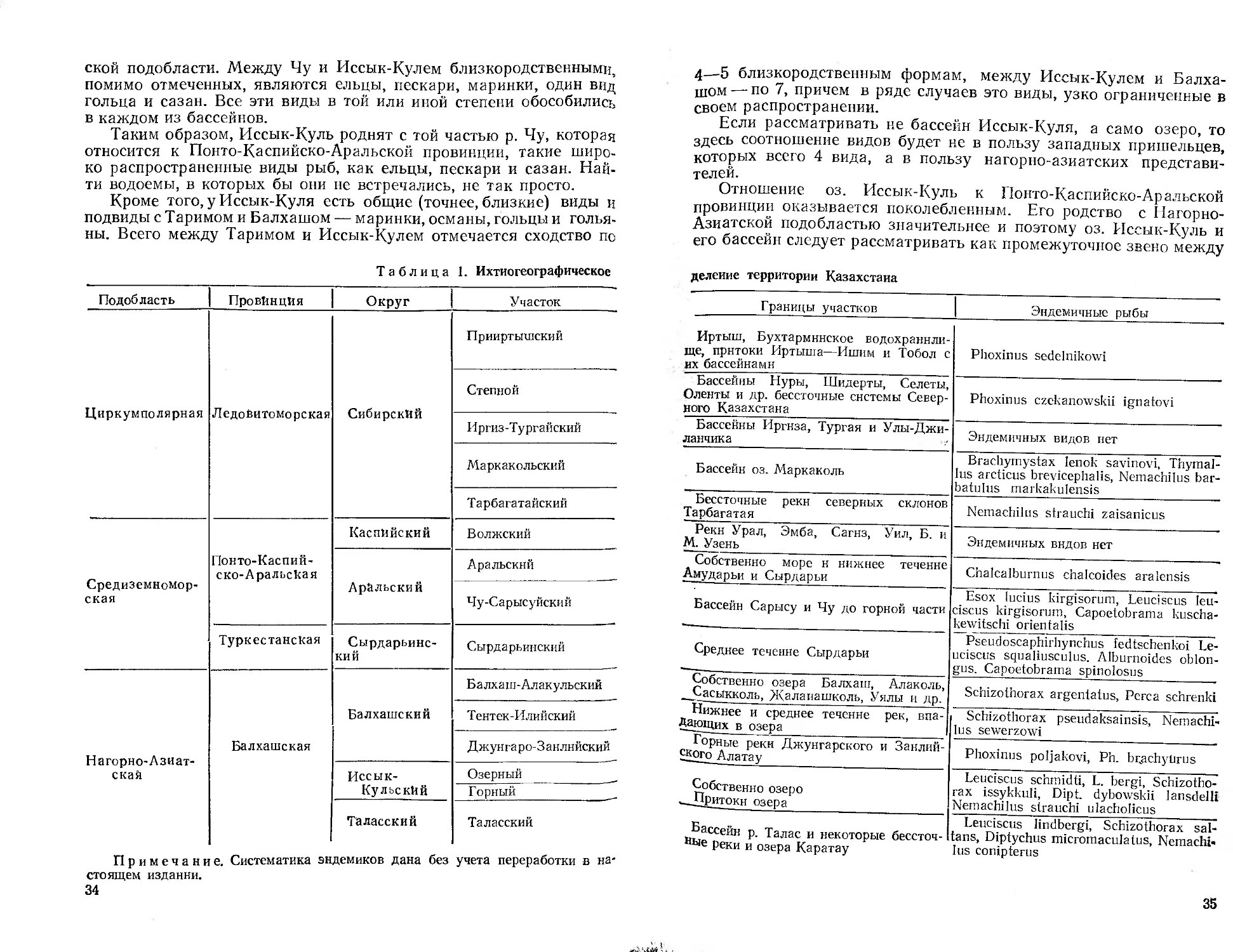
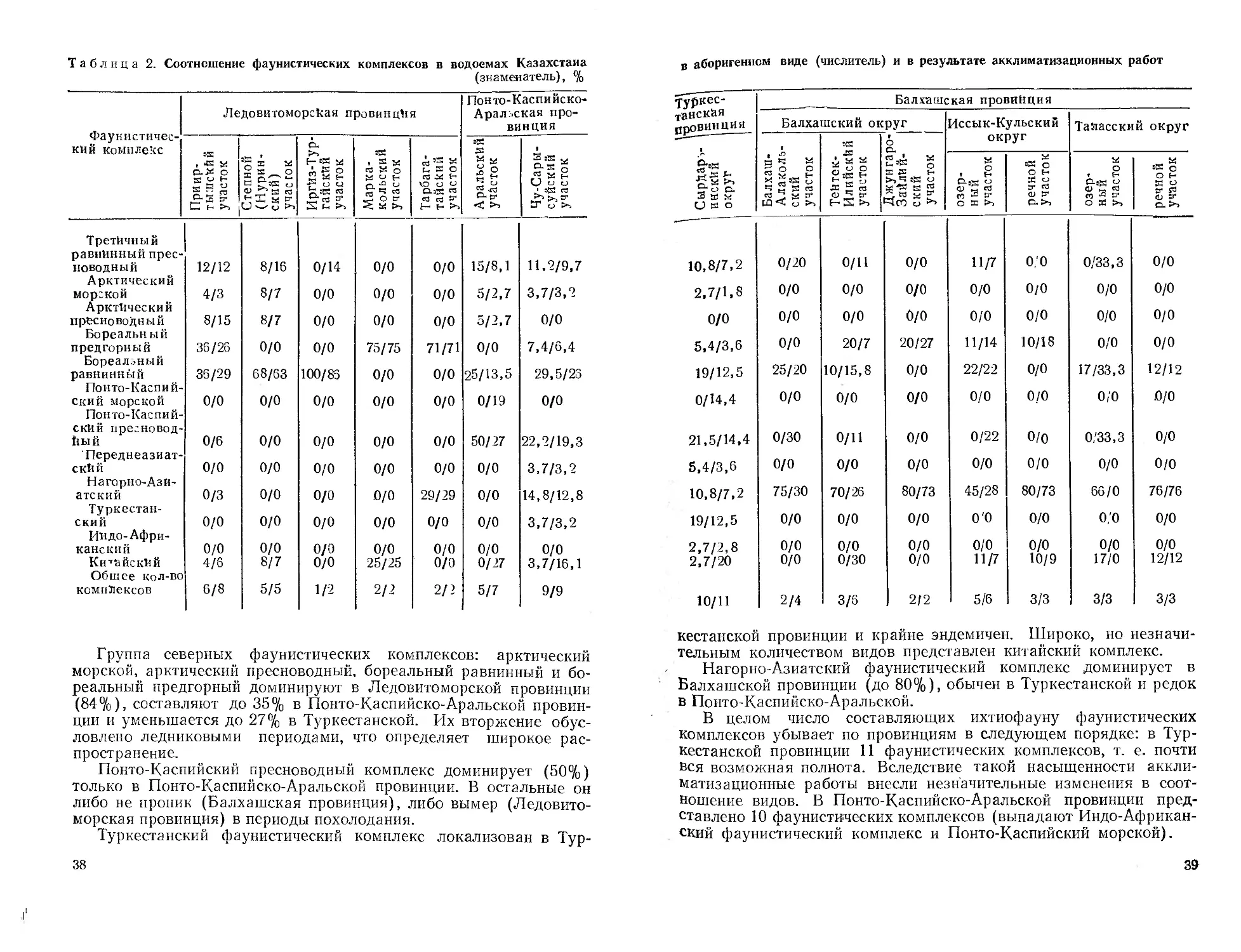
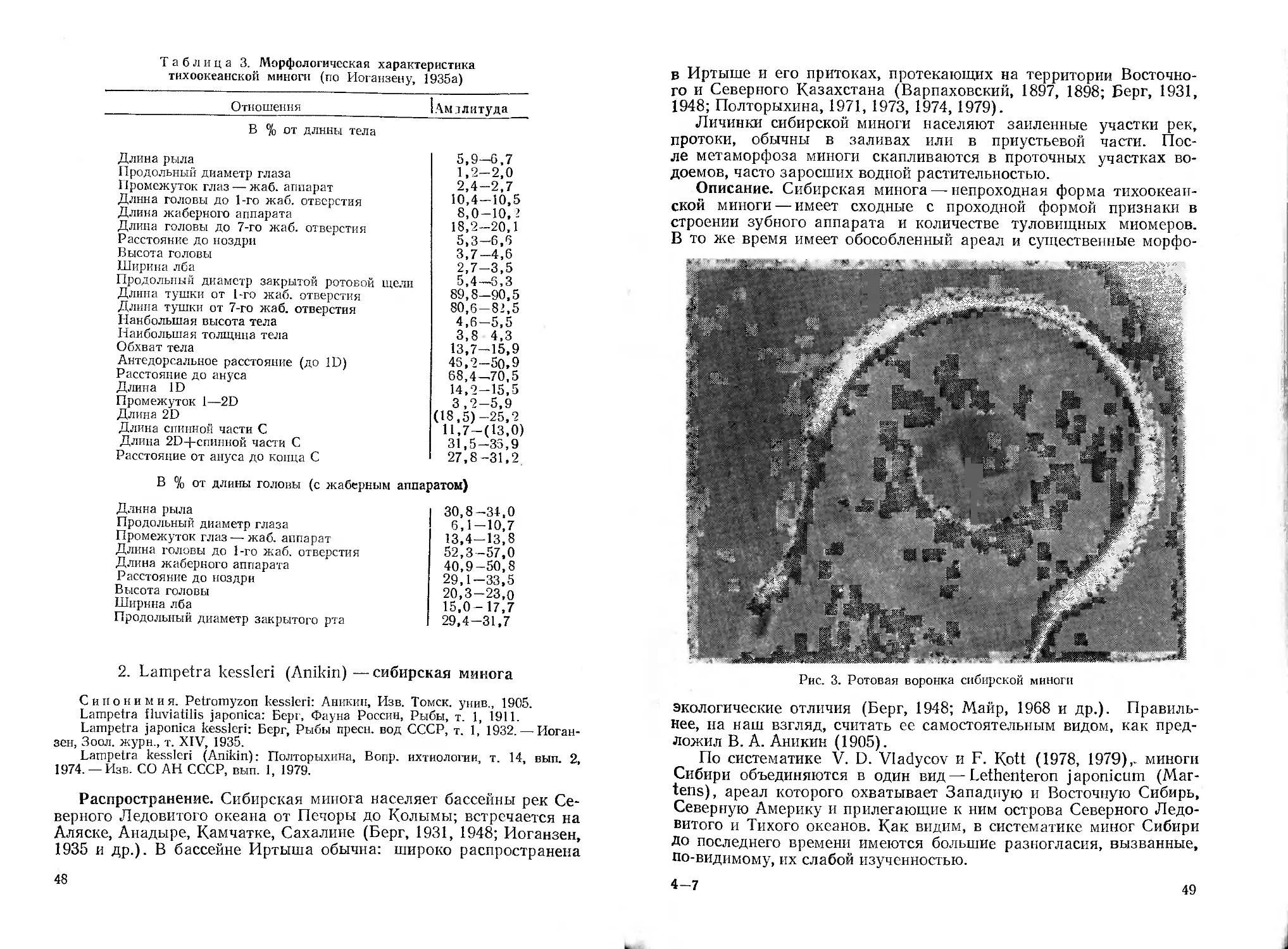
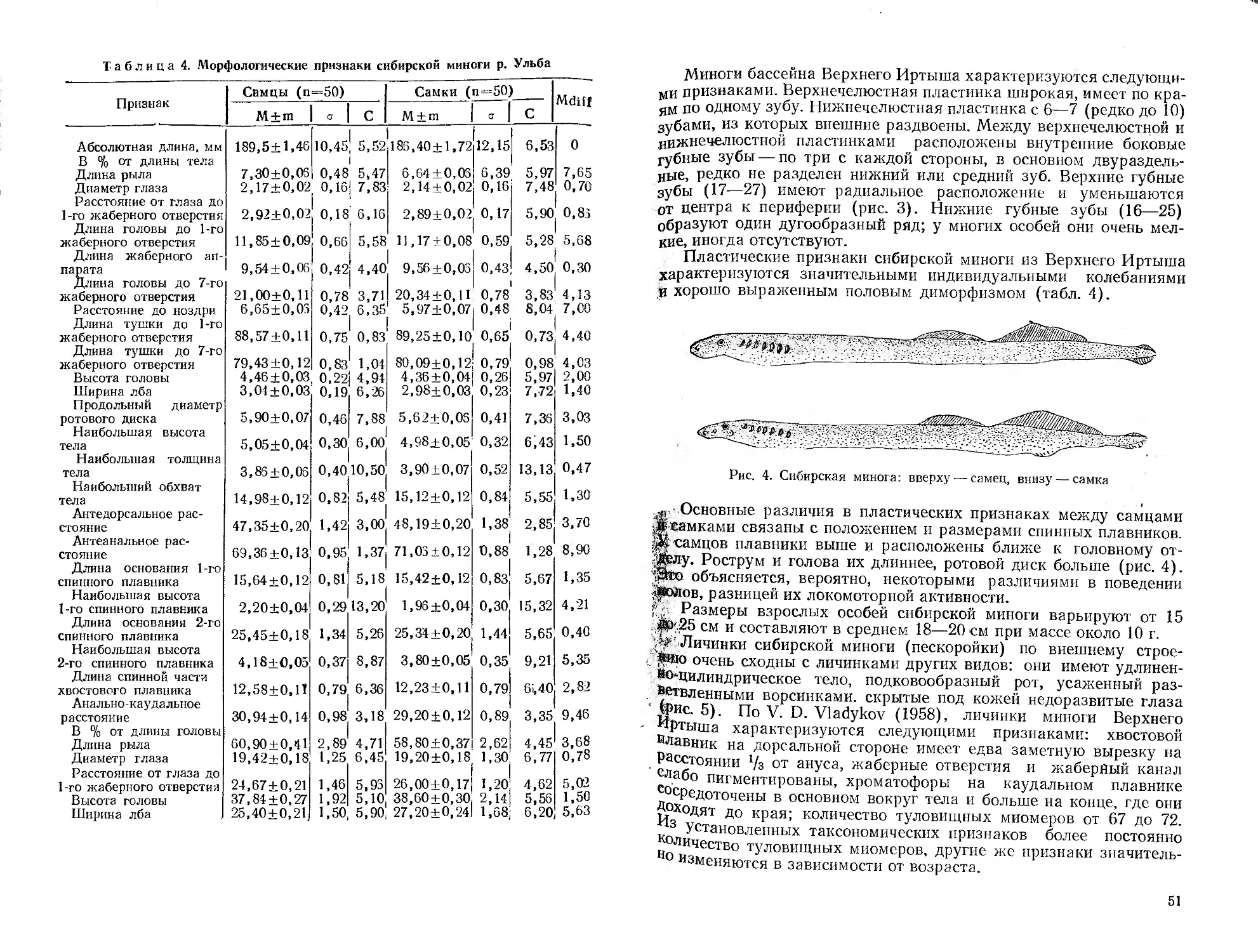
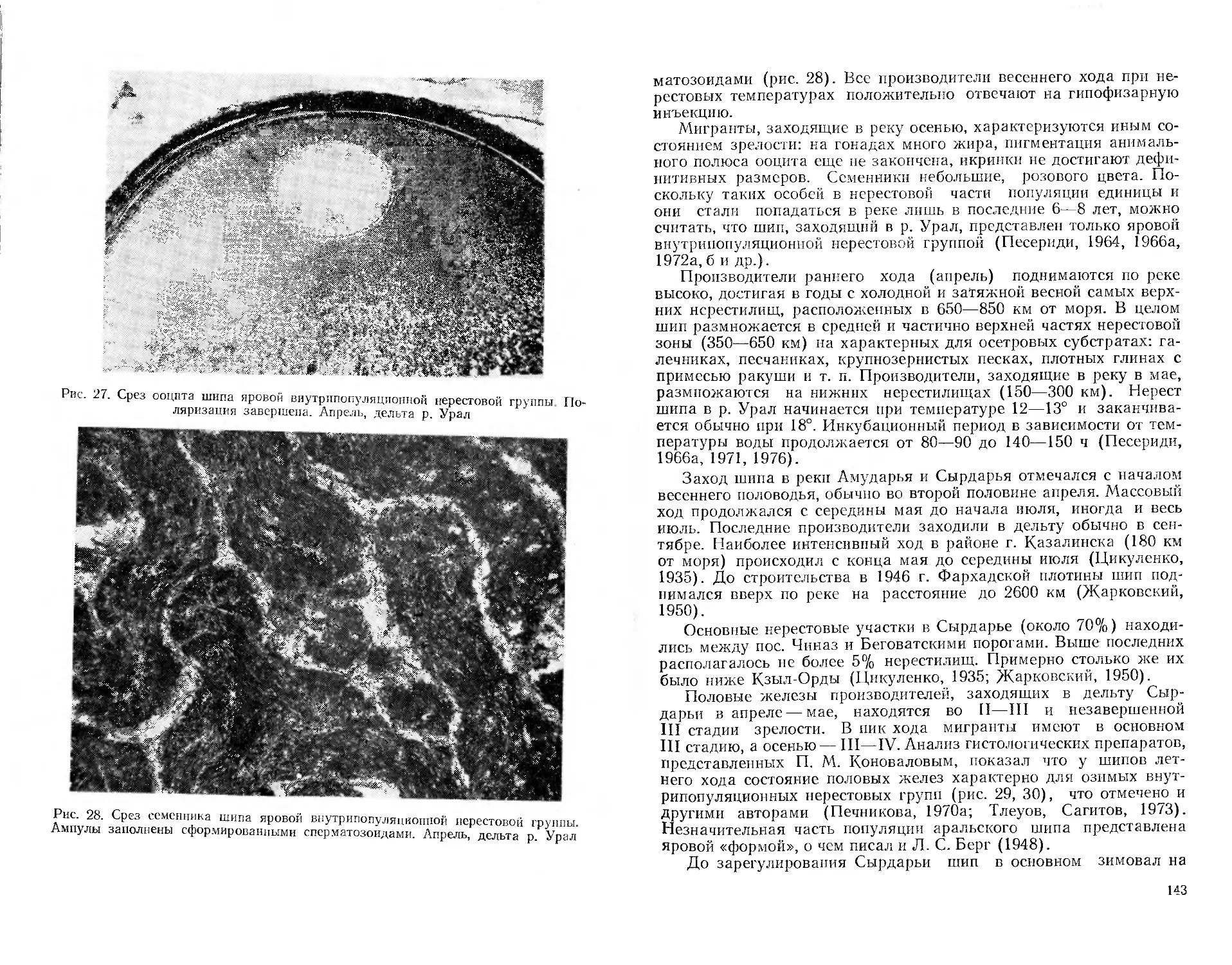
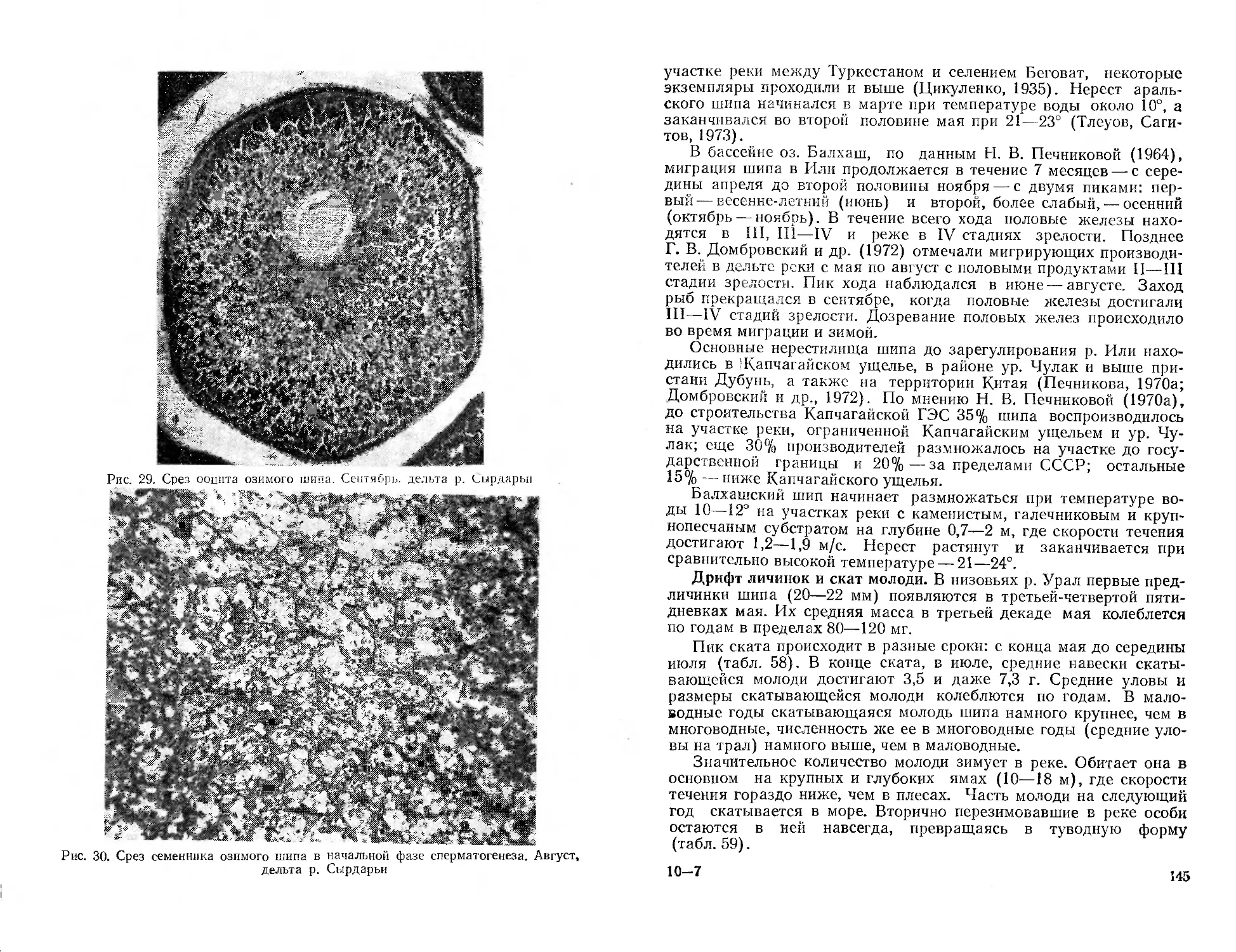
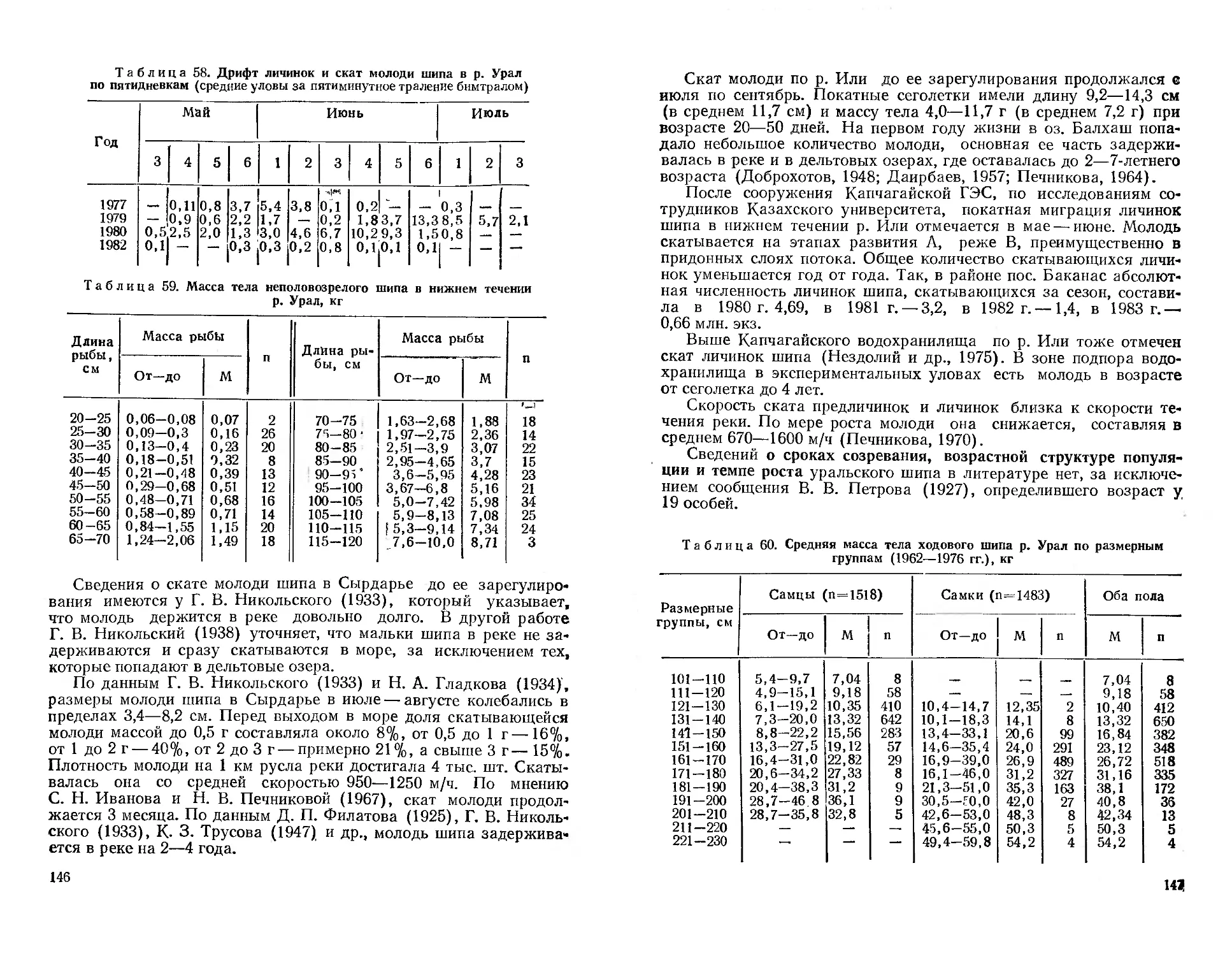
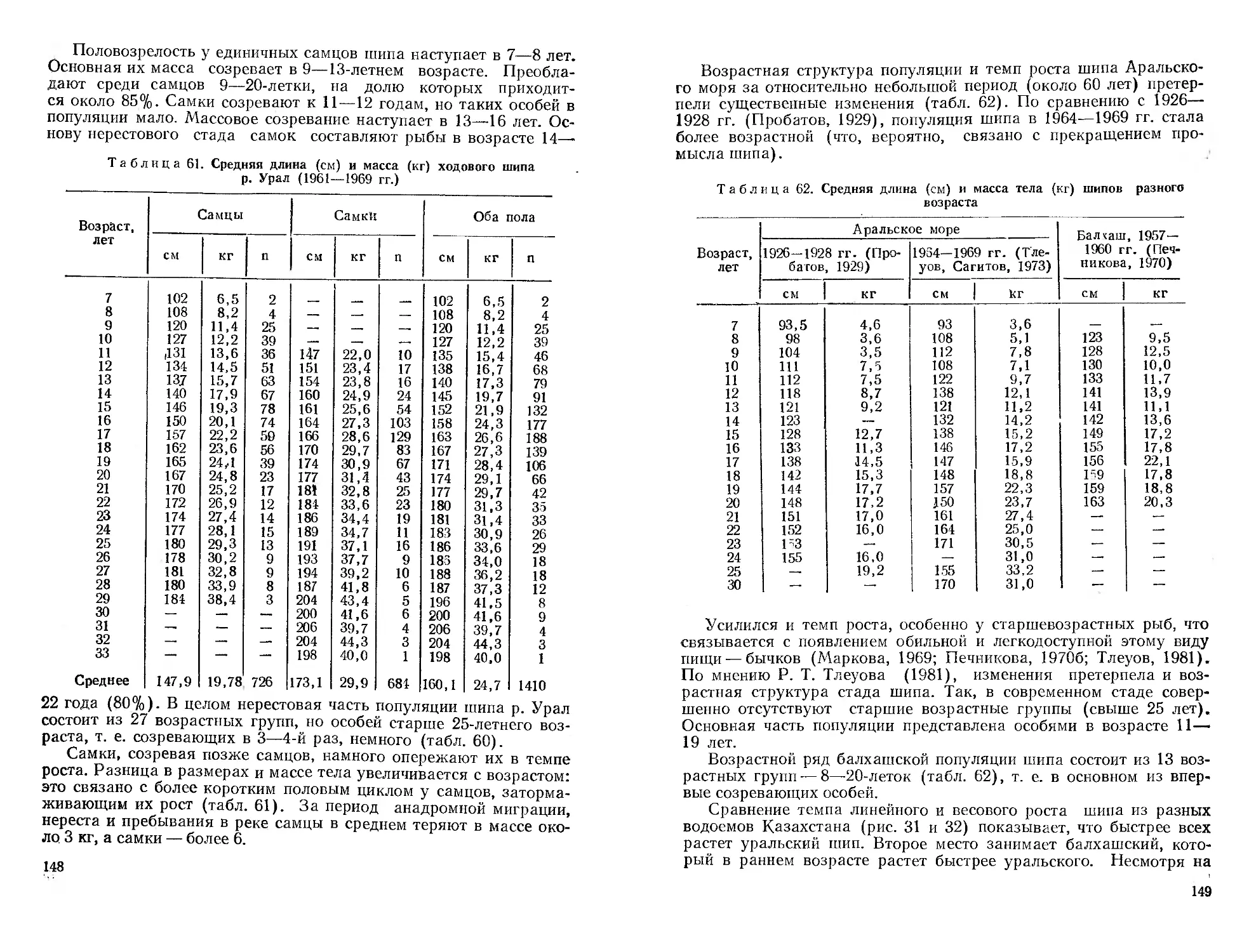
/


























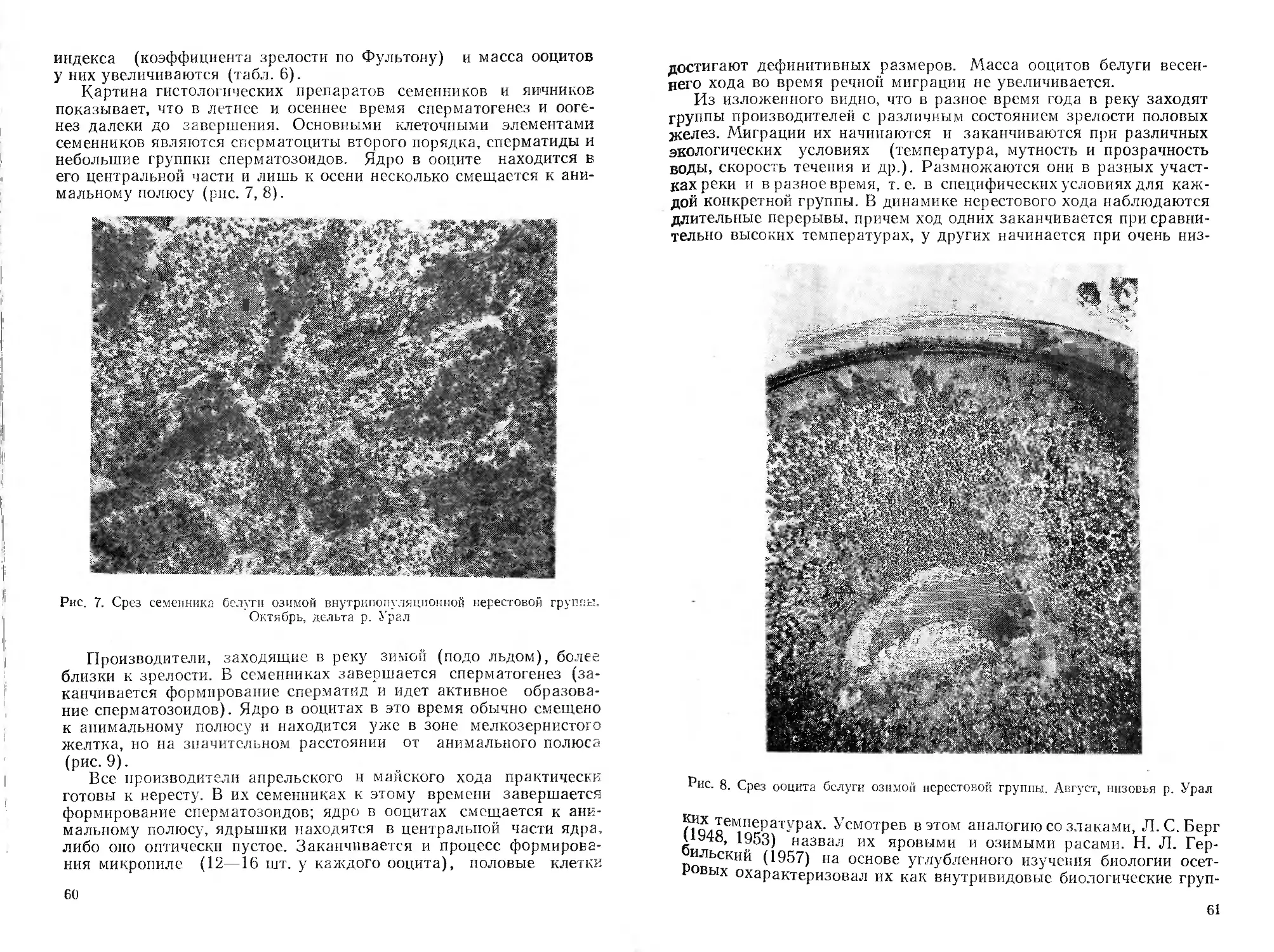
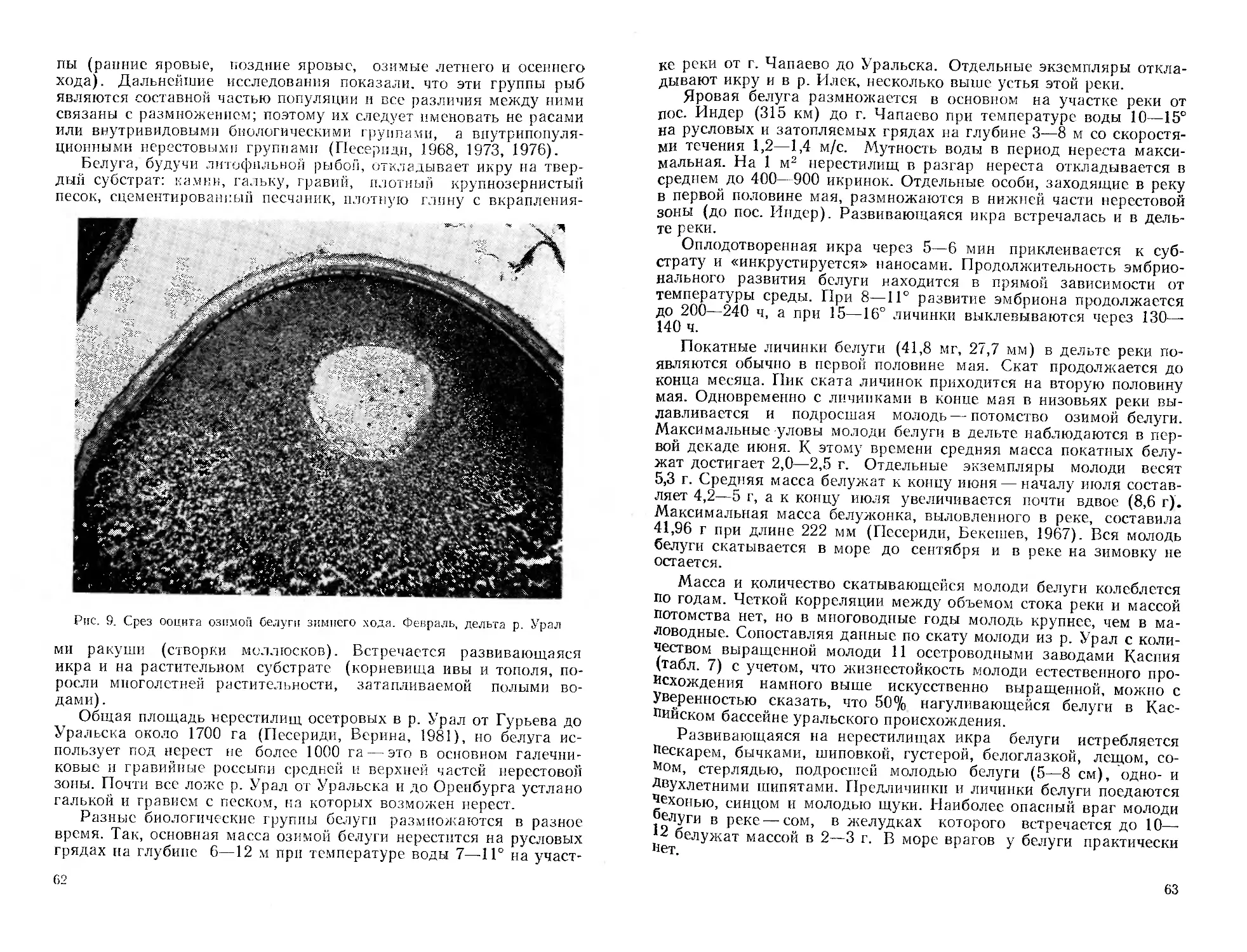
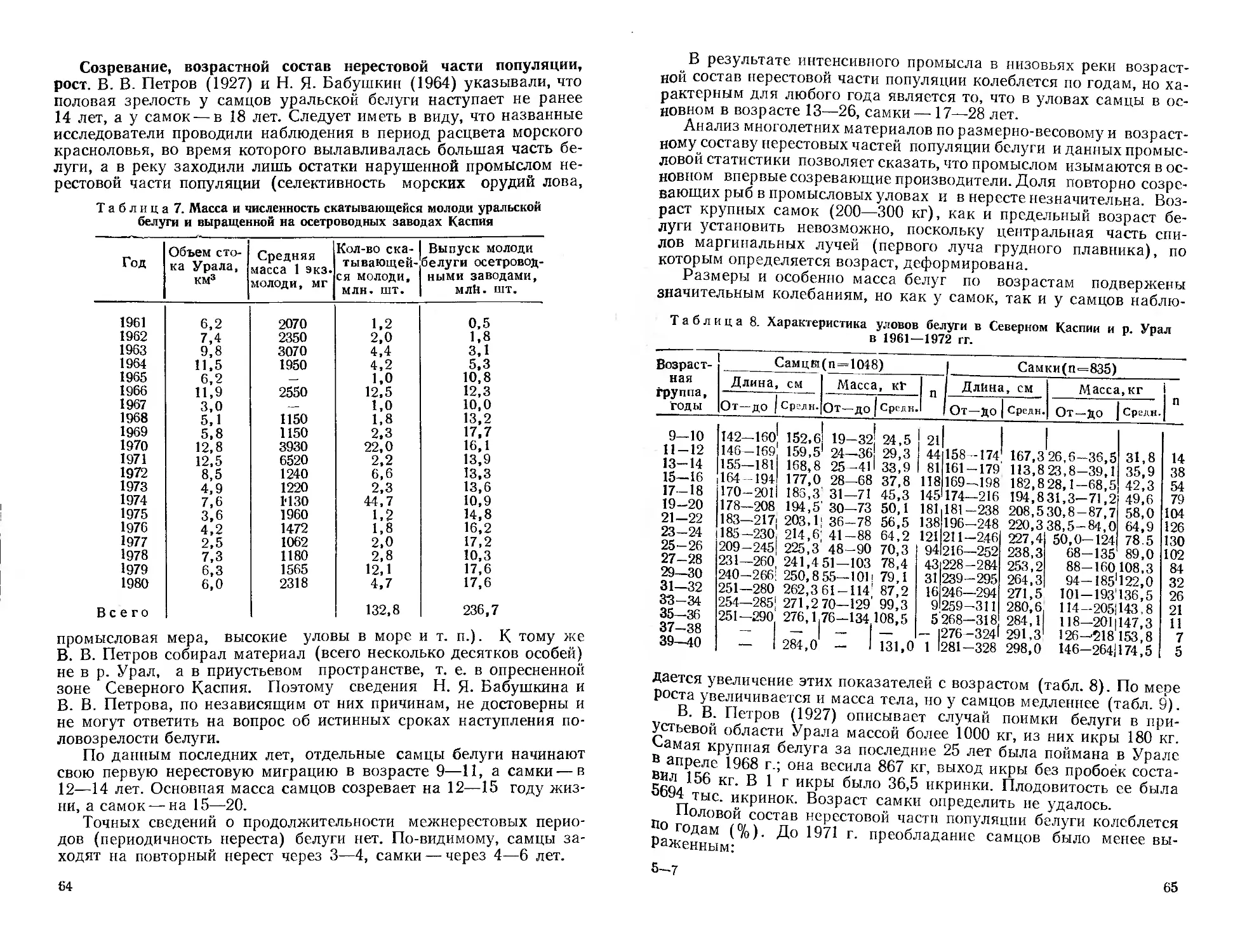























































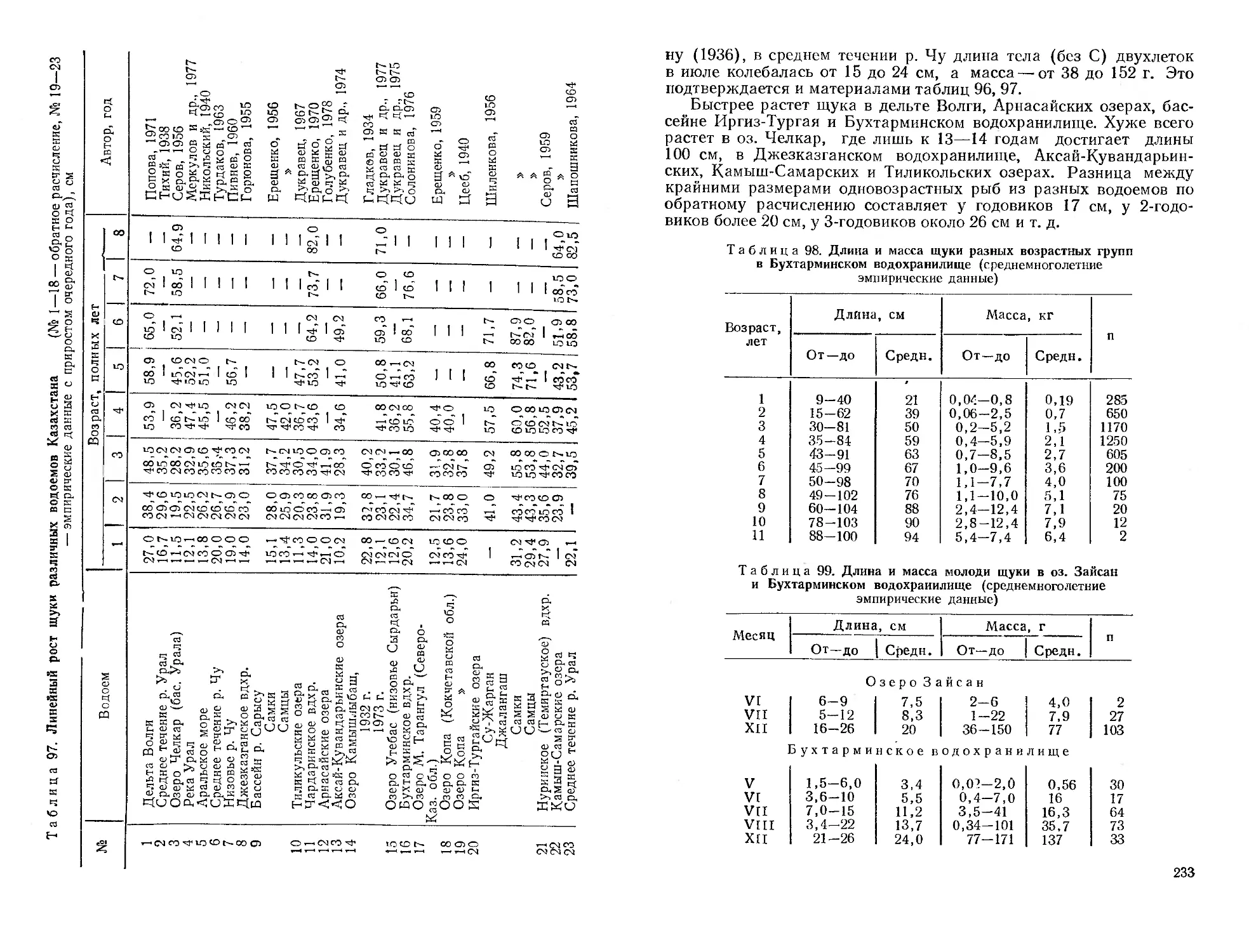


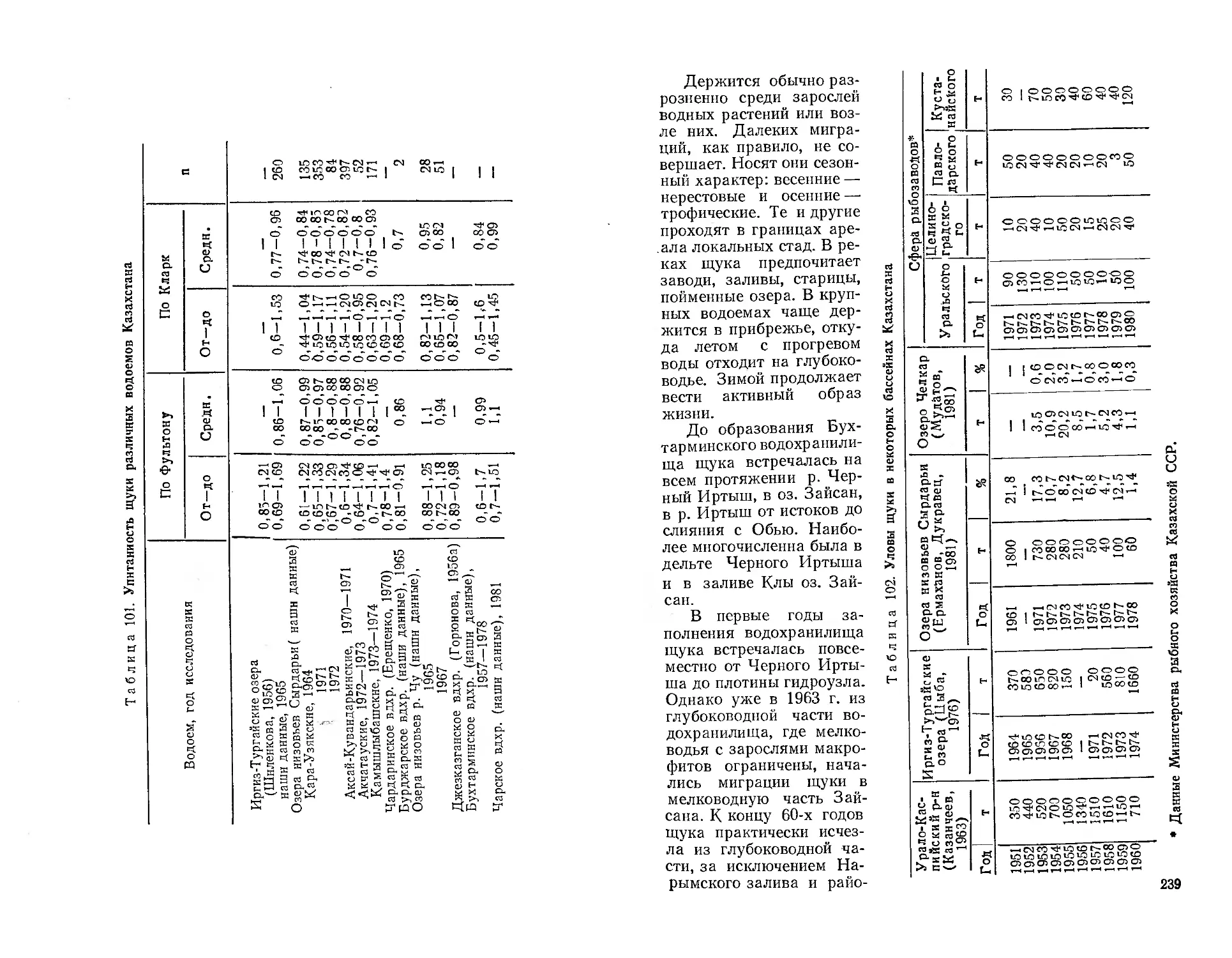
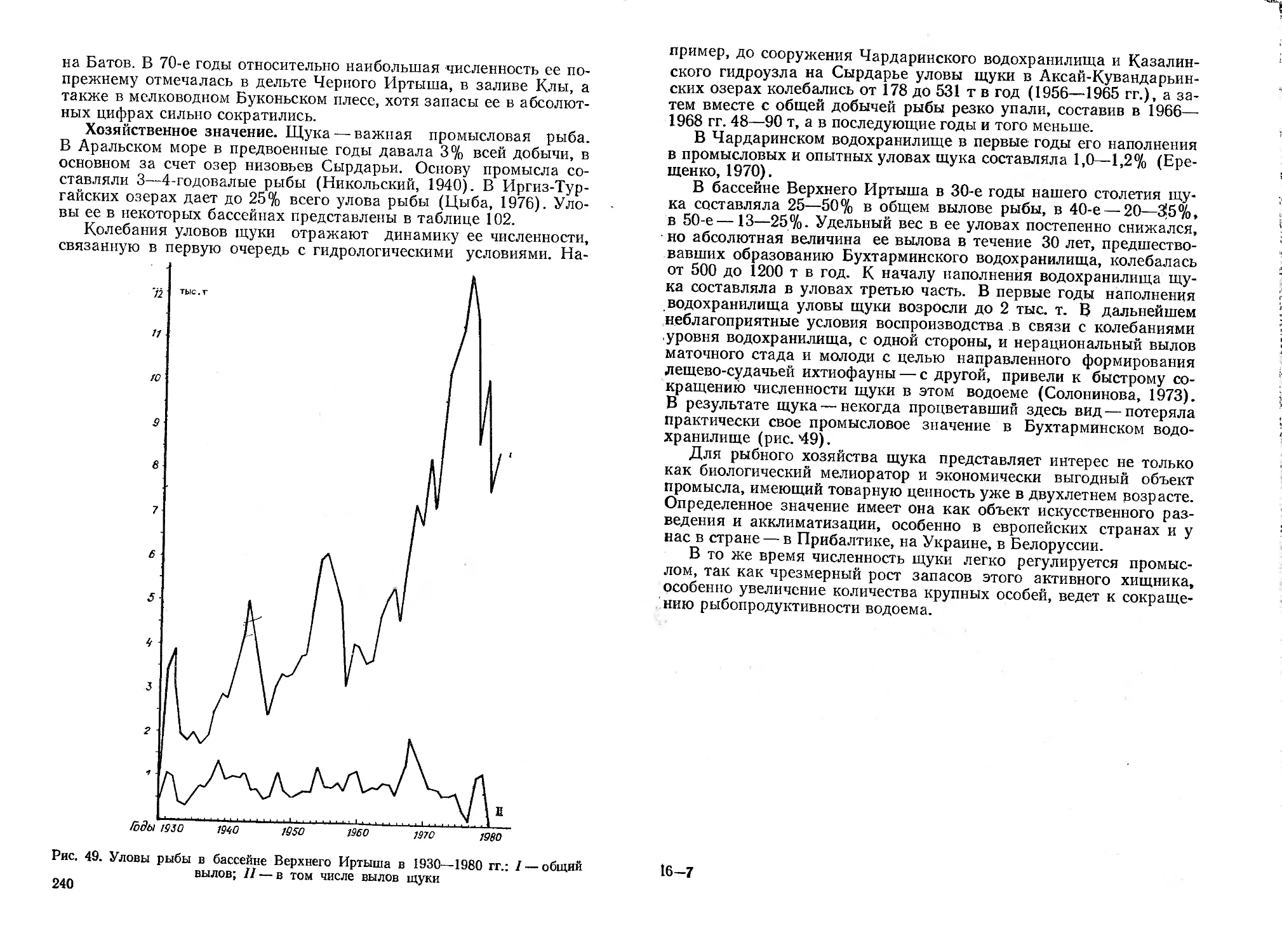
















Автор: Митрофанов В.П. Дукравец Г.М. Песериди Н.Е.
Теги: зоология ихтиология рыбное хозяйство
Год: 1986




Текст
—
АКАДЕМИЯ НАУК КАЗАХСКОЙ ССР
ИНСТИТУТ зоологии
КАЗАХСТАНА
ТОМ 1
МИНОГОВЫЕ, ОСЕТРОВЫЕ, СЕЛЬДЕВЫЕ,
ЛОСОСЕВЫЕ, ЩУНОВЫЕ
Издательство «Н А У К А» Казахской ССР
АЛМА-АТА- 1986
УДК 597
Рыбы Казахстана: В 5-ти т./Митрофанов В. П., Дукравец Г. М., Пе-
сериди Н. Е. и др. — Алма-Ата: Наука, 1986. Т. 1. Миноговые, Осетровые,
Сельдевые, Лососевые, Щуковые. — 272 с.
Обобщаются сведения по ихтиофауне пресных вод республики. Опи-
сываются история изучения рыб водоемов Казахстана и особенности фор-
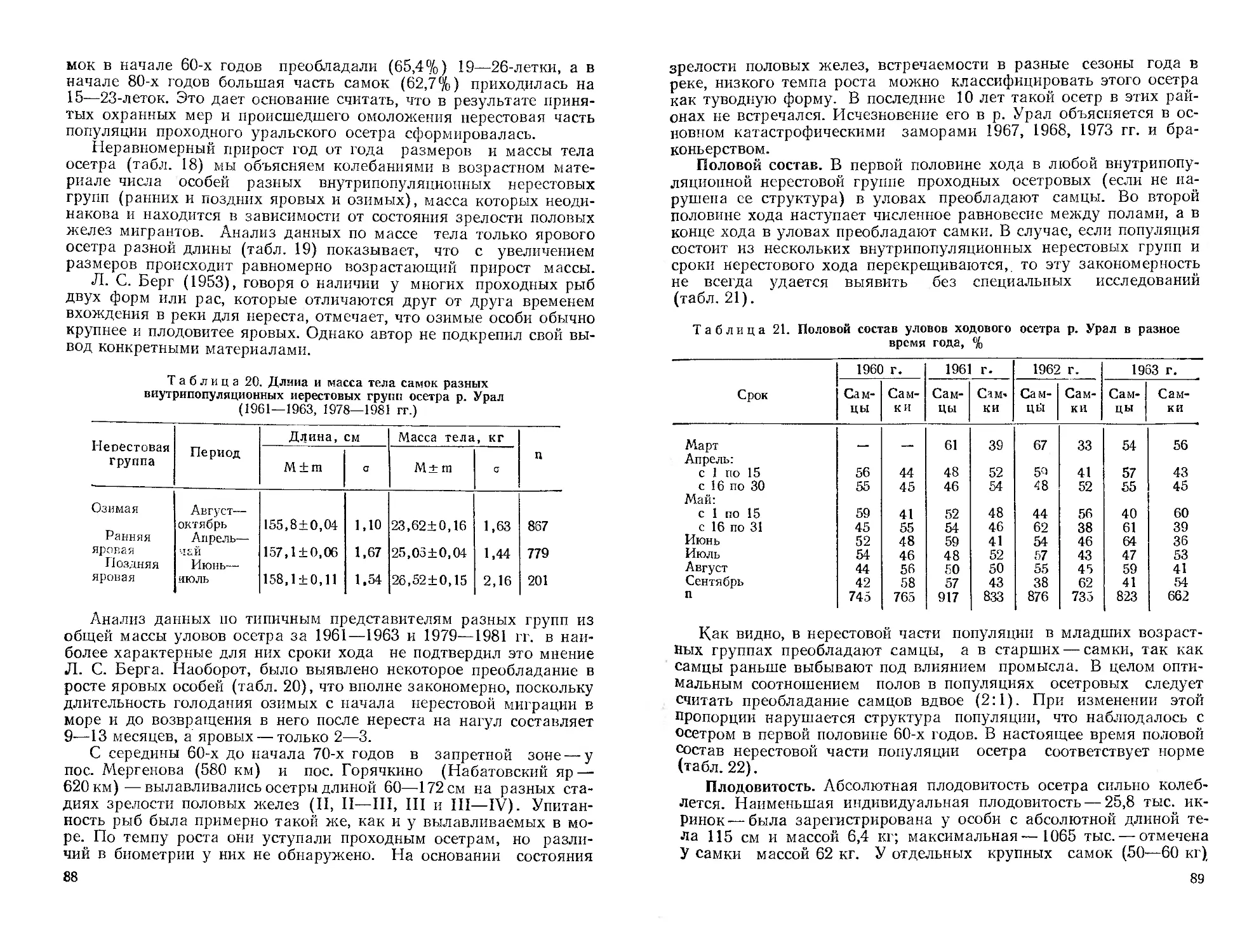
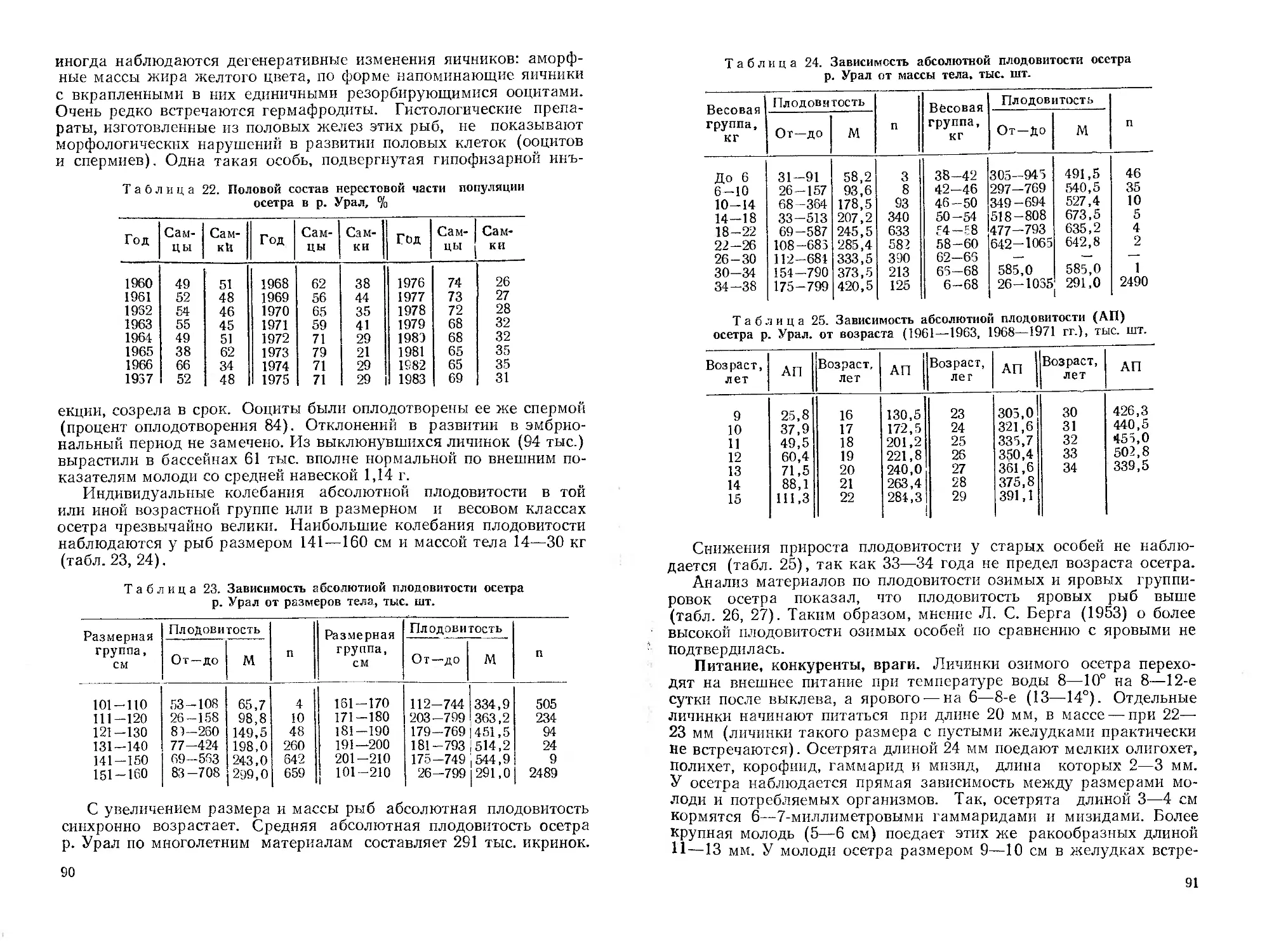
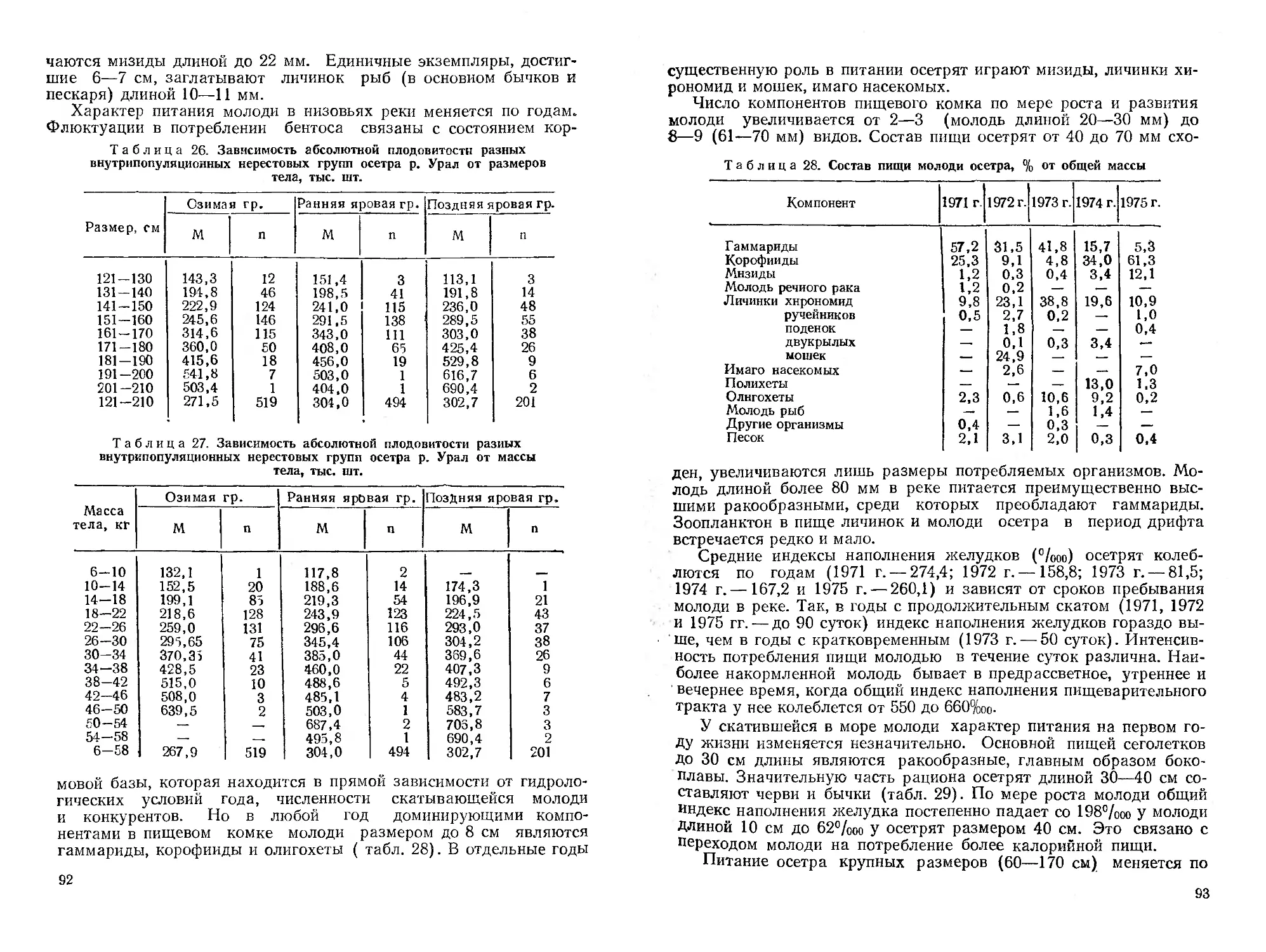
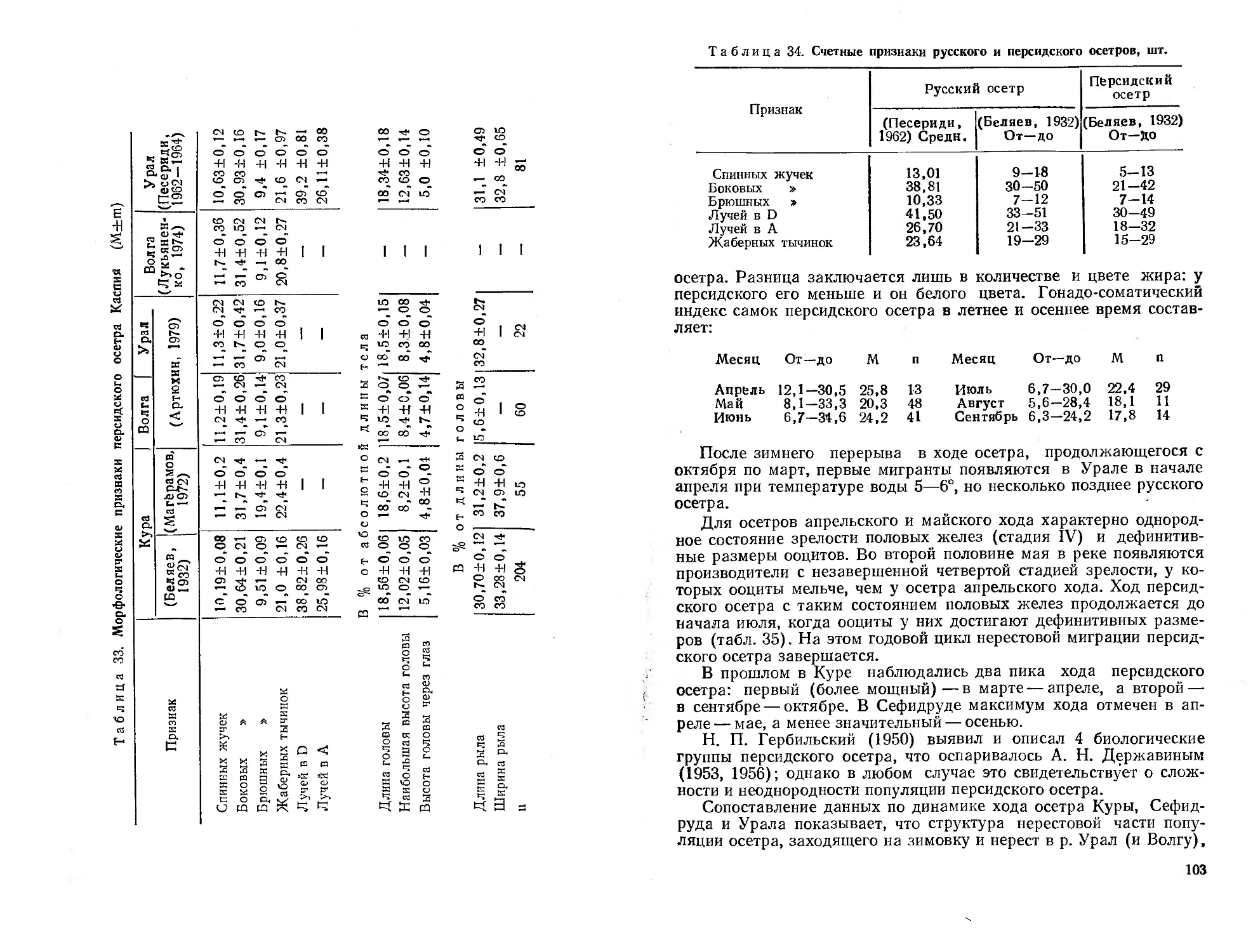
мирования его современной ихтиофаувы. Дается ихтиогеографическое
районирование республики. Приводятся оригинальные и литературные ма-
териалы о систематическом положении, морфологии, биологии, распростра-
нении, численности и хозяйственному значению представителей рыбообраз-
ных (миноги), а также осетровых, сельдевых, лососевых и щуковых рыб.
Книга рассчитана на воологов, ихтиологов, преподавателей биологии,
специалистов в области рыбного хозяйства, воспроизводства и охраны рыб-
ных запасов, а также на широкий круг любителей природы.
Библиогр. 546 назв. Ил. 49. Табл. 102,
Ответственные редакторы
академик АН КазССР Е. В. ГВОЗДЕВ,
доктор биологических наук профессор В. П. МИТРОФАНОВ
..2005000000—072 „ „
Р~ 407(05)—86 7686
© Издательство «Наука> Казахской ССР, 1986
ПРЕДИСЛОВИЕ
В последние годы огромное значение придается охране и ра-
циональному использованию природных ресурсов, в том числе жи-
вотных. В этой связи становится необходимым иметь более полные
сведения о рыбах, населяющих водоемы. .. ।
Изучение ихтиофауны водоемов, выявление рыбных ресурсов, и
разработка мер охраны и рационального использования запасов
промысловых рыб — насущная задача казахстанских ихтиологов.
В развитии этих исследований большое значение должна иметь
подготавливаемая к изданию фундаментальная пятитомная сводка
«Рыбы Казахстана», включающая все имеющиеся сведения о
морфологии, распространении, биологии и поведении рыб. Отсут-
ствие подобной сводки в значительной степени сдерживало реше-
ние ряда практических и научных выводов и разработок. Напри-
мер, составление в свое время «Красной книги Казахской ССР»
показало полное отсутствие сведений о многих редких видах рыб,
обитающих в водоемах республики. Из-за неполных данных ряд
редких рыб не попал в первое издание этой книги.
Видовой состав ихтиофауны различных бассейнов, прежде
всего Арала и Балхаша, претерпел такие изменения, что теперь
получить сведения о некоторых аборигенных видах можно только
по литературным данным, а их далеко не достаточно.
В течение последних десятилетий непромысловые, так называ-
емые малоценные и сорные, рыбы почти не изучались. Поэтому
достаточно полные сведения можно найти в литературе лишь о
некоторых из них. Большинство же сорных аборигенных видов
остается мало исследованными.
Под сомнением сейчас находится оправданность самостоятель-
ности ряда таксономических единиц — видов, подвидов, экологи-
ческих рас. Решить вопрос о праве их на существование в этих
рангах возможно только после тщательной ревизии отдельных ви-
дов и родов.
3
Планируемые дальнейшие коренные изменения гидрографиче-
ской сети Казахстана и продолжающиеся акклиматизационные
работы требуют ответа на многие вопросы, связанные с иссле-
дованиями ихтиофауны казахстанских водоемов. Все это еще более
остро ставит задачу подготовки и издания наиболее полной свод-
ки по рыбам Казахстана.
Приступая к ее созданию, авторы вполне отдают себе отчет в
том, что многие видовые очерки целиком компилятивны, во мно-
гих случаях описание сводилось к пометке «Нет сведений». И тем
не менее сводка выявит степень изученности отдельных видов и
послужит определенной вехой, точкой отсчета новых исследований
по изучению ихтиофауны республики. Обобщение накопленных
сведений поможет в планировании дальнейших исследований, из-
бавит от повторения допущенных промахов как в организации
науки, так и в методах ведения рыбного хозяйства.
В этой связи в сводке выделен раздел, посвященный акклима-
тизационным работам. Пора, наконец, иметь полное представление
° уом, к чему привели внушительные затраты на реконструкцию
ихтиофауны. Казахстанские водоемы фигурируют в литературе в
качестве наиболее убедительных примеров торжества идеи аккли-
матизации. Но если сделать простой подсчет, сколько и какой
рыбы вылавливалось до акклиматизационных работ и после них,
TQ результат получится неотрадный.
. Ихтиофауна Казахстана, сложившаяся в суровых условиях
.изолированных водоемов с непостоянным гидрологическим режи-
мом, исторически была адаптирована к ним и отличалась высоким
(Эндемизмом и своеобразием. В настоящее время это своеобразие
полностью утеряно и его вряд ли можно восстановить в полном
объеме. Поэтому создание заповедных водоемов стало назревшей
необходимостью не только и не столько для ценных промысловых
рыб, сколько для видов эндемичных.
Понимая всю сложность взятых на себя обязательств, авторы
.все же считают возможным провести ревизию некоторых родов,
поскольку концепция полиморфности вида им более близка и в
ряде случаев представляется единственно приемлемой для объяс-
нения происходящих изменений в морфологии, экологии и биоло-
гии близкородственных номинальных видов.
Структура сводки такова: в первом томе, кроме двух общих
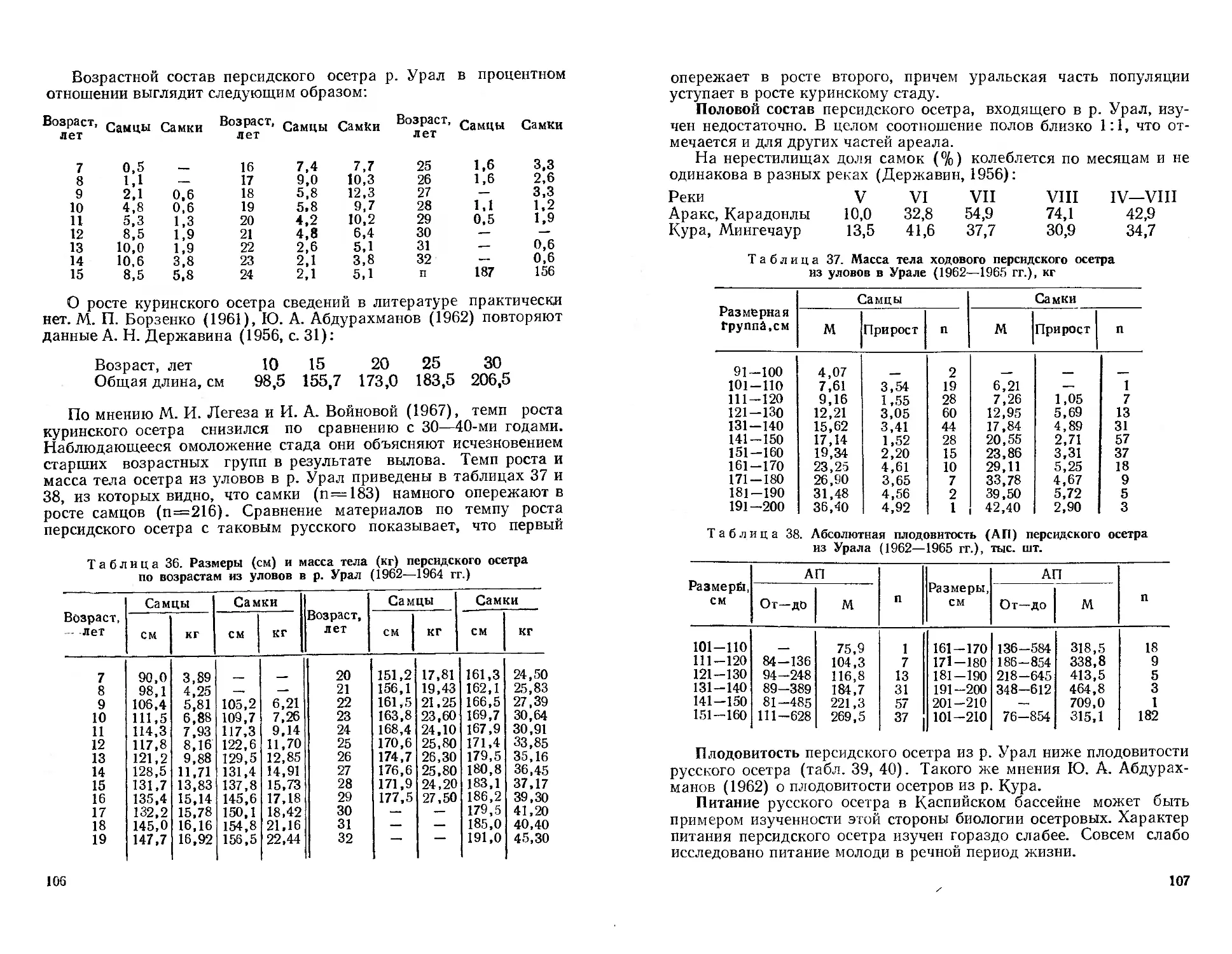
глав, приводится, описание аборигенных и переселявшихся в пре-
делах республики представителей семейств миноговых, осетровых,
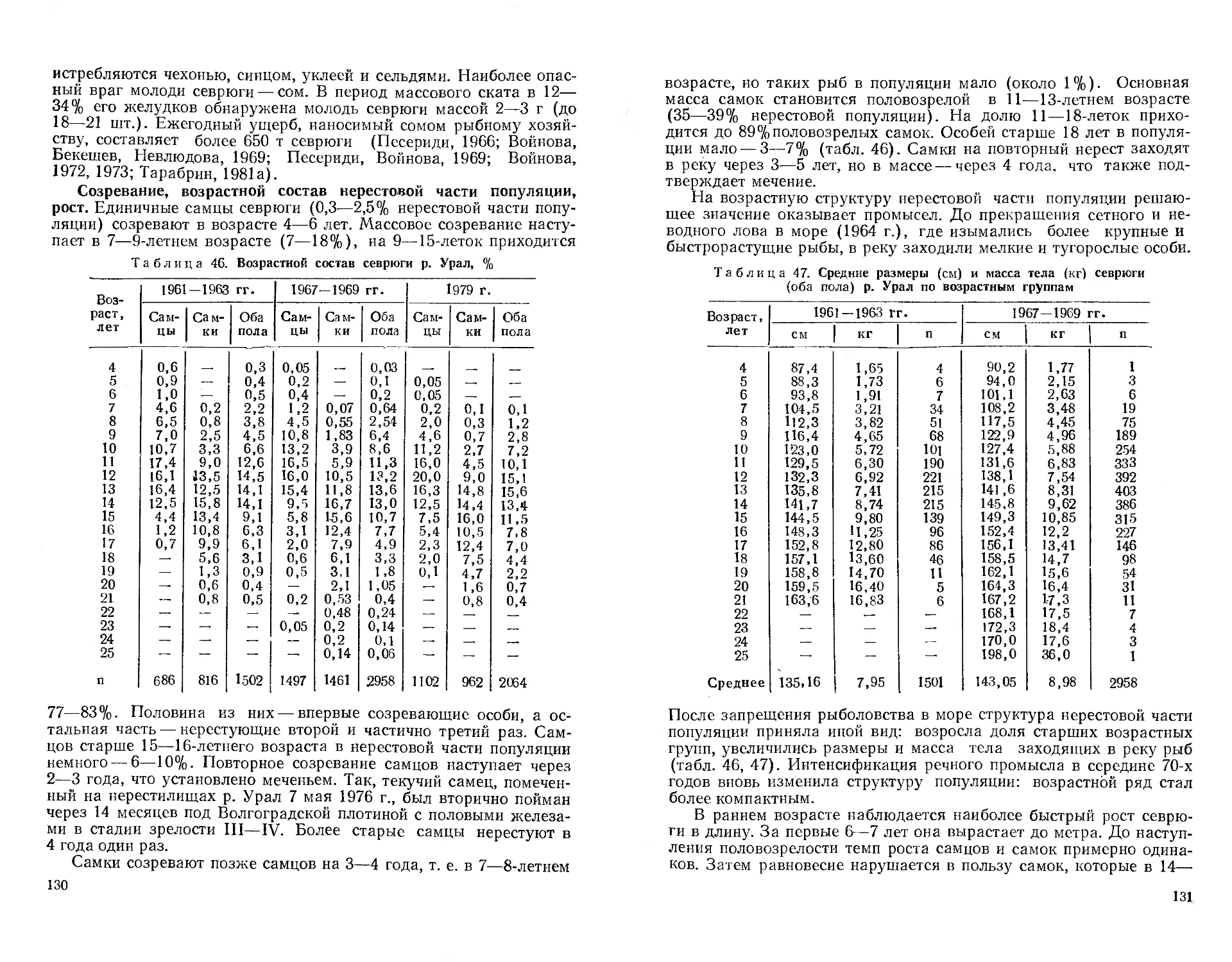
(Сельдевых, лососевых и щуковых рыб; второй и третий тома по-
священы аборигенным карповым рыбам; в четвертом томе будут
описаны представители остальных семейств, а в пятом сделан ана-
лиз акклиматизационных работ и дано описание новых для Казах-
стана натурализовавшихся видов. Рыбы, обитающие только в
Каспийском море и не заходящие в пресные воды, не описываются,
я В первом томе предисловие, глава 1 и вводная часть главы 3
написаны Г. М. Дукравцом и В. П. Митрофановым, глава 2 —
В П Митрофановым; круглоротые описаны А. Н. Полторыхиной;
осетровые (кроме лжелопатопоса и сибирского осетра) — Н. Е. Пе-
сеоиди; сырдарьинский лжелопатонос, сельдевые, аральский и кас-
пийский лососи, белорыбица и щука —Г. М. Дукравцом; сибир-
ский осетр и нельма — В. И. Ерещенко; таймень, ленок и хариус —
В П Митрофановым, он же составил и определительные таблицы.
В написании некоторых очерков в качестве соавторов принимали
участие: но белуге —С. С. Захаров, по шипу — В. А. Мельников
(аральская и балхашская популяции), по щуке — Л. Н. Солонино-
ва (бассейн р. Иртыш, преимущественно Бухтарминское водохра-
нилище), в доработке материалов по сибирскому осетру и нельме
и подготовке очерков об этих видах — И. В. Орлова; в работе над
очерком по нельме и в редактировании очерков по круглоротым и
осетровым — Г. М. Дукравец. Ряд замечаний и дополнений к гла-
ве 1, преимущественно по рыбоводству и гидропаразитологии, сде-
лала А. И. Горюнова. Научная редакция тома проведена
проф. В. П. Митрофановым.
Рисунки рыб и ротовых воронок миног выполнены в основном
на основе иллюстраций к сводкам Л. С. Берга (1948), Г. В. Ни-
кольского (1971), Т. 3. Захидова и др. (1971). Рисунки осетровых
рыб (кроме сибирского осетра и лопатоноса), микрофотографии их
половых желез и срезов маргинальных лучей стерляди сделал
Н. Е. Песериди, фотографии сибирской миноги (рис. 3—5) —
А. Н. Полторыхина, графики — авторы соответствующих очерков.
Остальные иллюстрации, в том числе и оригинальный рисунок
ленка из оз. Маркаколь, выполнил И. В. Митрофанов.
Авторы надеются, что эта книга будет полезна труженикам
рыбного хозяйства и научным работникам — ихтиологам, рыбово-
дам, зоологам, а также преподавателям, студентам и широкому
кругу любителей природы. Такая обширная сводка едва ли может
быть свободной от недостатков, и авторы будут признательны всем
за высказанные в этом отношении критические замечания и поже-
лания.
.’4
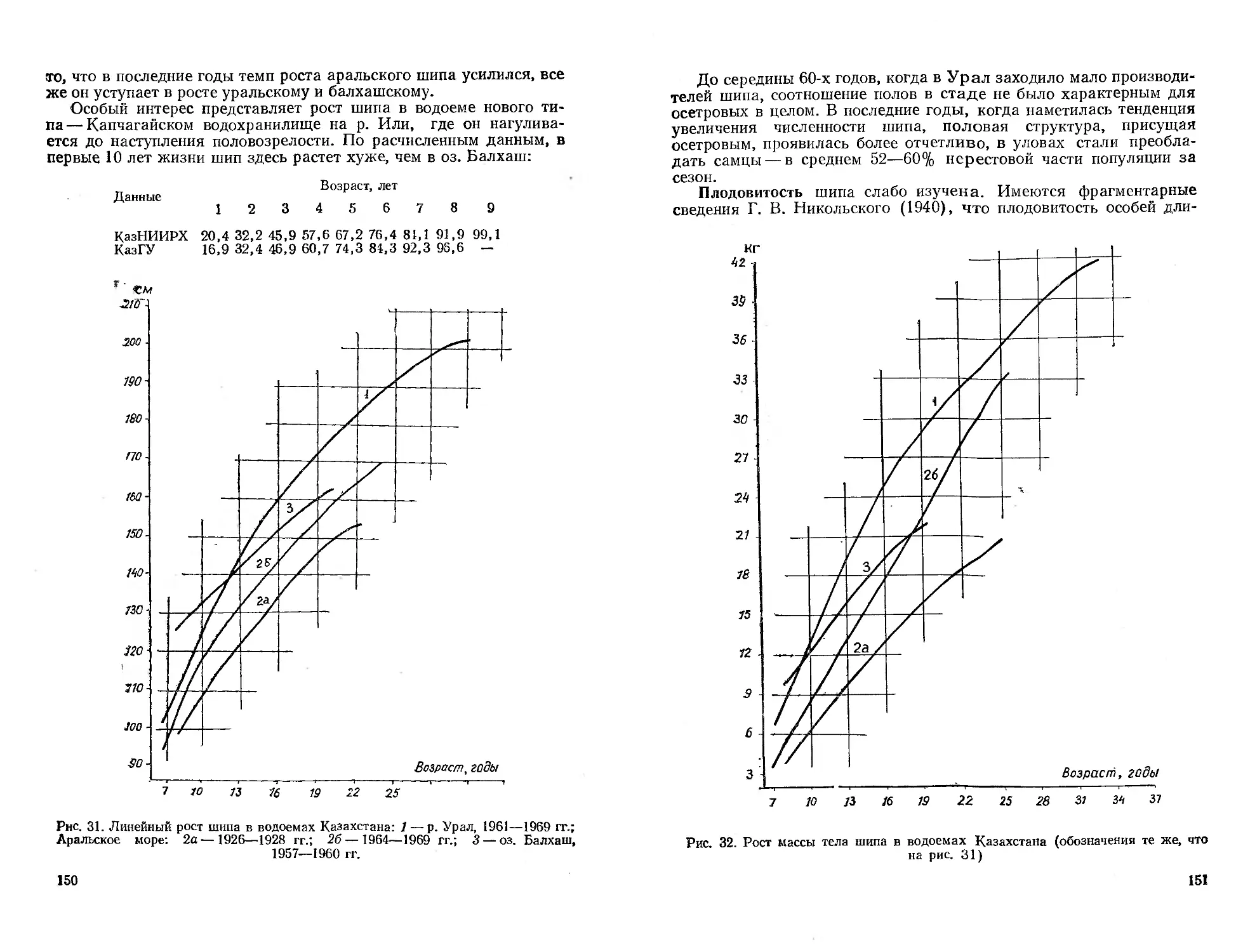
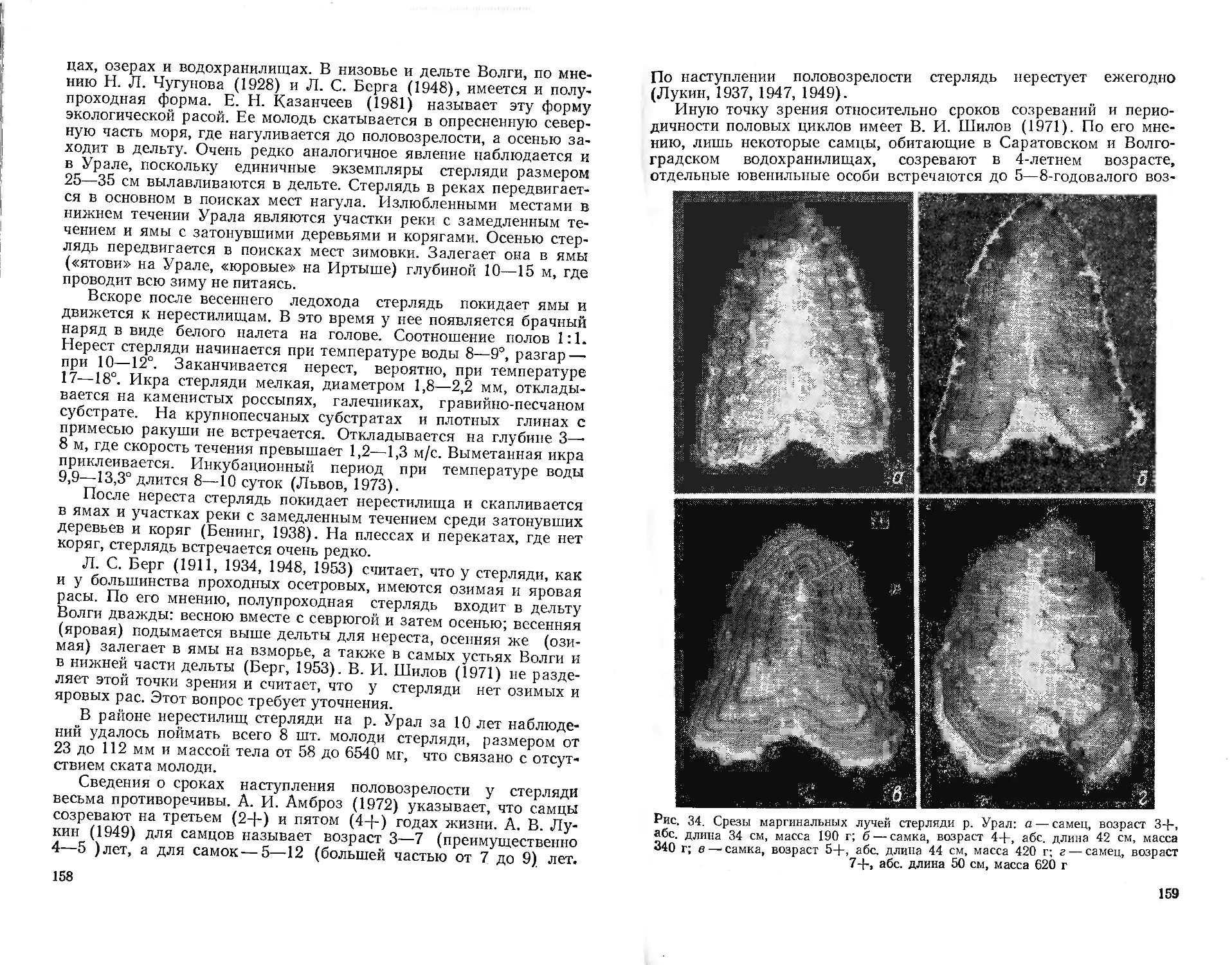
Глава 1
ИСТОРИЯ ИЗУЧЕНИЯ ИХТИОФАУНЫ КАЗАХСТАНА
Начало серьезным исследованиям рыб водоемов, расположен-
ных на территории Казахстана в его современных границах, по-
ложил один из крупнейших натуралистов XVIII в. Петр Симон
Паллас (1741—1811). В 1768—1773 гг. он совершил путешествие
от Петербурга до Забайкалья, исследовав по пути реки Волгу,
Урал, Терек, Иртыш, Обь, Енисей и Каспийское море.
В опубликованном на немецком языке 5-томном труде «Путе-
шествия по разным провинциям Российского государства»
П. С. Паллас дал, в частности, первое описание уральского рыбо-
ловства, которое «нигде в России столь хорошо не распоряжено и
законом не ограничено, как в здешнем месте» (цит. по П. Г. Бори-
сову, 1964). В этой же книге подробно описываются и отдельные
виды рыб, среди которых впервые — севрюга из р. Урал. В 1811—
1814 гг. вышел на латинском языке труд П. С. Палласа «Zoogra-
phia Rosso-Asiatica», в третьем томе которого описано около
300 видов морских и пресноводных рыб России, в том числе и мно-
гие водящиеся в Казахстане.
Первые научные сведения о рыбах Аральского моря дал
Г. Мейендорф — участник русского посольства в Бухару и Хиву в
1820 г. Он же привез экземпляр шипа, который в 1828 г. был опи-
сан А. Ловецким как новый вид. Некоторых рыб Арала и низовьев
Сырдарьи указывает А. И. Бутаков в отчете об эспедиции 1848—
1849 гг. (Володкин, 1961). Первые сведения об ихтиофауне Бал-
хаш-Илийского и Алакольского бассейнов получены А. И. Шрен-
ком в 1840 г. (Доброхотов, 1948).
Во второй половине XIX — начале XX в. изучение ихтиофауны
Казахстана велось более интенсивно в связи с освоением этого
края и благодаря усилиям ряда замечательных русских путеше-
ственников и ученых.
В 1853—1857 гг. в бассейне Каспийского моря работала боль-
шая экспедиция по изучению жизни рыб и рыболовства. Руководил
6
экспедицией академик К. Э. Бэр (1792—1876), вопросами рыбо-
ловства занимался Н. Я. Данилевский (1822—1885). Результаты
экспедиции вошли в многотомный труд «Исследования о состоя-
нии рыболовства в России», который йвился сводкой ценных дан-
ных о биологии и промысле рыб, сохраняющей научное значение
до настоящего времени.
Своими исследованиями К- Э. Бэр и Н. Я- Данилевский зало-
жили основы теории динамики численности рыб и биологической
продуктивности водоемов. Их важнейшей заслугой является опре-
деление путей и методов приложения ихтиологии к решению
практических запросов рыбного промысла, разработка четких
принципов рационального рыбного хозяйства. Кроме того, <
Н. Я. Данилевский дал подробное описание состояния уральского
рыболовства, список и некоторые данные по биологии рыб, водя-
щихся в бассейне Урала, включая оз. Челкар, Камыш-Самарские
и Кушумские озера.
* Познанию ихтиофауны Казахстана в значительной степени
способствовали сборы ученых-путешественников Н. А. Северцова
(1827—1885) и А. П. Федченко (1844—1873). Н. А. Северцов в
1857—1879 гг. совершил 7 путешествий в Казахстан и Среднюю
Азию: в 1857—1858 гг. побывал в низовьях Сырдарьи, где собрал
первую коллекцию рыб (около 17 видов) этой реки, в 1860—
1862 гг. — в бассейне Урала, в 1864 г. — в междуречье Чу — Сыр-
дарья, в 1865—1868 гг. путешествовал по Тянь-Шаню, в 1879 г.
посетил Семиречье. А. П. Федченко в 1869—1871 гг. совершил
4 путешествия по Средней Азии, результатом которых были сведе-
ния о рыбах среднего течения Сырдарьи и бассейна р. Или (Мур-
заев, 1956).
Сборы Н. А. Северцова по Сырдарье обработал профессор
К. Ф. Кесслер (1815—1881) и опубликовал в труде «Ихтиологиче-
ская фауна Туркестана» (1872). Это первые подробные сведения о
рыбах данного региона. В 1874 г. К- Ф- Кесслер опубликовал еще
одну работу, где описал 11 видов рыб, на основе обработки сбо-
ров А. П. Федченко, переданных в Зоомузей Московского универ-
ситета.
В 1874—1876 гг. К. Ф- Кесслер участвовал в работе Арало-Кас-
пийской экспедиции, снаряженной Петербургским обществом
естествоиспытателей. Материалы экспедиции вместе с результата-
ми обработки сборов А. И. Шренка, Н. А. Северцова и А. П. Фед-
ченко вошли в сводку К- Ф. Кесслера «Рыбы, водящиеся и встре-
чающиеся в Арало-Каспийско-Понтийской ихтиологической об-
ласти» (1877). В этом труде описано 279 видов рыб, в том'числе
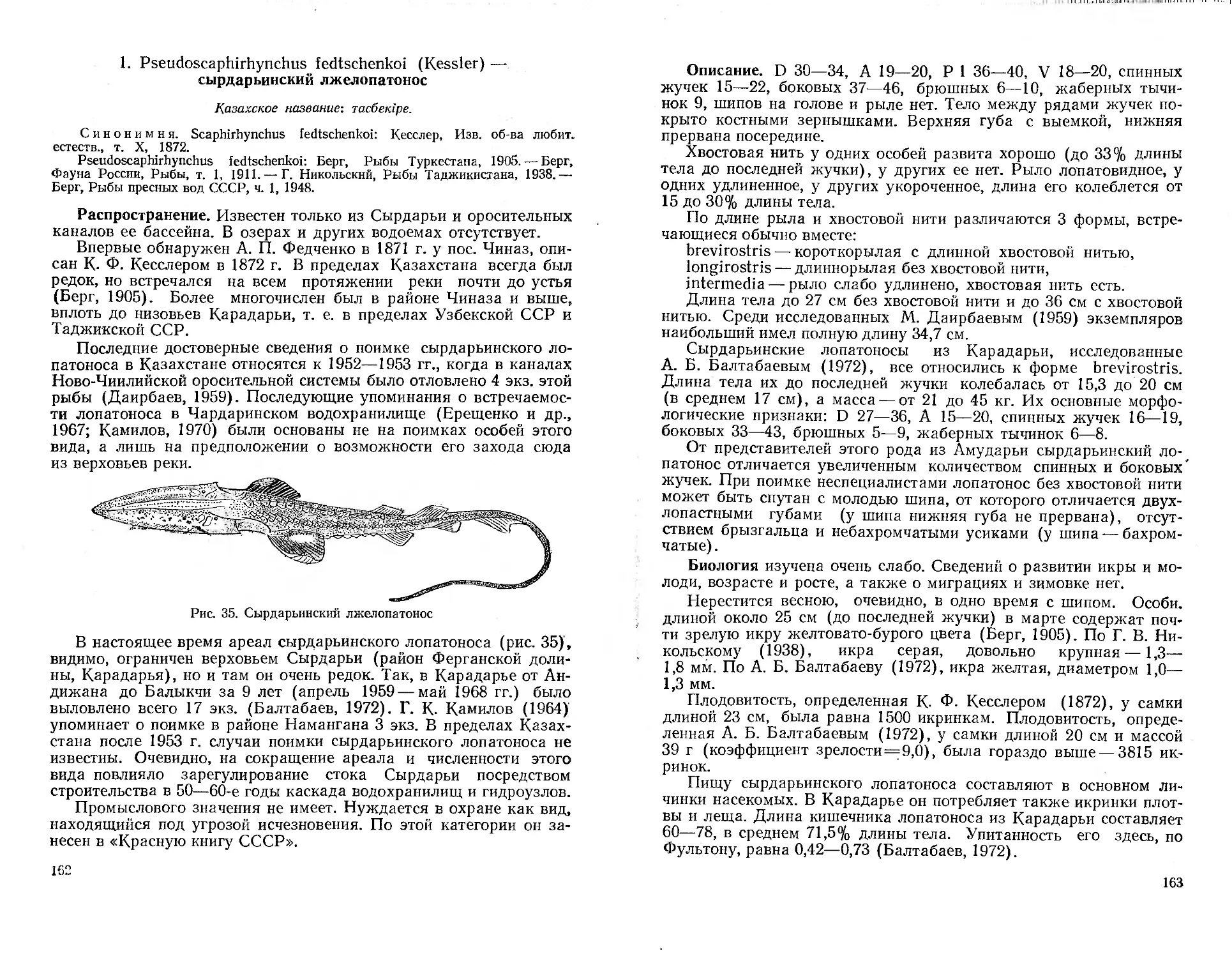
76 — новых для науки. Среди последних — обнаруженный в 1871 г.
А. П. Федченко сырдарьинский лжелопатонос и туркестанские
язь, пескарь и усач, балхашская маринка, голый осман, жерех-
лысач, остролучка, колюшка, балхашский окунь и др. К. Ф. Кес-
слер разработал первую экологическую классификацию рыб, подраз-
делив их на морские, солоновато-водные, разноводные, проходные,
7
полупроходные и пресноводные виды. Он стал основополож-
ником ихтиогеографического районирования нашей страны, пред-
ложив объединить три южных морских бассейна в одну общую
Арало-Каспийско-Понтийскую ихтиологическую область.
Свой вклад в изучение рыб Казахстана сделал и Н. М. Прже-
вальский (1839—1888), собравший коллекцию рыб в бассейне
р. Или во время Лобнорско-Джунгарского путешествия в 1876—•
1877 гг. Сборы его затем обработал и описал С. М. Герценштейн
(1854—1894) в труде «Научные результаты путешествий
Н. М. Пржевальского по Центральной Азии. Рыбы» (1888—1891),
где, в частности, впервые дал описание илийской маринки.
В 1877 г. коллекцию рыб из р. Тентек и озер Алаколь и Сасык-
коль собрал Г. И. Поляков. В 1879—1882 г. на Алакольских озерах
изучал рыб О. Финш. В Балхаш-Илийском бассейне следующие по
времени материалы собрали А. М. Никольский в 1884 г. и
Л. С. Берг в 1903 г.
В 1882 г. О. Финш и А. Брэм побывали в Горном Алтае, дали
первые сведения о рыбах оз. Маркаколь. Вслед за ними водоемы
Алтая и Джунгарии посетили известные натуралисты А. М. Ни-
кольский (1884), С. Абрамович (1888), А. Суворцев (1888),
П, Г. Игнатов (1897), В. В. Сапожников (1900), Н. И. Яблонский
(1907), И. М. Пахомов (1909) и др. Они с разной степенью точ-
ности сообщали об ихтиофауне водоемов, расположенных на пути
их следования, обращая в первую очередь внимание на «обилие
рыбы». Но уже в то время состояние запасов ценных промысловых
рыб вызывало беспокойство, как следствие «неразумного их ис-
пользования» (А. С., 1926).
В бассейне р. Урал в конце XIX в. работал Н. А. Бородин, кото-
рый дал материалы об оз. Челкар (Чархал), привел сведения о его
гидрофауне, впервые описал чархальскую селедочку и чархаль-
скую воблу (Серов, 1956). Кроме того, Н. А. Бородин изучал со-
стояние уральского рыболовства и биологию осетровых рыб
(1897а), впервые описал персидского осетра (18976).
Последний дореволюционный период в изучении ихтиофауны
Казахстана связан в основном с именем Л. С. Берга (1876—
1950)—крупнейшего советского ихтиолога и географа. Еще в
1898 г. он вместе с П. Г. Игнатовым (1874—1902) обследовал со-
леные озера Омского уезда (ныне Кокчетавская и Павлодар-
ская обл.) Кызылкак, Селетытениз и Теке. Рыбы в этих озерах не
было, но в р. Селеты обнаружены гольяны и караси.
В 1899 г. Л. С. Берг был назначен заведовать рыбными про-
мыслами на Арале и Сырдарье, а П. Г. Игнатов продолжил изуче-
ние озер Северного Казахстана, в частности Кургальджино и
Тенгиз в низовье р. Нура. В последнем ему удалось обнаружить
колюшку. В 1902 г. во время обследования озер Кокчетавской об-
ласти П. Г. Игнатов умер. Коллекции рыб, собранные им в Север-
ном Казахстане, в основном остались необработанными
(Р. Л. Берг, 1955).
Л С. Берг с 1899 по 1907 г. занимался изучением Аральского
моря' и р. Сырдарьи, в том числе ихтиофауны этого бассейна. Он
собрал богатейший материал по систематике, биологии и промыслу
рыб опубликованный в сводке «Рыбы Туркестана» (1905). Летом
1903 г. Л. С. Берг по поручению Туркестанского отдела Русского
географического общества провел краткосрочное обследование
оз. Балхаш и низовьев р. Или, где обнаружил 2 вида маринок,
гольца и балхашского окуня.
В 1908 г. Л. С. Берг впервые описал аральского лосося, а в
1912 г. опубликовал работу «О рыбах рек Сарысу и Нуры в Акмо-
линской области», где дал первые списки и описания рыб этих
рек. Конечно, списки были еще далеко не полными. Например,
для Сарысу описано лишь 6 видов, а для Нуры — 3 вида по коллек-
ции, собранной М. А. Балыклейским.
Из других ученых, прямо или косвенно способствовавших по-
знанию рыб Казахстана до Великого Октября, можно назвать еще
П, П. Семенова-Тян-Шанского, совершившего два путешествия в
Семиречье в 1856 и 1857 гг.; Г. Н. Потанина, путешествовавшего в
1863—1864 гг. по Тарбагатаю, Зайсану, Черному Иртышу; В. Грюн-
берга, опубликовавшего заметки о жизни аральского шипа
(1905); А. С. Юлу сова, дополнившего сведения об аральском лососе
(1910); А. С. Покровского, изучавшего рыб и рыболовство на
Аральском море (1916); А. Н. Седельникова, собравшего интерес-
ные сведения об истории рыболовства на озерах Зайсан (1910) и
Маркаколь (1914); В. Городецкого, сообщившего о появлении са-
зана в р. Или (1916) и др.
Во. время поездок в Западный Тянь-Шань собрали первые кол-
лекции рыб бассейна р. Талас Абрамов в 1905 г. (цит. по
Ф. А. Турдакову, 1955) и В. В. Чернавин в 1913 г. По бассейну
р., Урал наиболее полный список рыб и краткие сведения о их
биологии и распространении опубликовал И. П. Навозов (1912).
В первые годы Советской власти основные усилия ихтиологов'
были направлены на комплексное изучение ихтиофауны и состоя-
ния рыбных запасов крупных водных бассейнов республи-
ки — Урало-Каспийского, Арало-Сырдарьинского и Балхаш-Йлий-
ского.
Уже в 1920—1921 гг. на Аральском море и в дельтах Сырдарьи
и Амударьи работала научно-промысловая экспедиция под руко-
водством Ф. А. Спичакова (1922). В отчетах этой экспедиции и в
статьях ее участников Д. П. Филатова и С. Н. Дуплакова описаны
распределение, миграции, нерест, характер питания и некоторые
Другие стороны биологии многих промысловых рыб бассейна.
В 1925 г. на Арале побывала экспедиция отдела прикладной их-
тиологии Института опытной агрономии во главе с Л. С. Бергом,
который затем опубликовал сводку, посвященную анализу состоя-
ния аральского рыболовства и рыбных запасов бассейна (Волод-
Кин, 1961).
В последующие годы здесь работали А. Я. Недошивин, изучав-
8
9
ший рыболовство на озерах низовьев Сырдарьи; А. И. Рабинерсон,
опубликовавший сведения о биологии аральского усача; А. Г. Ка-
гановский, исследовавший аральского леща; М. Д. Ильин, изучав-
ший перспективы рыбоконсервного дела на Арале и собравший
некоторый материал по биологии рыб; М. И. Маркун и А. Н. Про-
батов, опубликовавшие ряд статей по систематике и биологии ос-
новных промысловых рыб и по динамике рыболовства.
В 1929 г. Всесоюзным научно-исследовательским институтом
рыбного хозяйства в г. Аральске была организована стационарная
рыбохозяйственная станция для систематического изучения гидро-
логии, гидрофауны и рыбного хозяйства Аральского моря.
В дальнейшем здесь в разные годы работали известные советские
исследователи, внесшие свой вклад в изучение гидрофауны бас-
сейна: А. Л. Бенинг, Э. А. Бервальд, А. В. Володкин, Н. А. Глад-
ков, А. Ф. Карпевич, П. М. Коновалов, А. П. Макеева, Н. П. Мо-
розова, В. Я. Панкратова, О. Д. Романычева, К- 3. Трусов,
М. А. Фортунатов, Н. 3. Хусаинова, Н. И. Чугунова, Е. А. Яблон-
ская и др.
Большой вклад в изучение ихтиофауны Аральского бассейна и
других водоемов Казахстана внес член-корр АН СССР Г. В. Ни-
кольский (1910—1977). Еще студентом он в 1929 г. принял участие
в составе геологической партйи в обследовании бассейна р. Чу, где
собрал материал для дипломной работы «Рыбы среднего и нижне-
го течения р. Чу», опубликованной в 1931 г. После окончания уни-
верситета он проводил в 1930 г. рыбохозяйственное обследование
озер Северного Казахстана.
С 1931 по 1940 г. Г. В. Никольский работал на Аральской ры-
бохозяйственной станции. В это время им изучена систематика, био-
логия, динамика численности многих рыб бассейна. Результаты
исследований изложены во многих статьях и обобщены в моно-
графии «Ихтиофауна бассейна Аральского моря», состоящей из
двух разделов: «Рыбы Таджикистана» (1938) и «Рыбы Аральского
моря» (1940).
Кроме того, Г. В. Никольский вместе с А. Л. Бенингом в 1932 г.
обследовал низовые озера р. Сарысу и добавил к списку рыб этого
бассейна еще 5 видов (1933). В 1937 г. он побывал на р. Или и
опубликовал затем (совместно с Н. А. Евтюховым) сводку «Рыбы
равнинного течения р. Или» (1940).
В области систематики казахстанских рыб Г. В. Никольский
изучил морфологию быстрянок и голого османа бассейна Сыр-
дарьи, описал самую мелкую пресноводную рыбу в СССР — голь-
ца Северцова, исследовал закономерности внутривидовой измен-
чивости пескарей, сазана, остролучки и других рыб. В области
экологии образцом видового очерка можно назвать работу Г. В. Ни-
кольского «Аральский сазан. Материалы по систематике, биоло-
гии и промыслу» (1934).
Первая крупная советская рыбохозяйственная экспедиция в
бассейне Балхаша работала в 1928—1931 гг. под руководством
П Ф Домрачева. Экспедиция выявила значительные запасы
оыб в озере, обосновала перспективность их промышленной экс-
плуатации, отметила низкую биомассу кормовых ресурсов рыб.
В 1933 г. для проведения стационарных исследований на Балхаше
было открыто отделение ВНИОРХ, где затем в разные годы пло-
дотворно работали Е. В. Бурмакин, Г. В. Домбровский, П. А. Дря-
гин Б. Ф- Жадин, С. Н. Иванов, В. А. Максунов, В. В. Покров-
ский, Н. О. Савина и другие исследователи.
В 1936 г. озера Зайсан и Маркаколь посетила экспедиция
Пермского государственного университета в составе А. О. Таусон,
М. И. Меньшикова, Е. Нельзиной и Л. Масленниковой, опублико-
вавших затем статьи по гидробиологии и ихтиологии этих озер.
В частности, М. И. Меньшиков (1938) описал новые подвиды пес-
каря и гольца и дал полный список ихтиофауны оз. Маркаколь.
В 1939—1945 гг. Н. Г. Некрашевич проводил подробные иссле-
дования ихтиофауны Алакольских озер. Их результаты вошли в
кандидатскую диссертацию «Рыбы Алакольских озер (системати-
ка, биология, промысел)», защищенную в 1946 г. и послужили ос-
нованием для включения озер в сферу деятельности рыбной про-
мышленности.
Первые сведения об ихтиофауне бассейна р. Талас в советское
время получил Д. Н. Кашкаров в 1926 г., который опубликовал в
работе «Экологический очерк района озер Бийлю-куль, Ак-куль
и Аще-куль Аулиеатинского уезда» (1928) сведения о трех ви-
дах рыб.
В 1933 г. рыбохозяйственное обследование этой реки от г. Аулие-
ата (Джамбул) до верховьев провел А. П. Занин (1936) в составе
Киргизской комплексной экспедиции АН СССР. Он описал 8 ви-
дов рыб.
Комплексное изучение ряда озер
Казахстана провели в 30-е годы П. И.* Усачев, М. П. Шеина,
Д. А. Ярошевский, Я. Я- Цееб, Н. 3. Хусаинова, П. Ф. Домрачев.
Я- Я- Цееб опубликовал в 1940 г. работу, в которой привел список
рыб этих озер и некоторые данные по их биологии. В конце 30-х —
начале 40-х годов Н. 3. Хусаинова и Т. И. Синица изучали гидро-
фауну высокогорных озер Тянь-Шаня.
В бассейне р. Урал в 20—30-е годы работали и опубликовали
результаты своих исследований ихтиологи |В. В. Перов, А. Н. Про-
батов, М. И. Маркун, К. А. Киселевич (цит. по Г. X. Шапошни-
ковой, 1964). Они изучали главным образом биологию и промысел
рыб среднего и нижнего течения реки, пойменных и старичных
°зер, а также оз. Челкар — реликтового водоема Понто-Каспий-
ского бассейна. Познанию ихтиофауны этого региона в немалой
степени способствовало и открытие в 1923 г. в г. Гурьеве стацио-
нарного наблюдательного пункта.
Наиболее полные исследования ихтиофауны Среднего Урала
провел ВНИОРХ в 1932—1933 гг. в связи с предполагавшимся
гидростроительством. Результаты их опубликованы М. И. Тихим
Центрального и Северного
10
11
в 1938 г. В 1931—1933 гг. в Северном Каспии проводила исследо-
вания Всекаспийская научная рыбохозяйственная экспедиция,
главной задачей которой было определение путей реконструкции
рыбного хозяйства, рыбных запасов, степени их использования и
перспектив на будущее, а также изучение миграций рыб.
В 1934—1937 гг. сотрудники ВНИРО под руководством
Г. Н. Монастырского с помощью научно-промысловой разведки
Северного Каспия провели, по существу, впервые, большую ис-
следовательскую работу по вобле, охватив и казахстанскую часть
ее ареала. Результаты этих исследований изложены в сборнике
«Вобла Северного Каспия» (ч. 1, 1939; ч. 2, 1940).
Следующий этап в изучении ихтиофауны республики наступил
после окончания Великой Отечественной войны, когда в Институте
зоологии АН КазССР образовался сектор, а затем отдел ихтиоло-
гии и гидробиологии, усиленный в дальнейшем Аральским (дирек-
тор—А. В. Володкин) и Балхашским (директор — Н. П. Серов)
отделениями и Алтайской базой (заведующий — В. И. Ерещенко).
С 1948 по 1959 г. указанным сектором (отделом) института за-
ведовал И. К. Иванов (1906—1968), в последние годы жизни
(с 1962 г.) он был директором КазНИИРХ. Ученый много сил отдал
организации ихтиологических и .гидробиологических исследований
в Казахстане; он — один из пионеров внедрения в республике био-
логического метода борьбы с кровососущими насекомыми путем
вселения в водоемы гамбузии.
Стационарные исследования в первые послевоенные годы про-
водились в трех основных рыбопромысловых районах — Аральском,
Балхашском и Зайсанском; основное внимание было направлено
на изучение запасов ценных промысловых видов рыб и рациональ-
ную организацию промысла. Назрела необходимость и более де-
тально изучить гидрофауну многочисленных в республике изоли-
рованных водных систем. От кратковременного их посещения уче-
ные переходят к длительным наблюдениям, проводимым обычно
совместно гидробиологами и ихтиологами. Так, под руководством
члена-корреспондента АН СССР В. А. Догеля, а затем —
В. И. Доброхотова в 1943 г. были начаты исследования на оз. Зай-
сан и р. Или, в которых принимали участие А. И. Горюнова,
К. В. Смирнова, Л. В. Таронова (Даришева), Е. В. Гвоздев,
П. А. Дианов, П. Ф. Мартехов и другие исследователи.
В конце 40-х — начале 50-х годов Н. П. Серов и С, К. Тютень-
ков исследовали бассейн р. Нуры с Кургальджинскими озерами,
систему Камыш-Самарских и Кушумских озер, оз. Челкар в бас-
сейне Урала. В 1949 г. на озерах Бийликоль и Акколь (бассейн
Таласа) побывали А. И. Горюнова, П. Ф. Мартехов и А. Ф. Си-
дорова, сделавшие описание биологии здешнего сазана (1956) и,
в частности, явление массового уродства его (Горюнова, 19566).
В 1951—1953 гг. А. И. Горюнова и А. С. Малиновская изучали
Джезказганское (Кенгирское) водохранилище и (совместно с
П. Ф. Мартеховым) Алакольские озера.
ГОДЫ "Г Г-- —.
----М. М. Даирбаев (1959), а в бассейне Иргиз — Тургая, остав-
---------------------------------------------------- — • •тлтттгуг О п Л/-ХГГ»С1
VIСЯ ним in nv — *----— - ------------------- . .
А. Ф. Сидорова, исследовавшая язя и карасей (1956, 1959),
, изучавшая щуку и окуня (1956, 1959) и
" '«ГЧГ-Г'Х ТЛ ЧГХГГ 1 ПСС —
В 1952__1953 гг. В. И. Ерещенко детально обследовал бассейн
Сарысу, дал список обитающих здесь рыб с кратким описанием
их морфологии, биологии и промыслового значения (1956). В эти
же годы ихтиофауну ирригационных систем бассейна Сырдарьи
изучал М. М. Даирбаев (1959), а в бассейне Иргиз — Тургая, остав-
шемся почти не изученным в ихтиологическом отношении, работа-
ли А. Ф. Сидорова, исследовавшая язя и карасей (1956, 1959),
А. К. Шиленкова, изучавшая щуку и окуня (1956, 1959) и
j/ Г. Сидоров, описавший паразитов рыб (1956). В 1955—1956 гг.
В. П- Митрофанов и В. А. Тэн изучали гидрофауну оз. Маркаколь,
а в 1956—1959 гг. А. П. Чабан и В. А. Киселева — гидрофауну Усть-
Каменогорского водохранилища.
. • В изучении ихтиофауны республики в это время принимали
участие не только казахстанские ученые. В 1947, 1951—1953 гг.
член-корреспондент АН КиргССР Ф. А. Турдаков (1899—1968)
собирал материалы в бассейне Таласа и в 1955 г. опубликовал
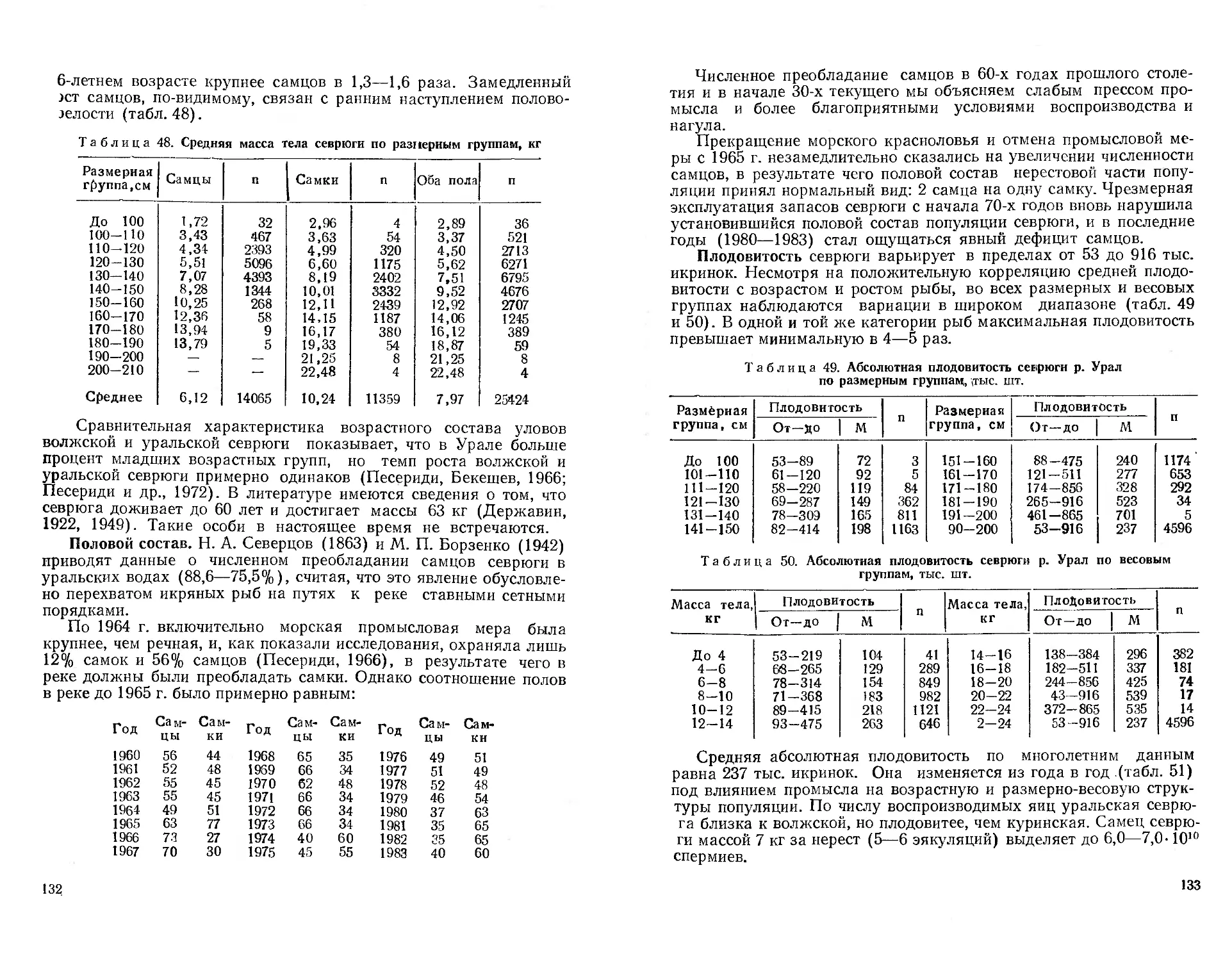
«Очерк ихтиофауны Таласского бассейна», где содержится наибо-
лее полное описание ихтиофауны, рассматриваются особенности
ее систематического состава и источники формирования. В 50-е
годы в бассейне р. Чу и на озерах бассейна р. Талас работал со
студентами доцент Киргизского университета И. А. Пивнев.
В 1949—‘1954 гг. в бассейне р. Урала проводила исследования
Г. X. Шапошникова (1964).
В ходе перечисленных и других подобных исследований были
в значительной степени уточнены и расширены сведения о ихтио-
фауне Казахстана, которые затем были обобщены в многочислен-
ных статьях и в серии монографических работ по отдельным вод-
ным системам.
В середине 50-х годов в связи с освоением целинных земель
возникла необходимость в рыбохозяйственном обследовании (бо-
нитировке) малых водоемов республики, сведения о которых были
крайне скудны и отрывочны. В то же время со всей очевидностью
встал вопрос о развитии прудового рыбоводства, оно было пред-
ставлено единственным в республике Алма-Атинским прудхозом.
Нужно было изучить сырьевую базу развития прудовых и озер-
ных рыбных хозяйств в различных климатических зонах Казах-
стана.
Экспедиционные отряды Института зоологии АН КазССР,
Института ихтиологии и рыбного хозяйства (КазНИИРХ), органи-
зованного в 1959 г. в системе республиканской Академии наук и
в 1962 г. переданного в систему Минрыбхоза; Института водного
хозяйства АСХН КазССР, Казахского государственного универ-
ситета им. С. М. Кирова, Казахского женского педагогического
института провели бонитировку водоемов большинства областей
Республики.
В- И. Ерещенко и А. С. Малиновская обследовали Северо-Ка-
захстанскую область, А. И. Горюнова — Кустанайскую и Целино-
радскую, Л. В. Таронова и М. М. Даирбаев — Кокчетавскую,
12
13
А. Ф. Сидорова и В. М. Обухова — Павлодарскую и Восточно-Ка-
захстанскую, В. П. Митрофанов и В. А. Тэн — Чимкентскую и
Джамбулскую. В 1960—1962 гг. В. П. Митрофанов изучал водо-
емы Алма-Атинской, Талды-Курганской и Карагандинской облас-
тей, а Г. М. Дукравец — Джамбулской области.
Дальнейшее расширение и углубление ихтиологических и гидро-
биологических исследований в республике сдерживалось нехваткой
специалистов. Поэтому в 1961 г. в КазГУ была открыта кафедра гид-
робиологии и ихтиологии во главе с инициатором ее создания про-
фессором Н. 3. Хусаиновой (1911—1971). Сотрудники и аспиран-
ты кафедры В. П. Митрофанов, А. Ф. Сидорова, Г. М. Дукравец,
А. П. Фаломеева, Т. И. Збарах и другие вместе со студентами ак-
тивно включились в решение научных и рыбохозяйственных проб-
лем. В дальнейшем из выпускников кафедры сформировались
мощные коллективы специалистов в самом университете и в
КазНИИРХ, Минрыбхозе КазССР, Казахрыбводе, прудовых хо-
зяйствах республики.
В 1963—1968 гг. экспедиционные отряды КазГУ провели бони-
тировку водоемов семи областей: Павлодарской — в 1963 г.
(А. Ф. Сидорова, А. А. Баимбетов, Р. X. Мамилова), Кзыл-Ордин-
ской — в 1964 г. (Г. М. Дукравец, А. А. Баимбетов, Р. X. Мамило-
ва), Актюбинской (А. А. Баимбетов, Г. М. Дукравец, А. П. Фа-
ломеева) и Чимкентской (Г. М. Дукравец, Р. X. Мамилова)—в
1965 г., Алма-Атинской — в 1966 г. (В. П. Митрофанов, Г. М. Дук-
равец) .Семипалатинской— в 1967 г. (Г. М. Дукравец, А. П. Фа-
ломеева) и Уральской — в 1968 г. (они же и Е. А. Меркулов).
Многие водоемы республики были обследованы экспедициями
КазНИИРХ: в 1963—1966 г.— под руководством И. К. Иванова в
Северо-Казахстанской, Целиноградской, Карагандинской и Ураль-
ской областях (И. А. Амиргалиев, Н. Б. Воробьева, В. В. Селез-
нев, О. Тленбеков, А. П. Чабан и др.); в 1969—1971 гг. — под ру-
ководством И. А. Пивнева в Джамбулской (В. С. Башунов,
Г. М. Сечной), Северо-Казахстанской и Кокчетавской областях
(И. Ф. Лысенко, М. И. Сироткин, А. С. Фокина); в 1971—1974 гг.—
под руководством Д. А. Амангалиева в Уральской области
(Л. А. Аксаментова, С. А. Попова, Г. С. Трофимов); в 1971—
1974 гг. — под руководством и при участии Г. М. Дукравца в
Кзыл-Ординской (А. И. Мачулип, Л. Н. Сумбаева, Л. И. Шарапова
и др.), Карагандинской, Целиноградской, Кокчетавской и Северо-
Казахстанской областях (Ю. А. Бирюков, Л. О. Пичкилы,
Л. И. Шарапова и др.). Экспедиции КазНИИРХ в 1972—1975 гг.
в Актюбинскую область (К. П. Цыба) и в 1974—1975 гг. в Тургай-
скую область (Н. Ф. Ефимова, Г. С. Трофимов) завершили бонити-
ровку водоемов республики.
Эта работа, растянувшаяся более чем на 20 лет, результаты
которой изложены в многочисленных отчетах и статьях, выявила
прежде всего непостоянство водного фонда республики. Повтор-
ные обследования ряда областей показали отставание реализации
рыбохозяйственных мероприятий от проводимой реконструкции
водных систем. Вместе с тем были получены основные гидрохими-
ческие, гидробиологические и ихтиологические характеристики
большинства водоемов, раскрыты некоторые связи изменения ви-
дового состава ихтиофауны с колебаниями уровня воды. В част-
ности, отмечено (Горюнова, 1962), что по мере усыхания степных
озер и ухудшения условий обитания в них в однополой (самки)
популяции серебряного карася появляются самцы.
По материалам бонитировок были даны рекомендации о про-
ведении рыбоводно-мелиоративных мероприятий на озерах, пред-
ложены, схемы ведения рациональных рыбных хозяйств и'комп-
лексного использования озер. Разработаны биологические обос-
нования на организацию озерно-товарных хозяйств: в Целино-
градской области — Майбалыкского и Ерментауского (А. И. Го-
рюнова), в Кокчетавской — Имантауского (Л. И. Фролова), в
Кустанайской —Аккабакского (К. П. Цыба) и Токтасского
(А. И. Горюнова), в Северо-Казахстанской — Петропавловского
(Н. Ф. Лысенко), в Кзыл-Ординской — Камышлыбашского
(Г.М. Дукравец), в Талды-Курганской — Канбактинского (А.И. Го-
рюнова) и др. При этом хозяйства южной зоны Казахстана были
ориентированы на выращивание карповых рыб, а северной зоны —
сиговых.
Бонитировочные работы вскоре получили новое направление в
связи с широко развернувшейся с 1957 г. акклиматизацией рыб в
казахстанских водоемах с целью повышения их рыбопродуктив-
ности и улучшения качества продукции. Начали давать результа-
ты и акклиматизационные мероприятия, проведенные до 1950 г.:
интродукция сазана из Балхаша в оз. Зайсан, шипа, усача и ле-
ща из бассейна Арала в оз. Балхаш, севрюги — из Волги и Урала
в Сырдарью и др.
Опыт акклиматизации нуждался в изучении, анализе и обоб-
щении. На этом в той или иной мере сосредоточили внимание их-
тиологи, гидробиологи и ихтиопаразитологи республики. Так,
Н. Е. Быков (1961, 1963, 1964, 1968 и др.) и Е. Л. Маркова (1962,
1964, 1975 и др.) изучали акклиматизацию рыб в Аральском море;
П. Ф. Мартехов (1959, 1961 и др.), В. И. Ерещенко (1966, 1968,
1972 и др.), X. К- Исмуханов (1976, 1978 и др.) —в бассейне Верх-
него Иртыша; Н. П. Серов (1963, 1972, 1975 и др.) —в Балхаше.
Монографические исследования отдельных видов рыб, натура-
лизовавшихся в Балхаше, провели: по вобле — В. С. Башунов
(1974), по жереху — Н. Н. Башунова (1974), по лещу — К. П. Цы-
ба (1975), по сому — Н. Ф. Лысенко (1976). Л. И. Фролова
(1974) изучала биологию пеляди и рипуса и возможность их раз-
ведения в водоемах Северного Казахстана.
Г. М. Дукравец в кандидатской диссертации и серии статей
проанализировал результаты реконструкции ихтиофауны в озерах
бассейна р. Талас (1962, 1964, 1965, 1966 и др.), затем изучал бело-
го амура в бассейне р. Или (1967а, 1968, 1972а, 1975 и др.), вместе
15
14
с А. И. Мачулиным — змееголова, вселенного попутно с раститель-
ноядными рыбами в бассейн Сырдарьи (1974, 1975, 1978 и др.).
А. Ф. Сидорова с группой выпускников университета в 1964 г.
взялась за внедрение радужной форели в горные водоемы Юго-
Восточного Казахстана и изучение особенностей ее акклиматиза-
ции здесь (Сидорова, Курмангалиева, 1968; Сидорова, Буранбае-
ва, 1970; Сидорова, 1973, 1978 и др.). С 1975 г. подобные иссле-
дования параллельно ведутся ею (вместе с Ю. А. Бирюковым и
сотрудниками кафедры ихтиологии МГУ) и по камчатской мики-
же, завезенной в Казахстан.
В ряде самостоятельных и совместных публикаций В. П. Митро-
фанов и Г. М. Дукравец описали некоторые закономерности
морфо-биологической изменчивости различных видов рыб при ак-
климатизации в водоемах Казахстана (Дукравец, Митрофанов,
1972, 1975; Дукравец, 1977, 1982 и др.), провели анализ акклима-
тизационных работ в некоторых южных бассейнах республики
(Митрофанов, Дукравец, 1963, 1966; Митрофанов, 19716; Митро-
фанов и др., 1980 и др.).
А. А. Баимбетов J1975, 1977), М. Н. Батраева (1970, 1975, 1981),
В. В. Селезнев (1972а, 1974) и другие исследователи изучали так
называемых «сорных» рыб, натурализовавшихся в бассейне Бал-
хаша. Это перечисление можно было бы продолжить.
Параллельно выполнялись и другие интересные исследования.
Так, В. В. Кормилин изучил размножение и развитие балхашской
маринки(1972), А. А. Баимбетов — морфо-биологическую измен-
чивость маринок бассейна Балхаша (1974), А. С. Стрельников —
рыб Алакольских озер (1974), М. А. Абдильдаев (1975) и
В. А. Мельников (1977) — возможность использования рыб в
борьбе с комарами и т. п.
Рост водохранилищ в республике, в том числе появление таких
крупных, как Бухтарминское, Чардаринское, Капчагайское и др.,
необходимость их рационального рыбохозяйственного освоения в
условиях комплексного использования водных ресурсов на базе
научных рекомендаций привели к созданию в 1975 г. в КазНИИРХ
лаборатории водохранилищ под руководством В. И. Ерещенко.
Однако и до этого основные водохранилища тщательно изуча-
лись в ихтиологическом отношении по мере их наполнения. На во-
дохранилищах Верхнего Иртыша, например, были организованы
постоянные наблюдения Алтайским отделением КазНИИРХ.
В результате были выполнены, в частности, монографические
исследования о рыбах Усть-Каменогорского водохранилища (Ча-
бан, 1965), о сазане (Федотова, 1973), щуке (Солонинова, 1974),
леще и судаке (Исмуханов, 1981) Бухтарминского водохранили-
ща; о формировании ихтиофауны водохранилищ канала Иртыш—
Караганда (Аббакумов, 1977); путях интенсификации рыбного
хозяйства на Бугуньском водохранилище (Кузнецова-Короткова,
1981). Подробное изучение ихтиофауны Чардаринского водохра-
нилища в первые годы его накопления провел с группой сотруд-
16
в В И- Ерещенко (1970). Кроме того, рекогносцировочное об-
НИК°ование малых водохранилищ Южного Казахстана выполнила
СЛТоВ4__1965 гг. экспедиция Казахского университета (Дукравец,
Митрофанов, 1970).
Успехи первых лет акклиматизации вселяли радужные надеж-
ы на резкое увеличение добычи рыбы в естественных и искусст-
венных водоемах. Однако уже к середине 60-х годов стало ясно,
что обойтись без интенсивных форм ведения рыбного хозяйства
невозможно. Поэтому значительная часть исследователей пере-
шла к изучению вопросов искусственного воспроизводства рыбы в
естественных водоемах (Н. С. Бабаев, Е. Л. Галактионова), био-
техники прудового и садкового рыбоводства, генетики и селекции
прудовых рыб.
Изучением особенностей выращивания рыб в прудах система-
тически начали заниматься в Казахстане с момента организации в
1962 г. в структуре КазНИИРХ рыбоводной группы (руководи-
тель—А. И. Горюнова), преобразованной в 1969 г. в лабораторию
прудового рыбоводства (с 1980 г. заведующий — Ж. Г. Сарсем-
баев).
Достигнуты значительные успехи в разработке биотехники за-
водского получения личинок карпа и растительноядных рыб, в
изучении закономерностей искусственного воспроизводства и вы-
ращивания рыб дальневосточного комплекса (белого амура, бело-
го и пестрого толстолобиков), в исследованиях экологических ос-
нов выращивания прудовых рыб (Г. М. Агапова, М. Н. Батраева,
А. И. Горюнова, О. М. Кан, В. В. Кормилин, М. Д. Линчевская,
С. А. Попова, Ж- Г. Сарсембаев и др.).
В качестве одного из интенсификационных мероприятий пру-
дового рыбоводства с 1975 г. начаты работы по разведению и вы-
ращиванию в прудах разных рыбоводных зон Казахстана трех ви-
дов буффало — новых для республики рыб (М. Н. Батраева,
Е. К- Саурская).
Большое научное и практическое значение представляют иссле-
дования по улучшению породы карпа и растительноядных рыб,
начатые с 1970 г. в рамках лаборатории прудового рыбоводства, а
с 1980 г. продолженные в самостоятельной лаборатории селекции и
генетики рыб КазНИИРХ (заведующий — Р. М. Цой).
Доказана перспективность использования химических мута-
генов в рыбоводстве, возможность сокращения срока создания от-
дельных линий карпа с помощью индуцированного гиногенеза,
раскрыты коррелятивные связи между морфологическими и фи-
зиологическими признаками прудовых рыб (Ю. Ф. Голодов,
Т. С. Зак, В. В. Кормилин, Н. X. Мусина, Р. М. Цой, С. Ф. Шус-
това и др.).
Первые опыты по садковому выращиванию радужной форели и
Карпа, в том числе на теплых водах Ермаковской ГРЭС, были про-
ведены в 1974—1976 гг. Казгосуниверситетом (Ф. Ф. Гибадулин,
А. Тлеубердин). Результаты показали принципиальную возмож-
2-7 и
ность и перспективность такого способа получения товарной рыбы,
В дальнейшем работа в этом направлении была продолжена со-
трудниками КазНИИРХ, которые разработали, в частности, тех-
нологическую схему товарного выращивания форели в садках на
Капчагайском водохранилище (Ж- Г. Сарсембаев, X. А. Тлеубер-
дин, П. И. Пак, М. М. Шарипова).
К изучению закономерностей биологической продуктивности
водоемов первыми в Казахстане приступили сотрудники КазГ^
под руководством сначала И. 3. Хусаиновой, а затем — В. П. Мит-
рофанова.
В 1966—1970 гг. они в рамках Международной биологической
программы (МБП) изучали особенности формирования биопро-
дукции на разных трофических уровнях в оз. Караколь бассейна
р. Или (Chusainova е. а., 1970; Мамилова, 1970; Шарапова, 1971
и др.). Результаты исследований получили высокую оценку спе-
циалистов. Признанием определенных достижений в этом направ-
лении явилось открытие в 1974 г. в университете проблемной лабо-
ратории по изучению биопродуктивности степных и полупустын-
ных водоемов Казахстана (заведующий — Г. М. Дукравец).
Широкое развитие орошаемого земледелия и ирригационных
систем в Южном Казахстане явилось стимулом для ихтиологов
КазГУ к развитию еще одного направления исследований — изу-
чению пространственно-временной структуры распределения рыб
в водоемах и на этой основе разработке (совместно с сотрудниками
ИЭМЭЖ АН СССР) экологических способов защиты рыб от по-
падания в различного типа водозаборные сооружения (Джарде-
малиев и др., 1978; Орлов, 1980; Нездолий, 1982 и др.).
В настоящее время постоянные ихтиологические наблюдения
на оз. Балхаш и Алакольских озерах проводит КазНИИРХ (ди-
ректор— Д. А. Амангалиев), на Аральском море и озерах низовьев
Сырдарьи в условиях изменяющегося гидрорежима ведут исследо-
вания сотрудники Аральской рыбохозяйственной лаборатории (ра-
нее— отделение) КазНИИРХ (до 1983 г. возглавляла Е. Л. Мар-
кова, а затем 3. Ермаханов). Для проведения исследований в Се-
верном Казахстане организована комплексная лаборатория этого
института в г. Петропавловске. В г. Усть-Каменогорске работает
Алтайское отделение КазНИИРХ. В бассейне р. Урала изучение
ихтиофауны, и в первую очередь осетровых, успешно проводит
Урало-Каспийское отделение ЦНИОРХ под руководством
Н. Е. Песериди.
Большой вклад в изучение ихтиофауны республики вносят ка-
захстанские гидробиологи, изучающие питание и пищевые взаи-
моотношения рыб. Среди них М. Я- Ветышева, Н. Б. Воробьева,
В. А. Киселева, Ш. Г. Курмангалиева, Э. В. Логиновских, А. С. Ма-
линовская, Р. X. Мамилова, Р. Е. Садуакасова, Т. С. Стуге,
С. К. Тютеньков, В. А. Тэн, А. П. Фаломеева и др.
С ихтиологическими исследованиями неразрывно связано изу-
чение паразитофауны рыб, начало которому в Казахстане поло-
паботы В. А. Догеля и Б. Е. Быховского (1934) на Араль-
ЖИ"м море. Регулярный и планомерный характер оно принимает
Сосле организации в 1943 г. специальной лаборатории при секторе
"° пгип Казахского филиала АН СССР под руководством
В А Догеля (с 1947 г. — Е. В. Гвоздева), а затем (в 1962 г.) —ла-
йппатории болезней рыб в КазНИИРХ, которой заведовали сна-
п!>ля К В. Смирнова, в 1974—1981 гг. — А. И. Агапова, а с
1981 г. — Н. С. Жук.
Изучение паразитов рыб получило широкий размах и охвати-
ло большинство бассейнов республики, а также прудовые хозяй-
ства. Исследования осуществлялись преимущественно в плане ре-
гистрации ихтиоценотических изменений, вызываемых акклимати-
зацией рыб и кормовых беспозвоночных, особенностей формирования
паразитофауны акклиматизантов. Анализ и обобщение
фаунистического материала позволили дать зоогеографическую ха-
рактеристику паразитофауны рыб водоемов Казахстана (Смирно-
ва, 1944, 1972, 1974; Догель, Гвоздев, 1945; Гвоздев, 1950; Добро-
хотова, 1953; Агапова, 1956, 1963, 1966; Сидоров, 1956, 1959; Бра-
гина, 1970, 1974; Тленбекова, 1980 и др.).
В последние годы развиваются новые направления гидропара-
зитологических исследований: изучение биологии и циклов раз-
вития гельминтов, биоценотических отношений и роли водных бес-
позвоночных в циркуляции гельминтозной инвазии рыб и рыбояд-
ных птиц (Гвоздев и др., 1978; Сысолятина, 1981; Сидоров,
1983 и др.).
Закономерная последовательность в познании ихтиофауны во-
доемов Казахстана, прослеживаемая в течение полутора столе-
тий,— от усилий натуралистов-одиночек до плановых перспектив-
ных исследований Академии наук Казахской ССР, Казахского
государственного университета им. С. М. Кирова и Казахского ин-
ститута рыбного хозяйства дала свои результаты. И тем не менее
в последние годы стали особенно видны те пробелы, которые обра-
зовались в результате незавершенности разных этапов исследо-
ваний.
Первой попыткой подведения итогов этого нелегкого пути по-
знания явилась монография В. П. Митрофанова «Карповые рыбы
Казахстана» (1973), защищенная как докторская диссертация.
Его опыт использован в работе над настоящей сводкой.
18
Глава 2
ФОРМИРОВАНИЕ СОВРЕМЕННОЙ ИХТИОФАУНЫ
КАЗАХСТАНА И ИХТИОГЕОГРАФИЧЕСКОЕ
РАЙОНИРОВАНИЕ
Современная аборигенная ихтиофауна отдельных водных си-
стем Казахстана образовалась в результате длительного процесса
эволюции и перемещений рыб различных фаунистических комплек-
сов. Чтобы понять пути ее формирования, необходимо проследить
в общих чертах смену теократических и талассократических фаз
развития территории Казахстана до середины третичного периода,
и более подробно — с конца третичного до современного.
В конце палеозоя в Казахстане произошли крупные сдвиги ко-
ренных пород. К этому времени относят появление основных гор-
ных систем: Мугоджар, Каратау, Тянь-Шаня, Джунгарии, Тарбага-
тая, Алтая и Центрального Казахстана. Казахстан превратился в
основном в континентальную страну, и некоторые местности впо-
следствии уже не подвергались трансгрессиям моря. С большой
долей вероятности это можно сказать о Центрально-Казахстан-
ском плато и Зайсанской котловине (Берг, 1908; Наливкин, 1930;
Терлецкий, 1931; Герасимов, 1933; Кассии, 1947; Синицин, 1962
и др.). Об этом свидетельствуют и палеонтологические находки.
Так, амии — типичные представители озерно-болотных ландшаф-
тов известны из района Зайсана, начиная с верхнемеловых отло-
жений, и просматриваются до верхнего миоцена (Стоянов, 1915;
Хисарова, 1967), хотя для Европы В. Н. Яковлев (1961) отмечает
вымирание амий в нижнем миоцене.
В конце мезозоя и в палеоцене произошло погружение суши
на огромных площадях, достигшее своего максимума в эоцене.
В это время море проникло далеко на восток по южному краю
Казахстана, достигнув Ташкентской впадины и образовав заливы
по Чуйской долине до Таласского Алатау и Киргизского хребта.
Активно проходило наступление моря и в севера. Через Йргиз-
Тургайский прогиб южное море соединилось с северным, затопив-
шим Западно-Сибирскую низменность. Именно к этому времени
(юра — мел) приурочивается происхождение карпообразных
20
'Никольский, 1950) и карповых (эоцен — Никольский, 1950; па-
леоцен — Яковлев, 1961).
Если принять точку зрения Г. В. Никольского (1938) о проис-
жлении карповых от индийских рыб, то именно к этому времени
^палеоцену — эоцену) следует отнести миграции их на север и
Распространение в Азии. Сопоставляя время возникновения кар-
повых и их расселение в Азии и Европе (Яковлев, 1961), нетрудно
сделать вывод, что свое эволюционное развитие карповые прохо-
дили в основном на территории нынешней Средней Азии и Казах-
стана. Этому способствовала и обильная озерно-речная система,
определявшая длительное время облик этих местностей.
Наиболее важно выделить те участки, которые не подвергались
трансгрессии моря со времени обособления современных предста-
вителей пресноводных семейств: осетровых, лососевых, щуковых,
карповых и окуневых. Большинство авторов (Никольский, 1950;
Яковлев, 1961; Берг, 1962; Синицин, 1962; Дарлингтон, 1966) отно-
сят их появление к эоцену — олигоцену. Во всяком случае, все ис-
следователи едины во мнении, что к нижнему миоцену сформиро-
вался родовой состав пресноводной фауны. Исключение составля-
ют лишь молодые филогенетические ветви расщепобрюхих карпо-
вых, обособившиеся в середине миоцена. Ф. А. Турдаков (1963)
выдвигает гипотезу об автохтонном происхождении нагорно-азиат-
ской фауны в противоположность точке зрения Г. В. Никольского
об ее индийском происхождении.
Вопрос о происхождении нагорно-азиатской фауны очень бли-
зок к проблеме возникновения карповых или одной из ветвей этой
группы, поэтому нет особых противоречий в выводах Г. В. Николь-
ского и Ф. А. Турдакова, так как определение «глубины» автох-
тонности может быть достаточно различным. В то же время нет
оснований сомневаться в проникновении ряда форм из Индии на
север и нашедших здесь новую родину.
Климатические и гидрологические условия в неогене позволили
расселиться так называемому третичному пресноводному фаунис-
тическому комплексу очень широко. Входящие в него виды разных
семейств обнаружены в ископаемом состоянии почти по всей Евро-
пе, Западной и частично Восточной Сибири, Средней Азии и Ка-
захстане, многие найдены и в Америке. Это позволило исследовате-
лям говорить о единой третичной пресноводной ихтиофауне. Судя
по времени проникновения отдельных видов в различные участки
Палеарктики, местом их возникновения считается море Тетис и
образовавшиеся на его месте озерные системы.
В Казахстане с середины третичного периода морским транс-
грессиям не подвергались следующие участки: Центральный Ка-
захстан, или Казахский мелкосопочник, регион современного Бал-
хаша и Зайсанская котловина. Остальные площади претерпевали
периодическое наступление моря с запада и севера. Однако все
эти изменения имеют еще весьма отдаленное отношение к совре-
менному составу ихтиофаун, поскольку распространение видов
21
было очень широким. Важно отметить лишь один факт — очень
древнее обособление Балхашской впадины, происшедшее раньше,
чем образовались современные семейства. Некоторые исследова-
тели (Кассин, 1947) предполагают, что Балхаш развивался изоли-
рованно с палеозоя.
О широком распространении основных родов пресноводных рыб
в южных, центральных и восточных частях Казахстана в олигоце-
не и миоцене свидетельствуют многочисленные палеоихтиологиче-
ские находки. Наряду с родами Rutilus, Leuciscus, Aspius, Scardi-
nius, Tinea, Chondrostoma, Gobio, Alburnus, Alburnoides, Abramis,
Cyprinus весьма обычны были окуневые — Lucioperca (—Stizoste-
dion), щуковые — Esox, сомовые — Silurus.
Эти находки типично пресноводных рыб заставляют с уверен-
ностью заключить, что большая часть территории Казахстана
(кроме западной) не подвергалась с этого времени трансгрессиям
моря. Все древнейшие трансгрессии — сарматская, меотическая,
понтическая, акчагыльская, апшеронская и новейшие — бакинская,
хозарская, хвалынская — сказались только на формировании фау-
ны Арала и Каспия. Формирование же ихтиофауны других водо-
емов шло своим путем.
Следовательно, исходная ихтиофауна середины третичного
периода была одного типа для всех бассейнов Казахстана, за
исключением, может быть, Балхаша. На дальнейшем формирова-
нии ее в отдельных системах и обособлении сказались тектониче-
ские процессы, прошедшие в конце третичного и продолжающиеся
в четвертичном периодах. Тогда и поднялись вновь древние систе-
мы Тянь-Шаня, Джунгарии, Тарбагатая, Алтая, некоторые из них
более чем на километр. Меньше повысились Казахское нагорье и
Мугоджары.
Собственно, с того времени, когда гидрографическая система Ка-
захстана приобрела очертания, близкие к современным, и следует
искать следы связей и образования современных ихтиофаун.
Хорошо известны изменения наших южных бассейнов — Каспия
и Арала. Их связи со Средиземноморьем и друг с другом неодно-
кратно возобновлялись и обусловили появление в фауне атланти-
ческих элементов. Полное отсутствие морских форм в Арале мож-
но объяснить двояко. Более распространен взгляд, что Арал пол-
ностью и неоднократно высыхал и от него сохранялись только ре-
ки, явившиеся резервациями (убежищами) пресноводных видов.
На этот счет хочется сделать некоторые критические замечания.
Полному высыханию Арала обязательно должно было предшест-
вовать исчезновение пойменных и придаточных озер рек Амударьи
и Сырдарьи, которые в первую очередь зависят от величины стока
рек. Убедительный пример исчезновения пойменных водоемов в
результате уменьшения стока дают Сырдарья и Иртыш в наше
время. Оставшаяся в Арале пресноводная фауна должна обладать
в таком случае следующими показателями: реофильностью, способ-
ностью размножаться в русле рек, а также питаться и зимовать
рсли же предположить, что от Арала (на месте его ложа)’
хпанялись озера какой-либо величины, то они должны быть по-
вышенной солености и способствовать сохранению не пресновод-
ной, а морской фауны.
В Казахстане имеются примеры подобных превращении в ми-
ниатюре, свидетельствующие, что в результате высыхания низовых
озер в реках сохраняется только реофильная фауна и погибает
лимнофнльная, хотя она и пресноводная. В Арале же значительная
часть видов — типичные лимнофилы, достаточно указать на саза-
на, леща, язя, шемаю.
Согласно второй точке зрения, разъединение Арала и
Каспия произошло раньше, чем в Каспий проникли понтиче-
ские элементы. Осуществлявшаяся позднее связь Аральского
бассейна с Каспием через Узбой не могла служить мостом
для проникновения таких видов, как Syngnatus, и в то же время
явилась хорошей водной артерией для расселения проходных рыб
бореального комплекса, например аральского лосося.
Вторая гипотеза более приемлема, хотя обе страдают больши-
ми пробелами. Трудно объяснить обедненность Арала осетровыми.
Ни первая, ни вторая гипотезы не дают на это ответа.
Последним общим бассейном, объединяющим Каспий и Арал,
был, по всей видимости, Апшеронский. Этой точки зрения придер-
живается Н. 3. Хусаинова (1959). Но в это время фауна осетровых
(как, впрочем, и других пресноводных семейств) была вполне со-
временной по своему видовому составу. Происходившие до апше-
ронскбй трансгрессии неоднократные теократические фазы развития
Аральской впадины не могли не отразиться на современном видо-
вом составе рыб и ответ следует искать в истории развития Арала
с самого конца третичного времени. Но почему в Арале не могло
сохраниться с конца третичного периода (или даже с начала чет-
вертичного) большинство каспийских видов осетровых? Объясне-
ние, приведенное Н. 3. Хусаиновой (1959) относительно ряда форм
беспозвоночных, данный случай мало проясняет. Н. 3. Хусаинова
предполагает, что Арал являлся как бы окраинной частью Апше-
ронского бассейна со своеобразным гидрохимическим и гидрогео-
логическим режимом, куда не могли проникнуть типичные морские
виды. Но для проходных осетровых это наиболее вероятные места
обитания или, во всяком случае, не препятствующие их распро-
странению.
Более вероятна третья точка зрения — об опреснении вод Ара-
ла в недавнем геологическом прошлом под воздействием усилив-
шегося притока вод Амударьи.
Л. С. Берг (1962) также допускает подобную возможность
Для Каспийского моря. Он пишет, что обмен между Курой и Сефид-
рУдом (и обратно) облегчался в те эпохи, когда воды Каспия
становились менее солеными, например, во время последней круп-
ой четвертичной трансгрессии—хвалынской, когда абсолютная
высота Каспия достигала 50 м; эту эпоху предположительно при-
22
23
урочивают к последнему, вюрмскому, оледенению. Подобны^
взгляд вполне применим и к Аралу. О том факте, что в Аральске^
море с севера некогда впадала мощная река, свидетельствуют ц
геологи и палеонтологи.
Одни считают, что это был Иртыш, повернутый наступающим
ледником (Кассии, 1947), другие пишут о крупной мощной реке
без конкретизации ее принадлежности. Наиболее четко выражена
эта идея П. Л. Пирожниковым (1937). Он считает, что в Арало-
Каспийскую впадину были повернуты не только реки бассейна
Иртыша, но и все реки Западной Сибири. В его представлении в
начале четвертичного периода Арал сделался проточным водоемом.
Доказанность этих предположений помогла бы ихтиологам решить
многие вопросы взаимопроникновения фаун, например, объяснить
сохранение в Арале эвритермных рыб, так как при опреснении
Арала в подобных условиях неизбежно должно произойти охлаж-
дение его водных масс.
Этим можно объяснить исчезновение таких видов, как пред-
ставители семейства Clupeidae. При просмотре палеоихтиологи-
ческих коллекций Института зоологии АН КазССР совместно с
сотрудниками лаборатории палеонтологии были определены ры-
бы, весьма близкие с Caspialosa caspia. Сборы сделаны М. Д. Би-
рюковым * в скальных породах середины третичного периода се-
вернее Аральского моря на несколько десятков километров.
Наряду с этими проходными видами сельдей здесь встречены
остатки рыб семейств Percidae, Siluridae, Esocidae, Cyprinidae и
рода Lota. Особенно интересно нахождение остатков налима, что
непосредственно указывает на вторжение северных элементов че-
рез Иргиз — Тургай.
В переотложенных песках Северного Приаралья весьма обыч-
ны зубы акул (коллекция Института зоологии АН КазССР, а так-
же Л. С. Гликман, 1964), встречающиеся одновременно с остатка-
ми носорогов, бобров и других млекопитающих, что дало основание
М. Д. Бирюкову и В. В. Кузнецову высказать мнение о пресновод-
ности ископаемых акул Приаралья. По-видимому, это не так.
Находки рыб рода Caspialosa указывают на морской характер вод
Арала в середине третичного периода. Морской облик имела и
его ихтиофауна, продержавшаяся, вероятно, до четвертичного
периода.
Четвертичный период ознаменовался для Арала усиленным
притоком северных вод, последующим опреснением (иначе чем
объяснить такую разницу в солености Арала и Каспия) и некото-
рым охлаждением вод. Представители морской понтическо-каспий-
ской ихтиофауны в Арале погибли, но в нем сохранились пресно-
водные, эвритермные рыбы и позднейшие вселенцы с севера, про-
1 * М. Д. Бирюкову, Б. С. Кожамкуловой, В. В. Кузнецову и Г. Хисаровсй
автор признателен за возможность ознакомления с коллекциями ископаемых рыС
: и палеонтологические консультации.
ие через Узбой. После потери притока северных вод началось
новое осолонение Арала, продолжающееся до наших дней.
Н В целом история развития и изменения ихтиофауны Аральского
бассейна представляется как процесс замещения одних комплексов
ЛОУгими, с включением некоторых видов от прежних комплексов.
Наиболе'е древними «останцами» являются роды Pseudoscaphir-
hvnehus, Aspiolucius и Acipenser, сохранившиеся здесь с третичного
времени. Затем следуют представители средиземноморского комп-
лекса, неоднократно наступавшие на конечные участки бассей-
на__горные потоки и отступавшие назад под воздействием наи-
более молодых групп нагорно-азиатских видов. Этот процесс про-
ходил в течение всего четвертичного периода и продолжается до
настоящего времени.
В ледниковый период сюда проникли представители ледовито-
морского комплекса — бычки и лососевые. Гидрологические изме-
нения отсортировали и сохранили из этих рыб типично пресновод-
ные формы, а поднятие горных систем привело к обозначению
четких границ между нагорно-азиатскими и понто-каспийско-
аральскими видами. Те и другие стремятся к расширению своих
ареалов в случае свободных стаций даже в мало свойственных им
условиях. Нагорно-азиатские виды спускаются в долинные водо-
емы, осваивают озерные акватории, но отступают при появлении
типичных лимнофилов. Лимнофилы продвигаются при увеличении
численности в предгорные участки рек.
В результате наиболее полные, богатые видами ихтиоценозы об-
разовались в срединных течениях рек Сырдарьи и Амударьи.
В меньшей степени подобная картина выражена в других реках
этого бассейна. Современное расхождение в видовом составе рыб
нижнего и среднего течения бессточных рек Южного и Централь-
ного Казахстана есть результат позднейших гидрогеологических
изменений и уменьшения стока. Эти изменения мало отразились
на видовом составе рыб горной части рек, что является прямым
следствием происхождения и разнородности комплексов, состав-
ляющих их ихтиофауну.
Перейдем к формированию ихтиофауны в северных водоемах
Казахстана. Отношение современного бассейна Иртыша к Ледо-
витоморской провинции в прошлом было не столь уж явным. Про-
никновение карповых рыб в древнезайсанскую котловину могло
происходить только по речным системам, а не через морские прос-
торы, так как пресноводность и даже заболоченность древнезай-
санской котловины и ее развитие без талассократической фазы с
начала третичного периода, если не с конца мезозоя-—доказанный
Фаот. Среди ископаемых рыб морские виды не встречены.
Смена же пресноводных форм от мела до современности про-
слеживается отлично. Вначале это были амии, которые обитали
196?Ь °Т мела до миоцена включительно (Лебедев, 1959; Хисарова,
И ® палеоцене к ним добавляются судак, окунь, щука,
сленно увеличивающиеся в миоцене (Хисарова, 1967). Значи-
25
24
тельно позже, вероятно, в середине миоцена, сюда проникают
карповые, близкородственные современным формам. В это время
наблюдалось наиболее широкое развитие озерной сети в Тургае,
который, по всей видимости, и был тем мостом, через который
понто-каспийско-аральские формы проникали в Сибирь и распро-
странялись далее к востоку от бассейна Оби.
К началу четвертичного периода в водоемах бассейна древнего
Иртыша, включая север Казахстана, как и по большинству сибир.
ских водоемов, карповые были представлены всеми основными ро-
дами этого семейства. Общими являются Acipenser, Esox, Ruti-
lus, Leuciscus, Aspius, Alburnus, Alburnoides, Abramis, Chondrosto-
ma, Gobio, Carassius, Cyprinus, Silurus, Perea, Lucioperca (=Sti-
zostedion). Обращает на себя внимание полное отсутствие нагор-
но-азиатских форм в этом списке, что также свидетельствует о
сравнительно позднем их возникновении (по Яковлеву — в конце
миоцена, по Турдакову — несколько позже) и большой самобыт-
ности.
Быстрее расселялись озерно-речные формы, могущие выдержи-
вать известное осолонение и приуроченные к низинному рельефу,
такие, как сазан, лещ и им подобные. Медленнее, но столь же
успешно расселялись рыбы, обитающие в реках с умеренным тече-
нием (пескарь, подуст), которые не могли преодолевать большие
пространства мезогалинных озер. И, наконец, группа рыб — оби-
тателей горных потоков, расселялась через горные системы (марин-
ка) и задержалась в своем движении в горной части Азии.
Сокращение ареалов большинства теплолюбивых видов проис-
ходило уже в плейстоцене. Из состава ихтиофаун выпали жерех,
уклея, быстрянка, лещ, подуст, сазан, судак. В это время произошли
разрывы в ареалах многих видов, разделение водных систем и об-
разовались современные ихтиофауны в Средней Азии, Казахстане
и Сибири. Их слагаемыми в Казахстане оказались средиземно-
морские, бореальные и нагорно-азиатские фаунистические элемен-
ты. Преобладание того или иного фаунистического комплекса яв-
ляется результатом позднейших гидрологических и климатических
условий.
История Балхаш-Алакольского бассейна освещена геологами
менее полно, чем, например, Каспия, Арала и Зайсана. Данные па-
леонтологии тоже чрезмерно скудны. Неоднократные колебания
уровня Балхаша — от огромного озера, объединяющего всю систе-
му Алакольских озер (по Синицину, 1962, например, это синхро-
низируется с хозарской трансгрессией), до, возможно, полного вы-
сыхания— привели к полному исчезновению его прежней ихтио-
фауны и замене ее выходцами из горных рек. А. М. Никольский
(цит. по Н. 3. Хусаиновой, 1955) считает Балхаш остатком древ-
него Ханхайского водоема и относит образование его к третичному
периоду. Подобным образом трактует этот вопрос и В. А. Обручев
(1940). Он считает, что Прабалхаш был большим единым озером,
объединяющим, может быть, современные Балхаш, Алаколь, Эби-
26
Н п и Тели-Нур с начала и до конца третичного периода. Это мнение
Н лучает некоторые палеонтологические доказательства. В районе
П° Андреевка (Талды-Курганская обл.) палеонтологи Института
С ологии АН КазССР обнаружили остатки амии, захороненные
одновременно с тапиром и датируемые эоценом.
В четвертичный период Балхаш усыхал, и большинство иссле-
дователей считают, что в среднем и верхнем плейстоцене и в го-
лоцене между Балхашем и Алаколем не существовало связи (Кос-
тенко, 1946; Сваричевская, 1952; Галицкий, 1957). К ним присо-
единяются некоторые ихтиологи (Серов, 1961) и гидробиологи
(Малиновская, 1959). Наиболее вероятно, что Балхаш потерял
связь с Алаколем в послеледниковое время, т. е. сравнительно не-
давно. К. В. Курдюков (1951) отмечает наличие озерных террас к
северу'от Алаколя на высоте около 400 м над современным уров-
нем озера. На основе этого он считает доказанным факт связи
Балхаша с Алаколем, но отрицает существование в прошлом ёди-
ногеЙЙссейна с Эби-Нуром.
Действительно, озерные террасы наблюдаются во многих окру-
жающих Балхаш горных системах и на отдельных скалистых
останцах. Они встречаются в хребтах Джунгарского Алатау в доли-
не р. Или и в Северном Прибалхашье более чем в 150 км от со-
временной береговой линии. Их абсолютная высота над уровнем
морЯлВ пределах 700—800 м. Это, конечно, не означает, что уро-
вень' Прабалхаша находился в этих отметках, так как сами тер-
расы могли подняться, что соответствует общему орогенезу дан-
нойгместности. Но то, что уровень озера был выше современного
на сотни метров, не вызывает сомнений.
Л; С. Берг (1958) считает, что Балхаш полностью высыхал в
сравнительно недавнем времени. Он утверждает, что Балхаш в
его-теперешнем виде — сравнительно молодое озеро; котловина
его раньше была сухой. Это положение Л. С. Берг связывает с
послеледниковой эпохой *.
С этой точки зрения современная ихтиофауна Балхаша должна
считаться одной из самых молодых в Казахстане. Проникновение
в нее окуня до сих пор остается загадкой. Наиболее аргументиро-
ванно рассмотрел этот вопрос Б. Ф. Жадин (1949), предположив-
ши®, что окунь проник в бассейн Балхаша из Зайсанской котлови-
ны в верхнетретичное время. Проникновение окуня в позднетретич-
ное время и переживание им полного высыхания Балхаша,
согласно гипотезе Л. С. Берга, трудно совместимы. Биология бал-
хашского окуня позволяет предположить возможность его су-
ществования в небольших заболоченных водоемах, но это доджен
быть не кратковременный период, а долгий путь эволюции в сто-
* Полное высыхание какого-либо водоема в некоторый период очень «удоб-
но» объясняет многие особенности его ихтиофауны. В основе фактов о пересы-
хании озер и их возобновлении лежит, как правило, степень их минерализации,
иногда наличие русел рек в современном ложе озер. Как указывалось выше, рас-
солоиеиие возможно как результат увеличения стока в ледниковый период.
27
рону образования тугорослых, карликовых форм или приспособ
ленности вида к речному образу жизни. И то и другое вряд
привело к столь сильному обособлению балхашского окуня от рец
ного. В его облике прослеживается больше древних черт, харак-
терных для предков окуневых (Жадин, 1949). Следует обратит;
внимание на развитие сенсорных каналов, свойственных в основ
ном рыбам соленых вод и больших водных пространств. Такщ
образом, строение и биология балхашского окуня не соответству.
ют предположению Л. С. Берга о полном высыхании Балхаша j
послеледниковый период.
По-видимому, этот вопрос останется спорным до появлешц
достоверных палеоихтиологических находок. Пока же можнс
предложить еще одну гипотезу проникновения окуня в бассещ
Балхаша. При обследовании верховьев р. Нуры и ее основной
притока Шерубай-Нуры (Чурбай-Нуры) было обнаружено, чтс
разграничить бассейны этих рек от бассейна р. Токрау, северной
притока Балхаша, ныне не доходящего до озера, порой очень за-
труднительно. В одних и тех же долинах ручьи текут в обе сторо-
ны, часто изменяют направление с широтного на меридиональное
и обратно, иногда заканчиваются слепо. Расстояние между водото-
ками не превышает 1 км.
В ихтиофауне Нуры вплоть до верховьев обычен окунь, состав-
ляющий в некоторых участках основной по численности вид. По-
скольку этот район не заливался морем в интересующий нас пе-
риод и не был в то же время выше современного уровня, то можно
представить (и это было почти наверное), что некоторые притоки
Нуры и Токрау меняли ориентацию и между ними осуществлялся
обмен ихтиофауной. Этот путь еще более вероятен для гольянов,
которые проникли в бассейн Балхаша явно позже маринки и
гольцов.
Нагорно-азиатские элементы в бассейне Балхаша также до-
вольно поздние вселенцы. Путь их вторжения хорошо охарактери-
зовал Ф. А. Турдаков (1963). Это бесспорный, очевидный путь
через долину Сан-Таш, расположенную между хребтами Терскей-
Алатау и Кунгей-Алатау.
В недалеком прошлом, по мнению Ф. А. Турдакова, здесь су-
ществовало озеро, которое могло иметь переменный сток в оба
бассейна —Илийский и Иссык-Кульский. Не исключена возмож-
ность и смены направления стока рек Тюп и Ирису, что, несомнен-
но, способствовало обмену ихтиофауной между Иссык-Кулем и
Балхашем. Это предположение не вызывает сомнений, но не совсем
понятно, почему ихтиофауна Иссык-Куля выводится от выходцев
из Балхаша, а не наоборот. Так, Ф. А. Турдаков пишет: «На запад
от перевала Сан-Таш в оз. Иссык-Куль обитает близкий вид
Sch. issykkuli, заведомо происшедший от предыдущего-
(Sch. pseudaksaiensis. — В. 7И.).
К «восточным» иммигрантам Иссык-Куля, т. е. проникшим из
бассейна Балхаша, Ф. А. Турдаков относит и другие виды рыб—-
Ph issykkulensis, D. dybowskii, N. dorsalis и N. strauchi. При этом
" мечает, что восточные иммигранты изменились в Иссык-Куле
больше, чем западные. Из шести видов, общих для бассейна Или и
Иссык-Куля, лишь три (D. gymnogaster, D. dybowskii, N. strauchi)
успели дать в последнем подвиды. Далее Ф. А. Турдаков расселяет
N dorsalis и маринку до р. Курагаты (приток р. Чу), а голого
осмаиа — до Р- Чирчика, отмечая, что происхождение чешуйчатых
османов и N. dorsalis в бассейне Сыра, а также N. kafirnigani в
Аму —иное.
Таким образом, Ф. А. Турдаков вполне сознательно выводит
упомянутые нагорно-азиатские виды из бассейна Балхаша. Совер-
шенно справедливо отмечает, что этот путь был закрыт в обрат-
ном направлении (из Иссык-Куля в Балхаш) для долинных форм
(ельцов, сазана, пескаря) в связи с крутизной стока р. Тюп
ниже Сан-Таша. Но для горных и даже предгорных видов такое
направление миграций возможно, однако Ф. А. Турдаков не до-
пускает подобных предположений. Точку зрения Ф. А. Турдакова о
проникновении маринок и османов из Балхаша в Иссык-Куль кос-
венно поддерживает А. А. Баимбетов (1974), который считает, что
в бассейн Балхаша нагорно-азиатские элементы проникли через
верховья р. Или из рек Нарын и Тарим. Такой путь возможен, но
поскольку из этого региона нет проверенных данных, он может
рассматриваться только предположительно.
Если Л. С. Берг прав, и Балхаш действительно высыхал в после-
ледниковый период, то тем более он не может быть исходным во-
доемом для таких видов, как маринка и гольцы. Возникает во-
прос, откуда же взялись эти представители нагорно-азиатской
фауны в Балхаше? Возможный путь их проникновения лежит че-
рез бассейн Иссык-Куля, иначе мы вынуждены будем выводить
всех гольцов от N. strauchi, а османов и маринок — от D. dybowskii
из рек Кандысу и Уласты бассейна Зайсана. Несостоятельность
подобного взгляда очевидна. Вероятно, Ф. А. Турдаков оказался в
плену исторической последовательности изучения ихтиофаун
Балхаша и Иссык-Куля и это автоматически было перенесено им
на пути расселения рыб. Поскольку Ф. А. Турдаков отмечает
«молодость» восточных вселенцев в Иссык-Куле, с полным правом
можно применить это положение, доказанное им вполне обосно-
ванно в отношении недавнего проникновения нагорно-азиатских
видов из Иссык-Куля в Балхаш.
Исходным видом маринок Балхаша следует считать Sch. pseu-
daksaiensis, обитающую в Иссык-Куле и придаточной системе
Балхаша. Балхашскую маринку — Sch. argentatus —надо .рас-
сматривать как производную форму, образовавшуюся в озерных
Условиях. Проведенные исследования позволяют рассматривать
ее не как вид, а как экологическую расу — Sch. pseudaksaiensis
argentatus.
Итак, заселение бассейна Балхаша нагорно-азиатскими вида-
Ми—маринками и гольцами — происходило из бассейна Иссык-
28
29
Куля. Это наиболее вероятный путь их появления в Балхаше. Все-
ление их следует отнести к межледниковым периодам, так как
часть из них (или только исходные формы, что вероятнее) успели
проникнуть в бассейн Алаколя. Окунь появился в бассейне Бал-
хаша раньше других своих современников из бассейна Иртыша.
Непосредственный путь его проникновения спорен.
Гольяны, наиболее молодые вселенцы этого бассейна, относят-
ся явно к послеледниковому периоду. К моменту их проникновения
и расселения Балхаш и Алаколь были разъединены и между ними
могли возникать только временные связи по речным системам.
Вселение гольянов могло происходить только через Токрау из
бассейна Нуры. Об этом со всей очевидностью свидетельствует их
современный ареал. Появиться в р. Чу гольяны могли либо из
Сырдарьи, либо из Иссык-Куля. Но в бассейне Сырдарьи гольяны
не обнаружены (Турдаков, 1963). Упоминание Л. С. Берга (1949)
о гольянах в р. Угам не подтвердилось. Следовательно, остается
один путь — через Иссык-Куль, куда они попали из Балхаша. Это
как раз тот случай, когда Ф. А. Турдаков полностью прав, выводя
иссык-кульских и чуйских гольянов из балхашских.
ИХТИОГЕОГРАФИЧЕСКОЕ РАЙОНИРОВАНИЕ
История формирования ихтиофаун водоемов Казахстана есть
процесс смешения разнообразных фаунистических комплексов.
К остаткам верхнетретичной ихтиофауны уже в четвертичном пе-
риоде подключились южные, западные, северные и восточные им-
мигранты. Ледниковый период сказался на водоемах Казахстана
в меньшей степени, чем в более северных широтах, и привел к об-
разованию современного состава ихтиофауны. Наибольшие изме-
нения в составе рыб происходили в Аральском и Зайсанском бас-
сейнах, каждый из которых долгое время был опорным пунктом
в расселении карповых, щуковых, окуневых и других пресновод-
ных семейств.
Многие замкнутые ныне бессточные системы Центрального
Казахстана претерпевали неоднократное вторжение то разных, то
одних и тех же видов с юга и с севера, и современный видовой со-
став рыб отражает эту картину (Нура, Иргиз, Тургай, Сарысу).
В стороне от путей расселения рыб оказался лишь бассейн Бал-
хаша, собравший виды бореального и нагорно-азиатского фау-
нистического комплексов. Проникновение северных вселенцев
происходило в этом бассейне, вероятно, через речные системы Ну-
ра—Токрау, а нагорно-азиатских — через бассейн Чарына из
Иссык-Куля.
Ихтиогеографическое деление Палеарктики, проведенное
Л. С. Бергом (1962), отражает описанную картину в том смысле,
что именно на территории Казахского мелкосопочника сходятся
границы трех подобластей — Циркумполярной, Средиземномор-
ской и Нагорно-Азиатской. Исторический подход к ихтиогеографи-
30
скому районированию в условиях развития акклиматизационных
чебот обретает особый смысл, так как помогает правильнее оце-
нивать исходный материал для акклиматизации.
Н Появившееся в последнее время стремление некоторых ихтио-
логов строить ихтиогеографическое районирование на основе ак-
климатизированных видов следует рассматривать критически,
р то же время нельзя отрицать влияние акклиматизантов (точнее,
деятельности человека) на изменение облика водоемов и состава
ихтиофауны. Достаточно вспомнить исчезновение аборигенных
видов под действием акклиматизированных в Балхаше. Однако
учет этих факторов как биогеографических приведет нас к потере
ретроспективы естественного формирования фауны.
Вслед за Л. С. Бергом ихтиогеографическое районирование не-
которых водоемов Казахстана проводили Ф. А. Турдаков (1963),
Б. Г. Иоганзен (1951) и Н. П. Серов (1961).
ф. А. Турдаков, обстоятельно аргументируя свою точку зрения
и применяя исторический подход к формированию фауны, пере-
смотрел некоторые положения Л. С. Берга относительно градации
Понто-Каспийско-Аральской провинции, выделил Туркестанскую
провинцию, уточнил границы Нагорно-Азиатской подобласти. Ос-
новной идеей Ф. А. Турдакова является несовместимость ихтиогео-
графических границ со всем течением реки. Как единственное
исключение он указывает на бассейн Балхаша, строго вписываю-
щийся в Балхашскую провинцию. Предложения Ф. А. Турдакова
поддержал Л. С. Берг, во всяком случае, в их основной части — в
выделении Туркестанской провинции.
Н. П. Серов (1961а, б) рассмотрел положение Иргиз — Тур-
гая и предложил дробное деление Балхашской провинции. Он
считает возможным отказаться от включения Иргиз — Тургая в
Понто-Каспийско-Аральскую провинцию и рассматривать эти во-
доемы как участок Западно-Сибирского округа Циркумполярной
провинции*.
Предполагая, что вся современная ихтиофауна Тургая и Иргиза
проникла из бассейна Иртыша, Н. П. Серов ищет возможные пу-
ти проникновения и находит их, ссылаясь на И. А. Долгушина
(in lift), в верховьях бассейна Ишима и р. Каратургай. Он отме-
чает, что Иргиз — Тургай не имел длительной водной связи с
Аральским морем в прошлом и поэтому здесь нет таких видов, как
аральский лосось, аральская плотва, туркестанский язь, аральский
Жерех, усач, аральская шемая и т. д. Свое мнение Н. П. Серов под-
крепляет ссылкой на морфологический анализ «отдельных пред-
ставителей ихтиофауны Иргиз — Тургая», сделанный А. Ф; Сидо-
ровой (1959) и А. К. Шиленковой (1959), который показывает
.Лйзость иргиз-тургайских рыб к иртышским, но не аральским
формам. Предложение Н. П. Серова ранее отвергалось (Митрофа-
Ов, 1971а), однако более подробное изучение вопроса заставляет
Вероятно, Н. П. Серов имел в виду Ледовнтоморскую провинцию.
31
согласиться с ним. Ихтиофауна Иргиз — Тургая на 100% состоит
из представителей бореально-равнинного фаунистического комп-
лекса, составляющего основу Ледовитоморской провинции Цир.
кумполярной подобласти.
Н. П. Серов (1961) предложил также подробное разделение
Балхашской провинции на округа и участки. Предложенное им
разделение Балхашской провинции на два округа — Балхашский и
Алакольский — вполне убедительно, как и разделение Балхашско-
го округа на участки, но это не единственно возможный вариант.
На основе работ упомянутых авторов и дальнейших исследова-
ний предлагается следующее разделение территории Казахстана
на провинции, округи и участки, детализирующее ранее существо-
вавшую градацию и вносящее некоторые изменения в установив-
шиеся положения ихтиогеографии.
Зоогеографические границы провинций, проходящие по терри-
тории Казахстана, в основном охарактеризованы Л. С. Бергом и
Ф. А. Турдаковым. Однако и здесь не все ясно.
Граница Ледовитоморской провинции может быть проведена
между Балхашской провинцией по водоразделу хребтов Саур и
Тарбагатай*. Далее к западу она проходит по водоразделам
между реками Аягуз, Баканас и Токрау бассейна Балхаша и меж-
ду реками Чаган, Тундык и Нура, относящимися к бассейну Ир-
тыша, т. е. вначале идет по хр. Чингизтау. С запада сюда примы-
кает бассейн Сарысу, относящийся уже к Понто-Каспийско-Араль-
ской провинции.
Далее на запад Ледовитоморская провинция граничит уже с
Понто-Каспийско-Аральской провинцией. Граница между ними
проходит в широтном направлении через весь Казахский мелкосо-
почник, разделяя бассейны Кургальджинских озер и оз. Тенгиз от
притоков Сарысу. Здесь она вплотную подходит к истокам р. Тур-
гай и, по Л. С. Бергу, резко поворачивает на северо-запад, огибает
бассейн р. Тургай с севера, оставляя северо-восточнее бассейн
Ишима (р. Терсакан). Далее она снова делает поворот на запад,
несколько южнее истоков р. Убаган, и четкая граница теряется,
так как проходит через озерную страну. Временные озера Куста-
найский области только с большой натяжкой можно отнести к
Ледовитоморской провинции. Тогда граница пройдет южнее
оз. Аксуат, обогнет озерную область и выйдет в верховья р. Тобол.
Отсюда она вновь приобретает четкость, поворачивает резко на
север, разделяя бассейны Урала и Тобола.
Если следовать Н. П. Серову (1961), то от истоков р. Кара-
кенгир, притока р. Сарысу, границу Ледовитоморской провинции
* Не следует относить реки Уласты и Кандысу к Балхашской провинции, как
это сделал Л. С. Берг. Правильнее точка зрения Б. Г. Иоганзена (1951), относя-
щего эти реки к Призайсанскому участку Западно-Сибирского округа Ледовито-
морской провинции. С таким же успехом можно отнести, например, верховья
р. Урала к Ледовитоморской провинции на основании встречи в них таких имми-
грантов, как хариус и налим.
32
следует проводить не на северо-запад, а на юго-запад по хр. Улу-
Tav захватывая верховья бассейнов рек Тургай и Джиланчик.
5 районе р. Байконур граница поворачивает на запад, оставляя
севернее низовья Тургая и Иргиза, и выходит на водораздел Юж-
ных Мугоджар. Здесь четко отграничены бассейны рек Иргиз и
Эмба, а затем — Иргиз и Орь (приток р. Урала). От водораздела
Тобол — Урал и далее граница идет по описанию Л. С. Берга.
Итак, отграничение Ледовитоморской провинции от Балхаш-
ской на востоке и Понто-Каспипско-Аральскоп на западе просле-
живается довольно хорошо.
Разделение Балхашской и Понто-Каспийско-Аральской про-
винции сложнее, хотя границы и здесь могут быть определены
конкретно. Стык трех провинций из трех разных подобластей рас-
положен в центре Казахского мслкосопочника, где от широтно
идущей границы Ледовитоморской провинции перпендикулярно на
юг отходит граница между Балхашской и Понто-Каспийско-
Аральской провинциями. Наметить ее здесь можно от пос. Байна-
зар до крайней западной точки оз. Балхаш. Отсюда она вполне
реально проходит по водоразделу Чу-Илийских гор, затем перехо-
дит на хр. Кендык-Тас, с него на хр. Жетпжол в районе перевала
Курдай и далее на водораздел Заилийского Алатау. Затем пере-
ходит на хр. Кунгей-Алатау, с него через перевал Сан-Таш на
Терскеп-Алатау и далее по водоразделам рек Чарын, Текес и
Кунгес.
На запад от Боомского ущелья, где протекает р. Чу, граница
Нагорно-Азиатской подобласти с Понто-Каспийско-Аральской
подобластью четко описана Ф. А. Турдаковым (1963).
Изолированным «вклиниванием» в территорию Нагорно-Азиат-
ской подобласти является оз. Иссык-Куль, относимое Л. С. Бергом
и Ф. А. Турдаковым на правах участка к Понто-Каспийско-Араль-
ской провинции. Бассейн Иссык-Куля разделяет две провинции
Нагорно-Азиатской подобласти и является тем мостом, через ко-
торый шло расселение нагорно-азиатских видов на север, в Бал-
хашскую провинцию. Ф. А. Турдаков отмечает, что «западные»
Бселенцы — сазан, пескарь и др. в Иссык-Куле более молоды, чем
«восточные» — османы, маринки и гольцы. Л. С. Берг (1962) и
Ф. А. Турдаков (1963) вряд ли правомерно выводят маринок, ос-
танов и гольцов в Иссык-Куле из бассейна Балхаша. Расщепо-
брюхие карповые, наоборот, проникли из Иссык-Куля в Балхаш.
_ Более правильно отнести бассейн Иссык-Куля, как это сделал
Ф. А. Турдаков в отношении притоков озера, к Нагорно-Азиатской
Подобласти на правах Иссык-Кульского округа Балхашской про-
винции.
Сравним, для примера, ихтиофауны окружающих Иссык-Куль
бассейнов Тарима, Чу и Балхаша. Общими для них являются два
ВиДа рода Diptychus и один вид рода Nemachilus. В бассейне
р. Чу эти виды обитают только в ее верхнем течении, которое от-
°сится, и по Л. С. Бергу и по Ф. А. Турдакову, к Нагорно-Азиат-
3"7 33
ской подобласти. Между Чу и Иссык-Кулем близкородственными,
помимо отмеченных, являются ельцы, пескари, маринки, один вид
гольца и сазан. Все эти виды в той или иной степени обособились
в каждом из бассейнов.
Таким образом, Иссык-Куль роднят с той частью р. Чу, которая
относится к Понто-Каспийско-Аральской провинции, такие широ-
ко распространенные виды рыб, как ельцы, пескари и сазан. Нац.
ти водоемы, в которых бы они не встречались, не так просто.
Кроме того, у Иссык-Куля есть общие (точнее, близкие) виды и
подвиды с Таримом и Балхашем — маринки, османы, гольцы и голья-
ны. Всего между Таримом и Иссык-Кулем отмечается сходство по
Таблица 1. Ихтиогеографическое
Подобласть Провинция Округ Участок
Циркумполярная Ледовитоморская Сибирский Прииртышский
Степной
Иргиз-Тургайский
Маркакольский
Тарбагатайский
Средиземномор- ская Понто-Каспий- ско-Аральская Каспийский Волжский
Аральский Аральский
Чу-Сарысуйский
Туркестанская Сырдарьинс- кий Сырдарьинский
Нагорно-Азиат- ская Балхашская Балхашский Балхаш-Алакульский
Тентек-Илийский
Джунга ро-3 анлнйский
Иссык- Кульскйй Озерный
Горный
Таласский Таласский
Примечание. Систематика эндемиков дана без учета переработки в на-
стоящем издании.
34
4__5 близкородственным формам, между Иссык-Кулем и Балха-
шем— по 7, причем в ряде случаев это виды, узко ограниченные в
своем распространении.
Если рассматривать не бассейн Иссык-Куля, а само озеро, то
здесь соотношение видов будет не в пользу западных пришельцев,
которых всего 4 вида, а в пользу нагорно-азиатских представи-
телей.
Отношение оз. Иссык-Куль к Понто-Каспийско-Аральской
провинции оказывается поколебленным. Его родство с Нагорно-
Азиатской подобластью значительнее и поэтому оз. Иссык-Куль и
его бассейн следует рассматривать как промежуточное звено между
деление территории Казахстана
Границы участков Эндемичные рыбы
Иртыш, Бухтармннское водохранили- ще, притоки Иртыша—Ишим и Тобол с их бассейнами Phoxinus sedelnikowi
Бассейны Нуры, Шидерты, Селеты, Олеиты и др. бессточные системы Север- ного Казахстана Phoxinus czckanowskii ignatovi
Бассейны Иргнза, Тургая и Улы-Джи- ланчика Эндемичных видов нет
Бассейн оз. Маркаколь Brachymystax lenok savinovi, Thymal- lus arcticus brevicephalis, Nemachilus bar- batulus markakulensis
Бессточные реки северных склонов Тарбагатая Nemachilus strauchi zaisanicus
Реки Урал, Эмба, Сагнз, Уил, Б. и М. Узень Эндемичных видов нет
Собственно море н нижнее течение Амударьи и Сырдарьи Chalcalburnus chalcoides aralensis
Бассейн Сарысу и Чу до горной части Esox lucius kirgisorum, Leuciscus leu- ciscus kirgisorum, Capoetobrama kuscha- kewitschi orientalis
Среднее течение Сырдарьи Pseudoscaphirhynchus fedtschenkoi Le- uciscus squaliusculus. Alburnoides oblon- gus. Capoetobrama spinolosus
Собственно озера Балхаш, Алаколь, . Сасыкколь. Жаланашколь, Уялы и др. Schizothorax argentatus, Perea schrenki
Нижнее и среднее течение рек, впа- дающих в озера Schizothorax pseudaksainsis, Nemachi- lus sewerzowi
1 орные реки Джунгарского и Занлий- 9кого Алатау Phoxinus poljakovi, Ph. br.achyUrus
Собственно озеро ^_Дритокн озера Leuciscus schmidti, L. bergi, Schizotho- rax issykkuli, Dipt, dybowskii lansdellt Nemachilus strauchi ulacholicus
Бассейн р. Талас и некоторые бессточ- ные реки и озера Каратау Leuciscus lindbergi, Schizothorax sal- tans, Diptychus micromaculatus, Nemachi- lus conipterus
35
бассейном Тарима и Балхаша. Не совсем ясно, к какой из окру-
жающих его провинций Нагорно-Азиатской подобласти можно от-
нести Иссык-Куль. С балхашской фауной его родство как будто
бы больше, чем с таримской, но, учитывая слабую изученность
таримской ихтиофауны, этого нельзя утверждать с полной основа-
тельностью.
Можно предложить, возможно, как временное решение вопроса,
объединение Иссык-Куля с Балхашской провинцией, с выделением
в ней Иссык-Кульского округа.
Крайнюю точку зрения высказали В. А. Догель и Е. В. Гвоздев
(1945), предложившие отнести Балхашскую, Таримскую и Тибет-
скую провинции к Аральскому округу Понто-Каспийско-Аральской
провинции, а Западно-Монгольскую — к Ледовитоморской провин-
ции, упразднив тем самым Нагорно-Азиатскую подобласть.
Опровергая их, Л. С. Берг высказал мнение, что Нагорно-Азиат-
ская подобласть в большей степени характерна тем, чего там нет,
а не тем, что есть. Говоря об основных чертах Нагорно-Азиатской
подобласти, он отмечает, что она отличается своей бедностью,
сильно выраженным видовым эндемизмом и оригинальностью ро-
дов. Для нее характерно присутствие родов Schyzothorax, Schyzo-
pygopsis, Dyptychus, Aspiorhynchus, а также большое количество
видов Nemachilus.
Таким образом, эта подобласть характеризуется прежде всего
расщепобрюхими карповыми, которых Ф. А. Турдаков считает ее
автохтонами. Поэтому довольно странно выглядит Западно-Мон-
гольская провинция в составе этой подобласти, с которой ее ниче-
го не роднит. Л. С. Берг (1962) сам отмечает, что с соседними
провинциями Нагорно-Азиатской подобласти Западно-Монгольская
не имеет ни одного общего вида (добавлю от себя: н ни одного
близкородственного!). Ее ихтиофауна несет на себе основной отпе-
чаток Ледовптоморской провинции. И, вероятно, в этом случае
В. А. Догель и Е. В. Гвоздев были правы: возможно, этот район
может войти в Ледовитоморскую провинцию на правах округа.
Предлагаемое ихтиогеографическое разделение территории,
смежной с подобластями Палеарктики, представлено в таблице 1.
Балхашская провинция. Балхашский округ распадает-
ся на три четко выраженных части: озера, крупные реки в их до-
линном течении и горные реки. Каждый из этих участков характе-
ризуется определенным ихтиофаунистическим составом, приуро-
ченностью к резко контрастным условиям оз. Балхаш и горных
потоков Тянь-Шаня и Джунгарии.
В то же время разница между ихтиофауной озер Балхаш-Ала-
кольской группы очень незначительна, как и между крупными ре-
ками, достигающими этих озер. Это позволило несколько упрос-
тить деление Балхашского округа.
Иссык-Кульский округ возможно разбить на два участка: озер-
ный и речной. Разница их ихтиофауны очевидна и хорошо показа-
на Ф. А. Турдаковым (1963).
36
Понто-Каспийско-Аральская провинция. Каспий-
ский округ принимается полностью по Л. С. Бергу.
Аральский округ, за исключением из его состава бассейна Ис-
сык-Куля, также принимается по Л. С. Бергу. Здесь следует выде-
лить такие участки: 1) Аральский, включающий собственно Арал
и нижнюю часть Сырдарьи, до границ с Туркестанской провинцией;
2) Чуйско-Сарысуйский, захватывающий весь бассейн р. Сарысу,
нижнее и среднее течение р. Чу.
Туркестанская провинция. Сырдарьинский округ
включает один участок — Сырдарьинский. Эндемичные виды Pseu-
doscaphirhynchus kafirnigani, Alb. oblongus и др. Границы в пре-
делах, указанных Ф. А. Турдаковым для этой реки *.
Л ед овито морская провинция. Сибирский округ при-
нимается по Л. С. Бергу (1962). На территории Казахстана в его
составе могут быть выделены следующие участки: 1) Прииртыш-
ский, включающий все течение р. Иртыша и его притоков Ишим,
Тобол и т. д.; 2) Степной, охватывающий все бессточные озерные
системы Северного Казахстана, бассейны Шидерты, Оленты, Се-
леты, Нуры и т. д.; 3) Иргиз-Тургайскип, включающий весь бас-
сейн одноименных рек и, возможно, являющийся переходным меж-
ду Понто-Каспийско-Аральской и Ледовитоморской провинциями;
4) Тарбагатайскпй—бессточные реки северных склонов Тарбага-
тая; 5) Алтайский (по Иоганзену, 1951); 6) Маркакольскип— бас-
сейн оз. Маркаколь.
Как видно, и здесь дробность деления увеличивается при при-
ближении к горам. Алтай, так же как и другие горные системы,
является местом, где происходила изоляция фаун зачастую в не-
больших бассейнах, что привело к образованию эндемичных форм
(Световидов, 1936; Митрофанов, 1958) и своеобразию видового
состава.
Для обоснования выдвинутых предложений рассмотрим соот-
ношение видов в приведенных границах провинций, округов и
участков в пределах Казахстана по их отношению к фаунистиче-
ским комплексам в характеристике Г. В. Никольского (1953).
В таблицу 2 включены данные по изменению видового состава в
результате акклиматизационных работ. Это позволяет со всей
наглядностью убедиться в их влиянии на облик ихтиофауны, при-
водящем к потере специфичности практически всех водных систем
Казахстана. Третичный равнинный пресноводный комплекс свой-
ствен всем провинциям. Это ранее широко распространенные ви-
ды по всей Палеарктике и они не могут служить основой для дроб-
ного биогеографического районирования. Этот комплекс нигде не
является доминирующим по численности видов. Максимальное ко-
личество его представителей (15%) сохранилось в Понто-Каспий-
ско-Аральской провинции.
* Бассейн р. Талас, ранее относимый к Туркестанской провинции (Митрофа-
нов, 1971а), по преобладанию в его аборигенной ихтиофауне видов Нагорно-
азиатского комплекса следует включить в состав Балхашской провинции.
37
Таблица 2. Соотношение фаунистических комплексов в водоемах Казахстана
(знаменатель), %
Фаунистичес- кий комплекс Ледовитоморская провинция Понто-Каспийско- Аральская про- винция
Приир- тыпе’кйй участок Степной (Нурин- ский) участок ИргНз-Тур- гайский участок Марка- кольский участок Тарбага- тайский участок Аральский учйсток Чу-Сары- суйский участок
Третичный равнинный прес- новодный 12/12 8/16 0/14 0/0 0/0 15/8,1 11.2/9,7
Арктический моргкой 4/3 8/7 0/0 0/0 0/0 5/2,7 3,7/3,2
Арктический пресноводный 8/15 8/7 0/0 0/0 0/0 5/2,7 0/0
Бореальн ый предгорный 36/26 0/0 0/0 75/75 71/71 0/0 7,4/6,4
Бореальный равнинней 36/29 68/63 100/83 0/0 0/0 25/13,5 29,5/26
Понто-Каспий- ский морской 0/0 0/0 0/0 0/0 0/0 0/19 0/0
Понто-Каспий- ский пресновод- ный 0/6 0/0 0/0 0/0 0/0 50/27 22,2/19,3
Переднеазиат- скйй 0/0 0/0 0/0 0/0 0/0 0/0 3,7/3,2
Нагорно-Ази- атский 0/3 0/0 0/0 0/0 29/29 0/0 14,8/12,8
Туркестан- ский 0/0 0/0 0/0 0/0 0/0 0/0 3,7/3,2
Индо-Афри- кане кий 0/0 0/0 0/0 0/0 0/0 0/0 0/0
Китайский 4/6 8/7 0/0 25/25 0/0 0/27 3,7/16,1
Общее кол-во комплексов 6/8 5/5 1/2 2/2 2/2 5/7 9/9
Группа северных фаунистических комплексов: арктический
морской, арктический пресноводный, бореальный равнинный и бо-
реальный предгорный доминируют в Ледовитоморской провинции
(84%), составляют до 35% в Понто-Каспийско-Аральской провин-
ции и уменьшается до 27% в Туркестанской. Их вторжение обус-
ловлено ледниковыми периодами, что определяет широкое рас-
пространение.
Понто-Каспийский пресноводный комплекс доминирует (50%)
только в Понто-Каспийско-Аральской провинции. В остальные он
либо не проник (Балхашская провинция), либо вымер (Ледовито-
морская провинция) в периоды похолодания.
Туркестанский фаунистический комплекс локализован в Тур-
38
в аборигенном виде (числитель) и в результате акклиматизационных работ
CbipJiap',- Iggl ИНСКЙЙ Is g о округ § ' И Балхашская провинция
Балхашский округ Иссык-Кульский округ Таласский округ
Балхаш- Алаколь- ский участок Тейтек- Илийскйй участок джунгаро- Зайлпй- ский участок
озер- ный участок речной участок озер- ный участок речной ; участок 1
10,8/7,2 0/20 0/11 0/0 И/7 0'0 0/33,3 0/0
2,7/1,8 0/0 0/0 0/0 0/0 0/0 0/0 0/0
0/0 0/0 0/0 0/0 0/0 0/0 0/0 0/0
5,4/3,6 0/0 20/7 20/27 11/14 10/18 0/0 0/0
19/12,5 25/20 10/15,8 0/0 22/22 0/0 17/33,3 12/12
0/14,4 0/0 0/0 0/0 0/0 0/0 0/0 0/0
21,5/14,4 0/30 0/11 0/0 0/22 о/о 0'33,3 0/0
5,4/3,6 0/0 0/0 0/0 0/0 0/0 0/0 0/0
10,8/7,2 75/30 70/26 80/73 45/28 80/73 66/0 76/76
19/12,5 0/0 0/0 0/0 0'0 0/0 0/0 0/0
2,7/2,8 0/0 0/0 0/0 0/0 0/0 0/0 0/0
2,7/20 0/0 0/30 0/0 И/7 10/9 17/0 12/12
10/11 2/4 3/6 2/2 5/6 3/3 3/3 3/3
кестанской провинции и крайне эндемичен. Широко, но незначи-
тельным количеством видов представлен китайский комплекс.
Нагорно-Азиатский фаунистический комплекс доминирует в
Балхашской провинции (до 80%), обычен в Туркестанской и редок
в Понто-Каспийско-Аральской.
В целом число составляющих ихтиофауну фаунистических
комплексов убывает по провинциям в следующем порядке: в Тур-
кестанской провинции 11 фаунистических комплексов, т. е. почти
вся возможная полнота. Вследствие такой насыщенности аккли-
матизационные работы внесли незначительные изменения в соот-
ношение видов. В Понто-Каспийско-Аральской провинции пред-
ставлено 10 фаунистических комплексов (выпадают Индо-Африкан-
ский фаунистический комплекс и Понто-Каспийский морской).
39
В Ледовптоморской провинции 7 фаунистических комплексов
(выпадают Понто-Каспийский морской, Понто-Каспийский пресно-
водный, Индо-Африканский и Туркестанский фаунистические
комплексы).
В Балхашской провинции 5 фаунистических комплексов, если
включить в нее бассейн Иссык-Куля, и только 2 комплекса в са-
мом бассейне Балхаша. В этом случае акклиматизационные рабо-
ты внесли коренные изменения в ихтиофауну, и в ее современном
составе доминируют Понто-Каспийско-Аральский и Третичный
пресноводный фаунистические комплексы.
Рассмотрение ихтпогеографического районирования с точки
зрения преобладания того или иного фаунистического комплекса,
специфичности и эндемизма фауны представляется достаточно объ-
ективным решением спорных вопросов. В этой главе мы пытались
показать сложность подобного подхода, не скрывая возникающих
при этом проблем. Сугубо теоретические исследования по харак-
теру формирования и современного распределения ихтиофауны
имеют прямое прикладное значение в реконструкции наших
водоемов.
Глава 3
ОПИСАНИЕ ВИДОВ ИХТИОФАУНЫ КАЗАХСТАНА
Видовое разнообразие пресноводных, проходных и солоновато-
водных аборигенных рыб * Казахстана включает представителей
9 отрядов, 16—17 семейств и более 50 родов. В основном это пред-
ставители Понто-Каспийско-Аральского и Бореально-равнинного
фаунистических комплексов. Однако значительное место занимают
останцы Третичного пресноводного и молодого Нагорно-Азиатского
комплексов.
Наиболее многочисленны в видовом отношении карповые ры-
бы, составляющие около 50% всех видов.
Указать достоверное число видов, а тем более подвидов, ихтио-
фауны Казахстана можно будет только по завершении рабо-
ты над настоящей сводкой. Сложность заключается не в сборе ли-
тературных источников. Такая работа проводится. Но подход их-
тиологов к описанию новых видов и подвидов в разное время
был различным. Многие виды описывались из эфемерных, ныне
не существующих, водоемов, по единичным образцам. Поэтому го-
ворить о их реальности без тщательного анализа конкретных ус-
ловий можно лишь с большими оговорками. Взять хотя бы такие
примеры, как описание ащикольскоп маринки Ф. А. Турдаковым и
алакольекой маринки Н. А. Некрашевичем.
Кроме того, следует иметь в виду, что исключение лишь одного
вида из состава ихтиофауны республики (например, вследствие
уточнения его ареала) может изменить число родов и даже се-
мейств. Последнее, в частности, зависит от туркестанского сомика,
границы ареала которого пока не ясны.
^Акклиматизация значительно пополнила список отрядов, се-
мейств, родов и видов рыб, населяющих водоемы Казахстана. По-
явились даже представители нового для ихтиофауны СССР рода
из отряда Cyprinodontiformes, систематическое положение которых
еЩе требует уточнения. Подробно обо всем этом будет сказано в
соответствующем разделе сводки.
Без круглоротых.
41
Пока же ориентировочно можно принять, что без рыб, обитаю-
щих только в Каспийском море и не заходящих в пресные воды, в
Казахстане обитает около 100 видов аборигенных рыб и около
20 акклиматизированных видов, натурализовавшихся в естествен-
ных водоемах.
В данном издании описание ведется по системе, принятой в
книге «Жизнь животных» (М., 1983, 2-е изд.). Все описываемые
животные относятся к типу Chordata — хордовые и подтипу Сга-
niata — черепные или Vertebrata — позвоночные. Рыбообразные
относятся к надклассу Agnatha — бесчелюстные, классу Cyclosto-
mata— круглоротые и подклассу Petromyzones — миноги. Все ка-
захстанские рыбы относятся к надклассу Gnathostomata — че-
люстноротые, классу Osteichthyes — костные рыбы, подклассу Acti-
nopterygii — лучеперые рыбы.
Таксономия от отряда и ниже приводится в очерках, которые
построены по схеме: а) латинское, русское, казахское (иногда и
местное) названия; б) синонимия; в) распространение; г) описа-
ние (морфологическое); д) биология (размножение, рост и воз-
раст, питание и упитанность, миграции и пр.); е) численность и
хозяйственное значение. По синонимам приводится не вся литера-
тура, а лишь основная или относящаяся к Казахстану.
Внутри родов виды пронумерованы арабскими цифрами, под-
виды обозначены, кроме того, и буквенной индексацией (1а, 16, 2а,
26), морфа обозначена 1а'.
ОТРЯД PETROMYZONIFORMES —МИ НОГООБРАЗНЫЕ
Семейство PETROMYZONIDAE — МИНОГОВЫЕ
Круглоротые — наиболее древние и просто организованные
позвоночные животные. От челюстноротых (Gnathostomata), в
частности рыб, они отличаются не только примитивным анатоми-
ческим строением, но и своеобразным эмбриональным и постэмбри-
ональным развитием.
У миног голое угреобразное тело. В их скелете нет костной
ткани и в течение всей жизни сохраняется хорда. Тела позвонков
отсутствуют. Парных плавников нет. Из непарных имеются только
2 спинных (реже один) и хвостовой. Мешковидные жабры открыва-
ются по бокам семью жаберными отверстиями, отчего и произошло
народное название миноги — семидырка. Челюсти отсутствуют,
рот присасывательный, в виде воронки, окружен кожистой бахро-
мой и вооружен множеством роговых образований, называемых
зубами. Они располагаются на верхнечелюстной и нижнечелюстной
пластинках, на поверхности ротовой воронки и на языке. Форма и
количество зубов являются основным таксономическим призна-
ком (Кесслер, 1870; Аникин, 1905; Берг, 1932, 1948; Иоганзен,
1935а, б и др.). Носовое отверстие непарное: оно расположено на
42
верхней стороне головы и сообщается с назо-гипофизарной по-
лостью. Имеется третий теменной глаз; он не содержит хрусталика,
В миноги воспринимают им только световые раздражения. Разме-
ры миног в зависимости от вида, к которому они относятся, варьи-
руют от 20 до 90 см.
Жизненный цикл миноги характеризуется длительным личиноч-
ным периодом. Личинки миног — пескоройки по своему строению, а
также и образу жизни значительно отличаются от взрослой формы.
Долгое время их принимали за самостоятельный род Ammocoetes.
В отличие от взрослых они имеют недоразвитые, скрытые под ко-
жей глаза; подковообразный, несмыкающийся рот, который уса-
жен разветвленными ворсинками; есть расположенные в борозде
жаберные отверстия. Тело личинок окаймлено сверху, сзади и сни-
зу непарной плавниковой складкой, которая с возрастом разделя-
ется на отдельные плавники.
Обитают пескоройки на заиленных участках рек, протоков и
ручьев. Большую часть времени они проводят зарывшись в ил, ко-
торым и питаются. Личиночный период у большинства видов длит-
ся от 3 до 6 лет, после чего происходит метаморфоз и личинка
превращается во взрослую форму (Иоганзен, 1935а, б; Берг, 1948;
Абакумов, 1964; Полторыхина, 1971 и др.).
Размножаются миноги в пресной воде. Среди них встречаются
крупные проходные виды, которые после метаморфоза скатыва-
ются в море и ведут паразитический образ жизни. Другие же по-
стоянно обитают в пресных водоемах и во взрослом состоянии не
питаются. После нереста миноги погибают.
Всего известно 20—24 вида миног, относящихся к 7 родам.
Из них в водах Казахстана обитают представители двух родов —
Caspiomyzon и Lampetra. Последний представлен как проходной
(Lampetra japonica), так и непроходной, туводной формой
(L. kessleri).
Род CASPIOMYZON BERG, 1906
1. Caspiomyzon wagneri (Kessler) —каспийская минога
Казахское название-, минога.
Синонимия. Petromyzon wagneri: Кесслер, Тр. СПб. об-ва ест., т. 1, 1870.
Caspiomyzon wagneri: Берг, Фауна России. Рыбы, т. 1, 1911. — Навозов,
Нести. рыбопром., № 8—10, 1912. — Лукаш, Рыбы Камы, 1920. — Абдурахманов,
Рыбы пресных вод Азербайджана. Баку, 1962. — Гинзбург, Вопр. нхтнол., т. 10,
вып. 4, 19/0.—Агамалиев, Труды Всесоюзн. научио-нсслед. ин-та морск. рыбн.
хоз-ва и океанографии, т. 86, 1971.
Распространение. Каспийская минога — единственный пред-
ставитель рода Caspiomyzon. Распространена в бассейне Каспий-
ского моря, по всему побережью Азербайджана и Северного Кас-
пия. Из моря входит в реки Волгу, Урал, Куру, Терек и их притоки.
Ло строительства гидроэлектростанций поднималась в Волге до
43
Рис. 1. Ротовая воронка каспий-
ской миноги
г. Калинина, в Каме — до Чусовой, в Оке — до Москвы-реки, в Те-
реке— до устья Баксана, в Куре — до г. Мцхета.
Теперь выше плотины Волжской ГЭС не встречается (Кузне-
цов, 1979). По р. Урал прежде поднималась до Оренбурга (Наво-
зов, 1912).
Описание. Строение зубного аппарата ротовой воронки кас-
пийской миноги характеризуется следующими признаками: на
верхней челюстной пластинке находится один зуб; нижняя че-
люстная пластинка имеет по 5 зубов, редко 4—6, из которых
крайние бывают двухраздельными; губные зубы располагаются ра-
диальными рядами; бока ротово-
го отверстия окружены 10—И
внутренними губными зубами
(рис. 1). Все зубы тупые. Взрос-
лые особи имеют однородную
темно-серую окраску (Берг, 1948;
Абдурахманов, 1962).
Биология. Размножение и ли-
чиночный период каспийской ми-
ноги проходят в реке. После пре-
вращения она скатывается в море,
где живет до наступления по-
ловой зрелости, после чего совер-
шает миграции на нерест. В пе-
риод нереста у миноги укорачи-
вается тело, увеличивается высо-
та спинных плавников и умень-
шается расстояние между ними.
Биология каспийской миноги из р. Урал не изучена. В литера-
туре имеются лишь некоторые сведения о ее распространении, сро-
ках нереста и питании личинок (Навозов, 1912).
По данным Л. С. Берга (1948), каспийская минога образует
две формы: крупную, достигающую 37—55 см длины, и мелкую,
которая становится половозрелой при длине 19—31 см. Как отме-
чает А. А. Абдурахманов (1962), основную массу миног, совершаю-
щих миграции на нерест в р. Кура составляют особи длиной от 41
до 46 см и массой от 45 до 204 г; преобладают особи массой от 100
до 140 г.
Ход миноги на нерест происходит с конца сентября до середины
декабря (Волга) и с ноября по февраль в реках Кура, Терек и их
притоках. В реки миноги входят с хорошо развитыми половыми
продуктами (III и III—IV стадии зрелости). В мае гонады дости-
гают IV стадии зрелости; их масса по отношению к массе тела
самки составляет около 17%. Плодовитость варьирует от 14,3 до
38 тыс. икринок. Размеры икринок к моменту нереста достигают в
среднем 1,15 мм.
Размножается минога на песчаных и галечных грунтах с хоро-
шей проточностью воды. В Волге и Урале нерест происходит с
44
лая по июнь. Так, по данным Н. А. Навозова (1912), нерест мино-
ги в притоках Урала у г. Оренбурга происходил в начале июня,
g бассейне Куры нерест каспийской миноги происходит в конце
апреля —начале мая, при температуре воды 21—23° (Агамалиев,
1971а); в Волге, в подплотинном пространстве Волгоградской
ГЭС— в конце мая — начале июня, при температуре воды 16°
(Гинзбург, 1970). Таким образом, размножение каспийской мино-
ги в различных водоемах происходит с апреля по июнь. Икра от-
кладывается в гнезда. Во время икрометания самка присасывается
к камню, а самец, присосавшись к верхней части ее головы, изви-
вается вокруг ее тела. Выделение икры и молок происходит одно-
временно.
Эмбриональное развитие миноги с момента оплодотворения до
выклева личинки при температуре воды от 18 до 20° длится 8—
,9 суток. При искусственном оплодотворении при температуре во-
ды 20—22° выклев личинок происходит через 7 дней. Выклюнув-
шиеся личинки похожи на мелких червей. Они прозрачны и имеют
желточный мешок. Вначале они прячутся между камнями, где на-
ходятся в течение нескольких дней, а затем, на 7—9-й день, сплы-
вают вниз по течению и закапываются в грунт на заиленных
участках рек со слабым течением, часто заросших водной расти-
тельностью. К этому времени у личинок заканчивается первичное
формирование кишечника и они начинают питаться детритом и
Мельчайшими диатомовыми водорослями (Агамалиев, 1971а).
^Указание Н. А. Навозова (1912) о том, что пескоройки питаются
^личинками водных насекомых, не было подтверждено дальнейшими
Исследованиями и, вероятно, ошибочно (Абакумов, 1964).
.й? Только что выклюнувшиеся из икринок личинки имеют длину
$йг 3,8 до 4,2 мм, в 20-дневном возрасте их длина достигает в
среднем 9,5 мм, а к концу месяца—10,3 мм; в возрасте 70 дней
Дичинка вырастает до 20 мм, а 8-месячная — до 35 мм (Абдурах-
манов, 1962; Гинзбург, 1970; Агамалиев, 1971 а, б). Опыты по вы-
ращиванию личинок в искусственных условиях, проведенные
, А. С. Агамалиевым (1971а), дают основание считать продолжи-
тельность личиночного периода каспийской миноги не менее 4 лет.
За это время пескоройки достигают длины 12 см.
Метаморфоз каспийской миноги в бассейне Куры начинается в
Августе, когда появляются личинки с расширенными кровеносны-
ми сосудами. В начале сентября у превращающихся личинок уже
Хорошо заметны глаза, а в январе метаморфоз, вероятно, уже за-
канчивается, так как появляются особи с ороговевшими зубами
^Агамалиев, 1971а, б).
После превращения минога скатывается в море, где она'живет,
видимо, около двух лет, после чего возвращается в реку, на не-
рест.
R О питании каспийской миноги в море почти ничего не известно.
Большинство авторов (Берг, 1948; Агамалиев, 19716 и др.) склон-
ны считать, что каспийская минога, имея тупые зубы, не может
45
паразитировать на рыбах; в кишечнике ее находили остатки водо-
рослей. В реке она не питается.
Численность и хозяйственное значение. Мясо каспийской ми-
ноги обладает прекрасными вкусовыми качествами и большим
(31—34%) содержанием жира. В прошлом минога занимала вид-
ное место в каспийском рыболовстве. В 1936—1941 гг. ее уловы ко-
лебались от 10 до 530 т, а средний годовой улов за это время со-
ставлял 350 т (Никольский, 1971). В последние годы наблюдается
резкое снижение уловов. Данных о промысловом значении миноги
в р. Урал не имеется.
Для воспроизводства численности миноги в условиях гидро-
строительства на реках Каспия необходимы организация ее искус-
ственного разведения на рыбоводных заводах и охранно-регули-
рующие мероприятия по ее естественному размножению.
Род LAMPETRA GRAY, 1851
1. Lampetra japonica (Martens)—тихоокеанская минога
Синонимия. Petromyzon fluviatilis (non L.): Лепехин, Путешествие
(1772 г.), т. IV. СПб., 1805. — Данилевский, Иссл. сост. рыболов, в России, т. VI,
1862.
Lampetra fluviatilis japonica: Берг, Фауна России, Рыбы, т. 1, 1911. — Сол-
датов, Рыбы Печоры, 1924.
Lampetra japonica septentrionalis: Берг, Ежегодн. Зоол. муз. Акад, наук,
т. XXXII, 1931. — Берг, Рыбы пресных вод СССР, т. 1, 1932. — Иоганзен, Зоол.
жури., т. XIV, 1935.
Lampetra japonica (Martens): Берг, Рыбы пресн. вод СССР, т. 1, 1948.
Lampetra japonica septentrionalis Berg: Абакумов, Жизнь животных, т. 1,
1970. — Полторыхнна, Изв. СО АН СССР, вып. 1, 1979.
Распространение. Тихоокеанская минога распространена в
бассейне Тихого океана — от Кореи на юге до р. Юкон (Аляска)
на севере, а также в морях Северного Ледовитого океана — от Оби
на востоке до северного побережья Норвегии на западе. В реках Вос-
точной Сибири отсутствует. Таким образом, тихоокеанская минога
имеет прерывистый ареал, что послужило основанием для выделе-
ния проходной миноги бассейна Северного Ледовитого океана в
подвид — Lampetra jap. septentrionalis (Berg)—ледовитоморская
(Берг, 1931). Впоследствии Л. С. Берг (1948) вновь отнес ее в си-
ноним тихоокеанской. Однако в последнее время наличие подвида
L. jap. septentrionalis (Berg) признается многими авторами
(Иоганзен, 1935а, б; Абакумов, 1970; Никольский, 1971; Полторы-
хнна, 1979) и может считаться вполне обоснованным.
В Обь-Иртышском бассейне минога распространена от Обской
губы до верховьев (Тура, Тобол, Иртыш, Томь). По Оби ледовито-
морская минога поднимается до Барнаула (Иоганзен, 1935а)-
В литературе нет данных о распространении этого вида в реках
Иртыш, Тобол, Ишим, протекающих по территории Северного и
Восточного Казахстана. В бассейне Верхнего Иртыша ледовито-
46
морская минога не встречается. Указание Н. А. Варпаховского
(1897, 1898) о наличии миноги в Иртыше у Семипалатинска, ско-
рее всего может быть отнесено к мелкой туводной форме —сибир-
ской миноге (см. далее).
Описание. По данным Б. Г. Иоганзена (1935а) и Л. С. Берга
(1948), L. jap. septentrionalis (Berg) характеризуется следующи-
ми меристическими признаками: верхнегубные зубы имеются у
всех особей (рис. 2); верхнечелюстных всегда 2; нижнечелюстных —
6—8, крайние всегда двураздель-
ны. петь нижнегуоные и краевые
зубы. Внутренних боковых губ-
ных зубов по три; они раздвоены,
Внешних боковых губных зубов
нет. Зубы ротовой воронки вне
времени икрометания острые, в
период икрометания — тупые.
Другим таксономическим при-
знаком, применяемым за послед-
нее время в систематике миног,
является число туловищных мио-
меров, расположенных между по-
следним жаберным отверстием и
анусом (Vladykov, 1958). Коли-
чество туловищных миомеров у
ледовнтоморской миноги, уста-
новленное при исследовании экс-
понатов лаборатории ихтиологии
Зоологического музея АН СССР,
равняется 66—72 (Полторыхина,
1979). Показатели пластических
Рис. 2. Ротовая воронка тихоокеан-
ской миноги
признаков приведены в таблице 3. Хромосомный набор: 2п=168
(Рыбы МНР, 1983).
Биология. По характеру питания L. jap. septentrionalis являет-
ся преимущественно паразитом: присасывается к рыбам и питает-
. ся их соками. Питание миноги, мигрирующей в Обь и Иртыш, в
основном происходит в Обской и Тазовой губах. Иногда она пита-
ется, видимо, и в реке, судя по ее острым зубам.
Размеры миног, совершающих миграции в реки Сибири, по дан-
ным Л. С. Берга (1931, 1948) и Б. Г. Иоганзена (19356), достигали
35—43 см, в р. Мезень-—до 54 см.
О нересте ледовитоморской миноги в Иртыше и других реках
Сибири ничего неизвестно. Нет данных и по другим вопросам
биологии этого вида.
47
Таблица 3. Морфологическая характеристика
тихоокеанской миноги (по Иоганзену, 1935а)
Отношения 1ам тлитуда
В % от длины тела
Длина рыла
Продольный диаметр глаза
Промежуток глаз — жаб. аппарат
Длина головы до 1-го жаб. отверстия
Длина жаберного аппарата
Длина головы до 7-го жаб. отверстия
Расстояние до ноздри
Высота головы
Ширина лба
Продольный диаметр закрытой ротовой щели
Длина тушки от 1-го жаб. отверстия
Длина тушки от 7-го жаб. отверстия
Наибольшая высота тела
Наибольшая толщина тела
Обхват тела
Антедорсальное расстояние (до 1D)
Расстояние до ануса
Длина 1D
Промежуток 1—2D
Длина 2D
Длина спинной части С
Длина 2О-{-спинной части С
Расстояние от ануса до конца С
5,9-6,7
1,2-2,0
2,4-2,7
10,4-10,5
8,0-10, 2
18,2-20,1
5,3-6,6
3,7-4,6
2,7-3,5
5,4—6,3
89,8-90,5
80,6-82,5
4,6-5,5
3,8 4,3
13,7-15,9
45,2-50,9
68,4-70,5
14,2-15,5
3 ,2-5,9
(18,5) -25,2
11,7—(13,0)
31,5-35,9
27,8-31.2
В % от длины головы (с жаберным аппаратом)
Длина рыла
Продольный диаметр глаза
Промежуток глаз — жаб. аппарат
Длина головы до 1-го жаб. отверстия
Длина жаберного аппарата
Расстояние до ноздри
Высота головы
Ширина лба
Продольный диаметр закрытого рта
30,8-34,0
6,1-10,7
13,4-13,8
52,3-57,0
40,9-50,8
29,1—33,5
20,3-23,0
15,0-17,7
29,4-31,7
2. Lampetra kessleri (Anikin)—сибирская минога
Синонимия. Petromyzon kessleri: Аникин, Изв. Томск, унив., 1905.
Lampetra fluviatilis japonica: Берг, Фауна России, Рыбы, т. 1, 1911.
Lampetra japonica kessleri: Берг, Рыбы пресн. вод СССР, т. 1, 1932. — Иоган-
зен, Зоол. журн., т. XIV, 1935.
Lampetra kessleri (Anikin): Полторыхина, Вопр. ихтиологии, т. 14, вып. 2,
1974. — Изв. СО АН СССР, вып. 1, 1979.
Распространение. Сибирская минога населяет бассейны рек Се-
верного Ледовитого океана от Печоры до Колымы; встречается на
Аляске, Анадыре, Камчатке, Сахалине (Берг, 1931, 1948; Иоганзен,
1935 и др.). В бассейне Иртыша обычна: широко распространена
48
в Иртыше и его притоках, протекающих на территории Восточно-
го и Северного Казахстана (Варпаховский, 1897, 1898; Берг, 1931,
1948; Полторыхина, 1971, 1973, 1974, 1979).
Личинки сибирской миноги населяют заиленные участки рек,
протоки, обычны в заливах или в приустьевой части. Пос-
ле метаморфоза миноги скапливаются в проточных участках во-
доемов, часто заросших водной растительностью.
Описание. Сибирская минога — непроходная форма тихоокеан-
ской миноги — имеет сходные с проходной формой признаки в
строении зубного аппарата и количестве туловищных миомеров.
В то же время имеет обособленный ареал и существенные морфо-
Рис. 3. Ротовая воронка сибирской миноги
экологические отличия (Берг, 1948; Майр, 1968 и др.). Правиль-
нее, на наш взгляд, считать ее самостоятельным видом, как пред-
ложил В. А. Аникин (1905).
По систематике V. D. Vladycov и F. Kott (1978, 1979),- миноги
Сибири объединяются в один вид—Lethenteron japonicum (Mar-
tens), ареал которого охватывает Западную и Восточную Сибирь,
Северную Америку и прилегающие к ним острова Северного Ледо-
витого и Тихого океанов. Как видим, в систематике миног Сибири
До последнего времени имеются большие разногласия, вызванные,
по-видимому, их слабой изученностью.
4—7
49
Таблица 4. Морфологические признаки сибирской миноги р. Ульба
Признак Самцы (п=50) Самки (п=50 ) с Mdiff
М + ш 1 ° 1 с М±т °
Абсолютная длина, мм 189,5±1,46 10,45 ! 5,52 186,40 + 1,72 12,15 6,53 0
В % от длины тела Длина рыла 7,30±0,05 0,48 1 5,47 6,64 + 0,03 6,39 5,97 7,65
Диаметр глаза 2,17±0,02 0,16 I 7,83 2,14 + 0,02 0,16 7,48 0,70
Расстояние от глаза до 1-го жаберного отверстия 2,92±0,02 0,18 6,16 2,89±0,02 0,17 5,90 । 0,83
Длина головы до 1-го жаберного отверстия 11,85 + 0,09 0,66 5,58 11,17 + 0,08 1 0,59 5,28 5,68
Длина жаберного ап- парата 9,54 + 0,06 0,42 4,40 9,56±0,03 0,43 4,50 0,30
Длина головы до 7-го жаберного отверстия 21,00±0,11 0,78 3,71 20,34±0,11 0,78 3,83 4,13
Расстояние до ноздри 6,65±0,03 0,42 6,35 5,97±0,07 0,48 8,04 7,00
Длина тушки до 1-го жаберного отверстия 88,57 + 0,11 0,75 1 0,83 89,25±0,10 0,65 0,73 4,40
Длина тушки до 7-го жаберного отверстия 79,43+0,12 0,83 1,04 80,09 + 0,12 0,79 0,98 4,03
Высота головы 4,46 + 0,03. 0,22 4,94 4,36±0,04 0,26 5,97 2,00
Ширина лба 3,04±0,03 0,19 6,26 2,98+0,03 0,23 7,72 1,40
Продольный диаметр ротового диска 5,90±0,07 0,46 7,88 5,62±0,03 0,41 7,36 3,03
Наибольшая высота тела 5,05+0,04 0,30 6,00 4,98 + 0,05 0,32 6,43 1,50
Наибольшая толщина тела 3,86+0,06 1 0,4010,50 3,90+0,07 0,52 13,13 0,47
Наибольший обхват тела 14,98+0,12 0,82 5,48 15,12 + 0,12 0,84 5,55 1,30
Антедорсальное рас- стояние 47,35±0,20 1,42 3,00 48,19±0,20 1,38 2,85 3,70
Антеанальное рас- стояние 69,36 ±0,13 0,95 1,37 71,03 + 0,12 0,88 1,28 8,90
Длина основания 1-го спинного плавника 15,64+0,12 0,81 5,18 15,42 + 0,12 0,83 5,67 1,35
Наибольшая высота 1-го спинного плавника 2,20+0,04 0,29 13,20 1,96±0,04 0,30 15,32 4,21
Длина основания 2-го спинного плавника 25,45±0,18 1,34 5,26 25,34 ±0,20 1,44 5,65 0,40
Наибольшая высота 2-го спинного плавника 4,18+0,05 0,37 8,87 3,80±0,05 0,35 9,21 5,35
Длина спинной части хвостового плавника 12,58 + 0,11 0,79 6,36 12,23±0,11 0,79 61,40 2,82
Анально-каудальное расстояние 30,94 ±0,14 0,98 3,18 29,20 + 0,12 0,89 3,35 9,46
В % от длины головы Длина рыла 60,90 + 0,41 2,89 4,71 58,80±0,37 2,62 4,45 3,68
Диаметр глаза 19,42+0,18 1,25 6,45 19,20±0,18 1,30 6,77 0,78
Расстояние от глаза до 1-го жаберного отверстия 24,67+0,21 1,46 5,93 26,00±0,17 1,20 4,62 5,02
Высота головы 37,84 + 0,27 1,92 5,10 38,60 + 0,30 2,14 5,56 1,50
Ширина лба 25,40 + 0,21] 1,50, 5,90 27,20±0,24i 1,68 6,20 5,63
Миноги бассейна Верхнего Иртыша характеризуются следующи-
ми признаками. Верхнечелюстная пластинка широкая, имеет по кра-
ям по одному зубу. Нижнечелюстная пластинка с 6—7 (редко до 10)
зубами, из которых внешние раздвоены. Между верхнечелюстной и
нижнечелюстной пластинками расположены внутренние боковые
губные зубы — по три с каждой стороны, в основном двураздель-
ные, редко не разделен нижний или средний зуб. Верхние губные
зубы (17—27) имеют радиальное расположение и уменьшаются
от центра к периферии (рис. 3). Нижние губные зубы (16—25)
образуют один дугообразный ряд; у многих особей они очень мел-
кие, иногда отсутствуют.
Пластические признаки сибирской миноги из Верхнего Иртыша
характеризуются значительными индивидуальными колебаниями
И хорошо выраженным половым диморфизмом (табл. 4).
Рис. 4. Сибирская минога: вверху — самец, внизу — самка
.^Основные различия в пластических признаках между самцами
леамками связаны с положением и размерами спинных плавников,
^самцов плавники выше и расположены ближе к головному от-
,делу. Рострум и голова их длиннее, ротовой диск больше (рис. 4).
объясняется, вероятно, некоторыми различиями в поведении
•ролов, разницей их локомоторной активности.
Ж Размеры взрослых особей сибирской миноги варьируют от 15
.Jwp'25 см и составляют в среднем 18—20 см при массе около 10 г.
^'Личинки сибирской миноги (пескоройки) по внешнему строе-
/ 1вйо очень сходны с личинками других видов: они имеют удлинен-
*о* *цилиндрическое тело, подковообразный рот, усаженный раз-
ветвленными ворсинками, скрытые под кожей недоразвитые глаза
ЙИс- 5)- По V. D. Vladykov (1958), личинки миноги Верхнего
Иртыша характеризуются следующими признаками: хвостовой
Илавник на дорсальной стороне имеет едва заметную вырезку на
Расстоянии 1/3 от ануса, жаберные отверстия и жаберный канал
слабо пигментированы, хроматофоры на каудальном плавнике
°средоточены в основном вокруг тела и больше на конце, где они
доходят до края; количество туловищных миомеров от 67 до 72.
3 установленных таксономических признаков более постоянно
оличество туловищных миомеров, другие же признаки значитель-
изменяются в зависимости от возраста.
51
Пластические признаки пескороек имеют существенные колеба-
ния, обусловленные размерно-возрастными изменениями. Срав-
нительный анализ трех размерных групп пескороек показывает
(табл. 5), что с увеличением их абсолютной длины, особенно на
первых этапах постэмбрионального развития, соотношение отдель-
ных частей тела резко меняется. Из 19 исследованных признаков
(в % к длине тела), реальные различия наблюдаются по 10 индек-
Рис. 5. Личинка сибирской миноги на первых стадиях метаморфоза
сам, из которых одни с ростом личинок увеличиваются, а другие,
наоборот, уменьшаются. К признакам, увеличивающимся с дли-
ной, относятся: длина тушки до 1 и 7-го жаберных отверстий, дли-
на спинной части стебля хвостового плавника, анально-каудальное
расстояние. К признакам, уменьшающимся с размером личинок,
относятся: длина головы до 1 и 7-го жаберных отверстий, ширина
лба, длина жаберного аппарата, антедорсальное и антеанальное рас-
стояние и, кроме того, длина основания 1-го спинного плавника
при сравнении личинок II и III размерных групп. У особей длиной
менее 7—8 см плавники дифференцированы слабо и их размеры в
таблице 5 не приводятся.
Изменения пластических признаков, объясняются, видимо, тем,
что скорость роста отдельных компонентов, составляющих индек-
сы, в течение всего периода развития миноги неодинакова. Как
известно, головной отдел животного, связанный с расположением
наиболее жизненно важных органов (нервная система, дыхатель-
ная, сосудистая), развивается быстрее и достигает своего оптимума
значительно раньше других отделов тела (Иванов, 1937). Осо-
бенно интересные данные получаются при сравнении антедорзаль-
ного, антеанального и анально-каудального расстояний, определя-
ющих положение плавников. У личинок сибирской миноги на раз-
ных этапах развития (I возрастная группа), анально-каудальное
расстояние значительно меньше, чем у личинок средней группы
(Mdiff = 14,5), в то время как при сравнении II и III размерных
групп это различие заметно сглаживается (Mdiff=3,5), хотя и
остается достоверным. Аналогичные изменения наблюдаются и в
52
положении спинного плавника (антедорсальное расстояние), ко-
торый у личинок младшего возраста располагается ближе к хвос-
товому отделу. Изменение расположения спинных плавников на
теле миноги с увеличением ее размеров может быть вызвано удли-
нением хвостового отдела.
Окраска личинок носит приспособительный характер, поэтому
часто ее вариации связаны с изменениями условий внешней сре-
ды. Как показали наблюдения, цвет тела пескороек сибирской ми-
ноги, обитающих в темном грунте, намного темнее, нежели у осо-
бей из светлого песчаного биотопа. Аналогичные результаты полу-
чены при выдерживании животных в аквариуме с разным затемне-
нием.
Биология. Размножается сибирская минога в водоемах Казах-
стана в конце мая — начале июня. Так, в протоке Иртыша, близ
г. Усть-Каменогорска нерест миноги происходил: в 1965 г. — с
24 мая по 3 июня, в 1966 г. — с 27 мая по 3 июня при температуре
воды 14—15°, в 1967 г. — с 23 мая по 2 июня при 13,5—15°, в
1969 г. — с 3 июня по 10 июня при 13,5—15° (Полторыхина,
1973).
Для нереста миноги поднимаются в те же реки и протоки, где
происходило их эмбриональное развитие. По своему характеру не-
рестовые миграции сибирской миноги сходны с миграциями про-
ходных миног, но они значительно короче. Наблюдения за мигра-
циями миноги в р. Ульба показали, что более крупные особи про-
ходят на нерест раньше и поднимаются в верхние участки реки, а
мелкие идут на нерест позднее и размножаются в нижних, при-
устьевых участках. Соотношение полов во время размножения не-
одинаково, но во всех нерестовых популяциях преобладают самцы.
Нерестовое поведение сибирской миноги сходно с поведением дру-
гих непроходных миног (Абакумов, 1964 и др.). Размножаются
миноги в неглубоких речках и протоках с песчано-галечным грун-
том и сравнительно быстрым течением.
Для откладки икры миноги строят гнезда-ямки, обычно у выхо-
да грунтовых вод или родников, обеспечивающих более постоян-
ный кислородный и температурный режим, лучший водообмен,
необходимый для развития эмбрионов. Строить гнездо начинает
самец. Перед этим он выбирает место, иногда вступая в борьбу с
другими самцами. К самцу присоединяются 2—3 самки, образуя
нерестовую «семью» и принимая участие в дальнейшей постройке
гнезда. Перекладывая гальку своими присосками, они углубляют
ямку. Позднее к некоторым семьям присоединяются еще один са-
мец и две-три самки. В основном «семьи» речной миноги состоят
из 3—4, реже 6—8 особей.
Вся икра у миноги в период размножения одинаковой зрелости,
но мечет ее самка в несколько приемов. Икра в гнезде, несмотря
на небольшое количество, распределяется также отдельными пор-
циями — «кладками». Каждую только что отложенную порцию
икры самец закладывает галькой. После нереста миноги поги-
54
бают. Перед этим они уходят в тихие глубокие места, заросшие
растительностью.
Эмбриональный период сибирской миноги длится около 10 дней
при температуре воды 14—19°. Через 10—12 дней после нереста в
гнездах обнаруживаются мелкие прозрачные личинки длиной 3—
5 мм, питающиеся за счет желтка.
Вначале они неподвижны, затем приобретают подвижность и
течением сносятся вниз, где в более тихих, заиленных участках
оседают на дно и зарываются в ил. Дрейф личинок продолжается
15—20 дней. К этому времени их длина увеличивается до 8—10 мм.
Размерный состав и плотность распределения личинок в раз-
личных биотопах неодинаковы и зависят в основном от мощности
иловых отложений и состава грунта. Мелкие личинки (до 3 см)
встречаются, как правило, в мелкозернистом, мягком грунте на
глубине 1—4 см. В этих же местах, но значительно глубже, могут
находиться и более крупные особи. Пескоройки длиной 5—15 см
предпочитают открытые, ровные места и расселение их по сравне-
нию с первыми более равномерное и плотное (до 10—15 экз. на
1 м2). Крупные личинки (15—25 см) населяют сильно заиленные
участки, удаленные от берега, иногда заросшие водной раститель-
ностью, их плотность не более 3—5 экз. на 1 м2.
Растут пескоройки медленно, достигая к концу первого лета
всего 20—23 мм, а к началу метаморфоза — 200—250 мм. В сред-
нем за год их размеры увеличиваются на 30—40 мм. Растут в ос-
новном летом, а зимой рост почти прекращается. В разные годы и
в разных водоемах бассейна Верхнего Иртыша рост пескороек ме-
няется незначительно, однако эти отклонения в совокупности с ин-
дивидуальными колебаниями роста существенно изменяют размер-
ный состав старших возрастных групп и в дальнейшем — нересто-
вых популяций. Длина личинок последней возрастной группы мо-
жет варьировать от 160 до 280 мм.
Питаются личинки сибирской миноги детритом и водорослями.
Соотношение органических остатков и водорослей в их питании
непостоянно и меняется в зависимости от возраста и сезона года.
Самые мелкие личинки кормятся мельчайшими планктонными орга-
низмами и пелогеном, оседающим на грунт и подводные предметы.
Пескоройки старших возрастных групп усваивают органические
вещества из ила, в котором попадают зеленые, сине-зеленые и
Диатомовые водоросли.
Продолжительность личиночного периода сибирской миноги не
Менее 5—6 лет, после чего происходит превращение личинки во
взрослую форму.
Начинается метаморфоз сибирской миноги в конце июля — на-
чале августа, при температуре воды 16—18°. Первые стадии мета-
морфоза протекают у личинок довольно быстро. Уже в первой
половине августа встречались пескоройки, которые имели откры-
тые глаза и округлое ротовое отверстие. Дальнейшее превращение
личинок во взрослую особь замедляется. Постепенно у пескороек
55
исчезают личиночные ворсинки и на их месте в виде бугорков
появляются зачатки зубов. На краях ротового отверстия образу-
ется оторочка из кожистых выростов, преназальпая часть головы
удлиняется, тело становится темным, серебристым, происходит
формирование плавников. На этих стадиях метаморфоза изменя-
ется и внутреннее строение животного: передняя часть кишечника
разделяется на два самостоятельных отдела — трубчатый пищевод
и жаберный отдел, открывающиеся в ротовую полость. С измене-
нием строения ротового и дыхательного аппаратов меняется и
поведение личинок: они становятся более активными и способными
присасываться к подводным предметам.
Самые последние стадии метаморфоза сибирской миноги харак-
теризуются дальнейшим обособлением и увеличением ротовой
воронки, ростом и ороговением зубов. С завершением ороговения
зубов метаморфоз миноги, как указывает П. П. Балабай (1958),
заканчивается. Таким образом, превращение личинок сибирской
миноги проходит в короткий период (август — ноябрь). В конце
ноября он полностью завершается.
В имагинальный период у миноги происходят укорачивание
тела, сближение спинных плавников и сморщивание кожи. Эти из-
менения вызываются, вероятно, созреванием половых продуктов и
голоданием в преднерестовый период. В среднем размеры песко-
роек перед метаморфозом больше взрослых особей на 25—30 мм.
Взрослые миноги проводят зиму под валунами, подводными
камнями, где обнаружить их очень трудно, почти невозможно, по-
этому о зимнем периоде жизни миноги в литературе нет сведений.
В это время миноги не питаются. Их кишечник постепенно атро-
фируется и превращается к моменту нереста в тонкую нить.
Миногами питаются щука, окунь, а также птицы-ихтиофаги.
Однако употребление взрослой миноги и ее личинок в пищу други-
ми животными носит чаще всего случайный характер и отмечается
в литературе крайне редко. Вполне возможно, что в природе си-
бирская минога служит кормом для других животных значительно
чаще, чем об этом известно, так как отсутствие костных образова-
ний в ее скелете способствует быстрому перевариванию и затруд-
няет тем самым их обнаружение среди прочих компонентов пищи.
В то же время нельзя не признать, что скрытый образ жизни ми-
ног защищает их от более сильных врагов. Добычей они становят-
ся, вероятно, только тогда, когда выходят из грунта или укрытий и
совершают незначительные миграции в поисках лучших мест оби-
тания. Особого внимания в этой связи заслуживают исследования
О. Ф. Гриценко (1968) и других авторов, отмечающих важное
значение бентических организмов, в частности личинок миног, в
аэрации грунтов, например, в местах эмбрионального разви-
тия лососевых рыб. Употребляя в пищу ил и прокладывая много-
численные ходы, пескоройки облегчают доступ кислорода в более
глубокие слои грунта, улучшая тем самым его химический состав
и структуру.
56
Хозяйственное значение. Промыслового значения сибирская
минога не имеет. Обычно пескоройки, выбираемые из ила, зачерп-
нутого лопатой, используются рыбаками для наживки при ловле
ценных промысловых рыб. В небольшом количестве миног вылав-
ливают специальными вилками или сачком для корма домашней
птицы.
ОТРЯД ACIPENSERIFORMES — ОСЕТРООБРАЗНЫЕ
Семейство ACIPENSER1DAE — ОСЕТРОВЫЕ
Пресноводные или проходные рыбы, обитающие в Европе, Се-
верной Азии и Северной Америке. В Казахстане водятся предста-
вители 3 родов.
Род HUSO BRANDT, 1869 — БЕЛУГА
Включает 2 вида: Н. dauricus (Georgi) — калуга в бассейне
Амура и Н. huso (Linne) — белуга в бассейнах юго-восточных мо-
рей Европы.
1. Huso huso (Linne)—белуга
Казахское название: кортпа
Синонимия. Acipenser huso: Linne, Syst nat, ed. X, 1758.
Huso huso: Берг, Фауна России, Рыбы, т. I, 1911.
Н. Я. Бабушкин (1942) предложил считать типом Huso huso дунайскую бе-
лугу, а каспийскую—классифицировать как подвид. Л. С. Берг (1948) указы-
вает, что типом следует считать каспийскую белугу и что для таксономии дунай-
ской белуги как типа достаточных оснований нет.
Распространение. Белуга (рис. 6) — самая крупная проходная
рыба, обитает в бассейнах Каспийского, Черного и Азовского мо-
рей, единичные экземпляры встречаются в восточной части Адриа-
тического моря, отдельные особи заходили в реки Анатолии. Наи-
более многочисленное «стадо» сохранилось в бассейне Каспия.
В пределах Казахстана обитает только в р. Урал, по которой дохо-
дит до устья р. Илек. Отдельные экземпляры заходят в эту реку и
поднимаются вверх по Уралу выше ее устья на 50—100 км.
Описание. Тело белуги торпедообразной формы. Рыло короткое,
заостренное, слегка вздернутое, мягкое сверху и с боков, не при-
крытое костяными щитками. Рот нижний, в закрытом состоянии
серпообразный, в приоткрытом — полулунный, не переходящий на
бока головы; нижняя губа прервана. Усики сплющены с боков и
имеют листовидные придатки. Половой диморфизм у неполовозре-
лых особей не выражен, но самку от самца можно отличить по
большому и более мягкому брюшку во время захода в реку на не-
рест, когда половые железы достигают III—IV и IV стадии зре-
лости.
57
D 58—75, в среднем 65,8: А 25—40, в среднем 32,4; спинных жу-
чек 10—15, в среднем 13,5; боковых — 40—51, в среднем 46,5;
брюшных — 9—11, в среднем 10,3; жаберных тычинок 19—30, в
среднем 22,9. Хромосом 60 (Серебрякова, 1974), а по последним
данным, условно 116±4 (Васильев и др., 1980).
Максимальная масса тела выловленной белуги за последние
25 лет равна 865 кг при длине 434 см.
Рис. 6. Белуга
Биология. Нерестовая миграция белуги начинается в море вда-
ли от «родной» реки. У созревающих особей миграционный им-
пульс появляется при определенном состоянии организма и дей-
ствии раздражителей, воспринимаемых извне (Гербильский, 1957;
Баранникова, 1975), когда половые железы одних (озимых) осо-
бей находятся в III и III—IV стадиях зрелости, а у других (яро-
вых) — в IV.
Началом нерестового хода следует считать период, когда в реку
начинают заходить первые половозрелые особи с развитыми поло-
выми железами. Это, по-существу, начало нового биологического
года. У уральской белуги он начинается в июле и лишь в исключи-
тельных случаях (в годы с очень ранней весной) —во второй поло-
вине июня при температуре воды 19—24°. Прозрачность речной
воды в это время бывает почти максимальной (0,5—0,8 м по диску
Секки), а расход воды и скорость течения близки к минимальным.
В августе ход белуги усиливается, максимума достигает в конце
месяца, но чаще в сентябре. С начала октября уловы в низовьях
реки начинают быстро падать. В конце этого месяца и особенно в
начале ноября в уловах нижних (лицевых) тоней белуга встре-
чается очень редко. В осеннее время по Яицкому рукаву и проры-
тому рыбоходному каналу в р. Урал не заходит из-за малой глу-
бины. Заход белуги в реку заканчивается при температуре 7—6°, а
в некоторые годы (1964 г.) —при 17°. Продолжительность летне-
осеннего хода белуги 35—90 суток, но в большинстве случаев —
60—75. Движется белуга летом со скоростью 15—20 км в сутки.
Осенью скорость перемещения мигрантов снижается и не превы-
шает 9—12 км в сутки.
С конца октября — начала ноября по декабрь включительно
(2 месяца) белуга в реку не заходит. При появлении прочного
льда и крепких морозов (конец декабря — начало января) белуга
58
вновь появляется в реке. В этот период в уловах встречаются лишь
единичные экземпляры, к концу января ход постепенно усиливает-
ся, достигая максимума к середине или концу февраля, а затем
затухает. Эта закономерность характерна для любого года, не-
смотря на неодинаковую интенсивность хода в разные годы, за
исключением суровых зим, когда толщина льда достигает 0,8—1 м,
а содержание растворенного в воде кислорода падает до нижних
пороговых величин (1,7 мг/л у пос. Махамбет—150 км от
взморья)*, что обусловливает скат ослабевших осетровых с зимо-
вальных ям нижнего течения р. Урал. Подо льдом белуга передви-
гается со скоростью 8—12 км в сутки. Перед таянием льда ход
белуги ослабевает или прекращается.
После короткого (15—25 дней) зимне-весеннего перерыва пер-
вые ходовые производители белуги вновь появляются в дельте
реки при температуре воды 2—3° (конец марта — начало апреля).
Массовый весенний ход обычно начинается в тот период, когда
верхние участки реки и значительные площади восточной части
Северного Каспия еще покрыты льдом. Белуги, зашедшие в реку
осенью и зимовавшие на ямах, в это время еще покоятся, ожидая
таяния льда и потепления воды. Почти вся белуга весеннего хода
до начала паводка заходит в р. Урал только через судоходный
канал. По рыбоходному каналу и Яицкому рукаву она мигрирует
в небольших количествах только в половодье.
Разгар весеннего хода белуги обычно наблюдается во второй
декаде апреля (6—8°), т. е. за 15—30 дней до пика паводка.
В маловодные и близкие к средней водности годы с ранней и теп-
лой весной пик хода совпадает с пиком паводка. Весенний ход бе-
луги в реку заканчивается при температуре воды 14—15° (пример-
но середина мая). После чего наступает третий длительный (пол-
тора-два месяца) летний перерыв в ходе белуги.
Производители белуги, заходящие в реку летом и осенью, дале-
ки от зрелости. По мере приближения нерестового периода (от ле-
та к осени, затем к зиме и весне) показатели гонадо-соматического
Таблица 6. Состояние зрелости половых желез и масса ооцитов
белуги р. Урал, мг
Сезоны года Гонадо-соматический индекс Масса ооцитов п
Самцы Самки От—S.0 М
От—до 1 М От—до М
Лето— осень 0,83—7,2 3,2 6,8-26,4 14,8 12,4-36,4 25,1 315
Зима 1,30-9,4 3,4 7,6-27,4 16,2 13,6-38,3 27,2 237
Весна 1,61-9,9 3,7 9,2-30,3 17,8 14,6-44,1 28,5 512
За биол. год 0,83-9,9 3,6 6,8-30,3 16,6 12,4-44,1 26,8 1064
* В дальнейшем изложении расстояния будут приводиться от взморья до
Указанного места по реке.
59
индекса (коэффициента зрелости по Фультону) и масса ооцитов
у них увеличиваются (табл. 6).
Картина гистологических препаратов семенников и яичников
показывает, что в летнее и осеннее время сперматогенез и ооге-
нез далеки до завершения. Основными клеточными элементами
семенников являются сперматоциты второго порядка, сперматиды и
небольшие группки сперматозоидов. Ядро в ооците находится в
его центральной части и лишь к осени несколько смещается к ани-
мальному полюсу (рис. 7, 8).
Рис. 7. Срез семенника
белуги озимой внутрипопуляционной нерестовой группы.
Октябрь, дельта р. Урал
Производители, заходящие в реку зимой (подо льдом), более
близки к зрелости. Б семенниках завершается сперматогенез (за-
канчивается формирование сперматид и идет активное образова-
ние сперматозоидов). Ядро в ооцитах в это время обычно смещено
к аиимальному полюсу и находится уже в зоне мелкозернистого
желтка, но на значительном расстоянии от анимального полюса
(рис. 9).
Все производители апрельского и майского хода практически
готовы к нересту. В их семенниках к этому времени завершается
формирование сперматозоидов; ядро в ооцитах смещается к ани-
мальному полюсу, ядрышки находятся в центральной части ядра,
либо оно оптически пустое. Заканчивается и процесс формирова-
ния микропиле (12—16 шт. у каждого ооцита), половые клетки
60
достигают дефинитивных размеров. Масса ооцитов белуги весен-
него хода во время речной миграции не увеличивается.
Из изложенного видно, что в разное время года в реку заходят
группы производителей с различным состоянием зрелости половых
желез. Миграции их начинаются и заканчиваются при различных
экологических условиях (температура, мутность и прозрачность
воды, скорость течения и др.). Размножаются они в разных участ-
ках реки и в разное время, т. е. в специфических условиях для каж-
дой конкретной группы. В динамике нерестового хода наблюдаются
длительные перерывы, причем ход одних заканчивается при сравни-
тельно высоких температурах, у других начинается при очень низ-
Рис. 8. Срез ооцита белуги озимой нерестовой группы. Август, низовья р. Урал
^температурах. Усмотрев в этом аналогию со злаками, Л. С. Берг
б 1953) назвал их яровыми и озимыми расами. Н. Л. Гер-
-ЬСКИЙ (1957) на основе углубленного изучения биологии осет-
Р ЫХ охарактеризовал их как внутривидовые биологические груп-
61
пы (ранние яровые, поздние яровые, озимые летнего и осеннего
хода). Дальнейшие исследования показали, что эти группы рыб
являются составной частью популяции и все различия между ними
связаны с размножением; поэтому их следует именовать не расами
или внутривидовыми биологическими группами, а внутрипопуля-
ционными нерестовымп группами (Песеридп, 1968, 1973, 1976).
Белуга, будучи литофильной рыбой, откладывает икру на твер-
дый субстрат: камни, гальку, гравий, плотный крупнозернистый
песок, сцементированный песчаник, плотную глину с вкрапления-
Рис. 9. Срез ооцита озимой белуги зимнего хода. Февраль, дельта р. Урал
ми ракуши (створки моллюсков). Встречается развивающаяся
икра и на растительном субстрате (корневища ивы и тополя, по-
росли многолетней растительности, затапливаемой полыми во-
дами) .
Общая площадь нерестилищ осетровых в р. Урал от Гурьева до
Уральска около 1700 га (Песеридп, Верина, 1981), но белуга ис-
пользует под нерест не более 1000 га — это в основном галечни-
ковые и гравийные россыпи средней и верхней частей нерестовой
зоны. Почти все ложе р. Урал от Уральска и до Оренбурга устлано
галькой и гравием с песком, на которых возможен нерест.
Разные биологические группы белуги размножаются в разное
время. Так, основная масса озимой белуги нерестится на русловых
грядах на глубине 6—12 м при температуре воды 7—11° на участ-
62
ке реки от г. Чапаеве до Уральска. Отдельные экземпляры откла-
дывают икру и в р. Илек, несколько выше устья этой реки.
Яровая белуга размножается в основном на участке реки от
рос. Индер (315 км) до г. Чапаево при температуре воды 10—15°
на русловых и затопляемых грядах на глубине 3—8 м со скоростя-
ми течения 1,2—1,4 м/с. Мутность воды в период нереста макси-
мальная. На 1 м2 нерестилищ в разгар нереста откладывается в
среднем до 400—900 икринок. Отдельные особи, заходящие в реку
в первой половине мая, размножаются в нижней части нерестовой
зоны (до пос. Индер). Развивающаяся икра встречалась и в дель-
те реки.
Оплодотворенная икра через 5—6 мин приклеивается к суб-
страту и «инкрустируется» наносами. Продолжительность эмбрио-
нального развития белуги находится в прямой зависимости от
температуры среды. При 8—11° развитие эмбриона продолжается
до 200—240 ч, а при 15—16° личинки выклевываются через 130—
140 ч.
Покатные личинки белуги (41,8 мг, 27,7 мм) в дельте реки по-
являются обычно в первой половине мая. Скат продолжается до
конца месяца. Пик ската личинок приходится на вторую половину
мая. Одновременно с личинками в конце мая в низовьях реки вы-
лавливается и подросшая молодь — потомство озимой белуги.
Максимальные уловы молоди белуги в дельте наблюдаются в пер-
вой декаде июня. К этому времени средняя масса покатных белу-
жат достигает 2,0—2,5 г. Отдельные экземпляры молоди весят
5,3 г. Средняя масса белужат к концу июня — началу июля состав-
ляет 4,2—5 г, а к концу июля увеличивается почти вдвое (8,6 г).
Максимальная масса белужонка, выловленного в реке, составила
41,96 г при длине 222 мм (Пессриди, Бекешев, 1967). Вся молодь
белуги скатывается в море до сентября и в реке на зимовку не
остается.
Масса и количество скатывающейся молоди белуги колеблется
по годам. Четкой корреляции между объемом стока реки и массой
потомства нет, но в многоводные годы молодь крупнее, чем в ма-
ловодные. Сопоставляя данные по скату молоди из р. Урал с коли-
чеством выращенной молоди 11 осетроводными заводами Каспия
(табл. 7) с учетом, что жизнестойкость молоди естественного про-
исхождения намного выше искусственно выращенной, можно с
Уверенностью сказать, что 50% нагуливающейся белуги в Кас-
пийском бассейне уральского происхождения.
Развивающаяся на нерестилищах икра белуги истребляется
Пескарем, бычками, шиповкой, густерой, белоглазкой, лещом, со-
мом, стерлядью, подросшей молодью белуги (5—8 см), одно- и
Двухлетними шипятами. Предличинкп и личинки белуги поедаются
чехонью, синцом и молодью щуки. Наиболее опасный враг молоди
белуги в реке — сом, в желудках которого встречается до 10—
*2 белужат массой в 2—3 г. В море врагов у белуги практически
Нет.
63
Созревание, возрастной состав нерестовой части популяции,
рост. В. В. Петров (1927) и Н. Я- Бабушкин (1964) указывали, что
половая зрелость у самцов уральской белуги наступает не ранее
14 лет, а у самок — в 18 лет. Следует иметь в виду, что названные
исследователи проводили наблюдения в период расцвета морского
красноловья, во время которого вылавливалась большая часть бе-
луги, а в реку заходили лишь остатки нарушенной промыслом не-
рестовой части популяции (селективность морских орудий лова,
Таблица 7. Масса и численность скатывающейся молоди уральской
белуги и выращенной на осетроводных заводах Каспия
Год Объем сто- ка Урала, км3 Средняя масса 1 экз. молоди, мг Кол-во ска- тывающей- ся молоди, млн, шт. Выпуск молоди белуги осетровод- ными заводами, млН. шт.
1961 6,2 2070 1.2 0,5
1962 7,4 2350 2,0 1,8
1963 9,8 3070 4,4 3,1
1964 11,5 1950 4,2 5,3
1965 6,2 — 1.0 10,8
1966 11,9 2550 12,5 12,3
1967 3,0 — 1,0 10,0
1968 5,1 1150 1,8 13,2
1969 5,8 1150 2,3 17,7
1970 12,8 3930 22,0 16,1
1971 12,5 6520 2,2 13,9
1972 8,5 1240 6,6 13,3
1973 4,9 1220 2,3 13,6
1974 7,6 1'130 44,7 10,9
1975 3,6 1960 1,2 14,8
1976 4,2 1472 1,8 16,2
1977 2,5 1062 2,0 17,2
1978 7,3 1180 2,8 10,3
1979 6,3 1565 12,1 17,6
1980 6,0 2318 4,7 17,6
Всего 132,8 236,7
промысловая мера, высокие уловы в море и т. п.). К тому же
В. В. Петров собирал материал (всего несколько десятков особей)
не в р. Урал, а в приустьевом пространстве, т. е. в опресненной
зоне Северного Каспия. Поэтому сведения Н. Я. Бабушкина и
В. В. Петрова, по независящим от них причинам, не достоверны и
не могут ответить на вопрос об истинных сроках наступления по-
ловозрелое™ белуги.
По данным последних лет, отдельные самцы белуги начинают
свою первую нерестовую миграцию в возрасте 9—И, а самки — в
12—14 лет. Основная масса самцов созревает на 12—15 году жиз-
ни, а самок — на 15—20.
Точных сведений о продолжительности межнерестовых перио-
дов (периодичность нереста) белуги нет. По-видимому, самцы за-
ходят на повторный нерест через 3—4, самки — через 4—6 лет.
64
В результате интенсивного промысла в низовьях реки возраст-
ной состав нерестовой части популяции колеблется по годам, но ха-
рактерным для любого года является то, что в уловах самцы в ос-
новном в возрасте 13—26, самки — 17—28 лет.
Анализ многолетних материалов по размерно-весовому и возраст-
ному составу нерестовых частей популяции белуги и данных промыс-
ловой статистики позволяет сказать, что промыслом изымаются в ос-
новном впервые созревающие производители. Доля повторно созре-
вающих рыб в промысловых уловах и в нересте незначительна. Воз-
раст крупных самок (200—300 кг), как и предельный возраст бе-
луги установить невозможно, поскольку центральная часть спи-
лов маргинальных лучей (первого луча грудного плавника), по
которым определяется возраст, деформирована.
Размеры и особенно масса белуг по возрастам подвержены
значительным колебаниям, но как у самок, так и у самцов наблю-
Таблица 8. Характеристика уловов белуги в Северном Каспии и р. Урал
в 1961—1972 гг.
Возраст- ная группа, годы Самцы(п=1048) Самки(п=835)
Длина, см Масса, кг п Длина, см Масса, кг п
От-до | Средн. От—до | Средн. От—до Средн. От-до Средн.
9—10 11-12 142—160 146—169 152.6 159,5 19-32! 24—36 24,5 29,3 21 44 158-174 167,3 26,6-36,5 31,8 14
13-14 155—181 168,8 25-411 33,9 81 161-179 113,8 23,8-39,1 35,9 38
15—16 164-194 177,0 28—68 37,8 118 169-198 182,8 28,1-68,5 42,3 54
17-18 170-201 185,3 31—71 45,3 145 174-216 194,831,3-71,2 49,6 79
19-20 178—208 194,5 30-73 50,1 181 181 -238 208,530,8-87,7 58,0 104
21-22 183—217 203,1 36-78 56,5 138 196-248 220,3 38,5 - 84,0 64,9 126
23-24 185-230 214,6 41-88 64,2 121 211-246 227,4 50,0-124 78.5 130
25-26 209- 245 225,3 48-90 70,3 94 216—252 238,3 68-135 89,0 102
27-28 231—260 241,4 51—103 78,4 43 228-284 253,2 88-160 108,3 84
29—30 240-266; 250,8 55—101! 79,1 31 239 -295 264,3 94-185'122,0 32
31—32 251—280 262,3 61-114' 87,2 16 246—294 271,5 101—193 136,5 26
33-34 254—285 271,2 70—129 99,3 9 259-311 280,6 114-2051143,8 21
35—36 251—290 276,1 76-134, 108,5 5 268-318 284,1 118—2011147.3 11
37-38 — - — 276-324 291,3 126-218 153,8 7
39—40 — 284,0 - 1 131,0 1 281-328 298,0 146-264)174,5 5
Дается увеличение этих показателей с возрастом (табл. 8). По мере
роста увеличивается и масса тела, но у самцов медленнее (табл. 9).
В. В. Петров (1927) описывает случай поимки белуги в при-
устьевой области Урала массой более 1000 кг, из них икры 180 кг.
Самая крупная белуга за последние 25 лет была поймана в Урале
в апреле 1968 г.; она весила 867 кг, выход икры без пробоек соста-
вил 156 кг. В I г икры было 36,5 икринки. Плодовитость ее была
6694 тыс. икринок. Возраст самки определить не удалось.
Половой состав нерестовой части популяции белуги колеблется
Во годам (%). До 1971 г. преобладание самцов было менее вы-
раженным:
Год Самцы Самки Год Самцы Самки
1961 73,8 26,2 1971 55,5 44,5
1962 61,0 39,0 1972 78,0 22,0
1963 67,7 32,3 1973 77,5 22,5
1964 58,1 41,9 1974 71,0 29,0
1965 62,0 38,0 1975 72,5 27,5
1966 54,2 45,8 1976 81,8 18,2
1967 57,0 43,0 1977 85,4 14,6
1968 60,5 39,5 1978 87,6 12,4
1969 58,7 41,3 1979 85,4 13,6
1970 65,7 34,3 1980 82,7 17,3
В последующие годы число самцов в уловах резко увеличилось,
что связано, во-первых, с изъятием большого количества самок в
60-х годах и, во-вторых, вступлением в промысел урожайных поко-
лений благоприятных по водности 1961—1966 гг. (самцы созревают
быстрее самок и раньше их начинают заходить в реку). В ближай-
шие годы, как показывают и исследования в море (здесь соотно-
шение полов близко 1:1), ожидается вступление в промысел со-
зревающих самок.
Таблица 9. Масса тела половозрелой белуги р. Урал
по размерным группам (длина абсолютная)
Размер- ные Группы, см Самцы Самки Оба пола (п-4193)
Средняя масса 1 экз., кг п Средняя масса 1 экз,, кг п Срёдняя масса 1 экз,, кг п 1 '
141-150 21,3 9 21,3 9
151—160 28,6 15 — — 28,6 15
161—170 31,0 67 — — 31,0 67
171—180 33,6 152 —. — 33,6 152
181-190 38,8 348 — — 38,8 348
191-200 47,2 480 42,9 17 47,1 497
201—210 53,1 615 54,0 55 53,1 670
211-220 59,3 567 66,3 75 60,1 642
221-230 59,3 345 89,5 99 71,8 444
231—240 79,1 158 91,8 130 84,8 288
241—250 92,8 123 104,2 110 98,3 233
251-260 104,5 95 118,7 122 113,1 217
261-270 116,2 52 138,1 111 134,1 163
271-280 136,8 53 153,6 105 147,8 158
281-290 151,2 17 170,8 72 167,3 89
291 -300 179,4 17 188,4 73 186,7 90
301-310 193,5 5 214,5 31 213,3 36
311-320 203,0 4 232,6 13 229,0 16
321 -330 212,0 3 253,5 13 246,5 17
331-340 — — 268,4 13 268,4 13
341-350 — — 308,5 13 308,5 13
351 - 360 —. — 325,8 10 325,8 10
361-370 — — 335,8 4 335,3 4
371-380 — — 344,0 2 344,0 2
66
Плодовитость белуги колеблется в широком диапазоне: от 89,6
до 5694 тыс. икринок (средняя — 825 тыс.). Несмотря на незначи-
тельные индивидуальные колебания, в целом наблюдается законо-
мерное возрастание плодовитости по мере увеличения размеров
и массы самок (табл. 10).
Таблица 10. Абсолютная плодовитость белуги р. Урал в связи с длиной
тела (тыс. шт.)
Размеры, см Плодовитость п Раз- меры, см Плодовитость п
От-до | Средн. От—до Средн.
141-150 118-158 138 2 261—270 495-1785 863 120
151-160 — 89,6 1 271-280 464-1824 966 121
161-170 — — — 281-290 525-1938 1004 80
171-180 244-319 291 3 291-300 709-2340 1276 64
181-190 217-317 280 3 301-310 787—1817 1310 30
191-200 197-402 288 8 311-320 938-2254 1557 14
201-210 191-718 318 41 312-330 1120-2316 1617 9
211-220 263-1006 496 63 331-340 1711-2140 1856 4
221—230 328-1097 560 99 341-350 1416-2352 1832 9
231-240 303-1147 625 120 351-360 1148-2610 1920 9
241-250 333-1134 682 129 361-370 1105—2790 1689 3
251-260 356-1424 748 132 141-370 89,6-2790 825 1064
Рассматривая вариации массы ооцитов белуг разного размера
без учета внутривидовой дифференциации, Н. Я. Бабушкин (1964)'
пришел к выводу, что с увеличением размеров рыб возрастает и
масса их икринок: с 22—25,2 мг у рыб размером 160—199 см до
30,7—37 мг у особей длиной 320—359 см (158 проб). Анализ раз-
меров и массы ооцитов 540 самок, у которых они достигли дефини-
тивных размеров, позволил установить лишь незначительные раз-
личия массы ооцитов у разноразмерных белуг. Так, средняя масса
икринок у белуг до 120 кг длиной до 260 см (259 проб) составила
26,9 мг, а у рыб свыше 120 кг (281 проба) и более 260 см — 28,8 мг.
Средний самец белуги массой 50—60 кг за один нерестовый
период эякулирует до 1,2—1,6 л семенной жидкости. В 1 см3 спер-
мы содержится до 10 млрд, сперматозоидов, в среднем 2—3 млрд.
Следовательно, средний самец за нерест выметывает до 2,8—-
4,2- Ю12 спермиев, что в 3,3—4,9 млн. раз больше количества ооцитов,
выметываемых одной средней самкой.
Питание, распределение в море. Скатывающаяся молодь белуги
(1,5—7 г) в реке питается бокоплавами, мизидами, личинками хи-
Рономид, поденок, ручейников, а также олигохетами, икрой и ли-
чинками осетровых и других рыб. Крупные мальки белуги поедают
севрюжат и осетрят. Для молоди белуги характерен каннибализм.
Состав пищевого рациона меняется по мере ее роста. Индексы на-
полнения желудков очень высокие — 280—4ОО°/ооо (Зачетнова, 1973;
Стыгар, Песериди, 1981).
С момента попадания молоди в опресненные участки моря она
пачинает расселяться вдоль мелководных берегов и постепенно
67
уходит в более глубокие осолоненные участки. Меньшая часть мо-
лоди белуги уральского происхождения нагуливается на участках
моря, расположенных в междуречье Волги и Урала. Основная
ее масса устремляется к Уральской бороздине и на восток Север-
ного Каспия.
В первый год жизни сеголетки белуги на морских пастбищах
продолжают питаться ракообразными (гаммариды, мизиды), мол-
люсками (метилястер, дрейссена, кардиум) и рыбами (бычки, пу-
головка, килька, молодь сельди и карповых). На втором году
жизни белужата почти полностью переходят на хищный образ
жизни. Доля рыб в пищевом комке достигает 98%.
Характер пищи белуги меняется по сезонам года и в зависи-
мости от мест нагула. В Среднем и Южном Каспии белуга пита-
ется в основном бычками, пуголовками, килькой, сельдью. В Се-
верном Каспии она поедает воблу, леща, сазана и бычков. В пого-
не за косяками воблы и леща нагуливающиеся неполовозрелые
белуги иногда заходят в нижнюю часть дельты р. Урал. Белуга на
морских пастбищах не прекращает питаться в течение всего года,
но в зимнее время интенсивность питания гораздо ниже. У 60—
70% белуг, выловленных зимой, желудки бывают пустыми.
В желудках отдельных крупных особей, выловленных весной и
осенью в дельте реки, встречается до 40—50 вобл. В желудке од-
ной самки белуги массой 280 кг, пойманной в дельте, был обнару-
жен 9-летний самец осетра массой 9,3 кг, 8 вобл и I лещ, а у другой
самки — 2 сазана (2,6 и 3,8 кг), 14 вобл и 3 леща, а также кости
скелета крупного судака. Такие случаи не единичны. В период
зимовки в реке белуга не прекращает питаться. По мере наступле-
ния половой зрелости перед началом нерестовой миграции интен-
сивность питания ее резко снижается или прекращается.
Неполовозрелая белуга нагуливается до июля повсеместно в
междуречье вплоть до западных границ восточной половины Се-
верного Каспия (казахстанские воды). В это время за 100 сетко-
часов здесь ловится до 4—8 неполовозрелых особей размером
66—122 см и массой 1,35—10,4 кг. В июле и первой половине ав-
густа белуги в этих районах моря нет. Наиболее высокие кон-
центрации белуги в междуречье и на свале Уральской бороздины
наблюдаются до середины сентября (до 9 шт. за 100 сетко-часов).
В конце октября — первой половине ноября белуга в этой части
моря отсутствует.
Вдоль северо-восточных и частично восточных берегов Север-
ного Каспия до конца мая на глубине 1,5—2 м нагуливается гораз-
до больше белуг (до 16 шт. за 100 сетко-часов), чем в между-
речье. Наибольшее количество белуги в июне (1970 г.) встречается
восточнее п-ова Пешной (до 30 шт. за 100 сетко-часов). В период
максимального прогрева воды (июль — первая половина августа)
белуга откочевывает в центральные районы Северного Каспия
(Уральская бороздина). На восточных мелководьях в это время
нагуливается небольшое количество белуги (не более 3—4 шт. за
68
100 сетко-часов). По мере спада жары она вновь появляется в
мелководной зоне, но встречается здесь гораздо меньше (9—10 шт.
за 100 сетко-часов), чем весной. В течение всего вегетационного
периода самые низкие концентрации белуги наблюдаются на мел-
ководье у Мертвого Култука и в районе островов Дурнева.
В октябре начинается второй, уже тотальный отход молоди и
нагуливающихся особей из мелководной зоны в глубокие районы
Северного Каспия на зимовку. В зимнее время белуга на мелко-
водных участках (до глубины 2—3 м), за исключением при-
устьевой области Урала, не встречается. Зимует в основном на
Уральской бороздипе и в Мангышлакском заливе (Киппер и др.,
1972; Песериди, 1972).
После зимовки на Уральской бороздине неполовозрелая часть
популяции белуги уральского происхождения как бы делится на
три группы, одна из них (большая часть) распределяется по всей
акватории Северного Каспия, другая (самая меньшая) направля-
ется к западным берегам Северного и Среднего Каспия, где она
встречается с белугой волжского происхождения, а третья, вместе
с белугой, зимовавшей в Казахском заливе, мигрирует вдоль вос-
точных берегов Среднего Каспия на юг моря. Здесь она нагулива-
ется с малочисленным стадом белуги куринского происхождения.
Излюбленными местами нагула белуги, где она образует срав-
нительно плотные скопления, является Уральская бороздина, Ка-
захский залив, восточное побережье Среднего Каспия и районы
моря у о. Огурчинского. В летний период белуга обитает в поверх-
ностных слоях преимущественно над глубинами 30—50 м. В зим-
нее время опускается до 100-метровой изобаты. Повторно созре-
вающие нагуливающиеся особи в восточной половине Северного
Каспия встречаются крайне редко. Сведений о возвращении непо-
ловозрелой белуги из южных районов моря в Северный Каспий и
откочевки ее обратно на юг моря (имеется в виду двух-трехкрат-
ные перемещения) нет.
Не исключено, что из особей, нагуливающихся в южных районах
моря (почти круглогодичное питание), формируется в основном
яровая часть популяции, а из зимующих в северной части Среднего
и в Северном Каспии (замерзающем) —озимая. Не исключено
также, что непрерывающееся питание белуги на юге бассейна и
прекращение его на севере обусловливает образование в одной и
той же возрастной группе резко различающихся между собой по
размерам и массе рыб и растягивает созревание особей одного и
того же года рождения на несколько лет (3—5).
Численность и хозяйственное значение белуги зависит, от мно-
гих факторов: условий воспроизводства и обитания, количества и
качества производителей, участвующих в нересте, обеспеченности
кормом, мер использования запасов и т. п. Величина отдельных
поколений белуги варьирует в диапазоне от 30 до 400 тыс. шт., о
чем можно судить по числу скатывающейся молоди в море в тот
Или иной год с учетом коэффициента промыслового возврата.
69
Общая численность белуги уральского происхождения в Кас-
пийском море, по данным о скате молоди из р. Урал, исчисляется
примерно в 2,5—3 млн. шт. В связи с тем, что созревание особей
одного поколения происходит не одновременно, а растягивается
на много лет, резких флюктуаций добычи по годам не наблюдается.
Основными факторами, лимитирующими численность этого
ценного вида при существующем биологически обоснованном ре-
жиме рыболовства в р. Урал (периодическое прекращение промыс-
ла на 2—3 дня в период нерестовой миграции), являются: возрас-
тающее безвозвратное водопотребление, слабая охрана производи-
телей на местах зимовки и нереста, отсутствие искусственного
воспроизводства.
До 1964 г. в связи с интенсивным промыслом в осеннее время
почти все озимые мигранты вылавливались в низовьях реки (Песе-
риди, 1964, 1966), лишь единичным экземплярам удавалось про-
рваться за пределы промысловой зоны. После введения в осеннее
время свободных от рыболовства дней (Песериди, 1963, 1964, 1967)
существенная часть озимой белуги стала проходить к местам зи-
мовки. Большая часть зимовальных ям (35 и 52 с глубинами до
20 м и более) расположена между пос. Горячкино (600 км) и
Уральском (850 км). Выше имеется не более 6—8 ям с глубинами
8—12 м. В последние годы единичные особи озимой белуги встре-
чаются у устья р. Илек (1155 км). Белуга, зашедшая в реку зимой,
доходит только до Уральска, яровая — до г. Чапаеве (650 км).
В летнее и осеннее время в реку в среднем за последние 15 лет
заходит около 4 тыс. производителей (до 33% нерестовой популя-
ции), зимой — 1,5, а весной — 7 тыс. шт. (55%). Общая числен-
ность нерестовой популяции колеблется по годам от 7 до 21,4 тыс.
рыб, а пропускаемой на зимовку и нерест — от 3 до 13 тыс.
По уловам среди осетровых Урало-Каспия белуга занимает
второе место (после севрюги), а по Каспийскому бассейну — третье
(после осетра и севрюги). Значение белуги в экономическом ба-
лансе определяется не только объемом добычи, но и ценностью из-
готавливаемых продуктов питания.
В первую пятилетку после прекращения морского красно-
ловья (1960—1964 гг.) уловы (т) белуги были несколько ниже,
чем в последующие три пятилетки:
Годы Улов, т
1960—1964 1564,1
1965—1969 2278,1
1970—1974 2039,0
1975—1979 2088,1
1980—1982 1367,9
Данные по численности нагуливающейся белуги показывают,
что добыча ее в Урало-Каспии к концу века достигнет 1 тыс. т.
В том случае, если в ближайшие годы будет осуществлена пере-
броска части стока Волги в Урал, вступит в строй запроектиро-
ванный осетровый завод, усилится охрана производителей на мес-
70
тах зимовки и нереста, улучшится защита молоди от гибели в во-
дозаборных сооружениях и ирригационных системах, уловы
белуги могут быть до 1,5 тыс. т в год.
Род ACIPENSER LINNE, 1758 — ОСЕТР
Включает 18 видов, из которых в Казахстане водится 5. Отличается
от рода Huso тем, что у его представителей жаберные перепонки
прикреплены к межжаберному промежутку, а не сращены между
собой; усики в разрезе цилиндрические, а не сплющенные.
1. Acipenser giildenstadti Brandt — русский осетр
Казахское название-, бексре.
В пределах Казахстана известны 2 подвида.
la. Acipenser giildenstadti giildenstadti
Brandt—русский осетр
Синонимия. Acipenser giildenstadti: Brandt, Med. Zool., t. 2, 1833.—<
Берг, Фауна России, Рыбы, т. 1, 1911.
Распространение. Обитает в бассейнах Черного, Азовского и
Каспийского морей. Более многочислен в Каспии, откуда заходит
на нерест в Волгу, Урал и Терек. По Уралу отдельные особи
поднимаются до устья р. Сакмара (1350 км). В р. Илек
редок. В последние годы в результате увеличения численности и
расширения ареала стал заходить в Куру (Магерамов, 1972).
Описание. Тело осетра веретенообразное, покрыто пятью ряда-
ми крупных острых жучек с радиально-зернистыми полосками
(рис. 10). У некоторых особей между боковыми и брюшными жуч-
ками наблюдаются зачатки дополнительных неполных рядов
Рис. 10. Русский осетр
мелких жучек, характерных для персидского подвида. Все тело
покрыто звездчатыми пластинками — бляшками, верхняя часть
головы — мощными радиально-зернистыми щитками. Нижняя гу-
ба прервана. Рот полулунный, промежуток между лопастями не-
большой, при питании образует хоботок. Усики без бахромы не до-
стают до рта, но достигают конца рыла, которое у части особей
короткое и тупое, у других заостренное. Выделить у осетра остро-
71
Таблица 11. Биометрическая характеристика русского осетра бассейна
Каспийского моря (М±т)
Волга Кура Волга 1 Урал
Признак (Белаев, 1932) (Магера- мов, 1972) (Артюхин, 1979) (Песериди, 1962)
Спинных жучек 12,11 ±0,09 12,7±0,2!12,19±0.16 13,4 + 0,21 13,01 ±0,10
Боковых жучек 39,01 ±0,26:36,8±0,537,5-с0,51 38,4 + 0,43 37,81 + 0,18
Брюшных жучек 9,84±0,09 10,2+0,2 10,0+0,22 10,0± 0,27110,33 + 0,13
Жаберных тычинок 23,5 ±0,16 23,1 ±0,4 23,9 + 0,45 24,1+0,38 23,64 ±0,24
Лучей в D 40,98±0,26 - — — 41,50+0,36
Лучей в А 25,81±О,15 — — 26,70±0,31
В % абс. длины тела (L.)
Длина головы Наибольшая высота го- — 19,7±0,1 17,4±0,17 17,3±0,14 17,20±0,21
ЛОВЫ — 8,9±0,1 8,9±0,06 9,1±0,12 9,20=0,10
Высота головы через
глаз — 5,1 ±0,1 5,0±0,03 5,1 + 0,06 5,40±0,08
п 204 45 60 22 106
возможным из-за
Таблица 12. Гонадо-соматический
индекс самцов русского осетра р. Урал
(по данным 1961—1963, 1968—1971
и 1979—1981 гг.)
Месяц От—до Средн. п
Апрель 0,3-11,2 4,05 1423
Май 0,3-9,6 3,29 565
Июнь 0,3-9,4 3,26 905
Июль 0,4-10,6 4,07 1042
Август 0,7-11,9 4,73 968
Сентябрь 0,9-10,4 5,23 371
Октябрь 0,8-9,7 4,83 101
За сезон 0,3-11,9 4,18 5375
рылую и тупорылую формы не представляется
множества вариаций. Цвет спины от серого до золотисто-коричне-
вого, брюшко желтое или светло-желтое с белизной.
По основным меристическим и пластическим признакам осетр
из разных рек Каспия существенно не различается (табл. 11).
Ранее считалось, что у осетра 130 хромосом (Серебрякова, 1974).
Исследования последних лет
показали, что у него ориен-
тировочно 249+5 хромосом
(Васильев и др., 1980).
Биология. Первые особи
ходового осетра III и III—
IV стадии зрелости появля-
ются в дельте Урала в кон-
це апреля — мае при темпе-
ратуре воды 8—12° в пик па-
водка. Пигментация ооцитов
еще не завершена, и яичник
имеет серый с желтоватым
оттенком цвет, а семенник —
розовый либо красновато-
невооруженным глазом раз-
малиновый. Анимальный полюс яйца
личается с трудом. Икринки мелкие: две трети полости в 1 г —
130—277 шт. Яичник и семенник занимают 2/3 полости тела, не до-
стигая перикардиальной перегородки и конца каудальной части
полости. Гонадо-соматический индекс как самок, так и самцов низ-
кий (табл. 12, 13). Жировые накопления значительны.
72
Слизистая оболочка пищевода, желудка и кишечника в это
время находится в инактивном состоянии. Лишь у немногих
мигрантов обнаружены створки моллюсков и измягченные кости
рыб, что свидетельствует о прекращении питания до захода в реку.
Как видно, осетры, заходящие в реку в конце апреля — мае,
далеки от готовности к нересту. Это подтверждает и анализ гисто-
логических препаратов. Полярность ооцитов на них не выражена,
они симметричны. Некрупное ядро с ядрышками в 2—3 ряда
(рис. 11) расположено в центре яйца (рис. 12). Все оболочки
Рис. 11. Ядро озимого осетра с ядрышками по периферии окружено глыбками
крупнозернистого желтка
ооцита (студенистая, первая и вторая желточные) толстые, хоро-
шо видна радиальная исчерченность желточных оболочек (рис. 13)'.
Зона мелкозернистого желтка тонким слоем расположена по пери-
ферии ооцита. В ампулах семенника лежат многочисленные деля-
щиеся сперматогонии, интерстициум тонкий. Клеточные элементы
следующих фаз сперматогенеза представлены сперматоцитами
первого порядка (рис. 14). Рыб с таким состоянием половых же-
лез мы относим к озимой внутрипопуляционной нерестовой группе
(II тип мигрантов по Н. Л. Гербильскому, 1957).
С конца мая ход озимого осетра начинает постепенно усили-
ваться. Пик хода в отдельные годы наблюдается в июле, но в ос-
новном приходится на август (рис. 15); в маловодные годы заре-
гистрирован в сентябре. Обычно со второй половины этого месяца
74
Рис. 12. Срезы ооцитов осетра озимой внутрипопуляционной нерестовой группы.
Июнь, дельта р. Урал
Рис. 13. Оболочки ооцита осетра озимой нерестовой группы
ход осетра затухает и в октябре прекращается. Температура воды
в это время не превышает 6—8°, прозрачность — 50—60 см.
С начала захода озимого осетра и до конца его осеннего хода
происходит постепенное дозревание половых продуктов, о чем
свидетельствует увеличение размеров ооцитов и показателей гона-
до-соматического индекса. Гистологические препараты показы-
вают, что к этому времени заканчивается поляризация ооцита.
Рис. 14. Срез семенника осетра озимой нерестовой группы в начальной фазе спер-
матогенеза. Июнь, дельта р. Урал
Крупное ядро смещается в сторону анимального полюса и нахо-
дится либо на границе крупнозернистого и мелкозернистого желт-
ков, либо окружено последним. Наличие «шлейфа» — свободной от
желтка зоны цитоплазмы — свидетельствует о движении ядра к
анимальному полюсу. На некоторых срезах удается обнаружить
микропиле. Клетки фолликулярного эпителия, как и их ядра, упло-
щаются (рис. 16). К осени приближается к концу и формирование
сперматозоидов. Большинство ампул семенника заполняется сфор-
мированными сперматозоидами, и лишь в некоторых имеются
группки сперматид (рис. 17).
Жира на гонадах у осетров осеннего хода намного меньше, чем
у заходящих в реку в летнее время, но тешка у них толстая, с
большой жировой прослойкой. Яичники и семенники в это время
почти полностью заполняют полость тела. Заход озимого осетра в
реку продолжается в течение 5,0—5,5 месяца. Мигранты раннего
хода за лето и осень успевают подняться высоко по реке и зимуют
76
Рис. 15. Динамика хода осетра в р. Урал (шт. за одно притонение речного
невода)
Рис. 16. Срез ооцита осетра озимой нерестовой группы на завершающей фазе
поляризации. Сентябрь, дельта р. Урал
Рис. 17. Срез семенника осетра озимой нерестовой группы. Завершение сперма-
тогенеза. Октябрь, дельта р. Урал
на ямах выше Уральска, достигая верхних границ ареала. Осетры
летнего и осеннего хода зимуют в реке ниже Уральска.
Движется озимый осетр со скоростью 8—10 км в сутки, придер-
живаясь участков реки с замедленным течением. Во время мигра-
ции он останавливается на отдых в заводях и небольших ямах.
После зимнего перерыва первые годовые самцы появляются в
дельте реки в конце марта — начале апреля при температуре воды
3—4°. Самки в реке появляются позже. Пик весеннего хода обыч-
но наблюдается в конце второй — начале третьей декады апреля,
т. е. до паводка, когда температура воды достигает 6—9°. К сере-
дине мая ход осетра затухает, но отдельные производители с хо-
рошо развитыми половыми продуктами встречаются в уловах
вплоть до конца первой декады июня.
Осетры, заходящие в реку в апреле, чрезвычайно близки к зре-
лости. Брюшко самок мягкое, отвислое, генитальное отверстие вос-
палено. Мощный яичник темно-серого или черного цвета запол-
Рис. 18. Срез ооцита осетра ранней яровой нерестовой группы. Апрель, дельта
р. Урал
няет всю полость тела. Ооциты имеют округлую форму и дости-
гают дефинитивных размеров. Анимальный полюс приобретает
характерный рисунок: светлое полярное пятно окружено темным
пигментным кольцом; между ним и вторым пигментным кольцом,
находящимся на границе вегетативной области, имеется светлый
ободок. У некоторых ооцитов встречается еще одно дополнитель-
79
Рис. 19. Оболочки анимального полюса ооцита осетра ранней яровой нерестовой
группы. Видны два микропиле, закрытые клетками фолликулярного эпителия.
Апрель, дельта р. Урал
Рис. 20. Срез семенника осетра ранней яровой нерестовой группы после заверше-
ния формирования сперматозоидов. Апрель, низовья р. Урал
ное кольцо. Гонадо-соматический индекс достигает предельных
величин. У отдельных особей масса яичника составляет 33—37%
массы тела. Семенник, заполняющий всю полость тела, белого
цвета, на его разрезе появляются капельки спермы; при разбавле-
80
Рис. 21. Срез ооцита русского осетра поздней яровой нерестовой группы. Сере-
дина мая, дельта р. Урал
Рис. 22. Срез семенника русского осетра поздней яровой нерестовой группы в
разгаре сперматогенеза. Май, дельта р. Урал
6-7
нии ее водой сперматозоиды приобретают активность. На гонадах
и в тешке жира почти нет.
На гистологических препаратах видно, что ядро целиком на-
ходится в зоне мелкозернистого желтка вблизи утонченных оболо-
чек анимального полюса. Сформированные микропиле закрыты
замыкательными клетками (рис. 18,19). Все ампулы семенника за-
полнены сформированными сперматозоидами (рис. 20). Рыб с та-
ким состоянием половых желез мы относим к ранней яровой внут-
рипопуляционной нерестовой группе (I тип мигрантов по
Н. Л. Гербильскому), продолжительность хода которой в среднем
2 месяца.
Ранний яровой осетр мигрирует по реке с большей скоростью,
преодолевая 650—700 км за 20—30 дней. Дальше середины верх-
ней части нерестовой зоны (пос. Коловертное, 720 км) он не под-
нимается.
Одновременно с производителями раннего ярового осетра в ре-
ку с конца апреля начинают заходить производители, отличающие-
ся рядом характерных признаков от описанных двух внутрипопу-
ляционных нерестовых групп. Их яичники и семенники заполняют
всю полость тела, однако ооциты у них мельче, чем у ранних яро-
вых, но намного крупнее озимых. Показатели гонадо-соматическо-
го индекса занимают промежуточное положение между озимым и
яровым. Поляризация ооцита у них еще не закончена: ядро, как
правило, находится на границе мелко- и крупнозернистого желт-
ков либо несколько смещено к анимальному полюсу (рис. 21).
У самцов сперматогенез не завершен: лишь отдельные ампулы се-
менника заполнены сформированными сперматозоидами. Пре-
обладающие клеточные элементы в семенниках представлены по-
ловыми клетками более ранних фаз сперматогенеза — сперматоци-
тами II порядка и сперматидами (рис. 22). Гаметогенез у таких
осетров происходит во время речной миграции и завершается
(в зависимости от температуры воды) примерно через месяц.
Производителей с таким состоянием половых желез мы относим к
поздней яровой внутрипопуляционной нерестовой группе.
Отличить раннего ярового осетра от позднего ярового по внеш-
ним признакам довольно трудно. Надежными отличительными кри-
териями являются масса ооцитов, гонадо-соматический индекс и
ответная реакция на гипофизарную инъекцию. Наиболее простой
способ в полевых условиях — это взятие пробы икры с помощью
щупа и определение положения ядра в ооците.
Заход позднего ярового осетра в реку заканчивается обычно в
середине июня, но отдельные особи встречаются в дельте до сере-
дины июля. Поднимается осетр до верхних границ средней части
нерестовой зоны (пос. Чапаеве, 650 км). Доля позднего ярового
осетра в нерестовой части популяции мала,и не превышает 6, яро-
вого— 31, а озимого — около 63%. В отдельные годы обнаружить
позднего ярового осетра в общей массе уловов визуально не уда-
ется.
82
Осетр является типичным представителем лито- и псаммофилов.
Нерестится озимый осетр летнего хода при температуре воды 9—•
12° на участках реки, удаленных от моря на 900—1200 км. Озимый
осетр осеннего хода размножается на нерестилищах, расположен-
ных между пос. Кожехарово и г. Уральском (670—850 км) при
этой же температуре. Излюбленным его нерестовым субстратом
являются галечники и гравелисто-песчаные гряды, находящиеся
на глубине 4—7 м. Нерест раннего ярового осетра начинается при
12—13° и заканчивается при 18—19°. Осваивает он нерестилища
центральной части нерестовой зоны (320—650 км от моря), где
ложе реки устлано гравелисто-песчаной смесью либо крупнозер-
нистым плотным песком. Нерест позднего ярового осетра проте-
кает при температуре 18—24° примерно на тех же нерестилищах,
где откладывает икру ранний яровой осетр. Инкубационный пе-
риод озимого осетра длится 200—240, раннего ярового — НО—-170,
а позднего ярового — 65—120 ч.
Таблица 14. Распределение скатывающихся личинок осетра
в нижнем течении р. Урал в 1979—1981 гг., % от общего ската
Горизонт Пологий берег Фарватер Крутой берег п
1 м от поверхности — 13,0 — 1309
В толще воды —- 12,4 •—- 1257
У дна 13,6 51,2 9,6 7522
В 1367 7750 971 10088
Дрифт личинок и скат молоди. Ранние личинки осетра появля-
ются в низовьях р. Урал обычно в середине мая. Длина их в это
время колеблется от 25 до 31 мм (средняя 27 мм), а масса — от
90 до 150 мг (средняя 115 мг). Личинки неравномерно распределе-
ны в толще воды. Основная их масса скатывается у дна и по фар-
ватеру (табл.14).
В третьей декаде мая средние размеры личинок увеличиваются
до 34 мм (колебания 21—56 мм), масса—136 мг (колебания 70—
540 мг). Обычно в последних числах мая в дельте встречается в
небольшом количестве и сравнительно крупная молодь массой
1—1,3 г. В первой половине июня наряду с мелкоразмерной мо-
лодью (потомство раннего ярового осетра) скатывается и молодь
массой 1,7—2 г (потомство озимого осетра). К концу второй дека-
ды июня средняя масса молоди в отдельные годы достигает 2—
2,5 г (колебания 0,25—5,85 г). В последних числах июня этот по-
казатель снижается до 0,8—0,9 г (колебания 0,25—3,24 г), что
обусловлено наличием в уловах молоди двух-трехнедельного воз-
Раста (потомство позднего ярового осетра). Средняя навеска ска-
тывающейся молоди в первой половине июля вновь увеличивается,
Достигая в разные годы 3,5—5,5 г (Песериди, 1966; Пёсериди, Бе-
83
кешев, 1967). Неравномерно протекает дрифт личинок и скат мо-
лоди в разное время суток. Большая часть молоди скатывается по
фарватеру, но в ночное время ее здесь, как у пологого берега,
меньше (табл. 15).
Таблица 15. Суточная динамика ската молоди осетра в нижием течении
р. Урал, % от общего ската
Место Время ската, ч
13 17 21 1 5 9 13 п
Крутой берег 0,3 0,6 1,2 3,8 0,7 1,7 1.2 104
Фарватер 9,1 8,1 7,8 4,0 6,4 6,6 12,7 597
Пологий берег 9,6 5,0 6,7 2,2 5,2 3,4 2,7 387
п 209 152 172 108 135 130 181 1088
Первые скатывающиеся личинки появляются в дельте реки
обычно в середине мая. Массовый скат наблюдается здесь с кон-
ца мая до середины июня. Завершается скат в конце июня — нача-
ле июля. Численность скатывающейся молоди осетра с момента
появления ее в дельте реки нарастает быстро, вторая половина
ската несколько растянута (Песериди, 1966, 1967; Невлюдова и др.
1969, Тарабрин,1981).
Таблица 16. Количественные и качественные показатели
ската молоди осетра р. Урал в разные по водности годы
Год Объем сто- ка р. Урал, км® Средний улэв молоди на трал, шт. Кол-во ска- тившейся молоди в море, млн. шт. Средняя на- веска моло- ди, мг
1961 6,24 5,3 11,2 1481
1962 7,44 4,5 14,9 1632
1963 9,80 10,5 20,9 2285
1964 11,52 4,0 10,5 1878
1966 11,16 10,6 30,1 1235
1967 3,01 14,9 6,2 1246
1969 5,83 25,0 18,3 1475
1970 12,83 17,9 29,4 2795
1971 12,50 5,9 -18,5 1079
1972 8,48 60,9 156,0 370
1973 4,87 46,6 50,0 370
1974 7,64 45,9 193,0 389
1975 3,56 22,3 8,5 385
1976 4,20 32,8 41,0 403
1977 2,54 36,5 20,0 290
1978 5,30 63,3 102,0 405
1979 6,25 51,6 135,0 675
1980 6,03 17,3 17,0 1244
1981 9,22 2,9 2,0 2510
1982 6,00 12,2 20,0 618
84
В маловодные годы, когда прогрев воды идет быстро и нере-
стовые температуры наступают рано, производители не только
осетра, но и севрюги и шипа не успевают дойти до исконных мест
нереста, размножаются на нижних, менее благоприятных нерести-
лищах. Скат молоди в такие годы начинается раньше, он компакт-
нее. Покатная молодь имеет мелкие размеры. В многоводные годы,
наоборот, молодь намного крупнее, скат продолжительный и рав-
номерный (табл. 16).
Определение возраста осетра по спилам луча грудного плавни-
ка затруднено из-за наличия дополнительных колец. У старше-
возрастных рыб отдельные дополнительные кольца схожи с годо-
выми. Образование, количество годовых и дополнительных колец,
вероятно, связано со сложной структурой популяции проходного
осетра, т. е. наличием у него озимых, ранних и поздних яровых
внутрипопуляционных нерестовых групп и туводных форм с разной
динамикой питания.
Таблица 17. Возрастной состав нерестовой части популяции
осетра р. Урал, %
Возраст, лет Самцы Самки
1961 — 1963 гг. 1969— 1970 гг. 1974— 1976 гг. 1980— 1982 гг. 1961 — 1963 гг. 1968- 1970 гг. 1974— 1976 гг. 1980— 1982 гг.
7 — —. 0,6
8 0,1 0,3 — 1,9 —. -
9 1,4 1,1 0,3 3,0 0,4 0,3 0,2 1 6
10 1,6 1,6 0,9 3,7 0,9 0,6 0,2 2’0
11 2,1 2,0 1,5 5,3 0,9 0,8 0,4 2’6
12 2,6 2,4 3,8 8,2 1,2 1,1 0,8 1’8
13 2,8 3,0 7,5 9,2 1,2 1,4 0,8 3’5 4’3 5’9 5’8
14 4,2 4,5 9,3 Ю,1 1,8 2,6 3,2
15 5,4 5,8 12,6 10,8 2,4 2,7 3,7
16 10,6 7,3 13,2 9,7 3,0 4,5 6,7
17 11,0 10,4 12,1 8,8 3,7 5,1 7,3 6’4 6 8
18 14,0 12,5 11,4 6,8 4,8 6,3 8,8
19 9,9 10,8 9,2 6,5 6,1 6,9 10,7 8;е 9 4
20 7,9 8,5 7,2 5,2 7,5 9,5 12,1
21 7,1 7,4 3,6 3,1 8,5 7,9 11,1 7,3 6,3 6,2 4,6 3,7 3,4 3,1 1,8 1,4 1,7 0,6 0,6 0,4 0,4 1389
22 5,2 5,8 3,8 2,6 8,6 7,1 9,9
23 3,4 4,3 1,6 1,3 8,6 6,8 7,6
24 25 26 27 28 29 30 31 32 33 34 п 3,1 2,4 1,4 0,7 0,5 0,2 0,5 0,2 0,5 0,2 423 4,0 3,0 2,1 1,2 0,9 0,4 0,4 0,2 0,1 854 0,8 0,4 0,3 0,4 1,0 1185 1,2 0,3 0,5 0,3 0,4 0,3 0,1 0,1 1432 10,5 8,1 7,5 4,0 2,5 3,2 1,4 0,9 1,2 0,9 347 6,5 6,0 6,3 5,8 4,9 3,1 1,7 0,9 0,6 0,3 0,3 648 6,0 2,9 2,3 1,8 1,2 0,9 0,5 0,3 0,6 640
85
Из-за наслоения дополнительных колец при определении воз-
раста осетра не исключены ошибки на 1—2 года в обе стороны.
При большом числе проб эти ошибки нивелируются и в целом
определенный возрастной состав нерестовой части популяции
близок к истинному.
Половая зрелость отдельных самцов осетра наступает в воз-
расте 7—8, у самок — в 9—10 лет (табл. 17), но таких особей еди-
ницы. Нерестовая часть популяции осетра (оба пола) состоит из
24 28 возрастных групп. Их значение непостоянно и колеблется
по годам, что зависит от характера эксплуатации запасов.
Анализ возрастного состава нерестовой части популяции за
20-летнии период (см. табл. 17) показывает, что со времени пре-
кращения морского лова, отмены промысловой меры и объявления
запрета на его промышленный лов произошло перемещение наи-
более многочисленных возрастных групп самцов из «средних»
возрастов (15 22-летки) к более молодым (12—17-летки). У са-
Таблипа 18. Средние размерно-весовые показатели осетра
р. Урал по возрастным группам (1961—1963, 1968—1970 гг.)
Возраст, лет Самцы ( п=1277) Самки 'п-995) Эба пола (п=2272
Длина, Масса, Длина, Масса, Длина, Масса,
см КГ СМ КГ СМ КГ
7 82,0 3,13 82,0 3 13
8 89,7 4,03 — 8),7 4,03
9 93,6 4,37 101,4 5,84 97,3 5,17
10 98,7 4,49 106,8 6,37 103,7 5 81
П 102,4 4,85 110,2 6,98 106,6 6’23
12 103,5 6,58 113,7 8,88 108,5 7,78
13 106,2 7,06 114,9 10,11 110,9 8 37
14 111,7 7,93 116,8 10,36 114,9 9,14 10,5
15 116,5 8,81 119,5 11,22 116,8
16 118,4 9,89 122,5 12,71 119,9 11,6
17 119,0 10,33 128,6 13,83 123,6 12Л
18 19 122,3 125,8 11,15 12,21 130,8 142,9 16,22 18,63 127,6 133,1 13,6 14,9
20 127,5 13,40 144,6 18,99 136,7 16,5
21 130,3 14,80 147,6 20,81 140,9 18,2
22 131,6 15,60 154,3 22,88 145,4 19М
23 133,5 16,30 158,7 23,75 148,2 90 1
24 141,9 16,80 162,1 24,90 153,8 22,3
2э 144,0 17,40 164,6 27,85 156,9 23,4
26 148,7 18,10 167,9 28,45 1 ',9,5 23,9
27 152,3 22,20 166,8 27,84 163,2 25,1
28 157,5 25,10 169,8 32,00 165,8 29,9
29 155,6 24,30 171,1 30,87 166,5 29,1
30 155,0 24,20 170,3 31,5 164,8 28 4
31 162,0 23,70 185,0 38,1 177,3 33,3
32 165,4 26,50 176,0 35,5 170,8 32 5
33 168,0 28,40 178,-5 37,6 174,3 32^9
34 175,1 33,4 175,1 33,4
86
мок в начале 60-х годов преобладали (65,4%) 19—26-летки, а в
начале 80-х годов большая часть самок (62,7%) приходилась на
15—23-леток. Это дает основание считать, что в результате приня-
тых охранных мер и происшедшего омоложения нерестовая часть
популяции проходного уральского осетра сформировалась.
Неравномерный прирост год от года размеров и массы тела
осетра (табл. 18) мы объясняем колебаниями в возрастном мате-
риале числа особей разных внутрипопуляционных нерестовых
групп (ранних и поздних яровых и озимых), масса которых неоди-
накова и находится в зависимости от состояния зрелости половых
желез мигрантов. Анализ данных по массе тела только ярового
осетра разной длины (табл. 19) показывает, что с увеличением
размеров происходит равномерно возрастающий прирост массы.
Л. С. Берг (1953), говоря о наличии у многих проходных рыб
двух форм или рас, которые отличаются друг от друга временем
вхождения в реки для нереста, отмечает, что озимые особи обычно
крупнее и плодовитее яровых. Однако автор не подкрепил свой вы-
вод конкретными материалами.
Таблица 20. Длина и масса тела самок разных
внутрипопуляционных нерестовых групп осетра р. Урал
(1961—1963, 1978—1981 гг.)
Непестовая группа Период Длина, см Масса тела, кг п
М±т а М± m а
Озимая Август— октябрь 155,8±0,04 1,10 23,62±0,16 1,63 867
Ранняя яровая Апрель— чай 157,1 + 0,06 1,67 25,03 + 0,04 1,44 779
Поздняя яровая Июнь— июль 158,1 + 0,11 1,54 26,52+0,15 2,16 201
Анализ данных по типичным представителям разных групп из
общей массы уловов осетра за 1961—1963 и 1979—1981 гг. в наи-
более характерные для них сроки хода не подтвердил это мнение
Л. С. Берга. Наоборот, было выявлено некоторое преобладание в
росте яровых особей (табл. 20), что вполне закономерно, поскольку
длительность голодания озимых с начала нерестовой миграции в
море и до возвращения в него после нереста на нагул составляет
9-—13 месяцев, а яровых — только 2—3.
С середины 60-х до начала 70-х годов в запретной зоне — у
пос. Мергенова (580 км) и пос. Горячкино (Набатовский яр —
620 км)—вылавливались осетры длиной 60—172 см на разных ста-
диях зрелости половых желез (II, II—III, III и III—IV). Упитан-
ность рыб была примерно такой же, как и у вылавливаемых в мо-
ре. По темпу роста они уступали проходным осетрам, но разли-
чий в биометрии у них не обнаружено. На основании состояния
88
зрелости половых желез, встречаемости в разные сезоны года в
реке, низкого темпа роста можно классифицировать этого осетра
как туводную форму. В последние 10 лет такой осетр в этих рай-
онах не встречался. Исчезновение его в р. Урал объясняется в ос-
новном катастрофическими заморами 1967, 1968, 1973 гг. и бра-
коньерством.
Половой состав. В первой половине хода в любой внутрипопу-
ляционной нерестовой группе проходных осетровых (если не на-
рушена се структура) в уловах преобладают самцы. Во второй
половине хода наступает численное равновесие между полами, а в
конце хода в уловах преобладают самки. В случае, если популяция
состоит из нескольких внутрипопуляционных нерестовых групп и
сроки нерестового хода перекрещиваются, то эту закономерность
не всегда удается выявить без специальных исследований
(табл. 21).
Таблица 21. Половой состав уловов ходового осетра р. Урал в разное
время года, %
Срок 1960 г. 1961 г. 1962 г. 1953 г.
Сам- цы Сам- ки Сам- цы Сам- ки Сам- цй Сам- ки Сам- цы Сам- ки
Март Апрель: — — 61 39 67 33 54 56
с 1 по 15 56 44 48 52 50 41 57 43
с 16 по 30 Май: 55 45 46 54 48 52 55 45
с 1 по 15 59 41 52 48 44 56 40 60
с 16 по 31 45 55 54 46 62 38 61 39
Июнь 52 48 59 41 54 46 64 36
Июль 54 46 48 52 57 43 47 53
Август 44 56 50 50 55 45 59 41
Сентябрь 42 58 57 43 38 62 41 54
п 745 765 917 833 876 735 823 662
Как видно, в нерестовой части популяции в младших возраст-
ных группах преобладают самцы, а в старших — самки, так как
самцы раньше выбывают под влиянием промысла. В целом опти-
мальным соотношением полов в популяциях осетровых следует
считать преобладание самцов вдвое (2:1). При изменении этой
пропорции нарушается структура популяции, что наблюдалось с
осетром в первой половине 60-х годов. В настоящее время половой
состав нерестовой части популяции осетра соответствует норме
(табл. 22).
Плодовитость. Абсолютная плодовитость осетра сильно колеб-
лется. Наименьшая индивидуальная плодовитость — 25,8 тыс. ик-
ринок— была зарегистрирована у особи с абсолютной длиной те-
ла 115 см и массой 6,4 кг; максимальная— 1065 тыс. — отмечена
У самки массой 62 кг. У отдельных крупных самок (50—60 кг)
89
иногда наблюдаются дегенеративные изменения яичников: аморф-
ные массы жира желтого цвета, по форме напоминающие яичники
с вкрапленными в них единичными резорбирующимися ооцитами.
Очень редко встречаются гермафродиты. Гистологические препа-
раты, изготовленные из половых желез этих рыб, не показывают
морфологических нарушений в развитии половых клеток (ооцитов
и спермиев). Одна такая особь, подвергнутая гипофизарной инъ-
Таблица 22. Половой состав нерестовой части популяции
осетра в р. Урал, %
Год Сам- цы Сам- ки Год Сам- цы Сам- ки Год Сам- цы Сам- ки
1960 49 51 1968 62 38 1976 74 26
1961 52 48 1969 56 44 1977 73 27
1932 54 46 1970 65 35 1978 72 28
1963 55 45 1971 59 41 1979 68 32
1964 49 51 1972 71 29 1989 68 32
1965 38 62 1973 79 21 1981 65 35
1966 66 34 1974 71 29 1982 65 35
1937 52 48 1975 71 29 1983 69 31
екции, созрела в срок. Ооциты были оплодотворены ее же спермой
(процент оплодотворения 84). Отклонений в развитии в эмбрио-
нальный период не замечено. Из выклюнувшихся личинок (94 тыс.)
вырастили в бассейнах 61 тыс. вполне нормальной по внешним по-
казателям молоди со средней навеской 1,14 г.
Индивидуальные колебания абсолютной плодовитости в той
или иной возрастной группе или в размерном и весовом классах
осетра чрезвычайно велики. Наибольшие колебания плодовитости
наблюдаются у рыб размером 141—160 см и массой тела 14—30 кг
(табл. 23, 24).
Таблица 23. Зависимость абсолютной плодовитости осетра
р. Урал от размеров тела, тыс. шт.
Размерная группа, см Плодовитость п Размерная группа, см Пл одовитость п
От—до М От—до М
101-110 53-108 65,7 4 161-170 112-744 334,9 505
111-120 26-158 98,8 10 171-180 203-799 363,2 234
121-130 89-260 149,5 48 181-190 179-769 451,5 94
131-140 77-424 198,0 260 191—200 181-793 514,2 24
141-150 69-563 243,0 642 201-210 175-749 544,9 9
151-160 83 -708 299,0 659 101-210 26-799 291,0 2489
С увеличением размера и массы рыб абсолютная плодовитость
синхронно возрастает. Средняя абсолютная плодовитость осетра
р. Урал по многолетним материалам составляет 291 тыс. икринок.
90
Таблица 24. Зависимость абсолютной плодовитости осетра
р. Урал от массы тела. тыс. шт.
Весовая группа, кг Плодовитость п Вёсовая группа, кг Плодовитость п
От—до М От—До М
До 6 31-91 58,2 3 38-42 305-945 491,5 46
6-10 26-157 93,6 8 42-46 297-769 540,5 35
10-14 68-364 178,5 93 46-50 349 -694 527,4 10
14-18 33-513 207,2 340 50-54 518-808 673,5 5
18-22 69-587 245,5 633 54—58 477-793 635,2 4
22-26 108-685 285,4 582 58-60 642-1065 642,8 2
26-30 112-681 333,5 390 62-65 — -— —
30-34 154-790 373,-5 213 65—68 585,0 585,0 1
34-38 175-799 420,5 125 6-68 26-1035 291,0 2490
Таблица 25. Зависимость абсолютной плодовитости (АП)
осетра р. Урал, от возраста (1961—1963, 1968—1971 гг.), тыс. шт.
Возраст, лет АП Возраст, лет АП Возраст, лег АП Возраст, лет АП
9 25,8 16 130,5 23 305,0 30 426,3
10 37,9 17 172,5 24 321,6 31 440,5
И 49,5 18 201,2 25 335,7 32 455,0
12 60,4 19 221,8 26 350,4 33 502,8
13 71,5 20 240,0 27 361,6 34 339,5
14 88,1 21 263,4 28 375,8
15 111,3 22 284,3 29 391,1
Снижения прироста плодовитости у старых особей не наблю-
дается (табл. 25), так как 33—34 года не предел возраста осетра.
Анализ материалов по плодовитости озимых и яровых группи-
ровок осетра показал, что плодовитость яровых рыб выше
(табл. 26, 27). Таким образом, мнение Л. С. Берга (1953) о более
высокой плодовитости озимых особей по сравнению с яровыми не
подтвердилась.
Питание, конкуренты, враги. Личинки озимого осетра перехо-
дят на внешнее питание при температуре воды 8—10° на 8—12-е
сутки после выклева, а ярового—на 6—8-е (13—14°). Отдельные
личинки начинают питаться при длине 20 мм, в массе — при 22—
23 мм (личинки такого размера с пустыми желудками практически
не встречаются). Осетрята длиной 24 мм поедают мелких олигохет,
полихет, корофиид, гаммарид и мпзид, длина которых 2—3 мм.
У осетра наблюдается прямая зависимость между размерами мо-
лоди и потребляемых организмов. Так, осетрята длиной 3—4 см
кормятся 6—7-миллиметровыми гаммаридами и мизидами. Более
крупная молодь (5—6 см) поедает этих же ракообразных длиной
Н—13 мм. У молоди осетра размером 9—10 см в желудках встре-
91
чаются мизиды длиной до 22 мм. Единичные экземпляры, достиг-
шие 6—7 см, заглатывают личинок рыб (в основном бычков и
пескаря) длиной 10—11 мм.
Характер питания молоди в низовьях реки меняется по годам.
Флюктуации в потреблении бентоса связаны с состоянием кор-
та б л и ц а 26. Зависимость абсолютной плодовитости разных
внутрипопуляционных нерестовых групп осетра р. Урал от размеров
тела, тыс. шт.
Размер, см Озимая гр. Ранняя яровая гр. Поздняя яровая гр.
М п М п М п
121-130 143,3 12 151,4 3 113,1 3
131-140 194,8 46 198,5 41 191,8 14
141-150 222,9 124 241,0 115 236,0 48
151-160 245,6 146 291,5 138 289,5 55
161-170 314,6 115 343,0 111 303,0 38
171-180 360,0 50 408,0 65 425,4 26
181-190 415,6 18 456,0 19 529,8 9
191-200 541,8 7 503,0 1 616,7 6
201-210 503,4 1 404,0 1 690,4 2
121-210 271,5 519 304,0 494 302,7 201
Таблица 27. Зависимость абсолютной плодовитости разных
внутрипопуляционных нерестовых групп осетра р. Урал от массы
тела, тыс. шт.
Масса тела, кг Озимая гр. Ранняя яровая гр. Поздняя яровая гр.
М п М п М п
6-10 132,1 1 117,8 2
10-14 152,5 20 188,6 14 174,3 1
14-18 199,1 85 219,3 54 196,9 21
18—22 218,6 128 243,9 123 224,5 43
22-26 259,0 131 296,6 116 293,0 37
26-30 295,65 75 345,4 106 304,2 38
30-34 370,35 41 385,0 44 369,6 26
34-38 428,5 23 460,0 22 407,3 9
38-42 515,0 10 488,6 5 492,3 6
42-46 508,0 3 485,1 4 483,2 7
46-50 639,5 2 503,0 1 583,7 3
50-54 — —. 687,4 2 705,8 3
54-58 — .—- 495,8 1 690,4 2
6-58 267,9 519 304,0 494 302,7 201
мовой базы, которая находится в прямой зависимости от гидроло-
гических условий года, численности скатывающейся молоди
и конкурентов. Но в любой год доминирующими компо-
нентами в пищевом комке молоди размером до 8 см являются
гаммариды, корофииды и олигохеты ( табл. 28). В отдельные годы
92
существенную роль в питании осетрят играют мизиды, личинки хи-
рономид и мошек, имаго насекомых.
Число компонентов пищевого комка по мере роста и развития
молоди увеличивается от 2—3 (молодь длиной 20—30 мм) до
8—9 (61—70 мм) видов. Состав пищи осетрят от 40 до 70 мм схо-
Таблица 28. Состав пищи молоди осетра, % от общей массы
Компонент 1971 г. 1972 г. 1973 г. 1974 г. 1975 г.
Гаммариды 57,2 31,5 41,8 15,7 5,3
Корофииды 25,3 9,1 4,8 34,0 61,3
Мнзиды 1.2 0,3 0,4 3,4 12,1
Молодь речного рака 1.2 0,2 — <— —
Личинки хнрономид 9,8 23,1 38,8 19,6 10,9
ручейников 0,5 2,7 0,2 — 1,0
поденок — 1,8 — — 0,4
двукрылых — 0,1 0,3 3,4 —
мошек — 24,9 —. — —
Имаго насекомых 2,6 — —k 7,0
Полихеты — — —- 13,0 1.3
Олигохеты 2,3 0,6 10,6 9,2 0,2
Молодь рыб —• — 1,6 1,4 —
Другие организмы 0,4 — 0,3 — —
Песок 2,1 3,1 2,0 0,3 0,4
ден, увеличиваются лишь размеры потребляемых организмов. Мо-
лодь длиной более 80 мм в реке питается преимущественно выс-
шими ракообразными, среди которых преобладают гаммариды.
Зоопланктон в пище личинок и молоди осетра в период дрифта
встречается редко и мало.
Средние индексы наполнения желудков (%оо) осетрят колеб-
лются по годам (1971 г. — 274,4; 1972 г. — 158,8; 1973 г. — 81,5;
1974 г.— 167,2 и 1975 г. — 260,1) и зависят от сроков пребывания
молоди в реке. Так, в годы с продолжительным скатом (1971, 1972
и 1975 гг. — до 90 суток) индекс наполнения желудков гораздо вы-
ше, чем в годы с кратковременным (1973 г. — 50 суток). Интенсив-
ность потребления пищи молодью в течение суток различна. Наи-
более накормленной молодь бывает в предрассветное, утреннее и
’ вечернее время, когда общий индекс наполнения пищеварительного
тракта у нее колеблется от 550 до 660°/ооо-
У скатившейся в море молоди характер питания на первом го-
ду жизни изменяется незначительно. Основной пищей сеголетков
До 30 см длины являются ракообразные, главным образом боко-
плавы. Значительную часть рациона осетрят длиной 30—40 см со-
ставляют черви и бычки (табл. 29). По мере роста молоди общий
индекс наполнения желудка постепенно падает со 198°/оОО у молоди
Длиной 10 см до 62%оо у осетрят размером 40 см. Это связано с
Переходом молоди на потребление более калорийной пищи.
Питание осетра крупных размеров (60—170 см) меняется по
93
сезонам, годам и месту обитания. Наиболее разнообразно оно в
восточных районах Северного Каспия, где в пищевом комке в раз-
ное время года встречается от 5 до 9 компонентов. В апреле почти
половину пищевого комка (48,12%) составляют ракообразные
(гаммариды и корофниды). Около четверти пищи приходится на
долю моллюсков и 18,5% —на долю рыб. В жаркое время года
(июнь — август), когда осетр откочевывает в глубокие участки мо-
ря, в его пище преобладают моллюски (17,7—37,43%) и рыба
(23,5—34,41%), а значение ракообразных падает до 12,52—17,7%.
В октябре половину пищевого комка составляет рыба, четвертую
часть — ракообразные.
Таблица 29. Состав пищи сеголетков осетра в Северном
Каспии, % по массе
Компонент Длина молоди, см
10 20 30 40
Амфиподы 84,7 48,7 29,3 31,8
Мизиды 11,3 38,3 29,7 1,0
Прочие ракообразные 0,2 1.0 1,3 7,1
Нереис — 4,2 17,8 21,7
Рыба — 4,4 5,0 438,4
Хирономиды 3,8 3,4 16,9 —
Средняя масса молоди, г Общий индекс наполнения же- 4,3 14,7 65,7 239,0
лудка, °/ооо 198 166 93 .62
У восточных берегов Среднего Каспия в феврале осетр питается
ракообразными (главным образом корофиидами и гаммаридами)
и рыбой. В апреле более половины пищевого комка приходится на
долю рыбы. Осенью осетр вновь усиленно потребляет ракообраз-
ных. Наиболее узкий спектр питания у осетра в восточной полови-
не Южного Каспия: здесь основную часть его пищи составляют
рыбы. Анализ характера питания молоди, ювенильных особей и
взрослых рыб позволяет сказать, что осетр — типичный эврифаг.
Главными пищевыми конкурентами скатывающейся молоди
осетра в реке являются пескарь, бычок-песочник, пуголовка, бело-
глазка и молодь леща. Индекс пищевого сходства (СП-коэффи-
циент) показывает, что характер питания неполовозрелого осетра
в Северном Каспии наиболее сходен с севрюгой (25,9%) и наиме-
нее— с воблой (10,5), у севрюги — соответственно с осетром
(25,9) и воблой (5,4), у леща — с осетром (21,7) и воблой (14,8).
Отложенную икру озимого осетра, размножающегося в верхней
части нерестовой зоны при низкой температуре воды, поедают в
основном белоглазка и крупные экземпляры пескаря. Икрой ран-
него ярового и позднего ярового осетров, которые нерестуют ниже
94
по течению реки и при более высокой температуре, питается гораз-
до большее количество видов рыб: белуга, шип, стерлядь, пескарь,
белоглазка, лещ, вобла, щиповка и др. Основным потребителем ли-
чинок и молоди осетра являются синец, чехонь, сельди, крупная
молодь белуги и главным образом сом. Отдельные некрупные осо-
би осетра встречаются в желудках белуги. Других врагов у осетра
в море нет (Желтенкова, 1964; Тарвердиева, 1965, 1968; Песериди,
1966; Полянинова, 1972, 1979; Зачетнова, 1972; Стыгар, Песериди,
1981; Стыгар, 1981).
Миграции и распределение в море. Молодь осетра в приустье-
вом пространстве не задерживается и быстро рассел'яется в вос-
Таблица 30. Возрастной состав уловов осетра в восточной половине
Северного Каспия
Возраст, лет Самцы Сам- ки Оба пола Возраст, лет Сам- цы Сам- ки Оба пола
До 2 1 1 17-18 5 16 21-
3-4 2 <— 2 19-20 3 11 14
5-6 2 1 3 21-22 2 5 7
7-8 5 3 8 23-24 .— 1 1
9-10 6 7 13 25—26 2 1 3
11-12 12 10 23 27-28 1 — 1
13-14 17 17 34 29-30 — 1 1
15-16 17 21 38 п 75 95 170
Северного Каспия. Большая часть ее мигрирует
материковой отмели. Незначительное количество
точной половине
вдоль восточной
сеголетков — на свале Уральской бороздины.
Численность осетра в восточной половине Северного Каспия
характеризуется снижением уловов от распаления льда к середине
лета в десять раз. После спада жары наблюдается второй подход
осетра к материковой отмели. Максимальные уловы наблюдались
вблизи залива Комсомолец в середине сентября. В этом же районе
моря зарегистрированы самые высокие уловы в середине мая.
Средние уловы осетра вдоль восточных берегов Северного Каспия
колеблются по периодам и годам от 2 до 6,6 особи за 100 сетко-ча-
сов, а в междуречье Волга — Урал — 1,3—1,8 шт. Минимальная
глубина, на которой был выловлен осетр в Северном Кас-
пии, — 0,6 м.
На глубине свыше 1,5 м в течение всего вегетационного периода
нагуливаются осетры всех возрастных категорий (табл. 30).
Часть популяции осетра, нагуливающаяся в восточной полови-
не Северного Каспия, зимует на Уральской бороздине и впадине
Мангышлакского залива. В феврале незначительное количество
осетра встречается вдоль казахстанского побережья Среднего Кас-
пия от мыса Тюб-Караган до залива Кендерли. Основная масса
осетра, вероятно, зимует в более теплых районах моря.
Наиболее высокие концентрации осетра в апреле наблюдались
95
у залива Кендерли (Пискунов, 1965; Легеза, 1970). В июне и ав-
густе значительные скопления осетра вдоль материковой отмели
Среднего и Южного Каспия встречались южнее п-ова Мангышлак и
у о. Огурчинского. Высокие концентрации осетра в октябре отмече-
ны на обширных площадях до 100-метровой глубины на траверзах
мыса Сагындык и залива Кендерли. Скопление осетра в октябре
на участках моря вдоль казахстанского побережья, не отличаю-
щегося высокой кормностью, можно объяснить встречей откочевы-
вающих рыб из северных районов моря на юг Каспия на зимовку с
нерестовой частью популяции будущего года, мигрирующей с юга
Каспия. У о. Огурчинского в это время года наблюдается разре-
женность концентрации осетра на весьма ограниченных участках
моря (Пискунов, 1965).
Таблица 31. Уловы осетра и севрюги в Урало-Каспийском
рыбопромысловом районе, тыс. ц
Год Улов Год Улов Год Улов
Осетр Севрюга Осетр Севрюга Осетр Севрюга
1932 19,9 14,1 1948 0,2 0,9 1954 2,5 21,9
1933 20,0 11,6 1949 0,2 0,8 1965 1,9 29,5
1934 14,9 8,3 1950 0,1 0,7 1966 0,6 27,7
1935 21,8 10,7 1951 0,2 1,8 1967 0,6 40,5
1935 24,0 9,5 1952 0,3 2,0 1968 0,8 41,8
1937 17,8 7,6 1953 0,9 2,8 1969 1,2 50,9
1938 8,8 3,8 1954 0,8 7,3 1970 0,4* 45,7
1939 7,8 3,5 1955 0,4 6,4 1971 0,1 44,9
1940 5,2 3,6 1956 0,4 15,9 1972 0,3 54,3
1941 1,9 3,2 1957 0,3 9,5 1973 0,5 56,3
1942 0,9 1,5 1958 1,1 18,2 1974 0,3 70,4
1943 1,1 1,1 1959 1,0 14,7 1975 0,1 80,2
1944 0,6 1,4 1960 0,8 13,6 1976 0,4 85,7
1945 0,4 1,1 1961 0,9 13,5 1977 0,6 98,7
1946 0,1 0,2 1962 2,6 29,1 1978 0,6 78,6
1947 0,3 0,9 1953 1,7 20,4 1979 0,7 84,1
* С 1970 г. установлен запрет на вылов осетра. В таблице 31 после этого
года приведены уловы с контрольных тоней для научно-исследовательских целей.
Численность и хозяйственное значение. Достоверные статисти-
ческие данные по добыче осетра в Урало-Каспийском рыбопро-
мысловом районе имеются с 1932 г. (Гуревич, Лопатин, 1962). До
1940 г. доля осетра в общем вылове красной рыбы была высокой
(61,6—67,7%). К началу 40-х годов запасы осетровых были по-
дорваны и общий вылов красной рыбы в Урало-Каспии упал до
436 ц в 1946 г., из которого доля осетра составила лишь 100 ц
(табл.31).
Упорядочение красноловья в конце 40—50-х годов (общее огра-
ничение морского лова, запрещение крючковой снасти, калады,
96
дрифтерного лова и распоров) привело к восстановлению числен-
ности осетровых (Песериди, 1966). К концу 50-х годов их уловы в
Урало-Каспии достигли 21,3 тыс. ц, но на долю осетра приходи-
лось 1 тыс. ц. На этом же уровне добыча осетра оставалась до
конца 60-х годов. Небольшая «вспышка» уловов в 1962—1965 гг.
(1,7—2,6 тыс. ц) произошла за счет вылова урожайных поколений
многоводных (1946—1948) лет.
К началу 70-х годов запасы осетра упали до катастрофически
малых величин — 0,4—0,-1 тыс. ц. В то же время уловы севрюги
достигли небывалых размеров за всю историю красноловья.
Как могло случиться, что в одной и той же незарегулирован-
ной реке запасы одного вида (осетр) находятся на грани исчез-
новения, а численность другого-—севрюги (как и осетра в Волге)
неуклонно возрастает (в Волго-Каспийском районе добыча осетра
в 1971—1975 гг. составила 78,0—100,1 тыс. ц)?
Изучение биологии обоих видов, условий обитания и режима
рыболовства на протяжении многих лет позволили выявить при-
чины возникшего феномена и разработать мероприятия по вос-
становлению запасов угнетенного вида и сохранению численности
процветающего.
Подрыв запасов уральского осетра обусловил ряд факторов:
1) гибель огромного числа его молоди и неполовозрелых рыб в пе-
риод морского лова; 2) низкая промысловая мера; 3) вылов боль-
шей части нерестовой популяции на путях подхода к реке; 4) чрез-
вычайно интенсивный промысел в осенний пик хода озимой
нерестовой группы, когда в реку заходит до 70% нерес-
товой части популяции; 5) почти поголовное изъятие производите-
лей раннего ярового осетра; 6) истребление браконьерами мигри-
рующих производителей позднего ярового и озимого осетра; 7) по-
головный вылов курилкинским садком-ловушкой производителей,
прошедших интенсивно облавливаемый участок нижней части
промысловой зоны в осеннюю межень; 8) отсутствие свободных от
рыболовства дней в разгар весеннего и осеннего хода осетра;
9) уничтожение отложений икры на нерестилищах лотовыми цепя-
ми во время буксировки барж; 10) выедание икры и истребление
личинок и молоди рыбами — икроедами и хищниками; 11) чрез-
вычайно суровые условия зимовки в реке в результате ее загряз-
нения стоками промышленных предприятий и горюче-смазочными
материалами судов, что неоднократно приводило к катастрофиче-
ским заморам в 1950—1973 гг.; 12) возрастающее безвозвратное
недопотребление; 13) выход из строя более 40% нерестилищ осет-
ровых в результате их заиления, а также разработка перекатов с
целью обеспечения судоходства и забора гальки из ложа реки;
14) гибель огромного количества личинок и молоди в водозабор-
ных сооружениях, ирригационных системах и во время дноуглуби-
тельных работ на перекатах нерестовой зоны; 15) отсутствие ры-
бохозяйственной мелиорации и искусственного воспроизводства.
Сопоставление биологии, условий обитания и воспроизводства с
7-7
97
учетом степени эксплуатации запасов севрюги и осетра показало,
что последний в значительно большей степени подвергается воз-
действию неблагоприятных факторов, чем севрюга, поскольку
большая часть его нерестовой популяции представлена озимой
«формой», жизненно важные процессы и воспроизводство которой
совпадают с периодом наибольшего воздействия различных отри-
цательных факторов на его популяцию. Несомненно, часть пере-
численных факторов оказывает неблагоприятное влияние и на
воспроизводство севрюги, но в меньшей степени, поскольку 93—
95% ее нерестовой части популяции приходится на яровые формы,
воспроизводящиеся в более благоприятных условиях (Песериди,
1962; Песериди и др., 1969, 1972, 1981; Бекешев, Песериди, 1972;
Верина, Песериди, 1979; Стыгар и др., 1979; Тарабрин, 1981а, б;
Песериди, Верина, 1981).
Для ограничения воздействия отрицательных факторов на эф-
фективность воспроизводства и восстановление запасов осетра
р. Урал после полного запрета морского лова был предложен ряд
мероприятий, большинство из которых уже реализовано. Со-'
кращена протяженность нерестовой зоны с 45 до 15 км, соответ-
ственно уменьшилось количество тоней с 25 до 10; осуществлен
концентрированный лов с периодическим прекращением промысла
на 3 дня через каждые 5 дней в апреле и октябре и на 2 дня через каж-
дые 3 дня в мае, августе и сентябре; отменена промысловая мера;
с 1970 г. объявлен круглогодичный запрет на промышленный лов
осетра; прекращен лов рыбы курилкинским садком-ловушкой;
Таблица 32. Пропуск осетра в р. Урал к местам зимовки и нереста
Расчетный пропуск осетра по промысловым уловам до запрета Расчетный пропуск осетра по по- штучным уловам верхней контрольной тони после запрета
Год Тыс. шт.| Год j Тыс. шт. Год I Тыс. шт.| Год | Тыс. шт.
1958 2,81 1964 6,38 1970 8,36 1976 23,16
1959 2,55 1965 4,85 1971 6,24 1977 17,70
1960 2,04 1966 1,53 1972 12,71 1978 59,91
1961 9,32 1967 1,53 1973 13,34 1979 35,28
1962 6,63 1968 2,04 1974 20,95 1980 33,28
1963 4,37 1969 3,06 1975 48,81 1981 30,90
запрещено применение лотовых цепей и судоходство в пределах
нерестовой зоны в период нереста и ската личинок осетровых
(с 1 мая по 15 июля), а также сокращен объем перевозок на судах
на 50% в пред- и посленерестовый периоды; прекращены дноуглу-
бительные работы на перекатах в районе нерестилищ, а в судо-
ходном Урало-Каспийском канале — в период нереста и ската мо-
лоди; не производится забор гальки и песчано-гравийной смеси с
ложа реки; наиболее продуктивная часть нерестовой зоны (от
устья р. Барабастау до Татарского яра Гурьевской обл.) объявле-
98
на заповедной; сокращена численность сома путем облова его
мест зимовки; разработаны биотехнические нормативы по искусст-
венному разведению осетра; разработан проект аэрации нижнего
течения р. Урал и др.
В результате этих мероприятий численность осетра стала уве-
личиваться, о чем можно судить по данным пропуска производи-
телей к местам зимовки и нереста до запрета на его лов и после
(табл.32).
16. Acipenser gCldenstadti persicus Borodin —
персидский (куринский) осетр
Синонимия. Acipenser persicus: Бородин, Вести, рыбопр., 1897. — Boro-
din, Ann. Mag. Nat. Hist. v. 9, N 20, 1926.
Acipenser giildenstadti: Каврайский, Осетровые Кавказа. Тифл., 1907,—
Берг, Фауна России, Рыбы, т. 1, 1911. — Беляев, Бюл. Всекасп. рыбохоз. эксп.
1932 г., № 5—6, Баку, 1932.
Acipenser giildenstadti persicus: Берг, Рыбы пресных вод СССР, т. 2, 1933;
там же, т. 1, 1948.
Распространение. Персидский осетр — проходная рыба, оби-
тающая в основном на юге Каспия, откуда входит главным обра-
зом в р. Кура. В гораздо меньшем количестве заходит на зимовку
и нерест в реки Волгу и Урал. В реках дагестанского (Самур, Су-
дак, Терек) и азербайджанского (Ленкоранка и Астара) побере-
жий малочислен.
В прошлом в р. Сефидруд (Иран) заходил в большом количе-
стве в течение всего года; после зарегулирования реки встречается
в ней только во время паводка (Rostami, 1961).
Рис. 23. Персидский осетр
Описание. Персидский осетр (рис. 23) впервые описан
Н. А. Бородиным (1897) в качестве вида. Позже Л. С. Берг (1911)
не согласился с этим мнением и предложил отнести персидского
осетра в качестве подвида русского осетра. Впоследствии Н. А. Бо-
родин (1926) считал возможным рассматривать персидского осет-
ра как подвид.
Эти точки зрения затем находили своих сторонников. Некото-
рые авторы осторожно обходили вопрос о систематической принад-
лежности указанного осетра, давая ему русские названия: пер-
сидский, куринский, южнокаспийский (Беляев, 1932; Державин,
1939; Борзенко, 1961 и др.). Вслед за Л. С. Бергом многие авторы
99
(Абдурахманов, 1962; Чугунов, Чугунова, 1964; Легеза, Войнова,
1967; Легеза, 1970) диагностировали его как подвид или даже как
нацию. Г. В. Никольский (1971) о персидском осетре не упоминает.
В последнее время первоначальную точку зрения Н. А. Боро-
дина поддержал В. И. Лукьяненко (1973, 1974), который на осно-
ве изучения степени антигенной дифференциации сывороточных
белков установил, что так называемый поздний яровой осетр Вол-
ги (по Баранниковой, 1957), или летненерестящийся (по Трусову,
1967), является не внутрипопуляционной нерестовой группой рус-
ского осетра, а персидским осетром. Проведя морфометрический
анализ и основываясь на некоторых сторонах биологии летнене-
рестящегося осетра, автор заключает, что «мы имеем дело
с двумя хорошими видами, полностью отвечающими тройному
критерию: генетическая (изолированы), морфологическая (хиа-
тус) и экологическая (имеют самостоятельный ареал) самостоя-
тельность. В соответствии с этим, вслед за Н. А. Бородиным
(1897), мы предлагаем именовать впредь южнокаспийского осетра
персидским (Acipenser persicus Borodin), а северокаспийского
осетра — русским...» (Лукьяненко, 1973, с. 54).
Е. П. Артюхин (1974) на основе анализа 4 меристических и
8 пластических признаков осетров, а также гистологического ана-
лиза пришел к выводу, что «поздний яровой, или летненерестящий-
ся осетр Волги представлен персидским подвидом (Acipenser persi-
cus Borodin); озимый же осетр морфологически полностью соот-
ветствует типичному русскому осетру» (с. 8—9). Позднее
Е. П. Артюхин (1979) отобрал в Урале весной из нерестовой части
популяции 22 особи озимого русского осетра и типичных предста-
вителей персидского осетра (оба пола) и, проведя анализ, сделал
заключение, что поздний яровой осетр Урала представлен только
персидским осетром и что описанной Н. Е. Песериди (1966, 1971)
малочисленной поздней яровой внутрипопуляционной нерестовой
группы русского осетра в Урале нет.
Из изложенных воззрений В. И. Лукьяненко и Е. П. Артюхина
следует, что персидский осетр, заходящий в Волгу и Урал, явля-
ется видом и представлен только одной поздней яровой группой,
размножающейся «летом на средних и нижних участках нерестовой
зоны при температуре 20—22° с конца июня по август» (Артюхин,
1979, с. 106). Авторы заостряют внимание на том, что персидский
осетр является типично летненерестящейся «формой», в чем и за-
ключается его главное отличие от русского осетра (ранней яровой
и озимой). Это дает им основание говорить о его генетической изо-
лированности.
Позднее Е. П. Артюхин и А. Е. Андронов (1981) пишут, что
«с самого начала хода создается впечатление совместной миграции
двух групп персидского осетра, различных по состоянию гонад и
численности относительно друг друга. Аналогию можно найти в
миграции персидского осетра р. Куры, когда в первой декаде мая
на конец хода близкого к зрелости раннего ярового осетра начи-
100
нает «накладываться» ход значительно менее зрелого позднего
ярового осетра, нерест которого происходит в сентябре» (с. 16).
Е. П. Артюхин (1983) указывает, что в дельте Волги количество
«зрелых» рыб максимально в начале хода, к лету увеличивается
число «незрелых» персидских осетров, но они не превышают ко-
личества более «зрелых»; осенью отмечались единично особи с
III стадией зрелости гонад и типичным «предзимним» состоянием
гипофиза — «озимые» (с. 9).
Из процитированного можно понять, что для персидского осет-
ра, как и для русского, характерна сложная структура популяции
и она представлена не одной поздней яровой группой (летненерес-
тящимся осетром), а несколькими. Подтверждают сказанное дан-
ные Л. А. Путилиной (1983, 1983а), позволяющие сделать предпо-
ложение о наличии озимого персидского осетра в р. Волге.
В последнем издании «Жизнь животных» (1983) персидский
осетр описан как вид без обоснования такой трактовки.
Таким образом, мнения разных авторов по таксономии и био-
логии персидского осетра чрезвычайно противоречивы. Что же
представляет собой персидский осетр?
Тело персидского осетра веретенообразное, прогонистое, утон-
чающееся к хвосту, покрыто пятью полными рядами крупных жу-
чек с радиально зернистыми полосками и в части случаев двумя
(между боковыми и брюшными) неполными рядами более мелких
жучек. На теле имеются мелкие звездчатые бляшки. Наибольшая
высота тела составляет '/в его длины (у русского—’/е). Нижняя
губа прервана. Рот полулунный, но уже, чем у русского. Рыло
мощное, длинное, несколько спущено вниз («клюворылый»).
Усики не имеют бахромок, далеко не хватают до рта, но достигают
конца рыла. Глаза с серебристо-белой радужной оболочкой. Груд-
ные плавники короче и уже, чем у русского. Цвет спины серо-го-
лубой, серебристо-белый или стальной; брюхо белое, светло-жел-
тое, серебристо-белое.
Сопоставление морфологических показателей персидского осет-
ра, по данным разных авторов, указывает на несущественные раз-
личия в средних величинах (табл. 33). Его отличия от русского
осетра в счетных признаках показаны в таблице 34.
Очевидно, характеризовать персидского осетра как самостоя-
тельный вид нет достаточных оснований, поскольку он не отвечает
тройному критерию современной концепции вида: генетически не
изолирован и экологически не самостоятелен. Различий по некото-
рым счетным и пластическим признакам достаточно только для
того, чтобы классифицировать его как подвид, что и сделал
Л. С. Берг (1934).
Биология. Особи ходового персидского осетра с половыми про-
дуктами в стадии зрелости III, III-—IV появляются в Урале в
Конце мая — первой половине июня и отмечаются до конца июля.
По состоянию половых желез мигранты июньского и июльского
хода практически не отличаются от ходового озимого русского
101
Таблица 33. Морфологические признаки персидского осетра Каспия (М±т)
СЧ О > (Песериди, 1962-1964) 10,63±0,12 30,93±0,16 9,4 ±0,17 21,6 ±0,97 39,2 ±0,81 26,11 ±0,38
СЧ «= С сс (Лукьянен- ко, 1974) 11,7±0,36 31,4±0,52 9,1 ±0,12 20,8±0,27
Урал I (Артюхин, 1979) 11,3±0,22 31,7±0,42 9,0±0,16 21,0±0,37
Волга | 11,2±0,19 31,4±0,26 9,1 ±0,14 21,3±0,23
сЗ О. 'Магёрамов, 1972) 11,1±0,2 31,7±0,4 19,4±0,1 22,4±0,4
(Беляев, ( 1932) 1 1О,19±0,08 30,64 ±0,21 9,51 ±0,09 21,0 ±0,16 38,82± 0,26 25,98±0,16
Признак Спинных жучек Боковых » Брюшных » Жаберных тычинок Лучей в D Лучей в А
СО о о ю
W— Tf СО
ООО о о
+1 +1 +1 +1 +1 га
со „
со со о — со
да ci ю — еч
— — со со
III III
Таблица 34. Счетные признаки русского и персидского осетров, шт.
Признак Русский осетр Персидский осетр
(Песериди, 1962) Средн. (Беляев, 1932) От—до (Беляев, 1932) От—ЙО
Спинных жучек 13,01 9-18 5-13
Боковых » 38,81 30—50 21-42
Брюшных » 10,33 7-12 7-14
Лучей в D 41,50 33-51 30-49
Лучей в А 26,70 21-33 18—32
Жаберных тычинок 23,64 19-29 15-29
тела 18,5±0,15 8,3±0,08 4,8±0,04
со
2 о о
&
Д- о со о
к +1 -н -н
ч ю
со СО
• ’
о СМ .
о
О о
г* +1 +1 о
2 со см -Н
г- ь 00
00 со
о
о
хо со ю со
W о о о
я, г.
f-l о о о
о +1 +1 +1
со СМ со
ио о «
о"- А
00 см ю
га т—< * 1
сч
о
+1 1 см сч
со
*
сч
со
со
л
CQ о
о 1 о
Ч -н со 1 со
О
ю
S сч со
и о о
я -н -н СО
с; О) СП ю
•* •»
со со
о
СЧ tJ*
о" №
о о
га +1 +1 S
о со см
О)
*
о со СО СО
осетра. Разница заключается лишь в количестве и цвете жира: у
персидского его меньше и он белого цвета. Гонадо-соматический
индекс самок персидского осетра в летнее и осеннее время состав-
ляет:
Месяц От—до М п Месяц От—до М п
Апрель 12,1-30,5 25,8 13 Июль 6,7-30,0 22,4 29
Май 8,1-33,3 20,3 48 Август 5,6-28,4 18,1 11
Июнь 6,7-34,6 24,2 41 Сентябрь 6,3-24,2 17,8 14
После зимнего перерыва в ходе осетра, продолжающегося с
октября по март, первые мигранты появляются в Урале в начале
апреля при температуре воды 5—6°, но несколько позднее русского
осетра.
Для осетров апрельского и майского хода характерно однород-
ное состояние зрелости половых желез (стадия IV) и дефинитив-
ные размеры ооцитов. Во второй половине мая в реке появляются
производители с незавершенной четвертой стадией зрелости, у ко-
торых ооциты мельче, чем у осетра апрельского хода. Ход персид-
ского осетра с таким состоянием половых желез продолжается до
начала июля, когда ооциты у них достигают дефинитивных разме-
ров (табл. 35). На этом годовой цикл нерестовой миграции персид-
ского осетра завершается.
В прошлом в Куре наблюдались два пика хода персидского
осетра: первый (более мощный)—в марте—апреле, а второй —
в сентябре — октябре. В Сефидруде максимум хода отмечен в ап-
реле — мае, а менее значительный — осенью.
Н. П. Гербильский (1950) выявил и описал 4 биологические
группы персидского осетра, что оспаривалось А. Н. Державиным
(1953, 1956); однако в любом случае это свидетельствует о слож-
ности и неоднородности популяции персидского осетра.
Сопоставление данных по динамике хода осетра Куры, Сефид-
руда и Урала показывает, что структура нерестовой части попу-
ляции осетра, заходящего на зимовку и нерест в р. Урал (и Волгу),
103
не повторяет структуру южнокаспийской популяции. Это, вероят-
но, обусловлено тем, что р. Урал является окраиной ареала, а
р. Кура и р. Сефидруд — ее центром.
Персидский осетр в р. Урал размножается на тех же участках и
в те же сроки, что и русский. Производители, зашедшие в реку ле-
том и осенью с половыми железами в стадии зрелости III—IV,
зимуют на ямах верхней части нерестовой зоны совместно с ози-
мым русским осетром и нерестятся в первой половине мая при
температуре воды 9—12°. Самки по завершении нереста сразу
покидают нерестилища и скатываются вниз по течению со ско-
ростью до 60 км в сутки. Самцы задерживаются на нерестилищах
до 2 месяцев и более.
Таблица 35. Соотношение ооцитов разного размера персидского осетра
Куры (банк, — Державин, 1956) и Урала (низовья, 1962—1964 гг.), % от числа
исследованных рыб
Месяц Кура Урал Кура Урал Кура Урал Кура Урал Кура Урал
7-14 мг 14., 1- 16 мг 16,1- 18 мг 18,1-22 -27 мг
Март — 10,0 - 25,0 65,0 — 20
Апрель —- 7,7 13,4 7,7 6,6 23,1 80,0 61,5 30 13
Май 5,0 6,0 5,0 4,0 10,0 52,0 80,0 38,0 20 48
Июнь 11,3 7,3 24,8 12,2 30,8 46,4 33,1 34,1 169 41
Июль 7,5 17,5 17,5 27,5 40,0 20,6 35,0 34,4 40 29
Август 1,7 7,1 11,8 37,4 20,4 36,4 66,1 9,1 168 11
Сентябрь — — — — — 71,4 100 28,6 14 14
Часть «озимых» самцов после ската самок остается на тех же
нерестилищах, а другая медленно сплывает вниз. По мере подхода
к нерестилищам самок, которые зашли в реку ранней весной со
стадией зрелости IV, IV—V («яровые»), возможно их спаривание
с самцами «озимого» и подошедшего «ярового» осетров. Аналогич-
ное скрещивание самцов «раннего ярового» осетра, возможно,
происходит с самками «позднего ярового», т. е. летненерестящегося.
Не исключено также скрещивание персидского и русского осетров,
поскольку сроки их размножения и места нереста одни и те же.
О беспрерывном нересте персидского осетра с середины апреля
до октября в р. Мпнгечаур свидетельствует А. Н. Державин
(1947). Среднемесячная температура, по данным этого автора, на
нерестилищах родных рек персидского осетра колеблется от 13
до 26°,
Из приведенного можно понять, что персидский осетр не ограж-
ден от скрещивания не только со своим ближайшим сородичем —
русским осетром, но и другими осетровыми, о чем свидетельствует
наличие помесей между разными видами осетровых, описанных
Л. С. Бергом (1949) и М. И. Легеза (1971). Наиболее многочис-
104
ленной из восьми вариантов помесей, по данным Т. С. Зарбалие-
вой (1979), на юге западного побережья Среднего Каспия была
помесь белуга X осетр (36,6%).
Обнаружить помеси между персидским и русским осетрами и
отличить их от исходных форм по меристическим и пластическим
признакам практически невозможно из-за совпадающей вариа-
бельности этих признаков, а также потому, что у обоих подвидов
имеется большое количество тупорылых, толсторылых, прогонис-
тых, высокоспинных с пластинками между рядами жучек и без
пластинок (Берг, 1948) и других рыб с различными оттенками
окраски тела.
Длительность инкубационного периода персидского осетра
примерно такая же, как и у русского.
В период эмбрионального и на ранних этапах постэмбриональ-
ного развития потомство персидского осетра практически не отли-
чимо от русского. Но поскольку сроки и места нереста обоих осет-
ров в Урале совпадают, очевидно, сроки ската их потомства оди-
наковые.
В р. Кура, где русского осетра меньше, первые личинки появ-
ляются в дельте во второй половине апреля (Маилян, Махмудбе-
ков, 1966, 1970). Личинки и молодь в возрасте 5—20 суток ловятся
в низовьях реки в июне, июле и августе (Захарян, 1971, 1972, 1974),
но всегда раньше севрюги (Захарян, 1981). Подросшая молодь
массой до 3—5 г встречается до глубокой осени, незначительная
часть остается на зимовку в реке (Маилян, Махмудбеков, 1970).
Созревание, возраст и рост. Половозрелость самцов персидского
осетра наступает в 7—8 лет, но таких особей в популяции едини-
цы, что для Куры отметили М. И. Легеза и И. А. Войнова (1967).
Отдельные самки, по нашим материалам, созревают несколько
позже: в 9—10-летнем возрасте. М. И. Легеза и И. А. Войнова
указывают наступление половозрелости самок в 11—12 лет. Уловы
осетра (оба пола) представлены 24 возрастными группами (8—-
32 года). В Куре известны производители до 48 лет (Бабушкин,
Борзенко, 1951).
Основную массу уловов самцов осетра в р. Урал составляют
рыбы в возрасте 10—21 года (84,7%) с модальной группой 12—
17лет (54%). На долю самцов до 10-летнего возраста приходится
всего 3,7%. Некоторые самцы в возрасте 13—14 лет, видимо, не-
рестуют второй раз, но основная часть повторно нерестующих сам-
цов представлена 15—18-летками. Самцов старше 21 года мало —
11,6%.
Большая часть уловов самок осетра в Урале представлена
особями 15—24-летнего возраста (76,4%). Ядро уловов составляют
17—20-летки (42,5%). На долю младших возрастных групп
(9—13 лет) приходится 6,3%, а более старых (свыше 25 лет) —
13,5%). Некоторые рыбы в возрасте 14—17 лет, вероятно, нерес-
туют вторично. Незначительное количество в уловах старшевозра-
стных рыб говорит об интенсивном промысле.
105
Возрастной состав персидского осетра р. Урал в процентном
отношении выглядит следующим образом:
Возраст, лет Самцы Самки Возраст, лет Самцы Самки Возраст, лет Самцы Самки
7 0,5 16 7,4 7,7 25 1,6 3,3
8 1,1 — 17 9,0 10,3 26 1,6 2,6
9 2,1 0,6 18 5,8 12,3 27 — 3,3
10 4,8 0,6 19 5,8 9,7 28 1.1 1.2
11 5,3 1,3 20 4,2 10,2 29 0,5 1,9
12 8,5 1,9 21 4,8 6,4 30 — —-
13 10,0 1,9 22 2,6 5,1 31 — 0,6
14 10,6 3,8 23 2,1 3,8 32 — 0,6
15 8,5 5,8 24 2,1 5,1 п 187 156
О росте куринского осетра сведений в литературе практически
нет. М. П. Борзенко (1961), 10. А. Абдурахманов (1962) повторяют
данные А. Н. Державина (1956, с. 31):
Возраст, лет 10 15 20 25 30
Общая длина, см 98,5 155,7 173,0 183,5 206,5
По мнению М. И. Легеза и И. А. Войновой (1967), темп роста
куринского осетра снизился по сравнению с 30—40-ми годами.
Наблюдающееся омоложение стада они объясняют исчезновением
старших возрастных групп в результате вылова. Темп роста и
масса тела осетра из уловов в р. Урал приведены в таблицах 37 и
38, из которых видно, что самки (п=183) намного опережают в
росте самцов (п=216). Сравнение материалов по темпу роста
персидского осетра с таковым русского показывает, что первый
Таблица 36. Размеры (см) и масса тела (кг) персидского осетра
по возрастам из уловов в р. Урал (1962—1964 гг.)
Возраст, • лет Самцы Самки Возраст, лет Самцы Самки
см КГ СМ кг СМ кг СМ кг
7 90,0 3,89 - 20 151,2 17,81 161,3 24,50
8 98,1 4,25 — 21 156,1 19,43 162,1 25,83
9 106,4 5,81 105,2 6,21 22 161,5 21,25 166,5 27,39
10 111,5 6,88 109,7 7,26 23 163,8 23,60 169,7 30,64
И 114,3 7,93 117,3 9,14 24 168,4 24,10 167,9 30,91
12 117,8 8,16 122,6 11,70 25 170,6 25,80 171,4 33,85
13 121,2 9,88 129,5 12,85 26 174,7 26,30 179,5 35,16
14 128,5 11,71 131,4 14,91 27 176,6 25,80 180,8 36,45
15 131,7 13,83 137,8 15,73 28 171,9 24,20 183,1 37,17
16 135,4 15,14 145,6 17,18 29 177,5 27,50 186,2 39,30
17 132,2 15,78 150,1 18,42 30 —. — 179,5 41,20
18 145,0 16,16 154,8 21,16 31 — — 185,0 40,40
19 147,7 16,92 156,5 22,44 32 — — 191,0 45,30
106
опережает в росте второго, причем уральская часть популяции
уступает в росте куринскому стаду.
Половой состав персидского осетра, входящего в р. Урал, изу-
чен недостаточно. В целом соотношение полов близко 1:1, что от-
мечается и для других частей ареала.
На нерестилищах доля самок (%) колеблется по месяцам и не
одинакова в разных реках (Державин, 1956):
Реки V VI VII VIII IV—VIII
Араке, Карадонлы Кура, Мпнгечаур 10,0 32,8 54,9 74,1 42,9
13,5 41,6 37,7 30,9 34,7
Таблица 37. Масса тела ходового персидского осетра
из уловов в Урале (1962—1965 гг.), кг
Размерная Группа,см Самцы Самки
М Прирост п М Прирост п
91-100 4,07 2
101-110 7,61 3,54 19 6,21 — 1
111-120 9,16 1,55 28 7,26 1,05 7
121-130 12,21 3,05 60 12,95 5,69 13
131-140 15,62 3,41 44 17,84 4,89 31
141-150 17,14 1,52 28 20,55 2,71 57
151-160 19,34 2,20 15 23,86 3,31 37
161-170 23,25 4,61 10 29,11 5,25 18
171 — 180 26,90 3,65 7 33,78 4,67 9
181-190 31,48 4,56 2 39,50 5,72 5
191-200 36,40 4,92 1 42,40 2,90 3
Таблица 38. Абсолютная плодовитость (АП) персидского осетра
из Урала (1962—1965 гг.), тыс. шт.
Размерй, см АП п Размеры, см АП п
От—до М От—до М
101-110 75,9 1 161-170 136-584 318,5 18
111-120 84-136 104,3 7 171-180 186-854 338,8 9
121-130 94 -248 116,8 13 181-190 218-645 413,5 5
131-140 89-389 184,7 31 191-200 348-612 464,8 3
141-150 81-485 221,3 57 201-210 — 709,0 1
151-160 111-628 269,5 37 101-210 76-854 315,1 182
Плодовитость персидского осетра из р. Урал ниже плодовитости
русского осетра (табл. 39, 40). Такого же мнения Ю. А. Абдурах-
манов (1962) о плодовитости осетров из р. Кура.
Питание русского осетра в Каспийском бассейне может быть
примером изученности этой стороны биологии осетровых. Характер
питания персидского осетра изучен гораздо слабее. Совсем слабо
исследовано питание молоди в речной период жизни.
107
Скатывающаяся молодь осетра (16—73 мм) в р. Кура, по дан-
ным Г. Б. Захаряна (1974), питается мизидами, хирономидами и
гаммаридами; общий индекс наполнения желудков 195—1ООО%оо-
Молодь (2—3-летки) и неполовозрелые особи (до 7 лет) размером
35—75 см, выловленные в юго-восточной части Каспия в 1966—
1968 гг., питались попеременно крабами и рыбой. С возрастом зна-
чение рыбной пищи (в основном, бычки) увеличивается до 78%:
рациона.
Таблица 39. Абсолютная плодовитость (АП) персидского
осетра по возрасту из Урала (1962—1965 гг.), тыс. шт.
Возраст, лет АП Возраст, лет АП Возраст, лет АП
9 75,9 17 159,8 I 25 355,0
10 89,0 18 194,5 26 371,2
11 85,0 19 216,8 27 375,9
12 72,3 20 236,7 28 394,2
13 76,5 21 271,3 29 379,0
14 94,2 22 296,8 I 30 —
15 116,5 23 316,7 31 645,0
16 124,3 24 343,1 1 Средн. 315,1
В Северном Каспии молодь персидского осетра (1 экз. длиной
41 см) потребляла гаммарид (26%оо), а взрослая особь (130 см)
питалась рыбой (30,4%оо). В Среднем Каспии у дагестанских бе-
регов питание молоди осетра длиной до 40 см было разнообразнее.
При питании нереисом индекс наполнения был низким — 6,7%оо.
При смешанном питании рыбой (пуголовки), ракообразными (ми-
зиды) и нереисов он достигал 46,7%оо- Особи длиной 41—80 см
были менее накормленными: при питании кумовыми индекс не
превышал 1,4%оо, а при потреблении моллюсков — 20,2°/000. В от-
дельные годы в желудках было обнаружено много балянуса (39%)
и кильки (61%). Осетр размером 81—120 см в основном потреблял
рыбу (53,4%), остальная часть рациона приходилась на балянуса
с ракушей.
У казахстанских берегов Среднего Каспия накормленность не-
половозрелых особей персидского осетра (до 80 см) была
высокой — 99,8—106,3%оо. Питались они здесь в основном гам-
маридами и бычками. Более крупные рыбы накормлены слабее —
14,2°/ООо-
В целом рацион питания персидского осетра неустойчив и ши-
роко варьирует в пределах одного района. Четкой зависимости ра-
ционов от кормовой базы нагульного ареала нет (Зарбалиева,
Деткина, 1981). По морю интенсивность питания неодинакова.
Наиболее накормленные особи размером 41—80 см: общий ин-
декс наполнения желудков 51,2°/00о (Зарбалиева, 1981). В течение
суток осетр питается неравномерно, наиболее активно в дневное и
ночное время (Филиппов, 1979).
108
Распределение в море. Персидский осетр неравномерно распре-
делен по Каспийскому морю. В результате падения его числен-
ности и сокращения ареала на нагульных площадях Северного
Каспия он встречается очень редко (Магерамов, 1972). В мелко-
водной зоне восточной половины Северного Каспия в 1972—1975 гг.
обнаружить его не удалось. Отдельные особи зимуют у Мангыш-
лакского залива (Селивестрова, 1969; Легеза, 1970).
Излюбленное место обитания персидского осетра — материко-
вая отмель Среднего Каспия и юга моря. Только летом он в массе
покидает воды Южного Каспия. Некоторые исследователи (Сели-
вестрова и др., 1979) считают, что нефтеносные площади Южного
Каспия являются местом нагула осетровых и имеют рыбохозяйст-
венное значение.
Численность и хозяйственное значение. Уловы персидского
осетра в реках Урал и Волга статистикой не учитывались, по-
скольку его доля в общих уловах составляла ничтожную часть.
Наиболее высокие его уловы (вместе с шипом и русским осетром)
в водах Азербайджана наблюдались в 1936 г. —1540 т. К началу
50-х годов они упали до 630—650 т, а к 60-м годам сократились до
310—340 т в год. После прекращения морского красноловья
(1964 г.) запасы персидского осетра не восстановились и находятся
на крайне низком уровне в Азербайджане.
Между тем в иранских морских водах уловы осетра ставными
сетями с 250 т в начале 50-х годов достигли 800—920 т к началу
60-х (Богородицкий, 1969). Высокие уловы осетровых в иранских
водах (по статистическому ежегоднику ФАО) наблюдаются и в
последнее десятилетие, а доля осетра среди них на юге Каспия
составляет 48,4—83,2% (Легеза, 1972):
Год АзССР Иран Год АзССР Иран Год АзССР Иран
1971 30 2400 1975 140 1500 1979 300 1500
1972 70 2200 1976 190 1500 1980 133 1500
1973 100 1500 1977 190 1500 1981 157 1500
1974 90 1500 1978 215 1500 — —.
В целях восстановления запасов персидского осетра необходи-
мо установить запрет на его лов во всех реках, впадающих в Кас-
пийское море, в течение всего года, а также форсировать масшта-
бы его искусственного разведения на Куре, Волге и Урале. Био-
технические нормативы по его искусственному разведению
разработаны (Маилян, Алекперов, 1976; Артюхин, 1978; Маилян,
1981).
2. Acipenser baeri Brandt — сибирский осетр
Местные названия: у русских — осетр, костерь (мелкие неполовозрелые осет-
ры); у казахов— бекре.
Синонимия. Acipenser baeri: Brandt, Med. biol. Acad. Petersburg, v. VII,
1869.
Acipenser stenorrhynchus: А. Никольский, Ежегодн. Зоол. музея АН, т. I,
1896.
Acipenser baeri: Берг, Фауна России, Рыбы, т. I, 1911. — Исаченко, Рыбы Ту-
руханского края, 1912.—Меньшиков, Уч. зап. Пермск. ун-та, т. II, вып. 1, 1936.—
Боган, Уч. зап. Пермск. ун-та, т. III, вып. 2, 1938.
Распространение. Ареал сибирского осетра (рис. 24) охваты-
вает площадь от Оби до Колымы и от северных районов Китая до
морей Советского Заполярья. В Казахстане обитает в Среднем и
Верхнем Иртыше, есть в Усть-Каменогорском и Бухтарминском
водохранилищах, по Черному Иртышу поднимается в территори-
альные воды Китая.
В бассейнах крупных рек стадо осетра обычно представлено
двумя формами: полупроходной и туводной, ареалы которых пере-
крываются. В бассейне Оби П. А. Дрягин (1948) указывает на
существование двух стад. Кроме того, в Верхней Оби, по данным
Рис. 24. Сибирский осетр
А. И. Петкевича (1952), имеется жилая форма осетра, ареал кото-
рой полностью покрывается миграционным путем проходного. По-
мимо реофильных форм (проходная, туводная) у сибирского
осетра известны озерные формы, для которых местом обитания
служат крупные озера, а речные миграции ограничиваются перио-
дом размножения. К таким озерным формам принадлежат
байкальский осетр и зайсанское стадо.
В Среднем и Верхнем Иртыше, как и во всем Обь-Иртышском
бассейне, стадо осетра было биологически неоднородным.
Ф. Е. Боган (1938) высказал предположение о наличии в Иртыше
двух самостоятельных биологически обособленных форм: типичной
полупроходной (обь-иртышская) и типичной туводной (зайса-
но-черноиртышская). Последующие исследования (Иоганзен,
1948, 1951; Вотинов, 1958, 1963; Ерещенко, 1966, 1969, 1970 и др.)
подтверждают наличие в Иртыше, в том числе и в Верхнем, оби-
тание двух форм — полупроходного обского («низовой») и жилого
осетра. Жилая форма была приурочена к Верхнему Иртышу, озеру
110
Зайсан и Черному Иртышу. Четких границ между двумя формами
не установлено.
После перекрытия Иртыша плотинами ГЭС верхнеиртышское
стадо оказалось изолированным и перестало пополняться за счет
многочисленного обь-иртышского. В настоящее время оно разделе-
но на две части, которые соответственно водохранилищам назы-
ваются усть-каменогорское и бухтарминское.
Описание. Морфологическое описание проводится по трем
пунктам: 1) нижний бьеф Усть-Каменогорской ГЭС (в пробе сме-
шанный материал жилой и проходной форм осетров); 2) Усть-
Каменогорское водохранилище (также две формы осетра); 3) пе-
рекаты в ур. Джамбай, расположенные вблизи истока Белого Ир-
тыша из оз. Зайсан (осетр локального зайсано-чернопртышского
стада).
D 36—48; А—(16)22—26(32); спинных жучек 12—16, боковых
жучек 39—49, брюшных жучек 7—13; жаберных тычинок 29—
38(51). Длина головы в среднем составляет 18,46% абсолютной
длины рыбы; длина рыла 36,75, ширина — 37,46% от длины голо-
вы; диаметр глаза 6,48%; заглазье 58,93%; наибольшая длина
усика 23,78%) от длины головы. Наибольшая высота тела 11,49,
наименьшая —3,27%), длина хвостового стебля 10,28% от абсолют-
ной длины рыбы; антедорсальное расстояние 62,49, антеанальное —
66,18%; длина D 11,28, длина А 5,0, длина Р 12,45, длина
V 7,53% от абсолютной длины рыбы. Другие пластические призна-
ки приведены в таблице 40.
Рот небольшой, поперечный. Жаберные перепонки приращены
к межжаберному промежутку и не образуют под ним складку.
Спинной плавник расположен далеко сзади, над анальным.
Окраска спины серовато-желтая, желто-серая и темно-грязного
цвета. Спинной и хвостовой плавники цвета спины. Грудной и
брюшной плавники светло-желтые, с разнообразными оттенками.
Брюхо светлое. Анальный плавник цвета брюшной части тела.
Окраска тела для сибирского осетра не является существенным
критерием, так как в одном водоеме можно встретить особей от
абсолютно черных до оранжевых с различными переходами цвета.
При сравнении пределов и средних показателей морфологиче-
ских признаков усть-каменогорского осетра с зайсанским обнару-
живается: у первого больше лучей в D и спинных жучек, меньше
жаберных тычинок. У усть-каменогорского осетра длиннее хвос-
товой стебель. Показатель наибольшей и наименьшей высоты тела
выше, тело толще, голова длиннее, рыло длиннее и шире, усики
короче. Есть и другие различия в средних показателях признаков
описываемых осетров из трех районов Верхнего Иртыша. Видимо,
такое явление закономерно для данного вида. А. Н. Петкевич и др.
(1950), А. Н. Петкевич (1953) и другие исследователи, специально
изучавшие морфологию осетра Средней и Верхней Оби, пришли к
заключению, что счетные и пластические признаки у него подвер-
жены большой возрастной и половой изменчивости, поэтому опери-
Ш
Таблица 40. Морфометрическая характеристика сибирского осетра бассейна р. Иртыш
Признак Нижний бьеф Усть- Каменогорской ГЭС; 1957-1958 гг. ;п =21 (Чабан, 1961) Усть-Каменогорское вдхр; 1959г. ; п = 19 (Чабан, 1961) Иртыш, р-н Джамбай; 1961 г. ; п-=12
От-до М±ш От—до М±т От —до М±т
1 2 з 4 5 6 7
Абсолютная длина, см Длина тела до конца средних лучей, см Длина тела до корней средних лучей, см Масса тела, кг Лучей в D Лучей в А Жаберных тычинок Спинных жучек Боковых жучек слева справа Брюшных жучек слева справа 41-46 24-25 12-15 38-50 9-12 66,06 43,11±0,58 24,25±0,22 13,38± 0,31 43,54±0,81 10,23± 0,27 35-51 22-27 22-35 12-16 39-49 36-50 9—12 9-12 68,75 43,34± 1,04 24,57 ±0,39 31,95± 0,46 14,14±0,28 43,06 ±0,73 43,47± 0,71 10,42± 0,24 10,53±0,22 50,5-107,0 42,1-94,5 39,0 - 88,5 0,411-7,400 36-48 (16)22-26(32) 29-38(51) 12-16 39-49 39-48 7-13 9-13 90,33 + 4,50 78,98±4,17 73,98±3,95 3,719± 0,512 41,42±0,79 24,70±0,34 33,06± 0,87 13,60+0,46 45,14± 0,93 44,50± 0,82 11,00± 0,49 10,5 ±0,52
В % от длины тела
Длина хвостового стебля —. —. 17,3-34,9 29,55±0,54 8,21 -11,39(15,88) 10,28±0,28
Наибольшая высота тела 11,9-12,8 12,4 11,4-15,6 13,05+0,24 10,46-12,50 11,49+0,15
Наибольший обхват тела —. — — — 30,99-43,22 37,10± 0,45
Наименьшая высота тела 1,6-3,8 3,2 1,9-4,6 3,44± 0,14 2,93-3,54 3,27±0,04
Наибольшая толщина тела 8,4-10,6 9,3 9,7-14,0 11,66± 0,30 9,01-11,97(14,35) 10,75+0,26
Высота головы у затылка —, — —- 8,11-11,58 10,22± 0,23
Высота головы через середину глаза —-' — 4,87-6,03 5,45± 0,07
Длина рыла Ширина рыла Диаметр глаза Длина головы Заглазничный отдел головы 6,4-8,7 7,3 7,3-11,0 9,29±0,22 5,78-7,61(10,09) 6,70± 0,15
8,0-9,2 1,4-1,6 18,6-19,9 8,3 1,5 18,9 7,5-9,2 0,8-1,4 18,4-22,1 8,38±0,13 1,17 + 0,04 19,92+0,26 7,85-9,12 1,05-1,58 16,21-20,49 8,35± 0,09 1,23±0,04 18,46± 0,26
10,2-11,1 10,3 8,4-11,3 9,53± 0,15 8,71-12.02 10,82± 0,24
Наибольшая ширина головы Ширина лба — 9,1-12,0 Ю,55±0,17 10,82-12,68 5,41-6,93 11,32 + 0,11 6,29± 0,10
Антедорсальное расстояние 52,0-62,8 57,7 56,0—62,5 60,28±0,45 56,03-68,31 62,49±0,59
Антевентральное расстояние 52,3-69,6 57,6 48,2-55,8 51,86± 0,39 50,09-60,52 53,29±0,85
Аитеанальное расстояние 56,5-60,0 62,9 63,5-68,0 65,55± 0,35 59,34-68,82 66,18± 0,76
Длина основания D 9,1-12,7 10,4 9,2-11,8 10,5910,09 10,62-12,08 11,28 + 0,09
Высота D 6,5-7,1 6,9 5,8-8,0 6,8810,14 5,85-8,71 6,89±0,23
Длина основания А 3,6-6,3 5,2 4,1-6,1 4,810,12 4,56-5,63 5,09+0,07
Высота А 4,2-7,5 6,2 6,1-7,8 6,8510,10 5,95-7,25 6,54± 0,09
Длина Р 12,9-15,1 14,6 9,7-14,7 12,5510,32 9,90—16,43 12,45± 0,43
Длина V — — 5,44-8,71 7,53 + 0,23
Расстояние между Р и V 31,7—37,0 35,8 30,2-34,8 33,0210,34 31,28-36,47 34,75±0,40
Расстояние между V и А —, — — — 11,39-16,66 14,74± 0,38
Расстояние от конца рыла до губы 6,9-10,1 8,0 8,1-11,9 10,2410,23 6,84-11,48 8,14± 0,39
Расстояние от конца рыла до хрящевого
свода рта — 5,89-10,49 7,18+0,39
Расстояние от конца рыла до средних
усиков 2,8-4,7 3,7 4,4-7,2 5,8110,22 2,84 -4,65,(6,53) 3,75± 0,13
Расстояние от основания усиков до хря-
щевого свода рта 4,4-5,2 4,5 3,2-6,8 5,3910,10 3,02-5,74 3,70± 0,24
Наибольшая длина усиков 3,5—5,9 4,3210,13 3,59-5,83 4,58+0,18
Ширина рта 5,9—6,3 6,0 4,2-7,0 5,0410,15 3,85-9,22 4,80± 0,28
В % о т длины головы
Длина рыла 34,35-43,70 39,1 40,5-51,3 46,5310,79 31,73-40,00 36,35± 0,60
(50,00)
Диаметр глаза 7,9-8,6 8,2 4,0-7,6 5,8610,21 5,52-8,25 6,48± 0,24
Заглазничный отдел головы 50,9—58,6 56,1 41,5-51,1 47,6310,77 52,17-64,93 (AQ 1о\ 58,93± 1,02
Ширина рта 31,7—33,7 32,4 20,0-32,2 25,0110,75 21,26 — 35,26 26,35+ 1,14
Наибольшая ширина головы — — 45,1 -58,9 52,9310,82 54,10—66,66 / 7 f) Л7\ 60,00± 1,00
Ширина рыла 43,2-49,6 45,8 36,4-49,2 53,7910,69 (/ 2 f и/ J 91,73-41,84 /г1 оо\ 37,46±0,90
Межглазничное пространство — — 26,6-97,2 91,5710,57 ^01,23) 34,59—49,01 43,77± 1,17
Наибольшая длина усика 24,7 -25,6 25,0 16,4-30,0 22,1110,46 19,02-30,62 23,78± 0,76
рование только одними средними показателями может быть оши-
бочным.
Длина рыла — самый неустойчивый признак и подвержен силь-
ному варьированию и изменению с возрастом. Различия у осетра в
форме рыла отмечаются на всех стадиях индивидуальной жизни.
Среди молодых особей у одних оно длиннее и сильно заострено, у
других — короткое, тупое или слегка заострено, но короткое. Меж-
ду этими крайними формами рыла наблюдаются различные пере-
ходы. У взрослых рыло округляется, однако остается у одних ко-
ротким, у других узким и вытянутым в длину. Очевидно, что изме-
нение признака длины рыла нарушает пропорцию всех остальных
частей головы.
На основании анализа морфобиологических признаков осетра
Обь-Иртышского бассейна многие исследователи приходят к за-
ключению о широкой приспособляемости этого вида к разным эко-
логическим условиям, что ведет к распаду его на дробные морфо-
логические вариации.
Половая и возрастная изменчивость верхнеиртышского осетра
не изучена. А. Н. Петкевич (1953) отмечает, что у обского осетра
количество спинных, боковых, брюшных жучек, лучей в D и А воз-
растной изменчивости почти не подвержено; жаберных тычинок с
возрастом становится больше. Длина головы, хвостового стебля и
рыла в процентах от длины тела с возрастом уменьшается. Увели-
чиваются наибольшая высота тела, антедорсальное и антевент-
ральное расстояния и наименьшая высота тела.
Максимальная длина осетра свыше 2 м, масса тела около
200 кг (Никольский, 1971). Мельников (цит. по Бергу, 1948) сооб-
щает о поимке в Зайсане осетра массой 192 кг. Ф. Е. Боган (1938)'
отмечает, что в Черном Иртыше и Зайсане нередко были особи
массой до 50—70 кг, чего не отмечается для низовьев Иртыша и
Оби.
Масса осетра Бухтарминского водохранилища, по сборам
1962—1965 гг., максимальная 31,7, средняя 9,3 кг. В Усть-Камено-
горском водохранилище минимальная длина 30,3, максимальная
163, средняя 112,5 см. Минимальная масса 0,4 кг, максимальная
26,5 кг, средняя 11,2 кг. Возраст рыб — от 4 до 24 лет (Ерещенко,
1970).
В р. Иртыш ниже Усть-Каменогорской ГЭС линейный размер
осетров колебался от 30 до 100, средний составил 66,6 см (Чабан,
1962). По исследованиям, проведенным в 1972—1973 гг., в Иртыше
от г. Усть-Каменогорска до г. Павлодара изредка встречались
осетрята (костери) массой 2—5 кг, крупных особей не обнару-
жено.
Биология. Наступление половой зрелости у сибирского осетра
в Нижнем Иртыше Ф. Е. Боган (1938) определяет для самцов в
возрасте 11, самок—-14—15 лет. В Оби, по исследованиям
Б. Г. Иоганзена и Ф. И. Вовк (1952), половозрелости самцы дости-
гают в возрасте 9 лет, по П. А. Дрягину (1949), — в 7—8 лет.
114
По нашим наблюдениям, в речных условиях Верхнего Иртыша
самцы созревают в возрасте 8—9 лет, редко — в 6 лет. В водохра-
нилище большинство самцов в возрасте 7 лет имеют зрелые поло-
вые продукты. По самкам таких наблюдений нет, но сокращение
миграционных путей и лучшие условия нагула в водохранилищах,
очевидно, будут способствовать более раннему наступлению поло-
возрелое™ и у самок (Ерещенко, 1969). Нерестовая популяция
осетра Усть-Каменогорского водохранилища представлена семью
возрастными группами самцов и только пятью возрастными груп-
пами самок (16—19 лет и 24 года).
Данные о соотношении полов сибирского осетра в казахстан-
ской части его ареала отсутствуют. Н. П. Вотинов (1955, 1963) от-
мечает, что в Иртыше, в нерестовой части полупроходного обского
стада, самцов гораздо больше, чем самок, в то время как на местах
нагула они встречаются примерно в равных количествах.
В Верхнем Иртыше (от оз. Зайсан до Семипалатинска) до
гидростроительства имелись обширные нерестовые площади, обес-
печивающие воспроизводство осетра в этом районе. Нерестился он
и в нижних участках рек Уба, Ульба, Бухтарма, Курчум.
Главные нерестовые площади находились от устья р. Шульба до.
г. Усть-Каменогорска. Этот участок Иртыша богат каменистыми и.
песчано-галечными россыпями с умеренными скоростями течения.
Участок Иртыша от Усть-Каменогорска до устья Бухтармы, но-;
сивший название Быстрый Иртыш, служил для осетра главным
образом местом зимовки. Для нереста этот участок не имел сущест-
венного значения. По Белому Иртышу (участок от истока Иртыша до
устья Бухтармы) нерестилища тянулись сплошной цепью от Бух-
тармы до Букони и далее вверх с разрывами до Джамбая.
Производители зайсанского локального стада частично подни-
мались по Черному Иртышу на нерестилища, расположенные пре-'
имущественно за пределами СССР. Другая часть нерестовой по-
пуляции, наоборот, мигрировала в Белый Иртыш, рассеиваясь по
нерестилищам от истока до пос. Кумашкино (Ерещенко, 1966,
1969, 1970).
В Тихом Иртыше, в пределах Павлодарской области, гидроло-
гические условия, по-видимому, были благоприятными для нереста
осетра (Боган, 1938; Вотинов и др., 1975).
Гидростроительство резко ухудшило условия воспроизводства
осетра в Иртыше. В связи с постройкой Усть-Каменогорской ГЭС
проходной обский осетр оказался изолированным от большинства
нерестилищ в Верхнем Иртыше. Выше зоны выклинивания Усть-
Каменогорского водохранилища условия воспроизводства осетра
оставались оптимальными до 1960 г. С созданием Бухтарминского
водохранилища все нерестилища, расположенные по Белому и
Быстрому Иртышу, попали в зону затопления. В Усть-Каменогор-
ском водохранилище для нереста пригоден был только речной уча-
сток водохранилища; длина его в зависимости от уровня воды ко-
лебалась от 5 до 10 км. С сооружением Бухтарминской ГЭС нерес-
115
тилища осетра в Усть-Каменогорском водохранилище утратили
свое значение из-за ухудшения гидрологических и термических усло-
вий. Из Бухтарминского водохранилища в нижний бьеф (Усть-
Каменогорское водохранилище) поступает холодная вода, и в рай-
оне расположения мест икрометания она не прогревается даже до
8—10°, изменилась и скорость течения воды.
В Бухтарминском водохранилище для воспроизводства осетра
сохранились только нерестилища по Черному Иртышу, причем
основные площади находятся за пределами страны (Ерещенко, 1960,
1969; Вотинов, 1965, 1975 и др.).
Иртыш от Усть-Каменогорска вниз до устья р. Убы из-за изме-
нения гидрологических, термических условий, связанных с гидро-
строительством, а главным образом санитарных, в последние годы
не посещают производители осетра (Ерещенко, 1966). Оставшиеся
нерестилища, расположенные по Иртышу от устья Убы до Семи-
палатинска, также потеряют свое значение с завершением строи-
тельства и вводом в эксплуатацию Шульбинской ГЭС.
Таким образом, в новых экологических условиях водоемов
Верхнего Иртыша естественное воспроизводство сибирского осет-
ра оказалось предельно ограниченным. В результате гидрострои-
тельства осетр лишился обширных нерестовых угодий в Верхнем
Иртыше.
В то же время установлено, что гаметогенез у сибирского осетра
может нормально протекать в условиях зарегулированного стока;
следовательно, скат молоди осетра в Обскую губу и последующая
анадромная миграция производителей не обязательны для осу-
ществления процессов гаметогенеза. Это подтверждает присут-
ствие в водохранилищах самок с различным состоянием ооцитов
периода трофоплазматического роста. Наличие половозрелых сам-
цов из генерации, появившейся уже после закрытия плотины Усть-
Каменогорской ГЭС, является доказательством того, что и сперма-
тогенез в условиях водохранилищ проходит нормально. Следова-
тельно, есть основания говорить о возможности создания в
водохранилищах Иртыша промысловых стад осетра. Никакого био-
логического барьера, в форме стерильности взрослых особей, при за-
регулировании стока не возникает (Ерещенко, 1970).
Нерестовые миграции осетра начинаются со вскрытием льда в
водоемах. Поднимаясь с зимовальных ям, производители продви-
гаются к местам нереста. К нерестилищам приближаются при
прогреве воды до 9—11°.
Нерестилища располагаются на глубине от 1 до 9 м (наиболее
оптимальная 2—6 м), грунт гравийный, галечный, песчано-галеч-
ный, реже каменистый; скорость течения воды 2—4 км/ч; чистая
вода с хорошим насыщением ее кислородом; оптимальная нересто-
вая температура 14—16°, может происходить нерест и в диапазоне
12—18°. Сроки нереста — с половины мая, июнь, иногда продолжа-
ется и в июле. Осетр — требовательная рыба к условиям нереста,
нарушение одного из перечисленных экологических факторов при-
116
водит к отрицательным последствиям. Этот вид обладает очень
высокой чувствительностью к качеству воды. По наблюдениям
Н. П. Вотинова и др. (1975), при малейшем загрязнении ее в рай-
оне нерестилищ сточными водами бытовых и промышленных
предприятий, остатками лесосплава (кора, щепа и др.) производи-
тели не подходят к местам икрометания.
Плодовитость одноразмерного осетра из разных участков Ир-
тыша существенных различий не имеет. С увеличением возраста,
особенно размеров тела, индивидуальная плодовитость возрастает.
Корреляция плодовитости с длиной и массой тела всегда положи-
тельная. У самок из Усть-Каменогорского и Бухтарминского водо-
хранилищ массой от 15 до 30 кг индивидуальная плодовитость ко-
леблется от 163,2 до 390,4 тыс., средняя 290 тыс. икринок.
По данным М. И. Меньшикова (1947), индивидуальная плодо-
витость осетра в р. Иртыше равнялась 299 тыс. икринок при коле-
баниях от 174,1 до 419,8 тыс., относительная плодовитость на 1 кг
живой массы рыбы 12,6 (7—17).
Индивидуальная плодовитость иртышского осетра, по данным
Ф. Е. Богана (1938), равняется 211,2 тыс. икринок (146,4—
303,8 тыс.), относительная — 12,4 (9,0—15,9) при длине рыб от
137 до 146 см и массе от 14 до 19 кг. У самки, выловленной в 1891 г.
в оз. Зайсан, весившей 192 кг, масса икры равнялась 64 кг (Берг,
1948). Плодовитость ее, по мнению Н. П. Вотинова (1963), была
не менее 3—3,5 млн. икринок.
Икрометание у осетра единовременное. Икра после оплодотво-
рения опускается на дно и приклеивается к грунту. Зрелая икрин-
ка около 3 мм в диаметре, форма ее шарообразная пли яйцевид-
ная, цвет довольно различен — от светло-серого (иногда голубова-
того) до темно-коричневого.
Сроки развития икры зависят от температуры воды: при 13—15°
инкубация продолжается 7—9 суток. Выклюнувшиеся личинки
имеют длину около 10 мм, в первые дни жизни это слабые мало-
подвижные существа. Задняя часть тела личинки окружена тон-
кой прозрачной каймой, плавников нет. Большой желточный ме-
шок затрудняет активное плавание. Личинки время от времени
поднимаются в толщу воды на несколько сантиметров и затем пас-
сивно опускаются на дно. Во время подъемов они током воды раз-
носятся на значительные расстояния. Через 5—7 дней желточный
мешок рассасывается примерно на 2/з, появляются грудные плав-
ники, личинки становятся более подвижными и начинают захваты-
вать пищу. Полный переход на внешнее питание происходит на
12—15-й день после выклева. К этому времени оформляются спин-
ной, хвостовой, анальный плавники (Вотинов, 1958).
Скат молоди происходит частично в первый год жизни, но зна-
чительная ее часть задерживается в реке до 2—3, а отдельные
особи — до 5 лет.
Линейный и весовой рост сибирского осетра широко освещен в
научной литературе. По темпу роста осетр из Верхнего Иртыша
117
существенно не отличается от нижнеиртышского и обского. Досто-
верных различий в темпе роста у рыб, выловленных в разных
участках бассейна, установить нельзя, поскольку исследователи
имеют дело с единой проходной формой, к которой примешивается
определенный процент жилой, туводной и локальной. Все эти фор-
мы осетра по внешним признакам не различаются, и выводы де-
лаются на смешанном материале, что обусловливает большие ко-
лебания по длине и массе в одной возрастной группе. Крайние ва-
рианты в пределах одной возрастной группы отклоняются от
Таблица 41. Темп линейного роста сибирского осетра из бассейна Оби
и Байкала, см
Район Возраст, лет
1+ 1 2+ | 3± | 4+ | 5+ 1 6+ | 7+
Усть-Каменогорское вдхр (Чабан, 1962) 38,9 62,5 79,5 86,5 101,0 109,4 126,4
Иртыш у плотины Усть-Ка- меногорской ГЭС - 48,5 64,5 69,7 96,6 ——
Иртыш, Уватский р-н (Мень- шиков, 1936) 27,5 34,4 39,8 55,0 86,0
Обская губа (Винокуров, — цит. по Чабану, 1962) 40,5 50,4 57,9 60,6 63,9 71,5
Средняя Обь (Петкевич и др. 1950) 23,3 25,4 32,9 38,9 43,5 45,0 61,0
Байкал (Егоров, 1947) 22,7 33,2 42,2 47,9 54,8 58,7 61,3
Ангара (Боброва, — цит. по Чабану, 1962) — 66,5 80,5 88,2 89,7 95,5
средней на 10% и более. По исследованиям Н. П. Вотинова (1963),
у рыб в возрасте 27 лет размеры колеблются от 93 до 134 см, у
33-летних особей — от 102 до 147 см.
Нет строгой зависимости между размерами и массой осетра.
Например, размеры рыб массой от 9 до 10 кг колеблются от 90 до
114 см, а масса рыб длиной.в 115 —120 см изменяется от 11 до
18 кг. Существенно меняется соотношение между размерами и
массой рыб в зависимости от сезона, а также физиологического
состояния и пола рыбы.
В водохранилищах сибирский осетр растет очень интенсивно,
что детально прослежено на усть-каменогорском стаде. Быстрый
рост осетра в этом водоеме отмечен только на молодой его популя-
ции, т. е. у рыб, начавших свой жизненный цикл в водохранилище
или попавших сюда на нагул в раннем возрасте.
В целом темп роста усть-каменогорского осетра значительно
превышает таковой осетра не только из других участков Иртыша,
но и других водоемов Сибири (табл. 41). Подобная картина отме-
чается и по росту массы тела (табл. 42). Даже у быстрорастущего
осетра из Ангары в 7-летнем возрасте масса в два раза меньше,
чем у усть-каменогорского.
118
Высокий темп линейного и весового роста сибирского осетра в
Усть-Каменогорском водохранилище связан с благоприятными
условиями нагула этого вида, осваивающего площадь до предель-
ных глубин.
Питание и упитанность. По данным Г. А. Богданова (1959), в
Усть-Каменогорском водохранилище пища осетра состоит главным
образом из личинок хирономид и их куколок (соответственно 56,9 и
17,70% по массе, 100 и 50% по частоте встречаемости).
Таблица 42. Рост массы тела осетра из бассейна Оби и Байкала, кг
Район * Возраст, лет
1 + 2+ 34- 44- | 54- 64- 74-
Усть-Каменогорское вдхр. 0,24 1,40 2,50 3,90 5,80 6,50 -
Иртыш у плотины Усть-Каменогорской ГЭС — — .— 1,26 1,54 2,45 4,80
Обская губа — 0,26 0,57 1,02 1,07 1,18 1,78
Средняя Обь 0,06 0,11 0,13 0,24 0,41 0,48 1,27
Байкал 0,04 0,13 0,28 0,44 0,68 0,85 1,01
Ангара — — 1,20 2,30 2,80 3,30 —
* Данные тех же авторов, что и в таблице 41.
Значительную роль в питании осетра играют личинки ручейни-
ков, составляющие по массе 6,16%, встречаемости — 10%• Личинки
поденок, веснянок и стрекоз большого значения в питании не име-
ют. Наполнение кишечников высокое, средний индекс равен 193°/оо-
В оз. Зайсан в желудках взрослых осетров нередко встречается
рыба. Важное место в питании осетра занимают личинки комара-
толкунца.
Не выяснены вопросы о питании осетра зимой в водоемах
Верхнего Иртыша. В других участках Обь-Иртышского бассейна
молодь до наступления половозрелости интенсивно питается зимой.
По данным Н. П. Вотинова (1958, 1963), Ф. И. Боган (1938), в
Иртыше во время зимовки особи III и IV стадий зрелости не пита-
ются. Взрослые рыбы I и II стадий зрелости, зимующие после не-
реста в реке, а также молодь осетра питаются довольно интенсив-
но. В период нерестовой миграции и нереста производители не
едят. Наиболее интенсивно нагуливаются осетры летом и осенью.
Коэффициенты упитанности сибирского осетра Усть-Каменогор-
ского водохранилища колеблются в следующих пределах: по Фуль-
тону, от 0,42 до 0,68; по Кларк, от 0,30 до 0,52 (Чабан, 1962).
Места обитания, миграции, зимовка. В водохранилищах сибир-
ский осетр в нагульный период распространен повсеместно, захо-
дит в затоны, устья рек, встречается и на мелководных участках.
Будучи придонной рыбой, осетр в поисках пищи осваивает всю
площадь до предельных глубин: в Бухтарминском — 70, Усть-Ка-
меногорском— 42 м.
119
Нерестовые миграции происходили в мае—-июне, половозрелые
особи образовывали значительные концентрации. В настоящее
время ярко выраженного хода не наблюдается из-за малочислен-
ности производителей. В Бухтарминском водохранилище нересто-
вые миграции осуществляются в Черный Иртыш, незначителен
подход в устье р. Курчум.
Условия воспроизводства осетра в р. Курчум полностью нару-
шены. В Усть-Каменогорском водохранилище производители миг-
рируют вверх в зону подпора на речной участок. В силу изменив-
шихся гидрологических и термических условий нерест осетра в по-
следние годы здесь не наблюдается.
В озерной части Бухтарминского водохранилища осетр зимует
наямахумысов Бархот, Бакланий и Волчий. В Черном Иртыше —у
пос. Ордынка и у Алкабекской заставы. В горной части этого во-
доема места зимовок осетра неизвестны. По Белому и Быстрому
Иртышу до зарегулирования стока располагалось 10 зимовальных
ям, в настоящее время они попали в зону затопления.
В Усть-Каменогорском водохранилище известны две зимоваль-
ные ямы — в районе Семи Братьев и Лога Грохолова. Зимоваль-
ная яма в районе пос. Огневка у скалы Петух из-за загрязнения
отходами Белогорской обогатительной фабрики потеряла свое зна-
чение. Рыбы в ямах занимают определенное положение: рылом в
сторону течения воды. Старшие возрастные группы — впереди ста-
да, младшие сзади. Уход осетра из ям происходит в момент таяния
льда или несколько раньше (Чабан, 1962).
В Иртыше от Усть-Каменогорска до Семипалатинска имею-
щиеся зимовальные ямы из-за загрязнения реки потеряли свое
назначение (Ерещенко, 1969а). В Среднем Иртыше, по данным
многих исследований, зимовальных ям было мало; этот участок ре-
ки служил преимущественно миграционным путем для осетра.
Численность и хозяйственное значение. Статистические данные
по вылову осетра в Среднем Иртыше и до Усть-Каменогорска от-
дельно не приводятся. Имеющиеся литературные материалы осве-
щают вылов осетра в целом для Иртыша или Иртыша без ка-
захстанской части.
Начало промысла осетровых в Верхнем Иртыше от Малокрас-
ноярки вверх, включая оз. Зайсан, относится к 1822 г. В первой по-
ловине XIX в. зайсано-черноиртышское стадо осетровых было
довольно многочисленным. В 1825 г. здесь было выловлено около
3 тыс. осетров массой от 48 до 96 и даже 125 кг. Однако в резуль-
тате интенсивного лова запасы осетра быстро истощились.
В 1900—1909 гг. уловы их колебались от 53,7 до 6,3 ц. В 30-х годах
вылов осетра по оз. Зайсан с дельтой Черного Иртыша не превы-
шал 23 ц, с годами сокращаясь до 3—1 ц (Ерещенко, 1970).
По понятным причинам приведенные данные весьма приблизи-
тельно отображают фактическую добычу осетра. Например, по
официальной статистике, вылов осетра в Верхнем Иртыше, вклю-
кая оз. Зайсан и Черный Иртыш, в 1935 г. составил 11,8 ц.
120
Ф. В. Боган (1938) на основе тщательного анализа вылов осетра
только в Черном Иртыше за 1935 г. определяет в 30 ц. В после-
военные годы промышленный лов осетра в Верхнем Иртыше был
запрещен. Однако потребительский, браконьерский лов продолжа-
ется по настоящее время. Длительные миграции, относительная
легкость вылова разнообразными орудиями, высокая ценность до-
бычи делают осетра излюбленным объектом лова.
Сокращение численности зависит от ряда причин: интенсивного
промысла, сопровождающегося массовым выловом неполовозре-
лых рыб в местах их нагула и производителей на зимовальных
ямах и во время нерестовых миграций, потери нерестовых площа-
дей в результате гидростроительства на Иртыше, загрязнения бас-
сейна промышленными и хозяйственно-бытовыми сточными во-
дами.
В настоящее время численность осетра в Бухтарминском и
Усть-Каменогорском водохранилищах и речном участке Верхнего
Иртыша крайне ограничена, запасы его сильно подорваны. Назре-
вает необходимость реакклиматизации сибирского осетра в каскаде
водохранилищ Верхнего Иртыша.
Для сохранения этой ценной промысловой рыбы необходимо
проведение мероприятий по охране молоди на местах нагула, про-
изводителей на местах зимовки, миграционных путях и нерестили-
щах. Основной путь восстановления запасов осетра, поддержания
высокой численности его в бассейне Иртыша — искусственное раз-
ведение.
К искусственному разведению сибирского осетра приступили с
1956 г. Соответствующая биотехника разработана (Петкевич, 1957;
Бетинов, 1957, 1963а, б, 1975; Злоказов, 1972 и др.). В настоящее
время в Западной Сибири в широких масштабах получают молодь
осетровых на рыбоводных пунктах и заводах. В 1962—1965 гг.
сотрудники КазНИИРХ провели опыты по отработке биотехники
искусственного разведения осетра в условиях Усть-Каменогорско-
го и Бухтарминского водохранилищ; они дали положительные
результаты.
В плане рыбохозяйственного освоения Бухтарминского водо-
хранилища уловы осетра предусматривались в размере 3,4 тыс. ц
в год. При этом 96%промыслового стада должно было создавать-
ся за счет искусственного воспроизводства. Водохранилище осваи-
вается промыслом уже третье десятилетие, однако практически ни-
чего еще не сделано для воспроизводства стада осетра.
3. Acipenser stellatus Pallas — севрюга
Казахское название: шоцыр.
Синонимия. Acipenser stellatus: Pallas, Reise, v. I, 1771.
Acipenser helops: Pallas, Zoogr. Rosso—’Asiat, v. Ill [1814].
Acipenser stellatus: Берг, Фауна России, Рыбы, т. I, 1911. — Борзенко, Бюл.
Всекасп. рыбохоз. эксп., № 5—6, 1932. — Берг, Рыбы пресных вод, т. I, 1948.
Распространение. Обитает в бассейнах Каспийского, Черного и
Азовского морей. Наиболее многочисленное стадо в настоящее
время населяет бассейн Каспийского моря. На нерест идет глав-
ным образом в Урал, Волгу, Куру, Терек, Сулак и Самур. Отдель-
ные особи встречаются в Эмбе, Сумгаите, Астаре и Сефид-Руде.
В 30-х и 50-х годах были попытки акклиматизировать севрюгу в
Аральском море (Коновалов, 1966), но она там не прижилась.
Описание. Спинных жучек 9—16 (в среднем 13), боковых 26—
43 (в среднем 34), брюшных 9—16 (в среднем 11,2). Среднее чис-
ло жаберных тычинок на первой дуге 25,3. D 40—54 (в среднем
42,4), А 22—35 (в среднем 23). Хромосом 60 (Серебрякова, 1974).
Рис. 25. Севрюга
Тело севрюги (рис. 25) торпедообразно-стреловидной формы,
вытянутое, окраска спины до боковых жучек почти черная. В от-
дельные сезоны года в реку заходят особи, у которых черная
окраска в виде узкой полосы или черных пятен вдоль спинных
жучек, остальная часть верхней половины тела светло-серая,
брюшко белое. Рыло удлиненное, мечевидное, конец его слегка
вздернут. Длина рыла составляет 60% и более от длины головы.
Рот небольшой, в закрытом состоянии щелеобразный, при захва-
тывании пищи образует небольшой «хоботок», загнутый книзу.
Максимальная масса тела севрюги, выловленной в Урале
(1967 г.), равна 43,4 кг при длине 219 см.
Биология. Нерестовую миграцию из Южного и Среднего Кас-
пия севрюга начинает в мае, когда ее половые железы достигают
II и III—IV стадий зрелости. К этому времени, накопив достаточ-
ное количество энергетического материала (жирность IV—V бал-
лов), который откладывается в основном в тешке, на гонадах и в
мышцах, севрюга прекращает питаться и направляется на поиски
«родной» реки.
Первые мигранты с наименее развитыми половыми железами
появляются в дельте Урала на спаде половодья, обычно в середине
июня, при температуре воды 22—25°. В июне в низовьях реки за
122
сутки одной тоней вылавливается не более 1—3 мигрантов. Брюш-
ко у них твердое, половые железы не заполняют всю полость тела.
Ооциты мелкие, неправильной формы, окружены жировой тканью.
Анимальный полюс ооцитов («шапочка») невооруженным глазом
трудно различим. В 1 г икры насчитывается 123—-250 икринок.
Генеративная часть семенника плотная, розового цвета, сплошь
покрыта жиром, от которого ее трудно отделить. Гонадо-сомати-
ческий индекс самок низкий — 9,3. На гистологических препаратах
видно, что полярность ооцита не выражена, ядро с ядрышками по
периферии находится в его центре. Вокруг ядра имеется свобод-
ная от желтка зона цитоплазмы. Узкий слой мелкозернистого
желтка расположен по периферии под оболочками.
Ампулы семенника заполнены многочисленными делящимися
сперматогониями, клеточные элементы последующих фаз спермато-
генеза встречаются редко. На препаратах видны каналы сравни-
тельно крупных кровеносных сосудов. Производители с описанным
состоянием половых желез составляют озимую внутрипопуляцион-
ную нерестовую группу.
В период максимального прогрева воды в приустьевой области
моря и реке (26—28° — вторая половина июля) заход озимой сев-
рюги в реку ослабевает или прекращается. С августа ход озимой
севрюги усиливается и достигает максимума в конце этого месяца
или в первой половине сентября. Обычно он приходится на период,
когда уровни и расходы воды в реке падают до минимума, а про-
зрачность ее бывает максимальной. Температура воды в разгар
хода колеблется в пределах 26—23°. Заход озимой севрюги с на-
чала октября начинает ослабевать и во второй половине месяца
прекращается.
Состояние половых желез мигрантов озимой севрюги осеннего
хода значительно отличается от летнего. Так, анимальный полюс
ооцитов севрюг, заходящих в реку в сентябре и октябре, хорошо
различается невооруженным глазом, «шапочка» икринки приоб-
ретает характерный рисунок (чередование темных и светлых ко-
лец с белым центром), ооциты становятся округлыми. Генератив-
ная часть семенников составляет более половины гонады. Жира в
ястыках осенних мигрантов хотя и много, но меньше, чем в летнее
время (жирность III—IV и IV балла). Величина гонадо-соматиче-
ского индекса из-за трудности отделения жира от гонад имеет не-
которую погрешность (табл. 43).
Существенные изменения наступают и во внутреннем строении
клеток и структурных элементах половых желез. На гистологиче-
ских препаратах видно, что к осени происходит активное передви-
жение ядра в сторону анимального полюса, на котором накапли-
вается мелкозернистый желток. Ядро (с ядрышками по периферии
или в его центре) находится на границе крупно- и мелкозернистого
желтков. К этому времени заканчивается процесс формирования
микропиле, которые закрыты замыкательными клетками фоллику-
лярного эпителия. Ампулы семенников почти достигают конечных
123
размеров. Интерстициум растянут. Основная масса мужских по-
ловых клеток представлена сперматоцитами и сперматидами. Лишь
в отдельных ампулах семенника встречаются небольшие группки
сформированных сперматозоидов. К концу осеннего хода (октябрь)
половые клетки производителей озимой севрюги еще более близки
к зрелости, но еще далеки от конечных фаз спермато- и ооге-
неза.
Таблица 43. Состояние зрелости половых желез и масса ооцита у севрюги
р. Урал (М±т)
Д1есяц Средний гонадэ-соматический индекс Масса ооцита, мг
Самцы | п Самки п Средн. п
Июль 2,37±0,28 107 20,20±0,20 167 11,90+0,03 481
Август 3,05±0,07 533 13,27±0,01 324 9,33 + 0,02 198
Сентябрь 3,81±0,16 1104 14,15+0,06 449 9,68 + 0,06 199
Октябрь 3,30+0,10 228 16,86±0,16 147 11,10+0,22 40
Апрель 5,43 + 0,03 3059 22,65±0,01 1305 12,54 + 0,03 875
Май 5,25+0,03 3815 22,88±0,01 4600 12,12+0,01 2568
Июнь 4,47±О,08 1176 23,55±0,04 2455 11,83± 0,06 801
В среднем за год 3,95 + 0,02 10032 20,38 ±0,05 9447 11,91± 0,01 5162
Половые железы производителей, выловленных перед ледоста-
вом на зимовальных ямах средней части нерестовой зоны (500—•
600 км от устья), находятся примерно в таком же состоянии, как и
у мигрантов позднего осеннего хода в низовьях реки, однако жи-
ра как в тешке (в мышцах), так и в гонадах севрюг из зимоваль-.
ных ям меньше, чем у производителей, выловленных в дельте. Это
свидетельствует о том, что во время миграции продолжается про-
цесс дозревания половых желез, и накопленный энергетический
материал, отложенный в разных частях тела, интенсивно расхо-
дуется.
При снижении температуры воды в реке до 6—-7° заход севрюги
в реку прекращается. Таким образом, озимая севрюга начинает
входить в реку при сравнительно высоких скоростях течения еще
мутной воды и почти максимальных температурах. Заканчивается
ее ход при совершенно противоположных условиях: минимальных
скоростях течения, максимальной прозрачности и низкой темпера-
туре воды, что дает основание характеризовать ее как эврибионт-
ный вид.
Сокращение объема стока за последние 15 лет сместило начало
захода производителей яровой севрюги на более ранние сроки.
Первые ходовые производители севрюги в низовьях Урала после
длительного зимнего перерыва (5,0—5,5 месяца) появляются в
дельте в последних числах марта (единицы) —первой декаде ап-
реля при температуре воды 3—4°, т. е. задолго до начала поло-
124
водья, чего не наблюдалось в 60-х годах, когда объем безвозврат-
ного водопотребления и численность севрюги были гораздо
меньше.
С момента появления первых мигрантов в низовьях реки ход
севрюги быстро усиливается и через 15—25 дней достигает своего
максимума. В последние 10 лет пик хода севрюги в низовьях реки
наблюдается в конце второй-третьей декады апреля. В начале
хода в уловах преобладают самцы, а в разгаре и во второй поло-
вине весеннего хода — самки. Аналогичное явление наблюдается и
во время летне-осенней миграции. Ход яровой севрюги в ослаблен-
ном виде продолжается до конца июня. Во второй половине этого
месяца ходовых самцов в низовьях реки практически нет. В нача-
ле июля в уловах встречаются лишь единичные самки.
Состояние половых желез мигрантов весеннего хода резко от-
личается от такового летнего и осеннего хода. Так, внутриполост-
ного жира у самок и самцов нет, тешка тонкая. Икринки, достиг-
шие дефинитивных размеров, легко отделяются от стромы яични-
ка, имеют округлую или полуовальную форму, анимальный полюс
различим невооруженным глазом. Семенник имеет молочно-белую
или бледно-розовую окраску; на его разрезе выделяются капельки
спермы, при активизации водой сперматозоиды приобретают ак-
тивность. Анальное отверстие у отдельных самок (особенно во вто-
рой половине мая) воспалено.
На гистологических препаратах видно, что оптически пустое
или с ядрышками в центре ядро находится в зоне мелкозернистого
желтка анимального полюса, вблизи оболочек у микропиле, обо-
лочка ядра имеет неправильную форму, из ядра выфильтровывает-
ся кариоплазма. Ампулы семенников сплошь заполнены сформи-
рованными сперматозоидами. После гипофизарной инъекции почти
100% производителей переходят в текучее состояние. Таких произ-
водителей мы относим к ранней яровой внутрипопуляционной не-
рестовой группе (Песериди, 1966).
Сравнение массы ооцитов ранней яровой севрюги, выловленной
в дельте реки и на нерестилищах, показывает, что увеличения раз-
меров яйцеклеток в период речной миграции не происходит (Песе-
риди, Исламгазиева, 1967).
Характерно, что севрюги, заходящие в реку в апреле, имеют
темную окраску спины. Начиная с середины мая в дельте реки
появляются особи со светлой окраской спины. Брюшко у них твер-
дое, генитальное отверстие не воспалено, в тешке имеется неболь-
шая прослойка жира. При изгибе тела самцов выделения спермы
не происходит.
Ооциты хотя и округлой формы, но они мельче, чем у самок
ранней яровой группы. На срезах видно, что ядро с ядрышками по
периферии находится на границе мелкозернистого и крупнозернис-
того желтков. В ампулах семенников наблюдаются завершающие
фазы сперматогенеза. Основными клеточными элементами являют-
ся сперматиды и небольшие группки сформированных сперматозои-
125
дов. По состоянию зрелости половых клеток они схожи с особями
озимой севрюги, заходящими в реку глубокой осенью или зимовав-
шими в реке. Мы их относим к поздней яровой севрюге. Заход ее
в реку начинается либо в пик паводка, либо вскоре после него при
температуре воды 12—14° и продолжается обычно до начала июля,
а в маловодные годы — вплоть до его середины. Ход поздней
яровой севрюги не имеет ясно выраженного пика и происходит на
спаде хода ранней яровой, отдельные производители которой, за-
ходящие в реку в мае, также имеют светлую окраску спины.
Основная масса озимой севрюги, как и яровой, до начала па-
водка заходит в р. Урал через судоходный канал и Золотой рукав.
В пик осеннего хода в Яицком рукаве встречаются лишь единичные
экземпляры. С начала подъема воды в реке и массового весеннего
хода уловы севрюги в Яицком рукаве возрастают и достигают
максимума в пик половодья. В это время наблюдается заход сев-
рюги и через рыбоходный канал западной части дельты. По мере
спада паводка значение Яицкого рукава как миграционной трассы
падает. Доля озимой и поздней яровой севрюги от всей нерестовой
части популяции в сумме составляет не более 10—15%.
Озимая севрюга с момента захода в дельту и до нереста пре-
бывает в реке в течение 7—11 месяцев. Движется она медленно —
7—11 км в сутки. По пути к местам зимовки часто останавливается
на отдых в ямах, заводях и суводях, где пребывает по нескольку
(3—5) дней. Зимует в основном в глубоких ямах верхней части не-
рестовой зоны — от г. Чапаеве до г. Уральска (650—800 км от
взморья). Мигранты осеннего хода зимуют в ямах средней части
нерестовой зоны —от пос. Красные Яры до г. Чапаеве (370—
600 км). Единичные экземпляры вылавливались нами на ямах ниж-
него течения реки у пос. Махамбет (Бакланья яма —130 км).
После вскрытия реки озимая севрюга покидает ямы и продолжает
свою миграцию в поисках нерестилищ (Песериди, 1971). Подни-
мается она вверх по реке до г. Уральска (850 км). Отдельные
особи доходят до пос. Рубежка (950 км).
Продолжительность пребывания ранней яровой севрюги в реке
с момента ее захода в дельту и до нереста не превышает 2,0—
2,5 месяца. Доходит она до середины верхней части нерестовой
зоны — пос. Кушум (770 км). Отдельные особи встречаются у
г. Уральска. Движется ранняя яровая севрюга вверх по реке
сравнительно быстро —15—30 км в сутки, избегая сильных пото-
ков и придерживаясь малых глубин. Самцы движутся быстрее
самок и раньше их достигают мест нереста. Мелкие особи мигри-
руют в прибрежной зоне, крупные —ближе к фарватеру и в тол-
ще воды, периодически поднимаясь в верхние слои. По мере при-
ближения нерестового периода самки все чаще выскакивают из
воды («бьют свечи»), ударяясь брюхом о воду.
Передвигаясь вверх по реке, яровая севрюга делает частые
остановки. Максимальное время пребывания в потоке со ско-
ростью течения более 1 м/с не превышает 3 ч (Сливка, 1974). Оста-
126
навливается она на отдых в основном в дневное время в заводях
и суводях, где накапливается в большом количестве. Наиболее ин-
тенсивная плавательная активность у севрюги проявляется в су-
мерках и ночью. В дельте реки это явление не наблюдается.
Характер миграционного поведения поздней яровой севрюги
примерно такой же, как и ранней яровой, за исключением того, что
она не поднимается так высоко вверх по реке, достигая лишь
г. Чапаеве (650 км), и делает более продолжительные остановки
во время миграции. Продолжительность пребывания поздней яро-
вой севрюги в реке не более 2 месяцев. Самки ранней и поздней
яровой севрюги нерестуют «сходу», а самцы находятся на нерести-
лищах в течение почти всего нерестового периода (1,5—2 месяца).
В результате в разгар нереста на одну самку приходится 6—8 сам-
цов, в конце нереста соотношение полов достигает 1:12—1:15.
Самцы эякулируют сперму до 5—6 раз за сезон, у самок икроме-
тание тотальное. Дольше всех в реке задерживаются самцы ози-
мой и ранней яровой севрюги. Первые скрещиваются как с самка-
ми озимой, так и ранней яровой севрюги, а самцы ранней яровой —
со своими «соплеменницами» и самками поздней яровой севрюги.
Озимая севрюга нерестится при температуре воды 13—17° в
пик паводка или в начале его спада при скоростях течения 1,3—
1,7 м/с и предельной мутности воды. Нерест ее начинается на всем
участке (от г. Чапаеве до пос. Рубежка 650—950 км) одновременно
и продолжается 10—15 дней. Численность севрюги, размножаю-
щейся выше г. Уральска невелика. Размножается озимая севрюга
на самых эффективных нерестилищах, ложе которых устлано галь-
кой и гравием с примесью крупнозернистого песка. Икру отклады-
вает на обширных затопляемых грядах на глубине 6—10 м. Плот-
ность кладок икры невысокая — 50—70 шт/м2, а на некоторых
нижних нерестилищах — 10—15 шт/м2.
После завершения нереста озимой севрюги (обычно через 15—
25 дней) на грядах верхней (а в основном средней и нижней)'
части нерестовой зоны начинается нерест ранней яровой севрюги.
Размножается она при диапазоне температур 15—25°. Нерест
продолжается с мая до начала июля. Откладывает икру на
всех видах нерестового субстрата на глубине 3—7 м — га-
лечниках, гравийно-песчаном грунте (верхняя часть нерестовой
зоны), плотных песках, сцементированных песчаниках, плот-
ных неразмываемых глинах с вкраплениями створок моллюсков
(средняя часть нерестовой зоны), плотных незаиливаемых мелко-
зернистых песках и на песчаных плесах с порослью ивы и даже
на временно не действующих промысловых тонях (нижняя часть
нерестовой зоны и дельта). Плотность кладок икры на верхних
нерестилищах колеблется от 50 до 800 шт/м2, на нерестилищах в
районе г. Чапаеве и пос. Индер — 0,5—2,5 тыс. шт/м2. Средняя
плотность кладок в разгар нереста 1000 шт/м2 (Песериди, Верина,
1981).
Верхняя граница размножения поздней яровой севрюги нахо-
127
дится в 650 км от взморья (г. Чапаеве). Нерестится она при тем-
пературе воды 20—26° в основном на русловых грядах на спаде
половодья при скоростях течения 1,2—0,8 м/с. Запоздалые самки
нерестятся на самых нижних грядах вплоть до дельты. Плотность
кладок икры поздней яровой севрюги невысокая — 50—100 шт/м2.
Наблюдениями установлено неоднократное использование од-
ной и той же гряды за один сезон, что обусловлено растянутостью хо-
да и постепенным подходом производителей к нерестилищам. Эм-
бриональное развитие при наиболее низких температурах (15°)
длится около 6 суток. При температуре 25° эмбрионы выклевыва-
ются через 55—60 ч.
Отнерестовавшие самки севрюги в дельте реки появляются
обычно в середине мая. Пик ската в июне, отдельные покатные
самцы встречаются в уловах до конца июля. Скат проходит по
фарватеру со скоростью до 75 км/сутки, головой против течения.
Наибольшее истощение наблюдается у крупных самок, которые
теряют до 30% и более массы тела. Посленерестовая смертность
незначительна.
Дрифт личинок и скат молоди. Сроки, количественные и качест-
венные показатели дрифта предличинок, личинок и ската молоди
севрюги колеблются по годам и находятся в прямой зависимости
от времени и мест нереста, термического и гидрологического ре-
жимов реки, численности пропускаемых производителей к местам
размножения и т. п. Осредненная схема сроков ската потомства
разных внутрипопуляционных нерестовых групп выглядит сле-
дующим образом.
Первые покатные предличинки и личинки севрюги (5—17 мм) в
дельте Урала появляются во второй или третьей декаде мая. Од-
новременно с личинками вылавливается и молодь, перешедшая на
активное питание (43 мм). К концу мая в отдельные годы в уловах
встречается молодь размером 29—56 мм и массой 160—525 мг.
В первой декаде июня средняя длина колеблется в пределах 21 —
43 мм при средней массе 88—307 мг. Массовый скат обычно идет
весь июнь. Средняя масса молоди к концу месяца достигает 1,2—
1,3 г. В конце июня наряду с крупной молодью (до 5,5 г) вылавли-
ваются и личинки (первые — потомство озимых, вторые — поздних
яровых). В начале июля, в конце ската молоди озимой и яровой
севрюги, средние навески несколько снижаются за счет прилова
потомства поздней яровой севрюги. В динамике ската наблюдаются
два явно выраженных пика: первый — первая и вторая декады
июня, второй — конец июня — первая декада июля (табл. 44).
Наиболее крупные экземпляры молоди (184—192 мм и 15,6—22 г)
скатываются в конце июля — августе. Незначительная часть моло-
ди остается на зимовку в реке (Бекешев, Песериди, 1966; Песери-
ди, Бекешев, 1967; Тарабрин, 1981).
Дрифт личинок происходит по всему поперечнику реки. Под-
росшая молодь сплывает по стержню, периодически выходя на заи-
ленные плесы или скапливаясь на ямах и участках реки с замед-
128
ленным течением. В многоводные годы (свыше 7 км3) скатываю-
щаяся молодь имеет более крупные размеры (табл. 45), чем в
маловодные, что определяется изменением условий покатной миг-
рации.
Яйца севрюги (до 4 мм по продольной оси) гораздо мельче,
чем у других осетровых. Поэтому они доступны щиповке, бычкам,
белоглазке, густере, лещу, сому, стерляди, а также молоди белуги,
шипа и осетра. Особое место в истреблении развивающейся икры
севрюги занимает пескарь (до 15 икринок в желудке), численность
которого в Урале колоссальна. Предличинки и личинки севрюги
Таблица 44. Динамика ската личинок и молоди севрюги р. Урал
по пятидневкам (средние уловы на одно пятиминутное траление
бимтралом), шт.
Год Май Июнь Июль
3 4 | 5 j 6 • 2 3 4 5 3 1 1 2 3
1977 12 91 2282 2293 1354 214 148 3 5,о' 1 2,0 3
1978 —• 91 147 711 345 33 ПО 131 188 97,0 81 3,5 2
1979 62 99 197 32 7 7 10 21 64 9,0 16 35,0 5
1980 —• 59 111 157 328 55 41 89 100 68,0 10 15,0 1
1982 6 1 19 44 99 97 78 90 19 0,4| 0,3 — —.
Таблица 45. Количественные и качественные показатели
ската молоди севрюги р. Урал в разные по водности годы
Год Эбъем стока р. Урал, км® Средний улэв мол ЭДИ на трал, шт. Кол-во ска- тившейся молоди в мо- ре, млн. шт Средняя на- веска моло- ди, мг
1961 6,24 47,0 129 1125
1962 7,44 43,2 130 1170
1963 9,80 26,1 141 1194
1964 11,52 17,9 137 1645
1965 6,24 50,0 169 1250
1966 11,16 16,7 123 1156
1967 3,01 250,0 124 405
1968 5,10 127,0 183 652
1969 5,83 124,8 270 596
1970 12,80 21,0 151 1422
1971 12,50 27,3 197 930
1972 8,48 129,7 409 330
1973 4,87 80,0 105 270
1974 7,64 49,6 258 310
1975 3,56 374,2 170 270
1976 4,20 134,8 213 270
1977 2,54 559,1 377 260
1978 5,30 216,6 422 270
1979 6,25 48,8 222 313
1980 6,03 103,0 389 350
1981 9,22 37,6 106 1100
1982 6,00 38,4 94,8 212
9—7
/
129
истребляются чехонью, синцом, уклеей и сельдями. Наиболее опас-
ный враг молоди севрюги — сом. В период массового ската в 12—
34% его желудков обнаружена молодь севрюги массой 2—3 г (до
18—21 шт.). Ежегодный ущерб, наносимый сомом рыбному хозяй-
ству, составляет более 650 т севрюги (Песериди, 1966; Войнова,
Бекешев, Невлюдова, 1969; Песериди, Войнова, 1969; Войнова,
1972, 1973; Тарабрин, 1981а).
Созревание, возрастной состав нерестовой части популяции,
рост. Единичные самцы севрюги (0,3—2,5% нерестовой части попу-
ляции) созревают в возрасте 4—6 лет. Массовое созревание насту-
пает в 7—9-летнем возрасте (7—18%), на 9—15-леток приходится
Таблица 46. Возрастной состав севрюги р. Урал, %
Воз- раст, лет 1961—1963 гг. 1967-1969 гг. 1979 г.
Са bi- ll ы Сам- ки Оба пола Сам- цы Са м- ки Оба пола Сам- цы Сам- ки Оба пола
4 0,6 , 0,3 0,05 0,03 __
5 0,9 0,4 0,2 — 0,1 0,05 —. —
6 1.0 — 0,5 0,4 —- 0,2 0,05 —. —
7 4,6 0,2 2,2 1,2 0,07 0,64 0,2 0,1 0,1
8 6,5 0,8 3,8 4,5 0,55 2,54 2,0 0,3 1,2
9 7.0 2,5 4,5 10,8 1,83 6,4 4,6 0,7 2,8
10 10,7 3,3 6,6 13,2 3,9 8,6 11,2 2,7 7,2
11 17,4 9,0 12,6 16,5 5,9 11,3 16,0 4,5 10,1
12 16,1 13,5 14,5 16,0 10,5 13,2 20,0 9,0 15,1
13 16,4 12,5 14,1 15,4 11,8 13,6 16,3 14,8 15,6
14 12,5 15,8 14,1 9,5 16,7 13,0 12,5 14,4 13,4
15 4,4 13,4 9,1 5,8 15,6 10,7 7,5 16,0 11,5
16 1,2 10,8 6,3 3,1 12,4 7,7 5,4 10,5 7,8
17 0,7 9,9 6,1 2,0 7,9 4,9 2,3 12,4 7,0
18 — 5,6 3,1 0,6 6,1 3,3 2,0 7,5 4,4
19 — 1,3 0,9 0,5 3,1 1,8 0,1 4,7 2,2
20 —- 0,6 0,4 — 2,1 1,05 — 1,6 0,7
21 — 0,8 0,5 0,2 0,53 0,4 — 0,8 0,4
22 — — — — 0,48 0,24 .— —. -
23 —. —. — 0,05 0,2 0,14 — —. —
24 — —. —- — 0,2 0,1 —- —
25 — — — — 0,14 0,06 — — —
п 686 816 1502 1497 1461 2958 1102 962 2064
77—83%. Половина из них — впервые созревающие особи, а ос-
тальная часть — нерестующие второй и частично третий раз. Сам-
цов старше 15—16-летнего возраста в нерестовой части популяции
немного — 6—10%. Повторное созревание самцов наступает через
2—3 года, что установлено меченьем. Так, текучий самец, помечен-
ный на нерестилищах р. Урал 7 мая 1976 г., был вторично пойман
через 14 месяцев под Волгоградской плотиной с половыми железа-
ми в стадии зрелости III—IV. Более старые самцы нерестуют в
4 года один раз.
Самки созревают позже самцов на 3—4 года, т. е. в 7—8-летнем
130
возрасте, но таких рыб в популяции мало (около 1%)- Основная
масса самок становится половозрелой в И—13-летнем возрасте
(35—39% нерестовой популяции). На долю 11—18-леток прихо-
дится до 89% половозрелых самок. Особей старше 18 лет в популя-
ции мало — 3—7% (табл. 46). Самки на повторный нерест заходят
в реку через 3—5 лет, но в массе — через 4 года, что также под-
тверждает мечение.
На возрастную структуру нерестовой части популяции решаю-
щее значение оказывает промысел. До прекращения сетного и не-
водного лова в море (1964 г.), где изымались более крупные и
быстрорастущие рыбы, в реку заходили мелкие и тугорослые особи.
Таблица 47. Средние размеры (см) и масса тела (кг) севрюги
(оба пола) р. Урал по возрастным группам
Возраст, лет 1961—1963 Гг. 1967-1969 гг.
см кг п см КГ п
4 87,4 1,65 4 90,2 1,77 1
5 88,3 1,73 6 94,0 2,15 3
6 93,8 1,91 7 101,1 2,63 6
7 104,5 3,21 34 108,2 3,48 19
8 112,3 3,82 51 117,5 4,45 75
9 116,4 4,65 68 122,9 4,96 189
10 123,0 5,72 101 127,4 5,88 254
11 129,5 6,30 190 131,6 6,83 333
12 132,3 6,92 221 138,1 7,54 392
13 135,8 7,41 215 141,6 8,31 403
14 141,7 8,74 215 145,8 9,62 386
15 144,5 9,80 139 149,3 10,85 315
16 148,3 11,25 96 152,4 12,2 227
17 152,8 12,80 86 156,1 13,41 146
18 157,1 13,60 46 158,5 14,7 98
19 158,8 14,70 11 162,1 15,6 54
20 159,5 16,40 5 164,3 16,4 31
21 163,6 16,83 6 167,2 17,3 И
22 —. — .— 168,1 17,5 7
23 —. — —- 172,3 18,4 4
24 — — — 170,0 17,6 3
25 — — — 198,0 36,0 1
Среднее 135,16 7,95 1501 143,05 8,98 2958
После запрещения рыболовства в море структура нерестовой части
популяции приняла иной вид: возросла доля старших возрастных
групп, увеличились размеры и масса тела заходящих в реку рыб
(табл. 46, 47). Интенсификация речного промысла в середине 70-х
годов вновь изменила структуру популяции: возрастной ряд стал
более компактным.
В раннем возрасте наблюдается наиболее быстрый рост севрю-
ги в длину. За первые 6—7 лет она вырастает до метра. До наступ-
ления половозрелости темп роста самцов и самок примерно одина-
ков. Затем равновесие нарушается в пользу самок, которые в 14—
131
6-летнем возрасте крупнее самцов в 1,3—1,6 раза. Замедленный
jct самцов, по-впдимому, связан с ранним наступлением полово-
эелости (табл. 48).
Таблица 48. Средняя масса тела севрюги по размерным группам, кг
Размерная г0уппа,см Самцы п Самки п Оба пола п
До 100 1,72 32 2,96 4 2,89 36
100-110 3,43 467 3,63 54 3,37 521
110-120 4,34 2393 4,99 320 4,50 2713
120-130 5,51 5096 6,60 1175 5,62 6271
130-140 7,07 4393 8,19 2402 7,51 6795
140-150 8,28 1344 10,01 3332 9,52 4676
150-160 10,25 268 12,П 2439 12,92 2707
160-170 12,36 58 14,15 1187 14,06 1245
170-180 13,94 9 16,17 380 16,12 389
180-190 13,79 5 19,33 54 18,87 59
190-200 — — 21,25 8 21,25 8
200-210 — — 22,48 4 22,48 4
Среднее 6,12 14065 10,24 11359 7,97 25424
Сравнительная характеристика возрастного состава уловов
волжской и уральской севрюги показывает, что в Урале больше
процент младших возрастных групп, но темп роста волжской и
уральской севрюги примерно одинаков (Песериди, Бекешев, 1966;
Песериди и др., 1972). В литературе имеются сведения о том, что
севрюга доживает до 60 лет и достигает массы 63 кг (Державин,
1922, 1949). Такие особи в настоящее время не встречаются.
Половой состав. Н. А. Северцов (1863) и М. П. Борзенко (1942)
приводят данные о численном преобладании самцов севрюги в
уральских водах (88,6—75,5%), считая, что это явление обусловле-
но перехватом икряных рыб на путях к реке ставными сетными
порядками.
По 1964 г. включительно морская промысловая мера была
крупнее, чем речная, и, как показали исследования, охраняла лишь
12% самок и 56% самцов (Песериди, 1966), в результате чего в
реке должны были преобладать самки. Однако соотношение полов
в реке до 1965 г. было примерно равным:
Год Са м- цы Сам- ки Год Сам- цы Сам- ки Год Сам- цы Сам- ки
1960 56 44 1968 65 35 1976 49 51
1961 52 48 1969 66 34 1977 51 49
1962 55 45 1970 62 48 1978 52 48
1963 55 45 1971 66 34 1979 46 54
1964 49 51 1972 66 34 1980 37 63
1965 63 77 1973 66 34 1981 35 65
1966 73 27 1974 40 60 1982 35 65
1967 70 30 1975 45 55 1983 40 60
132
Численное преобладание самцов в 60-х годах прошлого столе-
тия и в начале 30-х текущего мы объясняем слабым прессом про-
мысла и более благоприятными условиями воспроизводства и
нагула.
Прекращение морского красноловья и отмена промысловой ме-
ры с 1965 г. незамедлительно сказались на увеличении численности
самцов, в результате чего половой состав нерестовой части попу-
ляции принял нормальный вид: 2 самца на одну самку. Чрезмерная
эксплуатация запасов севрюги с начала 70-х годов вновь нарушила
установившийся половой состав популяции севрюги, и в последние
годы (1980—1983) стал ощущаться явный дефицит самцов.
Плодовитость севрюги варьирует в пределах от 53 до 916 тыс.
икринок. Несмотря на положительную корреляцию средней плодо-
витости с возрастом и ростом рыбы, во всех размерных и весовых
группах наблюдаются вариации в широком диапазоне (табл. 49
и 50). В одной и той же категории рыб максимальная плодовитость
превышает минимальную в 4—5 раз.
Таблица 49. Абсолютная плодовитость севрюги р. Урал
по размерным группам, ,тыс. шт.
Размерная группа, см Плодовитость п Размерная группа, см Плодовитость п
От—до М От—до М
До 100 53-89 72 3 151-160 88-475 240 1174'
101-110 61-120 92 5 161-170 121-511 277 653
111-120 58-220 119 84 171-180 174-856 328 292
121-130 69-287 149 362 181-190 265-916 523 34
131-140 78—309 165 811 191-200 461-865 701 5
141-150 82-414 198 1163 90-200 53-916 237 4596
Таблица 50. Абсолютная плодовитость севрюги р. Урал по весовым
группам, тыс. шт.
Масса тела, кг Плодовитость п Масса тела, кг Плодовитость п
От—до | М От—до М
До 4 53-219 104 41 14-16 138-384 296 382
4-6 68-265 129 289 16-18 182-511 337 181
6-8 78-314 154 849 18-20 244-856 425 74
8-10 71-368 183 982 20-22 43-916 539 17
10-12 89-415 218 1121 22-24 372- 865 535 14
12-14 93-475 263 646 2-24 БЗ—916 237 4596
Средняя абсолютная плодовитость по многолетним данным
равна 237 тыс. икринок. Она изменяется из года в год (табл. 51)
под влиянием промысла на возрастную и размерно-весовую струк-
туры популяции. По числу воспроизводимых яиц уральская севрю-
га близка к волжской, но плодовитее, чем куринская. Самец севрю-
ги массой 7 кг за нерест (5—6 эякуляций) выделяет до 6,0—7,0-1010
спермиев.
133
Питание, конкуренты. Переход личинок севрюги на активное
питание начинается на 5—9-е сутки после выклева при длине
18—19 мм. В пищу используются бентосные организмы на самых
ранних стадиях развития, размеры которых 1—2 мм, олигохеты,
полихеты, корофииды, гаммариды, мизиды, личинки и куколки хи-
рономид (в нерестовой зоне). Придонные формы циклопов в пище
личинки севрюги встречаются незначительно. На одну личинку
севрюги длиной 20—22 мм приходится в среднем 2,0—4,3 молоди
гаммарид, 0,2—0,6 экз. личинок хирономид, 0,2—1,6 олигохет и
0,08 циклопов.
Содержание пищеварительных трактов молоди севрюги (сред- няя длина 63 мм) из нижнего течения Урала состоит из личинок хирономид Таблица 51. Средняя абсолютная (44%), МИЗИД (22%), ПОде- плодовитость (АП) севрюги р. Урал нок (19%) и корофиид в разные годы, тыс. ШТ. (8%). Второстепенной пи-
Год АП Год АП Год щей служат имаго насеко- Ап мых (3%), личинки стрекоз
1961-1962 1963 1964 1965 1966 1967 1968 размера- фиидамг ков длин 211 213 218 208 195 209 211 ИИ МО , гамл ой до 1969 1970 1971 1972 1973 1974 1975 чоди гаридг 10 см. 197 225 228 222 208 228 I потр 1МИ, м 1976 1977 1978 1979 1980 1981 1982 ебляе изида 218 тех или иных групп организ- 236 мов (18 компонентов) в пи- 251 тании молоди на разных ста- ^55 циях неодинакова. Индексы 253 наполнения пищеваритель- 259 ных трактов молоди высо- кие — 104—133°/оо- Четкая прямолинейная связь между мыми ими ракообразными (коро- ми) проявляется только у сеголет-
По мере роста молоди интенсивность питания возрастает. Сред-
ний индекс наполнения желудка достигает 162°/000 (колебания от
2,5 до 640). В первый месяц пребывания молоди в море она питает-
ся в основном бокоплавами (89% массы пищи). Рацион молоди
крупнее 10 см состоит главным образом из мизид (56—70%). У мо-
лоди севрюги выражена суточная периодичность питания: в реке
она наиболее интенсивно питается в ночное время (22—24 ч), а в
-море — днем (12—16 ч).
Характер питания неполовозрелой севрюги меняется по мере
роста, по сезонам года и местам нагула. В восточной части Север-
ного Каспия в апреле севрюга размером до 80 см питается в основ-
ном корофиидами (до 39% массы пищи), кумацеями (30%) и не-
реис (28%). У более крупной севрюги (80—100 см) спектр корма
примерно такой же, но роль нереис падает до 6%. Половозрелая
севрюга (крупнее НО см), ранней весной поедает также корофиид
(26%), кумацей (36%), гаммарид и мизид (17%). В августе и
осенью основу пищи особей до ПО см составляют корофииды
134
(33—100%), гаммариды и мизиды (60%), частично — крабы
(до 4%).
Созревающая и половозрелая севрюга у Мангышлакского по-
бережья питается в основном ракообразными (корофииды — 53%,
гаммариды 13%), рыбами (27%). На долю хирономид, моллюсков
и других объектов питания приходится 7%. В последние годы сев-
рюга размером 50—ПО см в восточной части Каспия стала уси-
ленно питаться крабом. В желудках 25% севрюги встречается краб
в значительных количествах (до 30—50% массы пищевого комка).
У восточных побережий Среднего и Южного Каспия севрюга
питается в течение всего года, но зимой интенсивность питания
ослабевает. Перед началом нерестовой миграции, накопив необхо-
димое количество жира, она прекращает прием пищи. По заверше-
нии нереста и ската в море вновь усиленно питается.
Основные конкуренты молоди севрюги в реке — пескарь, бычок-
песочник, пуголовка, молодь леща, густеры и воблы. Степень пи-
щевого сходства с пескарем, бычком-песочником и пуголовкой в
период активного ската молоди достигает 50—93%• В Северном
Каспии степень пищевого сходства севрюги с осетром равна 26,
воблой — 5,4, лещом — 17,6% (Желтенкова, 1964, 1967; Тарвер-
диева, 1968; Зачетнова, 1972, 1976; Полянинова, 1972; Полянинова,
Кашенцева, Маленникова, 1981; Стыгар, 1981; Стытар, Песериди,
Батина, 1981).
Миграции и распределение в море. Скатившись в море, молодь
севрюги непродолжительное время пребывает в опресненном при-
устьевом пространстве, а затем начинает расселяться по восточной
половине Северного Каспия. Незначительная часть молоди мигри-
рует вдоль северной границы Уральской бороздины на запад Се-
верного Каспия, придерживаясь илистых и песчано-илистых грун-
тов, богатых гаммаридами и корофиидами. Обойдя Уральскую
бороздину с севера, часть молоди направляется к западным бере-
гам Северного Каспия, а другая устремляется в сторону п-ова Буза-
чи, Казахского залива и восточных берегов Среднего Каспия.
Основная часть молоди уральского происхождения из опреснен-
ного приустьевого пространства мигрируют вдоль свала глубин с
восточной стороны Уральской бороздины, направляясь к п-ову Бу-
зачи и Казахскому заливу. У побережий междуречья Волга — Урал
подрастающая молодь встречается редко. На Уральской бороздине
ювенильная севрюга появляется лишь во второй половине лета.
Обогнув Казахский залив, часть подрастающей севрюги направ-
ляется вдоль границы Северного и Среднего Каспия к западным
берегам моря, но основная масса нагуливается у казахстанских
берегов Среднего Каспия, двигаясь вдоль побережья на юг.
Изложенная схема миграции не означает, что все подрастаю-
щее поколение севрюги покидает Северный Каспий и нагуливается
только на юге моря. В Северном Каспии в течение всего года раз-
новозрастная, в том числе созревающая и нагуливающаяся сев-
рюга встречается повсеместно (табл. 52).
135
Отнерестовавшая севрюга мигрирует по морю, вероятно, по
тем же маршрутам, что и нагуливающаяся. Так, 2 севрюги, поме-
ченные у устья Урала 13 августа и 2 сентября 1977 г., были пой-
маны иранскими рыбаками у порта Пехлеви 2 марта 1978 г. Метки
нам возвратил директор Иранского научно-исследовательского ин-
ститута организации рыбного хозяйства X. Эмади.
Третья помеченная севрюга была поймана у г. Ленкорань
АзССР через 9 месяцев 19 дней. Она преодолела расстояние по
морю не менее 1300 км, передвигаясь со средней скоростью 5—7 км
в сутки.
Таблица 52. Состав траловых уловов севрюги
в Северном Каспин (Павлов, Захаров, 1971)
Возраст, лет Длина тела см Масса тела, кг п %
1+ 39.6 0,30 22 1,96
2+ 55,2 0,56 47 4,14
3+ 69,6 1,03 125 11,06
4+ 75,3 1,11 157 13,9
5+ 82,5 1,26 82 7,23
6+ 88,3 1,87 97 8,56
7+ 102,6 2,9-2 99 8,73
8+ 108,8 3,52 68 6,00
9+ 108,9 3,53 ПО 9,70
104- 111,7 4,15 116 10,27
11 + 121,2 5,92 58 5,11
12+ 128,7 6,67 71 6,26
13+ 132,5 8,2 20 1,76
14+ 130,3 7,26 25 2,20
15+ 142,5 11,03 5 0,44
16+ 132,5 9,6 8 0,70
17+ 147,8 9,3 19 1,68
18-Ь 161,3 14,1 4 0,30
Наиболее высокая концентрация севрюги наблюдается во вто-
рой половине апреля и в начале мая в районе восточных мелководий
(на глубине менее 2 м) вокруг устья Урала и приустьевой области
р. Эмба. В южных районах восточных мелководий, примыкающих
к заливу Комсомолец, в это время концентрация севрюги ниже.
Еще меньше она в районе Суендыковских, Балашовских и Кол-
хозных шалыг. К середине лета на восточных мелководьях севрю-
га встречается в очень небольшом количестве. Начиная с августа
ее численность здесь несколько увеличивается, а к концу сентября —
началу октября падает до минимума. В распределении севрюги на
мелководьях усматривается аналогия с динамикой нерестового хо-
да в реке. Основная масса встречающейся здесь севрюги (до 80%)
представлена половозрелыми рыбами-мигрантами, которые на
мелководьях пребывают очень недолго. С середины сентября на-
гуливающаяся севрюга в Северном Каспии направляется к местам
136
предстоящей зимовки — Уральской и Мангышлакской бороз-
динам.
В Среднем Каспии по мере охлаждения воды севрюга откоче-
вывает в глубь моря, концентрируясь в основном на траверсе зали-
ва Кандерли на глубинах от 50 до 100 м, а в Южном — у о. Огур-
чинского и вдоль Иранского побережья на этих же глубинах (Бор-
зенко, 1964; Легеза, 1970; Пискунов, 1970).
Численность и хозяйственное значение. Организованное рыбо-
ловство в р. Урал имеет почти четырехвековую историю. П. С. Пал-
лас (1809) характеризует Урал как севрюжью реку. В 30—40-х
годах XIX в. на долю севрюги вместе с белугой приходилось до
половины улова (Данилевский, 1858). Н. А. Северцев (1863), как
и П. С. Паллас, называет Урал севрюжьей рекой. В начале XX в.
в Урало-Каспии вновь преобладал осетр (Кевдин, 1915), как и в
1936—1940 гг., когда Урал занимал третье место по добыче севрю-
ги (тыс. шт.) среди рек Каспия (Борзенко, 1942):
Река 1936 г. 1937 г.
1938 г. 1939 г. 1940 г.
Кура 121,6 93,1 71,3 93,2 63,7
Волга 51,8 61,5 43,5 32,9 25,8
Урал 15,1 24,8 28,5 9,4 12,5
Безудержная эксплуатация осетровых в море всевозможными
средствами, а также концентрация большого количества орудий
лова в реке подорвали в 30-е годы запасы осетровых настолько,
что в 1946 г. в Урало-Каспии было выловлено всего 43,6 т красной
рыбы, т. е. в 112 раз меньше, чем в 1932 г.
Таблица 53. Добыча осетровых в р. Урал,
тыс. т
Год Прогноз План Фактичес- кий ВЫЛОВ Перелов
1971 4,0 5,10 4,95 0,95
1972 4,3 <4,32 5,84 1,54
1973 4,5 4,75 5,92 1,42
1974 5,2 4,85 7,49 2,29
1975 6,2 7,50 8,25 1,95
1976 6,2 8,20 9,00 2,70
1977 7,1 8,90 10,41 3,11
1978 7,1- 7,60 8,47 1,17
1979 7,1 8,10 8,92 1,62
1980 8,0 8,45 8,10 0,10
1981 7,5 7,73 7,64 0,14
1982 7,2 7,50 7,11 —0,09
Всего 74,4 83,00 92,10 17,70
Ограничение промысла в 50-е годы и его совершенствование в
последующем (см. очерк по русскому осетру) способствовали
быстрому восстановлению запасов осетровых и в первую очередь
137
севрюги, уловы которой с 1,36 тыс. т в 1960 г. (в море и реке) под-
нялись до 5 тыс. т в 1971 г. только в реке. Возросшая численность
севрюги могла стабильно обеспечить уловы в пределах 7—8 тыс. т
до конца века. Однако интенсификация промысла (нарушение ре-
жима рыболовства, открытие тоней в запретной зоне) в 70-х годах
в результате установления завышенных планов по сравнению с
научно обоснованными прогнозами и стремления промышленности
перевыполнить план привели к тому, что за 9 лет (1971—1979 гг.)-
было переловлено сверх прогнозируемого около 18 тыс. т севрюги
(табл. 53), которую надо было пропустить на нерест.
Перелов такого количества рыбы в реке за короткий срок не-
замедлительно сказался на численности популяции и ее качестве:
увеличилась средняя длина и навеска рыб за счет преобладания в
уловах старшевозрастных самок (табл. 54), возросла плодови-
та б л и ц а 54. Качественный состав севрюги р. Урал
(по уловам лицевой тони)
Год Самки Самцы Оба пола
Длина, с м Масса, кг Длина, см Масса, кг Длина, см Масса, кг
1974 146,1 11,0 128,8 6,3 135,4 8,6
1975 149,0 11,5 124,0 6,2 133,6 8,7
1976 150,1 11,8 131,7 6,5 140,2 8,9
1977 149,9 11,5 132,0 6,5 142,0 9,2
1978 151,3 11,9 132,7 6,7 142,1 9,4
1979 151,2 11,5 131,8 6,9 142,2 9,3
1980 153,6 12,1 133,3 6,8 144,5 10,0
1981 156,0 12,6 134,4 6,9 145,6 9,9
1982 154,8 12,2 132,8 6,5 145,6 9,8
тость, нарушился половой состав; в уловах в последние годы пре-
обладают самки, ощущается острый дефицит самцов на нерестили-
щах, на зимовку и нерест пропускается гораздо меньше производи-
телей, чем в прошлые годы (табл. 55), сократилась численность
севрюги в море. В результате этого уловы ее имеют тенденцию к
снижению.
Следует отметить, что при разработанном режиме рыболовства
Таблица 55. Пропуск севрюги к местам зимовки
и нереста, тыс. шт.
Год Пропуск Год Пропуск Год Пропуск Год Пропуск
1967 412 1971 393 1975 345 1979 182
1968 331 1972 441 1976 311 1980 175
1969 281 1973 436 1977 231 1981 177
1970 463 1974 349 1978 320 1982 221
138
к местам размножения гарантированно может подходить 35—40%
нерестовой части популяции (Песериди и др., 1972; Захаров, Песе-
риди, 1981). На самом же деле выше промысловой зоны в послед-
ние годы проходит гораздо меньше производителей из-за вылова
севрюги браконьерами как в низовьях реки, так и на местах зимов-
ки и нереста.
С целью восстановления и увеличения численности севрюги и
доведения стабильных уловов до 8—9 тыс. т в год необходимо:
1) провести мелиорацию нерестилищ; 2) усилить охрану севрюги в
море и на миграционной трассе в реке, а также в зимовальных ямах
и на нерестилищах; 3) наладить искусственное воспроизводство
этой ценной рыбы и соблюдать лимиты вылова.
4. Acipenser nudiventris Lovetzky — шип
Казахское называние — пиилай.
Синонимия. Acipenser nudiventris: Ловецкий, Нов. магаз, ест. ист., 1828.
— Берг, Фауна России. Рыбы, т. I, 1911. — Филатов, Дуилаков, Бюл. Среднеаз.
унив., № 15, 1927. — Г. Никольский, Рыбы Аральского моря, 1940.
Распространение. Относится к Понто-каспийско-аральскому
фаунистическому комплексу. Распространен в бассейнах Черного и
Азовского, Каспийского и Аральского морей. В реках этих бас-
сейнов—Дунае, Доне, Днепре, Кубани и Риони в настоящее время
практически не встречается. Наиболее многочислен в бассейнах
Каспийского, Аральского морей и оз. Балхаш (в последнем аккли-
матизирован). Из Каспийского моря заходит главным образом в
реки Кура и Урал.
В Урале отдельные экземпляры проходного шипа доходят до
Оренбурга. Основной район его обитания — нижнее течение реки
до Уральска (850 км). Здесь имеется немногочисленное «стадо»ту-
водного шипа (Песериди, 1971). В Волге вылавливались единич-
ные экземпляры (Книпович, 1923; Берг, 1948). Сведений о заходе
шипа в реки Дагестанского побережья (Терек, Самур, Сулак) и в
р. Эмба нет.
В Аральском море распространен повсеместно, основной нерес-
товой рекой была Сырдарья, по которой шип поднимался до
пос. Чиназ и Беговатских порогов (Узбекская ССР, 1800 км) и вы-
ше (Цикуленко, 1935; Жарковский, 1950). В Амударье встречался
в меньшем количестве, чем в Сырдарье, и поднимался до Баумана-
бада. В настоящее время на севере Аральского моря и р. Сырдарье
практически не встречается.
В Балхаш был завезен в 1933—1934 гг. и распространился по-
всеместно. По р. Или поднимается выше государственной границы.
Описание. Тело шипа (рис. 26) торпедообразное. Рыло конусо-
видное. Жаберные перепонки прикреплены к межжаберному про-
межутку. Рот нижний, поперечный, превышает половину ширины
139
рыла, вытягивается в хоботок. Характерный признак — крупная
первая жучка (за что и назван шипом) и сплошная, не прерванная
посередине нижняя губа. По данным Д. П. Филатова (1925),
Р. Т. Тлеуова и Н. И. Сагитова (1973), у 3—4% шипов с юга
Аральского моря нижняя губа раздвоена. В р. Урал особей с пре-
рванной нижней губой не зарегистрировано.
Рис. 26. Шип
Усики бахромчатые, почти достигают конца рыла. Тело кофей-
ного или коричневатого цвета, окраска схожа со стерлядью и от-
личается от других осетровых. Брюхо белое. Пять рядов крупных
светлых жучек. Морфометрическая характеристика шипа р. Урал
(оба пола) и Аральского моря приведены в таблице 56.
Таблица 56. Морфологические признаки шипа
Признак Аральское море (Никольский, 1940) Река Урал, 1962-1964 гг.
От-до М От—до | М±т
Количество
Спинных жучек 11 — 16 13,8 11-16 13,4± 0,21
Боковых » 52-72 63,1 51-74 60,4 + 0,15
Брюшных * » 12-17 13,8 11-17 13,3±0,И
Лучей в D 45-57 50,3 40-53 46,6+0,28
Лучей в А •23-37 29,4 24-34 29,2+0,19
Жаберных тычинок 24-36(42) — 30-43 37,7±0,34
В % от общей длины тела **
Длина головы 19-25 22,0 19-22 20,3+0,14
Антедорсальиое расстояние 71-80 74,4 61-73 67,4+ 0,24
Аитевентралыюе расстояние 61-66 63,5 53-60 57,5+ 0,16
Антеанальное расстояние 75-86 79,9 67-75 71,9+0,31
Наибольшая высота тела 9-16 12,3 12-16 14,4+0,36
В % от длины гол овы
Длина рыла 36-45 39,9 35-43 38,0+0,10
от конца рыла
до усиков 21-31 26,2 19-28 23,7+ 0,31
от усиков до рта 18-23 20,4 12-18 14,4±0,08
* Брюшные жучки у крупных особей частично или совсем отсутствуют.
** У Г. В. Никольского (1940) расчет сделан по старой промысловой длине:
от конца рыла до конца средних лучей хвостового плавника.
140
Между аральским и каспийским шипами имеются существен-
ные различия в числе боковых жучек, лучей в спинном плавнике,
длине головы, наибольшей высоте тела, характеризующие большую
прогонистость аральского шипа. По последним данным, из 23 уч-
тенных морфологических признаков аральского шипа изменилось
более 15 признаков, особенно сильно —10, в том числе и меристи-
ческие, что связывают с изменением характера питания шипа и
ухудшением гидрологического режима бассейна (Тлеуов, 1981).
Сравнив данные по небольшому количеству шипов из Араль-
ского, Каспийского и Азовского морей, Л. С. Берг (1911) заключил,
что в этих водоемах обитает один вид. У каспийской популяции
отмечается более массивное туловище и хвостовой стебель, мень-
ший диаметр глаза и большее заглазничное расстояние (Борзенко,
1950). Л. С. Берг (1911) и М. И. Легеза (1971) описали помеси
шипа с белугой, осетром и стерлядью. Помеси других осетровых в
реках, где шип не обитает, обычно также именуются «шипами».
Биология. В дельте р. Урал шип появляется вскоре после осво-
бождения реки и взморья от льда, при температуре воды 3—4° (на-
чало апреля). Сначала, как правило, преобладают самцы. К раз-
гару хода, приходящемуся обычно на середину апреля, с колеба-
ниями от второй до пятой пятидневки, устанавливается численное
равновесие между полами. К началу мая ход шипа обычно зату-
хает, а к третьей декаде прекращается. Летом шип в р. Урал не
заходит, осенью встречается очень редко. Основная масса шипа
движется в р. Урал через судоходный канал и рукав Золотой.
Особенностью производителей шипа, заходящих в р. Урал в
апреле и мае, является сравнительно однородное состояние зре-
лости половых желез, несмотря на значительные колебания массы
ооцитов (табл.57).
Таблица 57. Колебания массы ооцнтов у шипа
Месяц Масса ооцитов, мг М±т п
до 10 -12 -14 -16 -18 —20>20
Апрель 16 215 399 168 62 22 10 13,3±0,04 892
Май 14 54 48 10 6 6 3 12,6±0,11 141
Осень 28 16 4 1 — —. — 10,1 ±0,10 49
Гонадо-соматический индекс (ГСП) у самок в апреле равен в
среднем 20, в мае —19,4, осенью—15,7; у самцов в апреле в сред-
нем 5,8, в мае — 4,5. У всех производителей апрельского и майского
хода на гонадах отсутствует жир, икра с характерной пигмента-
цией, легко отделяется от стромы яичника. Ооциты почти дости-
гают дефинитивных размеров, поляризация закончена (рис. 27)
или находится на грани завершения. Оптически пустое ядро или с
ядрышками, перемещающимися к его центру, целиком находится
в зоне мелкозернистого желтка вблизи от оболочек анимального
полюса. Ампулы семенников заполнены сформированными спер-
141
Рис. 27. Срез ооцита шипа яровой виутрипопуляционной нерестовой группы. По-
ляризация завершена. Апрель, дельта р. Урал
Рис. 28. Срез семенника шипа яровой виутрипопуляциошюй нерестовой группы.
Ампулы заполнены сформированными сперматозоидами. Апрель, дельта р. Урал
матозоидами (рис. 28). Все производители весеннего хода при не-
рестовых температурах положительно отвечают на гипофизарную
инъекцию.
Мигранты, заходящие в реку осенью, характеризуются иным со-
стоянием зрелости: на гонадах много жира, пигментация анималь-
иого полюса ооцита еще не закончена, икринки не достигают дефи-
нитивных размеров. Семенники небольшие, розового цвета. По-
скольку таких особей в нерестовой части популяции единицы и
они стали попадаться в реке лишь в последние 6—8 лет, можно
считать, что шип, заходящий в р. Урал, представлен только яровой
виутрипопуляционной нерестовой группой (Песериди, 1964, 1966а,
1972а, б и др.).
Производители раннего хода (апрель) поднимаются по реке
высоко, достигая в годы с холодной и затяжной весной самых верх-
них нерестилищ, расположенных в 650—850 км от моря. В целом
шип размножается в средней и частично верхней частях нерестовой
зоны (350—650 км) на характерных для осетровых субстратах: га-
лечниках, песчаниках, крупнозернистых песках, плотных глинах с
примесью ракуши и т. п. Производители, заходящие в реку в мае,
размножаются на нижних нерестилищах (150—300 км). Нерест
шипа в р. Урал начинается при температуре 12—13° и заканчива-
ется обычно при 18°. Инкубационный период в зависимости от тем-
пературы воды продолжается от 80—90 до 140—150 ч (Песериди,
1966а,1971, 1976).
Заход шипа в реки Амударья и Сырдарья отмечался с началом
весеннего половодья, обычно во второй половине апреля. Массовый
ход продолжался с середины мая до начала июля, иногда и весь
июль. Последние производители заходили в дельту обычно в сен-
тябре. Наиболее интенсивный ход в районе г. Казалинска (180 км
от моря) происходил с конца мая до середины июля (Цикуленко,
1935). До строительства в 1946 г. Фархадскон плотины шип под-
нимался вверх по реке на расстояние до 2600 км (Жарковский,
1950).
Основные нерестовые участки в Сырдарье (около 70%) находи-
лись между пос. Чиназ и Беговатскими порогами. Выше последних
располагалось не более 5% нерестилищ. Примерно столько же их
было ниже Кзыл-Орды (Цикуленко, 1935; Жарковский, 1950).
Половые железы производителей, заходящих в дельту Сыр-
дарьи в апреле—мае, находятся во II—III и незавершенной
III стадии зрелости. В пик хода мигранты имеют в основном
III стадию, а осенью— III—IV. Анализ гистологических препаратов,
представленных П. М. Коноваловым, показал что у шипов лет-
него хода состояние половых желез характерно для озимых внут-
рипопуляционных нерестовых групп (рис. 29, 30), что отмечено и
другими авторами (Печникова, 1970а; Тлеуов, Сагитов, 1973).
Незначительная часть популяции аральского шипа представлена
яровой «формой», о чем писал и Л. С. Берг (1948).
До зарегулирования Сырдарьи шип в основном зимовал на
143
Рис. 29. Срез ооцита озимого шипа. Сентябрь, дельта р. Сырдарьи
Рис. 30. Срез семенника озимого шипа в начальной фазе сперматогенеза. Август,
дельта р. Сырдарьи
участке реки между Туркестаном и селением Беговат, некоторые
экземпляры проходили и выше (Цикуленко, 1935). Нерест араль-
ского шипа начинался в марте при температуре воды около 10°, а
заканчивался во второй половине мая при 21—23° (Тлеуов, Саги-
тов, 1973).
В бассейне оз. Балхаш, по данным Н. В. Печниковой (1964),
миграция шипа в Или продолжается в течение 7 месяцев — с сере-
дины апреля до второй половины ноября — с двумя пиками: пер-
вый — весенне-летний (июнь) и второй, более слабый, —осенний
(октябрь — ноябрь). В течение всего хода половые железы нахо-
дятся в III, III—IV и реже в IV стадиях зрелости. Позднее
Г. В. Домбровский и др. (1972) отмечали мигрирующих производи-
телей в дельте реки с мая по август с половыми продуктами II—III
стадии зрелости. Пик хода наблюдался в нюне — августе. Заход
рыб прекращался в сентябре, когда половые железы достигали
III—IV стадий зрелости. Дозревание половых желез происходило
во время миграции и зимой.
Основные нерестилища шипа до зарегулирования р. Или нахо-
дились в ‘Капчагайском ущелье, в районе ур. Чулак и выше при-
стани Дубунь, а также на территории Китая (Печникова, 1970а;
Домбровский и др., 1972). По мнению Н. В. Печниковой (1970а),
до строительства Капчагайской ГЭС 35% шипа воспроизводилось
на участке реки, ограниченной Капчагайским ущельем и ур. Чу-
лак; еще 30% производителей размножалось на участке до госу-
дарственной границы и 20%—за пределами СССР; остальные
15%) —ниже Капчагайского ущелья.
Балхашский шип начинает размножаться при температуре во-
ды 10—12° на участках реки с каменистым, галечниковым и круп-
нопесчаным субстратом на глубине 0,7—2 м, где скорости течения
достигают 1,2—1,9 м/с. Нерест растянут и заканчивается при
сравнительно высокой температуре — 21—24°.
Дрифт личинок и скат молоди. В низовьях р. Урал первые пред-
личинки шипа (20—22 мм) появляются в третьей-четвертой пяти-
дневках мая. Их средняя масса в третьей декаде мая колеблется
по годам в пределах 80—120 мг.
Пик ската происходит в разные сроки: с конца мая до середины
июля (табл. 58). В конце ската, в июле, средние навески скаты-
вающейся молоди достигают 3,5 и даже 7,3 г. Средние уловы и
размеры скатывающейся молоди колеблются по годам. В мало-
водные годы скатывающаяся молодь шипа намного крупнее, чем в
многоводные, численность же ее в многоводные годы (средние уло-
вы на трал) намного выше, чем в маловодные.
Значительное количество молоди зимует в реке. Обитает она в
основном на крупных и глубоких ямах (10—18 м), где скорости
течения гораздо ниже, чем в плесах. Часть молоди на следующий
год скатывается в море. Вторично перезимовавшие в реке особи
остаются в ней навсегда, превращаясь в туводную форму
(табл.59).
10-7
145
Таблица 58. Дрифт личинок и скат молоди шипа в р. Урал
по пятидневкам (средние уловы за пятиминутное траление бимтралом)
Год Май Июнь Июль
3 4 5 6 1 2 3 4 5 6 1 2 3
1977 — 0.11 0,8 3,7 5,4 3,8 0,1 0,2 - 0,3
1979 — 0,9 0,6 2,2 1,7 0,2 1,83,7 13,3 8,5 5,7 2,1
1980 0,52,5 2,0 1,3 3,0 4,6 6,7 10,29,3 1,50,8 —. —
1982 °-' - —• 0,3 |0,3 0,2 0,8 0,1|0,1 0,1 — —
Таблица 59. Масса тела неполовозрелого шипа в нижнем течении
р. Урал, кг
Длина рыбы, см Масса рыбы п Длина ры- бы, см Масса рыбы п
От—до М От—до М
20-25 0,06-0,08 0,07 2 70-75 1,63-2,68 1,88 18
25-30 0,09-0,3 0,16 26 75-80 1,97-2,75 2,36 14
30-35 0,13-0,4 0,23 20 80-85 2,51-3,9 3,07 22
35-40 0,18-0,51 9,32 8 85—90 2,95-4,65 3,7 15
40-45 0,21-0,48 0,39 13 90-95’ 3,6-5,95 4,28 23
45-50 0,29-0,68 0,51 12 95-100 3,67—6,8 5,16 21
50-55 0,48-0,71 0,68 16 100-105 5,0-7,42 5,98 34
55-60 0,58-0,89 0,71 14 105-110 5,9-8,13 7,08 25
60-65 0,84-1,55 1,15 20 110-115 ) 5,3-9,14 7,34 24
65-70 1,24-2,06 1,49 18 115-120 7,6-10,0 8,71 3
Сведения о скате молоди шипа в Сырдарье до ее зарегулиро-
вания имеются у Г. В. Никольского (1933), который указывает,
что молодь держится в реке довольно долго. В другой работе
Г. В. Никольский (1938) уточняет, что мальки шипа в реке не за-
держиваются и сразу скатываются в море, за исключением тех,
которые попадают в дельтовые озера.
По данным Г. В. Никольского (1933) и Н. А. Гладкова (1934),
размеры молоди шипа в Сырдарье в июле — августе колебались в
пределах 3,4—8,2 см. Перед выходом в море доля скатывающейся
молоди массой до 0,5 г составляла около 8%, от 0,5 до 1 г—16%.
от 1 до 2 г — 40%, от 2 до 3 г — примерно 21%, а свыше 3 г— 15%.
Плотность молоди на 1 км русла реки достигала 4 тыс. шт. Скаты-
валась она со средней скоростью 950—1250 м/ч. По мнению
С. Н. Иванова и Н. В. Печниковой (1967), скат молоди продол-
жается 3 месяца. По данным Д. П. Филатова (1925), Г. В. Николь-
ского (1933), К- 3. Трусова (1947) и др., молодь шипа задержива-
ется в реке на 2—4 года.
146
Скат молоди по р. Или до ее зарегулирования продолжался с
июля по сентябрь. Покатные сеголетки имели длину 9,2—14,3 см
(в среднем 11,7 см) и массу тела 4,0—11,7 г (в среднем 7,2 г) при
возрасте 20—50 дней. На первом году жизни в оз. Балхаш попа-
дало небольшое количество молоди, основная ее часть задержи-
валась в реке и в дельтовых озерах, где оставалась до 2—7-летнего
возраста (Доброхотов, 1948; Даирбаев, 1957; Печникова, 1964).
После сооружения Капчагайской ГЭС, по исследованиям со-
трудников Казахского университета, покатная миграция личинок
шипа в нижнем течении р. Или отмечается в мае — июне. Молодь
скатывается на этапах развития А, реже В, преимущественно в
придонных слоях потока. Общее количество скатывающихся личи-
нок уменьшается год от года. Так, в районе пос. Баканас абсолют-
ная численность личинок шипа, скатывающихся за сезон, состави-
ла в 1980 г. 4,69, в 1981 г. —3,2, в 1982 г, —1,4, в 1983 г.—
0,66 млн. экз.
Выше Капчагайского водохранилища по р. Или тоже отмечен
скат личинок шипа (Нездолий и др., 1975). В зоне подпора водо-
хранилища в экспериментальных уловах есть молодь в возрасте
от сеголетка до 4 лет.
Скорость ската предличинок и личинок близка к скорости те-
чения реки. По мере роста молоди она снижается, составляя в
среднем 670—1600 м/ч (Печникова, 1970).
Сведений о сроках созревания, возрастной структуре популя-
ции и темпе роста уральского шипа в литературе нет, за исключе-
нием сообщения В. В. Петрова (1927), определившего возраст у
19 особей.
Таблица 60. Средняя масса тела ходового шипа р. Урал по размерным
группам (1962—1976 гг.), кг
Размерные группы, см Самцы (п=1518) Самки (п=1483) Оба пола
От—до м п От—до М п М п
101-110 5,4—9,7 7,04 8 . 7,04 8
111-120 4,9-15,1 9,18 58 — — — 9,18 58
121-130 6,1-19,2 10,35 410 10,4-14,7 12,35 2 10,40 412
131-140 7,3-20,0 13,32 642 10,1-18,3 14,1 8 13,32 650
141-150 8,8-22,2 15,56 283 13,4-33,1 20,6 99 16,84 382
151-160 13,3-27,5 19,12 57 14,6—35,4 24,0 291 23,12 348
161-170 16,4-31,0 22,82 29 16,9-39,0 26,9 489 26,72 518
171-180 20,6-34,2 27,33 8 16,1-46,0 31,2 327 31,16 335
181-190 20,4-38,3 31,2 9 21,3-51,0 35,3 163 38,1 172
191-200 28,7-46 8 36,1 9 30,5—50,0 42,0 27 40,8 36
201-210 28,7-35,8 32,8 5 42,6-53,0 48,3 8 42,34 13
211-220 — — — 45,6—55,0 50,3 5 50,3 5
221-230 — — — 49,4-59,8 54,2 4 54,2 4
1Я
Половозрелость у единичных самцов шипа наступает в 7—8 лет.
Основная их масса созревает в 9—13-летнем возрасте. Преобла-
дают среди самцов 9—20-летки, на долю которых приходит-
ся около 85%. Самки созревают к 11—12 годам, но таких особей в
популяции мало. Массовое созревание наступает в 13—16 лет. Ос-
нову нерестового стада самок составляют рыбы в возрасте 14—-
Таблица 61. Средняя длина (см) и масса (кг) ходового шипа
р. Урал (1961—1969 гг.)
Возраст, лет Самцы СамкЯ Оба пола
СМ КГ п СМ КГ п СМ КГ п
7 102 6,5 2 102 6,5 2
8 108 8,2 4 — —. —, 108 8,2 4
9 120 11,4 25 — — — 120 11,4 25
10 127 12,2 39 —. —— 127 12,2 39
11 Д31 13,6 36 147 22,0 10 135 15,4 46
12 134 14,5 51 151 23,4 17 138 16,7 68
13 137 15,7 63 154 23,8 16 140 17,3 79
14 140 17,9 67 160 24,9 24 145 19,7 91
15 146 19,3 78 161 25,6 54 152 21,9 132
16 150 20,1 74 164 27,3 103 158 24,3 177
17 157 22,2 50 166 28,6 129 163 26,6 188
18 162 23,6 56 170 29,7 83 167 27,3 139
19 165 24,1 39 174 30,9 67 171 28,4 106
20 167 24,8 23 177 31,4 43 174 29,1 66
21 170 25,2 17 18! 32,8 25 177 29,7 42
22 172 26,9 12 184 33,6 23 180 31,3 35
23 174 27,4 14 186 34,4 19 181 31,4 33
24 177 28,1 15 189 34,7 11 183 30,9 26
25 180 29,3 13 191 37,1 16 186 33,6 29
26 178 30,2 9 193 37,7 9 183 34,0 18
27 181 32,8 9 194 39,2 10 188 36,2 18
28 180 33,9 8 187 41,8 6 187 37,3 12
29 184 38,4 3 204 43,4 5 196 41,5 8
30 -— —. —. 200 41,6 6 200 41,6 9
31 —• — — 206 39,7 4 206 39,7 4
32 — —. —- 204 44,3 3 204 44,3 3
33 — — — 198 40,0 1 198 40,0 1
Среднее 147,9 19,78 726 173,1 29,9 684 160,1 24,7 1410
22 года (80%)- В целом нерестовая часть популяции шипа р. Урал
состоит из 27 возрастных групп, но особей старше 25-летнего воз-
раста, т. е. созревающих в 3—4-й раз, немного (табл. 60).
Самки, созревая позже самцов, намного опережают их в темпе
роста. Разница в размерах и массе тела увеличивается с возрастом:
это связано с более коротким половым циклом у самцов, заторма-
живающим их рост (табл. 61). За период анадромной миграции,
нереста и пребывания в реке самцы в среднем теряют в массе око-
ло 3 кг, а самки — более 6.
148
Возрастная структура популяции и темп роста шипа Аральско-
го моря за относительно небольшой период (около 60 лет) претер-
пели существенные изменения (табл. 62). По сравнению с 1926—
1928 гг. (Пробатов, 1929), популяция шипа в 1964—1969 гг. стала
более возрастной (что, вероятно, связано с прекращением про-
мысла шипа).
Таблица 62. Средняя длина (ем) и масса тела (кг) шипов разного
возраста
Возраст, лет Аральское море Балхаш, 1957— 1960 гг. (Печ- никова, 1970)
1926-1928 гг. (Про- батов, 1929) 1934—1969 гг. (Тле- уов, Сагитов, 1973)
см | кг см | кг см | кг
7 93,5 4,6 93 3,6 — -—
8 98 3,6 108 5,1 123 9,5
9 104 3,5 112 7,8 128 12,5
10 111 7,5 108 7,1 130 10,0
11 112 7,5 122 9,7 133 11,7
12 118 8,7 138 12,1 141 13,9
13 121 9,2 121 11,2 141 11,1
14 123 — 132 14,2 142 13,6
15 128 12,7 138 15,2 149 17,2
16 133 11,3 146 17,2 155 17,8
17 138 14,5 147 15,9 156 22,1
18 142 15,3 148 18,8 159 17,8
19 144 17,7 157 22,3 159 18,8
20 148 17,2 150 23,7 163 20,3
21 151 17,0 161 27,4 — .—
22 152 16,0 164 25,0 —.
23 Г’З —- 171 30,5 — —
24 155 16,0 —. 31,0 —. —
25 —. 19,2 155 33,2 — —
30 —- — 170 31,0 — —
Усилился и темп роста, особенно у старшевозрастных рыб, что
связывается с появлением обильной и легкодоступной этому виду
пищи-—бычков (Маркова, 1969; Печникова, 19706; Тлеуов, 1981).
По мнению Р. Т. Тлеуова (1981), изменения претерпела и воз-
растная структура стада шипа. Так, в современном стаде совер-
шенно отсутствуют старшие возрастные группы (свыше 25 лет).
Основная часть популяции представлена особями в возрасте 11—
19 лет.
Возрастной ряд балхашской популяции шипа состоит из 13 воз-
растных групп —8—20-леток (табл. 62), т. е. в основном из впер-
вые созревающих особей.
Сравнение темпа линейного и весового роста шипа из разных
водоемов Казахстана (рис. 31 и 32) показывает, что быстрее всех
растет уральский шип. Второе место занимает балхашский, кото-
рый в раннем возрасте растет быстрее уральского. Несмотря на
149
то, что в последние годы темп роста аральского шипа усилился, все
же он уступает в росте уральскому и балхашскому.
Особый интерес представляет рост шипа в водоеме нового ти-
па— Капчагайском водохранилище на р. Или, где он нагулива-
ется до наступления половозрелости. По расчисленным данным, в
первые 10 лет жизни шип здесь растет хуже, чем в оз. Балхаш:
Возраст, лет
Данные
12345678 9
КазНИИРХ 20,4 32,2 45,9 57,6 67,2 76,4 81,1 91,9 99,1
КазГУ 16,9 32,4 46,9 60,7 74,3 84,3 92,3 95,6 —
Рнс. 31. Линейный рост шипа в водоемах Казахстана: 1 —р. Урал, 1961—1969 гг.;
Аральское море: 2а—1926—1928 гг.; 26 — 1964—1969 гг.; 3— оз. Балхаш,
1957—1960 гг.
150
Др середины 60-х годов, когда в Урал заходило мало производи-
телей шипа, соотношение полов в стаде не было характерным для
осетровых в целом. В последние годы, когда наметилась тенденция
увеличения численности шипа, половая структура, присущая
осетровым, проявилась более отчетливо, в уловах стали преобла-
дать самцы — в среднем 52—60% нерестовой части популяции за
сезон.
Плодовитость шипа слабо изучена. Имеются фрагментарные
сведения Г. В. Никольского (1940), что плодовитость особей дли-
Рис. 32. Рост массы тела шипа в водоемах Казахстана (обозначения те же, что
на рис. 31)
151
ной 126—150 см в возрасте 16—19 лет составляет 162—342,3 тыс.
икринок. Литературных сведений о плодовитости шипа р. Урал нет.
Наши материалы характеризуют плодовитость шипа р. Урал в
зависимости от возраста, размера и массы тела. Самая низкая
плодовитость — 101,4 тыс. икринок — была зарегистрирована у
особи размером 133 см, массой 14,1 кг, возраст которой 15 лет
(табл. 63, 64); наиболее высокая—1032 тыс. — была у рыбы дли-
ной 188 см и массой 47,4 кг.
Таблица 63. Плодовитость шипа р. Урал по размерным
группам, тыс. шт.
Размер, см От—до М п Размер, см От—до М п
121-130 175 175 1 171-180 270-811 499 269
131-140 101-258 314 5 181-190 277—1032 593 144
141-150 165-470 314 74 191 -200 547-1005 749 21
151-160 169-672 342 226 201-210 632-955 893 8
161-170 237—784 425 343 121-210 101-1032 451 1082
Таблица 64. Плодовитость шипа р. Урал по массе тела,
тыс. шт.
Масса, кг От—до М п М'асса, кг От—до М п
12-16 101-270 219 6 36-40 309-866 555 89
16-20 158 —464 304 62 40-44 460-904 652 28
20-24 166-522 332 164 44-48 562-1032 777 25
24-28 183-589 389 263 48-52 718-1005 868 И
28-32 258 - 672 454 265 52—56 771-955 866 7
32-36 270-811 532 156 56-60 7.54-913 842 3
Средняя плодовитость впервые созревающих рыб размером от
131 до 150 см составляет 314 тыс. икринок. Наибольший размах
колебаний плодовитости наблюдается у многочисленных размер-
ных категорий — от 151 до 190 см. Аналогичное явление отмечено
и с увеличением массы рыбы. Плодовитость достигает максимума
у особей массой от 40 до 48 кг. Равномерный прирост плодовитости
наблюдается у рыб до 28-летнего возраста. В целом шип весьма пло-
довитый среди осетровых вид.
Питание, конкуренты, враги. Питание взрослого шипа р. Урал
изучено недостаточно. Это обусловлено тем, что уральский шип
во время анадромной миграции, зимовки, нереста и поката не кор-
мится. Питание молоди изучено лучше.
Личинки шипа в р. Урал переходят па активное питание при
длине 20 мм (Стыгар, 1981). Качественный состав пищи молоди
шипа в разные годы почти не меняется, но в количественном соот-
ношении наблюдаются значительные колебания. Так, в различные
годы (1971—1973) в пище шипят (до 70 мм) гаммариды составля-
152
ли 38—69% (по массе), корофинды •— 22—30, личинки ручейников —
12—13, хирономид— до 13%. Средние индексы наполнения желуд-
ков шипят в 1971—1975 гг. колебались от 66,9 до 246%оо, индексы
наполнения всего пищеварительного тракта — от 224 до 411°/ооо-
Особи с пустыми желудками составляли не более 2%. Качествен-
ный состав пищи молоди шипа, осетра и севрюги весьма сходен.
Наибольшая степень пищевого сходства у шипа с осетром, наи-
меньшая— с белугой. Отдельные шипята размером более 70 мм
питаются и личинками осетровых (Бережной, 1971; Стыгар, 1981;
Стыгар, Песериди, 1981).
А. Н. Державин (1949), М. П. Борзенко (1950), М. В. Желтен-
кова (1964), Г. М. Филиппов (1972) указывают, что в море шип
питается преимущественно рыбой: бычками, пуголовками, атери-
ной, килькой (до 97%). Единично в его пище встречаются ракооб-
разные, моллюски.
О характере питания молоди шипа в Сырдарье сведений нет. По
данным Н. В. Печниковой (1972), молодь шипа на первом году
жизни в заливе Аббас Аральского моря питалась вначале мелки-
ми, а с конца июня — преимущественно крупными личинками хи-
рономид. В оз. Муйнак она поедала бычка-бубыря, а в заливе
Сарбас и морских участках дельты, кроме того, — личинок и куко-
лок хирономид.
В 30-х годах шип в Аральском море питался главным образом
моллюсками (Никольский, 1940). После натурализации в Араль-
ском море бычков шип с первого года жизни почти полностью пе-
решел на питание ими (Печникова, 1964; Тлеуов, Сагитов, 1973).
В желудках молоди шипа встречалось от 8 до 70 экземпляров быч-
ков-бубырей, а у взрослых особей — до 250 шт. бычков-песочников.
Ранняя молодь шипа в р. Или питается преимущественно мел-
кими формами хирономид. Подросшая молодь в период ската по-
мимо личинок хирономид (64,5% содержимого желудка) потреб-
ляет и икру карповых рыб (26%), куколок хирономид и кулицид
(6—47%). Общий индекс наполнения желудков молоди шипа ко-
леблется от 83 до 944%Оо-
В зависимости от мест обитания молоди шипа состав ее пищи
меняется. Так, в р. Или она преимущественно питается личинками
поденок—100% встречаемости. По данным С. К. Тютенысова
(1953), молодь шипа массой 19—400 г в р. Или питается пауками,
личинками веснянок, двукрылых, жуками и их личинками. Шипы
в возрасте 5—7 лет в р. Или потребляли одноцветного и пятнистого
губачей, мелкого окуня (Домбровский и др., 1972). В дельтовых
озерах в пище молоди преобладали личинки хирономид, встреча-
лись рыбы (губач, окунь, молодь маринки), насекомые и др. (Печ-
никова, Воробьева, 1961).
В оз. Балхаш шип питался губачом, окунем, гольцами. В желуд-
ках крупных экземпляров шипов встречалось до 6—15 гольцов,
29 окуньков общей массой 0,35—1,4 кг. С 1967 г. в желудках ши-
пов стали встречаться раковины вселенного моллюска — монодак-
153
ны. Численность малоценных рыб в Западном Балхаше после мас-
сового развития судака резко упала, и шип перешел на питание
моллюсками и мизидами (Печникова, Воробьева, 1961; Домбров-
ский и др., 1972).
В Капчагайском водохранилище 4—5-летние шипы питаются
преимущественно этими же видами кормовых организмов. Более
крупные едят преимущественно рыбу (амурский бычок, лжепес-
карь).
Упитанность аральского и уральского шипа во время нересто-
вой миграции колеблется в пределах 0,70—0,75 по Фультону.
В оз. Балхаш упитанность молоди, не перешедшей на хищное пи-
тание, низкая (0,34—0,46 по Кларк). Упитанность самок до нерес-
та— 0,73. Самцы обычно упитаннее самок. Наивысший коэффи-
циент упитанности— 0,87 — наблюдается в июне — июле (Печнико-
ва, 1964).
Конкурентами в питании молоди шипа являются пескарь, бело-
глазка, густера, вобла и молодь сазана, леща, белуги и осетра.
Взрослые шипы конкурируют с белугой и частично с судаком и
жерехом. Конкурентами шипа в Арале являются усач, судак, лещ,
сазан и вобла.
В оз. Балхаш наибольшие коэффициенты сходства пищи с саза-
ном, лещом, судаком (Воробьева, 1968). Большое количество мо-
лоди шипа в речной период жизни истребляют сом и частично су-
дак. В желудках крупных сомов массой 15—20 кг, выловленных в
зимовальных ямах среднего течения р. Урал, встречалось до Зэкз.
двухлеток шипа. По мнению Н. В. Печниковой (19706), молодь
шипа в Аральском море выедают большой лопатонос, судак, же-
рех и сом.
Численность и хозяйственное значение. В Каспийском море
шип распределен неравномерно. В западной половине Северного
Каспия и вдоль западных берегов Среднего, вплоть до северной
границы Азербайджана, шип практически не встречается. Южнее
(Худат-Апшеронский п-ов) доля шипа в уловах не превышает
0,04—0,05% от всего числа выловленных осетровых. В южной час-
ти Среднего Каспия шип в уловах встречается чаще — 0,9% На
восточных мелководьях Северного Каспия шип чрезвычайно редок.
Наибольшие скопления его в Северном Каспии наблюдаются во-
круг уральской приустьевой области моря, где он концентрируется
весной перед заходом в р. Урал. В волжской приустьевой области
моря его нет.
Миграционные пути шипа пролегают вдоль казахстанских берегов
Среднего Каспия. Обогнув п-ов Тюб-Караган, эта рыба вдоль вос-
точного свала Уральской бороздины движется на север в сторону
п-ова Пешной. Шип в море встречается на глубинах 11—25 м при
температуре воды 2,7—14,6° (Пискунов, 1965; Легеза, 1966, 1967,
1973; Селиверстова, 1968).
В оз. Балхаш концентрации шипа обнаружены между островами
Ультарахты, Кержук, в заливе Красный Октябрь, в районах Ту-
154
рангалык, Аджартас, с восточной стороны Мынаральских
островов.
До последнего времени шип в р. Урал фиксировался рыбозаго-
товителями под общим названием «осетр». В начале 30-х годов
здесь вылавливалось до 200 шипов в год (Державин, 1947); в на-
чале 60-х годов — более 500 (Песериди, 1963). К концу 60-х — на-
чалу 70-х годов в уловах появилось большое количество молодых
особей, а к середине 70-х годов численность шипа еще больше
возросла. В 1973—1975 гг. к местам размножения пропускалось
более 3,0—3,5 тыс. особей шипа, чего не наблюдалось и не наблю-
дается ни в одной реке. К началу 80-х годов запасы уральского
шипа увеличились по сравнению с серединой 70-х примерно в
1,5 раза. Сопоставление данных по уловам и пропуску шипа в Ку-
ре и Урале показывает, что к местам размножения в последнем
пропускается в 15—20 раз больше производителей, чем в Куре —
материнском водоеме шипа Каспия (Захарян, 1972; Овсепьян, Пе-
сериди, 1976).
По данным промысловой статистики, сравнительно стабильные
уловы (300—400 т) шипа в Аральском море наблюдали с 1928 по
1935 г. (табл. 65).
Таблица 65. Уловы шипа в бассейне Аральского моря
(Тлеуов, Сагитов, 1973), т
Год У лов, т Год Улов, т Год Улов, т Год Улов, т
1928 300 1936 620 1953 4,3 1962 1,6
1929 350 1937 417 1954 2,2 1963 6,5
1930 308 1938 2.7 1955 2,5 1964 5,9
1931 338 1939 5,4 1956 2,5 1965 9,3
1932 399 1940 2,2 1957 0,3 196S 8,7
1933 349 1943 0,1 1959 0,2 1967 9,7
1931 298 1951 0,2 1960 0,9 1938 6,5
1935 324 1952 6,0 1961 0,9 1969 7,2
В разгар эпизоотии (1936—1937 гг.) добыча и прием больного
и ослабевшего шипа возросли. После гибели шипа и установления
полного запрета на его отлов во всем бассейне Аральского моря
(с 1 июня 1940 г.) добыча упала до минимума, но не прекратилась,
так как разрешался его «прилов», достигший 6,0—4,3 т в 1952—
1953 гг. Наиболее высокий прилов отмечен в 1963—1969 гг. — 5,9—
9,1 т (Жарковский, 1950; Маркова, 1969; Тлеуов, Сагитов, 1973).
После зарегулирования стока Сырдарьи шип на севере Арала
практически не встречается. Прилов его на юге Арала у устья
Амударьи продолжается и в настоящее время. Как и прежде (Ни-
кольский, 1940), один из факторов, лимитирующих численность
шипа Аральского моря, — браконьерство. Фактический вылов его
значительно превышает данные официальной статистики.
Специализированный лов шипа в Балхаш-Илийском бассейне
155
запрещен, но разрешается прилов. По данным КазНИИРХа, вели-
чина прилова не стабильна и колеблется по годам:
Годы 1955 1962 1963 1976 1977 1978 1980 1981 1982
Улов, т 20,5 1,3 0,1 8,7 32,0 23,1 31,5 15,4 5,6
Расчеты показывают, что даже при минимальной средней.на-
веске, равной 15 кг, в Балхаш-Илийском бассейне в последние го-
ды (1976—1982) вылавливается по 370—1550 мерных особей в год.
По данным Г. В. Домбровского и др. (1972), в 22—23,5 тыс. сетей,
выставляемых в Западном Балхаше и в водоемах дельты Или,
ежегодно гибнет не менее 9—11 тыс. экз. молоди шипа, достигших
таких размеров, когда их естественная смертность не превышает
4—5%. Как видно, количество погибающих особей превышает
официальный прилов в 7-—30 раз, что по существу и является
главной причиной низкой численности шипа оз. Балхаш. Наряду с
этим существенный урон воспроизводству шипа оз. Балхаш на-
несло и строительство Капчагайского гидроузла, преградившего
путь шипу к основным местам его воспроизводства.
Заключая изложенное, можно сказать, что основными причи-
нами, лимитирующими численность шипа в бассейнах Арала и Бал-
хаша, являются гидростроительство, возрастающее из года в год
безвозвратное недопотребление, биологически и экономически не-
обоснованный и нерациональный промысел и браконьерство.
В настоящее время запасы шипа Аральского бассейна находят-
ся в крайне неблагоприятном состоянии и не исключена возмож-
ность гибели его здесь в ближайшем будущем. Несколько лучше
положение в Балхаше, где есть надежда на сохранение запасов
шипа.
В целях сохранения и восстановления численности шипа в бас-
сейне Арала и Балхаша необходимо: а) форсировать искусствен-
ное разведение аральского шипа на Нукусском осетроводном заводе;
б) построить в нижнем бьефе Капчагайского водохранилища
осетроводный завод мощностью до 10 млн. шт. молоди шипа в год;
г) разработать и внедрить в Балхаш-Илийском бассейне режим
рыболовства, сокращающий гибель подрастающего поколения
шипа.
5. Acipenser ruthenus Linne — стерлядь
Казахское название — суир1к
Синонимия. Acipenser ruthenus: Linne, Syst. nat., ed. X, 1758. — Берг,
Фауна России, Рыбы, т. 1, 1911. —Берг, Рыбы пресных вод СССР, ч. 1, 1948.
Распространение. Наиболее широко распространенный вид ро-
да осетров. Обитает в бассейнах Северного Ледовитого океана,
Черного, Азовского, Каспийского, Балтийского морей (в послед-
нем редок). В пределах Казахстана водится в реках Урал, Иртыш,
Тобол, но везде численность этого вида невелика.
156
Основная масса уральской популяции обитает в средней и
верхней частях нижнего течения реки. Выше Оренбурга стерляди
нет. Редко она ловится в дельте реки и устьях Урала и Эмбы
(Меньшиков, 1929; Меньшиков, Букиреев, 1934; Пробатов, 1935;
Цеханович, 1937; Берг, 1948; Борзенко, 1951; Jankovic, 1958; Абду-
рахманов, 1962; Демин, 1963; Егоров, 1963; Зырянова, 1963; Федо-
ров, Филиппова, 1963).
Описание. Сведений по морфологии стерляди из водоемов Ка-
захстана нет. Л. С. Берг (1948), ссылаясь на М. И. Меньшикова,
приводит следующие данные по меристическим признакам стерля-
ди. D 39—49, в среднем 44,1; А 20—30, в среднем 24,7. Спинных
жучек 12—16, чаще 13—14, в среднем 13,6; боковых 58—71, чаще
64—66, в среднем 64,3; брюшных 12—16, в среднем 13,4. Жаберных
тычинок в первой дуге (15) 16—21, в среднем 17,8; они не вееро-
образные. По А. В. Лукину (1949), количество спинных жучек ко-
леблется в пределах (10) 12—17, боковых (52) 57—71, брюшных
10—19. Жаберных тычинок 15—26 (31). Лучей в D 37—54, в
А 19—31. По В. И. Шилову (1971), длина головы (% от общей
длины тела) 19,7 (колебания 15—28), длина рыла (% от длины го-
ловы) 29—50, в среднем 38,6, от хрящевого свода до конца рыла
36—61, в среднем 47,9, от усиков до конца рыла 17—44, в среднем
28,2; высота головы 34—60, в среднем 45,6; заглазничное расстоя-
ние 38—65, в среднем 53,1 и ширина рта 11—31, в сред-
нем 23,8.
Рис. 33. Стерлядь
Тело стерляди (рис. 33) торпедообразное, покрыто очень мел-
кими гребенчатыми зернышками. Рыло умеренно вытянутое. По
длине рыла различают острорылую (типичная) и тупорылую фор-
мы. Некоторые исследователи (Шмидтов, 1939; Берг, 1948; Лукин,
1977) тупорылую форму относят к озимой расе, а острорылую-—к
яровой. Е. Н. Казанчеев (1963), высказывая оригинальную точку
зрения, считает, что тупорылые — это старые или хорошо расту-
щие рыбы. Остальные же — молодые особи или рыбы, живущие в
неблагоприятных условиях откорма. Нижняя губа посреди прерва-
на. Усики бахромчатые. Тело обильно покрыто слизью, светло-ко-
ричневого или кофейного цвета. Жучки светлые. Брюхо белое. По-
ловой диморфизм не выражен.
Биология. Стерлядь типично придонная пресноводная (тувод-
йая) рыба, в большинстве своем постоянно живет в реках, стари-
157
цах, озерах и водохранилищах. В низовье и дельте Волги, по мне-
нию Н. Л. Чугунова (1928) и Л. С. Берга (1948), имеется и полу-
проходная форма. Е. Н. Казанчеев (1981) называет эту форму
экологической расой. Ее молодь скатывается в опресненную север-
ную часть моря, где нагуливается до половозрелости, а осенью за-
ходит в дельту. Очень редко аналогичное явление наблюдается и
в Урале, поскольку единичные экземпляры стерляди размером
25—35 см вылавливаются в дельте. Стерлядь в реках передвигает-
ся в основном в поисках мест нагула. Излюбленными местами в
нижнем течении Урала являются участки реки с замедленным те-
чением и ямы с затонувшими деревьями и корягами. Осенью стер-
лядь передвигается в поисках мест зимовки. Залегает она в ямы
(«ятови» на Урале, «юровые» на Иртыше) глубиной 10—15 м, где
проводит всю зиму не питаясь.
Вскоре после весеннего ледохода стерлядь покидает ямы и
движется к нерестилищам. В это время у нее появляется брачный
наряд в виде белого налета на голове. Соотношение полов 1:1.
Нерест стерляди начинается при температуре воды 8—9°, разгар —
при 10—12°. Заканчивается нерест, вероятно, при температуре
17—18°. Икра стерляди мелкая, диаметром 1,8—2,2 мм, отклады-
вается на каменистых россыпях, галечниках, гравийно-песчаном
субстрате. На крупнопесчаных субстратах и плотных глинах с
примесью ракуши не встречается. Откладывается на глубине 3—•
8 м, где скорость течения превышает 1,2—1,3 м/с. Выметанная икра
приклеивается. Инкубационный период при температуре воды
9,9—13,3° длится 8—10 суток (Львов, 1973).
После нереста стерлядь покидает нерестилища и скапливается
в ямах и участках реки с замедленным течением среди затонувших
деревьев и коряг (Бенинг, 1938). На плессах и перекатах, где нет
коряг, стерлядь встречается очень редко.
Л. С. Берг (1911, 1934, 1948, 1953) считает, что у стерляди, как
и у большинства проходных осетровых, имеются озимая и яровая
расы. По его мнению, полупроходная стерлядь входит в дельту
Волги дважды: весною вместе с севрюгой и затем осенью; весенняя
(яровая) подымается выше дельты для нереста, осенняя же (ози-
мая) залегает в ямы на взморье, а также в самых устьях Волги и
в нижней части дельты (Берг, 1953). В. И. Шилов (1971) не разде-
ляет этой точки зрения и считает, что у стерляди нет озимых и
яровых рас. Этот вопрос требует уточнения.
В районе нерестилищ стерляди на р. Урал за 10 лет наблюде-
ний удалось поймать всего 8 шт. молоди стерляди, размером от
23 до 112 мм и массой тела от 58 до 6540 мг, что связано с отсут-
ствием ската молоди.
Сведения о сроках наступления половозрелости у стерляди
весьма противоречивы. А. И. Амброз (1972) указывает, что самцы
созревают на третьем (2+) и пятом (4-J-) годах жизни. А. В. Лу-
кин (1949) для самцов называет возраст 3—7 (преимущественно
4—5 )лет, а для самок — 5—12 (большей частью от 7 до 9) лет.
158
По наступлении половозрелости стерлядь нерестует ежегодно
(Лукин, 1937, 1947, 1949).
Иную точку зрения относительно сроков созреваний и перио-
дичности половых циклов имеет В. И. Шилов (1971). По его мне-
нию, лишь некоторые самцы, обитающие в Саратовском и Волго-
градском водохранилищах, созревают в 4-летнем возрасте,
отдельные ювенильные особи встречаются до 5—8-годовалого воз-
Рис. 34. Срезы маргинальных лучей стерляди р. Урал: а — самец, возраст 3+,
абс. длина 34 см, масса 190 г; б — самка, возраст 4-р, абс. длина 42 см, масса
340 г; в — самка, возраст 5+, абс. длина 44 см, масса 420 г; г — самец, возраст
74-, абс. длина 50 см, масса 620 г
159
раста, в массовом количестве в стадо производителей они вступа-
ют с 6-годовалого. Среди самцов старше 8 лет в весенних уловах
подавляющее большинство составляют особи со зрелыми семенни-
ками. Отдельные самки созревают в 7—8-годовалом возрасте, а в
массовом количестве (относительно одновозрастных рыб) начи-
нают принимать участие в нересте с 13—14 лет; самцы повторно
смогут принять участие в нересте через 2 года, а для.повторного
созревания самок необходим промежуток не менее 4—5 лет.
Сведений о плодовитости стерляди из водоемов Казахстана не-
достаточно. По р. Урал известна плодовитость 2 самок стерляди: у
первой (46,3 см, 490 г, 6 лет) 8,42 тыс. икринок, у второй (50 см,
1350 г, 9 лет) — 12,76 тыс. икринок, что соответствует данным по
другим водоемам. Известная максимальная плодовитость —
145 тыс. (Львов, 1973).
Определять возраст стерляди легко, поскольку годовые кольца
у нее выражены отчетливо (рис. 34). По материалам, собранным на
нерестовом участке р. Урал между пос. Индер и г. Уральском,
самцы стерляди становятся половозрелыми в 4—5-летнем возрасте,
а самки — на 1—2 года позже. Темп роста самцов и самок стерля-
ди до 6—7-летнего возраста одинаков (табл. 66).
Таблица 66. Основные показатели стерляди р. Урал (1961—1973 гг.)
Воз- раст, лет Самцы Самки
Средняя длйна, см Средняя масса, г Стадия зрелости п Средняя длина, см Средняя масса,г Стадия зрелости п
2 29,3 87 I—II 3 30,5 98 I—II 2
3 35,5 193 II 4 38,5 255 II 4
/ 40,0 300 II—III 4 40,7 314 II 3
5 —- — — 44,0 420 II—III 1
6 41,5 325 II—III 2 46,3 490 III—IV 3
7 48,5 560 п,V 2 —. .—, —- —
8 —• —- — —. 51,0 915 П-ПТ 1
9 57,0 1260 V 1 50,0 1350 III—IV 1
10 — — — .—. 53,0 1100 п—пт 1
11 51,5 980 nr-iv 2 — —, —
12 59,0 1380 V 1 58,0 1260 II—III 1
Рост стерляди в разных водоемах различен; уральская растет
быстрее и несколько упитаннее волжской и камской, но уступает
по всем показателям роста стерляди из р. Вычегды. Соотношение
полов у стерляди в большинстве водоемов близко 1:1. Личинки
стерляди переходят на активное питание на пятые сутки. Питание
молоди размером 2—13 см однотипно. Основную массу пищевого
комка (90—94% по весу) в низовьях реки составляют гаммариды.
Мизиды, кумовые личинки насекомых и хирономиды играют не-
значительную роль в питании скатывающейся молоди. Интенсив-
ность питания сеголетков во время ската высокая. В июне общий
индекс наполнения пищеварительного тракта стерлядок до 0,5 г
160
составлял 165°/ооо, в июле у молоди средней массой 1,3 г— 164%оо,
а в августе (6,4 г) — 105%оо- В июле молодь стерляди потребляет
в основном гаммарид. Мизиды, кумовые и хирономиды в желудках
стерлядок в это время лета встречаются нерегулярно. Максималь-
ное наполнение пищеварительного тракта (225°/ООо) наблюдается
в вечерние часы.
К осени спектр питания сеголетков становится разнообразнее.
Основу пищи молоди длиной до 20 см в это время составляют хи-
рономиды (52% по весу и 73% по встречаемости). Стерлядки раз-
мером 20—30 см питаются в равной степени кумовыми и хироно-
мидами, составляющими соответственно 48 и 38% по весу. Боль-
шая часть пищи стерляди размером 30—40 см приходится на
долю ручейников и речного скорпиона (77% по весу). Общий ин-
декс наполнения желудочно-кишечного тракта в октябре состав-
ляет 71°/ооо (Полянинова, 1973).
Половозрелая стерлядь ведет в основном ночной образ жизни
и кормится главным образом по ночам или на рассвете. По ночам
же иногда покидает свои постоянные места обитания (ямы) и под-
ходит в поисках корма к песчаным отлогим берегам. Во время
роения поденок («метлицы») поднимается в верхние слои воды.
В целом стерлядь — прожорливая рыба. При изобилии пищи она
наедается настолько, что становится похожа на икряную. В середине
июня у одной самки стерляди из низовьев Урала в желудке оказалось
около 31 тыс. икринок севрюги общей массой 420 г, что описал, по
нашему сообщению, Е. Н. Казанчеев (1981). У 31% стерлядей, вы-
ловленных в период нереста осетровых, в желудках была обнару-
жена их икра. В пище стерляди встречается и рыба — пескари,
бычки, щиповки.
Основные конкуренты стерляди — молодь всех видов осетровых,
пескарь, белоглазка, густера, сазан, вобла, лещ и, вероятно, другие
бентофаги. Врагов у взрослой стерляди немного — это сом и, ве-
роятно, белуга.
Численность и хозяйственное значение. Стерлядь — самый мало-
численный вид из осетровых, обитающих в р. Урал. В промысло-
вой зоне (низовья и дельта Урала) встречается исключительно
редко. В статистике сведений по уловам стерляди в р. Урал нет.
Проведенные еще в 1895 и 1899 гг. пересадки стерляди из Волги в
Урал (600 и 750 шт.) не были результативными.
Поскольку основная масса стерляди обитает в пределах нерес-
товой зоны и питается отложенной икрой осетровых, вряд ли це-
лесообразно заботиться об увеличении ее численности.
Род PSEUDOSCAPHIRHYNCHUS A. NIK-, 1900 — ЛЖЕЛОПАТОНОС
Относится к древнему верхнетретичному пресноводному комплек-
су. Включает 3 вида, два из которых — Р. kaufmanni (Bogdanov) и
Р. hermanni (Kessler) —водятся в Амударье и один — Р. fedtschen-
koi (Kessler) —в Сырдарье.
11-7
161
1. Pseudoscaphirhynchus fedtschenkoi (Kessler) —
сырдарьинский лжелопатонос
Казахское название: тасбек1ре.
Синонимия. Scaphirhynchus fedtschenkoi: Кесслер, Изв. об-ва любит,
естеств., т. X, 1872.
Pseudoscaphirhynchus fedtschenkoi: Берг, Рыбы Туркестана, 1905. — Берг,
Фауна России, Рыбы, т. 1, 1911. — Г. Никольский, Рыбы Таджикистана, 1938.—
Берг, Рыбы пресных вод СССР, ч. 1, 1948.
Распространение. Известен только из Сырдарьи и оросительных
каналов ее бассейна. В озерах и других водоемах отсутствует.
Впервые обнаружен А. П. Федченко в 1871 г. у пос. Чиназ, опи-
сан К. Ф. Кесслером в 1872 г. В пределах Казахстана всегда был
редок, но встречался на всем протяжении реки почти до устья
(Берг, 1905). Более многочислен был в районе Чиназа и выше,
вплоть до низовьев Карадарьи, т. е. в пределах Узбекской ССР и
Таджикской ССР.
Последние достоверные сведения о поимке сырдарьинского ло-
патоноса в Казахстане относятся к 1952—1953 гг., когда в каналах
Ново-Чиилийской оросительной системы было отловлено 4 экз. этой
рыбы (Даирбаев, 1959). Последующие упоминания о встречаемос-
ти лопатоноса в Чардаринском водохранилище (Ерещенко и др.,
1967; Камилов, 1970) были основаны не на поимках особей этого
вида, а лишь на предположении о возможности его захода сюда
из верховьев реки.
Рис. 35. Сырдарьинский лжелопатонос
В настоящее время ареал сырдарьинского лопатоноса (рис. 35)',
видимо, ограничен верховьем Сырдарьи (район Ферганской доли-
ны, Карадарья), но и там он очень редок. Так, в Карадарье от Ан-
дижана до Балыкчи за 9 лет (апрель 1959 — май 1968 гг.) было
выловлено всего 17 экз. (Балтабаев, 1972). Г. К. Камилов (1964)
упоминает о поимке в районе Намангана 3 экз. В пределах Казах-
стана после 1953 г. случаи поимки сырдарьинского лопатоноса не
известны. Очевидно, на сокращение ареала и численности этого
вида повлияло зарегулирование стока Сырдарьи посредством
строительства в 50—60-е годы каскада водохранилищ и гидроузлов.
Промыслового значения не имеет. Нуждается в охране как вид,
находящийся под угрозой исчезновения. По этой категории он за-
несен в «Красную книгу СССР».
162
Описание. D 30—34, А 19—20, Р 1 36—40, V 18—20, спинных
жучек 15—22, боковых 37—46, брюшных 6—10, жаберных тычи-
нок 9, шипов на голове и рыле нет. Тело между рядами жучек по-
крыто костными зернышками. Верхняя губа с выемкой, нижняя
прервана посередине.
Хвостовая нить у одних особей развита хорошо (до 33% длины
тела до последней жучки), у других ее нет. Рыло лопатовидное, у
одних удлиненное, у других укороченное, длина его колеблется от
15 до 30% длины тела.
По длине рыла и хвостовой нити различаются 3 формы, встре-
чающиеся обычно вместе:
brevirostris — короткорылая с длинной хвостовой нитью,
longirostris — длиннорылая без хвостовой нити,
intermedia — рыло слабо удлинено, хвостовая нить есть.
Длина тела до 27 см без хвостовой нити и до 36 см с хвостовой
нитью. Среди исследованных М. Даирбаевым (1959) экземпляров
наибольший имел полную длину 34,7 см.
Сырдарьинские лопатоносы из Карадарьи, исследованные
А. Б. Балтабаевым (1972), все относились к форме brevirostris.
Длина тела их до последней жучки колебалась от 15,3 до 20 см
(в среднем 17 см), а масса — от 21 до 45 кг. Их основные морфо-
логические признаки: D 27—36, А 15—20, спинных жучек 16—19,
боковых 33—43, брюшных 5—9, жаберных тычинок 6—8.
От представителей этого рода из Амударьи сырдарьинский ло-
патонос отличается увеличенным количеством спинных и боковых’
жучек. При поимке неспециалистами лопатонос без хвостовой нити
может быть спутан с молодью шипа, от которого отличается двух-
лопастными губами (у шипа нижняя губа не прервана), отсут-
ствием брызгальца и небахромчатыми усиками (у шипа -— бахром-
чатые) .
Биология изучена очень слабо. Сведений о развитии икры и мо-
лоди, возрасте и росте, а также о миграциях и зимовке нет.
Нерестится весною, очевидно, в одно время с шипом. Особи,
длиной около 25 см (до последней жучки) в марте содержат поч-
ти зрелую икру желтовато-бурого цвета (Берг, 1905). По Г. В. Ни-
кольскому (1938), икра серая, довольно крупная — 1,3—
1,8 мм. По А. Б. Балтабаеву (1972), икра желтая, диаметром 1,0—
1,3 мм.
Плодовитость, определенная К. Ф. Кесслером (1872), у самки
длиной 23 см, была равна 1500 икринкам. Плодовитость, опреде-
ленная А. Б. Балтабаевым (1972), у самки длиной 20 см и массой
39 г (коэффициент зрелости = 9,0), была гораздо выше —3815 ик-
ринок.
Пищу сырдарьинского лопатоноса составляют в основном ли-
чинки насекомых. В Карадарье он потребляет также икринки плот-
вы и леща. Длина кишечника лопатоноса из Карадарьи составляет
60—78, в среднем 71,5% длины тела. Упитанность его здесь, по
Фультону, равна 0,42—0,73 (Балтабаев, 1972).
163
ОТРЯД CLUPEIFORMES — СЕЛ ЬД ЕОБРАЗ НЫ Е
Семейство CLUPEIDAE — СЕЛЬДЕВЫЕ
В Казахстане представители этого семейства обитают только в
бассейне Каспийского моря, где выделяют 2 рода — Clupeonella
(тюлька, килька) и Alosa (сельдь). Боковая линия на теле отсут-
ствует. Брюхо покрыто шиповидными килевыми чешуйками. Хвос-
товой плавник с выемкой.
Род CLUPEONELLA KESSLER, 1877 —ТЮЛЬКА (КИЛЬКА)
Относится к понтическому морскому фаунистическому комплек-
су. Включает 4 вида мелких рыб; три из них обитают в бассейне
Каспия. Анчоусовидная и большеглазая тюльки населяют только
море, преимущественно его среднюю и южную части, где при соле-
ности воды не менее 8%о проходит их жизненный цикл. Каспийская
обыкновенная тюлька шире распространена в бассейне.
Название «килька» по отношению к каспийским представите-
лям рода Clupeonella неправильно. Оно укоренилось среди про-
мысловиков и населения из-за внешнего сходства с настоящими
кильками (род Sprattus), которых нет в Каспии.
1. Clupeonella delicatula (Nordmann) —
обыкновенная тюлька
Обитает в бассейнах Черного, Азовского и Каспийского морей.
Образует 2 подвида. В Казахстане встречается один с пресновод-
ной морфой.
la. Clupeonella delicatula caspia Svetovidov —
каспийская обыкновенная тюлька (килька)
Синонимия. Clupea delicatula: Кесслер, Рыбы Арало-Касп.-Понт. ихтиол,
обл., 1877.— Суворов, Тр. Касп. эксп. 1904, т. II, 1908.
Harengula delicatula: Берг, Мат. к позн. русск. рыбол., т. II, вып. 3, 1913:
Рыбы пресных вод России, 1916.
Clipeonella delicatula: Берг, Рыбы пресных вод СССР, т. 1, 1932. Киселевич,
Сельди Сев. Каспия, 1937.
Clupeonella delicatula caspia: Световидов, Зоол. журн., т. 22, вып. 4, 1943.—
ДАН СССР, нов. сер., т. 46, № 5, 1945. — Ловецкая, Касп. кильки и их промы-
сел, 1951. — Казанчеев, Рыбы Касп. моря, 1963.
Распространение. Каспийский подвид обыкновенной тюльки
(рис. 36) встречается в пелагиали по всему морю, но наиболее
густо населяет прибрежье и мелководье. Заходит в низовья Вол-
ги, Урала и Терека, иногда поднимаясь довольно высоко по тече-
нию. В реки Среднего и Южного Каспия не входит. В море обра-
зует локальные стада. Одно из них размножается в Северном Кас-
пии, другое постоянно обитает в Южном Каспии, где и мечет икру.
Эвригалинная и эвритермная рыба: переносит широкий диапа-'
зон солености воды (от пресной до 36%0) и температуры (от ме-
нее 2,5° до 27,6°).
164
Описание (Световидов, 1945; Берг, 1948; Ловецкая, 1951; Ка-
занчеев, 1981): D III—IV 11—13, в среднем 11,9; А III 14—18, в
среднем 16,2; V 1(6) 7, расположен под передней третью спинного
плавника; жаберных тычинок 50—63, в среднем 57,8; позвонков
42—43, в среднем 42,5; по Т. С. Рассу («Жизнь животных», 1971),
позвонков 41—45.
В процентах длины тела по Смитту (до конца средних лучей
хвостового плавника): наибольшая высота тела 18—24,2, в среднем
20,6; наименьшая высота 5,6—7,8, в среднем 7,0; длина головы
20,9—24,2, в среднем 22,5; высота головы 14,8—17,4, в среднем 16,0;
грудные плавники заостренные, длина их 15,6—18,7, в среднем 16,9;
длина брюшных плавников 8,6—10,5, в среднем 9,5. Глаза срав-
нительно небольшие: 5,3-—6% длины тела, или 22,8—29,1% длины
головы, в среднем 25,9%.
Рис. 36. Каспийская обыкновенная тюлька
Брюхо сильно сжатое с боков, покрытое на всем протяжении
киля от горла до анального отверстия 24—30 (в среднем 27—28)
шиповидными чешуйками. В анальном плавнике два последних
луча удлинены. Рот верхний, беззубый. Нижняя челюсть выдается
вперед, верхнечелюстная кость едва достигает переднего края гла-
за. Чешуя легко опадающая. Окраска типичная для пелагических
рыб: спина и верхняя часть головы серо-зеленые или сине-зеленые,
бока и брюхо серебристо-белые, иногда золотистые.
Длина тела до 14—15 см, масса до 20 г. Средние размеры в про-
мысловых уловах 8—10 см и около 10 г. Самки несколько крупнее
самцов.
Другие виды рода Clupeonella в пределах Казахстана встречаются
редко, живут только в море и внешне отличаются от обыкновенной
тюльки следующими признаками: у большеглазой тюльки более
крупная голова (в среднем 25% длины тела по Смитту) и глаза
(в среднем 28,6% длины головы); у анчоусовидной тюльки более
низкое вальковатое тело с нерезко выраженным килем.
От представителей рода Alosa в бассейне Каспия обыкновенная
тюлька отличается наличием удлиненных лучей в задней части
анального плавника, отсутствием удлиненных крыловидных че-
шуек в основании хвостового плавника и вырезки на середине
верхней челюсти. У предличинок тюльки, в отличие от сельдей, в
желточном мешке есть жировая капля.
В пресных водах (оз. Челкар бассейна Урала, затоны и ильмени
165
низовьев Волги и Урала, водохранилища Цимлянское, Куйбышев-
ское, Волгоградское) образует морфу tscharchalensis (см. ниже).
Биология (источники те же, а также Чаянова, 1951; Приходько,
1952, Никольский, 1971; Коблицкая, 1981).
Половозрелость у тюльки наступает рано: большинство самцов
созревают в годовалом возрасте, основная масса самок — на вто-
ром году жизни.
Икрометание происходит в Северном Каспии почти повсеместно
(особенно в Волжском придельтовом районе, в Урало-Эмбенском
районе и у п-ова Мангышлак), а также на глубине до 30 м в при-
брежных участках Среднего и Южного Каспия. Часть тюльки за-
ходит на нерест в Волгу и Урал.
Нерест в Северном Каспии проходит с середины апреля до
июля, разгар его обычно приходится на май. Оптимальные условия
для нереста: температура воды 11—20°, соленость воды 0,02—8%о-
Однако нерест отмечается и в воде соленостью более 2О°/оо, а живые
оплодотворенные икринки тюльки находили в воде до 15%о-
Соотношение полов у тюльки весной в Северном Каспии близко
1:1. Плодовитость колеблется от 9,5 до 60 тыс. икринок, в среднем
31,2 тыс. Икрометание порционное. Икра пелагическая, прозрач-
ная. Диаметр икринки, по А. А. Ловецкой (1951), равен 0,148—
1,46 мм, по Е. И. Казанчееву (1963),—0,7—1,28 мм. Икринка со-
держит жировую каплю фиолетового цвета размером около трети
диаметра яйца, благодаря чему легко удерживается в толще
воды.
Развитие оплодотворенной икры продолжается при средней
температуре воды 14,5° — 3 суток, при 17,6° около 30 ч. Предличин-
ки вылупляются при длине тела 1,3—2,7 мм. Первое время молодь
держится возле мест нереста, а затем рассредоточивается. К сере-
дине июня она достигает длины 16—18 мм.
Каспийская обыкновенная тюлька — короткоцикличный вид.
Возраст ее не превышает 6 полных лет. В уловах обычно преобла-
дают 2—4-годовалые рыбы. Темп линейного роста ее наиболее ин-
тенсивен до наступления половозрелости. Длина сеголетков в нояб-
ре равна 55—60 мм, средняя длина годовалых рыб — 57—66, двух-
годовалых — 70—80, трехгодовалых — 83—95, четырехгодовалых —
92—-110 мм.
Питается тюлька преимущественно зоопланктоном, в Северном
Каспии главным образом — веслоногими рачками. Кроме того, по-
требляет мизид, личинок хирономид и моллюсков, мальков рыб.
На местах нереста в ее желудках находили собственную икру. Наи-
более интенсивно питается летом. Питание в основном происходит
в светлое время суток: наибольшие индексы наполнения желуд-
ков относятся к середине дня. Ночью питание прекращается или
снижается до минимума, так как ориентация на пищу у нее в ос-
новном зрительная.
Сама тюлька служит важным пищевым объектом сельдей, ло-
сося, белорыбицы, сома, судака, других рыб и тюленя.
166
Коэффициенты упитанности тюльки невелики. Так, в 40-е годы
ее упитанность, по Фультону, весной в Среднем Каспии колебалась
от 0,58 до 1,01, при этом самки были упитаннее самцов, а мелкие
рыбы (7—9 см) упитаннее крупных (10—11 см). В то же время
тюлька очень жирна: в ее теле содержится до 12% жира.
Зимует тюлька обычно в Среднем и Южном Каспии. Весной
мигрирует к северу от мест зимовки и подходит для нереста в при-
брежные мелководья. В Северном Каспии часть ее идет на нерест
в пресные воды, поднимаясь по Волге и Уралу на десятки кило-
метров. У берегов Мангышлака косячный ход начинается в сере-
дине марта при температуре воды 4—5°, максимум его приходит-
ся на середину апреля при температуре воды 7—10°, а конец хо-
да— в первых числах мая. В дельте Волги начало хода во второй
половине апреля, массовый ход — в начале мая при температуре
воды 12—14°.
После икрометания тюлька уходит из Северного Каспия. При
этом в районе Мангышлака появляется в июне. Летом тюлька
обычно избегает поверхностных прогретых слоев воды и держит-
ся преимущественно в слое температурного скачка (15—30 м).
Осенью постепенно продвигается дальше на юг.
В период с июня по октябрь тюлька совершает многократные
миграции к берегу и от берега, связанные с соответствующими пе-
ремещениями ее кормовых организмов (зоопланктона), вызывае-
мыми резкими, но относительно кратковременными изменениями
гидрологического и термического режима прибрежной зоны под
влиянием сгонно-нагонных ветров. При этих миграциях тюлька
формируется в многочисленные и крупные косяки, на подходах ко-
торых к берегу был основан ее летний промысел малыми кошель-
ковыми неводами.
Численность и хозяйственное значение. Начало килечного про-
мысла на Каспии относится к 1925 г. Первыми орудиями были
мелкоячеистые волокуши и самодействующие ловушки — скипас-
ти; затем стали применяться ставные сети, ставные невода и коль-
цевые аламаны. В 1948 г. началось внедрение лова на электро-
свет конусными сетями, а затем и рыбонасосами, при котором ис-
пользовался резко выраженный положительный фототаксис
этих рыб.
До 1950 г. промыслом осваивалась в основном обыкновенная
тюлька, добыча которой доходила до 16,7 тыс. т в год. Освоение
лова на свет привело к увеличению ее добычи почти вдвое —
30,3 тыс. т в 1956 г. Однако дальнейшее развитие промысла затор-
мозилось по ряду причин. Так, береговой лов тюльки приводит к
значительному вылову молоди сельдей и поэтому был запрещен.
Летний лов обыкновенной тюльки аламанами вытеснен более до-
бычливым промыслом анчоусовидной тюльки, уловы которой на
свет достигли в 1961 г. 204 тыс. т.
Кроме того, численность обыкновенной тюльки заметно сокра-
тилась в связи с уменьшением кормности Северного Каспия из-за
167
уменьшения поступления биогенных элементов в результате заре-
гулирования Волги (Осепян, 1972).
Таким образом, в настоящее время она добывается в основном
как прилов к двум другим видам тюлек. В 1974—1979 гг. этот при-
лов составил 0,2—0,8% общего вылова тюлек, или 300—2500 т.
Динамика улова обыкновенной каспийской тюльки за 1950—1961 гг.
такова:
Год Улов, тыс. т Год Улов, тыс. т
1950 20,7 1956 30,3
1951 24,6 1957 20,0
1952 27,2 1958 14,4
1953 24,2 1959 1,1
1954 28,0 1960 1,1
1955 21,8 1961 2,4
Несмотря на сокращение численности и снижение уловов, за-
пасы обыкновенной каспийской тюльки, по мнению специалистов
(Борисов, 1951; Никоноров, 1963; Казанчеев, 1981), еще достаточ-
но велики. Для добычи ее нужно разрабатывать и внедрять новые
способы лова.
В пределах Казахстана основной район летне-осеннего промыс-
ла тюльки расположен у п-ова Мангышлак.
la'. Clupeonella delicatula caspia morpha
tscharchalensis (Borodin)—чархальская селедочка
Синонимия. Clupea cultriventris var. tscharchalensis: Бородин, Ежегодник
Зоол. музея АН, т. I, 1896.
Harengula tscharchalensis: Берг, Мат. к позн. русск. рыболовства, т. II, вып. 3,
1913; Рыбы пресных вод России, 1916.
1 1932е°Пе^а ^е^са^и а тогР^а tscharchalensis: Берг, Рыбы пресных вод СССР,
Clupeonella delicatula caspia morpha tscharchalensisi: Световидов, ДАН
СССР, т. 46, № 5, 1945; Сельдевые, Фауна СССР, Рыбы, т. II, вып. 1, 1952.—
Казанчеев, Рыбы Касп. моря, 1963.
Clupeonella delicatula caspia natio tscharchalensis; Серов, Сб. работ по их-
тиол. и гидробиол. АН КазССР, вып. 1, 1956.
Распространение. Пресноводная и солоноватоводная форма
обыкновенной каспийской тюльки (рис. 37). Обнаружена в оз. Чел-
кар (бассейн Урала), где соленость воды равна 3—7%о, а также
в пойменных озерах и затонах Среднего и Нижнего Урала, в зато-
нах и ильменях низовьев Волги (Световидов, 1945; Берг, 1948; Се-
ров, 1956; Казанчеев, 1963; Шапошникова, 1964). Появилась в во-
дохранилищах волжского каскада и в Цимлянском (Лапицкий,
1970; Шаронов, 1971).
Описание (по указанным выше источникам и сборам С. М. Му-
датова в оз. Челкар в 1980 г.). От типичной формы отличается
лишь меньшим числом жаберных тычинок — 40—50, в среднем 47,4
168
(у типичной 50—63, в среднем 57,8), более высоким телом и более
длинной головой (табл. 67). Некоторые авторы отмечают более
прямой профиль спины. Однако Л. С. Берг (1948) считает, что это
свойственно лишь относительно крупным особям; у мелких селе-
дочек (до 5,5 см) профиль спины не отличается от типичной
формы.
Рис. 37. Чархальская селедочка
Учитывая указанные отличия и обособленный ареал селедочки
из оз. Челкар, Н. П. Серов (1956) предложил выделить ее в само-
стоятельное племя (natio). Однако это вряд ли целесообразно, по-
скольку morpha tscharchalensis обитает не только в оз. Челкар, но
и в бассейне Волги, где ее ареал соприкасается с ареалом типичной
формы. Кроме того, обыкновенная каспийская тюлька (килька)
очень легко образует экологические формы.
Таблица 67. Морфологические признаки чархальской селедочки
(числитель — пределы, знаменатель — среднее)
Кол-во жаберных тычинок Длина тела по Смитту, мм В процентах к длине тела Автор, год
наибольшая высота длина голо- вы
41-50 40-46 51,5 30-70 25,6 18,6-26,5 26,2 24-29 Берг, 1948 Серов, 1956
22,9 25,9
41-50 21,5-28,8 Казанчеев, 1963 Шапошникова, 1964 Мудатов, 1981
47,4 40-46 41—48 32-70 32-65 27,4 24,3* 25,0* 25,4-30
* Длина тела 70 мм.
Так, К- А. Киселевич (1926) сообщает, что в дельте Волги за-
пертая в ильмене в 1925 г. каспийская «килька», перезимовав, к
весне следующего года совершенно изменила свою внешность и
Имела вид чархальской. Н. И. Лапицкий (1970) отмечает, что
IV
169
каспийская тюлька (килька), попав в 1959 г. в Цимлянское водо-
хранилище, через несколько лет образовала здесь, наряду с ти-
пичной формой, и крупную, высокотелую, очень схожую с чархаль-
ской.
Длина тела чархальской селедочки до 110 мм.
Биология почти не изучена. Имеющиеся сведения относятся
лишь к селедочке из оз. Челкар (Серов, 1956; Мудатов, 1981).
Половозрелость наступает на третьем году жизни. Нерест про-
ходит в мае — начале июня в открытой части озера. Икра выметы-
вается в толщу воды двумя порциями. Плодовитость колеблется от
3,8 до 11,65 тыс. икринок. Икринки мелкие: в 1950 г. — 0,15—
0,24 мм в диаметре (Серов, 1956), в 1980 г. — 0,21—0,35 мм.
Возраст известен до 4 лет. Рост более замедленный, чем у ти-
пичной формы, но стабильный, практически не изменившийся за
30 лет (табл. 68).
Таблица 68. Рост чархальской селедочки
в оз. Челкар
Показатель 1950 г. 1980 г.
Возраст, лет 0+ 1 + 2+ з+ 0+ 1 + 2+
Длина, мм 33 45 59 65 30 43 61
Масса, г 0,5 0,9 1,7 2,4 0,4 0,9 1,6
Питается селедочка планктоном: ветвистоусыми и веслоногими
рачками, коловратками, их яйцами, диатомовыми водорослями.
Обитает преимущественно в открытой части озера. К берегу в
массе подходит лишь в апреле, когда на мелководье начинается
интенсивное развитие фито- и зоопланктона. В мае снова отходит
в открытую часть озера, где нерестится, нагуливается и зимует.
В 1950 г. запасы селедочки в оз. Челкар оценивались как доста-
точно большие. Тогда в апреле ее вылавливалось до 50 кг за одно
притонение 15-метровой волокушей. Однако стабильный промысел
налажен не был и вылавливалась она лишь для нужд местного
населения.
В настоящее время промыслового значения не имеет в связи с
сокращением запасов из-за ухудшения гидрологического режима
озера. Служит объектом питания для хищников — жереха и су-
дака.
Род ALOSA CUVIER, 1829 —СЕЛЬДЬ
От рода Clupeonella морфологически отличается в основном
более крупными размерами, наличием вырезки на середине верх-
ней челюсти и удлиненных чешуй в основании хвостового плавни-
ка, отсутствием удлиненных лучей в задней части анального плав-
ника. У предличинок сельдей, в отличие от тюльки, жировой кап-
ли нет.
170
Представители рода Alosa широко распространены в Мировом
океане. В Каспийском море насчитывают 5—6 полиморфных ви-
дов, большинство из которых всю жизнь проводит в море и никог-
да не заходит в реки. Все они относятся к понтическому морскому
фаунистическому комплексу.
Жизненный цикл представителей двух видов: A. caspia (Eich-
wald)—пузанок и A. kessleri (Grimm)—проходная сельдь в
разной степени связан с реками и предустьевыми опресненными
участками моря. Морфологически эти виды отличаются от оби-
тающих только в море видов увеличенным числом тычинок на пер-
вой жаберной дуге: соответственно 55—170 и 20—48.
1. Alosa caspia (Eichwald)—пузанок
Обитает в Черном, Азовском и Каспийском морях. Образует
несколько подвидов, из которых в пресные воды казахстанской
части Каспия заходит лишь северокаспийский пузанок.
la. Alosa caspia caspia (Eichwald)—северокаспийский
(среднекаспийский) пузанок
Казахское название: ^арынсау
Синонимия. Clupea caspia: Eichwald, Bull. Soc. Nat. Moscou, v. XI, 1838;
Fauna caspio-caucasica, 1841.
Alosa caspia: Кесслер, Tp. СПб., об-ва ест., т. I, 1870.
Alosa caspica: Яковлев, Tp. об-ва ест. Казанск. унив., т. 1, 1871.
Clupea caspia: Кесслер, Тр. СПб. об-ва ест., т. V, № 1, 1874.
Clupea (Alosa) caspia: Суворов, Тр. Касп. эксп. 1904, т. I, 1907.
Clupeonella caspia: Берг, Мат. позн. русск. рыбол., т. II, № 3, 1913.
Caspialosa caspia: Берг, там же, т. IV, № 6, 1915.—Берг, Рыбы пресн.
вод России, 1916.
Caspialosa caspia caspia: Аверинцев, Сыч, Тр. Астрах, научн. рыбохоз. ст.,
' т. VIII, № 3, 1930, —Световидов, ДАН СССР, 1932.
Alosa caspia caspia: Световндов, Сельдевые, Фауна СССР, Рыбы, т. II, № 1,
1952. — Казанчеев, Рыбы Касп. моря, 1963.
Распространение. Наиболее широко распространенный подвид
пузанков (рис. 38). Встречается по всему морю. Размножается на
мелководьях Северного Каспия, наиболее интенсивно — в при-
устьевых районах Волги и Урала. В небольшом количестве захо-
дит в низовья этих рек (прежде заходил и в Эмбу), но выше дель-
ты обычно не поднимается.
Описание (Берг, 1948; Световидов, 1952; Казанчеев, 1963):
D III—IV 13—15, в среднем 13,7; AIII(IV) 15—20, в среднем 18,3;
позвонков 46—52, обычно 50. Жаберные тычинки тонкие и длин-
ные, в 1,5—2 раза длиннее жаберных лепестков. Число тычинок
на первой жаберной дуге, по Световидову — 68—135, в среднем
116; по Бергу — 70—149, в среднем 121,5; по Казанчееву — 90—
150, в среднем 120. Чешуй по боку тела 49—54. Зубы очень сла-
бые, на нижней челюсти обычно не заметны даже на ощупь. Пило-
171
рических придатков 30—59, в среднем 40,8. В основании хвостово-
го плавника имеются удлиненные крыловидные чешуйки.
Тело высокое, сжатое с боков, с характерным для пузанка «от-
висшим» брюшком. Глаза крупные. В процентах длины тела по
Смнтту: наибольшая высота тела 22,8—33,1, чаще 25—28; наи-
меньшая 6,8—8,9, в среднем 7,8; длина головы 20,4—29, чаще 25,5—
26,5; высота головы 18,8—22,0, в среднем 20,5; диаметр глаза
19,5—26, в среднем 22,2—23,4.
Рис. 38. Северокаспийский пузанок
Окраска спины и верхней части головы темная, по бокам пере-
ходящая в светло-зеленоватый цвет. За жаберной крышкой почти
всегда имеется темное пятно, иногда за ним на боках тела еще до
7 пятен.
Длина тела до 28 см, чаще 18—22 см. Масса до 220 г, средняя
100- 120 г. Самки обычно крупнее самцов.
Биология (источники те же, а также Остроумов, 1948; Ни-
кольский, 1971; Казанчеев, 1981). Половой зрелости основная
масса пузанка достигает в 3—4 года, но самцы созревают несколько
раньше самок: чаще в 2—3 года. В течение жизни нерестится до
4 раз, обычно 2—3 раза. Однако нерестовое стадо в основном со-
стоит из пополнения, т. е. впервые нерестующих рыб, число кото-
рых, например, в 1957—1961 гг. составляло 69,2—88,7%.
Икрометание происходит с конца апреля до середины июня, в
основном в мае, в пресной или слегка осолоненной воде Северного
Каспия с содержанием солей до 2°/оо, при температуре воды от 14
до 24°, в основном при 18—22°, на глубине от 1 до 3 м, редко
до 6 м.
Икра созревает порционно и выметывается обычно в три прие-
ма с промежутками 7—12 дней. Абсолютная плодовитость от 116
до 218, в среднем 148 тыс. икринок. Диаметр зрелой неоплодо-
творенной икринки 1,1-—1,4 мм, оплодотворенной разбухшей —
1,9—2,9 мм.
Икра полупелагическая, развивается в придонных слоях воды.
Инкубационный период при температуре около 20° длится 42—52 ч,
172
при 18,4° — 44—58 ч. Небольшое количество развивающихся икри-
нок встречается в воде соленостью до 6—8%о- Только что выклю-
нувшиеся личинки имеют длину около 4 мм.
Молодь нагуливается вблизи нерестилищ, достигая здесь наи-
большей концентрации к июлю. В конце лета начинает мигриро-
вать на юг, в основном вдоль западного берега. Производители
отходят на юг моря еще раньше. Таким образом, поздней осенью
пузанок в Северном Каспии уже почти не встречается.
Летние скопления пузанка в Среднем Каспии обычно невелики.
Держится он в это время в толще воды около зоны температурного
скачка на глубине 15—30 м. Осенью и особенно весной подни-
мается ближе к поверхности, в наиболее прогретые слои воды.
Зимует северокаспийский пузанок в Южном, реже в Среднем
Каспии, образуя наибольшие скопления в местах постоянных ни-
сходящих течений на глубине 30—100 м.
Нерестовые миграции начинаются в марте, когда основная
масса пузанков сосредоточивается близ западного побережья, по-
степенно продвигаясь на север. Вдоль восточного берега мигри-
руют разреженные косяки.
В северной части моря первые особи появляются в конце мар-
та — начале апреля при температуре воды 5—7°. Во второй поло-
вине апреля происходит усиление хода, в это время пузанок появ-
ляется у берегов Мангышлака. В начале мая, при температуре во-
ды 17—23°, наблюдается разгар хода с образованием крупных
скоплений рыб. Основные косяки идут в волжские предустьевые
пространства. В приуральские воды их проходит значительно
меньше. Ход в основном заканчивается во второй половине мая на
меляках, в реки обычно идут единичные экземпляры.
Таблица 69. Процентное соотношение
генераций северокаспийского пузанка в уловах
(Казанчеев, 1981)
Максимальный известный возраст пузанка — 9 полных лет.
Основу промысловых и экспериментальных уловов составляют
обычно 3—4-годовалые рыбы. В последнее время в уловах относи-
тельно возросло количество старшевозрастных рыб (табл. 69), что,
видимо, связано с прекращением морского лова пузанка.
Линейный рост в первые три года жизни довольно интенсивен:
средняя длина годовиков — 12, двухгодовиков—-17, трехгодови-
173
ков—19,2, четырехгодовиков — 20,8, пятигодовалых рыб — 22 см.
В 1934—1936 гг. средние размеры трех-четырехгодовалых рыб бы-
ли еще выше: соответственно 20,1 и 22,3 см. О росте массы тела
пузанка дают представление следующие цифры (в среднем): в
2 года — 64 г, в 3 года—-88, в 4 года—117, в 5 лет—146 г.
Кормится пузанок в основном планктонными ракообразными:
веслоногие рачки составляют в его пище более 70, мизиды — около
20% - Интенсивно питается во время преднерестовых и послене-
рестовых миграций, не прекращает питания и во время нереста.
Одна из самых жирных каспийских сельдей: весной содержит
до 13% жира (в Северном Каспии — 6,3—10,3%), осенью — до
18% жира.
Численность и хозяйственное значение. Северокаспийский пу-
занок был одной из важнейших промысловых рыб бассейна. Его
численность и уловы по годам сильно колебались в зависимости от
урожайности поколений и интенсивности промысла: в 1936—
1939 гг. от 5,6 до 75 тыс. т. В 1956—1961 гг. в общих уловах кас-
пийских сельдей он составил 40—77Не-
основная масса его добывалась у западного побережья моря во
время нерестовой миграции и в Северном Каспии на местах не-
реста. В Казахстане основные районы лова располагались у
п-ова Мангышлак и в приуральских водах.
Прекращение морского промысла привело к резкому сокраще-
нию уловов пузанка. В 1974—1979 гг. его контрольные среднегодо-
вые уловы колебались от 400 до 1000 т. При возобновлении мор-
ского лова добыча пузанка может возрасти в 5—6 раз.
2. Alosa kessleri (Grimm)—черноспинка
Обитает в Черном, Азовском и Каспийском морях. Образует
3 подвида. Из них в Казахстане встречаются два.
2а. Alosa kessleri kessleri (Grimm) — черноспинка
(бешенка, залом)
Казахское название: крражон, царацаптал балык;
Синонимия. Clupea alosa (поп Linne): Pallas, Reise, v. I, 1771.
Clupea kessleri: Гримм, Астраханская селедка, СПб., 1877.
Clupeonella kessleri: Берг, Мат. позн. русск. рыбол., т. II, № 3, 1913.
Caspialosa kessleri: Берг, Мат. позн. русск. рыбол., т. IV, № 6, 1915. — Рыбы
пресных вод России, 1916. — Киселевич, Промысл, рыбы Волго-Касп. района.
Астрах., 1926. — Берг, Рыбы пресных вод СССР, т. I, 1932, 1948.
Caspilosa caspia kessleri: Киселевич, Изв. отд. рыбол. и научно-пром, исслед.,
т. I, 1918; Касп.-волжские сельди, т. 1, 1923. — Световидов, ДАН СССР, 1932.
Caspialosa kessleri kessleri: Световидов, Зоол. жури., т. XXII. № 4, 1943;
Промысл, рыбы СССР, 1949.
Alosa kessleri kessleri: Световидов, Сельдевые, Фауна СССР, Рыбы, т. 11,
№ 1, 1952. — Казанчеев, Рыбы Касп. моря, 1963.
Распространение. Черноспинка (рис. 39) в Каспии встречается
174
чаще в западной части. Проходная форма, идущая для икромета-
ния, главным образом в Волгу и в небольшом количестве в Урал.
Описание (Берг, 1948; Световидов, 1952; Казанчеев, 1963,
1981). По многим признакам близка к волжской сельди, от которой
отличается главным образом признаками, связанными с питанием.
Рис. 39. Сельдь черноспинка
Так, количество жаберных тычинок у черноспинки гораздо мень-
ше—57—95 (среднее по разным выборкам 71,9—78,2); они часто
короче жаберных лепестков, у крупных рыб толстые и грубые,
частью обломанные, у некоторых рыб тычинки имеют боковые ши-
пики.
Довольно хорошо развиты у черноспинки зубы, всегда замет-,
ные на ощупь. Больше у нее и пилорических придатков: 36—62, в
среднем 48,2. Обычно много у нее жировых отложений в полости
тела и в спинной мускулатуре.
Голова наиболее широкая из всех форм вида: межглазничный
промежуток 18,4—23,0, в среднем 20,3% длины головы. Чешуя си-
дит плотнее, чем у волжской сельди.
Сведения о количестве позвонков у черноспинки противоречивы.
По Бергу, их 51—54, чаще 53; по Казанчееву, — 50—54, в среднем
52,6, что почти совпадает с числом позвонков у волжской сельди
(48—54, чаще 52—53). Однако, по Световидову, позвонков у черно-
спинки заметно меньше — 47—50, в среднем 48,9.
Окраска спины и верхней части головы темно-фиолетовая, часто
почти черная. Грудные плавники темные. Особенно сильно пиг-
ментированы конец рыла и основание грудных плавников. Неред-
ко за жаберной крышкой сверху есть темное пятно. Во время не-
реста спина и бока становятся зеленовато-серыми, а на жаберных
крышках появляются желто-бронзовые пятна.
Самая крупная из каспийских сельдей. Абсолютная длина те-
ла до 52 см, масса до 1800 г. Средняя длина половозрелых рыб око-
ло 40 см, средняя масса 950 г.
Биология (источники те же). Половой зрелости достигает в
4—5-годовалом возрасте, редко в 3—6-годовалом. Раньше считали,
что черноспинка нерестится только один раз в жизни, после чего
погибает. Теперь установлено, что это не так. Большинство самок
175
действительно гибнет после нереста от истощения. Однако часть
их и многие самцы выживают и через год нерестятся повторно, а
некоторые размножаются до трех раз в жизни, что подтверждает-
ся и нерестовыми марками на чешуе. Повторно нерестующие рыбы
составляют до 20% нерестового стада.
Черноспинка, в отличие от волжской сельди, входит в реки с
недоразвитыми половыми продуктами (обычно на III стадии зре-
лости), которые дозревают во время хода. В Урале она поднимает-
ся до Оренбурга, по Волге прежде доходила до г. Горького, входи-
ла в Каму и Оку. Теперь ход заканчивается у плотины Волгоград-
ской ГЭС, ниже которой и расположены оставшиеся нерестилища.
Нерест происходит на значительном протяжении реки, как и у
волжской сельди, не только в местах с замедленным течением (за-
водях, затонах), но и на стрежне. Начало нереста определяется
прогревом воды до 14—15° и спадом весеннего паводка, что обыч-
но наступает во второй половине мая — начале июня. Разгар не-
реста — с середины июня до середины июля при температуре воды
18—24°.
Икра созревает и выметывается несколькими порциями. Плодо-
витость 135—312, в среднем 218 тыс. икринок-—больше, чем у
других каспийских сельдей. Зрелые икринки имеют размеры 1,31—
1,51 мм, оплодотворенные и набухшие — 2,62—3,68 мм (по Казан-
чееву, 1981, — 2,5—4,1 мм). Выклев личинок при средней темпера-
туре 19,6° происходит через 49 ч, при 20,4° — через 42 ч.
Молодь в реке проводит около 2 месяцев. В Северном Каспии
она начинает появляться в конце июля. Наиболее интенсивный
скат молоди происходит в августе — сентябре. А в октябре начи-
нается откочевка молоди на юг моря, в основном вдоль западного
берега. В Южном Каспии черноспинка живет, не совершая мигра-
ций на север, до наступления половозрелости.
Известный максимальный возраст — 7 полных лет. В низовьях
Волги и Урала уловы состоят преимущественно из 4—6-годовалых
рыб.
Темп роста самый быстрый среди каспийских сельдей. В сред-
нем за 1968—1972 гг. он составил: в годовалом возрасте 9,8 см, в
2 года—18,6, в 3 года — 27,8, в 4 года — 32,5, в 5 лет —34,3,
в 6 — 35,5 см.
Взрослая черноспинка — хищник, питающийся преимуществен-
но мелкой рыбой (тюлька, атерина, молодь судака и др.), реже ра-
кообразными (мизиды, бокоплавы) и личинками насекомых. Ос-
новная пища сеголетков — веслоногие и ветвистоусые рачки. Во
время нерестовой миграции почти не питается, расходуя накоплен-
ные во время нагула энергетические запасы.
В море черноспинка — самая жирная из каспийских сельдей.
Содержание жира у рыб, выловленных в Среднем Каспии, дости-
гает 19%, в Северном Каспии и дельте Волги—11—18, в районе
нерестилищ до икрометания — около 7, после нереста — не более 3,
а в среднем 1,2%.
176
Зимует на юге Каспия, не образуя плотных скоплений. Точных
данных о распределении ее в море в это время нет. Весеннюю миг-
рацию начинает во второй половине марта. Большей частью подхо-
дит к западному берегу, вдоль которого и движется на север. Близ
восточного берега мигрирует немного черноспинки.
Ход происходит при температуре воды 6—13°. В Северный Кас-
пий входит в начале апреля. Здесь ее ход продолжается до конца
мая. В дельте Волги появляется обычно в середине апреля. Пер-
выми мигрируют более крупные рыбы, мелкие — позднее, в середи-
не мая. Соотношение полов в нерестовых косяках в целом пример-
но равное.
Численность и хозяйственное значение. Наиболее ценная по пи-
щевым качествам сельдь Каспийского моря. Добывается весной во
время нерестовых миграций. Величина годовых уловов сильно
колеблется. В начале столетия добывалось до 30 тыс. т. После
зарегулирования Волги уловы сильно сократились: в 1956—
1961 гг. — от 0,56 до 4,89, в 1974—1978 гг. — от 0,2 до 1,2 тыс. т
в год.
Необходима строгая регламентация вылова этой сельди.
26. Alosa kessleri volgensis (Berg) —
волжская многотычинковая сельдь
Казахское название: машиабац, Волга майшабагы
Синонимия. Alosa caspia (non Eichw.): Яковлев, Tp. Об-ва естествознан.
Казанск. унив., т. 1, № 2, 1871.
Clupea caspia (non Eichw.): Гримм, Астраханская селедка, СПб., 1887.—
Арнольд, Тр. Касп. эксп. 1904 г., т. 1, 1907.
Clupeonella caspia volgensis: Берг, Мат. позн. русск. рыбол., т. II, № 3, 1913.
Caspialosa caspia volgensis: Берг, Рыбы пресных вод России, 1916. — Кисе-
левич, Касп.-волжск. сельди, т. I, 1923.—Световидов, ДАН СССР, 1932.
Caspialosa volgensis: Киселевич, Промыс. рыбы Волго-Касп. района, Астрах.,
1926. — Берг, Рыбы пресных вод СССР, т. I, 1932.
Caspialosa kessleri volgensis: Световидов, Зоол. журн., т. XXII, № 4, 1943.
Alosa kessleri volgensis: Световидов, Сельдевые, Фауна СССР, Рыбы, т. II,
вып. 1, 1952. — Казанчеев, Рыбы Касп. моря, 1963.
Распространение. Проходная форма. В Каспийском море оби-
тает повсеместно. В массе заходит в Волгу, значительно меньше —
в Урал, единично — в Терек, в низовьях которых, а также перед
устьями, нерестится. Прежде по Волге сельдь (рис. 40) поднима-
лась до г. Горького, теперь ее ход ограничен плотиной Волгоград-
ской ГЭС. В Урале доходит до с. Котельникове (около 300 км от
устья).
Описание (Берг, 1948; Световидов, 1952; Казанчеев, 1981):
D III—IV 12—15, чаще 14; А III 15—21, чаще 18; поперечных ря-
дов чешуй 55, позвонков 48—54, чаще 51—53; пилорических при-
датков 30—56, в среднем 44,3; жаберных тычинок 87—158, в сред-
нем по разным выборкам 121,3—125,1. Тычинки длинные и тонкие,
12-7
177
обычно длиннее жаберных лепестков. Зубы развиты слабо, иногда
почти незаметны на ощупь. На переднем конце нижней челюсти
есть небольшой бугорок снизу.
Тело удлиненное, слегка вальковатое, наибольшая высота его
19,2—26,8, в среднем 23—24% длины. Голова довольно большая,
широкая, но низкая; длина ее обычно больше высоты тела: 21,5—
28,6, в среднем 24,8% длины тела, высота головы 15,8—18,5, в
среднем 17,4% той же длины. В процентах длины головы: межглаз-
ничный промежуток 14,9—19,6, в среднем 17,5; диаметр глаза
17,2—21,8, в среднем 19,7.
Рис. 40. Волжская сельдь
Жировых отложений в полости тела обычно нет или очень мало.
Окраска спины оливково-зеленая. Темных пятен на боках тела
обычно нет.
Длина тела до 40 см. Средние размеры половозрелых рыб в ре-
ке 26—31 см. Масса до 600 г, в среднем около 300 г. Самки крупнее
самцов.
Биология (источники те же, а также В. С. Танасийчук, 1948;
Н. П. Танасийчук, 1948). Зимует на юге Каспия. Миграции на се-
вер начинает рано — в феврале при температуре воды около 8°,
продвигаясь как по западной, так и по восточной части моря, в от-
далении от берега.
В Северном Каспии появляется в конце марта — начале апреля,
наибольший подход наблюдается при температуре воды 10—13°.
В предустьевое пространство Волги подходит в середине апреля, в
приуральских водах появляется обычно в конце апреля. Разгар
хода приходится на середину мая при температуре воды 14—17°,
конец хода отмечается во второй половине июня.
В реки входит в начале мая, двигаясь стрежневой частью рус-
ла и днем, и ночью, со среднесуточной скоростью около 20 км.
Массовый ход продолжается не более двух недель. В начале хода
обычно идет более крупная сельдь, в конце хода — более мелкая.
Нерест проходит в низовьях Волги и Урала на течении или в
затонах и заводях. Часть рыб мечет икру еще перед входом в дель-
ту, но в пресной воде. Начало нереста при температуре воды около
13°, разгар его при 15—19° во второй половине мая.
178
Основная масса сельди мечет икру вечером, после 16 ч. Ночью
1ерест не отмечался. Икрометание порционное, с интервалами
7—10 дней. Плодовитость 100—281 тыс. икринок, в среднем
179,4 тыс. Икринки полупелагические, развивающиеся во взвешен-
ном состоянии во время сноса в придонных слоях воды. Длитель-
ность инкубационного периода 44 ч при температуре воды 19,6—
21,5°.
Молодь сельди, скатываясь по реке, в светлое время держится у
поверхности, ночью — в толще воды. С конца июня до сентября
она скапливается в Северном Каспии, а затем откочевывает на юг,
преимущественно вдоль западного берега моря.
Массовой гибели отнерестившихся производителей не проис-
ходит, хотя часть особей погибает. Кроме того, ослабевшие покат-
ники становятся легкой добычей хищников. Скатившиеся в море
производители без задержки отходят на юг, не образуя больших
скоплений.
Половой зрелости волжская сельдь достигает в основном в
3—4-годовалом возрасте, реже к 5 годам. Самцы обычно созрева-
ют на год раньше самок и часть их нерестится уже в 2 года. Судя
по нерестовым маркам на чешуе, сельдь размножается до 4 раз в
жизни. Однако нерестовые косяки состоят в основном из впервые
созревающих особей. Повторно нерестующие составляют до 25%!
нерестового стада.
Продолжительность жизни волжской сельди — семь полных
лет. Основную массу уловов обычно составляют 3—5-годовалые
рыбы. Линейный рост характеризуется следующими цифрами:
средняя длина годовиков—10,6 см, в 2 года—18,6, в 3 — 24,2, в
4 — 27,6, в 5 лет — 30,5 см.
Питается волжская сельдь ракообразными (веслоногие рачки,
мизиды, амфиподы и др.) и рыбами. В районе п-ова Мангышлак в
ее пище были атерина, тюлька, бычки. Во время нерестовой мигра-
ции в реке питания не прекращает. Жирность ранней весной до-
стигает 10%, у ходовой в дельте Волги снижается до 6—7%.
Численность и хозяйственное значение. Одна из наиболее цен-
ных промысловых рыб бассейна. В прежние годы вместе с пузанком
составляла основу каспийского сельдяного промысла. Уловы ее в
30-е годы достигали 70 тыс. т в год.
В последнее время численность волжской сельди резко сократи-
лась, по-видимому, в связи с ухудшением условий размножения и не-
рациональным промыслом. В 1956—1961 гг. ее уловы колебались
от 2,8 до 6,3 тыс. т, а в последние годы составляли лишь 5—10 т.
Ловится главным образом весной во время нерестовых миграций.
Сохранение запасов связано с мелиорацией нерестилищ и рег-
ламентацией промысла.
179
ОТРЯД SALMONIFORMES — ЛОСОСЕОБРАЗНЫЕ
Семейство SALMONIDAE — ЛОСОСЕВЫЕ
Род SALMO LINNE, 1758 —ЛОСОСЬ
Положение лососей рода Salmo в системе фаунистических
комплексов пока не достаточно ясно. Так, Г. В. Никольский отнес
их сначала к арктическому пресноводному комплексу (1953), а
потом — к бореальному предгорному (1980).
Вопрос о систематическом положении лососей южных морей
СССР, в том числе Каспия и Арала, дискутировался в ихтиологи-
ческой литературе около 100 лет. Сейчас можно считать, что он,
наконец, решен благодаря проведенным в последние годы эмбрио-
логическим, кариологическим, остеологическим и гематологиче-
ским исследованиям, показавшим генетическую близость этих ло-
сосей к виду Salmo trutta—-кумжа и четкое отличие их от вида
Salmo salar — атлантический лосось (Никольский, 1971; Кудер-
ский,1974).
1. Salmo trutta Linne— кумжа (проходной лосось)
Вид распространен от Пиренеев до Исландии и р. Печора. Об-
разует проходные и жилые формы в Средиземном, Балтийском,
Белом, Черном, Каспийском и Аральском морях, выделяемые в
подвиды и морфы. В Казахстане встречаются представители двух
подвидов.
la. Salmo trutta caspius Kessler — каспийский лосось
Казахское название: албырт, Каспий албырты.
Синонимия: Salmo caspius: Кесслер, Рыбы Арало-Касп.-Понт. ихтиол.
Обл., 1877.
Salmo trutta caspius: Каврайский, Лососевые Кавказа, т. 2, 1897. — Берг,
Ежегодник Зоол. музея АН, т. 13, 1908.
Salmo trutta labrax: Берг, Рыбы пресных вод России, 1916.
Salmo caspius: Державин, Бюл. Всекасп. н.-рыбохоз. эксп., № 5—6, 1932.
Salmo trutta caspius: Берг, Рыбы пресных вод СССР, ч. 1, 1948. — Казан-
чеев, Рыбы Каспийского моря, 1963. — Коблицкая, Определитель молоди пресно-
водных рыб, 1981.
Распространение. Подвид кумжи (Salmo trutta Linne), насе-
ляющий Каспийское море, главным образом его юго-западную
часть, откуда входит в реки, стекающие с Кавказского хребта.
В Северном Каспии малочислен. В Казахстане редок (рис. 41).
Еще в начале XX столетия в небольшом количестве заходил в
Волгу и Урал (Берг, 1928; Державин, 1941). В последние годы све-
дений о поимках лосося в этих реках нет (Коблицкая, 1981). За-
несен в «Красную книгу Казахской ССР» (1978).
180
Описание (Берг, 1948; Державин, 1949). Самый крупный пред-
ставитель рода лососей. Отдельные особи его куринского стада
прежде достигали массы более 50 кг. Обычная масса половозре-
лых особей 2—7 кг, длина до 100 см.
D III IV 8—10, А II—III 7—9, Р I 11—12, V I (7) 8, 1.1. 115—
132, жаберных лучей 9—11, жаберных тычинок на первой дуге
16—22, позвонков 57—60.
Рис. 41. Каспийский лосось
Длина головы у взрослых лососей составляет 17—22, верхне-
челюстной кости 5,4—8,2, нижнечелюстной 10,3—12,8% длины те-
ла до конца средних лучей хвостового плавника. Наименьшая
высота тела обычно укладывается более 2 раз в длине хвостового
стебля. Задний конец верхнечелюстной кости заметно заходит за
глаз. Расстояние от жирового плавника на верхней части хвостово-
го стебля до первых лучей хвостового плавника равно высоте
анального плавника или меньше ее.
Бока тела у молоди покрыты большими овальными темными
пятнами, у взрослых рыб — обычно многочисленными х-образными
темными пятнышками. У взрослых во рту довольно крупные зубы:
на верхней челюсти их 9—14, на нижней 10—14, на языке 8—10.
Весьма близок к черноморскому лососю (Salmo trutta labrax
Pallas), от которого отличается более низким хвостовым стеб-
лем. У обоих подвидов, как и у кумжи, диплоидное число хромо-
сом (2 п) равно 80 (Дорофеева, 1965).
Биология каспийского лосося из казахстанской части ареала
не известна. Другие его популяции изучены довольно хорошо
(Берг, 1948; Державин, 1941, 1949; Казанчеев, 1963, 1981; Николь-
ский, 1971 и др.).
Обычно это проходная рыба, размножающаяся в реках с галеч-
никовым дном. Там она проводит первые годы жизни, а затем ска-
тывается в море, где растет и созревает. Но есть у нее и карлико-
вые самцы, созревающие в реке и не выходящие в море.
Каспийский лосось образует местные стада (популяции), свя-
занные с отдельными реками (Кура, Терек, Араке, Самур и др.) и
различающиеся размерами и морфо-экологическими признаками.
Так, рыбы куринского стада относительно более крупные (средняя
181
навеска прежде была 13 кг, в последние годы, после зарегулирова-
ния реки, около 10 кг при длине тела до 130 см), нерестятся один
раз в жизни в возрасте 5—9 лет. Лососи из стад, приуроченных к
небольшим речкам, мельче (до 3 кг), созревают раньше (на треть-
ем году) и нерестятся 5—6 раз в жизни.
Нерест проходит с октября до января при температуре воды
3—13°. Плодовитость от 1,5 тыс. до 45 тыс. икринок, в том числе у
куринского лосося прежде—16—30 тыс. икринок, в среднем около
23 тыс., в последние годы — около 10 тыс. икринок. Икра донная,
неклейкая, зарывается в гальку, диаметр ее 5—6,5 мм.
При температуре воды 10—11° икра развивается 26—27 суток,
а при более низких температурах до 90 суток. Покатной стадии ло-
сось частью (10—2О°/о) достигает уже на первом году жизни в ре-
ке и скатывается в море в период с декабря по апрель. Основная
же масса его остается в реке на второй год.
Максимальный известный возраст каспийского лосося 10 лет.
В последние годы особи старше 6 лет встречаются единично. Ос-
новной рост лосося происходит в море, где представители курин-
ского стада, например, к 4 годам достигали прежде средней длины
86—97 см и массы 7—11 кг. Сейчас рост лосося замедлился: 4-лет-
ки имеют среднюю длину тела 82 см и массу 6,9 кг, а 6-летки —
92 см и 10,1 кг.
Молодь питается личинками водных и воздушных насекомых,
бокоплавами; взрослые — рыбой (килька, атерина, сельдь), мизи-
дами, креветками, в реке не питаются.
Численность каспийского лосося никогда в нашем столетии не
была особенно велика. Однако в 1936—1939 гг. его уловы достигали
620 т. В последние десятилетия вылов лосося на юго-западе Каспия
сильно сократился. В других частях ареала лосось ловится еди-
нично. В 1970 г. его учтенный вылов не превышал 5 т.
Резкое сокращение численности лосося связано в первую оче-
редь с нарушением условий его естественного воспроизводства в
связи с зарегулированием стока рек и нерациональным промыслом.
Восстановление запасов этой ценной промысловой рыбы зави-
сит в основном от масштабов ее искусственного разведения с вы-
ращиванием молоди до покатной стадии (смолт).
16. Salmo trutta aralensis Berg —аральский лосось
Казахское название: албырт, Арал албырты.
Синоним и я. Salmo trutta aralensis: Берг, Ежегодник Зоол. музея АН,
т. 12, 1908.— Г. Никольский, Рыбы Аральского моря, 1940. — Берг, Рыбы прес-
ных вод СССР, ч. 1, 1948.
Распространение. Самая восточная форма кумжи, проникшая в
бассейн Арала из Каспия по древней системе Узбоя (Берг, 1961).
Известна только из Аральского моря и низовьев Амударьи (до
Тортколя). Была очень малочисленна: в год вылавливалось не
более 10 экз., в основном в южной части моря (Никольский, 1940).
182
В последние годы случаи поимки этого подвида не известны.
Есть основания считать, что он находится под угрозой исчезнове-
ния вследствие зарегулирования и сокращения стока Амударьи,
падения уровня Аральского моря. Внесен в «Красную книгу СССР»
по категории видов, находящихся под угрозой исчезновения.
Описание (Никольский, 1940; Берг, 1948): D III 10—11,
А II—III 8—9, 1.1. 117—120, жаберных тычинок на первой дуге
18—22, позвонков 53 (без уростиля).
Длина головы составляет 23—26% длины тела до конца чешуй-
ного покрова*, наибольшая высота тела 24—28, наименьшая —
9-—10, длина хвостового стебля 18—24, длина Р 13—16, длина V
10—12,5, антедорсальное расстояние 40—46, антевентральное—
49—56%. Длина рыла составляет 27,5—33,6% длины головы, дли-
на верхнечелюстной кости 40—47,5, длина нижней челюсти
62—69%.
Тело вальковатое, спина темная, бока серебристые, покрытые
многочисленными Х-образными черными пятнами, которые вместе
с жировым плавником служат хорошим отличительным признаком
этой рыбы. Длина тела до 100 см, масса до 13,5 кг (рис. 42).
Рис. 42. Аральский лосось
От каспийского лосося отличается в основном меньшим числом
позвонков, более крупной головой и длинными челюстями. От аму-
дарьинской форели, в первую очередь,-—меньшим числом позвон-
ков и большим количеством жаберных тычинок.
Биология. Для размножения входил в Амударью. Возможно,
отдельные особи поднимались на нерест до Кафирнигана и Вахша
(Максунов, 1968). Весной обычно ловились особи с незрелой икрой.
Единственный экземпляр, добытый осенью, имел почти зрелую ик-
ру (Никольский, 1940). Известны взрослые самцы с крючкообраз-
но загнутой нижней челюстью («брачный наряд»?).
Других сведений о биологии аральского лосося нет. Методы
его искусственного разведения не разработаны.
* Так у Г. В. Никольского (1940).
183
Род HUCHO GUNTHER, 1866 —ТАЙМЕНЬ
Представители этого рода относятся к непроходным лососевым
рыбам. Обитают в горных притоках крупных рек, совершая недлин-
ные кочевки. Отличаются от близкородственных родов Salvelinus и
Brachymystax строением скелета головы, расположением зубов на
сошнике и челюстях. Верхнечелюстная кость длинная, заходит за
вертикаль заднего края глаза.
Из 4 видов этого рода, выделенных Л. С. Бергом (1948), в Ка-
захстане обитает один.
1. Hucho taimen (Pallas) —таймень
Казахское название: таймень.
Синонимия. Salmo taimen: Pallas, Reise, v. II, 1773.
Salmo fluviatilis: Pallas, Zoogr. Rosso-Asiat., v. Ill [1814].
Salvelinus taimen: Берг, Рыбы бассейна Амура, 1909. — Навозов, Вести, ры-
бопр., 1912.
Hucho taimen: Берг, Рыбы пресных вод России, 1916.
Распространение. Таймень (рис. 43) —наиболее крупный пред-
ставитель бореально-предгорного фаунистического комплекса —
распространен от левобережных притоков Волги на западе до
Индигирки на востоке. Характерен по всему течению сибирских
рек, но наиболее многочислен в их верховьях. Обычен в бассейне
Амура, реках и озерах Монголии. В Казахстане встречается в пра-
вобережных притоках Иртыша, откуда на зиму уходит в основное
русло и водохранилища.
Рис. 43. Таймень
Описание. D III—V 9—12, А III—V 8—10, Р I—-III 12—16, V 1—II
8—10, позвонков 64—72, жаберных лучей 9—13, жаберных тычи-
нок 12—18. В боковой линии исследователи указывают различное
число чешуи. По Л. С. Бергу (1949), 193—242 поперечных ряда, по
Ф. Н. Кириллову (1972), — 190—242 в боковой линии, по «Рыбам
МНР» (1983), — 111—134, по данным В. И. Ерещенко * — 125—141.
Вероятно, как указывала Г. X. Шапошникова (1968) для ленка,
* Для написания очерка использованы материалы по 15 тайменям из р. Ир-
тыш, переданные нам В. И. Ерещенко.
184
происходит методическая неуточненность. В одних случаях указы-
вается число поперечных рядов чешуй (ио пишется — в боковой
линии), в других приводится число прободенных чешуй. Общий
диапазон оказывается очень широким —111—242. Для использо-
вания этого признака в систематических целях необходимо прове-
дение специальных исследований, так как систематика рода тай-
меней основана на количестве поперечных рядов чешуй: около 200
у Н. taimen и Н. hucho; около 150 у Н. ishikawai; около 100—120 у
Н. perryi. Нетрудно заметить, что по этому признаку описываемого
тайменя (Н. taimen) можно отнести к любому из указанных видов.
В таком же методически неуточненном аспекте находится и
второй систематический признак — число жаберных тычинок. По
их количеству разделяются виды Н. taimen и Н. hucho.
У первого 11—12 жаберных тычинок, у второго—16. Однако
внутри ареала для Н. taimen указывают значительно большее ко-
личество тычинок — до 21 (с зачаточными бугорками). Так, по
Ф. Н. Кириллову (1972), у тайменя из р. Лены и р. Оленек жа-
берных тычинок 10—16, с зачаточными—14—18; по данным
В. И. Ерещенко, у тайменя из р. Иртыш 14—21 жаберных тычинок.
Видимо, Л. С. Берг при разделении рода Hucho на виды исполь-
зовал не ключевые признаки. В описании Н. taimen он указывает:
жаберных тычинок (9) 11—12, считая с зачаточными — до 14—46.
Признак неточный, и последующие исследования внесли в него су-
щественные коррективы. Предложенной Л. С. Бергом определи-
тельной таблицей рода Hucho пользоваться практически нельзя, и
этот род ждет своих исследований.
В настоящее время нам представляется более верным рассмат-
ривать род Hucho как монотипический, с одним полиморфным ви-
дом Н. hucho Linne, как это сделано с ближайшими родственны-
ми родами — Salvelinus и Brachymystax. В известной мере такая
работа начата. К. Hensel и J. Holcik (1983) предложили объеди-
нить виды Н. hucho и Н. taimen, с чем, на наш взгляд, необходимо
согласиться.
Морфологическая характеристика тайменя из бассейна р. Ир-
тыш, добытого 29—30 апреля 1959 г. у подходного канала Усть-
Каменогорской ГЭС и 17—18 сентября 1961 г. в р. Курчум:
D IV—V 9—11, А IV 8—9, Р I—II 11—16, V I—II 8—10, жаберных
лучей 10—12. Остальные признаки представлены в таблице 70.
Интересной особенностью тайменя из р. Иртыш является боль-
шая изменчивость счетных признаков и значительно меньшая из-
менчивость пластических. Так, по количеству лучей в спинном
плавнике у 15 особей отмечены 5 различных сочетаний: IV — 9,
IV —10, IV—11, V — 9 и V—10. Столько же сочетаний в лучах
грудного плавника и четыре — в брюшных плавниках. В признаках
по числу чешуй наблюдаются большие хиатусы в ряду распреде-
ления (возможно, в связи с малочисленностью материала), а по
количеству жаберных тычинок выявляются две группы: 15—16 ты-
чинок (40%) и 19 тычинок (33%).
185
Таблица 70. Морфологическая характеристика тайменя
из р. Иртыш
Признак От-до Средн. (М + ш)
Длина рыбы, см 57,1—93,5 69,09
по Смитту, см 55,5-90,0 66,67
тела, см 52,2-85,5 62,33
Полная масса, кг 2,1—8,2 3,83
Масса без внутренностей, кг В % длины по Си 1,9-7,4 /1ИТТУ 3,47
Длина головы 21,8-23,7 22,82 + 0,13
рыла 5,8-6,6 6,07 + 0,06
верхней челюсти 7,6-9,1 8,4±0,12
Ширина верхней челюсти 2,1—2,7 2,44 + 0,04
Длина нижней » 12,5-14,1 13,53+0,11
Заглазничный отдел головы 13,2-15 14,24-0,11
Диаметр глаза 2,5-3,2 2,83 + 0,05
Ширина головы 9,7-14,4 12.33-0,34
Ширина лба 5,8-6,7 6.35+0,08
Высота головы у затылка 11,4-15,3 13,04+0,29
Высота головы через глаз 8,2-9,8 8,97 + 0,11
Наибольшая высота тела 17,1—22 20.05-0,35
Наименьшая » » 6,1-7,6 6,8+0,09
Толщина тела 11,8—15,6 13,37-0,29
Длина хвостового стебля . 10,8-14,2 13,01±0,2
грудного плавника 8,25-12,4 11,27 + 0,25
брюшного » 8,5-10,2 9,56 + 0,11
Антедорсальное расстояние 45—49 46,94 + 0,29
Постдорсальное » 35,5—40,5 37,53 L0,26
Антеанальное » 73-81 76,45 + 0,52
Антевентральное » 53-62 57,18 t0,57
Длина основания D 8,9-11,2 10,02 + 0,16
Высота D 9,05-10,2 9,58+0,1
Длина основания А 6 3-8,1 7,37±0 21
Высота А 9-10,8 10,01 ±0,14
Расстояние Р—V 31,5-35,5 34,0±0,21
Расстояние V—А 17—20,2 18,64 + 0,25
Длина верхней лопасти С 11-12,5 11,9±0,22
Длина нижней лопасти С 11-12,5 11 ,а7 + 0,26
Счетные признаки
Чешуи в 1. 1
Чешуи над 1. 1.
Чешуи под 1. 1.
Чешуи по боку хвостового стебля
Жаберных тычинок на 1-й дуге
125-141
27-38
22-34
24-30
14-21
1130,4±1,21
33,0±0,78
30,15-0,87
26,73 ±0,54
17,4±0,54
Половой диморфизм в счетных признаках не обнаружен. Воз-
растная изменчивость не изучена.
Биология. Размножается таймень весной вскоре после стаивания
льда. Судя по состоянию гонад особей, добытых в русле Иртыша в
конце апреля, в это время встречаются самки, готовые к откладке ик-
186
ры, и текучие самцы. Для нереста заходит в горные реки довольно
высоко по течению в поисках мест с крупным галечниковым грун-
том без иловых наносов.
После нереста задерживается в горных притоках почти на все
лето, лишь к осени скатываясь на зимовку в русло Иртыша *. По
левые продукты в сентябре достигают III стадии зрелости. Плодо-
витость тайменя из водоемов Казахстана неизвестна, так же как
и сроки развития икры и молоди. В пределах ареала для тайменя
указывается абсолютная плодовитость от 7 до 34 тыс. икринок
(Берг, 1949; Кириллов, 1972).
Неизвестны рост и питание тайменя из рек Казахстана. В пре-
делах ареала отмечаются большие различия в росте даже в одной
реке между популяциями из верховьев и низовьев (Кириллов,
1972). На первых годах жизни таймень растет медленно, достигая,
например, в р. Алдан 0,5 кг на четвертом году жизни. Обычная
масса тела тайменей в уловах по сибирским рекам и Амуру от 3—4
до 15 кг. Есть сообщение о таймене в 80 кг (Москаленко, 1960,—
цит. по Кириллову, 1972). В питании тайменя преобладает рыба, но
в целом спектр потребляемых организмов очень широк — от личи-
нок ручейников и хирономид до ондатр, белок, крыс, уток.
Численность и хозяйственное значение. Как крупный хищник,
обитающий обычно в небольших горных реках, таймень не образует
значительных скоплений, собираясь лишь на время нереста неболь-
шими косяками. В остальное время года держится разрозненно.
В водоемах Казахстана никогда не имел промыслового
значения ввиду малочисленности. В настоящее время отлавливает-
ся только рыбаками-любителями.
Таймень представляет собой несомненно одну из наиболее цен-
ных по вкусовым качествам рыбу и в будущем может стать объек-
том внимания рыбоводов. Для сохранения тайменя в казахез анских
реках необходимо ввести ограничения на его лов.
Род BRACHYMYSTAX GUNTHER, 1866 —ЛЕНОК
В последние годы в связи с ревизией систематики рыб род лен-
ков изучают многие ихтиологи (Шапошникова, 1968, Беседнов, Ку-
черов, 1972; Кифа, 1976; Мина, 1977; Васильева, 1978; Васильева,
Мина, 1980 и др.). Однако о количестве видов в роде пока нет еди-
ного мнения.
Одной из причин начала дискуссии было выделение подвида
Вг. lenok savinovi из оз. Маркаколь (Митрофанов, 1959).
М. И. Кифа (1976) нашел возможным выделить эту форму до ви-
дового ранга (Br. savinovi — для бассейна Амура). Е. Д. Василье-
ва и М. Н. Мина другого мнения: они подчеркивают, что в настоя-
щее время вообще трудно решить, один или более видов включает
род Brachymystax; можно лишь утверждать, что если этот род мо-
* На Иртыше существует народная примета: стали опадать листья у топо-
ля — начался осенний ход тайменя.
187
нотипический, то его единственный вид представляет собой комп-
лекс форм.
Основной причиной расхождения мнений послужило наличие
двух форм лепка — длиннорылой и короткорылоп, известных с
давних пор. Первооппсание сделано П. С. Палласом по экземпляру
из р. Енисея по длиннорылой форме (Кифа, 1976). Т. Мори (1930) эту
же длиннорылую форму описал как новый вид Br. tumensis из рек
Ялу и Тумень-Ула (Корея). Л. С. Берг (1909) отнес расхождения в
форме рыла к половому диморфизму (нами не обнаружен) и со-
хранил единый вид — Br. lenok. К этому мнению присоединились
Г. В. Никольский (1956), Г. X. Шапошникова (1968), Е. Д. Ва-
сильева (1978), М. В. Мина (1977) и др. К другой группе авторов,
считающих возможным разделение вида, относятся Ли Сы-чжун
(1966), Л. Н. Беседнов и А. Н. Кучеров (1972), М. И. Кифа (1976).
Для решения этого вопроса многими авторами, начиная с
Г. X. Шапошниковой (1968), подробно изучен скелет головы у
длиннорылых и тупорылых ленков. Однако и эти исследования не
привели, как было отмечено выше, к единому мнению. Трудность
заключается в том, что, во-первых, в ряде водоемов эти формы
обитают совместно, во-вторых, форма черепа неоднозначно корре-
лирует с такими признаками, как число жаберных тычинок, чешуй
в боковой линии (или число поперечных рядов) и другими. В свя-
зи с этим при описании вида Br. lenok его систематика рассматри-
вается на имеющемся материале возможно подробнее.
1. Brachymystax lenok (Pallas)—ленок
Синонимия: Salmo lenok: Pallas, Reise, v. Ill, 1776.
Salmo coregonoides: Pallas, Zoogr. Rosso-Asia!., v. Ill [1814] Финш и Брэм,
Путешествие в Западную Сибирь, И., 1882.
Brachymystax coregonoides: Варпаховский, Краткие данные об ихтиофауне
Азиатской России. Зап. Акад, наук, т. 59, кн. 1, 1889. — Игнатов, По Южному
Алтаю. Землеведение, т. 4, кн. 1—2, 1897.
Brachymystax lenok: Берг, Рыбы бассейна Амура, 1909. — Меньшиков, Рыбы
бассейна Оби. Докт. дне., 1948. Фонды библ. нм. В. И. Ленина.
Brachymystax tumensis: Mori Т., On the freshwater fishes from the Tumen
River, Korea, with descriptions of new species. J. Chosen Nat. Hist, soc., № 11, 1930.
Brachymystax lenok savinovi: Митрофанов, К систематике ленка из оз. Мар-
ка-куль. Сб. работ по ихтиологии и гидробиологии, вып. 2, Алма-Ата, 1959.
Brachymystax lenok swetowidowi: Кириллов, Ихтиофауна бассейна р. Вилюя.
Тр. Ин-та биол. Якут. фил. СО АН СССР, вып. 8, М., 1962.
Brachymystax lenok tsinlingensis: Li Sze-chung, On a new subspecies of
freshwater trout Brachymystax lenok tsinlingensis, from Taipaishan, Shansi, China.
Acta zootaxon, sinica, vol. 3, N 1, 1966.
Brachymystax savinovi: Кифа, Морфология двух форм ленка (род Brachy-
mystax, сем. Salmonidae) из басе. Амура и их систематическое положение. Зоо-
география и систематика рыб. ЗИН АН СССР. Л., 1976.
Распространение. Населяет водоемы от бассейна Оби до тихо-
океанского побережья. Встречается на островах, имеющих горные
реки (Шантарские); на юге распространен до Корен, включая.
188
весь бассейн Амура. Водится в горных озерах Монголии и Китая.
В Казахстане встречается в горных водоемах Южного Алтая.
Описание. В пределах ареала ленок (рис. 44) характеризуется
следующими признаками: D (II) III—IV (V) 8—12, А II—V 8—12,
1.1. 103—186, жаберных тычинок на первой жаберной дуге 17—33;
число позвонков 50—63; жаберных лучей 10—13, рядов чешуй над
боковой линией 26—37, под боковой линией 23—31; чешуй от задне-
го края анального плавника 17—-30, чешуй от заднего края жиро-
вого плавника 17—30.
Рис. 44. Ленок из оз. Маркаколь
Хромосомный набор ленка (Kang, Park, 1973, — цит. по «Ры-
бам МНР», 1983) соответствует 2п=90, при 26 метацентрических,
субтелоцентрических и 64 акроцентрических хромосомах. Число
NF=116.
Высокая вариабельность ленка по всем счетным признакам при-
водит к мысли о неоднородности вида. Однако многие признаки
варьируют столь же широко и в одном водоеме — оз. Маркаколь.
При этом кривые распределения за некоторым исключением одно-
вершинны и не оставляют сомнений в целостности популяции лен-
ка в озере:
8— 9—10—11—12 — количество мягких лучей в спинном плавнике,
1 16 52 30 1 —частота в °/о‘,
8— 9—10—11 — количество мягких лучей в А,
5 22 49 24 0 — частота в °/о;
50—51—52—53—54—55—56—57—58—59—60—61 —кол-во позвонк.
1 1 5 3 614 3119 9 8 1 1 —частота в %;
17—18—19—20—21—22—23—24 — количество жаберных тычинок,
1 4 10 28 31 14 9 2 — частота в % (п = 100), в т. ч.:
•— — 6 30 42 11 11—генерация 1951 г. (п = 17),
3,5 7 29 39 11 7 3,5 — генерация 1952 г. (п = 28),
13 9 30 26 13 4,5 4,5 — генерация 1953 г.(п=23);
число чешуй в боковой линии (пределы=103—186):
100—105—110—115—120—125—130—135—140—145—150—155—
160— 165— 170— 175— 180—185—190
1 26 27 15 9796604
0 3 2 0 1 1
189
Различия в числе жаберных тычинок Е. Д. Васильева и
М. В. Мина (1980) в разных выборках у одной и той же популя-
ции ленка из р. Керулен объясняют не субъективным критерием
(влияние оператора), а тем, что генеральные совокупности, из ко-
торых брались выборки, имели разное число жаберных тычинок.
Этот вопрос можно решить и проще, разложив одну выборку
(один оператор) на генерации. Результат получается следующий:
при одинаковых пределах колебаний числа тычинок в смежных ге-
нерациях наблюдается сдвиг модальной величины, а между неко-
торыми генерациями есть различия и в изменении пределов. Об-
щая закономерность — вариабельность этого признака убывает с
возрастом. Таким образом, число жаберных тычинок есть величи-
на достаточно переменная в одной и той же популяции, вероятно,
зависящая от фактора температуры в период развития икры. Раз-
ница в количестве тычинок между популяциями в значительной
мере объясняется типом питания (Митрофанов, 1977).
Интересен такой показатель, как число чешуй в боковой линии.
Г. X. Шапошникова (1968) выразила сомнение в правильности под-
счета различными исследователями прободенных чешуй и предло-
жила оперировать числом поперечных рядов чешуй. Вероятно,
этот признак у ленка более стабильный, чем число прободенных че-
шуй, и вариабельность должна снизиться по нижней границе при-
знака (число прободенных чешуй не может быть меньше количества
рядов, но последнее может быть много больше). При подсчете этого
признака ошибка оператора должна одинаково отразиться на ле-
вой и правой частях кривой распределения. Тем не мепее кривая
распределения резко асимметрична, растянута в правую сторону,
имеет тенденцию к двувершинности и «зубчатости» в правой части.
Возможно, это результат недостаточности материала по оз. Марка-
коль (п=100) при огромном размахе изменчивости, когда на
каждый класс вариационного ряда приходится от 0 до 8 особей при
интервале—1, или от 0 до 27 особей с интервалом 5. Модальная
величина соответствует 119 чешуям (п=8), модальный класс при
интервале 5=115—119 чешуям. Средняя величина ряда —
130 чешуй — не соответствует наибольшей встречаемости. В преде-
лах 113—124 чешуй расположено 48% особей, в пределах
113—148 чешуй — 85%. Менее 113 чешуй встречено у 3% осо-
бей, более 148 — у 11%. Непрерывное распределение встречается
от 113 до 140 чешуй (за исключением 125), но и в этом диапазоне
наблюдается неравномерность и асимметрия.
Неравномерность распределения чешуи прослеживается и по
боку хвостового стебля. Вариации здесь от 17 до 30 чешуй:
17—18—19—20—21 —22—23—24—25—26—27—28—29—30
3 1 1 5 16 9 9 10 13 21 5 1 3 2
Это распределение коррелирует с длиной хвостового стебля,
имеющего возрастную и половую изменчивость (r= —J-0,24 при сте-
пени свободы, равной 70, что обеспечивает Р=0,05). Длина хвосто-
вого стебля, более короткого у самок (13,58%), по сравнению с
190
самцами (14,8%) и неполовозрелыми особями (14,45%), дает дву-
вершинную кривую распределения числа чешуй по боку хвостового
стебля. Этот признак у самцов и самок отдельно дает одновершин-
ную кривую у самцов с модой 24—26 чешуй и двухвершинную кри-
вую у самок с модой 21 п 26 чешуй, что позволяет думать о
неоднородности стада. При этом средние значения числа чешуй по
боку хвостового стебля у самцов и самок не различаются (у сам-
цов — 23,8, у самок — 23,6).
Мы остановились на этом признаке столь подробно потому, что
подсчет числа чешуй по боку хвостового стебля более точен, чем
по всей рыбе и практически исключает ошибку. Не располагая ма-
териалом по остеологии черепа ленка и считая проведенные иссле-
дования в этом направлении вполне достаточными (Шапошникова,
1968; Васильева, 1978; Васильева и Мина, 1980 и др. работы), от-
метим большое разнообразие в строении скелета головы, особенно
у ленков из оз. Маркаколь. Во время наших исследований неодно-
кратно встречались особи с резко укороченным рылом (нижняя
челюсть выдается вперед), которые, к сожалению, не были изуче-
ны. Мопсообразная форма головы в ряде случаев сочеталась с ис-
кривлением позвоночника в хвостовом и туловищном отделах в
дорсовентральном направлении. Такие уродства наблюдались
преимущественно в районе впадения в озеро р. Тихушка, имеющей
отличный от всех остальных рек температурный и гидрологиче-
ский режим (что отражено в ее названии). Мы относим подобные
отклонения от нормы к действию повышенной температуры разви-
тия икры. Это предположение в отношении числа костных элемен-
тов у ленка в зависимости от условий развития икры высказывают
Ф. Н. Кириллов (1972) и другие авторы. Вероятно, многие спорные
вопросы его систематики определяются именно экологически-
ми условиями размножения и питания, а также жизнью в условиях
различного воздействия течения.
Несомненным остается разделение популяций ленка по числу
жаберных тычинок, что, на наш взгляд, свидетельствует о широте
спектра его питания.
Полиморфный вид Вг. lenok — типичный представитель боре-
ального предгорного комплекса в пределах своего обширного
ареала, вероятнее всего, образует несколько подвидов или эколо-
гических рас. Однако недостаточная изученность ленка из разных
мест обитания не позволяет утверждать это с большой убежден-
ностью. Выделение подвида Br. lenok savinovi из оз. Маркаколь не
отрицают большинство исследователей. Но по разделению вида на
два подвида: Br. lenok lenok — западносибирский и Br. lenok swe-
towidowi — восточносибирский возражении больше (Шапошникова,
1968 и др.). В литературе совершенно недостаточно сведений по
озерным популяциям этого вида, например из оз. Косогол (Хубсу-
гул). Способность ленка образовывать озерные расы гораздо вы-
ше, чем у тайменя, его ближайшего родственника. По этой способ-
ности ленок приближается к хариусам, имея практически совпа-
191
дающий ареал с сибирским, байкальским, монгольским и
косогольским видами хариусов. Экологические и, как следствие,
систематические параллели между родами Brachymystax и Thy-
mallus очевидны. Вероятно, в дальнейшем следует искать возмож-
ности разделения этого вида по образу жизни: речной и озерной
форм, поскольку ленки, обитающие в озерах и реках с замедлен-
ным течением, обладают большей вариабельностью призна-
ков.
la. Brachymystax lenok savinovi Mitrofanov —
маркакольский ленок
Местное название: ускуч.
Распространение. Озеро Маркаколь и впадающие в него реки.
Описание ленка из оз. Маркаколь: D II—IV 8—12, А II—III
8—11,5; 1.1. 103—186; рядов чешуй от заднего края жирового плавни-
ка 17—30; чешуй от заднего края анального плавника 17—30; жабер-
ных тычинок 17—25; жаберных лучей 10—13; позвонков 50—61, в
среднем 56; вершину D образуют 1—4 мягких луча; чаще 2—3;
вершину Р образуют 1—3, чаще 2; вершину V образуют 1—3, чаще
1—2; вершину А образуют 1—4, чаще 2—3.
Форма тела прогоиистая, вальковатая, несколько сжатая с бо-
ков. Рот конечный, верхняя челюсть слегка выдается над нижней.
Хвост слабо выемчатый, с возрастом выемка уменьшается. Окрас-
ка сильно варьирует: от темно-зеленой, почти черной, до серебрис-
то-белой, но тогда обязательно с пятнами. Преобладает темная
окраска. Пятнистость не обязательна у темноокрашенных рыб, хо-
тя в большинстве случаев она имеется, особенно у некрупных осо-
бей. Пятна, темно-зеленые или черные, покрывают спину, бока и
голову, величина их различна; иногда со светлыми пятнышками
посередине. Горло и брюхо всегда белые, без пятен. Плавники
окрашены различно: спинной и жировой под цвет спины, но спин-
ной светлее; грудные и брюшные серые. Брачная окраска парных и
анального плавников проявляется не всегда.
У самок в среднем 56 позвонков, 1.1. 130, мягких лучей в
D 10,58, в А 10,25; тычинок на первой жаберной дуге 20,74; жабер-
ных лучей 11,44; чешуй от заднего края А 24,02; чешуй от заднего
края жирового плавника 26,26.
В пластических признаках ленка (табл. 71) заслуживает вни-
мания разница в длине хвостового стебля. У неполовозрелых осо-
бей эта величина занимает промежуточное положение (14,45%)
между самцами и самками. С наступлением половозрелости у са-
мок относительная длина хвостового стебля становится меньше.
Показатель наибольшей высоты сильно варьирует в зависимости
от степени наполнения желудка и развития гонад. Разница в отно-
сительной величине глаза объясняется меньшими абсолютными
размерами самцов по сравнению с самками.
192
Таблица 71. Пластические признаки ленка из оз. Маркаколь (М±т),
% длины тела по Смитту
Признак Неполовоз- релые двух- и трехлетки (36 экз.) | Половозре- лые самки (25 &кз.) Половозре- лые самцы (25 экз.)
Масса тела (пределы), г 77-390 400-1650 300—1100
средняя 137,7 925 415
Длина по Смитту (пределы), см 19-30 33,8 -53,5 31-47
средняя 24 42,3 34,2
Длина всего тела 105,01 +0,29 105,61 +0,31
Длина тела без С 93,22 + 0,22 92,53 + 0,2 93,45+0,28
Наибольшая высота 16,65 + 0,23 20,52+0,19 17,82 + 0,19
Наименьшая » 6,86 + 0,08 7,38+0,08 7,06 + 0,05
Антедорсальное расстояние 43,08 t0,24 44,74 + 0,32 43,73 + 0,27
Постдорсальное » 39,95 + 0,29 39,71+0,37 40,23+0,22
Антевентральное » 52,42+0,26 53,05+0,44 51,89 + 0,25
Антеанальное » 71,45+0,4 73,07 t0,35 72,63 + 0,36
Длина хвостового стебля 14,45 + 0,27 13,58+0,17 14,8 +0,14
Расстояние Р—V 30,79+0,46 31,74 г0,29 31,9 ±0,18
Длина основания D 10,51+0,11 10,53 + 0,1 10,36 + 0,09
Высота D 12,67 + 0,15 11,38+0,17 11,94+0,18
Длина основания А 8,33+0,13 8,55+0,11 8,15 + 0,13
Высота А 12,55+0,16 12,54+0,17 12,94 + 0,21
Длина Р 14,6 +0,19 14,6 +0,2 14,74 + 0,19
V 11,03 + 0,14 10,64 + 0,16 11,02 + 0,19
верхней лопасти С 14,64 + 0,18 15,14+0,12
нижней лопасти С . 15,56+0,18 16,06 + 0,19
средних лучен С — 6,61 + 0,1 6,34+0,11
головы 20,88+0,15 20,26 + 0,15 20,35+0,17
рыли Диаметр глаза Заглавье Высота головы у затылка Высота головы через глаз Ширина лба Длина верхней челюсти 5,6 +0,115 4,58 t0,l 5,68+0,07 3,51+0,07 5,64 + 0,07 3,82 + 0,06
10,75 + 0,15 5,55 + 0,08 6,48+0,12 10,38 + 0,08 12,71 + 0,14 9,35+0,09 5,67+0,06 6,29 + 0,06 10,17 + 0,11 12,47+0,15 9,22+0,12 5,41+0,09 6,26 + 0,07
нижней челюсти . . 10,04 + 0,09 10,04 + 0,11
первой жаберной дуги 13 14+0 19 12,92+0,26
наибольшей жаберной
тычинки 1,98+0,07 2,01 + 0,05 2,02 + 0,04
По сравнению с самками самцы обладают более длинными пар-
ными плавниками, большей высотой непарных плавников и мень-
шей длиной их оснований. Все признаки в своем сочетании прида-
ют самцам более стройную форму тела с выделяющимися плавни-
ками. Разница в длине рыла между самцами и самками не
обнаружена. Возрастная изменчивость во многих случаях сходна с
половой и проявляется более четко, чем половой диморфизм.
Изменчивость в счетных признаках обнаружена между генера-
циями по числу жаберных тычинок, числу позвонков и чешуй в
боковой линии, проявляется в сдвиге модальных величин и сокра-
щении размаха варьирования у старших возрастных групп, что,
13-7
193
вероятие, связано с малочисленностью материала. Средние значе-
ния, за отдельными исключениями, остаются в пределах ошибки.
Биология. Нерест ускуча в Маркаколе приурочен к вскрытию
озера. Время ледохода и последующая за ним неделя — время ин-
тенсивного икрометания. Однако протяженность всего нерестового
периода значительно больше. Нерест начинается в середине мая,
когда все притоки озера уже вскрыты, а на озере образуются
большие предустьевые полыньи. В это время наблюдается заход в
реки крупных особей обоих полов. Вслед за вскрытием озера, что
обычно происходит в третьей декаде мая, в реки на нерестилище
заходит основная масса рыбы. С начала июня заход рыбы в реки
начинает сокращаться, но процесс икрометания идет интенсивно
до середины июня. К июлю нерест заканчивается, и в реках встре-
чаются только одиночные мелкие особи производителей.
Таблица 72. Встречаемость ленка с гонадами различной стадии
зрелости в течение года, %
Представление о сроках созревания половых продуктов дает
таблица 72. Рыба отлавливалась в разных участках озера и в ре-
ках на нерестилищах.
В преднерестовый месяц—апрель — основная масса полово-
зрелых особей подготовлена к нересту; половые продукты в
IV стадии зрелости; намечается переход в V стадию. В первой по-
ловине мая готовых к нересту особей становится больше за счет
почти полного исчезновения особей с III стадией зрелости гонад.
Единичные особи нерестуют. Во второй половине мая картина за-
метно изменяется. Наибольшее количество рыбы будет нересто-
вать в ближайшие десять дней или уже нерестует. В то же время
встречаются особи с IV стадией зрелости половых продуктов.
В уловах появляются и крупные отнерестовавшие особи. Первая
половина июня характерна массовым нерестом. Количество отне-
рестовавших и готовых к нересту особей одинаково. Во второй по-
ловине июня нерест протекает слабо; нерестуют только мелкие
особи и не на всех нерестилищах. Основная часть половозрелого
стада уже отнерестилась и находится в стадии выбоя. В июле резко
возрастает количество особей со II стадией зрелости икры и молок.
194
В начале месяца нередки экземпляры в стадии выбоя. В августе
наблюдения не проводились, но ход гаметогенеза легко устано-
вить.по состоянию гонад в сентябре. В это время половые железы
у большинства рыб переходят из И в III стадию зрелости. В конце
сентября появляются особи с половыми продуктами, очень близки-
ми к IV стадии. Октябрь интересен тем, что опять увеличивается
количество экземпляров со II стадией развития гоиад. В данном
случае это объясняется вливанием в нерестовую популяцию нового
пополнения. Большинство отнерестовавших в этом году особей в
октябре достигает IV стадии зрелости гонад. В течение зимних
месяцев изменений в гаметогенезе почти не замечается. В апреле
особи, готовящиеся к первому нересту, догоняют по развитию го-
над повторнонерестующих рыб. По состоянию гонад ленка в раз-
личные сезоны года нельзя сделать вывода о его неежегодном
нересте.
Таблица 73. Изменение абсолютной плодовитости лейка
в зависимости от массы тела
Показатель
Пределы АП
Средняя АП
п
Масса, г (пределы; в скобках — средняя)
250—600 (427) 601-900 (762,25) 901 -2400 (1168)
551-2684,5 1347 47 1125-3801 2377,5 23 2047.7-5059 3599,8 21
Таблица 74. Изменение абсолютной плодовитости ленка
в зависимости от размера рыбы
Показатель Длина по Смитту, мм (пределы; в скобках — средняя)
310-380 (349,2) 381 -425 (393) 426-570 (455,6)
Пределы АП Средняя ЛП п 551—2684,5 1328,5 38 894--3801 1845 27 2047,7-5059 3449,5 26
Для определения плодовитости был использован материал в
сто с лишним самок IV—V стадии зрелости, масса тела которых
колебалась в пределах 250—2400 г, длина по Смитту — от 310 до
570 мм и возраст — от 3 до 7 лет. Минимальная плодовитость — 551,
максимальная — 5059 икринок; средняя абсолютная плодови-
тость — 2004,8 икринки.
Абсолютная плодовитость (ЛП) закономерно изменяется в за-
195
висимости от возраста рыбы, размера и массы ее тела
(табл. 73—75).
Резкое увеличение массы икринок у 5-годовиков сменяется при
дальнейшем росте менее резкими скачками у 6—7-годовиков. От-
сутствие более взрослых рыб в сборах не позволяет найти грань
стабилизации икринок в одной величине.
Таблица 75. Изменение массы яичников и абсолютной
плодовитости ленка в зависимости от возраста
Показатель Возраст, лет
3 4 5 6 7-9
Средняя масса яичиика, г 26,3 50,4 103,9 176 248
Икринок в яичнике 904,3 1745,2 2822 4064,5 5021,2
Икринок в 1 г яичника 34,4 34,6 27,1 23,1 20,2
п 29 34 12 7 2
Относительная плодовитость остается почти неизменной у рыб
массой до 1 кг и резко снижается при массе свыше 2 кг, что опре-
деляется увеличением размеров икринок. На 1 г массы тела прихо-
дится 3,15 икринок при массе 427 г (п=47); 3,12 — при 762 г
(п=23); 3,08 —при 1168 г (п= 19); 2,16 —при 2324 г (п=2).
Относительная плодовитость продолжает увеличиваться при
достижении рыбой длины 60 см и, возможно, выше: при длине
36,8 см число икринок (на 1 см длины) достигает 35 (п=38), при
41,4 см — 46 (п=27), при 48,6 см — 71 (п=24), при 59,4 см — 82
(п = 2).
Низкая плодовитость ленка в оз. Маркаколь объясняется ран-
ней его половозрелостью. Самки, достигшие 3 полных лет, в боль-
шинстве своем половозрелы. Половозрелые самцы встречаются
массой 180—280 г при длине тела 315—340 см.
Соотношение полов в уловах характеризуется более чем двой-
ным преобладанием самок (табл. 76).
Таблица 76. Соотношение полов у лейка оз. Маркаколь, %
Самки
Самцы
и
69
31
104
77
23
94
72
28
128
44
56
145
84
16
163
35 36
49 164
83
17
97
79
21
32
67
33
38
66
34
17
70 722
30 309
- 1031
Диаметр зрелой овариальной икринки ленка составляет в сред-
нем 3,91 мм с колебаниями от 3,66 до 4,21 мм. Примерно такие же
данные приводят Д. Д. Дулькейт и Г. У. Линдберг (1929) —3,8—
196
4,2 мм. К. И. Мишарин (1942) для ангарского ленка дает предель-
ные размеры икринок в 4—4,5 мм, но измерял он оплодотворенную
набухшую икру. П. А. Дрягин (1949) указывает предельные раз-
меры икринок из гонад 4—4,5 мм, диаметр развивающихся икри-
нок после набухания—]5,5—6 мм. Инкубация икры ангарского
ленка при температуре 4,5° продолжается 50 суток, при темпера-
туре 4,6—4,9° — от 45 до 49 суток, общее количество градусодней
220—230 (Мишарин, 1942). П. А. Дрягин (1949) определяет сумму
температур для развития икры в 174°.
Опыты по искусственному размножению маркакольского ленка
(Вотинов, 1963) проводились с применением гипофизарных инъ-
екций и сбора икры от текучих самок. Средняя рабочая плодови-
тость оказалась равной 2000 икринкам, что полностью совпадает с
нашими данными по абсолютной плодовитости. Опыты проводи-
лись в условиях колебаний среднесуточной температуры воды от
7,2 до 14,8°, а абсолютной температуры — от 3,6 до 16,2°. Инкубация
икры продолжалась от 19 до 22 суток, а количество градусодней
колебалось от 176 до 265. Выклев личинок растягивался на не-
сколько дней. Отход икры в опытах был велик, в среднем 65,3%.
Отход при транспортировке оплодотворенной икры составил
16—27%.
Личинки после выклева малоподвижны и обычно держатся у
дна аппаратов (опыты проводились в естественных условиях Мар-
каколя). Активное питание зарегистрировано с 12—15 дня после
выклева при наличии желточного мешка, который рассасывается
на 20—25-е сутки. Личинки ленка проявили устойчивость к дефи-
циту кислорода. Гибель наступала при содержании кислорода
2,3—2,8 мг/л при температуре 10—11°. Верхняя летальная темпера-
тура развития личинок до месячного возраста около 28°.
Эти исследования Н. П. Вотинова подтвердили наше предполо-
жение (Митрофанов, 1959), что развитие икры и личинок ленка в
оз. Маркаколь происходит при более высоких температурах, чем
указывают К. И. Мишарин (1942) и П. А. Дрягин (1949). Выска-
занное ранее предположение, что именно температурный режим в
период развития определяет морфологические особенности марка-
кольского ленка находит, таким образом, подтверждение.
Например, в р. Тополевке (наиболее холодный приток) темпе-
ратура воды в нерестовый и инкубационный периоды была: 24 мая
в 11 ч 5°, 1 июня в 15 ч 40 мин 6,5°, 14 июня в 12 ч 30 мин 7,5”,
20 июня в 18 ч 12,5° и 24 июня в 13 ч 20 мин 12°. В одном из теплых
притоков озера, в р. Тихушке, температура менялась следующим
образом: 23 мая в 19 ч 30 мин 11,5°; 24 мая в 15 ч 8°; 1 июня в
13 ч 10°; 14 июня в 17 ч 12,5°.
Нерестилища ленка в оз. Маркаколь находятся в коротких горных
притоках. В озеро впадает более двух десятков мелких речек и масса
ключей. Последние ленок не использует. Основных притоков у озе-
ра четыре: реки Тополевка, Тихушка, Глухова и Мостовая. Они и
являются основными нерестилищами. Наиболее важное значение
197
имеет р. Тополевка. Она впадает в озеро почти десятью рукавами,
образуя помимо этого много разветвлений в среднем и предустье-
вом течении. Весной площадь нерестилищ здесь велика. Верхняя
граница захода ленка в реки обусловлена наличием подходящих
нерестилищ, которыми служат песчаные отмели, так называемые
«шивера», мелкогалечниковые небыстрые протоки, широкие пере-
каты. Нерестилища расположены на глубине, не превышающей
метра. Обычно такие места ленок находит в 10—12 км от устья, а
зачастую много ближе.
Ленок держится на нерестилищах небольшими стайками, ведет
себя осторожно, и при малейшем звуке или движении прячется в
более глубокие места: ямы под водопадами, подмывы под берега-
ми, под завалы реки деревьями и т. п.; обычно избегает протоков
с очень быстрым течением и охотно заходит в старицы; при необ-
ходимости проходит быстрые участки реки, не задерживаясь в них.
Вообще ленок предпочитает затененные участки русла реки или
озера.
Интересным доказательством отрицательного фототаксиса
ленка служит лов его во время вскрытия озера. Местные рыбаки
всегда ставят сети на дно. Обязательное условие для лова рыбы
днем — закрытие сетей льдом. Если сети льдом не закрыты, ни
один рыбак не поедет проверять пх в течение дня; но если сети
закрыты льдом хотя бы на один час, то рыбаки выезжают на их
проверку и берут хороший улов. Основной же лов ленка сетями в
озере и ловушками в реках производится ночью, когда рыба наибо-
лее активна.
Никаких приготовлений к откладке икры ни самец, ни самка не
делают. Самка становится головой к течению, прилегает ко дну, и,
производя боковые движения телом, откладывает икру. Самец на-
ходится несколько выше самки в толще воды и несколько ниже
по течению. Выметанная икра сносится по течению и забивается в
неровности грунта, где и проходит ее дальнейшее развитие.
Врагов у ленка на нерестилищах нет. Пескари, которых в озере
огромное количество, на нерестилища ленка не заходят. Гольцы
нерестуют в реках, но значительно ниже ленка, и количество их
невелико.
Хариус конкурирует с ленком только в самых верхних участках
нерестилищ последнего, в основной же массе нерестует еще выше.
Пернатых и четвероногих врагов ленок не имеет.
Наибольший урон выметанной икре наносит сам ленок, кото-
рый поедает свою икру в течение всего нерестового периода. В по-
жирании участвуют самцы и самки. На нерестилищах в р. Топо-
левке в середине июня из 70 вскрытых желудков ленка наполнен-
ными икрой были 56 (Тэн, 1959).
Ряд авторов (Никольский, 1956) отмечают икроядность ленка
на нерестилищах кеты и горбуши. Однако некоторые исследовате-
ли приходят к выводу, что ленок поедает вымытую из гнезд икру и,
таким образом, не является врагом этих видов.
198
Сразу после нереста ленок скатывается с нерестилищ в озеро.
Возраст и темп роста ленка изучались по жаберным крышкам
и позвонкам. Распределение взятой для анализа рыбы по длине и
массе тела представлено в таблице 77. Данные по темпу роста — в
таблице 78.
Таблица 77. Характеристика возрастного материала ленка
Показатель Возраст, лет
1 + 2-г з+ 4+ 5+ 6 7 8-Ь 9+ ю-ь
Абсолютная длина, мм 142 243 294 359 406 420 437 575 600 605
Масса, г 27,5 113 210 452 677 727 773 1900 2400 2400
п 2 10 17 41 38 21 17 2 1 1
Таблица 78. Линейный рост ленка из оз. Маркаколь
(обратное расчисленне), мм
Показатель Возраст, полных лет
1 2 3 4 5 6 7 8 9 п
Позвонки ЖаберНые 75 138 209 280 346 397 444 517 566 70
Крышки 93 162 238 299 356 395 441 505 583 51
Самцы 76 139 209 276 340 382 438 490 532 21
Самки 75 137 209 283 349 416 456 525 600 38
В течение всего года самцы в уловах мельче самок. Однако
сравнение их роста не дает существенных различий. Самки обго-
няют в росте самцов после наступления половой зрелости. Мень-
шие размеры самцов в уловах объясняются возрастным составом
половозрелого стада (табл. 79).
Таблица 79. Возрастной состав половозрелого стада
ленка, %
Пол рыб Возраст, лет
2+ 3+ |4+ 5+ 6+ |7Ь 8-10 Средн.
Самки — 25 43 26 4 2 Единичные особи 4,2
Самцы 3 37 38 16 4 2 То же 3,6
Оба пола I 29 41 23 4 2 — 4,1
199
Возрастной состав стада ленка свидетельствует о равномернос-
ти изъятия промыслом самцов и самок и о значительной интенсив-
ности промысла на оз. Маркаколь в 1950—1956 гг. Следовательно,
преобладание самок над самцами можно объяснить тем, что на
половой состав стада влияет преобладание на нерестилищах
впервые созревающих производителей с мелкой икрой (Митрофа-
нов, 1970).
Сравнение роста ленков из различных водоемов позволяет от-
метить некоторые свойства, характерные, по-видимому, для всего
рода Brachymystax — большие линейные и особенно весовые при-
росты на восьмом или девятом году жизни. Объясняется это явле-
ние, на наш взгляд, сменой питания в этот период, усилением хищ-
ничества.
Упитанность ленка с возрастом увеличивается независимо от
времени года. Осенью он более упитан, чем весной: коэффициент
по Кларк соответственно у 2—3-годовалых—1,07 и 0,96; 4—5-го-
довалых—1,3 и 1,06; у 6—7-годовалых—1,31 и 1,11. Определение
упитанности ленка (по Кларк) из разных участков озера дает
картину ее закономерной изменяемости в связи с кормовыми за-
пасами: у восточного берега—1,3, у северного—1,31, у южного—-
1,34 (для 4—5-годовалых рыб).
Незначительно изменяется упитанность в результате нереста,
что объясняется интенсивным питанием ленка во время икро-
метания.
Питание ленка изучала В. А. Тэн (1959) в течение всего года.
Исследовано содержимое более 300 желудков. По частоте встре-
чаемости в них преобладала рыба (33%), гаммарусы (23%) и
икра ленка (19%). В остальных желудках содержались пиявки,
олигохеты, клещи, моллюски, наземные клопы и жуки, муравьи,
бабочки, личинки стрекоз, веснянок, поденок, ручейников, хироно-
мид, тендепедпд и мух, а также растительная пища, что свидетель-
ствует о широте спектра питания, практически полном освоении
кормовых ресурсов озера. Предпочитает ленок крупных беспозво-
ночных, проявляя высокую элективность питания по отношению
к моллюскам, личинкам мух и ручейников, жукам, клопам, гамма-
русам. Растительная пища встречена в 22% желудков. Всего в
пище зарегистрировано 36 видов животных. Слабее других групп
ленок использует олигохет и личинок хирономид. Последних отме-
чено в пище 16 видов из 65, обитающих в озере.
Изменения в составе пищи ленка хорошо прослеживаются по
сезонам года, местам его концентрации и доступности (наличию)
того или иного вида пищи (табл. 80). Рыба и гаммарусы состав-
ляют основу его питания в зимовальный, преднерестовый и нересто-
вый периоды, меняясь лишь по весовому соотношению. В нагульный
период на первое место выступает рыба и моллюски, а в нерес-
товый большое значение приобретает собственная икра. В это вре-
мя наблюдается и наибольшее разнообразие питания.
Из рыб в пище ленка отмечены пескарь и голец. Первый чис-
200
ленно преобладает (результат колоссального количества его в
озере и крайне редкой встречаемости гольцов). Тем нс менее по
отношению к гольцу ленок проявляет повышенную избиратель-
ность. Это можно объяснить различиями в экологии гольцов и пес-
карей. Последние в озере долгое время были поголовно заражены
лигулезом. Огромные их стаи держатся в толще воды и у ее поверх-
ности. Голец ведет типично придонный образ жизни. У дна преиму-
щественно держится и ленок, лишь в крайних случаях поднимаю-
щийся в толщу воды.
Таблица 80. Состав пищи ленка по периодам года (Тэн, 1959)
Компоненты пищи Нагульный (сентябрь- октябрь) Зимоваль- ный (ноябрь- декабрь) Преднерес- товый (апрель— 15 мая) Нерестовый (15 мая— июнь)
1 2 1 2 1 2 1 2
Растения 30 5,4 21 2,2 22 2,4 20 2,4
Черви 6 2,6 — 18 2,8
1 аммарусы 20 1,9 46 19 1 58 62,8 30 23,9
Веснянки —. — — 13 0,4
Клопы — —. — ,—. 11 1,8
Жуки — — — —. — .—. 9 0,3
Ручейники 23 1,4 11 0,6 5 0,2 28 4,9
Хирономиды —. — 18 0,1 11 0,1 48 2,2
Двукрылые (без хпрономид) 3 0,8 — — — — 31 0,3
Муравьи — —. — — — •— 10 0,7
Моллюски 27 14,2 7 0,2 3 0,2 3 0,7
Рыбы 38 72,9 57 77.8 29 34,3 28 37,2
Икра — — — — — — 41 20,9
Примечание. 1 — частота встречаемости; 2 — значение ио массе.
Сопоставление питания и упитанности ленка по сезонам года
показывает полную обеспеченность его кормами круглогодично.
Нет значительного снижения упитанности и в результате нереста.
После нереста не гибнет.
Численность и хозяйственное значение. Добыча ленка в
оз. Маркаколь в промысловых масштабах ведется с середины
прошлого века. Это совершенно уникальное промысловое стадо
ленка, нигде больше не достигающего столь значительной числен-
ности. Ориентировочно его вылов в начале нашего века был равен
1,5 тыс. т в год (Яблонский, 1907), что в 15—20 раз больше, чем
вылавливалось в 50-х и в 1981 —1982 гг. В настоящее время вылов
100 т ленка в год является рекордным.
Организация заповедника на оз. Маркаколь определяет особые
условия эксплуатации запасов ленка. Полностью отказаться от его
вылова было бы неразумно: создаваемую продукцию надо исполь-
зовать, но с соблюдением рекомендуемых мероприятий. Добыча
ленка может быть разрешена и в весенний период, и в районе
р. Тополевка, но при условии организации его искусственного
201
воспроизводства в той же реке и сокращения заготовок икры. Нача-
тые работы по искусственному разведению ленка необходимо про-
должить, как и изучение состояния его запасов.
Семейство COREGONIDAE — СИГОВЫЕ
Род STENODUS RICHARDSON, 1836 — БЕЛОРЫБИЦА, ИЛИ НЕЛЬМА
Включает лишь один вид, распространенный в бассейне Север-
ного Ледовитого океана — от Белого моря до рек Северной Аме-
рики, а также в бассейне Каспийского моря. Самые крупные пред-
ставители семейства сиговых рыб. Относятся к арктическому
пресноводному комплексу.
1. Stenodus leucichthys (Giildenstadt)—нельма
В водах СССР, в том числе в пределах Казахстана, водятся
2 подвида этого вида: нельма — St. leucichthys nelma (Pallas) —
в бассейне Ледовитого океана и белорыбица — St. leucichthys leu-
cichthys (Giildenstadt)—в бассейне Каспия, различающиеся меж-
ду собой некоторыми остеологическими и пластическими призна-
ками.
la. Stenodus leucichthys nelma (Pallas) —нельма*
Казахское название: at] бальщ, сылан.
Синонимия. Salmo nelma: Pallas, Reise, v. II, 1773.
Salmo leucichthys: Pallas, Zoogr. Rosso-Asiat., v. Ill, [1814].
Salmo mackenzii: Richardson, Franklin’s Journ., 1823.
Coregonus lucius: Nilsson, Scandin. fauna, v. IV, 1853.
Stenodus nelma: Smitt, Salmonider, 1886.
Stenodus mackenzii: Jordan and Evermann, Fish. N. America, v. I, 1896.
Coregonus leucichthys: Иохельсон, Землеведение, 1898.
Stenodus leucichthys nelma: Берг, Рыбы пресных вод России, 1916. — Мень-
шиков, Изв. Пермск. бнол. ин-та, т. X, вып. 1—2, 1935. — Берг, Рыбы пресных
вод СССР, т. I, 1948.
Распространение. Нельма (рис. 45) широко распространена в
Ледовитоморской провинции, вплоть до ее южных районов; встре-
чается в некоторых бассейнах Тихоокеанской провинции. На запа-
де Ледовитоморской провинции границей распространения служат
реки Северная Двина, Онега, на востоке — Макензи и Юкон (Се-
верная Америка). В Обь-Иртышском бассейне встречается повсе-
местно (Березовский, 1930; Меньшиков, 1935; Смирнова, 1945).
В Казахстане нельма (Берг, 1948; Дрягин, 1948; Иоганзен,
1951; Конева, 1972) обитает только в водоемах Среднего и Верхне-
го Иртыша. По реке Черный Иртыш поднимается в пределы КНР.
В Бухтармипском водохранилище распространена преиму-
* Мелких, неполовозрелых, на Иртыше называют «кукча».
202
щественно в озерной части, в преддельте и дельте Черного
Иртыша. В других участках водохранилища встречается очень
редко. С годами ареал нельмы в водохранилище сокращается,
сдвигаясь в его верховье и р. Черный Иртыш. В Усть-
Каменогорском водохранилище на девятнадцатом году его су-
ществования нельма выпала из состава ихтиофауны (Ерещенко,
1972). Стала встречаться в водохранилищах канала Иртыш—-Ка-
раганда, куда попадает, видимо, при закачке речной воды. В 1973—
1975 гг. единичные экземпляры пойманы в водохранилищах 1, 5 и
6-го гидроузлов.
Рис. 45. Нельма
Описание. П. А. Дрягин (1948) отмечает подверженность
обской нельмы биотоппческой изменчивости: наряду с полупроход-
ной существует туводная форма, ареал которой приурочен к вер-
ховьям Обь-Иртышского бассейна. Нельма легко приспосаблива-
ется к новым условиям обитания и в пределах ареала образует
туводные формы, отличающиеся от полупроходной биологическими
особенностями. Биологическую разнокачественность обь-иртыш-
ской нельмы отмечает и И. А. Петрова (1972).
В настоящее время в р. Иртыш в пределах Казахстана обитают
2 формы нельмы: полупроходная, поднимающаяся из низовьев Ир-
тыша и Оби, н жилая — туводная.
До зарегулирования стока реки в Верхнем Иртыше, включая
оз. Зайсан, обитали 2 формы: туводная «зайсанка» и полупроходная
«иртышанка». Сейчас в Бухтарминском водохранилище обитает
только жилая форма нельмы, так как пути полупроходной обской
и нижнеиртышской отрезаны плотинами Усть-Каменогорской и
Бухтармипской ГЭС.
А. И. Березовский (1930) считает, что зайсанская нельма —
это местная форма, свойственная только оз. Зайсан и Черному Ир-
тышу. По проведенным К. В. Смирновой (1945) исследованиям,
основу стада в Зайсане в 1944 г. составляла туводная форма. Мне-
ние о существовании зайсанской локальной формы подтверждает и
тот факт, что сравнительно скоро после перекрытия реки плотиной
Усть-Каменогорской ГЭС в отчлененном отрезке Иртыша до
оз. Зайсан нельма исчезла.
Далее морфометрическая характеристика дается по 18 экз. из
оз. Зайсан, исследованным в 1958—1963 гг. D III—V 9—13, А IV—V
203
13—15, чешуй в боковой линии 99—108, над ней 11—14, под ней
10—13; жаберных лучей 9—10, тычинок на 1-й жаберной дуге
20—24; позвонков 64—69. Пластические признаки представлены в
таблице 81.
Сравнение этих данных с материалами прежних лет показывает
Таблица 81. Морфометрическая характеристика нельмы
Верхнего Иртыша
Признак От—до Среди.
Полная длина, см 56,0-91,0 72,28
Длина по Смитту, см 52,8-86,0 67,68
Длина без С, см 50,3-81,0 63,70
Полная масса, кг 1,5-5,5 3,36
В % длины тела поСмитту
Длина головы 10,76-24,19 22,21
рыла 4,26-5,68 4,92
верхнечелюстной кости 5,68-7,81 7,05
Ширина верхнечелюстной кости 1,51-2,36 1,99
Длина нижней челюсти 9,85-11.59 10,60
Заглавье 12,83-14,96 14,01
Диаметр глаза 2,99-3,87 3,32
Высота головы у затылка 10,79-14,07 12,52
Наибольшая высота тела 20,0-23,93 21,91
Наименьшая » » 5,95-7,70 6,79
Длина хвостового стебля 10,38-15,12 12,29
Длина Р 12,62-15,34 13,64
Длина V 11,76-14,11 12,87
Антедорсальное расстояние 43,94-49,37 47,52
Постдорсальное » 32,33-40,63 37,14
Антеанальное » 70,00—76,07 72,79
Антевентральное » 47,94-52,27 49,63
Длина основания D 6,12-11,58 9,97
Высота D 10,63—14,84 12,51
Длина основания А 10,91—13,89 12,28
Высота А 9,84-13,48 10,89
Р—V 24,29-30,26 26,31
V—А 21,31-26,69 23,31
В % длины головы
Длина рыла 19,1-24,7 22,23
Длина верхнечелюстной кости 26,1—37,0 31,71
Ширина верхнечелюстной » 6,8-11,9 8,96
Длина нижнечелюстной » 43,0-51,4 47,84
Заглавье 60,0-65,2 62,83
Диаметр глаза 12,6-17,0 14,95
Ширина лба 16,7-24,0 19,41
Высота головы у затылка 47,0-69,3 56,48
204
уменьшение у нельмы длины головы, наибольшей и наименьшей
высоты тела и увеличение диаметра глаза: в среднем, по
К. В. Смирновой (1945), было соответственно 23,17; 23,4 и 7,37%
длины тела по Смитту и 14,1 % длины головы.
Зайсанская нельма отличается от полупроходной иртышской
более длинной головой, большими высотой тела и диаметром гла-
за, длинными грудными плавниками и челюстными костями.
Рот у нельмы большой, конечный. Нижняя челюсть выдается
вперед, сочленена с черепом за задним краем глаза. Зубы мелкие,
расположены на челюстях, сошнике, небных костях и языке.
Окраска спины серовато-пепельная, бока серебристые, брюхо
белое. Спинной и хвостовой плавники светло-пепельные, с грязно-
ватым оттенком, остальные плавники светлые.
Половой диморфизм проявляется только в пластических при-
знаках и окраске: самцы обычно темнее и более прогонисты, брюш-
ные плавники их смещены назад; анальный плавник, верхняя и
нижняя челюсти длиннее, чем у самок. Размерно-возрастная из-
менчивость нельмы, обитающей в водоемах Верхнего Иртыша, не
изучалась.
Нельма — крупная рыба длиной свыше 1 м и массой до 40 кг
(Титеньков, 1961). В водоемах Верхнего Иртыша популяция этого
вида представлена сравнительно мелкими особями. К- В. Смирно-
ва (1945) отмечает, что за 3 года работы на оз. Зайсан ни разу
не встречались особи длиной более 100 см и массой свыше 7,5 кг.
В Бухтарминском водохранилище в первые годы его существо-
вания отлавливали нельм массой до 8 кг. В последующем эта по-
пуляция заметно помельчала. Так, по контрольным анализам рыб
из ставных неводов, в мае 1973 г. в районе промысловых участков
Байтугас, Бесчаны, Тююк, а затем в августе при опытно-промыш-
ленном облове ставными сетями северо-западной запретной зоны
оз, Зайсан, длина тела нельмы колебалась от 27 до 67 см (в сред-
нем 49—51,5 см), масса — от 0,2 до 5,3 кг (в среднем 1,8—2,1 кг),
В водохранилищах канала Иртыш—Караганда в 1973—1974 гг.
ловились экземпляры массой до 2,5 кг при длине тела (без хвосто-
вого плавника) до 65 см. В Иртыше в пределах Семипалатинской
и Павлодарской областей нельмы массой свыше 4—5 кг в настоя-
щее время не встречаются.
Биология. Половозрелость у нельмы оз. Зайсан наступала по
достижении длины тела 56—65 см в возрасте 5—6 лет (Смирнова,
1945), т. е. раньше, чем у обской полупроходной формы.
В Бухтарминском водохранилище самцы нельмы становятся
половозрелыми на пятом году жизни (4+), самки на шестом (5+),
реже на седьмом. В последние годы отмечается омоложение не-
рестовой популяции. Так, в 1955 г., который характеризовался
сильным снижением численности стада нельмы в Зайсане, нересто-
вая популяция при общей ее малочисленности состояла из 6 воз-
растных групп с преобладанием в ней 8—10-летних рыб. В 1964 г.
на контрольном запоре, установленном в устье Черного Иртыша,
205
среди покатных нельм, нерестующих в пределах СССР, не обнару
жены особи старше 6 лет.
Современное нерестовое стадо бухтарминской нельмы состоит
из одной возрастной группы самок — шестилеток (5+) и двух воз-
растных групп самцов — пяти-шестилеток. Повторно нерестящиеся
рыбы встречаются единично.
С середины и до конца мая особи, впервые достигшие полово-
зрелое™, поднимаются по Черному Иртышу. Среди этой мигри-
рующей части популяции встречаются и неполовозрелые экземпля-
ры. Мигрирующая весной нельма проходит на дальние нерестили-
ща, расположенные в пределах Китая. В реку весной идет нс вся
нерестовая популяция, часть ее продолжает нагуливаться в озере.
В июле-—августе нельма концентрируется в районе мысов Бархот
и Бакланий, затем постепенно продвигается к устью Черного и
Старого Иртыша, а в конце августа — сентябре начинает подни-
маться вверх по Черному Иртышу. Миграция продолжается в от-
дельные годы почти до конца октября. Осенняя нерестовая мигра-
ция не имеет ярко выраженного пика и очень растянута во вре-
мени.
Незначительное количество рыб задерживается на ближних
нерестилищах, расположенных на территории СССР, в районе
Мын-Арала, Бурана, основная же часть стада поднимается по реке
выше. Верхние границы миграционных путей нельмы по Черному
Иртышу в пределах КНР не известны, так же как и условия не-
реста, время ската молоди.
Обширные нерестилища нельмы, располагавшиеся рапсе по
Иртышу от Семипалатинска до Усть-Каменогорска, в настоящее
время не используются ввиду сильного загрязнения реки (Ере-
щенко, 1969). Основные нерестовые угодья нельмы по Быстрому
и Белому Иртышу попали под затопление. На каменисто-галечных
перекатах реки в пределах Павлодарской области воспроизводство
нельмы сохранилось.
В наших водах нельма нерестится во второй половине октября,
реже в начале ноября, при температуре воды 4—6°. Икру мечет на
каменисто-галечниковых грунтах, на глубине 1—3 м. После нерес-
та производители скатываются в озеро.
Судя по нахождению сеголеток и двухлеток нельмы в озерной
части водохранилища, нерест этого вида происходит ежегодно, но с
разной результативностью. О соотношении полов на нерестилищах
и продолжительности нереста сведений нет.
Абсолютная плодовитость нельмы в Белом Иртыше, по данным
А. В. Подлесного (цит. по Смирновой, 1945), составляла в среднем
107,38 тыс. икринок (пределы 98,5—120,25 тыс.). В Верхнем Ир-
тыше и Бухтарминском водохранилище опа несколько ниже: 92—
116,2, в среднем — 98,7 тыс. икринок. Инкубационный период дли-
тельный, продолжительностью 5—6 месяцев, с конца октября —
начала ноября до апреля.
Развитие икры и молоди нельмы в казахстанских водоемах не
206
изучено. Предположительно выклев ее начинается в первой поло-
вине апреля. Личинки скатываются в водохранилище, где и нагули-
ваются до наступления половой зрелости.
Растет нельма быстро. Сеголетки в сентябре — октябре дости-
гают длины 15—22 см и массы тела 50—100 г. Средняя длина
двухлеток (1+) составляет 31,7 см, масса — 320 г; трехлеток —
соответственно 41 см и 800 г; четырехлеток—50 см и 1000—1300 г.
Рост половозрелых особей показан в таблицах 82, 83.
Таблица 82. Темп роста нельмы в оз. Зайсан
(обратное расчленение), см
Время сбора 1 Возраст, полных лет
2 3 4 5 1 6 | 7 8 | 9
Зима, 1942 * 16,2 26,6 35,9 44,0 51,459,1:66,1 73,879,9
Лето, 1942 * 16,4 27,9 38,1 47,0 54,662,369,975,9 82,2
1962—1965 гг. 18,2 29,4 42,0 48,0 54,2 61,869,4 75,2 83,2 till
* Данные К. В. Смирновой (1945).
Как видно, в первые годы жизни наиболее интенсивен линейный
рост, а по достижении половозрелости—-рост массы тела. Самки
растут быстрее самцов. Расхождения в средних показателях одно-
возрастных рыб (5-J-, 6Ц-) составляют по длине тела 1—3 см, а по
массе — 500—800 г.
Таблица 83. Линейный рост и увеличение массы тела
нельмы оз. Зайсан (Смирнова, 1945; эмпирические данные)
Возраст, лет Средняя длина тела, см Прирост, см Средняя масса тела, г Прирост, г п
о-f- 59,10 6,20 2420 550 277
6+ 65,25 6,15 2875 455 174
7+ 71,10 5,85 4250 1375 33
8+ 75,35 4,25 53035 785 14
Известный предельный возраст нельмы из казахстанской части
ее ареала —10 лет. В настоящее время ее популяция здесь пред-
ставлена молодыми особями. Рыбы старше 8 лет в последние годы
не встречаются. Основную массу в опытных уловах составляют
3—5-годовики.
Нельма — типичный хищник. К активному питанию ес личинки
переходят уже на 10-й день после выклева. Молодь длиной от 14
до 25 мм и массой от 10 до 100 мг питается преимущественно зоо-
планктоном. Мальки длиной от 11'5 до 160 мм и массой от 12 до
35 г поедают уже молодь окуня, плотвы, язя, ельца и других рыб.
207
Обычно переход сеголетков нельмы на питание рыбой происхо-
дит в мае — июне с появлением в водоеме личинок и молоди других
видов рыб. Однако в рационе нельмы до 3 лет крупные формы бес-
позвоночных играют заметную роль. Взрослые особи питаются
преимущественно плотвой, ельцом, молодью язя. Зимой в их ра-
ционе встречаются ерш и окунь. По исследованиям 1973 г., в озер-
ной части Бухтарминского водохранилища нельма питалась пре-
имущественно молодью окуня и плотвы. В мае по частоте встре-
чаемости первое место занимала плотва (100%), второе — окунь
(75%). В августе плотва встречена только в 5% желудков, а
окунь — во всех. С акклиматизацией леща в водоемах Верхнего
Иртыша отмечены случаи поедания нельмой молоди переселенца.
Селективности в питании нельмы не установлено. Обычно она
поедает некрупных рыб наиболее массовых видов. В период нерес-
тового хода и нереста не кормится. С августа интенсивность пита-
ния возрастает и остается высокой в течение всей зимы. Различий
в характере питания самцов и самок нет.
Как большинство пресноводных хищников нельма не образует
больших стай, нагуливается и к местам нереста идет разрозненно.
Миграции ее носят сезонный характер: весенние и осенние нересто-
вые— вверх по р. Черный Иртыш. Нагуливаются и зимуют взрос-
лые особи и молодь в озерной части водохранилища. Зимой нель-
ма продолжает вести активный образ жизни.
Полупроходная нельма с Оби и Нижнего Иртыша совершает
нерестовые миграции, поднимаясь до плотины Усть-Каменогорской
ГЭС. Взрослые особи полупроходной формы после нереста медлен-
но скатываются в низовья, молодь их год-два задерживается на
верхних участках Иртыша. У жилой нельмы жизненный цикл про-
ходит в реке, нерестовые и трофические миграции короткие.
Численность и хозяйственное значение. Статистических данных
об объеме вылова нельмы в р. Иртыш в пределах ^Казахстана нет.
По оз. Зайсан в общей добыче рыбы нельма никогда не занимала
значительного места. По качеству же является одной из лучших и
наиболее ценных промысловых рыб Верхнего Иртыша.
В предвоенные годы в оз. Зайсан добывалось от 11,8 до 37,8, в
среднем около 17 т нельмы в год (Смирнова, 1945). Затем добыча
ее несколько возросла, достигнув в 1950—1954 гг. в среднем 31,2 т.
В дальнейшем уловы резко упали (до 2 т в год и ниже), а с 1957 г.
промысел нельмы был запрещен (Ерещенко, 1970). Причина резко-
го падения запасов нельмы — нерационально организованный про-
мысел, который был перенесен из озера на миграционные пути в
р. Черный Иртыш.
Тотальный лов нельмы запорными сетями и перетягами на не-
рестовых путях привел к омоложению стада, изъятию его полово-
зрелой части и в конечном счете к сокращению воспроизводитель-
ной способности, падению численности, подрыву запасов. Ската
нельмы с нерестилищ, расположенных по Черному Иртышу в пре-
делах КНР, в последние годы практически нет.
208
В итоге стадо нельмы в Бухтарминском водохранилище сведе-
но к минимуму. Рассчитывать на увеличение численности нельмы
в водохранилищах канала Иртыш — Караганда не приходится из-
за отсутствия здесь благоприятных условий для ее воспроизвод-
ства.
В целях сохранения этой ценной промысловой рыбы необходимо:
принять срочные меры по охране ее маточного стада и молоди на
местах нагула и зимовки; создать условия для естественного раз-
множения; обеспечить беспрепятственный пропуск производителей
на нерестилища и их обратный скат; осуществлять строгую охрану
основных нерестилищ нельмы вплоть до запрета лова на них дру-
гой рыбы.
Наряду с охранными мероприятиями необходимо ускорить ра-
боты по искусственному воспроизводству нельмы, биотехника
которого детально отработана (Злоказов, 1972 и др.).
16. Stenodus leucichthys leucichthys (Giildenstadt)—белорыбица
Казахское название: ак, балык;
Синонимия. Salmo leucichthys: Giildenstadt, Novi Comment. Petropol.,
v. XVI, 1772. — Pallas, Zoogr. Rosso-Asiat, v. Ill [1814].
Coregonus leucichthys: Кесслер, Tp. Сиб. об-ва, ест., т. I, 1870.
Stenodus leucichthys: Smitt, Salmonider, Ke 16, 188. — Берг, Рыбы пресных
вод России, 1916. — Подлесный, Белорыбица, Тр. Сиб. отд. ВНИРО,. т. VII,
вып. 1. Красноярск, 1947.
Stenodus leucichthys leucichthys. Смольянов, Тр. ИМЖ АН СССР, вып. 20,
1957.— Г. Никольский, Частная ихтиология, 1971.
Распространение. Белорыбица (рис. 46) — ближайший родствен-
ник нельмы, населяющий бассейн Каспийского моря. По едино-
душному мнению ихтиологов (Берг, 1928, 1948; Подлесный, 1947;
Никольский, 1971 и др.), она проникла сюда из бассейна Северно-
го Ледовитого океана, вероятнее всего, через Камско-Вычегодский
водораздел (р. Северная Кельтма в бассейне Печоры и р. Южная
Кельтма в бассейне Камы).
Рис. 46. Белорыбица
14-7
209
Проходная, холодолюбивая рыба. Обитает преимущественно в
Северном и Среднем Каспии, но встречается и южнее широты
Баку. Для размножения мигрирует в реки, в основном в Волгу.
Прежде поднималась до Ржева и заходила в Шексну, в Оке встре-
чалась до Рязани. Особенно многочисленной была в бассейне Камы
(реки Уфа, Белая, Вятка и др.). Единичные экземпляры изредка
входили в р. Терек.
В пределах Казахстана известна из Северного Каспия от Ман-
гышлака до Волги и из р. Урал, где, по мнению М. И. Тихого
(1938), поднималась выше Оренбурга. По данным А. В. Подлесно-
го (1947, 1949), в Урал белорыбица мигрировала как исключение,
в небольшом количестве, и уже возле Уральска ловилась единично
и не ежегодно.
Описание (Берг, 1948; Подлесный, 1947, 1949): D II—VI 9—13,
чаще III—IV 10-11; А II—IV 9—15, чаще II—III 12—13; 1.1. 99—
120, в среднем 109,3; жаберных тычинок 19—26, чаше 21—25; жа-
берных лучей 8—11, позвонков 65—68, пилорических придатков
191—193.
В процентах длины тела по Смитту в среднем: длина головы
19,1; высота головы 11,8; длина верхнечелюстной кости 5,5; длина
нижней челюсти 8,9; наибольшая высота тела 17,5; наименьшая —
5,6; длина хвостового стебля 14; антедорсальное расстояние 46,1;
постдорсальное — 37,5; расстояние Р — V 27,4; V — А 26. В про-
центах к длине головы (в среднем): длина рыла 25; ширина лба
23,2; горизонтальный диаметр глаза 10,5.
Тело удлиненное, слегка веретенообразное (рис. 46). Голова
коническая, рот большой, конечный. Сочленена нижняя челюсть с
черепом позади заднего края глаза; верхнечелюстная кость не до-
ходит до заднего края глаза. На челюстях, сошнике, небных кос-
тях, языке есть очень маленькие зубы. Затылочная часть головы и
спина сизые, бока серебристо-белые, брюхо белоснежное, блестя-
щее. Пятен на теле нет.
Прежде отдельные особи достигали длины 155 см и массы 32 кг.
В последние годы рыбы свыше 100 см и 11 кг не встречаются. Сред-
няя длина, по Смитту, равна 94—96% полной длины.
Половой диморфизм проявляется в том, что самцы в среднем
несколько мельче самок, и в нерестовый период у них на голове и
по бокам брюха появляются эпителиальные бугорки.
С возрастом у белорыбицы относительно уменьшаются длина го-
ловы и челюстей, диаметр глаза, увеличивается высота тела.
Биология (Подлесный, 1947; Берг, 1948; Смольянов, 1957; Ка-
занчеев, 1963, 1981; Никольский, 1971; Летичевский, 1981). От
малькового состояния до половозрелости, а также между нересто-
выми миграциями белорыбица живет и нагуливается в Каспийском
море на глубинах 10—50 м. Летом держится в глубоководных час-
тях моря, а осенью и зимой концентрируется в мелководном Се-
верном Каспии, где интенсивно питается.
Половозрелость у белорыбицы наступает в возрасте 5 лет (4+),
210
когда впервые нерестуют 23% самцов и 3% самок. Основная же
масса рыб созревает в 6 лет (5+). В течение жизни размножается
1—3, обычно 2 раза с интервалом в 2 года.
До зарегулирования стока Волги у белорыбицы к началу не-
рестовой миграции формировались в море два косяка: западный и
восточный (меньший по численности). Западный косяк начинал
входить в Волгу в сентябре, а большинство его представителей
(82—96%) мигрировали в декабре—марте, когда восточный ко-
сяк находился еще на подходе к Волге, так как совершал более
далекий путь от Мангышлака вдоль северного берега Каспия, ми-
мо устья Урала. В апреле нерестовый ход производителей в основ-
ном заканчивается.
Основные нерестилища белорыбицы находились в бассейне
р. Камы, где на речных перекатах с каменисто-галечным грунтом'
на глубине 1,5—2 м при температуре воды 0,2—6° с середины ок-
тября до конца ноября происходил нерест.
После сооружения на Волге каскада электростанций основные
нерестилища белорыбицы оказались для нее недоступными и есте-
ственное воспроизводство резко сократилось. Правда, часть произ-
водителей, скапливаясь ниже Волгоградского гидроузла, достигает
здесь половой зрелости и с наступлением в ноябре нерестовых тем-
ператур начинает нереститься. Однако эффективность этого раз-
множения ничтожна из-за недостатка соответствующего субстрата
и ряда гидрологических факторов (большая скорость течения, рез-
кие суточные колебания уровня и др.).
Сроки нерестовой миграции не изменились. Несколько снизи-
лась индивидуальная плодовитость самок, составляющая теперь в
среднем 100 тыс. икринок. Прежде плодовитость колебалась от
160 до 400 тыс. икринок. В среднем у 6-летних самок 198,5 тыс., у
7-леток 261,2 тыс., у 8-леток 296,2 тыс. икринок светло-желтого
цвета, диаметром 2,2—2,4 мм, при набухании — до 3,1 мм, массой
7,6—8 мг. Коэффициент зрелости был равен 26—28%.
Во время нерестовой миграции, вплоть до достижения нерести-
лищ, половые продукты производителей находятся на III стадии
зрелости. Дальнейшее созревание идет уже на нерестилищах, а пе-
реход в «текучее» состояние совершается быстро — в течение су-
57K30/^hC4CHHo НЭ неРестилищах преобладают самцы — в среднем
Икрометание единовременное. Выметанная икра слабоклейкая,
прикрепляется к камням, что, видимо, обеспечивает лучшее осеме-
нение, но через 3—4 дня отклеивается и забивается течением под
камни, где и развивается до выхода личинок.
Развитие икры продолжается более 6 месяцев при среднесуточ-
ной температуре воды около 1°. Массовый выход личинок происхо-
дит в конце апреля — начале мая. Размеры их 11—12 мм. При по-
вышенной температуре срок инкубации значительно сокращается.
Так, в эксперименте при температуре воды 3—6° выход жизнеспо-
собных личинок происходил через 83—101 день, т. е. вдвое быстрее.
211
На смешанное питание, которое продолжается около 3 недель,
личинки переходят на 6—10-й день после выклева.
В течение лета молодь белорыбицы скатывается в Каспий, дер-
жась преимущественно в верхних слоях потока. На зиму в реке,
как правило, не остается. Производители после нереста, потеряв
до 90% своей преднерестовой жирности (в начале хода — 21—26%,
после нереста— 1,5—2% жира в мясе), тоже скатываются в море.
Среднесуточная скорость ската молоди и производителей составля-
ет 40—50 км.
Стадо ходовой белорыбицы, как в дельте Волги, так и на нерес-
тилищах, прежде обычно состояло из 6 возрастных групп: от 4-ле-
ток (3+) до 9-леток (8+), но основу (82,3—85,4%) всегда состав-
ляли 6- и 7-летки. В 1970—1979 гг. численность последних стала
относительно меньше, зато возросло количество более старых и
более молодых производителей.
Так, в прошлом десятилетние (9+) особи встречались единич-
но, а старше не были известны. Теперь же в уловах попадаются и
11-летки (10—|—), а в целом 9—11-летние самки в 1970—1979 гг. со-
ставили 11,7% всех учтенных производителей. Максимальный воз-
раст самцов остался прежним — 8+-
Большая растянутость и равномерность возрастного ряда ходо-
вых производителей в современных условиях связаны, очевидно,
со значительным сокращением миграционных путей.
Растет белорыбица быстрее нельмы, достигая к 4 годам длины
тела около 80 см и массы 4—5 кг. После зарегулирования Волги
рост ее почти не изменился (табл. 84, 85).
Таблица 84. Рост белорыбицы до зарегулирования Волги (Подлесный, 1947)
Показатель Молодь (Каспий; п=22) Производители (р. Уфа; п=1892)
1 2 3 Пол 4+ 5+ 6+ 7+ 84-
Средняя Длй- на, см 27,6 47,5 58,2 Самцы Самки 81,9 89,3 86,6 91,1 90,8 94,4 93,7 100,3 101,1
Средняя мас- са, кг 0,21 1,13 2,19 Самцы Самки 4,5 5,7 5,7 6,3 6,4 7,6 6,8 9,1 10,5
Белорыбица — хищник, питающийся мелкой рыбой: сельдями,
атериной, бычками, молодью воблы, судака и других частиковых
рыб. Во время нерестового хода в реке почти не питается. Мальки
во время ската начинают хищничать уже в 2-месячном возрасте.
В море обеспеченность кормом молоди и производителей достаточ-
но хорошая. Так, на Среднем Каспии индексы наполнения желуд-
ков 4—8-годовалых рыб колеблются от 100 до 223 % о.
Численность и хозяйственное значение. Белорыбица — жирная
рыба. В дельте Волги ее жирность достигает 17—21%. В связи с
212
этим ее промысловое значение определяется не столько величиной
вылова, сколько высокой пищевой ценностью.
Максимальные уловы белорыбицы в Северном Каспии и дельте
Волги достигали в 30-х годах 1460 т. С зарегулированием стока
Волги, когда естественное воспроизводство белорыбицы практиче-
Таблица 85. Рост белорыбицы из Нижней Волги
в 1970—1979 гг. (Летнчевский, 1981)
Пол Воз- раст, лет Длина тела, см Масса тела, кг п, %
От—до М От—До М
Самки 5 77-82 80,0 4,6-6,0 5,5 2,8
(п=П32) 6 81-88 84,6 6,4-7,4 6,8 35,2
7 87-92 89,6 7,3—8,0 7,6 31,5
8 90-95 92,0 8,0-9,0 8,4 22,0
9 92-100 97,0 9,0—10,2 9,6 6,5
10 95-103 100,0 9,9-11,0 10,2 2.0
Самцы 4 68-73 70,0 3,8-4,2 4,0 7,7
(п=2103) 5 72-80 76,0 5,0-5,9 5,4 59,0
6 79-85 82,0 6,0-6,8 6,4 28.5
7 84-90 87,5 7,6-8,0 7,7 3,8
8 87-91 89,0 8,6-8,9 8,4 1,0
по данным
т, возникла
ски прекратилось, уловы ее резко упали. В 1959 г.,
М. А. Летичевского (1975), вылов составил всего 0,4
угроза полного уничтожения вида, и на лов был установлен запрет.
В настоящее время очень малочисленная популяция белорыби-
цы поддерживается лишь за счет незначительных нерестовых пло-
щадей Волги (ниже Волгоградской ГЭС) и Урала, а главным об-
разом за счет искусственного воспроизводства; биотехника его
известна с 1910 г., в условиях зарегулированного стока Волги деталь-
но разработана М. А. Летичевским (1963) и др. В 1961—1980 гг.
рыбзаводы дельты Волги вырастили на естественной кормовой ба-
зе и выпустили в реку 72,4 млн. экз. покатной молоди. За это же
время на Нижней Волге учтено 81 тыс. экз. ходовых производите-
лей, размеры которых в целом такие же, как были в прошлые годы.
Увеличение численности белорыбицы и возрождение ее промыс-
лового значения возможны при условии дальнейшего расширения
масштаба заводского выращивания и улучшения условий естествен-
ного воспроизводства путем сооружения мелиоративных каменисто-
галечных нерестилищ в сохранившемся нерестовом ареале.
Белорыбица занесена в «Красную книгу» Международного
союза охраны природы (МСОП).
213
Семейство THYMALLIDAE — ХАРИУСОВЫЕ
Род THYMALLUS CUVIER, 1829 —ХАРИУС
Центр происхождения хариусов, по А. Н. Световидову (1936),
находится на Алтае. В настоящее время в Европе и на севере Азии
водится 4 вида, из них 3 — в водоемах СССР (Берг, 1948).
В Казахстане обитает лишь 1 вид — Th. arcticus (Pallas). Евро-
пейский хариус (Th. thymallus), встречающийся в верхнем течении
р. Урал, в пределы Казахстана не заходит.
1. Thymallus arcticus (Pallas)—сибирский хариус
Казахское название: хариус.
Синонимия. Salmo arcticus: Pallas, Reise, v. Ill, 1776.
Thymallus vulgaris (non Nilss.): Финш и Брэм, Путешествие в Зап. Сибирь,
1882. — Абрамович, Вести, рыбопром., т. III, вып. 8—9, 1888.— Варпаховский,
Зап. ИАН, т. 59, кн. 1, СПб, 1889.
Thymallus nikolskyi: Кащенко, Рез-ты Алтайской зоол. эксп. 1898 г. Позво-
ночные. Томск, 1899.
Thymallus arcticus: Берг, Ежегодн. Зоол. музея АН, т. XII, 1908.—Световп-
дов, Тр. Зоол. ин-та АН СССР, т. III, 1936.
Thymallus arcticus brevicephalus: Митрофанов, Тр. конф, по рыбн. хоз-ву
респ. Ср. Азии и Каз-на. Фрунзе, 1961.
Распространение сибирского хариуса (рис. 47) довольно широ-
кое: от бассейна Оби на восток до Тихого океана. Водится на Кам-
чатке, в Анадыре, Амуре, на северо-западе Монголии, обилен на
Алтае. В Казахстане его распространение локализовано в правобе-
режных горных притоках Иртыша, где обитает подвид Th. arcticus
arcticus, и в оз. Маркаколь (подвидTh. arcticus brevicephalus).
Рис. 47. Сибирский хариус из р. Иртыш
Сведений по систематике и биологии западносибирского под-
вида из водоемов Казахстана не имеется. Дальнейшее описание
дается по маркакольскому хариусу.
214
la. Thymallus arcticus brevicephalus Mitrofanov —
маркакольский хариус
Описание. D VI—X 10—14, всего 19—22; A III—IV 8—11; 1.1.
78—96, чаще 80—84, над 1.1. 6—9, под 1.1. 5—9; позвонков 49—58,
чаще 54—56; жаберных тычинок на первой дуге 11—20, чаще 13—
17; жаберных лучей 5—10, обычно 8. Голова маленькая: короткая
и невысокая.
Таблица 86. Морфологические признаки сибирского
хариуса (М±т)
Признак Оз. Марка- коль Зап. Сибирь (Светови- дов, 1936}
Длина тела по Смитту, мм 236,4+6,24 233,5 + 3,54
В % длины тел а п< Длина тела без С т Смитту 94,26 + 0,21 94,32 + 0,097
Наибольшая высота тела 18,81 + 0,23 21,7 + 0,382
Наименьшая высота тела 6,65 + 0,06 7,47 + 0,083
Антедорсальное расстояние 33,58 + 0,26 32,22 ±0,264
Постдорсальное » 40,64+ 0,24 —-
Антевентральное » 46,27 + 0,21 44,75+0,189
Антеанальное » 70,25+0,30 69,42+0,212
Длина хвостового стебля 14,68+0,11 17,07+0,157
Пектровентральное расстояние 28,38 + 0,14 27,88 + 0,171
Вентроанальное » 22,65 + 0,13 25,48 + 0,221
Длина основания D 19,72+0,19 23,27 + 0,289
Наибольшая высота D 12,41+0,15 14,77 + 0,546
Длина основания А 7,96 + 0,12 9,41 ±0,124
Наибольшая высота А 12,4+0,10 12,05 + 0,159
Длина Р 15,24 + 0,14 16,56 + 0,149
V 13,34+0,13 16,09 + 0,143
ГОЛОВЫ 15,71+0,08 17,71+0,136
рыла 2,94 + 0,04 3,47 + 0,051
Продольный диаметр глаза 3,89+0,03 5,02 + 0,091
Заглазничнын отдел головы 8,02+0,05 9,03 г 0,076
Высота головы у затылка 11,69±0,11 14,44 + 0,146
Ширина лба 4,22 + 0,04 5,14+0,065
Длина верхней челюсти 4,7 + 0,04 5,82+0,068
Ширина верхней челюсти 1,72+0,03 1,69+0,021
Длина нижней челюсти 8,16 + 0,08 9,22 + 0,102
Счетные приз Общее число лучей в D н а к и 20,38 + 0,13 21,77 + 0,212
Разветвленных лучей в А 9,62+0,12 9,22 + 0,084
Чешуй в 1-1 83,79 + 0,28 81,52+0,569
Тычинок на первой жаберной дуге 14,6+0,33 17,69+0,173
Жаберных лучей 7,82 + 0,13 9,0 + 0,011
Позвонков 54,33 + 0,40 —
215
Окраска серебристая, с поперечными темными полосами у мо-
лодых особей и с глазчатыми пятнами у взрослых. Известные раз-
меры хариуса в озере до 30 см и 300 г массы. У сибирского хариу-
са несколько подвидов, образование которых связано с обособле-
нием популяций в озерах (Байкал, Косогол, Маркаколь), крупных
реках (Амур) или значительным географическим расстоянием
(Обь, Камчатка). Они различаются между собой количеством жа-
берных тычинок, чешуй в боковой линии, вариациями в размерах и
положении спинного, брюшного и грудного плавников.
Хариус из оз. Маркаколь — Th. arcticus brevicephalus — явля-
ется одним из наиболее уклонившихся подвидов сибирского ха-
риуса. По ряду признаков близок к западносибирскому подвиду, но
и отличается существенно. У него очень короткое основание спин-
ного плавника, роднящее его с байкальскими хариусами, так же
как длина и соотношение грудного и брюшного плавников. Основ-
ное его отличие от всех подвидов сибирского хариуса — маленькая
голова. В меристических признаках у хариуса из оз. Маркаколь
следует отметить наименьшее количество жаберных тычинок, жа-
берных лучей среди известных подвидов сибирского хариуса и
близкое к минимальному количество чешуй в боковой линии, лучей
в спинном плавнике, число позвонков (табл.86).
Выяснение полового диморфизма показало различие между
самцами и самками в 6 признаках (табл. 87). Все они связаны с
размерами плавников, которые у самцов лучше развиты.
Таблица 87. Половой диморфизм хариуса оз. Маркаколь
(М±т)
Признак Самки Самцы Mdlff
Высота А И.63±0,16 10,8±0,11 4,3
D И ,92±0,14 13,02 + 0,31 3,2
Длина основания D 19,23±0,24 20,35 ±0,30 3,0
V 12,92 + 0,13 13,8 + 0,27 2,9
основания А 7,68 1:0,13 8.3с+0,22 2,4
Р 14,97±0,12 15,53 _:.0,30 1,7
Биология. Половозрелым хариус в оз. Маркаколь становится
на третьем году жизни. В 3-годовалом возрасте нерестует, видимо,
вся рыба этой возрастной группы. Соотношение полов, по данным
за весь год, близко 1:1 (самцов 46,7, самок 53,3;%).
Нерест происходит ранней весной. Хариус собирается к устьям
рек и входит в них до вскрытия озера. В конце апреля в предустье-
вых пространствах наблюдается большое его скопление. Икромета-
ние начинается примерно в середине мая, когда озеро еще не
вскрылось. Ко времени вскрытия озера хариус уже интенсивно
нерестует в верховьях всех его притоков. Интенсивный нерест про-
216
должается до середины июня, а затем постепенно затихает. Во вто-
рой половине июня на нерестилищах встречаются единичные осо-
би с невыметанными половыми продуктами. В июле хариус спу-
скается вниз по рекам и выходит в озеро. В зто время он уже имеет
половые продукты в III и II—III стадиях зрелости. В сентябре
гонады переходят в стадии III—IV и IV. Зимует хариус с половыми
продуктами в IV стадии зрелости.
Плодовитость определялась у 47 самок в возрасте от 2 до 4 пол-
ных лет. Наименьшая — 966 икринок, наибольшая — 1989, сред-
няя—1467. Масса тела исследованных на плодовитость особей
колеблется от 90 до 212 г, длина по Смитту, — от 195 до 265; масса
зрелых яичников — от 8,6 до 18,9 г, относительная плодовитость в
среднем на 1 г массы составляет 9,9 и на 1 см длины — 61,4 ик-
ринки.
Икра у хариуса донная, свободно скатывающаяся по течению.
Средний диаметр неоплодотворенной икринки 2,6 мм. Развитие
икры при температуре 4—12° продолжается около 200 градусо-
дней. Через неделю после выклева мальки начинают пи-
таться.
Таблица 88. Питание хариуса оз. Маркаколь (Тэн, 1959), %
Компонент пищи Весна Лето Осень
1 2 1 2 1 2
Hirudinea 13 16,3 — —. —
Gammarus 29 13,3 — — —. —
Hydracarina 8 2,2 10 4,9 12 7,9
Hemiptera (наземные) 10 2,9 16 7,1 12 24,6
Coleoptera 10 9,3 52 24,1 24 16,9
Piecop tera 3 2,1 64 19,8 — —
Trichoptera (личинки) — — дб 27,1 —- —
Odonaia 10 7,8 — —- —- —-
Formicidae 6 2,4 28 2,3 20 22,7
Tendipedidae 55 5,2 12 8,6 — —
Pisces 13 — — — — —
Растительная пища 16 4,9 8 6,1 8 27,9
Примечание. 1 — частота встречаемости; 2 — значеи; ie гг., массе.
По линейному росту хариус из оз. Маркаколь относится к груп-
пе быстрорастущих. Темп роста хариусов (обратное расчисление)
в возрасте 1, 2, 3, 4 лет: у западносибирского (Световидов, 1936) —
соответственно 9,3, 14,6, 19, 23,6, у маркакольского—11,1, 17,7,
23,9, 28,5 см (Митрофанов, 1971 в). Как и у прочих форм хариуса,
самцы здесь несколько крупнее самок — соответственно размеры
одновозрастных групп 19,5—28,5, в среднем 24,4 см и 19—26,5, в
среднем 23 см.
217
Масса тела хариуса, по нашим данным, составляет в возрасте
1 год 18 г, в 2 года—88, в 3 года — 130, в 4 года—-183 г. (Митро-
фанов, 1971в). Для анализа использовались рыбы, выловленные
весной и не имеющие значительного прироста в текущем году.
Хариус в оз. Маркаколь питается преимущественно донными
беспозвоночными и наземными насекомыми. В зависимости
от мест обитания и сезонов года состав пищи заметно меняется
(табл. 88).
В весенний период питание хариуса наиболее интенсивно и раз-
нообразно, что связано с его обитанием в реках. В это время ос-
нову его пищи составляют рыбы (33,6% по массе) и гамма-
русы (13,2%). С потеплением и развитием наземной энтомофауны
(июнь) хариус переключается на питание жуками (24,1% по мас-
се), личинками веснянок (19,8%) и ручейников (27,1%). В летние
месяцы интенсивность питания хариуса падает и несколько подни-
мается осенью, когда увеличивается доля растительной пищи. По
сравнению с питанием хариусов из других водоемов следует отме-
тить высокий процент хищничества в весенний период и низкую
интенсивность питания летом.
Упитанность, по Кларк, определялась для весенне-летнего
(1,09) и осеннего периодов (1,18).
Численность и хозяйственное значение. В оз. Маркаколь хариус
образует довольно большие стада. Весной местами скоплений, как
указывалось выше, служат предустьевые пространства, в июле —
августе много хариуса попадается на песчаных отмелях и неглубо-
ких местах (до 5—6 м) с галечниковым грунтом. Много его в это
время и в низовьях рек, где наблюдается медленное течение.
Осенью он рассредоточивается по всему озеру и попадает в сети
только как прилов. Зимой также держится небольшими косяками
на галечниковом грунте. В это время нередки его уловы до 3—5 кг
на сеть.
Заходя весной в реки, хариус держится довольно разрозненно,
не образуя значительных стай. Местами обитания его служат
различного рода затиши, хотя спасается он от преследования на
стремнинах. Верхняя граница захода хариуса в реки трудно опре-
делима, так как в верховьях рек, по всей вероятности, обитает
речной хариус, не спускающийся в озеро. Вполне возможно, что эта
речная форма мало чем отличается от западносибирского хариуса.
По словам старожилов, в самых верхних участках реки встречают-
ся очень крупные хариусы—до килограмма и больше. Все сказанное
относится только к крупным притокам озера, имеющим больше
20 км длины, как реки Тополевка, Тихушка и др. В озере крупные
экземпляры хариуса (более 300 г) нами не встречены.
Врагов у хариуса на нерестилищах нет. Он — единственный
вид рыбы, нерестующей в верхнем течении рек, где встречаются
только единичные особи крохалей. Млекопитающих, кормящихся
рыбой, нет. Промысловое значение хариуса в оз. Маркаколь неве-
лико.
218
Семейство ESOC1DAE — ЩУКОВЫЕ
Род ESOX LINNE, 1758 —ЩУКА
Род Esox ведет свое происхождение из водоемов Восточного
Казахстана (Сычевская, 1976). В настоящее время в Евразии, в
том числе и в СССР, обитают 2 вида этого рода. Один из них —
Е. reicherti Dybowski — водится только на Дальнем Востоке (бас-
сейн Амура, оз. Ханка, Сахалин), а другой — Е. lucius Linne, от-
личающийся от первого в основном более крупной чешуей, имеет
очень широкое распространение.
1. Esox lucius Linne— щука
Казахское название: шортан.
Синонимия. Esox lucius: Linne, Syst. Nat. edition, v. X, 1758. — Кесслер,
Изв. об-ва любит, естеств., т. X, вып. 1, 1872. — Кесслер, Рыбы Арало-Касп,-
Понт. ихт. обл., 1877. — Берг. Рыбы Туркестана, 1905. — Никольский, Ежегод.
Зоол. музея АН СССР, т. 32, вып. 2, 1931. — Бенинг, Никольский, Тр. Аральского
отд. ВНИМОРХ, т. 2, 1933. — Никольский, Рыбы Аральского моря, 1940. — Берг,
Рыбы пресных вод СССР, ч. 1, 1948. — Серов, Сб. работ по ихт. и гидробиол.,
вып. 1, 1956. — Шиленкова, Сб. работ по ихт. и гидробиол., вып. 1, 1956.
Esox lucius kirgisorum: Турдаков, Пискарев, Тр. Ин-та зоол. и паразитол.
Кирг. ФАН СССР. Фрунзе, вып. 1, 1954.
Распространение. Относится к бореальному равнинному фауни-
стическому комплексу.
В Казахстане щука (рис. 48) распространена почти повсемест-
но, населяя как пресные, так и солоноватые воды. В Северном Кас-
пии и в Аральском море встречается в основном в прибрежной
зоне, преимущественно вблизи устьев рек; в открытом море обыч-
но не держится (Никольский, 1940; Казанчеев, 1963). Водится в ре-
Рпс. 48. Шука
ках Урал, Кушум, Большой и Малый Узень, Мухор, Уил, Сагиз,
Хобда, Эмба, в их притоках, водохранилищах, пойменных и ни-
зовых озерах, в том числе и в оз. Челкар (Тихий, 1938; Берг, 1948;
Серов, 1956, 1959; Дукравец, 1971; Меркулов и др., 1977).
Многочисленна в Иргиз-Тургайском бассейне (Шиленкова,
1956; Цыба, 1976). В Сырдарье — на всем протяжении, преимуще-
ственно в озерах и водохранилищах (Кесслер, 1872; Берг, 1905;
219
Рылов, Гладков, 1934; Никольский, 1938; Тютеньков, 1948; Дукра-
вец, 1978; Голубенко, 1978). Есть, по нашим данным, в бассейне
р. Арысь (приток Сырдарьи). В р. Чу известна от низовьев до Ток-
мака в Киргизии (Никольский, 1931; Дрягин, 1936; Пивнев, 1960;
Дукравец, Митрофанов, 1966). Обычна в бассейнах Сарысу, Нуры,
Улы-Жыланшика (Бенинг, Никольский, 1933; Серов, 1950, 1953;
Ерещенко, 1956; Дукравец, 1967, 1974; Селезнев, 19726; Дукравец,
Бирюков, 1976).
В Северном и Восточном Казахстане в значительном количестве
населяет бассейны Иртыша, Ишима, Тобола, Убагана, в том числе
озера, связанные хотя бы периодически с реками. Особенно много-
численна в бассейне Верхнего Иртыша (Седельников, 1910; Ефи-
мова, 1949; Доброхотов, 1959; Мартехов, 1959; Сидорова и др.,
1966; Чабан, 1966; Чабан, Дюсенгалиев, 1967; Солонинова, 19746).
По нашим данным и материалам Я- Я. Цееб (1940), В. И. Ере-
щенко (1959), М. М. Даирбаева (1966), Н. Ф. Лысенко и др.
(1972), В. В. Селезнева (19726), Л. Н. Солониновой (1981), щука
встречается в ряде изолированных, но не пересыхающих озер: Зе-
ренде, Щучьем, Большом и Малом Чебачьем, Айдабуле, Имантау,
Копа и др. в Кокчетавской области; Малом Тарангуле, Курейном,
Глубоком, Круглом, Карасевом в Северо-Казахстанской области;
Алаколе, Койбагаре, Карасоре и др. в Кустанайской области.
В 1970—1975 гг. щука вселена в водоемы Васильевского нако-
пителя шахтных вод Соколовско-Сарбайского комбината в Куста-
найской области, где широко распространилась и вышла на вто-
рое место в уловах (Цыба, 1978).
Во многих изолированных озерах Центрального и Северного
Казахстана щуки нет. Не водится она в Балхаш-Илийском и Та-
ласском бассейнах. Не обнаружена в бассейне р. Бугунь (Чимкент-
ская обл.), где ее обитание вполне вероятно, учитывая наличие
подходящих биотопов (Бугуньское водохранилище, Шошкаколь-
ские озера и др.) и периодическую связь с Сырдарьей.
Описание (в целом по республике; источники указаны в табл. 89).
D IV—X 12—19, A III—VIII 9—15, Р I 9—15, V I—II 8—12; че-
шуя мелкая — в боковой линии 105—147, над ней 11—17, под ней
11—19, число пор (прободенных чешуй) в боковой линии 43—57;
жаберных лучей 13—15, позвонков 56—63; жаберных тычинок на
первой дуге у щуки из Бухтарминского водохранилища 34—39, у
щуки из р. Чу — по переднему краю 13—21, по заднему 16—24.
Голова длинная, составляет 24—36% длины тела без хвостового
плавника; рыло вытянутое, сплющенное, длина его 10,7—18%
(около половины длины головы), диаметр глаза 2,1—6, заглавье
10,8—17,7%; наибольшая высота тела от 10% в Камыш-Самар-
ских озерах до 22,3% в оз. Челкар, наименьшая — 5—9%; анте-
дорсальное расстояние 65—80, длина хвостового стебля 10,2—17,4,
длина Р 9,3—17, длина V 10,3—16,8%. Другие пластические при-
знаки исследованы лишь в бассейнах Иргиз-Тургая и Чу, а также
в Бухтарминском водохранилище (табл. 90). В последнем мср-
220
Таблица 89. Счетные признаки щуки из различных водоемов Казахстана (числитель — пределы,
знаменатель — среднее или чаще встречающееся)
Бухтар- минское Bftxp. 5-10 6-7 13-19 15-16 3-8 6 11-15 13 1-П 9-11 121-147 132 11-17 14-15 11-17 14-15 44-57 49-50 59-63 09
Камыш- Самар- at/ua Лоа. • со о Ф 1С 2 еа CQ d 6-8 13-15 С£ 11-13 1 12-14 1 -П 8-10 120-131 13-14 61-62
Челкар (Серов, 1956) 6-8 13-17 V 12-15 I 12-15 120-132 14-15 60-62
ИргПз- Тургай (Шилен- йова, 1956) 7-9 8 12-16 13,6-14,5 6-8 7 9-14 11-12,2 119-144 СО CD 7 2 03 14-15 11-19 с 3 0 13-15 56-62 I 9'62
Чу (Пнв- нев, 1960) 4-8 13-17 13-15 3-6 10-14 со 03 lQ >—< —СО 1 1 IT О ОСО 1 110-115 12-15 13-14 13-19 14-15 45-56 ю 1 1 1 о ..
CXQJ и щен ко, I 1956) ! 6-9 13-17 СО 1 1 Ю Ю 03 оо 2 1 17 119 47-52 49,6 56-62 58
Арал (Николь- ский, 1940) 5-8 13-17 15,2 3-5 11-14 12,6 114-126 120 43-50 48 58-60
й <U Qi оо 2 Ю 2 § 1 1 1 1 1 1 1 2 X CD СО ’Ф 03 LQ 1
Признак Лучей в D неветвистых лучей в и ветвистых Лучей в А неветвистых Лучей в А ветвистых Лучей в Р Лучей в V Чешуй в 1. 1. Чешуй над 1. 1. Чешуй под 1. 1. Пор в 1. 1. Жаберных лучей XI D * D И СО D
Таблица 90. Пластические признаки щуки из различных водоемов Казахстана
(числитель — пределы, знаменатель — среднее или чаще встречающееся),
% от длины тела без С
Признак Арал (Ни- кольский, 1940) Иргиз-Тур- гай (Ши- ленкова, 1956) Чу (Пив- йев, I960) Бухтармин- ское вдхр.
Длина головы 29-35 24,1-32,8 29-36 25,1-30,1
32,0 28,6-29,8 32-33 28,6
Длина рыла 12-18 10,7-15,1 11,5-16 10,7-13,5
14,6 12,1 — 13,3 13-14 12,3
Диаметр глаза 3-6 2,7-5,6 2,7-5 2,1-3,7
4,5 3,2-3,8 3,5-4,5 3,0
Заглазье 12-17 10,8-17,7 12,6-17,5 11,7-14,0
13,7 12,3-13,5 13,5-15,5 13,1
Высота головы у затылка 9,6-18,3 10,5-15 10,0-14,6
12,6-12.9 12-13 12,6
Наибольшая высота тела 12-17 14,5-21,0 13,5-21 14,6—22,1
14,5 16,8-17,1 16-18 19,0
Наименьшая высота тела 5-7 5,3-8,7 5,5-9 5,9-7,4
6,2 6,3-6,8 7-8 6,6
Антедорсальное расстояние 70-76 65,0—76,0 70-80 70-74
73,2 70,8-72,9 73-76 72
Постдорсальное расстояние 11,9-19,5 12,5-17 12,6-16,5
14,2-14,7 14-16 14,6
Длина хвостового стебля 12-16 10,2-17,4 11,5-16 11,1-15,6
13,7 12,5-13.6 12-14 13,4
Длина D 11,2-17,8 11-16 11,7-15,3
13,2-14,3 13-15 13,6
Высота D 9,7-15,2 10-14 10,5-13,2
12,0-12,8 11,5-13 11,9
Длина А 8,2-14,0 9-12,5 9,1-12
10,7-11,1 9,5-11,5 10,4
Высота А —. 8,2-15,4 10-14 10,5-13,2
Длина Р 10-15 12,7 11-14 12,4 11,7-12,2 11,1-17,3 11,5-13,5 10-13 11,6 9,3-14,4
Длина V 12,9-13,8 10,3-16,8 11-12 10,5-13 12,1 8,7-13,6
12,2-14,3 11-12 11,7
222
фометрическому анализу в 1961 г. подвергли 25 экз., длина тела
которых (без С) была равна 39—85 см (в среднем 57,3 см), а пол-
ная масса — 0,8—6,3 кг (в среднем 2,2 кг).
Анализ таблиц 89 и 90 показывает, что в целом щука бассейнов
Урала (Челкар, Камыш-Самарские и Кушумские озера) и Иртыша
морфологически близка к европейско-сибирской. В то же время
щука из водоемов Аральского бассейна (Арал, Сырдарья, Чу, Са-
рысу) заметно отличается меньшим числом чешуи в боковой линии,
меньшим числом позвонков и жаберных тычинок, более длинной
головой и увеличенным антедорсальным расстоянием. В связи с
этим она, очевидно, может быть выделена в подвид Esox lucius
aralensis. Вряд ли целесообразно выделение особого подвида —
Е. lucius kirgisorum — только в р. Чу, как это предложили
Ф. А. Турдаков и К. В. Пискарев (1954).
Щука Иргиз-Тургайского бассейна занимает по этим при-
знакам промежуточное положение и, кроме того, имеет увеличен-
ный размах колебания большинства признаков. На географиче-
скую (широтную) изменчивость щуки указывал в свое время и
М. И. Меньшиков (1951).
Диплоидное число хромосом (2п) у щуки равно 50 (Николь-
ский, Васильев, 1973).
Половая и возрастная изменчивость детально не изучена. Обыч-
но половозрелые самки несколько крупнее самцов за счет более
интенсивного роста. Для аральской щуки отмечено относительное
уменьшение с возрастом длины головы, диаметра глаза и длины
парных плавников (Никольский, 1940). То же для длины головы
отметил Н. П. Серов (1956) у щуки из оз. Челкар: у молодых —
32—34,8, у взрослых — 27,6—32,8% длины тела.
Рот большой, вооружен острыми зубами, сидящими на многих
костях ротовой полости — межчелюстных, сошнике, небных,
нижней челюсти и на языке. Нижняя челюсть выдается вперед.
Задний край верхней челюсти свободный. Жаберные перепон-
ки не сращены между собой и свободны от межжаберного проме-
жутка. Щеки покрыты чешуей, а жаберная крышка — лишь в верх-
ней части. Спинной плавник расположен далеко сзади, над аналь-
ным.
Тело вальковатое, серо-зеленого, серо-желтоватого или серо-
бурого цвета; спина темнее, бока и брюхо светлее. На боках круп-
ные темно-оливковые пятна, иногда располагающиеся в попереч-
ibie полосы. Плавники, особенно непарные, тоже покрыты темны-
ми пятнами.
Максимальные размеры щуки достигают 1,5 м и 35 кг (Берг,
1948; Никольский, 1971; «Жизнь животных», 1971 и др.). В Казах-
стане самые крупные щуки (до 1,1 м и 11—16 кг) встречались в
(50-е годы в бассейне Иргиз-Тургая и в оз. Челкар. Правда,
Я- Я- Цееб (1940), исследовавший в 1933 г. озера Кокчетавской
области Копа, Имантау и др., называет еще более крупные раз-
меры щуки из этих озер — 16—26 кг. Однако эти сведения осно-
ваны лишь на опросных данных.
223
В последующие годы наиболее крупные щуки — длиной около
100 см и массой тела 9,1—12,4 кг —ловились в Бухтарминеком во-
дохранилище на Иртыше и в Арнасайских озерах бассейна Сыр-
дарьи. Последние территориально относятся к Узбекистану, но
расположены рядом с Чардаринским водохранилищем, за счет ко-
торого формируется их гидрологический режим и ихтиофауна.
Обычные размеры щуки в уловах везде до 60 см и до 2 кг
(табл. 91).
Размножение. Щука относится к рыбам с полицикличным и
единовременным икрометанием (Дрягин, 1949). Половозрелой в во-
доемах Казахстана она становится в 2—4 года, причем на юге —
на год раньше, чем на севере, а самцы на год раньше самок. Так, в
среднем течении Сырдарьи (Кайраккумское, Фархадское, Чарда-
ринское водохранилища, Арнасайские озера) и в бассейне р. Чу
она созревает, как правило, при длине тела (без С) около 30 см,
Таблица 91. Максимальные размеры щуки из водоемов Казахстана
Водоем Длина тела, см Масса тела, кг Автор, год
Бассейн р. Чу 100 6—7 Турдаков, 1963
Джезказганское вдхр. 48 1,06 Горюнова, 1956а
Бассейн р. Сарысу — 3,6 Ерещенко, 1956
Озера низовьев Сырдарьи 78 6,9 Рылов, Гладков, 1934
Чардаринское вдхр. 85 5,5 Ерещенко, 1970
Арнасайские озера 95 9,1 Голубенко, 1978
Бассейн Иргиз-Тургая НО 11,0 Шиленкова, 1956
Озеро Челкар НО 16,0 Серов, 1956
Камыш-Самарские и Кушумские
озера 96 5,8 Серов, 1959
56,5 1,5 Меркулов и др., 1977
Река Урал 54 1,35 »
Кушумские вдхр. 75 3,7 Малиновская, Тэн, 1983
Бухтарминское вдхр. 104 12,4 Солонинова, 1976
в том числе самцы — на втором-третьем году, а самки —на треть-
ем-четвертом (Турдаков, 1963; Максунов, 1968; Ерещенко, 1970;
Голубенко, 1978); в бассейне Сарысу— в 2—3 года, по достижении
длины 30—35 см (Ерещенко, 1956), в озерах Иргиз-Тургая — боль-
шинство в 3 года (Щиленкова, 1956); в оз. Челкар, Камыш-Самар-
ских и Кушумских озерах самцы созревают на третьем, самки —
на четвертом году жизни (Серов, 1956, 1959), в низовьях Волги —
самцы в 3 года, самки в 4 (Берг, 1948); в озерах Северного Казах-
стана самцы в массе (82,5%) созревают в возрасте 3 лет, самки — 3
(43%) —4 (57%) лет (Ерещенко, 1959).
В Бухтарминеком водохранилище часть самцов (около 30%) впер-
вые нерестится в годовалом возрасте, большинство созревает на вто-
ром-третьем году жизни при длине тела не менее 23 см, а самки
224
15—7
S
С4
О
СЗ
S
ч
Н
впервые нерестятся в 2—4 года (в массе-—в 3 года) при длине те-
ла не менее 32,5 см и массе 330 г (табл. 92).
В озерах низовьев Сырдарьи, судя по нерестовым маркам на
чешуе, щука впервые нерестится в возрасте 2 года (15—36%), ос-
новная масса — в 3 года (57—69%) и часть — в 4 года (7—15%)
по достижении длины тела 23—36 см, отдельные особи— 15—41 см.
Соотношение полов в уловах в различных озерах в разные се-
зоны и годы может сильно меняться. Например, в Арнасайских
озерах в среднем за год оно близко 1:1, но зимой в связи с более
высокой интенсивностью питания самок они в уловах преобла-
дают— 4:1 (Голубенко, 1978). На нерестилищах в Иргиз-Тургай-
Таблица 93. Соотношение полов в уловах щуки водоемов
Казахстана (оригинальные данные), %
Водоем, год исследования Сзм- ки Сам- цы
Озера низовьев р. Чу, 1959, 1965 50,0 50,0
1967 41,0 59,0
Тиликольские озера (бас. Сарысу), 1964 66,6 33,4
Озера низовьев Иргиз—Тургая, 1965 64,6 35,4
в том числе оз. Байтак 34,0 66,0
Бугункуль 79,6 20,4
7Каркуль 64,5 35,5
Малайдар 25,0 75,0
Кока рал 20,0 89,0
Кожакуль 66,6 33,4
Камыш-Самарские озера, 1968 Озера низовьев Сырдарьи 62,3 37,7
Караузякские, 1964, 1971—1972 50,0 50,0
Аксай-Кувандарьинские, 1970— 1971 50,0 50,0
Акчатауские, 1972—1973 80,0 20,0
Камышлыбашские, 1973—1974 76,0 24,0
ских озерах численно (59,8%) преобладают самцы (Шиленкова,
1956).
В Бухтарминском водохранилище в нерестовый период соотно-
шение полов близко 1:1, в другое время в уловах в 1,5 раза преоб-
ладают самки. В Джезказганском водохранилище на р. Кенгир
осенью 1951 г. самки составляли 55,5, самцы — 45,5% (Горюнова,
1956а); в Чарском водохранилище в Семипалатинской области
самок более 60%. В оз. Имантау Кокчетавской области их в 3,5 ра-
за больше, чем самцов. Таким образом, в целом в водоемах респуб-
лики чаще преобладают самки (табл. 93).
Нерестовые миграции щуки начинаются с таянием льда. Нерест
проходит на юге (Чардаринское водохранилище, Арнасайские озе-
ра) в конце февраля — начале марта (Ерещенко, 1970; Голубенко,
1978); на Арале и в бассейне р. Чу — в марте — начале апреля (Ни-
кольский, 1940; Турдаков, 1963); в Камыш-Самарских и Кушум-
ских озерах, оз. Челкар, бассейне р. Сарысу — в конце марта —
226
апреле (Ерещенко, 1956; Серов, 1956, 1959); в р. Нура — во второй
половине апреля (Серов, 1950); в Иргиз-Тургайских озерах — с се-
редины апреля до середины мая (Шиленкова, 1956); в дельте Вол-
ги— в апреле—мае (Берг, 1948; Монастырский, 1949); в это же
время нерестится щука и в Бухтарминском водохранилище (Соло-
нинова, 1969).
В оз. Зайсан и пойме р. Иртыш щука прежде нерестилась в кон-
це марта —начале апреля, в первые годы наполнения Бухтармин-
ского водохранилища — на неделю позже. С 1966 г. сроки икроме-
тания сдвинулись еще на 2—3 недели, продолжительность нереста
увеличилась (табл.94).
Таблица 94. Сроки нереста щуки в Бухтарминском водохранилище
Год Срок нереста Примечание
начало разгар конец
1961 31.111 — 15.IV До 4 мая встречались единичные самки с икрой IV стадии развития
1962 2.IV 8-10.IV 15.IV
1965 6.IV 15-19.IV 22.IV До 2 мая встречались единичные самки с икрой IV стадии развития
1966 16.IV 28.1 V—5 V 8.V До 11 мая встречались единичные самки С икрой IV стадии развития
1969 24. IV 18-20.V Не было До июля встречались многочислен- ные самки с невыметанной резорби- рующейся икрой
1970 12.IV —- Не было »
1971 12.1V 18-23.IV 26. IV До 9 мая встречались единичные самки с икрой IV стадии развития
1972 21. IV Не было В июне вылавливалось много самок с резорбирующейся икрой
Отдельные особи начинают икрометание в прибрежных прота-
линах, когда на водоеме еще держится ледовый покров. Так, в
Бухтарминском водохранилище «текучие» самки ловились при
температуре воды 0,8—1,8° подо льдом толщиной до 80 см. Массо-
вый нерест обычно длится 10—15 дней при температуре воды на
юге 4—12°, в бассейне Иргиз-Тургая — при 9—14°, на Бухтарме —
при 8—12°. Однако в зависимости от погодных условий нерест мо-
жет растягиваться и до 2,5 месяца (Голубенко, 1978; Коблицкая,
1981). В низовьях р. Б. Узень (бас. Урала) отдельные текучие осо-
би встречались даже в первой половине июля (Серов 1959). Обыч-
но же при температуре воды 18—20° икрометание угнетается, а
при более высокой температуре икра в яичниках перерождается.
Нерест происходит на вечерних и утренних зорях. Икра откла-
дывается единовременно в прибрежье или на разливах на глубине
до 1 м, преимущественно на прошлогодние растения, но иногда и
прямо на дно.
227
В Бухтарминеком водохранилище наиболее ценные нерестили-
ща щуки и других видов рыб — это дельта Черного Иртыша, Бу-
коньский плес и залив Клы. Прежде Черный Иртыш при впадении
в оз. Зайсан образовывал дельту площадью более 50 тыс. га, изо-
биловавшую протоками, рукавами, старицами, заливами и озера-
ми, разграниченными зарослями тростника и плавучими островами
отмершей растительности. Нерестовым субстратом для щуки в
этом лабиринте служили промытые корневища тростника и рогоза.
Кладки икры располагались узкой полосой в 5—10 см от поверх-
ности воды. На протяжении 1 м параллельно урезу воды насчиты-
валось до 25 икринок.
В зону затопления Буконьского плеса вошли р. Иртыш с об-
ширной поймой. Здесь заливались низины со степной, полупустын-
ной и вновь вегетирующей луговой растительностью, служившей
нерестовым субстратом для щуки. На свежезалитых кустиках зла-
ков или полыни насчитывалось до 15 икринок. Для икрометания
щука иногда выходит на такие мелководья, что над поверхностью
воды выступает ее спина.
В оз. Зайсан до образования водохранилища и в первые годы
его наполнения щука нерестилась также в заливах, образованных
длинными мысами-волнорезами (мыс. Бархот — протяженностью
8 км, Бесчаны — 4, Тополев — более 2 км). С подъемом уровня мы-
сы частично размылись, ушли под воду, растительность разбило
волной, в результате Тарбагатайское и Курчумское побережья по-
теряли значение в воспроизводстве рыбных запасов.
Первыми начинают нерест наиболее старые, крупные, особи,
последними выходят на нерестилища впервые созревающие щуки,
о чем свидетельствуют их средние линейные размеры в дельте Чер-
ного Иртыша по пятидневкам (1965 г.): 15—49.IV — 52 см, 20—
24.IV —49, 25—29.IV —47, 30.IV—4.V — 37, 5—9.V —28 см.
Икрометание у щуки единовременное, но содержимое яичника
выметывается не в один прием. Вероятно, это связано со ступенча-
тым дозреванием икры от каудальной части к краниальной. В пе-
риод нереста попадаются особи, у которых каудальная часть яич-
ника свободна от икры, а краниальная заполнена икрой IV стадии
развития. Часто это связано с резким похолоданием в период не-
реста.
Ход нереста и его эффективность зависят от характера весен-
него паводка и температуры воды. Наибольшее отрицательное
влияние на естественное воспроизводство щуки оказывают колеба-
ния уровня водоема. При низком, но постоянном уровне урожай-
ность молоди может быть высокой. Снижают эффективность не-
реста резкие суточные колебания температуры воды, а также
несовпадение благоприятного для икрометания уровенного и терми-
ческого режимов.
Абсолютная плодовитость щуки в разных бассейнах колеблется
в значительных пределах: от 6 тыс. икринок у двухгодовалой самки
массой 450 г из Бухтарминского водохранилища до 270,6 тыс. ик-
228
Таблица 95. Абсолютная плодовитость щуки водоемов Казахстана,
тыс. икринок
Водоем ДлПна без С,см Масса тела, кг Плодовитость Автор, год
От—до М
Джезказганское вдхр. * 32-46 0,55-0,9 17,0-40,3 26,7 Горюнова, 1956а
Бассейн Сарысу 40-62 —. 17,1-55,7 36,2 Ерещенко, 1956
Озеро Челкар 36-58 —' 12,9-39,8 — Серов, 1956
Кушумские озера 38-61 — 8,4-45,7 — Серов, 1959
Иргиз-Тургайские озе- 42-96 1,5-9,5 26,5-270,6 199,6 Шиленкова,
ра 10,0-18,0 1956
Среднее течение р. Чу — 0,4-0,65 — Турдаков, 1963
Чардаринское вдхр. 41-78 0,76-4,83 29,6-124,9 — Ерещенко, 1970
Арнасайские озера — — 16,3-89,1 44,6 Г олубенко,
Низовья р. Чу 37,5—50 0,56-1,1 32,9-61,4 50,0 1978 Наши дан-
Низовья Иргиз-Тургая Низовье р. Сырдарьи 53-69 1,4-2,75 34,8-93,7 54,1 ные. 1967 Наши дан- ные, 1965
Караузякские озера Озеро Кандарал 41-42 0,6-0,7 29,6-38,4 34,0 Наши дан- ные, 1964
41,0 0,68 50,2 50,2 »
Озеро Камышлыбаш Наши дан-
44,5 0,85 26,6 26,6 ные, 1973
Озеро Утебас — 2,0-2,8 68-168,6 106,0 Дукравец и др., 1975
ринок у самки массой 9,5 кг из озер низовьев Иргиз-Тургая. Обыч-
но же она не превышает 90 тыс. икринок, а в среднем составляет
26,6—54,1 тыс. (табл. 95).
С возрастом и увеличением размеров самок плодовитость их
возрастает. Так, в дельте Волги самки размером 36—40 см имели
среднюю плодовитость 16,6 тыс. икринок, размером 46—50 см—•
43,7 тыс., размером 56—60 см — 79,7 тыс. икринок (Берг, 1948).
В Джезказганском водохранилище плодовитость четырехлеток
(3+) была равна в среднем’22,8 тыс. икринок, пятилеток (4Д-) —
28,2 тыс. (Горюнова, 1956а). Подобное же отмечено у щуки из ни-
зовьев Иргиз-Тургая (Шиленкова, 1956), из Арнасайских озер (Го-
лубенко, 1978).
В Бухтарминеком водохранилище у впервые нерестящихся осо-
бей длиной тела 35—50 см и массой 0,3—1,5 кг в 2—3-годовалом
229
возрасте абсолютная плодовитость чаще составляет 15—20 тыс.
икринок, у повторно нерестящихся увеличивается вдвое. В каж-
дой размерно-возрастной группе плодовитость колеблется в широ-
ких пределах, часто максимальное значение превосходит мини-
мальное в 4—5, иногда в 10 раз. Основу маточного стада состав-
ляют особи с длиной тела 50—60 см, их плодовитость 30—35 тыс
икринок (Солопинова, 1975).
Относительная плодовитость колеблется от 12 до 81, а чаще от
20 до 45 икринок на 1 г массы тела. Она не столь явно, как абсо-
лютная плодовитость, зависит от длины тела и возраста щуки.
Диаметр зрелых икринок 2,1—3 (Монастырский, 1949; Серов,
1956), в бассейне Иргиз-Тургая несколько меньше — 2,19—2,25 мм
(Шиленкова, 1956). Цвет икринок светло-желтый.
У щуки Бухтармпнского водохранилища масса яичников, диа-
метр икринок, коэффициент плодовитости увеличиваются с ростом
рыбы. Среднее значение коэффициента зрелости повышается от
6,3 до 15,4 у рыб с длиной тела до 80 см и массой до 6 кг; у более
крупных особей оно снижается до 14,'5. Средний коэффициент пло-
довитости, рассчитанный на общую массу рыбы, колеблется от 1,4
до 4,6, а на массу без внутренностей — от 0,9 до 3,7. Диаметр икры
щуки в IV стадии зрелости изменяется от 1,8 до 2,9 мм (в среднем
2,4 мм). Масса яичников увеличивается пропорционально массе
рыбы и обычно составляет около 15%. У впервые нерестящихся са-
мок с длиной тела 30—50 см и массой до 1,5 кг масса яичников ко-
леблется от 50 до 200 г, у повторно нерестящихся особей повыша-
ется втрое, у самок размером более 70 см и массой выше 5 кг
достигает 1 кг. ?4аксимальная масса гонад—-1,3 кг — зафиксирова-
на у самки длиной 100 см, массой 10 кг. У промысловой части ста-
да с линейными размерами 50—60 см она составляет 200—300 г.
Выметанная икра обладает слабой клейкостью и в первые дни
развивается, приклеившись к растениям. Затем клейкость пропа-
дает, икра осыпается и дальше развивается на дне', возможно, по
П. А. Дрягину (1949), в полупелагическом состоянии, плавая око-
ло дна.
Развитие икры и молоди щуки в казахстанских водоемах почти
не изучено. Есть лишь сведения о том, что в Бухтарминском водо-
хранилище икра щуки развивается 10—13 дней при температуре
воды 9—12°. Выклюнувшиеся личинки имеют длину 6,2 мм и массу
9 мг. В европейской части СССР икра развивается 10—14 дней при
температуре воды 8—14° (Шамардина, 1957). Таков же инкуба-
ционный период у щуки Фархадского водохранилища на р. Сыр-
дарье в Таджикистане (Максунов, 1968).
Личинки на весь период эндогенного питания прикрепляются к
водным растениям. Через 8—10 суток, достигнув длины 12—13 мм,
они переходят на смешанное питание. Через 15 дней после вылуп-
ления, при длине тела около 15 мм, желточный мешок резорбирует-
ся и щука полностью переходит на экзогенное питание (Монастыр-
ский, 1949; Шамардина, 1957).
230
Во время смешанного питания молодь держится в прибрежье
небольшими стайками. Полностью переходя на внешнее питание,
щука начинает вести индивидуальный образ жизни, образуя замет-
ные концентрации лишь в нерестовый период.
Рост и возраст. Растет щука обычно очень быстро. В первые го-
ды жизни у нее наиболее интенсивен линейный рост, а после до-
стижения половозрелости — рост массы тела. Мальки в мае в Иргиз-
Тургайскпх озерах имеют длину 6,4 см (Шиленкова, 1956), в июле
в бассейне р. Чу—11 см (Дрягин, 1936), длина сеголетков осенью
в дельте Волги в среднем 19,6 см (Монастырский, 1949).
В то же время известно, что у щуки, особенно в первые годы
жизни, очень сильны колебания в темпе линейного роста, связан-
ные в первую очередь с конкретными условиями питания (Попова,
1971). Так, А. Ф. Коблицкая (1981) указывает, что размеры сего-
летков в сентябре в различных водоемах колеблются от 11 до 33 см,
т. е. в пределах 22 см. В Чардаринском водохранилище размеры
одновозрастных щук различаются у годовиков на 13—15 см, у
двухгодовиков — на 10—15 см (Ерещенко, 1970). По П. А. Дряги-
Т а б л и ц а 96. Линейный рост щуки различных водоемов Казахстана
(обратное расчисление), см
Водоем, год исследований Воз* раст, лет 1. 1в 1» 14 15 И
Бурдж а рское вдх р., 1965 3+ 16,8 27,1 36,2 40,0 2
Старица в р-не Турке-
стана, 1965 3-1—4+ 12,5 21,0 29,4 37,0 — — 3
Караузякскпе озера, 2-1—5+ 15,0 24,4 31,9 39,5 — 154
1964 2+-7+ 9,2 17,9 23,0 32,9 38,5 41,9 302
1971 2+-7+ 9,4 16,2 22,6 29,1 34,5 38,3 78
1972
Аксай-Кувандарьян- 1 + -7+ 11,0 20,5 29,4 35,3 40,8 48,9 263
ские озера, 1971
Акчатаускпе озера. 3+-6+ 10,7 20,8 30,1 37,5 42,9 46,6 50
1972—1973
Камбашскне озера, 3+-6+ 11,9 22,7 32,3 41,0 47,5 57,5 66
1973—1974 в т. ч. Раим, 1973
СЕМКИ 3+—5+ 12,3 22,8 31,7 40,4 45,4 — 56
самцы 3-1—5+ 12,0 21,9 30,6 38,1 42,9 — 24
Камбаш, 1974 самки 4+-6+ 11,3 22,3 32,9 42,6 50,1 83
самцы 4+ -5+ 11,5 23,2 32,9 41,0 46,1 — 9
Озера низовьев р. Чу,
1965 3+-6 + 10,0 18,9 27,1 34,8 39,7 45,9 28
1967 34—6+ 9,2 16,3 24,4 29,5 33,0 42,0 47
Иргиз-Тургайские озе- ра, 1965 1 + -7+ 14,3 24,6 34,2 41,0 45,4 51,2 258
Озера низовьев р. Ну-
ра, 1974 5+-7+ 12,1 23,2 30,3 38,1 49,7 58,9 11
231
Oi
ea
tl
К
4
KO
C3
E-
ну (1936), в среднем течении р. Чу длина тела (без С) двухлеток
в июле колебалась от 15 до 24 см, а масса — от 38 до 152 г. Это
подтверждается и материалами таблиц 96, 97.
Быстрее растет щука в дельте Волги, Арнасайских озерах, бас-
сейне Иргиз-Тургая и Бухтарминеком водохранилище. Хуже всего
растет в оз. Челкар, где лишь к 13—14 годам достигает длины
100 см, в Джезказганском водохранилище, Аксай-Кувандарьин-
ских, Камыш-Самарских и Тиликольских озерах. Разница между
крайними размерами одновозрастных рыб из разных водоемов по
обратному раскислению составляет у годовиков 17 см, у 2-годо-
виков более 20 см, у 3-годовиков около 26 см и т. д.
Таблица 98. Длина и масса щуки разных возрастных групп
в Бухтарминеком водохранилище (среднемноголетние
эмпирические данные)
Возраст, лет Длина, см Масса, кг И
От—до Средн. От — до Средн.
1 9-40 21 0,04—0,8 0,19 285
2 15-62 39 0,06-2,5 0,7 650
3 30-81 50 0,2-5,2 1,5 1170
4 35—84 59 0,4—5,9 2,1 1250
5 43-91 63 0,7-8,5 2,7 605
6 45-99 67 1,0-9,6 3,6 200
7 50-98 70 1,1-7,7 4,0 100
8 49-102 76 1,1-10,0 5,1 75
9 60—104 88 2,4—12,4 7,1 20
10 78-103 90 2,8-12,4 7,9 12
11 88-100 94 5,4—7,4 6,4 2
Таблица 99. Длина и масса молоди щуки в оз. Зайсан
и Бухтарминеком водохранилище (среднемноголетние
эмпирические данные)
Месяц Длина, см Масса, г п
От—до Средн. От—до Средн.
Озеро Зайсан
VI VII XII 6-9 5-12 16-26 □ухтарми 7,5 8,3 20 некое в 2-6 1-22 36-150 одохрани 4,0 7,9 77 л и щ е 2 27 103
V 1,5-6,0 3,4 0,01-2,0 0,56 30
VI 3,6-10 5,5 0,4—7,0 16 17
VII 7,0-15 11,2 3,5-41 16,3 64
VIII 3,4-22 13,7 0,34-101 35,7 73
XII 21-26 24,0 77-171 137 33
233
У щуки Бухтарминского водохранилища, по эмпирическим
данным, эта разница еще более значительна (табл. 98, 99).
В Бухтарминском водохранилище линейные размеры и масса
молоди щуки стали больше, чем прежде в оз. Зайсан и р. Иртыш в
те же календарные сроки, несмотря на то, что нерест здесь проис-
ходит на 2—3 недели позже. Более интенсивный рост молоди сви-
детельствует о том, что в водохранилище условия нагула благо-
приятнее, чем в исходных водоемах (Солонинова, 1966). Несколько
различается рост сеголетков по участкам водохранилища: напри-
мер, в августе 1972 г. быстрее росли они у Тарбагатайского побе-
режья (длина тела 22 см и масса НО г), в дельте Черного Иртышг
(18 см п 60 г) и в Нарымском заливе (20 см и 70 г). Замедленны!
рост наблюдался и в заливе Клы (8 см и 5 г). На остальных участ
ках средние длина и масса сеголетков были 15 см и 40 г.
Различия в темпе роста особей отдельных локальных стад ха
рактерны и для старшевозрастных групп щуки и связаны ш
столько с разной обеспеченностью пищей, сколько с термическим!
условиями при нагуле. В Бухтарминском водохранилище более вы
сокий темп роста щуки отмечен в участках, где летом вода на глу
бине 10 м прогревается до 15—20°, а замедленный рост — в мелко
водной зайсанской части, где вода от поверхности до дна прогре
вается до 22—26° (Вакулко, 1969).
Таблица 100. Рост массы тела щуки из водоемов Казахстана с возрастом, кг
Озера Возраст, лет Автор, год
2 3 4 । 5 1 6 1 7 1 8
Челкар Кушумские Камыш-Самарские Су-Жарган Джалангаш Самки Самцы 1 | 0,320,530,80 0,13 0,39 0,64 0,140,330,48 0,73.1,1111,62 0,95'1,942,43 0,881,602,23 1 1 1 1 1,19,1,632,63 1,14'1,532,01 0,65'1,074,71 2,73 3,845,61 4,43:6,69; — 3,91)5,50, - 3,54 2,38 2,23 11,0 Серов, 1956 Серов, 1959 » Ши ленкова, 1956 &
Замедленный рост щуки в некоторых водоемах объясняется их
неудовлетворительным гидрологическим режимом и, как следствие,
плохими условиями питания. По таблицам можно проследить ухуд-
шение роста щуки по годам исследования в бассейнах рек Чу, Са-
рысу, Сырдарья.
А. И. Ефимова (1949) считает, что рост щуки в водоемах с раз-
витой поймой происходит быстрее, чем в водоемах, где пойма раз-
вита слабее. Материалы Г. X. Шапошниковой (1964) по бассейну
Урала это подтверждают: в верховьях бассейна, где пойма развита
слабее, щука растет медленнее, чем в среднем течении.
Масса тела у щуки особенно интенсивно растет, начиная с пя-
того года жизни (табл. 100).
В Арнасайских озерах щука в возрасте 6—8 лет ежегодно при-
234
бавляет почти по 1,5 кг (Голубенко, 1978)'. Самки везде растут
лучше самцов (и по длине, и по массе тела). Так, на Бухтарме го-
довики самцов уступают самкам по длине лишь на 2 см, а по массе
различий нет; у 4-годовиков расхождение увеличивается по длине
до 8 см, по массе до 0,9 кг; у 7-годовиков — до 20 см и 2,5 кг.
Известный максимальный возраст щуки из водоемов Казахстана
равен 18 годам. Такой экземпляр исследовал Н. П. Серов в 1949—
1951 гг. на оз. Челкар. К сожалению, размеры этой щуки он не
указал. Там же 15-летняя щука имела длину 1,1 м и массу тела —
16 кг. В последние годы самые старые особи — 8—11 полных лет —
ловятся в Чардаринском водохранилище (Ерещенко, 1970), Арна-
сайских озерах (Голубенко, 1978) и Бухтарминском водохранили-
ще. Однако такие щуки встречаются единично. Основу промысло-
вых и экспериментальных уловов везде составляют 3—5-годовалые
рыбы.
Питание и упитанность. Щука — типичный хищник. Лишь очень
непродолжительное время после рассасывания желточного мешка
она питается преимущественно беспозвоночными: личинками хи-
рономид, поденок, ветвистоусыми и веслоногими рачками, пиявка-
ми и т. п. При длине тела около 2 см она уже может захватывать и
молодь рыб (Суворов, 1948; Шамардина, 1957; Махотин, 1977). По
достижении длины тела около 5 см щука почти полностью пере-
ключается на питание рыбой. Жертву обычно подстерегает среди
зарослей, настигая ее стремительным броском.
Видовой избирательности в пище нет. Питается обычно наи-
более многочисленными и доступными в конкретных обстоятельст-
вах видами. Поэтому там, где состав ихтиофауны богаче, ра-
цион щуки разнообразнее, и наоборот. Так, в дельте Волги в состав
ее пищи входят 15 видов рыб (Терентьев, 1937; Фортунатова, 1949).
Столько же видов, но, естественно, в несколько ином наборе, вхо-
дят в ее рацион в Чардаринском водохранилище с Арнасайскими
озерами (Ерещенко, 1970; Голубенко, 1978). В озерах низовьев
Сырдарьи, по нашим исследованиям 1970—1975 гг., щука питается
рыбами 14 видов в Камыш-Самарских и Кушумскях озерах, по
данным Н. П. Серова (1956, 1959),— 12 видов. В бассейне Верх-
него Иртыша за 1957—1978 гг. в пище щуки зарегистрировано рыб
14 видов и — единично — минога (Солонинова, 1972).
В то же время в бассейне р. Сарысу в пище щуки представлены
лишь 8 видов рыб (Ерещенко, 1956; Малиновская, 1956), столько
же, по нашим данным 1959—1967 гг., в низовьях р. Чу. В Иргиз-
Тургайских озерах, по А. К. Шиленковой (1956) и нашим исследо-
ваниям в 1965 г., щука использовала в пищу все 6 видов ихтио-
фауны этого бассейна.
Всего в пище щуки водоемов Казахстана обнаружено 36 видов
рыб, но основу ее питания везде составляют плотва (все подвиды),
елец, красноперка, язь, жерех, лещ, караси (2 вида), сазан, окунь,
ерш и щука. Размеры жертв увеличиваются по мере роста хищни-
ка, а максимальные составляют 40—60,% его длины в любой раз-
235
мерно-возрастной группе (Терентьев, 1937; Фортунатова, 1949).
В наших сборах наибольший размер добычи щуки равен 47 см
(без С): такой длины особь своего вида заглотила в Бухтармин-
ском водохранилище щука, собственная длина которой была 79 см.
В других водоемах республики наибольший размер жертвы щуки
равен 33 см (язь, жерех, судак и щука). Размеры других рыб-жертв
не превышают 25 см. Обычно же длина жертв составляет 5—15 см.
Количество рыб в одном желудке зависит от их размера и доступ-
ности; обычно оно не превышает 10 шт., а максимум — 19 мальков
окуня — был отмечен у щуки Бухтарминского водохранилища.
Свойственный щуке каннибализм (питание особями своего ви-
да) проявляется даже при благоприятных условиях питания, а
при недостатке другого корма является важным фактором выжи-
вания, чему способствует разноразмерность одновозрастных щук.
Среди нерыбных объектов в пище щуки нередко встречаются
креветки, бокоплавы, личинки стрекоз. Последних, размером 1,5—
3 см, мы насчитывали до 80 штук в одном желудке. Изредка обна-
руживаются в желудках щук лягушки (Пивнев, 1960) и наземные
позвоночные: ящерицы, мыши, землеройки (Ерещенко, 1970).
В Бухтарминском водохранилище было изучено питание личи-
нок и мальков щуки размером от 1,8 до 21 см (Солонинова, 1974а).
В мае — июне зоопланктон занимает существенное место по часто-
те встречаемости и в весовом отношении в пище мальков длиной
до 5 см (соответственно 70 и 12,4%); при этом ветвистоусые рачки
преобладают над веслоногими по массе, а по частоте встречаемос-
ти они находятся в равном соотношении. С увеличением размеров
щуки потребление зоопланктона сокращается, а в июле, по дости-
жении мальками длины 10 см, он совсем выпадает из их рациона.
Зообентос также имеет большой удельный вес в питании личи-
нок и мальков длиной от 2 до 10 см в мае и июне; в июле и августе
у мальков размером 10—20 см он встречается редко. Наиболее
часто сеголетки питаются личинками хирономид и поденок. Не-
редки в пищевом комке личинки жуков и стрекоз. Олигохеты обна-
ружены у сеголетков с длиной тела 2—4 см. Нематоды, пиявки, бо-
коплавы, личинки ручейников встречались единично и существен-
ного значения в питании не имели.
Личинки рыб входят в состав пищи щуки по достижении ею
длины 1,8 см. Сначала это в основном личинки ельца (57% по час-
тоте встречаемости и 47% —по массе). У более крупной молоди
рацион пополняется мальками плотвы, язя, окуня. Первостепенное
значение в рыбном рационе щуки длиной более 5 см по частоте
встречаемости приобретает плотва, несколько уступают ей елец,
язь и окунь. В весовом отношении доминирующее положение зани-
мает язь, меньший удельный вес имеют плотва, елец и окунь. Реже
встречается молодь ерша и леща, а такие виды, как щука, сазан,
карась, пескарь, шиповка и судак в пище щуки единичны.
Об интенсивности и периодичности питания щуки сведения
противоречивы. А. К. Шиленкова (1956) считает, что в Иргиз-Тур-
236
гайских озерах щука во время нереста практически не кормится,
поскольку 97,5% исследованных в это время желудков щуки были
пусты. В дельте Волги, по данным В. Терентьева (1937), щука пи-
тается и во время нереста, а интенсивность питания колеблется в
течение года. В Чардаринском водохранилище, по мнению
В. И. Ерещенко (1970), щука кормится круглый год без существен-
ных колебаний интенсивности. В Арнасайских озерах, непосред-
ственно связанных с Чардаринским водохранилищем, интенсив-
ность питания щуки, о которой судили по относительному количе-
ству кормящихся особей, наибольшая весной (50%) и осенью
(54,5%); зимой она вдвое слабее (23,3%), причем самки в это
время питаются интенсивнее самцов, а летом, с повышением темпе-
ратуры воды более 25°, щука вообще перестает есть (Голубенко,
1978). Подобное чередование периодов интенсивного и ослабленно-
го питания, а также прекращения приема пищи, свойственно и
щуке оз. Челкар (Серов, 1956). Сходно распределяется по сезонам
годовой рацион щуки в дельте Волги: весной — 23,9, летом — 20,6,
осенью — 35,9, зимой—19,6% (Казанчеев, 1981).
Наши материалы подтверждают данные о колебании интенсив-
ности питания щуки по сезонам года. Так, количество ее желудков,
содержащих пищу, на озерах низовьев Сырдарьи, Чу и Иргиз-Тур-
гая колебалось в сборах от 17—28% летом, до 42—47% весной и
осенью. В Бухтарминском водохранилище щука питается более
равномерно в течение года: весной в уловах бывает в среднем 50%
питающихся особей, летом — 40, осенью — 33, зимой-—54%. Прав-
да, самки здесь прекращают питаться в период нереста. По-види-
мому, периодическое снижение интенсивности питания у щуки свя-
зано и со сменой зубов.
Мясо щуки тощее, содержит 2—3% жира и является диетиче-
ским продуктом («Жизнь животных», 1971). Упитанность ее в це-
лом невысока: средний коэффициент по Фультону колеблется око-
ло 1 (табл. 101).
С возрастом упитанность понижается. Зависит она и от гидро-
логических условий в водоеме, как это видно на примере озер в
низовьях р. Чу и Караузякских, которые в годы исследований ин-
тенсивно усыхали. На Бухтарме отмечено некоторое снижение
упитанности в течение года, от весны к зиме (по Фультону в сред-
нем — весной 1,07, летом 1,01, осенью 0,94, зимой 0,89). Различия в
упитанности по полу не наблюдаются.
Места обитания, миграции, численность. Щука предпочитает
медленно текущие водотоки и озера с чистой прохладной водой.
Быстрого течения избегает. Может жить в «кислой» воде (pH до
4,8), но заморных водоемов не выносит. В водохранилищах числен-
ность ее обычно невелика, очевидно, в связи с колебаниями уровня,
отражающимися на воспроизводстве, высокими летними темпера-
турами воды и малочисленностью макрофитов. В Казахстане осо-
бенно многочисленна в дельтах Волги, Урала, в низовьях Сыр-
дарьи, Иргиза и Тургая, в бассейне Верхнего Иртыша.
237
Таблица 101. Упитанность щуки различных водоемов Казахстана
Держится обычно раз-
розненно среди зарослей
водных растений или воз-
ле них. Далеких мигра-
ций, как правило, не со-
вершает. Носят они сезон-
ный характер: весенние —
нерестовые и осенние —
трофические. Те и другие
проходят в границах аре-
ала локальных стад. В ре-
ках щука предпочитает
заводи, заливы, старицы,
пойменные озера. В круп-
ных водоемах чаще дер-
жится в прибрежье, отку-
да летом с прогревом
воды отходит на глубоко-
водье. Зимой продолжает
вести активный образ
жизни.
До образования Бух-
тарминского водохранили-
ща щука встречалась на
всем протяжении р. Чер-
ный Иртыш, в оз. Зайсан,
в р. Иртыш от истоков до
слияния с Обью. Наибо-
лее многочисленна была в
дельте Черного Иртыша
и в заливе Клы оз. Зай-
сан.
В первые годы за-
полнения водохранилища
щука встречалась повсе-
местно от Черного Ирты-
ша до плотины гидроузла.
Однако уже в 1963 г. из
глубоководной части во-
дохранилища, где мелко-
водья с зарослями макро-
фитов ограничены, нача-
лись миграции щуки в
мелководную часть Зай-
сана. К концу 60-х годов
щука практически исчез-
ла из глубоководной ча-
сти, за исключением На-
рымского залива и райо-
G3
Д'
к
хо
(Я
Н
Данные Министерства рыбного хозяйства Казахской ССР.
на Батов. В 70-е годы относительно наибольшая численность ее по-
прежнему отмечалась в дельте Черного Иртыша, в заливе Клы, а
также в мелководном Буконьском плесе, хотя запасы ее в абсолют-
ных цифрах сильно сократились.
Хозяйственное значение. Щука — важная промысловая рыба.
В Аральском море в предвоенные годы давала 3% всей добычи, в
основном за счет озер низовьев Сырдарьи. Основу промысла со-
ставляли 3—4-годовалые рыбы (Никольский, 1940). В Иргиз-Тур-
гайских озерах дает до 25% всего улова рыбы (Цыба, 1976). Уло-
вы ее в некоторых бассейнах представлены в таблице 102.
Колебания уловов щуки отражают динамику ее численности,
связанную в первую очередь с гидрологическими условиями. На-
Рис. 49. Уловы рыбы в бассейне Верхнего Иртыша в 1930—1980 гг.: I— общий
вылов; II — в том числе вылов щуки
240
пример, до сооружения Чардаринского водохранилища и Казалин-
ского гидроузла на Сырдарье уловы щуки в Аксай-Кувандарьин-
ских озерах колебались от 178 до 531 т в год (1956—1965 гг.), а за-
тем вместе с общей добычей рыбы резко упали, составив в 1966—
1968 гг. 48—90 т, а в последующие годы и того меньше.
В Чардаринском водохранилище в первые годы его наполнения
в промысловых и опытных уловах щука составляла 1,0—1,2% (Ере-
щенко, 1970).
В бассейне Верхнего Иртыша в 30-е годы нашего столетия щу-
ка составляла 25—50% в общем вылове рыбы, в 40-е — 20—35%,
в 50-е— 13—25%. Удельный вес в ее уловах постепенно снижался,
но абсолютная величина ее вылова в течение 30 лет, предшество-
вавших образованию Бухтарминского водохранилища, колебалась
от 500 до 1200 т в год. К началу наполнения водохранилища щу-
ка составляла в уловах третью часть. В первые годы наполнения
водохранилища уловы щуки возросли до 2 тыс. т. В дальнейшем
неблагоприятные условия воспроизводства в связи с колебаниями
уровня водохранилища, с одной стороны, и нерациональный вылов
маточного стада и молоди с целью направленного формирования
лещево-судачьей ихтиофауны — с другой, привели к быстрому со-
кращению численности щуки в этом водоеме (Солонинова, 1973).
В результате щука — некогда процветавший здесь вид — потеряла
практически свое промысловое значение в Бухтарминском водо-
хранилище (рис. *49).
Для рыбного хозяйства щука представляет интерес не только
как биологический мелиоратор и экономически выгодный объект
промысла, имеющий товарную ценность уже в двухлетнем возрасте.
Определенное значение имеет она как объект искусственного раз-
ведения и акклиматизации, особенно в европейских странах и у
нас в стране — в Прибалтике, на Украине, в Белоруссии.
В то же время численность щуки легко регулируется промыс-
лом, так как чрезмерный рост запасов этого активного хищника,
особенно увеличение количества крупных особей, ведет к сокраще-
нию рыбопродуктивности водоема.
16-7
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
ОПРЕДЕЛИТЕЛЬ СЕМЕЙСТВ РЫБ,
ВСТРЕЧАЮЩИХСЯ В ПРЕСНЫХ ВОДАХ КАЗАХСТАНА*
(2). Челюстей нет, жаберных отверстий 7 пар. Парные плавники отсут-
ствуют..................................................Petromyzonidae.
(1). Челюсти есть, жаберных отверстий одна пара. Парные плавники
имеются.
(4). На теле 5 рядов костных жучек: 1 спинной, 2 боковых и 2 брюшных
........................................ .. . Acipenseridae.
(3). Костных жучек нет. Тело покрыто чешуей, шнпиками, костными пласти-
нами или голое.
(6). Тело покрыто кольцевыми костными пластинами. . . . Syngnathidae.
(5). Тело покрыто чешуей или голое.
(8). Брюшные плавники превращены в колючки. Перед спинным плавником
свободные колючки.......................................Gasterosteidae.
(7). Брюшные плавники не в виде колючек.
(10). Брюшные плавники слиты, образуют присоску. . . . Gobiidae.
(9). Брюшные плавники обычной формы.
(12). Брюшиые плавники впереди грудных, на подбородке один усик. .
................................................................Gadidae.
(11). Брюшные плавники позади грудных или под ними. Усики, если есть,
парные.
(20). Жировой плавник есть.
(15). Есть усики..............................................Sisoridae.
(14). Усиков нет.
(17). В спинном плавнике более 17 лучей.....................Thymallidae.
(16). В спиниом плавнике менее 16 лучей.
(19). В боковой линии обычно более 120 чешуй. Есть suprapreoperculum и иет
dermosphenoticum.............................................Salmonidae.
(18). В боковой линии обычно менее 120 чешуй. Нет suprapreoperculum и есть
dermosphenoticum.............................................Coregonidae
(13). Жирового плавника нет.
(22). В спиииом плавнике около 50 лучей.................Ophiocephalidae.
(21.) В спинном плавнике (или двух плавниках) менее 40 лучей.
(24). Спинной плавник отнесен далеко назад, расположен над анальным. Ры-
ло длинное, уплощенное, на челюстях мощные зубы. . . Esocidae.
(23). Спиииой плавник (или плавники) расположен примерно посередине тела.
(28). Спииных плавников два. Передний только с нераздвоенными лучами.
(27). Жаберные крышки зазубрены. На челюстях зубы. Чешуя ктенондная.
...............................................................Percidae.
(26). Жаберные крышки ие зазубрены. Зубов на челюстях нет. Чешуя цик-
лоидная. ................................................. Atherinidae.
(25). Спиииой плавник один.
(32). Тело голое или чешуя скрыта в коже.
(31). Анальный плавник очень длинный, в нем более 70 лучей...............
....................................................... Siluridae.
(30). Анальный плавник значительно короче. В нем не более 6 мягких лучей.
............................................................. Cobitidae.
(29). Тело покрыто чешуей, иногда не по всей поверхности.
(36). Челюсти с мелкими зубами, боковой линии нет. Жаберные перепонки
свободные.
(35). На голове чешуи нет.....................................Clupeidae.
(34). Голова очешуена...................................Cyprinodontidae.
(33). Челюсти без зубов, жаберные перепонки соединены межжаберным про-
межутком. Есть глоточные зубы.......................Cyprinidae.
* Включая временно заходящих из Каспия и акклиматизированных рыб.
242
ОПРЕДЕЛИТЕЛЬ РОДОВ РЫБ, ОПИСАННЫХ В ТОМЕ 1
1
2
1
2
3
4
1
2
1
2
3
4
1
2
Сем. Petromyzonidae
(2). На верхнечелюстной пластинке 2 зуба по краям . . . Lampetra.
(1). На верхнечелюстной пластинке 1 зуб или 2, соприкасающиеся основа-
ниями .................................................Caspiomyzon.
Сем. Acipenseridae
(4). Брызгальце есть.
(3). Жаберные перепонки сращены и образуют свободную складку. Рот боль-
шой, полулунный...............................................Huso.
(2). Жаберные перепонки приращены к межжаберному промежутку. Сво-
бодной складки нет. Рот небольшой, поперечный . . . Acipenser.
(1). Брызгальца нет..........................Pseudoscaphirhynchus.
Сем. Clupeidae
(2). По середине верхней челюсти заметная вырезка. Сочленение нижней
челюсти позади вертикали заднего края глаза..................Alosa.
(1). На верхней челюсти нет вырезки или она едва заметная. Сочленение
нижней челюсти впереди вертикали заднего края глаза. . • Clupeonella.
Сем. Salmonidae
(4). Верхнечелюстная кость достигает заднего края глаза. Сочленение ниж-
ней челюсти позади вертикали заднего края глаза. Рот большой.
(3). Сошник удлиненный, рукоятка его всегда имеет зубы. Жаберных тычи-
нок больше 16 (16—22)........................................Salmo.
(2). Сошник короткий, в его задней части (на рукоятке) иет зубов. Зубы
на нёбных костях и сошнике образуют подковообразную полосу. Жабер-
ных тычинок 12—21, чаще 14—17................................Hucho.
(1). Верхнечелюстная кость не достигает заднего края глаза, сочленение
нижней челюсти впереди вертикали заднего края глаза. Рот малень-
кий...................................................Brachymystax.
Сем. Coregonidae
(2). Жаберных тычинок не более 30. Есть придаточная кость иа ннжией че-
люсти в месте сочленения articulare и dentale.............Stenodus.
(1) . Жаберных тычинок более 35. Нет придаточной кости на нижней че-
люсти..................................................Coregonus.
(Вселенцы. Описание см. в т. 5).
Сем. Thymallidae
....................................... . . . Thymallus.
Сем. Esocidae
Esox.
243
ОПРЕДЕЛИТЕЛЬ ВИДОВ И ПОДВИДОВ РЫБ, ОПИСАННЫХ В ТОМЕ 1
Род Lampetra
1 (2). Зубы на верхнечелюстной пластинке одновершинные................
................................L. japonica—тихоокеанская минога.
2 (1). Зубы на верхнечелюстной пластинке раздвоены....................
...................................L. kessleri — сибирская минога.
Род Caspiomyzon
...................................С. wagneri — каспийская минога.
Род Huso
,.....................................................Н. huso — белуга.
Род Acipenser
1 (2). Нижняя губа посередине не прервана.................................
............................................ A. nudiventris — шип.
2 (1). Нижняя губа посередине прервана,
3 (4). Рыло мечевидное, более 60% длины головы............................
.......................... ... A. stellatus — севрюга.
4 (3). Рыло коническое, менее 60% длины головы.
5 (6). Боковых жучек более 50. Усики бахромчатые..........................
,.............................................A. ruthenus — стерлядь.
6 (5). Боковых жучек менее 50. Усики не бахромчатые или малобахромчатые.
7 (8). Концы жаберных тычинок расщеплены. Жучкн слабозернистые.
...............................................A. baeri — сибирский осетр.
8 (7). Жаберные тычинки одновершинные. На жучках явные радиальные по-
лосы.
9 (10). Боковых жучек 30—50, в среднем 39.................................
...................................A. giildenstadti — русский осетр.
10 (9). Боковых жучек 21—42, в среднем 3'0,6..............................
....................................A. g. persicus — персидский осетр.
Род Pseudoscaphirhynchus
......................Ps. fedtschenkoi — сырдарьинский лжелопатонос.
Род Alosa
1 (2). Жаберных тычинок обычно более 90. Наибольшая высота тела более
25% его длины.........................................A. caspia — пузанок.
2 (1). Жаберных тычинок обычно менее 80. Наибольшая высота тела менее
25% его длины.
3 (4). Жаберные тычинки жесткие, не длиннее жаберных лепестков. .
........................ ......................A. kessleri — черноспинка.
4 (3). Жаберные тычинки мягкие, тонкие в полтора раза длиннее жаберных
лепестков...........................A. kessleri volgensis — волжская сельдь.
Род Clupeonella
1 (2). Высота тела 18—24% его длины. Жаберных тычинок 50—63'. .
......................................Cl. delicatula caspia — каспийская тюлька.
2 (1). Высота тела 23—27% его длины. Жаберных тычинок 40—50. .
. Cl. delicatula caspia m. tscharchalensis — чархальская селедочка.
244
Род Salmo
1 (2). На боках тела вдоль боковой линии у взрослых рыб идет широкая крас-
ная полоса. . . . S. gairdneri — стальноголовый лосось (в Казах-
. ставе акклиматизирована его пресноводная форма — радуж-
.......................................... ная форель, см. т. 5).
2 (1). На боках нет красной полосы...................S. trutta—-кумжа.
3 (4). Длина головы у взрослых самцов более 23% длины тела, обычно 23—
26%.................................S. trutta aralensis — аральский лосось.
4 (3). Длина головы у взрослых самцов менее 23% длины тела, обычно 17—
22%.................................S. trutta caspius — каспийский лосось.
Род Hucho
..........................................Н. hucho — таймень.
Род Brachymystax
...............................................Br. lenok — ленок.
1 (2). Длина головы в среднем 21—22% длины тела. Жаберных тычинок 19—•
27, в среднем 24—25................................Br. lenok lenok — ленок.
2 (1). Длина головы в среднем 20% длины тела. Жаберных тычинок 17—24,
в среднем 20—21. . . . Br. lenok savinovi — маркакольский ленок.
Род Stenodus
......................................St. leucichthys — белорыбица.
1 (2). В анальном плавнике 2—4 неразветвленных луча, чаще 2—3. Чешуй в
боковой линии 99—120......................St. 1. leucichthys—белорыбица.
2 (1). В анальном плавнике 4—5 неразветвленных лучей. В боковой линии 99—
108 чешуй......................................... St. 1. nelma — нельма.
Род Thymallus
...................................Th. arcticus — сибирский хариус.
1 (2). Жаберных тычинок 14—22, чаще 17—18; жаберных лучей 8—12. .
.............................Th. arcticus arcticus — западносибирский хариус.
2 (1). Жаберных тычинок 11—20, чаще 14—15; жаберных лучей 5—10, обыч-
но 8......................Th. arcticus brevicephalus — маркакольский хариус.
Род Esox
Е. lucius — щука.
A
С
D
P
V
1. 1.
1
Q
q
n
aD
pD
aA
aP
aV-
P—V
V—A
Коэффициент
по Фультону
Коэффициент
по Кларк
М (или х)
m
CV
ОЧ-
И-
СЛИСОК ТЕРМИНОВ, ОБОЗНАЧЕНИИ
И УСЛОВНЫХ СОКРАЩЕНИЙ В ТЕКСТЕ
-pinna analis — анальный (подхвостовой)' плавник
» caudalis — хвостовой плавник
» dorsalis — спинной плавник
» pectoralis — грудной плавник
» ventralis — брюшной плавник
» linea lateralis — боковая линия
- длина тела рыбы без хвостового плавника (без С)
11, 1г, 1з и т. д. » » в возрасте 1 год, 2 года,
3 года и т. д.
- масса тела рыбы
- масса рыбы без внутренностей (масса тушки,
порки)
- количество исследованных рыб
- антедорсальное расстояние
- постдорсальное »
антеанальное »
- антепектор альное »
- антевентральное »
- расстояние между Р и V (пектовентральное)
- расстояние между V и А (вентроанальное)
- упитанность, вычисленная по полной массе тела
- упитанность, вычисленная по массе тела без внут-
ренностей
- среднее арифметическое (вариационное) значение
признака
- средняя статистическая ошибка для средней ариф-
метической (М)
- коэффициент вариации (в однородном биологиче-
ском материале обычно не превышает 10—12%)
- сеголетки, т. е. рыбы, прожившие одно лето
-двухлетки, т. е. рыбы, прожившие более одного
года и т. д.
ЛИТЕРАТУРА
Абакумов В. А. Систематика и экология европейской ручьевой миноги Lam-
petra planeri (Bloch.). — Вопросы ихтиологии, 1964, т. 4, вып. 3(32), с. 423—432.
Аббакумов В. П. Биологические основы формирования ихтиофауны и рыбо-
хозяйственное освоение водохранилищ канала Иртыш—Караганда. Автореф.
дис. ... канд. биол. наук. Л., 1977. 22 с. _
Абдильдаев М. Использование Aplocheilus latipes (Temmink and Schlegel)
для борьбы с личинками кровососущих двукрылых иа юго-востоке Казахстана.
Автореф. дис. ... канд. биол. наук. Алма-Ата, 1975. 20 с.
Абдурахманов Ю. А. Рыбы пресных вод Азербайджана. Баку, 1962. 406 с.
Абрамович С. Озеро Марка-Куль. — Вести, рыбопромышленности, 1888, т. 3,
вып. 8—9, с. 195—197.
Агамалиев А. С. Личиночный период жизни каспийской миноги Caspiomy-
zon wagneri (Kessler). — Тр. Всесоюз. н.-и. ин-та морск. рыби. хоз-ва и океаногр.,
1971а, т. 86, с. 139—148.
Агамалиев А. С. Возрастные изменения в строении каспийской миноги Cas-
piomyzon wagneri (Kessler). — Тр. Всесоюз. и.-и. ин-та морск. рыбн. хоз-ва и
океаногр., 19716, т. 86, с. 149—157.
Агапова А. И. Паразиты рыб водоемов Западного Казахстана. — Тр. Ин-та
зоол. АН КазССР, 1956, т. 5, с. 5—60.
Агапова А. И. Изменения паразитофауны у рыб, акклиматизированных в Ка-
захстане.—В кн.: Акклиматизация животных в СССР: (Мат. конф. Фрунзе, 10—
15 мая 1963 г.). Алма-Ата, 1963, с. 336—337.
Агапова А. И. Паразиты рыб водоемов Казахстана. Алма-Ата, 1966. 342 с.
Аникии В. П. Описание новых азиатских пород рыб. — Изв. Импер. Томск,
ун-та, 1905, кн. 27, с. 1—17.
А. С. Озеро Марка-Куль.—Охотник и пушник Сибири, 1926, We 12, с. 13.
Артюхин Е. П. О положении позднего ярового осетра в Волге: (Тез. отчета,
сессии ЦНИОРХ). Астрахань, 1974, с. 7—9.
Артюхии Е. П. Инструкция по разведению волжской популяции персидского
Осетра. М, 1978. 12 с.
Артюхин Е. П. Персидский осетр в реках Северного Каспия и перспективы
его использования в осетровом хозяйстве. — В ки.: Биологические основы разви-
тия осетрового хоз-ва в водоемах СССР. М.; 1979, с. 105—114.
Артюхин Е. П. Структура нерестового контингента персидского осетра в Вол-
ге в связи с задачами воспроизводства. Автореф. дис. ... канд. биол. наук. Л.,
1983. 16 с.
Артюхин Е. П., Андронов А. Е. О внутривидовой дифференциации персидско-
247
го осетра Волги. — В кн.: Рацион, основы ведения осетрового хоз-ва: (Тез. докл.
конф.). Волгоград, 1981, с. 15—16.
Бабушкин Н. Я. К систематике каспийской белуги. — Изв. Азерб. н.-и. ры-
бохоз. ст., 1942, вып. 7, с. 131—134.
Бабушкин Н. Я. Биология и промысел каспийской белуги. — Тр. ВНИРО,
1964, т. 52, вып. 1, с. 183—258.
Бабушкин Н. Я., Борзенко М. П. Осетровые рыбы Каспия. М., 1951, с. 25—44.
Баимбетов А. А. Морфолого-экологическая изменчивость маринок водоемов
бассейна Балхаша. Автореф. дис. ... канд. биол. наук. Алма-Ата, 1974. 28 с.
Баимбетов А. А. Морфологическая характеристика амурского чебачка Капча-
гайского водохранилища. — В кн.: Биологические науки: Алма-Ата, 1975, вып. 8,
с. 48—55.
Баимбетов А. А. Морфо-биологическая изменчивость короткоцикличных рыб,
акклиматизированных в Капчагайском водохранилище (бассейн оз. Балхаш).—
В кн.: Советско-американское сотрудничество в области исследования Мирового
океана: (Симпозиум по реакции водных экосистем па вселение новых видов). М.,
1977, с. 4—6.
Балабан П. П. Метаморфоз миноги. Киев, 1958. 63 с.
Балтабаев А. Б. Сырдарьинский лопатонос Ps. fedtschenkowi (Kessl) реки
Карадарьи. — Вопросы ихтиологии, 1972, т. 12, вып. 6(77), с. 1118—1119.
Баранникова И. А. Биологическая дифференциация стада волго-каспийского
осетра: (в связи с задачами промышленного осетроводства в дельте Волги).—
Уч. зап. ЛГУ, 1957, № 228. Сер. биол., вып. 44, с. 57—72.
Баранникова И. А. Функциональные основы миграции рыб. Л., 1975. 207 с.
Батраева М. Н. К биологии амурского чебачка. — В кн.: Биология водоемов
Казахстана: (Сб. работ Казахстанского фил. ВГБО). Алма-Ата, 1970, с. 55—60.
Батраева М. Н. О воспроизводительной способности амурского чебачка в
условиях прудовых хозяйств предгорной зоны Казахстана. — В кн.: Рыбные ре-
сурсы водоемов Казахстана и их использование. Алма-Ата, 1975, вып. 9,
с. 67—69.
Батраева М. Н. Приспособительные особенности амурского чебачка и разра-
ботка мер борьбы с ним. — В кн.: Биология водоемов Казахстана. Алма-Ата, 1981,
вып. 3, с. 78—82.
Башуиов В. С. Биология и рыбохозяйственное значение воблы, акклиматизи-
рованной в озере Балхаш. Автореф. дис. ... канд. биол. наук. Л., 1974. 16 с.
Башуиова Н. Н. Биология и рыбохозяйственное значение жереха, акклима-
тизированного в озере Балхаш. Автореф. дис. ... канд. биол. наук. Л., 1974. 20 с.
Бекешев А. Б., Песериди Н. Е. Скат молоди осетровых реки Урал. — В кн.:
Биологические основы рыбного хоз-ва на водоемах Сред. Азии и Казахстана. Ал-
ма-Ата, 1966, с. 49—51.
Беляев В. Н. Осетр (Acipenser giildenstadti Br): Некоторые предварительные
данные. — Бюл. Всекасп. научн.-рыбохоз. экспедиции, 1932, № 5—6, с. 66—76.
Беиинг А. Л. Материалы по гидробиологии р. Урала. — В кн.: Большая Эмба,
1938, т. 2, с. 153—258.
Беиинг А. Л., Никольский Г. В. Материалы по рыбному хозяйству озер ни-
зовьев Сарысу. — Тр. Аральского отд. ВНИМОРХ, 1933, т. 2, с. 103—148.
Берг Л. С. Рыбы Туркестана. — Изв. Турк. отд. РГО, 1905, т. 4, Науч, рез-ты
Арал. эксп. вып. 6. СПб. 262 с.
Берг Л. С. Ежегодник Зоологического музея Академии наук (1907), 1908,
т. 12.
Берг Л. С. Аральское море. Опыт физико-географической монографии. — Изв.
Турк. отд. РГО, 1908, т. 5. Науч, рез-ты Арал, эксп., вып. 9. СПб. 580 с.
Берг Л. С. Рыбы бассейна Амура. — Зап. Российск. Акад, наук, 1909, т. 24,
№ 9. 270 с.
Берг Л. С. Фауна России. Рыбы. СПб., 1911, т. 1. 250 с.
Берг Л. С. О рыбах рек Сары-Су и Нуры в Акмолинской области. — Еже-
годник Зоол. музея АН, 1912, т. 17, Ns 1, с. 53—55.
Берг Л. С. О происхождении северных элементов в фауне Каспня. — ДАН
СССР. Сер. А, 1928, № 7, с. 107—112..
248
Берг Л. С. Обзор мнног Северного полушария. — Ежегодн. Зоол. музея АН
СССР, 1931, вып. 1, с. 87—116.
Берг Л. С. Яровые и озимые расы у проходных рыб. — Изв. АН СССР, 1934,
№ 5, с. 711—732.
Берг Л. С. Рыбы пресных вод СССР и сопредельных стран. М.; Л., 1948,
ч. 1. 468 с.
Берг Л. С. Рыбы пресных вод СССР и сопредельных стран. М.; Л., 1949,
ч. 2, с. 469—926.
Берг Л. С. Рыбы пресных вод СССР и сопредельных стран. М.; Л., 1949,
ч. 3, с. 927—1382.
Берг Л. С. Яровые и озимые расы у проходных осетровых. Очерки по общим
вопросам ихтиологии. М.; Л-, 1953, с. 242—260.
Берг Л. С. Изменения рельефа со времени засушливой послеледниковой эпо-
хи.— В кн.: Избр. труды: В 5-ти т. М., 1958. Т. 2, с. 76—85.
Берг Л. С. О лососе Аральского моря. — В кн.: Избр. труды: В 5-ти т. М.,
1961а. Т. 4, с. 622—628.
Берг Л. С. Опыты искусственного вывода севрюги на Урале. — В кн.: Избр.
труды: В 5-ти т. М., 19616. Т. 4, с. 629.
Берг Л. С. Зоогеография пресноводных рыб Передней Азии. — В кн.: Избр.
труды: В 5-ти т. М., 1962. Т. 5, с. 211—237.
Берг Р. Л. По озерам Сибири и Средней Азии: Путешествия Л. С. Берга
(1898—1906 гг.) и П. Г. Игнатова (1898—1902 гг.). М., 1955. 320 с.
Бердический Л. С., Иоганзен Б. Г., Соин С. Г. Г. В. Никольский и его роль
в развитии современной ихтиологии. — В кн.: Современные проблемы ихтиологии.
М.; 1981, с. 5—22.
Бережной В. И. Сезонная динамика питания молоди шипа в р. Урал. — В кн.:
Материалы объединен, научн. сессии ЦНИОРХ и АзНИИРХ. Астрахань, 1971,
с. 14—29.
Березовский А. М. Организация рыбного хозяйства на озере Зайсан. — Бюл.
рыбн. хоз-ва, 1930, № 11—12.
Беседнов Л. Н., Кучеров Н. Н. К систематическому положению ленков рода
Brachymystax р. Иман. — В кн.: Зоологические проблемы Сибири: (Материалы
IV совещания зоологов Сибири). Новосибирск, 1972, с. 220—221.
Боган Ф. Е. К биологии сибирского осетра (Acipenser baeri Brandt) бассейна
р. Иртыша. — Уч. зап. Пермского гос. ун-та, 1938, вып. 2, с. 145—162.
Богданов Г. А. Питание некоторых видов рыб Усть-Каменогорского водо-
хранилища.— В кн.: Сборник работ по ихтиологии и гидробиологии. Алма-Ата,
1959, вып. 2, с. 234—244.
Богородицкий П. А. Динамика промысловых уловов осетровых и кефалей в
районах южного побережья Каспия. — В кн.: Материалы конф., посвящ. 100-летню
осетроводства. Астрахань, 1969, с. 26—30.
Борзенко М. П. Каспийская севрюга. — Изв. Азерб. н.-и. рыбохоз. станции.
Баку, 1942. 114 с.
Борзенко М. П. Материалы по систематике, биологии и промыслу куринско-
го шипа. — Тр. Каспийск, бас. филиала ВНИРО. Астрахань, 1950, т. 2, с. 9—48.
Борзенко М. П. Распространение н рост стерляди в Каспийском море. —•
Рыбное хоз-во, 1951, № 2, с. 44.
Борзенко М. П. Современное состояние запасов и промысла осетровых в
Азербайджане и пути его рационализации. М., 1961. 35 с.
Борзенко М. П. Современное состояние и прогноз изменений запасов севрю-
ги в Каспийском море при зарегулированном стоке. — Тр. BHHfPO, 1964, т. 52,
с. 259—286.
Борисов П. Г. Состояние и перспективы лова каспийской кильки на электри-
ческий свет. — Рыбное хоз-во, 1951, № 11, с. 9—19.
Борисов П. Г. Научно-промысловые исследования на морских и пресных во-
доемах. М., 1964. 260 с.
Бородин И. А. Материалы к биологии осетровых. — Тр. Отдела ихтиологии
АН, 1897а, т. 2, с. 261—272.
Бородин Н. А. Отчет об экскурсии с зоологической целью летом 1895 года
249
на крейсере «Уралец» в северной части Каспийского моря. — Вести, рыбопромыш-
ленности, 18976, № 1, с. 17—24.
Брагина Е. В. Материалы по паразитофауне рыб водоемов Кустанайской
области. — В кн.: Рыбные ресурсы водоемов Казахстана и их использование. Ал-
ма-Ата, 1970, вып. 6, с. 290—297.
Брагина Е. В. Паразитофауна некоторых видов карповых и окуневых рыб
Бухтарминского водохранилища. — В кн.: Биологические основы рыбн. хоз-ва
респ. Ср. Азии и Казахстана. — Ашхабад, 1974, с. ПО—111.
Быков Н. Е. Материалы по акклиматизации в Аральском море рыб и кормо-
вых беспозвоночных. — Тр. конф, по рыбн. хоз-ву респ. Ср. Азии и Казахстана.
Фрунзе, 1961, с. 45—50.
Быков Н. Е. Роль акклиматизированных объектов в питании хищных рыб
Аральского моря. — В кн.: Акклиматизация животных в СССР. Алма-Ата, 1963,
с. 221—223.
Быков Н. Е. О случайном вселении бычков и атерины в Аральское море и
их взаимоотношении с акклиматизированными и местными видами рыб. — Бюл.
Моск, об-ва испыт. природы. Отд. биол., 1964, т. 69, вып. 1, с. 51—58.
Быков Н. Е. Акклиматизация балтийской салаки (Clupea harengus membras
L.) в Аральском море. М., 1968. 83 с.
Вакулко <П. П. Некоторые черты температурного режима Бухтарминского во-
дохранилища.— В кн.: Ихтиология и гидробиология. Душанбе, 1969, с. 6—19.
Варпаховский Н. А. Краткие данные об ихтиофауне Азиатской России. —
Зап. Акад. наук. СПб., 1889, т. 59, вып. 1, с. 1—21.
Варпаховский Н. А. Данные по ихтиологической фауне бассейна реки Оби. —
Ежегодн. Зоол. музея Акад. наук. СПб., 1897, с. 241—271.
Варпаховский Н. А. Рыболовство в бассейне реки Оби. — В кн.: Департ. зем-
лед. СПб., 1898, с. 159—201.
Васильев В. П., Соколов Л. И., Серебрякова Е. В. Кариотип сибирского
осетра (A. baeri Brandt) реки Лены и некоторые вопросы эволюции кариотипов
осетрообразных.— Вопросы ихтиологии, 1980, т. 20, вып. 6, с. 814—822.
Васильева Е. Д. Остеологическая характеристика ленка (род Brachymystax:
Salmoniformes, Salmonidae) озера Марка-Коль и р. Кальджир. — Зоол. журн.,
1978, т. 57, вып. 4, с. 555—561.
Васильева Е. Д., Мина М. В. Сравнительный анализ морфологических приз-
наков ленков из разных частей ареала рода Brachymystax (Salmoniformes, Sal-
monidae).— Зоол. журн., 1980, т. 59, вып. 1, с. 79—90.
Вобла Северного Каспия /Под ред. Монастырского Г. Н. (Тр. Всесоюз. н.-и.
ин-та морского рыбного хоз-ва и океанографии, т. 10). М.; Л., 1939, ч. 1. 184 с.
Вобла Северного Каспия /Под ред. Монастырского Г. Н. Там же, т. И. М.:
Л., 1940, ч. 2. 172 с.
Войнова И. А. О питании сома в условиях р. Урал. — В ки.: Состояние
сырьевых ресурсов, воспроизводства и рациональное использование рыбных за-
пасов Урало-Каспия: (Тез. докл.). Гурьев, 1972, с. 41—43.
Войиова И. А. Биология н промысел сома р. Урал. Автореф. дис. ... канд.
биол. наук. Баку, 1973. 23 с.
Войнова И. А., Бекешев А. Б., Невлюдова Г. И. Влияние хищников иа чис-
ленность скатывающейся молоди осетровых реки Урала. — В кн.: Материалы
конф., посвящ. 100-летию осетроводства. Астрахань, 1969, с. 38—39.
Володкии А. В. О развитии рыбохозяйственной науки иа Аральском море. —
В кн.: Сборник работ по ихтиологии и гидробиологии. Алма-Ата, 1961, вып. 3,
с. 10—44.
Воробьева Н. Б. Бентос и его значение в питании рыб озера Балхаш. Авто-
реф. дис.... канд. биол. наук. Алма-Ата, 1973. 22 с.
Вотинов И. П. Осетровые рыбы Обского бассейна. Тюмень, 1958. 43 с.
Вотииов Н. П. Состояние запасов осетра в Обь-Иртышском бассейне и меро-
приятия по их увеличению. — В ки.: Осетровое хозяйство в водоемах СССР. М.,
1963, с. 177—181.
Вотинов Н. П. Биологические основы искусственного воспроизводства обско-
250
го осетра. — В кн.: Искусственное разведение осетровых и сиговых рыб в Обь-
ИртЫшском бассейне. Тюмень, 1963, с. 5—102.
Вотинов Н. П. Маркакульский ленок как объект акклиматизации. — В кн.:
Акклиматизация животных в СССР. Алма-Ата, 1963, с. 232—233.
Вотинов Н. П. Условия воспроизводства сибирского осетра и лососевых рыб
в водохранилищах Верхнего Иртыша. — В кн.: Теоретические основы рыбоводст-
ва. М., 1965, с. 109—112.
Вотинов Н. П., Злоказов В. Н., Касьянов В. И., Сецко Р. И. Состояние за-
пасов осетра в реках Сибири и мероприятия по их увеличению. Свердловск,
1975. 96 с.
Вотинов Н. П., Петкевич А. Н., Сецко Р. И. Опыт искусственного разведения
осетровых рыб в Обь-Иртышском бассейне. Новосибирск, 1957. 14 с.
Галицкий В. В. Палеогндрография и неотектоника Восточного Прибалхашья.
— Изв. АН КазССР. Сер. геол., 1957, вып. 3(28), с. 40—69.
Гвоздев Е. В. Материалы по паразитофауне рыб озера Марка-Коль. — Изв.
АН КазССР. Сер. паразитол., 1950, вып. 8, с. 208—225.
Гвоздев Е. В., Османов С. О., Агапова А. И. Состояние перспективы развития
гидропаразитологических исследований в Средней Азии и Казахстане. — В кн.:
Биол. основы рыбн. хоз-ва водоемов Ср. Азии и Казахстана. Фрунзе, 1978,
с. 468—471.
Герасимов И. П. Каспийское море в четвертичный период. — Тр. Советской
секции Международной ассоциации по изучению четвертичного периода, 1933,
ч. 3.
Гербильский Н. Л. Биологические группы куринского осетра и основания
для их заводского воспроизводства. — Докл. АН СССР, 1950, т. 71, № 4,
с. 785—788.
Гербильский Н. Л. Пути развития внутривидовой биологической дифферен-
циации, типы анадромных мигрантов и вопрос о миграционном импульсе у осет-
ровых.— Уч. зап. ЛГУ, 1957, № 228, вып. 44, с. 11—32.
Герценштейн С. М. Научные результаты путешествий Н. М. Пржевальского
по Центральной Азии. — В кн.: Рыбы. СПб., 1888—1891. Т. 3, ч. 2. 262 с.
Гинзбург Я. И. Размножение миноги Caspiomyzon wagneri (Kessler) ниже
Волгоградской плотины и развитие ее личинок. — Вопросы ихтиологии, 1970,
т. 10, вып. 4, с. 655—665.
Гладков Н. А. Опыт перевозки производителей шнпа. — Рыбное хоз-во СССР,
1934, № 3, с. 43—45.
Гликмаи Л. С. Акулы палеогена и их стратиграфическое значение. М.; Л.,
1964. 229 с.
Голубенко Ю. А. Биология щуки в Арнасайской системе озер. — В кн.: Биол.
основы рыбн. хоз-ва водоемов Ср. Азии и Казахстана. Фрунзе, 1978, с. 285—288.
Городецкий В. О появлении сазана в р. Или. — Изв. Турк. отд. геогр. об-ва,
1916, т. 12, вып. 1, с. 240.
Горюнова А. И. Ихтиофауна Джезказганского водохранилища и пути ее ре-
конструкции.— Изв. АН КазССР. Сер. биол., 1955, вып. 9, с. 111—121.
Горюнова А. И. Формирование ихтиофауны Джезказганского водохранилища.
— В кн.: Сб. работ по ихтиологии и гидробиологии. Алма-Ата, 1956а, вып. 1,
с. 31—73.
Горюнова А. И. К вопросу об уродствах рыб.—Там же, 19566, с. 261—268.
Горюнова А. И. Периодические изменения ихтиофауны в озерах и реках Це-
линного края. — Вопросы ихтиологии, 1962, т. 2, вып. 4(25), с. 577—580.
Горюнова А. И., Мартехов П. Ф., Сидорова А. Ф. Биология сазана в озерах
Бийли-Куль и Ак-Куль Джамбулской области.— В кн.: Сб. работ по ихтиологии
и гидробиологии. Алма-Ата, 1956, вып. 1, с. 252—260.
Гриценко О. Ф. К вопросу об экологическом параллелизме между миногами
и лососями. — Изв. ТИНРО, Южно-Сахалинск, 1968, т. 65, с. 131—134.
Грюнберг В. Заметки о жизни аральского шипа в реке Сыр-Дарье. — Рус-
ское судоходство, 1905, № 7, с. 86—91.
Гуревич Г., Лопатин С. Добыча рыбы и морского зверя в Каспийском бас-
сейне.— Статистический справочник. Астрахань, 1962. 186 с.
251
Даирбаев М. Влияние ирригационных систем на скат молоди проходных я
некоторых жилых рыб Сыр-Дарьи. — Изв. АН КазССР. Сер. биол., 1957, вып. 2,
с. 37—42.
Даирбаев М. М. Формирование, состав и распределение ихтиофауны в раз-
личных типах водоемов ирригационной системы реки Сыр-Дарьи. — В кн.: Сб.
работ по ихтиологии и гидробиологии. Алма-Ата, 1959, вып. 2, с. 286—299.
Даирбаев М. М. Рыбохозяйственное использование водоемов Северо-Казах-
станской и Кокчетавской областей. — В кн.: Биол. основы рыбн. хоз-ва на водо-
емах Ср. Азии и Казахстана. Алма-Ата, 1966, с. 187—189.
Данилевский Н. Я. Краткий очерк Уральского рыбного хозяйства. — Вести.
Рус. геогр. об-ва, 1858, т. 22, с. 56—74.
Дарлиигтои Ф. Зоогеография. М., 1966. 519 с.
Демин Д. 3. Воспроизводство осетровых рыб Каспия в реках Дагестана. —
В кн.: Осетровое хозяйство в водоемах СССР. М., 1963, с. 102—107.
Державин А. Н. Севрюга: (Биол. очерк). Баку, 1922. 293 с.
Державин А. Н. Куринское рыбное хозяйство и Мингечаурская проблема.
Баку, 1939. 96 с.
Державин А. Н. Воспроизводство запасов каспийского лосося. Баку, 1941.
74 с.
Державин А. Н. Воспроизводство запасов осетровых рыб. Баку, 1947. 243 с.
Державин А. Н. Каспийский лосось. — В кн.: Промысловые рыбы СССР. Опи-
сания рыб. М., 1949, с. 178—180.
Державин А. Н. Севрюга. — Там же, с. 72—75.
Державин А. Н. Шип. — Там же, с. 49—52.
Державин А. Н. Методы получения зрелой икры осетровых и лососевых рыб.
— Тр. Всесоюз. конф, по вопр. рыбн. хоз-ва, 1953, вып. 1, с. 266—290.
Державин А. Н. Животный мир Азербайджана: Куринское рыбное хозяйство.
Баку, 1956. 428 с.
Джардемалиев К. К., Митрофанов В. П., Терещенко А. М., Мудатов С. М.
Выяснение возможности эксплуатации и повышения эффективности работы экс-
периментального воздушно-пневматического рыбозаградителя на Тасмурунском
оросительном канале р. Или. — В кн.: Биол. основы рыбн. хоз-ва водоемов Ср.
Азии и Казахстана. Фрунзе, 1978, с. 293—295.
Доброхотов В. И. Успехи акклиматизации некоторых промысловых видов
рыб в водоемах Казахстана. — Изв. АН КазССР. Сер. зоол., 1948, № 63, вып. 8,
с. 3—13.
Доброхотов В. И. Озеро Зайсан и его обитатели. — Сельское хозяйство Ка-
захстана, 1959, № 11, с. 76—80.
Доброхотова О. В. Паразиты рыб оз. Зайсан в связи с реконструкцией его
ихтиофауны. Автореф. дис. ... канд. биол. наук. Алма-Ата, 1953. 11 с.
Догель В. А., Быховский Б. Е. Фауна паразитов рыб Аральского моря.—
В кн.: Паразитологич. сб. Зоологич. ин-та АН СССР. М.; Л., 1934, т. 4,
с. 241—346.
Догель В. А., Быховский Б. Е. Паразиты рыб Каспийского моря. М- Л.,
1939. 121 с.
Догель В. А., Гвоздев Е. В. О желательности упразднения особой Нагорно-
азиатской подобласти в зоогеографии рыб. — Изв. АН КазССР Сер зоол 1945
№ 4, с. 49—52.
Домбровский Г. В., Серов Н. П., Диканский В. Я. Биология и промысел ши-
па Acipenser nudiventris в Балхаш-Илийском бассейне. — В кн.: Осетровые СССР
и их воспроизводство. М., 1972, с. 146—148.
Дорофеева Е. А. Кариологическое обоснование систематического положения
каспийского и черноморского лососей (Salmo trutta caspius Kessler, S. trutta
labrax Pallas).— Вопросы ихтиологии, 1965, т. 5, вып. 1(34), с. 38—45.
Драгомиров Н. И., Шмальгаузеи О. И. Эколого-морфологические особенно-
сти личинок лопатоноса. — Докл. АН СССР, 1952, т. 85, № 6, с. 1399—1402.
Дрягии П. А. Рыбы реки Чу и рыбохозяйственное использование этой ре-
ки,—В кн.: Рыбное хоз-во КиргАССР: (Тр. Кирг. компл. эксп. 1932—33 гг.).
М.; Л., 1936, т. 3, вып. 1, с. 49—87.
252
Дрягин П. А. Промысловые рыбы Обь-Иртышского бассейна. — Изв.
ВНИОРХ, 1947, т. 25, вып. 2.
Дрягин П. А. Половые циклы и нерест рыб. — Изв. ВНИОРХ, 1949, т. 28,
с. 3—113.
Дрягии П. А. Биология сибирского осетра, его запасы и рациональное исполь-
зование.— Изв. ВНИОРХ, 1949, т. 29.
Дукравец Г. М. Некоторые результаты акклиматизации судака в озерах
Джамбулской области и дальнейшее рыбохозяйственное их использование. —
Вести, сельскохоз. науки. Алма-Ата, 1962, № 6, с. 58—61.
Дукравец Г. М. К биологии размножения судака, акклиматизированного в
оз. Бийли-Куль.— Вести, сельскохоз. науки. Алма-Ата, 1964, № 9, с. 135—136.
Дукравец Г. М. Результаты акклиматизации рыб в озерах бассейна реки
Талас. Автореф. дис. ... канд. биол. наук. Алма-Ата, 1965. 20 с.
Дукравец Г. М. О реконструкции ихтиофауны водоемов равнинной части
бассейна р. Талас. — Вопросы ихтиологии, 1966, т. 6, вып. 1(38), с. 14—19.
Дукравец Г. М. Некоторые данные о распространении, систематике и биоло-
гии белого амура в Илийском бассейне. — В кн.: Биол. основы рыбн. хоз-ва респ.
Ср. Азии и Казахстана: (Тез. докл.). Балхаш, 1967а, с. 116—118.
Дукравец Г. М. Материалы по ихтиофауне Тиликульских озер. — В кн.:
Биол. основы рыбн. хоз-ва респ. Ср. Азии и Казахстана: (Тез. докл.). Балхаш,
19676, с. 115—116.
Дукравец Г. М. К систематике белого амура в бассейне р. Или, — Вести. АН
КазССР, 1968, № 7, с. 65—67.
Дукравец Г. М. Состояние ихтиофауны и перспективы рыбоводства в малых
водоемах Уральской области. — В кн.: Биол. науки, Алма-Ата, 1971, вып. 2,
с. 148—155.
Дукравец Г. М. Белый амур в бассейне р. Или. — Изв. АН КазССР. Сер.
биол., 1972а, № 1, с. 52—57.
Дукравец Г. М. Змееголов в бассейне р. Сырдарьи. — В кн.: Акклиматиза-
ция рыб и беспозвоночных в водоемах СССР. Фрунзе, 19726, с. 126—127.
Дукравец Г. М. Тиликульские озера и их ихтиофауна. — В кн.: Биология
водоемов Казахстана: Сб. работ Казах, фил. ВГБО, вып. 2. Рукопись дел.
ВИНИТИ, № 2922—74, 1974, с. 78—84.
Дукравец Г. М. Систематика и биология белого амура бассейна р. Или. —
В кн.: Биол. науки. Алма-Ата, 1975, вып. 9, с. 58—66.
' Дукравец Г. М. Морфо-биологическая изменчивость промысловых рыб при
акклиматизации в водоемах Казахстана. — В кн.: Советско-американское со-
трудничество в области исследования Мирового океана: (Симпозиум по реакции
водных экосистем на вселение новых видов). М., 1977, с. 38—39.
Дукравец Г. М. Динамика состава и состояния ихтиофауны в усыхающих
озерах Южного Казахстана. — Изв. АН КазССР. Сер. биол., 1978, вып. 4,
с. 21—27.
Дукравец Г. М., Бирюков Ю. А. Ихтиофауна бассейна р. Нуры в Централь-
ном Казахстане. — Вопросы ихтиологии, 1976, т. 16, вып. 2(97), с. 309—314.
Дукравец Г. М., Буранбаева Г. Я., Мачулин А. И. Краткая характеристика
ихтиофауны оз. Утебас в низовье р. Сырдарьи. — В кн.: Биол. науки. Алма-Ата,
1975, вып. 8, с. 42—45.
Дукравец Г. М, Мачулин А. И. Состояние и перспективы Аксай-Кувандарь-
инской системы озер и ее ихтиофауны. — В кн.; Рыбн. ресурсы водоемов Казах-
стана и их использование. Алма-Ата, 1974, вып. 8, с. 111—113.
Дукравец Г. М., Мачулии А. И. Морфологическая характеристика змееголо-
ва (Ophiocephalus argus warpachowskii Berg) бассейна Сырдарьи. — Вопросы
ихтиологии, 1974, т. 14, вып. 1, с. 179—181.
Дукравец Г. М., Мачулин А. И. Морфология и экология амурского змеего-
лова— Ophiocephalus argus warpachowskii Berg, акклиматизированного в бассей-
не Сырдарьи. — Вопросы ихтиологии, 1978, т.18, вып. 2(109), с. 222—228.
Дукравец Г. М., Митрофанов В. П. Биологические основы ведения рыбного
хозяйства на водохранилищах юга Казахстана. — В кн.: Биология водоемов Ка-
захстана: (Сб. работ Казах, фил. ВГБО). Алма-Ата, 1970, с. 73—77.
253
Дукравец Г. М., Митрофанов В. П. Некоторые закономерности морфологи-
ческой изменчивости рыб, акклиматизированных в Казахстане. — В кн.: Акклима-
тизация рыб и беспозвоночных в водоемах СССР. Фрунзе, 1972, с. 127—129.
Дукравец Г. М., Митрофанов В. П. О закономерностях морфологической из-
менчивости рыб при акклиматизации. — Изв. ГосНИОРХ, 1975, т. 103, с. 79—84.
Дукравец Г. М., Пичкилы Л. О., Шарапова Л. И., Дягилев Ю. В., Сумбае-
ва Л. Н., Тайпаков М. А., Бирюков Ю. А. Биологическое обоснование организа-
ции товарного рыбного хозяйства на Камышлыбашских озерах. — Изв. АН
КазССР. Сер. биол., 1977, вып. 1, с. 31—37.
Дулькейт Д. Д., Линдберг Г. У. Материалы по рыбам Шантарского моря.—
Изв. Тихоокеанской научно-пром, станции, 1929, т. 3, вып. 1.
Егоров А. Г. Состояние и перспективы развития осетрового хозяйства в сис-
теме Байкала и р. Ангары.— В кн.: Осетровое хозяйство в водоемах СССР. М.,
1963, с. 188—195.
Ерещенко В. И. Ихтиофауна бассейна р. Сары-Су.— В ки.: Сб. работ по
ихтиологии и гидробиологии. Алма-Ата, 1956, вып. 1, с. 94—123.
Ерещенко В. И. К биологии промысловых рыб озер Северного Казахстана. —
В ки.: Сб. работ по ихтиологии и гидробиологии. Алма-Ата, 1959, вып. 2,
с. 208—233.
Ерещенко В. И. Состояние рыбного хозяйства на водохранилищах Верхнего
Иртыша и мероприятия по его развитию. — В кн.: Биол. основы рыбн. хоз-ва на
водоемах Ср. Азии и Казахстана. Алма-Ата, 1966, с. 280—283.
Ерещенко В. И. Промысловое освоение леща, акклиматизированного в Кап-
чаганском водохранилище. — В кн.: Тезисы докл. конф, по вопросам рыбн. хоз-ва
респ. Ср. Азии и Казахстана. Фруизе, 1968, с. 63—64.
Ерещенко В. И. Влияние гидростроительства на воспроизводство осетра в
Верхнем Иртыше. — В кн.: Биологическая продуктивность водоемов Сибири. М.,
1969, с. 180—185.
Ерещенко В. И. Влияние загрязнения Иртыша и зарегулирования его стока
на условия воспроизводства рыб. — В кн.: Охрана и рациональное использование
живой природы водоемов Казахстана. Алма-Ата, 1969, с. 68—70.
Ерещенко В. И. Состояние стада сибирского осетра в водохранилищах Верх-
него Иртыша и пути его воспроизводства. — В кн.: Осетровые СССР и их воспро-
изводство. М., 1970, с. 158—163.
Ерещенко В. И. Разработка биологических основ рыбохозяйственного освое-
ния Чардаринского, Каратомарского, Сергеевского, Вячеславского водохранилищ:
(Отчет о НИР по теме № 3/19. КазНИИРХ Минрыбхоза СССР). Балхаш, 1970.
292 с.
Ерещенко В. И. Итоги и перспективы акклиматизации рыб в бассейне Верхне-
го Иртыша. — В кн.: Акклиматизация рыб и беспозвоночных в водоемах СССР.
Фрунзе, 1972, с. 37—40.
Ерещенко В. И. Изменения в составе ихтиофауны Усть-Каменогорского во-
дохранилища.— В кн.: Зоологические проблемы Сибири. Новосибирск 1972
с. 238—239.
Ерещенко В. И., Аксаментова Л. А., Сергеева Н. Г., Лопарева Т. В. Предва-
рительные итоги рыбохозяйственного обследования Чардаринского водохранили-
ща. — В кн.: Биологические основы рыби. хоз-ва респ. Ср. Азии и Казахстана:
(Тез. докл.). Балхаш, 1967, с. 128—130.
Ерещенко В. И., Малиновская А. С., Мусина Н. X., Серов Н. П., Селез-
нев В. В., Тэи В. А. Формирование промысловой ихтиофауны Капчагайского во-
дохранилища. — В кн.: Экология гидробионтов водоемов Казахстана. Алма-Ата,
1973, с. 58—84.
Ермаханов 3., Дукравец Г. М. Влияние водохозяйственных мероприятии на
ихтиофауну и рыбный промысел озер низовьев Сырдарьи и перспективы их рыбо-
хозяйственного использования. — В кн.: Рыбохоз. основы территориального пере-
распределения водных ресурсов. М., 1981, с. 154—162.
Ефимова А. М. Щука Обь-Иртышского бассейна. — Изв. ВНИОРХ, 1949,
т. 28, с. 114—174.
Жарковский А. А. К вопросу об охране рыбных запасов бассейна Аральско-
254
го моря в связи с гидростроительством. — В кн.: Материалы по ихтиофауне и
режиму бассейна Аральского моря. Нов. сер. М., 1959, вып. 19, с. 21—43.
Жадин Б. Ф. О происхождении балхашского окуня. — Докл. АН СССР, 1949,
т. 66, № 3, с. 499—502.
Желтенкова М. В. Питание осетровых рыб южных морей. — Тр. ВНИРО. М.,
1964, т. 54, с. 9—48.
Желтенкова М. В. Питание и использование кормовой базы бентосоядными
рыбами Каспийского бассейна. — В кн.: Осетровые СССР и их воспроизводство.
М., 1967, т. 1, с. 122—131.
Жизнь животных /Под ред. Т. С. Расса. М., 1971. Т. 4, ч. I. Рыбы. 656 с.;
изд. 2-е, перераб. М., 1983. Т. 4. Ланцетники, Круглоротые, Хрящевые рыбы.
576 с.
Занин А. П. Материалы к естественно-исторической и рыбохозяйственной ха-
рактеристике Таласского бассейна. — В кн.: Рыбное хоз-во КиргАССР: (Тр.
Кирг. компл. эксп. 1932—33 гг.). М.; Л., 1936, т. 3, вып. 1, с. 89—113.
Зарбалиева Т. С. Питание осетровых рыб в Южном Каспии.— В кн.: Материа-
лы объедин. научн. сессии ЦНИОРХ и АзНИИРХ. Астрахань, 1971, с. 31—32.
Зарбалиева Т. С. К питанию русского и персидского осетров у северных бе-
регов Азербайджана. — В кн.: Некоторые вопросы рыбного хозяйства Азербайд-
жана. Баку, 1972, с. 52—55.
Зарбалиева Т. С. Питание помесей осетровых в Каспийском море. — В кн.:
Осетровое хозяйство внутренних водоемов СССР; (Тез. и реф. II Всесоюз. со-
вещ.). Астрахань, 1979, с. 84—85.
Зарбалиева Т. С. Накормленность осетровых в современных условиях Кас-
пийского моря. — В кн.: Рациональные основы ведения осетрового хозяйства:
(Тез. докл. н.-практ. коиф.). Волгоград, 1981, с. 83—84.
Зарбалиева Т. С., Деткина Т. И. Характеристика питания персидского осетра
по районам Каспийского моря. — Там же, с. 84—85.
Захаров С. С., Песериди Н. Е. Основные закономерности миграции осетровых
р. Урал в зависимости от гидрологических условий реки. — Там же, с. 85—87.
Захарян Г. Б. Динамика ската молоди осетровых с естественных нерестилищ
в бассейне р. Куры. — В кн.: Материалы к объединенной науч, сессии ЦНИОРХ
и АзНИИРХ. Астрахань, 1971, с. 32—34.
Захарян Г. Б. Материалы к изучению условий естественного размножения
курииских осетровых. — В кн.: Осетровые СССР и их воспроизводство: (Тр.
ЦНИОРХ, № 4). М., 1972, с. 67—77.
Захаряи Г. Б. Характеристика питания скатывающейся молоди осетровых
рыб в р. Куре.— В кн.: Тез. отчетной сессии ЦНИОРХ. Астрахань, 1974, с. 51—52.
Захаряи Г. Б. Естественное размножение осетровых бассейна Куры в совре-
менных условиях. — В кн.: Рациональные основы ведения осетрового хозяйства:
(Тез. докл. н.-практ. конф.). Волгоград, 1981, с. 87—88.
Захидов Т. 3., Мекленбурцев Р. И., Богданов О. П. Природа и животный
мир Средней Азин. Ташкент, 1971. Т. 2. Позвоночные животные.
Зачетиова Т. И. К изучению бентоса низовьев р. Урал. — В ки.: Тез. отчетной
сессии ЦНИОРХ. Астрахань, 1972, с. 62—64.
Зачетиова Т. И. Сезонная динамика бентоса дельты р. Урал. — В кн.: Тез.
отчетной сессии ЦНИОРХ. Астрахань, 1973, с. 33—35.
Зачетиова Т. И. Изменения в питании молоди осетровых р. Урал в 1971—
1974 гг. — В кн.: Тез. отчетной сессии ЦНИОРХ. Гурьев, 1976, с. 57—58.
Злоказов В. Н. Опыт искусственного разведения полупроходных рыб бас-
сейна р. Оби.— В кн.: Зоологические проблемы Сибири. Новосибирск, 1972,
с. 240—241.
Зырянова Н. И. О размножении стерляди р. Вятки. — В кн.: Осетровое хо-
зяйство в водоемах СССР. М., 1963, с. 108—114.
Иванов П. П. Развитие миноги. — Общая и сравнительная эмбриология. М.;
Л., 1937, с. 520—537.
Иванов С. И., Печникова И. В. Наблюдения за скатом молоди шипа Acipen-
ser nudiventris Lovetzkii в низовьях Аму-Дарьи. — Вопр. ихтиол., 1967, т. 7,
вып. 4(45), с. 660—668.
255
Игнатов П. Г. По Южному Алтаю. СПб., 1897, т. 4, кн. 1—2.
Иоганзен Б. Г. Морфолого-биологические особенности круглоротых Сибири.
— Зоол. журн., 1935а, т. 14, вып. 2, с. 353—370.
Иоганзен Б. Г. Морфолого-биологические особенности круглоротых Сибири.
— Зоол. жури., 19356, т. 14, вып. 3, с. 501—522.
Иоганзен Б. Г. К ихтиогеографии северной части Казахстана. — В кн.: Во-
просы географии Сибири. Томск, 1951, сб. 2, с. 259—268.
Иоганзен Б. Г., Вовк Ф. И. Какую рыбу, когда, где и как лучше ловить. Но-
восибирск, 1952.
Исламгазиева Р. Б., Песериди И. Е. Итоги рыбоводного освоения осетровых
р. Урал. — В кн.: Биол. основы рыбн. хоз-ва на водоемах Ср. Азии и Казахстана.
Алма-Ата, 1966, с. 55—57.
Исмуханов X. К. Прогноз уловов хищных рыб в связи с акклиматизацией
судака в Бухтарминском водохранилище. — В кн.: Биол. основы рыбн. хоз-ва Ср.
Азии и Казахстана. Душанбе, 1976, с. 296—298.
Исмуханов X. К- Промысловые акклиматизанты Бухтарминского водохрани-
лища и обоснование к вселению новых видов. — В кн.: Биол. основы рыбн. хоз-
ва респ. Ср. Азии и Казахстана. Фрунзе, 1978, с. 314—316.
Исмуханов X. К. Морфо-экологическая изменчивость и промысловое значе-
ние леща и судака, акклиматизированных в Бухтарминском водохранилище. Ав-
тореф. дис. ... канд. биол. наук. Л., 1981. 16 с.
Казанчеев Е. И. Рыбы Каспийского моря. М., 1963. 180 с. 2-е изд. М., 1981,
168 с.
Кайымов К. Балыктар элемшде: (В мире рыб). Алматы, 1981. 172-6.
Камилов Г. К. Определитель рыб Узбекистана. Ташкент, 1964. 64 с.
Кассин И. Г. Материалы по палеогеографии Казахстана. Алма-Ата, 1947.
258 с.
Кашкаров Д. Н. Экологический очерк района озер Бийлю-Куль, Ак-Куль и
Аще-куль Аулиеатинского уезда. Ташкент, 1928. 54 с. (Тр. САГУ. Сер. VIII—а.
Зоология, вып. 2).
Кащенко И. Ф. Результаты алтайской зоологической экспедиции 1898 г. Поз-
воночные. Томск, 1899. 158 с.
Кевдин В. А. Современное рыболовство России. М., 1915, с. 64—77.
Кесслер К. Ф. Ихтиологическая фауна Туркестана. — Изв. об-ва любит, есте-
ствозн., 1872, т. 10, вып. 1, с. 47—79.
Кесслер К. Ф. Рыбы, водящиеся и встречающиеся в Арало-Каспийско-Пон-
тийской ихтиологической области. СПб., 1877. 360 с. (Тр. Арало-Касп. эксп.,
вып. 4).
Киппер 3. М., Песериди Н. Е., Марти Ю. Ю., Гоптарев Н. П. Отчленение
мелководной восточной части Северного Каспия. — В кн.: Состояние ресурсов,
воспроизводства и рациональное использование рыбных запасов Урало-Каспия.
Гурьев, 1972, с. 60—62.
Кириллов Ф. Н. Ихтиофауна бассейна р. Вилюя. — Тр. Ин-та биол. Якут,
фил. СОАН СССР, 1962, вып. 8.
Кириллов Ф. Н. Рыбы Якутии. М., 1972. 330 с.
Киселевич К. А. Промысловые рыбы Волго-Каспийского района, их привычки
и особенности. Астрахань, 1926. 52 с.
Кифа М. И. Морфология двух форм ленка (род Brachymystax, сем. Salmoni-
dae) из бассейна Амура и их систематическое положение. — В кн.: Зоогеография
и систематика рыб. Л., 1976, с. 142—155.
Книпович Н. М. Каспийское море и его промыслы. Берлин, 1923. 51 с.
Коблицкая А. Ф. Определитель молоди пресноводных рыб. М., 1981. 208 с.
Кожин Н. И. Осетровые СССР и их воспроизводство. — В кн.: Осетровые
южных морей Советского Союза: (Тр. ВНИРО, т. 52, сб. 1). М., 1964, с. 21—58.
Конева Л. А. Нельма верхнего бьефа плотины Новосибирской ГЭС. Автореф.
дис. ... канд. биол. наук. Томск, 1972. 21 с.
Коновалов П. М. Усовершенствование перевозки икры и личинок осетровых
рыб. — В кн.: Рыбные ресурсы водоемов Казахстана и их использование. Алма-
Ата, 1966, с. 62—75.
256
Кормилин В. В. Размножение и развитие балхашской маринки. Автореф.
дис. ... канд. биол. наук. Алма-Ата, 1972. 28 с.
Короткова В. М. Биологические основы интенсификации рыбного хозяйства
на ирригационных водохранилищах. Автореф. дис. ... канд. бнол. наук. Л.,
1981. 20 с.
Костенко Н. Н. К истории Балхаша. — Изв. КазФАН СССР. Сер. географ.,
1946, № 8(26), с. 96—104.
Кострова О. И., Песериди Н. Е. Сравнительная характеристика качественной
структуры нерестовой части популяции осетра р. Урал. — В кн.: Рациональные
основы ведения осетрового хозяйства. Волгоград, 1981, с. 113—116.
Кудерский Л. А. О происхождении лососей и форелей (S. trutta L). в бассей-
нах Аральского, Каспийского и Черного морей. — В кн.: Вопросы форелеводства.
Л., 1974, т. 97, с. 187—216.
Кузнецов В. А. Особенности воспроизводства рыб в условиях зарегулирова-
ния стока реки. Казань, 1979. 15 с.
Куленов К., Куленова С. Балык шаруашылыгынын. орысша-цазацша тус!нд1р-
ме сездт: (Русско-казахский толковый словарь по рыбному хозяйству). Алматы,
1980. 256-6.
Курдюков К. В. О колебаниях уровня озера Ала-Куль. — Вопросы географии,
1951, вып. 24, с. 117—133.
Лапицкий Н. И. Направленное формирование ихтиофауны и управление чис-
ленностью популяций рыб в Цимлянском водохранилище. — Тр. Волгоградок, отд.
ГосНИОРХ, 1970, т. 4. 278 с.
Лебедев В. Д. Неогеновая фауна пресноводных рыб Зайсанской впадины
и Западно-Сибирской низменности. — Вопросы ихтиологии, 1959, вып. 12,
с. 28—69.
Легеза М. И. Закономерности распределения осетровых рыб в Каспийском
море: (Тез. докл. отчетн. сессии ЦНИОРХ). Астрахань, 1966, с .48—50.
Легеза М. И. Влияние температурного фактора на распределение куринских
осетровых. — В кн.: Разработка биологических основ и биотехники развития
осетрового хозяйства в водоемах СССР. Астрахань, 1967, с. 27—33.
Легеза М. И. Закономерности распределения и формирование численности
осетровых в Каспийском море. — В кн.: Материалы научи, сессии ЦНИОРХ, по-
свящ. 100-летию осетроводства. Астрахань, 1969, с. 99—102.
Легеза М. И. Количественное распределение осетровых рыб в Каспийском
море. — Тр. ЦНИОРХ. М„ 1970, т. 2, с. 57—63.
Легеза М. И. О помесях каспийских осетровых. — В кн.: Осетровые СССР и
их воспроизводство. — Тр. ЦНИОРХ. М., 1971, т. 3, с. 196—206.
Легеза М. И. О добыче осетровых на юге Каспийского моря.-—В кн.: Неко-
торые вопросы рыбного хозяйства Азербайджана. Баку, 1972, с. 86—96.
Легеза М. И. Современное распределение осетровых рыб (сем. Acipenseridae)
в Каспийском море. — Вопросы ихтиологии, 1973, т. 13, вып. 6(83), с. 1008—1015.
Легеза М. И., Войнова И. А. Современное состояние запасов осетровых Кас-
пийско-К''ринского района. — В кн.: Осетровые СССР и их воспроизводство: (Тр.
ЦНИОРХ, т. 1). М„ 1967, с. 26—33.
Летичевский М. А. Воспроизводство белорыбицы в условиях зарегулирован-
ного стока Волги. М., 1963. 173 с.
Летичевский М. А. Опыт определения численности белорыбицы и эффектив-
ность ее заводского разведения в условиях дельты и Нижней Волги. — Вопросы
ихтиологии, 1975, т. 15, вып. 4, с. 630—635.
Летичевский М. А. Перспективы расширения заводского и естественного
воспроизводства белорыбицы в нижнем течении и дельте Волги. — Вопросы
ихтиологии, 1981, т. 21, вып. 5, с. 924—929.
Ловецкая А. А. Каспийские кильки и их промысел. М., 1951. 48 с.
Лукаш Б. С. Рыбы нижнего течения р. Вятки. — Тр. Вятск. ин-та краевед,
1933, т. 6, с. 13.
Лукин А. В. Наблюдения над биологией стерляди на Тетюшинском нерести-
лище «Черемта» летом 1934 г. — Тр. Об-ва естествоисп. при Казанск. гос. ун-те,
1937, т. 55, вып. 1—2, с. 143—163.
17-7
257
Лукин А. В. Основные черты экологии осетровых в Средней Волге, ч. 1.—
Тр. об-ва естественен, при Казанск. гос. ун-те., 1947, т. 57, вып. 3—4, с. 72—104.
Лукин А. В. Стерлядь.— В кн.: Промысловые рыбы СССР. М., 1949, с. 53—56.
Лукьяненко В. И. Внутривидовая дифференциация осетровых и ее значение
для рационального ведения осетрового хозяйства. — В кн.: Тез. отчетной сессии
ЦНИОРХ. Астрахань, 1973, с. 53—56.
Лукьяненко В. И. Итоги и перспективы физиолого-биологических исследова-
ний осетровых рыб. — В кн.: Рациональные основы ведения осетрового хозяйст>
ва. Волгоград, 1981, с. 151—154.
Лукьяненко В. И., Дубинин В. Н., Каратаева В. Б., Терентьев А. А. О видо-
вой принадлежности так называемого позднего ярового или летненерестящегося
осетра на Волге. — В кн.: Тез. отчета, сессии ЦНИОРХ. Астрахань, 1974,
с. 92—94.
Лукьяненко В. И., Каратаева Б. Б., Алтуфьев Ю. В., Умеров Ж. Б. Популя-
ционная структура каспийских осетровых в связи с проблемой их заводского
воспроизводства. — В кн.: Биол. ресурсы Каспийского моря: (Тезисы конф.).
Астрахань, 1972, с. 105—106.
Лукьяненко В. И., Умеров Ж. Г., Каратаева Б. Б. Южнокаспийский осетр —
самостоятельный вид рода Acipenser. — Изв. АН СССР. Сер. биол., 1974, № 5,
с. 736—738.
Львов Л. Ф. Результаты разведения стерляди Волги. — В кн.: Тез. отчет,
сессии ЦНИОРХ. Астрахань, 1973, с. 63—64.
Лысенко Н. Ф. Биология и рыбохозяйственное значение сома, акклиматизи-
рованного в оз. Балхаш. Автореф. дис. ... канд. биол. наук. Л., 1976. 22 с.
Лысенко Н. Ф., Сироткин М. Н., Григорьева Э. Н., Фокина А. С. К вопросу
о рыбохозяйственном освоении некоторых озер Кокчетавской области. — В кн.:
Рыбные ресурсы водоемов Казахстана и их использование. Алма-Ата, 1972,
вып. 7, с. 55—58.
Магерамов Ч. М. Появление волжского осетра в р. Куре. — В кн.: Осетровые
и проблемы осетрового хозяйства. М., 1972, с. 289—293.
Маилян Р. А. Современное состояние воспроизводства запасов куринских
осетровых.— В кн.: Тез. отчетн. сессии ЦНИОРХ. Астрахань, 1974, с. 94—95.
Маилян Р. А. Осетроводство — надежный способ регулирования численно-
сти каспийских осетровых. — В кн.: Рациональные основы ведения осетрового
хозяйства. Волгоград, 1981, с. 157—158.
Маилян Р. А., Махмудбеков А. А. Естественные нерестилища и скат молоди
куринских осетровых. — В кн.: Осетровые СССР и их воспроизводство: (Тр.
ЦНИОРХ, т. 2). М„ 1970, с. 95—104.
Маилян Р. А., Алекперов А. П. О росте и развитии осетровых. — В кн.: Те-
зисы отчет, сессии ЦНИОРХ по результатам работ в IX пятилетке (1971—
1975 гг.). Гурьев, 1976, с. 28—30.
Майр Э. Зоологический вид и эволюция. М., 1968. 600 с.
Максунов В. А. Промысловые рыбы Таджикистана. Душанбе, 1968. 100 с.
Малиновская А. С. Материалы по питанию рыб Джезказганского водохра-
нилища. — В кн.: Сборник работ по ихтиологии н гидробиологии. Алма-Ата,
1956, вып. 1, с. 74—93.
Малиновская А. С. Кормовая база Алакольских озер и ее использование ры-
бами.— Там же, 1959, вып. 2, с. 116—144.
Мамилова Р. X. Биомасса и продукция макрозообентоса озер Каракуль и
Соркуль (бассейн р. Или). Автореф. дис. ... канд. биол. наук. Алма-Ата, 1970. 26 с.
Маркова Е. Л. Некоторые данные о распространении новых каспийских все-
ленцев в Аральском море. — Бюл. МОИП. Отд. биол., 1962, № 5, с. 130—132.
Маркова Е. Л. Черный амур в Аральском море. — Рыбоводство и рыболов-
ство, 1964, № 1, с. 15—19.
Маркова Е. Л. О современном состоянии стада аральского шипа. — В кн.:
Материалы научн. сессии ЦНИОРХ, посвящ. 100-летию осетроводства. Астра-
хань, 1969, с. 119—120.
Маркова Е. Л. Результаты акклиматизационных работ, проведенных в Араль-
ском море за период с 1963 по 1971 г. — Изв. ГосНИОРХ, 1975, т. ЮЗ,
с. 164—168.
258
Мартехов П. Ф. Биологические основы создания стада ценных промысловых
рыб на Зайсане в связи с образованием Бухтармино-Зайсанского водохранили-
ща.— В кн.: Биологии, основы рыбн. хоз-ва: (Тр. Всесоюз. совещ. по биол.
основам рыбного хоз-ва 18—22.09.1956 г.). Томск, 1959, с. 191—200.
Мартехов П. Ф. К акклиматизации леща в водоемах Иртыш-Зайсанского бас-
сейна. — В кн.: Тр. конф, по рыбн. хоз-ву респ. Ср. Азии и Казахстана. Фрунзе,
1961, с. 63—72.
Махотин Ю. М. Эффективность нереста рыб в Куйбышевском водохранили-
ще и определяющие ее факторы. — Вопросы ихтиологии, 1977, т. 17, вып. 1(102),
с. 28—38.
Мачулин А. И., Дукравец Г. М. Возрастной диморфизм амурского змееголо-
ва в бассейне р. Сырдарьи. — В кн.: Рыбные ресурсы водоемов Казахстана и их
использование. Алма-Ата, 1975, вып. 9, с. 103—105-
Мельников В. А. Использование рыб для борьбы с комарами в аридной зоне
Казахстана. Автореф. дис. ... канд. биол. наук. Алма-Ата, 1977. 24 с.
Меньшиков М. И. Рыбы реки Камы и ее долины в окрестностях г. Перми. —
Изв. Биол. н.-и. ин-та при Пермск. гос. ун-те, 1929, т. 6, вып. 8, с. 377—407.
Меньшиков М. И. Материалы по систематике и биологии нельмы низовьев
р. Иртыш. — Изв. Пермск. биол. ин-та, 1935, т. *10, № 1—2, с. 1—28.
Меньшиков М. И. К биологии сибирского осетра и стерляди в р. Иртыш. —
Уч. зап. Пермск. гос. ун-та, 1936, вып. 1.
Меньшиков М. И. Об ихтиофауне оз. Марка-Куль. — Уч. зап. Пермск. гос.
ун-та, 1938, т. 3, вып. 2, с. 119—144.
Меньшиков М. И. Рыбы бассейна Оби. Автореф. дис. ... докт. биол. наука
(Фонд библ. им. В. И. Ленина), М., 1948.
Меньшиков М. И. Некоторые закономерности возрастной и географической
изменчивости рыб. — Тр. Карело-Финского отд. ВНИОРХ, 1951, т. 3, с. 292—306.
Меркулов Е. А., Митрофанов В. П., Дукравец Г. М. Ихтиофауна левобереж-
ных и правобережных водоемов реки Урал. Рукопись деп. ВИНИТИ, 1977,
№ 773—77. 42 с.
Мина М. В. Данные по морфологии и систематике ленка (род Brachymystax)
из рек Онон и Керулен (МНР). — Бюл. Моск, об-ва испыт. природы. Отд. биол.,
1977, т. 82, вып. 5, с. 58—65.
Митрофанов В. П. К систематике ленка из оз. Марка-Куль.—-В кн.: Сб. ра-
бот по ихтиологии и гидробиологии. Алма-Ата, 1959а, вып. 2, с. 267—275.
Митрофанов В. П. О размножении маркакульского ленка. — Там же, 19596,
с. 276—285.
Митрофанов В. П. Рыбы оз. Марка-Куль. — Тр. конф, по рыбному хоз-ву.
Фрунзе, 1961, с. 51—61.
Митрофанов В. П. Соотношение полов в популяциях рыб. — В кн.: Биоло-
гия водоемов Казахстана: (Сб. работ Каз. филиала ВГБО). Алма-Ата, 1970,
с. 96—104.
Митрофанов В. П. К вопросу ихтиогеографии узлового района Палеаркти-
ки.— В кн.: Биол. науки. Алма-Ата, 1971а, вып. 2, с. 158—165.
Митрофанов В. П. Некоторые вопросы акклиматизации рыб в Казахстане. —
В кн.: Материалы науч. конф, проф.-преп. состава, посвящ. XXIV съезду КПСС.
Изд. КазГУ, Алма-Ата, 19716, с. 92—95.
Митрофанов В. П. Хариус оз. Марка-Куль. — В кн.: Биол. науки. Алма-Ата,
1971b, вып. 1, с. 104—112.
Митрофанов В. П. Карповые рыбы Казахстана. Автореф. дис. ... докт. биол.
наук. Фрунзе, 1973. 36 с.
Митрофанов В. П. Экологические основы морфологического анализа рыб:
(Учебное пособие для студентов-ихтиологов). Алма-Ата, 1977. 35 с.
Митрофанов В. П., Дукравец Г. М. К вопросу реконструкции ихтиофауны
бассейна Таласа. — В кн.: Акклиматизация животных в СССР. Алма-Ата, 1963,
с. 271—273.
Митрофанов В. П., Дукравец Г. М. Состояние рыбных запасов малых водое-
мов бассейнов рек Чу и Талас и пути их рыбохозяйственного освоения. — В кн.:
259
Рыбные ресурсы водоемов Казахстана и их использование. Алма-Ата, 1966,
вып. 5, с. 303—316.
Митрофанов В. П., Дукравец Г. М., Дмитриев В. М., Никитин О. Л. Анализ
акклиматизационных работ в бассейне Балхаша и перспективы формирования
его ихтиофауны. — В кн.: Итоги и перспективы акклиматизации рыб и беспозво-
ночных в водоемах СССР. М., 1980, с. 76—77.
Мишарин К. И. Биология икры и молоди некоторых промысловых рыб из
Байкала и р. Ангары. — Тр. Вост.-Сиб. ун-та, 1942, т. 2, вып. 3.
Монастырский Г. Н. Щука. — В кн.: Промысловые рыбы СССР. Описание
рыб. М., 1949, с. 301—303.
Морозова А. В. Материалы по биологии и систематике тихоокеанской мино-
ги.— Вопросы ихтиологии, 1956, вып. 7, с. 149—157.
Мудатов С. М. Пути повышения рыбопродуктивности и улучшения рыбохо-
зяйственного использования оз. Челкар: (Отчет о НИР № ГР 79006344).
КазНИИРХ Минрыбхоза КазССР и Уральский ПИ Минпроса КазССР, 1981. 93 с.
Мурзаев Э. В далекой Азии: (Очерки по истории изучения Средней и Цент-
ральной Азии в 19—20 веках). М., 1956. 224 с.
Навозов Н. П. Материалы по ихтиофауне р. Урала. — Вести, рыбопромышл.,
1912, № 8—10, с. 253.
Наливкин Д. П. Палеогеография Средней Азии в Палеозое. — Тр. III съезда
геологов. М., 1930, т. 2.
Недошивин А. Я., Ильин Б. С. Уловы красной рыбы в водах южного побе-
режья Каспия в 1913—1915 гг. — Изв. Отд. прикладн. ихтиол., 1927, т. 7, с. 63.
Нездолий В. К. Покатная миграция молоди рыб р. Или в начальный период
зарегулирования стока. Автореф. дис. ... канд. биол. наук, М., 1982. 24 с.
Нездолий В. К., Митрофанов В. П., Дукравец Г. М., Терещенко А. М. Распре-
деление молоди промысловых и сорных рыб в Капчагайском водохранилище. —
В кн.: Биол. науки. Алма-Ата, 1975, вып. 9, с. 128—134.
Некрашевич Н. Г. Материалы по ихтиологии Алакольских озер. — В кн.:
Алакольская впадина и ее озера: (Вопросы географии Казахстана, вып. 12). Ал-
ма-Ата, 1965, с. 236—268.
Никольский А. М. Путешествие в Алтайские горы летом 1882 г. — Тр. СПб.
об-ва естествознания, 1884, т. 14, вып. 2, с. 150—218.
Никольский Г. В. Рыбы среднего и нижнего течения р. Чу. — Ежегодник
зоол. музея АН СССР, 1931, т. 32, вып. 2, с. 227—268.
Никольский Г. В. Обследование мест нереста шипа в районе Чиназа. — Тр.
Аральск, научн. рыбохоз. ст., 1933, т. 1, с. 95—НО.
Никольский Г. В. Аральский сазан. Материалы по систематике биологии и
промыслу. — Тр. Аральского отд. ВНИМОРХ, 1934, т. 3, с. 5—75.
Никольский Г. В. Рыбы Таджикистана. М.; Л., 1938. 217 с.
Никольский Г. В. Рыбы Аральского моря. — Бюл. МОИП. Нов. серия. Отдел,
зоол., вып. l(XVI). М., 1940. 216 с.
Никольский Г. В. Строительство Главного Туркменского канала и вопросы
рыбного хозяйства. — Рыбное хоз-во, 1950, № 12, с. 24—26.
Никольский Г. В. О биологической специфике фаунистических комплексов
и значение их анализа для зоогеографии. — В кн.: Очерки по общим вопросам
ихтиологии. М.; Л., 1953, с. 65—76.
Никольский Г. В. Рыбы бассейна Амура. М.; Л., 1956. 551 с.
Никольский Г. В. Частная ихтиология. М., 1950; изд. 3-е, испр. и доп. М.,
1971. 472 с.
Никольский Г. В. Структура вида и закономерности изменчивости рыб. М.,
1980. 182 с.
Никольский Г. В., Васильев В. П. О некоторых закономерностях в распреде-
лении числа хромосом у рыб. — Вопросы ихтиологии, 1973, т .13, вып. 1(78),
с. 3—22.
Никольский Г. В., Евтюхов Н. А. Рыбы равнинного течения р. Или. — Бюл.
МОИП. Отд. биол., 1940, т. 49, вып. 3/4, с. 187—200; вып. 5/6, с. 57—70.
Никоноров И. В. Лов рыбы на свет. М., 1963. 166 с.
260
Обручев В. А. Пограничная Джунгария. Т. 3: (Геологический очерк). М.;
Л., 1940. 292 с.
Овсепьян Г. П., Песериди Н. Е. Результаты наблюдений за динамикой хода
и составом нерестовой популяции шипа в р. Урал (1971 —1974 гг.). — В кн.: Тез.
отчета, сессии ЦНИОРХ по результ. работ в IX пятилетке. Гурьев, 1976,
с. 30—31.
Орлов А. В. Распределение ранней молоди промысловых рыб в прибрежье
Капчагайского водохранилища. — В кн.: Распределение и экологические способы
защиты молоди рыб. М., 1980, с. 89—96.
Осепян К. Г. Причины колебаний численности обыкновенной кильки Север-
ного Каспия. — Тр. ВНИРО, 1972, т. 83, с. 194—202.
Остроумов А. А. О составе нерестового стада каспийского пузанка. — Тр.
Касп. басе, филиала Всесоюзн. н.-и. нн-та морск. рыбн. хоз-ва и океаногр. Астра-
хань, 1948, т. 10, с. 97—106.
Павлов А. В., Захаров С. С. Распределение, качественный состав и числен-
ность осетровых в Северном Каспии в 1967 г. — В кн.: Осетровые СССР и их
воспроизводство. М., 1971, с. 235—269.
Паллас П. С. Путешествия. СПб., 1809, т. 1. 424 с.
Пахомов И. М. Рельеф местности вокруг оз. Марка-Куль. — Зап. Семипала-
тинского подотдела Западно-Сибирского отд. РГО, 1909, т. 4, с. 1—10.
Песериди Н. Е. Задачи по разведению осетровых рыб в дельте р. Урал. —
В кн.: Материалы конф, работников рыбн. пром. КазССР. Алма-Ата, 1961,
с. 56—60.
Песериди Н. Е. К вопросу о воспроизводстве осетровых р. Урал. — В кн.:
Вопросы рыбного хозяйства КазССР: (Тр. Ин-та ихтиол, и рыбн. хоз-ва, т. 4).
Алма-Ата, 1963, с. 33—47.
Песериди Н. Е. Меры по обеспечению естественного воспроизводства осет-
ровых рыб р. Урал. — Изв. ГосНИОРХ, 1964, т. 57, с. 127—134.
Песериди Н. Е. Состояние гонад производителей осетра в период нерестовой
миграции. — В кНЛ Рыбн, ресурсы водоемов Казахстана и их использование.
Алма-Ата, 1966а, вып. 5, с. 10—31.
Песериди Н. Е. К методике изготовления и просветления спилов лучей осет-
ровых рыб. — Там же, 19666, с. 146—155.
Песериди Н. Е. Регулирование промысла — одно из главных условий управ-
ления численностью осетровых рыб. — В кн.: Тез. докл. на отчета, сессии
ЦНИОРХ. Астрахань, 1966в, с. 69—75.
Песериди Н. Е. Некоторые данные по размножению осетровых и использо-
ванию ими нерестилищ р. Урал. — В кн.: Биол. основы рыбного хоз-ва на водое-
мах Ср. Азии и Казахстана. Алма-Ата, 1966 г., с. 46—48.
Песериди Н. Е. Причины падения запасов промысловых рыб Урало-Каспий-
ского района и пути увеличения их численности. — В кн.: Биол. основы рыбного
хоз-ва республ. Ср. Азии и Казахстана: (Тез. докл. конф.). Балхаш, 1967а,
с. 212—214.
Песериди Н. Е. Промысловая характеристика сома р. Урал и его влияние на
запасы осетровых. — Там же, 19676, с. 214—215.
Песериди Н. Е. Ступенчатый запрет и его роль в воспроизводстве осетро-
вых р. Урал.— Там же, 1967в, с. 215—217.
Песериди Н. Е. Нерестовые популяции осетра и севрюги р. Урал и мероприя-
тия по их воспроизводству. Автореф. дис. ... канд. биол. наук. Астрахань, 1968.
27 с.
Песериди Н. Е. О потенциальных возможностях осетровых р. Урал в увели-
чении численности этих рыб Каспийского моря. — Материалы научн. сессии
ЦНИОРХ, посвящ. 100-летию осетроводства. Астрахань, 1969, с. 144—146.
Песериди Н. Е. Сезонная динамика хода осетровых в низовьях р. Урал. —
В кн.: Осетровые СССР и их воспроизводство: (Тр. ЦНИОРХ, т. 3). М., 1971,
с. 355—358.
Песериди Н. Е. Роль р. Урал в воспроизводстве рыб Каспия. — В кн.: Биол.
ресурсы Каспийского моря. Астрахань, 1972а, с. 118—121.
261
Песериди Н. Е. О влиянии промысла на запасы осетровых р. Урал. Тез.
отчетн. сессии ЦНИОРХ. Астрахань, 19726, с. 126—128.
Песериди Н. Е. О необходимости развития осетроводства в низовьях р. Урал.
В кн.: Состояние сырьевых ресурсов, воспроизводства и рациональное использо-
вание рыбных запасов Урало-Каспия. Гурьев, 1972в, с. 69—72.
Песериди Н. Е. К вопросу диагностики внутрипопуляционных группировок
проходных осетровых. — В кн.: Тез. отчетн. сессии ЦНИОРХ. Астрахань, 1973,
с. 86—88.
Песериди Н. Е. О внутривидовой и внутрипопуляционной неоднородности
осетровых. — В кн.: Тез. отчетн. сессии ЦНИОРХ по результ. работ в IX пяти-
летке. Гурьев, 1976, с. 33—35.
Песериди Н. Е. К вопросу управления численностью осетровых Урало-Кас-
пия. — В кн.: Осетровое хозяйство внутренних водоемов СССР. (Тез. и рефер.
II Всесоюз. совещ. по осетр, хоз-ву). Астрахань, 1979, с. 196—198.
Песериди Н. Е., Бекешев А. Б. Возрастной состав нерестовой части популя-
ции осетровых р. Урал. — В кн.: Тез. докл. на отчетн. сессии ЦНИОРХ. Астра-
хань, 1966, с. 66—68.
Песериди Н. Е., Бекешев А. Б. Характеристика динамики ската покатной мо-
лоди осетровых р. Урал. — В кн.: Осетровые СССР и их воспроизводство. М.,
1967, с. 116—121.
Песериди Н. Е., Верина И. Л. О приемной мощности нерестилищ осетровых
р. Урал. — В кн.: Рацион, основы ведения осетр, хоз-ва: (Тез. докл. конф). Вол-
гоград, 1981, с. 113—115.
Песериди Н. Е., Войнова И. А. К вопросу управления численностью осетро-
вых и сома. — В кн.: Материалы научн. сессии ЦНИОРХ, посвящ. 100-летию
осетроводства. Астрахань, 1969, с. 146—147.
Песериди Н. Е., Едаменко В. Н., Бекешев А. Б. О влиянии судоходства на
запасы рыбы р. Урал. — Там же, с. 144—151.
Песериди Н. Е., Захаров С. С., Исламгазиева Р. Б., Шестых А, И., Жда-
нов А. П. Характеристика режима рыболовства на р. Урал и пути его дальней-
шего совершенствования. — В кн.: Рацион, основы ведения осетрового хоз-ва.
Волгоград, 1981, с. 113—115.
Песериди Н. Е., Исламгазиева Р. Б. Вариации веса овоцитов из разных
отделов яичника севрюги. — В кн.: Биол. основы рыбн. хоз-ва респ. Ср. Азии и
Казахстана: (Тез. докл. на конф.). Балхаш, 1967, с. 217—219.
Песериди Н. Е., Исламгазиева Р. Б., Бекешев А. Б., Шестых А. И., Войно-
ва И. А., Едаменко В. Н., Есжанов С. Е, Тарабрин А. Г. Оценка эффективности
нового режима рыболовства в низовьях р. Урал. — В кн.: Тез. отчетн. сессии
ЦНИОРХ. Астрахань, 1972, с. 131—133.
Песериди Н. Е., Соколова Т. К. Характеристика ската и питание молоди
осетровых р. Урал. — В кн.: Биол. основы рыбного хоз-ва респуб. Ср. Азии и Ка-
захстана: (Тез. докл. на конф.). Балхаш, 1967, с. 221—222.
Песериди Н. Е., Шестых А. И., Бекешев А. Б., Тлепов Б. Т. К вопросу о
воспроизводстве озимых осетровых р. Урал.—В кн.: Тез. отчетн. сессии ЦНИОРХ.
Астрахань, 1972, с. 128—130.
Песериди Н. Е., Шубина Т. Н., Сливка А. П. Сравнительная характеристика
возрастного состава волжской и уральской севрюги. — В кн.: Тез. докл. отчетн.
сессии ЦНИОРХ. Астрахань, 1972, с. 130—131.
Петкевич А. Н., Башмаков В. Н., Башмакова А. Я. Осетр Средней и Верхней
Оби. — В кн.: Тр. Барабинского отд. ВНИОРХ. Новосибирск, 1950, т. 4, с. 3—54.
Петкевич А. Н. Биология и воспроизводство осетра в Средней и Верхней
Оби в связи с гидростроительством. — Тр. Томского гос. ун-та, 1952, т. 119,
с. 39—64.
Петкевич А. Н. К морфологии сибирского осетра. — Тр. Барабинского отд.
ВНИОРХ. Новосибирск, 1953, т. 6, вып. 2, с. 3—16.
Петкевич А. Я. Состояние промысла осетра в бассейне Оби и его перспекти-
вы. Новосибирск, 1957, с. 1—7.
Петров В. В. Материалы по изучению роста и возраста каспийских осетро-
вых. М., 1927, т. 6, вып. 2, с. 156—215.
262
Петрова Н. А. К вопросу о разнокачественности полупроходной нельмы Обь-
Иртышского бассейна. — В кн.: Зоологические проблемы Сибири. Новосибирск,
1972, с. 265—266.
Печникова Н. В. Результаты акклиматизации аральского шипа (A. nudiven-
tris) в оз. Балхаш. — Вопросы ихтиологии, 1964, т. 4, вып. 1(30), с. 142—152.
Печникова Н. В. Шип Аральского моря и оз. Балхаш. Автореф. дис. ... канд.
биол. наук. Калининград, 1970а. 17 с.
Печникова Н. В. Изменение морфологии шипа (A. nudiventris) Аральского
моря и озера Балхаш в связи с акклиматизацией рыб и кормовых животных.—
Зоол. журн. 19706, т. 49, вып. 1, с. 96—105.
Печникова Н. В. Биология молоди аральского шипа на первом году жизни.
— В кн.: Осетровые СССР и их воспроизводство: (Тр. ЦНИОРХ, т. 4), М., 1972,
с. 139—145.
Печникова Н. В., Воробьева Н. Б. Питание шипа в бассейне оз. Балхаш. —
Вести. Каракалп. фил. АН УзбССР. Нукус, 1961, № 4(6), с. 68—73.
Пивнев И. А. Некоторые данные о морфологии и биологии щуки из бассейна
реки Чу. — Тезисы докл. IX науч. конф. Кирг. гос. ун-та, 1960, с. 47—50.
Пивнев И. А. Акклиматизационная изменчивость у северокаспийской воблы.
— В кн.: Рыбные ресурсы водоемов Казахстана и их использование. Алма-Ата,
1972. вып. 7, с. 8—14.
Пирожников П. Л. К вопросу о происхождении северных элементов в фауне
Каспия, —Докл. АН СССР. Нов. сер., 1937, т. 15, № 8, с. 513—516.
Пискунов А. И. Распределение осетровых в Каспийском море. — В ки.: Изме-
нение биол. комплексов Касп. моря за последние десятилетия. М., 1965,
с. 213—233.
Пискунов А. И. Материалы по биологии осетра и севрюги Каспия в морской
период жизни. — В кн.: Осетровые СССР и их воспроизводство. М., 1970,
с. 74—85.
Подлесный А. В. Белорыбица — St. leucichthys Gtild: (Биоэкологич. очерк).
— Тр. Сиб. отд. ВНИОРХ, Красноярск, 1947, т. 7, вып. 1. 184 с.
Подлесный А. В. Белорыбица. — В кн.: Промысловые рыбы СССР. М., 1949,
с. 215—217.
Покровский А. С. Рыболовство на реках Сыр-Дарье и Аму-Дарье и на Араль-
ском море. —В кн.: Материалы к познанию русского рыболовства. Пг., 1916,
т. 5, вып. 1.
Полторыхина А. Н. Рост и развитие сибирской миноги (Lampetra kessleri)
в реках Восточного Казахстана. — Изв. АН КазССР. Сер. биол., 1971, № 1,
с. 40—46.
Полторыхина А. Н. Метаморфоз сибирской речной миноги в Верхнем Ирты-
ше.—Вопросы ихтиологии, 1971, т. 11, вып. 2, с. 354—358.
Полторыхина А. Н. О размножении сибирской речной миноги Lampetra japo-
nica kessieri (Anikin) в бассейне Верхнего Иртыша. — В кн.: Проблемы эколо-
гии. Томск, 1973, т. 3, с. 167—174.
Полторыхина А. Н. Морфологические особенности и изменчивость сибирской
миноги Lampetra japonica kessleri (Anikin) водоемов Верхнего Иртыша. — Воп-
росы ихтиологии, 1974, т. 14, вып. 2, с. 218—230.
Полторыхина А. Н. К вопросу о систематическом положении, распростране-
нии и происхождении сибирской речной миноги Lampetra kessleri (Anikin). — Изв.
СО АН СССР. Сер. биол., 1979, вып. 1, с. 68—72.
Полянинова А. А. Особенности питания сеголетков стерляди в р. Волге и
Северном Каспии. — В кн.: Тез. отчетн. сессии ЦНИОРХ. Астрахань, 1972,
с. 137—138.
Полянинова А. А. Питание и пищевые отношения молоди осетровых, выра-
щенной на волжских заводах. Автореф. дис. ... канд. биол. наук. М., 1972. 27 с.
Полянинова А. А. Суточный ход питания и рационы осетра и севрюги в Се-
верном Каспии. — В кн.: Биол. основы развития осетрового хоз-ва в водоемах
СССР. М„ 1979, с. 170—180.
Полянинова А. А., Кашенцева Л. Н., Малеиникова И. А. Динамика пищевых
263
отношений осетровых в Северном Каспии. — В кн.: Рациональные основы веде-
ния осетрового хозяйства: (Тез. докл. конф.). Волгоград, 1981, с. 200—201.
Попова О. А. Биологические показатели щукн и окуня в водоемах с различ-
ным гидрологическим режимом и кормностыо. — В кн.: Закономерности роста и
созревания рыб. М., 1971, с. 102—152.
Правдии И. Ф. Руководство по изучению рыб. М., 1966. 375 с.
Приходько Б. И. Влияние среды на летние миграции кильки (Clupeonella
delicatula caspia Svetovidov) в северо-восточной части Среднего Каспия. — Тр.
Касп. бассейнового фил. ВНИРО. Астрахань, 1952, т. 12, с. 3—20.
Пробатов А. Н. О возрасте аральского шипа. — Изв. отд. прикладн. ихтиол.,
1929, т. 9, вып. 2, с. 55—77.
Пробатов А. Н. К изучению биологии Камской стерляди. — Изв. Биол. н.-и.
ин-та при Пермск. гос. ун-те, 1935, т. 10, вып. 1—2, с. 29—40.
Путилина Л. А. Морфологическая характеристика персидского осетра в
р. Волге. — В кн.: Комплексное использование биол. ресурсов Касп. и Азовск.
морей: (Тез. докл. Всесоюз. конф, молодых ученых и специалистов). М., 1983,
с. 70—71.
Путилина Л. А. Количественная и качественная оценка нерестовой популя-
ции персидского осетра в р. Волге. — Там же, с. 71—72.
Рыбы Монгольской Народной Республики /Под. ред. В. Е. Соколова, М.;
1983. 278 с.
Рылов В. М., Гладков Н. А. Рыбохозяйственное обследование Камышлыбаш-
ских озер. — Тр. Аральского отд. ВНИМОРХ, 1934, т. 3, с. 77—152.
Сапожников В. В. Доклад о путешествии 1899 г. — Протоколы об-ва естест-
воиспытателей и врачей за 1898—1899 гг. Томск, 1900, с. 1—3.
Сваричевская 3. А. К истории Балхаш-Алакульской впадины. — Вести. ЛГУ,
1952, № 7, с. 107—112.
Световидов А. Н. Европейско-азиатские хариусы. — Тр. ЗИН АН СССР, 1936,
г. 3, с. 183—301.
Световидов А. Н. О каспийских и черноморских сельдевых из рода Caspialo-
sa и Clupeonella и об условиях их формообразования.—Зоол. журн., 1943, т. 22,
вып. 4, с. 222—232.
Световидов А. Н. О каспийской и черноморско-азовской тюльке (Clupeonella
delicatula (Nordmann). — Докл. АН СССР. Нов. сер., 1945, т. 46, № 5, с. 226—228.
Световидов А. Н. Сельдевые. — В кн.: Фауна СССР. Рыбы, т. 2, вып. 1, М.;
Л., 1952. 331 с.
Световидов А. Н. Типы видов рыб, описанных П. С. Палласом в «Zoographia
Rosso—Asiatics». Л., 1978. 36 с.
Северцов Н. А. Жизнь красной рыбы в уральских водах. — Журн. Мин. го-
суд. имущества, 1863, т. 83, вып. 2, с. 92
Седельников А. Н. Озеро Зайсан. — Записки Зап.-Сиб. отд. Русск. геогр.
об-ва, 1910, т. 35, с. 253.
Седельников А. Н. Озеро Марка-Куль: (Предварит, отчет). — Ивв. Зап.-Сиб.
отд. РГО, 1914, т. 2, вып. 1—2, с. 1—6.
Селезнев В. В. Представители амурской ихтиофауны в Капчагайском водо-
хранилище. — В кн.: Акклиматизация рыб и беспозвоночных в водоемах СССР:
(Тез. докл. конф.). Фрунзе, 1972а, с. 75—77.
Селезнев В. В. К вопросу рыбохозяйственного использования водоемов Цент-
рального Казахстана. — В кн.: Биол. основы рыбн. хоз-ва республик Ср. Азии и
Казахстана. Ташкент; Фергана, 19726, с. 248—250.
Селезнев В. В. Малоценные и сорные виды рыб китайского комплекса в Кап-
чагайском водохранилище. — В кн.: Рыбные ресурсы водоемов Казахстана и их
использование. Алма-Ата, 1974, вып. 8, с. 143—148.
Селиверстова Н. Я. О видовом составе прилова осетровых в сельдяном райо-
не Азербайджана. — В кн.: Материалы отчета, сессии ЦНИОРХ. Баку, 1968,
с. 79—81.
Селиверстова Н. Я. Материалы по распределению молоди осетровых в Кас-
пийском море. — В кн.: Материалы научн. сессии ЦНИОРХ, посвящ. 100-летию
осетроводства. Астрахань, 1969, с. 172—174.
264
Селиверстова Н. Я. Особенности биологии осетровых, обитающих у северных
берегов Азербайджана. — В кн.: Осетровые СССР н их воспроизводство: (Тр.
ЦНИОРХ, т. 4). М., 1972, с. 78—94.
Селиверстова Н. Я., Аветисов К. Б., Аксенов В. И. Рыбохозяйственная ха-
рактеристика нефтеносных площадей Южного Каспия. — В кн.: Осетровое хозяй-
ство внутренних водоемов СССР: (Тез. и рефераты II Всесоюз. совещания).
Астрахань, 1979, с. 231—232.
Серебрякова Е. В. Изучение хромосомных комплексов и цитологии спермато-
генеза гибрида осетровых рыб. — Изв. ГосНИОРХ, 1964, т. 57, с. 279—285.
Серов Н. П. Ихтиофауна Нуринского водохранилища. — Изв. АН КазССР.
Сер. зоол., 1950, вып. 9, с. 87—95.
Серов Н. П. Рыбы и рыбный промысел Кургальджинских озер Акмолинской
области. — Тр. Ин-та зоол. АН КазССР, 1953, т. 2, с. 105—111.
Серов Н. П. Рыбы озера Челкар. — В кн.: Сб. работ по ихтиологии и гидро-
биологии. Алма-Ата, 1956, вып. 1, с. 278—320.
Серов Н. П. Ихтиофауна Камыш-Самарских и Кушумских озер. — В кн.:
Сб. работ по ихтиологии и гидробиологии. Алма-Ата, 1959, вып. 2, с. 152—175.
Серов Н. П. К вопросу об ихтиогеографическом положении бассейна Иргиз-
Тургая.— Тр. конф, по рыбн. хоз-ву респ. Ср. Азии и Казахстана. Фрунзе, 1961а,
с. 213—214.
Серов Н. П. Опыт разделения Балхашской ихтиологической провинции. —
Там же, 19616, с. 201—211.
Серов Н. П. Успехи акклиматизации рыб в бассейне Балхаша. — В кн.: Ак-
климатизация животных в СССР. Алма-Ата, 1963, с. 290—291.
Серов Н. П. Акклиматизация рыб в бассейне Балхаша. — В кн.: Акклимати-
зация рыб и беспозвоночных в водоемах СССР. Фрунзе, 1972, с. 77—79.
Серов Н. П. Акклиматизация рыб в бассейне озера Балхаш. — Изв.
ГосНИОРХ, 1975, т. 103, с. 172—174.
Сидоров Е. Г. Паразиты рыб в водоемах Иргиз-Тургайского бассейна. —
В кн.: Сб. работ по ихтиол, и гидробиол. Алма-Ата, 1956, вып. 1, с. 232—251.
Сидоров Е. Г. Паразитофауна рыб водохранилищ Центрального Казахстана.
—Тр. совещ. по болезням рыб. М.; Л., 1959, вып. 9, с. 134—137.
Сидоров Е. Г. Природная очаговость описторхоза. Алма-Ата, 1983. 240 с.
Сидорова А. Ф. Караси водоемов Иргиз-Тургайского бассейна. — В кн.: Сб.
работ по ихтиол, и гидробиол. Алма-Ата, 1956. вып. 1, с. 172—214.
Сидорова А. Ф. Язь водоемов Иргиз-Тургая. — В кн.: Сб. работ по ихтиол,
и гидробиол. Алма-Ата, 1959, вып. 2, с. 191—207.
Сидорова А. Ф. Акклиматизация радужной форели в водоемах Южного Ка-
захстана. — В кн.: Чтения памяти акад. Е. Н. Павловского. Алма-Ата, 1973,
с. 44—58-
Сидорова А. Ф. Закономерности линейного роста радужной форели при ак-
климатизации в горных водоемах. — В кн.: Биол. основы рыбн. хоз-ва респ. Ср.
Азии и Казахстана. Фрунзе, 1978, с. 400—403.
Сидорова А. Ф., Баимбетов А. А., Махмутова Р. X. Рыбохозяйственная оцен-
ка водоемов Павлодарской области. — В кн.: Биол. основы рыбн. хоз-ва на во-
доемах. Ср. Азии и Казахстана. Алма-Ата, 1966, с. 184—187.
Сидорова А. Ф., Буранбаева Г. Я. Условия размножения радужной форели
в водоемах бассейна р. Чилик. — В кн.: Биол. основы рыбн. хоз-ва респ. Ср.
Азии и Казахстана. Алма-Ата; Балхаш, 1970, с. 107—410.
Сидорова А. Ф., Курмаигалиева Ш. Г. Питание радужной форели в оз. Куль-
сай (Кунгей-Алатау). — В кн.: Тезисы докл. коиф. по вопросам рыбн. хоз-ва
респ. Ср. Азии и Казахстана. Фрунзе, 1968, с. 136—138.
Синицин В. М. Палеография Азии. М.; Л., 1962. 268 с.
Сливка А. П. Миграции осетровых в дельте и нижнем течении р. Волги. Ав-
тореф. дис. ... канд. биол. наук. М., 1974. 27 с.
Смирнова К- В. Паразитофауна рыб Алакульских озер. — Изв. КазФАН АН
СССР. Сер. зоол., 1944, № 3, с. 49—80.
265
Смирнова К. В. Нельма озера Зайсан. — Изв. АН КазССР. Сер. зоол., 1945,
№ 5, с. 79—93.
Смирнова К. В. Влияние акклиматизации на паразитофауну рыб. — В кн.:
Акклиматизация рыб и беспозвоночных в водоемах СССР. Фрунзе, 1972,
с. 267—269.
Смирнова К. В. Материалы по паразитофауне рыб — случайных вселепцев в
Балхаш-Илийский бассейн. — В кн.: Рыбные ресурсы водоемов Казахстана и их
использование. Алма-Ата, 1974, вып. 8, с. 192—193.
Смольянов И. И. Развитие белорыбицы St. leucichthys Giiid., нельмы St. 1.
nelma Pall, и сига-нельмушки Coregonus lavaretus nelmuschka Pravdin.. — Tp.
ИМЖ АН СССР, 1957, вып. 20, с. 232—294.
Солонинова Л. Н. Об урожайности и биологии молоди весенне-нерестующих
рыб Бухтарминского водохранилища. — В кн.: Биол. основы освоения, рацион,
использования и воспроизводства рыбных запасов в водоемах Ср. Азии и Казах-
стана. Алма-Ата, 1966, с. 283—286.
Солонинова Л. Н. К вопросу о размножении весенне-нерестующих рыб в
Бухтарминском водохранилище. — В кн.: Гидробиология и ихтиология. Душан-
бе, 1969, с. 164—178.
Солонинова Л. Н. Малоценные и сорные рыбы — основные компоненты пи-
тания щуки и окуня в Бухтарминском водохранилище. — В кн.: Рыбные ресурсы
водоемов Казахстана и их использование. Алма-Ата, 1972, вып. 7, с. 38—43.
Солонинова Л. И. Состояние сырьевых запасов щуки в Бухтарминском водо-
хранилище. — В кн.: Водоемы Сибири и перспективы их рыбохоз. использования.
Томск, 1973, с. 112—114.
Солонинова Л. Н. Питание молоди щуки в Бухтарминском водохранилище•
В кн.: Рыбные ресурсы водоемов Казахстана и их использование. Алма-Ата,
1974а, вып. 8, с. 149—152.
Солонинова Л. Н. Биология и хозяйственное значение щуки в Бухтармии-
ском водохранилище. Автореф. дис. ... канд. биол. наук. Казань, 19746. 23 с.
Солонинова Л. Н. Плодовитость щуки в Бухтарминском водохранилище. —
В кн.: Рыбные ресурсы водоемов Казахстана и их использование. Алма-Ата, 1975,
вып. 9, с. 109—110.
Солонинова Л. Н. О росте щуки в Бухтарминском водохранилище. — Гидро-
биол. журн., 1976, № 6, с. 58—66.
Солонинова Л. Н. О регулировании добычи сиговых на водоемах Кокчетав-
ской области. — В кн.: Тез. докл. V Всесоюз. совещ. лимнологов. Иркутск, 1981,
вып. 3, с. 141—143.
Спановская В. Д. О зоогеографических исследованиях Г. В. Никольского в
Московском университете. — В кн.: Современные проблемы ихтиологии. М., 1981,
с. 65—74.
Спичаков Ф. А. Аральская научно-промысловая экспедиция. — Рыбное хоз-во,
1922, № 1, с. 143—152.
Стояиов А. А. Об остатках амий из третичных отложений системы хребта
Монрак в Зайсанском уезде Семипалатинской области. — Изв. ГК, 1915, т. 34,
№ 4, с. 487—508.
Стрельников А. С. Рыбы и биологические основы рыбного хозяйства Ала-
кольских озер. Автореф. дис. ... канд. биол. наук. Томск, 1974. 20 с.
Стыгар В. М. О начале активного питания личинок осетровых р. Урал. —
В кн.: Рацион, основы ведения осетрового хоз-ва. Волгоград, 1981, с. 221—222.
Стыгар В. М., Песериди Н. Е. Пищевые отношения между молодью осетро-
вых и других рыб в р. Урал. — Там же, с. 222—224.
Стыгар В. М., Тарабрин А. Г., Песериди Н. Е. Особенности ската молоди
шипа в р. Урал. — Там же, с. 225—227.
Стыгар В. М., Песериди Н. Е., Спановская В. Д., Дума В. В. Питание ураль-
ского пескаря в период нереста осетровых. — В кн.: Осетровое хоз-во внутренних
водоемов СССР. Астрахань, 1979, с. 250—251.
Суворов Е. К. Основы ихтиологии. М., 1948. 433 с.
Суворцев А. Рыболовство на оз. Марка-Куль. — Вестник рыбопромышленно-
сти, 1888, т. 3, вып. 8—9, с. 189—195.
266
Сысолятина Н. А. Структура природного очага дилепидоза на оз. Балыкты-
коль. — В кн.: Биол. основы рыбн. хоз-ва водоемов Ср. Азии и Казахстана: (Тез.
докл. XVII науч. конф.). Фрунзе, 1981, с. 537—539.
Сычевская Е. К. Ископаемые щуковидные СССР и Монголии. М., 1976. 116 с.
Танасийчук Н. П. Нерестовые миграции волжской многотычинковой сельди.
— Тр. Касп. басе, филиала Всесоюзн. н.-и. ин-та морск. рыбн. хоз-ва и океаногр.
Астрахань, 1948, т. 10, с. 3—82.
Танасийчук В. С. Ход и нерест проходных сельдей в р. Урал.— Там же,
с. 93—96.
Тарабрии А. Г. Динамика ската личинок осетровых и распределение их в
русле р. Урал (по уловам икорных сеток в 1979—1980 гг.). — В кн.: Рациональ-
ные основы ведения осетров, хоз-ва. Волгоград, 1981а, с. 233—235.
Тарабрин А. Г. Качественный состав молоди севрюги р. Урал. — Там же,
19816, с. 235—236.
Тарвердиева М. И. Роль акклиматизированных организмов в питании осетра
и севрюги Каспийского моря в 1962 г.— В кн: Изменения биол. комплексов Кас-
пийского моря за последи, десятилетие. М., 1965, с. 234—255.
Тарвердиева М. И. Питание осетровых рыб в Каспийском море. Автореф.
дис. ... канд. биол. наук. М., 1968. 28 с.
Терентьев В. Влияние щуки и окуня на запасы промысловых рыб Волго-Кас-
пийского района.— Рыбное хоз-во, 1937, № 9, с. 34—40.
Терлецкий В. К. Балхаш-Алакульская впадина. — Тр. Геологоразв. упр.,
1931, вып. 105.
Титеньков И. С. Кубенская нельма. М., 1961. 52 с.
Тихий М. И. Использование и экология рыб р. Урала в связи с проектом ре-
гулирования реки. — В кн.: Большая Эмба: (Тр. Казах, фил. АН СССР, вып. 11).
М„ 1938, т. 2, с. 259—366.
Тленбекова И. К. Паразиты рыб бассейнов оз. Балхаш и Алакольской груп-
пы озер в связи с реконструкцией ихтиофауны. Автореф. дис. ... канд биол. наук.
Алма-Ата, 1980. 27 с.
Тлеуов Р. Т. Влияние антропических факторов на состояние популяций рыб
и рыбное хозяйство Аральского моря. Автореф. дис. ... докт. биол. наук. М.,
1981. 47 с.
Тлеуов Р. Т., Сагитов Н. И. Осетровые рыбы Амударьи. Ташкент, 1973. 155 с.
Трусов В. 3. Биологические и экспериментальные основы мероприятий по
воспроизводству запасов аральского шипа. — Тр. лаборат. основ рыбоводства.
Л., 1947, т. 1.
Трусов В. 3. Биологическое обоснование рыбоводных работ с летненерестя-
щимся (поздним яровым) осетром. — В кн.: Осетровые СССР и их воспроизвод-
ство: (Тр. ЦНИОРХ, т. 1). М., 1967, с. 168—180.
Турдаков Ф. А. Очерк ихтиофауны Таласского бассейна. — Уч. зап. биофака
КирГУ, 1955, вып. 5. Зоология, с. 128—163.
Турдаков Ф. А. Рыбы Киргизии. Фрунзе, 1963. 284 с.
Турдаков Ф. А., Пискарев К. В. Систематическое положение щуки из р. Чу. —
Тр. Ин-та зоол. и паразитол. Кирг. ФАН СССР. Фрунзе, 1954, вып. 1, с. 131—136.
Тютеньков С. К. Материалы по питанию некоторых промысловых рыб пой-
менных водоемов Кзыл-Ординской области. — Изв. АН КазССР, 1948, № 63, Сер.
зоол., вып. 8, с. 168—175.
Тютеиьков С. К., Исмуханов X. К., Козляткин А. Л. Акклиматизация рыб и
беспозвоночных в Бухтарминском водохранилище. — В кн.: Советско-американ-
ское сотрудничество в области исследования Мирового океана. — В кн.: Симпо-
зиум по реакции водных экосистем на вселение новых видов. ВНИРО. М., 1977,
с. 141—142.
Тэн В. А. Материалы по питанию ленка озера Марка-Куль. — В кн.: Сб. ра-
бот по ихтиол, и гидробиол. Алма-Ата, 1959, вып. 2, с. 256—261.
Тэн В. А. Питание хариуса из оз. Марка-Куль. — Там же, с. 262—266.
Федоров А. В., Филиппов И. А. Состояние гонад стерляди и перспективы
осетроводства в Верхнем Дону. — В ки.: Осетровое хоз-во в водоемах СССР. М.,
1963, с. 167—170.
267
Федотова Л. А. Биологическая характеристика сазана и пути рационального
использования его запасов в Бухтарминском водохранилище. Автореф. дис. ...
канд. биол. паук. Алма-Ата, 1973. 30 с.
Филатов Г. М. Питание осетровых восточной части Южного Каспия. — В кн.:
Тез. отчетн. сессии ЦНИОРХ. Астрахань, 1972, с. 167—168.
Филатов Д. П. К биологии и морфологии аральских рыб: (Предварительное
сообщение).—-Русек, зоол. жури., 1925, т. 5, вып. 1—2, с. 36—52.
Филиппов Г. М. Суточный ход питания персидского осетра в юго-восточном
районе Каспийского моря. — В кн.: Осетровое хоз-во внутренних водоемов СССР.
Астрахань, 1979, с. 263—264.
Фииш О., Брэм А. Путешествие в Западную Сибирь. СПб., 1882. 566 с.
Фортунатова К. Р. Некоторые данные по биологии питания хищных рыб в
дельте р. Волги. — Зоол. журн., 1949, т. 28, вып. 5, с. 453—460.
Фролова Л. И. Биология сиговых и их разведение в разнотипных водоемах
Северного Казахстана. Автореф. дис. ... канд. биол. наук. Л., 1974. 20 с.
Хисарова Г. Д. Палеоихтиологическая находка. — Изв. АН КазССР. Сер.
биол., 1967, № 5. 93 с.
Хусаинова Н. 3. К акклиматизации кормовых (для рыб) беспозвоночных в
Балхаше. — Уч. зап. КазГУ, 1955, т. 17, с. 118—130.
Хусаинова Н. 3. Генезис донной фауны Аральского моря. — В кн.: Сб. работ
по ихтиол, и гндробиол. Алма-Ата, 1959, вып. 2, с. 3—33.
Цееб Я. Я. К познанию фауны озер Северного Казахстана в связи с их ры-
бохозяйственной оценкой. — Уч. зап. Орловского гос. пед. ин-та. Сер. естествозн.
и химии, 1940, вып. 1, с. 37—125.
Цеханович Ю. В. Рыбы Урала и их ужение. Свердловск, 1937, с. 37—45.
Цикулеико П. В. К биологии шипа н о мерах его охраны в Сыр-Дарье. —
Тр. Аральск, отдел, н.-и. ин-та морского рыбн. хоз-ва н океанографии, 1935, т. 4,
с. 115—126.
Цыба К. П. Биология восточного леща озера Балхаш. Автореф. дне. ... канд.
биол. наук. Томск, 1975. 18 с.
Цыба К. П. Рыбохозяйственное значение озер Нижне-Тургайской и Иргиз-
ской систем и пути повышения их рыбопродуктивности. — В кн.: Биол. основы
рыбн. хоз-ва Ср. Азии и Казахстана. Душанбе, 1976, с. 383—385.
Цыба К. П. Обоснование организации озерно-товарного хозяйства на озерах
Васильевского накопителя. — В кн.: Биол. основы рыбн. хоз-ва водоемов Ср.
Азии и Казахстана. Фрунзе, 1978, с. 439—441.
Чабан А. П. Рыбохозяйственное освоение Усть-Каменогорского водохранили-
ща.— Вести, сельскохоз. науки. Алма-Ата, 1962, № 7, с. 80—86.
Чабан А. П. Рыбы Усть-Каменогорского водохранилища и биологические
основы его рыбохозяйственного освоения. Автореф. дис. ... канд. биол. наук.
Томск, 1965. 20 с.
Чабан А. П. Рыбохозяйственное использование Усть-Каменогорского водо-
хранилища. — В кн.: Рыбные ресурсы водоемов Казахстана и их использование.
Алма-Ата, 1966, вып. 5, с. 294—302.
Чабан А. П., Дюсеигалиев Т. Д. Биологические основы ведения рыбного хо-
зяйства на водоемах Кустанайской области. — В кн.: Биол. основы рыбн. хоз-ва
республик Ср. Азии и Казахстана. Балхаш, 1967, с. 300—302.
Чаянова Л. А. Питание кильки (Clupeonella delicatula caspia) в Северном
Каспии. — В кн.: Рыбы Каспийского моря: (Тр. Всесоюз. н.-и. ин-та морского
рыбн. хоз-ва и океанографии, т. 18). М., 1951, с. 245—255.
Чериавин В. В. Отчет о поездке в Западный Тянь-Шань летом 1913 г.—
Ежегодник Зоол. музея ИАН, 1914, т. 19, № 4, с. 138—168.
Чугунов Н. Л. Биология молоди промысловых рыб Волго-Каспийского райо-
на.— Тр. Астраханск. научн. рыбохоз. ст., 1928, вып. 4. 100 с.
Чугунов Н. Л., Чугунова Н. И. Сравнительная промыслово-биологическая
характеристика осетровых Азовского моря. — В кн.: Осетровые южных морей Со-
ветского Союза: (Тр. ВНИРО, т. 52, сб. 1), М., 1964, с. 87—182.
Шамардина И. П. Этапы развития щуки. — Тр. ИМЖ АН СССР. М., 1957,
вып. 16, с. 237—298.
268
Шапошникова Г. X. Биология и распределение рыб в реках уральского типа.
М., 1964. 176 с.
Шапошникова Г. X. Сравнительно-морфологическое изучение тайменя и лен-
ка.— Вопросы ихтиологии, 1968, т. 8, вып. 3(50), с. 440—464.
Шарапова Л. И. Зоопланктон озер Каракуль и Соркуль (бассейн р. Или) и
его продукция. Автореф. дис. ... канд. биол наук. Алма-Ата, 1971. 32 с.
Шаронов И. В. Расширение ареала некоторых рыб в связи с зарегулирова-
нием Волги. — В кн.: Материалы I конф, по изучению водоемов бассейна Волги.
Куйбышев, 1971, с. 226—232.
Шиленкова А. К. Материалы по биологии щуки Иргиз-Тургайских озер. —
В кн.: Сб. работ по ихтиол, и гидробиол. Алма-Ата, 1956, вып. 1, с. 215—231.
Шилеикова А. К. Материалы по систематике и биологи)! окуня озер Иргиз-
Тургайской системы. — В кн.: Сб. работ по ихтиол, и гидробиол. Алма-Ата, 1959,
вып. 2, с. 176—190.
Шилов В. И. О расах, росте, созревании и повторности нереста стерляди
Волгоградского водохранилища. — В кн.: Осетровые в Волгоградском и Саратов-
ском водохранилищах: (Тр. Саратовск. отд. ГосНИОРХ, т. 2). Саратов, 1971,
с. 112—153.
Юлусов А. С. Аральский лосось. — Вести, рыбопром., 1910, т. 25, с. 282.
Яблонский Н. И. Озеро Марка-Куль. — Природа и охота, 1907, № 10, с. 1—
14; № 11, с. 1—8; № 12, с. 1—10.
Яковлев В. Н. Распространение пресноводных рыб неогена Голарктики и зоо-
географическое районирование. — Вопросы ихтиологии, 1961, т. 1, вып. 2(19).
Berg L. S. Acipenser giildenstadti persicus, a sturgeon from the south Caspian
Sea. — Ann. Mag. Hist., London, 1934, 10 ser., v. 13, p. 317—318.
Borodin N. Acipenser persicus, a sturgeon from the Caspian Sea. — The An-
nals and Magazine of Nat. Hist., 1926, N XX, v. 9, p. 26—28.
Chusainova N. S., Sharapova L. I., Mamilova R. Kh., Kriusenko T. G., Mitro-
fanov V. P. On Biological Productivity of theKarakul Lake. — Preliminary papers
for UNESCO-1BP Symposium on Productivity Problems of Freshwaters. Warsza-
wa, 1970, v. 1, p. 209—215.
Dukravets G. M. Regularities of morpho-biological variability of fishes in Ka-
zakhstan. — Abstracts. Fourth congress of european ichthyologists. Hamburg,
Fed. Rep. of Germany, 1982, p. 76.
iankovis D. Ekologija dunavske kecige (Acipenser ruthenus L.). — Bioloski
Institut N. R. Serbije: Posebna izd. Beograd, 1958.
Li S. T. On a new subspecies et fresh-water trout Brachymystax lenok tsinlin-
gensis from Taipaishan, Shensi, China. — Acta zootaxonomica sinica, 1966, v. 3,
N 1, p. 92—94.
Mori T. On the freshwater fishes from the Tumen river with description of new
species. — J. Chosen Nat. Hist, soc., 1930, N 11.
Pallas P. S. Reise. V. 3. St. P„ 1776.
Pallas P. S. Zoographia Rosso-Asiatica. T. 3. Petropolis, 1814. 428 p.
Hensel K., Holcik I. On the identity of Hucho hucho and Hucho taimen (Pisces,
Salmonidae). — Folia zool., 1983, v. 32, N 1, p. 67—83.
Rostami I. Biologi et expiation de esturgions (Acipenser kaspiens). Imprimerie
Comte-jacqu et Bar-le-Dul (Maise). France, 1961, p. 221.
Vladykov V. D. The taxonomy of European ammocoetes (Petromyzonidae).—
XV th International Congress of Zoology. London, 1958, Sect. II, p. 38.
Vladykov V. D., Kott F. A new nonparasitic Species of Zethenteron Greaser
and Hubbs 1922 (Petromyzonidae) from Hokkaido, Japan.— Canad. Zook, 1978,
v. 56, N 8, p. 1792—1800.
Vladykov V. D., Kott F. List of Northern Hemisphere Lampeys (Petromyzoni-
dae) and Their Distribution. — Department of Fisheries and Oceans. Ottawa, 1979.
269
ОГЛАВЛЕНИЕ
Предисловие ............................................... 3
Глава 1. История изучения ихтиофауны Казахстана ... 6
Глава 2. Формирование современной ихтиофауны Казахста-
на и ихтиогеографическое районирование..................20
Глава 3. Описание видов ихтиофауны Казахстана ... 41
Отряд Petromyzoniformes — Миногообразные...................42
Семейство Potromyzonidae — Миноговые...................42
Род Caspiomyzon Berg, 1906 43
1. Caspiomyzon wagneri (Kessler)—каспийская ми-
нога ..............................................43
Род Lampetra Gray, 1851..............................46
1. Lampetra japonica (Martens) — тихоокеанская
минога ............................................46
2. Lampetra kessleri (Anikin)—сибирская минога 43
Отряд Acipenseriformes — Осетрообразные....................57
Семейство Acipenseridae — Осетровые....................57
Род Huso Brandt, 1869 — Белуга.......................57
1. Huso huso (Linne) — белуга.....................57
Род Acipenser Linne, 1758 —Осетр.....................71
1. Acipenser giildenstadti Brandt— русский осетр . . ц
la. Acipenser giildenstadti giildenstadti Brandt — рус-
ский осетр.........................................71
16. Acipenser giildenstadti persicus Borodin — персид-
ский (куринский) осетр.............................99
2. Acipenser baeri Brandt — сибирский осетр . . . 100
3. Acipenser stellatus Pallas — севрюга . 122
4. Acipenser nudiventris Lovetzky — шип .... 139
5. Acipenser ruthenus Linne — стерлядь .... 156
Род Pseudoscaphirhynchus A. Nik, 1900 —Лжелопа-
тонос ..............................................161
1. Pseudoscaphirhynchus fedtschenkoi (Kessler) —
сырдарьинский лжелопатонос........................162
Отряд Clupeiformes — Сельдеобразные ...... 164
Семейство Clupeidae — Сельдевые.......................154
Род Clupeonella Kessler, 1877 —Тюлька (килька) . . 164
1. Clupeonella delicatula (Nordmann)—обыкновен-
ная тюлька........................................164
la. Clupeonella delicatula caspia Svetovidov — кас-
пийская обыкновенная тюлька (килька) .... 164
270
la'. Clupeonella delicatula caspia morpha tscharcha-
lensis (Borodin)—чархальская селедочка . . . 168
Род Alosa Cuvier, 1829 — Сельдь...................170
1. Alosa caspia (Eichwald)—пузанок .... 171
la. Alosa caspia caspia (Eichwald)—северокаспий-
ский (среднекаспийский) пузанок ............... 171
2. Alosa kessleri (Grimm) — черноспинка . . . 174
2a. Alosa kessleri kessleri (Grimm) — черноспинка
(бешенка, залом)................................174
26. Alosa kessleri volgensis (Berg)—волжская много-
тычинковая сельдь...............................177
Отряд Salmoniformes — Лососеобразные....................180
Семейство Salmonidae — Лососевые....................180
Род Salmo Linne, 1758 — Лосось....................180
1. Salmo trutta Linne — кумжа (проходной лосось) 180
la. Salmo trutta caspius Kessler — каспийский лосось 180
16. Salmo trutta aralensis Berg—аральский лосось 182
Род Hucho Gunther, 1866 — Таймень.................184
1. Hucho taimen (Pallas) —таймень...............184
Род Brachymystax Gunther, 1866 — Ленок .... 187
1. Brachymystax lenok (Pallas)—ленок .... 188
la. Brachymystax lenok savinovi Mitrofanov — марка-
кольский ленок ................................ 192
Семейство Coregonidae — Сиговые.....................202
Род Stenodus Richardson, 1836 — Белорыбица, или
Нельма............................................202
1. Stenodus leucichthys (Giildenstadt)—нельма . . 202
la. Stenodus leucichthys nelma (Pallas) —нельма 202
16. Stenodus leucichthys leucichthys (Gfildenstadt) —
белорыбица......................................209
Семейство Thymallidae — Хариусовые..................214
Род Thymallus Cuvier, 1829 — Хариус...............214
1. Thymalius arcticus (Pallas)—сибирский хариус 214
la. Thymallus arcticus brevicephalus Mitrofanov —
маркакольский хариус............................215
Семейство Esocidae —Щуковые.........................219
Род Esox Linne, 1758 — Щука.......................219
1. Esox lucius Linne — щука....................219
Определитель семейств рыб, встречающихся в пресных водах
Казахстана...........................................242
Определитель родов, видов и подвидов рыб, описанных в
1 томе....................... .......................243
Список терминов, обозначений и условных сокращений в
тексте ................... ..........................246
Литература........................ .....................247
271
Валерий Петрович Митрофанов,
Геннадий Михайлович Дукравец,
Николай Евстафьевич Песериди,
Александра Николаевна Полторыхина и др.
РЫБЫ КАЗАХСТАНА
Том 1
Миноговые, Осетровые, Сельдевые, Лососевые, Щуковые
Утверждено к печати Ученым советом Института зоологии
Академии наук Казахской ССР
Рецензенты: доктор биол. наук А. Ф. Ковшарь,
кандидат биол. наук А. Ф. Сидорова
Зав. редакцией Д. М. Глазырина
Редактор А. Н. Ведерникова
Художественный редактор Н. Ф. Чурсин
Оформление художника Л. Г. Мироненко
Технический редактор В. М. Муромцева
Корректоры Е. С. Латыпова, А. Н. Утебаева
ИБ № 2003
Сдано в набор 29.11.85. Подписано в печать 29.04.86. УГ12042.
Формат 60Х90‘/1б- Бум. тип. № 1. Литературная гаппитчоа. Высокая печать.
Усл. п. л. 17. Усл. п. кр.-отт. 18. Уч.-изд. л. 21,7. Тираж 3700.
Заказ 7. Цена 2 р. 20 к.
Издательство «Наука» Казахской ССР.
480100, Алма-Ата, Пушкина, 1111113.
Типография издательства «Наука» Казахской ССР.
480021, Длма-Атц, Шевченшэ, 28.