/







































































































































































































Автор: Березина Н.А. Афанасьева Н.Б.
Теги: экология растений этология растение и окружающая среда ботаника экология
ISBN: 978-5-7695-5161-1
Год: 2009




Текст
Н. А. Березина
Н. Б. Афанасьева
экология
РАСТЕНИЙ
Ф
Ф
3
CD
ВЫСШЕЕ ПРОФЕССИОНАЛЬНОЕ ОБРАЗОВАНИЕ
Н. А. БЕРЕЗИНА, Н. Б. АФАНАСЬЕВА
ЭКОЛОГИЯ
РАСТЕНИЙ
Допущено
Учебно-методическим объединением
по классическому университетскому образованию
в качестве учебного пособия для студентов
высших учебных заведений, обучающихся
по специальности «Экология» и по направлению
«Экология и природопользование»
ACADEMA
Москва
Издательский центр «Академия»
2009
УДК SHI.5(075.8)
Ы»К 28.58я73
Б484
Рецензенты:
доктор географических наук, профессор МГУ им. М. В.Ломоносова
Г. Н. Огуреева',
доктор географических наук, старший научный сотрудник
Института географии РАН А.А. Гольева
Березина Н.А.
Б484 Экология растений: учеб, пособие для студ. высш. учеб,
заведений / Н.А.Березина, Н.Б.Афанасьева. — М. : Изда-
тельский центр «Академия», 2009. — 400 с.
ISBN 978-5-7695-5161-1
В учебном пособии рассмотрены вопросы взаимодействия растений и
их совокупностей со средой, а также факторы, влияющие на эти процес-
сы. Изложены основы устойчивости растительных организмов и типы их
экологической гетерогенности. Особое внимание уделено фундаменталь-
ным исследованиям в области экологии растений.
Для студентов высших учебных заведений. Может быть рекомендова-
но аспирантам, преподавателям вузов и специалистам, использующим в
своей работе данные о жизни и экологии растений.
УДК 581.5(075.8)
ББК 28.58я73
Учебное издание
Березина Наталья Александровна, Афанасьева Наталья Борисовна
Экология растений
Учебное пособие
Редактор Н.А. Соколова. Технический редактор Е.Ф. Коржуева
Компьютерная верстка: Л. А. Смирнова. Корректоры В. А. Жилкина, Г. Н. Петрова
Изд. № 101110266. Подписано в печать 27.06.2008. Формат 60x90/16.
Гарнитура «Таймс». Печать офсетная. Бумага офсетная № 1. Усл. печ. л. 25,0.
Тираж 3000 экз. Заказ № 6429
Издательский центр «Академия», www.academia-moscow.ru
Санитарно-эпидемиологическое заключение № 77.99.02.953.Д.004796.07.04 от 20.07.2004.
117342, Москва, ул. Бутлерова, 17-Б, к. 360.
Тел./факс: (495) 334-8337, 330-1092.
Отпечатано с электронных носителей !i.uvn<Jii*cnia.
ОАО "Тверской полиграфический комбинат", 170024. г. Turpi., нр-г Ленина, 5.
Телефон: (4822) 44-52-03,44-50 34, Геле<|и.и,/<|»ак< (4822) 44-42-15
Home page - www.tverpk.ru Электрон:*» почта (E-mail) sales»* verpk.ru
1
Оригинал-макет данного тдания является собственностью
Издательского центра «Академия», и его воспроизведение любым способом
без согласия правообладателя запрещается.
© Березина Н.А., Афанасьева Н.Б., 2009
iQniv отв « тао« ciAi 1 © Образовательно-издателнекий центр «Академия», 2009
ISBN 978-5-7695-5161-1 © оформление. Издательский центр «Академия», 2009
ПРЕДИСЛОВИЕ
Экология (от греч. oikos — дом, жилище, местообитание и lo-
gos — слово, учение) — наука об отношениях живых организмов
и образуемых ими сообществ между собой и с окружающей сре-
дой. С середины XX в. в связи с многократным усилением воз-
действия человека на природу экология получила особое значе-
ние как научная основа охраны природы и рационального приро-
допользования, а термин «экология» приобретает сейчас все бо-
лее широкий смысл, выходя за рамки науки. Под влиянием средств
массовой информации экология часто трактуется как сугубо при-
кладное знание о состоянии среды обитания человека. Хотя это и
имеет важнейшее практическое значение и решение проблемы
улучшения качества среды невозможно без экологических зна-
ний, задачи экологии намного шире: объяснение строения и фун-
кционирования биосферы, роли живого в круговороте химиче-
ских элементов и процессах превращения энергии; изучение за-
кономерностей взаимосвязи разных биологических объектов между
собой и со средой, распределения их в пространстве и изменения
во времени. Особое место экология занимает в системе современ-
ных биологических знаний. Она стала очень важной областью
науки, с которой связаны большие надежды человечества. Эколо-
гия — одна из главных точек междисциплинарного синтеза в ес-
тественно-научном познании. И именно в экологическом аспекте
традиционные биологические науки (ботаника, зоология и др.),
исследующие природу в ее естественном состоянии, наиболее вос-
требованы современным обществом.
В последнее десятилетие в соответствии с запросами времени в
системе экологического образования появились новые специаль-
ности, факультеты и вузы экологического профиля. Они созданы
на базе географических, инженерно-экономических, сельскохо-
зяйственных, строительных, педагогических, юридических и дру-
гих подразделений институтов и университетов для подготовки
специалистов, компетентных в вопросах рационального природо-
пользования. Учебные программы этих специальностей предпо-
лагают обязательное изучение основ классической экологии. Эко-
логия растений — один из ее разделов. Данное учебное пособие
адресовано студентам разных экологических специальностей, в
том числе тем, в чьи учебные планы не входит специальное изу-
чение курса таких классических университетских дисциплин, как
анатомия, морфология, физиология, география растений, фито-
испили! ин. псе э и аспекты важны для изложения материала об
жологии растений, поэтому им в книге уделено особое внима-
ние. Учебное пособие ставит своей целью дать будущему экологу
систематизированное и эмпирически аргументированное представ-
ление об автотрофном блоке экосистем.
Экология растений описывает процессы взаимодействия рас-
тений и их совокупностей со средой, а также факторы, влияющие
на эти процессы. На воздействия среды реагируют ботанические
объекты разного уровня — от клеточного до ценотического. По-
этому экологическая ботаника рассматривает явления на уровне и
клетки (цитоэкология), и организма (аутэкология), и разных со-
вокупностей растений (синэкология). Она также обсуждает воп-
росы экологической физиологии растений, экологии видов, эко-
логии популяций и сообществ растений. При этом в соответствии
с традициями биологического экологического образования основ-
ным остается аутэкологический уровень (уровень особей), на ко-
тором рассмотрены эффекты действия факторов среды. Но в при-
роде растения не встречаются поодиночке, поэтому в предлагае-
мой книге освещаются вопросы демэкологии и фитоценологии
(экологии популяций и растительных сообществ).
В основу пособия положена программа курса «Экология расте-
ний», читаемого на кафедре геоботаники биологического факуль-
тета Московского государственного университета им. М. В. Ломо-
носова и введенного в учебный план основателем кафедры
В. В. Алехиным еще в 1923 г. Каждый из преподавателей кафедры,
читавших этот курс (В. В. Алехин, Н.Я.Кац, М.С.Двораковский,
И.М.Культиасов, Н. А. Березина), вносил свои изменения, уточ-
нения и усовершенствования в программу. Менялись понятия,
концепции, подходы, но в своей основе курс опирался в первую
очередь на учение об экологических факторах. Читавшийся на
кафедре курс в разные годы был отражен в университетских учеб-
никах (В.В.Алехин, 1950; М.С.Двораковский, 1978; И.М.Куль-
тиасов, 1982).
Авторы признательны всем коллегам биологического факуль-
тета МГУ, помогавшим в работе над рукописью, а также выража-
ют искреннюю благодарность доктору биологических наук, про-
фессору В. Г. Онипчснко, доктору биологических наук, профессо-
ру Н. И. Шориной, доктору географических наук, профессору
Г. Н. Огуреевой и доктору географических наук, старшему науч-
ному сотруднику А. А. Гольевой за ценные советы по ее улучше-
нию. Будем благодарны студентам, преподавателям и специалис-
там разных направлений за конструктивные критические замеча-
ния и пожелания.
ИСТОРИЯ ЭКОЛОГИЧЕСКОГО ИЗУЧЕНИЯ
РАСТЕНИЙ
Историю науки по экологическому изучению растений обычно
связывают с первыми научными публикациями, появившимися в
основных центрах цивилизации на разных континентах. Так, пер-
вые экологические сведения были запечатлены на древнехеттских
глиняных табличках, возраст которых более 3 тыс. лет. Накопле-
ние научных сведений об экологических особенностях растений
исторически тесно связано с развитием ботаники и географии.
1.1. История экологии растений до XX в.
Первые научные обобщения, касающиеся экологии растений,
содержатся в трудах древнегреческого естествоиспытателя, уче-
ника Аристотеля, Теофраста (372 — 287 гг. до н.э.), одного из пер-
вых ботаников древности. В своих трудах «Естественная история»
(10 томов) и «О причинах растений» (8 томов) он рассматривал и
вопросы распространения растений, приуроченности их к мес-
тообитаниям, отмечал изменение внешнего вида и формы расте-
ний при произрастании на разных почвах. Римский ученый Пли-
ний Старший (23 — 79 гг. н.э.) в «Естественной истории»
(37 книг) — энциклопедии естественно-научных знаний антич-
ности — излагал результаты эколого-ботанических исследований.
В период раннего Средневековья ботанические знания в Евро-
пе накапливались в основном в монастырях. Изучение природы,
в том числе растений, сочеталось с суевериями, верой в магиче-
ские силы растений и колдовство. Средневековье — время не толь-
ко накопления знаний, но и оформления научной этики, когда
изучение природы сочеталось с бескорыстным поиском истины.
Обязательное знание латыни, обмен научной информацией при-
вели к формированию науки как интернационального явления и
бурному ее развитию в Западной Европе в последующие века.
В период позднего средневековья (XII —XIII вв.) развитие науки
во многих европейских странах привело к открытию светских школ
и первых университетов.
Сведения о растениях можно найти в трудах Альберта Велико-
го (1193—1280), немецкого философа и теолога, члена монаше-
ского ордена доминиканцев, в ведении которого была инквизиция
и борьба с ересью. Он преподавал в Кёльне и Париже и в своем
трактате о растениях касался разных экологических вопросов:
причин зимнего покоя растений, влияния качества почвы на жизнь
растения, зависимости роста и развития растений от солнечного
тепла. Знание природы в его трудах неизбежно сочеталось с пред-
рассудками, характерными для его времени (например, он изла-
гал представления о взаимном превращении видов, связанные с
изменением почв: ячменя — в пшеницу, дуба — в виноград).
Великие географические открытия дали огромный ботаниче-
ский материал и возможность его обобщения. В ботанических са-
дах с познавательными целями стали создаваться коллекции рас-
тений из разных стран. Много сведений экологического характе-
ра опубликовано в трудах ботаников XVI—XVII в. Итальянский
врач и ботаник А.Чезальпино (1519—1603) описал функции от-
дельных частей растений, разработал систему растительного цар-
ства. Французский ботаник и путешественник Ж.Ф.Турнефор
(1656—1708) исследовал распределение растений в горах. Англий-
ский ботаник Д. Рей (1627 —1705) в своем труде «Historia plantarum»
специальную главу посвятил описанию местообитаний растений.
Он указал, что на распространение растений влияют географиче-
ская широта и высота над уровнем моря, показал, что растения
имеют различную экологическую амплитуду по отношению к фак-
торам местообитания.
К XVIII в. накопилось большое количество сведений о расте-
ниях. Ботанические исследования в этом веке в первую очередь
связаны с К.Линнеем (1707—1778). Он не только ввел в научный
обиход бинарную номенклатуру и создал основы систематики орга-
низмов в «Системе природы» (1735), но и провел экологические
исследования. К. Линней описывал растительный покров гор, тундр
и болот, предложил типологию местообитаний растений. В его
трудах присутствует также представление об «экономии приро-
ды» — упорядоченности естественных процессов, направленных
на поддержание равновесия в природе. Эта упорядоченность по-
нималась им с точки зрения креационизма: Творец специально
создал разные группы организмов для исполнения определенных
ролей, поэтому растения служат пищей травоядным, которым
хищники не позволяют чрезмерно размножаться. К. Линней был
первым президентом Шведской академии наук и в 1739 г. был
избран почетным членом Петербургской академии наук.
Значительный вклад в разработку ботанических и экологиче-
ских знаний внесли академики Петербургской академии наук:
С. П. Крашенинников (1711 — 1755) при описании Камчатки пред-
ставил элементы ее ботанической географии; И. Г. Гмелин (1709—
6
1755) изучал растительной покров Западной и Восточной Сиби-
ри; П.С. Паллас (1741 — 1811) исследовал растения и раститель-
ные сообщества Крыма; И.И.Лепехин (1740—1802) описал рас-
тительный покров пустынь, тропиков, умеренной зоны, указал на
зависимость растений от изменения климата.
Во второй половине XVIII в. начинается отступление от креа-
ционистского понимания природы. От внешнего описания при-
роды исследователи переходят к выявлению внутренних связей,
определяющих ее развитие. Так, И.Кант в прочитанных в Кениг-
сбергском университете лекциях по физической географии под-
черкнул необходимость целостного описания природы, учитыва-
ющего взаимодействие физических процессов с деятельностью
живых организмов.
Начало XIX в. ознаменовалось публикацией трудов А. Гумбольд-
та: «Идеи о физиономичности растений» (1806) положили начало
изучению жизненных форм, «Идеи о географии растений» (1807)
легли в основу ботанической географии. А.Гумбольдт (1769—1859)
предпринимал экспедиции в Центральную и Южную Америку, на
Урал, в Сибирь, исследовал разные страны Европы. Его ботани-
ко-географические исследования носили и экологический харак-
тер: он не только выявлял различия в распределении разных групп
растений, но и объяснял их климатическими причинами. А. Гум-
больдту принадлежит заслуга в том, что от изучения отдельных
растений он перешел к познанию растительного покрова как це-
лостности. Заложив основы географии растений, исследования
А. Гумбольдта повлекли за собой целый ряд подобных работ.
В начале XIX в. во Франции Ж. Б. Ламарк предложил свою кон-
цепцию круговорота веществ на Земле, в которой живым орга-
низмам отводилась очень важная роль. Он полагал, что только
деятельность организмов, приводящая к созданию сложных со-
единений, способна противостоять естественным процессам рас-
пада. Концепция Ламарка была умозрительной, не всегда соот-
ветствовавшей химическим данным, но в ней был предугадан ряд
положений о биосфере, получивших развитие много позже.
XIX в. отмечен большим количеством основополагающих ра-
бот по экологии растений. Так, английский ботаник X. Уотсон в
1833 г. предложил субординированную типологию экологических
факторов, включающих тепло, увлажнение, условия освещения,
экспозицию склонов, механические и химические свойства почв
и др., и при этом отмечал важность сочетания факторов. Круп-
нейший французский ботаник О. П. Декандоль (1778 —1841) в книге
«Очерки начальной географии растений» (1820) дает определения
и подчеркивает различие понятий «местонахождение» и «место-
обитание». В 1832 г. он обосновал выделение особой науки «эпир-
реологии», соответствующей современному пониманию аутэко-
логии. Среда определялась им как совокупность условий, воздей-
7
ствующих на растения. Выясняя роль разных факторов в распре-
делении растительности, О. Декандоль подчеркивал также важность
конкуренции между видами за ресурсы. Его сын А.Декандоль
(1806— 1893), продолжив исследования отца, в книге «География
растений» (1855) классифицировал местообитания, основываясь
на таких факторах внешней среды, как температура, свет, влага,
почва. А. Декандоль отмечал ограничивающее влияние на рас-
пространение растений климатических факторов, в первую оче-
редь температуры периода вегетации (особенно периода цвете-
ния), изучал связь растений с почвами и указал на высокую пла-
стичность растений из-за их «прикрепленности» и невозможно-
сти «уйти от неблагоприятных факторов». Позднее, в 1872 г.,
выходит капитальный труд А. Гризебаха (1814—1879), где впер-
вые дано описание основных растительных сообществ всего зем-
ного шара.
Весомый вклад в развитие экологии растений внесли ученые,
занимавшиеся агрохимией. Заложивший ее основы Ж. Б. Буссенго
показал, что все растения нуждаются в почвенном азоте. Он так-
же выявил, что для успешного завершения развития растениям
необходимо разное количество тепла, которое можно учесть, ежед-
невно суммируя температуры в вегетационный период. В 1840 г.
Ю. Либих сформулировал «Закон минимума», не потерявший сво-
его значения и в современной экологии. Он показал, что необхо-
димые растению химические элементы незаменимы. В современ-
ной экологии выявленная им закономерность учитывается при
изучении факторов, ограничивающих распределение и численность
организмов.
Эволюционное учение и публикация в 1959 г. книги Ч. Дарви-
на (1809—1882) «Происхождение видов путем естественного от-
бора» оказали большое влияние на развитие биологических наук.
Представления о единстве организма и среды, изменчивости орга-
низмов, наследовании признаков и естественном отборе стали
основополагающими и для экологии растений. Эволюционная
теория созревала в умах разных естествоиспытателей (Э. Дарвин,
А. Р. Уоллес и др.) и до выхода в свет основных трудов Ч. Дарвина.
В России А. Н. Бекетов в работе «Гармония в природе» (1858) из-
лагал сведения об изменении растений в различных условиях оби-
тания, о борьбе за существование. Однако только после 1859 г.
эволюционное мышление стало господствующим среди естест-
воиспытателей. Так, в русле эволюционных идей проводил ис-
следования по экспериментальной экологии растений профес-
сор Казанского университета Н.Ф.Леваковский. Его диссерта-
ция «О влиянии некоторых внешних условий на форму корней»
была написана в 1868 г. А последователь и пропагандист дарви-
низма немецкий ученый Э. Геккель в 1869 г. ввел в науку термин
«экология».
8
Экология растений фактически оформляется в самостоятель-
ное научное направление в конце XIX в. после работ датского
ботаника Е. Варминга (1842—1924), а официально признается раз-
делом ботаники в 1910 г. на III Всемирном ботаническом конг-
рессе в Брюсселе. Е.Варминг в 1895 г. возрождает забытый уже
геккелевский термин «экология» и в труде «Plantesamfund» (1895)
систематизирует и обобщает основную экологическую информа-
цию того времени. Он дает обзор и классификацию всех абиоти-
ческих и биотических факторов, влияющих на жизнь растений,
вводит в научный обиход представления о гидрофитах, ксерофи-
тах, мезофитах и галофитах. Книга Е. Варминга сыграла большую
роль в развитии экологии растений в разных странах. В России
она была переведена и издана дважды — в 1901 и 1902 гг. В после-
днем издании под названием «Распределение растений в зависи-
мости от внешних условий (экологическая география растений)»
в приложении была дана обобщающая сводка Г. И.Танфильева о
растительном покрове России. В своих работах Г. И.Танфильев
особое внимание обращал на роль почвенного покрова в распре-
делении растений и растительных сообществ. Продолжением «мор-
фолого-биологического» направления в экологии растений, ос-
нованного Е.Вармингом, явилась работа немецкого ботацика и
фитогеографа О.Друде «Экология растений» (1913), где он пред-
ставил систему жизненных форм растений (55 основных групп со
многими подразделениями).
Основателем экологии растений наряду с Е. Вармингом можно
считать и немецкого ботаника А. Шимпера (1856—1901). В 1898 г.
вышла его книга «География растений на физиологической осно-
ве», где обобщена вся западная фитогеографическая, экологиче-
ская и фитоценотическая литература конца XIX в., послужившая
основой для эколого-физиологического направления исследова-
ний. Особое внимание А.Шимпер уделял трем экологическим
факторам: теплу, гидрометеорологическим условиям (в том числе
ветру) и почве. Отмечалась им и приуроченность различных ви-
дов растений к определенным экотопам, обращалось внимание
на эколого-физиологические особенности растений. В это время
экология рассматривается учеными как ответвление физиологии,
исследующее природу непосредственно и акцентирующее внима-
ние на факторы среды, действующие на организм.
Подытоживая историю развития биологии в XIX в., К. А.Тими-
рязев (1908) подчеркнул ее бурное развитие и назвал это столетие
веком наук, когда возникли и развились новые естественные на-
уки и направления. Подчеркнув значение эволюционных идей для
развития науки, он назвал XIX в. веком Ч. Дарвина. К. А. Тимиря-
зев также полагал, что «рядом с морфологией описательной на-
рождается морфология экспериментальная» и наметилось «сбли-
жение задач морфологии и физиологии».
9
1.2. Современный этап развития экологии растений
В XX в. развитие экологии растений в нашей стране продол-
жил Б. А. Келлер (1901, 1903, 1907 и др.). Его работа (совместно с
почвоведом Н.А.Димо) «В области полупустыни» (1907) и труды
по экологии галофитов (1912) являются классическими. С усовер-
шенствованием полевых экологических приборов появилось эко-
лого-физиологическое направление, продолженное в работах
Л.А. Иванова (влияние света), Н.А. Максимова (засухоустойчи-
вость), И. И.Туманова (морозостойкость), П.А. Генкеля (солеус-
тойчивость). Ряд работ был посвящен экологии фотосинтеза. В эко-
логии растений оформилось особое направление — функциональ-
ная, или физиологическая, экология.
Большой вклад в развитие экологии растений внесли фитоце-
нологи, изучающие сообщества растений. С начала XX в. призна-
ние мировой научной общественности завоевали экологические
работы, выполненные американскими ботаниками. В США раз-
вивалось индикационное направление, связанное с изучением
растений, указывающих на характер почв. Были выделены рас-
тения-индикаторы карбонатных, песчаных, гипсовых, засолен-
ных почв и субстратов, содержащих отдельные химические эле-
менты (труд Ф.Клементса «Растения-индикаторы» (1920), работа
Дж. Е. Уивера и Ф. Клементса «Экология растений» и многие дру-
гие).
Важную роль в формировании экологической проблематики и
ее методологии сыграло представление о сукцессии. Так, в США
Г. Коулс детально описал последовательность изменения расти-
тельности песчаных дюн около оз. Мичиган. Разновозрастность
дюн позволяла отыскать там растительные сообщества разного
возраста (от бедных видами разреженных травяных фитоценозов
начинающих зарастать песков до леса на старых дюнах). Концеп-
цию сукцессии детально разработал Ф. Клементс. Растительное
сообщество он считал целостным образованием, подобным орга-
низму и претерпевающим изменения, подобные онтогенезу.
В России ботаники тоже уделяли большое внимание синэко-
логическим исследованиям. Так, С. И. Коржинский, изучая гра-
ницу между лесом и степью, отмечал важность воздействия самих
растений на среду, их способность делать ее более пригодной для
произрастания других видов. Большое значение для развития эко-
логии растительных сообществ имели труды В. В. Алехина, В. Н. Су-
качева, Л. Г. Раменского, А. П. Шенникова и других геоботаников.
В. Н. Сукачев предложил свою классификацию сукцессий, экспе-
риментально исследовал конкуренцию, изучал историю раститель-
ности. Он внес весомый вклад в учение о растительных сообще-
ствах, которые рассматривал как целостные структуры, образо-
ванные в результате коэволюции растений. В 1940-е гг. он разра-
10
ботал учение о биогеоценозе, включающем наряду с раститель-
ным сообществом почву, климат, воды, животных, микроорга-
низмы и т.д.
В 1920—1940 гг. формируется теоретическая основа современ-
ной экологии, предлагаются первые математические модели, вы-
рабатывается методология. Оформляются два главных подхода
современной экологии: популяционный, акцентирующий внима-
ние на динамике численности организмов и их распределении в
пространстве, и экосистемный, концентрирующийся на кругово-
роте веществ и трансформации энергии (А. М. Гиляров, 2003).
Главной целью популяционного подхода было выявление за-
кономерностей динамики численности популяций. Для этого ис-
пользовались простые математические модели, в которых форму-
лы отражали связь между разными характеристиками состояния
популяции (рождаемостью или у растений появлением всходов,
смертностью, скоростью роста, плотностью и др.). Они позволя-
ли проверять последствия разных допущений, выявляя условия
для реализации определенных вариантов популяционной динами-
ки. Популяционный подход в экологии долгое время развивался
преимущественно зоологами. Ботаники больше исследовали сооб-
щества, которые трактовали как целостные образования, доволь-
но четко отграниченные друг от друга. Однако уже в 1920-е гг.
были высказаны противоположные взгляды о том, что виды рас-
тений — «не рота солдат, шагающих в ногу», что они неодинаково
реагируют на факторы среды и по-разному распределены в ней.
Исходя из этого, растительные сообщества должны рассматри-
ваться как условные образования с размытыми границами. Этот
новый взгляд на растительное сообщество в 1924 г. сформулиро-
вал Л. Г. Раменский, подчеркнув экологическую индивидуальность
видов и непрерывность растительного покрова, зависимость его
от множества факторов. Неизменными он считал только законы
сочетаемости растений, которые и следовало изучать. В те же годы
в США сходные взгляды независимо развивал Г.Глизон. В его
«индивидуалистической концепции», выдвинутой в противовес
представлениям Ф.Клементса (о сообществе как аналоге организ-
ма), также подчеркивалась независимость распределения видов
друг от друга и непрерывность растительного покрова. Но широ-
ко популяционные исследования растений развернулись только в
1960-е гг. В СССР лидерами этого направления были Т.А.Работ-
нов и А.А.Уранов, в Великобритании — Д. Харпер.
Другое направление экологических исследований XX в. — эко-
системное. Термин «экосистема» был предложен в 1935 г. англий-
ским ботаником А.Тенсли для обозначения естественного комп-
лекса живых организмов и физической среды, в которой они оби-
тают. В 1930-х гг. в СССР Л.Л. Россолимо предложил «балансо-
вый подход», уделяющий основное внимание круговороту веществ
11
и трансформации энергии. В рамках его Г. Г. Винберг разработал
метод исследования создания растениями органического вещества,
позволявший судить о продуктивности фотосинтеза по количе-
ству выделившегося кислорода. Позднее в США Г. Райли и Р. Лин-
деманом были сделаны аналогичные работы, в которых предло-
жена общая схема трансформации энергии в экосистеме. В част-
ности, было продемонстрировано, что при переходе энергии с
одного трофического уровня на другой (от растений к травояд-
ным животным и т.д.) организмам каждого следующего уровня
достается не более 10 % энергии предыдущего уровня. Рассматри-
валась роль растений в экосистемных процессах.
Во второй половине XX в. завершается становление экологии
как самостоятельной науки, имеющей собственную теорию и ме-
тодологию, круг проблем и подходы к их решению. В связи с
широким применением математических подходов для решения
различных экологических проблем развилась количественная эко-
логия. Математические модели становятся более реалистичными,
их предсказания проверяются в эксперименте и наблюдениях в
природе. А эксперименты и наблюдения все чаще проводятся для
тестирования теоретически разработанных гипотез. Работы 1960—
1970-х гг. были ориентированы на выяснение общих принципов
устройства любых сообществ, а в 1980-е гг. основное внимание
перенесено на механизмы формирования их структуры. Напри-
мер, при изучении вытеснения одного вида другим экологи преж-
де всего стали интересоваться процессом этого вытеснения и осо-
бенностями видов, предопределяющими исход взаимодействия.
Так, выяснилось, что при конкуренции растений за элементы
минерального питания (азот, фосфор) победителем часто оказы-
вается не тот вид, который в принципе (без дефицита ресурсов)
может расти быстрее, а вид, способный поддерживать хотя бы
минимальный рост при низкой концентрации этого элемента.
Особое внимание стало уделяться эволюции жизненного цик-
ла и стратегии выживания растений. Поскольку возможности орга-
низмов всегда ограничены, а за каждое эволюционное приобрете-
ние приходится расплачиваться, между признаками возникают
отрицательные корреляции. Например, растение не может очень
быстро расти и одновременно много тратить на защиту от фито-
фагов. Изучение таких корреляций позволяет выяснить, как в
принципе достигается возможность существования организмов в
определенных условиях.
В современной экологии сохраняет актуальность и ряд про-
блем, имеющих давнюю историю исследований: установление за-
кономерностей динамики численности организмов, оценка роли
факторов, ограничивающих рост популяций, выяснение причин
колебаний численности. В этой области достигнут значительный
прогресс: для многих конкретных популяций выявлены механиз-
12
мы регуляции численности, продолжаются исследования конку-
ренции и взаимовыгодного сотрудничества видов. Большой про-
гресс достигнут в изучении круговорота веществ и потока энер-
гии. Примером может быть дистанционное (со спутников) опре-
деление содержания хлорофилла в поверхностных водах, позво-
ляющее составить карты распределения фитопланктона для всего
Мирового океана и оценить сезонные изменения его продукции.
Как продолжение идей XIX в. развивалась эволюционная эко-
логия — теоретическая основа для понимания экологических за-
кономерностей эволюционного процесса и эволюционного ста-
новления современных экологических взаимоотношений. В XX в.
в связи с возникновением и развитием палеоботанических мето-
дов в сочетании с использованием результатов исследований смеж-
ных и гуманитарных наук накопился обширный материал по ис-
тории природы, в том числе о роли антропогенных факторов в
формировании современного растительного покрова.
В настоящее время большое внимание стало уделяться методо-
логии экологических исследований. Было обращено внимание на
то, что многие фундаментальные экологические теории, сформу-
лированные в XX в., содержали «мифологическую» (романтиче-
скую) компоненту, опирались на признание слишком высокой
целостности экологических явлений и существование «жестких» свя-
зей между средой и биологическими объектами (А. М. Гиляров,
2001; Б. М. Миркин, Л. Г. Наумова, 2005 и др.). Была подвергнута
сомнению незыблемость ряда установившихся экологических кон-
цепций (правило 10 %, модель «хищник—жертва», положения «ос-
тровной биогеографии», принцип конкурентного исключения и
др.). Выявление множества исключений из установленных ранее
принципов и правил экологии привело к необходимости очер-
тить рамки их применимости, внести коррективы, смягчающие
«жесткость» законов. Стала разрабатываться идея шкалирования
биологического времени и пространства (признание необходимо-
сти выделять разные закономерности для объектов разного разме-
ра и продолжительности жизни). Было показано, что нестабиль-
ность в малых масштабах может быть условием стабильности в
больших: так, устойчивость коренного разновозрастного леса свя-
зана с динамическими процессами нарушения (вывалы, порой
и т.п.) и восстановления в отдельных его локусах. Таким образом,
как и во многих науках, в это время в экологии наметился отход
от универсализации экологических закономерностей, внимание к
разнообразию, частным случаям (case studies).
Значение экологических исследований в современном мире
возрастает. Кризисное состояние биосферы ставит проблемы ох-
раны природы и восстановления биогеоценозов в число важней-
ших проблем современности. Накоплен огромный фактический
материал, требующий обобщения и выхода в практику. Необхо-
13
димость правильного взаимоотношения человека и природы для
обеспечения устойчивого жизнеобеспечения на планете должна
быть осознана не только экологами, но и широкими массами на-
селения, а также государственными руководителями и деловым
сообществом. Для последних десятилетий XX — начала XXI в.
характерен процесс экологизации мышления (в первую очередь
исследователей разных специальностей), распространение эколо-
гических знаний по разным каналам коммуникации. Поскольку
экологические проблемы приняли планетарный масштаб, разви-
вается международное сотрудничество в согласовании экологиче-
ских программ. В 1960-е гг. биологи разных стран объединились
для решения задачи оценки продуктивности биосферы в рамках
Международной биологической программы (МБП). В 1970-е гг.
была организована международная программа «Человек и био-
сфера» для выявления и характеристики важнейших экологиче-
ских проблем Земли. Ныне международное сотрудничество про-
должается в рамках всемирных научных программ «Биоразнооб-
разие», «Изменение климата» и др. При этом во многих случаях
для решения конкретных экологических проблем требуется не
столько дальнейшее накопление знаний, сколько совокупность
грамотных управленческих решений и волевых усилий по внедре-
нию рекомендаций экологов.
ГЛАВА 2
ОСНОВНЫЕ МЕТОДЫ ЭКОЛОГИИ
РАСТЕНИЙ
В арсенале современной экологии растений существуют раз-
ные методы исследования, среди которых можно выделить три
основные группы: полевые наблюдения, эксперименты в поле и
лаборатории и моделирование (В. Д. Федоров, Т. Г. Гильманов,
1980).
2.1. Наблюдение и эксперимент
Метод наблюдения подразумевает невмешательство (или ми-
нимально возможные нарушения, вызываемые процессом наблю-
дения) исследователя в природную систему. Он представляет со-
бой наиболее важный и исторически первый прием экологичес-
кого исследования.
Сравнительные эколого-географические исследования прошли
долгий путь от красочных описаний картин природы, сделанных
натуралистами прошлых веков, до современных комплексных
программ экологических наблюдений и изменений с помощью
точной аппаратуры при длительных стационарных исследованиях.
Широкое применение в экологии растений получил экспери-
ментальный метод. Особенность эксперимента в том, что иссле-
дователь изучает систему, в которой специально изменены опре-
деленные параметры (в том числе и достаточно сильно, что не
характерно для ее естественного состояния). Методики экспери-
ментальной работы постоянно совершенствуются, и корректность
получаемых в эксперименте данных серьезно обсуждается.
Возможные воздействия экспериментатора на растение и его
окружение очень разнообразны, поэтому очень разнообразны и
экспериментальные подходы в экологии растений. Например,
эксперименты различаются по степени контроля над объектом
исследования. Так, большинство полевых экспериментов в есте-
ственных условиях относятся к неконтролируемым, так как вна-
чале воздействуя на экосистему или ее отдельные объекты, далее
экспериментатор следит только за изменениями, развертывающи-
15
мися на фоне всевозможных, часто непредвиденных и нежела-
тельных, с точки зрения задач эксперимента, внешних воздей-
ствий (например, погодных). В лабораторных условиях обычно
имеются условия для проведения в разной степени контролируе-
мого эксперимента (почти полный контроль за множеством эко-
логических факторов возможен только в специальных сложней-
ших установках типа фитотронов). Большую роль в экологии ра-
стений играют эксперименты по специальному культивированию
растений в лабораторных условиях и на делянках под открытым
небом.
Если в науках о неживой природе (физике прежде всего) рас-
пространен классический однофакторный эксперимент, когда изу-
чается влияние одного избранного фактора при фиксированных
значениях всех остальных, то в экспериментировании с биологи-
ческими объектами, поведение которых зависит от целого комп-
лекса взаимосвязанных природных факторов, этот подход оказал-
ся менее эффективным. Поэтому в экологии растений наряду с
однофакторным получили распространение многофакторные эк-
сперименты, когда в каждом варианте из серии опытов экспери-
ментатор изменяет не один, а сразу несколько факторов, значе-
ния» которых комбинирует особым способом, в результате чего
при математической обработке получается многофакторное опи-
сание изучаемого явления.
Экологи растений также постоянно имеют дело с результатами
стихийных «экспериментов», которые ставит сама природа (засу-
ха, наводнение, резкое похолодание и т.д.). Им остается в нуж-
ный момент и подходящим способом установить результат этих
экспериментов измерениями (Г. Вальтер, 1974). Широкую извест-
ность получили также непредвиденные последствия преобразова-
ния человеком экосистем и отдельных их компонентов, а также
интродукции видов. Анализ таких непредвиденных «эксперимен-
тов» внес заметный вклад в развитие теории экологии.
Среди специальных методов экологии растений особо выделя-
ются сравнительные эколого-географические наблюдения и по-
левые эксперименты. Для сравнительного изучения отношения
растений к среде широко используется разработанный Б.А. Кел-
лером (1908) метод экологических рядов. Экологический ряд со-
ставляют конкретные местообитания вида, располагаемые в по-
рядке постепенного изменения какого-либо экологического фак-
тора (влажность, богатство почвы, pH, степень и состав засоле-
ния, освещенность и пр.). Таким образом выясняются экологиче-
ские амплитуды вида по отношению к факторам среды, границы
оптимумов и пессимумов. Кроме естественных местообитаний
экологические ряды могут составлять экспериментальные участ-
ки (с различным количеством вносимых удобрений, влаги и пр.).
В экологическом ряду меняются морфология, анатомия, темпы
16
онтогенеза, химический состав, гидратура и температура отдель-
ных растений и другие характеристики. При изучении распреде-
ления растений в зависимости от экологических условий можно
учитывать нескольких совместно действующих факторов.
2.2. Моделирование
Одно из плодотворных методов естественных наук представля-
ет собой моделирование, т.е. построение, проверка и исследова-
ние моделей, интерпретация полученных с их помощью результа-
тов. При моделировании реальная природная система замещается
другой более простой системой, являющейся ее неполным подо-
бием. Модель — это упрощенный образ оригинала, при котором
сознательно огрублены отдельные черты изучаемого объекта или
процесса.
В зависимости от характера упрощения для одного и того же
оригинала можно получить несколько различных моделей, в том
числе удобные для изучения тех или иных свойств оригинала.
Стратегия моделирования заключается в стремлении путем упро-
щения получить модель, свойства и поведение которой можно
было бы эффективно изучать, но которая в то же время остава-
лась бы достаточно сходной с оригиналом, чтобы результаты это-
го изучения были применимы к оригиналу. Обратный переход от
модели к оригиналу — интерпретация модели. Эта процедура все-
гда неоднозначна в силу принципиально неполного соответствия
модели и моделируемой системы.
В зависимости от особенностей объекта изучения и задач ис-
следования в экологии растений применяются модели разных ти-
пов. Прежде всего модели подразделяют на реальные (натураль-
ные, аналоговые) и идеальные (знаковые).
Реальные модели отражают оригинал по своей физической при-
роде. Например, аквариум со всем его населением воспроизводит
некоторые черты естественных водоемов. Главная проблема рабо-
ты с такими моделями — трудность установления степени адек-
ватности модели оригиналу и обоснование возможности приме-
нения результатов моделирования к исходной системе — ориги-
налу.
Знаковые модели представляют собой условное описание ориги-
нала с помощью символов и операций над ними. Полученные
результаты при использовании кода символического языка интер-
претируются и сопоставляются с реальными природными образо-
ваниями. Наибольшее значение в экологии имеют две разновид-
ности знаковых моделей — математические и концептуальные.
Концептуальная модель — более формализованный вариант
традиционного естественно-научного описания изучаемого объекта
17
или процесса, состоящий из научного текста, сопровождаемого
схемами, графиками и другим иллюстративным материалом. На-
значение этих моделей — дать ясное обобщенное и достаточно
полное выражение знаний об изучаемом явлении в рамках опре-
деленной концепции. Например, в русле концепции «соматичес-
кой редукции» предложены схемы эволюции жизненных форм
растений, а в рамках «энергетической» концепции построены схемы
потока вещества и энергии в биогеоценозе, которые сопровожда-
ются поясняющим текстом, таблицами и графиками. Концепту-
альные модели широко применяются в науке благодаря своей уни-
версальности, гибкости, богатству средств выражения и др. Но им
свойственны также высокая неоднозначность интерпретации и
определенная статичность, затрудняющая описание постоянно
изменяющихся природных объектов.
Математические модели используют разного рода абстракт-
ные знаковые построения. Они дают возможность формально оп-
ределять взаимодействия отдельных компонентов экосистем при
их количественном выражении.
ГЛАВА 3
ЭКОЛОГИЧЕСКИЕ ФАКТОРЫ
Влияющие на организмы элементы окружающей среды назы-
вают экологическими факторами. Совокупность всех факторов,
действующих на растения в данном месте, объединяют в понятие
местообитание. Так как комплекс условий в конкретном месте
сильно изменен живыми организмами, принято различать поня-
тия «экотопа» и «биотопа». Экотоп — это первичный комплекс
факторов физико-географической среды, а биотоп (от греч. bios —
жизнь и topos — место) — среда, уже видоизмененная средообра-
зующей деятельностью организмов (Б. М. Миркин и др., 1989).
3.1. Типы экологических факторов.
Фитоиндикация
Классификации экологических факторов. В любое время среда
действует на растение как единое целое, поэтому выделение от-
дельных ее факторов в значительной степени условно. Однако
для удобства изучения экологические факторы выделяют и объе-
диняют в группы.
Традиционно по типу воздействующего объекта экологические
факторы принято делить на 2 группы: абиотические (факторы
неорганической среды) и биотические (связанные с влиянием
живых организмов). К абиотическим факторам относят климати-
ческие (свет, тепло, воду, воздух), эдафические (совокупность всех
свойств почвы) и орографические (условия рельефа). Среди био-
тических факторов обычно выделяют фитогенные (влияние рас-
тений) и зоогенные (влияния животных). Многогранную деятель-
ность человека, воздействующего как непосредственно на расте-
ния и их сообщества, так и на все параметры местообитания, рас-
сматривают как антропогенные факторы.
Возможны и другие основания для классификации экологи-
ческих факторов. Г. Вальтер, обратив внимание на их взаимодей-
ствие, предложил делить факторы среды на первичные и комп-
лексные (рис. 3.1). К первичным он отнес тепло, воду, свет, хи-
19
Первичные
Экологические факторы
Тепло
Комплексные
Вода
Свет
Химизм — —
- I-Механические
Климатические -]->
n J
Орографические
Эдафические -I
Биотические
Рис. 3.1. Классификация экологических факторов по их действию
(по Г. Вальтеру, I960)
мизм и механические факторы, а к комплексным — климатиче-
ские, орографические, эдафические и биотические факторы. Обе
группы факторов тесно связаны. Например, тепловые условия ме-
стообитания зависят от климата региона и микроклимата фитоце-
ноза, а также от рельефа и характера поверхности почвы. Одно-
временно взаимодействуют сами первичные и комплексные фак-
торы. Так, тепло связано с обеспеченностью местообитания све-
том, водой, а также с гранулометрическим составом почв и отча-
сти их химизмом (например, богатые гумусом почвы темнее и
лучше прогреваются), а такой комплексный фактор, как эдафи-
ческий, определяется и климатом, и рельефом, и влиянием жи-
вых организмов и т. п.
Еще одна классификация делит экологические факторы по характеру
их действия. В этом случае их разделяют на прямодействующие и кос-
венно действующие. Прямодействующие непосредственно влияют на
обмен веществ и развитие организмов, а косвенно действующие влияют
на них через изменение других факторов. Примером прямодействующих
факторов могут быть тепло, вода, состав и движение воздуха, сенокоше-
ние, поедание животными, пожары, перенос семян и плодов и т.п. При-
мер фактора, действующего косвенно, — рельеф (высота над уровнем
моря и экспозиция склона), который влияет на растения, перераспреде-
ляя тепло и воду на поверхности Земли. К косвенно действующим фак-
торам относятся также географическая широта и удаленность от океана,
материнская горная порода и гранулометрический состав почвы, влия-
ние соседних растительных сообществ и т.д. Все эти факторы действуют
и на растения, и на другие компоненты экосистемы, которые, в свою
очередь, опосредованно влияют на ботанические объекты. Бывает и так,
что один и тот же фактор в одних условиях действует косвенно, а в дру-
гих прямо. Например, карбонатные породы действуют на кальцефобные
растения (плохо переносящие этот химический элемент) прямо, а на
многие другие виды косвенно (через изменение плодородия почв, теп-
лового и водного режима). Косвенные факторы важны не менее прямо-
действующих, так как нередко именно через них растение может полу-
чить комплекс условий, обеспечивающий ему возможность жизни на
данной территории.
20
В современной экологии принято выделять факторы-ресурсы
и факторы-условия. Экологические ресурсы — факторы, которые
организмами в процессе жизнедеятельности потребляются и рас-
ходуются (А. М. Гродзинский и др., 1991). В сообществах фонд
экологических ресурсов делится между сообитающими видами.
За ресурсы между растениями происходит конкуренция. К эколо-
гическим ресурсам Т. А. Работнов (1998) отнес свет, воду, элемен-
ты минерального питания, диоксид углерода вокруг фотосинтези-
рующих органов, кислород в почвенном воздухе и воде. Для орга-
низмов другие организмы (живые и мертвые) имеют значение как
пищевые ресурсы. Ресурсом является и физическое пространство,
так как все другие ресурсы занимают какую-то территорию (так,
для питания растению нужны определенная освещаемая площадь
и некоторый объем почвы). При высокой плотности популяции
нехватка пространства (как вместилища ресурсов) влияет на от-
ношения между особями и их состояние (повышает смертность,
вызывает изменения внешнего вида и физиологии). Иногда де-
фицит пространства возникает вне связи с ресурсами питания
(например, выталкивают друг друга растущие луковицы). А вот
тепло, влажность воздуха, кислотность среды, соленость и ско-
рость течения воды, содержание неиспользуемых в питании за-
грязняющих веществ ресурсами не являются. Это экологические
условия, которые не расходуются организмами, и те не могут
сделать их более или менее доступными для других. Хотя конку-
ренции за факторы-условия не происходит, они существенно вли-
яют на способность растений конкурировать за ресурсы.
Фитоиндикация. Оценка свойств местообитаний (среды) по
растениям называется фитоиндикацией. Для получения инфор-
мации о действующих в данном месте экологических факторах
могут использоваться виды растений, их внутривидовые подраз-
деления и тераты (формы с аномальными особенностями), а так-
же растительные сообщества — наиболее надежные и хорошо за-
метные показатели. Возможна фитоиндикация и по состоянию
отдельных растений. Благодаря быстроте ответной реакции на
клеточном, тканевом и органном уровне этот вариант индикации
удобен при оценке деталей антропогенного воздействия.
Индикационная связь может быть прямой и косвенной (через
промежуточное звено). Прямые индикаторы надежнее косвен-
ных, но они более редки. В аридных районах примером прямых
индикаторов подземных вод могут служить сообщества с господ-
ством растений, постоянно связанных корнями с грунтовыми во-
дами, например чий (Achnatherum splendens), виды верблюжьей
колючки (Alhagi). А в песчаных пустынях Средней Азии заросли
аристиды (Aristida pennata) служат косвенным индикатором скоп-
лений верховодки, так как это растение является пионером-псам-
мофитом, указывающим на слабую закрепленность песка, кото-
21
рая обусловливает хорошую аэрацию и проникновение осадков,
благоприятствующих образованию верховодки.
По степени географической устойчивости индикационных связей
индикаторы могут быть разделены на панареальные, региональные и ло-
кальные. Связь первых с индицируемым параметром сходна на всем аре-
але. Так, тростник {Phragmites australis) является панареальным инди-
катором повышенной влажности субстрата в пределах развития своей
корневой системы (его корни и корневища могут располагаться и на
поверхности донных отложений озера, и на глубине более Юму зеркала
грунтовых вод в аридных местообитаниях). Панареальные индикаторы
немногочисленны и обычно относятся к прямым. Значительно более рас-
пространены региональные индикаторы, имеющие индикационное зна-
чение в определенной физико-географической области, и локальные, со-
храняющие индикационное постоянство лишь в определенном физико-
географическом районе. Локальные и региональные индикаторы обыч-
но считаются косвенными.
Параметры среды, определяемые с помощью ботанических по-
казателей, очень разнообразны. Ими могут быть различные типы
природных объектов (почва, горные породы, подземные воды
и т.п.) и их отдельные свойства (гранулометрический состав, за-
соленность, трещиноватость и др.), процессы в среде (эрозион-
ные, карстовые, тектонические, заболачивание и т.п.) и ее от-
дельные свойства (например, климат). Индицировать по растени-
ям можно и антропогенные изменения, происходящие при мели-
орации, воздействии промышленных предприятий, строительстве,
добыче полезных ископаемых и т. п.
3.2. Закономерности действия экологических факторов
Закон оптимума. Закон оптимума указывает, что любой эколо-
гический фактор имеет определенные пределы положительного
влияния на живые объекты. При отклонении от них наступает
угнетение организмов, а по достижении критических значений —
гибель. Например, растения плохо переносят и сильную жару, и
сильные морозы, для большинства из них оптимальны средние
температуры. Одинаково неблагоприятны для них засухи и посто-
янные проливные дожди. Закон оптимума указывает на важность
меры воздействия каждого фактора на жизнеспособность орга-
низма.
Графически эта зависимость выражена оптимальной (колоко-
ловидной) кривой, где по горизонтали указывается интенсивность
воздействия фактора, а по вертикали — степень благоприятности
его воздействия (рис. 3.2). В центре под такой кривой располага-
ется зона оптимума. При оптимальных значениях фактора на-
22
блюдаются наилучшие показатели жизнедеятельности растений:
они активно растут, питаются, размножаются. Чем больше откло-
нение фактора от этих показателей, тем менее это благоприятно
для растений. На схеме по обе стороны от зоны оптимума распо-
лагаются зоны пессимума (или зоны угнетения). В месте пересе-
чения кривой с горизонтальной осью находятся две критические
(кардинальные) точки — минимума и максимума. Они соответ-
ствуют крайним значениям фактора, за пределами которых насту-
пает смерть организма. Расстояние между критическими точками
показывает степень выносливости организмов по отношению к
изменениям данного фактора, или его толерантность. И опти-
мумы, и пессимумы по конкретному фактору могут быть смеще-
ны действием других факторов. Так, прогрессивная агротехника
может существенно расширить зону оптимума и пределы толе-
рантности (экологическую амплитуду) возделываемого вида. Ус-
ловия, соответствующие точкам, близким к критическим, особенно
тяжелы для выживания и их называют экстремальными. Для ра-
стений важна дозировка экстремальных проявлений фактора —
сила, продолжительность и повторяемость его воздействия. Так,
кратковременные и редкие морозы и засухи обычно не ограничи-
вают возможность распространения вида в данном районе, но осо-
бенно жестокая засуха или очень сильный мороз приводят к нео-
братимым повреждениям тканей.
В экологии растений принято выделять два типа оптимумов
(или экологических ареалов) вида — аутэкологический (физио-
логический, потенциальный) и синэкологический (фитоценоти-
ческий, фактический). Первый подразумевает те условия, кото-
рые вид потенциально может занять при отсутствии конкуренции
с другими видами. Он свойствен виду, отражает его физиологи-
3 фактора
Рис. 3.2. Схема действия экологического фактора на растение:
1 — зона оптимума; 2 — зоны пессимума; 3 — пределы толерантности; max, min —
кардинальные точки
23
Рис. 3.3. Схема положения аутэкологического и синэкологических
оптимумов сосны обыкновенной {Pinus sylvestris):
1 — аутэкологический оптимум; 2, 3 — синэкологический оптимум сосны обык-
новенной
ческие возможности, соответствует оптимальным значениям абио-
тических факторов, при которых может наблюдаться наилучший
рост растения (наибольшее накопление биомассы и т. п.). Аутэколо-
гический оптимум может быть выявлен в одновидовых группиров-
ках-(например, в посевах или посадках). Условия среды, в кото-
рых вид играет наибольшую роль в естественном растительном
покрове, конкурируя при этом с соседями, — оптимум синэколо-
гический. Он формируется под влиянием всего комплекса взаи-
модействий в фитоценозе и выявляется в естественных сообще-
ствах, а из-за зависимости от конкурентной обстановки не явля-
ется для вида неизменным. Синэкологический оптимум, как пра-
вило, уже аутэкологического. Поэтому в ботанических садах и
других специально поддерживаемых посадках многие растения
растут далеко за пределами своих естественных ареалов.
Соотношение аут- и си нэкологического оптимумов зависит от жиз-
ненных стратегий видов и используется при описании их экологических
ниш (см. разд. 5.6). У некоторых растений эти оптимумы совпадают, а у
некоторых нет. Например, у сосны обыкновенной {Pinus sylvestris) аут-
экологический оптимум приходится на умеренно влажные почвы, а фи-
тоценотический находится в совершенно других условиях — в области
очень сухих и очень влажных почв (рис. 3.3). Так, в Европейской России
сосна на умеренно влажных и достаточно богатых почвах вытесняется
более конкурентоспособной в этих условиях елью {Picea abies). А на
сухих песках и сфагновых болотах она играет большую роль в раститель-
ном покрове, господствует в древесном ярусе, образуя сосняки.
Правило экологической индивидуальности видов. Часто то, что
оптимально для одного вида, вызывает угнетение другого. Разно-
образие видов проявляется и в положении критических точек эко-
логических кривых. Они могут быть по-разному смещены от цен-
24
тра диапазона значения фактора, а также тесно сближены или
широко расставлены. Поэтому, с одной стороны, можно выде-
лить виды, приспособленные к жизни при разной интенсивности
действия фактора (например, светолюбивые и тенелюбивые).
С другой стороны, ряд видов может жить только в очень стабиль-
ных условиях, а другие выдерживают широкие колебания значе-
ний фактора. Для обозначения первых употребляется приставка
стено-, а вторых — эври-. Так, если рассматривается тепловой
фактор, то виды, живущие в узком диапазоне температур, — сте-
нотермы, а в широком — эвритермы (рис. 3.4).
Индивидуальные наборы кривых по всем факторам, которыми
могут быть охарактеризованы все виды, отражают сформулиро-
ванное Л. Г. Раменским (1924) правило экологической индивиду-
альности видов: виды экологически индивидуальны, имея инди-
видуальный набор приспособлений к среде, особым образом бу-
дучи распределены в ней, занимая уникальное место в жизни со-
обществ, по-своему приходя и уходя из него. Генетическое разно-
образие организмов обусловливает то, что нет двух видов, иден-
тичных по адаптивным возможностям. Более того, каждая особь
эколого-генетически индивидуальна, и поэтому возможности ре-
агирования на факторы среды и возможности адаптации к ним
различны и у всех видов, и у всех особей.
Закон совместного действия факторов. Правило совместного
(совокупного) действия факторов было установлено в 1909 г. не-
мецким агрохимиком и физиологом растений А. Митчерлихом и
названо им законом эффективности факторов (или законом
физиологических взаимосвязей). Он подчеркнул, что урожай сель-
скохозяйственных культур всегда зависит от действия многих фак-
торов. При этом имелось в виду, что можно по отдельности изу-
Степень
благо-
приятности
фактора
Рис. 3.4. Схема действия температурного фактора на растения разных
типов:
I — эвритермный вид со средними требованиями к теплообеспеченности; 2 —
эвритермный холодолюбивый вид; 3 — эвритермный теплолюбивый вид; 4 —
стенотермный холодолюбивый вид; 5 — стенотермный теплолюбивый вид
25
чить влияние разных факторов и потом суммировать результаты,
т.е. сначала не учитывалось взаимодействие факторов и разные
влияния их в определении жизнеспособности растений.
Но экологические факторы взаимосвязаны и действуют всегда
совокупно. При этом изменение одного из них часто ведет к из-
менению других (например, иссушение обычно возрастает с рос-
том температуры). Именно совместное действие факторов, вза-
имное их усиление или ослабление определяют эффект воздей-
ствия на организм и успешность его жизни. Б. Бауле (1918), под-
робно разрабатывавший идею совместности действия факторов
среды, назвал это законом совокупного действия (или законом
совместного действия факторов). Дополнил и развил его А.Тине-
ман, и теперь имена всех этих исследователей часто включают в
название закона. Формула Митчерлиха —Бауле была первым ма-
тематическим выражением явления взаимодействия факторов, и
тем самым работы А. Митчерлиха и его последователей стимули-
ровали изучение многофакторных зависимостей в экологии, сель-
ском хозяйстве и смежных дисциплинах.
Опыты показывают, что факторы среды часто взаимодейству-
ют таким образом, что при экстремальных значениях одного из
них Диапазон устойчивости к другим факторам сокращается. Так,
при экстремальной температуре организм обычно выживает в бо-
лее узких пределах влажности, чем при оптимальной. Часто имеет
значение последействие факторов, которые влияли на растение
ранее. Например, жизнь побегов данного года сильно зависит от
того, каковы были условия предыдущих сезонов, когда заклады-
вались почки, формировались защитные структуры, образовыва-
лись семена. Таким образом, при изменении одних экологиче-
ских факторов меняется относительное влияние других (изменя-
ются зоны толерантности, положение оптимума, масштабы и фор-
мы реагирования на каждый фактор). Поэтому методической ос-
новой факторальной экологии служат комплексные подходы, ис-
пользующие приемы многофакторного эксперимента и многомер-
ного математического анализа.
Принцип ограничивающих факторов и закон толерантности. На
организм действует много факторов, но в совокупном давлении
среды часто выделяют факторы, сильнее всего ограничивающие
успешность его жизнедеятельности. Идея о том, что в конкрет-
ных условиях выносливость организма определяется «самым сла-
бым звеном» его экологических возможностей, впервые была обо-
снована немецким агрохимиком Ю. Либихом в 1840 г. Рисуя бочку
с отверстиями, он указывал, что уровень жидкости в ней опре-
деляет только нижняя дыра. Ю.Либих исследовал в основном
влияние химических веществ, находящихся в минимуме. Поэто-
му выявленная закономерность была названа им законом мини-
мума.
26
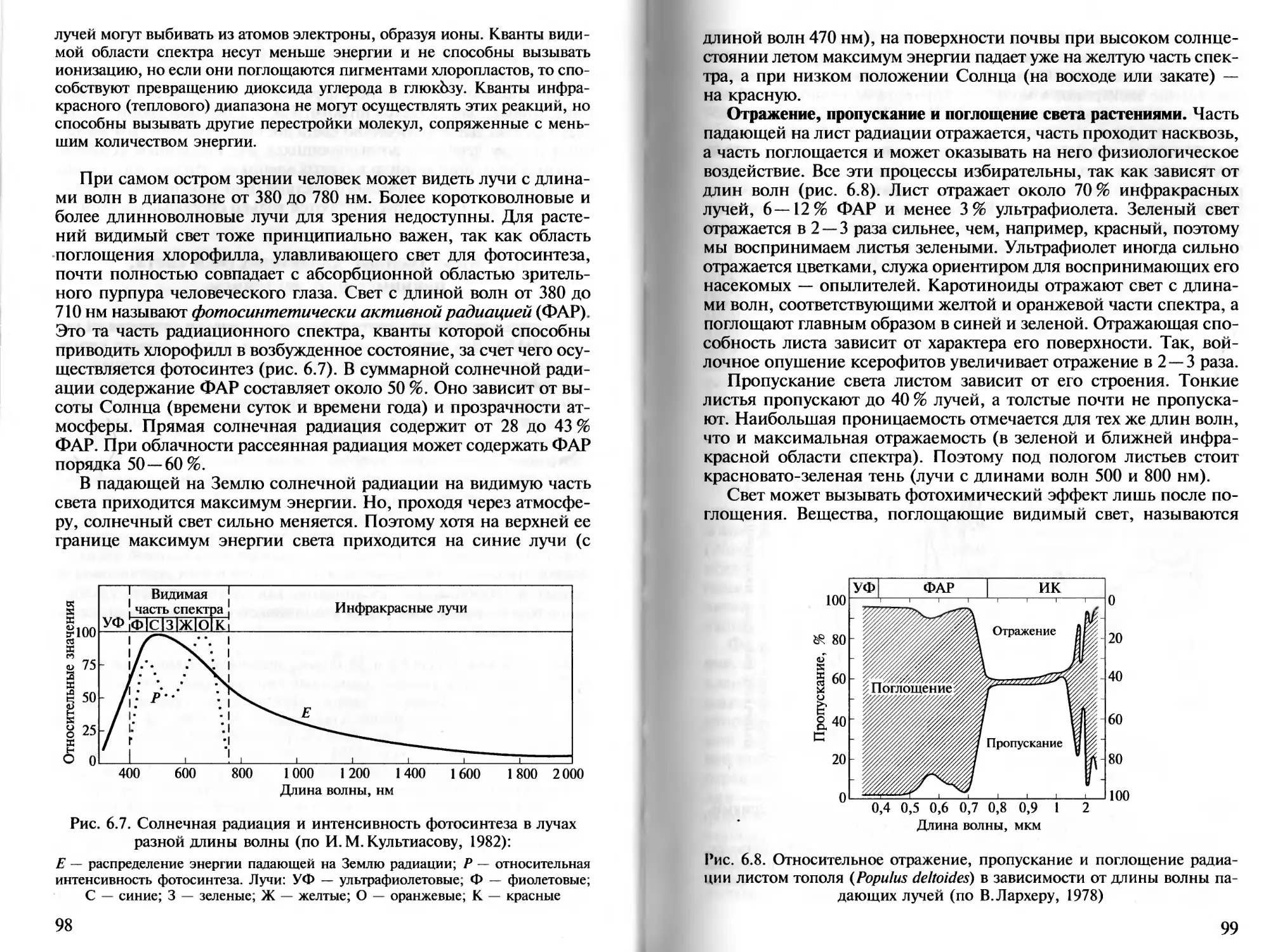
Развивая идею Ю. Либиха, Ф. Блэкман (1909) предложил прин-
цип ограничивающих факторов. Он гласит, что для организма
наиболее значим фактор, больше всего отклоненный от оптималь-
ных для него значений. Именно эти ограничивающие, или «ли-
митирующие», факторы среды, имеющие в конкретных условиях
значения, близкие к критическим, и ограничивают возможность
существования вида в этих условиях, несмотря на оптимальные
значения других факторов. Так, в приполярных районах для рас-
тений лимитирующим фактором обычно бывает тепло, а в тропи-
ческой пустыне — вода. В 1913 г. ту же закономерность сформу-
лировал в виде закона толерантности американский специа-
лист по экологии водных животных В. Шелфорд. Он акцентиро-
вал внимание на том, что жизнедеятельность лимитируют и ми-
нимальные, и максимальные значения фактора, а диапазон меж-
ду ними определяет величину выносливости (толерантности) орга-
низма к данному фактору.
Впоследствии закон толерантности был дополнен. Указывалось,
что неоптимальные условия по одному фактору могут снижать
толерантность по другим, и конкурентные отношения также мо-
гут мешать использованию оптимальных условий среды. С другой
стороны, организмы могут иметь очень разные диапазоны толе-
рантности по разным факторам. Так, стеногалинные виды (на-
пример, солянки — Salsola) с узкими диапазонами по параметру
засоления могут иметь широкие диапазоны выносливости по от-
ношению к теплу (эвритермность) и т.д. При этом виды с широ-
кими диапазонами по всем факторам (эврибионты) наиболее
распространены. Узкий же диапазон толерантности — проявле-
ние специализации видов (стенобионтов).
Компенсаторные закономерности. Поскольку любой фактор
может быть лимитирующим, все они важны для существования
организма. Положение о незаменимости фундаментальных фак-
торов связывают с именем В. Р. Вильямса, который в 1949 г. жес-
тко сформулировал правило: полное отсутствие в среде фунда-
ментальных экологических факторов (свет, вода, биогены и т.д.)
не может быть заменено другими факторами. Так, на Крайнем
Севере недостаток тепла полностью ничем не заменить.
Хотя ни один фактор не способен вполне заменить другой, воз-
можна частичная компенсация. Это отражено в законе компенса-
ции (эффект взаимозаменяемости) факторов, сформулированном
в 1930 г. Э. Рюбелем. Отмечалось, что на севере влияние на резуль-
таты фотосинтеза продолжительного летнего освещения частично
компенсирует недостаток там тепла, а в теплице недостаток света
отчасти может быть компенсирован обилием диоксида углерода.
Многочисленные данные по компенсаторным закономерностям были
получены при изучении минерального питания растений. К. А. Куркин
27
(1976) показал, что на лугах при уменьшении влажности почвы повыша-
ется содержание в ней нитратов, а при высокой обеспеченности азотом
растения лучше переносят засуху. Экспериментально обнаружено, что
потребность в некоторых ионах может зависеть от снабжения растения
другими соединениями. Отмечено также, что хотя азот как элемент не-
заменим, для большинства растений взаимозаменяемы источники азот-
ного питания — ионы аммония и нитрат-ионы.
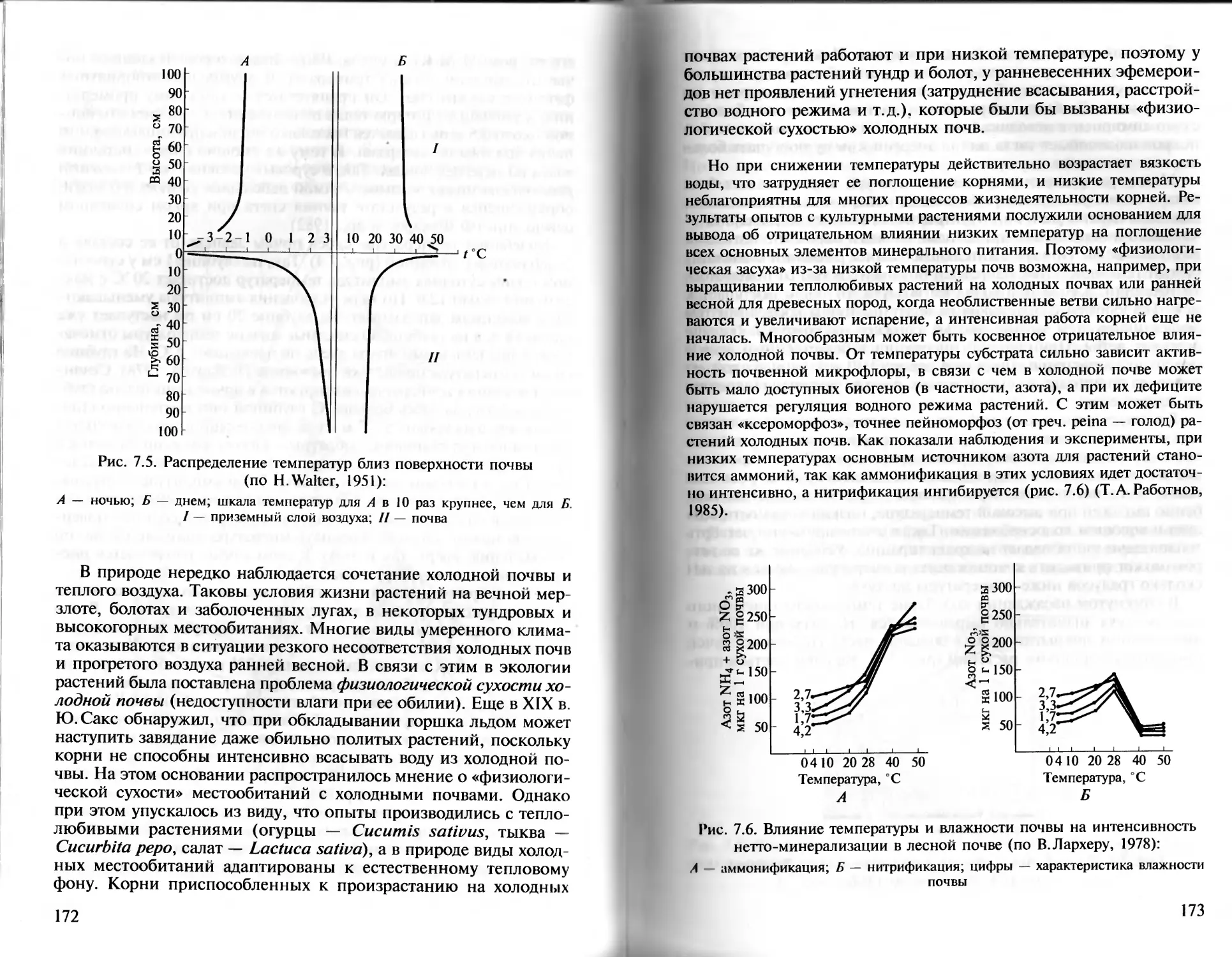
Правило относительного постоянства местообитания. Компен-
саторный эффект отражен также в известном правиле постоян-
ства местообитания (или правиле замещения), сформулирован-
ном в начале XX в. В. В. Алехиным. Он подчеркивал, что в приро-
де одни условия среды могут до некоторой степени быть замеще-
ны другими, что позволяет одним и тем же растениям произрас-
тать в довольно разных местообитаниях. Сходное экологическое
действие может оказывать разная совокупность факторов. Напри-
мер, степные растения, продвигаясь на север, переходят на юж-
ные склоны или на теплые известковые почвы.
Близко по смыслу и ботанико-географическое правило пред-
варения В. В. Алехина, которое гласит, что плакорный вид (и био-
ценоз) предваряется на юге или на севере в соответствующих био-
топах (там условия существования для него меняются мало в свя-
зи с меньшей или большей инсоляцией склонов разной экспози-
ции). Так, в ряде случаев формируются фрагменты растительнос-
ти, соответствующие более северным или южным зонам (напри-
мер, степные участки в тайге или участки леса в степной зоне)
(рис. 3.5). Однако эти экстразональные сообщества обычно бед-
нее видами.
Правило селективности действия фактора. Эффект действия
экологических факторов будет неодинаковым при различных со-
стояниях организма. Ослабленные и угнетенные особи сильнее
страдают в неблагоприятных условиях. Уязвимы также растения,
находящиеся в старческом состоянии или на начальных периодах
онтогенеза, так как с возрастом меняется структура тканей (на-
пример, развиваются дополнительные защитные слои покровов),
мощность и расположение органов. Ю.Одум (1986) в этой связи
указывает, например, что взрослый кипарис может расти и с по-
стоянно погруженными в воду корнями, и на сухом холме, но для
Рис. 3.5. Схема правила предварения (по В. В. Алехину, 1950).
Северное влаголюбивое растение (/) переходит на юге на северные склоны и
дно балок, южное растение (2) при движении к северу переходит на лучше на-
греваемые южные склоны
28
развития его проростков необходима влажная, но незаливаемая
почва.
Необходимо учитывать также генетическую неоднородность
популяций. Экологические факторы могут по-разному действо-
вать на вегетативно и генеративно размножающиеся растения.
Неодинаково они действуют и на различные функции организма.
Так, оптимум для одних процессов (например, дыхания) — не
есть оптимум для других (например, фотосинтеза).
Выносливость организмов может зависеть и от их пола. Так,
мужские особи обычно страдают и элиминируются больше, но у
них также отмечается и более частое появление мутантных форм,
в том числе перспективных с точки зрения выработки новых адап-
таций к ужесточившимся условиям. В экологическом мониторин-
ге на учете пыльцевых зерен (мужских заростков), отклоняющих-
ся от нормы, основан один из методов контроля состояния окру-
жающей среды (О. Ф. Дзюба и др., 2005 и др.).
ГЛАВА 4
УСТОЙЧИВОСТЬ РАСТЕНИЙ
И ИХ РЕАКЦИИ НА ДЕЙСТВИЕ
НЕБЛАГОПРИЯТНЫХ ФАКТОРОВ
Растения, как и все другие организмы, существуют в сложных
и изменчивых условиях среды. Они способны поддерживать свою
структуру и физиологические процессы (гомеостаз). Общие видо-
вые свойства, а также особенности, возникшие в результате при-
способления растений к конкретным экологическим условиям,
обусловлены наследственностью. Характерная черта растений —
экологическая пластичность, т.е. способность к изменчивости на
всех уровнях своей организации под влиянием условий жизни.
Пределы изменчивости растений в разных условиях предопреде-
ляет генотип.
Благодаря фенотипической пластичности растения выживают
в среде, изменившейся в неблагоприятном направлении, а также
могут распространяться на новые места обитания с иными эколо-
гическими условиями.
4.1. Общие вопросы устойчивости растений
Пути стабилизации живых систем. Устойчивость клеток и тка-
ней к факторам среды и способность многоклеточных систем к
восстановлению во многом обусловлены их функциональной и
структурной гетерогенностью, а также регуляцией размножения
отдельных групп клеток. Системы стабилизации, предотвращаю-
щие возможные нарушения, описываются рядом принципов —
избыточности, гетерогенности и др. (Л. Г. Косулина и др., 1993).
Избыточность структур и функциональных возможностей —
один из основных способов обеспечения надежности систем.
Резервирование в живых объектах имеет разные проявления. Яр-
кий пример — избыток клеточных органелл. Так, для обеспече-
ния растения продуктами фотосинтеза достаточно лишь части име-
ющихся хлоропластов (А.Л. Курсанов, 1976). А на Крайнем Севе-
ре повышенное число митохондрий в клетках мезофилла важно
для процессов репарации (восстановления) и адаптации. Резерви-
рование проявляется и в накоплении предшественников для био-
30
синтеза многих соединений. Запасенные вещества способствуют
успешности прорастания семян, роста проростков, цветения, а
также для репарации и адаптации в неблагоприятных условиях.
Например, запасание углеводов помогает ряду болотных расте-
ний переносить существенный дефицит кислорода в почве (так
как в этих условиях для усиления гликолиза нужны углеводы). На
организменном уровне принцип избыточности выражается в обра-
зовании большого количества гамет и семян; на популяционном —
в большом числе особей, на биоценотическом — популяций.
Для надежности систем организма большое значение имеет
компартментация — структурное разобщение разных процессов,
основанное на мембранной организации клетки. Пространствен-
ное разобщение поддерживает локальные концентрации веществ
(промежуточных соединений обмена, конечных продуктов, фермен-
тов), pH среды и др. Это обеспечивает оптимальные условия для
координации физиологических процессов, способствует поддержа-
нию внутриклеточного гомеостаза и минимизации повреждений.
Для достижения устойчивости организма большое значение
имеет также регенерация, обеспечиваемая образовательными тка-
нями. Устойчивость растений к повреждениям во многом связана
с устойчивостью их образовательных тканей, наличием спящих
почек и способностью к образованию вторичных меристем. Важ-
ный резерв восстановления — покоящиеся клетки меристем с
повышенной устойчивостью. У многих растений они выходят из
периода покоя только при гибели значительного числа активно
делящихся клеток. Присутствие у растений постоянных меристем
во многом определяет устойчивость организма и длительность его
жизни. А их гибель стимулирует возникновение вторичных мери-
стем из дифференцированных клеток. Так, широко известно об-
разование новых побегов и корней после повреждения их верху-
шек. При этом новые органы могут возникать, например, из эпи-
дермы и т.п.
Основные приспособления растений к действию неблагоприят-
ных факторов. Выделяют несколько основных способов приспособ-
ления организмов к действию неблагоприятных факторов (А. Г. Во-
ронов, 1963). Первый связан с избеганием неблагоприятных воз-
действий благодаря особенностям поведения. Этот вид реагиро-
вания у неподвижных растений встречается редко. Среди приме-
ров — закрывание цветков в дождь и складывание листочков стыд-
ливой мимозы (Mimosa pudica) при прикосновении к ним. К дан-
ному типу реагирования можно отнести движения некоторых ра-
стений и их органов от источника воздействия (отрицательные
тропизмы и таксисы).
Второй способ избегания организмами неблагоприятных усло-
вий — перестройка цикла развития для переживания неблагопри-
ятного периода в покоящемся состоянии. Примеры — зимний
31
покой растений и развитие эфемеров и эфемероидов пустынь толь-
ко после весенних дождей.
Третий тип приспособлений к действию неблагоприятных фак-
торов — расширение экологической амплитуды вида. Приобрете-
ние организмом выносливости компенсирует невозможность ухо-
да в более благоприятные условия. Выживать в суровых условиях
позволяют таким организмам специфические анатомо-морфоло-
гические и физиолого-биохимические свойства. При этом расши-
рение экологической амплитуды часто идет путем образования
внутривидовых групп, приспособленных к обитанию при разных
значениях фактора.
Четвертый способ реагирования растений на воздействия сре-
ды — создание относительного постоянства благоприятных усло-
вий внутренней среды. Так, регулируя движение устьиц, растения
до некоторой степени могут поддерживать свою оводненность и
температуру.
Стрессовые реакции растений. Ответ растений на действие эк-
стремальных факторов среды включает неспецифические реакции
и процессы специализированной адаптации. Биологическое зна-
чение неспецифических реакций заключается в срочной мобили-
зации защитных систем для сохранения жизнеспособности в не-
благоприятных условиях. А специализированная адаптация обес-
печивает сохранение высокого уровня продуктивности растений
в данных условиях.
Важное свойство живых объектов — способность поддерживать
гомеостаз — относительное постоянство внутренней среды при
действии внешних факторов. При действии на растение неблаго-
приятных факторов (стрессоров) в нем возникает напряженное
состояние, отклонение от нормы. Ответные реакции, индуцируе-
мые в организме любыми внешними воздействиями, часто называ-
ют «адаптационным синдромом» или «стрессом» (от англ, stress —
напряжение). В физиологии стресс — это комплекс неспецифи-
ческих изменений, возникающих в организме в ответ на дей-
ствие неблагоприятных факторов (В. В. Полевой и др., 2001).
Реакция клеток на неблагоприятные воздействия сходна: повыша-
ется проницаемость мембраны, отмечаются изменения в ядре, бел-
ки денатурируются, цитоплазма коагулирует и т.п. Неспецифич-
ность этих признаков выражается в том, что они сопутствуют раз-
ным повреждениям и наблюдаются у клеток любых тканей и однокле-
точных организмов. Этот комплекс неспецифических физико-хи-
мических признаков повреждения был назван паранекротическим
(«вблизи смерти»). При нормализации условий эти изменения мо-
гут полностью исчезнуть, если повреждение не было радикальным.
В соответствии с созданной в 1930-е гг. канадским физиологом Г. Се-
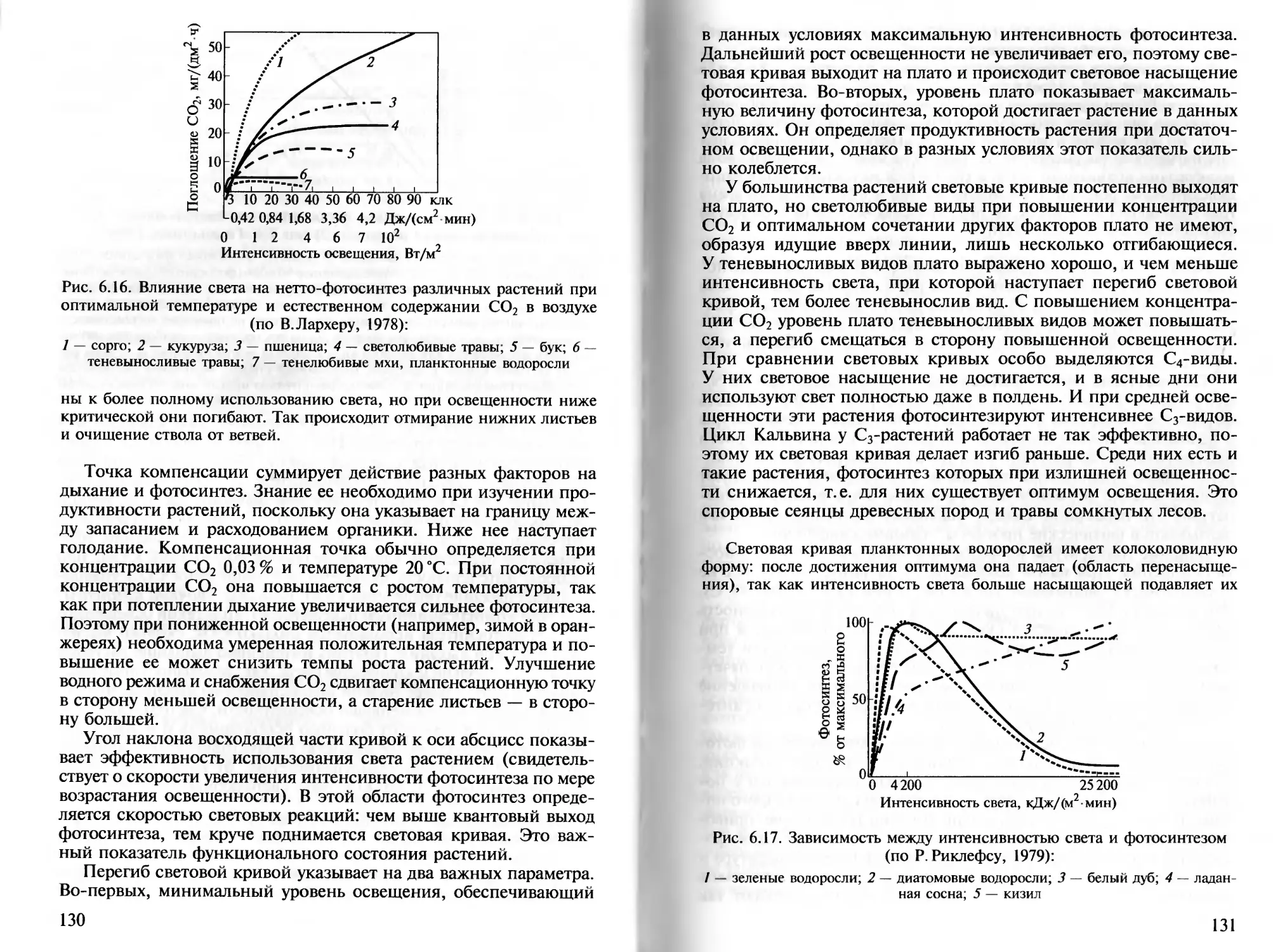
лье теорией общего адаптационного синдрома (стресса), у растений можно
32
выделить три фазы ответной реакции на действие стрессора: первичную
стрессовую реакцию, адаптацию и истощение ресурсов надежности. Так,
листья проростков фасоли (Phaseolus), обдуваемые феном (+38 °C), че-
рез 12 — 30 мин опускаются из-за завядания (фаза первичной реакции),
а затем, несмотря на продолжение обдува, поднимаются (фаза адапта-
ции) (Л. Г.Косулина и др., 1993). Можно также выделить три группы
факторов, вызывающих стресс у растений (В. В. Полевой, 1989): 1) физи-
ческие с । рессоры - недостаточная или избыточная влажность, освещен-
ность, темпера гура, радиоактивное излучение, механические воздействия;
2) химические стрессоры — некоторые соли, газы и другие вещества
(в том числе разные ксенобиотики — пестициды, промышленные отхо-
ды и др ); 1) биологические стрессоры — возбудители болезней, живот-
ные — вредители, конкуренты.
Любые стрессоры вызывают общие изменения процессов об-
мена веществ. У растительных организмов в отличие от животных
реакция на стресс проявляется не в активации метаболизма, а,
наоборот, в снижении функциональной активности. Активность
клеток тормозится в ре вульгате действия ингибиторов и переклю-
чения HicpieiiriccKiix ресурсов на преодоление неблагоприятных
сдвигов. Очень \арлк1грны торможение синтеза гормонов роста и
усиление обрл юнлпия иш нбн торов, вызывающие замедление ро-
ста pacieiiiin через снижение скорости деления и роста клеток.
При cipcrcc у рл» И нин 1лкже ымегно подавляются энергетичес-
кие прощт » i.i < ipr< < IHIIJI! о!|к*1 спя tan и с нарушением функци-
онирования мсмг»р.1п oim< член я чю у высших растений увели-
чивается нч пропинаемо» и. нроисхо ин деполяризация илазма-
леммы. При ном на t|M»iie ш iiiB.iiiiiii ( Hine ia специальных стрес-
совых белков и виноыгн ч общее снижение синтетических про-
цессов. ()дно1»||1 менно у< и.. процессы гидролиза и закис-
ление ниloii'iа 1мы (В В Полевой и др., 2001). Все эти изменения
развиваюн я > лг к.к i ллные процессы, позволяя мобилизовать ре-
зервы । in fn.it । pin о общею ответа на действие раздражителя.
В невысо* нч ло1л\ повторяющиеся стрессы способствуют закали-
ванию Opl ЛИИ IM.I
У видов, устойчивых и неустойчивых к стрессу, выработались
разни» । омненелгорные механизмы, обеспечивающие жизнеспо-
собн»м и. организма в экстремальных условиях. Наиболее общими
НОКЛ1.1Н* 1ями жишенного состояния на организменном уровне
ЯВИЯ1О11 я биомасса и скорость ее накопления и соответственно у
мллоу» юнчивых к стрессу растений в неблагоприятных условиях
ум» । напас гея площадь ассимилирующей поверхности, а устойчи-
вые виды могут увеличивать листовую площадь, приходящуюся
и.। с шпицу массы (поэтому они дольше сохраняют и меньше сни-
жаю! онюсительную скорость роста).
11рисн()собления растительного мира к стрессовым воздействи-
ям протекают на разных уровнях организации — клеточном, орга-
33
низменном, популяционном и биоценотическом. В пределах од-
ного генотипа проявляется существенная фенотипическая плас-
тичность: возможны несколько морфологических, физиологиче-
ских и поведенческих состояний. Организмы изменяют активность
синтеза определенных соединений, включают физиологическую
пластичность, формируют определенные анатомо-морфологиче-
ские образования. Популяции используют поливариантность он-
тогенеза, корректируют параметры участия в сообществе, особен-
ности возобновления, стратегии выживания и т.п.
Соотношение специфических и неспецифических ответных ре-
акций. В ответных реакциях на повреждающие факторы выделя-
ют элементы неспецифической устойчивости (включающиеся в
самых разных стрессовых ситуациях) и специфические процессы,
инициируемые в растении только определенным типом стрессо-
вых воздействий. На формирование неспецифических элементов
устойчивости (синтез белков теплового шока, полиаминов) тре-
буется гораздо меньше времени, чем для прохождения специфи-
ческих адаптивных реакций (синтез белков-антифризов, переклю-
чение фотосинтеза на CAM-путь и др.) (С. С. Медведев, 2004).
Специфическое реагирование на действие экстремальных факто-
ров .контролируется генетическими механизмами через работу бе-
локсинтезирующего аппарата. В основе неспецифического реаги-
рования лежит физиологическая пластичность (изменение струк-
туры и активности клеточных белков, пластичность мембранных
компонентов и пр.).
Неспецифические приспособления считаются основными и ярко
проявляются на молекулярном и клеточном уровнях. Но уже на
них можно зафиксировать и зависимость изменений от характера
повреждающего агента, что служит основой специфических приспо-
соблений. Они происходят на всех уровнях организации, но более
выражены на высоких (уровень тканей, органов, организмов). При
специфических реакциях неспецифический ответ на низших уров-
нях организации корректируется высшими ступенями.
Соотношение специфических и неспецифических ответных
реакций в значительной степени зависит от длительности дей-
ствия фактора. При кратковременном воздействии высокой дозы
неблагоприятного фактора в основном наблюдаются нсспецифи-
ческие реакции. При длительном же действии срабатывает боль-
шее число метаболических звеньев, часть которых специфична
для данного организма. Постепенное нарастание действия стрес-
сора приводит к включению процессов специализированной адап-
тации, обеспечивающих надежность внутриклеточных процессов.
Соотношение специфических и неспецифических ответов зависит от
биологических особенностей объекта. Например, у огурца (Cucumis
sativus) при низких температурах в основном задействованы определяе-
34
мне геномом специфические ответные реакции, так как стабильные тем-
пературы на его родине в Юго-Восточной Азии не обеспечили форми-
рования пластичности обмена. А у пшеницы (Triticum) становление рода
шло на фоне существенных колебаний температуры и других факторов,
поэтому на своем обширном ареале (от Северного полярного круга до
юга Африки и Америки) она хорошо приспособлена к очень разным
условиям. Залог ее широкого распространения — развитая система спе-
цифического реагирования, подкрепленная механизмами неспецифичес-
кой устойчивости. Эволюция пшеницы шла путем выработки механиз-
мов лабильности мембранных компонентов, пластичности регуляторных
механизмов, подвижности структуры и функции внутриклеточных бел-
ков, что и позволило этому важному хлебному злаку иметь такой широ-
кий ареал (Л. Г.Косулина и др., 1993).
Во всех случаях провести резкую границу между специфичес-
кими и нсспецифическими реакциями невозможно. Неспецифич-
пость многих признаков повреждения не абсолютна. На фоне од-
нотипных нсспецифических реакций обычно выявляются и спе-
цифические, поэтому, вероятно, имеет место не тождество явле-
ний, а ли...сходство. В целом для всех организмов характерна
способность в ответ на внешние воздействия эффективно соче-
i,in. |Х‘акции исснсцифичсскис (в значительной мере не завися-
щие «и прпро лы по действующего фактора) со специфическими.
A'HiiiiiHiiiMr 1111ци111о-11|)иснособителы1ые реакции растений.
В iipoin < < 4* .Ы.1Н1.1ННИ ирешеходит «подгонка» свойств живых объек-
р)П ► у 1О1ЦН1М i унпт Hioii.iiiiHi, 4)(кч'испивающая непрерывное по-
г |ум п ни» и» оЬм» ।ими\ р4ч vpi он Адаптация и гомеостаз — взаимо-
||||||(»||ни|о||Н1<* чн н ния I’riyuhi.ii многих адаптивных стратегий —
Ho/i/irpA.iiiiii' IOMCO4 она по при ном во многих случаях поддер-
ФПВ.1ГЦ .Я не (огюинпг (. грукгур, а их функция. Так, при действии
icMiicpaiyp нязкоегь биомембрап изменяется, но таким образом,
чюбы могли норма»....о функционировать связанные с ними фер-
мсигы. грапснортныс системы, рецепторные молекулы и др.
Временные параметры адаптации варьируют в широких преде-
1Л\ oi длительных периодов, необходимых для эволюционных
и 1МСПСПИЙ в аминокислотных последовательностях, до долей се-
кунды, ta которые изменяется активность уже присутствующих в
клетке ферментов (Л. Г. Косулина и др., 1993). Чем больше време-
ни для изменений, тем больше выбор возможных адаптивных стра-
тегий. Эволюционная (генетическая) адаптация связана с пере-
стройкой генома. Это длительный процесс, затрагивающий мно-
гие поколения. Он зависит от приобретения новой генетической
информации и определяет новые признаки фенотипа, в том чис-
ле позволяющие организму осваивать ранее недоступные место-
обитания. Данные изменения связаны с накоплением мутаций:
изменением числа хромосом (геномные мутации), изменением их
структуры (хромосомные мутации), изменением отдельных генов
35
(генные мутации), мутациями во внеядерном генетическом мате-
риале пластид и митохондрий. Кроме того, мобильные генетичес-
кие элементы, перемещающиеся в геноме или между ними, а так-
же взаимодействующие с растениями вирусы и бактерии могут
изменять их наследственные свойства. В частности, заражение
агробактериями (Agrobacterium tumefaciens) индуцирует образо-
вание корончатых галлов, что вызывает перенос бактериальных
ДНК в геном растений.
В ответ на действие среды могут происходить и более быстрые
ненаследственные модификации признаков, расширяющие воз-
можность вида успешно произрастать в разнообразных условиях.
К такому типу адаптаций относятся модификации на уровне от-
дельных организмов, органов и тканей при средней скорости от-
вета (в течение месяцев и годов), а также на уровне клетки (моле-
кул, органоидов) с еще более быстрым ответом (дни, часы, мину-
ты). Примером реакции средней по времени ответа может быть
акклиматизация — приспособление к изменяющимся условиям
среды при выращивании растений в непривычных климатических
условиях. Таковы, например, изменения сезонного развития рас-
тений при интродукции. Этот тип адаптации формируется от не-
скольких часов до нескольких месяцев и завершается при жизни
организма. Приспособление в этом случае обеспечивает уже со-
державшаяся в геноме информация.
Быстрая адаптация осуществляется путем изменения активно-
сти уже имеющихся ферментов и представляет собой «первую
линию защиты» организма от неблагоприятных факторов среды.
Пример — таксисы (направленные перемещения по отношению
к источнику действия). Так, хлоропласты имеют отрицательный
фототаксис к яркому свету и положительный — к слабому.
В адаптивные процессы вовлечены все уровни организации жи-
вого. Так, многие растения избегают перегрева под пологом других
видов (биоцснотичсский уровень адаптации). Охлаждение путем
транспирации — адаптация на уровне организма. Пристеночное
положение хлоропластов при сильном освещении — клеточная адап-
тация. А высокая тсрмоустойчивость белков у термофильных орга-
низмов — проявление адаптации на уровне молекулярном. Благо-
даря многообразию приспособительных реакций в процессе эво-
люции сформировалось большое разнообразие растений, заселив-
ших разные регионы и местообитания. Приспособление к небла-
гоприятным условиям привело к тому, что в ряде случаев они ста-
ли необходимы для жизни этих организмов. Например, растения-
галофиты лучше развиваются на засоленном субстрате.
В целом специализированная адаптация обеспечивает жизне-
деятельность растений в неблагоприятных условиях за счет мно-
гих процессов: изменения экспрессии генов и новообразования
макромолекул (стрессовых белков и др.), повышения уровня про-
36
текторных соединений (пролина, полиаминов и др.), изменения
гормонального баланса и ростовых процессов (торможение роста,
регенерационная замена поврежденных органов, рост почек и
новых корней, образование аэренхимы и др.), стабилизации ре-
зервных возможностей организма при изменении уровня и на-
правленности физиологических процессов, за счет повышения спо-
собности не допускать или ликвидировать повреждения, обеспе-
чения разнообразных связей между клетками, тканями и органа-
ми в целом организме (Л. Г. Косулина и др., 1993).
Репаративные способности растений. Действие повреждающих
факторов всегда усиливает расщепление сложных соединений, но
это вызывает или усиливает реакцию ресинтеза. Поэтому уже во
время действия повреждающего фактора (и особенно после него)
репарационные биосинтетические процессы усиливаются: идет
приспособительный синтез структурных и ферментных белков,
восстанавливаются запасы энергии, расходуемые во время дей-
ствия неблагоприятных факторов. Репаративные способности —
важная составляющая устойчивости организмов. Именно за счет
их растения могут переносить воздействие многих экстремальных
условий. Выживаемость растений и способность сохранять про-
дуктивность в экстремальных условиях во многом зависят от на-
дежности их репаративных систем.
Репарация — восстановление, ликвидация повреждений. Сис-
темы восстановления работают на разных уровнях — молекуляр-
ном, клеточном, организменном, популяционном и биоценоти-
ческом. Репаративные процессы, связанные с постоянным вос-
созданием нарушенных структур, идут с затратой энергии и мате-
риалов, поэтому как только обмен веществ прекращается, пре-
кращается и репарация. В экстремальных условиях особенно важ-
но сохранение дыхания, дающего энергию для репарации.
Количественно репаративные способности характеризуются ско-
ростью восстановления поврежденной функции и величиной ре-
паративной зоны (диапазоном действия фактора, вызывающим
обратимые изменения). Чем сильнее повреждение, тем медленнее
репарация. Например, после 5-минутного прогрева при 45 °C дви-
жение цитоплазмы в клетках эпидермы листа колокольчика вос-
станавливается на второй день, а после прогрева при 50 °C — на
пятый. Скорость репарации у разных объектов неодинаковая. Раз-
мер репаративной зоны зависит от объекта и длительности дей-
ствия фактора. Так, для листьев традесканции (Tradescantia) при
5-минутном нагреве минимальная температура прекращения дви-
жения цитоплазмы составляет 45 °C, а максимальная, обратимо
останавливающая ее движение, достигает 52 °C, т.е. репаративная
зона составляет 7 °C. А у клеток эпидермы листа шелковицы
(Morus) при 5-минутном прогреве репаративная зона 5°C (от 46
до 51 °C), а при 640-минутном — всего 1,3 °C (от 42,7 до 44 °C).
37
Репаративные зоны разных физиологических процессов могут су-
щественно отличаться (Л. Г. Косулина и др., 1993).
Репаративные способности клеток адаптированных организмов
во многом определяются устойчивостью их белков. Большое зна-
чение имеют слабые связи, нарушаемые при денатурации. Так, ус-
тойчивость спор и семян к экстремальным температурам обычно
объясняется снижением денатурации их белков после обезвожива-
ния. А поскольку основным поставщиком энергетического мате-
риала у растений является фотосинтез, то от устойчивости фото-
синтетического аппарата и его способности восстанавливаться после
повреждений сильно зависят энергообеспечение клетки и связан-
ные с ним процессы, в том числе репаративные. В крайних услови-
ях растения активизируют системы, обеспечивающие синтез ком-
понентов мембран тилакоидов, торможение автоокисления липи-
дов, поддержание и восстановление ультраструктуры пластид.
Сопряженная устойчивость. Во многих случаях закаливание
по отношению к одному неблагоприятному фактору способству-
ет повышению устойчивости организма и к некоторым другим
(кросс-устойчивость или кросс-адаптация) (С. С. Медведев, 2004).
П. А. Генкель (1982) сформулировал это в положении о сопряжен-
ной* устойчивости. Она может быть положительной и отрица-
тельной. Например, предпосевная обработка хлоридом кальция
семян проса (Panicum), повышающая его жароустойчивость, мо-
жет сопровождаться повышением его солеустойчивости (положи-
тельная сопряженная устойчивость). А увеличивающая засухоус-
тойчивость обработка семян табака (Nicotiana) понижает его ус-
тойчивость к хлоридам (отрицательная сопряженная устойчивость),
но увеличивает сульфатоустойчивость (положительная сопряжен-
ная устойчивость).
Сходство в реакциях растений на ряд неблагоприятных факто-
ров среды (например, холод и тепло), наличие положительной
сопряженной устойчивости свидетельствуют о том, что резистен-
тность к разным экстремальным воздействиям может контроли-
роваться одними и теми же внутренними факторами. Существен
вклад в этот эффект множества неспецифических приспособи-
тельных реакций. И специфическое реагирование на разные вне-
шние воздействия осуществляется по единому типу генетической
регуляции физиологических процессов. Так, сходство ответных
реакций растений на температурный, водный и солевой стресс,
по-видимому, определяется тем, что в этих условиях в клетках
создается водный дефицит, который может быть устранен с по-
мощью однотипных защитно-приспособительных процессов.
Ответные реакции растений на разные факторы носят колебатель-
ный характер. Например, при действии высокой температуры вязкость
цитоплазмы сначала уменьшается, а затем увеличивается. Скорость дви-
38
жсния цитоплазмы и выход веществ из клетки также меняются волно-
образно: сначала наблюдается усиление этих процессов, а затем они за-
медляются. В зависимости от силы повреждающего воздействия меняется
характер этих колебаний (амплитуда, длина волны, время пуска ответной
реакции). Колебательный характер клеточных процессов при действии раз-
дражителей отражает сложную природу ответных реакций, которые име-
ют разную направленность: одни имеют деструктивный характер, а другие
направлены на сохранение внутриклеточных структур и процессов.
Приспособительные изменения растений на организменном и
популяционном уровнях. На организменном уровне к клеточным
механизмам адаптации добавляются новые, отражающие взаимо-
действие органов. В неблагоприятных условиях растения создают
и сохраняют минимум генеративных органов, но в достаточной
мере обеспеченных необходимыми веществами для нормального
созревания, достижения обычных размеров и качества. Так, более
половины заложившихся завязей в соцветиях культурных злаков
и в кронах культурных плодовых деревьев могут редуцироваться и
опасть. В неблагоприятных условиях резко ускоряются процессы
старения и опадения нижних листьев. При этом нужные растени-
ям вещества перемещаются в молодые органы. Идет замена и ре-
генерация поврежденных или утраченных частей растения, акти-
визируется рост пазушных почек.
В процессах защиты, адаптации и репарации растений участвуют
все системы их регуляции (генетическая, гормональная, электро-
физиологическая и др.). Так, при действии неблагоприятных ус-
ловий в растении резко возрастает содержание этилена и абсци-
зовой кислоты, которые снижают обмен веществ, тормозят росто-
вые процессы, способствуют старению и опадению органов, пе-
реходу растения в состояние покоя. Одновременно в тканях сни-
жается содержание ауксина, цитокинина и гиббереллинов. Эта
стереотипная реакция на экстремальные условия, при которой
ведущую роль играют гормоны, тормозящие функциональную
активность, очень характерна для растений (В. В. Полевой, 1989).
При этом изменения затрагивают весь организм.
При длительном и сильном стрессе быстрее гибнут те растения,
у которых уже норма реакции на экстремальные факторы. В резуль-
тате семенное потомство образуют лишь генетически более устой-
чивые к стрессу растения и общий уровень устойчивости в популя-
ции возрастает. Таким образом, на популяционном уровне в стрессо-
вую реакцию включается отбор, приводящий к появлению более
приспособленных организмов. Предпосылкой этому служит внутри-
популяционная изменчивость устойчивости по разным факторам.
Уровень устойчивости популяции (сорта) к тому или иному
фактору или их группе определяется устойчивостью составляю-
щих популяцию организмов. Наиболее устойчивые сорта имеют в
своем составе ряд биотипов, обеспечивающих хорошую продук-
39
тивность даже в неблагоприятных условиях. Приспособительные
изменения растений связаны также с разными типами гетероген-
ности популяций — возрастной, половой и др.
4.2. Реакции клеток растений на действие
неблагоприятных факторов
Антиокислительная система растительной клетки. Накоплен боль-
шой экспериментальный материал о важности окислительных про-
цессов в мембранах при разных внешних воздействиях (экстремаль-
ном освещении и температуре, дефиците влаги и др.). Во многих
стрессовых ситуациях они развиваются на ранних этапах патологи-
ческих процессов. Из-за перекисного окисления липидов у чув-
ствительных к стрессу растений могут разрушаться хлоропласты,
подавляться фотосинтез и гибнуть клетки. Окислительная деграда-
ция полиненасыщенных жирных кислот, хлорофиллов и накопле-
ние токсичных продуктов перекисного окисления липидов отмеча-
ется у неадаптированных форм растений при интенсивном осве-
щении, переохлаждении и т.п. Инфекция тоже активирует пере-
кисное окисление липидов клетки, в результате чего патоген лока-
лизуется и его распространение ограничивается.
В растительных клетках существует эффективная антиокисли-
тельная система, обеспечивающая защиту клетки при действии
многих факторов среды. Устойчивость растений к повреждающим
агентам во многом зависит от детоксикации активных форм кис-
лорода и свободных радикалов. Обезвреживание осуществляет ряд
ферментов. Так, концентрацию супероксидных радикалов эффек-
тивно понижает супероксиддисмутаза, обнаруженная практиче-
ски во всех тканях цветковых растений, у некоторых голосемен-
ных, мхов и водорослей. Активность ее выявлена в цитозоле, ми-
тохондриях, пероксисомах, хлоропластах. В ряде случаев подтвер-
ждена связь между ее активностью и устойчивостью клеток к фо-
топовреждению. А каталаза и пероксидазы препятствуют накоп-
лению токсичных перекисей.
Важные антиоксиданты — витамины Е, С и каротиноиды. Во-
дорастворимый витамин С и жирорастворимый Е имеют слабые
О—Н-связи и поэтому являются активными ловушками радика-
лов. В присутствии супероксиддисмутазы они предотвращают вы-
цветание пигментов и переокисление липидов. Каротиноиды же
благодаря своей относительной стабильности выступают в каче-
стве защитных пигментов, предохраняя светочувствительные хло-
рофиллы от фотоокислительных разрушений.
Роль состава мембран в устойчивости клеток. Большую роль в
адаптации организмов играют клеточные мембраны. Характерная
их черта — способность к структурным изменениям, которые мо-
40
гут переключать клетку из одного метаболического состояния в
другое. Мембраны определяют активность и чувствительность ре-
цепторов, а также через свои перестройки влияют на изменение
метаболизма клеток.
Свойства мембран во многом обусловлены взаимодействием
белковых компонентов с липидным матриксом, обеспечивающим
вязкость мембран. Количественные соотношения разных типов
липидов и их жирно-кислотный состав определяют стабильность
и устойчивость мембран. Уровень подвижности углеводородных
цепей в липидах важен и для функционирования ферментных
систем и рецепторов, обеспечения проницаемости мембран.
Для большинства растений зафиксировано увеличение доли нена-
сыщенных жирных кислот при снижении температуры, что прида-
ет мембранам повышенную текучесть, препятствуя повреждению
растений холодом и морозом. Вообще экстремальные температуры
приводят к избыточному затвердению или разжижению жирных
кислот (при повышении температуры увеличивается доля туго-
плавких насыщенных жирных кислот, а при переохлаждении —
наоборот) и в некоторых случаях это определяет тсрмоустойчивость
клеток.
Клетки способны адаптивно изменять состав жирных кислот
в липидах и соотношение липидных фракций в ответ на измене-
ние условий среды. Это может достигаться разными путями: из-
менением скорости их синтеза и распада, изменением подбора
включаемых в мембрану жирных кислот, разной активностью фер-
ментов, катализирующих образование двойных связей (Л. Г. Косу-
лина и др., 1993). В целом гибкость мембран определяется соста-
вом и количеством ненасыщенных жирных кислот, а также нали-
чием белков с высокой подвижностью. Благодаря своей эластич-
ности мембраны сохраняют способность к обратимому сокраще-
нию и набуханию в широком диапазоне действия факторов.
Биохимическая адаптация клеток растений. Сохранение кле-
точного гомеостаза при постоянных изменениях условий окружа-
ющей среды определяется адаптивными возможностями клеток.
Процессы биохимической адаптации выполняют в клетке следу-
ющие функции: 1) поддержание структурной целостности макро-
молекул при их функционировании в специфических условиях;
2) достаточное снабжение клетки «энергетической валютой» (АТФ),
восстановителями и предшественниками для синтеза белков и
нуклеиновых кислот; 3) поддержание систем, регулирующих ме-
таболизм в соответствии с потребностями организма и их измене-
ниями при изменении условий среды.
Основные направления адаптации клетки, связанные с макро-
молекулами, следующие (по Л. Г. Косули ной и др., 1993): 1) изме-
нение количества или типов макромолекул, имеющихся в клетке;
2) регулирование функциональной активности макромолекул;
41
3) изменение среды, позволяющее защитить макромолекулы от
действия неблагоприятных факторов. Комбинация этих механиз-
мов позволяет настраивать клеточный метаболизм так, чтобы орга-
низм имел возможность непрерывно получать необходимые про-
дукты и энергию. В целом качественный и количественный со-
став белков организма определяется его геномом, а интенсивность
их синтеза — условиями среды (рис. 4.1). При этом состояние
ферментных систем и изменение концентрации субстратов и про-
дуктов реакций регулируют расходование и синтез белков клетки.
Синтез ряда белков часто ограничивается репрессированием уча-
стков генома или усиливается в результате индукции или дереп-
рессии, вызванных повышением активности клетки или повреж-
дающим воздействием. Изменение активности ферментов, а так-
же усиление их синтеза могут быть вызваны усилением расщепле-
ния (и расходования) веществ при изменении концентрации про-
дуктов обмена и условий существования.
Повреждающие факторы различно действуют на ферменты. При
их активизации усиливается расходование источников энергии, а
освободившиеся из клеточных структур белки подвергаются рас-
щеплению. Расщепляться могут и сами ферменты. Это приводит
к возрастанию концентрации метаболитов, которые нередко слу-
жат активаторами генетического аппарата. Они способствуют от-
щеплению от гена репрессоров, ограничивавших синтез белков, и
запускают их биосинтез. В результате усиливается построение
структурных и ферментных белков, восстанавливаются клеточные
Рис. 4.1. Расщепление и синтез белков при действии повреждающих
факторов среды (по Н. Н.Яковлеву, 1986).
Утолщенные стрелки — синтез, тонкие — влияние пути синтеза белков, штрихо-
вые — их расщепление
42
структуры. При этом может увеличиваться количество фермен-
тов, готовых функционировать в усиленном режиме, число кле-
точных структур (например, митохондрий) и в целом возрастать
функциональная мощность клетки, органа и организма в целом.
Очень важна активация синтеза стрессорных белков при одно-
временном ослаблении синтеза других, образуемых в нормальных
условиях. Синтез стрессовых белков в ответ на сигналы внешней
среды — универсальный механизм быстрой адаптации организмов.
Так, у многих растений выявлены белки теплового шока. У ку-
курузы (Zea mays), например, их синтез индуцируется температу-
рой +45 °C. Полупериод жизни этих белков около 20 ч, в течение
которых клетка сохраняет терморезистентность. Некоторые из них
уже существовали в цитоплазме и в условиях стресса активирова-
лись. В ядре клетки белки теплового шока связывают матрицы
хроматина, необходимые для нормального метаболизма, а по окон-
чании стрессового состояния эти матрицы освобождаются и вновь
начинают функционировать. Еще одна важная функция стрессо-
вых белков — структурная: они стабилизируют мембраны и со-
держимое клеток в условиях стресса.
Приспособительный рост синтеза белков и увеличение содер-
жания их в клетке связаны прежде всего с деятельностью генети-
ческого аппарата. Но ряд свойств белков изменяется уже после
синтеза их первичной структуры. При этом отдельные аминокис-
лоты превращаются в производные, в геноме не кодируемые (на-
пример, пролин окисляется в оксипролин). Изменяется также
форма (и свойства) белков при трансформации их под влиянием
сдвигов во внутренней среде организма (изменение pH, концент-
рации ионов, субстратов и продуктов реакций, действие актива-
торов и ингибиторов). Большое значение имеет активирование
протеинкиназ, которые, присоединяя фосфат, изменяют структу-
ру и активность белков, в свою очередь активирующих другие
ферменты. Биологический смысл этого каскада активации в том,
что он многократно усиливает первоначальный сигнал, индуци-
рующий целый комплекс защитно-приспособительных реакций
(Л. Г. Косулина и др., 1993).
При неблагоприятных условиях в клетках возрастает также со-
держание протекторных веществ (углеводы, аминокислоты), уча-
ствующих в защитных реакциях и стабилизирующих состояние
цитоплазмы. Например, при водном дефиците и засолении у яч-
меня (Hordeum), шпината (Spinacia), хлопчатника (Gossypium)
концентрация пролина в цитоплазме возрастает в 100 раз и более.
Благодаря гидрофильным группам он хорошо растворяется и спо-
собен связываться с поверхностными гидрофильными остатками
белков, что повышает их растворимость и защищает от денатура-
ции. Накопление осмотически активного пролина благоприятствует
удержанию веществ в клетке (В. В. Полевой, 1989).
ГЛАВА 5
ЭКОЛОГИЧЕСКАЯ ГЕТЕРОГЕННОСТЬ
РАСТЕНИЙ
Каждое растение индивидуально в отношении требовательно-
сти к совокупности экологических факторов. Нет двух видов с
абсолютно одинаковыми стратегиями жизни, каждый приспосаб-
ливается к экологическим условиям по-своему. Однако несмотря
на признанную биологами экологическую индивидуальность, орга-
низмы со сходной экологией (жизненными формами, ареалами и
др.) можно объединить в группы. Они характеризуются некото-
рыми общими существенными свойствами, хотя все экологиче-
ское своеобразие при этом не учитывается. Это удобно для эколо-
гических обобщений, поэтому обычно каждый экологический
признак растений характеризуется именно такими упрощенными
системами типов, сводящими реальное многообразие до некото-
рого количества условных классификационных единиц, между
которыми неизбежны переходы.
5.1. Экологические группы растений
Основные подходы к изучению экологии видов. Поскольку от-
ношение разных видов к факторам среды индивидуально, то наи-
более последовательным методом изучения экологии видов долж-
на быть ординация — упорядочение по положению их оптимумов
вдоль осей изменения экологических факторов. Этот вариант оцен-
ки экологии видов базируется на признании экологической ин-
дивидуальности и континуума в экологических свойствах видов.
Континуальные результаты ординационного анализа часто пре-
образуются в более простую и удобную систему дискретных еди-
ниц, какими являются экологические группы.
Экологическая группа — это группа растений, сходно реаги-
рующих на действие какого-либо фактора среды (освещенность,
влажность и т.п.). При выделении экологических групп обычно
выбирают ведущий (наиболее важный в данных условиях) фактор
и весь градиент его разбивают на ступени. В экологические же
группы объединяют виды, наиболее часто встречающиеся в выде-
44
ленном диапазоне условий. При этом не важно, насколько широ-
ко они вообще распределены по градиенту (живут только в таких
условиях или и во множестве других). Важно лишь то, что обычно
они встречаются при данном значении фактора. К одинаковым
условиям растения могут приспосабливаться по-разному, поэто-
му одна экологическая группа может объединять организмы раз-
ных жизненных форм (например, среди растений засушливых мест
есть и деревья, и травы, и полукустарники). Классические представ-
ления об экологических группах, разрабатывавшиеся в экологи-
ческой ботанике более полутора столетий (от А. Гризебаха до
А. П. Щенникова, Г. Вальтера и др.), оказались очень плодотвор-
ными для понимания жизни растений и их разнообразия.
В целом главный способ оценки экологии видов состоит в учете
их распределения по разным местообитаниям. А для выявления
нюансов экологических свойств растений можно провести допол-
нительные физиологические исследования. Полезные сведения дает
определение типа фотосинтеза, осмотического давления клеточно-
го сока, суточных циклов транспирации и оводненности тканей, а
также учет анатомических свойств, помогающих экономить влагу
(редукция листа, суккулентность, развитость эпидермы) и др.
Экологические группы и экологические шкалы. Экологические
шкалы представляют собой таблицы с экологическими характе-
ристиками видов. По описанию растительности они позволяют
определить условия произрастания растений. Эти шкалы нашли
применение в решении разных экологических задач (биоиндика-
ция, экологическое картирование и др.). В России наиболее изве-
стны экологические шкалы Л. Г. Раменского (1956), разработан-
ные для экологической оценки сельхозугодий. Они содержат све-
дения более чем по 1,5 тыс. видов растений европейской части
СССР. При составлении каждой шкалы подбиралось большое
количество геоботанических описаний площадок из местообита-
ний, характеризующихся разными значениями данного экологи-
ческого фактора. На оси изменения фактора устанавливались ди-
апазоны распределения видов при разном их обилии, а также оп-
тимальные для видов части градиента (где их участие в сообще-
ствах наибольшее). Последователи Л. Г. Раменского составили ана-
логичные шкалы для других регионов (Средняя Азия, Урал, Си-
бирь, Кавказ, Памир, Карпаты, Балканы).
В экологических шкалах Л. Г. Раменского экология каждого вида
оценена по пяти факторам среды: увлажнению, богатству почвы,
пастбищной дигрессии, аллювиальности и переменности увлаж-
нения. По этим осям ординации автор использовал большое чис-
ло ступеней: 120 ступеней для увлажнения, по 30 для богатства и
засоления почвы, 20 для переменности водного режима и по 10
для аллювиальности и пастбищной дигрессии. Экологические груп-
пы Л. Г. Раменский не выделял, но охарактеризованные его шка-
45
лами виды можно разбить на группы по приуроченности их диапа-
зона распределения (при максимальном обилии) к разным частям
градиента. Так, по оси увлажнения Л. Г. Раменский объединил 120
ступеней в 12 более крупных градаций, соответствующих место-
обитаниям с разным типом увлажнения, и по приуроченности к
ним можно выделить экологические группы (например, ступени
18 — 30 соответствуют полупустынному увлажнению и гиперксеро-
фитам, а ступени 94—103 — болотному увлажнению и гигрофи-
там). Экологические шкалы показывают также, что единично рас-
тение может встречаться в разных условиях, но массово — только в
узком (близком к оптимальному) диапазоне значения фактора.
Для Центральной Европы экологические шкалы, основанные
на многолетних наблюдениях и экспериментах, были составлены
Г.Элленбергом. В них охарактеризовано более 2 700 видов и внут-
ривидовых таксонов растений по отношению к освещению, теп-
ловому режиму, континентальности климата, увлажнению, кис-
лотности почвы, засолению и обеспеченности доступным азотом.
Отношение видов к увлажнению оценивается 12 баллами, осталь-
ных параметров — 9. Отдельно отмечено отношение растений к
переменности увлажнения и затоплению. В 1991 г. Г.Элленберг с
соавторами опубликовал экологические шкалы для мохообразных
и лишайников.
Сходно составлены и популярные в Европе экологические
шкалы Э.Ландольта (1977). Но в них на осях факторов среды вы-
деляется небольшое число градаций и виды сразу объединяются в
экологические группы. Так, градиенты увлажнения и богатства
почвы азотом имеют по пять ступеней, а у шкалы засоления всего
две ступени. Отдельно составлены шкалы для кислотности по-
чвы, температурного режима, континентальности климата, осве-
щенности, структурности почвы и ее гумусированности.
5.2. Жизненные формы растений
Подходы к пониманию жизненных форм растений. В природе на
растение всегда воздействует сумма факторов, взаимодействую-
щих и модифицирующих проявления друг друга. Приспосаблива-
ясь к условиям жизни, растения эволюционно выработали вне-
шний облик, внутреннее строение, физиологические и биологи-
ческие особенности, наиболее отвечающие окружающей их об-
становке. В результате в природе сформировались группы расте-
ний, соответствующие разнообразным вариантам условий среды.
Для их обозначения датский ботаник Е. Варминг в 80-х гг. XIX в.
ввел понятие жизненная форма.
Идея классифицировать растения по внешнему виду берет на-
чало от А. Гумбольдта, который еще в начале XIX в. обратил внима-
46
ние на связь ландшафтов и внешнего облика господствующих в
них растений. Е. Варминг разрабатывал эту идею в онтогенетиче-
ском аспекте (растение от прорастания семени до отмирания име-
ет форму, гармонирующую со средой). В дальнейшем под жизнен-
ной формой стали понимать выраженное во внешнем облике и
внутреннем строении организмов приспособление их ко всему ком-
плексу условий, складывающихся в определенных типах местооби-
таний. Изучение же жизненных форм растений шло в основном в
трех направлениях: 1) выявление и классификация их многообра-
зия; 2) исследование связи жизненных форм с условиями среды;
3) построение морфогенетических рядов жизненных форм.
Существуют два основных подхода к пониманию жизненных
форм. В одном случае акцент делается на морфологическом ас-
пекте, а в другом жизненные формы понимаются шире — как
морфолого-физиологические типы растений. Структурные при-
знаки в качестве главных критериев приспособленности растения
к среде рассматриваются во многих системах жизненных форм, и
иногда само понятие жизненной формы определяют как «струк-
турный тип растений». Однако они не отражают всех сторон адап-
тации растений к условиям местообитания, так как приспособле-
ние к среде включает также адаптивные изменения физиологи-
ческих и биохимических процессов, ритмов развития и т.д.
Эколого-физиологический аспект в процессе формирования
жизненных форм выражен очень ярко, но предпочтение струк-
турным признакам часто отдается из-за большей доступности их
наблюдения в природе по сравнению с физиолого-биохимичес-
кими параметрами. Кроме того, внешний вид растений нередко
определяет облик растительных сообществ и целых ландшафтов,
поэтому отражающие его типы жизненных форм востребованы в
фитоценологических, географических и общеэкологических ис-
следованиях.
Жизненные формы и систематические подразделения растений.
Жизненные формы растений, как правило, не связаны с их систе-
матическим положением, так как влияние одних и тех же условий
па представителей различных систематических групп может дать
одинаковый внешний эффект. Например, кактусовидная жизнен-
ная форма (покрытый колючками сочный стебель) характерна для
жарких пустынь, и в сходных условиях ее выработали представи-
тели совершенно неродственных семейств (Кактусовые, Молочай-
ные, Ластовневые и др.) (рис. 5.1). Иногда только во время цвете-
ния можно увидеть, что столь сходные внешне растения относят-
ся к разным семействам. При этом во всех этих семействах есть и
растения с другими жизненными формами. Так, к кактусовым
относятся и несуккулентные облиственные деревья, кустарники,
нодушковидные формы с нормально развитыми широкими лис-
тьями (например, Pereskia).
47
Рис. 5.1. Кактусовидная жизненная форма у растений разных семейств
(по В. В.Алехину, 1950):
1 — Echinocereus blankii (семейство Кактусовые); 2 — Heumia peuzigii (семейство
Ластовневые); 3 — Echinopsis cereiforme (семейство Кактусовые); 4 — Euphorbia
blaumeriana (семейство Молочайные)
Лишь изредка жизненные формы оказываются связанными с систе-
матическими подразделениями растений. Так, в семействе Нимфейные
все представители — водные растения, имеющие округлые плавающие
листья (кубышка — Nuphar, кувшинка — Nymphaea, виктория — Victoria
и др.). Но обычно такого совпадения не бывает, и к одной жизненной
форме относятся растения разных систематических групп.
Хорошо отражают приспособление к среде не любые свойства
организма. Еще в XIX в. предлагалось делить признаки растений
на «организационные» и «приспособительные». Первые (число ча-
стей околоцветника, листорасположение и т.п.) прочно закрепле-
ны в генотипе и не подвергаются быстрым изменениям под влияни-
ем среды. Немного экологической информации несут и признаки,
унаследованные от предковых форм, отражающие приспособление
к условиям прошлых эпох (например, вечнозеленость некоторых
растений ельников). Для выделения жизненных форм более важны
пластичные «приспособительные» признаки, быстро реагирующие
на изменение среды (ритм сезонного развития, защищенность ме-
ристем, вегетативное размножение и т.п.). Именно они в сходной
среде делают похожими систематически отдаленные растения.
48
С другой стороны, как в систематике принадлежность к одной
таксономической единице означает общность происхождения, так
и в экологии отнесение к одной жизненной форме означает оди-
наковый путь приспособления к среде. В целом аналогично тому,
как систематика в качестве основных единиц имеет дело с вида-
ми, жизненные формы неоднократно предлагалось считать основ-
ными единицами в экологии растений, подчеркивая этим адек-
ватность отражения ими связи растений со средой.
Классификации жизненных форм растений. В XIX—XX вв. в
экологической ботанике была предложена серия экологических
классификаций растений, акцентировавших внимание на разных
признаках растений. И. Г.Серебряков (1962) разделил их в соот-
ветствии с разными подходами к классификации жизненных форм
на эколого-физиономические и морфолого-биологические. Исто-
рически они развивались независимо.
Эколого-физиономические классификации жизненных форм
при выделении групп растений опираются только на их внешний
вид. Так, австрийский ботаник А. Кернер в 1863 г. создал систему
«Основных форм растений» умеренных широт, где описал 12 таких
типов: лазящие, вьющиеся, дернистые, деревья, кустарники и т.п.
В 1872 г. А. Гризебах издал труд по растительности земного шара,
где предложил экологическую классификацию растений, призван-
ную отразить климат и историю регионов. Он выделил 7 «основ-
ных форм» растений (деревянистые, сочные, вьющиеся, эпифи-
ты, травы, злакообразные и бессосудистые), подразделив их на 54
«растительные формы». В начале XX в. свою физиономическую
систему форм растений предложил О.Друде. Он описал 55 групп
растений (включая низшие), отдельно рассмотрев обитателей раз-
ных сред жизни (аэрофиты, гидрофиты) и впервые обратившись
к вопросам эволюции жизненных форм. В 1930-е гг. подробную
экологическую систему растений предложил Г.Э.Дю Рие, исполь-
зующую множество разнородных признаков и жестко не связан-
ную с адаптацией растений к среде.
В нашей стране в рамках эколого-физиономического направле-
ния была предложена целая серия классификаций жизненных форм
растений для разных природных зон. Для таежных лесов ее разра-
ботал В. Н. Сукачев, для европейских степей — В. В. Алехин, для
степей, пустынь и полупустынь — Б. А. Келлер. Ряд систем жиз-
ненных форм был описан для пустынных видов (Е. П. Коровин,
Д. Н. Кашкаров, А. В. Прозоровский, Л. Е. Родин, Н.Т. Нечаева).
Морфолого-биологическое направление связано с использо-
ванием для классификации жизненных форм разных биологиче-
ских особенностей растений (длительность жизни, ритм разви-
тия, способ питания, структура побега и т.п.). Первую такую клас-
сификацию представил в 1818 г. швейцарский ботаник О. П.Де-
кандоль, который выделил 8 групп растений (одно-, дву- и мно-
49
голетники, моно- и поликарпики и т.д.), подразделив их по еже-
годности плодоношения, отмиранию побегов и т.п. Предложен-
ные им термины используются до сих пор. Большое значение имела
классификация жизненных форм, разработанная Е.Вармингом
(1896). Он поставил своей целью отразить видимыми классифи-
кационными признаками сущностные процессы, происходящие в
растениях. Е.Варминг опирался на время жизни побега и всего
растения, количество периодов цветения, способность к вегетатив-
ному разрастанию, тип корневой системы, характер зимнего по-
коя. К морфолого-биологическому направлению относится и са-
мая известная система жизненных форм растений, созданная
К. Раункиером.
Морфолого-биологическое направление не антагонистично
эколого-физиономическому, так как многие важные биологичес-
кие признаки растений находят отражение в их внешнем облике
и многие свойства растений в онтогенезе существенно меняются.
Так, начальные стадии развития разных групп растений часто
имеют много общих черт, поэтому классификация жизненных
форм строится по взрослым особям с полностью выраженными
биологическими особенностями. Определяя жизненную форму
растений, надо учитывать и пластичность в развитии многих их
структур. Так, А. П. Шенников отметил, что гемикриптофиты (из
системы К.Раункиера) иногда зимуют как хамефиты, а геофиты
зимуют с надземными побегами; что есть виды (горошек забор-
ный — Vicia sepium, чина луговая — Lathyrus pratensis, льнянка
обыкновенная — Linaria vulgaris и т.п.), одновременно являю-
щиеся геофитами и гемикриптофитами. Изменчивость жизнен-
ной формы вида может зависеть и от фитоценотических условий.
Например, Ю.Е. Алексеев (1996) указывает, что для осок очень
важны изменения освещенности и влажности и при экстремаль-
ном их значении у них может меняться направление роста побега
и длина междоузлий. Так, у ряда кочкообразующих видов могут
появляться корневищеподобные побеги (в условиях повышенной
влажности у Carex elata, С. canescens, С. echinata, С diandra, а
при затенении у Carex alba, С. rhizina, С. remold).
Система жизненных форм растений К.Раункиера. Первый ва-
риант своей знаменитой системы датский ботаник К. Раункиер
предложил в 1905 г. Автору удалось свести все разнообразие жиз-
ненных форм растений в небольшое число типов, выделенных все-
го по одному критерию — способу защиты почек возобновления в
неблагоприятный сезон (холодный или сухой). К. Раункиер указал,
что положение почек возобновления по отношению к поверхности
почвы (или воды) зимой помогает объединить самые разные расте-
ния в небольшое число типов (рис. 5.2), которые позволяют делать
ботанико-географические и экологические обобщения. Защита
меристем обеспечивает выживание растения в условиях постоянно
50
Рис. 5.2. Жизненные формы растений по системе К. Раункиера:
/ — фанерофиты (тополь — Populusy, 2— хамефиты (черника — Vdccinium myrtillusy,
3 — гемикриптофиты (лютик — Ranunculus, одуванчик — Taraxacum, злаки); 4 —
геофиты (ветреница — Anemone, тюльпан — ТиНраУ, 5 — терофиты (фасоль —
Phaseolus). Зимующие части (внизу) отделены пунктиром
изменяющейся среды. Избранный К. Раункиером частный признак
коррелирует с множеством других (в том числе физиологических),
и потому отражает приспособление растений к комплексу факто-
ров местообитания. Но так как система К. Раункиера не физионо-
мична, в одну группу в ней часто собраны внешне непохожие рас-
тения (например, эпифиты, лианы и деревья).
На систему К. Раункиера опирались авторы многих других си-
стем жизненных форм (Ж. Браун-Бланке, И.Шмитхюзен и др.).
Они дополняли его группы, разделяли их по дополнительным при-
знакам. Современное развитие этого подхода представлено в сис-
теме жизненных форм Д. Мюллер-Дембуа и Г.Элленберга
(D. Mueller-Dombois, H.Ellenberg, 1974).
Рассмотрим основные категории жизненных форм растений в систе-
ме К. Раункиера.
Фанерофиты (от греч. phaneros — видимый, открытый, явный) име-
ют зимующие почки, расположенные высоко над землей. Сюда относят-
ся в основном деревья и кустарники. К.Раункиер подразделил фанеро-
фиты на 15 подтипов по размеру и длительности жизни листьев, харак-
теру почечных покровов. Среди них есть мегафанерофиты — очень вы-
сокие деревья (выше 30 м), мезофанерофиты — деревья от 8 до 30 м,
микрофанерофиты — большей частью кустарники, нанофанерофиты (от
греч. nanus — очень маленький) — мелкие кустарники. Отдельно выде-
лен подтип стеблесуккулентных растений (древесные кактусы, молочаи
и т.п.). К фанерофитам отнесены также растения, совершенно непохо-
жие на деревья, однако имеющие почки возобновления высоко над зем-
лей (например, растущие на высоких растениях эпифиты). Удаленность
от земли предполагает перезимовку почек без защиты снегом или иным
предохраняющим от замерзания покровом. Поэтому наиболее хорошо
эта жизненная форма представлена во влажных тропиках. Фанерофиты
51
обычны также в достаточно суровых условиях лесной зоны умеренного
пояса, однако их видовое разнообразие здесь небольшое.
Хамефиты (от греч. chamai — наземный) — невысокие растения с
почками возобновления, находящимися невысоко над поверхностью
почвы (не выше 20 — 30 см). Почки этих растений зимуют под защитой
почечных чешуй, снега, отмерших органов. Среди хамефитов много ку-
старничков (брусника — Vaccinium vitis-idaea, черника — Vaccinium
myrtillus, вереск — Calluna vulgaris, линнея — Linnaea borealis и др.),
широко распространенных в таежной зоне. Хорошо представлены в этой
группе и полукустарнички с отмирающими в неблагоприятный сезон ча-
стями побегов. Их особенно много в средиземноморском климате с вы-
раженным засушливым летом. К хамефитам относятся и некоторые тра-
вы, сохраняющие зимой почки на лежащих и приподнимающихся побе-
гах или в пазухах отмерших листьев (например, барвинок — Vinca minor,
звездчатка ланцетовидная — Stellaria holostea и др.). Хамефитами явля-
ются и растения-подушки с крайне ограниченным приростом в высоту.
Гемикриптофиты (от греч. hemi — половина и kryptos — скрытый)
располагают почки непосредственно на поверхности почвы, под под-
стилкой. Это травянистые многолетники, надземные органы которых (или
большая их часть) к концу вегетации отмирают. Почки возобновления
их защищены отмершими листьями, подстилкой и снегом. Гемикрипто-
фиты широко распространены во внетропических областях. Среди них
много обычных растений степей, лугов, лесов. Группа эта очень разно-
образна. Розеточные гемикриптофиты имеют укороченные побеги, ко-
торые могут зимовать на уровне почвы (одуванчик — Taraxacum, при-
мула — Primula). Перед перезимовкой ось розеточного побега, как пра-
вило, втягивается вглубь вплоть до почки, остающейся на поверхности.
М. Вит. Марков (1990) указывает, что розеточный побег не только слу-
жит для успешной перезимовки, но еще и способствует экономному рас
ходованию энергии, поэтому такие растения могут раньше начать рост,
эффективно использовать весеннюю влагу и при этом избегать конку-
ренции с более сильными видами. Протогемикриптофиты — расте-
ния с обычными удлиненными надземными побегами, ежегодно отми-
рающими до основания, где и находятся почки возобновления. В благо-
приятных условиях у них иногда сохраняются и воздушные побеги с
почками, например у крапивы (Urtica dioica) и чистеца лесного (Stachys
sylvatica).
Криптофиты (от греч. kryptos — скрытый) имеют «скрытые», спря-
танные почки возобновления. Это многолетние травянистые растения,
во множестве представленные в степях, пустынях, водоемах. К. Раунки-
ер подразделил их на три подгруппы: геофиты — сухопутные растения с
подземными почками возобновления, гелофиты — болотные растения,
воздушные побеги которых находятся.над водой, а почки под водой, и
гидрофиты — водные растения, полностью находящиеся под водой. У гео-
фитов на зиму отмирают не только надземные органы, но и часть под-
земных. Зимующие почки находятся под землей. В соответствии со стро-
ением зимующих органов геофиты делят на луковичные (тюльпаны —
Tulipa, луки — Allium), клубневые (картофель — Solanum tuberosum,
чистяк — Ficaria), корневищные (ландыш — Convallaria, пырей ползу-
52
чий — Elytrigia repens), корнеотпрысковые (бодяк полевой — Cirsium
arvense) и т.д. Почва хорошо предохраняет меристемы от повреждения
и холодом, и засухой, поэтому геофиты распространены в разных климати-
ческих условиях. Сюда относятся и многие обычные лесные травы, и
весенние эфемероиды пустынь, степей и лиственных лесов. В подзем-
ных органах эфемероидов накапливаются питательные вещества, что по-
зволяет растениям быстро расти и рано зацветать. Эфемероидам пустынь
это дает возможность быстро завладеть кратковременно влажной поч-
вой. Многие геофиты развивают утолщенные втягивающие корни, кото-
рые, высыхая, сокращаются и втягивают луковицу или клубень в почву,
и растения через несколько лет оказываются на значительной глубине.
У гидрофитов листья (плавающие или погруженные) на зиму отмирают,
а почки возобновления зимуют на дне водоема. Они могут располагаться
на многолетних корневищах (кубышка — Nuphar, кувшинка — Nymphaea)
или турионах, опускающихся на дно осенью и всплывающих к весне
(ряска — Lemna, рдест — Potamogetori). Благодаря постоянной зимней
температуре в придонном слое почки гидрофитов зимуют в условиях
еще более защищенных, чем у геофитов.
Терофиты (от греч. theros — лето) — однолетники, переживающие
неблагоприятный период в виде семян или спор, имеющих хорошую
морфологическую (плотные покровы) и физиологическую защиту (по-
кой, исключающий несвоевременное прорастание при кратковременном
возврате благоприятных условий). Важнейшая черта этой жизненной
формы — способность быстро проходить цикл развития, используя крат-
ковременные благоприятные условия. Однако они не достигают боль-
ших размеров и не оказывают сильного средообразующего воздействия.
Терофиты плохо представлены в умеренном и холодном климате, но очень
разнообразны в аридных районах. Особенно характерны они для пус-
тынь с коротким влажным периодом. К терофитам относятся, напри-
мер, пустынные и степные весенние эфемеры, которые успевают завер-
шить цикл развития до знойного лета. Некоторые из них, например ве-
роника весенняя — Veronica verna, вегетируют всего несколько недель.
Однако эфемеры встречаются и в достаточно увлажненных районах, где
есть местообитания с очень кратким периодом, благоприятным для ве-
гетации. Например, в низовьях Амура на неоднократно затопляемых в
течение года берегах в сухие периоды развивается большая группа ви-
дов, проходящих жизненный цикл за 1 — 1,5 мес. Терофиты активно за-
хватывают также антропогенно нарушенные территории.
Статистически сопоставив соотношения разных жизненных
форм в регионах с разным типом климата, К. Раункиер обнару-
жил, что каждая климатическая зона имеет особый биологиче-
ский спектр жизненных форм. Поэтому климатические регионы
он предложил классифицировать по представленности в них ха-
рактерных жизненных форм растений. Спектры жизненных форм
сильно изменяются также при подъеме в горы.
В целом для районов тропической зоны с обилием тепла и
осадков закономерно господство фанерофитов («климат фанеро-
фитов») (рис. 5.3). В субтропической зоне с зимними дождями
53
Рис. 5.3. Биологический спектр лесов по К. Раункиеру
(по Т. К. Горышиной, 1979):
А — дождевой тропический лес; Б — листопадный лес умеренных широт; / —
эпифиты; 2 — фанерофиты; 3 — хамефиты; 4 — гемикриптофиты; 5 — геофиты;
6 — терофиты. Цифры над колонками — проценты видов соответствующих групп
во флоре
обильны терофиты («климат терофитов»). Обычно это эфемеры,
заканчивающие развитие до жары и засухи. Согласно Т. К. Горы-
шиной (1979), они могут быть хорошим показателем аридности:
во флоре Прованса на юге Франции терофитов 29 %, в семиарид-
ных областях Марокко на севере Африки — 56%, а в аридных
районах на границе с Сахарой — от 75 до 91 %. Гемикриптофита-
ми богата умеренная зона, где этим растениям помогает суще-
ствовать снежный покров. В холодных полярных областях тоже
много гемикриптофитов, а на втором месте по распространенно-
сти — хамефиты. В этих суровых условиях их тоже защищает снег,
а перезимовка всей вегетативной части помогает эффективно ис-
пользовать короткий вегетационный период («климат гемикрип-
тофитов» характерен для большей части умеренной и холодной
зон, а «климат хамефитов» — для холодных районов).
Для биологически значимых особенностей климата показатель-
но также участие жизненных форм в отдельных семействах на
сравниваемых территориях. Так, в тропиках невысока представ-
ленность травянистых растений даже в таких семействах, как Слож-
ноцветные и Фиалковые (в умеренных широтах все они исключи-
тельно травянистые виды). Биологические спектры жизненных
форм отражают и частные особенности условий жизни растений
в разных типах растительности одного района. Так, на северо-
западе России показательно преобладание в хвойных лесах при-
способленных к суровым условиям хамефитов, а на лугах и в ли-
стопадных лесах — гемикриптофитов.
Система жизненных форм растений Д. Мюллер-Дембуа и Г. Эл-
ленберга. Система жизненных форм растений Д. Мюллер-Дембуа
и Г.Элленберга (D. Mueller-Dombois, H.Ellenberg, 1974) развивает
54
подход К. Раункиера. Она детализирует классификацию, доведя
ее иерархию до пяти ступеней. В этой системе на разных уровнях
используется 12 групп признаков, что ведет к огромному числу
потенциально возможных типов (29 млн), из которых описано
около 10 тыс. Однако если отбросить маловероятные сочетания
параметров, то число типов жизненных форм для наземных рас-
тений будет около 90, и благодаря четкой структурированности
ориентироваться в этой системе оказывается несложно.
Основными параметрами, на которые опирается система Д.Мюллер-
Дембуа и Г.Элленберга, являются: 1) тип питания (автотрофные, полу-
автотрофные — полупаразиты, гетеротрофные — паразиты и сапротрофы);
2) анатомическое строение (листостебельные и слоевищные); 3) харак-
тер автономности стебля и тип укоренения (укорененные в земле и с
автономными стеблями, лианы, полуэпифиты, эпифиты, водные расте-
ния); 4) общая архитектоника растений (с одним главным стеблем, с
многочисленными стеблями, растения-подушки, розеточные, полурозе-
точные, с ползучими неукореняющимися стеблями, с ползучими укоре-
няющимися стеблями, цепляющиеся); 5) степень одревеснения тканей:
древесные, полудревесные, травы; 6) сезонный ритм (вечнозеленые, зим-
не-зеленые, летне-зеленые, весенне-зеленые); 7) характер органов веге-
тативного размножения (со столонами, корневищами, луковицами, поч-
ками возобновления на стержневом корне); 8) наличие специфических
тканей (суккуленты, галоморфные растения).
Отечественные исследователи экологической морфологии рас-
тений. В российских системах жизненных форм всегда уделялось
большое внимание детальной морфологии растений (Г. Н. Высоц-
кий, 1915; В. В.Алехин, 1930; Б.А. Келлер, 1933; Е. М.Лавренко,
1935 и др.). Многими исследователями были предложены частные
системы для отдельных групп растений или регионов (В. Р. Вильям-
сом и А. М. Дмитриевым — для луговых трав, А. И. Мальцевым —
для сорняков, В. В. Скрипчинским и М.В.Сеняниновой-Корча-
гиной — для Ленинградской области, Г. Н. Высоцким, М.С. Ша-
лытом, В. Н. Голубевым — для степей и др.). Во всех этих систе-
мах в основу был положен принцип разделения растений на дере-
вья, кустарники, полукустарники, кустарнички, травы, которые
далее делились по типу строения надземных и подземных орга-
нов. Разработанные специально для растительного покрова конк-
ретных регионов, эти системы подробны, корректны и удобны.
Некоторые авторы при определении жизненных форм включили
в рассмотрение также позицию растения в сообществе.
Обобщив работы, сделанные в первой половине XX столетия,
свою систему жизненных форм растений в 1960-е гг. предложил
основатель отечественной школы экологической морфологии ра-
стений И. Г. Серебряков. Его система базируется на признании
исторически сложившегося соответствия структуры и функции
55
растений и нацеливает на рассмотрение эволюции жизненных форм.
Так, от одноствольного предка могут быть выведены разные вари-
анты деревьев (лиановидные, суккулентные, стелющиеся, древес-
ные подушки) и далее прослежены эволюционные линии к кустар-
никам, травянистым поликарпикам и монокарпикам.
Система И. Г. Серебрякова строго иерархична. Все жизненные
формы растений он разбивает на 4 отдела: древесные, полудревес-
ные, травы наземные и водные. Каждый отдел разделен на типы
(например, древесные — на деревья, кустарники и кустарнички,
полудревесные — на полукустарники и полукустарнички); каждый
тип — на множество дробных единиц (всего 8 рангов подразделе-
ний). Поскольку, как и в системе К.Раункиера, эти жизненные
формы характеризуют адаптацию растений к переживанию небла-
гоприятных условий, их соотношение во флоре различных зон не-
одинаково. Так, для влажных тропических районов характерны пре-
имущественно деревья и кустарники, для районов с холодным кли-
матом — кустарники и травы, а в жарком и сухом климате много
однолетников и т.д. Высокая экологическая информативность си-
стемы И. Г. Серебрякова способствует широкому использованию ее
в о^щеэкологических и фитоценотических исследованиях.
Вопросы эволюции жизненных форм растений. Эволюционные
построения, касающиеся жизненных форм растений, появились
намного позже утверждения дарвинизма, к началу XX в. При этом
ботаников давно волновал вопрос о филогенетических отноше-
ниях между деревьями и травами. Еще в XIX в. А. Н. Красновым
была высказана мысль о первичности древесных форм. Он также
предположил, что однолетние травы являются новейшими обра-
зованиями, произошедшими при «вырождении» древесных типов,
и связывал эти процессы с тектоническими и палеоклиматиче-
скими событиями, особенное значение придавая похолоданию.
Позднее о первичности древесной жизненной формы у покрыто-
семенных писали многие ботаники (М. П. Голенкин, А.А. Грос-
сгейм, Б. М. Козо-Полянский, А.Л.Тахтаджян, Р. Веттштейн и др.).
В 1905 г. Г.Галлир описал эволюционную цепочку от древней-
ших магнолиевых к лианам, кустарникам, многолетним травам
и однолетникам. В 1930-х гг. Дж. Корнер изложил по этому по-
воду «теорию дурьяна» (по названию растения Durio zibethinus).
А в 1940-е гг. М. Г. Поповым был предложен термин «соматиче-
ская редукция», вошедший в название теории, описывающей про-
исхождение от деревьев более мелких жизненных форм.
К. Раункиер также считал, что климат времени начала эволю-
ции покрытосеменных был равномерно теплым и влажным, по-
добным влажно-тропическому, в котором господствуют фанеро-
фиты. Поэтому древесные растения следует поставить в начало
эволюционного пути жизненных форм покрытосеменных. В су-
хом и холодном климате рост крупных деревьев затруднялся, раз-
56
меры их уменьшались и через кустарники они преобразовывались
в низкорослые хамефиты. При дальнейшем ухудшении климата
возникли формы с более защищенными (снегом, подстилкой) поч-
ками — гемикриптофиты, а далее и полностью скрывающие поч-
ки криптофиты. В пустынях же появились предельно сократив-
шие жизненный цикл терофиты.
С этих позиций главным содержанием процесса эволюции
жизненных форм у покрытосеменных было сокращение продол-
жительности жизненного цикла и соответствующие анатомо-мор-
фологические изменения побегов. Для древовидных растений эво-
люцию жизненных форм подробно рассмотрели И. Г. Серебряков
и его последователи. Ботанико-географические и филогенетичес-
кие соображения нашли подтверждение в палеоботанических ма-
териалах. Так, самые древние порядки покрытосеменных состоят
только из древесных форм, и произошли они от голосеменных,
среди которых травы отсутствуют. Среди травянистых покрытосе-
менных водные формы произошли от наземных. Вторичными в
большинстве случаев являются и однолетники, продолжающие
приспособление к засушливым условиям. Таким образом, рекон-
струированная в середине XX в. эволюция жизненных форм покры-
тосеменных растений, вероятно, в основном шла таким путем:
деревья — кустарники — травы многолетние — травы однолетние
и водные. При этом редукционная соматическая эволюция разде-
лилась на две основные линии — на криофильную (в холодных
условиях) и ксерофильную (в аридных условиях) (рис. 5.4).
Рис. 5.4. Эволюционные изменения древесной формы роста
(по А. П. Хохрякову, 1973).
В криофильных условиях: Ли Б — образование стлаников и подушек; В — нанизм.
В ксерофильных условиях. Г и Д — образование суккулентных и кустарниковых форм
57
Концепция соматической редукции как основного направле-
ния эволюции жизненных форм растений была принята большин-
ством ботаников и хорошо аргументирована данными сравнитель-
ной анатомии, морфологии, палеоботаники, ботанической гео-
графии. При этом конкретные пути эволюции жизненных форм в
пределах основного направления были разнообразны в разных
регионах и в разные эпохи. Так, по мнению ряда авторов, эволю-
ция древесных форм сначала шла по линии увеличения размеров,
а затем под влиянием ухудшения климата во внетропических об-
ластях — в сторону уменьшения их и потери одревеснейности. Но
в некоторых случаях мог быть другой ход развития. Так, у полы-
ней (Artemisia) при аридизации климата происходил переход от
мезофильных многолетних трав к ксерофильным полукустарнич-
кам, у которых почки приподняты над почвой (вероятный уход от
обжигающего действия песка в пустынях и полупустынях).
5.3. Эколого-фитоценотические стратегии
растений
Фитоценотипы и эколого-фитоценотические стратегии. Фито-
ценотипы — это группы видов, выделенные по роли в организа-
ции сообществ. К. А. Куркин (2002) определяет их как типы пози-
ций, которые виды занимают в фитоценозе, ролей, которые они в
них играют, и свойств, которые эти позиции и роли предопреде-
ляют. Сначала к выделению фитоценотипов подходили с количе-
ственных позиций, отделяя преобладающие по массе виды (доми-
нанты) от видов, обладающих меньшей биомассой, но потом ста-
ли рассматриваться и качественные критерии с акцентом на сре-
дообразующую роль видов.
Наиболее популярная система фитоценотипов принадлежит
В. Н. Сукачеву и Г. И. Поплавской. В ней особое внимание уделе-
но эдификаторам (от лат. aedificator — строитель) — средообра-
зующим видам. Эдификаторы формируют среду сообщества и во
многом определяют существование в нем других видов. Соэдифи-
каторы и ассектаторы (соучастники) — подчиненные малообиль-
ные виды, не вносящие существенного вклада в фитоценотиче-
ские отношения, они должны к этой среде приспосабливаться.
Система В. Н.Сукачева хорошо применима к лесам, где деревья
во многом определяют состав нижних ярусов. Термины эдификатор-
ность и доминантность не являются синонимами. Эдификатор —
частный случай доминанта, так как нет прямой связи между фи-
томассой и средообразующей ролью вида. Не всякий доминант
становится эдификатором и так сильно влияет на среду, что обус-
ловливает состав прочих видов. Например, эфемероиды весенне-
го леса — доминанты, но не являются эдификаторами, неправо-
58
мочно считать эдификаторами и доминантов тундровых и пус-
тынных сообществ.
Эколого-фитоценотические стратегии — это способы вы-
живания популяций растений в сообществах и экосистемах
(Б. М. Миркин и др., 1989). J.P. Grime (2001) пишет в этом случае
о группах сходных генетически определенных признаков, кото-
рые часто повторяются среди видов или популяций растений,
проявляясь в экологическом сходстве. Фитоценотипы и эколого-
фитоценотические стратегии — дополняющие друг друга поня-
тия. К.А. Куркин (2002) подчеркивает, что фитоценотипы показыва-
ют отношение растений к фитоценотическому стрессу (создавае-
мому средообразователями), а типы жизненных стратегий отражают
способы адаптации растений к любому стрессу (вызываемому и
абиотическими, и биотическими факторами). Таким образом,
понятие «фитоценотип» менее универсально, так как может исполь-
зоваться только для оценки поведения видов внутри сообществ с
высоким уровнем взаимосвязей между растениями. А эколого-фи-
тоценотические стратегии (или типы поведения) — наиболее обоб-
щенные характеристики видов, отражающие способ выживания в
самых разных условиях (в разных экотопах, сообществах, при раз-
ных режимах нарушений). Они позволяют объяснить реакцию лю-
бых видов в любых экотопах на стрессы и нарушения любого типа.
Системы типов эколого-фитоценотических стратегий. Первый
вариант системы жизненных стратегий растений в конце XIX в.
предложил Дж.Маклиод. Использовав социально-политические
аналогии, он разделил растения по способу выживания на «капи-
талистов» и «пролетариев». Первые затрачивают основную энер-
гию на поддержание взрослых особей и зимуют с «капиталом» из
многолетних тканей (стволы, ветви, корневища, клубни и пр.).
А «пролетарии», не имея «капитала» многолетних органов, зиму-
ют в виде семян, которых образуют очень много. Таким образом,
первая система типов стратегий растений была одномерна и ос-
новывалась на учете одного свойства — роли репродуктивного
усилия в выживании вида.
В середине XX в. эти типы стратегий были фактически переот-
крыты П. МакАртуром и Э. Уилсоном (1967), которые описали их
как результат двух типов отбора. В одном случае эволюция орга-
низмов идет в направлении увеличения затрат на размножение
(r-отбор), а во втором — на поддержание жизни взрослого организ-
ма (К-отбор). Хотя эти стратегии были разработаны для живот-
ных, они хорошо соответствовали стратегиям растений-«пролета-
риев» и «капиталистов». В 1970-е гг. эти эколого-эволюционные
идеи развил Э. Пианка. При этом он специально подчеркнул, что
в природе чаще встречаются организмы с переходными типами
стратегий (например, луговые травы и часто образуют довольно
много семян, и имеют зимующие многолетние органы).
59
Двумерный вариант типов стратегий отражает отношение рас-
тений к двум факторам и показывает возможность выживания их
в условиях разной степени благоприятности и нарушенности.
Первоначально в 1938 г. Л. Г. Раменский разделил все виды на три
«ценобиотических типа» — виолентов, патиентов и эксплерентов.
Виоленты (от лат. violentia — насилие, сила) — конкурентно мощ-
ные растения, энергично развивающиеся, захватывающие терри-
торию и удерживающие ее за собой, подавляя соперников энер-
гией жизнедеятельности и полнотой использования среды. Па-
тиенты (от лат. patientia — терпеливость, выносливость) выжи-
вают благодаря выраженной выносливости к крайне суровым ус-
ловиям. А эксплеренты (от лат. explere — наполнять), имея низ-
кую конкурентоспособность, могут очень быстро размножаться и
расселяться, захватывая освобождающиеся территории, однако они
так же легко вытесняются потом более сильными видами. Л. Г. Ра-
менский уподобил виоленты — львам, патиенты — верблюдам,
эксплеренты — шакалам.
В 80-е гг. XX в. подобные стратегии выделил Дж.Ф. Грайм,
сделав акцент на функциональном аспекте жизни растений. Он
подробно обосновал выделение двух важнейших факторов диф-
ференциации местообитаний растений — стресса и нарушений.
Под стрессом Дж.Ф. Грайм понимал набор явлений, ограничи-
вающих фотосинтетическую продукцию (недостаток света, воды,
минерального питания, неоптимальные температуры). Наруше-
ния же связаны с частичным или полным разрушением биомас-
сы растений, возникающим в результате деятельности фитофа-
гов, патогенов, человека, ветра, мороза, засухи и т.п. Реакции
представителей трех основных типов жизненных стратегий на
стресс фундаментально отличаются: рудералы (от лат. ruderis —
мусорный) формируют хотя бы минимальное число семян, кон-
куренты (от англ, competitor — конкурент) усиливают поглоще-
ние ресурсов, а стресс-толеранты (от англ, stress-tolerant —
устойчивый к стрессу) обеспечивают лучшую сохранность уже
поглощенных ресурсов. Нарушения губительны для стресс-толе-
рантов и конкурентов, но создают возможность для вспышки
численности рудералов.
Графически эта система типов стратегий часто изображается в
виде треугольника. Стороны его лежат на векторах, показываю-
щих усиление / ослабление нарушений и ухудшение / улучшение
условий (рис. 5.5). Углы треугольника соответствуют крайним
возможным типам местообитаний и обозначены буквами С, R, и
S в соответствии с первичными типами стратегий. Верхний угол
показывает исходно наиболее благоприятные и наименее нару-
шенные местообитания (соответствуют С-видам). Левый нижний
угол соответствует наиболее нарушенным местообитаниям (мес-
тообитания R-видов), а правый нижний угол свидетельствует об
60
Рис. 5.5. Треугольник Грайма
(по Б. М. Миркину, Л. Г. Наумовой, 1998).
Первичные типы стратегии: С — конкуренты; S —
сгресс-толеранты; R — рудералы; переходные типы
стратегий — CR, SC, RS, CSR. Сплошная стрелка —
усиление нарушения; пунктирная стрелка — ухуд-
шение условий
исходно неблагоприятных условиях (местообитания S-видов).
Области внутри треугольника, обозначенные сочетаниями букв,
отмечают вторичные (переходные) типы стратегий. Отдельные
виды — это точки на плоскости данного треугольника, и чем бли-
же точка располагается к какому-то углу, тем ярче выражена эта
стратегия у данного вида.
Сочетание у одного вида ярко выраженных признаков разных
типов стратегий невозможно: за совершенствование одних показа-
телей приходится расплачиваться другими. Здесь в скрытой форме
присутствует одна из организующих идей современной эволюци-
онной экологии — идея компромисса (трейдоффа), указывающая
на отрицательную корреляцию между отдельными признаками
жизненных циклов. С 1980-х гг. в экологии широким фронтом ве-
дутся работы, в которых в основном методами оптимизационного
моделирования исследователи пытаются установить, какие комби-
нации признаков жизненных циклов организмов приближаются к
оптимальным, а какие маловероятны или недопустимы.
В последние десятилетия XX в. в Великобритании проведено масш-
табное экспериментальное определение типов экологических стратегий
двух сотен видов растений на основе показателей морфологии и относи-
тельной скорости роста, а также изучены физиологические, биохими-
ческие и экологические особенности тестируемых видов британской
флоры (J.P. Grime et al., 1988; J. Р. Grime, 2001). Многие параметры ока-
зались тесно скоррелированы друг с другом. Эти материалы позволили
прогнозировать изменение растительности Британских островов при уве-
личении глобальной температуры и возможную реакцию растений на
увеличение концентрации СО2 в атмосфере (J. Р.Grime et al., 1996 и др.).
Однако корректное определение в полевых условиях многих из этих па-
раметров для большого числа видов затруднительно. В. И. Пьянков и
Л. А. Иванов предложили более доступные методы идентификации жиз-
ненных стратегий растений бореальной зоны. Акцент был ими сделан на
морфологическом и химическом аспектах, а особое внимание обращалось
на различия растений разных стратегических групп по биомассе, вкладу
в нее основных органов, содержанию N и соотношению C/N. Этот под-
ход может быть полезен при массовой идентификации типов эколого-
фитоценотических стратегий в естественных условиях (В. И. Пьянков,
61
Л.А. Иванов, 2000, 2001). Их инвентаризация в реальных ландшафтах
даст полезные материалы для экологического мониторинга и построе-
ния прогнозов реакции растительного покрова при глобальных измене-
ниях климата и разнообразном антропогенном вмешательстве.
Содержание первичных типов эколого-фитоценотических стра-
тегий Л. Г. Раменского — Дж.Ф.Грайма. Рассмотрим содержание
первичных типов эколого-фитоценотических стратегий Л. Г. Ра-
менского — Дж.Ф.Грайма, опираясь на изложение Б. М. Миркина
и Л. Г. Наумовой (1998).
Виолент — силовик, «лев» растительного мира. Это мощные
растения, чаще деревья, кустарники или высокие травы. Они ча-
сто имеют запасающие органы, позволяющие быстро разверты-
вать листья в наиболее благоприятный для фотосинтеза период.
Для них характерна относительно высокая скорость роста и быс-
трый ростовой отклик на обогащение ресурсами отдельных учас-
тков пространства, быстрое отрастание после частичной дефоли-
ации и невысокий вклад ассимилятов в защитные вещества и струк-
туры. Виоленты произрастают в благоприятных условиях (полное
обеспечение водой, элементами питания, теплом), формируя там
большую биомассу и площадь листовой поверхности, распростер-
тые кроны или корневища. За счет этого они держат под контро-
лем условия среды и почти полностью используют обильные ре-
сурсы таких местообитаний. Виоленты в фитоценозах — явные
доминанты. Таковы, например, крапива двудомная (Urtica dioica),
иван-чай (Chamaenerion angustifolium), тростник (Phragmites
australis) в дельтах рек. Однако они могут процветать только при
значительном количестве ресурсов, а при ухудшении условий (вы-
сыхание и обеднение почвы, засоление, затенение и т.д.) погиба-
ют, не имея специальных приспособлений для их переживания.
При нарушениях же (вырубка, рекреационные нагрузки, пожары
и т.д.) они также выпадают из сообществ, так как не образуют
семенного запаса в почве, а потому не могут конкурировать с оби-
лием быстро прорастающих эксплерентов.
Патиенты — «выносливцы», «верблюды» растительного мира,
обитающие в стрессовых условиях, ограничивающих образование
биомассы. Неблагоприятность условий может быть абиотической
(засуха, засоление, дефицит света или ресурсов минерального
питания, холодный климат) или биотической (затенение в лесу и
т.д.). Поэтому патиенты подразделяют на 2 подтипа. Экотопиче-
ские патиенты приспособлены к жизни в исходно бедных мес-
тообитаниях с постоянным абиотическим стрессом. Это растения
тундр, пустынь, солончаков, высокогорий и пещер, характерная
особенность которых — жесткая экономия и обеспечение жизне-
деятельности за счет самых ограниченных ресурсов. А фито-
ценотические патиенты растут в исходно благоприятных мес-
62
гах, где стресс создают виоленты (например, копытень — Asarum
еигораеит под сомкнутым пологом широколиственных деревь-
ев). М. Вит. Марков (1990) отмечает, что в ряде случаев выжива-
ние их обусловлено не столько толерантностью к влиянию эди-
фикаторов, сколько избеганием конкуренции с ними.
Для переживания стресса у патиентов развита система многочислен-
ных специальных приспособлений. Характерные черты их — низкая от-
носительная скорость роста, нечастое цветение, низкая морфологиче-
ская пластичность, отсутствие резко выраженной сезонной динамики
биомассы побегов и т.п. Дж.Ф.Грайм (2001) также отмечает адаптиро-
ванность их к временной пульсации доступности ресурсов, которая ха-
рактерна для бедных местообитаний и подчеркивает плохую поедаемость
животными. С химической точки зрения по сравнению с растениями
других типов стратегий у патиентов в биомассе отмечается минимальная
концентрация азота и максимальная углерода. Большое количество уг-
лерода связано с накоплением лигнина, целлюлозы и гемицеллюлоз в
клеточных стенках, сильно развиваемых при защите от разного рода стрес-
сов. А малое количество азота коррелирует с невысокой активностью
ферментов фотосинтеза и соответственно его невысокой интенсивнос-
тью и невысокими скоростями роста (В. И. Пьянков, Л.А. Иванов, 2001).
Патиентам свойственна также отрицательная реакция на нарушения, не-
высокое репродуктивное усилие и невысокий запас жизнеспособных се-
мян в почве (В.Г.Онипченко и др., 1991). Дж.Ф.Грайм (2001) еще отме-
чает природоохранный аспект: эвтрофикация, прекращение экстенсив-
ных форм сельского землепользования, усиление нарушений — все эти
формы антропогенного воздействия снижают участие патиентов, пред-
ставляющих значительную долю редких и исчезающих видов растений.
Эксплерент — рудерал, «бродяга», «шакал» растительного мира.
Эти растения замещают виолентов в исходно благоприятных ус-
ловиях среды при нарушениях, когда разрушается уже образован-
ная биомасса, или используют ресурсы стабильных местообита-
ний в период, когда они не востребованы доминантами. Они харак-
теризуются непродолжительным периодом жизни, высокой мор-
фологической пластичностью, способностью образовывать в усло-
виях стресса карликовые формы. Большинство эксплерентов —
однолетники или малолетники с высоким репродуктивным уси-
лием. Они обычно образуют много мелких семян, формируют
высокий запас жизнеспособных зачатков в почве и могут бурно
прорастать после нарушений местообитания, как, например, марь
белая (Chenopodium album). Многие из них имеют эффективные
приспособления для распространения плодов и семян (летучки у
одуванчика — Taraxacum и бодяка — Cirsium, прицепки у лопуха —
Arctium и череды — Bidens). К интенсивному семенному размно-
жению нередко добавляется вегетативное, например корневища-
ми и корневыми отпрысками (бодяк полевой — С. arvense и др.).
63
Эксплеренты — характерные растения побережий, обочин до-
рог, гарей, пустынные однолетники, мусорные виды вытаптывае-
мых участков, многие культурные растения и сорняки. Обычно
они первыми начинают восстанавливать растительность после
нарушений: семена одних видов уже имелись в почвенном банке,
а другие быстро доставляются ветром или другими агентами.
К эксплерентам относятся и виды, которые периодически дают
вспышки обилия в стабильных фитоценозах без нарушений. Это
происходит, во-первых, при обильных ресурсах, когда временно
ослаблено конкурентное влияние виолентов (например, весенние
эфемероиды развиваются до облиствения деревьев). Во-вторых,
это бывает в суровых экотопах при постоянно ослабленном режи-
ме конкуренции и внезапно увеличивающемся количестве ресур-
са, который патиенты не могут освоить. Например, пустынные
эфемеры после дождей быстро образуют сплошной покров, не
испытывая конкуренции с разреженным пологом кустарников.
Эксплеренты также населяют регулярно изменяющиеся, пятнис-
тые местообитания.
В биомассе эксплерентов большая доля приходится на стеблевую часть
и репродуктивные органы (относительная масса последних в три раза
больше, чем масса репродуктивных органов у растений других страте-
гий), и очень небольшая часть массы приходится на корни (в шесть раз
меньшая, чем у виолентов и патиентов) (В. И. Пьянков, Л. А. Иванов, 2000).
Они характеризуются также максимальной долей ассимилирующих тка-
ней, что позволяет развивать наибольшую относительную скорость рос-
та и развития. С химической точки зрения у эксплерентов отмечается
минимальная концентрация углерода и максимальная азота, что, воз-
можно, связано с увеличенным количеством ферментов фотосинтеза (око-
ло 75% азота листьев находится в белках хлоропластов) (В. И. Пьянков,
Л. А. Иванов, 2001).
Переходные эколого-фитоценотические стратегии. Пластичность
стратегий. Три первичных типа стратегий связаны множеством
переходных типов, которые называют вторичными. Видов с вто-
ричными стратегиями больше, они занимают основную площадь
треугольника Грайма. К.А. Куркин (2002) отмечает, что конку-
рентоспособность вида увеличивается при сочетании виолентно-
сти с другими стратегиями. Например, виолент осока дернистая
(Carex caespitosa) проявляет себя и как патиент (вынослива к
резкой перемене влажности и затенению). В Барабинской лесо-
степи на слабоосушенных тростниковых займищах она поселяет-
ся под полог тростника (Phragmites australis) и вытесняет его,
формируя почти чистые дернисто-осочники. Кострец безостый
(Bromopsis inermis) соединяет стратегию-виолента с патиентнос-
тью (вынослив к отложению наилка) и эксплерентностью (быст-
ро захватывает свежие аллювиальные наносы), поэтому в поймах
64
рек с активно откладывающимся аллювием он формирует моно-
доминантные кострецовые луга.
Переходными стратегиями характеризуются многие широко
распространенные растения (Б. М. Миркин и др., 2001). Приме-
ром вида со стратегией виолент-патиент (SC) является сосна обык-
новенная (Pinus syluestris), которая может формировать высокие
древостои на бедных песчаных почвах, а также все виды ели, ко-
торые произрастают в холодном климате на бедных кислых (но
хорошо увлажненных) почвах. Стратегию виолент-рудерал (CR)
имеет разрастающаяся на вырубках ольха серая (Alnus incana).
Виды со стратегией рудерал-патиент (RS) растут в пустыне на
вытоптанных площадках вокруг колодцев. А многие луговые рас-
тения представляют смешанный тип стратегии (CSR), сочетаю-
щий у разных видов в разном соотношении черты виолентности,
патиентности и эксплерентности (например, у пырея ползучего —
Elytrigia repens повышена доля эксплерентности), причем в раз-
ных условиях эти доли могут изменяться. В связи с этим В. Г. Онип-
ченко предлагает от этикетных определений стратегий переходить
к относительным.
Многие виды также обладают пластичностью стратегии. На-
пример, виолетность дуба черешчатого (Quercus robur) у границ
ареала, где он представлен кустарниковой формой, существенно
снижена, а патиентность повышена. Тростник, в дельте рек де-
монстрирующий стратегию виолента, становится патиентом на
засоленных почвах. На олиготрофных сфагновых болотах вырас-
тают специфические болотные формы сосны, например f. pumila
Рис. 5.6. Болотная форма сосны (Pinus sylvestris f. pumila)
(по P. И.Аболину из В. В.Алехина, 1950).
Пунктиром обозначена поверхность болота
65
в столетнем возрасте имеет высоту около метра, короткую хвою и
мелкие шишки (рис. 5.6).
Т.А.Работнов (1992) подчеркивал, что особи разных возраст-
ных состояний могут различаться по своим жизненным стратеги-
ям и это важно учитывать при общей оценке стратегии выжива-
ния вида. Так, по Ю. Э. Романовскому (1989), виолентами могут
быть только взрослые особи, а юные индивидуумы тех же видов —
патиенты. При этом чем выше их выносливость (патиентность),
способность выживать при ограниченных ресурсах, тем выше об-
щая виолентность вида. Еще больше различаются эколого-фито-
ценотические стратегии спорофитов и гаметофитов высших спо-
ровых растений.
5.4. Экологическая гетерогенность популяций
растений
Популяции и ценопопуляции. Слово «популяция» (от лат. рори-
los — народ, население) означает население вида на какой-то тер-
ритории (или совокупность особей одного вида, обладающих еди-
ным генофондом и занимающих некоторую территорию). Термин
«популяция» был введен в науку в 1903 г. датским генетиком
В. Иогансоном для обозначения генетически неоднородной груп-
пы особей вида (в отличие от однородной, именуемой «чистой
линией»). Изначально понятие «популяция» возникло примени-
тельно к животному миру, но потом приобрело общебиологиче-
ское значение.
Территорию, на которой встречаются представители данного
вида, называют ареалом. В пределах ее встречаются как места
более пригодные, так и менее пригодные или непригодные для
жизни вида. Поэтому возникает некоторая отграниченность од-
ной популяции от другой. Соседние популяции обычно сообща-
ются друг с другом в процессе расселения и переноса частей орга-
низмов. У некоторых видов такая связь постоянна (как у многих
луговых и рудеральных видов), у других — эпизодична (у популя-
ций некоторых озерных растений). С отграниченностью популя-
ций могут быть связаны их размеры и четкость границ. Так, одни
виды состоят из множества мелких популяций (например, горные
эндемики), а другие — из небольшого числа крупных (распрост-
раненные древесные лесные виды и злаки, растущие на обшир-
ных равнинах). Но и у одного вида могут образовываться разные
по размеру популяции. Так, В. Грант (1991), описывая популяции
Мамонтова дерева (Sequoiadendron giganteum) на западных скло-
нах гор Сьерра-Невада в Калифорнии, отмечает, что они изменя-
ются от изолированных небольших рощиц на севере их области
распространения до прерывистого лесного пояса на юге.
66
В разных частях ареала условия жизни могут сильно отличать-
ся. Папоротник орляк (Pteridium aquilinum) во Флориде, на юге
Африки, в Австралии, Индокитае и средней полосе России живет
в условиях совершенно разного климата, в разных типах сооб-
ществ, вместе с разными симбионтами, паразитами и конкурен-
тами. Каждая популяция приспосабливается к своим условиям
обитания. Поэтому любой широко распространенный вид оказы-
вается экологически неоднороден. А параметры конкретной по-
пуляции обусловлены взаимодействием ее генетического потен-
циала и сопротивления среды, которая включает локальный ком-
плекс всех абиотических и биотических факторов.
В середине XX в. в науке сложилось два понимания популяции —
генетическое и экологическое. Генетическая популяция — это
совокупность особей одного вида, связанных отношениями пан-
миксии (свободного обмена генами при равновероятном скрещи-
вании особей внутри этой совокупности). Такая популяция имеет
общий генофонд. Генетические популяции называются также еще
менделевскими. Несмотря на то что о закономерностях их форми-
рования и динамики написано очень много, установить в приро-
де генетическую популяцию практически невозможно (Б. М. Мир-
кин и др., 2001). Поэтому в экологии чаще используется другое
понимание популяции — локальное. Локальная популяция — это
совокупность особей одного вида в пределах однородного экото-
па. Поскольку такой экотоп обычно занят одним фитоценозом,
границы локальных популяций совпадают с границами фитоце-
ноза. Такие локальные популяции растений называются ценопо-
пуляциями или фитоценотическими популяциями. Размер цено-
популяции зависит от размеров однородного экотопа и соответ-
ственно занимающего его фитоценоза. Для описания ценопопу-
ляций используется много признаков, из которых главные — плот-
ность (количество особей на единицу площади) и гетерогенность
(наличие в ее составе особей, различающихся по возрасту, жиз-
ненности и т.д.).
Популяции клональных растений. Клональными называют рас-
тения, способные к вегетативному разрастанию. Структурными
элементами их популяций служат два типа образований — генеты
и раметы. Генеты — особи, выросшие из семян или спор. Раме-
ты — вегетативные побеги, в определенной степени автономные
от материнского растения, представляющие собой части генет (на-
пример, надземные побеги корневищного растения). Основную
роль в захвате и удержании ресурсов у клональных растений игра-
ют раметы.
Несмотря на относительную автономность рамет, между ними
складывается своеобразное «разделение труда», взаимосогласован-
ное развитие. По мере улучшения условий развития клона число
рамет и их дифференциация по размерам возрастают. Обычно
67
плотность генет и число рамет, приходящихся на одну генету,
связаны обратной зависимостью. Их соотношение — индикатор
состояния популяции, а оптимизация этого соотношения являет-
ся одной из задач агрономии.
Клональные растения различаются по степени вегетативной
подвижности. Выделяют виды-фалангисты и виды-партизаны.
Первые собраны в плотные скопления и хорошо удерживают тер-
риторию. Например, вейник тростниковидный (Calamagrostis
arundinaceae) разрастается крайне мало и почти никакие виды не
допускает в свою дернину. Т. Herben (1993) назвал такие виды сидя-
щими в крепости «сиднями». А виды-партизаны (ожика волосис-
тая — Luzula pilosa, седмичник — Trientalis еигораеа, орляк —
Pteridium aqudinum), имея менее скученное размещение, плохо
удерживают территорию и довольно быстро перемещаются. Сте-
пень мобильности вида зависит от стратегии роста и неоднородно-
сти среды.
Клональные виды — важные компоненты многих широко рас-
пространенных сообществ — лугов, степей, саванн и др. В сомк-
нутых лесах часто вегетативно размножаются растения нижних
ярусов, у которых в этих условиях ослаблено семенное возобнов-
ление (рис. 5.7). Вегетативное размножение хорошо представлено
не только среди трав, но и среди кустарников. Оно повышает
устойчивость их к выгоранию, помогает выжить на подвижных
субстратах (приморские пляжи, дюны). Роль клоновости обычно
возрастает в неблагоприятных условиях. Клоновые растения очень
важны в сменах растительности, так как вегетативное размноже-
ние позволяет им дольше удерживаться в сообществах. Виды, со-
Рис. 5.7. Вегетативное размножение при помощи подземных побегов
у растений елового леса (по В. В. Алехину, 1950):
А — вороний глаз (Paris quadrifolia)\ Б — седмичник (Trientalis еигораеа)', В —
кислица (Oxalis acetosella)
68
метающие семенное и вегетативное размножение, чаще имеют
более обширные ареалы (Б. М. Миркин, Л. Г. Наумова, 1998).
Регулирование плотности популяций растений. Существует два
основных типа регулирования плотности популяций растений: за-
висимостью от смертности и пластичностью. В первом случае
происходит избирательное самоизреживание, и с возрастом число
особей в популяции уменьшается до определенного порога. При
л ом еще в конце XIX — начале XX в. Г. Ф. Морозов и В. Н. Сукачев
показали, что зависимая от плотности смертность связана с усло-
виями жизни (возрастает при их улучшении). Она направлена про-
тив неограниченного роста популяции и стабилизирует ее числен-
ность в пределах, близких к оптимальному для данных условий.
Пластичность растений обусловливает полиморфизм размеров
особей: при увеличении плотности популяции он уменьшается.
При разной плотности популяции размер особей может разли-
чаться в тысячи раз. Миниатюризация особенно характерна для
малолетников (М. Вит. Марков, 1990). Она часто встречается, на-
пример, у сорных и рудеральных видов. Но при высокой плотно-
сти посадки растения не представляют собой лишь уменьшенные
копии с меньшим числом модулей. Внутривидовая конкуренция
влияет не только на скорость роста, но и на скорость развития и
созревания, на распределение биомассы по частям растения. Так,
Дж. Харпер показал, что конкуренция в посевах кукурузы (Zea
mays) гораздо сильнее влияет на продукцию семян (массу почат-
ков), чем на рост побегов. В результате при более плотном посеве
выход семян с единицы площади снижается (М. Бигон и др., 1989).
При повышении плотности популяции изменяется рост расте-
ний, хуже развиваются генеративные органы. При этом у ряда ви-
дов (например, однолетних) падение семенной продуктивности
способно обеспечить регуляцию плотности популяции. Плотность
популяции оказывает влияние и на скорость развития растений.
Как показал В. Н. Сукачев (1941), при увеличении плотности попу-
ляции развитие однолетников ускоряется, а многолетников — за-
медляется. В ряде случаев комплекс регуляторных процессов по-
зволяет компенсировать уменьшение плотности популяции увели-
чением роста особей, в результате чего при изменениях плотности
популяции растения могут сохранять общую биомассу довольно
постоянной. Такое «постоянство конечного урожая» Д. Харпер (1977)
показал для древесных пород и многих однолетников.
В экстремальных условиях (сильное затенение, засоление и т. п.)
главные типы регулирования плотности популяций дополняются
жотопической регуляцией. В этом случае плотность популяции
зависит от степени неблагоприятности среды и патиентности вида
(приспособленности к этим условиям). В природе у большинства
видов плотность популяций регулируется одновременно несколь-
кими способами. Так, при повышении плотности и усилении стрес-
69
са процессы самоизреживания и миниатюризации часто идут па-
раллельно.
Формы гетерогенности популяций растений. Разнообразие —
важное свойство природных популяций, обеспечивающее лучшую
адаптированность и более полное использование ими ресурсов.
Различающиеся особи популяций могут быть разделены на груп-
пы по размерам, возрасту, темпам развития и другим показате-
лям, реагирующим на изменяющуюся среду. Среди параметров,
наиболее важных для функционирования популяций растений, —
характеристики ассимиляционного аппарата (количество, разме-
ры, форма и характер расположения листьев), размеры побегов,
биомасса особей и вклад в нее органов репродукции, количество
зачатков генеративного и вегетативного размножения, показатели
роста особей. Вариации этих признаков определяются как гене-
тически, так и фенотипически. При этом по геному различаются
и растения одной популяции.
Б. М. Миркин и Л. Г. Наумова (1998) выделяют следующие ос-
новные формы разнообразия особей внутри популяции растений:
морфогенетическую (наличие генет и рамет), возрастную (разно-
возрастность особей), генетическую (наличие генетически различ-
ных групп), размерную (наличие особей разного размера), интег-
рационную (отражает степень скоррелированности признаков). Для
характеристики гетерогенности популяций используют два основ-
ных подхода: изучение возрастного состава и анализ морфологи-
ческой гетерогенности, отражающей их жизнеспособность.
Есть много причин формирования особей разных размеров. Большое
влияние оказывает разнокачественность семян. Так, всходы из крупных
семян лучше обеспечены питанием, быстрее развиваются, что позволяет
им сразу стать конкурентоспособными. Растения могут вырасти разны-
ми и из-за разных условий прорастания семян. Для возобновления раз-
ных видов бывают благоприятными разные условия, причем важны не
только абиотические, но и биотические факторы (напряженность кон-
куренции, деятельность гетеротрофов и др.). Так, поражение всходов фи-
тофагами во многом случайно, но бывают более предпочитаемые живот-
ными экотипы, обладающие некоторыми биохимическими особеннос-
тями. «Невкусные» всходы при этом получат преимущества. Разница в
темпах роста растений результирует влияние многих факторов и может
также зависеть от генотипа растения, так как разные группы одной по-
пуляции могут различаться по скорости прохождения фаз онтогенеза.
Итог же влияния всех факторов — разная конкурентоспособность. Луч-
ше развитое растение выигрывает в конкуренции и его преимущество
будет нарастать.
Разнокачественность особей популяций может меняться при
возрастных преобразованиях. Возможны четыре варианта онтоге-
нетической тактики (Ю.А. Злобин, 1989): стабилизация (уровень
70
неоднородности сохраняется), конвергенция (уровень неоднород-
ности снижается), дивергенция (уровень неоднородности повы-
шается), неопределенность (уровень неоднородности варьирует).
1>. М. Миркин и Л. Г. Наумова (1998) отмечают, что, вероятно, бо-
лее распространена тактика дивергентная, так как возрастание
инутрипопуляционного разнообразия позволяет полнее исполь-
ювать ресурсы местообитания. На онтогенетические тактики раз-
вития популяций влияют способы регулирования их плотности.
Регулирование зависимостью от смертности предрасполагает к
дивергентной тактике, а конвергентная чаще встречается в попу-
ляциях эксплерентов, где идет миниатюризация растений.
Онтогенетические состояния растений и типы ценопопуляций по
возрастному спектру. В онтогенезе растения испытывают возраст-
ные физиологические изменения, морфологически выражающие-
ся в размерах тела, а также в смене структуры корневых и побего-
вых систем, соотношении вегетативных и генеративных органов,
способности к вегетативному размножению и др. (рис. 5.8). Из-за
постоянной смены органов и разрушения старых частей абсолют-
ный возраст растений обычно определяется с трудом. Так, воз-
раст, подсчитанный по годичным приростам корневища, обычно
соответствует лишь возрасту наиболее молодой, живой части рас-
тения. Кроме того, особь семенного происхождения может рас-
пасться на несколько дочерних. При этом каждая часть (парпги-
кула) и несет на себе печать возраста материнской семенной осо-
би, и может быть значительно омоложена (таковы особи, вырос-
шие из спящих почек корневищ, корневые отпрыски). Таким об-
разом, дочерние особи проходят собственный жизненный цикл,
начинающийся уже не с прорастания семени, а с момента отделе-
ния от материнского растения.
Рис. 5.8. Схема состава ценопопуляции многолетнего травянистого
растения (по В. В. Петрову и др., 1994):
/ — семена; 2 — проростки; 3 — молодые растения; 4 — взрослые растения, не
образующие генеративных органов; 5 — генеративные растения; 6 — старческие
особи
71
У трав, быстро сменяющих побеги, каждый из них проходит
жизненный цикл от развертывания почки до отмирания надзем-
ной части («малый цикл»). Поэтому отдельно выделено понятие
«большого жизненного цикла», включающего весь онтогенез рас-
тения — от возникновения зародыша в семени до смерти всех вегета-
тивных потомков особи. Большой жизненный цикл складывается
из малых циклов разного масштаба (отдельных побегов, парциаль-
ных кустов и т.д.). У вегетативно неподвижных растений границы
особи и клона явны, а у вегетативно подвижных расплывчаты и на
поздних этапах большого жизненного цикла часто неопределимы.
Т.А. Работнов (1950) предложил разделять онтогенез растения
на четыре периода, специфичные по морфологии, физиологии и
функции в жизни популяции. Латентный период (от лат. latens —
скрытый, невидимый) представляют покоящиеся семена. Вирги-
нильный период (от лат. vitginatus — девственность) длится от про-
растания до первого цветения. В этом периоде часто выделяют
три возрастных состояния растений.
Проростки появляются из семян и сохраняют еще зародышевые
органы (семядоли и остатки эндосперма). Ювенильные растения (от
лат. juvenilis — юношеский) еще несут семядоли и следующие за ними
мелкие листья, иногда отличающиеся по форме от листьев взрослых осо-
бей. Имматурные (от лат. immaturus — незрелый) — полувзрослые осо-
би, потерявшие ювенильные черты, но еще недооформившиеся.
Генеративный период (от лат. generate — порождать, произво-
дить) отсчитывают от первого до последнего цветения. По оби-
лию цветущих побегов, их размерам, соотношению живых и мер-
твых подземных частей различают молодые, зрелые и старые ге-
неративные особи. Для монокарпиков полный онтогенез закан-
чивается этой стадией. Сенильный (от лат. senilis — старческий)
период — от потери способности к цветению до отмирания.
Онтогенетические состояния связаны с абсолютным (кален-
дарным) возрастом, но не всегда могут быть точно привязаны к
нему. Скорость индивидуального онтогенеза у отдельных орга-
низмов не совпадает, и конкретные особи могут достигать опре-
деленного возрастного состояния в разные календарные сроки.
Однако функционально особи каждого онтогенетического состо-
яния сходны. Морфологические признаки разных возрастных групп
для разных жизненных форм неодинаковы, хотя физиологичес-
кие границы основных возрастных периодов универсальны (про-
растание, первое цветение, последнее цветение, отмирание). Се-
нильные травянистые растения по внешнему виду часто похожи
на ювенильные, и только анализ подземных органов выявляет их
глубокое различие. В каждом возрастном состоянии растение об-
ладает специфическими приспособительными чертами, которые
не остаются постоянными. Так, ювенильные растения часто бо-
72
лес тенелюбивы и гигроморфны, чем взрослые, что помогает им
выжить под пологом взрослых особей.
По соотношению представителей разных возрастных групп
Т. А. Работнов предлагает различать три типа ценопопуляций. Ин-
вазионные ценопопуляции (от лат. invasio — нашествие) состоят в
основном из молодых (прегенеративных) растений. Они харак-
терны для начальных этапов освоения видом экотопа. Нормаль-
ные ценопопуляции включают все (или почти все) возрастные со-
стояния. Они способны к самоподдержанию и при этом не зави-
сят от внешнего поступления зачатков. Такие ценопопуляции ус-
тойчиво существуют в сообществе. В достигших равновесия нор-
мальных ценопопуляциях характерный возрастной спектр опре-
деляется особенностями биологии вида (продолжительностью он-
тогенеза, длительностью возрастных состояний, способами само-
поддержания, способностью формировать банк зачатков и др.).
Регрессивные ценопопуляции (от лат. regressus — возвращение)
представлены в основном сенильными особями. Таким образом,
возрастной состав ценопопуляции во многом определяет ее со-
стояние и роль в сообществах.
Т. А. Работнов дополнительно выделил еще ложноинвазионные цено-
популяции. Их образуют растения, имеющие почвенный банк семян или
вегетативных зачатков. Обычно их особи в фитоценозе отсутствуют, но
если подействует фактор, способный вывести часть зачатков из состоя-
ния покоя, то происходит массовое развитие этих растений. Так, на ме-
сте вывалов в лесу быстро развиваются ложноинвазионные ценопопуля-
ции рудералов. Б. М. Миркин и Л. Г. Наумова (1998) отмечают важную
роль этих популяций, так как со вспышек их обилия часто начинается
восстановление сообществ.
Поливариантность развития растений. Растениям свойственна
поливариантность развития. Неоднократно отмечалась высокая
пластичность их популяций, основные причины которой — моду-
лярная организация растений, неограниченное нарастание, при-
крепленный образ жизни, возможность разных специализаций
меристем. Эти свойства реализуются у растений благодаря боль-
шому набору структурных единиц (модулей), их разным сочета-
ниям и скорости формирования у разных биоморф, разной дли-
тельности жизни. Поэтому в любом возрасте у растений суще-
ствует большой диапазон изменчивости структурной организации,
жизненности, способности размножаться, темпов развития в пре-
делах генетической программы онтогенеза вида.
В основе поливариантности развития растений лежит генети-
ческий полиморфизм и модификационная изменчивость. На базе
одного генотипа наследуются разные программы развития и вы-
бор их определяется воздействием среды. Провоцирующие и сиг-
нальные факторы, объединяясь и преломляясь через индивиду-
73
альные особенности организмов, направляют онтогенез, но жест-
кой детерминации при этом не существует. Один фактор, дей-
ствуя на разные по генотипу особи, включает разные программы
онтогенеза. Способность к формированию нескольких феноти-
пов на базе одного генома возникла в результате длительного от-
бора, позволяя популяциям эффективно и адаптивно маневриро-
вать при изменении среды.
У растений можно выделить пять типов поливариантности, объеди-
ненных в две группы (Л. А.Жукова, 2001 и др.). Структурная полива-
риантность включает размерную, морфологическую и связанную с раз-
множением. Размерный тип (дополненный учетом анатомо-морфологи-
ческих признаков) позволяет судить о степени процветания популяции
и ее перспективах. Морфологический вариант связан с формированием
в ценопопуляции биоморфологических групп, обусловленных разнооб-
разными воздействиями (эдафическими, антропогенными, биотически-
ми и др.). Нередко важным показателем некоторых воздействий служит
повышенная тератность (уродства, аномалии) организмов. Например, у
валерианы (Valeriana officinalis) в придорожных условиях до 80 % осо-
бей имеют морфологические отклонения, а у подорожника большого
(Plantago major) при воздействии гербицидов и патогенных грибов фор-
мируется двурозеточность и метельчатость соцветий. Поливариантность
размножения проявляется в добавлении к обычным формам воспроиз-
ведения иных вариантов в определенных условиях. Например, кроме
семенного размножения иногда корневыми отпрысками может размно-
жаться одуванчик (Taraxacum officinale), подземными столонами — лютик
ползучий (Ranunculus repens), а ежа (Dactylis glome rata) — надземными
корневищами.
Динамическая поливариантность может быть разделена на ритмо-
логический тип и тип, связанный с темпами размножения. Первый обус-
ловлен асинхронностью в наступлении сроков фенофаз у растений од-
ной ценопопуляции или ценопопуляций из разных мест. Например, в
городах у березы повислой (Betula pendula) зрелые генеративные особи
имеют сближенное время цветения, а молодые и старые генеративные —
растянутое время цветения. Что касается темпов развития, то при мони-
торинге большого числа видов выявлено, что в природе средними тем-
пами развития характеризуются лишь около 20 % растений. Впервые о
существовании внутрипопуляционных групп, отличающихся по темпам
развития, писал еще В. Н. Сукачев (1941). В этой связи был выделен ряд
вариантов. Во-первых, замедленное развитие или его задержка обычны
у многолетников (например, часто встречается у чистотела — Chelidonium
majus). Вторичный покой на разных стадиях онтогенеза характерен, на-
пример, для подорожников большого и ланцетного (Plantago major,
Р. lanceolata). Другой вариант — ускорение развития, например неоте-
ния, — цветение ювенильных и имматурных особей (у подорожника боль-
шого, некоторых крестоцветных и др.) (Т.А. Работнов, 1978; А. А.Чистя-
кова, 1982 и др.). Причины такого развития могут быть разными — засуха,
конкуренция, воздействие удобрений и гербицидов. Возможен и вариант
омоложения, когда зацветают субсенильные особи. Сильное омоложе-
74
нис происходит при активизации спящих почек. Последовательность пе-
рехода растений из одного возрастного состояния в другое может нару-
шаться и формировать сложные поливергентные сети, когда отдельные
возрастные состояния выпадают и время пребывания растения в них
меняется.
Поли вариантность развития растений — важный адаптацион-
ный механизм популяционного уровня, способствующий устойчи-
вости ценопопуляции. Она создает большие адаптационные воз-
можности для растений в самых разных экологических ситуациях.
Половая форма гетерогенности популяций растений. Экологи-
ческие различия мужских и женских особей иногда настолько су-
щественны, что разделение полов способствует дифференциации
экологических ниш и более полному использованию экологичес-
ких ресурсов. Большинству видов растений свойственна либо обое-
полость цветков, либо однодомность; раздельнополые (двудом-
ные) цветковые растения составляют не более 5 % от общего чис-
ла современных видов. Анализ полового состава популяций про-
водят для двудомных растений. У таких видов раздельнополые
особи могут отличаться не только строением цветков, но и осо-
бенностями вегетативного тела, сроками наступления генератив-
ного периода. У мужских растений цветение часто наступает рань-
ше, поэтому при развитии популяции сначала в молодом генера-
тивном состоянии могут преобладать мужские особи, а среди стар-
ших генеративных растений соотношение полов выравнивается
или несколько сдвигается в сторону преобладания женских осо-
бей. У некоторых видов часть особей в начале генеративного пе-
риода продуцирует только женские органы, а к возрасту макси-
мального плодоношения они становятся двуполыми (Л. Б.Зауголь-
нова и др., 1988).
В природе нередко можно встретить не только мономорфные муж-
ские или женские экземпляры, но и особи, промежуточные по половому
составу, у которых наряду с раздельнополыми цветками встречаются и
обоеполые. Такие растения называют многодомными. Их много среди
кленов (Acer), гречишных и др. Оказалось, что очень распространена
женская двудомность — сочетание экземпляров с женскими и обоепо-
лыми цветками (А. Е. Васильев и др., 1988). Указывается, что у многих
губоцветных популяции могут существенно отличаться друг от друга по
соотношению особей с женскими и обоеполыми цветками и только с
обоеполыми цветками, а популяции гравилата речного (Geum rivale) от-
личаются по долям растений с мужскими и обоеполыми цветками, толь-
ко с мужскими и только с обоеполыми (А. В.Яблоков, 1987). В итоге
можно говорить о континууме по половой принадлежности у растений и
половой структуре их популяций.
По половому составу могут различаться и популяции в разных
условиях существования. Например, у ариземы японской (Arisaetna
75
Рис. 5.9 Ареал (сплошной и
точечный) женских растений
белокопытника (Petasites hyb-
ridus) в Великобритании (по
А. В. Яблокову, 1987)
japonica) решающий фактор в опреде-
лении пола — масса клубней: только
самые крупные клубни дают растения
с женскими цветками (А. В. Яблоков,
1987). А у клональных видов возможна
ситуация, когда популяции целого рай-
она состоят из особей только одного
пола. Так, белокопытник гибридный
(Petasites hybridus) встречается практи-
чески по всей территории Великобри-
тании, однако лишь в центре и частич-
но на юге страны в популяциях этого
вида есть и мужские, и женские особи.
По всей остальной территории его по-
пуляции сформированы только муж-
скими растениями, возобновляющими-
ся вегетативно (рис. 5.9).
Соотношение полов в популяциях
некоторых двудомных растений в зна-
чительной степени определяется раз-
личной конкурентоспособностью и
устойчивостью мужских и женских осо-
бей. Замечено, что при большой норме
высева мужские экземпляры часто бывают мельче женских и
имеют меньшую продолжительность жизни. Эти отличия особей
разного пола особенно хорошо проявляются у однолетников.
А наблюдаемая иногда у многолетников разная численность осо-
бей разного пола бывает связана с особенностями устойчивости
женских и мужских экземпляров к неблагоприятным факторам
среды.
В. Н. Сукачев (1941) приводит результаты опыта, в котором у конопли
(Cannabis) в загущенном посеве на удобренной почве ко времени цвете-
ния погибло 44% мужских и лишь 13% женских особей. Меньше эти
различия у двулетников (Trinia glauca) и часто совсем не проявляются у
многолетников (ивы корзиночной — Salix viminalis, дремы — Silene dioica
и др.). В. Н. Сукачев объяснял это тем, что у однолетников более раннее
отмирание мужских особей, выполнивших свое назначение, полезно для
женских экземпляров, так как ослабляет конкуренцию за ресурсы. У дву-
летников оно имеет уже меньшее значение.
Мужские и женские особи могут отличаться биохимическим соста-
вом, важным для гетеротрофных консортов. Так, плотность пилильщи-
ков на женских и мужских экземплярах ивы (Salix lasiolepis) отличается
в 1,5 — 3 раза, что объясняется разным содержанием в их листьях биоге-
нов и фенолов (И. А. Шилов, 2001).
Генетическая неоднородность популяций растений. Генетическая
структура популяций определяется соотношением частот аллелей
76
и генотипов, а также характером подразделенности популяции на
группы генетически близких организмов и характером связи меж-
ду этими группами в пространстве и времени (потоком аллелей).
На генетическую структуру популяций растений накладывает от-
печаток разнообразие способов их размножения в сочетании с
неподвижным образом жизни.
Популяции многих растений состоят из вегетативных потомков не-
многих особей. Так, у секвойи вечнозеленой (Sequoia sempervirens) из
пневой поросли образуются новые группы особей, сроки жизни которых
исчисляются тысячелетиями. В природных популяциях в эти группы
входит до 80 % особей (проросткам удается выжить только на периферии
популяции). Клональные группы деревьев хорошо известны у осин и
тополей (род Populus).
Так как наследуются не сами признаки, а спектр возможнос-
тей развития в зависимости от условий среды (норма реакции ге-
нотипа) и на реализацию каждого признака в онтогенезе влияют
многие гены, возможности быстрого определения генетической
структуры популяций по учету разового соотношения фенотипов
ограничены. В 1970-е гг. генетическую структуру популяций ста-
ли описывать с использованием маркеров генотипического соста-
ва. Ими могут быть либо выявляемые при электрофорезе особен-
ности белковых молекул, либо фены — дискретные морфологи-
ческие признаки, моно- или олигогенная природа которых экс-
периментально доказана. В генетической структуре популяций и
внутрипопуляционных групп растений описано много различий,
фиксируемых по разным маркерам (от деталей строения цветка и
других частей растения до различий хромосомного аппарата и
биохимических особенностей). Иногда их удается связать с давле-
нием отбора в определенной обстановке. Так, в Европе картиро-
вание межпопуляционных различий клевера ползучего (Trifolium
repens) по такому генетически контролируемому признаку, как
содержание цианидов, показало зависимость их от средней ян-
варской температуры (что нашло потом объяснение в биоценоти-
ческих исследованиях).
По серии морфологических признаков и цитологическим мар-
керам у множества видов удалось надежно выделять внутрипопу-
ляционные пространственно-генетические группы разного ранга.
М.В.Придня (1984) по специфической комбинации наследствен-
но закрепленных фенотипических признаков описал у кавказ-
ской пихты (Abies nordmanniana) семьи, состоящие из несколь-
ких сотен особей. Аналогичные данные получены В. и К. Грантами
(1980) по опунции (Opuntia lindheimeri), Р. Ботмер (1975) по луку
(Allium bourgeaui) и т.п. Показано также, что разные ценопопу-
ляции вида обычно характеризуются резкими перепадами частот
ряда признаков.
77
Существование в популяциях растений генетически разнород-
ных групп может быть объяснено также распространением авто-
гамии (самооплодотворение), агамоспермии (образование семян
без оплодотворения), полиплоидии, анеуплоидии и естественной
гибридизации. Например, к полиплоидии склонны многие широ-
ко распространенные растения: лапчатка гусиная (Potentilla
anserina), клевер луговой (Trifolium pratense), мятлик луговой (Роа
pratensis) и др.
5.5. Экотипы растений
Экотипы и их варианты. Еще в 20-е годы XX в. Г.Турессон
установил адаптивную природу многих расовых признаков расте-
ний. Он также показал, что особи одного вида, происходящие из
разных климатических областей или несходных местообитаний,
при одинаковых условиях в культуре в целом сохраняют первона-
чальный облик. Эти наследственно закрепленные расы, обитаю-
щие в природе в разных экологических условиях, и были названы
экотипами. Среди них было выделено несколько вариантов —
климатические, эдафические и биотические.
Экотипы представляют собой группы близкородственных осо-
бей, приуроченные к некоторым условиям и характеризующиеся
общими наследственными признаками, соответствующими им.
Экотипы — самые крупные экологические внутривидовые под-
разделения, т.е. вид — это совокупность экотипов, отличающих-
ся друг от друга рядом внутренних и внешних признаков, имею-
щих значение в выживании вида. Экотипы не совпадают с подви-
дами и разновидностями, выделяемыми таксономистами. Они
могут различаться морфологически, но это не обязательно. Так, у
полиморфных видов разным местообитаниям часто соответству-
ют экотипы, отличающиеся формой роста, размерами листьев, опу-
шением и другими признаками, но у большинства растений эко-
логически различающиеся формы внешне не отличаются. Поэто-
му экотипы устанавливают в эксперименте.
Экологами собран значительный материал по микроэволюции эко-
типов, который свидетельствует о быстром приспособлении растений
при изменении условий произрастания. Так, на Ротамстедской биостан-
ции (Великобритания) в опытах по длительному применению на лугу
удобрений было отмечено появление экотипов душистого колоска
(Anthoxanthum odoratum), различающихся морфологически и экологи-
чески. Растения, выросшие из семян, собранных с делянок, на которые
в течение 40 — 60 лет вносили известь, лучше росли на карбонатных по-
чвах, а выросшие из семян с делянок без известкования лучше развива-
лись на кислой почве. О скорости возникновения экотипов могут свиде-
тельствовать и наблюдения Т. А. Работнова (1998) над полевицей собачей
78
(Agrostis canina) и овсяницей овечьей {Festuca ovina): растения, произ-
растающие на пастбище непосредственно под оцинкованной проволо-
кой изгороди, сооруженной 25 лет назад, характеризовались повышен-
ной устойчивостью к цинку. Аналогичные данные есть о возникновении
у луговых растений экотипов, устойчивых к меди, свинцу и другим тя-
желым металлам.
Чем шире ареал вида, тем больше местообитаний встречается в
нем и тем разнообразнее набор его экотипов. Как правило, наи-
большее число экотипов отмечается в центральных частях ареала,
а в экстремальных условиях у его границ их число падает. Разные
виды в одинаковых условиях могут образовывать сходные экоти-
пы. Например, щавелек {Rumex acetosella), полынь полевая
{Artemisia campestris) и ряд других видов в хорошо освещенных
сухих местообитаниях образуют полевой экотип с распростерты-
ми побегами, на дюнах — песчаный экотип, на засоленных по-
чвах — солевой, в высокогорьях — горный.
Среди экотипов наиболее известны климатические. Преобла-
дающее большинство экотипов, изученных в классических рабо-
тах Г.Турессона, относится именно к ним (у гравилата — Geum,
калужницы — Caltha, тысячелистника — Achillea и др.). Клима-
тические экотипы — это группы ценопопуляций, занимающие
определенную часть ареала и сформировавшиеся главным обра-
зом под влиянием климатических факторов. Воздействие климата
ясно выступает в расовых различиях по срокам цветения. Так,
при совместном выращивании северные расы многих видов (земля-
ники лесной — Fragaria vesca, колокольчика круглолистного —
Campanula rotundifolia, гравилата речного — Geum rivale и др.)
зацветают раньше южных. Нередко экотипы ограничены частью
видового ареала и тяготеют к специфическим местообитаниям.
В лесоводстве и сельском хозяйстве климатические экотипы на-
зывают расами разного географического происхождения и давно
учитывают при подборе саженцев и семенного материала. Клима-
тические экотипы свойственны многим хозяйственно важным ро-
дам: сосне, ели {Picea), тимофеевке {Phleum), овсянице {Festuca),
люцерне {Medicago), бруснике {Vaccinium vitis-idaed) и др.
Лесоводам давно известно, что сосна обыкновенная {Pinus sylvestris)
состоит из ряда климатических экотипов, которые сменяют друг друга в
широтном и долготном направлениях и в родных регионах способны к
наилучшему развитию (деревья северных рас, к примеру, плохо растут в
южных районах). Изучение выращенных в одинаковых условиях сосен
разного происхождения показало различие в их способности к росту,
холодо- и засухоустойчивости и др. Обычно эти растения отличаются и
морфологически (длина игл, форма кроны, строение ствола и т.д.). Но
все эти расовые признаки склонны к модификации и не имеют значе-
ния для выделения вида.
79
У травянистых растений нередко образуются эдафические эко-
типы с разными требованиями к почвам. Часто эндемичные расы
(и виды) образуются на отложениях гипса или выходах известня-
ка. Описаны также эдафические расы, связанные с почвами, со-
держащими тяжелые металлы, например у полевицы тонкой
(Agrostis tenuis) на почвах со свинцом. Из деревьев эдафические
экотипы хорошо известны у сосны (Pinus sylvestris). Так, типич-
ный эдафоэкотип — меловая сосна, растущая на известняковых
обнажениях, иногда даже описывавшаяся как особый вид. Мор-
фологически существенно отличаются сосны, растущие на верхо-
вом болоте и в лесу. Н. В. Гришиной (1985) было показано, что
из-за разных сроков фенологического развития вероятность ус-
пешного перекрестного опыления таких группировок сосны со-
ставляет всего 0,7 —1,5 %, что свидетельствует о реальной биоло-
гической изоляции этих экотипов.
Уже из работ Г. Бонье (вторая половина XIX в.) известно раз-
личие растений, выросших на равнине и в горах. Замечено, что
вид, произрастая в горных условиях, формирует более укорочен-
ные междоузлия, более мелкие надземные органы и т.п., причем
эти признаки растения в основном сохраняют при выращивании
в измененной среде (рис. 5.10). В этой связи были выделены рав-
нинные и горные экотипы. Так, при культивировании равнин-
ных форм в горных питомниках растения хотя и видоизменяются
(мельчают, формируют более узкие листья, меньшее количество
цветков при более яркой окраске), однако достаточно резко отли-
чаются от растущих рядом горных форм. Аналогично ведут себя
горные формы в культуре на равнине. Подобно климатическим
экотипам, расы лапчатки (Potentilla glandulosa) и тысячелистни-
ка (Achillea millefolium) из альпийского пояса гор Сьерра-Невада
зацветают на экспериментальных участках раньше, чем расы, оби-
тающие в тех же горах на средних высотах.
Фитоценотические экотипы, или ценоэкотипы, формируют-
ся главным образом под влиянием условий фитоценоза. Так, выде-
ляют луговой и лесной экотипы у ежи сборной (Dactylis glo-
merata). А Р.Теркингтон (1979) экспериментально изучил био-
типы клевера ползучего (Trifolium repens), адаптированные к про-
израстанию вместе с некоторыми растениями (в совместном посе-
ве с ними дающие большую биомассу). В Малой Азии Е. Н.Син-
ская (1931) описала две биотические расы Brassica nigra — встре-
чающуюся в естественных условиях и в посевах. Более специа-
лизирована раса, распространение которой ограничивается по-
лями. Ее особенности позволяют успешно расти, образовывать
полноценные семена и распространять их вместе с посевным
видом. Аналогично некоторые расы рыжика (Camelina sativa)
представляют собой специализированные сорняки льняных по-
лей (форма linicola).
80
Рис. 5.10. Изменение облика солнцецвета (Helianthemum vulgare) в связи
с изменением внешних условий (по В. В. Алехину, 1950):
А — на равнине; Б, В — в горах (А, Б — в одном масштабе, В — увеличен)
Различия между расами могут быть связаны с опылением. Так, расо-
вая дифференциация по отношению к животным-опылителям обнару-
жена у нескольких видов семейства Синюховые. В. Грант (1984) отмеча-
ет, что различия в строении цветка между расами Gilia splendens корре-
лируют с опылением их преимущественно жужжалами — насекомыми с
длинными хоботками и колибри. А у Eriastrum densifolium и G. achilleifolia
есть расы с широким зевом венчика, произрастающие в Калифорнии
вдоль береговой линии и опыляемые крупными видами пчел, а есть расы
с маленьким зевом, опыляемые мелкими пчелами и произрастающие на
удаленных от моря хребтах.
В ряде случаев можно выделить экотипы, формирующиеся под
влиянием регулярно повторяющихся воздействий. При длитель-
ном выпасе скота (а также диких животных) сформировались па-
стбищные экотипы ежи, клевера, плевела (Lolium), кульбабы
(Leontodori) и др. Например, тимофеевка луговая (Phleum pratense)
дает стелющуюся форму на пастбищах и прямостоячую на сено-
косах. Существуют сенокосные (сезонные) экотипы клевера, ов-
сяницы, погремков (Rhinanthus). Например, у клевера лугового
(Trifolium pratense) двуукосная форма формируется в южной Ев-
ропе, а одноукосная (более морозоустойчивая) — в северной. А в
субальпийских лугах Высоких Татр произрастает горный подвид
погремка (R. major montanus), характеризующийся растянуты -
81
ми сроками цветения. С внедрением здесь сенокоса в середине
лета этот подвид дифференцировался на две формы: с весенним
цветением {R.m. vernalis) и осенне-летним (R.m. aestivalis).
Когда в конце августа стали практиковать покос отавы, сфор-
мировалась третья форма, отличающаяся поздним цветением
{R.m. polycladus). Появление в субальпике полей ржи {Secale
cereale) вызвало проникновение погремка в ее посевы, где он
закрепился как постоянный сорняк. У этой формы плодоноше-
ние начинается одновременно с плодоношением ржи, семенные
коробочки перестали раскрываться, а у семян редуцировались
приспособления к полету. В результате семена не выносятся вет-
ром и не отвеиваются от зерен ржи, что обеспечивает постоян-
ное подсеивание погремка на полях. Так сформировалась еще
одна форма — R. т. apterus.
Экоклины и смены биотопов. Наряду с наследственно закреп-
ленными экотипами в экологически контрастных местообитани-
ях существуют ненаследуемые модификации, внешне от них не
отличимые. Так, наземные формы водных растений представляют
собой исключительно модификации. Например, горец земновод-
ный {Polygonum amphibium) образует в воде, на влажной почве и
на сухих песчаных дюнах три разные ненаследуемые формы. То
же в ряде случаев касается карликовости, а также равнинных и
горных форм, у которых функциональные и наследственно зак-
репленные адаптации удается различать только в эксперименте с
пересадкой.
Г. Вальтер (1982) приводит пример сивца лугового {Succisa pratensis),
образующего на приморских лугах небольшие плотные розетки и невы-
сокие соцветия, а на влажных лугах обитают растения крупных размеров
с обильно ветвящимися соцветиями. Когда сивец выращивают в саду,
где все растения находятся в одинаково благоприятных условиях, лишь
часть экземпляров с приморских лугов сохраняет низкорослость, другие
же пышно разрастаются. Таким образом, карликовость была наследствен-
но закреплена не у всех особей, у некоторых это была модификация,
определяемая местообитанием.
На территории Северной Америки для тысячелистника {Achillea
millefolium) было установлено, что для горного типа наследствен-
но закрепленных рас характерны плавные переходы, тем более
постепенные, чем ближе одно к другому располагались места взя-
тия проб (рис. 5.11). Таким образом, можно говорить не о четко
обособленных экотипах, а о непрерывном ряде экотипов, называ-
емом экоклином. По ряду морфологических и биохимических при-
знаков экоклины специально прослежены в США для можже-
вельника виргинского {Juniperus virginiana) и губастика длинно-
цветкового {Mimulus longiflorus) (В. Грант, 1984). Клинальная из-
82
менчивость присуща и обычным видам среднерусской флоры,
например калужнице (Calthapalustris), лютику едкому (Ranunculus
acris), таволге вязолистной (Filipendula ulmaria), колокольчику
круглолистному (Campanula rotundifolid), ястребинке зонтичной
(Hieracium umbellatum) и др. (Г. Вальтер, 1982). Она характерна
для многих широко распространенных деревьев умеренной зоны.
Вероятно, существование экотипов часто лишь кажущееся, когда
число проб мало или они не отобраны в переходных зонах между
разными местообитаниями.
Приуроченность вида к разным местообитаниям в разных час-
тях ареала далеко не всегда объясняется возникновением новых
экотипов. В краевых частях ареала часто можно обнаружить сме-
ну биотопа данным видом. Так, многие травы восточноевропей-
Рис. 5.11. Схематический профиль от Тихого океана до Большого Бассей-
на в Америке с местами сбора семян тысячелистника (Achillea), культи-
вировавшихся затем в Станфорде (по Г. Вальтеру, 1974).
Изображены средние растения и вариационные кривые
83
ских степей в Центральной Европе встречаются только на скло-
нах южной экспозиции с благоприятным для них температурным
режимом. И довольно часто виды континентального климата, ко-
торые в центре своего ареала не приурочены к каким-либо по-
чвам, начинают предпочитать известковые почвы на его перифе-
рии. А многие лесные растения Центральной Европы, произрас-
тающие на легких или умеренно влажных почвах, в сухих облас-
тях юга России встречаются только в заболоченных и пойменных
лесах (Ф.Фукарек и др., 1982). Такая смена биотопов связана с
тем, что, компенсируя климатические изменения, вид меняет ме-
стообитание и в пределах ареала обеспеченность местообитаний
вида теплом и водой остается сравнительно постоянной.
5.6. Экологические ниши растений
Самые разнообразные экологические сведения о видах сводят-
ся воедино в понятии «экологическая ниша». Она характеризует
все стороны жизни вида: границы выносливости по отношению к
разным факторам, характер связи с другими видами, распределе-
ние в пространстве, потребление ресурсов и т.д. Понятие ниши
двойственно: с одной стороны, она является свойством самого
вида, определяемым его эколого-фитоценотической стратегией, а
с другой — это то экологическое место, которое вид занимает в
экосистеме. Таким образом, экологическая ниша представляет со-
бой и свойство самого вида, и совокупность экологических фак-
торов. В этом понятии зафиксированы все обстоятельства того,
как вид живет и использует местообитание.
Ю. Одум (1986) называл экологическую нишу «профессией» вида.
Б. М. Миркин и Л. Г. Наумова (1998) образно развивают это сравнение:
экологическая ниша — это все то, что определяет «работу по профессии»
(совокупность занимаемого видом пространства, потребностей в ресур-
сах и временного ритма их использования). Если рассматривать экосис-
тему как совокупность условий среды и организмов, в результате дея-
тельности которых производится органика, то «профессия» видов опре-
деляется тем, из какого материала, в какое время, на какой территории и
какое органическое вещество они производят.
Совокупность требований вида к среде и реальное положение
популяции в экосистеме могут существенно различаться. В связи
с этим были введены понятия «фундаментальной» и «реализован-
ной» ниши. Фундаментальная ниша — это экологическая ниша,
которую может занять популяция взрослых растений при отсут-
ствии конкуренции, а реализованная ниша — это часть фунда-
ментальной ниши, которую реально занимает популяция при кон-
куренции с другими видами. Фундаментальные ниши обычно очень
84
широки, поэтому в садах и парках при отсутствии конкуренции
успешно выращиваются очень разные растения, которые, однако,
нуждаются в защите от аборигенных видов.
Различия фундаментальной и реализованной ниш хорошо выявились
в опытах со степными растениями, проведенными в 30-е гг. XX в.
А. В. Прозоровским. В чистом посеве при поливе ковыли (Stipa) и тип-
чак (Festuca sulcata) давали урожай выше, чем на сухом черноземе, т.е.
при отсутствии луговых видов степняки продуктивнее при хорошем увлаж-
нении. Фундаментальная ниша этих «сухотерпцев» намного шире реа-
лизованной и включает очень разные по увлажнению местообитания.
Но реализованная ниша охватывает именно засушливые условия, где
выжить им помогает мощная корневая система, узкие листья и т. п. Без
конкуренции степняки лучше бы росли при большем увлажнении, но в
природе в таких условиях они не выдерживают конкуренции с луговыми
травами. Аналогичные результаты были получены в многочисленных опы-
тах по удобрению естественных лугов и чистых посевов из тех же видов
(Т.А. Работнов, 1985; Б. М. Миркин, Л. Г. Наумова, 1998 и др.).
Соотношение объемов фундаментальной и реализованной
ниш — важная составляющая стратегии растений. Так, виоленты
прочно удерживают свою фундаментальную нишу, и чем выше
виолентность, тем большую ее долю занимает ниша реализован-
ная. При этом у чистых виолентов формируются практически од-
новидовые сообщества. Пример тому — заросли тростника
(Phragmites ausrtalis) в низовьях рек юга России. Наиболее раз-
личаются фундаментальная и реализованная ниши у эксплерен-
тов, которые могут иметь очень широкие пределы толерантности
по любому фактору среды, но при появлении виолентов из сооб-
щества вытесняются. Повышение эксплерентности вида увеличи-
вает расхождение его фундаментальной и реализованной ниш, а
поскольку эксплерентность свойственна большинству видов, фун-
даментальная и реализованная ниши редко совпадают. В есте-
ственных сообществах очень узкую реализованную нишу имеют
культурные растения, поэтому без защиты от сорняков они быст-
ро выпадают из сообществ (Б. М. Миркин, Л. Г. Наумова, 1998).
Различиями фундаментальных и реализованных ниш объясняют-
ся многовершинные распределения видов вдоль экологических
градиентов: они возникают при оттеснении конкурентно слабых
видов к периферии их фундаментальных ниш (см. подразд. 3.2.1).
ГЛАВА 6
СВЕТ КАК ЭКОЛОГИЧЕСКИЙ ФАКТОР
Подавляющее большинство растений — фотоавтотрофы. Бла-
годаря энергии Солнца они создают первичное органическое ве-
щество — основу жизнеобеспечения почти всех обитателей Зем-
ли. Перехватывая солнечные лучи, растения используют для фо-
тосинтеза лишь малую их часть. Свет действует на растение и как
источник энергии для фотохимических реакций, и как регулиру-
ющий развитие раздражитель, а также может оказывать повреж-
дающее действие. Кроме участия в питании, формообразовании и
развитии растений он определяет ряд особенностей их географи-
ческого распространения и топографического размещения. В про-
цессе эволюции растения приобрели способность по-разному адап-
тировать свой обмен, строение и развитие к тому количеству и
качеству света, которое преобладает в месте их произрастания.
6.1. Количественные характеристики принимаемого
растением света
Общее поступление солнечной энергии к земной поверхности.
Энергия солнечного излучения постоянным потоком поступает
на планету благодаря термоядерным реакциям, протекающим в
недрах Солнца. Мощность излучения Солнца — 3,8 • 1020 МВт.
Пройдя 150 млн км, одна двухмиллиардная часть этой энергии
попадает на Землю, и именно благодаря ей на нашей планете про-
исходит круговорот воды (на испарение ее тратится треть посту-
пившей энергии), дуют ветры, развивается жизнь. Вся энергия,
скрытая в горючих ископаемых, таких, как нефть, уголь, торф,
природный газ, — изначально это тоже энергия Солнца.
Полный поток солнечного излучения, приходящийся на еди-
ницу земной поверхности, изменяется от 1,31 до 1,4 кВт/м2 в за-
висимости от расстояния до Солнца. Стандартное его значение
(для среднего расстояния между центрами Солнца и Земли) назы-
вается солнечной постоянной и равно 1,353 кВт/м2 (или 1,94 кал
на 1 см2 в 1 мин, или 4 871 кДж на 1 м2 в 1 ч). Солнечная постоян-
86
ная определяется количеством лучи-
стой энергии, поступающей в 1 мин
на 1 см2 площади, перпендикуляр-
ной к солнечным лучам и находя-
щейся над верхней границей атмос-
феры при среднем расстоянии Зем-
ли от Солнца (1,495* 108 км) (Фо-
тосинтез и биопродуктивность: ме-
тоды определения, 1989). При не-
которой ритмичности в геологичес-
ком масштабе времени эта величи-
на обладает удивительным посто-
янством. Солнечная постоянная, за-
висящая преимущественно от мас-
сы Солнца и радиуса орбиты пла-
неты, создает космическую основу
поддержания на Земле благоприят-
Рис. 6.1. Ослабление солнечной
радиации при прохождении че-
рез атмосферу (по В.Лархеру,
1978)
ного для жизни климата.
Солнечная постоянная измерена за пределами атмосферы, а до
земной поверхности доходит обычно не более 61 % этой энергии,
но и это при ясной погоде. Обычно же из потока общей солнеч-
ной радиации около 34 % отражается обратно (25 % облаками и
9 % рассеянными молекулами газов, каплями паров, кристаллами
льда, частицами пыли). Еще 19 % ее поглощается (10 % облаками
и 9 % аэрозолями). Таким образом, в среднем лишь около 47 %
поступившей к Земле солнечной энергии может быть доступно
обитателям биосферы (рис. 6.1). Эта радиация лишь наполовину
состоит из прямых солнечных лучей, беспрепятственно прони-
завших толщу атмосферы. Остальная часть — это рассеянный свет,
подвергшийся рассеянию облаками и частицами воздуха. Рассе-
янная радиация характеризуется особым спектральным составом,
а количество ее варьирует в зависимости от высоты стояния сол-
нца и облачного слоя, и даже в ясные дни она составляет 10—
30 % от полной солнечной радиации. Дошедшая до земной повер-
хности совокупность прямой и рассеянной солнечной радиации
называется суммарной радиацией, и в средних широтах на уров-
не моря горизонтальная поверхность получает ее в полдень до
900 Вт/м2
Изменчивость получаемой радиации. В разных точках планеты
обеспеченность солнечной радиацией сильно зависит от геогра-
фической широты, высоты над уровнем моря, рельефа, облачнос-
ти. Поэтому на Земле наблюдаются большие региональные и ло-
кальные различия радиационного довольствия.
Тропические области высоких давлений, приходящиеся на пу-
стыни Африки и Австралии, получают очень много света. Эти
малооблачные сухие пространства принимают в среднем не 47 %,
87
a 70 % радиации, поступившей на верхнюю поверхность атмосфе-
ры. Экологически важно, что в аридных условиях из-за малого
количества водяных паров интенсивность света гораздо выше. Это
позволяет пустынным растениям, резко сократив испаряющую
поверхность, все же нормально ассимилировать. Однако для нужд
фотосинтеза свет в аридных пустынях избыточен — он не может
быть полностью утилизирован растениями из-за высоких темпе-
ратур и недостатка воды.
Суммарная интенсивность радиации велика также в высокого-
рьях, так как там потери энергии уменьшаются при прохождении
ее через более тонкий и чистый слой атмосферы. Так, для пер-
пендикулярной к лучам поверхности при подъеме со 100 до 4000 м
над уровнем моря инсоляция увеличивается в 2 раза. Большую
роль играет также экспозиция склона. Усиление инсоляции в
высокогорье имеет немаловажное значение для альпийских рас-
тений.
Напротив, в гумидных областях, где постоянна облачность и
водяные пары экранизируют свет, значительно снижается и осве-
щенность. Это происходит во влажных тропических районах, по-
ясах туманов в горах, областях сырого океанического климата.
Все они в среднем получают меньше солнечной радиации.
Немалое значение в обеспечении светом местообитаний имеет
рельеф (угол наклона поверхности и экспозиция склонов). Изме-
нение угла падения солнечных лучей влияет на интенсивность
радиации: чем ближе угол падения лучей к прямому, тем больше
света получает участок (рис. 6.2). Параллельно нарастает и темпе-
ратурное различие. Поэтому локальные топографические вариа-
ции силы света могут иметь существенное экологическое значе-
ние. Это относится к большому разнообразию растительности в
горах, но особенно актуально для приполярных территорий: в
Рис. 6.2. Суточный ход потока прямой солнечной радиации на перпен-
дикулярную (7) и горизонтальную (2) поверхность для Павловска в июле
и январе (по И. А. Шульгину, 1967)
88
высоких широтах именно различия в интенсивности освещения
южных и северных склонов во многом объясняют разницу в их
растительности. И в средних широтах растения, растущие совсем
недалеко, но на разных элементах рельефа, могут оказаться в ус-
ловиях освещенности, различие которых сравнимо с зональными.
Так, южный склон крутизной 20° на широте Санкт-Петербурга
(60° с.ш.) в летний день получает больше солнечной радиации,
чем горизонтальная поверхность на широте Харькова (50° с.ш.).
Это находит ботаническое отражение в правиле предварения
В. В.Алехина, по которому при движении от зоны к зоне харак-
терные для них растения и сообщества предваряются на соответ-
ствующих склонах предыдущих зон (в соответствии с меньшей
или большей инсоляцией склонов разной экспозиции). В некото-
рых случаях в рельефе могут создаваться особые местообитания,
когда растения попадают в условия особенно сильного затенения
(пещеры, расщелины скал, нависающие глыбы).
Существенно влияет на количество получаемого растениями света
запыленность атмосферы, вызванная ее загрязнением. Это наблюдается
при естественных катастрофических явлениях (вулканизм, пыльные бури),
которые на некоторых территориях бывают более часты. А широко об-
суждавшаяся модель «ядерной зимы», предложенная группой академика
Н. Н. Моисеева, прогнозирует, что результатом ядерной войны может быть
сильное запыление атмосферы и глобальное похолодание из-за подня-
тия в воздух огромных масс твердых частиц. Аналогичный эффект дает
падение крупных небесных тел. Так, расчеты показывают, что падение
10-километрового астероида обеспечивает естественную «ядерную зиму»,
когда под непроницаемым покровом пыли и пепла даже на экваторе
температура полгода может держаться ниже 0°. Возможно, в прошлом
падение крупных небесных тел могло вызывать длительное запыление
атмосферы с соответствующим падением температуры и эволюционны-
ми последствиями. Например, рядом ученых предполагалась именно та-
кая причина, спровоцировавшая на границе мела и палеогена вымира-
ние крупных рептилий. В наше время зафиксировано уменьшение сол-
нечной радиации, проходящей через запыленную атмосферу крупных
индустриальных центров.
Растения открытых мест получают дополнительный свет, отра-
женный от поверхности почвы, воды, снега, других организмов.
Эта величина, называемая нижним светом, сильно зависит от
свойств отражающих поверхностей. Отношение отраженной ра-
диации к падающей называется альбедо. Оно зависит от цвета
субстрата, его структуры, влажности и выравненное™ поверхнос-
ти. У почв, покрытых растительностью, оно зависит также от осо-
бенностей растений, цвета листьев и стеблей. Альбедо сильно раз-
личается у разных субстратов. Так, свежевыпавший снег отражает
около 80% падающей энергии, песчаные дюны — до 60% света,
89
поэтому лист над сухим песком может получить энергии на 20 %
больше. Выходы мела отражают до 70 % падающего света, а влаж-
ный чернозем — всего 8%. Альбедо орошаемых участков на 5 —
11 % ниже, чем альбедо сухих (И. С. Кауричев и др., 1982). Доля
отраженного света существенна в околоводных местообитаниях,
особенно при низком стоянии солнца (утром и вечером). Так,
при высоте солнца над горизонтом 2 ° альбедо воды составляет
около 78 %. Очень интенсивен нижний свет на южных склонах
берегов: в солнечный день он может составлять до 85 % от пря-
мого.
Растения как приёмники солнечной радиации. Фоторецепто-
ры — светочувствительные и световоспринимающие образования,
способные генерировать физиологический сигнал в ответ на по-
глощение кванта света. Светочувствительным элементом фоторе-
цептора служит мембрана, содержащая пигменты. У растений фо-
торецепторами являются пластиды. У высших растений эффек-
тивно организованы расположение и работа световоспринимаю-
щих органов — листьев.
Лист как оптическая система отличается сложной структурой.
На уровне тканей ее элементами являются различные слои и сис-
темы клеток, межклетники с разными оптическими свойствами;
на уровне клетки — хлоропласты, другие органеллы и вещества,
способные к перемещениям. На уровне хлоропласта — система
тилакоидов, гран, пигментов. Это структурное разнообразие и
обилие межфазных границ создает большие возможности для внут-
рилистового рассеяния и отражения света. Поэтому лист погло-
щает значительно больше энергии, чем слой раствора хлорофилла
соответствующей концентрации. Кроме того, структурированность
Рис. 6.3. Схема поступления прямой и рассеянной солнечной радиации
к растениям с горизонтальными, вертикальными и различно ориентиро-
ванными листьями (по И. А. Шульгину, 1967).
Расположение листьев: А — горизонтальное, Б — вертикальное, В — различное.
S — прямая радиация; D — рассеянная радиация; S2 — поступление к листьям
прямой радиации; 7,2 — листья с разными углами наклона
В Часы дня
90
листа позволяет реагировать на различия освещенности путем
перестройки на разных уровнях организации.
Кроме собственно оптических свойств листа на количество
получаемого света влияет его положение в пространстве. Большое
значение имеет угол наклона листовой пластинки. При той же
плотности светового потока горизонтально расположенный лист
поглощает квантов больше, чем лист наклоненный. Существенно
отличается и радиационный режим растений с горизонтально и
вертикально расположенными листьями. Например, у розеточных
листьев поступление прямой радиации в первую половину дня
нарастает, а у прижатых к прямостоячему стеблю или опущенных
вдоль него — падает (рис. 6.3). К горизонтально ориентирован-
ным листьям рассеянного света поступает больше и нарастание
этого поступления идет быстрее. У эффективно использующих
свет сельскохозяйственных растений верхние листья обычно на-
клонены, а нижние находятся в горизонтальном положении, что
обеспечивает максимальное поглощение света всем посевом.
Поскольку растения получают свет от одного главного источ-
ника, важно положение световоспринимающих поверхностей по
отношению к нему. Наилучшим вариантом были бы листья, по-
ворачивающиеся вслед за солнцем. Так у подсолнечника (Helian-
thus annuus) вместе с соцветием поворачиваются листочки обер-
тки. Но это встречается редко. Обычно ориентация листьев на
растении диффузна и наибольшее количество света получают то
одни из них, то другие. Поэтому растение сравнивают с непод-
вижной локаторной системой, у которой часть приемников все-
гда оптимально ориентирована (Т. К.Горышина, 1979). Но в не-
которых случаях листовые пластинки расположены строго направ-
ленно. Это бывает, во-первых, при одностороннем освещении в
условиях общего дефицита света (листья лесных трав близ опуш-
ки или окна в древесном пологе повернуты в сторону просвета).
Во-вторых, это характеризует растущие на открытых местах ком-
пасные растения (например, латук — Lactuca располагает лис-
тья в вертикальной плоскости, ориентируя их узкой стороной на
юг).
Для обеспечения фотосинтеза все листья должны быть доста-
точно освещены. Это хорошо достигалось бы при расположении в
один слой, когда они не затеняли бы друг друга, как побеги ряски
(Lemna) на воде. Но такая фотосинтезирующая система исполь-
зовала бы не весь падающий свет, поэтому количество и площадь
световоспринимающих элементов увеличиваются: растет число
листьев на растении, появляется ярусность фитоценоза. Однако
при этом большая часть листьев оказывается затенена. Частично
это компенсируется особым расположением листовых пластинок
в кроне. Так, у вяза (Ulmus) верхние листья часто располагаются
почти вертикально, лежащие глубже — наклонно, а нижние, по-
91
лучаюшие в основном рассеянный свет, — горизонтально. Боль-
шое значение имеют форма и густота кроны. Подобные законо-
мерности организации световоспринимающих поверхностей есть
и в растительных сообществах.
Световое довольствие. Характеристикой условий освещения
служит относительное световое довольствие. Оно показывает,
какую часть полного солнечного света получает растение в дан-
ном местообитании, и выражается дробью или процентами от пол-
ной освещенности. В 1907 г. А. Визнер предложил измерять све-
товое довольствие скоростью почернения фотобумаги (в дроби
числитель 1, а знаменатель показывает число секунд, необходи-
мых для почернения фотобумаги). Сейчас освещенность измеря-
ют люксметрами.
Растения различаются по кардинальным точкам светового до-
вольствия. Так, большинство лесных видов избегает полного ос-
вещения и характеризуется разным переносимым максимальным
световым довольствием (Lmax). Виды сильно отличаются и по
минимальному световому довольствию (Lmin) — наименьшей от-
носительной освещенности, при которой у них еще выражен по-
ложительный баланс веществ. Минимальное световое довольствие
определяет минимальную для данного вида потребность в свете.
От нее зависит, например, густота крон: чем меньше эта величи-
на, тем больше сомкнутость крон. Недостаток света препятствует
закладке новых и существованию уже сформированных листьев,
поэтому у светолюбивых видов (березы — Be tula, лиственницы —
Larix, сосны — Pinus sylvestris) с минимальным световым до-
вольствием 10 —20 % кроны ажурны, а у теневыносливых деревь-
ев (пихты — Abies, ели — Picea, бука — Fagus) с минимальным
световым довольствием 1 — 3 % — плотные. Чем хуже общие усло-
вия существования, тем больше света необходимо растению, по-
этому минимальное световое довольствие повышается при низ-
ких температурах, на бедных почвах и т. п. Например, для ореш-
ника (Corylus avellana) на бедных почвах Lmin = 5,3 —4,3 %, а на
богатых — 2 %. Леса на бедных почвах обычно имеют меньшую
сомкнутость крон (требуется больше света).
Световое довольствие в 1 —2 % обычно считается нижним пре-
делом для жизни высших зеленых растений. Ниже начинается так
называемая «мертвая тень», при которой могут встречаться толь-
ко гетеротрофы. А. Визнер называл «мертвой лесной тенью» осве-
щенность ниже ’/70 от полной, однако кислица (Oxalis acetosella)
и ряд папоротников могут существовать при освещенности */юо
от полной. Некоторые наземные водоросли сохраняются в плохо
освещенных местообитаниях до Lmin= 0,1 %. Но в природе есть
виды, приуроченные к местообитаниям с экстремально низкой
освещенностью. Это в первую очередь пещерные растения. Напри-
мер, мох Schistostega osmundacea живет при освещенности 'Лоо,
92
улавливая свет особыми линзовидными светособирающими клет-
ками. У папоротника Asplenium trichomones максимальная по-
требность в свете в обычных условиях лежит в пределах а в
пещерах он переносит снижение освещенности до 1/\ 380. Папо-
ротник венерин волос (Adianthum capillis veneris) обнаружен в
пещерах Средиземноморья и при освещенности У^оо- К крайне
теневыносливым фотоавтотрофам относятся некоторые цианобак-
терии и зеленые водоросли, живущие в трещинах скал и камней,
а также входящие в состав накипных лишайников. При очень сла-
бом освещении живут многие водные растения. А некоторые водо-
росли и цианобактерии (например, Gloeocapsa и Protococcus) вы-
живают некоторое время при освещенности до 1/25оо (И.М. Куль-
тиасов, 1982).
Поглощение света в фитоценозе. Индекс листовой поверхности.
Растительные сообщества имеют разную вертикальную структуру,
их растения по-разному отражают и пропускают свет, поэтому
световой режим разных фитоценозов неодинаков. В многоярус-
ном сообществе поступающая радиация используется почти пол-
ностью. Благодаря повторному отражению и последовательному
поглощению до почвы доходит так мало света, что световое до-
вольствие составляет там проценты от полной освещенности. Ос-
лабление света проявляется также в травостоях и мохово-лишай-
никовом покрове. Градиент освещенности зависит от густоты об-
листвения и расположения листьев.
Густоту облиствения количественно выражают с помощью ин-
декса листовой поверхности (ИЛП). Он показывает отношение
общей площади листьев к площади поверхности почвы, над кото-
рой они находятся. Чтобы охарактеризовать степень перекрыва-
ния фотосинтетически активных слоев в водоемах, обычно ука-
зывают содержание хлорофилла в заполненном планктоном стол-
бе воды под 1 м2 водной поверхности. Обычно ИЛП выражают в
квадратных метрах площади листьев на 1м2 поверхности почвы.
Отмечено относительное постоянство ИЛП в сомкнутых фитоце-
нозах независимо от видового состава (за исключением хвойных с
особым строением фотосинтезирующих органов). Чаще всего ИЛП
равен 5 — 6. Вероятно, столько слоев листьев оптимально и при
этом продуктивность фитоценоза максимальна. При таком зате-
нении почвы лесные травы, например, обычно проходят полный
цикл развития.
Оптимальная поверхность листьев — важное приспособление
к улавливанию света. При малой плотности листвы световое до-
вольствие отдельных растений было бы больше, но при пересчете
на площадь почвы несомкнутый растительный покров продуци-
рует сухого вещества меньше, чем сомкнутый. Однако если расте-
ния расположены слишком близко друг к другу и листья много-
кратно перекрываются, то в самых затененных местах для постоян-
93
ного поддержания положительного баланса СО2 не хватает света
и урожай с единицы поверхности снижается. Затенение вызывает
также недоразвитие механической ткани, что может повлечь по-
легание растений. В реальных условиях рост ИЛП имеет предел и
потому, что листья не только ассимилируют СО2, но и испаряют
влагу. С ростом ИЛП растет транспирация, поэтому при недоста-
точном водоснабжении растения не могут увеличивать листовую
поверхность.
Густота облиствения растений и расстояние между ними не
только являются важными факторами продуктивности, но и сами
зависят от условий, влияющих на продуктивность. В благоприят-
ной среде ИЛП зависит от интенсивности освещения. Оптималь-
ное его значение наблюдается тогда, когда теневые листья, распо-
ложенные в нижней части крон, имеют положительный баланс
вещества. При недостаточном обеспечении водой и минеральным
питанием растениям не хватает материала для построения боль-
шого листового аппарата и ИЛП не достигает оптимального зна-
чения. Так, в Германии во влажное лето ИЛП в посевах подсол-
нечника (Helianthus annuus) равнялся 9, а при той же агротехни-
ке в сухое лето был в 2 раза меньше (Г. Вальтер, 1968). На экстре-
мально засушливых, переобводненных и чрезвычайно бедных суб-
стратах, в областях с очень коротким вегетационным периодом
всегда создаются слабосомкнутые сообщества с небольшим ИЛП.
Другой важный показатель, влияющий на градиент освещен-
ности в фитоценозе, — расположение листьев. Интенсивность
света, проходящего через разные сообщества, снижается с разной
скоростью. Для полей хлебных злаков и лугов с листьями, рас-
положенными под большим углом (более 3/4 листьев выше 45°),
на среднем уровне травостоя освещенность уменьшается не более
Относительное световое
довольствие, %
чем вдвое по сравнению с открытым
местом (злаковый тип светового гра-
диента). А в сообществах с горизон-
тально расположенными широкими
листьями, например на клеверном поле
или высокотравном лугу, уже на по-
ловине высоты травостоя поглощается
2/3—3/4 падающего света (двудольный
тип светового градиента) (рис. 6.4).
Высокое светопоглощение характерно
Рис. 6.4. Уменьшение освещенности в тра-
вяных сообществах (по В.Лархеру, 1978):
/ — высокотравный разнотравный луг; 2 — вы-
сокозлаковый луг; 3 — клеверное поле; 4 — низ-
козлаковый луг
94
1,5
о
------1----1____I____I
0 12 3 4
Время, мин
Рис. 6.5. Колебания интенсивности света (длина волн 400—700 нм) на
солнечном блике под пологом тропического леса (по О. Bjorkman,
М. Ludlow, 1972)
для знаменитого дальневосточного высокотравья, где травы дос-
тигают 5,5 м высоты. В них поглощается максимально возможное
количество солнечной энергии (85 — 90%) при минимальном от-
ражении и пропускании (0,9 —5,1 %). При этом важное значение
имеют оптимальная архитектоника сообществ и размеры листо-
вой поверхности (Г.А. Белая, В.Л. Морозов, 2002). Леса с боль-
шой сомкнутостью крон уже в верхнем ярусе поглощают так мно-
го света, что на почву радиации попадает очень мало. В них гра-
диент освещенности оказывается таким же или более крутым, чем
под двудольными травами. А в несомкнутых древостоях и лесах из
светолюбивых пород (берез — Betula, сосен — Pinus sylvestris,
эвкалиптов — Eucalyptus) градиент освещенности пологий, как и
в злаковниках.
В течение суток растения испытывают влияние постоянно ме-
няющейся освещенности. Это связано с положением солнца, со-
стоянием атмосферы, движением воздуха, а также с изменением
просветов в верхних ярусах. Так, под пологом леса приборы пери-
одически фиксируют «световые удары», которым подвергаются
травы из-за движения световых бликов в кронах (рис. 6.5). Блики
и затененные участки создают в лесу сложный режим мелькаю-
щего света.
Сезонные изменения светообеспеченности растений. Свет — один
из самых динамичных факторов среды. Сезонные изменения све-
тообеспеченности связаны и с астрономическим положением Зем-
ли около Солнца, и с сезонным ритмом климата (сезоны дождей
и т.д.). На разных широтах сезонная вариабельность светообеспе-
чснности выражена по-разному: от почти полного отсутствия в
зоне экватора до сильно дифференцированных полярных режи-
мов (с полярной ночью зимой и полярным днем летом). Наблю-
дается сезонная изменчивость и качественного состава света.
95
Рис. 6.6. Схема сезонной динамики относительной освещенности под
пологом елового леса (7), дубравы (2) и посева хлебных злаков (3)
(по Т.К. Горышиной, 1979).
За 100% принята освещенность на открытом месте
В растительных сообществах сезонная динамика световых ус-
ловий зависит и от фенологического состояния видов-средообра-
зователей. Облиствение изменяется на протяжении года, поэтому
в листопадных лесах и травяных сообществах характерный гради-
ент радиации устанавливается только после завершения роста
побегов. В лиственных лесах умеренного пояса зимой до почвы
доходит 50—70% света, а при полном облиствении деревьев —
менее 10 %. Вегетационный период здесь имеет два световых мак-
симума — весенний и осенний, а все лето в этих лесах травы
растут при сильном затенении (рис. 6.6). Это особенно характер-
но для лесов с самыми теневыносливыми средообразователями
(буком — Fagus и липой — Tilia).
Растения приспособились к сезонным изменениям светового
режима. Ряд из них (кислица — Oxalis acetosella, фиалки — Viola
и др.) при затенении листвой деревьев снижает интенсивность
дыхания. У других видов газообмен не перестраивается, и летом
они голодают, имея отрицательный баланс органического веще-
ства, а осенью, после осветления древесного полога, их ассимиля-
ция достигает положительных величин. К этой группе принадле-
жат многие лесные виды, развивающиеся летом, — земляника
лесная (Fragaria vesca), звездчатка жестколистная (Stellaria
holostea), зверобой продырявленный (Hypericum perforatum), гру-
шанка круглолистная (Pyrola rotundifolia) и др. При большей до-
ступности света у опушки они развиваются значительно лучше.
К периодам лучшей освещенности приурочено цветение цело-
го ряда трав. Весенние эфемероиды широколиственных лесов цве-
тут ранней весной до облиствения древесного яруса. В европей-
ских лиственных лесах есть также травы, цветущие осенью. Иног-
да молодые древесные растения, например сеянцы дуба и бука,
сбрасывают листву гораздо позднее, чем взрослые деревья, при-
способившись таким образом использовать благоприятные осен-
96
ние условия освещенности. А в агрофитоценозах есть пожнивные
сорняки, развивающиеся после уборки урожая, получая в это вре-
мя максимум света.
Под кронами вечнозеленых деревьев в течение всего года све-
товое довольствие остается низким. Так, в дождевом тропическом
лесу максимальное количество света достается опушечным расте-
ниям и ряду деревьев, возвышающихся над сплошным верхним
ярусом. Постоянно темно и в густых ельниках, что сказывается на
обеднении их флористического состава (при максимальном раз-
витии крон в нижнем ярусе остаются только споровые растения).
6.2. Качественные характеристики света,
принимаемого растением
Спектральный состав света и фотосинтетически активная ради-
ация (ФАР). Для оценки светового довольствия растения кроме
количественных характеристик важно знать и качественный со-
став света — соотношение лучей разной длины, или спектраль-
ный состав света. Солнечная радиация представляет собой элек-
тромагнитное излучение в широком диапазоне волн. Световые вол-
ны составляют непрерывный спектр с длинами от 200 до 4 000 нм.
Ультрафиолетовые лучи короче 290 нм губительны для живого,
но они поглощаются озоновым слоем в верхних слоях атмосферы.
Из лучей, достигающих поверхности земли, 10 % составляют уль-
трафиолетовые (с длиной волн от 290 до 400 нм), около 45 % —
видимый свет (400 — 750 нм) и еще 45% — инфракрасная радиа-
ция (750 — 4000 нм). В диапазоне с длинами волн более 4000 про-
исходит тепловое излучение Земли. Спектральный состав света в
разных местообитаниях неодинаков. Так, интенсивность ультра-
фиолетового излучения резко увеличивается в горах на высотах
больше 2 000 м.
Как показал в начале XX в. М. Планк, энергия излучения может быть
представлена в виде порций, названных квантами (для электромагнит-
ного излучения — фотоны). Энергия квантов прямо пропорциональна
частоте излучения: чем больше длина волны, тем ниже частота и тем
меньше энергия квантов. Так, квант ультрафиолетового излучения обла-
дает большей энергией, чем квант синего света, а он в свою очередь
несет больше энергии, чем квант красного света. При соударении с мо-
лекулой квант лучистой энергии может быть поглощен ею, в результате
чего молекула переходит в возбужденное состояние и способна вступать
в реакцию, которая была для нее невозможна, когда она находилась на
более низком энергетическом уровне. Чтобы вызвать определенную хи-
мическую реакцию, квант должен обладать энергией, превышающей
некую критическую величину, характерную для данной реакции. На-
пример, кванты рентгеновских и коротковолновых ультрафиолетовых
97
лучей могут выбивать из атомов электроны, образуя ионы. Кванты види-
мой области спектра несут меньше энергии и не способны вызывать
ионизацию, но если они поглощаются пигментами хлоропластов, то спо-
собствуют превращению диоксида углерода в глюкбзу. Кванты инфра-
красного (теплового) диапазона не могут осуществлять этих реакций, но
способны вызывать другие перестройки молекул, сопряженные с мень-
шим количеством энергии.
При самом остром зрении человек может видеть лучи с длина-
ми волн в диапазоне от 380 до 780 нм. Более коротковолновые и
более длинноволновые лучи для зрения недоступны. Для расте-
ний видимый свет тоже принципиально важен, так как область
поглощения хлорофилла, улавливающего свет для фотосинтеза,
почти полностью совпадает с абсорбционной областью зритель-
ного пурпура человеческого глаза. Свет с длиной волн от 380 до
710 нм называют фотосинтетически активной радиацией (ФАР).
Это та часть радиационного спектра, кванты которой способны
приводить хлорофилл в возбужденное состояние, за счет чего осу-
ществляется фотосинтез (рис. 6.7). В суммарной солнечной ради-
ации содержание ФАР составляет около 50 %. Оно зависит от вы-
соты Солнца (времени суток и времени года) и прозрачности ат-
мосферы. Прямая солнечная радиация содержит от 28 до 43 %
ФАР. При облачности рассеянная радиация может содержать ФАР
порядка 50—60%.
В падающей на Землю солнечной радиации на видимую часть
света приходится максимум энергии. Но, проходя через атмосфе-
ру, солнечный свет сильно меняется. Поэтому хотя на верхней ее
границе максимум энергии света приходится на синие лучи (с
Рис. 6.7. Солнечная радиация и интенсивность фотосинтеза в лучах
разной длины волны (по И. М. Культиасову, 1982):
Е — распределение энергии падающей на Землю радиации; Р — относительная
интенсивность фотосинтеза. Лучи: УФ — ультрафиолетовые; Ф — фиолетовые;
С — синие; 3 — зеленые; Ж — желтые; О — оранжевые; К — красные
98
длиной волн 470 нм), на поверхности почвы при высоком солнце-
стоянии летом максимум энергии падает уже на желтую часть спек-
тра, а при низком положении Солнца (на восходе или закате) —
на красную.
Отражение, пропускание и поглощение света растениями. Часть
падающей на лист радиации отражается, часть проходит насквозь,
а часть поглощается и может оказывать на него физиологическое
воздействие. Все эти процессы избирательны, так как зависят от
длин волн (рис. 6.8). Лист отражает около 70% инфракрасных
лучей, 6—12% ФАР и менее 3% ультрафиолета. Зеленый свет
отражается в 2 — 3 раза сильнее, чем, например, красный, поэтому
мы воспринимаем листья зелеными. Ультрафиолет иногда сильно
отражается цветками, служа ориентиром для воспринимающих его
насекомых — опылителей. Каротиноиды отражают свет с длина-
ми волн, соответствующими желтой и оранжевой части спектра, а
поглощают главным образом в синей и зеленой. Отражающая спо-
собность листа зависит от характера его поверхности. Так, вой-
лочное опушение ксерофитов увеличивает отражение в 2 — 3 раза.
Пропускание света листом зависит от его строения. Тонкие
листья пропускают до 40 % лучей, а толстые почти не пропуска-
ют. Наибольшая проницаемость отмечается для тех же длин волн,
что и максимальная отражаемость (в зеленой и ближней инфра-
красной области спектра). Поэтому под пологом листьев стоит
красновато-зеленая тень (лучи с длинами волн 500 и 800 нм).
Свет может вызывать фотохимический эффект лишь после по-
глощения. Вещества, поглощающие видимый свет, называются
Рис. 6.8. Относительное отражение, пропускание и поглощение радиа-
ции листом тополя {Populus deltoides) в зависимости от длины волны па-
дающих лучей (по В.Лархеру, 1978)
99
пигментами. Какие именно длины волн он будет поглощать, за-
висит от числа и расположения двойных связей и ароматических
колец в его молекуле. В результате поглощения кванта света рас-
пределение электронов в молекуле пигмента изменяется и он пе-
реходит в «активированную» форму. Лист как поглощающая сис-
тема очень эффективно останавливает определенные виды волн.
Так, 95 —98 % ультрафиолета задерживает эпидермис, около 70 %
ФАР поглощают пигменты хлоропластов. Спектр поглощения хло-
рофилла имеет максимумы в синих и красных лучах, которые наи-
более активны в фотосинтезе. В поглощении света участвуют так-
же желтые пигменты — каротиноиды, которые наряду с хлоро-
филлом в большом количестве содержатся в хлоропластах.
Красные, бурые водоросли и цианобактерии наряду с хлоро-
филлом включают большое количество пигментов из группы фи-
кобилинов (фикоэритрин, фикоцианин, аллофикоцианин и др.),
а также специфические каротиноиды (фукоксантин, перидинин
и др.). По сравнению с зелеными растениями спектр поглощения
света у этих организмов заметно отличается (рис. 6.9). Фикоэрит-
Рис. 6.9. Спектры поглощения света водорослями и цианобактериями
(по Е. Gabrielsen, 1960; А. Гэлстону и др., 1983):
А — зеленая водоросль Ulva taeniatcr, Б — бурая водоросль Coilodesme califomiccr,
В — красная водоросль Porphyra naiadum, Г — спектры поглощения пигментов
цианобактерий; 1 — хлорофилл а и каротиноиды; 2 — фикоэритрин; 3 — фико-
цианин; 4 — аллофикоцианин; 5 — хлорофилл а
100
рин хорошо поглощает радиацию в сине-зеленой области спектра
и потому выглядит красным, а фикоцианин и аллофикоцианин
наиболее интенсивно поглощают ее в желтом и красном диапазо-
нах и соответственно окрашены в синий или зеленый цвета. По-
глощение света фикобилинами хорошо заполняет разрыв в той
области, где хлорофилл поглощает свет слабо. Есть также водо-
росли, у которых соотношение разных фикобилинов зависит от
спектрального состава света. Они меняют окраску при освещении
светом разных длин волн. Это позволяет глубоководным водорос-
лям, несмотря на иное количество и качество там света, погло-
щать достаточно необходимой для фотосинтеза энергии.
Спектр поглощения листьев в целом примерно соответствует сумме
спектров поглощения фотосинтетических пигментов, однако значитель-
ное количество света в оранжевой области поглощают еще органические
соединения, не участвующие в фотосинтезе. Показательно, что ближняя
инфракрасная радиация (700 —3 000 нм) почти не поглощается: лист рас-
сеивает большую ее часть. Роль этого явления состоит в том, чтобы умень-
шить тепловую нагрузку от волн тех длин, которые не используются в
фотосинтезе. Однако дальняя инфракрасная радиация (тепловая) пере-
хватывается почти полностью (97 %). А поскольку хорошие поглотители
служат хорошими источниками излучения, листья способны эффектив-
но рассеивать избыток тепла в длинноволновой области спектра (Фото-
синтез и биопродуктивность: методы определения, 1989).
Листья разных типов имеют разные спектры поглощения
(рис. 6.10). Так, толстые, сильно пигментированные листья ин-
жира (Ficus carica) поглощают 85 % зеленого света, а листья табака
(Nicotiana) — только 50%. В целом вследствие избирательного
поглощения растениями свет, проникающий под полог высоко-
сомкнутого сообщества (например, леса), оказывается специфи-
чески изменен. Он относительно богат в зеленой и инфракрасной
частях спектра, но беден в красно-оранжевой и голубой.
Физиологическое действие частей солнечного спектра на расте-
ния. Физиологические эффекты радиации зависят от поглощения
квантов разной энергии (света разных длин волн). Кванты погло-
щают разные рецепторы. Радиацию для фотосинтеза поглощают
хлорофиллы и дополнительные пигменты. Фитохромной систе-
мой растения регулируются такие фотоиндуцируемые процессы,
как прорастание семян, образование цветков, листопад, сезонное
чередование активности и покоя. Это относится и к фотоморфо-
ш — зависимым от света дифференцировкам тканей и внешним
формообразовательным процессам. Направленный рост (фото-
тропизм) регулируется рецепторами синего света типа флаво-
протеидов или каротиноидов.
По действию лучей на растение солнечный спектр можно раз-
делить на 4 физиологические зоны. Волны длиной 300—520 нм
101
500 1000 1500 2000
Длина волны, нм
юо
80
60
40
20
400 500 600 700
Длина волны, нм
В
Рис. 6.10. Поглощение лучистой энер-
гии листьями (по И. А. Шульгину, 1967,
1973):
А — «средний» лист (обобщенные данные по
измерениям на 1 400 видах); Б — листья ра-
стений разных экологических типов: 1 — ме-
зофиты, 2 — ксерофитьГй растения с ксеро-
морфными листьями, 3 — суккуленты; В —
диапазон измерений поглощения в пределах
одного вида: 4 — световые листья, 5 — тене-
вые листья
поглощаются пигментами, фермен-
тами, клеточными оболочками, раз-
ными частями цитоплазмы, но боль-
шая их часть поглощается хлорофил-
лом. Волны длиной 520 — 700 нм
(оранжево-красные лучи) поглоща-
ются исключительно хлорофиллом и
имеют решающее значение для мно-
гих физиологических процессов (фо-
тосинтез, рост, развитие и т.д.). Вол-
ны длиной 700—1050 нм (инфра-
красные лучи) играют незначитель-
ную роль в жизни растений («абио-
тическая радиация»). Волны длиной
выше 1 050 нм (2 000 нм) — инфракрасные лучи дальнего крас-
ного света, поглощающиеся цитоплазмой и водой, важны для
теплового режима растений и играют регуляторную роль в об-
менных процессах. Инфракрасные лучи обычно вызывают на-
гревание листа, но в некоторых условиях они полезны и для
фотосинтеза. Так, при низких температурах эти лучи частично
поглощает хлорофилл и их устранение приводит к снижению
продуктивности.
В области видимого света хлорофилл имеет два максимума по-
глощения. Наиболее интенсивно лист поглощает оранжево-крас-
ные и красные лучи (600 — 680 нм). Меньший максимум погло-
щения находится в сине-фиолетовых лучах (460—490 нм). Эти
характеристики чрезвычайно стабильны: они проверены на мно-
гих сотнях растений разных по экологии, географии и системати-
ческому положению. Поэтому можно говорить о листе как уни-
версальном устройстве для поглощения солнечной радиации. Ло-
кализация главного максимума поглощения листа эволюционно
объяснима: на красно-оранжевые лучи падает максимум энергии
в спектре солнечной радиации, поэтому именно зеленый цвет листа
обеспечивает лучшее энергоснабжение растения. Минимум по-
глощения света приходится на желто-зеленые лучи (550—575 нм).
102
Непосредственно на фотосинтез они влияют слабо, но, видимо,
служат источником сильного светового возбуждения.
В ультрафиолетовой части спектра действует несколько групп
лучей. Как известно, жесткий ультрафиолет губительно воздей-
ствует на живое. Но волны короче 280 нм, сильно повреждающие
генетический материал, в биосферу не проникают. Более длин-
ные УФ-лучи проходят в растение неглубоко, так как их эффек-
тивно задерживает эпидермис, который сам может отмереть, но
глубжележащие клетки ассимиляционной ткани обычно оказыва-
ются неповрежденными. На хлорофилл УФ-лучи почти не дей-
ствуют, но стимулируют его образование у выросших в темноте
бледных растений. Эффект действия на клетки ультрафиолетовых
волн с длинами около 300 нм зависит от дозы облучения (при
малых дозах — стимуляция, при больших — угнетение).
Повреждающее действие света. Свет может вызывать повреж-
дения эпидермы, а также изменять химический состав более глу-
боких тканей. Обилие света особенно опасно для растений, испы-
тывающих недостаток воды или минерального питания. У не за-
щищенных от избытка света организмов это вызывает глубокие
нарушения обмена веществ и может приводить к гибели. Расте-
ния бедных минеральными веществами местообитаний (верховые
болота, песчаные почвы и др.) развивают ряд приспособлений,
защищающих их от избытка света (опушение, утолщенная кути-
кула, выработка экранирующего свет пигмента антоциана).
Поглощение света в присутствии кислорода может нарушать
многие структуры и функции клеток растений (фотодинамиче-
ское действие). На клеточном уровне фотодинамическое действие
вызывает стимуляцию или торможение деления клеток, мутаген-
ные эффекты, бактерицидные проявления, повреждение биомем-
бран. Этому действию подвержены многие физиологические и био-
химические процессы — фотосинтез, дыхание, окислительное фос-
форилирование. В основе многих эффектов лежит повреждение
белков при окислении входящих в них аминокислот. Наследствен-
ный аппарат клетки страдает от разрушения светом азотистых
оснований. Избыток света вызывает также изменения в хлоро-
филле, накопление токсичных активных форм кислорода и про-
дуктов перекисного окисления липидов.
Хорошо известно повреждающее действие УФ-лучей. У орга-
низмов, произрастающих в условиях сильной УФ-нагрузки, вы-
работался ряд специфических особенностей. У их листьев изме-
няются цвет и блеск, уменьшается их площадь и увеличивается
толщина (за счет развития покровной и столбчатой тканей). Од-
нолетники могут превращаться в многолетники, появляется розе-
гочность и т. п. В ряде случаев клеточные структуры надежно эк-
ранированы от ультрафиолета опушением, а также накапливаю-
щимся в эпидерме антоцианом. Этот пигмент придает характер-
103
ный красноватый оттенок ряду высокогорных растений и видов
верховых болот, а также некоторым молодым листьям древесных
растений.
В условиях удовлетворительного питания растений сильный
видимый свет, попадая на клетки после действия УФ-лучей, умень-
шает повреждающее действие ультрафиолетового излучения. Это
явление называется фотореактивацией. Она возникла как за-
щита от губительного воздействия ультрафиолета и является од-
ной из важнейших форм репарации, предохраняющей от повреж-
дений генетический аппарат (см. разд. 4.1.6). Для разных жизнен-
ных процессов (синтез гормонов, дифференциация меристем и
др.) характерна неодинаковая степень фотореактивации.
Состав и использование растениями рассеянного света. Спект-
ральный состав прошедшего через облака солнечного света обед-
нен рассеивающимися коротковолновыми ультрафиолетовыми и
сине-фиолетовыми лучами. В рассеянном свете по сравнению с
прямым оказывается больше полезных для фотосинтеза оранже-
во-красных лучей и до 60% ФАР (рис. 6.11). Кроме того, по-
скольку хлорофилл лучше всего воспринимает лучи красной и
синей частей спектра, то слабый свет, поступающий к растениям
при облачности и низком стоянии солнца, усваивается ими пол-
нее. В прямом же свете важных для фотосинтеза лучей обычно
лишь около трети. А прямые лучи большой интенсивности для
растений опасны.
Длина волны
Рис. 6.11. Относительное распределение энергии в спектре солнечной
радиации при высоте солнца 30° (по Ю.Д.Янишевскому, 1957; О. Авасте
и др., 1962):
1 — прямая радиация; 2 — рассеянная радиация. Длина волны дана в логариф-
мическом масштабе
104
В ясные дни рассеянный свет составляет 10—15% от общей
радиации, а в пасмурные — 100 %. Но соотношение между рассе-
янной и прямой солнечной радиацией, между интенсивностью
света и его спектральным составом неодинаково на разных широ-
тах, при разной континентальности, океаничности, на разной
высоте над уровнем моря. Оно зависит от состояния атмосферы,
рельефа, характера растительности и т.д. Эти соотношения от-
личаются в разные часы дня, в разные сезоны и разные годы.
Как правило, утром и вечером из-за низкого стояния Солнца
больше рассеянного света с преобладанием длинноволновой радиа-
ции, а в полдень преобладает свет прямой, богатый короткими
волнами.
Коэффициент использования рассеянного света, характеризующего-
ся в среднем */10 интенсивности прямых лучей, велик. Поэтому в высо-
ких широтах преобладающего рассеянного света вполне достаточно для
активного фотосинтеза и продуктивность растений здесь ограничивает-
ся не светом, а низкими температурами. Например, в Крыму количество
солнечной энергии, приходящейся в течение года на единицу площади,
в пять раз больше, чем на Шпицбергене, и в два раза больше, чем в
Санкт-Петербурге, но если учесть рассеянный свет, которого на севере
больше, за 4 мес вегетационного периода эти различия сильно сглажи-
ваются. Так, если иметь в виду только световое довольствие, то с 1 га на
Шпицбергене можно было бы собрать столько же продукции, сколько с
1 га в средней полосе России. Но из-за недостатка тепла в Арктике зна-
чительная часть света оказывается недоиспользованной.
Обычно отмечается, что при движении от экватора к полюсам
поглощение солнечной энергии атмосферой увеличивается и ее
поступает на поверхность почвы все меньше. Однако одновре-
менно увеличивается и количество рассеянного света. Поэтому в
умеренном поясе летом суммарной солнечной энергии получает-
ся больше, чем на экваторе, причем падает и содержание УФ-
лучей.
Световой режим водоемов и распространение в них растений.
Особый световой режим складывается в водоемах. Он зависит от
общей освещенности над водой, степени отражения водной по-
верхностью, поглощения и рассеяния света в воде. На него силь-
но влияют гидродинамические показатели — турбулентность, про-
зрачность, цветность и мутность воды. Во многих водоемах эти
показатели характеризуются сезонной изменчивостью. Например,
в холодное время промерзшие грунты меньше эродируются река-
ми, мутность воды падает и наблюдаются вспышки обилия ряда
видов фитопланктона.
При высоком стоянии солнца гладкая водная поверхность в
среднем отражает 6 % падающего света, при сильном волнении —
около 10%. При низком стоянии Солнца отражение значительно
105
увеличивается и большая часть света в воду не проникает. Поэто-
му под водой день короче, чем на суше.
Вода ослабляет радиацию намного сильнее атмосферы. Длин-
новолновые тепловые лучи поглощаются уже в верхних милли-
метрах водяного слоя, инфракрасная радиация — в верхних сан-
тиметрах. УФ-лучи проникают на глубину от нескольких деци-
метров до метра. Но ФАР достигает больших глубин. С глубиной
интенсивность радиации убывает экспоненциально. Свет погло-
щается и рассеивается водой, растворенными веществами, взве-
шенными частицами и планктоном. В мутных водах уже на глу-
бине около 0,5 м количество света может уменьшиться до 7 %
(как под пологом темного елового леса). Разные глубины характе-
ризуются и разной освещенностью, и разным спектральным со-
ставом света (рис. 6.12). Красное излучение, как правило, не про-
никает ниже 34 м, желтая часть спектра задерживается на глубине
около 177 м, а зеленая — на глубине 322 м. На больших глубинах
царит слабое зеленовато-голубоватое освещение. Глубже 500 м не
проникают даже синие и фиолетовые лучи.
Слой воды ниже границы обитания автотрофных растений на-
зывается дисфотической зоной, а выше ее — эуфотической.
В прозрачных озерах 1 % падающей ФАР может достигать глуби-
ны 5—10 м, поэтому высшие растения могут встречаться там на
пятиметровой глубине, а прикрепленные ко дну водоросли и на
глубинах до 20—130 м. В открытом море граница эуфотической
зоны лежит глубже. Так, в прибрежной полосе Средиземного моря
1 % радиации проникает до глубины 60 м, а в прозрачных водах
океана — до 140 м.
Разные группы водорослей неодинаково относятся к спектраль-
ному составу достигающего их света. В процессе фотосинтеза одни
из них (зеленые, бурые, диатомовые) преимущественно использу-
ют красные и в меньшей степени синие лучи спектра. Другие во-
Рис. 6.12. Ослабление в воде цветных
лучей (по G. Clarke, 1957).
Лучи: УФ-Ф — ультрафиолетовые — фио-
летовые, С, Г — синие и голубые, 3 — зеле-
ные, Ж — желтые, О — оранжевые, К —
красные. Шкала по оси абсцисс логарифми-
ческая
106
доросли (красные и цианобактерии), наоборот, больше использу-
ют синие и в меньшей степени красные лучи. Они активно фото-
синтезируют благодаря присутствию красного пигмента фико-
эритрина. Представители красных водорослей доходят в морях и
океанах до самых больших глубин, а распространение большин-
ства зеленых водорослей, имеющих фотосинтетические пигмен-
ты, поглощающие ту же часть спектра, что и наземные растения,
ограничено поверхностными водами. Т. Энгельман назвал это яв-
ление хроматической комплементарной адаптацией водорослей.
Некоторые организмы (например, цианобактерии) изменяют
свою окраску под влиянием цвета освещающих лучей и интен-
сивнее усваивают СО2 в дополнительных к их окраске лучах спек-
тра. Это явление связано с развитием других пигментов, особенно
фикобилинов и каротиноидов. Так, если эти организмы выращи-
вать на красном свету, у них будет преобладать фикоцианин, а
при освещении зеленым светом главное место займет фикоэрит-
рин. Способность к хроматической адаптации обнаружена не у
всех водорослей. Некоторые выживают на глубине благодаря син-
тезу большего количества пигмента.
К количеству света водоросли в целом очень нетребовательны.
Обычно они довольствуются очень слабым освещением, гораздо
меньшим, чем наземные растения. Проникающий свет лимитирует
глубину расселения водорослей, как правило, не превышающей
150 м. Так, в прозрачной морской воде основная масса видов
фитопланктона размещается на глубине 40 — 70 м, иногда опус-
каясь до 100 —120 м. В озерах они сосредоточены на глубине 10—
15 м, а в водах с малой прозрачностью — в поверхностном слое
толщиной не более 3 м.
Распространение в глубину фитобентоса часто зависит не только
от количества проникающего света, но и от возможностей его
использования. Степень использования света зависит от сопря-
женного действия целого ряда факторов: температуры, характера
грунта, содержания в воде минеральных и органических веществ.
Так, интенсификации фотосинтеза способствует постоянное об-
новление водных масс. Косвенно это подтверждает тот факт, что
бентосные водоросли наиболее пышно развиваются именно в ме-
стах с интенсивным перемешиванием воды (в реках и ручьях —
на перекатах, где вода течет с большой скоростью, в морях — в
проливах с течениями, в прибрежной прибойной зоне). Глубина,
на которой разрастаются бентосные растения, в разных водоемах
неодинакова. Дно глубоких бассейнов свободно от фотоавтотроф-
ных водорослей. Глубина, на которую они опускаются в разных
морях, неодинакова. Например, в северном регионе граница встре-
чаемости бентосных растений лежит на незначительных глуби-
нах, около 40 — 50 м, у берегов Флориды морские водоросли опус-
каются до глубины 100 м, а в отличающемся прозрачностью Сре-
107
диземном море они растут на глубине 130 и даже 180 м (Л.Л. Ве-
ликанов и др., 1981).
6.3. Свет и функционирование растений
Световой фактор оказывает значительное влияние на рост, раз-
витие и размещение растений и их сообществ. Действие света на
растения можно подразделить на фотосинтетическое, регулятор-
но-фотоморфогенетическое и тепловое. Свет определяет многие
фотобиологические явления: фотопериодизм, фотоморфогенез,
фототаксисы, фототропизмы, фотонастии и др. Формообразую-
щее действие света сказывается на облике растительного покрова
разных областей Земли. Состав и структура растительных сооб-
ществ тесно зависит от требований растений к свету.
Влияние интенсивности освещения на рост и развитие растений.
Зеленое растение — не просто механизм, работающий на солнеч-
ной энергии: сама структура этого механизма частично определя-
ется количеством и качеством получаемой лучистой энергии
(А. Гэлстон и др., 1983). Зависимые от света процессы роста и
развития растения, определяющие его форму и структуру, назы-
ваются фоупоморфогенезом. В ходе его растения оптимизируют
свою структуру для поглощения света в конкретных условиях.
Известно действие света на рост растений через фотосинтез:
при слабой освещенности они растут плохо. При недостатке света
проростки гибнут, израсходовав запас питательных веществ семе-
ни. Так, всходы ели (Picea) часто погибают под кроной ее взрос-
лых особей. Затенение — один из существенных факторов повы-
шенной смертности и медленного развития молодых растений на
лугах (отчасти это связано со снижением при затенении устойчи-
вости к паразитам) (Т. А. Работнов, 1985). Для фотосинтеза требу-
ется довольно высокий уровень энергии, но воздействие на рост и
развитие растений возможно и при недостаточной для фотосин-
теза интенсивности света. В этом случае играет роль сигнальное
действие света, запускающее физиологические реакции.
Проростки некоторых растений появляются глубоко в почве,
куда свет не проникает. У них формируются светлые тонкие быс-
тро вытягивающиеся (этиолированные) побеги с искривленным
подсемядольным коленом и маленькими листьями. Адаптивный
смысл этого — в экономном и быстром росте таких проростков:
уязвимая меристема верхушки побега защищена изгибом, а на рост
листьев энергия не тратится, так как в этих условиях они не могут
функционировать. Когда же проросток достигнет поверхности
почвы, изгиб стебля выпрямляется, скорость удлинения умень-
шается, листья увеличиваются и зеленеют. Рост затененных лис-
товым пологом растений сходен с ростом этиолированных проро-
108
стков под землей (вытянутые тонкие стебли и неразвитые лис-
тья). Эти важные для выживания особенности регулируются све-
том через изменение фоторецепторов при разной освещенности.
Большая освещенность, напротив, ингибирует вытягивание
междоузлий, тоже влияя на форму и размеры формирующегося
побега. В тени листья вырастают крупнее, чем на свету. Сильная
коротковолновая радиация в горах интенсифицирует заложение
новых почек, активизирует их рост и развитие, ускоряет отложе-
ние запасных веществ. Поэтому там формируются многочислен-
ные подушечные формы с сильно укороченными стеблями. Ис-
кусственное экранирование ультрафиолетовой части спектра у ряда
растений ведет к ускорению развития на несколько дней, умень-
шению роста боковых побегов, увеличению длины стеблей, а так-
же числа и величины листьев.
У древесных растений на разных стадиях онтогенеза освещен-
ность может оказывать разное воздействие, но в целом рост в
высоту почти не меняется при изменении интенсивности света от
50 до 90 % от полной освещенности. Однако в более темных усло-
виях проявляется карликовость. Уменьшение освещенности при-
водит к снижению прироста. В зависимости от теневыносливости
критическое значение, при котором баланс органических веществ
становится равен нулю и прирост останавливается, у разных рас-
тений варьирует. Теневыносливые виды страдают в меньшей сте-
пени. Так, прирост прекращается у осины (Populus tremula) при
интенсивности света 4,5 %, у клена (Acer), дуба (Quercus robur) и
ясеня (Fraxinus excelsior) — при 3 %, у липы (Tilia cordata) —
при 2 %. Аналогично реагируют на недостаток света споровые ра-
стения. Так, мох Pleurozium schreberi, перекрытый лопастями ли-
шайника Peltigera aphtosa, уменьшает продукцию фотосинтеза,
не образует спорогонов, желтеет и отмирает.
Недостаточное освещение подавляет цветение. Поэтому при
высокой облачности вегетативные органы, развиваясь за счет ре-
продуктивных, могут давать больший урожай. Однако успех плодо-
ношения зависит от многих условий, и в целом для влажного мор-
ского климата увеличение числа ясных дней полезно, а для сухого
континентального предпочтительнее увеличение облачности. На
цветение большое влияние оказывает также продолжительность
длины дня (см. разд. 6.6.1).
Действие на растения красного и синего света. Хлорофилл и
вспомогательные пигменты фотосинтеза в процессах фотоморфо-
генеза не играют главной роли. Эти реакции индуцируются све-
том, включающим особые фоторецепторы. В растениях обнару-
жены две пигментные системы фоторецепторов, связанные с мем-
бранами клеток, — фитохром, поглощающий красный свет, и крип-
тохром, поглощающий синий свет. Содержание этих пигментов в
растениях чрезвычайно мало, и энергия, необходимая для насы-
109
щения соответствующих фотопроцессов, на несколько порядков
ниже, чем при фотосинтезе. А.А.Красновский (1975) отмечает,
что данные пигменты поглощают ничтожную долю падающего сол-
нечного излучения, которая используется для переключения ме-
таболических путей по сигнальному типу. Так, если при фотосин-
тезе необходимо 8—10 квантов света для высвобождения всего
одной молекулы О2, то это же число квантов на одну клетку мо-
жет полностью определить репродуктивную судьбу растения или
направление роста всего побега.
Фитохром — фикобилиновый пигмент, состоящий из не-
большого поглощающего свет хромофора и крупного бесцветного
белка. После поглощения света определенной длины волны фор-
ма хромофора изменяется, что меняет и форму белкового компо-
нента. В растениях фитохром существует в двух взаимно-обрати-
мых формах. Одна образуется под действием красного света (Фк),
а другая — дальнего красного (ФДК). Синтезируется фитохром в
форме Фк (и этиолированные проростки содержат его), а облуче-
ние превращает большую его часть в физиологически активную
Фдк-форму. Соотношение красного и дальнего красного света важ-
но для роста ряда растений в закрытых сообществах.
Превращение Фк в Фдк при кратковременном воздействии крас-
ного света влияет на биохимические реакции растений в течение
нескольких последующих часов темноты. Стойкость эффекта крат-
кого облучения зависит от того, что Фдк, будучи «активной» фор-
мой фитохрома, относительно стабилен в темноте. При +25 °C
период полужцзни его в этиолированной ткани составляет около
2 ч (т. е. за 2 ч содержание Фдк снижается до половины его перво-
начального количества), а в ткани на свету достигает 8 —
12 ч. В некоторых опытах с растущими на свету тканями актив-
ность Фдк можно обнаружить и спустя 72 ч после облучения крас-
ным светом. В отличие от этого активированные формы других
пигментов растений очень нестабильны (у возбужденного хлоро-
филла, например, период полужизни составляет всего несколько
миллисекунд).
Фитохром находится во всех клетках растения, но больше все-
го его в меристемах. Он участвует в регуляции многих видов
жизнедеятельности растений: прорастании светочувствительных
семян, развитии гипокотиля проростков, развертывании семядо-
лей, дифференциации эпидермиса и устьиц, формообразовании
тканей и органов, ориентации хлоропластов, синтезе антоциана и
хлорофилла, контроле метаболизма фитогормонов. Фитохром оп-
ределяет фотопериодическую реакцию растений, регулирует на-
чало цветения, опадение листьев, клубнеобразование, старение и
переход в состояние покоя. Красный свет при этом тормозит де-
ление и способствует удлинению клеток: растения вытягиваются,
становятся тонкостебельными.
ПО
Синий свет также регулирует многие фотоморфогенетические
и метаболические реакции растений. Фоторецепторы синего све-
та — флавины и каротиноиды — имеются в клетках всех тканей.
Желтый пигмент, рецептирующий синий (ближний ультрафиоле-
товый свет, называемый криптохромом), присутствует у всех ра-
стений. В ультрафиолетовой части спектра, вероятно, работает еще
одна воспринимающая система.
Рецепторы претерпевают превращения, быстро передавая элек-
троны другим акцепторам. Фототропизм растений определяется
рецепторным комплексом стеблевого апекса, включающего крип-
тохром и каротиноиды.
Синий свет влияет на многие процессы: открывание устьиц,
движение цитоплазмы и хлоропластов, развитие листьев, угнетает
прорастание семян и др. Он вызывает фототропический изгиб
проростка и других осевых органов. Синие и фиолетовые лучи
стимулируют деление, но задерживают растяжение клеток, спо-
собствуя формированию мелкоклеточности и карликовости рас-
тений. При недостатке синего света в загущенных посевах и по-
садках растения вытягиваются и полегают. Это случается и в теп-
лицах, стекла которых задерживают синие и сине-фиолетовые лучи.
Влияние света на транспирацию растений. Высокая интенсив-
ность света сказывается и на повышении транспирации. Свет сти-
мулирует открывание устьиц, повышает чувствительность мемб-
ран.
При хорошем водоснабжении устьица открываются на свету
тем шире, чем больше интенсивность освещения (фотоактив-
ное открывание).
Краткое яркое освещение действует на устьица по-разному в
зависимости от теневыносливости растений. Так, устьица тене-
выносливого бука (Fagus) открываются значительно быстрее, чем
у светолюбивого тюльпанного дерева (Liriodendron tulipifera). Кро-
ме того, на разных сторонах листа устьица отличаются разной
чувствительностью к свету. Так, у табака (Nicotiana) закрывание ус-
тьиц на верхней поверхности листа вызывается освещенностью в
10 раз большей, чем на нижней поверхности. При иссушении те-
невые растения под воздействием света закрывают устьица быст-
рее, чем световые. Таким образом, устьица листьев растений зате-
ненных местообитаний реагируют на повышение освещенности
быстрее, и их реакция меньше ограничена низкой освещеннос-
тью, чем у растений открытых мест. Это дает им возможность
четче регулировать фотосинтез при значительных колебаниях ос-
вещенности.
Влияние света на прорастание семян. Свет сказывается на про-
растании семян неоднозначно. Для разных растений его воздей-
ствие может быть прямо противоположным. В одних случаях он
способствует ему, в других тормозит.
111
Свет ускоряет прорастание семян моркови (Daucus carota), конского
щавеля (Rumex confertus), ели (Picea), сосны (Pinus). Световая стимуля-
ция требуется семенам посевного салата (Lactuca sativa) и коровяка
(Verbascum). Много света необходимо для прорастания мятликов (Роа).
Только на свету прорастают семена осоки песчаной (Carex arenaria) и
ряда горных осок (С. albonigra, С. ebenea) (Ю.Е.Алексеев, 1996). На
свету летом дозревают семена бутеня (Chaerophyllum tainturieri), выйдя
из состояния характерного для них неглубокого покоя, и если не будет
достаточного количества света и высокой температуры (20 —35 °C), до-
развитие их зародыша не начнется и, осенью семена снова впадут в со-
стояние покоя (М. Г. Николаева, 2002). Напротив, многие лилейные,
тыквенные, примулы (Primula) и однолетний костер (Bromus tectorum)
лучше прорастают в темноте. А для семян табака (Nicotiana) достаточно
побыть освещенными в течение 0,01 с.
Способность семян прорастать на свету лучше, чем в темноте,
со временем меняется. Потребность в световом влиянии на про-
растание семян может быть изменена разными воздействиями (тем-
пература, кислород, азот, pH). Например, семенам многих сорня-
ков для прорастания свет не нужен, но потребность в нем возни-
кает при помещении их в почву. У многих растений семена ста-
новятся чувствительными к свету при смачивании. Набухнув и
получив необходимую дозу света, они могут высыхать и хорошо
прорастать потом и будучи покрытые почвой. Полагают, что это
явление связано с изменением состояния фитохрома. В сухом со-
стоянии он дегидратируется и не способен к взаимопревращени-
ям Фк и Фдк. А во влажной почве фитохром семян частично гид-
ратируется и происходит его реверсия, которая у хранящихся се-
мян служит эффективным механизмом выживания.
Свет, проходя через полог зеленых листьев и в результате из-
бирательного поглощения хлорофиллом волн определенной дли-
ны, относительно обогащается лучами дальней красной области
по сравнению с красной. Это препятствует прорастанию чувстви-
тельных к свету семян после развития листового полога. Поскольку
прорастание в среде с недостаточным освещением приводило бы
к плохому росту, подавление прорастания, обусловленное отсут-
ствием Фдк, имеет значение для выживания. В начале следующего
вегетационного сезона перед развитием листового полога семена
смогут прорасти и проростки будут хорошо развиваться (А. Гэл-
стон и др., 1983).
Влияние света на движение растений и их органов. Побеги и
корни многих растений растут, поворачиваясь в соответствии с
положением источника света. Ростовые изгибы органов растений
под влиянием одностороннего освещения называются фототро-
пизмом. У стеблей он положителен, т.е. они растут в сторону
источника света. Особенно ярко это проявляется у светолюбивых
видов. А корни обычно обладают положительным геотропизмом
112
и отрицательным фототропизмом и растут от источника света (ис-
ключение составляют дыхательные корни некоторых тропичес-
ких растений илистых субстратов). А у многих листьев, распола-
гающихся перпендикулярно к направлению света, фототропизм
поперечный. Предполагают, что в рецепции света для этих про-
цессов принимают участие каротиноиды и флавины. Сам же на-
правленный рост органов вызывается перераспределением гормо-
нов роста (ауксинов), вырабатываемых апексами стебля и корня.
Фототропические реакции подчиняются «закону количества раз-
дражения», т.е. важны не сами по себе интенсивность и продол-
жительность освещения, а доза полученного света.
Фототропизм связан с торможением роста на освещенной сто-
роне органа, что приводит к его повороту или деформации. При
равномерном освещении листья обычно располагаются более или
менее горизонтально. Однако у ряда растений (эвкалипт —
Eucalyptus, гладиолус — Gladiolus, ирис — Iris, многие злаки)
расположение листьев близко к вертикальному. Положение лис-
тьев «компасных растений» позволяет им избегать полуденной пе-
реосвещенности и перегрева и максимально использовать более
мягкий утренний и вечерний свет. Экологическое значение раз-
нообразных фототропических проявлений заключается в том, что
для растения важно сориентировать ассимилирующие органы та-
ким образом, чтобы они получали оптимальное количество света.
Способность подвижных организмов перемещаться к источни-
ку света называется фототаксисом. Он характерен, например,
для планктонных водорослей. Другой его пример — перемещение
хлоропластов под влиянием света: в темноте они располагаются
более или менее равномерно, при слабом освещении концентри-
руются на освещенной стороне клетки, а при сильном прямом
свете переходят к боковым стенкам. Подобные перемещения и
здесь связаны с достижением оптимального освещения. Еще иногда
отмечают фотокинезис — ненаправленное увеличение или умень-
шение подвижности организма в ответ на изменение степени ос-
вещенности.
Фотонастии — это движения органа растения в зависимости
от изменения интенсивности света. В этом случае направление
роста стеблей, листьев, лепестков вызывается неравномерным ос-
вещением и обусловлено в первую очередь строением органа.
Например, направление роста боковых побегов шалфея (Salvia)
зависит от интенсивности освещения: чем она сильнее, тем гори-
зонтальнее они расположены, но при этом побеги растут во все
стороны, т.е. фототропизма нет. Здесь реакция вызвана светом,
но не его направлением.
У хризантем (Chrysanthemum) побеги на свету растут вертикально,
но если прикрыть конуса нарастания, то они займут горизонтальное по-
113
ложение. А листья у них в темноте растут вертикально (так как сильнее
вытягиваются клетки нижней поверхности листа — гипонастия), а на
свету переходят в горизонтальное положение (т. е. энергичнее растет вер-
хняя сторона — эпинастия). Аналогично у подорожника среднего
(Plantago media) при умеренном освещении листья поднимаются вверх,
а при сильном освещении изгибаются книзу. Подобное явление отмече-
но также у фасоли (Phaseolus) и др.
С влиянием света связаны и никтинастические движения. На-
пример, многие цветки и соцветия (одуванчик — Taraxacum, мать-
и-мачеха — Tussilago farfara) открываются утром и закрываются
к ночи. Некоторые растения раскрывают цветки в определенное
время суток: мак (Рараиег) — на рассвете, лен (Ыпит) — на не-
сколько часов позднее, дурман (Datura stramonium) — вечером.
Аналогичные явления отмечены и у листьев (кроме света оказы-
вает влияние и изменение тургора). Так, у томата (Lycopersicon)
листья, горизонтально расположенные днем, ночью занимают более
вертикальное положение, листочки клевера (Trifolium) на ночь
складываются вдоль средней жилки кверху, складываются и по-
никают при сильном освещении листочки кислицы (Oxalis aceto-
sella). Все эти движения стебля и листьев под влиянием света
тоже связаны с экологической оптимизацией функционирования
растений.
6.4. Свет и фотосинтез
Поглощенная солнечная энергия любых длин волн в конечном
счете приводит к нагреванию главным образом атмосферы и по-
верхности планеты, а также частично обратно излучается в кос-
мическое пространство. Фотосинтез же — уникальный процесс,
позволяющий запасти эту энергию, сделать ее доступной для со-
вершения работы живыми организмами. В процессе фотосинтеза
участвуют вызываемые светом фотохимические реакции, не нуж-
дающиеся в свете ферментативные (темновые) процессы и про-
цессы диффузии, при которых происходит обмен диоксидом уг-
лерода и кислородом между хлоропластами и наружным возду-
хом. Каждый из этих процессов находится под влиянием разных
факторов, которые могут ограничить фотосинтез.
В целом фотосинтез заключается в преобразовании и запаса-
нии солнечной энергии (477 кДж на каждый ассимилированный
моль СО2), в результате чего из простых неорганических синтезиру-
ются сложные органические вещества и выделяется молекуляр-
ный кислород. В общем виде этот процесс можно описать уравне-
нием
Н2О + СО2= О2 +’/6 (С6Н12О6)
114
Условия этой реакции — свет и кислородовыделяющие фото-
синтезирующие организмы. Обратная реакция (окисление орга-
нических веществ с образованием воды и диоксида углерода) пред-
ставляет собой процесс дыхания.
Фотосинтез — очень распространенный в биосфере химиче-
ский процесс, но зеленые растения используют в нем солнечную
энергию малоэффективно. В среднем по всей земной поверхно-
сти на фотосинтез ежегодно затрачивается всего лишь около
138 Дж/см2, т.е. около 1/2ооо наличной энергии. Правда, суще-
ственная часть солнечной радиации падает на Землю там, где ра-
стительности нет. Поэтому если включить в расчет только то коли-
чество света, которое действительно поглощается зелеными расте-
ниями, то эффективность фотосинтеза (отношение запасенной
лучистой энергии к поглощенной) окажется более высокой — около
нескольких процентов (А.Гэлстон и др., 1983).
Зеленый лист поглощает в среднем 75 % падающей на него энер-
гии Солнца, но коэффициент использования ее на фотосинтез
довольно мал. Выход фотосинтеза часто лимитируется сильным
освещением, пониженным поступлением СО2, уменьшением ак-
тивности ферментов в процессе развития (например, при старе-
нии) и т.д. Поэтому в природных условиях КПД использования
света в фотосинтезе невысок. При интенсивном освещении на
фотосинтез тратится примерно 1—2%, а при более низкой осве-
щенности — до 10 % поглощенной листом лучистой энергии. Ос-
тальные 90—99 % ее переходят в тепловую энергию, которая тра-
тится на транспирацию и другие процессы.
Масштабы фотосинтетического преобразования и запасания
энергии огромны: каждый год на Земле за счет фотосинтеза обра-
зуется более 200 млрд т биомассы, что эквивалентно энергии, рав-
ной 30,2 • Ю20 Дж (А. Гэлстон и др., 1983). При этом фотосинтез —
единственный биологический процесс, протекающий с увеличе-
нием свободной энергии, а все остальные процессы в организмах
идут за счет химической энергии, накопленной в них. Следова-
тельно, практически вся живая материя на Земле представляет
собой прямой или отдаленный результат фотосинтетической дея-
тельности. В ближайшие миллиарды лет растения служат посред-
никами между неиссякаемым источником энергии (Солнцем) и
живым миром нашей планеты. Основанный на окислении воды
фотосинтез стал мощным поставщиком энергии в биосферу, уси-
лил биоэнергетические процессы благодаря включению в метабо-
лизм кислорода, защитил живое от разрушительного действия
космической радиации сформировавшимся озоновым экраном. Все
это индуцировало мощное развитие жизни, приведшее в конеч-
ном счете к появлению современных сложных организмов и со-
обществ, освоению огромных пространств суши и атмосферы.
В настоящее время фотосинтез лежит в основе биологического
115
круговорота на Земле, от масштабов которого зависит жизнь на
планете, ее разнообразие и возможности.
Типы фотосинтеза
С3~ и С4-пути ассимиляции диоксида углерода. У большинства
растений темновая фаза фотосинтеза осуществляется в цикле ре-
акций, получившем по имени его открывателя название «цикла
Кальвина». В нем акцептором СО2 служит пентозофосфат, а про-
дукт реакции, содержащий 6 атомов углерода, сразу распадается
на 2 трехуглеродные молекулы (3-фосфоглицериновой кислоты).
Поэтому такой способ ассимиляции СО2 называют пентозофос-
фатным или «С3-путем». Далее полученные молекулы восстанав-
ливаются (до 3-фосфоглицеринового альдегида) и используются
для синтеза различных углеводов. Это происходит у большинства
растений. Однако в 60-х гг. XX в. при сравнительном изучении
продуктивности фотосинтеза было установлено, что у ряда видов,
особенно у злаков тропического происхождения (сахарный трос-
тник — Saccharum officinarum, кукуруза — Zea mays, сорго —
Sorghum) и некоторых двудольных (например, амарант —
Amaranthus), достигаются чрезвычайно высокие показатели, при-
чем эти растения используют и свет наибольшей интенсивности.
Всемирную известность этот тип метаболизма получил как «цикл
Хэтча — Слэка», названный по именам австралийских исследова-
телей, предложивших законченную схему цикла темновых реак-
ций у С4-растений.
Во второй половине XX в. количество обнаруженных С4-видов
приблизилось к тысяче. Они относятся главным образом к по-
крытосеменным растениям примерно из 200 родов двух десятков
семейств. Наиболее богаты этими растениями злаковые (около
половины видов), осоковые, щирицевые, маревые, гвоздичные,
молочайные, сложноцветные, аизовые, портулаковые, фитолак-
ковые, губоцветные. Среди С4-форм отмечен также ряд низших
фотоавтотрофов (например, Anacystis из цианобактерий). Все эти
организмы обладают очень высокой фотосинтетической способ-
ностью и часто обитают в аридных областях. В подавляющем боль-
шинстве этот механизм фотосинтеза встречается у короткоднев-
ных растений тропического происхождения.
Фиксация СО2 в мезофилле у С4-растений осуществляется дру-
гим акцептором — фосфоэнолпируватом (ФЕП). Фермент, ката-
лизирующий эту реакцию, обладает очень большим сродством к
диоксиду углерода. Поэтому он работает даже при очень низких
ее концентрациях (активность фосфопируваткарбоксилазы в 50—
60 раз выше, чем у растений С3-пути) (А. Г. Юсуфов, 1995). В ре-
зультате карбоксилирования образуется щавелевоуксусная кисло-
116
та, которая может перейти в аспарагиновую (у аспартатной груп-
пы С4-растений) или восстановиться до яблочной (у малатной груп-
пы). Эти кислоты являются дикарбоновыми и имеют по 4 атома
углерода, что и определило название этого пути (путь дикарбоно-
вых кислот, или «С4-путь»).
Первичные продукты фиксации СО2 перерабатываются в сахара не в
той же клетке, а транспортируются в специализированные для этого об-
кладочные клетки проводящих пучков. В этом случае СО2 переносится
на фермент (РДФ-карбоксилазу) более эффективно, чем из атмосферы,
и она работает в условиях насыщающих концентраций СО2, что пред-
ставляет одно из главных преимуществ С4-фотосинтеза. В клетках об-
кладки дикарбоновые кислоты перерабатываются с высвобождением СО2,
используемой далее по циклу Кальвина. Другой продукт реакции — пи-
руват — возвращается в обычные клетки мезофилла, где регенерирует
снова до фосфоэнолпирувата, главного акцептора СО2 в этом цикле.
Регенерация акцепторов СО2 создает возможность непрерывного функ-
ционирования циклов. Таким образом, у С4-растений два типа карбок-
силирования оказываются пространственно разобщены.
Так как при С4-фотосинтезе реакция карбоксилирования про-
исходит дважды, КПД фотосинтеза повышается. Высокая интен-
сивность фотосинтеза позволяет растению в единицу времени
фиксировать больше СО2, быстро создавая запасы углеводов. Эта
сложная комбинация синтеза дикарбоновых кислот с С3-циклом
дает преимущество С4-растениям в использовании СО2, так как
многообразие акцепторов обеспечивает более полную его утили-
зацию и утилизацию восстановителей. Чрезвычайно высокое срод-
ство к диоксиду углерода фермента, катализирующего его присо-
единение к акцептору, позволяет этим растениям немедленно ре-
утилизировать дополнительные ее порции, образующиеся при
фотодыхании. Имея низкую точку компенсации по СО2, С4-рас-
тения лучше используют недостаточные ресурсы диоксида угле-
рода. Ферментативная фиксация СО2 у С4-растений приводит к
снижению его концентрации внутри листа до гораздо более низ-
ких значений, чем у С3-растений. Скорость выделения водяного
пара от этого не меняется, а скорость диффузии диоксида углеро-
да внутрь листа возрастает. Следовательно, эффективный фото-
синтез может происходить у них при значительно сниженной от-
крытости устьиц, вызванной, например, водным стрессом. При-
чем они продолжают фотосинтезировать даже при закрытых усть-
ицах, высоких температурах и интенсивном освещении.
Таким образом, для С4-растений характерен процесс вторич-
ного карбоксилирования, являющийся первой ступенью челноч-
ного механизма, развившегося как способ доставки атмосферного
СО2 к местам фотосинтетического восстановления углерода в хло-
ропластах клеток обкладки сосудистых пучков. Особенности ана-
117
томии и биохимии, использование клеток мезофилла как «биохи-
мической помпы» диоксида углерода, стабильно высокая продук-
тивность определяют С4-путь фотосинтеза как дополнение к стан-
дартному С3-пути, увеличивающее эффективность фиксации СО2
(Ю. В. Гамалей и др., 1992). Однако хотя в пересчете на площадь
листвы эффективность фотосинтеза у С3-растений ниже, именно
эти растения создают большую часть фотосинтетической продук-
ции мира. Причиной этого может быть то, что они более конку-
рентоспособны в смешанных сообществах, где растения затеняют
друг друга и где освещенность, температура и другие факторы
ближе к средним значениям, чем к предельным. А при ослаблен-
ной освещенности и невысокой температуре С3-растения эффек-
тивнее С4-видов (Ю.Одум, 1986).
С4-виды — доминанты сообществ тропических и субтропиче-
ских пустынь, где летний максимум тепла и света связан с высо-
кой полуденной высотой Солнца и малооблачной погодой. Ряд
С4-видов имеет пантропические ареалы, но исходные формы свя-
заны с зонами экстраинсоляции. Показательно, что в горных пу-
стынях сумма эффективных температур недостаточна для форми-
рования С4-растений. Там С4-метаболизм не характерен даже для
злаков из триб с его доминированием (просовые, тростниковые,
бородачовые). И на лесных территориях с муссонным климатом
нет эндемичных С4-растений, так как там из-за большой облач-
ности инсоляция недостаточна.
С4-виды обладают выраженной способностью эффективно ис-
пользовать воду. По количеству связанного углерода на единицу
израсходованной при транспирации воды они превосходят С3-ра-
стения примерно в два раза. Многие из С4-растений произраста-
ют в жарких аридных условиях. Так, Ю.Одум (1986) отмечает, что
в США летом среди пустынных однолетников С4-виды составля-
ют 100%. Показательно, что немногие С4-виды, достигающие
умеренных широт, например Spartina, встречаются в морских или
иных обогащенных солями местообитаниях, где осмотические
условия могут предоставлять растениям, эффективно использую-
щим воду, особые преимущества. С4-видов много также среди
трудноискоренимых сорняков (свинорой — Cynodon dactylon, щи-
рица — Amaranthus, щетинник — Setaria, куриное просо —
Echinochloa crusgalli и др.), процветающих на хорошо прогревае-
мых засушливых нарушенных участках.
Однако мнения исследователей относительно экологических преи-
муществ С4-видов неоднозначны. Распространена точка зрения, что
С4-путь — это адаптация к режиму с малой влажностью, высокой темпера-
турой и интенсивным освещением, хотя наличие С4-пути не всегда обес-
печивает наибольшую скорость роста. Другие авторы считают С4-синд-
ром адаптацией только к высокой температуре, указывая, что С4-фото-
118
синтез эффективен и при высокой влажности. Кроме того, были обнару-
жены виды и популяции с переходным типом фотосинтеза. А П. А. Ген-
кель (1982) показал, что в крайне жестких условиях пустынь у некоторых
растений хлорофиллоносные ткани служат субстратом для различного
типа метаболизма. Эколого-флористические исследования тоже не по-
зволяют делать однозначных выводов о преимуществах С4-видов в жар-
ких условиях. Например, во флоре Каракумов встречается лишь несколько
С4-видов, причем максимальная интенсивность фотосинтеза некоторых
С3-видов в этой пустыне даже несколько выше таковой у С4-растений, а
по положению верхних температурных границ фотосинтеза они почти
не отличаются (Л.Г.Косулина и др., 1993).
Главные пищевые растения человечества (пшеница — Triticum
aestivum, рис — Oryza sativa, картофель — Solanum tuberosum,
большая часть овощей) относятся к С3-видам, поскольку эти куль-
туры были выведены преимущественно в умеренной зоне Север-
ного полушария. Но культуры тропического происхождения (куку-
руза — Zea mays, просо — Panicum, сорго — Sorghum, сахарный
тростник — Saccharum officinarum) являются С4-видами. Они же
известны и как одни из наиболее продуктивных культур. Так,
интенсивность фотосинтеза у кукурузы — 85 мг СО2 на 1 дм2/ч,
сорго — 55 мг на 1 дм2/ч, тогда как у пшеницы всего 31 мг на 1 дм2/ч
(Н.И.Якушкина, 1993). Поэтому в сельском хозяйстве существу-
ет потребность в выведении новых С4-сортов растений для возде-
лывания в орошаемых пустынях тропиков.
Фиксация диоксида углерода суккулентами (САМ-фотосинтез).
В 30-х гг. XX в. был обнаружен особый суточной ритм изменения
кислотности клеточного сока и особый путь метаболизма у негало-
фитных суккулентов из семейства Толстянковые (Crassulaceae). Его
назвали САМ-путь (crassulacean acid metabolism). Этот способ асси-
миляции свойствен суккулентам, живущим преимущественно в
жарких и сухих местообитаниях (около 300 видов почти из двух
десятков семейств). Они хорошо представлены в семействах Ли-
лейные, Бромелиевые, Орхидные, Кактусовые, Аизовые и Ластов-
невые. Среди них есть также папоротники и голосеменные.
Фиксация поглощенной СО2 у САМ-видов, как и у С4-расте-
ний, происходит с использованием фосфоэнолпирувата (ФЕП), и
в результате тоже образуется яблочная кислота (наряду с этой ре-
акцией идут и другие, в которых образуются органические кисло-
ты). Пространственное разделение этапов кислотного метаболиз-
ма между мезофиллом и обкладочными клетками, отмеченное для
С4-растений, у растений с САМ-путем заменено разделением во
времени. Они поглощают СО2 при широко раскрытых устьицах
ночью (когда водный стресс спал), а фотосинтетически перераба-
тывают его только на следующий день (для расщепления образо-
вавшихся кислот необходим свет). Для суккулентов, обитателей
сухих мест, такой путь ассимиляции диоксида углерода, разводя-
119
щий во времени его фиксацию (ночью при открытых устьицах) и
переработку (днем — при закрытых), экологически очень выго-
ден. При дневном водном дефиците это спасает их от чрезмерной
потери и позволяет получить достаточное количество углерода через
широко раскрытые устьица ночью.
Образующиеся ночью органические кислоты отводятся и накаплива-
ются в клеточном соке, повышая его кислотность. Утром они переходят
из вакуолей обратно в пластиды и, как у С4-растений, декарбоксилиру-
ются с освобождением СО2. Он же при закрытых днем устьицах исполь-
зуется в цикле Кальвина. По мере расходования кислот, накопленных в
вакуолях, кислотность клеточного сока снова уменьшается, демонстри-
руя характерную ритмичность. Таким образом, крупные вакуоли сукку-
лентов служат не только для запасания воды, но и содержат резерв угле-
рода для фотосинтеза, делая его на некоторое время независимым от
поступления СО2 из внешней среды.
При отсутствии дефицита воды фиксация СО2 у этих растений
происходит по С3-пути. К переключению метаболизма на САМ-тип
часто приводит длительная засуха или засоление. Например, пус-
тынный однолетник Mesembryanthernum crystallinum во влажный
период формирует крупные листья и использует С3-путь, а в пери-
од засухи его рост приостанавливается и он переходит на САМ-
путь (А. Г. Юсуфов, 1995). В эксперименте удается стимулировать
САМ-обмен переносом некоторых растений в раствор NaCl. САМ-
обмен характерен также для некоторых горных растений, посто-
янно испытывающих температурные стрессы.
У многих видов обмен пластичен и соотношение между С3-, С4- и
САМ-метаболизмом может значительно меняться в онтогенезе при ва-
рьировании условий роста и минерального питания. Часть растений при
ухудшении условий переходит с С3- на С4-путь, а другие, например пор-
тулак (Portulaca), при дефиците воды, коротком дне и низкой ночной
температуре С4-путь переключают на САМ-метаболизм. Если сравни-
вать пути метаболизма, то в целом среди покрытосеменных преобладает
С3-путь, хотя у суккулентов — САМ-метаболизм. Возможно, С4-путь и
САМ-путь — полифилетически возникшие модификации обычного С3-
пути (А. Г. Юсуфов, 1995). При этом САМ-путь представляет собой ме-
ханизм переживания условий специфических именно для суккулентов.
Возможно также, что С4-путь возник как следующий шаг эволюции ра-
стений с САМ-метаболизмом.
Фотосинтез и продуктивность
Фотодыхание. Фотодыхание (световое дыхание) — процесс
поглощения кислорода и выделения диоксида углерода, происхо-
дящий в фотосинтезирующих клетках при участии света. Фотоды-
хание увеличивается на свету и прекращается в темноте (рис. 6.13).
120
При фотодыхании окисляется промежуточный продукт цикла
Кальвина — гликолат, поэтому фотодыхание зависит от интен-
сивности фотосинтеза, в котором он образуется. Возникший в хло-
ропластах гликолат перемещается в пероксисомы, где происходит
первый этап фотодыхания: гликолат окисляется кислородом. За-
тем продукт превращается в глицин, который покидает перокси-
сому и поступает в митохондрию. Там две молекулы глицина со-
единяются, высвобождая при этом СО2. Продукт возвращается в
пероксисому, где через цепь реакций превращается в фосфогли-
цериновую кислоту. Она переходит в хлоропласт и включается в
цикл Кальвина, замыкая круг. В целом в фотодыхании участвуют
продукты фотосинтеза, поглощается кислород и выделяется ди-
оксид углерода. В процессе участвуют три типа органелл — хло-
ропласты, пероксисомы и митохондрии.
С3- и С4-растения имеют разный метаболизм гликолата. С4-
растения образуют его меньше, они также сразу используют вы-
деляемый при фотодыхании диоксид углерода. Поэтому фотоды-
хание у С4-растений незаметно. У С3-растений фотодыхание со-
провождается значительной потерей углерода из цикла Кальвина
из-за снижения способности РДФ-карбоксилазы фиксировать СО2
и повышения ее активности окислять субстрат (РДФ) в присут-
ствии кислорода.
В отличие от обычного темнового дыхания фотодыхание уси-
ленно реагирует на изменение концентрации О2 (обычное дыха-
ние почти нечувствительно к изменению содержания кислорода
от 2 до 100%). Окисление гликолата при фотодыхании сильно
ускоряется повышенным содержанием кислорода и прекращается
при снабжении им в концентрации менее 2%. Рассчитано, что
если содержание кислорода в воздухе понизить в 10 раз и исклю-
чить фотодыхание, то поступление СО2 в фотосинтезирующие
органы увеличится на 50 %. В естественных же условиях продук-
Рис. 6.13. Фотосинтез и дыхание листь-
ев подсолнечника (Helianthus annuus)
при 21 % О2, 0,003 % СО2 и t 22 °C при
возрастающей интенсивности освеще-
ния (по ВЛархеру, 1978):
Фн — нетто-фотосинтез; Дт — темновое ды-
хание; Дсв — световое дыхание
освещения, клк
121
тивность фотосинтеза понижена примерно на 20—50 % из-за по-
терь СО2 на фотодыхание. Вычисляя степень использования энер-
гии и экономичность фотосинтеза, В. Лархер предлагает исходить
из того, что у С3-растений при хорошей освещенности фотодыха-
ние в 1,5 —3,5 раза интенсивнее, чем темновое дыхание.
Таким образом, процесс фотодыхания связан с потерей угле-
рода и метаболической активностью, сопряженной со значитель-
ными затратами энергии. Причины, по которым такой расточи-
тельный процесс сохранился в эволюции, представляют большой
интерес не только с теоретической, но и с практической точки
зрения. В настоящее время общепризнанных представлений о
физиологической роли фотодыхания нет. Многие исследователи
полагают, что фотодыхание выполняет роль поставщика более
широкого ассортимента продуктов фотосинтеза. Действительно,
гликолатный путь приводит к синтезу аминокислот (глицина и
серина), и в многочисленных опытах прослежена связь фотоды-
хания с азотным обменом. Кроме того, фотодыхание поставляет
энергетически ценные продукты (например, восстановленный
НАДФ). Другая концепция предполагает важность защитной роли
фотодыхания: сброс избыточной энергии позволяет предотвратить
разрушение фотосистем, избежать прекращения фотохимических
процессов при высокой освещенности и температуре, закрытых
при водном дефиците устьицах (Н. Н.Третьяков и др., 2000). Тео-
ретические проблемы, касающиеся фотодыхания, имеют важное
практическое значение, так как ставят под вопрос предпринима-
ющиеся попытки уменьшить величину фотодыхания путем отбо-
ра мутантных форм. Если процесс фотодыхания случаен, то этим
путем повысить продуктивность растений можно. Однако если он
действительно играет существенную роль (прежде всего предох-
ранительную), эти работы к положительному результату не при-
ведут.
Характеристики результатов фотосинтеза. Часть ассимилятов,
выработанных в процессе фотосинтеза, постоянно расходуется на
дыхание. Обычно днем при фотосинтезе СО2 потребляется боль-
ше, чем выделяется при дыхании. Количество ассимилятов, в це-
лом выработанное растением (в том числе для покрытия расходов
на дыхание), называют истинным, или брутто-фотосинтезом.
А видимый, наблюдаемый, или нетто-фотосинтез, — это раз-
ность между количеством диоксида углерода, ассимилированного
листом, и выделенного в процессе дыхания.
Другая характеристика результатов фотосинтеза — продукция,
под которой понимают массу органических веществ, производи-
мых организмом (или сообществом) за определенный отрезок вре-
мени. А интенсивность фотосинтеза (продуктивность) — это ко-
личество СО2, поглощенное за единицу времени единицей повер-
хности листьев или единицей массы (сухой или сырой). Это ос-
122
новной показатель в оценке накопления растениями органиче-
ского вещества, хотя прямой зависимости урожая от интенсивно-
сти фотосинтеза в разных условиях не наблюдается. Если фото-
синтез рассчитывают на единицу площади листа, то при этом ха-
рактеризуют эффективность ассимиляционной деятельности ос-
вещаемой поверхности. Эта величина дает возможность судить об
эффективности усвоения солнечной энергии и СО2 данным орга-
ном. Однако она сильно зависит от анатомо-морфологических
особенностей листа и поэтому требуется учитывать характер ас-
симилирующей поверхности (складчатость, выпуклость и т.д.).
Фотосинтетическая способность — это интенсивность нет-
то-фотосинтеза при данном состоянии растения (развитии, ак-
тивности), естественном содержании СО2 и оптимальных величи-
нах прочих внешних факторов. Эта величина, определяемая в стан-
дартных условиях, показывает потенциальную интенсивность
фотосинтеза. Ее используют для характеристики видов, сортов,
экотипов и жизненных форм растений. Растения сильно различа-
ются по фотосинтетической способности. Наивысшие показатели
характерны для С4-видов, которые ассимилируют почти вдвое
интенсивнее, чем большинство сельскохозяйственных культур, и
в 30 раз активнее, чем мхи, лишайники и водоросли. За С4-вида-
ми идут селекционные сельскохозяйственные С3-культуры, среди
которых наиболее продуктивны рис (Oryza sativa), пшеница
(Triticum aestivum), картофель (Solanum tuberosum), подсолнеч-
ник (Helianthus annuus). Минимальная фотосинтетическая спо-
собность характерна для галофитов.
Отмечено, что у видов с коротким вегетационным периодом нетто-
фотосинтез обычно выше, чем у длительно вегетирующих. У большин-
ства трав интенсивность наблюдаемого фотосинтеза выше, чем у деревь-
ев. Из древесных форм максимальная интенсивность фотосинтеза на-
блюдается у листопадных деревьев и кустарников умеренной зоны. Сре-
ди вечнозеленых растений нет видов с интенсивностью фотосинтеза выше
25 мг СО2 на 1 г сухой массы в 1 ч. Теневыносливые формы усваивают
углерод в 2 — 3 раза слабее растений солнечных местообитаний. Виды, у
которых ассимилирующие органы развивают небольшую поверхность
(злаки со свернутыми в трубку листьями, кустарнички с эрикоидными
листьями, хвойные, безлистные кустарники и суккуленты), довольно слабо
используют свет и характеризуются умеренной фотосинтетической спо-
собностью. У безлистных опробковевших побегов фотосинтезирует лишь
хлоренхима коры, однако этот процесс малоэффективен и играет неко-
торую роль лишь в компенсации потерь на дыхание. Ряд растений отли-
чается очень слабой фотосинтетической активностью. Так, у папоротни-
ков, мхов и лишайников интенсивность фотосинтеза не превышает 4 мг
СО2 на 1 г сухой массы в 1 ч. Очень мало связывают СО2 водные расте-
ния. Так, фитопланктон фиксирует 2 —3 мг СО2на 1 г сухой массы в 1 ч
и очень слабая продуктивность выявляется у погруженных в воду выс-
123
ших растений (около 7 мг СО2 на 1 г сухой массы в 1 ч). Существенной
причиной этого В.Лархер (1978) считает то, что СО2 очень медленно
диффундирует в воде и погруженные водные растения хуже снабжаются
СО2, чем наземные виды.
Фотосинтетическая способность изменяется в ходе онтогенеза.
На стадии распускания листья еще не способны эффективно свя-
зывать СО2, поэтому фотосинтез в это время незначителен и ды-
хание превосходит его по интенсивности. Молодые побеги дре-
весных растений в фазе растяжения тоже еще должны снабжаться
углеводами из других частей растения. С развертыванием листьев
фотосинтез быстро интенсифицируется и полностью развитая
молодая листва обладает наивысшей продуктивностью. Через не-
сколько недель фотосинтетическая способность ослабевает, осо-
бенно быстро у однолетних органов. У вечнозеленых растений из
районов с холодной зимой фотосинтетическая способность лис-
тьев снижается после каждой зимы. Так, у ели (Piceа) уже на 3 —
4 год хвоя имеет отрицательный баланс ассимилятов. Дополни-
тельно ослабляет ее новый прирост хвои, поэтому если ее уда-
лить, старая хвоя будет жить дольше и фотосинтезировать актив-
нее. Зависимость фотосинтетической способности от стадии раз-
вития растения связана с несколькими причинами. У очень моло-
дых листьев еще невелика поверхность, поэтому они поглощают
недостаточно света. К тому же они очень интенсивно дышат и
часто бедны хлорофиллом. Кроме того, процессы развития связа-
ны с изменением контролируемой фитогормонами активности фер-
ментов. Незадолго до отмирания листа хлорофилл разрушается и
фотосинтетическая способность падает до нуля.
Для растений в конкретном местообитании важны не единич-
ные максимальные фотосинтетические показатели, а среднее по-
глощение СО2. При этом даже в благоприятных условиях в уме-
ренной зоне средние показатели нетто-фотосинтеза составляют
50 — 60% от наивысшего за день значения (30 — 50% максималь-
ной фотосинтетической способности). Для накопления углерода
решающее значение имеет продолжительность фотосинтеза в пе-
риод облиствения в светлые часы суток (если не препятствуют
мороз, жара или засуха). Из суммарного поглощения углерода
вычитается выделенное в виде СО2.
Суточный баланс — это чистая продуктивность фотосинтеза
за 24 ч, а сумма суточных балансов дает годовой баланс углерод-
ного газообмена. Суточный баланс положителен, если дневной
приход больше ночного расхода, и он тем выше, чем благоприят-
нее для фотосинтеза условия днем и чем короче и прохладнее
ночь. Если поглощение СО2 непродолжительно или неэффектив-
но, то чистый выход продуктов ассимиляции невелик. Это проис-
ходит в засушливые периоды, в сухих местообитаниях и после
124
морозов. В глубокой тени суточный баланс обычно слабоположи-
тельный (а при теплых ночах иногда отрицательный), что может
приводить к голоданию растений. Таким образом, зависимость
распространения определенных групп растений от минимума све-
тового довольствия имеет эколого-физиологическое объяснение.
Для определения продуктивности важно содержание нефото-
синтезирующих тканей. Их расходы на дыхание особенно суще-
ственны, когда масса стеблей, корней, цветков и плодов значи-
тельна по сравнению с листовой массой. Так, у взрослых деревьев
фотосинтетически активные органы составляют не более 5 % фи-
томассы. У вересковых кустарничков и пустынных растений —
10 — 20%, у злаков — 30—50%, а у споровых растений тундр —
более 95% (В.Лархер, 1978). Углерод, не расходуемый на дыха-
ние, может быть использован на прирост и запасы. Скорость при-
роста обычно увеличивается пропорционально фотосинтетиче-
ской способности. Так, С4-злаки дают в 2—4 раза больший при-
рост, чем С3-травы, и в 100 раз больший, чем САМ-растения.
Прирост зависит и от жизненной формы растений: у трав он при-
мерно в 10 раз больше, чем у древесных видов, а у лиственных
деревьев больше, чем у хвойных.
Среди культурных растений одну из самых больших продук-
ций сухой массы дает сахарная свекла (Beta vulgaris) — 18 — 20 т/га
(Г. Вальтер, 1982). Но и она не достигает своей потенциальной
продуктивности, так как в мае и начале июня листовая поверх-
ность еще слишком мала, а осенью из-за старения листьев фото-
синтетическая активность падает. Образование сухого вещества в
основном ограничено 2,5 месяцами — с середины июня по конец
августа (потери на дыхание составляют 30 — 50%) (рис. 6.14).
Как для углеродного баланса отдельного растения, так и для
годового урожая в сообществах большое значение имеет продол-
жительность вегетационного периода. Обычно в умеренно-теп-
Рис. 6.14. Анализ продукции сахарной
свеклы (Beta vulgaris) (по Г. Вальтеру,
1982).
Общая площадь — максимальная теоретиче-
ская брутго-продукция: 1 — потери из-за ды-
хания листьев и корнеплодов; 2 — потери из-
за неполностью развитой листовой поверхно-
сти весной и ранним летом; 3 — потери из-за
пониженной продуктивности фотосинтеза в
стареющих листьях; 4 — реальная продукция;
утолщенная линия — максимальная первич-
ная продукция, теоретически возможная при
полностью развитых листьях в течение всего
вегетационного периода и при отсутствии их
старения (в 1,5 раза больше фактической)
Месяцы
125
лых зонах при умеренной интенсивности фотосинтеза, но суще-
ственной его продолжительности органики накапливается много.
Но если ассимиляция возможна только в течение короткого пе-
риода, то эти показатели остаются низкими, даже если растения
обладают большой фотосинтетической способностью и экономно
используют ассимиляты, как это происходит в высокогорьях, Арк-
тике и засушливых областях.
Длительность благоприятного для эффективного фотосинтеза перио-
да зависит от жизненной формы растений. Он максимален у вечнозеле-
ных растений теплых влажных областей (например, видов дождевых тро-
пических лесов). Меньше его величина у вечнозеленых растений с пере-
рывом продуктивного периода, который может быть связан с холодным
временем года (таежные хвойные деревья) или сухим сезоном (среди-
земноморские жестколистные кустарники). Таким образом, выраженное
преимущество вечнозелености — в удлинении продуктивного для фото-
синтеза времени, особенно заметное там, где вегетационный период со-
кращен из-за летней засухи или долгой зимы (горы, лесная зона, грани-
ца леса). Там вечнозеленые растения могут преобладать над листопад-
ными. Меньше благоприятный период для эффективного фотосинтеза у
листопадных растений, фотосинтезирующих короткое время. Среди них
выделяют виды с полным использованием периода облиствения в бога-
тых осадками областях (лиственные деревья умеренной зоны), с частич-
ным использованием периода облиствения из-за недостатка света (рас-
тения нижнего яруса в лиственных лесах) и с частичным использовани-
ем периода облиствения из-за засухи (деревья в лесостепи). Очень недол-
гое время продуктивного фотосинтеза у растений, фотосинтезирующих
в короткие благоприятные периоды между длинными неблагоприятны-
ми. К ним относятся высшие растения в пустынях с нерегулярным вы-
падением осадков (до 30—40 благоприятных дней), высшие растения
Арктики и высокогорий (60 — 90 благоприятных дней), мхи, лишайники
и наземные водоросли, фотосинтезирующие в короткие периоды после
увлажнения. В холодных морях продуктивный период ограничен поляр-
ным летом, когда в течение нескольких недель возможен фотосинтез без
ночного перерыва.
Коэффициент полезного действия фотосинтеза показыва-
ет, сколько процентов поглощенной световой энергии запасено в
форме энергии химических связей при превращении диоксида
углерода в углеводы. В процессе фотосинтеза на 1 г полученного
углевода используется 25,9 кДж энергии. Умножение этого коэф-
фициента на интенсивность брутто-фотосинтеза дает количество
связанной энергии (величины пересчитываются на одинаковую
поверхность и одинаковый промежуток времени). КПД фотосин-
теза обычно выражают по отношению либо к падающей, либо к
поглощенной растениями ФАР. В большинстве случаев листья
работают с КПД меньше 5 — 10 %, но у некоторых видов в особо
благоприятных условиях он достигает 15 % (у С4-злаков 24 %).
126
Несмотря на высокую эффективность начальных фотофизи-
ческих и фотохимических стадий (около 95 %), в урожай перехо-
дит лишь несколько процентов солнечной энергии. Потери обус-
ловлены лимитированием процесса на биохимическом и физио-
логическом уровнях, а также неполным поглощением света. Во
всех сообществах получаемая солнечная радиация используется
неэффективно. К факторам, ограничивающим первичную продук-
цию на суше, относятся: 1) недостаток воды, ограничивающий
скорость фотосинтеза; 2) нехватка элементов минерального пита-
ния, замедляющая скорость образования ассимилирующей ткани
и снижающая эффективность фотосинтеза; 3) неблагоприятная
для роста температура; 4) попадание большой части радиации мимо
фотосинтезирующих органов (из-за сезонного опадения листьев,
действия фитофагов и паразитов); 5) низкая эффективность фо-
тосинтеза в листьях (даже при идеальных условиях в наиболее
продуктивных сельскохозяйственных системах она редко бывает
выше 10 % ФАР). При изменяющихся условиях ассимиляции КПД
растительных сообществ суши в среднем ниже 1 — 3%. Макси-
мальная эффективность фотосинтеза интенсивных зерновых куль-
тур при идеальных условиях 3—10%. А если рассматривать пла-
нету в целом, то КПД падающей ФАР составляет всего около 0,2 %
(Н. И.Якушкина, 1993).
Первичная продукция водных сообществ. В водной среде пер-
вичную продукцию чаще всего лимитируют свет, наличие эле-
ментов минерального питания и интенсивность выедания живот-
ными. Среди минеральных веществ наибольшее значение имеют
азот и фосфор. При высокой их концентрации формируются са-
мые высокопродуктивные сообщества.
Озера сильно различаются по степени эвтрофикации. В них
необходимые минеральные вещества поступают при выветрива-
нии горных пород, эрозии почв, с дождевой водой. Неоднократно
отмечалась обратная зависимость между первичной продукцией
озера и географической широтой. Частично это связано с уси-
лением освещенности и удлинением вегетационного периода в
низких широтах. Но там более высока также скорость минера-
лизации и в озерах накапливается больше биогенов. Однако чем
богаче ими вода, тем меньше эуфотическая зона, так как там боль-
ше поглощающего свет фитопланктона (аналогично затенению
верхним ярусом леса). Поэтому если фитопланктона много, то
даже в мелководных водоемах заросли макрофитов не развива-
ются.
В морях и океанах первичная продукция высоких показателей
гоже достигает в обогащенных биогенами местах, связанных с
прибрежной шельфовой зоной эстуариев и океаническими апвел-
лингами (места подъема глубинных вод). В первом случае высо-
кая продукция объясняется относительно высокой прозрачнос-
127
тью воды, обеспечивающей значительную толщину эуфотической
зоны. Ближе к берегу концентрация биогенов выше, но вода мут-
нее, поэтому продукция падает. Наименее продуктивные зоны
находятся на внешнем шельфе и в открытом океане, несмотря на
высокую прозрачность воды и большую глубину эуфотической
зоны. Концентрации биогенов там очень низкие.
Океанические апвеллинги, характеризующиеся высоким содержани-
ем минеральных веществ, наблюдаются на континентальном шельфе в
тех местах, где ветры регулярно дуют параллельно или под небольшим
углом к берегу. При этом поверхностные слои отгоняются, а на их место
поднимаются холодные воды с накопленными в ходе осаждения мине-
ральными веществами. Мощные апвеллинги возникают также около
подводных хребтов и в районах с сильными течениями. Там, где эти
богатые биогенами воды выходят на поверхность, продукция фитоплан-
ктона резко увеличивается.
Сезонные изменения первичной продукции водоемов хорошо
прослеживаются в умеренных и высоких широтах. Неблагоприят-
ные погодные условия и перемешивание водных масс вынуждают
планктон погружаться на глубину, где затраты на дыхание уже
плохо компенсируются. При увеличении температуры и светово-
го дня, снижении ветрового перемешивания продукция фитоплан-
ктона повышается. Чем ближе к полюсу, тем позднее начинается
этот процесс. Снижение продукции из-за ухудшения погодных
условий и истощения биогенов тоже происходит сначала в высо-
ких широтах. Но в ряде озер и морей умеренных широт иногда
наблюдается два сезонных пика биомассы. Возможно, это связано
с тем, что биогены, летом лимитирующие фотосинтез, осенью при
перемешивании поднимаются с глубинными водами к поверхно-
сти. А поскольку погодные условия еще благоприятны для фото-
синтеза, то это может дать второй пик продукции фитопланктона
(М. Бигон и др., 1989).
Экология фотосинтеза
Световая кривая фотосинтеза. Световая кривая графически
выражает зависимость интенсивности фотосинтеза от освещен-
ности (рис. 6.15). Она характеризует весь газообмен диоксида уг-
лерода на свету (показывает нетто-фотосинтез). Для световых кри-
вых характерна гиперболическая форма: поднявшись вверх, они
выходят на плато, и дальнейшее усиление освещенности почти не
влияет на газообмен. Ряд параметров этой кривой имеют важный
экологический смысл.
Точка пересечения световой кривой с осью ординат находится
ниже нулевой отметки по оси абсцисс. Пока интенсивность света
128
Рис. 6.15. Световые кривые фотосинтеза для светолюбивых (7)
и теневыносливых растений (2) (по Т. К. Горышиной, 1979):
аь а2 — ниже оси абсцисс: растения в темноте, фотосинтез отсутствует, идет
только дыхание; бн б2 — «компенсационные точки» (фотосинтез уравновешива-
ет дыхание): вь в2 — перегиб светой кривой фотосинтеза, она выходит на плато
насыщения; г(, г2 — проекция точек в(, в2 на ось абсцисс, характеризует «насы-
щающую» интенсивность света, выше которой свет не повышает интенсивность
фотосинтеза; дь д2 — проекция точек вь в2 на ось ординат — наибольшая интен-
сивность фотосинтеза для данного вида в данном местообитании; а — углы на-
клона световой кривой к абсциссе, отражающей степень увеличения фотосинтеза
при возрастании радиации (в области сравнительно низкой интенсивности света)
мала, идет выделение СО2, а не поглощение его, так как фотосин-
тез не компенсирует дыхание. При некотором значении освещен-
ности световая кривая пересекает линию абсцисс. Это значит, что
количество поглощенного в фотосинтезе диоксида углерода рав-
но количеству СО2, выделенному при дыхании. За этой точкой
уже преобладает фотосинтез, т.е. с повышением освещенности
потребление СО2 будет превышать его выход при дыхании. Ми-
нимальное освещение (при нормальном содержании СО2), при
котором интенсивность фотосинтеза (поглощения СО2) уравно-
вешивает дыхание (выделение СО2), называют точкой компенса-
ции (или компенсационным освещением). При такой освещенно-
сти для фотосинтеза используется столько СО2, сколько высво-
бождается при дыхании. Поэтому интенсивно дышащие растения
нуждаются в большем количестве света. Так, теневыносливые виды
характеризуются более низкой интенсивностью дыхания, что по-
зволяет им расти при меньшей освещенности. Светолюбивые ра-
стения лучше используют большую освещенность и в этих усло-
виях имеют гораздо более высокую продуктивность фотосинтеза.
Поэтому сельскохозяйственные растения, от которых требуется
высокая урожайность, должны быть светолюбивыми (рис. 6.16).
Компенсационная точка варьирует даже у разных листьев одной кро-
ны. Выросшие в тени дышат слабее световых, поэтому компенсация у
них наступает при меньшем освещении. Теневые листья лучше исполь-
зуют слабый свет, и насыщение у них наступает очень рано (примерно
при 10 клк). Растения или органы с низкой точкой компенсации способ-
129
О 1 2 4 6 7 102
Интенсивность освещения, Вт/м2
Рис. 6.16. Влияние света на нетто-фотосинтез различных растений при
оптимальной температуре и естественном содержании СО2 в воздухе
(по В.Лархеру, 1978):
1 — сорго; 2 — кукуруза; 3 — пшеница; 4 — светолюбивые травы; 5 — бук; 6 —
теневыносливые травы; 7 — тенелюбивые мхи, планктонные водоросли
ны к более полному использованию света, но при освещенности ниже
критической они погибают. Так происходит отмирание нижних листьев
и очищение ствола от ветвей.
Точка компенсации суммирует действие разных факторов на
дыхание и фотосинтез. Знание ее необходимо при изучении про-
дуктивности растений, поскольку она указывает на границу меж-
ду запасанием и расходованием органики. Ниже нее наступает
голодание. Компенсационная точка обычно определяется при
концентрации СО2 0,03 % и температуре 20 °C. При постоянной
концентрации СО2 она повышается с ростом температуры, так
как при потеплении дыхание увеличивается сильнее фотосинтеза.
Поэтому при пониженной освещенности (например, зимой в оран-
жереях) необходима умеренная положительная температура и по-
вышение ее может снизить темпы роста растений. Улучшение
водного режима и снабжения СО2 сдвигает компенсационную точку
в сторону меньшей освещенности, а старение листьев — в сторо-
ну большей.
Угол наклона восходящей части кривой к оси абсцисс показы-
вает эффективность использования света растением (свидетель-
ствует о скорости увеличения интенсивности фотосинтеза по мере
возрастания освещенности). В этой области фотосинтез опреде-
ляется скоростью световых реакций: чем выше квантовый выход
фотосинтеза, тем круче поднимается световая кривая. Это важ-
ный показатель функционального состояния растений.
Перегиб световой кривой указывает на два важных параметра.
Во-первых, минимальный уровень освещения, обеспечивающий
130
в данных условиях максимальную интенсивность фотосинтеза.
Дальнейший рост освещенности не увеличивает его, поэтому све-
товая кривая выходит на плато и происходит световое насыщение
фотосинтеза. Во-вторых, уровень плато показывает максималь-
ную величину фотосинтеза, которой достигает растение в данных
условиях. Он определяет продуктивность растения при достаточ-
ном освещении, однако в разных условиях этот показатель силь-
но колеблется.
У большинства растений световые кривые постепенно выходят
на плато, но светолюбивые виды при повышении концентрации
СО2 и оптимальном сочетании других факторов плато не имеют,
образуя идущие вверх линии, лишь несколько отгибающиеся.
У теневыносливых видов плато выражено хорошо, и чем меньше
интенсивность света, при которой наступает перегиб световой
кривой, тем более теневынослив вид. С повышением концентра-
ции СО2 уровень плато теневыносливых видов может повышать-
ся, а перегиб смещаться в сторону повышенной освещенности.
При сравнении световых кривых особо выделяются С4-виды.
У них световое насыщение не достигается, и в ясные дни они
используют свет полностью даже в полдень. И при средней осве-
щенности эти растения фотосинтезируют интенсивнее С3-видов.
Цикл Кальвина у С3-растений работает не так эффективно, по-
этому их световая кривая делает изгиб раньше. Среди них есть и
такие растения, фотосинтез которых при излишней освещеннос-
ти снижается, т.е. для них существует оптимум освещения. Это
споровые сеянцы древесных пород и травы сомкнутых лесов.
Световая кривая планктонных водорослей имеет колоколовидную
форму: после достижения оптимума она падает (область перенасыще-
ния), так как интенсивность света больше насыщающей подавляет их
Рис. 6.17. Зависимость между интенсивностью света и фотосинтезом
(по Р. Риклефсу, 1979):
/ — зеленые водоросли; 2 — диатомовые водоросли; 3 — белый дуб; 4 — ладан-
ная сосна; 5 — кизил
131
фотосинтетическую активность (рис. 6.17). Поэтому наибольшая интен-
сивность фотосинтеза при высоком стоянии солнца отмечается не у са-
мой поверхности, а несколько глубже (в озерах на глубине — 2—5 м, в
открытом море — 10—15 м) в зависимости от силы света над водой и ее
мутности. Вблизи поверхности воды свет избыточен, и здесь, особенно в
солнечные дни, может происходить ингибирование фотосинтеза. Веро-
ятно, это обусловлено в основном тем, что скорость поглощения радиа-
ции пигментами так высока, что ее энергия не может быть использована
водорослями нормальным путем и расходуется на деструктивные фото-
окислительные реакции. В пасмурные же дни и в бедное светом время
года торможения фотосинтеза светом в водоемах нет и область оптимума
смещается до самой поверхности.
Параметры световых кривых специфичны для разных по отно-
шению к свету экологических групп растений, они значительно
меняются в зависимости от действия других экологических фак-
торов, стадии развития растения, площади и расположения лис-
тьев. Таким образом, световые кривые несут много важной эко-
логической информации.
Температурная кривая фотосинтеза. Температура влияет на об-
мен веществ, изменяя скорость химических реакций и активность
ферментов. Ускорение фотосинтеза при повышении температуры
в целом идет экспоненциально, опережая возрастание скорости
неферментативных реакций. При низких температурах это уско-
рение более существенно, так как в этих условиях лимитируют
процесс обычно ферментативные реакции, а при высоких темпе-
ратурах это нарастание слабее, поскольку лимитирующими там
оказываются физические процессы, например диффузия.
Влияние температуры на фотосинтез зависит от интенсивнос-
ти освещения. При малом количестве света фотосинтез от нее не
зависит (идет с одинаковой скоростью и при 15 °C, и при 25 °C).
Это связано с тем, что при низкой освещенности интенсивность
фотосинтеза лимитируется скоростью световых реакций, а при
высокой освещенности она определяется уже протеканием тем-
новых реакций, и влияние температуры становится очень отчет-
ливым. Так, для подсолнечника (Helianthus annuus) повышение
температуры от 9 до 19 °C увеличивает интенсивность фотосинте-
за в 2,5 раза (Н. И.Якушкина, 1993).
При достаточном освещении действие температуры на фото-
синтез связано в основном с влиянием на темновые реакции.
И поглощение, и восстановление СО2 обычно ускоряются с по-
вышением температуры вплоть до достижения ею некоторого оп-
тимального диапазона. А при очень высокой температуре, приво-
дящей к инактивации ферментов и рассогласованию реакций, фо-
тосинтез быстро прекращается. Аналогично влияет температура и
на дыхание: с ее повышением оно быстро растет, но при очень
высоких температурах биохимические процессы протекают так
132
быстро, что снабжение субстратом и метаболитами не поспевает
за обменом веществ, и интенсивность дыхания быстро падает.
Итоговый результат зависит от разности между скоростями асси-
милирования и выделения диоксида углерода при той же темпе-
ратуре. На общий итог влияет также зависимость от температуры
ширины открывания устьиц. В целом повышение температур сна-
чала усиливает нетто-фотосинтез, а потом тормозит его.
Зависимость нетто-фотосинтеза от температуры характеризу-
ется температурной кривой, которая имеет колоколовидную фор-
му. Кривая асимметрична: с постепенным подъемом и довольно
быстрым падением. Нижняя температурная точка довольно по-
стоянна для разных видов и обычно совпадает с замерзанием тка-
ней. У большинства растений она приходится на диапазон от -10
до -2 °C. У весенних эфемероидов, высокогорных и многих веч-
нозеленых растений этот показатель составляет от -5 до -7 °C, а
лишайники могут поглощать СО2 даже тогда, когда их талломы
уже частично замерзли (некоторые при температурах до -25 °C).
Фотосинтез же тропических растений начинается при положи-
тельных температурах (от 5 до 7 °C). Зимние температурные ми-
нимумы близки у многих растений.
При перегреве продуктивность фотосинтеза резко падает и од-
новременно высвобождается много СО2 при повышении интен-
сивности дыхания. Верхняя температурная кардинальная точка
фотосинтеза лежит на 2 —12 °C ниже температуры тепловой смер-
ти и соответствует условиям, при которых отмечается равновесие
между ингибированием фотосинтеза и повышением скорости ды-
хания. Поэтому температурный максимум нетто-фотосинтеза мож-
но рассматривать как тепловую точку компенсации, которая дос-
тигается тем раньше, чем чувствительнее к теплу фотосинтез и
чем круче подъем дыхания.
Усиливающийся расход СО2 на дыхание может быть покрыт
фотосинтезом при более высокой интенсивности света. Поэтому
если освещенность повышается, то максимальная температура
фотосинтеза тоже увеличивается. При постоянной же освещенно-
сти восходящая кривая дыхания и нисходящая фотосинтеза пере-
секаются в верхней точке. Верхняя температурная граница нетто-
фотосинтеза подвержена сезонным и адаптивным изменениям
(вызываемым жарой и засухой). Она тем выше, чем южнее ареал
вида. Так, для растений высоких широт она лежит около 30 °C,
для видов умеренной зоны — в диапазоне от 37 до 44 °C, для сре-
диземноморских растений — в диапазоне 42—48 °C, а для видов
жарких пустынь составляет 55 °C. У некоторых тропических зла-
ков и маревых из чрезвычайно жарких местообитаний тепловые
точки компенсации близки к абсолютному пределу переносимос-
ти температур (58 — 60 °C), однако обычно такие показатели в ес-
тественных условиях не наблюдаются (И. М. Культиасов, 1982).
133
Колоколообразная форма кривой зависимости фотосинтеза от тем-
пературы иногда полностью объясняется соотношением между фотосин-
тезом и дыханием, нарастающим с повышением температуры. Однако
необходимо принимать во внимание и многие другие факторы. Более
точной считается схема В.Лархера (1978), учитывающая еще транспира-
цию и содержание СО2 в воздухе при разных температурах (рис. 6.18).
В этой схеме брутто-фотосинтез возрастает благодаря температурной ак-
тивации участвующих в этом процессе ферментов до тех пор, пока недо-
статочное поступление СО2 и тормозящие эффекты (связанные, напри-
мер, с ферментами) не приведут к замедлению фотосинтеза. Дыхание
экспоненциально возрастает в более широком диапазоне температур и
быстро прекращается при высоких, повреждающих цитоплазму. Разность
между брутто-фотосинтезом и дыханием определяет уровень нетто-фо-
тосинтеза, положение его оптимума и кардинальных точек. Кроме того,
абсолютный температурный оптимум фотосинтеза может быть получен
только при устранении ограничивающего влияния других факторов. Но
в этом случае проявится потенциальный аутэкологический оптимум, а
синэкологический реализуется только при совместном влиянии всех
факторов с учетом конкуренции. Этот реальный температурный опти-
мум фотосинтеза всегда смещен в область более низких температур.
Оптимальной температурной областью фотосинтеза считают
температуру, при которой наблюдаемый фотосинтез достигает 90 %
максимально возможной величины. При этом температурный
оптимум нетто-фотосинтеза уже оптимального интервала темпе-
ратур для активности фотосинтетических ферментов, так как в то
время, когда брутто-фотосинтез близок к максимуму, дыхание
усиливается, снижая чистую продуктивность.
Рис. 6.18. Взаимосвязь между фотосинтезом, дыханием, транспирацией
и содержанием СО2 (по В.Лархеру, 1978).
I — брутто-фотосинтез при оптимальном содержании СО2; II — брутто-фотосин-
тез при естественном содержании СО2
134
Рис. 6.19. Зависимость от температуры и положение кардинальных точек
нетто-фотосинтеза при световом насыщении у разных типов растений
(по В.Лархеру, 1978)
У С3-растений кардинальные точки температурной кривой фо-
тосинтеза в основном зависят от теплового режима местообита-
ния. По В.Лархеру (1978), растущие в тени растения нагреваются
меньше и имеют оптимум фотосинтеза между 10 и 20 °C. Анало-
гично это наблюдается для раннецветущих, высокогорных и дру-
гих растений, растущих в такое время года или в таких местах, для
которых характерны низкие средние температуры. Так, мох
Rhacomitrium lanuginosum господствует на приатлантических бо-
лотах и скалах в горах с резко выраженным океаническим клима-
том. Оптимальная величина нетто-ассимиляции отмечается у него
при температуре 13 —15 °C (при температуре ниже 10 °C или выше
20°С интенсивность его фотосинтеза незначительна).
Растения теплых местообитаний наивысшей продуктивности
достигают при более высоких температурах (20 —30 °C). У С4-рас-
тений оптимум продуктивности лежит при температурах выше 30 °C
(рис. 6.19), а у некоторых даже при 50 °C. У CAM-растений днев-
ная температура почти не имеет значения для фотосинтеза, а тем-
новая фиксация СО2 хорошо согласуется с низкими ночными тем-
пературами (оптимум между 5 и 15 °C, выше 25 °C процесс пре-
кращается).
Оптимальная температура фотосинтеза повышается с увеличе-
нием освещенности. При меньших освещенностях оптимумы сме-
щаются в область более низких температур. Менее значительные
изменения температурного оптимума наблюдаются при измене-
нии концентрации СО2, колебания которой в естественных усло-
виях незначительны. При низкой интенсивности света и нормаль-
ном содержании СО2 растения сильнее фотосинтезируют при бо-
лее низких температурах, а при высокой освещенности наблюда-
ется обратное соотношение. Однако в этом случае усиливается
дыхание, поэтому растение проигрывает в нетто-фотосинтезе.
135
Температурный оптимум нетто-фотосинтеза при слабой интен-
сивности света лежит в пределах низких температур, а при высо-
кой освещенности повышается. Эти закономерности экологиче-
ски важны, так как малая освещенность часто связана с низкими
температурами (например, в холодный период года или утром и
вечером). В полярных областях это имеет значение во время по-
лярных ночей в конце лета, когда освещенность падает. Однако
полярные растения используют и эту малую интенсивность света
для образования ассимилятов при невысоких температурах. В связи
с коротким вегетационным периодом в полярных районах почти
круглосуточный фотосинтез играет большую роль, так как расте-
ния здесь должны развиваться быстро.
Влияние на фотосинтез содержания диоксида углерода и кис-
лорода. Основной источник углерода для фотосинтеза — СО2 ат-
мосферы. Содержание его сильно влияет на ход фотосинтеза. Нор-
мальная концентрация СО2 около 0,03 % и его снижение вызывает
уменьшение интенсивности фотосинтеза, а увеличение (до опреде-
ленного предела) ведет к усилению фотосинтеза (см. разд. 9.2.2).
Разные растения неодинаково используют одни и те же кон-
центрации углекислоты. С4-виды обладают большей способнос-
тью к связыванию СО2 благодаря более высокой активности осу-
ществляющего процесс фермента. У них процесс фотосинтеза
может происходить при содержании СО2 от 0,0005 %, в то время
как для С3-растений необходимая концентрация СО2 в 10 раз боль-
ше. Повышение содержания диоксида углерода до 1,5% прямо
пропорционально возрастанию интенсивности фотосинтеза у зер-
новых. Для других растений это наблюдается в основном до кон-
центрации СО20,1 %.
Концентрация СО2, после достижения которой интенсивность
фотосинтеза начинает снижаться, имеет примерно десятикратное
Концентрация СО2, об. %
Рис. 6.20. Продуктивность нетто-фо-
тосинтеза при повышенном снабже-
нии растений СО2 (по В.Лархеру, 1978):
1 — Cucumis\ 2 — Lycopersicunr, 3 — Fagus
(световые листья); 4 — Ginkgo (зрелые ли-
стья); 5 — Ginkgo (молодые листья); 6 —
Fagus (теневые листья)
136
Рис. 6.21. Зависимость фотосинтеза кисли-
цы (Oxalis acetoselld) от освещенности и
содержания СО2 в воздухе (по Н. Walter,
1960):
1 - 0,09 % СО2; 2 - 0,06 % СО2; 3 - 0,03 % СО2
Освещенность, %
превышение нормального значения (например, для бука — Fagus —
по достижении 0,2 % СО2 в воздухе) (рис. 6.20). Избыточное со-
держание СО2 (5 — 20%) фотосинтез ингибирует. Есть растения
более чувствительные к повышению концентрации СО2, и у них
ассимиляция тормозится раньше. Депрессия фотосинтеза вызы-
вается разными причинами: при увеличении содержания СО2 за-
крываются устьица, ингибируются темновые реакции фотосинте-
за. В природе, где содержание СО2 в воздухе редко поднимается
выше 0,04 %, растения связывают обычно не более 80 мг СО2 на
1 дм2 в 1 ч. Малое содержание диоксида углерода может ограни-
чивать возрастание фотосинтеза, особенно при ярком освещении,
когда лимитирующими являются темновые реакции. Кроме того,
днем содержание СО2 в воздухе вокруг растений понижается (над
полем кукурузы — Zea mays — в три раза).
В нижних ярусах леса при нормальном содержании СО2 уже
при 10%-й освещенности (с учетом бликов) фотосинтез обычно
достигает максимальных значений (световая кривая переходит на
плато). Использование более интенсивного света возможно только
при повышении концентрации СО2 в воздухе, так как усиление
освещенности ведет к повышению уровня насыщения (рис. 6.21).
В широколиственном лесу в жаркий день в Подмосковье содер-
жание СО2 в приземном слое на порядок выше, чем в среднем по
атмосфере, за счет интенсивности микробиологических процес-
сов.
При искусственно повышенном (до 0,3—1 %) содержании СО2 в воз-
духе, сильном освещении, хорошем водоснабжении, благоприятной тем-
пературе (т.е. в оптимальных условиях) травы способны усваивать за 1 ч
до 150 мг СО2 на 1 дм2 листовой поверхности. При этом достигается
предел «потенциального фотосинтеза». Возможность повышения про-
дуктивности фотосинтеза путем «удобрения диоксидом углерода» исполь-
зуют в теплицах. Так, выращивая растения при концентрации СО2 в
воздухе 0,1 %, можно повысить интенсивность фотосинтеза зерновых в
два раза, листовых овощей, томатов (Lycopersicori) и огурцов (Cucumis
137
sativus) в четыре раза, а прирост их — в два раза. В полевых условиях
углекислое удобрение может частично достигаться за счет усиления ды-
хания почвы при внесении органических удобрений. А особенно эффек-
тивны такие подкормки в закрытом грунте (Н.И.Якушкина, 1993).
Несмотря на то что кислород — один из продуктов фотосинте-
за, при полном анаэробиозе этот процесс прекращается. Это свя-
зано с торможением дыхания и накоплением продуктов неполно-
го окисления, в частности органических кислот, отчего резко сни-
жается pH. В кислой среде влияние анаэробиоза более резко. Обыч-
ная концентрация кислорода превышает оптимальную для фото-
синтеза величину. У растений с высоким уровнем фотодыхания,
например у бобов (Vicia faba), уменьшение концентрации кисло-
рода с 21 до 3 % фотосинтез усиливает, а у кукурузы (Zea mays)
(с низким уровнем фотодыхания) это изменение на интенсив-
ность фотосинтеза не влияет. Повышенная концентрация кисло-
рода (25 — 30%) также тормозит фотосинтез (эффект Варбурга).
Особенно сильно это сказывается при интенсивном освещении.
Фотосинтез и водообеспеченность. Фотосинтез необходимо рас-
сматривать во взаимосвязи с водным режимом растений, поскольку
и на поглощение диоксида углерода, и на испарение воды влияет
работа устьичного аппарата (транспирация фактически становит-
ся побочным процессом открытости устьиц при газообмене). Вода,
как и СО2, — необходимый субстрат для фотосинтеза, но лимити-
рует его лишь та вода, которая нужна для поддержания необходи-
мого набухания цитоплазмы. От этого зависят процессы клеточ-
ного метаболизма. Уменьшение степени набухания тормозит фо-
тосинтез, так как ослабляет и фотохимические, и темновые реак-
ции, снижает активность ферментов.
У слоевищных растений степень набухания цитоплазмы изме-
няется вместе с влажностью воздуха и соответственно содержания
в нем водяного пара. При обезвоживании фотосинтез и дыхание
снижаются. Важное экологическое значение при изменении влаж-
ности имеет точка компенсации. Фотосинтетический аппарат на-
земных водорослей, лишайников и мхов адаптирован к частым и
сильным колебаниям содержания воды. Высохшие слоевища бы-
стро возобновляют фотосинтез после увлажнения, что определя-
ет, например, возможность жизни лишайников в пустыне. Для
фотосинтеза они используют там раннее утро, поглощая росу и
влагу воздуха до восхода солнца. Пока цитоплазма набухает, они
несколько часов фотосинтезируют, а потом высыхают и фотосин-
тез прекращается.
У высших растений при сильном обезвоживании цитоплазмы
фотосинтез тоже снижается. Он ограничивается водным дефици-
том и косвенно (из-за его влияния на устьица). При обезвожива-
нии тургор устьичных клеток ослабевает, поэтому прекращается
138
приток СО2. На это разные листья реагируют неодинаково: при
напряженном водном балансе теневые листья устьица закрывают,
а световые — нет. Интенсивность дыхания тоже падает. В целом
чем чувствительнее вид к недостатку воды и чем сильнее засуха,
тем в течение дня раньше сократится фотосинтез. У САМ-расте-
ний дневной фотосинтез идет и при закрытых устьицах, так как
они используют диоксид углерода, получающийся из яблочной
кислоты, накопленной за ночь.
Обычно для фотосинтеза оптимально не полное насыщение
водой тканей, а небольшой водный дефицит (около 5 — 20% от
полного насыщения) («эффект Бриллиант») (рис. 6.22). Это связа-
но, видимо, с тем, что в местообитаниях наземных растений обычно
наблюдается подобное состояние недонасыщения листьев водой.
Больший дефицит воды ингибирует фотосинтез, а при 50%-м со-
держании воды от полного насыщения он прекращается.
У деревьев суточный ход нетто-ассимиляции листьев в целом анало-
гичен изменениям транспирационного режима, так как оба процесса
связаны с состоянием устьичного аппарата. При оптимальной влаго-
обеспеченности нетто-ассимиляция зависит от интенсивности освеще-
ния: наибольшая ее величина отмечена на более освещенной части дере-
ва, наименьшая — в наиболее затененных частях кроны. У очень высо-
ких деревьев нетто-ассимиляция на вершине снижается из-за трудности
подачи воды на большую высоту.
Сухость воздуха усиливает значение фактора воды, фотосинтез тене-
вых листьев снижается, поскольку у них преждевременно закрываются
устьица. Если сухость воздуха усугубляется сухостью почвы, то устьица и
у световых листьев смыкаются, что ограничивает транспирацию. При
таких погодных условиях больше всего вещества продуцируется в пред-
полуденные часы. В пасмурные дни, но при светлой облачности бук
Рис. 6.22. Зависимость интенсивности фотосинтеза от водного дефицита
ассимиляционной ткани у картофеля (Solatium tuberosum) (по В. М. Иван-
ченко, 1974)
139
(Fagus), например, продуцирует не меньше вещества, чем в ясные теп-
лые дни (Г. Вальтер, 1968).
Фотосинтез и минеральное питание. Если почва недостаточно
богата необходимыми солями, продуктивность растений будет
низкой вне зависимости от солнечности погоды, обилия влаги и
устойчивости температуры. «Сколько не имей энергии, а расте-
ние строит свой организм не из нее, а из вполне «осязаемых»
веществ и элементов», — пишут в «Теоретической экологии эко-
систем» Г.Орген и Э. Боссат (1996). Они отмечают, что в традици-
онных моделях круговорота веществ основное внимание уделяет-
ся углероду, но «продукция растений — это не только углеводы,
но и белки», поэтому все большее внимание уделяется доступно-
сти минеральных веществ, особенно азота и фосфора.
Элементы минерального питания могут влиять на интенсив-
ность углеродного обмена и прямо, и косвенно, через обмен ве-
ществ и рост. Прямое действие связано с тем, что они входят в
состав ферментов, пигментов или непосредственно участвуют в
фотосинтезе как активаторы. Например, Мп служит активатором
фотолиза, К связан с переносом энергии через мембраны тилако-
идов, N и Mg входят в состав хлорофилла, Fe, Со и Си содержатся
в различных ферментах, а фосфат — в нуклеотидах.
Недостаток минеральных веществ или нарушение соотноше-
ния между ними может повлиять на количество, размеры, ультра-
структуру хлоропластов и содержание хлорофилла, притом даже
если эти элементы не входят в состав молекулы хлорофилла (на-
пример, Fe). При недостатке N и Fe возникают хлорозы, приво-
дящие к сильному снижению поглощения СО2. Аналогичные по-
следствия возможны при недостатке Mg. Из-за низкого содержа-
ния хлорофилла световые растения уже не могут использовать
сильный свет и ведут себя как теневые. Кроме того, минеральное
питание сказывается на газообмене, влияя на реакции устьиц,
анатомическое строение, размеры и продолжительность жизни
листьев. При недостатке N листья развиваются мелкие, с менее
подвижным устьичным аппаратом, а избыток его сильно повы-
шает интенсивность дыхания, снижая этим нетто-фотосинтез.
Из почвенных биогенов на продуктивность растений наиболь-
шее влияние имеют соединения азота. Поэтому, вероятно, нет
сельскохозяйственной культуры, которая не отреагировала бы на
азотное удобрение повышением первичной продукции. По-види-
мому, это справедливо и для естественной растительности. Вне-
сение азотных удобрений в почву леса обычно стимулирует при-
рост древесины. Из-за недостатка других элементов продукция
сообщества также может быть гораздо ниже теоретически возмож-
ного уровня. Например, в Южной Австралии при существенном
дефиците фосфора и цинка экономически оправдано выращива-
140
ние сосняков из Pinus radiata только при искусственном внесе-
нии в почву этих биогенов (М. Бигон и др., 1989).
Фотосинтез и болезни растений. Многие грибные болезнетвор-
ные организмы не приводят к быстрой гибели хозяев, однако ин-
тенсивность фотосинтеза больных растений обычно значительно
ниже нормы (рис. 6.23). Это происходит в основном из-за нару-
шения структуры фотосинтетического аппарата клеток. Так, у
пораженных ржавчиной листьев овса (Avena) и ячменя (Hordeum)
резко падают числа хлоропластов на единицу площади листа, объем
хлоропластов, концентрация хлорофилла и соотношение хлоро-
филлов а и Ь.
Отрицательное влияние на фотосинтез бактериальных инфек-
ций связано прежде всего с интенсивным разрушением структуры
хлоропласта, что обусловлено активным выделением бактериями
токсинов в ткани хозяина. Например, табтоксин, образуемый
Pseudomonas tabaci, подавляет активность РДФ-карбоксилазы в
листьях табака (Nicotiana).
Вирусное воздействие на фотосинтез растений может быть двух
типов. В первом случае, например у тыквы (Cucurbita pepo), ин-
фекция ведет к снижению числа хлоропластов, но на интенсив-
ность фотосинтеза влияет мало. В другом случае (у китайской ка-
пусты) вирусное воздействие подавляет фотофосфорилирование
и фотосинтез, не оказывая заметного влияния на структуру мезо-
филла листа.
На общий результат продуктивности
растений влияет также изменение пато-
генами дыхания растений. Обычно оно
повышается у инфицированных особей.
Это может сочетаться с высокой долей
анаэробного дыхания, а также с наруше-
нием сопряженного с окислением про-
цесса фосфорилирования из-за ингиби-
рующего действия продуктов жизнедея-
тельности патогена. В результате снижа-
ется эффективность дыхания клеток рас-
тения и усиливаются гидролитические
процессы. Отмечено также отрицатель-
ное влияние на ферментативный аппа-
рат метаболитов патогена.
Рис. 6.23. Влияние заражения мучнистой ро-
сой листьев ячменя (Hordeum) на нетто-фо-
тосинтез (Л), дыхание (Б) и фотодыхание (В)
(по Н.Н. Третьякову, 2000).
Контроль — неокрашенные символы
141
У иммунных сортов инфекция, наоборот, индуцирует образо-
вание и специфических ферментов, и новых рибосом, и митохон-
дрий. У них активирование дыхания, усиление окислительного
фосфорилирования, использование дополнительной энергии на
синтез ферментативных и структурных белков успешно противо-
стоит влиянию токсинов и иногда даже повышает эффективность
запасания энергии (Н.Н.Третьяков и др., 2000).
Совокупное влияние факторов на фотосинтез. В природе эколо-
гические факторы действуют совместно, а газообмен растений
отражает взаимодействие множества внутренних и внешних фак-
торов. Какой-то из них может принимать экстремальное значе-
ние и в течение некоторого времени лимитировать фотосинтез,
но влияние оказывают и остальные факторы. Например, с повы-
шением интенсивности света оптимальная температура и верхняя
тепловая граница нетто-фотосинтеза сдвигаются на несколько
градусов вверх. Для растения это выгодно, так как сильное осве-
щение всегда связано с нагревом. При перегреве и связанного с
ним недостатка воды ассимиляция СО2 и продуктивность снижа-
ются.
Влияние на фотосинтез света, диоксида углерода и температу-
ры осуществляется в сложном взаимодействии. Особенно тесно
связаны свет, действующий на скорость фотохимических реак-
ций, и температура, контролирующая скорость ферментативных
процессов. При высокой интенсивности света и низких темпера-
турах (5—10°C), когда главным фактором, лимитирующим ско-
рость всего процесса, становятся контролируемые температурой
ферментативные реакции, скорость комплекса сопряженных с
восстановлением углерода энзиматических реакций при повы-
шении температуры на 10 °C возрастает более чем в четыре раза
(Qio > 4). При более высоких значениях температурный коэффи-
циент Q|0 снижается до 2. При низких же интенсивностях света
Qio = 1, т.е. фотосинтез относительно независим от температуры,
Рис. 6.24. Зависимость фотосинтеза кар-
тофеля (Solatium tuberosum) от темпера-
туры, света и содержания СО2 в воздухе
(по И. М. Культиасову, 1982):
7, 2 — при низкой концентрации СО2 и сла-
бом освещении; 3 — на полном свету и при
нормальной концентрации СО2; 4 — на пол-
ном свету и при повышенной концентрации
СО2
142
так как в данном случае его скорость ограничивается фотохими-
ческими реакциями (В. В. Полевой, 1989).
Если концентрация СО2 и сила света низка, то фотосинтез силь-
но ослабевает, а при повышенном содержании СО2 и полном све-
те интенсивность его резко подскакивает при большей темпера-
туре, но и быстро падает уже при небольшом ее превышении в
связи с перегревом и недостатком влаги (рис. 6.24). В целом в
природе благоприятное сочетание факторов — большая редкость,
поэтому фотосинтез редко достигает максимальных величин. По
В.Лархеру (1978), наивысшая интенсивность поглощения СО2 за
день обычно в среднем составляет 70—80% фотосинтетической
способности.
6.5. Приспособления растений к световому режиму
На Земле существует огромное разнообразие местообитаний,
отличающихся по световому режиму (интенсивность света, его
спектральный состав, продолжительность освещения, распреде-
ление света в пространстве и времени). Растения живут и при
очень сильном освещении в пустынях и высокогорьях, и в тем-
ных пещерах и подводных глубинах. В процессе длительной эво-
люции растительные организмы приобрели эффективные приспо-
собления к успешной жизни при световом режиме их местооби-
таний.
Экологические группы растений по отношению к свету. В. Н. Лю-
бименко предложил выделять три группы растений по отноше-
нию к свету: гелиофиты (от греч. helios — солнце), или световые
(светолюбивые), сциофиты (от греч. skia — тень), или теневые
(тенелюбивые), и теневыносливые. Они отличаются прежде все-
го положением светового оптимума и кардинальных точек свето-
вого довольствия (рис. 6.25). У гелиофитов оптимальное световое
довольствие LonT находится в области почти полного освещения,
Рис. 6.25. Экологические оптимумы
растений по отношению к свету (по
Т.К.Горышиной, 1979):
/ — тенелюбивый вид; 2 — светолюби-
вый вид; 3 — теневыносливый вид
143
и сильное затенение их угнетает. Световое довольствие этих рас-
тений близко к 100%, они растут на открытых местах, где инсо-
ляция велика и растительный покров изрежен. К этой группе от-
носятся виды жарких пустынь, тундр, высокогорий, скал и каме-
нистых осыпей, рудералы пустырей и обочин дорог, большинство
культурных растений открытого грунта и сорняков, многие при-
брежные и водные растения с надводными листьями, деревья пер-
вого яруса и ранневесенние травы листопадных лесов. Гелиофи-
тами являются, в частности, сосна обыкновенная (Pinus sylvestris),
береза повислая (Betulapendula), мать-и-мачеха (Tussilagofarfara),
очиток едкий (Sedum acre) и др.
Однако границы между дискретными экологическими группа-
ми условны: гелиофиты образуют непрерывный ряд по измене-
нию светолюбия. Действительно, некоторые из них могут расти в
слабозатененных условиях. Поэтому различают облигатные ге-
лиофиты, не выносящие затенения, и факультативные, хорошо
растущие и при небольшом затенении. Кроме того, их экологи-
ческие предпочтения пластичны в онтогенезе и конкурентной
среде. С хозяйственной точки зрения, поскольку полевые сорня-
ки большей частью светолюбивы, действенное средство борьбы с
ними — затенение. Засорены же в первую очередь поля с зерно-
выми, дающими мало тени (Г. Вальтер, 1982).
Теневыносливые растения имеют широкую экологическую ам-
плитуду по отношению к свету, и их точнее называть свето- и
теневыносливыми. Максимальное световое довольствие (Lmax) их
равно 100%, а минимальное (Lmin) разнообразно и характеризует
способность углубления в тень. Как правило, световая кривая их
асимметрична: они лучше растут при большой освещенности, но
хорошо адаптируются и к слабому свету. Экспериментально по-
казано, что многие растения затененных мест при предоставле-
нии им несколько большего освещения развиваются лучше, т.е.
они именно теневыносливы. Это распространенная и очень плас-
тичная группа. Сюда относятся, например, многие лесные травы.
А в соответствии с континуальным изменением данного признака
у лесоводов давно существуют шкалы теневыносливости древес-
ных пород.
Сциофиты имеют оптимум в области слабой освещенности и
не выносят сильного света. Он подавляет их, так как интенсивно
разрушает хлорофилл, а они медленно его производят. Кроме того,
теневые растения при сильной инсоляции так быстро теряют вла-
гу при транспирации, что вынуждены закрывать устьица, что при-
водит к торможению фотосинтеза и голоданию. Поэтому сцио-
фиты не растут на полном свету (у них всегда Lmax < 100 %). Также
они заходят в более глубокую тень, чем теневыносливые виды, и
их минимальное световое довольствие всегда ниже. Сциофиты
предпочитают ослабленное освещение.
144
К тенелюбивым видам принадлежат растения сильно затенен-
ных мест. При этом исходно затененные местообитания (пещеры,
расщелины скал, водные глубины и т.п.) сравнительно редки.
В основном в экосистемах затенение — результат перехвата света
более высокими растениями. Поэтому среди сциофитов много
лесных видов нижних ярусов (мхи, папоротники, ряд трав, на-
пример грушанки — Pyrola, и др.). Оказавшись на вырубке, эти
растения желтеют из-за разрушения хлорофилла и вскоре погиба-
ют. К тенелюбам относятся также многие комнатные растения,
являющиеся выходцами из травяного покрова или эпифитов тро-
пических лесов. По способности расти на свету среди сциофитов
тоже можно выделить факультативные и облигатные виды.
Степень теневыносливости или светолюбия не является строгим ви-
довым признаком. Внутривидовые анатомо-морфологические и физио-
логические различия выражены у особей, растущих в разных условиях
освещенности, — в глубине леса и на опушке, в густом травостое и при
одиночном произрастании, на северной и южной стороне стволов и т.д.
Дозируя интенсивность и продолжительность освещения в период за-
кладки и формирования листа, из саженцев можно получить особи и со
световыми, и с теневыми листьями. Так, листья вырастающих в тени
сеянцев эвкалиптов {Eucalyptus) сохраняют черты теневыносливости
(Т. К. Горышина, 1979). Существуют различия световых кривых фото-
синтеза у глубоководных и мелководных форм водорослей одного вида.
Есть различия в структуре и физиологии листьев разных частей кроны: с
ослаблением света в нижних и внутренних частях кроны они приобрета-
ют теневой характер. Так же на опушке сторона дерева, обращенная к
лесу, имеет листья теневого типа, а на открытой стороне преобладают
листья световые.
Способность растений выносить недостаток света в большой
степени зависит от комплекса других экологических факторов.
В оптимальных климатических и почвенных условиях теневынос-
ливость вида обычно выше. Этим объясняются закономерные гео-
графические изменения по отношению к свету широко распрост-
раненных видов. Например, в северных частях ареала виды сред-
ней полосы более светолюбивы, чем в южных. Световое доволь-
ствие увеличивается и при подъеме в горы, что отмечено на при-
мере дикого ячменя (Hordeum): на высоте 180 м он выносит ’/9
полного освещения, а на 2 500 м ему требуется не менее */5 (Т. К. Го-
рышина, 1979). Возрастание светолюбия при понижении темпе-
ратуры по мере продвижения растений к полюсу — одна из при-
чин смены местообитаний некоторых видов на более осветлен-
ные. Так, лесная черника (Vaccinium myrtillus) в тундре хорошо
растет на открытых местообитаниях, а сныть (Aegopodium podagra -
ria), один из наиболее теневыносливых видов дубрав, на севере
России обитает на опушках и лугах. Аналогично отношение рас-
145
тений к свету и при подъеме в горы. Особо меняется светолюбие
у растений арктической зоны, где летом освещение круглосуточ-
но: в холодные ночи они лучше используют свет слабой интен-
сивности (как тенелюбы), а днем хорошо используют интенсив-
ный свет (как светолюбы).
Формообразование растений под действием света. Растения и
их части, развивающиеся на полном свету, анатомо-морфологи-
чески отличаются от теневых. В связи с этим выделяют черты
гелиоморфоза (или фотоморфоза) — формообразования под дей-
ствием света. Некоторые из них — видовые, другие развиваются
лишь в определенных условиях. Часто эти черты сопряжены с
ксероморфными признаками, что связано с напряженным водо-
снабжением в сильно освещаемых местах. У теневых же листьев
обычно проявляются мезо- и гигроморфные признаки.
В целом побеги гелиофитов обычно более толстые, с хорошо
развитой ксилемой и механической тканью. Корни их более длин-
ные и разветвленные, а у бобовых на них развивается больше клу-
беньков и они крупнее. Большинство светолюбивых древесных
пород — анемохоры с мелкими семенами, и всходы их лучше вы-
живают на оголенной почве или под изреженным покровом низ-
ких трав. В ажурных кронах этих деревьев свет ослабевает посте-
пенно, а сильное развитие осевого побега у некоторых видов при-
водит к пирамидальности кроны. В сомкнутом сообществе ниж-
ние ветви этих деревьев рано отмирают, стволы очищаются и кроны
располагаются высоко. Для гелиофитов характерны также выра-
женный фототропизм и возникновение асимметричности при од-
ностороннем освещении.
Прямой яркий свет тормозит рост в длину. Междоузлия трав-
светолюбов нередко укорочены, что ведет к образованию розе-
точности. А сочетание укорачивания междоузлий с сильным вет-
влением приводит к подушковидности (рис. 6.26). В связи с мед-
Рис. 6.26. Схема продольных разрезов через растения-подушки
(по В. В.Алехину, 1950):
А — проломник гельвейский (Androsace Helvetica); Б — камнеломка (Saxifraga sp.)
146
ленным ростом растения-подушки по долговечности сравнимы с
деревьями.
При недостатке света у многих растений (особенно у гелиофи-
тов с запасающими органами) стебли аномально вытягиваются,
приобретают бледную окраску и мелкие листья. Утолщение и од-
ревеснение их в затенении отстает от нарастания в длину. Развив-
шиеся в темноте побеги вытянутые, бледные и не способные под-
держивать себя в вертикальном положении, называют этиолиро-
ванными (по франц. — «хилые»). Элементы этиолированности
часто встречаются у растений при совместном произрастании — в
лесу, на лугах, в загущенном посеве. Так, у клевера (Trifolium) в
густом травостое междоузлия удлинены и листья уже по сравне-
нию с экземплярами, выросшими при меньшей сомкнутости.
Многие теневые растения закрепили некоторую этиолированность
в своей жизненной форме. Они нередко становятся стелющимися
или ползучими, а некоторые имеют полегающие стебли, легко
укореняющиеся при достаточной влажности. Если при этом силь-
ное вытягивание стеблей может сочетаться с недоразвитием лис-
тьев, образуются столоны.
Особенности листьев световых и теневых растений. На распо-
ложение, форму и строение листьев влияет множество факторов,
но решающее значение в связи со специализацией листа как при-
емника света и поверхности транспирации имеют световой ре-
жим и водообеспеченность.
Размер листьев — экологически очень важный параметр. Дей-
ствительно, одно из наиболее наглядных различий внешнего об-
лика растений в разных световых условиях — неодинаковая вели-
чина их листовых пластинок. От этого в большой степени зависит
количество радиации, получаемое растением. Этот признак во
многом наследственно детерминирован, но также связан и с мо-
дификациями в разных условиях светообеспеченности (и обеспе-
ченности водой).
Уже простые модели, основанные на оптимизации использова-
ния воды листом (количество граммов СО2, ассимилированного на
1 г израсходованной воды), предсказывают существование несколь-
ких размерных классов листьев (рис. 6.27): например, крупных у
растений затененных влажных теплых мест и мелких у растений
холодных или более теплых и одновременно солнечных местооби-
таний (Э.Пианка, 1981). При одинаковом водообеспечении у све-
толюбивых растений листья обычно мельче, чем у тенелюбивых.
Это хорошо видно при сравнении родственных видов, живу-
щих при разном освещении. Так, мелколистные колокольчики
(поникший Campanula patula и круглолистный С. rotundifolia)
растут на лугу, а колокольчики с широкими листьями (крапиво-
листный С. trachelium и широколистный С. latifolia) обитают в
лесу. Лесные злаки и осоки тоже обычно представлены видами с
147
Рис. 6.27. Схема распределения размеров листьев в зависимости
от условий обитания растений (по Э. Пианка, 1981).
Заштрихованный участок соответствует преобладающим в природе условиям
более широкой листовой пластинкой (овсяницы лесная и гигант-
ская — Festuca sylvatica, F. gigantea, осока волосистая — Carex
pilosa), чем луговые. Отмечен и сезонный диморфизм размеров
листьев: у одной и той же особи медуницы (Pulmonaria obscura)
весенние побеги, развивающиеся при сильном освещении в еще
необлиственном лесу, несут мелкие сидячие листья, а летом в тени
крон широколиственных пород их сменяют крупные листья с
широкой листовой пластинкой. Крупнолистные травы под поло-
гом тенистых широколиственных лесов даже называют широко-
травъем.
Однако все же затенение по-разному может влиять на размеры листа.
У светолюбивых видов любое затенение ведет к редукции площади лис-
тьев и максимальных размеров они достигают только при самом силь-
ном освещении (например, у подсолнечника — Helianthus annuus). Ли-
стья же теневыносливых видов, как это было показано на недотроге
(Impatient noli-tangere), при умеренном затенении становятся крупнее,
но при сильном затенении мельчают. Увеличение листовых пластинок
при затенении до некоторой степени компенсирует снижение интенсив-
ности фотосинтеза ростом фотосинтезирующей поверхности. При этом
еще уменьшается относительная масса неассимилирующих органов и
соответственно падает расход органики на дыхание. Однако чрезмерное
увеличение площади листьев вредно, поскольку нарушает оптимальное
соотношение между поверхностями листьев и корней, что может поста-
вить растение под угрозу при ухудшении условий среды (дефиците водо-
снабжения, усилении инсоляции и т.д.).
С затенением может быть связано уменьшение числа листьев
на побеге (например, у сеянцев дуба — Quercus robur и многих
148
тропических деревьев). Ослабление освещения вызывает редук-
цию листьев как за счет слабого облиствения главных побегов,
так и за счет ослабления формирования боковых. У разных побе-
гов одной кроны размер листьев при затенении меняется по-раз-
ному, но в целом суммарная площадь листьев на растении при
умеренном затенении оказывается больше, чем у незатененных
экземпляров, а при сильном затенении общая площадь листьев
все же падает. Отмечается также, что по форме листовые плас-
тинки сциофитов обычно более простые, чем у светолюбивых ра-
стений, которые чаще развивают рассеченные листья. Рассечение
увеличивается при росте растения на свету.
Морфологической адаптацией является и разное расположе-
ние листовых пластинок в пространстве в условиях избытка или
недостатка света. У гелиофитов листья обычно ориентированы
так, чтобы избежать избыточного полуденного света. Их листо-
вые пластинки расположены вертикально или под большим уг-
лом к горизонту, поэтому днем листья получают лишь скользя-
щие лучи. Среди древесных пород такое положение листьев очень
характерно для эвкалиптов (Eucalyptus). Пластинки листочков
сложных листьев мимоз (Mimosa) и акаций (Acacia) тоже де-
монстрируют такое расположение. Среди трав свойство повора-
чивать листья сообразно падающим лучам имеют многие степ-
ные виды.
Другая адаптация, связанная с необходимостью ослабления
получаемой радиации, встречается у компасных растений. Их ли-
стовые пластинки расположены в одной плоскости, которая ори-
ентирована с севера на юг, поэтому в полдень приход радиации к
листу минимален. Типичное компасное растение — дикий латук
(Lactuca serriola), растущий в разных открытых местообитаниях.
На сильно освещенных местах тенденцию к компасному располо-
жению листьев приобретает и ряд обычных луговых трав (пижма
обыкновенная — Tanacetum vulgare, тысячелистник обыкновен-
ный — Achillea millefolium и др.). Однако в тени компасность
теряется и ориентация листьев становится беспорядочной (Т. К. Го-
рышина, 1979).
У теневыносливых растений, растущих при ослабленном осве-
щении, напротив, листья направлены так, чтобы максимизиро-
вать поглощение света. Поэтому листья растений нижних ярусов
леса часто ориентированы по направлению к просветам в пологе
древостоя. Если же просветов в густом пологе нет, то листья трав
и подроста расположены почти горизонтально, улавливая рассе-
янный свет. То же отмечается в густых луговых травостоях и ниж-
них частях плотных крон. Известным приспособлением, умень-
шающим затенение, является листовая мозаика, получающаяся
при расположении листьев в одной плоскости (рис. 6.28). Она
характерна для подроста и нижних ветвей многих широколиствен-
149
Рис. 6.28. Листовая мозаика у плюща (Hedera) (по А. Е. Васильеву и др.,
1988)
ных деревьев (клен — Acer, вяз — Ulmus, липа — Tilia). Подобное
расположение листьев встречается и у трав сильно затененных
лесов.
Некоторые теневые растения способны менять положение ли-
стовых пластинок при попадании яркого света. Так, избегая све-
тового повреждения, оказавшаяся на вырубке кислица (Oxalis
acetosella) складывает листья так, что их доли приобретают вер-
тикальное положение.
Многие гелиофиты имеют белесоватую окраску, что предохра-
няет их от чрезмерного светового и теплового воздействия. При
этом поверхность органов часто имеет особенности, способству-
ющие отражению лучей. Она может быть блестящей, покрытой
восковым налетом, опушенной светлыми волосками. Характерно,
что у дорсовентральных листьев отражение от нижней стороны
почти в 3 раза выше, чем от верхней. Это защищает нежную губ-
чатую паренхиму и устьица от перегрева в случае поворачивания
листа ветром. Сильно ослабляют свет густое опушение, толстая
кутикула и утолщенные наружные стенки клеток эпидермиса. Так,
эпидермис световых растений пропускает не более 15% падаю-
щего света, а эпидермис теневых — до 98 %. В эпидермисе неко-
торых гелиофитов, растущих на мелах, обнаруживаются кристал-
лические включения, экранирующие световые лучи. То же иногда
отмечается у липы (Tilia) при избытке освещения. Защитную роль
играет и гиподерма хвои сосны (Pinus) и ели (Picea).
Внутреннее строение листа гелиофитов отличается рядом черт
«световой структуры» ассимиляционной ткани. Это прежде всего
хорошо развитый столбчатый мезофилл (рис. 6.29), составляю-
щий около 60 % от общей поверхности фотосинтезирующей тка-
ни (Л. А. Иванов, В. И. Пьянков, 2002). Он состоит из узких длин-
ных клеток, лежащих иногда в 2 — 3 ряда. Вдоль них по межклет-
никам свет эффективно проникает в глубь листа, достигая хло-
ропластов, постенно расположенных в узких клетках. Многослой-
ная столбчатая паренхима световых листьев позволяет гелиофи-
150
там поглощать большие порции света. Многим гелиофитам свой-
ственны также изопалисадные листья, в которых столбчатая ткань
хорошо развита на обеих сторонах (рис. 6.30). Это явление часто
наблюдается у растений с вертикальным положением листьев и
видов на меловых выходах, отражающих падающий на них свет.
В связи с развитием мощной столбчатой ткани возрастает общая тол-
щина листа, поэтому световые листья часто толще теневых. Обычно еди-
ница площади световых листьев имеет значительно большую массу, чем
теневых, т. е. у них меньше удельная поверхность листа (площадь, при-
ходящаяся на единицу массы). В более толстом листе гелиофитов гуще и
сеть жилок, а клетки обычно расположены компактно, без крупных меж-
клетников. Для листьев гелиофитов типична и мелкоклеточность, также
обусловливающая меньшие размеры их листьев. Устьица их обычно бо-
лее мелкие и многочисленные.
Характерно, что свет прежде всего влияет на меристематиче-
скую ткань, поэтому листья развивают структуру, соответствую-
щую освещенности прошлого сезона, когда в почках закладыва-
лись их зачатки. В целом структура светового листа обеспечивает
и достаточный перехват солнечной радиации, и хорошо защища-
ет лист от чрезмерного облучения.
У растений затененных местообитаний листья имеют черты
«теневой» структуры. Листовые пластинки их более тонкие, с боль-
шей удельной поверхностью, что способствует проникновению
Рис. 6.29. Поперечный срез листь-
ев сирени (Syringa) (по И. С. Михай-
ловской, 1977):
А — световой лист; Б — теневой лист
Рис. 6.30. Поперечный срез листа
олеандра (Nerium oleander) (по
И. С. Михайловской, 1977):
1 — двуслойный эпидермис с кутику-
лой; 2 — гиподерма; 3 — изопалисад-
ный мезофилл; 4 — углубление ниж-
ней стороны листа с устьицем и во-
лосками
151
слабого света в глубь листа. Клетки теневых листьев, в том числе
устьичные, обычно довольно крупные. Устьица редкие, рассеян-
ные и в отличие от световых листьев расположены только на ниж-
ней стороне листа. Для теневых листьев характерны также изви-
листые очертания клеток эпидермиса. Мезофилл теневых листьев
или совсем не дифференцирован на столбчатую и губчатую па-
ренхиму, или клетки верхнего слоя лишь несколько отличаются
от нижних (не бывают узкими и длинными). А губчатая паренхи-
ма в теневых листьях всегда хорошо выражена. В ней много меж-
клетников, что обеспечивает полное поглощение света из-за мно-
гократного рассеивания его здесь.
На уровне хлоропластов световые и теневые листья тоже имеют раз-
личия. Листья гелиофитов мелкоклеточные и потому содержат хлоро-
пластов относительно немного. Однако на единицу объема клетки у них
приходится хлоропластов больше, чем у теневыносливых видов с их бо-
лее крупными клетками, хлоропластами и межклетниками. Кроме того,
у световых листьев существенно большая толщина мезофилла. В итоге в
условиях сильной освещенности число хлоропластов, приходящееся на
единицу площади листа, оказывается в несколько раз больше, чем у ра-
стений в затенении. Так, очень много хлоропластов на 1 см2 листьев у
ксерофитов аридных пустынь, обитающих в условиях высокой интен-
сивности света: у астрагала хивинского {Astragalus chiwensis) — 89 млн,
у осоки вздутой {Carex physodes) — 72 млн (А. Г. Юсуфов, 1985). При
этом в густо наполненном хлоропластами световом листе не все пласти-
ды получают сильный свет из-за взаимного затенения. Сами хлороплас-
ты у гелиофитов более мелкие и светлые (с меньшим содержанием хло-
рофилла), а у сциофитов — крупные и темные. Так, содержание хлоро-
филла в одном хлоропласте теневыносливых растений в 5 —10 раз боль-
ше (А. Г. Юсуфов, 1985). То же характерно для низших растений. У водо-
рослей подводных гротов хроматофоры значительно крупнее, чем у осо-
бей с мелководий. При этом общая поверхность хлоропластов оказыва-
ется довольно велика, например у бука {Fagus) она почти в 200 раз пре-
вышает площадь листьев. Такая организация пластидного аппарата те-
невых листьев способствует лучшему освещению каждого хлоропласта и
лучшему обеспечению светом ассимилирующих тканей. Повышение ин-
тенсивности света в диапазоне слабого освещения ведет к увеличению
величины хлоропластов, но число их в клетке не меняется. По достиже-
нии пороговой интенсивности размеры хлоропластов становятся макси-
мальными. Дальнейшее увеличение интенсивности света способствует
увеличению числа хлоропластов в клетке и уменьшению их размеров.
Таким образом, поскольку интенсивность фотосинтеза генетически оп-
ределена числом хлоропластов, активностью единичного хлоропласта и
скоростью оттока метаболитов из него, эффективное жизнеобеспечение
у разных типов растений достигается за счет изменения их количествен-
ных соотношений.
Хорошо известна и способность хлоропластов к изменению ориенти-
ровки и перемещениям в клетке (рис. 6.31). Она особенно выражена у
152
Рис. 6.31. Схема изменения положения хлоропластов и хроматофоров
в клетках растений под влиянием света (по Т. К. Горышиной, 1979):
А — ряска тройчатая (Lemna trisulca)\ Б — зеленая водоросль Mougeotia sp.; В —
селягинелла (Selaginella martensii), 1 — слабое освещение; 2 — сильное освеще-
ние; 3 — темнота; стрелки показывают направление освещения
световых растений как защитная реакция на избыточный свет. На силь-
ном свету в течение 15 — 20 мин хлоропласты занимают постенное поло-
жение и становятся узкой стороной к направлению лучей. При слабом
же освещении они распределяются в клетке диффузно. Изменение ори-
ентировки в зависимости от силы и направления света отмечены также у
хроматофоров водорослей.
Физиолого-биохимические адаптации растений к световым ус-
ловиям. Основные физиологические адаптации к свету касаются
фотосинтеза. Изменение концентрации хлорофилла в листьях свя-
зано с приспособлением для улавливания света. При оптималь-
ном содержании хлорофилла световые растения создают больше
сухого вещества, чем теневые. При изменении освещенности у
гелиофитов количество пигментов нередко падает, причем как при
увеличении, так и при уменьшении интенсивности света. В ясные
солнечные дни у световых растений содержание хлорофилла сни-
жается, и минимальное его количество отмечается при наиболее
сильном дневном освещении.
Классические работы по изучению физиологических адапта-
ций растений к условиям освещенности проделал В. Н.Любимен-
ко еще в начале XX в. Изучив более 600 видов растений, он при-
шел к выводу, что световые листья лучше работают при более
сильном свете, а теневые — при слабом. Причем активный фото-
синтез тенелюбов при слабом освещении связан с более высокой
концентрацией хлорофилла (у них более темная окраска в тени).
Это различие хорошо заметно при сравнении содержания хлоро-
филла в единице массы листа. Так, у светолюбов листья содержат
153
1,5 — 3 мг хлорофилла в 1 г листа, а у теневых видов — 4—6 мг
(до 7 — 8 мг). В. Н.Любименко также показал, что если светолю-
бивые растения реагируют на затенение снижением содержания
хлорофилла, то для теневыносливых типично возрастание его кон-
центрации.
От условий освещенности зависит не только количество, но и
состав пигментов. Так, хлорофилл представлен в листе рядом форм,
отличающихся структурой и спектром поглощения. Форма а зна-
чительно преобладает над формой b в хорошо освещенных место-
обитаниях, а в затенении доля хлорофилла а падает. Пигменты
незеленого цвета тоже содержатся в определенном отношении к
хлорофиллу в разных местообитаниях. Так, Т.К. Горышина (1979)
отмечает, что у сильно освещенных высокогорных растений от-
ношение зеленых пигментов к желтым обычно выше, чем у расте-
ний с более низких высот, и намного выше, чем у растений зате-
ненных местообитаний. Особенно ярко адаптивное изменение
состава пигментов проявляется у водорослей при изменении спек-
трального состава света на разных глубинах. Так, у красных водо-
рослей отношение содержания фикоэритрина к хлорофиллу тем
больше, чем глубже они обитают. Закономерное изменение со-
става пигментов водорослей на разной глубине, существенно сдви-
гающее спектральную кривую поглощения света, называется хро-
матической адаптацией.
Антоциан — дополнительный красный пигмент, снижающий повреж-
дающее действие яркого света. Он локализован в поверхностных клет-
ках, экранирует избыточный свет и защищает от ультрафиолета глубже-
лежащие ткани. С другой стороны, красные пигменты хорошо отражают
нагревающие длинноволновые лучи. Поэтому повышение содержания
антоциана предохраняет растение и от перегрева. И. М. Культиасов (1982)
приводит данные о том, что у плодов под красными пятнами отмечалась
температура на 22 °C ниже, чем под зелеными. У некоторых видов коли-
чество антоциана прямо пропорционально интенсивности света, при ко-
торой развивается растение в данных условиях.
При резком недостатке света своеобразной формой физиоло-
гической адаптации может служить потеря растением пигментов,
участвующих в фотосинтезе, и переход к миксотрофному (сме-
шанному) питанию. Иногда в связи с полной потерей хлорофил-
ла переход к гетеротрофности становится безвозвратным. Таковы
ряд вольвоксовых и хлорококковых водорослей, а также орхид-
ных в еловых лесах (например, гнездовка — Neottia nidus-avis).
Факультативными гетеротрофами становятся некоторые циано-
бактерии, обитающие в почве (на поверхности ее они сохраняют
способность к фотосинтезу).
Другой важный способ физиологической адаптации к услови-
ям освещенности состоит в том, что у растений сильно- и слабо-
154
освещенных местообитаний зависимость фотосинтеза от света
неодинакова. Поэтому и световые кривые фотосинтеза, графи-
чески отражающие это, различаются. У световых растений точка
компенсации соответствует высокой освещенности и лежит в пре-
делах 500—1000 лк и более. С одной стороны, это объясняется
трудностью проникновения света через их более плотные листья.
С другой стороны, это часто связано со значительным количе-
ством у них неассимилирующих тканей, которые дышат, но не
фотосинтезируют. Поэтому у светолюбов даже при небольшом
затенении потери на дыхание начинают преобладать, и баланс
органического вещества становится отрицательным. Это хорошо
видно на примере саженцев различных по теневыносливости дре-
весных пород, выращенных на разном световом фоне. Высокую
световую компенсационную точку имеют и молодые листья, по-
скольку у них тоже высока интенсивность дыхания.
У растений тенелюбивых и теневыносливых точка компенса-
ции лежит в области более низкой освещенности — около 1/юо~
Умо от полной (от 50 до 100 — 200 лк). Это связано с низкой ин-
тенсивностью дыхания теневых листьев. Ослабленное дыхание,
увеличение ассимилирующей площади, уменьшение относитель-
ной массы нефотосинтезирующих тканей — компенсация слабого
фотосинтеза, позволяющая теневыносливым растениям даже при
сильном затенении сохранять положительный баланс органиче-
ского вещества. Однако если сравнить положение точки компен-
сации с интенсивностью света, при которой теневые виды (май-
ник — Majanthemum, кислица — Oxalis и др.) реально существу-
ют в лесу, то окажется, что при минимальном световом доволь-
ствии они находятся на границе голода. На нижней границе све-
тового довольствия свет уже является лимитирующим фактором.
При увеличении света фотосинтез у гелиофитов нарастает мед-
ленно (угол наклона кривой невелик), и выход кривой на плато
происходит при высокой освещенности, составляющей от */з Д°
1/2 и более от полной. А очень светолюбивые виды совсем не име-
ют «плато насыщения», и фотосинтез у них возрастает с увеличе-
нием освещенности до максимально возможных величин. Угол
наклона световой кривой у сциофитов более крутой, что свиде-
тельствует о более эффективном использовании доступного им
слабого света. У теневыносливых растений кривая выходит на плато
насыщения при низкой освещенности, но максимальная интен-
сивность фотосинтеза при насыщающей освещенности ниже, чем
у светолюбивых видов.
При естественном сильно меняющемся освещении растения
открытых мест приспосабливаются к довольно широкому диапа-
зону изменчивых условий освещения, а растения сильно затенен-
ных местообитаний адаптированы к более узким оптимумам. При
полном же освещении у многих тенелюбов наступает депрессия
155
фотосинтеза (световая кривая опускается ниже плато) (Т.К. Го-
рышина, 1979) (рис. 6.32). Например, это наблюдается у сныти
(Aegopodium podagraria), ряда папоротников {Polypodium vulgare,
Dtyopteris austriaca) и др.
Световые кривые фотосинтеза бывают довольно изменчивы и
у одного вида, отражая функциональную перестройку фотосинте-
тического аппарата в соответствии с условиями освещенности и
стадиями онтогенеза растения. Так, у весенних эфемероидов све-
товые кривые фотосинтеза характерно изменяются в течение года.
Например, у лука медвежьего (Allium ursinum) весной в необли-
ственном лесу точка компенсации фотосинтеза лежит при 650 лк,
а в летней тени снижается до 200 лк. У осоки волосистой (Carex
pilosa) ранней весной в дубняке молодые листья имеют плато свето-
вого насыщения фотосинтеза при 20—25 тыс. лк, при летнем затене-
нии приобретают способность более эффективно использовать
слабый свет, а после перезимовки эти же листья под пологом без-
листного весеннего леса вновь обнаруживают световые черты.
Ранневесенняя вегетация дает эфемероидам возможность избе-
жать затенения под пологом леса: они прорастают и начинают
цвести сразу после таяния снега, ко времени полного развития
листвы на деревьях теряют надземные органы и период глубокого
затенения переживают в виде покоящихся подземных органов. Их
способ адаптации к временному затенению основывается на сдвиге
годичного цикла вегетации на светлый отрезок благоприятного
сезона.
Другой тип сезонной адаптации к режиму освещенности под поло-
гом листопадных лесов наблюдается у длительно вегетирующих трав,
сезонное развитие которых начинается еще в безлистном лесу и конча-
ется глубокой осенью. Это сныть (Aegopodium podagraria), копытень
(Asarum europae ит), медуница (Pulmonaria obscura) и др. У них основ-
ная фотосинтетическая деятельность приурочена к концу весны — нача-
лу лета, когда под пологом леса еще достаточно света (скрытая эфеме-
роидность). За этот период растения накапливают основную массу орга-
ники. Летом же их фотосинтез в тени идет на таком низком уровне, что
Освещенность, тыс. лк
Рис. 6.32. Световые кривые фотосин-
теза растений из различно освещен-
ных местообитаний (по Т. К. Горыши-
ной, 1979):
7 — калужница арктическая (Caltha arctica)
(тундра); 2 — Eurotia ceratoides (Памир);
3 — пролеска сибирская (Scilla sibirica)
(ранневесенний эфемероид дубовых лесов);
4 — типчак (Festuca sulcata) (степь); 5 —
кислица (Oxalis acetosella)\ 6 — сныть
(Aegopodium podagraria) (дубрава, лето)
156
не всегда покрывает расходы на дыхание. Параметрами отношения к
свету могут отличаться иногда и листья одной особи. Так, весенние ли-
стья у медуницы и сныти световые, а летние — теневые (Т. К. Горышина,
1979). И в этом случае развитие растений в основном идет за счет весен-
него периода, поэтому глубокая летняя тень для них не губительна. Та-
ким образом, длительно вегетирующие лесные травы с такой сезонной
динамикой в целом теневыносливы и занимают в лесах наиболее зате-
ненные местообитания.
Различное отношение листьев к свету иногда носит возрастной
характер. Как правило, первые листья побега имеют более тене-
вую структуру, более теневыносливы всходы и молодые растения.
Например, несмотря на то что дуб черешчатый (Quercus robur)
является слаботеневыносливым видом, его сеянцы способны не-
сколько лет переносить затенение всего в 1,2 —1,5 % от полного
света. Часто подрост под пологом леса выносит сильное затене-
ние материнскими растениями, так как у него есть ряд теневых
приспособлений. То же отмечается для всходов и молодых расте-
ний в травяных сообществах. Повышенную теневыносливость
растений в начале онтогенеза следует рассматривать как адапта-
цию к воспроизведению в сомкнутом сообществе.
6.6. Фотопериод и фотопериодические реакции
растений
Фотопериодическая реакция растений. Важнейшей характерис-
тикой светового режима местообитания является фотопериод —
продолжительность светлого времени суток, обусловленная вра-
щением Земли вокруг своей оси. Он неодинаков в течение года и
на разных широтах. Чем ближе к полюсу, тем длиннее там день в
летнее время (рис. 6.33). В низких широтах фотопериод в течение
года меняется мало, но при удалении от тропиков соотношение
светлого и темного периодов суток все больше различается. В сред-
них широтах зимой день вдвое короче, чем летом, а за Полярным
кругом наступает долгая полярная ночь зимой после долгого по-
лярного дня летом. Ближе к полюсам интенсивность света ослаб-
ляется, но освещенность становится продолжительнее и преобла-
дает длинноволновый рассеянный свет. При движении к экватору
день укорачивается, но интенсивность света становится выше, а
он богаче коротковолновым излучением. Таким образом, в высо-
ких широтах растения растут в условиях длинного дня, а в низких —
короткого дня.
Растения приспособились к жизни при чередовании опреде-
ленной продолжительности дня и ночи. Способность организмов
реагировать на суточный ритм освещения (соотношение светлого
157
I II III IV V VI VIIVIII IX X XI XII
Месяцы
Рис. 6.33. Сезонное изменение длины дня на разных широтах Северного
полушария (по А. С. Данилевскому, 1961)
и темного периодов) называется фотопериодической реакцией.
Выражается она в изменении процессов роста и развития при из-
менении длины дня и обеспечивает адаптацию онтогенеза расте-
ний к сезонным особенностям климата. Длина дня используется
растением как часы, показывающие лучшее время для перехода к
цветению или для подготовки к неблагоприятному сезону.
Основным критерием фотопериодической реакции считается
стимулирование некоторой продолжительностью дня перехода к
генеративной фазе развития. Иногда достаточно всего несколь-
ких суток с нужным соотношением света и темноты, чтобы расте-
ние зацвело {фотопериодическая индукция). Так, дурнишник
(Xanthium strumarium) не цветет целое лето при неподходяще длин-
ном дне, но зацветает уже после одного короткого дня. Оказа-
лось, что растения способны воспринимать продолжительность
дня с большой точностью. Так, для хризантемы (Chrysanthemum)
критическая длина дня, обеспечивающая цветение, составляет 14 ч
40 мин, но уже при 13 ч 50 мин бутоны не образуются. А у дур-
нишника пенсильванского (X. pennsylvanicum) критическая дли-
на дня лежит между 15 ч и 15 ч 30 мин (Т.К.Горышина, 1979).
Для растений важен также характер изменения дня (укорачи-
вающийся или удлиняющийся). Так, у ряда многолетних злаков
переход к цветению стимулируется удлинением дня весной, а со-
кращающийся день той же продолжительности в конце лета этого
эффекта не вызывает.
Фотопериодическая индукция обнаруживается при намного
более слабой интенсивности света, чем фотосинтез. Например,
дурнишник реагирует на длину дня при освещении 100-ваттной
158
лампой на расстоянии 150 м. При такой чувствительности расте-
ний к слабому свету при определении критической длины дня
требуется учитывать также светлую часть сумерек.
Как указывает И. М. Культиасов (1982), фотосинтез отмечен даже при
лунном свете. Интенсивность света в полнолуние в безоблачную ночь на
широте 50° иногда достигает 0,3 лк, а в тропических областях — в 3 раза
больше. Низкая ночная освещенность (около 0,1 лк) у некоторых расте-
ний может влиять на фотопериодический счет времени, в то время как
другие виды выработали адаптивные механизмы, препятствующие влия-
нию лунного света на фотопериодизм. Рецепторами фотопериода слу-
жат листья, поэтому изменение их положения ночью может влиять на
количество принимаемого листовой поверхностью света. Так, у бобовых
(соя — Glycine, клевер — Trifolium, земляной орех — Arachis hypogaed)
сонные движения листьев ночью приводят их в вертикальное положе-
ние. Это снижает интенсивность падающего на лист света на 85 —95 %,
т.е. до уровня, уже не влияющего на счет времени. У некоторых расте-
ний (Albizzia, Cassia) листья не только ориентируются ночью верти-
кально, но и поворачиваются вокруг своей оси так, что парные листочки
складываются вместе, затеняя друг друга и подставляя свету менее чув-
ствительную нижнюю сторону. Некоторые растения лучше цветут при
слабом ночном освещении (около 0,5 лк), а не в полной темноте. У этих
видов ночной свет увеличивает количество цветков, образующихся при
коротком дне.
Круг явлений, контролируемых длиной дня, гораздо шире, чем
только индукция зацветания. От фотопериода в значительной мере
зависят продуктивность растений, устойчивость к инфекции, спо-
собность к сожительству с другими организмами, анатомия веге-
тативных органов, степень ветвления, дольчатость листьев, зимо-
стойкость, прорастание и сезонное развитие. Так, к перенесению
неблагоприятного периода организмы готовятся заблаговремен-
но. Задолго до наступления холода начинается пожелтение ли-
ствы у древесных растений, одревеснение и опробковение побе-
гов. Многолетние травы и озимые культуры при закаливании на-
капливают защитные вещества. У водных растений зимующие
почки опускаются на дно. Сигнал, запускающий все эти процес-
сы, — сокращение фотопериода. Уменьшенная длина дня стано-
вится именно сигнальным фактором, так как сама по себе не вредна
для растения, а лишь указывает на близящееся понижение темпе-
ратуры.
Экспериментально доказано, что именно осенний фотопериод
вызывает листопад, формирование морозостойкости, вызревание
древесных побегов и другие формы подготовки к зимнему покою.
Так, южные растения, перенесенные на север, не становятся мо-
розостойкими и по причине того, что длинный день не позволяет
им закончить к осени ростовые процессы, накопить в тканях ве-
159
щества, повышающие морозоустойчивость. Если искусственным
затенением увеличить для них продолжительность ночи на не-
сколько часов, то морозоустойчивость их сильно повышается.
Такие факты получены при изучении яблонь {Malus domestica)
на Урале (А. Г. Воронов, 1963).
Типы растений по фотопериодической реакции. В процессе эво-
люции сформировались три основные группы растений с различ-
ной фотопериодической реакцией (рис. 6.34).
Растениям короткого дня для перехода к цветению нужно
менее 12 ч света в сутки. При длине дня более 12 ч цветение не
наступает или задерживается. Это большей частью растения низ-
ких широт (а в умеренных областях — ближе к субтропикам).
Перенесенные в высокие широты (с более длинным днем, чем
критический фотопериод), они имеют ненормальное развитие:
сильно вытягиваются, развивают мощную вегетативную массу, но
позднее зацветают или не цветут. Нормальное развитие их можно
обеспечить искусственным затенением на несколько часов в день.
Характерно, что чем короче критический период, тем короче жиз-
ненный цикл этих растений. Например, соя {Glycine) при 12-ча-
совом фотопериоде зацветает на 100-й день, а при 5-часовом — на
37-й.
Культурными короткодневными растениями являются соя, рис {Oryza
sativa), просо {Panicum), бобы {Vicia faba), лук {Allium сера), свекла
{Beta vulgaris), хлопчатник {Gossypium), конопля {Cannabis sativa), та-
бак {Nicotiana).
Растения длинного дня зацветают только тогда, когда про-
должительность дня достаточно велика — обычно более 12 ч. В
условиях короткого дня эти растения дают большую вегетатив-
ную массу, но не цветут. У них часто укорачиваются междоузлия,
А
100- -------
80-
60
40
20
J
0 4 8 12 16 20 24
Число часов света в сутки
Б
0 4 8 12 16 20 24
Рис. 6.34. Типы фотопериодической реакции у растений
(по Б. С. Мошкову, 1961):
А — короткодневный (у периллы — Perilla)-, Б — длиннодневный (у шпината —
Spinacid); В — нейтральный (у горчицы — Sinapis)
160
они могут приобретать розеточную форму. Чем выше широта ме-
стности, тем продолжительнее день, что позволяет растениям се-
верных областей проходить полный цикл развития в более корот-
кие сроки.
К длиннодневным относятся многие дикорастущие виды приполяр-
ных и умеренных широт (вереск обыкновенный — Calluna vulgaris, мо-
лодило — Sempervivum tectorum, белена — Hyoscyamus niger), а также
широко известные культурные растения (рожь — Secale cereale, пшени-
ца — Triticum aestivum, ячмень — Hordeum, овес — Avena, редис —
Raphanus sativus, картофель — Solanum tuberosum, шпинат — Spinacia,
мак — Papaver, хризантема — Chrysanthemum).
Нейтральные растения безразличны к длине фотопериода.
Их цветение не зависит от длины дня и наступает при любой
продолжительности освещения, кроме очень короткой, ведущей
к голоданию. Но количество цветков и урожай плодов у них все
же зависят от соотношения длины дня и ночи.
К этой группе относятся широко распространенные виды, например,
одуванчик (Taraxacum officinale), а также ряд культурных растений (гре-
чиха — Fagopyrum esculentum, горох — Pisum sativum, томаты —
Lycopersicon, некоторые сорта хлопчатника — Gossypium).
Иногда выделяют еще растения с фотопериодической реакци-
ей промежуточного типа, например, гваюла (Parthenium argenta-
tum). Их цветение наступает при узком диапазоне фотопериода —
не длиннее и не короче определенных критических величин.
В целом каждому виду (и сорту) присущ свой критический фото-
период и за пределами определенной продолжительности дня они
не развиваются. Для большинства видов этот период лежит в пре-
делах 12 —14 ч.
Элементарной единицей, обладающей однородной фотопери-
одической реакцией, является биотип. Накоплено много данных
о генетической обусловленности фотопериодической реакции, ее
полигенной природе и передаче потомству при гибридизации.
Обычно гены, определяющие короткодневную реакцию, ведут себя
как рецессивные. Длиннодневные формы при скрещивании до-
минируют над нейтральными. Нередко спонтанное изменение
фотопериодической реакции связано с полиплоидией (А. Г. Юсу-
фов, 1995).
Существует четкая связь между географическим распростране-
нием растений и типом их фотопериодической реакции. Так, виды
и сорта, распространенные в высоких широтах, в основном длин-
нодневные. Среди видов средних широт имеются как растения
длинного дня, так и короткого. Из них те, что цветут летом, обычно
длиннодневные, а цветущие весной или осенью — короткоднев-
161
ные. Соотношение растений этих групп меняется: чем ближе к
Полярному кругу, тем короткодневных растений меньше. Виды
тропиков и субтропиков преимущественно короткодневные или
нейтральные. Таким образом, растения обычно не выходят за пре-
делы той географической широты, которая обеспечивает необхо-
димую для половой репродукции длину дня. Однако есть расте-
ния, цветущие с весны до осени, нейтральные по отношению к
длине дня. Это потенциальные космополиты. Но у этих видов с
обширными ареалами отдельные географические популяции раз-
личаются по значению критического фотопериода. В этом случае
фотопериодическая реакция выступает как механизм, подгоняю-
щий свойства растений к разнообразию условий на обширном
ареале. Фотопериодическая реакция культурных растений часто
соответствует региону происхождения. Так, абиссинские сорта
пшеницы (Triticum aestivum) короткодневные, а сибирские —
длиннодневные.
ГЛАВА 7
ТЕПЛО КАК ЭКОЛОГИЧЕСКИЙ ФАКТОР
У большинства растений активная жизнь протекает в довольно
узких температурных пределах, в среднем от 0 до 50 °C. Темпера-
турные минимумы для многих растений лежат в интервале 5 —
15 °C, оптимумы — при 15 —30 °C, максимумы — при 37 —50 °C.
Но вообще температурные пределы существования растений весьма
широки: у побегов опунции (Opuntia) зарегистрирована темпера-
тура 65 °C, в Якутии при температуре до -68 °C существуют леса, а
некоторые покоящиеся споры и семена переносят температуру
кипения воды и близкую к абсолютному нулю.
Растения — это пойкилотермные организмы, т.е. температура
их тел определяется температурой среды. Но днем их органы час-
то нагреваются выше температуры воздуха, а когда физиолого-
биохимические процессы идут с большой интенсивностью (на-
пример, в цветущем початке арума — Arum или в больших скоп-
лениях растений), температура их может существенно превысить
окружающую. При этом отдельные ткани и органы растений име-
ют различные кардинальные температурные точки. Например, у
пшеницы (Triticum aestivum) прорастание семян начинается при
0 °C, рост листьев — при 6 °C, а тычиночных нитей — при 15 °C
(Л. Г. Косулина и др., 1993). По-видимому, у вида закрепляется
связь онтогенеза с характерным ходом температур.
Роль тепла в жизни растений многообразна. Оно влияет прак-
тически на все процессы жизнедеятельности — фотосинтез, дыха-
ние, транспирацию, прорастание семян, рост побегов, цветение и
многие другие. Разные виды нуждаются в тепле неодинаково, по-
грому разнообразие тепловых условий на планете во многом оп-
ределяет границы ареалов, топографическое размещение, а также
«шальную структуру растительного покрова.
7.1. Тепловой режим местообитаний
Баланс радиации на земной поверхности. Как уже отмечалось,
па верхней границе атмосфера получает инсоляцию в объеме сол-
163
нечной постоянной (около 1,353 кВт/м2), но далеко не весь пря-
мой поток радиации доходит до поверхности почвы и формирует
тепловой режим местообитаний (часть энергии отражается, рассеи-
вается или поглощается элементами атмосферы) (см. разд. 6.1).
Длинноволновую тепловую часть излучения сильно поглощают
диоксид углерода и непостоянные составляющие атмосферы (пыль,
водяные пары, газовые примеси). Ночью «парниковый эффект»
удерживает тепло у поверхности почвы (иначе температура в это
время опускалась бы намного ниже нуля). Так атмосфера смягча-
ет суточные температурные колебания.
Среднее количество солнечной энергии, падающей на едини-
цу площади земной поверхности, зависит от многих условий. Оно
прямо связано с географической широтой, что объясняется, во-
первых, тем, что в высоких широтах лучи входят в атмосферу не
под прямым углом и их энергия распределяется на большую пло-
щадь земной поверхности. Во-вторых, лучи, проникающие в ат-
мосферу не под прямым углом, проходят через больший слой воз-
духа и при этом значительная часть их отражается. Имеет значе-
ние также облачность. Поэтому с возрастанием широты отмечает-
ся снижение среднегодовых температур. Количество солнечного
тепла, достигающего земной поверхности, меняется и по сезонам:
его больше летом, когда лучи падают отвеснее. И в ясные дни
почва принимает тепла больше, чем в пасмурную погоду.
Одновременно с прогревом идет и отдача тепла земной поверх-
ностью. Во-первых, тепло проникает в глубь почвы. Оно зависит
от теплопроводности: чем она больше, тем меньше поверхность
нагревается. Большое значение имеют верхние слои почвы. На-
пример, из-за малой теплопроводности подстилки весной почва
остается холодной. У сухой почвы теплопроводность невысока,
поэтому ночью после засухи почва нередко становится холоднее
воздуха. На проникновение тепла в почву сильно влияет и снеж-
ный покров: зимой на определенной глубине температура почвы
выше воздушной.
Другой путь отдачи тепла почвой — излучение, происходящее
в основном в инфракрасном диапазоне. Оно сильно поглощается
атмосферой (это называется противоизлучением атмосферы),
приводя к прогреву нижних слоев воздуха. Чем больше противо-
излучение атмосферы, тем меньше общая величина земного излу-
чения. Этот параметр называют эффективным излучением зем-
ной поверхности и определяют как разность между излучением
земной поверхности и энергией, получаемой ею обратно от про-
тивоизлучения атмосферы. Из-за возрастания противоизлучения
при облачности, тумане и задымлении эффективное излучение в
эти периоды сильно падает. Поэтому при заморозках сады пыта-
ются спасать дымом. Эффективное излучение велико при сухом
воздухе и безоблачном небе в пустынях, потому для них характер-
164
пы холодные ночи. Эффективное излучение усиливается и при
подъеме в горы, так как противоизлучение сильно уменьшается с
истончением слоя воздуха над поверхностью. Поэтому в альпий-
ском поясе выражена контрастность микроклимата (интенсивная
радиация днем и сильная потеря тепла ночью из-за возросшего
эффективного излучения). В лесу эффективное излучение умень-
шается, так как кроны увеличивают противоизлучение (И. М. Куль-
тиасов, 1982).
Третий путь отдачи тепла почвой — теплообмен. Он имеет боль-
шое значение для нагрева припочвенного воздуха, который эф-
фективно прогревается прямой теплоотдачей с поверхности суб-
страта. Нагретый воздух легче и он поднимается, а холодные плот-
ные воздушные массы размещаются внизу. В местообитании воз-
дух постоянно перемещается, поэтому температура нижнего слоя
очень изменчива.
Тепло теряется и при испарении, поэтому температура почвы
зависит от влажности. Тепло, затрачиваемое на испарение с влаж-
ных почв, может составлять до 30 % теплооборота. К тому же теп-
лопроводность сырых почв выше, что ведет к еще большему их
охлаждению. Поэтому при равной инсоляции поверхность сырой
почвы холоднее. Отдача полученного при поглощении радиации
тепла неодинакова на разных поверхностях. Так, воздух над мо-
рем почти не получает от него тепла. Мало отдают тепла песок и
скалы. А сфагновый покров, снег и особенно листовая подстилка
передают воздуху большую часть поступившего тепла (листовой
опад более 90%) (Г. Вальтер, 1982) (рис. 7.1).
Баланс радиации, которую получает поверхность за определен-
ное время, складывается из баланса коротковолновой радиации и
длинноволнового теплового излучения. Баланс коротковолновой
радиации показывает поступление энергии в местообитание и
состоит из прямой солнечной радиации, рассеянного света неба
за минусом отраженной коротковолновой части. Длинноволно-
вый баланс показывает в основном потерю энергии и складывает-
Поглощение тепла
Отдача в воздух
ВШШ^ОЙМоре (подвижное)
Покров ИЗ сфанговых MXOB^j
СнегЦ
Листовая подстилка |[
] Спокойная вода
Рис. 7.1. Реализация получаемого тепла при поглощении радиации
различными видами земной поверхности (по Г. Вальтеру, 1982)
165
Рис. 7.2. Потоки коротковолновой (Ф5)
и длинноволновой (Ф/) лучистой энер-
гии между листом и его окружением
(по кн.: Фотосинтез и биопродуктив-
ность: методы определения, 1989)
ся из длинноволнового обратного излучения атмосферы за выче-
том теплового излучения тел. Все это влияет на потоки энергии
между растением и его окружением (рис. 7.2). Общий баланс ра-
диации положителен, если коротковолновая его часть превышает
длинноволновую (приход радиации больше потери энергии). Это
характерно для фазы инсоляции, которая имеет место днем и за-
канчивается перед заходом солнца. Баланс становится отрицатель-
ным, если потерю энергии перестает компенсировать ее поступ-
ление. Это происходящая ночью фаза излучения. Избыток энер-
гии из этого баланса идет на фотосинтез, испарение, нагрев рас-
тений, почвы и воздуха.
Влияние рельефа на теплообеспеченность местообитаний. На
склонах разной экспозиции разница температур может быть зна-
чительной. Она во многом зависит от географической широты,
формы рельефа, крутизны склона, сезона и др. На экваторе в пол-
день экспозиция почти не имеет значения, но перед полуднем
больше тепла получают восточные склоны, а после полудня —
западные. В Северном полушарии по мере движения к полюсам
все больше тепла получают южные склоны, а в Южном — север-
ные. В полярных областях прямая инсоляция значительно умень-
шается, а диффузное излучение увеличивается, и разницы темпе-
ратур, обусловленные экспозицией, становятся несущественны-
ми, но для организмов они имеют большое значение, поскольку
температура здесь является лимитирующим фактором. С увеличе-
нием высоты над уровнем моря диффузное излучение уменьшает-
ся и экспозиция склонов, особенно в умеренных широтах, приоб-
ретает немаловажное значение. Особенную роль играет экспози-
ция весной, когда растения начинают вегетацию. Так, в среднеев-
ропейских горах даже в холодный период года на южных склонах
могут зацвести некоторые виды, в то время как на северных chci
лежит до половины лета.
В Северном полушарии южные склоны, на которые солнечные
лучи падают под большим углом, нагреваются больше и при этом
менее увлажнены. В результате растительность их обычно имеет
166
более южный, ксерофитный характер по сравнению с раститель-
ностью ровных пространств. Северные же склоны более влажны
и в сочетании с пониженными температурами на них создаются
условия, характерные для более северных зон. Таким образом, за
пределами зоны под влиянием факторов, действующих косвенно,
формируются экологически равноценные им местообитания. Так,
в пределах лесной зоны на склонах южных экспозиций встреча-
ются степные растения, а в степной зоне на склонах северной
экспозиции растут леса (см. разд. 3.2.6). Так же в горах Северного
полушария лес в среднем поднимается по южному склону выше,
чем по северному.
Количество солнечной радиации, получаемое поверхностью,
сильно зависит от крутизны склона, так как связано с углом паде-
ния солнечных лучей. Например, летом в полдень (при высоте
солнца около 68° над горизонтом) наиболее освещен и согрет склон
с уклоном в 22°, весной и осенью — в 45°, а зимой (при высоте
солнца около 22°) — в 68°. При этом даже в Арктике поверхность,
на которую солнечные лучи падают почти перпендикулярно, мо-
жет нагреваться до 50 °C (Ф.Фукарек и др., 1982).
В горных странах отмечается большая пестрота термических
условий. С увеличением высоты температура воздуха понижает-
ся, т.е. в горах существует высотный температурный гради-
ент. Он меняется по сезонам, зависит от экспозиции и положе-
ния на склоне, но в среднем составляет около Г на 180 м высо-
ты. Это вызывает изменение длительности фенофаз и сокраще-
ние вегетационного периода. Так, в Альпах на высоте 1 800 м он
продолжается 5 мес, а на высоте 3 000 м — 1,5. И есть высоты в
горах, где растений нет, так как круглый год там лежит снег.
При движении от полюсов к экватору граница вечных снегов
поднимается: на Шпицбергене она находится на высоте 460 м, а в
приэкваториальных областях — на высоте 5 000 м. Падение тем-
пературы с подъемом в горы обусловливает соответствующие
и 1менения климатических условий. С высотой закономерно сме-
няются тепловые пояса — от жарких и умеренно-теплых у под-
ножия до холодных в зоне вечных снегов. Закономерно изменя-
ются и факторы увлажнения (количество осадков, облачность,
влажность и плотность воздуха). Эти изменения сильно отража-
ются на размещении растительности: в горах формируется вы-
сотная поясность. Высотный пояс — это сравнительно широ-
кая горизонтальная полоса растительности в горах, где на дан-
ной высоте господствует определенный ее тип. Дифференциа-
ция почв и растительных сообществ перерастает в вертикальную
поясность при колебаниях высот более 200 м. Но уже в условиях
центра и юго-востока Русской равнины возвышенности отлича-
ются более холодным климатом и более коротким вегетацион-
ным периодом по сравнению с таковыми низменностей. Горные
167
массивы также во многом формируют природные условия на
прилегающих к ним равнинах.
Тепловой режим почвы. Большое значение для растений имеет
тепловой режим почвы. От него зависят поступление в растение
воды и минеральных веществ, рост, дыхание корней, жизнеспо-
собность почвенных организмов, разложение органических остат-
ков и т. п. А сам температурный режим почвы зависит от темпера-
туры воздуха и свойств почвы. В глубину изменение температуры
распространяется очень медленно, и ее колебания в почве значи-
тельно меньше, чем в воздухе. В целом в зависимости от характе-
ра почвенного покрова в одной климатической зоне условия теп-
лообеспеченности растений могут быть очень разными.
Тепловой режим почвы во многом зависит от ее теплоемко-
сти — свойства поглощать тепло. Она выражает количество теп-
ла, затрачиваемое на нагревание 1 г или 1 см3 сухой почвы на 1 °C.
Теплоемкость почвы зависит от минералогического и грануломет-
рического состава, влажности, содержания органики. По мере по-
вышения влажности теплоемкость почвы возрастает. Так, глини-
стые почвы считают холодными, так как они более влагоемки,
больше расходуют тепла на испарение и поэтому весной медлен-
нее прогреваются. А песчаные и супесчаные почвы прогреваются
быстрее, поэтому их называют теплыми. Теплоемкость почв тес-
но связана с гидрофильностью ее коллоидов, количеством и каче-
ством гумуса. Более теплоемки почвы с большим содержанием
гидрофильных коллоидов и более гумусированные. Рыхлые, по-
ристые почвы также более теплоемки, чем плотные.
Теплопроводность почвы (способность проводить тепло) и теп-
лообмен с воздухом сильно зависят от структуры и порозности
почвы, ее химического и гранулометрического состава, влажнос-
ти, плотности и температуры. Чем крупнее почвенные частицы,
тем больше теплопроводность. Например, при одинаковой влаж-
ности и пористости у крупнозернистого песка она в 2 раза боль-
ше, чем у крупной пыли. Теплопроводность твердых частиц в 100
раз выше теплопроводности воздуха, поэтому рыхлая почва имеет
более низкий коэффициент теплопроводности, чем плотная. Так,
при изменении плотности почвы с 1,1 до 1,6 г/см3 теплопровод-
ность ее возрастает в 2—2,5 раза, а при увеличении пористости с
30 до 70% она уменьшается в 6 раз (И.С.Кауричев и др., 1982).
Теплопроводность почвы зависит и от ее температуры: с ее повы-
шением увеличивается теплопроводность почвенного воздуха и,
следовательно, почвы в целом. Прямое влияние на теплопровод-
ность оказывает степень увлажнения почвы. По данным А. И. Гу-
пало, при повышении влажности пахотного горизонта южного чер-
нозема до 25 % теплопроводность возрастает в 5 раз. В целом чем
почва суше, чем больше в ней воздуха, тем меньше ее теплопро-
водность, сильнее прогревается ее поверхность и тем большую
168
часть поглощенного тепла она отдает прилегающему слою возду-
ха. Но запасают тепла за день такие почвы меньше, поэтому но-
чью их поверхность охлаждается сильнее, что вызывает большее
охлаждение прилегающего воздуха.
Большое значение имеет глубина промерзания и скорость оттаива-
ния почвы, которые зависят от снежного покрова и характера субстрата.
Например, сильно гумусированные почвы, особенно торфянистые, от-
таивают очень медленно. Так, в Центральной Европе богатая гумусом
почва оттаивает на месяц позже, чем базальтовый щебень (рис. 7.3).
Декабрь1939 г. Январь Февраль Март 1940 г.
1 15 30.1 15 30.1 15 29.1 15 30.1
Рис. 7.3. Промерзание (изотерма 0 °C) различных типов почв
и их оттаивание весной (по Г. Вальтеру, 1974)
169
Поэтому вечнозеленые виды чаще растут там на каменистых осыпях и
песках (Г. Вальтер, 1974).
На теплообмен почвы и прилегающего воздуха сильно влияет
ее цвет. Например, светлые почвы отражают много тепла, а тем-
ные поглощают его почти полностью. В связи с этим теплообмен
можно изменить применением органических удобрений. Так, без-
морозный период на светлом субстрате можно удвоить, «утеплив»
Месяцы
Б
Рис. 7.4. Изменение температуры песчаной почвы на разной глубине
около г. Павловска (по В.Лархеру, 1978):
А — суточный ход температур (в мае); Б — годовой ход температур. Цифры па
кривых — глубина почвы, см
170
его торфом (И. М. Культиасов, 1982). Зимой хорошей защитой по-
чве от слишком низких температур и других неблагоприятных
факторов служит снег. Он препятствует ее глубокому промерза-
нию и уменьшает потерю тепла от излучения. Слой снега толщи-
ной около 0,5 м оказывается настолько теплонепроницаемым, что
почва под ним не замерзает. К тому же тающий снег — источник
влаги на мерзлых почвах. Так, в суровых условиях гор Loiseleuria
procumbens может всасывать зимой небольшое количество влаги,
образующейся в результате таяния снега при ярком солнечном
освещении (Ф.Фукарек и др., 1982).
Колебания температуры слоев почвы зависят от ее состава и
ослабляются с глубиной (рис. 7.4). Так, на глубине 1 см у супесча-
ной почвы суточная амплитуда температур достигает 20 °C с мак-
симумом около 12 ч. По мере углубления амплитуды уменьшают-
ся, а максимум запаздывает: на глубине 20 см он наступает уже
около 18 ч, а на глубине 40 см самые низкие температуры отмеча-
ются в полдень и амплитуда здесь не превышает 2 °C. На глубине
80 см температура почвы уже постоянна (В.Лархер, 1978). Сезон-
ные колебания температур фиксируются в почве значительно глуб-
же и амплитуды здесь больше. С глубиной они постепенно сгла-
живаются, а на уровне 5 —7 м из-за запаздывания тепловой волны
ход температур становится обратным (летом эти слои холоднее).
Но эти значения зависят от выраженности климатических сезо-
нов. Так, в тропиках даже на глубине 0,5 м амплитуда не превы-
шает 2 °C. Таким образом, в полдень наивысшая температура на-
блюдается на границе между воздухом и нагретой солнцем повер-
хностью почвы. От этого уровня температура понижается как по
направлению вверх, так и вниз. В этом случае наблюдается рас-
пределение температур типа инсоляции (рис. 7.5).
Ночью припочвенный теплообмен полностью меняется: скла-
дывается распределение температур типа излучения. В это
время поверхность почвы имеет самую низкую температуру и
она возрастает как по направлению вверх, так и вниз. Хотя излу-
чение Земли как космического тела в 204 раз слабее излучения
Солнца, оно играет большую роль в формировании климата при-
ночвенного слоя воздуха, где обитают растения (И. М. Культиа-
сов, 1982). Это излучение особенно велико в ясные ночи и у
почв, имеющих низкую теплопроводность. Поэтому, например,
гс иловой режим осушенных торфяников зимой очень неблаго-
приятен, так как из-за плохой теплопроводности поток тепла из
глубоких слоев в них затруднен. Одним из следствий ночного
выхолаживания поверхности почвы является конденсация водя-
ного пара и формирование росы летом и инея зимой. Им объяс-
няются также существенные перепады между дневными и ноч-
ными температурами в горах, а также раннеосенние и поздневе-
ссннис заморозки там.
171
Рис. 7.5. Распределение температур близ поверхности почвы
(по Н.Walter,’ 1951):
А — ночью; Б — днем; шкала температур для А в 10 раз крупнее, чем для Б.
I — приземный слой воздуха; // — почва
В природе нередко наблюдается сочетание холодной почвы и
теплого воздуха. Таковы условия жизни растений на вечной мер-
злоте, болотах и заболоченных лугах, в некоторых тундровых и
высокогорных местообитаниях. Многие виды умеренного клима-
та оказываются в ситуации резкого несоответствия холодных почв
и прогретого воздуха ранней весной. В связи с этим в экологии
растений была поставлена проблема физиологической сухости хо-
лодной почвы (недоступности влаги при ее обилии). Еще в XIX в.
Ю.Сакс обнаружил, что при обкладывании горшка льдом может
наступить завядание даже обильно политых растений, поскольку
корни не способны интенсивно всасывать воду из холодной по-
чвы. На этом основании распространилось мнение о «физиологи-
ческой сухости» местообитаний с холодными почвами. Однако
при этом упускалось из виду, что опыты производились с тепло-
любивыми растениями (огурцы — Cucumis sativus, тыква —
Cucurbita pepo, салат — Lactuca sativa), а в природе виды холод-
ных местообитаний адаптированы к естественному тепловому
фону. Корни приспособленных к произрастанию на холодных
172
почвах растений работают и при низкой температуре, поэтому у
большинства растений тундр и болот, у ранневесенних эфемерои-
дов нет проявлений угнетения (затруднение всасывания, расстрой-
ство водного режима и т.д.), которые были бы вызваны «физио-
логической сухостью» холодных почв.
Но при снижении температуры действительно возрастает вязкость
воды, что затрудняет ее поглощение корнями, и низкие температуры
неблагоприятны для многих процессов жизнедеятельности корней. Ре-
зультаты опытов с культурными растениями послужили основанием для
вывода об отрицательном влиянии низких температур на поглощение
всех основных элементов минерального питания. Поэтому «физиологи-
ческая засуха» из-за низкой температуры почв возможна, например, при
выращивании теплолюбивых растений на холодных почвах или ранней
весной для древесных пород, когда необлиственные ветви сильно нагре-
ваются и увеличивают испарение, а интенсивная работа корней еще не
началась. Многообразным может быть косвенное отрицательное влия-
ние холодной почвы. От температуры субстрата сильно зависит актив-
ность почвенной микрофлоры, в связи с чем в холодной почве может
быть мало доступных биогенов (в частности, азота), а при их дефиците
нарушается регуляция водного режима растений. С этим может быть
связан «ксероморфоз», точнее пейноморфоз (от греч. peina — голод) ра-
стений холодных почв. Как показали наблюдения и эксперименты, при
низких температурах основным источником азота для растений стано-
вится аммоний, так как аммонификация в этих условиях идет достаточ-
но интенсивно, а нитрификация ингибируется (рис. 7.6) (Т.А.Работнов,
1985).
-а 300
О ”
Z g250
о «
5 ^200
^150
Н S 100
< I 50
Температура, °C
Б
Рис. 7.6. Влияние температуры и влажности почвы на интенсивность
нетто-минерализации в лесной почве (по В.Лархеру, 1978):
Л — аммонификация; Б — нитрификация; цифры — характеристика влажности
почвы
173
Тепловой режим растительного покрова. Для характеристики
теплового режима растительного покрова необходимо знать уро-
вень поступления, потребления и излучения энергии, параметры
теплообмена. В общем энергообмене растений доля метаболиче-
ского компонента невелика (около 1 — 2 %) и тепло растительный
покров накапливает тогда, когда энергии к нему поступает боль-
ше, чем отводится.
Между растениями и средой температура выравнивается бла-
годаря теплопроводности, конвекции, испарению и конденсации
водяного пара. В растении и фитоценозе проведение тепла незна-
чительно и перенос его происходит главным образом с потоками
воздуха и воды. При положительном балансе радиации конвекци-
онный тепловой поток направлен большей частью от поверхнос-
ти растений, а если она холоднее воздуха, то тепло поступает к
ней. Теплообмен с окружающим воздухом путем конвекции тем
эффективнее, чем мельче листья, сложнее их контур и больше
скорость ветра. При сильной инсоляции в безветренный день у
поверхности растения образуется перегретый воздушный слой, из
которого поднимается теплый воздух, а ветер ускоряет теплоотда-
чу, сдувая его.
Благодаря устьичному регулированию транспирации на тепло-
вой режим растительного покрова существенно влияют физиоло-
гические процессы. Если растения транспирируют, то происхо-
дит их охлаждение, а если на листьях конденсируется роса или
иней, — то потепление. Охлаждающий эффект испарения осо-
бенно выражен при высокой температуре, низкой влажности воз-
духа и хорошем водоснабжении. Так, в степи примерно четверть
теплоотдачи листа падает на транспирацию. Усиление же ее вет-
ром может приводить к понижению температуры листьев на не-
сколько градусов ниже температуры воздуха.
В сомкнутом насаждении колебание температуры приземного
слоя воздуха значительно выравнивается. На лугу при 100%-м
проективном покрытии почвы большая часть солнечных лучей
поглощается органами растений (рис. 7.7). На слой листьев при-
Рис. 7.7. Поглощение радиации травостоем луга (в Джоулях)
(по И.М.Культиасову,1982)
174
ходится максимальное нагревание, а нагрев почвы снижается,
поэтому кривые распределения температур сильно изгибаются. Ход
ночных температур в слое травянистых растений тоже имеет спе-
цифику: излучение идет и с почвы, и из травостоя. Весной при
небольшой высоте растений их неплотный слой не препятствует
опусканию холодного воздуха и почва остается более холодной.
Но по мере подрастания растений температурный минимум пере-
мещается в их слой. Поэтому колебания температур на почве ста-
новятся меньше, чем на открытой поверхности.
В травяных фитоценозах формируется особый фитоклимат,
зависящий от сомкнутости и высоты надземных органов. Специ-
фический тепловой режим, создаваемый травостоем, важен для
растений. Так, стабилизируя температуру припочвенного воздуха,
заросли иван-чая (Chamaenerion angustifolium) благоприятству-
ют развитию проростков деревьев на гарях. В плотных кустах и
кочках обычно тоже создаются благоприятные тепловые условия.
Например, в Гренландии в куртинах растений часто фиксируются
температуры на 8—10°C выше температуры воздуха. Кочки пу-
шицы влагалищной (Eriophorum vaginatum) высотой 20 см рань-
ше освобождаются от снега, и длительность периода вегетации на
них может быть больше на 5 —10 %. Внутри кочки субстрат оттаи-
вает быстрее, температура раньше достигает летнего максимума, она
выше на 6 —8 °C и более устойчива, чем между кочками (Т. А. Ра-
ботнов, 1985). Повышенные температуры наблюдаются и внутри
плотных растений-подушек, распространенных в холодных высо-
когорных пустынях и приполярных областях.
В лесу до 80 % солнечной радиации перехватывают кроны де-
ревьев, поэтому под ними складывается особый микроклимат.
Поскольку радиация поглощается в верхних частях крон, там от-
Месяцы
Рис. 7.8. Снижение суточных колебаний температур в лесных насажде-
ниях по сравнению с открытой местностью (по И. М. Культиасову, 1982):
1 — буковый лес; 2 — еловый лес; 3 — сосновый лес
175
мечаются и самые высокие температуры. Из-за сильного энерго-
обмена там же наблюдаются и большие их суточные колебания.
А вблизи почвы температура воздуха обычно остается постоянно
невысокой. В высокоствольном насаждении интенсивность про-
никающей радиации быстро падает и обратное излучение тоже
эффективно задерживается. В результате лес значительно снижа-
ет суточные колебания температур. Например, летом в жаркие
дни температура тут ниже, а ночью выше, чем на открытом месте.
И летние почвенные температуры в лесу ниже, а зимой выше, чем
вне него (поэтому весенние заморозки заканчиваются раньше, а
осенние начинаются позже). Суточные амплитуды температур тем
меньше, чем больше сомкнутость крон. Зависит это и от характе-
ра кроны лесообразующей породы. Например, в июле колебания
температур под лесным пологом в сосняке уменьшаются на 3 °C, а
в буковом лесу — на 4,5 °C (рис. 7.8).
7.2. Температура растений
Особенности температуры растений. Растения — пойкилотер-
мные организмы, и их температура следует за температурой окру-
жающей среды. Однако это соответствие неполное. В результате
разных причин температура растений может значительно отли-
чаться от температуры воздуха. Поэтому в экологии наряду с теп-
ловыми характеристиками среды необходимо знать и температуру
растений, так как именно она представляет истинный тепловой
фон для физиологических процессов.
Температура растения определяется соотношением поглоще-
ния и отдачи им энергии, а эти величины зависят от многих свойств
среды (приход радиации, температура воздуха, ветер) и самих ра-
стений (величина и расположение листьев, окраска, блеск, опу-
шение и т. д.). В результате действия всего комплекса причин тем-
пература растений обычно отличается от температуры воздуха их
местообитаний, и иногда значительно. В целом в жарких место-
обитаниях температура надземных частей растений ниже, а в хо-
лодных — выше температуры воздуха. Совпадение же температу-
ры растений с температурой окружения встречается гораздо реже.
Это бывает в условиях, исключающих сильное нагревание и ин-
тенсивную транспирацию (например, в нижнем ярусе тенистых
лесов и на открытых местах в пасмурную погоду).
О. Ланге предложил выделять группы растений по соответствию
температуры их и воздуха местообитаний: 1) супратемператур-
ные виды с температурой выше температуры окружающего воздуха;
2) субтемпературные растения с температурой ниже температу-
ры воздуха; 3) растения с температурой, очень близкой к темпера-
туре среды.
176
Супратемпературные виды растений. Супратемпературные рас-
тения встречаются довольно часто в разных условиях. При силь-
ной инсоляции отдача тепла путем конвекции и транспирации
часто оказывается недостаточной, чтобы уравнять температуру
побега с температурой воздуха, и листья нагреваются на 10 —15 °C
выше температуры окружающей среды. Так, у кожистых листьев
кофейного дерева (Coffea) фиксируются температуры до 45 °C при
30—35 °C воздуха (И. М. Культиасов, 1982).
У массивных органов с плохим теплообменом (листья и стебли
суккулентов, плоды и стволы деревьев) разность температур с воз-
духом может достигать 20 °C. Сильно прогреваются слабоиспаря-
ющие мясистые стебли кактусов и утолщенные листья толстянко-
вых. Так, в жарких пустынях при температуре воздуха 40—45 °C
кактусы нагреваются до 55 — 60 °C. Летом в умеренном поясе лис-
тья очитков (Sedum) нередко имеют температуру 45° С, а у розет-
ки молодила горного (Sempervivum montanum) при температуре
воздуха 22 °C зафиксирована температура 51 ° (В. Лархер, 1978) (рис.
7.9).
Сильно нагреваются мясистые плоды. Так, спелые томаты
(Lycopersicon) и арбузы (Citrullus lanatus) на 10—15 °C теплее
воздуха, а температура спелых плодов аронника (Arum maculatum)
доходит до 50 °C. Существенное превышение температуры отме-
45°С 42°С
Рис. 7.9. Температурный профиль суккулентов при сильной инсоляции
и отсутствии ветра (по В.Лархеру, 1978):
А — кактус (Ferocactus wislizenii) на высоте 900 м в Аризоне (температура воздуха
*2 *С); F — молодило (Sempervivum montanum) на высоте 2 200 м в Альпах (темпе-
ратура воздуха 22 °C)
177
чено внутри цветка с полузакрытым околоцветником, сохраняю-
щим тепло, выделяемое при дыхании или нагретой почвой. Это
одно из адаптивных приспособлений ряда лесных эфемероидов
(пролески — Scilla, хохлатки — Corydalis и др.), сохраняющих их
цветки ранней весной, когда температура воздуха близка к О °C
(Т. К. Горышина, 1979).
Сильно нагревается поверхность стволов одиночно стоящих
деревьев (а весной и осенью и в лиственных лесах). С юга темпе-
ратура камбия бывает на 10—20 °C выше, чем его температура на
северной стороне, а при температуре воздуха около 0 °C на сол-
нечной стороне может достигать 30 °C (И. М. Культиасов, 1982).
В жаркие дни темные стволы ели (Piced) нагреваются до 55 °C,
что может приводить к ожогам камбия. Внедренные в кору тер-
мопары показывают, что стволы разных пород защищены в раз-
ной степени. Так, у березы (Betula) температура камбия быстро
следует колебаниям температуры воздуха, а у сосны (Pinus sylvestris)
она более постоянна из-за теплозащитных свойств коры. Стволы
хорошо аккумулируют тепло, поэтому весеннее нагревание дере-
вьев в безлистном лесу существенно влияет на микроклимат
(Т. К. Горышина, 1979).
Превышение температуры растений над температурой воздуха
бывает и в холодных местообитаниях. Этому часто способствуют
оптические свойства растений, увеличивающие поглощение ра-
диации (например, темная окраска в результате накопления анто-
циана в эпидерме) и снижающие транспирацию анатомо-морфо-
логические особенности. А в стеблях травянистых растений Арк-
тики нередко редуцируется сердцевина, они становятся полыми,
и температура внутри полостей может превышать температуру
воздуха на 20 °C.
Многократно показано существенное нагревание органов арк-
тических растений. Так, нижние листья Novosiversia glacialis на
Таймыре прогреваются до 19,4 °C, составляя градиент с вышеле-
жащими участками стебля в 6 °C (Б. А.Тихомиров, 1963) (рис. 7.10).
А у арктической ивы (Salix arctica) на Аляске листья в ночные
часы теплее воздуха на 1 —3 °C, а днем — на 2 —11 °C. Изучение
распределения температур в арктических растениях показывает,
что наиболее нагретые органы расположены около почвы (розет-
ки листьев, стебель). А в Антарктиде летом даже под слоем снега
более 30 см температура темноокрашенных лишайников бывает
положительной, благодаря чему они имеют положительный ба
ланс углеродного обмена.
Супратемпературность растений важна и в холодные сезоны в
умеренной зоне. Так, по наблюдениям Т. К. Горышиной (1961),
ранней весной температура цветка и листьев только что вышед-
шей из-под снега пролески превышает температуру воздуха на
7 —8 °C (рис. 7.11). Зимой значительно нагревается хвоя и даже в
178
Рис. 7.10. Температурный профиль
Novosiversia glacialis в таймырской
тундре (по Б. А. Тихомирову, 1962,
1963). Температура воздуха +15,5 °C
Рис. 7.11. Температурный профиль
пролески сибирской {Scilla sibirica)
в дубраве (по Т. К. Горышиной,
1961)
мороз возможно превышение ее температуры над температурой
воздуха на 9—12 °C. Экспериментально показано, что при силь-
ном освещении и отрицательной температуре (-5...-6°C) листья
могут нагреться до 17—19°C и фотосинтезировать. Такое повы-
шение температуры растения экологически очень важно, так как
физиологические процессы при этом получают некоторую неза-
висимость от окружающего теплового фона.
Субтемпературные виды растений. Снижение температуры рас-
тений по сравнению с окружающим воздухом обычно отмечается
в сильно освещенных и прогретых местообитаниях (степи, пусты-
ни) у видов с редуцированной листовой поверхностью и усилен-
ной транспирацией (эффект «гидротерморегуляции»). У интен-
Рис. 7.12. Отличие температуры листьев хлопчатника (Gossypium) от тем-
пературы воздуха при разном водоснабжении (в Средней Азии) (по
I .Вальтеру, 1982). Тонкая кривая — плохое водоснабжение, толстая —
нормальное
179
сивно транспирирующих видов снижение температуры листьев
достигает 15°, причем охлаждение уже на 3 — 4 °C может предох-
ранить их от смертельного перегрева (он начинается при затруд-
нении водоснабжения) (рис. 7.12). Именно перегрев быстро при-
водит пустынные растения к гибели при смазывании вазелином
нижней поверхности листьев (с устьицами).
В пустынях Средней Азии у некоторых кустарников и полуку-
старников, сбрасывающих листья в жаркий период, например у
Astragalus paucijugus, а также у травянистых облиственных мно-
голетников (эфемероидов) температура ассимилирующих органов
всегда держится ниже температуры воздуха. Максимальная раз-
ница (8,1 °C) отмечена у туркестанского ревеня (Rheum turkesta-
nicum). Интересно, что у пустынных деревьев и кустарников с
редуцированными листьями (саксаулы — Haloxylon, песчаная ака-
ция — Ammodendron) утром температура ассимилирующих орга-
нов обычно выше (на 1,6 °C) окружающей температуры, но когда
температура воздуха становится выше 25 °C, температура расте-
ний опускается ниже нее.
Температура листа во многом зависит от его толщины и кон-
систенции (рис. 7.13). У тонких листьев теплоемкость невысока и
они сильнее реагируют на колебания инсоляции, чем мясистые
суккуленты, у которых из-за небольшой поверхности меньше ин-
тенсивность теплообмена и ниже транспирация. Температура тон-
кого листа быстро повышается при внезапном освещении и быс-
тро падает при затенении.
Рис. 7.13. Изменение температуры листа в зависимости от консистен-
ции при внезапном освещении (по И. М. Культиасову, 1982):
I — Papaver discolor, II — Magnolia sphenocarpa, I — температура листа;
2 — температура воздуха
180
7.3. Влияние температуры на рост и развитие растений
Температурные показатели роста растений. Рост растений во
многом зависит от температурных условий. Кривая зависимости
роста от температуры колоколовидна. Минимальные температу-
ры роста обычно лежат чуть выше точки замерзания тканей, а
максимальные находятся на несколько градусов ниже показателя
тепловой смерти. Процессы деления и дифференцировки клеток
могут проходить, хотя и очень медленно, даже зимой при низких
температурах. Например, они идут в почках плодовых деревьев.
Однако для активного митоза нужны более высокие температуры
(примерно на 5° выше, чем для процесса растяжения). При этом
кардинальные температурные точки для роста могут изменяться в
зависимости от температурной адаптации, фазы развития, сезона
и времени суток.
Есть и географические аспекты этих зависимостей. В целом
чем выше температуры естественных местообитаний, тем выше
лежат и кардинальные точки температурной кривой роста. Так,
рост побегов большинства растений умеренной зоны начинается
при температуре на несколько градусов выше О °C, а у тропиче-
ских растений — только при 12 —15 °C. Минимальная температу-
ра роста для северных культурных растений (гороха — Pisum
sativum, ржи — Secale cereale, пшеницы — Triticum aestivum)
составляет 2 —5 °C, а для южных видов (дыня — Melo sativus,
сорго — Sorghum) — 15—18°C. У растений холодных областей,
раноцветущих и горных видов ростовые процессы начинаются уже
около О °C. В районах с низкими температурами, лимитирующи-
ми жизнедеятельность, уже небольшое их повышение сильно ска-
зывается на росте. Поэтому у арктических и высокогорных расте-
ний температурная кривая роста круто поднимается вверх и они
быстро растут уже при температурах, близких к О °C. Особенность
арктических растений в том, что рост их может начинаться при
отрицательных температурах, а интенсивное перемещение угле-
водов из подземных органов в побеги поддерживает рост при низ-
ких температурах. Для арктических видов типично и быстрое сезон-
ное развитие, чему способствуют зимнезеленость и высокая ско-
рость фотосинтеза с пиком при низких температурах (А. П.Тыр-
гиков, 1980). Большое значение имеет антоциановая окраска, со-
здающая условия для повышения температуры органов.
Отличаются у разных географических групп видов и оптималь-
ные температуры для процессов роста. Для роста побегов расте-
ний умеренной зоны наиболее благоприятны температуры от 15
до 25 °C, а для растений тропиков и субтропиков — от 30 до 40 °C.
Однако есть виды, которые проходят весь цикл развития в райо-
нах с постоянно высокой или низкой температурой. Так, многие
арктические и северные морские растения живут в местообитани-
181
ях с температурами чуть выше О °C, а водоросли горячих источни-
ков всю жизнь могут существовать при температурах выше 60 °C
(В.Лархер, 1978).
Температурные границы роста корней обычно имеют широкую амп-
литуду. У древесных растений умеренной зоны нижний предел этого
процесса лежит между 2 и 5 °C, поэтому корни начинают расти еще до
распускания почек и заканчивают рост поздней осенью (рис. 7.14). Кор-
ни растений теплых регионов предъявляют к температуре более высокие
требования. Так, у лимона (Citrus limori) они растут только при темпера-
турах выше 10 °C, но в пределах его естественного ареала температура
почвы даже в самое холодное время не опускается ниже. Однако культи-
вирование цитрусовых в других местах этот фактор ограничивает, так
как плоды у них созревают зимой и потому именно в холодное время им
требуется обильное снабжение водой и элементами минерального пита-
ния. Клетки корней арктических растений делятся даже в слое много-
летней мерзлоты, что подтверждено опытами в фитотроне. При низких
температурах удлинение корней замедляется, но они значительно утол-
щаются, что снижает полезное соотношение их поверхности и объема.
Но при очень низких температурах в полярных пустынях уменьшается и
толщина корней (А.П.Тыртиков, 1964).
На разных фазах развития растений требуется определенная
температура. Например, пониженная температура способствует
кущению злаков и образованию боковых побегов у бобовых.
У молодых растений оптимальная температура роста выше, чем у
взрослых (Т.А. Работнов, 1985). Иногда имеет значение продол-
жительность действия неблагоприятных температур, причем ско-
Месяцы Месяцы
д — начало растяжения почек v — начало пожелтения почек
Рис. 7.14. Прирост в длину побега и корней различных древесных
пород в течение года (по В.Лархеру, 1978).
- Прирост корней зачернен
182
рость роста может значительно меняться и от кратковременного
температурного шока.
Температурные параметры цветения и плодоношения. Для ус-
пешной жизнедеятельности вида необходимы подходящие усло-
вия для генеративного размножения. Температурный диапазон
образования и функционирования генеративных органов, созре-
вания зачатков, прорастания всходов и жизни проростков часто
иной, чем для роста вегетативных органов. Так, для созревания
плодов и семян тепла требуется больше, чем для окончания роста
побегов и корней. Поэтому в местообитаниях с коротким и про-
хладным вегетационным периодом распространены виды, способ-
ные к эффективному вегетативному размножению.
Цветение индуцируется в довольно узком интервале темпера-
тур, а для развития и распускания цветков благоприятны иные тем-
пературы. Озимые, однолетние и двулетние растения для нормаль-
ного цветения нуждаются в холодовом воздействии. Это относится
и к почкам некоторых древесных растений, например персика
(Persica vulgaris). Они готовы к цветению только в том случае,
если несколько недель подвергались воздействию температур меж-
ду -3 и +13 °C (лучше от 3 до 5 °C), прошли яровизацию. Но если
холодный период короток или наступает несвоевременно, преры-
вается подъемом температуры выше 15 °C, то цветение отсутствует.
Летом в Арктике может выпадать снег, заносящий цветущие
растения. При этом они обычно оледеневают, становятся хрупки-
ми, но при оттаивании продолжают развитие. Е. АТихменев (1980)
обнаружил, что цветение камнеломок (Saxifraga) на о. Врангеля
происходит уже при температурах 3,4 —6,3 °C. Для этого региона
характерно также продолжительное время (до 3 лет) между заложе-
нием первых цветочных бугорков в почках и зацветанием, что при-
водит к превращению однолетников в многолетники (И. Г.Сереб-
ряков, 1962).
В суровых температурных условиях (Крайний Север, высоко-
горья) многие виды плодоносят только в годы с особенно теплым
вегетационным сезоном. Существуют данные и о приуроченности
возобновления растений на верхней границе леса в горах к пери-
одам улучшения термического режима. Так, на Алтае возрастная
структура лиственичников из Larix sibirica показывает увеличен-
ную долю деревьев, появившихся и выживших в благоприятные
по температуре годы. При этом максимальные годичные прирос-
ты ствола коррелируют и с максимальным появлением новых де-
ревьев (П.А. Моисеев, 2002). Аналогичные материалы поучены
С. Г. Шиятовым (1967) для деревьев на верхней границе леса на
Полярном Урале.
Температурные параметры прорастания семян. На прорастание
семян температура влияет двояко. С одной стороны, она опреде-
ляет скорость прорастания, а с другой — температурное воздей-
183
ствие может снять состояние покоя. При этом температурные кар-
динальные точки для прорастания спор и семян должны соответ-
ствовать тем условиям, при которых возможно дальнейшее разви-
тие молодых растений.
Скорость прорастания семян увеличивается с повышением тем-
пературы. Так, семена видов умеренных и приполярных широт
прорастают в теплое время года, а при еще низких температурах
весной этот процесс идет очень медленно. Это имеет важный адап-
тивный смысл, так как на холодном субстрате не смогут нормаль-
но развиваться проростки. Поэтому пока почва не прогреется
(обычно выше 10 °C), семена не трогаются в рост, но как только
температура субстрата станет благоприятной, они развиваются
очень быстро. Характерно, что семена большей части тундровых
растений лучше всего прорастают при температурах 20—30 °C, что
увеличивает надежность выживания проростков (А. П.Тыртиков,
1980). В тундрах отмечается также очень медленный рост всходов.
Например, в Восточной Гренландии Oxyria digyna в первое лето
развивает всего два листа по 2,5 мм. Из-за короткого вегетацион-
ного сезона семена многих тундровых растений опадают не до-
зрев и дозревают под снегом (А. Б.Тихомиров, 1963).
У некоторых растений существуют механизмы, препятствующие
прорастанию семян в неблагоприятное время. Так, семена многих
видов умеренных и холодных широт прорастают легче, если при
набухании некоторое время подвергались воздействию низких тем-
ператур (прошли стратификацию). Например, Ю.Е.Алексеев
(1996) экспериментально показал, что без стратификации не мо-
гут прорасти семена некоторых осок (Carex elongata, С. paupercula,
С. loliacea, С. curaica). Семена яблони (Malus), рябины (Sorb us),
ряда видов клена (Acer), характеризующиеся глубоким покоем,
нуждаются в длительной стратификации холодом (1,5 — 3 мес при
0 — 3 °C), причем никакие другие воздействия ее не заменят
(М. Г. Николаева, 2002). А для полного проращивания семян та-
кого субарктического вида, как морошка (Rubus chamaemorus),
необходимо воздействие на набухшие семена низких (около 5 °C)
температур в течение 9 мес.
В противоположность северным растениям виды из областей с
мягкой зимой используют долгий вегетационный период прорас-
тая уже зимой при более низких температурах и в более узких
температурных границах. Так, семена многих растений умерен-
ной зоны начинают прорастать при температуре около 8 °C. При
этом, например, у ясеня (Fraxinus) чем дальше отодвинута от по-
люса граница ареала вида, тем больше размер его зародыша (он
будет быстрее дозревать) и тем менее глубок его покой (М. Г. Ни-
колаева, 2002). Температурный интервал прорастания семян и спор
также коррелирует с широтой ареала вида, и у космополитов он
обычно очень широк.
184
Рис. 7.15. Влияние температуры на ско-
рость прорастания (время до прорас-
тания) свежесобранных семян гвоздич-
ных (по В.Лархеру, 1978):
/ — Silene secundiflorcr, 2 — S. viscosa;
3 — Coronaria flos-cuculi
Температура, °C
Эти закономерности хорошо прослеживаются в семействе Гвоздич-
ные, чьи представители имеют разные типы ареалов. Так, у средиземно-
морских видов (например, Silene secundiflora) семена прорастают зимой
(7 °C). Виды из Средней и Восточной Европы (S. viscosa, Coronaria flos-
cuculi) прорастают летом при температуре 12—17°C. А континенталь-
ный вид смолевки (S. viscosa) обладает самым широким температурным
оптимумом прорастания (14—28 °C) (В.Лархер, 1978) (рис. 7.15).
Для прорастания семян ряд растений нуждается в повышен-
ных температурах. Семена тропических растений дают наиболь-
ший процент всхожести при температурах от 15 до 30 °C. У ряда
видов умеренных широт (крупка — Draba verna, ежовник —
Echinochloa tumerana и др.) неглубокий покой семян снимается
в течение нескольких недель температурой около 15 °C, а актив-
ный выход из покоя наблюдается при 28 °C (20—35°). Неглубокий
покой, нарушаемый высокой температурой, встречается в жарких
областях (прорастание семян там связано также с водообеспечен-
ностью) (М. Г. Николаева, 2002). А семена некоторых видов мож-
но простимулировать к прорастанию кратким (менее 1 мин) воз-
действием высоких температур. Это характерно для вересковых,
растущих в местообитаниях, подверженных частым пожарам, на-
пример для вереска обыкновенного (Calluna vulgaris), эрики пе-
пельной (Erica cinered) и других пирофитов. А у некоторых видов
прорастание семян стимулирует перепад температур.
Экологические группы растений по отношению к теплу. В зави-
симости от особенностей температурных кривых жизнедеятель-
ности и положения их кардинальных точек различают экологи-
ческие группы растений по отношению к температуре. У тепло-
любивых форм (термофильных, мегатермных) оптимум лежит в
области повышенных температур. Они обитают в тропиках и суб-
тропиках, а в умеренных поясах — в сильно прогреваемых место-
обитаниях. Для холодолюбивых (криофильных, микротермных)
растений оптимальны низкие температуры. К ним принадлежат
виды, живущие в полярных и высокогорных областях или зани-
мающие холодные местообитания. Можно выделить также про-
185
межуточную группу мезотермных растений. Различие этих групп
проявляется по положению оптимума на температурной кривой
фотосинтеза, а положение точек минимума и максимума этой кри-
вой указывает на их выносливость к экстремальным температурам.
Зависимостью распределения растений от температуры ботаники и
географы интересовались давно. В 1874 г. А. Декандоль предложил выде-
лить шесть групп растений, связанных с климатическими поясами. Ну-
левая группа — мегистотермы, нуждающиеся в среднегодовой темпе-
ратуре выше 30 °C. Это растения каменноугольного периода, вымершие
при похолодании климата. Первая группа — мегатермы, которым необ-
ходимы высокая температура и постоянная влажность. Они не перено-
сят морозов и соответствуют влажным тропикам и субтропикам со сред-
ней годовой температурой более 18 °C. Вторая группа — ксеротермы,
приспособленные к климату сухих субтропиков с высокими температу-
рами и сухим периодом в течение нескольких месяцев. Хорошо перено-
сят высокие температуры и низкую влажность. Третья группа — мезо-
термы. Это растения умеренно теплого климата с холодным периодом,
который не прерывает вегетацию (например, средиземноморские). Обыч-
но это чувствительные к морозу вечнозеленые виды или растения с го-
лыми почками. Четвертая группа — микротермы, главным образом ра-
стения умеренной зоны, приспособленные к прохладному лету и про-
должительной морозной зиме. Пятая группа — гекистотермы — расте-
ния полярного пояса и высокогорий, существующие в условиях мини-
мального тепла и удовлетворяющиеся коротким вегетационным перио-
дом. Для роста древесных форм тут условий нет.
Современную шкалу экологических групп растений по отноше-
нию к теплу предложил в 70-е гг. XX в. Г.Элленберг. Он тоже вы-
делил шесть групп: Т-1 — крайне морозостойкие виды; Т-2 — холо-
достойкие растения, редко выходящие за северную границу леса;
Т-3 — среднехолодостойкие, в основном виды смешанных лесов;
Т-4 — теплолюбивые растения южных склонов и «теплых» почв;
Т-5 — очень теплолюбивые виды, крайне чувствительные к моро-
зу; Т-0 — растения индифферентные, безразличные к теплу, имею-
щие широкую амплитуду приспособленности к нему. Группы Г.Эл-
ленберга применимы в основном для условий Средней Европы.
7.4. Действие экстремальных температур на растения
За долгий период эволюционного освоения планеты растения
приспособились к воздействию разных температур. Но эти адап-
тации не защищают их абсолютно. Экстремальные температуры
нарушают жизнедеятельность и ограничивают распространение
растений. Действие жары и холода зависит от их интенсивное ги,
продолжительности, периодичности, а также от состояния расгс-
186
ний. Многие виды обладают выраженной устойчивостью к дей-
ствию экстремальных температур, но обычно она не является их
постоянным свойством, а формируется в процессе воздействия
некоторых тепловых условий. При настройке адаптационных си-
стем растительные организмы приобретают способность перено-
сить экстремальную теплообеспеченность.
Термостресс и тепловые границы жизни растений
Температурные границы функционирования и распространения
растений в районах экстремальных температур. Высшие наземные
растения обычно более эвритермны (растут в более широком ди-
апазоне температур), чем водные. В большинстве случаев их ак-
тивная жизнедеятельность протекает при температурах от 5 до
55 °C, причем их продуктивность не выходит за пределы 40 °C.
Среди водных растений, особенно низших, немало стенотермов,
приспособленных к очень узким температурным диапазонам.
Иногда они становятся экстремофилами. Например, зеленые и
диатомовые водоросли полярных и высокогорных снегов и льдов
растут только вблизи точки замерзания. Так, Chlamydomonas
nivalis, живущая в оснеженных высокогорьях, существует в интер-
вале от -5 до 5 °C и имеет температурный оптимум около 0 — 1 °C.
А фототрофная цианобактерия Oscillatoria, живущая на о. Ява в воде
с температурой 64°C, погибает при 68°C уже через 5—10 мин.
Для экологических целей необходимо знать тепловые потреб-
ности видов и ряд температурных границ функционирования ра-
стений. Температурные границы жизни — самые низкие и самые
высокие температуры, которые выдерживает данное растение.
Различают латентные и летальные границы. После перехода через
латентную границу жизненные процессы снижаются до мини-
мального уровня, однако этот процесс обратим. При достижении
летальной границы возникают необратимые повреждения и расте-
ние погибает. Наиболее узок температурный диапазон жизни у вод-
ных растений (от -1 — 0 до 25 — 40 °C), а наиболее широк у С4-видов
сухих субтропиков (от -5 до 45 —60°C), горных (от -4...-10 до
38 — 52 °C) и средиземноморских растений (от -5...-7 до 48 — 54 °C)
(В.Лархер, 1978).
Только под водой и глубоко в почве температуры всегда оста-
ются в безопасных для растений пределах. Почти во всех других
местах при суточных или сезонных колебаниях теплообеспечен-
ности создаются опасные для них условия. Есть и постоянно су-
ровые местообитания. Особенно высокие температуры воздуха в
тропиках. В пустынях Ливии и Мексике отмечены абсолютные
максимумы в 57 —58 °C. Около четверти суши может иметь сред-
негодовой максимум температуры воздуха выше 40 °C. Но и вне
187
этих областей на скалах и других открытых местообитаниях воз-
можен прогрев до 70 °C. Большинство цветковых растений гибнет
при 45 —50 °C, но растения скал выдерживают нагревание выше
60 °C. А пустынный лишайник манна (Lecanora esculenta) в су-
хом виде нагревается без повреждения и выше 70 °C.
При крайне высоких температурах из фотоавтотрофов развиваются
только цианобактерии. Например, Synechococcus живет в горячих гейзе-
рах при температурах от 45 до 82 °C (оптимум 70 °C). Эукариотические
водоросли выдерживают высокие температуры (65 —75 °C) только при
полном освещении, а в затененных местах не выносят температур выше
50 —55 °C. Видовое богатство водорослей горячих источников превыша-
ет 200 видов, но с повышением температуры число видов, способных ее
переносить, заметно уменьшается. Большинство термофильных фотоав-
тотрофов живет при температуре 35—40°, а при 85 —90 °C обнаружено
всего два вида (Л.Л. Великанов и др., 1981).
В умеренных, приполярных и горных областях растения еже-
годно в течение нескольких месяцев подвергаются действию низ-
ких температур. Кроме того, в отдельных районах и в более теп-
лые сезоны они могут испытывать кратковременные воздействия
пониженных температур во время заморозков. И есть местооби-
тания, где вся жизнь растений проходит в суровых холодных ус-
ловиях (высокогорья, приполярные снежные поля и морские воды).
Ниже 0 °C температур не бывает только на трети суши. На 42 %
поверхности Земли среднегодовой минимум температуры воздуха
опускается ниже -20 °C. В Гренландии и Восточной Сибири ми-
нимальные температуры воздуха доходят до -68 °C (при этом Ой-
мякон окружает лиственничная тайга). Самая же низкая темпера-
тура на Земле (около -90 °C) была зарегистрирована в Антаркти-
де, но в тех местах растений нет.
Тепло для растений необходимо, но на поверхности Земли фактиче-
ски нет территории, где какие-нибудь растения не могли бы жить имен-
но из-за недостатка тепла. Так, в поверхностных слоях снега и льда (на-
пример, в горах, в Заполярье) обитает криофитон, представленный бо-
лее чем 100 видами снежных водорослей. Значительна и продукция этих
организмов: из 1 м3 льда можно выделить около 1 кг сырой биомассы
(Л.Л. Великанов и др., 1981). При массовом размножении эти водоросли
окрашивают снег в разные цвета. Фазы их активной жизнедеятельности
обычно проходят на оттаявшей поверхности при температуре около 0 °C.
А пределы устойчивости к низким температурам колеблются у них от •
-34...-36 (Sphaerella nivalis, Chlamydomonas nivalis) до -40...-60°C
(Pediastrum borianum, Phormidium flaccidum). Так же велика холодостой
кость фитопланктона полярных морей, нередко зимующего в корке льда.
Кроме абсолютных температур для растений очень важен ре -
жим изменения теплообеспеченности. Эпизодические морозы.
188
даже несильные, обычно очень опасны, так как могут застать ра-
стения неподготовленными, в уязвимой фазе развития. К сезон-
ным же экстремальным температурам организмы эволюционно
подготовлены путем закалки.
Термостресс и термонарушения клеток растений. Тепловой
стресс — необычная для организма нагрузка, вызывающая в нем
характерную общую реакцию. При термострессе цитоплазма сна-
чала отвечает быстрым усилением метаболизма. Резко интенси-
фицируется дыхание, что отражает попытку исправить появив-
шиеся дефекты и создать предпосылки для приспособления к новой
ситуации. Если этот результат не достигается, возможности клет-
ки истощаются и она гибнет. Если температура переходит крити-
ческую точку, клеточные структуры могут повреждаться, и прото-
пласт быстро отмирает. Так внезапная гибель клеток происходит,
например, в заморозки. Но повреждения могут возникать и по-
степенно. Отдельные функции все сильнее угнетаются, наруша-
ются, что приводит к разбалансировке процессов жизнедеятель-
ности и гибели клеток.
К температуре очень чувствительны слабые связи макромоле-
кул (водородные, ионные, гидрофобные, вандерваальсовы). От них
швисят третичная и четвертичная структуры белка, надмолеку-
лярные связи его в мембранах, структура нуклеиновых кислот,
поды, поэтому температура сильно влияет на метаболический ап-
парат клеток. Для клетки эти связи очень важны, так как придают
биологическим структурам лабильность и способность быстро из-
меняться под действием разных агентов. Различные типы этих
взаимодействий неодинаково реагируют на повышение и пони-
жение температуры, поэтому влияние ее на систему со слабыми
связями частично зависит от того, какие из них играют в данном
случае главную роль.
Для многих внутриклеточных структур (белков, нуклеиновых
кислот и др.) важно сохранение определенного уровня их кон-
формационной гибкости, которая, например, определяет степень
устойчивости макромолекул к температурной денатурации. Кон-
формационная гибкость возрастает с повышением температуры и
понижается при ее падении. Поэтому эволюционно выработан-
ные адаптации поддерживают соответствие между температурой
существования организма и конформационной гибкостью макро-
молекул (она не должна быть ни избыточной, ни недостаточной)
(Л. Г. Косулина и др., 1993).
Различные жизненные процессы растений неодинаково чув-
ствительны к неоптимальной температуре (рис. 7.16). Довольно
быстро повреждаются хлоропласты, нарушается фотосинтез. А при
нарушении дыхания заметно падает интенсивность движения ци-
топлазмы, которая зависит от энергоснабжения клетки. Дыхание
как источник энергии очень важно в адаптации организмов к ус-
189
Выпадение
функции
Нарушение
функции
Рис. 7.16. Нарушение клеточных функций под действием холода
и жары (по В.Лархеру, 1978)
ловиям среды. Повреждающие температуры вызывают у неустой-
чивых растений прогрессирующее уменьшение интенсивности
поглощения кислорода и снижение эффективности дыхания.
У поврежденных холодом или жарой растений уровень дыхания
сильно колеблется и часто остается повышенным и после возвра-
щения температуры к умеренным показателям.
В конечной стадии термонарушений утрачивается полупрони-
цаемость мембран, разрушаются клеточные структуры (особенно
активно тилакоиды пластид) и клеточный сок выходит в меж-
клетники.
Действие на растения низких температур
Повреждение растений холодом и морозом. В экологии рас ге-
ний принято различать действие холода (низкой положительной
температуры) и мороза (отрицательных температур). Негативное
190
влияние холода зависит от диапазона понижения температур и
продолжительности их воздействия. Уже неэкстремальные низ-
кие температуры неблагоприятно сказываются на растениях, по-
скольку тормозят основные физиологические процессы (фотосин-
тез, транспирацию, водообмен и т.д.), снижают энергетическую
эффективность дыхания, изменяют функциональную активность
мембран, приводят к преобладанию в обмене веществ гидролити-
ческих реакций. Внешне повреждение холодом сопровождается
потерей листьями тургора и изменением их окраски из-за разру-
шения хлорофилла. Резко замедляются рост и развитие. Так, лис-
тья огурца (Cucumis sativus) теряют тургор при 3 °C на 3-й день,
растение завядает и гибнет из-за нарушения доставки воды. Но и
в насыщенной водяными парами среде пониженные температуры
неблагоприятно влияют на обмен веществ растений. У ряда видов
усиливается распад белков и накапливаются растворимые формы
азота.
Основная причина повреждающего действия низкой положи-
тельной температуры на теплолюбивые растения — нарушение
функциональной активности мембран из-за перехода насыщен-
ных жирных кислот из жидкокристаллического состояния в гель.
В результате, с одной стороны, повышается проницаемость мем-
бран для ионов, а с другой — увеличивается энергия активации
ферментов, связанных с мембраной. Скорость реакций, катали-
зируемых мембранными ферментами, снижается после фазового
перехода быстрее, чем скорость реакций, связанных с раствори-
мыми энзимами. Все это приводит к неблагоприятным сдвигам в
обмене веществ, резкому возрастанию количества эндогенных ток-
сикантов, а при длительном действии низкой температуры — к
гибели растения (В. В. Полевой, 1989). Так, при снижении темпе-
ратуры до нескольких градусов выше О °C гибнут многие растения
тропического и субтропического происхождения. Отмирание их
идет медленнее, чем при вымерзании, и является следствием рас-
стройства биохимических и физиологических процессов в орга-
низме, оказавшемся в несвойственной обстановке.
Выделено множество факторов, губительно действующих на
растения при отрицательных температурах: потеря тепла, разрыв
сосудов, обезвоживание, льдообразование, повышенные кислот-
ность и концентрация клеточного сока и т.п. Гибель клеток от
мороза обычно связывают с дезорганизацией обмена белков и нук-
леиновых кислот, а также с не менее важным нарушением прони-
цаемости мембран и прекращением тока ассимилятов. В результа-
те процессы распада начинают преобладать над процессами синте-
за, накапливаются яды, нарушается структура цитоплазмы.
Многие растения, не повреждаясь при температурах выше О °C,
гибнут от образования льда в тканях. В обводненных незакален-
ных органах лед может образовываться в протопластах, межклет-
191
никах и клеточных стенках. Г. А.Самыгин (1974) выделил три типа
вымерзания клеток, зависящие от физиологического состояния
организма и его готовности к перезимовке. В первом случае клет-
ки гибнут после быстрого образования льда сначала в цитоплаз-
ме, а потом в вакуоле. Второй тип вымерзания связан с обезвожи-
ванием и деформацией клетки при образовании межклеточного
льда (рис. 7.17). Третий тип гибели клеток наблюдается при соче-
тании межклеточного и внутриклеточного льдообразования.
При замерзании, как и в результате засухи, протопласты отда-
ют воду, сжимаются и содержание растворенных в них солей и
органических кислот возрастает до токсичных концентраций. Это
вызывает инактивацию ферментных систем, участвующих в фос-
форилировании и синтезе АТФ. Перемещение воды и замерзание
продолжаются до тех пор, пока не установится равновесие сосу-
щих сил между льдом и водой протопласта. А оно зависит от тем-
пературы: при температуре -5 °C равновесие наступает при 60 бар,
а при -10 °C уже при 120 бар (В.Лархер, 1978).
При длительном действии мороза кристаллы льда вырастают
до значительных размеров и могут сжимать клетки и повреждать
плазмалемму. Процесс образования льда зависит от скорости по-
нижения температуры. Если вымерзание идет медленно, лед об-
Внеклеточное образование льда
______________I______________
' Механический Осмотический
Обезвоживание етпе.с.с. стпеее
Денатурация мембранных Изменение взаимодействия
белков липид/белков
Инактивация калиевого Разобщение
и сахарного насосов ** фосфорилирования
Пассивная утечка К и сахаров из клетки i
1 Инфиль- трация ткани I Потеря тургора Замена Са2 + в мембране наК+ Набухание цитоплазмы Наруш в митохо и хлорог 1 ения ндриях шастах
1 Нехватка кислорода 1 Разрушение мембранной системы
t
Гибель клетки
Рис. 7.17. Схема повреждений клетки, вызванных внеклеточным
льдообразованием и оттаиванием (по Дж.П.Палту, П.Х.Ли, 1983)
192
разуется вне клеток, и при оттаивании они остаются живыми.
Когда же температура падает быстро, вода не успевает проник-
нуть сквозь клеточную стенку и замерзает между нею и протопла-
стом. Это вызывает разрушение периферических слоев цитоплаз-
мы, а потом и необратимое повреждение клетки. При очень быс-
тром падении температуры вода не успевает выйти из протоплас-
та и кристаллы льда быстро распространяются по клетке. Следо-
вательно, клетки быстро замерзают в том случае, если вода из них
не успела оттечь. Поэтому важен быстрый транспорт ее в меж-
клетники, чему способствует поддержание высокой проницаемо-
сти мембран, связанное с большим содержанием в их составе не-
насыщенных жирных кислот (В. В. Полевой, 1989). У закаленных
растений при отрицательных температурах мембраны «не засты-
вают», сохраняя функциональную активность. Морозоустойчивость
клетки также повышается, если вода прочно связана со структу-
рами цитоплазмы.
Мороз может сильно нарушать структуру мембран. Мембран-
ные белки дегидратируются и денатурируют, что инактивирует
важные системы активного транспорта сахаров и ионов. Сверты-
вание белков под действием мороза особенно характерно для
южных растений, отмирающих до образования льда. А морозный
распад липидных компонентов мембран сопровождается гидро-
лизом фосфолипидов и образованием фосфорной кислоты. В итоге
поврежденные мембраны теряют полупроницаемость, потеря воды
клетками усиливается, тургор падает, межклетники заполняются
водой, и из клеток интенсивно вымываются необходимые ионы.
Мороз повреждает и пигментную систему растений. Причем
действие температурного стресса зимой часто сочетается с повреж-
дением ассимилирующих органов светом. Так, в хлоропластах хвои
повреждается электрон-транспортная цепь, но эти повреждения
обратимы. У зимующих растений увеличивается содержание ка-
ротиноидов, защищающих хлорофилл от повреждения светом.
Сохранение пигментов и фотосинтеза важно для устойчивости
растений и осенью, когда при низких положительных температу-
рах синтезируются протекторные соединения, и для перезимовки
растений. При отрицательных температурах у озимых злаков за
счет фотосинтеза происходит частичная компенсация затрат на
поддержание жизнеспособности в стрессовых условиях (Л. Г.Ко-
сулина и др., 1993).
Мороз может вызвать и механические повреждения раститель-
ных организмов. В этом случае особенно страдают стволы деревь-
ев и крупные ветви. Зимой при сильном ночном охлаждении ствол
быстро теряет тепло. Кора и наружные слои древесины охлажда-
ются быстрее, чем внутренняя часть ствола, поэтому в них возни-
кает значительное напряжение, которое при быстром изменении
температуры приводит к вертикальному растрескиванию дерева.
193
Кроме того, возможны тангентальные трещины и отслойки коры.
Морозобойные трещины при активной работе камбия закрыва-
ются, но если новые слои древесины образоваться не успевают,
трещины распространяются по радиусу внутрь ствола. В них по-
падает инфекция, которая, проникая в соседние ткани, нарушает
работу проводящей системы и может привести дерево к гибели.
Морозобойные повреждения возникают и днем. При длительных
морозах, особенно в солнечную погоду, возвышающиеся над снегом ча-
сти растений могут пересыхать от дисбаланса транспирации и поглоще-
ния воды из холодной почвы (имеет значение также сжатие клеток при
обезвоживании и образовании льда, замораживание клеточного сока).
У древесных растений в районах с солнечной зимой (Восточная Сибирь,
Северный Кавказ, Крым и др.) даже отмечаются зимне-весенние «ожо-
ги» на южной стороне ветвей и молодых незащищенных стволов. Ясны-
ми зимними и весенними днями у неопробковевших частей растений
клетки нагреваются, теряют морозостойкость и не выдерживают после-
дующих морозов. А в лесотундре морозобойные повреждения могут об-
разовываться и летом во время заморозков. Особенно им подвержен мо-
лодой подрост. Его камбий быстро охлаждается, так как еще не сформи-
ровался достаточный теплоизолирующий слой коры, и поэтому невели-
ка теплоемкость тонких стволов. Особенно опасны эти воздействия в
середине лета, когда активность камбия максимальна (М.А. Гурская,
С.Г.Шиятов, 2002).
Уплотнение и растрескивание замерзшей почвы приводит к
механическому повреждению и разрыву корней. Так же может
действовать и морозное «выпирание» растений, которое вызыва-
ется неравномерным замерзанием и расширением почвенной влаги.
При этом возникают силы, выталкивающие растение из почвы.
В результате выворачиваются дернины, оголяются и обрываются
корни, вываливаются деревья. Суммируя данные о зимних по-
вреждениях растений, кроме собственно холодостойкости и мо-
розостойкости, отражающих способность переносить прямое
действие низких температур, в экологии выделяют еще зимо-
стойкость — способность к перенесению всех неблагоприятных
зимних условий (замерзание, выпревание, выпирание и т.п.). При
этом специальных морфологических приспособлений, защищаю-
щих только от холода, у растений нет и в холодных местообита
ниях защита осуществляется от всего комплекса неблагоприят-
ных условий (ветры, иссушение, холод и т.д.)
Холод воздействует на растение не только прямо (через термонару
шения), но и косвенно, через физиологическую «зимнюю засуху». При
зимнем интенсивном освещении и потеплении температура воздуха мо
жет превысить температуру почвы. Надземные части растений усилива
ют транспирацию, а поглощение воды из холодной почвы замедлено.
194
В результате в растении повышается осмотическое давление, наступает
водный дефицит. При длительных холодах и интенсивной инсоляции
эго может привести даже к летальным повреждениям. Иссушающее дей-
ствие холода усугубляют усиливающие транспирацию зимние ветры.
А уменьшает зимнее иссушение сокращение транспирирующей поверх-
ности, что и происходит при осеннем сбрасывании листьев. Очень силь-
но транспирируют зимой зимне-зеленые растения. Р.Трен (1934) опре-
делил, что в окрестностях Гейдельберга безлистные побеги черники
(Vaccinium myrtillus) транспирировали в три раза интенсивнее, чем хвоя
елей (Pice а) и сосен (Pinus). В 20 раз интенсивнее была транспирация
вереска (Calluna vulgaris). А сохраняющиеся живыми до зимы на стенах
домов побеги льнянки (Linaria cymbalaria) и Parietaria ramiflora испа-
ряли в 30—50 раз интенсивнее древесных видов. В некоторых местооби-
таниях зимняя засуха может быть существенно ослаблена. Например,
растения, находящиеся под снегом или в расселинах стен, значительно
меньше расходуют влаги на транспирацию и во время оттепелей могут
восполнять дефицит воды.
Формообразующее действие холода. Явным морфологическим
проявлением действия холода на растения считается нанизм (от
греч. nannos — карлик). Это свойство характерно для многих тра-
вянистых многолетников, кустарничков и кустарников полярных
и высокогорных областей. В некоторых родах выработался целый
ряд карликовых видов и форм (например, среди ив полярная —
Salix polaris, арктическая — S. arctica, травяная — S. herbacea).
Эти растения часто имеют высоту не более нескольких сантимет-
ров, сближенные междоузлия, мелкие листья. Рост в толщину их
гоже незначителен (например, у можжевельника — Juniperus тол-
щиной 8 см было насчитано 544 годичных кольца). Высота карли-
ковых растений часто соответствует глубине снежного покрова,
под которым они зимуют, так как выступающие над снегом орга-
ны вымерзают и высыхают. Но в образовании карликовых форм
сметную роль играют также торможение фотосинтеза низкими
температурами и бедность почв в результате подавления холодом
микрофлоры. Независимо от причины формирования карлико-
вость дает растениям преимущество в условиях низкой темпера-
туры. Они находятся в припочвенном слое, наиболее прогревае-
мом летом, а зимой защищенном снегом и получающем неболь-
шой дополнительный приток тепла из почвы.
Хорошо адаптированы к холоду стелющиеся растения. Пере-
ходя к горизонтальному росту, древесные растения образуют стла-
никовые формы. Стланики часто встречаются в областях, где дре-
весные породы находятся на границе своего распространения,
например на границе леса. Они известны у сосен (кедровый стла-
ник — Pinus pumila), можжевельников (Juniperus communis, J.
sibirica, J. turkestanica), рябины (Sorbus) и др. Ветви их расплас-
гапы по земле и не поднимаются выше снежного покрова. Иног-
195
да это результат отмирания ствола и разрастания нижних ветвей (у
ели — Picea), а иногда это рост лежащего дерева с укоренившим-
ся во многих местах стволом и приподнятыми ветвями (кедровый
стланик) (рис. 8.6, 7.18). В ряде случаев стланиковая форма явля-
ется видоизменением обычной формы роста древесного вида в
экстремальном местообитании. Так, туркестанский можжевель-
ник (J. turkestanica) в лесном поясе гор имеет обычную древо-
видную форму роста, а в субальпийском поясе принимает форму
стланика. Но есть и стланики, выделившиеся в самостоятельный
вид, например кедровый стланик — родственник горной сосны
(Р. mugo) (Т. К. Горышина, 1979). В экстремально холодных усло-
виях стелющимися могут стать разные кустарники, кустарнички
и даже лишайники. Так, на скалах Антарктиды стелющиеся слое-
вища образуют кустистые виды лишайников.
К числу жизненных форм, способствующих выживанию расте-
ний в холодных местообитаниях, принадлежит и подушковидная
форма, образующаяся в результате усиленного ветвления и край-
не замедленного роста побегов. У этих растений мелкие листья и
цветки расположены по периферии, а между ветвями скапливает-
ся мелкозем, в результате чего некоторые подушки (смолевка бес-
стебельная — Silene acaulis, проломник — Androsace Helvetica,
Azorella и др.) приобретают большую плотность. Благодаря ком-
пактной структуре они успешно противостоят холодным ветрам.
Поверхность подушки нагревается почти так же, как почва. Отме-
чены и случаи существенного повышения температуры внутри них
благодаря аккумуляции тепла в своеобразном «парнике».
Например, у лапчатки двуцветковой {Potentilla biflora) на Алтае сред-
няя температура внутри подушки оказывается выше средней температу-
Рис. 7.18. Стелющаяся форма ивы сетчатой {Salix reticulata) в тундре
(по В. В. Алехину, 1950)
196
ры воздуха на 2 °C. После морозной ночи температура внутри ее круп-
ных подушек составляла 7,5 °C. А у распространенного в высокогорьях
Тянь-Шаня Dryadanthe tetrandra зафиксирована температура 23 °C при
тепловом фоне воздуха 10 °C. Подушковидность существенно снижает
колебания температуры между ветвями. Так, на глубине 10 см в подуш-
ках лапчатки двуцветковой и остролодочника (Oxytropis tragacanthoides)
температурные колебания составили 3 — 3,3 °C, в то время как темпера-
тура воздуха изменилась на 12—13,7°C (И.В.Волков, А.С.Ревушкин,
2000).
Морфолого-анатомические признаки, свойственные арктиче-
ским и альпийским видам (нанизм, мелколистность, подушко-
видность, стланиковость и др.), способствуют перенесению рез-
ких колебаний температур. А признаки, присущие многим расте-
ниям холодных областей, развиты и у видов, произрастающих в
сухих и жарких местах (образование мощной пробки, выделение
смол, низкорослость, сильное опушение, плотные почечные че-
шуи, глянцевые листья и т.д.). Эти признаки — в основном, след-
ствие экономного водного режима, а не собственно экстремаль-
ных температур. В арктических условиях гибель частей растений,
возвышающихся над снегом, происходит не только от холода или
снежной корразии, но и от зимнего или весеннего иссушения,
усиливающегося действием ветра. Аналогично и у низших расте-
ний нет специальных морфологических приспособлений к пере-
несению экстремальных температур. Формообразующее действие
тепла практически невозможно вычленить из действия всего ком-
плекса природных факторов.
Стимулирующее воздействие на растения низких температур.
Холод оказывает стимулирующее влияние на развитие растений,
переход их к росту и репродукции. Необходимая сумма низких
температур варьируется в зависимости от вида растения и его
физиологического состояния.
Холодовое воздействие необходимо для развития почек боль-
шинства древесных растений умеренного климата (для выхода их
из глубокого покоя). Пониженная температура инициирует се-
юнные изменения ультраструктуры клеток апикальной меристе-
мы почек. Как показали опыты с тополем (Populus euroamericana),
в середине января — начале февраля (после выхода из состояния
покоя) в пластидах этих клеток начинают активно формироваться
нроламеллярные тельца. А в оранжерее этих изменений у расте-
ний нет и почки у них в срок не распускаются. Таким образом,
возобновление роста, связанное с формированием проламелляр-
ных ультраструктур, зависит от стимулирующего влияния низкой
температуры.
Низкие температуры — необходимое условие и для развития
уже сформированных многолетних органов. Они могут вызывать
процессы, не связанные с формированием механизмов устойчи-
197
вости клеток к зимним условиям. Так, длительное воздействие
низких температур — пусковой фактор зимней стадии биосинтеза
хлорофилла и обновления фотосинтетического аппарата. Осеннее
похолодание стимулирует деление хлоропластов, в результате чего
число их увеличивается (Е. А. Мирославов, Н. К.Котеева, 2002).
Некоторые виды нуждаются в низких температурах для того,
чтобы перейти к цветению и полностью завершить жизненный
цикл. Холод часто необходим для стимуляции образования цве-
точных почек. Стимулирующее влияние его сказывается также на
начальных этапах развития озимых. Например, высеянная весной
озимая пшеница (Triticum aestivum) не зацветет и может погиб-
нуть при весенних морозах. Но если ее влажные наклюнувшиеся
семена подвергнуть воздействию температур, близких к замерза-
нию, они и при весеннем посеве дадут неплохой урожай. Озимым
растениям необходим короткий период воздействия холода после
прорастания семян. Такой тип влияния низкой температуры на-
зывается яровизацией или вернализацией (приведением «в весен-
нее состояние»). Но для человека это может быть и нежелатель-
ным: двулетники после поздних заморозков могут завершить жиз-
ненный цикл в первый год, и капуста (Brassica oleracea), напри-
мер, не завяжет кочана.
В действии низких температур для энергичного прорастания
нуждаются семена некоторых растений. Этот процесс называется
стратификацией. Семена ряда древесных пород (лиственница —
Larix, липа — Tilia, ясень — Fraxinus и др.) могут прорасти толь-
ко после нее. Стратификация осуществляется во влажном состоя-
нии при близких к 0 °C положительных температурах. У некото-
рых видов рост зародышевого корешка стимулируется холодом
первой (после прорастания) зимы, а рост эпикотиля — второй.
Стимулирующее воздействие низких температур используется
при интродукции растений. Если вид продвинуть далеко в высо-
кие широты, он рано попадает под охлаждение, и его рост может
возобновиться в еще опасный по температуре период. А будучи
перенесен далеко в низкие широты, он может подвергнуться ох-
лаждению только в конце зимы, и его развитие задержится.
Зимний покой растений. В умеренном поясе годовой ход темпе-
ратур имеет четко выраженный холодный период, когда активная
жизнедеятельность невозможна. Соответственно у пойкилотерм-
ных организмов в годичном цикле чередуются фазы активности и
покоя. Однолетние растения переживают холодный период в виде
семян, а у многолетников многолетние органы переходят в состо-
яние покоя. Таким образом, у растений два основных физиологи-
ческих состояния — вегетирующее и зимостойкое (Л.Г.Косулипа
и др., 2003). Активный рост и зимостойкость несовместимы. Вхож
дение растений в состояние зимнего покоя — одно из условий
формирования их устойчивости.
198
Покоем принято называть такое состояние растений, при ко-
тором у них отсутствует видимый рост. Покоящиеся органы ды-
шат, в них идет превращение запасных веществ, но скорость про-
цессов очень мала. В период покоя растения пребывают в таком
физиологическом состоянии, которое позволяет им с наимень-
шими потерями пройти через неблагоприятный период. Так, ли-
стопад и отмирание надземных неодревесневших органов сокра-
щают расходы на дыхание и спасают растения от зимней засухи,
уменьшая транспирирующую поверхность. А для того чтобы у
зимующих органов максимально сократились отдача воды и об-
мен веществ, в период зимнего покоя их устьица находятся в зак-
рытом состоянии, в клетках уменьшаются содержание свободной
воды и проницаемость цитоплазмы, сильно снижается интенсив-
ность дыхания и скорость окислительно-восстановительных про-
цессов, резко падает количество ростовых веществ, инактивиру-
ется фотосинтетический аппарат.
В период зимнего покоя накопленное органическое вещество
почти не расходуется, поэтому, например, долгую полярную ночь
северные растения переносят без ущерба. В целом у зимующих
растений существенно понижен обмен веществ и ограничено вза-
имодействие со средой, что делает их менее чувствительными к
повреждающим воздействиям. Поэтому в покое растения облада-
ют повышенной устойчивостью к действию разных экстремаль-
ных факторов (холод, жара, сухость и др.). В состоянии покоя они
хорошо адаптированы и к перенесению долгих и сильных моро-
юв. В холодных районах почки деревьев и хвоя успешно пережи-
вают суровые морозы ниже -40 °C. В опытах покоящиеся семена
и споры выдерживают краткое охлаждение до -197 °C, а ветви де-
ревьев — до -200 °C. Но весной, когда состояние покоя прекра-
щается, устойчивость растений резко падает. Так, в Европе у ли-
ственных пород зимующие почки легко переносят температуры
до -30 °C, а распускающиеся могут погибнуть при -3 °C. Поэтому
гак опасны заморозки поздней весной.
В покоящихся растениях происходят существенные изменения.
В первом периоде никакие воздействия не заставят растение на-
чать развитие. Если осенью или в начале зимы поставить в воду
срезанную ветку дерева или выкопать травянистый многолетник
(например, луковицу тюльпана — Tulipa) и поместить их в тепло,
го почки не раскроются очень долго (у некоторых вообще не рас-
кроются). В это время растения не вывести из состояния покоя и
другими воздействиями — химическими, механическими и др.
1акой период жизни организмов называется периодом органи-
ческого (глубокого) покоя. Однако с конца декабря — начала ян-
варя покой становится менее глубоким и почки быстрее трогают-
ся в рост, а с конца января они распускаются уже через 2 — 3 дня.
Таким образом, на втором этапе зимы глубокий покой заканчива-
199
ется, и растения готовы к возобновлению развития, но из-за низ-
ких температур оно пока невозможно. Поэтому вторую фазу зим-
него покоя называют вынужденным покоем. Фаза глубокого по-
коя выражена не у всех растений. Некоторые весь холодный пе-
риод находятся в состоянии вынужденного покоя. Так, ряд трав
продолжает развитие теплой осенью и при перенесении в теплое
помещение зимой сразу начинают рост. Обычно это теплолюби-
вые растения, и от осеннего возобновления вегетации их предох-
раняют высокие температурные пороги начала развития.
Для разных видов и сортов требуется различная длительность
покоя. Однако сопоставление сроков наступления вынужденного
покоя с ходом температур показывает, что у большинства расте-
ний умеренных широт глубокий покой заканчивается уже в сере-
дине зимы, когда стоят самые сильные холода. Это экологически
оправдано: глубокий покой необходим в начале холодного перио-
да, когда температурный режим неустойчив и потепления могут
спровоцировать рост и развитие растений, которые будут обрече-
ны на гибель при неизбежном дальнейшем похолодании. Поэто-
му адаптивный смысл глубокого покоя заключается в защите рас-
тений от преждевременного развития. А в середине зимы, когда
устанавливаются устойчивые холода, необходимость в глубоком
покое отпадает, так как теперь от возобновления развития расте-
ния надежно предохранены низкими температурами. Поэтому у
большинства растений умеренных широт переход от глубокого
покоя к вынужденному совпадает со временем установления ус-
тойчивых низких температур.
Подготовка к состоянию покоя происходит заблаговременно.
Задолго до холодов начинаются пожелтение и сбрасывание ли-
ствы, одревеснение и опробковение побегов, закаливание; у вод-
ных растений зимующие почки (гибернакулы) опускаются на дно.
Сигнал к запуску этих процессов — сокращение длины дня. Вы-
ход же из состояния покоя (начало сокодвижения, раскрывание
почек, отрастание побегов, бутонизация и т.д.) совпадает с дей-
ствием весеннего повышения температуры, т.е. отмечается гораз-
до позже окончания периода глубокого покоя. Сигнальным фак-
тором к окончанию покоя служит повышение температуры, а нс
удлинение фотопериода, так как большинство растений в это время
находится в безлистном состоянии, а фотопериодическая инфор-
мация воспринимается листьями.
Действие на растения высоких температур
Реакция растений на высокую температуру тесно связана с со-
стоянием их обводненности и включает изменение многих об-
менных процессов. В онтогенезе растений отмечаются периоды,
200
более чувствительные к высоким температурам и водному дефи-
циту. Как правило, они совпадают с периодом наиболее интен-
сивного роста и формирования гамет.
Перегрев может вызывать водный дефицит и характерные для
него изменения у растений, но высокая температура и сама по
себе приводит к гибели из-за повреждения мембран, инактива-
ции и денатурации белков. При перегреве в растении обычно па-
дает активность биосинтеза и усиливаются гидролитические про-
цессы. Даже если из строя выходят немногие термолабильные
ферменты, то это ведет к гибели клеток от расстройства обмена
нуклеиновых кислот и белков. Растворимые азотистые соедине-
ния накапливаются в таких концентрациях, что диффундируют
из клеток. Образуются ядовитые продукты распада, которые больше
не обезвреживаются: например, могут гидролизоваться белки до
аммиака, вызывающего сильное отравление клеток и гибель рас-
тений (П.А. Генкель, 1982). Но при этом у жаростойких видов
увеличивается также содержание органических кислот, связыва-
ющих избыточный аммиак.
Характерно, что большинство растений повреждается темпе-
ратурами немного выше 30 °C, хотя ферменты и органеллы, выде-
ленные из растений, при таких температурах обычно не повреж-
даются. Вероятно, при перегреве быстро распадаются белково-
липидные комплексы и изменяется структура мембран. Чувстви-
тельность мембран клеток и органелл к изменениям температуры
объясняется плавлением или затвердеванием жирных кислот в их
фосфолипидах. Известно, что при высоких температурах расте-
ния синтезируют больше насыщенных жиров с более высокой тем-
пературой плавления (Л. Г. Косули на и др., 1993).
Другая причина повреждения клеток при перегреве состоит в
гом, что при повышенных температурах некоторые биологически
активные вещества — необходимые для роста витамины, амино-
кислоты, гормоны — быстро разрушаются или не образуются в
нужном количестве. Поэтому, зная химическую природу инду-
цированного температурой замедления роста, можно улучшать
состояние растений внесением нужных веществ (А.Гэлстон и др.,
1983).
Высокие температуры существенно изменяют энергетический
обмен, нарушая баланс ассимиляции и усиливая дыхание при сни-
жении фотосинтеза. Многие растения переносят потепление хуже,
чем похолодание, так как холод приостанавливает вегетацию, а
гспло принуждает растение к иному, неприемлемому балансу. Так,
в ботанических садах субтропические растения, чей оптимум тем-
ператур около 15 °C, легко переносят похолодание до 10 °C и даже
до 5 °C, но температура в 20 °C может привести к их чрезвычайно-
му угнетению. По С. М. Разумовскому (1999), в эволюции биосфе-
ры потепление зимы могло быть тем катастрофичным фактором,
201
который привел к гибели многие субтропические растения и со-
кратил площадь древнейших лавровых лесов.
На Земле многие виды обитают в климатических условиях с
высокими температурными показателями. Пройдя в этих местах
долгий эволюционный путь, они приспособились к такой среде,
и высокие температуры во многих случаях их существенно не по-
вреждают. Растения же, обычно обитающие в более мягких, теп-
ловых условиях, жарой повреждаются сильнее. В умеренном кли-
мате от высоких температур иногда страдают сочные плоды (ви-
нограда — Vitis v in if era, яблони — Matus domestica), получающие
«солнечные ожоги». У деревьев умеренной зоны жарой могут по-
вреждаться сеянцы, иногда испытывающие ожоги коры (особен-
но весной). Растения, выросшие в тенистых условиях, страдают
от жары сильнее, чем растения светлых, сухих и прогреваемых
местообитаний. «Солнечные ожоги» зимой и весной могут быть у
хорошо освещенных стволов деревьев. Так, прямыми наблюдени-
ями установлено, что при температуре воздуха около 0° темпера-
тура камбия на солнечной стороне ствола может повышаться до
30 °C. Характерно, что виды из мест с более холодным климатом
повреждаются уже сравнительно невысокими температурами. Так,
органы растений умеренной зоны обычно должны повреждаться
при температуре воздуха между 40 и 55 °C, которая в этой зоне
наблюдается редко, однако термальная гибель листа в умеренном
климате — нередкое событие, так как если транспирационное
охлаждение по каким-либо причинам снижено, он довольно быс-
тро нагревается и при более низких температурах (И. М. Культиа-
сов, 1982).
Растения получают тепловые повреждения и при пожарах. Кро-
ме обычных ожогов, огонь может затронуть корневую шейку и
вызвать отмирание поверхностных корней из-за перегрева почвы.
Термостойкость растений
Составляющие термостойкости растений. Приспособительные
механизмы, обеспечивающие противостояние неблагоприятным
температурам, могут действовать на всех уровнях организации —
молекулярном, клеточном, организменном и популяционном.
Термостойкость — это способность организма переносить жару
или холод без необратимого повреждения. Она складывается из
двух составляющих. Во-первых, это выносливость (или толеран-
тность) — способность цитоплазмы переносить экстремальные тем-
пературы в силу собственных физико-химических свойств. А во-
вторых, это избегание — комплекс имеющихся у растения защит-
ных приспособлений, которые снижают вредное действие факто-
ра, замедляют или предотвращают развитие повреждений. Термо-
202
стойкость изменяется в онтогенезе. Проростки, растущие весен-
ние побеги не способны закаливаться (повышать свою термостой-
кость) и поэтому чрезвычайно чувствительны к неоптимальным
температурам. Но в онтогенезе организм приобретает способность
изменять метаболизм в ответ на изменения температуры.
Длительное и регулярное воздействие крайне неблагоприят-
ных температур растения выдерживают только тогда, когда жаро-
или морозоустойчива сама цитоплазма. Это свойство связано с
синтезом белка и ряда протекторных веществ и обусловлено гене-
тически, поэтому у разных видов и сортов выражено в разной
степени, а также зависит от репрессирования и активирования
генов (рис. 7.19). Исходную термотолерантность у растений фор-
мируют конститутивные реакции биосинтеза, происходящие как
при экстремальных, так и при нормальных температурах. Они
определяются генетическими возможностями организма. Инду-
цированные же ответные реакции проявляются только под дей-
ствием индуктора (гипо- или гипертермии) и вызывают измене-
ние термотолерантности у менее устойчивых растений. Процессы
индукции тоже контролируются геномом и зависят от функцио-
нирования регуляторных механизмов (Л. Г. Косулина и др., 1993).
Растения с развитыми механизмами избегания перегрева или
холода обычно менее выносливы к морозу и жаре по сравнению с
видами, не имеющими этих приспособлений. Компоненты тер-
мостойкости могут компенсировать и дополнять друг друга. На-
пример, жаровыносливость зависит от интенсивности транспира-
ционного охлаждения: чем оно ниже, тем цитоплазма жаровы-
носливее. Так, у средиземноморских растений с разной интен-
сивностью транспирации различие жаровыносливости достигает
12°.
При экстремальных температурах для растений очень важно
сохранить достаточный уровень энергообеспечения роста и репара-
ции возможных повреждений, что определяется интенсивным
дыханием. На примере памирских растений (горец живородя-
Рецепторы
।
I
Индуктор
Репрессор
Температура
Геном
иРНК
• • * Полирибосомы
Внутриклеточные структуры
Макромолекулы
Биохимические реакции
Конститутивный
синтез
Индуцированный
синтез
Рис. 7.19. Влияние температуры на внутриклеточные процессы
и системы (по Л. Г. Косулиной и др., 1993)
203
щий — Polygonum viviparum, первоцвет памирский — Primula
pamirica и др.) показано, что в высокогорьях интенсивное дыха-
ние сохраняется и после действия температур от -6 до -10° или
выше 40 °C. При этом все причины, позволяющие сохранять оп-
тимальную интенсивность дыхания, способствуют выживанию
растений. Показано, что низкие положительные температуры,
индуцирующие рост Холодовыносливости у теплолюбивых расте-
ний, вызывают у них перестройку дыхательной системы. Включе-
ние альтернативного пути метаболизма приводит к расширению
адаптивных возможностей. У устойчивых растений и при сниже-
нии интенсивности поглощения кислорода эффективность дыха-
ния долго сохраняется. Вероятно, это связано с повышенным со-
держанием в мембранах митохондрий ненасыщенных жирных
кислот, позволяющих сохранять их эластичность и при низких
температурах.
Важный фактор защиты растений от неблагоприятной темпе-
ратуры — возможность сохранять достаточный уровень биосинте-
тических процессов. Так, для многих холодостойких организмов
характерна адаптация фотосинтеза к низким температурам. Она
выражается в изменении липидного состава хлоропластов и росте
термоустойчивости ферментов. Усиление фотосинтеза в этих ус-
ловиях способствует синтезу защитных веществ, помогающих стой-
ким растениям переносить холод. У таких растений изменяется
также азотный обмен и рост клеток (Л. Г. Косулина и др., 1993).
На организменном уровне защита проявляется в опадении или
недоразвитии ряда плодоэлементов и в ускоренном старении ниж-
них листьев. На популяционном уровне термозащита реализуется
в выживаемости особей с более широкой нормой реакции на эк-
стремальный фактор.
Термостойкость разных органов растений. Действия холодов и
жары обычно ограничены по времени, и растения нередко избе-
гают их воздействия, сбрасывая чувствительные части. В ряде слу-
чаев они редуцируют тело до защищенных подземных органов, а
когда неблагоприятный сезон проходит, вновь образуют надзем-
ные части.
Разные органы и ткани растений сильно отличаются по термо-
стойкости. Например, у кукурузы (Zea mays) и гречихи (Fagopyrum
esculentum) от холода быстрее всего отмирают стебли, у риса (Oryza
sativa) — листья, у сои (Glycine) — черешки листьев, а у арахиса
(Arachis hypogaea) — корневая система. Обычно особенно чув-
ствительны к холоду репродуктивные органы, а зародышевый
мешок может погибнуть и тогда, когда завязь еще не повреждена.
Степень устойчивости цветков и их зачатков, семян, а также мо-
лодых растений, как правило, определяет границы расселения вида.
Очень чувствительна меристема конуса нарастания, поэтому боль
шое значение имеет защита почек. А среди тканей побега наибо
204
лее устойчив в закаленном состоянии камбий, который может ос-
таться живым и зимой, когда древесина уже отмирает, и весной,
когда при сильной инсоляции получают ожог кора и хвоя (Г. Валь-
тер, 1974).
Довольно чувствительны к холоду и жаре подземные органы
многих растений. У древесных форм устойчивость одревесневших
участков корневой системы, прежде всего корневой шейки, имеет
решающее значение для выживания. Так, в умеренной зоне ран-
ней весной могут наблюдаться ожоги корневой шейки молодых
дубков (Quercus robur). У травянистых растений лиственных ле-
сов умеренного пояса морозостойкость малозащищенных надзем-
ных почек и перезимовывающих листьев часто гораздо выше, чем
у закрытых подземных органов (корневищ, корней и др.) (рис.
7.20). Разница может достигать 11,5° (Г. Вальтер, 1982).
Разной термостойкостью обладают неодинаково нагреваемые
органы. Например, у сильно транспирирующего листа хлопчат-
ника (Gossypium) предел устойчивости 46°, а у коробочек 50°, что
на несколько градусов выше температуры воздуха. А у медуницы
(Pulmonaria obscura) отмечено различие теплоустойчивости ве-
сенней и летней генерации листьев: у весенних органов она за-
метно ниже. Ветви наружных частей крон сильнее других частей
древесных растений охлаждаются при излучении тепла и развива-
ют более высокую холодостойкость, чем внутренние ветви в кро-
не и находящийся под пологом древостоя подрост. Это относится
и к растениям кустарникового и травяного ярусов, у которых по-
беги и почки становятся более холодостойкими по мере удаления
от почвы и подстилки. Так, в Центральной Европе морозостой-
кость листьев вечнозеленых видов на высоте 3 — 5 см составляет
от -11,5 до -14,5°С, а на высоте 10 — 20 см — от -13 до -20 °C.
Морозоустойчивость почек травянистых растений под подстил-
О Листья о Бутоны ЛИСТЬЯ
Рис. 7.20. Сезонные изменения морозостойкости у печеночницы
(Hepatica) (по Г. Вальтеру, 1982)
205
кой от -7 до -11,5°, а над ней — от -15 до -19,5 °C. Границы же
морозостойкости почек деревьев и кустарников лежат в пределах
от -21 до -40 °C. Устойчивость почек важна для оценки послед-
ствий действия холода: потеря листвы не так опасна, если сохра-
нены почки. Из них наиболее устойчивы спящие и при их вы-
нужденном пробуждении деревья начинают ветвиться.
Термоустойчивость особей разных возрастных стадий неоди-
накова. Так, ткани проростков часто очень чувствительны к экст-
ремальным температурам, что мешает возобновлению деревьев на
гарях и вырубках. Например, проростки бука (Fagus) погибают
при повреждении основания стебля повышенной температурой
оголенной почвы (Г. Вальтер, 1982). Обычно молодые активно ра-
стущие особи с обводненными органами менее устойчивы, чем
старые. Так, у каменного дуба (Quercus ilex) мороз при -4° может
уничтожить однолетний подрост, а наступающие каждую зиму
морозы от -8 до -10 °C исключают его возобновление, хотя взрос-
лые деревья от этих температур не страдают. Для взрослого древо-
стоя губительны только морозы ниже -20°, причем для того чтобы
холод глубоко проник в толстые стволы, необходимо достаточно
продолжительное его действие.
Группы растений по стойкости к охлаждению. По отношению к
жаре и холоду виды растений обладают разной чувствительнос-
тью и неодинаковой закаливаемостью. Мерой устойчивости слу-
жит летальная температура (ТЬ5о), при которой погибает по-
ловина взятых растений. В зависимости от степени и характера
стойкости к охлаждению выделяют три группы растений.
Нехолодостойкие (теплолюбивые) растения серьезно повреж-
даются уже при температурах выше точки замерзания. Обычно
они погибают при положительных температурах ниже 10 °C. Осо-
бенно губительны для них температуры до 4—5 °C, при которых
наблюдается самое плотное состояние воды, ведущее к наруше-
нию обмена. Минимальная температура их прорастания от 10—
14 °C, а всходы появляются при 12—15 °C (Л.Г.Косулина и др.,
1993). К этой группе относятся и водоросли теплых вод, многие
тропические и субтропические виды. Так, у водорослей тропичес-
ких морей нижняя температурная граница лежит в пределах 5 —
14 ° (для сравнения: у водорослей арктических морей верхняя теп-
ловая граница 16 °C). Невысокими плюсовыми температурами по-
вреждаются некоторые сельскохозяйственные растения — выход-
цы из тропиков: рис (Oryza sativa), хлопчатник (Gossypium), фа-
соль (Phaseolus), сорго (Sorghum), арахис (Arachis hypogaea), арбуз
(Citrullus lanatus), дыня (Melo sativus), огурцы (Cucumis sativus),
тыква (Cucurbita pepo) и др. Саженцы тропических древесных
пород гибнут при 3 — 5 °C.
Холодостойкие, но неморозостойкие (умеренно теплолю-
бивые) растения погибают при образовании в тканях льда. Эти
206
виды повреждаются, но не погибают при кратковременных лег-
ких заморозках (до -3 °), и понижение температуры ниже 5 ° пе-
реносят без значительных повреждений. Их всходы появляются
при 8—12°C. От повреждения они защищены только средства-
ми, замедляющими замерзание. В холодное время года у них по-
вышается концентрация осмотически активных веществ, что пре-
дотвращает или замедляет образование льда при температурах
примерно до -7 °C (а при постоянном переохлаждении и до бо-
лее низких температур). В период вегетации неморозостойки все
листостебельные растения. К этой группе относятся многие сель-
скохозяйственные культуры: томаты (Lycopersicon), картофель
(Solanum tuberosum), гречиха (Fagopyrum esculentum), просо
(Panicum), кукуруза (Zea mays), соя (Glycine) и др. В районах их
произрастания возможны кратковременные понижения темпе-
ратур. Чувствительны к образованию льда также глубоководные
водоросли холодных морей и некоторые пресноводные виды, ра-
стения умеренно теплых районов, ряд тропических и субтропи-
ческих древесных пород.
Морозостойкие (льдоустойчивые) растения в холодное вре-
мя года переносят внеклеточное замерзание воды и связанное с
ним обезвоживание. Они выдерживают заморозки до -8...-10 °C.
Семена их прорастают уже при 5 — 6 °C. К ним относятся конопля
(Cannabis sativa), горчица (Sinapis), яровые пшеница (Triticum
aestivum) и ячмень (Hordeum), овес (Avena sativa), горох (Pisum
sativum), люпин (Lupinus), подсолнечник (Helianthus annuus),
свекла (Beta vulgaris), капуста (Brassica oleracea) (Л. Г. Косулина
и др., 1993).
Устойчивыми к образованию льда становятся некоторые на-
емные и пресноводные водоросли, водоросли приливной зоны,
многолетние наземные сосудистые растения областей с холодной
шмой и все мхи. Так, лиственница (Larix) хорошо выдерживает
замерзание тканей и понижение температуры воздуха до -62 °C.
Некоторые водоросли, многие лишайники и древесные растения
способны к чрезвычайно сильному закаливанию, и их можно ох-
лаждать даже до температуры жидкого азота.
Группы растений по жаростойкости. Действие тепла на расте-
ния существенно зависит от его дозы (силы воздействия и про-
должительности): умеренная жара при большой продолжительно-
сти оказывает такое же повреждающее действие, как и кратковре-
менная сильная жара. Поэтому жароустойчивость характеризуют
переносимостью некоторых температур при их получасовом воз-
действии. Если бы вместо этого высокая температура поддержи-
валась в течение часа, то пределы устойчивости лежали бы на 1 —
?’ ниже.
По жаростойкости различают ряд групп растений. Нежаро-
стойкие виды повреждаются уже при 30—40 °C. Это водоросли и
207
подводные листостебельные растения, лишайники в набухшем
состоянии и большинство нежестколистных сосудистых растений.
Они заселяют лишь те местообитания, где не подвергаются боль-
шому перегреву, а более сильную жару выносят только при эф-
фективном транспирационном охлаждении.
Жаровыносливые растения — эукариотические организмы сол-
нечных и сухих местообитаний, как правило, с высокой способ-
ностью к закаливанию. Они переносят получасовое нагревание
до 50 —60 °C. Температура выше 60 °C обычно является границей
жизнеспособности для эукариотических клеток. Наибольшей ус-
тойчивостью к прогреву поверхности субстрата обладают лишай-
ники, особенно эпилитные, что связано с их высокой обезвожен-
ностью. Температурный порог их составляет 92— 101 °C (Л.Г.Ко-
сулина и др., 1993).
Жаростойкие виды — термофильные прокариоты (из фотоав-
тотрофов — цианобактерии). Они обладают особенно устойчивы-
ми нуклеиновыми кислотами и белками. Некоторые из них пере-
носят чрезвычайно высокие температуры (более 80 °C). Но среди
высших растений термофилов нет.
Устойчивость к жаре — очень специфичное свойство, и даже
близкородственные виды заметно различаются по этому призна-
ку. Различия эти можно связать с условиями существования рас-
тений в естественных ареалах. Так, стойкость к жаре листьев мно-
гих тундровых видов составляет 42 °C, таежных 44 °C, а видов жар-
ких полупустынь 47 °C. Также чем суше местообитание, тем выше
температурный максимум растений. Кроме того, жаростойкость,
как и морозостойкость, связаны с обводненностью растения: чем
она меньше, тем жаростойкость выше.
У некоторых видов жаростойкость подвержена сезонным изменени-
ям. Характерно, что у видов умеренных и приполярных регионов она
часто выше зимой и обычно коррелирует со степенью зимнего покоя.
А жаростойкость видов тех регионов, где нет выраженного зимнего по-
коя (средиземноморские растения), выше летом. Так, летом в Арктике
брусника (Vaccinium vitis-idaea) и толокнянка (Arctostaphylos uva-ursi)
переносят нагрев до 48...52°C; вечнозеленые растения умеренной зоны
(ель европейская — Picea abies и пихта белая — Abies alba) переносят
летом температуры до 46°C, а маслина европейская (Olea еигораеа) в
Средиземноморье — до 55 °C. Жаростойкость также хорошо коррелирует
со стадией развития растения.
Так, молодые активно растущие ткани менее устойчивы, чем старые
и покоящиеся. Для многих растений высокие температуры особенно
опасны в период цветения, так как вызывают стерильность цветков и
опадение завязей. Например, у пшеницы (Triticum aestivale) высокая
температура в фазе кущения тормозит дифференциацию колосков, что
приводит к уменьшению числа цветков в колоске и колосков в колосе
(Л. Г. Косулина и др., 1993).
208
Изоляция растений от неблагоприятных тепловых
воздействий
Покровы, экранирующие растения от неблагоприятных тепло-
вых воздействий. К основным приспособлениям, защищающим
растение от тепловых повреждений, относят покровные образо-
вания, задерживающие горячие лучи и изолирующие глубжележа-
щие ткани. Волоски, чешуйки и толстая кутикула предохраняют
мезофилл листа, стебли и почки, слои пробки — флоэму и кам-
бий стволов, пробка или аэренхима — корневую шейку пустын-
ных растений, а зимнее засмоление защищает почки хвойных.
Эти приспособления при опасной инсоляции не пропускают лучи
к тканям. Важны они и как защита от иссушения.
Хвойные деревья с толстой коркой лучше выдерживают высо-
кие температуры пожаров. Так, весьма устойчивы сосны (Pinus),
которые не страдают от таких пожаров, в которых погибают дру-
гие виды лесных растений. А возраст отдельных секвой (Sequoia),
несмотря на регулярные пожары в местах их обитания, насчиты-
вает тысячи лет. Характерно, что многие из этих приспособлений
защищают растения и от мороза, и от засухи.
Во избежание температурного повреждения почки возобнов-
ления закрываются почечными чешуями, смолой, располагаются
в глубине кроны, прячутся под снегом, подстилкой, почвой, во-
дой. В густых кронах и подушках почки, листья и цветки, находя-
щиеся в глубине и ближе к земле, меньше подвергаются опаснос-
ти замерзнуть при отдаче тепла путем излучения. Признак защи-
щенности почек возобновления лег в основу самой известной си-
стемы жизненных форм растений.
В растительном мире есть и примеры морфолого-поведенческих адап-
таций, направленных на кратковременную изоляцию уязвимых тканей и
сохранение тепла в отдельных частях растений. Так, в высокогорьях Во-
сточной Африки и Южной Америки у гигантских розеточных деревьев
(Lobelia, Espeletia) выработалась оригинальная защита от частых ноч-
ных морозов: ночью листья розетки закрываются, защищая конус нара-
стания. При этом у некоторых видов листья снаружи опушены, а у дру-
гих в розетке скапливается выделяемая растением жидкость (ночью за-
мерзает лишь самый верхний ее слой, и конус нарастания защищен от
мороза в своеобразной ванне) (Т. К. Горышина, 1979).
Избегание перегрева путем уменьшения поглощения радиации.
В целом анатомо-морфологические черты, предотвращающие пе-
регрев, в основном те же, что ослабляют приток света к тканям
наземных частей растений, так как при сильной инсоляции ре-
ально опасно повышение температуры растений. Перегреву хоро-
шо противодействуют отражательные свойства поверхности и
уменьшение площади, поглощающей радиацию.
209
В жарких и сухих местах часто проявляется адаптивная редук-
ция листовой поверхности, сокращающая и испаряющую и на-
греваемую площадь. Афилльные формы широко распространены
среди разных групп растений (саксаул — Haloxylon, эфедра —
Ephedra и т.п.). У многих трав и древесных форм элементы по-
бега редуцировались до колючек (акации — Acacia и др.). Умень-
шает поверхность, воспринимающую радиацию, свертывание ли-
стовых пластинок, хорошо выраженное, например, у ряда злаков
(ковыль — Stipa и др.).
Эффективно противодействует перегреву и такое расположе-
ние листьев, когда лучи не могут падать на них перпендикулярно.
Этому способствует вертикальная и меридиональная ориентация
листьев. Например, ряд древесных бобовых при температуре выше
35 °C отвесно складывают листочки сложных листьев и при от-
личном водоснабжении, благодаря чему поглощение радиации со-
кращается вдвое. Не случайно у многих деревьев саванн (мимозо-
вые, цезальпиниевые, протейные и др.) распространены перис-
тые листья. У ряда видов осота (Sonchus) и клевера (Trifolium) в
течение суток изменяется угол наклона листьев, а у тополей (Popu-
lus diversifolia, Р. pruinosa), растущих в поймах среднеазиатских
рек, во время солнцепека листовые пластинки повернуты к сол-
нечным лучам ребром, а утром и вечером — широкой стороной.
Н. Н.Слемнев (2000) описывает поведение в пустыне Гоби травянис-
того многолетника парнолистника (Zygophyllum potanii), избегающего
перегрева в знойные дни путем настических движений: ночью и утром
его побеги распростерты по земле, с повышением температуры почвы
они поднимаются и в полдень располагаются под углом 25 — 30°, а в
самое жаркое время (с 15 до 18 ч) нагретые почти до 50 °C и утратившие
тургор листья складываются.
Защитную роль играет также усиленное отражение падающих
лучей, чему способствует светлая окраска органов и опушения,
блестящие поверхности.
Избегание перегрева охлаждающим действием транспирации. На
испарение воды затрачивается энергия, поэтому этот процесс мо-
жет понижать температуру испаряющего тела. В связи с этим транс-
пирация служит эффективной защитой растений от перегрева.
В опытах неоднократно показано, что листовая пластинка с высо-
кой транспирирующей способностью снижает нагрев многих рас-
тений. Но этот механизм работает только при хорошем водоснаб-
жении. Если же одновременно с высокими температурами насту-
пает водный дефицит, то транспирация тормозится и начинаются
тепловые повреждения растений (В.Лархер, 1978).
Влияние транспирации можно оценить, ограничив отдачу воды рас-
тением. Если прервать доступ воды (перерезав черешок листа или обра-
210
ботав растение тормозящим транспирацию веществом), то через несколько
минут температура листьев повысится. Так, в пустыне у политого арбуза
(Citrullus lanatus) листья почти на 10°C холоднее воздуха. Но у нетран-
спирирующего срезанного листа температура менее чем за час подска-
кивает (на 25°) и становится выше температуры воздуха (на 10 °C), дос-
тигая уровня, при котором наступает повреждение тканей (И. М.Куль-
тиасов, 1982). Таким образом, выжить в жарких местообитаниях ряду
нежаростойких видов помогает усиленная транспирация.
Охлаждение при транспирации способствует тому, что темпе-
ратура листьев пустынных и степных растений всегда на 4 — 6 °C
холоднее окружающего воздуха (иногда и на 15 °C). Из-за силь-
ной транспирации средиземноморских подушечников температу-
ра их поверхности обычно не только ниже температуры воздуха,
но и внутри их создается особый микроклимат (например, влаж-
ность держится на уровне 70 — 80% при влажности наружного
воздуха 30 %).
Роль транспирации в предотвращении тепловых повреждений
может существенно различаться у растений, обитающих на од-
ной территории. Характерные количественные результаты были
получены в Средиземноморье О. и Р.Ланге (1962). При изуче-
нии травянистых растений (Heliotropium europaeum, Convolvulus
althaeoides, Psoralea bituminosa) и орошаемых культур (Solanum
melongea, Melo, Citrullus) оказалось, что интенсивность их транс-
пирации иногда достигает 50 мг/дм2 в мин, что в 5 — 10 раз боль-
ше, чем у местных древесных растений. Из-за транспирацион-
ного охлаждения температура листьев этих трав по сравнению с
температурой нетранспирирующих листьев (смазанных вазели-
ном или отрезанных) оказывается существенно ниже: на 5,7 °C
у Psoralea, на 8,5 °C у Heliotropium, на 13,9 °C у Convolvulus, на
15,7 °C у Solanum. Благодаря этому температура листьев не дос-
тигает 40 °C, что очень важно, так как граница их устойчивости
лежит в пределах 44 —47 °C.
Избегание неблагоприятных условий в покоящемся состоянии.
В период активного функционирования все растения чувствитель-
ны к температурам и переход в покоящееся состояние позволяет
им избегать экстремальных условий при заторможенных процес-
сах жизнедеятельности. В неактивном состоянии избегают холода
и жары подземные органы многолетников, семена и споры, ткани
стволов под толстой коркой.
Экстремальные температуры растение переносит легче при
малом содержании в нем воды, что характерно для многих поко-
ящихся стадий. Так, сухие семена переносят без вреда и отрица-
тельные температуры в -200 °C, и нагрев до 120 °C. Но набухшие
погибают уже при нескольких градусах мороза. Высохшие мхи и
лишайники успешно переносят температуры до 60 °C и более, но
211
устойчивость их резко снижается при увлажнении. Деревья зимой
могут выносить понижения температуры до -35 °C, когда в их поч-
ках и ветвях очень мало воды. Но если даже небольшие морозы
наступят при распускании почек весной, когда в них много влаги,
они погибнут.
Своеобразное физиологическое приспособление к температурам
среды, превышающим адаптивные возможности организмов, —
анабиоз. В этом состоянии растение обезвоживается и почти пол-
ностью приостанавливает жизнедеятельность. Хорошо известна
способность мхов и лишайников переносить длительное промер-
зание в анабиотическом состоянии. Т. К. Горышина (1979) приво-
дит пример опыта, в котором лишайник Cladonia замораживали
более двух лет при -15 °C. После оттаивания фотосинтез и рост у
него возобновились. В крайне холодных условиях у лишайников
периоды анабиоза удлиняются, а активная жизнедеятельность
наступает лишь в краткое благоприятное время и не каждый год.
Однако анабиоз — крайняя мера в борьбе с холодом и жарой, так
как приостановка жизненных процессов приводит к резкому сни-
жению продуктивности. Кроме того, далеко не все эукариоты спо-
собны на обратимую радикальную перестройку своих сложноор-
ганизованных клеток.
Избегать неблагоприятные условия в покоящемся состоянии
способствуют особенности годичного цикла развития эфемеров и
эфемероидов-геофитов. У них вся вегетация может сдвигаться на
время с более благоприятными температурами. Так, в раститель-
ном покрове ряда пустынь и степей есть группы видов, начинаю-
щие вегетацию очень рано весной и завершающие ее до наступле-
ния жары. Летний неблагоприятный период они переживают в
состоянии покоя: эфемеры-однолетники в виде семян (крупка
весенняя — Draba иета, бурачок пустынный — Alyssum desertorum
и др.), а эфемероиды-многолетники в виде подземных луковиц,
клубней, корневищ (тюльпаны — Tulipa, крокусы — Crocus, мят-
лик луковичный — Роа bulbosa и др.).
Прятать от неблагоприятных условий почки в защищенных
местах помогает растениям (пролеске — Scilla sibirica и др.) и
углубление покоящихся частей в почву, в том числе с помощью
сократительных корней. Подсыхая, они сильно сокращаются в
длину (что внешне заметно по их поперечной морщинистости) и
втягивают в почву корневища, луковицы, зимующие почки во-
зобновления. Сократительные корни встречаются у многих расте-
ний высокогорий, тундр, степей. В холодных местообитаниях они
позволяют также противостоять морозному выпиранию из почвы.
А если растение полегло, такие корни еще и ориентируют почку
возобновления перпендикулярно поверхности. Глубина втягива-
ния у разных растений и на разных почвах варьирует, достигая
иногда нескольких десятков сантиметров.
212
Термотолерантность растений
Белки теплового шока. Белки теплового шока (БТШ) синтези-
руются в клетке в ответ на действие экстремальных температур
при одновременном ослаблении синтеза обычных белков. Их син-
тез — пример быстрой адаптации растительных клеток. БТШ со-
ставляют основу индуцированной теплоустойчивости, выполняя
важные защитные, регуляторные и структурные функции. При
большей гипер- или гипотермии, когда БТШ уже не могут обес-
печить сохранность клеточных структур, разрушаются жизненно
важные биополимеры.
На сое (Glycine), рисе (Oryza sativa) и куколе (Agrostemma)
было показано, что при термострессе (как при охлаждении, так и
при нагревании) быстро активируются (или ингибируются) одни
и те же генные локусы. При этом синтезируются одинаковые бел-
ки. Синтез БТШ обнаруживается уже через 10—15 мин после на-
чала перегрева или переохлаждения. Это свидетельствует о быст-
ром изменении синтеза РНК, а именно об образовании специфи-
ческой мРНК, способной транслировать несколько полипепти-
дов. Максимальный синтез стрессовых белков в растительных клет-
ках, как правило, наблюдается в течение 1 — 3 ч. При увеличении
действия экстремальных температур их производство снижается
и восстанавливается синтез белков, происходящий в нормальных
условиях. Полупериод жизни БТШ составляет около 20 ч, в тече-
ние которых клетки сохраняют терморезистентность. Очевидно,
эти белки необходимы на начальных этапах стресса для запуска
адаптивных клеточных процессов (синтез протекторов, измене-
ние компонентов мембран и др.).
Обнаружено много форм БТШ, как высокомолекулярных, так
и низкомолекулярных. Большинство их кодируется в ядре, но есть
и белки, синтезирующиеся в цитоплазме и органеллах (пластиды,
митохондрии и др.). Вместе с мРНК они образуют в клетках рас-
тений стрессовые гранулы (гранулы теплового шока). Вероятно,
роль БТШ в составе гранул теплового шока заключается в защите
от повреждения временно нетранслируемых мРНК, необходимых
для синтеза нормальных белков. Предполагается, что гранулы теп-
лового шока предохраняют внутриклеточные биополимеры от
разрушения. Так, они могут защищать РНК от действия нуклеаз,
активность которых при высокой температуре возрастает. В ядре и
ядрышке БТШ образуют гранулы, связывающие матрицы хромати-
на, необходимые для нормального метаболизма. БТШ, ассоцииро-
ванные с митохондриями, участвуют в регуляции их активности,
способствуя увеличению сопряженности дыхания и фосфорилиро-
вания, а также интенсивности окислительных процессов. Хлоро-
пластные БТШ участвуют в сборке субъединиц ферментов и в адап-
тивных изменениях фотосинтетической активности.
213
БТШ еще могут временно регулировать активность рецептор-
ных молекул, участвуют в нормализации процессов жизнедеятель-
ности клеток в стрессовых условиях, выборочно разрушают на-
капливающиеся при стрессе денатурированные белки (Л. Г. Косу-
лина и др., 1993).
Осморегуляция и изменение температурных показателей физио-
логических процессов как механизм толерантности. Особую роль в
устойчивости растений к неблагоприятным факторам играет обес-
печивающая высокий водный потенциал осмотическая регуляция,
которую многие исследователи относят к механизмам толерант-
ности. В качестве осморегуляторов, которые накапливаются за счет
ассимиляции, часто выступают сахара и аминокислоты.
Один из механизмов биохимической адаптации связан с на-
коплением пролина, содержание которого резко возрастает в ус-
ловиях жары. Он препятствует развитию осмотического шока, так
как поддерживает высокую степень гидратации биополимеров.
У некоторых культур (ячмень — Hordeum, хлопчатник —
Gossypium) концентрация пролина в таких условиях возрастает
более чем в 100 раз. Уровень этого осморегулятора прямо пропор-
ционален интенсивности засухи, поэтому этот показатель можно
использовать для сравнительной оценки засухоустойчивости сор-
тов. Накопление пролина, низкомолекулярных гидрофильных
белков и моносахаров обеспечивает водоудерживающую способ-
ность клеток. Благодаря осмотической регуляции при жаре и за-
сухе у растений сохраняется тургор, и потому возможен нормаль-
ный рост, открытость устьиц и высокая активность синтетичес-
ких процессов. При стрессовых воздействиях уровень осморегу-
ляции видо- и сортоспецифичен и зависит от их напряженности
(В. В. Полевой и др., 2001). В экстремально аридных районах при
суровой жаре и жесткой засухе значение осморегуляции в обеспе-
чении устойчивости растений падает. В этих условиях толерант-
ность зависит в основном от стабильности клеточных мембран
(Д.Левитт, 1980).
Изменение температурных оптимумов и границ физиологиче-
ских процессов — важная адаптация растений к неблагоприятным
по теплообеспечен ности местообитаниям. Так, большое значение
в адаптации растений к холоду имеет сохранение нормальной
жизнедеятельности путем снижения температурных оптимумов
физиологических процессов и нижних температурных границ, при
которых эти процессы возможны. Эти явления хорошо выражены
у растений холодных местообитаний. Например, у альпийских и
антарктических лишайников для фотосинтеза оптимальна темпе-
ратура около 5 °C, а заметный фотосинтез обнаруживается уже
при -10 °C. Оптимум фотосинтеза арктических и высокогорных
растений, ранневесенних эфемероидов также лежит при низких
температурах. Зимой при отрицательных температурах к фото-
214
синтезу способны многие хвойные. Факты повышения темпера-
турных оптимумов известны при адаптации растений к высоким
температурам.
У разных популяций одного вида температурные оптимумы
фотосинтеза часто связаны с изменением условий. Например, у
горных экотипов травянистых многолетников (Oxyria digyna,
Thalictrum alpinum и др.) они более низкие, чем у равнинных.
Показательно также сезонное смещение оптимума фотосинтеза
по мере сезонного изменения температуры. Так же важны и ана-
логичные изменения других физиологических процессов.
Жароустойчивость растений. На тепловой стресс растения реа-
гируют быстрой индуктивной адаптацией. Молекулярный меха-
низм приспособления цитоплазмы к жаре основан прежде всего
на конформационных изменениях и стабилизации структуры бел-
ков. Сочетание прочности и гибкости позволяет им поддерживать
структуру и функциональную активность в крайних условиях. Так
же быстро происходят сдвиги температурного оптимума активно-
сти важнейших ферментов. Благодаря этому у устойчивых расте-
ний высокая скорость ресинтеза белка и стабильные мембраны.
После прекращения жары репарация обеспечивается в первую
очередь генетическими системами. Для этого у растений суще-
ствуют механизмы защиты ДНК (так, она частично выводится из
активного состояния путем блокирования гистонами и стрессо-
выми белками).
Устойчивость растений к экстремальным температурам тесно
связана с их оводненностью. Резкое повышение температуры до
повреждающих значений приводит к обезвоживанию тканей. На-
пример, при действии температуры 45 °C содержание воды в се-
мядольных листьях огурца (Cucumis sativus) уже через сутки сни-
жается вдвое, сопровождаясь изменениями фракционного соста-
ва воды. Так, при действии высокой закаливающей температуры
(40 °C) в листьях томата (Lycopersicon) содержание свободной воды
уменьшается, а связанной — увеличивается. Таким образом, при
закаливании снижение подвижности воды способствует повыше-
нию устойчивости растения. Однако некоторое количество под-
вижных форм воды растение сохраняет для обеспечения транспи-
рации и метаболизма. Разные экологические группы растений
содержат неодинаковое количество такой воды: покрытосемен-
ные ксерофиты — 18 —20, а мезофиты — 35 —50 % (П. А. Генкель,
1982).
У устойчивых к жаре растений существенно изменяется ли-
пидный состав мембран: накапливаются нейтральные липиды,
богатые энергией вещества и фосфолипиды, количественные со-
отношения которых определяют мембранную проницаемость. Для
оптимального функционирования мембран при высоких темпе-
ратурах важно увеличение в составе липидов доли насыщенных
215
жирных кислот. Они влияют на текучесть мембран, имеющую
важное значение для осуществления в клетке катализа, транспор-
та, рецепции и других функций. Поэтому у жароустойчивых рас-
тений изменяется либо число двойных связей в молекулах жир-
ных кислот, либо число типов жирных кислот, включаемых в
фосфолипиды. Чем меньше двойных связей, тем выше может быть
температура, при которой сохраняется необходимая вязкость мем-
бран. В слишком жидком липидном бислое липидная фаза мемб-
ран менее упорядочена, что может приводить к дезинтеграции
структуры мембран и инактивации мембранно-связанных фермен-
тов.
Жаростойкие растения отличаются высокой вязкостью цито-
плазмы и содержанием специальных защитных веществ (слизи,
соли, органические кислоты), препятствующих ее свертыванию и
нейтрализующих токсичные продукты гидролиза. Имеет значе-
ние состав и состояние белков в клетке. По Д. Левитту, при быс-
тром тепловом воздействии изменяется конформация макромо-
лекул: частичное развертывание белков, а потом свертывание в
более термостабильную форму, обогащенную более прочными при
высокой температуре гидрофобными связями. При долговремен-
ных адаптациях к высокой температуре изменяются число и тип
слабых взаимодействий: синтезируются более термостабильные
белки, у которых за счет уменьшения водородных связей (ослаб-
ляемых при повышении температуры) увеличено число гидрофоб-
ных взаимодействий. В результате белок принимает одну из энер-
гетически выгодных конформаций, делающую молекулу более тер-
моустойчивой. Большое значение в формировании жаростойкос-
ти принадлежит также белкам теплового шока.
При экстремально высоких температурах и засухе в раститель-
ных организмах существенно перестраивается гормональная сис-
тема: увеличивается содержание ингибиторов роста (АБК и эти-
лена) и уменьшается содержание его активаторов (ауксина, гиб-
береллина, цитокинина). В целом к жаре и засухе растения адап-
тируются путем последовательной перестройки метаболизма, в
результате чего интенсивность процессов синтеза и гидролиза ус-
танавливается на более низком уровне. Чем выше устойчивость
вида или сорта, тем выше этот уровень и тем быстрее происходит
адаптация.
Обеспечение толерантности растений к низким температурам.
Механизм устойчивости растений к низким температурам обес-
печивается уменьшением обезвоживания и механической дефор-
мации протопласта. Он включает в себя предотвращение образо-
вания льда внутри клетки и повышение устойчивости к межкле-
точному льду. Растворенные вещества понижают точку замерза-
ния растворов, поэтому повышение концентрации клеточного сока
(главным образом за счет растворимых углеводов) — приспособ-
216
ление, препятствующее замерзанию растений. Этому способству-
ет и повышение доли коллоидно-связанной воды. Клеточный сок
обычно замерзает при температурах от -1 до -5 °C, но клетки в
тканях замерзают при более низкой температуре, так как на по-
нижение точки замерзания действуют и другие силы, связываю-
щие воду.
Ряд растений способен сохранять жизнеспособность в промерзшем
состоянии. Есть виды (мокрица — Stellaria media, маргаритка — Bellis
perennis), замерзающие осенью в фазе цветения и продолжающие цвести
весной после оттаивания. Бутоны ложечной травы (Cohlearia arctica,
рис. 7.21) выдерживают зимой температуру до -46 °C и после стаивания
снега нормально развиваются. Ранневесенние лесные эфемероиды не-
однократно переносят ночные заморозки, при которых цветки и листья
покрываются инеем, но уже через 2 —3 ч после восхода солнца возвра-
щаются в нормальное состояние. Лепестки цветков ряда высокогорных
видов при сильном охлаждении ночью иногда тоже замерзают, а днем
без последствий оттаивают. Отмечено, что в подобных случаях возможна
витрификация', клетки переходят в стекловидное состояние, в котором
способны переносить очень низкие температуры (до -200 °C). Витрифи-
кацию обеспечивают криопротекторы, связывающие воду и затрудняю-
щие кристаллизацию. В стекловидном состоянии согласованность реак-
ций обмена сохраняется, но скорость их снижается почти до остановки
Рис. 7.21. Ложечная трава (Cochlearia arctica) (по В. В.Алехину, 1950)
217
(клетки переходят в состояние анабиоза). Обратный процесс (девитри-
фикация) должен проходить очень быстро, иначе внутриклеточная кри-
сталлизация погубит клетку (Л.Г.Косулина и др., 1993).
Вода в клетках способна к переохлаждению: она может охлаж-
даться ниже точки замерзания без образования льда. Однако пе-
реохлажденное состояние неустойчиво и редко сохраняется доль-
ше нескольких часов. Обычно оно возможно лишь при несколь-
ких градусах ниже 0° и очень редко доходит до температур ниже
-20 °C (как в древесинной паренхиме некоторых лиственных де-
ревьев). Вследствие глубокого переохлаждения воды внутрикле-
точного замерзания избегают сердцевинные лучи древесины, за-
чатки цветочных почек у плодовых культур (яблоня — Malus
domestica, груша — Pyrus communis), зачаточные побеги озимых
(Л.Г.Косулина и др., 1993).
Возможность образования переохлажденной воды в клетках
обусловлена разными причинами: особенностями их субмикро-
скопической структуры в закаленном состоянии (гелирование ци-
топлазмы), частичным обезвоживанием, отсутствием центров кри-
сталлизации.
В период вегетации понижение точки замерзания дает единст-
венную надежную защиту от мороза. Она достаточна даже зимой
для вечнозеленых листьев субтропических растений. Закончив-
шим дифференцировку листьям опасность замерзания обычно не
угрожает до температур -3...-5 °C, а к зиме в связи с накоплением
осмотически активных веществ температура замерзания в вечно-
зеленых листьях снижается еще на 2—5 °C. Но молодые листья,
цветки и сочные плоды замерзают уже при температуре -1 ...-2 °C.
У морозоустойчивых растений при действии низких темпера-
тур существенно изменяется функциональная активность мито-
хондрий: режим их работы становится менее интенсивным, но
более устойчивым.
Включение этого аварийного механизма имеет важное биоло-
гическое значение, так как позволяет обеспечить энергией реак-
ции, необходимые для репарации холодовых повреждений. А ча-
стичное разобщение окислительного фосфорилирования при пе-
реходе митохондрий в этот режим приводит еще и к генерации
тепла, освобождающегося при окислении субстратов. В результа-
те температура побегов морозоустойчивых видов оказывается выше
температуры среды. Митохондрии же неустойчивых к морозу ра-
стений либо вообще не переходят в низкоэнергетическое состоя-
ние, либо переходят после длительного действия низких темпера-
тур. У них при значительном охлаждении (ниже -18 °C) ультра-
структура митохондрий нарушается, интенсивность дыхания рез-
ко снижается и фосфорилирование прекращается (Л.Г.Косулина
и др., 1993).
218
Закаливание растений
Формирование морозостойкости растений. В областях с сезон-
ным климатом растения осенью способны переносить образова-
ние льда в тканях. Подготовка к перенесению низких темпера-
тур {закаливание) наступает только после замедления роста. Мо-
розоустойчивость повышают факторы, увеличивающие способ-
ность цитоплазмы не терять жизненных свойств при обезвожива-
нии и механических повреждениях. Весной растения эту способ-
ность утрачивают и могут быстро вымерзнуть в заморозок. Таким
образом, морозостойкость растений сезонно колеблется между ми-
нимальной величиной в период вегетации и максимумом зимой
(рис. 7.22).
Закаливание — временная адаптация цитоплазмы, определяю-
щая меру ее устойчивости к последующим низкотемпературным
воздействиям, и чем ниже температуры закаливания, тем сильнее
морозостойкость. Формирование морозостойкости — сложная
обратимая метаболическая и структурная перестройка растений
на разных уровнях. Способность к закаливанию меняется в онто-
генезе: она тем меньше, чем ближе растение к репродуктивной
фазе. Закаливание — генетически закрепленный признак, реали-
зуемый только при определенных условиях. При этом большую
роль играет жизненное состояние растения: оно может не начать-
ся или быть слабым (рис. 7.23), если задерживался рост или при-
останавливался жизненный цикл под действием засухи, затопле-
ния, холодной весны.
24.1 16.111 7.IV 24.VI ll.VIll 1.Х 23.XI 2.1 12.11 27.111
Рис. 7.22. Сезонные изменения морозостойкости альпийских растений
(по Г. Вальтеру, 1982).
Вверху — кривая температурных минимумов
219
Зима
Лето
Рис. 7.23. Устойчивость органов Loiseleuria procumbens к низким
температурам (по В.Лархеру, 1978).
Обведенные цифры — уровень морозостойкости хорошо закаленных растений,
необведенные — значения морозостойкости растений со слабой закалкой
Процесс закаливания растений обычно делят на два этапа.
Первый проходит на свету при низких положительных темпера-
турах, а второй — при медленном охлаждении при отрицательных
температурах. Закаливание начинается многодневным воздействи-
ем температур чуть выше 0° и первыми заморозками. Низкие тем-
пературы и сокращение фотопериода — сигнал к торможению
роста и стимул для накопления его ингибиторов. Одновременно
уменьшается обводненность клеток и снижается активность фер-
ментов. Однако в первую фазу закаливания идут важные синтети-
ческие процессы, образуются необходимые для перестройки клетки
макроэргические соединения. В этот период наиболее выражен
процесс образования криопротекторов (сахара, белки, нуклеино-
вые кислоты, липиды, гемицеллюлозы и др.).
Благодаря криопротекторам клетка становится более защищенной от
формирования внутриклеточного льда и обезвоживания. Они связывают
воду, образуя гидратные оболочки вокруг молекул, или обволакивают
кристаллы льда и тормозят их рост. Клетки становятся беднее свободной
водой, а центральная вакуоль распадается на множество мелких, и в них
повышается осмотическое давление клеточного сока. Благодаря этому
снижается точка замерзания тканей и цитоплазма оказывается подго-
товленной к следующей фазе закалки.
Вторая фаза закаливания проходит при регулярных слабых
морозах от -3 до -5° (-8 °C). Под воздействием отрицательных
температур лед сначала образуется в межклетниках, куда вода тран-
220
спортируется из клеток. При этом ультраструктуры и ферменты
цитоплазмы перестраиваются так, что клетки получают возмож-
ность переносить обезвоживание. Только после этого растения
могут завершать закаливание, когда вода клеток превращается в
лед. При этом сначала практически прекращается фотосинтез, а
потом, когда вся легкозамерзающая вода превратится в лед, тор-
мозится и дыхание.
Заканчивается закаливание при продолжительном заморажи-
вании или температурах от -10 до -30'С и ниже. В это время
появляется устойчивость к морозному высыханию и ткани без вреда
можно помещать в жидкий азот. При этом замерзшие органы не
погибают при температурах ниже -40 °C и после оттаивания фо-
тосинтез и дыхание у них полностью восстанавливаются. Расте-
ния, прошедшие закалку, значительно легче переносят и морозы,
и другие неблагоприятные условия.
Низкие температуры сами по себе менее опасны для расте-
ний, чем сезоны с аномальным ходом температур (например,
мягкие зимы и весны с поздними заморозками). Оттепель, осо-
бенно в конце зимы, вызывает быстрое снижение морозоустой-
чивости растений. И в середине зимы после нескольких дней с
умеренной или повышенной температурой (от 10 °C) растения в
значительной мере закалку теряют и потом сильно страдают от
морозов. В период оттепелей они утрачивают типичные для вто-
рой фазы закаливания свойства и уровень их морозостойкости
падает. В этом случае цитоплазма опять переходит в первую фазу
закаливания, но пока растения остаются в состоянии покоя, они
при понижении температуры вновь могут дозакалиться.
По окончании зимнего покоя степень закалки и способность к
закаливанию быстро утрачиваются. Весеннее снижение морозо-
стойкости, вызываемое повышением температуры и обусловлен-
ное процессом, обратным закалке, называется изнеживанием.
Молодые листья очень чувствительны к холоду и погибают уже
при небольшой отрицательной температуре, поэтому поздние за-
морозки, наступающие после распускания почек, наиболее опас-
ны. Летом растения к закалке не способны, однако летнее сни-
жение холодостойкости не опасно, так как отрицательных тем-
ператур в это время, как правило, не бывает. Более того, это
снижение холодостойкости необходимо для успешной жизнеде-
ятельности, так как она связана с уменьшением активности ци-
топлазмы.
Тепловая закалка и тепловая настройка растений. Жароустой-
чивость растений подвержена некоторым сезонным изменениям,
но с меньшей амплитудой, чем морозоустойчивость. Они регули-
руются в основном процессами развития растений и немного за-
висят от температуры воздуха. Многие виды умеренной зоны наи-
большую жароустойчивость приобретают в период зимнего по-
221
коя. Наряду с этим встречается дополнительное летнее или толь-
ко летнее повышение жароустойчивости, а у некоторых растений
сезонных колебаний теплоустойчивости нет.
Водоросли отличаются динамичной теплоустойчивостью, об-
ратимо приспосабливая ее к температуре воды. Теплоустойчивость
водорослей максимальна в конце лета, а ниже всего зимой. Годо-
вая амплитуда ее тем шире, чем больше сезонная разница темпе-
ратур воды.
Происходит такая адаптация очень быстро — в течение не-
скольких часов. Например, у литоральных Fucus уже во время
отлива теплоустойчивость повышается по сравнению с теплоус-
тойчивостью во время прилива.
Такую модификацию теплоустойчивости называют тепловой
настройкой.
Другой путь приведения теплоустойчивости клеток в соответ-
ствие с изменившимися условиями характерен для сосудистых
растений. Их теплоустойчивость стабильна, если температура
близка к оптимуму, но повышается при кратковременном дей-
ствии высоких температур. Явление повышения теплоустойчи-
вости к супероптимальным температурам называют тепловой за-
калкой. Благодаря ей растения переносят самые жаркие дни. Ве-
личина закалки измеряется разностью температур, характеризу-
ющих теплоустойчивость объекта до и после нее. Тепловая за-
калка зависит от дозы нагрева. Дающая эффект закалки темпе-
ратура тем выше, чем больше исходная теплоустойчивость. На-
пример, максимальный закаливающий эффект для огурцов (Сиси-
mis sativus) дает температура 38 °C и 43 °C — для хлопчатника
(Gossypium).
Динамика теплоустойчивости хорошо согласуется с ходом тем-
ператур. Так, показано, что в Туркмении у ряда злаков (напри-
мер, у Aristida) и древесных пород (например, у Catalpa) в ре-
зультате тепловой закалки теплоустойчивость повышается в жар-
кие летние месяцы. В жарких пустынях максимальная теплоус-
тойчивость растений проявляется летом после наступления зной-
ных дней. Закаливание к жаре может осуществляться за несколько
часов, поэтому в жаркие дни теплоустойчивость после полудня
выше, чем утром. Но обычно она быстро исчезает, если стано-
вится прохладнее. Однако у акклиматизированных к жаре эко-
типов цитоплазма остается жароустойчивой, если они развива-
лись при постоянно повышенной температуре (Л.Г.Косулина и
др., 1993).
Жароустойчивость может повыситься и под действием отрица-
тельных температур — в тундре и лесотундре. При этом увеличи-
вается и обезвоживание цитоплазмы, и стабильность белковых
молекул, что обусловливает, по-видимому, повышение устойчи-
вости клеток и к жаре, и к холоду.
222
7.5. Термопериодизм и фенологические особенности
действия теплового фактора
Термопериодизм. Важным условием жизни растений являются
периодические изменения факторов среды, которые имеют раз-
личную амплитуду и могут оказывать как субстратное, так и регу-
ляторное воздействие. Периодические изменения климатических
факторов связаны с суточными и сезонными изменениями сол-
нечной радиации. Росту и развитию многих растений благопри-
ятствует смена температур. Термопериодизм — это реакция рас-
тений на смену повышенных и пониженных температур, выража-
ющаяся в изменении процессов роста и развития. Это общая ре-
акция пойкилотермных организмов, способствующая выживанию
и успешному прохождению онтогенеза. Чем ближе к полюсу, тем
ярче выражена термопериодическая приспособленность растений
и тем большие колебания температур становятся потребностью
для многих видов.
Различают термопериодизм суточный и сезонный. Многими
исследованиями отмечена полезность для растений чередования
повышенной дневной и пониженной ночной температур. Восход
и заход солнца вызывают ритмические изменения факторов сре-
ды: утром уменьшается влажность воздуха, увеличиваются осве-
щенность и температура, а после полудня идут обратные измене-
ния. Большинство растений приспособилось к регулярным суточ-
ным сменам температур, и они стали необходимы для их нор-
мального развития.
Переменные температуры стимулируют многие физиологиче-
ские процессы. Так, томаты (Lycopersicon) быстрее и лучше цве-
тут, завязывают более крупные плоды, если дневная температура
26,5 сменяется ночной около 17...20°C. При постоянной темпе-
ратуре 26,5 °C они плодоносят гораздо хуже (Т. К. Горышина, 1979).
Семена многих луговых видов тоже гораздо лучше прорастают при
переменных температурах.
Чередование высоких и низких температур, как и фотоперио-
дизм, регулирует «внутренние часы» растений. Возможно, низкие
температуры повышают активность ферментов, гидролизующих
крахмал в листьях, а образующиеся растворимые формы углево-
дов потом передвигаются в корни и боковые побеги (Н.Н. Треть-
яков и др., 2000). Относительно низкие ночные температуры ус-
коряют рост боковых побегов и корневой системы томатов, повы-
шают урожай клубней картофеля (Solanum tuberosum), сахарис-
тость корнеплодов сахарной свеклы (Beta vulgaris) (Н.И.Якуш-
кина, 1980).
По В.Лархеру (1978), для тропических растений оптимальная разни-
ца между дневными и ночными температурами составляет 3—6 °C, для
223
Рис. 7.24. Области оптимальных температур для роста и развития
различных растений (по В.Лархеру, 1978):
1 — Ceiba pentandra’, 2 — Pinus ponderosa’, 3 — Saintpaulia ionantha’, 4 — Coffea,
5 — Zea mays’, 6 — Zinnia’, 7 — Pinus taeda’, 8 — Mathiola', 9 — Ageratum’, 10 —
Papaver, 11 — Bellis’, 12 — Viola
большинства растений умеренного пояса — 5 — 7° (рис. 7.24), а растения
континентальных областей развиваются лучше, если ночь на 10—15°
холоднее дня. Но это закреплено не у всех растений. Так, сахарный тро-
стник {Saccharum officinarum) и арахис {Arachis hypogaea) хорошо рас-
тут без суточного изменения температуры. А узамбарская фиалка
{Saintpaulia ionantha) и североамериканская желтая сосна {Pinus
ponderosa) предпочитают более высокую ночную температуру.
В процессе эволюции большое значение имела смена времен
года. Она вызывает у растений закономерное чередование перио-
дов активного функционирования и покоя, в которые даже в тро-
пиках периодически погружаются многие растения. С сезонной
сменой температур скоррелированы многие клеточные процессы.
Так, зимой и в начале весны, когда складывается стрессовая ком-
бинация пониженной температуры и яркого света, фотосинтети-
ческий аппарат реорганизуется в сторону светового типа, что су-
щественно снижает вероятность его повреждения.
На многочисленных опытах было показано, что растения уме-
ренных широт для нормального развития нуждаются в понижен-
ных осенних температурах. Продолжительность действия холода
должна быть в среднем не менее трех месяцев, а температура при
этом не выше 3 — 5 °C. В природе растение получает необходимую
дозу холода в начале зимы и, вероятно, это служит сигналом для
перехода их из глубокого покоя в вынужденный. Таким образом,
потребность растений умеренных широт в пониженных осенних
224
и повышенных весенних температурах означает, что для них име-
ет значение не столько общий уровень получаемого тепла, сколь-
ко чередование холодных и теплых сезонов года (сезонный тер-
мопериодизм).
Однако внутрисезонные колебания температуры могут разбаланси-
ровать ход развития растений. Так, чередование морозов и оттепелей
может снять морозостойкость. Поэтому действие переменных темпера-
тур может быть для растений и положительным, и отрицательным в за-
висимости от амплитуды колебаний, их продолжительности и фона воз-
действия.
Длительность вегетационного периода и ритм вегетации. Для
развития растений большое значение имеет длительность перио-
да с температурами, при которых возможно прохождение полного
цикла развития. Вид хорошо акклиматизируется в данном место-
обитании, если полностью использует вегетационный период и
не получает повреждений при наступлении неблагоприятного се-
зона. Если ритм вегетации вида не совпадает с ритмом вегетаци-
онного периода, то это ограничивает его развитие.
Обычно наступление любой фазы развития растений, особен-
но в первой половине вегетационного периода, связано с перехо-
дом температур через некоторый уровень, т.е. оно становится воз-
можным, когда температура воздуха и почвы не опускается регу-
лярно ниже определенного значения. Эти уровни для раноцвету-
щих и высокогорных растений лежат ниже, чем для поздноцвету-
щих видов и видов более теплых мест.
Для большинства древесных пород температурный порог раскрыва-
ния почек находится в пределах 5 — 10 °C, а весенние лесные эфемерои-
ды зацветают при прогреве почвы до 2 —6 °C. В северном полушарии
наступление у вида одной фенофазы при движении на север в среднем
запаздывает на 4 дня на каждый градус широты (примерно на 111 км).
Но на эти сроки часто влияют местные условия. Под Санкт-Петербур-
гом, например, облиствение многих растений начинается тогда, когда
среднесуточные температуры не опускаются ниже 10°, а для Москвы эта
температура составляет 13,8 °C. Листопад начинается примерно при тех
же температурах. Развитие растений часто задерживается облачностью,
обилием осадков, низкими температурами, высотой местности над уров-
нем моря.
Для умеренных широт вегетационным периодом обычно счи-
тают отрезок времени, когда ежедневные средние температуры
превышают 10 °C. Но виды более холодных районов начинают
вегетацию при более низких температурах, а более теплых —
при более высоких. Во второй половине вегетационного перио-
да для наступления фенофаз уже более важны фотопериод и сум-
225
мы температур (продолжительность действия тепла). Для успеха
размножения растений нередко оказывается очень важен и ход
температур.
Так, у ели европейской (Picea abies) в Подмосковье успешное семя-
ношение связано с погодными условиями многих месяцев (М.Д. Мерз-
ленко, В. А. Брынцев, 2000). Сначала это преобладание антициклона в
феврале и марте предшествующего года (холодный февраль и сухой теп-
лый март), когда при большом количестве солнечных дней ростовым
процессам в почках способствует их прогрев. Потом сырой май с мини-
мальными при этой погоде заморозками, сухой и теплый июнь, когда
включаются процессы образования генеративных почек. И далее с июля
предшествующего года влияют все факторы, изменяющие количество и
качество будущего урожая (заморозки, сырость, фитофаги и т. п.).
На сезонные изменения растений влияют и эндогенные фак-
торы. В ряде случаев продолжительность фенофаз оказывается
почти одинаковой независимо от внешних условий. Например,
на обширных территориях лесной зоны и субарктических райо-
нов длительность созревания плодов некоторых растений практи-
чески постоянна (малины — Rubus idaeus — 38 — 40, черники —
Vaccinium myrtillus — 46 — 49, брусники — V. vitis-idaea — 57 —
60 дней). Проявлением эндогенного сезонного ритма являются и
случаи сохранения его в неизменной или неритмично меняющей-
ся среде. Например, в тропиках у яблони (Malus domestica) со-
храняется листопадность. Выращивание пустынных эфемеров и
эфемероидов в районах с умеренным и холодным климатом при
достаточном количестве летних осадков показало, что среди них
также есть виды с эндогенной ритмичностью, в любых условиях
сохраняющие короткую вегетацию и длительный покой (но есть
и виды, становящиеся длительно вегетирующими в благоприят-
ных условиях увлажнения). В целом благодаря сочетанию экзо-
генных и эндогеных факторов сезонный ритм растений синхро-
низируется с климатическим.
Продолжительность фенофаз зависит от современных условий
произрастания и происхождения растений. Виды могут иметь раз-
ные циклы сезонного развития. По этому признаку их разбивают
на группы феноршпмотипов. Параметры, описывающие фено-
ритмотип, — сроки начала и конца вегетации, длительность веге-
тационного периода, периодичность развития листьев, зимнее со-
стояние, время цветения. В районах, отличающихся динамикой
климатических факторов, один и тот же вид может иметь разный
ход сезонного развития и, следовательно, принадлежать к разным
феноритмотипам. Например, в западноевропейских лесах звезд-
чатка ланцетовидная (Stellaria holostea) и ясменник пахучий
(Asperula odorata) вечнозеленые, а в восточноевропейских дубра-
226
нах — весенне-летне-осеннезеленые. Состав феноритмотипов раз-
ных фитоценозов связан с климатом и почвами района, особен-
ностями фитоклимата сообществ и историей формирования рас-
тительного покрова. Так, в тундрах преобладают вечно- и зимне-
зеленые виды, в тайге чаще встречаются летнезеленые и вечнозе-
леные, в пустынях — эфемеры и эфемероиды, а в широколиствен-
ных лесах много летнезеленых растений и весенних эфемероидов.
Фенологическое расхождение растений по времени активной вегета-
ции может служить важным механизмом сосуществования видов. На-
пример, в зарослях таволги (Filipendula ulmarid) может произрастать те-
невыносливый пролесник (Mercurialis perennis), но весной там же раз-
вивается ветреница (Anemone nemorosa), а осенью и весной на опаде
растет мох Brachythecium rutabulum. На известковых лугах сначала раз-
виваются более требовательные к влажности злаки, а в более засушливый
период — глубокоукореняющиеся двудольные (J.P. Grime, 2001). Разно-
временное цветение — тоже проявление снижающей конкуренцию диф-
ференциации экологических ниш.
ГЛАВА 8
ВОДА КАК ЭКОЛОГИЧЕСКИЙ ФАКТОР
По наиболее распространенной в естествознании точке зрения,
жизнь зародилась в океане. Около 3,5 млрд лет назад в воде появи-
лись автотрофные прокариотические организмы. Сначала, вероят-
но, это были хемоавтотрофы, получающие органику за счет энер-
гии окисления химических веществ, потом фотоавтотрофные бак-
терии, способные к бескислородному фотосинтезу, и затем выде-
ляющие кислород цианобактерии (синезеленые водоросли). 2 млрд
лет назад уже сформировалось разнообразие современных циано-
бактерий, среди которых были и многоклеточные нитчатые формы
(с разными типами клеток, ветвящиеся). Около 1,5 млрд лет назад
возникли эукариоты, а 1 млрд лет назад они стали многочислен-
ными. Около 600 млн лет назад появились многоклеточные слое-
вищные растения, давшие начало разным типам водорослей, без
больших изменений сохранившихся до наших дней. И примерно
400 млн лет назад в сложных экологических условиях прибрежных
экотопов сформировались растительные организмы, которые ста-
ли осваивать сушу. Таким образом, жизнь, фотосинтез, многокле-
точность, эукариотичность и растения зародились в воде.
В процессе эволюции живые организмы постепенно освоили
почти всю поверхность Земли. Водные местообитания, где расте-
ния не испытывают затруднений в водоснабжении, очень разно-
образны, но создают для растений ряд экологических проблем,
связанных с тепловым режимом, газообменом, химизмом, осве-
щением и др. На суше достаточно света, кислорода, минеральных
веществ, но критическим фактором при ее освоении была вода.
Процессы водоснабжения — центральная экологическая пробле-
ма наземных растений.
8.1. Вода в растении
Содержание воды в растительных клетках и ее типы. Растения
на 50 — 98% состоят из воды. Даже сухие части их в состоянии
анабиоза (почвенные водоросли, семена, споры) содержат воду.
Без воды невозможен ни один физиологический процесс.
228
Вода в растительных клетках содержится в разных формах: хи-
мически связанная конституционная вода поддерживает состоя-
ние набухания цитоплазмы и других структур, вода в виде раство-
ров присутствует в клеточном соке вакуолей и проводящей систе-
ме растений. Наименьшее количество воды, при котором расте-
ние способно поддерживать постоянство внутренней среды (го-
меостаз), называется гомеостатической водой. У растений тех
или иных экологических групп уровень гомеостатической воды
разный: у видов засушливых мест — 25 — 27, у растений средних
условий увлажнения — 45 — 60, у организмов в условиях обильно-
го увлажнения — 65 — 70% от массы сырого вещества (П. А. Ген-
кель, 1982; Л.Г.Косулина и др., 1993 и др.).
Для характеристики степени насыщенности водой цитоплаз-
мы растительных клеток и целого организма введено понятие гид-
ратуры (оводненности) (Н. Walter, 1931). При насыщенной водя-
ными парами атмосфере гидратура клетки или целого растения
составляет 100%. Она снижается при понижении степени насы-
щения воздуха парами воды или при повышении концентрации
осмотически активных веществ в клеточном соке.
Пойкилогидричность и гомойогидричность растений. В процессе
эволюции у растений возникли разные приспособления к измен-
чивому водному режиму окружающей среды. По отношению к
воде Г. Вальтер предложил разделить растения на две группы: пой-
килогидрические (от греч. poikilos — различный) — переменно-
увлажняющиеся и гомойогидрические (от греч. homois — одина-
ковый) — постоянно увлажненные, способные поддерживать от-
носительное постоянство обводненности тканей (рис. 8.1).
Пойкилогидрические растения способны выносить сильное и
длительное обезвоживание. Устойчивость к нему имеет генети-
ческую основу и служит главным общим адаптивным признаком
пойкилогидрических растений. Соответственно у них имеются ре-
парационные механизмы, восстанавливающие жизнедеятельность
клеток при последующем увлажнении. Способность переносить
обезвоживание более свойственна низшим растениям (эпифит-
ным и почвенным водорослям, лишайникам) и мохообразным.
Продолжительное высыхание выносят даже некоторые постоян-
но обитающие в воде морские водоросли. Но в основном дли-
тельное обезвоживание способны переносить те растения, кото-
рые приспособились к водному режиму своих естественных мес-
тообитаний. Это морские водоросли приливной полосы, обитате-
ли периодически пересыхающих водоемов, эпифитные, эпилит-
ные и почвенные водоросли. Из высших растений периодическое
обезвоживание переносят некоторые виды мхов и папоротников,
а также виды цветковых растений в жарких пустынях.
Водоснабжение пойкилогидрических растений осталось на до-
вольно низком эволюционном уровне: они не регулируют водо-
229
Рис. 8.1. Изменение водного режима растений в связи с приспособлени-
ем их к наземному образу жизни; переход от пойкилогидричности к го-
мойогидричности (по Н.Walter, 1967)
обмен, гидратура их клеток полностью зависит от внешних усло-
вий. Многие виды эпигейных, эпилитных и эпифитных мхов в
разных ботанико-географических зонах и горных поясах настоль-
ко приспособились к неустойчивому водному режиму своих мес-
тообитаний, что пересыхание стало необходимым условием для
их выживания. Так, эпигейный мох, обычный обитатель смешан-
ных и лиственных лесов умеренной зоны Rhytidiadelphus triquetrus,
погибает, если его в течение длительного времени держать сырым
при температуре 37 °C. Способность к быстрому высыханию —
хорошая защита от опасной для него высокой температуры: обез-
воженный мох становится термоустойчивым (Р. Библь, 1965). За-
сушливый период легче переносят также куртинки мхов, а не еди-
ничные особи: медленное высыхание и капиллярное поднятие
влаги в куртинах препятствуют иссушению.
К группе пойкилогидрических растений относятся также не-
которые папоротники и отдельные виды покрытосеменных рас-
тений, способные переносить сильное обезвоживание цитоплаз-
мы и не имеющие морфологических приспособлений для защиты
от иссушения. Эта своеобразная особенность цитоплазмы неко-
торых цветковых растений — вновь приобретенный адаптивный
признак, свидетельствующий о большой экологической пластич-
ности этой группы растений. Так, водное растение Chamaegigas
inrepidus из семейства Норичниковые, живущее в мелких водо-
емах среди раскаленных гнейсовых холмов на юго-западе Афри-
230
Рис. 8.2. Осока пустынная
(Carex physodes)
(по А. П. Шенникову, 1950)
ки, может месяцами находиться в су-
хом состоянии (листья сложены, клет-
ки деформированы), а затем оживать в
течение нескольких минут после сма-
чивания дождем. У пустынной осоки
илак (Carexphysodes) (рис. 8.2), укоре-
ненной в поверхностном слое песка в
Каракумах, листья при иссушении ста-
новятся ломкими (их можно растереть
в порошок), но после смачивания они
приобретают эластичность и зеленеют.
Гомойогидрические организмы свой
водный режим регулируют. Подавляю-
щее большинство наземных растений,
в основном покрытосеменных, в про-
цессе эволюции приспособились к под-
держанию высоких значений гидратур
в нестабильных условиях увлажнения.
Выработаны разные механизмы поддер-
жания гомеостаза метаболических фун-
кций. Адаптации растений к резким
изменениям водного режима имеют генетическую основу и осу-
ществляются на разных уровнях: молекулярном, клеточном, орга-
низменном, биоценотическом. По Р. Библю (1965), суть гомойо-
гидричности растений состоит в их анатомо-морфологических и
физиологических особенностях, защищающих цитоплазму от силь-
ного обезвоживания и создающих для нее свою собственную сре-
ду с более высоким, чем в атмосфере, водным потенциалом.
Экологическое значение осмотического давления. У высших ра-
стений, в первую очередь покрытосеменных, оводненность ци-
топлазмы сбалансирована с оводненностью клеточного сока и
осмотическое давление его может служить мерой гидратуры ци-
топлазмы. Границы гидратуры определяются оптимальным (ти-
пичным для вида) и максимальным значениями осмотического
давления клеточного сока. Чем шире амплитуда между оптималь-
ным и максимальным значениями, тем больше приспособитель-
ных возможностей изменения водного режима имеет растение.
Виды с широким диапазоном между оптимальным и максималь-
ным осмотическим давлением клеточного сока получили назва-
ние эвригидрические. Они могут существовать при значительных
изменениях водного режима или в резко отличающихся условиях
обводненности. Растения с небольшим диапазоном между опти-
мальным и максимальным осмотическим давлением называются
стеногидрическими. Они не выносят резких колебаний условий
увлажнения, возможности расселения их ограничены и для них
характерно локальное распространение.
231
Растения, способные поддерживать свою гидратуру, называ-
ются гидростабильными. Гидростабильность поддерживается раз-
ными путями: развитием мощной корневой системы или путем
ограничения расходов воды на транспирацию. Один из путей ее
достижения — запасание воды в тканях (у суккулентов). Виды с
малыми возможностями поддержания осмотического давления,
меняющимся осмотическим давлением, называются гидролабиль-
ными. Наиболее засухоустойчивы эвригидрические гидролабиль-
ные виды. Засухоустойчивыми могут быть и стеногидрические
гидролабильные виды. Стеногидрические гидролабильные виды
также часто обитают в местах с постоянно высоким увлажнением
(водные, болотные, растения влажных лесов).
Осмотическое давление — индикатор водного режима расте-
ний. Оно подвержено многим изменениям в результате колеба-
ний содержания воды и осмотически активных веществ (колло-
идно-химических изменений) в цитоплазме. Годовой ход осмоти-
ческого давления у разных видов, особенно гидролабильных, чет-
ко отражает меняющиеся условия увлажнения. Осмотическое дав-
ление растет по мере увеличения сухости. При оптимальной влаж-
ности почвы и воздуха гидратура поддерживается беспрепятственно.
Однако затруднение в водоснабжении обычно ведет к пассивному
повышению осмотического давления, затем к изменению струк-
туры цитоплазмы, активному поднятию осмотического давления,
повышению засухоустойчивости на клеточном уровне и далее к
усилению ксероморфной структуры новых органов.
Минимальное, оптимальное и максимальное значения осмо-
тического давления — кардинальные точки гидратуры, свойствен-
ные отдельным видам растений и являющиеся их экологической
характеристикой. Для сравнения осмотических свойств видов и
сообществ разных местообитаний Г. Вальтер (1960) предложил
составлять осмотические спектры (рис. 8.3). Измерение диапа-
зонов осмотического давления клеточного сока листьев показало
низкие значения осмотического давления у кустарников и эпи-
фитов влажных тропических лесов (от 3 до 11 бар). Высоких зна-
чений достигает осмотическое давление у галофитов. У растений
пустыни Каракумы оно в пределах 13,6—58,9 бар. В период засу-
хи уровень максимального осмотического давления повышается.
Наибольшая его амплитуда отмечена у. пустынных эфемероидов.
Осмотическое давление закономерно меняется при онтогене-
тическом и сезонном развитии растений. В благоприятных усло-
виях молодые листья имеют, как правило, более низкие его зна-
чения, что оптимально для ранних стадий развития. По мере раз-
вития листа осмотическое давление постепенно повышается и
достигает значений, оптимальных для вида. Способность подни-
мать осмотическое давление к зиме при накоплении в клетках
сахаров способствует продвижению растений к полюсам. Выше
232
Осмотическое давление, бар
О 10 20 30 40 50
ушшжжжял Водные растения
ушхшжяь Гсофиты
Луговые растения
>
Культурные растения
ушщщшт Сорняки
юяпял Травы влажных лесов
Травы сухих лесов
Хвойные деревья
Лиственные деревья и кустарники
Средиземноморские
древесные растения
'/SSS////SS//7/, 77Л fSSSSSSSSS/SSSs
ДО 60
до 60
Кустарники
Степные растения
тшшШ/шА Пустынные растения
Суккуленты
Мангровые растения
.1 Растения солончаков
|Гало-
। фиты
Рис. 8.3. Осмотические спектры (диапазоны осмотического давления
клеточного сока) листьев растений разных эколого-ценотических групп
(по Н.Walter, 1960):
/ — горец змеиный {Polygonum bistorta)-, 2 — одуванчик {Taraxacum officinale)-, 3 —
подмаренник мягкий {Galium mollugo)-, 4 — тысячелистник {Achillea millefolium)-,
5 — козлобородник луговой {Tragopogon pratensis)-, 6 — мятлик луговой {Роа
pratensis); 7 — дрема белая {Melandrium album)-, 8 — плевел многолетний {Lolium
регеппе); 9 — райграс высокий {Arrhenatherum elatius)
оптимального осмотическое давление может повыситься в суро-
вую засуху.
У разных видов максимальное значение осмотического давле-
ния неодинаково. В пределах вида у закаленных и засухоустойчи-
вых экземпляров его значение выше. При недостатке воды и го-
лодании в случае снижения или прекращения фотосинтеза осмо-
тическое давление может опуститься ниже оптимального. Это
вызывает сильную перестройку цитоплазмы, потребление раство-
ренных запасных веществ и еще большее снижение осмотическо-
го давления. При сильном его снижении растение может погиб-
нуть.
Формообразующее значение гидратуры. Между гидратурой рас-
тений, осмотическим давлением клеток и морфологической струк-
турой их органов существует тесная связь. Морфология листа,
in «атомическое строение и физиологические особенности расте-
ний, выросших в оптимальных условиях (высокая гидратура), от-
нимаются от растений, находящихся в условиях затрудненного
водоснабжения (низкая гидратура). При затрудненном водоснаб-
жении у растений возникают признаки ксероморфизма'.
233
1. Мелкоклеточность, уменьшение размеров органов (как стебля,
так и площади листовой пластинки) и всего организма (нанизм).
2. Устьица мелкие, но на единицу площади листа их много
(обеспечение высокой интенсивности транспирации на единицу
поверхности листа при благоприятном водоснабжении).
3. Утолщение листовой пластинки, при этом сильное развитие
столбчатой ткани, что связано с интенсивным фотосинтезом в
благоприятных условиях; губчатый мезофилл развит слабее.
4. Клетки ксилемы мелкие, в них много механических воло-
кон, общая площадь сосудисто-волокнистых пучков относитель-
но велика. Сеть жилок густая, промежутков между ними немного.
5. Сильное развитие опушения, кутикулы, утолщение клеточ-
ных стенок, что в засуху при закрытых устьицах исключает кути-
кулярную транспирацию.
6. Развитая корневая система, превосходящая по массе надзем-
ную часть.
Растения с ксероморфными признаками устойчивы к завяда-
нию, отличаются ранним цветением и плодоношением, долговеч-
ностью. Признаки ксероморфизма конвергентно развиваются у
разных растений, приспособившихся к засушливым местообита-
ниям. Для видов аридных районов такие структуры генетически
обусловлены. Нов онтогенезе они возникают и у нексерофитных
растений в пределах одного организма или органа.
В начале XX в. В. Р. Заленский (1904) установил закономер-
ность, получившую название «правило Заленского»: выше распо-
ложенные листья побега, развивающиеся в условиях менее благо-
приятного водоснабжения, обладают более ксероморфными ана-
томо-морфологическими признаками. Верхние листья часто мельче
и более рассечены по сравнению с нижними, число устьиц и жи-
лок у них на единицу площади больше. Правило Заленского вер-
но даже в пределах одного листа: верхушка его имеет больше ксе-
роморфных признаков, чем основание. И в пределах кроны дли-
тельно освещенные листья обычно имеют более ксероморфную
структуру, чем затененные.
Как правило, водный дефицит представляет собой результат
совокупного действия факторов. Сходные анатомо-морфологичес-
кие и физиологические реакции, составляющие основу устойчи-
вости растений, могут возникать не только из-за водного дефици-
та, но и по другим причинам: высокая температура, недостаток
кислорода или минерального питания и др.
Поглощение воды корнем. Растение получает воду в основном
из почвы с помощью корневой системы. Масса корней составляет
от 20 до 90% от общей фитомассы. Кончик корня ежесуточно
растет примерно на 5—10 мм. В гумидных зонах рост корней нс
прекращается в течение вегетационного периода, в аридных райо-
нах он прерывается летом лишь на период засухи. Корни облада-
234
куг положительным гидротропизмом', они сами «отыскивают» но-
вые влажные участки почвы. Огромная всасывающая поверхность
корневых волосков, постоянный рост корней, а также микориза
обеспечивают поступление воды из большого объема почвы.
Основные силы, вызывающие поступление воды в растение и
ее передвижение, — транспирация и корневое давление, в резуль-
тате чего возникает разница водных потенциалов и вода передви-
гается осмотическим путем до проводящих элементов ксилемы.
Корневое давление — сила, вызывающая в растении односторон-
ний ток воды с растворенными веществами, не зависящий от транс-
пирации. Корневое давление связано с осмотической и метаболи-
ческой активностью корней. Особенное значение для растений
подача воды с помощью корневого давления имеет в отсутствие
транспирации ранней весной.
На формирование корневой системы и поступление воды в
растение влияют многие экологические факторы (влажность по-
чвы, температура, обеспечение корней кислородом, минеральное
питание растений).
1. Распределение корней в почве в значительной степени зави-
сит от характера увлажнения почвенных горизонтов. Кончики кор-
ней постоянно растут в направлении влажных слоев, активно
отыскивая воду. В аридных районах есть растения с разными кор-
невыми системами (рис. 8.4). У одних она глубокая, достигающая
А
Рис. 8.4. Корневые системы растений аридных мест
(по Т. К. Горышиной, 1979):
I — корневая система верблюжьей колючки (Alhagi camelorum)', Б — корневая
система аристиды (Aristida karelini)\ Z — в профиль; II — в плане
235
зеркала грунтовых вод (солодка — Glycyrrhiza, верблюжья колюч-
ка — Alhagi camelorum). Другие, наоборот, развивают поверхност-
ную корневую систему, которая быстро перехватывает атмосфер-
ные осадки (кактусы, осока илак — Carexphysodes, злак Aristida).
Часто растения с такими корневыми системами растут в одном
фитоценозе, формируя подземную ярусность. В заболоченных ме-
стообитаниях недостаток кислорода приводит к плохому разви-
тию корневых систем. У болотных растений, приспособившихся
к недостатку кислорода, система межклетников составляет еди-
ную вентиляционную систему растения от листьев до корней (см.
разд. 9.1).
2. Рост корней, формирование корневых систем и корневое
питание зависят от температуры. Оптимальная температура для
роста и деятельности корней несколько ниже по сравнению с
оптимальной температурой роста надземных органов, но сильное
понижение температуры почвы тормозит рост корней.
3. Рост корней, как и других органов, зависит от обеспеченно-
сти растения минеральным питанием. Показано, что внесение
фосфорных удобрений способствует росту и углублению корне-
вых систем, а добавка азотных удобрений — их усиленному ветв-
лению (Н.И.Якушкина, 1993).
Корневые системы отличаются большой пластичностью, изме-
няясь в зависимости от условий местообитания (рис. 8.5).
Транспирация. Траты воды, поступившей в растение через кор-
невую систему, прошедшей через тело растения и, наконец, от-
данной в атмосферу в процессе транспирации, очень велики. 97 —
99 % воды, поступившей в растение, идет на транспирацию.
В основном испарение воды происходит активно — через устьица
и некоторая ее часть уходит путем внеустьичной (кутикулярной)
транспирации. На это расходуется большая часть энергии, полу-
чаемой растением. Транспирация тесно связана с анатомо-мор-
фологическими и физиологическими особенностями растений.
Огромные количества воды, которые растения пропускают через
себя при транспирации, обеспечивают непрерывный ток ее от
корней к листьям, связывающий органы и ткани в единое целое.
С транспирационным током передвигаются минеральные и отча-
сти органические питательные вещества. Чем интенсивнее транс-
пирация, тем быстрее идет этот процесс.
Транспирация снимает опасность перегрева, который может
грозить растению на прямом солнечном свету (см. разд. 7.2). Тем-
пература интенсивно транспирирующего листа бывает на несколько
градусов ниже температуры нетранспирирующего завядающего
органа. Такая регуляция способствует созданию оптимальных ус-
ловий для фотосинтеза. Благодаря интенсивной транспирации
растения переносят не только повышенную температуру, но и
атмосферную засуху.
236
Процесс транспирации подчиняется физическим законам ис-
парения и.зависит от внешних условий, но устьичная регулиров-
ка смягчает это влияние. Транспирация тем выше, чем суше ат-
мосфера; при затрудненном корневом водоснабжении она умень-
шается. Движение устьичных клеток зависит от температуры сре-
ды: чем она выше, тем быстрее открываются устьица. Однако при
температуре выше оптимальной они закрываются. При отрица-
тельной температуре устьица также не открываются. Закрывание
устьичных щелей может быть и при переполнении водой окружа-
ющих устьица паренхимных клеток, которые могут механически
сдавливать замыкающие клетки. Это бывает после обильных дож-
дей или поливов и приводит к торможению фотосинтеза.
Транспирация и получение растением диоксида углерода для
фотосинтеза идет одним и тем же путем, через устьица. При хоро-
Рис. 8.5. Корневая система сосны {Pinus sylvestris) (по А. П.Шенникову,
1950):
А — в лесу; Б — на сфагновом болоте
237
шем водоснабжении устьица открыты тем шире, чем сильнее ос-
вещенность. Фотоактивная реакция устьичного аппарата связана
с процессом фотосинтеза: у большинства растений (кроме сукку-
лентов с САМ-путем фотосинтеза) устьица открываются на свету
и закрываются в темноте. Растение постоянно «стоит на грани
голода и жажды»: при недостатке влаги оно должно закрывать
устьица, что сокращает и поступление СО2, а следовательно, и
фотосинтез. При открытых устьицах усиливается транспирация и
поступает СО2, но чрезмерное испарение опасно при недостатке
влаги. Измерения транспирации показали, что это пульсирующий
процесс: увеличение ее приводит к уменьшению содержания воды
в растении, что вызывает сокращение испарения, а это в свою
очередь приводит к увеличению количества влаги в растении, и
транспирация возрастает и т.д.
Транспирацию характеризуют различными показателями. Ин-
тенсивность транспирации — количество испаряемой растени-
ем воды с единицы площади листа за единицу времени (измеря-
ется в граммах на 1 дм2 в час). Эта величина колеблется в преде-
лах 0,15—1,47 г/дм2 ч (Н.И.Якушкина, 1993). Транспирацион-
ный коэффициент — количество воды (в граммах), испаряемой
растением при накоплении 1 г сухого вещества. У засухоустойчи-
вых растений транспирационный коэффициент невысок. Так, у
пшеницы (Triticum aestivum) в зависимости от экологических усло-
вий он колеблется от 217 до 755 (т.е. для накопления 1 г сухого
вещества ей необходимо пропустить через себя 200—750-кратное
количество воды), а для более засухоустойчивого проса (Panicum)
он значительно ниже — 162—447 (Н. И.Якушкина, 1993). Продук-
тивность транспирации — величина, обратная транспирацион-
ному коэффициенту, показывающая количество сухого вещества
(в граммах), накопленного за период транспирации 1кг воды.
Гигроскопические движения растений. Разная гигроскопичность
отмерших и одревесневших тканей может приводить к удлине-
нию или сокращению отдельных органов и их частей. Ветви мно-
гих деревьев и кустарников (ив — Salix, тополей — Populus, кон-
ского каштана — Aesculus, кленов — Acer, вязов — Ulmus, ли-
ственниц — Larix, берез — Betula, ольх — Alnus, можжевельни-
ков — Juniperus, сосен — Pinus, елей — Picea, дубов — Quercus и
др.) способны к значительным движениям этого типа. По анатоми-
ческому и химическому строению древесина верхних и нижних
сторон ветвей отличается, а расположение фибрилл в клеточных
стенках имеет разное направление. При высыхании или набуха-
нии клеточных стенок происходит продольное сокращение или
удлинение древесины. Амплитуда движения ветвей тем больше,
чем больше разница в потере влаги верхней и нижней сторонами.
Гигроскопические движения ветвей имеют важное экологичес-
кое значение для перезимовки растений в суровых условиях тунд-
238
Рис. 8.6. Кедровый стланик (Pinus pumila)
(по Б. А.Тихомирову из А. П. Щенникова, 1950)
ры и лесотундры, а также в крайне засушливых областях. Многие
растения аридных регионов скручиваются или сжимаются в засуху
и разворачиваются в сырую погоду. Благодаря этим движениям стла-
никовые формы благополучно перезимовывают под снегом, избе-
гая воздействия низкой температуры и сильного ветра. Так, у сте-
лющейся формы березы тощей (Betula exilis) при понижении тем-
пературы до -17 °C ветви пригибались на 22 — 25 см. Ветви кедро-
вого стланика (Pinus pumila) (рис. 8.6) зимой прижаты к поверхно-
сти почвы, но летом вершины его побегов поднимаются над яру-
сом трав и кустарничков. Кроме того, с наступлением вегетацион-
ного периода стланики могут распрямляться и выносить ассимиля-
ционную поверхность на высоту до 4—5 м (Г. Э. Гроссет, 1959).
Гигроскопические движения обеспечивают разбрасывание се-
мян при резком раскрывании плодов у многих бобовых, кресто-
цветных и др. Они же способствуют зарыванию в почву зерновок
ковылей (Stipa). А плоды овсюга (Avena fatua) могут самостоя-
тельно передвигаться за счет гигроскопического скручивания и
раскручивания остей. К этому способны также споры хвощей, снаб-
женных гигроскопическими нитями — элатерами.
8.2. Водный режим местообитаний
Атмосферные осадки и коэффициент увлажнения. Для назем-
ных растений основной источник воды — атмосферные осадки.
Количество их зависит от общеклиматических условий, распреде-
239
ление — от рельефа, а конкретные условия увлажнения — еще и
от почвы. Форма атмосферных осадков может быть разная (дождь,
снег, роса и т.д.). Важен также характер распределения их по се-
зонам. Некоторое количество влаги в почве возникает и при не-
посредственной конденсации паров воды из воздуха. Конденса-
ционная влага — одно из важных условий существования расте-
ний в песчаных пустынях. Значительную роль в водоснабжении
растений играют также грунтовые воды: корни многих видов рас-
тений достигают зеркала грунтовых вод.
Очень важна для жизни растений относительная влажность
воздуха (процентное отношение реального давления водяного пара
к давлению насыщенного пара при той же температуре). Она по-
казывает долю (процент) от того максимального количества вла-
ги, которое могло бы содержаться при полном насыщении данно-
го объема воздуха при данной температуре. Растения страдают от
недостатка атмосферной влаги при низких показателях относи-
тельной влажности воздуха. При длительной засухе и суховеях
возникает резкий воздушный дефицит влаги, вызывающий по-
вреждение в первую очередь листьев культурных растений (так
называемый запал).
Снег и град — это вода в твердой фазе, прямо физиологически недо-
ступная для растений, но оказывающая большое косвенное воздействие
на них. Снег особенно воздействует на водный режим территорий в ве-
сенний период. Снеготаяние определяет поемность рек, что влияет на
состав, рост и развитие луговых растений. Тающий снег пополняет вод-
ные запасы почвы. Леса и болота задерживают до 85 % зимних осадков и
медленно их расходуют в течение вегетационного периода, нивелируя
недостаток и неравномерность поступления осадков летом. При инфиль-
трации снеговой воды в почву пополняется уровень грунтовых вод. Из-
морозь и иней также способствуют накоплению воды в почве, так как
при их таянии часть влаги попадает в почву. Отрицательное воздействие
на растения оказывает град. Особенно страдают виды с широкими лис-
тьями, получающие серьезные механические повреждения.
Роса, туман, изморозь и иней в отличие от дождя и снега относятся к
неизмеряемым атмосферным осадкам. Они появляются в результате кон-
денсации водяных паров и тоже могут играть большую роль в жизни
растений. Теплый воздух, как правило, содержит некоторое количество
влаги. При охлаждении его относительная влажность увеличивается. Когда
она достигает 100% {«точка росы»), начинается конденсация паров в
виде капель. Выпадение осадков в виде росы может достигать 10—20%
от годовой суммы осадков. В безводных пустынях за счет этого суще-
ствуют лишайники, поддерживается водный режим листьев ряда видов.
Во влажных тропических лесах это явление важно для существования
многочисленных эпифитов. В ряде горных систем в «туманных лесах»
роса служит источником увлажнения листьев и почвы. В районах, харак-
теризующихся неустойчивыми климатическими характеристиками, она
сглаживает неравномерность выпадения дождей и уменьшает опасность
240
I \
высыхания растений за счет восприятия влаги листьями. Так, после жар-
кого сухого дня прохладная ночь с росой спасает растения от обезвожи-
вания, в утренние часы препятствует быстрому нагреву и задерживает
начало интенсивной транспирации, увеличивая общее содержание воды
в надземной части растений. В центральных областях России продолжи-
тельность выпадения росы составляет около 1000 ч в год. Вместе с тем
систематические обильные росы способствуют заражению растений па-
тогенными грибами.
Величины осадков в разных ботанико-географических зонах и
в разных орографических условиях значительно отличаются. Так,
в восточной части Сахары дождей не бывает по 10 лет, в пустынях
Южной Африки выпадает 25 — 30 мм осадков в год, в сухом кон-
тинентальном климате Приаралья и Прибалхашья — 80 мм. А во
влажном тропическом лесу среднее количество осадков составля-
ют около 2 500 мм в год. В отдельных местах тропической зоны
эти показатели достигают рекордных значений: в прибрежной
полосе на западном побережье Северной Гвинеи выпадает 4000—
5000 мм осадков («Гвинейский климат»), а у подножия гор — до
10000 мм. Каждый миллиметр осадков имеет огромное значение:
I мм осадков означает, что 1 м2 площади получает 1 л воды, т.е.
на 1 га приходится 10 т (!) воды (А. Г. Исаченко, А. А. Шляпников,
1989).
Для характеристики водного режима местности указание толь-
ко количества осадков недостаточно. Так, территории с очень раз-
ными условиями для существования растений, такие как острова
Северного Ледовитого океана, тайга Восточной Сибири, пустыни
Средней Азии, получают примерно одинаковое количество осад-
ков (около 200 мм в год). Поэтому важным экологическим пока-
зателем служит характеристика гидротермических условий. На
каждой территории связь водного и теплового режимов выража-
ется с помощью коэффициента увлажнения, получающегося от
деления среднегодовой суммы осадков на величину испаряемости
с открытой водной поверхности в данных условиях. Территории,
где коэффициент увлажнения больше 1, получили название гу-
мидных, а области с коэффициентом увлажнения менее 1 называ-
ются аридными. В гумидных областях коэффициенты увлажне-
ния разнообразны: в тундрах России этот показатель равен 1,5 —
2,5, на большей части лесной зоны (кроме Восточной Сибири) —
1,33, а во влажных тропиках у подножия гор на побережье Гви-
нейского залива — 5. В аридной же степной зоне он составляет
0,66, а в пустынях и полупустынях Средней Азии — 0,12—0,33.
Зона широколиственных лесов и лесостепь имеют коэффициент
увлажнения около 1.
Неравномерное поступление воды создает для растений ряд
экологических проблем. Так, обильное разовое выпадение осад-
241
ков может вызвать повреждения растений. Последствиями поле-
гания травостоя после сильного ливня могут быть снижение уро-
жая, создание условий для развития грибных заболеваний и т.п.
Неравномерность поступления атмосферных осадков бывает се-
зонной. Так, в умеренной зоне северного полушария основные
осадки обычно поступают весной. Во влажных тропиках, где сред-
ние значения коэффициента увлажнения достигают 3 — 5, выпа-
дение осадков имеет муссонный ритм: в течение 3—4 мес коэф-
фициент увлажнения здесь меньше 1 (до 0,25). Для атмосферных
осадков характерно и неравномерное распределение их по повер-
хности. Так, огромную роль в перераспределении атмосферной
влаги играет рельеф, что особенно важно в районах с дефицитом
влаги.
Водный режим почвы и показатели ее влажности. Вода почвы —
основной источник влаги для растений, зависящей от общекли-
матических условий (поступление атмосферных осадков, конден-
сация паров воды воздуха) и от капиллярной связи с грунтовыми
водами.
Атмосферные осадки богаты кислородом, но бедны минеральными
солями, а грунтовые воды минерализованы, но обеднены кислородом.
Для растений важно, что значительная часть их водоснабжения связана
с грунтовыми водами. Они образуются как за счет атмосферных осадков,
так и за счет глубинных подземных вод. Грунтовые воды обеспечивают
резерв почвенной влаги, устраняющий перебои в поступлении воды. На
почвах с небольшим резервом влаги случаются резкие колебания водо-
снабжения. Так, в таежной зоне на почвах, сформировавшихся на одно-
родных кварцевых песках, лесообразователем может быть только непри-
хотливая сосна обыкновенная {Pinus sylvestris). Для растений важна и
капиллярная влага, удерживаемая в порах почвы: при интенсивном по-
треблении воды растением она перемещается, пополняя запас почвен-
ной влаги.
Водный режим почвы — это совокупность всех поступлений,
перемещений, удержания и расхода влаги за определенный пери-
од. Расход почвенной воды включает испарение, потребление воды
растениями {десукция) и сток (поверхностный, внутрипочвенный,
грунтовый). В России основные типы водного режима почв сле-
дующие (по А. А. Роде).
1. Мерзлотный тип\ почва в течение вегетационного периода
оттаивает на сравнительно небольшую глубину. Под талым слоем
сохраняется постоянно мерзлый водоупорный горизонт. Почвы
обильно увлажнены за счет талого слоя и надмерзлотной верхо-
водки из атмосферной влаги. На мерзлотных почвах успешно раз-
вивается лиственничная тайга.
2. Промывной тип: почвенно-грунтовая толща ежегодно про-
мачивается атмосферными и талыми водами до грунтовых вод.
242
Грунтовые воды, подпитываясь атмосферными осадками, получа-
ют водорастворимые соединения из почвенного профиля. Нисхо-
дящий ток воды преобладает над восходящим. Этот тип водного
режима характерен для гумидных областей и, например, свой-
ствен подзолистым и болотно-подзолистым почвам.
3. Периодически промывной тип', в разные годы водный ре-
жим складывается то по промывному, то по непромывному типу.
Формируется в областях, где коэффициент увлажнения примерно
равен 1, но соотношение между поступающими атмосферными
осадками и испаряемостью очень неустойчиво. Грунтовые воды
располагаются глубоко (за пределами корнеобитаемого слоя). Та-
кой тип водного режима имеют серые лесные почвы широколи-
ственных лесов и лесостепные черноземы.
4. Непромывной тип', почва промачивается на относительно
небольшую глубину. Характерен для аридных областей глубоким
залеганием грунтовых вод. Вся влага, инфильтрованная в почву,
возвращается в атмосферу при испарении и десукции. Ниже го-
ризонта промачивания обычно накапливаются водорастворимые
соединения (гипс, карбонат кальция и др.). Такой режим характе-
рен для степных черноземов и каштановых почв.
5. Десуктивно-выпотной тип', возвращение влаги в атмос-
феру происходит в основном благодаря транспирации. Возникает
в областях с коэффициентом увлажнения меньше 1, где испаряе-
мость значительно превышает сумму осадков, а залегание грунто-
вых вод неглубокое, и капиллярная кайма входит в почвенный
слой. Почвенно-грунтовые воды обычно сильно минерализова-
ны. Этот тип водного режима свойствен лугово-черноземным и
лугово-каштановым почвам.
6. Выпотной тип', влага непосредственно физически испаря-
ется. Возникает в аридных областях при очень близком залегании
почвенно-грунтовых вод (верхняя граница капиллярной каймы
достигает поверхности почвы). При постоянном восходящем токе
воды и испарении содержащиеся в грунтовых водах соли выпада-
ют в основном в поверхностном слое почвы. Такой водный ре-
жим имеют солончаки.
Влажность почвы — динамичная величина. Количество воды в
почве меняется в течение суток, по сезонам и по годам. Постоян-
но меняются и формы воды в почве. Твердая фаза почвы со-
ставляет 40—70% ее объема, а оставшееся пространство может
быть заполнено жидкостью и газами. Вода в почве может нахо-
диться во всех агрегатных состояниях (в виде льда, жидкости, пара)
(рис. 8.7). Кроме того, в почве выделяют следующие формы воды:
I) химически связанную, 2) парообразную, 3) сорбционно-свя-
laimyio (гигроскопическую и пленочную), 4)свободную воду (ка-
пиллярную и гравитационную). Основная форма почвенной вла-
|п, используемая растениями, — капиллярная. Химически свя-
243
Поверхность
Рис. 8.7. Вода в почве
(по Г. В.Добровольскому, 1976):
1 — частицы почвы; 2 — гравитационная вода осадков,
просачивающаяся в почву; 3 и 5 — гигроскопическая (пле-
ночная и коллоидальная) вода; 4 — почвенный воздух с
парами воды; 6 — зона, в которой часть пор заполнена
воздухом, часть — капиллярной водой; 7 — вода капил-
лярная; 8 — все поры почвы заполнены капиллярной
водой; 9 — уровень грунтовой воды; 10 — грунтовая вода
занная, парообразная и сорбционно-связанная
вода для растений недоступна.
Количество воды, удерживаемое почвой,
называется влагоемкостью. Полная влагоем-
костъ — наибольшее количество воды, кото-
рое может быть в почве при заполнении всего
свободного почвенного пространства. Полевая
влагоемкость — это максимально возможный
запас влаги, который удерживает почва после
оттока гравитационной воды и значительного уменьшения скорос-
ти ее нисходящего движения (т.е. количество воды, удерживаемое
почвой против силы тяжести). У разных почв показатели полевой
влагоемкости различаются: от 10 % в легких почвах до 30 % — в
тяжелых.
Резкий почвенный дефицит влаги опасен для растений (особенно
культивируемых, сохраняющих мезофитную структуру). Он получил на-
звание «захвата»: после сравнительно влажных весны и начала лета, ког-
да растения развили большую листовую поверхность, резкая засуха вы-
зывает дефицит водоснабжения, завядание и глубокие нарушения осмо-
тических свойств клеток и обмена веществ. После восстановления гид-
ратуры у «захваченных» растений ферментативное равновесие полнос-
тью не восстанавливается, что отрицательно сказывается на ростовых
процессах и урожае.
При длительной почвенной засухе завядание может быть не-
обратимым. Тот процент влажности почвы, при котором проис-
ходит необратимое завядание, называется влажностью устой-
чивого завядания (или коэффициентом завядания). Помимо ус-
ловий увлажнения она зависит от гранулометрического состава
почвы, ее плотности, засоленности и др. Эта величина наимень-
шая у песков, а наибольшая — у глин. Коэффициент завядания
различен для разных экологических групп (засухоустойчивых, вла-
голюбивых), а также отличается у растений одного вида и даже у
отдельного организма в разные фазы онтогенеза.
Для растений очень важна подвижность воды в почве, так как
с ней связано обеспечение корней воздухом (постоянно движу-
244
щаяся почвенная вода, обеспечивающая растения влагой и кисло-
родом, играет роль «наружного кровообращения растений»). При
медленном опускании почвенно-грунтовых вод корни остаются
обеспечеными водой, так как активно «отыскивают» увлажненные
участки. Но слишком быстрое заглубление вод для растений опасно
из-за отрыва корнеобитаемого слоя от увлажненного горизонта.
Водный режим фитоценозов. В растительных сообществах со-
здается особый фитоклимат. Так, в лесных фитоценозах суще-
ственно перераспределяются атмосферные осадки. Летом они ча-
сто не доходят до поверхности почвы, так как значительная их
часть задерживается кронами (до 75 %) и затем испаряется. Чем
гуще лес, тем больше атмосферной влаги расходуется на смачива-
ние растений. Очень гигроскопична лесная подстилка, впитыва-
ющая большое количество талой воды. Около 85 % зимних осад-
ков, задержанных лесом, попадает на почву и просачивается, по-
полняя грунтовые воды. Лес выделяет большое количество паров
воды в результате транспирации, при этом повышается влажность
лесного воздуха и понижается температура. При пересыщении
воздуха влагой пар конденсируется на ветвях. Повышенная влаж-
ность обусловливает существование лесных трав-гигрофитов.
Лесные массивы влияют на климат обширных территорий.
Благодаря перераспределению влаги внутри лесной зоны обле-
сенные районы всегда относительно более богаты осадками. Кон-
векционные токи из леса выносят воздух более холодный и влаж-
ный по сравнению с безлесной площадью, и над лесными площа-
дями осадков выпадает больше, чем над безлесными.
Способствуя медленному таянию снега, лес сильно уменьшает
наземный сток, препятствуя эрозии почв. При сведении его на
обширных площадях происходит бурное снеготаяние и, как след-
ствие, сильные и краткие разливы рек с ранним наступлением
межени (низкого стояния воды в реках). При уничтожении есте-
ственного растительного покрова и усилении поверхностного стока
возникают проблемы, связанные с эрозией почв. Так, по исследо-
ваниям в Вологодской области старопахотные почвы на склонах
потеряли уже 50—60 см верхнего слоя. Почвенная эрозия способ-
ствует оврагообразованию, размыву речных берегов, усилению
выноса песка и ила в дельты, расширению и обмелению русел,
(’ведение леса в горах приводит не только к увеличению поверх-
ностного стока, но и к изменению климата долин, пересыханию
юрных источников, повышению лавиноопасности.
В луговых травостоях тоже формируется свой микроклимат.
Для одних лугов единственный источник увлажнения — атмос-
ферные осадки, для других — еще грунтовые и (или) полые воды.
Чем больше сомкнут травостой и чем больше его масса, тем боль-
ше он может задержать атмосферных осадков и тем больше влаж-
ность внутри него и над ним. Это создает благоприятные условия
245
для паразитических грибов (ржавчинных и др.), а также для фил-
лосферных азотфиксирующих бактерий.
Особый гидрологический режим формируется на болотах. Боль-
шой гигроскопичностью отличаются сфагновые мхи. При накоп-
лении торфяных толщ, для которых обычна высокая влажность и
недостаток кислорода, формируются внутриболотные водотоки и
особый гидрологический горизонт болотных вод, обусловливаю-
щий увлажнение не только самого болота, но и обширных приле-
гающих территорий. Мелкие болота способствуют формированию
влажного и прохладного микроклимата, обширные — влияют на
климат регионов. Уничтожение болот на больших территориях
ведет к усилению континентальности климата, понижению уров-
ня грунтовых вод и влажности воздуха, изменению паводково-
го режима рек и газового состава атмосферы на прилегающих про-
странствах.
8.3. Экологические группы растений по отношению
к водному режиму
В конце XIX в. А. Шимпер и Е. Варминг предложили различать
три экологические группы сухопутных растений, облик которых
определяется водным режимом их местообитаний: гигрофиты,
мезофиты, ксерофиты. Позже Е. Варминг добавил группу водных
растений. Это подразделение легло в основу последующих более
дробных эколого-морфологических классификаций растений по
водному режиму. По А. П. Шенникову (1950), выделенные группы
представляют собой морфологическое проявление коллоидно-хи-
мических проблем растений.
Гигрофиты. Гигрофиты (от греч. hygros — влажный) — расте-
ния влажных местообитаний. Количество осадков, увлажнение
почвы, относительная влажность воздуха обеспечивают им срав-
нительно благополучный водный режим. У гигрофитов нет необ-
ходимости развивать приспособления, ограничивающие расход
воды. У них слабо развита корневая система, крупные листовые
пластинки, постоянно открыты немногочисленные устьица, тон-
кая кутикула. Преобладает не устьичная, а кутикулярная транс-
пирация. Клетки крупные, с тонкими оболочками, осмотический
потенциал их невысок, механические ткани слабо развиты.
Типичные, или теневые, гигрофиты обитают в сырых теплых
и тенистых тропических лесах. Растения там не испытывают не-
достатка воды в почве и плохо переносят даже колебания влажно-
сти воздуха. В умеренно-холодном климате теневые гигрофиты
развиваются под пологом лесов, где в течение вегетационного се-
зона сохраняется постоянная повышенная влажность приземного
слоя воздуха. У этих растений тонкие листья, слабая способность
246
к регуляции транспирации, они быстро вянут в сухом воздухе.
Таковы кислица (Oxalis acetosella), недотрога (Impatiens noli-
tangere), адокса (Adoxa moschatellina) и др.
Световые гигрофиты растут на почвах, насыщенных или за-
литых водой, в местообитаниях с высокой влажностью приземно-
го слоя воздуха, подвергающегося эпизодическому высыханию.
К светолюбивым тропическим гигрофитам относятся рис (Oryza
sativa), болотная пальма (Phoenixpaludosa) и др. Папирус (Суperus
papyrus), например, образующий на болотах Африки густые за-
росли, испаряет так много воды, что понижает уровень Нила.
Временное подсыхание почвы или воздуха вызывает снижение
жизнедеятельности и завядание гигрофитов. Их корни и стебли
снабжены аэренхимой, слабо развиты механическая ткань, эпи-
дермис и кутикула. Таковы калужница болотная (Caltha palustris),
сердечники (Cardamine pratensis и др.), подмаренник болотный
(Galium palustre) и др.
Среди гигрофитов болотные растения обычно выделяют в от-
дельную группу гелофитов (от греч. helos — болото). Местооби-
тания их характеризуются обильным застойным увлажнением, где
остро не хватает кислорода. Болотные растения — многолетние
травы с хорошо развитой системой межклетников и воздушных
полостей. Нормальный газообмен в листьях компенсирует недо-
статок кислорода в корнях: выделяющийся в результате фотосин-
теза кислород распределяется по межклетникам вплоть до кончи-
ков корней. Таковы белокрыльник (Calla palustris), вех ядовитый
(Cicuta virosa), омежник (Oenanthe aquatica), вахта трехлистная
(Menyanthes trifoliate) и др. Сплетения корней и укороченных
корневищ некоторых гелофитов (осока дернистая — Carex
cespitosa, осока омская — С. omskiana и др.) образуют кочки,
высота которых зависит от уровня воды в вегетационный сезон и
необходимости выноса узла кущения на воздух. Эти растения —
обитатели низинных эвтрофных болот, в которых увлажнение со-
здается выходящими на поверхность грунтовыми водами, богаты-
ми минеральными веществами.
Растения верховых (олиготрофных) болот составляют своеоб-
разную экологическую группу ацидогигрофитов (Б.А. Быков,
1983). Кроме того, что эти растения страдают от недостатка кис-
лорода в застойной воде, они испытывают постоянный голод, так
как болотные воды здесь отличаются крайней бедностью мине-
ральными солями, а также подвергаются влиянию повышенной
кислотности из-за выделений сфагновых мхов. Для верховых бо-
лот типичны кустарнички из семейств Вересковые и Брусничные:
багульник (Ledum palustre), болотный мирт (Chamaedaphne
calyculata), подбел (Andromedapolifolia), клюква (Oxycoccus). Они
отличаются вечнозеленостью, низкорослостью, мелкоклеточнос-
тью и своеобразной структурой листьев. Ряд черт морфологии этих
247
растений напоминает приспособления к недостатку воды («черты
ксероморфоза»). Однако болотные кустарнички, являясь мико-
симбиотрофами, не испытывают недостатка в воде и не страдают
от физиологической сухости. Своеобразное устройство листьев этих
кустарничков (рис. 8.8) обусловлено необходимостью защититься
от света на открытых болотных пространствах при резком недо-
статке минерального питания (в первую очередь азота).
Защищаясь от света, ослабив фотосинтез, транспирацию и дыхание,
растение снижает уровень метаболизма и обеспечивает устойчивость об-
менных процессов в этих специфических условиях. Листья болотных кус-
тарничков имеют утолщенные наружные стенки верхнего эпидермиса,
сильно развитую кутикулу. Для листьев багульника характерно сильно
развитое войлочное опушение на нижней стороне, погруженные устьица.
Рис. 8.8. Строение листьев болотных кустарничков
(по А. П. Шенникову, 1950; И. С. Михайловской, 1977):
1 — багульник {Ledum palustre)-, 2 — вереск {Calluna vulgaris)-, 3 — мирт болотный
{Chamaedaphne calyculata)-, 4 — подбел {Andromedapolifolid)', 5 — клюква {Oxycoccus)
{a — взрослый однолетний лист; б — прошлогодний лист)
248
Рис. 8.9. Строение стебля и листа сфагнума {Sphagnum) (по Ю. К.Дун-
дину из кн. «Водоросли, лишайники и мохообразные СССР», 1978):
Л — стебель с ретортовидными клетками (а); Б — строение листа в плане: / —
живые фотосинтезирующие клетки, 2 — гиалиновые клетки с кольцевидными
утолщениями стенок (5) и отверстиями в стенках (4)
У багульника и болотного мирта отмечена повышенная концентрация
антоцианов в клеточном соке верхнего эпидермиса, защищающая мезо-
филл от избытка света. Нижняя сторона поднятых листьев подбела, обра-
щенная к свету, покрыта белым восковым налетом, отражающим свет.
Г. Вальтер (1982) своеобразную морфологию обитателей олиготрофных
болот предложил считать проявлением пейноморфоза (от греч. peina —
голод).
Если у болотных кустарничков в олиготрофных условиях стали сни-
жаться обменные процессы, то у других болотных растений появилась
иасекомоядность. На олиготрофных болотах России часто встречаются
росянки {Drosera), липкие волоски на их листьях выделяют протеолити-
ческие ферменты, переваривающие прилипших насекомых.
К ацидогигрофитам относятся и другие растения верховых бо-
лот: пушица влагалищная {Eriophorum vaginatum), осока топяная
{Carex limosa), а также сфагновые мхи {Sphagnum), зеленые во-
доросли и цианобактерии. Сфагнумы поддерживают высокий уро-
вень увлажнения, как свой собственный, так и всего местообита-
ния благодаря своеобразным гиалиновым клеткам в их листьях и
ре тортовидным клеткам в коре стеблей (рис. 8.9). Засасывая воду
через поры этих клеток, сфагновые мхи в состоянии удерживать
воды в 20 раз больше собственной массы. Произрастание плотны-
ми коврами позволяет им минимизировать испарение воды, так
как плотно расположенные верхушки изолируют нижнюю часть
растений в засушливую погоду и длительное время поддерживают
общее высокое увлажнение.
249
Гидрофиты. Гидрофиты (от греч. hydror — вода) — растения
водных местообитаний. В широком смысле гидрофиты — это и
водоросли, эволюция и существование которых постоянно связа-
ны с водой, и высшие растения, которыми водная среда была
освоена вторично. Однако по традиции в группу гидрофитов объе-
диняют макрофиты, т. е. высшие водные растения. В морях мак-
рофитов мало, а пресноводная флора их очень разнообразна.
Высшие водные растения вынуждены адаптироваться к осо-
бенностям минерального питания, газового и светового режимов,
движению воды в водоемах. Г. И. Поплавская (1948) предложила
подразделять их на несколько групп.
1. Гидатофиты — полностью погруженные растения (над во-
дой могут быть лишь цветки). Среди них различают: а) неукоре-
няющиеся, или взвешенные (пузырчатки — Utricularia, роголис-
тники — Ceratophyllum и др.), и б) укореняющиеся (водяные люти-
ки — Batrachium, уруть — Myriophyllum spicatum, валлиснерия —
Vallisneria spicata и др.).
2. Аэрогидатофиты — растения с плавающими листьями.
В этой группе также можно выделить: а) неукореняющиеся (водо-
крас — Hydrocharis morsus-ranae, ряски — Lemna и др.) и б) уко-
реняющиеся (кувшинки — Nymphaea, кубышки — Nuphar, во-
дяные лютики и рдесты — Potamogeton natans с плавающими
листьями).
3. Собственно гидрофиты (гидрофиты в узком смысле) име-
ют листья, расположенные над водой. Они обычны по берегам
водоемов (стрелолист — Sagittaria sagittifolia, частуха — Alisma
plantago-aquatica, камыш озерный — Scirpus lacustris и др.).
Б
Рис. 8.10. Гетерофиллия (по А. Е. Васильеву и др., 1988):
А — водяной лютик {Batrachium)-, Б — стрелолист (Sagittaria sagittifolia). Листья:
1 — подводные; 2 — плавающие; 3 — воздушные
250
Многие виды характеризуются гетерофиллией — образуют листья
разного строения в зависимости от среды их образования (рис.
8.10). У некоторых гидрофитов (например, стрелолист, поручей-
ник — Sium latifolium и др.) одновременно развиваются воздуш-
ные, плавающие и погруженные в воду листья, отличающиется
морфологически, анатомически и функционально.
Для гидрофитов, испытывающих затруднения с газообменом,
характерны листья с увеличенной поверхностью, часто сильно
рассеченные. Нитевидные доли имеют «листья-жабры» роголист-
ника, урути, пузырчатки, водных лютиков и др. Тесьмовидность
листьев ряда водных растений увеличивает их поверхность и пре-
дохраняет их от повреждения текучей водой. А некоторые гидрофи-
ты (рдесты, валлиснерия и др.) имеют очень тонкие листовые
пластинки. Предельно тонки листья элодеи (Elodea canadensis) —
всего два слоя клеток.
Для гидрофитов характерно слабое развитие механических тка-
ней, центральное положение которых в стебле обеспечивает эла-
стичность и прочность в условиях постоянного изгибания теку-
щей водой. В них развита система межклетников, обеспечиваю-
щих снабжение кислородом погруженных в воду органов. Кроме
того, эти полости обеспечивают плавучесть стеблей и листьев.
Для многих гидрофитов характерно слабое развитие корневой
системы, что связано со способностью всасывать воду всей повер-
хностью тела. Основная роль корней — закрепление растений в
грунте, а насасывание ими растворов идет слабо. Поэтому элодея,
например, может развивать корни, а может обходиться без них.
Корневища некоторых гидрофитов (кувшинки, кубышки и др.)
играют роль запасающих органов и обеспечивают вегетативное
размножение.
Недостаток в водоемах минеральных веществ и света, затрудняющий
фотосинтез, способствует возникновению растений-хищников, питающихся
мелкими водными организмами. В России и сопредельных государствах в
олиготрофных водоемах распространены 8 видов пузырчаток и альдрованда.
Пузырчатки получили свое название по ловчим пузырькам, имеющим
«ротовое» отверстие с волосками и щетинками по краям. От верхнего края
пузырька отходит эластичный клапан, выделяющий приманки для мел-
ких животных (личинок насекомых, рачков, дафний, циклопов, червей,
инфузорий). Если жертва попала в пузырек, клапан захлопывается. Про-
дукты пищеварения поглощаются всасывающими клетками. Род пузыр-
чатка насчитывает около 200 видов, в основном тропических, среди кото-
рых есть болотные и эпифитные. Альдрованда принадлежит семейству
Росянковые, которое насчитывает около 100 видов растений-хищников,
растущих на болотах и в водоемах. Ловчий аппарат ее — две полукруглые
половинки листовой пластинки, в центре которой множество пищевари-
тельных железок и чувствительных волосков. Микроскопические обита-
тели водоемов, задевая волоски, вызывают смыкание листьев-ловушек.
251
Гидрофиты могут накапливать в своем теле разные минераль-
ные вещества, концентрация которых иногда в тысячи раз боль-
ше, чем в окружающей воде (например, элодея накапливает в клет-
ках углекислый кальций). Такое избирательное накопление изме-
няет среду обитания (кислотность, солевой состав и т.п.) и может
приводить к смене экосистем.
Для растений, жизнь которых связана с обводненными место-
обитаниями, характерна гидрохория — распространение плодов и
семян с помощью воды. Кувшинки, виктория (Victoria), эвриала
(Euryale ferox) имеют на семенах губчатые мешковидные выросты,
играющие роль поплавков. Лишь после их разрушения семена опус-
каются на дно и прорастают. Крупные плавучие плоды и семена
имеют многие обитатели тропических морских побережий и манг-
ровых зарослей. Семена некоторых лютиков (Ranunculus lingua,
R. sceleratus), растущих на болотах и в ручьях, имеют под эпидер-
мой плавательный пояс из крупных опробковевших воздухонос-
ных клеток. У болотных осок (Carex vesicaria, С. rostrata, С. rhyn-
chophysa) семя заключено в заполненные воздухом вздутые мешоч-
ки. У калужницы болотной (Caltha palustris) семена приобретают
плавучесть при разбухании. А плоды кокосовой пальмы (Cocos
nucifera) содержат обеспечивающий плавучесть воздух и запас воды
для зародыша, что позволяет им разноситься морскими течениями
на тысячи километров, не теряя всхожести в течение месяцев.
Ксерофиты. Ксерофиты (от греч. xerox — сухой) — растения,
приспособившиеся к жизни в засушливых местах. Благодаря пла-
стичности своей организации они сумели адаптироваться к усло-
виям неустойчивого увлажнения и безводным пространствам, до-
вольствуясь минимальным количеством воды. Освоение засушли-
вых местообитаний шло по-разному, поэтому обширная группа
ксерофитов экологически очень неоднородна и по морфологи-
ческим особенностям подразделена на несколько подгрупп.
Л. Бриггс и Г. Шантц (1914) разделили растения засушливых мес-
тообитаний (ксерофиты в широком смысле) на три группы (при-
способления первых двух дают возможность сохранять мезофит-
ную организацию в засушливых местообитаниях).
1. Избегающие засухи (escaping) — пустынные и степные виды
с коротким, обычно весенним периодом развития. Безводный
период эфемеры переживают в виде семян (например, пустын-
ный мак павлиний — Papaver pavonicum), а эфемероиды — в
виде подземных органов (например, тюльпаны — Tulipa, при мас-
совом цветении покрывающие степь яркими коврами).
2. Уклоняющиеся (evading) — виды с мощной поверхностной
корневой системой, быстро поглощающей дождевую влагу или
развивающие корни, достигающие грунтовых вод (фреатофиты).
Например, у верблюжьей колючки (Alhagi camelorum) они про-
никают на глубину более 15 м (см. рис. 8.4).
252
3. Выдерживающие засуху (enduring) — настоящие ксерофиты
(ксерофиты в узком смысле).
По анатомо-морфологическому строению и характеру адапта-
ций к засухе настоящие ксерофиты делят на суккуленты и скле-
рофиты. Суккуленты (от лат. succulentus — сочный) — многолет-
ние сочные растения с сильно развитой водозапасающей парен-
химой. Коллоиды их цитоплазмы отличаются высокой водоудер-
живающей способностью, расход воды у них экономный, рост
очень медленный. Это обитатели аридных областей, где периоди-
чески наступает влажный период, и их поверхностная корневая
система способна быстро перехватывать выпадающие осадки, а
также росу. В засушливый период боковые корни их отмирают,
но способны быстро отрастать при увлажнении. Листья и стебли
покрыты толстой кутикулой, количество устьиц небольшое. Фи-
зиолого-морфологическая организация позволяет суккулентам
длительно поддерживать высокую гидратуру, а обезвоживания они
не переносят.
Если запас воды концентрируется в листьях, то это листовые сукку-
ленты. К ним относятся многочисленные виды рода агава (Agava) из
засушливых областей Центральной Америки, алоэ (Aloe) из аридных ре-
гионов Африки и Мадагаскара. В нашей стране на сухих песчаных про-
греваемых местообитаниях встречаются листовые суккуленты семейства
Толстянковые: несколько видов рода очиток и молодило (Sedum,
Sempervivum, Jovibarba). Среди них есть известные лекарственные рас-
тения (очиток розовый — Sedum rosea, или золотой корень, каланхоэ —
Kalanchoe daigremontiana, К. pinnata и др.).
Стеблевые суккуленты отличаются сильным развитием водозапаса-
ющей ткани в коре и сердцевине стебля. Листья их редуцированы или
превращены в колючки. Стеблевыми суккулентами является большин-
ство представителей семейства Кактусовые (Opuntia, Се re us, Cactus и др.),
характерного для американских пустынь, и семейства Молочайные из за-
сушливых областей Африки. Сходство в строении и функциях органов
африканских молочаев и американских кактусов — классический пример
конвергенции: результат одинаково направленных эволюционных про-
цессов в сходных экологических условиях. Кактусовые имеют разнооб-
разные по форме и размерам сочные зеленые безлистные стебли, покры-
тые колючками и (или) волосками, которые не только защищают расте-
ния от поедания животными, но и способствуют конденсации влаги. Стенки
эпидермиса стеблей утолщены, покрыты толстым слоем кутикулы. Ребри-
стая поверхность, характерная для некоторых видов, способствует равно-
мерному распределению света и тени. Устьица погружены в гиподерму
(слой клеток под эпидермисом). Днем они закрыты и транспирация очень
незначительна. Расход воды очень медленный, так как клеточный сок
наряду с органическими кислотами и сахарами содержит слизь, способ-
ную набухать и удерживать воду. Кактусы накапливают огромные запасы
воды: колонновидный Cereus высотой 10 м запасает до 3 000 л воды,
50-килограммовый кактус содержит около 45 кг воды.
253
Корневые суккуленты развивают водозапасающие ткани в подзем-
ной части растения. В засушливых местах Африки встречаются низко-
рослые многолетники из семейства Молочайные, имеющие довольно
крупные подземные органы, запасающие воду. А небольшое дерево сейба
мелколистная (Ceiba parvifolia) из баобабовых, растущее в Мексике, имеет
на корнях вздутия диаметром до 30 см, губчатая паренхима которых на-
капливает влагу. Эти «резервуары» могут функционировать много лет.
Другая подгруппа настоящих ксерофитов — склерофиты (от
греч. scleros — твердый). Это растения с сухими жесткими листь-
ями, имеющими толстую кутикулу и развитые механические тка-
ни. Даже в периоды полной обеспеченности водой они мало об-
воднены, а в периоды засух способны без заметного ущерба те-
рять 25 — 75% влаги (гигрофиты и мезофиты вянут, потеряв 1 —
2 % воды). Растения этой группы тем более засухоустойчивы, чем
мельче их клетки и вакуоли. В этом случае опасность деформаций
при высыхании и набухании клеток минимальна. Другое их при-
способление к крайне засушливым условиям — образование ме-
таболической воды за счет дыхания.
Важное свойство склерофитов — высокое осмотическое давле-
ние клеточного сока, увеличивающее сосущую силу корней и по-
зволяющее добывать воду из сухих почв. К тому же в пустынных
сообществах масса их корней намного превышает надземную фи-
томассу. Но для склерофитов характерна не только способность
добывать воду, но и активно использовать ее, если позволяют об-
стоятельства. Так, эти растения могут интенсивно транспириро-
вать в условиях достаточного водообеспечения. Н.А. Максимов
(1952) показал, что при благоприятном водоснабжении мезофиты
обычно расходуют 3—4 мг воды в 1 ч на 1 см2 листовой поверхно-
сти, а ксерофиты — 8 — 14 мг воды. Снижающие испарение при-
Рис. 8.11. Поперечные срезы листьев ковыля (JStipa) и типчака (Festuca
sulcata) (по А. П. Шенникову, 1950):
1 — лист ковыля; 2 — лист типчака; 3 — схема строения листа ковыля в развер-
нутом виде; 4 — сочленовные клетки
254
способления (скрученные листья, погруженные устьица и др.,
рис. 8.11) имеют большое значение при затрудненном водоснаб-
жении, но влагу дождей склерофиты используют очень эффек-
тивно. Поэтому А. П. Шенников подчеркивал, что они не «сухо-
любы», а «сухотерпцы» (влажная среда для них благоприятнее, в
условиях достаточного водоснабжения они развиваются лучше).
Склерофиты фотосинтезируют интенсивнее мезофитов. Их хлоро-
пласты хорошо удерживают воду, что позволяет им сохранять вы-
сокий уровень фотосинтеза в засуху. Так, даже при очень сильном
обезвоживании, когда оводненность клеток падает на 75 %, овод-
ненность хлоропластов уменьшается меньше, чем наполовину.
Мезофиты. Мезофиты обитают в условиях умеренного увлаж-
нения, и приспособление большей части растений связано имен-
но с ним. К данной группе относится большинство лиственных
древесных пород, луговые и многие лесные травы, сорняки, боль-
шинство культурных растений. Эту группу составляют разнород-
ные растения как с «типичным» мезоморфным строением, так и с
разными уклонениями в сторону гигро- или ксероморфной орга-
низации. Соответственно А. П. Шенников предлагает различать
типичные мезофиты, ксеромезофиты, гигромезофиты, психроме-
зофиты и т.д.
Для обеспечения оптимальной гидратуры растениям необхо-
димы определенные тепловые условия, достаточное минеральное
питание и аэрация. Обычно мезофиты одновременно представля-
ют собой мезотермофиты, мезотрофы и мезоаэрофиты. Такие ра-
стения характеризуются интенсивным обменом веществ, большой
скоростью роста, крупными размерами листьев и всего растения
и большой продуктивностью.
В связи с обитанием в разных условиях А. П. Шенников (1950)
выделил несколько групп мезофитов.
1. Вечнозеленые мезофиты влажных тропических лесов — это глав-
ным образом деревья и кустарники, круглый год обеспеченные влагой,
элементами минерального питания и теплом, что позволяет им постоян-
но расти. У них крупные мезоморфные листья, большей частью теневые
(но верхние листья крупных деревьев имеют световое строение с приспо-
соблениями к уменьшению транспирации). В теплом сыром воздухе влаж-
ного тропического леса обычна интенсивная гуттация (от лат. gutta —
капля) — пассивное выделение растворов через специальные комплексы
клеток (гидатоды, или «водяные устьица»). Это наиболее древний тип
мезофитов, из которого при изменениях климата развились остальные
группы мезофитов и ксерофиты.
2. Зимнезеленые деревянистые мезофиты распространены в кон-
тинентальных частях тропической и субтропической зоны, где за теплой
влажной зимой следует засушливое лето. Деревья и кустарники теряют
листву на время летней засухи (поэтому А. Шимпер назвал их тропофи-
тами). В период вегетации они обеспечены влагой и имеют мезоморф-
255
ное строение листьев, а в весенний листопад освобождаются от излиш-
ней испаряющей поверхности.
3. Летнезеленые деревянистые мезофиты — деревья и кустарники
умеренных зон, сбрасывающие листья на холодный период. Отличаются
большой изменчивостью признаков.
4. Летнезеленые многолетние травянистые мезофиты — в ос-
новном обитатели лугов. Строение листьев и физиологические особен-
ности растений этой группы разнообразны, с уклонами в гигро- или
ксероморфность.
5. Эфемеры и эфемероиды сохраняют мезоморфность в засушливых
местообитаниях пустынь и степей. Ряд исследователей относят их к ксе-
рофитам (в широком смысле).
8.4. Экологическое действие на растения снега и льда
Защитное действие снежного покрова. Снег — хороший тепло-
изолятор, эффективно задерживающий отдачу тепла почвой. Под
снежным покровом сохраняется температура, которая может быть
значительно выше атмосферной. Измерения показали, что при
высоте его в 40 см и температуре воздуха -20 °C температура под
снегом может быть лишь ненамного ниже 0 °C или нулевая. Снеж-
ный покров также хорошо предотвращает зимнее иссушение рас-
тений. Однако сама поверхность снега и тонкий прилежащий к
ней слой воздуха охлаждаются очень сильно. Поэтому в тундре и
горах возвысившиеся над снежным пологом части растений гиб-
нут, формируя ровную (как бы подстриженную) поверхность рас-
тительного сообщества.
Снег хорошо защищает зимнезеленые растения (зеленчук —
Galeobdolon luteum, копытень — Asarum еигораеит и др.). У ряда
лесных осок (Carex pilosa, С. digit at а, С. rhizina, С. sylvatica и
др.) сохранение фотосинтезирующих прошлогодних листьев спо-
собствует прохождению репродуктивных фаз до распускания но-
вых листьев (Ю. Е. Алексеев, 1996). Только под снегом перезимо-
вывают зимующие побеги летнезеленых и полузимнезеленых кус-
тарничков, например черники (Vaccinium myrtillus). А в Средней
Европе многие вечнозеленые растения, например плющ (Hedera),
часто отмерзают до уровня снега.
Снеговой покров позволяет выживать ряду древесных расте-
ний на границе существования леса в горах. Так, в Снежных
горах на юго-востоке Австралии границу произрастания деревь-
ев отмечает кустарниковый эвкалипт (Eucalyptus niphophila). Там
он защищен толстым снежным покровом 6 месяцев в году и по-
тому морозы переносит легко. Но попытки культивировать его в
Центральной Европе окончились неудачей, так как здесь без до-
статочной снежной защиты его побеги вымерзают (Г. Вальтер,
1974).
256
От высоты снежного покрова зависит видовой состав тундровых ли-
шайников (Ф.Фукарек и др., 1982). Так, в Евразии ягельники с Cladonia
и Cetraria islandica расположены преимущественно в западной Аркти-
ке, а для Восточной Сибири характерны тундры с Alectoria ochroleuca и
Cetraria cucullata, которые могут расти в условиях бесснежных зим и
переносить жесткость погоды, губительную для ягельников. Ягельникам
же требуется для перезимовки глубокий снежный покров, поэтому они и
распространены в районах, находящихся под влиянием влажных ветров
с Атлантики. Характерно, что на крайнем востоке Азии ягельные тундры
появляются опять и тут они снова связаны с увеличением снежного по-
крова.
Выпревание и вымокание растений под снегом. Относительно
высокая температура под снегом представляет для растений боль-
шую опасность. В этих условиях озимые гибнут от истощения.
Находясь под снегом при температуре около О °C в очень влажной
среде и почти в полной темноте, растения, сохранившие на зиму
зеленые листья, тратят на дыхание значительное количество за-
пасных веществ, а фотосинтетического восполнения энергии в
таких условиях не происходит. Расходуя запасы (содержание са-
харов уменьшается в 5—10 раз), растения истощаются и могут
погибнуть от голода. Это явление получило название выпрева-
ния. Гибель растений от него наблюдается преимущественно в
теплые зимы с большим снеговым покровом. Выпревание наибо-
лее часто наблюдается у неуспевших пройти закалку растений,
сохранивших под снегом зеленые листья.
Влажный снег способствует также развитию паразитических грибов.
Так, молодые сеянцы хвойных в этих условиях часто поражаются «снеж-
ной плесенью» (кедровая сосна — Pinus sibirica страдает от гриба
Lophodermium pinastri, а сеянцы ели — Picea от Herpotrichia nigra). По
наблюдениям на лугах Финляндии зимой злаки часто поражаются
Typhula, Fusarium nivale, Sclerotinia borealis. Бобовые, особенно клевер
луговой (Trifolium pratense) и люцерна посевная (Medicago sativа), стра-
дают от Sclerotinia trifoliorum. Даже устойчивая к зимней инфекции ти-
мофеевка в некоторые годы от нее погибает (Т.А. Работнов, 1985).
При избыточно увлажненной почве под снеговым покровом
растениям опасно также вымокание. Оно проявляется в период
таяния снега, преимущественно в пониженных местах. Реже вы-
мокание приходится на время длительных оттепелей, когда на
поверхности замерзшей почвы накапливается талая вода, которая
не впитывается и может затопить растения. В этом случае причи-
ной их гибели служит резкий недостаток кислорода в воде и по-
чве. У растений, оказавшихся под слоем воды, нормальное дыха-
ние нарушается, а анаэробное усиливается, в результате чего об-
разуются токсичные вещества. В условиях избытка влаги в почве
257
появляются вредные для растений закисные соединения, а ряд
элементов минерального питания переходит в неусвояемое состо-
яние. В итоге растения погибают от истощения и отравления.
Весной при низких температурах выходящие из-под снега расте-
ния к затоплению относительно устойчивы, но с повышением
температуры устойчивость их резко снижается. Так, для многих
трав повышение температуры воды до 10° на сутки приводит к
снижению урожая на треть, на 2 суток — наполовину, а при со-
хранении повышенной температуры в течение 8 суток урожай
практически будет уничтожен (Н. Н. Третьяков и др., 2000).
Экологическое действие почвенного льда и ледяной корки. Пре-
вращаясь в лед, вода увеличивается в объеме, поэтому подземные
органы растений, оказавшиеся в почве, насыщенной замерзаю-
щей водой, испытывают значительное давление. Выпирание рас-
тений наблюдается тогда, когда осенью морозы наступают в бес-
снежье или при осенней засухе, а также при оттепелях, когда сне-
говая вода успевает впитаться в почву. Замерзание воды тогда на-
чинается не с поверхности почвы, а на некоторой глубине, где
есть влага. Образующаяся там прослойка льда за счет продолжаю-
щегося поступления по капиллярам воды утолщается и поднима-
ет верхние слои почвы вместе с растениями. Это приводит к об-
рыву корней, проникших на значительную глубину. После оттаи-
вания почвы такие растения погибают от иссушения, если они
вторично не укореняются. Особи разных видов, возрастных и жиз-
ненных состояний отличаются по устойчивости к выпиранию, что
связано с морфологией и анатомией корней (прочностью, спо-
собностью к растяжению), развитостью корневых систем (в част-
ности, количеством придаточных корней). Молодые особи со сла-
боразвитой корневой системой более подвержены выпиранию, чем
взрослые (Т. А. Работнов, 1971).
При замерзании воды, застаивающейся на поверхности почвы,
появляется ледяная корка, вызывающая массовое отмирание трав,
почвенных животных и бактерий. Она образуется при наступле-
нии морозов до формирования снегового покрова, а также зимой
во время сильных оттепелей или ранней весной при заморозках
после стаивания снега. Особенно страдают растения в районах,
где частые оттепели сменяются сильными морозами. В местах,
где ледяная корка возникает регулярно, происходит отбор видов
и экотипов, устойчивых к ней. Например, широкое распростра-
нение лисохвостовых лугов в центральных частях многих пойм,
возможно, обусловлено также устойчивостью лисохвоста лугового
(Alopecuruspratensis) к ледяной корке. Т. А. Работнов (1985) отме-
чает, что ледяная корка воздействует на растения многосторон-
нее: действует механически, создает неблагоприятные тепловые
условия, изменяет состав почвенного воздуха. Она также косвен-
но влияет на растения, воздействуя на другие группы организмов.
258
Механическое воздействие ледяной корки связано с расширением воды
при замерзании (происходит разрыв корней). Вторая причина ее отри-
цательного действия связана с большей теплопроводностью льда, в ре-
зультате чего сильно понижается температура среды, окружающей под-
земные органы. Степень повреждения зависит от мощности ледяной
корки, температуры воздуха и длительности воздействия льда. Поздней
осенью, когда растения уже находятся в состоянии покоя или входят в
него, действие льда менее опасно, чем весной, когда они перешли в
активное состояние. Третья причина повреждения растений при образо-
вании ледяной корки связана с малой газопроницаемостью льда. Пре-
кращение газообмена приводит к снижению содержания в почве кисло-
рода и накоплению диоксида углерода. Устойчивые к гипоксии болот-
ные виды и растения, находящиеся в глубоком покое, от ледяной корки
страдают гораздо меньше.
ГЛАВА 9
ВОЗДУХ КАК ЭКОЛОГИЧЕСКИЙ ФАКТОР
Газы — важнейшие прямодействующие условия жизни расте-
ний. Состав основных компонентов атмосферного воздуха почти
постоянен: в сухом состоянии он содержит N2 — 78,1 %, О2 —
21 %, СО2 — 0,032 %, Аг — 0,9 % и др. В воздухе всегда имеется
непостоянное количество примесей: пыль, микроорганизмы, спо-
ры, пыльца, мелкие семена, газообразные выделения организ-
мов, отходы производств и т. п. Кроме того, в воздухе всегда есть
водяной пар, иногда в значительном количестве. Такой состав
воздуха во многом определяет возможность на Земле жизни.
Биосфера коренным образом трансформировала атмосферу: она
почти в тысячу раз уменьшила содержание в ней СО2 (с 35 %) и
в двести раз увеличила количество О2 (с 0,1 %) (К. С. Лосев, 1985),
что изменило окислительно-восстановительные свойства и теп-
лопроводность атмосферы. Суша покрылась растительностью и
стала более аккумулятивной в отношении тепла и влаги. По-
требляя и выделяя газообразные вещества, изменяя свойства зем-
ной поверхности, растения и сейчас оказывают большое влия-
ние на атмосферу.
Кислород, диоксид углерода, водяные пары и ряд примесей
воздуха существенно влияют на жизнь растений. Так, кислород
необходим для дыхания, а диоксид углерода — для углеродного
питания. Воздух воздействует на наземные растения и как физи-
ческая среда, их окружающая. Менее плотный, чем вода, он выз-
вал необходимость формирования у наземных растений механи-
ческих тканей, поддерживающих над землей их тела. Атмосфера
задерживает губительное ультрафиолетовое излучение, сглажива-
ет колебания температуры, меняет распределение тепла и света.
Существенное экологическое значение для растений имеет и дви-
жение воздуха. Горизонтальное перемещение воздушных масс
оказывает механическое и иссушающее влияние, служит важным
агентом распространения диаспор. Вертикальные конвекционные
потоки способствуют переносу пыльцы и легких семян, а также
во многом определяют тепловой режим территорий (перемешива-
ние, стекание холодных масс в депрессии и т.д.).
260
9.1. Экологическое значение кислорода воздуха
В атмосфере Земли почти весь кислород накоплен в результате
деятельности цианобактерий и зеленых растений. Цианобактерии,
развив тип фотосинтеза, побочным продуктом которого стало
выделение О2, начали кардинальное изменение атмосферы, сде-
лавшее возможным эволюцию аэробной жизни. В конечном счете
кислород воздуха происходит из расщепленной при фотосинтезе
воды, и почти вся вода Земли (около 1,5 млрд км3) примерно за
2 млн лет проходит через цикл фотосинтез / дыхание. Выделив-
шийся при фотосинтезе кислород поступает в атмосферу и, по
расчетам П. Клауда и А.Джибора (1972), примерно за 2000 лет
проходит через живое вещество. Сейчас атмосфера Земли содер-
жит около 1,2 • 1015 т кислорода. Растения при фотосинтезе повы-
шают его содержание (производя 70 • 109 т в год) по отношению к
диоксиду углерода, а дыхание всего живого его понижает (В. Л ар-
хер, 1978). Конечный баланс СО2 и О2 в атмосфере во многом
зависит от жизнедеятельности организмов, и если растения по-
гибнут, то свободный кислород на Земле быстро исчезнет.
Кислород в клетке как главный окислитель жизненно необхо-
дим для существования эукариотических организмов и многих
прокариот. Их клетки получают основную энергию в процессе
кислородного дыхания. Ряд организмов разлагает органику в ана-
эробных условиях (брожение), но такой метаболизм удовлетворя-
ет энергетические потребности менее эффективно. Так, при аэроб-
ном окислении глюкозы образуется в 18 раз больше энергии, чем
при ее анаэробном сбраживании. Однако роль свободного кисло-
рода в организмах двойственна. Атомарный кислород и озон яв-
ляются клеточными ядами, и эволюция жизни была бы невоз-
можна без развития систем защиты от их прямого действия. В ат-
мосфере чистого кислорода дыхание растений снижается, а при
длительном его действии они гибнут. Это связано с усилением
свободнорадикальных реакций, повреждением мембран вследствие
окисления их липидов и в конечном итоге — с нарушением мно-
гих процессов обмена веществ (см. разд. 4.2.1).
Кислород и дыхание растений. Кислород воздуха служит конеч-
ным акцептором водорода при дыхании. Дыхание относится к
процессам диссимиляции, при которых происходит распад орга-
нических субстратов организма. При этом от них отщепляется
водород и освобождается некоторое количество энергии. Однако
максимальный ее выход связан с переносом водорода на конеч-
ный акцептор, который получает его через дыхательную цепь в
митохондриях или глиоксисомах. Этот процесс связан с цепью
переноса электронов.
В дыхательной цепи благодаря фосфорилированию образуется
универсальная энергетическая «валюта» клеток — АТФ. Еще вы-
261
сокоэнергетические соединения образуются при фосфорилирова-
нии на уровне субстратов. При брожении водород присоединяет-
ся к органическим соединениям, способным далее восстанавли-
ваться, а при анаэробном дыхании — к некоторым неорганичес-
ким акцепторам водорода (нитратам, сульфатам). Однако между
водородом и этими окислителями разность потенциалов меньше,
чем между ним и кислородом. Поэтому кислородное дыхание,
при котором конечным акцептором водорода служит О2, дает зна-
чительно больший выход энергии (КПД 30—40%).
Школой А.Л.Курсанова была показана перспективность использова-
ния в сельскохозяйственной практике экзогенного нитрата как терми-
нального акцептора электронов в бескислородных условиях. Азотные
удобрения на анаэробных почвах предоставляют растению дополнитель-
ные акцепторы электронов, что положительно сказывается на урожае.
В воздухе кислорода достаточно для дыхания наземных частей
растений. Газы попадают в их тела через устьица, растворяются в
жидкой фазе клеточных стенок и оттуда проникают в клетку. Этот
процесс требует сохранения влажности контактирующих с возду-
хом оболочек и мембран. Водоемы всегда беднее кислородом, чем
атмосфера. В самых благоприятных условиях (при 0 °) в 1л воды
может раствориться не более 14,7 мг О2. С повышением темпера-
туры этот показатель падает, и при 25 °C растворимость кислоро-
да уже в 2 раза меньше. Кроме того, в погруженные в воду органы
(там нет устьиц) он проникает только путем диффузии через стенки
клеток, а это происходит очень медленно (скорость диффузии
кислорода в воде в 104 раз меньше, чем в воздухе). Ситуация усу-
губляется тем, что в воде О2 диффундирует медленнее СО2. По-
этому для находящихся в почве и воде корней, семян и аэробных
микроорганизмов кислород часто становится лимитирующим фак-
тором и его недостаток сильно сказывается на жизни растений
(Т. В. Чиркова, 1988 и др.). Еще кислород активно расходуется на
дыхание микрофлоры при разложении в воде органики, причем
при высоких температурах этот процесс ускоряется. Такое «био-
химическое потребление кислорода» особенно характерно для сто-
ячих вод, где накапливаются органические загрязнители.
Аэрация почвы. Большое значение в жизни растений имеет аэра-
ция почвы. От нее зависят скорость проникновения в почву кис-
лорода и удаления из нее диоксида углерода, интенсивность на-
копления анаэробно образующихся соединений (сероводорода,
метана и др.), потеря и образование микроорганизмами необхо-
димых растениям веществ (например, нитратов). Газообмен ну-
жен для поддержания в почве достаточной концентрации кисло-
рода, поскольку без притока из атмосферы его запасы летом, на-
пример, могут быть исчерпаны за 20—100 ч. Аэрация обеспечива-
262
сгся системой почвенных пор, заливание которых препятствует
проникновению туда кислорода.
Содержание О2 в почвенном воздухе зависит от многих при-
чин: погоды, физико-химических свойств почвы (объем порового
пространства, размер пор, дренированность и др.), биологичес-
ких параметров (интенсивность дыхания почвенных организмов
и корней), типа угодий (пашня, лес, луг). В почвах нормального
увлажнения содержание кислорода, как правило, уменьшается от
верхних горизонтов к нижним. На пашне состав воздуха связан с
агротехникой и возделываемой культурой. Например, на паровом
поле в почвенном воздухе кислорода становится больше, а диок-
сида углерода меньше. На количество О2 в почве сильно влияют
температура и влажность, но эта связь неоднозначна. Так, при
оптимальной влажности с повышением температуры содержание
О2 в почвенном воздухе падает, но в летнюю жару и засуху кон-
центрация его в почве может быть максимальная (И. С. Кауричев
и др., 1982).
При нехватке кислорода изменяется отношение растений с
почвенными организмами: увеличивается поражаемость парази-
тическими грибами, снижается активность микоризообразовате-
лей (Т.А. Работнов, 1985). Недостаток кислорода в заболоченной
почве существенно изменяет деятельность почвенной микрофло-
ры. Так, в затопленном субстрате повышается активность анаэ-
робных аммонифицирующих и денитрифицирующих бактерий, что
ведет к значительным потерям азота. С другой стороны, избыточ-
ное увлажнение заметно снижает интенсивность разложения гу-
мусовых веществ аэробными микроорганизмами, что замедляет
переход их в доступные для растений минеральные формы. Осо-
бенно ослабляется деятельность нитрифицирующих бактерий,
обогащающих почву нитратами: они активны лишь в хорошо аэри-
руемых богатых органикой незакисленных почвах.
Избыточно увлажненные почвы часто обедняются также со-
единениями Са, К, Mg и основаниями, так как в составе их гу-
муса преобладают фульвокислоты, которые образуют с ними ра-
створимые соли, мигрирующие вниз с просачивающейся влагой
(И.С. Кауричев и др., 1982). Растения страдают здесь от мине-
рального голодания еще и потому, что даже если корни и не от-
мирают при нехватке кислорода, они могут прекращать всасыва-
ние элементов минерального питания (М.Бигон и др., 1989). Не-
хватка О2 усиливает также токсическое действие некоторых про-
дуктов разложения, увеличивает кислотность почвы, ускоряет
оподзоливание и оглеение, образование вредных для растений
шкисных соединений. Замедленное разложение остатков организ-
мов при дефиците кислорода препятствует образованию гумуса и
формированию благоприятной для корней комковатой структуры
почвы.
263
Потребности корней в кислороде удовлетворяются не только
за счет почвенного воздуха. Часть его диффундирует вниз от по-
бегов. По межклетникам воздух, обогащенный выделяющимся при
фотосинтезе кислородом, поступает в корни. Велика и роль лис-
тьев в снабжении растений кислородом, поэтому при лишении
света и дефолиации у растений могут обнаруживаться все симп-
томы анаэробного обмена, что было показано на тростниковых
сообществах Южного Казахстана при использовании их как сено-
косов (Т. А. Работнов, 1985). И опытами установлено, что при по-
мещении корней в анаэробные условия листья начинают погло-
щать кислорода больше, особенно у гигрофитов (у них больше
устьиц и слабее развита кутикула). Однако кислород в подземных
органах расходуется быстро, поэтому возможность снизить его
потребление способствует устойчивости растений в таких услови-
ях (Т. А. Работнов, 1985). Направленная вниз диффузия кислоро-
да, по-видимому, обычное явление, особенно у видов, корни ко-
торых содержат много пустот.
Воздух может выделяться из корневых окончаний в почву и аэриро-
вать прикорневой участок, поддерживая на поверхности корней комп-
лекс аэробных организмов. У разных растений этот процесс идет с раз-
ной интенсивностью: количество кислорода, выделяемого единицей по-
верхности корней в единицу времени, у пушицы многоколосковой
(Eriophorum polystachyon) в 13, а у вахты (Menyanthes trifoliata) — в 16
раз больше, чем у молинии (Molinia) (М. Бигон и др., 1989). К. А. Куркин
(1976) отмечает выдающуюся способность к аэрированию болотной по-
чвы у тростника (Phragmites australis), обладающего сильно развитой
воздухоносной тканью. Выделение кислорода из корней в почву, нахо-
дящуюся в состоянии анаэробиоза, часто сопровождается окислением
закисного железа, в результате чего устраняется его токсическое дей-
ствие на растения. Вокруг корней формируются гидроокиси железа, и
по их мощности судят об окисляющей силе корней. Так, выявлено, что
болотные растения способны выделять кислорода больше, чем корни
растений, приуроченных к менее влажным почвам. Например, одно ра-
стение вахты может окислить в день от 6,3 до 45,5 мг оксида железа (II),
а побег молинии — 1,24—3,4 мг.
Приспособление растений к гипоксии и аноксии. Явления ги-
поксии (недостаток кислорода) и аноксии (отсутствие кислорода)
широко распространены в природе. В условиях кислородной не-
достаточности очень часто оказываются озимые, ячмень (Hordeum),
соя (Glycine), рис (Oryza sativa), хлопчатник (Gossypium), неко-
торые древесные растения (ель — Picea, сосна — Pinus, береза —
Betula) и др. Одни растения при переувлажнении погибают очень
быстро, другие способны переносить затопление и связанную с
ним аноксию гораздо дольше. Поскольку все растения аэробы, в
условиях аноксии они могут пребывать лишь временно. Воздей-
264
етиие гипоксии и аноксии на растение не идентично. При гипок-
сии отключаются только системы, обладающие низким сродством
к кислороду (например, окислительные системы микросом), а
электрон-транспортная цепь митохондрий продолжает работать,
хотя и с ограничениями. При аноксии электрон-транспортная цепь
отключается полностью (если нет соединений, заменяющих кис-
лород как акцептор электронов) (В. В. Полевой и др., 2001).
Устойчивость растений к гипоксии связана прежде всего с воз-
можностью сохранять близкое к нормальному содержание кисло-
рода в тканях. Этому способствуют разнообразные анатомо-мор-
фологические и физиологические особенности (поверхностная
корневая система, аэренхима, способность листьев к поглощению
дополнительного кислорода и т. п.). В случае же их недостаточно-
сти растения используют специальные метаболические приспо-
собления (Т. В. Чиркова, 1988) (рис. 9.1). В связи с этим школой
А.Л. Курсанова была разработана концепция двух стратегий адап-
тации растений в условиях анаэробного стресса. Первая — ис-
тинная толерантность, или адаптация на молекулярном уров-
не, реализуемая в условиях полного исключения кислорода из
среды обитания. Вторая стратегия — кажущаяся толерантность,
или адаптация на уровне целого организма, при которой клетки
не имеют механизма резистентности к аноксии и избегают анаэ-
робиоза благодаря транспорту кислорода из аэрированных частей
в органы, локализованные в бескислородной среде.
Метаболические приспособления растений к недостатку кис-
лорода связаны с различными звеньями обмена веществ, из них
ведущие — приспособления дыхания. Так, устойчивые к гипок-
сии растения демонстрируют низкую интенсивность дыхания даже
при нормальной аэрации. Экономное потребление кислорода дает
им возможность стабилизировать дыхание, сохраняя его почти
неизменным и при дефиците О2. У этих растений отмечается и
качественное своеобразие дыхательного метаболизма: меняется
соотношение разных способов дыхания, что необходимо для об-
разования достаточного количества высокоэнергетических соеди-
нений и веществ для биосинтеза (В. В. Полевой и др., 2001).
При усилении гипоксии и при аноксии повышается доля гли-
колиза, который при умеренной интенсивности и длительном
функционировании играет определяющую роль в устойчивости
растения. Для поддержания высокой скорости гликолиза необхо-
димо непрерывное реокисление восстанавливающихся в ходе его
коферментов. При аноксии, когда электрон-транспортная цепь
митохондрий отключена, НАДН может окисляться в конечных
|какциях молочнокислого и спиртового брожения, но при этом
накапливаются токсичные продукты этих процессов, особенно
лиловый спирт (этанол). У приспособленных к гипоксии расте-
ний его накапливается меньше. Этанол выделяется в ризосферу,
265
ПУТИ ПРИСПОСОБЛЕНИЯ
РАСТЕНИЙ К ГИПО- И АНОКСИИ
Рис. 9.1. Схема путей приспособления растений к гипо- и аноксии
(по Т. В. Чирковой, 1984)
что понижает его содержание в тканях растений. Но у неустойчи-
вых видов, несмотря на более интенсивное выделение спирта, его
концентрация и в тканях, и в ризосфере нарастает. У приспособ-
ленных к гипоксии растений низкое накопление продуктов бро-
жения связано также с частичным предотвращением их образова-
ния при переключении реакций на синтез нетоксичных промежу-
точных продуктов. Меньшее накопление этанола в тканях устой-
чивых растений связано и с его частичной переработкой. Таким
образом, есть несколько механизмов, избавляющих растения от
высоких концентраций спирта, и способность растений к деток-
сикации продуктов анаэробного обмена играет важную роль в ус-
тойчивости их к гипо- и аноксии.
266
Несмотря на то что при аноксии электрон-транспортная сеть мито-
хондрий не работает, у приспособленных растений ее компоненты дли-
тельно сохраняют потенциальную активность. Поэтому использование
при аноксии других акцепторов электронов у устойчивых растений тоже
сохраняется дольше. Так, в анаэробных условиях оказываются важными
процессы аноксического эндогенного окисления, в ходе которого элект-
роны переносятся на такие вещества, как нитраты (нитратное дыхание),
двойные связи ненасыщенных соединений (жирные кислоты, каротино-
иды). Перенос электронов на подобные акцепторы играет роль запасно-
го выхода и способствует продлению жизни растений.
Приспособления обмена веществ устойчивых к гипоксии и
аноксии растений не ограничиваются особенностями дыхания.
К метаболическим компенсаторным изменениям можно отнести
и осуществляющийся в бескислородной среде синтез белков (боль-
шая часть их — ферменты анаэробного обмена), идущий несмот-
ря на преобладающие процессы белкового распада. Большое зна-
чение имеют стрессовые белки, появляющиеся при аноксии и
исчезающие в аэробных условиях. Индукция их синтеза при ана-
эробном стрессе играет существенную роль в защите мембран.
Влияние аэрации почвы на находящиеся в ней органы растений.
Кислород почвы имеет большое значение для прорастания семян.
У некоторых видов погребенные в почве семена долго сохраняют
всхожесть, пребывая в условиях высокой концентрации СО2, не-
достатка кислорода, влажности и света, но, будучи вынесены на
поверхность, успешно прорастают. Семена ряда бобовых начина-
ют прорастать только после механического нарушения (скарифи-
кации) плотной кожуры, из-за которой зародыш не мог дышать.
Однако у некоторых видов семена прорастают и при очень низ-
ких концентрациях кислорода. В целом на начальных стадиях
онтогенеза требовательность к кислороду у разных растений от-
личается, но общая закономерность заключается в угнетении про-
растания семян в бескислородной среде. Вероятно, кислород сти-
мулирует механизмы ростовых процессов семян, а в бескислород-
ных условиях накапливаются ингибиторы роста (А. Г. Юсуфов,
1995). Недостаток кислорода в почве угнетает и рост появивших-
ся из семян проростков.
Оптимальные условия для развития корней растений создают-
ся при содержании кислорода в почвенном воздухе около 20%
(И.С.Кауричев и др., 1982). При 2%-м содержании О2 корни всех
растений еще развиваются нормально, если содержание СО2 в
почвенном воздухе не превышает 30 — 50%. При концентрации
кислорода менее 0,5 % корневые системы многих видов тоже мо-
гут развиваться, но медленно и в течение ограниченного времени.
В плохо дренированной почве корни большинства растений кон-
центрируются в поверхностном горизонте и под слой грунтовых
вод, где содержание кислорода очень мало, их корни не проника-
267
ют. Но ряд видов на ухудшение аэрации практически не реагиру-
ет: корни рогоза (Typha), камыша (Scirpus), болотного кипариса
(Taxodium} и некоторых ив (Salix) могут расти ниже уровня грун-
товых вод, где нет свободного кислорода.
Развитие корней при анаэробиозе зависит от температуры: чем
выше температура почвы, тем больше концентрация кислорода,
необходимая для нормального развития корней. Так, при кон-
центрации кислорода в почве около 3 % угнетение корней начи-
нается при 18°, а при 10%-й концентрации скорость роста корней
снижается только при 30 °C. В теплых почвах тропиков недоста
ток кислорода может образоваться при большом его потреблении
почвенными организмами и при этом могут возникать анаэроб-
ные повреждения корней.
Снижение концентрации О2 по-разному влияет на активность кор-
ней разных видов. Например, при недостатке кислорода ряд луговых
трав быстро образует расширенные межклетники и газовые полости,
новые придаточные корни. К. весеннему затоплению более устойчивы
корни сосны (Pinus sylvestris), менее устойчивы корни березы пушистой
(Betula pubescens), еще менее — ели (Picea abies). Для древесных расте-
ний очень важна аэрированность среды вокруг корней. Так, по данным
А.Я.Орлова (1974), в южной тайге минимальная концентрация кислоро-
да в почвенной воде, которая обеспечивает возможность роста корней
сосны, ели и березы пушистой, колеблется около 1 —2 мг/л. В условиях
анаэробиоза их рост обычно прекращается, но может быть восстанов-
лен, если продолжительность пребывания в бескислородной среде нс
превышала 3 — 5 дней. При больших сроках корни прекращают активное
поглощение, отмирают их верхушечная меристема и ткани зоны растя-
жения. Гибель корневых окончаний в период интенсивного роста отме-
чена при концентрации О2 в 0,5 мг/л. Однако в состоянии покоя корни
выносят длительный анаэробиоз хорошо.
В ряде случаев (во время ливней, паводков, образования ледя-
ной корки) воздуха может не хватать также надземным органам и
даже целым растениям (низкорослым и молодым). В этом случае
в критической ситуации по отношению к О2 оказывается листо-
вой аппарат.
Формообразующее действие недостатка кислорода. В заболо-
ченных почвах содержание кислорода иногда настолько мало, что
растения вынуждены перестраивать свою морфологию и анатоми-
ческое строение. Некоторые из них приобрели способность обра-
зовывать кочки и размещать корни вне анаэробных условий, напри-
мер на сырых лугах это наблюдается у многих осок (Carex cespitosa,
С. juncella и др.) и злаков (луговик дернистый — Deschampsia
cespitosa, вейник Лангсдорфа — Calamagrostis langsdorffii).
У ряда болотных и водных видов развивается аэренхима -
воздухоносная ткань с увеличенными межклетниками и полостя-
268
ми (рис. 9.2), протягивающимися из надземных или надводных
частей растений в органы, находящиеся в заболоченной почве или
воде. Иногда в основании стебля разрастаются рыхлые ткани.
В корнях выраженность воздухоносной ткани усиливается с их
возрастом и заглублением. По Г.Элленбергу (1978), развитую аэрен-
химу в корнях имеют более 13 % сосудистых растений Централь-
ной Европы.
Воздухоносная ткань характерна и для видов, приспособленных к
переменному увлажнению. У них также формируется толстостенная оп-
робковевшая экзодерма, что служит приспособлением к периодическо-
му пребыванию корней в увлажненной почве. К этой группе растений
относятся хвощ полевой (Equisetum arvense), осока высокая (Carex elata),
кровохлебка лекарственная (Sanguisorba officinalis), сухопутная форма
горца земноводного (Polygonum amphibium). Сюда же можно отнести
•эксплеренты вымокания», в частности лютик ползучий (Ranunculus
repens), преобладающий на пойменных лугах в местах застоя полых вод.
Ряд растений хорошо аэрируемых почв формирует аэренхиму, оказав-
шись в условиях недостаточной аэрации. Это свойственно даже ксеро-
фильному кострецу прямому (Bromopsis erecta), а также мезофитам —
еже сборной (Dactylis glomerata), овсянице луговой (Festuca pratensis) и
др. При ухудшении аэрации многие виды образуют придаточные корни
с аэренхимой.
Рис. 9.2. Участки поперечных срезов листьев (А) и стеблей (Б) расте-
ний из обводненных местообитаний (по И.С.Михайловской, 1977):
/ рдест плавающий (Potamogeton natans)', 2 — рдест курчавый (Р. crispusy, 3 —
► уншинка белая (Nymphaea alba)\ 4 — воздушный лист стрелолиста (Sagittaria
\agutifoHa)’, 5 — камыш озерный (Scirpus lacustris)-, 6 — уруть (Myriophyllum)\
а — воздухоносные полости
269
Для приморских тропических растений мангровых зарослей
очень характерны пневматофоры (дыхательные корни). Во вре-
мя прилива над водой возвышаются только кроны мангровых де-
ревьев, а во время отлива обнажаются стволы и система особых
корней — ходульных и дыхательных. Пневматофоры необходимы
мангровым растениям, так как ил, в котором они растут, почти не
содержит кислорода. Они имеют возвышающиеся над субстратом
части, содержащие отверстия, через которые воздух поступает в
воздушные полости и проходит в подземные части растений. Дыха-
тельные корни могут быть разного типа. У одних видов (Вги-
guiera) они коленчато выступают из ила, у других (Xylocarpus) —
ползучие змеевидные, у третьих — вертикально выходят из полужид-
кого субстрата (Avicennia, Sonneratia) (рис. 9.3). А у Rhizophora
дыхательную функцию берут на себя выступающие из воды части
ходульных корней. Кроме мангровых растений на почвах со слабой
аэрацией дыхательные корни развивают некоторые пальмы, панда-
нусы (Pandanus), сахарный тростник (Saccharum officinarum) и др.
Из анатомо-морфологических изменений, вызываемых недо-
статком аэрации почвы, отметим еще утончение клеточных сте-
нок в корнях, ослабление корневого ветвления и образования кор-
невых волосков, формирование придаточных корней в основании
стебля. Ризосфера при этом обычно имеет меньший объем, корни
укорачиваются, занимая более поверхностное положение, масса
надземной части растения снижается, площадь листьев и содер-
жание в них хлорофилла падают, иногда наступает хлороз. У вод-
ных растений с целью максимизации поступления кислорода обыч-
Рис. 9.3. Дыхательные корни Sonneratia alba в мангровых зарослях во
время отлива (по В. В.Алехину, 1950)
270
но увеличивается поверхность подводных органов, что часто дос-
тигается сильным удлинением, истончением и рассечением листьев
(см. разд. 8.3.2).
9.2. Экологическое значение диоксида углерода
воздуха
Диоксид углерода очень важен для растений, так как они стро-
ят органические вещества своего тела за счет его ассимиляции
при фотосинтезе. Для образования 1 г глюкозы требуется погло-
тить 1,47 г (или 0,75 л) СО2 (учитывая его концентрацию, такое
количество СО2 извлекается из 2 500 л воздуха). Диоксид углерода
имеет большое значение и в химическом изменении минеральной
части почв: насыщенный им почвенный раствор растворяет мно-
гие соединения почвы. Растворяющее действие СО2 на карбонаты
при благоприятных условиях обеспечивает их вынос из верхних
горизонтов почвы или, наоборот, подтягивание из нижних гори-
юнтов (И. С. Кауричев и др., 1982).
Диоксид углерода и запасы углерода на Земле. Общий запас
углерода на Земле составляет более 26 • 1015 т (В. Лархер, 1978). По
оценке Межправительственной комиссии по вопросам измене-
ния климата в атмосфере, его около 730 млрд т, еще 2 011 млрд т
содержится в почве и 466 млрд т — в наземной растительности.
Но больше всего углерода растворено в Мировом океане или вхо-
дит в состав планктона — 38 000 млрд т. Пока человек не вмеши-
вался в круговорот углерода в природе, наземные растения еже-
годно поглощали 120 млрд т его, но он почти весь возвращался за
счет дыхания организмов и разложения остатков их тел. Эта гло-
бальная система вышла из равновесия в XIX в. С начала промыш-
иенной революции в атмосферу с диоксидом углерода попало по-
•I ги 500 млрд т углерода (2/3 за счет сжигания ископаемого топли-
ва и % из-за разрушения естественных сообществ — вырубки ле-
сов, распашки пустошей и т.п.). Сейчас в результате хозяйствен-
ной деятельности в атмосферу попадает около 6,3 млрд т углерода
в год. Еще около 1,6 млрд т ежегодно выделяется при лесных
пожарах. При этом около 2,3 млрд т СО2 ежегодно растворяется в
Мировом океане и примерно столько же поглощает естественная
растительность. Важнейшую роль в поглощении СО2 на Земле
hi рают акватория океанов в южном полушарии и область тайги и
смешанных лесов — в северном.
Основная масса углерода на Земле содержится в неорганиче-
ских соединениях, и только 0,05% — в органических (В. Лархер,
1978). Запасы неорганического углерода откладываются большей
частью в осадочных горных породах (около 2,5 • 10,6т). Воды со-
держат около 0,14 % углеродного запаса в виде карбонатов, бикар-
271
бонатов, растворенного диоксида углерода. При этом основная
его масса находится в глубинах океана, а поверхностные слои, где
живут растения, содержат только 1/60 запаса неорганического угле-
рода гидросферы. В атмосфере углерода примерно столько же,
сколько в поверхностных водах океана, и между ними идет посто-
янный обмен СО2. В среднем по атмосфере концентрация СО2
составляет 0,03 %, но в промышленных районах она может повы-
шаться до 0,05 % из-за сжигания ископаемого топлива (угля, не-
фти, газа). При этом в атмосферу возвращается СО2, забранный
из нее миллионы лет назад фотосинтезировавшими тогда организ-
мами.
Органический углерод находится в биосфере и верхних слоях лито-
сферы. В живых и отмерших организмах его содержится около 3,5 • 10,2т.
Порядка 64 % находится в ископаемых отложениях, 32 — в органических
остатках в почве и водоемах и около 4 % — в живых организмах. Наи-
большая часть биомассы приходится на наземные растения, при этом
главные резервы органического углерода сосредоточены в лесах (углерод
древесины составляет 3/4 всего углерода наземных растений) (Н. И. Ба-
зилевич, Л. Е. Родин, 1971) (рис. 9.4). А в почве и на ее поверхности
наибольшие запасы углерода образуются в тундре и северных лесах, где
органические остатки разлагаются медленно.
Содержание диоксида углерода в воздухе и его изменения. Отно-
сительное постоянство содержания СО2 в атмосфере поддержива-
ется благодаря равновесию всех составляющих биогеохимического
круговорота углерода и буферной роли Мирового океана. Однако
концентрация СО2 в воздухе постоянно увеличивается. В начале
XXI в. количество СО2 в атмосфере эквивалентно 7,3 • 1011 т угле-
рода, в то время как в 1980 г. оно соответствовало 7,1 • 10м т, а до
1860 г. атмосфера содержала лишь 6,1 • 10й т. Это почти 20%-е уве-
личение связывают прежде всего с появлением дополнительного
источника СО2 при интенсивном сжигании ископаемого топлива.
Большая часть углерода содержится в виде карбонатов в оса-
дочных породах, живых и отмерших организмах, и даже в Миро-
вом океане в десятки раз больше углерода, чем в атмосфере, что
связано с высокой растворимостью СО2 в воде. Можно было бы
предположить, что дополнительное поступление СО2 в результате
сжигания ископаемого топлива незначительно и не должно при-
водить к заметному увеличению его концентрации в атмосфере.
Однако быстрый обмен углерода с атмосферой (за 6 — 7 лет) осу-
ществляется в верхних слоях океана, содержащих лишь 1,5 % ра-
створенного углерода, а для установления равновесия с глубин-
ными слоями океана требуется несколько тысячелетий. Вследствие
этого промышленное сжигание ископаемого топлива способству-
ет накоплению СО2 в атмосфере (с 0,027 % в доиндустриальную
эпоху до 0,035 % в настоящее время). Ряд расчетов показывает.
272
Рис. 9.4. Распределение фитомассы на Земле (в тоннах сухого вещества
на 1 га) (по Н. И. Базилевич, Л. Е. Родину, 1971)
□ < 10 т/га
□3 10-50 т/га -
ШИШ 50—150 т/га
НИМ 150 — 400 т/га
I < 400 т/га
ч го к 2035 г. содержание СО2 в атмосфере может удвоиться и со-
ставить около 0,06 %.
Сезонные колебания концентрации атмосферного диоксида
углерода связаны с фотосинтезом и влияющими на почвенную
микрофлору изменениями увлажнения, аэрации и температуры
почвы. Так, в высоких широтах после продолжительных дождей
•ктивность аэробных почвенных микроорганизмов падает и со-
держание СО2 в приземном слое воздуха снижается. А в аридных
областях то же вызывается засухой. Годичные колебания СО2 в
воздухе особенно сильны в северном полушарии, где преобладает
суша. Весной в северных областях потребление диоксида углеро-
да развивающимися растениями значительно превышает поступ-
ке пие его из почвы. Как указывает И. М. Культиасов (1982), с апреля
по сентябрь атмосфера северного полушария к северу от 30-й па-
раллели теряет почти 3 % запаса СО2 (3 млрд т углерода). Учи-
। ывая, что одновременно она пополняется СО2 за счет разложения
органического вещества, в целом в этот период в северном полу-
шарии из атмосферы усваивается до 6 млрд т углерода (’Д—’/5
годовой продуктивности суши). Общие цифры сезонных колеба-
ний атмосферного углекислотного баланса СО2 таковы: в север-
ном полушарии содержание его в воздухе в течение вегетацион-
ного периода ниже, чем зимой, на 0,00008%, а в южном — на
0,00002 %.
('уточные колебания содержания СО2 более выражены, чем
се юпные. В фитоценозах обычно наблюдается связанное с фото-
273
синтезом дневное уменьшение концентрации СО2. Например, уг-
ром в лесу с началом фотосинтеза содержания СО2 падает. К по
лудню температура повышается, дыхание усиливается и количе-
ство диоксида углерода в кронах отчасти выравнивается. После
захода солнца фотосинтез прекращается, начинает преобладать
дыхание и концентрация СО2 вблизи почвы повышается. В без-
ветренную погоду она может достигнуть 0,4 %, что объясняется
«дыханием почвы» и отсутствием конвекционных токов. Суточ-
ные колебания содержания диоксида углерода в густых сообще-
ствах могут достигать 25 % от средних величин (И. М. Культиасов,
1982).
Дыхание почвы. СО2 выделяется при извержении вулканов (65
млрд т в год), сжигании горючих веществ, некоторую прибавку
дает разложение карбонатных горных пород, однако главный ис-
точник СО2 в сообществах суши — дыхание почвы. Под ним по-
нимают выделение эдафоном (совокупностью живых организмов
почвы) СО2 в прилегающий к почве слой воздуха. В этом процес-
се на долю дыхания корней приходится до 10—30 %, дыхание даже
крупных животных очень мало и основное количество СО2 обра-
зуется за счет микробиологической переработки остатков отмер-
ших организмов.
В почвах с нормальным увлажнением содержание СО2, как
правило, увеличивается от верхних горизонтов к нижним и растет
с повышением температуры. Но при летней жаре и влажности,
близкой к влажности завядания, в почве наблюдаются самые низ-
кие концентрации диоксида углерода. Пик дыхания почвы обыч-
но приходится на лето, но годовая динамика концентрации СО2 в
почве очень изменчива и зависит от возможностей газообмена,
связанных с многими факторами (увлажнение почвы, температу-
ра и др.) (И.С. Кауричев и др., 1982). При некоторых условиях
содержание СО2 в почве может превысить содержание его в ат-
мосфере в 50 раз. Этому способствует активное разложение орга-
ники при ее обильном поступлении, слабокислой или слабоще-
лочной реакции почвы, достаточном доступе О2 и воды и при
оптимальной температуре. Влияние оказывают также диффузия и
движение припочвенного воздуха. Так, при одинаковой темпера-
туре увлажнение усиливает дыхание почвы, но переувлажнение
его снижает. Имеют значение объем почвенных пор и степень
заполненности их водой. Суточные колебания содержания СО2 в
воздухе леса могут быть связаны также с большей турбулентнос-
тью воздуха днем (поэтому обогащение диоксидом углерода при-
почвенных слоев особенно ощущается ночью) (рис. 9.5).
Выделение диоксида углерода в течение суток из разных типов
почв может существенно отличаться: из черноземов — 100 кг/га,
серых лесных — 80, подзолистых — 50—80, каштановых — 40, из
пустынных — 5—10 кг/га (Т.К.Горышина, 1979). Интенсивность
274
Рис. 9.5. Суточные колебания содержания СО2 в воздухе сосняка
(бонитет II, возраст 75 лет, сомкнутость крон 0,7)
(по Г. Вальтеру, 1982).
Средние данные для июня —июля: 7 — в почве; 2 — на высоте 15 см; 3 — на
высоте 130 см; 4 — у основания крон; 5 — в кронах; 6 — в 2 — 3 м над кронами
дыхания почвы в разных сообществах так же неодинакова. По
I. Вальтеру (1960), на 1 м2 поверхности в 1 ч еловый лес на подзо-
лах выделяет 0,36 г СО2, ельник на известняке — 0,431 г, буковый
лес — 0,407 г, сырой мелкоосоковый луг — 0,551 г, сухой луг с
лисохвостом — 0,78 г СО2. Из-за небольшой скорости минерализа-
ции хвои дыхание почвы в хвойных лесах невелико. В кислой среде
верховых болот минерализация тоже слаба, так как здесь мало мик-
роорганизмов и слаборазложившиеся остатки образуют торф.
Для оценки местообитаний существенное значение имеет ло-
кальное содержание СО2. Так, открытые сообщества, как прави-
ло, имеют невысокие концентрации диоксида углерода, посколь-
ку ветер все время его удаляет. В приземном же слое лесного воз-
духа содержание его может в 6 раз превышать среднее, а над удоб-
ренной окультуренной почвой и в 10 раз. Интенсивность «по-
чвенного дыхания» бывает столь велика, что создает более благо-
приятные условия для фотосинтеза растений в приземном слое.
В лесу, например, минимум содержания СО2 приходится на слой
крон, где идет его интенсивное поглощение при фотосинтезе, а
максимум наблюдается в нижнем ярусе. Повышенная концентра-
ция СО2 в припочвенном воздухе леса частично компенсирует
рас тущим здесь растениям недостаток света. Это особенно важно
пня нижних ярусов влажного тропического леса. Так, на о. Ява в
М) см припочвенного воздуха отмечены концентрации СО2 днем
к> 0,09 %, а ночью до 0,26%, что позволяет достаточно хорошо
ра шиваться травам, до которых доходят лишь сотые доли света,
падающего на поверхность лесного полога.
Круговорот углерода и живые организмы. Углерод встречается
ни 1смле в разных формах: в виде твердых тел (организмы, почва,
275
литосфера), в растворенной форме (водоемы, почвенная и грун-
товая вода, жидкости организмов) и в виде газа (атмосфера, поры
почвы и межклетники). Между разными формами углерода по-
стоянно идет обмен. Литосферный углерод переходит в раствор,
растворенный СО2 уравновешивается с СО2 воздуха, а углеродный
обмен организмов приводит к постоянному круговороту углерода.
Круговорот углерода на Земле поддерживается прежде всего
благодаря биологическим процессам (рис. 9.6). Соединения угле-
рода составляют основу живого вещества биосферы. Они непре-
рывно синтезируются, передаются, изменяются и разлагаются
организмами. Основная миграция углерода на Земле идет от СО2
атмосферы в живые организмы и обратно. Атмосферный диоксид
углерода проходит этот цикл примерно за 300 лет. По сравнению
с биологическим циклом углерода геохимические его превраще-
ния ничтожны. Они включают выделение СО2 при извержении
вулканов, пожарах и промышленных выбросах. Вместе взятые, они
дают менее 10% выделяемого в год СО2.
Круговорот углерода начинается с фиксации атмосферной СО2
в процессе фотосинтеза, Годовое фотосинтетическое потребление
диоксида углерода растениями составляет около 6 — 7 % СО2 ат-
мосферы и поверхностных вод. Растения суши и фитопланктон
фиксируют примерно одинаковые количества углерода. В водных
сообществах водоросли поглощают диоксид углерода из воды, в
которой он находится в свободном или связанном состоянии (в
виде карбонатов и бикарбонатов). Основные потребители СО2 на
суше — леса и болота. Если не считать горючих ископаемых, дре-
весина — главный резервуар биологически связанного углерода.
В лесах запасено 400—500 млрд т углерода (примерно 2/3 атмос-
ферного запаса). Во влажном тропическом лесу на 1 м2 за год
фиксируется 1 — 2 кг СО2, что равно его количеству в столбе воз-
духа сечением 1 м2, доходящем до границ атмосферы. В тундрах
Свет
Фитомасса
(наземная)
СО 2 атмосферы
Фитофаги «ф Плотоядные I—► Плотоядные II
Растительный опад + Экскременты и трупы животных
------п---------f-----------------------------
Корни—Сапрофаги — Плотоядные I —- Плотоядные
о
со
£
Микроорганизмы почвы
Биомасса
почвы
Рис. 9.6. Схема пищевых цепей и круговорота СО2 в лесу
(по Г. Вальтеру, 1974)
276
же и аридных пустынях фиксируется лишь около 1 % этого коли-
чества (И.М.Культиасов, 1982).
При ассимиляции растениями СО2 образуются углеводы и выс-
вобождается кислород. Около трети полученного вещества исполь-
зуется растениями на дыхание, продукт которого — СО2 — воз-
вращается в атмосферу. При этом у растений дышат и нефото-
сиптезирующие ткани, которые существуют за счет деятельности
листьев. Например, у степных, пустынных и высокогорных расте-
ний 70 — 90% фитомассы приходится на подземные органы.
СО2 ассимилируется не только в процессе фотосинтеза. Ее использу-
ют также хемосинтетические автотрофные микроорганизмы. Часть энер-
гии, выделяющейся при окислении ими неорганических соединений,
идет на восстановление СО2 и синтез органики. В 1914 г. А. Ф. Лебедев
предположил, что и гетеротрофы могут частично ассимилировать угле-
род из СО2. Впоследствии гетеротрофная фиксация СО2 была экспери-
ментально подтверждена. Оказалось, что гетеротрофы усваивают СО2,
связывая его с кетокислотами. Гетеротрофное усвоение СО2 свойствен-
но также корням высших зеленых растений. Диоксид углерода почвы
при этом присоединяется к пировиноградной кислоте, образующейся
при расщеплении в корнях сахаров. Получившиеся органические кисло-
1Ы перемещаются в листья, где содержащийся в них углерод использует-
си для биосинтеза. Активно усваивают СО2 и клубеньки азотфиксирую-
пшх растений. Таким образом, СО2 почвы — важный дополнительный
источник углеродного питания растений. Но при этом следует подчерк-
нуть, что автотрофы могут синтезировать органику полностью за счет
неорганических соединений (Н2О и СО2), а гетеротрофы делают это лишь
имея в распоряжении готовые органические соединения (например, пи-
ровиноградную кислоту) (В.Л.Кретович, 1986).
Почти вся органика, оставшаяся после использования части ее
на дыхание растений, становится пищей для гетеротрофных орга-
шпмов (лишь ’Лоос доля ассимилированного углерода выбывает
и । круговорота при погребении растительных остатков). Дыха-
ние, брожение и гниение, происходящие в организмах, высво-
бождают СО2 и пополняют ее общий запас. Отмирающие ткани
растений и животных разлагаются (минерализуются) микроорга-
низмами. При этом углерод окисляется до СО2 и через «дыхание
почвы» возвращается в атмосферу. Деятельность почвенных мик-
роорганизмов в круговороте углерода имеет очень большое значе-
ние. В эволюции биосферы вместе с развитием автотрофного блока,
производящего органику, сопряженно, но с опозданием шло со-
вершенствование редуцентов, минерализующих органические ос-
ники и способствующих более полному замыканию биологиче-
4 кого круговорота. Так, известны мощные слои битумов из циано-
бакгерий в докембрийских отложениях, накопление которых в
4 («временных условиях (с обилием разнообразных редуцентов)
277
невозможно. Так же количество почвенных грибов, разлагающих
древесину, нарастало постепенно. В девоне их было еще немного,
поэтому с тех времен остались огромные запасы недоразложив-
шейся древесины. СО2 атмосферы связывалась тогда растениями
заболоченных лесов и депонировалась в торфяных остатках, ко-
торые метаморфизировались в каменный уголь. Со временем роль
сапротрофов росла, активизировался почвообразующий процесс,
и растения расселялись на ранее малодоступные субстраты.
Циркуляция воздушных масс в атмосфере связывает наземные
экосистемы с атмосферным резервуаром СО2. Морские течения
перемешивают поверхностные воды океанов и медленно вырав-
нивают различия поверхностных и глубинных вод. Относительно
самостоятельные круговороты углерода в атмосфере и океане свя-
заны через обмен СО2 между воздухом и водоемами, особенно в
ветреную погоду. СО2 хорошо растворима в воде, и по расчетам,
почти весь диоксид углерода атмосферы мог бы раствориться в
океане за 5—10 лет. Но притом, что в год в океане растворяется
до 100 млрд т атмосферного СО2, почти столько же за это время и
выделяется из него в атмосферу. Таким образом, для всемирного
круговорота углерода Мировой океан представляет собой глобаль-
ный буфер.
Поступление диоксида углерода в растение. Газообмен у расте-
ний происходит путем диффузии, и на пути от атмосферного воз-
духа до хлоропластов СО2 преодолевает ряд препятствий, состав-
ляющих диффузное сопротивление. Кроме константы диффузии
сюда относятся сопротивление на поверхности раздела фаз, дли-
на пути диффузии и его поперечное сечение. В целом сопротив-
ление транспорту СО2 создают: сопротивление пограничного слоя
(воздушной пленки около листа), физиологически регулируемое
устьичное сопротивление, диффузное сопротивление в системе
межклетников, сопротивление при растворении и транспорте СО?
в жидкой фазе клеточной стенки и в цитоплазме, а также сопро-
тивление в процессе карбоксилирования.
Сначала тормозить диффузию может пограничный слой между
листом и окружающим воздухом. Толщина его зависит от величи-
ны и расположения листьев, их опушенности и силы ветра. В без-
ветренную погоду он имеет толщину в несколько миллиметров, а
сильный ветер полностью его сдувает. Густое и высокое опушс
ние увеличивает пограничный слой, поэтому СО2 быстрее пере
ходит в лист, чем притекает извне, и воздушная оболочка вокру|
листа становится беднее СО2 Но в целом сопротивление погра
ничного слоя ниже, чем последующие устьичное и мезофиллы юс
сопротивление.
У наземных растений СО2 проникает в лист через устьица. Ус-
тьичное сопротивление — главное препятствие для свободною
поступления диоксида углерода в растение. Закрытые устьица
278
практически полностью перекрывают поток СО2. При недостатке
света, когда устьичные щели суживаются, устьичное сопротивле-
ние становится фактором, ограничивающим диффузию СО2. Но
при широко раскрытых устьицах это сопротивление ниже сопро-
тивления мезофилла. Изменяя ширину устьичных щелей, расте-
ние контролирует поступление СО2 и одновременно потерю воды.
Количество, распределение, размеры, форма, подвижность усть-
иц видоспецифичны, но в некоторой степени зависят от условий
местообитаний и индивидуальных особенностей растений. Для
поступления СО2 в растение решающее значение имеет площадь
отверстий устьиц. С ее уменьшением устьичное сопротивление
диффузии экспоненциально возрастает. Суммарная площадь ус-
тьичных щелей, или относительная площадь пор, определяется
умножением густоты расположения устьиц (числа устьиц на 1 мм2
листовой поверхности) на максимальную площадь устьичной щели.
У большинства растений отверстия устьиц занимают 0,5—1,5%
поверхности листа. Из-за слабой открытости минимальная пло-
щадь устьичных щелей отмечена у листьев вечнозеленых жестко-
шстных растений в засушливом маквисе и кустарничковых пус-
юшах.
Максимальная диффузия газов в лист достигается при максимальном
открытии устьиц, которое зависит от формы замыкающих клеток и осо-
бенностей строения их стенок. Степень открытости устьиц зависит так-
же от состояния растения и внешней среды. Из экологических факторов
наиболее значимы свет, температура, влажность воздуха и водообеспе-
•iriiiiocTb. Из внутренних факторов сильнее всего на поступление диок-
i ида углерода влияют парциальное давление в межклетниках, гидрата-
ция растения, баланс ионов и фитогормонов. На состояние устьиц вли-
яю г также возраст листьев, стадия развития растения и эндогенные су-
точиые ритмы. При хорошем водоснабжении устьица открываются тем
шире, чем интенсивнее освещение (фотоактивное открывание). Хемоак-
। пиная регуляции устьичных движений связана с содержанием в воздухе
( ()?. Так, при снижении его содержания ниже критического уровня они
шкрываются и в темноте. Решающее влияние концентрации СО2 на от-
крытость устьиц прослеживается у суккулентов с САМ-фотосинтезом.
VipoM, когда при расщеплении малата в межклетниках накапливается
< <)_>, их устьица хемоактивно закрываются, а ночью, когда СО2 активно
расходуется на образование органических кислот и ее содержание в меж-
кис i никах остается низким, они открыты (см. разд. 6.4.2.2). Реакцию
м н.пц индуцирует и ряд других веществ (SO2, озон, сульфат алюминия
и ip ). На состояние устьиц воздействуют также токсины паразитов (на-
пример, выделения гриба Hehninthosporium maydis) и вещества, приме-
няемые для борьбы с ними (например, фунгицид фенилмеркурацетат).
1емнсратура влияет на скорость открывания устьиц через выработку
шергии для их движения. При более высокой (но неэкстремальной)
н мнературе энергии у клетки больше и устьица открываются быстрее
' модельных растений при невысоких положительных температурах (ниже
279
5 °C) устьица открываются очень медленно и не полностью, а при отри-
цательных температурах они остаются закрытыми (В.Лархер, 1978). При
сверхоптимальных температурах устьица также открываются не полнос-
тью, однако у ряда видов в жару (40 — 45 °C) ширина устьиц вновь дости-
гает максимума.
Важный фактор регуляции состояния устьиц — водообеспеченность.
Устьичная щель закрывается при потере воды и падении тургора замы-
кающих клеток, которые могут транспирировать сильнее других клеток
эпидермиса. Недостаток водоснабжения в устьичном аппарате становится
заметен раньше, поэтому в сухом воздухе реакция закрывания устьиц
наступит раньше, чем растение начнет испытывать серьезный недоста
ток воды.
Совокупное влияние внешних факторов обычно обусловлива-
ет не максимальную, а среднюю степень открывания устьиц, так
как все условия, способствующие этому, совпадают редко. Но и
полное закрытие устьичных щелей вызывают тоже только экстре-
мально неблагоприятные условия. В целом регуляция устьичного
сопротивления газообмену описывается как система с различны-
ми обратными связями.
Прошедший через устьичную щель диоксид углерода попадает
в межклетники, где пополняется СО2, выделившимся при дыха-
нии клеток. Сопротивление диффузии в системе межклетников
зависит от строения листа. Обычно газопроводность теневых ли-
стьев выше, чем световых, а в мягких листьях она лучше, чем в
жестких органах. Сопротивление диффузии в межклетниках при-
мерно равно сопротивлению при переходе в жидкую фазу в кле-
точных стенках и сопротивлению в цитоплазме и хлоро пластах.
В клеточных стенках СО2 растворяется, и этот процесс сильно
тормозит диффузию. Внутри клетки растворенный СО2 очень мед-
ленно (по сравнению с газообразной фазой) движется к хлоро-
пластам, где присоединяется к акцептору. Карбоксилирование мо-
жет отставать от световых реакций фотосинтеза и лимитировать
скорость процесса в целом. Сопротивление карбоксилирования
зависит от эффективности работы ферментов, отличающихся в
разных группах растений. Так, из-за того, что ФЕП-карбоксилаза
катализирует связывание углекислоты эффективнее, чем РДФ-
карбоксилаза, сопротивление карбоксилирования у С4-видов зна-
чительно меньше, чем у С3-растений.
9.3. Экологическое значение непостоянных
компонентов воздуха
Непостоянные компоненты воздуха. Примеси в воздухе содер-
жатся в атмосфере разных местообитаний в разных количествах и
имеют большое значение для растений, особенно в промышлен-
280
них центрах. Наиболее вредные компоненты промышленных га-
зов — оксиды серы, азота, соединения фтора и хлора. Основной
источник дымовых газов — сжигаемое топливо. Среди атмосфер-
ных загрязнителей для растений особенно опасны продукты сго-
рания, содержащие сернистый ангидрид (SO2), который вызывает
разрушение хлорофилла, недоразвитие пыльцы, нарушает работу
проводящей системы и др. Выхлопные газы содержат токсично
действующий на растения свинец. Вблизи металлургических пред-
приятий неблагоприятно влияют на растения соединения метал-
пов (меди, цинка, кобальта, никеля, кадмия и др.), поступающие
как из почвы, так и из воздуха.
Газообразные примеси абсорбируются почвой и подстилкой,
соединяются с водой и могут образовывать кислоты. Они повы-
шают кислотность почвы и уменьшают насыщенность ее основа-
ниями, что снижает биохимическую и микробиологическую ак-
тивность почвы. Кислоты могут также переводить в растворимую
форму ряд веществ, токсичных для растений.
Пыль снижает прозрачность воздуха и, уменьшая освещенность,
ограничивает углеродное питание растений. Осевшие на поверх-
ности листьев пылевые частицы действуют как экран, сокращаю-
щий доступ ФАР и усиливающий поглощение тепла. Они могут
привести также к закупориванию устьичных щелей. Все это ухуд-
шает условия фотосинтеза и водообмена. Особенно вредна це-
ментная пыль, во влажной среде образующая на листьях плотную
корку. Токсичны для растений и взвешенные в воздухе частицы
моющих средств.
Пыль и дымовые газы могут существенно модифицировать
микроклимат местообитаний. В промышленных зонах нередко
повышается температура и падает влажность воздуха. А в аридных
районах нередко случаются пыльные бури, способствующие силь-
ному иссушению воздуха.
Проникновение токсичных газов в клетки растений. Многие ток-
сичные вещества проникают в растение из воздуха при газообме-
не, с дождем, при осаждении тумана и пыли. Основная часть пе-
реносимых по воздуху загрязняющих веществ перехватывается
нистьями. Они адсорбируются и выводятся из обмена кутикулой,
к пс точными стенками, срединными пластинками, вакуолями.
Первая преграда на пути проникновения токсинов из атмос-
феры в растение — кутикула. Она представляет собой гетероген-
ную мембрану, состоящую из тонкого внешнего слоя (препятству-
ющего попаданию веществ из среды) и внутреннего сорбционно-
и» благодаря им она выполняет двойную роль: барьера и погло-
I тельной емкости. Вещества кутикулы способны ковалентно свя-
ii.iiurrb загрязнители и сохранять их в растении, не подвергая де-
м»кснкации, но эти соединения могут высвободиться в пищева-
ршеньном тракте животных и человека или при разложении под-
281
стилки (М. Riederer, Е. Schonherr, 1993). Кутикулярный барьер
обычно не пропускает токсичные газы. Поэтому основная масса
их попадает в листья при газообмене через открытые устьица п
чечевички. При этом нарушается регуляторная функция замыка-
ющих клеток, токсичные вещества накапливаются в межклетни-
ках и через клеточные стенки проникают к клеткам мезофилла.
Многие токсичные газы растворяются водой клеточной стенки
(она составляет около 60 % массы листа) и реагируют с ней, обра-
зуя кислоты. При этом содержание подвижной воды в клетке па-
дает, что снижает скорость движения цитоплазмы, нарушает внут-
риклеточный транспорт и приводит к плазмолизу. Чем больше
растворимость загрязнителя в воде (а газообразные соединения
фтора, серы и азота имеют очень высокую растворимость), тем
выше скорость его поглощения и сильнее воздействие на орга-
низм.
Газы, образующие кислоты, снижают буферную емкость кле-
точных стенок, благодаря которой они были способны к много-
кратной адсорбции и десорбции ионов. При понижении pH сте-
пень адсорбции ряда ионов падает, что может быть причиной
вымывания из листьев ионов Са2+ и Mg2+ у растений, произрас-
тающих в условиях хронического загрязнения воздуха кислыми га-
зами. Атак как CaF2, образующийся при действии HF, в воде прак-
тически нерастворим, он выводит ионы кальция из метаболизма,
поэтому связанный кальций накапливается в листьях, подвергших-
ся действию фторидов (Р.Гудериан, 1979). Осаждение CaF2 в стен-
ках ведет также к увеличению сопротивления току воды в них.
Далее токсичные соединения атакуют плазмалемму и прони-
кают в цитоплазму. При этом часто нарушаются структура и фун-
кциональная активность клеточных мембран, благодаря чему со-
здаются условия для быстрого проникновения токсинов внутрь
клетки и нарушения обмена веществ.
Действие токсичных газов на растения. Вредное влияние ток-
сичных газов на растения происходит путем прямого их воздей-
ствия на ассимиляционный аппарат и косвенно — через почву.
Первое приводит к ухудшению роста, отмиранию органов, сни-
жению количества и качества урожая, а второе уменьшает плодо-
родие почвы, вызывает гибель полезной микрофлоры, отравление
корневой системы, нарушение минерального питания и как след-
ствие — снижение урожая. Токсичные газы могут также усили-
вать накопление мутаций. У видов одно и то же вещество вызыва-
ет разные типы и разную степень повреждений. С другой сторо
ны, сходные нарушения могут быть вызваны различными веще-
ствами. Токсическое повреждение растений зависит от получен
ной ими дозы.
Дымовые газы действуют на растения комплексно. Сначала они
вызывают большой спектр реакций, типичный для растений в
282
с трессовой ситуации, а затем нарушают процессы жизнедеятель-
ности клетки. При больших концентрациях ядовитых примесей в
воздухе растения могут получить обширные ожоги и погибнуть, а
длительное воздействие низких концентраций газообразных ток-
синов ведет к накоплению хронических нарушений. При этом у
пораженных растений снижается устойчивость к засухе, холоду,
вредителям. При больших концентрациях газообразных токсинов
клетки деформируются, клеточный сок закисляется, повреждают-
ся хлорофилл и хлоропласты, происходит сбой движения цито-
плазмы и работы ферментных систем. Все это приводит к разру-
шению важных органических соединений и последующему отми-
ранию клеток (С. П. Васфилов, 2003).
Изменение фотосинтетической активности листьев служит чув-
ствительным показателем их повреждения загрязняющими веще-
ствами, так как одна из первых мишеней их действия — фотосин-
тетические реакции. Попав внутрь листа, токсичные газы запус-
кают на свету процессы фотоокисления белков, аминокислот и
других важных веществ. В клетках мезофилла накапливаются пе-
роксид водорода и кислородные радикалы, ингибируются одни
ферменты (каталаза, кислая фосфатаза) и активизируются другие
(например, пероксидаза, разрушающая гормон — индолилуксус-
ную кислоту), деградируют тилакоиды, набухают и разрушаются
х поропласты. В результате у сосны {Pinus sylvestris), например,
фотосинтез может снижаться более чем вдвое, причем ингибиру-
с гея он задолго до появления видимых повреждений.
Изменяются и другие физиологически важные процессы. Так,
интенсивность дыхания сначала возрастает, а потом сильно пада-
ет. Токсичные атмосферные примеси даже в невысоких концент-
рациях могут в 1,5 — 2 раза снизить интенсивность транспирации.
11анример, деревья в промышленных зонах часто начинают Сухо-
ве ршинить, так как на верхушке кроны транспирация падает осо-
(к’нно быстро, а из-за нарушения движений устьичных клеток
л нс иной ход транспирации у поврежденных деревьев становится
Гюлсс изменчивым.
Нарушая ход важнейших физиологических и биохимических процес-
« он, тормозя рост и развитие и вызывая падение продуктивности, соеди-
нения токсичных газов сильно ослабляют растения. Например, одна из
ш рных реакций хвойных на загрязнение — снижение устойчивости к
пи 1ким температурам. Атмосферные токсиканты сбивают естественную
/ишамику вхождения деревьев в зимний покой, нарушая состав сахаров.
Например, сосны на загрязненных территориях позже входят в состоя-
ние покоя, который имеет меньшую глубину, и раньше из него выходят.
Часн> по завершившие переход к зимнему покою растения оказываются
не югоны к суровым условиям зимы и сильно повреждаются морозом.
1ак, на Кольском полуострове при сильном загрязнении сернистым га-
юм морозоустойчивость хвои сосны падает в 2 —4 раза. А выбросы мед-
283
но-никелевого комбината привели там к гибели лесов на 40 тыс. га
(А.Л.Федорков, 2002).
Токсины заметно изменяют внутриклеточные и анатомические
структуры в растениях. Так, усиление окислительно-восстанови-
тельных процессов приводит к лавине неуправляемых реакций и
нарушению компартментов клетки, повреждению органелл. В дре-
весных побегах развиваются лизигенные межклетники, склери-
фицируются первичная кора и флоэма, уменьшается доля вторич-
ных тканей. В ксилеме увеличивается количество сосудов, но уко-
рачиваются длина и диаметр их члеников. Так как деятельность
камбия подавляется, то уменьшается растяжимость клеток и со-
кращается время камбиальной активности. У разных растений
выраженность этих признаков неодинакова. Например, у осины
(Populus tremula) работа камбия нарушается сильнее, чем у дру-
гих тополей (Р. nigra, Р. alba) (К.Ф.Хмелев, В.Н.Хватова, 2003).
Разные органы и ткани растений неодинаково чувствительны
к атмосферным загрязнителям. При действии дымовых газов преж-
де всего сильно нарушается углеродный и азотный обмен в асси-
милирующей ткани. Поэтому листья растений быстрее стареют.
Проводящие ткани страдают меньше, так как одревесневшие клет-
ки почти не изменяются. Однако живые клетки флоэмы повреж-
даются довольно сильно (например, существенно подавляется их
растяжение). Промышленные газы неблагоприятно действуют и
на корневые системы. Значительно уменьшаются общая масса кор-
ней и число активных сосущих корневых окончаний.
Отрицательные последствия загрязнения окружающей среды
широким спектром поллютантов разной природы находят свое
выражение и в увеличении скорости мутационных и рекомбина-
ционных процессов, увеличении генетического груза и падении
биоразнообразия. Таким образом, повреждения растений в заг-
рязненной атмосфере проявляются на разных уровнях, и общая
устойчивость организма складывается за счет разных форм адап-
таций.
Токсичность промышленных выбросов в разных условиях дей-
ствует неодинаково. Так, ядовитость оксида серы зависит от тем-
пературы и светового режима. Поэтому самые сильные поврежде-
ния растения получают в полдень при максимальных освещенно-
сти и температуре, а минимальные — ночью. На открытом месте
повреждения также будут существеннее, чем в затенении. Силь-
ное влияние на токсичность газов оказывает влажность воздуха.
Например, при высокой концентрации сернистого газа увеличе-
ние влажности среды приводит к образованию сернистой, а затем
серной кислот, в результате чего токсичное действие усугубляет-
ся. Усилить отрицательное действие газов при их высокой кон-
центрации может безветрие. Отмечен также синергетический эф-
284
фект, при котором загрязнители (например, соединения серы и
азота) усиливают действие друг друга (Р. Гудериан, 1979).
Газочувствительность и газоустойчивость. Чувствительность и
устойчивость к токсичным газам — разные аспекты реагирования
растений. Газочувствительность отражает скорость и степень
проявления у растений патологической реакции при воздействии
газа. Обычно она снижается при усилении ксероморфности, а
древесные растения в целом менее чувствительны к загазованно-
сти, чем травянистые. Наиболее чувствительные виды служат ин-
дикаторами загрязнения.
Газоустойчивость определяют как способность растения про-
тивостоять действию газов, сохраняя нормальный рост и разви-
тие. Например, ель (Picea abies) чувствительна к сернистому газу,
сосна (Pinus sylvestris) еще чувствительнее к ней, но она одновре-
менно и более устойчива, чем ель. Из-за слабой кутикулы очень
чувствительна к вредным газам лиственница (Larix), но ее же ус-
тойчивость к ним обусловлена ежегодным сбрасыванием хвои.
Сезонное выключение газовоспринимающего аппарата (устьиц)
позволяет ей неплохо сохраняться в городах. Определение газоус-
тойчивости важно в озеленительной практике, особенно при ис-
пользовании вечнозеленых растений, листва которых долго под-
вержена воздействию повреждающих газов.
Реакция одних и тех же растений на различные газы неодинако-
ва. Например, кукуруза (Zea mays) очень чувствительна к высокой
концентрации HF и устойчива к SO2. Фасоль (Phaseolus vulgaris)
и томаты (Lycopersicori) чувствительны к воздействию азотсодер-
жащих газов и относительно устойчивы к воздействию HF, а капус-
та (Brassica oleracea), тыква (Cucurbita pepo) и лук (Allium сера)
устойчивы к азотсодержащим газам и неустойчивы к HF и SO2.
По газоустойчивости отмечено большое разнообразие расте-
ний. Она зависит от состояния ассимиляционного аппарата и кор-
невой системы растений, фазы их развития и возраста, интенсив-
ности роста и экологической пластичности видов, эколого-гео-
графического происхождения и положения в филогенетической
системе (В.Т.Ярмишко, 1997 и др.). Так, вблизи химических за-
водов нередко полностью гибнут сосняки, а березняки в этих же
условиях признаков угнетения практически не обнаруживают. Для
хвойных часто вредны обычные городские условия: в загрязнен-
ном воздухе городов отмечается их неправильное ветвление, от-
мирают верхушки деревьев и хвоя. Из хвойных пород сильнее
всего подавляется рост у лиственницы в охвоенном состоянии. Из
лиственных пород к дымовым газам очень чувствительна осина
(Populus tremula), а липа (Tilia cordata) с кленом (Acerplatanoides)
н малочувствительны, и устойчивы к сернистому газу.
Газоустойчивость реализуется благодаря разным биологическим
особенностям растений. На основании этого Ю.З. Кулагин (1973)
285
выделил несколько типов газоустойчивости. Анатомическая га-
зоустойчивостъ связана со структурными особенностями расте-
ний, ограничивающими газообмен и затрудняющими проникно-
вение газов в ткани. Ею обладает ряд суккулентов и многие расте-
ния с жесткими листьями (Rhododendron, самшит — Buxus и др.).
Физиологическая газоустойчивость основана на особенностях
взаимодействия внутренних тканей с окружающим воздухом. Био-
химическая газоустойчивость базируется на защите систем об-
мена веществ и ферментных комплексов. Особую роль в этом иг-
рает окисляемость клеточного содержимого (газоустойчивость тем
выше, чем она меньше). Габитуальная газоустойчивость умень-
шает возможности контакта листьев и цветков с токсичными га-
зами из-за высоты растения, особенностей ветвления, образова-
ния стланиковых и подушковидных форм и т.д. Феноритмичес-
кая газоустойчивость определяется несовпадением во времени
воздействия газа и критических периодов вегетации. Анабиоти-
ческая газоустойчивость связана с периодами покоя растений.
Регенерационная газоустойчивость обусловлена способностью
быстро восстанавливать поврежденные органы благодаря повтор-
ному облиствению и побегообразованию (этот вариант газоустой-
чивости развила, например, бузина — Sambucus). Популяцион-
ная газоустойчивость зависит от разных возможностей и по-
требностей особей разных возрастных состояний в популяции. Фи-
тоценотическая газоустойчивость связана с вертикальной и го-
ризонтальной неоднородностью фитоценоза, препятствующей про-
никновению газов.
Газоустойчивость часто характерна для определенных родов и
семейств растений. Это определяется их анатомо-морфологиче-
ской и физиолого-биохимической спецификой. Например, окис-
ляемость клеточного сока зависит от систематического положе-
ния видов (очень газоустойчивы жимолостные, пасленовые, ки-
парисовые). С другой стороны, у ряда жизненных форм и эколо-
гических групп свойства, способствующие устойчивости к ток-
сичным газам, связаны с анатомической и габитуальной устойчи-
востью (ксерофиты, деревья).
С эволюционной точки зрения, газы промышленных выбросов
начали действовать на растения совсем недавно. Выработать к ним
специфические адаптации виды еще не успели, поэтому нельзя
выделить специальные анатомо-морфологические или физиоло-
гические признаки, связанные исключительно с выносливостью
к загрязнениям. Исходя из этого, газоустойчивость должна опре-
деляться теми свойствами, которыми растения уже располагали в
естественной среде. Это отражено в разработанной Ю.З. Кулаги-
ным преадаптационной концепции. Противостоя токсичным га-
зам, растения мобилизуют уже существовавшие средства устойчи-
вости к другим неблагоприятным влияниям (холоду, засухе и пр.).
286
Поэтому в ряде случаев устойчивыми к загрязнению оказываются
виды, выносливые и к другим экстремальным факторам. Напри-
мер, более ксерофильные дуб (Quercus robur) и белая акация
(Robinia pseudacacia) газоустойчивее, чем мезофилльные липа
(Tilia cordata) и конский каштан (Aesculus). Более стойки также
растения засушливой зоны, происходящие из районов с карбонат-
ными и слабозасоленными почвами. Вероятно, как и при адаптаци-
ях галофитов, это связано с повышенным содержанием у них ионов
кальция, калия и натрия, которые могут эффективно связывать
поглощенные анионы. Стойки к загрязнению и многие сорняки,
например пикульник (Galeopsis tetrahit), чему способствуют осо-
бенности листьев (восковой налет, защищенные устьица и др.).
Фитоиндикация загрязнения атмосферы. Возможность возник-
новения синергетических и антагонистических эффектов факто-
ров совместного действия обусловливает некорректность оценок
опасности поллютантов, основанных исключительно на данных
химико-аналитического контроля. Поэтому во многих случаях
перспективен биологический мониторинг, позволяющий получать
интегрированную оценку последствий действия суммы загрязняю-
щих веществ. Очень чувствительны к внешним воздействиям гене-
ративные органы, поэтому оценивать опасность загрязнения эффек-
тивнее по параметрам репродуктивной способности растений (се-
менная продуктивность, масса и размеры семян, их всхожесть, а
также морфологические нарушения у семян и проростков).
У высших растений физиологические повреждения атмосфер-
ными токсикантами часто сопровождаются ожогами, некротичес-
кими пятнами, побурением листьев и отмиранием их краев, фор-
мированием уродливых форм (смятых, скрученных и др.). У дре-
весных растений хорошими индикаторными признаками могут
служить также явления, связанные с нарушением водного обмена
(опадение листьев и суховершинность). Рост поврежденных дере-
вьев ослабляется, кольца в древесине становятся более узкими.
При больших дозах токсических веществ может произойти пол-
ная дефолиация и гибель растений.
В определении степени загрязнения воздуха используется ряд
особо чувствительных высших растений. Например, отдельные
сорта гладиолусов (Gladiolus) и тюльпанов (Tulipa) служат хоро-
шими индикаторами загрязнения фтористым водородом. Хоро-
шим биоиндикатором служит и распространенный мятлик одно-
летний (Роа annua), чувствительный к токсическим газам. Пока-
штелями загрязнения воздуха могут быть и некоторые древесные
растения. Так, у лещины (Corylus), платана (Platanus), катальпы
(Catalpa) листья повреждаются при хроническом действии даже
небольших концентраций SO2. А при фитоиндикации утечки газа
из трубопроводов фиксируют признаки отравления и факты ис-
чезновения определенных видов растений.
287
Хорошими показателями чистоты воздуха являются лишайни-
ки. Они эффективно накапливают минеральные катионы, извле-
кая их из дождевой воды, стекающей по субстрату, капающей с
растений и др. На этом факте основан принцип лихеноиндика-
ции. Выделены виды лишайников, устойчивые к городской среде
и неустойчивые. Выявлена корреляция между загрязнением воз-
духа (серный диоксид, оксиды азота, соединения фтора и т.п.) и
видовым разнообразием лишайников: чем выше загрязнение ат-
мосферы, тем беднее лихенофлора. Предложены специальные ли-
хенометрические индексы и формулы расчета атмосферного заг-
рязнения, учитывающие состав, число видов и встречаемость раз-
ных групп лишайников. Периодическое картирование эпифит-
ных лишайников позволяет вести мониторинг атмосферного заг-
рязнения. Для оценки степени загрязнения воздуха разработан
также метод трансплантации лишайников в места промышлен-
ных выбросов. В этом случае оценивается повреждение слоевищ,
выставленных вместе с кусочками коры, на которых они росли.
По сравнению с другими методами лихеноиндикация позволяет
учитывать кумулятивное действие длительных нагрузок и прово-
дить сравнения через большие промежутки времени.
Многократно показано, что распространение лишайников в городах
хорошо отражает степень загрязнения атмосферы. Лихеноиндикацион-
ные карты составлены для многих промышленных центров (рис. 9.7). На
них вокруг загрязняющего предприятия обычно располагается «лишай-
никовая пустыня», где лишайники совсем или почти совсем отсутству-
ют. Вокруг этой территории располагается «зона борьбы», где могут при-
сутствовать немногие виды, обычно с малым покрытием и в угнетенном
состоянии. Дальше лишайники становятся обильнее, но сдвиг в видовом
составе еще указывает на токсическую нагрузку.
Наряду с высокочувствительными растениями в качестве био-
индикаторов часто используют и газоустойчивые виды. Благодаря
своей выносливости они могут накапливать вредные вещества,
поэтому их относят к аккумулирующим индикаторам. Обнару-
живают действие некоторых токсикантов (SO2, HF и НС1) на ра-
стения по аккумулирующим индикаторам с помощью химическо-
го анализа на содержание определенных элементов в органах ин-
дикаторных растений. Так, капуста возле промышленных пред-
приятий накапливает в листьях Fe, S, Pb, а вблизи алюминиевых
заводов в таких индикаторах сильно возрастает концентрация А1.
Во многих случаях важна ранняя диагностика начавшегося по-
глощения вредных веществ. Морфологические повреждения еще
могут отсутствовать, но уже обнаруживаются химические, функ-
циональные и некоторые структурные изменения. Ранней диаг-
ностике начавшегося повреждения могут служить повышение кон-
288
Рис. 9.7. Карта распространения лишайников в г. Мюнхене
(по PJurging, 1975):
1 — «лишайниковая пустыня»; 2 — «зона борьбы»; 3 — зона нормального роста
центрации вредных веществ в растении, сдвиг pH на поверхности
органов и в тканях, изменение активности ферментов, распад хло-
рофилла, показатели депрессии фотосинтеза, изменения белко-
вого обмена и мобилизации вторичных метаболитов, нарушение
роста, снижение урожая, изменение проницаемости клеток, флу-
оресценция и др. Так, чувствительным способом обнаружения в
воздухе примеси сернистого ангидрида служит определение элек-
тропроводности экстракта из коры ели (В. В. Полевой и др., 2001).
При этом картина газовых повреждений многообразна и неспе-
цифична: одно и то же вещество у тех или иных растений может
вызывать разные эффекты, а сходные симптомы способны вы-
звать разные вещества.
9.4. Влияние на растения перемещения
воздушных масс
Ветер. Представляет собой перенос воздушных масс вследствие
различий в давлении воздуха. В некоторых районах он оказывает
определяющее влияние на климат (тропические области пассатов
и муссонов). А в ряде мест, перенося частицы субстрата, ветер
создает особые формы рельефа (барханы и дюны), заселяемые
только теми растениями, которые могут переносить и обнажение
ветром корневой системы, и засыпание стеблей. Связанный с пе-
реносом тепла и воздушных масс, ветер играет важную роль в
тепло- и газообмене растений, формировании их облика и в рас-
пространении. Например, на Фолклендских островах из-за силь-
289
ных и частых ветров деревья не растут. В подобных условиях ве-
тер оказывает настолько сильное влияние на растительный по-
кров, что, например, безлесные субантарктические острова были
названы А. Шимпером «ветровыми пустынями». Во многих рай-
онах Арктики отдельные деревья или лесные сообщества встреча-
ются только по берегам рек, где они более защищены от ветра.
Действие ветра на организмы может быть прямым и косвен-
ным. При прямом влиянии он вызывает механические деформа-
ции растений, обусловленные торможением движущегося возду-
ха, переносит их диаспоры. Как механический фактор ветер обычно
эффективен лишь там, где дует с особой силой (прежде всего в
горах и на морских побережьях). При косвенном действии он из-
меняет режимы других экологических факторов. Так, раскачива-
ние крон меняет световой режим для лесных растений. Даже лег-
кий ветер оказывает влияние на тепловой баланс сообществ. При
сильной инсоляции он понижает температуру растений из-за ак-
тивизации теплообмена, но может также подводить теплый воз-
дух с более сильно нагретых участков. В степях и полупустынях
переносящие горячий и сухой воздух суховеи существенно повы-
шают испарение и транспирацию, что отрицательно сказывается
на урожае. В городах ветер распространяет загрязнения, поэтому
розу ветров учитывают при благоустройстве и планировании эко-
логического мониторинга.
Ветер действует на растения не только как экстремальный кли-
матический фактор, хорошо известно его положительное влияние
на размножение и расселение растений. У многих видов он обес-
печивает перенос пыльцы, спор, семян, плодов, частей растений
и даже целых организмов.
Деструктивное механическое влияние ветра на растения (ветро-
валы, буреломы, ветровое полегание растений). Сильные порывы
ветра могут ломать деревья (вызывать бурелом) и вырывать их с
корнем (ветровал). Ветровальные нарушения — распространен-
ное явление на лесных территориях. Так, по Н. Г. Улановой (2004),
в разновозрастных ельниках Центрально-лесного заповедника они
занимают 7—13% площади. Массовый ветровал ведет к полной
гибели древостоя и резкому изменению экологических условий, в
первую очередь микроклимата (освещенность, влажность). Раз-
ные виды растений неодинаково страдают от ветров. Это зависит
от характера их корневой системы и анатомии ствола (зимой еще
имеет значение хрупкость промерзших стволов).
Ветровалу особенно подвержены крупные деревья с большой
парусной поверхностью и поверхностной корневой системой.
Например, на подзолистых почвах средней полосы России имею-
щая поверхностную корневую систему ель (Picea) — типичная
ветровальная порода. Но на более богатой почве, где укоренение
глубже, она хорошо выдерживает напор ветра даже на открытых
290
местах. В искусственных же посадках ель часто страдает от ветро-
вала в чистых одновозрастных насаждениях: деревья формируют
там тонкие высокие стволы с небольшой кроной, и порывы ветра,
сильно раскачивая их, вырывают с корнем или ломают. Опас-
ность возрастает также при рубках и на лесосеках, нарушающих
сомкнутость крон. Сосна (Pinus sylvestris) на сырых глинистых
почвах, наоборот, укоренена довольно глубоко, и потому при силь-
ных ветрах вывала не происходит. Но при очень сильном ветре у
сосны может сломаться ствол, а при резких его порывах иногда
отрывается крона. Однако на неглубоких почвах (заболоченных,
при близком залегании грунтовых вод или многолетней мерзло-
ты) ветровалу бывает подвержена и она.
В целом сосна и пихта (Abies), в норме имеющие довольно
глубокие корневые системы, являются «буреломными породами»,
ветер чаще не может вывернуть их с корнем и переламывает ствол.
К буреломной породе из-за непрочной древесины относится и
осина (Populus tremula). А вот липа (Tilia) от бурелома страдает
меньше, так как имеет развитый луб. Ветроустойчивы же породы
с глубокой и разветвленной корневой системой — дубы (Quercus),
сосна горная (Pinus mugo), сосна сибирская (Р. sibiricd), секвойя
(Sequoia), тутовое дерево (Morus) и др. Сильно подвержены вет-
ровалу некоторые вечнозеленые деревья (лавр — Laurus, олива —
Olea, мирт — Myrtus). Из летнезеленых пород таковы ивы (Salix),
березы (Betula), боярышник (Crataegus), бук (Fagus) и многие
тополя (Populus) (Т. К. Горышина, 1979 и др.).
В сомкнутых фитоценозах ветер встречает большое сопротив-
ление, поэтому сильно страдают от ветровала разреженные на-
саждения и оставшиеся на вырубке одиночные деревья. Ветровал
обычно открывает путь проникновению ветра в глубь насажде-
ния, что вызывает новые ветровалы. В северных лесах это иногда
создает непроходимые завалы на десятках гектаров. И восстанов-
ление леса после возобновляющихся здесь ветровалов может идти
очень долго.
Ветер вызывает также полегание многих травянистых расте-
ний. Молодые особи могут выпрямиться за счет более сильного
роста боковых частей стебля, но в конце периода вегетации этого
уже не происходит. Полегание сильно затрудняет уборку зерно-
вых и ведет к большим потерям урожая хлебных злаков, кукурузы
(Zea mays), сахарного тростника (Saccharum officinarum) и др.
Скорость ветра возрастает с высотой над поверхностью почвы
и различается с разных сторон препятствия (например, она мак-
симальна в кроне дерева с наветренной стороны). Растительный
покров значительно снижает скорость ветра. Поэтому борьбе с
ветровалом помогает создание перпендикулярных направлению
господствующих ветров ветроустойчивых опушек из лиственных
деревьев и кустарников с плотной кроной. Препятствуют ветру и
291
Рис. 9.8. Изменение скорости ветра в растительных сообществах
(по Р. Гейгер, 1960; О. Stocker, 1933; J. Wilson, 1959):
А — в лесу (7 — 1,5 м/с; 2 — 2,5 м/с; 5 — 4 м/с); Б — в степи, В — в тундре
морфологические особенности лесообразователей. Так, при ско-
рости ветра на поле 2,8 м/с в ельнике на расстоянии 40 м от
опушки она достигает лишь 1 — 1,5% от исходной. И травяной
покров может эффективно останавливать движение воздуха.
Т. К. Горышина (1979) указывает, что даже при штормовых ветрах
приземистые облиственные растения редко подвергаются действию
ветра со скоростью более чем 10 м/с (рис. 9.8).
Формообразующее действие ветра на растения. Ветры могут
оказывать сильное воздействие на внешний вид растений. Это
наблюдается в местах, где они постоянно дуют в одном направле-
нии (на перегибах горных хребтов, морских побережьях и т.д.).
Рис. 9.9. Флаговые формы крон деревьев (по Т. К. Горышиной, 1979).
А — ель (Picea) на верхней границе леса в Высоких Татрах; Б — сосна (Pinus) на
Балтийском побережье
292
Рис. 9.10. Деформирующее влияние
господствующих ветров на радиаль-
ный прирост ствола лиственницы
(Larix) на Таймыре (по Н.В.Лове-
лиусу, 1973):
/ — величина годичного прироста ство-
ла (в мм) по румбам; 2 — повторяемость
ветров по направлениям (дней в году).
Преобладают ветры С, СВ и В румбов, с
наветренной стороны прирост мини-
мальный
Такие ветры нарушают нормальный рост деревьев, вызывая не-
равномерный прирост древесины, наклон ствола, однобокость
кроны. Некоторые деревья формируют плоскую крону, другие —
флагообразную (вытянутую в подветренную сторону) (рис. 9.9).
Образование таких форм вызвано отмиранием почек с наветрен-
ной стороны. Иногда меняется даже строение ствола, и древесина
на срезе принимает асимметричное строение (рис. 9.10).
Так как у земли сила ветра из-за трения становится меньше,
ветровое действие на кустарниках сказывается слабее, чем на де-
ревьях. Но и у них при постоянно сильных ветрах образуются
ветровые формы. Так, на Южном берегу Крыма встречаются вет-
ровые формы можжевельника (Juniperus excelsd), представляю-
щие собой стланики с ветвями, обращенными в противополож-
ную от господствующих ветров сторону (рис. 9.11). А на Шпиц-
бергене отмечено одностороннее отмирание растений-подушек с
наветренной стороны.
Губительным последствием ветрового влияния может быть ох-
лестывание деревьев собственными ветвями или кроной сосед-
него дерева. Так, сильно охлестывает своими длинными ветвями
береза (Betula) соседние деревья. Поэтому растущая с ней рядом
ель (Picea) может иметь деформированную крону. Внутрикроно-
вое охлестывание может снижать густоту крон, а межкроновое —
Рис. 9.11. Ветровая стланиковая форма можжевельника (Juniperus
excelsa) на Южном берегу Крыма (по В. В. Алехину, 1950)
293
уменьшать их сомкнутость. Знание этих фактов полезно при пла-
нировании лесохозяйственных мероприятий.
Причины отмирания ветвей под действием ветра могут быть
чисто механическими: обламывание изгибающихся побегов, об-
рывание листьев и т.д. Однако более губительно иссушающее дей-
ствие ветра. Под его влиянием ветви могут испарять влаги намно-
го больше, чем обычно, в результате чего клетки теряют воду и
погибают. Ветровое воздействие приобретает абразивные свойства,
когда потоки воздуха несут твердые частицы (песок, соль, снег).
В пустынях, например, ветер с песком сечет листья, обтачивает
кору и стволы. Особенно страдают крупнолистные деревья.
Растения морских побережий деформируются во многом под дей-
ствием солей, переносимых ветром (Г.Вальтер, 1982). Капли соленой воды
разбрызгиваются, развеиваются ветром и осаждаются на твердых повер-
хностях, что было экспериментально показано для наветренных сторон
деревьев и кустарников. Осажденная соль проникает в листья через мел-
кие трещины кутикулы, вызывая гипертрофию клеток, а при более вы-
сокой концентрации — отмирание листьев. Поэтому на морских побере-
жьях деревья с флаговидной кроной встречаются особенно часто. Видов
же, плохо переносящих соли, здесь вообще нет; например, ель европей-
ская {Picea abies) и сосна обыкновенная {Pinus sylvestris) не растут на
западных морских побережьях Европы.
В суровых приполярных и высокогорных условиях сильный
ветер часто несет острые снежные частицы, сдирая дернину и со-
стригая ветви кустарников на небольшой высоте. Это явление
называют снежной корразией (от лат. corrado — скоблю, соскре-
баю). Под ее воздействием зимой гибнут все органы растений,
оказавшиеся выше снежного покрова. В результате летом древес-
ные виды в этих местах выглядят подстриженными. Так, в высо-
когорьях описаны столовые формы деревьев и кустарников с низ-
Рис. 9.12. Столовые формы древесных пород (по Е. Fries, 1913;
P.Plesnik, 1971, из Т.К.Горышиной, 1979):
А — береза {Betula) на северной границе леса; Б — ель {Picea) на склоне Белан
ских Татр. Пунктир — граница снегового покрова
294
Рис. 9.13. Ветвление ели (Piced) в местообитаниях с сильными ветрами
на верхней границе леса в Высоких Татрах (по Р. Plesnik, 1971, из Т. К. Го-
рышиной, 1979)
кой и очень плоской кроной (рис. 9.12). Своеобразные «юбочные
кроны» (рис. 9.13) могут образовываться у ели в горах и тундрах.
Нижние ветви одиноко стоящих деревьев, защищенные зимой
снежным покровом, остаются неповрежденными, а средняя часть
ствола, выступающая над снегом и открытая воздействию пере-
носимого ветром снега, оголяется из-за отмирания боковых вет-
вей. Выше боковые ветви опять появляются, так как тяжелые кри-
сталлы льда их не достигают, и крона тут развивается, хотя и де-
формированная.
Другой вариант формообразования при механическом и иссу-
шающем действии ветра выработался у деревьев саванн. Для них
характерны уплощенные зонтиковидные кроны, подставляющие
ветру минимальную поверхность. Самый известный пример —
зонтиковидная акация (Acacia spinocarpa и др., рис. 9.14). При
сходных условиях зонтиковидные кроны образуются у деревьев
разных семейств (Caesalpiniaceae, Myrtaceae, Burseraceae).
Карликовость арктических и горных растений отчасти тоже фор-
мируется под влиянием ветра. Растения, развивающиеся под постоян-
ным его воздействием, из-за иссушения имеют пониженную обвод-
ненность, сниженный тургор. В этих условиях в период роста и растя-
жения клеток они не вытягиваются до нормальных размеров и стано-
вятся карликовыми. Деревья здесь и при значительном возрасте могут
едва достигать высоты обычного кустарника. Благодаря приземистости
карликовые растения избегают влияния сильного ветра, заметно ослаб-
295
Рис. 9.14. Зонтиковидные акации (Acacia spinocarpa) в восточноафри-
канской саванне (по В. В.Алехину, 1950)
ляющегося у земли. Для них имеет значение и более высокая темпера-
тура почвы по сравнению с воздухом. Защищает такие растения от вет-
ра и микрорельеф.
Иссушающее действие ветра на растения. Ветер способствует
испарению даже во влажной атмосфере из-за усиления конвек-
ции. Обычно он повышает транспирацию, сдувая с поверхности
листа сырой воздух. Особенно интенсивно этот процесс идет у
мелких листьев. Однако транспирация не пропорциональна уве-
личению скорости ветра. У разных видов он может изменять транс-
пирацию по-разному: у одних уже несильный ветер резко повы-
шает водоотдачу, а у других на ветру она не меняется или даже
снижается. Эффект ветрового иссушения связан с состоянием ус-
тьиц, чья регулирующая роль при ветре обычно усиливается. При
сильной ветровой нагрузке устьица закрываются и влага теряется
только при слабой кутикулярной транспирации. Скручивая лис-
тья, ветер также сжимает межклетники, усиливая выдавливание
водяного пара через устьица.
Особенно сильно ветер иссушает растение в холодную пого-
ду. Так он действует на ветви листопадных деревьев и кустарни-
ков зимой. И весной, когда надземные органы заметно нагрева-
ются, а почвенная влага еще малодоступна, ветровое иссушение
может быть губительным. Вредно оно и в летнюю жару, когда
почва содержит мало доступной влаги и дует сухой ветер. При
этом транспирация резко возрастает, а корни не успевают по-
дать нужное количество воды, и растения вянут. А суховей в
конце цветения — начале созревания злаков влечет за собой сла-
бое развитие колоса, малочисленность и маловесность зерен. Это
может наблюдаться даже при культивировании растений в усло-
296
виях полной обеспеченности водой (например, у риса — Oryza
sativa).
В особенно ветреных условиях (на островах, в высокогорьях) у
растений прослеживаются признаки ксероморфоза и угнетенного
водного режима, в частности повышение осмотического давле-
ния. Эти изменения особенно заметны на наветренной стороне
растений и у высокорослых форм.
К сильным ветрам хорошо приспособлены высокогорные по-
душечники, и иногда ветер способствует образованию подушек у
растений, в норме не подушковидных (у них отмирают верхушки
побегов и усиливается ветвление). Плохо ослабляют ветер и силь-
но страдают от иссушения афилльные ксерофиты. У пустынных
саксаулов при сильных ветрах годичный прирост снижается в 2 —
3 раза, а содержание воды в ветках при скорости ветра 13 м/с
падает на четверть.
Анемофилия. Анемофилия (от греч. anemos — ветер) — опы-
ление с помощью ветра — очень распространенный способ дос-
тавки пыльцы. Среди покрытосеменных около 10% видов ане-
мофильны. Ветроопыляемые растения преобладают среди голо-
семенных и однодольных. Опыляется ветром большинство лесо-
образующих пород лесной зоны (сосна — Pinus sylvestris, ель —
Picea, береза — Betula, дуб — Quercus robur, осина — Populus
tremula), однако под пологом леса анемофилов мало. Анемофи-
лы — основные доминанты пустынь, саванн, степей и лугов (по-
лыни — Artemisia, солянки — Salsola, осоки — Carex, злаки).
В лесах умеренного климата к ним относятся около 20 % видов
(А. Е. Васильев и др., 1988). Этот процент снижается в тропиках,
но возрастает в полярных широтах и районах, где господствуют
злаковники (степи, саванны). На океанических островах, где обыч-
ны сильные ветры, до трети видов ветроопыляемы. Анемофилов
также обычно много и в тех условиях, где насекомые немногочис-
ленны (в Арктике).
Существует ряд характерных приспособлений к ветроопыле-
нию. Анемофильные растения часто обладают раздельнополыми
соцветиями. Цветки их обычно мелки, невзрачны, голые или с
чашечковидным околоцветником, лишены запаха. Характерны
длинные перистые рыльца, особой сложности и разнообразия до-
стигающие в семействе Крапивные. Пыльники анемофилов обычно
сидят на длинных, легко раскачивающихся тычиночных нитях (как
у злаков и конопли — Cannabis). Раскачиваются ветром и сереж-
ки. А тычиночные нити крапивы (Urtica) с силой раскручиваются
и выбрасывают пыльцу. Пыльца анемофилов мелкая, сухая, с тон-
кой нескульптурированной оболочкой или облегчающими массу
воздушными мешками (рис. 9.15). Облегчает перенос пыльцы вет-
ром и раннее цветение растений. Так, ряд древесных и кустарни-
ковых пород пылит до распускания листьев.
297
2 3
5 6
Рис. 9.15. Формы пыльцевых зерен ветроопыляемых
и насекомоопыляемых растений (по В.Л. Комарову, 1949):
А — ветроопыляемые; Б — насекомоопыляемые; I — сосна (Pinus sylvestris); 2 —
береза (Betula)', 3 — конопля (Cannabis)', 4— мальва (Malva)', 5— тыква (Cucurbita
pepo)', 6 — цикорий (Cichorium)
Пыльца анемофилов образуется в огромных количествах. Например,
у кукурузы (Zea mays) в мужской метелке содержится до миллиона пы-
линок. А рогоз узколистный (Typha angustifolia) образует так много пыль-
цы, что, например, маори, живущие в Новой Зеландии, собирают ее,
разводят водой и едят как кашу (В. В. Петров, 1981).
Обычно считается, что анемофилия менее надежна и более
расточительна, чем, например, энтомофилия, поскольку у пыль-
цы меньше шансов попасть на рыльце того же вида растения и в
нужное время. Поэтому анемофильные растения часто растут боль-
шими группами и вырабатывают очень много пыльцы. Однако в
процессе эволюции растения, захватывающие пыльцу из воздуха,
трансформировались, чтобы лучше функционировать в окружаю-
щей аэродинамической обстановке. При этом изменялись и пара-
метры пыльцевых зерен, и морфология воспринимающих пыльцу
частей растений.
Так, особые аэродинамические условия создаются в шишках хвой-
ных. Потоки воздуха, проходящие между чешуями, отклоняют движение
пыльцы наподобие водоворота, направляя ее преимущественно к семя-
почкам. Опытами по улавливанию пыльцы разных видов было показано,
что при этом собирается преимущественно пыльца своего вида, облада-
ющая определенными характеристиками. Цветковые чешуи тоже спо-
собны создавать завихрения вокруг цветка, способствующие улавлива-
нию пыльцы. Имеет значение и устройство соцветий. Например, сильно
сжатое соцветие щетинника (Setaria geniculata) ведет себя подобно шиш-
298
ке, и треть пыльцы улавливается им на подветренной стороне. А у име-
ющей рыхлое соцветие полевицы (Agrostis hiemalis) преобладает инер-
ционное столкновение, и больше собирается пыльцы на наветренной
стороне (К. Niklas, 1985). Важно и поведение осей растения под действи-
ем ветра. Так, щетинник обычно колеблется параллельно потоку возду-
ха, а движение полевицы имеет еще перпендикулярный компонент. По-
этому, проигрывая в сборе пыльцы в статичных условиях, она компен-
сирует это волнообразными движениями стебля, увеличивающими воз-
можность столкновения с пыльцой. Возможно, поэтому растениям с плот-
ными соцветиями выгоднее расти в тесных скоплениях, а со свободны-
ми — в разреженных группах.
Легкая пыльца анемофилов может переноситься ветром на сотни
километров. Так, пыльцевые зерна сосны улавливаются в Атлан-
тике более чем за 100 км от берега. Пыльца ряда деревьев была
обнаружена в воздухе пустынь (Каракумы, Сахара), а также на
Шпицбергене за многие десятки километров от мест произраста-
ния. Но дальний занос обычно не обеспечивает опыление, так
как пыльца анемофилов легко высыхает, теряя жизнеспособность.
Анемохория. Анемохория — расселение с помощью ветра —
один из наиболее эффективных способов распространения расте-
ний, иногда на огромные расстояния. Так, в Арктике шторм мо-
жет унести семена на 2 000 км (В. Грант, 1991). Анемохоры обыч-
но первыми заселяют свободные субстраты.
Исследованием Л. И. Абрамовой (1968) зарастания выработанных тор-
фяников Мещерской низменности после завершения торфодобычи вы-
явлено, что 65% растений, поселяющихся здесь в первые 2 — 5 лет, —
анемохоры. Среди них есть древесные растения (березы — Betula, ивы —
Salix, осина — Populus tremula), сорные и сорно-луговые травы (иван-
чай — Chamaenerion angustifolium, кульбаба — Leontodon automnalis,
крестовник обыкновенный — Senecio vulgaris), водно-болотные виды
(рогоз — Typha latifolia, тростник — Phragmites australis, пушицы —
Eriophorum). Аналогичные материалы получены во многих регионах мира.
Те части растений, которые разносятся ветром, обладают ря-
дом сходных черт. Во-первых, они часто очень мелки и легки.
К анемохорам принадлежат споровые растения, так как споры
легко подхватываются ветром, а также некоторые покрытосемен-
ные, имеющие очень мелкие семена или плоды. Минимальная
масса семян достигается за счет предельной редукции запасаю-
щей ткани и зародыша (иногда до нескольких клеток). В резуль-
тате у орхидных, вересковых, грушанковых, росянковых, зарази-
ховых, саррацениевых, раффлезиевых возникают пылевидные се-
мена (масса их редко превышает 0,002 г).
У более крупных плодов и семян существуют различные «аэро-
динамические» приспособления, придающие им парусность и ле-
299
тучесть (рис. 9.16). Так, у многих анемохоров есть крыловидные
выросты. Их, например, имеют многие пустынные растения и
древесные лесные виды. Форма диаспор и эффективность их рас-
сеивания в этом случае весьма разнообразны.
Плоды или семена, планирующие в воздухе, симметричны, их цент-
ры тяжести лишь слегка смещены вперед. Некоторые из них имеют по
2 боковых крыла, как плоды берез. У других видов лишь одно огибаю-
щее крыло, удачным примером чего служит семя тропической лианы
Macrozanonia из семейства Тыквенные. Эти семена легкие (0,25 г), а
крыло их достигает 15 см, поэтому уже при очень слабых движениях
воздуха они эффектно планируют, улетая на большие расстояния. Осо-
бенно богато видами с планирующими семенами семейство Bignoniaceae.
Сходным образом устроены плоды и семена с крылом в виде диска (пло-
ды вязов — Ulmus, санталового дерева — Pterocarpus и др.). Они облада-
ют хорошей летучестью и могут быть далеко отнесены ветром. Меньшие
расстояния преодолевают крутящиеся плоды и семена. У них одно кры-
ло и центр тяжести расположен сбоку. Таковы плоды клена (Acer), ясеня
(Fraxinus), многих хвойных (ель — Picea, пихта — Abies, сосна обыкно-
венная — Pinus sylvestris). Их семена и плоды разносятся ветром на рас-
стояния до 5 км.
Третья группа анемохоров включает плоды и семена с летуч-
ками из волосков, увеличивающими поверхность и позволяющи-
ми снизить скорость падения. Центр тяжести этих диаспор нахо-
дится внизу, поэтому полет осуществляется как с парашютом. По-
мимо летучести волоски помогают семенам пружинить на повер-
хности снега или песка. Так, у джузгуна (Calligonum) плоды пред-
ставляют собой легкие каркасы из упругих, образующих ажурное
сплетение волосков и выростов. Опушенные плоды и плоды с
волосовидными отростками типичны для растений песчаных пус-
Рис. 9.16. Плоды анемохорных растений пустыни
(по А. В. Кожевникову, 1951):
1 — Smimowia turkestana (тип дирижабля); 2, 3, 5 — разные виды Calligonum (плоды
с выростами и крылатками); 4 — Ammodendron conollyi (тип пропеллера); 6 —
Aristida pennata (тип парашюта)
300
Рис. 9.17. Кермек (Statice gmelinii) (по В. В. Алехину, 1950):
А — в период цветения; Б — в период плодоношения
тынь. Эффективному распространению с помощью ветра служат
также длинные ости, как у ряда злаков.
Летучкой с пучком волосков обладают плоды многих злаков (напри-
мер, тростник — Phragmites, пушица — Eryophorum, астрагалы —
Astragalus, кипрей — Epilobium), большинства сложноцветных и др. По-
добные хохолки есть у семян ив (Salix), тополей (Populus) и др. Зонти-
ковидными летучками снабжены плоды ворсянковых, валериановых,
многих сложноцветных (одуванчика — Taraxacum, козельца — Scorzonera
и козлобородника — Tragopogon). Летучки обеспечивают эффективное
разнесение плодов и семян по воздуху. Так, Г. Вальтер, изучая заселение
гари близ Берлина, обнаружил, что среди видов, появившихся в первые
годы, около 86 % составили анемохоры, а среди них примерно половина
размножалась очень мелкими семенами, половина имела плоды или се-
мена с хохолками или зонтиковидными летучками. Аналогично шло за-
селение растениями острова Кракатау после извержения в 1885 г. вулка-
на, уничтожившего все живое.
Распространению ветром служат также вздутые оболочки пло-
дов и семян некоторых растений (смирновия — Smirnoivia, осока
вздутая — Carex physodes, физалис — Physalis и др.). В пустынях
эти шары легко перекатываются под действием ветра или как воз-
душные шары медленно передвигаются в приземном слое в пото-
ках нагретого воздуха (Carex bohemica, С. duriuscula и др.).
У ряда других анемохоров приспособления к переносу семян
или плодов ветром выражаются в облике и форме роста всего ра-
301
стения. Это встречается у жизненной формы «перекати-поле».
В основном это растения шаровидного облика, который достига-
ется обильным ветвлением и отгибанием веточек книзу (рис. 9.17).
Подобный тип рассеивания семян выработался независимо в раз-
ных семействах: к перекати-поле относятся петросимония (Petrosi-
monia) из маревых, клоповник (Lepidium) из крестоцветных, со-
лончаковая астра (Aster tripolium) из сложноцветных, резак
(Falcarid) из зонтичных и т.п. Растут эти растения на открытых
ветру пространствах — в степях, полупустынях и пустынях, на
приморских пляжах. К концу вегетационного периода они часто
подсыхают, и ветер их выдергивает или обламывает. Ко времени
рассеивания семян представители большинства таких видов ста-
новятся более или менее шарообразными, что увеличивает их па-
русную поверхность и способствует перемещению. Будучи очень
легкими, эти шары перекатываются, рассеивая семена. При этом
для распространения семян на большей площади выработался ряд
приспособлений к их постепенному разбрасыванию: отверстия
коробочек периодически становятся узкими и внутри них разви-
ваются волоски, затрудняющие высыпание семян.
ГЛАВА 10
ПОЧВА КАК ЭКОЛОГИЧЕСКИЙ ФАКТОР
Основоположник современного почвоведения В. В. Докучаев
сформулировал понятие о почве как «самостоятельном естествен-
но-историческом теле, которое является продуктом совокупной
деятельности грунта, климата, растительных и животных организ-
мов, возраста страны, а отчасти и рельефа местности». Позже к
факторам почвообразования добавили деятельность человека.
Наибольшее экологическое значение для всех почв имеют клима-
тические факторы. Они обеспечивают условия теплообеспечен-
ности и увлажнения, а также связанные с ними воздушный и со-
левой режимы. С климатическим зонированием связаны зоны почв
и растительности. Многообразие почв в значительной степени
обусловливает почвообразующая порода (минеральный субстрат,
на котором развиваются почвы). От нее зависит не только мине-
ральный состав почв, но и их химические особенности, а также
механические, водно-физические и многие другие свойства. Гор-
ные породы, из которых формируются почвы, очень разнообраз-
ны. В процессе выветривания происходят глубокие изменения их
)цементного и минерального состава: они превращаются в кору
выветривания (глинистую, песчаную и др.). В верхней части ее
при участии организмов формируется почва.
10.1. Основные свойства почвы и их экологическое
значение
Возраст почвы. Возраст почвы можно рассматривать как осо-
бый экологический фактор. Стечением времени меняются ее свой-
с i на: вымываются карбонаты, накапливаются органические веще-
< 1ва, щелочная реакция может смениться на кислую. По мере
формирования почвы образуется характерный вертикальный по-
чвенный профиль с горизонтами, создающими разные экологи-
ческие условия для жизни разнообразных почвенных организмов.
Oi возраста во многом зависит мощность почвы и, следователь-
303
но, глубина залегания подстилающей материнской породы, что
обусловливает глубину распространения корней.
Почвы на Земле имеют разный возраст. Древние латеритные
почвы находятся в тропиках. В них почвообразованием затронуты
слои в несколько метров. В дождевых тропических лесах за долгое
время существования биогены перешли в биомассу, которая при
отмирании быстро минерализуется и вновь вовлекается в цикл
производства органики. Поэтому под этими богатейшими лесами
почвы содержат мало элементов минерального питания. В горных
системах распространены почвы молодые: постоянные процессы
смыва и переотложения ведут к их обновлению. Молодые почвы
формируются у подножия ледников, в речных долинах, на зарас-
тающих промышленных отвалах.
Результаты определения абсолютного возраста почв радиоуглеродным
методом на Русской равнине, в Западной Сибири, Северной Америке и
Западной Европе показали, что время образования профиля современ-
ных почв составляет от нескольких сотен до нескольких тысяч лет
(Г. В. Добровольский, 1989). Многие из них сформировались на отложе-
ниях плейстоцена, в течение которого в результате мощной тектоничес-
кой деятельности значительно увеличились площадь суши и ее абсолют-
ные высоты, а неоднократные похолодания (ледниковые периоды) при-
вели к распространению в этих регионах холодостойких и засухоустой-
чивых растений. На слабо сформированных почвах складывались откры-
тые фитоценозы и светолюбивые растения могли тогда далеко мигриро-
вать на те территории, куда их не пустили бы в другое время сомкнутые
сообщества. В это время во внеледниковых районах ксеротермические
(засушливые и жаркие) эпохи сменялись плювиальными (влажными).
От этого времени в почвах остались погребенные реликтовые горизон-
ты: твердая латеритная корка от ксеротермических периодов и гумуси-
рованные погребенные слои от плювиальных.
Почвенный режим как экологический фактор. Ряд параметров,
характеризующих почву, переменчив, поэтому для экологии рас-
тений необходимы данные о режимах их изменения. Водный ре-
жим характеризуется большой изменчивостью (см. разд. 8.2.2).
Особенно резко она выражена в аридных областях. Помимо соб-
ственно содержания воды, для растений важно, что с почвенной
влагой связано их минеральное питание. Вода в почве — это все-
гда почвенный раствор, в котором находятся необходимые им ми-
неральные вещества. Растения приспособились к перемене вод-
ного режима своих местообитаний. В экологических шкалах для
характеристики резко переменного водного режима введены спе-
циальные обозначения и можно выделить растения, индицирую-
щие переменный водный режим (полевица белая — Agrostis alba,
горец земноводный — Polygonum amphibium и др.).
304
Тепловой режим почвы в основном определяется климатом,
но важную роль в его формировании играет еще рельеф, в частно-
сти экспозиция склона. Тепловой режим почвы обусловливает мно-
гие физиологические и анатомо-морфологические адаптации ра-
стений, включая корневые системы, а также влияет на структуру
растительного покрова в целом.
Воздушный режим почвы в основном зависит от ее водного
режима. Повышенное увлажнение почвы часто связано с недо-
статком в ней воздуха (см. разд. 9.1). При затоплении или забола-
чивании почвенный воздух вытесняется водой, а кислород диф-
фундирует в ней значительно медленнее, чем в газовой среде.
В торфяно-болотных почвах количество кислорода составляет ме-
нее 1 %, а ниже уровня грунтовых вод падает до нуля (Т. В. Чирко-
ва, 2002). Периодическую гипоксию испытывают растения мно-
гих почв гумидных областей. Иссушение же почвы, обеспечивая
ее аэрацию, связано с опасной для растений почвенной засухой.
Гранулометрический состав почвы как экологический фактор.
Гранулометрический (механический) состав почвы — следствие
процессов выветривания и дальнейшей биологической переработки
материнской породы. Он характеризуется процентным соотноше-
нием минеральных частиц разного размера: от крупных обломков
горной породы до коллоидов размером в сотые доли микрона.
Промежуточные по размеру образования, формирующие почвы, —
гравий и хрящ (более 3 мм), песок (0,05 — 3 мм), глинистые час-
тицы (мельче 0,01мм), пылеватые частицы и ил (менее 0,001 мм).
В зависимости от соотношения этих фракций различают песча-
ные, супесчаные, суглинистые и другие типы почв. Грануломет-
рический состав во многом обусловливает физические свойства
почвы. От него зависит ее плотность (и возможность проникно-
вения корней), порозность и связанные с ней водо- и воздухопро-
ницаемость, водоподъемность, гигроскопичность, поглотительная
способность и др.
Гранулометрический состав почвы существенно влияет на ее
тепловой и водный режимы. Во влажном прохладном климате гли-
нистые почвы считаются «холодными и тяжелыми», так как они
более теплопроводны и медленно прогреваются. Песчаные же
почвы — «легкие и сухие». В области многолетней мерзлоты они
летом протаивают быстрее и глубже. Но сильное нагревание их
поверхности днем связано с резкими суточными колебаниями тем-
пературы. В глинистых почвах составляющие их мелкие частицы
плотно «упакованы», для них характерна большая сила молеку-
лярного сцепления и водоудерживающая способность, часто де-
лающая воду недоступной для растений. Эти почвы менее водо-
проницаемы и слабо аэрируемы, в них могут создаваться анаэ-
робные условия с накоплением диоксида углерода, сероводорода,
метана и др. Хорошо выраженный капиллярный подъем воды
305
вызывает пересыхание глинистых почв на большую глубину.
В песчаных же почвах капиллярное поднятие влаги ограничено,
водоудерживающая сила мала, а за счет промывания и слабого
поглощения пески обеднены минеральными солями.
В аридных зонах песчаные почвы влажнее глинистых, так как
они просыхают на меньшую глубину, и в некоторых случаях в них
образуется горизонт с конденсационной влагой. В результате рас-
тительность песчаных пустынь оказывается намного богаче пус-
тынь глинистых. А каменистые почвы промачиваются еще лучше,
удерживая влагу на глубине. Но на каменистых осыпях и скалах
создаются жесткие тепловые, водные и ветровые условия в соче-
тании с недостатком минерального питания. Растения там при-
способились закрепляться в щелях и трещинах, прижиматься к
камням. Это растения-литофиты (от греч. lithos — камень), от-
личающиеся карликовостью, ползучим искривленным ростом и
цепкой корневой системой (рис. 10.1).
Виды, приспособившиеся к жизни на песках, называются псам-
мофитами (от греч. psammos — песок). В связи с обычно небла-
гоприятным водным режимом здесь они ксероморфны. На под-
вижных песках растения приспособились к непрерывному пере-
носу частиц ветром. Ряд их при засыпании образует придаточные
корни (рис. 10.2). Другое приспособление к выдуванию субстрата
и оголению корней — защитные футляры из пробковой ткани
или из песчинок, склеенных корневыми выделениями. Кроме видов
песчаных пустынь, они характерны также для растений примор-
ских и приречных песков в средних широтах (овсяница Беккера —
Festuca beckeri и др.).
Рис. 10.1. Литофит остролодоч-
ник углубленный (Oxytropis
immersd) в Западном Тянь-Шане
(по И.М. Культиасову, 1955)
Гранулометрический состав почв
во многом обусловливает химические
свойства почвы. Малый размер час-
тиц, образующих глинистые и суг-
линистые почвы, способствует боль-
шему растворению минеральных ве-
ществ и переходу их в почвенный
раствор. Их мелкие частицы в сум-
ме имеют огромную поверхность, ко-
торая удерживает растворы мине-
ральных солей, препятствуя их вы-
мыванию. Поглотительная способ-
ность глинистых почв высока и на-
долго задерживает элементы мине-
рального питания в зоне распрост-
ранения корней. А низкая поглоти-
тельная способность песчаных почв
слабее удерживает минеральные соли,
в том числе и внесенные удобрения.
306
Рис. 10.2. Приспособления растений к жизни в песчаных пустынях
(по М. В. Петрову, 1950):
А — образование придаточных корней при засыпании кустарника движущимся
барханом; Б — корневые футляры из сцементированных песчинок у Aristida karelini
Органическое вещество почвы как экологический фактор. Орга-
ническая часть почвы представлена в основном гумусом (пере-
гноем), состоящим из продуктов неполного разложения остатков
организмов (большей частью растительных). Гумус представляет
собой комплекс темноокрашенных высокомолекулярных органи-
ческих соединений почвы: гуминовые кислоты, гумин и другие
вещества полимерной природы, состоящие из полифенолов,
углеводов, азотистых гетероциклических соединений. Кроме С, Н,
О, N, в нем находится значительное количество S, Fe, Са, К,
микроэлементов. От содержания гумуса зависит плодородие по-
чвы, ее водные и тепловые свойства.
Количество гумуса связано с процессами накопления органи-
ческих остатков и их минерализацией. Накопление его во многом
зависит от продуктивности растительности, ежегодного опада и
массы отмирающих корней. Величины ежегодного опада в разных
типах растительных сообществ колеблются от 25 т/га во влажном
тропическом лесу до 1 т/га в пустынях и тундрах. В ельниках эта
величина составляет 3,5 —5,5 т/га, в дубравах — около 6 т/га, в
луговых степях — до 4 т/га. Отмершая фитомасса в процессе гумифи-
кации превращается в гумус в результате деятельности микроорга-
низмов-редуцентов, минерализующих органические остатки. Их
активность и соответственно скорость гумификации зависят от про-
должительности вегетационного периода и запаса влаги в почве.
Наиболее активно процесс минерализации идет в почвах влаж-
ных тропиков в условиях высоких температур, влажности и дос-
таточной аэрации. Эти почвы бедны гумусом, так как продукты
гумификации, не накапливаясь, быстро усваиваются растениями.
Масса гумуса в степных почвах намного превышает общую фито-
массу одной генерации растений, и его накопление формирует
307
здесь плодороднейшую почву — чернозем. Довольно богаты орга-
никой почвы тундр, так как при их высокой влажности и недо-
статке кислорода, низких температурах и коротком вегетацион-
ном периоде активность редуцентов тут слабая. Поэтому остатки
организмов здесь накапливаются, не разлагаясь. В высокогорных
районах в условиях пониженной температуры и хорошего дрена-
жа продуктивность растительных сообществ может быть высокой,
а процессы гумификации заторможены, в результате чего также
накапливается мощный слой гумуса. Под болотной растительнос-
тью деятельность сапротрофов ослаблена из-за недостатка кислоро-
да при застойном увлажнении и, как правило, кислой реакции сре-
ды. Это способствует накоплению многометровых толщ торфа —
недоразложившейся органической массы.
При отмирании и разложении растений полученные ими ми-
неральные вещества возвращаются в почву. Накопление коллои-
дов обусловливает ее водоудерживающую способность, а количе-
ство гумуса определяет ее структуру (склеивание минеральных
частиц ведет к образованию структурных агрегатов разного раз-
мера и прочности). Например, в богатых гумусом черноземах фор-
мируется благоприятная для корней зернистая или комковато-зер-
нистая структура, а подзолистые почвы содержат мало гумуса и
имеют непрочную структуру, где условия для жизни растений
намного хуже. Плодородие почв поддерживают длительные цик-
лы накопления органики, гумификации и минерализации.
Экологическое значение живого населения почвы. По В. И. Вер-
надскому, почва — биокосное тело, так как кроме неживых со-
ставляющих она содержит живые организмы. Растения (высшие и
низшие), бактерии, грибы, животные (от микроскопических до
крупных) — важные компоненты почв. Их бесчисленные поколе-
ния проделывают огромную геохимическую работу.
Водоросли (зеленые, желтозеленые, диатомовые) и цианобак-
терии в значительных количествах присутствуют почти во всех
типах почв (табл. 10.1).
Таблица 10.1
Количество водорослей в некоторых почвах
(Э. А. Штина, 1960) (тыс. клеток в 1 г почвы)
Почвы Циано- бактерии Зеленые Диато- мовые Всего
Подзолистые 0-2 3-25 2-7,5 5-30
Дерново- подзолистые 2-24 10-128 10-76 12-220
Черноземы 5-50 10-85 8-35 25-120
Т емно-каштановые 660-2000 6-35 86-116 800-2160
Бурые сухостепные 43 37 15 96
308
В лесных почвах преобладают зеленые водоросли, в степных
увеличивается доля цианобактерий, в заболоченных и болотных
почвах помимо этих групп значительно участие диатомовых. Мак-
симальное развитие водорослей связано с влажными сезонами.
Создавая в процессе фотосинтеза органику, почвенные водоросли
обогащают почву. Они влияют и на состав почвенного воздуха,
поглощая диоксид углерода и выделяя кислород. Образуя слизи,
водоросли способствуют структурированию почвы, благоприятно
влияющему на ее воздушно-водный режим. Выделяя в почву биоло-
гически активные вещества, они воздействуют на состав и числен-
ность почвенных микроорганизмов. В целом, по мнению Т. А. Ра-
ботнова (1984), несмотря на конкуренцию с другими растениями
за элементы минерального питания почвенные водоросли, напри-
мер для луговодства, имеют положительное значение.
Высшие растения часто накапливают в почве большую под-
земную биомассу. Среди естественных фитоценозов по массе кор-
ней на первом месте стоят широколиственные и влажные тропи-
ческие леса (табл. 10.2). Так, в дубравах корни расположены яру-
сами и распространяются на значительные глубины. Очень боль-
шая доля от общей биомассы (до 90 %) приходится на корни в
степях. А наименьшая масса корней отмечена в пустынях, хотя у
ксерофитов она часто значительно превосходит массу надземных
органов.
Слой почвы толщиной 2 — 3 мм близ поверхности корней, со-
держащий многочисленные организмы, составляет ризосферу. Осо-
бенно много их у корневых волосков. За счет корневых выделе-
ний развиваются эккрисотрофы. Состав их зависит от выделений
Таблица 10.2
Запасы корней в почвах природных зон
(по Н. И. Базилевич, Л.Е.Родину, 1968)
Природная зона Запас корней
ц/га % от фитомассы
Арктические тундры 6-80 70
Кустарниковые тундры 200-300 80-85
Леса хвойные 300-800 21-25
Леса лиственные 250-950 15-33
Степи, прерии, луга 100-200-250 80-90
Пустыни 3-30 40-85
Пампы и саванны 200-400 30-60
Влажные тропические леса 1000 20
309
растений. Ризосферные организмы продуцируют многие вещества,
важные для высших растений (стимуляторы роста, антибиотики и
др.). Здесь же некоторые свободноживущие микроорганизмы осу-
ществляют азотфиксацию.
В кислых почвах ризосферными организмами обычно являют-
ся грибы. Их мицелий развивается на поверхности корней выс-
ших растений (эктомикориза) или в клетках корня (эндомикори-
за). При микосимбиотрофии микориза обеспечивает развитие ра-
стений на почвах с обилием слаборазложившихся органических
остатков. Преимущественно грибами разлагается в лесу хвойный
опад. При этом образуются органические кислоты, увеличиваю-
щие кислотность среды. В опаде лиственных лесов с кислотно-
стью, близкой к нейтральной, грибов значительно меньше (пре-
обладают сапротрофные бактерии).
Общая численность почвенных микроорганизмов может коле-
баться, но измеряется миллиардами в 1 г почвы. В поверхностном
слое почвы (до 25 см) их биомасса достигает 3 — 7 т/га (0,1 —0,2 %
массы почвы) и резко убывает с глубиной (Г. В. Добровольский,
1989). Бактерии-сапротрофы играют огромную роль в разложе-
нии органических веществ. Разные группы прокариот специали-
зированы на разрушении разных типов органических соединений.
Нитрифицирующие бактерии окисляют аммиак до азотной кис-
лоты, и таким образом образуется до 300 кг ее солей на 1 га.
В результате деятельности в поверхностной части почвы серобак-
терий накопление сульфатов достигает 200 — 250 кг/га. Велико зна-
чение свободноживущих и симбиотрофных азотфиксирующих
прокариот, которые переводят азот атмосферы в доступные для
растений соединения. Азот органики, недоступный высшим рас-
тениям, переводится в усвояемую форму бактериями-аммонифи-
каторами и другими группами, разлагающими азотсодержащие
вещества.
Почвенные животные создают зоомассу от единиц до тысяч кг/га.
Простейшие (инфузории, амебы, жгутиконосцы) содержатся в
количестве более миллиона особей в 1 г почвы, насекомые и их
личинки — в тысячах экземпляров на 1 м2. В некоторых почвах
число позвоночных (змеи, ящерицы, грызуны, насекомоядные и
пр.) достигает нескольких тысяч на 1 га. Однакб основная зоо-
масса создается почвенной мезофауной', членистоногими, нема-
тодами, дождевыми червями. Количество беспозвоночных в по-
чве зависит от обеспечения ее теплом и влагой. Особенно много
их в лесах умеренного климата, где при благоприятных гидротер-
мальных условиях ежегодное поступление опада хорошо обеспе-
чивает их пищей. Наибольшая масса и разнообразие беспозво-
ночных приходится на серые лесные почвы широколиственных
лесов умеренного климата. Сырая зоомасса достигает там 600 —
2 000 кг/га (Г. В. Добровольский, 1989). В дубовых лесах лесостепи
310
их биомасса оценивается в 81,2 г/м2 (Г. Вальтер, 1974). В умерен-
ном климате на 1 м2 лесной почвы может содержаться более 10
млн особей нематод и простейших, 100 тыс. коллембол и клещей
и 50 тыс. прочих беспозвоночных (M.Begon et al., 1986). На 80 —
90 % они представлены сапротрофами, деятельность которых ус-
коряет разложение опада в 3 — 8 раз (Т.А.Работнов, 1993).
Из почвенных животных необходимо особо отметить дождевых чер-
вей. Они широко распространены в природе и входят с состав биогеоце-
нозов разных природных зон. На территории России и сопредельных
стран отмечено более 80 видов этих животных. На некислых луговых и
лесных почвах их содержится до 1 млн особей на 1 га, и они могут со-
ставлять до 90 % и более почвенной зоомассы. Для них благоприятны
достаточно влажные почвы, но без застоя воды, засоления и повышен-
ной кислотности, поэтому много дождевых червей в почвах широколи-
ственных лесов (до 500 на 1 м2) и луговых степей (свыше 100 на I м2).
Здесь они в период от 30 до 200 лет полностью перерабатывают 20-сан-
тиметровый слой почвы (Т.А.Работнов, 1993). На одного червя в год
приходится до 400 г заглатываемой смеси органических остатков и ми-
неральных частиц (И. М. Культиасов, 1982). Ч. Дарвин отмечал, что вряд
ли есть «другие животные, которые заняли бы в истории земной коры
столь видное место». Они не только перерабатывают опад, но и оказыва-
ют существенное прямое и косвенное влияние на все почвенные компо-
ненты. Пронизывая почву ходами, улучшая ее аэрацию, водопроницае-
мость и влагоемкость, дождевые черви создают благоприятные условия
для развития и растений, и разных почвенных организмов, участвующих
в разложении органического вещества. Питаясь отмершими органами
растений и экскрементами животных, дождевые черви заглатывают так-
же много бактерий, грибов, простейших и нематод. Участвуя в разложе-
нии экскрементов скота на пастбищах, они частично переносят их в
глубь почвы, обогащая эти слои. Стенки их ходов пропитываются выде-
лениями червей, содержащими аммиак и мочевину; так что общее коли-
чество поступившего в почву азота колеблется от 18 до 50 кг/га. А выде-
ляемые дождевыми червями копролиты представляют достаточно водо-
стойкие агрегаты, способствующие созданию комковатой структуры по-
чвы. Все это улучшает условия жизни растений, что неоднократно было
доказано многими опытами.
Большое и разнообразное влияние оказывают на почву рою-
щие животные (сурки, суслики, кроты, полевки и др.). Они изме-
няют микрорельеф, увеличивают площадь соприкосновения по-
чвы с воздухом, способствуют проникновению в нее осадков, улуч-
шают условия минерализации органики. Все это благоприятно
сказывается на растениях. Перерывая почву, землерои выносят из
глубины на поверхность отличающийся по свойствам субстрат.
Рыхлые выбросы заселяются пионерными растительными груп-
пировками с большим участием однолетников, семена которых
сохранились в почве. Поэтому на лугах, например, деятельность
311
кротов нередко приводит к пятнистости растительного покрова, а
растительность «сурчин» долго выделяется на общем фоне степи.
Роющая деятельность животных обычно сочетается с фитофа-
гией. При этом органы растений повреждаются и при поедании
их животными, и при прокладывании ходов и устройстве нор, а
также при погребении растений под выбросами земли. Поэтому в
местах скопления землероев сомкнутость растительного покрова
может снижаться. В ряде случаев это сказывается и на эрозион-
ных процессах. Погребение выбросами земли приводит многие
растения к гибели. Гибнут напочвенные мхи, лишайники, а также
не размножающиеся вегетативно травы. Особенно страдают сла-
бые молодые и угнетенные особи. В противоположность им длин-
нокорневищные и корнеотпрысковые растения часто положитель-
но реагируют на прикрытие почвой. При этом имеет значение
хорошая аэрация, небольшая плотность рыхлой почвы, а также
ослабление конкуренции с другими травами. У этих растений также
распространена способность образовывать новые побеги при трав-
мировании подземных органов.
Т.А. Работнов (1993) отмечает, что в годы массового размноже-
ния землероев фиксируется заметное увеличение обеспеченности
растений элементами минерального питания (в частности, азо-
том). Создаются благоприятные условия для семенного и вегета-
тивного размножения тех видов, которые долго их не имели. Омо-
лаживается популяция клональных растений, а также появляются
условия для существования однолетников. В целом землерои —
характерный компонент многих естественных биогеоценозов, и в
процессе коэволюции растения приобрели способность к сосуще-
ствованию с ними. Если численность роющих животных не пре-
вышает некоторого предела, общая продукция растительного со-
общества не снижается. Разные виды растений по-разному реаги-
руют на деятельность землероев, и многие из них получают пре-
имущество в годы с определенной их численностью. Порой по-
чвенные ходы и выбросы разнообразят условия среды, и этим зем-
лерои помогают созданию динамически устойчивых богатых ви-
дами фитоценозов.
10.2. Экологическое значение химических свойств
почвы
Почвенный раствор играет настолько важную роль в жизни
растений, что Г. Н. Высоцкий (1925) образно назвал его «кровью
почвы». Корневая система приспособлена для извлечения дефи-
цитного минерального питания: она постоянно растет в поисках
минеральных веществ, выделяя соединения, способствующие пе-
реходу необходимых питательных элементов в почвенный раствор.
312
Для живых организмов требуется большинство химических эле-
ментов, обычно содержащихся в земной коре. Несколько десят-
ков так называемых биогенных элементов, необходимых для их
нормального функционирования, — постоянные компоненты их
химического состава. Большинство этих элементов (кроме С и О)
растения получают из почвы. Еще 7 извлекаемых из почвы мак-
роэлементов (N, Р, К, Са, Mg, Fe, S) являются основными, без
них невозможно успешное развитие растений. Si, Na, Cl, Al также
относятся к макроэлементам и в почве их обычно достаточно.
Растения постоянно нуждаются и в микродозах микроэлемен-
тов (Мп, В, Sr, Си, Ti, Zn, Li, Ba, Br, F, Rb, Sn, Ni, As, Mo, Co,
Se, J и многие другие). В незначительных количествах растениям
требуются ультрамикроэлементы (Ge, Pb, Hg, Ag, Au, Ra), хотя
некоторые виды способны их накапливать (например, кукуруза —
Zea mays и хвощ — Equisetum могут концентрировать в тканях
заметные количества золота). Микроэлементы, постоянно содер-
жащиеся в организмах, включаются в обмен веществ в составе
биологически активных соединений и являются незаменимыми.
При недостатке или избытке многих из них нарушаются рост и
развитие растений.
Экологическое значение кислотности почвы
Показатели кислотности почв и ее формирование. Находящиеся
в почвенном растворе водородные ионы обусловливают актив-
ную кислотность почв, а поглощенные — потенциальную. Для
растений наиболее важна первая. Она обозначается pH и пред-
ставляет собой отрицательный десятичный логарифм концентра-
ции ионов водорода в растворе. По pH почвы делятся на сильно-
кислые (pH 3 — 4), кислые (pH 4 — 5), слабокислые (pH 5 — 6),
нейтральные (pH 6,5 — 7), слабощелочные (pH 7 — 7,5), щелочные
(pH 7,5 —8,5) и сильнощелочные (pH 8,5 и более). Нейтральную
реакцию имеют черноземы, кислую — подзолистые, серые лес-
ные и болотные почвы, щелочную — каштановые почвы и серозе-
мы пустынь, сильнощелочную — солонцы. От концентрации во-
дородных ионов в почвенном растворе зависят и существование
высших растений, и микробиологические процессы, а также весь
ход почвообразования. Как правило, в кислых почвах хлориды,
сульфаты и карбонаты отсутствуют, а в щелочных накапливаются.
В гумидных районах при обилии осадков и низких температурах мед-
ленное разложение растительных остатков сопровождается образовани-
ем большого количества органических кислот. Так, под хвойными леса-
ми почвы обычно сильнокислые и кислые (среди продуктов преимуще-
ственно грибного разложения многолетней хвои много фульвокислот).
313
Торф верховых болот имеет pH 3,5 — 4 из-за выделений сфагновых мхов.
В аридных зонах почвы обычно щелочные и в них присутствуют карбо-
наты Са и Na, так как при недостатке осадков и сильном испарении
затруднено их промывание и выщелачивание. При pH более 8,3 почвы
насыщены щелочами и для них характерно присутствие свободных ионов
Са. При нейтральной реакции среды почти все минералы достаточно
хорошо растворимы и доступны растениям. В гумидной зоне щелочная
или нейтральная реакция почв встречается нечасто и связана с особен-
ностями почвообразующих пород или близостью грунтовых вод с повы-
шенным содержанием извести.
Кислотность и щелочность почв формируются разными путя-
ми. Если почвообразующие породы содержат кислые соединения
(граниты, безызвестковые ледниковые наносы и др.), реакция
почвы будет кислая. Кислотность увеличивается при активизации
почвенных организмов, выделяющих СО2. Она значительно уси-
ливается при формировании подстилки из опада хвойных пород
и вересковых кустарничков. Сильно подкисляют среду выделения
мхов (сфагновые, кукушкин лен — Polytrichum commune и др.).
Существенно повышают кислотность почв промышленность и
транспорт, загрязняя атмосферу продуктами сгорания каменного
угля и нефтепродуктов. Присутствующие в них газы, главным
образом диоксид серы, а также оксиды азота образуют так назы-
ваемые кислотные дожди.
Значения pH субстратов изменчивы. В почвенном профиле
верхние горизонты обычно более кислые, нижние — более ще-
лочные. Поэтому виды с разным отношением к кислотности по-
чвы могут находиться в одном фитоценозе: виды с «кислыми»
рН-оптимумами (ацидофилы) располагают корни близ поверхно-
сти почвы, а растения с более «щелочными» оптимумами (бази-
филы) заглубляют корневую систему. В водоемах обычен суточ-
ный ритм кислотности: ночью она повышается из-за увеличения
концентрации СО2 в результате дыхания, а днем снижается из-за
потребления его при фотосинтезе.
Почва обладает буферностью — способностью сохранять кис-
лотность при некотором (обычно в естественных условиях неболь-
шом) добавлении кислот или щелочей. Буферность почв зависит
в основном от состава поглощенных оснований. При действии
кислот на почву с нейтральной реакцией происходит обмен их на
Н+ кислоты и раствор остается нейтральным. Таким образом, ионы
водорода изымаются из раствора, адсорбируются почвой и общая
концентрация их существенно не меняется.
Влияние кислотности почвы на растения. Помимо прямого вли-
яния водородных ионов, с кислотностью среды связан целый ряд
побочных явлений, на которые реагируют организмы. На кислых
почвах возрастает содержание токсичного свободного иона алю-
миния, на что многие растения реагируют отрицательно. В кис-
314
лой почве переходят в раствор, становясь токсичными, ионы Fe,
Мп, Zn, Си.
К реакции среды чувствительны почвенные бактерии. При pH
ниже 6 или выше 7,7 затормаживаются процессы нитрификации
и азотфиксации, снижается деятельность редуцентов. В кислых
почвах практически отсутствуют дождевые черви. При низких
значениях pH почвенного раствора образуются труднораствори-
мые фосфаты алюминия и железа и фосфор становится менее
доступен растениям. Все это обусловливает недостаток фосфора и
азота в питании растений на кислых почвах и может быть причи-
ной неодинакового распределения растений на почвах с разным
значением pH.
В сельском хозяйстве широко применяется снижающее кис-
лотность почв известкование, поскольку культурные и луговые
растения предпочитают слабокислую или нейтральную среду. Это
приводит к изменению набора видов в фитоценозе. А щелочные
почвы подкисляют до нейтральных или слабокислых, используя
сульфаты, торф и другие удобрения.
Растения значительно отличаются по реакции на pH почвен-
ного раствора. Экологические шкалы Г.Элленберга (1974) содер-
жат информацию об отношении к кислотности почв среднеевро-
пейских сосудистых растений и мхов.
Группа Rt — растения, предпочитающие сильнокислые почвы. Ин-
дикаторы кислых почв: щавелек (Rumex acetosella), пушица влагалищ-
ная (Eriophorum vaginatum), клюква (Oxyccocus), голубика (Vaccinium
uliginosum), черника (V. myrtillus), брусника (V. vitis-idaea), вереск обык-
новенный (Calluna vulgaris), водяника черная (Empetrum nigrum), ку-
кушкин лен обыкновенный (Polytrichum commune), дикранум метло-
видный (Dicranum scoparium), сфагнумы верховых болот.
Группа R2 — растения кислых почв, но встречающиеся на слабокис-
лых и нейтральных: сурепка (Raphanus raphanistrum), чистец полевой
(Stachys arvensis), луговик извилистый (Lerchenfieldia fiexuosa), марь-
янник луговой (Melampyrum pratense), плаун годичный (Lycopodium
annotinum), грушанка (Pyrola sp.), орляк (Pteridium aquilinum), верони-
ка лекарственная (Veronica officinalis), ястребинка зонтичная (Hieracium
umbellatum), ландыш майский (Majanthemum bifolium), сфагнум Гир-
гензона (Sphagnum girgensohnii), кукушкин лен можжевельниковый
(Polytrichum juniperinum).
Группа R3 — растения слабокислых почв: ромашка лекарственная
(Matricaria recutita), ожика волосистая (Luzula pilosa), кислица (Oxalis
acetosella), бор развесистый (Milium effusum), звездчатка жестколистная
(Stellaria holostea), подмаренник лесной (Galium sylvaticum), ветреница
дубравная (Anemone nemorosa), щитовник мужской (Dryopteris filixmas),
мицелис (Mycelis muralis), купена многоцветковая (Polygonatum
multifiorum), мниум растопыренный (Mnium rostratum).
Группа R4 — растения, предпочитающие слабокислые почвы, но встре-
чающиеся и на щелочных: горчица полевая (Sinapis arvensis), дымянка
315
(Fumaria officinalis), коротконожка лесная (Brachipodium sylvaticum), гра-
вилат городской (Geum urbanum), печеночница (Hepatica triloba), лю-
тик золотистый (Ranunculus auricomus), волчье лыко (Daphne mezereum),
чистец лесной (Stachys sylvatica), вороний глаз (Paris quadrifolia), осока
пальчатая (Carex digitata).
Группа Rs — растения в основном щелочных почв (но могут встре-
чаться и на нейтральных): адонис летний (Adonis aestivalis), чина клуб-
невая (Lathyrus tuberosus), сокирки полевые (Consolida regalis), копы-
тень (Asarum еигораеит), пролесник многолетний (Mercurialisperennis),
колокольчик крапиволистный (Campanula trachelium), чина весенняя
(Lathyrus vernus), ветреница лютичная (Anemone ranunculoides), хох-
латка полая (Corydalis cava), лук медвежий (Allium ursinum), водосбор
обыкновенный (Aquilegia vulgaris), володушка длиннолистная (Bupleurum
longifolium).
Группа Rq — индифферентные к значениям pH растения: василек
синий (Centaurea cyanus), пастушья сумка (Capsella bursa-pastoris), лютик
ползучий (Ranunculus repens), горец птичий (Polygonum aviculare), бо-
дяк полевой (Cirsium arvense), незабудка полевая (Myosotis arvensis),
молиния голубая (Molinia coerulea), золотая розга (Solidago virgaurea),
костяника (Rubus saxatilis), полевица обыкновенная (Agrostis vulgaris),
земляника лесная (Fragaria vesca), ритидиадельфус (Rhytidiadelphus
triquetrus), бриум волосовидный (Bryum capillare).
Распределение луговых растений по таким экологическим груп-
пам осложнено особенностями водного режима. Так, индикато-
рами кислых почв в сухих условиях являются белоус (Nardus
stricta), луговик извилистый (Lerchenfieldia flexuosa), полевица
собачья (Agrostis canina), а во влажных — ситник нитевидный
(Juncus filiformis), арника горная (Arnica montana), пушица влага-
лищная (Eriophorum vaginatum) и др. Отдельные виды могут быть
индикаторами кислотности только в конкретных регионах. Напри-
мер, широко распространенный сорняк щавелек (Rumex acetosella)
в средней полосе Европы принято считать указателем сильно кис-
лых почв, но, попав в Австралию, он широко распространился и
на щелочные почвы. Результаты исследований по приуроченнос-
ти видов к pH почвы в разных регионах редко совпадают, более
надежный индикатор кислотности — растительные сообщества.
Экологическое значение обеспеченности почвы азотом
Азот — главный элемент минерального питания, требующийся
для нормального роста и развития растений. Он входит в состав
почти всех важнейших органических веществ (нуклеотидов и
нуклеиновых кислот, аминокислот и белков, АТФ, алкалоидов,
липидов и др.), участвует в формировании хлорофилла, фермен-
тов, витаминов, ростовых веществ и др. Цитоплазма в значитель-
ной степени состоит из азотсодержащих соединений.
316
Фиксация атмосферного азота. Атмосфера на 78 % состоит из
азота, но инертный молекулярный азот воздуха для растений не-
доступен. Они усваивают его в виде минеральных солей аммония
(NH4), нитритов (NO2) и большей частью нитратов (NOj) и ами-
нокислот. Растения поглощают азот из почвы и (или) получают
его от свободноживущих или симбиотически связанных с ними
азотфиксирующих организмов. Фиксация атмосферного азота
прокариотами — основной источник пополнения почвы этим эле-
ментом. После отмирания микроорганизмов соединения азота
остаются в почве.
Группа несимбиотических азотфиксаторов включает ряд аэроб-
ных и анаэробных видов из родов Azotobacter, Aerobacter,
Beijerinckia, Mycobacterium, Methylobacteria, Apirillum, Bacillus,
Enterobacter, Klebsiella, Rhodopseudomonas, Pseudomonas,
Clostridium, Mechanobacterium и др. Для фиксации азота они
используют энергию, содержащуюся в органическом веществе
почвы (сапротрофы) или в прижизненных выделениях автотро-
фов (эккрисотрофы) (Т.А.Работнов, 1979). Эккрисотрофы обна-
ружены в ризосферах многих растений, поэтому такая азотфикса-
ция еще именуется «ассоциативной» (М.М. Умаров, 1986). В ус-
ловиях умеренного климата общее количество азота, фиксируе-
мого свободноживущими почвенными организмами, колеблется
от нескольких килограммов до 40 — 50 кг/га в год. Особенно зна-
чительная фиксация азота отмечена в ризосферах С4-растений,
поскольку с сокращением затрат на фотодыхание больше углево-
дов поступает в ризосферу с корневыми выделениями. Предпола-
гается также, что фиксация азота ризосферными бактериями сти-
мулируется микоризообразующими грибами (И. М. Рыжова,
М.М.Умаров, 1979 и др.). Свободноживущие азотфиксаторы зат-
рачивают значительно больше энергии на фиксацию азота, чем
симбиотически связанные с бобовыми: в среднем 100 г глюкозы
на 1 г фиксируемого азота (W. Larcher, 1984). При этом до 80%
усвоенного ими азота выделяется в среду и становится доступным
для растений, остальная часть поступает в почву после отмирания
азотфиксаторов (Т.А. Работнов, 1992).
Азотфиксирующие микроорганизмы находятся и на листьях (в фил-
лоплане). К ним относятся Azotobacter, Azotococcus, Aerobacter, Klebsiella,
Beijerinckia и др. В США для пшеницы {Triticum aestivum), костра безо-
стого {Bromopsis inermis), мятлика лугового {Роа pratensis) установлено,
что в их филлоплане фиксируется до 10 кг/га азота в год. Для филлоп-
ланных азотфиксаторов большое значение имеет хорошее обеспечение
водой (с этим, возможно, связано высокое содержание азота в болотных
и водных растениях) (Т.А.Работнов, 1992). Фиксация азота в филлопла-
пе, вероятно, способствует выделению ими в окружающую среду боль-
ших количеств органических соединений. Для этого типа азотфиксации
важно также снабжение его энергетическим материалом, содержащимся
317
в выделениях листьев (в наибольших количествах это происходит тогда,
когда ассимиляты, образующиеся при фотосинтезе, не полностью ис-
пользуются для синтеза более сложных соединений из-за недостатка азо-
та). На способности листьев усваивать нитраты основана агрохимичес-
кая практика внекорневой подкормки азотными удобрениями.
Большое значение в накоплении азота в почве имеет симбио-
тическая азотфиксация. Симбиотрофы получают энергию от ав-
тотрофов, с которыми находятся в симбиотических взаимоотно-
шениях. Такие отношения азотфиксирующих клубеньковых бак-
терий (Rhizobium, Azorhizobium, Bradyrhizobium, Sinorhizobium
и др.) с большинством видов семейства Бобовые широко распро-
странены в фитоценозах разных зон (рис. 10.3). Клубеньки на
корнях, образованные азотфиксирующими бактериями, обнару-
жены и у травянистых растений других семейств. Например, на
корнях злака лисохвоста лугового (Alopecuruspratensis) они обра-
зованы азотфиксирующей бактерией Bacillus alopecuri. Для всех
типов азотфиксирующих клубеньков общим является развитие их
из зачатков, возникающих во внутренних тканях корня под дей-
ствием микробных сигналов. Клубеньки, составляя до 5 % массы
растения, выполняют комплекс взаимосвязанных функций, обес-
печивающих вместилище для микроорганизмов, структурную ос-
нову для обмена партнеров метаболитами и контроля над числен-
ностью и физиологической активностью бактерий. Клубеньки на
корнях — важное приспособление для работы фермента азотфик-
сации (нитрогеназы): в них микроорганизмы находятся в среде со
сниженным содержанием О2, благоприятными кислотностью и
температурой, с постоянным обеспечением энергетическим мате-
Рис. 10.3. Клубеньки на корнях бобовых (по А.П. Шенникову, 1950):
1 — клевер (Trifolium)’, 2 — фасоль (Phaseolus); 3 — соя (Glycine)', 4 — люпин
(Lupinus)
318
риалом. При этом расход энергии на фиксацию азота намного
меньше, чем у свободноживущих азотфиксаторов.
Значительную роль в фиксации атмосферного азота играют
актиномицеты (актинобактерии) из рода Frankia. Они формиру-
ют на корнях автотрофных сосудистых растений многолетние де-
ревянистые клубеньки. Актиноризу образуют более 200 видов ра-
стений из 8 семейств (Березовые, Розоцветные, Крушиновые, Ло-
ховые, Казуариновые, Восковниковые, Кориариевые, Датисковые)
(Н.А. Проворов и др., 2002). Все они — покрытосеменные, дву-
дольные, обычно деревья или кустарники. Распространены акти-
норизные растения на всех материках — от субарктики до тропи-
ков. Для фитоценозов умеренного климата Евразии большое зна-
чение имеют симбиотические отношения актиномицетов с оль-
хой (Abuts) и облепихой (Hippophae*), для Северной Америки — с
Ceanothus, для Австралии — с Casuarina, а в Новой Зеландии —
с Coriaria. Многим из этих растений мутуалистические взаимоот-
ношения с актиномицетами дали возможность поселяться в бед-
ных азотом местообитаниях. Среди них есть растения-пионеры,
заселяющие свежий аллювий и песчаные дюны, сильно эродиро-
ванные почвы, каменистые россыпи, ледниковые морены. В от-
личие от бобовых они способны произрастать в суровых припо-
лярных и высокогорных районах (куропаточья трава — Dryas), а
также на кислых и заболоченных почвах (ольха, восковник —
Myrica). Как отмечает Т.А. Работнов (1992), по числу видов эта
группа растений значительно уступает бобовым, но если учиты-
вать только дикорастущие виды, их биомасса и масса их клубень-
ков не меньше. И фиксация азота актиномицетами может проис-
ходить так же энергично, как и клубеньковыми бактериями бобо-
вых. Общее количество фиксируемого в год азота составляет для
Myrica gale — 10—30 кг/га, Casuarina angustifolia — 60—70 кг/га,
Alnus rugosa — 85—160 кг/га, A. glutinosa — 210 — 225 кг/га.
Большое значение в азотфиксации играют цианобактерии. Роль
их особенно велика на начальных стадиях зарастания обнажен-
ных субстратов, а также для фитоценозов в экстремальных усло-
виях (Арктика, Антарктика, жаркие пустыни, олиготрофные бо-
лота), где деятельность других микроорганизмов затруднена. Ци-
анобактерии хорошо фиксируют азот, поселяясь на (или внутри)
мохообразных. На субарктических болотах они связывают 0,2 —
12 г/м2 азота в год. Меньше (65— 104 мг/м2 в год) фиксируют его
в тундровых осоковых фитоценозах, но здесь цианобактерии обес-
печивают 65 — 80% от поступающего в систему азота. На олиго-
трофных сфагновых болотах азота фиксируется около 5 кг/га в
год (А. В. Тюрин, 1956). Особенно благоприятны для азотфикса-
ции условия гипновых болот, где обеспечивается устойчиво высо-
кое увлажнение, грунтовые воды богаты фосфором, всегда хоро-
шее освещение и реакция среды близка к нейтральной (Т.А. Ра-
319
ботнов, 1993). Азот также интенсивно фиксируется цианобакте-
риями филлоплана, поселяющимися на болотных и водных рас-
тениях. В водоемах цианобактерии повышают эвтрофность по-
верхностных слоев.
Очень распространен симбиоз цианобактерий с автотрофными орга-
низмами (печеночники, сфагновые мхи, водный папоротник Azol 1а, са-
говники, ряд видов покрытосеменных). У печеночников {Blasia,
Clavicularia) они живут в слизистых полостях таллома, и растение реа-
гирует на их присутствие образованием тонких нитей, увеличивающих
контакт между симбионтами. У сфагновых мхов цианобактерии могут
обитать и снаружи, и внутри их воздухоносных клеток. В Юго-Восточ-
ной Азии Azolla, живущий в симбиозе с цианобактерией Anabaena,
широко культивируется на рисовых полях. Количество фиксируемого в
таких условиях азота может превышать 300 кг/га в год. Для возделыва-
ния риса {Oryza sativa) большое значение имеют также свободноживу-
щие цианобактерии, обитающие в воде рисовых полей. Общее количе-
ство фиксируемого ими азота исчисляется 7 млн т в год (Т. А. Работнов,
1993). Из семенных растений все виды саговниковых имеют на корнях
коралловидные вздутия, образованные цианобактериями, способными
фиксировать достаточно много азота. А из покрытосеменных симбиоз с
цианобактериями образуют виды рода Gunnera {Nostoc punctiforme об-
разует характерные вздутия у основания приземных листьев G. scabra).
Некоторые цианобактерии обнаружены в ризосфере цветковых расте-
ний и клубеньках клевера {Trifolium alexandrinum). А в симбиозе с гри-
бами азотфиксирующие цианобактерии образуют лишайники. Напоч-
венные лишайники родов Stereocaulon, Peltigera, Nephroma широко рас-
пространены в Субарктике и играют важную роль в жизни тундровых
фитоценозов, являясь постоянным источником доступного азота.
Пополнение почвы азотом при минерализации органических ос-
татков. В почвенном растворе естественная концентрация нитра-
тов обычно низкая (не более 2 % от общего запаса азота в почве),
так как они быстро потребляются растениями. Но количество азота
быстро пополняется при минерализации органических остатков.
В почве азот содержится преимущественно в органическом веще-
стве (гумус, отмершие организмы). Его минерализация, осуще-
ствляемая микроорганизмами, сначала проходит этап аммонифи-
кации, когда получающийся аммиак сразу соединяется с почвен-
ными кислотами, образуя соли аммония. Аммонификация связа-
на с разными неспециализированными организмами. Процесс этот
идет в довольно разных условиях среды, но медленно. Во влаж-
ных плохо аэрируемых почвах (тундры, болота) продукты аммо-
нификации могут накапливаться и играть основную роль в азот-
ном питании растений.
Второй этап минерализации органического вещества почвы —
нитрификация. При этом аммонийные соли в результате дея
тельности почвенных микроорганизмов преобразуются в нитриты
320
и нитраты. Нитрификация идет тем интенсивнее, чем больше в
почве аммония. Под действием нитрозобактерий в присутствии
О2 аммоний превращается в нитриты. Нитриты токсичны, но в
почве не накапливаются, так как очень быстро переводятся в нит-
раты нитробактериями. Интенсивность нитрификации снижает-
ся с увеличением сухости почвы, а высокая влажность ингибирует
этот процесс из-за ухудшения аэрации. Для процесса нитрифика-
ции оптимальная температура составляет около 30°, а ниже 5° и
выше -ИО °C она резко снижается.
Часть азота почвы теряется, вымываясь или выделяясь в ат-
мосферу в результате процесса денитрификации, при котором из
нитритов и нитратов образуются газообразные формы азота (N2,
N2O и аммиак). Важнейшие денитрификаторы — Pseudomonas
denitrificans, Micrococcus denitrificans, Thiobacillus denitrificans.
Этот процесс характерен для почв гумидного климата, где микро-
организмы используют нитриты и нитраты в качестве окислите-
лей (вместо О2).
В лесных фитоценозах, где ежегодно в почву и на ее поверх-
ность поступает большое количество опада, в минерализации орга-
нических остатков велико значение сапротрофных грибов. Отли-
чаясь от бактерий более развитой ферментативной системой, они
принимают основное участие в разрушении ряда трудно разлага-
ющихся соединений, в частности целлюлозы, лигнина, суберина
и кутина. Грибы способствуют аммонификации, а на очень кис-
лых почвах — нитрификации (Т. А. Работнов, 1993).
Влияние на растения обеспеченности субстрата азотом. Виды
растений используют разные источники азота, и необходим он
им в разных количествах. Некоторые растения существуют лишь
в местообитаниях с повышенным содержанием доступных форм
азота (нитрофилы), другие мирятся с бедными им почвами. А для
многих видов, сформировавшихся в олиготрофных условиях, эти
местообитания необходимы для благополучного существования:
они более приспособлены к перенесению неблагоприятных для
большинства растений условий — недостатку минерального пита-
ния. Виды с широкой экологической амплитудой адаптировались
к обитанию в местах с разной обеспеченностью азотом.
Недостаток азота подавляет фотосинтез. Растения (в том числе
их листовые пластинки и стебли) измельчаются, возникают при-
знаки, сходные с ксероморфозом (пейноморфоз). Эти черты воз-
никают у растений-мезофитов, в том числе культурных, в случае
азотного голодания. Однако наиболее ярко черты пейноморфоза
выражены у растений тундр, верховых болот, бедных песчаных и
каменистых почв.
При обилии азота в почве изменяется морфология, анатомия
растений, их химический состав. Увеличиваются надземная био-
масса, количество и размеры листьев, содержание в них хлоро-
321
филла и воды. Меняется соотношение подземной и надземной
биомассы: масса надземных органов возрастает больше, чем под-
земных. У некоторых видов при обилии азота увеличение вегета-
тивной массы сочетается с задержкой цветения и плодоношения.
Улучшение обеспеченности азотом способствует не только уве-
личению общей продуктивности растений, но и более экономно-
му расходованию воды. При этом снижается потребление воды на
единицу урожая для всего сообщества, включая расход на транс-
пирацию и испарение. Азот может также служить акцептором элек-
тронов при дыхании корней (нитратное дыхание) в обстановке
нехватки кислорода в почве (см. разд. 9.1.3). Внесение азотных
удобрений на заболоченные луга и верховые болота приводит к
мезофитизации растительности (уменьшению черт гигроморфиз-
ма, увеличению доли мезофитов) и повышению урожайности.
Под влиянием улучшения обеспечения азотом изменяется
отношение растений-мезофитов к теплообеспеченности, повы-
шается теневыносливость растений, что частично связано с уве-
личением содержания хлорофилла. Таким образом, лучшее
снабжение азотом расширяет экологический ареал растений в
отношении обеспечения их теплом, водой, светом и др. (Т. А. Ра-
ботнов, 1979).
Экологическое значение обеспечения почвы другими
элементами минерального питания
Экологическое значение обеспечения почвы кальцием. Кальций
во многом определяет важнейшее свойство почвы — ее плодоро-
дие. Растения получают его из почвенного раствора, и он требует-
ся им в значительных количествах как элемент минерального пи-
тания. Са необходим для деления клеток, роста и развития моло-
дых органов. Его соединения стабилизируют мембраны, снижают
их проницаемость, способствуют устойчивости растений в усло-
виях стресса. Кальций участвует в регуляции активности фермен-
тов. Значительные количества его присутствуют в составе пекти-
новых веществ клеточной стенки и срединной пластинки. Много
Са расходуется на нейтрализацию щавелевой кислоты — продукта
обмена растений. Нерастворимые в клеточном соке кристаллы
оксалата кальция накапливаются в клетках всех тканей растений
и особенно много в старых органах.
От недостатка Са страдают меристематические ткани и кор-
невая система, нарушаются процессы деления клеток. При дефи-
ците Са увеличивается проницаемость мембран, нарушаются
сшивки между соединяющими клетки пектиновыми веществами.
В результате клеточные стенки ослизняются, корни и части стеб-
лей загнивают, края листьев белеют, затем листья чернеют, скру-
322
чиваются и отмирают. На почвах с дефицитом Са многие расте-
ния не могут существовать.
Кроме прямого действия как необходимого химического био-
гена, Са оказывает на растения значительное косвенное влияние,
формируя актуальную кислотность почвы. В присутствии гумуса
он способствует созданию прочной комковатой структуры почвы,
улучшая ее водно-воздушный, тепловой режимы и плодородие.
Соли Са способствуют переводу труднорастворимых фосфатов в
более доступные для растений формы.
Углекислый кальций (СаСО3) формирует материнские породы
(мел, известняк), на которых развиваются почвы, характеризую-
щиеся своеобразными условиями для жизни растений. Тепловой,
водный и воздушный режимы этих почв более благоприятны для
растений, чем эти же режимы на бескальциевых субстратах. Фор-
мируется там и особый световой режим. На меловых почвах встре-
чаются крайне стенотопные виды, отличающиеся своеобразными
анатомо-морфологическими и физиологическими особенностями:
шиверекия подольская (Schivereckia podolica), норичник мело-
вой (Scrophularia cretacea), солнцецвет (Helianthemum сапит),
волчеягодник Софии (Daphne sophia), иссоп меловой (Hyssopus
cretaceus) и др. На меловых горах растет своеобразная меловая
форма сосны обыкновенной (Pinus sylvestris f. cretacea). Для этих
растений характерна ксероморфная световая структура. Анатоми-
ческая особенность их листьев — расположение основной фото-
синтезирующей ткани (столбчатой) не только около верхнего эпи-
дермиса, но и около нижнего (рис. 10.4). Это приспособление
возникло для восприятия отраженного (нижнего) света (Р.П.Ба-
рыкина, Н.В.Чубатова, 2005). Меловые выходы являются син-
экологическим, но не аутэкологическим оптимумом для этих рас-
Рис. 10.4. Строение листьев растений меловых обнажений
(по А.П. Шенникову, 1950):
А — лист иссопа (Hyssopus officinalis) типичной формы; Б — лист иссопа
(Я. officinalis) меловой формы; В — лист смолевки меловой (Silene cretacea)
323
тений. Они — патиенты, вытесненные на неудобные местообита-
ния более сильными конкурентами.
Богатые кальцием почвы могут формироваться и на других ма-
теринских породах: в гипсе (CaSO4- 2Н2О), доломите (CaMg(CO3)2),
мергеле. В почве присутствуют кальциевые соли фосфорной, крем-
ниевой, органических кислот. Многие растения с глубокой кор-
невой системой, извлекая Са из глубоких слоев почвы, способ-
ствуют его накоплению в верхних горизонтах. К таким накопите-
лям относятся бобовые и некоторые деревья (например, липа —
Tilia, ясень — Fraxinus).
По отношению к содержанию Са в почве выделяют три эколо-
гические группы растений.
1. Кальций-постоянные виды — нуждающиеся для нормаль-
ного развития в богатых известью субстратах.
2. Калъцефилы — растения, предпочитающие почвы с кальци-
ем: лиственница сибирская (Larix sibirica), бук (Fagus), ясень
(Fraxinus), люцерна серповидная (Medicago falcata), костер реч-
ной (Bromopsis riparia), ветреница лесная (Anemone sylvestris),
венерин башмачок (Cypripedium calceolus).
3. Калъцефобы — избегающие извести обитатели кислых почв
(белоус — Nardus stricta, сфагнумы, вересковые). Они обходятся
минимальным количеством Са, необходимым для обменных про-
цессов.
Многие виды, произрастая на юге лесной зоны и в степи на
разнообразных почвах (вплоть до кислых и безызвестковых), к
северу становятся кальций-постоянными. Таковы, например, дуб
пушистый (Quercuspubescens), бук лесной (Fagus sylvatica), кос-
тер прямой (Bromus erectus). На северном пределе распростране-
ния некоторые виды становятся кальцефилами, так как известко-
вые почвы суше, теплее, лучше проницаемы для воздуха. Таким
образом, понятия кальцефильности и кальцефобности условны.
Экологическое значение обеспечения почвы калием. Калий —
один из самых важных для растений элементов. В их тканях К+
значительно больше, чем других катионов. Калий участвует в важ-
ных электрофизиологических системах регуляции растений (транс-
порт через мембраны, регуляция устьичного аппарата и др.).
От него зависит гидратация клеточных коллоидов, он активирует
более 60 ферментных систем, обеспечивает работу проводящих
тканей, участвует в транспорте ассимилятов к органам запасания.
Растения поглощают ион К+ из почвенного раствора. В боль-
шинстве почв он присутствует в достаточных количествах в соста-
ве полевых шпатов, слюд, глинистых минералов. Исследования
показали, что концентрация ионов калия в клетках корней гороха
(Pisum sativum) в 75 раз выше, чем в почвенном растворе, а вакуо-
ли клеток корней брюквы (Brassica napus var. napobrassica) со-
держат в 10 000 раз больше калия, чем наружный раствор (П. Рейвн
324
и др., 1990). Если калия недостаточно, растения приобретают черты
ксероморфности (пейноморфоза), как и при недостатке других
элементов. При калиевом голодании листья желтеют с краев, затем
края и верхушки их буреют и засыхают (листья выглядят обо-
жженными), отмирают верхушечные почки. При этом активирует-
ся рост боковых побегов и растение становится низкорослым и
кустистым. Значительные нарушения возникают в водно-минераль-
ном обмене, развитии и функционировании проводящих тканей.
Разным видам растений необходимы разные количества калия.
Так, его много накапливают осока лисья (Carex vulpina), ежа сбор-
ная (Dactylis glomerata), люцерна хмелевидная (Medicago
lupulina), подмаренник болотный (Galium palustre), из культур-
ных растений — свекла (Beta vulgaris) и картофель (Solanum
tuberosum). Малым количеством калия обходятся душистый ко-
лосок (Anthoxanthum odoratum), нивяник (Leucanthemum vulgare),
белоус (Nardus stricta), таволга вязолистная (Filipendula ulmaria),
Черноголовка (Prunella vulgaris).
Экологическое значение обеспечения почвы фосфором. Фосфор
необходим растениям в значительно меньших количествах, чем
Са, К, N (по сравнению с азотом его требуется на порядок мень-
ше). По содержанию в растениях фосфор занимает среди других
элементов одиннадцатое место, однако роль его в метаболических
процессах огромна. Фосфор принимает участие в энергетическом
обмене, является компонентом фосфолипидов и нуклеиновых
кислот, участвует в регуляции активности белков (реакции фос-
форилирования). Он необходим для синтеза белка, образования
мембран, процессов репродукции, фотосинтеза и анаэробного
дыхания.
Обеспеченность почв фосфором отличается в разных зонах, а в
пределах зоны зависит от почвообразующей породы и гидрологи-
ческого режима. Так, подзолистая почва содержит около 0,1%
Р2О5, мощные и обыкновенные черноземы — 0,18 %, сероземы —
0,12 %. В суглинистых и глинистых почвах фосфора больше, чем в
песчаных. В почвах фосфор содержится в их минеральной части,
почвенном растворе, гумусе, организмах, но доступных для расте-
ний форм его мало (они образуются, например, при минерализа-
ции отмерших организмов). Большая часть фосфора находится в
почве в виде труднорастворимых солей ортофосфорной кислоты
(с Са, Al, Fe). Растения поглощают фосфор в виде ионов этой
кислоты, поэтому доступность его зависит от количества этих
ионов, перешедших в почвенный раствор. Виды растений разли-
чаются по способности поглощать фосфор из почвы. Например,
гречиха (Fagopyrum) и горчица (Sinapis) способны эффективно
поглощать фосфор из труднодоступных фосфатов.
Фосфорное питание растений в значительной степени связано
с микосимбиотрофией. Экспериментально показано, что грибной
325
компонент микоризы улучшает минеральное питание растений и
в первую очередь обеспечение фосфором. Это связано с тем, что
ионы ортофосфорной кислоты в почве малоподвижны, поэтому
вокруг корней на длину корневых волосков создаются зоны исто-
щения доступного фосфора (шириной 1 — 1,5 мм). А внешние гифы
гриба микоризы отходят от корня на значительно большие рас-
стояния (20—30 мм и более), и по ним фосфор поступает в ко-
рень, как это установлено в опытах с мечеными атомами (Т. А. Ра-
ботнов, 1993). При этом, сильно разветвляясь, гифы «обсасыва-
ют» большой объем почвы. Подсчитано, что увеличение всасыва-
ющей поверхности корней за счет микоризы выгоднее непосред-
ственного роста корневой системы (энергетические затраты на по-
строение 1 см корня в тысячи раз больше, чем на построение 1 см
намного более простых гифов).
На бедных фосфором почвах образующаяся микориза повы-
шает скорость роста растения — хозяина и количество в нем фос-
форосодержащих соединений. Т. А. Работнов (1993) отмечает так-
же, что в некоторых опытах установлена способность микориз-
ных растений использовать фосфор фосфоритов. Показано, что
микоризные корни интенсивнее продуцируют фосфатазу, способ-
ствующую растворению труднорастворимых фосфатов. Возможен
также синэргизм микоризных грибов с бактериями, растворяю-
щими фосфорные соединения. Таким образом, грибной компо-
нент существенно повышает эффективность использования фосфо-
ра микоризными растениями. Для растительных организмов, ис-
пользующих эту микоризу как источник фосфатов, симбиоз обычно
является «экологически облигатным», т.е. прохождение онтоге-
неза возможно и без него, но он необходим растению для выжи-
вания в типичных для него экологических условиях (Н.А. Прово-
ров и др., 2002). На почвах же, богатых доступными фосфатами,
микориза может стать для растения обременительной тратой.
Растения, испытывающие недостаток фосфора и приспособив-
шиеся к низкому содержанию его в почве, приобретают, как и
при азотном голодании, «ксероморфные» анатомо-морфологичес-
кие признаки (явление пейноморфоза).
Экологическое значение обеспечения почвы прочими биогенами.
Железо выполняет каталитические функции, активизирует ряд
ферментов. Несмотря на содержание в почве, оно может быть в
дефиците, особенно в карбонатных почвах (соединения Fe плохо
растворимы в щелочной среде). Переводу труднорастворимых со-
единений железа в более доступные для растений формы способ-
ствуют органические кислоты, выделяемые корнями.
Сера входит в состав аминокислот, участвует в построении ком-
понентов хлоропластов. В почве она встречается в виде сульфи-
дов и сульфатов, а также высвобождается при распаде органичес-
ких соединений. Ряд почвенных микроорганизмов могут окис-
326
лять или восстанавливать серу. Корни растений поглощают ее
главным образом в виде сульфатов. Листья могут усваивать сер-
нистый газ из атмосферы, но большая часть его попадает в почву,
где биологическим путем окисляется до серной кислоты. Сульфа-
ты легко вымываются из дренированных почв, а обогащены ими
почвы аридных областей с близким к поверхности стоянием суль-
фатных грунтовых вод (это вызывает сульфатное засоление и по-
селение растений-галофитов). Своеобразны растения гипсовых
(CaSO4-2H2O) почв с pH 7,5 — 8: в основном это кальцефилы.
Много серы содержится в растениях семейств Зонтичные и Крес-
тоцветные (особенно в горчичных маслах), в семенах бобовых.
Цинк необходим растениям в составе ферментов, обеспечива-
ющих окислительно-восстановительные процессы. Он участвует
в углеводном, фосфорном и белковом обмене. В почве Zn нахо-
дится в составе фосфатов, карбонатов, сульфидов, силикатов, окис-
лов. Если его много в субстрате, он может накапливаться в кор-
нях и побегах растений, формируя галмейную флору, состоящую
из растений не только толерантных к высокой концентрации цин-
ка, но и успешно развивающихся в этих условиях. Так, ярутка
альпийская (Thlaspi alpestre) может содержать цинка в листьях до
25 000 мг/кг сухой массы (при норме 120) (В.Лархер, 1978). Как
правило, для таких местообитаний характерно повышенное со-
держание и других тяжелых металлов (Си, РЬ), где цинк усугубля-
ет их токсическое действие. На почвах с обилием цинка деревья и
кустарники обычно не развиваются, а травы представлены вида-
ми, приобретшими устойчивость к этому неблагоприятному фак-
тору (Viola calaminaria, Minuartia verna и др.).
Магний входит в состав хлорофилла, а также участвует в регуля-
ции набухания коллоидов. В почве он находится в виде карбона-
тов (магнезит, доломит), силикатов (оливин, серпентин и др.),
сульфатов и хлоридов. На некоторых кислых почвах легкого грану-
лометрического состава отмечается недостаток соединений маг-
ния и для растений характерны хлороз и ослабленный рост. Избы-
ток магния в почвах создает своеобразные условия для растений:
эти почвы бедны основными элементами минерального питания,
но лучше обеспечены теплом благодаря темной окраске. На них
формируется серпентинная флора из эндемичных видов (форзи-
ция европейская — Forsythia еигораеа, костенец — Asplenium
cuneifolium, бурачок — Alyssum bertolonii и др.), подвидов и форм
обычных растений, накапливающих в тканях большие количества
Mg. У таких растений развитая корневая система, но надземная
их часть, в том числе генеративные органы, развиваются слабо.
Кадмий (Cd2+) и хром (Сг2+) — активаторы нескольких фер-
ментов в живых организмах. Мышьяк в малых дозах стимулирует
жизненные процессы и повышает урожай сельскохозяйственных
культур (В. Б. Ильин, 1991).
327
Экологическое значение почвенных
токсикантов
Токсиканты и токсикофиты. Ядовитые для организмов веще-
ства называют токсикантами. Угнетающие растения вещества очень
разнообразны, а совместное действие их усиливает отрицатель-
ный эффект. Но среди растений встречаются и токсикофиты —
устойчивые к высоким концентрациям токсичных веществ. От
вредных веществ, накопившихся в тканях, растения освобожда-
ются только с отмиранием и опадением своих частей, поэтому
смена листьев у деревьев, отмирание наземных побегов трав —
необходимый выделительный процесс.
Некоторые микроэлементы присутствуют в ряде почв в ано-
мально больших концентрациях. Это может быть вызвано есте-
ственными причинами (нахождение близ рудных месторождений)
или техногенным загрязнением. Необходимые в микродозах Cd,
Pb, Hg, As, Zn, Mo, Co, Ni, Си, Сг становятся токсикантами при
высоких концентрациях.
Элементы с атомной массой более 50 называют тяжелыми
металлами (В. В. Ковальский, 1974). В естественном состоянии
почвы с большой концентрацией их — редкое явление. Загрязне-
ние почв тяжелыми металлами обычно связано с деятельностью
промышленных предприятий (особенно металлургических), руд-
ников, тепловых электростанций, автотранспорта, коммунально-
го хозяйства и т.п. В загрязненных почвах тяжелые металлы при-
сутствуют в обменной форме, в виде карбонатов, соединений с
Fe, Мп, связаны с органическими веществами почвы.
Для многих растений высокие концентрации тяжелых метал-
лов токсичны, но некоторые виды накапливают их в значитель-
ных количествах. Например, листья букашника горного (Jasione
montana) могут концентрировать мышьяк до 6 200 мг на 1 кг су-
хой массы (при норме 0,2 мг/кг), а минуарция весенняя (Minuartia
verna) способна накапливать в листьях свинец до 11 400 мг/кг (на
почвах с нормальным содержанием РЬ — 12 мг/кг) (В.Лархер,
1978). Характерна флора почв, богатых кобальтом и медью (мед-
ные флоры на соответствующих месторождениях). К купрофи-
там (способным накапливать ионы меди) относятся смолевка
обыкновенная (Silene vulgaris), качим (Gypsophila patrim), виды
рода шпажник (Gladiolus), некоторые злаки и мхи.
Токсичные соединения ртути широко используются как пес-
тициды, при производстве бумажной массы, для получения поли-
винилхлорида. Со сточными водами они попадают в почву, грун-
товые воды, водоемы и циркулируют в наземных и водных пище-
вых цепях. Растения, вынужденные поглощать соединения ртути,
способны освобождаться от нее: восстановленная ртуть поступает
в атмосферу в очень ядовитом элементарном виде.
328
Промышленные сточные воды и моющие средства обусловли-
вают повышенное содержание мышьяка в грунтовых водах и во-
доемах. Попав в водоросли, он накапливается затем в телах жи-
вотных, занимающих более высокое положение в пищевых цепях.
Водоросли тропических водоемов, поглощая растворенные арсе-
наты, восстанавливают их в арсениты и затем через ряд химичес-
ких реакций связывают с фосфолипидами. В связанном виде As
сохраняется в жировых каплях цитоплазмы и (или) клеточных
мембранах. Таким образом, тропические водоросли в значитель-
ной степени предотвращают опасность отравления мышьяком. Во
внетропических водоемах водоросли не способны обезвреживать
арсенаты, которые аккумулируются в клеточных белках, убивая
растения (В.Эйхлер, 1993). В верхних горизонтах незагрязненных
почв содержание мышьяка колеблется от 0,35 до 8,5 мг/кг воз-
душно-сухой почвы (растения выносят концентрацию As от 15 до
60 мг/кг) (В. Б. Ильин, 1991).
Выхлопные газы автомобилей и отходы ряда производств при-
водят к свинцовому загрязнению среды. Например, в поверхнос-
тных горизонтах незагрязненных почв Западной Сибири соеди-
нений свинца содержится от 2,8 до 20,2 мг на 1 кг воздушно-
сухой почвы (в дозе до 50 мг/кг он не вызывает отрицательного
биологического эффекта) (В. Б. Ильин, 1991). Но близ автострад
количество РЬ возрастает на порядок. Почвенные микроорганиз-
мы способны переводить его нерастворимые в воде соединения в
доступную для растений форму. Свинец накапливается в стеблях,
листьях, корнях (в меньшей степени в плодах и семенах). Поэто-
му нельзя употреблять в пищу растения, собранные на 50-метро-
вой полосе близ шоссе, а земли близ промышленных предприя-
тий должны быть выведены из сельскохозяйственного использо-
вания.
Очень чувствительны к свинцовому загрязнению низшие растения:
например, некоторые виды лишайников не выживают в крупных горо-
дах и близ трасс с интенсивным движением автотранспорта. Они часто
служат индикаторами загрязнения среды. Из высших растений довольно
чувствительны к свинцовому загрязнению мохообразные, а у ряда видов
найдены отложения свинца в ядрах, пластидах, вакуолях, митохондриях
и плазмодесмах. У цветковых растений система детоксикации более со-
вершенна. Например, у рдеста (Potamogeton) свинец в основном изоли-
руется в клеточной стенке. Деревья депонируют его в коре вне живых
клеток. Но при сильном загрязнении тяжелые металлы обнаруживаются
и в живых тканях, где концентрируются в вакуолях. Так, в листьях с 1 га
лиственного леса близ Вены обнаружено 376 г РЬ. Ежегодный опад спо-
собствует накоплению его на поверхности почвы (В.Эйхлер, 1993).
Источник загрязнения почв кадмием — промышленные газо-
образные выбросы (он используется в качестве катализатора при
329
производстве поливинилхлорида; 1т угля, сжигаемого в котель-
ных, содержит около 2 г Cd). Этот элемент легко поглощается
корнями растений, поэтому загрязнение почвы сульфатом кад-
мия приводит к снижению урожайности сельскохозяйственных
культур. В исключительно больших количествах накапливают кад-
мий грибы.
Сильно токсичны и широко используемые в хозяйстве пестициды
(от лат. pestis — зараза и cid — убивать). Это разные химические соеди-
нения, используемые для уничтожения или сокращения численности бак-
терий {бактерициды), грибов {фунгициды), нематод {нематоциды), на-
секомых {инсектициды), клещей {акарициды), позвоночных {зооциды),
сорных растений {гербициды). Все пестициды являются биоцидами.
Самоочищение почвы и защита растений от избытка вредных
веществ в ней. Загрязненная тяжелыми металлами и (или) пести-
цидами почва в определенной степени способна к самоочищению
(детоксикации) — выведению токсикантов из круговорота веществ.
При небольшом загрязнении они могут перейти в малоактивную
форму, что способствует сохранению почвенной биоты, фитоце-
ноза и не мешает сельскохозяйственным культурам. К такому са-
моочищению способны в первую очередь глинистые почвы, имею-
щие большую адсорбционную поверхность, высокие значения pH,
достаточное количество органического вещества, способного к
комплексообразованию. Верхний слой почвы толщиной в несколь-
ко сантиметров, состоящий из глинистых и органических колло-
идов, создает на 1 га миллионы квадратных метров активной ад-
сорбирующей поверхности. Адсорбция происходит в силу физи-
ческих причин (силы Ван-дер-Ваальса), межмолекулярного взаи-
модействия, а также химического связывания. Связь пестицидов
с почвенными коллоидами со временем становится все более проч-
ной, и химические вещества теряют биологическую активность.
Однако защитные возможности почв ограничены. Особенно малы
они у малогумусных кислых почв легкого гранулометрического
состава.
Некоторые растения приобрели наследственную устойчивость
к неблагоприятному минеральному составу местных почв. Они
вынужденно поглощают избыточное количество токсичных эле-
ментов, но обезвреживают их путем осаждения или изоляции в
вакуолях, клеточных стенках, кутикуле. Важную роль в защите
растений от вредных веществ играют корни. Они препятствуют
поглощению избыточных ионов и токсичных соединений, в ре-
зультате чего в надземных органах обеспечивается приемлемая их
концентрация. Эффективным барьером, контролирующим поступ-
ление веществ в проводящую систему, являются пропускные клетки
эндодермы корней. Например, в опыте с поглощением корнями
330
кукурузы (Zea mays) раствора с ионами лантана эндодерма пре-
дотвратила поступление этого элемента в глубь корня: он был
обнаружен только в стенках клеток коры (П.Рейвн и др., 1990).
Но при высоких концентрациях токсикантов в почве корни не
могут полностью предотвратить их попадание в ксилему. В этом
случае растения используют другой способ защиты — инактива-
цию попавших ядов. Они концентрируются вдали от метаболи-
ческих центров в клеточных стенках и межклетниках, в малопод-
вижных соединениях. Защитные механизмы обеспечивают мини-
мальное поступление тяжелых металлов и пестицидов в плоды и
запасающие органы (клубни, корнеплоды). Но в зоне большого
загрязнения деревья и кустарники отмирают, травяной покров
изреживается, мохово-лишайниковый ярус исчезает и возникают
маловидовые сообщества из растений, приспособленных к новым
экологическим условиям (В. Б. Ильин, 1991).
Экологическое значение засоления
Типы засоленных почв. Соли необходимы для жизнедеятельно-
сти растений. Вредное их действие начинает сказываться на неко-
торых, особенно культурных, не приспособленных к засолению
растениях уже при концентрации 0,1 % от массы сухой почвы.
Засоленными считаются почвы, если в каком-либо горизонте со-
средоточено более 0,25 % солей от массы почвы. Если почва со-
держит 0,5 % и более количества от массы почвы легкораствори-
мых солей, тогда определяющим экологическим свойством мес-
тообитания становится засоление. Повышенная концентрация лег-
корастворимых и, следовательно, осмотически сильно действую-
щих солей (NaCl, Na2SO4, Na2CO3 и др.) характерна для почв мно-
гих естественных местообитаний. Засоление широко распростра-
нено в аридных областях (пустыни, степи), а также по берегам
морей и соленых озер (Баскунчак, Чаны и др.). Засолены около
25 % почв земного шара.
Засоленные почвы разнообразны по составу солей и степени
засоленности. Почву считают слабосолончаковатой, если соли
составляют более 0,25 % от ее массы и находятся лишь на глубине
80—150 см. Если такая концентрация солей встречается на глуби-
не 30—80 см, почву называют солончаковатой. В солончаковой
почве повышенное содержание солей отмечается в пределах 5 —
30 см от поверхности. А солончак в самом верхнем горизонте имеет
не менее 1 % солей. Концентрация солей в солончаках может до-
стигать нескольких десятков процентов. Они постоянно и сильно
увлажнены солеными водами и лишь в середине лета подсыхают
и покрываются корками и выцветами солей. Для них характерна
особая галофитная растительность. Из солончаков по мере вымы-
331
вания солей и их заглубления (при понижении грунтовых вод или
увлажнении климата) возникают солонцы. Соли по мере их вы-
мывания остаются в подсолонцовом горизонте. В процессе осо-
лонцевания и освобождения от солей верхних горизонтов меня-
ются растительные сообщества (солянковые на полынные и зла-
ковые). Солонцы и солончаки — основные типы засоленных почв.
Участки их часто располагаются вперемешку, создавая солонцо-
во-солончаковые комплексы почв и растительности. При даль-
нейшем развитии процесса рассоления и понижения грунтовых
вод возникают солоди, а процесс вымывания и заглубления солей
называют осолодением. Особенно интенсивно он идет в пониже-
ниях рельефа, где застаивается влага.
Рассолению и заглублению солей способствует зимнее промер-
зание сильноувлажненных почв. Это природное явление издавна
учитывается земледельцами и целенаправленно используется в
сельском хозяйстве степного Поволжья и юга Западной Сибири
(зимний полив, вызывающий промерзание и заглубление почвен-
ных солей).
Характер засоления почв определяется по преобладающим ани-
онам: хлоридное, сульфатно-хлоридное, хлоридно-сульфатное,
сульфатно-содовое, изредка встречается нитратное засоление и др.
В солончаках преобладают соли натрия, которые находятся не
только в растворе, но и насыщают коллоиды почвенного погло-
щающего комплекса. Кроме преобладающих NaCl и Na2SO4 там
присутствуют и другие высокорастворимые соли (СаС12, MgCl2,
MgSO4, Na2SO4, Na2CO3, NaHCO3 и др.). При хлорид но-сульфат-
ном засолении реакция почвенного раствора близка к нейтраль-
ной, а при содовом достигает 9—11.
Особо негативное воздействие на растения оказывает хлорид-
ное засоление. Повышенная концентрация хлористых солей мо-
жет разобщать окисление и фосфорилирование, нарушать снаб-
жение растений макроэргическими фосфорными соединениями.
Na препятствует накоплению других катионов, в том числе необ-
ходимых для жизни растений К и Са. При обилии NaCl в почве
нарушается азотный обмен, накапливается аммиак и другие ядо-
витые вещества, нарушается обмен соединений серы.
В условиях сильного засоления многие виды (мезофиты) не
дают всходов. Но соленый раствор не могут потреблять и взрос-
лые растения, так как осмотическое давление почвы оказывается
выше, чем в клетках корней. Большинство видов обладает резер-
вом устойчивости к засолению (некоторые приспосабливаются
переносить довольно высокое засоление — до 3 — 5 г/л). Вредное
влияние высокой концентрации легкорастворимых солей на рас-
тения, не адаптированные к этим условиям, связано с поврежде-
нием мембранных структур, в результате чего возрастает их про-
ницаемость и теряется способность к избирательному пропуска-
332
нию веществ. При этом соли поступают в клетки пассивно вместе
с транспирационным током воды. В аридных зонах, где транспи-
рация активна, солей в растение поступает много.
Растения-галофиты. Растения, приспособившиеся к жизни на
почвах с высоким содержанием солей (более 5 % от сухой массы
почвы), получили название галофитов (от греч. hats — соль).
Некоторые из них развивают огромное осмотическое давление.
Так, у выжатого сока лебеды (Atriplex confertifolia) осмотический
потенциал равен 200 бар, и в 1 л его содержится 67,33 г хлоридов
(у большинства растений осмотический потенциал составляет 5 —
30 бар, а у растений пресных водоемов — около 1 бар) (Н. И.Якуш-
кина, 1993). Адаптации растений к засолению разнообразны и
затрагивают разные уровни (молекулярный, клеточный, организ-
менный, популяционный, фитоценотический).
Галофиты могут быть факультативными, выносящими повы-
шенное содержание солей, но лучше развивающимися при низ-
ком их содержании. Факультативным галофитом, например, яв-
ляется хлопчатник (Gossypium). Для облигатных галофитов по-
вышенное засоление — благоприятный фактор, стимулирующий
их рост и жизнедеятельность. Растения, выносящие разные кон-
центрации солей, называются эвригалинными, например частуха
подорожниковая (Alisma plantago-aquatica), растущая в пресных
водоемах, однако выносящая pH от 2,0 до 12,0 и концентрацию
солей до 3 % (Т. К. Горышина, 1979). Растения с узкой экологи-
ческой амплитудой по отношению к засолению называются сте-
ногалинными.
Среди галофитов есть растения разных семейств, по-разному
приспособившиеся к избытку солей в почве. По способам адапта-
ции к засолению, анатомо-морфологическим и физиологическим
особенностям П.А. Генкель объединил галофиты в несколько
групп.
1. Эугалофиты — «соленакапливающие» растения. Осмоти-
ческое давление клеточного сока у них очень высокое, и содержа-
ние солей может быть в несколько раз больше, чем в почве. Так,
в прежние века солянки (Salsola kali, S. soda) и поташник
(Kalidium) служили источником сырья для получения соды и по-
таша. Эти растения могут расти на мокрых солончаках в аридных
областях, по берегам соленых озер и морским побережьям. К ним
относятся многие виды семейства Маревые (из родов солянка —
Salsola, шведка — Sueda, солерос — Salicornia), выдерживающие
засоление до 30—50 % и более. Нередко они имеют черты ксеро-
фитов, отличаются суккулентностью, безлистностью, членистос-
тью стебля и ветвей (рис. 10.5). Доминантами в засоленных райо-
нах становятся и многочисленные виды полыней (Artemisia). Для
эугалофитов повышенная концентрация солей в почве — необхо-
димый и благоприятный фактор. Опыты с культивированием со-
333
iPnc. 10.5. Солерос (Salicomia herbacea)
Л ль (по А. П. Шенникову, 1950):
t КЗ? — целое растение; Б — окончание ветви (увеличено)
лероса и подорожника морского (Plantago
| w maritima) показали, что для них оптимальна
л 2—3%-я концентрация поваренной соли в
ь растворе, а пресная вода эти растения угне-
| тает. В пустынях эугалофиты существуют в
i 4 условиях засоленности почв, постоянной
14 S почвенной и атмосферной сухости, перегре-
П ва, резких суточных колебаний температур,
с интенсивной инсоляции, сильных ветров.
\ VSW" 0? Такие условия другие растения не выносят.
г Jjf 'l- Криногалофиты — «солевыделяющие»
растения. Таковы виды родов кермек
I (Statice), армерия (Armeria), тамариск
I (Tamarix), франкения (Frankenia) и др. Ци-
топлазма их клеток легкопроницаема для
солей, и эти растения пропускают через себя
соляной раствор. Избыток солей выделяется
f / в виде раствора через солевыделительные
f Н \ железки. Выделившаяся соль лежит на по-
верхности листьев, осыпается, сдувается вет-
л ром, смывается дождем. При большом из-
бытке солей растения могут сбрасывать лис-
тья или годичные приросты. Из золы чилийского кустарника
Frankenia berteroana аборигены добывали поваренную соль. Мно-
гие криногалофиты по строению листьев близки к мезофитам.
3. Гликогалофиты — «соленепроницаемые» растения. Это эв-
ригалинные виды, потребность которых в солях сравнительно
невелика. Значительное осмотическое давление у них достигается
не высокой концентрацией солей в вакуолях, а повышенным со-
держанием органических веществ, особенно углеводов. Такие адап-
тации к засолению известны у некоторых видов полыней и пус-
тынных злаков. К этой группе относится свекла (Beta vulgaris),
дикий предок которой — прибрежно-морская форма (Beta vulgaris
subsp. maritima), произрастающая на засоленных морских побе-
режьях Южной и Западной Европы.
Кроме того, выделяют группу псевдогалофитов, которые, про-
израстая на засоленной почве, избегают засоления благодаря очень
глубокой корневой системе, достигающей малозасоленных овод-
ненных горизонтов. Таков тростник (Phragmites australis), расту-
щий и в пресных водоемах, и в пустынях, кендырь сибирский
(Apocinum sibiricum) и др. А растения, не имеющие приспособле-
ний к засолению почв, называют гликофитами.
334
Экологические условия торфяных почв
Торфяные болота широко распространены в разных зонах, за-
нимая 4,4% территории Земли (Торфяные ресурсы мира, 1988).
В России общая площадь оторфованных земель составляет при-
мерно пятую часть ее территории (С. Э. Вомперский, 1994). Осо-
бенно много болот в лесной зоне. Растительные сообщества тор-
фяных болот сложены большей частью гигрофитами или растени-
ями с широкой экологической амплитудой, приспособившимися
к обитанию в условиях застойного увлажнения.
Торф — органическая масса, образовавшаяся в результате отмирания
и неполного разложения растений в условиях повышенной влажности и
недостатка кислорода. Он состоит из недоразложившихся растительных
остатков, гумуса и минеральных веществ. Естественная влажность торфа
86—95 %. В разных видах его в зависимости от характера питающих бо-
лото вод может содержаться от 2 до 50 % минеральных вешеств.
На торфяниках грунтового и речного питания (болота низин-
ного типа) обитают растения, требующие повышенного содержа-
ния минеральных веществ и формирующие эвтрофную раститель-
ность. Химический состав питающих вод может быть разным, и в
связи с этим формируются или маловидовые монодоминантные
зеленомоховые и травяные (осоковые, злаковые) сообщества, или
богатые по видовому составу фитоценозы из растений разных
жизненных форм. При обильном и разнообразном минеральном
питании формируются облесенные болота с деревьями (ольха —
Alnus, береза — Betula), кустарниками (виды ив — Salix), травами
(осоки — Carex и болотное разнотравье: вахта трехлистная —
Menyanthes trifoliata, белокрыльник — Calla palustris, сабель-
ник — Comarum palustre, вех ядовитый — Cicuta virosa и др.),
зелеными мхами (виды Drepanocladus, Calliergon, Aulacomnium
и др.). На торфяниках атмосферного питания (верховой тип бо-
лот) образуется растительный покров из олиготрофных видов:
болотных форм сосны (Pinus sylvestris), кустарничков из семейств
Вересковые (багульник — Ledum palustre, болотный мирт —
Chamaedaphne calyculata, подбел — Andromeda polyfolia) и Брус-
ничные (голубика — Vaccinium uliginosum, клюква — Oxycoccus),
сфагновых мхов. Зольность торфа тут низкая (2,5 —5 %).
Низинные и верховые торфяные болота — стадии единого бо-
лотообразовательного процесса: по мере накопления торфа рас-
тительность все больше изолируется от влияния грунтовых вод, и
в гидрологическом режиме болота атмосферные воды становятся
преобладающими. Низинные и переходные стадии развития бо-
лот могут различаться по длительности. Верховые болота обычно
начинали свое развитие с низинных стадий.
335
Верховые болота — своеобразные в экологическом отношении
местообитания: их сильная обводненность связана с особеннос-
тями строения сфагновых мхов и сфагнового торфа (наличие гиа-
линовых клеток, обладающих высокой влагоемкостью, а также
большое содержание в субстрате гидрофильных коллоидов).
В торфе затруднено передвижение воды: вертикальная фильтра-
ция составляет около 0,02 м в сутки, горизонтальная — около
0,6—0,7 м (С.Н.Тюремнов, 1976). Минерализация вод низкая (40—
70 мг/л). Источник минеральных веществ на верховых болотах —
атмосферная пыль: в промышленных районах на поверхность бо-
лот выпадает до 2 т/га пыли (Н.И.Пьявченко, 1967). Атмосфер-
ная пыль (в том числе загрязняющие вещества) передвигается в
сторону пониженной температуры (явление термофореза) и осе-
дает на поверхности болот. Она частично утилизируется растени-
ями, а остальная ее часть выводится из круговорота, захороняясь
в торфе. Это одна из важных экологических функций болот, по-
зволяющих считать их биогеохимическим барьером природы, спо-
собствующим очищению атмосферы и вод.
Недостаток минерального азота на верховых болотах воспол-
няется деятельностью азотфиксирующих цианобактерий, которые
живут среди листьев и стеблей сфагновых мхов, а некоторые рас-
тения (например, росянки — Drosera) обеспечивают себя азотом,
поедая мелких животных. Основных минеральных веществ расте-
ниям верховых болот не хватает, и олиготрофность обусловливает
их специфическую морфологию (пейноморфоз).
Застойность болотных вод вызывает анаэробность среды, по-
давляет деятельность редуцентов. Недостаток кислорода вынуж-
дает растения располагать корни в самом верхнем 30-сантиметро-
вом слое (деятельном горизонте), где существует слабое движе-
ние воды. Органы даже жестких вечнозеленых кустарничков име-
ют воздухоносные полости. Постоянный вертикальный прирост
сфагновых мхов (от 0,5 до 2,5 см в год) (С.Н.Тюремнов, 1976)
вынуждает растения к постоянному приросту побегов и развитию
придаточных корней (у болотных кустарничков, рис. 10.6), пере-
мещению вверх зимующей почки (у росянки), изгибанию и росту
корней вверх, выше корневой шейки (у сосны).
Своеобразен тепловой режим торфяной почвы: поверхность
торфяника может сильно нагреваться (до 30—40°), однако внутри
торфяной толщи температура понижена (4 °C). Теплоизоляцион-
ные свойства торфа таковы, что в его толще прослойки льда могут
сохраняться в течение всего лета. Микосимбиотрофия болотных
растений способствует их бесперебойному снабжению водой, не-
смотря на большой температурный градиент. Явление «физиоло-
гической сухости» в таких условиях отсутствует (см. разд. 7.1.3).
Растения верховых болот, растущие в условиях олиготрофнос-
ти, недостатка О2, резких температурных перепадов, находятся
336
Рис. 10.6. Погребенный мхом куст багульника (Ledum palustre)
(по А.А. Ниценко, 1967):
/ — поверхность современного мохового покрова; 2 — уровень корневой шейки
(прежняя поверхность мохового покрова); 3 — придаточные корни
здесь в режиме синэкологического (фитоценотического) оптиму-
ма, где развиваются вне конкуренции с другими видами. Будучи
слабыми конкурентами, в иных сообществах в сочетании с други-
ми видами они не встречаются. Опыты с болотными кустарнич-
ками показали, что их аутэкологические (физиологические) оп-
тимумы по многим показателям сильно отличаются от синэколо-
гических: при отсутствии конкурентов они положительно реаги-
ровали на внесение азотных удобрений, повышение аэрации и
улучшение тепловых условий.
10.3. Экологическая полифункциональность почв
Почва — важная составная часть биогеоценоза, выполняющая
множество функций. Все экологические функции почв взаимо-
связаны и все вместе во многом определяют условия существова-
ния организмов. Г. В. Добровольский и Е.Д. Никитин (1986) мно-
гочисленные экологические функции почв объединили в ряд групп.
I. Функции почвы, обусловленные ее физическими свойствами.
1. Жизненное пространство. С почвой связано существование ог-
ромного числа разнообразных видов, в том числе подавляющей части
цветковых растений. Жизненное пространство почв может характери-
зоваться как горизонтальное (распространение по поверхности суши) и
вертикальное (распределение по почвенному профилю). Оно также мо-
337
жет быть временным (связанным с изменениями погодных условий, се-
зонными явлениями, изменчивостью по годам, климатическими рит-
мами).
2. Жилище и убежище. Почва предоставляет организмам разнооб-
разные убежища. Она предохраняет их от переохлаждения и перегрева.
Температура и влажность воздуха в почве подвержены менее резким ко-
лебаниям, чем на поверхности земли. Эта особенность важна в периоды
резких изменений погоды, в экстремальных условиях тундр и пустынь.
3. Опора. В почве растения заякориваются с помощью корней, что
помогает им сохранять вертикальное положение и быть устойчивыми к
ветру.
4. Депо семян и других зачатков. Благодаря «консервирующей» спо-
собности, зависящей от режима тепла и влаги, повышенного содержа-
ния диоксида углерода и отсутствия света, в почвах длительно сохраня-
ются семена, споры и другие зачатки многих организмов.
II. Функции почвы, связанные преимущественно с ее хими-
ческими и биохимическими свойствами.
1. Источник элементов минерального питания растений. Полу-
чение и усвоение минеральных веществ из почвы зависит от концентра-
ции — благоприятного соотношения доступных элементов, pH среды,
гранулометрического состава и других характеристик почв.
2. Депо элементов минерального питания, энергии и влаги. Почва
имеет резерв полезных компонентов и играет роль буфера, помогающе-
го устранять перебои в снабжении ими при исчерпании наиболее дос-
тупных запасов.
3. Стимулятор и ингибитор биохимических и других процессов.
В почву поступают разнообразные продукты метаболизма, которые мо-
гут стимулировать или угнетать жизнедеятельность организмов. Значи-
тельную роль во взаимоотношениях растений играет взаимовлияние их
через корневые выделения. Известно также самоугнетение растений («поч-
воутомление») в посевах культурных растений без севооборота и в неко-
торых древостоях.
III. Функции почвы, определяемые в основном физико-хими-
ческими свойствами.
1. Сорбция веществ, поступающих из атмосферы и с грунтовы-
ми водами. Поверхность мелкозема и коллоиды почвы адсорбируют газы
и жидкости, содержащие молекулы и ионы, поступающие в почву. По-
глотительная способность почв обеспечивает удержание элементов ми-
нерального питания, а также адсорбцию загрязняющих веществ.
2. Сорбция микроорганизмов, обитающих в почве. Благодаря этой
функции микроорганизмы защищены от выноса нисходящим током воды
за пределы почвенного профиля. Различные свойства поверхностей по-
чвенных микрозон создают условия для сорбции разных организмов. Эта
способность связана с разным гранулометрическим и минералогическим
составом, кислотностью, особенностями формирования почв. Почвы
338
тяжелого механического состава сорбируют микроорганизмов больше,
так же как черноземы по сравнению с дерново-подзолистыми и серыми
лесными почвами. От процессов адсорбции зависят жизнедеятельность,
развитие и размножение почвенных микроорганизмов.
IV. Информационные биогеоценотические функции почвы.
1. Сигнал для сезонных и других биологических процессов. Эта фун-
кция связана с периодически изменяющимися режимами почвы. Темпе-
ратура почвы служит сигналом начала или прекращения сезонных цик-
лов жизнедеятельности организмов, определяет протекание физиологи-
ческих процессов. В аридных зонах начало или задержка вегетации обус-
ловлена динамикой водного режима почв. От сезонных колебаний ак-
тивности разных почвенных микроорганизмов зависит обеспеченность
растений элементами минерального питания, а от температуры и влаж-
ности почв — развитие насекомых (в том числе фитофагов).
2. Регулятор состава и структуры биогеоценоза. Почва — компо-
нент биогеоценоза, от которого во многом зависит его состав и структу-
ра. Свойства ее существенно влияют на продуцентов и связанных с ними
консументов и редуцентов.
3. Пусковой механизм сукцессий. Процессы заболачивания, засоле-
ния, остепнения, вытаптывания и другие, даже если они возникли под
влиянием внешних причин, протекают как стадии преобразования по-
чвы, вызывая соответствующие сукцессии биогеоценозов.
4. «Память» биогеоценоза. Почва хранит большое количество ин-
формации. Согласно концепции В.О.Таргульяна и И. А. Соколова (1976),
она состоит из комплекса устойчивых свойств, возникших в ходе ее раз-
вития (почва-память), и совокупности изменчивых свойств и процес-
сов в момент наблюдения (почва-момент).
V. Интегральные биогеоценотические функции почвы.
1. Трансформация вещества и энергии. При почвообразовании ис-
ходные вещества (материнские породы и продукты, поступающие с пы-
лью, осадками, поверхностными и грунтовыми водами, растительными
остатками) превращаются в субстрат, пригодный для формирования био-
геоценоза. Освобождение энергии в ходе разложения органических ос-
татков имеет не меньшее значение, чем образование органических со-
единений при фотосинтезе.
2. Санитарная функция. Деструкция микроорганизмами и беспоз-
воночными поступающих в почву продуктов обмена и органических ос-
татков предохраняет ландшафты от самозагрязнения. Микробное насе-
ление почвы обладает антисептическими свойствами, подавляет разви-
тие болезнетворных микроорганизмов. Почвенный покров эффективно
защищает грунтовые воды от проникновения бактериальных и химичес-
ких загрязнений. Но некоторые болезнетворные микроорганизмы со-
храняются в почве длительное время.
3. Защитный и буферный биогеоценотический экран. Сформиро-
вавшиеся в ходе длительной эволюции биогеоценозы отличаются устой-
339
чивостью к воздействию внешних факторов, способностью к гомеоста-
тическому регулированию, поддержанию своих свойств. Почвы способ-
ны в значительной степени нивелировать перепады влажности и темпе-
ратуры в биогеоценозах. Буферная роль почв проявляется в поддержа-
нии плодородия, противостоянии водной и ветровой эрозии, химичес-
кому загрязнению, в способности к восстановлению нарушенных биоге-
оценозов за счет запаса семян.
4. Плодородие — способность обеспечивать условия для жизни рас-
тений. Это интегральная функция почв, выражение всех ее свойств.
ГЛАВА 11
БИОТИЧЕСКИЕ ЭКОЛОГИЧЕСКИЕ
ФАКТОРЫ
Совокупность влияний одних организмов на другие относится
к биотическим экологическим факторам. Они — важная состав-
ляющая природной среды, существенно влияющая на жизнедея-
тельность растений и функционирование растительных сообществ.
11.1. Типы отношений растений с другими организмами
Прямые и косвенные отношения растений с другими организма-
ми. Прямые (контактные) отношения между организмами проис-
ходят при непосредственном контакте. Их подразделяют на меха-
нические и физиологические. Примеры механического влияния
на растения — поселение на них других организмов и охлестыва-
ние кронами. Прямые физиологические отношения растений очень
разнообразны. Условно среди них выделяют связи, дающие орга-
низмам обоюдную пользу и приносящие пользу одностороннюю.
Примеры первых — связи растений с симбиотическими азотфик-
сирующими прокариотами и микоризными грибами. Примеры
прямых физиологических отношений, приносящих односторон-
нюю пользу, — паразитизм и хищничество.
Прямым взаимоотношениям противопоставлены косвенные,
осуществляемые через посредство других объектов. Косвенные
отношения подразделяют на трансабиотические и трансбио-
тические. Первые заключаются во влиянии одних организмов на
другие через изменение абиотической среды. Такой тип взаимо-
отношений широко распространен в природе: влияние на расте-
ния часто осуществляется путем изменения организмами свойств
почвы, воды, воздуха, условий освещения. К трансабиотическим
отношениям можно отнести и конкуренцию, которая возникает
при совместном использовании ресурсов. Косвенные трансбио-
тические взаимодействия происходят при посредстве других орга-
низмов. Такая форма взаимоотношений наблюдается, например,
на лугах при предпочтении скотом некоторых видов, когда не-
поврежденные растения разрастаются и угнетают соседей. Другой
341
пример: желтоцветковая ястребинка (Hieracium piloselloides) опы-
ляется лучше, когда растет рядом с ярко-оранжевой Н. auranticum.
Или некоторые виды отрицательно влияют на другие, являясь
промежуточными хозяевами их паразитов. Например, в средневе-
ковой Европе обсаживание межей барбарисом (Berberis) небла-
гоприятно сказывалось на урожае, так как на этом кустарнике
развивается одна из стадий ржавчинных грибов, поражающих хлеб-
ные посевы.
Классификация отношений растений с другими организмами по
действию на жизнеспособность партнера. Классификация взаимо-
отношений организмов по действию на жизнеспособность парт-
нера («биологическому эффекту») делит их на шесть типов: му-
туализм (взаимополезные отношения), антагонизм (полезно-
вредные отношения), комменсализм (полезно-нейтральные от-
ношения), конкуренция (взаимовредные отношения), аменсализм
(вредно-нейтральные отношения) и нейтрализм (безразличные
отношения) (Ю.Одум, 1975).
Если мутуалистические отношения не обязательны для суще-
ствования видов, такой тип взаимоотношений называют прото -
кооперацией. Пример — распространение муравьями семян лес-
ных растений. А если присутствие партнера становится обязатель-
ным условием жизни, то такие связи видов в этой классификации
традиционно называют симбиозом. Однако использование этого
термина в данном случае неудачно, так как буквально «симбиоз»
(от греч. simbiosis — совместная жизнь) означает сожительство,
без указания на вред или пользу (см. разд. 11.2). Обычно приво-
димые примеры симбиоза — микориза (взаимоотношения гриба с
высшим растением) и лишайник (сожительство гриба и водорос-
ли). Однако при тщательном рассмотрении здесь часто наблюда-
ется тонко подогнанная взаимоэксплуатация.
Форма взаимоотношений, при которых один вид получает пре-
имущество, не принося другому ни вреда, ни пользы, широко
распространена в природе. Например, крупные млекопитающие
служат разносчиками плодов и семян с крючочками, не получая от
этого сами ни существенного вреда, ни пользы. Комменсализм —
это одностороннее использование одного вида другим без нанесе-
ния ему ущерба. Связи комменсалов могут быть условно разделе-
ны на несколько вариантов. С участием растений может происхо-
дить сотрапезничество — потребление разных частей одного ре-
сурса. Пример — отношения между почвенными бактериями-сап-
ротрофами, перерабатывающими органику гниющих остатков, и
высшими растениями, потребляющими образовавшиеся при пере-
гнивании минеральные соли. Другой вариант комменсализма —
квартирантство (использование других видов в качестве жили-
ща) — широко распространен: большая группа эпифитов живет
на других растениях.
342
Антагонистические отношения хищник —жертва и паразит—
хозяин — это, по существу, все варианты пищевых связей, основ-
ные отношения в экосистемах, так как любой гетеротрофный орга-
низм может существовать лишь за счет других организмов. Хищ-
никами обычно называют организмы, питающиеся другими, ко-
торых они ловят и умерщвляют. Такое охотничье поведение ха-
рактерно для насекомоядных растений. Паразитизм — это ос-
лабленный вариант хищничества, когда организм, испытывающий
вредное влияние (хозяин), обычно не погибает сразу, а некоторое
время используется паразитом (Э. Пианка, 1981). Довольно часто
под влиянием паразита у хозяина проявляется токсикоз. При па-
разитизме живой организм хозяина обычно служит для паразита
не только источником пищи, но и местом постоянного или вре-
менного обитания, прикрепления. Поэтому паразиты обычно мень-
ше хозяина и часто узкоспециализированы. Однако в природе
существует много форм отношений организмов, не укладываю-
щихся в жесткую схему. Например, Э. Пианка (1981) предлагает
относить к паразитизму питание растительноядных животных.
Конкуренция возникает, когда необходимого ресурса не хватает
для всех особей сообщества. В результате каждое растение, исполь-
зуя дефицитный ресурс, снижает обеспечение им других (Т.А. Ра-
ботнов, 1998). Конкуренция чрезвычайно широко распростране-
на в природе, если виды со сходными потребностями обитают
совместно. Она отрицательно сказывается на росте и выживании
обоих конкурентов и может привести к вытеснению менее при-
способленного (конкурентное исключение).
При аменсализме один вид испытывает угнетение, в то время
как другой не получает ни вреда, ни пользы (например, взаимо-
действие светолюбивых трав и затеняющего их дерева). О нейт-
рализме же говорят в том случае, если виды непосредственно не
влияют друг на друга и, уживаясь в одном сообществе, связаны
только через цепь других видов (например, птицы и напочвенные
грибы). Однако в природе истинный нейтрализм или очень ре-
док, или его нет, поскольку между всеми видами возможны опос-
редованные взаимодействия, незафиксированные и неизученные.
То же относится и к другим проявлениям «нейтральности». Так,
деревья сильно затеняют травы, но и испытывают их влияние,
вступая в корневую конкуренцию.
Другой вариант классификации взаимоотношений организмов по
результату взаимодействия делит эти связи на антибиоз, нейтрализм и
симбиоз (И. А. Шилов, 2001). Антибиоз — крайнее выражение конкурен-
ции, при которой один вид полностью препятствует поселению другого
в пределах своей зоны влияния. Он чаще связан с химическим воздей-
ствием и наиболее свойствен грибам и прокариотам. Обычный пример
его — сильное подавление водных растений выделениями цианобакте-
343
риального планктона. Нейтрализм — тип отношений, при котором у парт-
неров исключены значимые прямые взаимодействия и они не оказывают
друг на друга заметного воздействия (например, виды несмежных трофи-
ческих уровней). А симбиозом в этой классификации считается система
отношений, при которой формируются тесные функциональные взаимо-
действия. Если они выгодны обоим видам, — это мутуализм, а если толь-
ко одному, — комменсализм (переходный вариант к нейтрализму).
11.2. Симбиоз
В настоящее время складывается учение о симбиозе, где взаи-
модействия организмов рассматриваются в самых разных аспек-
тах (генетическом, биохимическом, популяционном, эволюцион-
ном, экологическом и т. п.). Симбиоз — особая стратегия адапта-
ции, достигающаяся путем объединения с неродственными орга-
низмами. Она отличается от индивидуальных адаптаций, проис-
ходящих за счет физиологических или поведенческих реакций
свободноживущего организма. Поэтому симбиозу как стратегии
выживания путем объединения должен противопоставляться не
паразитизм, а автобиоз — стратегия выживания своими силами.
Разная трактовка симбиоза. В современной экологии устанав-
ливается понимание симбиоза как стратегии кооперативной адап-
тации неродственных организмов, объединяющихся в стабильную
надорганизменную систему (А. Е. Douglas, 1994). Однако термин
«симбиоз» широко используется и в других смыслах. Основопо-
ложник учения о симбиозе фитопатолог Де Бари в XIX в. предла-
гал считать главным его признаком длительность взаимодействия,
сопоставимую с продолжительностью жизни партнеров. Иногда
встречается очень широкое понимание, когда его отождествляют
с любыми взаимодействиями, включая отношения природы и че-
ловека. Иногда принципиальным для симбиоза считают длитель-
ность взаимодействия, специфичность или образование специ-
альных органов. В России распространено узкое понимание сим-
биоза как взаимовыгодного взаимодействия.
Узкое определение симбиоза пришло в науку в связи с изуче-
нием паразитарных систем (К. И. Скрябин, 1991). Паразитизм стали
выделять как однозначно вредный для хозяина тип отношений,
противопоставив ему симбиоз. Но оправданный с точки зрения
медицинской и сельскохозяйственной практики, этот подход стал
противоречить накапливающимся наблюдениям. Во-первых, ока-
залось, что отношения партнеров могут переходить от антагонис-
тических к мутуалистическим и обратно в зависимости от усло-
вий среды.
Например, зеленая водоросль Chlorella улучшает развитие пресно-
водной гидры на свету и в отсутствие соединений углерода, когда по-
344
ставляемые автотрофным симбионтом продукты фотосинтеза — един-
ственные источники питания хозяина. Но в темноте развитие гидры во-
дорослью подавляется, так как фотосинтез прекращен, и эндосимбионт
потребляет метаболиты хозяина (А. Е. Douglas, 1994). Другой пример —
семена орхидных, в природе прорастающие только при воздействии гриб-
ного симбионта. Они очень мелки, практически лишены запасных ве-
ществ, и микориза снабжает зародыш питанием. Гриб стимулирует разви-
тие проростков, но в ряде случаев агрессивность его повышается и на на-
чальном этапе взаимоотношений он может подавить зародыш (И. В. Ка-
ратыгин, 1993).
Во-вторых, эффект зависит от генотипов особей. Например,
клубеньковые бактерии могут превращаться в паразитов в резуль-
тате единичной мутации, нарушающей синтез нитрогеназы или
поверхностных полисахаридов. Некоторые их штаммы вырабаты-
вают токсины, вызывающие у растений хлороз. В-третьих, отно-
шения партнеров могут по-разному проявляться на уровне инфи-
цированного органа, организма или популяции хозяина. Напри-
мер, спорынья (Claviceps), проникая в завязь цветка злака, разру-
шает ее и в период развития склероция потребляет растительные
метаболиты. Но после созревания склероция питание оставшихся
зерновок продолжается, и продуктивность растения обычно не
снижается. А поскольку продуцируемые грибом алкалоиды ток-
сичны для животных, питающихся семенами, общая приспособ-
ленность популяции растения в результате инфицирования мо-
жет существенно возрасти.
Эти примеры показывают, что польза не может быть надеж-
ным основанием для выделения симбиоза. Поэтому с учетом ме-
таболических и регуляторных процессов, экологических особен-
ностей сложилось представление о нем как о надорганизменной
системе, в которой осуществляются разные варианты взаимодей-
ствий, включая мутуализм и антагонизм (Н.А. Проворов, 2001).
Типы симбиозов. Наиболее простая классификация симбиозов
основана на морфологических параметрах. При проникновении
одного партнера внутрь другого говорят об эндосимбиозе. Обыч-
но эндосимбионты сильно влияют на партнеров и иногда образу-
ют особые структуры (клубеньки у бобовых, талломы лишайни-
ков). Довольно широко распространен внутриклеточный эндосим-
биоз, при котором водоросли поселяются в клетках животных при
сохранении самостоятельности питания (Л.Л. Великанов и др.,
1981).
Водоросли Zoochlorella и Zooxanthella обнаружены в клетках многих
фораминифер, инфузорий, губок, некоторых червей, иглокожих, мол-
люсков и кишечнополостных. Они защищены телом животного от не-
благоприятных воздействий, диоксид углерода и азот получают из выде-
лений животных клеток, а благодаря прозрачности тел хозяев для них
345
возможен и фотосинтез. Животные же получают от водорослей раство-
римые углеводы и кислород. Такие объединения водорослей и животных
образуют в богатых минеральными веществами водах популяции очень
высокой плотности, которые эффективно связывают СО2 и минераль-
ные вещества. В них зафиксирована самая высокая для одноклеточных
скорость образования органики. В некоторых случаях автотрофный ком-
понент в таких комплексах содержится постоянно и при размножении
животных передается потомкам. В этом случае водоросль приспосабли-
вается к сожительству только с определенными видами животных. При-
мер такого симбиоза — обитающие в пресных водоемах зеленая гидра
(Hydra viridis) и водоросль Chlorella (М. Бигон и др., 1989). В опытах
свободноживущие хлореллы воспринимаются гидрами как пищевые
объекты, а выделенные из гидр не поедаются. Они по одной окружаются
вакуолями и перемещаются к основанию пищеварительных клеток, где
размножаются. Рост эндосимбионта и хозяина взаимосогласованы. Гид-
ра на свету получает от водоросли углеводы и кислород, а также как
животное может использовать и поступающую извне органику. В морях
подобные мутуалистические связи с беспозвоночными образуют диноф-
лагелляты. Экспериментально показано, что хозяин может ускорять рост
и выделение метаболитов у водоросли. Важная экологическая роль при-
надлежит симбиотическим комплексам динофлагеллят с коралловыми
полипами. Водоросли не только обеспечивают их углеводами, но и в
качестве побочного эффекта вызывают осаждение карбоната кальция,
что позволяет кораллам строить скелеты, составляющие основу рифов.
При эктосимбиозе (ассоциативном симбиозе) проникновения
партнеров не наблюдается. К нему относят, например, эпифит-
ные ассоциации и эктопаразитические системы. Иногда эффект
эктосимбиоза бывает очень силен. Так, ризосферные бактерии
могут существенно улучшать развитие растений благодаря азот-
фиксации, выработке фитогормонов и подавлению патогенов.
Поселяющиеся на животных водоросли имеют тесный наружный кон-
такт с ними при сохранении автономности питания. Как правило, они
тяготеют к определенной группе животных, являясь их комменсалами.
Нередко наблюдаются характерные водорослевые обрастания раковин
моллюсков, панцирей ракообразных и т.д. Так, зеленая водоросль
Basicladia, нуждаясь для развития в кератине, растет только на панцире
пресноводных черепах. А на желобчатых волосах южноамериканского
ленивца зеленые водоросли иногда развиваются так обильно, что обес-
печивают ему зеленоватую окраску и дополнительную защиту.
Более сложная классификация симбиозов учитывает характер
взаимодействия участников. По биологическому эффекту симби-
озы делят на мутуалистические, антагонистические и коммен-
сальные. Характерно, что у водорослей и ряда разных групп бес-
позвоночных выработались отношения, включающие одновремен-
но мутуализм, паразитизм и хищничество. Животные научились
346
переваривать водоросли, сохраняя в своих тканях способные к
фотосинтезу хлоропласты. Такие отношения особенно распрост-
ранены между сифоновыми водорослями и брюхоногими моллюс-
ками. Хлоропласты в животном остаются активными более двух
месяцев и, пока не потеряли способности к фотосинтезу, защи-
щены от переваривания (М. Бигон и др., 1989).
По степени зависимости партнеров симбиозы делят на факуль-
тативные и облигатные. Это деление также условно, и проявле-
ния его зависят от особенностей среды. Например, в отсутствие
азота бобовые выживают только за счет симбиоза с азотфиксиру-
ющими бактериями, но в присутствии его способны и к симбио-
трофному, и автотрофному азотному питанию.
По характеру трофических взаимодействий симбиозы подраз-
деляют на биотрофные (источник питания — живые клетки парт-
нера, а используют их и патогены и мутуалисты), некротрофные
(источник питания — специально умерщвленные клетки) и сап-
ротрофные (источник питания — естественно погибшие клетки).
Эта классификация изначально применялась для грибных симби-
онтов растений (Ю.Т. Дьяков, 1980).
Часто симбиозы разделяют на специфические и неспецифи-
ческие. В первом случае во взаимодействие вступают таксоно-
мически узкие группы, а во втором симбиоз формируется раз-
ными неродственными организмами. Специфическими симбио-
зами часто являются паразитарные системы, а мутуалистически-
ми — наоборот (иногда для одной из групп). Например, везику-
лярно-арбускулярная микориза встречается у многих групп расте-
ний, но микобионты принадлежат одному порядку Зигомицеты
(Giomales).
Функциональные концепции симбиоза. Применительно к пара-
зитарным системам на основе анализа механизмов взаимодействия
организмов были выработаны три концепции симбиоза — мета-
болическая, экологическая и регуляторная. Теперь они широко
используются и для характеристики мутуалистических отноше-
ний.
Метаболическая концепция основывается на представлении
о том, что партнеры тесно связаны интенсивным переносом ис-
точников питания и энергии. При этом во многих случаях тесные
трофические связи сводятся к передаче неспецифических метабо-
литов, а эндосимбионты могут и сами синтезировать вещества,
получаемые от хозяина. В других случаях один из партнеров при-
обретает новую функцию, которую он не может выполнять само-
стоятельно. Такие отношения очень характерны для взаимодей-
ствия эукариот с прокариотами. Обладающие более высокой мор-
фологической организацией эукариоты обеднены биохимически-
ми функциями, которые восполняются за счет симбионтов. При
этом не просто передаются полезные вещества, а формируются
347
скоординированные биохимические пути. У неподвижных расте-
ний зависимость от этих взаимодействий сильнее, чем у живот-
ных, поэтому у них более развита способность к метаболической
интеграции с микробами — донорами новых функций.
Экологическая концепция симбиоза также появилась в русле
паразитологии при рассмотрении внутренней среды хозяина как
новой экологической ниши для паразита, где складывается пара-
зитоценоз с разными отношениями между эндопаразитами. Сей-
час эти аспекты рассматриваются и при изучении мутуализма. Так,
грибы-эндофиты используют хозяина сначала как место, позво-
ляющее выжить в неблагоприятных условиях, и только после ги-
бели хозяина питаются его тканями. С точки зрения этой концеп-
ции, симбиозы про- и эукариот основаны на том, что эукариоты
предоставляют структурно более простым прокариотам ряд офор-
мленных вместилищ (внутриклеточные компартменты, межкле-
точные пространства, полости).
Регуляторная концепция позволяет рассматривать симбиоз как
результат взаимного контроля партнерами численности, интен-
сивности метаболизма и репродуктивной активности. Наиболее
важен контроль активности эндосимбионта, у которого потенци-
альная скорость размножения обычно выше, чем у хозяина. Эта
концепция оформилась из патофизиологии паразитизма, где клю-
чевая роль отводится защитным системам хозяина, ограничиваю-
щим патоген, и факторам вирулентности, позволяющим паразиту
преодолевать эту защиту. Теперь активно изучается контроль раз-
множения эндосимбионтов в мутуалистических системах. Его ме-
ханизмы разнообразны: регуляция активности генов симбионта,
ограничение его питания, частичный лизис и выброс в среду
(А. Е. Douglas, 1994). Например, бобовые контролируют числен-
ность клубеньковых бактерий и на уровне общего числа клубень-
ков, и на уровне числа бактерий в клубеньке. В первом случае
контроль системный (с участием веществ, выработанных в над-
земных органах), а во втором — локальный (с участием тканей
клубенька).
Регуляция мутуалистических микробно-растительных отноше-
ний основана на тех же механизмах, что и защита от фитопато-
генов. Так, транскрипция генов клубеньковых бактерий, конт-
ролирующих формирование клубенька, индуцируется раститель-
ными флавоноидами, многие из которых идентичны защитным
веществам. Синтез каллозы, пероксидаз и других защитных ве-
ществ в небольших количествах происходит и в процессе нор-
мального развития клубеньков. Таким образом, растения обла-
дают универсальными системами регуляции отношений с мик-
роорганизмами, которые могут быть мутуалистическими или
антагонистическими в зависимости от симбионта, среды, гено-
типов. У растений, чья адаптивная способность сильно зависит
348
от симбиоза с микробами, защитные системы организованы про-
ще, чем у животных (нет фагоцитоза и выработки антител, спе-
циальных защитных структур). Их системные защитные механиз-
мы по отношению к партнерам малоспецифичны. По Н.А. Про-
ворову (2001), это позволяет растениям поддерживать широкий
круг эндосимбионтов, обеспечивающий высокую экологическую
пластичность растений.
Интеграция партнеров симбиоза. Симбиотические взаимодей-
ствия, как и другие адаптивные функции организма, находятся
под контролем генных систем. Генетическая интеграция партне-
ров симбиоза осуществляется в форме перекрестной регуляции
генов, обмена продуктами их действия (белки, РНК), дублирова-
ния (амплификации) участвующих во взаимодействии генов.
К тому же длительные и тесные контакты партнеров определяют
возможность спорадического переноса между ними генов. На уров-
не фенотипа интеграционные процессы проявляются в форме
сигнальных взаимодействий, объединения метаболических сис-
тем партнеров и развития симбиотических органов.
Сигнальные процессы определяют формирование симбиоти-
ческих систем и взаимную регуляцию деятельности партнеров.
Их результатом становится перекрестная регуляция генов (в от-
личие от трофических взаимодействий, сводящихся к обмену ис-
точниками питания или энергии). Сигнальные процессы в хозяи-
не контролируют также численность эндосимбионтов, при избытке
которых мутуалистическое взаимодействие переходит в антагони-
стическое.
Сигнальные взаимодействия хорошо изучены для бобовых и клубень-
ковых бактерий. Так, семена и корни этих растений секретируют флаво-
ноиды, которые активируют транскрипцию бактериальных генов, под кон-
тролем которых синтезируются вещества, запускающие образование клу-
беньков. Сигнальное взаимодействие включает не только синтез, перенос
и рецепцию сигнальных молекул, но и их видоизменение. Например, у
клубеньковых бактерий есть системы расщепления секретируемых бобо-
выми флавоноидов, а выделяемые бактериями вещества модифицируются
ферментами растений. При нарушении сигнального контроля у бобовых
образуется очень много клубеньков, но биомасса растений-мутантов рез-
ко снижается из-за перерасхода энергии на их образование.
Для многих симбиозов характерна глубокая метаболическая
интеграция организмов. Она выражается либо в установлении
между ними тесных трофических связей, либо в приобретении
одним из партнеров новых биохимических функций. Так, пере-
крестная регуляция генов эндосимбионта и хозяина, осуществля-
емая посредством сигнального взаимодействия, приводит к выбо-
рочной работе тех генов, функционирование которых обеспечи-
вает симбиоз.
349
Например, в растительных клетках клубеньков синтезируются осо-
бые формы ферментов азотного обмена, которые обусловливают пере-
ключение бобовых на усвоение продуктов азотфиксации. Они синтези-
руют также специфичные изоферменты углеродного обмена, благодаря
которым микросимбионт снабжается наиболее доступными для него ис-
точниками углерода. Эти изоферменты кодируются специальными рас-
тительными генами, работа которых многократно усиливается или даже
индуцируется в клубеньках. Например, в промоторах генов легоглобина
(обеспечивающего в клубеньке необходимые для азотфиксации бескис-
лородные условия) выявлены участки, гомологичные бактериальным,
которые могут быть мишенями для сигналов, поступающих от бактерий.
Выделены также бактериальные ДНК-связывающие белки, взаимодей-
ствующие с промоторами гена легоглобина. Параллельно в бактероидах
синтезируется ряд компонентов, не характерных для свободноживущих
бактерий (например, нитрогеназы и белки, обеспечивающие их созрева-
ние и работу).
Кроме перекрестной регуляции генов, этой универсальной стра-
тегии метаболической интеграции при всех симбиозах, интегра-
цию симбионтов могут обеспечивать еще как минимум два про-
цесса: перенос макромолекул и генов. Перенос макромолекул
(РНК, белков и др.) происходит, например, между цитоплазмой
растительной клетки, хлоропластами и митохондриями. Но эука-
риотическая клетка — продукт очень давнего симбиоза, компо-
ненты которой уже во многом утратили автономность. Для менее
интегрированных симбиозов перенос сложных молекул не обна-
ружен (например, в клубеньках бобовых нет передачи белков и
нуклеиновых кислот между бактероидами и растительной цитоп-
лазмой). Но перенос генов выявлен при симбиозе растений с агро-
бактериями. А в клубеньках бобовых ряд бактериальных генов (гены
некоторых ферментов и белков цитоскелета) имеют столь близ-
ких гомологов у эукариот, что их появление у бактерий в резуль-
тате горизонтального переноса генов весьма вероятно (Н.А. Про-
воров, 2001).
Интеграция партнеров симбиоза может проявляться и в обра-
зовании симбиотических морфологических структур. Для многих
мутуалистических симбиозов характерно формирование хозяином
специализированных органов, содержащих микробные клетки. Они
выполняют три тесно связанные функции: 1) вместилище для сим-
бионта; 2) структурная основа для обмена партнерами метаболи-
тов и регуляторных факторов; 3) локализация симбионта, которая
облегчает контроль над его численностью и активностью. Все эти
функции выполняют, например, клубеньки. При внутриклеточ-
ных симбиозах микробы часто находятся в клетке-хозяине внутри
особых мембранных структур — симбиосом (А. Е. Douglas, 1994).
Их образование характерно, например, для кораллов с водорос-
лью Symbiodinium, растений с азотфиксаторами (ризобии, Frankid)
350
и эндомикоризными грибами (Glomus). При этом часть системы
контроля морфогенеза этих образований вынесена за пределы ге-
нотипа хозяина. Так, в клубеньках бобовых большая часть мате-
риала мембран образована клеткой растения, но часть белков имеет
бактериальное происхождение.
Развитие растительно-микробных симбиозов отличается тем,
что оно может базироваться на дедифференцировке специализи-
рованных клеток, которая обычно происходит лишь при повреж-
дениях. Однако в целом наблюдается большое сходство процес-
сов формирования обычных органов и симбиотических. Таким
образом, морфогенез симбиотических органов генетически свя-
зан с морфогенезом прочих органов, но многие его этапы специ-
фичны и могли возникнуть только при длительном взаимодей-
ствии партнеров.
В целом ключевые процессы симбиоза (сигнальные взаимо-
действия, метаболическая интеграция партнеров и развитие спе-
циальных органов) показывают, что симбиоз необходимо рассмат-
ривать как генетический феномен, основанный на формирова-
нии надорганизменных генных систем.
Коэволюция симбионтов. Формирование симбиоза — результат
сопряженной эволюции взаимодействующих организмов. Коэво-
люция рассматривается как процесс обоюдной адаптации, проис-
ходящей в результате взаимодействия их популяций и существен-
но влияющий на эволюцию видов и надвидовых таксонов. Коэво-
люция участников симбиоза основана на преадаптациях и осу-
ществляется под действием различных форм отбора.
Симбиотические системы имеют разное происхождение. Так,
у бобовых симбиоз возник парафилетически (параллельно в раз-
ных таксонах). Способные к симбиозу виды у них сосредоточены
в эволюционно продвинутых таксонах, а не способные — в при-
митивных. Это позволяет предположить наличие у предка бобо-
вых преадаптаций, обеспечивших появление симбиоза. Ими мог-
ли быть сформированные на корнях клубеньковоподобные струк-
туры, первоначально служившие для запасания крахмала, и осо-
бенности защитных систем, которые позволяли растениям разли-
чать полезные и вредные бактерии (Н.А. Проворов, 1998). Пре-
адаптациями предков клубеньковых бактерий могли быть способ-
ность к азотфиксации и особенности клеточной поверхности,
позволявшие избегать действия защитных систем растений на ран-
них стадиях симбиоза и стабильно существовать в растительных
клетках позднее.
Кроме преадаптаций в эволюции симбиозов существенную роль
играют факторы среды. Так, эволюция бобово-ризобиального сим-
биоза происходила под давлением отбора в пользу растений, спо-
собных развиваться при дефиците связанного азота. Бобовые воз-
никли в меловой период, когда почвы были бедны азотом и не
351
могли обеспечить полноценное питание растений. У клубенько-
вых бактерий эволюция симбиотических признаков происходила
под давлением отбора, который обусловил уход их от конкурен-
ции с почвенными сапротрофами, а также использование про-
дуктов растительного фотосинтеза.
По Н.А.Проворову (2001), стратегия эволюции симбиотических мик-
роорганизмов включает две составляющие. Во-первых, это упрощение
генома при облигатном симбиозе (утрата генов, необходимых для само-
стоятельного существования, перенос необходимых для взаимодействия
генов в геном хозяина). Во-вторых, это усложнение генома при факуль-
тативном симбиозе (повышение его пластичности, дифференциация на
системы контроля свободноживущей и симбиотической стадий жизнен-
ного цикла). Например, у клубеньковых бактерий это усложнение выра-
жается в повышении рекомбинационной активности, накоплении по-
вторяющихся и мобильных элементов, разделении генома на группы
сцепления, появлении больших «молчащих» областей генома.
Микосимбиотрофия. Микосимбиотрофия (питание с помощью
грибов) среди высших растений распространена гораздо шире (ох-
ватывает около 225 тыс. видов), чем другие симбиотические ассо-
циации (И. В. Каратыгин, 1993). По И. А. Селиванову (1981), око-
ло 80 % видов флоры бывшего СССР — микоризообразователи, в
том числе все голосеменные, 77,5 % покрытосеменных, 60 % спо-
рофитов высших споровых. Облигатно микосимбиотрофны гаме-
тофиты плауновидных и 90 % хвощевидных. Как правило, мико-
трофны виды таких больших семейств, как Лилейные, Лютиковые,
Сложноцветные, Бобовые. Микориза обнаружена у растений раз-
ных жизненных форм и экологических групп, почти во всех при-
родных зонах, горных поясах и типах наземной растительности.
Наиболее хорошо она развивается в теплых условиях при умерен-
ном увлажнении в сочетании с хорошей аэрацией. Выраженность
микоризы зависит от вида растения, его онтогенетического со-
стояния и условий произрастания.
Отсутствует микориза у видов, сформировавшихся в условиях обиль-
ного обеспечения фосфором, водно-болотных (в воде фосфаты находят-
ся в растворе, и потому отсутствует стимул для симбиоза с грибом), па-
разитных и насекомоядных (получающих элементы минерального пита-
ния от растений-хозяев и животных-жертв), а также у накапливающих
вредные для микоризообразующих грибов вещества (алкалоиды, высо-
кие концентрации солей и др.) и сформировавшихся в неблагоприятной
для них среде (низкие температуры, недостаток воды, щелочность и за-
соление почвы). Наибольшее число безмикоризных видов отмечено сре-
ди крестоцветных (87 %), осоковых (74%), маревых (61 %), ситниковых
(56 %), а также рдестовых, шейхцериевых, ежеголовниковых, рогозовых,
частуховых. Безмикоризны древесные виды большого южнополушарно-
го семейства Proteaceae (И. В. Каратыгин, 1993).
352
Большое значение в процессе микоризообразования имеют ус-
ловия освещения. Экспериментально установлено, что максималь-
ное образование микориз отмечено при интенсивности освеще-
ния 25 — 50% от полного. Понижение интенсивности света при-
водит к уменьшению развитости микоризы и замене симбионтов,
а при 6%-м освещении микоризообразование не наступает. Недо-
статочное освещение влияет и на взаимоотношения симбионтов:
при неблагоприятных условиях освещения гриб ведет себя агрес-
сивно, вызывая повреждения корней. Свет влияет на грибной сим-
бионт через фотосинтез: слабое освещение снижает его интенсив-
ность и, следовательно, отток ассимилятов в корни, что лимити-
рует развитие микоризных грибов. Микосимбиотрофия встреча-
ется при разной кислотности почв (от 2,9 до 8,0). Небольшое за-
соление ей не препятствует. Наиболее распространена она на поч-
вах, богатых органикой, но бедных доступными минеральными
веществами (особенно N и Р). На богатых азотом и фосфором
почвах микориза часто отсутствует, хотя образование ее угнетает-
ся и низким содержанием элементов минерального питания, сдер-
живающим синтетические процессы у растения (И. А. Селиванов,
1981). Выраженность микоризы может зависеть и от влияния дру-
гих видов растений. Например, брюква (Brassica napobrassica)
отрицательно влияет на образование микоризы у лука (Allium сера),
а душистый колосок (Anthoxanthum odoratum) — у клевера пол-
зучего (Trifolium repens). Некоторые грибы приобрели способ-
ность соединять гифами разные растения, получающие при этом
возможность обмениваться продуктами фотосинтеза (А. М. Гиля-
ров, 2003).
Взаимоотношения между растением и грибным компонентом
в большинстве случаев разносторонни и взаимовыгодны. Обще-
известно, что растение обеспечивает гриб создаваемыми при фо-
тосинтезе углеводами, а тот содействует минеральному питанию
растения. Зависимость грибных компонентов микоризы от расте-
ний обусловлена тем, что они в отличие от сапротрофных грибов
обычно не могут использовать лигнин и целлюлозу. Поэтому все,
что снижает фотосинтез (сокращение фотопериода, уменьшение
интенсивности освещения и т.п.), ухудшая обеспечение гриба уг-
леводами, сказывается на выраженности микоризы. Все, что сни-
жает поступление углеводов в корни (объедание листвы фитофа-
гами, скашивание травы и стравливание ее скотом), тоже отрица-
тельно сказывается на микоризе.
Грибы микориз влияют на организмы ризосфер. Отмечена меньшая
поражаемость микоризных растений почвенными паразитическими гри-
бами и бактериями. По мнению Б.Зака (1971), грибной чехол затрудняет
проникновение инфекции в корень. Грибной компонент выделяет в по-
чву антибиотики и, используя углеводы, лишает патогенные организмы
353
энергии. Микоризные грибы, являясь, вероятно, ослабленными в про-
цессе эволюции паразитами, приспособились к совместной жизни с ра-
стениями, но паразитические свойства в какой-то мере сохранили. По-
этому, поселяясь на корнях, они стимулируют действие защитных меха-
низмов и предохраняют корни от инфекции (И.А. Селиванов, 1981).
К настоящему времени получено много данных о полифунк-
циональности микориз: их грибы не только обеспечивают расте-
ния элементами минерального питания, но и улучшают снабже-
ние их водой, повышают устойчивость к патогенам, увеличивают
проницаемость мезофилла для СО2, увеличивают концентрацию
хлорофилла, уменьшают поступление тяжелых металлов, увели-
чивают устойчивость к засолению, благоприятно изменяют струк-
туру почв и динамику почвенных микроорганизмов (увеличивают
число азотфиксаторов), выделяют ростовые вещества, способствуют
утилизации лигнина и целлюлозы, участвуют в горизонтальном
переносе генов, передают вещества между растениями (Т.А. Ра-
ботнов, 1993; Каратыгин, 1993; Б.М. Миркин, Л. Г. Наумова, 1998
и др.). Важно, что, обладая неодинаковыми ферментными систе-
мами, растение и гриб вырабатывают разные биологически ак-
тивные вещества и могут обмениваться ими. По М. М. Камшилову
(1979), гриб — вставочное звено, ускоряющее миграцию биоген-
ных элементов за счет «быстроты их оборачиваемости» через ми-
целий, т. е. микоризу можно рассматривать как механизм интен-
сификации круговорота биогенов в экосистемах, повышающий
качество жизни растений.
11.3. Фитофагия и защита растений от нее
Фитофаги и фитопаразиты. Растения составляют основу тро-
фической системы и потому отношения с фитофагами и фитопа-
разитами особенно значимы в экосистемах. Существует множе-
ство форм фитофагии: сокращение при съедании поверхности
поедаемого органа, выедание полостей внутри него (минирова-
ние) или высасывание содержимого, образование галлов (защи-
щающих ткани растения от поступления метаболитов эндобион-
та) и др. В некоторых случаях деятельность фитофага сразу на
растении может и не отражаться. Например, тли прокалывают
своими стилетами ткани растения и высасывают сок из ситовид-
ных трубок, которые в это время не прекращают активной работы
(М.Бигон и др., 1989). Но во всех случаях, используя ткани расте-
ния, фитофаг ослабляет его.
Среди фитофагов есть моно-, олиго- и полифаги (питающиеся
одним, немногими или многими видами). Некоторые животные
специализируются на поедании отдельных органов и тканей рас-
354
тений. Соответственно выделяют семяедов, ризофагов, ксилофа-
гов и др. Большинство трофических отношений между фитофага-
ми и поедаемыми растениями можно отнести к варианту хищни-
чества. Однако эти взаимоотношения значительно отличаются от
типичных отношений в системе хищник—жертва. Растения не
могут прятаться от нападающего животного или убегать. Поэтому
в процессе коэволюции у них сформировался ряд анатомо-мор-
фологических и физиолого-биохимических приспособлений, по-
зволяющих уменьшать наносимый животными вред.
Фитопаразитами обычно называют тех фитофагов, которые
обитают внутри тел хозяев, и потому не только получают от ра-
стений питание, но и выделяют в них свои метаболиты, чем от-
личаются от хищников. Фитопаразиты могут быстро приводить
растение к гибели или существенно подрывать его конкуренто-
способность. Другую точку зрения относительно паразитизма и
хищничества в связи с растениями представляет Р. Риклефс (1979).
Он называет хищниками тех растительноядных животных, кото-
рые поедают растения целиком (обычно семена или проростки), а
объедающих отдельные части растений он относит к паразитам.
Последствия влияния растительноядных организмов зависят от
того, в какой период жизни и какие органы растения подверглись
нападению. Разные способы воздействия фитофагов (обкусыва-
ние листьев, высасывание сока, поедание меристем, повреждение
цветков или плодов, подгрызание корней) также неодинаково дей-
ствуют на растения. Последствия лишения листа для проростка
будут совсем иными, чем для взрослой особи. Результат зависит и
от реакции растения. Оно может перераспределить вещества в
организме, изменить уровень обмена, сформировать дополнитель-
ные защитные образования. Отмечается также, что фитофаги чаще
повышают чувствительность растений к факторам смертности,
нежели сами убивают их (М.Бигон и др., 1989).
Для фитофагов потенциальная пищевая ценность растений
может быть снижена выработкой разных защитных приспособле-
ний: физических и химических, анатомо-морфологических и фи-
зиологических, поведенческих и биоценотических. На формиро-
вание способов защиты растение расходует ресурсы и при этом
делает недоступными для фитофагов свои органы. Цель этих при-
способлений — помочь выжить и оправиться после нападения.
М. Бигон, Дж. Харпер, К Таундсен (1989) отмечают, что эволюция
защищенности растений влияет на эволюцию потребляющих их
животных, развивающих способы обхода этой защиты. Это не-
прерывное эволюционное взаимодействие авторы уподобляют
противоборствующим армиям, когда одна из них разрабатывает
все новые средства нападения, а другая — новые средства защиты.
Эволюция каждого взаимодействующего таксона зависит от эво-
люции другого. Взаимодействие такого типа называют сопряжен-
355
ной эволюцией или коэволюцией. Как отмечают Б. М. Миркин и
Л. Г. Наумова (1998), эти коадаптации обычно носят коллектив-
ный характер, что обеспечивает возможность быстрого переклю-
чения фитофагов на питание другим видом. При этом если у рас-
тения выработалась способность выделять вещества, отпугиваю-
щие большинство хищников, а у кого-то из них появился способ
справляться с этим соединением, такое животное получает источ-
ник пищи, недоступный конкурентам, и специализируется на
питании этим типом растений.
Другим эволюционным аспектом развития фитофагии можно
назвать формирующееся биоразнообразие. С. Стенли (1976), ана-
лизируя проблемы макроэволюции, пишет, что появление более
высокого трофического уровня способно повысить разнообразие
на предыдущем уровне. Коэволюция растений и растительнояд-
ных насекомых — важнейший механизм, обеспечивающий рост
видового богатства обеих этих групп на протяжении всей их эво-
люции. Так, около 600 млн лет назад появление первых расти-
тельноядных организмов привело к выеданию монокультур водо-
рослей и могло спровоцировать мощнейшую за всю историю Земли
вспышку эволюционной активности, называемую эволюционис-
тами «кембрийским взрывом», когда всего за несколько милли-
онов лет резко повысилось биоразнообразие. А расцвет цветко-
вых растений совпал с появлением чешуекрылых. Химическая
дивергенция средств защиты растений понижает эффективность
питания полифагов и приводит к ускорению эволюции стенофа-
гов. Отмечено также, что поскольку родственные виды содержат
сходные метаболиты, эволюция растительноядных насекомых,
особенно бабочек, идет параллельно филогении растений-хозяев
(Э.Пианка, 1981).
Фитофаги (и паразиты) обычно уничтожают и повреждают лишь
часть особей фитоценоза, но при этом могут сильно изменять
конкурентную обстановку в сообществах. Эти важные посредни-
ки во многом определяют исход конкуренции растений: они мо-
гут настолько ослабить вид, что под влиянием конкурентов он
будет вытеснен из сообщества.
Последствия взаимодействия с паразитами сильно зависят от эволю-
ционной коадаптированности партнеров. Например, в США растения
природных пастбищ поражаются грибами в 3 — 5 раз меньше, чем посе-
вы люцерны (Medicago) и клевера (Trifolium). Особенно большой вред
наносят паразиты, привнесенные из других регионов. По этой причине,
например, в течение 50 лет из широколиственных лесов Северной Аме-
рики практически выпал распространенный доминант — зубчатый каш-
тан (Castanea dentata), составлявший 40 % биомассы лесов в районе Ап-
палачей (Ю.Одум, 1986). Это произошло после того, как в 1904 г. из
Китая вместе с интродуцированными деревьями был завезен поражаю-
щий кору гриб Endothia parasitica. По Б. Небелу (1993), в результате
356
этой крупнейшей ботанической катастрофы погибло около 3,5 млрд де-
ревьев. Гибель каштана ускорялась тем, что ослабленные паразитом ра-
стения проигрывали в конкуренции другим доминантам.
Анатомо-морфологические приспособления растений, препятству-
ющие доступу фитофагов. Растения не тратят энергию на убегание
от хищника, поэтому они вкладывают довольно много ресурсов в
«оборонительные сооружения», препятствующие доступу фитофа-
гов к пище. Защите растений от поедания животными хорошо
служат густое опушение, развитые механическая и покровная тка-
ни, колючие выросты (могут так же отпугивать животных).
Из избытка углеводов растения легко формируют целлюлозные и
лигниновые образования, которые могут служить основой прочных обо-
ронительных покровов для важных тканей (меристем, зародышей и т. п.),
с которыми связаны перспективы роста и размножения. Чешуи и твер-
дые околоплодники, обычно содержащие мало белка и дефицитных био-
генов (N, Р, К и др.), обходятся растению сравнительно недорого. Но
толстая скорлупа или волокнистая шишка существенно увеличивают время
добывания пищи, в результате чего число поедаемых семян уменьшает-
ся. При этом давно было замечено, что семена, имеющие толстые и твер-
дые покровы, обычно не ядовиты. Прочность внешней защиты делает
излишней трату энергии на другие способы защиты (D. Н Janzen, 1981).
Наружные покровы с разнообразными трихомами и их произ-
водными используются многими растениями как защита. В гус-
том покрове волосков насекомое может запутаться, колючки мо-
гут его проткнуть, а плотные ткани делают недоступными лежа-
щие под ними съедобные образования. Роль этих средств зашиты
показана в многочисленных наблюдениях и экспериментах. Так,
дубовый коконопряд не трогает колючих листьев падуба (Ilex), но
при удалении колючек охотно ими питается. Меристемы в кону-
сах нарастания побегов обычно защищены от мелких фитофагов
чешуями, покрытыми кутикулой или одревесневшими. Но круп-
ные потребители эту защиту легко обходят. Например, снегири
весной легко склевывают бутоны плодовых деревьев. Иногда поч-
ки могут быть защищены шипами, находящимися на стволах и
крупных ветвях.
Только что созревшие семена, еще сосредоточенные на материнских
растениях, в значительной степени беззащитны, а рассеянные находятся
уже в относительной безопасности. Большинство культурных растений
человек селекционировал так, чтобы они семена не рассеивали (соби-
рать их с земли трудно), поэтому у большинства сельскохозяйственных
культур они стали удобными неподвижными мишенями для семяедов.
Дикие же предки этих растений, быстро рассеивавшие семена после со-
зревания, рисковали намного меньше. Легкость рассеивания семян у
растений с плодам и-коробочкам и зависит от размера отверстий, через
357
которые выпадают семена. Поэтому у диких маков (у Papaver rhoeas,
Р. dubium) с коробочками, имеющими большие отверстия, которые от-
крываются сразу после созревания семян, плоды голые. А у видов с ко-
робочками, имеющими мелкие отверстия (Р. agremone, Р. hybridum),
рассеивающих семена месяцами, плоды снабжены шипиками (рис. 11.1).
Иногда Papaver agremone образует формы с гладкими коробочками — и
они страдают от птиц больше всего. А культурный мак (Р. somniferum)
выводился с расчетом на то, чтобы семена его не рассеивались, поэтому
птицы причиняют его посевам огромный вред, легко используя его скон-
центрированные семена (М. Бигон и др., 1989 и др.).
Постоянно повреждаемые растения обычно усиливают защи-
ту, что ослабляет эффект последующих нападений. Так, шипы на
ежевике (Rubus caesius), которую объедает крупный рогатый скот,
отрастают более длинными и острыми, чем на неповрежденных
растениях. А у лиственницы (Larix) после сильной дефолиации
лиственничной молью в следующие годы образование хвои за-
медляется, повышается ее жесткость, возрастает концентрация смол
и волокон, понижается содержание азота. Нередко хорошо защи-
щенные растения возвышаются на стравленных скотом пастби-
щах. Таковы колючие сложноцветные и можжевельник (Juniperus)
на европейских лугах. А в Средиземноморье скот, поедая поросль
лиственных деревьев, не трогает жесткие побеги склерофитов,
особенно лавра (Laurus), чем способствует повышению конку-
рентоспособности этих растений (Г. Вальтер, 1974).
Особой анатомо-морфологической реакцией растения на фи-
тофаги может быть образование галлов, отграничивающих от тка-
ней растения личинки животных (минирующие мухи, галлицы,
осы-орехотворки). Для запуска этой морфогенетической реакции
достаточно бывает откладки одного яйца и не обязательно при-
сутствие потом личинок (возможно, паразиты влияют на генети-
ческую трансформацию клеток растения через передачу им плаз-
мид). Чрезмерное разрастание клеток растения, образование клу-
Рис. 11.1. Коробочки маков {Papaver spp.) различных видов
(по М. Бигон и др., 1989):
1 — Р. rhoeas, 2 — Р. lecoqiv, 3 — Р. dubiunr, 4 — Р. argemone\ 5 — Р. hybridum
358
беньков и других деформаций могут вызывать также нематоды
(М. Бигон и др., 1989).
Способность растений к регенерации как защитное приспособ-
ление от поедания животными. В связи с ограниченными возмож-
ностями избегать нападения животных у растений широко рас-
пространена способность к регенерации. Обладая открытой сис-
темой роста и модульным строением, растительные организмы в
благоприятных условиях быстро образуют новые вегетативные
органы взамен съеденных и поврежденных.
Отрастание после стравливания животными обычно включает
и продолжение роста поврежденных побегов, и образование но-
вых. Способность к восстановлению зависит от возраста расте-
ний, времени воздействия, окружающей обстановки. У видов с
растянутым периодом облиствения потеря молодых листьев ком-
пенсируется появлением новых. Иногда образование такой гене-
рации листьев отражается в формировании дополнительного го-
дичного кольца древесины. При повреждении верхушечных по-
чек деревья усиливают ветвление, образуют шарообразные и зон-
тиковидные кроны (и в озеленительной практике стрижка спо-
собствует усилению побегообразования). При сильном объедании
некоторые деревья приобретают форму кустарников (Т. А. Работ-
нов, 1993).
При восстановлении растений часто перераспределяется ток
ассимилятов: он направляется или к наиболее поврежденным орга-
нам, или к наименее пострадавшим. Часто особи с поврежденны-
ми корнями, недополучающие воду и минеральное питание, на-
правляют больше ассимилятов на восстановление корней, а особи
с поврежденными побегами и пониженным из-за этого фотосин-
тезом большую часть биогенов тратят на восстановление стеблей
и листьев. Возможно также локальное перераспределение асси-
милятов при компенсации небольших повреждений: например,
соя {Glycine) компенсирует потерю плодов увеличением массы
отдельных семян.
Защита растений от поедания животными путем выделения ток-
сических веществ. Растения содержат немало вторичных метабо-
литов, вероятно, не участвующих в основном обмене веществ. Они
очень разнообразны: от простых соединений (щавелевая и синиль-
ная кислоты) до довольно сложных (гликозиды, алкалоиды, тер-
пеноиды, флавоноиды и др.). Некоторые из них обладают токси-
ческим действием на животных, и в ряде случаев с их помощью
растения могут эффективно защищаться от фитофагов. Так, ток-
сичные («необычные») аминокислоты, присутствующие в семе-
нах тропических бобовых, вызывают гибель фитофагов, вероятно,
вследствие того, что включаются в белки вместо нормальных ами-
нокислот (А. Гэлстон и др., 1983). Некоторые токсичные вещества
образуются у растений при повреждении клеток.
359
В опытах Дж. Харпера (1977) выяснено, что некоторые популяции кле-
вера ползучего (Trifolium repens) при нарушении целостности тканей
выделяют синильную кислоту. Оказалось, что эта способность определя-
ется двумя парами аллелей: одна ответственна за наличие или отсутствие
гликозида линамарина, а другая — фермента бета-глюкозидазы. При
повреждении тканей происходит катализируемая бета-глюкозидазой ре-
акция, в которой от линамарина отщепляется ядовитый цианистый во-
дород. Листья цианогенных форм моллюски только пробуют, но не едят,
а растения, лишенные этого фермента или гликозида, улитками и слиз-
нями поедаются, поэтому они проигрывают в конкуренции цианоген-
ным формам.
В ряде случаев в зависимости от содержания токсических ве-
ществ растения либо вовсе не поедаются животными, либо у них
поедаются только те органы и в то время, когда содержание ток-
синов незначительно. Обычно одновременно у животных возни-
кают приспособления к детоксикации содержащихся в растениях
токсинов и ответная реакция растений, синтезирующих новые
токсические соединения. Защитная реакция растений против по-
едания в периоды, наиболее важные для их роста, проявляется в
сезонном изменении содержания нежелательных для фитофагов
веществ. Это особенно широко распространено у растений пус-
тынь и полупустынь (полыней — Artemisia, солянок — Salsola),
которые поедаются животными в основном после созревания се-
мян поздней осенью и зимой, когда значительная часть солей и
эфирных масел вымывается и изменяется состав веществ, препят-
ствующих поеданию.
И. А. Шилов (2001) собрал интересные примеры приспособле-
ния фитофагов к растениям с ядовитыми выделениями. В некото-
рых случаях животные научились избегать такие растения, в дру-
гих случаях у них формируется устойчивость к вторичным мета-
болитам растений даже при их высокой токсичности. Многие на-
секомые выработали способность к детоксикации ядов. А живот-
ные, которым содержащиеся в поедаемых растениях яды не при-
носят вреда, могут накапливать их и использовать для самозащи-
ты. Фитофаги, преодолевшие химическую защиту растения, иногда
потом специализируются на его потреблении, не испытывая кон-
куренции со стороны других претендентов на этот пищевой ре-
сурс.
Гусеницы молочайного бражника имеют особые железы, с помощью
которых яды, попавшие в тело из пищи, выделяются на поверхность
кутикулы.
В Калифорнии Larrea tridentata, содержащая значительное количе-
ство феноловых смол и потому не поедаемая большинством млекопита-
ющих, зимой составляет 75 % рациона древесной крысы, обладающей
повышенной устойчивостью к этим веществам.
360
Лемуры обычно избегают пищу, содержащую цианиды, но мадагас-
карский Hapalemur aureus, поедая верхушки бамбука (Cephalostachyum
viguieri), потребляет в день 12-кратную смертельную дозу цианидов.
Гусеницы бабочки данаиды кормятся на ваточниках (Asclepias), со-
держащих сердечные гликозиды. Накапливая эти вещества, гусеницы
передают их взрослой стадии насекомого. Поскольку для позвоночных
эти гликозиды ядовиты, бабочки становятся для птиц и млекопитающих
несъедобными.
На Большом Барьерном рифе распространена водоросль Chlorodesmis
fascigiata, концентрирующая токсичный хлородесмин и практически не
поедаемая рыбами. Однако краб Caphyra rotundifrons питается исклю-
чительно ею и прячется в ее зарослях. Питается ею и моллюск элизия,
накапливающий хлородесмин и становящийся несъедобным для рыб.
В семенах пастернака (Pastinaca sativd) содержится фуранокумарин,
опасный для большинства бабочек, но пастернаковая моль выработала
против него противоядие. Однако в ответ на это растение приобрело
способность образовывать два типа семян: бесплодные с небольшим ко-
личеством яда и с зародышем — образующие много яда. Интересно так-
же, что фуранокумарин вырабатывается у представителей 10 семейств
растений, и в ряде этих семейств видовой состав более разнообразен
именно в родах, вырабатывающих этот яд. И в семействах насекомых
(Oecophoridae, Papilionidae) видовой состав более разнообразен в родах,
в которых гусеницы выработали устойчивость к этому веществу. Устой-
чивость к кумаринам расширила кормовую базу и снизила уровень кон-
куренции среди фитофагов.
В целом за миллионы лет выработалось своеобразное равнове-
сие между защищающимися растениями и нападающими живот-
ными. В эволюционных масштабах взаимные приспособления
растений и фитофагов привели к усилению адаптивной радиации
и тех и других.
Отрицательное действие на фитофагов могут оказывать и ве-
щества растений, не являющиеся ядами, но попадающие в орга-
низм в неподходящее время. Так, пихта бальзамическая (Abies
balsamea) продуцирует аналог ювенильного гормона насекомых,
приводящий к невозможности завершения личиночной стадии
развития и гибели личинок. А декоративное растение Ageratum,
наоборот, образует вещества, подавляющие у насекомых секре-
цию ювенильного гормона и вызывающие преждевременное пре-
вращение неполовозрелых личинок в аномальную взрослую фор-
му (крошечных стерильных имаго) (А.Гэлстон и др., 1983). Эти
вещества, влияющие на насекомых и безвредные для человека,
перспективны как инсектициды.
Защита растений от поедания животными путем выделения де-
террентов. Ряд видов вырабатывает вещества-детерренты (терпе-
ны, танины, каучуки, лигнин, целлюлоза, ингибиторы протеоли-
тических ферментов), понижающие перевариваемость и усвояе-
мость съеденного фитофагами корма. Так, танины химически свя-
361
зываются белками и делают содержащие их ткани (например, ду-
бовые листья) трудно перевариваемыми. Имеет значение и несба-
лансированность веществ, необходимых для нормальной жизне-
деятельности (в частности, малое содержание белков и аминокис-
лот). Так, у ряда видов выработались приспособления против низ-
кого стравливания животными, заключающиеся в значительном
снижении кормовой ценности их нижних частей. Это ведет к бо-
лее высокому обкусыванию растений, способствуя их лучшему
отрастанию.
Характерное явление отмечено при взаимодействии с фитофа-
гами дуба (Quercus): с повышением концентрации танина в лис-
тьях рост гусениц зимней пяденицы замедляется (М. Бигон и др.,
1989). Пищевая ценность листьев из-за большого содержания та-
нинов понижается, так как они препятствуют перевариванию бел-
ков. Такая химическая защита эффективна против гусениц, но
личинки жуков, минирующие дубовые листья, преодолевают ее.
Пробуравливая лист, они поедают только его внутренние ткани,
избегая наполненных танином частей, расположенных под эпи-
дермисом (Р. Риклефс, 1979).
Семена большинства бобовых тоже содержат вещества, подав-
ляющие действие пищеварительных ферментов насекомых. На-
пример, бобы сои (Glycine) устойчивы к большинству долгоноси-
ков. А личинки большинства зерновок гибнут вскоре после про-
никновения под семенную кожуру бобов.
У ряда растений (например, берез — Betula, ив — Salix) после
нападения фитофагов концентрация защитных веществ повыша-
ется, а выделение пораженными особями сигнальных молекул (эти-
лен) усиливает образование веществ, сдерживающих поедание и у
тех растений, которые еще не подверглись нападению фитофагов.
Защита растений от поедания животными путем выделения ре-
пеллентов. Ряд растений выделяет отпугивающие животных ве-
щества — репелленты. Это соединения по какой-то причине (вкус
или чаще запах) неприятные для них. Так, скот на лугу обычно
обходит зверобой (Hypericum). Этот эффект может быть сопря-
жен с токсичностью содержащихся в зверобое веществ. А некото-
рые ароматические вещества подавляют развитие полезных сим-
бионтов животных (например, в рубце жвачных), чем ослабляют
фитофагов.
У растений роль репеллентов играют многочисленные изопре-
ноидные соединения, в частности терпены и эфирные масла.
А ядовитые цианогенные гликозиды разлагаются при поврежде-
нии ткани с выделением цианистого водорода, что делает их мощ-
ными репеллентами. Агентом, отпугивающим травоядных, часто
служат танины, поэтому животные иногда поедают только моло-
дые части растений и не трогают старые, содержащие танины и
другие репелленты. К другим соединениям, защищающим расте-
362
ния от фитофагов, относят эфирные масла, смолы, алкалоиды,
терпены и терпеноиды, причем последние два класса веществ
имеют резкие запах и вкус. Показательно, что дикие животные,
эволюция которых шла параллельно эволюции ядовитых расте-
ний, их не едят, но домашний скот многие ядовитые травы поеда-
ет (при этом сесквитерпены для овец смертельны).
Известны факты отпугивания ядовитыми растениями животных, по-
едающих совместно произрастающие с ними съедобные виды растений.
Так, овсяница красная {Festuca rubra) меньше поедается скотом, если
растет рядом с лютиком луковичным {Ranunculus bulbosus). А клевер
земляничный (Trifolium fragiferum) в смешанном посеве с клевером пол-
зучим (Т. repens) перестает поедаться зайцами.
Защита растений от поедания животными путем привлечения
хищников. Защита растений путем привлечения хищников отно-
сится к группе трансбиотических взаимоотношений (с участием
посредника). Ряд растений выделяет аттрактанты — вещества,
привлекающие насекомых-«защитников». Так, поврежденные гу-
сеницами совок растения выделяют в среду сигнальные вещества,
привлекающие животных, питающихся этими фитофагами. Такие
«охранники» обнаружены у нескольких десятков видов растений,
в том числе у культурных (томаты — Lycopersicon, огурцы —
Cucumis sativus, кукуруза — Zea mays).
Часто в зависимости от того, какой вид фитофага вызвал по-
вреждение, растения выделяют разные сигнальные вещества (для
привлечения разных зоофагов). Некоторые растения, особенно с
экстрафлоральными нектарниками, привлекают хищников и па-
разитов, которые уничтожают фитофагов, поедающих не только
данный вид, но и другие, совместно с ним произрастающие. То
же наблюдается при поселении на растении тлей, выделяющих
сахаристую жидкость. Есть мнение, что и сам нектар выработался
не для питания зоофильных видов, а для привлечения ос и других
врагов галлообразующих и листогрызущих насекомых (И. А. Ши-
лов, 2001).
Связь растения с насекомым-защитником может быть эволю-
ционно закреплена и таким образом, что химические изменения,
приводящие к образованию аттрактантов, могут начаться задолго
до того, как растительная ткань будет повреждена. Так, листья
вяза (Ulmus) начинают выделять привлекающие хищников веще-
ства еще до того, как из отложенных на них яиц выведутся гусе-
ницы-бабочки.
Хорошие примеры привлечения хищников, являющихся есте-
ственными врагами насекомых-вредителей, демонстрирует ряд
тропических растений, страдающих от муравьев. Испытывая вред
от одних видов, растения привлекают для защиты другие виды
363
муравьев. Так, в некоторых регионах тропической Америки мура-
вьи-листорезы могут тотально уничтожить листья ряда видов.
Привлечение же других агрессивных видов муравьев позволяет
растениям справиться с листорезами. В случае мирмекофилии на-
блюдается обоюдная польза от сожительства растений с муравья-
ми: насекомые поселяются в растениях и получают от них место
для устройства гнезда и пищу, а сами защищают их от вредите-
лей. Экспериментально показано, что у акаций (Acacia), остав-
ленных без защиты муравьями-помощниками, из-за повреждений
растительноядными насекомыми резко сокращался рост побегов.
Взаимополезные отношения между растениями и муравьями ши-
роко распространены в тропиках. Отмечено 258 видов из 19 се-
мейств таких «муравьиных деревьев». Многие из них уже утрати-
ли способность самостоятельно противостоять фитофагам.
Растения выработали ряд приспособлений, полезных для муравьев.
У некоторых акаций (Acacia cornigera) при основании листьев имеются
большие колючки с деревянистой оболочкой и рыхлой сердцевиной. За-
кладывая колонию, муравьиная матка Pseudomyrmex ferruginea пробу-
равливает у основания колючки отверстие и извлекает часть сердцеви-
ны, освобождая место для потомства (D.H. Janzen, 1967). По мере роста
колонии осваиваются и другие колючки. Пищей же муравьям служат
богатые белками округлые тельца на концах листочков и сахаристая жид-
кость, выделяемая из нектарников при основании черешков. В свою оче-
редь муравьи убивают всех растительноядных насекомых, пытающихся
напасть на акацию, отпугивают крупных млекопитающих, щиплющих
листья, и даже уничтожают появляющиеся у ее ствола проростки расте-
ний-конкурентов. Развитие мутуалистических вза-
имоотношений сопровождается формированием у
партнеров приспособлений, повышающих эффек-
тивность ассоциации. Так, Pseudomyrmex актив-
ны круглые сутки (что не характерно для муравь-
ев), чем обеспечивают непрерывную защиту ака-
ции. A Acacia cornigera сохраняет листву круглый
год, непрерывно снабжая муравьев пищей (боль-
шинство акаций в засушливый период листья сбра-
сывают). У видов рода Cecropia муравьи поселя-
ются в полых стволиках (рис. 11.2), пробуравли-
вая утонченные места, заметные на стебле в виде
округлых углублений. Пищей им служат специ-
альные тельца, находящиеся при основании че-
решков. Можно установить ряд градаций приспо-
Рис. 11.2. Верхняя часть стебля Cecropia
(по В. В. Алехину, 1950):
А — стебель с вскрытой полостью; Б — мюллеровы тельца
при основании черешков
364
собления растений к защищающим муравьям. Так, если у Acacia мура-
вьи пробуравливали стенки колючек, то у Cecropia есть утончения на
стебле, а у ряда видов (Humboldtia laurifolia, Ficus inaequalis и др.) и
готовые отверстия. Особый тип мирмекофилов представляют эпифит-
ные Myrmecodia и Hydnophytum, живущие на островах Малайского ар-
хипелага. У них многочисленные сообщающиеся полости имеются в клуб-
нях. Кроме растений с полостями в органах стеблевого типа существует
немало мирмекофильных форм, у которых муравьи живут в полостях
прилистниковых колючек. Вероятно, для привлечения их могут служить
и экстрафлоральные нектарники, расположенные на вегетативных орга-
нах растений 39 семейств. Они не связаны с обеспечением перекрестно-
го опыления, но помогают растениям содержать защитников.
Защита от фитофагов путем изменения участия растений в фи-
тоценозе. Губительного воздействия фитофагов и их размножения
растения могут избегать путем цикличности своего доминирова-
ния (или значительного участия) в фитоценозах. Сдерживать вли-
яние фитофагов удается и благодаря уходу вегетативно подвиж-
ных видов с участка, на котором размножаются фитофаги. Такое
избегание (в пространстве и времени) мест массового скопления
фитофага характерно, например, для взаимоотношения на лугах
ряда бобовых с клеверной нематодой. При этом клевер ползучий
(Trifolium repens) и чина луговая (Lathyrus pratensis) смещаются
с мест локализации червей, развивая органы вегетативного раз-
множения. А клевер луговой (Trifolium pratense) сдерживает раз-
множение клеверной нематоды, циклично изменяя свое участие в
фитоценозе: так называемые «клеверные годы» сменяются вре-
менным отсутствием в травостое его наземных побегов, что на
некоторый срок лишает фитофага кормовой базы (Т. А. Работнов,
1993).
Редкие и эфемерные виды, малодоступные животным, выраба-
тывают разнообразные простые в химическом отношении спосо-
бы защиты, которые представляют собой эффективные эволюци-
онные барьеры на пути влияния неадаптированных полифагов,
наиболее вероятных врагов этих растений. Многочисленные же и
часто встречающиеся растения травоядным легкодоступны. У них
развились более дорогостоящие способы обороны (жесткие лис-
тья с низким содержанием питательных веществ, большое коли-
чество неспецифических соединений, например танинов). Эти
способы обеспечивают серьезный экологический барьер, но в эво-
люционном плане представляют слабую помеху за исключением
тех случаев, когда есть качественные химические средства защи-
ты. У некоторых растений имеются оба типа защиты (Э. Пианка,
1981).
Хорошее средство от семяедов — периодичность плодоношения.
Это относится к типичным зоохорам (дубы — Quercus, буки —
Fagus, каштаны — Castanea, сибирская сосна — Pinus sibirica) и
365
в меньшей степени к анемохорам, семена которых тоже поедают-
ся животными (ели — Picea, сосны — Pinus sylvestris, пихты —
Abies, лиственницы — Larix). Отсутствие синхронности плодоно-
шения и его периодичность у растений, чьи семена служат кор-
мом животным, в неурожайные годы приводит к снижению чис-
ленности семяедов и неполному уничтожению ими семян в сле-
дующие годы обильного плодоношения.
11.4. Отношения растений с паразитическими
организмами и устойчивость к инфекции
Характеристики инфекционных агентов. Инфекционные болез-
ни растений вызываются вирусами, бактериями, грибами, нема-
тодами, паразитическим растениями. Наибольшие потери урожая
обычно вызывают грибные инфекции, что связано с очень боль-
шим количеством видов грибных паразитов (более 10000).
Паразиты характеризуются патогенностью, вирулентностью и
агрессивностью. Патогенность — способность вызывать заболе-
вание. Вирулентность отражает способность поражать разных хо-
зяев: разные формы паразита могут быть вирулентны по отноше-
нию к разным растениям. Вирулентность меняется в результате
генетических изменений паразита и почти не зависит от среды.
А агрессивность патогена выражает степень поражения им вос-
приимчивого растения. Она определяется скоростью роста пара-
зита и факторами среды. Вирулентность и агрессивность отража-
ют качественную и количественную характеристику патогенности
(В. В. Полевой, 1989).
Различают несколько групп фитопатогенов. По характеру пи-
тания (и воздействию на растения) различают некротрофных
паразитов, сначала убивающих ткани хозяина, а затем питаю-
щихся ими, и биотрофных — питающихся живым хозяином. Не-
кротрофы поселяются на предварительно убитой ими ткани. Для
этого служат токсины разного действия (мембранолитические,
нарушающие азотный и энергетический обмен). Они также вы-
деляют в среду различные ферменты (пектиназы, целлюлазы, фос-
фатазы, кутиназы и др.), с помощью которых разрушают мемб-
раны, клеточную стенку растений и срединную пластинку, вы-
зывают мацерацию тканей. С их помощью патогены превраща-
ют содержимое убитых клеток в доступную для усвоения форму.
Эти особенности некротрофов позволяют им поражать широкий
круг хозяев (они обычно не имею! узкой специализации). Не-
кропаразитов можно рассматривать как пионеров детритофагии,
опережающих конкурентов благодаря способности умерщвлять
хозяина, который слабо сопротивляется их воздействию и мало-
устойчив к ним.
366
Обычная реакция зараженных растений — сбрасывание пораженных
листьев. Так поступает, например, люцерна (Medicago), инфицирован-
ная грибом Pseudopeziza medicaginis. Другая реакция — образование изо-
лирующих барьеров. Например, на картофеле (Solatium tuberosum) в от-
вет на поражение актиномицетом Actinomyces scabies возникают твер-
дые струпья, изолирующие паразита и препятствующие его распростра-
нению (М. Би гон и др., 1989). Если хозяин — модул ьно устроенный орга-
низм (побеговая система, состоящая из повторяющихся элементов), не-
кропаразит часто убивает его не целиком, а отдельными модулями, и
растения наращивают потом новые органы. Некропаразиты могут также
ускорять естественное отмирание отдельных модулей растения, оставляя
другие живыми. Многие некропаразиты, особенно грибы рода Pythium,
вызывают быструю гибель проростков — наиболее уязвимую стадию раз-
вития растений (Л.Л. Великанов и др., 1981).
Биотрофы питаются только живыми тканями, некоторое вре-
мя сосуществуя с живыми клетками хозяина. Они проникают в
них, минуя защитные образования и не выделяя вредных для хозяи-
на токсинов. Часто грибы-биотрофы обитают в межклетниках,
добывая питание из клетки с помощью гаусторий. Биотрофы тесно
связаны с особенностями обмена хозяина и сильно от него зави-
сят. Поэтому по крайней мере на первых этапах заражения они не
сильно травмируют хозяина (растение обычно повреждается пос-
ле спороношения паразита). Эта группа патогенов отличается уз-
кой специализацией по отношению к хозяевам. Биотрофных па-
разитов делят на 2 группы: сбалансированные паразиты способ-
ны длительно питаться живыми тканями хозяина, не вызывая их
гибели, и деструктивные паразиты, так же питающиеся живы-
ми тканями хозяина, но вызывающие их быструю гибель.
Большинство видов выработали устойчивость к действию высокоспе-
циализированных биотрофов. Например, клетки растения при проник-
новении ржавчинных и мучнисторосяных грибов остаются живыми.
В ряде случаев в них нет и серьезных изменений, кроме некоторого умень-
шения продолжительности жизни зараженных клеток, изменения в них
скорости метаболизма и потери воды. В других случаях паразит приво-
дит к глубоким нарушениям обмена веществ хозяина. Характерно также,
что, поражая листья, многие биотрофы стимулируют формирование вокруг
инфицированной зоны области повышенной активности фотосинтеза.
Сюда направляются метаболиты из незараженных частей организма.
Биотрофные паразиты могут вызывать у хозяев и морфологические из-
менения. Например, гриб Plasmodiophora brassicae стимулирует деле-
ние и увеличение размеров клеток корня капусты (Brassica oleracea),
расширяя объем тканей для заселения (М. Бигон и др., 1989).
По месту локализации паразитические грибы условно делят
на экто- и эндопаразитов. Эктопаразиты, например мучнисто-
росяные грибы, распространяются по ассимилирующим частям
367
растений, внедряя в клетки хозяина только гаустории. Спороно-
шение их происходит на поверхности растения. Эндопаразиты
тоже выносят свои спороношения наружу, проходя через устьи-
ца или прорывая растительные ткани, но таллом их остается
внутри хозяина (в межклетниках, клеточных стенках, внутри
клеток).
Паразиты отличаются и специализацией по отношению к хо-
зяевам. Облигатные паразиты не могут существовать без хозяев
и обычно высокоспецифичны по отношению к ним. К этой груп-
пе относятся многие вирусы и грибы, выработавшие способность
проникать в ткани хозяина, минуя защитные барьеры. Некоторые
из них, например мучнисторосяные грибы, не могут развиваться
на искусственных питательных средах. Другие, например ржав-
чинные, растут на этих средах и даже образуют там типичные
спороношения. Это экологически облигатные паразиты, так как
они развиваются как сапротрофы при отсутствии конкуренции со
стороны других организмов, но в естественных условиях суще-
ствуют только как паразиты на живых тканях, недоступных для
конкурентов (Л.Л. Великанов и др., 1981). К облигатно биотроф-
ным грибам относятся важнейшие с экономической точки зрения
патогены, вызывающие массовые поражения растений (панфи-
тотии), например ржавчина злаков (Puccinia graminis). Во мно-
гих случаях они повреждают хозяина незначительно (это пример
эволюционно сложившегося компромисса: чтобы выжить, хозяева
должны постоянно совершенствовать способы защиты, а парази-
ты — возможности получения достаточного питания).
Факультативные паразиты обычно развиваются как сапротрофы и
живут на мертвых растениях, но могут паразитировать и на ослабленных
живых особях или невегетирующих органах, например плодах. Такие
грибы обычно мало специализированы и, поражая живые ткани, выжи-
вают после гибели хозяина, а также легко культивируются на искусст-
венных питательных средах. К этой группе относятся многочисленные
сумчатые грибы, образующие во время паразитирования бесполое спо-
роношение, а на погибшем хозяине — половое (например, возбудитель
парши яблони — Venturia inaequalis\
Факультативные паразиты близки к некротрофам, не способ-
ным поражать здоровые ткани. Сначала грибы поселяются на от-
мерших участках хозяина, а затем расширяют область распростра-
нения, отравляя прилегающие части растения. Иногда их токси-
ны переносятся по сосудам хозяина, поражая далеко отстоящие
от места внедрения органы (например, некроз листьев после вне-
дрения в основание стебля). Между факультативными паразита-
ми и некротрофами есть переходы (Э. Мюллер, В. Леффлер, 1995).
В эволюции, вероятно, неоднократно происходил переход к пара-
зитизму от сапротрофности через некрофитизм, хотя некоторые
368
микологи стоят на противоположной позиции (J. Kohlmeyer, 1975;
V. Demoulin, 1985).
Фитоиммунитет и специфическая устойчивость растений к дей-
ствию патогенных организмов. Растения, постоянно живя в среде
с другими организмами, выработали защиту от неблагоприятных
биотических факторов. Устойчивость к инфекции представляет
собой способность предотвращать, ограничивать и задерживать
ее развитие. Основоположник учения об устойчивости растений
Н. И. Вавилов выделял устойчивость неспецифическую (видовую)
и специфическую (сортовую). Видовую устойчивость еще называ-
ют фитоиммунитетом. Он представляет собой полную невосп-
риимчивость (возбудитель не поражает данный вид растения). На-
пример, бобовые не поражаются болезнями пасленовых. Неспе-
цифический иммунитет защищает растения и от множества сапро-
трофных организмов, в окружении которых они постоянно нахо-
дятся. Благодаря ему растения поражаются лишь небольшим чис-
лом возбудителей. Поэтому иммунитет в природе — правило, а
восприимчивость — исключение (Л. Г. Косулина и др., 1993). Но в
процессе эволюции некоторые патогены сумели преодолеть видо-
вой иммунитет и в разной степени стали поражать данный вид
растений. Они стали паразитами, специфичными для этого вида,
и в отношении их правильнее говорить только об устойчивости.
На такие патогены приходится 90 % потерь урожая сельскохозяй-
ственных культур.
Очень важны генетические аспекты устойчивости растений к
действию патогенных организмов. Обусловленная генами устой-
чивость накладывается на биохимические пути базовой устойчи-
вости, ограничивающей рост патогена после заражения. Генети-
ческий анализ отношений растений и фитопатогенов показал
высокую специфичность этого взаимодействия («ген-на-ген»).
Белковые продукты генов устойчивости полифункциональны,
и сами эти гены имеют разное происхождение. Гены устойчивос-
ти происходят от генов, кодировавших необходимые для развития
белки, участвующие в эндогенных сигнальных системах. Напри-
мер, растения риса (Oryza sativa) для обнаружения патогена мо-
билизуют белки, определяющие межклеточные связи с участием
внеклеточного компонента. Некоторые белки, связанные с ини-
циацией защитного отклика, подобны белкам, вызывающим за-
программированную смерть клетки.
Исследователи отмечают приобретенный растениями специ-
фичный механизм быстрой эволюции генов устойчивости, отсут-
ствующий у животных, которые имеют развитую соматическую
иммунную систему. Для растений характерна быстрая диверген-
ция этих генов (хотя и разная в разных семействах: у Злаков —
быстрее, у Пасленовых — медленнее). При этом коэволюция рас-
тений и паразитов определила связанную изменчивость генов ви-
369
рулентности паразитов и генов устойчивости хозяев, называемую
в иммунологии «гонкой вооружений», аналогичной для подобных
экологических систем (С. Н. Шамрай, 2003).
Устойчивость растений к возбудителям болезней могут изме-
нять внешние условия (погода, удобрения, возраст растения и
органа, биотическое окружение). Например, у растений зафикси-
ровано появление устойчивости ко второму возбудителю после
предшествующей инфекции или ослабление устойчивости к па-
тогену при заболевании, вызванном другим возбудителем.
Механизмы защиты от патогенов и теории устойчивости. Уже
первые исследователи устойчивости растений к инфекции связы-
вали это свойство с особенностями химического состава тканей,
исходя из того, что паразиты положительно хемотропичны (пере-
мещаются или растут по направлению к питательным веществам).
Первая теория устойчивости, названная хемотропичной, исходи-
ла из того, что растения поражаемы, если содержат привлекаю-
щие паразитов вещества, диффундирующие через клеточную стен-
ку, а если таких веществ нет, то растение устойчиво. Однако было
показано, что споры грибов могут прорастать не только через жи-
вую ткань, но и через клетки эпидермиса, лишенные содержимо-
го. Кроме того, они одинаково прорастают через клеточную стен-
ку как восприимчивых, так и устойчивых растений, а заражение
зависит от процессов, развивающихся при взаимодействии про-
дуктов патогена с мембраной клетки. Дальнейшее изучение роли
химического состава растений в устойчивости к патогенным орга-
низмам привело к возникновению еще двух теорий. Тормозящая
теория акцентирует внимание на том, что устойчивые растения
содержат вещества, тормозящие развитие патогена. А питатель-
ная теория (теория «неполной среды») связывает устойчивость
растения с отсутствием в нем необходимых паразиту веществ.
В настоящее время защита растений от патогенов рассматри-
вается как комплексное явление, включающее механизмы кон-
ституционные (присутствовавшие в тканях хозяина до инфек-
ции) и индуцированные (возникшие в ответ на контакт с парази-
том или с его выделениями). Конституционные механизмы вклю-
чают: 1) особенности клеток и тканей, обеспечивающие механи-
ческий барьер для проникновения инфекции; 2) способность вы-
делять антибиотические вещества; 3) создание в тканях недостат-
ка важных веществ для роста и развития паразита. Индуцирован-
ные механизмы устойчивости характеризуются реакцией расте-
ния на инфекцию через: 1) усиление дыхания и энергетического
обмена; 2) накопление веществ, обеспечивающих общую неспе-
цифическую устойчивость (фитонциды, фенолы и продукты их
окисления); 3) создание дополнительных механических барьеров;
4) возникновение реакции сверхчувствительности; 5) быстрый
синтез низкомолекулярных защитных веществ — фитоалексинов.
370
Общая стратегия защиты растения состоит в недопущении дей-
ствия паразита на свои клетки или в локализации инфекции и
гибели паразита.
Реакция растений на поражение некротрофами и биотрофами
отличается. Устойчивость к биотрофам создается с помощью ме-
ханизмов распознавания паразита, включения реакции сверхчув-
ствительности и последующего уничтожения патогена в некроз-
ной зоне с участием синтезированных в ответ на инфекцию фито-
алексинов. Устойчивость же к некротрофам обеспечивается с по-
мощью: 1) детоксикации токсинов паразита; 2) потери устойчи-
выми растениями чувствительности к специализированным ток-
синам; 3) присутствия на плазмалемме хозяина рецептора, связы-
вающего токсин; 4) инактивации экзоферментов неспецифичес-
кими ингибиторами; 5) задержки синтеза экзоферментов парази-
та устранением или маскировкой субстратов (например, усиление
суберинизации и лигнификации клеточных стенок в месте пора-
нения маскирует пектиновые вещества, необходимые для начала
синтеза пектиназ паразита); 6) повреждения клеточных стенок
паразита ферментами растения (хитиназой, глюконазой); 7) син-
теза белков-антиферментов в ответ на гидролитические фермен-
ты паразита (В. В. Полевой, 1989).
Механическая защита растения от патогена. Взаимодействие с
паразитом начинается на поверхности растения. Патоген или его
споры должны сначала удержаться на ней. Препятствовать этому
может восковой слой, делающей эпидерму гладкой и плохо сма-
чиваемой водой, необходимой для прорастания спор. Микроорга-
низмы преодолевают этот барьер через устьица, чечевички и раны.
Грибы могут еще разрушать кутикулу своими выделениями. По-
кровные ткани служат не только механическим барьером, но еще
и до заражения содержат антибиотические вещества.
Инфекция вызывает и активное ответное развитие механичес-
ких барьеров. Так, широко распространено усиление одревесне-
ния клеточных стенок в ответ на заражение. Оно резко затрудняет
проникновение паразита, так как лигнин могут расщеплять не-
многие микроорганизмы. При поражении лигнифицируются даже
стенки тех клеток, в которых лигнин до того не откладывался.
Это повышает механическую прочность стенок, ограничивает рас-
пространение токсинов паразита и приток питания к нему, защи-
щает стенку клетки от действия ферментов паразита. Лигнифици-
роваться может и клеточная стенка грибов-паразитов, что оста-
навливает их рост (В. В. Полевой, 1989).
Эффективным механическим барьером между очагом инфек-
ционного некроза и живой тканью становится и образующаяся
перидерма. Она препятствует распространению паразита, затруд-
няет приток питательных веществ в пораженную зону и защи-
щает здоровые клетки от токсинов гибнущих клеток. А механи-
371
ческой преградой для патогенов, распространяющихся по сосу-
дам растения, становятся образующиеся в них тилы — покрытые
пектиновым чехлом выпячивания содержимого соседних парен-
химных клеток. Распространение паразита при этом задерживает-
ся, и в закупоренном сосуде он повреждается антибиотическими
веществами.
Питательная теория устойчивости растений. Питательная тео-
рия (теория «неполной среды») связывает устойчивость растения
с отсутствием в нем необходимых паразиту веществ. Она опирает-
ся на работы по мутантным грибам, неспособным самостоятельно
синтезировать необходимые для роста и развития вещества. Такие
грибы не поражают растения, которые не содержат данных ве-
ществ, но при искусственном введении их устойчивость растения
к паразиту теряется.
Питательная теория применима к объяснению взаимодействия
растений с зависимыми от них высокоспециализированными био-
трофами. Примером может быть устойчивость пшеницы (Triticum
aestivum) к фузариозу. Растение теряет устойчивость к грибу
Fusarium graminearum во время цветения, так как заражение про-
исходит через пыльники, в которых содержатся азотсодержащие
метилированные спирты, входящие в состав его фосфолипидов.
Экспериментальное добавление этих веществ усиливает инфек-
ционность гриба, но не влияет на другие патогены, не нуждаю-
щиеся в них. И в целом круг растений, поражаемых этим грибом,
ограничен видами, содержащими высокие концентрации данных
спиртов. Реже питательной теорией можно объяснить сортовую
устойчивость. Так, некоторые сорта риса (Oryza sativa) устойчи-
вы к Xanthomonas oryzae. Эта бактерия зависит от наличия в сре-
де аминокислот, содержащих серу, так как не может использовать
неорганические ее источники. Добавление выделенного из злака
цистеина стимулирует рост паразита и повышает его патогенность.
Поэтому сорта, не содержащие этого вещества (или бедные им),
устойчивы к данной бактерии (Л.Г.Косулина и др., 1993).
С точки зрения индуцированной устойчивости формирование
«неполной среды» может происходить в ответ на заражение. При
этом растение исключает из обмена необходимые паразиту веще-
ства. Так, К. Т. Сухоруков (1952) показал, что при поражении про-
водящей системы хлопчатника (Gossypiuni) возбудителем вилта
Verticillium в тканях устойчивых сортов понижается содержание
биотина и пантотеновой кислоты. Таким образом, устойчивые
растения в отличие от неустойчивых устраняют из очага пораже-
ния необходимые грибу вещества, создавая неблагоприятную сре-
ду для его роста.
Л. В. Метлицкий и О.Л.Озерецковская (1985) обнаружили, что устой-
чивые к фитофторозу сорта картофеля (Solatium tuberosum) лишают
372
Phytophthora infestans стеринов, источником которых для гриба служат
клетки растения-хозяина (способность к синтезу их фитофтора утратила
в коэволюции с растением). Бета-ситостерин необходим патогену для
образования зооспор, но у устойчивых растений в месте инфицирования
он перестает синтезироваться, так как его предшественник быстро по-
требляется для начавшегося синтеза защитных веществ. При недостатке
бета-ситостерина паразит не размножается, а его мембраны поврежда-
ются и становятся чувствительнее к фитоалексинам.
Фитонциды. Важную роль в неспецифической защите расте-
ний от патогенов играют продуцируемые ими антибиотические
вещества — фитонциды. Эти соединения способны эффективно
задерживать развитие и убивать многие микроорганизмы. Они
играют у растений столь же важную и универсальную роль, как и
фагоцитоз у животных. Внутри клеток фитонциды часто накап-
ливаются в вакуолях. Летучие фитонциды, выделяемые при пора-
нении клеток, например у лука (Allium сера) и чеснока (A. sativum),
защищают растения от патогенов уже над поверхностью органа.
Нелетучие фитонциды содержатся в покровных тканях, участвуя
в создании поверхностных барьеров. После повреждения расте-
ний их содержание резко возрастает, предотвращая инфицирова-
ние раны. Специализированные паразиты довольно легко при-
спосабливаются к антибиотическим веществам своего хозяина, но
роль фитонцидов в неспецифическом фитоиммунитете достаточ-
но велика.
С химической точки зрения фитонциды — низкомолекуляр-
ные вещества разных групп. Среди них наиболее распространены
терпеноиды, алкалоиды, эфирные масла, пигменты, глюкозиды и
фенольные соединения. Защитные свойства ряда фитонцидов
проявляются при их превращениях. Так, при выходе из состояния
покоя луковиц лука в них активируется фермент аллииназа, под
влиянием которого основной компонент эфирных масел лука ал-
лиин превращается в обладающий фунгицидным действием алли-
цин. При повреждении патогеном окрашенных органов растений
антоциановые пигменты распадаются с образованием токсичного
для микроорганизмов антоцианидина.
Неоднократно показано, что в поврежденных и инфицирован-
ных клетках активируются процессы, приводящие к накоплению
фенольных соединений (Л.Г.Косулина и др., 1993). Была уста-
новлена также связь устойчивости мертвых тканей с содержанием
в них фенолов. Так, луковицы с сухими окрашенными чешуями
устойчивы к Botrytis allii, так как ингибирующие гриб фенолы
диффундируют в попавшую на чешую каплю с паразитом. При
удалении сухих чешуй луковица будет поражена. Фенольные со-
единения эффективно инактивируют экзоферменты патогенов, ог-
раничивают диффузию токсинов и питательных веществ, прида-
ют прочность механическим тканям, тормозят рост гифов.
373
Растениям фенолы необходимы еще для синтеза лигнина, который
экранирует клеточные стенки от атаки паразита, так как его не может
расщеплять большинство патогенов. В связи с этим высказывалось пред-
положение, что одревеснение наземных растений, приведшее к глобаль-
ным последствиям для планеты, могло стать побочной реакцией борьбы
растений с грибной инфекцией (И. В. Каратыгин, 1993).
Алкалоиды токсичны и для самих растений, поэтому не могут
накапливаться во всех тканях, но концентрирование их в отдель-
ных группах клеток играет предохраняющую роль. Например, в
корнях магонии (Mahonia trifoliata), устойчивой к грибу Phymato-
trichum omnivorum, отдельные клетки паренхимы содержат алка-
лоид берберин в концентрации в 79 раз более высокой, чем кон-
центрация, полностью останавливающая развитие паразитичес-
кого гриба в культуре. Другие же клетки заметных количеств это-
го алкалоида не содержат. Алкалоидам отводится важная роль в
защите от раневых патогенов. Например, в раневой перидерме,
образующейся в местах повреждения клубней картофеля (Solanum
tuberosum), накапливается много альфа-соланина и альфа-чако-
нина, которые помогают защитить клубень от Fusarium solani.
Терпеноиды, содержащиеся в смоле деревьев (особенно хвой-
ных), эффективно защищают древесину от дереворазрушающих
грибов. Например, монотерпены защищают хвойные от пузырча-
той ржавчины, а сесквитерпеновые ортохиноны препятствуют за-
ражению вязовых грибом Ceratocystis ulmi.
Глюкозиды сами по себе не токсичны для патогенов, но ток-
сичны накапливающиеся продукты их распада. В клетке они на-
ходятся в вакуоле, а разрушающие их ферменты — в цитоплазме,
поэтому только при повреждении клетки, когда тонопласт нару-
шается, глюкозиды поступают в цитоплазму, где становятся дос-
тупными для расщепления ферментами. Такая система защиты
эффективна против некротрофов, вызывающих повреждения за-
раженных клеток. Биотрофы же, оказывая мягкое действие на
клетки хозяина, не вызывают образования токсичных продуктов
гидролиза глюкозидов.
Фитоалексины. Особое место в химической защите растений
имеют фитоалексины — низкомолекулярные вещества, быстро
вырабатываемые растениями в ответ на контакт с фитопатогеном.
Они обнаружены у бобовых, пасленовых, маревых, мальвовых,
зонтичных, сложноцветных и многих других растений (но не най-
дены у злаков). Их выделяют погибающие от воздействия парази-
та клетки и их ближайшее окружение. В зоне поражения фито-
алексины накапливаются до концентраций, подавляющих пара-
зита, и таким образом способны предотвратить распространение
инфекции. Однако в отличие от иммунной реакции животных
этот ответ довольно неспецифичен (М. Бигон и др., 1989).
374
Вследствие разнообразия растений, патогенов и их взаимодей-
ствий велико и разнообразие фитоалексинов. У бобовых это изо-
флавоноиды, у пасленовых — сесквитерпеноидные вещества, у
сложноцветных — полиацетилены и т.д. Кроме того, в одном ра-
стении в ответ на инфекцию могут образовываться разные фито-
алексины и другие антибиотики. Это помогает бороться с патоге-
нами, легко приобретающими устойчивость к одному антибиоти-
ку, но гораздо труднее привыкающими к нескольким веществам.
Кроме того, у разных фитоалексинов часто неодинаков механизм
повреждающего действия на патоген.
В здоровых тканях фитоалексины отсутствуют. Они — конеч-
ные продукты измененного заражением метаболизма. От погиба-
ющих клеток поступает сигнал о необходимости синтеза фито-
алексинов, которые образуются в соседних живых клетках и пере-
мещаются в некротическую зону, где находятся паразиты. Фито-
алексины подавляют их рост и дезактивируют их ферменты.
Образование фитоалексинов стимулируют разные биотичес-
кие и абиотические агенты. Некоторые индукторы получены из
тел патогенов и их выделений. Обнаружены и эндогенные индук-
торы — соединения растительной клетки, освобождающиеся из
нее под действием патогена (например, фрагменты пектинового
полисахарида клеточной стенки, освобождаемые пектинолитичес-
кими ферментами патогена). Образование фитоалексинов может
индуцироваться не только паразитами и их метаболитами, но и
некоторыми химикатами (соли тяжелых металлов, ферментные
яды, антибиотики и др.). Но для этого требуется концентрация
химикатов на 5 —10 порядков выше, чем при использовании био-
тических индукторов, и даже при этом не удается образовать фи-
тоалексины в летальных для паразита концентрациях.
Фитоалексины играют важную роль в фитоиммунитете. Расте-
ние становится восприимчиво к патогену, если он не индуцирует
синтез фитоалексинов. При подавлении у растения способности
к образованию фитоалексинов оно также становится восприим-
чивым ко многим патогенам, ранее не поражавшим его. Но высо-
коспециализированные патогены могут преодолевать фитоалек-
синовый барьер. Некоторые из них выработали толерантность к
количествам фитоалексинов, накапливаемым в зараженных тка-
нях; другие разлагают их до менее токсичных соединений; третьи
тормозят их синтез, выделяя вещества, блокирующие индукторы
этого процесса (Л. Г. Косулина и др., 1993).
Реакция сверхчувствительности. Реакция сверхчувствительнос-
ти (СВЧ) — универсальная защитная реакция растений, лежащая
в основе большинства случаев проявления устойчивости к патоге-
нам. Суть ее в том, что в месте контакта с биотрофным паразитом
клетки устойчивых к нему растений быстро отмирают, создавая
зону некроза, останавливающую дальнейшее проникновение па-
375
разита, а затем некротическая зона окружается механическим барье-
ром из перидермы.
Реакция устойчивых растений основана на повышенной чув-
ствительности их к данной инфекции. Скорость этой реакции
велика. Так, при контакте возбудителя фитофтороза с листом кар-
тофеля (Solatium tuberosum) клетки отмирают уже через 30 мин.
Чем растение устойчивей, тем быстрее гибнут клетки в месте по-
ражения и тем меньше клеток затрагивается инфекцией. У неус-
тойчивых сортов клетки остаются живыми и паразит быстро рас-
пространяется по телу. Основная функция реакции СВЧ — по-
давление спороношения биотрофного паразита, происходящего
лишь при контакте с живыми клетками.
При заражении картофеля фитофторой выяснено, что генерация ак-
тивных форм кислорода наблюдается уже через час после контакта пато-
гена с растением, а максимум их образования совпадает с началом гибе-
ли клеток растения (через 4 —6 ч после заражения). Экспериментально
показано, что свободные радикалы образуются на плазматической мем-
бране и могут ее повреждать, индуцируя процессы переокисления липи-
дов, приводящие к нарушению липид-белковых взаимодействий в мем-
бране и повреждению мембранных белков. Однако не все работы это
подтверждают. Ряд исследователей считает также невыясненным, явля-
ется ли реакция СВЧ случаем неудачной защиты или запрограммиро-
ванным процессом (Л. Г. Косулина и др., 1993).
11.5. Содействие животных размножению
и распространению растений
Животные играют важную роль в распространении пыльцы,
спор, семян, плодов и вегетативных органов растений, влияя при
этом на их размножение и расселение. Деятельность животных
обычно мотивирована пищевыми потребностями, но использова-
ние ими в пищу семян, плодов и пыльцы для растений полезно,
так как способствует их распространению. Поэтому хотя живот-
ные и выступают как хищники, растения в этом случае вряд ли
можно рассматривать как жертвы.
В настоящее время типы опыления и распространения диас-
пор рассматриваются на вероятностной основе — как наиболее
часто реализуемый вариант (Б. М. Миркин и др., 2001). Так, насе-
комоопыление, ветроопыление, самоопыление и апомиксис у од-
них и тех же видов могут сочетаться и чередоваться в разные годы.
То же относится к способам распространения плодов и семян.
Так, кроме животных, сочные плоды черемухи (Padus avium) и
боярышника (Crataegus) могут транспортироваться водой, а се-
мена многих анемохорных, гидрохорных, эпизоохорных и авто-
376
хорных растений часто распространяются эндозоохорно, не теряя
всхожести при прохождении через пищеварительный тракт жи-
вотных, питающихся вегетативными частями растений.
Зоофилия. Во флоре Земли господствующее положение зани-
мают цветковые растения, а цветки большинства видов опыляют-
ся животными (чаще насекомыми, реже — птицами и млекопита-
ющими). Поэтому между зоофильными видами и опылителями
устанавливаются тесные взаимоотношения. К цветкам животных
привлекает пища, которую они здесь находят (пыльца, нектар), а
в отдельных случаях имитация цветками самок насекомых, с ко-
торыми самцы пытаются спариться (известна псевдокопуляция
самца наездника Lissapimpa excelsa с орхидеей Cryptostylis
subulata). Так как в основе зоофилии лежит трофическая привле-
кательность пыльцы, животные могут уничтожать ее в большом
количестве. Растениям это невыгодно, но в то же время эффек-
тивное перекрестное опыление обеспечивает необходимую гене-
тическую гетерогенность популяций. Поэтому формирование не-
ктарников в процессе эволюции можно рассматривать как «комп-
ромисс», при котором животные, потребляя высококалорийный
и легкоусвояемый нектар, одновременно осуществляют перенос
пыльцы.
У растений, приспособившихся к опылению животными, яр-
кие лепестки или чашелистики, привлекательный запах, содер-
жится нектар. Одни цветки, широко открытые, доступны для мно-
гих опылителей, другие, с длинной трубкой венчика, — только
для некоторых (с длинным хоботком или клювом, рис. 11.3). На-
секомые отыскивают цветки в первую очередь с помощью зрения,
и этому соответствуют приспособления растений. Показано, что
окраска цветков лежит в области наиболее чувстви-
тельной части цветового зрения опылителей. На-
пример, это касается опыления цветков разного
цвета у живокости (Delphinium nelsonil). Обычно
она имеет синие цветки, но встречаются и расте-
ния с белыми венчиками, которые дают на 20%
меньше семян. Это связано с тем, что шмели и пти-
цы посещают их на 24 % меньше, притом что не-
ктара они вырабатывают столько же, сколько и
обычные. При искусственном опылении белые
цветки производят такое же количество семян, что
и синие.
Рис. 11.3. Цветки тропических растений, опыляемые ноч-
ными бабочками (по В. В.Алехину, 1950):
А — Oxyanthus hirsutus, Б — Exostema floribundum А Б
377
Зоофилия часто приводит к коадаптации взаимодействующих
видов. Характерна синхронизация циклов развития растений и
опылителей. Так, виды, опыляемые ночными животными, откры-
вают цветки вечером и закрывают утром, имеют белую окраску и
сильный запах. И опылители подстраивают свой цикл к ритму
жизни опыляемых растений. А у насекомых с коротким временем
питания (например, чешуекрылых) жизненный цикл может быть
очень жестко скоординирован с цветением определенных видов
растений. Так взаимосвязаны репродуктивные циклы юкки (Yucca)
и юкковой моли {Tegeticula maculata). Моль опыляет цветки, от-
кладывает в их завязь яйца, и личинки регулируют число семян,
выедая их часть. Но в конце сезона самки откладывают яйца не в
цветки, а в плоды, так как из поздних цветков плоды часто не
образуются.
Но в некоторых случаях взаимоотбора партнеров по взаимовы-
годному сотрудничеству может и не произойти. Например, раз-
ные виды ваточника {Asclepias) опыляются шмелями одного вида.
Однако устройство их цветков таково, что пыльца пристает к раз-
ным частям тела насекомого, поэтому потом она попадает на рыль-
ца только соответствующего вида растения, что препятствует меж-
видовому опылению. В этом примере опылитель не специализи-
рован, но растения специализироваться вынуждены. К неспециа-
лизированным универсальным опылителям относятся и долгожи-
вущие животные (пчелы, летучие мыши, грызуны). В течение се-
зона они используют цветки разных видов, а при недоступности
нектара переходят на другие корма (М. Бигон и др., 1989).
Зоофильныс виды обычно встречаются только там, где есть их опы-
лители. Поэтому отсутствие соответствующего таксона растений в дан-
ной местности при прочих благоприятных условиях может быть объяс-
нено иногда лишь отношениями с опылителями. Так, северная граница
распространения нектарниц, главных птиц-опылителей Старого Света,
примерно соответствует северной границе орнитофильных растений
{Musa, Hibiscus, Loranthus, Bombax и др.). Связь между распростране-
нием этих растений и опыляющих их птиц вполне вероятна, хотя не
ясно, какой из партнеров ограничивает распространение другого (Ф.Фу-
карек и др., 1982).
Зоохория. Эпизоохорными (от греч. epi — на, над, zoon — жи-
вотное, choreo — распространяюсь) называются растения, диас-
поры которых переносятся благодаря приспособлениям, облегча-
ющим прикрепление их семян и плодов к шерсти, перьям и коже
животных. Растениям эпизоохория особенно выгодна: семенам не
грозит разрушение при разжевывании и переваривании, и время
пребывания их на переносчике часто не ограничено. Животное
часто не замечает прикрепившихся снаружи диаспор, поэтому за-
378
цепившиеся семена и плоды разносятся на большие расстояния.
Основные эпизоохорные разносчики семян — млекопитающие,
но значительную роль играют и птицы.
В природе выработалось множество приспособлений к эпизоохории
и число видов, расселяющихся таким способом, очень велико. Так, око-
ло 10% цветковых растений имеют прицепляющиеся плоды и семена
(А. Г. Воронов, 1963). Цепляющимися выростами снабжены липучка
{Lappula), репейник {Arctium), череда {Bidens), чертополох {Carduus), гра-
вилат {Geum), морковь {Daucus) и др. У некоторых растений (якорцы —
Tribulus, рогоплодник — Ceratocarpus) плоды впиваются шипами в лапы
животных. Ряд эпизоохорных видов (в родах линнея — Ыппаеа, ситник —
Juncus, кувшинка — Nymphaea, кубышка — Nuphar, лен — Linum, омела —
Viscum и др.) имеет семена и плоды с липким соком (Ф. Фукарек и др.,
1982 и др.).
Эпизоохорное расселение происходит и в тех случаях, когда у
растения нет специальных приспособлений для приклеивания или
прицепления. Например, у болотных и водных видов диаспоры
часто очень мелкие и вместе с влажным илом пристают к лапам,
перьям и клювам водоплавающих птиц, которые переносят их
между водоемами. Такая эпиорнитохория отмечена Ю.Е. Алексе-
евым (1996) для ряда осок {Carex vesicaria, С. rostrata, С. rhyn-
chophysa). Так же могут переноситься побеги ряски (Lemna) и
части тел разных водных растений.
Процесс распространения растений, при котором животные
съедают семена, а затем выделяют их неповрежденными и спо-
собными прорасти, называется эндозоохорией (от греч. endon —
внутри). Сочные плоды и семена часто входят в пищевой рацион
животных. При этом прохождение семян через пищеварительный
тракт нередко подготавливает семена к прорастанию и служит их
распространению. Эндозоохории часто способствуют млекопита-
ющие. Так, нередко можно наблюдать всходы черемухи (Padus
avium), малины {Rubus idaeus) и других ягодных растений в тех
местах, где были экскременты медведя. Но преимущественно эн-
дозоохория связана с птицами, и хотя семена в их пищеваритель-
ном тракте находятся всего несколько часов, этого достаточно,
чтобы они оказались перенесены на километры. Так распростра-
няются рябина {Sorb us), бузина (Sambucus), бересклет {Euonymus),
можжевельник {Juniperus) и др. Некоторые твердые семена про-
глатываются птицами в качестве тел, способствующих измельче-
нию пищи в желудке (таковы, например, семена рдестов {Potamo-
getori), проглатываемые утками). Большая часть таких семян ис-
тирается и погибает, но некоторые проходят через кишечный тракт,
не потеряв всхожести.
И.А. Шилов (2001) отмечает, что так как специализация на
питании животных семенами сопряжена с выработкой приспо-
379
соблений, повышающих эффективность их переваривания, роль
таких видов в распространении семян снижается. Например, ти-
пично зерноядные воробьи переваривают семена клевера
(Trifolium) полностью, а у более всеядных грачей треть съеденных
семян выводится пригодными для прорастания. В процессе эво-
люции растения реагировали на регулярное потребление семян
формированием более плотных оболочек, препятствующих пере-
вариванию. В ряде случаев адаптация к эндозоохории привела к
повышению всхожести таких семян после прохождения их через
кишечник животных. Например, всхожесть семян борщевика
(Heracleum laciniatum), взятых из экскрементов медведя-гризли,
в 1,5 — 2 раза превысила всхожесть тех же семян, собранных с
растений.
Синзоохория (от греч. syn — вместе) — распространение рас-
тений животными, запасающими плоды и семена или случайно
утаскивающими диаспоры вместе со строительным материалом
для жилищ. Запасы плодов и семян устраивают многие грызуны
(хомяки, бурундуки, лесные мыши и др.), и при этом ряд живот-
ных нередко забывает или отказывается от них при некоторых их
повреждениях. Добывая и транспортируя съедобные диаспоры, жи-
вотные нередко случайно их теряют. А часть запасов остается не-
использованной при гибели хозяина. Так, даже падая с большой
высоты, плоды бразильского ореха (Bertholletia sp.) не раскалы-
ваются, но их разгрызают имеющие крепкие резцы агути. Съесть
все семена зверек сразу не может, часть их он закапывает, а потом
забывает о запасах.
Некоторые запасенные семена прорастают в норах. Кроме того,
очищая свои убежища, животные выбрасывают из них семена и
плоды с мусором. Так, выбросы из нор европейского сурка отме-
чены «сурочьими садами». Аналогичные заросли можно наблю-
дать и около нор других зверей. По И. К. Пачоскому, именно из
мест с колониями землероев пришли на поля многие сорняки.
Животные очень важны для возобновления деревьев с тяжелы-
ми семенами и плодами. Так, возобновление кедра европейского
и сибирского (Pinus cembra, Р. sibirica) в значительной мере за-
висит от сбора семян кедровками. Зарывая кедровые орешки, они
потом их теряют, чем способствуют облесению гарей, лесосек и,
следовательно, расширению площади обитания кедров. При этом
область произрастания кедровых сосен (Pinus sibirica, Р. pumila)
хорошо совпадает с сибирским участком ареала основного рас-
пространителя их семян — кедровки. Так же ведут себя по отно-
шению к желудям сойки. Наблюдения показали, что собранные
ими желуди отличались лучшей всхожестью, и при этом птицы
запасают их больше, чем могут. А Н. Г. Холодный (1949) считал,
что в широколиственных лесах главный распространитель дуба че-
решчатого (Quercus robur) именно сойка, которая осенью и зимой
380
питается исключительно желудями. В расселении европейских ду-
бов еще участвуют грачи, дятлы и белки. Синзоохорно распростра-
няются также лещина (Corylas), грецкий орех (Juglans), настоя-
щий каштан (Castanea). Вероятно, участие животных сыграло глав-
ную роль в возвращении этих пород после отступления ледника.
11.6. Взаимоотношения между растениями
Взаимоотношения между растениями довольно разнообразны,
существенно влияя на жизнь растительных организмов и фитоце-
нозов. Наиболее часто используется классификация взаимоотно-
шений растений, предложенная В. Н. Сукачевым (1956). Он раз-
личал три основные формы влияния растений друг на друга при
их совместном произрастании: прямые и косвенные — трансаби-
отические и трансбиотические (см. разд. 11.1). В прямых (контак-
тных) отношениях растения действуют друг на друга непосред-
ственно, а в косвенных — через посредство другого агента. При
прямых отношениях растения могут механически давить, разру-
шать друг друга, поселяться друг на друге, питаться друг другом.
При косвенных посредником могут быть неживая среда (транс-
абиотические отношения) и другие организмы (трансбиотичес-
кие отношения). Среди трансабиотических отношений можно вы-
делить конкуренцию за ресурсы, создание фитосреды и влияние
через выделения (прижизненные и от разлагающихся остатков).
Все формы отношений растений проявляются одновременно, дей-
ствуют друг на друга, и поэтому в природе их влияние обычно
трудно разграничить.
Формы прямых (контактных) взаимоотношений растений.
В. Н. Сукачев (1956) различал две формы контактных взаимоотно-
шений растений: со срастанием и без него. Обе довольно широко
распространены в отношениях паразит-хозяин, лиана-опора, эпи-
фит-форофит. Особый вариант контактных взаимоотношений —
срастание корней, распространенное у древесных растений. Наи-
более часто срастаются корни одной особи, реже — особей одного
вида и еще реже — разных видов. С возрастом древостоя сраста-
ние корней существенно увеличивается. Экспериментально дока-
зана возможность переноса веществ (включая органику) через
сросшиеся корни.
При совместном произрастании нередко одни растения созда-
ют механические препятствия для нормальной жизнедеятельнос-
ти других. Например, перехват пыльцы снижает вероятность опы-
ления цветков. Плохо влияет на семенное размножение задержи-
вание плодов и семян мхами, лишайниками и надземными орга-
нами других растений. Сплошной мохово-лишайниковый покров
снижает также всхожесть семян и иногда приводит к гибели вехо-
381
дов, что связано с истощением запасов семени до достижения
корешком проростка почвы, с колебаниями влажности в слое мхов
и лишайников, с «выдергиванием» всходов при набухании увлаж-
ненных лишайников.
Плоды и пыльца, оседая на поверхности листьев, могут сокра-
щать их фотосинтезирующую поверхность. Такое воздействие плос-
ких плодов вяза (Ulmus), обильной пыльцы сосны (Pinus) и ели
(Picea) недолговременно. Однако растения могут надолго созда-
вать препятствия для перемещения подземных органов других
растений, которые вынуждены обходить препятствие или прони-
зывать его. Так, Т. А. Работнов (1993) отмечает прободение корне-
вищами пырея ползучего (Elytrigia repens) и свинороя (Cynodon),
корневищ щавеля конского (Rumex confertus), корней клевера лу-
гового (Trifolium repens) и клубней картофеля (Solanum tuberosum).
В этих взаимоотношениях один вид наносит другому вред, одна-
ко тратит на это много энергии.
Угнетение одних растений другими происходит и при пере-
движении особей в результате разрастания. Например, у белоуса
(Nardus stricta) молодые растения испытывают давление разрас-
тающихся мощных особей. И хотя всходы обычно гибнут от зате-
нения и перехвата взрослыми особями воды и минеральных эле-
ментов раньше их физического уничтожения передвигающимися
растениями, судьба молодых экземпляров может зависеть и от того,
где по отношению к более мощным соседям они возникли. Быс-
трорастущие водоросли часто механически вытесняют медленно-
растущие. А в некоторых случаях, например при гнездовом посе-
ве моркови (Daucus carota), может происходить «выпирание од-
них особей другими».
Особую форму взаимовлияния растений представляют меха-
нические воздействия, осуществляемые при действии других аген-
тов (ветер, текучая вода). Например, в ветреную погоду охлесты-
вающая крона одного дерева может повреждать листву соседнего
(береза — Betula и ель — Picea). К контактным взаимоотношени-
ям можно отнести также использование одними растениями дру-
гих как субстрата для поселения (укоренение в моховом покрове,
кочках и подушках растений).
Экология лиан и эпифитов, их взаимодействие с растениями-
опорами. Лианы и эпифиты реализуют особый вариант контакт-
ных отношений — топические, при которых одни организмы по-
селяются на других и при этом не используют растение-опору как
источник пищи. Но влияние лиан может быть весьма существен-
ным, почему их еще называют «структурными паразитами» или
«нетрофическими полупаразитами» (Т. А. Работнов, 1998).
К лианам относят растения со стеблем, нуждающимся в опоре
для роста. Они укореняются в почве, а стебли других видов ис-
пользуют как подпорки для поднятия вверх. Среди лиан есть дре-
382
весные и травянистые виды, представители разных групп высших
растений, в том числе папоротники и голосеменные (Gnetum).
Среди них встречаются бактерио- и микосимбиотрофы, насеко-
моядные растения (Nepenthes). Лианы возникли из обычных ви-
дов в результате приспособления к жизни в сомкнутых сообще-
ствах. В онтогенезе сначала они образуют прямостоячие всходы,
но, найдя опору, начинают расти очень быстро, цепляясь или об-
вивая ее. Конкурентное преимущество лианы получают вынося
ассимилирующие и генеративные органы к свету при сэкономлен-
ных затратах материала и энергии. Лиановидная жизненная форма
очень характерна для тропических лесов (около 2 000 видов).
Взаимодействия лиан с растениями-опорами часто рассматри-
ваются как комменсализм. Считается, что эти взаимоотношения
полезны для лиан и не приносят вреда их партнерам. Однако сре-
ди них есть и практически не воздействующие на опору, и ги-
бельные для хозяев (лианы-душители). Лианы влияют на исполь-
зуемые растения и непосредственно, и косвенно (изменяя среду).
Непосредственное влияние проявляется, например, в создании
нагрузки на стволы и ветви, что может приводить к их обламыва-
нию. Лианы часто оказывают формообразующее влияние на дере-
вья-опоры, которые приобретают однобокую форму. Если лиан
много, они изменяют микроклимат фитоценоза (световой режим,
влажность воздуха, опад). Поглощая СО2 и создавая тень, они могут
существенно влиять на питание растений. При этом особенно стра-
дают подрост и низкорослые особи. Лианы конкурируют со свои-
ми опорами и окружающими растениями и за минеральные ре-
сурсы почвы. В лесном хозяйстве тропиков они рассматриваются
как сорняки, засоряющие плантации древесных культур, и удале-
ние их (например, Bursera simurula в Коста-Рике) увеличивает
плодоношение деревьев (Т. А. Работнов, 1993).
Лианы прошли долгий путь взаимодействия с опорами, и в результа-
те сопряженной эволюции деревья приспособились к поселяющимся на
них местным лианам. Но при интродуцировании лиан из других регио-
нов они могут нанести большой ущерб. Так, распространенная в есте-
ственных лесах Флориды декоративная жимолость японская (Lonicera
japonica) стала наносить существенный вред местным деревьям и кус-
тарникам, чего не могла делать у себя на родине. А в Европейской Рос-
сии занесенная из Северной Америки Echinocystis lobata стала быстро
распространяться по приречным ивнякам и угнетать ивы (Salix).
К эпифитам относят поселяющиеся на растениях автотрофные
организмы, не поглощающие воду и элементы минерального пи-
тания из живых органов растений-опор. Форофитами (от греч.
phoros — несущий), на которых поселяются эпифиты, называют
виды с одревесневшими органами (от деревьев до полукустарни-
ков и кустарничков). Эпифиты происходят от наземных расте-
383
ний, приспособившихся избегать конкуренции за свет, поселяясь
достаточно высоко и не тратя при этом материалы на построение
массивных опорных органов. При этом сопряженная эволюция
минимизировала ущерб для форофитов. Различают эпифиты ко-
ровые и гумусные. Коровые прикрепляются к коре форофита, а
гумусные укореняются в скоплениях органического вещества на
нем. Иногда эта аэротопная почва используется и придаточными
корнями форофитов и тогда эпифиты конкурируют с ними за нее.
Обеспеченность водой и элементами минерального питания ко-
ровых и в значительной мере гумусных эпифитов зависит от со-
держания их в воздухе.
Деревья представляют собой гетерогенное местообитание для эпи-
фитов, так как микроклиматические условия изменяются от основания
стволов к верхним частям крон, а в пределах крон — от периферии к
центру. Микроклиматические условия существования эпифитов зависят
также от экспозиции и наклона стволов и ветвей. Большое значение
имеют поверхность коры, ее механические свойства и химический со-
став. Деревья с бороздчатой корой заселены обильнее, так как на них
лучше удерживаются пыль и зачатки эпифитов и тем самым создаются
условия для поселения большого числа видов. Деревья же с отслаиваю-
щейся чешуйчатой корой, как у сосны (Pinus sylvestris), эпифитами бед-
ны (J.J. Barkman, 1969). Важен и химический состав коры, особенно со-
держание в ней азота. Ряд эпифитов требовательны к кислотности коры,
но иногда и сами подкисляют среду.
У многих эпифитов возникли приспособления к эффективно-
му использованию воды атмосферных осадков. Они проявляются
в ксероморфной (иногда суккулентной) структуре, экономном
использовании воды, способности впитывать водяной пар и осад-
ки надземными органами (например, веламеном — пористой тка-
нью, покрывающей воздушные корни, рис. 11.4) и запасать воду в
специальных резервуарах. Эпифиты, особенно коровые, плохо
обеспечены элементами минерального питания, так как получают
их в основном с атмосферными осадками. Например, Т.А. Работ-
нов (1993) указывает, что зольный состав тилляндсии (Tillandsia
usneoides), обитающей на растениях и на телеграфных проводах,
почти не отличается. Характерная для многих эпифитов вечнозе-
леность позволяет длительно удерживать поглощенные элементы
минерального питания, а обычная редукция вегетативных орга-
нов способствует использованию значительной их части на гене-
ративное размножение.
Сосудистые эпифиты (около 30 тыс. видов из 68 семейств) — в
основном многолетние травы (кустарники представлены вереско-
выми и меластомовыми), распространенные почти исключитель-
но в условиях теплого и влажного климата. Богатство эпифитов в
384
Рис. 11.4. Воздушные корни у эпифитных орхидей
(по А. Е. Васильеву и др., 1988):
А — Oncidium на ветке дерева; Б — часть поперечного разреза воздушного корня
Dendrobiunr, 1 — веламен
тропических лесах обусловлено большим объемом среды обита-
ния (деревья достигают 50 м и более), ее гетерогенностью (боль-
шим видовым разнообразием форофитов, разнообразием крон
деревьев и местообитаний внутри их). В пределах одного дерева
иногда живут более 50 видов эпифитов. Изредка сосудистые эпи-
фиты встречаются в лесах умеренного климата. Так, В. В. Мазинг
(1966) отмечает, что в Эстонии как эпифиты могут расти недотро-
га мелкоцветковая (Impatiensparviflora), мокрица (Stellaria media),
бузина (Sambucus), рябина (Sorbus), клен (Acer) и береза (Betula).
В тропических лесах почти все лишайники, мхи и водоросли —
эпифиты. Эпифитизм — очень частое явление среди водорослей
(Л.Л. Великанов и др., 1981). Все их отделы имеют представите-
лей, ведущих эпифитный образ жизни. При этом продолжитель-
ность эпифитирования у разных видов неодинакова: у одних она
занимает почти весь жизненный цикл, а у других — только часть
его. Эпифитные лишайники и мхи широко распространены и в
лесах умеренного климата. Мохообразные обычно занимают бо-
лее влажные места в нижних частях стволов, лишайники подни-
маются до крон, а водоросли покрывают места, не занятые ли-
шайниками и мхами.
Воду и минералы они поглощают всем телом почти исключительно
из атмосферы. Т.А. Работнов (1993) отмечает, что способность бессосу-
дистых эпифитов поглощать воду очень высока (у мхов она достигает
1700% от сухой массы). Они характеризуются высоким осмотическим
385
давлением: у лишайников оно выше 1 000 бар, а у эпифитных мхов оно
обычно выше (20 — 90 бар), чем у наземных (6—12 бар). Это обеспечива-
ет им возможность поглощать водяной пар даже при невысокой влажно-
сти воздуха.
Эпифиты связаны с форофитами в основном топически, ис-
пользуя их как субстрат для поселения. Трофические связи их
проявляются в основном в форме потребления эпифитами ресур-
сов, содержащихся в отмерших тканях форофита (чаще — в коре).
Но отношения между эпибионтом и хозяином не всегда безраз-
личны для организма-опоры. Хотя эти взаимоотношения часто
относят к комменсализму, они могут быть благоприятны для хо-
зяина (в продуктах жизнедеятельности гумусных эпифитов могут
располагаться придаточные корни форофита), но в некоторых
случаях эпифиты явно отрицательно воздействуют на опору. При
массовом развитии они могут создавать значительную нагрузку на
ветви деревьев, что иногда приводит к их поломке. Так, на Тихо-
океанском побережье Америки в лесах из Pseudotsuga масса эпи-
фитных лишайников достигает 3 кг на стволах и 15 кг на ветвях.
Поэтому опыты по удалению эпифитов (биомасса сосудистых эпи-
фитов может достигать 10 т/га) приводили к улучшению состоя-
ния форофитов (Т.А.Работнов, 1998). Образуя достаточно сомк-
нутый покров, эпифиты снижают освещенность нижних ярусов,
изменяют температуру и влажность коры, ее химизм и физичес-
кие свойства. Поглощая стекающую по ветвям и стволам воду,
они уменьшают поступление ее в почву и препятствуют исполь-
зованию деревьями вымытых из крон веществ. Есть также наблю-
дения о выделении эпифитными лишайниками токсинов, прони-
кающих в ткани форофита. Эпифиты могут ускорять разрушение
коры дерева, создавая условия для поселения паразитов и фито-
фагов. Так, в опытах на о. Ява было показано, что микоризообра-
зующий симбионт ряда эпифитных орхидных и папоротников
паразитирует на деревьях-опорах (Т.А.Работнов, 1993). А у неко-
торых эпифилльных лишайников грибной компонент может па-
разитировать на листьях.
Растения-паразиты и полупаразиты. Паразитизм и полупарази-
тизм — варианты прямых физиологических отношений между
организмами разных трофических уровней. Автотрофный хозяин
при этом во всех случаях угнетается. Среди цветковых растений
имеется более 570 видов полных паразитов (голопаразитов), от-
носящихся к 50 родам и 8 семействам. Больше всего их встречает-
ся среди повиликовых, заразиховых, баланофоровых, раффлезие-
вых. Паразиты почти не имеют хлорофилла и поэтому являются
гетеротрофами, полностью питающимися за счет хозяев. Обычно
у них ярко выражена специфичность в выборе партнера, причем
чем сильнее зависимость от хозяев, тем ограниченнее их выбор.
386
Паразитический образ жизни накладывает отпечаток на анато-
мию и морфологию растений. Паразиты утрачивают зеленую ок-
раску, у них редуцируются стебли и листья. Корни их развиты
слабо или отсутствуют, вместо них формируются гаустории (при-
соски, внедряющиеся в ткани растения-хозяина и соединяющие-
ся с его проводящей системой). В тело жертвы гаустории выделя-
ют ферменты, разрушающие клеточные стенки. С помощью ра-
диоактивных меток было показано, что паразиты поглощают из
тела хозяина в основном сахарозу, глутаминовую и аспарагино-
вую кислоты и их амиды (В. В. Полевой, 1989).
Ярко выраженный паразит — раффлезия Арнольда (Rafflesia arnoldii),
питающаяся соками корней тропических лиан. Вегетативное тело ее,
напоминающее мицелий с гаусториями, все погружено в подземные орга-
ны хозяина. Снаружи располагается только цветок, самый крупный в
мире (диаметром около метра). Другой пример — повилика (Cuscuta)
(рис. 11.5). Подземных корней она не образует, а ее побеги обвивают
стебли жертв и присасываются к ним с помощью дисков, плотно приле-
гающих к коре хозяина. Группа клеток из центральной части диска вне-
дряется в проводящие ткани хозяина, откуда получает воду, минераль-
ные и органические вещества. Проростки повилики находят жертву, со-
вершая ростовые вращательные движения. При этом они хемотропичес-
ки реагируют на выделяемые хозяином вещества и на градиент влажно-
сти. Паразитируют повилики на побегах многих диких и культурных
растений (лен — Linum, клевер — Trifolium, люцерна — Medicago, ива —
Salix, крапива — Urtica и др.) (А. Е. Васильев и др., 1988). А у заразихи
(Orobanche) семена прорастают только под влиянием корневых выделе-
ний жертвы. Соприкоснувшись с корнем хозяина, кончик корня пророс-
тка паразита преобразуется в присоску, начинает выделять гидролазы, ра-
створяющие клеточные стенки, и активно внедряется в жертву. В период
роста заразиха поглощает из корней хозяина воду и большое количество
Рис. 11.5. Повилика (Cuscuta) (по А. Е. Васильеву и др., 1988):
А — общий вид растения на клевере; Б — стадии развития проростков; В — часть
стебля с присосками; Г — схема поперечного среза стебля растения-хозяина
с внедрившимися в его проводящие пучки отростками присосок повилики
387
углеводов, азотистых веществ и минеральных элементов (особенно фос-
фор). Так, в пораженных ею томатах (Lycopersicon) содержание белкового
азота снижается в 3 раза, а сахаров — в 16 раз (В. В. Полевой, 1989).
В природных фитоценозах массовой гибели хозяев от расте-
ний-паразитов обычно не наблюдается, так как эти виды прошли
длительный путь коэволюции. Но если сопряженной эволюции
паразита и хозяина не было, то результатом их совместного выра-
щивания может стать быстрая гибель растений-хозяев. Так про-
изошло в России в начальный период возделывания завезенного
из Америки подсолнечника (Helianthus annuus). Он был массово
поражен заразихой, паразитировавшей ранее на полынях
(Artemisia) и не приносившей им особого вреда. На родине у под-
солнечника такого врага не было и защитных свойств против та-
кого воздействия не выработалось (Т. А. Работнов, 1993).
Кроме настоящих паразитов, полностью утративших способ-
ность к фотосинтезу, имеются еще растения-полупаразиты (ав-
тотрофные паразиты), сохраняющие нормальные зеленые листья,
но заменяющие почвенное питание паразитическим. Они образу-
ют присоски, внедряющиеся в ткани корней или ветвей хозяина.
Имея хлорофилл, полупаразиты осуществляют нормальный фо-
тосинтез, а у хозяев берут в основном воду и элементы минераль-
ного питания. Потому в отличие от паразитов они неспецифичны
и могут иметь десятки видов хозяев. Среди цветковых растений
полупаразитов насчитывается более 2,5 тыс. видов из 11 семейств.
Особенно много их среди ремнецветных (почти 1 000 видов), но-
ричниковых (около 500 видов), санталовых (около 430 видов).
В экосистемах умеренной полосы полупаразиты распростране-
ны больше, чем паразиты. Среди луговых, лесных, а также болот-
ных и полевых трав полупаразиты часто представлены однолет-
ними норичниковыми (погремки — Rhinanthus, очанки —
Euphrasia, марьянники — Melampyrum) и многолетними мытни-
ками (Pedicularis). К полупаразитам относится известное расте-
ние иван-да-марья (Melampyrum nemorosum) и широко распрос-
траненный на лугах, верховых болотах и в лесах средней полосы
марьянник луговой (Melampyrum pratense). К ним принадлежит
и успешно растущий в Западной Европе кустарничек омела
(Viscum), поселяющаяся в кронах деревьев. Полупаразиты дере-
вьев проникают в заболонь хозяина с помощью гаустории, ветвя-
щейся уже в коре, или с помощью корней, оплетающих ветви
хозяина и внедряющих в них множество гаусторий. Неоднократно
было показано, что развитие полупаразитов существенно замедляет
рост растения-хозяина. Так, омела приводит к потере !/5— '/3 объе-
ма деловой древесины (М. Бигон и др., 1989).
Паразитические свойства у разных видов полупаразитов выра-
жены в разной степени. Факультативные полупаразиты имеют в
388
почве слабую корневую систему, которая присасывается к корням
растений-хозяев, и в этой группе есть виды с выраженной спо-
собностью к самостоятельному существованию. Типичные пред-
ставители — норичниковые (марьянники, мытники), паразитиру-
ющие на большом числе видов трав и некоторых древесных ви-
дах. Облигатные же полупаразиты не имеют своей корневой сис-
темы. Они присасываются к ветвям древесных растений (являют-
ся стеблевыми полупаразитами). Типичные представители этой
группы — ремнецветные, множество видов которых развивается в
дождевых тропических лесах.
Воздействуя в разной степени на разные виды, полупаразиты
изменяют их конкурентоспособность, влияя на взаимоотношения
между растениями в фитоценозах. Ослабленные организмы пора-
жаются в первую очередь. В особенно больших количествах одно-
летние полупаразиты разрастаются там, где травостои низкорос-
лы и растения-хозяева в силу разных причин угнетены. В благо-
приятных же условиях, когда образуются высокие сомкнутые тра-
востои, полупаразитов мало. Это в значительной степени опреде-
ляется их большой требовательностью к освещению, а также ус-
тойчивостью неугнетенных растений к присасыванию пророст-
ков полупаразитов. На постоянных пастбищах однолетние полу-
паразиты отсутствуют, так как не выносят длительного вытапты-
вания. Численность полупаразитов, размножающихся семенами,
зависит от возможности обсеменения. Поэтому эффективный
прием борьбы с ними — скашивание травы в срок, исключающий
обсеменение (Т. А. Работнов, 1984).
В луговых сообществах полупаразиты становятся фактором поддер-
жания высокого видового разнообразия. Это связано с тем, что они по-
давляют доминантов, высвобождая часть ресурсов для других видов. Од-
нако продуктивность сообществ при этом падает, так как при переходе с
одного трофического уровня на другой энергия теряется. По этой при-
чине массовое развитие на лугах погремка (до 100 и более особей на 1м2)
может снизить урожай сена на 20 — 25 %. Т. А. Работнов (1998) указывает,
что в опыте общий урожай трав (без учета погремка) снизился в 2 раза
(все злаки и многие виды разнотравья отрицательно прореагировали на
его внедрение, в 4 раза ухудшился рост бобовых). А урожай некоторых
видов (колокольчик скученный — Campanula glomerata, бодяк полевой —
Cirsium arvense, звездчатка злаковая — Stellaria graminea, тысячелист-
ник обыкновенный — Achillea millefolium) в присутствии полупаразита
не сократился. Они или не повреждались им, или повреждались мень-
ше других видов, что способствовало повышению их конкурентоспо-
собности.
Конкурентные взаимоотношения растений. Конкуренция возни-
кает, если какого-то ресурса не хватает для удовлетворения в нем
потребности всех совместно обитающих растений (Т. А. Работнов,
389
1993, 1998 и др.). Большинство фитоценологов считает ее основ-
ным механизмом взаимоотношений между растениями. Конку-
рентные отношения между видами зависят от условий произрас-
тания, количественных соотношений видов и воздействия других
организмов.
Большой материал о конкуренции получен в работах лесово-
дов и на сельскохозяйственных опытных станциях. Примерами
внутривидовой конкуренции могут быть изреживание подроста в
лесу и угнетение злаков в загущенном посеве. При этом снижает-
ся семенная продуктивность и сильнее выражена периодичность
плодоношения деревьев (Т.А.Работнов, 1993).
Внутривидовая конкуренция по-разному влияет на однолет-
ники и многолетники. Об этом свидетельствует «правило Сука-
чева»: в загущенном посеве развитие и переход в генеративное
состояние у однолетников ускоряется, а у многолетников замед-
ляется.
В загущенном посеве у однолетних растений обычно все выжившие
особи, несмотря на сильное угнетение, цветут и плодоносят. Так, у мари
белой {Chenopodium album) плодоношение наблюдалось даже на стадии
семядоли. Ускорение развития однолетников при усилении конкурен-
ции объясняется тем, что в эволюции они приобрели способность с ухуд-
шением условий произрастания давать потомство за более короткий срок.
Многолетники же в загущенном посеве замедляют развитие, чтобы на-
копить ресурсы для более мощных генеративных органов. Наблюдения
над многими монокарпическими двулетниками привели к выводу, что
за 2 года эти виды завершают жизненный цикл в питомниках только в
разреженных посевах. А в природе, где конкуренция между растениями
достаточно интенсивна, они живут значительно дольше (нередко до 10 и
более лет) (Т. А. Работнов, 1960 и др.).
Большое значение в определении конкурентных отношений
между особями вида имеет разнокачественность семян. Разные
размеры семян определяют неодинаковую обеспеченность всхо-
дов питанием. В экспериментах с клевером подземным (Trifolium
subterraneum) было показано, что выросшие из крупных семян
растения имели более длинные черешки и выносили листья выше,
чем растения, возникшие из мелких семян. Это позволило круп-
носемянным особям побеждать в конкуренции за свет.
У клональных растений конкуренция может возникать между
побегами, причем она тем сильнее, чем слабее конкуренция меж-
ду клонами (Б. М. Миркин, Л. Г. Наумова, 1998). Конкуренция при-
водит и к тому, что возрастная структура популяций может ме-
няться при изменении условий среды. Например, при внесении
азотно-фосфорных удобрений общее число побегов осоки песча-
ной (Carex arenaria) не изменилось, но соотношение побегов раз-
ного возраста поменялось резко. Так, на удобренных участках стали
390
преобладать молодые побеги, а более старые, многочисленные на
контрольных площадках, стали редки, так как не выдержали кон-
куренции (М. Бигон и др., 1989).
Виды в фитоценозе обычно представлены несколькими биотипами,
между которыми тоже существует конкуренция. В. Н. Сукачев (1953) в
экспериментах с клонированными растениями установил, что реакция
различных биотипов на степень густоты травостоев и древостоев неоди-
накова. Биотипы, более сильные в редких культурах, могут оказаться
более слабыми в густых. Победу биотипа часто определяют незначитель-
ные эколого-физиологические отличия. В опытах со многими культур-
ными растениями было показано, что в изменении со временем участия
отдельных сортов в составе сортосмеси большое значение имеет их нео-
динаковая устойчивость к вредителям и болезням, что определяет их
разную конкурентоспособность. Так же урожай сорта в монокультуре и в
смеси зависит от условий произрастания. Например, у клевера белого
(Trifolium repens) одни эдафические экотипы приурочены к кислым по-
чвам, а другие — к карбонатным. При выращивании в совместном посе-
ве на кислой почве более конкурентоспособным оказывался ацидофиль-
ный экотип, а на карбонатной — кальцефильный. Отбор признаков, свой-
ственных растениям разных клонов, происходил также под действием
поедающих их слизней и овец.
На остроту конкуренции между растениями влияет обеспе-
ченность местообитания условиями для жизни растений. Еще в
1920-е годы В. Н. Сукачев (1928) обнаружил в опытах, что при улуч-
шении обеспечения элементами минерального питания отмира-
ние особей в загущенном посеве возрастает. Это обусловлено уве-
личением мощности растений на удобренной почве и возрастани-
ем конкуренции за свет и воду. Результат этого опыта согласуется
с установленной лесоводами закономерностью: снижением плот-
ности взрослых деревьев по мере улучшения условий произраста-
ния. Д. Харпер (1977) предложил назвать это явление «эффектом
Сукачева». В определении конкурентоспособности вида очень важ-
на обеспеченность светом. Так, в опыте с лесными травами в сме-
шанном посеве на свету была более конкурентоспособна звезд-
чатка жестколистная (Stellaria holostea), а в тени — зеленчук
(Galeobdolon luteum) (в монокультуре оба вида росли лучше при
полном освещении). Конкурентоспособность видов может силь-
но изменяться в зависимости от уровня обеспечения водой. Так,
при двухлетнем совместном выращивании ежи сборной (Dactylis
glomerata) и щучки (Deschampsia cespitosa) на сухой почве уро-
жай ежи превышал урожай щучки почти в 30 раз, а на сырой
почве — только в 3 раза. О значении засоления Т.А. Работнов
(1993) предлагает судить по результатам вегетационного опыта с
овсяницей красной (Festuca rubra) и армерией морской (Armerid)
в монокультуре и смеси: в отсутствие засоления более конкурен-
391
I и
[II
IV
Увеличение богатства ресурсов почвы
Увеличение освещенности
Рис. 11.6. Основной комплексный градиент конкуренции
(по Д.Тильману из Б. М. Миркина, Л. Г. Наумовой, 1998):
I — конкуренция отсутствует; II — парциальная конкуренция за ресурсы почвы;
III — интегральная конкуренция за свет и ресурсы почвы; IV — парциальная
конкуренция за свет
тоспособна овсяница, а при засолении — армерия. Влияет на кон-
курентоспособность и кислотность почвы. Так, на сильнокислой
почве в смешанном посеве овес {Avena sativa) конкурентоспособ-
нее, чем ячмень {Hordeum), а на нейтральной почве — наоборот.
В некоторых случаях фактор кислотности оказывает косвенное
влияние на изменение конкуренции. Так, при совместном выра-
щивании клевера белого и лядвенца {Lotus) низкая конкуренто-
способность клевера на кислой почве обусловлена неспособнос-
тью его поглощать фосфор из-за высокого содержания в ней ра-
створимого алюминия, к которому лядвенец устойчив.
В связи с вопросом о зависимости остроты конкуренции от
обеспеченности ресурсами Д.Тильман (1988) сформулировал пред-
ставление о главном комплексном градиенте конкуренции, кото-
рый объединяет обеспечение растений светом и почвенным пита-
нием. На богатых почвах формируются сомкнутые фитоценозы с
высокой плотностью популяций. Растения там конкурируют в ос-
новном за свет, причем чем почвы богаче, тем конкуренция ост-
рее. А на бедных почвах (сухих, засоленных) фитоценозы разомк-
нуты и растения конкурируют за почвенные ресурсы. В централь-
ной части градиента конкуренция идет за оба фактора (рис. 11.6).
Влияние трансформации растениями экотона на другие расте-
ния. В результате экотоп жизнедеятельности организмов преобра-
зуется в биотоп. Средообразующее действие растений проявляет-
ся в выраженности действия основных экологических факторов
(свет, тепло, обеспечение водой и минеральным питанием, состав
воздуха). Биотическая трансформация экотопа может быть про-
слежена в макромасштабе (для совокупности многих фитоцено-
зов) и микромасштабе (на уровне отдельных растений). Фито-
392
генное поле (зона влияния) — сфера влияния растения на среду.
Особь может воздействовать на окружение через затенение, иссу-
шение, изменение концентрации минеральных веществ, толщину
подстилки, pH почвы и др. Размер фитогенного поля зависит от
жизненной формы растения (от нескольких сантиметров у трав
до нескольких метров у деревьев), возрастного состояния и жиз-
ненности вида в разных частях ареала (высокой в области эколо-
го-фитоценотического оптимума и низкой в экстремальных усло-
виях у границ распределения вида по градиентам среды).
Глубина преобразования экотопа может существенно отличать-
ся. Биотическое преобразование экотопа по типу «момент» по-
верхностно, и его результаты исчезают вместе с гибелью средооб-
разователей (В.О.Таргульян, И.А. Соколов, 1976). Этот вариант
трансформации экотопа обычно выражается в изменении напря-
женности режимов прямодействующих факторов (например, ос-
вещенности около дерева). А преобразования типа «память» на-
капливаются и сохраняются после гибели средообразователей. Они
проявляются в нарастании почвы на оголенном субстрате, вы-
щелачивании ее, отложении торфа, сапропеля и наилка. Такое ку-
мулятивное средообразование может зависеть от использования фи-
тоценоза человеком. Так, при скашивании травы и сборе урожая из
почвы регулярно изымаются минеральные элементы, способствуя
ее обеднению, а при внесении удобрений почва обогащается.
В результате влияния на среду разных растений в фитоценозе
создаются разнообразные микроместообитания, отличающиеся по
фитоклимату и эдафическим условиям. На изменение среды рас-
тениями накладывается влияние связанных с ними гетеротрофов.
Кроме того, фитогенные поля отдельных растений перекрывают-
ся. Поэтому фитоценоз в целом образует единую сферу средооб-
разующего действия.
В некоторых случаях биотическое преобразование экотопа мо-
жет затрагивать большие территории. Например, при теплой по-
годе и длительном застое воды в поймах можно наблюдать массо-
вое размножение нитчатых водорослей, которые оседают на по-
верхность почвы и при высыхании образуют корку толщиной до
1 см, трудно пробиваемую побегами трав. Т.А. Работнов (1984)
указывает, что в нижнем течении Иртыша и Оби в годы длитель-
ного затопления разрастание зеленой водоросли Cladophora fracta
на больших территориях ведет к снижению урожая кормовых трав
и ухудшению качества кормов.
Аллелопатия. Аллелопатия означает взаимовлияние растений
через изменение среды в результате выделения в нее продуктов
жизнедеятельности, не имеющих значения как источник энергии
и элементов питания. В широком смысле к аллелопатическим
веществам относят также соединения, образующиеся при разло-
жении опада. Роль их в природе может быть больше, чем прижиз-
393
ненных выделений, особенно в лесах, где продукты разложения
подстилки регулируют развитие проростков деревьев (Б. М. Мир-
кин, Л. Г. Наумова, 1998).
Количество и состав аллелопатических веществ зависят от вида
растения, органа, фенологического состояния и условий произра-
стания (Г.Грюммер, 1957). Среди них наиболее значимы следую-
щие: 1) неспецифические вещества, выделяемые в процессе транс-
пирации, дыхания и фотосинтеза (сахара, аминокислоты, органи-
ческие кислоты и др.); 2) специфические для отдельных видов
балластные соединения, от которых растение освобождается;
3) аттрактанты и репелленты, выделяемые для привлечения или
отпугивания животных; 4) вещества, способствующие выживанию
при высоких температурах, дефиците влаги и т.п.; 5) соединения,
образующиеся при поражении фитофагами и паразитами; 6) по-
глощаемые растением вещества, которые в данный момент не могут
полностью использоваться.
Большую роль аллелопатия может играть в отношениях расте-
ний-паразитов с хозяевами. Так, семена многих паразитных рас-
тений всходят лишь под влиянием на них корневых выделений
хозяев. Проростки, израсходовав питательные вещества семени,
присасываются к корню хозяина. В ряде случаев удалось раскрыть
формулу этих веществ и синтезировать их аналоги. В результате
спровоцированные этими веществами проростки заразихи
(Orobanche), не найдя рядом питающего корня хозяина, погиба-
ли (Б. М. Миркин, 1986).
Аллелопатические взаимодействия — часть эволюционно сло-
жившейся системы отношений в фитоценозах. Р.Уиттекер счита-
ет, что аллелопатия — побочный продукт биохимических взаимо-
действий с фитофагами. Сначала токсины могли служить для за-
щиты от насекомых (часто они продолжают выполнять эту функ-
цию), влияние же на другие растения — закрепленный отбором
побочный эффект. Растения, долго обитавшие совместно, лучше
адаптированы к выделениям друг друга, поэтому аллелопатиче-
ское отталкивание в наибольшей мере проявляется при интро-
дукции. Например, на Кавказе и в Северной Америке под эвка-
липтами (Eucalyptus) местные травы практически не растут, а на
их родине в Австралии травяной покров в эвкалиптовых лесах
развит хорошо. В естественных сообществах в процессе коэволю-
ции подбирались растения, устойчивые к токсическим метаболи-
там других компонентов биогеоценоза, создавались механизмы де-
токсикации. В результате сопряженной эволюции сформирова-
лись биоценозы, в которых обычно аллелопатические воздействия
или отсутствуют, или слабые (Б. М. Миркин и др., 2001).
ЗАКЛЮЧЕНИЕ
К настоящему времени накопился большой объем знаний,
объясняющих механизмы взаимосвязей отдельных элементов в
природных системах. Многочисленны и разнообразны факты вред-
ных последствий нарушений этих взаимосвязей в результате дея-
тельности людей. Исчезновение отдельных звеньев в природных
цепях вызывает глубокие нарушения в экосистемах. Погодные
аномалии, обмеление рек, вспышки болезней растений и живот-
ных — это не только общеэкологические проблемы, они тесно
связаны и с экологией растений. Проблема сохранения биоразно-
образия — в первую очередь проблема сохранения устойчивости
естественных биогеоценозов (экосистем), важнейшей частью ко-
торых (автотрофным блоком) являются растения.
Лавинообразно растет поток информации об экологических
нарушениях и сбоях, множатся факты природных катастроф, усу-
губляются их последствия. Увеличивается количество видов, став-
ших редкими, и наряду с «красными книгами» катастрофически
пополняются «черные книги» — списки исчезнувших уже в наше
время видов растений, грибов, животных. Сохранение отдельных
видов невозможно без сохранения среды их обитания. Современ-
ное состояние многих природных сообществ уже таково, что, по-
теряв часть компонентов, самостоятельно, без помощи человека,
они уже не могут восстановиться. Экологический кризис — ре-
альное состояние современной природы.
Необходимо расширять созидательную практику восстановле-
ния природных объектов, разрабатывать экологически оправдан-
ные схемы природопользования. Экологизация мышления и дея-
тельности людей необходима и во всех областях науки, и в поли-
тике, и в хозяйственной деятельности, и в культуре повседневной
жизни. Экологическое образование и воспитание, формирование
эксцентрического сознания — важнейшая современная социальная
задача. И во многом от ее ответственного решения в XXI в. будет
зависеть благополучие Земли.
СПИСОК ЛИТЕРАТУРЫ
Основная литература
Бигон М. Экология. Особи, популяции и сообщества: В 2 т. / М. Би-
гон, Дж.Харпер, К.Таундсен. — М.: Мир, 1989. — Т. 1. — 667 с.; Т. 2. —
477 с.
Ботаника: Морфология и анатомия растений / [А.Е. Васильев и др.]. —
М.: Просвещение, 1988. — 480 с.
Горышина Г. К. Экология растений. — М.: Высшая школа, 1979. — 368 с.
Ботаника: В 4 т.; на основе учебника Э.Страсбургера (и др.). Т. 4.
Экология / [П.Зитте и др.]. — М.: Издательский центр «Академия», 2007. —
256 с.
Косулина Л. Г. Физиология устойчивости растений к неблагоприят-
ным факторам среды / Л.Г.Косулина, Э.К.Луценко, В.А.Аксенова. —
Ростов-на-Дону: Изд-во Ростовского ун-та, 1993. — 240 с.
Кузнецов Вл. В. Физиология растений / Вл. В. Кузнецов,
Г.А. Дмитриева. — М.: Высшая школа, 2005. — 736 с.
Кулыпиасов И. М. Экология растений. — Изд-во МГУ, 1982. — 379 с.
Лархер В. Экология растений. — М.: Мир, 1978. — 384 с.
Миркин Б. М. Современная наука о растительности / Б. М. Миркин,
Л. Г. Наумова, А. И.Соломещ. — М.: Логос, 2001. — 264 с.
Одум Ю. Экология: В 2 т. — М.: Мир, 1986. — Т.1 — 328 с.; Т. 2 —
276 с.
Полевой В. В. Физиология растений. — М.: Высшая школа, 1989. —
464 с.
Работное Т.А. Фитоценология. — М.: Изд-во МГУ, 1992. — 352 с.
Дополнительная литература
Физиология растений / [Н.Д.Алехина и др.]. — Издательский центр
«Академия», 2005. — 640 с.
Барыкина Р. П. Большой практикум по ботанике. Экологическая ана-
томия цветковых растений / Р. П. Барыкина, Н. В.Чубатова. — М.: Т-во
науч, изданий КМ К, 2005. — 77 с.
Библь Р. Цитологические основы экологии растений. — М.: Мир,
1965. - 465 с.
Быков Б. А. Экологический словарь. — Алма-Ата: Наука, 1983. —
216 с.
Вальтер Г. Растительность земного шара. В 3 т. — М.: Прогресс. —
Т.1, 1968. - 552 с.; Т.2, 1974. - 424 с.; Т.З, 1975. - 430 с.
Веселова Т.В. Стресс у растений: Биофизический подход / Т. В. Ве-
селова, В.А. Веселовский, Д. С. Чернавский. — М.: Изд-во МГУ, 1993. —
144 с.
Виноградов Б. В. Растительные индикаторы и их использование при
изучении природных ресурсов. — М.: Высшая школа, 1964. — 324 с.
396
Биогеография с основами экологии / [А.Г.Воронов и др.]. — М.:
Высшая школа, 2002. — 392 с.
Гиляров А. М. Популяционная экология. — М.: Изд-во МГУ, 1990. —
191 с.
Гиляров А. М. Становление эволюционного подхода как объяснитель-
ного начала в экологии // Журнал общей биологии, 2003. — Т. 64. —
№ 1. - С. 3-22.
Гэрышина Т. К. Фотосинтетический аппарат растений и условия сре-
ды — Л.: Изд-во ЛГУ, 1989. — 203 с.
Биоморфология растений: иллюстрированный словарь / [П.Ю.Жмы-
лев и др.]. — М., 2002. — 240 с.
Ценопопуляции растений (очерки популяционной биологии) /
[Л. Б.Заугольнова и др.]. — М.: Наука, 1988. — 183 с.
Почвоведение / [И. С. Кауричев и др.]. — М.: Колос, 1982. — 496 с.
Кретович В.Л. Биохимия растений. — М.: Высшая школа, 1986. —
503 с.
Куркин К. А. Системные исследования динамики лугов. — М.: На-
ука, 1976. - 284 с.
Марков М. Вит. Популяционная биология растений. — Казань: Изд-
во Казанского гос. ун-та, 1986. — 108 с.
Медведев С. С. Физиология растений. — СПб.: Изд-во СПбГУ, 2004. —
336 с.
Метлицкий Л. В. Как растения защищаются от болезней / Л. В. Мет-
лицкий, О.Л.Озерецковская. — М.: Наука, 1985. — 189 с.
Миркин Б.М. Наука о растительности (история и современное со-
стояние концепций) / Б. М. Миркин, Л. Г. Наумова. — Уфа: Гилем, 1998. —
413 с.
Миркин Б. М. Словарь понятий и терминов современной фитоценоло-
гии / Б.М.Миркин, Г.С.Розенберг, Л.Г.Наумова. — М.: Наука, 1989. —
223 с.
Пианка Э. Эволюционная экология. — М.: Мир, 1981. — 400 с.
Проворов Н.А. Сравнительная генетика и эволюционная морфоло-
гия симбиозов растений с микробами-азотфиксаторами и эндомикориз-
ными грибами // Журнал общей биологии / Н.А.Проворов, А. Ю. Бори-
сов, И. А. Тихонович. - 2002. - Т. 63. -№ 6. - С. 451-472.
Работное Т.А. Актуальные вопросы экологии растений // Итоги на-
уки и техники. Ботаника, Т. 3. — М.: Изд-во ВИНИТИ, 1979. — 70 с.
Работное Т.А. Экология луговых трав. — М.: Изд-во МГУ, 1985. —
176 с.
Раменский Л. Г. Избранные работы. Проблемы и методы изучения
растительного покрова. — Л.: Наука, 1971. — 334 с.
Серебряков И. Г. Экологическая морфология растений. — М.: Выс-
шая школа, 1962. — 377 с.
Сукачев В.Н. Избранные труды. В 3 т. Т.1. Основы лесной типоло-
гии и биогеоценологии. — Л.: Наука, 1972. — 420 с.
Третьяков Н. Н. Физиология и биохимия сельскохозяйственных ра-
стений / [Н. Н.Третьяков и др.]. — М.: Колос, 2000. — 640 с.
Усманов И.Ю. Экологическая физиология растений / И. Ю. Усма-
нов, З.Ф. Рахманкулова, А. Ю. Кулагин. — М.: Логос, 2001. — 223 с.
397
Чиркова Т.В. Физиологические основы устойчивости растений. —
СПб.: Изд-во СПбГУ, 2002. - 238 с.
Шилов И. А. Экология. — М.: Высшая школа, 2001. — 512 с.
Штина Э.А. Экология почвенных водорослей / Э. А. Штина,
М.М.Голлербах. — М.: Наука, 1976. — 140 с.
Яблоков А. В. Популяционная биология. — М.: Высшая школа, 1987. —
303 с.
Якушкина Н.И. Физиология растений. — М.: Просвещение, 1993. —
335 с.
Biochemistry and Molecular Biology of Plants / Eds. B.B. Buchanan,
W.Gruissem and R.L. Jones. — American Society of Plant Physiologists, 2000. —
1408 p.
Etherington J.R. Environment and plant ecology. 2d ed. — Chichester
etc.: Wiley, 1982. - 487 p.
Grime J.P. Plant strategies and vegetation process. — Chichester etc.:
Wiley, 1979. - 222 p.
Harper J. L. Population biology of plants. — London; New York: Academic,
1977. - 892 c.
Larcher W. Physiological plant ecology. — N. Y.: Springer, 1995. — 506 p.
Leivitt J. Responses of plants to environmental stress. — N.Y.; L.: Acad.
Press, 1972. — 697 p.
Mohr H., Schopfer P. Plant Physiology. — Springer, 1995. — 629 p.
Plant ecology / Ed. By M. J. Crawley. — Oxford, London, Edinburg, Boston,
1986. - 496 p.
ОГЛАВЛЕНИЕ
Предисловие...................................................3
Глава 1. История экологического изучения растений.............5
1.1. История экологии растений до XX в........................5
1.2. Современный этап развития экологии растений.............10
Глава 2. Основные методы экологии растений...................15
2.1. Наблюдение и эксперимент................................15
2.2. Моделирование...........................................17
Глава 3. Экологические факторы...............................19
3.1. Типы экологических факторов. Фитоиндикация..............19
3.2. Закономерности действия экологических факторов..........22
Глава 4. Устойчивость растений и их реакции на действие
неблагоприятных факторов......................................30
4.1. Общие вопросы устойчивости растений.....................30
4.2. Реакции клеток растений на действие неблагоприятных
факторов..................................................40
Глава 5. Экологическая гетерогенность растений...............44
5.1. Экологические группы растений...........................44
5.2. Жизненные формы растений................................46
5.3. Эколого-фитоценотические стратегии растений.............58
5.4. Экологическая гетерогенность популяций растений.........66
5.5. Экотипы растений........................................78
5.6. Экологические ниши растений.............................84
Глава 6. Свет как экологический фактор.......................86
6.1. Количественные характеристики принимаемого растением
света........................................................86
6.2. Качественные характеристики света, принимаемого
растением....................................................97
6.3. Свет и функционирование растений........................108
6.4. Свет и фотосинтез......................................114
6.5. Приспособления растений к световому режиму..............143
6.6. Фотопериод и фотопериодические реакции растений.........157
Глава 7. Тепло как экологический фактор......................163
7.1. Тепловой режим местообитаний............................163
7.2. Температура растений....................................176
399
7.3. Влияние температуры на рост и развитие растений........181
7.4. Действие экстремальных температур на растения..........186
7.5. Термопериодизм и фенологические особенности действия
теплового фактора...........................................223
Глава 8. Вода как экологический фактор......................228
8.1. Вода в растении........................................228
8.2. Водный режим местообитаний.............................239
8.3. Экологические группы растений по отношению
к водному режиму............................................246
8.4. Экологическое действие на растения снега и льда........256
Глава 9. Воздух как экологический фактор....................260
9.1. Экологическое значение кислорода воздуха...............261
9.2. Экологическое значение диоксида углерода воздуха.......271
9.3. Экологическое значение непостоянных компонентов воздуха.280
9.4. Влияние на растения перемещения воздушных масс.........289
Глава 10. Почва как экологический фактор....................303
10.1. Основные свойства почвы и их экологическое значение....303
10.2. Экологическое значение химических свойств почвы.......312
10.3. Экологическая полифункциональность почв...............337
Глава 11. Биотические экологические факторы.................341
11.1. Типы отношений растений с другими организмами.........341
11.2. Симбиоз...............................................344
11.3. Фитофагия и защита растений от нее....................354
11.4. Отношения растений с паразитическими организмами
и устойчивость к инфекции ..................................366
11.5. Содействие животных размножению и распространению
растений....................................................376
11.6. Взаимоотношения между растениями......................381
Заключение..................................................395
Список литературы...........................................396