/








































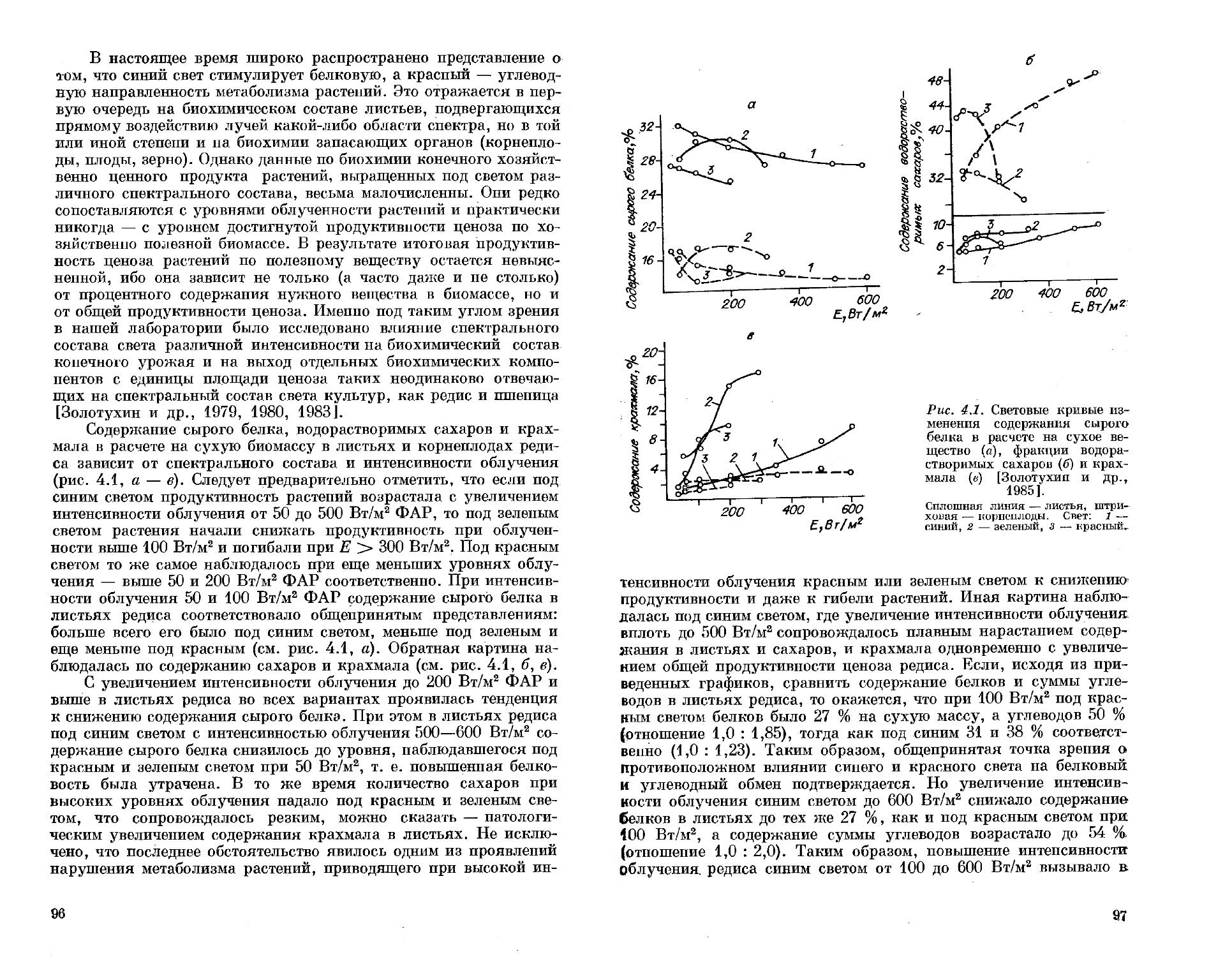






























Автор: Тихомеров А.А. Лисовский Г.М. Сидько Ф.Я.
Теги: физиология растений освещение светотехника растения биология ботаника
ISBN: 5-02-030057-8
Год: 1991




Текст
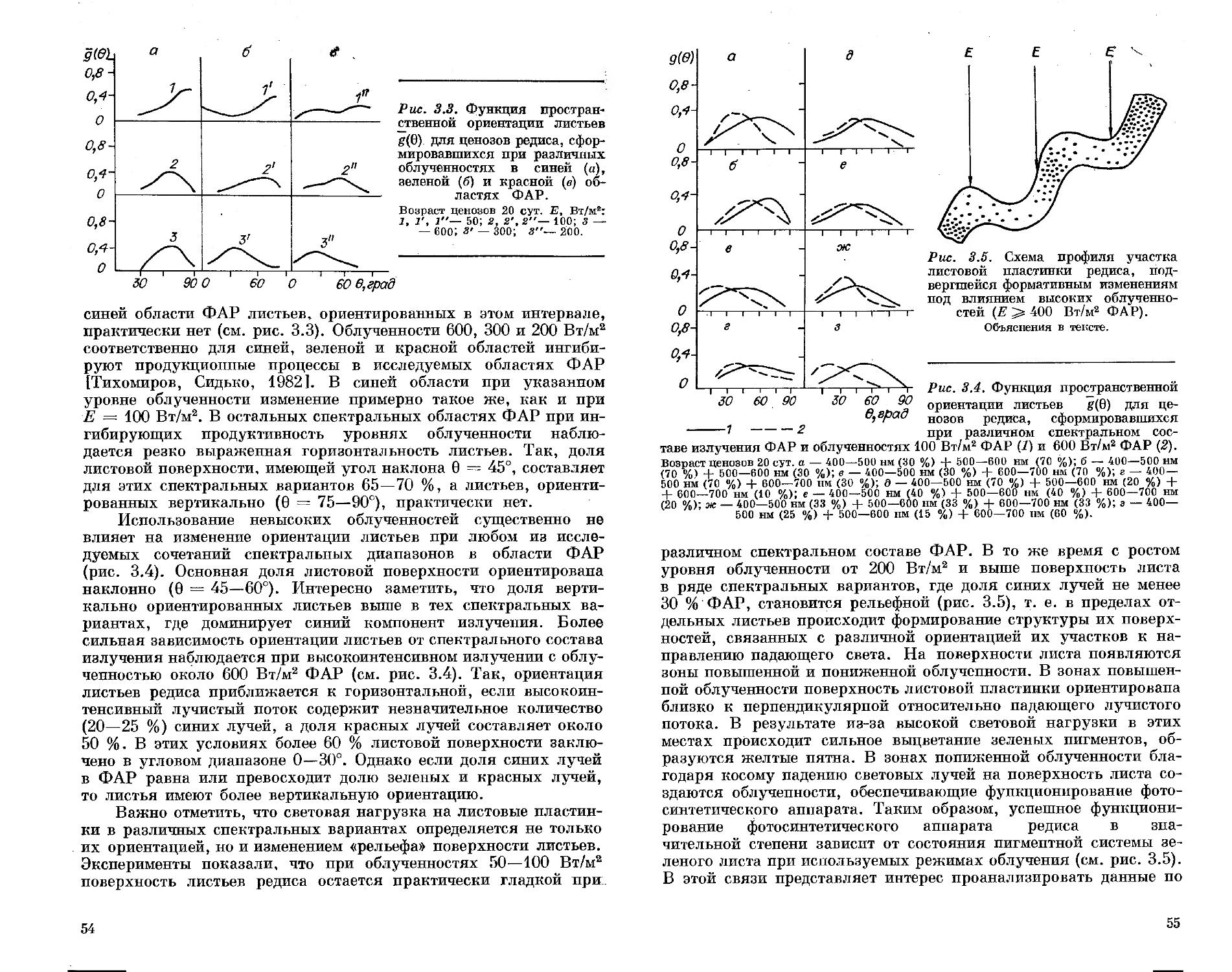
СПЕКТРАЛЬНЫЙ СОСТАВ СВЕТА
ПРОДУКТИВНОСТЬ РАСТЕНИЙ
А. А. ТИХОМИРОВ Г. М. ЛИСОВСКИЙ Ф. Я. СИДЬКО
АКАДЕМИЯ НАУК СССР
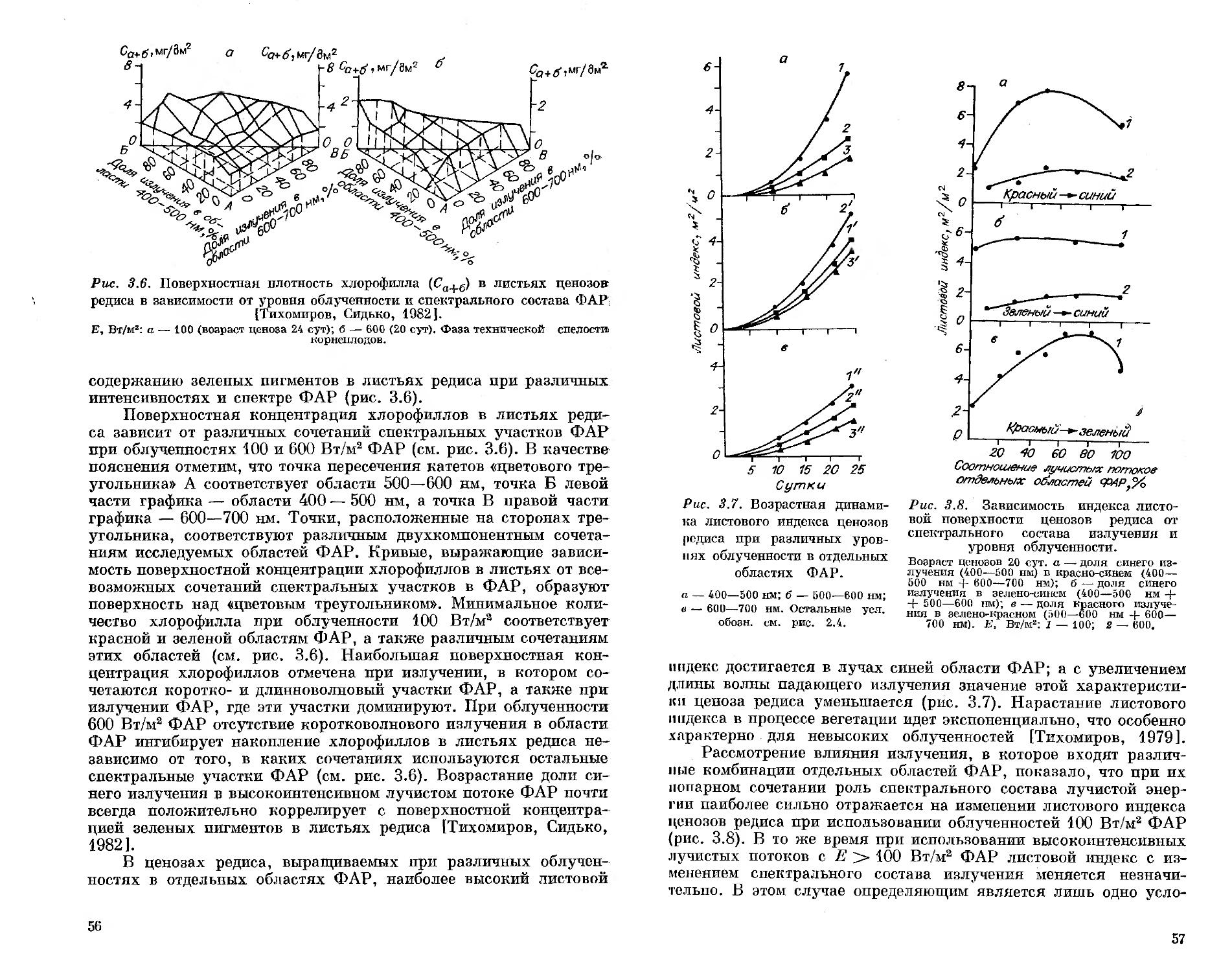
СИБИРСКОЕ ОТДЕЛЕНИЕ
ИНСТИТУТ БИОФИЗИКИ
А. А. ТИХОМИРОВ Г. М. ЛИСОВСКИЙ
Ф. Я. сидько
СПЕКТРАЛЬНЫЙ
СОСТАВ СВЕТА
И
ПРОДУКТИВНОСТЬ РАСТЕНИЙ
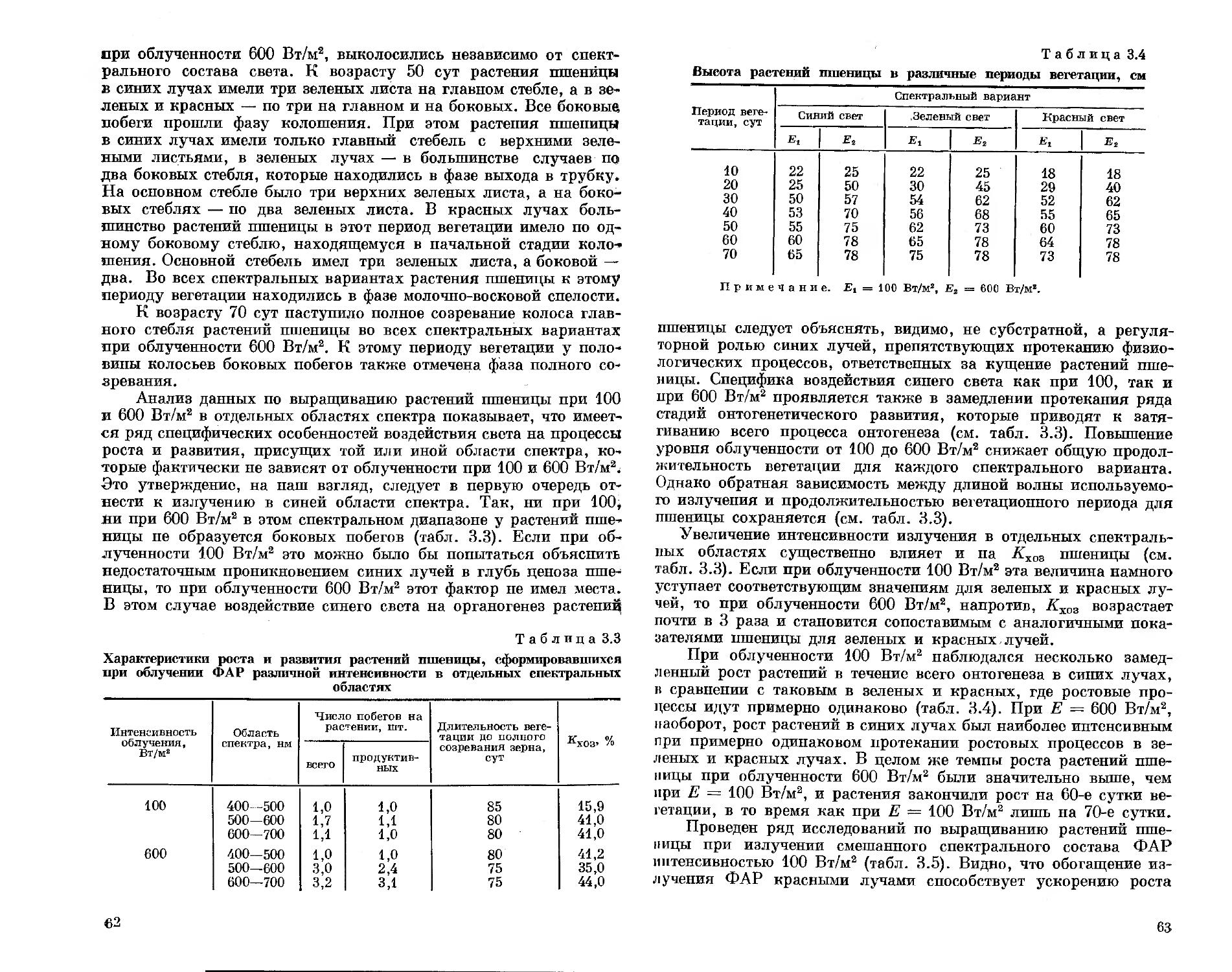
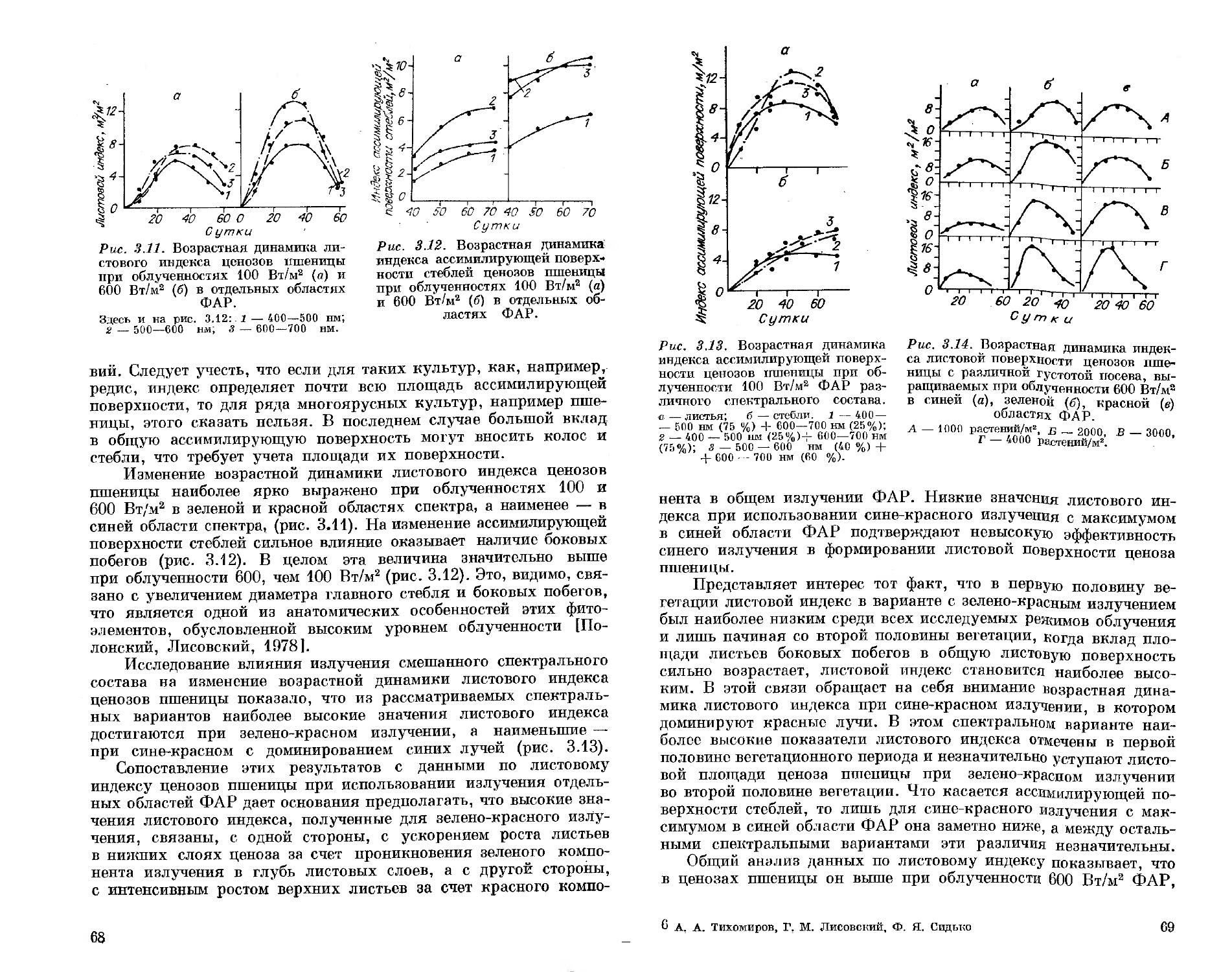
Ответственный редактор доктор биологических наук, профессор В. М. Гольд
в
НОВОСИБИРСК «НАУК А» СИБИРСКОЕ ОТДЕЛЕНИЕ 1991
.-ТЬИЫЙ РК -.ЕМП-W
УДК 581.14.035.4 : 628.981
Спектральный состав света и продуктивность растепий/Тихо-миров А. А., Лисовский Г. М., Сидько Ф. Я.— Новосибирск: Наука. Сиб. отд-ние, 1991.—168 с.
ISBN 5-02-030057—8.
В монографии рассмотрено длительное воздействие искусственного излучения различного спектрального состава и интенсивности фотосинтетически активной радиации (ФАР) на процессы, определяющие продуктивность растений: фотоморфогенез, рост и развитие, направленность биосинтеза, фотосинтез, накопление биомассы в контролируемых условиях среды, а также на состояние пигментного аппарата.
Книга рассчитана на биофизиков и биохимиков растений, светофизио-логов, светотехников, занимающихся вопросами разработки растениеводческих ламп, а также на специалистов в области светокультуры растений.
Табл. 33, Ил. 52. Библиогр.: 173 назв.
Рецензенты
доктор биологических наук | II. П. Воскресенская | кандидат биологических наук В. И. Полонский
Утверждено к печати
Институтом биофизики СО АН СССР
1906000000—155 _
Т—п/«ипо\—61—532—91 I полугодие © Издательство «Наука». 1991
ISBN 5—02—030057—8
ПРЕДИСЛОВИЕ
Продукционный процесс у растений исключительно сложен. Он складывается из разного рода взаимодействий и обратных связей в работе фотосинтетического аппарата, начиная от первичных процессов фотосинтеза и кончая ростовыми и оргапообразователь-ными процессами у растений в условиях фитоценоза, где формируется реальный урожай.
К настоящему времени установлен ряд фундаментальных положений о роли спектра и интенсивности фотосинтетически активной радиации (ФАР) в формировании наиболее важных составляющих продукционного процесса. Так, установлены общие закономерности воздействия излучения различных участков спектра ФАР на рост, развитие, направленность биосинтеза, фоторегуляцию и па другие процессы, влияющие на формирование конечной продукции. Исследования по этим вопросам отражены в монографиях советских ученых Н. А. Максимова [1955], А. Ф. Клешнина [1954], Н. П. Воскресенской [1965], Б. С. Мошкова [1966], В. М. Лемана [1971], И. А. Шульгина [1973] и др., а также зарубежных: Рабиновича [1951, 1953, 1959], Ван дер Вина и Мейер [1962] и др. Однако среди этих работ лишь монография Н. П. Воскресенской [1965] целиком посвящена спектральному составу света. В остальных работах изучению роли спектрального состава света отводилась только часть излагаемого материала. Более того, ни в одной из монографий не рассматривались последовательно вопросы формирования продуктивности растений в условиях различного спектрального состава ФАР. Внимание уделялось либо частным вопросам влияния излучения различных источников света на урожай растений, либо отдельным слагаемым продукционного процесса, таким как фотосинтез, рост и развитие и др. Что касается непосредственно исследований влияния спектрального состава света на продуктивность растений, то, как правило, опыты проводили на молодых отдельно стоящих растениях, не испытывающих влияния ценотических взаимодействий, а совокупность используемых спектральных комбинаций и уровней облученностей ФАР была крайне ограниченной. Систематический подход к оценке роли спектрального состава в продуктивности ценозов растений на фоне широкого варьирования спектрального состава света и интенсивности ФАР при
3
соблюдении единых методик в контролируемых условиях среды не был осуществлен ни отечественными, ни зарубежными авторами.
Это дает нам основания считать, что роль спектрального состава света в формировании продукционного процесса растений до сих пор исследована крайне недостаточно.
Несомненно, что углубленное изучение отдельных составляющих продукционного процесса при различном спектральном составе света будет последовательно и постепенно приближать нас к пониманию ответа фитоценоза как системы в целом на тот или иной спектральный режим облучения. Поэтому такое направление исследований имеет право на существование, оно создает определенные возможности детально разобраться в механизмах воздействия света различного спектрального состава на фотосинтетический аппарат растений на разных уровнях его организации. Однако понимание продукционного процесса в целом, прогнозирование конечной продуктивности растений при такой постановке исследований вряд ли достижимы.
Для решения этого вопроса необходимо, на наш взгляд, изучение воздействия спектрального состава света на высшем уровне организации фотосинтетического аппарата — цепотическом. При работе с ценозами растений выявление лимитирующих факторов, которые могут находиться на разных уровнях организации фотосинтетического аппарата, позволяет целенаправленно искать реальные пути повышения продуктивности растений. При этом можно с достаточной полнотой исследовать различные процессы, лежащие в основе жизнедеятельности растений, выявляя степень их связи с продуктивностью растений при заданных спектральных режимах облучения.
Последнее обстоятельство может быть корректно использовано, если информация получена в серии экспериментов, выполненных на общей методической основе, когда меняются только спектральные или энергетические характеристики ФАР. К сожалению, эти условия редко удается реализовать, и исследователи, имея информацию, полученную в разных условиях и при разном метрологическом обеспечении, часто приходят к неодинаковым выводам. Кроме того, ценность результатов резко снижается, когда в качестве источников света различного спектрального состава используются лампы без учета различий их инфракрасного и ультрафиолетового излучений, характера линейчатости спектра, пространственного распределения лучистого потока и других факторов, привносимых конструктивными особенностями тех или иных типов ламп. Использование в экспериментах серийных источников света не позволяет изучить влияние излучения отдельных спектральных областей ФАР и их дозированных сочетаний на те или иные стороны продукционного процесса, а использование светофильтров резко снижает уровни облученности ФАР и ввиду этого делает бессмысленными исследования по продуктивности растений.
4
Таким образом, подходы к оценке роли спектрального состава света в продуктивности растений нуждаются в совершенствовании.
С учетом изложенного целью настоящей книги является рассмотрение продукционного процесса в целом и ряда его составляющих (фотосинтез, рост и развитие, качество биомассы) при широком варьировании спектральных и энергетических характеристик лучистого потока. Для решения поставленной задачи были проведены комплексные исследования, выполненные по единой методике в контролируемых условиях среды при использовании универсальных светотехнических устройств, позволяющих менять спектральный состав и интенсивность ФАР при постоянстве всех других светотехнических характеристик.
Разумеется, книга не претендует на полноту охвата всей исследуемой проблемы. Основное внимание сосредоточено на рассмотрении спектральной эффективности действия ФАР для ряда вышеуказанных составляющих продукционного процесса, в то время как вопросы фоторегуляции, направленности фотосинтетического метаболизма и другие тонкие механизмы, лежащие в основе жизнедеятельности растений, в данной работе не рассматриваются.
На основе полученного экспериментального материала предложены физиологически обоснованные принципы оценки спектральной эффективности излучения ФАР в продуктивности растений и сформулированы предложения по созданию специализированных растениеводческих ламп с оптимизированным спектральным составом ФАР для выращивания определенных видов растений.
Разумеется, не все закономерности взаимодействия света различного спектрального состава с фотосинтетическим аппаратом растений, представленные в настоящей книге, удалось проанализировать достаточно глубоко, не всегда понятен механизм такого взаимодействия. Дальнейшее движение в этом направлении невозможно без привлечения физических, биофизических, биохимических и других современных методов исследования растений, комплексных усилий специалистов разных профилей. Изложенные в данной работе результаты помимо самостоятельного значения могут служить ориентирами в таком движении.
Подобные исследования, по нашему мнению, должны быть в первую очередь направлены на изучение вопросов оптимизации спектра и интенсивности ФАР в продукционном процессе растений на разных этапах их вегетации, а также на изучение возможностей использования света разного спектрального состава в организации направленного биосинтеза биологически ценных соединений различного назначения. Совершенно не исследованной в продуктивности растений остается пока спектральная эффективность действия ФАР, представленная линейчатыми спектрами разной структуры, ждут своего решения вопросы оценки роли инфракрасной радиации различного спектрального состава в жизнедеятельности растений и так далее.
5
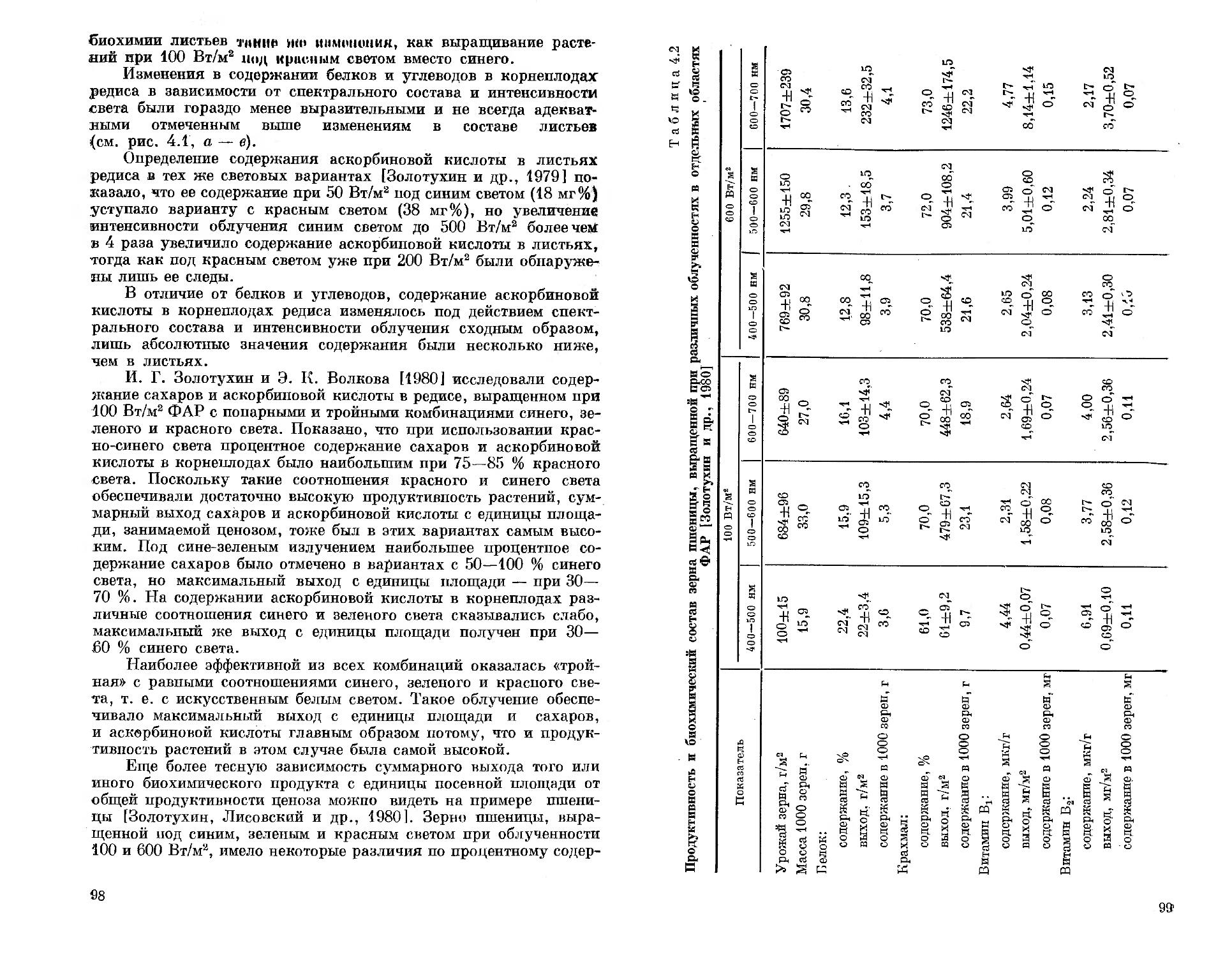
Авторы глубоко благодарны академику А. Т. Мокроносову и члену-корреспонденту АН СССР А. А. Ничипоровичу за внимание и поддержку при проведении исследований, доктору биологических наук |Н. П. Воскресенской) и кандидату биологических наук Н. Н. Протасовой за обсуждение результатов исследований и критические замечания, доктору технических наук Г. С. Сарычеву и кандидату технических наук Л. Б. Прикупцу, принимавшим участие в обсуждении прикладных аспектов исследуемой проблемы и предоставившим квазимонохроматические источники света собственной разработки, с использованием которых была выполнена большая часть фотобиологических экспериментов. Авторы искренне признательны научному сотруднику И. Г. Золотухину, совместно с которым проведены эксперименты и получены результаты, использованные в главах по росту и развитию, биохимии и продуктивности растений. Авторы благодарят всех коллег, принявших участие в обсуждении и оказавших содействие данной работе.
Глава 1
МЕТОДИКА ПРОВЕДЕНИЯ ЭКСПЕРИМЕНТОВ И ИСПОЛЬЗУЕМАЯ АППАРАТУРА
§ 1. Объекты исследования и условия их культивирования
Основными объектами исследования служили растения огурца и томата — культуры, широко используемые для выращивания в условиях искусственного освещения,— и редис как модельное растение многих физиологических экспериментов. Из зерновых культур, с которыми ведется селекционная работа в условиях искусственного освещения, была выбрана пшеница. Для решения некоторых конкретных вопросов использовали растения кукурузы, рапса, подсолнечника, смородины.
Из сортов огурца предпочтение было отдано сорту Московский тепличный, по которому имеется достаточный опыт культивирования в светокультуре [Курец и др., 1984; Кособрюхов, Чермных, 1986; и др. ], что облегчило задачу выбора условий его выращивания в планируемых экспериментах.
Из томатов был взят зарубежный сорт Старфайер, обстоятельно исследованный в условиях светокультуры в АФИ (г. Ленинград). Кусты томатов этого сорта отличаются компактностью, довольно высокой урожайностью [Мошков идр., 1984], что позволяет работать с ценозами этой культуры на весьма ограниченных посевных площадях в вегетационных шкафах.
Для работы с пшеницей был выбран специально выведенный для использования в условиях светокультуры короткостебельный и устойчивый к полеганию сорт 232 [Лисовский, 1973]. Ценоз этого сорта пшеницы обеспечивает определенную стабильность своей структуры в течение большей части вегетационного периода. Это, в частности, облегчило проведение запланированных оптических и фотосинтетических измерений без риска нарушить структуру ценоза. Сорт 232 имеет сравнительно короткую вегетацию, которая в условиях используемых нами радиационных режимов составляла около 65—75 сут.
Из сортов редиса выбран Вировский белый, который хорошо использует лучистую энергию при непрерывном облучении и способен эффективно продуцировать фитомассу в широком диапазоне облученностей [Тихомиров и др., 1976]. При этом техническая спелость корнеплодов у него наступает па 20—24 сут.
Среди указанных объектов Вировский белый имеет в 3 раза меньший период вегетации, чем все другие используемые в экспе
7
риментах виды растений. Поэтому ценоз редиса был выбран в качестве основной модельной культуры для проведения исследований по спектральной и энергетической эффективности ФАР в продукционном процессе.
Длительность периода вегетации при выращивании растений определялась наступлением технической спелости хозяйственно полезного урожая. При тех режимах облучения, когда рост и развитие растений были аномальными и получение хозяйственно полезного урожая было невозможным, длительность вегетации определялась моментом прекращения накопления сухого вещества растением, после чего деструкция фотосинтетического аппарата начинала преобладать над биосинтезом.
Все исследуемые культуры (как основные, так и дополнительные) выращивали при облученности (Е) 100 Вт/м2 ФАР различного спектрального состава. Такой выбор был связан с тем, что при этой или близких облученностях обычно достигается максимальная эффективность фотосинтеза растений. Такие уровни облученности часто используются при выращивании как отдельных растений, так и фитоценозов в условиях светокультуры [Лисовский и др., 1987]. Поэтому определение фотосинтетической продуктивности растений при разном спектральном составе ФАР для такой облученности может иметь не только научный, но и практический интерес.
В целом выбор уровней облученностей диктовался особенностями реакции фитоценозов на излучения разного спектрального состава и имеющимися техническими возможностями. Например, при выборе нижней границы облученности учитывали наиболее распространенные в практике светокультуры уровни облученности, при которых выращиваются растения, что соответствует примерно Е = 50 Вт/м2 ФАР [Сидько и др., 1977]. Такое значение облученности и было выбрано как минимальное при изучении реакции ценозов большинства исследуемых культур на спектр излучения различных областей ФАР. В то же время верхний уровень облученности ценоза редиса, на котором эксперименты выполнены наиболее подробно, определялся достижением полного ингибирования его продукционных процессов и составлял около 200 и 300 Вт/м2 соответственно для красной и зеленой областей ФАР. Для синей области ФАР технически возможным оказалось получить Е ~ 600 Вт/м2, при которой наблюдалось начало ингибирования продукционных процессов. Для ценозов пшеницы верхним уровнем облученности была Е = 600 Вт/м2 ФАР, поскольку ценозы этого сорта пшеницы имеют весьма высокие уровни светового насыщения продукционного процесса [Полонский, 1978]. В то же время максимальный КПД фотосинтеза пшеницы достигается при облученности 80—100 Вт/м2 ФАР [Лисовский и др., 1987]. Поэтому для пшеницы было целесообразно взять нижний уровень Е — 100 Вт/м2 видимой радиации в отличие от Е = 50 Вт/м2, которую использовали для других исследуемых культур.
8
Для ценозов огурца и томатов основным был уровень облученности 100 Вт/м2 ФАР как наиболее распространенный в интенсивной светокультуре [Лисовский, Долгушев, 1086; Лисовский и др.., 1987], в условиях которой наиболее целесообразно выращивать эти виды растений [Александрова, Радомысльская, 1980; Мошков п др., 1984; Кособрюхов, Чермных, 1986],
При выборе продолжительности суточного облучения учитывался значительный опыт, накопленный при выращивании цено,-зов исследуемых культур в условиях светокультуры как в Институте биофизики СО АН СССР, так и в других научных учреждениях. Многочисленными экспериментами было показано, что. для исследования сортов пшеницы и редиса наиболее приемлемо непрерывное облучение, при котором сокращаются сроки вегетационного периода и сохраняется высокий урожай хозяйственно полезной части [Лисовский и др., 1987]. Для ценозов огурца и томатов был выбран фотопериодический режим облучения 14 и 16 ч соответственно, который на основании ряда исследований можно считать оптимальным [Мошков и др.,. 1984; Кособрюхов, Чермных, 1986]. Густота стояния растений также выбиралась, исходя из данных этих опытов. Для редиса она составляла 180 растений/м2 [Тихомиров и др., 1976], а для пшеницы 1000 растений/м2 [Тихомиров, 1979]. В некоторых опытах с целью повышения эффективности усвоения падающей энергии проводили загущение ценозов пшеницы. Загущение ценозов редиса не проводили, так как в ряде случаев это может снизить выход хозяйственно полезной биомассы [Тихомиров и ДР-, 1976].
Как показали специальные эксперименты [Сидько и др., 1983], для обеспечения наиболее эффективного продукционного процесса ценоз огурца целесообразно формировать в виде одпо-стеблевого посева высотой около 1 м и с густотой 10 растений/м2, что и было использовано в настоящей работе. Ценозы томата формировали с густотой 6 растений/м2, с 2—3 цветочными кистями на растении, как это рекомендовано рядом исследователей [Рекомендации ..., 1986].
При культивировании фитоценозов поддерживались такие условия среды, которые по возможности не являлись бы лимитирующими при использовании высоких облученностей. В связи с этим питательным раствором служил раствор Кнопа тройной концентрации [Полонский, Лисовский, 1978], полив растений которым осуществлялся при Е 400 Вт/м2 ФАР через каждые 3 ч, а при Е <400 Вт/м2 — через каждые 6 ч [Там же]. Содержание СО3 в атмосфере колебалось в пределах 0,1—0,5 % в зависимости от уровня облученности ФАР. Температура воздуха поддерживалась па уровне 20 ± 1 °C круглосуточно для редиса и пшеницы [Гительзон и др., 1975], 25 °C [Курец и др., 1984] и 28 СС [Мошков, 1966] днем и 20 СС [Курец и др., 1984] и 25 °C [Мошков, 1966] ночью для огурца и томатов соответственно.
9
Относительная влажность воздуха составляла 70—80 %, а температура корнеобитаемого слоя керамзита — 16—17 °C [Полонский, Лисовский, 19781 для всех исследуемых культур.
§ 2. Установки для выращивания растений
Для реализации выбранных условий выращивания ценозов растений наиболее удобны терморегулируемые герметизируемые вегетационные шкафы. С этой целью в экспериментах использовались вегетационные камеры, разработанные и изготовленные в Институте биофизики СО АН СССР. В зависимости от масштаба эксперимента объем камер колебался от 340 до 3000 л. Вегетационная камера (рис. 14) представляет собой сваренный из листовой нержавеющей стали полый параллелепипед (7) с потолком из органического стекла и герметическим люком в одной из стенок. Для устранения повреждающего действия ультрафиолетового излучения источников света на органическое стекло укладывали силикатные стекла. Для уменьшения доли инфракрасной радиации источники света располагались над потолком со слоем проточной воды 35—50 мм.
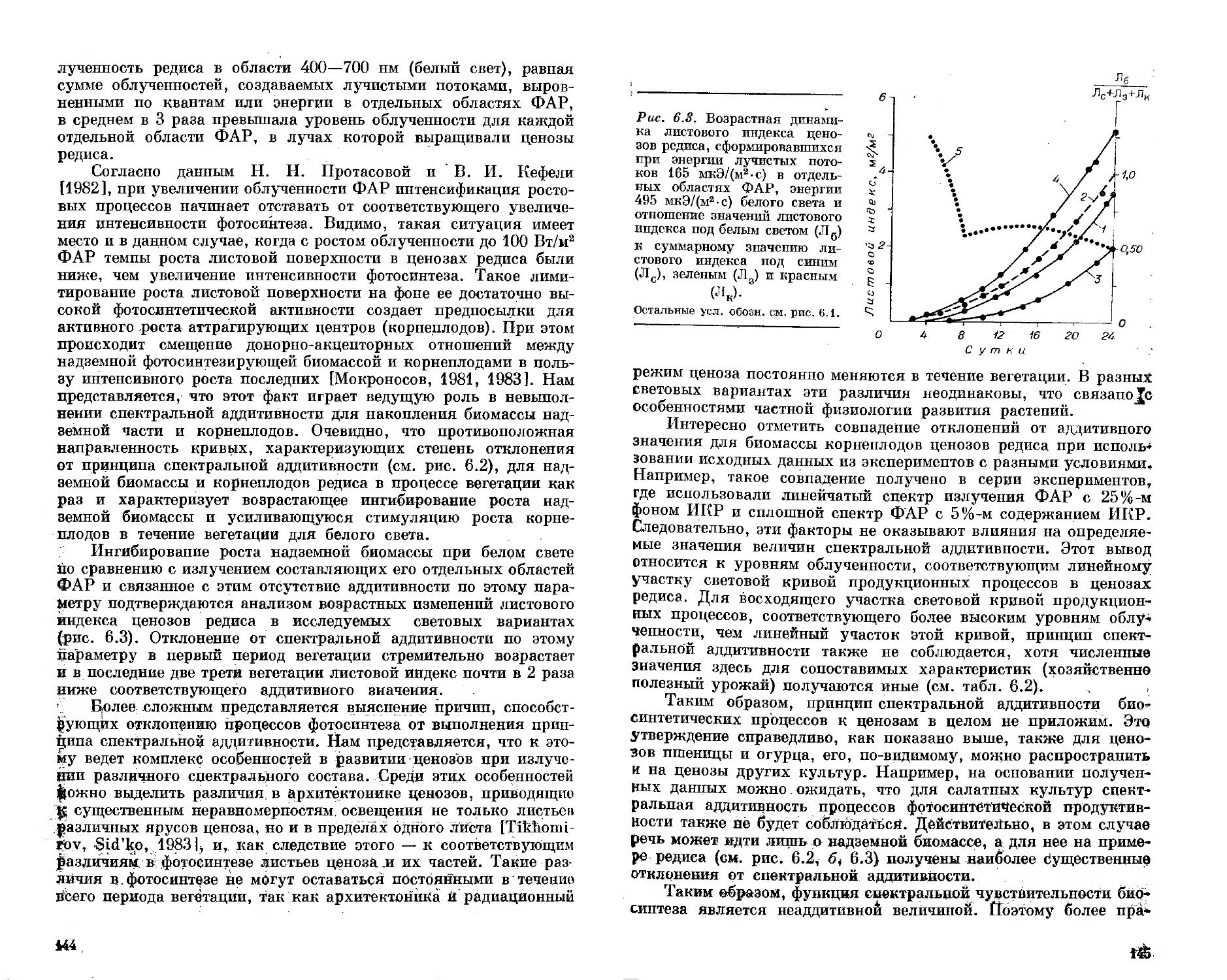
Рис. 1.1. Схема установки для культивирования фитоценозов.
Пояснения в тексте.
40
Субстратом для корнеобитаемой зоны служил слой керамзита (2) толщиной 20—30 см, питательный раствор периодически закачивался в него из соединенного с вегетационной камерой гормотического бака (5) с помощью насоса (4). К камере подключили непрерывно регистрирующий концентрацию углекислоты газон пал изатор АО-5501 или «Инфралит-4» (ГДР) со шкалой 0— (1,5 % СО2 (5). Содержание углекислоты поддерживалось в заданных пределах. Изменение давления воздуха в камере при добавке углекислоты уравновешивалось компенсатором с переменным объемом. Образующийся в ходе фотосинтеза избыток кислорода удалялся один раз в несколько суток разгерметизацией и провет-р и ванием к амеры.
Температура воздуха в вегетационных камерах автоматически поддерживалась на заданном уровне с помощью системы, включающей 12-канальный самописец, термисторы, расположенные в нескольких точках над ценозом, исполнительный механизм и вентилятор. При нагреве воздуха в вегетационной камере выше заданной температуры в нее с помощью вентилятора поступал охлажденный воздух, прошедший через радиатор (6), по которому циркулировала холодная водопроводная вода. При снижении температуры воздуха в камере до заданных пределов подача холодного воздуха автоматически прекращалась. Воздух в вегетационной камере нагревался за счет падающего лучистого потока ламп (7) и электронагревателя. Температура воздуха в камере регистрировалась на диаграммную ленту самописца. Аналогично регистрировали температуру корнеобитаемого слоя. Температурные показатели терморегулирующей системы периодически све^ ряли с показаниями контрольного термометра, находящегося в камере.
Уровень облученности фитоценозов регулировали с помощью изменения высоты подвеса поддопа с растениями (2) или светильников с лампами. Интенсивность лучистого потока, падающего па ценоз, регистрировали пиранометром Янишевского. Отклонения облученности по отдельным точкам над ценозами не превышали ±15 % от ее среднего значения.
§ 3. Световое обеспечение растений
В экспериментах использовали источники излучения, способные создавать лучистые потоки различной интенсивности и спектрального состава.
В ряде опытов удобными источниками излучения оказались ксеноновые лампы ДКсТВ-6000 [Маршак и др., 1963; Вассерман, 1989]. испытания которых в условиях светокультуры показали возможность создания уровней облученностей порядка солнечных и выше [Тихомиров и др., 1976; Полонский, Лисовский, 1978] и благоприятное влияние спектра этих ламп на выращивание растений различных видов [Леман, 1971; Протасова, Добровольский, 1973; Тихомиров и др., 1976]. Обладая практически равно-
11
Рис. 1.2. Спектры пропускания (Z, %) широкополосных свето-
фильтров, используемых для выделения отдельных областей ФАР.
Спектры пропускания фильтров: 1 — синего, 2 — голубого, 3 — желтого, 4 — красного. Заштрихована область спектра, выделенная при комбинировании голубого и желтого фильтров для получения зеленого излучения с помощью ламп ДКсТ В-6000.
.1
Рие. 1.3. Схема установки для получения белого света, практически не содержащего ИКР.
1 — лампа ДКсТВ-6000; 2 — водяные
фильтры; 3 — фильтр, выделяющий область 400—-600 нм и поглощающий ИКР;
4 — лампы ЛКБ-40; 5 — стеклянный теплозащитный светофильтр; 6—молочное стекло; 7 — растения в вегетационном шкафу; 8 — керамзит. Е — поток лучистой энергии.
энергетическим спектром в области ФАР, эти лампы могут успешно использоваться как источники высокоинтенсивпых лучистых потоков разного спектрального состава в сочетании с широкополосными фильтрами. Такая возможность была реализована в серии экспериментов по изучению выполнимости принципа спектральной аддитивности для биосинтетических процессов в фито-ценозах [Тихомиров, Сидько, 1987]. Лампы ДКсТВ-6000 использовали в сочетании с синим, голубым или желтым светофильтрами (рис. 1.2).
Для получения излучения в области 400—500 нм использовали синий и голубой фильтры, через которые пропускали излучение лампы ДКсТВ-6000. В данном случае голубой фильтр применяли для исключения инфракрасной радиации (ИКР) лампы ДКсТВ-6000. Излучение в области 500—600 нм выделяли из лучистого потока лампы ДКсТВ-6000 с помощью комбинации голубого и желтого светофильтров. В этих, а также в ряде других экспериментов использовали красные безртутиые люминесцентные лампы ЛКБ-40 [Волкова и др., 1982], излучение которых сосредоточено в красной (600—700 им) области спектра. Как правило, лампа ЛКБ-40 применялась в сочетании с красным светофильтром для предотвращения подсветки фоновым и естествен-йым излучением.
В этой же серии экспериментов получали «равноквантовый» спектр ФАР комбинацией лучистого потока сине-зеленой области (400—600 нм), выделяемой путем пропускания излучения лампы
12
ДНс i B tiiMJO через голуоой светофильтр, с дооавками излучения лампы JIKB-40. При этом суммарный поток белого света формировали таким образом, чтобы его энергия равнялась сумме энергий квантовых потоков используемых спектральных областей ФАР, каждый из которых составлял 165 мкЭ/(м2-с), что соответствовало уровням облученности 30, 36, 45 Вт/м2 соответственно в красной, зеленой и синей областях спектра. Тогда уровень облучен пости белым светом составлял 496 мкЭ/(м2-с).
Для сведения к минимуму влияния инфракрасной радиации во всех световых вариантах использовали стеклянные теплозащитные фильтры и водяные экраны. В частности, ИКР в излучении ламп ДКсТВ-6000 практически полностью срезает голубой фильтр, погруженный в воду. ИКР в излучении ламп ЛКБ-40 устраняли с помощью прозрачных теплозащитных фильтров, погруженных в слой воды (рис. 1.3). Доля ИКР в этой серии экспериментов не превышала 5 % интегрального лучистого потока.
При проведении фотобиологических исследований по оценке <'центральной эффективности различных серийных источников света кроме вышеуказанных ламп использовали ЛБ-40, КГ-1000, ДРЛФ-400, ДРФ-1000, ДНаТ-400, ДРИ-2000-6, ДМЗ-ЗООО, ДМ4-3000, ДРОТ-2000.
Основными источниками узкополосного излучения в области ФАР при проведении опытов по оценке спектральной эффективности ее различных диапазонов в продукционной деятельности растений служили экспериментальные металлогалогенные лампы, разработанные и изготовленные во ВНИСИ [Прикупец и др., 1973]. Достоинством этих ламп является способность давать высокоинтенсивные лучистые потоки с максимумами излучения в отдельных областях ФАР. Опыт использования ламп этого типа в условиях светокультуры показал, что они являются удобным инструментом для исследования специфики воздействия отдельных областей ФАР на различные физиолого-биохимические процессы высших растений [Терентьев и др., 1973; Павловский и др., 1976; Сидько и др., 1977]. Характер распределения энергии в спектре излучения этих ламп дает возможность использовать светофильтры, повышающие «чистоту» используемых лучистых потоков в отдельных областях ФАР без существенного снижения их мощности.
Так, для получения излучения в области 400—500 нм (условно синяя область ФАР) фильтром служило синее органическое стекло, что позволило повысить долю излучения ртутно-галлиево-ипдиевой лампы в этом спектральном диапазоне до 97 %, а примесь излучения в области 500—600 нм снизить до 3 %. Источником излучения в области 500—600 нм (зеленая область ФАР) служила ртутио-таллиевая лампа, излучение которой пропускали через желто-зеленый светофильтр из силикатного стекла, что дало возможность повысить долю излучения в области 500— 600 нм до 98 %, в то время как на долю синих лучей приходилось не более 2 % от общего излучения лампы. Красная область спект-
13
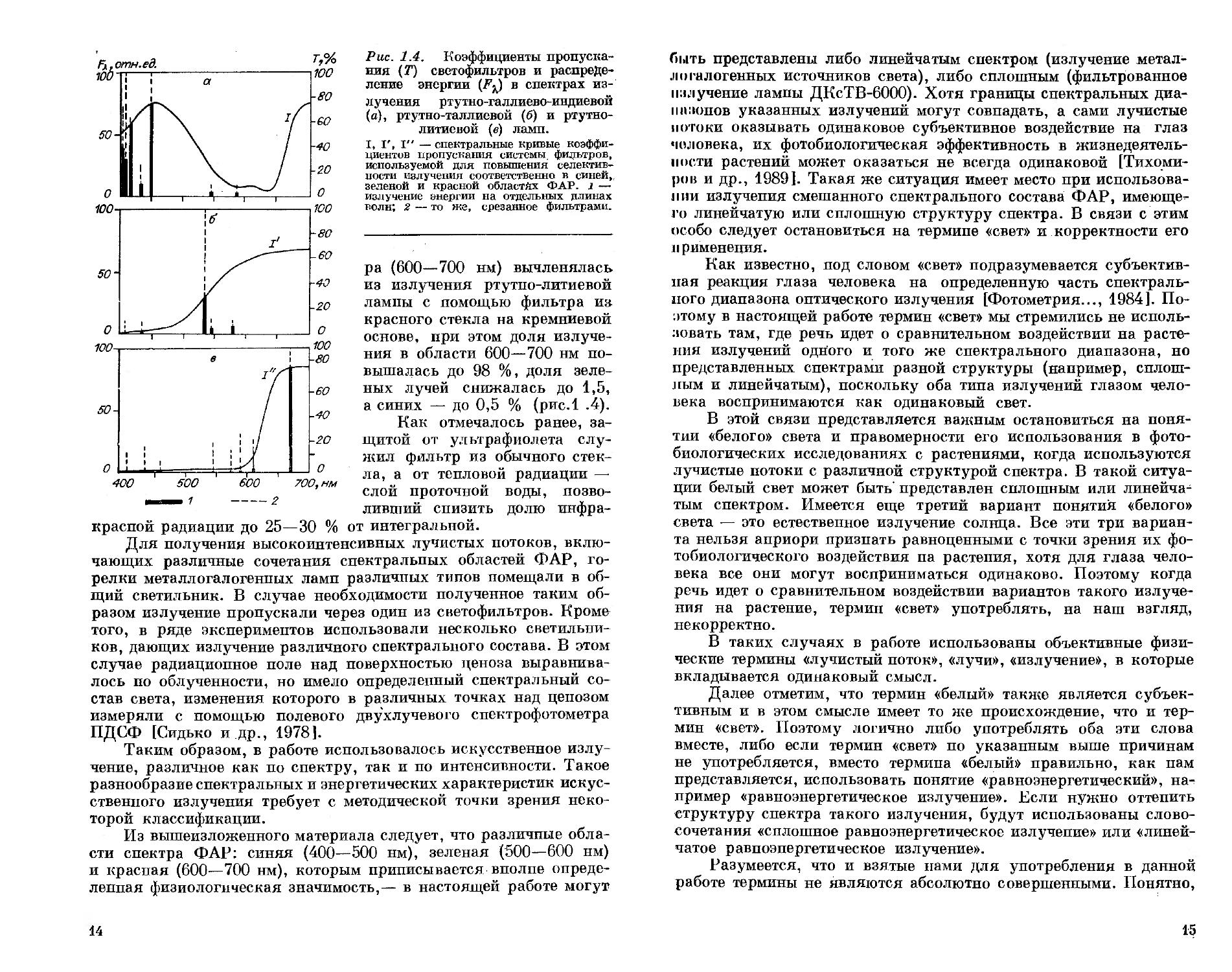
Рис. 1.4. Коэффициенты пропускания (Т) светофильтров и распределение энергии (/у) в спектрах излучения ртутно-галлиево-индиевой (а), ртутно-таллиевой (б) и ртутно-литиевой (в) ламп.
I, I', I" — спектральные кривые коэффициентов пропускания системы фильтров, используемой для повышения селективности излучения соответственно в синей, зеленой и красной областях ФАР. 1 — излучение энергии на отдельных длинах волн; 2 — то же, срезанное фильтрами.
ра (600—700 нм) вычленялась из излучения ртутно-литиевой лампы с помощью фильтра и» красного стекла на кремниевой основе, при этом доля излучения в области 600—700 нм повышалась до 98 %, доля зеленых лучей снижалась до 1,5, а синих — до 0,5 % (рис.1 .4).
Как отмечалось ранее, защитой от ультрафиолета служил фильтр из обычного стекла, а от тепловой радиации — слой проточной воды, позволивший снизить долю инфра-
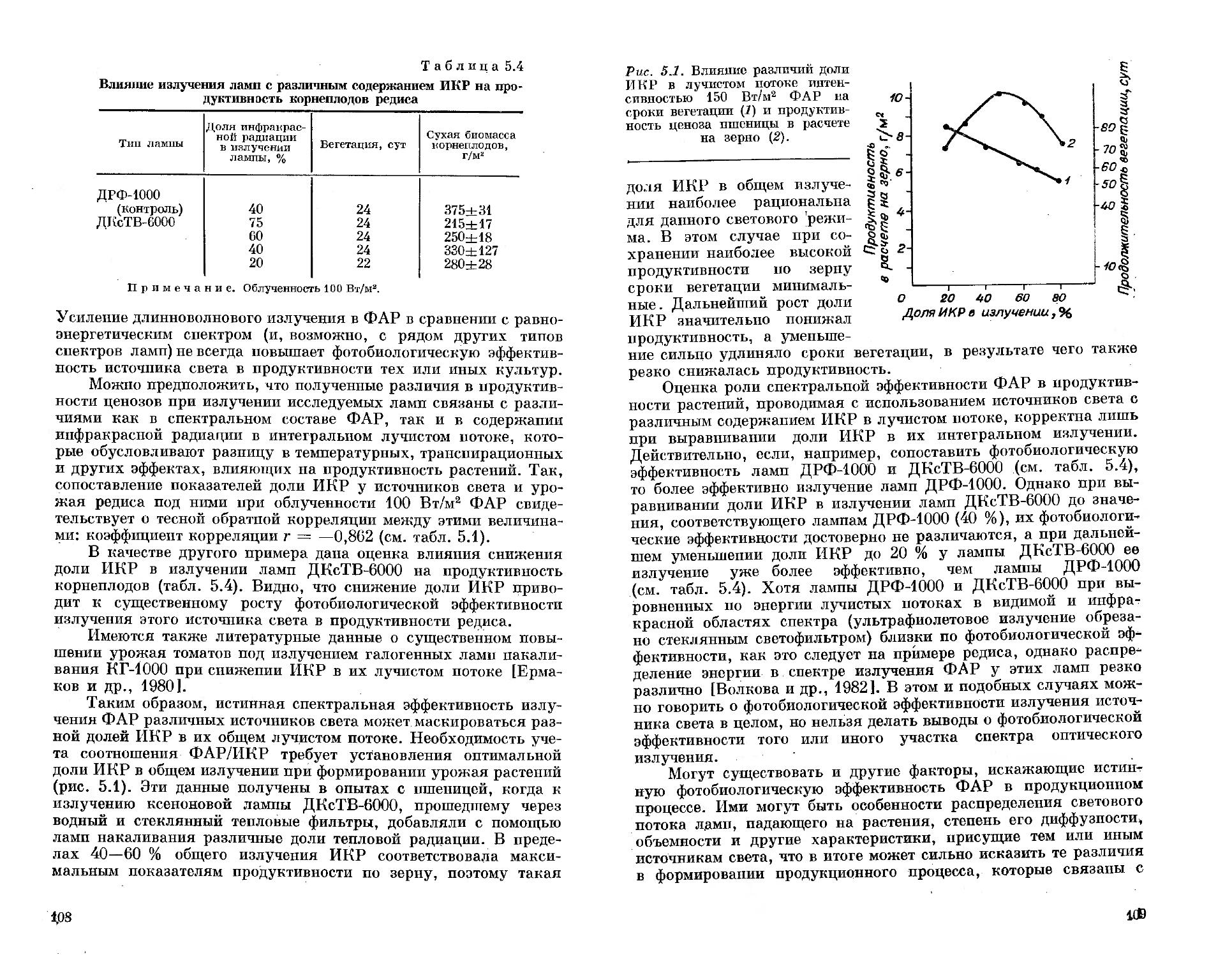
красной радиации до 25—30 % от интегральной.
Для получения высокоинтенсивных лучистых потоков, включающих различные сочетания спектральных областей ФАР, горелки металлогалогенных ламп различных типов помещали в общий светильник. В случае необходимости полученное таким образом излучение пропускали через один из светофильтров. Кроме
того, в ряде экспериментов использовали несколько светильников, дающих излучение различного спектрального состава. В этом случае радиационное поле над поверхностью ценоза выравнивалось по облученности, но имело определенный спектральный со
став света, изменения которого в различных точках над ценозом измеряли с помощью полевого двухлучевого спектрофотометра ПДСФ [Сидько и др., 1978].
Таким образом, в работе использовалось искусственное излучение, различное как по спектру, так и по интенсивности. Такое разнообразие спектральных и энергетических характеристик искусственного излучения требует с методической точки зрения некоторой классификации.
Из вышеизложенного материала следует, что различные области спектра ФАР: синяя (400—500 нм), зеленая (500—600 нм) и красная (600—700 нм), которым приписывается вполне определенная физиологическая значимость,— в настоящей работе могут
14
быть представлены либо линейчатым спектром (излучение металлогалогенных источников света), либо сплошным (фильтрованное излучение лампы ДКсТВ-6000). Хотя границы спектральных диапазонов указанных излучений могут совпадать, а сами лучистые потоки оказывать одинаковое субъективное воздействие на глаз человека, их фотобиологическая эффективность в жизнедеятельности растений может оказаться не всегда одинаковой [Тихомиров и др., 1989]. Такая же ситуация имеет место при использовании излучения смешанного спектрального состава ФАР, имеющего линейчатую или сплошную структуру спектра. В связи с этим особо следует остановиться на термине «свет» и корректности его н рименения.
Как известно, под словом «свет» подразумевается субъективная реакция глаза человека на определенную часть спектрального диапазона оптического излучения [Фотометрия..., 1984]. Поэтому в настоящей работе термин «свет» мы стремились не использовать там, где речь идет о сравнительном воздействии на растения излучений одного и того же спектрального диапазона, но представленных спектрами разной структуры (например, сплошным и линейчатым), поскольку оба типа излучений глазом человека воспринимаются как одинаковый свет.
В этой связи представляется важным остановиться на понятии «белого» света и правомерности его использования в фотобиологических исследованиях с растениями, когда используются лучистые потоки с различной структурой спектра. В такой ситуации белый свет может быть' представлен сплошным или линейчатым спектром. Имеется еще третий вариант понятия «белого» света — это естественное излучение солнца. Все эти три варианта нельзя априори признать равноценными с точки зрения их фотобиологического воздействия па растения, хотя для глаза человека все они могут восприниматься одинаково. Поэтому когда речь идет о сравнительном воздействии вариантов такого излучения на растение, термин «свет» употреблять, на наш взгляд, некорректно.
В таких случаях в работе использованы объективные физические термины «лучистый поток», «лучи», «излучение», в которые вкладывается одинаковый смысл.
Далее отметим, что термин «белый» также является субъективным и в этом смысле имеет то же происхождение, что и термин «свет». Поэтому логично либо употреблять оба эти слова вместе, либо если термин «свет» по указанным выше причинам не употребляется, вместо термина «белый» правильно, как нам представляется, использовать понятие «равноэнергетический», например «равноэнергетическое излучение». Если нужно оттенить структуру спектра такого излучения, будут использованы словосочетания «сплошное равноэнергетическое излучение» или «линейчатое равпоэиергетическое излучение».
Разумеется, что и взятые нами для употребления в данной работе термины не являются абсолютно совершенными. Понятно,
15
например, что термин «линейчатое равноэнергетическое излучение» является весьма условным, так как здесь имеется в вйду лишь равенство по энергии излучения для отдельных физиологически значимых областей ФАР — синей, зеленой и красной, представленных рядом резонансных линий излучения.
Есть также основания считать, что и сплошной равноэнергетический спектр ФАР, полученный от источников искусственного излучения, нельзя признать во всем идентичным сплошному равноэнергетическому излучению солнца. Последнее обстоятельство указывает на то, что некорректно в общем случае при работах с различным по спектральному составу искусственным излучением брать в качестве контроля естественное излучение. Более того, если в качестве контроля взято искусственное равноэнергетическое по спектру излучение, то оно должно быть «составлено» из отдельных областей ФАР, имеющих ту же структуру спектра, что и лучистые потоки этих областей ФАР или их сочетаний, которые используются в опытных вариантах. Поэтому во всех наших экспериментах по изучению воздействия на растения лучистых потоков ФАР, представленных двух- и трехкомпонентными сочетаниями отдельных областей, использованы потоки, составленные только на основе «исходных» излучений этих спектральных областей с фиксированной структурой линейчатого или сплошного спектра. Эти принципы заложены и в формирование равноэнергетического спектра ФАР, который является в рамках данного подхода частным случаем трехкомпонентного лучистого потока ФАР.
§ 4. Аппаратура и устройства для измерения оптических и фотосинтетических характеристик растений
Для проведения экспериментов по снятию световых кривых фотосинтеза и спектров действия фотосинтеза зеленых листьев, адаптированных к длительному пребыванию на свету различного спектрального состава, использовали листовую камеру из органического стекла, разработанную А. А. Тихомировым (рис. 1.5).
Листовая камера выполнена из оргстекла, имеет круглую форму с диаметром рабочей части 25 мм. Камера состоит из двух половин, одна из которых представляет крышку с резьбой (7), которая закручивается па корпус камеры (2). Герметичность здесь достигается за счет использования уплотнительного резинового кольца либо нанесением тонкого слоя вакуумной замазки па соприкасающиеся поверхности. Лист располагается на внутренней стороне крышки (3). Чтобы исключить подвядание листа, внутренняя поверхность крышки выстлана слоем влажного поролона. В центре внутренней поверхности крышки камеры находится пластилиновый выступ, на который крепится лист. Освещение листа осуществляется через кювету с проточной водой (4). Для обеспечения эффективного перемешивания и создания достаточного однородного газового потока над поверхностью листа в камере-
16
Рис. 1.5. Листовая камера для измерения фотосинтеза участков зеленого листа.
а — крышка камеры; б — корпус камеры (вид сверху); в — то же (вид снизу). Остальные усл. обозн. см. текст.
предусмотрено вводное отверстие большого диаметра (5), через которое газ поступает в замкнутое пространство для перемешивания. Затем газ поступает в пространство, где находится лист, через несколько отверстий (б), обеспечивающих равномерную протяжку газового потока над поверхностью листа. Аналогично устройство листовой камеры на выходе газового потока. Для стабилизации температурных условий в листовой камере входящий в пее воздух предварительно пропускали через змеевик, омываемый водой, температура которой поддерживалась постоянной с помощью термостата. Для предотвращения перегрева поверхности листа за счет падающего лучистого потока последний пропускали через кювету с проточной водой, температуру которой также стабилизировали с помощью термостата. Однородность освещения поверхности листовой камеры достигалась за счет регуляции оптической системы осветительного устройства.
В качестве осветительного устройства использовали оптическую систему диапроектора «Свитязь» с ксеноновой лампой ДКСТ-150 либо диапроектор «Киев-66 универсал» с лампой КГМ 24-250. В последнем случае при необходимости приближения спектра излучения лампы КГМ 24-250 к равпоэнергетическому использовали корректирующие светофильтры. Различные уровни облученности листа создавали! за счет использования металлических сеток, служивших в качестве ослабителей света. В остальном придерживались методики, описанной в работе Маккри [McCree, 1972а I и Инада [Inada, 1976, 19771.
Вышеописанная листовая камера и осветительная установка эксплуатировались в комплексе с газоанализатором «Инфралит-4» (ГДР) со шкалой на СО2 — 0—0,1 %.
2 А. А. Тихомиров, Г. М. Лисовский, Ф. Я. Сидько
17
§ 5. Методы оценки фотометрических и биологических характеристик растений и фитоценозов
Для оценки поглощательной способности зеленого листа, «формировавшегося при длительном воздействии излучения ФАР различного спектрального состава, использовали листья редиса, пшеницы и огурца.
Коэффициенты отражения R и пропускания Т для отдельных листьев определяли на спектрофотометре СФ-18 по известной методике [Брандт, Тагеева, 1967], после чего находили соответствующие значения коэффициентов поглощения А, используя формулу
А = 1 - (Т + R). (1.1)
При вычислении А для колосьев и стеблей полагали Т = 0. Средние значения коэффициентов поглощения для лучистых потоков различного спектрального состава вычисляли по формуле
j=k
------• а-г»
3=г
где A^ik — среднее значение коэффициента поглощения для спектрального интервала, ограниченного длинами волн и lft;
— спектральная плотность излучения; АХ = 10 нм.
Изучение ослабления радиации и изменения ее спектрального •состава проводилось в основном на ценозах редиса и пшеницы, имеющих разную архитектонику и выращиваемых при излучении различного спектрального состава и в широком диапазоне облученностей: 100—600 Вт/м2 ФАР. Средние значения коэффициентов ослабления для лучистых потоков различного спектрального состава вычисляли по формуле
где Jo — интенсивность лучистого потока, упавшего на ценоз;
— интенсивность лучистого потока, дошедшего до уровня ценоза на глубине Н.
Некоторые оптические характеристики сформировавшихся ценозов измеряли с помощью прибора ФАР-74 [Хазанов, 1975].
При изучении ориентации листьев в фитоценозах определяли функцию пространственной ориентации листьев — gL (z, 0, ср), где z — высота ценоза, 0 — угол наклона нормали листа, отсчитываемый от вертикальной оси, <р — азимут нормали листа [Росс, Нильсон, 1966, 1968; Росс, 1972; Тоомипг, 1977 ],,
При этом считали, что азимутальная ориентация листьев примерно равномерная [Росс, 1975; Тоомипг, 1977 ]. Интервал углов
18
наклона составлял 15° [Росс, Нильсон, 1968; Тооминг, 19771. В связи с этим по оси ординат откладывали величину jr/12g*(6), характеризующую долю листовой поверхности, ориентированную в исследуемом угловом диапазоне [Тооминг, 1977 ]. Согласно этой работе, введем обозначение n/12g*(0) = g(0). Из определения функции пространственной ориентации листьев следует, что она носит вероятностный характер, поэтому
О <£(0) < 1.
Для мопослойных ценозов редиса расчет g(0) проводили для всего слоя ценоза, а для многоярусных ценозов пшеницы и огурца g(0) рассчитывали как для всего ценоза, так и для верхней и нижней его половин [Тихомиров, 1980].
При анализе пигментного аппарата исследуемых растений определяли содержание зеленых пигментов в листьях и стеблях по методике Шлыка [1968]. Спектры поглощения растворов пигментов записывали на спектрофотометре СФ-18. Концентрацию пигментов пересчитывали на единицу площади фитоэлементов.
Воздействие излучения различного спектрального состава и интенсивности на жизнедеятельность растений оценивалось по продуктивности и эффективности использования лучистой энергии в расчете на накопление общей или хозяйственно полезной биомассы.
Под продуктивностью растений понималось количество общей (общая продуктивность) или хозяйственно полезной (хозяйственно полезная продуктивность) биомассы (сухой или сырой), накопленной за определенный период вегетации и отнесенной к единице посевной площади и единице времени (г/(м2 • сут)). В ряде случаев при одинаковом периоде вегетации растений сравнение разных вариантов вели по общему или хозяйственно полезному урожаю.
Под энергетической эффективностью излучения в продукционном процессе понимали эффективность использования излучения ФАР того или иного спектрального состава ценозами растений. Это понятие фактически совпадает с понятием коэффициента полезного действия (КПД) фитоценозов, предложенного Тоомингом [1977] для условий естественного произрастания. Ряд исследователей называет эту величину «КПД фотосинтеза ценоза» [Тооминг, 1977]. Энергетическую эффективность излучения в продукционном процессе (ц) вычисляли по формуле [Там же]
где q — калорийность растений (ккал/г); М — сухая биомасса растений (общая или хозяйственно полезная), накопленная за вегетационный период с единицы площади (г/м2); ^<2$— сумма фотосинтетически активной радиации за рассматриваемый вегета
2*
19,
ционный период (ккал) в расчете на единицу площади фитоценоза (м2).
Под спектральной эффективностью излучения понимали относительную величину, характеризующую отношение интенсивности протекания процесса (фотосинтез, рост, развитие и т. д.) при излучении в исследуемой области спектра AXj, к интенсивности протекания этого процесса /д^ при излучении в сравниваемой (контрольной) области спектра ДХ2:
Площади листовой поверхности растений, за исключением пшеницы, вычисляли весовым методом [Росс, 1975], а площадь листьев пшеницы по формуле [Тихомиров, 1979]
s = Zc/iZ,
где s — площадь листа пшеницы; h — максимальная ширина листа; I — длина листа по средней жилке; к = 0,8 — экспериментально определенный поправочный коэффициент.
При вычислении площади поверхности стеблей форма последних принималась цилиндрической.
Пробы, взятые на каждый анализ, составляли по 3—5 растений редиса, огурца или томата и по 15—20 растений пшеницы. В каждой биологической повторности анализы для растений редиса брали в среднем через 3—5 сут, а для растений пшеницы, огурца и томата — через 10—12 сут.
Представленные на рисунках и в таблицах данные являются средними из трех биологических повторностей опытов с отклонением не более 10—15 % при 95 % вероятности с использованием коэффициента Стьюдента.
Глава 2
ФОТОСИНТЕЗ И СПЕКТРАЛЬНЫЙ состав света
Введение
Фотосинтез является одной из важнейших составляющих продукционного процесса. Оптимизация спектрального состава света для фотосинтеза в значительной степени сказывается на ходе продукционного процесса. Поэтому исследование спектральной эффективности фотосинтеза — один из путей к решению проблемы оптимизации световых условий для протекания продукционного процесса. Такие исследования проводятся на различных уровнях организации фотосинтетического аппарата. Анализ этих исследо-
20
пиний дает основания выделить следующие основные уровни: пер-ппчиых механизмов фотосинтеза, или молекулярный, листовой, ненотический. При кратковременном (несколько минут и менее) гистоном воздействии лучистого потока того или иного спектрального состава на фотосинтетический аппарат фотосинтез на любом ri указанных уровней — ценотическом, листовом и даже тканевом фактически не изменяется. В то же время при длительном (многочасовом, мпогосуточпом) воздействии света на фотосинтетический аппарат фотосинтез зависит от взаимодействия быстрых и медленных реакций фоторегуляции, связанных с особенностями формирования фотосинтетического аппарата, морфогенеза, роста и развития, а также других медленно протекающих во времени процессов в растительном организме.
Первые работы, где обращалось серьезное внимание на возможности фоторегуляции фотосинтеза за счет варьирования спектральным составом света, появились в первой половине XX столетия и принадлежат советским исследователям [Данилов, 1935, 1936, 1940; и др.; Мальчевский, 1938, 1940; и др.].
Современные представления о роли быстрых и медленных реакций фоторегуляции в фотосинтезе стали интенсивно развиваться с 70-х годов нашего столетия в работах Н. П. Воскресенской [1975, 1987; и др.] и других исследователей [Белл, 1972; Петров и др., 1978]. Однако сама «наработка» информации о спектральной эффективности излучения отдельных областей видимого спектра в процессе фотосинтеза была начата еще в XIX в. К. А. Тимирязевым [1956]. В этот период такие исследования ограничивались воздействием кратковременного излучения того пл н иного спектрального состава и, как правило, низких уровней облученности на фотосинтетический аппарат листа. Такая ограниченность вполне закономерна и была связана с низким уровнем экспериментальной техники, не позволявшим расширять эти исследования с использованием более мощных и длительно действующих лучистых потоков разного спектрального состава. Последнее, правда, в крайне ограниченных масштабах стало возможно лишь с 20-х годов нашего столетия. Начиная с этого времени все чаще появляются исследования, связанные с изучением длительного (многосуточного) воздействия спектрального состава света различной интенсивности на фотосинтез растений [Мальчевский, 1940; и др. ]. Корректное проведение таких исследований требует мощных источников света с различным спектральным составом излучения, достаточно сложной аппаратуры для непрерывной длительной регистрации фотосинтеза растений и ценозов. Именно отсутствие примерно до 60-х годов нашего столетия таких возможностей предопределяло сравнительно недавнее начало исследований по оценке спектральной эффективности фотосинтеза при длительном воздействии света.
При оценке спектральной эффективности фотосинтеза при постоянном выращивании растений в условиях искусственного освещения логично использовать данные по длительному воздействию
21
спектральных потоков ФАР па фотосинтез растений и фи-тоцепозов.
Однако в этом вопросе пет единства, поскольку ряд исследователей считает, что для оценки спектральной эффективности действия ФАР в продукционном процессе достаточно пользоваться данными по спектрам действия фотосинтеза зеленого листа, получение которых связано с кратковременным (минутным) воздействием спектральных потоков ФАР низкой интенсивности па фотосинтетический аппарат. Дискуссия по этому вопросу проводилась в журнале «Светотехника» (1971—1973, 1978—1981 гг.), однако единого мнения выработано не было. Нам представляется, что одной из причин этого был недостаток экспериментальных данных по обсуждавшимся вопросам, поскольку спорная ситуация может быть разрешена лишь на основании экспериментально обоснованного заключения. Оно может быть получено после анализа данных по фотосинтезу при кратковременном и длительном воздействии излучения на фотосинтетический аппарат растения. Результат такого анализа может дать ответ на вопрос об адекватности или принципиальных различиях этих подходов в оценке спектральной эффективности фотосинтеза при постоянном выращивании растений в условиях искусственного облучения.
Поиску ответов на эти вопросы и посвящена настоящая глава.
§ 1. Спектры действия и спектральная эффективность фотосинтеза листа
Исследования спектральной эффективности фотосинтеза имеют длительную историю. Еще во второй половине XIX в. К. А. Тимирязев [1956] показал зависимость между интенсивностью фотосинтеза листа и спектром видимого излучения. В дальнейшем это направление работ по фотосинтезу было развито многочисленными исследователями. Обзор таких работ на период до середины 60-х годов дан в монографиях Е. Рабиновича [1953] и Н. П. Воскресенской [1965]. В дальнейшем наиболее значительные работы по этой тематике выполнены в 70-е годы Маккри [МсСгее, 1972а] и Инада [Inada, 1976, 1977]. В этих работах на листьях растений различных систематических групп были получены примерно одинаковые по структуре спектры действия фотосинтеза (рис. 2.1). Четкий методический подход и достаточно высокий уровень экспериментальной техники окончательно подтвердили мнение о том, что у зеленых листьев растений различных видов структура спектра действия фотосинтеза листа может быть принята как универсальная.
Общность структуры таких спектров для различных видов зеленых растений указывает на соответствующую общность некоторых характеристик фотосинтетического аппарата листьев. Среди них в первую очередь следует отметить, что у всех зеленых растений основные максимумы поглощения находятся в синей и красной областях спектра, а минимум — в желто-зеленой. Как пра-
22
Рис. 2.1. Кривые спектральной эффективности процессов. / - поглощение света; 2 — спектр дейстнин фотосинтеза в расчете на поглощенные кванты; 3 — спектр действия фотосинтеза в расчете на падающую энергию [МсСгее, 1972а].
пило, такой характер поглощения света листом обеспечивается и основном за счет хлорофиллов и «желтых» пигментов. Характер криной поглощения света листом и структура его спектра действия фотосинтеза в целом имеют общую тенденцию (см. рис. 2.1). <)тсюда можно предположить, что спектр поглощения хлорофиллов в значительной степени определяет спектр действия фотосинтеза листа. Однако в листьях растений кроме зеленых пигментов (хлорофиллов) имеются еще дополнительные пигменты, имеющие свои спектры поглощения, отличные от таковых у хлорофиллов. Ввиду того что дополнительные пигменты находятся в листе и имеют максимумы поглощения, находящиеся в области широких < иних полос поглощения хлорофиллов, их изучение in vivo оптическими методами затруднено. Тем не менее многообразие пигментов в зеленом листе указывает и на многообразие соответствующих спектров действия фотобиологических процессов, которые регулируют те или иные процессы жизнедеятельности растений в вносят свой вклад в формирование продукционного процесса. Разумеется, что среди этих процессов фотосинтезу отводится очень важная роль. Вполне закономерно, что получению и анализу спектров действия фотосинтеза зеленого листа на протяжении длительного времени уделялось большое внимание [Gabrielsen, 1948; Stoy, 1955; Bulley et al., 1969; МсСгее, 1972a; Inada, 1977; и др.]. Поэтому не случайно, что в литературе появились предложения использовать такие спектры действия фотосинтеза для оценки спектральной эффективности этого процесса в реальных условиях выращивания растений [Свентицкий, 1960, 1977].
В связи с этим остановимся более подробно па таких спектрах действия и возможности их использования для оценки спектральной эффективности фотосинтеза в реальных световых условиях выращивания растений. При получении спектров действия фотосинтеза необходимым условием является использование низких уровней облученности, когда между интенсивностью фотосинтеза и уровнем облученности сохраняется линейная зависимость. Это условие связано с необходимостью отделить световые стадии фотосинтеза от темновых. Последние лимитируют фотосинтез, когда зависимость между фотосинтезом и уровнем облученности находится за пределами линейного участка световой кривой
23
фотосинтеза. Если выдержать это методическое условие, то можно, сопоставив спектры поглощения пигментов зеленого листа й спектр действия фотосинтеза, сделать заключение о роли пигмента в фотосинтезе листа. Именно с учетом этой точки зрения записаны и проанализированы спектры действия фотосинтеза зеленого листа [Литвин, Хэ-И-Тань, 1967; Санадзе, Баазов, 1982; Баазовг Санадзе, 1984а, б]. Так, в работе Ф. Ф. Литвина и Хэ-И-Таня [1967] по измерениям спектров действия фотосинтеза взрослых и зеленеющих листьев показано, что в фотосинтезе участвуют несколько различных форм хлорофилла а, причем относительная доля их участия на разных стадиях фотосинтеза может быть различной. В этой же работе проведено исследование спектра действия эффекта Эмерсопа па листьях высших растений. Сопоставляя данные Ф. Ф. Литвина и Хэ-И-Таня [1967 ] с собственными исследованиями, Г. А. Санадзе и Д. И. Баазов [1982] пришли к выводу об универсальности структуры и функции фотохимических пигментов систем у высших растений и водорослей. Продолжая работы в этом направлении, исследователи на листьях тополя получили спектры действия, аналогичные ранее известным для других растений и зеленых водорослей [Баазов, Санадзе, 1984а, б]. В этих же работах подтверждено участие в фотосинтезе тех же групп пигментов, о которых говорилось в более ранних работах других авторов, и дана дальнейшая детализация исследований по оценке структуры и функции пигментов, участвующих в фотосинтезе зеленого листа, где на основании анализа длинноволнового пика в спектре действия фотосинтеза в красной области спектра предпринята попытка оценить активность разных форм хлорофиллов, поглощающих свет в этом спектральном диапазоне.
Таким образом, различными исследователями в той или иной форме подтверждается универсальность кривых спектров действия фотосинтеза зеленых листьев разных видов растений, в основе которых, очевидно, лежит общность структурных и функциональных характеристик зеленых пигментов, обеспечивающих фотосинтез листа. Однако в условиях длительного (многосуточного) выращивания растений их фотосинтез определяется не только особенностями структуры и функции пигментного аппарата, но и другими параметрами, среди которых важную роль играют темновые реакции фотосинтеза. Их влияние на формирование структуры спектра действия фотосинтеза может быть учтено, если сами спектры действия фотосинтеза будут записываться при достаточно высоких уровнях облученности, выходящих за линейный участок световой кривой фотосинтеза.
Попытка построения спектров действия фотосинтеза при использовании высоких (насыщающих фотосинтез) уровней облученности, когда фотосинтез зависит не только от световых, но и темновых реакций, предпринята Маклеодом [McLeod, 1961]. В этом случае спектр действия фотосинтеза имел более уплощенную форму по сравнению с обычным, однако в принципиальном отношении его структура пе претерпела изменений. К сожалению,
эта работа выполнена только на водорослях. Возможно, что на зеленых листьях высших растений результат такого исследования может оказаться иным. Е. И. Ефимцев с соавторами [1978] предложили и обосновали методику построения спектров действия фотосинтеза при использовании высоких уровней облучения. Авторы этой работы убедительно показали, что в таких случаях спектр действия фотосинтеза корректно измерять по обратным значениям интенсивностей света, необходимых для поддержания постоянной скорости при всех длинах волн. Реализовать предложенную методику снятия спектров действия фотосинтеза при пс,пользовании высоких облученностей авторы работы смогли на хлорелле, а листья растений не исследовали.
Таким образом, достаточно полных данных о влиянии высоких уровней облученности на формирование структуры спектра действия фотосинтеза в литературе нами не обнаружено. Однако косвенные данные, полученные на хлорелле [McLeod, 1961], укалывают, что такие спектры действия в целом сохраняют свою структуру, которая делается более уплощенной, поскольку пики и синей и красной областях становятся более размытыми, чем у спектров действия фотосинтеза, полученных при использовании монохроматических потоков низкой интенсивности.
Уплощение структуры спектра действия фотосинтеза с ростом уровня облученности согласовывается с ранее широко распространенным мнением, что спектральная эффективность действия излучения в фотосинтезе выравнивается при высоких облученностях [Мошков, 1966; Cathey, Campbell, 1977 ]. Такой вывод был распространен этими исследователями на длительные условия выращивания растений. Последнее указывает на то, что данные по спектрам действия фотосинтеза переносились на оценку его спектральной эффективности при длительном выращивании без должного анализа корректности такого переноса. В данном случае не учитывалось влияние адаптации фотосинтетического аппарата листа к длительному воздействию света того или иного спектрального состава. Дело в том, что при спятии обычных спектров действия фотосинтеза зеленый лист экспонируется в лучах монохроматического потока ФАР той или иной длины волны около 3—5 мин IMcCree, 1972а; Inada, 1977]. Причем фотосинтетический аппарат листа в этих случаях предварительно формировался либо при солнечном, либо при близком к равпоэпергетическому искусственном освещении. Таким образом, при снятии классических спектров действия фотосинтеза зеленого листа никак не учитывался фактор длительного пребывания листа на свету различного спектрального состава и интенсивности.
Попытка оценить спектральную эффективность фотосинтеза листа при длительном пребывании на высокоинтенсивном краспом или синем свету была предпринята в исследованиях Н. П. Воскресенской [1965]. В этой работе показано, что наиболее эффективным может быть как красный, так и синий свет в зависимости от вида растения. В отличие от этих исследований, где использо-
24
25
вался широкополосный свет, В. М. Гольд [1969, 1975] изучал фотосинтез листьев разных видов при высокоинтенсивном излучении отдельных резонансных линий в коротковолновой области ФАР и широкополосном излучении в красной области спектра. Им получены результаты, сходные с данными работы Н. П. Воскресенской [1965]. В отмеченных работах длительность воздействия излучения на лист колебалась от 20—40 мин до нескольких часов. Эти исследования показали, что спектральная эффективность высокоиптенсивного излучения даже при таком относительно небольшом времени адаптации к нему фотосинтетического аппарата листа уже неодинакова в различных областях ФАР. В данном случае, а также при дальнейшем изложении материала мы будем понимать под адаптацией всю совокупность характеристик фотосинтетического аппарата растительного объекта (листа, растения, фитоценозов), обеспечивающих поддержание его жизнедеятельности в тех или иных измененных режимах облучения.
Логично предположить, что такого рода адаптивные изменения фотосинтетического аппарата листа, которые произошли за 1—2 ч, связаны с перестройкой работы механизмов тонкой регуляции фотосинтеза и не могут идти в сравнение с более глубокими адаптационными перестройками в структуре и функции фотосинтетического аппарата, которые связаны с процессом фотоморфогенеза и происходят при многосуточном выращивании растений в лучах того или иного спектрального состава ФАР. При длительном воздействии излучения на фотосинтетический аппарат регистрируемый фотосинтетический ответ подготовлен работой всей фоторегуляторной системы, включающей в себя реакции, присущие длительному воздействию излучения определенного спектрального состава на фотосинтетический аппарат растений. Особенности протекания таких реакций в конечном итоге оказывают решающее воздействие на величину фотосинтеза, либо ингибируя последний, например, при длительном воздействии красного света, либо поддерживая его на стационарном уровне, например, при длительном воздействии синих лучей [Тихомиров и др., 1983, 1987].
Различия по воздействию кратковременного и длительно действующего излучения различного спектрального состава на растения легко объяснимы, если учесть, что при длительном воздействии излучения на фотосинтетический аппарат ответная реакция растений есть результат работы всей фоторегуляторной системы, включающей реакции быстрой и медленной фоторегуляции, которые и определяют метаболические процессы, влияющие на специфику обмена веществ, а через него на особенности морфогенеза и фотосинтеза [Воскресенская, 1975]. Ничего подобного не происходит при кратковременном (минутном) воздействии излучения того или иного спектрального состава (в том числе и красных лучей). В этом случае регистрируется лишь чисто фотосинтетический ответ, а все метаболические, морфологические и другие эффекты, способные оказать на него влияние, отсутствуют.
26
Рис. 2.2. Обобщенный спектр дей-< п1ия фотосинтеза листьев редиса, сформировавшихся в лучах отдельных областей ФАР различной интенсивности.
I - для облученности при предвари-le.ui.noM выращивании при 50 Вт/м2 ФАР < нинасыщающий уровень ФАР); 2 — при Вт/м2 ФАР (световое насыщение фото-i-цптгаа); 3 — спектр действия фотосинтезе по данным Маккри [МсСгее, 1972] и Инада [Inada, 1976, 1977].
Поскольку для выращивания растений необходимо длительное воздействие излучения определенного спектрального состава, то представляется интересным построить спектры действия фотосинтеза с учетом тех механизмов фоторегуляции, которые имеют место при длительном выращивании растений в лучах различных длин волн. При построении таких спектров использовали листья, сформировавшиеся в синих (400—500 нм), зеленых (500—600 нм) н красных (600—700 нм) лучах [Тихомиров и др., 1987]. Первоначально для этих трех групп листьев сняли спектры действия фотосинтеза по обычной методике с использованием интерференционных фильтров, имеющих полуширину 20 нм. На основании этих спектров действия был составлен «обобщенный» спектр действия фотосинтеза, для чего из каждого первоначально полученного спектра действия была взята только та часть кривой, которая соответствовала спектральному диапазону излучения, в котором сформировался данный лист. Иными словами, в «обобщенном» спектре действия фотосинтеза участок кривой в диапазоне 100—500 нм был взят от спектра действия фотосинтеза листа, выросшего при излучении, сосредоточенном в этом спектральном интервале, участок кривой в диапазоне 500—600 нм — от спектра действия фотосинтеза листа, сформировавшегося при излучении спектрального диапазона 500—600 нм. Аналогично было сделано и для участка кривой в диапазоне 600—700 нм. Таким образом был построен «обобщенный» спектр действия фотосинтеза для всего диапазона ФАР с учетом длительного воздействия излучения отдельных областей видимого спектра па фотосинтетический аппарат листа. При таком методическом подходе характер кратковременных фотосинтетических ответов обусловлен структурными и функциональными изменениями, происшедшими в листовом аппарате при длительном воздействии излучения той физиологически активной области спектра, в пределах которой ищется фотосинтетический ответ.
Для листа редиса построен «обобщенный» спектр действия фотосинтеза листа, адаптированного к излучению отдельных спектральных областей ФАР (рис. 2.2). Критический анализ рис. 2.2 может вызвать возражение, что представленный на нем «обобщенный» спектр действия, полученный по вышеуказанной методике, представляет собой совокупность трех спектров дейст
27
вия для отдельных областей ФАР и трех разных листьев, сформировавшихся при излучении этих спектральных областей. Однако объединяет такие спектры действия то, что они получены на листьях, имеющих общую генетическую и физиологическую основу, и различия в фотосинтезе этих листьев, обусловленные соответствующими различиями в структуре и функции фотосинтетического аппарата и индуцированные под влиянием света различного спектрального состава, носят обратимый характер. Такая обратимость для красного и синего света была проверена Н. П. Воскресенской [1975], а также нами в отдельных экспериментах, когда у растений (возраст 10 сут), переставленных с излучения одного спектрального состава света на другой, листья через 2— 3 сут приобретали все основные характеристики, присущие листьям, постоянно растущим на данном свету. Обратимый характер воздействия излучения исследуемых спектральных диапазонов на организацию фотосинтетического аппарата и дает основание объединить совокупность фотосинтетических ответов в отдельных областях ФАР в единый спектр действия фотосинтеза листа для всего диапазона ФАР.
С увеличением уровня облученности, при котором формировался исследуемый растительный материал, происходит трансформация структуры такого спектра действия фотосинтеза листа, в частности, максимум фотосинтеза перемещается из красной в синюю область спектра (см. рис. 2.2). Это связано с тем, что с ростом уровня облученности спектральная эффективность синих лучей для фотосинтеза редиса растет, а красных падает [Tikhomirov, Sid’ko, 1982; Тихомиров и др., 1983]. Таким образом, анализ возможностей использования спектров действия фотосинтеза, полученных в работах Маккри [МсСгее, 1972а] и Инада [Inada, 1977] (см. рис. 2.1) для оценки спектральной эффективности этого процесса в условиях долговременного выращивания растений, показал, что имеются причины, которые могут исказить эту оценку. Среди них следует назвать в первую очередь кратковременность воздействия и низкие уровни используемых облученностей, а также отсутствие длительной адаптации к спектру и интенсивности лучистых потоков. Все это характерно для классических спектров действия фотосинтеза, и поэтому кривые таких спектров не могут характеризовать спектральную эффективность фотосинтеза при длительных (многосуточных) режимах выращивания растений.
Максимально приближенным к длительным условиям выращивания является «обобщенный» спектр действия фотосинтеза зеленого листа (см. рис. 2.2). Он построен с учетом длительного воздействия излучения определенного спектрального состава и интенсивности на фотосинтетический аппарат листа, и с изменением этих параметров структура такого спектра действия фотосинтеза может меняться. В силу этого есть основание считать, что форма «обобщенного» спектра действия фотосинтеза может в целом отражать спектральную эффективность этого процесса для данного листа при длительных режимах его выращивания.
28
§ 2. Фотосинтез листа при длительном воздействии излучения различного спектрального состава и интенсивности
Попытки оцепить влияние света различного спектрального состава на фотосинтез растений в условиях их длительного выращи-IUI в ня при искусственном освещении предпринимались почти сразу после появления возможности использовать для этих целой электрический свет. Однако отсутствие в то время достаточно надежных приборов для измерения света и регистрации фотосинтеза в значительной степени обесценивало результаты проводимых исследований. Эти трудности стали заметно преодолеваться лишь в послевоенный период.
Многочисленные исследования длительного влияния спектрального состава света на фотосинтез растений связаны с изучением фотосинтеза листа при излучении различных источников спета. Обзор ранних работ по этому вопросу дан в монографии В. М. Лемана [1971]. В. М. Леман провел ряд таких исследований, используя наиболее распространенные в то время источники для искусственного освещения растений. Измеренные им значения фотосинтеза листьев растений, длительно выращиваемых под люминесцентными, неоновыми, ртутными, ксеноновыми лампами и лампами накаливания, показали, что наиболее высокие значения фотосинтеза дают ксеноновые лампы, несколько хуже — люминесцентные и лампы ДРЛ, а остальные источники света дают значительно худшие результаты [Леман, 19711. К сожалению, по этим данным можно лишь ориентировочно оценить спектральную эффективность действия излучения отдельных областей ФАР и их сочетаний. Это связано, с одной стороны, с отсутствием в указанной работе данных по выравниванию используемых для измерения фотосинтеза интенсивностей света и температуры, а с другой — с различиями у использованных источников света спектральных характеристик излучения не только в видимой, но и в инфракрасной и ультрафиолетовой областях спектра, что также может повлиять на интенсивность фотосинтеза.
Более поздние исследования по изучению длительного воздействия света различного спектрального состава на фотосинтез листьев растений уже позволяют сделать выводы об эффективности тех или иных лучей спектра ФАР в фотосинтезе при их длительном воздействии на лист. Такие работы в большинстве выполнены либо на проростках, либо на молодых растениях. Так, в работе В. Г. Гапонепка с соавторами [1979] на листьях 18-дневных растений кукурузы показано, что фотосинтез на белом свету люминесцентных ламп и зеленом свету ртутно-таллиевых ламп при облученностях 50—60 Вт/м2 ФАР фактически одинаков. В другой работе на 7—8-дневных растениях ячменя исследовали влияние излучения синих, зеленых и красных люминесцентных ламп на фотосинтетическую активность листьев растений [Гапо-ненка и др., 1982]. Показано, что интенсивность фотосинтеза уменьшалась в порядке синий — зеленый — красный свет. К со-
29
жалению, в данной работе использовали весьма низкие облученности порядка нескольких единиц облученности ФАР, которые в 5—10 раз ниже, чем в реальных условиях выращивания растений. С другой стороны, синий свет, на котором получена максимальная фотосинтетическая активность листьев ячменя, имел примесь как зеленых, так и красных лучей, что могло завысить его фотосинтетическую эффективность.
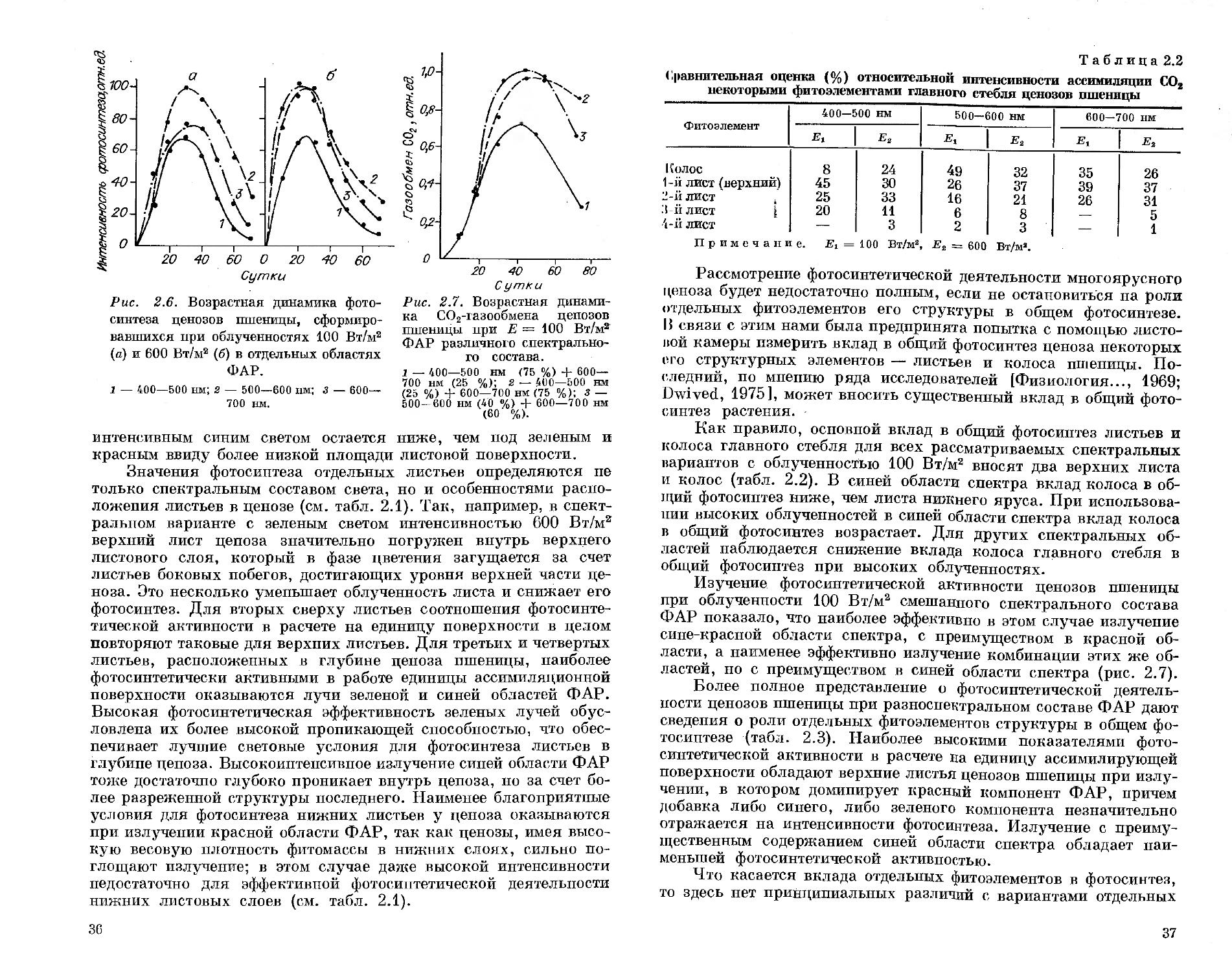
Интересные данные о спектральной эффективности белого, зеленого, красного и синего света в фотосинтезе зеленого листа бадана толстолистого и серпухи венценосной опубликованы в работе Р. А. Карначук [1987]. У световых кривых фотосинтеза листьев этих культур, выращенных при таких спектральных режимах облучения, плато светового насыщения понижалось в направлении синий — красный — белый — зеленый свет. Это было справедливо как для молодых, так и для взрослых листьев. Интересен вывод относительно зеленого света. Фотосинтез на нем в расчете на хлоропласт был наибольшим, однако ввиду малой концентрации хлоропластов на единицу поверхности листа фотосинтез последнего оставался на низком уровне. В работе сделан оригинальный вывод о важной роли зеленого света, регулирующего взаимосвязь роста и фотосинтеза, что обосновывает необходимость присутствия излучения этого спектрального диапазона в потоке ФАР для обеспечения нормальной регуляции сопряжения функций роста и фотосинтеза [Карначук, 1987].
В работе Н. Н. Протасовой [1987] на растениях перца оценивалась эффективность в фотосинтезе излучения синих, зеленых, красных, сине-красных и белых люминесцентных ламп. Для листьев, сформировавшихся при излучении этих ламп, записывали световые кривые фотосинтеза на белом свету. Такие измерения показали максимальный фотосинтез для листьев, выращенных на синем свету, с последовательным его уменьшением после выращивания на сине-красном (25 % синего света), белом, красном и зеленом свету.
Важное значение для понимания роли соотношения синих и красных лучей в смешанном спектральном составе ФАР в обеспечении высокого фотосинтеза листьев растений имеют работы И. С. Дроздовой с соавторами [1987] и А. В. Кумакова [1988]. Эти работы рассматривают механизмы фоторегуляции процесса фотосинтеза при разном соотношении синего и красного света. Показано, что повышение содержания синего света более 35 % в сине-красном излучении ФАР вряд ли целесообразно в интенсификации фотосинтеза зеленого листа. Этот интересный вывод, позволяющий глубже понять эффекты фоторегуляции фотосинтеза на свету различного спектрального состава, не может быть, к сожалению, в полной мере использован для оценки спектральной эффективности фотосинтеза в реальных условиях выращивания растений. Это связано с тем, что используемые в экспериментах облученности ФАР были весьма низкими, а само деление области ФАР пополам без учета роли зеленого света весьма спор-
но, особенно если речь идет о продуктивном фотосинтезе, где должно быть учтено скоординированное взаимодействие функций роста и фотосинтеза, в значительной мере зависящее от оптимального содержания зеленых лучей в потоке ФАР [Карначук, 1987]. ,
Общей особенностью вышеуказанных работ по оценке спектральной эффективности фотосинтеза зеленого листа является оценка фотосинтеза в условиях, не соответствующих нативным условиям жизнедеятельности листа, поэтому полученные результаты скорее характеризуют потенциальные, чем реальные, возможности фотосинтетического аппарата растений, сформировавшихся в данных спектральных условиях облучения. Это затрудняет использование таких данных по фотосинтезу растений для оценки его спектральной эффективности в условиях длительного выращивания.
Среди немногих работ, в которых приведены данные по спектральной эффективности фотосинтеза листьев растений, когда фотосинтез измеряли на том же спектральном свету, на котором росли растения, являются работы японских исследователей Ко, Ябуки [Ко, Yabuky, 1974] и Ко [Ко, 1982]. Используя разное по спектральному составу излучение люминесцентных ламп, они показали, что наиболее фотосинтетически активна красная область ФАР, а наименее — зеленая.
При оценке спектрально!! эффективности фотосинтеза зеленого листа важно знать, влияет ли на нее интенсивность излучения исследуемого спектрального интервала. Весьма ограниченная попытка дать такую оценку сделана в работе Ко [Ко, 1982]. Ее ограниченность связана с использованием низких уровней облученности 12 и 24 Вт/м2 ФАР, которые соответствуют линейному участку световой кривой фотосинтеза листа. Поэтому не удивительно, что принципиальных изменений в спектральной эффективности фотосинтеза зеленого листа для красной области спектра в этой работе в сравнении с данными работы Ко и Ябуки [Ко, Yabuky, 1974] не получено. Однако оценить роль интенсивности красного света в длительной фотосинтетической активности листа представляется весьма важным. В связи с этим нами было продолжено исследование длительного влияния красного света на фотосинтез листьев растений огурца в сравнении с излучением других участков спектра при облученностях 50 и 100 Вт/м2. В работе исследовали листья растений огурца сорта Московский тепличный, которые 25 сут выращивали при облученностях 50 и 100 Вт/м2 в синей, зеленой и красной областях спектра, создаваемых квази-монохроматическими лампами в сочетании с цветными светофильтрами (см. гл. 1, рис. 1.4).
При сопоставлении этих данных с данными, полученными японским исследователем Ко [Ко, 1982] при работе с низкими облученностями (рис. 2.3, а, б), видно, что спектральная эффективность фотосинтеза зеленого листа огурца может измениться с ростом уровня облученности, при которой предварительно выращива-
30
31
Рис. 2.3. Фотосинтез листьев огурца, сформировавшихся в синих (с), зеленых (з) и красных (к) лучах различной интенсивности. Е, Вт/м» ФЛР: а — 12; б — 24. По: Но [Ко, 1982]; в — 50; г — 100. По: [Тихомиров п др., 1983].
ли листья, вплоть до противоположной. Интересно, что в последнем случае возрастает эффективность синего света. Это указывает на то, что при достаточно высоких облученностях происходит реализация потенциальной эффективности синего света. Однако при низких
уровнях облученности, как показали, в частности, данные работы Ко [Там же], такая потенциальная фотосинтетическая эффективность синего света может и не быть реализована, а максимальный фотосинтез в этих случаях, как правило, наблюдается в красных лучах. Таким образом, расхождения в интенсивности фотосинтеза в нативных и иных условиях измерения могут носить не только количественный, но и качественный характер, связанный со смещением области максимальной спектральной эффективности фотосинтеза. Главная причина возникающих различий связана, на наш взгляд, с характером предыстории формирования растительного материала, являющегося объектом исследования. Если при измерении потенциального фотосинтеза листа используются спектральные режимы облучения, отличающиеся от стационарных условий выращивания растений, то при прямых измерениях фотосинтетический аппарат листа формировался при длительном излучении той спектральной области, в пределах которой исследовали фотосинтетический ответ.
Из сказанного ясно, что надежная оценка спектральной эффективности фотосинтеза листа может быть получена лишь при проведении прямых измерений фотосинтеза непосредственно в тех световых условиях, в которых сформировался фотосинтетический аппарат листа. В этой связи любая экстраполяция может сильно исказить истинную фотосинтетическую активность листа в тех или иных световых условиях его произрастания.
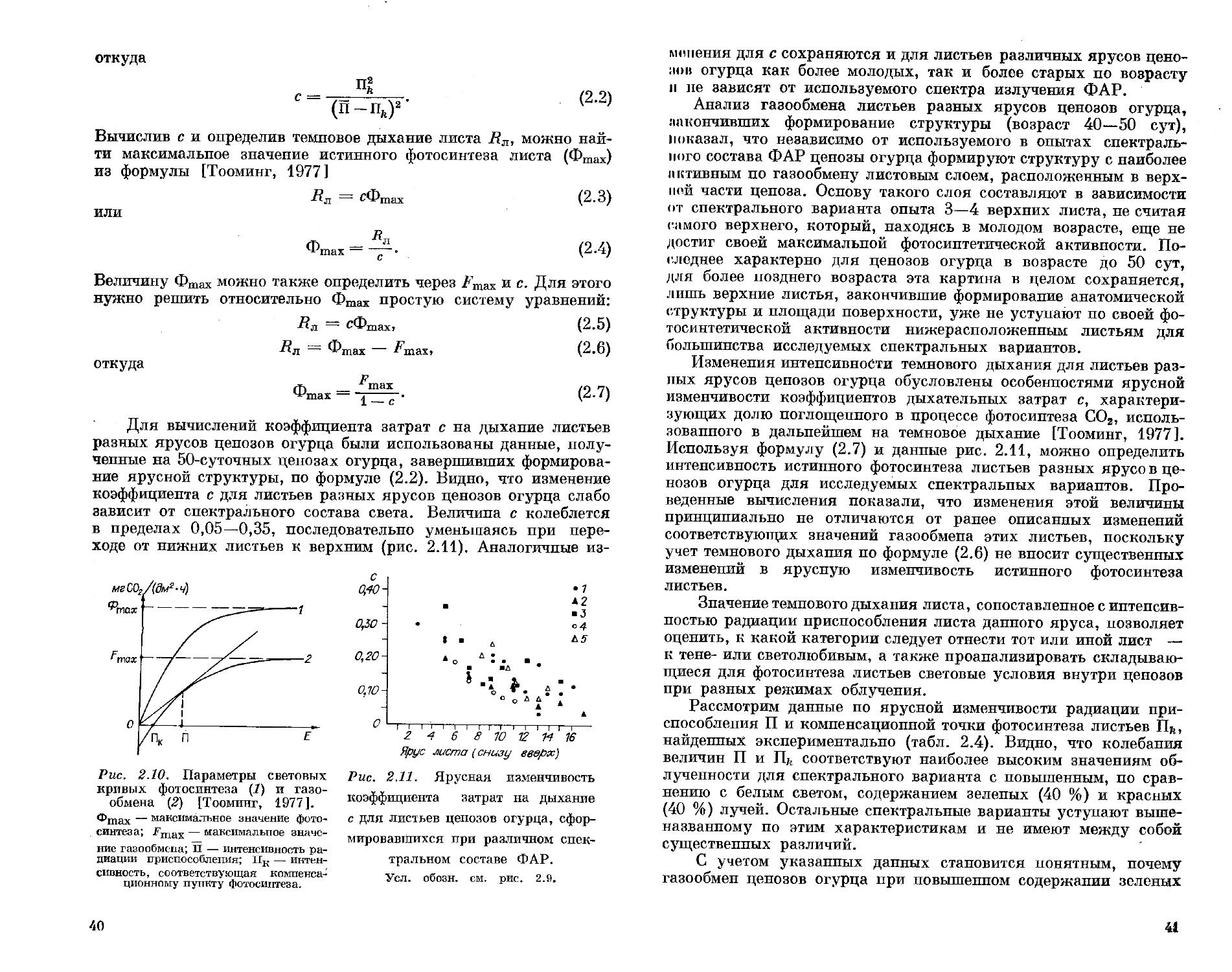
Однако, даже зная спектральную эффективность фотосинтеза для листа растения, нельзя гарантировать ее адекватный перенос на ценоз. В лучшем случае такой перенос возможен для отдельно стоящих растений. Конечно, обеспечив оптимальный по спектральным и энергетическим характеристикам режим облучения отдельных листьев, можно добиться их максимального фотосинтеза. Однако таким способом невозможно обеспечить максимальный фотосинтез всех листьев ценоза, где формируется реальный урожай. X. Г. Тоомипг [1984] отмечает, что в полноценном ценозе листья
32
работают не с максимальной интенсивностью фотосинтеза, а с оптимальной интенсивностью, соответствующей зоне интенсивности радиации приспособления (ИРП) и максимальному КПД. Отсюда напрашивается вывод, что механический перенос данных по с, центральной эффективности фотосинтеза отдельных листьев или их частей на ценоз без учета его структурных особенностей не корректен.
Поэтому вполне обоснованной выглядит необходимость оценивать спектральную эффективность фотосинтеза растений для условий их длительного выращивания, проводя прямые эксперименты на фитоценозах, используя световые режимы облучения с теми спектральными и энергетическими характеристиками, которые реально пригодны для условий формирования полноценного урожая.
§ 3. Спектральная эффективность фотосинтеза ценозов растений
Систематических исследований по влиянию излучения различного спектрального состава на фотосинтез ценозов растений в литературе нами не обнаружено. Встречаются лишь отдельные работы, где оценивается фотосинтез ценозов при излучении отдельных источников света различного спектрального состава [Ни-ловская, 1973; Чучалин, 1980; Александрова, Радомысльская, 1980; и др. ]. В то же время для решения данной задачи требуется комплексная оценка воздействия излучения как отдельных спектральных областей ФАР, так и их различных комбинаций на фотосинтетическую активность ценозов на протяжении всего вегетационного периода при константных режимах выращивания.
Исследования в этом направлении проведены в Институте биофизики СО АН СССР. Условия выращивания растений описаны выше (см. гл. 1, § 1—4).
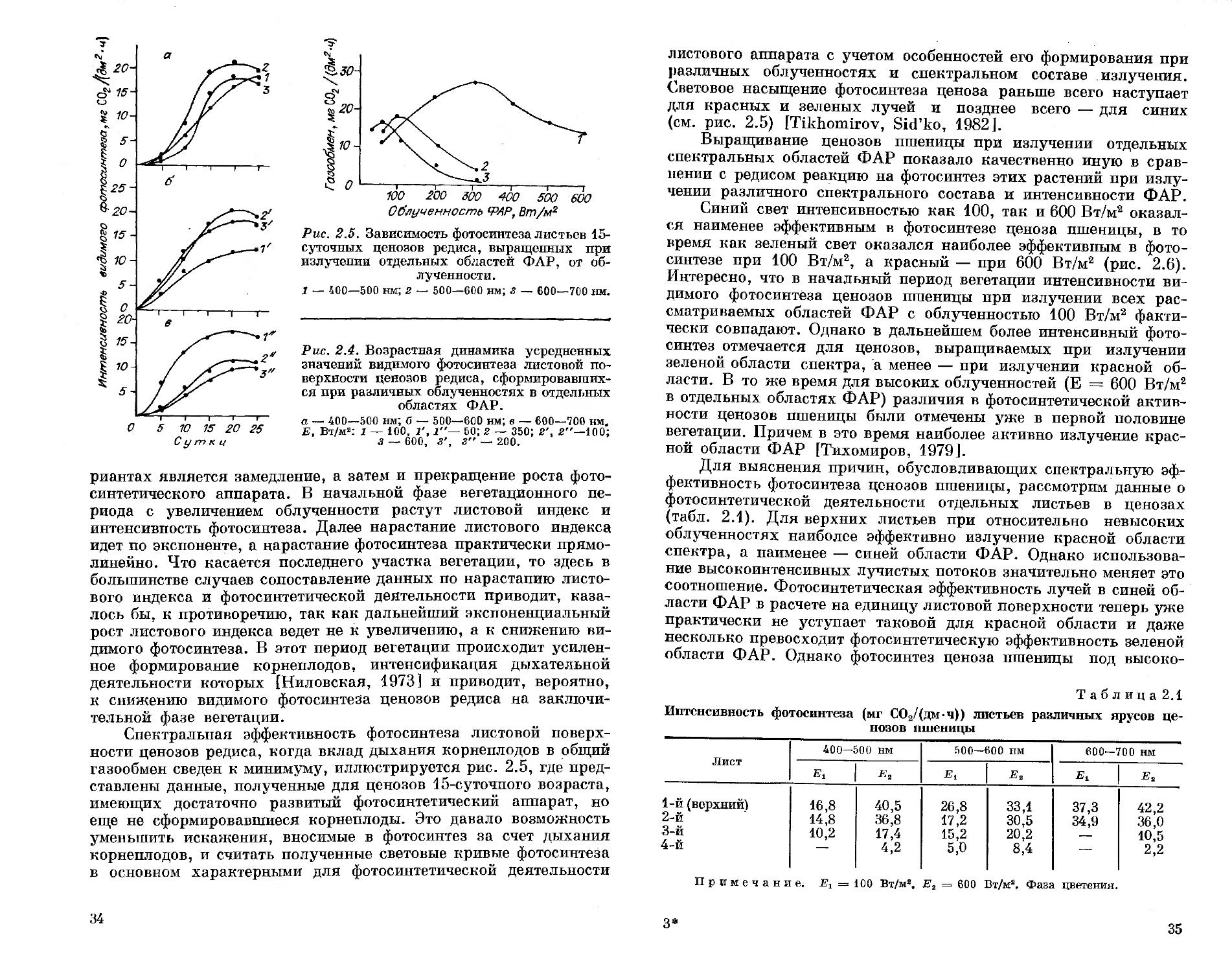
С повышением облученности в синей области ФАР возрастные кривые фотосинтеза ценозов редиса приближаются к 5-образной кривой, причем с ростом облученности увеличивается период лаг-фазы (рис. 2.4). В зеленой и красной областях ФАР ход возрастных кривых фотосинтеза имеет более плавный характер, а сама фотосинтетическая активность ценозов ингибируется при значительно меньших облученностях, чем в синей области спектра [Тихомиров, 1977].
Общим для всех спектральных вариантов является нарастание фотосинтеза ценозов в первой половине вегетационного периода. В этот период кривая зависимости видимого фотосинтеза от возраста ценоза характеризуется прямолинейным участком. Общность хода возрастных кривых фотосинтеза ценозов редиса в различных спектральных вариантах может быть объяснена соответствующей общностью протекания ряда ростовых и органообразовательных процессов. Действительно, общей чертой действия нарастающих интенсивностей в различных спектральных ва-
3 А. А. Тихомиров, Г. М. Лисовский, Ф. Я. Сидько
33
риантах является замедление, а затем и прекращение роста фотосинтетического аппарата. В начальной фазе вегетационного периода с увеличением облученности растут листовой индекс и интенсивность фотосинтеза. Далее нарастание листового индекса идет по экспоненте, а нарастание фотосинтеза практически прямолинейно. Что касается последнего участка вегетации, то здесь в большинстве случаев сопоставление данных по нарастанию листового индекса и фотосинтетической деятельности приводит, казалось бы, к противоречию, так как дальнейший экспоненциальный рост листового индекса ведет не к увеличению, а к снижению видимого фотосинтеза. В этот период вегетации происходит усиленное формирование корнеплодов, интенсификация дыхательной деятельности которых (Ниловская, 1973] и приводит, вероятно, к снижению видимого фотосинтеза ценозов редиса на заключительной фазе вегетации.
Спектральная эффективность фотосинтеза листовой поверхности ценозов редиса, когда вклад дыхания корнеплодов в общий газообмен сведен к минимуму, иллюстрируется рис. 2.5, где представлены данные, полученные для ценозов 15-суточпого возраста, имеющих достаточно развитый фотосинтетический аппарат, но еще не сформировавшиеся корнеплоды. Это давало возможность уменьшить искажения, вносимые в фотосинтез за счет дыхания корнеплодов, и считать полученные световые кривые фотосинтеза в основном характерными для фотосинтетической деятельности
34
листового аппарата с учетом особенностей его формирования при различных облученностях и спектральном составе излучения. Световое насыщение фотосинтеза ценоза раньше всего наступает для красных и зеленых лучей и позднее всего — для синих (см. рис. 2.5) [Tikhomirov, Sid’ko, 1982].
Выращивание ценозов пшеницы при излучении отдельных спектральных областей ФАР показало качественно иную в сравнении с редисом реакцию на фотосинтез этих растений при излучении различного спектрального состава и интенсивности ФАР.
Синий свет интенсивностью как 100, так и 600 Вт/м2 оказался наименее эффективным в фотосинтезе ценоза пшеницы, в то время как зеленый свет оказался наиболее эффективным в фотосинтезе при 100 Вт/м2, а красный — при 600 Вт/м2 (рис. 2.6). Интересно, что в начальный период вегетации интенсивности видимого фотосинтеза ценозов пшеницы при излучении всех рассматриваемых областей ФАР с облученностью 100 Вт/м2 фактически совпадают. Однако в дальнейшем более интенсивный фотосинтез отмечается для ценозов, выращиваемых при излучении зеленой области спектра, а менее — при излучении красной области. В то же время для высоких облученностей (Е = 600 Вт/м2 в отдельных областях ФАР) различия в фотосинтетической активности ценозов пшеницы были отмечены уже в первой половине вегетации. Причем в это время наиболее активно излучение красной области ФАР [Тихомиров, 1979].
Для выяснения причин, обусловливающих спектральную эффективность фотосинтеза ценозов пшеницы, рассмотрим данные о фотосинтетической деятельности отдельных листьев в ценозах (табл. 2.1). Для верхних листьев при относительно невысоких облученностях наиболее эффективно излучение красной области спектра, а наименее — синей области ФАР. Однако использование высокоинтенсивных лучистых потоков значительно меняет это соотношение. Фотосинтетическая эффективность лучей в синей области ФАР в расчете на единицу листовой поверхности теперь уже практически не уступает таковой для красной области и даже несколько превосходит фотосинтетическую эффективность зеленой области ФАР. Однако фотосинтез ценоза пшеницы под высоко-
Т а б лица 2.1
Интенсивность фотосинтеза (мг СО2/(дм-ч)) листьев различных ярусов ценозов пшеницы
Лист 400—500 нм 500—600 нм 600—700 нм
к2 Е, Е2 Е, Е,
1-й (верхний) 16,8 40,5 26,8 33,1 37,3 42,2
2-й 14,8 36,8 17,2 30,5 34,9 36,0
3-й 10,2 17,4 15,2 20,2 — 10,5
4-й — 4,2 5,0 8,4 — 2,2
Примечан! е. Е, = 100 Вт/м2. Е2 = 600 Вт/м2, Фазг цветения.
3*
35
Рис. 2.6. Возрастная динамика фотосинтеза ценозов пшеницы, сформировавшихся при облученностях 100 Вт/м2 (я) и 600 Вт/м2 (б) в отдельных областях ФАР.
1 — 400—500 нм; 2 — 500—600 нм; 3 — 600— 700 нм.
ка СО2-газообмена ценозов пшеницы при Е = 100 Вт/м2 ФАР различного спектрального состава.
1 — 400—500 нм (75 %) + 600— 700 нм (25 %); 2 — 400—500 нм (25 %) + 600—700 нм (75 %); 3 — 500—600 нм (40 %) + 600—700 нм (60 %).
интенсивным синим светом остается ниже, чем под зеленым и красным ввиду более низкой площади листовой поверхности.
Значения фотосинтеза отдельных листьев определяются не только спектральным составом света, но и особенностями расположения листьев в ценозе (см. табл. 2.1). Так, например, в спектральном варианте с зеленым светом интенсивностью 600 Вт/м2 верхний лист цепоза значительно погружен внутрь верхнего листового слоя, который в фазе цветения загущается за счет листьев боковых побегов, достигающих уровня верхней части ценоза. Это несколько уменьшает облученность листа и снижает его фотосинтез. Для вторых сверху листьев соотношения фотосинтетической активности в расчете на единицу поверхности в целом повторяют таковые для верхних листьев. Для третьих и четвертых листьев, расположенных в глубине ценоза пшеницы, наиболее фотосинтетически активными в работе единицы ассимиляционной поверхности оказываются лучи зеленой и синей областей ФАР. Высокая фотосинтетическая эффективность зеленых лучей обусловлена их более высокой проникающей способностью, что обеспечивает лучшие световые условия для фотосинтеза листьев в глубине ценоза. Высокоинтенсивное излучение синей области ФАР тоже достаточно глубоко проникает внутрь цепоза, но за счет более разреженной структуры последнего. Наименее благоприятные условия для фотосинтеза нижних листьев у ценоза оказываются при излучении красной области ФАР, так как ценозы, имея высокую весовую плотность фитомассы в нижних слоях, сильно поглощают излучение; в этом случае даже высокой интенсивности недостаточно для эффективной фотосинтетической деятельности нижних листовых слоев (см. табл. 2.1).
36
Таблица 2.2
Сравнительная оценка (%) относительной интенсивности ассимиляции СО2 некоторыми фитоэлементами главного стебля ценозов пшеницы
Фитоэлемент 400—500 нм 500—600 нм 600—700 нм
К, Ег Et Ег К, Ег
Колос 8 24 49 32 35 26
1-й лист (верхний) 45 30 26 37 39 37
2-й лист 25 33 16 21 26 31
3 й лист [ 20 11 6 8 — 5
4-й лист — 3 2 3 — 1
Примечание. Et = 100 Вт/м2, Е2 == 600 Вт/м2.
Рассмотрение фотосинтетической деятельности многоярусного цепоза будет недостаточно полным, если не остановиться на роли отдельных фитоэлементов его структуры в общем фотосинтезе. В связи с этим нами была предпринята попытка с помощью листовой камеры измерить вклад в общий фотосинтез ценоза некоторых его структурных элементов — листьев и колоса пшеницы. Последний, по мнению ряда исследователей [Физиология..., 1969; Dwived, 1975], может вносить существенный вклад в общий фотосинтез растения.
Как правило, основной вклад в общий фотосинтез листьев и колоса главного стебля для всех рассматриваемых спектральных вариантов с облученностью 100 Вт/м2 вносят два верхних листа и колос (табл. 2.2). В синей области спектра вклад колоса в общий фотосинтез ниже, чем листа нижнего яруса. При использовании высоких облученностей в синей области спектра вклад колоса в общий фотосинтез возрастает. Для других спектральных областей наблюдается снижение вклада колоса главного стебля в общий фотосинтез при высоких облученностях.
Изучение фотосинтетической активности ценозов пшеницы при облученности 100 Вт/м2 смешанного спектрального состава ФАР показало, что наиболее эффективно в этом случае излучение сине-красной области спектра, с преимуществом в красной области, а наименее эффективно излучение комбинации этих же областей, по с преимуществом в синей области спектра (рис. 2.7).
Более полное представление о фотосинтетической деятельности ценозов пшеницы при разпоспектралыюм составе ФАР дают сведения о роли отдельных фитоэлементов структуры в общем фотосинтезе (табл. 2.3). Наиболее высокими показателями фотосинтетической активности в расчете па единицу ассимилирующей поверхности обладают верхние листья ценозов пшеницы при излучении, в котором доминирует красный компонент ФАР, причем добавка либо синего, либо зеленого компонента незначительно отражается на интенсивности фотосинтеза. Излучение с преимущественным содержанием синей области спектра обладает наименьшей фотосинтетической активностью.
Что касается вклада отдельных фитоэлементов в фотосинтез, то здесь нет принципиальных различий с вариантами отдельных
37
Таблица 2.3
Сравнительная фотосинтетическая активность фитоэлементов главного стебля ценозов пшеницы при различных спектральных режимах
Фитоэлемент с - 75 %, к - 25 % с - 25 %, к - 75 % 3 — 40 %, к-60 %
1 2 1 2 1 2
Колос — 28 - 38 — 45 -
1-й лист (верхний) 26,5 37 39,4 32 38,8 26
2-й лист 19,8 20 26,5 22 30,1 19
3-й лист 11,5 11 12,2 7 14,4 8
4-й лист 3,3 4 — — 6,4 2
Примечание. 1 — интенсивность фотосинтеза, мг СО2/(дмг-ч), 2 — доля усвоения СО2, %, с — 400—500 нм; в — 500—600 нм; к — 600—700 нм. Фаза цветения.
областей ФАР, если сравнивать их с соответствующими вариантами, где доминирует одна из областей ФАР.
Следует заметить, что если сравнивать вариант с синим светом и сине-красным, где доминирует синий, то можно отметить, что в последнем случае роль колоса возрастает почти в 3 раза, а эффективность фотосинтеза верхних листьев — в среднем в 1,5 раза. Тем не менее отсутствие боковых побегов, что характерно для структуры ценоза пшеницы при синем свете, не снимается 25%-й добавкой красного компонента.
Данные по фотосинтезу ценозов огурца при излучении отдельных областей ФАР представлены на рис. 2.8. Анализ этого рисунка показывает, что наиболее активно фотосинтезирующими вплоть до 20-суточного возраста являются растения огурца, выращиваемые в красных лучах, а наименее активно — в зеленых и синих. Однако в дальнейшем картина меняется. Фотосинтез ценозов под красным светом резко уменьшается, и к возрасту 30— 40 сут растения погибают вследствие полного выцветания зеленых пигментов листьев. Примерно аналогичная ситуация характерна и для спектрального варианта с зеленым светом, правда, здесь ингибирование фотосинтеза и отмирание растений растягиваются на более продолжительный срок. В то же время длительное выращивание растений огурца в синих лучах не приводит к ингибированию фотосинтеза. В этом спектральном варианте наблюдается монотонный рост фотосинтеза ценоза огурца в процессе вегетации вплоть до 50-суточного возраста, а дальнейшее некоторое снижение фотосинтеза связано с постепенным старением фотосинтетического аппарата.
Рис. 2.8. Возрастная динамика газообмена ценозов огурца Московский тепличный при облучении в отдельных областях ФАР интенсивностью 100 Вт/м2.
1 — 400—500 нм; 2 — 500—600 нм; 3 — 600—700 нм.
38
Рис. 2.9. Динамика газообмена ценозов огурца Московский тепличный, сформировавшихся при различном спектральном составе ФАР. « — спектральные режимы облучения с одним максимумом излучения в области ФАР: 1 — 400— 500 нм (20 %) + 500—600 нм (20 %) + 600—700 нм (60 %); 2 — 400—500 нм (20 %) + 500— 600 нм (60 %) + 600—700 нм (20 %); 3 — 400—500 нм (60 %) + + 500—600 нм (20%) + 600— 700 нм (20 %).
б — спектральные режимы облучения с двумя максимумами излучения в области ФАР и «белым» светом: 4 — 400—500 нм (20 %) + 500—600 нм (40 %) + 600—700 нм (40 %); 5 — 400—500 нм (34 %) + 500—600 нм (33 %) + 600—700 нм (33 %); 6 — 400—500 нм (40 %) + 500—600 нм (20 %) + 600—700 нм (40 %); 7 — 400—500 нм (40 %) + 500— 600 нм (40 %) + 600—700 нм (20 %).
Изучен газообмен целого ценоза огурца для режимов облучения с различными комбинациями отдельных спектральных областей в общем потоке ФАР (рис. 2.9). Видно, что наибольших значений газообмен достигает при излучении, имеющем максимум в красной (рис. 2.9, а) и зелено-красной (рис. 2.9, б) областях спектра. Существенная доля зеленых лучей в трехкомпонентном излучении ФАР не только не оказывает ингибирующий эффект на газообмен ценоза огурца, но и обусловливает его весьма высокие значения, особенно если зеленый свет дается в присутствии значительной доли красных лучей. В то же время повышенное содержание как синих, так и сине-зеленых лучей не способствует высокому газообмену ценозов огурца (см. рис. 2.9, а, б).
Рассмотренные различия в спектральной эффективности фотосинтеза ценозов зависят от комплекса факторов, среди которых важную роль играет фотосинтетическая активность листьев различных ярусов. Оценка этого параметра может быть проведена путем измерения и анализа световых кривых фотосинтеза отдельных листьев растений. Такой способ оценки привлекателен тем, что он позволяет достаточно комплексно оценить фотосинтетическую активность листа, определив как его фотосинтетические, так и дыхательные характеристики. Кроме того, можно оценить и световые условия, в которых находится лист в ценозе.
Анализ световой кривой газообмена листа с целью определения комплекса таких параметров, характеризующих фотосинтетическую и дыхательную функции листа, достаточно подробно разработан Тоомингом [1977] (рис. 2.10).
Взяв за основу экспериментальные данные, которые получены при снятии и анализе световых кривых листьев разных ярусов ценозов огурца при различных спектральных режимах их предварительного выращивания, можно вычислить ряд других важных характеристик фотосинтетической деятельности листа. Так, можно найти коэффициент затрат на дыхание с, пользуясь формулой [Тооминг, 1977 ]
П„ - П(УС7С>, (2.1)
39
откуда
Вычислив с и определив темновое дыхание листа 7?л, можно найти максимальное значение истинного фотосинтеза листа (Фтах) из формулы [Тооминг, 1977 ]
= сФтах (2-3)
ИЛИ
Фтах = 4^' <2Л)
Величину Фтах можно также определить через Fmax и с. Для этого
нужно решить относительно Фтах простую систему уравнений:
Rr — сФтах» (2.5)
= Фтах — ^тах, (2-6)
откуда
Ф™, - 4^- (2-7)
Для вычислений коэффициента затрат с на дыхание листьев разных ярусов ценозов огурца были использованы данные, полученные на 50-суточных ценозах огурца, завершивших формирование ярусной структуры, по формуле (2.2). Видно, что изменение коэффициента с для листьев разных ярусов ценозов огурца слабо зависит от спектрального состава света. Величина с колеблется в пределах 0,05—0,35, последовательно уменьшаясь при переходе от нижних листьев к верхним (рис. 2.11). Аналогичные из-
• 1
М
I . й Л5
I I 1 I I I I I I I I I I J Г”
2 4 6 8 10 12 14 16
Ярус листа (снизу вверх)
с 0,40-
0,10 -
0,20-
0,10-
Рис. 2.10. Параметры световых кривых фотосинтеза (7) и газообмена (2) [Тооминг, 1977].
Фтах — максимальное значение фотосинтеза; Гтах — максимальное значение газообмена; П — интенсивность радиации приспособления; Пк — интенсивность, соответствующая компенсационному пункту фотосинтеза.
Рис. 2.11. Ярусная изменчивость коэффициента затрат на дыхание с для листьев ценозов огурца, сформировавшихся при различном спектральном составе ФАР.
Усл. обозн. см. рис. 2.9.
40
лишения для с сохраняются и для листьев различных ярусов ценозов огурца как более молодых, так и более старых по возрасту и не зависят от используемого спектра излучения ФАР,
Анализ газообмена листьев разных ярусов ценозов огурца, закончивших формирование структуры (возраст 40—50 сут), показал, что независимо от используемого в опытах спектрального состава ФАР ценозы огурца формируют структуру с наиболее активным по газообмену листовым слоем, расположенным в верхней части ценоза. Основу такого слоя составляют в зависимости от спектрального варианта опыта 3—4 верхних листа, не считая самого верхнего, который, находясь в молодом возрасте, еще не достиг своей максимальной фотосинтетической активности. Последнее характерно для ценозов огурца в возрасте до 50 сут, для более позднего возраста эта картина в целом сохраняется, лишь верхние листья, закончившие формирование анатомической структуры и площади поверхности, уже не уступают по своей фотосинтетической активности нижерасположенным листьям для большинства исследуемых спектральных вариантов.
Изменения интенсивности темнового дыхания для листьев разных ярусов ценозов огурца обусловлены особенностями ярусной изменчивости коэффициентов дыхательных затрат с, характеризующих долю поглощенного в процессе фотосинтеза СО2, использованного в дальнейшем на темновое дыхание [Тооминг, 1977]. Используя формулу (2.7) и данные рис. 2.11, можно определить интенсивность истинного фотосинтеза листьев разных ярусо в ценозов огурца для исследуемых спектральных вариантов. Проведенные вычисления показали, что изменения этой величины принципиально не отличаются от ранее описанных изменений соответствующих значений газообмена этих листьев, поскольку учет темнового дыхания по формуле (2.6) не вносит существенных изменений в ярусную изменчивость истинного фотосинтеза листьев.
Значение темнового дыхания листа, сопоставленное с интенсивностью радиации приспособления листа данного яруса, позволяет оценить, к какой категории следует отнести тот или иной лист — к тене- или светолюбивым, а также проанализировать складывающиеся для фотосинтеза листьев световые условия внутри ценозов при разных режимах облучения.
Рассмотрим данные по ярусной изменчивости радиации приспособления П и компенсационной точки фотосинтеза листьев П&, найденных экспериментально (табл. 2.4). Видно, что колебания величин П и Щ соответствуют наиболее высоким значениям облученности для спектрального варианта с повышенным, по сравнению с белым светом, содержанием зеленых (40 %) и красных (40 %) лучей. Остальные спектральные варианты уступают вышеназванному по этим характеристикам и не имеют между собой существенных различий.
С учетом указанных данных становится понятным, почему газообмен ценозов огурца при повышенном содержании зеленых
41
Таблица 2.4
Характеристики радиационного режима для листьев ценозов огурца при облучении ФАР различного спектрального состава
Спектральный вариант Диапазон изменений Hfc, Вт/мг Диапазон изменений П, Вт/м2
40 сут 50 сут 40 сут 50 сут
С — 2б, з — 60, к — 20 10—18 5-15 30—90 15-75
с — 60, з — 20, к — 20 5-10 5-15 15-45 17—60
с — 40, з — 20, к — 20 7—18 10-15 25-55 30-70
с — 34, з — 33, к — 33 5—25 5-15 20—70 15—60
с — 20, з — 40, к — 40 8-18 15—30 25—80 40—90
с — 20, з — 20, к — 60 9—15 7-15 25—60 20—65
Примечание, с — 400—500 нм; а — 500—600 нм; к — 600—700 нм. Возраст ценозов 40 и 50 сут. Остальные уел. обозн. см. текст.
лучей (40 %) на фоне достаточного обеспечения красным светом (40 %) (см. рис. 2.9, б) выше, чем для других спектральных вариантов. Для этого спектрального варианта характерно формирование ценоза с листьями, имеющими, как правило, более высокие значения интенсивности радиации приспособления и, следовательно, находящимися при более высоких облученностях и более эффективно работающими, чем листья соответствующих ярусов в других спектральных вариантах (см. табл. 2.4).
Таким образом, оценка фотосинтетической активности листьев разных ярусов в ценозах огурца путем анализа световых кривых фотосинтеза таких листьев позволяет на ценотическом уровне определить причину повышения или понижения газообмена ценоза как целой системы при изменении спектрального режима облучения.
Изложенные выше результаты исследований показывают, что различная эффективность излучения отдельных участков ФАР в обеспечении фотосинтеза ценоза растений в условиях длительного выращивания сильно зависит от таких факторов, как интенсивность света, вид растений, а также фаза вегетации. В начальный период вегетации практически всегда эффективность красных лучей в фотосинтезе ценоза растений выше, чем зеленых и синих. Однако в дальнейшем в зависимости от вида растений картина может измениться. В этом случае растет эффективность синих лучей, а красных — падает. Максимальная спектральная эффективность фотосинтеза ценоза с ростом уровня облученности, как это было видно на примере редиса, характеризуется смещением в синюю область спектра. В то же время для ценозов пшеницы увеличение интенсивности света со 100 до 600 Вт/м2 ФАР не меняет наибольшей в фотосинтезе эффективности красных лучей. Правда, различия между фотосинтетической активностью ценозов пшеницы при синем и красном свете интенсивностью 600 Вт/м2 уже не такие большие, как при облученности 100 Вт/м2 ФАР.
Сильно зависит от вида растений и от архитектоники образуемого ими ценоза эффективность фотосинтеза на зеленом свету.
42
Если для фотосинтеза монослойного ценоза редиса зеленый свет оказывает ингибирующее воздействие уже при облученностях выше 100 Вт/м2, то для пшеницы повышение облученности от 100 до 600 Вт/м2 способствует активному фотосинтезу ценоза. Обращает на себя внимание роль зеленого света в фотосинтезе ценоза огурца: при облученности 100 Вт/м2 в зеленой области спектра фотосинтез постепенно падает и растения погибают, а при той же облученности, но с добавками к зеленому свету синих и красных лучей наблюдается устойчивый и достаточно высокий фотосинтез. Это позволяет сделать вывод, что высокая эффективность зеленых лучей в фотосинтезе ценозов огурца может быть достигнута лишь при наличии в светопотоке определенного количества синих и красных лучей.
Таким образом, спектральная эффективность фотосинтеза для ценозов растений имеет видовую специфику.
§ 4. Специфика реакций растений разных видов на качество света
при оценке спектральной эффективности фотосинтеза
В исследованиях по длительному выращиванию растений в потоках ФАР различного спектра нередко можно отметить специфику в реакции по фотосинтезу у растений разных видов на излучения различного спектрального состава и интенсивности [Карначук, Постовалова, 1986]. Особенно ярко эта специфика проявлялась при длительном выращивании растений в лучах отдельных спектральных областей ФАР в широком диапазоне облученностей [Тихомиров и др., 1987].
Актуальность исследований по выявлению специфики реакций растений разных видов на различный спектральный состав ФАР связана как с расширением общих представлений о роли спектра ФАР в фотосинтетической продуктивности ценоза, так и с возможностями использования результатов таких исследований для систематизации представлений о многообразии реакций растений на спектр и интенсивность ФАР. Такая систематизация десьма полезна для объединения близких по реакции на спектр и интенсивность ФАР видов растений в отдельные группы, поскольку индивидуальный подбор спектра и интенсивности для обеспечения оптимальной фотосинтетической продуктивности для каждого вида растений, незначительно различающихся по спектральной чувствительности к фотосинтезу, вряд ли целесообразен.
Как известно, наибольшую физиологическую активность имеют синяя и красная области спектра, поэтому выявление реакций растений на спектр излучения ФАР должно включать оценки действия на них в первую очередь излучений этих областей. В связи с этим следует заметить, что при длительном воздействии излучения синей области спектра на растения последние хотя и могут в ряде случаев иметь аномалии в ростовых и органо-образовательных процессах, но тем не менее не гибнут [Тихоми-
43
ров и др., 1983]. В то же время длительное воздействие излученид красной области спектра на растения некоторых видов нередко приводит к ингибированию фотосинтеза и их гибели [Там же],
В этой связи красный свет может выступать как своеобразны! индикатор раннего выявления спектральной специфичности различных видов растений на начальных этапах проведения таких исследований.
Для выявления видовой специфики растений к спектральному составу света в отношении их фотосинтетической деятельности были проведены эксперименты по анализу состояния фотосинтетического аппарата после смены спектрального режима облучения. Для этого растения первоначально выращивали на белом свету, на котором происходил сбалансированный процесс формирования структуры и функции фотосинтетического аппарата, а затем осуществляли перевод части растений на красный свет.
Для анализа были выбраны растения огурца сорта Московский тепличный, которые при длительном пребывании на красном свету погибают, растения томатов сорта Старфайер и кукурузы сорта Минусинка, которые хорошо растут на красном свету, а также растения подсолнечника сорта Енисей, особенностью которых являлась быстрая реакция на присутствие красного света, проявляющаяся в разных морфофизиологических изменениях, наступающих уже через 4—5 сут после перестановки с белого на красный свет, в то время как у других исследуемых растений никаких внешних изменений еще не наступало. Растения выращивали в вегетационных шкафах при облученности 100 Вт/м2 «белого» света, получаемого с помощью лампы ДРИ 2000-6 (контроль), и «красного» (600—700 нм) света, выделяемого из излучения той же лампы с помощью красного светофильтра (см. рис. 1.2). Фотопериод во всех световых вариантах составлял 14 ч, темновой период — 10 ч, температура воздуха днем поддерживалась в пределах 25 ± 1 °C, ночью — 18 ± 1 °C, относительная влажность воздуха 70—80 %, концентрация СО2 атмосферная.
При оценке изменений фотосинтетической активности листьев растений, обусловленной их видовой спецификой к спектру ФАР, анализировали световые кривые фотосинтеза листьев этих растений, сформировавшихся при переходных спектральных режимах облучения. Для этого растения огурца, подсолнечника и кукурузы выращивали при облученности 100 Вт/м2 ФАР «белого» света ламп ДРИ 2000-6, а через 9—14 сут вегетации, в зависимости от появления первого сформировавшегося взрослого листа, половину исследуемых растений переводили под «красный» свет той же интенсивности, получаемый вышеуказанным способом. Оставшиеся под «белым» светом растения служили контролем. Снятие световых кривых фотосинтеза в контрольных и опытных вариантах проводили на взрослых листьях растений с помощью газоанализатора «Инфралит-4» со специально изготовленной терморегулируемой листовой камерой (см. гл. 1).
Анализ световых кривых фотосинтеза (рис. 2.12), как показано рядом исследователей [Воскресенская и др., 1968], позволяет вы-44
/•iii 12. Световые кривые фо-Ци питсла листьев огурца (а), Ии цпишечника (б) и кукурузы (и), «•формировавшиеся под белым спетом (1) и переставленные г полого света па красный (2). Л измерения, проведенные по истечении сут после перенесения растений г iic.iHiro света на красный; Б — то же, iini .ir более длительного пребывания pin гений на красном свету (пояснения в тексте;.
Облученность ФАР, Вт/м2 после перестановки опытных рас-
•шить возможные причины, нодущие к тем или иным физиологическим изменениям в листьях, сформировавшихся при излучениях различного спектрального состава. Первый цикл измерений сделан по истечении двух суто!
тений с белого на красный свет (см. рис. 2.12). В этом случае ход световых кривых фотосинтеза и их выход на плато существенно не различаются. В дальнейшем ход этих кривых как на линейном участке, так и выход на плато сильно зависят от вида растений (см. рис. 2.12). В частности, у огурца эти различия четко проявляются через 7 сут пребывания на красном свету, а у подсолнечника — уже через 5 сут. В то же время у кукурузы таких различий не наступает.
В работе Н. П. Воскресенской с соавторами [1968] показано, что более высокое расположение плато светового насыщения фотосинтеза зеленого листа может быть связано с благоприятным влиянием длительного воздействия спектра излучения (синий свет) на реакции переноса электрона в фотосинтезе, сопряженного с фотофосфорилированием. Кривые замедленной флуоресценции листьев огурца, полученные Т. В. Нестеренко [Нестеренко и др., 1985], указывают на то, что при длительном воздействии красного света на фотосинтетический аппарат происходит замедление скорости электронного транспорта, уменьшение активности фотосистемы II. Последнее обстоятельство уже конкретно указывает па тот факт, что в более низком расположении плато световых кривых фотосинтеза листьев, длительное время произраставших на красном свету, по сравнению с листьями, находившимися все время на белом свету, несомненна важная роль фотохимических реакций.
Следует отметить, что представленные на рис. 2.12 данные по специфичности реакций для разпых видов растений на длительное воздействие красного света совсем не проявляются при кратковременном (минутном) воздействии света, которое можно зафиксировать в виде фотосинтетического ответа. Как отмечалось ранее, такой ответ всегда однозначен: фотосинтез зеленого листа максимален в красных лучах. Этот результат и нашел свое отражение в спектрах действия фотосинтеза, которые имеют максимум
45
в красной области ФАР независимо от вида зеленого растения [МсСгее, 1972а; Inada, 1977] и спектрального режима предварительного выращивания [Тихомиров и др., 1983]. Это вполне объяснимо, если учесть, что при длительном воздействии излучения определенного спектрального состава и интенсивности в формировании фотосинтеза участвует совокупность как быстро, так и медленно протекающих во времени фоторегуляторных механизмов. Именно совокупность этих эффектов решающим образом определяет видовую специфику растений в их реакции на свет разного спектрального состава и интенсивности. Если исключить влияние этих эффектов, вычленив лишь чисто фотосинтетический кратковременный ответ на излучение данного спектрального состава, то следует ожидать получение близких по структуре кривых спектров действия фотосинтеза зеленых листьев различных видов растений. Именно такой результат и был получен в работах Маккри [МсСгее, 1972а] и Инада [Inada, 1977].
Таким образом, длительное воздействие на растения излучения определенного спектрального состава и интенсивности, затрагивающее структуру и функцию фотосинтетического аппарата, может изменить и саму спектральную эффективность фотосинтеза (см. рис. 2.2, 2.5, 2.6, 2.8). В то же время такие изменения, затрагивающее структуру и функцию фотосинтетического аппарата, не реализуются при кратковременном воздействии излучения определенного спектрального состава, что, в частности, видно при сопоставлении данных по фотосинтезу, полученных с учетом длительного и кратковременного воздействия излучения определенного спектрального состава на фотосинтез зеленого листа (см. рис. 2.2, 2.3). Оценка кратковременного воздействия излучения различного спектрального состава на фотосинтез зеленого листа в виде спектра действия фотосинтеза отражает универсальные характеристики фотосинтетического аппарата, присущие зеленым листьям растения независимо от их видовой принадлежности. Под такими характеристиками мы имеем в виду наличие одних и тех же зеленых пигментов, определяющих сходство в спектрах поглощения энергии листом, общих закономерностей миграции энергии к реакционным центрам, единых принципов работы фотосистем и ряд других общих черт строения фотосинтетического аппарата, определяющих быстротекущие реакции фоторегуляции. Получение и анализ таких спектров действия принесли несомненную пользу в понимание общих закономерностей, связывающих структуру и функцию фотосинтетического аппарата с поглощением излучения ФАР различного спектрального состава. Однако энергетическое обеспечение продукционного процесса в условиях искусственного освещения связано с длительным воздействием излучения определенного спектрального состава на це-позы растений. Здесь как единое целое участвуют быстрые и медленно протекающие во времени фоторегуляторные механизмы. Именно они в существенной степени обусловливают особенности в направленности биосинтеза, ведущие к глубоким структурным изменениям фотосинтетического аппарата, проявляющимся в особенностях морфогенеза и определяющим фотосинтез растений.
46
Глава 3
ВЛИЯНИЕ СПЕКТРАЛЬНОГО СОСТАВА СВЕТА НА РОСТ И РАЗВИТИЕ РАСТЕНИИ
Введение
В настоящее время достаточно четко установлено, что на продукционный процесс оказывает влияние тесная взаимосвязь фотосинтеза с ростом и развитием [Мокроносов, 1981]. А. Т. Мокро-носов [1983] отмечает, что фотосинтетическая функция сама контролируется процессами онтогенеза и формирование урожая детерминировано прежде всего эпигенетическими событиями. В этой связи понимание особенностей формирования урожая растений без рассмотрения характера протекания ростовых и органообразовательных процессов нельзя признать достаточно полным.
Первые попытки исследования характеристик роста и развития растений при использовании искусственного облучения относятся к концу XIX — началу XX в. и отмечены в монографии А. Ф. Клешнина [1954]. Однако крайне низкие уровни облученности, неблагоприятный спектр излучения и ненадежность работы источников света в тот период времени позволяют рассматривать результаты таких работ лишь как первые попытки дать оценку росту и развитию растений в условиях искусственного освещения, действию искусственного освещения и его спектрального состава на ростовые и органообразовательные характеристики растений. Достаточно обстоятельные исследования в этом направлении стали возможными лишь начиная с середины 50-х годов нашего столетия, когда появились источники света, способные обеспечить выращивание растений вплоть до получения конечного урожая в отдельных областях ФАР.
В экспериментальном отношении исследование процессов роста и развития растений в зависимости от спектра и интенсивности излучения удобно проводить при использовании искусственных источников света. Поэтому вполне естественно, что многие исследователи, занимающиеся этими вопросами, использовали в работе искусственное излучение с различными спектральными и энергетическими характеристиками. Практически во всех монографиях по использованию искусственного света для выращивания растений в той или иной степени затронуты вопросы их роста и развития при различной интенсивности и спектральном составе света. Многочисленные материалы, представленные в этих работах, получены при использовании разнообразных источников света при отличающихся условиях выращивания растений многих видов. Воздействие излучения различного спектрального состава на растения не всегда было длительным, а тем более постоянным, спектральные источники излучения часто использовали лишь для досветки растений [Свет и морфогенез..., 1978]. Ввиду того что
47
часто растения выращивали при излучении источников света разного спектрального состава без учета содержания в них ультрафиолетовой и инфракрасной радиации, резко снижалась достоверность выявленных закономерностей влияния тех или иных спектральных областей ФАР на процессы роста и развития. Так, например, долгое время ускорение ростовых и органообразова-тельпых процессов у ряда культур приписывали оранжево-красным лучам, и лишь после появления возможности раздельного исследования излучения для этой области спектра и инфракрасного излучения выяснилось, что этот эффект принадлежит инфракрасным лучам [Шульгин, 1971].
Несмотря на это определенные успехи были достигнуты начиная с 60-х годов нашего столетия. Главным среди них следует, видимо, признать установление общих закономерностей фотоморфогенеза, а также прогресс в детальных исследованиях этапов органогенеза различных видов растений и дифференциации верхушечной меристемы [Куперман, 1984]. Эти успехи в значительной мере способствовали становлению морфофизиологии растений как науки о закономерностях органогенеза и морфофизиологической изменчивости различных жизненных форм высших растений. Вместе с тем, как справедливо отмечалось Ф. М. Куперман [1984], отдельные разделы морфофизиологии растений еще недостаточно разработаны. Последнее объясняется различием методов и приемов исследований в морфофизиологии.
Нам представляется, что одним из таких недостаточно исследованных вопросов является морфофизиологическая изменчивость растений в онтогенезе при длительном воздействии излучения различного спектральпого состава и интенсивности ФАР. Изучение этого вопроса становится особенно важным для понимания особенностей формирования урожая растений в условиях искусственного облучения в связи с поиском его оптимальных спектральных и энергетических характеристик. Процессы роста и развития могут быть показателями реакции растений на действие света определенного спектрального состава, характеризующими тот или иной уровень продуктивности растений.
Среди относительно немногочисленных исследований, выполненных по изучению роста и развития растений в условиях длительного воздействия излучения ФАР различного спектрального состава, эксперименты, как правило, проводили с индивидуально растущими растениями [Клешнин, 1954; Леман, 1971; Шульгин, 1971; Протасова, Кефели, 1982; и др.]. В то же время для поиска более корректных взаимосвязей между параметрами роста и развития растений и реальным продукционным процессом важно оценивать процессы морфогенеза и развития с учетом ценоти-ческого взаимодействия между растениями. Нам представляется целесообразным анализировать как параметры роста и развития отдельных растений и их фитоэлементов, сформировавшихся в ценозе, так и параметры, характеризующие по возможности особенности поведения всего ценоза как целостной системы при тех или
48
иных спектральных режимах облучения. Но в ряде случаев целесообразно проанализировать эти процессы и на более низком уровне организации фотосинтетического аппарата. В качестве поиска такого комплексного подхода мы попытались оценить на ряде культур особенности воздействия излучения различного спектрального состава и интенсивности на формирование анатомической структуры листа.
Таким образом, была определена задача проведения исследований по изучению особенностей роста и развития на разных уровнях организации фотосинтетического аппарата: тканевом, листовом, организменном и ценотическом.
§ 1. Влияние света и интенсивности ФАР на морфологические характеристики растений
При выращивании растений в условиях искусственного облучения была отмечена их различная видовая реакция на спектральный состав излучения, что, разумеется, отразилось на их ростовых и органообразовательных характеристиках. В параграфе приводятся результаты исследований по редису, огурцу, томатам и пшенице, т. е. культурам, у которых в достаточной мере проявляются такие различия.
Для томатов оценка ростовых и органообразовательных процессов на ценотическом уровне не проведена. Это связано с тем, что структура ценоза растений томата во второй половине вегетации, когда проявлялись различия между спектральными вариантами, претерпевала систематические изменения, связанные с вмешательством в структуру ценозов при проведении агротехнических мероприятий, требуемых по методике выращивания этих растений в условиях интенсивной светокультуры.
Для редиса не рассматриваются данные по генеративному развитию, поскольку растения выращивали только до стадии формирования корнеплодов.
Для оценки ростовых и органообразовательных процессов в ценозах растений использовали такие известные характеристики, как функция пространственной ориентации листовой поверхности g(Q) (см. гл. 1, § 5), листовой индекс ценоза, распределение фотосинтезирующей поверхности по вертикали ценозов. Эти характеристики, являясь усредненными ценотическими показателями, позволяют оценить ростовые и органообразовательные процессы, происходящие непосредственно в ценозах растений. Они тесно связаны с формированием реального урожая в ценозах. На такие связи указывают исследователи, изучавшие эти цено-тические характеристики. Так, например, показано, что более вертикальная ориентация листьев способствует повышению плато светового насыщения такого ценоза [Полонский, 1978]. Однако работ по изучению ориентации листовой поверхности в ценозах
4 А. А. Тихомиров, Г. М. Лисовский, Ф. Я. Сидько
49
крайне мало и среди них не встречается исследований по изучению ориентации листовой поверхности в зависимости от спектрального состава ФАР. В немногочисленных работах этого направления объектом исследования, как правило, служили отдельные растения, часто молодого возраста [Virgin, 1962; Inada, 1969; Kimura, 1974].
Поэтому имеющиеся данные не позволяют достаточно полно представить ответную реакцию ценоза как фотосинтезирующей системы на спектральный состав падающего излучения, заключающуюся в перестройке его структуры за счет изменения ориентации листовых пластинок. Большое внимание при изучении взаимосвязи организационной структуры и продуктивности ценоза придается листовому индексу [Monsi, Saeki, 1953]. Показано, что при его оптимальных значениях достигается максимальная продуктивность фитоценоза. В зависимости от культуры листовые индексы могут колебаться от 1,0—1,05 у ценозов, приближающихся к монослою [Тарчевский, 19771, до 30 у ряда тропических растений [Plant..., 1971]. Большинство таких исследований выполнено для естественных условий произрастания. Для условий искусственного облучения растений показана принципиальная возможность формирования высокопродуктивных ценозов с большим листовым индексом при использовании высокоинтенсивного облучения [Полонский, 1978]. Однако роль спектрального состава ФАР в формировании листового индекса в ценозах растений практически не исследовалась.
Листовой индекс, отражая общий характер изменения площади листовой поверхности, не позволяет оценить формообразовательные процессы, происходящие внутри ценоза в зависимости от тех или иных воздействующих факторов. В этом отношении весьма информативно изучение распределения листовой поверхности в ценозах в зависимости от стадии вегетации при различном спектре и интенсивности ФАР. Немногочисленные исследования в этом направлении ранее выполнялись только для условий естественного освещения [Ничипорович, 1961; Росс, 1975; Тооминг, 1977; и др. ].
Таким образом, вопросы роста и развития растений в ценозах при длительном воздействии излучения различной интенсивности и спектра ФАР изучены еще недостаточно. Излагаемый ниже материал призван хотя бы частично восполнить этот пробел.
Редис. Влияние светового режима на рост и развитие редиса исследовалось наиболее активно с начала 60-х годов нашего столетия. Было установлено, что спектральный состав как видимой, так и инфракрасной радиации может играть решающую роль в формировании корнеплодов [Шульгин, 1971, 1973; Куперман, 1984; и др.]. Увеличение интенсивности ФАР также может оказывать очень сильное влияние на ход органообразовательных процессов редиса, в том числе и на образование корнеплодов [Шульгин, 1973; Тихомиров и др., 1976; Протасова и др., 1982]. Поэтому, очевидно, что варьирование спектральным составом и ин-
50
тенсивностыо ФАР является мощным фактором в изучении разнообразия ростовых и органообразовательных процессов у редиса. Для реализации этих условий требуется комплексное изучение влияния различных сочетаний спектра и интенсивности ФАР в широком диапазоне их изменений на рост и развитие редиса. Работ, выполненных в этом направлении, крайне недостаточно. Среди этих немногочисленных исследований следует отметить работы Шульгина с соавторами [1963а, б], в которых изучался органогенез редиса при досвечивании синими, красными или инфракрасными лучами. Морфогенез редиса при постоянном выращивании на свету разного спектрального состава изучали И. С. Дроздова с сотрудниками [1987]. Использовалось излучение от люминесцентных ламп низкой интенсивности (30—70 Вт/м2 ФАР). Было показано, что нормальный морфогенез растений редиса может быть обеспечен присутствием уже 10 % синего света в сине-красном излучении низкой интенсивности. Длительное воздействие разноспектральных потоков ФАР интенсивностью 100 Вт/м2 и более на морфогенез редиса практически не изучалось. Следует отметить, что наиболее хорошо исследованный диапазон уровней облученности, где оценивали роль спектрального состава света в процессе развития растений (в том числе и редиса), ограничивался во многих работах облученностью 50 Вт/м2 ФАР или ниже [Клешнин, 1954; Шульгин, Мерцалов, 1963; и др.] В экспериментах было показано, что развитие редиса может ускоряться синими, а задерживаться оранжево-красными лучами.
Для расширения представлений о спектральной эффективности действия ФАР различной интенсивности в ростовых и органообразовательных процессах растений редиса нами были проведены эксперименты по выращиванию этой культуры в течение всей вегетации в диапазоне облученностей от 50 до 600 Вт/м2 ФАР при различном спектральном составе (рис. 3.1).
Синие лучи ускоряют процесс развития у редиса, оцениваемый по стрелкованию стебля, особенно при относительно низких уровнях облученности. В то же время из этого рисунка следует, что с ростом уровня облученности рассматриваемые различия во влиянии исследуемых спектральных потоков на развитие редиса все более сглаживаются, а затем вообще исчезают. Таким
Рис. 3.1. Зависимость стрелкования растений редиса сорта Вировский белый от интенсивности ФЛР в отдельных спектральных областях [Сидько и др., 1977]. 1 — 600—700 нм; 2 — 500—
600 нм; 3 — 400—5ОО нм.
4*
51
Таблица 3.1
Зависимость ростовых процессов целых растений и листовых пластинок редиса от спектра и интенсивности ФАР
400- -500 им 500- 600 нм 600- -700 нм
Д и Линейные и • Линейные га Линейные
л размеры ли- д размеры ли- д размеры ли-
£.8 н о стовой пла- S о стовой пла- Я о стовой пла-
5ИВ1 Вт/ сб и а стинки, см О в Е? стинки, см С ее S стинки, см
к - о ь О о
KG о и Я е; о S Длина Ширина о о И М О S F Длина Ширина о S И R о S Длина Ширина
50 40 7 13 8 23 5 12 6 21 5 12 6
100 31 6 12 7 22 4 9 6 15 4 6 4
200 28 6 11 6 19 4 7 4 11 4 4 3
300 25 5 10 6 И 4 4 3 — — — —
400 16 5 8 5
500 И 4 7 4
600 7 4 6 4
Примечание.
Возраст ценоза 24 сут.
образом, можно добиться отсутствия стрелкования у растений редиса при его выращивании во всех исследуемых спектральных диапазонах ФАР, однако уровни облученности при этом будут разные. Наименьшие затраты энергии для исключения стрелкуто-щих форм редиса требуются при использовании красных лучей (100 Вт/м2), в то время как при использовании зеленых лучей уровень облученности необходимо увеличить в 2 раза по сравнению с красным светом, при использовании синих — в 6 раз (см. рис. 3.1).
С ростом уровня облученности высота растений редиса уменьшается во всех спектральных вариантах опыта (табл. 3.1). Растения редиса, выращенные в синих лучах, были всегда выше, чем соответственно в зеленых и красных при использовании одинако вых уровней облученности. Такая же закономерность просматривается и для линейных размеров листовых пластинок редиса Действительно, с увеличением длины волны падающей радиации усиливается ингибирующее действие излучения па рост и формирование листового аппарата редиса, что проявляется в уменьшении числа листьев и линейных размеров листовых пластинок Этот факт является общим для всех рассматриваемых участкоь ФАР, но достигается он при различных облученностях. Следуй Г отметить, что с увеличением облученности количество листьев достигнув четырех, в дальнейшем не уменьшается даже при очень высоких уровнях облученности. На фоне определенного постоянства количества листовых пластинок увеличение облученности влияет на их размеры, значительно уменьшая скорость роста. Так, например, в синей области ФАР увеличение облученности с 50 до 600 Вт/м2 сопровождается уменьшением линейных размеров листовых пластинок более чем в 2 раза, а для зеленых
52
Рис. 3.2. Зависимость коэффициента хозяйственной эффективности (КуоВ) ценозов редиса Вировский белый от интенсивности ФАР в отдельных спектральных областях [Тихомиров, 1979].
I — 400—500 нм; 2 — 500—600 нм; 3 — 600—700 нм.
и красных лучей этот эффект достигается при изменении облученности от 50 до 300 Вт/м2.
Торможение роста листового аппарата редиса при высоких облученностях часто сопровождается усиленным ростом корнеплодов [Шульгин, 1973; Тихомиров и др., 1976; Протасова, Ке-фели, 1982]. Распространенным показателем, характеризующим распределение ассимилятов между листовой поверхностью и корнеплодами, является коэффициент хозяйственной эффективности но накоплению биомассы (Ах03). Значения А”х03 для различных спектральных режимов выращивания редиса неодинаковы (рис. 3.2). Наиболее высокие значения Кхоз могут быть достигнуты при излучении красной и синей областей спектра. Однако если в первом случае для этого требуется облученность 100 Вт/м2, то во втором — в 5 раз большая.
Следует иметь в виду, что одинаковый Кх03 может достигаться при различной как общей, так и хозяйственно полезной биомассе. Нами показано, например, что резкое увеличение интенсивности синего [Тихомиров, 1979] и белого [Тихомиров и др., 1976] света сопровождается одновременным ростом Zfx03 и продуктивности растений, в то время как в зеленых и красных лучах с ростом уровня облученности Кх03 может возрастать, а накопление биомассы падать.
Для регуляторных процессов, определяющих увеличение Ах03 и подавляющих стрелкование растений, наиболее эффективен в энергетическом отношении красный свет, а наименее — синий. Зеленый свет занимает промежуточное положение (см. рис. 3.1, 3.2).
При невысоких облученностях (Е = 50 Вт/м2) в отдельных областях ФАР в интервале углов наклона 0 = 75—90° находятся 40—50 % всей листовой площади для синей и зеленой областей спектра (рис. 3.3). При переходе к направлениям с 6 = 45—60° наблюдается примерно одинаковое уменьшение величины g(6) для всех спектральных интервалов. Листья, ориентированные близко к горизонтали (0 = 0—30°), практически отсутствуют для спектрального варианта с излучением в синей области. Увеличение облученности до 100 Вт/м2 приводит к смещению максимума функции в сторону более горизонтальных направлений. Однако горизонтальность листьев еще слабо выражена, так как на угловой интервал 0—30° приходится около 20 % листовой поверхности для красной области, примерно 10 % для зеленой, а для
53
Рис. 3.3. Функция пространственной ориентации листьев g(0) для ценозов редиса, сформировавшихся при различных облученностях в синей (а), зеленой (б) и красной (в) областях ФАР.
Возраст ценозов 20 сут. Е, Вт/м2: 1, 1', 1"— 50; 2, 2', 2"— 100; з — — 600; 3' — 300; 3"— 200.
синей области ФАР листьев, ориентированных в этом интервале, практически нет (см. рис. 3.3). Облученности 600, 300 и 200 Вт/м2 соответственно для синей, зеленой и красной областей ингибируют продукционные процессы в исследуемых областях ФАР [Тихомиров, Сидько, 1982]. В синей области при указанном уровне облученности изменение примерно такое же, как и при Е = 100 Вт/м2. В остальных спектральных областях ФАР при ингибирующих продуктивность уровнях облученности наблюдается резко выраженная горизонтальность листьев. Так, доля листовой поверхности, имеющей угол наклона 0 — 45°, составляет для этих спектральных вариантов 65—70 %, а листьев, ориентированных вертикально (0 = 75—90°), практически нет.
Использование невысоких облученностей существенно не влияет на изменение ориентации листьев при любом из исследуемых сочетаний спектральных диапазонов в области ФАР (рис. 3.4). Основная доля листовой поверхности ориентирована наклонно (0 = 45—60°). Интересно заметить, что доля вертикально ориентированных листьев выше в тех спектральных вариантах, где доминирует синий компонент излучения. Более сильная зависимость ориентации листьев от спектрального состава излучения наблюдается при высокоинтенсивном излучении с облученностью около 600 Вт/м2 ФАР (см. рис. 3.4). Так, ориентация листьев редиса приближается к горизонтальной, если высокоинтенсивный лучистый поток содержит незначительное количество (20—25 %) синих лучей, а доля красных лучей составляет около 50 %. В этих условиях более 60 % листовой поверхности заключено в угловом диапазоне 0—30°. Однако если доля синих лучей в ФАР равна или превосходит долю зеленых и красных лучей, то листья имеют более вертикальную ориентацию.
Важно отметить, что световая нагрузка на листовые пластинки в различных спектральных вариантах определяется не только их ориентацией, но и изменением «рельефа» поверхности листьев. Эксперименты показали, что при облученностях 50—100 Вт/м2 поверхность листьев редиса остается практически гладкой при
54
Рис. 3.5. Схема профиля участка листовой пластинки редиса, подвергшейся формативным изменениям под влиянием высоких облученностей (Е 400 Вт/м2 ФАР).
Объяснения в тексте.
Рис. 3.4. Функция пространственной ориентации листьев g(0) для ценозов редиса, сформировавшихся при различном спектральном составе излучения ФАР и облученностях 100 Вт/м2 ФАР (7) и 600 Вт/м2 ФАР (2).
Возраст ценозов 20 сут. а — 400—500 нм (30 %) + 500—600 нм (70 %); б — 400—500 нм (70 %) + 500—600 нм (30 %); в — 400—500 нм (30 %) + 600—700 нм (70 %); г — 400 — 500 нм (70 %) + 600—700 нм (30 %); д — 400—500 нм (70 %) + 500—600 нм (20 %) + + 600—700 нм (10 %); е — 400—500 нм (40 %) + 500—600 нм (40 %) + 600—700 нм (20 %); не — 400—500 нм (33 %) + 500—600 нм (33 %) + 600—700 нм (33 %); з — 400— 500 нм (25 %) + 500—600 нм (15 %) + 600—700 нм (60 %).
различном спектральном составе ФАР. В то же время с ростом уровня облученности от 200 Вт/м2 и выше поверхность листа в ряде спектральных вариантов, где доля синих лучей не менее 30 % ФАР, становится рельефной (рис. 3.5), т. е. в пределах отдельных листьев происходит формирование структуры их поверхностей, связанных с различной ориентацией их участков к направлению падающего света. На поверхности листа появляются зоны повышенной и пониженной облученности. В зонах повышенной облученности поверхность листовой пластинки ориентирована близко к перпендикулярной относительно падающего лучистого потока. В результате из-за высокой световой нагрузки в этих местах происходит сильное выцветание зеленых пигментов, образуются желтые пятна. В зонах пониженной облученности благодаря косому падению световых лучей на поверхность листа создаются облученности, обеспечивающие функционирование фотосинтетического аппарата. Таким образом, успешное функционирование фотосинтетического аппарата редиса в значительной степени зависит от состояния пигментной системы зеленого листа при используемых режимах облучения (см. рис. 3.5). В этой связи представляет интерес проанализировать данные по
55
Рис. 3.6. Поверхностная плотность хлорофилла (Са+б) в листьях ценозов редиса в зависимости от уровня облученности и спектрального состава ФАР; [Тихомиров, Сидько, 1982].
Е, Вт/м2: а — 100 (возраст ценоза 24 сут); б — 600 (20 сут). Фаза технической спелости корнеплодов.
содержанию зеленых пигментов в листьях редиса при различных интенсивностях и спектре ФАР (рис. 3.6).
Поверхностная концентрация хлорофиллов в листьях редиса зависит от различных сочетаний спектральных участков ФАР при облученностях 100 и 600 Вт/м2 ФАР (см. рис. 3.6). В качестве пояснения отметим, что точка пересечения катетов «цветового треугольника» А соответствует области 500—600 нм, точка Б левой части графика — области 400 — 500 нм, а точка В правой части графика — 600—700 нм. Точки, расположенные на сторонах треугольника, соответствуют различным двухкомпонентным сочетаниям исследуемых областей ФАР. Кривые, выражающие зависимость поверхностной концентрации хлорофиллов в листьях от всевозможных сочетаний спектральных участков в ФАР, образуют поверхность над «цветовым треугольником». Минимальное количество хлорофилла при облученности 100 Вт/ма соответствует красной и зеленой областям ФАР, а также различным сочетаниям этих областей (см. рис. 3.6). Наибольшая поверхностная концентрация хлорофиллов отмечена при излучении, в котором сочетаются коротко- и длинноволновый участки ФАР, а также при излучении ФАР, где эти участки доминируют. При облученности 600 Вт/м2 ФАР отсутствие коротковолнового излучения в области ФАР ингибирует накопление хлорофиллов в листьях редиса независимо от того, в каких сочетаниях используются остальные спектральные участки ФАР (см. рис. 3.6). Возрастание доли синего излучения в высокоинтенсивном лучистом потоке ФАР почти всегда положительно коррелирует с поверхностной концентрацией зеленых пигментов в листьях редиса [Тихомиров, Сидько, 1982].
В ценозах редиса, выращиваемых при различных облученностях в отдельных областях ФАР, наиболее высокий листовой
56
Рис. 3.7. Возрастная динамика листового индекса ценозов редиса при различных уровнях облученности в отдельных областях ФАР.
Соотношение лучистых потоков отдельных областей
Рис. 3.8. Зависимость индекса листовой поверхности ценозов редиса от спектрального состава излучения и уровня облученности.
Возраст ценозов 20 сут. а — доля синего излучения (400—500 нм) в красно-синем (400— 500 нм + 600—700 нм); б—доля синего излучения в зелено-синем (400—500 нм + + 500—600 нм); в — доля красного излучения в зелено-красном (зОО—600 нм + 600— 700 нм). Е, Вт/м2; 1 — 100; 2 — 600.
« — 400—500 нм; б — 500—600 нм; в — 600—700 нм. Остальные усл.
обозн. см. рис. 2.4.
индекс достигается в лучах синей области ФАР; а с увеличением длины волны падающего излучения значение этой характеристики ценоза редиса уменьшается (рис. 3.7). Нарастание листового индекса в процессе вегетации идет экспоненциально, что особенно характерно для невысоких облученностей [Тихомиров, 1979].
Рассмотрение влияния излучения, в которое входят различные комбинации отдельных областей ФАР, показало, что при их попарном сочетании роль спектрального состава лучистой энергии наиболее сильно отражается на изменении листового индекса ценозов редиса при использовании облученностей 100 Вт/м2 ФАР (рис. 3.8). В то же время при использовании высокоинтенсивных лучистых потоков с Е > 100 Вт/м2 ФАР листовой индекс с изменением спектрального состава излучения меняется незначительно. В этом случае определяющим является лишь одно усло-
57
Таблица 3,fi
Листовые индексы ценозов редиса, сформировавшихся при излучении смС«-шанного спектрального состава
Интенсивность облучения, Вт/м2 Спектральный вариант
с-70, s-20, к—10 с—40, 3—40, к—20 с-ЗЗ, з-ЗЗ, к—33 с—20, 3—50, к—30 с—20, 3—40, к—40 с—20, 3—20, к—60
100 4,4 4,0 5,0 6,0 5,4 4,5
600 1,7 2,3 1,6 2,0 1,1 1,2
Примечание, с — 400—500 нм, з — 500—600 нм, к — 600—700 нм.
вне — входит ли излучение синей области ФАР в интегральный лучистый поток. Обогащение синими лучами ФАР способствует увеличению листового индекса ценозов редиса, а отсутствие этих лучей в высокоинтенсивном лучистом потоке приводит к быстрой гибели ценоза из-за интенсивного выцветания зеленых пигментов (см. рис. 3.6).
Спектральные варианты опытов по изучению листового индекса ценозов редиса выбраны таким образом, чтобы одна или две области ФАР представляли преимущественную долю в излучении ФАР (табл. 3.2). Это позволяет оценить с достаточной степенью уверенности роль каждой отдельной области, входящей в ФАР, при формировании листового индекса ценоза редиса. Наиболее высокие значения листового индекса ценоза редиса отмечены для излучения, обогащенного зелеными лучами при Е — 100 Вт/м2 ФАР, и излучения с преимущественным содержанием синих и зеленых лучей при Е — 600 Вт/м2 ФАР (см. табл. 3.2).
Увеличение листового индекса ценозов редиса при обогащении ФАР зелеными лучами отмечалось нами и ранее, когда мощность лучистого потока составляла около 100 Вт/м2 ФАР. Однако листовые индексы такого же порядка достигаются и при использовании двухкомпонентного сине-красного излучения с максимумом в красной области. Однако процесс формирования ценоза в этих спектральных вариантах протекал по-разному. При использовании сине-красного излучения скорость нарастания листовой поверхности значительно опережала таковую при использовании ФАР, обогащенной зелеными лучами. На заключительных этапах вегетации повышенная доля зеленых лучей способствовала интенсивному росту не только верхних, но и нижних листьев за счет улучшения световых условий внутри ценоза и поэтому общий листовой индекс в этом случае превзошел таковой для синекрасного излучения. Что касается высокоинтенсивного излучения, то лишь при преимущественном излучении в зелено-красной области ФАР листовой индекс ценозов редиса заметно уменьшается. Этот факт вполне объясним, если учесть влияние отдельных областей ФАР на листовой индекс. Именно при зеленом и красном высокоинтенсивпом излучении ценоз редиса погибал.
58
20 40 О 20 40 О 20 40
Плои^адь листьев в единице объема ценоза, м2у
Рис. 3.9. Распределение площади листьев по вертикальному профилю ценозов редиса, сформировавшихся при различных облученностях в синей (а), зеленой (б) и красной (в) областях ФАР.
Площадь листьев в единице объема ценоза, мг/м3
Рис. 3.10. Распределение площади листьев по вертикальному профилю ценозов редиса при облученностях 100 (А) и 600 Вт/ма ФАР (Б).
Усл. обозп. спектральных вариантов см. рис. 3.4.
Е, Вт/м2: 1 — 100; I' — 350; Г — 600; г — 100; г' — 200; 2” — 300; 8 — 50; з’ — юо; з"— 200.
Поэтому вполне закономерно, что повышение доли излучения этих областей в высокоинтенсивпом излучении ФАР отрицательно сказывается на листовом индексе [Тихомиров, 1979].
Уровни облученности, превышающие 200 и 300 Вт/м2 соответственно для красной и зеленой областей, являются летальными для ценоза редиса, поэтому рассмотрение особенностей его структуры, сформировавшейся при излучении этих спектральных интервалов, ограничивается указанными облученностями, а для синей области, в силу тех же обстоятельств, взяты облученности до 600 Вт/м2 (рис. 3.9).
С ростом уровня облученности в синей области спектра характер распределения листовой поверхности по вертикальному профилю ценоза является более равномерным, чем при высоких уровнях облученности в других исследуемых областях ФАР (см. рис. 3.9). Лишь при облученности 100 Вт/м2 это распределение аналогично таковому для зеленой и красной областей ФАР этого же уровня облученности.
59
Указанная особенность структуры ценоза редиса, выращен-* него при высоких уровнях облученности в синей области спектра, связана с тем, что здесь не происходит отмирания листьев. Воз-* действие синего света на листовой аппарат проявляется в характерном скручивании листовых пластинок, замедлении увеличения их площади, что в итоге приводит к более равномерному распределению листьев по вертикали ценоза [Тихомиров, Сидько, 1982].
Для получения кривых вертикального распределения листовой поверхности в ценозах редиса при различных комбинациях отдельных областей ФАР из всевозможного разнообразия их сочетаний выбраны наиболее характерные, отражающие тенденцию в изменении листовой поверхности ценоза редиса при Е = 100 и 600 Вт/м2 (рис. 3.10). Так, при рассмотрении попарных сочетаний синей, зеленой и красной областей ФАР взяты варианты с преимущественным излучением одной из спектральных областей и некоторой добавки другой. Сочетание зеленой и красной областей здесь не рассматривается ввиду летальности такого излучения для ценоза редиса при Е = 600 Вт/м2. При рассмотрении сочетания всех трех исследуемых областей ФАР сопоставляются спектральные варианты, в которых одна или две области ФАР доминируют. Ведущим фактором, определяющим распределение листовой поверхности по вертикали в ценозах редиса при использовании трехкомпонентных спектральных потоков, является интенсивность облучения, так как при любом сочетании спектральных участков ФАР плотность листьев в единице объема ценоза заведомо ниже при Е = 600, чем при Е — 100 Вт/м2 (см. рис. 3.10).
При облученности 100 Вт/м2 максимумы вертикального распределения листовой поверхности приходятся в большинстве спектральных вариантов на верхнюю часть ценоза. Наиболее равномерное распределение площади листьев отмечено для варианта с сине-красным излучением, где красный компонент доминирует, а наименее равномерное — для излучения ФАР, обогащенного зелеными лучами (см. рис. 3.10). Степень неоднородности кривой вертикального распределения листьев в ценозе обусловлена их площадью и количеством, а также характером ориентации.
При Е = 600 Вт/м2 листовая поверхность ценоза редиса в большинстве спектральных вариантов сосредоточена в верхней части ценоза, что связано с близким к горизонтальному расположением листовых пластинок. Лишь в варианте с излучением, где доминирует синяя область ФАР, листья расположены более вертикально, что создает и более однородное распределение площади листьев по вертикали ценоза.
Пшеница. Подробный обзор по физиологии роста, развития и онтогенеза пшеницы дан в работе Ф. М. Куперман [1969]. Однако вопросам влияния спектрального состава и интенсивности искусственного излучения на эти процессы уделено мало места. Первые достаточно подробные исследования в этом направлении касались действия подсветки дополнительных к солнечному свету излучений в различных областях спектра на морфогенез и тем
60
пы развития пшеницы [Куперман, 1969, 1984]. Эти исследования проводили, как правило, с использованием маломощных источников света, дающих спектральные потоки низкой интенсивности.
В настоящее время в связи с проблемой ускорения селекционного процесса и рядом других задач выращивание пшеницы при интенсивном искусственном освещении получило довольно широкое распространение [Леман, 1975; Лисовский, Долгушев, 1986; Мошков, 1987; и др.]. В этой связи знания особенностей роста н развития пшеницы при длительном выращивании в условиях интенсивной светокультуры при разнообразных спектральных н энергетических режимах облучения крайне важны. Однако таких исследований выполнено недостаточно, а данные по росту и развитию пшеницы при длительном выращивании в лучах отдельных спектральных областей фактически отсутствуют.
В Институте биофизики СО АН СССР были проведены исследования по выращиванию пшеницы при искусственном излучении ФАР различной интенсивности и спектрального состава.
Выращивание пшеницы в течение всего вегетационного периода проводили в синих, зеленых и красных лучах при интенсивностях 100 и 600 Вт/м2.
При выращивании пшеницы в лучах отдельных областей ФАР интенсивностью 100 Вт/м2 растения к возрасту 10 сут имели по три листа и практически между собой не различались. К возрасту 20 сут в разных спектральных вариантах растения пшеницы имели па главном стебле по пять зеленых листьев. Наличие боковых побегов не отмечено. К возрасту 30 сут растения пшеницы под сипим, зеленым и красным светом имели шесть листьев, причем в последнем варианте зарегистрирована фаза выхода в трубку. К возрасту 40 сут растения вступили в фазу колошения и имели под синим светом четыре, а под зеленым и красным — по пять зеленых листьев на главном стебле. Примерно каждое третье растение под зеленым светом имело слаборазвитый боковой побег с двумя листьями. К возрасту 60 сут растения пшеницы находились в фазе молочно-восковой спелости и имели под синим и красным светом три, а под зеленым — четыре зеленых листа на главном стебле. Полное созревание растений пшеницы при облученности 100 Вт/м2 наступало в синих лучах через 85 сут вегетации, в зеленых и красных — через 80.
При использовании высокоинтенсивного излучения с Е = = 600 Вт/м2 в отдельных областях спектра молодые растения пшеницы, выращенные в синих и красных лучах, в возрасте 10 сут практически не имели видимых различий, а у растений пшеницы, выращенных в зеленых лучах, было отмечено появление зачатка бокового побега. К возрасту 30 сут растения в этом спектральном варианте имели один-два боковых побега, в красных лучах большинство растений имело по одному боковому побегу, а в синих боковых побегов пе образовалось. Во всех спектральных вариантах при 600 Вт/м2 растения пшеницы находились в фазе выхода в трубку. К возрасту 40 сут растения пшеницы, выращиваемые
61
при облученности 600 Вт/м2, выколосились независимо от спектрального состава света. К возрасту 50 сут растения пшеницы в синих лучах имели три зеленых листа на главном стебле, а в зеленых и красных — по три па главном и на боковых. Все боковые побеги прошли фазу колошения. При этом растения пшеницы в синих лучах имели только главный стебель с верхними зелеными листьями, в зеленых лучах — в большинстве случаев по два боковых стебля, которые находились в фазе выхода в трубку. На основном стебле было три верхних зеленых листа, а на боковых стеблях — по два зеленых листа. В красных лучах большинство растений пшеницы в этот период вегетации имело по одному боковому стеблю, находящемуся в начальной стадии коло-шения. Основной стебель имел три зеленых листа, а боковой — два. Во всех спектральных вариантах растения пшеницы к этому периоду вегетации находились в фазе молочно-восковой спелости.
К возрасту 70 сут наступило полное созревание колоса главного стебля растений пшеницы во всех спектральных вариантах при облученности 600 Вт/м2. К этому периоду вегетации у половины колосьев боковых побегов также отмечена фаза полного созревания.
Анализ данных по выращиванию растений пшеницы при 100 и 600 Вт/м2 в отдельных областях спектра показывает, что имеется ряд специфических особенностей воздействия света на процессы роста и развития, присущих той или иной области спектра, которые фактически не зависят от облученности при 100 и 600 Вт/м2. Это утверждение, на наш взгляд, следует в первую очередь отнести к излучению в синей области спектра. Так, ни при 100* ни при 600 Вт/м2 в этом спектральном диапазоне у растений пшеницы не образуется боковых побегов (табл. 3.3). Если при облученности 100 Вт/м2 это можно было бы попытаться объяснить недостаточным проникновением синих лучей в глубь ценоза пшеницы, то при облученности 600 Вт/м2 этот фактор не имел места. В этом случае воздействие синего света на органогенез растений
Таблица 3.3
Характеристики роста и развития растений пшеницы, сформировавшихся при облучении ФАР различной интенсивности в отдельных спектральных областях
Интенсивность облучения, Вт/м2 Область спектра, нм Число побегов на растении, шт. Длительность вегетации до полного созревания зерна, сут ^Х03> %
всего продуктивных
100 400—500 1,о 1,0 85 15,9
500—600 1,7 1,1 80 41,0
600—700 1,1 1,0 80 41,0
600 400—500 1,0 1,0 80 41,2
500—600 3,0 2,4 75 35,0
600—700 3,2 3,1 75 44,0
Таблица 3.4
Высота растений пшеницы в различные периоды вегетации, см
Период вегетации, сут Спектральный вариант
Синий свет .Зеленый свет Красный свет
К, К2 К, к2 К,
10 22 25 22 25 18 18
20 25 50 30 45 29 40
30 50 57 54 62 52 62
40 53 70 56 68 55 65
50 55 75 62 73 60 73
60 60 78 65 78 64 78
70 65 78 75 78 73 78
Примечание. £, = 100 Вт/м2, Es = 600 Вт/м£.
пшеницы следует объяснять, видимо, не субстратной, а регуляторной ролью синих лучей, препятствующих протеканию физиологических процессов, ответственных за кущение растений пше-ницы. Специфика воздействия синего света как при 100, так и при 600 Вт/м2 проявляется также в замедлении протекания ряда стадий онтогенетического развития, которые приводят к затягиванию всего процесса онтогенеза (см. табл. 3.3). Повышение уровня облученности от 100 до 600 Вт/м2 снижает общую продолжительность вегетации для каждого спектрального варианта. Однако обратная зависимость между длиной волны используемого излучения и продолжительностью вегетационного периода для пшеницы сохраняется (см. табл. 3.3).
Увеличение интенсивности излучения в отдельных спектральных областях существенно влияет и на Кхов пшеницы (см. табл. 3.3). Если при облученности 100 Вт/м2 эта величина намного уступает соответствующим значениям для зеленых и красных лучей, то при облученности 600 Вт/м2, напротив, Кх,.а возрастает почти в 3 раза и становится сопоставимым с аналогичными показателями пшеницы для зеленых и красных лучей.
При облученности 100 Вт/м2 наблюдался несколько замедленный рост растений в течение всего онтогенеза в синих лучах, в сравнении с таковым в зеленых и красных, где ростовые процессы идут примерно одинаково (табл. 3.4). При Е = 600 Вт/м2, наоборот, рост растений в синих лучах был наиболее интенсивным при примерно одинаковом протекании ростовых процессов в зеленых и красных лучах. В целом же темпы роста растений пшеницы при облученности 600 Вт/м2 были значительно выше, чем при Е — 100 Вт/м2, и растения закончили рост на 60-е сутки вегетации, в то время как при Е — 100 Вт/м2 лишь на 70-е сутки.
Проведен ряд исследований по выращиванию растений пшеницы при излучении смешанного спектрального состава ФАР интенсивностью 100 Вт/м2 (табл. 3.5). Видно, что обогащение излучения ФАР красными лучами способствует ускорению роста
62
63
Таблица 3.5
Характеристики растений пшеницы, сформировавшихся при облучении ФАР различного спектрального состава интенсивностью 100 Вт/м2
Распределение энергии в области ФАР, % Число побегов на растении, шт. кхоз> % Длительность вегетации до полного созревания, сут
всего продуктивных
с — 34, з — 33, к — 33 1,5 1,0 42,0 80
с — 75, з — 25 1,0 1,0 34,0 85
с — 25, з — 75 1,28 1,0 44,0 83
с — 25, к — 75 1,8 1,38 35,0 78
с — 15, к — 85 1,75 1,40 41,0 75
с —75, к —25 1,36 1,0 36,0 83
и развития пшеницы, что в итоге сокращает сроки вегетации. До-Аминирование красных лучей в спектре ФАР независимо от того, синие или зеленые лучи в этом случае дополнительные, приводит к образованию продуктивных боковых побегов. Наиболее высокие значения Кх03 соответствуют спектральным режимам, для кото-' рых характерно доминирование зеленых или красных лучей в ФАР, а также трехкомпонентному сочетанию этих областей в равноэнергетическом излучении (табл. 3.5). Доминирование синих лучей в ФАР приводит к затягиванию вегетационного периода, а красных — к его ускоренному протеканию. Это согласуется с ролью синих лучей в ростовых и органообразовательных процессах, отмеченных выше.
Таким образом, повышенное содержание синих лучей в ФАР независимо от того, зеленые или красные лучи являются в этом случае дополнительными, приводит к замедлению ростовых и органообразовательных процессов у растений пшеницы. На основании этого можно считать, что при доминировании в ФАР синих лучей сохраняются в основном все закономерности роста и развития пшеницы, присущие ей при выращивании на чистом синем свету.
Ориентация листовой поверхности в ценозах пшеницы при искусственном облучении рассматривалась в работах В. И. Полонского [1978], где показано, что вертикальная ориентация листьев ценозов пшеницы — один из важнейших факторов адаптаций к условиям высокоинтенсивного излучения. Влияние спектрального состава излучения на отклонение первого листа у пророст--ков пшеницы показано в работе Кимура [Kimura, 1974]. Листья освещали синим, зеленым, красным и белым светом. Наиболее сильное отклонение листа в сторону горизонтальной ориентации наблюдалось при синем свете, далее следовали соответственно белый, зеленый и красный свет. В то же время в работе Виржин [Virgin, 1962] отмечено влияние красного света на отклонение листьев пшеницы. К сожалению, результаты последних двух ра
бот получены при различных по интенсивности воздействия лучистых потоках, что затрудняет их сопоставление и в ряде случаев придает им противоречивый характер. Другим недостатком этих работ является выполнение исследований на отдельных растениях, часто молодого возраста. Поэтому имеющиеся данные не позволяют достаточно полно представить ответную реакцию фитоценоза на спектральный состав падающего излучения, заключающуюся в перестройке его структуры за счет изменения ориентации листовых пластинок.
Наши данные по изменению ориентации листовой поверхности ценозов пшеницы в различные периоды вегетации при облученности 100 Вт/м2 в отдельных областях ФАР подтверждают, что у 10-суточных ценозов пшеницы горизонтальная ориентация листьев практически отсутствует и более половины листовой поверхности ориентировано с 9> 45°. Для ценозов такого возраста разделение по слоям не представлялось целесообразным. Наиболее четко выраженная вертикальная ориентация листовой поверхности наблюдается в ценозах молодых растений пшеницы при использовании излучения синей области. Для 20-суточных ценозов пшеницы независимо от спектрального состава используемого излучения функция g(0), рассчитанная на весь ценоз, имеет два максимума, причем больший максимум, находящийся в угловом интервале 75—90°, характеризует хорошо выраженную вертикальность листьев, расположенных в верхнем слое ценоза, а меньший связан с близкой к горизонтальной ориентацией листьев нижнего слоя ценоза.
Для 30-суточных ценозов пшеницы наблюдается исчезновение максимума в области малых углов для всех спектральных вариантов. Это связано с тем, что, хотя листья нижнего слоя ценоза пшеницы по-прежнему ориентированы в окологоризонтальных направлениях, однако ввиду уменьшения их фотосинтезирующей поверхности за счет подсыхания вклад последних в общую листовую площадь уменьшается, в то время как площадь листьев верхнего слоя, преимущественно ориентированных в околовертикаль-пых направлениях, продолжает возрастать. Последнее обстоятельство характеризует наличие максимумов в угловом интервале 75—90°. Однако при переходе в фазу цветения (40 сут) этот максимум перемещается в сторону меньших углов и ориентация листьев становится более горизонтальной.
На заключительных этапах вегетации интенсивно идет процесс старения и отмирания листьев, что отражается на изменении функции g(0). Так, для красной области спектра, где процессы старения листьев идут наиболее интенсивно, максимум g(0), определенный для нижних слоев ценоза, находится в угловом интервале 40—50°. Однако, как и в течение всего вегетационного периода, главный максимум функции распределения g(0) для всего цепоза определяется листовой поверхностью, сосредоточенной в верхней половине ценоза, где доминируют листья с околовер-тикалькой направленностью.
6 А. А. Тихомиров, Г, М. Лисовский, Ф. Я. Сидько 65
64
В первые 10 сут ценозы пшеницы имеют резко выраженную вертикальность листьев независимо от спектрального состава используемого излучения. В последние 10 сут вертикальность листьев становится несколько менее выраженной, максимум g(0) для ценоза пшеницы сдвигается в сторону меньших углов (0= — 60—75°). После 10 сут вегетации начинают проявляться различия в формировании структуры ценозов пшеницы при излучении различного спектрального состава. Это связано в первую очередь с появлением боковых побегов у- ценозов, выращиваемых при зе-> леном и красном излучении. Ввиду этого в нижней части ценоза появляются листья, ориентированные в угловых интервалах, близких к вертикали. Последнее обстоятельство нашло отражений в смещении максимума функции g(0) для нижней половины ценоза в сторону меньших углов. В более поздние периоды вегетации эта тенденция усиливается, в результате чего максимумы g(0), определенные для верхней и нижней половин ценоза, оказываются в одном или смежных угловых интервалах, при этом максимумы g(6) сдвигаются в сторону больших углов. Это указывает на то, что в нижних слоях ценоза возрастает доля листовой поверхности, ориентированная в сторону вертикальных направлений, что связано как с образованием новых, так и с ростом взрослых листьев на боковых побегах. Однако суммарная g(0), рассчитанная для всего ценоза, имеет тенденцию к сдвигу в сторону меньших углов, поскольку верхние листья, вносящие основной вклад в общую листовую поверхность, с возрастом отклоняются в горизонтальном направлении.
Сходный характер изменения g(0) с возрастом ценоза отмечен и для пшепицы, выращиваемой в синей области спектра. Но поскольку боковых побегов здесь не образуется, причины изменений ориентации листьев несколько иные. Так, максимум g(0), рассчитанный для нижней половины ценоза, сдвигается с возрастом ценоза в сторону меньших углов из-за активного отмирания горизонтально расположенных листьев. Этот процесс протекает значительно активнее, чем в других спектральных областях ФАР, так как из-за более разреженной структуры (отсутствие боковых побегов) горизонтально ориентированные нижние листья испытывают повышенную световую нагрузку и интенсивно отмирают.
С ростом уровня облучеппости доля вертикальных листьев в общей листовой площади ценоза пшеницы возрастает независимо от спектрального состава излучения исследуемых областей ФАР. Это характерно для молодых, интенсивно растущих ценозов пшеницы вплоть до фазы цветения. В этот период большая часть листовой поверхности (70 %) ориентирована в угловом интервале 60—90°. В то же время при облученности 100 Вт/м2 в исследуемых спектральных областях ФАР распределение площади листьев в угловом диапазоне 0° g(0) <1 90° более равномерное.
Отметим, что на заключительных этапах вегетации в ценозах пшеницы, выращиваемых при облученности 600 Вт/м2 происходит
66
существенное изменение ориентации листовой поверхности в сторону горизонтальных углов, на что указывает сдвиг в сторону меньших углов. При использовании Е = 100 Вт/м2 таких резких изменений не наблюдается.
В ориентации листовой поверхности ценозов пшеницы при использовании излучения смешанного спектрального состава принципиальных различий пе наблюдается. Основную роль в изменении g(0) играют возрастные различия структуры ценозов. Следует лишь отметить более значительный сдвиг в сторону больших углов максимума g(0), рассчитанной для нижней половины ценозов, выращиваемых при сине- и зелено-красном излучении с доминированием красного компонента излучения, по сравнению с вариантом, где используется сине-красное излучение с доминированием синего компонента. Это указывает на возрастание доли листовой поверхности, ориентированной вертикально в нижних слоях ценоза, и связано с появлением боковых побегов в двух первых спектральных вариантах.
Рядом исследователей показано, что синие лучи способствуют сильному отклонению первого листа в проростках пшеницы вплоть до горизонтального положения, в то время как в зеленых и красных лучах лист по ориентации близок к вертикальному. Сопоставление этих данных с нашими (см. рис. 3.11, 3.12) показывает, что таких резких различий не наблюдается. Очевидно, это связало с тем, что в наших опытах растения пшеницы составляют ценоз и изменения ориентации листьев в нем весьма ограничены и зависят от архитектоники и оптической плотности ценоза. Поэтому спектральные эффекты указанного типа могут нивелироваться структурой ценоза.
В то же время рост облученности от 100 до 600 Вт/м2 в различных спектральных областях ФАР приводит к заметному изменению ориентации листовой поверхности. Общей тенденцией в этом случае является увеличение доли вертикально ориентированной поверхности листьев в ценозах пшеницы, независимо от спектрального состава используемого излучения. Этот факт может служить проявлением адаптационной перестройки в структуре ценоза пшеницы при действии высокоинтенсивных лучистых потоков.
Для оценки особенностей роста листовой поверхности пшеницы рассмотрим ход листового индекса в зависимости от используемых режимов облучения в течение вегетации. Систематических исследований по оценке листовых индексов пшеницы при искусственном излучении различного спектрального состава и интенсивности света нам в литературе обнаружить не удалось. Среди немногочисленных трудов по этому вопросу представляет интерес работа В. И. Полонского [1978], где показано, в частности, что при высокоинтенсивном искусственном излучении пшеница может формировать ценозы с листовым индексом, достигающим 30, т. е. в 5—7 раз превосходит эту величину для естественных усло-
5*
67
Рис. 3.12. Возрастная динамика индекса ассимилирующей поверхности стеблей ценозов пшеницы при облученностях 100 Вт/м2 (а) и 600 Вт/м2 (б) в отдельных областях ФАР.
Рис. 3.11. Возрастная динамика листового индекса ценозов пшеницы при облученностях 100 Вт/м2 (а) и 600 Вт/м2 (б) в отдельных областях ФАР.
Здесь и на рис. 3.12: 1 — 400—500 нм; 2 — 500—600 нм; 3 — 600—700 нм.
вий. Следует учесть, что если для таких культур, как, например, редис, индекс определяет почти всю площадь ассимилирующей поверхности, то для ряда многоярусных культур, например пшеницы, этого сказать нельзя. В последнем случае большой вклад в общую ассимилирующую поверхность могут вносить колос и стебли, что требует учета площади их поверхности.
Изменение возрастной динамики листового индекса ценозов пшеницы наиболее ярко выражено при облученностях 100 и 600 Вт/м2 в зеленой и красной областях спектра, а наименее — в синей области спектра, (рис. 3.11), На изменение ассимилирующей поверхности стеблей сильное влияние оказывает наличие боковых побегов (рис. 3.12). В целом эта величина значительно выше при облученности 600, чем 100 Вт/м2 (рис. 3.12). Это, видимо, связано с увеличением диаметра главного стебля и боковых побегов, что является одной из анатомических особенностей этих фитоэлементов, обусловленной высоким уровнем облученности [Полонский, Лисовский, 1978].
Исследование влияния излучения смешанного спектрального состава на изменение возрастной динамики листового индекса ценозов пшеницы показало, что из рассматриваемых спектральных вариантов наиболее высокие значения листового индекса достигаются при зелено-красном излучении, а наименьшие — при сине-красном с доминированием синих лучей (рис. 3.13).
Сопоставление этих результатов с данными по листовому индексу ценозов пшеницы при использовании излучения отдельных областей ФАР дает основания предполагать, что высокие значения листового индекса, полученные для зелено-красного излучения, связаны, с одной стороны, с ускорением роста листьев в нижних слоях ценоза за счет проникновения зеленого компонента излучения в глубь листовых слоев, а с другой стороны, с интенсивным ростом верхних листьев за счет красного компо-
68
Рис. 3.13. Возрастная динамика индекса ассимилирующей поверхности ценозов пшеницы при облученности 100 Вт/м2 ФАР различного спектрального состава, с — листья; б — стебли. 1 — 400— — 500 нм (75 %) -I- 600—700 нм (25%); 2 — 400 — 500 нм (25%)4- 600—700 нм (75%); 3 — 500 — 600 нм (40 %) + + 600 — 700 нм (60 %).
Рис. 3.14. Возрастная динамика индекса листовой поверхности ценозов пшеницы с различной густотой посева, выращиваемых при облученности 600 Вт/м8 в синей (а), зеленой (б), красной (в) областях ФАР.
А - 1000 растений/^, Б _ 2000, В — 3000, Г 4000 растений/м2.
нента в общем излучении ФАР. Низкие значения листового индекса при использовании сине-красного излучения с максимумом в синей области ФАР подтверждают невысокую эффективность синего излучения в формировании листовой поверхности ценоза пшеницы.
Представляет интерес тот факт, что в первую половину вегетации листовой индекс в варианте с зелено-красным излучением был наиболее низким среди всех исследуемых режимов облучения и лишь начиная со второй половины вегетации, когда вклад площади листьев боковых побегов в общую листовую поверхность сильно возрастает, листовой индекс становится наиболее высоким. В этой связи обращает на себя внимание возрастная динамика листового индекса при сине-красном излучении, в котором доминируют красные лучи. В этом спектральном варианте наиболее высокие показатели листового индекса отмечены в первой половине вегетационного периода и незначительно уступают листовой площади ценоза пшеницы при зелено-красном излучении во второй половине вегетации. Что касается ассимилирующей поверхности стеблей, то лишь для сине-красного излучения с максимумом в синей области ФАР она заметно ниже, а между остальными спектральными вариантами эти различия незначительны.
Общий анализ данных по листовому индексу показывает что в ценозах пшеницы он выше при облученности 600 Вт/м2 ФАР
6 А, А. Тихомиров, Г, М. Лисовский, Ф. Я. Сидько
69
чем при Е = 100 Вт/м2 ФАР. При этом существенное влияние оказывает спектральный состав излучения.
Для более полного выявления действия спектрального состава лучистой энергии на листовой индекс представляет интерес рассмотреть формирование площади листьев в ценозах с разной густотой посева. Для этого проведены эксперименты на ценозах пшеницы с различной густотой посева: 103, 2-Ю3, 3-103, 4-103 расте-ний/м2 в каждом спектральном варианте опытов при уровне облученности 600 Вт/м2 в исследуемых областях ФАР. Проведенные эксперименты позволяют оценить максимальные значения листовых индексов ценозов пшеницы при различных спектральных составах высокоинтенсивного излучения и их возрастные изменения (рис. 3.14). Наибольший листовой индекс — 16 соответствует главным образом излучению зеленой области ФАР при нескольких значениях «стартовых» густот ценозов пшеницы, причем тем быстрее, чем более загущен ценоз. Это указывает на то, что существует определенный предел в условиях данного режима облучения, при котором дальнейшее увеличение листового индекса ценоза невозможно ввиду достижения критического значения оптической плотности ценоза, так как образование дополнительной фитоплощади невозможно из-за недостатка лучистой энергии. Таким образом, ценоз в этом случае ведет себя как саморегулирующая система. Максимальные значения листовых индексов различны и зависят от спектра излучения. Наиболее эффективно в этом отношении излучение зеленой области ФАР, а наименее — синей.
На рис. 3.15 представлены данные по вертикальному распределению листовой поверхности в ценозах пшеницы в процессе вегетации при облученности 100 Вт/м2 в отдельных областях ФАР. Наиболее резкая выраженность максимумов для кривых вертикального распределения поверхности листьев в многоярусном ценозе пшеницы соответствует одному и тому же возрастному периоду ценозов для спектральных вариантов с облученностью 100 Вт/м2 в синей и красной областях спектра и приходится примерно на 30-е сутки вегетации. В то же время для излучения зеленой области характер распределения листовой поверхности в ценозе в этот период вегетации более равномерный, а в возрасте 40 сут имеет небольшой пик в нижней части ценоза, что связано с появлением к этому времени листьев боковых побегов. К концу вегетации максимумы в рассмотренных спектральных вариантах становятся менее выраженными в связи с пожелтением листьев (рис. 3.15, А).
Несколько ипой характер возрастной динамики вертикального распределения листовой поверхности в ценозах пшеницы наблюдается при облученности 600 Бт/м2 в изучаемых спектральных областях ФАР (рис. 3.15, Б). Высокоинтенсивное излучение исследуемых спектральных диапазонов оказывает сходное воздействие на распределение листовой поверхности по вертикали ценоза в различные периоды вегетации. В отличие от уровня об-
70
1--1--Г ГП--1-Г I—Г——I—г
20 О 2020 О 2020 О 20
Площадь сритомассы в единице объема ценозаf м2 /м3
----2 — . j
Рис. 3.15. Возрастные изменения вертикального распределения фитоплощади в единице объема ценоза пшеницы при облученностях 100 Вт/м2 (Л) и 600 Вт/м2 (В) в синей («), зеленой (б) и красной (в) областях ФАР.
Возраст ценоза, сут: I — 10; II — 20; III — 30; IV—40; V — 50; VI — 60, VII — 70. 1 —листья; 2 — стебли; з — листья + стебли.
в
лученности 100 Вт/м2, максимумы кривых вертикального распределения листовой поверхности в ценозах пшеницы, формирующихся при высокоинтенсивпом излучении исследуемых спектральных интервалов ФАР, имеют тенденцию к концентрации в нижней части ценоза, особенно во второй половине вегетации (см. рис. 3.15), что не характерно для ценозов пшеницы, выращиваемых при облученности 100 Вт/м2. Это связано с тем, что мощность лучистого потока, проникающего в глубь ценозов молодых растений, довольно длительное время (около 20 сут) остается весьма высокой, что создает условия для дополнительного формирования в нижних слоях ценоза фитомассы в виде боковых побегов.
6*
71
Как отмечалось ранее, они появляются при высокоинтенсивном излучении зеленой и красной областей спектра, в то время как в синей области ФАР, несмотря на достаточное количество лучистой энергии, боковых побегов не образуется.
Общим для всех представленных спектральных вариантов является наличие явно выраженных максимумов в распределении фитоплощади, которые находятся в большинстве случаев в верхней части ценозов. Очевидно, такая структура ценоза биологически более целесообразна, поскольку в падающем на ценоз лучистом потоке содержится высокая доля лучей, сильно поглощаемых хлорофиллоносными структурами.
Действительно, если рассмотреть 40-суточные ценозы, закончившие формирование листьев, то наиболее выраженная неоднородность в распределении площади листьев по вертикали характерна для ценоза пшеницы, выращиваемого в сине-красных лучах, с доминированием красного компонента. У такого ценоза почти 70 % листовой поверхности сосредоточено в верхней части ценоза, что в 1,5 раза больше, чем в остальных спектральных вариантах (рис. 3.16). В то же время для зелено-красного излучения максимум вертикального распределения фитоплощади сдвинут в более глубокие слои ценоза, чем для сине-красного. В этом случае, наряду с высоким поглощением красного компонента излучения верхним слоем ценоза, лучи зеленого компонента, проникая в более глубокие слои, создают условия для дополнительного формирования фитомассы в нижних слоях ценоза.
Огурец. Изучение влияния спектральных режимов облучения на рост и органогенез огурца ведется уже более 30 лет [Клешнин, 1954; Баханова, Львова, 1961; Шульгин и др., 1963; Львова, 1978; и др. ]. Одним из первых наиболее значительных исследований в этом направлении явилось изучение ростовых и органообразовательных процессов растений огурца при излучении лю-
Рис. 3.16. Возрастные изменения в вертикальном распределении площади фитоэлементов в единице объема ценоза пшеницы при облученности 100 Вт/ма ФАР различного спектрального состава.
а _ 400—500 нм (75 %) + 600—700 нм (25 %); б — 400—500 нм (25 %) + 600—700 нм (75 %); в — 500—600 нм (40 %) + 600—700 нм (60 %). Остальные усл. обозн. см. рис.
О 4 ц
72
минесцептных ламп разного спектра [Бахапова, Льиона, 19(11 |. В этой работе, в частности, было показано, что синий свит способ-ствовал развитию пестичных цветков, а при красном спито преобладали тычиночные цветки. В работах И. А. Шульгина с соавторами [1963] и И. Н. Львовой [1978] установлены различия во влиянии спектрального состава света на некоторые сорта огурца при разных фотопериодах. В работе Ф. Я. Сидько с соавторами [1983] показаны возможности проведения целенаправленных формообразовательных процессов растений огурца при различных режимах облучения для увеличения хозяйственно полезного урожая.
В задачу настоящего исследования входило изучение некоторых закономерностей ростовых и органообразовательных процессов растений огурца при излучении различного спектрального состава. Растения выращивали при излучении отдельных спектральных областей ФАР интенсивностью 50 и 100 Вт/м2, а также при излучении ФАР смешанного спектрального состава интенсивностью 100 Вт/м2.
При использовании излучения отдельных областей спектра ФАР первые различия у растений огурца по органообразовательным процессам (образование листьев) стали проявляться к 10-суточному возрасту. Так, па 10-е сутки вегетации в варианте с синим светом при интенсивности 50 Вт/м2 отмечено наличие одного настоящего листа, а при интенсивности 100 Вт/м2 зарегистрировано наличие двух листьев. В спектральном варианте, где использовали излучение зеленой области спектра интенсивностью 50 и 100 Вт/м2, у растений огурца к возрасту 10 сут отмечено появление третьего листа. В красных лучах в этот период вегетации растения огурца имели при облученности 50 Вт/м2 два, а при облученности 100 Вт/м2 — лишь один настоящий лист.
Таким образом, к возрасту 10 сут процесс образования листьев наиболее активно проходил при излучении зеленой области спектра при интенсивности как 50, так и 100 Вт/м2. В синих лучах интенсивностью 50 Вт/м2 этот процесс протекал менее энергично, чем в красных, в то время как при интенсивности 100 Вт/м2 наблюдалась обратная картина. Полученные результаты дают основание считать, что в этот период вегетации световое насыщение по такому органообразовательному процессу у растений огурца, как появление листьев, выше всего расположено для синих лучей (за пределами облученности 100 Вт/м2), находится на плато светового насыщения в диапазоне 50—100 Вт/м2 для зеленых лучей и ингибируется в диапазоне 50—100 Вт/м2 для красных лучей.
К возрасту 30 сут при облученности 50 Вт/м2 наиболее интенсивно образование листьев шло по-прежнему в зеленых лучах (7 листьев), медленнее — в синих и красных (соответственно 6 и 5 листьев). Однако при интенсивности 100 Вт/м2 наибольшая скорость образования листьев отмечена в синих лучах (7 листьев на растение), в то время как в зеленых и красных лучах образовалось по 6 листьев на растение огурца.
73
Рис. 3.17. Возрастная динамика хлорофиллов (Л + Б) первых настоящих листьев ценозов огурца под излучением отдельных областей ФАР интенсивностью 100 Вт/м2.
Усл. обозн. см. рис. 3.11.
К возрасту 30 сут четко обозначились различия по старению и отмиранию листьев растений огурца в исследуемых спектральных вариантах. Наиболее интенсивно эти процессы шли под зеленым и красным светом. При облученности 50 Вт/м2 в красной области спектра растения огурца имели три нижних сухих листа, а растения, сформировавшиеся при излучении зеленой и синей областей спектра, сухих листьев не имели. При облученности 100 Вт/м2 растения огурца, растущие в красных лучах, имели четыре сухих нижних листа, в зеленых — два, а в синих лучах все листья были живыми.
Отставание в скорости образования новых листьев у растений огурца под красным светом к возрасту 30 сут усугублялось активным отмиранием листовых пластинок. С учетом процесса отмирания листовых пластинок огурца наиболее благоприятным к 30-суточному периоду вегетации оказался синий спектральный вариант, в котором функционировали все образовавшиеся листья огурца при интенсивности 50 и 100 Вт/м2.
Отмирание листовых пластинок растений огурца при исследуемых режимах облучения тесно связано с состоянием их пигментной системы. В синих лучах наблюдается наиболее интенсивное накопление зеленых пигментов на единицу поверхности листа как при Е = 50, так и при Е — 100 Вт/м2 (рис. 3.17). При длительном выращивании огурца в зеленых или красных лучах происходит интенсивное разрушение пигментных систем листьев огурца (Нестеренко и др., 1985].
При дальнейшем выращивании растений различия по содержанию пигментов в листьях для этих спектральных вариантов еще больше возрастают. Это приводит к гибели растений огурца при использовании красного света интенсивностью 50 и 100 Вт/м2 в возрасте 40 и 35 сут соответственно, так как к этому периоду во гетации у растений отмирает большая часть фотосинтезирующей листовой поверхности. К аналогичному результату приводит выращивание растений огурца в зеленых лучах при интенсивности 100 Вт/м2, когда гибель растений наступает на 45-е сутки вегета
74
ции. В остальных спектральных вариантах растения огурца при дальнейшем выращивании не погибли и вступили в стадию репродуктивного развития.
Так, в возрасте 30 сут, раньше, чем в других спектральных вариантах, начался процесс образования завязей у растений огурца в зеленых лучах при облученности 50 Вт/м2, в 38—40 сут у них наблюдалось массовое цветение. К возрасту 60 сут у. растений этого спектрального варианта продукционная деятельность резко упала ввиду активного отмирания листьев и выращивание растений было прекращено. К этому периоду вегетации растения имели по пять зеленцов.
Так же до возраста 60 сут выращивали растения огурца в синих лучах интенсивностью 50 Вт/м2. Массовое цветение огурца наступило в 45—50 сут, однако плодов фактически не образовалось. Более длительное выращивание, до 70 сут, проводили для растений огурца в синих лучах при интенсивности 100 Вт/м2. Лишь к этому периоду вегетации стало наблюдаться прогрессирующее отмирание фотосинтетического аппарата, который снабжал асспмилятами в среднем 10 зеленцов на каждом растении. Их налив к этому периоду вегетации прекратился.
Сравнительный анализ особенностей репродуктивного развития огурца при рассмотренных спектральных режимах облучения показывает, что уровень облученности ФАР может иметь принципиальное значение в этом процессе в зависимости от спектра излучения. Особенно контрастные различия в этом отношении дает изменение интенсивности излучения от 50 до 100 Вт/м2 в зеленой области спектра. Так, если для зеленых лучей при интенсивности 50 Вт/м2 обеспечивается прохождение растением огурца всех стадий репродуктивного развития вплоть до плодоношения, то при 100 Вт/м2 растения погибают, не вступив в стадию репродуктивного развития. Для синей и красной областей спектра повышение уровня облученности в рассматриваемых пределах принципиально не меняет, а лишь усиливает характер воздействия на те или иные стороны роста и развития растений.
Таким образом, увеличение уровня облученности с 50 до 100 Вт/м2 в исследуемых спектральных областях может не только повлиять на рост и развитие растений огурца при использовании того или иного спектрального состава света, но и изменить сравнительную спектральную эффективность действия излучения исследуемых областей ФАР. Так, если при облученности 50 Вт/м2 темпы развития были наибольшими в зеленых лучах, то при облученности 100 Вт/м2 более эффективными для обеспечения процессов развития оказались синие лучи, преимущество которых становится ощутимым к 30-суточному возрасту растений.
Специфика воздействия спектрального состава света на ростовые и оргапообразовательные процессы растений огурца резко изменяется при добавлении к излучению рассмотренных отдельных областей ФАР излучения других спектральных диапазонов при выровненных уровнях облученности. Исключение составляло лишь двухкомпонентное зелено-красное излучение ФАР.
75
Выращивание растений в потоке двухкомпонентного зелено-^ красного излучения интенсивностью 100 Вт/м2 показало, что процессы роста и развития принципиально не отличаются от их хода при излучении зеленой или краспой областей спектра той же интенсивности и не зависят от соотношения этих лучей в общем излучении. Растения погибали в возрасте 35—40 сут. При всех остальных сочетаниях отдельных областей спектра в ФАР летального воздействия излучения на фотосинтетический аппарат растений огурца при исследуемых интенсивностях излучения не наблюдается.
Эксперименты с двухкомпонентными сочетаниями исследуемых отдельных областей ФАР интенсивностью 100 Вт/м2 показали, что добавление синих лучей к красным в соотношении 1 : 4 значительно изменяло габитус растений в сравнении со спектральным вариантом, где использовался один синий свет. При этом уменьшалась длина междоузлий и высота растений примерно на 1/3. Дальнейшее увеличение доли красного компонента в синекрасном излучении вплоть до 60 % фактически не изменяло габитус растения огурца — длина междоузлий и высота практически не изменялись, растения к возрасту 30 сут имели по пять листьев.
Сине-зелепое излучение, где доминируют зеленые лучи (80 %), обеспечивало в первые 20 сут вегетации фактически такие же темпы образования листьев на растении, как и чисто зеленое излучение той же интенсивности. Однако в дальнейшем образование листьев при использовании двухкомпонентного излучения ФАР резко увеличивалось: к возрасту 30 сут у растений было по 8— 9 листьев (против 6 листьев на зеленом свету). Растения зацвели и вегетировали до возраста 80 сут, дав урожай плодов огурца, чего пе было при использовании зеленого излучения той же интенсивности.
Обратное соотношение синих и зеленых лучей в сине-зеленом излучении (80 % синих лучей) приводило к замедлению темпов развития растений огурца в сравнении с двухкомпонентным излучением, где доминируют зеленые лучи. Фактически 20 % добавки зеленых лучей к синим не изменяли скорости появления листьев, сроков цветения и лишь незначительно увеличивали урожай плодов огурца в сравнении со спектральным вариантом, где использовано излучение синей области спектра. Последнее обстоятельство дает основание полагать, что для сине-зеленого излучения более эффективно такое соотношение синих и зеленых лучей, когда доминируют зеленые лучи. Излучение, где доминируют зеленые лучи, обеспечивает наиболее высокую площадь листовой поверхности и хорошие соотношения между биомассой листьев и общей биомассой надземной части растений (табл. 3.6).
Таким образом, увеличение доли зелепых лучей в сине-зеленом излучении от 20 до 80 % ускоряло рост растений огурца.
При использовании трех компонентного излучения ФАР для накопления зеленых пигментов в листьях растений огурца нет
76
Таблица 3.6 Ростовые характеристики растений огурца, сформировавшихся при излучении с различным соотношением синих и зеленых лучей
Соотношение синих и зеленых лучей, % Биомасса надземной части растений огурца, г Биомасса листовых пластинок растений огурца, г Суммарная площадь листьев одного растения, см3 Масса листьев ty0 Надземная биомасса
Синие 100 Синие 80, зеленые 20 Синие 20, зеленые 80 Зеленые 100 Примечание. Об.1 4,00 2,30 6,5 2,2 тученность 100 2,4 1,7 4,5 1,4 Вт/м® ФАР. I 1357 458 1381 255 возраст растет 60 74 69 63 «й 70 сут.
существенной разницы, излучение какой области спектра должно доминировать в общем потоке ФАР. Однако такие различия становятся весьма существенными при анализе роста и органообразовательных процессов. Так, при использовании трехкомпонентного излучения ФАР с различным сочетанием синих, зеленых и красных лучей наиболее ранние различия (в возрасте 10 сут) по ростовым и органообразовательным процессам начинают проявляться лишь в ряде спектральных вариантов, особенно там, где доминирует какая-либо отдельная область ФАР. К возрасту 10 сут растения огурца в спектральных вариантах с доминированием синих, зеленых или красных лучей (60 % от 100 Вт/м2 ФАР) имели по три настоящих листа. Рост листовых пластинок наиболее активно шел при излучении, где доминировали красные лучи, листья, формирующиеся при излучении, где доминируют зеленые лучи, имели наиболее короткие междоузлия. К возрасту 20 сут определялось отставание в темпах образования листьев у растений огурца, формирующихся при излучении, где доминируют синие лучи. К возрасту 30 сут эти различия еще более возросли. Так, растения огурца, растущие при излучении ФАР, где доминируют красные и зеленые лучи, имели в этом возрасте по девять, а где синие — по восемь листьев. Кроме того, в первых двух спектральных вариантах наступило цветение, в то время как при излучении, где доминируют синие лучи, цветения еще не было. Наибольшая длина междоузлий — 4,2 см зарегистрирована у растений огурца при излучении, где доминируют красные лучи, в двух других спектральных вариантах различия по этому параметру практически отсутствовали и длина междоузлий составляла в среднем 3,1 см.
Таким образом, ростовые и органообразовательные процессы в ценозах огурца существенно зависят от соотношения синих, зеленых и красных лучей в общем потоке ФАР.
При использовании излучения синей области спектра интенсивностью 100 Вт/м2 около 70 % листовой поверхности находится в интервале углов наклона 15—30° (рис. 3.18, с). При излучений,
77
Рис. 3.18. Функция пространственной ориентации листьев д(6)«' для ценозов огурца, сформировавшихся при излучении отдельных областей ФАР («) и при излучении смешанного спектрального состава ФАР (б).
Облученность 100 Вт/м2 ФАР. Возраст ценозов 30 сут. 1 — 400—500 нм; 2 — 500—600 нм; з — 600—700 нм; 4 — 400—500 нм (60 %) + 500—600 нм (20 %) + 600—700 нм (20 %); 5 — 400—500 нм (20 %) + 500—600 нм (60 %) + 600—700 нм (20 %); 6 — 400—500 нм (20 %) -4- 500—600 нм
(20 %) + 600—700 нм (60 %).
в котором отсутствуют синие лучи, максимум функции g(6) еще больше сдвигается в сторону малых углов, и в этом случае около 80 % листовой поверхности расположено почти горизонтально.
Такая роль синего света прослеживается и при анализе ориентации листовой поверхности при выращивании ценозов огурца при излучении смешанного спектрального состава ФАР (рис. 3.18, б). Спектральные варианты, которые характеризуются преимущественно содержанием синих лучей, отличаются наличием резко выраженного максимума функции g(&), указывающего, что более половины всей листовой поверхности находится в угловом диапазоне 0—15° (см. рис. 3.18, б). С уменьшением доли синих лучей в потоке ФАР такой максимум в большинстве спектральных вариантов сохраняется, но становится менее выраженным и смещается в сторону отрицательных углов. Последнее обстоятельство указывает, что возрастает доля листовой поверхности, ориентированной в нижнюю полусферу. Такой характер ориентации листьев увеличивается не только с уменьшением доли синих лучей в потоке ФАР, но и с возрастом ценозов внутри каждого спектрального варианта.
Решающее влияние на увеличение вертикальной ориентации листовой поверхности в ценозах огурца оказывает доля синих лучей в общем потоке ФАР.
Листовой индекс в ценозах огурца с увеличением длины волны падающего излучения уменьшается. Причем возрастная динамика этой характеристики имеет свои особенности для излучения исследуемых областей ФАР.
В первой половине вегетации наиболее интенсивно увеличение листового индекса идет под зеленым и красным светом, а наименее — под синим. Однако параллельно с этим наблюдается постепенное выцветание зеленых пигментов в листьях ценозов огурца при использовании красного и зеленого света. После 20-суточного возраста эти процессы способствуют снижению листового индекса, причем это снижение идет более интенсивно при красном свете, чем при зеленом (рис. 3.19).
При синем свете возрастная динамика листового индекса имеет обширное плато, связанное с длительным функционированием листового аппарата ценоза огурца.
78
Листовой
Рис. 3.19. Возрастная динамика листового индекса ценозов огурца, сформировавшихся при излучении отдельных областей ФАР интенсивностью 100 Вт/м2.
1 — 400—500 нм; 2 — 500—600 нм; 3 — 600—700 нм.
Рис. 3.20. Динамика листового индекса ценозов огурца, сформировавшихся при различном спектральном составе ФАР интенсивностью 100 Вт/м2.
Усл. обозн. см. рис. 2.9.
При излучении смешанного спектрального состава света формирование листового индекса ценозов огурца в первые 30 сут происходит наиболее интенсивно с доминированием в зеленых лучах (рис. 3.20), когда листовой индекс в этом спектральном варианте в 1,5 раза превышает значения листовых индексов для ценозов огурца, сформировавшихся при излучении, где доминируют синие или красные лучи. В дальнейшем увеличение листового индекса идет опережающими темпами при излучении, где доминируют красные лучи, и уже к 40 сут вегетации листовой индекс достигает максимального значения — 3,2 против 2,6 для варианта, где доминируют зеленые лучи, и 2,2 — где синие. Причем в двух последних случаях максимальные значения листовых индексов соответствуют 50-суточным возрастам ценозов.
Нарастание листового индекса в первые 30 сут вегетации идет наиболее быстро в зеленых лучах или потоках ФАР, где эти лучи занимают доминирующее положение (см. рис. 3.19, 3.20). Этот результат может найти применение при выращивании рассады огурца, а также в интенсивной светокультуре при комбинированном облучении растений разноспектральными потоками ФАР в разные периоды вегетации.
Распределение листовой площади по вертикали для 30-суточных ценозов огурца имеет один максимум, который находится в верхней половине ценоза (рис. 3.21). В этой части ценоза сосредоточена подавляющая часть листовой поверхности, причем наи-
79
op 1p 2,0 Зро,о 1,0 2p зр Площадь листьев
в единице объема ценоза, м2/м3
Рис 3.21. Распределение площади листьев в единице объема ценоза огурца при облученностях 50 (Л) и 100 (Б) Вт/м2 в различных областях спектра,
а — 400—500 нм; б — 500—600 нм; в — 600—700 нм. Возраст ценоза 30 сут.
большая ее плотность в единице объема отмечается для спектрального варианта с зеленым светом, а наименьшая — с красным.
Томаты. Исследования по влиянию света на рост, развитие и органогенез томатов показывают, что развитие этих растений значительно ускоряется при доминировании оранжево-красной и инфракрасной радиации [Мошков, 1966; Куперман, Русу, 1971; и др.]. Эти данные получены в экспериментах с использованием невысоких облученностей, создаваемых различными цветными люминесцентными лампами [Куперман, Русу, 1971], либо при достаточно высоких уровнях облученности (Е = 150—200 Вт/м2 ФАР) для интенсификации процессов роста и развития [Мошков, 1966], но при очень ограниченных вариациях спектрального состава ФАР. Систематические исследования процессов роста и развития томатов при достаточно интенсивном излучении ФАР (Е — — 100—150 Вт/м2 и выше) в отдельных спектральных областях и их различных сочетапий вообще не проводились, а имеющиеся данные весьма фрагментарны [Александрова, Клочкова, 1983; Протасова, 1987; и др.].
С учетом этого нами были проведены систематические исследования по оценке ряда показателей роста и развития томатов при излучении как отдельных спектральных областей, так и смешанного спектрального состава ФАР.
При работе с лучистыми потоками отдельных областей спектра использовали уровни облученности 50 и 100 Вт/м2. Использование таких режимов облучения для выращивания томатов показало, что наиболее активно ростовые и оргапообразовательные процессы идут при излучении красной области спектра, а наименее активно — синей. Промежуточное положение по воздействию на ростовые и оргапообразовательные процессы растений томата занимает излучение зеленой области спектра. Эффективность ее воздействия на эти процессы по характеру ближе к красной области спектра, чем к синей (табл. 3.7). Увеличение интенсивности излучения с 50 до 100 Вт/м2 в исследуемых областях спектра наиболее заметно сказывается на ускорении процесса начала цвете-
80
Таблица 3.7
Некоторые показатели роста и развития томатов сорта Старфайер
Область спектра, нм Облученность, Вт/м2 Число листьев на 1 растение при возрасте растений, сут Высота растений в возрасте 30 сут, см Начало цветения, сут
10 20 30
400—500 50 1 4 6 30 52
100 2 5 7 25 47
500—600 50 3 4 8 40 44
100 3 5 9 33 35
600—700 50 2 4 9 45 42
100 1 4 10 35 34
ния, практически не меняя временного разрыва по этому показателю между спектральными вариантами опыта.
Морфологический анализ взрослых растений томатов, сформировавшихся при излучении отдельных областей ФАР интенсивностью как 50, так и 100 Вт/м2, показал, что к концу вегетации наибольшую листовую массу имели томаты при излучении синей области спектра, а наименьшую — красной области. Однако появление и развитие репродуктивных органов шло более интенсивно в красных лучах, а наименее интенсивно — в синих. В отличие от этих характеристик, слабо зависящих от интенсивности излучения, такой показатель, как линейный рост томатов, существенно зависел от спектра ФАР и уровня облученности в ее отдельных областях. Заметные различия по росту стебля в зависимости от спектра наблюдались при облученности 50 Вт/м2, когда наиболее интенсивный рост стебля был зарегистрирован в красных лучах, а наименее интенсивный — в синих. Однако двухкратное увеличение уровня облученности резко сократило эти различия (см. табл. 3.7).
При анализе процессов генеративного развития томатов весьма важной характеристикой является начало цветения. Существенные различия по срокам наступления цветения у томатов, выращиваемых в лучах отдельных областей ФАР, наблюдались при облученности 50 Вт/м2. Самое раннее цветение (иа 42-е сутки вегетации) наступает в красных лучах, наиболее позднее (на 52-е сутки вегетации) — в синих лучах, а для томатов, выращиваемых в зеленых лучах, начало цветения приходится на 47-е сутки вегетации. При повышении уровня облученности до 100 Вт/м2 в отдельных областях ФАР сроки наступления цветения у растений томатов сократились во всех спектральных вариантах, причем для спектральных вариантов, где используются зеленые и красные лучи, различия в наступлении цветения практически исчезли.
Процесс плодообразования у растений томата существенно различался во всех спектральных вариантах опыта. Замедленно
81
он шел в синих лучах, а наиболее интенсивно — в красных лучах. Так, при облученности 100 Вт/м2 к возрасту 60 сут растения томатов, сформировавшиеся в красных лучах, имели около поло-, вины созревших плодов, сформировавшиеся в зеленых лучах,— 20 %, а в синих лучах все плоды томатов оставались в этот период вегетации зелеными. Подобная же картина наблюдалась и при облученности 50 Вт/м2 в исследуемых спектральных областях, однако она сдвигалась к более позднему периоду вегетации (75 сут).
Резюмируя вышеизложенное, отметим, что несмотря на различия в росте и развитии при излучении отдельных областей ФАР при облученности как 50, так и 100 Вт/м2 растения томатов прошли все стадии онтогенеза и дали урожай плодов.
Использование двухкомпонентных сочетаний различных областей ФАР для выращивания томатов показало, что доминирование синих лучей в излучении замедляет процессы органообразования, цветения и в конечном итоге плодоношение томатов. Это происходило независимо от того, какая область спектра — зеленая или красная использовалась в сочетании с синей областью.
Эксперименты по выращиванию растений томатов при различных сочетаниях зеленых и красных лучей показали, что получаемые параметры по росту и развитию находились в диапазона значений, соответствующих отдельному использованию зеленых и красных лучей.
Для растений томата, сформировавшихся при излучении трехкомпонентных сочетаний исследуемых спектральных областей в ФАР, сохранялась общая закономерность специфики воздействия излучения той области спектра, которая доминирует в общем потоке ФАР (табл. 3.8). При излучении, где доминируют синие лучи, растения томата имели меньшую высоту стебля, меньшее количество листьев на стебле, более позднее наступление цветения, чем при излучении с преобладанием зеленых или красных лучей (см. табл. 3.8). Следует отметить, что спектральный вариант с белым светом дает средние показатели по исследуемым параметрам роста и развития. Наиболее высокие значения этих показателей соответствовали спектральным вариантам, где доминируют красные лучи.
Таблица 3.8
Показатели роста 30-суточных растений и сроки зацветания томатов сорта Старфайер при облученности ФАР 100 Вт/м2 смешанного спектрального состава
Соотношение спектральных областей в ФАР, % Высота растений, см Числе листьев на 1 растении Начало цветения, сут
с — 60, з — 20, к — 20 20 7 40
с — 20, з — 60, к — 20 25 9 35
с — 20, з — 20, к — 60 30 10 30
с — 34,з — 33, к — 33 30 8 33
82
Изложенный материал позволяет сделать ряд обобщений по спектральной эффективности процессов роста и развития ценозов исследуемых культур.
Так, за исключением ценозов огурца, спектральная эффективность роста листовой поверхности ценозов в течение вегетационного периода фактически не претерпевает резких качественных изменений. В то же время для ценозов огурца спектральная эффективность роста листовой поверхности смещается в синюю область (см. рис. 3.19). Однако вряд ли обоснованно считать, что ростовая функция в данном случае инициируется синими лучами более активно, чем зелеными (или красными). Здесь, как нам представляется, следует иметь в виду, что при длительном воздействии на фотосинтетический аппарат огурца зеленых или красных лучей ростовая функция подвергается существенному негативному воздействию со стороны ряда других составляющих продукционного процесса. Среди них важная роль должна быть отведена фотосинтезу. Известно, что подавление фотосинтеза листьев за счет различных нарушений метаболизма приводит к замедлению, а затем и остановке роста листовой поверхности [Мокроно-сов, 1981, 1983]. Именно такие нарушения выявлены нами для листьев огурца при их длительном пребывании на зеленом и красном свету [Тихомиров и др., 1987]. Это, вероятно, и предопределило замедление, а затем и остановку роста листовой поверхности ценозов огурца в зеленых и красных лучах, а поддержание ростовой функции листьев ценоза огурца оказалось возможным лишь в синих лучах. Таким образом, смещение спектральной эффективности процессов роста листовой поверхности ценозов огурца из зеленой в синюю область ФАР связано, вероятно, не с переключением каких-то тонких механизмов фоторегуляции, усиливающих эффективность использования синих лучей в процессах роста, а с «выключением» зеленых и красных лучей из фотоконтроля за ростовой функцией.
Однако такой контроль сохраняется, если используется смешанное излучение ФАР, когда присутствие синих лучей в общем излучении препятствует возникновению отмеченных выше причин блокировки фотосинтеза листа, а через них — ростовой функции.
Поскольку рост листовой поверхности ценозов в принципе обеспечивается лучистыми потоками всех исследуемых спектральных областей ФАР, можно полагать, что фотоконтроль этой составляющей продукционного процесса осуществляется в синих, зеленых и красных лучах, а фоторецепторы, ответственные за этот процесс, имеются в каждой из названных спектральных областей. Важно отметить, что в ряде случаев добавка синих лучей к красным и зеленым принципиально важна для обеспечения длительного роста и функционирования листовой поверхности, как это имело место, например, для ценозов огурца.
Необходимость добавки синих лучей к зеленым и красным для обеспечения устойчивого роста листовой поверхности может
83
оказаться крайне важной с увеличением уровня облученности. Для ряда культур (огурец, редис) это условие обязательно, так как предотвращает остановку роста и деструкцию фотосинтетического аппарата {Тихомиров, Сидько, 19821, в то время как для таких культур, как пшеница и томаты, это не имеет кардинального значения, а лишь может способствовать в большей или меньшей степени усилению роста и увеличению длительности функционирования ассимилирующей поверхности ценозов. В свете изложенного подтверждается мнение Н. П. Воскресенской [1987 ], что синий свет активизирует сенсоры, чувствительные к высоким уровням облученности. Однако, несмотря па достаточно большое количество гипотез и отдельных экспериментальных фактов, роль синего света в высокоинтепсивном излучении ФАР с точки зрения обеспечения ростовой функции остается во многом неясной. Пока лишь можно утверждать, что высокая спектральная эффективность процессов роста листовой поверхности ценозов в зеленых и красных лучах может быть гарантированно реализована на большем или меньшем (что зависит от вида растений) фоне синих лучей. Следует полагать, что синие лучи в зтом случае «ослабляют или снимают» отрицательные обратные связи со стороны других составляющих продукционного процесса (фотосинтез, фотосинтетический метаболизм и т. д.) на ростовую функцию и создают условия для более сбалансированной работы всех звеньев фотосинтетического аппарата.
Процессы развития растений разных видов в ценозах имеют резко выраженную видовую специфику реакций к спектру и интенсивности излучения различных областей ФАР. В этом плане растения можно разделить на две группы: характеризующиеся нарушением процесса развития на определенном этапе онтогенеза (огурец, редис); способные пройти все этапы онтогенеза и дать урожай независимо от используемого спектрального состава излучения исследуемых областей ФАР (томаты, пшеница). Наличие такой закономерности весьма важно, так как принципиальная возможность использовать излучение той или иной области ФАР для выращивания растений до получения хозяйственно полезного урожая определяется тем, обеспечивает ли данный спектр излучения ФАР прохождение всего процесса онтогенеза растения.
На основании изложенного материала можно заключить, что роль излучения отдельных областей ФАР в обеспечении прохождения онтогенеза растений разных видов нельзя признать однозначной. Такая неоднозначность обусловлена, с одной стороны, наличием видовой специфики в реакции растений на различный спектр излучения ФАР [Тихомиров и др., 1987], а с другой стороны, зависимостью спектральной эффективности излучения исследуемых областей ФАР в отношении различных процессов роста и развития от уровней облученности ФАР в исследуемых спектральных областях.
Среди исследованных нами видов растений наименее критичной в отношении прохождения всех этапов онтогенеза при излу-
84
чении отдельных спектральных областей оказалась пшеница, для которой это было подтверждено экспериментально как при Е = = 100, так и при Е = 600 Вт/м2 ФАР, причем в последнем случае темпы прохождения этапов органогенеза и продуктивность растений были выше. Этот факт позволяет считать, что указанные спектральные потоки ФАР обеспечат прохождение всех этапов, онтогенеза у пшеницы и при еще более высоких уровнях облученности ФАР, чем достигнутые в наших экспериментах.
Для всех исследованных отдельных областей ФАР в более ограниченном диапазоне уровней облученности (50—100 Вт/м2} показана возможность прохождения всех этапов онтогенеза для растений томатов. Нами также предпринимались попытки выращивания томатов при Е > 200 Вт/м2 в отдельных областях ФАР. Хотя в этих экспериментах синие, зеленые и красные лучи обеспечивали прохождение томатами всех этапов органогенеза, однако общая продуктивность растений в зеленых и особенно красных лучах снижалась. Это позволяет считать, что диапазон уровней облученности для излучения отдельных областей ФАР, обеспечивающих прохождение всех этапов онтогенеза, для томатов значительно уже, чем для пшеницы.
Такой диапазон уровней облученности оказался еще более узким для редиса, поскольку уже при Е 200 Вт/м2 ФАР происходит гибель растений вначале в красных, а затем в зеленых лучах.
Самыми критичными из исследуемых видов растений к спектральному составу света, обеспечивающему прохождение всех этапов органогенеза, оказались растения огурца, для которых лишь зеленые лучи при Е — 50 Вт/м2 ФАР и синие лучи при Е = = 100 Вт/м2 обеспечивали прохождение всех этапов онтогенеза до получения плодов.
На примере растений огурца, а также редиса особенно отчетливо видно, какую важную роль играет уровень облученности узкоспектральных потоков ФАР в обеспечении прохождения этапов онтогенеза. При этом вовсе не обязательно, чтобы неэффективность излучения той или иной области спектра в обеспечении прохождения этапов органогенеза была связана с гибелью растения на одной из его стадий, как это отмечалось выше для растений редиса и огурца при действии зеленых и красных лучей. Например, неэффективность синих лучей интенсивностью 50 Вт/м2: для растений огурца связана с нарушением плодообразования, хотя сами растения при этом не погибают. Гибель растений, наступающая из-за деструкции пигментного аппарата, наблюдалась, как это было видно из вышеприведенных материалов, в зеленых и красных лучах у растений огурца и редиса. Не исключен такой процесс и для синих лучей, но интенсивности здесь будут много выше.
Разумеется, что и для таких растений, как пшеница и томаты, имеются критические уровни облученности для излучения отдельных областей ФАР, когда растения должны погибнуть,
7 А. А. Тихомиров, Г.. М. Лисовский. Ф. Я. Сидько
85
не успев пройти все стадии онтогенеза, но такие уровни облученности весьма высоки и находятся за пределами тех интенсивностей ФАР, которые реально используются в условиях светокультуры.
Важно отметить, что понимание роли отдельных областей ФАР в рассмотренных процессах роста и развития на разных уровнях организации фотосинтетического аппарата лишь отчасти и в определенных пределах позволяет прогнозировать общий характер протекания этих процессов при использовании излучения смешанного спектрального состава, где доминирует та или иная область ФАР. Такой прогноз оказался справедлив для различных вариантов сочетаний зеленых и красных лучей в зеленокрасном излучении, которые действительно оказались бесперспективными при использовании такого спектрального состава ФАР для обеспечения успешного протекания всех этапов органогенеза у культур, рост и развитие которых ингибируется по отдельности в зеленых и красных лучах (например, огурец, редис). В то же время синий свет, который среди излучений отдельных областей ФАР был вполне благоприятным для прохождения онтогенеза, при доминировании в смешанном потоке ФАР снижал темпы онтогенеза. Это выражалось, как правило, в затягивании всего онтогенеза в сравнении с вариантами, где в ФАР доминировали другие ее области. Доминирование синих лучей в ФАР может быть оправдано для успешного прохождения онтогенеза лишь при весьма высоких уровнях облученности (Е > > 200 Вт/м2 ФАР), как это было установлено на примере ценозов редиса. В то же время при использовании уровней облученности до 100—150 Вт/м2 наиболее сбалансированные процессы роста и развития у большинства растений разных видов обеспечиваются излучением, где доля синих лучей весьма невелика (20—25 %) при некотором доминировании красных или равном соотношении зеленых и красных лучей. В ряде случаев в качестве альтернативы может быть использовано и равноэнергетическое излучение ФАР либо иные спектральные сочетания на фоне пониженной доли синего компонента.
§ 2. Спектральный состав света и формирование мезоструктуры листьев различных растений
Процессы роста как на уровне целого растения, так и на тканевом, клеточном и других, более низких уровнях организации фотосинтетического аппарата взаимосвязаны и в значительной мере определяют формирование продуктивности растений. Среди них весьма важным представляется изучение 'различных показателей мезоструктуры листа, которая весьма чувствительна к спектральному составу света, что может быть охарактеризовано показателями роста и клеточной дифференцировки ткани листа. Эти показатели часто использовали для изучения их зависимости от спектрального состава света [Кахнович, 1980; Карначук и др.,
86
1988]. В частности, было установлено, что клеточный рост наиболее активен на синем свету, а наименее — на красном [Карна-чук и др., 1988]. Причем на красном свету формируются листья более тонкие, с мелкими хлоропластами [Власова, Воскресенская, 1973]. *
В последнее время значительное внимание уделяется роли зеленого света в формировании мезоструктуры листа. Сделан весьма важный вывод, что на синем, зеленом и красном свету фотосинтез в расчете на единицу поверхности хлоропласта имеет несущественные различия в сравнении с фотосинтезом на единицу поверхности листа [Карначук, 1987]. С другой стороны, отмечается видовая специфика ростовых процессов на тканевом уровне для листа, сформировавшегося при различных спектральных режимах облучения [Карначук, 1987; Карначук и др., 1988]. Таким образом, ростовые процессы, происходящие на тканевом уровне организации фотосинтетического аппарата, весьма важны для оценки фотосинтетической активности листа, а следовательно, и продукционной деятельности растения при тех или иных спектральных режимах облучения.
В связи с этим мы специально исследовали особенности формирования мезоструктуры листьев различных видов растений при излучении разного спектрального состава [Шалаева и др., 1991].
Наиболее тонкие листья формировались в красных лучах, а наиболее толстые — в синих. При этом толщина слоя палисадной паренхимы в сравнении с губчатой паренхимой была в 2—3 раза меньше при любом спектре излучения ФАР (табл. 3.9)?.
Повышение уровня облученности от 50 до 100 Вт/м2 способствует увеличению большинства рассмотренных параметров мезоструктуры листа огурца также независимо от спектра излучения отдельных областей.
С другой стороны, ряд характеристик мезоструктуры листа более отзывчив на рост уровня облученности, чем на изменение спектра ФАР. Так, с ростом уровня облученности наиболее существенным, слабо зависящим от спектра ФАР является увеличе-
Таблица 3.9
Параметры мезоструктуры настоящего листа растения огурца
Параметр 400—500 нм 500—600 нм 600—700 нм
К, е2 Е. Eq Ei к2
Общая толщина листа, мкм 210 240 150 205 80 155
Толщина палисадной паренхимы, мкм 60 55 40 70 30 45
Толщина губчатой паренхимы, мкм Количество клеточных слоев палисад- 150 185 ПО 135 50 ПО
ной паренхимы, шт. Средняя высота клеток палисадной па- 1—2 1 1 1 1 1
репхимы, мкм Средпий размер клеток губчатой па- 60 55 40 70 30 45
репхимы, мкм 20 20 23 30 15 20
Примечание. В, — 50, Е2 — 100 Вт/м2. Возраст растений 10 сут.
87
Eiie толщины листовой пластинки, а также увеличение толщины палисадной и губчатой паренхим. В то же время средние размеры палисадных и особенно губчатых клеток возросли незначительно. Рост облученности в отдельных областях ФАР не привел к увеличению числа клеточных слоев в губчатой и палисадной паренхимах.
По мнению ряда исследователей, роль палисадной и губчатой тканей в фотосинтетической деятельности листа не однозначна. Существуют данные, что основной вклад в фотосинтез листа обеспечивает палисадная ткань [Мокроносов и др., 1973]. Такой вывод в этой работе был сделан для условий выращивания листьев растений на естественном белом свету. В связи с этим представляет интерес установить, сохраняется ли такая закономерность при выращивании растений в разпоспектральных потоках для отдельных областей ФАР. Нам представляется целесообразным провести такую оценку в первую очередь для излучения красной области спектра, поскольку именно для него наблюдаются наиболее контрастные различия в фотосинтетической деятельности зеленых листьев разных видов растений. Так, например, растения огурца при облученности 50 Вт/м2 в красных лучах погибают через 30—40 сут [Нестеренко и др., 1985], то же самое происходит с редисом, но при более высоких интенсивностях красного света [Тихомиров, Сидько, 1982], в то время как пшеница и томаты хорошо растут на красном свету в широком диапазоне облученностей. Подсолнечник занимает промежуточное положение, так как после стрессового состояния адаптируется к красному свету. С учетом этого листья указанных растений были выбраны для оценки влияния красного света в сравнении с белым на образование палисадной ткани (табл. 3.10). Красный свет резко снижает образование палисадной ткани у всех этих растений, независимо от их фотосинтетической активности при его длительном воздействии. Поэтому можно предположить, что изменение фотосинтетической активности листа на красном свету может быть связано не обязательно с замедлением образования палисадной ткапи, по и с рядом других причин.
Нельзя исключить, что в качестве таких причин могут выступать другие параметры мезоструктуры листа. С этой целью на примере разновозрастных листьев растений огурца и подсолнеч-Таблица 3.10 Число слоев палисадной паренхимы в листьях некоторых растений, сформировавшихся при красном и белом (контроль) свете интенсивностью 100 Вт/м2
Культура Спектральный вариант Культура Спектральный вариант
Красный свет Белый свет Красный свет Белый свет
Огурец 1 2 Томаты 1 2
Подсолнечник 1 2—3 Пшеница 1 2—3
Редис 2 3—4
88
Таблица 3.11
Количественные характеристики анатомической структуры листьев огурца разного возраста, сформировавшихся при белом и красном свете интенсивностью 100 Вт/м2
Спектральный состав света Возраст растения, сут Количество клеток, тыс./см2 Количество хлоропластов в одной клетке Количество хлоропластов, млн/см2 Общее количество клеток, тыс./см2 Доля палисадных клеток от всех, %
П* п г
Белый 10 196,8 169,2 96,3 76,3 29,2 366,0 53,8
30 144,1 81,8 102,0 80,2 21,3 225,9 63,8
Красным 10 190.0 160,1 58,9 50,6 19,2 350,1 54,3
30 34,0 56,6 17,6 19,5 1,7 91,0 37,5
* П — палисадная паренхима.
** Г — губчатая паренхима.
дика, имеющих различную реакцию на красный свет, рассмотрим более подробно поведение ряда количественных характеристик мезоструктуры листьев этих растений, сформировавшихся на белом (контроль) и красном свету (табл. 3.11).
У листьев растений огурца при длительном воздействии красного света уменьшается количество клеток и содержание в них хлоропластов как в палисадной, так и в губчатой ткани. При этом снижается и число хлоропластов в расчете на одну клетку палисадной и губчатой паренхим, где эти процессы шли неодинаково, поскольку количество клеток в палисадной ткани листа огурца с возрастом уменьшалось сильнее, чем в губчатой ткани. В контроле (белый свет) за рассматриваемый период (20 сут) зарегистрировано уменьшение количества клеток палисадной и губчатой паренхим и уменьшение общего количества хлоропластов. Однако скорость уменьшения этих параметров в контроле была примерно в 2 раза ниже, чем для соответствующих параметров ме-Эоструктуры листа огурца, растущего в красных лучах. В то же время количество хлоропластов в расчете на одну клетку как палисадной, так и губчатой паренхим для белого света, в отличие от красного, не уменьшалось с возрастом листа. В целом после 20-суточного пребывания в красных лучах доля палисадных клеток и содержащихся в них хлоропластов у листьев огурца уменьшилась в 1,5—2 раза, в то время как в контроле за этот период времени эти показатели мезоструктуры не только не уменьшились, но даже несколько увеличились (см. табл. 3.11). Таким образом, для листа огурца после 20 сут пребывания на красном свету наблюдается уменьшение всех рассматриваемых параметров мезоструктуры.
Этого не происходит для листьев подсолнечника, выравниваемых при аналогичных условиях облучения (табл. 3.12). Так, например, число клеток столбчатой паренхимы и количество в них
89
Таблица 3.12
Количественные характеристики анатомической структуры листьев подсолнечника разного возраста, сформировавшихся при белом и красном свете интенсивностью 100 Вт/м2
аст ра-1Й, сут Спектральный вариант П, 10’ кл./см2 Г, 103 кл./см2 Количество хлоропластов в палисадной клетке Количество хлоропластов в губчатой клетке Число слоев П Толщина листа, мм П+Г 100 %
Возр 3 *
1Е Белый 283 367 54,1 39,3 2 0,26 43,5
Красный 161 208 46,9 48,0 1 0,19 43,6
ЗС Белый 298 351 49,1 23,3 2 0,29 45,9
Красный 179 356 38,3 45,8 1 . 0,24 33,4
40 Белый 290 328 50,2 31,6 2 0,29 46,9
Красный 136 248 21,9 37,3 1 0,20 35,4
Примечание. Усл. обозн. см. табл. 3.11.
хлоропластов уменьшается до 20-суточного возраста листа подсолнечника, а затем выходит на стационарный уровень.
Примерно аналогичная ситуация имеет место и для содержания хлоропластов в расчете на клетку палисадной ткани. Что касается губчатой ткани, то здесь наблюдалось даже возрастание числа клеток в расчете на единицу поверхности листа подсолнечника в период 15—20 сут и их дальнейшая стабилизация (см. табл. 3.12).
Под белым светом листья подсолнечника сохраняют практически стабильными рассматриваемые параметры мезоструктуры в; исследуемый временной период (см. табл. 3.12). Таким образом, как у листьев огурца, так и у листьев подсолнечника длительное воздействие красного света приводит к резкому уменьшению палисадной ткани, однако если у листьев огурца этот процесс идет непрерывно и приводит к полной деструкции фотосинтетического аппарата, то для листьев подсолнечника происходит стабилизация анатомической структуры листа и, следовательно, ее адаптация к красному свету. Интересно, что губчатая ткань листа подсолнечника практически не страдает от красного света, в то время как у листьев огурца здесь, как и для палисадной ткани, наблюдается сильная деструкция (см. табл. 3.11).
Дальнейшие исследования показали, что существуют культуры, анатомическая структура листьев которых вообще не испытывает деструктивного влияния длительного воздействия красного света. В качестве примера можно привести данные по анатомической структуре листа томата (табл. 3.13).
Конечно, общие закономерности, связанные с формированием на красном свету большего количества и более мелких по размерам хлоропластов, чем, например, в синей области спектра, от-
90
Таблица 3.13
Количественные характеристики анатомической структуры листьев томата разного возраста, сформировавшихся при облученности 100 Вт/ма в отдельных областях ФАР
Область спектра Возраст, сут П, 103 кл./см2 Г, 103 кл./см2 Количество хлоропластов Общее количество клеток па 1 см2 поверхности листа, 10s п+г 100 %
п г
400—500 20 368 311 50 16 679 54.2
30 421 328 43 27 749 56,2
500-600 20 216 194 33 28 410 52,7
30 238 174 37 22 412 57,8
«600—700 20 228 211 48 49 439 51,9
30 334 266 46 50 600 55,7
Примечание.
Усл. обозн. см. табл. 3.11.
меченные ранее рядом исследователей [Власова, Воскресенская, 4973], остаются справедливыми и для томатов (см. табл. 3.13). Однако длительное воздействие красного света на анатомическую ^структуру листа не способствует в данном случае деградации хлоропластов и не снижает их количества как в палисадной, так и в губчатой паренхиме.
Таким образом, анализ длительного воздействия света различных участков спектра на формирование анатомической структуры листьев разных видов показал в ряде случаев их различную реакцию на спектр излучения. Если в отношении синих лучей такие различия не были значительными, то в отношении красного света такие различия проявились достаточно четко. Диапазон их может быть прослежен от деструкции анатомической структуры и гибели зеленого листа (огурец), первоначальной деструкции и последующей стабилизации анатомической структуры (подсолнечник) и до фактически неизменных параметров анатомической •структуры листа томата после 20 сут и более пребывания на красном свету.
Наблюдаемые деструктивные изменения под действием красных лучей на лист идут в основном за счет палисадной паренхимы (см. табл. 3.11), в то время как губчатая ткань страдает меньше, а в ряде случаев (например, с подсолнечником) именно она берет на себя основную роль в функционировании клеточной «структуры листа. Эта особенность губчатой паренхимы проявляется не только при воздействии неблагоприятного по спектру излучения ФАР, но и при воздействии достаточно высоких интенсивностей ФАР (Е — 200 Вт/м2 и выше). Увеличение интенсивности ФАР в более низких диапазонах изменения мощности лучистого потока приводит к практически одинаковым изменениям в пали-
91
© 6 0 ogo
Рис. 3.22. Анатомическое строение-листьев редиса Вировский белый,, выращенных при облученностях 200 (а), 300 (б), 600 (в) Вт/м2 ФАР/ m — общая толщина листа, п — толщина-палисадной паренхимы, k — толщина губ-, чатой паренхимы, t — толщина слоя палисадной паренхимы, лишенной хлорофиллов (точки — зеленые хлоропласты в клетках палисадной и губчатой паренхим).
садной и губчатой паренхимах, связанных с уменьшением параметров мезоструктуры в расчете на единицу поверхности листа. Лишь начиная с уровня облученности 200 Вт/м2 ФАР и выше в анатомической структуре листа некоторых видов растений появляются указанные различия в дифференцировке структуры палисадной и губчатой паренхим [Тихомиров, Сидько, 1984]. Рассмотрим такие изменения на примере листа редиса.
При облученности 200 Вт/м2 ФАР верхний слой клеток палисадной паренхимы практически лишен хлорофиллов (рис. 3.22, а), при 300 Вт/м2 ФАР лишены хлорофилла два слоя палисадных клеток (рис. 3.22, б), а при 600 Вт/м2 ФАР уже практически весь слой палисадной паренхимы не содержит зеленых пигментов (рис. 3.22, в). Лишь клетки губчатой паренхимы имеют зеленую окраску во всех исследуемых световых вариантах.
Размеры клеток как палисадной, так и губчатой паренхимы, а также общая толщина листа уменьшаются с ростом уровня облученности (табл. 3.14).
Таблица 3.14
Количественные характеристики анатомического строения листьев редиса Вировский белый, выращенных при различных уровнях ФАР, мкм
Облученность, Вт/м2 Общая толщина листа, m Толщина паренхимы Слой клеток, лишенных хлорофилла, t
палисадной, п губчатой, k
200 460 203 257 68
300 378 186 192 126
600 196 153 43 153
Примечание. Усл. обозп. см. рис. 3.22.
92
Содержание хлорофиллов (а + 6) в листьях редиса Вировский белый в зависимости от используемого уровня облученности (200, 300 и 600 Вт/м2 ФАР) составляло соответственно 3,94; 2,15 и 1,45 мг/дм2. Видно, что с ростом уровня облученности содержание хлорофиллов в листьях уменьшается.
Толщина зеленого слоя листа при 600 Вт/м2 ФАР меньше, чем при 200 Вт/м2 ФАР, в 3,5 раза, а содержание хлорофиллов — в 5 раз (см. рис. 3.22, табл. 3.14). Это позволяет заключить, что при облученности 600 Вт/м2 ФАР усредненное по объему зеленого слоя листа распределение хлорофиллов менее плотное, чем при 200 Вт/м2 ФАР. Для облученности 300 Вт/м2 ФАР эти величины являются промежуточными.
Поскольку с ростом уровня облученности толщина клеточного слоя, лишенного хлорофиллов, возрастает (см. рис. 3.22), то это позволяет предположить, что при высоких облученностях верхние слои столбчатых клеток листа играют защитную роль, снижая световую нагрузку на нижерасположенные слои фотосинтезирующей ткани, где в основном сосредоточены зеленые пигменты.
В зависимости от уровня облученности меняются анатомические и оптические характеристики листа редиса, выращиваемого при высокоинтенсивном облучении. Эти изменения регулируют световое обеспечение фотосинтезирующих клеток листа, что позволяет поддерживать жизнеспособность растений при облученностях порядка 600 Вт/м2 ФАР. При этом важную роль играет как ориентация листа, так и защитная функция верхних, лишенных хлорофиллов клеток палисадной ткани листа, причем с ростом уровня облученности роль последних возрастает. Все это указывает па широкие приспособительные возможности листьев растений к высокоинтенсивному излучению и к фотосинтетической деятельности при облученностях порядка солнечных и выше.
Таким образом, и спектральный состав и интенсивность ФАР могут оказывать существенное влияние на анатомическую структуру листа. При ряде экстремальных световых условий — резком увеличении уровня облученности или сосредоточении излучения в одной из спектральных областей ФАР — возможны адаптивные изменения в анатомическом строении листовой пластинки, главной чертой которых является перераспределение функциональной активности между палисадной и губчатой паренхимой. В этом случае основную роль в фотосинтетической активности листа начинает играть губчатая паренхима, а палисадная выполняет в значительной мере защитную роль. Следует полагать, что указанные адаптивные изменения анатомической структуры листа к спектру и интенсивности ФАР не должны рассматриваться как универсальная реакция растений различных видов. Экспериментально доказанные различия реакции растений разных видов на спектр и интенсивность ФАР [Тихомиров и др., 1987] дают основания считать, что это распространяется на тканевой уровень организации фотосинтетического аппарата, что способствует адаптации и успешной продукционной деятельности растений.
93
В заключен ih> пнстошцой главы отметим, что различное отношение рассматри и немых видов растений к излучению отдельных спектральных областей ФАР в отношении процессов роста и развития связано с неодинаковым характером изменчивости исследуемых параметров как на ценотическом, так и на организменном листовом и тканевом уровнях организации фотосинтетического аппарата. При тех спектральных режимах облучения, которые оказываются благоприятными для прохождения растениями этапов органогенеза до получения конечного урожая, наблюдается своеобразная «настройка» исследуемых параметров фотосинтетического аппарата на данный радиационный режим.
На ценотическом уровне это, в частности, проявляется в бо~ лее рациональном распределении листовой поверхности по вертикали ценоза, как это наглядно видно, например, при выращивании пшеницы в зеленых лучах, на уровне целого растения и листа — в ориентации листьев и характерных морфофизиологи-ческих изменениях их поверхности (у редиса), на тканевом уровне — в отсутствии процессов деградации составляющих анатомической структуры при длительном излучении отдельных областей ФАР (воздействия красного света на мезоструктуру листа томата). В качестве интересного примера процесса адаптации анатомии ческой структуры листа к длительному воздействию красного света следует отметить перестройки мезоструктуры листа подсолнечника, связанные с переходом основной фотосинтетической, функции от палисадной к губчатой паренхиме. Это явление, вероятно, носит достаточно универсальный характер не только при адаптации к спектру, но и к высоким интенсивностям ФАР, как это было показано на примере листьев редиса.
Глава 4
СПЕКТРАЛЬНЫЙ СОСТАВ СВЕТА И БИОХИМИЯ РАСТЕНИЙ
Спектральный состав света, особенно при длительном его воздействии на растения, может оказывать влияние на синтез и накопление самых различных биохимических продуктов.
Еще в 40-х годах было показано, что синтез основного пигмента фотосинтеза — хлорофилла имеет ярко выраженную спектральную зависимость и наиболее интенсивно осуществляется в синей и красной областях спектра [Frank, 1946]. В то же время желтые пигменты листа интенсивнее синтезируются на зеленом свету [Окунцов и др., 1964; Симонова, 1965], при этом этиолированные проростки сильнее реагируют на качественный состав света, чем зеленые. Содержание хлорофилла [Воскресенская, Нечаева, 1967 ] в первом листе растений ячмепя в период его активного роста было наибольшим на синем, средним — на красном.
94
Таблица 4.1
Содержание РБФ-карбоксилазы в листьях разного возраста у растений ячменя, мг/г сырой массы
Свет Возраст листа, дни
4 9 10 13 15
Синий 4,29 3,08 3,39 2,45 0,83
Красный Отношение сиций/крас- 3,28 2,01 1,95 1,12 0,31
ный 1,31 1,53 1,74 2,19 2,86
а наименьшим — на зеленом свету, но при старении листа оно выравнивалось во всех трех вариантах. Исследуя зависимость накопления фотосинтезирующих пигментов в листьях растений и от спектрального состава, и от интенсивности освещения, В. М. Терентьев с соавторами [1974] отмечает, что при низкой освещенности максимальное накопление пигментов характерно для зеленой области спектра, а при высокой — для красной (у капусты) или синей (у пшеницы).
Определенные количественные различия в пигментной системе листьев растений, зависящие от спектрального состава света, показаны в работе С. М. Кочубей с соавторами [1987]. Однако эти исследователи подчеркивают, что набор пигментных форм в каждой из фотосистем остается неизменным на красном и синем свету и совпадает с набором форм у растений, выращенных на белом свету.
Можно отметить, что помимо пластидных пигментов спектральный состав света влияет и на образование пигментов клеточного сока — антоцианинов [Mancinelli et al., 1985]. Наряду с пигментами, ответственными за фотосинтез, отмечено влияние спектрального состава света па содержание фермента РБФ-карбоксилазы, играющего ведущую роль в самом процессе фотосинтеза [Poyarkova et al., 1973; Кумаков, 1987]. Так, по данным А. В. Кумакова [1987], в листьях ячменя содержание РБФ-карбоксилазы на синем свету в течение всего периода их активности было больше, чем на красном, и хотя оно с возрастом листа снижалось, отношение синий/красный даже возрастало (табл. 4.1).
Влияние спектрального состава света на общую биохимическую направленность фотосинтеза проявляется уже на ранних этапах этого процесса.
Как показала Н. П. Воскресенская [Воскресенская, 1965; Voskressenskaya, 1972], растения, предварительно выращенные па белом свету, уже через несколько секунд после экспозиции под синим светом усилили синтез неуглеводных продуктов: аминокислот (алапин, аспартат) и органических кислот (малат, цитрат), а под красным светом — растворимых углеводов. Через несколько минут экспозиции под сипим светом интенсифицировался синтез белков, а под красным — крахмала.
95
В настоящее время широко распространено представление о том, что синий свет стимулирует белковую, а красный — углеводную направленность метаболизма растений. Это отражается в первую очередь на биохимическом составе листьев, подвергающихся прямому воздействию лучей какой-либо области спектра, но в той или иной степени и на биохимии запасающих органов (корнеплоды, плоды, зерно). Однако данные по биохимии конечного хозяйственно ценного продукта растений, выращенных под светом различного спектрального состава, весьма малочисленны. Они редко сопоставляются с уровнями облученности растений и практически никогда — с уровнем достигнуто!! продуктивности ценоза по хозяйственно полезной биомассе. В результате итоговая продуктивность ценоза растений по полезному веществу остается невыясненной, ибо она зависит не только (а часто даже и не столько) от процентного содержания нужного вещества в биомассе, но и от общей продуктивности ценоза. Именно под таким углом зрения в нашей лаборатории было исследовано влияние спектрального состава света различной интенсивности на биохимический состав конечного урожая и на выход отдельных биохимических компонентов с единицы площади ценоза таких неодинаково отвечающих на спектральный состав света культур, как редис и пшеница [Золотухин и др., 1979, 1980, 1983].
Содержание сырого белка, водорастворимых сахаров и крахмала в расчете на сухую биомассу в листьях и корнеплодах редиса зависит от спектрального состава и интенсивности облучения (рис. 4.1, а — в). Следует предварительно отметить, что если под синим светом продуктивность растений возрастала с увеличением интенсивности облучения от 50 до 500 Вт/м2 ФАР, то под зеленым светом растения начали снижать продуктивность при облученности выше 100 Вт/м2 и погибали при Е >> 300 Вт/м2. Под красным светом то же самое наблюдалось при еще меньших уровнях облучения — выше 50 и 200 Вт/м2 ФАР соответственно. При интенсивности облучения 50 и 100 Вт/м2 ФАР содержание сырого белка в листьях редиса соответствовало общепринятым представлениям: больше всего его было под синим светом, меньше под зеленым и еще меньше под красным (см. рис. 4.1, а). Обратная картина наблюдалась по содержанию сахаров и крахмала (см. рис. 4.1, б, в).
С увеличением интенсивности облучения до 200 Вт/м2 ФАР и выше в листьях редиса во всех вариантах проявилась тенденция к снижению содержания сырого белка. При этом в листьях редиса под синим светом с интенсивностью облучения 500—600 Вт/м2 содержание сырого белка снизилось до уровня, наблюдавшегося под Красным и зеленым светом при 50 Вт/м2, т. е. повышенная белковость была утрачена. В то же время количество сахаров при высоких уровнях облучения падало под красным и зеленым светом, что сопровождалось резким, можно сказать — патологическим увеличением содержания крахмала в листьях. Не исключено, что последнее обстоятельство явилось одним из проявлений нарушения метаболизма растений, приводящего при высокой ин-
96
а
б
Е,Вг/мг
Рис. 4.1. Световые кривые изменения содержания сырого белка в расчете на сухое вещество (а), фракции водорастворимых сахаров (б) и крахмала (в) [Золотухин и др., 1985].
Сплошная линия — листья, штриховая — корнеплоды. Свет: 1 — синий, 2 — зеленый, 3 — красный.
тенсивности облучения красным или зеленым светом к снижению продуктивности и даже к гибели растений. Иная картина наблюдалась под синим светом, где увеличение интенсивности облучения вплоть до 500 Вт/м2 сопровождалось плавным нарастанием содержания в листьях и сахаров, и крахмала одновременно с увеличением общей продуктивности ценоза редиса. Если, исходя из приведенных графиков, сравнить содержание белков и суммы углеводов в листьях редиса, то окажется, что при 100 Вт/м2 под красным светом белков было 27 % на сухую массу, а углеводов 50 % (отношение 1,0 : 1,85), тогда как под синим 31 и 38 % соответственно (1,0 : 1,23). Таким образом, общепринятая точка зрения о противоположном влиянии синего и красного света па белковый и углеводный обмен подтверждается. Но увеличение интенсивности облучения синим светом до 600 Вт/м2 снижало содержание белков в листьях до тех же 27 %, как и под красным светом при 100 Вт/м2, а содержание суммы углеводов возрастало до 54 %, (отношение 1,0 : 2,0). Таким образом, повышение интенсивности облучения, редиса синим светом от 100 до 600 Вт/м2 вызывало в
97
биохимии листьев тмине »мо инмопония, как выращивание растений при 100 Вт/м2 иод красным светом вместо синего.
Изменения в содержании белков и углеводов в корнеплодах редиса в зависимости от спектрального состава и интенсивности света были гораздо менее выразительными и не всегда адекватными отмеченным выше изменениям в составе листьев (см. рис. 4.1, а — в).
Определение содержания аскорбиновой кислоты в листьях редиса в тех же световых вариантах [Золотухин и др., 1979] показало, что ее содержание при 50 Вт/м2 под синим светом (18 мг%) уступало варианту с красным светом (38 мг%), но увеличение интенсивности облучения синим светом до 500 Вт/м2 более чем в 4 раза увеличило содержание аскорбиновой кислоты в листьях, тогда как под красным светом уже при 200 Вт/м2 были обнаружены лишь ее следы.
В отличие от белков и углеводов, содержание аскорбиновой кислоты в корнеплодах редиса изменялось под действием спектрального состава и интенсивности облучения сходным образом, лишь абсолютные значения содержания были несколько ниже, чем в листьях.
И. Г. Золотухин и Э. К. Волкова [1980] исследовали содержание сахаров и аскорбиновой кислоты в редисе, выращенном при 100 Вт/м2 ФАР с попарными и тройными комбинациями синего, зеленого и красного света. Показано, что при использовании красно-синего света процентное содержание сахаров и аскорбиновой кислоты в корнеплодах было наибольшим при 75—85 % красного света. Поскольку такие соотношения красного и синего света обеспечивали достаточно высокую продуктивность растений, суммарный выход сахаров и аскорбиновой кислоты с единицы площади, занимаемой ценозом, тоже был в этих вариантах самым высоким. Под сине-зеленым излучением наибольшее процентное содержание сахаров было отмечено в вариантах с 50—100 % синего света, но максимальный выход с единицы площади — при 30— 70 %. На содержании аскорбиновой кислоты в корнеплодах различные соотношения синего и зеленого света сказывались слабо, максимальный же выход с единицы площади получен при 30— ВО % синего света.
Наиболее эффективной из всех комбинаций оказалась «тройная» с равными соотношениями синего, зеленого и красного света, т. е. с искусственным белым светом. Такое облучение обеспечивало максимальный выход с единицы площади и сахаров, и аскорбиновой кислоты главным образом потому, что и продуктивность растений в этом случае была самой высокой.
Еще более тесную зависимость суммарного выхода того или иного биохимического продукта с единицы посевной площади от общей продуктивности ценоза можно видеть на примере пшеницы [Золотухин, Лисовский и др., 1980]. Зерно пшеницы, выращенной под синим, зеленым и красным светом при облученности 100 и 600 Вт/м2, имело некоторые различия по процентному содер-
98
99-
жанию белка, крахмала, витаминов Вх и В2 (табл. 4.2). Варианты под красным и под зеленым светом при облученности 100 и 600 Вт/м2 были довольно сходны между собой почти по всем исследованным биохимическим компонентам. Достаточно существенно отличалось зерно, выращенное под сипим светом. При интенсивности облучения 100 Вт/м2 процентное содержание белка, витаминов BjH В2 в нем было примерно в 1,5 раза выше, чем в зерне, выращенном под красным и зеленым светом, а содержание крахмала заметно ниже. При облученности 600 Вт/м2 между всеми спектральными вариантами различия в содержании белка и крахмала сгладились, содержание витамина Bt под синим светом понизилось, а В2 — повысилось.
Сопоставляя процентное содержание биохимических компонентов в зерне, выращенном при 100 Вт/м2 в разных областях спектра, следует обратить внимание па то, что отмеченные выше особенности биохимии зерна в варианте с синим светом главным образом определяются плохим его наливом, выразившемся в сниженной массе 1000 зерен. Поэтому при высоких относительных (процентных) показателях содержания белка и витаминов Вг и В2 в варианте с синим светом абсолютное содержание этих компонентов в расчете на 1000 зерен не превышало, а по белку даже уступало аналогичным показателям в вариантах с красным и зеленым светом. Если сравнивать спектральные варианты по выходу того или иного биохимического компонента с единицы площади, то во всех случаях этот выход определялся не относительным (процентным) содержанием компонента в зерне, а урожаем зерна с единицы площади. Сказанное остается справедливым и для разных спектральных вариантов, выращенных при интенсивности облучения 600 Вт/м2. Хотя в этом случае пониженное содержание витамина Вг и повышенное — В2 в варианте с синим светом не было связано с плохим наливом зерна, а зависело, по-видимому, непосредственно от спектрального состава света, суммарный выход этих витаминов с единицы площади все равно в конечном счете определялся большей или меньшей общей продуктивностью растений.
Вместе с тем даже с практической точки зрения не следует, видимо, отдавать приоритет только суммарному выходу нужного биохимического компонента с единицы площади, ибо далеко не всегда количественная сторона в получении продукта оказывается важнее качественной. Так, например, при получении растительного лекарственного сырья процентное содержание в нем необходимого алкалоида или другого продукта вторичного обмена может быть важнее общего сбора этого вещества. Известно, что спектральный состав света оказывает влияние на синтез гормонов растений, алкалоидов, эфирных масел и т. д. [Шульгин, 1969; Кузнецова и др., 1978; Карначук, 1989], однако это влияние неоднозначно. Так, накопление соласодина в листьях паслена дольчатого на синем свету идет активнее, чем на красном [Кузнецова и др., 1978], а содержание стероидного соединения экдистерона
100
в листьях и подземных органах левзеи, выращенной под красным светом, было в несколько раз больше, чем под зеленым и синим [Карначук, 1989]. Таким образом, спектральный состав света в сочетании с необходимой интенсивностью облучения можно рассматривать как один из факторов управления и количественной, и качественной стороной продукционного процесса у растений.
Глава 5
СПЕКТРАЛЬНЫЙ СОСТАВ СВЕТА И СКОРОСТЬ НАКОПЛЕНИЯ РАСТИТЕЛЬНОЙ БИОМАССЫ
§ 1. Спектральный состав излучения искусственных источников света и продуктивность растений
Принципиальная возможность выращивать растения «от семени до семени» при искусственном освещении была установлена еще в 20-х годах нашего столетия Гарвеем [Harvey, 1922] в США и независимо от него Максимовым [1925] в СССР. Однако серьезные исследования зависимости формирования урожая растений от спектрального состава света стали возможны лишь начиная с конца 40-х — начала 50-х годов. В решающей степени это связано с тем, что до этого времени не было технических возможностей создавать источники света, имеющие разнообразные спектральные характеристики в видимой области оптического излучения. Конечно, нельзя сказать, что до начала 50-х годов исследования по изучению влияния спектрального состава света на урожай растений вообще не проводились. Даже имея скудный набор источников света с разными спектральными характеристиками, исследователи предпринимали попытки дать оценку роли того или иного спектрального состава света на формирование урожая растений.
Обзор исследований за этот период изложен в монографии А. Ф. Клешнина [1954] и дает основание считать, что для синтеза биомассы растений и формирования урожая более благоприятным, как правило, является излучение, где доминируют оранжево-красные лучи либо для некоторых растений — синие. Однако следует помнить, что такого рода выводы были сделаны при использовании источников света с сильно различающейся структурой в спектрах излучения исследуемых физиологических областей. В частности, под оранжево-красными лучами подразумевалось как излучение натриевой лампы низкого давления, излучающей в области ФАР практически па одной длине волны 587 нм, так и излучение лампы накаливания, имеющей сплошное излучение в оранжево-красной области спектра. Часто источники света, ис
101
пользовавшиеся исследователями в тот период, излучая преимущественно в одной физиологически значимой области ФАР (синей, зеленой или красной), имели при этом разную «примесь» излучения других спектральных областей, что снижало достоверность результатов фотобиологических исследований, трактуемых как воздействие той или иной области спектра на продукционный процесс.
Из сказанного следует, что исследователи не имели достаточных возможностей для ведения целенаправленных работ но оценке влияния спектрального состава света на продуктивность растений. Различия в светотехнических характеристиках источников света, отсутствие надежных и объективных методов и приборов для измерения уровней облученности и спектрального состава падающего на растения излучения, отсутствие подходявщх светофильтров с нужными спектральными характеристиками пропускания света зачастую сводили на нет попытки оценить роль излучения того или иного спектрального состава в формировании урожая растений.
В конце 40-х — начале 50-х годов стали все шире использоваться люминесцентные лампы, которые за счет введения в них определенных типов люминофоров дали возможность варьировать спектральный состав излучения этих источников света в очень широких пределах. Такая возможность стимулировала широкое использование люминесцентных ламп различного спектрального состава для исследований роли спектра ФАР в различных сторонах жизнедеятельности растений и, в частности, в продукционном процессе. В СССР эти исследования были активно начаты А. Ф. Клешниным [1954]. Им было установлено, что среди люминесцентных ламп более эффективны при выращивании длинно-дневных растений для получения урожая те, в спектре излучения которых преобладают оранжево-красные лучи, а для короткодневных растений — лампы, где преобладают синие лучи. Люминесцентные лампы, в излучении которых преобладают зеленые лучи, оказались наименее пригодными для выращивания растений.
Исследованием влияния спектра излучения люминесцентных ламп на урожай растений длительное время занимался В. М. Леман [1971 ]. Им также было показано, что среди люминесцентных ламп наиболее эффективными оказались те, в спектре излучения которых была «усилена» красная область. За счет меньшего содержания инфракрасной радиации излучение люминесцентных ламп зачастую было более эффективно в формировании урожая растений, чем излучение источников света предшествующих типов, главным образом ламп накаливания. Однако общий анализ периода, когда в исследовательской практике широко использовались люминесцентные лампы в качестве инструмента для оценки роли спектрального состава в продукционном процессе, показывает, что надежного и всестороннего решения этой проблемы не было. Это, во-первых, связано с тем, что, несмотря на принци-
402
щтальную возможность широкого варьирования спектра излучения люминесцентных ламп, их серийные промышленные образцы имели, как правило, лишь несколько градаций по спектру ФАР, определявшихся в основном светотехническими задачами, не связанными с выращиванием растений. Это ограничивало возможности спектральных фотобиологических исследований. Ограниченная мощность люминесцентных ламп в большинстве случаев не позволяла работать с уровнями облученности выше 50—60 Вт/м2 ФАР, что часто не соответствует максимальному КПД по использованию растениями лучистой энергии в расчете на хозяйственно полезный урожай [Полонский, Лисовский, 1980]. Правда, были предприняты попытки создать несколько типов люминесцентных ламп специально для выращивания растений [Справочная книга..., 1983], однако они не сняли большинства из указанных выше ограничений использования люминесцентных ламп для всестороннего исследования влияния спектрального состава света на продукционный процесс растений.
В конце 50-х — начале 60-х годов появились и начали успешно использоваться для выращивания растений ксеноновые лампы. Важнейшей особенностью этих ламп по сравнению с ранее известными источниками света является возможность использования высокоинтенсивных лучистых потоков со спектром, близким к равноэпергетическому в области ФАР [Маршак и др., 1963; Вассерман, 1989]. В. М. Леман [1971], используя ксеноновые лампы (ДКсТВ-6000), получил для ряда овощных растений значения продуктивности, в 2—4 раза превосходящие таковые, полученные при использовании люминесцентных ламп, выровненных по установленной удельной мощности с лампами ДКсТВ-6000. В работах Тихомирова с соавторами [1976], Протасовой и Кефе-ли [1982] при использовании ксеноновых ламп также получены весьма высокие показатели продуктивности для различных видов растений. Появление ксеноновой лампы способствовало разработке высокоэффективных режимов облучения растений самых различных видов. Это позволило, в частности, успешно выращивать с применением ксеноновых ламп пе только овощные, но и зерновые культуры, что.дало возможность разработать и осуществить основы световой технологии для селекции различных культур в условиях искусственного облучения [Лисовский, Долгушев, 1986], интенсивной светокультуры [Лисовский и др., 1987]. Универсальность излучения ксеноновых ламп в отношении получения достаточна высоких урожаев за сравнительно небольшие периоды вегетации для растений различных систематических групп явилась основанием для выбора ксеноновых ламп (ДКсТВ-6000) в качестве источников света для успешного выращивания растений в замкнутых системах жизнеобеспечения [Гительзон и др., 1975]. Плодотворным явилось использование ксеноновых ламп для научных исследований роли интенсивности света в процессах роста, развития и продуктивности растений [Леман, 1971; Протасова и др., 1972; Тихомиров и др., 1976; и др.]. В то же время исполь
103
зование ксеноновых ламп для исследования роли спектрального состава ФАР в жизнедеятельности растений возможно лишь при наличии специальных цветных светофильтров [Тихомиров, Сидь-ко, 1987 ], что технически реализовать удается далеко пе всегда. .Поэтому использование ксеноновых ламп в такого рода экспериментах весьма ограничено и существенно не продвинуло вперед эти исследования.
Качественно новый этап в развитии исследований по влиянию спектрального состава света на продукционный процесс растений произошел в начале 60-х годов. Он связан с появлением мощных ртутных ламп высокого давления, дающих высокоинтенсивное излучение преимущественно в синей и зеленой областях ФАР. Многочисленные исследования на этом этапе характеризовались фотобиологическими испытаниями таких источников света [Леман, 1971]. Особенностью этих исследований являлось то, что такие лампы наряду с ксеноновыми позволяли создать необходимые для формирования полноценного урожая уровни облученности ФАР. Это позволяло исключить лимитирующую роль интенсивности света в формировании урожая и получить более высокие абсолютные значения, чем при использовании маломощных ламп предыдущих поколений. Кроме того, появилась возможность более корректно оценить роль спектрального состава света, ограниченного, правда, в основном сине-зеленой областью ФАР, в формировании урожая при использовании высокоинтенсивных лучистых потоков, что не представлялось возможным ранее при использовании большинства источников света предыдущих поколений.
Дальнейшее расширение этих исследований связано с разработкой на базе ртутных ламп высокого давления металлогалогенных ламп с добавками иодидов металлов, что позволило создать мощные лампы с различным спектром ФАР. Первой из металлогалогенных ламп явилась лампа ЛОР-ЮОО [Сулацков и др., 19711. Важнейшей особенностью спектра излучения лампы ЛОР-ЮОО была достаточно высокая доля излучения в красной области спектра (600—700 им), на которую приходилось около 40 % излучения в области ФАР (излучение резонансной линии лития на длине волны около 670 нм).
Результаты экспериментов, проведенных на ряде овощных и зерновых культур, показали, что излучение лампы ЛОР-ЮОО более эффективно в формировании урожая, чем излучение всех других, использованных в этих экспериментах источников света [Свентицкий, 1982]. Несомненно, что такой результат связан с достаточно высоким содержанием красных лучей в излучении лампы ЛОР-ЮОО.
О необходимости присутствия в светопотоке красных лучей для нормального протекания жизненно важных процессов в организме растений известно еще со времен К. А. Тимирязева. До 60-х годов нашего столетия, пожалуй, единственно доступным источником света для выращивания растений, имеющим высокое
104
содержание красных лучей в светопотоке достаточно высокой интенсивности, являлась лампа накаливания. Однако это ее преимущество перед другими источниками света часто нивелировалось высоким содержанием тепловых лучей, отрицательно влиявших па процессы роста и развития и в конечном итоге на продуктивность растений. Тем не менее работы Б. С. Мошкова [1966] показали, что если снять с помощью водяного экрана большую часть тепловой радиации, то излучение лампы накаливания по своей спектральной эффективности для ряда культур ие уступает многим известным в то время источникам света. Успешное использование галогенных ламп накаливания с водным экраном для выращивания овощных растений отмечается в работе К. С. Арбузовой с соавторами [1983]. Целесообразность добавки излучения ламп накаливания к излучению источников света, практически не имеющих красного компонента ФАР, нашла свое отражение в создании комбинированной лампы ДРВ-750, совмещающей ртутную горелку, дающую преимущественное излучение в синей (400— 500 нм) области спектра, и вольфрамовую спираль, излучающую в видимой области преимущественно красные лучи [Справочная книга..., 1983]. Однако все эти технические устройства, имея существенные светотехнические и эксплуатационные недостатки, ие нашли длительного и широкого применения в светофизиоло-гии. В связи с этим создание лампы ЛОР-ЮОО (в более поздней модификации лампа ДРФ-1000) с повышенным содержанием красных лучей в потоке ФАР было, несомненно, крупным шагом в технической реализации спектральных световых условий обеспечения продукционного процесса растений.
Достаточно высокая фотобиологическая эффективность излучения такой лампы явилась одним из стимулов для создания других типов металлогалогенных ламп, в которых конструкторы-светотехники попытались «усилить» оранжево-красную часть излучения. Среди таких ламп следует отметить в первую очередь лампы типа ДМ (ДМ 3-3000, ДМ 4-3000, ДМ 4-6000) [Левин, 1986], ДРОТ-2000 [Васильев и др., 1982], фотобиологическая эффективность которых в ряде случаев сравнима с таковой для лампы ДРФ-1000 [Квашин и др., 1983]. Достаточно успешно идет внедрение в практику светокультуры ламп типа ДРИ-2000-6 и натриевых ламп высокого давления ДНаТ-400 [Федоров и др., 1986].
Таким образом, были подтверждены научные представления о повышенной фотобиологической эффективности излучения ФАР, обогащенного красными лучами, в продукционном процессе растений. В настоящее время, на наш взгляд, наибольший научный интерес представляет решение вопроса о том, каковы для растения и в конечном счете урожая критичные уровни красного компонента в излучении ФАР, каким должно быть соотношение различных спектральных областей ФАР для оптимального протекания продукционного процесса, какие для этого пужны уровни облученности, каким должен быть фон инфракрасной радиации и т. п.
® А. А. Тихомиров, Г. М. Лисовский, Ф. Я. Сидько
105
Работы в этом направлении могут вестись двумя методическими путями. Первый — проведение и анализ фотобиологических экспериментов с источниками света, имеющими разнообразные спектральные и энергетические характеристики. Второй — проведение аналогичных исследований на базе единого светотехнического устройства, позволяющего варьировать спектр и интенсивность ФАР при стабилизации всех других светотехнических параметров.
Первый путь в практическом отношении представляется более доступным, и поэтому не случайно, что вопросы оптимизации спектрального состава света в продуктивности растений многие исследователи пытаются решить на основании использования ламп, дающих излучение различного спектрального состава. Рассмотрим, какие возможности и ограничения могут ожидать исследователя на таком пути. Для этого необходимо проанализировать влияние спектрального состава различных источников света на продуктивность растений. В табл. 5.1 приведены данные по фотобиологической оценке эффективности излучения различных источников при облученности 100 Вт/м2 ФАР. Учитывая, что люминесцентные лампы часто используются при более низких облученностях, их фотобиологическая эффективность дополнительно оценивалась и при Е = 50 Вт/м2 ФАР, а для газоразрядных ламп, способных создавать более высокие уровни облученности, представлялось целесообразным дополнительно провести их фотобиологическую оценку при Е = 200 Вт/м2 ФАР, т. е. при облученности, которую можно использовать в условиях практической светокультуры [Полонский, Лисовский, 1980].
Таблица 5.1
Влияние непрерывного излучения различных источников света на продуктивность редиса Вировский белый (по корнеплодам) [Волкова и др., 1982]
Тип источника излучения Доля ИКР в интегральном излучении, % Уровень облученности, Вт/м2 Сырая биомасса корнеплодов, . к г/м2 Сухая масса корнеплодов, г/м2 Длительность вегетации» сут
ДКсТВ-6000 75 100 4,3+0,34 215±17 24
75 200 5,4+0,52 300+29 - 22
ДРФ-1000 40 100 7,5+0.62 375+31 24
40 200 8,2±0.69 455+38 .. . 22
ДНаТ-400 60 100 5,2+0,40 260±20 24
60 200 7,0+0,61 420+34 22
ЛБ-40 15 50 2,6+0,19 125+9 30
15 90 5,6±0,44 282+22 24
ЛКБ-40 15 50 5,0±0,36 240+17 28
15 90 7,1±0.50 355±25 22
ДМЗ-ЗООО 40 100 8,0±0.9 400+45 24
КГ-1000 90 100 2,0±0,4 100+20 24
106
Таблица 5.2
Влияние непрерывного излучения различных источников света на продуктивность (по зерну) ценоза пшеницы 232 селекции Г. М. Лисовского [Волкова и др., 1982]
Тип источника света Уровень облученности, Вт/м2 Абсолютно сухая масса зерна, г/м2 Масса 1000 зерен, г Среднесуточная продуктивность, г/(м2-сут)
ДКсТВ-6000 100 650+74 26,1±2,2 Ю,0±1,1
ДРФ-1000 100 694±68 30,5+2,1 8,7±0,8
ДМЗ-ЗООО 100 818±66 31,2+1,8 10,9+0,8
ДНаТ-400 100 551±62 25,3+2,0 7,9±0,9
ДРИ-2000 100 432±48 22,8+1,9 5,8+0,6
ЛКБ-40 80 708+74 35,0+3,1 9,4+1,0
Для продуктивности редиса при Е — 90—100 Вт/м2 ФАР наиболее эффективно излучение ламп ДРФ-1000 и ЛКБ-40, а наименее — ДКсТВ-6000 й КГ-1000. Для уровней облученности 50 Вт/м2 ФАР, создаваемых маломощными источниками света, сравнивали эффективность излучения красных безртутных люминесцентных ламп ЛКБ-40 и широко известных белых люминесцентных ламп ЛБ-40 (см. табл. 5.1). Более эффективным оказалось излучение ламп первого типа, под светом которых продуктивность корнеплодов редиса была в 2 раза выше.
В повышении продуктивности пшеницы наиболее эффективным оказалось излучение ламп ДМЗ-ЗООО, ДКсТВ-6000 и ЛКБ-40 (табл. 5.2).
Сравнительно высокая фотобиологическая эффективность одних типов ламп относительно других не является постоянной для различных культур. Так, например, относительно высокая фотобиологическая эффективность лампы типа ДМЗ-ЗООО в сравнении С лампой ДКсТВ-6000, полученная в отношении продуктивности редиса (см. табл. 5.1), может измениться на противоположную при выращивании некоторых других видов растений (табл. 5.3).
Таблица 5.3
Продуктивность гороха Солянский (по семенам) и ячменя сорта Унион (по Верну), выращенных при непрерывном излучении различных искусственных источников света интенсивностью 80—100 Вт/м2 ФАР [Лисовский, Долгу-шев, 1986]
Культура ДКсТВ-6000 ДМЗ-ЗООО
Хозяйственно полезный урожай, г/м2 Среднесуточная продуктивность, г/(м2-сут) Хозяйственно полезный урожай, г/м2 Среднесуточная продуктивность, г/(м2-сут)
Горох, сорт Солянский Ячмень, сорт Унион 671 909 11,8 17,2 444 756 7,8 11,5
107
Таблица 5.4
Влияние излучения ламп с различным содержанием ИКР на продуктивность корнеплодов редиса
Тип лампы Доля инфракрасной радиации в ивлучении лампы, % Вегетация, сут Сухая биомасса корнеплодов, г/м2
ДРФ-1000
(контроль) 40 24 375+31
ДКсТВ-6000 75 24 215+17
60 24 250±18
40 24 330±127
20 22 280±28
Примечание. Облученность 100 Вт/м2.
Усиление длинноволнового излучения в ФАР в сравнении с равноэнергетическим спектром (и, возможно, с рядом других типов спектров ламп) не всегда повышает фотобиологическую эффективность источника света в продуктивности тех или иных культур.
Можно предположить, что полученные различия в продуктивности ценозов при излучении исследуемых ламп связаны с различиями как в спектральном составе ФАР, так и в содержании инфракрасной радиации в интегральном лучистом потоке, которые обусловливают разницу в температурных, транспирационных и других эффектах, влияющих на продуктивность растений. Так, сопоставление показателей доли ИКР у источников света и урожая редиса под ними при облученности 100 Вт/м2 ФАР свидетельствует о тесной обратной корреляции между этими величинами: коэффициент корреляции г — —0,862 (см. табл. 5.1).
В качестве другого примера дана оценка влияния снижения доли ИКР в излучении ламп ДКсТВ-6000 на продуктивность корнеплодов (табл. 5.4). Видно, что снижение доли ИКР приводит к существенному росту фотобиологической эффективности излучения этого источника света в продуктивности редиса.
Имеются также литературные данные о существенном повышении урожая томатов под излучением галогенных ламп накаливания КГ-1000 при снижении ИКР в их лучистом потоке [Ермаков и др., 1980].
Таким образом, истинная спектральная эффективность излучения ФАР различных источников света может маскироваться разной долей ИКР в их общем лучистом потоке. Необходимость учета соотношения ФАР/ИКР требует установления оптимальной доли ИКР в общем излучении при формировании урожая растений (рис. 5.1). Эти данные получены в опытах с пшеницей, когда к излучению ксеноновой лампы ДКсТВ-6000, прошедшему через водный и стеклянный тепловые фильтры, добавляли с помощью ламп накаливания различные доли тепловой радиации. В пределах 40—60 % общего излучения ИКР соответствовала максимальным показателям продуктивности по зерну, поэтому такая
1.08
Рис. 5.1. Влияние различий доли ИКР в лучистом потоке интенсивностью 150 Вт/м2 ФАР на сроки вегетации (?) и продуктивность ценоза пшеницы в расчете на зерно (2).
доля ИКР в общем излучении наиболее рациональна для данного светового режима. В этом случае при сохранении наиболее высокой продуктивности по зерну сроки вегетации минимальные. Дальнейший рост доли ИКР значительно понижал
продуктивность, а уменьшение сильно удлиняло сроки вегетации, в результате чего также резко снижалась продуктивность.
Оценка роли спектральной эффективности ФАР в продуктивности растений, проводимая с использованием источников света с различным содержанием ИКР в лучистом потоке, корректна лишь при выравнивании доли ИКР в их интегральном излучении. Действительно, если, например, сопоставить фотобиологическую эффективность ламп ДРФ-1000 и ДКсТВ-6000 (см. табл. 5.4), то более эффективно излучение ламп ДРФ-1000. Однако при выравнивании доли ИКР в излучении ламп ДКсТВ-6000 до значения, соответствующего лампам ДРФ-1000 (40 %), их фотобиологические эффективности достоверно не различаются, а при дальнейшем уменьшении доли ИКР до 20 % у лампы ДКсТВ-6000 ее излучение уже более эффективно, чем лампы ДРФ-1000 (см. табл. 5.4). Хотя лампы ДРФ-1000 и ДКсТВ-6000 при выровненных по энергии лучистых потоках в видимой и инфракрасной областях спектра (ультрафиолетовое излучение обрезано стеклянным светофильтром) близки по фотобиологической эффективности, как это следует на примере редиса, однако распред-деление энергии в спектре излучения ФАР у этих ламп резко различно [Волкова и др., 1982]. В этом и подобных случаях можно говорить о фотобиологической эффективности излучения источника света в целом, но нельзя делать выводы о фотобиологической эффективности того или иного участка спектра оптического излучения.
Могут существовать и другие факторы, искажающие истин-? ную фотобиологическую эффективность ФАР в продукционном процессе. Ими могут быть особенности распределения светового потока ламп, падающего на растения, степень его диффузпости, объемности и другие характеристики, присущие тем или иным источникам света, что в итоге может сильно исказить те различия в формировании продукционного процесса, которые связаны с
1®
особенностями спектрального распределения энергии в видимой области этих источников света.
Разумеется, все указанные светотехнические характеристики источников света, являющиеся дополнительными факторами и влияющие на продуктивность растений, вряд ли играют важную роль, если источник света имеет крайне неблагоприятное распределение энергии в спектре ФАР, предопределяющее гибель растения или низкие значения урожая, а также в случае, если эксперименты ведутся в неконтролируемых условиях среды. В то же время если источник света не имеет указанных недостатков, а выращивание растений проводится в контролируемых, оптимизированных по несветовым параметрам условиях, что в комплексе обеспечивает поддержание продукционного процесса на достаточно высоком уровне, то отмеченные дополнительные факторы могут оказывать существенное влияние на формирование урожая растений, маскируя тем самым истинную фотобиологическую эффективность спектра ФАР.
Отсюда следует вывод, что излучение ламп с достаточно разным распределением энергии в спектре ФАР может обеспечить показатели продукционного процесса, которые в ряде случаев перекрываются. К такому же выводу пришли некоторые зарубежные исследователи [Ilieva и др., 1985].
Такая ситуация затрудняет оценку «в чистом виде» роли того или иного спектрального распределения ФАР в продукционном процессе, если оценка дается при использовании различных источников света.
Подводя итоги исследованиям роли спектрального состава света в продукционном процессе на основании использования данных по фотобиологическому испытанию различных источников света (как старых, так и современных конструкций), следует отметить, что этот этап работы, дав ряд интересных результатов по влиянию спектрального состава света на продукционный процесс растений, тем не менее не позволил достаточно четко сформулировать требования растений к спектру ФАР в связи с оптимизацией их продукционной деятельности. Нам представляется, что главная причина этого заключается в следующем: проводя исследования с использованием разнообразных серийных и не серийных источников света, исследователи в принципе не могут в достаточной степени корректно сопоставлять полученные результаты. Это, на наш взгляд, связано в первую очередь с тем, что каждая лампа, имея присущие только ей характеристики светотехнического или чисто эксплуатационного характера, может привносить свои особенности в оценку фотобиологической эффективности излучения. Из этих соображений эксперименты необходимо унифицировать в том смысле, что источники света должны быть одинаковы во всех отношениях, кроме спектрального состава излучения. В частности, при изучении влияния спектра ФАР на продукционный процесс ультрафиолетовая и инфракрасная области спектра должны быть одинаковы, а меняться может лишь спектр излучения в области ФАР.
МО
Попыткой имитации такой ситуации является создание серии источников света, которые обладали бы близкими светотехническими параметрами, но различались бы спектральными характеристиками в исследуемой области оптического диапазона излучения.
В рамки такого подхода укладывается серия экспериментальных работ минских светофизиологов [Федюпькин и др., 1988], В качестве источников света они использовали серию квазимоно-хроматических металлогалогенных ламп, изготовленных во ВНИСИ (Москва) [Прикупец и др., 1973], дающих интенсивное узкополосное излучение в различных спектральных областях ФАР. Поскольку результаты этих исследований оказались свободными от методических недостатков, связанных с использованием различных серийных источников света, они позволили с достаточной степенью уверенности установить наличие ряда общих принципиальных закономерностей воздействия спектрального состава ФАР на накопление биомассы молодых растений. Среди таких закономерностей следует отметить зависимость воздействия спектрального состава света различных областей ФАР на продукционный процесс от интенсивности облучения, а также от вида растений [Павловский и др., 1976]. К сожалению, эти исследования не свободны от ряда недостатков. В частности, выводы о влиянии излучения отдельных областей ФАР на различные стороны жизнедеятельности разных видов растений (в том числе и на продуктивность), полученные в работах Федюнькина с соавторами [1988] и Павловского с соавторами [1976], нельзя признать достаточно точными, поскольку в лучистых потоках, которые авторы называют синим, зеленым и красным, присутствовали значительные (до 15—20 %) примеси излучений других спектральных областей ФАР. Например, когда при оценке воздействия па растения наиболее физиологически значимых синей и красной областей спектра в «красном» излучении содержалось до 20 % и более синих лучей, это может принципиально исказить воздействие красных лучей на изучаемые физиологические процессы в растении [Тихомиров, Сидько, 1982; Дроздова и др., 1987]. Кроме того, недостаточно широко в этих работах представлены уровни облученности 100—150 Вт/м2 в области ФАР, которые являются физиологически обоснованными «рабочими» при выращивании растений на урожай при полном искусственном освещении [Лисовский и др., 1987]. Исследования проводились в большинстве случаев для облученности около 50 Вт/м2 ФАР преимущественно на зеленных или овощных культурах [Федюпькин и др., 1988], Эти обстоятельства также ограничивали возможности оценки оптимальности спектрального состава ФАР при выращивании растений на урожай при полностью искусственном освещении, когда целесообразно использовать облученности 100 Вт/м2 ФАР и выше [Тихомиров и др., 1983]. Оценка влияния спектрального состава света на конечный урожай растений определенного вида не всегда может быть распространена на другие виды растений,
что будет показано ниже. Не представляется обоснованным и проведение исследований по влиянию спектрального состава света на конечный урожай растений по оценке реакции на этот фактор одиночных растений, не говоря уже о кратковременной фотосинтетической реакции отдельных листьев. Это связано с тем, что световое насыщение фотосинтетической продуктивности последовательно возрастает при переходе от листового к организменному и ценотическому уровням. А именно ценоз, являясь продуктивным слоем, где формируется урожай, и должен лежать в основе исходного объекта исследований по влиянию спектрального состава света на продукционный процесс. К сожалению, этот момент редко принимался во внимание и ценотических исследований в этом направлении проведено крайне мало. В то же время экспериментально показано, что разрыв между цепотическим и более низкого порядка уровнями исследований зачастую имеет качественный характер [Мурей, 1978] и «стыковка» здесь может оказаться чрезвычайно затруднительной, что признается рядом исследователей [Цельникер, 1978; Тооминг, 1984].
Достаточно четкие и надежные сведения о влиянии спектрального состава света на продуктивность растений могут быть получены при использовании светотехнических установок, позволяющих менять как спектральный состав, так и интенсивность ФАР в достаточно широких пределах, а в качестве объекта исследования должны быть использованы ценозы растений разных видов при их выращивании на урожай в течение всей вегетации в контролируемых условиях. Такая постановка задачи легла в основу наших исследований влияния спектрального состава света на продукционный процесс.
§ 2. Продуктивность ценозов растений при различном спектральном составе и интенсивности искусственного облучения
Поиск, оптимального спектрального состава излучения для облучения растений проводили путем изучения воздействия различных участков ФАР на формирование ценозов растений и их продуктивность при застабилизированном содержании ультрафиолетового и инфракрасного излучения в интегральном лучистом потоке. При этом уровни облученности растений изменяли в широких пределах, поскольку в зависимости от этого может меняться физиологическая эффективность того или иного участка ФАР [Тихомиров и др., 1983]. Эксперименты проводили в строго контролируемых условиях, где изменяли только световой фактор, а остальные параметры среды стабилизировали на нелимитирующих уровнях (минеральное питание, температура, концентрация СО2 и др.). Опыты проводили на ценозах различных растений: редисе, пшенице, огурце и др.
Редис. Выращивание редиса в отдельных областях ФАР показало, что продукционный процесс в большой степени зависел от
112
Рис. 5.2. Зависимость среднесуточной продуктивности ценозов редиса в расчете па общую сухую биомассу (а) и корнеплоды (б) от облученности в отдельных областях ФАР.
1 — 400—500 нм; 2 — 500—600 нм; 3— 600—700 нм.
0;
5
J 20-
|--------* *-----------•'< в
% 10-______________•—
rJ lz w
S Зеленый —*- красный
0 -----1---1——-т----т-----1
20 40 60 80 100
Соотношение лучистых потоков отдельных областей ФАР,%
Рис. 5.3. Зависимость среднесуточной продуктивности ценозов редиса от облученности и соотношения спектральных областей в красно-синем (а), зелено-синем (б) и красно-зеленом (в) излучении ФАР.
1 — общая сухая биомасса при облученности 100 Вт/м2; 2 — то же, при 600 Вт/м2; 3 —• масса корнеплодов при облученности 100 Вт/м2; 4 — то же, при 600 Вт/м2.
спектрального состава и от интенсивности лучистого потока. В коротковолновой области ФАР ингибирование среднесуточной продуктивности по общей биомассе начиналось при Е > 500 Вт/м2, а максимум продуктивности отмечен при Е — 500 Вт/м2 (рис. 5.2). Аналогичная закономерность отмечена и для корнеплодов. В то же время использование желто-зеленых лучей сильно снижало плато насыщения по продуктивности, оно соответствовало Е = 100 Вт/м2. Дальнейший рост облученности до 200 Вт/м2 сопровождался уменьшением продуктивности, а при Е = 300 Вт/м2 продукционные процессы полностью ингибировались. Рассматривая световую зависимость среднесуточной продуктивности по сухой биомассе редиса в длинноволновой области ФАР, следует отметить четко выраженный максимум этой величины при Е = 50 Вт/м2. Дальнейшее повышение облученности сопровождалось резким ингибированием продукционных процессов. Световые зависимости среднесуточной продуктивности по корнеплодам имели аналогичный характер в зеленой и красной областях ФАР. Отсутствие единого плато для спектральных световых кривых продуктивности биомассы редиса связано, очевидно, с различным характером протекания фотосинтетических и сопряженных с ними ростовых процессов. В частности, устойчи-
113
600-
1г -
* 5 400-
I -
200
1
I I I I I ’
100 150 200
Облученность ФАР, Вт/ мг
Рис. 5.4. Световые кривые продуктивности корнеплодов ценозов редиса Вировский белый при трехкомпонентном сочетании отдельных областей ФАР.
Длительность вегетации — 21 сут. 1— с — 60 %, 3 — 20 %, к — 20 %; 2 — с — 34 %, з — 33 %, к — 33 %; 3 — с — 40 %, 3 — 20 %, к — 40 %; 4 — с — 20 %, 3 — 20 %, к — 60 %.
вость ценозов редиса к высокоиптенсивному излучению синего света в немалой степени связана с близкой к вертикальной ориентацией листьев в посеве, характерным скручиванием листовых пластинок и рядом других адаптационных перестроек, предохраняющих фотосинтетический аппарат от ингибирующего воздействия высоких облученностей. Этого не наблюдалось у растений в ценозах редиса, культивируемых в условиях облучения зеленой и красной областями ФАР [Тихомиров, Сидько, 1982]. С увеличением длины волны падающей радиации усиливалось ингибирующее действие света на рост и формирование листового аппарата редиса, что проявлялось в уменьшении числа листьев и их линейных размеров (см. гл. 3).
Полученный материал дает основание считать, что специфичность воздействия отдельных областей ФАР на продукционную •деятельность ценозов редиса выражается в том, что для каждой из них области светового насыщения по накоплению биомассы и ряду других физиологических показателей находятся в различных интервалах облученности. Наиболее быстрое световое насыщение по всем рассмотренным физиологическим характеристикам наступает на красном свету, в то время как достижение такого эффекта в коротковолновой области требует почти десятикратного увеличения облученности.
Исследование влияния на продуктивность ценозов редиса попарного сочетания отдельных областей ФАР при Е = 100 и 600 Вт/м2 показало, что наиболее интенсивно биосинтез шел в сине-зеленых и сине-красных лучах (рис. 5.3). Зелено-красное излучение оказалось наименее эффективным в продукционной деятельности ценозов редиса при Е = 100 Вт/м2, а при Е — = 600 Вт/м2 оказывало летальный эффект.
Последнее обстоятельство указывает на то, что для обеспечения интенсивной продукционной деятельности ценоза редиса лучи коротковолновой области ФАР должны обязательно входить в состав двухкомпонентного излучения. При этом с ростом облученности должна увеличиваться доля синих лучей в двухкомпонентном излучении ФАР независимо от того, является ли вторым компонентом зеленая или красная область спектра.
На рис. 5.4 представлены данные по продуктивности редиса в расчете на корнеплоды при использовании трехкомпопеитных
114
-
разноспектральных потоков ФАР. С ростом уровня облученности все более эффективным становилось излучение, в котором возрастала доля синих лучей, а эффективность излучения, где доминировали красные лучи, снижалась с ростом уровня облученности. Такое поведение спектральных световых кривых вполне объяснимо (см. рис. 5.2). В красных лучах насыщение по продуктивности редиса наступало быстрее всего, а в синих — наиболее медленно. Поэтому вполне логично, что для излучения, где доминируют красные лучи, световое насыщение для продуктивности редиса по накоплению биомассы произойдет довольно быстро (см. рис. 5.4, кривая 4), а добавка к такому излучению небольшого количества синих лучей за счет уменьшения доли красного света замедлит процесс светового насыщения по накоплению биомассы корнеплодов (см. рис. 5.4, кривая 3).
Еще более эффективным в продуктивности корнеплодов редиса для диапазона 100—200 Вт/м2 ФАР было равноэнергетическое излучение ФАР (см. рис. 5.4, кривая 2). Более высокое расположение плато световой кривой 2 по накоплению биомассы по сравнению с кривой 4 связано с резким увеличением содержания синих и уменьшением доли красных лучей в лучистом потоке. Более высокое расположение плато световой кривой 2 по сравнению с кривой 3 может быть объяснено увеличением доли зеленых лучей за счет уменьшения доли красных в первом случае по сравнению со вторым. Энергетическая эффективность зеленых лучей при 100—200 Вт/м2 ФАР выше, чем красных, поэтому вполне логично' предположить, что частичная замена красных лучей на зеленые может сдвинуть выход на плато насыщения по продуктивности редиса для такого излучения в сторону более высоких облученностей. Это мы и наблюдаем, сравнивая кривые 2 и 3 (см. рис. 5.4).
Излучение, где доминируют синие лучи (см. рис. 5.4, кривая 1) имело сравнительно низкую эффективность при облученности 100 Вт/м2 ФАР, однако в диапазоне 150—200 Вт/м2 эффективность этого излучения резко возрастала. Очевидно, что при высоких облученностях синие лучи наиболее эффективны для синтеза биомассы (см. рис. 5.2). Такое предположение подтверждается данными по продуктивности ценозов редиса при высокоинтенсивном излучении ФАР с уровнем облученности 600 Вт/м% (табл. 5.5).
В продуктивности ценозов редиса эффективность излучения ФАР с равным содержанием (40 %) синих и зеленых лучей существенно выше, чем для всех других представленных спектральных-вариантов. Интересно сопоставить данные по продуктивности ценозов редиса для облученности 100 и 600 Вт/м2 различного спектрального состава ФАР. При 100 Вт/м2 световая зависимость продуктивности ценозов редиса соответствует восходящему, а при 600 Вт/м2 — нисходящему участку световой кривой. Наименее эффективным в продукционном процессе ценозов редиса было излучение, представленное одной из исследуемых областей ФАН (см. рис. 5.2—5.4, табл. 5.5). Более высокие значения продукт
11£
Таблица 5.5
Продуктивность ценозов редиса при Е — 600 Вт/м2 ФАР различного спектрального состава
Номер варианта Спектральный состав ФАР, % Абсолютно сухое вещество, г/мг
Общая биомасса Биомасса корнеплодов
1 с — 70, з — 15, к — 15 471,1±51,7 283,3±6,6
2 с — 55, з — 30, к — 15 621,1^74,9 355,4± 53,3
3 с — 40, з — 40, к — 20 780,8±61,3 387,7+81,2
4 с — 40, з — 20, к — 40 528,2±54,2 331,б±27,4
5 с — 34, з — 33, к — 33 577,9± 55,4 376,2+28,2
6 с — 20, з — 50, к — 30 500,9± 20,0 293,8±19,8
7 с — 20, з — 20, к — 60 459,3±23,4 234,2±10,6
Примечание, с— 400—500 нм; з — 500—600 нм; к — 600—700 нм.
Продолжительность вегетации 20 сут.
тивпости цепоза редиса могут быть достигнуты при двухкомпо-нептном излучении ФАР. Однако наиболее высокие показатели продуктивности для ценозов редиса отмечены при использовании трехкомпопентного потока ФАР. Эти выводы справедливы для облученности 100 и 600 Вт/м2 ФАР.
Интересно также сравнить данные по продуктивности ценозов редиса под действием равноэнергетического излучения, полученного с помощью комбинаций равных по энергии долей исследуемых областей ФАР, с соответствующими данными при использовании трехкомпонентного излучения ФАР различного спектрального состава. Продуктивность по корнеплодам для ценоза редиса при облученности 100 Вт/м2 равпоэнергетического излучения ФАР достоверно не превышала лучший из полученных в других спектральных вариантах. Однако при облученности 600 Вт/м2 равноэнергетического излучения урожай ценоза редиса уступал таковому, полученному при использовании излучения с максимумом в сине-зеленой области ФАР. Существенно, что если для Е = 100 Вт/м2 спектральный сдвиг в излучении ФАР без снижения продуктивности цепоза редиса возможен в длинноволновую область, то при Е — 600 Вт/м2 — в коротковолновую. Эти различия, очевидно, связаны с особенностями морфофизиологических перестроек в структуре цепоза редиса с увеличением облученности (см. гл. 3).
Пшеница. Влияние отдельных областей ФАР с Е = 100 и 600 Вт/м2 па продуктивность ценозов пшеницы иллюстрирует табл. 5.6.
Анализ этих данных показывает, что при облученностях 100 Вт/м2 ФАР наименее эффективным в продуктивности пшеницы был свет коротковолнового участка ФАР. Так, при более затянутом вегетационном периоде общая сухая биомасса в этом свето, вом варианте была в 2,5 раза, а масса зерна — в 6 раз ниже-116
Таблица 5.6
Продуктивность пшеницы 232 при различных облученностях в отдельных областях ФАР
Облученность ФАР, Вт/м2 Область спектра, нм Абсолютно сухая масса 1000 зерен, г Среднесуточная продуктивность, г/(м2-сут) Длительность вегетации, сут
по общей биомассе по зерну
100 400—500 15,9 7,4 1,2 85
500—600 33,0 20,6 8,6 80
600—700 27,0 19,3 8,0 80
600 400—500 30,8 23,3 9,6 80
500-600 29,8 47,1 16,7 75
600-700 30,4 51,8 22,8 75
чем при свете зеленого и красного участков ФАР. Следует отметить, что в последних двух спектральных вариантах как общая продуктивность, так и продуктивность по зерну достоверных различий не имели.
Примерно одинаковые значения продуктивности для зеленого и красного света при Е = 100 Вт/м2 связаны с тем, что при меньшей фотосинтетической активности единицы листовой поверхности пшеницы в варианте с зеленым светом общий листовой индекс был выше, чем в варианте, где использовали излучение длинноволновой области ФАР (см. табл. 5.6). Это способствовало выравниванию продукционных возможностей посевов в этих световых вариантах.
Повышение интенсивности излучения до 600 Вт/м2 ФАР активизировало продукционные процессы во всех исследуемых интервалах ФАР. Эффективность синей области спектра в продукционном процессе и в этом случае была ниже по сравнению с зеленым и красным светом, по различия по продуктивности менее существенны, чем в опытах с Е = 100 Вт/м2 ФАР. Кроме того, высокоинтенсивный красный свет оказался наиболее эффективным для ценоза пшеницы. Это, очевидно, можно связать с тем, что высокоинтенсивный красный свет, способствуя образованию продуктивных боковых побегов (см. табл. 3.3) и имея большую фотосинтетическую эффективность в расчете на единицу ассимиляционной поверхности (см. табл. 2.1), обеспечил более высокие темпы накопления биомассы, чем свет зеленой и синей областей спектра.
Среди спектральных комбинаций с Е = 100 Вт/м2 наиболее эффективны в обеспечении продукционного процесса ценозов пшеницы варианты с доминированием в ФАР красных лучей, а наименее эффективны — с преобладанием синих независимо от того, какая область ФАР является дополнительной — зеленая или красная. В последнем случае снижаются темпы накопления общей биомассы и зерна на фоне увеличения вегетационного периода (табл. 5.7). В большинстве спектральных вариантов продуктив-
117
Таблица 5.7
Продуктивность ценоза пшеницы при Е = 100 Вт/м2 ФАР различного спектрального состава
Номер варианта Распределение энергии в области ФАР, % Про до л жи-, тельность вегетации, сут Сухая биомасса, г/№ Среднесуточная продуктивность по зерну, г/(м2-сут)
Общая Зерно
1 с — 34, з — 33, к — 33 75 1635,4+164,3 659,3+67,8 8,8+2,2
2 с — 75, з — 25 80 1210,6+133,1 420,0+52,3 5,3±1,7
О с — 25, з — 75 75 1495,4+179,4 668,5+77,4 8,9+1,0
4 с — 75, к - 25 78 1593,3± 143,4 573,2+55,4 7,3+0,71
5 с — 50, к — 50 75 1974,3±185,5 803,3+93,8 10,7+1,3
6 с — 25, к — 75 70 2457,5+247,4 868,4+82,3 12,4+1,2
7 с — 15, к — 85 70 1624,2±194,9 668,3+77,8 9,5±1,1
8 з — 40, к — 60 70 2274,4+272,8 881,1+90,1 12,6+1,3
ный по зерну стеблестой был сформирован только за счет главных стеблей. Однако в спектральных вариантах, где наблюдалась наибольшая продуктивность, определенный вклад внесли и боковые побеги (№ 6, 8).
Ценозы огурца. За первые 10 сут вегетации при излучении интенсивностью 50 и 100 Вт/м2 темпы накопления сухого вещества были выше при излучении красной области спектра в обоих случаях, однако в дальнейшем более высокое накопление сухого вещества происходило при излучении синей области спектра (рис. 5.5). Наиболее контрастные различия в характере накопления сухого вещества ценозами огурца наблюдались между зеленым и красным спектральными вариантами* с одной стороны, и синим спектральным вариантом — с другой. Так, если в зеленых лучах при 50 Вт/м2 и в красных при 50 и 100 Вт/м2 ФАР происходило замедление, а затем и прекращение накопления сухого вещества, ведущее к гибели ценозов огурца, то в варианте с синим светом при обоих уровнях облученности процесс накопления сухого вещества неуклонно продолжался все 60 сут эксперимента (см. рис. 5.5).
Сопоставление данных рис.- 5.5 с- данными по накоплению зелёных пигментов (см. рис. 3.20) и данными по фотосинтезу (см. рис. 2.8) показывает, что прекращение биосинтеза биомассы в ценозах огурца в зеленых и красных лучах связано с выцветанием зеленых пигментов в листьях этих растений и прогрессирующим падением фотосинтеза ценозов огурца, в то время как в синих лучах процессы накопления хлорофиллов в листьях ценозов огурца и фотосинтез последних не подвергались ингибированию.
Лишь при облученности 50 Вт/м2 зелёного света эти процессы Протекали сравнительно замедленно, в результате чего в этом спектральном варианте сформировался нёбольИгой хозяйственно
118
160
Сутки
Рис. 5.5. Возрастная динамика накопления сухой вегетативной биомассы ценозов огурца Московский тепличный при облученностях 50 (я) и 100 (б) Вт/м2 в отдельных областях ФАР.
Остальные уел. обозн. см. рис. 5.2.
полезный урожай (0,9 кг/м2). В остальных вариантах с использованием зеленых или красных лучей растения погибли, не успев вступить в стадию плодоношения. В синих лучах урожай плодов огурца составил 0,6 кг/м2 при облученности 100 Вт/м2, а при 50 Вт/м2 процесс плодоношения вообще не наступил.
Эти результаты указывают на то, что излучение ни одной из рассматриваемых отдельных областей ФАР не может быть признано эффективным для формирования урожая огурца. Такая непригодность излучения зеленой и красной областей, с одной стороны, и синей — с другой, обусловлена разными причинами. Если в первом случае это связано с нарушениями биосинтеза зеленых пигментов, влекущими за собой гибель ценозов огурца, то во втором — с нарушением процессов плодоношения.
Исследовано влияние различных сочетаний синих и красных лучей на накопление биомассы в ценозах огурца 30-суточного
119
Доля красных лучей е сине-красном излучении, %
Рис. 5.6. Зависимость накопления сухого вещества ценозами огурца от соотношения спектральных областей ФАР в сине-красном излучении интенсивностью 100 Вт/м2.
Возраст ценозов 30 сут.
Рис. 5.7. Временная динамика сбора урожая плодов томатов, сформировавшихся при облученности 100 Вт/м2 в отдельных областях ФАР.
1 — 400—500 нм; 2 — 500—600 нм; 3 — 600—700 нм.
возраста (рис. 5.6). Видно, что увеличение доли красных лучей в сине-красном излучении до примерного равенства с долей синих способствовало увеличению биомассы растений. Дальнейшее увеличение доли красных лучей замедляло темпы накопления биомассы растений вплоть до их гибели, происходящей при полной замене синих лучей красными, что отмечалось ранее. Такое замедление темпов накопления биомассы растений связано, вероятно, с усилением процессов выцветания зеленых пигментов в листьях ценозов огурца при резком возрастании доли красных лучей в сине-красном излучении.
Исследование различных сочетаний синих и зеленых лучей в сине-зеленом излучении интенсивностью 100 Вт/м2 показало, что наиболее эффективным в формировании урожая был лучистый поток, где доминировали синие лучи (80 %), а наименее эффективным — с преобладанием зеленых лучей (80 %). В первом случае урожай огурцов составил 3,95 кг/м2, а во втором — 1,22 кг/м2.
Эксперименты по изучению воздействия различных двухкомпонентных сочетаний зеленых и красных лучей на формирование урожая в ценозах огурца показали, что любое их сочетание в зелено-красном излучении (интенсивность 100 Вт/м2 ФАР) приводило к выцветанию зеленых пигментов и гибели ценозов огурца.
Из трехкомпонентных сочетаний исследуемых спектральных областей ФАР интенсивностью 100 Вт/м2 наиболее эффективными, обеспечивающими высокие показатели продуктивности для огурца, были лучистые потоки, где доминировало зелено-красное излучение (табл. 5.8). Анализ последовательности спектральных вариантов показывает, что с ростом доли синих лучей в потоке ФАР продуктивность ценозов огурца уменьшалась. При этом
120
Таблица 5.8
Продуктивность ценозов огурца сорта Московский тепличный при облучении различного спектрального состава ФАР интенсивностью 100 Вт/м2 [Тихомиров и др., 1991]
Номер варианта Соотношение спектральных областей в ФАР, % Урожай плодов огурца, кг/м2 Продолжительность вегетации, сут Среднесуточная продуктивность в расчете на урожай плодов, кг/(м2-сут)
1 с — 20, з — 40, к — 40 27,5±1,3 70 392,9±22,9
2 с — 34, з — 33, к — 33 22,5±1,9 75 300,0±29,3
3 с — 30, з — 20, к — 50 22,1±1,5 75 299,74-20,0
4 с — 40, з — 20, к — 40 20,8±1,6 80 260,04-20,0
5 с — 60, з — 20, к — 20 46,1±1,3 90 178,8±14,4
Примечание, с — 400—500 нм, з — 500—600 нм, к — 600—700 нм.
снижался урожай плодов огурца и увеличивалась продолжительность вегетации.
Ценозы томатов. Накопление вегетативной биомассы и хозяйственно полезный урожай томатов зависят от излучения отдельных областей ФАР разной интенсивности (табл. 5.9). Для 30-суточных растений томатов, вступающих в фазу цветения, наиболее эффективным в накоплении биомассы было излучение красной области спектра независимо от используемых уровней облученности, тогда как синие и зеленые лучи по своей спектральной эффективности в накоплении биомассы растений одинаково уступали красным и не имели достоверных различий между собой по урожаю плодов томатов за 100 сут вегетации. Синие лучи хотя и уступали красным, но оказались заметно эффективнее зеленых и при 50, и при 100 Вт/м2 ФАР. Это позволяет предположить, что спектральная эффективность излучения в продукционном процессе в целом не всегда может быть достоверно определена на основании данных, полученных на молодых растениях.
На рис. 5.7 показана временная динамика сбора плодов томатов под излучением отдельных спектральных областей ФАР интенсивностью 100 Вт/м2. Видно, что процесс формирования и созре-
Таблица 5.4>
Влияние излучения отдельных областей ФАР на накопление сухого вещества растений томатов за 30 сут и урожая плодов за 100 сут вегетации
Область спектра, нм Сухая биомасса 30-дневного растения, г Сырая биомасса плодов томатов, кг/м2
К, Ег Ei Ег
400-500 1,0±0,1 4,7±0,2 3,3±0,2 7,7±0.8
500-600 1,3±0,1 4,5±0,2 2,5±0,1 5,5±0,4
600—700 3,5±0,2 5,7±О,3 5,8±0.6 8,6±0,8
Примечаг и е. Et = 50, Е 2 = 100 Вт/м2,
9 А. А. Тихомиров, Г. М. Лисовский, Ф, Я. Сидько
121
Таблица 5.10
Продуктивность томатов сорта Старфайер при облучении ФАР различного спектрального состава интенсивностью 100 Вт/м2 (совместно с И. Г. Золотухиным)
Номер варианта Соотношение спектральных областей в ФАР, % Период вегетации, сут Урожай томатов, кг/м8 Среднесуточная продуктивность в расчете на урожай плодов, г/(м8-сут)
1 с — 10, з — 15, к — 75 100 18,5±1,2 185,0± 12,0
2 с — 20, з — 20, к — 60 100 16,9+1,2 169,0± 12,0
3 с — 20, з — 40, к — 40 по 15,5±0,9 140,9+8,2
4 с — 40, з — 20, к — 40 120 15,7+1,0 130,8+8,3
5 с — 34, з — 33, к — 33 ПО 15,4±1,1 140,0+10,0
6 с — 20, з — 60, к — 20 120 11,1+0,7 92,5+5,9
7 с — 60, з — 20, к — 20 130 13,2+0,9 101,5+6,9
Примечание, с — 400 — 500 нм; 3 — 500 — 600 нм; к — 600 — 700 нм.
вания плодов наиболее интенсивно протекал при излучении красной области спектра, где раньше всего стали образовываться спелые плоды. Дружное созревание плодов в синих лучах обеспечило в конце вегетации урожай плодов существенно больший, чем под зеленым светом. Длительность вегетации томатов под излучением отдельных спектральных областей составляла 100 сут, после чего выращивание растений прекращали. Поэтому последний сбор плодов томатов в конце вегетации включал в себя как спелые, так и зеленые плоды.
Наиболее быстрое созревание урожая плодов томатов при смешанном спектральном составе ФАР происходило при использовании спектра, обогащенного красными лучами, составляющими 60—75 % в общем излучении ФАР (табл. 5.10, варианты 3, 7). Весьма эффективным в формировании урожая плодов томатов было равноэнергетическое излучение ФАР, а также излучение с небольшими отклонениями от него по спектру в синюю или красную область (см. табл. 5.10, варианты 4—6). Однако сроки вегетации в трех последних вариантах оказались более продолжительными, а следовательно, продуктивность ниже, чем в лучших по урожаю вариантах.
§ 3. Эффективность использования фитоценозами излучения ФАР различного спектрального состава и интенсивности (КПД фитоценозов)
Спектральная эффективность излучения в продукционном процессе может быть оценена с помощью величины, получившей название энергетической эффективности фитоценозов [Ничипоро-вич, 1956], или КПД фитоценозов [Тооминг, 1977]. Такая оценка в условиях искусственного облучения представляет большой ин-
122
Рис. 5.8. Эффективность усвоения (КПД) излучения отдельных областей ФАР различной интенсивности ценозами редиса в расчете на падающую (а) и поглощенную (б) энергию.
Усл. обозн. см. рис. 5.2.
терес, так как позволяет определить экономичность использования излучения ФАР, имеющей различные спектральные и энергетические характеристики в продукционном процессе. Вначале рассмотрим КПД ценозов при излучении отдельных областей ФАР.
КПД использования излучения отдельных областей ФАР (в расчете на падающую энергию) ценозами редиса зависит от уровня облученности ФАР (рис. 5.8, а). Наиболее высокие значения КПД соответствуют низким уровням облученности для всех исследуемых спектральных областей ФАР. Для синего света в отличие от зеленого и особенно красного эта зависимость выражена наименее резко. Для расчетов использования энергии фитоценозами в ряде случаев считается более целесообразным вести расчеты на поглощенную ценозом радиацию ФАР [Тооминг, 19771 (рис. 5.8, б).
При расчете КПД для разноспектральных потоков ФАР такой подход может оказаться целесообразным в связи с различным поглощением ФАР для отдельных ее областей (например, зеленых лучей по сравнению с синими и красными).
Принципиальных различий в зависимости КПД от уровня облученности в отдельных областях ФАР как при расчете на падающую, так и поглощенную энергию нет (см. рис. 5.8, а, б). Для КПД при расчете на падающую энергию характерно лишь некоторое уменьшение их абсолютных значений в сравнении с соответствующими, рассчитанными на поглощенную энергию, в то время как взаимное расположение и форма, кривых (см. рис. 5.8, а, б) остались практически неизменными. Это объясняется тем, что вариации спектрального режима облучения мало влияют на общую оптическую плотность ценоза.
Однако при интегральной, примерно одинаковой, поглощательной способности фитоценозов для лучей различных спектральных областей ФАР распределение лучистой энергии по ярусам ценоза может быть различным и зависит-от спектрального
9*
123
Таблица 5.11
Средний за вегетацию КПД ценозов пшеницы в расчете на общую биомассу (тр, %) и зерно (ц2, %) при различной густоте посадки и интенсивности облучения 600 Вт/м2 в отдельных областях ФАР
Густота посадки, растений/м2 400—500 нм 500—600 нм 600—700 нм
41 Па 41 42 41 4г
1000 (контроль) 0,77 0,32 1,46 0,52 1,6 0,7
2000 1,44 0,68 2,2 0,86 2,67 0,97
3000 1,43 0,61 2,68 1,04 2,75 1,1
4000 1,45 0,49 3,24 1,17 2,67 1,07
состава используемого излучения. Это подтверждается прямыми измерениями, проведенными нами для многоярусных ценозов пшеницы [Тихомиров, 1977; Tikhomirov, Sid’ko, 1983]. Отсюда можно сделать вывод, что при общем, примерно равном поглощении радиации разного спектрального состава оптически плотных ценозов верхние слои, наиболее ответственные за формирование урожая, поглощают радиацию разного спектрального состава неодинаково [Тихомиров, 1979]. Это дает основание предполагать, что даже для полнопоглощающих ценозов можно повысить эффективность использования ими лучистой энергии (КПД) за счет загущения. В этом случае основную роль будет играть спектральный состав радиации, неодинаково поглощаемый верхними слоями ценоза, а следовательно, по разному влияющий на его продуктивность.
С этой целью нами на основании вычисления КПД рассмотрена целесообразность степени загущения многоярусных ценозов при использовании высокоинтенсивных лучистых потоков различного спектрального состава (табл. 5.11).
Во всех световых вариантах отмечается рост КПД ценозов по мере их загущения в расчете на общую биомассу. При этом наиболее высокий прирост КПД отмечается при загущении ценозов, выращиваемых при зеленом излучении, а наименьший — при синем. Однако, чтобы оценить эффективность использования излучения в формировании урожая зерна, о целесообразности характера загущения лучше судить по КПД в расчете па хозяйственно полезную часть биомассы (зерно). Такая оценка показывает, что целесообразность загущения может быть обоснована при использовании высокоинтенсивного красного и особенно зеленого излучения, в то время как для синей области ФАР более чем двухкратное загущение по сравнению с контролем снижает КПД в расчете на зерно. В то же время ухудшение радиационного режима в ценозах пшеницы при их загущении с использованием длинноволнового излучения ФАР в меньшей степени снижает фотосинтетическую деятельность отдельных растений в ценозе. Поэтому в последнем случае за счет загущения удается повысить урожай, а следовательно, и КПД ценоза. Однако в наиболее вы
124
годном положении при загущении находятся ценозы, выращиваемые в лучах зеленой области ФАР. За счет более высокой проникающей способности зеленых лучей в глубь ценоза даже при сильном загущении световые условия для фотосинтеза фитоэле-мептов в этом случае более благоприятны, чем в других. Ввиду этого и рост КПД ценозов с увеличением их оптической плотности здесь выше, чем в остальных световых вариантах.
Ценозы пшеницы во всех спектральных вариантах независимо от степени загущения были практически полнопоглощаю-щими структурами. Поэтому можно полагать, что рост КПД таких ценозов обусловлен спецификой поглощения света разного спектрального состава верхними слоями посевов.
Таким образом, учет спектрального состава лучистой энергии при выборе степени загущения ценозов может существенно повысить эффективность использования радиации на построение хозяйственно полезной части фитомассы.
Примерное равенство поглощательной способности ценозов, сформировавшихся при различном спектральном составе ФАР, наблюдается при использовании излучения смешанного спектрального состава. В этом случае поглощательная способность ценозов, имеющих равные стартовые густоты посадки растений на единицу площади, не ниже, а в большинстве случаев выше, чем при излучении отдельных спектральных областей ФАР. Возникающие при этом морфогенетические изменения в структуре ценозов под воздействием излучения различного смешанного спектрального состава слабо влияют на их общую поглощательную способность. Поэтому расчеты КПД по падающей энергии для различных спектральных вариантов не будут принципиально отличаться от аналогичных расчетов по поглощенной ценозами ФАР. Поскольку оценка КПД по падающей энергии более проста, а для расчетов затрат энергии и более удобна, мы в дальнейшем воспользуемся этим способом оценки КПД.
Для оценки зависимости КПД фитоценоза от спектрального состава используемой радиации ФАР представляет интерес проанализировать такую зависимость на примере ценозов, выращенных при режимах облучения, различающихся плавным изменением спектральных характеристик облучения. В качестве таких режимов целесообразно взять двухкомпонентные сочетания излучения отдельных спектральных областей ФАР, для которых в экспериментальном отношении не представляет затруднений достаточно плавный переход от излучения, где доминирует одна из двух спектральных областей, к излучению, где доминирует другая спектральная область ФАР (рис. 5.9). Существенную роль в изменении эффективности использования падающей на ценозы энергии играют различия как в ее спектральном составе, так и в используемых уровнях облученности. Максимальные значения таких КПД при сине-краспом и зелено-красном излучении, когда используются облученности 100 Вт/м2 ФАР, более чем в 5—6 раз превосходят максимальные значения КПД при облученности
125
Рис. 5.9. Эффективность усвоения (КПД) красно-синего (о), зелено-синего (б) и красно-зеленого (в) излучения ФАР интенсивностью 100 и 600 Вт/м2 ценозами редиса.
Усл. обозн. см. рис. 5.3.
600 Вт/м2 ФАР для тех же комбинаций отдельных областей ФАР (см. рис. 5.9).. Сближение этих значений КПД наблюдается лишь при сопоставлении наиболее неблагоприятных в отношении формирования урожая сочетаний синих и красных или синих и зеленых лучей при облученности 100 Вт/м2 ФАР и наиболее благоприятном для формирования урожая редиса сочетании этих же лучей при интенсивности 600 Вт/м2 ФАР. Однако сопоставимые значения получаются в этом случае лишь при расчете КПД усвоения падающей энергии в расчете на хозяйственно полезный
урожай, в то время как при расчете КПД на общую биомассу эти различия остаются весьма существенными.
Если оценивать влияние спектрального состава света на КПД в пределах одного из используемых уровней облученности, то при 100 Вт/м2 ФАР неблагоприятное сочетание синих и красных или синих и зеленых лучей в двухкомпонентном излучении может снизить КПД в расчете на общий урожай в 2,5, а для сочетания красных и зеленых лучей — в 2 раза. Примерно такая же картина наблюдается и при использовании облученностей 600 Вт/м2 ФАР, за исключением зелено-красного излучения, когда растения редиса погибают независимо от соотношения зеленых и красных лучей в общем двухкомпонентном потоке (см. рис. 5.9).
В условиях искусственного освещения КПД фотоценозов зависит от количества накопленного сухого вещества, уровня облученности, длительности фотопериода и вегетационного периода. Влияние спектрального состава света на КПД является опосредованным и может влиять на него через накопление биомассы и длину вегетационного периода, а также за счет изменения поглощательной способности ценоза.
12В
§ 4. Некоторые особенности воздействия спектрального состава света на продукционный процесс растений
Изучение роли излучения отдельных областей ФАР в продукционном процессе показало неоднозначность влияния разноспектральных потоков ФАР как на накопление общей биомассы растений разных видов, так и на формирование хозяйственно полезного урожая. При этом излучение синей области спектра в отличие от зеленой и красной обеспечивает накопление общей биомассы на протяжении всего периода вегетации для полного набора исследуемых культур. В то же время не все эти культуры в синих лучах формируют хозяйственно полезный урожай или он весьма низок.
Контрастной оказалась реакция растений разных видов на излучение красной области спектра. Это проявилось в том, что при длительном выращивании редиса, огурца в красных лучах через 20—30 сут вегетации эти культуры гибли. В то же время ценозы пшеницы, томатов, кукурузы способны не только поддерживать структурную и функциональную активность фотосинтетического аппарата, но и давать хозяйственно полезный урожай в красных лучах [Тихомиров и др., 1987].
Что касается зеленых лучей, то их влияние на формирование продукционного процесса в качественном отношении сходно с действием красной области спектра. Для зеленых лучей, так же как и для красных, характерно либо негативное воздействие на формирование фотосинтетического аппарата, приводящее к гибели те же виды растений, что при красном свете, либо позитивное, обеспечивающее формирование ценоза, способного дать урожай (например, пшеницы, томатов), который в большинстве случаев ниже, чем под красным светом. Зная положительное или отрицательное воздействие зеленых лучей па продукционный процесс у того или иного вида растений, можно ожидать, что качественно в этом же направлении будет характеризоваться воздействие красных лучей на продукционный процесс растений данного вида.
Эффективность излучения отдельных спектральных областей в формировании продукционного процесса сильно зависит от его интенсивности. Такая зависимость наиболее подробно изучена на ценозах редиса и отчасти на ценозах пшеницы. Ряд экспериментов в этом направлении выполнен для ценозов огурца и томата. Общий вывод, который следует из этих экспериментов, указывает на возрастающую спектральную эффективность (но не КПД) синих лучей в сравнении с красными в продукционном процессе с ростом уровня облученности. Однако увеличение эффективности синих лучей в продуктивности ценозов растений разных видов с ростом уровня облученности происходит неодинаково. Так, например, если для ценозов редиса максимальная эффективность синих лучей в накоплении биомассы находится в диапазоне 400—
127
500 Вт/м2 ФАР (см. рис. 5.2), то для пшеницы, образующей многоярусный оптически более плотный ценоз, облученность должна быть выше. Несомненно, что плато насыщения по накоплению биомассы под синим светом (так же, как под лучами других спектральных областей) для ценозов разных культур может быть достигнуто при различных интенсивностях облучения. Это связано с тем, что при высоких облученностях деструктивное влияние синего света на фотосинтетический аппарат растений сказывается в меньшей степени, чем зеленого и красного. Такая устойчивость фотосинтетического аппарата к высокоинтенсивному синему свету может быть связана, с одной стороны, с более замедленным, чем в зеленых и красных лучах, старением фотосинтетического аппарата, в первую очередь хлоропластов [Власова, 1975], а с другой стороны — с активным синтезом в синих лучах каротиноидов, флавинов и других веществ белковой природы, имеющих защитную функцию к воздействию высокоинтенсивного излучения [Воскресенская, 1987].
При использовании смешанного потока ФАР, в котором представлены в тех или иных пропорциях синяя, зеленая и красная области, формирование урожая сельскохозяйственных культур более эффективно, чем при излучении той же интенсивности, сосредоточенном в одной или двух из названных спектральных областей. Таким образом, оптимальный для продукционного процесса спектр ФАР зависит от уровня облученности, с одной стороны, и вида используемой культуры — с другой. Поэтому выбор спектральных и энергетических режимов облучения растения необходимо вести с учетом конкретно поставленной задачи. Например, с точки зрения максимального КПД при выгонке рассады огурцов оптимальным будет, согласно нашим данным, уровень облученности 50—60 Вт/м2 ФАР [Лисовский и др., 1987], в то же время для формирования хозяйственно полезного урожая (томаты, пшеницы) уровни облученности должны находиться в диапазоне 100—200 Вт/м2 ФАР [Полонский, Лисовский, 1980; Александрова, Клочкова, 1983]. При выращивании растений в условиях, когда энергетические затраты не являются лимитирующим фактором, можно искать оптимум, например, для продуктивности при весьма высоких облученностях порядка солнечной и выше [Тихомиров и др., 1976; Полонский, 1978]. При столь разных уровнях облученности оптимальный спектр ФАР для продуктивности одних и тех же растений может оказаться разным.
С другой стороны, при одинаковых уровнях облученности ФАР для ценозов различных культур оптимальный спектр излучения тоже неодинаков. Действительно, если сравнить данные по оптимальному в накоплении урожая растений распределению энергии в спектре ФАР интенсивностью 100 Вт/м2 для ценозов редиса, пшеницы, огурца и томатов, то все они различны (см. § 2). Так, например, необходимая доля синих лучей, обеспечивающая при облученности 100 Вт/м2 ФАР оптимум по продуктивности, выше для ценоза редиса, но ниже для ценоза огурца.
128
Существенную роль играет в оптимальном спектре ФАР зеленый компонент излучения, как это было показано, например, на ценозах пшеницы и огурца. Одиако варьирование синим и зеленым компонентами ФАР для обеспечения высокой продукционной деятельности фитоценоза с учетом его оптической плотности и вида растений может быть успешным лишь на фоне достаточного обеспечения красным светом. На основании анализа собственного материала и литературных данных нам представляется, что наращивание доли излучения красной области ФАР для обеспечения максимальной продуктивности ценоза растений должно быть тесно увязано со знанием потребностей такого ценоза в синих и зеленых лучах. Как это следует из ранее представленных материалов, чрезмерное увеличение в ФАР доли красных лучей (вплоть до «чистого» красного света) может привести к гибели некоторые виды растений. В то же время резкое уменьшение или полное исключение красных лучей из ФАР никогда не вызывает гибели ценозов растений, хотя и может существенно снизить их продуктивность.
В этой связи возникает необходимость установить те предельные значения доли красного света в ФАР, которые бы соответствовали максимальной продукционной деятельности фитоценозов. Несомненно, что для разных культур такие значения будут отличаться.
Результаты наших исследований по длительному воздействию излучения красной области спектра на ценозы различных видов растений (табл. 5.12) позволяют предположить сугцествование двух групп растений, различающихся по реакции на красный свет: первая группа характеризуется нарушением метаболизма и падением продуктивности, вторая — отсутствием аномальных изменений в метаболизме и достаточно хорошей продуктивностью.
В практическом плане такое подразделение растений на две группы по реакции на красный свет предполагает, что растения
Таблица 5.12
Влияние облученности 100 Вт/м2 ФАР в синей (отс) и красной (тк) областях спектра на накопление биомассы растений
Растение, сорт Возраст, сут Общая сухая биомасса 1 растения, г
тс nic/™K
Огурец Московский тенлич- 3,34
ный 30 4,01 1,2 *
Редис Вировский белый 20 1,61 0,75 * 2,15
Подсолнечник Енисей 20 12,1 ** 7 14 ** 1,7
Кукуруза Минусинка 20 2,8 ** 6,4 ** 0,44
Пшеница 232 30 0,25 0,25 1,00
Томаты Старфайер 30 0,95 1,63 0,58
* При дальнейшем выращивании растения постепенно погибают.
•• До конца вегетации растения не выращивали.
129
второй группы для обеспечения эффективного продукционного процесса должны выращиваться при излучении ФЛР с большим содержанием красных лучей, нежели растения первой группы. Например, для обеспечения наибольшей продукционной деятельности томатов в условиях интенсивной светокультуры (Е — 100— 150 Вт/м2) доля красных лучей в ФАР должна составлять 65— 75 %, а для огурца только 40—50 % (но не менее 40 %).
Таким образом, рассмотренный материал по изучению роли красного компонента в общем излучении ФАР показывает достаточную сложность и неоднозначность его влияния на повышение продуктивности ценозов растений разных видов как в качественном, так и в количественном отношении.
Этот вывод подтверждает предположение о том, что простым увеличением доли излучения в красной области спектра проблема оптимизации в распределении энергии излучения растениеводческих ламп полностью не решается. Отсутствие в настоящее время серийных источников света с достаточно высокой долей красных лучей в ФЛР позволяет считать первым этапом создания эффективной растениеводческой лампы для выращивания растений на урожай при полностью искусственном освещении повышение доли излучения лампы в красной области ФАР до 40— 50 % с сохранением примерного равенства синих и зеленых лучей. Дальнейшим резервом в повышении спектральной эффективности излучения представляется создание специализированных по группам растений и задачам выращивания типов ламп с физиологически обоснованным распределением энергии излучения в синей, зеленой и красной областях ФАР на основании данных фотобиологических экспериментов с ценозами различных растений.
Глава 6
ОПТИМИЗАЦИЯ СПЕКТРАЛЬНЫХ И ЭНЕРГЕТИЧЕСКИХ ХАРАКТЕРИСТИК ФАР В СВЕТОКУЛЬТУРЕ РАСТЕНИЙ
Проблема повышения эффективности использования искусственных источников света при выращивании растений в защищенном грунте весьма актуальна.
Еще 20—25 лет назад эта проблема не стояла так остро, поскольку тогда главным потребителем источников света для выращивания растений являлась сравнительно небольшая сеть тепличных хозяйств, которые довольствовались 2—3 типами маломощных и по современным оценкам малоэффективных источников света.
В настоящее время интенсивная светокультура растений приобретает все более широкое распространение в овощеводстве се
130
верных районов страны, а в многочисленных селекционных центрах и исследовательских учреждениях построены фитотроны и тепличные комплексы для ускоренного выведения новых сортов растений и интенсификации исследований по физиологии растений, генетике и др. Расширение масштабов использования светокультуры растений и необходимость внедрения энергосберегающих технологий придают проблеме эффективного использования искусственных источников света в этой области серьезное народнохозяйственное значение.
Одним из перспективных путей решения этой проблемы является оптимизация спектрального состава излучения и уровней облученности растений.
§ 1. Методы оценки спектральной эффективности действия ФЛР в условиях светокультуры
К настоящему времени высказано две точки зрения по оценке спектральной эффективности ФАР для выращивания растений в условиях искусственного освещения.
Первая из них базируется на том, что белый солнечный свет является лучшим для растений, поскольку филогенетическое развитие растений происходило при нем, а следовательно, растения к нему наиболее приспособлены.
Простое постулирование наибольшей эффективности солнечного спектра для выращивания растений в искусственных условиях породило и основанную на этом оценку физиологической эффективности источников света по следующим формулам [Рождественский, Клешнин, 1980]:
С«Ф —
Л'сф-ЮО % г/ *ИКБ
---Т----- ЛЦНБ — 7-----
ксф кИКБ
*4з-100% ^жз
7 > _ *икд ^икд — 7----
'ИКД
(6-1)
/Сок’ЮО %
где /с' — коэффициент, характеризующий физиологическую эффективность источника излучения по сравнению с коэффициентом к, соответствующим прямой солнечной радиации (эталону), падающей на землю для различных областей ФАР: сф — сине-фиолетовой, ок — оранжево-красной, жз — желто-зеленой; /сИкб, &икб, /сикд> ^икд — коэффициенты, характеризующие соответственно доли потоков ИКР в участках 720—1200 нм и выше 1200 нм от интегрального потока излучения для сравниваемого источника и солнечного света; ZHKB, £Икд — отношения интенсивностей соответственно в участках спектра ИКР 720—1200 нм и выше 1200 нм.
131
Формулы (6.1) даны для случая, когда интенсивности ФАР источника и эталона равны.
Однако авторы такой концепции не учли, что многие растения адаптированы не просто к солнечному свету, а к тому спектральному составу света, который доходит до них в условиях их экологической ниши.
Действительно, в естественной обстановке, когда имеется единственный источник света — солнце, для огромного множества растений, произрастающих при различных температурных режимах, влагообеспеченности, плодородии почвы, экологических условиях, освещение существенно различается как по интенсивности, так и по спектральному составу света. Хорошо известно, например, что в ФАР, проникшей в глубь прозрачной океанской воды, доминируют синие лучи [Иванов, 1975], а ФАР в горных районах обогащена сине-фиолетовыми лучами [Шульгин, 1973]. Зеленая область спектра существенна в освещении растений, обитающих, например, под пологом леса.
Таким образом, близкий к равноэнергетическому спектр солнечного излучения скорее всего можно признать наиболее универсальным для энергетического обеспечения различных видов растений в разных экологических нишах. Однако принцип универсализации не соответствует принципу максимальной эффективности и поэтому вполне понятно, что белый свет не может быть признан максимально эффективным по своему спектральному составу для обеспечения наивысшей продукционной деятельности любого растения.
Другой принцип оценки спектральной эффективности искусственной ФАР для продукционной деятельности растений основывается на поиске и установлении эффективных потоков излучения для различных видов растений.
Принцип оценки спектральной эффективности излучения зародился и получил свое развитие в физике, в частности, для оценки спектральных характеристик приемников оптического излучения. В биологии этот подход был успешно использован при изучении глаза как приемника оптического излучения и разработке в связи с этим методов оценки спектральной эффективности излучения для человеческого гйаза.
Прежде чем перейти к обсуждению возможностей использования принципов оценки спектральной эффективности ФАР для растений, напомним основные физические характеристики, которые для этого потребуются.
Так как у нас приемником излучения является биологический объект растительного происхождения, то мы должны ввести понятие спектральной чувствительности такого биологического приемника g^:
, <1ф'..
<6-2>
где (1Ф}— эффективно преобразованный биологическим прием-
132
ником монохроматический лучистый поток с длиной волны Л. (т. е. тот поток излучения, энергия которого затрачена растительным организмом на многогранные процессы жизнедеятельности: фотосинтез, рост, развитие, формирование урожая и т. д.); —
монохроматический лучистый поток, падающий на биологический приемник от источника света; <рх — спектральная плотность падающего монохроматического лучистого потока в интервале длин волн dk.
Тогда, если известно спектральное распределение энергии в потоке излучения источника света <р(Х), можно определить эффективный поток излучения как
Ф' = I' (p(X)g'(X)JX, (6.3)
где Xj и Z2 — границы участка спектра, в пределах которого заключен поток излучения, падающего на биологический приемник излучения.
Для того чтобы определение эффективного потока по формуле (6.3) было правомерным при оценке эффективности излучения по его действию на биологический приемник, необходимо выполнение условий, суть которых сводится к следующему [Юров, 1958]:
1. Выбранный критерий равенства разноспектральных излучений должен однозначно соответствовать эффекту, на основе которого строится система эффективных величин.
2. Метод сравнения разноспектральных излучений должен быть стандартизирован.
3. Должно учитываться изменение спектральных характеристик биологического приемника под влиянием адаптации к длительному воздействию излучений разного спектрального состава и при разных условиях реакции.
4. Аддитивность действия разноспектральных излучений должна быть изучена при длительном действии излучения.
5. Необходима экспериментальная проверка соответствия расчетной и экспериментальной оценки эффективности излучений.
Рассмотрим, что означают эти условия относительно использования принципа оценки эффективности излучения для светокультуры растений.
Во-первых, должен быть выбран критерий равенства действия разноспектральных потоков ФАР. Для светокультуры таким критерием может быть либо общая биомасса растений (при выращивании рассады), либо хозяйственно полезный урожай. В качестве измеряемого эффекта может быть взято изменение подходящего параметра растительного объекта, который должен коррелировать с выбранным критерием. В качестве такого параметра предпринимаются попытки использовать газообмен, некоторые оптические характеристики растительного объекта.
133
Второе условие предполагает сравнение разноспектральпых излучений по едипому эффекту, третье — учет изменений спектральных характеристик биологического приемника при длительном воздействии лучистого потока того или иного спектрального состава. Это чрезвычайно важно для фотосинтетического аппарата растений, который, как известно, имеет разную степень адаптации к длительному воздействию излучения того или иного спектрального состава [Воскресенская, 1975; Тихомиров и др., 1987].
Четвертое условие предполагает, что общий эффект от спектрального состава равен сумме эффектов, получаемых от воздействия отдельных спектральных потоков, независимо от их мощности.
Пятое условие требует экспериментальной проверки выработанного принципа оценки спектральной эффективности действия излучения.
Попытка использовать понятия эффективного потока для оценки спектральной эффективности ФАР при выращивании растений была предпринята более 30 лет назад и изложена в работе И. И. Свентицкого [1960]. Суть ее сводилась к тому, что для искусственного излучения, используемого при выращивании растений как на общую биомассу, так и на хозяйственно полезный урожай, существует универсальный спектр ФАР, наиболее благоприятный для фотосинтеза абсолютного большинства зеленых растений.
Такой подход к оценке спектральной эффективности ФАР при выращивании растений в условиях искусственного излучения вызвал бурную дискуссию (см., например, журнал «Светотехника» за 1971—1973 и за 1978—1981 гг.). Поскольку такой подход, имея энергичных сторонников, особенно среди агрономов, светотехников, специалистов лампового производства, получил определенное распространение, целесообразно рассмотреть его более подробно.
Идея использования универсального эффективного потока, определяемого формулой (6.3), в 70-е годы получила дополнительный импульс. Это связано с появлением работ Маккри [МсСгее, 1972а] и Инада [Inada, 1976, 1977], в которых на листьях растений различных систематических групп были получены примерно одинаковые спектры действия фотосинтеза зеленых листьев.
Универсальность таких спектров действия фотосинтеза для листьев различных видов растений, высокий уровень экспериментальной техники, использованный при выполнении этих работ, сразу же привлекли к себе внимание, и сторонники введения эффективного потока для выращивания растений заменили функцию спектральной чувствительности g'(K), основанную на спектре поглощения среднего листа [Свентицкий, 1960], функцией, основанной на спектре действия фотосинтеза g(X) [Свентицкий, 1977], полученной по данным работ Маккри [МсСгее, 1972а] и Инада [Inada, 1976, 1977]. С учетом этого формула (6.3) может быть за-
1о4
писана как
Х=700 НМ
Ф- f <pWg(X)A. (6.4) Х=400 НМ
В таком виде понятие эффективного потока (Ф), который получил название «фотосинтезный», отстаивается его сторонниками и в настоящее время.
Наиболее привлекательным в этом подходе по оценке спектральной эффективности действия ФАР является его универсальность, из которой следует наличие единого эффективного потока излучения, т. е. излучения ФАР, спектральное распределение энергии которой наиболее оптимально для всех видов растений и условий их выращивания. Разумеется, несомненны достоинства такого подхода при его практической реализации. Однако постановка вопроса о практической реализации такого подхода правомерна лишь тогда, когда проведен научный анализ обоснованности выделения универсального эффективного потока излучения при выращивании растений в условиях искусственного освещения.
Нам представляется, что в качестве научного подхода здесь должно выступать физиологическое обоснование выполнимости тех принципов, которые заложены в условия определения эффективного потока [Юров, 1958].
Действительно, реальным критерием равенства действия разноспектральных потоков ФАР для выращивания растений при искусственном освещении может быть признан лишь критерий по накоплению растительной биомассы — либо общей, либо хозяйственно полезной.
При введении эффективного «фотосинтезного» потока отмечалось, что оценка разноспектральных потоков основана на измерении фотосинтеза, причем не реального, имеющего место при длительном выращивании растений при излучении того или иного спектрального состава, а «потенциального» [Свентицкий, 1977], который измеряется при кратковременной (3—5 мин) экспозиции на монохроматическом потоке того или иного участка ФАР с относительно низкой (15—30 Вт/м2) облученностью.
Разумеется, что для условий выращивания растений при интенсивном искусственном освещении такая оценка эффективности разноспектральных потоков ФАР не дает объективного представления о продуктивности. В таком случае необходима тесная корреляция между интенсивностью отмеченного выше «потенциального» фотосинтеза и продуктивностью растений. Необходимость такого сопоставления диктуется жестким требованием — конечная цель выращивания растений в светокультуре — это формирование урожая растений.
Поэтому, хотя в основу понятия эффективного фотосинтезного потока положена интенсивность «потенциального» фотосинтеза, реально в светокультуре сравнение разноспектральных
135
потоков ФАР ведется на основе данных по накоплению биомассы растений. Как сторонникам, так и оппонентам эффективного фо-тосинтезного потока такое сравнение приходится вести при длительном воздействии излучения того или иного спектрального состава на образование биомассы растений с контрольным вариантом, где спектральное распределение энергии излучения подобно кривой спектра действия фотосинтеза зеленого листа, полученной в работах Маккри [МсСгее, 1972а] и Инада [Inada, 1976, 1977] при кратковременных воздействиях монохроматического излучения на лист.
Таким образом, для первого условия, выполнение которого необходимо при введении понятия эффективного потока для биологического приемника, в данном случае можно отметить подмену критерия равенства разноспектральных излучений, основанного на кратковременном фотосинтезе листа, критерием равенства по накоплению биомассы.
Допустима ли такая замена критериев? Известно, что фотосинтез и накопление биомассы при длительном воздействии излучения смешанного спектрального состава коррелируют далеко не всегда [Протасова, Кефели, 1989]. Что же касается «потенциального» фотосинтеза, то обсуждение его корреляции с накоплением биомассы, которое происходит лишь при достаточно длительном воздействии света на растения, дает отрицательные выводы, поскольку в этом случае приходится сопоставлять кратковременный минутный эффект (фотосинтез) и длительный многосуточный эффект, оцениваемый по конечному результату (накопление биомассы).
Даже в рамках одного процесса — фотосинтеза — кратковременное и длительное воздействие спектрального состава света на растительный организм может оказаться несовпадающим (см. гл. 2). Так, например, действие красного света на лист огурца при кратковременном воздействии дает максимальный фотосинтез по сравнению с синими и зелеными- лучами, а при длительном приводит зеленый лист к гибели [Тихомиров и др., 1987]. Последнее обстоятельство экспериментально доказывает, что четкой и устойчивой корреляции между «потенциальным» фотосинтезом и биомассой растений не может быть.
Отсюда следует вывод, что недопустимо подменять критерий равенства разноспектральных потоков по интенсивности потенциального фотосинтеза критерием такого равенства на основе накопления биомассы, что фактически вынуждены делать сторонники введения эффективного фотосинтезного потока. Иными словами, первое условие введения понятия эффективного потока для искусственного облучения растений в рамках обсуждаемой концепции нарушено.
Что касается второго условия, то оно также строго не выполняется. Действительно, стандартизация измерений в данном случае основана на использо'вании классических спектров действия фотосинтеза, которые по ряду причин (см. гл. 2) неприменимы
133
к реальным условиям, в которых формируется урожай сельско-хо зяйственных культур.
Невыполненным следует считать и третье условие введения эффективного фотосинтезного потока. Оно требует изучения изменения спектральных характеристик листа при длительном воздействии спектрального состава света на лист. Мы уже достаточно подробно рассматривали влияние длительного воздействия спектра излучения ФАР на адаптационные характеристики листа (см. гл. 2, 3). В частности, показано, что при длительных условиях пребывания на свету, лишенном синего компонента ФАР, листья ряда растений как биологические приемники излучения могут вообще прекратить свое существование. Таким образом, адаптация листа как приемника оптического излучения к разноспектральным потокам ФАР не учитывалась при введении эффективного фотосинтезного потока.
Одним из необходимых условий корректности формулы (6.4), определяющей эффективный фотосинтезный поток, является соблюдение для функции спектральной чувствительности листа закона спектральной аддитивности. Функция спектральной чувствительности подчиняется закону спектральной аддитивности, когда соблюдается следующее условие:
Кп
£=j’g(X)dX, (6.5)
Ч
где 3 — функция спектральной чувствительности листа для всего спектрального диапазона, ограниченного длинами волн и
В работе Маккри [МсСгее, 1972b] для низких облученностей порядка 10—15 Вт/м2 в отдельных участках спектра ФАР экспериментально показано, что кратковременные фотосинтетические ответы могут быть представлены как независимые и аддитивные. Аналогичный вывод при кратковременном облучении листа разноспектральными потоками ФАР был сделан в работе В. С. Хазанова, Г. К. Кузнецовой [1973а]. Анализ двух последних работ показывает, что функция & определена для отдельных листьев при облученностях 10—30 Вт/м2 и АХ 20 нм. Выполнимость принципа спектральной аддитивности фотосинтеза (формула (6.5)) при Е > 30 Вт/м2 не исследовалась. Кроме того, если использовать формулу (6.5) для оценки эффективного потока ФАР в реальных условиях выращивания растений, как это и имеет место в условиях интенсивной светокультуры, где целесообразно использовать уровни облученности 100—150 Вт/м2 ФАР [Лисовский и др., 1987], то спектральные эффекты могут проявляться на не--линейном участке световой кривой исследуемого процесса (фото-.* синтез, общая биомасса, урожай и т. д.). Поэтому необходимым условием применимости формулы (6.4) при определении эффективного потока Ф является выполнимость принципа спектральной аддитивности для функции 3 в диапазоне облученностей 100 <
Ю А. А. Тихомиров, Г, М. Лисовский, Ф, Я. Сидько
137
< Е 150 Вт/м2 ФАР, которые реально используются в условиях интенсивной светокультуры [Там же]. Этот вопрос также до сих пор не изучался (см. гл. 6, § 2).
Что касается последнего — пятого — условия, то следует отметить, что строгая экспериментальная проверка оценки эффективности излучений, основанной на спектре действия фотосинтеза листа при разных уровнях облученности, весьма проблематична. Это связано с тем, что, исходя из принципа построения эффективного фотосинтезного потока, такую проверку нужно осуществлять по интенсивности фотосинтеза в условиях светокультуры при том или ином используемом спектре и интенсивности излучения. Однако это, как уже отмечалось выше, противоречит задачам выращивания растений в условиях светокультуры. Поэтому такая проверка как сторонниками, так и оппонентами эффективного фотосинтезного потока проводилась не по интенсивности фотосинтеза, а по анализу формирования урожая растений, как правило, его хозяйственно полезной части.
В рамках рассматриваемой задачи достаточно подробно зависимость урожая растений от спектра и интенсивности ФАР изложена в гл. 5. Здесь мы лишь отметим, что спектральная эффективность действия ФАР сильно зависит от интенсивности последней. Общей тенденцией является усиление эффективности синих лучей и ослабление эффективности красных с ростом уровня облученности ФАР. Этот факт указывает, что универсальность эффективного фотосинтезного потока для разных уровней облученности отсутствует, что также указывает на нарушение данного пункта условий при введении принципа оценки спектральной эффективности действия ФАР по обсуждаемой концепции.
§ 2. Выполнимость принципа спектральной аддитивности для биосинтетических процессов в фитоценозах
Если известны показатели исследуемого биологического процесса как при использовании отдельных монохроматических потоков, так и при их совместном воздействии на растительный организм, то выполнимость принципа спектральной аддитивности можно оценить следующим соотношением:
(г=п \ £; 2
i=1-~ > (6-6)
2 bi (^ М »=1
где bj(E\ — значения исследуемого процесса (газообмена, биосинтеза биомассы и т. д.) при облученности Е (в дальнейшем для краткости функции типа Ь(Е; л) будем обозначать Ь(Е-Л), где Е} — энергия излучения на данной длине волны X), Bj (Е; 2^-i) — значения этого же процесса при воздействии лучистого потока, по интенсивности и спектральному составу равного сумме падающих
138
монохроматических (или излучений в узких спектральных областях потоков, т. е. имеющих суммарную облученность
г=п
Е = 2 Еч. (6.7)
г=1
При этом в общем случае (и АХг) могут быть не равны друг другу, однако во многих случаях желательно, чтобы
Еыл = Е^2 = .. • = (6.8)
при АХг = const.
Облученности Е и Е^, как правило, выражаются в единицах квантового потока (эйнштейнах, Э) или в энергетических единицах (ВТ), приходящихся на единицу площади. Спектральная аддитивность выполняется, если, согласно (6.6), А = 1 и отсутствует, если А =£ 1.
При вычислении по формуле (6.6) необходимо иметь в виду, что при интенсивностях монохроматических потоков Е^, соответствующих линейному участку световой кривой исследуемого процесса, уровень облученности Е, определяемый формулой (6.7), может оказаться насыщающим или даже ингибирующим данный процесс. Поэтому выбор Е^ и Е для вычисления по формуле (6.6) должен определяться тем, на каком участке световой кривой данного процесса представляется целесообразным оценивать выполнимость принципа спектральной аддитивности.
При такой оценке для значений исследуемых процессов, соответствующих восходящему участку световой кривой, но лежащему за пределами ее линейной части, предпочтительно проводить вычисления при условии, когда данный процесс Ь; поддерживается на постоянном уровне. В этом случае оценка выполнимости принципа спектральной аддитивности может быть проведена по соотношению следующего вида:
где bj (E^i) — const при различных лг- за счет варьирования уровня спектральной облученности Е^:
Ь> (Л.,.,) - bi (Еч) - ... - (6.10)
— значение исследуемого процесса при совместном действии монохроматических потоков, суммарная интенсивность которых ослаблена в п раз:
(6.Н)
Разумеется, что по формуле (6.9) можно вести вычисления и для линейного участка световой кривой исследуемого процесса,
ю*
139
как это было сделано, например, в работе В. С. Хазанова и Г. К. Кузнецовой [19736 J.
Рассмотрим последовательно выполняемость принципа спектральной аддитивности на примере ценозов редиса для фотосинтеза и накопления биомассы в различные периоды вегетации. Для такого анализа нам представлялось в экспериментальном отношении более удобным провести оценку выполнимости принципа спектральной аддитивности по формуле (6.6). С учетом методических особенностей эксперимента формулу (6.6) перепишем в следующем виде:
(6.12)
г=1
где —значения исследуемых в данной ра-
боте процессов (динамика газообмена, накопления надземной и подземной биомассы, листового индекса); Е^ = 165 мкЭ/(м2-с); Е — 495 мкЭ/(м2-с); п = 3, причём AZt = 400—500 нм; Д/.2 — i=3
— 500-600 нм; ДХ3 = 600-700 нм; 2 ДМ = 400 — 700 нм.
i=l
Для вычисления выполнимости принципа спектральной аддитивности по фотосинтезу для ценозов редиса в различные периоды вегетации можно воспользоваться данными по возрастной динамике фотосинтеза ценоза редиса в различных участках видимого спектра и по выполнимости принципа спектральной аддитивности по этому параметру, вычисленными с помощью формулы (6.12) (рис. 6.1). Видно, что для фотосинтеза в первую половину вегетации, примерно до 12 сут, можно допустить выполнение закона спектральной аддитивности с удовлетворительной погрешностью.
Рассмотрим выполнимость принципа спектральной аддитивности для процессов накопления биомассы ценозами редиса
Рис. 6.1. Возрастная динамика видимого фотосинтеза ценозов редиса, сформировавшихся под излучением 165 мкЭ/(м2-с) в синей (/), зеленой (2) и красной (5) областях ФАР и под излучением 495 мкЭ/(м2 • с) в области 400—700 нм (4) и отношение фотосинтеза при белом (Фб) к суммарному фотосинтезу под синим (Фс), зеленым (Ф3) и красным (Фк) светом (5).
140
Рис. 6.2. Возрастная динамика накопления биомассы ценозами редиса, сформировавшимися под излучением 165 мкЭ/(м2 • с) в синей, зеленой и красной областях ФАР и под излучением 495 мкЭ/(м2 • с) в области 400— 700 нм и отношения значений биомассы, полученной под белым светом к соответствующим суммарным значениям биомассы, полученной под синим (Бс), зеленым (Ба) и красным (Бк) светом.
Остальные уел. обозн. см. рис. 6.1.
(рис. 6.2). Для корнеплодов отклонение от аддитивности с возрастом ценоза увеличивается и к концу вегетации в 1,55 раза превышает аддитивное значение (рис. 6.2, а).
Сутки
Лишь в короткий период вегетации, примерно соответствующий 10— 16-суточным ценозам редиса, можйо считать, что закон спектральной аддитивности выполняется с удовлетворительной погрешностью. Для процесса накопления надземной биомассы еще более четко про-
слеживается возрастание отклонения от спектральной аддитивности с возрастом ценозов. Здесь примерное соблюдение принципа спектральной аддитивности для молодых (8-суточных) ценозов редиса быстро исчезает с возрастом и к концу вегетации надземная биомасса составляет менее половины от соответствующей аддитивной (рис. 6.2, б). -
Есть периоды вегетации, когда аддитивность проявляется (с допустимой погрешностью). Эти периоды приходятся на первую Половину вегетации, они более-продолжительны для фотосинтеза, Нем для накопления биомассы надземной части и корне1 плодов. Анализ рис. 6.1 и 6.2 показывает, что ориентировочно ДЛЯ 9—11-суточных ценозов редиса принцип спектральной аддитивности соблюдается с допустимой погрешностью как для процессов фотосинтеза, так и для процессов накопления надаем1 ной биомассы и корнеплодов. Однако в целом по рассмотренным характеристикам он не работает.
Для проверки этого вывода можно использовать данные некоторых наших работ [Сидько и др., 1977; Тихомиров и др/, 1985J. В этих работах речь также идет о ценозах редиса Виров-ский белый, выращенных при указанных выше облученностях И других аналогичных условиях эксперимента, но с использованием источников света с линейчатым спектром. Рассмотрим, кай выполняется принцип спектральной аддитивности на примере
141
надземной биомассы и хозяйственно полезного урожая для ценозов редиса в этой серии экспериментов (табл. 6.1). Проведенные расчеты показали, что конечный урожай корнеплодов ценоза редиса в 1,6 раза превышал аддитивную величину, а надземная биомасса составляла 0,42 от соответствующего аддитивного значения, что полностью соответствует данным предыдущего эксперимента.
Теперь перейдем к рассмотрению выполнимости принципа спектральной аддитивности по методике, в основе которой лежит постоянство исследуемого фактора. В качестве такого фактора используем величину хозяйственно полезного урожая ценоза редиса. Для расчетов используем формулу (6.9), которую для данного случая запишем в виде
(6.13)
где bK — хозяйственно полезная биомасса ценоза редиса, сформировавшаяся при облученности Е^; ВК(Е) — то же, сформировавшаяся при облученности Е, определенной по формуле (6.11); i = 1, 2, 3; значения АЛ,-, те же, что в формуле
(6.12).
Значения параметров, необходимых для вычисления коэффициента А по формуле (6.13), представлены в табл. 6.2. Пользуясь данными табл. 6.2 и формулой (6.13), получаем А — 2,55. Таким образом, и такой способ вычисления указывает на отсутствие спектральной аддитивности.
Работы, проведенные на других культурах, также дают отрицательные результаты по соблюдению принципа спектральной аддитивности по хозяйственно полезной биомассе. Так, для огурца при излучении 50—100 Вт/м2 в рассмотренных отдельных областях ФАР урожай формируется только в синих лучах при 400 Вт/м2 и он крайне низкий. Это указывает на то, что ни по фотосинтезу, ни по урожаю принцип спектральной аддитивности не выполняется и для ценозов огурца, даже если сделать поправку на равноквантовые потоки. Аналогичный вывод справедлив для ценозов пшеницы. Как показали эксперименты, урожай зерна ценоза пшеницы, выращенного при облученности 45 Вт/м2 в синей области спектра, равен нулю. То же самое получено для зеленых и красных лучей, выровненных с синими по квантам. Это позволяет утверждать, что для белого света интенсивностью 400 Вт/м2 принцип спектральной аддитивности для ценозов пшеницы не соблюдается.
Попытаемся проанализировать возможные причины, ведущие к невыполнению принципа спектральной аддитивности рассмотренных физиологических процессов.
Известно, что увеличение уровня облученности у редиса тормозит рост надземной биомассы и стимулирует увеличение массы корнеплодов независимо от спектрального состава ФАТ’ (Тихомиров и др., 1983]. В ряде наших экспериментов общая об-
142
я
табл. 6.1
Характеристика ценозов редиса Вировский белый, сформировавшихся при облученностях в отдельных областях ФАР по всей области ФАР [Тихомиров, Сидько, 1987]
SW/I •( III «5 сч CD ей tr
'Ёс <ч Я R 'С
‘ Лп(Едц)- г/м’ 50 130 255 1 F О Й E c К s о и й и ф rt Н
м д Рч < н к "к Я) | 1 | 408 ft о 1 к Ч S
% 77 7г < 'ЁС 14 140 90 25 1 'ft A m n c E5 E Ф E P c
С? м Д ф к я ч 1 1 1 495 rt о о ей E о s ю ей и о
с4 Jri и 2 S 165 165 165 1 1 < н у Та cq
Область спектра, им 400-500 i 500-600 600-700 400-700 «Т • К А Q S а
Спектральный интервал тМ «4 << О ст << < << MS й е о S © й
X
F-
Ф
© Я д
©
/-Ч.
и Я < . . 1 °О о
я 111 *<г*
X
*.
ф t£ S.
Е- к
© <4
©
*
в:
ф
©
№
Я £
а.
в® о с о ,
to << со О О 1
С <
ы
й к
л Л к
А
я о
R
2 «Г
© 02
я. 2
Я S
N
05 И д *
U £ X е4 Я““ = 6н н И 1118
fcj
Св Л. tt
в 5
s h
S
а. ч
2 to >& о £
“ 5S р
ф m ООО |
’5 § Я Jr СО Ш 1
Ч о <
£в KI
ЗЯК|
5 s
М Я. S&
Л = ей ©
Яеь к ООО о
а!5 мм о ООО о
С ю со t>. г—
ceG > С нм III 1
ф ООО о
н я Б й ООО о Ю © vf*
& £ О
» о
©
«
о
я
ф
я-
ЭК
ка в ган-
е ф ё * £ а »-i еа М << <<<<.< <J
ф Еч & ь ф£ в s < м
и и
©
р.
©
и
Величина Е' получена по формуле (6.11), Усл, обозн, см. табл. 6.1.
143
лученность редиса в области 400—700 нм (белый свет), равная сумме облученностей, создаваемых лучистыми потоками, выровненными по квантам или энергии в отдельных областях ФАР, в среднем в 3 раза превышала уровень облученности для каждой отдельной области ФАР, в лучах которой выращивали ценозы редиса.
Согласно данным Н. Н. Протасовой и В. И. Кефели [1982], при увеличении облученности ФАР интенсификация ростовых процессов начинает отставать от соответствующего увеличения интенсивности фотосинтеза. Видимо, такая ситуация имеет место и в данном случае, когда с ростом облученности до 100 Вт/м2 ФАР темпы роста листовой поверхности в ценозах редиса были ниже, чем увеличение интенсивности фотосинтеза. Такое лимитирование роста листовой поверхности на фоне ее достаточно высокой фотосинтетической активности создает предпосылки для активного роста аттрагирующих центров (корнеплодов). При этом происходит смещение донорно-акцепторных отношений между надземной фотосинтезирующей биомассой и корнеплодами в пользу интенсивного роста последних [Мокроносов, 1981, 1983]. Нам представляется, что этот факт играет ведущую роль в невыполнении спектральной аддитивности для накопления биомассы надземной части и корнеплодов. Очевидно, что противоположная направленность кривых, характеризующих степень отклонения от принципа спектральной аддитивности (см. рис. 6.2), для надземной биомассы и корнеплодов редиса в процессе вегетации как раз и характеризует возрастающее ингибирование роста надземной биомассы и усиливающуюся стимуляцию роста корнеплодов в течение вегетации для белого света.
Ингибирование роста надземной биомассы при белом свете по сравнению с излучением составляющих его отдельных областей ФАР и связанное с этим отсутствие аддитивности по этому параметру подтверждаются анализом возрастных изменений листового индекса ценозов редиса в исследуемых световых вариантах (рис. 6.3). Отклонение от спектральной аддитивности по этому параметру в первый период вегетации стремительно возрастает и в последние две трети вегетации листовой индекс почти в 2 раза ниже соответствующего аддитивного значения.
Более сложным представляется выяснение причин, способствующих отклонению процессов фотосинтеза от выполнения принципа спектральной аддитивности. Нам представляется, что к этому ведет комплекс особенностей в развитии ценозов при излучении различного спектрального состава. Среди этих особенностей |щжно выделить различия, в архитектонике ценозов, приводящие существенным неравномерностям освещения не только листьев различных ярусов цепоза, но и в пределах одного листа [Tikhomirov, Sid’ко, 1983 ]т и, как следствие этого — к соответствующим различиям в фотосинтезе листьев ценоза .и их частей. Такие различия в. фотосинтезе не могут оставаться постоянными в течение всего периода вегетации, так как архитектоника й радиационный
Ш .
Рис. 6.3. Возрастная динамика листового индекса ценозов редиса, сформировавшихся при энергии лучистых потоков 165 мкЭ/(м2-с) в отдельных областях ФАР, энергии 495 мкЭ/(м2-с) белого света и отношение значений листового индекса под белым светом (Лб) к суммарному значению листового индекса под синим (Лс), зелёным (Л3) и красным (Лк).
Остальные уел. обозн. см. рис. 6.1.
режим ценоза постоянно меняются в течение вегетации. В разных световых вариантах эти различия неодинаковы, что связапо^с особенностями частной физиологии развития растений.
Интересно отметить совпадение отклонений от аддитивного значения для биомассы корнеплодов ценозов редиса при исполь* зовании исходных данных из эксперимептов с разными условиями. Например, такое совпадение получено в серии экспериментов, где использовали линейчатый спектр излучения ФАР с 25 %-м фоном ИКР и сплошной спектр ФАР с 5%-м содержанием ИКР. Следовательно, эти факторы не оказывают влияния па определяемые значения величин спектральной аддитивности. Этот вывод относится к уровням облученности, соответствующим линейному участку световой кривой продукционных процессов в ценозах редиса. Для восходящего участка световой кривой продукционных процессов, соответствующего более высоким уровням облу-ценности, чем линейный участок этой кривой, принцип спектральной аддитивности также не соблюдается, хотя численные значения здесь для сопоставимых характеристик (хозяйственно полезный урожай) получаются иные (см. табл. 6.2).
Таким образом, принцип спектральной аддитивности биосинтетических процессов к ценозам в целом не приложим. Это утверждение справедливо, как показано выше, также для ценозов пшеницы и огурца, его, по-видимому, можно распространить и на ценозы других культур. Например, на основании полученных данных можно ожидать, что для салатных культур спектральная аддитивность процессов фотосинтетической продуктивности также пё будет соблюдатьсй. Действительно, в этом случае речь может идти лишь о надземной биомассе, а для нее на примере редиса (см. рис. 6.2, б, 6.3) получены наиболее существенные отклонения от спектральной аддитивности.
Таким образом, функция спектральной чувствительности биосинтеза является неаддитивной величиной. Поэтому более пр а-
вильной будет ее запись в виде
(i—n \ 2 >-< . 1=1 /
То есть функция спектральной чувствительности биосинтеза зависит не просто от длины волны Хг-, обособленной внутри спектрального диапазона ФАР, а от Xz, которая рассматривается как составная часть всего исследуемого спектрального диапа-1—71
зона 2 ^г-1=1
Если мы рассматриваем зависимость функции спектральной чувствительности биосинтеза от отдельной величины Хг вне ее расположения внутри спектрального диапазона ФАР, то вместо (n X
У, Xi I функция спектральной чувствительности приобре-i=i /
тает вид В (Е^,кг). Ввиду отсутствия спектральной аддитивности справедлива запись:
: п \ п
в\Е^ у Xi =#2 Ь(Е^,Ы, \ 1=1 / 1=1
причем
B(^;Xi) = b(EXi;Xi).
Важно также отметить, что хотя функция В (Е^, 2^i) неаддитивна, а следовательно, ее нельзя выразить через интеграл, это еще не говорит о том, что понятие эффективного потока нельзя использовать для оценки спектральной эффективности излучения ФАР в светокультуре.
Сам факт существования функции В (ZiXi; 2^1)> хотя это и неаддитивная величина, указывает, что оптимальное распределение ФАР для выращивания растений может быть описано покой кривой, формула которой зависит от функции В (Е^', У Xi). В связи с неаддитивностью рассматриваемой функции спектральной чувствительности возникает необходимость более подробно рассмотреть вопрос оценки спектральной эффективности потоки ФАР, поскольку ранее известные подходы к этому вопросу оспе вывались на аддитивности функции спектральной чувствительности фотосинтеза.
§ 3. Эффективные потоки и возможности их использования для оптимизации излучения в светокультуре
Рассмотренный выше материал показал, что использование традиционного понятия эффективного потока для строгой оценки спектральной эффективности ФАР при длительном выращивали и растений в условиях светокультуры с научной точки зрения но-
446
корректно. Если в качестве критерия оценки в этом случае взять тестовую величину потенциального фотосинтеза, как предлагается в работе И. II. Свентицкого [1977], то фактически нарушаются все фотометрические требования в определении эффективного потока ФАР, указанные в § 1.
Несомненно, что идеальным при решении вопроса оптимизации оптического излучения в растениеводстве было бы нахождение единого эффективного потока излучения, т. е. излучения со спектральным распределением энергии, оптимальным для всех видов растений и условий их выращивания. Однако результаты исследований по фотосинтезу и продуктивности растений, проведенные па ценозах при использовании различного спектрального состава и интенсивности ФАР, показывают, что у растений разных видов различаются требования к оптимальному сочетанию спектральных и энергетических характеристик светового режима их выращивания. -Это вносит ряд существенных усложняющих особенностей при введении и использовании основных терминов, необходимых для корректного определения самого понятия эффективного потока ФАР применительно к реальным условиям выращивания растений при искусственном освещении. Среди этих терминов исключительно важное значение имеет функция спектральной чувствительности S? биологического приемника излучения. Здесь следует отметить, что в качестве такого приемника в данном случае наиболее корректно рассматривать ценоз растений, а не отдельно взятый зеленый лист. Это связано с тем, что всевозможные характеристики, снятые па листовом и ценотическом уровнях, могут существенно разниться.
Ценотичёское взаимодействие растений в посеве оказывает большое влияние на формирование урожая [Мурей, 1978; Тоо-минг, 1984]. Сам урожай растений есть результат сложного взаимодействия различных процессов в растительных организмах, начиная с фотофизических и кончая биохимическими. Поэтому в обобщенном виде вместо функции спектральной эффективности фотосинтеза листа введем функцию спектральной чувствительности биосинтеза ценоза растений, которая зависит не только от Спектрального состава излучения, но и от другого светового фактора — уровня облученности. С учетом этого по аналогии с функцией g(%) введем функцию «относительная спектральная чувствительность для важнейших процессов биосинтеза» bj определив ее соотношением
^Лпах (^тах
(6.14)
где. tij (Ех) и ц;тах (^Wx) — эффективности влияния излучения С длинами волн % и Хишх на образование данного продукта; 7,1пах — длина волны излучения, при которой достигается максимальная эффективность-биосинтеза; / — исследуемый процесс биосинтеза! образование биомассы, фотосинтез, хозяйственно полезный урожай, и др.
Следует отметить, что формулу (6.14) нельзя признать «работающей» при выращивании растений в различных условиях искусственного освещения. Условия длительности и непрерывности воздействия спектральных потоков ФАР на растения, имеющие место при реальных условиях выращивания, характеризуются помимо других особенностей использованием не отдельных длин волн X, а некоторого спектрального диапазона АХ, который будет представлять достаточно широкую область спектра. Это связано с тем, что излучение с узким спектральным диапазоном может не дать возможность урожаю сформироваться. Разумеется, что границы АХ не должны заходить за пределы спектральной области, излучению которой приписывается принципиально иное значение по воздействию на растение (фотосинтетическое, регуляторное и т. д.). С учетом этого формулу (6.14) запишем в виде
bi (АЛ; Е) = Ь, (Еы) _ г.
6’гпах \ д^-тах/
Кроме того, в реальных условиях выращивания растений длительная работа с монохроматическими потоками излучения в широком интервале облученностей в настоящее время технически неосуществима. Но и в перспективе физиологические особенности растений ограничат возможности полноценного протекания физиологических процессов, ответственных за формирование конечного биологического продукта (зерно, корнеплоды и т. д.) при использовании очень узких спектральных интервалов. Поэтому уменьшение ширины спектрального интервала ограничено определенными пределами. С другой стороны, нелимитированное увеличение ширины такого спектрального интервала, как отмечалось выше, может привести к выходу за рамки физиологической значимости области спектра (синей, зеленой, красной). Поэтому речь может идти об оптимальных размерах АХ.
Необходимо также иметь в виду, что современные источники света, используемые в светокультуре, в своем большинстве имеют не сплошной, а линейчатый спектр. Если в первом случае поток излучения (Fc) источника света можно представить в виде
Fc = f (p(X)dX, (6.15)
Ч
где Хх и Хп — границы области ФАР; <р(Х) — спектральная плотность излучения, то для источника света с линейчатым спектром роток излучения (Ал) записывается, в виде -
^л= 2ф'(Х)ДХй (6.16)
г=1
Где ср'(^) _ спектральная плотность излучения отдельной i-й резонансной линии; т — количество резонансных линий. При этом должно выполняться условие п т в границах одного и того жо
№
спектрального диапазона. С учетом линейчатости спектра излучения плотности потоков излучения <р'(А,г) Для отдельных длин воли могут быть весьма велики и даже соответствовать не только насыщению, но даже ингибированию отдельных фотобиологических процессов, фоторецепторы которых имеют максимум поглощения на данной длине волны Хг. Такая ситуация может иметь место даже в том случае, если общий лучистый поток ФАР данного источника света еще не достигает насыщающих значений по фотосинтезу или продуктивности. Действительно, если, пользуясь формулами (6.15) и (6.16), поток излучения Fn сравнить с потоком Fo от сплошного источника света, то можно записать три следующих случая при Fn = Fc:
ф'(М » Л); <PW > ф'(М « ф(^)-
Эффект светового насыщения или перенасыщения на отдельных длинах волн может наблюдаться при использовании линейчатого спектра в первом и втором случаях. Напрашивается вывод, что в реальных условиях выращивания растений мы можем иметь дело с эффектами насыщения для отдельных фотобиологических процессов при работе с далекими от насыщения в формировании урожая облученностями. Учитывая, что в условиях реальной светокультуры используются достаточно интенсивные лучистые потоки, даже при использовании источников света со сплошным спектром ряд фоторецепторов, обеспечивающих протекание важных для формирования урожая биологических процессов, может находиться либо в условиях нелинейной зависимости от света, либо в условиях светового насыщения. Отсюда традиционное представление о диапазоне уровней облученности, при которых реакция биологического приемника излучения зависит от интенсивности падающего излучения, в данном случае будет нарушено. Такое представление основано на зависимости характеристик биологического приемника от спектра падающего излучения без учета структуры спектра. Ранее было показано также, что спектральные потоки ФАР разной интенсивности при длительном воздействии на биологический приемник (ценоз растений) могут сильно изменить его спектральную чувствительность (зависимость формирования урожая от спектра ФАР).
Формирование урожая в разноспектральных потоках ФАР есть результат работы комплекса фотореакций и фотопроцессов, имеющих фоторецепторы с максимумами поглощения в различных областях ФАР. Такая множественность процессов и, как отмечалось выше, возможная нелинейность световых кривых для некоторых из них указывают, что вряд ли для такой ситуации существует универсальный спектр действия, как это постулируется для эффективного фотосинтезного потока [Свентицкий, 1977, 1982; и др.].
Согласно нашим представлениям, физиологически обоснованной будет такая запись эффективного потока ФАР, когда учтены свойства фитоценоза как адаптивного приемника оптического
149
г—п \
ф(Х)ЙХ, (6.17)
1=1 /
излучения, длительно функционирующего в условиях светокультуры. Учет этих свойств фитоценоза может быть выполнен при использовании соответствующей функции спектральной чувствительности — В!Еъ, 2 а сама запись формулы для эффек-тивного потока примет вид
(i—n \
Е^\ 2 ^1) — функция спектраль-i=i /
ной чувствительности биологического процесса; ф(к) — спектральная плотность излучения.
Если известно, что функция спектральной чувствительности биосинтеза может быть представлена определенной кривой, то в (г=п \
Е^; 2 1 можно записать ее аналити-
г=1 /
ческий вид. Здесь важно иметь банк данных таких эффективных кривых, которые затем могут быть объединены в две-три группы сходных кривых. Можно утверждать, что одной общей группы, а следовательно, и общей универсальной кривой эффективного потока, аппроксимирующего достаточно точно отдельные кривые, не будет. Об этом свидетельствуют весьма контрастные требования растений к спектру ФАР для обеспечения интенсивной продукционной деятельности (см. гл. 5), особенно с ростом уровня облученности и выходом на использование мощных лучистых потоков, соответствующих интенсивной светокультуре. В связи с этим нам представляется, что в основу распределения энергии по спектру ФАР необходимо положить функцию спектральной чувствительности по хозяйственно полезному урожаю 2^-*)-Вид функции должен быть обобщен для-группы растений разных видов, близких по требованию к спектру ФАР при интенсивной продукционной деятельности. В этом случае становится возможным на практике придерживаться нужных соотношений физиологически значимых областей ФАР в видимом излучении для выращивания данной культуры или группы культур, имеющих близкие значения 2? (£\; 2 ^)-
В качестве примера практической реализации данного подхода рассмотрим возможности оценки эффективного потока растениеводческих ламп. Для этого используем способ, в основе которого лежит спектрозональный метод, достаточно широко применяемый в светотехнике для оценки цветопередачи источников света [Лампы разрядные..., 1978]. Используя такой метод для оценки эффективного потока растениеводческих ламп, необходимо, разумеется, учитывать те особенности растений, которые они проявляют как адаптирующиеся к длительному воздействию спектра и интенсивности ФАР приемники излучения;
150
G учетом этого доля (Д) эффективного потока для каждой спектральной зоны излучения ФАР растениеводческих ламп может быть представлена как
fr
?—400НМ
где ср (Z) — относительная спектральная плотность излучения растениеводческой лампы; В (£\; 2 М) — относительная функция спектральной чувствительности того биологического процесса, который является целью выращивания растений (общий урожай, хозяйственно полезный урожай, фотосинтез и т. д.); —
длины волн, соответствующие границам i-й спектральной зоны.
Поскольку в числителе (6.18) стоит значение эффективного потока Ф/ в спектральном диапазоне, ограниченном длинами волн Xi и Zj, а в знаменателе — значение эффективного потока Ф для всего спектрального диапазона ФАР, то Фг и Ф связаны соотношением
г=?г Ф = 2 Фг, г=1
a fi можно определить как
Таким образом, предлагаемый метод оценки спектральных характеристик растениеводческих ламп включает в себя оценку эффективности потока этих ламп в соответствии с требованиями, предъявляемыми к спектру излучения того или иного типа растениеводческой лампы. Несомненно, что такой метод оценки должен базироваться на физиологически обоснованных данных. В качестве примеров рассмотрим, как можно использовать такой метод для оценки спектральных характеристик источников света, предназначенных для выращивания огурца и томата.
В первую очередь необходимо определить относительное распределение эффективного потока по спектральным зонам ФАР. В светотехнике в аналогичной ситуации используется 8-зонная система [Лампы разрядные..., 1978]. Что касается растениеводческих ламп, то здесь нужно основываться на наличие, как минимум, трех физиологически значимых участков спектра: синего (400—500 нм), зеленого (500—600 нм) и красного (600—700 нм)< Разумеется,'что при этом физиологически значимые области спектра могут быть разбиты и на более узкие спектральные зоны. Однако такое сужение спектральных диапазонов приводит к ря-
151
ду трудностей биологического и технического характера, связанных с оценкой в реальных условиях эффективности излучения этих участков спектра в формировании урожая растений. Поэтому, пока эти трудности не преодолены, представляется целесообразным ограничиться разбиением спектрального диапазона ФАР на три вышеуказанных участка. Тогда долю /г (в процентах) эффективного потока для каждого такого участка спектра можно определить, пользуясь формулой (6.18), где интегрирование заменяем суммированием: п
2 ЧЦХ) Я ( £дХ. 2 ДХН ДХг
/• = 7=55=-------j-----ГТ!---га- «О * • (0-19)
2 Ф(Х)В ЕД) ; 2 Л>-( м Х=400нм \ г=1 /
где обозначение <р(Х) то же, что и в формуле (6.18); В Е^, 1=3 \
2 АХ; — относительная функция спектральной чувствительности i=i I
по хозяйственно полезному урожаю; i = 1, 2, 3; АХ; = 100 нм; <=з
2 АХ, = 400 — 700 нм; X, и X; — длины волн, соответствующие ,i=l t
границам j-й спектральной зоны, где при t = 1, Х^ = 400 нм, Xj = 500hm, при i — 2, Х2 = 500 нм, Х2 = 600нм, при i = 3, Х8 = = 6С0 нм, Х3 = 700 нм.
Таким образом, можно конкретизировать формулу (6.18) для записи эффективных потоков в каждой спектральной зоне. Эталонные значения /г, как это вытекает из рапее полученного материала (см. гл. 5), составляют для огурца: Д = 20 %,f2 = 40 %, /з = 40 %, а для томатов = 20 %, /2 = 15 %, /8 = 65 %.
Особое значение при выращивании огурца и томата на урожай следует придавать значениям /8. Колебания /8 как в сторо-’ ну уменьшения, так и в сторону увеличения более существенно, чем колебания Д и /2, влияют на продуктивность растений. В частности, /8 40 % для огурца может не только резко снизить урожай, но и привести к гибели растений [Тихомиров и др., 1987]. Если провести аналогию между величиной /3 и методом определения «красного соотношения» источников света, согласно ГОСТ 23198-78 [Лампы..., 1978], то величина /8 как раз й выражает «красное соотношение». Нам представляется, что если бы спектральных зон, в которых определяются эффективные потоки, согласно формуле (6.18), было бы значительно больше, чем 3, то метод оценки красного излучения приобрел бы для контроля спектрального состава растениеводческих ламп самостоятельное значение. Не исключено, что дальнейшие исследования в этом направлений позволят реализовать вышесказанное предположение.
§ 4. Перспективы создания растениеводческих ламп с оптимальным спектром излучения
Рассмотренный в настоящей главе экспериментальный материал показал, что различные виды растений и одни и те же виды на разных возрастных этапах развития могут иметь весьма контрастные требования к спектру ФАР для максимизации продукционной деятельности. Эти требования могут существенно зависеть и от уровня облученности в области как ФАР, так и ИКР. Разумеется, что ИКР оказывает определенное влияние на ход продукционного процесса, влияя на него через морфогенез и ряд регуляторных механизмов (фитохром, гормональная регуляция и др.), что требует специального изучения и является дальнейшим резервом в повышении эффективности излучения искусственных источников света в светокультуре растений. Если считать, что доля ИКР в лучистых потоках современных металлогалогенных ламп составляет 40—50 % и, как правило, не зависит от варьирования спектром в области ФАР, то такое содержание ИКР в излучении растениеводческих ламп можно считать удовлетворительным (см. гл. 5).
Что касается ультрафиолетовой радиации (УФР), то ее содержание в оптическом излучении, как известно [Дубров, 1963], может также существенно влиять на продуктивность растений. Однако поскольку колбы современных металлогалогенных ламп одинаково ограничивают пропускание УФР, то влияние последней на продуктивность растений в светокультуре мы здесь не рассматриваем. УФР, как и ИКР, является предметом специальных исследований и может также рассматриваться как резерв в дальнейшем повышении продуктивности растений.
В настоящее время ближайшей, а поэтому наиболее реальной перспективой повышения эффективности излучения растениеводческих ламп является оптимизация спектрального состава их ФАР. Это связано, с одной стороны, с достаточно высоким уровнем светотехнических разработок, позволяющих варьировать в широких пределах спектр излучения ФАР различных по мощности металлогалогенных ламп [Справочная книга..., 1983], а с другой стороны, с накопленным фактическим материалом по физиологическому воздействию излучения различных участков ФАР на разнообразные стороны жизнедеятельности высших растений [Кахнович, 1980; Протасова, 1987; Воскресенская, 1987; Федюнькин и др., 1988; и др.], включая продукционный процесс, исследованный на ценотическом уровне [Тихомиров и др., 1983]. Именно исследования па ценотическом уровне являются необходимым заключительным звеном, позволяющим перейти к формулировке физиологически обоснованных требований по спектральному составу ФАР растениеводческих ламп.
Ранее мы уже отмечали, что ввиду отсутствия единых требб-ваний растений к спектру ФАР в отношении их продукционной деятельности необходимо создание нескольких типов универсаль-
Н А. А. Тихомиров, Г. М. Лисовский, Ф. Я. Сидько 153
ных и специализированных ламп для растениеводства. Это прежде всего один-два типа ламп, являющихся универсальными для различных видов растений. Такая универсализация должна гарантировать достаточно высокий (но не всегда наибольший) выход по хозяйственно полезному продукту и являться резервным вариантом при отсутствии необходимых типов специализированных ламп. Обобщение фактического материала по роли интенсивности и спектра ФАР в формировании урожая растений позволяет считать, что для выращивания растений на выгонку рассады, когда уровни облученности ФАР не превышают 40—50 Вт/м2, в качестве универсального можно рекомендовать квантовый поток ФАР, где доминируют красные лучи (около 50 %), а синих и зеленых содержится поровну. Для условий интенсивной светокультуры, где целесообразно использование уровней облученности в диапазоне 100—200 Вт/м2 ФАР [Полонский, Лисовский, 1980], в качестве универсального представляется источник света с равноэнергетическим спектром ФАР. Единицы квантового потока достаточно широко используются в научной литературе, однако их использование часто дается безотносительно к мощностям лучистых потоков в отдельных областях спектра.
Нетрадиционными представляются рекомендации по созданию и использованию специализированных источников света. В этой связи следует отметить, что дифференциация требований растений к спектру ФАР усиливается с ростом уровня облученности. Поэтому наиболее актуальным в использовании специализированных источников света является режим интенсивной светокультуры. Учитывая; что в СССР самыми распространенными видами растений, выращиваемыми в условиях искусственного освещения, являются огурцы и томаты, рассмотрим в первую очередь на их примере создание специализированных растениеводческих ламп.
Что касается растений огурца, то наиболее оптимален для него спектр ФАР с высоким содержанием зеленых и красных лучей (по 40 %) и 20%-й добавкой синих лучей (см. гл. 5). Разумеется, возможны некоторые отклонения от этих соотношений в отдельных областях ФАР. Так, дополнительные эксперименты показали, что увеличение доли красных лучей в ФАР без снижения продуктивности растений еще возможно до 50 %, а дальнейшее наращивание ведет к прогрессирующему падению продуктивности растений. Понижение доли красных лучей в ФАР для ценозов огурца ниже 40 % также ведет к падению продуктивности.
Для интенсивной светокультуры томатов оптимальным является сцектр ФАР с ярко выраженным максимумом в красной области. Доля красных лучей в ФАР может колебаться в диапазоне 60—70 % с примерно равными дополнительными долями синих и зеленых лучей. Незначительное уменьшение урожая плодов томатов происходит при выравнивании доли красных и синих лучей (цо 40 %) без изменения доли зеленого излучения в ФАР, однако при этом удлиняются сроки вегетации, что ведет к падению продуктивности.
154
Таким образом, создание специализированных растениеводческих ламп для огурца и томатов связано с усилением излучения в зеленой (огурец) и красной областях спектра (огурец, томаты).
Для интенсивной светокультуры редиса при облученности 100 Вт/м2 ФАР его оптимальный спектр приближается к равноэнергетическому. Поэтому специализированный источник света для этой культуры создавать не нужно, поскольку спектр его излучения совпадает со спектром излучения ламп, рекомендуемых нами как универсальный для интенсивной светокультуры. С другой стороны, нам представляется, что вряд ли следует создавать специализированный источник света для каждой овощной культуры, выращиваемой в условиях искусственного облучения. Экономически это может быть целесообразно для огурца и томата, которые нашли массовое использование в светокультуре. Вероятно, что, имея по спектральным характеристикам три типа растениеводческих ламп (для светокультуры огурца, томатов и источник света с равноэнергетическим спектром ФАР), можно выбрать из них наиболее близкий по оптимуму спектрального распределения и для других овощных культур.
В настоящее время достаточно широкое распространение получило выращивание различных видов растений в селекционных центрах при искусственном свете. Здесь также необходимы исследования по выработке физиологически обоснованных рекомендаций для создания искусственных источников света с оптимальным спектром излучения для выращивания тех или иных культур. Так, наши исследования показали, что некоторым зерновым культурам (пшеница, кукуруза) требуется для высокой продуктивности значительно большее содержание красных и зеленых лучей в излучении ФАР, чем представителям масличных культур (подсолнечник).
Создание новых источников света для выращивания растений в условиях искусственного освещения является непростой светотехнической задачей, которая осложняется тем, что большинство источников света на фоне различного соотношения синих и зеленых лучей должно иметь повышенное содержание красных лучей в спектре ФАР. Решение такой задачи требует создания новых технологий изготовления материалов электродов и оболочек ламп, стабилизации плазменного шнура в горелке ламп, приготовления самих наполнителей, обеспечивающих излучение с нужным распределением энергии в спектре ФАР, а также решения ряда других достаточно сложных технических вопросов. К сожалению, без этого серьезный прогресс в создании высокоэффективных растениеводческих ламп трудно представить. В то же время важной особенностью, облегчающей светотехнические разработки таких растениеводческих ламп, является возможность маневра при выборе спектрального состава излучения ФАР, что позволяет вести поиск по этому параметру в пределах указанных спектральных зон, руководствуясь наиболее оптимальным сочетанием светотехнических и экономических показателей разрабатываемых источников света.
11* 155
В связи с этим использование спектральной кривой, имитирующей структуру классических спектров действия фотосинтеза зеленого листа, может рассматриваться как частный случай в наборе физиологически обоснованных спектральных вариантов. Но такой частный случай, выданный за общий и рекомендованный как основа для создания источников света с данным распределением энергии в спектре ФАР [Свентицкий, 1977], объективно лишает разработчиков-светотехников физиологически обоснованных возможностей варьировать спектр ФАР, исходя из тех или иных технических и экономических особенностей при создании растениеводческих ламп. Такое положение дел может привести ряд спектральных разработок к отбраковке, хотя они в действительности могут обеспечить высокоэффективный поток ФАР для продукционной деятельности тех или иных растений.
Разумеется, в нашу задачу не входит дать конкретные пред- ложения технического характера по созданию того или иного типа растениеводческих ламп. Высказанные здесь предложения по перспективам оптимизации спектральных и энергетических характеристик ламп для светокультуры растений имеют целью показать, каковы на сегодня биологические предпосылки для движения в этом направлении.
СПИСОК ЛИТЕРАТУРЫ
Алексеев В. А. Некоторые вопросы оптических свойств леса // Проблемы экологии и физиологии лесных растении.— Л.: Изд-во ЛТА, 1963.— Вып. 1.- С. 47—80.
Александрова Л. С., Клочкова М. П. Зависимость продуктивности томатов от интенсивности излучения газоразрядных ламп // Светотехника.— 1983.— № 2.— С. 10—11.
Александрова Л. С., Радомысльская Т. М. Динамика фотосинтеза томатных растений в условиях интенсивного освещения // Проблемы светокультуры растений.— Симферополь: Крымский СХИ, 1980.— С. 97—99.
Анисимова И. И., Глуховский Б. М. Фотоэлектронные умножители.— М.: Сов. радио, 1974.— 64 с.
Арбузова К. С., Жарков Д. Г., Малышев В. В. Об использовании галогенных ламп накаливания при выращивании растений // Светотехника.— 1983.— № 7.— С. 11—12.
Баазов Д. И., Санадзе Г. А. Спектр действия фотосинтеза у листьев тополя // Изв. АН СССР. Сер. биол.— 1984а.— Т. 10, № 2.— С. 137—139.
Баазов Д. И., Санадзе Г. А. Спектр действия фотосинтеза I фотосистемы // Материалы межфакультетской конф. Тбил. ун-та по естественным наукам: химия, биология, география, геология; май, 1984.— Тбилиси, 19846.— С. 153—165.
Баханова С. Г., Львова И. Н. Влияние качества света на рост и развитие растений огурца // Морфогенез растений,— М.: Изд-во МГУ, 1961.— Т. II.— С. 79—83.
Белл Л. И. Качество света как фактор, регулирующий энергетику фотосинтеза // Теоретические основы фотосинтетической продуктивности.— М.: Наука, 1972.— С. 50—59.
Брандт А. Б., Тагеева С. В. Оптические характеристики растительных организмов.— М.: Наука, 1967.— 301 с.
Ван Дер Вин Р., Мейер Г. Свет и рост растений.— М.: Сельхозиэдат, 1962.—‘ 183 с.
Васильев В. И., Вассерман А. Л., Квашин Г. Н. и др. Тепличный облучатель ОТ-2000 И Светотехника.— 1982.— № 10.— С. 13—14.
/Вассерман А. Л. Ксеноновые трубчатые лампы и их применение.— М.: Энергоатомиздат, 1989.— 88 с.
Власова М. П. Действие света разной интенсивности и качества на организа-цию фотосинтетического аппарата: Автореф. дис. ... канд. биол. наук/Ин-т физиологии растений АН СССР.— М., 1975.— 32 с.
Власова М. П., Воскресенская Н. П. Тонкая структура хлоропластов нормальных и мутантных растений гороха, выращенных на свету различи ного спектрального состава // Физиология растений.— 1973.— Т. 20, № 1.— С. 96—101.
Волкова Е. Б., Золотухин И. Г., Лисовский Г. М. и др. Фотобиологическая эффективность некоторых источников света в условиях светокультуры // Светотехника.— 1982.— № 9.— С. 1—3.
Воскресенская Н. П. Фотосинтез и спектральный состав света.— М.: Наука, 1965.— 311 с.
Воскресенская Н. П. Принципы фоторегулпрования метаболизма растений
W
и регуляторное действие красного и синего света на фотосинтез // Фоторегуляция метаболизма и морфогенеза растений.— М.: Наука, 1975.— С. 16—36.
Воскресенская Н. П. Фоторегуляторные реакции и активность фотосинтетического аппарата // Физиология растений.— 1987.— Т. 34, вып. 4.— С. 669—684.
Воскресенская Н. П., Нечаева В. П. Действие синего, красного и зеленого света на содержание белка, нуклеиновых кислот и хлорофилла в молодых растениях ячменя // Там же.— 1967.— № 2, вып. 14.— С. 299— 307.
Воскресенская Н. П., Ошарова И. С., Крылов Ю. В. Влияние длительного п кратковременного действия синего света на фотосинтез растений гороха И ДАН СССР.— 1968.— Т. 182, № 6.— С. 1443.
Гапоненка В. Г., Балева Е. Ф., Шаучук С. М. Фотосинтез i асдмгляцыйныя л!к! раслшн кукурузы, вырашчаных на вяленым i белым святле // Весц1 акадэмп навук БССР.— 1979.— № 4.— С. 52—55.
Гапоненка В. Г., Николаева Г. Н., Куперман И. М. Метаболизм хлорофилла, интенсивность фотосинтеза и ассимиляционные числа у растений, выросших на зеленом свету И Ботаника. Исследования.— М., 1982,— С. 176—183.
Гительзон И. И., Ковров Б. Г., Лисовский Г. М. и др. Экспериментальные экологические системы, включающие человека // Проблемы космической биологии.— М.: Наука, 1975.— Т. 28.— 312 с.
Гольд В. М. Некоторые эколого-физиологические особенности характера спектра действия фотосинтеза при световом насыщении // Физиология растений.— 1969.— Вып. 4.— С. 594—602.
Гольд В. М. Синий свет и фотосинтез: Автореф. дне. ... докт. биол. наук.— Иркутск, 1975.— 51 с.
Данилов А. Н. Качество света как фактор, определяющий пути использования лучистой энергии в процессе фотосинтеза // Советская ботаника.— 1935.— Т. 4.— С. 3-8.
Данилов А. Н. Своеобразие биологического действия световых лучей разной длины волны// Архив биологических наук.— 1936.— Т. 42, вып. 2/3.— С. 365—372.
Данилов А. Н. Изменение внутренних факторов фотосинтеза под действием света И Экспериментальная ботаника.— М.; Л., 1940.— Вып. 4.— С. 42—52,
Дроздова И. С., Бондарь В. В., Воскресенская Н. П. Совместное действие фоторегуляторных реакций, вызываемых красным и спним светом, на фотосинтез и морфогенез растений редиса // Физиология растений.— 1987.— Т. 34, вып. 4.— С. 786—794.
Дубров А. П. Действие ультрафиолетовой радиации на растения.— М.: Изд-во АН СССР, 1963.— 124 с.
Ермаков Е. И., Медведева И. В., Черноусов И. Н. Эффективные фотосинтез-ные единицы и близкая инфракрасная радиация // Светотехника.— 1980,— № 9.— С. 14—16.
Ефимцев Е. И., Бойченко В. А., Ефимцева Е. Г., Литвин Ф. Ф. Полярографический метод исследования спектров действия и кинетики газообмена О2 при фотобиологических процессах // Физиология растений.— 1978.— Т. 25, вып. 4.— С. 860—868.
Золотухин И. Г., Волкова Э. К. Спектральный состав излучения и качество растительной продукции // Проблемы светокультуры растений: Тез. Всесоюз. науч. конф.— Симферополь, 1980.— С. 123—125.
Золотухин И. Г., Лисовский Г. М., Баянова Ю. И. Действие света различного спектрального состава и интенсивности на биосинтез аскорбиновой кислоты в растениях // Физиология и биохимия культ, растений.— 1979.— Т. 11, № 2.— С. 141—146.
Золотухин И. Г., Лисовский Г. М., Волкова Э. К. Продуктивность и биохимический состав редиса, выращенного под излучением различной интенсивности и спектрального состава // Физиология растений,— 1983.— Т. 30, вып. 4.— С. 647—652.
15$
Золотухин И. Г., Лисовский Г. М., Трубачев И. Н., Волкова Э. К. Биохимический состав зерна пшеницы, выращенной при различных облученностях в отдельных областях ФАР // Физиология и биохимия культ, растений.— 1980.— Т. 2, № 5.— С. 451—457.
Иванов А. П. Физические основы гидрооптики.— Минск: Наука и техника, 1975.— 526 с.
Карначук Р. А. Регуляторное влияние зеленого света на рост и фотосинтез листьев И Физиология растений.— 1987.— Т. 34, вып. 4.— С. 765— 773.
Карначук Р. А. Регуляторная роль света разного спектрального состава в процессах роста и фотосинтетической активности листа растений: Автореф. дис. ... докт. биол. наук.— М.: ТСХА, 1989.— 42 с.
Карначук Р. А., Протасова Н. Н., Головацкая И. Ф. Рост растений и содержание гормонов в зависимости от спектрального состава света // Рост и устойчивость растений.— Новосибирск: Наука. Сиб. отд-ние, 1988.—• С.‘ 71—81.
Карначук Р. А., Поставалова В. М. Роль света в жизнедеятельности и продуктивности растений // Пути рационального использования почвенных, растительных и животных ресурсов Сибири.— Томск, 1986.—• С. 126-130.
Кахнович Л. В. Фотосинтетический аппарат и световой режим,— Минск: Изд-во БГУ, 1980.— 144 с.
Квашин Г. Н., Косов Ю. А., Садовой А. Ф. Эффективность воздействия источников света на растения ячменя /7 Светотехника.— 1983.— № 2.— С. 12.
Кособрюхов А. А., Чермных Л. Н. Влияние световых условий выращивания на температурный оптимум и интенсивность фотосинтеза растений огурца // С.-х. биология.— 1986.— № 5.— С. 76—79.
Кочубей С. М., Ляшенко В. Н., Бухов Н. Г., Воскресенская Н. П. Влияние света различного спектрального состава на организацию пигментного аппарата хлоропластов // Изв. АН СССР. Сер. биол.— 1987.— № 6.— С. 902-911.
Кузнецова Г. К., Пушкина Г. П., Шайн С. С. О фотоактивации биосинтеза гликоалкалоидов у паслена дольчатого // Проблемы фотоэнергетики,— Алма-Ата, 1978.— С. 112—121.
Кулаков А. В. Участие синего света в регуляции фотосинтетического СОа-газообмена проростков ячменя: Автореф. дис. ... канд. бпол. наук.— М.: ИФР АН СССР, 1988.— 20 с.
Куперман Ф. М. Физиология развития, роста и органогенеза пшеницы // Физиология сельскохозяйственных растений.— М.: Изд-во МГУ, 1969.— Т. 4.— С. 7—203.
Куперман Ф. М. Морфофизиологии растений.— М.: Высш, шк., 1984.— 240 с. Куперман Ф. М., Русу Ф. О роли света в фотоморфогенезе томатов И Морфогенез овощных растений.— Новосибирск: Наука. Сиб. отд-ние, 1971.— С. 79—89.
Курец В. К., Кособрюхов А. А., Марковская Е. Ф. и др. Методические подходы к характеристике температурного оптимума продуктивности сорта // Термо резистентность и продуктивность сельскохозяйственных растений.— Петрозаводск, 1984.— С. 104—113.
Лампы разрядные. Методы измерения спектральных и цветовых характеристик. ГОСТ 23198—78.— М.: Государственный комитет СССР по стандартам.
лЛевин И. А. Высокоинтенсивные трехфазные источники света // Светотехника.- 1986,— № 10.— С. 8—10.
.Леман В. М. Культура растений при электрическом свете.— М.: Колос, 1971.— 320 с.
Леман В. М., Фанталов О. С., Власова О. П. Роль пространственной структуры светового поля в формировании урожая яровой пшеницы // Селекция и семеноводство.— 1975.— № 4.— С. 20—22.
Лисовский Г. М. Фототрофы как звено замкнутой экологической системы, включающей человека: Автореф. дис. ... докт. биол. наук.— Новосибирск, 1973.— 52 с.
159
Лисовский Г. М., Долгушев В. А. Очерки частной светокультуры растений.— Новосибирск: Наука. Спб. отд-ние, 1986.— 128 с.
Лисовский Г. М., Сидько Ф. Я., Полонский В. И. и др. Интенсивность и качество света как факторы, определяющие формирование ценоза и урожай растений в светокультуре // Физиология растений.— 1987.— Т. 34, вып. 4.—С. 636—643.
Литвин Ф. Ф., Хэ-И-Тань. Спектры действия фотосинтеза, эффект Эмерсона и индукционные явления у высших растений // Там же.— 1967.— Т. 14, вып. 2.— С. 219.
Львова И. Н. Влияние светового режима па морфогенез различных сортов огурца И Свет и морфогенез растений.— М.: Изд-во МГУ, 1978.— С. 113-136.
Максимов Н. А. Культура растений на электрическом свете и применение ее для семенного контроля и селекции // Науч.-агр. журн.— 1925.— Т. 2, № 7/8.— С. 395—404.
Максимов Н. А. Биологическая основа светокультуры растений // Тр. пн-та/Ин-т физиол. растений АН СССР.— 1955.— Т. 10.— С. 7—16.
Мальчевский В. П. Применение искусственного свота для повышения урожайности // Докл. ВАСХНИЛ.— 1938,— Вып. 1/2 (10/11).— С. 70—74.
Мальчевский В. П. Применение искусственного света для ускорения роста и развития сеянцев древесных пород // Тр. ин-та/Ин-т физиол. растений АН СССР.— 1946,- Т. 3, вып. 2.— С. 3—46.
Маршак И. С., Васильев В. И., Тохадзе И. А. Малогабаритная безбалласт-пая трубчатая ксеноновая лампа с водяным охлаждением // Светотехника.— 1963.— № 11,— С. 13—17.
Мокроносов А. Т. Онтогенетический аспект фотосинтеза.— М.: Наука, 1981.— 196 с.
Мокроносов А. Т. Фотосинтетическая функция и целостность растительного организма. 42-е Тимирязевские чтения.— М.: Наука, 1983.— 64 с.
Мокроносов А. Т., Багаутдинова Р. И., Бубнова Е. X. Фотосинтетический метаболизм в палисадной и губчатой тканях листа И Физиология растений.— 1973.— Т. 20, вып. 6.— С. 1191—1197.
\Мошков Б. С. Выращйвапие растений при искусственном освещении.— Л.: Колос, 1966.— 287 с.
Мошков Б. С. Актиноритмия растений, — М.: Агропромпздат, 1987.— 272 с.
Мошков Б. С., Дроздов В. Н., Клочкова М. П. Промышленное выращивание томатов при искусственном освещении в Заполярье // Докл. ВАСХНИЛ.— 1984.— № 4.— С. 13—15.
Мурей И. А. Количественный анализ роста растений в посеве: Автореф. дне. ... канд. физ.-мат. наук.— Красноярск: ИФ СО АН СССР, 1978.— 30 с.
Нестеренко Т. В., Тихомиров А. А., Золотухин И. Г., Волкова Э. К. Индукция флуоресценции хлорофилла а и некоторые характеристики листьев огурца, сформировавшихся при действии света разного спектрального состава и интенсивности I/ Физиология и биохимия культ, растений,— 1985.— Т. 17, № 1.— С. 11—16.
Нийлиск X. И. Спектральный радиационный режим посева кукурузы и расчет фотосинтетически активной радиации (ФАР) // Изв. АН ЭССР, сер. физ.-мат. наук и техн. наук.— 1969.— Т. 13, № 3.— С. 177—191.
Пяловская Н. Т. Изучение газообмена и продуктивности посевов в фитотронах: Автореф. дис. ... д-ра биол. наук.— Киев, 1973.— 48 с.
Ничипорович А. А. Фотосинтез и теория получения высоких урожаев // Тимирязевское чтение.— М.: Изд-во АН СССР', 1956.— С. 1-93.
Ничипорович А. А. О свойствах растений как оптической системы // Физиология растений.— 1961.— Т. 8, № 5.— С. 53б—546.
Окунцов М. М., Роньжина О. А., Симонова Е. И. Влияние спектрального’ состава света йа биосинтез ка ретиноидов в растениях // Работы проблемной лаборатории фотйстггсза при Каф. физиологии? и биотямин растений Томского г ос университета. — Томск, 1964. — Вып. 1.— С. 93—113.
160
Павловский В. И., Прикупец Л. Б., Сарычев Г. С. и др. Фотобиологические исследования с селективными металлогалогенными лампами высокого давления // Светотехника.— 1976.— № 9.— С. 3—6.
Петров В. Е., Лоскутников А. И., Лосева Н. Л. Реактивация фотосинтетической функции ассимилирующей клетки после действия экстремальных температур И Продуктивность фотосинтезирующих систем в экстремальных условиях.— Улан-Удэ: БФ СО АН СССР, 1978.— С. 78-81.
Полонский В. И. Продуктивность ценоза пшеницы при высоких интенсивностях ФАР в светокультуре: Автореф. дне. ... канд. бпол. наук.— М., 1978.— 24 с.
Полонский В. И., Лисовский Г. М. Анатомо-морфологическая характеристика растений пшеницы при высоких интенсивностях фотосинтетически активной радиации (ФАР) в светокультуре // Ботап. журн.— 1978.— Т. 63, № 2,— С. 263—269.
Полонский В. И., Лисовский Г. М. Оптимизация продукционной деятельности фитоценозов при искусственном освещении // Проблемы светокультуры растений: Тез. докл.— Симферополь: Крымский СХИ, 1980,— С. 70—72.
Прикупец Л. Б., Сарычев Г. С., Головин В. И. и др. Перспективы применения ртутно-галогенидиых источников излучения для фотобиологических исследований // Управление скоростью и направленностью биосинтеза v растений.— Красноярск: ИФ СО АН СССР, 1973.— С. 76— 77, 161—162.
Протасова Н. Н. Светокультура как способ выявления потенциальной продуктивности растений // Физиология растений.— 1987.— Т. 34, вып. 4.— С. 812—822.
Протасова Н. Н., Добровольский М. В. Биологическая эффективность и эксплуатационные характеристики различных модификаций ксеноновых ламп, применяемых для выращивания растений // Биологическая спектрофотометрия и фитоактинометрия.— Красноярск: ИФ
СО АН СССР, 1973.— С. 76—77.
Протасова Н. Н., Кефели В. И. Фотосинтез и рост высших растений, их взаимосвязи и корреляции // Физиология фотосинтеза.— М.: Наука, 1982.— С. 251-270.
Рабинович Е. Фотосинтез.— М.: Изд-во иностр, лит., 1951, 1953, 1959.— Ч. 1-3.
Рекомендации по проектированию теплиц со светонепроницаемым ограждением для выращивания сельскохозяйственных растений при интенсивной светокультуре для районов Крайнего Севера.— Орел: Гппро-нисельпром, 1986.— 158 с.
Рождественский В. И., Клешнин А. Ф. Управляемое культивирование растений в искусственной среде.— М.: Наука, 1980.— 199 с.
Росс Ю. К. Математическое моделирование фотосинтетической продуктивности растений // Вестп. АН СССР.— 1972.— № 12.— С. 99—104.
Росс Ю. К. Радиационный режим и архитектоника растительного покрова.— Л.: Гпдрометеоиздат, 1975.— 344 с.
Росс Ю. К., Бихеле 3. Расчет фотосинтеза растительного покрова // Фотосинтез и продуктивность растительного покрова.— Тарту: ИФА АН ЭССР, 1968.— С. 46—74.
Росс Ю. К., Бихеле 3. Расчет фотосинтеза растительного покрова И Фотосинтетическая продуктивность растительного покрова.— Тарту: ИФА АН ЭССР, 1969.— Ч. 2,— С. 5—43.
Росс Ю. К., Нильсон Т. Пространственная ориентация листьев в посевах и методика ее определения // Фотосинтезирующие системы высокой продуктивности,— М.: Наука, 1966.— С. 109—125.
Росс Ю. К., Нильсон Т. Расчет фотосинтетически активной радиации в растительном покрове // Режим солнечной радиации в растительном покрове.— Тарту: ИФА АН ЭССР, 1968.— С. 5—54.
Санадзе Г. А., Баазов Д. И. Эффект усиления фотосинтеза (Эмерсона) у листьев тополя // Физиология растений.— 1982.— Т. 29, вып. д.-c. 901-906.
161
Свентицкий И- И. К вопросу об оценке оптического излучения в растениеводстве // Вести, с.-х. науки.— 1960.— № 2.— С. 106—122.
Свентицкий И. И. Количественная оценка фотосинтетической эффективности оптического излучения // Проблемы практической фотобиологии.— Пущино, 1977.— С. 73—76.
Свентицкий И. И. Экологическая биоэнергетика растений и сельскохозяйственное производство.— Пущино: ОНТИ НЦБИ АН СССР, 1982.— 222 с.
Свет и морфогенез растений/Под ред. Ф. М. Куперман, Е. И. Ржа новой.— М.: Изд-во МГУ, 1978.— 188 с.
Сидько Ф. Я., Лисовский Г. М., Сарычев Г. С. и др. Действие света различной интенсивности и спектрального состава на продукционные процессы в ценозах редиса // Интенсивная светокультура растений.— Красноярск: ИФ СО АН СССР, 1977.— С. 3—14.
Сидько А. Ф., Филимонов В. С., Сидько Ф. Я. и др. Полевой двухлучевой дифференциальный спектрофотометр // Жури, прикл. спектроскопии.— 1978.— № 5,— С. 950—955.
Сидько Ф. Я., Шарупич Т. С., Шарупич В. П. Оптимизация организационной структуры ценозов огурца и режимов их облучения в светонепроницаемых теплицах/ИФ СО АН СССР.— Препр.— Красноярск, 1983,— 34 с.
Симонова Е. И. Влияние света разного спектрального состава на биосинтез пигментов в растениях: Автореф. дис. ... канд. биол. наук.— Томск, 1965.— 20 с.
Справочная книга по светотехнике/Под ред. Ю. А. Айзенберга.— М.: Энерго-атомиздат, 1983.— 472 с.
Сулацков В. Г., Свентицкий И. И., Неклеса Н. П. Лампа типа ЛОР-1000 для облучения растений // Светотехника.— 1971.— № 10.— С. 3—4.
Тарчевский И. А. Основы фотосинтеза.— М.: Высш, шк., 1977.— 253 с.
Терентьев В. М., Головнева Н. Б., Фёдюнькин Д. В. и др. Зависимость накопления фотосинтезирующих пигментов в листьях растений от спектрального состава и интенсивности света И Хлорофилл.— Минск: Наука и техника, 1974.— С. 340—349.
Терентьев В. М., Федюнькин Д. В., Головин В. Н. и др. О фотобиологической реакции молодых растений на облучение узкополосным светом И Управление скоростью и направленностью биосинтеза у растений.— Красноярск: ИФ СО АН СССР, 1973.— С. 172—173.
Тимирязев К. А. Солнце, жизнь и хлорофилл.— М.: Гос. изд-во с.-х. лит.— 1956,— 228 с.
Тихомиров А. А. Оптические характеристики фитоэлементов и фитоценозов при свете различной интенсивности в отдельных областях ФАР // Интенсивная светокультура растений.— Красноярск: ИФ СО АН СССР, 1977а,— С. 81—100.
Тихомиров А. А. Формирование структуры и фотосинтез ценозов при использовании света различной интенсивности в отдельных областях ФАР // Там же.— 19776.— С. 58—80.
Тихомиров А. А. Оптические свойства и продуктивность ценозов растений при использовании излучения различной интенсивности и спектрального состава: Автореф. дис. ... канд. биол. наук.— Красноярск:
ИФ СО АН СССР, 1979.— 25 с.
Тихомиров А. А. Некоторые особенности структуры радиационного поля в ценозах редиса и пшеницы, сформировавшихся при различных режимах облучения И Тез. докл. XI Всесоюз. совещания по актинометрии. Ч. VII: Фитоактинометрия.— Таллинн: Изд-во АН ЭССР, 1980.— С. 151—154.
Тихомиров А. А., Золотухин И. Г., Лисовский Г. М., Сидько Ф. Я. Специфика реакции растений разных видов на спектральный состав ФАР при искусственном освещении .// Физиология растений.— 1987.— Т. 34, вып. 4.— С. 774—785.
Тихомиров А. А., Золотухин И. Г., Лисовский Г. М. и др. Способ выращивания огурца при искусственном облучении: А. с. 1620062 СССР // Бюл. изобрет.— 1991.— № 2.
162
Тихомиров А. А., Золотухин И. Г., Сидько Ф. Я. Влияние световых режимов на продуктивность и качество урожая редиса // Физиология растений,— 1976.— Т. 23.— С. 502—507.
Тихомиров А. А., Сидько Ф. Я. Состояние пигментного аппарата и формирование структуры ценозов редиса в связи с их продуктивностью при различной интенсивности и спектре излучения // Там же.— 1982.— Т. 29, вып. 3,— С. 457—464.
Тихомиров А. А., Сидько Ф. Я. Некоторые особенности энергетики листа редиса в условиях интенсивной светокультуры // Биофизические методы исследования экосистем.— Новосибирск: Наука. Спб. отд-ние, 1984,— С. 91—98.
Тихомиров А. А., Сидько Ф. Я. Зависимость урожая редиса в светокультуре от спектра и интенсивности излучения // Плодоовощное хозяйство.— 1985.— № 7.— С. 35—36.
Тихомиров А. А., Сидько Ф. Я. К вопросу о спектральной аддитивности биосинтетических процессов в фптоценозах // Физиология растений.— 1987.— Т. 34, вып. 1.— С. 39—50.
Тихомиров А. А., Сидько Ф. Я., Лисовский Г. М. и др. Проблема оптимизации спектральных и энергетических характеристик излучения растениеводческих ламп.— Препр.— Красноярск: ИФ СО АН СССР, 1983.— 25 с.
Тооминг Г. X. Солнечная радиация и формирование урожая.— Л.: Гидро-метеоиздат, 197'7,— 200 с.
Тооминг X. Г. Экологические принципы максимальной продуктивности.— Л.: Гидрометеоиздат, 1984.— 264 с.
Федоров А. К., Мудрак Е. И., Юрьева Н. А., Феофидов Э. В. Светотехнические установки для овощеводства //. Плодоовощное хозяйство.— 1986.— № 8.— С. 39—42.
Федюпькин Д. В., Головнева Н. Б., Кошелева Л. Л., Бахнова К. В. Интенсивная культура растений в искусственных условиях,— Минск: Наука и техника, 1988.— 214 с.
Физиология сельскохозяйственных растений.— М.: Изд-во МГУ, 1969.— Т. 4.— 556 с.
Фотометрия. Термины и определения. ГОСТ 26148-84.— М.: Изд-во стандартов, 1984.— 24 с.
Хазанов В. С. Абсолютные измерения излучения с Помощью фарометра ВНИСИ ФАР-74 // Проблемы фотоэнергетики растений.— Киев: Наук, думка, 1975.— С. 102.
Хазанов В. С., Кузнецова Г. К. О спектральной эффективности излучения в обеспечении фотосинтеза // Науч. докл. высшей школы. Биол. науки,— 1973а.— № 6,— С. 84.
Хазанов В. С., Кузнецова Г. К. Об аддитивности действия разноспектральных излучений при фотосинтезе 7/ Там же.— 19736.— № 2.— С. 78.
Цельникер Ю. Л. Физиологические основы теневыносливости растений.— М.: Наука, 1978,— С. 215—216.
Чучалин А. И. Радиационный режим и использование ФАР ценозом пшеницы в условиях интенсивной светокультуры: Автореф. дис. ... канд. биол. наук.— Красноярск: ИФ СО АН СССР, 1980.— 25 с.
Чучалин А. И., Ерошин Н. С., Тихомиров А. А. и др. Содержание пигментов и оптические свойства листьев пшеницы в условиях интенсивной светокультуры растений // Изв, СО АН СССР. Сер. бпол. наук.— 1977.— Вып. 2.— С. 38—43.
Шалаева Е. Е., Лисовский Г. М., Тихомиров А. А. Спектральный состав света и некоторые особенности мезоструктуры листьев различных растений // Физиология растений.— 1991.— Т. 38, вып. 1.—С.55—62.
Шлык А. А. О спектрофотометрическом определении хлорофилла А + Б // Биохимия.— 1968.— Т. 33, № 2.— С. 275—295.
Шульгин И. А. Солнечная радиация и растение.— Л.: Гидрометеоиздат, 1967.— 180 с.
Шульгин И. А. О физиологическом действии дальней красной (ближней инфракрасной) радиации на развитие некоторых овощных растений //
163
Морфогенез овощных растений.— Новосибирск: Наука. Сиб. отд-ние, 1971.— С. 42—48.
Шульгин И. А. Растение и солнце.— Л.: Гидрометеоиздат, 1973.— 212 с.
Шульгин И. А., Клешнин А. Ф. О корреляции между оптическими свойствами и содержанием хлорофилла в листьях растений И ДАН СССР.— 1959.— Т. 125, № 6,— С. 1371—1373.
Шульгин И. А., Куперман Ф. М., Мерцалов В. М. Влияние спектрального состава, интенсивности радиации, продолжительности фотопериодов на развитие, рост и морфогенез растений // Вести, с.-х. науки.— 1963а.— № 4.— С. 21—33.
Шульгин И. А., Мерцалов В. М. Влияние продолжительности фотопериода на развитие редиса в зависимости от спектрального состава лучистой энергии // Экспериментальный морфогенез.— М.: Изд-во МГУ, 19636.— С. 68—70.
Юров С. Г. Общая схема построения системы фотометрических величин // Светотехника.— 1958.— № 9.— С. 10—17.
Bulley N. В., Nelson С. D., Tregunna Е. В. Photosynthesis: action spectra for leaves in normal and low oxygen /I Plant Physiol.— 1969.— Vol. 44j N 5,— P. 678-684.
Cathey H. M., Campbell L. B. Plant productivity: new approches to efficient sources and environmental control // Trans. ASAE.— 1977.— P. 360 — 366, 371.
Duncan W. G., Loomis B. S., Wiliams W. A., Hanau B. A Model for simulating photosynthesis in plant communiteis // Halgardia.— 1967.— Vol. 38, N 4.— P. 181—205.
Dwived S. B. Studies on the efficiency of energy. Conversion in photosynthetic parts of wheat plants. {Triticum aestivum L.) // Ann. Bot.— 1975.— Vol. 39, N 164.— P. 1077—1085.
Frank S. K. The effectiveness of the spectrum in chlorophyll formation // J. gen. Physiol.— 1946,— Vol. 29, N 3.— P. 157—179.
Gabrielsen E. K. Influence of light of different wave-lenght on photosynthesis in foliage leaves // Physiol. Plant.— 1948.— Vol. 1, N 1.— P. 113.
Gausman H. W., Allen W. A. Optical parameters of leaves of 30 plant species 7/ Plant Physiol.— 1973,— Vol. 52, N 1.— P. 57—62.
Harvey В. B. Growth of plants in artificial light from seed to seed // Sci.— 1922.—Vol. 56, N 1448.—P. 366—367.
Ilieva V., Andreichin B., Keechlibarov T. Efficiency of artificial light sources in plant growing // Int. Agrophys.— 1985.— N 1,— P. 115—120.
Inada K. Effect of blue light on the photonastic reaction of rice leaves // Plant Cell Physiol.— 1969.—Vol. 10.— P. 485.
Inada K. Acton spectra for photosynthesis in higher plants // Ibid.— 1976.— N 17,- P. 355—365.
Inada K. Effects of leaf color and the light quality applied to leaf developing period on the photosynthesis responce spectra fn crop plants // Jap. J. Crop Sci.— 1977.— Vol. 9.— P. 37—44.
Kimura K. Effect of light on leaf inclination of Triticum Aestirum // Ohala Inst. Landwirt. Biol. Okayama Univ.— 1974.— Vol. 16, N 2.— P. 47—56. • ,.
Ko B. Optical characteristics and spectral dependence of photosynthesis ef crop leaf developied under different light qnolities 7/ Environ. Cont-. rol. Biol.— 1982,— Vol. 20, N 1.—P. 1—7.
Ko B., Yabuki K. The dependence of the relation between light quality and photosynthesis on optical and anatomical characteristics of leaves (fj. The relation between light quality and photosynthesis of leaves irradiated from both sides // J. Agric. Meteorology.— 1974.— Vol. 2§, N 4.— P. 229—237. •
MancinelIi A. L. Light dependent anthocyanin synthesis // Bot. Hew.— 1985.— Vol. 51, N 1.— P. 107—155.
McCree K. J. The action spectrum, absorptance and quantum yield of photosynthesis in crop plants 7/ Agric. Meteorology.— 1972a.— Vol. 9.— P. 192-216.
164
McCree К. J. Significance of onhencement for calculations based on the action spectrum for photosynhtesis // Plant Physiol.— 1972b.— Vol. 49.— P. 704-700.
McLeod G. C. Action spectrum of light saturated photosynthesis // Ibid.— 1961.— Vol. 36, N 1.— P. 430—434.
Monsi M., Saeki T. Uber den Lightfaktor in den Pflansengesellschaften und seine Bedeutung fur die Stoffproduktion // Jap. J. Bot.— 1953.— Vol. 29, N 113.— S. 22—52.
Monteith I. L. Light distrabution and photosynthesis in field crops // Ann. Bot.— 1965.— Vol. 29, N 113.— P. 17—37.
Moss D. N. Optimum lighting of leaves H Crop Sci. (Madison).— 1964.— Vol. 4.— P. 131—136.
Osborne B. A., Raven J. A. Light absorption by plants and its implications for photosynthesis // Biol. Rev.— 1986.— Vol. 61.— P. 1—61.
Plant photosynthetic production. Manual and methods.— Hague.— 1971.— 800 p....................................................
Poyarkova N., Drozdova L, Voskresenskaya N. Effect of blue light on the activity of carboxylating enzymes // Photosynthetica.— 1973.— Vol. 7, N 1.— P. 58:
Stoy V. Action of different light qualities of simultaneous photosynthesis and nitrate assimilation in wheat leaves// Physiol. Plant.— 1955.— Vol. S.-Р. 963—986.
Tikhomirov A. A., Sid’ko F. Ya. Photosynthesis and structure of radish and wheat canopies and spectral composition // Photosynthetica.—,1982.— Vol. 16, N 2.— P. 191—195.
Tikhomirov A. A., Sid’ko F. Ya. Optical characteristics of individual plant elements and plant canopies growth under radiation regimes of different Spectral composition and intensity // Appl. Optics.— 1983.— Vol. .22, N 18.— P. 2874—2881..............................
Virgin H. L. Light induced unfolding of the grass leaf // Physiol. Plant.— 1962.— Vol. 15.— P. 380—389.
Voskresenskaya N. P. Blue light and carbon metabolism // Ann. Rev. Plant Physiol.— 1972.— Vol. 23.— P. 219—234.
ОГЛАВЛЕНИЕ
Предисловие ................................................. j 3
Глава 1. Методика проведения экспериментов и используемая аппаратура .............................................. .... s 7
§ 1. Объекты исследования и условия их культивирования —
§ 2. Установки для выращивания растений................... 10
§ 3. Световое обеспечение растений....................... 11
§ 4. Аппаратура и устройства для измерения оптических и фотосинтетических характеристик растений................. 16
§ 5. Методы оценки фотометрических и биологических характеристик растений и фитоценозов........................... 18
Глава 2. Фотосинтез и спектральный состав света.............. . 20
Введение.............................................. —
§ 1. Спектры действия и спектральная эффективность фотосинтеза листа............................................... 22
§ 2. Фотосинтез листа при длительном воздействии излучения различного спектрального состава и интенсивности ... 29
§ 3. Спектральная эффективность фотосинтеза ценозов расте- 33
ний................................................
§ 4. Специфика реакций растений разных видов на качество света при оценке спектральной эффективности фотосинтеза 43
Глава 3. Влияние спектрального состава света на рост и развитие растений......................................................... 47
Введение.................................................. —
§ 1. Влияние спектра и интенсивности ФАР на морфологические характеристики растений.............................. 49
§ 2. Спектральный состав света и формирование мезоструктуры листьев различных растений........................... 86
Глава 4.Спектральный состав света и биохимия растений............ 94
Глава 5. Спектральный состав света и скорость накопления растительной биомассы.................................................101
§ 1. Спектральный состав излучения искусственных источников света и продуктивность растений........................ —
§ 2. Продуктивность ценозов растений при различном спектральном составе и интенсивности искусственного облучения ................................................... 112
§ 3. Эффективность использования фитбценозами излучения ФАР различного спектрального состава и интенсивности (КПД фитоценозов)........................................ 122
§ 4. Некоторые особенности воздействия спектрального состава света на продукционный процесс растений . . . . 127
Глава 6. Оптимизация спектральных и энергетических характеристик ФАР в светокультуре растений..................................130
§ 1. Методы оценки спектральной эффективности действия ФЛР в условиях светокультуры............................... 131
§ 2. Выполнимость принципа спектральной аддитивности для биосинтетических процессов в фитоценозах................... 138
§ 3. Эффективные потоки и возможности их использования для оптимизации излучения в светокультуре.................. 146
§ 4. Перспективы создания растениеводческих ламп с оптимальным спектром излучения .............................. 153
Список литературы.................................................157
Научное издание
Тихомиров Александр Аполлинариевич Лисовский Генрих Михайлович Сидько Федор Яковлевич
СПЕКТРАЛЬНЫЙ СОСТАВ СВЕТА И ПРОДУКТИВНОСТЬ РАСТЕНИЙ
Редактор издательства Н. Л. Шварцкопп Художник Л. Л. Мордохович
Технический редактор Г. Я. Герасимчук Корректор Н* М, Горбачева
ИБ № 42802
Сдано в набор 21.11.00. Подписано к печати 02.08.91. Формат 60x90'/ie. Бумага кн.-журнальная. Обыкновенная гарнитура. Высокая печать. Усл. печ. л. 10,5. Усл. кр,-отт. 10,8. Уч.-изд. и. 11,2. Тираж 750 экз. Заказ № 503. Цена 2 р. 90 к.
Ордена Трудового Красного Знамени издательство «Наука», Сибирское отделение. 630099 Новосибирск, ул. Советская, 18.
4-я типография издательства «Наука». 630077 Новосибирск, ул, Станиславского, 25
2 р. 90 и.
[од
„НАУКА” СИБИРСКОЕ ОТДЕЛЕНИЕ