основного источника красными люминесцент-
ными лампами \ совершенно не имеющими коротковолновых ультра-
фиолетовых и сине-фиолетовых лучей, или подбирать такие растения,
которые не теряли бы своих декоративных качеств от определенного
расположения листьев и органов в пространстве, например, аспарагус
(Asparagus Sprengeri, A. plumosus) и др. Как бы то ни было, эту труд-
ность можно без особого труда преодолеть.
Нет необходимости говорить, что с помощью последнего приема —
выращивания растений исключительно при беспрерывном искусствен-
ном освещении — на станциях метрополитена можно зимою полностью
воспроизвести летние клумбы и цветники. Выбор растений здесь
Красные лучи, как известно, не вызывают фототропического изгиба.
IX. Светокультура декоративных растений
389
значительно разнообразнее и богаче, чем в первых двух случаях,
когда приходится пользоваться исключительно теневыносливыми расте-
ниями. Для озеленения 1 м2 достаточно при этом 300—450 вт, т. е.
10—15 люминесцентных ламп, если пользоваться трубками мощностью
в 30 вт.
В связи с тем, что совершенно отсутствует литература по культуре
растений с декоративными целями
внутри помещений, куда не прони-
кает естественный солнечный свет,
в 1951 г. мы начали специальное
исследование в этом направлении.
В качестве объектов служили по-
левица {Agrostis stolonijera), тимо-
феевка {Phleum pratense), райграсе
{Lolium регеппе), гвоздика {Dian-
thus Нeddewigii), львиный зев {An-
tirrhinum majus папит), нигелла
{Nigella damascena), лобелия {Lo-
belia erinus), анютины глазки {Vio-
la tricolor), бархатцы {Tagetes pa-
tula), плюмбаго {Plumbago capen-
sis), фуксия {Fuchsia hybrida), ки-
тайский розан {Jlibiscus rosa-sinen-
sis), ay куба {Aucuba japonica), гла-
диолус {Gladiolus sp.), драцена
{Dracaena sp.), колумнея {Kolum-
nea kewensis), панданус {Pandanus
veitchii), эпифиллум {Epiphyllum
hibridum), филлокактус {Phylloca-
ctus hibridus), традесканции {Zeb-
rina pendula, Tradescantia virgi-
nica, Spironema jragrans), пей-
реския {Peireskia sp.), солянум
{Solanum Hendersonii), лилия {Li-
lium tigrinum), амариллис {Ama-
ryllis hybrida), кливия {Clivia sp.),
пеларгония {Pelargonium zonale),
перец черный {Piper nigrum), гор-
тензия {Hydrangea hortensis), бего-
нии {Begonia rex, B. semperjlorens),
ахирантес {Achyranthes sp.), дуб
{Quercus sp.), лимон {Citrus limon),
ель {Picea sp.), лиственница {Larix
sp.) и другие. Первые восемь
Фиг. 176. Схема создания на станциях
метрополитена ложного окна, декориро-
ванного растениями
видов высевались семенами, а ос-
тальные высаживались в виде отводков, луковиц или черенков. Расте-
ния выращивались с помощью люминесцентных ламп, расположенных
в междурядьях растений непосредственно на уровне почвы. Лампы нахо-
дились на расстоянии 2,5 см друг от друга (600 вт/м2), повышая темпе-
ратуру окружающего воздуха на 2—3° выше температуры окружающей
среды (телшература в камере колебалась от 20 до 25°). Растения росли
исключительно при освещении, создаваемом с помощью люминесцентных
ламп белого света, получая свет не сверху, как в естественных условиях,
а снизу (фиг. 174). Опыт производился с 1 января по 10 июня 1951 г.
390
Частная светокультура растений
Исследования показали, что при нижнем освещении могут расти все
перечисленные растения, однако для декоративных целей годятся лишь
некоторые из них. Для создания .клумб и газонов пригодны тимофеевка,
райграс, полевица, лобелия, гвоздика, герань, ахирантес, бегония месяч-
ная и другие. Особенно хороша лобелия, которая, почти полностью
закрывая лампы, зацветает через 50 дней после посева и беспрерывно
цветет в течение нескольких месяцев. Беспрерывное цветение наблюдается
также у бегонии {Begonia semper flor ens). Гвоздика цветет через 60 дней;
после срезки она снова дает побеги. Нигелла цветет в возрасте 60 дней,
плюмбаго цветет каждые 40 дней, а пеларгония
Метеор — каждые 70 дней. Тимофеевка, рай- /ИГх
Фиг. 17/. Схема выращивания растений
в нише на станциях метрополитена
А — вид сбоку; Б — вид в плане
Фиг. 17<S. Схема де-
ки]) и рования вьющимися
растениями колонн
грасс и полевица нуждаются в стрижке через каждые 30—35 дней.
Очень красивые и мощные растения образуют бархатцы.
Исключительно эффектны также панданус (фиг. 165) и спиронема
{Spironema {Tradescentia) fragrans). На фоне других растений можно
культивировать также ель, дуб и другие древесные, которые в условиях
беспрерывного освещения становятся вечнозелеными.
Таким образом, для газонов в условиях станций метрополитена можно
рекомендовать смеси злаков (райграсе, полевица, тимофеевка и другие)
с лобелией. На фоне газонов в качестве второго яруса можно создать раз-
личные сочетания из бархатцев, пандануса, бегонии месячной, гвоз-
дики, ахирантеса, солянума (фиг. 169), пеларгонии, дуба, ели, спиро-
немы {Spironema fragrans), традесканции {Tradescantia virginica, фиг. 170)
и других.
Совсем непригодны для названных целей бегония королевская {Bego-
nia rex), львинный зев и другие широколиственные культуры, листья
которых под влиянием нижнего освещения имеют тенденцию сильно
закручиваться (фиг. 179). Это происходит в силу стремления листьев
располагаться так, чтобы верхняя сторона листовой пластинки рас-
полагалась перпендикулярно направлению лучей.
IX. Светокультура декоративных растений
391
В силу тех же причин при нижнем освещении нельзя культивировать
китайский розан (Hibiscus rosa-sinensis) (фиг. 179), фуксию и другие.
Плюмбаго (Plumbago capeusis) в этом случае растет не в высь, а в стороны,
располагая свои побеги в горизонтальном направлении. Все перечислен-
ные растения, так же как и гладиолусы, пеларгонию, солянум, панданус,
спироиему, пейрсскию (фиг. 179) и другие, следует рекомендовать для
культуры в нишах (фиг. 180).
Фиг. '79. Растения, выращенные при нижнем освеще-
нии люминесцентными лампами (600 вт/м2). Рено
видно опускание и скручивание листьев
1 — пейреския (Peireskia sp.); 2 — роза китайская (Hibiscus
rosa-sim.-7>.sis); 3—яблоня (Malus sp.)
Для декорирования карнизов по схеме, представленной на фиг. 173
(без верхнего освещения), пригодны панданус, традесканция (Tr. rir-
ginica'), амариллис и другие. В качестве ампельного растения при нижнем
освещении очень эффектна для декорирования карнизов зебрина (Zeb-
rina penduld) (фиг. 171).
В 1952 г. опыты, преследующие цель озеленения станций метрополитена,
были продолжены. С этой целью была оборудована небольшая камера-
ниша (80X70X70 см), освещаемая люминесцентными лампами белого
света мощностью 15 вт. 11 ламп располагались в виде горизонтальной
решетки под потолком на расстоянии 70 см от пола, 10 ламп находились
сбоку (по 5 с каждой стороны) в виде вертикальной решетки с вертикаль-
ным расположением ламп и 5 ламп — у задней стенки также в верти-
кальной плоскости, но с горизонтальным расположением ламп. При
общей удельной мощности 700 вт/м2 в камере создавалась интенсивность
физиологической радиации в 25 тыс. эрг/см2-сек (около 7000 лк).
Все лампы защищались специальными козырьками (фиг. 180) и были
совершенно не видны. Объектами являлись бегония (Begonia semperjlo-
392
Частная светокультура растений
reus, В. melallica, В. lawiana, В. Feastii), комнатная роза (фиг. 156 и 157),
китайская роза (Hibiscus rosa-sinensis\ фиг. 164), колеус (Coleus &р.-, фиг.
158), глоксиния (Gloxinia sp.), колокольчик (Campanula тауг. фиг. 161),
пеларгония Метеор (Pelargonium zonale, фиг. 166), ель (Picea sp.), лист-
венница (Larix sp.), ипомея (Ipomoea sp.), бархатцы (Tagetes patula-,
фиг. 155), гладиолус (Gladiolus sp.; фиг. 159), кипарис (Cupressus sp.),
Фиг. 180. Общий вид камеры-ниши, освещаемой
люминесцентными лампами
крестовник (Senecio articulatin', фиг 168), рейнекия (Reineckia сагпеа:
фиг. 167), кротон (Croton salicijolius\ фиг. 162), драцена (Dracaena dere-
mensis-, фиг. 163), пальма (Trachycarpus Fortunei), биота (Biota orienta-
lis\ фиг. 160). пеперомия (Peperomia argyraea.-, фиг. 160), белокрыльник
(Calla aethiopica, фиг. 162), фуксия (Fuchsia hybrida), панданус (Pandanus
oeitchii), кактус (Opuntia sp.), узамбарская фиалка (Saintpaulia ionantha)
и другие.
Опыт, проводившийся с 25 ноября по 1 июля, показал, что все перечис-
ленные растения прекрасно чувствуют себя в камерах-нишах при выра-
щивании целиком на люминесцентном освещении, нормально растут,
нормально цветут и выглядят весьма декоративно. Бегония (Begonia
semper] lor ens), фуксия, узамбарская фиалка и колокольчик цветут бес-
прерывно в течение нескольких месяцев; нормально цветут бархатцы,
рейпекия, бегонии (Begonia lawiana и В. Feastii)-, пеларгония зацветает
через 133 дня после черенкования и цветет каждые 50—60 дней; комнат-
IX. Светокультура декоративных растений
393
ная роза начинает цвести через 42 дня и цветет каждые 20—50 дней
(6—26 ноября, 14—31 марта и 20 апреля—5 мая), гладиолус зацветает
через 90 дней и цветет в течение 10 дней (5—15 марта). Очень эффектно
выглядят различные формы колеуса. образующие сильно ветвящиеся
растения с ярко окрашенными листьями; у зеленых пестролистных форм
развивается ярко выраженная пестролистность.
Одновременно было выяснено, что изолома (1 soloma hirsutum}, аню-
тины глазки (Viola tricolor}, синнипгия (Sinningia sp.), астры (Calliste-
phus chinensis} и некоторые другие растения в условиях наших опытов
(при смешанном верхнем и боковом освещении) или растут очень медленно
(1 soloma}, или обнаруживают сильные признаки этиоляции (Calhste-
phus}. В специальных опытах, проведенных (• лампами, расположенными
в горизонтальной плоскости непосредственно над растениями (на высоте
10 см), было показано, что все перечисленные растения при одном верх-
нем освещении растут нормально. В этом случае для астр необходима
интенсивность физиологической радиации порядка (20—40 тыс. эрг/см2-сек,
для изоломы и синнингии — около 25 тыс эрг/см2-сен. Попутно было
установлено, что бегония месячная (Begonia semperflorens) лучше всего
развивается и быстрее зацветает (через 70 дней после черенкования) при
интенсивности 23 тыс. эрг/см2-сек, колокольчик (Campanula mayi} —
при 25 тыс. эрг/см2-сек, а колеус наиболее мощно растет при интенсив-
ности порядка 18 тыс. эрг/см2-сек. Астры, перенесенные в период буто-
низации в камеру-нишу, нормально зацветают, находятся в цветущем
состоянии в течение 15—20 дней и при люминесцентном освещении выгля-
дят значительно эффектнее, чем в естественных условиях.
Эти предварительные исследования, находящиеся в самом начале,
показывают полную возможность озеленения станций метрополитена
с помощью современных люминесцентных ламп. Вопрос заключается
лишь в подборе растений, разработке их агротехники и размещении
ламп.
11. выводы
Таким образом, подводя итоги литературным данным по дополнитель-
ному освещению цветочных (декоративных) растений, следует заметить,
что этот прием можно применять как для выгонки рассады ранней вес-
ной, так и для культуры растений в зимнее время.
Хорошо разработан вопрос о культуре цветочных растений зимою.
Литература по этому вопросу представлена сравнительно большим чис-
лом работ, большинство которых принадлежат советским, скандинавским,
голландским, немецким и американским исследователям. При этом бро-
сается в глаза своеобразное разделение труда: исследования советских
и североевропейских авторов касаются главным образом тенелюбивых
или теневыносливых оранжерейных и луковичных растений, а исследо-
вания североамериканских авторов — светолюбивых однолетних и мно-
голетних растений цветников. Сказанное находится в связи с резкими
климатическими различиями между СССР и северной Европой, с одной
стороны, и Северной Америкой — с другой.
В этой же связи находятся и различия между интенсивностями допол-
нительного освещения, которые рекомендуют европейские и амери-
канские авторы. Последние обычно применяют освещенности порядка
300—600 лк, так как в дневные часы растения находятся в условиях
сравнительно высокой естественной освещенности. Подобные величины,
достаточные для удлинения дня с целью вызвать определенный фото-
394 Частная светокультура растений
периодический эффект, совершенно недостаточны для культуры цветоч-
ных растений в зимние месяцы в условиях умеренной полосы СССР и
стран, расположенных в северной Европе.
Поэтому лишь на юге СССР при культуре цветочных растений в зим-
ние месяцы можно ограничиться интенсивностью дополнительного осве-
щения в 2000—6000 эрг/см2-сек (300—1000 лк), а в умеренной полосе сле-
дует рекомендовать интенсивность порядка 30—60 тыс. эрг/см2-сек
(5000—10 000 лк).
При высоте подвеса ламп 50 см и употреблении зеркальных глубоко-
излучателей в первом случае необходима удельная мощность 25—75 вт/м2,
а во втором — 200—500 вт/м2. В случае применения обычных эмалирован-
ных широкоизлучателей эти величины возрастают до 40—100 вт/м2 и
300—750 вт/м2.
Опыт показывает, что длительность дополнительного освещения в зим-
ние месяцы должна равняться 8—10 часам и постепенно, по мере удлине-
ния естественного дня, снижаться с приближением весны. При этом сле-
дует исходить из прочно установленного факта, что для большинства рас-
тений оптимальным является 16—17-часовоп день. Одновременно сле-
дует помнить, что дополнительное освещение короткодневных растений
возможно лишь в пределах короткого 10—12-часового дня. Освещение
в ночные часы в данном случае можно применять лишь в том случае,
если необходимо задержать цветение и получить мощный вегетативный
рост.
Что касается выбора источника радиации, то в настоящее время пра-
ктически можно говорить лишь о лампах накаливания (особенно зеркаль-
ных), хотя для культуры декоративных и цветочных растений они наи-
менее пригодны, вызывая перегрев и вытягивание растений. С физиологи-
ческой точки зрения наиболее пригодными следует признать неоновые
и люминесцентные лампы, позволяющие выращивать мощные, призе-
мистые растения.
Нет необходимости подробно останавливаться на том, что для успеш-
ного выращивания цветочных растений необходимо строго поддерживать
необходимую для каждой культуры температуру и тщательно избегать
перегрева растений, одновременно создавая оптимальный режим в смысле
подбора почвы и удобрения, полива и подкормки углекислотой.
В случае соблюдения всех перечисленных условий с помощью допол-
нительного ночного или дневного освещения в зимние месяцы можно
вырастить и заставить цвести любое цветочное растение.
Почти совершенно отсутствует литература по применению дополни-
тельного освещения для получения рассады декоративных растений.
Па практике этот метод применяется чрезвычайно редко.
Г л а г. а X
СВЕТОКУЛЬТУРА ПОЛЕВЫХ РАСТЕНИИ
(Светокультура в селекции и семеноводстве)
Светокультура сельскохозяйственных половых растений представ-
ляет большой интерес для селекции и семеноводства. Позволяя выращи-
вать растения зимою в условиях оранжерей, этот прием дает возможность
вести круглогодичную селекцию и быстрое размножение ценных сортов.
При этом следует иметь в виду, что для селекции пригодны лишь те
источники радиации, спектр которых максимально приближается к спектру
естественных солнечных лучей. В противном случае растения, выращи-
ваемые в условиях светокультуры, будут резко отличаться по своим внеш-
ним и внутренним признакам от растений, выращиваемых в естествен-
ных полевых условиях. Это обстоятельство не только затрудняет отбор,
но может значительно исказить этот процесс. Наиболее совершенными и
пригодными источниками • следует . поэтому признать люминесцентные
лампы дневного света «ДС-ЗО» или «ДС-15». От применения других источни-
ков следует воздержаться. К сожалению, большинство работ проведено
с лампами накаливания, поэтому в дальнейшем нам придется рассматри-
вать материал, полученный почти исключительно с названным типом
ламп и имеющий значение лишь для биологической характеристики рас-
тений (светолюбив, длина дня и др.). Для целей семеноводства, напро-
тив, наиболее пригодны источники радиации, богатые оранжево-крас-
ными лучами: неоновые лампы, лампы накаливания и люминесцентные
лампы красного света, под влиянием которых растения быстро про-
ходят световую стадию и дают максимальый урожай плодов и семян.
В настоящей главе мы рассмотрим растения полевых культур —
зерновые, бобовые, травы, технические культуры, имея в виду возмож-
ности селекции и семеноводства перечисленных растений в зимнее время
в оранжереях. Заметим, кстати, что мы не будем касаться вопроса о тем-
пературном режиме растений, предполагая, что читатель знаком с тео-
рией стадийного развития и знает, что озимые и двухлетние растения для
прохождения стадии яровизации нуждаются в низких температурах.
1. ЗЕРНОВЫЕ
Гречиха (Fagopyrum esculentum)
Нейтральные и короткодневные формы. Оптимальная длина дня для
ростовых процессов — 17 часов [667]. Длина вегетационного периода —
70—85 дней.
39G
Частная светокультура растений
В опытах Рэмэли [697] зимою гречиха цвела одновременно (через
70 дней) как при освещении (160—320 лк; с 17 до 22 час.), так и в конт-
роле; в опытах Адамса 1365] — сперва в контроле, а затем при
освещении.
При дополнительном ночном освещении растения имеют больший
вес и высоту. По данным Адамса [365], при освещении с помощью ламп
накаливания в 300—500 вт (18-часовой день) сырой вес 1 растения рав-
нялся 188 г, а в контроле (12-часовой день) — 120 г. В другом опыте
высота растений в возрасте 17 дней равнялась при освещении 16,4, а
в контроле •— 13,9 см.
Хороший эффект дает прерывистое ночное освещение с помощью ламп
накаливания в 500 вт (табл. 114). В этом случае при культуре зимою-
сухой вес 2-недельного растения равняется 0,42 г, а контрольного расте-
ния — 0,25 г [383, 387].
Таким образом, при дополнительном освещении гречиху можно куль-
тивировать зимой. Для нейтральных форм освещение дается в ночные
часы, а для короткодневных — в пределах короткого дня (12 часов).
Для дополнительного освещения, невидимому, будет достаточно
удельной мощности порядка 500 вт/м2.
Кукуруза (Zea mays)
Исключительно светолюбивое растение. Относится к группе коротко-
дневных или нейтральных растений и успешно растет лишь при темпера-
туре более 17°. Оптимальная длина дня для ростовых процессов равна
16 часам [703, 704]. Длина вегетационного периода — 85—140 дней.
Несмотря на то что опыты по выращиванию кукурузы зимою с помо-
щью дополнительного освещения начались давно [470, 692, 812], литера-
тура по светокультуре этого растения очень невелика.
Дополнительное ночное освещение в 160—320 лк в большинстве слу-
чаев задерживает или полностью устраняет цветение короткодневных
форм [618, 697, 710]. Нейтральные формы цветхт одновременно с контро-
лем [715, 716, 862].
Культура кукурузы зимою при дополнительном освещении удается
на юге. В умеренной полосе, несомненно, необходимо мощное дополни-
тельное дневное освещение.
Овес (Avena sativa)
Длиннодневное растение (фиг. 143, 1) с длиною вегетационного периода
от 93 до 110 дней.
В зимние месяцы в большинстве случаев овес приступает к плодоно-
шению лишь при дополнительном освещении; в условиях короткого дня
он или не цветет совсем, или значительно запаздывает с цветением [101,
266, 376, 379, 501, 535, 716, 754, 771, 837]. Особенно сильный эффект
дополнительное освещение производит при одновременном удобрении
углекислотой (табл. 119 [379]). При дополнительном освещении коло-
шение овса наступает через 40—90 дней, а без освещения — на 28—75 дней
позднее (через 70—130 дней) или совсем не происходит; в последнем слу-
чае овес продолжает бесконечно куститься, совершенно не переходя
в трубку. К первой группе следует отнести сорта из Палестины, Малой
Азии, Шведский, Уинтер Тэрф и другие, а ко второй — Шатиловский,
Победа, Финляндия, A. byzantina и другие. По данным В. И. Разумова
X. Светокультура полевых растений
397
[266], в условиях короткого дня колосятся лишь южные сорта, однако,
как видно из приведенного выше списка сортов, это наблюдается не
всегда.
Для дополнительного освещения овса обычно применяют лампы нака-
ливания. Однако лучший эффект получается под солнечными лампами
(табл. 90 [5091]. В опытах Смита [771] овес зацветал быстрое всего при
дополнительном освещении лампами накаливания; слабее влияли нео-
новые лампы, а натриевые лампы ускорения зацветания не дали.
В. И. Разумов [266] и В. М. Катунский [101] показали, что наибольшее
ускорение колошения наблюдалось в условиях беспрерывного освещения
(фиг. 143, 1). Однако в этом случае происходит значительное уменьшение
сухого веса растений (фиг. 144, 7; табл. 119). При длине дня в 10, 16,
18 и 24 часа вес растений соответственно равнялся 16,5; 12,34; 9,8 и
8,26 г [101]. Одновременно в результате удлинения дня может значи-
тельно уменьшиться и высота растений [501, 771]. По данным Смита
[771], оптимальная длина дня равна 18 часам. В этом случае полу-
чается наибольшие вес растений, кущение (фиг. 145, 2) и урожай
семян.
Для перехода некоторых сортов к репродуктивному состоянию доста-
точно дать дополнительное освещение порядка 10—50 лк [101, 266],
однако для полного зацветания лучше пользоваться освещенностью
в 300—500 лк. Названные величины пригодны для культуры овса зимою
в южных районах СССР. В умеренной зоне с низкими естественными осве-
щенностями зимою необходимо дополнительное освещение порядка
8000 лк [379].
Просо (Panicum miliaceum)
Короткодневпые и нейтральные светолюбивые растения с длиною
вегетационного периода 50—110 дней.
Большинство исследований по дополнительному освещению проса
проведены летом [62, 101, 173, 266]. Как правило, в результате искус-
ственного удлинения короткого дня наблюдается значительное запазды-
вание плодоношения растений (фиг. 143, 5). При беспрерывном освеще-
нии, по данным В. И. Разумова [266] и В. М. Катунского [101], задержка
равняется 10—60 дням. Если на коротком дне колошение наступает через
13—35 дней, то в условиях беспрерывного освещения — через 36—87 дней.
Сказанное относится к сортам: Бухарскому, Памирскому, Иркутскому,
Украинскому, Воронежскому, Западно-Сибирскому, Безенчукскому,
Тулунскому, Саратовскому и другим. Сорта Афганское, Персидское,
Корейское, Китайское, Монгольское и другие на коротком дно (8—10
часов) колосятся через 24—32 дня, а при беспрерывном освещении не
колосятся совсем.
При дополнительном освещении проса значительно увеличивается
сухой вес растений. Последний в опытах В. М. Катунского [101] с просом
Саратовское при длине дня 10, 16, 18 и 24 часа соответственно равнялся
5,1; 1э,83; 17,99 и 19,22 г. Сухой вес растений достигает максимума при
длине дня 18—24 часа.
При культуре проса дополнительное освещение можно давать лишь
в пределах короткого 10—12-часового дня. В противном случае может
наступить значительное запаздывание колошения. У южных сортов
это наблюдается уже при освещенности 0,5 лк, а обычно вызывается
освещенностью 50—100 лк [101, 266].
398
Частная светокультура растений
Пшеница (Triticum)
Длинподневные светолюбивые растения. Длина вегетационного пери-
ода 85—110 дней. В настоящее время по светокультуре этого растения
имеется сравнительно большая литература [1, 3, 12, 142, 160, 174,
267, 284, 309, 310, 313, 334-336, 345, 365, 366, 376, 379, 450, 482, 493,
501,535,559—561, 577,619,620, 632, 697, 716, 756, 759, 771, 811, 830,
837], причем охвачено большое число видов и сортов пшеницы (Tr. vul-
gare, озимые сорта: Пооператорна, Харьковская, Украинка, Московская
(№ 3251 и № 2411), Белоколоска, Дюрабль, Гостианум 237, Лютесценс
329, Гарвест Куин, Мингарди, Гарди Нортерн, Турка, Ред уэйв, Мин-
турки, Конрад и другие; яровые сорта: Гирка, Эритроспермум 341, Лютес-
ценс 62, Цезиум 111, Маркиз, Престон, Кота, Пурин Строу, Руби, Санора,
Тайнее Блустем, Гард Федерэйшн, Прелюд и другие; Tr. durum: Кубанка,
Арнаутка, Гордеиформе 10, Меланопус 69 и другие; Tr. dicoccum, Tr.
compactum, Tr. spella и другие).
Большинство исследований проведено с сортами озимой пшеницы
Tr. aestivum (Tr. vulgare). Известно, что этим сортам для прохождения
стадии яровизации необходима низкая температура. Дополнительное осве-
щение в этот период имеет значение лишь для усиления вегетативного
роста растений. На переход к цветению оно влияет только при прохо-
ждении световой стадии. В зависимости от условий яровизации, темпера-
туры и других факторов озимая пшеница зимою колосится без освеще-
ния через 144—199 дней, а при освещении — через 131—154 дня,
ускоряя колошение на 17—66 дней [365, 501, 560, 577]. Интересно,
что эффективность дополнительного освещения проявляется относительно
тем сильнее, чем в меньшей степени пройдена стадия яровизации [309.
310, 620].
Особенно сильное влияние дополнительное освещение производит
на яровые формы Tr. vulgare, многие из которых (Лютесценс 62, Маркиз,
Гайнес Блустем, Санора, Гард Федерэйшн, Цезиум 111, Эритроспермум 341,
Ветвистая кахетинская и другие) не колосятся в условиях естественного
зимнего или искусственного летного короткого дня (8—12 часов), а прп
дополнительном ночном освещении колосятся через 32—78 дней [3, 160,
334, 336, 620], В других случаях — у сортов Престон, Кота, Прелюд и
других — колошение наступает при освещении через 32—75 дней, а без
освещения — через 43—184 дня [365, 560, 620].
Хорошая реакция на дополнительное освещение наблюдается и
у яровых пшениц Tr. durum, Tr. dicoccum, Tr. compactum, Tr. spella и дру-
гих [266, 535, 620]. Следующие цифры характеризуют число дней до коло-
шения у названных видов при дополнительном освещении и в контроле:
Tr. durum—31—35 и 37—63, Tr. compactum—-35—41 и 50—79, Тг.
dicoccum — 26—35 и 36—55 и Tr. spelta — 42—47 и 51—78.
Опыты показывают, что у пшениц максимальное ускорение развития
наблюдается в условиях беспрерывного освещения (фиг. 143, 12:
[3, 266, 620, 759, 771]); в этом случае пшеница колосится в наиболее ранние
сроки — через 32—67 дней, ускоряя наступление этой фазы развития по
сравнению с 14-часовом днем на 6—38 дней, а по сравнению с 16—18-часо-
вым днем — на 0—19 дней.
Однако, несмотря на то что беспрерывное освещение является опти-
мальным для дифференциации и роста зачаточного колоса [3, 334—336,
345], названный вариант не является наиболее оптимальным для нормаль-
ного развития пшеницы. Дело в том, что при беспрерывном освещении
с помощью ламп накаливания растения настолько быстро начинают
X. Светокультура полевых растений
399
выходить в трубку и выколашиваться, что значительно снижается уро-
жай как соломы, так и зерна (фиг. 144, 10; табл. 112 [334—336, 560, 759,
771]). Для получения более мощных и нормальных растении необходимо
избегать беспрерывного освещения и пользоваться 17—18-часовым днем
(фиг. 143, 12; фиг. 144, 10; фиг. 145, 4 и фиг. 146. 7). Можно также перво-
начально, в течение 30—40 дней, создавать короткий 10—12-часовой
день, а затем помещать растения в условия беспрерывного освещения
[334—336]. В этом случае пшеница запаздывает с колошением, но дает
более высокий урожай.
В зимние месяцы пшеница хорошо реагирует на совместное действие
дополнительного освещения и удобрения <10., (табл. 119 [379]).
Для того чтобы вызвать колошение пшеницы, достаточно дополни-
тельное освещение в 250—1000 лк. При высоте подвеса ламп 50 см и упо-
треблении обычного глубокоизлучателя эта освещенность создается при
удельной мощности 25—100 вт/м'2.
Однако для получения нормальных растений в зимние месяцы в уме-
ренной зоне этой удельной мощности далеко не достаточно. По данным
М. А. Бассарской и П. В. Алехина [12] нормальная культура пшеницы с по-
мощью ламп накаливания возможна лишь при дополнительном беспрерыв-
ном или ночном освещении при удельной мощности 2000 вт/м'2. М. Я. Тре-
губевко [313] показал, что нормальное развитие пшеницы (7 г. vulgare
Лютесценс 62 и Tr. durum Меланопус 69) с образованием колоса в
12—15 зерен возможно лишь при дневном освещении (16 часов) при удель-
ной мощности 1000 вт/м2 и дополнительном ночном освещении (8 часов)
при удельной мощности 200 вт/м'2. В этом случае при посеве 17 октября
выход в трубку совершается через 30, колошение — через 58, цвете-
ние — через 67, а восковая спелость — через 108 дней (6 февраля). Однако
лучший эффект при зимней культуре пшеницы дают люминесцентные
лампы [160].
В этом случае получаются нормальные растения с нормальным
циклом развития (цветение через 65—71 день, созревание — через 107—
112 дней) и нормальным урожаем при удельной мощности 300—600 вт/м'2,
длине дня 16 часов и интенсивности дополнительного ночного и дневного
освещения 5000—8000 лк.
Рис (Oryza sativa)
Короткодневные или нейтральные светолюбивые растения с длиною
вегетационного периода 90—130 дней. Дополнительное ночное освещение
задерживает или полностью устраняет цветение [581, 794]. При освеще-
нии с помощью ламп накаливания в 200 вт (24-часовой день) колошение
у одних сортов не наступает совсем, у других же начинается через 123—
142 дня, а без освещения (12-часовой день) — через 66—93 дня.
Рожь (Secale cereale)
Длиннодневные, в большинстве случаев озимые растения. Для про-
хождения стадии яровизации нуждаются в низких температурах. До-
полнительное освещение, удлиняющее естественный короткий день, имеет
смысл лишь после прохождения процесса яровизации.
Работ по дополнительному освещению ржи имеется немного [1, 310,
334—336, 365, 501, 535, 716, 771, 811, 837], причем лишь некоторые из
них относятся к зимнему периоду [365, 501, 716, 811], остальные же каса-
ются дополнительного освещения летом.
400
Частная светокультура растений
В условиях естественного или искусственного короткого дня рожь
или нс цветет совсем [334, 837] или зацветает с большим запозданием;
колошение в этом случае наступает через 130—175 дней [365, 001],
а в условиях беспрерывного освещения — через 64—120 дней (после
предварительной яровизации, а у яровых форм — даже через
29—42 дня).
У ржи, как и пшеницы, беспрерывное освещение не является наи-
более благоприятным режимом, несмотря на наиболее раннее колошение
(фиг. 143, 8). Более мощные и урожайные растения формируются в усло-
виях 17—18-часового дня (фиг. 144, 8; фиг. 145, 3 и фпг. 146,2) или после
предварительной культуры в условиях короткого дня (10—12 часов)
в течение 30—40 дней и последующего переноса на беспрерывное осве-
щение [1, 334—336],
Обычно применяется дополнительное освещение порядка 300—1000 лк.
В умеренной полосе для получения нормальных растений в зимнее время
этих величин, конечно, недостаточно. Здесь, невидимому, необходим
тот же режим, что и для пшеницы.
Ячмень (Hordcum vulgare)
Длиннодневные растения с длиною вегетационного периода 85—100
дней.
Современная литература ио светокультуре ячменя представлена
небольшим числом работ [1, 101, 267, 334—336, 376, 379, 393, 428, 468, 482,
483, 488, 501, 511, 535, 754, 771, 837]. Лишь немногие работы проведены
по дополнительному зимнему освещению [428, 501, 535, 771], большин-
ство же проведено летом.
В условиях искусственного короткого дня или без дополнительного
освещения зимою ячмень или не цветет [101, 334, 468, 535] или цветет
с большим запозданием, выколашиваясь через 64—146 дней [267, 468,
488, 501]. При дополнительном освещении в условиях беспрерывного
дня колошение наступает через 35—83 дня (фиг. 143, 2).
Дополнительное освещение в зимнее время дает особенно сильный
эффект при одновременном удобрении углекислотой (табл. 119,. [379]).
Опыты В. М. Катунского [101] показывают, что оптимальной длиной
дня для ячменя следует признать 18 часов. В этом случае наблюдается
максимальный вес растений. То же самое отмечают Артур (табл. 119,
[379]) и Смит [771]. По данным Чайлахяна [335, 336], беспрерывное осве-
щение дает несколько худший результат, чем нормальный летний день.
В. И. Разумов [267] нашел, что колошение ячменя Винер наступает
одновременно как в условиях беспрерывного освещения, так и на 14-часо-
вом дне. В свете этих данных следует признать, что для культуры ячменя
наиболее благоприятен 18-часовой день (фиг. 143, 2; фиг. 144, 2;
фиг. 146, 4).
Что касается интенсивности дополнительного освещения, то Фа-
биан [488] и Деиффер [468] нашли, что ускорение колошения вызывает уже
освещенность 20 лк, однако для полного эффекта необходима освещенность
100—150 лк (фиг. 142, 10 и фиг. 152).
По данным Фабиан [488], утреннее освещение (с 1 до 8 час.) дает
несколько лучший эффект, чем вечернее (с 18 до 1 час.).
Особенно мощные растения ячменя получаются при культуре сперва
в условиях короткого дня (около 30—40 дней), а затем в условиях длин-
ного дня или беспрерывного освещения [335, 336].
X. Светокультура полевых растений
401
2. БОБОВЫЕ
Бобы (Vicia faba)
Длиннодневные или нейтральные растения с длиною вегетационного
периода 93—102 или 133—142 дня.
Зимою можно культивировать при дополнительном ночном освещении
[697, 812]. Растения зацветают через 71—75 дней.
Горох (Pisum sativum)
Длиннодневиое растение с длиною вегетационного периода от 60 до
100 дней. Может культивироваться и в закрытом грунте.
Культура гороха зимою с помощью дополнительного освещения впер-
вые была осуществлена еще в конце прошлого столетия [398, 466, 467,
470, 692, 754, 756].
Сименс [754], пользуясь вольтовой дугой и экраном из обычного стекла,
получил зрелые семена гороха 16 февраля (при посеве 23 октября). В опы-
тах Бэйли [398] также с вольтовой дугой горох при дополнительном осве-
щении цвел и плодоносил раньше контроля на 7 дней. Асо и Мураи [393]
установили, что при дополнительном ночном освещении (лампы накали-
вания 100 вт) горох растет быстрее и зацветает раньше, чем в контроле.
Бос [428] показал, что при дополнительном освещении в 7000 лк (лампы
накаливания в 200 вт па высоте 40 см) и длине дня 17 часов горох цветет
и плодоносит в декабре—январе.
По данным В. М. Катунского [101], горох Бисмарк лучше всего раз-
вивается в условиях беспрерывного освещения. В этом случае полу-
чается максимальный сухой вес и наиболее раннее цветение (фиг. 143, 3;
фиг. 144, 3 и фиг. 146, 5). В опытах Смита [771] лучший результат наблю-
дался в условиях 18-часового дня. В этом случае цветение наступает одно-
временно с беспрерывным освещением, а сухой вес растений достигает
максимальной величины.
Гелин и Бюрстрём [509], сопоставляя различные источники излуче-
ния, нашли, что для гороха наиболее пригодны солнечные лампы (табл. 90).
По данным Клифтона [447], с помощью дополнительного освещения
(150 вт/м2) в течение зимы (октябрь—май) можно получить два дополни-
тельных урожая гороха.
Соя (Soja hispida)
Исключительно светолюбивое растение, но начинающее бутонизиро-
вать уже при интенсивности физиологической радиации в 6000 эрг/см2-сек
и полностью (100% растений) бутонизирующее при 8000 эрг/см2-сек
(фиг. 134, 2 [528]). Большинство сортов относится к группе растений
короткого дня (Билокси, Авуалль и другие), а некоторые к нейтральным
(Баторовка, Мандарин, Виргинская и другие). Длина вегетационного
периода колеблется от 100 до 170 дней.
Исследований по дополнительному освещению сои сравнительно
много. Однако большинство их относится к изучению световой стадии
[70, 101, 170, 171, 173, 174, 256—260, 289, 365, 366, 379, 394, 420, 497,
500, 528, 642, 656, 715, 716, 822, 855].
В силу того что соя относится к типичным короткодневным расте-
ниям [500] и в большинстве случаев совершенно не цветет в условиях
длинного дня (фиг. 143, 7), дополнительным освещением в ночные часы
26 А. Ф. Клешнин
402
Частная светокультура растений
обычно пользуются для предотвращения или задержки цветения [170,171,
173, 174, 497]. Полное устранение цветения у короткодневных форм имеет
место при 14—18-часовом дне [101, 420, 642]. В некоторых случаях длин-
ный день вызывает быстрое пожелтение и опадение листьев [258] и задер-
живает все фазы развития от всходов до появления плодов, но ускоряет
процесс созревания [257]. Для устранения цветения достаточно добавоч-
ного освещения в 65—85 лк [101]. По данным Бортвика и Паркера [419],
задержка цветения наступает уже при 10 лк (табл. 106). Оптимальная
длина дня для короткодпевпых сортов, при которой наблюдается макси-
мальное количество цветущих растений и наибольшее число цветков и
стручков, равна 10—12 часам, а для нейтральных форм— 14—16 ча-
сам [420]. У последних начиная с 14-часового дня цветочные почки за-
кладываются тем выше по стеблю, чем больше длина дня. Следовательно,
дополнительное освещение сои с целью получения максимального урожая
семян следует производить в пределах короткого 10—-12- пли 14—16-
часового дня.
Как растение чрезвычайно светолюбивое, соя исключительно снльнорс-
агирует на дополнительное освещение. В опытах Артура и его сотрудников
[379] с соей Токио, Мандарин, Пекин и Билокси ежедневное дополнитель-
ное 6-часовое освещение при интенсивности радиации 0,45 г-кал/см2>мин
(8190 лк) увеличивало вес опытных растений по сравнению с кон-
тролем в 23/2—5 раз (табл. 119). В результате дополнительного освещения
имеет место увеличение веса и высоты растений, числа узлов и площади
листьев [70, 254, 260, 420, 822].
По данным В. М. Катунского [101], оптимальным для вегетативных
ростовых процессов является 16-часовой день (фиг. 144, 7). Паркер и
Бортвик [656], однако, нашли, что лучший результат дает беспрерыв-
ное освещение. Впрочем, в их опытах разница в весе растений 16-часо-
вого дня и беспрерывного освещения была сравнительно невелика (105,3 и
107,1 г). Практически оптимальным следует поэтому признать 16-
часовой день.
Для нормального роста и развития соя нуждается в большой интен-
сивности дополнительного освещения. Обычно пользуются лампами нака-
ливания в 1000 вт 170, 822], создавая освещенность порядка 6500—
8000 лк.
Как показывают опыты Уитроу [855], лучшее накопление сухой массы
у сои наблюдается под обычными лампами накаливания (табл. 89).
Фасоль (Phaseolus)
Большинство сортов фасоли относятся к видам Ph. vulgaris и Ph.
coccineus (Ph. multi floras). Среди первых преобладают короткодневные,
а среди вторых — длиннодневные сорта. В естественных условиях длина
вегетационного периода фасоли колеблется в пределах 100—120 дней.
В условиях закрытого грунта урожай стручков составляет 3—6 кг/м2.
Положительные результаты были получены уже в первых опытах
с искусственным освещением [466, 467, 470, 692, 754, 755]. Позднее
дополнительным освещением фасоли занимались Тинкер [806, 807, 810],
Роденбург 1717], Линдонбейн [580, 603], Леман [160] и другие.
Гэйден и Штейпметц [544], пользуясь лампами накаливания в 500 вт,
висевшими на расстоянии 40 см друг от друга на высоте 90 см, показали,
что в зимнее время (декабрь—февраль) при беспрерывном горении ламп
(7000 лк) фасоль бутонизнрует через 21 день, а через 55 дней дает первый
урожай. Контрольные растения урожая не дают, а бутоннзируют через
36 дней. В опытах Тьеббеса и Упгофа [811] фасоль Шеврт.е, посеянная
А. Сие/покулынура полевых растений
403
14 января под пампами накаливания в 200 вт, находившимися на высоте
20—30 см и горевшими с 22 до 6 час., зацвела 28 февраля, а 14 марта
дала прекрасно развитые стручки (длиной 10—12 см). У контрольных
растений размеры стручков не превышали нескольких ^миллиметров.
По данным Шуара [441], при дополнительном освещении в течение 7 ча-
сов (300—500 лк) цветение и плодоношение ускоряются на 23—35 дней,
а урожай удваивается ио сравнению с контролем. При посеве 21 января
цветение фасоли при освещении наступило 7 марта, а плодоношение —
3 апреля; соответствующие сроки в контроле — И апреля и 26 апреля.
Оден [646] установил, что Ph. nanus (сорт Энко^инг) при дополнительном
освещении интенсивностью 0,2 г-кал/см2-млн в условиях 15-часового
дня зацветает через 35 дней и даст нормальные стручки. Другой сорт
1’нзбринкен в тех же условиях в зимние месяцы зацветает через
25—30 дней. По данным Лпндснбейна [603], дополнительное освещение
зимою ускоряет цветение на 12—16 диен.
В тех же опытах Оден показал, что коротковолновые лучи (при пере-
кале ламп накаливания) вызывают опадение цветков, а длинноволновая
радиация благоприятствует плодоношению.
В. М. Катунский [101] специальными опытами показал, что для
фасоли Золотая гора и Осборн наиболее благоприятен 16—18-часовой
день (фиг. 143, 4 и фиг. 144, 4). Хотя цветение раньше всего наступает
в условиях беспрерывного освещения (через 29—33 дня), но максималь-
ный вес растений получается при 16-часовом дне.
Н. А. Максимов [188] и О. В. Тагеева [307] установили, что развитие
фасоли без дополнительного освещения в условиях Ленинграда и Москвы
возможно лишь в период с 10 февраля по 20 октября. В остальное время
фасоль нуждается в дополнительном ночном освещении (фиг. 139). С. В.
Тагеева [307] нашла, что в зимние месяцы лучшее накопление органиче-
ского вещества наблюдается иод лампами накаливания в 1000 вт при вы-
соте подвеса 60 см. Лампы в 500 вт дают значительно худший результат.
В опытах II. А. Максимова [188] фасоль в декабре под лампами накали-
вания (1000 вт, с 14 до 10 час.) росла так же хорошо, как она обычно растет
в июне или августе. По данным С. В. Танеевой [307], кустовые сорта
фасоли (Триумф) под влиянием ламп накаливания становятся вьющимися.
Сильное вытягивание фасоли иод лампами накаливания отмечает и
Н. А. Максимов [188].
По данным Клифтона [447], при дополнительном освещении (150 вт/м2)
зимою (октябрь—май) можно получить еще один урожай фасоли.
Фасоль в зимние месяцы (январь—март) хорошо развивается под люми-
несцентными лампами (300—600 вт/м?) в условиях 16-часового дня при
интенсивности дополнительного освещения 5000—8000 лк. Сорт Лат-
вия 800 в этом случае зацветает через 31 и созревает через 97 дней, образуя
нормальные семена (абсолютный вес 362 г) с повышенной всхоже-
стью [160].
.3. ТЕХНИЧЕСКИЕ КУЛЬТУРЫ
Гваюла (Parthenium argentatum)
Длиннодневное растение. Цветет при длине дня более 13 часов. При
дополнительном ночном освещении люминесцентными лампами белого
света (220 лк) максимальное количество цветущих растений (80%) наблю-
дается в условиях беспрерывного освещения, а максимальный вес семян
(388 мг) — при 15-часовом дне [850]. Увеличение веса семян по сравнению
26*
404
Частная светокультура растений
с контролем1 равняется 83%. По данным Б. С. Мошкова [212], беспрерыв-
ное освещение дает отрицательный эффект. В его опытах наиболее мощные
растения и 100% цветения наблюдались в условиях 15-часового дня.
Горчица (Sinapis alba)
Длиннодневное растение. Длина вегетационного периода 80—120 дней.
Литература но дополнительному освещению очень невелика [365, 468,
496, 512, 697]. Зимою при дополнительном освещении (160—320 лк;
с 17 до 22 час.) за в,встает через 48, а без освещения — через 90 дней [697].
Клещевина (Ricinus communis)
Короткодневное растение. На дополнительное освещение реагирует
усилением вегетативного роста, увеличивая сырой вес [365].
Кок-сагыз (Taraxacum kok-saghyz)
Длинподневное растение. При дополнительном освещении с помощью
ламп накаливания в 500 лк растет и развивается зимой, причем макси-
мальное число бутонизирующих растений (48?zo) наблюдается в условиях
16-часового дня [422]. Опыты В. А. Давганя и М. II. Карася [60] по
влиянию дополнительного освещения на кок-сагыз показали, что осве-
щение на ранних фазах развития практически не влияет на последующее
развитие и урожай корней кок-сагыза.
Конопля (Cannabis sativa)
Короткодневные или нейтральные растения. Первые в условиях корот-
кого 10-часового дня цветут через 24—31 день, на 16-часовом дне через
34—41 день, а при 18—24-часовом дне не цветут совсем (фиг. 143, 6 [101]).
Сухой вес растений увеличивается вместо с длиной дня и достигает ма-
ксимума при 18—24-часовом дне (фиг. 144, 6 [101]). По данным
Адамса [365], сырой вес при дополнительном освещении (20-часовой день)
меньше, чем в контроле (15-часовой день), на 17,5%, а цветение запазды-
вает на 4—10 дней.
Для устранения цветения достаточно дополнительного освещения
в 65—80 лк [101].
Крым-сагыз (Taraxacum megalorrhizon, Т. gymnanthum),
Короткодневпое растение. По данным Н. Ф. Соколовой [299], растения
второго-третьего года в условиях беспрерывного освещения лампами нака-
ливания в 500 вт уходят в покой, совершенно сбрасывая листья через
15 дней. Отрастание розеток (выход из состояния покоя) начинается уже
при 19-часовом дне. Максимальное количество листьев образуется в усло-
виях 19-часового дня, а максимальное количество цветков — на корот-
ком 9-часовом дне. Запаздывание в цветении на длинном дне составляет
50 дней.
Растения первого года лучше развиваются на коротком дне; в этом
случае образуется больше листьев, увеличивается диаметр розетки и
формируется толстый и короткий стержневой корень. На длинном дне
при дополнительном освещении корни сильно ветвятся.
1 Длина дня в течение опыта падала от 14 часов 45 минут до 9 часов .30 минут.
X. Светокультура полевых растений
405
Кунжут (Scsamum indicum)
Короткодневное растение, задерживающее цветение при дополнитель-
ном освещении. В условиях Краснодара одни сорта (Кубанец 55) дают
лучший прирост при беспрерывном освещении, а другие (Японский) —•
одинаковый как па нормальном дне, так и при беспрерывном освеще-
нии [294].
Леи (Linum usitatissimum)
Длинно дневное растение. Зимою можно культивировать с помощью
дополнительного освещения [365, 428, 811]. В этом .случае цветение насту-
пает через 61—84 дня, а зрелые семена — через 75 дней.
Для культуры льна необходима длина дня около 18 часов и дополни-
тельное освещение порядка 4500 лк. В условиях 18-часового дня наблю-
дается минимальный вегетационный период и максимальный урожай
семян (фиг. 143, 21); наибольший вес и высота растений имеют место
в условиях беспрерывного освещения (фиг. 144, 22; фиг. 146, 7 [759]).
„ Монарда (Monarda)
Многолетние нейтральные тенелюбивые эфирномасличные растения,
цветущие в июле—августе и слабо реагирующие на дополнительное осве-
щение. М. didyma имеет тенденцию к длииподвсвности, а М. punctata —
к короткодневности [372].
Подсолнечник (Helianthus annuus)
Светолюбивое растение. При дополнительном освещении в условиях
длинного дня цветет позднее, но дружнее, увеличивает вегетативную
массу и слабее поражается заразихой [11, 75,365, 366]. При беспрерыв-
ном освещении цветение менее дружно, чем в условиях 16-часового дня.
Рапс (Brassica napus var. oleifera)
Длиннодневнос растение. По данным Чайлахяна [335—337], яровые
формы при дополнительном освещении в условиях длинного дня бутони-
зируют через 26, а цветут через 35 дней. Беспрерывное освещение менее
благоприятно, чем 17-—18-часовой день, вызывая уменьшение высоты
растений, снижая их сухой вес и уменьшая урожай семян.
Сун (Crotalaiia juncea)
Короткодневное растение, цветущее через 40—50 дней в условиях ко-
роткого дня (8—12 часов), и через 70—90 дней — па длинном дне (15—18
часов; фиг. 143,10). Плодоношение на коротком дне наступает через 70—80,
а на длинном — через 100—105 дней. Высота растений, сухой и сырой
листовая поверхность, длина волокон и урожай
вместе с увеличением длины дня и достигают макси-
вес, числолистьев
волокна'^врастают
мума при беспрерывном освещении. Максимальное ветвление (фиг. 145, 5)
и максимальный урожай семян получаются в условиях 12-часового
дпя [758—761].
406
Частная светокультура растении
Табак (Nicotiana tabacum)
В большинстве случаев короткодневные или нейтральные растения.
Встречаются и длиннодневные сорта (Дюбек, Гаванский, Варатик,
Самсун и другие).
Дополнительное освещение может применяться как для выгонки рас-
сады [68, 579], так и для культуры в зимние месяцы [376, 379, 500, 715,
716, 742]. По данным Г. А. Евтушенко [68], рассада короткодневных сор-
тов (Гаванна 1708, Виргиния 1459, Самсун 27, Трано.зунд 93, 709 и 1272,
Тык-кулак, Висконсин, Мэрйлэнд и другие) летом развивается лучше
в условиях короткого дня, а длиннодневных сортов (Дюбек, Варатик) —
при беспрерывном освещении с помощью ламп накаливания (500 вт/м2;
около 2000 лк).
Понятно, что при культуре зимою длиниодпевные сорта можно осве-
щать в ночные часы, а короткодневные — лишь в пределах короткого
12-часового дня. В опытах Робертса и Стракмсйера [715] мощные растения
Мэрйлэнд Маммот и Гаваппа развивались при дополнительном освеще-
нии в 300—800 лк, а в опытах Артура [379] — при 8200 лк. В последнем
случае вес 1 растения в возрасте 38 дней равнялся при освещении 1,23 кг,
а в контроле — 0,87 кг (табл. 119). По данным французских авторов
[610, 613], с помощью люминесцентных ламп дневного света (30 вт) при
освещенности 3000 лк зимою можно выращивать такие же табачные расте-
ния, что и летом.
Невидимому, освещенность порядка 300—800 лк следует рекомендо-
вать для юга, а 3000—8000 лк— для умеренной полосы.
Фенхель (Foeniculum vulgare)
Длинно дневное, многолетнее, культивируемое как двулетник эфирно-
масличное растение. При дополнительном освещении (с 17 до 22 час.;
160—320 лк) в зимние месяцы цветет через 102, а в контроле — через
122 дня [697].
Хлопчатник (Gossipium hirsutum)
Короткодневные и светолюбивые растения. В условиях короткого дня
(9—12 часов) цветут через 50—100 дней, а на длинном дне — на 5—30 дней
позднее или совсем не цветут. Дополнительное освещение (300 вт/м2),
по данным Н. Н. Константинова [126], перемещает заложение первой
плодовой ветви с 9-го узла на 34—40-й и вызывает сильную задержку
в наступлении цветения. При дополнительном освещении в 400 лк цвете-
ние не наступает совсем, при меньшей освещенности (100 лк) бутонизация
имеет место через 150 дней вместо 37—42 дней в условиях 9-часового
дня. В опытах Лысенко [174] многолетний хлопчатник 1632 плодо-
носил лишь на коротком дне. При беспрерывном освещении он не цвел
совсем.
По данным Беркли [407], высота растений хлопчатника растет вместе
с увеличением длины дня и достигает максимума при беспрерывном осве-
щении. Нормальный рост имеет место в условиях 16-часового дня; на корот-
ком дне формируются приземистые, а при беспрерывном освещении —-
сильно вытянувшиеся растения.
X. Светокультура полевых растений
407
4. КЛУБНЕПЛОДЫ II КОРНЕПЛОДЫ
Земляная груша (Helianthus tuberosus)
Максимальный вес наземной массы и высота растений наблюдаются
в условиях длинного дня (17 часов), а максимальный урожай клубней —
на коротком дне (12 часов [806]).
Картофель (Solanum tuberosum)
В большинстве случаев длинподновное растение, ускоряющее
наступление цветения при дополнительном освещении. Зимою легко
культивируется при дополнительном освещении и обычно цветет [172,
289, 290, 366, 379, 402, 446, 471, 501, 511, 610, 613, 715, 787, 845—847],
а в контроле не цветет. Цветение наступает примерно через 40 дней,
а количество цветущих растений, в зависимости от сорта, колеблется
в пределах 20—70% [172, 501, 787]. Освещаемые растения отличаются
большой мощностью, высотой и весом. В опытах Адамса [366] растения
в возрасте 49 дней имели высоту при освещении 31.7 см, а в контроле
23,3 см. По данным Отевепсопа л Кларка [787], освещаемые растения
отличались исключительной мощностью и через 61 день достигли высоты
134 см, а без освещения были значительно слабее (61 см). Аналогичное
увеличение высоты отмечает также Мак Клилланд [618]. По его данным,
в условиях 10-часового дня высота растений равняется 18—19 см, а при
дополнительном освещении (15-часовой день) — 29—60 см. Французские
авторы [610, 613] нашли, что в условиях северной Франции (Париж)
картофель зимою вообще нс растет, а при дополнительном освещении
с помощью люминесцентных ламп дневного света (3000 лк) развивается
так же, как и в естественных условиях весной (в мао).
Вес растений при дополнительном освещении, в зависимости от усло-
вий, возрастает на 20—80% (табл. 119 [379, 501)], а в некоторых слу-
чаях — в несколько раз. Так, в опытах Мак Клилланда [618], проведен-
ных в летние месяцы, сорта Ириш Коббер, Лукаут и Ред Блисс при 10-ча-
совом дне имели вес наземной массы 23—78 г, а при дополнительном осве-
щении (15-часовой день) — 237—506 г.
Дополнительное освещение, удлиняющее естественный короткий день,
как правило, приводит к значительному снижению клубнеобразования
[172, 501, 618], хотя в ряде случаев клубни хорошо образуются и в усло-
виях длинного дня, в частности, при дополнительном освещении зимою
[379, 787]. Лысенко и Долгушин [172] в опытах с сортами Эпикур, Вольт-
ман и другими установили, что клубнеобразование зависит не столько
от длины дня, сколько от стадийного состояния растений. У стадийно
молодых растений клубнеобразование есть результат задержки стадий-
ного развития; в этом случае клубни образуются лучше в условиях корот-
кого дня, задерживающего процесс перехода к цветению. У стадийно
старых, отцветающих растений клубни образуются раньше и обильное
в условиях беспрерывного освещения.
Оптимальная длина дня для картофеля равна 16—18 часам (фиг. 147).
В этом случае наблюдается максимальный вес растений [402], макси-
мальное количество цветков и плодов [446] и образуется максимальное
количество семян [471]. Если в условиях 9—13-часового дня урожай
семян с 1 растения равняется 107—129 мг, то при 17-часовом дне он дости-
гает 382 мг. В опыте Уорнера [847] по культуре картофеля зимою лучший
результат наблюдался при беспрерывном освещении. В этом случае
на растении образовалось по 12 цветков и созрело по 11 плодов, причем
408
Частная светокультура растений
вес одного плода равнялся 79 г, а воздушносухой вес семян— 1,15 г.
Соответствующие данные для 18-часового дня были: 7 цветков, 6 плодов,
41 и 0,55 г.
Для культуры картофеля зимою необходима интенсивность дополни-
тельного освещения порядка 8000 лк [379]. Опыты показывают, что опти-
мальные условия создаются при освещении лампами накаливания в 500 вт;
более мощные лампы (1000 вт) дают тот же эффект [787]. В опытах Уер-
нера [847] эффективность возрастала с увеличением освещенности от 600
до 5000 лк. В последнем случае наблюдалось максимальное количество
цветков (15—18) и плодов (11—15) и максимальный вес плода (88—112 г)
и семян (1,24—1,60 г).
Для получения нормальных растений наиболее пригодны, невиди-
мому, люминесцентные лампы белого или дневного света [820].
Свекла (Beta vulgaris)
Длиннодневные двухлетние и однолетние растения, цветущие лишь
при длине дня более 15 часов. Для нормального роста необходима осве-
щенность в 8000—12 000 лк и температура 10—23° [246].
Дополнительное освещение свеклы может быть применено для зимней
культуры однолетней свеклы [399, 655, 715, 716, 789] и для выращивания
семенников в зимние месяцы [136, 271, 293, 505, 651, 811].
Однолетняя сахарная и столовая свекла в зимние месяцы стрелкуется
только при дополнительном ночном освещении в 250—800 лк в условиях
17—18-часового дня при температуре порядка 12° [715, 789]. 1 Го данным
Нэйлора [637], наиболее быстрое цветение — через 20 дней — наступает
у однолетней свеклы в условиях беспрерывного освещения люминесцент-
ными лампами (800—1000 лк). При длине дня менее 10 часов свекла
остается в вегетативном состоянии.
В. В. Селихов [293] пользовался дополнительным освещением в ноч-
ные часы для получения семян двухлетней сахарной свеклы на первом
году вегетации. Яровизируя семена и высевая их в поло 13 апреля, автор
нашел, что при ночном освещении в 1000 лк лампами накаливания
(100 вт/м2) растения стрелкуются через 25—36 дней после появления
всходов (20—31 мая), бутонизируют через 36—49 дней (31 мая—13 июня),
зацветают через 52—58 дней (16—22 июня), а зрелые семена дают
через 64—68 дней (28 июня—4 июля). В конце июля каждое растение
дает 30—40 г крупных, хороших семян. Опыты показывают, что такие
сорта, как Верхнячка 770, Веселый подол 724, Романов 1044, Стандарт
2-й пятилетки, Ивановка 1009, Кальник, Первомайск 744, Немерча 722
и 1038, Льгов 778 и 1057, Хос-хос 739, Уладовка 755 и Романовка 763,
обычно дающие на первом году вегетации 0,2—0,9% цветухи, при бес-
прерывном освещении даже без яровизации увеличивают цветушность
до 8,3—83,3%, а после яровизации — до 64,3—100% (Уладовка 755
до 64,3%, Романовка— до 69,2%, а остальные перечисленные сорта —
до 100%). В опытах Толмачева [312] беспрерывное освещение молодых
растений, в зависимости от длительности освещения (30—90 дней), уве-
личивало цветушность с 23 до 58—100% у сорта Уладовка и с 8 до
42—100 % у сорта Степнячка. Для других сортов достаточно 40-днев-
ного беспрерывного освещения в зимние месяцы (с 10 февраля по
20 марта), чтобы вызвать полное цветение растеши! на первом
году [403],
II. М. Толмачев [311, 312] показал, что беспрерывное освещение
высадок ускоряет созревание семян на 3—4 недели по сравнению с нор-
X. Светокультура полевых растений
409
мальным летним днем. Дополнительное освещение в этом случае имеет
смысл лишь после прохождения стадии яровизации.
Дополнительное ночное освещение является хорошим методом для
культуры семенников в зимние месяцы. Для этой цели высадки сахарной
свеклы вначале культивируются при 10°, а после стрелкования — при 15°.
В этом случае при дополнительном освещении (100—200 вт/м2) стрелко-
вание начинается через 80—100, бутонизация — через 90—130, цвете-
ние — через 110—150, а образование семян — через 145—190 дней [246,
2931. Без освещения в зимнее время свекла не стрелкуется совсем [293]
или стрелкуется с запозданием [246, 811].
Ф. Э. Реймерс [271] нашел, что столовая свекла при температуре
15—20° днем и 10° ночью стрелкуется через 17—39 дней при беспрерыв-
ном освещении лампами накаливания в 500 вт (330 вт/м2) и через
35—68 дней в условиях 11-часового дня. Чем позже высаживаются
корнеплоды (2 ноября, 3 января и 10 марта), тем раньше начинается
стрелкование.
В опытах Смита [778] было установлено, что в зимние месяцы стрелко-
вание корнеплодов темнокрасной свеклы Детройт лучше всего совер-
шается при беспрерывном освещении (3000 лк). В этом случае при тем-
пературе 12—15° стрелкование всех растений наступает через 100 дней.
При интенсивности дополнительного освещения 6О0О лк полное стрелко-
вание наблюдается уже через 72 дня.
По данным Одена и Расмусена [651], для культуры семенников в зим-
ние месяцы необходимы лампы накаливания в 500—750 вт, создающие
интенсивность дополнительного освещения в 0,1 г-кал/см2* мин
(70 тыс. эрг/см2-сек). В этом случае зимою (7 января—16 марта) еже-
месячно затрачивается около 2,9 кг-кал/см2, причем 1,22 кг-кал/см2
(42%) приходится на естественный свет, а 1,67 кг-кал/см2 (58%) — на до-
полнительное искусственное освещение. Авторы нашли, что в зимние
месяцы при дополнительном освещении можно получить даже больший
урожай семян, чем при обычной летной культуре. В их опытах с 1 расте-
ния был получен следующий урожай: сахарная свекла — 60 г, кормовая
свекла Баррес — 38 г, красная свекла — 32 г и свекла Оборндорфер —
100 г. Летом эти же сорта дали урожай семян соответственно в 1, 11,
4 и 16 г. В. Д. Пинчук [246] рекомендует освещенность 5000—6000 лк
(200—300 вт/м2).
Цикорий (Cichorium intybus)
Длиннодневное растение с длиною вегетационного периода 120 дней.
Зимою можно получить мощные цветущие растения при дополнительном
освещении в 300—800 лк при температуре 21° [715].
5. ТРАВЫ
Бородач (Andropogon)
Длинно дневные или короткодневные многолетние кормовые растения.
Длиннодневный A. scoparius зацветает лишь при дополнительном осве-
щении (1000—1800 лк) в условиях длинного дня (более 13 часов). При
длине дня 14—15 часов выход в трубку наблюдается через 76—104, а ко-
лошение — через 110—184 дня [591]. Максимальный сухой вес и высота
растений в большинстве случаев получаются в условиях 15-часового-
дня [591].
410
Частная светокультура растений
Короткодневный A. furcatus цветет лишь в условиях короткого дня
при температуре 24° дном и 15° ночью; в этом случае цветение наступает
через 101 день [405]. Дополнительное освещение в 350 лк, удлиняющее
день до 18 часов, полностью предотвращает цветение. При дополнитель-
ном ночном освещении сухой вес растений увеличивается по сравнению
с контролем в 15—40 раз [405]. Увеличивается также и высота расте-
ний [751].
Бутелуя (Bouteloua)
Нейтральные, короткодневные и длиннодневные растения. Одни формы
В. curtipendula цветут лишь в условиях 13-часового дня, другие пред-
почитают 16—20-часовой день [653]. В условиях благоприятной длины
дня цветение наступает через 55—123 дня. Зимою В. curtipendula растет
значительно лучше при дополнительном освещении (16-часовой день;
300 вт/м2), чем в контроле [751],
В. gracilis в условиях естественного зимнего дня (10 час.) цветет через
72—93 дня, а при дополнительном ночном освещении (350 лк) на 18—•
20-часовом дне — через 100—113 диен; часто при дополнительном осве-
щении растения но цветут совсем [405]. Освещение увеличивает высоту
растений в 3—6 раз [751], а вес сухой массы — в 1/4—2/4 раза [405].
Вика (Vicia)
Длиннодневные яровые (К. sativa) или озимые (К. villosai) растения.
Зимою можно культивировать при дополнительном освещении порядка
300—800 лк [716]. По данным М. X. Чайлахяна [334—336], лучше
всего развивается в условиях 17—18-часового дня; в этом случае на-
блюдается максимальный вес растений и наиболее быстрый переход к цве-
тению (через 40 дней). Беспрерывное освещение, по ускоряя развития,
приводит к значительному снижению веса растений.
Донник (Melilotus)
Длиннодневные двулетние растения. М. dentatus хорошо развивается
в зимние месяцы при дополнительном освещении в 300—800 лк [715].
В этих условиях образуются мощные, высокие, а без освещения — низ-
кие, стелющиеся растения. М. officinalis при увеличении длины дня
с помощью дополнительного освещения с И —13 до 17 часов увеличивает
вес растений на 25—120%, а высоту — в 4 раза [779]. Корневая система
у М. albus, М. dentatus и М. ofjicinalis относительно лучше формируется
в условиях короткого дня [715, 779]; на длинном дне она составляет
8—15% , а на коротком дне — 22—64% общего веса растений.
Ежа сборная (Dactylis glomerate)
Длиннодневное растение. Весною колосится одновременно как при
дополнительном освещении лампами накаливания (200 вт; 3000—4000 лк),
так и в контроле; максимальная высота растений и максимальное куще-
ние наблюдаются в условиях 16-часового дня [373].
X, Светокультура полевых растений
411
Клевер (Trifolium)
Длиннодневные растения. 13 зимние месяцы культивируются с помощью
дополнительного освещения 1365, 376, 379, 571, 715, 716, 772]. Как пока-
зал Адамс [365], дополнительное освещение зимою в полтора раза увели-
чивает высоту растении (Tr. dubiuiri) и па 12 дней ускоряет наступление
цветения: при освещении цветение через 109 дней, а без освещения —
через 121 день. В опытах Артура [379] Tr. pratense цвел лишь при допол-
нительном освещении (6 часов; 7000—8000 лк), причем сухой вес расте-
ний под влиянием последнего увеличивался в 5—10 раз: с 0,6—1,1 г
до 5,9—6 г. Особенно хороший эффект наблюдался при одновременном
удобрении СО, (табл. 119). Tr. repens зимою хорошо развивается при
дополнительном освещении в 300—800 лк при температуре 10°, образуя
мощные растения [715].
Дополнительное освещение в ночные часы оказывает благоприятное
воздействие даже в летние месяцы, особенно в широтах с коротким лет-
ним днем [145, 162, 365, 496].
И. С. Крячков [145], пользуясь дополнительным освещением различ-
ных поздних и ранних сортов Tr. pratense (Сиверский, Башкирский,
Псковский), получал семена в условиях беспрерывного дня на первом
году вегетации. У позднеспелых сортов при освещении в течение 2—3 ме-
сяцев он получил с растения ио 5 головок и 0,5 г семян; с контрольных
растений было получено по 0,25 головки и 0,003 г семян. Аналогичные
результаты были получены Б. 11. Лисицыным [162], который высевал
Tr. pratense 17 апреля в оранжерее, а затем высаживал растения в грунт
(12 июня). В его опытах сорта Весь и Среднерусский цвели и дали семена
лишь при беспрерывном освещении, а Ленкорань и Памир ускорили
бутонизацию, цветение и созревание на 24—29 дней. При дополнитель-
ном освещении бутонизация началась через 110, цветение—через 116—119,
а созревание головок — через 125—136 дней после посева; соответ-
ствующие сроки для контроля — 129, 143 и 154 дня. При дополнитель-
ном освещении автор получил но 3—15,6 головки с куста и по 15,3—
51,2 семени с головки, а в контроле — по 0—4,5 головки и 0—14,7 семени.
По данным Смита [772], лучшим источником для дополнительного
освещения клевера служат неоновые лампы. По накоплению сухого
вещества они в 5,5—9 раз эффективнее, чем лампы накаливания,
и в 4,5 раза, чем натриевые лампы.
Келлер и Петерсон [571 ] исследовали влияние дополнительного осве-
щения лампами накаливания в 200 вт в зимние месяцы (ноябрь—февраль)
на культуру Tr. pratense в травосмеси с тимофеевкой. При сопоставле-
нии 10-, 14- и 18-часового дня авторы нашли, что клевер цветет лишь
при длине дня более 14 часов. Для накопления максимального сухого
веса клевера необходим 18-часовой день, а для максимального кущения
и образования наибольшего числа листьев — 10-часовой день. Относи-
тельное содержание клевера в травосмеси падает с увеличением длины
Дня.
Костер безостый (Bromus inermis)
Длиннодневное многолетнее растение. При дополнительном освеще-
нии (3000—4000 лк) ускоряет цветение на 21 день [373]. Урожай сена
в два раза выше при дополнительном освещении (650—800 лк) в условиях
длинного дня (18 часов), чем на коротком дне [838]. Зимою цветет лишь
412
Частная светокультура растений
при дополнительном освещении (200 вт/м2) в условиях 16-часового
дня [640].
Леспедеца (Lespedeza)
L. stipulacea ( Kiimrnerovia stipulacea) и L. striata — растения корот-
кого дня. При дополнительном освещении (2500 лк) в условиях длинного
дня (более 14 часов) названные виды не цветут [777].
Люцерна (Medicago)
М. sativa (синяя люцерна) и М. jalcata (желтая люцерна) относятся
к группе растений длинного дня. Зимою можно культивировать при до-
полнительном ночном освещении в 300—800 лк [644, 716, 819].
П. А. Лубенец [165] применял дополнительное освещение люцерны
в летние месяцы с целью получения 2—3 урожаев семян в год. В его опы-
тах раннеспелые сорта (Грим-Занкевича, Волховская, Ново-Зеландская
и Малоазиатская) при дополнительном освещении (20-часовой день;
400 вт/м2) бутонизировалп через 9—13, а в контроле (прп естественной
длине дня) — через 14—17 дней. Цветение в первом случае наступило'
через 19-—22 дня, а во втором — через 25—31 день. Соответствующие
сроки для наступления восковой зрелости — 55—61 день и 58—74 дня.
Аналогичные данные были получены и для позднеспелых сортов (Армян-
ская, Ладак, Дагестанская и Хивинская). Бутонизация здесь наступила:
при освещении — через 18—24 дня, без освещения — через 26—37 дней,
цветение — через 31—37 и 45—51 день, а восковая зрелость — через
64—69 и 74—84 дня. В условиях короткого дня (10 часов) восковая спе-
лость в обеих группах сортов не наступала.
Мятлик (Роа)
Р. conipressa, Р. pratensis и Р. bulbosa — длиннодневные или нейтраль-
ные растения.
Зимою можно культивировать при дополнительном освещении с по-
мощью ламп накаливания в 500 вт [666, 715, 716]. Кущению благоприят-
ствует короткий, а накоплению сухой массы — длинный день; макси-
мальное количество метелок и максимальный вес растений наблюдаются
в условиях 15—19-часового дня [487, 666].
Дополнительное освещение в летние месяцы эффекта не дает [373].
Полевица (Agrostis stolonifera)
Длиннодневное растение, цветущее лишь при длине дня порядка
16—18 часов [373].
Хорошо растет при нижнем освещении с помощью люминесцентных
ламп (1000 вт/м2). Можно рекомендовать в качестве газонной культуры
для станций метрополитена [120, 123].
Просо итальянское (Могар) (Setaria italica)
Короткодпевпое растение, цветущее в условиях короткого дня через 55,
а в условиях длинного дня — через 90—100 дней. Для задержки цвете-
ния достаточно уже 15 лк, но полный эффект наблюдается прп дополни-
тельном освещении в 200 лк (фиг. 142, 5 и фиг. 152 [488]). Более сильная
задержка наблюдается при освещении в утренние часы (с 1 до 8 час.).
X. Светокультура полевых растений
413
Пырей (Agropyrum Smithii)
Длиннодневное многолетнее кормовое растение, в зимние месяцы цве-
тущее лишь при дополнительном освещении (через 120—150 дней 1405]).
Без освещения растения в это время почти совершенно не растут [751].
Для перехода к цветению необходимо дополнительное освещение
порядка 350 лк.
Тимофеевка (Phleum pratense)
Длиннодневные или нейтральные растения. Зимою можно культиви-
ровать при дополнительном освещении в 300—800 лк [715, 716]. Допол-
нительное освещение с помощью ламп накаливания в 200 вт (3000—
4000 лк), удлиняющее естественный день до 16—18 часов, ускоряет на-
ступление колошения у самых различных форм и сортов на 2—53
дня [373, 486].
По данным Смита [772], оптимальным является 18-часовой день
(фиг. 143, 9; фиг. 144, 9 и фиг. 146, 6). В этом случае наблюдаются наи-
более быстрое цветение и максимальный вес и высота растений. Анало-
гичные данные получены в опытах Келлера и Петерсон [571] по дополни-
тельному освещению тимофеевки в чистом посеве и в смеси с клевером.
Относительное содержание тимофеевки в травосмеси возрастает вместе
с увеличением длины дня.
Тимофеевка хорошо растет при нижнем освещении с помощью люми-
несцентных ламп (600 вт/м2), образуя хороший газон. Можно рекомендо-
вать как газонную культуру для озеленения станций метрополитена
[120, 123].
6. ВЫВОДЫ
Таким образом, вопрос о светокультуре сельскохозяйственных поле-
вых растений, в сущности говоря, сводится к дополнительному ночному
и дневному освещению в зимние месяцы с целью получения зимних поко-
лений для ускорения процесса отбора и к дополнительному ночному
освещению лотом для ускорения получения семян. Последнее особенно
необходимо для длиннодневных растений в южных районах страны
Для этой цели вполне достаточно освещенности порядка 200—500 лк.
Эта же величина освещенности необходима при дополнительном ноч-
ном освещении зимою с целью вызвать цветение длиннодневных растений
или, напротив, задержать цветение растений короткого дня. В южных
районах страны с высокой естественной освещенностью в дневные часы
подобный режим, невидимому, будет достаточен для нормальной культуры
растений.
Что касается умеренной зоны СССР, то здесь необходимо прибегать
или к интенсивному дополнительном}' ночному освещению (порядка
8000 лк) или к интенсивному освещению в дневные часы. Иными словами,
в данном случае при употреблении ламп накаливания необходимо обеспе-
чить удельную мощность в 2000 вт/м2 в течение 12—16 часов и 200 вт/м2 —
в течение последующих 6—8 часов.
Предпочтение следует отдать люминесцентным лампам, которые раз-
мещаются в вертикальной плоскости в междурядьях растений и при удель-
ной мощности 300—600 вт/м2 дают освещенность 5000—8000 лк, позволяя
получать в зимние месяцы нормальные растения. Следует помнить при
этом, что длина дня в большинстве случаев не должна превышать
16—18 часов.
Глава XI
СВЕТОКУЛЬТУРА ДРЕВЕСНЫХ РАСТЕНИИ
Светокультура древесных растений в нас тоящее время, в связи с озе-
ленением городов, приобретает очень большое значение. Проблема прак-
тически сводится к ускоренному выращиванию саженцев древесных и зим-
ней выгонке цветочных кустарников. В настоящей главе мы рассмотрим
лишь первый вопрос, так как выгонка кустарников рассмотрена нами
в главе IX.
Ускоренное выращивание саженцев древесных возможно двумя спо-
собами: 1) с помощью беспрерывного исключительно искусственного осве-
щения или 2) с помощью ночного освещения, дополнительного к есте-
ственной дневной радиации. Первый метод мы рассмотрели в главе VI,
здесь же нам остается рассмотреть лишь метод дополнительного ночного
освещения. Этот прием дешевле и вполне осуществим как в зимние месяцы
в теплицах, так и в летний период непосредственно в питомниках. Он
заключается в том, что сеянцам древесных создаются условия беспрерыв-
ного освещения лампами, включаемыми в ночные часы. Подобное воздей-
ствие позволяет получить несколько приростов в течение вегетационного
сезона и в течение нескольких лет вырастить взрослые растения даже
медленно растущих пород. При этом световое воздействие производится
в течение одного сезона на первом или втором году вегетации, а даль-
нейший рост сеянцев происходит в условиях естественного дня. В резуль-
тате длительного последействия развитие и рост растений настолько
ускоряются, что позволяют значительно сократить время выращивания
саженцев древесных.
Акация (Acacia)
Вечнозеленые светолюбивые растения. .1. melanoxylon в условиях
беспрерывного освещения растет равномерно и несколько интенсивнее,
чем в естественных условиях: годовой прирост в первом случае равняется
119 см, а во втором— 108 см. Л. longtfoUa на беспрерывное освещение
реагирует отрицательно, давая годовой прирост в 45 см вместо Ю8 см
в контроле [531.
Белая акация (Robinia pseudacacia)
Хорошо реагирует на беспрерывное освещение, усиливая рост и пре-
вращаясь в вечнозеленое растение. В опытах Б. С. Мошкова [208, 2091
белая акация в результате дополнительного освещения прекратила листо-
XI. Светокультура древесных растений
415
пад и продолжала расти всю зиму. Прирост за 8 месяцев равнялся 100 см
вместо 25 см в контроле.
По данным Портера и Ли [678], усиление роста вызывает не только
беспрерывное освещение, но и ночное освещение в течение 4% часов.
Аналогичные данные получены также Крамером [584] при освещенности
1000—2000 лк. Необходима удельная мощность 100 вт/м2.
Апельсин (Citrus sinensis)
В опытах М. X. Чайлахяна и Л. Д. Азбукина [337] слабо реагировал
на беспрерывное освещение, в точение 40 дней дав прирост в 15 см вместо
13 см в контроле, т. е. в условиях естественного зимнего дня.
Береза (Betula verrucosa)
Растение, обычно дающее один прирост в год. Исключительно отзыв-
чива на беспрерывное освещение с помощью ламп накаливания и неоно-
вых ламп, сильно увеличивая прирост [53, 196]. В опытах В. П. Маль-
чевского даже 4—6-часовое дополнительное освещение в течение 2—3 ме-
сяцев увеличивает прирост до 40—45 см вместо 20—28 см в контроле [196].
Этот эффект сопровождается увеличением размеров листа и толщины колец
древесины и коры. В. 3. Гулисашвили [53] нашел, что в условиях бес-
прерывного освещения лампами накаливания в 1000 вт береза дает
за 12 месяцев прирост 62 см против 10 см в контроле.
По данным В. П. Мальчевского, для березы необходима интенсивность
дополнительного освещения в 4000 лк. Ежесуточный расход энергии —
0,8 квт-час на 1 м2 неоновыми лампами (200 вт/м2) пли 2,4 квт-час на 1 м2
лампами накаливания (400 вт/м2).
Зимою береза хорошо растет при дополнительном освещении люминес-
центными лампами (16 часов: 300—600 вт/м2; 5000—8000 лк), обнаружи-
вая сильную ветвистость [160].
Бирючина (Ligustrum)
По данным Одена [646], L. vulgare, и L. pyramidale не реагируют на осен-
нее дополнительное освещение; листопад начинается одновременно как
при освещении, так и в контроле.
Боярышник (Crataegus)
Литературных данных по культуре боярышника с помощью допол-
нительного освещения нет. Имеются лишь работы но нарушению зимнего
покоя с помощью ламп накаливания [764, 811]. Опыты с Crataegus sp.
и С. oxyacantha показывают, что для нарушения покоя нужна высокая
температура (21°), дополнительное же освещение влияет незначительно.
Распускание почек происходит через 10, а полное распускание листьев
через 17 дней.
Бузина (Sambucus sp.)
Дополнительное освещение в ночные часы (600 лк) применялось лишь
для зимней выгонки растений [547, 556, 700]. Освещение, применяемое
после тепловой ванны, вызывает цветение через 30—40 дней, ускоряя
распускание цветков на 9 дней. При тепловом воздействии в конце декабря
416
Частная светокультура растений
цветение наступает в начале февраля. По данным Гёстермана [5561, при
дополнительном освещении зимою (ноябрь—январь) в 300—900 лк бузина
образует больше цветков с более яркой окраской и более сильным запахом.
Бук (Fagus)
Растения с периодическим ростом, обычно дающие два прироста в год.
F. grandtfolia, по данным Крамера [584], значительно усиливает рост
и удлиняет период вегетации в результате дополнительного освещения
в ночные часы лампами накаливания в 200 вт (1000—2000 лк). F. silvatica
и F. orient alls на дополнительное освещение, удлиняющее естественный
короткий день, не реагируют [53, 646]. У этих видов годовой прирост
(И—12 см) не увеличивается, а листопад начинается одновременно с кон-
тролем.
Для нарушения зимнего покоя F, silvatica необходима теплая ванна
и дополнительное 12-часовое освещение в 60 г-кал/см2 в день. В этом
случае распускание листвы осуществляется через 30 дней [646].
Виноград (Vitis vinifera)
Светолюбивое растение. Сименс [754] с помощью вольтовой дуги полу-
чал зрелые гроздья винограда через 83 дня после начала освещения
(14 марта).
Я. И. Потапенко [251] в результате исследований над мичуринскими
и южными сортами винограда (Русский Конкорд, Сеянец Малерма, Силь-
ванер, Агадан, Семильон и Сеянец амурский) нашел, что в большинстве
случаев беспрерывное освещение не увеличивает прироста побегов, но
вызывает снижение сухого веса и числа листьев и сильно ухудшает вызре-
вание стебля. Последнее может привести к снижению морозостойкости
винограда.
Граб (Carpinus)
Медленно растущие растения, обычно дающие по одному приросту
в год. При беспрерывном освещении С. betulus растет в течение 335 дней
вместо 245 дней в контроле; соответствующие данные для С. orientalis —
328 и 247 дней [53].
Дерен (Cornus)
С. alba и С. mas после обработки ацетиленом нормально цветут зимою
при дополнительном освещении лампами накаливания [646, 811]. По дан-
ным Одена [646], при ежедневной затрате лучистой энергии 60 г-кал/см2
дерен зацветает через 20 дней (27 декабря).
При дополнительном освещении зимою С. stolonijera распускает листья
несколько раньше, чем в контроле [764].
Дуб (Quercus)
Медленно растущее растения, обычно дающие один прирост в год.
Дуб относится к группе растений, исключительно сильно реагирующих
на дополнительное, а в особенности на беспрерывное освещение. По дан-
ным Крамера [584], Q. stellata и (). alba при дополнительном освещении
с помощью ламп накаливания в 200 вт (1000—2000 лк), удлиняющем
XI. Светокультура древесных растений
417
день до 14% часов, усиливают интенсивность роста и удлиняют его пе-
риод, одновременно сокращая период покоя. Как в оранжерее, так и в пи-
томнике названные виды уходят в зимний покой значительно позднее,
чем в условиях естественного короткого дня. Лишь Q. borealis var. maxima
в условиях длинного дня сокращает как период роста, так и период
покоя. Зимою этот вид легко трогается в рост, как только попадет в усло-
вия дополнительного освещения. В. 3. Гулисашвили [53] нашел, что
Q. macranthera, растущий в естественных условиях в течение 184 дней
и дающий в этом случае один прирост в год, при беспрерывном освещении
увеличивает период роста до 284 дней, a Q. iberica, растущий обычно
в течение 187 дней (один прирост), в условиях беспрерывного освещения
дает четыре прироста в год, увеличивая прирост до 40 см вместо 8 см
в контроле.
Вечнозеленый Q. subur, дающий в естественных условиях один при-
рост в год (14 см), при дополнительном освещении (беспрерывный день)
увеличивает годовой прирост до 27 см.
Иными словами, несмотря на то, что различные виды дуба реагируют
на дополнительное ночное освещение по-разному, все они, тем не менее,
в условиях беспрерывного освещения дают значительно больший при-
рост, чем в условиях естественного короткого дня.
Для получения соответствующего эффекта необходима сравнительно
невысокая интенсивность дополнительного освещения — порядка 1000—
2000 лк.
По данным В. М. Лемана [160], зимою дуб хорошо растет при допол-
нительном освещении люминесцентными лампами (в течение 16 часов
300—600 вт/м2; 5000—8000 лк).
Ель (Picea sp.)
Теневыносливое медленно растущее растение. В опытах В. П. Маль-
чевского [196] в летние месяцы слабо реагировала на дополнительное
6-часовое освещение в 4000 лк, практически совершенно не увеличивая
прирост. В. М. Леман [196] выращивал ель зимою при дополнительном
ночном и дневном освещении люминесцентными лампами (16 часов;
300—600 вт/м2: 5000—8000 лк).
Жимолость (Loiiicera campcstre)
По данным Одена [646], при дополнительном освещении прекращает
листопад и становится вечнозеленой.
Ива (Salix)
Быстро растущие растения. Исключительно хорошо реагируют на до-
полнительное освещение [25, 208, 209, 616, 811]. 5. luminalis и S. phyli-
cijolia в условиях беспрерывного освещения (100 вт/м2) становятся вечно-
зелеными, вргетируют всю зиму и годовой прирост увеличивают
в 2—3 раза по сравнению с контролем; S. phylicifolia, кроме того, при
беспрерывном освещении зацветает на первом году, а в контроле не
цветет в течение 3 лет [208, 209]. Даже освещение уличных фонарей,
расположенных на расстоянии 1—1,5 м, значительно удлиняет вегета-
ционный период и вызывает более поздний листопад [616]. У S. jragilis
такое же влияние наблюдается уже при освещенности в 10 лк.
27 А. Ф. Клешнин
418
Частная светокультура растений.
Дополнительное освещение применяется также для нарушения зим-
него покоя ивы. S. viminalis, находящаяся в глубоком покое (ноябрь),
при освещении в 6000 лк распускается через 24—27 дней при беспре-
рывном освещении, через 24—31 день — при 18-часовом дне и через
76—85 дней в условиях естественного короткого дня (6—10 часов); осве-
щение в конце периода покоя (март) на распускание почек влияния не
оказывает, и листья распускаются через 10—20 дней как при освещении,
так и в контроле [25]. S. alba var. vitellina зимою распускается и цветет
лишь при дополнительном освещении [811].
Инжир (Ficus carica)
По данным В. М. Лемана [160], в зимние месяцы (январь—март) хо-
рошо растет при дополнительном ночном и дневном освещении люминес-
центными лампами (16 часов; 300—600 вт/м2; 5000—8000 лк), обнаружи-
вая сильное ветвление.
Калина (Viburnum opulus)
По данным Одена [646], при дополнительном освещении зимою стано-
вится вечнозеленой и цветет. Ранке [556, 700] также вызывала цветение
калины зимою с помощью дополнительного освещения, выводя ее из со-
стояния покоя. В опытах Скока [764] названное растение реагировало
на дополнительное освещение лишь в условиях низких температур (10°),
распуская листья прп освещении через 19 дней и не распуская в контроле;
при температуре 21° распускание происходило в обоих случаях одновре-
менно — через 10—11 дней.
Катальпа (Catalpa sp.)
Растение, обычно дающее один прирост в год. Прп беспрерывном
освещении период роста увеличивается с 241 дня до 288 дней, а годовой
прирост — с 13 до 22 см [53].
Каштан (Aesculus)
По наблюдениям Лауборта [592], A. pavia под влиянием уличных фона-
рей, горящих по 7—8 часов в сутки, задерживает листопад на 2—3 недели.
Дополнительное освещение в середине зимы (январь) не нарушает покоя
A. hippocastanum [811].
По данным Фейера [489], сильное уличное освещение сокращает период
роста и вызывает листопад.^
Кедр (Pinus sibirica)
В. М. Леман [158] показал, что кедр реагирует на дополнительное
(беспрерывное) освещение значительным усилением роста (на 80—100%)
и ускорением в развитии. В его опытах годовой прирост при освещении
составлял 9—10 см, а в контроле •— 5 см. Пучковая хвоя, обычно по-
являющаяся на 2—3-м году, под влиянием беспрерывного освещения
образовалась на первом году.
Опыт показывает, что дополнительное освещение с помощью лампы
накаливания в 500 вт (330 вт/м2; 1000—1500 лк) дает лучший эффект,
чем от лампы накаливания в 100 вт (67 вт/м2; 500—800 лк).
XI. Светокультура древесных растений.
419
Керрия (Kerria japonica)
По данным Клейна [579], названное растение хорошо реагирует на до-
полнительное освещение зимою, быстрее выходя из состояния покоя
и равномернее и дружнее зацветая.
Клен (Acer)
Теневыносливые растения, обычно дающие один прирост в год. В ре-
зультате дополнительного беспрерывного освещения становятся или
вечнозелеными или значительно увеличивают период вегетации [53, 646].
A. campestre, по данным В. 3. Гулисашвили [53], в естественных условиях
растет 245 дней, давая прирост 5 см, а при беспрерывном освещении
растягивает период роста до 331 дня и дает прирост 10 см. A. trautvetteri
также удлиняет период вегетации с 184 до 312 дней, но при этом умень-
шает прирост с 16 до 4 см. Повидимому, этот вид относится отрицательно
к беспрерывному освещению.
Для нарушения зимнего покоя /1. palmatum необходима интенсивность
радиации 0,08 г-кал/см2-мин (5—6 тыс. эрг/см2-сек) и длина дня 15—17 ча-
сов; в этом случае полное распускание происходит за 41 день [646].
Ликвидамбар (Liquidambar styraciflua)
При дополнительном освещении в 1000—2000 лк растения растут
значительно интенсивнее, причем в условиях 16-часового дня растут
почти всю зиму, а при беспрерывном освещении становятся вечнозеле-
ными. Дополнительное освещение нарушает зимний покой и значительно
сокращает цикл, состоящий из периода роста и периода покоя [584].
Лимон (Citrus limon)
Теневыносливое растение, зимою при дополнительном беспрерывном
освещении увеличивающее прирост побегов на 20—30% [337].
Хорошо растет при дополнительном ночном и дневном освещении
от люминесцентных ламп (16 часов; 300—600 вт/м2; 5000—8000 лк),
обнаруживая сильное ветвление [160].
Лиственница (Larix sp.)
Светолюбивое растение. Дополнительное освещение в летние месяцы
не влияет на его рост [196]. Зимою хорошо растет при дополнительном
освещении люминесцентными лампами [160].
Миндаль (Amygdalus sp.)
В зимние месяцы можно культивировать с помощью дополнительного
освещения от ламп накаливания в 200 вт при освещенности 300—900 лк
[556].
Ольха (Alnus glutinosa)
Зимою (январь) цветет при дополнительном освещении лампами нака-
ливания несколько раньше, чем в контроле [811].
27*
420
Частная светокультура растений
Орех (Corylus avellana)
Зимою (январь) зацветает одновременно как при освещении, так
и в контроле [811].
Осина (Populus tremula)
Быстро растущее растение.
Зимою из состояния глубокого покоя (ноябрь) выводится лишь с по-
мощью беспрерывного дополнительного освещения в 6000 лк; распуска-
ние почек в этом случае происходит через 40—96 дней. Дополнительное
освещение влияет слабее в период окончания периода покоя (март);
в данном случае распускание почек совершается через 11—13 дней,
а в контроле — через 47 дней [25].
Платан (Platanus)
Р. orientalis является растением, на котором впервые было уста-
новлено, что уличное освещение вызывает особенно мощное развитие
побегов [456]. Позднее Чил [437, 784] установил, что в этом случае зна-
чительно запаздывает листопад. Аналогичное явление было отмечено
у Р. acerijolia и Р. occidentalis [616].
Для задержки листопада достаточна интенсивность дополнительного
освещения в 10 лк. в
Персик (Pcrsica vulgaris)
По данным Сименса [754], хорошо реагирует па дополнительное ночное
освещение.
Пробковое дерево (Phellodendron amurense)
Ph. amurense при дополнительном освещении лампами накаливания
в 1000 вт растягивает период роста до 352 дней в году вместо обычных
275 дней в естественных условиях, увеличивая ежегодный прирост
до 12 см против 7 см в контроле [53].
Пуэрария (Pueraria hirsuta, Dolichos japonica)
Обычно осенью сбрасывает листья и уходит в покой. При дополни-
тельном освещении хорошо растет всю зиму [501 ].
Смородина (Ribes)
R. nigrum при дополнительном освещении зимою трогается в рост
на 29 дней раньше контроля и увеличивает прирост па 76% [366]. По дан-
ным Б. С. Мошкова [209], R. nigrum в условиях беспрерывного освещения
(100 вт/м2) становится вечнозеленой, увеличивает прирост в несколько
раз по сравнению с контролем и зацветает па первом году (обычно сморо-
дина цветет на третьем году).
R. rubrum зимою (январь) распускается одновременно как при осве-
щении, так и в контроле [811].
XI. Светокультура древесных растений
421
Сосна (Pinus)
Светолюбивое растение. Хорошо реагирует на дополнительное осве-
щение, усиливая рост и ускоряя развитие. Р. taeda в условиях беспрерыв-
ного освещения (1000—2000 лк) в зимние месяцы растет в несколько раз
интенсивнее, чем в контроле [584]. Р. pinea, обычно дающая один при-
рост в год (15 см), в условиях беспрерывного освещения дает годовой
прирост 56 см [53]. Р. silvestris при беспрерывном освещении увеличивает
прирост на 33—100% по сравнению с контролем, причем лампы накали-
вания в 500 вт (330 вт/м2; 1000—1500 лк) дают лучший результат, чем
лампы в 100 вт (67 вт/м2; 500—800 лк). В первом случае прирост дости-
гает 12 см, во втором — 8 см, а в контроле — 6 см. Максимальный при-
рост и максимальное число листьев у Р. silvestris имеют место в условиях
20-часового дня (фиг. 146, 10 [835]). По данным Уоринга [834, 835], осен-
нее удлинение дня до 15 часов с помощью ламп накаливания (150—200 лк)
значительно усиливает активность камбия у Р. silvestris, обычно прекра-
щающего свою деятельность в осенние дни. Как показывают опыты того же
автора [835], нарушение летнего покоя сосны возможно лишь с помощью
дополнительного освещения при длине дня 20—24 часа; 18-часовой день
дает значительно худшие результаты. Покой нарушается легко беспре-
рывным освещением в июле—сентябре, но с трудом и лишь частично —
в октябре. Интересно, что парная хвоя, которая обычно появляется
на 2—3-м году, при беспрерывном освещении формируется через месяц
[158].
В. П. Мальчевский [196] показал, что в летние месяцы дополнитель-
ное освещение в течение 6 часов даже при освещенности 4000 лк на рост
сосны не оказывает никакого влияния, если не считать появления более
крупной хвои. Неоновые лампы дают лучший эффект, чем лампы нака-
ливания.
Сумах (Rhus)
Светолюбивые растения. R. grabra и R. copallina при дополнительном
освещении становятся вечнозелеными или задерживают листопад на не-
сколько недель [501].
Тополь (Populus canadensis)
По наблюдениям Мацке [616], осенью под влиянием уличных фонарей,
расположенных даже на расстоянии 1,5 м, значительно запаздывает
с листопадом, причем для получения этого эффекта достаточно уже 10 лк.
Зимою под влиянием дополнительного освещения (6000 лк) легко
нарушает покой и трогается в рост. Состояние глубокого покоя (ноябрь)
при беспрерывном освещении нарушается через 13—70 дней, в условиях
18-часового дня — через 25—70 дней, а в контроле — через 37—74 дня.
Весною (март) дополнительное освещение на выход из состояния покоя
влияния не оказывает [25].
Тунг (Aleurites sp.)
По данным Б. С. Мошкова [209], при беспрерывном освещении увели-
чивает прирост в 2—3 раза по сравнению с естественным днем.
422
Частная светокультура растений
Тюльпанное дерево (Liriodendron tulipifera)
В условиях длинного дня значительно сокращает период покоя; при
беспрерывном освещении растет всю зиму. Из состояния покоя выходит,
как только подвергается действию дополнительного освещения [584]
Хинное дерево (Chinchona)
По данным Б. С. Мошкова [210], С. Ledgeriana и С. succirubra дости-
гают максимальных размеров в условиях короткого дня (10—12 часов).
Беспрерывное освещение (200 вт/м2) приводит к резкому снижению веса
растений — в 1,6—2,2 раза по сравнению с естественным днем и в 4,8—
6,0 раз по сравнению с 10—12-часовым днем.
Хорошо растет в зимние месяцы при дополнительном ночном и днев-
ном освещении люминесцентными лампами (в течение 16 часов; 300—
600 вт/м2; 5000—8000 лк [160]).
Шиповник (Rosa canina)
При дополнительном освещении лампами накаливания в летние ме-
сяцы растет лучше, чем в условиях естественного дня [196].
Зимою при переносе в оранжерею трогается в рост под лампами бы-
стрее, чем в контроле [811].
Ясень (Fraxinus)
По наблюдениям Крамера [584], F. americana и F. pennsylvanica. на до-
полнительное освещение (1000—2000 лк) не реагируют; во всяком случае
листопад при освещении начинается одновременно с контролем.
ВЫВОДЫ
Таким образом, дополнительное освещение является хорошим методом
для ускорения роста и нарушения летнего и зимнего покоя древесных
растений. Лучший результат наблюдается при этом в условиях беспре-
рывного дня. В большинстве случаев при беспрерывном освещении дре-
весные становятся вечнозелеными и усиливают прирост побегов в не-
сколько раз. В умеренной полосе под влиянием дополнительного ночного
освещения листопад наступает значительно позднее, а весною растения
раньше трогаются в рост.
У ряда пород эффект наблюдается уже при освещенности в 10 лк,
обычно же пользуются освещенностью порядка 500—1000 лк. При зимней
культуре древесных в большинстве случаев необходима освещенность
4000—6000 лк.
Для культуры древесных пригодны любые источники радиации,
если их излучение не содержит вредных ультрафиолетовых лучей. Пови-
димому, на усиление роста в высоту лучше влияют лампы накаливания,
которые имеют ряд преимуществ благодаря крайней простоте эксплуатации.
Кроме того, при нарушении состояния покоя не последняя роль принад-
лежит инфракрасным лучам, которыми так богаты лампы накаливания.
Для получения нормальных растений предпочтение следует отдать люми-
несцентным лампам, помещая их между растениями в вертикальной
плоскости (300—600 вт/м2).
ЗАКЛЮЧЕНИЕ
Светокультура растений — выращивание растений целиком на искус-
ственном свете или при дополнительном искусственном освещении в ноч-
ные или дневные часы — возникла в конце прошлого столетия, хотя
первые попытки применения искусственных источников радиации для
культуры растений имели место еще в середине столетия [319, 612]. В на-
стоящее время число исследователей, так или иначе связанных с пробле-
мой светокультуры, превышает 480 человек, а число публикаций по свето-
культуре растений — более 800.
Исследования по светокультуре растений развиваются главным
образом в странах, расположенных в умеренных и северных широтах
(СССР, Финляндия, Швеция, Норвегия, Голландия, Дания, Германия,
США и другие). Исследования в других странах носят случайный и спо-
радический характер. Постановка исследований в этом направлении
в СССР связана с именами Н. А. Максимова и Н. А. Артемьева, в Голлан-
дии — Роденбурга (Roodcnburg), в Швеции — Одена (Oden) и Оберга
(Aberg), в Норвегии — Смита (Smith), в Финляндии — Меурмана (Менг-
man), в Германии — Гёстермана (Hostermann) и Рейнгольда (Reinhold),
а в Соединенных Штатах Америки — Гарвея (Harvey), Артура (Arthur)
и других.
Светокультура растений представляет собою проблему, объединяю-
щую в себе вопросы биологии, физики и агротехники. Успешное решение
этой проблемы возможно лишь в результате совместных исследований
биологов, физиков и агротехников. Между тем, до сих пор она решалась
главным образом лишь физиологами растений при весьма незначитель-
ном участии физиков. Именно поэтому в настоящее время решены лишь
принципиальные физиологические стороны проблемы, а вопросы агро-
техники, частной светокультуры и светотехники ждут еще своего разре-
шения. Вопросом, настоятельно требующим своего решения, является
температурный режим в условиях светокультуры. Актуальна также
задача разработки специальных осветителей и рациональных источников
излучения. В ближайшие годы, несомненно, встанет задача зимней про-
изводственной культуры томатов, огурцов, земляники и других расте-
ний. Повидимому, возникнет и задача озеленения станций метрополитена.
Светокультура растений — проблема большого как теоретического,
так и практического значения. В результате почти столетних исследова-
ний многочисленных авторов установлено, что с помощью искусственных
источников радиации можно культивировать любое автотрофное расте-
ние — от водорослей до высших цветковых растений. Для этой цели,
в зависимости от конкретных задач культуры растений, пригодны раз-
ные источники радиации — лампы накаливания, ртутные, неоновые
424
Заключение
натриевые, дуговые, люминесцентные и даже керосиновые лампы, если
их излучение сосредоточено в области физиологической радиации
(400—700 mp„), если их мощность достаточно велика, чтобы обеспечить
необходимую интенсивность радиации, и если в спектре излучения от-
сутствуют вредные витацидные лучи (менее 300 ту.).
Практическое значение проблемы связано главным образом с допол-
нительным освещением растений при селекции, в овощеводстве и цвето-
водстве. В настоящее время, применяя дополнительное освещение в ноч-
ные или дневные часы, можно обеспечить выгонку рассады овощных
и цветочных растений к любому времени года, можно культивировать
цветочные растения зимою и вести круглогодичную селекцию и семено-
водство культурных растений.
Огромная заслуга в постановке и решении проблемы светокультуры
растений принадлежит исследователям СССР (Н. А. Максимов, Н. А. Ар-
темьев, Н. П. Красинский, В. П. Мальчевский, В. М. Леман, Б. С. Мош-
ков и другие). Советская литература по светокультуре растений содержит
свыше 300 названий и связана с исследованиями большого числа авторов
(около 130 человек). СССР является родиной камер искусственного кли-
мата (И. А. Артемьев, 1023), советские исследователи одни из первых
сумели вырастить растения «от семени до семени» целиком на искусствен-
ном свете (Н. А. Максимов, 1925); в Советском Союзе поставлена и ус-
пешно решается проблема светокультуры древесных (В. П. Мальчевский,
1946; В. М. Леман, 1946); советские исследователи в настоящее время
успешно работают над теорией светокультуры растений.
В СССР светокультура растений приобретает все более и более широ-
кое значение как метод ранней выгонки овощей и как способ продвижения
овощных и декоративных растений па север.
Несомненно, что в ближайшие годы по мере вступления в строй Волж-
ских гигантов-электростанций и в связи с появлением дешевой электро-
энергии светокультура растений будет играть еще большую роль. Задача
заключается в том, чтобы продвинуть южные растения на север, чтобы
обеспечить население СССР свежими овощами и фруктами на протяже-
нии всего года и чтобы широко продвинуть в быт цветочные и декоратив-
ные растения. Не подлежит сомнению, что советские ученые успешно
справятся с этой задачей.
Л ИТ Е Р А Г У Р А
В список литературы включены лишь исследования, использованные автором
и находящиеся в библиотеке Отделения биологических паук АН СССР, библиотеке
Сельскохозяйственной академии им. К. А. Тимирязева, Всесоюзной библиотеке
им. В. И. Левина, Государственной научной библиотеке Министерства высшего обра-
зования СССР, Библиотеке Московского общества испытателей природы, Централь-
ной политехнической библиотеке, Научной библиотеке им. А. М. Горького Москов-
ского университета им. М. В. Ломоносова, Центральной сельскохозяйственной библио-
теке, Библиотеке Академии Наук СССР (Ленинград) и Библиотеке Ботанического
института им. В. Л. Комарова (Ленинград). Сравнительно небольшое число работ,
отсутствующих в библиотеках СССР, включено по рефератам. В этом случае, помимо
основного источника, указано название реферативного источника.
Работы, не прямо относящиеся к светокультуре растений, но имеющие для этого
вопроса значение, всюду отмечены звездочкой.
Автор не претендует на абсолютную полноту списка. Всякое указание на отсут-
ствие того или иного исследования в этом списке он примет с благодарностью.
1. Ава к я н А. А. и '1' а г и - В а д е А. X. О так называемой «яровизации»
светом. Яровизация, ,№ 1, 65—107 (1935).
2. А в а к я и А. А. О биологии развития томатов. Яровизация, 2—3 (5—6),
34—66 (1936).
3. Алекперов С. А. Динамика формирования колоса в связи со стадийностью
растений. Изд. Наркомпроса АзССР. Баку (1939).
4. А л е к с а и д ров С. В. Опыты выращивания овощей и цветов на острове
Диксон. Советская Арктика, № 7, 68—73 (1938).
5. Александров С. В. Два года на острове Диксон. Плодоовощное хозяй-
ство, № 8-9, 40—43 (1938).
6. А и т о и ю к А. А. Использование тепловых отходов промышленности. Дости-
жения науки и передового опыта в сельском хозяйстве, № 10, 85—88 (1952).
7. Аптон ю к А. А. Опыт работы зеленного цеха завода. Сад и огород, № 9, 48—51
(1952).
8. Артемьев И. А. Проблемы энерговоздействия на рост растений. Изд.
ВАСХНИЛ, М. (1936).
9* . Б а л а ш и и с к и й Б. Л., С о к о л о в И. И. и Ф о м и и В. Н. Справоч-
ник по электрическим лампам накаливания. Изд. Бюро техн, информации МПСС
СССР. М. (1947).
10. Банников Н. А., Пищальников Д. Н., Пожидаев А. И.
и Рузин О. М. Ордена Ленина колхоз «Память Ильича». Сельхозгиз, М.
(1952).
11. Баранский Д. П. О биологическом анализе сортов подсолнечника при
различной длине дня. Яровизация, № 2—3 (5—6), 67—76 (1936).
12. Б а с с а р с к а я М. А. и А л е х и н 11. В. Влияние интенсивности и качества
света на развитие пшеницы. Яровизация, № 4—5 (19—20), 165—172 (1938).
13* . Белькпнд Л. Д. Электроосветительные приборы ближнего действия. Гос-
эпоргоиздат. М. (1945).
14* . Б е р л и н е р А. Курс физики. Ч. 2. рып. 2. Оптика. Гостехиздат. М. (1933).
15. Биологическое действие света при выращивании рассады овощей. Электрифика-
ция сельского хоз-ва, № 1, 49—50 (1937).
16. Битюков К. К. Растение и его жизнь. Сельхозгиз (1953).
17. Борисов В. Цветы для метро. Советский метрополитен, № 130 (2756) (1952).
18. Борисов М. А. Выращивать овощи в зимнее время. Рабочая Москва, № 262
от 14 ноября 1938 г.
19. Боровицкий II. II. Краткий справочник преподавателя естествознания.
Учпедгиз, \1. (1951).
426
Литература
20. Брызгалов В. А. Овощеводство защшценного групта северной полосы
СССР. Ленсельхозгиз, Л. (1934).
21. Брызгалов В. А. и Русин М. И. Влияние дополнительного электро-
освещения на развитие и урожай огурцов. Зап. Ленингр. плодоовощного ин-та,
вып. 1, 5—14 (1934).
22. Б р ы з г а л о в В. А. Осенне-зимняя культура овощей. Лениздат, Л. (1944).
23. Б у д з к о И. А. К вопросу о добавочном освещении в теплицах. Электрифика-
ция сельского хоз-ва, № 5/57—62 (1934).
24. Б у д з к о И. А. Новые опыты по искусственному освещению для воздействия
на развитие растений. Электрификация сельского хоз-ва, № 2, 79 (1936).
25. Булгакова 3. II. Влияние длины дня на распускание покоящихся почек
V древесных растений. Бот. журн. СССР, 22, № 5, 413—419 (1937).
26. В комиссии по светотехнике ОТН АН СССР. И.зв. АН СССР, ОТН, № 6, 964 (1949).
27. В лучах электроламп. Вечерняя Москва, № 2/1 (8785) от 17 ноября 1952 г.
28*. Вавилов С. И. О «теплом» и «холодном» свете. Изд-во АН СССР, М. (1949).
29. Вавилов С. И. Люминесценция и ее применение в светотехнике. Собр. соч.,
т. 2, 281—292 (1952).
30. В е р з и л и н И. М. Выращивание растений на электрическом свете. Биология
и химия в средней школе, № 4 (8), 8—14 (1935).
31. Верзилин II. М. Путешествие с домашними растениями. Дстгиз, М., 284—
287 (1949).
32. Веселовский И. А. Выращивание земляники на искусственном электро-
освещении. Плодоовощное хоз-во, № 2, 58—59 (1936).
33. Веселовский II. А. Выращивание овощей при электроосвещении. Докл.
ВАСХНИЛ, вып. 18, 17—18 (1940).
34. Власов И.А., Н ацентов Д. И. и др. Овощеводство. Ссльхозгпз, М. (1951).
35. Вобликова Т. В. Интенсивность фотосинтеза и дыхания листа в зависи-
мости от возраста. Докл. АН СССР. 33, Al 1, 76—77 (1941).
36. В о б л и к о в а Т. В. Фотосинтез и дыхание растений в условиях светокультуры
растений. Тр. Ин-та физиологии растений им. К. А. Тимирязева АН СССР, 8,
вып. 1, 184—209 (1953).
37. В о с к р е с е н с к а я Н.П. О влиянии спектрального состава света на соот-
ношение веществ, образующихся при фотосинтезе. Докл. АН СССР, т. 86, № 2,
429—432 (1952).
38. Воскресенская Н. II. Влияние условий освещения на состав продуктов
фотосинтеза. Тр. Ин-та физиологии растений им. К. А. Тимирязева АН СССР,
8, вып. 1, 42—56 (1953).
39. Временная инструкция по эксплуатации люминесцентных ламп. Изд. МЭЛЗ (1948).
40. Второе совещание по люминесценции. Вести. АН СССР, «№ 8, 84—88 (1948).
41. Г а в р и л о в Н. II. Новые задачи светокультуры. 1-я конференция молодых
ученых ТСХА. Вып. 28, 1—2 (1936).
42. Гаевская Н. С. Опыт применения газосветных ламп для культивирования
протококковых водорослей. Бголл. Моск, об-ва испыт. природы, биология, 57,
вып. 4, 35—41 (1952).
43. Г с и к е л ь И. А. и И с а к о в а О. В. II. А. Максимов. Изд-во АН СССР,
М. (1949).
44. Герасимов М. С. Растения и цветы выращены без солнца. Московская
правда, № 34 (9983) от 26 декабря 1952 г.
45*. Гер лих II. Фотоэлементы. Гостехиздат. М. (1948).
46. Г и л ь м е р Е. Опыты по применению света и тепла в садоводстве Восточной
Пруссии. Электрификация сельского хозяйства, <№ 5—6, 97—101 (1932).
47*. Гинзбург В. Л. Светофильтры. Гостехиздат (1936).
48*. Голдовский Е. М. Светотехника киносъемки. Госкнноиздат, М. (1944).
49. Голованова Е. А. Иаш опыт выращивания ранних овощей. Московский
колхозник, № 4, 28—29 (1953).
50. Гончаров Б. И. Получение раннего урожая томатов в Ленинградской области
путем выращивания рассады па искусственном свете. Изд. ВАСХИПЛ. Л. (1952).
51. Горновесов Г. В. и др. Электрификация сельского хозяйства Запорожья.
Сельхозгпз, М. (1939).
52. Горюнова С. В. Культивирование водорослей в лабораторных условиях.
Микробиология, 17. вып. 5, 402—408 (1948).
53. Г у л и с а ш в и л и В. 3. Нарушение покоя, периодичности вегетации и рит-
мики роста у некоторых древесных пород в условиях светокультуры. Природа,
№ 3, 63—66 (1948).
54*.Д а и и и г е р А. С. Электрическая лампочка. Гостехиздат, М. (1949).
55. Декоративное садоводство. Краткий словарь-справочник. Сельхозгиз, М., 400—
401 (1949).
Л итература
427
56. Джалилова Р. А. Агротехника ранних культур томатов в условиях защи-
щенного грунта. Конференция молодых ученых ТСХА, посвященная XVIII съезду
. ВКП(б) (1939).
57. Джапаридзе Г. Культура томат на искусственном свете. Вести, с.-х. ин-та
Грузии, 1—2, 155—163 (1933).
58. Д и л н г е н с к а я Н. Под лучами ламп (выращивание цветов). Вечерняя
Москва, № 34 (8548) от 9 февраля 1952 г.
59. Доброхотова С. И. Культура томатов на искусственном свете. Тр. Лабо-
ратории светофизиологии ФАИ, вып. 1, 51—73 (1938).
60. Довгань В. А. и Карась М. II. Влияние различной длины дня при пита-
нии аммиачным и нитратным азотом на урожайность и качественные изменения
кок-сагыза. Научи, тр. Ин-та физиологии растений и агрохимии АП Укр. ССР,
№ 3, 288—299 "(1951).
61. Долгушин Д. А. К вопросу о фотопериодическом последействии. Бюлл.
яровизации, № 1, 30—35 (1932).
62. Долгушин Д. А. О комплексе факторов, обусловливающих плодоношение
проса. Бюлл. яровизации, № 1, 36—38 (1932).
63. Дополнительное искусственное освещение растений. Электрификация сельского
хоз-ва, № 3, 116 (1936).
64. Дорохов Л. М. и Д е м у сен к о А. 11. Исследования стадийного развития
редиса. Научи, зап. Молдавск. плодоовощного ин-та, вып. 1, 150—197 (1939).
65. Е в р е и н о в М. Г. Научные проблемы во второй пятилетке. Электрификация
сельского хоз-ва, № 1, 12—19 (1932).
66. Е в р о и н о в М. Г. Применение электричества в сельском хозяйстве. Сельхоз-
гиз, М. (1948).
67. Е в р е и н о в М. Г. Электричество в биологических процессах сельского хозяй-
ства. Докл. ВАСХНИЛ, вып. 6, 98—103 (1949).
68. Евтушенко Г. А. Изменение длины вегетационного периода у различных
сортов табака в зависимости от выращивания рассады па различной длине дня.
Сборник работ по физиологии табака и махорки, 1, 31—45 (1939).
69* . Е п а н с ш и и ков М. М. и Соколов М. В. Электрическое освещение.
Госэнсргоиздат, М. (1950).
70. Еременко В. Г. Динамжа вуглевод’ж i азотних сполук у сои в звъязку
з фотоверюдизмом. Сб)рник роб!т з агроф!з1ологп, 1. 84—109 (1936).
71. Е р ми л о в Г. Б. Растения и свет. Сельхозгиз, М. (1952).
72 *. Е р м о л и н с к и и Н. II. Осветительные зеркальные лампы. Всстн. электро-
пром., № 1, 23—27 (1949).
73. Жуков Г. И. Ранние овощи на Севере. Архангельское пзд-во (1947).
74. 3 а х а р е в и ч Н. Теплично-парниковое хозяйство колхоза «Память Ильича».
Изд-во «Моск, большевик» (1945).
75. Захаров Б. С. Влияние длины дня на поражасмость подсолнечника зара-
зихой. Докл. АН СССР, 27, № Ь, 265—267 (1940).
76. Зацепин И. И. Опыт выращивания растений при электрическом свете. Се-
веро-Кавказский учитель, № 5, 40—43 (1936).
77. Из работ Всесоюзного института электрификации сельского хозяйства (ВИЭСХ).
Электрификация сельского хоз-ва, № 2, 55—56 (1933).
78* . И в а и о в А. П. Электрические источники света. Лампы накаливания. Гос-
техи.здат, М. (1938).
79* . Иванов А. П. Электрические источники света. Лампы газового разряда.
Госэнсргоиздат, М. (1948).
80* . Иванов Л. А. Новые приборы для измерения полной и физиологической
радиации. Метеорол. вести., № 3 (1925).
81. И ванов Л. А. О применении искусственных источников света для культуры
растений. Бот. жури. СССР, 23, № 1, 33—60 (1938).
82. И ванов С. И. Влияние фотопериода на морозоустойчивость цитрусовых расте-
ний. Докл. АН СССР, 28, № 8, 737—739 (1910).'
83. И г н а т ь е в а В. М. Действие лучистой энергии на культуру томатов. Электри-
фикация сельского хоз-ва, № 1, 29—35 (1937).
84*. И з в е к о в Р. Г. Ртутные лампы высокого давления («Игар»). Каталогиздат (1939).
85*. Ильина А. А. Спектры поглощения и отражения листьев растений.
Жури. физ. химии, 21. вып. 2, 145—159 (1947).
86*. Инструкция по эксплуатации ртутных ламп высокого давления типа «Игар-2».
Изд. МЭЛЗ (1949).
87*, Инструкция по эксплуатации ртутных ламп сверхвысокого давления. СВДЩ-250.
Изд. МЭЛЗ (1946).
88. Поффе А. Пути развития современной физики. Известия, № 80 (8996)
от 3 апреля 1952 г.
428
Литература
89*. Казарян В. О. О значении интенсивности света в фотопериодизме расте-
ний. Докл. АН Арм. ССР, 4, № 1, 15—20 (1947). Тр. Бот. ин-та АН Арм. ССР,
т. 6, 1—62 (1949).
90. Калинин М. С. Влияние электрического ноля высокого напряжения и иони-
зации на рост и развитие огурцов. Электрификация сельского хоз-ва, № 6, 40—51
(1933).
91. Кал и в и н М. С. Влияние электрического света различного состава на энер-
гию развития озимой пшеницы и томатов. Электрификация сельского хоз-ва,
№ 1, 43—55 (1934).
92. Кал и и и и М. С. Опыт выращивания шпината на электрическом свете разного
состава. Электрификация сельского хоз-ва, № 5, 34—39 (1934).
93*. Калит ин Н. 11. Лучи Солнца. Изд-во АН СССР. М. (1947).
94*. К а л и т и н Н. Н. Тепло и свет солнечных лучей. Гидромстсоиздат, Л. (1947).
95. Каменский К. В. Основы сельскохозяйственного семеноведения. Сельхоз-
гиз, 255—257 (1931).
96. Каневский С. Лимоны в комнате. Огонек, № 33 (1314), 28—29 (1952).
97. Карманов В. Г. Влияние мощности лучистого потока и температуры воздуха
на температуру листа растения. Докл. АН СССР, 77, № 5. 913—915 (1951).
98* . Касаткин А. С. ц 11 е р е к а л и и М. А. Электротехника. Энергоиздат,
М. (1947).
99. К а т у и с к и й В. М. Кратковременные периодические воздействия светом
как метод управления растительным организмом. Докл. АН СССР, 3 (12), № 6
(101), 301—303 (1936).
100. К а т у н с к и й В. М. О возможности «полива» светом как новый прием агро-
техники. Электрификация сельского хоз-ва, № 3, 96—99 (1936).
101. К а ту нс кий В. М. Об изменении фотосинтетической деятельности расте-
ний в процессе их роста и развития в связи с проблемой углекислотного пита-
ния. Изв. АН СССР, сер. биол., № 1, 85—102 (1939).
102. Киселев Г. Е. Комнатное цветоводство. Моск, рабочий (1948).
103. К и с е л е в Г. Е. Цветоводство. Сельхозгиз, М. (1949).
104* . Кислов В. П. Новая термоигла для измерения температуры в растениях.
Докл. ВАСХНПЛ, вып. 4, 41—45 (1940).
105* . Кислов В. II. Термоигла для тонких объектов. Жури. техн, физики, 11,
вып. 7, 674—679 (1941).
106. Китаев И. И. и Китаев С. Н. Тепличное овощеводство. Сельхозгиз,
М. (1947).
107. К и т а е в С. И. Выращивание овощей в теплицах и парниках. Сельхозгиз (1953).
108. К ла и г И. И. Оранжереи и парники в декоративном садоводстве. Изд. МКХ
РСФСР, М. (1950).
109* . Клешнин А. Ф. К вопросу о роли спектрального состава света в фото-
периодической реакции. Докл. АН СССР, 40, № 5, 237—240 (1943).
110* . Клешнин А. Ф. Значение спектрального состава света для фотопериоди-
ческого и формообразовательного процессов на различных фазах развития. Докл.
АН СССР, 52, № 9, 819—822 (1946).
111* . Клешнин А. Ф. К вопросу о значении спектрального состава света для
ростовых процессов. Докл. АН СССР, 53, № 2, 157—159 (1946).
112. К л е in и и н А. Ф. Без солнечного света. Молодой колхозник, № 1, 23 (1948).
113. Клешни н А. Ф. Люминесцентные лампы как источник радиации для свето-
культуры растений. Нзв. АН СССР, сер. физ., 13, ,№ 2, 294—300 (1949).
114. К ле in н и н А. Ф. Применение люминесцентных ламп для выращивания расте-
ний. Вести. АН СССР, № 4, 97 (1949).
115. Клешнин А. Ф. К вопросу о формативном действии различных областей
спектра физиологической радиации на растения. Докл. АН СССР, 67, № 3, 569—
571 (1949).
116. Клешнин А. Ф. Светокультура растений. Со. «Фотосинтез растений», вып. 2,
40—42 (1949).
117. К л е ш и и и А. Ф. Светокультура растений. Со. «Фотосинтез растений», вып. 3,
70—71 (1950).
118. Клешнин А. Ф. Значение различных областей спектра физиологической
радиации для роста и развития растений. Докл. АН СССР, 70, № 5, 891-—894
(1950). Реферат: Природа № 3. 59—60 (1951).
119. К л е ш и и н А. Ф. К вопросу о температуре листьев растений при искусствен-
ном освещении. Докл. АН СССР, 79, № 6. 1029—1032 (1951).
120. К л е in н и и А. Ф. Выращивание растений прп искусственном освещении.
Природа, № 10, 99 -103 (1952).
121. К л era и и и А. Ф. Выращивание растений с помощью люминесцентных ламп.
Естествознание в школе. № 6. 25—30 (1952).
Литература
429
122. Клешнин А. Ф. Электрическое «солнце». Пионерская правда, № 2 (3609)
от 6 января 1953 г.
123. Клешнин А. Ф. Теория и практика светокультуры растений. Тр. Ин-та фи-
зиологии растений им. К. А. Тимирязева АН СССР, 8, вып. 1, 131—163 (1953).
124* Клешнин А. Ф. Проблемы измерения лучистой энергии для физиологиче-
ских целей. Тр. Ин-та физиологии растений им. К. А. Тимирязева АП СССР,
8, вып. I, 219—228 (1953).
125. Коновалов И. Н. Несколько опытов по влиянию искусственного света
на развитие растений. Зап. Воронежск. с.-х. ин-та, 10, 167—172 (1928).
126. Константинов И. Н. Значение интенсивности освещения для развития
различных форм хлопчатника. Докл. АН СССР, 27, № 6, 607—610 (1940).
127. Корнилов А. А. Цветение периллы при непрерывном освещении. Докл.
АН СССР, 64, № 3. 401—403 (1949).
128. Корольков Е. Д. Культура огурцов при искусственном электрическом
освещении. Сад и огород, № 6, 5—7 (1929).
129. К о р о л ь к о в Е. Д. Энергетическая база тепличного овощеводства. Электри-
фикация сельского хоз-ва, № 11—12, 43—56 (1932).
130. Корольков Е. Д. Добавочное электрическое освещение. Электрификация
сельского хоз-ва, № 5, 50—57 (1934).
131. Красинский Н. П. Электросветокультура и удобрение углекислотой.
Изд. Акад. ком. хоз-ва. М. (1934).
132. К рас и и с к и й Н. II. Электросветокультура и удобрение углекислотой.
Зеленое строительство (орган треста зелен, строительства), № 2, 2 (1934).
133. Красинский Н. П. Электросветокультура и удобрение углекислотой.
За соц. рекопстр. городов, № 3, 48—52 (1934).
134. К р а с и н с к и й Н. II. и Князева Е. И. Электросветокультура и удобре-
ние углекислотой. Сон. город., № 7, 34—37 (1935).
135. Крас и и с к и й Н.П. и II а у к с е р Г. Электросветокультура. Сов. суб-
тропики, № 2 (6), 81—85 (1935).
136. Красинский Н. П. Методы ускоренной выгонки цветочных растений.
Ссльхозгиз. М. (1937).
137. Красинский Н.П. Удобрение углекислотой как прием повышения урожая
овощей. Ссльхозгиз, М. (1945).
138. К р а с и н с к и й Н. П. Итоги 15-летней работы по изучению действия углеки-
слотного удобрения на растения. Уч. заи. Горьк. ун-та, вып. 13, 223—242
(1947).
139. Красинский II. П. Электросветокультура. Декоративное садоводство.
Ссльхозгиз, М., 431 (1949).
140. К р а с и н с к и й Н. II. Удобрение углекислотой. Наука и жизнь, 18, № 12,
13—16 (1951).
141. Красинский И. П. Словарь «Русские ботаники». Т. 4. М., 451—454 (1952).
142. Крас и о о к И. II. Влияние света на онтогенез яровой пшеницы. Докл. АП
СССР, 28, А» 6, 541—543 (1940).
143. Кру ш и и с к и й С. Слово о мире. Правда, № 320 (11792) от 16 ноября 1950 г.
144. Крокер В. Рост растений. Изд-во иностр, лит-ры. М., 266—332 (1950).
145. К рячков И. С. Световая стадия у красного клевера. Селекция и семено-
водство, № 3, 15—20 (1936).
146. Куньщиков В. А. Использование отбросного тепла конденсационных
электростанций и добавочное облучение растений люминесцентными лампами
в теплицах в сельском хозяйстве. Изд. ТСХА, М. (1949).
147. Курбатов 11. М. и Васи л ь о в а 'Г. М. Выращивание овощных культур
в теплицах с добавочным электрическим светом. Электрификация сельского
хоз-ва, № 2, 36—40 (1936).
1 48. К у р с а нов А. Л. В союзе с практикой. Паука и жизнь, 19, № 10, 7—8 (1952).
149. К ю з И. II. и Б р ы з г а л о в В. А. Овощеводство. Ссльхозгиз, М. (1934).
150. Л а д ы ж ейская К. И. Лабораторная культура мохообразных. Природа,
№ 8, 38—43 (1939).
151*. Лазарев Д. И. Ультрафиолетовая радиация. Госэпергопздат. Л. (1950).
152*. Ландсберг Г. С. Оптика. Гостехиздат, М. (1940).
153. Л а и и н II. 11. Регулирование сроков цветения декоративных растений. Сов.
ботаника, № 1. 69—74 (1939).
154. Лема и В. М. Опыт практического использования светокультуры дуба. Докл.
АН СССР, 59, № 4. 777—780 (1948).
155. Леман В. М. О поведении сеянцев дуба при непрерывном электрическом
освещении. Докл. АН СССР, 60, № 7, 1261 —1264 (1948).
156. Л е м а н В. М. Светокультура как метод ускоренного выращивания сеянцев
древесных пород. Докл. ТСХА, вып. 9, 70—72 (1949).
430
Л итература
157. Леман В. М. Ускоренное выращивание сеянцев древесных при помощи элек-
трического света. Лесное хоз-во, № 1, 60—63 (1950).
158. Леман В. М. Об ускорении развития сеянцев некоторых хвойных пород при
непрерывном освещении. Докл. АН СССР, 71, № 1, 163—166 (1950).
159. Леман В. М. Светокультура сеянцев древесных пород. Естествознание
в школе, № 5. 27—32 (1950). Реферат: Anale Romano-Sovetice, 7, 115—116 (1951).
160. Леман В. М. Применение люминесцентных ламп для выращивания растений
в теплицах. Рефераты докладов ТСХА, вып. 15, 131—134 (1952).
161. Леман В. М. Под лампами дневного света. Наука и жизнь, № 2, 33—34 (1953).
162. Лисицын Б. 11. Ускорение плодоношения у красного клевера. Селекпия
и семеноводство, № 4, 55—60 (1936).
163. Листов П. Великие стройки коммунизма и электрификация сельского хозяй-
ства. Культурно-просв. работа, № 3, 6—10 (1951).
164. Л и х о н о с Ф. Д. Ранняя выгонка плодов и ягод. Сельхозгпз, М. (1949).
165. Лубенец И. А. Изучение стадийного развития сортов люцерны. Селекция
и семеноводство, № 12, 47—50 (1937).
166*. Л у г о в с к о й Б. II. Люминесцентное освещение зданий. Госэнергоиздат,
М. (1950).
167*. Луизов А. В. Люминесцентные лампы. Природа, № 8, 3—15 (1949).
168*. Лукьянов С. Ю. Фотоэлементы. Изд-во АП СССР, М. (1948).
169*. Лучистая энергия на производстве. Изд. Ин-та гигиены труда и профзаболе-
ваний, Л. (1938).
170. Лысо и к о Т. Д. Б вопросу о регулировании длины вегетационного периода
с.-х. растений. Бюлл. яровизации, № 1, 5—12 (1932).
171. Лысенко Т. Д. Присуще ли природе сельскохозяйственных растений требо-
вание фотоперподов. Бюлл. яровизации, № 2—3, 16—34 (1932).
172. Лысенко Т. Д. и Долгушин Д. А. Ускорение развития картофеля
в полевых условиях социалистического хозяйства. Бюлл. яровизации, № 2—3,
35—45 (1932).
173. Лысенко Т. Д. К вопросу яровизации кукурузы, проса, суданки, сорго
и сои. Бюлл. яровизации, № 2—3, 46—63 (1932).
174. Лысенко Т. Д. Основные результаты работ по яровизации сельскохозяй-
ственных растений. Бюлл. яровизации, № 4, 3—57 (1932).
175*. Л ы с е н к о Т. Д. Агробиология. Сельхозгпз, М. (1943).
176. Львов С. Д. Характеристика научной деятельности академика II. А. Макси-
мова. Бот. жури., 35, № 5, 544—555 (1950). (Н. А. Максимов. Пзбр. работы.
Изд-во АН СССР, М., 1—13 (1952).
177. Люминесцентные лампы. Реферат, сборник «Современная физика», сер. 2, вып. 5,
Люминесценция, 51—57 (1956s); вып. 9, Люминесценция, 112—122 (1952).
178*. М а й з о л ь С. О. Новые источники света. Изд-во «Правда», М. (1947).
179. М а к с и м о в Н. А. Культура растений па электрическом свете и применение
ее для семенного контроля и селекции. Иаучно-агроп. жури., 2, № 7-8, 395—404
(1925). Biol. Ztibl., 45, II. 10, 627—639 (1925).
180. Максимов Н. А. Культура растений па искусственном свете. Природа,
№ 5-6, 61—75 (1933).
181. Максимов II. А. Применение искусственного освещения для ускорения
селекционной работы. Семеноводство, № 4, 16—19 (1934).
182. Максимов Н. А. Применение искусственного света для ускорения роста
растений. Научи, конференции ТСХА. Докл. вып. 1, 14—16 (1945).
183. М а к с и м о в II. А. и Л о м а и В. М. Культура сеянцев древесных на элек-
трическом свете. Докл. ТСХА, вып. 3, 46—51 (1946).
184. М а к с и м о в И. А. и К л с ш и и п А. Ф. Люминесцентные лампы как источ-
ник радиации для светокультуры растений. Докл. АН СССР, 57, № 2, 20'1—204
(1947).
185. М а к с и м о в II. А. Очерк истории физиологии растений в России. Тр. Ин-та
истории естествознания, 1, 21—79 (1947).
186. Максимов II. А. Физиология растений. Сб. «Очерки по истории русской
ботаники». Изд. МОПП, М., 211—273 (1947).
187. Макс и м о в Н. А. Краткий курс физиологии растений. Сельхозгпз. М. (1948).
188. Максимов II. А. Опыт физиологической оценки условий роста растений
в оранжерее Ленинградского ботанического сада. Пзв. АН СССР, сор. биол.,
№ 4, 81—88 (1951).
189. Макс и м о в Н. А. Лаборатория климата. Огонск, 29, № 52 (1281), 28 (1951).
190. М а к с и м о в И. А. Лаборатория искусственного климата. Новый мир, № 5,
161—169 (1952).
191. М альчевс к и й В. П. Выступление на IV сессии ВАСХНПЛ (19—27 де-
кабря 1936 г.). Сб. «Достижения советской селекции», 71—72 (1937).
Л шпература
431
192. Мальчевский В. II. Применение искусственного света для повышения
урожайности. Докл. ВАСХНПЛ, вып. 1-2 (10—11), 70—74 (1938).
193. Мальчевский В. П. Действие некоторых лучей спектра на развитие расте-
ний. Тр. лабор. светофизиологии ФАИ, вып. 1, 4—50 (1938).
194. М а л ь ч е в с к и й В. И. и Доброхотова С. И. О причинах ускорения
роста и развития растений в условиях светокультуры и значение светокультуры
для практики. Тезисы докладов совещания по физиологии растений. Изд-во АН
СССР, М., 46—47 (1940).
195* . Мальчевский В. П. Новые приборы для измерения света в полевых усло-
виях. Тезисы докладов совещания по физиологии растений. Изд-во АН СССР,
М., 183—184 (1940).
196. Мальчевс к и й В. II. Применение искусственного света для ускорения
роста и развития сеянцев древесных пород. Тр. Ин-та физиологии растений
им. К. А. Тимирязева АН СССР, 3, вып. 2, 3—46 (1946).
197. Марков В. М. Овощеводство в Арктике. Плодоовощное хоз-во. № 11, 4—9
(1936).
198. Марков В. М. Что надо показать по овощеводству на Всесоюзной сельско-
хозяйственной выставке 1937 г. Плодоовощное хоз-во, № 7, 21—24 (1937).
199. Марков В. М. и .Маркова Л. И. Осенняя культура томатов при элек-
трическом освещении. Плодоовощное хоз-во, № 8, 32—34 (1937).
200* . Марков В. М. Рост овощеводства закрытого грунта. Плодоовощное хозяй-
ство, № И, 51—55 (1937).
201. Марков В. М. и Хаев М. К. Овощеводство. Сельхозгиз, М., 420—426 (1953).
202*. Масл а к о в е ц Ю. П. и М а к а р е в с к и й Н. И. Приборы для изме-
рения света в полевых условиях. Тр. лабор. светофизиологии ФАИ, вып. I,
101—109 (1938).
203* . Мешков В. В. и С околов II. II. Курс осветительной техники. Гос-
энергоиздат, М. (1948).
204. Мещерский М. II. Комнатная электротеплица. Естествознание в школе,
№ 1, 88 (1952).
205. Милославов Л. Культура огурцов и томатов при электрическом освещении.
Сад и огород, № 11, 7—11 (1929).
206. Молиш Г. Физиология растений как теория садоводства. Сельхозгиз, М.,
51—55 (1933).
207. Мошков Б. С. Фотопериодизм и морозоустойчивость многолетних растений.
Тр. по прикл. ботанике, генетике и селекции, сер. 3. № 6, 235—261 (1935).
208. М о ш к о в Б. С. Листопад у древесных растений. Сов. субтропики, № 12 (16),
73 (1935).
209. Мош к о в Б. С. Свет и листопад древесных растений. Сов. субтропики, № 7,
62—64 (1936).
210. Мошков Б. С. Повышение урожая хинного дерева фотопериодическим воз-
действием. Сов. субтропики, № 7 (35), 71—74 (1937).
211. Мошков Б. С. Фотопериодизм и засухоустойчивость многолетних растений.
Докл. АН СССР, 22, .М 4, 186—188 (1939)/
212. Мошков Б. С. О критических и оптимальных фотопериодах. Сов. ботаника,
№ 4, 32—45 (1940).
213. Мошков Б. С. Новые данные о потребности растения в лучистой энергии.
Вести. АН СССР, № 8, 77 (1949).
214. Мошков Б. С. Значение отдельных участков спектра физиологической
области излучения для роста и развития некоторых растений. Докл. АП СССР,
71, № 1, 171 — 174 (1950).
215. М о ш к о в Б. С. Влияние инфракрасной радиации на темновые процессы у расте-
ния короткого дня периллы масличной. Докл. АП СССР, 71. № 2, 391—394 (1950).
216. М о ш ко в Б. С. Вырагапваипе растений при искусственном освещении. Агро-
биология, № 2, 66—74 (1950).
217* . М о ш к о в Б. С. Влияние спектрального состава лучистой энергии на рост
земляники п бриофиллума. Докл. АН СССР, 76, № 4, 593—596 (1951).
218. Мошков Б. С. Влияние искусственного света на жизнедеятельность сельско-
хозяйственных растений. Сб. «Сельское хозяйство Помп АССР», Изд-во АН СССР,
155—167 (1951).
219. М о ш к о в Б. С. Воздействие на процессы связывания растениями лучистой
энергии. Естествознание в школе, № 6, 20—25 (1951).
220. М о in к о в Б. С. Характер использования лучистой энергии томатными расте-
ниями в зависимости от мощности лучистого потока и продолжительности еже-
суточного освещения. Докл. АН СССР, 84, № 2, 369—372 (1952).
221. Мошков Б. С., Карманов В. Б. и Гончаров Б. И. О выращи-
вании рассады томатов при электрическом освещении для получения ранних
432
Литература
урожаев. Сб. «Достижения науки по сельскохозяйственному производству. Ово-
щеводство и картофелеводство». Лениздат, 135—140 (1952).
222. Мошков Б. С. и Карманов В. Б. Значение температуры воздуха при
выращивании растений на электрическом освещении. Агробиология. № 4, 76—82
(1952).
223. Мошков Б. С. Некоторые вопросы светокультуры растений. Тр. Ин-та физио-
логии растений им. К. А. Тимирязева АН СССР, 8, вып. 1, 164—173 (1953).
224. Мошков Б. С. Выращивание растений на искусственном освещении. Сель-
хозгиз (1953).
225. Мэттьюс Р. Б. Электрокультура и светокультура. Электрификация сель-
ского хоз-ва, А!: 3, 66—79 (1931).
226. Мэттьюс Р. Б. Электрификация сельского хозяйства. Гостехиздат, М. (1931).
227* Народное хозяйство Мурманской области. Изд-во «Полярная Правда» Мурманск
228. Н а ц е н т о в Д. И. и Ж у к о в а Е. К. Организуем круглогодичное исполь-
зование парников и теплиц. Плодоовощное хоз-во, № 7, 48—50 (1935).
229. Нане н т о в Д. 11., Евсеев К. М., Шереметеве кий П. В. и
Боков В. В. Парники и теплицы. Сельхозгиз, М. (1936).
230. Н а ц е н т о в Д. И. Обеспечим ранний и высокий урожай овощей в парниках
и теплицах. Сад и огород, № 3, 29—35 (1949).
231. Немцов В. Л. Семь цветов радуги. «Молодая гвардия» (1950).
232. Н и к и т и н И. Н. О новых идеях в древоводстве и лесоводстве. Лесное хоз-во,
№ 5, 4—14 (1941).
233. Никитин И. Н. Возрастная стадийность и методы ускорения роста дуба
в первые годы его жизни. Лес и степь, № 2, 16—26 (1949).
234. Николаев А. Цветы, выращенные без солнца. Гудок, № 223 (8204)
от 19 сентября 1952 г.
235* Н и л о и д е р Р. А. Люминесцентные лампы и их применение. Госэиерго-
издат, М. (1948).
236. Н и ч и и о р о в и ч А. А. Фотосинтез как фактор урожайности. Пзв. АН СССР,
сер. биол., № 4, 3—30 (1952).
237. Н и ч и и о р о в и ч А. А. Продукты фотосинтеза и физиологическая роль
фотосинтетического аппарата. Тр. Ин-та физиологии растений им. К. А. Тимиря-
зева АН СССР, 8, вып. I, 3—41 (1953).
238. Новоселов А. А. Выращивание овощей в теплицах и парниках на Крайнем
Севере. Изд. Главсевморпуть, М. (1946).
239. Овощеводство защищенного грунта. Итоги VIII пленума секции плодоовощных
культур ВАСХНИЛ 12—18 января 1938 г. Изд. ВАСХНИЛ, М. (1939).
240* Оптика в военном деле. Т. I. Изд-во АН СССР, М. (1945).
241* О строительстве Куйбышевской гидроэлектростанции на реке Волге. Известия,
№ 199 (10348) от 22 августа 1950 г.
242* О строительстве Сталинградской гидроэлектростанции па р. Волге, об ороше-
нии и обводнении районов Прикасппя. Известия, № 207 (10356) от 31 августа 1950 г.
243* О строительстве Каховской гидроэлектростанции на реке Днепре, Южно-
Украинского канала, Северо-Крымского канала и об орошении земель южных
районов Украины и северных районов Крыма. Известия, № 225 (10374) от 21 сен-
тября 1950 г.
244. П а лило в И. Электричество в сельском хозяйстве. Леноблиздат. Л. (1935).
245* II а и ч е н к о В. Г. О тепловых и люминесцентных источниках света. При-
рода, № 3, 25—36 (1950).
246. II и н ч у к В. Д. Культура семенников сахарной свеклы в оранжерее. Свекло-
водство, т. 1, Киев, 810—821 (1940).
247. По боди мо ва Е. Г. Влияние электрического освещения ла развитие Stel-
laria media L. Пзв. Гл. бот. сада СССР, 28, вып. 1-2, 75—94 (1929).
248. П о бе дим о на Е. Г. Произвольное изменение длительности пмклов разви-
тия при электрическом свете. И.зв. Гл. бот. сада, 30, вып. 1-2, 147—188 (1932).
249. По л у я х то в. Предварительное сообщение о результатах электросветокуль-
туры в Свердловском комбинате зеленого строительства. Сборник работ Ураль-
ской опытной станции зеленого стр-ва, вып. 1, 136—137 (1939).
250. II о к р о в с к и й Н. С. Расчет электрического освещения для выращивания
растений на искусственном свете. Тр. Лабор. свстофизиологии ИМИ, вып. 1,
75—89 (1938).
251. Потапенко Я. И. и Захар о в а Е. И. Реакция винограда на световые
и температурные условия развития. Сб. «Селекция плодово-ягодных растений»,
82—106 (1937).
252* Прингсгейм II. и Фоге л ь М. Люминесценция жидких и твердых
тел и ее практическое применение. Изд-во иностр, л-ры, М. (1948).
Л итература.
433
253. Протасова И. Н. Применение искусственного досвечивания при выращи-
вании рассады огурцов и томатов. Изд-во АН СССР, М. (1952).
254. Протасова Н. П. Выращивание рассады в теплицах при различном осве-
щении. Сад и огород, № 12, 49—54 (1952).
255. Протопопов Б. «Зеленые цеха». Электрификация сельского хоз-ва, Л» 2,
60—65 (1932).
256. П с а р е в Г. М. О локализации фотопериодического цикла у сои. Сов. бота-
ника, № 3, 88—90 (1936).
257. Псарев Г. М. Влияние смены дня и ночи на характер изменения пофазного
ритма развития у сои. Докл. АН СССР, 17, № 8, 431—434 (1937).
258. Псарев Г. М. Влияние продолжительности освещения на явление пожелте-
ния и отмирания листьев у сои. Докл. АП. 25, № 8, 681—684 (1939).
259. Псаре в Г. М. Влияние длины дня на поперечный рост стебля растения.
Докл. АН СССР, 28, Оу 6, 538—540 (1940).
260. П cape в Г. М. и 11 е й м а и II. Ф. Влияние длины дня на активность кам-
биального роста. Докл. АП СССР, 29, Л» 7, 497—499 (1940).
26'1 *. П у т и л о в К. А. и Ф а б р и к а н т В. А. Курс физики. Г. 2. Гостохиздат,
М. (1945).
262. Р а в и ч 1V1. Б. и Сарычева А. II. Использование электросвета и продук-
тов горения для зимней культуры растений. Электричество, № 11, 76—78 (1938).
263. Р а в и ч М. Б. и Сарычева Л. П. Углекислотное удобрение растений
с использованием продуктов горения твердого топлива. Взв. Эперг. ин-та АН
СССР, 12, 111—119 (1944).
264. Р а в и ч М. Б. Поверхностное беспламенное горение. Изд-во А.П СССР, М.,
299—324 (1949).
265. Радченко С. 11. Температурный режим и рост растений. Пзв. Акад, педаг.
наук РСФСР, вып. 29, 57—74 (1950).
266. Разумов В. И. О вегетационном периоде у растений. Соц. растениеводство,
сер. А, № 15, 15—38 (1935).
267. Разумов В. П. Поведение отдельных растений чистой линии ла различной
длине дня. Яровизация, ..V 2 (11) (1937).
268. Растения при искусственном свете. Природа и люди, 24, ЛЬ 29, 462 (1913).
269. Реймерс Ф. Э. Стадийное развитие у разных биологических групп салата.
Докл. АП СССР, 20, № 7_8j 609—614 (1938).
270. Реймерс Ф. Э. Об изменении «знака фотопериодической реакции» у репы.
Докл. АН СССР, 20, № 7-8, 615—617 (1938).
271. Реймерс Ф. Э. Влияние длины дня на образование цветоносных побегов
у корнеплодов столовой свеклы. Докл. ВАСХНПЛ, вып. 2—3, 31—34 (1939).
272. Реймерс Ф. Э. Динамика сахаров и ферментов у растений лука, выращенных
на разной длине дня. Вести, с.-х. пауки. Овощи и картофель, вып. 4, 17—30(1940).
273. Рейнгольд И. Применение освещения лампами «Осрам» и неоновым све-
том при выгонке ранних овощей. Плодоовощное хоз-во, № 5, 79 (1936).
274*. Риль Н. Люминесценция. Гостехиздат, М. (1946).
275* . Родионов С. и Ф р а и к Г. Вопросы светофизиологии и измерения света.
Гостехиздат (1934).
276. Родников II. Влияние продолжительности дневного периода на развитие
некоторых огородных растений. Сад и огород, № 3, 23—30 (1929).
277. Ромаш онко в Д. Л. Влияние досвечивания рассады па урожай томатов.
Овощеводство, № 2-3, 23—24 (1939).
278. Рубинштейн Д. Л. Задачи электро биологии в сельском хозяйстве. Элек-
трификация сельского хоз-ва, № 5-6, 140—142 (1935).
279. Сааков С. Г. Культура некоторых декоративных растений при дополнитель-
ном электроосвещении. Рефераты работ учреждений ОБИ АП СССР за 1940 1’.,
44—45 (1941).
280. Сааков С. Г. Культура некоторых декоративных растений при дополнитель-
ном электрическом освещении. Интродукция растений и зеленое строительство,
вып. 1, 295—303 (1950).
281. Сазонов Н. А. За постепенное внедрение электроэнергии в производствен-
ные процессы сельского хозяйства. Электрификация сельского хоз-ва, № 1,
11—19 (1934).
282. Сазонов Н. А. Электричество в сельском хозяйстве СССР. Электричество,
№ И, 32—38 (1938).
283. Салтыковский А. Выращивание растений при искусственном освеще-
нии. Электрификация сельского хоз-ва, № 9—10, 100—103 (1932).
284. Самохина 3. Ф. и 3 и х е р м а и X. Я. Влияние температуры на ско-
рость развития яровой пшеницы после прохождения световой стадии. Докл.
ВАСХНПЛ, вып. 9, 7 —11 (1941).
28 А. Ф. Клешнин
434
Литература
285*. С а м ы г и н Г. А. Фотопериодизм растений. Тр. Ин-та физиологии растений
им. К. А. Тимирязева АН СССР, 3, вып. 2, 129—262 (1946).
286. С а м ы г п и Г. А. Фотопериодизм. Декоративное садоводство. Сельхозгиз,
М., 400—401 (1949).
287. Сарычева А. П. Удобрение углекислотой декоративных растений. Уч.
зап. естеств. ф-та МОП11, 2, 55—83 (1941).
288. Свекловодство, I. Биология, генетика и селекция сахарной свеклы. Гос. изд-во
колх. и совхозн. л-ры УССР, Киев, 202—232, 810—821 (1940).
289. Светокультура растений. Реферативный сборник «Фотосинтез растений», вып. 2,
43—54 (1949).
290. Светокультура растений. Реферативным сборник «Фотосинтез растений», вып. 3,
72—88 (1950).
291. Светофизиология и светокультура растений. Тр. Лабор. светофизиологии ФАИ,
вып. 1 (1938).
292* . Свечников Л. В. Исследование параметров люминесцентных ламп в зави-
симости от напряжения, температуры и частоты. Изв. Кмевск.. политехи, ин-та,
11, 320—331 (1952).
293. С е л и х о в Б. В. Стадийность развития и селекция сахарной свеклы. Научн.
зап. по сахари, пром-сти. Агрономический выпуск, № 3, 15—32 (1935).
294. С и н с к а я Е. Н. О типах взаимоотношения между развитием и ростом у ра-
стений «длинного» и «короткого» дня. Краткий отчет о научно-иссл. работе за
1950 г. Всес. научно-иссл. ин-та масличных культур, 154—162 (1951).
295. С к а з к и II Ф. Д. О постановке новых опытов по физиологии растений
(V класс). Биология и химия в средней школе, № 4 (8), 3—7 (1935).
296. С к а з к и н Ф. Д., Л о в ч и н о в с к а я Е. 11. и Красносельская
I. А. Практикум по физиологии растений. Изд-во «Сов. наука», М.,173—174 (1948).
297. С к у д и а я О. Д. Динамика накопления пигментов в растениях закрытого
грунта прп освещении люминесцентными лампами дневного света. Изд. Бело-
русск. ун-та, Минск (1952).
298* . С околов А. С. Автомобильные электрические лампочки. Воениздат, М. (1949).
299. Соколова Н. Ф. Влияние влажности почвы и длины дня на развитие крым-
сагыза. Тр. Гос. Никитск. бот. сада, 21, вып. 1, 61—76 (1935).
300. Стампа Г. Отепление растений на открытом воздухе. Сов. субтропики,
№ 4 (32), 102—105 (1937).
301. С т с л ь м а х о в и ч М. Л. и Викулина Л. А. Ускорение развития цве-
точных растений. Сборник работ Уральск, опытной станции зеленого строитель-
ства, вып. 1, 94—111 (1939).
302. Строгонов Б. II. Установка для выращивания растений в контролируемых
условиях газового режима. Тр. Пи та физиологии растений им. К. А. Тимиря-
зева АН СССР, 6, вып. 2, 255—259 (1949).
303* . Стронг Д. Практика современной физической лаборатории. Гостехиздат,
М. (1948).
304. Сурж и н а. Обогрев растений лучистой теплотой. Плодоовощное хоз-во,
№ 10, 73—74 (1936).
305. Сысина 11. А. Управление сроками цветения гладиолусов. Бюлл. Гл. бот.
сада, вып. 11, 63—66 (1952).
306. Танеева С. В. Обзор работ и перспективы электросветокультуры. Рефераты
научно-иссл. работ за 1944 г. ОБН АН СССР, 54—55 (1945).
307. Тагеева С. В. К познанию светового режима оранжерей г. Москвы. Изв.
АН СССР, № 4, 89—119 (1951).
308. Т и м и р я з е в К. А. Возможна ли культура растений на электрическом свете.
Сб. «Публичные лекции и речи». М. (1888). Сб. «Земледелие и физиология расте-
ний», М. (1906). Соч., т. 3, Сельхозгиз, М. 318—330 (1937).
309. Тимофеева М. Т. Морозоустойчивость озимых злаков в связи со стадий-
ным развитием и закаливанием растений. Докл. АН СССР/ 1, № 1, 61—64
(1935).
310. Т и м о ф о е в а М. Т. О роли термического фактора и фотопериодизма в раз-
витии озимых злаков. Тр. по прикл. ботанике, генетике и селекции, сер. А, 14,
177—183 (1935).
311. Толмачов П. М. и Ильинский Б. П. О продвижении сахарной
свеклы на север СССР. Зап. ио сахарной пром-сти, Агрономический вып. 11,
кн. 40, № 2, 18—33 (1934).
312. Толмачев И. М. Температура, свет, влажность и минеральное питание
как факторы развития сахарной свеклы. Свекловодство, 1,202—232, Киев (1940).
313. Трегуб о и к о М. Я. Температурно-световой режим для выращивания
пшениц в теплице в ноябре—январе. Селекция и семеноводство, № 11, 25—30
(1938).
Л итература
435
314. Треспе Г. Г. Выгонка цветов. Сельхозгиз, М. (1937).
315. Уайт Р. Возделывание сельскохозяйственных растений и окружающая среда.
Изд-во иностр, л-ры, М. (1949).
316. Ускоренное выращивание растений. Наука и жизнь, № 8, 13 (1952).
317*. Фабрикант В. А. Физика и техника люминесцентных ламп. Успехи
физ. наук, 27, вып. 2, 158—198 (1945).
318*. Фабрикант В. А. Газосветная лампа. БСЭ, 10, 79—82 (1952).
319. Фаминцин А. С. Описание аппарата для исследования действия искусствен-
ного света на растения. Тр. I Съезда русских естествоиспытателей и врачей.
СПб. (1868).
320. Федин II. Е. Температурные и световые условия выращивания сладких пер-
цев в Ленинградской области. Изд. ВИР, Л. (1953).
321*. Федоров Б. Ф. Общий курс светотехники. Госэнергоиздат, М. (1944).
322. Физиологическая лаборатория искусственного климата. Вести. АН СССР, № 4,
107—108 (1950).
323. Ф и л л и п и ш и н И. С. Электротеплофикация теплиц и парников. Сб. «Ово-
щеводство защищенного грунта», М., 53—63 (1939).
324. Финкельштейн 11. И. Электричество па службу овощеводству. Дости-
жения науки и передового опыта в сельском хозяйстве, № 10, 37—41
(1951).
325. Финкельштейн И. II. Лампы накаливания в движении как источник
света для светокультуры растений. Тр. Ин-та физиологии растений им. К. А. Ти-
мирязева АН СССР, 8, вып. 1, 175—183 (1953).
326. Ф о л ь г К. Электрическое освещение при выращивании томатов. Электри-
фикация сельского хоз-ва, № 5, 93—94 (1934).
327. Францев А. В. Опыт оценки гидробиологической производительности
москворецкой воды. Микробиология, 1, вып. 2, 112—130 (1932).
328. Хаев М. К., Чижов С. Г., Сукорцева К. Д. и Заостров-
ская Е. Н. Овощеводство. Сельхозгиз, М. (1943).
329. X а й к и и а Р. Я. Теплично-парниковый комбинат. Сад и огород, № 11, 47—51
(1951).
330. Хлебникова Н. А. и Московец К. Г. К биологии мари душистой.
Докл. АН СССР, 32, № 2, 156—157 (1941).
331. Цветники в метро. Пионер, № 5, 79 (1952).
332. Цветы в метро. Техника молодежи, <№ 9, 4 (1952).
333. Цицин Н. В. Крупнейшая лаборатория советской ботаники. Природа, № 6,
33—41 (1952).
334. Ч а й л а х я и М. X. Яровизация растений светом. Докл. АН СССР, .М 5,
224—229 (1933).
335. Чайлахян М. X. Исследования физиологической природы различий яро-
вых и озимых растений. Пзд-во АН СССР, Л. (1934).
336. Чайлахян М. X. Световое управление растением. Тр. Лабор. ПФР, 1,
149—184 (1934).
337. Чайлахян М. X. и Азбук и и Л. Д. Влияние светового режима па рост
многолетних растений. Тр. Ин-та физиологии растений им. К. А. Тимирязева
АН СССР, 2, вып. 2, 126—132 (1938).
338. Ч а й л а х я и М. X. и Некрасова Т. В. Влияние света люминесцентных
ламп на рост сеянцев лимона и апельсина. Докл. АП СССР, 78, № 4, 807—810
(1951).
339. Ч е л я д и п ц с в а А. П. Фотопериодическая реакция у Arachis hypogaea.
Изв. Научного ин-та им. Лесгафта, 20, вып. 2, 127—134 (1937).
340. Ч е л я д и н ц е в а А. Иг Влияние температуры на реакцию арахиса к длине
дня. Докл. АН СССР, 31, № 1, 55—57 (1941).
341. Чернявская II. М. Влияние интенсивности света и азотного питания
на фотосинтез и накопление сухого вещества в ранних фазах роста растений.
Изд-во АН СССР, М. (1952).
342* . Шаронов В. В. Таблицы для расчета природной освещенности и видимости.
Пзд-во АН СССР, М. (1945).
343. Ш е р с м е т е в с к и й П. В. Новое в агротехнике овощеводства защищенного
грунта. Сб. «Овощеводство защищенного грунта». М., 103—115 (1939).
344. Ш е р е и е т е в с к и й П. В. Овощеводство в парниках и теплицах. Сельхоз-
гиз, М. (1945).
345. Шестаков В. Е. и Смирнова А. Д. Температурная закалка и диффе-
ренциация зачаточного колоса у озимых пшениц при прохождении световой стадии
развития. Докл. АП СССР, 3 (12), № 8 (103), 399—403 (1936).
346. Шестаков В. Е. Морозоустойчивость ржано-пшеничных гиоридов в связи
со стадиями развития. Селекция п семеноводство, № 1, 14—16 (1936).
436
Литература
347. Шестаков В. Е. и Сергеев JI. II. Изменение морозостойкости и свойств
протоплазмы клеток у озимых пшениц при прохождении световой стадии. Бот.
жури., 22, № 4, 351—363 (1937).
348* . Ш и п ч и и с к и й А. В. Основы метеорологии и климатологии. Сельхозгпз,
М. (1933).
349* . 111 по льс кий Э. В. Спектр поглощения хлорофилла в растворе и в есте-
ственном состоянии. Нзв. АН СССР, сер. биол., № 3, 397—406 (1947).
350. Ш т а д т К. Э. Применение электричества в садоводстве Голландии. Электри-
фикация сельского хоз-ва, № 2, 76—79 (1932).
351. Л! у б о в И. А. Новое в тепличной технике. Голос стахановца (Орган РК ВК11(б)
и райсовета депутатов трудящихся Краснополянского района Московской
области) № 149 (257) от 8 сентября 1940 г.
352. Эдельштейн В. И. Продвижение овощей на север. Плодоовощное хоз-во,
№ 8, 11—13 (1932).
353. Эдельштейн В. 11. Пути развития овощеводства защищенного грунта
в третьей пятилетке. Сб. «Овощеводство защищенного грунта», М., 3—11 (1939).
354. Эдельштейн В. И. Овощеводство. Сельхозгпз, М. (1944).
355. Эдельштейн В. И. К вопросу о круглогодовом выращивании овощей.
Сад и огород, A's 1, 46—52 (1951).
356. Эйхфсльд. Комнатная культура цитрусовых в Хибинах. Плодоовощное
хоз-во, № 3, 38—39 (1937).
357. Электрический свет и семеноводство. Электрификация сельского хоз-ва, № 5,
95 (1934).
358. 10 д к и н Ф. М. Культура овощей в теплицах и парниках. Сельхозгпз, М. (1939).
359. Ю х и м ч у к Д. Печь для удобрения растений углекислотой. Плодоовощ-
ное хоз-во, № 8—9, 45—46 (1938).
360. Я к у ш е в а Е. И. Морозостойкость клевера и люцерны в связи с условиями
выращивания в предшествующий вегетационный период. Тр. Ин-та физиологии
растений им. К. А. Тимирязева АН СССР, 4, вып. 2, 147—159 (1945).
361* . Я и и га е в с к и й ТО. Д. Методы радиационных измерений для экологических
целей. Экспериментальная ботаника, № 5, 22—45 (194'1).
362. А а с h Н. G. Uber Waclistum und Zusammensetzung von Chlorella pyrenoidtma bei
unterschiedlichen Liclitstarkcn and Nitratmcngrn. Arch. f. Microbiol., 17, II. 3,
213—246 (1952).
363. A berg B. Vaxtodling i artificiell ljus (Kunstljuskultur) med sorskild hansyn
till tomat. Symbolae botanicao UpsalienscJ, 4, H. 5, 1—98 (1941).
364. A b e r g B. Physiologische und okologische Studien uber die Pflanzliche Photo"
morphose. Symbolae botanicae Upsalienses, 8, H. 1, 1—189 (1943).
365. A d a m s .1. Duration of light and growth. Ann. BoL., 38, N 151, 509—523 (1924).
366. A d a m s J. Some further experiments on the relation of light to growth. Amer.
Journ. Bot., 12, N 7, 398—412 (1925).
367. A 1 g e u s S. Glycocole as a source of nitrogen for Scene de smus obliquus. Physio-
logia plantarum, 1, Fasc. 1, 65—'84 (1948).
368. Algous S. Studies on the cultivation of algae in artificial light. Physiologia
plantarum, 4, Fasc. 4, 742—753 (1951).
369. Allard H. A. Flowering behavior of the hog peanut in response, to length day.
Jour. Agric. Research, 44, N 2, 127 — 137 (1932).
370. Allard H. A. Length of day in relation to the natural and artificial distribution
of plants. Ecology, 13, N 3, 221—234 (1932).
371. Allard H. A. Complete or partial inhibition of flowering in certain plant when
days are too short or too long. Jour. Agric. Research, 57, N 10, 775—789 (1938).
372. Allard H. A. Further studies of the photoperiodic behavior of some mints.
Jour. Agric. Research, 63, N 1, 55—64 (1941).
373. Allard H. A. and Evans M. W. Growth and flowering of some tame and
wild grosses in response to different photoperiods. Jour, zkgric. Research, 62, N 4,
193—228 (1941).
374. Allard H. A. a. Garner W. W. Responses of some plants to equal and
unaqual ratios of light and darkness in cicles ranging from I hour to 72 hours. Jour.
Agric. Research, 63, N 6 (1941).
375. A r t h u r J. M. Work to date of Bayce Thompson institute for plant research.
Trans. Ilium. Engin. Soc., 19, N 10, 995—997 (1924). Ref.: Licht u. Lampe, N 23,
802—804 (1925).
376. Arthur J. M. a. Guthrie J . D. Effect of light, carbon dioxide and tempera-
ture on flower and fruit production. Mem. llort. Soc. New York, 3, 73—74 (1927).
377. A r t h u r J. M. Artificial climate and plant growth. Tcchn. Engin News. N 2 (1928).
378. Arthur J. M. a. Guthrie J. D. Some effects of light and carbon dioxide on
the growth and flowering of plants. Physical Therapeutics, 46, N 3, 130—132 (1928).
Л итература
437
379. Arthur J. M., G u t h г i e J. D. a. Newell J. M. Some effects of arti-
ficial climate on the growth and chemical composition of plants. Amer. Journ. Bot.,
17, N 5, 416—482 (1930).
380. Arthur J. M. Light and the green plant. Scientific Monthly, .31, N 4 (181),
343—346 (1930).
381. Arthur J. M. Artificial light and plant growth. Boyce Thompson Inst. Prof.
Pap., 1, N 22, 212—221 (1932)1 Agric-Enginecring, 13, N 11, 288—291 (1932).
382. Arthur J. M. a. Steward W. D. Relative growth and dry weight produc-
tion of plant tissue under Mazda, neon, sodium and mercury vapor lamps. Contr.
Boyce Thompson Institute, 7, N 2, 119—130 (1935).
383. A r t h u r J. M. a. P а г к e r L. C. A new type of insulated green house heated
and lighted by Mazda lamps. Gontr. Boyce Thompson Institute, 7, N 2, 131—146
(1935)/
384. Arthur J. M. Plant growth in continoiis illumination. Biological effects of
Radiation, 2, 715—725 (1936).
385. Arthur J. M. a. llarvill E. K. Plant «growth under continous illumi-
nation from sodium vapor lamps supllemented by nierciir ark lamps. Contr. Boyce
Thompson Institute, 8, N 5, 433—443 (1937).
386. A ! t h u r J. M. Tageslange, Temperature, Belic.htung und Blumenzucht. 12 Inter-
nationale!’ Gartenbaukongross, Berlin, 1938, Section 17, 8 (1939).
387. A r t h u r J. M. a. H a r v i 1 1 E. K. Healing and lighting green house with
intermittent light. Contr. Boyce Tompson Institute, 10, N 1, 15—44 (1938).
388. Arthur J. M. a. II a r v i 1 1 E. К. Intermittent light and flowering of gla-
diolus and carnation. Contr. Boyce Thompson Institute, 11, N 2, 93—103. (1940).
389. A r t h u r J. M. a. H a r v i 1 1 E. K. Flowering in Digitalis purpurea initiated by
low temperature and ligth. Contr. Boyce Thompson Institute, 12, N 2, 11 1 —117(1941).
390. A shby E., Bolas B. D. a. Il e n d e r s о n T. Y. The interaction of fac-
tors in the growth of Lemna. I. Ann. Bot., 42, N 167, 771—782 (1928).
391. Ashby E. The interaction of factors in the growth of Lemna. TIT—IV. Ann.
Bot., 43, N 170, 333—354 (1929); 43, N 172, 805—816 (1929).
392. Ashby E. a. Oxley T. A. The interaction of factors in the growth ol Lemna.
VI. Am/ Bot., 49, N 194, 309—336 (1935).
393. A s о К. a. M urai U. An experiment on the promotion of plant groth by the
influence of electric light. Jour, sclent. Agric. Soc., 254, 31—36 (1924). Ref.: Bot.
Abstr., 15, N 2, 337 (N 2127) (1926).
394. A u c h t e r E. C. a. Harley С. P. Effect of various length of day on develop-
ment and chemical composition of same horticultural plants. Proc. Amer. Soc. Hort.
Sei., 21, 199—214 (1924).
395. Austin .1. P. Some effects of the photoperiod on development of Im pa liens
balsamina. Plant Physiology, 10, N 3, 545—552 (1935).
396. A ustin J. P. Minimum intensity of artificial illumination effective on supl-
lementing the normal photoperiod. Papers Michigan Acad. Sci., Arts and Letters,
22, 25—26 (1936).
397. Austin J. P. The influence of length of the photoperiod on the vegetative and
reproductive development of Rudbeckia bicolor superba, Delphinium ajacis, Cosmos
sulphurous and Jmpatiens balsamina. Amer. Journ. Bot., 28, N 3, 244—250 (1941).
398. Bailey L. 11. Some preliminary studies of the influence of the electric arc lamps
upon greenhouse plants. Cornell Univ Agric. Exper. Sta. Bull., 30, 83—122 (1891);
Ref.: Bicdermann s Zrbl. f. Agrikulturchemie, 21, 180—196 (1892). Forsch. auf
Gcbiete d. Agrikulturphysik, 16, 104—106 (1892).
399. Bailey L. 11. Second report upon Electrochorticulturc. Gornell Univ. Agric.
Exper. Sta. Bull. 42, 133—146 (1892). Ref.: Biedermann's Ztrb. f. Agrikultur-
chemie, 29, 105—106 (1894). Forsch. auf d. Gebiete Agrikulturphvsik, 16, 63—64
(1893).
400. Bail e у L. II. Greenhouse nots for 1890—1893. 1. Third report upon elcctrohor-
tikulture. Cornell Univ. Agric. Exper. Sta. Bull., 55, 147—157 (1893).
401. В a n durski R. G., Scott F. M., P f 1 u g M. a. W e n t F. W. The
effect of temperature on the color and anatomv of tomato leaves. Amer. Jour. Bot.,
40, N 1, 41—46 (1953).
402. Beaumant J. 11. a. W e a v e r J. S. Effect of light and temperature on
the growth and tuberisation of potato seedlings. Proc. Amer. soc. Hort Sci., 28,
285—290 (1931).
,403. Bell Ct. D. II. a. Bane r A. B. Experiments on growing sugar beet under
continuous illumination. Journ. Agric. Sci., 32, part 1, 112—141 (1942): 33, Part 2,
85—94 (1943).
404. В e n a r d A. Influence de la lumiere electrique sur la vegetation. Rev. gen.
Inst, Agronom., Louvain, 325—329 (1908).
438
Литература
405. Benedict Н. М. Effect of day length and temperature on the flowering
and growth of four species of grasses. Journ. Agric. Research., 61, N 9, 661—671
(1940).
406* . Benedict H. M. Growth of some range grasses in reduced light intensities
at Cheyenne Wyoming. Bot. gaz., 102, N 3, 582—589 (1941).
407. Berkley E. E. Studies of the effects of different length of day, with variations
in temperature, in vegetative growt and reproduction in cotton. Ann. Missouri Bot.
Garden, 18, N 4, 573—602 (1931).
408. Bestralungsanweisung fiir Gloxinien. Philips Pflanzenbcstrahler (1934).
409. Bewley W. F. a. Bolas B. D. The use of electric light in propagation
houses. Rep. Exper. and Research. Sta. Cheshunt. Rpt., 16, 84—86 (1931). Ref.:
Biol, abstr., 8, N 3, 732 (N 6623) (1934).
410. Bewley W. F. Heizung, Beleuchtung und Konstruktion von Clashausern
in England. 12 Internationaler Gartcnbaukom>ress, Berlin (1938), Section 18, 1
(1939).
411. В i e b e 1 J. P., Green L. a. Wit h г о w R. B. Flowering to temperature
in China aster. Proc. Amer. Soc. Hort. Sci., 33, 645—646 (1935).
412. В i e b e 1 J. P. Temperature, photoperiod, flowering and morphology in cosmos
and china aster. Proc. Amer. Soc. Hort. Sci., 34, 635—643 (1936).
413. В i g e a u 1 t E. Le chauffage et le for^age des plantes par panneaux a chaleur
rayonnante. Revue horticole, An. 107, 24, N 15, 372—374 (1935).
414. Bigeaul t E. Le probleme du chauffage et 1’utilisation des rayons calorifiques
obscures. Conference faite a la Societe Nationale d’Horticulture de France. Paris,
le 14 mars 1935.
415. Bold H. C. The cultivation of algae. Bot. Rev., 8, N 2, 69—138 (1942).
416. Bonnier G. Influence de la lumiere electrique continue et discontinue sur
la structure des arbres. C. R. Ac. Sc. Paris, 115, N 12, 447—450 (1892).
417. Bonnier G. Influence de la lumiere electrique sur la structure des plantes
lierbaceus. C. R. Ac. Sc. Paris, 115, N 214, 475—478 (1892).
418. Bonnier G. Influence de la lumiere electrique continue sur la forme et la struc-
ture des plantes. Rev. gen. Bot., 7, N 78—82, 241—257, 289—306,332—342,409—
419 (1895).
419. Bortwick H. A. a. Parker M. W. Photoperiodic perception in Biloxi
Soyabean. Bot. gaz., 100, N 2, 374—387 (1938).
420. В о r t w i с к H. A. a. Parker M. W. Photoperiodic reponses of several
verities of soybeans. Bot. gaz., 101, N 2, 341—365 (1939).
421. В о r L w i с к H. A., Parker M. W. a. H e i n z e P. H. Effect of photope-
riod and temperature on development of Barley. Bot. gaz., 103, N 2, 326—341 (1941).
422. В о r t w i с к H. A., Parker M. W. a. Scully N. Y. Effect of photo-
period and temperature on growth and development of Kok-Sa%hys. Bot. gaz., 105,
N 1, 100—107 (1943).
423. Bortwick H. A. Photoperiod response as a factor in choice of plants for testing
soil deficiencies. Soil Science, 62, N 1, 99—107 (1946).
424* . В о r t w i с к H. A., Hendricks S. B, a. Parker M. W. Action spec-
trum for the control of flowering in winter barley a long day plant. Amer. Jour.
Bot., 34, N 10, 398 (1947).
425* . В о r t w i с к H. A., Hendricks S. B. a. Parker M. W. Action spec-
trum for photoperiodic control of floral initiation of a long day plant. Bot. "az.,
110, N 1, 103—118 (1948).
426. В о r t w i с к H. A., Parker M. W. a. Hendricks S. B. Recent deve-
lopments in the control of flowering by photoperiod. American naturalist, 84, N 815,
117—134 (1950). ’
427. Bos H. Vergroeging van uien door lichtaanvulling Landbcuwkundig. Tijdschrift.
36, 273 (1924). Ref.: Ztschr. f. Pflanzenernahr. und Dungung. A. Wiss. Toil, 4,
526—527 (1925).
428. Bos H. Die Anwendung kiinstlicher Beleuchtung bei der Gartenechtheitspriifung
der Samen im Winter. Amgew. Bot., 1, II. 1, 25—53 (1929).
429. В о у e R. Amerikanische Versuche fiber electrische Bestrahlung von Blumen.
Tcchnik in der Landwirtschaft, 15, N 3, 61 (1934).
430. В г а с к e t t F. S. a. A b b о t C. G. Report on the division of radiation and
organism. Smithsonian. Report, 125—127 (1931).
431. Bremer A. H. Chinesischer Kohl oder Selleriekohl {Brassica pekinensis Rub.)
eine Langtags Pflanze. Gartenbauwiss., 11, N 4, 473—482 (1938).
432* . В finning E. Pbototropismus und Karotinoide. I—III. Planta, 26, II. 5,
719—737 (1937); 27, H. 5, 148—158 (1937); 27, H. 5, 583—610 (1938).
433. С a n n о r s С. H. Gesichtspunkte fiir Ziichten von Blumen in Gewachshausern.
12 Internationaler Gartenbaukongrcss. Berlin, 1938, Section 18, 3 (1939).
Литература
439
434. Central Bureau der V. D. E. N. Bevordering van der groei van planton door elec-
trische verlichting. Electrotechnick, Jg. 6, N 22, 72—83 (1928).
435. Chaminade R. et Boucher J. Influence de 1'eclairage artificiel sur
la floraison des plantes. C. R. Ac. Agric. France, 24, N 29, 964—970 (1938).
436. Chaumier M. Application do la lumiere electrique a la culture des plantes
d'appartement. Elektrizitiits Verwcrtung. Jg. 11, N 2/3, 62—64 (1936/1937).
437. Cheei H. Electric light and Vegetation. Australian Naturalist, 2, 117 (1912).
Ref.: Nature, 106, N 2674, 694 (1921).
438. Childers N. F. a. Brody H. W. An environment controlchamber for
study of photosynthesis, respiration and transpiration of horticultural plants.
Proc. Amer. Soc. Hort. Sei., 37, 384—390 (1939).
439. Chmelar Fr. a M о s t о v о j К. I. Je mozno rychlc rozlisosobati v labo-
ratoii ozime, jarni a piesivkove formy obilinin i bez umeleho osvelovani. (1st es
moglich Winter, Sommer und Wecbselformen von Getreide auch ohne kiinstliche
Boleuchtung im Laboratorium zu crkenne?) Vestnik ceskoslovenske Akademio zeme-
delske. B. 5, c. 1, 10—16 (1929).
440. C h о u a r d P. Chaleur lumiere et radiations. Revue horticale, An. 107, 24,
N 24, 579—583 (1935); An. 108, 25, N 1, 17—26 (1926).
441. Chouard P. Quelques effects de la lumiere sur la croissance, la floraison,
1’enraciment et le bourgeonnement de diverses plantes. С. R. Ac. Sci. Paris, 202,
N 17, 1454—1457 (1936).
442. Chouard P. Die gartnerische Verwcrtung und die Entdeckung neuer Verfahren
zur Einwirkung auf die Entwicklung von Pflanzcn. 12 Intcrnationaler Gartonbau-
kongress, Berlin, 1938, Section 18, 2—3 (1939).
443. Chouard P. Sur les reactions de croissance au photoperiodism. C. R. Ac. Sc.
Paris, 223, N 26, 1174—1176 (1946).
444. Claes II. Die Beteilung des disslmilatorischen Stoffwechsels an der photope-
riodischen Reaktion von Hyoscyamus niger. Ztschr. f. Naturforschung, 2b, 11. 1—2,
45—55 (1947).
445. Claes H. u. Lang A. Die Blutonbildiing von Hyosc.uamus niger in 48-stun-
digem Licht-Dunkelcyklen. Ztschr. f. Naturforscb. 2b, N 1—2, 56—62 (1947).
446. Clarke A. E. a. Lombard P. M. Relation of length of day to flower and
seed production in potato varieties. Amer. Potato Jour., 16, N 9, 236—244 (1939).
447. Clifton A. T. three seed crops a year with soil heating cable and lamps. Elec-
tricity on the Farm, 10, N 1, 15, 27 (1937).
448. Cochran H. L. Factors effecting flowering and fruit setting in the pepper.
Proc. Amer. Soc. Hort Sci., 29, 434—437 (1932).
449. Contributions from Boyce Thompson Institute for plant Research. 1, N 1, 1—58(1925).
450. Cooper H. P. The inheritance"of spring and winter growing habit in crosses
between tipical spring and tipical winter Wheats and the response of wheat plants
to artificial light. Jour. Amer. Soc. Agron., 15, N 1, 15—25 (1923).
451. Cooper J. Commercial uses of electricity in nurseries. Rural Electrification,
10, N 119, 372—373 (1935).
452. Corbet L. C. A study of the effect of incadescent gas light on plant growth.
West Virginia Agric. Expcr. Sta. Bull. 62, 77—110 (1899); Ref.: Just’s Bot. Jah-
resber. Jg. 28, Abt. 2, 287 (1900).
453. Crist J. W. a. Stout G. I. Relation between top and root size in herbaceous
plants. Plant Physiology, 4, N 1, 63—85 (1929).
454. Crocker W. Discussion of the work at Boyce Thompson Institute and its
application. Trans. Jllum. Engin. Soc., 19, N 10, 998—999 (1924).
455. Crocker W. Developments in plant research. Anglo-American Industrial Newslet-
ter, Jan. 1947, 23—26.
456. Cultivo electrico de platanos. Anales de la Souciedad cientifica Argentina, 44, 78
(1897).
457. Darrow G. M. a. Waldo G. F. The practical significance of increasing
the daily light period of winter for strawberry breeding. Science, 69, N 1793, 496—
497 (1929).
458. Darrow G. M. Tomatoes, berries and other crops under continuous light in
Alaska. Sciene, 78, N 2025, 370 (1933).
459. Darrow G. M. a. Waldo G. F. Responses of strawberry varieties and spe-
cies to duration of the daily light period. U. S. Depart. Agric. Washington Techn.
Bull., N 453, 1—31 (1934).
460. Darrow G. M. Interrelation of temperature and photoperiodism in the pro-
duction of fruitbuds and runners in the strawberry. Proc. Amer. Soc. Hort. Sci.,
34, 360—363 (1936).
461. Darrow G. M. Breaking the rest period of the strawberry by long day at high
temperature. Science, 85, N 2207, 391—392 (1937).
440
Литература
462. Davis Л. R. a. Hoagland D. R. An apparatus for the growth of plants
in a controlled environment. Plant physiology, 3, 277—292 (1928).
463. Davis A. R. a. H о a g 1 a n <1 D. R. Father experiments on the growth of
plants in a controlled environment. Amer. Jour. Bot., 15, N 10, 624 (1928).
464. Davy de V i r v i 1 1 e Ad. Influence' de la lumiere electrique discontinue
sur la flore d’une grotte. 1—5 (1928).
465. D e a t s M. The effects of the increase and decrease of the period of illumination
over that of the normal day period. Amer. Jour. Bot., 12, N 7, 384—392 (1925).
466. D e h e r a i n P. P. Influence de la lumiere electrique sur le dcveloppement des
vegetaux. Revue scientifique de la France et de I’etranger, Ser. 3, An. 1, Semestre 2,
28, N 7, 209—212 (1881); N 21, 649—653 (1881).
467. D eh e r a i n P. P. Experiences sur 1’influonce qu’excerce la lumiere electrique
sur le dcveloppement des vegetaux. Ann. agronomiques, 7, 551—575 (1881). Ref.:
Forsch. auf d. Gebjete d. Agricultnrphysik, 5, 488—490 (1882).
468. D e n f f e r D. Uher das Zusammenwirken von Keimstimmung und taglicher
Belichtungsdauer auf die Entwicklung von Sinapis alba und Hordeum. Jahrb. Wiss.
Bot., 88, II. 5, 759—815 (1939).
469. Discussion on the application of electric light to plants. Trans. Ilium. Engin. Soc.,
19, N 4, 326—335 (1924).
470. Dudgeon E. C. Growing crops and plants by electricity. London, 1911. Ref.:
Jour. Roy. Hort. Soc., 37, Part I, 15—20 (1911).
471. Edmundson W. C. Response of several varieties of potatoes to different
photoperiods. Amer. Potato Jour., 18, N 4, 100—112 (1941).
472. E d h о 1 m H. Ett nytt an vandningsomrade for elektriciteten: Drivning av
vaxter i drivhus och bankar. Meddel. fr. Svenska Motokulturforen, N 113, 1—35
(1926).
473. Ed ho Im II. Elektriciteten och Triidgardsodlingen. II. Den elcktriska driv-
banken. Meddel. fr. Kungl. Landbruksakad. Experimentalfalts Tradgardsavdelning,
N 24, 26—64 (1927).
474. E d h о 1 m H. Den elektriska drivbanken. Tekn. Meddel. fr. Kungl. Vattenfall-
styrelscn. Ser. E. N 13, 1—119 (1927).
475. Edwards J. K. Response of Adianthum cunealum to the photoperiod. Amer.
Jour. Bot., 25, N 10, 12 s. (1938).
476. E к b r a n t L. Belysningsforsok med begoniasticklingar. Viola, 35, N 7,45—51 (1929).
477. Electric light and plant growth. Rural electrification, 10, N 112, 134 (1934).
478. Electric light and seed production. Hural electrification, 9, N 98, 62 (1933).
479. Electric light for wheat growing. Rural Electrification, 5, N 68, 294 (1930).
480. Electricity in the garden. Rural Electrification, 10, N 112, 113—115 (1934).
481. E m s w e 1 1 e r S. L., Stuart N. W. a. Byrnes J. W. Using a short
interval of light during night to delay blooming of chrysanthemum. Proc. Amer.
Hort. Soc., 39, 391—392 (1941)
482. E n о m о t о N. On the physiological difference between the spring and winter
types in wheat and barley. Jour. Imper Agric. Exper. Sta. Nishigahara, Tokyo,
1, N 2, 136—138 (1929).
483. E n о m о t о N. On the growth period when the heading of spring barley is most
highly affected by night illumination. Proc. Crop. Sci. Soc. Japan, 3, 32—37 (1929).
Ref.: Japan Jour. Bot., 4, N 4, 81—82 (1929).
484. Erikson E. Forsok med elektrisk belysning vid forsokning av Begonia. Allman.
Svensk Trad gardstigning, 1, 61—63 (1929).
485. Erster Ausflug der Deutschen Gartenbaugesellschaft nach Gartnerleliranstalt in
Dahlem 22 Mai 1919. Gartenflora, 68, 146—154 (1919).
486. Evans M. W. a. Alla r. d H. A. Relation of length of day to growth of ti-
mothy. Jour. Agric. Research., 48, N 7, 571—586 (1934).
487. Evans M. W. a. Watkins I. N. The growth of Kentucky bluegrass and
Canada bluegrass in late spring and in automn as affected by the lenght of day.
Jour. Amer. Soc. Agron., 31, N 9, 767—774 (1939).
488. Fabian I. Beitrage ziim Lang und Kurztagsproblem. Zlschr. I. Bot.. 33, H. 8,
305—357 (1938).
489. Feh er I. Der Einfluss der Strassenbeleuchtung auf den Sommerbaubfall der
Rosskostanien. Botan. Kozlcmenyck, 29, 76 (1932). Ref.: Ztschr. 1. Pflanzencrn,
Toil B, Jg. 13, H. 4, 180 (1934).
490. Findlay H. Preliminary studies in the response of plants to artificial light.
Trans. Ilium. Engin. Soc., 19, N 4, 318—325 (1924).
491. Findlay II. Electric light in greenhause culture. Jour. Hort. Soc. New York,
4, N 1, 1—7 (1924).
492. F i s c h e r H. Bericht fiber 1928 und 29 ausgefiihrte Versuche mit kiinstlicher
Beleuchtung zu Pflanzen. Gartenflora, Jg. 80, 155—157, 191 —192 (1931).
Литература
441
493. Forste г Н. С., Tinker М. А. Н., V a s е у A. Y. a. Wadham S. М.
Experiments in England, Wales and Australia on the effect of length of day on
various cultivated varieties of Wheat. Ann. Appl. Biol., 19, N 3, 378—412
(1932).
494* . Fran к S. K. The effectiveness of the spectrum in chlovophyll formation, Jour,
gen. physiology, 29, N 3, 157—179 (1946).
495. Funke G. L. Over het Kweken van planten bij Neonlichet. I—II. Mededeelingen
der Landbouwhoogeschool en dor oproekingsstations van den Staat, Gent. Deel 4,
N 2, 67—90 (1936); Deel. 5, N 1, 30—40 (1937).
496. Funke G. L. Proeven over photoperiodiciteit bij verschillend gekleurd licht.
Dodonaca 3, 225—261 (1936); 4, 346—361 (1937); 5, 404—424 (1938); 6, 351—376
(1939).
497. Funke G. L. Observations on the flowering Photoperiodicity. Rec. Trav. Bot.
Neerlands, 40, 393—412 (1943—1945).
498. Fiirtaucr R. Untersuchungen iibcr die Beziohung zwischen Photoperiode,
Lichtintensitat sowie Temperatur und Plasmavererbung bei Epilobium.Jahrb.
f. wiss. Bot., 89, H. 3, 412—460 (1940).
499. G a b r i e 1 s о n E. K. Untersuchungen uber des Kohlenstoffhaushalt einer Ge-
wachshauspflanze in Winter bei Tageslicht rnit elcktrischer Zusalzbcleuchtung.
Planta, 22, H. 2, 180—189 (1934).
500. Garner W. W. a. Allard H. A. Effect of the relative length of day and
night and other factors of the environment on growth and reproduction in plants.
Jour. Agric. Research, 18, N 11, 553—606 (1920).
501. Garner W. W. a. Allard H. A. Further studies in photoperiodism: the
response of the plant to relative length of day and night. Jour. Agric. Research,
23, N 11, 871—920 (1923).
502. Garner W. W. a. Allard H. A. Photoperiodism in relation to hydrogen-
ion concentration of the cell sap and the carbohydrate content of the plant. Jour.
Agric. Research, 27, N 3, 119—156 (1924).
503. Garner W. W. a. Allard H. A. Localisation of the response in plants
to relative length of day and night. Jour. Agric. Research, 31, N 6, 555—566 (1925).
504. Garner W. W. a. Allard H. A. Effect of abnormally long and short alter-
nations of light and darkness on growth and development of plants. Jour. Agric.
Research, 42, N 10, 629—651 (1931).
505. Garner W. W. Plant growth by artificial light has possibilities. U. S. Depart.
Agric. Yearbook of Agriculture, A 1197, 1—4 (1931).
506. Garner W. W. Comparative response of long-day and short-day plants to rela-
tive lenght of day and night. Plant Physiology, 8, N 3, 347—356 (1933).
507. Garner W. W. Photoperiodism. Biological Effects of Radiation, 2, 677—713
(1936).
508. G e h e n i о P. M. and L u у e t B. J. Effect of light in inducing reproductive
processes in the Mycctozoon Physarella oblonga. Biodynamica, 7, N 136, 85—99
(1951).
509. G e 1 i n О. E. V. a. Burs triim 11. A study of artificial illumination of
greenhouse culture. Physiologia plantarum, 2, Ease. I, 70—77 (1949).
510. Gerlach A. Kiinstliches Wachstum der Pflanzen. Ertragssteigerung durch
Elektrizitat. Deutsch Landw. Presse, Jg. 51, N 40, 460 (1924).
511. Gerlach M. Versuch fiber die Einwirkung der Elektrizitat auf das Pflanzen
Wachstum. Mitteil. d. Keiser Wilhelm Institut, Bromberg, 4, 11. 4, 354 (1912).
Ref.: Biedermann’s Ztrbl. f. Agrikulturchemie, 41, II. 12, 820—821 (1912).
512. Gerlach M. Versuche fiber die Belichtung wachsender Pflanzen wahrend
der INacht. Mitteil. d. Keiser. Wilhelm Institut, Bromberg, 4, H. 4, 368—373 (1912).
Ref.: Biedermann’s Ztrb. f. Agrikulturchemie, 41, 11. 12, 819—820 (1912).
513* . Gilles E. Effcts des rayons ultraviolets sur les vegetaux superieures. Rev.
gen. bot., 51, 327—353 (1939).
514. G о о d w i n H. Schwedische Versuche mit kiinstlicher Beleuchtung im Gar-
tenbau. Gartonschonheit, N 10, 147—149 (1929).
515* . Goodwin R. H. a. Owens О. H. An action spectrum for inhibition of the
first internode of Avena by light. Bull. Torrey Bot. Club, 75, N 1, 18—21 (1948).
516* . Goodwin R. H. a. Owens О. H. The effectivness of the spectrum in Avena
internode inhibition. Bull. Torrey Bot. Club, 78, N 1, 11—21 (1951).
517* . Gordon N. T. Water cooling of incandescent lamps. Jour. Sci. Motion Picture,
Engin. 14, 332—343 (1930).
518. G о u 1 d e R. U. Die Anwendung der Elektrizitat im Hollandischen Gartenbau.
Elektrizitatswirtschaft. Jg. 34, H. 10, 214—219 (1935).
519. Gran O. Electrisk drit i Gartneriet. Norsk Gartnerforenings Tidskrift, N 16,
148 (1926).
442
Литература
520. Grass J. Beitrage zur Entwicklungsgeschichte der Protophyten. VII. Arch,
f. Protistenkunde, 73', N 2, 206—234 (1931).
521. Green L., Withrow R. B. a. Richman M. W. The response of green-
hause crops to electric light supplementing daylength. Purdue Univ. Agric. Exper.
Sta.^ Lafayette, Indiana Bull., 366, 1—20 (1932).
522. Haul G. och Norinder H. Nogra forsok med electrisk belysning sor driv-
ning av Kulturer i Vaxthus. Sveriges Tradgiirdsolding Arsbok, 108—116 (1925).
523. Haul S. och Norinder H. Elektrisk ljus vid driwning i vaxthus. Tcknisk
Tidskrift, Ag. 56, II. 6, Elektroteknik, 2, 26—29 (1926).
524. Haul S. och Norinder II. Elektrisk ljus vid driwning i vaxthus. Svensk
Land, N 11, 239—240 (1929).
525. Hagens II. W. Ein neuer photo thermostat fiir Mikroorganismen. Ztschr. f.
Naturforsch., 6b, H. 5, 277—278 (1951).
526. Hahn E. Intcressantes aus nordischen Betrieben. Gartenwelt, Jg. 36, N 25,
359—360 (1932).
527. Hamner К. C. a. Naylor A. W. Photoperiodic responses of dill (Anethum
graveolens) a vary sensitive long day plant. Bot. gaz., 100, N 4, 850—861 (1939).
528. Hamner К. C. Interelation of light and darkness in photoperiodic induction.
Bot. gaz., 101, N 3, 658—687 (1940).
529. II a m n e г К. G. Description of a chamber for growing plants under controlled
conditions. Bot. gaz., 105, N 4, 437—441 (1944).
530. Harder R., Keppler E. u. Reuss H. Beobachtungen iibor das Pflan-
zenwachstum und die Kohlensaure Assimilation bei Kohlensaurediingung und
nachtlicher Zusatzbeleuchtung. Gartenbauwissenschaft, 5, H. 5, 389—428 (1931).
531. Harder R., Fabian J. u. D e n f f e r D. Lichtinsensitat und Photo-
periodismus. Zuchter, 9, H. 2, 41—46 (1937).
532. Harder R. u. Witsch H. Wirkung von Photoperiodismus und Jarowisa-
tion auf die Blutenbildung von Kalanchoe blossfeldiana. Gartenbauwissenschaft,
15, If. 2, 225—246 (1940).
533. Harder R. u. Bode O. Uber die Wirkung von Zwischenbelichtungen wah-
rend der Dunkelperiode auf das Bliihen, die Verlaubung und die Blattsukkulenz
bei der Kurztagspflanze Kalancho'e blossjeldiana. Planta, 33, H. 4,469—504(1943).
534. Haring I. M. Plant growth under electric light. Brvologist, 33, 89—91 (1931).
Ref.: Bot. Ctrbl., 21 (163), H. 3/4, 78—79 (1932).
535. Harrington J. B. Growing wheat and barley hybrids in winter by means
of artificial light. Scientific Agriculture, 7, N 4, 125—130 (1926).
536. H a r t s e m a A. en L u у t e n J. Snelle bloei van hollandse irissen var. Impe-
rator. IV. Proc. Kon. Nederl. Akad. Wetcnsch., 56, N 1, ser. C, 81—105 (1953).
537. Hartmann H. T. a. McKinnon L. R. Environment control cabinets
for studying the interrelation of temperature and Photoperiod on the growth and
development of plants. Proc. Amer. Soc. Hort. Sci., 42, 475—480 (1943).
538. Hartmann H. T. Some effects of temperature and photoperiod on flower for-
mation and runner production in the strawberry. Plant physiology, 22, N 4, 407—
420 (1947).
539. Hartmann M. Untersuchungen fiber die Morphologic und Physiologic des
Formwechsels des Phytomonadinen III. Arch. f. Protistenkunde, 43, II. 1—2, 223—
286 (1921).
540. Harvey R. B. Growth of plants in artificial light from seed to seed. Science,
56, N 1448, 366—367 (1922).
541. Harvey R. B. Growth of plants in artificial light. Bot. gaz., 74, N 4, 445—-451
(1922). Ref.: Minnesota Agr. Exp. Sta. Ann. Bull., 31, 21 (1922).
542. Harvey R. B. Carbohydrate'production and growth in plants under artificial
light. Trans, ilium. Engin. Soc., 19, N 4, 311—317 (1924).
543. II a u t I. C. Photoperiodic response of sweet pea. Proc. Amer. Soc. Hort. Sci.,
27, 314—318 (1930).
544. Hayden J. L. u. Steinmetz С. P. Effect of artificial light on the growth
and ripening of plant. Gen. Electr. Rev., 21, N 3, 232—236 (1918).
545. H e 1 1 b о E. Nagra undersokningar rorande morfologiska egenskaper hos vid
artificiele belysning uppdragna ungplantar av vissa crucifer. (A study of morpho-
logical characters on the joung plant of some cruciferous species grown under arti-
ficial light). Meddelanden fran. Statens Centrala Frokontrollanstalt, N 10, 66—87
(1935).
546. Hendricks E. a. Harvey R. B. Growth of plants in artificial light.
II. Intensities of continuous light required for blooming. Bot. gaz., 77, N 3, 330—
334 (1924).
547. Herbatschek O. Elektrische Pflanzenbeleuchtung. Lichttechnik, Jg. 6,
II. 4—6, 33—40, 52—53, 61—64 (1929).
Литература
443
54 8'. Н i b b е n S. G. Influence of colored light on plant growth. Trans. Ilium.Engin.
Soc., 19, N 10, 1000—1010 (1924).
549. H i b b e n S. Greenhouse lighting installations to hasten plant growth. Ilium.
Engin. Bureau Westinghauso Co. March, II, 1—9 (1924).
550. Hilmer E. Ostpreussische Versuche mit Licht und Warme ini Gartenbau.
Elektrizitatswirtschaft. Jg. 30, N 16, 477—488 (1931).
551. H i о r t h G. Die Anwendung elektrischer Beleuchtung fiir Vererbungsversuche
mit Pflanzcn. Zuchter, Jg. 1, H. 7, 204—209 (1929).
552. Honeywell E. R. Aster culture. Purdue (Indiana) Univ. Agric. Exper. Sta.
Circular, "N 200, 1—20 (1933).
553. Hoover W. 11. The dependence of carbone dioxide assimilation in a higher
plant on wave length of radiation. Smthsonion Misc. collections, 95, N 21,1 —13 (1937).
554. H i> s t e r in a n n G. Versuche mit Nconlicht. Ber. d. Konigl. Gartnerlehranstalt,
Dahlem, 1916/1917, 76 (1918).
555. H о s t о r m a n n G. Kulturversuche mit elektrischem Licht. Landw. Jahrb. 57,
Erganzungsbd. I: Ber. d. hoh. Gartnerlehranstalt zu Dahlem, 1920/1921, 116—118
(1922).
556. H 6 s t c r m a n n G. Kulturversuche mit elektrischem Lucht. I. Gartenwelt,
Jg. 26, N 8, 74—75 (1922).
557. II о s t e r m a n n G. Pflanzenkultur versuche mit kiinstlichem Licht. Ztschr.
Vereins deutsch. Ingen., 66, N 21, 523 (1922).
558. Hume E. P. The response of plants to intermittent supplementary light. Proc.
Amer. Soc. Hort. Sci., 37, 1059—1066 (1939).
559. IIurd-Karrer A. M. The formative effect oi day length on wheat seedlings.
Jour. Maryland. Acad. Sci., 1, N 2, 115—126 (1930). Ref.: Biol. Abstr., 5, N 11,
2693 (N 27212) (1931).
560. H u r d - К a г г о r A. M. Comparative reponses of a spring and a winter wheat
to day length and temperature. Jour. Agric. Research., 46, N 10, 867—888 (1933).
561. Hurd-Karrer A. M. a. Dickson A. D. Carbohydrate and nitrogen
relations in wheat plants. Plant Physiology, 9, N 3, 533—565 (1934).
V562. I g g e n a M. L. Beobachtungen uber Wirkung des Lichtes auf das Wachstum
von Blaualgen und Griinalgen. Arch. Mikrobiol., 9, H. 2, 129—166 (1938).
563. Irradiation artificielle des plantes. Resultats obtenus en different pays. Genie
rural, An. 29, 28 (1936).
564. J. K. Elektervalgusture Kasutamisest Kasvuhoeneis taimekasvu soodustajna.
Eesti Pallumajandus, N 3, 183 (1948).
565. Jacobsen G. Elektrisitetens an Vendelse til opwarmning av drivbanker
og dens innflydelse pa Plante-Veksten Kunstig lys. Norsk Gartnerforenings Tid-
skritt, 16, 69—75, 130—133 (1926).
566. Johnston E. S. The seosonal march of climatic conditions of a greenhause
as related to plant growth. Agr. Exp. Sta. Maryland Bull., 245, 41—97 (1921).
567. Jones K. L. Studies on Ambrosia. Bot. gaz., 98, N 2, 296—306 (1936).
568. Jones L. H. Forcing gladiolus with the aid of artificial light. Massachusetts
Exper. Agric. Sta. Bull. 260, 343—344 (1930). Bull. 271, 246 (1931).
569. Joins M. J. Acetylene light for forcing plants. Cornell countryman, N 3, 183—
185 (1906).
570. J u 1 1 e r E.. Beitrage zur Entwicklungsgeschichtc der Protophyten. XIII. Arch,
f. Protistenkundc, 89, H. Im 55—93 (1937).
571. Keller E. R. a. Peterson M. L. Effect of photoperiod on red clower and
timothy strainsgrowth in association. Agron. Jour., 42, N 12, 598—603 (1950).
572. Ketchum В. H. a. Redfield A. C. A method for maintaining a conti-
nuous supply of marine diatomos by culture. Biol. Bull., 75, N 1, 165—169 (1938).
573. Keyes C. G. Bud formation, abscission and flower production of gardenia as
affected by light and temperature. Proc. Amer. Soc. Hort. Sci., 37, 1034—1036
(1939).
574. К i n d W. Praktische Anwendungsgcbiote der Elektrizitat im Gartenbau. Elek-
trizitatswirtschaft. Jg. 30, N 16, 472—476 (1931).
575. Kind W. Elektrisches Licht und Pflanzenwachstum. Umschau, Jg. 39, H. 3,
52—53, 55 (1935).
576. Kind W. Elektrische Heizung und Beleuchtung im deutschen Gartenbau. Elek-
trizitatswirtschaft, Jg. 34, H. 10, 219—220 (1935).
577. К 1 a ges R. H. Metrical attributes and physiology of hardy varieties of winter
wheat. Jour. Amer. Soc. Agron., 18, 7, 529—566 (1926).
578. Klebs G. Uber das Treiben der einheimischen Baume speziell der Buche.
Abhandl. Heidelberg Akad. Wiss. Math. Nat. KI. 3, 1 —116 (1914).
579. Klein G. Die Elektrizitat im Dienste des Gartenbaues. Festschr. Osterr. Gar-
tenbaues. 33—53 (1927). Beitrage zur Gartenbaukunden, 33—53 (1928).
444
Л итература
580. Koernicke М. u. Lindenbein W. Untersuchungen uber die Pflan-
zenphysiologischen Grundlagen der Vcrwendung von Electrolicht- und Warme
in ireibbeeten. Gartenbauwissenschaft, 10, H. 2, 151—183 (1936).
581. Kondo M., О к a m и r a S. u. Kasahara S. Untersuchungen fiber
Photoperiodismus der Reispflanzen. Ber. Ohara Inst. f. Landw. Forsch. in Ku-
rashiki, 5, 11. 2, 243—260 (1932).
582. Korda P. et Chouard P. L’eclairage d’appoint en horticulture. Revue
horticole, An. 108, 25, N 9, 259—264 (19,36).
583. К о t h P. a. H a m n e г К. C. Responses of Marchantia polymorpha to nutri-
entsupply and photoperiod. Bot. gaz., 102, N 1, 169—205 (1940).
584. Kramer P. J. Effect of variation in length of day on growth and dormancy
of trees. Plant Physiology, 11, N 1, 127—139 (1936).
585. К r a m e r P. ,1. Photoperiodic stimulation of growth by artificial ligth as a cause
of winter killing. Plant Physiology, 12, 11. 3, 881—883 (1937).
586. Van d e г К r a f t. De electricileit in den tuinbouw. Veldbodo, Jg. 28, N 1400,
44—48 (1929).
587. Kufferath A. Pflanzenzucbt bei Neonzusatzlicht. Nsturfrcund. I, 12—13
(1947).
588. L.-B., D. Untersuchungen fiber die Wirkung des Liclites auf das Wachstum tier
Pflanzen. Licht und Lampe, N 23, 802—804 (1925).
589. Lang A. u. Melchers G. Die Photoperiodische Reaktion von //yotcyamus
niger. Planta, 33, H. 5, 653—702 (1943).
590. Lang A. Beitrage zur Genetik des Photoperiodismus. II. Ztschr. f. Naturforsch.
2b, II. 1—2, 36—44 (1947).
591. Larsen E. G. Photoperiodic responses of geographical strains of Andropogan
scoparius. Bot. gaz., 109, N 2, 132—149 (1947).
592. L a и b e r t R. Eine Beobachtung fiber den Einfluss von Laternen auf Baume.
Gartenwelt, Jg. 12, N 15, 172—173 (1908).
593. Laurie A. Forcing aster with light. Amer. Florist, A 70, 7—8 (1928).
594. L a и r i e A. Photoperiodism. Practical application to greenhause culture. Proc.
Amer. Soc. Bort. Sci., 27, 318—322 (1930).
595. Laurie A. a. Poesch G. IL Photoperiodism: the value of supplementing
illumination and reduction of light on flowering plants in the greenhause. Ohio
Agric. Exper. Sta. Bull., 512, I—42 (1932).
596. Laurie A. a. Poesch G. H. The use of artificial light and reduction of the
daylight period for flowering plants in the greenhause. Ohio Agric. Exper. Sta.
Bull, 559, 1—43 (1935).
597. Laurie A. A retrospect looking back ten years on floriculture research. Proc.
Amer. Soc. Hort. Sci., 34, 669—676 (1936).
598. L aiirie A. a. К i p 1 i n g e r D. C. Commercial flower forcing. Philadelphia
(1945).
599. Lewis 11. a. W e n t F. W. Plant growth under controlled conditions. IV. Res-
ponse of California annuals to photoperiod and temperature. Amer. Jour. Bot.,
32, N 1, 1—2 (1945).
600. L i n d G. och P a 1 m g a r d A. Eleklriciteten och Tragard sodlingen. III. Med-
del. fr. Kungl. Landbruksakad. Experimentalfalts Tradsgardsa\delning, N 24,
65—84 (1927).
601. Lind G. Elektrisk Tradgardsodling. Handl, och Tidskr. Kungl. Lanbr.-akad., 78,
49—65 (1928). Medd. fr. Kungl. Landbr.-Akad. Experimentalfalts Triidgiirdsovdel-
ning, N 25, 1—17 (1928).
602. Lindback. Praktisko erfaringer Vedrorende eleklrisi telen 1 plante К ulturens
tieneste. Norsk Gartnerforenings 'lidskritt, N 16, 143—14(5 (1926).
603. Lindenbein W. Uber die Bedeulung der Tageszeit bei zusalzlicliein Kiin-
stlichtgaben im Winter. Gartenbauwiss., 13, H. 4, 587—597 (1939).
604. Link C. Further microchemical studies of flower bud differentiation in relation
to the Photopcriod. Proc. Amer. Soc. Hort. Sci., 35, 810—814 (1938).
605. Loschakowa N., Schick R., Scngbusch R. u. Vogl K. 1st
kiinstliche Beleuchtung bei Tomaten wirtschaflich3 Obst und Gemiisebau, Jg. 78,
H. 11, 169—170 (1932).
606. Lo w A. J. Fluorescent lichting for pot culture: an aid to winter research. Agri-
culture, 55, N 5, 210—211 (1948).
607. Ludwig. Kunstliches Licht im Gewachshaus. Blumen und Pflanzenbau, Jg. 45,
Sondernumer, 1—16 (1930).
608. Lund (). Eloktriciteten og gartnerne. Norsk Gartnerforenings Tidskr., 16,
129—130 (1926).
609. M adsen S. C. Influence of photoperiod on microsporogenesis in Cosmos sul-
phurous var. Klondyke. Bot. gaz., 109, N 2, 120—132 (1947).
Литература
445
610. М a g г о u J., М a n i g a u 1 t Р. et М а г i a t F. Sur un dispositif
d’eclairement artii’iciel des semis. C. R. Ac. Sc. Paris, 222, N 10, 521—523
(1946).
611. Magruder R. a. Allard H. A. Bulb formation in some american and
european varieties of onions as affected by length of day. Jour. Agric. Research,,
54, N 10, 719—752 (1937).
612. Mango n H. Production de la matiere verte des feuilles sous 1’influence de
la lumiere electrique. C. R. Ac. Sc. Paris, 53, N 6, 243—244 (1861).
613. Maniga ult P., Magrou I. et M a r i a t F. Utilisation au laboratoire
des lampes fluorescentes. Ann. Inst. Pasteur, 73, N 1, 89—93 (1947).
614. Massart J. L’action de la lumiere continue sur la structure des feuilles. Acad,
royale Belgique, Bull. cl. sc., Ser. 5, 6, 37—43 (1920).
615. Matthews R. B. Electro-farming. Intensive illumination on a Market garden.
Electr. Rev., 97, N 2501, 685—687 (1925).
616. Matzke E. B. The effect of street lights in delaying leaffall in certain trees.
Amer. Jour. Bot., 23, N 6, 446—452 (1936).
617. M с С 1 e 1 1 a n d T. B. The photoperiodism of Thcphrosia Candida. Jour. Agric.
Research, 28, N 5, 445—460 (1924).
618. McClelland T. B. Studies of the photoperiod ism e of some economic Plants.
Jour. Agric. Research, 37, N 10, 603—628 (1928).
619. McKinney H. H. a.w 8 a n d о W. Y. The behavior of Winter wheat in arti-
ficial environments. Science, 71, IN 1852, 668—669 (1930).
620. McKinney H. II. a. S a n d о W. Y. Earliness and seasonal growth habit
in wheat as influenced bv temperature and photoperiodism. Jour. Heredity, 24,
N 5, 169—179 (1933).
621. Men rm an O. Valtion puutarhakoeasemalla Noon. Kasvihuonelampula suori-
tetun alnstavan Kurkuntaimien Valaistuksen tulokset (Die Ergebnisse eines Vor-
laufige Bestrahlungsversuches mit Gurkenpflanzen, mil einer Neon-Gewachshau-
slampe ausgefiihrt an der Staatlichen Versuchsstation fiir Gartenbau). Valtion Maato-
louskoctoiminncn tiedonantoja, N 81 (1934).
622. M e и r in a n O. Untersuchungen iiber (lie Bedoutung des Neonlichtes fiir Ge-
wachshauskulturen. II. Versuchsergebnisse mit Gloxinien. Maatosoustieteellinen
Aikakauskirja Helsinki, Vussik 7, Vinko 4, 171—181 (1935).
623. M e и r m a n O. Tutkimuksia neon valon merkityksesta Kasvihuoneviljelyksisa.
Valtion maatolouskoetoiminnam julkaisuja, N 76, 70—73 (1936).
624. M e и r m a n O. Versuchc mit Neonlicht zur Fruhtreiberei untcr beson-
derer Beriicksichtigung der Tomatenkulturen. Gartenbauwirtschaft, 55, N 22,
19 (1983).
625. Mitchell J. W. Effects of carbon arc light on the chemical composition and
vegetative propagation of tomato plants growth with a limited supply of nitrogen.
Plant Physiology, 11, N 4, 833—841 (1936).
626. Mitchell J. W. Responses by tomato plants to artificial illumination. Bot.
gaz., 99, N 2, 412—419 (1937).
627. Mitchell K. J. High light intensities for experiments in crop physiology.
Nature, 166, N 4209, 35 (1950).
628. Mitchell K. J. Influence of light and temperature on the growth of Ryegras.
Physiologia plantarum, 6, Fasc. I, 21—46 (1953).
629. Molen S. van der. Photocultur in Groningen. Electrotechnick, Jg. 6, N 17,
17—20 (1928).
630. Montpellier J. A. Influence de 1’eclairage electrique sur les plantes. Rev.
scientifique, 49, N 11, 339—342 (1892).
631. Morton F. Piante verdi presso le lampadc dcll’illuminazione electrica nelle
Grotte di Postumia. Ее grotte d’Italia, Ser. 2, N 4, 23—28 (1941). Ref.: Bot. Ztrbl.,
36 (178), H. 8/10, 260—261 (1945).
632. Munerati O. Illuminazione artificiale notturna come mezzo per accelerate
le fasi del ciclo dei vegetali. Atti rcale. Accad. nazion. Linzei, ser. 5, Rendiconti
cl. sc. fis., mat. e naE, 33, Fasc., 10, 402—405 (1924).
633. Munerati O. Dal seme al seme in esclusiva luce artificiale. Atti reale Istituto
veneto Sc. letterc e arti, 83, Parte 2, 751—758 (1924).
634. Nakata S. Photoperiodic response of lespedeza. Plant physiology, 27, N 3,
644—647 (1952).
635. Naylor A. W. a. Gerncr G. Fluorescent lamps as a saurce of light for
growning plants. Bot. gaz., 101, N 3, 715—716 (1940).
636. Naylor A. W. The use of fluorescent light on experimental work. Trans. Illi-
nois. Acad. Sci., 34, N 2, 82—84 (1941).
637. Naylor A. W. Effects of some environmental factors on Photoperiodic induction
of beet and dill. Bot. gaz., 102, N 3, 557—575 (1941).
446
Литература
638. Naylor A. W. Effect of nutrition and age upon the rate of development of ter-
minal staminate inflorescences of Xanthium pensylvanicum. Bot. gaz., 103, N 2
342—353 (1941).
639. Naylor F. L. Effect of length of induction period on floral development of
Xanthium pensylvanicum. Bot. gaz., 103, N 1, 146—154 (1941).
640. Newell L. C. Controlled life cycles of Bromegrass, Bromus inermis used impro-
vement. Agr. Jour., 43, N 9, 417—424 (1951).
641. N i с о 1 a i W. Orchideen. Berlin (1939).
642. Nielsen Ch. S. Effects of photoperiod on microsporogenesis in Biloxi Soybean.
Bot. gaz., 104, N 1, 99—106 (1942).
643. N о m b 1 о t. Influence de Г eclairage artificial sur la floraison des plantes. C. R.
Ac. Agric. France, 24, N 29, 962—964 (1938).
644. Oakley R. A. a. Westover II. L. Effect of length of day on seedlings
of alf alfa varieties. Jour. Agric. Research, 21, N 8, 599—607 (1921).
645. Oden S. Elektriciteten och Tradgardsodlingen. I. Vaxterna och ljuset. Meddel.
------. fr. Kungl. Landbruksakad. Experimentalfalts Triidgardsavdelning, N 24, 1—25 (1927).
1646) Oden S. Viixtodling i elektrisk Ijus. Meddel. fr. Kungl. Landtbruksakad. Ve-
tenskaps. Stockholm, N 1, 1—161 (1929).
647. Oden S. Elektrohortikultur in der Schweiz. Beibl. f. Land und Forstwirtsch.
d. Neuc Zurich Ztg., N 21 und 23, 8 (1929).
648. Oden S. Ergcbnisse und Erfahrungen auf vier Jahren Elektro-Gartenbau beim
Experimentafolde. Handl, och Tijdskr. Kungl. Landbr. Akad., IV 2, 213 (1930).
Ref.: Fortschr. J.andwirtsch. Jg. 5, H. 20, 692 (1930).
649. Oden S., Kohler G. och Nilsson G. Den elektriska Vaxtb.elysningens
praktiska utforande och Kostnaderna Harfor. (Die praktische Ausfiihrung der elek-
trischen Pflanzenbeleuchtung und die Kosten dcrsclben). Meddel fr. Centralanstal-
ten for forsoksvasendet pa jordbruksomradet, N 393, Avdelningen for landbruks-
kemi, N 43, Stockholm (1931).
650. Oden S., Kohler G. a. Nilsson G. Plant cultuvation with the electric
light. Proc. 117. Internal. Ilium. Congress. 1931. 1298'—1326 (1932). Ref.: Rural
Electrification, 8, N 85, 24—26 (1932).
651. Oden S. och Rasmussen J. Frodling med elektrisk hjalpljus (Samem-
zuchtung mit elektrischer Hilfsbeleuchtung). Meddel. fr. Centralanstaltcn for
forsoksvasendet pa jordbruksomradet, N 424. Avdelningen for landbrukskemi,
N 50, Stockholm (1932).
652. Oden och Rasmussen J. Frodling med elektrisk Ijus. Handl. Kundl.
Landbruks. Akad., 70, N 80, 1094 (1932).
653. Olmsted Ch. E. Growth and development in range grosses. IV. Bot. gaz.,
106, N 1, 46—74 (1944).
654. 0 s t e r 1 i n d G. Growth cultivations of the algae Scenedesmus quadricaula.
Symb. Bot. Upsalienses, 10, N 23, 3—141 (1949).
655. О wen F. V., C a r s n e r E. a. S t о u t M. Phototermal induction of flo-
wering in sugar beets. Jour. Agric. Research., 61, N 2, 101—124 (1940).
656. P a г к с r M. W. а. В о r t w i с к II. A. Effect of photoperiod on development
and metabolism of the Biloxi soybean. Bot. gaz,, 100, N 3, 651—689 (1939).
657* . Parker M. W., Hendricks S. В., В о r t w i с к II. A. a. Scull y
N. J. Action spectrum for the photoperiodic control of floral initiation in Biloxi
soybean. Science, 102, N 2641, 152—155 (1945).
658. Parker M. W. Environmental factors and their control in plant experiment.
Soil Science, 62, N 1, 109'—119 (1946).
659* . Parker M. W., H e n d r i с к s S. B., Bor t w i с к H. A. a. Scull у
N. J. Action spectrum for the photoperiodic control of floral initiation of short
day plants. Bot. gaz., 108, N 1, 1—26 (1946).
660* . Parker M. W., II e n d r i с к s S. В., В о г t w i с к II. A. a. W e n t F. W.
Spectral sensitivities for leaf and stem growth of etiolated pea seddlings and their
similarity to action spectra for Photoperiodism. Amer. Jour. Bot., 36, N 2, 194—
204 (1949).
661. Parker M. W. a. Bortwick 11. A. Growth and composition of Biloxi
soybean grown in controlced environment with radiation from dificrcnt carbon-arc
sources. Plant Physiology, 24, N 3, 345—358 (1949).
662. Parke r M. W. а. В о r t w i с к II. A. A modified circuit for slimline fluo-
rescent lamps for plant growth chamber. Plant physiology, 25, N 1, 86—91 (1950).
663. Parker M. W. a. Bor t w i с к H. A. Influence of light on plant growth.
Annual Review of plant Physiology, 1, "43—58 (1950).
664* . Parker M. W., H e n d r i с к S. В. а. В о r t w i с к H. A. Action spec-
trum for the photoperiodic control of floral initiation of the long day plant Hyo-
scyamus niger. Bot. gaz., Ill, N 3, 242—252 (1950).
Литература
447
665. Pendred L. Plant life in Cheddar caves. Nature, 105, N 2649, 709 (1920).
666. Peterson M. L. a. Loomis W. E. Effects of photoperiod and temperature
on growth and flowering of Kentucky Blue grass. Plant physiology, 24, N 1, 31—42
(1949).
667. Pfeiffer N. E. Microchemical and morphological studies of Effect of light
on plants. Bot. gaz., 81, N 2, 173—196 (1926).
668. Pigorini L. Elektrisch light bij botanische onderzoekingen en toepassingen.
Illuminazione, Jg. 2, N 3, 17—20 (1929).
669. P i n к h о f M. Invloed van het Neonlicht op de ontwikkeling van planten. Han-
del. 22. Niderl. Nat. en Geneeskund. Congress, 155 (1929).
670. Pirschle K. u. We t is t ein F. Einige verlaufige Beobachtungen fiber
die Wirkung verschiedener Lichtintensitatcn und qualitatet auf hohere Pflanzen
untcr konstanter Bedingungen. Biol. Ztrbl., 60, II. 11/12, 628—658 (1940).
671. Pirschle K. Weitere Beobachtungen liber den Einfluss von langcwelliger
und mittelwelligcr U.-V.-Strahlung auf hohere Pflanzen. Biol. Ztrbl., 61, II. 9/10,
452—473 (1941).
672. Poclt H. Uber Entwicklungsboschleunigung und Aufzucht von Winterweizen
in Laboratorium. Landw. Forsch., 2, II. 1, 21—27. Ref.: Biol. Abstr., 25, N 11,
3058 (N 34472) (1951).
673. P о e s c h G. H. Forcing plants with artificial light. Proc. Amer. Soc. Hort. Sci.,
28, 402—406 (1931).
674. Poes ch G. H. Supplementary illumination from Mazda, mercury and neon
lamps on some greenhouse plants. Proc. Amer. Soc. Hort. Sci., 33, 637—638
(1935).
6/5. Poes ch G. II. Prolonging the flowering Period of Chrysanthemums with the
use of supplementary illumination. Proc. Amer. Soc. Hort. Sci., 34, 624—626 (1936).
676. Popp 11. W. Summary of literature on some phase of the effect of light on plant
growth. Trans. Ilium. Engin. Soc., 19, N 10, 981 — 994 (1924). Ref.: Licht mid
Lampe, N 23, 802—804 (1925).
677. Porter L. C. Light for Sight, plant growth and beauty. Gen. Electr. Rev.,
37, N 5, 239—242 (1934).
678. Porter L. C. a. Lee F. B. Stimulating the growth of plants by the use of
artificial light. Gen.. Electr. Co. Aug. (1934).
679. Porter L. C. Recent advances in the use of light for plant research. Gen. Electr.
Rev., 38, N 6, 268—271 (1935).
680. Porter L. C. Electrical floriculture. Scientific American, 154, N 3, 133—135
(1936).
681. Porter L. C, Stimulating plant growth with artificial light. Gen. Electr., Co.
(1937).
682. Portsmouth S. B. The effect of alternate periods of light and darkness of
short duration an the growth of the cucumber. Ann. Bot., 1, N 1, 175—189 (1937).
683. Post K. The effects of day length and light intensity on vegetative growth and
flowering of China aster (Callistephus chinensis). Proc. Amer. Soc. Hort. Sci., 32,
626—630 (1934). Ref.: Gartenbauwiss., 10, H. 5/6, 99 (1937).
684. Post K. Further responses of miscellaneous plants to temperature. Proc. Amor.
Soc. Hort. Sci., 34, 627—629 (1936).
685. Post K. The relationship of temperature to flower bud formation in chrysan-
themums. Proc. Amer. Soc. Hort. Sci., 37, 1003—1006 (1939).
686. Post K. a. We d d 1 e Ch. L. The effect of temperature and photoperiod on
the srrowth and flowering of miscellaneous annuals. Proc. Amer. Soc. Hort. Sci.,
37, 1037—1044 (1939).
687. Post K. Problems in forcing Easter lilies. Proc. Amer. Soc. Hort. Sci., 39,
415—418 (1941).
688. Post K. Effects of day length and temperature on growth and flowering of sonic
florist crops. Cornell Univ. Agric. Exper. Sta. Bull. 787 (1942).
689. Practical intensive illumination of plants. Rural Electrification, 6, N 67, 216—217
(1930).
690. Pringsheiin E. G. Kulturversche mit Chloropbyllfuhrenden Microorganis-
men. V. Bcitr. Biol. Pflanzen, 14, 283—312 (1926).
691. Pringsheim E. G. Pure culture of algae. Cambridge (1949).
692. Prist] ey J. II. Electricity in relation to horticulture. Jour. Roy. Hort. Sci.,
37, Part I, 15—25 (1911).
693* . R a b i d e a u G. S., French C. S. a. Holt A. S. The absorption spectra
of leaves, chloroplast suspension and chloroplast pigments as measured in an Ulbricht
sphere. Amer. Jour. Bot., 33, N 10, 769—777 (1946).
694. R a m a 1 e у F. Growth of plants under continous liuht. Science, 73, N 189!),
566—567 (1931).
448
Литература
695. R a m а 1 о у F. Some caryophyllaceous plants influenced in growth and struc-
ture by artificial illumination supplemental to daylight. Bot. gaz., 92, N 3, 311—319
(1931)
696. R a m a 1 e у F. A working bibliography of day length and artificial illumination
as effecting growth of seed plants. Univ. Colorado Studies, 20, N 2 and 3, 257—263
(1933). (Univ. G lorado Bull., voll. 33, N 2, Gen. Ser., N 322, 257—263 (1933).
697. Rama ley F. Influence of supplemental light on blooming. Bot. gaz., 96,
N 1, 165—174 (1934).
698. R a n e F. W. Electrohorticulture with the incandescent lamp. West Virginia
Agric. Exper. Sta. Bull., 37, 1—27 (1894).
699. R a n e F. W. Effect of acetylene gaslight on plant growth. New Hampshire Coll.
Agric. Exper. Sta. Techn. Bull., 4, 1—8 (1902).
700. Ranke A. Kulturversuche mit elektrischem Licht. IL Durchfiihrung und
Ergcbnissc der Versuche. Gartenwelt, Jg. 26, N 9 und 10, 90—91, 98—100 (1928).
701. R a ts e к J. C. Effects of day length on growth f orchid seedlings. Proc. Amer.
Soc. Hort. Sci., 29, 555—557 (1932).
702. R e d i n g t о n G. a. P r i s t I e у S. H. A study of the effect of diurnal perio-
dicity upon fibre production. Proc. Leeds Phil. Soc., N 1, 17—26 (1925).
703. R e d i n g t о n G. The effect of the duration of light upon the growth and deve-
lopment of the Plant. Biol. Rev. and biol. Proc. Cambridge Phill. Soc., 4, N 2,
180- 208 (1929). Ref.: Biol. Abstr., 5, N 6 — 7, 1624 (N 16716) (1931).
704. R e d i n g t о n G. A study of the effect of diurnal periodicity upon plant growth.
Trans. Roy. Soc. Edinburg, 56, part 2, 247—271 (1929).
705. R e i n a u E. Pflanzen und Licht. Jg. 4, II. 9 und 10, 161—165, 211—216 (1934).
706. Reinhold J. 1st Kiinstliche Belichtung zur Treibgurkenkultur wirtschaftlich?
Obst und Gemiisebau, Jg. 78, H. 12, 182—183 (1932).
707. Reinhold J. Osram und Neon Belichtungsversuche zu Friihgeniiisekulturon.
Gartenbauwiss., 9, N 6, 558—574 (1935).
708. Reinders-Gouwentak G. A. Het apweken van stoocktomaten met beh-
ulp van de hoogedruk-kwiklamp Philips «110-200». Mededeel Landbowhosch. Wage-
ningen, 50, N 4, 63—71 (1950). Ref.: Biol. Abstr., 25, N 10, 2824 (N 31990) (1951).
709. Reinders-Gou wentа к C. A. a. Sipkens J. The influence of
photoperiod dormancy breaking and growth hormone treatment on poplar cuttings.
Proc. Kon. Nedere. Akad. Wetensch., 56, N 1, Ser. C, 71—78 (1953).
710. Richey Fr. D. a. S p r a g u e G. F. Some factors effecting the reversal of sex ex-
pression in the tassels of Maize. Amer. Naturalist, 66, N 706, 434—443 (1932).
711. Richman M. W. Aster seedlings given additional light bloom quicker. Florist
Review, 16 Febr., 41—42 (1933).
712. R ingwald F. Neuere Anwendungen der Elektrizitat in der Landwirtschaft
und Verwandten Gebieten. Bull. Schweizer. Elektrotechn. Vercin, Jg. 16, N 5-bis,
307—324 (1925).
713. Rivera V. Illuminazione ed incremento di vegetazione. Mem. R. Staz. Patal.
Veg. Roma, 18 (1919).
714. Roberts La. Wadsworth S. E. The all-electric greenhouse. Washington,
Agric. Exper. Sta. Bull. 404, 1—22 (1941).
715. Roberts R. II. a. Struckmeyer В. E. The effects of temperature and
other environmental factors upon the photoperiodic responses of some of the higher
plants. Jour. Agric. Research, 56, N 9, 633—678 (1938).
716. Roberts R. H. a. Struckmeyer В. E. Futher studies of the effect
of temperature and other environmental factors upon the photoperiodic responses
of plants. Jour. Agric. Research, 59, N 9, 698—709 (1939).
717. Roodenburg J. M. Kunstlichtcultuur (Cultivation of plants in artificial
light). Mededeel. Lantbouwhoogeschool, Wagcningen, 34, N 14 (1930).
718. Roodenburg J. W. M. Kunstlichtkultur. Angew. Bot., 13, 162—166 (1931).
719. Roodenburg J. W. M. Bestraling van Komkommerplanten met Neonlicht.
Tuinderij, Jg. 12, N 40, 40—41 (1932).
720. Roodenburg J. W. M. Kunstlichtcultuur II. Over de noodzakelijke licht
intensiteit bij bestraling van planten en neonbelichting bij bloemcultures. Mede-
deel. Landbouwhoogeschool, Wageningen, 36, Verb. 2 (1932).
721. Roodenburg J. W. M. Zum Problem der Kiinstlichkultur. Gartenwelt,
Jg. 36, N 52, 736 (1932).
722. Roodenburg J. W. M. Pflanzenbestrahlung mit Neonlicht. Schweizer Elec-
trorundschau, Sondcrausgabc «Die Elektrizitat im Gartenbau», 181—186, Juli (1933).
723. Roodenburg J. W. M. L’irradiation, des plantes avec la lumiere au Neon.
Elektrisitate-Verwertung (L’Electrique), N 10/11, 283—287 (1933/1934).
724. Roodenburg J. W. M. De invloed op den lengtegroei van Kasplanten. Han-
del. 24. Nederland. Nat. en Geneeskund. Congres, 17—18 (1933).
Литература
449
725. R о о denburg J. W. M. Rotlicht fiir Treibhauspflanzen. Gartenztg. d. Osterr.
Gartenbau. Ges. Wien, 146—148 (1934). Ref.: Rot. Ctrbl., 26 (168), H. 9/10, 286
(1935).
726. Roodenburg J. W. M. Aardbein under neonlicht, Tuinderij, Jg. 16, 16/VI,
17—18 (1936).
727. Roodenburg J. W. M. Strawberries under Neonlicht. Jour. Roy. Hort,
Soc., 61, part 2, 504—509 (1936).
728. Roodenburg J. W. M. u. Z e c h о r G. Pflanzenbestrahlung mit Neon-
licht. Philips Techn. Tijdschrifts,1 1, N 7, 193—199 (1936).
729. Roodenburg J.W. M. Der Einfluss der Tageslange im Zusammenhang mit der
kiinstliche Pflanzenbeleuchtung im Winter. Ber. d. d. bot. Ges., 55, H. 1, 5—32 (1937).
730. Roodenburg J. W. M. Daglengte en Plantebestraling. Nederland. Kruid-
kund. Archief. Deel., 47, 113—114 (1937).
731. Roodenburg J. W. M. Die Pflanzenphysiologischen Bedingungen fiir die
Anwendung des kiinstlichen Lichtes im Gewachshaus. 12 Internationaler Garten-
baukongress, Berlin, 1938, Section 17, 8 (1939).
732. Roodenburg J. W. M. Het Vervroegen van aardbein met Kunstlicht (Das
Treiben von Erdbeeren mittels Zusatbeleuchtung). Landbauwkundig Tijdschrift,
Jg. 51, N 632, 806—832 (1939).
733. Roodenburg J. W. M. Das Verhallen von Pflanzen in Verschiedenfarbigem
Licht. Rec. trav. botan. neerl., 37, livr. 2, 301—376 (1940).
734. Rowland F. E. Stimulating plant growth. Electric. Rev., 117, N 3019, 437—
438 (1935).
735. R о w 1 e e W. W. Effect of electric light upon the tissues of leaves. Proc. Soc.
Prom. Agric. Sci., 17, N 1, 1—15 (1898).
i 736. Sakr E. S. a. Thompson H. C. Effect of temperature and photoperiod
on seedstalk development in carrots. Proc. Amer. Soc. Hort. Sci., 41, 343—346
(1942).
737. Sande-Bakhuyzen H. L., Van de. Studies of wheat growth under con-
stant condition. I—II. Plant physiology, 3, N 1, 1—30 (1928).
738. Sande-Bakhuyzen H. L., Van de. Studies on wheat growth under con-
stant conditions. Food Research Institute Stanford university, California (1937/.
739. Sartoris G. B. The behavior of sugarcane in relation to length of day. Sugar
News, 20, N 10, 421—424 (1939).
740. Schaffner J. H. The change of opposite to alternate phyllotaxy and repeated
rejuvenations in hemp by means of changed photoperiodicity. Ecology, 7, N 3,
315—325 (1926).
741. Schaffner J. H. Futher experiments in repeated rejuvenations in hemp and
their bearing on the general problem of sex. Amer. Jour. Bot., 15, N 1, 77—85 (1928).
742. Schappelie N. A. Effect of narrow ranges of wave lenght of radiant energy
and other factors on the reproductive growth of long-day and short-day plants. Cor-
nell univ. Agric. Exper. Sta. Mem., 185, 1—33 (1936).
743. Schmitz J. liber Beziehungen zwischen Blumenbildung im verschieden Licht-
Dunkel kombination. Planta, 39, H. 4, 271—308 (1951).
744. Schneider L. u. Vo gl K. Kiinstliche Beleuchtung fiir Pflanzen. Elektre-
zitatswirtschaft. Jg. 30, N 16, 458—464 (1931).
745. Schratz E. Einfluss kiinstlichor Beleuchtung auf hohere Pflanzen (Sammel-
referat). Zuchter, 3, H. 2, 45—57 (1931).
746. Scully N. J., Parker M. W. a. Bortwick H. A. Interaction of nitro-
gen nutrition and photoperiod as expressed in bulbing and flowerstalk development
of onion. Bot. gaz., 107, N 1, 52—61 (1945).
747. Scully N. J., Parker M. W. а. В о r t w i с к H. A. Relationship of
photoperiod and nitrogen nutrition to initiation of flower primordia in savbean
varieties. Bot. gaz., 107, N 2, 218—231 (1945).
748. Scully N. J. a. Domingo W. E. Effects of duration and intensity of light
upon flowering in several varieties and hybrids of castor bean. Bot. gaz., 108, N 4,
556—570 (1947).
749* . Seybold A. Uber die optischen Eigenschaften der Laubblatter. Planta,
16, 195 (1932); 18, 479 (1932); 20, 577—601 (1933); 21, 251—265 (1933).
750. Seybold A. u. Egle K. Lichtfeld und Blattfarbstoffe. I—II. Planta,
26, H. 3, 491—515 (1937); 28, H. 1, 87—123 (1938).
751. Shepherd H. R. Studies in breaking the rest period of grass plants by treat-
ments with potassium thyocyanate and stimulating growth with artificial light.
Trans. Kansas. Acad. Sci., 41, 139—153 (1938).
1 Имеются немецкое и английское издание — Philips technische Rundschau
и Philips technical Review.
29 А. Ф. Клешнин
450
Литература
752. Shih-Wey Loo. Preliminary experiments on the cultivation of Baeria chry-
sostoma under steril conditions. Amer. Jour. Bot., 33, N 5, 382—389 (1946).
753. Shirley H. L. The influence of light intensity and quality upon the growth
of plants. Amer. Jour. Bot., 16, N 5, 354—390 (1929).
754. Siemens C. W. On the influence of electric light upon vegetation and certain
physical principes involved. Proc. Roy. Soc. London, 30, N 202, 210—219 (1880).
Nature, N 541, 456—457 (1880). Ref.: Bot. Ctrbl., 8, N 5, 189—190 (1881). Fortschr.
auf d. Gebiete d. Agricultur-physik, 5, 486—488 (1882).
755. Siemens C. W. Some futher observations on the influence of electric light
upon vegetation. Proc. Roy. Soc. London, 30, N 202, 293—295 (1880).
756. Siemens G. W. On some applications of electric energy to agricultural and
horticultural purpose. Brit. Ass. Adv. Sci. Rep. 474—480 (1882).
757. S i g a f о о s M. M. Growth of certain plants under continous illumination from
fluorescent lamps. Ohio. State Univ. Doctoral Diss. 52, 275—281 (1946/47). Ref.:
Biol. Abstr. 22, N 9, 2247 (N 22289) (1948).
758. Singh' B. N., Singh S. N. a. Srivastava M. B. Photoperiodism
a factor in determing the manual efficiency and distribution of Crotalaria juncea.
Jour. Amer. Soc. Agron., 29, N 2, 123—133 (1937).
759. Singh B. N., К о p о о r G. Р. a. G h о u d h г i R. S. The light factor in
crop production. Proc. Indian. Acad. Sci. Sect. B., 7, N 3, 143—160 (1938).
760. Singh B. N. a. G h о и d h r i R. S. The influence of light on shoot elongation
and branching in Crotalaria juncea. Trop. Agric. Trinidad, 15, N 9, 202 (1938).
761. Singh B. N. a. Singh S. N. Photoperiodism and the phasic development
of Crotalaria juncea. Proc. Indian. Ac. Sci. Sect. B. 9, N 6, 323—330 (1939).
762. S i v о r i E. a. Went F. W. Photoperiodicity of Baeria chrysostoma. Bot.
gaz., 105, N 3, 321—329 (1944).
763. Skinner H. T. Factors effecting shoot growth and flower bud formation in
rhododendrons azalias. Proc. Amer. Soc. Hort. Sci., 37, 1007—1011 (1939).
764. S к о к I. Effect of length day and temperature on the opening of buds of dormant
twigs. Trans. Illinois Ac. Sci., 34, N 2, 76—77 (1941).
5. Smith F. Om plante dyrkning i kunstig lys. Norsk. Gartnerforenings Tidskrift,
Arg. 9, 87—90, 97—100, 103—106, 113—114, 127 (1919); Arg. 14, 86—89, 92—94
(1924).
766. Smith F. Om Lysreguleryng i plantekulturens tjeneste. Havedyrkningens
Venner. Medlemsskr., 4, 106—125 (1926).
767. Smith F. Om varmesumsbehavest avhengigheit av daglaengte, lysmaenyde,
lysintensitet og lyskvalitet. Norsk Landmansblad., 45, 232—235 (1926); 46, 257—
258, 267—270 (1927).
768. Smith F. Forel bige resultater angaaendc virkingen av forskielligc kunstige
lyslag paa plantene. Norsk. Landmansblad., 47, 168 (1928).
769. Smith F. Det kunstige lys i Plantekulturens tieneste Elektrisitetens uthytting
i Gartneri og havebruk. Oslo (1928).
770. Smith F. Om inflydelsen av kunstig ag naturlig lys paa plantene. Nordisk.,
lordbriigstorskning. H. 5, 337—344 (1930). Ref.: Gartenbauwiss. Referate, 5, H. 1,
3—4 (1931).
771. Smith F. Researches on the influence of natural and artificial light on plants.
I. On the influence of the length of day. Meldinger fra Norges Landbrukshoiskole,
13, N 1—5 (1933).
772. Smith F. Glodlampelys, neonlys og natriumdamplys som tilsetning til naturlig
dagslys. Norsk Gartnerforenings Tidskrift, Arg. 25, N 40 og 41, 329—332, 339—341
(1935).
773. Smith F. Glodlampsljus, neonljus,. och natriumangljus som tellskatt till det
naturliga dagsljuset. Allman Svensk Tradgardstidning, N , 13, 397—399
(1936).
774. Smith F. Daglengdens innflytelsc pa unge agurk-og tomatplanter. Norsk Gart-
nerforenings Tidskrift, Arg. 27, N 45, 437—442 (1937).
775. Smith F. Lysklima og vekst hos tomat planter. Norsk Gartnerforenings Tid-
skrift, Arg. 28, N 1 og 24, 3—7, 221—225 (1938).
776. Smith F. Fremskynding av vekst hos tomat planter ved glodelampelys. Norsk
Gartnerforenings Tidskrift, Arg. 28, N 48, 497—500 (1938); Arg. 29, N 49, 619—621
(1939).
777. Smith G. E. The effect of photoperiod on the growth of Lespedeza. Jour. Amer.
Soc. Agron., 33, N 3, 231—236 (1941).
778. Smith O. Premature seeding studies with beets. Proc. Amer. Soc. Hort. Sci.,
37, 793—798 (1939).
779. Smith T. J, Responses of biennial sweet clover to moisture, temperature and
length of day. Jour. Amer. Soc. Agron., 34, N 10, 865—876 (1942).
Л итература
451
780. Snyder W. Е. Effect of light and temperature on floral initiation in cocklebur
and Biloxi soybean. Bot. gaz., 102, N 2, 302—321 (1940),
781. Snyder W. E. Mechanism of the photoperiodic response of Plantago lanceolatus.
Amer. Jour. Bot., 35, N 8, 520—525 (1948).
782. S t a d t С. E., Van der. Versuche fiber Elektrizitats Anwendung im Hollandischen
Gartenbau. Elektrizitatswirtschaft, Jg. 30, N 16, 481—484 (1931).
783. S t a m p a G. Action de la chaleur rayonnantc et des radiations lumineuses sur
la vie des plantes. Chauffage et forcage des vegetaux a Fair libre. Bull, mensuel
renseignemcnt technique. Roma, An. 27, N 9 et 10, 339—348, 383—393 (1936).
784. Steel T. Electric light and Vegetation. Nature, 106, N 2674, 694 (1921).
785. Steinberg R. A. An apparatus for growing plant under controlled environ-
mental conditions. Jour. Agric. Research, 43, N 12, 1071—1084 (1931).
786. Steinberg R. A. a. Garner W. W. Response of certain plants to length
of day and temperature under controlled conditions. Journ. Agric. Research, 52,
N 12, 942—960 (1936).
787. Stevenson F. J. a. Clark C. F. Artificial light as an aid in Potato bree-
ding. Amer, potato Jour., 10, N 6, 103—107 (1933).
788. Steward F. G., Caplin S. M. a. Miller F. K. Investigation on growth
and metabolism of plant cells. Ann. Bot., 16, N 61, 57—77 (1952).
789. Stout M. Translocation of the reproductive stimulus in sugar beets. Bot. gaz.,
107, N 1, 86—94 (1945).
790. S t о u t e m e у e г V. T. a. Close A. W. Plant propagation under fluores-
cent lamps. U. S. Depart. Agric. Pamphlet, 1—15 (1946).
791. Stroud E. The experimental use of artificial light in connection with the gro-
wing of cucumbers in Denmark. Ilium. Engineer, 25, 123—125 (1932).
792. Stuart N. W. Controlling time of blooming of chrysanthemus by the use of
lights. Proc. Amer. Soc. Hort. Sci., 42, 600—606 (1943).
793. Taylor F. J. Oxidative assimilation of glucose by Scenedesmus quadricaula.
Journ. Exp. Bot., 1, N 3, 301—321 (1950).
794. Torao H. a. Katayama T. On the premature heading in Poddy rice.
Jour. Imp. Agric. Exper. Sta. 1, 25—40 (1929). Ref.: Japon Journ. Bot., 4, N 4,
105 (1928).
795. The electrical illumination of greenhouses. Rural Electrification, 5, N 57, 277—278
(1930).
796. Thelen O. Naturliches, kiinstliches und monochromatisches Licht in seiner Be-
deutung fiir die Entwicklung und die Stoffproduction einiger Kulturpflanzen.
Rostock (1910). Ref.: Bot. Ctrbl., B. 117, N 3, 56—57 (1911).
797. Thomas Fr. Uber durch elektrisches Licht hervorgerufene Vegetation. Verh.
Bot. Verein Brandenburg, Jg. 39, XCI—XCII (1897). Ref.: Bot. Ctrbl., 75, N 12,
389 (1898).
798. Thomas Fr. Uber Moosvegetation in elektrisch beleuchten Hohlcn. Verhaldl.
Bot. Verein Brandenburg, Jg. 45, 1903, XXIX. Ref.: Just’s Jahres ber. Jg. 31,
Abt. I, 567 (1905).
799. Thompson H. C. Seedstalk and bulb development in the onion {Allium сера).
Cornell Univ. Agric. Exper. Sta. Bull. 708, 1—21 (1938).
800. Thorsr und A. Technick im Gartenbau im Verhaltnis zum Betriebsgrosse-
und Form. 12 Internationaler Gartenbaukongress, Berlin, 1938, Section 18, 6—7
(1939).
801. Th waite В. II. Electric cultivation in relation to horticulture. Jour. Roy.
Hort. Soc., 33, Part. 2, 401—416 (1908).
802. T i e d j e n s V. Л. Stimulation of plant growth by means of electric lighting.
Massachusetts Agric. Exper. S.ta. N 31, 1924, 1—3. Trans. Ilium. Engin. Soc., 19,
N 10, 1011—1019 (1924).
803. Tiedjens V. A. Sex ratios im cucumber flowers as effected by different con-
ditions of soil and light. Jour. Agric. Research, 36, N 8, 721—746 (1928).
804. Tiedjens V. A. The relation of environment to shape of fruit im Cucucmis
sativus L. and its bearing on the genetic potentialities of the plant. Jour. Agric.
Research, 36, N 9, 795—809 (1928).
805. Tiedjens V. A. Controlling damping off with electric lamps. Science,
69, N 1782, 226 (1929).
806. T i n с к с r M. A. H. On the effect of length of daily period of illumination upon
the growth of plants. Jour. Roy. Hort. Soc., 54, Part. 2, 354—378 (1929).
807. T i n с к e r M. A. H. Response of some commongardenplants to the daily period
of light. Jour. Roy. Hort. Soc., 57, Part. 2, 321—325 (1932).
808. T i n с к e r M. A. H. The influence of the daily period of light and the supply
of potassium on the rate of growth of tubers and other storage organs. Jour. Roy.
Hort. Soc., 57, Part 2. 326—331 (1932).
29*
452
Литература
809. Tincker М. А. Н. The influence of the length of day on lettuces. Gardener’s
chronicle, 93, N 2424, 404—405 (1933).
810. T i n с к о r M. A. H. a. Darbshire F. V. Studies on the formations of
tubers and other storage organs. Ann. Bot., 47, N 185, 28—53 (1933).
811. T j e b b e s K. u. U p h о f J. C. Der Einfluss des elektrischen Lichtes auf das
Pflanzenwachstum. Landw. Jahrb., 56, 313—326 (1921).
812. T о 1 о in e i G. Sopra 1’azione della luce del magncsio sullo sviluppo dei vegetali.
Staz. Sperment. Agrar. Ital. Modena, 24, 377—386 (1893). Ref.: Just’s Bot. Jahres-
ber., 21, Abt. 1, 32 (1893). Biederman’s Ctrbl., 23, 106—107 (1894).
813. Tottingham W. E. A plant test of the sun lamp. Plant physiol., 7, N 3,
551—553 (1932).
814. Truffaut G. et T h u r n e s s e n G. Influence de la lumiere artificielle
sur la croissance dos plantos supcrieurcs. C. R. Ac. Sc. Paris, 188, N 5, 411—412
(1929). Jardinage, N 16, 163—165 (1929).
815. Truffaut G. Influence de la lumiere artificielle sur les plantes superieures.
Jardinage, N 16, 166—169 (1929).
816. T s c h e r m а с к E. Kiinstliche Bclichtung als Stimulationsmittel in Gewachs-
hausern in Dienste der Pflanzcnzuchtung. Anzeigcr Akad. Wiss. Wien. Math.-Nat.
KI. Jg. 68, N 24, 260—261 (1931).
817. T s c h e r m а с к E. Kiinstliche Bclichtung der Pflanzen. Gartenztg. Oster.
Gartcnbaues. N 1 und 2, 2—4, 18—20 (1932).
818. T u 1 u к d a r L. Photoperiodic behaviour of Hibiscus subdarijja. Nature, 170,
N 4324, 458—459 (1952).
819. T у s d a 1 H. M. Influence of light, temperature and soil moisture on the harde-
ning process in alfalfa. Jour. Agric. Research, 46, N 6, 483—515 (1933).
820. Ullrich H. Zur Frage des Entwicklung der Pflanzen bei ausschliesslich kiinst-
licher Belcuchtung. Ber. d. d. bot. Ges., 59, H. 6, 195—232 (1941); 60, H. 2, 152—
178 (1942).
821. Ullrich A. The influence of temperature and light factors on the growth and
development of sugar beets in controlled climatic environments. Agronomy Journal,
44, N 2, 66—73 (1952).
822. U 1 v i n G. B. Chlorophyll production under various environmental conditions.
Plant Physiology, 9, N 1, 59-—81 (1934).
823. Veen R., Van der. Wachstumsforderung von Tulpen mittels Kunstlicht. Philip’s
Techn. Rundschau i, Jg. 10, H. 9, 282—286 (1949).
824. Veen R., Van der. Die Aufbewarung von Saatkartoffeln in Kiinstlich Beleuch-
teten Kellern. Philip’s Techn. Rundschau1, Jg. 10, II. 10, 318—332 (1949).
825. Veen R., Van der. Der Einfluss von Licht auf Pflanzen. Philips’ Techn. Rund-
schau1, Jg. 11, H. 2, 47—53 (1949).
826. Veen R., Van dor. Trcibkastcn zur Untersuchung des .Pflanzenwachstum’s
unter reproduzierbaren Bedingungen. Philip’s Techn. Rundschau1, Jg. 12, N 1,
1—5 (1950).
827. Veen R., Van der. Influence of daylength on the dormansy of some species of
the genus Populus. Physiologia Plantarum, 4, Fasc. 1, 35—40 (1951).
828* . V о e г к e 1 S. H. Untersuchungen fiber die Phototaxis der chloroplasten. Planta,
21, H. 1, 156, 205 (1934).
829. V о g 1 K. Elektriscbe Beleuchtung bei Tomatenanzucht. Eloktrizitatswirtschaft,
Jg. 32, N 9, 194—196 (1933).
830. Voss J. Untersuchungen fiber Entwicklungsbeschleunigung und Anzucht von
Winterweizen im Warmhause. Pflanzenbau, 10, 321—337 (1934).
831. Voss J. Uber den Einfluss verschiedener Licht- und Strahlarten auf die Ent-
wicklung landwirtschaftlicher Kulturpflanzen. Angew. Bot., 18, II. 1, 43—75
(1936).
832. Wangermann E. a. Ashby E. Studies in Morphogenesis of leaves. VII.
New phytologist, 50, N 2,. 186—209 (1951).
833. Warburg O. Uber die Geschwindigkeit der Photochemischen Kohlensaurezer-
setzung in lebenden Zellen. Bioch. Ztschr., 100, 230—270 (1919).
834. W a r e i n g P. F. Photoperiodic control of leaf growth and cambial activity
in Pinus silvestris. Nature, 163, N 4150, 770—771 (1949).
835. W a r e i n g P. F. Growth studies in woody species. I—III. Physiologia plan-
tarum, 3, Fasc. 3, 258—276, 300—324 (1950); 4, Fasc. 1, 41—56 (1951).
836. W a r e i n g P. F. A new photoperiodic phenomenen in short day plants. Nature,
171, N 4353, 614—615 (1953).
837. W a r f D. J. Vernalisation of Marquis wheat and other spring cereals. Bot. gaz.,
101, N 2, 457—481 (1939).
1 См. сноску к № 728 (стр. 449).
Литература
453
838. Watkins J. М. The growth habits and chemical composition of bromo grass
Bromus inermis, as affected by different environmental condition. Jour. Amer. Soc.
Agron., 32, N 7, 527—538 (1940).
839. Weigel R. G. u. Knoll О. H. Lichtbiologische Beeinfhissung der Aufzucht
von Gemusepflanzen. Licht, 6, H. 10—11, 219—220, 237—242, 259—261 (1936).
840. W e i n a r d F. F. a. Decker S. W. Experiments in forcing Gladioli. Illinois
Agric. Exper. Sta. Bull., 357, 343—361 (1930).
841. W el lensiek S. J. De Vervanging van ZonJicht door kunstlicht bij Pootaar-
dappelbewa ring. (The substitution of sunlight by artificial light in seed-potato
storing). Tijdschrift over Planzenziekten Jg. 35, Afb. 9, 241—250 (1929).
842* . Went F. W. Effects of light on stem and leaf growth. Amer. Jour. Bot., 28,
N 2, 83—95 (1941).
843. Went F. W. Plant growth under controlled conditions. I—II. Amer. Jour. Bot.,
30, N 2, 157—167 (1943), 31, N 3, 135—150 (1944).
844. Went F. W. The Earhart plant research laboratory. Chronica Botanica, 12,
N 3, 89—108 (1950).
845. W e r n e r H. O. Effect on berry production of varied day length during the life
of two Triumph Potato strains. Amer. Potato Jour., 18, N 6, 174—179 (1941).
846. Werner H. O. Flower and berry Production by potatoes as influenced by two
ligth intensities and two midwinter planting date. Amer. Potato Jour., 18, N 12,
349—355 (1941).
847. Werner H. 0. Relation of length of Photoperiod and intensity of supplemental
ligth to the production of flowers and berries in the greenhouse by several varieties
of potatoes. Jour. Agr. Research, 64, N 5, 257—274 (1942).
848. Wettstein F. u. Pirschle K. Klima Kammern mit Konstanten Bedin-
gungen fur die Kultur hoherer Pflanzen. Naturwiss. Jg. 28, H. 34, 537—543 (1940).
849. White T. H. Studies in the forcing of Gladiolus. Amer. Soc. Hort. Sci., 27,
308—312 (1930).
850. Whitehead M. R. a. Mitchell J. W. Effects of nutrient, photoperiod
and high temperature on the development of guayule seeds. Bot. gaz., 105, N 1,
14—24 (1943).
851. Wilson A. L. Relation of hydrogen-ion concentration to the growth of onions.
N. J. (Cornell) Agric. Exper. Sta. Mem., 145, 1—59 (1932).
852. Wilson P. W. a. G e о r g i С. E. Methods for controlling the environment
of greenhouse plants. Bot. gaz., 94, N 2, 346—363 (1932).
853. Withrow A. P. Comparative effects of radiation and indolebuteric acid emul-
sion on tomato fruit production. Proc. Amer. Soc. Hort. Sci., 46, 329—335
(1945).
854. Withrow A. P. a. Withrow R. B. Plant growth with artificial sources
of radiant energy. Plant physiology, 22, N 4, 494—513 (1947).
855. Withrow A. P. a. Withrow R. B. Comparision of various lamp sources
for increasing growth of greenhouse crops. Proc. Amer. Soc. Hort. Sci., 49, 363—366
(1947).
856. Withrow A. P. a. Withrow R. B. Photoperiodic chlorosis in tomato.
Plant physiology, 24, N 4, 657—663 (1949).
857. Withrow R. B. a. Richman M. W. Artificial radiation as a means of
forcing greenhouse crops. Purdue (Indiana) Univ. Agric. Exper. Sta. Bull., 380,
1—20 (1933). Ref.: Rural Electrification, 10, N 112, 134 (1934).
858. Withrow R. B. Response of crops to electrical light. Electrical world, 102,
N 13, 411—412 (1933).
859. Withrow R. B. Plant forcing with electric lights. Purdue (Indiana) Univ.
Agric. Exper. Sta. Circular N 206, 1—12 (1934). Ref.: Biol. Abstr., 10, N 9, 2301
(N 21877) (1934).
860. Withrow R. B. The photoperiodic response of certain long and short day plants
to filtered visible radiation applied as a supplement to day-light. Proc. Amer. Soc.,
Hort. Sci., 33 , 501 (1935).
861. Withrow R. B. a. Benedict H. M. Photoperiodic responses of certain
greenhouse annuals as influenced by intensity and wavelength of artificial light
used to lenghten the day light period. Plant physiology, 11, N 2, 225—249 (1936).
862. Withrow R. В. a. В i e b e 1 J. P. Photoperiodic response of certain long
and short day plants to filtered radiation applied as supplement to day length. Plant
physiology, 11, N 4, 807—819 (1936).
863. Womeldorf D. Windowless greenhouse. Electricity on the Farm, 19, N 10,
7—8 (1946).
СОДЕРЖАНИЕ
Стр‘
От автора.............................................................. 3
Часть I
ФИЗИЧЕСКИЕ И ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ СВЕТОКУЛЬТУРЫ РАСТЕНИЙ
Введение .............................................................. 7
Глава I. Лучистая энергия и оптические свойства растений............... 9
1. Лучистая энергия . . ......................................... 9
2. Основные законы фотохимии...................................... И
3. Оптические свойства растений................................. 12
4. Выводы ....................................................... 15
Глава II. Физиологическая радиация.................................... 17
1. Формирование и движение листьев и пластид..................... 17
2. Фотосинтез.....................................................20
3. Развитие растений............................................. 21
4. Формообразование растений..................................... 23
5. Значение ультрафиолетовой радиации........................... 29
6. Значение инфракрасной радиации ............................... 29
7. Температура листьев........................................... 30
8. Значение температуры среды для физиологических процессов растения 37
9, Значение интенсивности физиологической радиации............... 37
10. Выводы ....................................................... 40
Глава III. Источники радиации......................................... 42
1. Энергетические и светотехнические характеристики источников ра-
диации ......................................................... 43
2. Общие законы теплового излучения.............................. 52
3. Лампы накаливания............................................. 57
4. Элсктродосвстные лампы........................................ 68
5. Ртутные лампы................................................. 73
6. Натриевые лампы .............................................. 80
7. Неоновые лампы................................................ 81
8. Гелиевые трубки.......................................... . 82
9. Криптоновые и ксеноновые лампы............................... 83
10. Цинковые и кадмиевые лампы................................... 86
11. Цезиевые лампы ............................................... 88
12. Люминесцентные лампы.......................................... 89
13. Влияние напряжения на светотехнические показатели ламп .... 95
14. Сравнительная характеристика физиологической эффективности ламп 96
Содержание
45
15. Размещение источников радиации................................. 103
16. Осветители (светильники)....................................... 108
17. Охлаждение источников радиации................................. 112
18. Выводы .......................................................
Глава IV. Солнечная радиация........................................... 117
1. Общая характеристика .......................................... 117
2. Спектральный состав солнечной радиации......................... 120
3. Интенсивность солнечной радиации............................... 124
4. Длина дня...................................................... 130
5. Количество лучистой энергии.................................... 133
6. Выводы ........................................................ 134
Глава V. Методы измерения лучистой энергии............................. 135
1. Интенсивность радиации ........................................ 135
2. Освещенность .................................................. 136
3. Переход от освещенности к интенсивности радиации............... 136
4. Применение светофильтров ..................................... 141
5. Измерительные приборы ......................................... 144
6. Определение количества квантов ........................ 149
7. Расчеты интенсивности радиации и освещенности.................. 150
8. Измерение температуры листьев.................................. 153
9. Выводы . . .................................................... 154
Часть II
ОБЩАЯ СВЕТОКУЛЬТУРА РАСТЕНИЙ
Глава VI. Выращивание растений исключительно при искусственной радиации 157
1. Культура растений при помощи дуговых ламп...................... 157
2. Культура растений при помощи ламп накаливания.................. 159
3. Значение инфракрасной радиации................................ 171
4. Значение ультрафиолетовой радиации............................. 173
5. Культура растений при помощи газосветных ламп. Значение различ-
ных областей спектра физиологической радиации..................... 177
6. Культура растений при помощи люминесцентных ламп. Значение раз-
личных областей спектра физиологической радиации.................. 180
7. Значение интенсивности физиологической радиации............... 201
8. Значение длиньГдня ........................................... 204
9. Значение прерывистого освещения............................... 210
10. Значение фотопериодических циклов............................. 211
11. Различные типы люмпностатов .................................. 212
12. Выводы ...................................................... 218
Глава VII. Выращивание растений при помощи дополнительного) искусствен-
ного освещения • . . .......................................•..... 220
1. Значение различных областей спектра физиологической радиации.
Сравнение различных источников радиации.......................... 224
2. Значение интенсивности дополнительного освещения.............. 241
3. Дополнительное дневное освещение.............................. 254
4. Значение длительности дополнительного освещения............... 255
5. Значение времени дополнительного освещения.................... 264
6. Значение прерывистого и кратковременного ночного дополнительного
освещения ....................................................... 264
7. Значение времени года для дополнительного освещения........... 269
8. Температурный режим при дополнительном освещении.............. 271
9. Удобрение углекислотой ....................................... 274
10., Выводы ..................................................... 279
456
Содержание
Часть III r
ЧАСТНАЯ СВЕТОКУЛЬТУРА РАСТЕНИЙ
Глава VIII. Светокультура овощных и садовых растений ............... 283
1. Овощные растения ........................................... 286
2. Садовые растения............................................ 306
3. Выводы ..................................................... 310
.Глава IX. Светокультура декоративных растений ..................... 312
1. Однолетники................................................. 312
2. Двулетники ................................................. 333
3. Многолетники ............................................... 334
•4. Декоративно-лиственные растения............................. 348
5. Клубневые и луковичные растения............................. 350
6. Оранжерейные и комнатные растения........................... 353
7. Дикорастущие растения....................................... 378
8. Лекарственные растения ..................................... 381
9. Комнатное цветоводство...................................... 382
10. Озеленение станций метрополитена ........................... 384
И. Выводы ...................................................... 393
Глава X. Светокультура полевых растений (светокультура в селекции и семе-
новодстве) ..................................................... 395
1. Зерновые.................................................... 395
2, Бобовые................•.................................... 401
3. Технические культуры ....................................... 403
4. Клубнеплоды и корнеплоды................................ 407
5. Травы....................................................... 409
6. Выводы .................................................... 413
Глава XI. Светокультура древесных растений.......................... 414
Заключение ......................................................... 423
Литература ......................................................... 425
Утверждено к печати
Институтом физиологии растений
Академии Наук СССР
Редактор издательства Ю. А. Самыгин Технический редактор Е. И. Зеленкова
Корректор А. К. Бессмертная
РИСО АН СССР №31-27 В Т-06938. Издат. № 148. Тип. заказ № 1152. Поди, к печ. ЗОТХ 1954 г.
Формат бум. 70x 108'',,. Бум. л. 14,25 + 2 вкл. Печ. л. 39,04 + 2 вкл. Уч.-издат. 38,7. Тираж 3500.
Цена по прейскуршту 1952 г. 28 р. во к.
1-я тип. Издательства Академии Наук СССР. Ленинград; В. О.. 9 лин., д. 12
С II И С О К И СПРАВ Л Е И 11 П И О 1 [ Е Ч А Т О К
Ct (гра- ниц а Строка Напечатано Должно оытъ
35 табл. 6, 1-я графа, строка 1. «ВС—15» «БС—15»
124 1 ся. фиг. 78, 2 и 3. фиг. 79, 2 п 3
128 Подпись под рис. 88, стр. 3 сп. Со Се
203. 24 св. 8 тые. 18 тыс.
244 23 сп. фиг. 142, 6—9 фиг. 142, 6—8
24!) 14 св. фиг. 142, 5, 10 и 11 фиг. 142, 5, 9-11
250 3 св. 60 и 75 тыс. 30 и 7,5 тыс.
334 20 сп. табл. 113 табл. 102
.‘СЗб 22 сп. ромашка василек
339 22 сп. (697) (695)
403 10 св. Энкорииг Эпкопинг
405 4 сп. вырастают возрастают
А. Ф. Клешнин.