/






















































































































Автор: Решетников Ю.С.
Теги: зоология систематика животных специальные зоологические науки атлас ихтиология рыбы рыбное хозяйство
ISBN: 5-02-002873-8
Год: 2003




Текст
АТЛАС
пресноводных
рыб России
Том 2
“НАУКА”
РОССИЙСКАЯ АКАДЕМИЯ НАУК
ИНСТИТУТ ПРОБЛЕМ ЭКОЛОГИИ
И ЭВОЛЮЦИИ им. А. Н. СЕВЕРЦОВА
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ
ИНСТИТУТ БИОЛОГИЧЕСКИХ
ПРОБЛЕМ СЕВЕРА
МИНИСТЕРСТВО ПРОМЫШЛЕННОСТИ, НАУКИ
И ТЕХНОЛОГИЙ РОССИЙСКОЙ ФЕДЕРАЦИИ
НАУЧНЫЙ СОВЕТ ПО ПРОГРАММЕ
“БИОЛОГИЧЕСКОЕ РАЗНООБРАЗИЕ”
МОСКОВСКИЙ ГОСУДАРСТВЕННЫЙ
УНИВЕРСИТЕТ им. М. В. ЛОМОНОСОВА
ATLAS
of Russian
Freshwater Fishes
In Two Volumes
Vol. 2
Edited by
Professor of Biology
Yu. S. RESHETNIKOV
Second edition
e
MOSCOW “NAUKA” 2003
АТЛАС
пресноводных
рыб России
В двух томах
Том 2
Под редакцией
доктора биологических наук
Ю. С. РЕШЕТНИКОВА
Второе издание
6
МОСКВА “НАУКА” 2003
УДК 59
ББК 28.693.32
А92
Авторы:
Ю.С. Решетников, О.А. Попова, Л.И. Соколов, Е.А. Цепкий,
В.Г. Сиделева, Е.А. Дорофеева, И.А. Черешнев,
К.И. Москалькова, Ю.Ю. Дгебуадзе, Г.И. Рубан, В.В. Королев
Рецензенты:
доктор биологических наук Ж А. Черняев,
доктор биологических наук Г.Г. Новиков
Атлас пресноводных рыб России: В 2 т. Т. 2 / Под ред. Ю.С. Решетникова. -
М.: Наука, 2003. - 253 с.: ил.
ISBN 5-02-002873-8
ISBN 5-02-002872-Х (т. 2)
Второй том "Атласа” продолжает краткое описание пресноводных рыб России и повторяет
структуру первого тома. Даются латинское и русское названия вида согласно последней ревизии,
рисунок вида, его краткое систематическое описание с указанием характерных видовых призна-
ков и числа подвидов в водах России. Приводится уточненный ареал с картой распространения,
максимальные размеры и возраст, кратко дается описание образа жизни и статус вида с указани-
ем его промысловой ценности и охранного статуса. “Атлас” снабжен указателем русских и ла-
тинских названий рыб. Для экологов, зоологов, ихтиологов, специалистов в области сохранения
биологического разнообразия и рыбного хозяйства, для студентов и аспирантов университетов и
ВУЗов, для рыболовов-любителей.
Atlas of Russian Freshwater Fishes: In 2 Vol. Vol. 2 / Ed. by Yu.S. Reshet-
nikov. - M.: Nauka, 2003. - 253 p.: il.
ISBN 5-02-002873-8
ISBN 5-02-002872-X (Vol. 2)
All the fish, including the agnathans, found in the freshwaters of Russia are described. The Atlas con-
tains 293 species representing 13 orders, 33 families, and 138 genera. The Latin and common Russian
names, as well the taxonomic changes made after the last revision of the “Annotated Check-list” (1998) are
presented. More precise distributional information, data on subspecies, some biological data, status of
fishery and conservation are given. List of scientific and Russian names is added. For ecologists, zoologists,
ichthyologists, and scientists in biological diversity conservation and fishery, University students and post-
graduates, and for fishermen.
ISBN 5-02-002873-8 © Российская академия наук, 2003
ISBN 5-02-002872-X (т. 2) © Издательство “Наука”
(художественное оформление), 2003
ПРЕДИСЛОВИЕ
Второй том “Атласа пресноводных рыб России” является продолжени-
ем первого тома и сохраняет его структуру. Том начинается с отряда сомо-
образных и кончается скорпенообразными. В отличие от “Аннотированно-
го каталога круглоротых и рыб континентальных вод России” (1998) в него
включены только пресноводные виды рыб, поэтому отсутствуют камбало-
образные и другие морские рыбы, которые заходят в низовья рек, но в пре-
сной воде не размножаются. Из завезенных видов включены только те, ко-
торые прижились в наших водах (американские сомики, медака, гамбузия,
тилапия).
По сравнению с “Аннотированным каталогом” внесены небольшие изме-
нения в таксономическое положение рыб. Сомообразные, карпозубообразные
и трескообразные даются практически без изменений. Систематика колюшко-
вых дана по В.В. Зюганову (1991), бычковых - по В.И. Пинчуку (1976, 1978,
1981, 1984; Пинчук, Савчук, 1982). Значительно переработана систематика
керчаковых, байкальских голомянок и широколобою В.Г. Сиделева дает опи-
сание новых родов и видов, приводит оригинальные рисунки этих рыб, а также
основные данные по их образу жизни.
Заканчивается “Атлас” списком литературы и указателями названий рыб
на русском и латинском языках.
Полагаем, что книга может быть полезна при составлении региональных
сводок и кадастров, а также для работников рыбного хозяйства и специалистов,
работающих по проблеме сохранения биологического разнообразия.
Авторы будут искренне благодарны лицам, которые пришлют свои крити-
ческие замечания, поправки и дополнения.
Ответственный редактор Ю.С.Решетников
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ
ВИДОВ
Группа PISCES - РЫБЫ
Класс II. OSTEICHTHYES - КОСТНЫЕ РЫБЫ
Отряд VII. SILURIFORMES - СОМООБРАЗНЫЕ
Сем. 16. SILURIDAE Cuvier, 1816 - Сомовые
Род 87. Parasilurus Bleeker, 1862 - дальневосточные сомы
189. Р. asotus (Linnaeus, 1758) - амурский сом.............. 11
Род 88. Silurus Linnaeus, 1758 - обыкновенные сомы
190. S. glanis Linnaeus, 1758 - обыкновенный (европейский) сом . 13
191. S. soldatovi Nikolsky et Soin, 1948 - сом Солдатова........ 16
Сем. 17. BAGRIDAE Regan, 1911 - Косатковые
Род 89. Leiocassis Bleeker, 1858 - косатки
192. L. brashnikowi (Berg, 1907) - косатка Бражникова........... 18
193. L. herzensteini (Berg, 1907) - косатка Герценштейна ....... 20
194. L. ussuriensis (Dybowski, 1872) - косатка-плеть, уссурийская ко-
сатка ......................................................... 22
Род 90. Mystus Scopoli, 1777 - мистусы
195. М. mica Gromov, 1970 - косатка-крошка...................... 24
Род 91. Pelteobagrus Bleeker, 1862 - косатки-скрипуны
196. Р. fulvidraco (Richardson, 1846) - косатка-скрипун......... 26
Сем. 18. ICTALURIDAE Taylor, 1954 - Икталуровые
Род 92. Ictalurus Rafinesque, 1820 - американские сомики
197.1, nebulosus (Lesueur, 1819) - американский сомик........... 28
198.1, punctatus (Rafinesque, 1818) - канальный сомик........... 30
Отряд VIII. CYPRINODONTIFORMES - КАРПОЗУБООБРАЗНЫЕ
Сем. 19. ORYZIATIDAE - Оризиевые
Род 93. Oryzias Jordan et Snyder, 1906 - оризии
199. О. latipes (Temminck and Schlegel, 1846) - медака.......... 32
Сем. 20. POECILIIDAE Bonaparte, 1838 - Пецилиевые
Род 94. Gambusia Poey, 1854 - гамбузии
200. G. holbrooki (Girard, 1859) - хольбрукская гамбузия........ 34
6
Отряд IX. GADIFORMES - ТРЕСКООБРАЗНЫЕ
Сем. 21. LOTIDAE Jordan et Evermann, 1898 - Налимовые
Род 95. Lota Oken, 1817 - налимы
201. L. lota (Linnaeus, 1758) - налим................................ 37
Отряд X. GASTEROSTEIFORMES - КОЛЮШКООБРАЗНЫЕ
Сем. 22. GASTEROSTEIDAE Bonaparte, 1832 - Колюшковые
Род 96. Gasterosteus Linnaeus, 1758 - трехиглые колюшки
202. G. aculeatus Linnaeus, 1758 - трехиглая колюшка ................ 40
Род 97. Pungitius Costa, 1848 - многоиглые колюшки
203. P. platygaster (Kessler, 1859) - малая южная колюшка ........... 43
204. P. pungitius (Linnaeus, 1758) - девятииглая колюшка............. 45
205. P. sinensis Guichenot, 1869 - китайская, или амурская, колюшка . 47
206. Р. tymensis (Nikolsky, 1889) - сахалинская колюшка ............. 49
Отряд XI. SYNGNATHIFORMES - ИГЛООБРАЗНЫЕ
Сем. 23. SYNGNATHIDAE Rafinesque, 1810 - Игловые
Род 98. Syngnathus Linnaeus, 1758 - иглы
207. S. nigrolineatus Eichwald, 1831 - черноморская игла............. 51
Отряд XII. PERCIFORMES - ОКУНЕОБРАЗНЫЕ
Подотряд PERCOIDEI
Сем. 24. PERCICHTHYIDAE Gill, 1860 - Перцихтовые
Род 99. Morone Mitchill, 1814 - лавраки
208. Morone saxatilis (Walbaum, 1792) - полосатый лаврак, полосатый
окунь.................................................... 53
Род 100. Siniperca Gill, 1863 - аухи
209. S. chuatsi (Basilewsky, 1855) - ауха, китайский окунь .......... 55
Сем. 25. CENTRARCHIDAE Bleeker, 1859 - Центрарховые
Род 101. Micropterus Lacepede, 1802 - черные окуни
210. М. salmoides (Lacepede, 1802) - большеротый окунь............... 57
Сем. 26. PERCIDAE Cuvier, 1816 - Окуневые
Род 102. Gymnocephalus Bloch, 1793 - ерши
211. G. acerinus (Giildenstadt, 1775) - донской ерш ................. 60
212. G. cemuus (Linnaeus, 1758) - обыкновенный ерш .................. 62
Род 103. Perea Linnaeus, 1758 - пресноводные окуни
213. Р. fluviatilis Linnaeus, 1758 - речной окунь ................... 64
Род 104. Percarina Nordmanrf, 1840 - перкарины
214. Р. demidoffi Nordmann, 1858 - перкарина......................... 67
Род 105. Stizostedion Rafinesque, 1820 - судаки
215. S. lucioperca (Linnaeus, 1758) - обыкновенный судак............. 69
216. S. volgense (Gmelin, 1788) - волжский судак, берш............... 72
7
Сем. 27. CICHLIDAE Bleeker, 1858 - Цихловые
Род 106. Sarotherodon Rtippell, 1852 - тилапии
217. S. mossambicus (Peters, 1852) - мозамбикская тилапия.............. 74
Подотряд GOBIOIDEI
Сем. 28. ELEOTRIDIDAE Regan, 1911 - Головешковые
Род 107. Micropercops Fowler et Bean, 1920 - микроперкопсы
218. M. cinctus (Dabry, 1872) - китайский элеотрис .................... 77
Род 108. Perccottus Dybowski, 1877 - головешки
219. P. glenii Dybowski, 1877 - головешка-ротан ....................... 79
Сем. 29. GOBIIDAE Bonaparte, 1832 - Бычковые
Род 109. Aboma Jordan et Starks, 1895 - абомы
220. A. lactipes (Hilgendorf, 1878) - японская абома................... 82
Род 110. Acanthogobius Gill,1859 - акантогобиусы
221. A. flavimanus (Temminck et Schlegel, 1845) - японский речной бы-
чок ................................................................ 83
Род 111. Benthophilus Eichwald, 1831 - пуголовки
222. В. baeri Kessler, 1877 - пуголовка Бэра........................... 85
223. В. casachicus Rahimov, 1978 - казахская пуголовка ................ 87
224. В. granulosus Kessler, 1877 - зернистая пуголовка................. 89
225. В. macrocephalus (Pallas, 1787) - каспийская пуголовка ........... 90
226. В. magistri Iljin, 1927 - азовская пуголовка, пуголовка магистра . 92
227. В. mahmudbejovi Rahimov, 1976 - пуголовка Махмудбекова............ 94
228. В. stellatus (Sauvage, 1874) - звездчатая пуголовка .............. 96
Род 112. Caspiosoma Iljin, 1927 - каспиосомы
229. С. caspium (Kessler, 1877) - каспиосома .......................... 98
Род 113. Chaenogobius Gill, 1858 - дальневосточные бычки
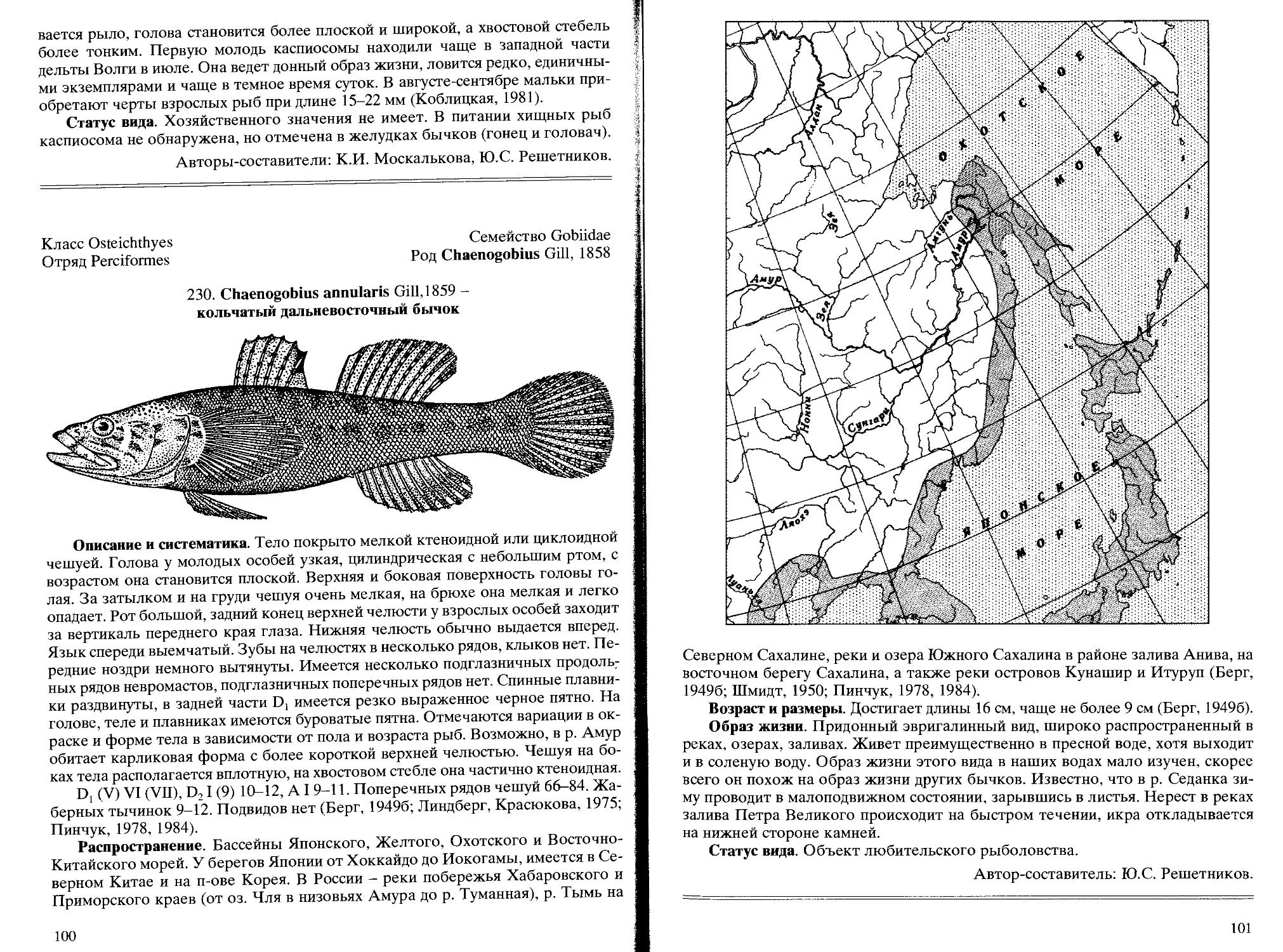
230. С. annularis Gill, 1859 - кольчатый дальневосточный бычок ....... 100
231. С. castaneus (O’Shaughnessy, 1875) - каштановый дальневосточный
бычок............................................................. 102
232. С. macrognathus (Bleeker, 1860) - большеротый бычок.............. 103
233. С. taranetzi Pinchuk, 1978 - дальневосточный бычок Таранца ...... 105
Род 114. Hyrcanogobius Iljin,1928 - гирканогобиусы
234. Н. bergi Iljin, 1928 - бычок Берга .............................. 107
Род 115. Knipowitschia Iljin, 1927 - бычки Книповича
235. К. caucasica (Berg, 1916) - бычок-бубырь......................... 108
236. К. longecaudata (Kessler, 1877) - длиннохвостый бычок Книповича .... 110
Род 116. Mesogobius Bleeker, 1874 - бычки-мартовики
237. М. batrachocephalus (Pallas,1814) - бычок-кнут, мартовик......... 112
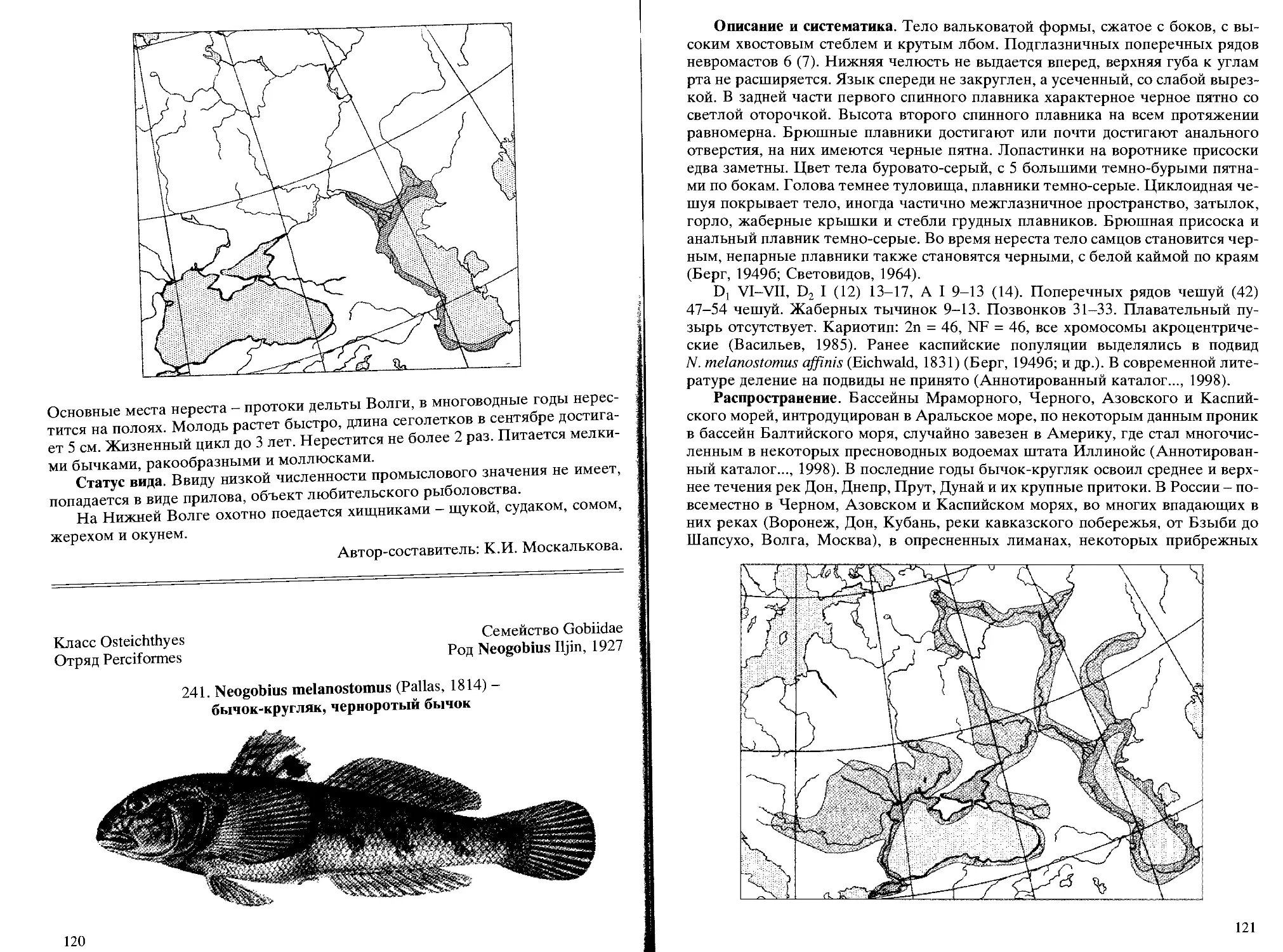
Род 117. Neogobius Iljin,1927 - черноморско-каспийские бычки
238. N. fluviatilis (Pallas,1814) - бычок-песочник..................... 114
239. N. gymnotrachelus (Kessler, 1857) - бычок-гонец ................. 117
240. N. iljini Vasiljeva et Vasiljev, 1996 - каспийский бычок-головач . 119
241. N. melanostomus (Pallas,1814) - бычок-кругляк, черноротый бы-
чок .............................................................. 120
242. N. platyrostris (Pallas,1814) - бычок-губан...................... 123
8
243- N. rhodioni Vasiljeva et Vasiljev, 1994 - речной бычок Родиона . 125
244. N. syiman (Nordmann,1840) - бычок-ширман ....................... 126
Род 118. Pomatoschistus Gill, 1864 - бычки-лысуны
245. P. marmoratus (Risso, 1810) - леопардовый лысун ................ 128
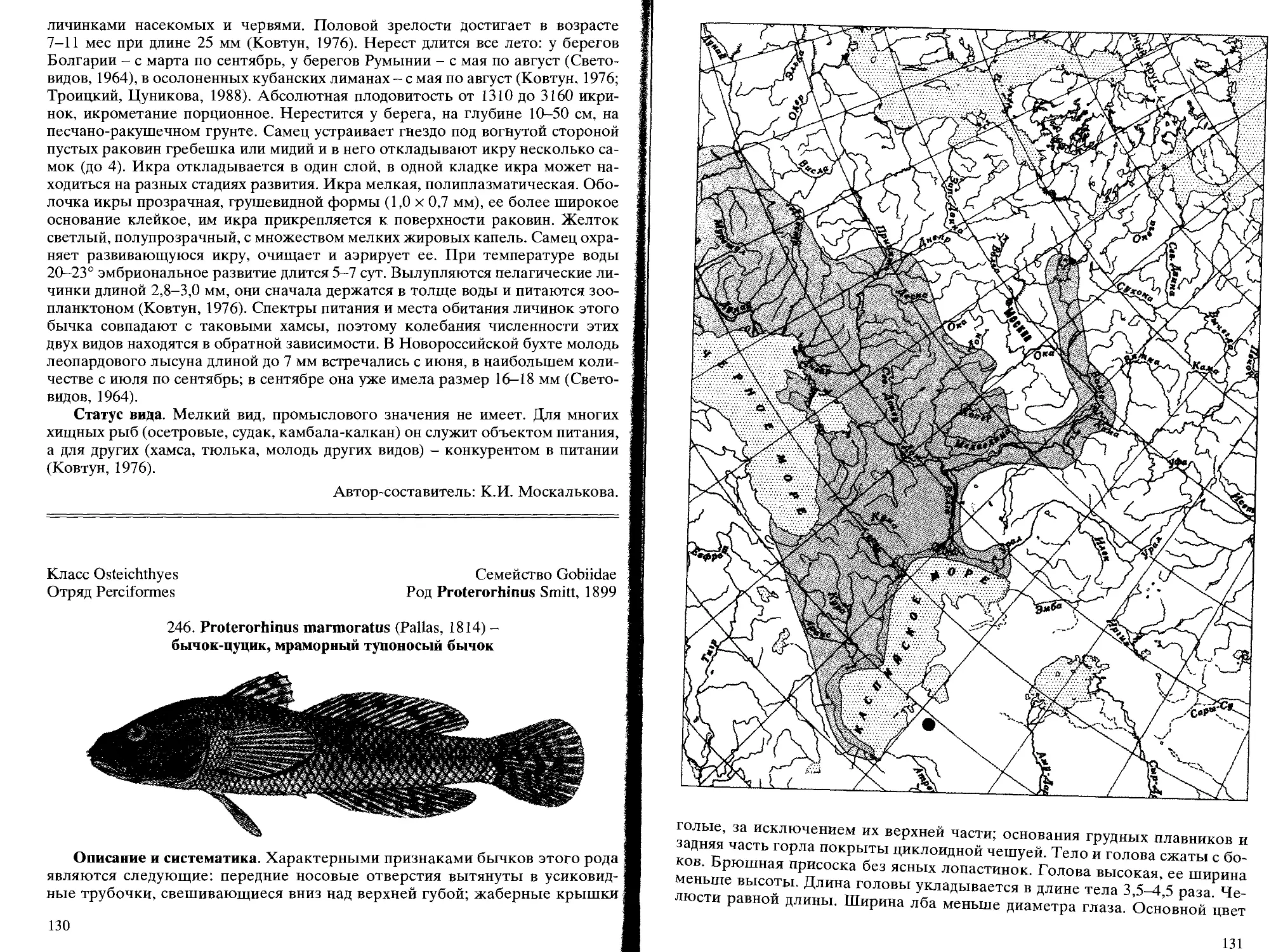
Род 119. Proterorhinus Smith,1899 - тупоносые бычки
246. P. marmoratus (Pallas,1814) - бычок-цуцик, мраморный тупоносый
бычок.......................................................... 130
Род 120. Rhinogobius Gill, 1859 - носатые бычки
247. R- brunneus (Temminck et Schlegek, 1845) - амурский бычок... 133
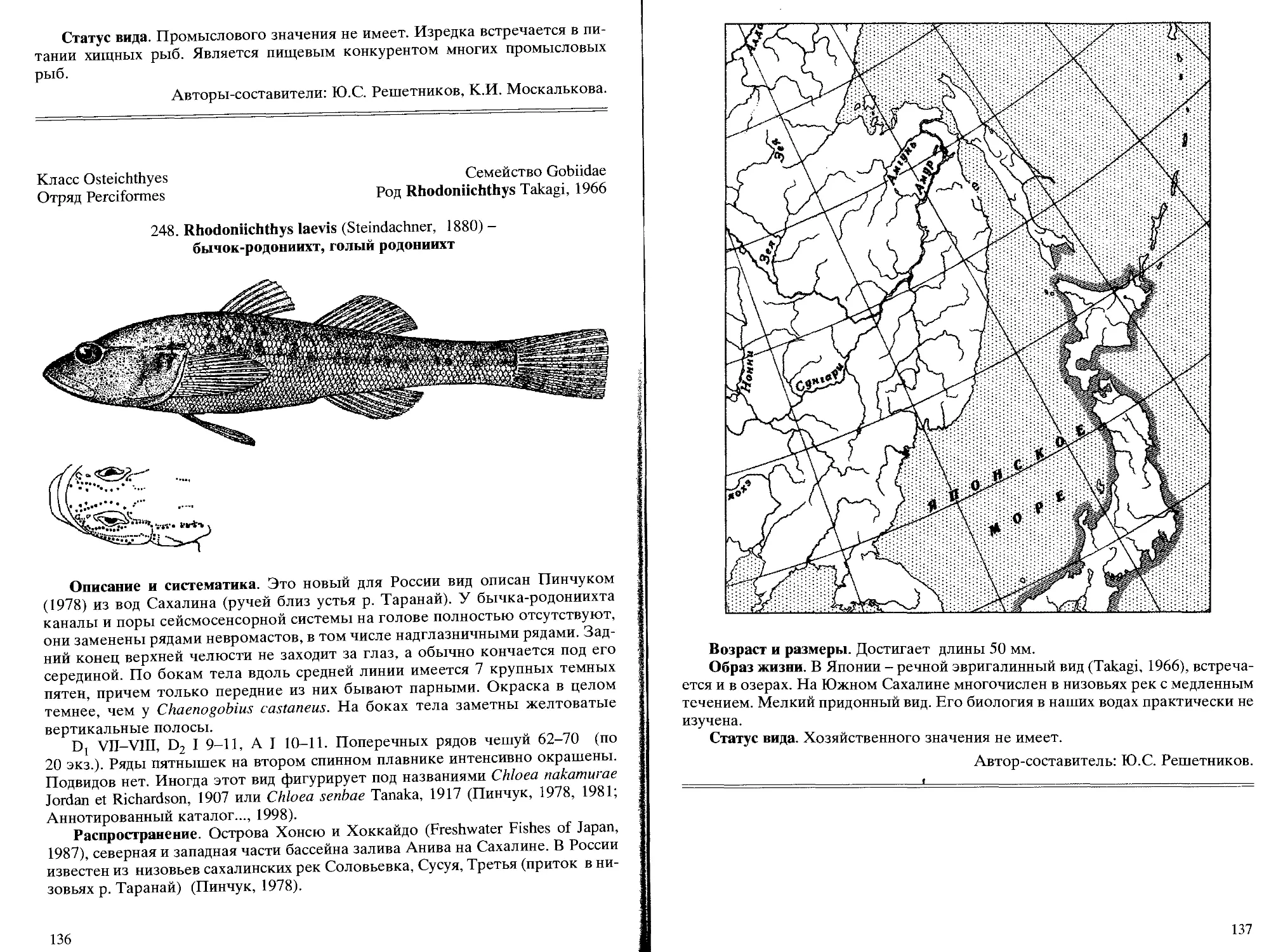
Род 121. Rhodonichthys Takagi, 1966 - бычки-родониихты
248. R. laevis (Steindachner, 1880) - бычок-родониихт, голый родониихт ... 136
Род 122. Tridentiger Gill,1858 - трехзубые бычки
249. Т. obscurus (Temminck et Schlegel, 1845) - темный трехзубый бычок 138
250. Т. trigonocephalus (Gill, 1858) - полосатый трехзубый бычок. 140
Подотряд CHANNOIDEI
Сем. 30. CHANNIDAE Berg, 1940 - Змееголовые
Род 123. Channa Scopoli, 1777 - змееголовы
251. С. argus (Cantor, 1842) - змееголов ............................ 141
Отряд XIII. SCORPAENIFORMES - СКОРОПЕНООБРАЗНЫЕ
Подотряд COTTOIDEI
Сем. 31. COTTIDAE Bonaparte, 1832 - Керчаковые
Род 124. Batrachocottus Berg, 1903 - большеголовые широколобки
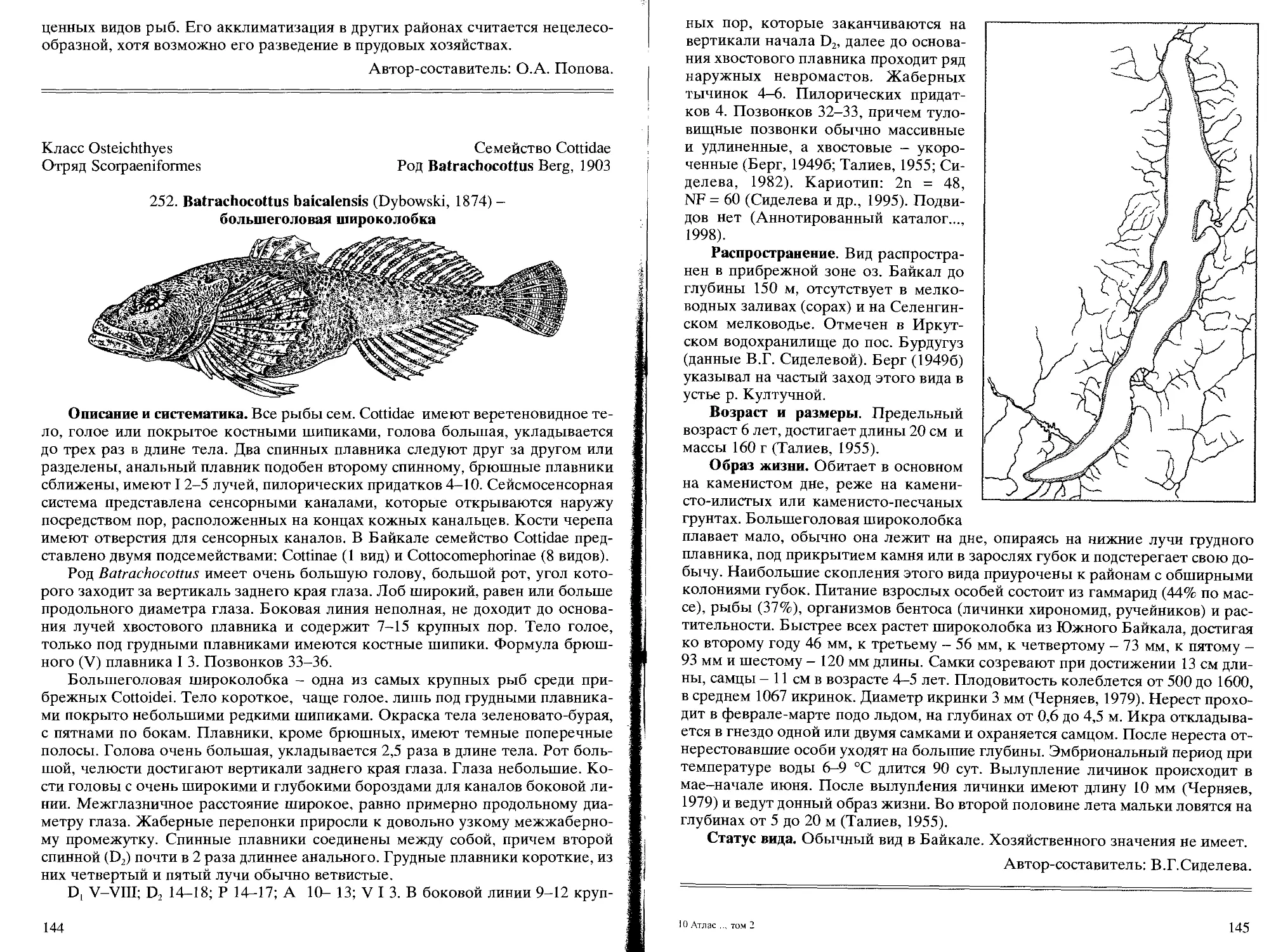
252. В. baicalensis (Dybowski, 1874) - большеголовая широколобка .... 144
253. В. multiradiatus Berg, 1907 - пестрокрылая широколобка ......... 146
254. В. nikolskii (Berg, 1900) - жирная широколобка.................. 147
255. В. talievi Sideleva, 1999 - широколобка Талиева................. 149
Род 125. Cottocomephorus Pellegrin, 1900 - желтокрылки
256. С. alexandrae Taliev, 1935 - северобайкальская желтокрылка.. 150
257. С. grewingkii (Dybowski, 1874) - желтокрылая широколобка ....... 152
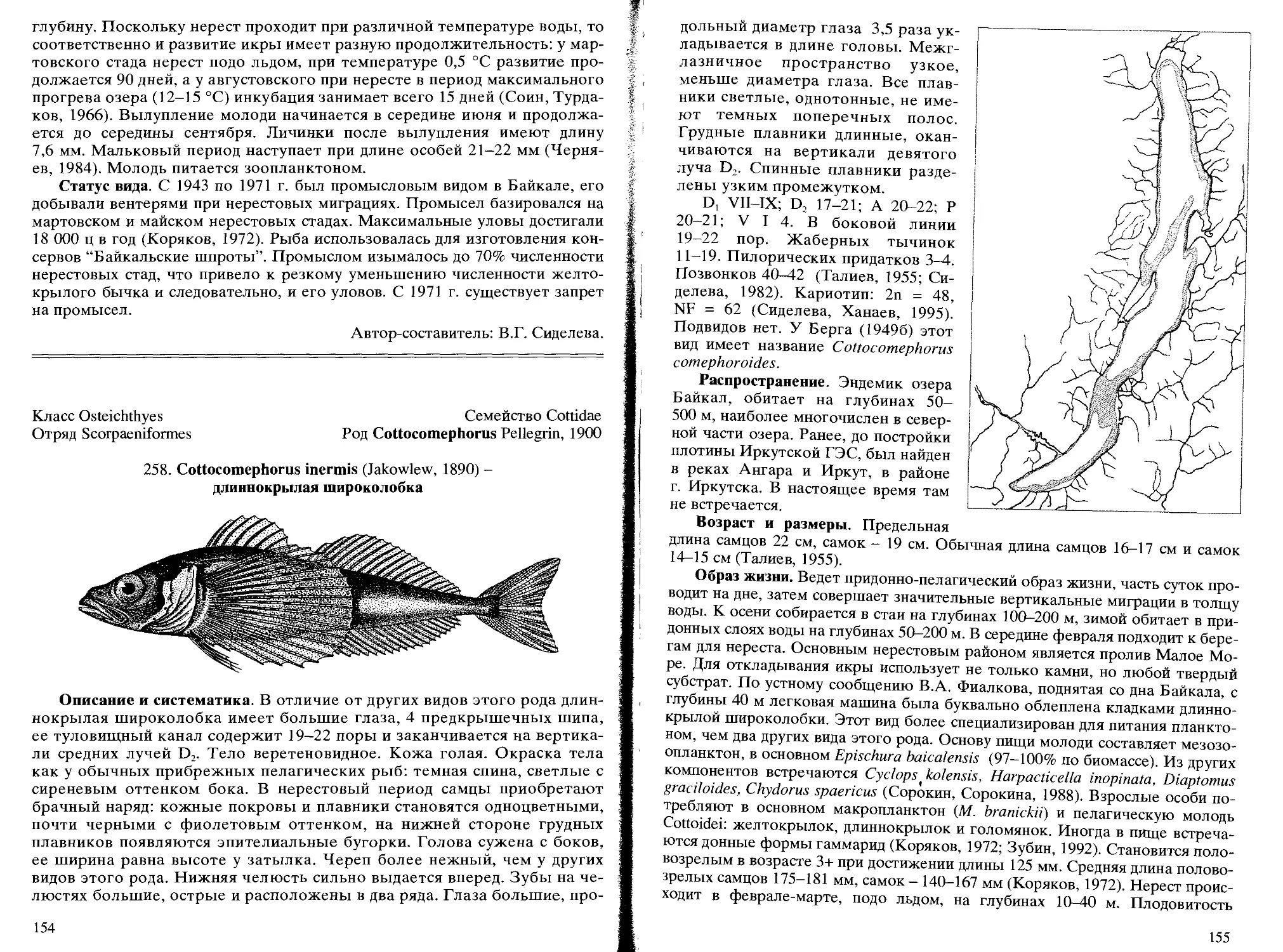
258. С. inermis (Jakowlew, 1890) - длиннокрылая широколобка...... 154
Род 126. Cottus Linnaeus, 1758 - подкаменщики
259. Cottus amblystomopsis Schmidt, 1904 - сахалинский подкаменщик ... 156
260. С. cognatus Richardson, 1836 - слизистый подкаменщик ........... 158
261. С. czerskii Berg, 1913 - подкаменщик Черского .................. 160
262. С. gobio Linnaeus, 1758 - обыкновенный подкаменщик.............. 162
263. С. hangiongensis Mori, 1930 - японский подкаменщик.............. 164
264. С. poecilopus Heckel, 1836 - пестроногий подкаменщик ........... 166
265. С. sibiricus Kessler, 1899 - сибирский подкаменщик.............. 168
Род 127. Leocottus Taliev, 1955 - песчаные широколобки
266. L. kesslerii (Dybowski, 1874) - песчаная широколобка ........... 170
Род 128. Mesocottus Gratzianow, 1907 - подкаменщики-широколобки
267. М. haitej (Dybowski, 1869) - амурская широколобка .............. 173
Род 129. Paracottus Taliev, 1949 - каменные широколобки
268. Р. knerii (Dybowski, 1874) - каменная широколобка............... 174
9
Род 130. Triglopsis Girard, 1851 - рогатки
269. T. quadricomis (Linnaeus, 1758) - четырехрогий бычок, рогатка. 177
Сем. 32. COMEPHORIDAE Bleeker, 1859 - Голомянковые
Род 131. Comephorus Lacepede, 1801 - голомянки
270. С. baicalensis (Pallas, 1776) - большая голомянка........... 179
271. С. dybowski Korotneff, 1905 - малая голомянка .............. 180
Сем. 33. ABYSSOCOTTIDAE Taliev, 1949 -
Глубоководные широколобки
Род 132. Abyssocottus Berg, 1906 - глубинные широколобки
272. A. elochini Taliev, 1955 - елохинская широколобка........... 182
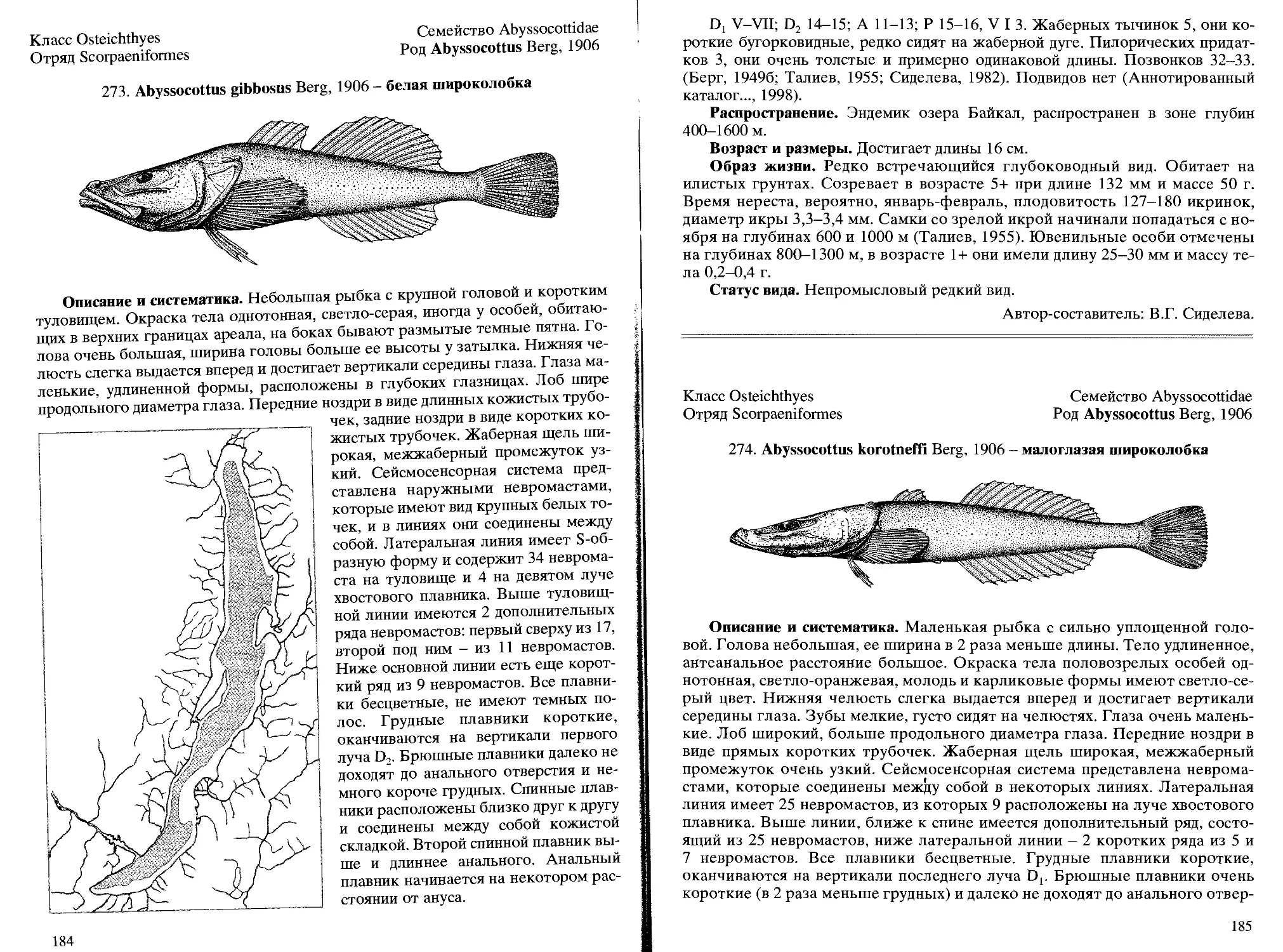
273. A. gibbosus Berg, 1906 - белая широколобка.................. 184
274. A. korotneffi Berg, 1906 - малоглазая широколобка .......... 185
Род 133. Asprocottus Berg, 1906 - шершавые широколобки
275. A. abyssalis Taliev et Korjakov, 1947 - глубоководная широколобка 187
276. A. herzensteini Berg, 1906 - шершавая широколобка Герценштейна 188
277. A. korjakovi Sideleva, 2001 - широколобка Корякова.......... 190
278. A. parmiferus Taliev, 1955 - панцирная широколобка.......... 191
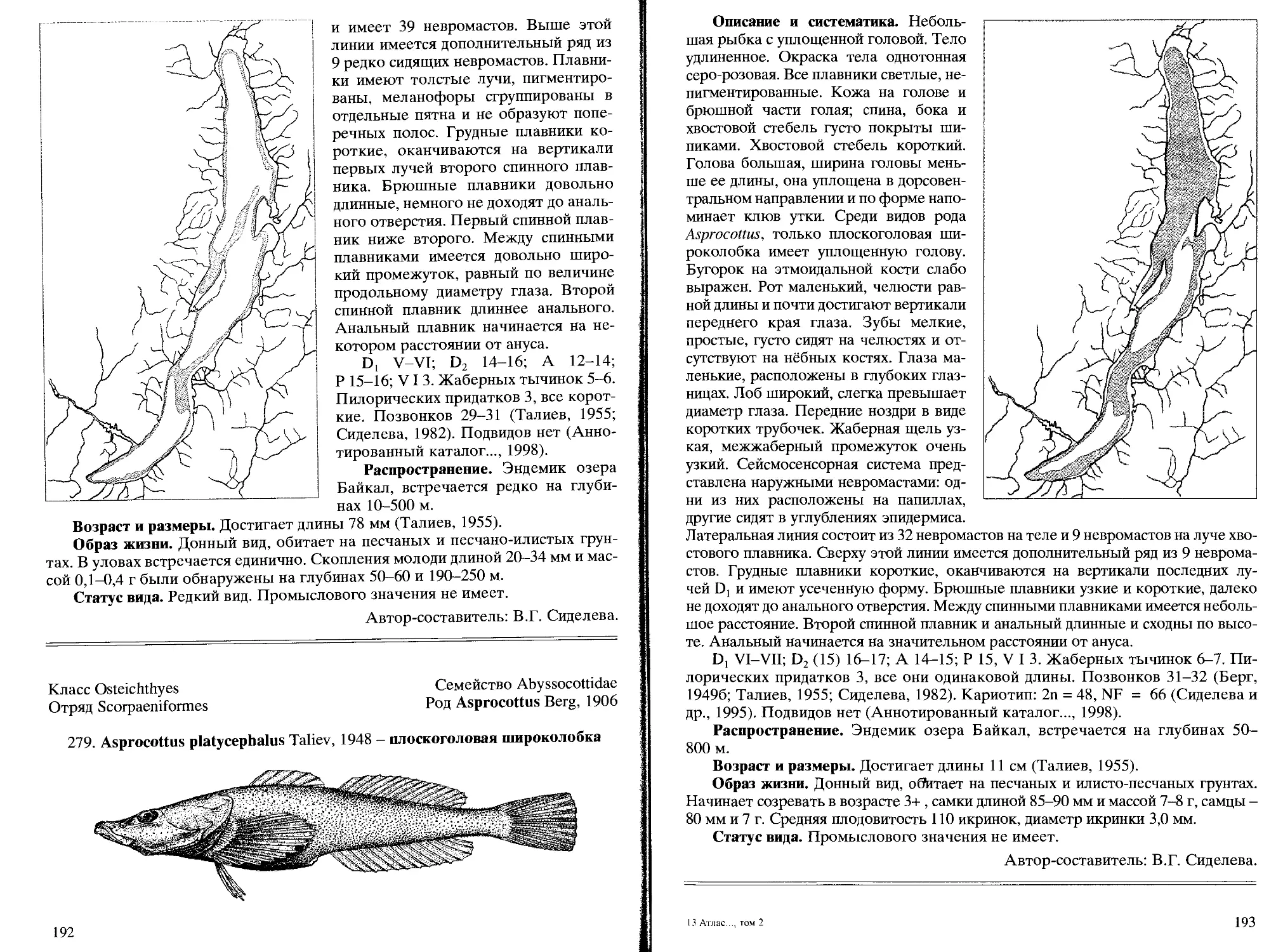
279. A. platycephalus Taliev, 1948 - плоскоголовая широколобка... 192
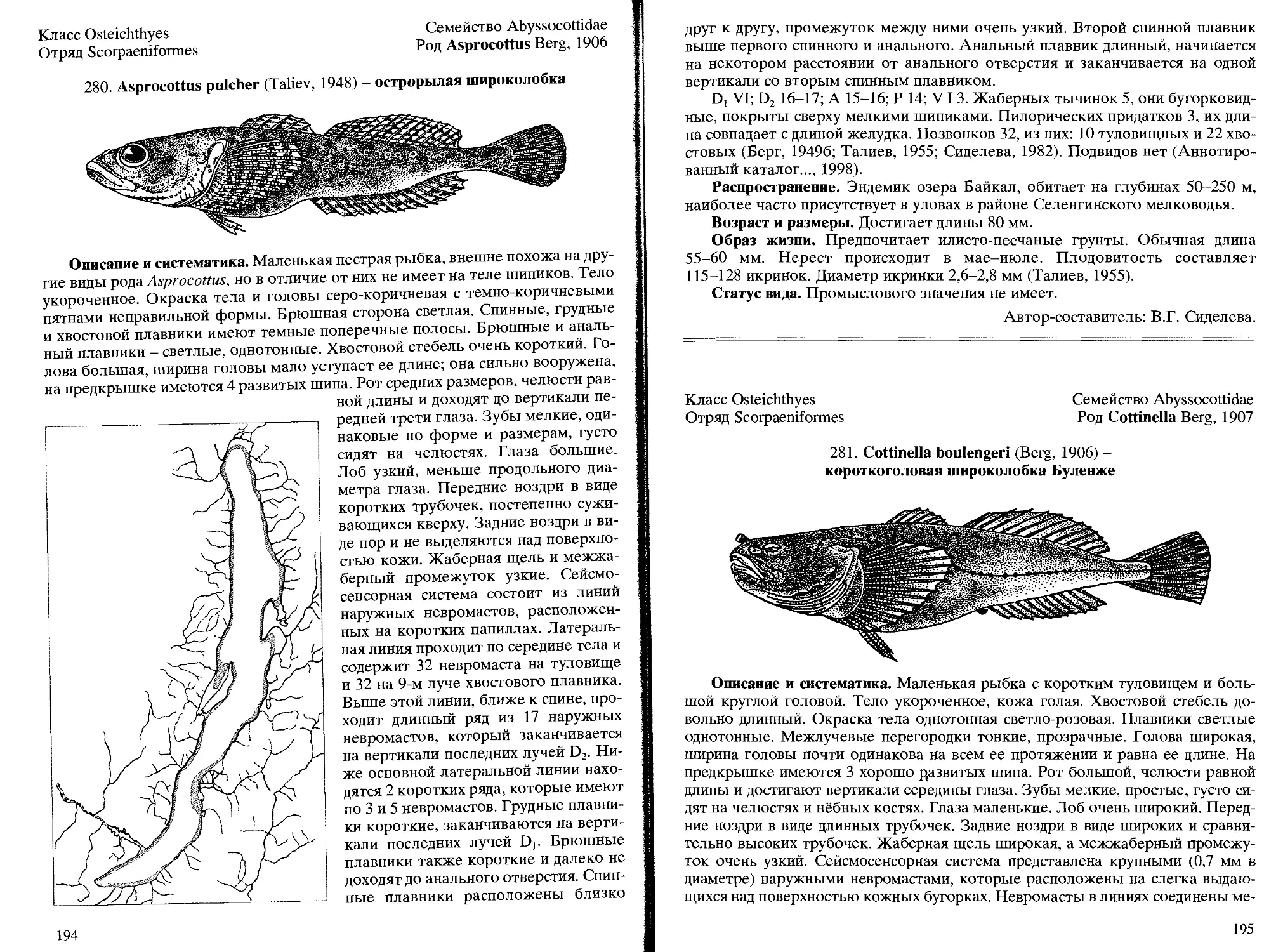
280. A. pulcher (Taliev, 1948) - острорылая широколобка ......... 194
Род 134. Cottinella Berg, 1907 - короткоголовые широколобки
281. С. boulengeri (Berg, 1906) - короткоголовая широколобка Буленже 195
Род 135. Cyphocottus Sideleva, 2001 - горбатые широколобки
282. L. eurystomus (Taliev, 1955) - широкорылая широколобка ..... 197
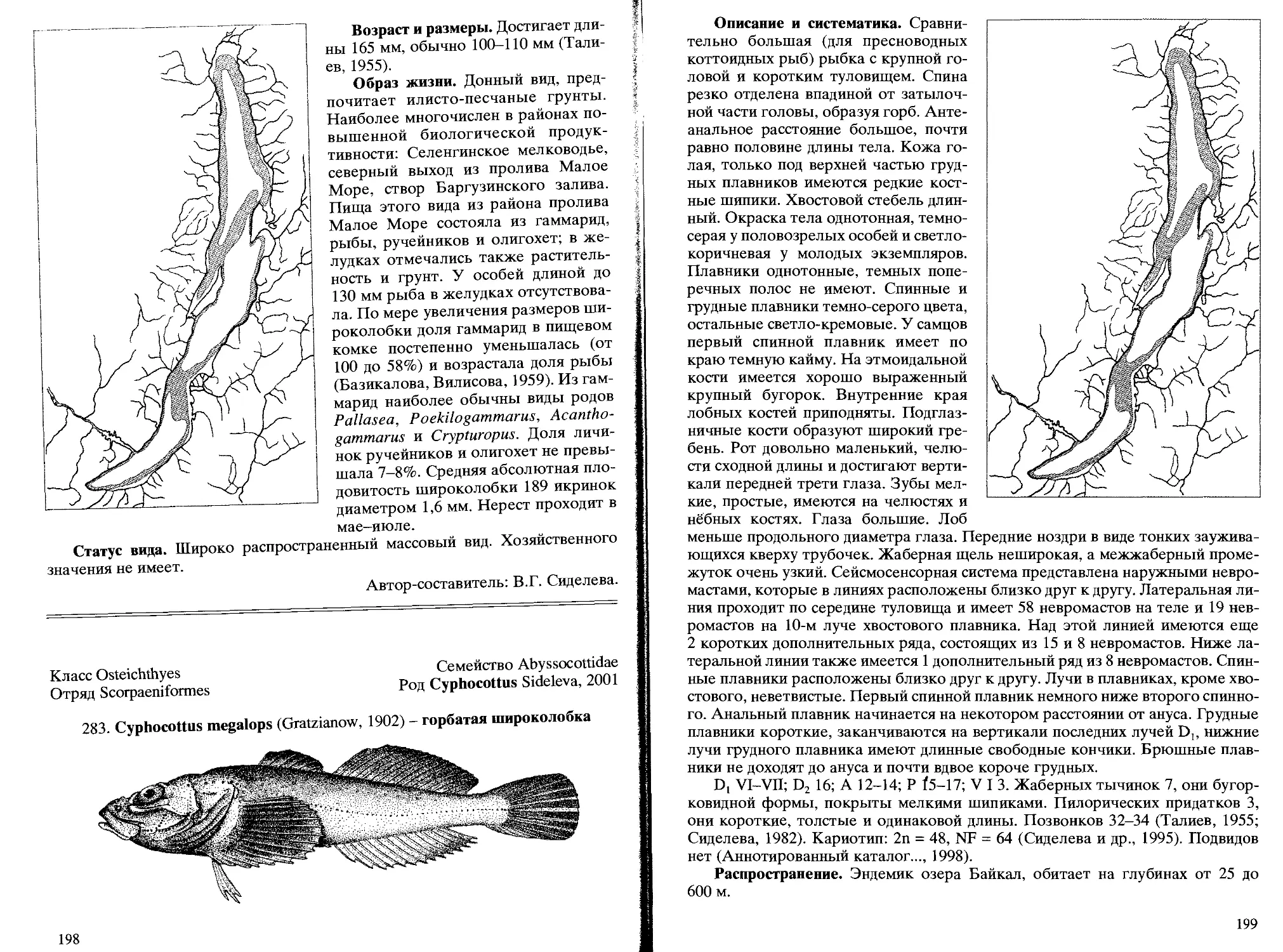
283. L. megalops (Gratzianow, 1902) - горбатая широколобка ...... 198
Род 136. Limnocottus Berg, 1906 - озерные широколобки
284. L. bergianus Taliev, 1935 - плоская широколобка............. 200
285. L. godlewskii (Dybowski, 1874) - крапчатая широколобка...... 202
286. L. griseus Taliev, 1948 - темная широколобка................ 203
287. L. pallidus Taliev, 1948 - узкая широколобка ............... 204
Род 137. Neocottus Sideleva, 1982-рыхлые широколобки
288. N. thermalis Sideleva, 2002 - тепловодная широколобка ...... 206
289. N. werestschagini (Taliev, 1935) - рыхлая широколобка....... 207
Род 138. Procottus Gratzianow, 1902 - красные широколобки
290. Р. gotoi Sideleva, 2001 - широколобка Гото ................. 209
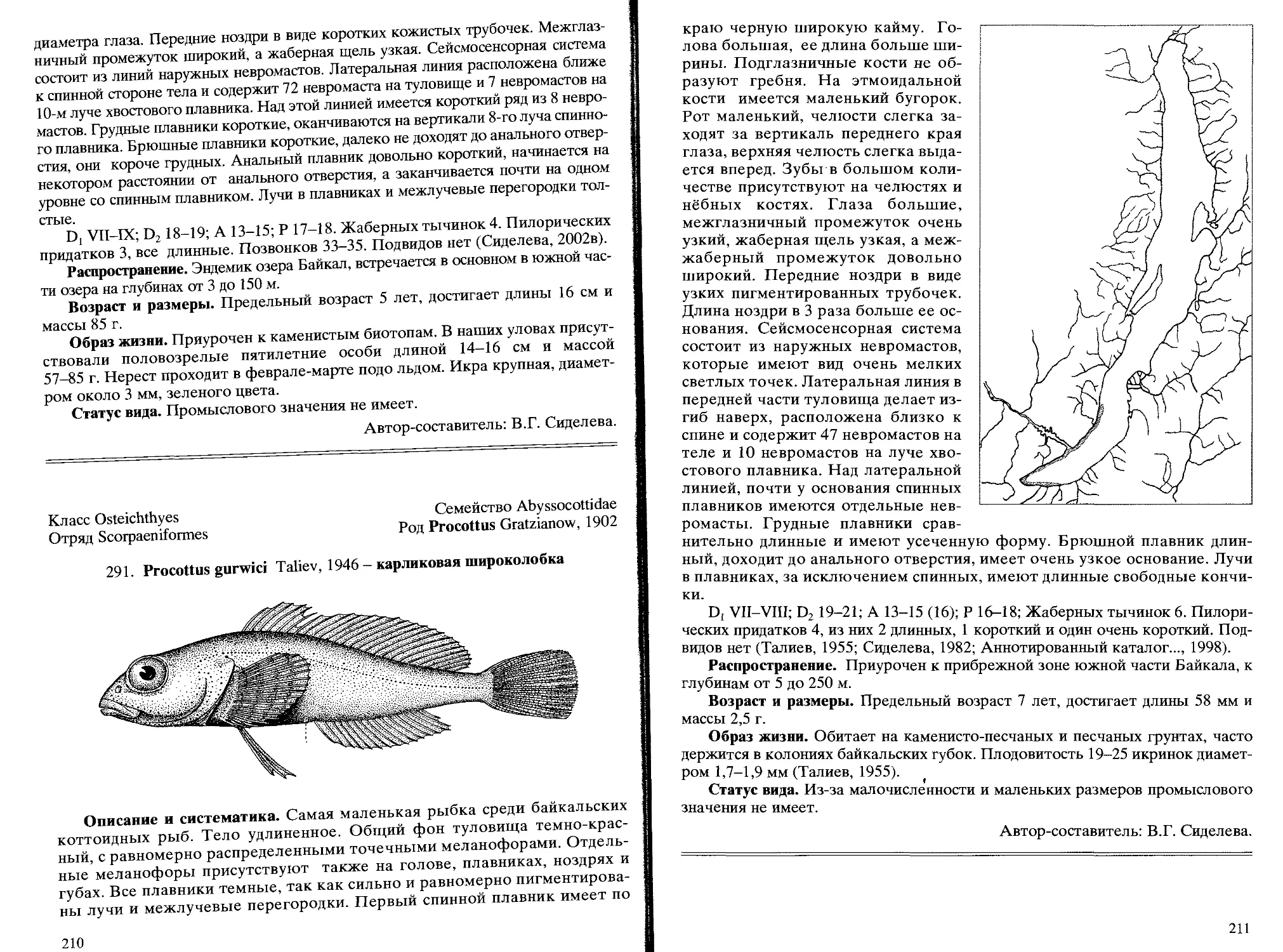
291. Р. gurwici Taliev, 1946 - карликовая широколобка............ 210
292. Р. jeittelesii (Dybowski, 1874) - красная широколобка ...... 212
293. Р. major Taliev, 1944 - большая широколобка................. 213
Класс Osteichthyes
Отряд Siluriformes
Семейство Siluridae
Род Parasilurus Bleeker, 1862
189. Parasilurus asotus (Linnaeus, 1758) - амурский сом
Описание и систематика. Более мелкий, чем сом Солдатова. Отличается
от него меньшими размерами головы (едва доходит до основания грудного
плавника) и тем, что нижняя челюсть доходит только до середины глаза.
У амурского сома 2 пары усиков: одна пара верхнечелюстных (заходят за ос-
нование грудного плавника) и две пары коротких нижнечелюстных, причем
задняя пара нижнечелюстных усиков имеется в зачаточном состоянии толь-
ко у молоди. Окраска тела оливково-зеленая или темно-серая, с расплывча-
тыми темными пятнами. Брюшко и брюшные плавники белые, остальные
плавники - темные. Колючий луч в грудном плавнике сильный, утолщенный
и сильно зазубрен по заднему краю. В отличие от рода Silurus есть анальная
папилла.
D 4-5, А (72) 75-84, V I 10-11. В боковой линии 63-70 пор. Жаберных ты-
чинок 8-14. Позвонков 60-63. Кариотип: 2п - 58, NF = 106, 100 (Берг, 1949а;
Никольский, 1956; Choi et al., 1990; Rab, Karakousis, Peios, 1994).
Распространение. Широко распространен в реках бассейнов Японского,
Желтого и Южно-Китайского морей. Населяет реки Китая от северо-восточ-
ного побережья до Гуаньчжоу на юге, водоемы Монголии (Онон, Керулен,
Халхин-Гол, оз. Буйр-Нур). Есть на Тайване, в Японии (кроме островов Хок-
кайдо и Рюкю), на п-ове Корея и во Вьетнаме (Берг, 1949а, Freshwater Fishes
of Japan, 1987; Choi et al., 1990). На российской территории широко распро-
странен в бассейне Амура, от верховьев (Ингода, Шилка, Онон, Аргунь,
оз. Кенон) до Амурского лимана, включая Сунгари, Уссури и оз. Ханка, и на
северо-западе Сахалина (Никольский, 1956). В 1932 г. вселен в оз. Шакша в
Забайкалье, откуда проник в бассейн Селенги, в Южный Байкал, Ангару и
Братское водохранилище, где широко распространился (Асхаев, 1958; Кара-
сев, 1987).
11
Возраст и размеры. В уловах встречаются особи длиной до 1 м и массой
6-8 кг в возрасте до 18 лет. Однако в большинстве водоемов его популяции со-
стоят из особей 8-10 возрастных групп, их основу составляют особи 4-6 лет
длиной до 60 см и массой 1,5-2,0 кг.
Образ жизни. Этот вид больше связан с придаточной системой и выходит в
русло реки при сильном падении уровня и промерзании придаточных водоемов.
Амурский сом ведет преимущественно ночной и сумеречный образ жизни, он
не является хищником-засадчиком, а активно отыскивает свою добычу. Сего-
летки потребляют беспозвоночных (личинок хирономид, поденок, водяных
клопов, моллюсков) и молодь рыб. Переход на полное питание рыбой начина-
ется со второго года жизни, уже с августа в их спектре питания появляется мо-
лодь 10 видов рыб. Пища взрослых особей - мелкие массовые виды рыб и мо-
лодь крупных видов, а также беспозвоночные и органические остатки. Размер
жертвы составляет 20% длины тела у молоди и 10-15% у взрослых рыб. Охо-
тится он чаще в одиночку, но иногда 15-30 особей объединяются в группу для
совместной охоты во время массового хода мелких видов рыб (корюшки).
В пище взрослого сома отмечено более 13 видов рыб. В годы высокого уровня
воды обеспеченность сомов кормом снижается. Амурский сом растет очень
медленно, достигая длины 13-15 см в годовалом и 65 см - в 10-летнем возрас-
те. Однако в ряде холодных водоемов с бедной кормовой базой такой длины он
достигает только к 17 годам (Никольский, 1956; Вронский, 1960). Полово-
зрелым становится в возрасте 3+ по достижении длины 40 см. Соотношение по-
лов на нерестилищах близко 1:1. Плодовитость колеблется в пределах
15,5-141,0 тыс. икринок в зависимости от возраста и размера самок. Нерест
происходит в теплые тихие вечерние часы с середины июня до середины июля
при температуре воды от 12-17 до 26 °C. Места нереста - густо заросшие тра-
вой прибрежные участки водоема с глубинами 60-80 см. Рыбы нерестуют по-
парно во время брачных игр. Амурский сом гнезд не строит и икру не охраня-
ет. Икра выметывается очень разрозненно, на большой площади и приклеива-
ется к субстрату. Икра зеленоватого цвета, с двумя оболочками; при этом на-
ружная очень толстая и студенистая. Диаметр икринок в ястыке 1,6-1,9 мм и
12
после вымета 3,7-5,2 мм за счет сильного разбухания наружной оболочки. Раз-
витие в икре продолжается 5-6,5 сут в зависимости от температуры. Выклю-
нувшиеся личинки имеют среднюю длину 7,25 мм. После вылупления они не-
надолго подвешиваются к растениям, а потом лежат на дне до исчезновения
желточного мешка (Крыжановский и др., 1951). На 8-9-е сутки при длине
9-10 мм молодь сома начинает активно питаться личинками рыб, молодью го-
ловастиков, крупным зоопланктоном, разлагающимися остатками водных жи-
вотных.
Статус вида. Ценный промысловый объект на всем протяжении Амура.
Благодаря высокой пластичности в питании амурский сом мало зависит от ко-
лебаний гидрологического режима и изменения кормовой базы, его числен-
ность стабильна, а уловы раньше постоянно составляли 0,4-2,0 тыс. т. в год.
В настоящее время уловы падают из-за снижения численности под влиянием
сильного промысла (использование глухих забоек и лов на нерестилищах). Вид
нуждается в охране.
Автор-составитель: О.А. Попова.
Класс Osteichthyes
Отряд Siluriformes
Семейство Siluridae
Род Silurus Linnaeus, 1758
190. Silurus glanis Linnaeus, 1758 -
обыкновенный (европейский) сом
Описание и систематика. Тело длинное, округлое в передней части и
сжатое с боков — в задней. Голова сильно сжата в дорсовентральном напра-
влении. Глаза маленькие, расположены ближе к затылочной части. Рыло
плоское, широкое, закругленное. Передние и задние ноздри далеко отстоят
друг от друга. Рот большой, нижняя челюсть длиннее верхней и чуть заги-
бается вверх. На челюстях и нёбе мелкие многочисленные зубы в виде ще-
ток. На верхней губе одна пара,длинных усиков, доходящих до конца груд-
ного плавника. На подбородке имеются 2 пары коротких усиков, причем пе-
редние в 3 раза короче задних. Окраска тела почти черная на спине, темно-
зеленая с пятнами по бокам и грязно-белая - на брюхе. В открытых проточ-
ных водоемах сом окрашен светлее, а в заросших с медленным течением во-
доемах он почти весь черный. Спинной плавник очень короткий, сильно
сдвинут к голове и расположен над грудными плавниками. Анальный очень
длинный, невысокий, соединен с хвостовым. В грудном плавнике наружный
13
луч превращен в колючку, иногда с зазубринами. Хвостовой плавник за-
кругленный. Тело голое, с мягкой кожей и большим количеством слизи-
стых желез. Жаберные перепонки спереди прикреплены к межжаберному
промежутку.
D 3-5; А 77-92 (в бассейне Арала 70-108); Р I 14-17; V I (9) 11-13 (14).
Жаберных тычинок 10-17. Позвонков 67-74. Кариотип: 2п = 60, NF = 120.
14
Подвидов нет (Берг, 1949а; Аннотированный каталог..., 1998; Rab et al.,
1991).
Распространение. Широко распространенный вид в водоемах Балтийско-
го Черного, Азовского, Каспийского и Аральского морей. В европейской час-
ти ареал сома простирается от Рейна и Дуная на западе до Урала на востоке,
есть в Боденском озере. Северная граница ареала сома проходит по 60° с.ш., по
югу Швеции и Финляндии, а южная доходит до берегов Черного и Эгейского
морей. Естественный ареал за пределами России в Азии - в бассейне Арала с
реками Сырдарья; Амударья и Сарысу. Позже проник в Каракумский канал, в
озера Сарыкамыш и Балхаш и в реки Чу, Или. В России сом обитает в бассей-
не Невы (озера Ладожское, Онежское, Ильмень), Днепра, Дона, Кубани, Тере-
ка, Волги и Урала. Южная граница проходит по рекам Северного Кавказа.
В водоемах Ледовитоморского бассейна сом отсутствует. Особенно многочис-
лен сом в дельтовых районах крупных рек (Волга, Дон, Днепр) и в водохрани-
лищах с замедленным стоком (Волгоградское, Куйбышевское, Саратовское). В
верховьях рек редок и малочислен.
Возраст и размеры. Известны максимальные размеры сома до 5 м длиной
и массой 300 кг (Берг, 1949а). В настоящее время встречаются особи длиной не
более 2,5 м и массой 150 кг в возрасте до 30 лет (р. Урал), средние же размеры
в большинстве водоемов 70-150 см и 5-50 кг в возрасте до 15 лет.
Образ жизни. На всем ареале распространена жилая форма сома, хотя в ни-
зовьях крупных рек он использует для нагула и мелководные морские пастби-
ща, прилегающие к дельтам. Больше связан с водоемами речной системы, хо-
тя есть в некоторых озерах и водохранилищах, предпочитая хорошо прогрева-
емые водоемы со слабым течением. Миграции его весьма ограничены, а места
нагула и нереста сближены. Зимуют сомы большими стаями на глубоких ямах,
прекращая питаться до весны. Сом - крупный хищник, ведущий малоподвиж-
ный придонный образ жизни в тихих захламленных участках водоемов, у обры-
вистых берегов, среди коряг или подмытых корней деревьев. Чаще охотится в
одиночку как типичный хищник-засадчик, коротким броском схватывая близ-
ко проплывающую жертву. Но иногда образует большие группы на мелково-
дье, в местах интенсивного хода или ската молоди рыб (в дельтовых протоках);
здесь сомы стоят против течения и заглатывают молодь рыб целыми стайками
(Фортунатова, Попова, 1973). Сомы питаются преимущественно в сумерках,
ориентируясь при поиске жертвы усиками и всей кожей на колебания воды.
Пищу молодых сомов составляют мизиды, личинки хирономид, веснянки, водя-
ные клопы и жуки, пиявки, моллюски, головастики, а также личинки и молодь
рыб. Рыба появляется в питании сеголетков 4 см длиной, а при достижении
12-13 см рыба становится основной пищей сома, ее дополняют крупные насе-
комые (саранча, медведка), раки, лягушки, грызуны и даже птицы. В пресных
водоемах взрослые сомы питаются в основном придонными видами рыб, кото-
рые наиболее многочисленны в данном районе - карповыми, окуневыми, быч-
ковыми, молодью осетровых; в ^орских водах и в волжских водохранилищах -
сельдевыми (килькой, тюлькой) (Фортунатова, Попова, 1973). Относительный
размер жертвы у взрослых сомов составляет 10-30%. Растет сом быстро, осо-
бенно в южных дельтовых участках рек: в дельте Волги и Урала к концу пер-
вого года жизни он достигает длины 35^15 см и массы 300-600 г, а к 10 годам -
125 см и 15-18 кг. В то же время в Аральском море годовалые особи сома ед-
ва достигают 19 см, а 10-летние - только 95 см. В связи с этим и время наступ-
ления половой зрелости в дельте Волги бывает в возрасте 3-5 лет при длине
15
55-65 см и массе 1,5-2,5 кг, а в Арале - в 4-6 лет при длине 60-70 см и массе
2,5-3,5 кг. Нерестится сом на юге (Днепр, Волга) с марта-апреля до июня, а на
севере ареала - в мае-августе при температуре воды 17-23 °C. Обычно у самок
в гонадах имеются две порции икры, нерест чаще порционный, но в холодных
водоемах выметывается только одна порция икры (вторая остается как резерв
следующего года или рассасывается). Диаметр икринок первой порции -
1,7-3,0, второй - 1,0-1,6 мм. Плодовитость колеблется от 11 до 900 тыс. икри-
нок в зависимости от размера и возраста самок. Перед нерестом самец строит
гнездо на глубине 30-70 см в виде округлой плоской ямки со стенками из рас-
тительности (Константинов, 1941). Икра клейкая, приклеивается ко дну или
стенкам гнезда и охраняется самцом. Развивается икра быстро, через 2,5-3 сут
вылупляются предличинки длиной 7-7,5 мм и первое время держатся в гнезде,
приклеиваясь к его стенкам. Через 5-6 сут после рассасывания желточного
мешка личинки длиной 12-15 мм уходят из гнезда и переходят на внешнее пи-
тание.
Статус вида. Ценный промысловый вид. Его уловы в ряде водоемов в пред-
военные годы достигали 1,2-А,2 тыс. т, но в последние годы снизились. Из-за
интенсивного промысла и браконьерского лова повсеместно упала числен-
ность сома, в большинстве водоемов он стал редким и нуждается в охране. За-
несен в “Красную книгу Карелии” (1995) как редкий вид.
Автор-составитель: О.А.Попова.
Класс Osteichthyes
Отряд Siluriformes
Семейство Siluridae
Род Silurus Linnaeus, 1758
191. Silurus soldatovi Nikolsky et Soin, 1948 - сом Солдатова
Описание и систематика. Окраска тела и размеры схожи с таковыми у ев-
ропейского сома. Спина и бока - коричнево-серые, с темными расплывчаты-
ми разводами. Брюшко светлое. Плавники темные. У молоди сома Солдатова
окраска тела значительно темнее, чем у молоди амурского сома. Боковая ли-
ния четкая, светлая. По сравнению с европейским сом Солдатова имеет более
крупную голову и более массивную нижнюю челюсть. Рот верхний, очень ши-
рокий, нижняя челюсть заходит за вертикаль заднего края глаза. Усов 3 пары,
но верхнечелюстные короче, чем у европейского сома, они доходят лишь до
грудных плавников. Передние нижнечелюстные усики длиннее задних. На
16
концах верхнечелюстных усов есть гроздевидные железки красноватого цвета
с большим количеством чувствительных клеток. В грудном плавнике первый
луч очень слабый и незазубренный по наружному краю.
D 6, А 83-90. Жаберных тычинок 13-16. Подвидов нет ( Никольский, 1956;
Аннотированный каталог..., 1998).
Распространение. Известен только из бассейна Амура. В российской части
Амура распространен от Благовещенска до устья, а также в Уссури, в оз. Хан-
ка и в Сунгари с ее притоками. Отмечались флуктуации численности и измене-
ния ареала в связи с ними.
Возраст и размеры. В 1930-е годы встречались особи более 3 м длиной.
В уловах 1950-х годов максимальная длина была 115-123 см и масса до 40 кг в
возрасте 8-9 лет. Основу уловов составляли рыбы 65-90 см 4-5-летнего
возраста.
Образ жизни. Встречается чаще в русле Амура. В придаточную систему
заходит в годы с низким уровнем воды и промерзанием зимовальных ям в ос-
новном русле (в 1950-е годы). Придерживается более глубоких мест, чем
амурский сом, избегает прибрежных мелководных участков. Питание моло-
ди не изучено. Пищу взрослых рыб составляют в основном взрослые караси
2 Атлас ... том 2
17
и молодь сазана, пятнистого коня, амурской щуки, а также колючий горчак и
другие виды, т.е. преимущественно рыбы придонных слоев - одного биотопа
с сомом Солдатова. Дополнительной пищей были монгольский краснопер,
амурский подуст, чебак, косатка-скрипун, косатка Бражникова, а также по-
лоз и серая крыса. Размер жертвы у этого вида больше, чем у амурского со-
ма: у молоди - 32%, а у взрослых рыб - 15-27% длины тела. Зимой не пита-
ется. Весной откорм начинается еще подо льдом (апрель), и максимальный
откорм приходится на июль-август. Кормится в придаточной системе Амура,
а зимует в самом русле. Питание молоди не известно. Растет сом Солдатова
быстро: в возрасте 1 года - 21-29 см, 2 лет - 42-47 см, 3 лет - 53-64 см и
8 лет - 115-118 см. Размножение не изучено. Рыбы в возрасте 3+ были еще
неполовозрелыми (Никольский, 1956).
Статус вида. Ценный промысловый вид, но в настоящее время немногочис-
лен и в промысле отдельно не учитывается (обычно идет вместе с мелким
амурским сомом). Уловы составляли от нескольких экземпляров до 10 т в год.
В некоторых частях ареала редок. Нуждается в охране.
Автор-составитель: О.А. Попова.
Класс Osteichthyes
Отряд Siluriformes
Семейство Bagridae
Род Leiocassis Bleeker, 1858
192. Leiocassis brashnikowi (Berg, 1907) - косатка Бражникова
Описание и систематика. Все косатковые в отличие от сомовых рыб име-
ют жировой плавник и сильную колючку в спинном плавнике. Тело прогони-
стое с длинным и тонким хвостовым стеблем, только впереди на боках по-
крыто мелкими ворсинками. Хвостовой плавник с глубокой выемкой и заост-
ренными лопастями. Спина лилово-серая или синевато-серая, брюхо светлое.
На боках на светло-желтом фоне неправильные темные пятна, не заходящие
на хвостовой плавник. У всех плавников темная оторочка по краю. Основа-
ние жирового плавника длиннее основания спинного, но много короче осно-
вания анального. Колючка спинного плавника мощная, сильно зазубрена и по
длине равна грудной. Грудная колючка с зазубринами изнутри и гладкая с
внешней стороны. Голова и глаза покрыты толстой кожей, у глаз векообраз-
ной складки нет, глаза крупные. Имеется 4 пары усиков: 1 пара у задних ноз-
дрей, 1 пара - на верхней челюсти и 2 пары на нижней. Носовые усики дохо-
18
дят до середины глаза, верхнечелюстные не достигают грудного плавника,
нижнечелюстные - короткие. Рот маленький, верхняя челюсть значительно
выдается над нижней. Губы толстые. Зубы на челюстях и на нёбе очень мел-
кие, густо сидящие.
D I 6-7; А 20-27; Р I 6; V I 6. Жаберных тычинок 9-12. Позвонков 34-40
(Берг, 1949а; Никольский, 1956).
Распространение. Встречается в нижнем и среднем течении Амура от
Благовещенска и до лимана. Много в Сунгари, Уссури и оз. Ханка. Вне бас-
сейна Амура не встречается (Никольский, 1956; Аннотированный ката-
лог..., 1998).
Возраст и размеры. Предельный возраст 6 лет, длина до 22 см, в промысло-
вых уловах чаще 8-19 см. Самцы крупнее самок.
Образ жизни. Этот вид те<^но связан с озерами придаточной системы
Амура. Летом обитает в озерах и протоках с медленным течением; в русле
равнинной части Амура только в местах с почти полным отсутствием тече-
ния. Зимой концентрируется в протоках, редко в русле. Взрослые особи пи-
таются личинками хирономид. Им свойствен узкий спектр питания (стенофа-
гия), но в разных местах доминируют разные кормовые объекты: в русле —
ручейники, в озерах - крупные личинки и куколки хирономид. Питание ры-
бой не отмечено. Самки мельче самцов, но их больше в уловах (соотношение
19
полов 60 : 40). Половозрелым становится в возрасте 2+ при длине 12 см. Пло-
довитость 1000-2700 икринок, размер икры с оболочкой 2,3 мм в диаметре.
Нерест порционный, в начале июля, в русле реки, на участках с тихим тече-
нием у обрывистого берега на глубине 0,6-1,0 м. Икра приклеивается к кор-
ням подмытой наземной растительности. Оболочка икринки инкрустируется
песчинками. Вылупление через 2,5-3 дня при длине 5,3 мм. Сначала органа-
ми дыхания является кровеносная система желточного мешка, но уже у
4-дневных личинок появляются дефинитивные жабры, покрытые жаберной
крышкой. Первое время личинки светобоязливы и прячутся на дне, вблизи
мест нереста. На 7-й день при длине 8,9 мм рассасывается желточный ме-
шок, и личинки начинают активное питание. Молодь питается пелагически-
ми ракообразными.
Статус вида. Хотя численность в некоторых местах велика, но из-за малых
размеров специального промысла не ведется. Используется только на тук. Ма-
лоценный вид и местами даже вреден, так как забивает сети, зацепляясь плав-
никовыми колючками.
Автор-составитель: О.А. Попова.
Класс Osteichthyes
Отряд Siluriformes
Семейство Bagridae
Род Leiocassis Bleeker, 1858
193. Leiocassis herzensteini (Berg, 1907) - косатка Герценштейна
Описание и систематика. Тело прогонистое, менее высокое, чем у косатки
Бражникова, спереди покрыто мелкими кожными ворсинками. Окраска одно-
тонная: спина зеленовато-серая, бока и брюхо - светлее, пятен нет. Окраска
плавников соответствует их положению на теле. Хвостовой стебель длинный,
тонкий. Хвостовой плавник слабовыемчатый, лопасти закруглены. Длина ос-
нования жирового плавника равна или больше основания анального. Спинная
колючка сильная, сзади слабо зазубрена и короче грудной колючки. Колючка
в грудном плавнике сильно зазубрена на внутренней стороне и гладкая на
внешней. Голова и глаза покрыты толстой кожей, векообразной складки нет.
Глаза маленькие. Носовые усики не доходят до середины глаз, верхнечелюст-
20
ные далеко не доходят до основания грудного плавника, так же как и нижнече-
люстные. Рыло тупое. Рот нижний, верхняя челюсть сильно выдается над ниж-
ней. D I 6-7; А III 15-17 (всего 18-20 ); Р I 6; V I 6 (Берг, 1949а; Никольский,
1956).
Распространение. Эндемик Амура. Встречается в бассейне его верхнего и
среднего течения (реки Онон, Шилка и Комиссаровка в бассейне оз. Ханка).
Сомнительно нахождение в р. Селемдже (притоке Зеи) (Никольский, 1956; Ка-
расев, 1987; Аннотированный каталог..., 1998).
Возраст и размеры. Предельная длина до 45 см, в уловах 11 -45 см.
Образ жизни. Живет в реках предгорного типа на быстром течении и в бо-
лее прозрачной воде, чем в основном русле Амура в равнинной зоне. В питании
преобладают личинки поденок, веснянок, хирономид, водяные клопы и молодь
рыб. Нерест в начале лета, характер и места нереста неизвестны, как и эмбрио-
генез.
Статус вида. Редкий вид.
Автор-составитель: О.А. Попова.
21
Класс Osteichthyes
Отряд Siluriformes
Семейство Bagridae
Род Leiocassis Bleeker, 1858
194. Leiocassis ussuriensis (Dybowski, 1872) -
косатка-плеть, уссурийская косатка
Описание и систематика. Тело сильно вытянуто в длину, особенно хво-
стовой стебель, в передней части покрыто короткими ворсинками. Окраска
обычно однотонная, желтовато-серая, на спине темнее, чем на брюхе. По
краю плавников темная оторочка. Хвостовой плавник слабовыемчатый,
длинный. Колючка в спинном плавнике сильная, сзади зазубрена, по своей
длине равна колючке в грудном плавнике. Колючка в грудном зазубрена
только на внутренней стороне, а снаружи гладкая. Голова и глаза покрыты
толстой кожей, векообразующей складки нет. Глаза маленькие. Усики не-
большие: носовые едва доходят до середины глаза, верхнечелюстные дале-
ко не доходят до основания грудного плавника, нижнечелюстные еще коро-
че и не достигают даже межжаберного промежутка. Верхняя челюсть не-
сколько длиннее нижней. Полоска зубов на нёбе посредине резко сужается.
Половые различия выражены сильнее, чем у других косаток: у самца длин-
нее тело, хвостовой стебель и анальный плавник, но меньше высота тела и
величина глаза.
DI 7; А (14) 17-21; РI 6-8; V15. В боковой линии 73-94 пор. Жаберные ты-
чинки длинные, тонкие, их 10-13 (14). Позвонков 47^19 (Берг, 1949а; Николь-
ский, 1956; Карасев, 1987; Choi et aL, 1990).
Распространение. Обитает в Северном Китае (до низовья Хуанхэ и Цзина-
ня), на п-ове Корея и в России, в бассейне Амура (Никольский, 1956; Choi et aL,
1990). В России она встречается по всему течению Амура, вниз до Сухановки.
Есть в низовьях Онона, в Ингоде, Зее, много в Уссури, Сунгари и в оз. Ханка
(Никольский, 1956).
Возраст и размеры. Л.С. Берг (1949а) указывает максимальную длину до
1 м, что сомнительно, так как в уловах и даже в ископаемых остатках в России
нет рыб более 55 см (Никольский, 1956); на п-ове Корея - более 50 см (Choi et
al., 1990). Предельный возраст 10 лет. Самцы живут дольше самок.
Образ жизни. В бассейне Амура, очевидно, существует несколько обособ-
ленных стад. Этот вид больше привязан к русловой части реки, в озера не за-
ходит даже на нагул. В русле Амура и в протоках держится на участках со сред-
ним течением или в прибрежье. Значительных миграций не совершает. Пита-
ется в течение всего года, хотя зимой и менее интенсивно. В реке основной
22
корм - личинки поденок и ручейников, в придаточной системе - личинки хиро-
номид, моллюски и водяные клопы. Летом поедает муравьев, майских жуков и
других насекомых; в оз. Ханка потребляет мизид. Молодью рыб питается неча-
сто (20% по встречаемости). Растет амурская косатка быстрее других видов: в
1-й год достигает 9-12 см, во 2-й - 14-17 см, в 3-й - 18-25 и к 10 годам выраста-
ет до 46-51 см. Половая зрелость наступает в возрасте 3+ и 4+ при длине самок
20 см и самцов 25-26 см. В период нереста соотношение полов близко 1:1, хо-
тя в обычное время самцов больше (вероятно, самки ведут более оседлый об-
раз жизни). Плодовитость самок длиной 27-32 см 3960-9260 икринок; икринки
диаметром 2 мм, более крупные, чем у других косаток. Вся икра выметывает-
ся в один прием, нерест с середины июня при температуре воды 19-23 °C. Сна-
чала нерестуют более крупные особи. Предположительно самец роет нору и
охраняет икру. Личинки в середине июля имели длину 10-13 мм, а мальки в
конце июля - 15-19 мм. Они питаются зоопланктоном и бентосом (Николь-
ский, 1956).
Статус вида. Второстепенный промысловый вид, в уловах среди косаток за-
нимает второе место после косатки-скрипуна, но обычно в виде прилова при
промысле других более ценных видов рыб. Из-за колючек и большого отхода
считается малоценным пищевым продуктом.
Автор-составитель: О.А. Попова.
23
Семейство Bagridae
Род Mystus Scopoli, 1777
Класс Osteichthyes
Отряд Siluriformes
195. Mystus mica Gromov, 1970 - косатка-крошка
Описание и систематика. Тело короткое, вальковатое. Окраска тела
одинаковая у самцов и самок, на желтом фоне темные полосы. На голове
сверху идут продольные полосы от конца рыла до затылка. Поперечная
широкая полоса соединяет основания грудных плавников и основание спин-
ного. От нее назад вдоль тела идут 3 темных полосы: по спине, вдоль боко-
вой линии до хвоста, разветвляясь на хвостовые лопасти, и от основания
грудного косо вниз до ануса. Анальный плавник короткий. Основание жи-
рового плавника длиннее, чем основание анального. Спинной плавник с ко-
лючкой, слабозазубренной с внутренней стороны и гладкой с внешней; ко-
лючка в спинном плавнике меньше, чем в грудном. Колючка в грудном
сильно развита, с внешней стороны она гладкая, с внутренней - с сильны-
ми, изогнутыми шипами. Хвостовой плавник с глубокой выемкой, его ло-
пасти закруглены. Голова маленькая, глаз крупный, покрыт кожей со сла-
бовыраженной векообразной складкой. Верхняя челюсть немного высту-
пает над нижней. На челюстях и нёбе мелкие шипиковидные зубы. Жабер-
ные тычинки тонкие и длинные. Жаберные перепонки не сросшиеся. Уси-
ки длинные, носовые достигают заднего края глаза или заходят за него,
верхнечелюстные и крайние нижнечелюстные заходят за основания груд-
ных плавников.
D I 7; А 13-16; Р I 4. Число зубчиков в грудной колючке 9-13. Жаберных
тычинок 9-13. Позвонков 32-34 (Громов, 1970).
Отнесение этого вида к южноазиатскому роду Mystus ставится под сомне-
ние (Аннотированный каталог..., 1998), этот род рыб не указывается для пре-
сных вод соседних стран (Nichols, 1943; Freshwater Fishes of Japan, 1987; Choi et
al., 1990). По некоторым признакам косатка-крошка близка к роду Pelteobagrus
(Аннотированный каталог..., 1998).
Распространение. Эндемик Амура. Вид описан из Среднего Амура у с. Ле-
нинское (оз. Омми) (Громов, 1970). В Хабаровском крае широко распростра-
нен везде в озерах средней части бассейна Амура (Хумми, Гасси, Падали).
Позже обнаружен в р. Мельгуновке из бассейна оз. Ханка, где многочислен
(Самуйлов, Свирский, 1976; Богуцкая, Насека, 1996; Аннотированный ката-
лог..., 1998).
Возраст и размеры. Максимальная длина пойманных экземпляров до
4,1 см, масса - до 1,4 г; предельный возраст неизвестен (Громов, 1970).
24
Образ жизни. Практически не изучен. Обитает в прибрежной части русла
Амура со слабым течением и песчаными или песчано-илистыми грунтами. Все
пойманные самки имели длину 3,4-4,1 см при массе 0,8-1,4 г, самцы - 3,0-3,4 см
и 0,4-0,7 г. Стадия зрелости гонад определена как III—IV при коэффициенте
зрелости 14-28% у самок и 1,2-2,4% у самцов. Очевидно, нерест порционный.
У трех самок абсолютная плодовитость составила в среднем 633 икринки с ко-
лебаниями от 570 до 700 икринок. Зрелые икринки зеленоватого цвета, около
1 мм в диаметре. Яичники парные, короткие, округлые, семенники у самцов
парные, белые и бахромчатые. В районе с. Ленинское в желудках косатки-
крошки найдены личинки хирономид, веслоногие и ветвистоусые ракообраз-
ные, остатки наземных насекомых, водяные клещи, личинки стрекоз и детрит
(Громов, 1970).
Статус вида. Малоизученныр вид.
Автор-составитель: О.А. Попова.
25
Семейство Bagridae
Род Pelteobagrus Bleeker, 1862
Класс Osteichthyes
Отряд Siluriformes
196. Pelteobagrus fulvidraco (Richardson, 1846) - косатка-скрипун
Описание и систематика. Тело умеренной длины, голое, у взрослых рыб
сплошь покрыто мелкими ворсинками. Хвостовой стебель короткий и высо-
кий. Спина зеленовато-черная, брюхо ярко желтое, на боках на желтом фоне
идут продольные прерывающиеся темные полосы от головы до лопастей хво-
стового плавника. Есть темные полосы и на других плавниках. Спинной плав-
ник находится в передней части спины, он короткий и с зазубренной колючкой.
Анальный удлиненный. Жировой плавник сзади свободный, его основание
меньше длины основания анального. Грудной плавник с более длинной колюч-
кой, чем спинной, она сильно зазубрена изнутри и имеет слабые шипики по на-
ружному краю. Хвостовой плавник с глубокой выемкой. Голова умеренной
длины, по ее средней части идет плоская борозда. Кожа на голове тонкая, гла-
за не покрыты кожей, имеют векообразную складку. Задние ноздри далеко от-
несены от передних. Имеется 4 пары усиков, одна пара у задних ноздрей (дли-
ной до конца глаза), вторая пара - на верхней челюсти (они длинные и доходят
до начала грудных плавников) и 2 пары на нижней челюсти. Рот большой, поч-
ти конечный; верхняя челюсть немного длиннее нижней. Губы тонкие. Имеют-
ся зубы на челюстях и сошнике.
D I 6-7; А 16-26; Р I 6; V I 5. Жаберных тычинок 13-16 (на п-ове Корея -
12-14). Позвонков 33^10 (Никольский, 1956; Choi et al., 1990). Кариотип у близ-
кого вида Р. nudiceps 2п = 56 (Васильев, 1985).
Распространение. Вид широко распространен в равнинной части рек Вос-
точной Азии, от Амура до юга Китая, севера и запада Корейского п-ова (реки
Ляохэ и Ялу) и Японии (юг о-ва Хондо и Сикоку) (Freshwater Fishes of Japan,
1983; Choi et al., 1990). В бассейне Амура встречается от устья Онона и Воскре-
сеновки вплоть до лимана, в бассейне Сунгари, Уссури, оз. Ханка и на северо-
западе Сахалина. В верховьях Амура обитает в реках Онон, Шилка, Ингода, но
здесь косатка-скрипун уже редка. В Монголии не отмечена (Никольский, 1956;
Карасев, 1987; Сафронов, Никифоров, 1995).
Возраст и размеры. Максимальная длина до 34,5 см, но в уловах обычно
длиной 11-32 см и массой до 400 г. Самцы крупнее самок (Никольский, 1956).
26
Образ жизни. Самая распространенная косатка в нижнем и среднем тече-
нии Амура. Летом держится преимущественно в местах с тихим течением, у ме-
дяков, в протоках и озерах, главным образом на участках с глинистыми и или-
стыми грунтами. На зиму скапливается на глубоких местах в протоках, в самом
русле Амура и в озерах. Косатка-скрипун - типичный эврифаг. Взрослые едят
бентосных, нектобентосных и воздушных насекомых (личинки хирономид, ру-
чейников, поденок), ракообразных, моллюсков (Planorbis, Viviparus), а также
молодь рыб (карася, малоротую корюшку, коня, чебака). Молодь не ест рыбу
и потребляет тех же беспозвоночных, что и взрослые особи косатки-скрипуна.
Растет косатка медленно, в возрасте 5+ достигает 22-27 см. Половозрелой ста-
новится в возрасте 2-3 лет, самцы - при более крупных размерах, чем сам-
ки. Нерест порционный, видимо, выметываются две порции икры. Икра откла-
дывается в конце июня-июле на мелководьях с песчаным или илистым грун-
том. Плодовитость косатки-скрипуна - 2,3-11,5 тыс. икринок диаметром
1,5-1,9 мм, со слизистой оболочкой - до 2,5 мм. Предварительно самец выка-
пывает норку кувшинообразной формы с отверстием 6-14 см, которое затем
суживается и опять расширяется до 14 см шириной и 16 см глубиной. Икра от-
кладывается самкой на дно и охраняется самцом. Самец в это время не питает-
ся, активно аэрирует икру и вышедших личинок до начала их активного пита-
ния (около 8 дней). Гнезда концентрируются в большие колонии с плотностью
13-14 гнезд на 1 м2. При температуре воды 24-26 °C вылупление происходит
через 2 сут при длине 5 мм. Личинки светобоязливы, они сначала лежат на дне
гнезда и машут хвостом, аэрируя желточный мешок, являющийся в это время
органом дыхания. В возрасте 7-8 сут при длине 7,8-8,0 мм желточный мешок
27
рассасывается, и личинки покидают гнездо. Молодь в больших количествах
держится в прибрежной зоне и активно питается. К осени сеголетки достигают
длины 4 см (Никольский, 1956).
Статус вида. Промысловый, но малоценный вид во всем Амуре. Многочис-
ленный вид, быстро восстанавливает свою численность после вылова.
Автор-составитель: О.А. Попова.
Класс Osteichthyes
Отряд Siluriformes
Семейство Ictaluridae
Род Ictalurus Rafinesque, 1820
197. Ictalurus nebulosus (Lesueur, 1819) - американский сомик
Описание и систематика. Тело вальковатое спереди и сжатое с боков в хво-
стовой части. Спина темно-коричневая или желтовато-коричневая, реже чер-
ная; бока светлее, с темными пятнами, брюхо белое или кремовое, плавники
темные. Голова крупная, уплощенная. Рот большой, конечный, челюсти оди-
наковой длины. На них многочисленные мелкие зубы в виде щеток. Усиков
4 пары: 2 пары на верхней части головы (одна из них длинная - в углах рта, вто-
рая - короткая, позади ноздрей) и две пары коротких усиков на подбородке
расположены в один ряд. Тело голое, без чешуи, с толстой кожей и снабжено
большим количеством слизистых желез. Боковая линия полная, хорошо замет-
ная от головы до хвоста. В отличие от обычного сома у американских сомиков
есть жировой плавник.
D I 6, А 18-24. Жаберных тычинок 13-14, из них 9 на верхней части и 4-5
на нижней части дуги. Позвонков 34-39 (Scott, Crossman, 1973). Кариотип:
2п = 60, NF = 76, 80 (Васильев, 1985). В Северной Америке описаны 3 подвида,
в Европу завезен номинативный подвид I. nebulosus nebulosus (Lesueur, 1819).
Распространение. Североамериканский вид, обитающий в бассейне Вели-
ких озер (кроме оз. Верхнее) и до Флориды и Дакоты на юге. Акклиматизиро-
ван на тихоокеанском побережье США, расселен по всему Западному полуша-
рию. Завезен в Новую Зеландию и в Европу, сначала в Англию, Румынию и
Германию, откуда проник в Польшу. В 1935 г. появился в водоемах Белоруссии
и Украины: бассейн р. Припяти, в Брестской и Волынской областях сначала
как объект прудоводства, оттуда проник в речные системы и сейчас широко
28
распространился в водоемах Белоруссии и Западной Украины. Возможно нахо-
ждение в прилегающих областях России.
Возраст и размеры. На родине сомик достигает максимальной длины 50 см
и массы 3 кг, в Европе - до 45 см и 2 кг (редко). В белорусских водоемах - до
30 см и 500 г в возрасте 8 лет. Чаще в уловах преобладают рыбы 15-20 см и
50-120 г в возрасте 3-6 лет (Жуков, 1965; Пенязь и др., 1973).
Образ жизни. Предпочитает медленнотекущие и стоячие водоемы с заи-
ленным дном и зарослями прибрежной растительности. Вынослив к высоким
температурам и низкому содержанию кислорода в воде подобно карасю и
вьюну. Может жить там, где другие виды рыб жить не могут. Очень непри-
хотлив в еде. Потребляет организмы бентоса (личинок хирономид, ручейни-
ков, стрекоз, ракообразных, моллюсков, водоросли и остатки высших водных
растений), а также мелких рыб и их икру. Зимой сомик не питается, с насту-
плением холодов забирается в глубокие ямы, откуда выходит только после
распаления льда. Растет сомик медленно: в возрасте 1 года он достигает
5-12 см и 4-10 г массы, 2 лет - 12-15 см и 20^15 г, в возрасте 4 лет - 18-24 и
65-190 г, в максимальном возрасте 8 лет - 30 см и 500 г. Рост зависит от корм-
ности и гидрологического режима водоема. Половозрелости достигает в воз-
расте 3^1 лет при длине 16-20 см. Нерест бывает в мае-июне при температу-
ре воды 17-20 °C в прибрежье, где есть затопленные деревья, песчаные или
каменистые грунты и много растительных остатков. Самка предварительно
готовит гнездо в виде ямки. Нерест осуществляется одной парой и сопровож-
дается брачными играми. Самка откладывает икру, слипшуюся в рыхлый ко-
29
мок. Плодовитость составляет 1200-5000 икринок размером 3^1 мм. Икра
бледно-кремового цвета. После окончания нереста самец остается у гнезда,
охраняя икру и выклюнувшуюся молодь. Инкубационный период длится
5-8 дней в зависимости от температуры воды. Вышедшие предличинки име-
ют длину 4-6 мм. Они сбиваются в плотные стайки в поверхностных слоях во-
ды и охраняются одним из родителей до рассасывания желточного мешка и
перехода на активное питание.
Статус вида. Объект прудового рыбоводства, особенно в теплых водах
электростанций. В естественных водоемах, где сомик прижился, приобретает
промысловое значение. Уловы в Брестской области составили 30 т в год. Бла-
годаря заботе о потомстве и повышенной степени выживания молоди он быст-
ро размножается и обладает высокой продуктивностью.
Автор-составитель: О.А. Попова.
Класс Osteichthyes
Отряд Siluriformes
Семейство Ictaluridae
Род Ictalurus Rafinesque, 1820
198. Ictalurus punctatus (Rafinesque, 1818) - канальный сомик
Описание и систематика. Это наиболее обычный вид из всех сомиков рода
Ictalurus. Тело удлиненное, торпедообразное. У молоди спина палево-голубая
или палево-оливковая, с серебристым отливом, брюшко серебристо-белое, бо-
ка с большим количеством мелких черных крапин. У взрослых особей спина и
верх головы серо-стальные, бока серые, брюхо и низ головы белые или сереб-
ристо-белые. Цвет плавников такой же, как цвет тела у их основания. Усы тем-
ные и с точками. Самцы в брачный период имеют голубой оттенок. Голова не-
большая. Рот конечный, рыло уплощено и заострено. Усиков 4 пары: 2 пары
на верхней части головы (одна пара длинная - в углах рта, вторая - короткая,
позади ноздрей) и две пары коротких усиков на подбородке расположены в
один ряд. Глаза крупные. Тело голое, гладкое. Боковая линия полная, хорошо
заметная. Есть жировой плавник. Спинной плавник высокий, его первый ко-
лючий луч не зазубрен. В анальном плавнике два первых луча зачаточные.
В грудном плавнике первый неветвистый луч имеет мелкие зазубрины. Хво-
стовой плавник длинный, острый, глубоко вырезанный.
D I 6, А 23-25, Р I 8-9. Жаберных тычинок 14-18 (9-12 на нижней части ду-
ги и 5-6 - на верхней). Позвонков 42^14 без Веберова аппарата (Scott,
Crossman, 1973). Кариотип: 2п = 56 или 58; NF = 92 или 94 (Rab. 1981). В Север-
30
ной Америке описаны 2 подвида, в Европу завезен номинативный подвид
/. punctatus punctatus (Rafinesque, 1818).
Распространение. В Северной Америке обитает от бассейна Великих озер
(кроме оз. Верхнее) и бассейна р. Миссисипи на севере до Флориды и Мекси-
канского залива на юге. Переселен в Европу, в частности в 1972 г. завезен в
пруды в бассейне Кубани (Виноградов, Ерохина, 1973а, б), откуда проник в реч-
ную систему не только Кубани, но и Дона, где встречается во всех равнинных
водоемах, но в небольшом количестве. В 1975-1976 гг. был посажен в водоем-
охладитель АЭС “Электрена” (Литва), где хорошо прижился и достигает высо-
кой численности. В России отмечено также самовоспроизводство в водоемах с
теплыми сбросными водами (при Черепетской ГРЭС, в Тульской области, на
Краснодарской ТЭЦ), а также разводится в прудовых хозяйствах Московской
и других областей (Кудерский, 1982; Емтыль, 1997; Аннотированный ката-
лог..., 1998).
Возраст и размеры. В Америке достигает максимальной длины 70 см и мас-
сы 6 кг в возрасте 12 лет (р. Миссисипи), есть сведения даже о рыбах 117 см
длиной и массой 12 кг в возрасте 40 лет, но обычно средние размеры 35—53 см
и 800-1600 г. В Европе размеры много меньше. Так, в Дону и Кубани ловятся
31
особи не более 5 кг при средней массе 350-600 г 2-8-летних рыб (Троицкий,
Цуникова, 1988).
Образ жизни. Обитает в озерах и крупных реках. Предпочитает чистые
глубокие места с песчано-галечниковым грунтом и проточной водой, а не при-
брежные участки, как I. nebulosus. Днем держится в укрытиях, в пещерах, под
камнями. Молодь в сумерках выходит на охоту, придерживаясь поверхностных
слоев воды. Часто заходит в протоки. Взрослые особи встречаются даже в оп-
ресненных участках моря, в устьях рек с соленостью 5-12%с. Питание разнооб-
разное. У молоди это личинки поденок, веснянок, хирономид, ракообразные,
головастики, лягушки, моллюски, водоросли и остатки высших растений (семе-
на деревьев). У взрослых особей (более 30 см длиной) - рыбы (основные мел-
кие виды в водоеме). Растет быстрее американского сомика, но темп роста за-
висит от водоема. В водоемах-охладителях он достигает 30 г в возрасте 1 год,
400 г - в 2 года и 1 кг - в 3 года. Созревает канальный сомик в возрасте 5-8 лет
при длине 27^10 см, а в некоторых водохранилищах даже в возрасте 2 лет. Не-
рестится поздней весной или в начале лета, когда температура воды достигнет
24—29 °C (оптимум 26,7°). Самец подготавливает гнездо - ямку среди камней,
коряг и растительности. Выделяя слизистый секрет, он скрепляет гнездо. Сам-
ка единовременно выметывает икру кучкой. Плодовитость в Америке равна
4,0-34,5 тыс. икринок, в наших водах - много меньше. Икра желтого цвета,
3,5-4,0 мм в диаметре. Самец охраняет гнездо, аэрирует его, чистит икру дви-
жениями тела и плавников. Икра развивается 5-9 дней при температуре
24—29 °C. Вылупившиеся предличинки имеют большой желточный мешок,
длину 10-11 мм и массу 20 мг. Они остаются на дне гнезда или около его сте-
нок 2-5 дней, а после рассасывания желточного мешка при длине 12-13 мм под-
нимаются к поверхности и начинают активно питаться. Самец первое время ох-
раняет стайки молоди. Мальки растут очень быстро: в возрасте 1 мес они име-
ют длину 5-10 см, а в возрасте 1 год - 9-19 см.
Статус вида. В Дону и Кубани встречается повсеместно, но в небольших ко-
личествах. Ценный объект рыбоводства. При выращивании на теплых водах
его рыбопродуктивность может достигать 400 кг/м3 (Кудерский, 1982;
Virbidkas, 1986).
Автор-составитель: О.А. Попова.
Класс Osteichthyes
Отряд Cyprinodontiformes
Семейство Oryziatidae
Род Oryzias Jordan et Snyder, 1906
199. Oryzias latipes (Temminck et Schlegel, 1846) - медака
Описание и систематика. Небольшая рыбка со сравнительно высоким телом.
По внешнему виду похожа на самок гамбузии, от которых отличается длинным ;
анальным плавником и тем, что у самцов анальный плавник не образует гонопо-
дия (не свернут в трубочку). Сдавленное с боков тело, сплющенная голова, не-
большой рот и большие глаза. Рыло меньше диаметра глаза. Спинной плавник
сдвинут назад, его начало над задней третью анального. Грудные плавники рас-
положены ближе к средней линии тела, брюшные - на середине тела и в сложен-
ном виде достигают анального. Чешуя крупная, циклоидная. Боковой линии нет.
Половой диморфизм у медаки хорошо выражен. Окраска спины варьирует от
желто-розовой до светло-коричневой, брюшко палевое. Имеется черная про-
дольная полоса на спине, от затылка до начала спинного плавника, и по бокам
тела - слабая темноватая полоска вдоль средней линии. Лучи брюшных и аналь-
ного плавников имеют черные крапинки. Брюшина черная. В роде Oryzias на-
считывается до 11 видов, распространенных от Западного Пакистана до Япон-
ских островов. Часто фигурировала под названием Aplocheilus latipes.
Для казахстанских популяций основные признаки имеют следующие пока-
затели. D 5-8, А 14-22, Р 7-11, V 5-7. Поперечных рядов чешуи (24) 25-36, жа-
берных тычинок 10-16, туловищных позвонков 9-16, хвостовых - (13) 15-19
(Рыбы Казахстана, 1992). Кариотип: 2и = 48, NF - 68 (Васильев, 1985).
Распространение. Естественный ареал медаки - п-ов Корея, Япония, Китай
(The Freshwater Fishes of China, 1982; Freshwater Fishes of Japan, 1987; Choi et al.,
3 Атлас.. . том 2
33
1990). В бывший СССР медака скорее всего попала из Китая при рыбоводных
работах. Впервые она зафиксирована в 1971 г. в бассейне р. Или (Казахстан),
сейчас стала массовым видом (Рыбы Казахстана, 1992; Аннотированный ката-
лог..., 1998). В Казахстане она прижилась в отдельных водоемах Семипалатин-
ской и Павлодарской областей, на рисовых чеках Кзыл-Ординской области,
натурализовалась в Чардаринском водохранилище и в р. Чу (Рыбы Казахстана,
1992). В России впервые отмечена в 1978 г. на Кубани, живет в пойменных озе-
рах р. Анапки (Емтыль, 1997). Вид завезен в Краснодарский край из Казахста-
на в 1970-х годах в целях борьбы с малярийным комаром.
Возраст и размеры. Длина тела до 5 см. Максимальный возраст не более
2 лет в Кубани и не более 5 лет в Казахстане (Рыбы Казахстана, 1992; Емтыль,
1997).
Образ жизни. Населяет самые мелководные биотопы, почти всегда при-
держивается одного места. Держится стайками. Основу питания составляют
личинки насекомых (чаще комаров), а также коловратки, рачки, олигохе-
ты, остракоды, водяные клещи. Растянутость нерестового сезона приводит
к значительной неравномерности роста, обычно за год медака достигает
длины 9-17 мм, на втором году - 14-20 мм и на третьем - 21-26 мм. Общая
продолжительность жизни медаки обычно не более 3 лет. Часть самок со-
зревает уже на первом году жизни, в июле-августе, а самцы даже в июне.
Размеры половозрелых рыб от 12 до 32 мм. Икрометание порционное, в ка-
ждой порции 15-50 икринок. Частота вымета икры зависит от температуры
воды, при 26 °C и выше самка мечет икру ежедневно. В Японии и Казахста-
не нерест с апреля по сентябрь, в среднем за сезон самка выметывает около
2000 икринок. Размножение происходит ночью или под утро необычным
способом. Сначала икра повисает за анальным отверстием самки в виде
грозди, где она оплодотворяется. Здесь проходят первые этапы ее развития,
затем самка рассеивает икринки по водным растениям. Развитие икры при
24-28° продолжается 10-15 сут. Вышедшие из икры личинки с небольшим
запасом желтка сразу же приступают к внешнему питанию (Рыбы Казахста-
на, 1992).
Статус вида. Промыслового значения не имеет. Считается перспективным
использование медаки в борьбе с кровососущими насекомыми, но достоверно
этот эффект не доказан. Медака часто используется как объект цитологиче-
ских, медицинских, генетических исследований и в генной инженерии, а также
как тест-объект для оценки загрязнения водоемов и выявления канцерогенов.
Автор-составитель: Ю.С. Решетников.
Класс Osteichthyes Семейство Poecilidae
Отряд Cyprinodontiformes Род Gambusia Роеу, 1854
200. Gambusia holbrooki (Girard, 1859) - хольбрукская гамбузия
Описание и систематика. Тело короткое, сжатое с боков. Рот направлен
кверху. На челюстях мелкие щетинковидные зубы. Боковая линия отсутству-
ет. Хвостовой плавник закругленный. Окраска тела обычно серо-зеленая с
темными пятнами на боках. Спинной и хвостовой плавники с рядами темных
пятнышек. Под глазом хорошо заметно темное пятно. Резко выражен половой
34
диморфизм: самцы значительно мельче и стройнее самок. У самцов имеется
совокупительный орган - так называемый гоноподий, образованный 3, 4 и 5-м
удлиненными свернутыми в трубочку лучами анального плавника; задний край
1-го луча в гоноподии резко зазубрен. D (6) 7-8 (9), А 8-11, Р 11-14. (Берг,
19496; Световидов, 1964; Рыбы Казахстана, 1992). Кариотип: 2п = 48, NF - 48
(Васильев, 1985).
Два близких вида этого рода - G. affinis (Baird et Girard, 1853) и G. holbrooki
(Girard, 1859) ранее рассматривались как два подвида одного вида: G. affinis affi-
nis (Baird et Girard, 1853) и G. affinis holbrooki (Girard, 1859). По последней реви-
зии они вновь восстановлены в ранг самостоятельных видов (Аннотированный
каталог..., 1998). В Узбекистане отмечали оба вида и возможные гибриды ме-
жду ними. В России обитает именно хольбрукская гамбузия (Световидов, 1964;
Аннотированный каталог..., 1998).
Распространение. Родина гамбузии - пресноводные водоемы Северной
Америки, от Нью-Джерси до Флориды и Алабамы. Широко расселена по стра-
нам света для борьбы с малярийным комаром. Завезена в Европу, Японию, Ки-
тай, Австралию, на Гавайские и Филиппинские острова. В СССР была приве-
зена в 1925 г. из Италии и выпущена в болотистые водоемы Абхазии, в районе
Сухуми. В последующем широко распространилась по югу территории бывше-
го СССР: она встречается на Украине, в Грузии, Азербайджане, в республиках
Средней Азии и Казахстана (Берг, 19496; Световидов, 1964; Рыбы Казахстана,
1992). На Каспии встречается от Хачмаса на юг до Куры, Ленкоранской низ-
менности, рек Ирана и до Атре^а (Туркмения). В России гамбузия отмечена в
Краснодарском крае (бассейн Кубани, реки Лаба и Ветта, сбросные каналы
Краснодарской ТЭЦ) и даже в центральных районах России на теплых водах
(Аннотированный каталог..., 1998). В 1960-е годы гамбузией буквально кише-
ли пруды-охладители тепловой электростанции в г. Электрогорске Москов-
ской области (Рыбы Подмосковья, 1988).
Возраст и размеры. Максимальные размеры самцов 4 см, самок - 8 см,
обычно значительно меньше. Предельный возраст не более 3 лет.
35
Образ жизни. Пресноводная и солоноватоводная рыба. Обычно обитает в
мелких водоемах, сильно заросших растительностью. Весьма неприхотлива,
не избегает даже очень загрязненных водоемов. Теплолюбивая рыба, предпо-
читает в наших водах температуру 20-30 °C , но выносит даже высокую тем-
пературу - пл юс 41,5 °C. Держится стайками 10-30 особей на глубине до 1,5 м
(Эланидзе, 1983; Емтыль, 1997). При температуре воды ниже 10 °C она зары-
вается в ил и впадает в спячку. В суровые зимы, когда водоемы скованы
льдом, в массе погибает. Отличается скороспелостью и высокой плодовито-
стью. Половое созревание наступает в возрасте 1—1,5 мес. Период размноже-
ния - с апреля по ноябрь при температуре воды выше 15°, оптимальная тем-
пература для размножения - 20-30 °C (Казанчеев, 1981; Эланидзе, 1983). Оп-
лодотворение внутреннее. Живородящая рыба. Период вынашивания икры
занимает 3^1 недели. Количество порций, выметываемых самкой за лето, мо-
жет достигать 5-6, обычно 3^1. В каждой порции в среднем около 70 мальков
длиной 8-10 мм. Они полностью сформированы, тотчас начинают плавать и
активно питаться. Максимальное количество детенышей, выметываемых од-
ной самкой за нерестовый сезон, в материнских водоемах составляет 315 экз., -
в Узбекистане - 266, в Казахстане - 169, в Грузии и Азербайджане -
100-150 экз. Для этого вида характерны очень резкие колебания численно-
сти: в отдельные годы в водоеме ее может быть очень много, затем она поч-
ти полностью исчезает. Только что родившиеся мальки гамбузии питаются
инфузориями, коловратками и другими мельчайшими организмами. Взрос-
лые рыбы поедают планктонных ракообразных, личинок водных беспозво-
ночных, растительность, икру рыб (Берг, 19496; Световидов, 1964; Эланидзе,
1983; Рыбы Казахстана, 1992).
Статус вида. Промыслового значения не имеет. Довольно эффективна в
борьбе с малярией, активно поедает личинок комаров. Так, в опытных водо-
емах Казахстана после ее вселения численность личинок малярийных комаров
снизилась с 128 до 32 экз./м2; в водоемах, где гамбузия отсутствовала, числен-
ность этих личинок доходила до 12 700 экз./м2 (Рыбы Казахстана, 1992). В то
36
же время гамбузия весьма нежелательна в рыбоводных хозяйствах, так как она
уничтожает икру и личинок рыб и является серьезным пищевым конкурентом
молоди промысловых рыб.
Авторы-составители: Ю.С. Решетников, Л.И. Соколов.
Класс Osteichthyes
Отряд Gadiformes
Семейство Lotidae
Род Lota Oken, 1817
201. Lota lota (Linnaeus, 1758) - налим
Описание и систематика. Единственный исключительно пресноводный вид
отр. Gadiformes. Тело удлиненное, невысокое, округлое в передней части и
сильно сжатое с боков - в задней. Голова уплощена, ее длина превышает мак-
симальную высоту тела. Глаз маленький. Рот большой, полунижний, верхняя
челюсть достигает вертикали заднего края глаза, нижняя челюсть короче верх-
ней. На челюстях и головке сошника имеются мелкие щетинковидные зубы, но
их нет на нёбе. На подбородке один усик (20-30% длины головы) и у передне-
го края ноздрей имеется по 1 короткому усику. Окраска тела довольно разно-
образная: чаще темно-бурая или черновато-серая, с большими светлыми пят-
нами на боках тела и непарных плавниках. Брюхо и плавники светлые. В зави-
симости от места обитания окраска может различаться: серая, коричнево-жел-
тая, оливково-черная. Могут варьировать форма и величина пятен. Спинных
плавников два, передний - короткий, задний - длинный. Анальный плавник
тоже длинный. D2 и А начинаются впереди вертикали середины тела и тянутся
до хвоста. Грудные плавники округлые. Брюшные расположены на горле, впе-
реди брюшных; второй луч брюшного плавника вытянут в длинную нить, снаб-
женную чувствительными клетками, как на усике. Хвостовой плавник округ-
лый и не соединяется с D2 и А. Чешуя циклоидная, очень мелкая, покрывает
все тело и часть головы сверху до ноздрей и жаберной крышки. Боковая линия
полная до начала хвостового стебля, далее к хвосту может прерываться. Обыч-
но она прямая, иногда волнистая.
Dj 10-15, D2 70-91, А 69-85, £ 16-23, V 6-8. Жаберных тычинок 4-10. Пи-
лорических придатков 21-67 в европейской части ареала и 42-180 - в азиат-
ской. Позвонков 61-66 (Берг, 19496; Световидов, 1948). Кариотип: 2п - 48,
NF = 74 (Раб, 1986).
Одни исследователи считают вид монотипическим (Кириллов, 1962; Нови-
ков. 1966; Scott, Crossman. 1973: McPhail. Lindsey, 1970), другие выделяют 2 или
3 подвида, в частности два в Евразии: L. lota lota (Linnaeus, 1758) - обыкновен-
37
ный налим (Европа и Азия до Лены), L. lota leptura Hubbs et Schultz, 1941 - тон-
кохвостый налим (северо-восток Сибири и Аляска). Третий подвид водится
только в Северной Америке (Световидов, 1948; Берг, 19496; Андрияшев, 1954;
Аннотированный каталог..., 1998).
Распространение. Широко распространен в пресных водах северных районов
Европы, Азии и Северной Америки. В Европе на западе встречается в Новой
Англии (нет в Шотландии и Ирландии), во Франции (в основном в бассейне Ро-
ны, изредка в верховьях Сены и Луары), в Италии (бассейн р. По), в западной
Швейцарии и повсеместно в бассейне Дуная и в водоемах бассейна Балтийского
моря. Нет его на западном побережье Скандинавии, на Пиренейском, Апеннин-
ском и Балканском полуостровах. В России повсеместно в водоемах арктической
и умеренной зон, в бассейнах Балтийского, Белого, Черного и Каспийского мо-
рей и в бассейнах всех сибирских рек от Оби до Анадыря на всем их протяжении.
В европейской части налима нет в Крыму, в Закавказье (кроме низовьев Куры и
Сефидруда), но есть на Северном Кавказе - в бассейне Кубани. Северная грани-
ца ареала - ледовитоморское побережье: есть на п-ове Ямал (кроме самых се-
верных речек), на Таймыре (бассейн рек Пясина и Хатанга, оз. Таймыр), на Но-
восибирских островах. На юге распространен в бассейне Обь-Иртышского бас-
сейна, от верховий (озера Телецкое и Зайсан) до Обской губы. Нет налима в
Средней Азии, в бассейне Аральского моря. В бассейне Енисея - в оз. Байкал и
его притоках повсеместно. В бассейне Селенги спускается на юг до Монголии
(озера Хубсугул и Буйр-Нур, верховья рек Онон, Керулен и Халхин-Гол; но нет
в системе озер Западной Монгольской провинции). Налим встречается во всем
бассейне Амура с его притоками Уссури, Сунгари. Есть в верховьях р. Ялу (бас-
сейн Желтого моря на границе Китая и п=ова Корея). На тихоокеанском побе-
режье налима нет в Восточном Приморье и на Камчатке, но есть на Сахалине и
Шантарских островах (Берг, 19496). Выходит в опресненные участки морей с со-
леностью до 12%с (Балтика, губы Оби и Енисея).
38
Возраст и размеры. Достигает длины 120 см и массы 24 кг, предельный воз-
раст - 24 года. Обычно в промысловых уловах до 60-80 см и 3-6 кг.
Образ жизни. Налим - холодолюбивая рыба, нерестится и нагуливается в
холодное время года. Он предпочитает холодные и чистые водоемы с камени-
стым иловатым дном и ключевой водой. Налим - очень хороший индикатор
чистоты воды. Летом при температуре воды выше 10-15 °C он становится вя-
лым и прячется в норы, ямы, под коряги, под обрывистыми берегами, впадая в
состояние оцепенения, очень мало питается, при температуре 27 °C погибает.
С наступлением осени и понижением температуры воды он начинает активно
передвигаться в водоеме и интенсивно откармливается перед нерестом. На-
лим - хищник с обонятельной и тактильной ориентацией (Малинин, 1971). Пи-
тается преимущественно ночью, максимальная двигательная и пищевая актив-
ность в 22-01 ч. В молодом возрасте он питается беспозвоночными: в первый
месяц - зоопланктоном, со 2-го мес - личинками водных насекомых, гаммари-
дами и другими ракообразными, икрой, личинками и молодью карповых рыб.
С годовалого возраста при длине 12-15 см налим начинает активно потреблять
рыбную пищу наряду с бентосом и только с 3-4 лет питается исключительно
рыбой. Состав пищи зависит от кормовой базы конкретного водоема. В сред-
ней полосе это преимущественно окуневые, карповые, корюшковые. В север-
ных водоемах к этим видам добавляются колюшки, молодь сиговых, подкамен-
щик (в Байкале - песчаная широколобка, гольян). У наиболее крупных особей
кроме рыб в пище встречаются лягушки (Световидов, 1948; Сорокин, 1976; Ре-
шетников и др., 1982).
Столь обширный ареал обусловливает большие различия в темпе роста на-
лима в водоемах с различным температурным режимом и разной кормностью.
Так, в Немане в 2-летнем возрасте налим достигает длины 36 см и массы 79 г,
а в 7 лет - 68 см и 2,4 кг; в водоемах Ямала в 2 года - 14,2 см и 20 г, а в 7 лет -
52 см и 1,46 кг. Половое созревание также наступает в разные сроки. В прибал-
тийских водоемах самцы созревают на 2-м году, самки - на 3-4 году при длине
35—40 см, в водоемах Крайнего Севера (Ямал) самцы - на 6-м году и самки - на
7-м году при длине 54—55 см. С наступлением зимнего похолодания налим вхо-
дит в мелкие реки на нерест, нерестилища располагаются в местах впадения
ручьев, где есть хорошая аэрация, вода прозрачная и температура более низ-
кая, чем в русле реки. Нерест происходит обычно сразу после ледостава, при
температуре воды около 0 °C в ноябре-декабре на севере или в декабре-фев-
рале - в умеренной зоне.
Икра полупелагическая, с жировой каплей, неклейкая, диаметром 0,75-
0,92 мм в ястыке и 1,05-1,15 мм уже в воде после вымета. Нерест на песчаном
или галечном грунте на глубинах 0,5-3,0 м. При слабом колебании воды икра
держится в подвешенном состоянии на нерестилищах или оседает на дно и за-
бивается под камни, а весной с увеличением скорости течения в реках посте-
пенно сносится вниз. Абсолютная плодовитость колеблется от 50 тыс. до 5 млн
икринок. Икра развивается при температуре воды, близкой к 0 °C, но выклев
происходит при температуре водк от 1,5 до 3-4 °C, что совпадает с распалени-
ем льда, через 60-90 сут после оплодотворения. Неоплодотворенная икра у на-
лима способна к партеногенетическому развитию (Крыжановский, 1949). В мо-
мент выклева личинки имеют длину 3,2-5,0 мм и сразу становятся подвижны-
ми, держатся вертикально, совершая постоянные движения вверх и вниз в тол-
ще воды в процессе ската по течению. Спустя 3-4 дня они начинают плавать
горизонтально у поверхности воды и опускаются на дно при опасности. Спустя
39
еще 2 недели при длине 8-9 мм они подходят к берегам, заросшим раститель-
ностью, с илистым грунтом и богатой кормовой базой и начинают активно пи-
таться.
Статус вида. Ценная промысловая рыба, однако отмечаются повсеместное
уменьшение средних размеров и снижение численности налима из-за ухудше-
ния условий существования (загрязнение воды и перелов).
Автор-составитель: О.А. Попова.
Класс Osteichthyes
Отряд Gasterosteiformes
Семейство Gasterosteidae
Род Gasterosteus Linnaeus, 1758
202. Gasterosteus aculeatus Linnaeus, 1758 - трехиглая колюшка
Описание и систематика. У представителей рода Gasterosteus задние отро-
стки тазовой кости образуют брюшной щит, брюшная колючка зазубрена,
спинные колючки расположены по одной линии, жаберные перепонки прира-
щены к межжаберному промежутку, тело голое или покрыто костными пла-
стинками. У трехиглой колюшки брюшная колючка без заметного бугорка у
основания. На спине перед плавником обычно три колючки, причем первые
две из них длиннее последней. Первая колючка расположена над основанием
грудных плавников и впереди брюшной колючки. Рыло короче заглазничного
отдела головы. По бокам тела 0-35 костных пластин, образующих на хвосте
более или менее ясный киль. Тело серебристо-белого цвета, верх головы и спи-
ны синие. В нерестовый период у самцов красное брюхо и голубые глаза.
D (II) III (IV) (9) 10-14; АI (7) 8-10; Р 10-11; VII. Жаберных тычинок 18-27.
Позвонков 29-36 (туловищных 14-15 и хвостовых 18-19) (Берг, 19496; Зюга-
нов, 1991). Кариотип: 2п = 42, NF = 54 (Chen, Reisman, 1970).
Подвидов нет, но отмечается большая изменчивость многих признаков:
число колючек, боковых пластин, количество позвонков и жаберных тычинок,
окраска тела и т.п. Вид представлен тремя формами (trachurus, leiurus и semiar-
matus), различающимися морфологически и по типу жизненного цикла. Часто-
та встречаемости форм в разных участках ареала различна, между ними воз-
можны переходы, в ряде случаев выявлена репродуктивная изоляция между
ними. Чаще их рассматривают вместе в ранге G. aculeatus complex (Scott,
Crossman, 1973; Зюганов, 1991).
Распространение. Вид широко распространен в бассейнах северной части
Атлантического и Тихого океанов. В европейской части России - от Кольско-
го полуострова и Новой Земли до Черного моря; обитает в бассейне Баренце-
во
ва, Белого (Имандра, все реки Карелии и Архангельской области, включая Се-
верную Двину), Балтийского (Ладожское и Онежское озера) и Черного морей
(Дон, устье Кубани). Однако в Печоре ее уже нет, отсутствует она в бассейне
Каспийского и Аральского морей и в реках Сибири. Вновь появляется в бассей-
не Тихого океана: по азиатскому побережью от Берингова пролива до Японии
и юга п-ова Корея, по американскому - от пролива до Калифорнии (Берг,
19496; Никольский, 1956; Зюганов, 1991; Аннотированный каталог..., 1998;
Scott, Crossman, 1973; Freshwater Fishes of Japan, 1987; Choi et al., 1990). В Рос-
сии - от Берингова пролива, Анадырского лимана, включая Камчатку и Ку-
рильские острова, а также все охотоморское побережье, Амур (только в лима-
не), Сахалин и Приморье до р. Туманная.
Возраст и размеры. Предельный возраст 5 лет, длина до 11 см, масса до 9 г
(Берг, 19496; Черешнев, 1996а).
Образ жизни. Имеются три экологические формы: морская, проходная и
пресноводная. Морская форма проводит весь свой жизненный цикл в море.
Так, в Баренцевом, Белом и Черном морях колюшки ведут пелагический образ
жизни, уходя довольно далеко в открытое море. В конце мая или в начале ию-
ня в Белом море они подходят на нерест к берегам, нерест в середине июня.
Размножается в море в прибрежных участках на мелководье при солености
20-25%с (Кандалакшский залив Белого моря). Проходная форма живет в море,
но на размножение идет в пресные воды (заход в Дон и кубанские лиманы из
Азовского моря); после нереста взрослые особи погибают или уходят обратно
в море. Молодь на первом году жизни также скатывается в море. Пресновод-
ная форма постоянно живет и размножается в пресной воде, в море не выхо-
дит. даже если этот водоем и связан с морем. Известно, что между морскими и
проходными колюшками нет генетических различий, но эти различия сущест-
вуют между проходной и жилой формами (Зюганов. 1991).
41
Трехиглая колюшка обитает в самых разнообразных водоемах: от ультра-
олиготрофных с минерализацией менее 10 г/л до супергалинных с соленостью
до 100%о. Температура также варьирует от 0° (и ниже зимой в северных морях)
до 32° в горячих источниках Камчатки (Зюганов, 1991; Черешнев, 1996а). На-
ходили колюшку в торфяных карьерах, где никакой другой рыбы нет, и даже в
артезианских колодцах (Берг, 19496).
Спектр питания довольно широк: от фито- и зоопланктона до бентоса (ли-
чинки насекомых, черви, ракообразные), в питании присутствуют также воз-
душные насекомые, икра, личинки и даже мальки рыб, поедает она и собствен-
ную молодь. В свою очередь колюшек охотно поедают водные насекомые, пи-
явки, лягушки, ужи, болотная черепаха, хищные рыбы, рыбоядные и хищные
птицы, млекопитающие (норка, выдра и даже медведь на Камчатке) (Макси-
мов, Долгов, 1983). В России основными хищными рыбами, которые питаются
трехиглой колюшкой, являются щука, окунь, арктический голец, мальма, сем-
га, кумжа, хариус, голавль, сом, судак, налим в пресных водах и треска, салака,
сельдь, бельдюга, маслюк, бычки и другие - в море (Зюганов, 1991).
Колюшка живет в среднем 2-3 года, максимум до 5 лет. Растет она быстро
и через 3—4 мес достигает размеров половозрелых рыб. Созревает рано, в воз-
расте 1 года. В некоторых популяциях после первого нереста отмечается мас-
совая гибель. Нерест зависит от географической широты водоема, температу-
ры и длины светового дня. Обычно на севере нерест летом, на Дону и в кубан-
ских лиманах - с апреля до конца июня. Нерест порционный (до 10 актов раз-
множения). Во время нереста появляется яркий брачный наряд. Самец строит
гнездо на дне из остатков растительности или другого материала, склеивая их
специальным клейким секретом. Размеры гнезда различны, обычно меньше
чайной чашки. После этого самец приглашает самку, которая откладывает
около 100 икринок и уходит. Плодовитость от 170 до 1000 икринок, икра свет-
ло-желтая, диаметром 1,8 мм. После оплодотворения этой икры самец может
пригласить другую самку. После заполнения гнезда икрой от 3—4 самок самец
становится агрессивным, охраняет кладку и аэрирует ее движениями своих
грудных плавников. Через 8-12 дней начинается выклев личинок, но и после
выхода всех личинок самец еще 5-7 дней охраняет их (Гомелюк, 1978; Неелов,
1987; Зюганов, 1991).
Как у всякого короткоциклового вида, у колюшки отмечены резкие коле-
бания численности. В отдельные годы колюшка составляла до 40-44% общего
вылова рыбы в прибрежной зоне Финского залива (1980-1981 гг.) и до 46% в
Куршском заливе (1970-1975 гг.). Пик ее численности отмечался в 1941 и
1948 гг. на Азовском море; в 1937-1939 и 1970-х годах - на Белом море; в
1979-1982 гг. - по всему восточному побережью Камчатки и в 1979-1982 гг. -
на Балтике (Рижский и Финский заливы). Только в р. Камчатку, по расчетам их-
тиологов, ежегодно заходили 500 млн рыб при биомассе 1500 т и при плотности
во время хода 1000 экз./м2 (Максимов, Долгов, 1983). В северных пресноводных
экосистемах ее биомасса может быть достаточно высокой и сопоставимой с
биомассой основных промысловых рыб. Так, в оз. Дальнем на Камчатке чис-
ленность колюшки колебалась от 1 до 6 млн особей при общей биомассе 2-12 т
(или 14-88 кг/га). Колюшка выедала до 1/3 биомассы зоопланктона, который
потреблялся всеми рыбами (колюшкой, гольцом и молодью красной); биомасса
же молоди красной была равна 15-30 т (110-220 кг/га) (Крохин, 1970). В водо-
емах южной Англии плотность колюшки составляла 3-15 экз./м2 (или около
6-30 кг/га), и она потребляла 10-100 г корма с 1 м2 площади озера в год.
42
Статус вида. Раньше существовал специальный промысел колюшки на Бе-
лом море, на Балтике (в Финском, Рижском и Куршском заливах), в Азовском
море и даже на Камчатке в связи со вспышкой ее численности. Рыбий жир и
кормовая мука из корюшки очень высокого качества. Во время блокады Ле-
нинграда жир колюшки с высоким содержанием каротиноидов (до 5%) с успе-
хом применяли в госпиталях при лечении ран и ожогов. Кроме того, колюшка
используется для откорма животных, приготовления искусственных кормов
для рыб и как удобрение для полей. В настоящее время хозяйственное значе-
ние колюшки невелико. Она всеядна, охотно поедает икру и личинок рыб, ока-
зывая вредное влияние на численность потомства ценных промысловых видов
(Гомелюк, 1978; Неелов, 1987; Зюганов, 1991).
Автор-составитель: Ю.С. Решетников.
Класс Osteichthyes
Отряд Gasterosteiformes
Семейство Gasterosteidae
Род Pungitius Costa, 1848
203. Pungitius platygaster (Kessler, 1859) - малая южная колюшка
Описание и систематика. Тело веретенообразное, покрыто не чешуей, а ма-
лозаметными костными пластинками (или ими покрыта только передняя часть
тела). Перед спинным плавником 8-11 отдельно посаженных колючек, накло-
ненных попеременно в разные стороны. В брюшном и анальном плавниках
имеется по одной колючке. Брюшные колючки зазубрены или нет. Отличает-
ся от Р. pungitius присутствием ряда костных пластин на теле, отсутствием ки-
ля на хвостовом стебле, меньшим числом позвонков и лучей в D и А, более вы-
соким и коротким телом и окраской. Хвостовой плавник округлой формы.
Плавательный пузырь замкнут. Спина темно-зеленая или оливково-зеленая,
бока светло-зеленые. В период нереста самцы чернеют.
D VIII-XI (6) 7-10; А I (6) 7-9; Р 10-11; V I 0-1. Жаберных тычинок 10-13.
Позвонков 30-31 (Берг, 19496; Казанчеев, 1981; Зюганов, 1991). Кариотип:
2п = 42, NF = 54 (70) (Васильев, 1985). Описаны 2 подвида: Р. platygaster platy-
gaster (Kessler, 1859) - малая южная колюшка - из бассейнов Черного, Азов-
ского и Каспийского морей и Р. platygaster aralensis (Kessler, 1877) - аральская
колюшка - из бассейна Аральского моря и водоемов, ранее с ним связанных
(Зюганов, 1991; Аннотированный каталог..., 1998).
Распространение. Номинативный подвид малой южной колюшки населяет со-
лоноватые участки Черного, Азовского и Каспийского морей, впадающие в них
реки (в России - Дон, Кубань, Сулак, Терек, Кума, Волга, Урал и др. ) и ряд озер
(Маныч-Гудило, Сарпинское, Банное, Чархал). В бассейне Волги встречается по-
43
всеместно в дельте и низовьях; после зарегулирования Волги и других рек быстро
расселяется в водохранилищах (отмечена в Волгоградском водохранилище и в
районе Рыбинской ГРЭС). Подвид аральская колюшка населяет бассейн Араль-
ского моря и водоемы, ныне не связанные с морем, - реки Чу, Сарысу, оз. Топья-
тан на Узбое, р. Нура и другие, но в Балхаше ее нет. В 1980-е годы аральская ко-
люшка проникала из р. Нура в р. Ишим (приток Иртыша) и стала встречаться в
России, в 1983 г. она спустилась до Усть-Ишима. Отметим, что немного южнее, в
400 км ниже по Иртышу, у г. Тобольска, проходит южная граница вида Р. pungitius
(Берг, 19496; Зюганов, 1991).
Возраст и размеры. Длина тела до 7 см, вес чуть более 1 г. Живет до 3 лет.
Образ жизни. Населяет заросшие погруженной водной растительностью
участки водоемов различной солености, иногда держится в опресненных участ-
ках Азовского моря и Таганрогского залива (Троицкий, Цуникова, 1988). Оби-
тает в солоноватых лиманах и заливах морей, а также в типично пресноводных
водоемах: реках, ручьях, озерах. В отличие от девятииглой колюшки не обра-
зует стай и скоплений, а ее численность невелика. Держится преимущественно
в неглубоких стоячих или полупроточных водоемах с хорошо развитой расти-
тельностью, склонна к скрытому образ}' жизни в зарослях. Питается зооплан-
44
ктоном и бентосом, причем бентос преобладает (личинки насекомых, прежде
всего мелкие личинки хирономид и др.), зарослевым планктоном, высшими ра-
кообразными, моллюсками, собственной икрой и растительностью. Созревает
на втором году жизни. Нерест в Нижней Волге с апреля по июнь, в водоемах
Азербайджана - в конце апреля, на Кубани - в мае. Икрометание порционное,
отмечено до 5-6 нерестовых актов за сезон, за один раз выметывается
60-90 икринок. Самец строит гнездо из обрывков растений и брачным танцем
привлекает самку. После нереста охраняет потомство. Плодовитость невели-
ка - 300-550 икринок (Казанчеев, 1981). Вид более устойчив к высоким темпе-
ратурам, чем Р. pungitius, икра малой южной колюшки способна развиваться
при температуре даже 28 °C, в то время как икра Р. pungitius при этой темпера-
туре гибнет (Зюганов, 1991). Молодь растет быстро.
Статус вида. Хозяйственного значения не имеет. Поедается окунем, суда-
ком и другими хищными рыбами.
Автор-составитель: Ю.С. Решетников.
Класс Osteichthyes
Отряд Gasterosteiformes
Семейство Gasterosteidae
Род Pungitius Costa, 1848
204. Pungitius pungitius (Linnaeus, 1758) - девятииглая колюшка
Описание и систематика. Передняя часть тела голая, без вертикальных ко-
стных щитков, или покрыта мелкими пластинками над грудными плавниками.
На хвостовом стебле хорошо выражен киль, покрытый маленькими костными
пластинками. Спинных колючек 8-10. Брюшные колючки не зазубрены. Боко-
вой тазовый отросток хорошо развит и достигает основания грудного плавни-
ка. Окраска меняется в зависимости от сезона: зимой спина и голова рыбки
темно-голубые, бока серебристо-белые с мелкими темными пятнышками, ле-
том нижняя часть головы с красным оттенком, брюхо светло-зеленое, с золо-
тистым отливом. В период нереста бока и брюхо самцов становятся черными,
брюшные колючки - белыми; у самок развивается “зеркальце” - блестящее
пятно на боку тела, не покрытое пигментом.
D (VI) VII-XII9-12; А 17-11; Р (9) 10; VI. Позвонков 30-35 (туловищных 14,
хвостовых - 18-19) (Зюганов, 19^1). Кариотип: 2п = 42, NF = 70 (Chen, Reisman,
1970). Ранее было 4 подвида, из которых амурская и сахалинская колюшки вы-
делены в самостоятельные виды, из двух оставшихся подвидов в России обита-
ет номинативный подвид Р. pungitius pungitius (Linnaeus, 1758), а подвид Р. pun-
gitius laevis (Cuvier, 1829) живет в Западной Европе (Зюганов, 1991; Аннотиро-
ванный каталог..., 1998).
Распространение. Циркумполярный вид, встречается в морях, реках и озе-
45
pax от бассейнов Северного, Балтийского, Белого и Баренцева морей в Евро-
пе, вдоль всего севера Сибири до Чукотки и Северной Америки. По Тихооке-
анскому бассейну от Чукотки до Японии по западному побережью и от Берин-
гова пролива до о-ва Кадьяк по восточному. В Америке встречается также
вдоль всего северного побережья, от Аляски до Гудзонова залива и по атлан-
тическому побережью до штата Нью-Джерси. Есть на юге Гренландии. В Рос-
сии обитает в реках Балтики, но обычно далеко вверх по ним не поднимается;
однако есть в Псковско-Чудском водоеме, в озерах Веряже и Ясы в бассейне
р. Великой, в Ладожском и Онежском озерах; в Карелии и Мурманской облас-
ти повсюду. В бассейнах рек Онеги и Мезени - до среднего течения, в Печоре -
до Усть-Цильмы. Отмечена на о-вах Колгуев и Новая Земля (губа Белушья).
Выше всего поднимается по Оби (до Иртыша у Тобольска). В Хатанге - до Бо-
ганиды, в Енисее - до Туруханска, в Лене - в нижней четверти реки. Есть изо-
лированные популяции в озерах Большое Чебачье и Майбалык. В Колыме из-
вестна до Верхнеколымска и ее много в озерах по Колыме. Повсеместно встре-
чается на Чукотке, в бассейне Анадыря, на Камчатке, в реках Охотского моря,
от Камчатки до устья Амура, есть на Сахалине, Шантарских, Беринга и Ку-
рильских островах (Берг, 19496; Зюганов, 1991; Черешнев, 1996; Аннотирован-
ный каталог..., 1998). В бассейне Каспийского моря и Волги ее не было. Непо-
нятно, как она была завезена в бассейн Верхней Волги, в последние годы
встречается в реках Самарской и Ульяновской областей (Евланов и др., 1998).
Возраст и размеры. Длина тела до 9 см. Продолжительность жизни 5 лет,
но в большинстве популяций 2-3 года (Зюганов, 1991).
Образ жизни. Девятииглая колюшка представлена как жилыми озерно-
речными, так и полупроходными формами, которые нагуливаются в опреснен-
ных участках моря, а нерестятся в солоноватых лагунах, заливах, эстуариях или
поднимаются на нерест в реки. Живет максимум 5 лет, но в большинстве попу-
ляций не более 2-3 лет и даже 1 год (Канада). По сравнению с трехиглой ко-
46
люшкой этот вид встречается в морской воде с соленостью до 32%с. более ус-
тойчив к дефициту кислорода и низким температурам (Зюганов, 1991). В Ка-
релии, Мурманской области и на Дальнем Востоке населяет самые разнообраз-
ные пресные водоемы (реки, ручьи, озера, озерки среди верховых болот) (Че-
решнев, 1996). В бассейне Верхней Волги она распространена в небольших ре-
ках и малых озерах с илисто-песчаным грунтом. Держится небольшими стая-
ми. Спектр питания довольно широк: зоопланктон, бентос, личинки хироно-
мид, моллюски, икра и молодь рыб (в том числе и своего вида). Половой зре-
лости достигает на второе лето после рождения. Порционный нерест бывает в
апреле-июле в зависимости от географической широты. Самец сооружает ша-
ровидное гнездо не на дне, а над грунтом среди зарослей водных растений,
склеивая кожной слизью куски стеблей и веточек. Самки откладывают икру
порциями по 60-160 икринок. Интервалы между актами 6^48 ч, за сезон раз-
множения наблюдается до 6-8 актов размножения у одной самки. Общая пло-
довитость самки от 350 до 960 икринок. Самец охраняет икру и выклюнувшую
молодь в течение 5-6 дней, причем для личинок он строит специальное второе
гнездо (“колыбельку”), располагающееся выше первого (Зюганов, 1991). Пос-
ле нереста проходные особи уходят зимовать в море, а пресноводные остают-
ся в своем водоеме.
Статус вида. Промысловое значение небольшое, вылавливается в водоемах
тундровой зоны Сибири (Якутия) на корм ездовым собакам и для приманки пуш-
ных зверей. В Финском заливе вместе с трехиглой колюшкой используется для
приготовления рыбной муки. Численность местами может быть значительной.
Объект питания многих ценных видов рыб. В бассейне Волги редкий вид.
Автор-составитель: Ю.С. Решетников.
Класс Osteichthyes
Отряд Gasterosteiformes
Семейство Gasterosteidae
Род Pungitius Costa, 1848
205. Pungitius sinensis (Guichenot, 1869) -
китайская, или амурская, колюшка
Описание и систематика. Бока тела с вертикальными костными боковыми
пластинками (32-36), образующими киль не только на хвостовом стебле, но и
на боках тела. Хвостовой стебель и брюшные колючки длиннее, чем у девяти-
иглой колюшки. Высота последней спинной колючки равна высоте предыду-
щих. Прозрачная пленка оторачивает сзади каждую колючку спинного плавни-
ка. Спина, бока тела, спинной и хвостовой плавники зеленовато-черные, груд-
ные и анальный плавники более светлые, брюхо светлое. Во время нереста са-
мец становится черным, а брюшные колючки - белыми.
47
D (VII) VIII-XI 8-13; A I (7) 8-11; P (9) 10; V I 1-2. Жаберных тычи-
нок 8-12. Позвонков 31-34 (Берг, 19496; Зюганов, 1991). Кариотип: 2п = 42,
NF = 50-54 (Muramoto et al., 1969). Многие европейские и американские ис-
следователи считают китайскую колюшку лишь подвидом девятииглой
Р. pungitius sinensis. В зонах контакта с Р. pungitius образует жизнеспособные
плодовитые гибриды, но так как они не имеют тенденции к полному слия-
нию, то считаются самостоятельными видами (Зюганов, 1991). На п-ове Ко-
рея и в Японии обитает самостоятельный подвид Р. sinensis kaibarae (Tanaka,
1915) (Choi et al., 1990).
Распространение. Встречается в Китае (до залива Б охай), на п-ове Корея
(восточное побережье), в Японии (Хондо, Хоккайдо), в оз. Долай-Нор, в бас-
сейне Амура и на Камчатке (Берг, 19496; Зюганов, 1991; Choi et aL, 1990).
В России отмечена на материковом побережье Западной Камчатки (к северу
от р. Большой), Курильских островах (Итуруп, Шикотан), на Сахалине, Шан-
тарских островах, в Амуре, от устья до рек Бира, Уссури и Сунгари; есть в
оз. Ханка и на всем приморском побережье от Амура до р. Туманная (Берг,
19496; Зюганов, 1991; Гриценко и др., 2002).
48
Возраст и размеры. В пресной воде достигает длины 65 мм, в морской -
90 мм. Максимальный возраст 2 года.
Образ жизни. Это чисто пресноводный вид (Зюганов, 1991), хотя есть ука-
зание и на обитание в море (Берг, 19496). В реках предпочитает затишные
места, особенно много ее в тихой зоне озер, на п-ове Корея - в водохранили-
щах. Биология изучена недостаточно. Отмечено, что в р. Димитрова на о-ве
Шикотан ведет скрытый образ жизни; днем прячется в зарослях растительно-
сти, а ночью выходит на участки со слабым течением (Гриценко и др., 2002).
В питании преобладают личинки хирономид, планктонные ракообразные, ди-
атомовые и зеленые водоросли, отмечены также воздушные насекомые, чер-
ви и собственная икра. Живет 1-2 года, достигая длины 55-65 мм. Созревает
при достижении длины 40 мм. Размножается только в пресных водах с февра-
ля по сентябрь. Нерест порционный. Самец строит гнездо среди растений над
грунтом. За один раз самка выметывает в среднем 80-150, максимально - 230
икринок.
Статус вида. Промыслового значения не имеет. Численность в реках не из-
вестна, в основной части ареала высокая, особенно в крупных озерах. Объект
питания хищных рыб и рыбоядных птиц (Черешнев, 1996а).
Автор-составитель: Ю.С. Решетников.
Класс Osteichthyes
Отряд Gasterosteiformes
Семейство Gasterosteidae
Род Pungitius Costa, 1848
206. Pungitius tymensis (Nikolsky, 1889) - сахалинская колюшка
Описание и систематика. Тело голое или покрыто 1-2 мелкими боковыми
пластинками в передней части. Киль на хвостовом стебле есть, состоит из 4-7
пластин, но выражен гораздо слабее, чем у Р. pungitius. Отличается от близких
видов тем, что боковые отростки тазовых костей слабо развиты, брюшные ко-
лючки короткие; спинные колючки очень низкие, а высота последней колюч-
ки в 2 раза больше высоты предыдущих; киль на хвостовом стебле слабо вы-
ражен, а сам хвостовой стебель короче и выше. Окраска темно-коричневая или
оливково-зеленая, иногда с пятнами. В нерестовый период самец чернеет, а его
брюшные колючки становятся белыми.
D VII-XII 9-12; А I 8-11; Р 10; V I. Жаберных тычинок 6-11. Позвонков
30-31 (Берг, 19496; Зюганов, 1991). Кариотип: 2п = 42, NF - 54 (Muramoto et
al.,1969). Подвидов нет. Некоторые авторы рассматривают эту колюшку как
подвид Р. pungitius (Линдберг, Легеза, 1965), другие - как самостоятельный вид
(Берг, 19496; Зюганов, 1991; Аннотированный каталог..., 1998).
4 Атлас . том 2
49
Распространение. В России вид встречается только на о-ве Сахалин - по-
всюду в пресноводных водоемах, от г.Оха на севере до залива Анива на юге.
В Японии отмечен только на о-ве Хоккайдо, в реках, впадающих в Тихий
океан, Японское и Охотское моря. На Курильских островах, в водах п-ова
Корея и о-ва Хонсю не отмечен (Берг, 19496; Зюганов, 1991; Freshwater
Fishes of Japan, 1987).
Возраст и размеры. Длина до 70 мм, в среднем 40-60 мм; в Японии - до
80 мм (Зюганов, 1991; Freshwater Fishes of Japan, 1987).
Образ жизни. Типично пресноводная форма. Предпочитает водоемы, густо
заросшие растительностью, со слабым течением воды. Не образует стай и из-
бегает открытой воды, ведет скрытый образ жизни в зарослях литоральной зо-
ны. Типичный бентофаг, питается личинками хирономид, моллюсками и чер-
вями. Нерест бывает в апреле-июле. По характеру размножения схожа с
Р. pungitius (Зюганов, 1991).
Статус вида. Промыслового значения не имеет.
Автор-составитель: Ю.С. Решетников.
50
Класс Osteichthyes
Отряд Syngnathiformes
Семейство Syngnathidae
Род Syngnathus Linnaeus, 1758
207. Syngnathus nigrolineatus Eichwald, 1831 - черноморская игла
Описание и систематика. Тело длинное, очень тонкое, с длинным хвосто-
вым стеблем, покрыто шестигранными кольцами из костных пластинок. Рыло
трубчатое и длинное (особенно у каспийских популяций), на его боках есть гре-
бешки. Жаберные крышки сильно выпуклые и только спереди с гребнем. На
темени есть слабый гребешок. Спинной плавник длинный и начинается перед
анальным отверстием, хвостовой очень маленький. Туловищных поясков
15-17, хвостовых - 36-41. Под спинным плавником 7-9 (10) поясков. Брюшных
плавников нет. Грудной плавник короче хвостового. Передний край сросшихся
внизу половин плечевого пояса заострен. Выводковая камера у самца защище-
на покровными пластинками и снабжена двумя длинными покровными склад-
ками, соединяющимися сзади и заходящими далеко за спинной плавник. Окра-
ска тела зеленовато-бурая или красновато-бурая, со светлыми поперечными
полосками посредине каждого пояска. Брюхо беловатое, а брюшной киль чер-
новатый. На спинном плавнике не бывает пятен.
D 30-43; А 3; Р 11-14 (Берг, 19496; Световидов, 1964). В Каспийском море
обитает свой подвид S. nigrolineatus caspius Eichwald, 1831, который переносит
совершенно пресную воду (Берг, 19496; Решетников и др., 1997; Аннотирован-
ный каталог..., 1998).
Распространение. По всем берегам Каспийского, Черного и Азовского
морей, входит в реки и связанные с ними озера. В Каспийском море обита-
ет во всех районах; в осолоненных заливах Мертвый Култук и Кайдак обна-
ружена карликовая форма. Встречается в пресной воде, в дельтах Волги,
Урала, Терека, низовьях Куры и в реках южного побережья моря (Берг,
19496; Световидов, 1964; Казанчеев, 1981). До зарегулирования Волги не
поднималась выше Астрахани. В последние десятилетия игла поднимается
вверх по Волге. Впервые обнаружена в Куйбышевском водохранилище в
1962 г., позднее отмечена также в Волгоградском, Саратовском и даже в
Рыбинском водохранилищах (Аннотированный каталог..., 1998; Евланов и
др., 1998). Возможно, идет саморасселение каспийского подвида из низовьев
Волги, или она сюда случайно завезена при акклиматизации мизид из Цим-
лянского водохранилища (типичный черноморский подвид). В бассейне
Азовского моря черноморская игла широко распространена в кубанских ли-
манах в воде с различной соленостью. В Кубани и ее притоках, а также в Се-
верском Донце раньше не отмечалась; после строительства Краснодарско-
го водохранилища заселила нижнее течение Кубани (Емтыль, 1997). В бас-
сейне Дона встречается в нижнем течении (Троицкий, Цуникова, 1988).
Имеется в оз. Палеостоми.
51
Возраст и размеры. Предельный возраст 6 лет, длина до 23 см, масса до 5 г.
Образ жизни. Эвригалинный вид, может жить как в пресных, так и в со-
леных водах (до 35%с). Держится в зарослях водных растений. Морская игла-
рыба в весеннее время заходит в реки и озера, поднимаясь иногда на значи-
тельные расстояния (в Днепре до 900 км). Пресноводная форма ведет тувод-
ный образ жизни в озерах, водохранилищах и старицах, придерживаясь одних
и тех же мест обитания в течение всей жизни (Долгий, 1993). Питается мо-
лодь только зоопланктоном, а взрослые - планктоном, крупными ракообраз-
ными, личинками насекомых, а иногда личинками и молодью рыб (Троицкий,
Цуникова, 1988; Долгий, 1993). В поисках жертвы ориентируется при помощи
зрения. Растет черноморская игла медленно, достигая в 5 лет длины 19 см и
массы 5 г. Весьма своеобразен процесс размножения. Нерест бывает в
мае-июне. После брачных игр с партнером самка обвивается вокруг него и
откладывает икру в выводковую камеру самца, расположенную на хвостовом
стебле. При этом икра оплодотворяется, после чего выводковая камера за-
крывается кожистой складкой. Плодовитость невелика, до 100 икринок. За
один раз самка может отложить до 20 икринок. Если выводковая камера у
самца не заполнена, то он может принять икру от другой самки. За сезон сам-
ка откладывает до трех порций икры (Долгий, 1993). Самцы с икрой и эмбри-
онами в выводковой камере встречались в мае-июле (Троицкий, Цуникова,
1988), в водохранилищах Волги - в июне-августе (Евланов и др., 1998). Икра
располагается в ячейках выводковой камеры и совершенно изолирована от
внешней среды. Она получает кислород из крови отца, притекающей к сли-
зистой оболочке выводковой камеры. В камере может быть 30-85 икринок;
у мелких самцов они располагаются в 2 ряда, у крупных - в 3 ряда. Самец вы-
нашивает икру, а затем личинок до конца августа. После выхода мальков сли-
зистая оболочка отделяется подобно плаценте млекопитающих. Молодь в
районе Новороссийска длиной 17 мм и более встречалась в мае-августе. Рас-
тет она довольно быстро, достигая к концу первого года жизни 10-12 см (Тро-
ицкий, Цуникова, 1988).
52
Статус вида. В волжских водохранилищах стал обычным (или даже много-
численным) видом на отдельных участках. Рыбохозяйственого значения не
имеет. Объект питания хищных рыб.
Автор-составитель: Ю.С. Решетников.
Класс Osteichthyes .
Отряд Perciformes
Семейство Percichthyidae
Род Morone Mitchill, 1814
208. Morone saxatilis (Walbaum, 1792) -
полосатый лаврак, полосатый окунь
Описание и систематика. Тело удлиненное, сжатое с боков, с двумя разде-
ленными спинными плавниками. Спина оливково-зеленая, спереди темно-си-
няя или черная, бока палевые, брюшко серебристо-белое. На теле 7-8 горизон-
тальных темных полос, совпадающих с рядами чешуи, причем 5 верхних идут
до хвоста, а 2 нижних - только до середины анального плавника. Такая окра-
ска появляется у молоди с 3-месячного возраста. Голова крупная, треугольной
формы. Рот большой, верхняя челюсть заходит за середину глаза, нижняя че-
люсть слегка выдвинута вперед. Зубы мелкие, щетинковидные и многочислен-
ные на челюстях и сошнике, на языке собраны в виде двух щеток. Спинные
плавники разделены, хотя и стоят близко. В анальном плавнике 3 колючих лу-
ча, последовательно увеличивающихся. Крупная ктеноидная чешуя покрывает
тело, жаберные крышки и щеки.
D, VII-XII; D21 8-14; A III 7-12; Р I 28-34; VI 5. В боковой линии 58-64 че-
шуй. Жаберных тычинок 20-29. Позвонков 24. Кариотип: 2п = 48, NF - 50. По-
двидов нет (Scott, Crossman, 1973).
Распространение. Естественный ареал вида - американское побережье Ат-
лантического океана, от р. Святого Лаврентия на севере до р. Сен-Джонс во
Флориде, и южное побережье Мексиканского залива от Флориды до Луизиа-
ны. В 1879 и 1882 гг. молодь полосатого окуня была выпущена на тихоокеан-
ском побережье в эстуарий р. Сан-Франциско, откуда он распространился на
север до Ванкувера и на юг до Калифорнийского залива. С 1996 г. широко ин-
тродуцирован в водохранилища бассейна рек Миссури (Озарк, Бул-Шоалс),
Сент-Франсис и Колорадо (Scott, Crossman, 1973). В 1965 г. молодь завезена в
СССР в водоемы бассейна Азовско-Черноморского и Каспийского морей для
искусственного разведения в питомнике “Горячий Ключ” на Кубани. Отсюда в
1973 г. полосатый окунь был пересажен в Шапсугское водохранилище, а в
53
1975 г. - в водоемы Литвы (VirbiCkas, 1986). Отмечался в самой Кубани, Про-
токе и в устьях других рек (Троицкий, Цуникова, 1988).
Возраст и размеры. На родине достигает максимальной длины 180-200 см
и массы 50 кг (Каролина). В северной части ареала размеры меньше - до 70 см
и 20 кг (р. Св. Лаврентия). В наших водах - обычно до 40-45 см и 2-5 кг.
Образ жизни. Морской и эстуарный прибрежный вид, населяющий опреснен-
ные участки взморья и заходящий в реки до 200 км от устья во время нерестовых
миграций. Переносит значительные колебания температуры и солености воды,
что и послужило основой для его акклиматизации в Азовском море. Молодь обыч-
но держится стайками, взрослые охотятся в одиночку. Личинки полосатого окуня
питаются мелким зоопланктоном. В возрасте 1 мес при длине 2-2,5 см молодь на-
чинает потреблять мелких личинок насекомых, мизид и отставших в росте собст-
венных собратьев. В возрасте 2 мес ( 5-6 см) полосатый окунь становится настоя-
щим хищником, потребляя молодь рыб, бокоплавов, червей, личинок хирономид,
креветок. Со второго года жизни при длине 8-12 см и позже питается исключи-
тельно рыбой и крупными ракообразными. Растет быстро, достигая в возрасте
2 лет 16^26 см, 3 лет - 27-36 см, 4 лет - 34-43 см и 9 лет - 48-68 см. Половозрелы-
ми самцы становятся в возрасте 3^4- лет, самки - 4-5 лет. Плодовитость от 14 тыс.
икринок у самок массой 1,2 кг до 3 220 тыс. икринок у самок массой 20 кг. Отме-
чены пропуски нереста. Икра перед нерестом имеет диаметр 1,0-1,35 мм, а после
попадания в воду разбухает до 3,6 мм. Она имеет большое перивителлиновое про-
странство и крупную жировую каплю. Икра неклейкая, полупелагическая, сносит-
ся течением. Нерест весной при температуре 16-19 °C, обычно в низовьях рек, на
течении (0,1-1,5 м/сек), в хорошо аэрируемых местах с содержанием кислорода
5-8 мл/л, обычно с постоянной величиной pH в диапазоне 6-10 и с хорошим осве-
щением. Развитие идет быстро, 1,5-3,5 сут в зависимости от температуры воды.
К моменту выклева икра сносится в эстуарный район с солоноватой водой
2-10%о. Однако полосатый окунь способен образовывать жилые формы в замк-
нутых пресных водоемах, что и послужило основой для использования этого ви-
54
да в рыбоводстве. Длина личинок при выклеве - 2,9-3,7 мм. Переход личинок на
активное питание происходит на 8-е сутки после выклева при длине 9—13 мм.
В течение первого года молодь держится стайками в прибрежных местах с пес-
чаным и галечным грунтом. Позже перемещается в прибрежные участки моря с
океанической соленостью (Дорошев, 1970; Scott, Crossman, 1973; Pflieger, 1991).
Еще в 1960 г. в СССР были начаты работы по перевозке из Америки поса-
дочного материала в виде разновозрастной молоди полосатого окуня в прудо-
вые хозяйства Ростовской области и Азербайджана (т.е. в районы, близкие по
климатическим условиям к естественному ареалу со среднегодовой температу-
рой воды 15 °C) для подращивания и получения маточного стада. В настоящее
время в России полностью разработана и освоена биотехника содержания про-
изводителей, получения от них икры, ее оплодтворения и инкубации, а также
выращивания всех возрастных групп от личинок до производителей. При ин-
тенсивном методе выращивания в прудах темп роста полосатого окуня высо-
кий: он достигает в возрасте 2 лет веса 0,5-1,0 кг; 3 лет - 0,8-1,5 кг; 4 лет -
1,0-1,8 кг; 5 лет - 2-3 кг и 6 лет - 3-6 кг (Стребкова, 1995).
Статус вида. Ценная промысловая рыба. Объект интенсивного рыбоводства.
Автор-составитель: О.А.Попова.
Класс Osteichthyes
Отряд Perciformes
Семейство Percichthyidae
Род Siniperca Gill, 1863
209. Siniperca chuatsi (Basilewsky, 1855) - ауха, китайский окунь
Описание и систематика. Тело высокое, сжатое с боков. Спина серая или
зеленовато-серая, бока зеленовато-желтые, с серебристым отливом. На теле
крупные черные сливающиеся п!ятна, есть пятна и на плавниках. Под первым
спинным плавником широкая поперечная темная полоса. От конца рыла через
глаз на спину идет узкая черная полоса. Голова крупная, ее длина равна высо-
те тела и около трех раз укладывается в длине тела. Глаза большие, подвиж-
ные. Рот очень большой, выдвижной. Зубы мелкие щетинковидные на челю-
стях, сошнике и нёбных костях. На челюстях есть еще крепкие клыки. Пред-
крышка зазубрена, жаберная крышка сзади имеет острый шип. Чешуя мелкая
55
циклоидная, покрывает все тело и бока головы (щеки и крышки). Два спинных
плавника соединены вместе, второй спинной выше первого; первый весь из ко-
лючек, второй - только ветвистые лучи. Колючки имеются также в брюшных
и анальном плавниках. Грудные и хвостовой плавники закруглены.
D, XI-XII; D2 13-14 ; Р I 5, A III 6-11. В боковой линии 108-120 чешуй. Пи-
лорических придатков 3, они ветвистые. Позвонков 28. Данных по кариотипу
этого вида нет, у близкого вида S. scherzeri из Японии; 2п = 48, NF = 52. Подви-
дов нет (Берг, 19496; Никольский, 1956; Аннотированный каталог..., 1998).
Распространение. Пресноводный вид. Встречается в реках Китая и
п-ова Корея (The Freshwater Fishes of Cina, 1982; Choi et al., 1990). В России - в
бассейне Амура, в его среднем и нижнем течении, в притоках Уссури и Сунга-
ри и в оз. Ханка, отмечен в водоемах северо-запада Сахалина (Никольский,
1956; Сафронов, Никифоров, 2002).
Возраст и размеры. Крупная рыба, достигает длины 65-70 см и массы
4-8,6 кг, максимальный возраст - 9 лет. В настоящее время в уловах встреча-
ются рыбы длиной до 53 (в среднем 30-35) см и массой 1 кг.
Образ жизни. Обитает как в русле крупных рек, так и в придаточных во-
доемах (озерах и протоках) с равномерным течением и прозрачной водой
вблизи зарослей растений. Хищник-засадчик со зрительной ориентацией.
Охотится в толще воды, где выслеживает добычу стоя неподвижно, или тихо
подбирается к ней, а затем коротким броском настигает ее и заглатывает. Ос-
нову пищи аухи составляют мелкие непромысловые виды рыб. В низовьях
Амура летом она кормится в придаточной системе, где ест в основном кара-
56
ся и горчаков, а осенью выходит в русло, где использует главным образом ко-
ней и пескарей. Зимует в русле крупных рек, почти не питаясь. В среднем те-
чении Амура летом ауха откармливается как в самом русле, так и в поймен-
ных озерах и заливах. Особи до 25 см длиной потребляют молодь пескарей и
креветок. Рыбы 25—30 см в русле поедают преимущественно молодь песка-
рей, косаток и подуста, а в заливах - молодь амурского чебачка, щуки и кара-
ся. Рыбы 30-60 см питаются большей частью косатками и чебаком (Соколов,
1962). Растет китайский окунь быстро, достигая к концу первого года
10-12 см, второго - 17-20 см, третьего - 23-26 см. Самцы в первые три года
растут быстрее самок, а в дальнейшем по темпу линейного и весового роста
самки обгоняют самцов. Обычно живут до 8 лет, достигая 45-50 см длины
(Никольский, 1956). Половая зрелость наступает у самцов в 2-3 года при до-
стижении 25-30 см, а у самок - на год позднее, при длине 32-34 см. Плодови-
тость 48-380 (в среднем 160) тыс. икринок. Икра пелагическая, с толстой обо-
лочкой, 1,9-2,2 мм в диаметре, с крупной жировой каплей (1,3 мм). Нерест
порционный (возможно, три порции), происходит в конце июня-начале июля
при температуре воды 20-26 °C на участках со спокойным течением и песча-
ным дном. Развитие идет быстро, через 36 ч выклевываются личинки, кото-
рые имеют длину 4,5-5,0 мм и сразу начинают активно питаться. Через
6-7 сут у личинок длиной 6-7 мм рассасывается желточный мешок, и они пе-
реходят в придаточную систему, где потребляют зоопланктон, мизид и личи-
нок карповых рыб. Размер добычи у личинок аухи может достигать 100%
длины тела, у взрослых рыб - 30-45% (Никольский, 1956).
Статус вида. Ценная промысловая рыба, но численность ее невелика. Еже-
годная добыча в бассейне Амура была не более 25 т в год. В настоящее время
вид крайне редок из-за вылова на нерестилищах и загрязнения водоемов, нахо-
дится на грани исчезновения. Занесен в “Красную книгу Российской Федера-
ции” (2001). Для увеличения численности рекомендовано прекратить вылов и
начать искусственное разведение.
Автор-составитель: О.А.Попова.
Класс Osteichthyes
Отряд Perciformes
Семейство Centrarchidae
Род Micropterus Lacepede, 1802
210. Micropterus salmoides (Lacepede, 1802) - большеротый окунь
57
Описание и систематика. Тело стройное, удлиненное, сжатое с боков.
Высота тела составляет 1/3 его длины. Верхняя часть головы и спина от тем-
но-оливкового до темно-зеленого цвета. Бока светлее, с серебристым отли-
вом и мелкими коричневыми пятнышками. Брюхо белое. Вдоль середины
тела от носа через глаз до хвоста идет широкая прерывистая черная полоса,
у молоди она четкая, с возрастом бледнеет. Старые особи мрачного серо-зе-
леного цвета. Голова большая, ее длина равна высоте тела. Большой рот,
верхняя челюсть заходит за вертикаль заднего края глаза. Нижняя челюсть
длиннее верхней, выступает вперед. Глаза крупные. Спинные плавники со-
единены друг с другом, но на границе между ними глубокая впадина. Хвост
широкий, с небольшой выемкой. Чешуя мелкая, покрывает все тело и бока
головы.
Dj X; D2 12-14; А 1П 10-12; Р 13-15; V I 5. В боковой линии 60-68 чешуй.
Жаберные тычинки длинные, их 8-10 штук. Пилорических придатков 24—28.
Позвонков 30-32 (Берг, 19496; Scott, Crossman, 1973). Кариотип: 2п = 46,
NF = 48 (Васильев, 1985).
Распространение. Акклиматизированный вид. Родина - Северная Аме-
рика, где естественный ареал охватывает территорию Канады и США в
пределах бассейнов Великих озер и р. Миссисипи с притоками. От атланти-
ческого побережья ареал простирается на восток и на юг до Мексиканско-
го залива (Флорида и северо-западная Мексика). Вид был интродуцирован и
в другие западные штаты США, Центральную и Южную Америку, Европу,
Южную Африку, Сянган (Гонконг), Филиппины. В Европу завезен в начале
XX в. в Англию, Шотландию, Германию, Францию, откуда попал в Данию,
Бельгию, Нидерланды, Польшу. В Россию завезен в озера Абрау и Лиман-
чик под Новороссийском и в подмосковные пруды. Однако большой чис-
ленности нигде не имеет, встречается единично (Виноградов, Ерохина,
1973а, б; Виноградов, 1995; Аннотированный каталог..., 1998; Scott,
Crossman, 1973; Lee et al., 1980).
58
Возраст и размеры. На родине часто достигает длины 65 см и массы
3,5^1 кг. Как редкость во Флориде отмечались длина 82 см и масса 8 кг. Мак-
симальный возраст до 15 лет. В Европе размеры крупных особей - до 52 см и
2,3 кг (Чехия), средние - 40-50 см и 1,5 кг.
Образ жизни. Обитает в неглубоких (до 6 м) тепловодных стоячих озерах
и прудах, в медленнотекущих речках, вблизи прибрежных зарослей. Держит-
ся часто в укрытии под корягами или в пещерах и норах под берегом. Круп-
ный хищник со зрительной ориентацией. Молодь до 3-5 см длиной питается
зоопланктоном, личинками поденок и других насекомых, бокоплавами; после
достижения длины 5-7 см поедает поденок, стрекоз, креветок, и впервые в ее
питании появляются личинки рыб. Взрослые особи питаются рыбой, кревет-
ками, лягушками, моллюсками. Утром и вечером ловит добычу у поверхно-
сти, а днем - в толще воды и у дна. Иногда охотится стаями вблизи берега (во
время ската молоди других видов рыб). Часто потребляет собственную мо-
лодь. Растет быстро, причем самки быстрее самцов. Половое созревание на-
ступает в 3-4 года у самцов и в 4—5 лет у самок при длине 25-35 см. Нерест
бывает в мае-июле при температуре воды 16-18 °C среди зарослей подводной
растительности или затопленных коряг и деревьев. Самец предварительно
строит гнездо, расчищая на глубине 0,2-2,0 м площадку диаметром 60-100 см
на песчаном или каменистом грунте, иногда окруженную валиком из расте-
ний. Нересту предшествуют брачные игры. Самец последовательно приводит
к гнезду несколько самок, которые, отложив икру, уходят. Однако одна сам-
ка может нереститься с несколькими самцами в разных гнездах. Плодови-
тость в зависимости от размеров составляет 2-109 тыс. икринок. Диаметр ик-
ринки 1,5-1,7 мм. После нереста у гнезда остается самец, который охраняет
икру и молодь. Выклев личинок происходит через 3-5 дней, длина личинок
3 мм. Они остаются в гнезде 6—7 дней, пока не рассосется желточный мешок
(при длине 5,6-6,3 мм), после этого они всплывают к поверхности, состаива-
ются и начинают питаться. Молодь светло-зеленого цвета еще около 30 дней
держится стайками и охраняется самцом. В дальнейшем она ведет одиночный
образ жизни. Рост в течение первого года жизни очень быстрый. Уже в октя-
бре в оз. Онтарио мальки имеют длину до 13 см, а в возрасте 1 года - 10-17 см.
В южных штатах США они растут еще быстрее, и уже в октябре сеголетки
имеют длину 18-23 см.
Статус вида. Ценный промысловый вид и излюбленный объект спортив-
ного рыболовства. Акклиматизация в Европе не имела успеха в естественных
водоемах из-за низких температур воды (оптимальная температура для вида
26-28 °C). Успешное разведение отмечено только в озерах Абрау и Лиман-
чик, у Новороссийска, где температуры воды близки к оптимальной. В пос-
ледние годы большеротый окунь в этих озерах встречается очень редко (Ем-
тыль, 1997).
Автор-составитель: О.А.Попова.
59
Класс Osteichthyes
Отряд Perciformes
Семейство Percidae
Род Gymnocephalus Bloch, 1793
211. Gymnocephalus acerinus (Goldenstadt, 1775) - донской ерш
Описание и систематика. У видов рода Gymnocephalus рот небольшой, вы-
движной, на челюстях щетинковидные зубы. Голова голая, на ней большие по-
лости сенсорной системы. Предкрышка с 8-13 шипами, жаберная крышка с
1 шипом. Жаберные тычинки короткие. Оба спинных плавника соединены.
У донского ерша тело удлиненное, несколько сжатое с боков, покрыто мелкой
чешуей. Высота тела составляет 15—20% длины тела. Рот небольшой, конеч-
ный. Рыло удлиненное, в два раза больше диаметра глаза. На предкрышке
6-7 шипов по заднему краю и 3 по нижнему. Шип на крышечной кости слабый.
Верхняя часть тела светло-желтая, брюхо беловатое. На боках тела и спинном
плавнике небольшие круглые черные пятнышки.
Dj XVII-XIX; D2 12-14; А II 5-6; Р 14-15; V14-5. В боковой линии 50-55 че-
шуй (Берг, 19496). Кариотип: 2п = 48, NF = 48 (Rab et al., 1987). Подвиды не опи-
саны. Близкий вид G. schraetser (Linnaeus, 1758) живет в Дунае.
Распространение. Встречается только в водоемах бассейнов Черного и
Азовского морей. В Днестре встречается до верховий, в Буге редко, в Днепре
обычен от низовьев до Дорогобужа и в его притоках (Припяти, Тетереве, Дес-
не, Соже). В России живет в верховьях Днепра и его притоков, в р. Дон и его
притоках на всем их протяжении (реки Северский Донец. Воронеж, Усмань,
Черная Калитва, Тихая, Сосна, Хопер, Медведица), а также в дельте Кубани
(Берг, 19496; Александрова, 1976; Троицкий, Цуникова, 1988; Емтыль, 1997;
Аннотированный каталог..., 1998).
Возраст и размеры. Достигает длины 21 см, массы - 150 г, но обычные раз-
меры в уловах 8-16 см и масса 9-30 г. Предельный возраст 10-11 лет.
Образ жизни. Чаще живет в русле рек на довольно быстром течении, но
встречается в протоках и реже в пойменных озерах. Летом держится на пере-
катах. В отличие от обыкновенного ерша предпочитает плотные песчаные
грунты. В реке держится небольшими стайками в придонных слоях воды.
Обычно активен в сумерках и начинает питаться в это время, но на ямах пи-
тается и днем. С сентября собирается в большие стаи и уходит на более глу-
бокие места. Зимует на ямах, где держится до распаления льда. Ведет осед-
лый образ жизни и больших миграций не совершает. Питается преимущест-
60
венно донными беспозвоночными (ракообразными, личинками насекомых,
мелкими моллюсками), реже мальками рыб, поедает также икру рыб. Растет
довольно медленно, причем самки растут быстрее самцов. Половозрелым
становится на 2-3-м году жизни при длине 12-16 см. Нерестится в апреле-мае
на участках с быстрым течением и галечным или песчаным грунтом. Плодо-
витость достигает 1,8-24,7 тыс. икринок в бассейне Дона и 5-30 тыс. икринок
в бассейне Днестра. Икра слабоклейкая, диаметром 1,0-1,1 мм, почти не раз-
бухает в воде. Икрометание порционное. Первый нерест бывает при темпе-
ратуре воды 6-8 °C, второй - при 12-14 °C. Нерестится небольшими стаями.
Инкубационный период длится 6-8 дней при температуре воды 14—16 °C. Вы-
клев при длине 4 мм. Личинки сначала неподвижно лежат на дне, в возрасте
9 сут с появлением грудных плавников всплывают в средние слои воды, сно-
сятся вниз по течению. На 12-е сутки при длине 6,5 мм переходят на активное
61
питание - охотятся в придонных прибрежных участках водоема (Федоров,
1960; Долгий, 1993; Емтыль, 1997).
Статус вида. Численность невелика. Второстепенный промысловый вид.
Является объектом любительского рыболовства.
Автор-составитель: О.А.Попова.
Класс Osteichthyes
Отряд Perciformes
Семейство Percidae
Род Gymnocephalus Bloch, 1793
212. Gymnocephalus cernuus (Linnaeus, 1758) - обыкновенный ерш
Описание и систематика. Окраска серо-зеленая на спине, с меланиновыми
бурыми пятнами на боках, а также на спинном и хвостовом плавниках. Тело
короткое, сжатое с боков, его высота составляет 20-30% длины тела. Рыло
тупое. Рот небольшой, нижний. На челюстях щетинковидные зубы, клыков
нет. Голова голая, на ней большие полости сенсорной системы. Предкрышка
на заднем крае имеет 5-10 шипов, на нижнем - 3. Крышечная кость снабжена
сильным шипом. Сильная колючка имеется в брюшных плавниках, и две силь-
ные колючки в анальном. Грудь часто не покрыта чешуей.
D, XI-XVI; D2 10-15; А II 4-7; Р 10-15; V I 4-6. Чешуй в боковой линии
33-Л2. Жаберные тычинки короткие (6-14). Позвонков 32-38. Кариотип:
2п = 48, NF = 48 (Берг, 19496; Решетников и др., 1997; Rab et al., 1987; Popova et
al., 1998). Подвиды не выделяются, но многие авторы отмечали высокую гео-
графическую и экологическую изменчивость ерша (Аннотированный ката-
лог..., 1998; Popova et al., 1998).
Распространение. Широко распространенный в Евразии вид от Англии и
Франции на западе до Колымы включительно на востоке. Северная граница
проходит почти по побережью Северного Ледовитого океана, кроме Северно-
го Таймыра и Северного Ямала. Имеется в водоемах по атлантическому побе-
режью Скандинавии, Северной Англии и Ирландии; повсеместно в водоемах
бассейнов Северного, Балтийского, Белого, Баренцева, Черного и Каспийско-
го морей в Европе, а в Сибири - в бассейнах рек Северного Ледовитого океа-
на. Ерша нет в Испании, Италии, Югославии, в Крыму, Закавказье. Ранее оби-
62
тал в бассейне Арала, теперь исчез. В Сибири южная граница распространения
ерша проходит по верховьям рек, текущих на север. Нет его в Байкале (но есть
в Ангаре), в бассейне Амура и по всему тихоокеанскому побережью, нет и на
Чукотке. В последние годы наблюдается расширение ареала ерша: он появил-
ся в Боденском озере, проник на север Англии и Шотландии, случайно был ин-
тродуцирован в Великие Озера Северной Америки, где быстро наращивает
свою численность (Решетников и др. 1997; Аннотированный каталог..., 1998;
Popova et al., 1998).
Возраст и размеры. В большинстве водоемов в уловах преобладает мелкий
ерш. Максимальная длина ерша - 18,5 см, масса - 208 г. Есть указания, что в
некоторых случаях он может достигать массы 500 г и длины 27 см при макси-
мальном возрасте 15 лет (Popova et al., 1998).
Образ жизни. Обитает в озерах, реках, водохранилищах, дельтовых рай-
онах рек и опресненных заливах морей. В водоемах держится в придонных го-
ризонтах как прибрежной зарослевой зоны, так и в профундали открытой зо-
ны озер, в водохранилищах поднимается и в пелагиаль. Ерш - типичный бен-
тофаг, очень пластичный в выборе корма. Излюбленной его пищей являются
личинки хирономид и гаммариды, но при их недостатке в водоеме он легко пе-
реключается на другие виды корма, тем более что ассортимент его кормовых
организмов включает все формы бентоса, зоопланктона и рыбную пищу (икру
и молодь). С возрастом увеличиЬаются размеры потребляемых им организ-
мов, наиболее крупные особи становятся хищниками. Растет ерш медленно, од-
нако в хороших условиях обитания (термического режима, кормовой базы)
темп его роста резко увеличивается. На юге ерш растет быстрее. В Днепре в
возрасте 1 года он достигает длины 10 см, а в возрасте 5 лет имеет максималь-
ный размер 15 см. В карельских озерах в возрасте 1 год ерш не достигает и 4 см
длины, а максимальный размер 13 см имеет в возрасте 13 лет. В большинстве
63
водоемов ерш - короткоцикловый вид. Большие различия в темпе роста опре-
деляют и различия в сроках созревания. Половая зрелость наступает в 2-^ го-
да при длине 9-12 см. Абсолютная плодовитость равна 2-104 тыс. икринок в за-
висимости от размера самок. Нерест продолжительный, порционный, с апре-
ля по июнь выметывается до 3 порций икры. Сроки нереста и температура во-
ды, при которой он происходит, варьируют в водоемах разных широт: 1 -я пор-
ция - при 4—9°, 2-я - при 11-13° и 3-я - при 18-20 °C. Нерест обычно происхо-
дит на песчаных и каменистых грунтах, иногда на растительности и корнях де-
ревьев на глубине 0,5-3,0 м. Инкубационный период занимает 5-6 сут при тем-
пературе 15-16 °C и до 4,5 сут при 20 °C. Размер личинок при выклеве -
3,8—4,3 мм, переход к активному питанию в придонном горизонте - в возрасте
11 сут при длине 5,5 мм.
Статус вида. В крупных водоемах озерного типа с высокой кормностью
ерш достигает большой численности и крупных размеров, подвержен резким
флуктуациям численности. Здесь он является второстепенным промысловым
видом и одновременно служит важным кормовым объектом для многих цен-
ных хищных видов рыб и конкурентом в питании ценных бентофагов. Высокая
воспроизводительная способность ерша при перелове крупных видов рыб не-
избежно вызывает вспышку его численности и нарушение баланса в водоеме.
Ерш очень чувствителен к загрязнению воды и эвтрофированию водоемов.
В последние годы его численность в водоемах европейской части России резко
снизилась.
Автор-составитель: О.А.Попова.
Класс Osteichthyes
Отряд Perciformes
Семейство Percidae
Род Perea Linnaeus, 1758
213. Perea fluviatilis Linnaeus, 1758 - речной окунь
Описание и систематика. Тело, сжатое с боков, покрыто мелкой ктеноид-
ной чешуей, щеки целиком в чешуе. Крышечная кость имеет один прямой шип,
предкрышка сзади зазубрена. Межчелюстные кости выдвижные. Щетинковид-
64
ные зубы расположены полосами во много рядов на челюстях, сошнике, нёб-
ных и внешнекрыловидных костях. Клыков нет. Жаберные перепонки не сра-
щены между собой. Тело зеленовато-желтое, на боках 5-9 поперечных черных
полос. Первый спинной плавник серый, на его конце черное пятно; второй
спинной - зеленовато-желтый, грудные плавники - желтые, иногда красные.
Два спинных плавника соприкасаются или слегка раздвинуты, причем первый
спинной выше второго.
D] XII-XVI, D2I-IV 12-17 А П-Ш 7-11, Р I 15-16, V I 4—6. В боковой линии
53-74 чешуй, она не переходит на хвостовой плавник. Число жаберных тычи-
нок 16-29. Позвонков 38—44. Подвидов нет, североамериканский желтый окунь
Р. flavescens Mitchill, 1814 теперь считается самостоятельным видом (Берг,
19496; Попова и др., 1993; Popova et al., 1997). В крупных озерах разные геогра-
фические и экологические формы иногда описывались как самостоятельные
единицы в ранге подвида или расы (Меньшиков, 1951; Покровский, 1951). На
самом деле это экологические формы окуня с разным способом питания и по-
ведения (Попова и др., 1993; Аннотированный каталог..., 1998). Кариотип:
2п = 48, NP = 48 (Васильев, 1985).
Распространение. Широко населяет равнинные водоемы Евразии - реки,
озера, прибрежные участки моря. Нет его на Пиренейском п-ове, на севе-
ре Англии, в Ирландии и на атлантическом побережье Скандинавии, в гор-
ных районах Закавказья, в Средней Азии, на юге Монголии, в бассейне
Амура, на Дальнем Востоке, Камчатке и Чукотке. В России северная гра-
ница распространения окуня проходит почти по побережью Северного Ле-
довитого океана, от р. Пасвик до Колымы, на юге — до Черного моря, Се-
верного Кавказа и до верховий сибирских рек. Его не было в Крыму, но в
1955 г. он был пересажен из Днепра в Альминское и Симферопольское во-
дохранилища, где хорошо прижился. По палеонтологическим данным, пре-
5 Атлас.. . том 2
65
жде окунь обитал и в бассейне Амура. В 1960-е годы был акклиматизиро-
ван в озерах Кенон и Иван в верховьях Амурского бассейна. Ареал его
расширился за счет вселения в водоемы Австралии, Новой Зеландии, Юж-
ной Африки и Азорских островов (Берг, 19496; Попова и др., 1993; Popova
et al., 1997).
Возраст и размеры. Максимальный возраст 17 лет, длина - 51 см и масса -
4,8 кг. Обычно в промысловых уловах преобладают особи длиной до 30 см, в
среднем 15-20 см и массой 200-300 г в возрасте 4-6 лет.
Образ жизни. Окунь - озерно-речной вид, приспособленный к жизни в
прибрежной зарослевой зоне водоема, где он питается зоопланктоном, бен-
тосными организмами и молодью разных видов рыб, которые сменяют друг
друга в рационе по мере его роста. Однако в разных водоемах пища окуня
значительно различается в связи с составом кормовой базы. В некоторых во-
доемах окунь в течение всей жизни потребляет зоопланктон, или остается
бентофагом, не переходя на хищничество. В крупных озерах и водохранили-
щах с богатой и разнообразной кормовой базой и обилием подходящих для
него биотопов окунь образует 2 или 3 экологических формы (расы), различа-
ющиеся местом обитания, составом пищи и темпом роста. Прибрежный мел-
кий окунь растет медленно и питается беспозвоночными, а глубинный растет
быстро и ведет преимущественно хищный образ жизни, питаясь молодью
разных видов рыб (главным образом карповых и окуневых). Темп роста и
сроки полового созревания на столь обширном ареале окуня сильно различа-
ются. В мелких и малокормных водоемах за первый год он едва достигает
5 см длины, а к 6 годам - 20 см. В крупных озерах и водохранилищах, в дель-
тах крупных рек годовалый окунь имеет длину 12 см, а пятилетний - 35 см.
В соответствии с этим и половая зрелость у него наступает в разные сроки и
при разной длине, обычно в возрасте 2-3 лет. Нерест бывает ранней весной,
после распаления льда: в феврале-марте - на юге, в мае-июне - на севере при
температуре воды 7-8 °C. Плодовитость колеблется от 12 до 300 тыс. икри-
нок. Икра в виде длинных сетчатых лент откладывается на прошлогоднюю
растительность. Икринки сильнообводненные, имеют диаметр 2,0-2,5 мм. Та-
кой способ откладки икры обеспечивает высокую выживаемость икры и ли-
чинок. Нерест однократный. Развитие длится 2 недели. Личинки при вылуп-
лении имеют длину около 6 мм и почти резорбированный желток, поэтому
сразу начинают активно плавать и охотиться за планктонными ракообразны-
ми. Молодь в первое лето питается зоопланктоном и бентосом, а в некоторых
водоемах при достижении длины 4 см - и молодью других видов рыб (Попо-
ва, 1971, 1979а).
Статус вида. Благодаря высокой численности является одним из основных
или второстепенных объектов промысла. Но при перелове в водоеме крупных
хищников окунь быстро наращивает свою численность, мельчает и становится
“сорным” видом, конкурирующим в пище с ценными рыбами-бентофагами.
В то же время мелкий окунь служит одним из основных объектов питания мно-
гих хищных видов рыб.
Автор-составитель: О.А.Попова.
66
Семейство Percidae
Род Percarina Nordmann, 1840
Класс Osteichthyes
Отряд Perciformes
214. Percarina demidoffi Nordmann, 1840 - перкарина
Описание и систематика. Тело сжато с боков, чешуя тонкая, легко опадаю-
щая. Голова голая или частично покрыта чешуей, на ней большие полости сен-
сорных органов. Рот выдвижной. Щетинковидные зубы на внешней и внутрен-
ней поверхностях челюстей, на сошнике и нёбных костях. Предкрышка снаб-
жена шипами по краю, причем нижние шипы направлены вперед. Предкрыш-
ка и жаберная крышка покрыты чешуей. Спинные плавники слабо соприкаса-
ются. Хвостовой плавник выемчатый, грудные длинные, доходят до конца
первого спинного. Брюшные под грудными, имеют сильную колючку. Окраска
от молочно-желтоватого цвета до серебристо-белого на боках, с перламутро-
вым переливом, спина сероватая, брюхо белое. Вдоль боковой линии ряд чер-
ных точек, а при основании спинного плавника 8-9 темных пятен и 1 темное
полулунное пятно на затылке.
Dj VIII-XI; D2 (I) II—III 9-13; A II 7-11; P 10; V I 5. Боковая линия не доходит
до хвостового плавника, в ней 33-37 чешуй. Жаберных лучей 7-8. Жаберные
тычинки длинные, их 22-28. Позвонков 33-35 (Берг, 19496; Световидов, 1964).
Кариотип: 2п = 48, NF = 68 (Suciu, Rab, 1991). Образует два подвида: Р. demidof-
fi demidoffi Nordmann, 1840 - черноморская перкарина, обитающая в лиманах
Днестра, Днепра, Южного Буга; в России живет Р. demidoffi maeotica Kuznetzov,
1888 - азовская перкарина (Берг, 19496; Световидов, 1964; Аннотированный ка-
талог..., 1998). У азовской перкарины в отличие от черноморской спинные плав-
ники почти не соприкасаются, в спинном плавнике меньше лучей, грудь покры-
та чешуей, меньше диаметр глаза и больше число чешуй в боковой линии.
Распространение. В России встречается подвид азовская перкарина, кото-
рый населяет северо-восточную и восточную части Азовского моря, в основ-
ном Таганрогский и Темрюкский заливы. В Керченском проливе, устье Дона и
в Кубанском лимане встречается редко, больше ее в Большом Ахтанизовском
лимане и в дельте Кубани (Берг, 19496; Троицкий, Цуникова, 1988; Аннотиро-
ванный каталог..., 1998).
Возраст и размеры. Небольшая рыба. Предельная длина - 11 см, обычно
самки - до 9,0-10,5 см, самцы - 6,0-7,2 см. Предельный возраст - 4 года у самок
и 2 (редко 3) - у самцов (Световидов, 1964; Долгий, 1993).
67
Образ жизни. Стайная рыба, ведет придонный образ жизни. Обитает в
осолоненных и пресных водах. В состав ее пищи входят организмы планк-
тона (Copepoda), нектобентоса (прежде всего мизиды), реже организмы
бентоса (черви, моллюски, личинки насекомых) и молодь рыб (бычки и
тюлька). Короткоцикловый вид, в популяции не более 3 возрастных групп.
Растет медленно, самки растут быстрее самцов. Годовики имеют длину
4—5 см, 2-годовики - 6 см и 3-годовики - 7-8 см. Созревает на втором году
жизни. Нерестится в наиболее опресненной части Таганрогского залива в
июне-июле, при температуре воды 24—25 °C. Плодовитость — до 3000 ик-
ринок. Икра донная, клейкая, мелкая, 0,7 мм в диаметре, выметывается
прямо на илистый грунт, она инкрустируется мелкими частицами ила и к
грунту не приклеивается. Взрослые особи после нереста выходят в Азов-
ское море. Развитие икры продолжается около двух суток. Выклюнувши-
еся личинки длиной 2,5 мм сначала лежат на дне, а через двое суток начи-
нают скачкообразными движениями вверх-вниз подниматься к поверхно-
сти воды и ведут пелагический образ жизни. При длине 6 мм начинают ак-
тивно питаться сначала зоопланктоном (мелкие ветвистоусые и веслоно-
гие ракообразные), при длине 16-40 мм — крупными веслоногими и мизи-
дами, а со второго года жизни основной пищей становятся бычок Knipowit-
schia longecaudata (Kessler, 1877) и тюлька размерами 18-30 мм (Светови-
дов, 1964).
Статус вида. Второстепенный объект промысла. При вылове выделяет
много слизи, поэтому ценность улова резко снижается. Конкурирует в пище с
хамсой и атериной, а также с молодью тюльки, судака, сельдей, чехони и быч-
ков. Сама перкарина служит пищей судаку, чехони и сому.
Автор-составитель: О.А.Попова.
68
Семейство Percidae
Род Stizostedion Rafinesque, 1820
Класс Osteichthyes
Отряд Perciformes
215. Stizostedion lucioperca (Linnaeus, 1758) - обыкновенный судак
Описание и систематика. Тело удлиненное, сжатое с боков. Спина и верх
головы зеленовато-серые, брюхо белое. На боках 8-12 буро-черных попе-
речных полос. На спинных и хвостовом плавниках ряды темных пятнышек,
расположенных на перепонках между лучами. Парные и анальный плавники
бледно-желтые. Рот большой, верхняя челюсть заходит за вертикаль заднего
края глаза. Зубы расположены узкими рядами на челюстях, сошнике и
нёбных костях; на челюстях и нёбных есть сильные клыки. Предкрышечная
кость сзади зазубрена, внизу с шипами. Щеки голые или только сверху по-
крыты чешуей.
D! XIII-XVII; D2 I-III 19-24; A II-III 10-13; Р I 14-18; V I 5. Два спинных
плавника слегка раздвинуты. Боковая линия заходит и на хвостовой плавник, в
ней 80-97 чешуй. Жаберных тычинок 10-16, они короткие в виде бугорков, гу-
сто усаженных зубчиками. Пилорических придатков 4-9. Позвонков 45-48
(Берг, 19496; Аннотированный каталог..., 1998). Кариотип: 2п = 48, NF = 48
(Васильев, 1985). Подвиды не описаны. В водохранилищах может образовы-
вать несколько самостоятельных устойчивых популяций.
Распространение. Естественный ареал судака охватывает все крупные реч-
ные и озерные водоемы бассейнов Балтийского (Эльба, Одер, Висла, Неман,
Западная Двина, Нева, речки Ботнического и Финского заливов), Черного
(Марица, Дунай, Днестр, Днепр, Дон, Кубань), Каспийского (Волга, Урал, Те-
рек, Кура, Сефидруд, Атрек) и Аральского (низовья Амударьи, вся Сырдарья,
Сарысу, Чу) морей, от верховий Дуная и Эльбы на западе до Уральских гор на
востоке. Его северная граница доходила до Северного полярного круга в Шве-
ции и Финляндии, до 63° с.ш. - в Карелии (озера Ладожское и Онежское) и до
61° с.ш. - в Вологодской области (Белое озеро). В России судак обитал только
в европейской части, от Карелии до Закавказья, его не было в Сибири. Не бы-
ло судака в Англии, Франции, Дании, в странах Средиземного моря (Берг,
19496; Кудерский, 1964; Pethon, 1^89).
После строительства многочисленных каналов и водохранилищ и акклима-
тизационных работ ареал судака значительно расширился. В Европе он был
акклиматизирован в Англии (р. Темза), во Франции (р. Рона), в Германии
(р. Рейн), в Южной Швеции (оз. Венерн), в бассейне Белого моря, в Карелии,
Вологодской и Архангельской областях и в Крыму (Кудерский, 1964; Зуянова,
1989; Делямуре, 1964; Аннотированный каталог..., 1998). Появился судак в
69
Средней Азии (в Каракумском канале, в озерах Саракамыш, Катташор, Бал-
хаш, Алаколь, Иссык-Куль и в реках Талас, Нура и во многих водохранили-
щах) (Рыбы Казахстана, 1989; Сальников, Решетников, 1991; Решетников, Ша-
кирова, 1993; Сальников, 1995).
Судак акклиматизирован во многих озерах Карелии (Сегозеро, Выгозеро,
Вядлозеро) и Вологодской области (Кубенское, Воже, Лача), принадлежащих
к бассейну Белого моря, где раньше его не было (Кудерский, 1964; Зуянова,
1989). Из оз. Воже судак проник в оз. Лача и р. Онегу, а из оз. Кубенское - в
Северную Двину (Новоселов, 2000). Акклиматизирован в Западной Сибири в
водохранилищах верховий Иртыша и в Новосибирском водохранилище
70
(р. Обь), откуда он спустился до низовий Оби, ее уральских притоков и до Об-
ской губы. Посажен в озеро Ханка, из которого он проник в Амур (Богуцкая,
Насека, 1996; Аннотированный каталог..., 1998). Новые места обитания судака
на карте обозначены штриховкой и точками.
Возраст и размеры. Достигает 130 см длины, массы до 18 кг и предельно-
го возраста 14 лет, но в уловах чаще встречаются особи длиной 40-60 см и мас-
сой 1-3 кг.
Образ жизни. Судак населяет как пресные, так и солоноватые воды, об-
разует полупроходные формы. Пелагический хищник, обитающий в откры-
той зоне озер и водохранилищ. Полупроходной судак откармливается в оп-
ресненных мелководных районах морей с соленостью 7-9%о, а нерестится в
реках. Периодически он появляется в Черном море вдоль кавказского побе-
режья, от Новороссийской бухты до Туапсе и Гудаут. Очень чувствителен
к содержанию кислорода в воде (до 13%). Молодь в первые месяцы жизни
питается зоопланктоном, который вскоре заменяется нектобентическими
ракообразными (мизидами, гаммаридами, изоподами) и молодью других
рыб. Пищу взрослого судака составляют мелкие массовые виды рыб: на се-
вере - молодь окуневых (окунь, ерш), корюшка, ряпушка, молодь сигов, на
юге - тюлька, хамса, перкарина, бычки (Фортунатова, Попова, 1973; Попо-
ва, 19796). Темп его роста сильно различается в пределах ареала в зависи-
мости от температурного режима и кормности водоема. Так, на севере
(Онежское озеро) в возрасте 1 года он имеет длину 11 см, в 14 лет - 60 см, а
на юге в дельте Кубани годовики достигают 17 см, а 8-летки — 70 см. Соот-
ветственно половое созревание у северных популяций наступает в возрасте
5-7 лет при длине 50 см, а на юге - в 3-4 года при длине 35-ЛО см. Плодови-
тость в зависимости от размеров рыб составляет 70-1180 тыс. икринок. Ик-
ра бледно-желтого цвета, диаметром 1,0-1,1 мм в ястыке и до 1,5 мм - при
попадании в воду. Нерест в Онежском озере идет при температуре воды
15-16 °C в середине июня, а в Азовском море — с середины апреля до нача-
ла июня при температуре 17-18 °C. Икра откладывается в гнезда, устраива-
емые самцом на мелководьях с песчаным грунтом или под подмытыми кор-
невищами тростника. Гнездо обычно имеет круглую или овальную форму
диаметром 0,5 м и глубиной 15-20 см. Икра откладывается кучно. Самец ак-
тивно охраняет гнездо и выклюнувшихся личинок, чистит от заиления и аэ-
рирует гнездо движениями грудных плавников. Инкубационный период
длится 5-6 сут при температуре 16-18° и 72-80 ч — при 20-22°. Выклюнув-
шиеся личинки имеют среднюю длину 4,6 мм, активное питание начинается
на 6-7-е сутки после вылупления при длине 6 мм (Берг, 19496; Фортунато-
ва, Попова, 1973).
Статус вида. Ценный объект промысла, особенно в крупных озерах, водо-
хранилищах, дельтовых районах рек и в опресненных заливах морей. В послед-
ние годы в связи с загрязнением воды его уловы в большинстве водоемов рез-
ко падают, поэтому необходимо поддерживать численность судака за счет раз-
ведения молоди и мелиорации нёрестилищ.
Автор-составитель; О.А.Попова.
71
Семейство Percidae
Род Stizostedion Rafinesque, 1820
Класс Osteichthyes
Отряд Perciformes
216. Stizostedion volgense (Gmelin, 1788) - волжский судак, берш
Описание и систематика. В отличие от обыкновенного и морского судаков
у берша нет клыков, щеки сплошь покрыты чешуей, лоб узкий (меньше диа-
метра глаза), верхняя челюсть доходит лишь до вертикали середины глаза. Го-
лова короче и выше, чем у судака. Окраска как у судака, но светлее и число по-
перечных полос на теле меньше (7-9). Спинные плавники соприкасаются, они
выше, чем у других видов; второй спинной плавник длиннее первого. Аналь-
ный плавник короткий.
Dj XII-XIX; D2 I—П 20-22; А II 9-10; Р 114-15; V I 5. Боковая линия короче,
чем у других судаков, в ней 70-83 чешуй. Жаберных тычинок 13-19. Число по-
звонков равно 43. Пилорических придатков 3, меньше, чем у других видов
(Берг, 19496; Казанчеев, 1981). Кариотип: 2п = 48, NF = 48 (Rab et al., 1987). По-
двиды не описаны (Аннотированный каталог..., 1998).
Распространение. Обитает только в пресных водоемах бассейна Волги (от
низовьев.до оз. Белое), Урала (вверх до Чапаевска), редок в Тереке, Сулаке и
Самуре, отсутствует в Куре. Активно заселяет водохранилища, где достигает
высокой численности, лучше растет и имеет длинный жизненный цикл. В бас-
сейне Черного моря имеется в Дону (до верховьев), Днепре (от устья до Кре-
менчуга), Буге (редок), Днестре, Дунае и Тисе. Акклиматизирован в Балхаше
вместе с судаком и воблой (Берг, 19496; Аннотированный каталог..., 1998).
В 1985 г. впервые обнаружен в Кубани, в настоящее время широко распростра-
нен во всех равнинных водоемах бассейна Кубани, от лимана до водохранилищ
и малых рек (Москул, 1994; Емтыль, 1997).
Возраст и размеры. По размерам меньше других судаков, достигает не бо-
лее 45 см длины и массы 1,2-1,4 кг. В реках живет до 6-8 лет. В водохранили-
щах живет до 12 лет и достигает длины 60 см и массы более 2 кг (Евланов и др.,
1998).
Образ жизни. Обитает только в пресной воде, в море не выходит, ведет
придонный образ жизни. Летом концентрируется в местах скопления корма:
молодь - ближе к берегу, взрослые - в открытой глубокой части русла реки,
озер и водохранилищ; зимой - в глубоких придонных слоях воды. В водохрани-
лищах Волги обитает как на небольших глубинах с медленным течением воды,
песчаным или каменистым дном, так и на заиленных русловых участках с глу-
бинами до 40 м. Более активен в вечерние и утренние часы (Евланов и др.,
1998). В дельте Волги совершает небольшие миграции, нечто подобное отме-
72
чено в Самарском и Куйбышевском водохранилищах. Питание у берша сме-
шанное: молодь длиной 1,6-2,5 см питается планктоном (циклопы, диаптому-
сы, дафнии, моины), у особей 2,5~i,0 см длиной планктон сменяется куколками
хирономид и личинками стрекоз. Берш длиной более 4 см начинает потреблять
и нектобентических ракообразных (мизид, гаммарид), речных раков и мелких
рыб (пескаря, щиповку, бычка-песочника, молодь окуневых рыб). В крупных
водохранилищах Волги и Дона с проникновением в них тюльки берш стал по-
треблять и ее. Растет берш гораздо медленнее, чем судак. В водохранилищах
рост интенсивнее, чем в этом же районе в речных условиях. В р. Дон годова-
73
лый берш имеет длину 7,7 см, а предельной длины 29 см достигает в 5 лет.
В Цимлянском водохранилище годовики имеют длину 9 см, здесь берш живет
до 12 лет, достигая 44,5 см. Темп роста в водохранилищах резко возрастает в
связи с улучшением условий откорма на всех этапах жизненного цикла. Созре-
вает берш на 3-4-м году жизни при длине 20-30 см. На нерест подходит в мел-
ководные места с песчаными косами, где строит гнезда на глубинах до 2 м, в ко-
торые откладывает икру на обмытые корневища растений. Гнезда берша час-
то располагаются вблизи от гнезд судака или с кладками окуня. Самец охраня-
ет икру (Дмитриева, 1973). Нерест начинается в апреле-мае при температуре
воды 10-22° и продолжается в течение месяца (выметываются две порции ик-
ры). Икра желтого цвета, мелкая, 0,75-0,84 мм в диаметре в первой порции и
0,52-0,65 мм - во второй. Плодовитость составляет 119-346 тыс. икринок в
Куйбышевском водохранилище, 64-472 тыс. - в Цимлянском и 44-254 тыс. ик-
ринок в Веселовском водохранилище. Инкубационный период длится 4—5 сут.
Длина личинок при выклеве - 4,5-5,0 мм, переход к внешнему питанию в воз-
расте 7 сут при длине 5-6 мм. Первая пища - коловратки, диатомовые водорос-
ли и зоопланктон (Константинов, 1957).
Статус вида. Ценный промысловый вид. В последние годы уловы берша
повсеместно упали в связи с загрязнением воды и осушением нерестилищ вес-
ной из-за колебаний уровня при зарегулировании стока. Занесен в “Красную
книгу Российской Федерации” (2001).
Автор-составитель: О.А.Попова.
Класс Osteichthys
Отряд Perciformes
Семейство Cichlidae
Род Sarotherodon Riippell, 1852
217. Sarotherodon mossambicus (Peters, 1852) - мозамбикская тилапия
Описание и систематика. Тело короткое, сжатое с боков и высокое (его вы-
сота укладывается 2,5-3 раза в длине). Голова большая. Окраска тела у моло-
ди и у самок - однотонная серебристая или сероватая. Есть темное пятно на
заднем конце жаберной крышки. У самцов окраска обычно зеленая, с метал-
74
лическим блеском, при возбуждении на боках появляются 4 белых пятна, беле-
ют нижняя губа и жаберная крышка. У половозрелых самцов - ярко красные
грудные плавники и оторочка спинных и хвостового плавников, а тело приоб-
ретает бархатно-черный цвет, с белыми крапинками на спинном и хвостовом
плавниках. Глаз крупный. Рот конечный, среднего размера, верхняя челюсть
доходит до переднего края глаза. Зубы на челюстях короткие, слабые, двух-
или трехвершинные. В наружном ряду нижней челюсти 60-100 зубов. Спинной
плавник один, длинный, начинается на уровне заднего края жаберной крышки,
а кончается у хвостового стебля. Лучи задней части спинного и анального плав-
ников длинные, доходят почти до конца хвостового плавника. Грудные и
брюшные плавники достигают анального. Чешуя крупная, циклоидная или
ктеноидная. Верх головы спереди покрыт голой кожей, остальная ее поверх-
ность, щеки и жаберные крышки покрыты чешуей. Боковая линия с переры-
вом: передняя ее часть от конца жаберной крышки идет ближе к спине, а зад-
няя - посредине тела. В боковой линии 30-33 чешуй. Кишечник очень длинный
(в 3,5-5 раз длиннее тела), с большим количеством петель.
D XI-XXIII 9-16; A III—IV (V,VI) 5-6. Позвонков 25-33 (Миронова, 1969).
Кариотип: 2п = 44, NF = 46 (Васильев, 1985).
Распространение. Родина тилапии - Восточная Африка от Занзибара до
Наталя, а также Израиль, Иордания и Сирия. В 1830-е годы началось ее бы-
строе распространение по странам Южной Азии и по тропическим архипела-
гам Индийского океана (Индонезия, Ява, Малайзия, Тайвань, Ямайка, Таити,
Гаваи, Филиппины и др.). В России появилась впервые как аквариумный объ-
ект. а с 1960-х годов как объект рыборазведения в прудовых хозяйствах юга и
в водоемах-охладителях ГРЭС, ТЭЦ, АЭС (Веригин, 1963; Краев, 1966). Часто
из рыбоводных хозяйств попадает в естественные водоемы, например в Крас-
нодарском крае (Емтыль, 1997).
Возраст и размеры. Максимальный возраст 11 лет, максимальные дли-
на 39 см и масса 700-800 г (единичные особи до 40 см и 2,5 кг) (Миронова,
1969).
75
Образ жизни. Эвригалинный вид, обитает в пресных и солоноватых во-
дах рек, эстуариев, а также в больших и малых тропических водоемах (озе-
ра, водохранилища, речки, лагуны, болота, каналы). В культуре использует-
ся в прудах, отстойных водоемах и рисовых чеках. Молодь питается планк-
тонными диатомовыми водорослями, а взрослые — водорослями (нитчатые,
зеленые), мягкой высшей растительностью и разлагающимися раститель-
ными остатками, а также мелкими беспозвоночными (личинками хироно-
мид и других водных насекомых), ракообразными, червями и мелкой рыбой
(гамбузией). В водоемах-охладителях поедает нитчатые, синезеленые и ха-
ровые водоросли, а также поденок, жуков. В культуре прудов — как естест-
венные, так и искусственные корма, включая кухонные остатки. Суточный
рацион в 3-месячном возрасте составляет 20-40% массы тела при питании
животным кормом и 30—60% - растительным; в 6 мес - 5-15 и 10-25% соот-
ветственно: у годовалых рыб - около 3% и в возрасте 6-7 лет - 1,0-1,5%. Ти-
лапия отличается большой экологической пластичностью, обитая в широ-
ком диапазоне температур. Темп роста и конечные размеры сильно варьи-
руют в зависимости от условий существования. Максимальных размеров -
36-38 см - она может достигать или за несколько месяцев, или за несколь-
ко лет. В холодных, малокормных или небольших водоемах она вообще
редко достигает 15 см.
Температурный режим и кормность водоема обусловливают как темп ро-
ста, так и темп полового созревания. Возраст полового созревания варьирует
от 2 до 5 мес, длина - от 6,5 до 18 см и масса от 2,5 до 69 г. По достижении по-
ловой зрелости рост замедляется, и рыбы начинают интенсивно размножать-
ся. Икрометание происходит в течение теплого сезона при температуре воды
20-35 °C, в тропиках - круглый год. Интервалы между икрометанием соста-
вляют 16-40 сут. За год одна самка может нереститься от 5 (Вьетнам) до 11
раз (Африка). Плодовитость от 80 икринок (у самок 8-9 см и 11-12 г) до
1000 икринок (у рыб 20 см и массой 150 г). Нерест в природе осуществляется
на гравийном грунте. Партнеры после непродолжительных брачных игр вы-
капывают ямку, самка откладывает туда икру, самец ее оплодотворяет. Икра
имеет грушевидную форму размером 2,25-2,95 х 1,55-2,30 мм. Через некото-
рое время самка прогоняет самца, а икру собирает в рот для дальнейшей ин-
кубации. Продолжительность вынашивания икры от 2-3 сут до 21 дня. Ли-
чинки при выклеве имеют длину 5-9 мм и еще несколько дней используют
рот матери как убежище при опасности. Благодаря такой заботе о потомстве
выживание молоди очень высокое, а при частом икрометании численность
тилапии быстро растет. В ряде прудовых хозяйств уже в возрасте 4,5 мес мо-
лодь достигает массы 250-380 г, а максимальная рыбопродуктивность -
14-18 т/га. Тилапия нетребовательна к содержанию кислорода в воде (до 1
мг/л) и загрязнению при уровне pH 6,0-6,5, что делает ее удобным объектом
для рыбоводства.
Статус вида. В естественном ареале - ценный объект рыболовства и интен-
сивного прудового хозяйства. В России - объект рыбоводства в прудах и в во-
доемах-охладителях, кроме того, в последних используется как биологический
мелиоратор (Веригин, 1963).
Автор-составитель: О.А.Попова.
76
Класс Osteichthyes
Отряд Perciformes
Семейство Eleotrididae
Род Micropercops Fowler et Bean, 1920
218. Micropercops cinctus (Dabry, 1872) - китайский элеотрис
Описание и систематика. Форма тела бычковидная. Тело вальковатое, сжа-
тое с боков, хвостовой стебель короткий. Голова также сжата с боков. Рот не-
большой, конечно-верхний. Нижняя челюсть выдается вперед, верхняя че-
люсть чаще достигает вертикали переднего края глаза. Зубов на сошнике и на
нёбных костях нет, на челюстях зубы щетинковидные в несколько рядов. Ры-
ло и межглазничное пространство голые, остальная часть головы (крышка и
предкрышка) и спина покрыты циклоидной чешуей. На теле чешуя ктеноид-
ная, средней величины. Предкрышка без шипа. Диаметр глаза составляет око-
ло 25% длины головы. Язык на переднем крае закруглен. На голове имеются
сейсмосенсорные каналы и поры. Окраска светло-серая, на теле (6)8-13 широ-
ких поперечных темных полос, некоторые из них могут раздваиваться. На спи-
не и боках много темных пятен неправильной формы. Под глазом начинается
темная полоска и идет за задний край нижней челюсти. Радужная оболочка
глаз голубая. Самцы несколько отличаются от самок по пластическим призна-
кам и по окраске, особенно во время нереста. Спинные плавники темные, раз-
делены небольшим промежутком. Анальный расположен под вторым спин-
ным, хвостовой и грудные плавники закруглены. Анальная папилла конусооб-
разная. Имеется плавательный пузырь.
D! VII-IX (X); D21-П 8-13; А 1-П 7-11; Р I (10) 12-13 (14); V I 5. Попереч-
ных рядов чешуй 31-36, чаще 33-35. Жаберных тычинок 11-13 (14). Позвонков
32-33 (35), в том числе туловищных 16-18 и хвостовых 14-17. Подвидов нет
(Берг, 19496; Никольский, 1956; Еловенко, 1981; Рыбы Казахстана, 1992; Ан-
нотированный каталог..., 1998).
Систематика головешковых разработана слабо, поэтому относительно ро-
дового названия единства нет: ранее его помещали в род Eleotris или
Hypseleotris (Берг, 19496; Никольский, 1956; Рыбы Казахстана, 1992; The
Freshwater Fishes of China, 1982; Choi et al., 1990), затем одни авторы помещали
его в род Perccottus, другие - в род Micropercops. Первоначально вид имел на-
звание Hypseleotris swinhonis (Gunther, 1873), однако по правилу приоритета вид
должен называться М. cinctus (Dabry, 1872) (Васильева, Макеева, 1988; Анноти-
рованный каталог..., 1998).
Распространение. Пресноводный вид, населяет воды Северного Вьетнама,
п-ова Корея и Китая к северу от Сянгана (Гонконга). Есть в реках Ляохэ (Те-
лии) и Хуньхэ (Мукден) (The Freshwater Fishes of China, 1982; Choi et al., 1990).
77
В России из бассейна Амура известны всего 2 экз., пойманные в р. Ялухэ, впа-
дающей в р. Нонни, приток Сунгари (Берг, 19496; Никольский, 1956). В респуб-
ликах бывшего СССР впервые обнаружен в 1960 г. в Туркмении, куда он попал
вместе с растительноядными рыбами. Потом он расселился в Сырдарье и ее
притоках на территории Узбекистана, в Арнасайских озерах, в Чардарьинском
водохранилище и ниже по течению этой реки. В р. Или обитает на всем ее про-
тяжении, включая оз. Балхаш, Капчагайское водохранилище. Обнаружен также
в реках Каратал, Чу и в Кировском водохранилище (Рыбы Казахстана, 1992).
Возраст и размеры. Длина до 5 см, изредка 7 см и масса 1,1г (Choi et al.,
1990). В аквариуме доживали до 6,8 см (Рыбы Казахстана, 1992). Максималь-
ный возраст 5 лет.
Образ жизни. Данные по биологии этого вида из вод Амура практически не
известны. В водоемах Казахстана предпочитает стоячие или медленнотекущие
воды, в реках придерживается затишных мест. На п-ове Корея встречается в
водоемах с медленным течением и особенно в прудах, где довольно обилен
(Choi et al., 1990). Питается мелким бентосом (чаще личинки хирономид), план-
ктонными ракообразными, поедает также икру и молодь карповых рыб. Рас-
тет медленно. Половой зрелости достигает на втором году жизни при длине
2,4-4,1 см. Количество зрелых икринок в гонадах самок 120-350 штук. Нерест
порционный и сильно растянут во времени: с конца мая до сентября с пиком в
июне-августе. Самка откладывает до 7 порций икры в укрытия из камней
78
(гнезда обычно располагаются на нижней поверхности камня) или на твердый
субстрат (камни, раковины моллюсков, коряги) в один ряд. Икринки мелкие,
овальной формы, с выростами оболочки на одном из полюсов, с помощью ко-
торых икра приклеивается к субстрату. В одной кладке бывает 35-150 икринок
диаметром 0,9-1,0 мм. Желток окрашен в светло-желтый цвет, имеются яркие
жировые капли. В нересте с одним самцом участвует несколько самок. Отме-
чены брачные игры. Гнездо охраняет самец, он обмахивает кладку грудными
плавниками. Во время охраны гнезда самец не питается. Развитие икры про-
должается 10-14 дней при температуре 17-20°С, при 25-28°С - 4-5 дней. Пер-
вое время после вылупления предличинки остаются прикрепленными к суб-
страту. После этого происходит заполнение воздухом плавательного пузыря, и
при длине 2,8-3,2 мм они переходят к пелагическому образу жизни, который
позволяет им широко расселяться в пределах речных систем. Экзогенное пита-
ние начинается на вторые сутки. К придонному образу жизни элеотрис перехо-
дит в возрасте 30-40 дней. Молодь на ранних этапах развития питается ветви-
стоусыми и веслоногими ракоообразными, потом в ее рационе появляются ли-
чинки хирономид и водоросли; икра встречается в питании рыб после достиже-
ния ими длины 1,3-1,5 см (Васильева, Макеева, 1988; Рыбы Казахстана, 1992).
Статус вида. Промыслового значения не имеет.
Автор-составитель: Ю.С. Решетников.
Класс Osteichthyes
Отряд Perciformes
Семейство Eleotrididae
Род Perccottus Dybowski, 1877
219. Perccottus glenii Dybowski, 1877 - головешка-ротан
Описание и систематика. Форма тела бычковидная. Тело не очень удлинен-
ное, спереди вальковатое, сзади сжатое. Брюшные плавники не соединены в
диск. Голова большая приплющенная, ее длина укладывается не более 3 раз в
длине тела. На голове нет сейсмосенсорных каналов и пор, но есть три ряда под-
глазничных невромастов. Рот конечный, большой и широкий. Нижняя челюсть
выдается вперед, верхняя челюсть доходит до заднего края глаза. Зубы на сош-
нике имеются, на челюстях они щетинковидные и несколько изогнутые, клы-
ковидных зубов нет. Чешуя умеренной величины, на боках скорее ктеноидная,
на спине - циклоидная. Голова покрыта чешуей вплоть до середины лба, бока
головы также покрыты чешуей. Предкрышка без шипа. Спина обычно черно-
79
вато-зеленая, бока желтовато-зеленые, на боках темно-бурые пятна непра-
вильной формы. От рыла через глаз к концу предкрышки идет узкая темная
полоса. В период нереста самцы становятся черными, на лбу появляется не-
большое вздутие. У самцов спинные плавники сближены, выше и ярче окраше-
ны, чем у самок. На теле и непарных плавниках появляются сверкающие зеле-
ные пятна. Спинные плавники без колючек. Имеется плавательный пузырь.
Dj VI-VIII; D21-II 9-11; A I-III 7-10; Р I (10) 12-13 (14); V I 5. Поперечных
рядов чешуй 36-43. Жаберных тычинок 11-13 (14). Кариотип: 2п = 44, NF = 44;
все хромосомы акроцентрические (Берг, 19496; Никольский, 1956; Еловенко,
1981). Подвидов нет (Аннотированный каталог..., 1998).
В соответствии с первоописанием правильное название вида - Perccottus
glenii Dybowski, 1877 (Голубцов, 1990; Аннотированный каталог..., 1998). Это
написание было неоправданно исправлено Н.А. Варпаховским и С.М. Герцен-
штейном (1887) на Р. glehni, что и закрепилось во многих русских публикациях
(Берг, 19496; Никольский, 1956; Еловенко, 1985; и др.).
Распространение. Ротан населяет пресные воды на северо-востоке п-ова
Корея, Северного Китая и Приморья, обычен в нижнем и среднем течении
Амура, его притоках Сунгари, Уссури и в оз. Ханка. На север от лимана Аму-
ра идет до р. Тугур, впадающей в Охотское море, на юг - до Владивостока и
рек Суйфун и Туманная (Берг, 19496; Никольский, 1956) и далее на юг до
п-ова Корея. Ротан завезен аквариумистами в европейскую часть России, где
он сейчас активно расселяется по мелким водоемам Московской, Ленинград-
ской, Калининградской и Нижегородской областей. В Волге отмечен от вер-
ховьев до Иваньковского (впервые в 1995 г.), Саратовского (в 1983 г.) и Куй-
бышевского водохранилищ, где истребляет аборигенные виды рыб (Еловенко,
1985; Неелов, 1987; Рыбы Подмосковья, 1988; Аннотированный каталог...,
1998; Евланов и др., 1998). В 2001 г. первые экземпляры ротана пойманы в
Онежском озере (Карелия).
80
Возраст и размеры. Длина до 25 см, масса до 300 г, но чаще 8-12 см.
Образ жизни. Предпочитает стоячие водоемы, пруды и болота. Очень не-
прихотлив к условиям среды, особенно к дефициту кислорода в воде. Выдер-
живает почти полное высыхание и промерзание водоемов, зарываясь в ил. Из-
бегает водоемов с быстрым и даже умеренным течением. Населяет пруды,
мелкие, зарастающие и заболоченные озера, старицы рек. Ведет оседлый об-
раз жизни, охотится как типичный хищник-засадчик, спрятавшись в густых за-
рослях подводных растений.
Интересное наблюдение по зимовке ротана в небольших придаточных во-
доемах в бассейне Амура проведено рыболовами-любителями (Соколов, 2001).
Ротан в конце декабря образует большие скопления в полостях среди льда, за-
полненных воздушно-ледовой влажной массой. При прозрачном льде эти ско-
пления легко просматриваются с поверхности: полость во льду, в которой зи-
муют ротаны, имеет вид полусферы диаметром от 20 см до 2 м, ее верхняя точ-
ка располагается в 30-60 см от верхней кромки льда. Температура в ней близ-
ка к 0-1 °C. Рыбы находятся в состоянии оцепенения и будучи вынутыми слег-
ка шевелятся. При помещении в воду они быстро отходят и начинают активно
плавать. В придаточных водоемах Амура ротан “входит” в лед в конце декабря
и просыпается от спячки в конце апреля (Соколов, 2001). Однако в Подмоско-
вье и в ряде других водоемов европейской части России ротан вообще не впа-
дает в спячку. На московских прудах рыболовы ловят его круглый год, в том
числе и зимой (Рыбы Подмосковья, 1988).
Питается животной пищей (бентосом и реже планктоном) доступного раз-
мера любого вида, в том числе поедает молодь рыб и икру. Несмотря на малые
размеры, ротан очень прожорлив и почти всеяден. Излюбленной пищей в на-
чале лета являются головастики. К концу первого года достигает 3—4 см и мас-
сы 0,8 г, второго года - 6-7 см и 4 г, третьего года - 9-11 см и 11-12 г, четвер-
того года - 12-13 см и 13-15 г. Половой зрелости достигает в возрасте 2-3 лет
при достижении длины около 6 см.
Плодовитость до 1000 икринок. Порционный нерест бывает в мае-июле
при температуре воды 15-20°С. Откладывает икру на донные предметы (кор-
невища растений, коряги, камни), на нижнюю поверхность плавающих в воде
предметов и листья водной растительности. Самец охраняет кладку и молодь.
Икра удлиненной формы, размером 3,8 х 1,3 мм. Как и у других бычковидных
рыб, на одном конце икринки имеются клейкие ворсинки, которыми она при-
клеивается к субстрату. Икринки откладываются в один ряд, обычно*около по-
верхности воды. Вылупившиеся личинки имеют размер 5,5 мм. Плавательный
пузырь наполняется воздухом еще у эмбрионов в оболочке, что предупрежда-
ет опускание личинок на дно, в неблагоприятные кислородные условия. Личин-
ки сначала ведут пелагический образ жизни. К активному питанию приступа-
ют на вторые сутки после выклева, потребляя мельчайшие планктонные орга-
низмы, а затем и более крупных беспозвоночных (Никольский, 1956; Рыбы
Подмосковья, 1988). г
Статус вида. Непромысловый вид, раньше в местах естественного обитания
его вылавливали на корм собакам. В городских и пригородных водоемах евро-
пейской части России ротан часто является единственным объектом любитель-
ского рыболовства, численность его здесь возрастает, а ареал расширяется.
Автор-составитель: Ю.С. Решетников.
6 Атлас..., том 2
81
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Aboma Jordan et Starks, 1895
220. Aboma lactipes (Hilgendorf, 1878) - японская абома
Описание и систематика. Голова и грудь голые, бока покрыты крупной че-
шуей, впереди спинного плавника отдельные чешуйки. Два спинных плавника.
У половозрелых самцов лучи первого спинного плавника волосовидные. Голо-
82
ва сжата с боков. Язык без выемки. Зубы простые, конические. Имеется под-
глазничный ряд невромастов. На боках тела темные пятна. Брюшная присоска
обычно светлая в середине и темная по бокам. Характерна окраска хвостового
плавника: на его верхней части несколько поперечных рядов темных пятны-
шек, которых нет внизу плавника.
D[ VIII—IX; D2 I 10-12; A I 10-12. Боковой линии нет, поперечных рядов че-
шуй 33^4-0. Позвонков 32-33. Плавательный пузырь имеется. Подвидов нет
(Берг, 19496; Линдберг, Красюкова, 1975). Кариотип: 2п = 40, NF = 40 (Василь-
ев, 1985).
Распространение. Мелкий эвригалинный донный вид тихоокеанского побе-
режья Азии, от Амурского залива до берегов Китая, п-ова Корея и Японии
(Choi et al., 1990). В России - от Амурского залива, к северу от линии Седанка -
мыс Песчаный, до залива Петра Великого. Обитает в бухтах залива Посьет, в
оз. Лебяжье, р. Киевке и в оз. Тунайча на восточном побережье Сахалина, но
нет в лагуне Буссе и Вавайских озерах (Пинчук, 1978).
Возраст и размеры. Достигает длины 90 мм.
Образ жизни. Населяет эстуарии с песчаным и илистым дном. Биология ви-
да в наших водах практически не изучена, описана для японских вод (Dotu, 1959).
Статус вида. Хозяйственного значения не имеет.
Автор-составитель: Ю.С. Решетников.
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Acanthogobius Gill, 1859
221. Acanthogobius flavimanus (Temminck et Schlegel, 1845) -
японский речной бычок
Описание и систематика. В отличие от других дальневосточных бычков нет
подглазничного дуговидного ряда( невромастов, вместо него косой ряд, идущий
от заднего края глаза вперед и вниз. Бока головы и ее верх покрыты чешуей,
вверху чешуя доходит до глаз. Жаберные крышки вверху покрыты мелкой че-
шуей, аналогично покрыто чешуей и горло. Рот большой, углы рта доходят до
переднего края глаза. Губы к углам рта не расширены. Глаза сидят высоко и
близко расположены друг к другу, ширина плоского лба меньше диаметра гла-
за. Передние ноздри вытянуты в очень короткие трубочки. Воротник брюш-
ной присоски без лопастинок. Тело желтоватое, вдоль боков тянется несколь-
83
ко темных пятен, особенно четко выделяется пятно в основании хвостового
плавника. На жаберной крышке есть пятнышки, на рыле от глаза ко рту косые
темные полоски. На спинных плавниках темные пятнышки, ряд зигзагообраз-
ных темных пятен есть в верхней части хвостового плавника.
Di VIII-IX, D21 12-14, All 1-13. Спинные плавники явно разделены проме-
жутком. Во втором спинном плавнике не менее 12 лучей. В спинном и грудном
плавниках нет волосовидных лучей. Хвостовой плавник закруглен. Грудной
плавник темноватый, при основании с рядами темных пятнышек. Брюшные
плавники не доходят до ануса. Поперечных рядов чешуй 47-54. Есть плава-
тельный пузырь. Подвиды не описаны (Берг, 19496; Линдберг, Красюкова,
1975). Кариотип: 2п = 44, NF = 44 (Васильев, 1985).
Распространение. Встречается в бассейнах Японского и Желтого морей, на
тихоокеанском побережье Японии, от Хоккайдо к югу, есть в Китае и на
п-ове Корея (Choi et al., 1990). В России обитает от Амурского залива, к севе-
ро-востоку от линии Седанка - устье р. Раздольной, до залива Петра Велико-
го, отмечен в реках Цимухэ (в Уссурийском заливе) и Суйфун (Берг, 19496;
Линдберг, Красюкова, 1975; Пинчук, 1978).
Возраст и размеры. В Японии достигает максимальной длины 25 см, в зали-
ве Петра Великого - 18,7 см.
84
Образ жизни. Мелкий эвригалинный донный вид, проникающий в низовья
рек, в больших и средних реках с медленным течением встречается значитель-
но выше устья. Биология описана в работах японских авторов (Miyazaki, 1940;
Dotu, Mito, 1955; Imamura, Hashitani, 1957).
Статус вида. В Японии - промысловый вид, в Амурском заливе и устьях
впадающих в него рек - объект любительского лова.
Автор-составитель: Ю.С. Решетников.
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Benthophilus Eichwald, 1831
222. Benthophilus baeri Kessler, 1877 - пуголовка Бэра
Описание и систематика. Тело несколько укороченное, его передняя
часть и особенно голова расширенные и толстые. Ширина головы почти рав-
на ее длине, а при отведенных жаберных крышках почти круглая. Жаберные
щели маленькие. Рыло закругленное, его длина меньше половины заглазнич-
ного пространства. Рот большой, челюсти почти одинаковой длины. На под-
бородке небольшой кожистый усик. По одному усику имеется позади угла
рта с каждой стороны на том месте, где у большинства пуголовок продолго-
ватые кожистые складки. Спинные мышцы частично покрывают череп. На
85
поверхности головы нет каналов и пор боковой линии; нет подглазничного
продольного ряда невромастов. На щеках имеются 6 подглазничных попереч-
ных рядов невромастов. Передние ноздри вытянуты в небольшие трубочки,
прилегающие к верхней губе. Нет плавательного пузыря. Длина брюшной
присоски немного меньше длины грудного плавника. На теле с каждой сторо-
ны по 3 ряда костных бугров: в спинном ряду их 12-17, в боковом - 5-12, в
брюшном - 10-14. Самые крупные бугры в спинном ряду, по бокам тела и на
верхней стороне головы. Половозрелые самцы бугры теряют и становятся
голыми. Спина, верхняя часть головы и бока имеют бледно-серый цвет с тем-
но-серыми крапинками, нижняя часть тела светлая. Грудные, второй спинной
и хвостовой плавники испещрены темно-серыми крапинками, а анальный и
брюшные плавники - бледные. D, 1-П, D216-8, А I 5-8 (Берг, 19496; Рагимов,
1985а). Подвидов нет.
Распространение. Вид встречается в Каспийском море, в основном у вос-
точных берегов, наиболее многочислен в южной зоне (Казанчеев, 1981; Раги-
мов, 1981). В России - в западной части Северного Каспия: севернее о-ва Че-
чень, в Бахтемирской бороздине, у юго-восточного побережья о-ва Тюлений
(Рагимов, 1981), по побережью Дагестана, есть в р. Сулак (Аннотированный
каталог..., 1998).
Возраст и размеры. Достигает длины 80 мм и массы 10 г; предельный воз-
раст- 1 год (Берг, 19496; Казанчеев, 1981).
Образ жизни. Биология плохо изучена. Мелкий прибрежный морской
вид, заходящий в пресную воду. В холодное время года мигрирует на глуби-
ны 60-70 м. В море питается ракообразными (52% - кумацеи, амфиподы) и
полихетой нереис (47%). Обычные размеры 2,9-5,0 см, масса 1-5 г (Казан-
чеев, 1981). Нерест происходит со второй половины мая по август, редко за-
тягивается до конца сентября. Икра откладывается на плотных илистых или
илисто-песчаных грунтах с примесью ракушки. В Северном Каспии размно-
жается в мелководной зоне на глубинах 5-10 м, а в Среднем и Южном Кас-
пии нерестовые скопления рыб отмечались на глубинах 10-15 м. Икромета-
ние порционное. В яичниках зрелых самок содержатся две порции икринок
86
диаметром 1,2-1,8 и 0,3-0,5 мм. В июле, а иногда и в августе ловятся самки
с икрой на стадии зрелости III-IV. Абсолютная плодовитость самок длиной
3,2-5,2 см и массой 1,2-5,3 г составляла 450-1860 икринок. Период инкуба-
ции не известен. Мальки в море в конце июня-июле достигают длины
1,9-2,4 см при массе 0,2-0,5 г; к концу ноября они уже имеют размеры
2,7-3,7 см и массу 0,5-1,7 г. При этом половые продукты у них находятся на
II и П-Ш стадиях зрелости. К февралю следующего года они вырастают до
4—6 см при массе 2,0-8,5 г, и их гонады переходят на III-IV стадию зрелости
(Рагимов, 1985а). .
Статус вида. Мелкий малоизученный вид. Хозяйственного значения не имеет.
Автор-составитель: К.И. Москалькова.
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Benthophilus Eichwald, 1831
223. Benthop hilus casachicus Rahimov, 1978 - казахская пуголовка
Описание и систематика. Передняя часть тела, особенно голова, расшире-
на. Ширина головы немного меньше ее длины. Имеется желобообразное уг-
лубление между глазами и позаДи них. Тело довольно высокое, высота тела
больше его ширины. Грудные плавники значительно длиннее хвостового, но
короче брюшных. Длина основания второго спинного плавника и высота его
лучей аналогичны тем же показателям анального плавника. Поверхность те-
ла и головы покрыта крупными и мелкими звездчатыми костяными буграми,
пластинками и зернышками с шипиками. По бокам тела крупные бугры об-
разуют 3 продольных ряда с каждой стороны: спинной ряд содержит 25-27
87
бугров и тянется от затылка до хвостового плавника, боковой из 9-17 бугров
начинается на уровне D( и не доходит до хвостового, брюшной из 18-21 бугра
начинается позади основания грудных плавников и достигает основания хво-
стового. Цвет тела бурый или беловато-серый. Поверхность головы и туло-
вища усеяна многочисленными темно-бурыми пятнышками. На спине име-
ются 3 крупных темно-бурых пятна: первое не сплошное, а в виде кольца рас-
полагается между затылком и второе - полукольцевидное охватывает
заднюю часть Dt; третье - кольцевидное охватывает заднюю часть D2. Кро-
ме них имеются более мелкие пятна на боках тела и у основания хвостового
плавника. У экземпляра, пойманного на большой глубине (46 м), тело про-
зрачное и пятна выделяются слабо. Спинные, грудные и хвостовой плавники
испещрены темно-серыми полосками, а анальный и брюшные плавники бе-
ловатого цвета и без пятен. Dj III; D2 I 8-9; A I 8-9 (Рагимов, 1978). Подвидов
нет. Первоначально был описан как подвид звездчатой пуголовки В. stellatus
casachicus Rahimov, 1978.
Распространение. Каспийское море: восточное побережье Среднего Кас-
пия, южнее Кендерлинской косы, и северная часть о-ва Огурчинского; в водах
России - дельта Волги у вододелителя (Рагимов, 1978; Васильева, 1983).
Возраст и размеры. Достигает длины 76 мм (Рагимов, 1978).
Образ жизни. Очень редкий неизученный вид, известный по единичным
экземплярам. Встречается в морской и пресной воде (Рагимов, 1978; Василь-
ева, 1983).
Статус вида. Хозяйственного значения не имеет.
Автор-составитель: К.И. Москалькова.
88
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Benthophilus Eichwald, 1831
224. Benthophilus granulosus Kessler, 1877 - зернистая пуголовка
Описание и систематика. Передняя часть тела расширена. Голова с отведен-
ными жаберными крышками, почти круглая, плоская, без продольного желобка
между глазами. Рыло закругленное и укороченное, его длина в 2,0-2,5 раза мень-
ше заглазничного пространства. Тело и голова сверху и с боков густо покрыты
мельчайшими костными зернышками; они имеются также на рыле и на нижней
стороне тела, а у молоди - и на нижней стороне головы. Бугры отсутствуют.
У половозрелых самцов зернышки теряются, и тело становится голым. На под-
бородке короткий кожистый усик. Углы рта доходят до вертикали середины гла-
за. Кожистая складка за углами рта развита слабо. Брюшная присоска широкая,
ее длина заметно больше длины грудного и хвостового плавников. Грудные
плавники не доходят до начала D2. Окраска тела в зависимости от места обита-
ния варьирует от серой до желтоватой. На спине и по бокам тела 3 крупных тем-
но-бурых пятна: первое в области Dt, второе охватывает кольцом заднюю поло-
вину основания D2 и третье - в виде полоски у основания хвостового плавника.
На спинных, грудных и хвостовом плавниках мелкие бурые пятнышки. Плава-
тельного пузыря нет. D, Ш-IV (V); D2 I (5) (6) 7—9; АI (5) 6-8 (Берг, 19496; Раги-
мов, 19856). Подвидов нет (Аннотированный каталог..., 1998).
Распространение. Эндемичный вид Каспийского моря, в прибрежье кото-
рого он очень широко распространен, но малочислен, есть перед устьями Вол-
ги и Урала. В России - все каспийское побережье и низовья Волги; вид обычен
в дельте и отмечен в самой Волге, севернее Астрахани (Казанчеев, 1981; Раги-
мов, 1981).
Возраст и размеры. Достигает длины 66 мм и массы 4 г. Предельный воз-
раст - 2 года (Казанчеев, 1981; Рагимов, 1966, 19856).
Образ жизни. В море населяет олигогалинную и мезогалинную зоны, захо-
дит в реки. Встречается во всех зонах дельты Волги. В теплое время года зер-
нистая пуголовка обитает в прибрежной зоне, где размножается, питается
предпочтительно ракообразными. В холодное время года она мигрирует на
глубины 60-70 м. Длина половозрелых особей в возрасте 1-2 года - 3,4-6,6 см.
В Северном Каспии нерест с конца апреля по май, преимущественно на мелко-
водьях среди растительности. Первые мальки появляются в мае и обитают в
придонных слоях. Икрометание в дельте Волги происходит с конца мая до кон-
ца июля. Молодь в этом районе появляется в середине июня, ловится редко,
единичными экземплярами (Казанчеев, 1981). Вылупившаяся из икры молодь
пуголовки имеет длину 4,5-5,0 мм и большой яйцевидный желточный мешок.
У нее короткое толстое тело, широкая голова, высокий и выпуклый лоб, ры-
89
ло тупое, рот нижний. Все плавники полностью сформированы. На голове,
спине и на боках тела 3 овальных пигментных пятна. Желточный мешок рас-
сасывается при длине 6,0-6,5 мм. При длине 12-15 мм голова сильно уплощает-
ся, лоб становится низким, глаза как бы “проваливаются”, передние ноздри вы-
тягиваются в трубочки, появляется кожистый усик на подбородке. В октябре
сеголетки достигают длины 25-36 мм и по строению не отличаются от взрос-
лых рыб; их тело покрыто мелкими костными зернышками (Коблицкая, 1981).
Статус вида. Мелкий вид, хозяйственного значения не имеет.
Автор-составитель: К.И. Москалькова.
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Benthophilus Eichwald, 1831
225. Benthophilus macrocephalus (Pallas,1787) - каспийская пуголовка
90
Описание и систематика. Голова большая, широкая и плоская, без продоль-
ного желоба; ширина головы почти равна ее длине. Каналов и пор сейсмосен-
сорной системы на голове нет. Затылок и темя густо покрыты шипиками. По
всему телу мелкие костные зернышки и ряды крупных звездчатых пластинок
(бугров). На боках тела бугры образуют по 3 продольных ряда: в спинном ря-
ду 23-25, в брюшном - 18-24 бугров, боковой ряд доходит до середины хвосто-
вого стебля. На темени и на затылке два ряда бугров являются как бы продол-
жением спинных. Горло, жаберные перепонки и брюхо голые. Половозрелые
самцы утрачивают зернышки и бугры и становятся совершенно голыми. На
подбородке кожистый усик, такие же усики в каждом углу рта. Брюшная при-
соска длиннее грудных плавников. Окраска однотонная, пепельно-серая, без
пятен. Плавательного пузыря нет. D[ III—IV; D2 I (II) 7-9 (10); A I (6) 7-9 (Берг,
19496; Казанчеев, 1981). Подвидов нет.
Распространение. Каспийское море от северного до южного побережья
(Энзели, Астрабадский залив): в Северном Каспии встречается повсеместно,
есть в устьях Волги и Урала; в Среднем Каспии держится вблизи западных бе-
регов, в Южном Каспии - до Астары, но довольно редка (Берг, 19496; Казан-
чеев, 1981). У берегов России - в Северном Каспии, есть в дельте Волги, встре-
чается даже в районе Астрахани, в Среднем Каспии - преимущественно в рай-
оне от о-ва Чечень до устья р. Сулак, отмечена в Новом устье Терека и в Са-
муре (Казанчеев, 1981; Васильева, 1983; Аннотированный каталог..., 1998).
Возраст и размеры. Среди пуголовок самая крупная: достигает максималь-
ной длины 12,6 см и массы 35 г, но в уловах преобладают особи длиной 5-8 см
и массой 5-16 г. Продолжительность жизни 1 год (Берг, 19496; Казанчеев,
1981; Рагимов, 1966, 1985а).
Образ жизни. Широко распространенный, но малочисленный вид. Почти
во все сезоны года держится в прибрежной полосе. В теплое время года кас-
91
пийская пуголовка обычна на глубине 0,5-10 м, зимой переселяется на глуби-
ны 20-25 м, но в Южном Каспии остается в прибрежной зоне (Рагимов, 1981,
1988). Питается в основном моллюсками (48%), рыбой (16%), полихетой нере-
ис и в меньшем количестве ракообразными. Из моллюсков предпочитает син-
десмию и кардиум, из рыб - мелкие виды бычковых (Рагимов, 1988). Во время
нереста образует небольшие скопления в прибрежных участках моря, где име-
ются плотные илистые или илисто-песчаные грунты с примесью ракуши. Не-
рестится в Южном и Среднем Каспии в апреле-мае, редко до середины июня, в
Северном Каспии - позднее. Икра откладывается в крупные пустые раковины
моллюсков. Икрометание порционное. В яичниках половозрелых самок ик-
ринки двух размерных групп: 2,5-3,0 и 0,3-0,6 мм. Абсолютная плодовитость
самок 5-8 см длиной и массой 5-16 г составляла 700-3180 икринок. Зрелая ик-
ра овальной формы, имеет размеры 5,0 х 1,75 мм (Рагимов, 1985а). Первая мо-
лодь, пойманная в восточной части Северного Каспия в мае-июне, имела дли-
ну 6-14 мм. У нее удлиненное тело, большая, широкая и плоская голова, хоро-
шо развита брюшная присоска. Тело интенсивно пигментировано, с тремя ря-
дами шипов с каждой стороны, заметны два темных пигментных пояска попе-
рек тела в области двух спинных плавников (Казакова, 1951). В июне молодь
уже имела длину 2,4-3,5 см и массу 0,3-1,0 г. К концу сентября сеголетки дос-
тигали длины 4-6 см и массы 1,5-6,2 г, а к концу ноября - 7-8 см и 10-15 г.
В феврале-марте следующего года они имели гонады на III-IV стадии зрело-
сти (Рагимов, 1985а).
Статус вида. Мелкий малочисленный вид. Хозяйственного значения не
имеет.
Автор-составитель: К.И. Москалькова.
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Benthophilus Eichwald, 1831
226. Benthophilus magistri Iljin, 1927 -
азовская пуголовка, пуголовка магистра
Описание и систематика. Тело удлиненное, его передняя часть и голова
расширены, но ширина головы заметно меньше ее длины. Поверхность голо-
вы плоская, без желобообразного углубления. Тело и голова покрыты мелки-
ми зернышками и костными пластинками (бугорками), мало выдающимися из
кожи. Бугорки образуют 3 слабо заметных продольных ряда: в спинном -
21-27, в боковом - 9-20, в брюшном - 15-26 пластинок. Более крупные пла-
92
стинки располагаются по бокам головы. Под подбородком есть небольшой ко-
жистый усик, округлые кожистые складки располагаются в углах рта. Брюш-
ная присоска расширена и длиннее грудных плавников. Окраска тела серая, без
пятен и полос. D, III-IV (V); D21 (7) 8—11; A I (6) 8-10 (Берг, 19496; Световидов,
1964; Казанчеев, 1981; Рагимов, 1985а).
Номинативный азовский подвид В. magistri magistri Iljin, 1927 обитает в
Азовском море, а в Каспийском море живут 2 других подвида. Северо-каспий-
ские популяции выделяются в подвид Абдурахманова В. magistri abdurahmanovi
Rahimov,1978; они отличаются от номинативного азовского подвида меньшим
числом лучей в спинном и анальном плавниках (Рагимов, 1978; Пинчук, Раги-
мов, 1979). Южнокаспийские популяции рассматриваются как ленкоранский
подвид В. magistri lencoranicus Rahimov, 1978; по числу лучей и бугров в боко-
вых рядах на теле они более сходны с номинативным подвидом, но отличают-
ся формой рыла и туловищных бугров (Рагимов, 1982).
Распространение. Азовский подвид населяет опресненные части Азовского
моря, главным образом Таганрогский залив, Миусский, Ейский и опресненные
приморские лиманы Кубани, Ахтанизовский лиман, на запад до Керченского
пролива (Световидов, 1964); в Сиваше и западной части Азовского моря его
нет; в совершенно пресной воде обнаружен в лимане Куркуй (р. Кубань). По-
двид В. т. abdurahmanovi обитает в Северном Каспии почти повсеместно: в при-
уральских водах, в устье Волги, у островов Тюлений и Чечень и на юг вдоль за-
падного побережья Среднего Каспия до Махачкалы (Рагимов, 1978; Казанче-
ев, 1981). Подвид В. т. lencoranicus встречается в Южном Каспии - в юго-вос-
точной части в районе Зеленого и Белого Бугров и вдоль ленкораньского по-
бережья (Рагимов, 1982). В России пуголовка магистра представлена 2 подви-
дами: азовская пуголовка - по побережью Азовского моря и подвид Абдурах-
манова - в Северном Каспии, у о-вов Тюлений и Чечень, на юг до Махачкалы,
93
в дельте Волги и Новом устье Терека (Световидов, 1964; Рагимов, 1981, 1982;
Казанчеев, 1981; Васильева, 1983; Аннотированный каталог..., 1998).
Возраст и размеры. Достигает максимальной длины 8,5 см. Продолжитель-
ность жизни - 1 год.
Образ жизни. Мелкий морской вид, проникающий в пресную воду, местами
весьма многочислен (особенно в Северном Каспии). Биология мало изучена.
В Азовском море придерживается предустьевых участков с илистыми грунта-
ми, в море - с кардиевым ракушечником, питается беспозвоночными. В Север-
ном Каспии половозрелые рыбы обнаруживаются на мелководьях с глубинами
3-5 м, на плотных илистых или песчаных грунтах с раковинами моллюсков.
Нерест проходит с мая по август, икрометание порционное. В яичниках поло-
возрелых самок содержится крупная (диаметром 1,1-1,6 мм) и мелкая
(0,3-0,5 мм) икра. Наименьшая плодовитость - 577 икринок - отмечена у сам-
ки длиной 4 см и массой 1,75 г, а наибольшая - 1750 икринок - у самки длиной
5,6 см и массой 5,3 г. В середине июня ловили молодь длиной 14-19 мм, в кон-
це сентября - 31—48 мм и массой 0,8-2,3 г (Рагимов, 1982, 1985а).
Статус вида. Малоизученный мелкий вид. Хозяйственного значения не имеет.
Автор-составитель: К.И.Москалькова.
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Benthophilus Eichwald, 1831
227. Benthophilus mahmudbejovi Rahimov, 1976 -
пуголовка Махмудбекова
Описание и систематика. Верхняя сторона головы и туловища покрыты от-
носительно мелкими костными буграми и пластинками со слабо развитыми
шипиками. Бугры на боковой поверхности головы значительно крупнее. Круп-
ные бугры по бокам тела образуют 3 продольных ряда с каждой стороны:
спинной из 20-27 бугров, боковой (9-17) и брюшной (17-23). Тело несколько
укороченное, его передняя часть и голова сильно сплющены. Высота головы
вдвое меньше ее ширины, а ширина головы всегда меньше ее длины. Между
глазами на поверхности головы нет углубления. Передние носовые отверстия
в виде коротких, прилегающих к губе трубочек. Позади подбородка неболь-
шой кожистый усик, а в углах рта по небольшой удлиненной кожистой склад-
ке. Челюсти равной длины. Длина хвостового стебля в 2 раза меньше длины го-
ловы. Лучи в D2 длиннее, чем в анальном. У половозрелых самцов костяные
бугры и пластинки спадают, тело становится голым, а плавники разрастаются,
щеки расширяются и становятся более вздутыми. Цвет тела сероватый или
желтовато-бурый. На спине и у основания хвостового плавника всегда имеют-
ся 3 темно-бурых пятна (перевязки). У рыб, пойманных на большой глубине
94
(40 м), тело прозрачное и перевязки на теле мало заметны. D, III—IV; D21 6-10;
A I 6-9 (Рагимов, 1976). Подвидов нет (Аннотированный каталог..., 1998).
Распространение. Каспийское море и Таганрогский залив Азовского моря.
В Северном Каспии вид встречается почти повсеместно, в Среднем и Южном
Каспии - только у восточных берегов (мысы Песчаный, Ракушечный, Кендыр-
линская коса, мыс Карасингир, п-ов Челекен, о-в Огурчинский) (Рагимов,
1976). В России - Таганрогский залив, Волга до Волгограда, ее притоки и дель-
та, каспийское побережье (Кизлярский залив, северное побережье о-ва Тюле-
ний) (Рагимов, 1976, 1981).
Возраст и размеры. Максимальная длина 6,6 см. Живет 1 год.
Образ жизни. Мелкий эвригалинный вид. Биология изучена слабо. В Се-
верном Каспии средняя длина самцов 51 мм, самок - 49 мм (Казанчеев, 1981).
Половозрелые рыбы в брачном наряде встречаются в Северном Каспии на
глубине 1-11 м, чаще - на 3-6 м; а в Среднем и Южном Каспии - на глубине
2(Р40 м. Нерест сильно растянут: в Северном Каспии он продолжается с мая по
август (иногда до конца сентября), а в Среднем и Южном - со второй полови-
ны апреля до ноября-декабря. Икрометание порционное. В гонадах половозре-
лых самок имеются икринки 3 размерных групп: первая - с диаметром
1,5-2,0 мм, вторая - 0,3-0,5 мм и третья - более мелкие. Наименьшая абсолют-
ная плодовитость около 100 икринок - зафиксирована у самки 1,8 см длиной и
массой 0,15 г, а наибольшая плодовитость - 518 икринок - у самки 4,7 см дли-
ной и массой 3,8 г. Молодь от раннего нереста к середине июля достигает дли-
ны 14-17 мм и массы 0,05-0,10 г, к концу сентября - 20-32 мм и к началу нояб-
ря 37—49 мм и массы 1-2 г. Многие особи к этому времени имели гонады на ста-
дии зрелости II и П-Ш (Рагимов, 1985а).
Статус вида. Мелкий вид. Хозяйственного значения не имеет.
Автор-составитель: К.И. Москалькова.
95
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Benthophilus Eichwald, 1831
228. Benthophilus stellatus (Sauvage, 1874) - звездчатая пуголовка
Описание и систематика. Тело сверху сильно уплощенное, без чешуи. Го-
лова плоская, широкая, без углубления между глазами. Жаберная щель не-
большая. Передние ноздри имеют вид коротких трубочек, а на подбородке
один маленький клиновидный усик. За углами рта с каждой стороны имеется
продольная кожистая складка. На голове и теле шиповатые или бугорчатые
костные пластинки. На теле они образуют 3 ряда: спинной, боковой и брюш-
ной. В спинном ряду 25-30 пластин, они сильно выдаются из кожи; в брюш-
ном - 20-25. Половозрелые самцы их теряют и становятся совершенно голы-
ми; голова у них приплющивается, щёки сильно раздуваются, грудные плав-
ники удлиняются (Троицкий, Цуникова, 1988). Окраска светло песочная или
серая. На теле 3 поперечных коричневых пятна, окаймляющие плавники:
первое - кольцевидное, спереди прерванное, охватывает Dt; второе - кольце-
видное, окаймляет заднюю часть D2 и третье - у основания хвостового плав-
ника. D1 III—IV; D2 I (6) (7) 8-9 (10); А I 7-9 (Берг, 19496; Световидов, 1964).
Кариотип: 2п = 46, NF = 46-48 (Васильев, 1985). Каспийские (более глубоко-
водные) популяции выделялись в особый подвид (Берг, 19496; Пинчук, Раги-
мов, 1979), который отличается сильным развитием шипов, однако система-
тическое отношение его к номинативному подвиду остается пока неясным
(Аннотированный каталог..., 1998).
Распространение. Бассейны Черного, Азовского и Каспийского морей:
населяет опресненные лиманы, заливы, реки и прибрежные озера (Светови-
дов, 1964; Miller, 1986). В Черное море не выходит, но по рекам поднимается
довольно высоко: Дунай - до Свищово, Днестр - до Бендер, Буг - до порога
Гард, Днепр - до порога Вольного, Дон - до Воронежа (Берг, 19496), но с
1950-х годов в Воронежской области не встречается (Федоров, 1960). В Кас-
пийском море звездчатая пуголовка известна из всех районов моря, но везде
сравнительна редка (Казанчеев, 1981). В России - Азовское море, Миусский,
Ахтанизовский и Ейский лиманы, заходит в р. Бейсуг, Керченский пролив,
Дон и Цимлянское водохранилище (Берг, 19496; Световидов, 1964); Каспий-
ское море, дельта Волги и Волга (Казанчеев, 1981; Васильева, 1983). Раньше
в Волге встречалась лишь на 30 км выше Астрахани, теперь известна во мно-
гих водохранилищах Средней Волги: сначала звездчатая пуголовка была об-
наружена в Куйбышевском водохранилище в 1970-х годах, в настоящее вре-
мя обычна в Куйбышевском, Саратовском и Горьковском (Евланов и др.,
1998). Возможно, этот вид вселен в волжские водохранилища вместе с кормо-
96
выми организмами, которые завозились на Среднюю Волгу из низовьев Вол-
ги и Дона (Евланов и др., 1998).»
Возраст и размеры. Достигает длины 13,5 см и массы более 23 г, обычно го-
раздо мельче, живет 1 год (Берг, 19496; Казанчеев, 1981).
Образ жизни. В море придерживается опресненных районов, высоко под-
нимается в реки. Предпочитает слабосолоноватые (до 8%о) и пресные воды.
Держится преимущественно в русле, на замедленном течении, на илистых или
заиленных грунтах. В водохранилищах Волги предпочитает биотопы с твер-
7 Атлас ... том 2
97
дым или илистым грунтом, часто встречается на значительной глубине. Пита-
ется мелкими донными беспозвоночными и моллюсками. В Куйбышевском во-
дохранилище весной потребляет личинок хирономид, а летом и осенью - мол-
люсков (дрейссену, вальвату, пизидиум) и бокоплавов. Короткоцикловый вид,
скорее всего большинство особей живут один год. Половозрелыми становятся
при длине 3,5 см. Нерестится в Азовском море с апреля по июль, нерест порци-
онный на заиленных песчаных грунтах с крупными раковинами, на которые
откладывается икра (Троицкий, Цуникова, 1988). Нерест в волжских водо-
хранилищах бывает в мае-июне. Икрометание порционное. После нереста
самки сразу погибают, самцы охраняют кладку и гибнут после выхода молоди
(Евланов и др., 1998). Плодовитость от 700 до 3500 икринок. Икра олигоплаз-
матическая, яйцевидной формы, высотой 4,0-4,5 мм и шириной 2,1-2,4 мм. Раз-
витие икры идет по прямому типу без стадии свободной личинки. Вылупивша-
яся молодь имеет длину 4,5-5,0 мм и желточный мешок. При длине 6,5-7,0 мм
мешок резорбируется, и мальки по всем признакам напоминают взрослых
рыб (Коблицкая, 1981). Мальки питаются в основном бентосными рачками,
позднее в питании появляются нереис и моллюски.
Статус вида. Мелкий вид. Хозяйственного значения не имеет. Стал обычным
видом в волжских водохранилищах, где входит в состав пищи ерша и судака.
Авторы-составители: Ю.С. Решетников, К.И. Москалькова.
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Caspiosoma Iljin, 1927
229. Caspiosoma caspium (Kessler, 1877) - каспиосома
Описание и систематика. Тело удлиненное, голое, без чешуи. Голова узкая,
приплюснутая. Межглазничное расстояние меньше диаметра глаза. Нет плава-
тельного пузыря. На поверхности головы нет каналов и пор системы боковой
линии, нет также подглазничного продольного ряда невромастов, на щеках
имеется 6 рядов подглазничных невромастов. Спинные мышцы сверху покры-
вают лишь половину черепа; передние ноздри в виде коротких трубочек. Хво-
стовой плавник закругленный. Окраска беловатая. Бурые окаймляющие поло-
сы имеются на спине перед началом D2 и в конце его, с обеих сторон хвостово-
го плавника по пятну. На щеках от глаз идет продольная полоска. Нерестую-
щие самцы темнее, задний конец D( и весь анальный плавник у них становятся
черными. D, (V) VI (VII), D2 I—II (10) 11-13, А I (7) 8-10. Подвидов нет (Берг,
19496; Световидов, 1964; Троицкий, Цуникова, 1988; Емтыль, 1997).
98
Распространение. Дельты рек, впадающих в северо-западную часть Черно-
го моря: в Дунае и Пруте не обнаружена; в бассейне Днестра обитает в Курчу-
ганском водохранилище, есть в низовьях Днепра, до Берислава, и в низовьях
Буга, в Днепровско-Бугском и Березанском лиманах. В России каспиосома
встречается в кубанских лиманах, в дельте Дона и в устьях других рек Азовско-
го моря (Световидов, 1964; Троицкий, Цуникова, 1988; Емтыль, 1997). В Кас-
пийском море обитает в его северной части и у западных берегов Среднего
Каспия, обычна в авандельте Волги, реже в Нижней Волге (Казанчеев, 1981).
Возраст и размеры. Мелкий вид. Достигает длины 5 см, но обычная длина
2-4 см и масса 0,2-1,0 г (Световидов, 1964; Казанчеев, 1981).
Образ жизни. Биология вида плохо изучена. Обитает в солоноватых и прес-
ных водах. Питается бентосом. До зарегулирования стока Волги каспиосома
размножалась только в дельте, в последние годы она распространилась
выше и размножается в Волго-Ахтубинской пойме. Икрометание в низовьях
Волги бывает с конца мая по конец июля, или даже до августа (Казанчеев, 1981;
Коблицкая, 1981). Икра откладывается в мелководной зоне, в проточных местах
в пустые раковины моллюсков. Плодовитость низкая, в кладке 20-60 ик-
ринок (Казанова, 1951). Икра крупная, олигоплазматическая, с прозрачной обо-
лочкой, удлиненной, эллипсовидной формы (длина 4,25-4,34 мм, наибольшая
ширина 1,75-1,80 мм). Верхний конец икринки заострен, нижний более тупой,
пучком клейких нитей прикреплен к раковине. Икринки плотно сидят одна к
другой. Оплодотворенную икру находили и в Северном Каспии (Казанчеев,
1981). Нерест в Краснодарском крае отмечен с июня по август в верховьях эсту-
арных частей и в низовьях рек (ЕМтыль, 1997). Развитие протекает по прямому
типу, без пелагической личинки. Вылупившаяся молодь имеет длину 4,5-5,0 мм,
тело короткое и высокое, с крупной головой, у нее сформированы все плавники,
но имеется большой яйцевидной формы желточный мешок. Вдоль всего тела
от глаз до хвостового плавника тянется темная полоса со сгущениями пиг-
мента под Dj и D2 и у основания хвостового плавника. Желточный мешок у
молоди рассасывается при достижении длины 6 мм. По мере роста у них вытяги-
99
вается рыло, голова становится более плоской и широкой, а хвостовой стебель
более тонким. Первую молодь каспиосомы находили чаще в западной части
дельты Волги в июле. Она ведет донный образ жизни, ловится редко, единичны-
ми экземплярами и чаще в темное время суток. В августе-сентябре мальки при-
обретают черты взрослых рыб при длине 15-22 мм (Коблицкая, 1981).
Статус вида. Хозяйственного значения не имеет. В питании хищных рыб
каспиосома не обнаружена, но отмечена в желудках бычков (гонец и головач).
Авторы-составители: К.И. Москалькова, Ю.С. Решетников.
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Chaenogobius Gill, 1858
230. Chaenogobius annularis Gill, 1859 -
кольчатый дальневосточный бычок
Описание и систематика. Тело покрыто мелкой ктеноидной или циклоидной
чешуей. Голова у молодых особей узкая, цилиндрическая с небольшим ртом, с
возрастом она становится плоской. Верхняя и боковая поверхность головы го-
лая. За затылком и на груди чешуя очень мелкая, на брюхе она мелкая и легко
опадает. Рот большой, задний конец верхней челюсти у взрослых особей заходит
за вертикаль переднего края глаза. Нижняя челюсть обычно выдается вперед.
Язык спереди выемчатый. Зубы на челюстях в несколько рядов, клыков нет. Пе-
редние ноздри немного вытянуты. Имеется несколько подглазничных продолы
ных рядов невромастов, подглазничных поперечных рядов нет. Спинные плавни-
ки раздвинуты, в задней части D, имеется резко выраженное черное пятно. На
голове, теле и плавниках имеются буроватые пятна. Отмечаются вариации в ок-
раске и форме тела в зависимости от пола и возраста рыб. Возможно, в р. Амур
обитает карликовая форма с более короткой верхней челюстью. Чешуя на бо-
ках тела располагается вплотную, на хвостовом стебле она частично ктеноидная.
Dj (V) VI (VII), D21 (9) 10—12, A19-11. Поперечных рядов чешуй 66-84. Жа-
берных тычинок 9-12. Подвидов нет (Берг, 19496; Линдберг, Красюкова, 1975;
Пинчук, 1978, 1984).
Распространение. Бассейны Японского, Желтого, Охотского и Восточно-
Китайского морей. У берегов Японии от Хоккайдо до Иокогамы, имеется в Се-
верном Китае и на п-ове Корея. В России - реки побережья Хабаровского и
Приморского краев (от оз. Чля в низовьях Амура до р. Туманная), р. Тымь на
100
Северном Сахалине, реки и озера Южного Сахалина в районе залива Анива, на
восточном берегу Сахалина, а также реки островов Кунашир и Итуруп (Берг,
19496; Шмидт, 1950; Пинчук, 1978, 1984).
Возраст и размеры. Достигает длины 16 см, чаще не более 9 см (Берг, 19496).
Образ жизни. Придонный эвригалинный вид, широко распространенный в
реках, озерах, заливах. Живет преимущественно в пресной воде, хотя выходит
и в соленую воду. Образ жизни этого вида в наших водах мало изучен, скорее
всего он похож на образ жизни других бычков. Известно, что в р. Седанка зи-
му проводит в малоподвижном состоянии, зарывшись в листья. Нерест в реках
залива Петра Великого происходит на быстром течении, икра откладывается
на нижней стороне камней.
Статус вида. Объект любительского рыболовства.
Автор-составитель: Ю.С. Решетников.
101
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Chaenogobius Gill, 1858
231. Chaenogobius castaneus (O’Shaughnessy, 1875) -
каштановый дальневосточный бычок
Описание и систематика. Внешне похож на кольчатого бычка. Имеет
почти цилиндрическое тело, узкий и сжатый с боков хвостовой стебель.
Цвет тела светлый, с желтовато-коричневым оттенком, поверх тела идут
нерегулярные полосы, имеется ряд черных точек по средней линии тела.
Отличительные признаки, по Пинчуку (1978, 1984), для бычков из вод Рос-
сии (р. Шишкевича и Чибисанское озеро) следующие. Каждый канал систе-
мы боковой линии на голове состоит из двух частей: надглазничной и за-
глазничной у верхне-заднего края орбиты; каждый канал открывается тре-
мя порами. Рот небольшой, задний конец верхнечелюстной кости доходит
лишь до середины глаза. Вдоль средней линии тела имеются некрупные
темные парные пятна.
D, VI-VII, D2110-11, AI 10-11. Поперечных рядов чешуй 63-71. Подви-
дов нет (Пинчук, 1978, 1984). У корейских популяций признаки таковы: Dj
VII-VIII; D2 I 8-11; A I 8-11; поперечных рядов чешуй 70-80 (Choi et al.,
1990).
Распространение. Бассейны Японского, Желтого и Охотского морей.
В Китае бассейн Желтого моря, весь п-ов Корея, в Японии — острова Кюсю,
Хонсю, Хоккайдо (Freshwater Fishes of Japan, 1987; Choi et al., 1990). В Рос-
сии - острова Шикотан и Сахалин (залив Анива, лагуна Буссе, низовья
р. Шишкевича, Вавайские озера) (Берг, 19496; Линдберг, Красюкова, 1975;
Пинчук, 1978, 1984).
Возраст и размеры. Достигает длины 5 см в России и 10 см на п-ове Корея
(Choi et al., 1990).
Образ жизни. Мелкий донный вид, населяющий моря, озера и устья рек.
102
В Японии встречается на рисовых полях. Биология в российских водах практи-
чески не изучена.
Статус вида. Хозяйственного значения не имеет.
Автор-составитель: Ю.С. Решетников.
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Chaenogobius Gill, 1858
232. Chaenogobius macrognathus (Bleeker, 1860) - большеротый бычок
103
Описание и систематика. Отличается от других видов этого рода тем,
что челюсти примерно одинаковой длины, голова уплощенная. Лучи Dj уд-
линены, особенно третий, сам плавник сверху заострен; последний луч в нем
отделен от предпоследнего примерно таким же промежутком, как проме-
жутки между остальными лучами; в задней части плавника нет черного пят-
на. Чешуя на боках тела располагается не вплотную, на хвостовом стебле
она ктеноидная.
D( (V) VI - VII; D21 11-12, AI 9-11. Поперечных рядов чешуй 49-53 (Пин-
чук, 1978, 1984). Подвидов нет. Систематическое положение бычков из России,
относимых к этому виду, особенно из оз. Чля, нуждается в уточнении (Николь-
ский, 1956; Пинчук, 1984). В Японии и на п-ове Корея часто идет под видовым
названием Ch. urotaenia (Линдберг, Красюкова, 1975; Freshwater Fishes of Japan,
1987; Choi et al., 1990).
Распространение. Населяет воды всего п-ова Корея, Китая, Японии и
нашего Приморья. В России известен от р. Туманная до Сахалина и ни-
зовьев Амура (оз. Чля), в реках залива Петра Великого (Седанка, Суйфун,
Сучан, Квандагоу и др.), отмечен на южных Курильских островах (Иту-
руп), в реках Южного и Северного Сахалина (Берг, 19496; Никольский,
104
1956; Линдберг, Красюкова, 1975). Но распространение этого вида в водах
России требует уточнения, так как даже его рисунки в прежних работах
(Берг, 19496; Линдберг, Красюкова, 1975) принадлежат другим видам
(Пинчук, 1984).
Возраст и размеры. Достигает длины 10 см.
Образ жизни. Населяет преимущественно пресные воды (среднее течение
рек), хотя выходит и в соленые. Много было его в р. Седанка, где он проводил
зиму в малоподвижном состоянии, зарывшись в листья. Нерест в реках залива
Петра Великого проходит на быстром течении, икра откладывается на нижней
стороне камней (Берг, 19496). Биология вида в наших водах практически не
изучена, описан образ жизни в японских водах (Dotu, 1955).
Статус вида. Хозяйственного значения не имеет.
Автор-составитель: Ю.С. Решетников.
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Chaenogobius Gill, 1858
233. Chaenogobius taranetzi Pinchuk, 1978 -
дальневосточный бычок Таранца
Описание и систематика. Тело веретенообразное, покрыто чешуей сред-
него размера. Голова несколько приплюснута сверху и спереди. Передние
ноздри с выступающими вверх краями. Рот небольшой, слегка косой. Ниж-
няя челюсть слегка выдается вперед. Верхняя губа не расширена по бокам.
Темя, затылок, жаберные крышки, горло и большая часть брюха голые. Во-
ротник брюшной присоски хорошо развит, но без лопастинок и бахромы,
брюшные плавники не достигают анального отверстия. Спинные плавники
разделены довольно большим промежутком. Это новый вид бычка, выде-
ленный Пинчуком (1978) и отличающийся от других видов этого рода следу-
ющими признаками. Каждый Kaijan системы боковой линии на голове состо-
ит только из одной части: нет надглазничной ветви, а имеется только часть
у верхнезаднего края орбиты, которая открывается двумя порами. Надглаз-
ничная часть канала заменена рядом невромастов. Задний край верхней че-
люсти достигает вертикали середины глаза. По бокам тела некрупные тем-
ные пятна. Сетчатый рисунок на спине и голове такой же, как и у вида
С/г. castaneus. Во время нереста у зрелых самок из р. Артемовки темнеет низ
головы и плавников.
105
D! VI-VIII, чаще VII; D21 9-11; A I 9-11. Поперечных рядов чешуй 53-71.
Подвидов нет. По существенным отличиям в строении каналов боковой линии
предлагается выделить в особый подрод Lindbergiana (Берг, 19496; Пинчук,
1978, 1984).
Распространение. Эндемик Приморского края и соседних районов. В При-
морском крае от р. Туманная до р. Чамочагоуза, впадающей в Уссурийский за-
лив, по-видимому, часть побережья п-ова Корея. В России - Амурский залив,
р. Артемовна (Уссурийский залив), оз. Хасан и Рязановское, залив Петра Вели-
кого (Пинчук, 1978, 1984).
Возраст и размеры. Достигает длины 83 мм, обычно 50 мм.
Образ жизни. Мелкий эвригалинный вид, обитает в устьях рек,
пресных озерах, умеренно опресненных и более солоноводных частях за-
ливов (Пинчук, 1978, 1984). В Амурском заливе с умеренно опресненной
водой встречается вдали от устьев рек. В заливе Петра Великого с более
солоноватой водой обитает в устьях рек, местами в их низовьях и даже в
совершенно пресных озерах. Занимая по морфологии промежуточное по-
ложение между видами Ch. castaneus и Rhodonichthys laevis, бычок Таранца
в Приморском крае фактически заселяет биотопы обоих видов благодаря
своей очень широкой эвригалинности.Образ жизни в наших водах мало
изучен.
Статус вида. Хозяйственного значения не имеет.
Автор-составитель: Ю.С. Решетников.
106
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Hyrcanogobius Iljin, 1928
234. Hyrcanogobius bergi Iljin, 1928 - бычок Берга
Описание и систематика. Этот род близок к р. Knipowitschia, от которого
отличается тем, что на поверхности головы в надглазничных каналах сейсмо-
сенсорной системы отсутствуют передние отделы, а задние отделы начина-
ются порами у заднего края глаз и продолжаются каналами на жаберной
предкрышке. Ширина лба равна 2/з диаметра глаза. Тело веретеновидное, го-
лова неприплюснутая, спинные плавники разделены. Спина голая до начала
Db далее тело покрыто ктеноидной чешуей. Хвостовой плавник симметрич-
ный, закругленный. На боках тела бурые крапинки, у самца образующие по-
лоски. У основания хвостового плавника нет темного пятна. Есть плаватель-
ный пузырь.
D] VI; D21 8-9; А I 8-9. Поперечных рядов чешуй около 30 (Берг, 19496).
Подвидов нет.
Распространение. Северная часть Каспийского моря, перед устьями рек
Эмба, Урал, Волга, в пресной воде озер нижнего течения р. Атрек. В России
107
встречается повсеместно в Северном Каспии, у берегов Дагестана, в пресно-
водных участках устья Волги редок (Берг, 19496; Казанчеев, 1981).
Возраст и размеры. Самая мелкая рыба в наших водах. Достигает длины
36 мм, но обычные размеры самцов - до 26,5 мм, самок - до 31 мм. Масса в сре-
днем 0,36 г. Продолжительность жизни 1 год.
Образ жизни. Мелкий полупелагический прибрежный морской вид, лишь
изредка встречается в пресной воде. Биология изучена слабо. Питается рако-
образными. Половозрелым становится при длине 21-22 мм. Во второй полови-
не мая в яичниках самок обнаружено 37СС-410 икринок; наряду с крупными зре-
лыми икринками диаметром 0,65-0,75 мм содержались и мелкие, диаметром
менее 0,3 мм. По-видимому, икрометание порционное. Развивающаяся икра
была найдена в юго-западной части Северного Каспия, на глубине 4 м, в мае.
Икра была приклеена к внутренней поверхности пустой раковины. Оболочки
икринок с хорошо сформированными эмбрионами имели грушевидную форму
высотой 1,1-1,4 мм и шириной 0,6-0,9 мм (Казанова, 1951). Из икринок вылуп-
ляются предличинки с остатками желточного мешка и незаполненным плава-
тельным пузырем длиной около 3 мм. При длине 3,4—3,5 мм пузырь заполняет-
ся воздухом, а при 4,2^1.6 мм полностью резорбируется желточный мешок и
начинается личиночный период.
Статус вида. Хозяйственного значения не имеет.
Автор-составитель: К.И. Москалькова.
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Knipowitschia Iljin, 1927
235. Knipowitschia caucasica (Berg, 1916) - бычок-бубырь
Описание и систематика. Тело веретеновидное, голова не приплюснута.
Глаза направлены в стороны, а не вверх. На боках тела ктеноидная чешуя,
но голова, передняя часть спины до второго спинного плавника, горло и
брюхо голые. Верхняя губа по бокам не расширена, язык усеченный. Лоб
выпуклый. На голове есть органы чувств системы боковой линии: каналы с
порами и ряды невромастов. Передние поры надглазничного канала распо-
ложены позади переднего края глаз. Грудные плавники широкие и длинные,
почти достигают анального плавника. Есть плавательный пузырь. Общая
окраска темно-серая, с желтоватым оттенком. Самцы отличаются от самок
более темной окраской тела, по бокам у них резкие темные пятна (иногда
108
поперечные полосы) и на заднем конце первого спинного плавника темное
пятно (у живых оно голубого цвета). Самки светлее, без полос (пятна непра-
вильной формы), Dj с многочисленными темными пятнышками. Подборо-
док черного цвета. Во время нереста темное пятно на первом спинном плав-
нике и черная пигментация подбородка исчезают, брюхо становится желто-
ватым.
Dj VI; D21 (6) 7-9; A I (6) 7-9. Поперечных рядов чешуй 30-37 (Берг, 19496;
Световидов, 1964; Эланидзе, 1983; Троицкий, Цуникова, 1988). Подвидов нет
(Аннотированный каталог..., 1998).
Распространение. Каспийское, Азовское, Черное, северные части Эгейско-
го и Адриатического морей, интродуцирован в Аральское море; встречается и
в совершенно пресной воде (озера Грузии) (Эланидзе, 1983; Miller, 1986). В Кас-
пии распространен по всему морю и в низовьях всех рек, включая болотные
участки. В России - приволжский участок Каспийского моря, авандельта и ни-
зовья Волги, бассейн нижнего течения Кумы (Рагимов, 1981; Казанчеев, 1981).
В Азовском море встречается в кубанских лиманах, в пресной и осолоненной
воде; в бассейне Дона не отмечен (Троицкий, Цуникова, 1988).
Возраст и размеры. Длина менее 50 мм. Продолжительность жизни не бо-
лее 1 года (Троицкий, Цуникова, 1988)
Образ жизни. Мелкий эвригалинный вид, встречается как в пресной, так и
в воде с соленостью до 23%о. Многочислен в низовьях Волги и в ее авандельте.
Предпочитает слаботекущие воды, заросшие водной растительностью. В от-
крытых частях моря встречается редко. Биология вида изучена недостаточно.
Питается в основном бокоплавами и личинками хирономид. В кубанских лима-
нах питается зоопланктоном, придонными ракообразными, личинками и взрос-
лыми насекомыми (Троицкий, Цуникова, 1988).
Половое созревание наступает при длине 22-25 мм. Самцы крупнее самок.
Размеры производителей на Каспии колеблются от 20 до 49 мм при средней
длине самок 30 мм и самцов - 33 мм. В низовьях Волги нерестится с конца ап-
реля до конца июня при температуре воды 12-25 °C, на мелководье, на глуби-
109
не 15-40 см. Самцы устраивают гнездо под пустыми створками крупных мол-
люсков, на нижнюю поверхность которых самки приклеивают икру. Икринки
могут откладываться и на растения. Икра в гнездо выметывается несколькими
самками (от 2 до 4), общее число икринок в кладке до 3000. Плодовитость от
210 до 780 икринок, в осолоненных лиманах Каспия, где рыбы мельче, плодо-
витость ниже (в среднем 295 икринок), в водах Грузии плодовитость колеблет-
ся от 100 до 460 икринок (Эланидзе, 1983). Оболочка икринки прозрачная, име-
ет удлиненную форму с размерами 1,7-2,6 х 0,7-0,9 мм. Узким концом она при-
клеивается к субстрату. Икра полиплазматическая. Самец охраняет кладку.
Процесс эмбриогенеза не изучен. Вылупившиеся личинки длиной 3,0-3,5 мм,
тонкие, прозрачные, с желточным мешком и плавательным пузырем. Они ве-
дут пелагический образ жизни и питаются зоопланктоном. При достижении
длины 9-11 мм они превращаются в мальков, переходят к донному образу жиз-
ни и питанию бентосом (Коблицкая, 1961). На зимовку бубырь отходит в глу-
бокие участки авандельты.
Статус вида. Хозяйственного значения не имеет. Встречается в питании мо-
лоди судака и других рыб (Троицкий, Цуникова, 1988).
Автор-составитель: К.И. Москалькова.
Класс Osteichthyes
Отряд Percifonnes
Семейство Gobiidae
Род Knipowitschia Iljin, 1927
236. Knipowitschia longecaudata (Kessler, 1877) -
длиннохвостый бычок Книповича
Описание и систематика. Тело веретеновидное, голова не приплюснутая,
спинные плавники разделены значительным промежутком. Голова, передняя
часть спины до второго спинного плавника и низ тела до анального плавника
голые, на боках тела ктеноидная чешуя. Хвостовой плавник несимметричный.
Брюшная присоска развита слабо. Глаза расположены близко друг к другу и
направлены вверх, ширина лба равна половине диаметра глаза. Надглазнич-
ные каналы сейсмосенсорной системы слиты на коротком протяжении, от ме-
ста их слияния назад отходит короткий каналец, оканчивающийся порой. Ок-
раска бледно-серая, спина в мелких буроватых крапинках. У основания хвосто-
вого плавника большое темное пятно неправильной формы. Во время нереста
у самцов все плавники, за исключением хвостового, темнеют, непарные удли-
няются, на боках тела появляются буроватые поперечные полосы. Есть плава-
тельный пузырь.
Dj (V) VI (VII), D21 7-9, AI 8-9. Поперечных рядов чешуй (36) 37-45 (Берг,
19496; Световидов, 1964; Троицкий, Цуникова, 1988). Подвидов нет, однако та-
ксономический статус некоторых популяций нуждается в уточнении. Описан-
ный Пинчуком (1978) самостоятельный вид К. georghievi не получил признания
других авторов (Miller, 1986; Аннотированный каталог..., 1998).
Распространение. Лагуны, лиманы и эстуарии Черного, Азовского и Кас-
пийского морей, а также устья впадающих в них рек. В России - по побережью
Азовского моря (многочислен в Таганрогском заливе, в опресненных лиманах,
редок в Керченском проливе), в устье Дона (вверх до ст. Кочетовской), во всех
манычских водохранилищах, почти по всей акватории Северного Каспия, у за-
падных берегов Среднего Каспия, от Махачкалы к югу, в низовьях дельты
Волги (Световидов, 1964; Рагимов, 1981; Казанчеев, 1981; Miller, 1986).
Возраст и размеры. Живет не более 1 года. Предельная длина самок 40 мм,
самцов - 50 мм, в среднем длина около 30 мм, масса - 0,6-0,7 г.
Образ жизни. Мелкий полупелагический вид, поэтому его окраска более
светлая. Обитает не на дне, а в толще воды над песчаными и илистыми грунта-
ми, в слабосоленой (не выше 5%о) или пресной воде. Питается придонными и
донными беспозвоночными: мизиды, бокоплавы, полихеты, личинки насеко-
мых (Москалькова, 1962). Этот бычок - самая раннесозревающая рыба в на-
шей фауне, часть популяции достигает половой зрелости в возрасте 2,5 мес при
длине тела 24 мм. Нерест в Таганрогском заливе длится со второй декады мая
по сентябрь включительно, с пиком в июле. Икрометание порционное. В ясты-
ках содержатся три порции икры разного диаметра. Диаметр икринок на V ста-
дии зрелости в среднем равен 0,7441,80 мм, на IV стадии - 0,5 мм, на III стадии -
0,25 мм. Абсолютная плодовитость самок длиной 22-33 мм колеблется от 270
до 800 икринок. Нерестится бычок на песчано-ракушечном грунте, недалеко
от берега, на глубинах 20-40 см. В Каспии кладка найдена на глубине 4 м на
створках раковины моллюска Mcmodacna. В Азовском море икру откладывает
в пустые раковины Cardium, причем часто по 2-4 самки выметывают икру в од-
ну раковину. В нерестовом стаде самок вдвое больше самцов. Оболочка отло-
111
женной икринки имеет удлиненную эллипсовидную форму. Узким концом она
приклеивается ко внутренней поверхности раковины. Самец энергичными дви-
жениями плавников и тела засыпает раковину песком, оставляя небольшой
вход под нее, куда и забирается, выставляя из гнезда только часть головы. Он
энергично атакует приближающихся врагов. Нерест начинают во второй поло-
вине мая самые крупные годовики, в июне к ним присоединяются более мел-
кие особи. После нереста все годовики погибают и к середине июля исчезают
из популяции. С июля по сентябрь нерестятся сеголетки. Первые из них от ран-
него нереста в мае созревают в возрасте 2,5 мес при длине 24 мм и вступают в
нерест в конце июля. Дочерние особи этих сеголетков созревают к следующей
весне, имея длину около 30 мм и возраст около 10 мес (Москалькова, 1960).
Оболочка оплодотворенной икринки тонкая, прозрачная, высотой 1,4 и шири-
ной 0,6-0,8 мм. Желток светло-желтого цвета с разными по размеру жировы-
ми каплями (Калинина, 1976). При температуре 23-26 °C на 7-8-е сутки вылу-
пляются предличинки длиной 3,0-3,2 мм. Пока у них нет воздуха в плаватель-
ном пузыре, они держатся у дна. С наполнением пузыря воздухом при длине
3,5—4,0 мм они всплывают в верхние слои воды. Основная масса сеголетков
обитает в толще воды. Самые маленькие личинки 3-9 мм длиной держатся у
поверхности, а мальки крупнее 20 мм опускаются в придонные слои. В Азов-
ском море молодь и, возможно, взрослые формы совершают вертикальные су-
точные миграции, поднимаясь ночью в верхние слои воды вслед за своим из-
любленным кормовым объектом Calanipeda aquaedulcis. Сеголетки питаются
планктонными формами ракообразных, преимущественно копеподами (Мос-
калькова, 1962).
Статус вида. Мелкий вид. Хозяйственного значения не имеет, но служит пи-
щей некоторых промысловых рыб и является конкурентом в питании их молоди.
Автор-составитель: К.И. Москалькова.
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Mesogobius Bleeker, 1874
237. Mesogobius batrachocephalus (Pallas, 1814) -
бычок-кнут, мартовик
Описание и систематика. Представители этого рода близки к бычкам
р. Neogobius, от которых отличаются тем, что затылок, темя, жаберные крыш-
ки и щеки не покрыты чешуей; сама чешуя мелкая или средней величины; во
втором спинном плавнике не менее 15 ветвистых лучей, а в анальном - не менее
112
12; подглазничных рядов невромастов 6-10; воротник брюшной присоски без
лопастинок. Характерные признаки мартовика: поперечных подглазничных ря-
дов невромастов 8-10, продольного ряда нет, поперечных рядов чешуй 65-84.
Тело удлиненное, сзади сжатое. Голова длинная, приплюснутая, нижняя че-
люсть заметно выдается вперед, а ее задний конец доходит до середины глаза.
Губы, особенно верхняя, мясистые. Межглазничное пространство меньше диа-
метра глаза. Брюшная присоска не достигает анального отверстия. Основной
цвет желтоватый или серый, на голове и на теле бурые пятна и перевязки. Хво-
стовой, спинные и грудные плавники с поперечными темными полосками, час-
то 1-2 темных пятна бывают в основании грудных плавников. Половой димор-
физм не выражен. Во время нереста окраска не меняется. Плавательный пу-
зырь отсутствует в течение всей жизни.
DjVI; D2 I 16-18 (19); A I 12-16 (17). Чешуя мелкая, поперечных рядов че-
шуй (65) 68-79 (84) (Берг, 19496; Световидов, 1964). Подвидов нет (Аннотиро-
ванный каталог..., 1998).
Распространение. Черное море по всем берегам, включая Босфор, есть в
Днестровском, Бугском и Днепровском лиманах. Изредка встречается в прес-
ных водах: в Буге до Вознесенска, в Днепре - до Херсона и Берислава, живет в
пресном озере у Констанцы (Румыния) и в Балта-лимане у Стамбула. В водах
России - Азовское море до устья Дона и кавказское побережье Черного моря
(Новороссийская бухта, Геленджик) (Берг, 19496; Световидов, 1964).
Возраст и размеры. Самый крупный из наших бычков: максимальная дли-
на 35 см и масса около 600 г. Продолжительность жизни 5-6 лет.
Образ жизни. Солоноватоводный, эвригалинный, донный вид, изредка за-
ходит в пресную воду, но предпочитает воды с соленостью 8-12%0. Ведет ма-
лоподвижный образ жизни, совершает небольшие миграции ранней весной,
во время подхода к берегам на нерест. Чаще обитает на песчаных и ракушеч-
ных грунтах. Зимой отходит на более глубокие места. Питается в основном
8 Атлас.... том 2
ИЗ
мелкой рыбой (атериной, хамсой, тюлькой, бычками, молодью ставриды),
которая составляет до 70% содержимого желудка, а также моллюсками, кре-
ветками, мелкими крабами и червями. Молодь питается ракообразными (ми-
зиды, бокоплавы) и полихетами (Nereis). Во время нереста желудки мартови-
ка бывают пустыми, но гибели бычков не отмечали (Световидов, 1964; Ем-
тыль, 1997). Самки немного крупнее самцов. Средние размеры самцов в
Азовском море - 19,2 см, самок - 21 см. Половой зрелости достигает на тре-
тьем году жизни при длине 15-19 см и массе 60-70 г. Нерестится раньше дру-
гих бычков, обычно в марте-апреле при температуре воды 6-9 °C. В это вре-
мя он образует скопления. Нерест дружный и единовременный. Плодови-
тость 5650-10150 икринок, в среднем 4 тыс. икринок в Черном море и 8 тыс. -
в Азовском (Калинина, 1976; Емтыль, 1997). Самцы устраивают гнезда под
камнями на песчано-галечном грунте, на глубине 0,5-1,5 м. Икринки яйцевид-
ные, очень крупные: высота 4—6 и ширина 1,8-2,5 мм. Более широким клей-
ким концом икринка прикрепляется к нижней поверхности камней. В одно
гнездо откладывают икру 1-2 самки, создавая кладки в виде однослойной ле-
пешки диаметром около 10 см. Оболочка икринок плотная, беловатая, полу-
прозрачная, по мере развития эмбриона она несколько растягивается. Жел-
ток ярко-желтый, диаметром около 2 мм, с мелкими жировыми каплями. При
температуре 8-15° развитие в оболочке длится 3-4 недели, оно включает и
личиночный период (поэтому нет свободноживущей личинки). В результате
вылупляются донные мальки длиной 7,5-8,5 мм с остатками желточного
мешка (Калинина, 1976).
Статус вида. Объект промысла и спортивного рыболовства. Включен в по-
следнее издание “Международной Красной книги” в статусе вида, данных по
которому недостаточно (Аннотированный каталог..., 1998).
Автор-составитель: К.И. Москалькова.
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Neogobius Iljin, 1927
238. Neogobius fluviatilis (Pallas, 1814) - бычок-песочник
Описание и систематика. Как и у всех представителей этого рода, тело
относительно крупное, удлиненное, сжатое с боков, нет продольного под-
114
глазничного ряда невромастов, поперечных подглазничных рядов 6. Нет
плавательного пузыря. Крупная олигоплазматическая икра. У бычка-пе-
сочника стройное тело с зауженным хвостовым стеблем и заостренным ры-
лом, второй спинной плавник кзади равномерно понижается. Брюшная при-
соска доходит до анального отверстия, ее лопастинки мало заметны. Перед-
ние ноздри в виде коротких трубочек. Межглазничное расстояние явно
меньше диаметра глаза. Нижняя челюсть выдается вперед. Верхняя губа к
углам рта не расширена. Циклоидная чешуя покрывает не только затылок,
но и темя, вдаваясь в промежуток между глазницами; чешуя частично по-
крывает жаберные крышки, заднюю половину горла и все остальное тело.
По сравнению с другими видами бычок-песочник окрашен в более светлые
тона. Тело полупрозрачное, желтовато-серого цвета с бледным буроватым
рисунком из сливающихся пятен (8-12) вдоль боков. Часто небольшое тем-
ное пятно бывает на верхней части основания грудных плавников. На спин-
ном и хвостовом плавниках имеются темные пятнышки. Имеются аберра-
ции зеленоватого тона. Во время нереста самцы становятся совершенно
черными, непарные плавники сильно удлиняются, и по их краям появляется
желтая бахрома.
Di VI, D21 (14) 15-17 (18), A112-15 (17). При жизни чешуя мало заметна, по-
перечных рядов чешуй (54) 58-65 (67) (Берг, 19496; Световидов, 1964; Казанче-
ев, 1981; Евланов и др., 1998). Кариотип: 2п = 46, NF = 46 (Васильев, 1985). Кас-
пийские популяции бычка-песочника выделяются в подвид N. fluviatilis pallasi
(Berg, 1916).
Распространение. Бассейны Черного, Азовского и Каспийского морей,
интродуцирован в Аральское море (Световидов, 1964; Miller, 1986). Изве-
стен по всему побережью Черного моря (кроме Румынии, где встречается
только в пресных водах), а также в бассейне Мраморного моря. Входит в ре-
ки, а также в связанные с ними прибрежные озера и лиманы. В России
встречается в верховьях Днепра (у Смоленска) и Десны (у Новгорода-Се-
верского) , населяет опресненные лиманы Азовского моря, Таганрогский
115
залив, бассейны Северского Донца, Дона с его притоками Медведица, Хо-
пер, Воронеж и Усмань; нижнее и среднее течение Кубани, есть в Керчен-
ском проливе и в речках черноморского побережья от Чемеса до Нечепсу-
хо и Дагомыса (у Сочи) (Берг, 19496; Федоров, 1960; Световидов, 1964; Тро-
ицкий, Цуникова, 1988; Емтыль, 1997). В Каспийском море обитает в Север-
ном Каспии и в опресненных участках западного побережья до Ирана (Ра-
гимов, 1981). В наших водах встречается в опресненных районах по всему
побережью, а также в озерах Нижнего Терека и в дельте Волги (Аннотиро-
ванный каталог..., 1998). После постройки плотин впервые отмечен сначала
в Волгоградском водохранилище (1975 г.), затем в Саратовском (1980-е го-
ды) (Евланов и др., 1998).
Возраст и размеры. Достигает максимальной длины 20 см и массы 50 г, но
обычно в пресных водах не более 10-16 см и 20-30 г. Живет не более 5 лет, а в
кубанских лиманах - не более 2 лет (Казанчеев, 1981; Троицкий, Цуникова,
1988; Емтыль, 1997).
Образ жизни. Эвригалинный прибрежный вид, предпочитает опреснен-
ные участки моря, но заходит и в пресные воды. Придерживается песчаных
и песчано-илистых грунтов. Далеких миграций не совершает, на зиму отко-
чевывает от берега, весной возвращается для нереста. Питается в основном
ракообразными (бокоплавами, мизидами, кумовыми), отчасти червями, ли-
чинками хирономид, моллюсками и рыбой. Но в Азовском море поедает
преимущественно моллюсков и полихету нереис. Половой зрелости дости-
гает на втором году жизни при длине 7-8 см и массе 5-9 г в бассейне Азов-
ского моря (Емтыль, 1997) и при 9-12 см - в бассейне Черного (Долгий,
1993). Нерест в мае-июле (разгар - май-июнь) при температуре воды
13-20 °C. В ястыках находят от 340 до 2800 икринок (Билько, 1968; Калини-
на, 1976). Нерестилища в море находятся в узкой опресненной прибрежной
зоне с песчаным дном и россыпями небольших камней. Самка за сезон вы-
метывает 2-3 порции икры. Самец устраивает гнездо в виде норки под кам-
нями, а самка, заходя в него, приклеивает икру на потолок в виде однослой-
ной лепешки. В одном гнезде в разное время могут нереститься до 4 самок.
Неоплодотворенная икра округлой формы, после оплодотворения и набуха-
ния оболочка удлиняется и становится яйцевидной, с заостренным свобод-
ным концом (ее высота до 4,0 мм и ширина - до 1,8 мм). Кладка всегда ох-
раняется самцом. В том случае, когда нет камней, икра откладывается на
любые предметы: на сваи мостов и шлюзов, отмершую растительность. Ик-
ра крупная, диаметр желтка 1,5-1,6 мм, но плазменного белка в ней мало.
Эмбриогенез включает и личиночное развитие, поэтому продолжитель-
ность инкубации достигает 14—16 дней. Молодь вылупляется длиной
6,5-7,8 мм, с костными лучами в плавниках, сформированной присоской, но
с еще достаточно большим желточным мешком. Она держится на дне и сра-
зу переходит на активное питание (Дмитриева, 1968).
Статус вида. Является объектом промысла и любительского лова. Молодь
и взрослый бычок поедаются хищными рыбами. Включен в последнее издание
“Международной Красной книги” в статусе вида, данных по которому недоста-
точно.
Автор-составитель: К.И. Москалькова.
116
Семейство Gobiidae
Род Neogobius Iljin, 1927
Класс Osteichthyes
Отряд Perciformes
239. Neogobius gymnotrachelus (Kessler, 1857) - бычок-гонец
Описание и систематика. Голова вальковатая, ее длина составляет
21-25% длины тела без хвостового плавника. Ширина головы немного бо-
лее ее высоты. Нижняя челюсть не выдается вперед. Губы, особенно верх-
няя, мясистые, верхняя губа к углам рта не расширена. Задний край верхней
челюсти заходит за вертикаль переднего края глаза. Межглазничное рас-
стояние в среднем вдвое меньше диаметра глаза. Поперечных подглазнич-
ных рядов невромастов 6. Темя, затылок, жаберные крышки, горло и стеб-
ли грудных плавников голые. Спина перед D] обычно покрыта циклоидной
чешуей. Изредка чешуя простирается на затылок, темя и верхнюю часть
жаберных крышек. В Dj концы лучей выдаются над перепонками. Брюш-
ная присоска без лопастинок на воротнике, обычно она доходит до анально-
го отверстия, но у крупных особей может и не доходить. Икра крупная. Пла-
вательного пузыря нет. Тело желтовато-серого цвета, на боках бурые пят-
на, иногда образующие поперечные полоски, косо направленные вперед.
На голове сверху и по бокам извилистые темные полоски. Три темные по-
лоски имеются на Dp иногда они бывают и при основании других непарных
плавников. Брюшная присоска и анальный плавник темно-серые (Берг,
19496; Световидов, 1964).
D, VI-VII, D2 I (14) 15-18, A I 12-15 (16). Поперечных рядов чешуй (54)
56-69. Ранее рассматривался в составе рода Mesogobius. Каспийские популяции
выделяются в особый подвид N. gymnotrachelus macrophthalmus (Kessler, 1877);
они имеют более крупные глаза и крупную чешую (47-57, чаще 53-54 попереч-
ных рядов чешуй) (Казанчеев, 1981; Аннотированный каталог..., 1998).
Распространение. Бассейны Черного, Азовского и Каспийского морей, где
населяет лиманы, прибрежные озера, водохранилища и предустьевые про-
странства. Обычно обитал в нижнем течении рек, но теперь поднимается вы-
ше, заселяя реки и водохранилища: Днепр и Каховское водохранилище, Буг,
Днестр с притоками и Дубоссар£кое водохранилище, Дунай до Бухареста, Кам-
чия, реки Турции, Риони. В России населяет опресненные лиманы Азовского
моря, Таганрогский залив, Дон, Донец, Аксай (до Новочеркасска) и Кубань
(Световидов, 1964; Емтыль, 1997). Бычок-гонец обычен в Северном Каспии,
особенно в его восточных районах; у западного побережья Среднего Каспия
чаще встречается вблизи устья р. Сулак (Рагимов, 1981). В Нижней Волге
чаще обитает в нижней зоне дельты.
117
Возраст и размеры. Достигает максимальной длины 26 см, но обычно сред-
ние размеры 7-13 см, а у каспийского подвида - 2,1-6,3 см и масса 0,6-2,2 г (Ка-
занчеев, 1981).
Образ жизни. В бассейнах Черного и Азовского морей обитает главным об-
разом в пресных водах низовьев рек, в озерах и водохранилищах, но проникает
и в солоноватоводные (до 5%о) участки моря и лиманы. В бассейне Каспия кро-
ме Волги в реках не обнаружен, населяет олигогалинную и мезогалинную зоны
моря. Придерживается песчаных и песчано-илистых грунтов, иногда встречает-
ся среди камней или в зарослях водорослей. Питается главным образом ракооб-
разными (Amphipoda, Mysidae, Cumacea), личинками хирономид и частично ры-
бой (Световидов, 1964). В других случаях пищей служат моллюски, черви, рако-
образные, икра и молодь рыб (Емтыль, 1997). В Каспии его рацион составляют
только ракообразные: Gammaridae, Corophiidae, Amphipoda, Mysidae (Казанчеев,
1981). Половой зрелости достигает на втором году жизни. Плодовитость самки
длиной 13 см и массой 32 г составила 1,5-2,0 тыс. икринок. Нерест в низовьях
Днепра происходит в апреле-мае при температуре воды 9-12 °C, в Каховском
водохранилище икрометание наблюдали при температуре воды 16-19 °C. Икра
выметывается двумя порциями с промежутком около трех недель. Оплодотво-
ренная икринка имеет яйцевидную форму и размеры 3,2 х 1,8 мм, оболочка
плотная, беловатая, с пучком клейких нитей, желток непрозрачный, светлый
(Билько, 1968; Калинина, 1976). Икра олигоплазматическая и полилецитальная.
Кладка икры развивается в гнезде под охраной самца. В нерестовый период
окраска самца не меняется. Минимум через 2 недели из оболочки вылупляется
молодь с признаками мальков, но с крупным желточным мешком. В июле-
августе в Северном Каспии ловятся уже совсем сформировавшиеся мальки
длиной 20-37 мм (Рагимов, 1981).
Статус вида. Хозяйственного значения не имеет в силу малочисленности, по-
падается в уловах при промысле других рыб. Включен в последнее издание “Ме-
ждународной Красной книги” в статусе вида, данных по которому недостаточно.
Автор-составитель: К.И. Москалькова.
118
Семейство Gobiidae
Род Neogobius Iljin, 1927
Класс Osteichthyes
Отряд Perciformes
240. Neogobius iljini Vasiljeva et Vasiljev,1996 -
каспийский бычок-головач
Описание и систематика. Вид отличается от N. kessleri (Gunther, 1861) карио-
типом и расположением чешуи. Тело сжатое с боков, хвостовой стебель корот-
кий и высокий. Голова приплюснута, ее ширина намного больше высоты. Весь
затылок, задняя часть горла, верхняя треть жаберных крышек и основания груд-
ных плавников покрыты циклоидной чешуей. Ктеноидная чешуя покрывает
только бока тела. Щеки выпуклые и округлые. Нижняя губа выдается, верхняя
слегка расширена и резко сужается к углам рта. Спинные плавники соединяют-
ся, второй спинной плавник в середине повышается. Брюшная присоска округ-
лой формы, далеко не достигает анального отверстия и имеет заостренные ло-
пастинки. Грудной плавник достигает анального отверстия. Хвостовой плавник
закругленный. Тело желтовато-бурого цвета, с мраморным рисунком, иногда по
средней линии имеется 4-8 пятен неправильной формы. Рисунок сетчатый или в
виде пятен заметен на щеках, верхней губе и у основания грудных плавников. На
спине 5 темных поперечных полос. Непарные плавники с рядами темных пятен.
В нерестовый период самцы темнеют, хотя и не становятся черными.
D, VI-VII, D2 I 17-19, А I (12) 13-15 (16). Диплоидный набор представлен
2 субтелоцентрическими и 44 акроцентрическими хромосомами: 2п = 46, NF = 46
(Васильев, 1985). Подвидов нет. Ранее рассматривался в составе черноморско-
го вида N. kessleri (Gunther, 1861), не встречающегося в водах России, иногда в
ранге его каспийского подвида N. kessleri gorlap (Берг, 19496; и др.); пресновод-
ные популяции зачастую неверно отождествлялись с кавказскими речными
бычками (Берг, 19496; Аннотированный каталог..., 1998).
Распространение. Каспийское море, в основном в прибрежных районах, низо-
вья Волги, мелкие речки Дагестана, Азербайджана и Северного Ирана, Кура до
Мингечаура, озера Нижнего Терека, оз. Ясхан, низовья Урала. В России - все по-
бережье Среднего Каспия (прибрежные морские и пресные воды), низовья Вол-
ги до Волгограда, (Vasil’eva, Vasil’ev, 1996; Аннотированный каталог..., 1998).
Возраст и размеры. Самый крупный из волжских бычков, достигает
длины 22 см.
Образ жизни. Эвригалинный вид, обитает в прибрежных районах моря и в
реках. Биология не изучена. По аналогии с близким видом предполагается, что
самки достигают половой зрелости при длине 5,0-5,5 см, самцы - при 5,5-6 см
на втором году жизни. Плодовитость 1300-6900 икринок. Нерест в апреле-мае.
119
Основные места нереста - протоки дельты Волги, в многоводные годы нерес-
тится на полоях. Молодь растет быстро, длина сеголетков в сентябре достига-
ет 5 см. Жизненный цикл до 3 лет. Нерестится не более 2 раз. Питается мелки-
ми бычками, ракообразными и моллюсками.
Статус вида. Ввиду низкой численности промыслового значения не имеет,
попадается в виде прилова, объект любительского рыболовства.
На Нижней Волге охотно поедается хищниками - щукой, судаком, сомом,
жерехом и окунем.
Автор-составитель: К.И. Москалькова.
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Neogobius Iljin, 1927
241. Neogobius melanostomus (Pallas, 1814) —
бычок-кругляк, черноротый бычок
120
Описание и систематика. Тело вальковатой формы, сжатое с боков, с вы-
соким хвостовым стеблем и крутым лбом. Подглазничных поперечных рядов
невромастов 6 (7). Нижняя челюсть не выдается вперед, верхняя губа к углам
рта не расширяется. Язык спереди не закруглен, а усеченный, со слабой вырез-
кой. В задней части первого спинного плавника характерное черное пятно со
светлой оторочкой. Высота второго спинного плавника на всем протяжении
равномерна. Брюшные плавники достигают или почти достигают анального
отверстия, на них имеются черные пятна. Лопастинки на воротнике присоски
едва заметны. Цвет тела буровато-серый, с 5 большими темно-бурыми пятна-
ми по бокам. Голова темнее туловища, плавники темно-серые. Циклоидная че-
шуя покрывает тело, иногда частично межглазничное пространство, затылок,
горло, жаберные крышки и стебли грудных плавников. Брюшная присоска и
анальный плавник темно-серые. Во время нереста тело самцов становится чер-
ным, непарные плавники также становятся черными, с белой каймой по краям
(Берг, 19496; Световидов, 1964).
D, VI-VII, D2 I (12) 13-17, A I 9-13 (14). Поперечных рядов чешуй (42)
47-54 чешуй. Жаберных тычинок 9-13. Позвонков 31-33. Плавательный пу-
зырь отсутствует. Кариотип: 2п = 46, NF = 46, все хромосомы акроцентриче-
ские (Васильев, 1985). Ранее каспийские популяции выделялись в подвид
N. melanostomus affinis (Eichwald, 1831) (Берг, 19496; и др.). В современной лите-
ратуре деление на подвиды не принято (Аннотированный каталог..., 1998).
Распространение. Бассейны Мраморного, Черного, Азовского и Каспий-
ского морей, интродуцирован в Аральское море, по некоторым данным проник
в бассейн Балтийского моря, случайно завезен в Америку, где стал многочис-
ленным в некоторых пресноводных водоемах штата Иллинойс (Аннотирован-
ный каталог..., 1998). В последние годы бычок-кругляк освоил среднее и верх-
нее течения рек Дон, Днепр, Прут, Дунай и их крупные притоки. В России - по-
всеместно в Черном, Азовском и Каспийском морях, во многих впадающих в
них реках (Воронеж, Дон, Кубань, реки кавказского побережья, от Бзыби до
Шапсухо, Волга, Москва), в опресненных лиманах, некоторых прибрежных
121
озерах, отдельных водохранилищах (Световидов, 1964). Раньше в Волге выше
Волгограда не поднимался, однако потом широко расселился по ее бассейну: в
1968 г. отмечен в Куйбышевском водохранилище, затем в Саратовском и Ры-
бинском, а также в Канале им. Москвы и в р.Москве (Рыбы Подмосковья,
1988; Евланов и др., 1998).
Возраст и размеры. Достигает максимальной длины 25 см и массы до 180 г,
но обычные размеры в уловах 6-10 см. Живет до 5 лет.
Образ жизни. Эвригалинный донный вид, предпочитающий солоновато-
водные прибрежные участки морей с глубинами 3-5 м, но обитает также в ре-
ках, водохранилищах и озерах. Отличается необычайно широким диапазоном
адаптационных возможностей (Москалькова, 1996). Выдерживает изменение
солености от 0 до 40,5%о. В морях встречается на глубинах от 0,2 м во время не-
реста до 10-15 м (Черное и Азовское моря) и даже до 70 м (Каспий) в период
зимних миграций. Живет в диапазоне температур от -1 до 30 °C. Устойчив к де-
фициту кислорода, обладает развитой способностью к кожному дыханию.
Предпочитает твердые каменистые грунты, но встречается и на ракушечно-пес-
чаном и даже илистом грунте, а в Среднем Каспии - среди донной раститель-
ности. Спектр питания очень широк, но основной пищей служат моллюски
(Cardium, Corbulomya, Mytilaster, Syndesmia, Dreissena), составляющие в Азов-
ском море до 80-90% рациона. Второстепенными объектами являются ракооб-
разные (преимущественно крабы), черви и рыба (Костюченко, 1960). В Волге
и ее водохранилищах основной пищей служат беспозвоночные (моллюски -
Dreissena и Pisidium, ракообразные, личинки насекомых, черви), икра и мальки
рыб, может он потреблять и водную растительность (Евланов и др., 1998). По-
ловая зрелость наступает на втором году жизни при длине 4—6 см. Нерестится
с апреля по август в Азовском море и по сентябрь - в Каспии при диапазоне
температур 10-30 °C. Размножение характеризуется асинхронным типом ви-
теллогенеза и многопорционным икрометанием (5-6 порций за сезон с переры-
вом в 17-20 дней). Приводимая обычно величина средней плодовитости около
1400 икринок (а для Каспия - около 700) (Берг, 19496; Казанчеев, 1981) отно-
сится лишь к одной порции. У самки длиной 116 мм в одной порции насчитыва-
лось 1540-1760 икринок, а общая плодовитость составляла 9770 икринок (Ку-
ликова, Фандеева, 1975).
Икрометание в море происходит в прибрежной зоне на глубине до 3 м, ино-
гда до 7-8 м. Икра откладывается на нижнюю поверхность камней, а при их от-
сутствии - на любые предметы на дне. Оплодотворенная икринка яйцевидной
формы, ее расширенная часть пучком клейких нитей прикрепляется к субстра-
ту. Икринки располагаются плотно друг к другу в виде однослойной лепешки,
в создании кладки могут принимать участие несколько самок. Самец охраняет
кладку, препятствует ее заилению и аэрирует движением грудных плавников.
Икра крупная (4x2 мм), олигоплазматическая, диаметр желточного мешка
около 1,8 мм. Большой запас питательных веществ и охрана самцом позволя-
ют удлинить развитие в оболочке, в которой проходит и личиночный период.
При температуре воды 19-26 °C через 14-20 сут вылупляется донная молодь
длиной 6,5-7,0 мм с дефинитивными органами локомоции, зрения, пищеваре-
ния и дыхания. Однако у вылупившихся особей сохраняется довольно круп-
ный желточный мешок (Москалькова, 1978). Уже на следующий день они ло-
вят науплиусов циклопов. Но часто на протяжении 4-7 дней еще не покидают
гнезда, прячась среди пустых оболочек, и самец продолжает их охранять, по-
ка не уменьшится желточный мешок, мешающий их передвижению. Затем
122
мальки рассеиваются по мелководью среди камней. Сообщения о вылуплении
бычка-кругляка на 4—7-й день (Рыбы Подмосковья, 1988; Рыбы Казахстана,
1989) следует считать ошибочными. Мальки до 10 мм длиной питаются круп-
ными формами зоопланктона, при длине 15-30 мм начинают потреблять мол-
люсков, которые уже составляют основу рациона.
Статус вида. Из всех бычковых самый многочисленный и наиболее ценный
объект промысла в Азовском море. К настоящему времени его запасы сильно
подорваны. Включен в последнее издание “Международной Красной книги”.
Автор-составитель: К.И. Москалькова.
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Neogobius Iljin, 1927
242. Neogobius platyrostris (Pallas, 1814) - бычок-губан
Описание и систематика. Характерными для этого вида являются следую-
щие признаки: боковые части верхней губы сильно раздуты, немного выдает-
ся нижняя челюсть, короткие брюшные плавники (не более 19% длины тела),
на первом спинном нет полос. Темя, затылок, спина, горло и стебли грудных
плавников покрыты циклоидной чешуей; жаберные крышки почти голые.
Голова спереди сплюснута, ее ширина в этом отделе больше высоты. Под-
глазничных поперечных рядов невромастов 6. Брюшная присоска с заострен-
ными лопастинками на воротнике не достигает анального отверстия. Хвосто-
вой стебель короткий. Плавательного пузыря у взрослых и молоди нет. Ок-
раска серо-бурая, с мелкими светлыми пятнами на боках; брюшная часть
светлее - желтоватая или беловатая. Первый спинной плавник темно-серый
и без полос, остальные плавники полосатые, у живых особей с темно-синим
оттенком. Половой диморфизм проявляется в разной высоте головы, в фор-
ме рта и щек.
D] VI, D2 I (15) 16-18 (20), АД (11) 12-14. Поперечных рядов чешуй (55)
58-78 (71) (Берг, 19496; Световидов, 1964; Емтыль, 1997).
Пинчук (1977) выделял 4 подвида: 1) номинативный морской восточночер-
номорский подвид; 2) N. platyrostris eurycephalus (Kessler, 1874), распространен-
ный в Азовском море и Керченском проливе; 3) N. platyrostris odessicus Pinchuk,
1977 из северо-западной части Черного моря и 4) N. platyrostris constructor
(Nordmann, 1840), представленный речными популяциями бассейнов Черного,
Азовского и Каспийского морей. В настоящее время N. р. eurycephalus и
123
N. p. odessicus рассматриваются в составе самостоятельного вида N. eurycepha-
lus (Kessler, 1874) (Miller, 1986). Подвид N. p. constructor, в понимании Пинчука,
в настоящее время рассматривается как совокупность нескольких самостоя-
тельных видов пресноводных бычков: У. constructor (Nordmann, 1840) из бассей-
нов рек, впадающих в Черное море южнее Бзыбского хребта (в водах России
не встречается), N. cyrius (Kessler, 1874) из верхнего и среднего течения Куры с
ее притоками (в водах России не встречается), а также новый вид N. rhodioni
Vasiljeva et Vasiljev, 1994 из бассейнов рек кавказского побережья, впадающих
в Черное море севернее Бзыбского хребта, и из бассейна Кубани (Аннотиро-
ванный каталог..., 1998).
Распространение. Черное море у берегов Крыма и Кавказа. Живет в по-
лигалинной зоне у открытых морских берегов, а в Керченский пролив и ли-
маны не заходит. В России обитает от Анапы на юг, вдоль черноморского по-
бережья Кавказа. По неопубликованным данным В.И. Пинчука, встречается
в связанным с морем озерце на мысе Большой Утриш (Аннотированный ка-
талог..., 1998). М.Х. Емтыль (1997) отмечает его присутствие в низовьях Ку-
бани.
Возраст и размеры. Достигает длины 225 мм и возраста 5 лет. У рыбы дли-
ной 160 мм масса тела 78 г.
Образ жизни. О биологии этого вида известно мало. Населяет россыпи
крупной гальки у открытых морских берегов. В Новороссийской бухте дер-
жится среди скал и камней в зарослях цистозиры, молодь - ближе к берегам
среди зеленых водорослей. Питается крабами, бокоплавами, полихетами,
молодью рыб и моллюсков (Емтыль, 1997). Половозрелость наступает в воз-
расте 2 года при длине тела 8 см и массе 5 г. Плодовитость от 225 до 915 ик-
ринок, икра крупная. Нерест в Краснодарском крае в апреле-мае (Емтыль,
1997), в других регионах более растянут (Световидов, 1964). Икрометание
порционное. У самки длиной 80 мм отмечено 91 крупных и 134 мелких икри-
124
нок, а у самки 110 см длиной соответственно 308 и 268 икринок. Можно пред-
положить, что, как и у других видов этой группы бычков, икра бычка-губа-
на также развивается под охраной самца не менее двух недель, после чего из
оболочки выходит относительно крупная донная молодь. Мальки длиной
18 см и более встречались в Новороссийской бухте с июня по сентябрь (Све-
товидов, 1964).
Статус вида. В морской прибрежной зоне обычен, в отдельных местах яв-
ляется объектом любительского лова.
Автор-составитель: К.И. Москалькова.
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Neogobius Iljin, 1927
243. Neogobius rhodioni Vasiljeva et Vasiljev, 1994 -
речной бычок Родиона
Описание и систематика. Отличается от других видов этого рода отсутст-
вием хромосомного полиморфизма и специфической окраской: спина перед
первым спинным плавником (D() однотонная, бока однотонные или мрамор-
ные. D, равномерно покрыт мелкими крапинками или полосами мелких пят-
нышек. Самцы во время нереста чернеют. D2 равномерно высокий или не-
сколько повышается в средней части. Хвостовой плавник закругленный.
Брюшная присоска округлой формы, реже продолговатая, не доходит до
анального отверстия. Лопастинки воротника остроконечные. Голова при-
плюснута, ее ширина заметно больше высоты. Верхняя губа расширена и
вздута к углам рта. Нижняя челюсть чуть выдается вперед. Тело и затылок
покрыты циклоидной чешуей. D! VI-VII; D2 I 17-19 (20); А I (12) 13-15 (16).
Поперечных рядов чешуй 57-84. Кариотип: 2п =46, NF = 46 (Васильева, Ва-
сильев, 1994). '
Распространение. Бассейны рек, впадающих в Черное море севернее Бзыб-
ского хребта, и бассейн р. Кубань. В России - бассейн Кубани и мелкие горные
речки кавказского побережья Черного моря (от р. Псоу до Нечепсуго) (Ва-
сильева, Васильев, 1994). Внутривидовой статус кубанских популяций требует
уточнения.
Возраст и размеры. Длина тела достигает 14 см.
125
Образ жизни. Обитает только в пресной воде, живет как в горных речках с
быстрым течением и каменистым грунтом, так и на широких плёсах с заилен-
ным дном и медленным течением. Биология практически не известна.
Статус вида. Мелкий пресноводный бычок. Хозяйственного значения не
имеет.
Автор-составитель: К.И. Москалькова.
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Neogobius Iljin, 1927
244. Neogobius syrman (Nordmann, 1840) - бычок-ширман
Описание и систематика. Отличительные признаки этого вида: стройное
тело, нижняя челюсть выдается вперед, лопастинки на брюшной присоске ту-
126
пые, длина хвостового стебля в 1,5 раза больше его высоты. Ширина головы
немного больше ее высоты. Диаметр глаза значительно больше ширины лба.
Рот большой, углы рта находятся под серединой глаза. Верхняя губа не расши-
рена к углам рта. Спина, затылок, темя, верхняя часть жаберных крышек, гор-
ло и брюхо покрыты циклоидной чешуей. Стебли грудных плавников голые
или также покрыты чешуй. Высота второго спинного плавника посредине не-
много повышается. Брюшная присоска не достигает анального отверстия.
Подглазничных поперечных рядов невромастов 6-8, обычно 7. Окраска тела
серая. На боках крупные светло-бурые пятна, расположенные в шахматном
порядке. От глаза к верхней губе тянется темная полоска. На первом спинном
плавнике имеются светлые и темные поперечные полосы, на втором спинном,
хвостовом и анальном - голубые полоски вдоль лучей.
Dj VI, D2 I (15) 16-18, A I (10) 12-14 (15). Поперечных рядов чешуй (56)
58-71 (78) (Берг, 19496; Световидов, 1964). Каспийские популяции выделялись
ранее в подвид N. syrman eurystomus (Kessler, 1877) (Берг, 19496; и др.). В совре-
менной литературе подвиды не выделяются (Пинчук, 1976; Miller, 1986; Анно-
тированный каталог..., 1998).
Распространение. Черное, Азовское и Каспийское моря, интродуцирован в
Аральское море. Опресненные лиманы, некоторые пресноводные озера и ни-
зовья ряда рек Черноморско-Азовского бассейна. В Каспии его много у Боль-
шой Жемчужной банки, в приуральских водах и у берегов Мангышлака (Ка-
занчеев, 1981). В России - Азовское море, а также Северный и Средний Кас-
пий (Световидов, 1964; Пинчук, 1976; Рагимов, 1981). В Дону отмечен единич-
но от устья до Ростова и ст. Аксай; в бассейне Кубани встречается в большом
количестве в осолоненных лиманкх, но есть и в пресной воде (Троицкий, Цуни-
кова, 1988). В бассейне Каспия обычен в море, в реках не встречается, в Север-
ном Каспии сравнительно немногочислен, но распространен широко.
Возраст и размеры. Максимальная длина тела 24,5 см, но в уловах домини-
руют особи длиной 12-14 см (Берг, 19496). В Каспии особи длиной 16-19 см
имели массу 88-181 г, причем в Северном Каспии преобладали более мелкие
особи длиной 5-12 см и массой 3-29 г (Казанчеев, 1981).
127
Образ жизни. Солоноватоводный донный вид, заходит в опресненные и пре-
сные воды, а участки с соленостью выше 13%с избегает. В Черном море живет у
берегов, до глубины 10 м, и предпочитает биоценоз кардиевого ракушечника.
В Азовском море держится разрозненно по всей акватории, более концентриро-
вано встречается в центральных районах моря на мягких илах, где часто наблю-
даются заморные явления. Однако ширман выдерживает падение кислорода в во-
де до 0,4 мг/л, поскольку у него сильно развито кожное дыхание (до 35% общего).
Молодь ширмана питается ракообразными (Harpacticoida, Cereopagis, Mysidae).
Взрослые особи - типичные хищники-ихтиофаги: тюлька, хамса, бычки и другие
рыбы составляют до 98% пищевого комка; из нерыбных объектов встречаются
ракообразные (крабы, изоподы) и моллюски (Казанчеев, 1981; Рагимов, 1988;
Троицкий, Цуникова, 1988). Половозрелым становится на втором году жизни.
Минимальная длина тела половозрелых особей 6 см (Трифонов, 1955). Массовый
нерест происходит в Каспии в апреле-мае (Рагимов, 1968; Трифонов, 1955), а
Азовском и Черном морях - в мае-июне (Световидов, 1964; Троицкий, Цуникова,
1988). Есть сведения, что он может длиться до августа (Емтыль, 1997), однако до-
стоверных сведений о порционности нереста не существует. Плодовитость шир-
мана в Черном море составляет 340-1450, в среднем 820 икринок (Трифонов,
1955), в Среднем Каспии самки длиной 14—22 см имели 3120-12040 икринок (Ра-
гимов, 1968). Клейкая икра откладывается на мелководье между камнями или под
ними и охраняется самцом. Форма икринок после оплодотворения яйцевидная
(1,9 х 4,2 мм), с крупным желточным мешком, олигоплазматическая. Развитие
икры длится не менее двух недель. Из оболочки вылупляется хорошо развитая
молодь длиной 7,1-7,3 мм с желточным мешком и сразу ведет донный образ жиз-
ни. К осени сеголетки достигают длины 5 см и массы 2,5 г.
Статус вида. Один из наиболее крупных видов бычков, является объектом
промысла. Включен в последнее издание “Международной Красной книги”.
Автор-составитель; К.И. Москалькова.
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Pomatoschistus Gill, 1864
245. Pomatoschistus marmoratus (Risso, 1810) - леопардовый лысун
Описание и систематика. Рыбы рода Pomatoschistus имеют плавательный
пузырь, хорошо развитую сейсмосенсорную систему на голове в виде кана-
лов, пор и рядов невромастов; межглазничный промежуток узкий, глаза на-
правлены косо вверх или в стороны, передние ноздри не вытянуты в виде
трубочек, бока тела покрыты ктеноидной чешуей, а голова, горло, брюхо и
основания грудных плавников голые. У леопардового лысуна на основании
128
хвостового плавника имеется неясное пятно, первый спинной плавник без
ряда резких пятен, глаза направлены косо вверх. Надглазничные каналы
сейсмосенсорной системы слиты более чем на половину своей длины, про-
дольный подглазничный ряд состоит из 20-23 невромастов, на щеках имеет-
ся 6-7 поперечных рядов невромастов. Есть межглазничная пора и канал над
жаберной крышкой. Окраска тела серовато-коричневая с многочисленными
пятнышками по всей поверхности (леопардовый рисунок). От глаза в перед-
не-нижнем направлении тянется черноватая полоска (Емтыль, 1997). Рого-
вица глаз сероватая. У самцов во время нереста на боках появляются попе-
речные темные полосы, а на первом спинном плавнике блестящее синее пят-
но (у живых рыб).
D; VI; D21 8-10; АI (7) 8-10. Поперечных рядов чешуй (35) 38-45 (48). Поз-
вонков 31 (Берг, 19496; Световидов, 1964; Емтыль, 1997) Подвидов нет.
Распространение. Восточная Атлантика (Пиренейский полуостров),
Средиземное, Черное и Азовское моря, Суэцкий канал; встречается в опрес-
ненных озерах и иногда заходит в устья рек; интродуцирован в оз. Кварун
(Египет) (Световидов, 1964; Miller, 1986). В России - повсюду у берегов Азов-
ского моря, в Новороссийской бухте, в кубанских лиманах и иногда в реке; в
бассейне Дона его нет (Пинчук, Савчук, 1982; Троицкий, Цуникова, 1988;
Емтыль, 1997).
Возраст и размеры. Достигает максимальной длины 8,0 см, но обычно не
более 5 см и массы 2 г. Продолжительность жизни около 2 лет (Ковтун,
1976).
Образ жизни. Держится в прибрежной зоне моря и в бухтах не глубже 10 м,
встречается при солености до 40%о, в солоноватых и опресненных озерах, мес-
тами заходит в устья рек (Световидов, 1964). Рыба небольшая, но многочислен-
ная. Взрослый бычок ведет малоподвижный донный образ жизни, стайная ры-
ба. Молодь питается планктонными ракообразными, а взрослые - мизидами,
9 Атлас..., том 2
129
личинками насекомых и червями. Половой зрелости достигает в возрасте
7-11 мес при длине 25 мм (Ковтун, 1976). Нерест длится все лето: у берегов
Болгарии - с марта по сентябрь, у берегов Румынии - с мая по август (Свето-
видов, 1964), в осолоненных кубанских лиманах - с мая по август (Ковтун. 1976;
Троицкий, Цуникова, 1988). Абсолютная плодовитость от 1310 до 3160 икри-
нок, икрометание порционное. Нерестится у берега, на глубине 10-50 см, на
песчано-ракушечном грунте. Самец устраивает гнездо под вогнутой стороной
пустых раковин гребешка или мидий и в него откладывают икру несколько са-
мок (до 4). Икра откладывается в один слой, в одной кладке икра может на-
ходиться на разных стадиях развития. Икра мелкая, полиплазматическая. Обо-
лочка икры прозрачная, грушевидной формы (1,0 х 0,7 мм), ее более широкое
основание клейкое, им икра прикрепляется к поверхности раковин. Желток
светлый, полупрозрачный, с множеством мелких жировых капель. Самец охра-
няет развивающуюся икру, очищает и аэрирует ее. При температуре воды
20-23° эмбриональное развитие длится 5-7 сут. Вылупляются пелагические ли-
чинки длиной 2,8-3,0 мм, они сначала держатся в толще воды и питаются зоо-
планктоном (Ковтун, 1976). Спектры питания и места обитания личинок этого
бычка совпадают с таковыми хамсы, поэтому колебания численности этих
двух видов находятся в обратной зависимости. В Новороссийской бухте молодь
леопардового лысуна длиной до 7 мм встречались с июня, в наибольшем коли-
честве с июля по сентябрь; в сентябре она уже имела размер 16-18 мм (Свето-
видов, 1964).
Статус вида. Мелкий вид, промыслового значения не имеет. Для многих
хищных рыб (осетровые, судак, камбала-калкан) он служит объектом питания,
а для других (хамса, тюлька, молодь других видов) - конкурентом в питании
(Ковтун, 1976).
Автор-составитель: К.И. Москалькова.
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Proterorhinus Smitt, 1899
246. Proterorhinus marmoratus (Pallas, 1814) -
бычок-цуцик, мраморный тупоносый бычок
Описание и систематика. Характерными признаками бычков этого рода
являются следующие: передние носовые отверстия вытянуты в усиковид-
ные трубочки, свешивающиеся вниз над верхней губой; жаберные крышки
130
голые, за исключением их верхней части; основания грудных плавников и
задняя часть горла покрыты циклоидной чешуей. Тело и голова сжаты с бо-
ков. Брюшная присоска без ясных лопастинок. Голова высокая, ее ширина
меньше высоты. Длина головы укладывается в длине тела 3,5-^,5 раза. Че-
люсти равной длины. Ширина лба меньше диаметра глаза. Основной цвет
131
буровато-серый или желто-серый. На боках имеется несколько поперечных
темных пятен. На рыле с каждой стороны темное пятно, окаймленное сза-
ди белой полоской. В основании хвостового плавника треугольное черное
пятно, окаймленное двумя белыми пятнами. На плавниках (кроме брюш-
ных) могут быть пятнышки. В нерестовый период самцы становятся темно-
коричневыми, их плавники темнеют и грудные плавники удлиняются, а по
краю грудных и второго спинного плавников появляется оранжевая кайма.
Плавательного пузыря нет.
Dj VI-VII, D2 I 14-17 (19), A I 11-15 (16). Поперечных рядов чешуй 36-48.
Жаберных тычинок 6-9. Позвонков 30-33 (Берг, 19496; Световидов, 1964). Ка-
риотип: 2п = 46, NF = 46 (Васильев, 1985). Монотипический род, подвидов нет
(Аннотированный каталог..., 1998).
Распространение. Побережья Мраморного, Эгейского, Черного,
Азовского и Каспийского морей, населяет лиманы, прибрежные озера и
впадающие в них реки; отмечен в оз.Ясхан на Узбое; интродуцирован в
Аральское море (Световидов, 1964). В бассейнах Дуная, Днестра, Буга и
Прута встречается повсеместно, исключая их горные участки. В России
водится в бассейне Азовского (бассейн Северского Донца и Дона с при-
токами, дельта Кубани) и Каспийского морей (у берегов Северного и
Среднего Каспия, бассейны Волги и Кумы, некоторые озера и водохра-
нилища) (Берг, 19496; Световидов, 1964; Казанчеев, 1981; и др.). В бас-
сейне Дона этот вид немногочислен в низовьях (Троицкий, Цуникова,
1988), но довольно обычен в Воронежской области, где встречается в
притоках Дона - Лукавка, Матыра, Воронеж, Икорца, Усмань, Тихая
Сосна (Федоров, 1960). В бассейне Кубани встречается только в лиманах,
отмечен также в р. Адагум и в верховьях Ей (Емтыль, 1997). Отмеча-
ется постоянное продвижение бычка-цуцика вверх по Волге: в 1982 г.
он впервые обнаружен в Саратовском водохранилище (Евланов и др.,
1998).
Возраст и размеры. Достигает максимальной длины 12-15 см. Обычно
преобладающая длина 2-7 см, масса 1-7 г. Продолжительность жизни 2-3 го-
да, лишь для Прута указан максимальный возраст 4 года (Казанчеев, 1981; Дол-
гий, 1993).
Образ жизни. Эвригалинный вид, в море держится у берегов, чаще все-
го в мелких заливах, в опресненных прибрежных участках. Многочислен в
лиманах и устьях рек, предпочитает зарослевые биотопы (Казанчеев,
1981; Троицкий, Цуникова, 1988; Евланов и др., 1998). Ведет малоподвиж-
ный донный образ жизни. Молодь питается зоопланктоном, а взрослые -
главным образом донными беспозвоночными (мелкими ракообразными и
моллюсками, червями, личинками насекомых), в пище также отмечены
остатки рыб, водная растительность, взрослые насекомые и детрит
(Троицкий, Цуникова, 1988; Долгий, 1993). Бычок-цуцик в Черноморском
бассейне крупнее (до 115 мм), чем в Каспийском (до 76 мм). В Черном
море размножается с апреля по август, в низовьях Дона, Кубани и
дельте Волги - с апреля по июль. Созревает в возрасте 1 года при дости-
жении длины 2,9 см. Нерестится в прибрежной зоне на глубине 20—150 см
на песчаном и песчано-каменистом дне. Икрометание порционное. Икра
откладывается в пустые створки раковин (мидий, устриц) или прямо
под камни. В гнезде обычно 2-3 кладки икры, каждая по 250-300 штук
(Калинина, 1976). Полагают, что каждая самка выметывает 200-800 икри-
132
нок, максимум 1080 икринок (Эланидзе, 1983; Емтыль, 1997). Икринки
яйцевидной формы, с острым верхним концом; на широком конце пучок
клейких нитей, которыми икра прикрепляется к субстрату. Гнездо охраня-
ет самец. Икра крупная, мезоплазматическая (плазма составляет 1/3 диа-
метра желточного мешка). Размер икринок 2,5 х 1,4 мм. При темпера-
туре 18-21 °C эмбриональное развитие длится не менее 8 сут. Развитие
без свободной личинки, вылупляется донная молодь длиной 5,2-5,4 мм
с довольно крупным остатком желточного мешка (Калинина, 1976).
Молодь в дельте Волги держится преимущественно в западной
части. Мальки ловились в Каспии при солености воды 24—31%о (Казанче-
ев, 1981).
Статус вида. Промыслового значения не имеет. Объект питания хищных рыб.
Автор-составитель: К.И. Москалькова.
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Rhinogobius Gill, 1859
247. Rhinogobius brunneus (Temminck et Schlegel, 1845) -
амурский бычок
Описание и систематика. В отличие от других бычков этого рода у
амурского бычка нет вертикальных рядов невромастов, но имеются ко-
роткий подглазничный продольный ряд и заглазничный и надкрышеч-
ный каналы сейсмосенсорной системы. Тело веретеновидное, слегка
сжатое с боков. Голова не приплюснутая, спинные и анальный плавники
короткие, хвостовой закруглен. Передние ноздри вытянуты в очень не-
большие трубочки. Глаза расположены высоко. Межглазничное про-
странство очень узкое. Рот коечный. Губы толстые, нижняя прервана
посредине. Спинные плавники разделены промежутком. Лучи первого
спинного плавника (Dj) заметно вытянуты (но не волосовидные), что де-
лает плавник высоким и остроконечным. На брюшной присоске хорошо
развитый свободный воротничок, есть лопастинки. Брюшные плавники
заметно короче грудных. Зубы простые, не трехраздельные. Язык без
выемки, закругленный. Окраска палево-желтая. На боках имеется 6-8
больших темных пятен, на спине подобные им бурые пятна. На хвосте
133
обычно бывает 5 темных полос, у многих особей в основании хвостового
плавника имеется яркое черное пятно. На боках головы нет зигзагооб-
разных темных полос. У самцов в брачном наряде основной тон тела ста-
новится буровато-оранжевым, а пятна на боках черными; у самок окра-
ска не меняется. У взрослых особей плавательного пузыря нет. По сред-
ней линии затылка имеется чешуя у типичного амурского подвида, и нет
ее у уссурийского.
Dj VI, D2 I 8-9, A I 8-9. Поперечных рядов чешуй 28-33 (35) (Берг,
19496; Никольский, 1956; Choi et al., 1990). Кариотип: 2n = 44, NF = 44
(Васильев, 1985). Ранее этот вид именовался у нас Rh. similis (Берг,
19496; Линдберг, Красюкова, 1975). Популяции бассейна Амура, Уссури,
оз. Ханка иногда выделяются в особый подвид Rh. brunneus lindbergi
Berg, 1933, который отличается наличием темного пятна между первы-
ми двумя лучами Dj. К номинативному подвиду Rh. brunneus brunneus
(Temminck et Schlegel, 1845) в России относятся популяции рек заливов
Уссурийского, Петра Великого и Амурского (реки Малая Седанка и
Черная) (Пинчук, 1974, 1978). Однако выделение подвидов принимается
не всеми ихтиологами. В Японии отмечается полиморфизм по окраске,
морфологии и экологии, некоторым формам предлагается придать ви-
довой статус (Аннотированный каталог..., 1998; Freshwater Fishes of
Japan, 1987).
Распространение. Бассейны рек Японского и Охотского морей, тихоокеан-
ского побережья Японии, островов Хоккайдо, Рюкю, Тайвань, реки п-ова Ко-
рея, материкового Китая и Филиппин (Берг, 19496; Линдберг, Красюкова,
1975; Freshwater Fishes of Japan, 1987; The Freshwater Fishes of China, 1988; Choi et
al., 1990). В России - бассейн Амура, от села Корфовского до Савинского, ре-
ки Сунгари и Уссури, оз. Ханка, реки Уссурийского залива и залива Петра Ве-
ликого, р. Туманная (Пинчук, 1974, 1978). Видимо, при перевозке растительно-
ядных рыб этот вид случайно попал в Южный Казахстан, где широко распро-
странился в озерах и реках Сырдарьинского и Балхаш-Илийского бассейнов
(Рыбы Казахстана, 1992).
Возраст и размеры. Достигает длины 10 см (в реках п-ова Корея - до 12 см),
обычно 4—5 см. Максимальный возраст - 4-5 лет.
Образ жизни. Мелкий донный вид, населяет реки, озера, а также усть-
евые и эстуарные участки с глубинами до 5 м. В Китае и на п-ове Корея на-
селяет верхние и средние участки рек с песчаным и каменистым грунтом
(The Freshwater Fishes of China, 1988; Choi et al., 1990). В Амуре держится как
в прибрежной зоне основного русла, так и в пойменных озерах. Дальних пе-
ремещений не совершает, взрослые бычки ведут оседлый образ жизни (Ни-
кольский, 1956). Расселение происходит главным образом за счет ската пе-
лагических личинок. Питается ветвистоусыми и веслоногими ракообраз-
ными и личинками хирономид, изредка олигохетами и водяными клопами.
Созревает на втором году жизни при длине тела 20-22 мм. Зафиксирован
нерест в оз. Медовом, у Елабуги, в июне при температуре воды 24 °C.
У самки длиной 38 мм и массой 1,03 г было 2258 икринок. Икра была отло-
жена на затонувшее бревно. Кладку охранял самец. Икра мелкая полиплаз-
матическая и удлиненная (длина 2,3-2,7 мм и ширина 0,6 мм). Выход из ик-
ры на шестые сутки, предличинка длиной 3,2 мм уже имеет заполненный
воздухом плавательный пузырь и сразу переходит к пелагическому образу
жизни. На 10-е сутки рассасывается желточный мешок (Крыжановский и
134
др., 1951; Никольский, 1956). В бассейне Сырдарьи в экспериментальных
условиях вылупившиеся предличинки длиной 3,3 мм придерживались дна, и
плавательный пузырь у них заполнялся воздухом через двое суток (Макее-
ва, 1972).
В Казахстане нерест порционный, длится с мая по август при температуре
воды 12-28°С (Рыбы Казахстана, 1992). Икрометание порционное. Плодови-
тость 700-1240 икринок. Икра приклеивается на нижнюю или боковую поверх-
ность камней, коряг и других предметов, образуя однослойные кладки. Кладка
икры формируется несколькими самками, имеет неправильную форму до 7 см
в поперечнике и охраняется самцом. Развитие икры длится около 5 сут. Вылу-
пившиеся личинки ведут пелагический образ жизни, совершая суточные верти-
кальные пищевые миграции, дне’м придерживаются дна. В Казахстане при до-
стижении длины 19 мм в июле они переходят к донному образу жизни и скап-
ливаются на мелководье. При этом у них редуцируется плавательный пузырь.
Молодь сначала питается планктонными ракообразными, а по достижении
длины 17-19 мм переходит на бентос, предпочитая личинок хирономид (Рыбы
Казахстана, 1992).
135
Статус вида. Промыслового значения не имеет. Изредка встречается в пи-
тании хищных рыб. Является пищевым конкурентом многих промысловых
рыб.
Авторы-составители: Ю.С. Решетников, К.И. Москалькова.
Класс Osteichthyes Семейство Gobiidae
Отряд Perciformes Род Rhodoniichthys Takagi, 1966
248. Rhodoniichthys laevis (Steindachner, 1880) -
бычок-родониихт, голый родониихт
Описание и систематика. Это новый для России вид описан Пинчуком
(1978) из вод Сахалина (ручей близ устья р. Таранай). У бычка-родониихта
каналы и поры сейсмосенсорной системы на голове полностью отсутствуют,
они заменены рядами невромастов, в том числе надглазничными рядами. Зад-
ний конец верхней челюсти не заходит за глаз, а обычно кончается под его
серединой. По бокам тела вдоль средней линии имеется 7 крупных темных
пятен, причем только передние из них бывают парными. Окраска в целом
темнее, чем у Chaenogobius castaneus. На боках тела заметны желтоватые
вертикальные полосы.
Dj VII-VIII, D2 I 9-11, A I 10-11. Поперечных рядов чешуй 62-70 (по
20 экз.). Ряды пятнышек на втором спинном плавнике интенсивно окрашены.
Подвидов нет. Иногда этот вид фигурирует под названиями Chloea пакатигае
Jordan et Richardson, 1907 или Chloea senbae Tanaka, 1917 (Пинчук, 1978, 1981;
Аннотированный каталог..., 1998).
Распространение. Острова Хонсю и Хоккайдо (Freshwater Fishes of Japan,
1987), северная и западная части бассейна залива Анива на Сахалине. В России
известен из низовьев сахалинских рек Соловьевка, Сусуя, Третья (приток в ни-
зовьях р. Таранай) (Пинчук, 1978).
136
Возраст и размеры. Достигает длины 50 мм.
Образ жизни. В Японии - речной эвригалинный вид (Takagi, 1966), встреча-
ется и в озерах. На Южном Сахалине многочислен в низовьях рек с медленным
течением. Мелкий придонный вид. Его биология в наших водах практически не
изучена.
Статус вида. Хозяйственного значения не имеет.
Автор-составитель: Ю.С. Решетников.
137
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Tridentiger Gill, 1858
249. Tridentiger obscurus (Temminck et Schlegel, 1845) -
темный трехзубый бычок
Описание и систематика. Форма тела как у всех бычков, оно покрыто
среднего размера чешуей. Цилиндрическое и вытянутое тело, сплющенная
голова. Зубы у бычков этого рода расположены в два ряда, зубы внешнего
ряда трехраздельные с более длинным средним зубом. Усиков нет. В отли-
чие от полосатого трехзубого бычка у этого вида нет темных полос, а чешуя
более крупная (менее 50 поперечных рядов). Чешуей покрыты голова свер-
ху и брюшко. Щеки толстые, голые. Язык закруглен. На голове есть каналы
и поры сейсмосенсорной системы, имеется подглазничный дуговидный ряд
невромастов, но нет вертикальных подглазничных рядов. Хвостовой плав-
ник закруглен. Лучи первого спинного плавника длинные, вытянутые в ни-
ти, особенно у самцов. Окраска тела темная. На голове имеются несколько
мелких светлых пятнышек. Первые лучи каждого спинного плавника с 4 ря-
дами темных пятен. Второй спинной и анальный плавники по краю белые.
У основания грудных плавников вертикальная белая полоса (у живых она
оранжевая) и темное пятно в верхней его части. Хвостовой плавник с черны-
ми перепонками.
Dj VI, D2 I 10-12, A I 9-11. Поперечных рядов чешуй (32) 34-37. Кариотип:
2п = 44, NF = 54 (Берг, 19496; Линдберг, Красюкова, 1975; Choi et al., 1990). По-
пуляции этого вида, населяющие пресные воды среднего течения рек и озер
Японии, выделяются в особый подвид Т. obscurus brevispinis Katsuyama, Arai et
Nakamura, 1972; они отличаются от номинативного подвида, обитающего глав-
ным образом в устьях рек и эстуариях, короткими лучами первого спинного
плавника, не вытянутыми нитевидно даже у самцов в период нереста. В водах
России этот подвид не обнаружен (Пинчук, 1978; Аннотированный каталог...,
1998).
Распространение. Бассейны Японского (от р. Ольги, впадающей в залив
Ольги, до Пусана на п-ове Корея, весь п-ов Корея, Китай, побережье Японии
138
от Хоккайдо до района Санин), Желтого (о-ва Чечжудо, Иньчхонь, Чжилий-
ский залив) и Южно-Китайского морей, тихоокеанское побережье Японии
(о-в Рюкю) (Линдберг, Красюкова, 1975; Choi et al., 1990). В России - р. Седан-
ка, низовья р. Ольгинки, Вавайские озера на Сахалине, р. Змейка (приток Ку-
рилки) на о-ве Итуруп (Пинчук, 1978).
Возраст и размеры. Достигает длины 135 мм.
Образ жизни. Речной эвригалинный вид. Населяет эстуарии, солоноватые
воды и связанные с ними водоемы с илистым дном. Средняя длина 7-10 см
(Dotu, 1958; Choi et al., 1990). Биология вида в наших водах практически не изу-
чена. Полагают, что длинные нитевидные лучи первого спинного плавника ха-
рактерны для бычков из эстуарных и устьевых участков. Однако в речках Ма-
лая и Большая Седанка выше водохранилища (среднее течение) зрелые самцы
имели такие же длинные нитевидные лучи, как и рыбы из устьев рек Седанка
и Ольгинка (Пинчук, 1978).
Статус вида. Хозяйственного «значения в России не имеет, в Японии - объ-
ект любительского рыболовства.
Автор-составитель: Ю.С. Решетников.
139
Класс Osteichthyes
Отряд Perciformes
Семейство Gobiidae
Род Tridentiger Gill, 1858
250. Tridentiger trigonocephalus (Gill, 1858) -
полосатый трехзубый бычок
Описание и систематика. В отличие от Tridentiger obscurus этот вид имеет
более мелкую чешую (более 50 поперечных рядов) и характерные две темные
полосы вдоль тела: одна идет по спине у основания спинных плавников, а вто-
рая - по середине бока, от конца рыла до основания хвостового плавника. У ста-
140
рых особей эти полосы плохо заметны. Все плавники, кроме брюшных, с ряда-
ми бурых полосок, идущих поперек лучей. На боках головы и внизу ее имеют-
ся мелкие светлые пятнышки на буром фоне. У основания грудных плавников
темная дуговидная полоса. Второй спинной и анальный плавники с белой кай-
мой по краю. Подглазничный ряд невромастов не доходит до середины глаза.
Dj VI, D2 I 12-14, А I 10-11. Поперечных рядов чешуй 50-54, у корейских
популяций - 50-60. Кариотип: 2п = 44, NF = 60 (Берг, 19496; Линдберг, Красю-
кова, 1975; Choi et al., 1990). Подвидов нет.
Распространение. Встречается по азиатскому побережью Тихого океана от
лимана Амура на севере до Южного Китая и Филиппинских островов на юге.
Бассейны Японского (от залива Петра Великого до Пусана на п-ове Корея, по-
бережье Японии, от мыса Сякотан до островов Цусима и Кюсю), Желтого
(Иньчхонь, Чжифу), Восточно-Китайского и Южно-Китайского морей, тихо-
океанское побережье Японии, Филиппинские острова. В России - бухта Чоми
в лимане Амура, Амурский залив, бухта Новгородская залива Посьет, устья и
низовья рек залива Петра Великого и Амурского залива (Берг, 19496; Николь-
ский, 1956; Линдберг, Красюкова, 1975; Пинчук, 1978; Аннотированный ката-
лог..., 1998; Choi et al., 1990).
Возраст и размеры. Достигает длины 83 мм в наших водах и 110 мм - в Японии.
Образ жизни. Мелкий морской эвригалинный вид, проникающий в устья и
низовья рек (Dotu, 1958; Choi et al., 1990 ). В местах обитания часто многочис-
лен. В малых речках с быстрым течением встречается лишь в устье (речки Се-
данка, Черная). В больших и средних реках с медленным течением он поднима-
ется значительно выше устья (р. Лянчиха). Биология в наших водах практиче-
ски не изучена.
Статус вида. Хозяйственного значения не имеет.
Автор-составитель: Ю.С.Решетников.
Класс Osteichthys
Отряд Perciformes
Семейство Channidae
Род Channa Skopoli, 1777
251. Channa argus (Cantor, 1842) - змееголов
Описание и систематика. Тело длинное, спереди полуцилиндрическое, позади
сжатое с боков. Его высота составляет 20% длины. Голова очень большая, упло-
щенная, ее длина в 2 раза больше высоты тела. Первый спинной и анальный
плавники очень длинные, без колючек. Рот конечный, большой. Верхняя че-
люсть далеко заходит за глаз. Глаза сдвинуты к переднему концу головы. Имеют-
141
ся зубы на челюстях, сошнике и нёбных костях. Спина темная или зеленовато-
черная. По бокам тела крупные расплывчатые пятна черновато-зеленого цвета.
Брюхо светлое. Брюшные плавники светлые, а спинной и хвостовой - темно-зе-
леные. На боках головы за глазами две узких темных полосы до заднего края жа-
берной крышки. В верхней части жаберной полости имеется наджаберный орган
для воздушного дыхания, который состоит из двух пластин. Боковая линия от
верхнего края жаберной крышки сначала идет в верхней части тела, затем на
уровне 15-20 луча спинного плавника изгибается вниз и идет до хвоста.
D 49-54, А 32-38, Р 15-19. Чешуя мелкая, покрывает все тело и голову.
В боковой линии 60-75 чешуй. Позвонков 50-51. Пилорических придатков 2.
Отмечены два подвида: С. argus argus (Cantor, 1842) - номинативный подвид,
который обитает в Китае, на п-ве Корея и в Японии, и С. argus warpachowskii
Berg, 1909 - амурский подвид, который живет только в бассейне Амура и отли-
чается от обычного большим числом чешуй в боковой линии и лучей в спин-
ном и анальном плавниках (Берг, 19496; Никольский, 1956; Аннотированный
каталог..., 1998).
Распространение. Вид распространен в России, Китае (от Амура на юг до
Хуанхэ), на п-ве Корея и в Японии (на о-ве Хонсю и южнее). В Южном Китае,
во Вьетнаме и странах Азии встречаются другие виды (The Freshwater Fishes of
China, 1982; Freshwater Fishes of Japan, 1987; Choi et al., 1990). В России обитает
в среднем и нижнем течении Амура (оз. Удыль), но ниже Комсомольска встре-
чается редко, а вверх по течению идет выше Благовещенска. Обычен в Уссу-
ри, Сунгари и оз. Ханка (Никольский, 1956; Богуцкая, Насека, 1996).
При акклиматизационных работах в 1960-х годах завезен из р. Янцзы (Ки-
тай) в водоемы Средней Азии, где из прудовых хозяйств проник в естественные
водоемы. Прижился и широко распространился в водоемах бассейна Амударьи
и Сырдарьи. В Амударье - повсеместно до низовьев, включая озера, водохра-
нилища, оросительные и сбросные каналы коллектора. Проник в Хаузханское
и Копетдагское водохранилища и распространился по всему Каракумскому ка-
налу (Сальников, Решетников, 1991; Решетников, Шакирова, 1993; Сальников,
1995). В Сырдарье известен от Чардарьинского водохранилища до приустье-
вых участков Аральского моря. Отмечен также в низовьях Сарысу, в Аксай-
Кувандарьинских, Камышлыбашских и других озерах, в бассейне рек Талас и
Чу (Рыбы Казахстана, 1992). Выпускался в низовья Волги, но не прижился.
Возраст и размеры. В естественном ареале достигает длины 80-85 см (на
п-ове Корея даже до 1 м) и массы 7 кг с максимальным возрастом 8+ (Берг.
19496; Choi et al., 1990). В бассейне Арала - до 80 см и массы 6-8 кг, изредка от-
мечались особи до 20 кг.
Образ жизни. Обитает в неглубоких, сильно заросших и хорошо прогревае-
мых водоемах, где есть дефицит кислорода. Периодически поднимается к поверх-
ности воды и с чавканьем заглатывает воздух. Для дыхания помимо жабр исполь-
зует наджаберный орган. Может вылезать на сушу и переползать по мокрой тра-
ве на значительные расстояния. На зиму уходит из придаточных водоемов в бо-
лее глубокие места, зимует небольшими группами в 5-10 экз. в норах под круты-
ми берегами. Это типичный хищник, ведущий малоподвижный образ жизни. Кор-
мится преимущественно утром и вечером, ловит свою жертву броском из засады.
Молодь змееголова питается ракообразными и личинками хирономид, пос-
ле достижения длины 5 см начинает потреблять рыбу, а особи свыше 20 см дли-
ной питаются исключительно рыбой. Размер жертвы составляет 20-40% дли-
ны хищника. Примерно такой же спектр питания и в новых местах обитания.
142
Растет змееголов быстро. Молодь в августе достигает 5-6 см, к ноябрю -
10-15 см. В возрасте 1 года ее длина 22-24 см, в возрасте 2 лет - 34—37 см. По-
ловозрелости достигает на 3-4-м, редко на 2-м годах жизни при длине 30 см.
Нерест растянут с середины мая до середины июля и проходит при температу-
ре воды 18-27 °C. В естественном ареале змееголов выметывает до 3 порций
икры, в бассейне Арала - от 1 до 3 порций. Он устраивает гнездо диаметром до
1 м из растительности в прибрежных районах с глубинами до 2,5 м. Плодови-
тость колеблется от 7 до 104 тыс., составляя в среднем 20-50 тыс. икринок. Ик-
ра пелагическая, желтого цвета с крупной жировой каплей, диаметром
1,2-1,60 мм, редко 2,0 мм. При изменении уровня воды часть икры вымывает-
ся из гнезда и сносится вниз по течению. Развитие икры при 23-25 °C продол-
жается около 2 сут (Соин, 1960). Самец охраняет гнездо. Личинки при выходе
из икры имеют длину около 7 мм и покрыты черным пигментом. Сначала они
спокойно лежат в гнезде, а к концу третьих суток при длине 7,3 мм уже способ-
ны к активному питанию и плавают плотными стайками. На 4-е сутки выходят
за пределы гнезда, образуя более крупные стайки, но они находятся под охра-
ной самца еще 2 недели. После этого у мальков начинает исчезать черный
цвет, к месячному возрасту они приобретают бурый цвет, их охрана прекраща-
ется, и они переходят к одиночному образу жизни. Молодь все лето питается
ракообразными, личинками водных насекомых, а по достижении 5 см длины
начинают потреблять икру и молодь рыб.
Статус вида. Ценная промысловая рыба. В самом Амуре уловы невелики
(2—40 ц), в оз. Ханка уловы были больше, но и там они в настоящее время за-
метно снизились (Никольский, 1956; Богуцкая, Насека, 1996). В бассейне Аму-
ра змееголов считается биомелиоратором, но в бассейне Арала поедает много
143
ценных видов рыб. Его акклиматизация в других районах считается нецелесо-
образной, хотя возможно его разведение в прудовых хозяйствах.
Автор-составитель: О.А. Попова.
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Cottidae
Род Batrachocottus Berg, 1903
252. Batrachocottus baicalensis (Dybowski, 1874) -
большеголовая широколобка
Описание и систематика. Все рыбы сем. Cottidae имеют веретеновидное те-
ло, голое или покрытое костными шипиками, голова большая, укладывается
до трех раз в длине тела. Два спинных плавника следуют друг за другом или
разделены, анальный плавник подобен второму спинному, брюшные плавники
сближены, имеют I 2-5 лучей, пилорических придатков 4-10. Сейсмосенсорная
система представлена сенсорными каналами, которые открываются наружу
посредством пор, расположенных на концах кожных канальцев. Кости черепа
имеют отверстия для сенсорных каналов. В Байкале семейство Cottidae пред-
ставлено двумя подсемействами: Cottinae (1 вид) и Cottocomephorinae (8 видов).
Род Batrachocottus имеет очень большую голову, большой рот, угол кото-
рого заходит за вертикаль заднего края глаза. Лоб широкий, равен или больше
продольного диаметра глаза. Боковая линия неполная, не доходит до основа-
ния лучей хвостового плавника и содержит 7-15 крупных пор. Тело голое,
только под грудными плавниками имеются костные шипики. Формула брюш-
ного (V) плавника I 3. Позвонков 33-36.
Большеголовая широколобка - одна из самых крупных рыб среди при-
брежных Cottoidei. Тело короткое, чаще голое, лишь под грудными плавника-
ми покрыто небольшими редкими шипиками. Окраска тела зеленовато-бурая,
с пятнами по бокам. Плавники, кроме брюшных, имеют темные поперечные
полосы. Голова очень большая, укладывается 2,5 раза в длине тела. Рот боль-
шой, челюсти достигают вертикали заднего края глаза. Глаза небольшие. Ко-
сти головы с очень широкими и глубокими бороздами для каналов боковой ли-
нии. Межглазничное расстояние широкое, равно примерно продольному диа-
метру глаза. Жаберные перепонки приросли к довольно узкому межжаберно-
му промежутку. Спинные плавники соединены между собой, причем второй
спинной (D,) почти в 2 раза длиннее анального. Грудные плавники короткие, из
них четвертый и пятый лучи обычно ветвистые.
D! V-VIII; D2 14-18; Р 14—17; А 10- 13; V I 3. В боковой линии 9-12 круп-
144
ных пор, которые заканчиваются на
вертикали начала D2, далее до основа-
ния хвостового плавника проходит ряд
наружных невромастов. Жаберных
тычинок 4-6. Пилорических придат-
ков 4. Позвонков 32-33, причем туло-
вищные позвонки обычно массивные
и удлиненные, а хвостовые - укоро-
ченные (Берг, 19496; Талиев, 1955; Си-
делева, 1982). Кариотип: 2п = 48,
NF = 60 (Сиделева и др., 1995). Подви-
дов нет (Аннотированный каталог...,
1998).
Распространение. Вид распростра-
нен в прибрежной зоне оз. Байкал до
глубины 150 м, отсутствует в мелко-
водных заливах (сорах) и на Селенгин-
ском мелководье. Отмечен в Иркут-
ском водохранилище до пос. Бурдугуз
(данные В.Г. Сиделевой). Берг (19496)
указывал на частый заход этого вида в
устье р. Култучной.
Возраст и размеры. Предельный
возраст 6 лет, достигает длины 20 см и
массы 160 г (Талиев, 1955).
Образ жизни. Обитает в основном
на каменистом дне, реже на камени-
сто-илистых или каменисто-песчаных
грунтах. Большеголовая широколобка
плавает мало, обычно она лежит на дне, опираясь на нижние лучи грудного
плавника, под прикрытием камня или в зарослях губок и подстерегает свою до-
бычу. Наибольшие скопления этого вида приурочены к районам с обширными
колониями губок. Питание взрослых особей состоит из гаммарид (44% по мас-
се), рыбы (37%), организмов бентоса (личинки хирономид, ручейников) и рас-
тительности. Быстрее всех растет широколобка из Южного Байкала, достигая
ко второму году 46 мм, к третьему - 56 мм, к четвертому - 73 мм, к пятому -
93 мм и шестому - 120 мм длины. Самки созревают при достижении 13 см дли-
ны, самцы - 11 см в возрасте 4-5 лет. Плодовитость колеблется от 500 до 1600,
в среднем 1067 икринок. Диаметр икринки 3 мм (Черняев, 1979). Нерест прохо-
дит в феврале-марте подо льдом, на глубинах от 0,6 до 4,5 м. Икра откладыва-
ется в гнездо одной или двумя самками и охраняется самцом. После нереста от-
нерестовавшие особи уходят на большие глубины. Эмбриональный период при
температуре воды 6-9 °C длится 90 сут. Вылупление личинок происходит в
мае-начале июня. После вылупЛения личинки имеют длину 10 мм (Черняев,
1979) и ведут донный образ жизни. Во второй половине лета мальки ловятся на
глубинах от 5 до 20 м (Талиев, 1955).
Статус вида. Обычный вид в Байкале. Хозяйственного значения не имеет.
Автор-составитель: В.Г.Сиделева.
10 Атлас .., том 2
145
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Cottidae
Род Batrachocottus Berg, 1903
253. Batrachocottus multiradiatus Berg, 1907 -
пестрокрылая широколобка
Описание и систематика. Сравнительно большая рыбка с коротким те-
лом и большой головой, которая примерно 3 раза укладывается в длине те-
ла. Окраска тела светло-коричневая, с темными пятнами на боках. Плавни-
ки, кроме брюшных и анального, имеют поперечные полосы. Угол рта до-
стигает переднего края глаза. Межглазничное пространство немного шире
диаметра глаза. Интересно отметить,
что пестрокрылая широколобка име-
ет очень крупные отолиты, по своим
размерам они сходны с таковыми у
трески и не встречаются у пресно-
водных коттоидных рыб. Кожа го-
лая, под грудными плавниками име-
ются густо сидящие костные шипи-
ки. Хвостовой стебель короткий.
Грудной плавник заканчивается на
вертикали второго луча второго
спинного плавника. Спинные плав-
ники соединены между собой низкой
кожистой складкой.
D, (VI) VII-VIII; D2 15-19; А (12)
13—15; Р 16-19; V I 3. В боковой линии
14-16 крупных пор. Жаберных тычи-
нок 5-6. Пилорических придатков 4.
Позвонков 35-36, из них туловищных
12-13, которые не увеличены в разме-
рах; хвостовые позвонки несколько
удлинены (Берг, 19496; Талиев, 1955;
Сиделева, 1982). Кариотип: 2п = 48,
NF = 68 (Сиделева и др., 1995). Подви-
дов нет (Аннотированный каталог...,
1998).
Распространение. Эндемик оз.
Байкал, распространен в большом ди-
апазоне глубин, от 50 до 900 м. Встре-
чается почти во всех районах озера,
предпочитая участки с илисто-песча-
146
ным и илистым дном. В притоках Байкала, в р. Ангара или мелководных сорах
этот вид не обнаружен.
Возраст н размеры. Предельный возраст 7 лет. Достигает длины 20,3 см,
обычно 11,5-13 см. Максимальная масса - 160 г.
Образ жизни. Предпочитает биотопы с илисто-песчаными грунтами, с вы-
соким содержанием детрита. Летом взрослые особи держатся на глубинах
200-400 м, молодые - на 50-200 м. Питается не только бентосными, но и при-
донно-пелагическими организмами. Состав пищи пестрокрылой широколобки
включает 4 основных компонента: гаммариды (частота встречаемости 72%),
рыба - 13% (малая голомянка и донные Abyssocottidae), икра донных коттоид-
ных рыб и детрит (Сиделева, Механикова, 1990). Растет медленно: в возрасте
1+ достигает длины 60 мм, в 2+ - 87 мм и 3+ - 109 мм. Созревает при достиже-
нии длины 115- 130 мм на 4-м году жизни. Самка имеет среднюю плодовитость
455 икринок. Икринки ярко-оранжевого цвета, диаметром 1,7 мм. Нерест про-
исходит в июне-июле на глубинах 180-200 м. Выклюнувшиеся личинки ведут
донный образ жизни. Длина сеголетков в сентябре-октябре 14-15 мм.
Статус вида. Обычный вид в Байкале. Промыслового значения не имеет.
Автор-составитель: В.Г.Сиделева.
Класс Osteichthyes Семейство Cottidae
Отряд Scorpaeniformes Род Batrachocottus Berg, 1903
254. Batrachocottus nikolskii (Berg, 1900) -
жирная широколобка
Описание и систематика. Этот вид отличается от других широколобок
тем, что имеет длинный хвостовой стебель, спинные плавники разделены
большим промежутком, очень низкий первый спинной плавник, однотонную
окраску тела и плавников и не имеет шипов на предкрышке. Туловище ко-
роткое, его высота у анальногб плавника почти в 1,5 раза меньше наиболь-
шей высоты тела. Окраска тела светлая, кремово-розовых оттенков. Все
плавники светлые, однотонные и не имеют темных поперечных полос. Голо-
ва более 3,5 раз укладывается в длине тела. Нижняя челюсть выдается впе-
ред. Вооружение на предкрышке полностью отсутствует. Межглазничное
пространство очень широкое, в 2,5 раза больше продольного диаметра гла-
за. Кожа на туловище голая, под грудными плавниками ниже латерального
147
канала имеются тупые шипики. Хво-
стовой стебель удлиненный. Все лучи
в грудном плавнике толстые, невет-
вистые. Брюшные плавники корот-
кие, в 1,5 раза короче грудных, дале-
ко не доходят до анального отвер-
стия. Спинные плавники разделены
между собой широким промежутком,
равным по величине ширине лба. Dj
очень низкий, вдвое ниже D2. Хвосто-
вой плавник имеет усеченную форму.
Di (V) VI—VIII; D2 14-18; А 11-14; Р
15-17; V I 3. В боковой линии 10-14
очень крупных пор. Жаберных тычи-
нок 6-7. Пилорических придатков 3-5.
Позвонков 35-37 (Берг, 19496; Талиев,
1955; Сиделева, 1982).
Распространеие. Эндемик озера
Байкал, обитает на глубинах от 100 до
1300 м.
Возраст и размеры. Максимальная
длина 268 мм при массе 280 г, средняя
длина половозрелых особей 220 мм и
масса 167 г.
Образ жизни. Самый глубоковод-
ный вид из семейства Cottidae. Предпо-
читает мягкие грунты с обилием дет-
рита. Жирная широколобка чаще
встречается на глубинах 800 и 1000 м
(Сиделева, 1999). Большие концентрации жирной широколобки обнаружены
на периферии района подводного гидровента в бухте Фролиха (Северный Бай-
кал), на глубине 430-450 м. Основной пищей В. nikolskii являются донные гам-
мариды и рыба. Часто (20% частота встречаемости) у взрослых особей в же-
лудках можно найти пелагических голомянок (Comephoridae). Как ловит
В. nikolskii голомянок, остается невыясненным, так как визуальные наблюде-
ния из подводных аппаратов “Пайсис” показали, что особи этого вида ведут ма-
лоподвижный образ жизни. Начинает созревать в возрасте 6+ при длине
203 мм. Плодовитость составляет в среднем 1100 икринок при колебаниях от
1000 до 1400. Икра крупная, светло-желтого цвета, диаметр икринки 3,2 мм.
Нерест происходит в июне-июле. Жирная широколобка получила свое русское
название в связи с тем, что в мышцах, печени и полости тела содержится боль-
шое количество жира. В связи с этим обстоятельством ее плотность (удельный
вес) составляет в среднем всего 1,028, этот показатель наименьший среди дон-
ных коттоидных рыб Байкала (Талиев, 1955).
Статус вида. Непромысловый вид.
Автор-составитель: В.Г.Сиделева.
148
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Cottidae
Род Batrachocottus Berg, 1903
255. Batrachocottus talievi Sideleva, 1999 - широколобка Талиева
Описание и систематика. Сравнительно большая рыбка с крупной голо-
вой и толстым укороченным туловищем. Окраска тела однотонная, темно-
коричневая на спине и боках и светлая на брюшной стороне. Голова боль-
шая, укладывается три раза в длине тела. Угол рта достигает середины
глаза. Межглазничное пространство широкое, равно продольному диамет-
ру глаза. Межжаберный промежуток узкий, в шесть раз меньше длины го-
ловы. Туловищный канал (боковая линия) заканчивается на вертикали пя-
того луча второго спинного плавника и содержит 17 пор. Тело короткое,
его высота у начала анального плавника в 1,4 раза меньше наибольшей вы-
соты, которая содержится 4 раза в длине тела. Кожа на боках, ниже туло-
вищного канала, покрыта костными шипиками, которые доходят до верти-
кали последнего луча первого спинного плавника. Хвостовой стебель ко-
роткий, содержится 6 раз в длине тела, его высота почти в 3 раза меньше
его длины. Все плавники однотонные и не имеют. поперечных полос.
В грудном плавнике шестой и седьмой лучи ветвистые. Спинные плавники
не соединены между собой и разделены узким промежутком. Анальный
плавник начинается на некотором расстоянии от анального отверстия.
Брюшные плавники короткие, далеко не доходят до анального отверстия,
расстояние от конца лучей брюшного плавника до анального отверстия
равно длине плавника.
D] (V) VI-VIII; D2 15-18; А 13-14; Р 16-18; V I 3. В боковой линии 16-17
очень крупных пор. Жаберных тычинок 5-6. Пилорических придатков 3. Поз-
вонков 35-36 (Сиделева, 1999).
Распространение. Эндемик озера Байкал, обитает на глубинах от 300 до
1300 м, половозрелые особи - в основном на глубинах 300-900 м. Максимум
численности приходится на зону глубин от 450 до 500 м.
Возраст и размеры. Максимальные размеры 223 мм при массе 150 г, обыч-
ная длина 170 мм. *
Образ жизни. Донный вид, предпочитает мягкие заиленно-песчаные
грунты с обилием детрита. В состав пищи входят гаммариды, рыба, детрит.
Гаммариды были обнаружены у всех исследованных широколобок и пред-
ставлены 15 видами. Наряду с донными и придонными формами гаммарид в
9% случаев в пище присутствовал пелагический бокоплав Macrohectopus
branickii. Рыба в пище была обнаружена у особей, пойманных на глубине
149
Класс Osteichthyes
Отряд Scorpaeniformes
900-950 м, и состояла из пелагиче-
ских голомянок (Comephoridae) и
донных широколобок (Abyssocotti-
dae). Из других компонентов присут-
ствовали свободноживущие немато-
ды (22% встречаемости) и детрит
(18%) (Сиделева, Механикова, 1990).
Созревает при длине 155 мм и массе
тела 80 г, в возрасте 6 лет. Нерест
происходит в июне-июле, вероятно,
на глубинах от 140 до 550 м. В районе
мыса Голого 11 июля 1987 г. на глу-
бине 480-540 м был пойман самец,
охраняющий гнездо, в желудке кото-
рого найдена развивающаяся икра из
кладки этого вида, находящаяся на
стадии начала гаструляции. Плодо-
витость колеблется в пределах 350-
760 икринок, диаметр икры 3,3-
3,7 мм (Сиделева, 1999).
Статус вида. Широко распростра-
ненный обычный вид в глубоководной
области Байкала. Встречается практи-
чески во всех траловых уловах. Про-
мыслового значения не имеет.
Автор-составителы В.Г. Сиделева.
Семейство Cottidae
Род Cottocomephorus Pellegrin, 1900
256. Cottocomephorus alexandrae Taliev, 1935-
северобайкальская желтокрылка
Описание и систематика. Все представители рода Cottocomephorus име-
ют сжатую с боков голову, характерную для пелагических рыб, выемча-
тый хвостовой плавник, большое число жаберных тычинок (8-16), лучей
150
во втором спинном (17-20) и анальном плавниках (20-23), позвонков (37-41).
Данный вид отличается от всех других представителей этого рода большим
числом пор боковой линии и большим числом жаберных тычинок, наличием
большого расстояния между спинными плавниками и недифференцированной
зубной пластинкой. Глаза маленькие, продольный диаметр глаза около 5 раз
умещается в длине головы. Межглазничное пространство широкое, в 1,5 раза
больше диаметра глаза. Все плавники, кроме грудных, не имеют темных попе-
речных полос. Грудные плавники длинные, более 2 раз в длине тела, они
заканчиваются на вертикали пятого луча D2, имеют светло-желтую окраску и
темные поперечные полосы, сходные с С. grewingkii. Брюшные плавники ко-
роткие, далеко не доходят до анального отверстия. Хвостовой плавник имеет
выемчатою форму.
Dt VIII—X; D2 17-19 (20); А 20-22; Р 18-21; V I 4. В боковой линии 18-23
мелких пор. Жаберных тычинок 18— 20. Пилорических придатков 2. Поз-
вонков 38-40 (Берг, 19496; Талиев, 1955). Д.Н. Талиев (1955) рассматривал
С. alexandrae в качестве подвида С. grewingkii. Согласно нашим представле-
ниям, С. alexandrae имеет уникальный комплекс признаков, а также отлич-
ные от других видов рода Cottocomephorus характеристики, что, позволило
поднять таксономический ранг подвида до ранга вида Cottocomephorus
alexandrae.
Распространение. Озеро Байкал,
вид наиболее многочислен в южной
части озера, проливе Малое Море и на
Селенгинском мелководье.
Возраст и размеры. Достоверных
данных о предельных размерах и воз-
расте нет.
Образ жизни. Биология вида изу-
чена слабо, так как его таксономи-
чес-кий статус долгое время был
спорным. Обитает в толще воды над
подводным склоном и в районах, ха-
рактеризующихся повышенной био-
логической продуктивностью. Со-
вершает значительные миграции при
нагуле и нересте. Пищу этого вида
составляют мезозоопланктон (55%
по биомассе), молодь Cottocomep-
horus (20%), пелагический бокоплав
М. branickii (14%) и донные гаммари-
ды (11%). Согласно данным Д.Н. Та-
лиева (1955), нерест у него происхо-
дит с февраля по апрель на глубине
5-10 м, в нагульный период северо-
байкальская желтокрылка обитает
глубже, от 10 до 400 м, но предпочи-
тает глубины 200-220 м. Мальки это-
го вида практически не отличаются
от мальков С. inermis.
151
Статус вида. Обычный вид в Байкале, но специального промысла нет. Слу-
жит пищей омуля, черного и белого хариусов (Талиев, 1955).
Автор-составитель: В.Г. Сиделева.
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Cottidae
Род Cottocomephorus Pellegrin, 1900
257. Cottocomephorus grewingkii (Dybowski, 1874) -
желтокрылая широколобка
Описание и систематика. Характерные признаки вида - маленькие
глаза, малое число пор в боковой линии (12—15) и меньшее, в сравнении с
другими видами рода, число жаберных тычинок (16-19), наличие в окра-
ске туловища и головы темного пигмента в виде пятен неправильной фор-
мы. Тело веретенообразное и вытянутое, его высота у начала анального
плавника в 1,5 раза меньше наибольшей высоты. Кожа голая. Голова и
спина темно-зеленые, бока светлые, желто-зеленого (оливкового) цвета,
с коричневыми пятнами неправильной формы, плавники светло-оливко-
вые. Первый спинной плавник окаймлен более темной зеленовато-жел-
той полосой. В нерестовый период у самцов грудные плавники окрашива-
ются в яркий лимонно-желтый цвет, на их внутренней стороне, на лучах
появляются эпителиальные бугорки. Спина и голова приобретают темно-
зеленую или почти черную окраску. Окраска самок в нерестовый период
практически не меняется, и эпителиальные бугорки на лучах не развива-
ются.
Голова сужена с боков. Имеются два предкрышечных шипа. Нижняя че-
люсть выдается вперед. Антеанальное расстояние большое, почти равно поло-
вине длины тела. Анальный плавник длинный, в нем более 19 лучей. Плавни-
ки, кроме брюшных и анального, имеют темные поперечные полосы. Спинные
плавники расположены близко друг к другу и соединены кожистой складкой.
Хвостовой плавник имеет посредине выемку.
Dj VII-IX; D2 17-20; А 19-22; Р 17-19; V I 4. Боковая линия неполная, туло-
вищный канал оканчивается у последних лучей DP В боковой линии 12-15 пор.
Жаберных тычинок 16-19 (21). Пилорических придатков 3-5. Позвонков
37^-0 (Берг, 19496; Талиев, 1955; Сиделева, 1982). Кариотип: 2п = 48, NF = 64
(Ханаев, Сиделева, 1995). Подвидов нет. Выделяемый ранее подвид С. grew-
ingkii alexandrae Taliev, 1935 теперь рассматривается нами как самостоятель-
ный вид.
152
Распространение. По всему Бай-
калу и в районе истока р. Ангары, до
поселка Никола. Вид наиболее при-
урочен к южной части озера.
Возраст и размеры. Предель-
ный возраст 5+ лет. Достигает длины
19 см, обычно меньше. Самки меньше
самцов.
Образ жизни. Придонно-пелаги-
ческий вид, в нагульный период рас-
пространен на глубинах от 20 до
350 м. Во время нереста близко под-
ходит к берегам. Состав пищи моло-
ди желтокрылки ограничен двумя
компонентами мезопланктона: Epis-
chura baicalensis и Cyclops kolensis.
В возрасте 2+ наряду с мезопланкто-
ном (85-94%) в пище появляется ма-
крозоопланктон, который представ-
лен видом Macrohectopus branickii
(3-11%), а также молодь желтокрыл-
ки и голомянок (1-6%), которые ста-
новятся основной пищей у особей
старших возрастных групп (Зубин,
1992). Половая зрелость наступает
на 2-3-м году жизни при длине
63-70 мм и массе 2,3—3,0 г (при пло-
довитости 465-565 икринок). На при-
мере желтокрылого бычка экспери-
ментально была показана роль половых феромонов в нерестовом поведе-
нии коттоидных рыб. Выявлено, что источником половых сигнальных ве-
ществ у самцов является мочевая жидкость, а у самок - овариальная жид-
кость. В преднерестовый период у самцов мочевой пузырь увеличивается в
10 раз и более. Половые феромоны действуют в ничтожно малых концент-
рациях: мочевая жидкость - в концентрации 1 х 10~16, овариальная жид-
кость - 1 х 10~12 (Остроумов, 1987). Обычно средняя длина и масса произво-
дителей таковы: самки - 104 мм и 13,4 г; самцы - 116 мм и 21,1 г. Абсолют-
ная плодовитость колеблется в пределах 465-2200, составляя в среднем 1850
икринок. Диаметр икринок 1,60-1,85 мм (Сиделева и др., 1990).
Желтокрылка образует три нерестовых стада, которые различаются
размерами производителей, сроками нереста, районами и глубинами рас-
положения нерестилищ. Названия стад соответствуют названию месяца,
на который приходится пик нереста: мартовское, майское и августовское.
После нереста самки отходят на глубины, а самцы охраняют икру до вы-
хода личинок. Самец может нереститься с несколькими самками, поэтому
в гнезде у желтокрылого бычка может быть от одной до двадцати кладок.
Охраняющий икру самец обмахивает ее плавными асинхронными движе-
ниями грудных плавников. При гибели охраняющего самца погибают и все
кладки икры от заиления или выедания их гаммаридами и планариями.
После вылупления личинок часть самцов погибает, а остальные уходят на
153
глубину. Поскольку нерест проходит при различной температуре воды, то
соответственно и развитие икры имеет разную продолжительность: у мар-
товского стада нерест подо льдом, при температуре 0,5 °C развитие про-
должается 90 дней, а у августовского при нересте в период максимального
прогрева озера (12-15 °C) инкубация занимает всего 15 дней (Соин, Турда-
ков, 1966). Вылупление молоди начинается в середине июня и продолжа-
ется до середины сентября. Личинки после вылупления имеют длину
7,6 мм. Мальковый период наступает при длине особей 21-22 мм (Черня-
ев, 1984). Молодь питается зоопланктоном.
Статус вида. С 1943 по 1971 г. был промысловым видом в Байкале, его
добывали вентерями при нерестовых миграциях. Промысел базировался на
мартовском и майском нерестовых стадах. Максимальные уловы достигали
18 000 ц в год (Коряков, 1972). Рыба использовалась для изготовления кон-
сервов “Байкальские шпроты”. Промыслом изымалось до 70% численности
нерестовых стад, что привело к резкому уменьшению численности желто-
крылого бычка и следовательно, и его уловов. С 1971 г. существует запрет
на промысел.
Автор-составитель: В.Г. Сиделева.
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Cottidae
Род Cottocomephorus Pellegrin, 1900
258. Cottocomephorus inermis (Jakowlew, 1890) -
длиннокрылая широколобка
Описание и систематика. В отличие от других видов этого рода длин-
нокрылая широколобка имеет большие глаза, 4 предкрышечных шипа,
ее туловищный канал содержит 19—22 поры и заканчивается на вертика-
ли средних лучей D2. Тело веретеновидное. Кожа голая. Окраска тела
как у обычных прибрежных пелагических рыб: темная спина, светлые с
сиреневым оттенком бока. В нерестовый период самцы приобретают
брачный наряд: кожные покровы и плавники становятся одноцветными,
почти черными с фиолетовым оттенком, на нижней стороне грудных
плавников появляются эпителиальные бугорки. Голова сужена с боков,
ее ширина равна высоте у затылка. Череп более нежный, чем у других
видов этого рода. Нижняя челюсть сильно выдается вперед. Зубы на че-
люстях большие, острые и расположены в два ряда. Глаза большие, про-
154
дольный диаметр глаза 3,5 раза ук-
ладывается в длине головы. Межг-
лазничное пространство узкое,
меньше диаметра глаза. Все плав-
ники светлые, однотонные, не име-
ют темных поперечных полос.
Грудные плавники длинные, окан-
чиваются на вертикали девятого
луча D2. Спинные плавники разде-
лены узким промежутком.
Dj VII-IX; D2 17-21; А 20-22; Р
20-21; V I 4. В боковой линии
19-22 пор. Жаберных тычинок
11-19. Пилорических придатков 3-4.
Позвонков 40-42 (Талиев, 1955; Си-
делева, 1982). Кариотип: 2п = 48,
NF = 62 (Сиделева, Ханаев, 1995).
Подвидов нет. У Берга (19496) этот
вид имеет название Cottocomephorus
comephoroides.
Распространение. Эндемик озера
Байкал, обитает на глубинах 50-
500 м, наиболее многочислен в север-
ной части озера. Ранее, до постройки
плотины Иркутской ГЭС, был найден
в реках Ангара и Иркут, в районе
г. Иркутска. В настоящее время там
не встречается.
Возраст и размеры. Предельная
длина самцов 22 см, самок - 19 см. Обычная длина самцов 16-17 см и самок
14-15 см (Талиев, 1955).
Образ жизни. Ведет придонно-пелагический образ жизни, часть суток про-
водит на дне, затем совершает значительные вертикальные миграции в толщу
воды. К осени собирается в стаи на глубинах 100-200 м, зимой обитает в при-
донных слоях воды на глубинах 50-200 м. В середине февраля подходит к бере-
гам для нереста. Основным нерестовым районом является пролив Малое Мо-
ре. Для откладывания икры использует не только камни, но любой твердый
субстрат. По устному сообщению В.А. Фиалкова, поднятая со дна Байкала, с
глубины 40 м легковая машина была буквально облеплена кладками длинно-
крылой широколобки. Этот вид более специализирован для питания планкто-
ном, чем два других вида этого рода. Основу пищи молоди составляет мезозо-
опланктон, в основном Epischura baicalensis (97-100% по биомассе). Из других
компонентов встречаются Cyclops kolensis, Harpacticella inopinata, Diaptomus
graciloides, Chydorus spaericus (Сорокин, Сорокина, 1988). Взрослые особи по-
требляют в основном макропланктон (Л/, hranickii) и пелагическую молодь
Cottoidei: желтокрылок, длиннокрылок и голомянок. Иногда в пище встреча-
ются донные формы гаммарид (Коряков, 1972; Зубин, 1992). Становится поло-
возрелым в возрасте 3+ при достижении длины 125 мм. Средняя длина полово-
зрелых самцов 175-181 мм, самок - 140-167 мм (Коряков, 1972). Нерест проис-
ходит в феврале-марте, подо льдом, на глубинах 10-40 м. Плодовитость
155
2,5-3,5 тыс. икринок, диаметр икринки 1,4-1,5 мм. Самец охраняет кладку ик-
ры. После нереста самки уходят на глубины свыше 100 м. Продолжительность
эмбриогенеза около 80 дней. Личинки после вылупления имеют длину
11,2-13,5 мм и ведут пелагический образ жизни. В дневное время они опуска-
ются на глубину 30-40 м, а ночью поднимаются к поверхности. Молодь пита-
ется зоопланктоном.
Статус вида. Промыслового значения не имеет, хотя может быть в прило-
ве. Ее молодь составляет пищу омуля, а взрослых особей поедают сиг, осетр и
нерпа.
Автор-составитель: В.Г. Сиделева.
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Cottidae
Род Cottus Linnaeus, 1758
259. Cottus amblystomopsis Schmidt, 1904 - сахалинский подкаменщик
Описание и систематика. Как у всех представителей рода Cottus, голова
слабо вооружена, на голове нет гребней, кожа голая или покрыта шипиками,
имеются зубы на челюстях и сошнике, жаберные перепонки приращены к
широкому межжаберному промежутку, боковая линия в один ряд, иногда не-
полная. Сахалинский подкаменщик похож по окраске и внешнему виду на
подкаменщика Черского, но отличается от него отсутствием нёбных зубов.
Это крупная для пресноводных коттоидных рыба с немного удлиненным ту-
ловищем. Кожа толстая, голая, только под грудными плавниками имеются
маленькие и редкие шипики. Окраска тела серая, на сером фоне рассеяны
черные мелкие пятна, которые у основания второго спинного плавника обра-
зуют темные скопления. Голова темно-серая, почти черная. Нижняя сторона
тела светлая. Плавники имеют темные поперечные полосы. На голове кожа
морщинистая. Хвостовой стебель короткий, его высота почти в 2 раза мень-
ше его длины. Голова большая и сильно сплюснутая. На предкрышке 2 тупых
шипа, скрытых в коже. Рот большой, нижняя челюсть слегка выдается впе-
ред и достигает вертикали середины глаза. Зубы мелкие, многочисленные,
одинаковые по форме и размерам. Глаза маленькие, межглазничный проме-
жуток широкий. Передние ноздри в виде маленьких коротких трубочек.
Грудные плавники короткие, заканчиваются на вертикали первых лучей вто-
рого спинного плавника. Брюшные плавники, особенно у самцов, очень длин-
156
ные, заходят за анальное отверстие и доходят до третьего луча анального
плавника. У нерестовых самцов на лучах брюшного плавника присутствуют
эпителиальные бугорки. Между спинными плавниками нет промежутка,
плавники следуют друг за другом. Второй спинной плавник длиннее анально-
го, но ниже его. Анальный плавник начинается сразу же за анальным отвер-
стием. Сейсмосенсорная система представлена узкими сенсорными каналами
с очень мелкими порами.
D! VIII-IX; D2 17-19; А 14; Р (14) 15-16; V I 4. Туловищный канал полный,
доходит до основания лучей хвостового плавника и содержит 38-40 пор. Жа-
берных тычинок 6. Позвонков 37 (Берг, 19496; Линдберг, Красюкова, 1987).
Подвиды отсутствуют (Аннотированный каталог..., 1998).
Распространение. Реки и опресненные участки тихоокеанского побережья
вдоль берегов России, п-ова Корея, Японии. В России известен из рек о-ва Са-
халин (Лютога, Александровка, Поронай, Тымь), Курильских островов, (Куна-
шира, Итурупа) и Приморья (Копи, Ботчи, Туманная). Встречается также в ре-
ках о-ва Хоккайдо (Берг, 19496; Линдберг, Красюкова, 1987; Freshwater Fishes
in Japan, 1987).
157
Возраст и размеры. Известная предельная длина 208 мм (Берг, 19496).
Образ жизни. Пресноводный вид, но встречается и в опресненных участках
моря. Биология этого вида изучена у рыб, обитающих в реках о-ва Хоккайдо.
Половозрелые особи обитают в нижнем и среднем течении рек. Днем они за-
рываются в гальку, а после захода солнца мигрируют к середине русла реки и
питаются плывущими по течению организмами дрифта (насекомыми и бентос-
ными организмами). Представители этого вида так же, как и другие пресновод-
ные коттоидные рыбы, строят гнезда. Ранней весной, когда уровень воды под-
нимается вследствие таяния снегов, половозрелые самцы занимают простран-
ство под большими плоскими камнями в речном русле, строят гнездо и “ведут”
в него готовых к нересту самок для выметывания икры. Икра мелкая, ее диа-
метр 1,6-1,9 мм. Плодовитость 600-3000 икринок в зависимости от размеров
самок. Икра откладывается на нижнюю сторону камней, и примерно в течение 4
месяца, до вылупления ее охраняет “самец-отец”. Выклюнувшиеся личинки
сносятся вниз по реке к морю. В море они держатся в пелагиали в течение трех |
недель, после чего они возвращаются в реку и ведут донный образ жизни (Goto, 1
Nakano, 1993). 1
Статус вида. Промыслового значения не имеет. 1
Автор-составитель: В.Г. Сиделева. а
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Cottidae
Род Cottus Linnaeus, 1758
260. Cottus cognatus Richardson, 1836 - слизистый подкаменщик
Описание и систематика. Небольшая рыбка с широкой, приплюснутой го- I
ловой, равномерно суживающимся к хвосту телом, большим ртом, мелкими зу- ]
бами на челюстях и сошнике (на нёбных костях зубов нет), короткими и густы- |
ми шипиками только под грудными плавниками, неполной боковой линией, од- |
ним (верхним) шипом на предкрышечной кости, двумя порами нижнечелюст- 1
ного канала сейсмосенсорной системы на подбородке. Общий фон тела тем- |
ный, брюшко и брюшные плавники желтоватые, без пятен и перетяжек. Ос- 1
тальные плавники с рядами мелких темных пятен, под вторым спинным плав- |
ником два крупных темных пятна. В период нереста у самцов по краю первого |
спинного плавника появляется заметная красная кайма. Спинные плавники |
почти соприкасаются, последний луч анального плавника обычно двойной. 1
158
D1 VII-IX; D2 15-18; A 9-12; P 12-15; V I 3-4. В туловищном канале боко-
вой линии 11-32 (15-25) пор. Жаберных тычинок 3-7, пилорических придатков
2-5. Позвонков 33-34 (Черешнев, 1976). Выделяются два подвида: С. cognatus
cognatus Richardson, 1836 - западный слизистый подкаменщик, большая часть
ареала которого расположена в районах северо-западной Америки, а также на
Аляске и на Чукотке; С. cognatus gracilis Heckel, 1840 - восточный слизистый
подкаменщик с атлантического побережья Северной Америки (Аннотирован-
ный каталог..., 1998).
Распространение. Широко распространен в северных районах Северной
Америки (Scott, Crossman, 1973; Lee et al., 1980). На Северо-Востоке России
встречается только на Восточной Чукотке - на арктическом побережье, к вос-
току от р. Амгуэма, и на беринговоморском, от Берингова пролива к югу до
р. Хатырка, есть в бассейне р. Анадырь от р. Белой до лимана, живет в оз. Эль-
гыгытгын (Берг, 19496; Черешнев, 1976, 1996а; Аннотированный каталог...,
1998).
Возраст и размеры. Предельный возраст 8 лет, достигает длины 13,8 см,
массы 20 г.
Образ жизни. Обитает в самых разнообразных водоемах - ручьях, реках,
горных озерах, лиманах, опресненных лагунах, термальных лужах, но избегает
159
заморных термокарстовых озер. Предпочитает чистые быстрые водотоки с
крупнокаменистым грунтом; молодь обычна на песчаных мелководьях. Ведет
малоподвижный, скрытный образ жизни, прячась под камнями и другими ук-
рытиями. Питается преимущественно бентосом, но иногда хищничает, поедая
молодь и икру рыб. Созревает на 3-4-м году жизни при длине тела 6-10 см и ве-
се 2-14 г. Размножается в июне-июле при температуре воды 3-11° С (опти-
мальная - 6°). Плодовитость равна 80-780 икринок. Самка откладывает икру в
виде лепешки на нижнюю поверхность крупного камня; эту кладку затем охра-
няет самец, аэрируя ее грудными плавниками. Число икринок в одной кладке
может достигать нескольких сот. Икра светло-желтого цвета, диаметром
2,2-2,7 мм. Нерестилища обычно располагаются в проточной воде, но могут
быть и в озерах, на мелководьях. Инкубационный период 28-29 дней при тем-
пературе воды 8-18° С. Выклюнувшиеся личинки имеют длину 5,7-6,3 мм
(Heufelder, 1982).
Статус вида. Обычный вид в водоемах Восточной Чукотки. Промыслом не
используется. Численность повсеместно высокая в предпочитаемых биотопах.
Служит объектом питания хищных гольцов, хариуса, налима.
Автор-составитель: И.А. Черешнев.
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Cottidae
Род Cottus Linnaeus, 1758
261. Cottus czerskii Berg, 1913 - подкаменщик Черского
Описание и систематика. Довольно крупная рыба с большой головой и
узким туловищем. Хвостовой стебель короткий. Характерные признаки это-
го вида: тело голое, некоторые экземпляры имеют под грудными плавника-
ми редкие мелкие костные шипики, зубы на нёбных костях хорошо развиты.
На предкрышке один направленный назад и загнутый вверх шип. Рот боль-
шой, верхняя челюсть слегка выдается вперед и доходит до вертикали задне-
го края глаза. Зубы мелкие, многочисленные, одинаковые по форме и раз-
мерам. Глаза маленькие, межглазничный промежуток довольно узкий, мень-
ше диаметра глаза. Передние ноздри в виде коротких прямых трубочек, зад-
ние - в виде пор. Межжаберный промежуток узкий. У крупных экземпляров
жаберные перепонки образуют складку поперек межжаберного промежут-
ка. Сейсмосенсорная система состоит из узких сенсорных каналов, которые
открываются наружу мелкими порами. Все плавники имеют на лучах тем-
160
ные пятнышки. Грудные плавники доходят до вертикали третьего луча вто-
рого спинного плавника. Брюшные плавники длинные и у самцов заходят за
анальное отверстие. У половозрелых особей на внутренней стороне брюш-
ных плавников имеются бугорки. Спинные плавники соединены друг с дру-
гом, второй спинной плавник длиннее анального, но ниже него. Анальный
плавник короткий, но высокий и начинается на некотором расстоянии от
анального отверстия.
D, VII-IX; D218-22; А 14-16; Р 15-16; VI 3-4. Туловищный канал, как пра-
вило, полный, доходит до начала лучей хвостового плавника и содержит 40-44
поры. Жаберных тычинок 6, пилорических придатков 5-7. Позвонков 39 (Берг,
19496). Подвидов нет (Аннотированный каталог..., 1998).
Распространение. Обитает в реках, впадающих в Японское море, от При-
морья до п-ова Корея. В России известен из рек Приморья: Сахобэ, Седанка,
Суйфун, Туманная, Кифая (Линдберг, Красюкова, 1987; Шедько, 2001).
1! Атлас,.., том 2
161
Возраст и размеры. Предельная длина 19,5 см.
Образ жизни. Ведет донный образ жизни. Данные по биологии отсутствуют.
Статус вида. Промыслового значения не имеет.
Автор-составитель: В.Г. Сиделева.
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Cottidae
Род Cottus Linnaeus, 1758
262. Cottus gobio Linnaeus, 1758 - обыкновенный подкаменщик
Описание и систематика. Тело голое или покрыто мелкими костными ши-
пиками, количество и место расположения которых на туловище варьирует у
разных популяций. Все плавники, кроме брюшных, покрыты рядами мелких
темных пятен. Брюшные плавники чаще лишены пигментации, или имеют пят-
на, которые при этом никогда не образуют полосатого рисунка, как у пестро-
ногого подкаменщика. В период нереста первый спинной плавник у самцов
имеет желтую или оранжевую кайму. Голова слабо вооружена, на предкрыш-
ке имеется один острый шип и два редуцирующихся. Брюшные плавники
обычно не достигают анального отверстия, но иногда, у половозрелых самцов,
доходят до него. Нижнечелюстной канал сейсмосенсорной системы открыва-
ется на подбородке, как правило, одной порой, изредка - двумя.
D] V-IX; D2(14) 15-19; А 10—15; Р 11-16; V 14. В туловищном канале боко-
вой линии насчитывается от 11 до 37 пор. Позвонков 30-35, обычно 33-34
(Берг, 19496; Koli, 1990; Королев, 1995, 2002). Кариотип: 2п = 52, NF = 72 (Ва-
сильев, 1985).
Вид состоит из двух подвидов: С. gobio gobio Linnaeus, 1758 - западноевро-
пейский подкаменщик и С. gobio koshewnikowi Gratzianov, 1907 - русский подка-
менщик (Koli, 1969, 1990; Королев, 1995; Аннотированный каталог..., 1998).
Однако по последним данным морфологические признаки подвида С. gobio
koshewnikowi, по которым он и был выделен, укладываются в пределы популя-
ционной изменчивости номинативного подвида С. gobio gobio. Согласно пос-
ледним данным, подвид С. gobio koshewnikowi является синонимом С. gobio
gobio.
Распространение. Широко распространен в водоемах (реках и озерах) Ев-
ропы, от Пиренейского и Апеннинского полуостровов до Уральских гор. Под-
каменщика нет в Ирландии, на севере Англии и в Норвегии. Населяет европей-
скую часть России, за исключением Кольского п-ова (Берг, 19496; Озера Каре-
162
лии, 1959; Сидоров, 1974; Koli, 1969.). В Карелии обычен на каменистой лито-
рали многих озер Карелии, в том числе и принадлежащих к бассейну Белого
моря (Сегозеро, Ондозеро, Топозеро, Пулозеро, Энозеро, Пяозеро и др.), поэ-
тому указание на то, что его нет в бассейне Белого моря (Павлов и др., 1994),
неверно. На юге Карелии является обычной рыбой многих озер (Онежского,
Сямозера, Святозера, Миккелъского и др.) (Озера Карелии, 1959). По северно-
му побережью его ареал доходит до Кары включительно. Он обычен в водо-
емах Архангельской области, в бассейнах рек Онега, Двина, Мезень, Волонга,
Печора, Кара (Новоселов, 2000). Далее за Уральским хребтом и в бассейне
Оби его нет. Отнесение к этому вида подкаменщика из бассейнов рек Иртыш
и Катунь нами ставится под сомнение (Решетников и др., 1997). Подкаменщик
из водоемов Ямала (Богданов и др., 1997) также скорее принадлежит к сибир-
скому виду С. sibiricus Kessler, 189^. На юг ареал обыкновенного подкаменщи-
ка простирается до низовьев Урала, Волги и Дона; в Кубани его уже нет, хотя
он встречается в Крыму и на юге Украины.
Возраст и размеры. Максимальные размеры взрослых особей 20 см. Дожи-
вает до 9 лет.
Образ жизни. Пресноводный вид, лишь изредка встречается в опресненных
морских заливах. Предпочитает небольшие речки с каменистым дном и сред-
ней скоростью течения, реже обитает в олиготрофных озерах. Большую часть
163
времени проводит под камнями, которые служат ему укрытием, местом пита-
ния и размножения. В реках держится обычно на перекатах, на небольшой глу-
бине. В нерестовый период самец охраняет свою территорию. Основу питания
составляют донные беспозвоночные: личинки поденок, веснянок, хирономид.
Изредка в желудках встречается молодь хариуса, гольяна, а также собственная
молодь; может поедать икру других рыб (форели, хариуса, гольяна). Растет
медленно и в возрасте 2-3 года достигает длины 5-6 см и массы 2-3 г. Половая
зрелость наступает в возрасте четырех лет при длине тела около 4 см. Самцы
численно преобладают над самками. Нерестится в зависимости от широты рас-
положения водоема в апреле-мае. Индивидуальная абсолютная плодовитость
колеблется в пределах 40-410 икринок. Икринки довольно крупные, разме-
ром 2,0-2,5 мм, желтовато-розового цвета. В одном гнезде в зависимости от ве-
личины самца может находиться от одной до пяти кладок, отложенных разны-
ми самками. Самец охраняет икру до выхода личинок из икры. Развитие икры
при температуре воды 10-15° С длится 2-4 недели.
Статус вида. Во многих водоемах обычный и многочисленный вид, при за-
грязнении водоемов отмечаются снижение численности и полное исчезнове-
ние. Многие авторы отмечали снижение численности при ухудшении качества
воды, особенно при наличии стока в реку отходов сельскохозяйственного про-
изводства. На севере европейской части России многочислен и занимает одно
из ведущих мест в питании хищных рыб (хариуса, щуки, окуня, налима), млеко-
питающих (выдры, норки) и птиц (оляпки, крохаля). Вид причислен к редким
видам рыб Европы (Павлов и др., 1994) и занесен в “Красную книгу Российской
Федерации” (2001). Подкаменщик р. Кары внесен в “Красную книгу Ямало-Не-
нецкого автономного округа” (1997). В связи с многочисленностью этого вида
на севере европейской части России вопрос о внесении его в разряд редких ви-
дов рыб, нуждающихся в охране, является спорным.
Авторы-составители: В.В. Королев, Ю.С. Решетников.
Класс Osteichthys
Отряд Scorpaeniformes
Семейство Cottidae
Род Cottus Linnaeus, 1758
263. Cottus hangiongensis Mori, 1930 - японский подкаменщик
Описание и систематика. Небольшая рыбка с удлиненным телом и сравни-
тельно маленькой головой. Кожа голая, только под грудными плавниками име-
ются очень мелкие шипики. Хвостовой стебель короткий. Длина головы равна
ее ширине и 4 раза укладывается в длине тела. Спина и бока тела серо-коричне-
164
вые, с черными мелкими пятнами, брюш-
ко светлое. Все плавники имеют темные
поперечные полосы, а грудной и первый
спинной плавники еще и светлую кайму по
краю. У корейских подкаменщиков тем-
ные пятна сливаются и образуют 5 тем-
ных поперечных полос по бокам тела. Го-
лова гладкая, без гребней. На предкрыш-
ке один хорошо развитый прямой шип.
Рот небольшой, верхняя челюсть слегка
выдается вперед и не достигает вертикали
середины глаза. Зубы очень мелкие, зуб-
ная пластинка не дифференцирована. Гла-
за маленькие, лоб узкий, меньше диаметра
глаза. Передние ноздри в виде коротких
кожистых трубочек. Межжаберный про-
межуток широкий, не образует попереч-
ной складки. Сейсмосенсорная система со-
стоит из узких сенсорных каналов, кото-
рые, кроме предкрышечно-нижнечелюст-
ного, соединены в единую систему. Поры
очень мелкие. Туловищный канал распо-
ложен ближе к спинной стороне тела, пол-
ный, доходит до основания лучей хвосто-
вого плавника. Лучи в плавниках простые, неветвистые. Спинные плавники со-
единяются друг с другом. Первый спинной плавник начинается позади вертика-
ли грудных плавников, второй - длинный, почти доходит до основания хвостово-
го. Грудные плавники короткие, оканчиваются на вертикали третьего луча D2.
Анальный плавник короткий, меньше второго спинного и начинается далеко по-
зади анального отверстия. Брюшные плавники короткие, не доходят до ануса и
имеют хорошо выраженные темные поперечные полосы.
Ц (VII) VIII-IX (X); D2 (16) 21-22 (24); А (13) 15-16; Р (13) 14; V 14 (в скоб-
ках приведены данные для С. hangiongensis из корейских и японских вод). В ту-
ловищном канале 41^43 поры. Жаберных тычинок 6-7, тычинки мелкие, бу-
горковидные (Шедько, 1999; Goto, 1984; Choi et al., 1990). Кариотип: 2n = 48,
NF = 54 (Abe, 1972).
Распространение. Обитает по восточному берегу п-ова Корея, на севере
Японии (о-в Хоккайдо и северная часть о-ва Хонсю) и на Кунашире (Goto, 1984;
Freshwater Fishes in Japan, 1987; Choi et al., 1990 ). В России найден в ручье о-ва
Кунашир (Шедько, 1999).
Возраст и размеры. Максимальная длина: самцы 170, самки 120 мм. Макси-
мальный возраст 5-6 лет (Goto, 1984).
Образ жизни. Взрослые особи обитают в нижнем и среднем течении рек на ка-
менистом дне. В пище доминируют водные насекомые (Goto, 1986). В нижнем те-
чении обитают более мелкие особи, которые созревают в возрасте 2-3 лет при
длине 70 мм. Обитатели среднего течения созревают позднее, в возрасте 4-5 лет и
длине 120 мм. Длина половозрелых самцов варьирует от 72 до 129 мм. Нерест
происходит с марта по май (Goto, 1987). Половой диморфизм выражается в том,
что самцы крупнее самок, имеют рот большего размера и более длинный аналь-
ный плавник (Goto, 1984). Обычно самец нерестится с 5 самками, и в одном гнезде
165
в среднем присутствует 5 кладок икры, которые различаются цветом, формой и
стадией развития. Более крупные самцы могут спариваться и с большим числом
самок. Икра некрупная, ее диаметр 2,0-2,4 мм. Личинки пелагические (Goto, 1984).
Статус вида. Промыслового значения не имеет.
Автор-составитель: В.Г. Сиделева.
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Cottidae
Род Cottus Linnaeus, 1758
264. Cottus poecilopus Heckel, 1836 - пестроногий подкаменщик
Описание и систематика. В отличие от обыкновенного подкаменщика
пестроногий отличается длинными брюшными плавниками, которые, как
правило, заходят за анальное отверстие и имеют резкие темные попереч-
ные полосы, за что вид и получил свое название. Основной цвет тела олив-
ково-зеленый, с бурыми пятнами по бокам (4—5 штук, у корейских особей
они образуют поперечные полосы), одно пятно у основания хвостового
плавника. Первый спинной плавник имеет оранжевую кайму. Туловище ве-
ретенообразное, равномерно удлиненное. Кожа голая, только под грудны-
ми плавниками имеются очень мелкие, редкие и скрытые в коже костные
шипики. Хвостовой стебель короткий и невысокий. Голова меньше, чем у
других коттоидных рыб, ширина головы почти равна ее длине. Челюсти
сходной длины. На предкрышке 2 шипа. Верхний шип хорошо развит и
скрыт в коже. Рот большой и почти доходит до вертикали заднего края гла-
за. Зубы очень мелкие, густо сидят на челюстях, одинаковые по форме и
размерам; на нёбных костях зубов нет. Глаза маленькие. Передние ноздри в
виде коротких, прямых трубочек. Межжаберный промежуток широкий.
Спинные плавники следуют друг за другом, но никогда не соединены кожи-
стой складкой. Второй спинной плавник (D2) длиннее анального, но одной с
ним высоты. Анальный плавник начинается на небольшом расстоянии от
анального отверстия. Грудные плавники короткие и заканчиваются на вер-
тикали первого луча D2. Брюшные плавники длинные, доходят до анально-
го отверстия, а у самцов заходят за него. Сейсмосенсорная система предста-
влена узкими каналами, которые открываются мелкими порами. На подбо-
родке имеются 2 поры, которые расположены близко друг к другу. Лате-
166
ральный канал проходит ближе к спинной стороне туловища, неполный, не
доходит до вертикали последних лучей второго спинного плавника.
Dj VII-IX; D2 16-19; А 12-15 (16); Р 12-14, V I 4. Канал боковой линии
содержит 28 мелких пор. Жаберных тычинок 3—9, пилорических придат-
ков 2-6. Позвонков 34—35. Кариотип: 2п = 48, NF = 56 (Берг, 19496; Кирил-
лов, 1972). В бассейне Тихого океана, южнее 48° с.ш., обитает подвид
С- poecilopus volki Taranetz, 1933 (Берг, 19496; Аннотированный каталог...,
1998), однако корейские ихтиологи подвидов не выделяют (Choi et al.,
1990).
Распространение. Широко распространен в бассейне Северного Ледови-
того океана от Скандинавии до Чаунской губы и в бассейне Тихого океана
от Камчатки и Охотского моря до п-ова Корея (реки Туманная, Ялу). Встре-
чается в верховьях Дуная, Днестра, а также в некоторых водоемах бассейна
Балтийского моря (территории государств: Дания, Германия, Польша, Лит-
ва, Норвегия, Швеция, Финляндия). В России обитает в бассейне Балтийско-
го моря, в том числе в Неве, Ладожском и Онежском озерах, в р. Суна, а так-
же на севере Карелии — оз. Паанаярви и р. Оланга. В Северной Двине и Пе-
чоре его нет. Отсутствует в бассейне Волги (Варпаховский, 1886, Зиновьев,
1963; Слынько и др., 2000). Неизвестно о его распространении в бассейне
Урала. Вновь появляется в Оби, Енисее, отмечен в Телецком озере и в
р. Бия; но его нет в бассейне Байкала. Обычен в бассейне рек Оленек и Ле-
на, особенно в ее верховьях (реки Киренга, Миня, Чая, Витим, Чара, Токко,
Олекма и озера в их бассейне, включая Ирбо, Леприндо, Даватчан, Кирял-
тинские и др.) (Кириллов, 1972; Богданов, 2000). Отмечен в реках Колымо-
Индигирской низменности, в самой Колыме редок, ловится чаще в ее верх-
нем и среднем течении (Новиков, 1966). В бассейне Анадыря его уже нет,
здесь его заменяет слизистый подкаменщик. Встречается на северо-восточ-
ном побережье Камчатки (залив Корфа) и на материковом побережье
167
Охотского моря (Черешнев, 1996). В Забайкалье его распространение свя-
зано с бассейнами Амура (в Шилке, Аргуни, Ингоде и Ононе, включая Мон-
голию) и Лены (Карасев, 1987; Рыбы МНР, 1983). В самом Амуре встреча-
ется от верховьев до лимана (Никольский, 1956), есть в речках Сихотэ-Али-
ня (от Сучана до Такема) (Берг, 19496).
Возраст и размеры. Максимальная длина - 145 мм. Продолжительность
жизни составляет 6-7 лет (Берг, 19496). Максимальная длина из водоемов Во-
сточной Сибири 116 мм и масса 16,5 г.
Образ жизни. Пресноводный вид, обитает в реках и озерах с холодной во-
дой, ведет скрытный донный и малоподвижный образ жизни на местах с галеч-
ным или песчаным грунтом, прячась под камнями, чем схож с обыкновенным
подкаменщиком. Может менять окраску под цвет грунта. В составе пищи пре-
обладают донные организмы: личинки насекомых (поденки, ручейники, вес-
нянки, хирономиды), черви, а также придонные ветвистоусые и ракушковые
ракообразные, икра и молодь рыб (Кириллов, 1972; Карасев, 1987; Черешнев,
1996; Богданов, 2000). Некоторые авторы указывают на присутствие в желуд-
ках водорослей. У молоди может встречаться и зоопланктон. В возрасте 1+ до-
стигает длины 40-50 мм, 2+ - 70 мм (Никольский, 1956). Как правило, живет не
дольше 5-6 лет. Половое созревание в возрасте 3-4 лет. Плодовитость состав-
ляет 108-336, в среднем - 197 икринок у речных популяций и всего 59-79, в сре-
днем 69 икринок в олиготрофном озере Леприндо (Забайкалье) и 45-149 икри-
нок в оз. Леприндокан (Богданов, 2000). У особей из р. Лены плодовитость вы-
ше: 194-4-74 икринок (Кириллов, 1972). Нерест происходит в весеннее время, а
на крайнем Северо-Востоке России - в июне-июле. Икра светло-желтая, диа-
метром около 2,3 мм. Личинки ведут донный образ жизни, так же как и взрос-
лые особи, сидят под камнями, но предпочитают участки рек с более быстрым
течением.
Статус вида. Промыслового значения не имеет. Является пищевым объек-
том для налима, тайменя, ленка, щуки и гольца. Занесен в “Красную книгу Ка-
релии” (1995).
Автор-составитель: В.Г. Сиделева.
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Cottidae
Род Cottus Linnaeus, 1758
265. Cottus sibiricus Kessler, 1899 - сибирский подкаменщик
168
Описание и систематика. От обыкновенного подкаменщика отличается
тем, что брюшные плавники доходят до анального отверстия и на них не быва-
ет поперечных темных полос, боковая линия полная и тянется посреди тела, на
предкрышке имеется хорошо развитый и загнутый вовнутрь шип. Тело удли-
ненное. Обычно кожа густо покрыта костными шипиками, они присутствуют
на голове, спине и боках выше боковой линии, особи из р. Иртыш имеют едва
заметные шипики. Окраска тела серая, с мелкими темными пятнами. Все плав-
ники имеют темные поперечные полосы. Голова довольно большая, ширина
головы немного уступает ее длине. Голова сильно вооружена, на предкрышке
3 шипа, верхний из них сильно развит, острый и загнут внутрь. Рот большой,
челюсти равной длины и достигают вертикали середины глаза. Зубы мелкие,
одинаковые по форме и размерам, густо сидят на челюстях. Глаза маленькие,
межглазничный промежуток широкий. Передние ноздри в виде коротких, за-
уживающихся кверху пигментированных трубочек. Межжаберный промежу-
ток широкий. Лучи в плавниках (кроме хвостового) неветвистые и толстые.
Грудные плавники короткие, оканчиваются на вертикали первых лучей D2.
Брюшные плавники длинные, доходят до анального отверстия. Спинные плав-
ники соединяются друг с другом, второй спинной плавник длиннее анального.
Сейсмосенсорная система представлена узкими каналами, которые открыва-
ются наружу мелкими порами. На подбородке имеется одна пора. Туловищный
канал проходит ниже середины тела, он полный, доходит до основания лучей
хвостового плавника.
D, VII-IX; D, 16-19; А 12-14 (16); Р 15-16; VI 4. Канал боковой линии име-
ет 40 пор. Жаберных тычинок 6. Пилорических придатков 3, все они сравни-
тельно короткие (Берг, 19496; Кириллов, 1972; Гундризер и др., 1981). Подви-
ды отсутствуют (Аннотированный каталог..., 1998).
Распространение. Реки и озера бассейна Северного Ледовитого океана на
территории Сибири от Оби до Яны (Обь, Иртыш, Телецкое озеро, Енисей, при-
токи Ангары, Лена, Яна). Восточнее Яны его нет, не обнаружен в реках Хро-
ма, Индигирка и Колыма. В Енисее и Яне встречается вплоть до устья (Берг,
19496; Андрияшев, 1954; Кириллов, 1972; Гундризер и др., 1981). Это обычный
вид в водоемах бассейна Верхней Лены (реки Киренга, Витим, Лена) и Ангары
(Иркут, Олха, Белая) и их притоках горного типа (Калашников, 1978; Карасев,
1987; Богданов, 2000). В бассейне Байкала сибирского подкаменщика нет.
Возраст и размеры. Предельный возраст 10 лет (Телецкое озеро, Лена), в
Ангаре - 9. Максимальная длина рыб отмечена для особей из р. Лены - 158 мм
и масса 61,8 г. Обычно в уловах попадаются рыбы в возрасте до 5 лет длиной
7 см и массой 8 г (Гундризер и др., 1981; Богданов, 2000).
Образ жизни. Литофильный вид, обитает на участках рек, дно которых
сложено крупными плоскими камнями. Для него, как и для других подкамен-
щиков, характерны непродолжительные миграции: весной - нерестовые, осе-
нью - зимовальные. В реках миграции обусловлены промерзанием в зимнее
время тех участков рек, к котором приурочены нерест и нагул рыб. В осенне-
зимний период рыбы перемещаются в более глубокие участки водоемов. Пи-
щей сибирского подкаменщика являются гаммариды, личинки веснянок, поде-
нок и ручейников. Основной способ охоты - подкарауливание добычи в укры-
тии. У молоди в пище представлены бентосные организмы, у половозрелых
особей в пищевом комке, помимо бентоса большое значение имеет рыба, поэ-
тому их можно характеризовать как факультативных хищников (Богданов,
2000). В местах симпатрического обитания сибирского подкаменщика и камен-
169
ной широколобки (Paracottus knerii) доля рыбной пищи возрастает за счет по-
требления молоди каменной широколобки. Растет медленно, достигая в воз-
расте 3+ - 4-5 см, 4+ - 6 см, 5+ - 7-8 см и 6+ - 9 см (Гундризер и др., 1981). В Те-
лецком озере доживает до 10 лет, достигает длины 120 мм и массы 43 г; в Ан-
гаре - 9 лет, 149 мм и 47 г. В бассейне р. Ангары сибирский подкаменщик на-
чинает созревать на 4-5-м году жизни, в бассейне Лены - на 5-6-м году. Пло-
довитость рыб в бассейне Ангары 126-664 икринок, в бассейне Лены - 209-614
икринок. Начало нереста обычно совпадает с периодом вскрытия рек и нача-
лом весеннего паводка. Как и у других видов, икра откладывается на нижнюю
поверхность камня, и самец охраняет кладку.
Статус вида. Непромысловый вид. Играет важную роль в питании налима
и тайменя (Богданов, 2000).
Автор-составитель: В.Г. Сиделева.
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Cottidae
Род Leocottus Taliev, 1955
266. Leocottus kesslerii (Dybowski, 1874) - песчаная широколобка
170
Описание и систематика. Отличается от всех прочих донных видов подка-
менщиков Палеарктики своим длинным анальным плавником (более 20 лу-
чей). От байкальских представителей семейства Cottidae отличается наличием
полного канала боковой линии, который заканчивается у основания лучей хво-
стового плавника и имеет более 40 мелких пор. Тело песчаной широколобки
равномерно удлиненное, хвостовой стебель короткий. Кожа покрыта очень
мелкими, редко сидящими шипиками. Окраска тела оливково-серая, с темны-
ми пятнами на боках. Длина головы почти равна ее ширине. Верхняя челюсть
немного выдается вперед. Рот маленький, угол рта достигает вертикали перед-
него края глаза. Межглазничное пространство узкое. Плавники, кроме брюш-
ных и анального, имеют поперечные полосы. У половозрелых самцов Dj со
светлой каймой. Песчаной широколобке в нерестовый период свойствен поло-
вой диморфизм. Самцы имеют более крупные размеры, массивные кости, вы-
сокие спинные и анальный плавники. Грудные плавники заканчиваются на вер-
тикали первого луча D2. Брюшные плавники длинные, доходят до анального
отверстия. Боковая линия полная и заканчивается у основания лучей хвостово-
го плавника.
Dt VII-IX; D2 (17) 18-20 (21); Р 17-19; А 20-22; V14. В боковой линии 36-42
мелких пор. Жаберных тычинок 4-6. Позвонков 38-39 (Берг, 19496; Талиев,
1955). Кариотип: 2п = 48, NF = 58 (Сиделева и др., 1995). Образует 2 подвида:
L. kesslerii arachlensis (Tarchova, 1962) - арахлейская песчаная широколобка,
обитает в оз. Арахлей (Читинская область) и L. kesslerii gussinensis (Tarchova,
1962) - гусиноозерская песчаная широколобка, живет в р. Селенге и оз. Гуси-
ном (Бурятия) (Тархова, 1962; Карасев, 1987). Все водоемы относятся к бассей-
ну Байкала.
Распространение. Обитает по всему побережью Байкала, заходит в мелко-
водные хорошо прогреваемые заливы-соры и крупные притоки озера (В. Ан-
гара, Селенга); имеется в озерах Кулинда, Гусиное, Арахлей, Цайдамские; рас-
пространена в водохранилищах Ангарского каскада. Нахождение в р. Лене и
Баунтовских озерах (Талиев, 1946; Берг, 19496) не подтвердилось (Кириллов,
1972, Скрябин, 1977; Сиделева, 1982).
Возраст и размеры. Предельный возраст 7 лет, достигает длины 14 см, при-
чем самцы крупнее самок.
Образ жизни. Песчаная широколобка обитает в основном на песчаных
грунтах, закапывается в грунт. Летом распространена от прибрежья Байка-
ла до глубин 50-150 м. Весной в мае перед нерестом концентрируется на
глубинах 3-5 м. Нерестится на каменистых биотопах, на глубинах от 0,1 до
6,0 м. Спектр ее питания весьма широк и насчитывает 9 компонентов: в ос-
новном бокоплавы, личинки хирономид и ручейников, меньше олигохеты,
рыба и икра рыб (чаще своего вида), моллюски, остракоды, растительные
остатки (Базикалова, Вилисова, 1959). Жизненный цикл составляет 7 лет, в
уловах чаще преобладают рыбы в возрасте 3^4 лет длиной 8-11 см. Растет
медленно, достигая в возрасте 1+ - 57 мм, 2+ - 80 мм, 3+ - 105 мм и 4+ -
118 мм. Созревает в возрасте 2+ при длине 54 мм. Нерестовое стадо состоит
в основном из особей в возрасте от 2+ до 4+. Плодовитость колеблется от
1400 до 10 000 икринок. Это самое высокое значение абсолютной плодови-
тости среди всех байкальских и других пресноводных коттоидных рыб. Диа-
метр икры 0,8-1,0 мм. Нерест проходит с конца мая до середины июля на
глубинах от 10 см до 6 м при температуре воды 4-5° С. На период нереста
песчаная широколобка меняет нагульный песчаный биотоп на каменистый.
171
При этом крупные самцы песчаной широколобки занимают центральную
часть каменистого биотопа, вытесняя на периферию более мелких самцов
каменной широколобки (Munehara, Sideleva, 1998). Самцы песчаной широ-
колобки оккупируют наиболее крупные и удобные для нереста камни и уст-
раивают гнезда. Несколько самок откладывают икру в одно гнездо и самец
сторожит ее. Иногда встречаются смешанные гнезда, состоящие из кладок
двух видов: песчаной широколобки и каменной широколобки или песчаной
широколобки и желтокрылой широколобки. В этих случаях гнездо сторо-
жит наиболее крупный и сильный самец песчаной широколобки (Munehara,
Sideleva, 1998). Среди байкальских коттоидных рыб песчаная широколобка
имеет в гнезде наибольшее число кладок и самую мелкую икру. В гнезде,
как правило, находится 3—4 кладки. Развитие продолжается 30-50 дней. Вы-
клюнувшиеся личинки длиной 5,2-5,4 мм (Черняев, 1977) ведут пелагиче-
ский образ жизни. Мальки длиной 19-22 мм начинают покидать пелагиаль
и опускаться в придонные слои воды. Взрослые особи являются типично
донными формами.
Статус вида. Промыслового значения не имеет. Песчаную широколобку
потребляют налим, омуль, осетр и нерпа (Талиев, 1955).
Автор-составитель: В.Г. Сиделева.
172
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Cottidae
Род Mesocottus Gratzianow, 1949
267. Mesocottus haitej (Dybowski, 1869) - амурская широколобка
Описание и систематика. Сравнительно крупная рыба с большой головой и
коротким туловищем. Кожа толстая, густо покрыта костными шипиками, ко-
торые отсутствуют только на брюшной стороне тела. Хвостовой стебель ко-
роткий. Спина и бока серого, иногда коричневатого цвета. Брюшко светлое, за
первым спинным плавником и у основания хвоста имеются 3 темных полоски,
которые доходят до боковой линии. Плавники, кроме брюшных и анального,
имеют поперечные темные полосы, грудные плавники короткие, оканчивают-
ся на вертикали первых лучей D2. Брюшные плавники также довольно корот-
кие, не доходят до анального отверстия. Между спинными плавниками проме-
жуток отсутствует. Анальный плавник короче D2, но одинаковой с ним высо-
ты и начинается сразу за анальным отверстием. Голова очень большая и широ-
кая, сильно вооружена, на предкрышке 4 хорошо развитых шипа, верхний шип
очень острый. Рот большой, челюсти равной длины и доходят до вертикали се-
редины глаза. Зубная пластинка слабо дифференцирована, зубы, расположен-
ные по середине челюстей, крупнее всех остальных. Глаза маленькие, межглаз-
ничный промежуток широкий. Передние ноздри в виде коротких, зауживаю-
щихся кверху трубочек, задние имеют вид пор. Межжаберный промежуток до-
вольно узкий. Сейсмосенсорная система представлена узкими сенсорными ка-
налами, которые открываются наружу мелкими порами. На подбородке име-
ются 2 поры. Латеральный канал полный и проходит посередине туловища.
Dj VIII-IX; D212-15; Р 16-17; А 10-13; V13-4. В боковой линии 35 пор. Жа-
берных тычинок 4—6, по форме они бугорковидные, покрыты мелкими шипи-
ками. Позвонков 32-33 (Берг, 19496; Никольский, 1956; Карасев, 1987). Подви-
дов нет.
Распространение. Встречается в России, Монголии (р. Онон), Северном
Китае и Северной Корее (р. Ялу). В России обитает в бассейне Амура от вер-
ховьев (реки Шилка, Аргунь, Онон, Ингода) до лимана, имеется в бассейнах
Уссури и Сунгари. Обитает в районе Амурского лимана (Берг, 19496; Николь-
ский, 1956; Рыбы МНР, 1983; Карасев, 1987; Аннотированный каталог..., 1998).
Возраст и размеры. Предельный возраст 5 лет, достигает длины 20 см.
Образ жизни. В основном речной вид, ведущий малоподвижный донный об-
раз жизни. Живет преимущественно в русле реки, придерживаясь галечного,
реже песчаного грунта. В состав пищи входят личинки поденок, веснянок, ру-
чейников и хирономид, мизиды, гаммариды и рыба (колюшка, гольяны и гор-
чаки). У молодых особей длиной 50-60 мм чаще встречаются хирономиды, у
более крупных экземпляров преобладают личинки поденок (Берг, 19496; Ни-
кольский, 1956). Из рыб в пище амурской широколобки в верховьях Амура до-
173
минируют пескари, горчак и молодь карася (Карасев, 1987). В возрасте 1+ име-
ет длину 65 мм, 2+ - 115 мм, 3+ - 145-150 мм и 4+ - 160 мм. Созревает на тре-
тьем году жизни при длине 10-11 см. Основная масса половозрелых особей
имеет размеры 10-13 см. Плодовитость, определенная по одному экземпляру,
составила 2822 икринки. Нерест проходит в мае-июне при температуре воды
10-16° С.
Статус вида. Промыслового значения не имеет. Вероятно, является пище-
вым конкурентом хариуса и ленка (Никольский, 1956).
Автор-составитель: В.Г. Сиделева.
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Cottidae
Род Paracottus Taliev, 1949
268. Paracottus knerii (Dybowski, 1874) - каменная широколобка
174
Описание и систематика. Представители этого рода имеют голое тело,
мелкие шипики располагаются лишь под грудными плавниками, широкий
межжаберный промежуток, слабое вооружение на голове, разрывы в сен-
сорных каналах головы. Голова составляет 1/4 длины тела, ее ширина при-
мерно равна ее длине. Меглазничный и межжаберный промежутки широ-
кие. Глаза выпуклые и приподняты над лбом. Костные шипики под грудны-
ми плавниками расположены ниже туловищного канала и образуют около
20 рядов. Хвостовой стебель высокий, его высота составляет половину его
длины. Окраска тела оливково-зеленая, по бокам с темно-зелеными и тем-
но-бурыми пятнами неправильной формы. Плавники не имеют попереч-
ных полос, только у половозрелых самцов на D, имеется черное пятно.
Спинные плавники сближены и соединены между собой низкой кожистой
складкой. D2 длиннее анального плавника. Анальный плавник начинается
на некотором расстоянии от ануса. Брюшные плавники светлые и не дохо-
дят до анального отверстия, их длина примерно равна длине хвостового
стебля.
Dj VII-IX; D2 15-19; А 12-14; Р 16-17; V I 4. Боковая линия обычно окан-
чивается в области второй половины D2 и имеет 11-17 мелких пор. Жаберные
тычинки бугорковидные, несут шипики, число тычинок 6-7. Пилорических
придатков 4. Позвонков 33-35 (Берг, 19496; Талиев, 1955). Кариотип: 2п = 48,
NF = 62 (Сиделева и др., 1995). Каменная широколобка из оз. Верхняя Агата
описана как особый путоранский подвид Р. knerii putorania Korjakov et Sidelev,
1976 (Коряков, Сиделев, 1976).
Распространение. Встречается в прибрежной зоне оз. Байкал до глуби-
ны 150 м, заходит во все его притоки, кроме р. Селенги. Отмечена в реках
Ангара и Енисей и их притоках, а также озерах Тувы и в оз. Верхняя Агата
(бассейн Енисея, плато Путорана). Имеется в озерах Грамнинских, Кулинда
и Верхнее Кичерское (бассейн Байкала) (Гундризер, 1975; Смирнов, 1980).
Нахождение этого вида в оз. Баунт (Талиев, 1955) не подтвердилось (Скря-
бин, 1977).
Возраст и размеры. Предельная длина 14,5 см, обычно 7-9 см. Самцы круп-
нее самок. Живет до 7 лет.
Образ жизни. В Байкале предпочитает каменистые грунты на глубинах
до 50-70 м, лишь единичные особи опускаются до 150 м. С конца лета, осе-
нью и зимой этот вид придерживается глубин более 10 м, а с момента ледо-
става весной и в первой половине лета переходит на меньшие глубины -
0,1-1,5 м, а также заходит в устьевые участки мелких притоков Байкала.
Молодь ведет донный образ жизни и питается придонными формами гамма-
рид и циклопов. Пища взрослых особей каменной широколобки из оз. Бай-
кал состоит из 12 компонентов, основу которых составляют донные гамма-
риды (37—100%). Каменная широколобка, обитающая в реках, потребляет
личинок хирономид, поденок, веснянок, ручейников и других насекомых
(Богданов, 2000). Растет медленно, достигая в возрасте 1н— 50 мм, 2н—
63 мм и 3+ - 70 мм. Созревает нА третьем году жизни при длине 6-7 см. Не-
рестовое стадо представлено особями в возрасте от 2+ до 4+. Абсолютная
плодовитость колеблется от 350 до 750 икринок. У рыб из р. Ангары, ее
притоков и оз. Верхняя Агата значения плодовитости колеблются от 100 до
750 икринок (Коряков, Сиделев, 1976; Богданов, 2000). Диаметр икринок
1,8-2,3 мм. Нерест проходит с конца мая до середины июля на глубинах от
10 см до 5 м при температуре воды 6-10° С. В качестве нерестового субстра-
175
та особи используют неподвижные валуны первого яруса, размером
20x30 см. Если камни лежат в несколько ярусов, то используются для нере-
ста камни второго и даже третьего ярусов значительно меньших размеров -
5x5 см. Если широколобка занимает камни из многоярусной толщи, то на
одном квадратном метре может находиться 60 кладок икры и более. На пер-
вом ярусе икра от одной или нескольких самок откладывается на нижнюю
сторону камня, на втором и последующих ярусах кладка может находиться
как на нижней, так и на верхней стороне камня. В каждом гнезде обычно
3-4 кладки икры, иногда встречаются гнезда с 6 кладками. После нереста
самки уходят на глубину, а самцы охраняют икру до вылупления личинок и
аэрируют ее движениями грудных плавников, а затем тоже уходят на глуби-
ну. Если отловить самца, то кладка погибает от заиления или от выедания
хищными беспозвоночными. Развитие в икре длится 28-32 дня. Личинки вы-
лупляются с большим запасом желтка и с хорошо сформированными луча-
ми в непарных плавниках. Они ведут донный образ жизни в прибрежной по-
лосе Байкала, на глубинах до 5-6 м.
Статус вида. Многочисленный, но непромысловый вид. Молодь камен-
ной широколобки поедается хариусами, сигом, тайменем, ленком, окунем и на-
лимом.
Автор-составитель: В.Г. Сиделева.
176
Класс Osteichthyes Семейство Cottidae
Отряд Scorpaeniformes Род Triglopsis Girard, 1851
269. Triglopsis quadricornis (Linnaeus, 1758) -
четырехрогий бычок, рогатка
Описание и систематика. Представители этого рода имеют голое тело и силь-
но вооруженную голову, жаберные перепонки образуют свободную складку попе-
рек межжаберного промежутка. Тело невысокое, впереди немного уплощенное.
Голова небольшая, низкая, ее длина составляет около 30% длины тела. На пред-
крышке имеются 4 хорошо развитых шипа. На голове у морских форм присутст-
вуют заглазничные и затылочные бугры, у пресноводных форм они или сглажены
или вообще отсутствуют. Глаза небольшие. Лоб довольно широкий, превышает
диаметр глаза. Окраска тела темно-серая, иногда с неясными пятнами на спине и
светлая на брюшной стороне. Плавники имеют черные поперечные полосы или
пятна. Второй спинной плавник сильно удлинен и заканчивается на одной верти-
кали с анальным плавником. Сейсмосенсорная система характеризуется развити-
ем фонтанелей в костях головы, которые сверху затянуты кожной мембраной. От
широких сенсорных каналов отходят кожные канальцы, которые заканчиваются
микроскопически малыми порами. Отдельные канальцы не имеют пор и заканчи-
ваются слепо. На подбородке имеется одна подбородочная пора. Туловищный ка-
нал часто неполный, не доходит до основания лучей хвостового плавника.
D, VI-X; D2 (12) 13-16; А (12) 13-17; Р 15-18; V I 3. В туловищном канале
от 28 до 48 пор. Жаберных тычинок 7-10, пилорических придатков 6-10. Поз-
вонков 37—42 (Берг, 19496; Адрияшев, 1954, Неелов, 1979; Неелов, Сиделева,
1979; Разнообразие рыб Таймыра, 1999).
Полиморфный вид, внутривидовая структура изучена слабо, хотя она дос-
таточно сложная. Иногда его рассматривают как комплексный вид. Ранее опи-
санные подвиды признано целесообразным не употреблять до подробного ис-
следования вида (Неелов, 1979; Аннотированный каталог..., 1998).
Распространение. Арктический циркумполярный вид. Прибрежные мор-
ские бычки, заходящие в сильно опресненные или пресные воды. Имеются ре-
ликтовые чисто пресноводные фо{5мы в ряде крупных озер Норвегии, Швеции,
Финляндии, России и Северной Америки. В России живет в озерах Карелии (Се-
гозеро, Остер, Куйто), в Ладожском и Онежском озерах, ряде озер п-ова Тай-
мыр (Андермей, Кета, Лабаз, Лама) и в бассейне рек (Пясина, Боганида, Хан-
тайка, в нижних участках рек Хатанга и Индигирка) (Михин, 1941; Кириллов,
1955). Обычен в Балтийском и Белом морях, встречается в реках Нева, Нарова
и в устьях рек, впадающих в Северный Ледовитый океан. Южнее Берингова
12 Атлас..., том 2
177
пролива известен только до Анадырского лимана. На карте ареала местонахо-
ждение пресноводных популяций четырехрогого бычка показано точками.
Возраст и размеры. Предельная длина морских форм 40 см, масса 500 г,
озерных - до 20-28 см. Живет более 11 лет (Берг, 19496).
Образ жизни. Обитает в прибрежной зоне северных морей, заходит в заливы
и в низовья рек, живет в морских, солоноватых и пресных водах. Образует чис-
то озерные формы. В летнее время питание рогатки состоит в основном из ры-
бы, в море - это корюшка, сельдь, трехиглая колюшка, камбала, сиг, навага.
В зимнее время большее значение в питании приобретают бентосные организ-
мы - ракообразные, многощетинковые черви, крабы, моллюски, а также водо-
росли и высшие растения (Андрияшев, 1954; Мухомедияров, 1967). В реках и озе-
рах - моллюски, личинки хирономид и молодь рыб, чаще сиговых (Разнообразие
рыб Таймыра, 1999), а также их икра. Рост ледовитоморской рогатки различен в
зависимости от места обитания. Пресноводная форма рогатки растет медленнее
морской. Так, рогатка из Онежского залива Белого моря в возрасте 1+ имеет
длину 69 мм, 2+ - 165 мм, 3+ - 180 мм, 4+ - 185 мм, 5+ - 215 мм, 6+ - 227 мм, 7+ -
230 мм (Мухомедияров, 1967). Обычно в возрасте 5-6 лет достигает длины
20-22 см, к 7-8 годам - 24-27 см. Средняя длина 22-24 см, масса 150-200 г и
возраст 6-7 лет. Половая зрелость у самок наступает на третьем и четвертом го-
ду, у самцов на год раньше. Соотношение полов в нерестовом стаде близко 1:1.
Массовый нерест происходит в декабре-январе при температуре -1 —1,2° С, на
глубинах 1-1,5 м подо льдом. Икра откладывается на камни среди бурых водо-
рослей и имеет цвет темно-оливковый, сходный с цветом водорослей. Плодови-
тость сильно варьирует в зависимости от возраста и соответственно размеров
рыбы. При длине 20-22 мм средняя индивидуальная плодовитость составляет
3013 икринок, у рыб длиной 26-28 мм - 7790 икринок, у крупных особей -
38^10 мм - 16 650 икринок. Диаметр икры 2 мм. Вылупление личинок происхо-
дит в мае, к августу они достигают длины 20-22 мм.
Статус вида. Самостоятельного промыслового значения ни в морских, ни в
пресных водах не имеет, является приловом при промысле массовых рыб. В
Белом море рогатка составляла всего лишь 0,5-1% промыслового улова (Му-
хомедияров, 1967).
Автор-составитель: В.Г. Сиделева.
178
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Comephoridae
Род Comephorus Lacepede, 1801
270. Comephorus baicalensis (Pallas, 1776) - большая голомянка
Описание и систематика. У всех видов этого семейства и единственно-
го рода, как у пелагических рыб, тело сплющено с боков и покрыто очень
нежной и голой кожей, тело удлиненное, голова приплюснута и лишена
вооружения. Плавниковые лучи очень тонкие, прозрачные, неветвистые;
брюшных плавников нет; в D2 более 28 лучей; в анальном - 27-35 гибких
лучей и иногда бывает колючка; грудные плавники очень длинные; хво-
стовой плавник усеченный или слегка выемчатый. Кости тела и головы
очень тонкие и пористые. В костях головы интенсивно развиты фонтане-
ли для сейсмосенсорной системы. Сама система представлена сенсорными
каналами, которые имеют вид обширных, последовательно соединенных
друг с другом полостей. Жаберные дуги длинные и тонкие. Жаберные от-
верстия очень широкие, а жаберные лепестки маленькие, жаберные ты-
чинки короткие булавовидные. Тело бесцветное, только у некоторых эк-
земпляров на спине, верхней части головы и у основания хвостового плав-
ника присутствуют редкие звездчатые меланофоры. Все виды рода живо-
родящие.
Отличительными признаками вида Comephorus baicalensis являются боль-
шие глаза (их диаметр равен ширине лба или больше ее), а боковая линия до-
ходит до середины D2. Тело удлиненное. Хвостовой стебель очень короткий,
лучи анального плавника доходят до краевых лучей хвостового. Рот большой,
имеет косое расположение, челюсти доходят до вертикали заднего края глаза.
Зубы мелкие, волосковидные, имеются на внутренней и внешней сторонах че-
люстей. Жаберные перепонки разделены между собой и не прикреплены к
межжаберному промежутку. Грудные плавники длинные и веерообразные.
Спинные плавники сближены, между ними узкий промежуток.
D, VI-VIII; D2 28-34; А 27-36; Р 10-15. Боковая линия доходит до середины
D, и имеет 26-30 пор. Жаберных тычинок 12-16. Пилорических придатков 2-4.
Позвонков 48-50 (Берг, 19496; Талиев, 1955). Кариотип: 2п = 48, NF = 56 (Ха-
наев, 1997). Подвидов нет (Аннотированный каталог..., 1998).
Распространение. Эндемик оз. Байкал, обитает в толще открытых вод и
встречается до глубины 1700 м.
Возраст и размеры. Предельная длина 21 см, обычно длина самок 19,8, сам-
цов - 13,4 см. Максимальный возраст 8 лет.
Образ жизни. Ведет пелагический образ жизни. Приурочен к открытым
районам озера. Зимой и в начале весны подо льдом держится на глубинах
25—400 м и совершает вертикальные миграции вслед за своим основным
кормовым объектом. В летний период встречается на глубинах от 300 до
179
Класс Osteichthyes
Отряд Scorpaeniformes
1700 м. С осени в ночное время под-
нимается в более верхние слои. Пи-
ща взрослых особей состоит из мак-
ропланктона (М. branickii), личинок
своего вида и малой голомянки. Пи-
ща молоди практически на 100% со-
стоит из Epischura baicalensis. В воз-
расте 1+ достигает длины 52 мм, 2+ -
95 мм, 3+ - 159 мм, 4+ - 175 мм,
5+ - 191 мм и 6+ - 210 мм. Обычная
длина большой голомянки в уловах
13-20 см. Большая голомянка явля-
ется чрезвычайно жирной рыбой, ее
удельный вес меньше удельного веса
воды, у взрослых самок жирность те-
ла достигает 40% сырого веса. Поло-
вая зрелость наступает в возрасте 3+.
Абсолютная плодовитость самок в
возрасте 3-6 лет в среднем 1200-
2675 икринок (Стариков, 1977). Для
видов этого рода характерно живо-
рождение. Массовый вымет личинок
происходит во второй половине ию-
ля или в августе и приходится на пе-
риод максимального прогрева воды.
Вымет растянут, самки с развиты-
ми эмбрионами встречаются круг-
лый год (Коряков, 1964). Личин-
ки большой голомянки держатся
днем на больших глубинах, а ночью поднимаются до глубины 10 м.
Статус вида. Многочисленный, но непромысловый вид, основной пищевой
объект нерпы, составляющий до 28% ее рациона (Гурова, Пастухов, 1974).
Автор-составитель: В.Г. Сиделева.
Семейство Comephoridae
Род Comephorus Lacepede, 1801
180
271. Comephorus dybowski Korotneff, 1905 - малая голомянка
Описание и систематика. Сохраняются общие признаки для рода голо-
мянок. Малая голомянка отличается от большой маленькими глазами (их
диаметр вдвое меньше ширины лба) и тем, что боковая линия не доходит до
начала D2. Тело удлиненное. Голова сужена с боков, ее длина в 3 раза боль-
ше ширины. Рот имеет косое расположение. Челюсти длинные, далеко за-
ходят за вертикаль заднего края глаза. Зубы мелкие, волосковидные, загну-
ты назад, присутствуют на внутренней и наружней сторонах челюсти. Жа-
берные перепонки разделены между собой и не прикреплены к межжабер-
ному промежутку. Жаберные дуги тонкие и длинные. Хвостовой стебель
короткий, анальный плавник почти доходит до хвостового. Грудные плавни-
ки очень длинные.
Dt VIII-IX; D2 30-34; А 31-36; Р 13-15. Туловищный канал боковой линии
короткий, оканчивается до начала D2 и имеет 12-17 пор, причем последняя
пора крупнее всех остальных. Жаберных тычинок 24-28. Пилорических при-
датков 2-4. Позвонков 48-50 (Берг, 19496; Талиев, 1955). Кариотип: 2п = 48,
NF = 52 (Мамонтов, Яхненко, 1981). Подвидов нет (Аннотированный ката-
лог..., 1998).
Распространение. Эндемик оз. Байкал, обитает в пелагиали от 130 до мак-
симальных глубин - 1700 м.
Возраст и размеры. Самцы имеют максимальный размер 135, обычно
80-90 мм; самки - 158, обычно 100-110 мм. Живут до 7 лет (Талиев, 1955).
Образ жизни. Обычно обитает в
толще вод открытого озера от по-
верхности (зимой) до максимальных
глубин. Летом и осенью этот вид
придерживается глубин более 100 м,
с момента ледостава переходит на
меньшие глубины. Пища молоди
практически на 100% состоит из
Epischura baicalensis. У взрослых
особей основу пищи составляет
М. branickii, иногда в желудках
встречается молодь своего вида и
большой голомянки. Как и у боль-
шой голомянки, у малой известны
суточные пищевые вертикальные
миграции. В возрасте 1+ самки име-
ли длину 45 мм, 2+ - 76 мм, 3+ -
105 мм, 4+ - 127 мм, 5ч— 137 мм и
6+ - 143 мм. Самцы растут медлен-
нее самок. Обычные размеры рыб
80-110 мм. Половая зрелость насту-
пает в возрасте 2+ и 3+. Абсолютная
плодовитость у особей в возрас-
те 2-5 лет в среднем 670-2160 икри-
нок, при этом диаметр икринки пра-
ктически остается постоянным —
1,02—1,03 мм. Оплодотворение внут-
реннее. Развитие эмбрионов от спа-
ривания до вымета личинок длится
181
90-100 дней (Стариков, 1977). Вымет личинок происходит в феврале-мар-
те подо льдом в верхних слоях воды. Личинки держатся рассеянно от по-
верхности до глубины 500 м, не совершая суточных миграций (Берг,
19496). У мальков, как и у взрослых рыб, появляются суточные вертикаль-
ные миграции (Талиев, 1955).
Статус вида. Многочисленный, но непромысловый вид, так как не образу-
ет скоплений. Молодь малой голомянки поедают омуль и другие виды рыб, а
взрослые особи - основной пищевой объект нерпы, который составляет в ее
рационе 61% (Гурова, Пастухова, 1974).
Автор-составитель: В.Г. Сиделева.
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Abyssocottidae
Род Abyssocottus Berg, 1906
272. Abyssocottus elochini Taliev, 1955 - елохинская широколобка
Описание и систематика. Глубоководные широколобки сем. Abyssoco-
ttidae внешне похожи на донных керчаковых рыб сем. Cottidae. Это в основ-
ном небольшие рыбки, часто с уплощенной в дорсовентральном направле-
нии головой, живут только в Байкале и на больших глубинах. От всех рыб
отряда Scorpaeniformes отличаются отсутствием сенсорных каналов на го-
лове, сейсмосенсорная система представлена только наружными неврома-
стами.
Рыбы рода Abyssocottus характеризуются совершенно голым телом, бо-
ковая линия в передней части тела состоит из нескольких рядов, средний
ряд из 30-60 невромастов, которые заходят на срединный луч хвостового
плавника. Голова уплощенная, большая. Глаза маленькие, расположены в
глубоких глазницах. Подглазничные кости черепа выдаются. Позвон-
ков 31-33.
Елохинская широколобка A. elochini Taliev имеет короткое тело, большую и
сильно уплощенную голову, ее ширина больше высоты у затылка. Окраска го-
ловы и верхней части туловища однотонная, темно-коричневая. Нижняя часть
туловища светлая. Рыло широкое, лопатообразной формы с бугорком на перед-
нем крае. Челюсти почти достигают вертикали середины глаза. Лоб широкий,
больше продольного диаметра глаза. Передние ноздри в виде длинных узких
182
пигментированных кожистых трубо-
чек, слегка зауживающихся кверху.
Задние ноздри имеют вид коротких,
широких пигментированных трубочек.
Жаберная щель небольшая, межжабер-
ный промежуток очень узкий. Сейсмо-
сенсорная система представлена на-
ружными невромастами, которые рас-
положены на тонких, нежных, волос-
ковидных папиллах, высотой до 1 мм,
частично пигментированных у основа-
ния. Латеральная линия состоит из
33 невромастов, из которых 2 располо-
жены на срединном луче хвостового
плавника. Выше латеральной линии
имеются 2 дополнительных ряда невро-
мастов: верхний, длинный, из 18 и ниже
его короткий, из 5 невромастов. Ниже
латеральной линии располагается еще
один дополнительный ряд из 8 невро-
мастов. Лучи всех плавников пигменти-
рованы, но не в равной степени. На лу-
чах спинных, грудных и хвостового
плавников пигмент формируется в тем-
ные полосы, а на лучах анального и
брюшных плавников - только в виде
отдельных меланофоров. Межлучевые
перегородки тонкие, непигментирован-
ные. Грудные плавники короткие,
оканчиваются на вертикали первого луча D2. Брюшные плавники не доходят до
анального отверстия, но сравнительно длинные, почти одинаковой длины с груд-
ными плавниками. Спинные плавники соединены кожистой складкой. Второй
спинной плавник выше анального, но имеет равную с ним длину.
D] V, D2 15, А 13, Р 16. Жаберных тычинок 4, они маленькие бугорковид-
ные, полностью покрыты шипиками, на жаберной дуге сидят редко. Пилори-
ческих придатков 3, они очень короткие и толстые. Позвонков 32 (10 туловищ-
ных и 22 хвостовых). Подвидов нет (Аннотированный каталог..., 1998).
Распространение. Эндемик озера Байкал. Среди подводных скал, на глуби-
не 250-300 м, бимтралом было поймано 3 экз.: один длиной 63 мм и два маль-
ка (Талиев, 1955). В настоящее время в ихтиологической коллекции Зоологи-
ческого института РАН присутствует в единственном экземпляре. После
1951 г. в наших уловах никогда не встречался.
Возраст и размеры. Достигает длины 6,3 см.
Образ жизни. Экземпляр A. Elochini был пойман на скальном грунте. Сведе-
ния по биологии отсутствуют.
Статус вида. Исключительно редкий вид, обитающий среди подводных
скал, т.е. на грунтах, не доступных для траления и постановки сетей. Хозяйст-
венного значения не имеет.
Автор-составитель: В.Г. Сиделева.
183
Семейство Abyssocottidae
Род Abyssocottus Berg, 1906
Класс Osteichthyes
Отряд Scorpaeniformes
273. Abyssocottus gibbosus Berg, 1906 - белая широколобка
Описание и систематика. Небольшая рыбка с крупной головой и коротким
туловищем. Окраска тела однотонная, светло-серая, иногда у особей, обитаю-
щих в верхних границах ареала, на боках бывают размытые темные пятна. Го-
лова очень большая, ширина головы больше ее высоты у затылка. Нижняя че-
люсть слегка выдается вперед и достигает вертикали середины глаза. Глаза ма-
ленькие, удлиненной формы, расположены в глубоких глазницах. Лоб шире
продольного диаметра глаза. Передние ноздри в виде длинных кожистых трубо-
чек, задние ноздри в виде коротких ко-
жистых трубочек. Жаберная щель ши-
рокая, межжаберный промежуток уз-
кий. Сейсмосенсорная система пред-
ставлена наружными невромастами,
которые имеют вид крупных белых то-
чек, и в линиях они соединены между
собой. Латеральная линия имеет S-об-
разную форму и содержит 34 неврома-
ста на туловище и 4 на девятом луче
хвостового плавника. Выше туловищ-
ной линии имеются 2 дополнительных
ряда невромастов: первый сверху из 17,
второй под ним - из 11 невромастов.
Ниже основной линии есть еще корот-
кий ряд из 9 невромастов. Все плавни-
ки бесцветные, не имеют темных по-
лос. Грудные плавники короткие,
оканчиваются на вертикали первого
луча D2. Брюшные плавники далеко не
доходят до анального отверстия и не-
много короче грудных. Спинные плав-
ники расположены близко друг к другу
и соединены между собой кожистой
складкой. Второй спинной плавник вы-
ше и длиннее анального. Анальный
плавник начинается на некотором рас-
стоянии от ануса.
184
D, V-VII; D2 14-15; A 11-13; P 15-16, V I 3. Жаберных тычинок 5, они ко-
роткие бугорковидные, редко сидят на жаберной дуге. Пилорических придат-
ков 3, они очень толстые и примерно одинаковой длины. Позвонков 32-33.
(Берг, 19496; Талиев, 1955; Сиделева, 1982). Подвидов нет (Аннотированный
каталог..., 1998).
Распространение. Эндемик озера Байкал, распространен в зоне глубин
400-1600 м.
Возраст и размеры. Достигает длины 16 см.
Образ жизни. Редко встречающийся глубоководный вид. Обитает на
илистых грунтах. Созревает в возрасте 5+ при длине 132 мм и массе 50 г.
Время нереста, вероятно, январь-февраль, плодовитость 127-180 икринок,
диаметр икры 3,3-3,4 мм. Самки со зрелой икрой начинали попадаться с но-
ября на глубинах 600 и 1000 м (Талиев, 1955). Ювенильные особи отмечены
на глубинах 800-1300 м, в возрасте 1+ они имели длину 25-30 мм и массу те-
ла 0,2-0,4 г.
Статус вида. Непромысловый редкий вид.
Автор-составитель: В.Г. Сиделева.
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Abyssocottidae
Род Abyssocottus Berg, 1906
274. Abyssocottus korotneffi Berg, 1906 - малоглазая широколобка
Описание и систематика. Маленькая рыбка с сильно уплощенной голо-
вой. Голова небольшая, ее ширина в 2 раза меньше длины. Тело удлиненное,
антеанальное расстояние большое. Окраска тела половозрелых особей од-
нотонная, светло-оранжевая, молодь и карликовые формы имеют светло-се-
рый цвет. Нижняя челюсть слегка выдается вперед и достигает вертикали
середины глаза. Зубы мелкие, густо сидят на челюстях. Глаза очень малень-
кие. Лоб широкий, больше продольного диаметра глаза. Передние ноздри в
виде прямых коротких трубочек. Жаберная щель широкая, межжаберный
промежуток очень узкий. Сейсмосенсорная система представлена неврома-
стами, которые соединены между собой в некоторых линиях. Латеральная
линия имеет 25 невромастов, из которых 9 расположены на луче хвостового
плавника. Выше линии, ближе к спине имеется дополнительный ряд, состо-
ящий из 25 невромастов, ниже латеральной линии - 2 коротких ряда из 5 и
7 невромастов. Все плавники бесцветные. Грудные плавники короткие,
оканчиваются на вертикали последнего луча DP Брюшные плавники очень
короткие (в 2 раза меньше грудных) и далеко не доходят до анального отвер-
185
стия. Спинные плавники сходны по
высоте и разделены довольно боль-
шим промежутком, D2 и анальный
плавники равны по высоте, но аналь-
ный плавник несколько длиннее D2.
Анальный плавник начинается на не-
котором расстоянии от ануса.
Dj V-VII; D2 13-16; А 12-14;
Р 15-16 (17); V I 3. Жаберных тычи-
нок 7, они бугорковидной формы и по-
крыты густыми мелкими шипиками.
Позвонков 33-35. Кариотип: 2п = 48,
NF = 72 (Берг, 19496; Сиделева, 1982,
1985; Сиделева и др., 1995). Подвидов
нет (Аннотированный каталог...,
1998).
Распространение. Эндемик озера
Байкал, обитает в зоне глубин
400-1600 м. Наибольшее количество
малоглазой широколобки попадалось
в уловах на Северном Байкале.
Возраст и размеры. Предельная
длина 126 мм. Максимальный воз-
раст 12 лет.
Образ жизни. Обитает на или-
стых грунтах, визуальные наблюде-
ния из подводных аппаратов “Пай-
сис” показали, что малоглазая ши-
роколобка способна закапываться в
ил. Удалось сфотографировать экземпляр, тело которого было спрятано
в иле, голова торчала снаружи. Пища A. korotneffi состоит из гаммарид.
Гаммариды представлены всего пятью видами: Р. petersi, Р. zienkowezi,
G. cabanisi, A. grewinki и О. margaritaceus (Сиделева, Механикова, 1990). В
желудках отмечен также детрит. Созревает в возрасте 5-8 лет при длине
86-109 мм и массе 5-12 г. Плодовитость 16-31 икринка, икра крупная, диа-
метром 3,5—3,7 мм. Этому виду свойственно наличие карликовой формы,
которая созревает в возрасте 4—6 лет, обитает вместе с неполовозрелыми
особями и составляет на разных глубинах от 7 до 69% общей численности.
Карликовые созревшие самцы имеют длину 45-59 мм и массу 0,8-2,1 г;
самки - 53-67 мм и 1,1—2,8 г, плодовитость последних колеблется в преде-
лах 5-12 икринок диаметром 2,4 мм (Сиделева, 1985). Нерест происходит в
мае-июле.
Статус вида. Самый многочисленный глубоководный вид из семейства
Abyssocottidae. Промыслового значения не имеет.
Автор-составитель: В.Г. Сиделева.
186
Семейство Abyssocottidae
Род Asprocottus Berg, 1906
Класс Osteichthyes
Отряд Scorpaeniformes
275. Asprocottus abyssalis Taliev et Korajakov, 1947 -
глубоководная широколобка
Описание и систематика. У видов рода Asprocottus (кроме A. pulcher) тело
покрыто мелкими костными шипиками. У всех видов голова хорошо вооруже-
на, на предкрышке имеется 4 острых шипа. Рот маленький, угол рта доходит
только до вертикали переднего края глаза. Наружные невромасты сидят на ко-
жистых папиллах.
A. abyssalis - небольшая рыбка с коротким туловищем. Кожа толстая, ту-
ловище полностью покрыто густыми
шипиками, но на голове они отсутству-
ют. Хвостовой стебель короткий. Ок-
раска тела однотонная, светло-розо-
вая. Все плавники светло-кремовые и
не имеют пигментации. Голова очень
большая и сильно вооружена. Нижняя
челюсть слегка выдается вперед и за-
канчивается вблизи вертикали перед-
него края глаза. Передние ноздри в ви-
де длинных трубочек, их длина равна
половине ширины лба. Зубы очень
мелкие, одинаковые по форме и раз-
мерам, сидят на челюстях очень густо
и на нижней челюсти образуют 6 не-
правильных рядов. Глаза большие,
лоб узкий, меньше продольного диа-
метра глаза. Жаберная щель и межжа-
берный промежуток узкие. Латераль-
ная линия проходит посередине туло-
вища и имеет 32-36 невромастов на те-
ле и 22-25 расположены по всей Длине
9-го луча хвостового плавника. В на-
чале латеральной линии, выше и ниже
нее находятся 2 ряда, имеющие по 7 и
3 невромаста. Грудные плавники ко-
роткие, заканчиваются на вертикали
последних лучей Db Брюшные плав-
187
ники также короткие и далеко не доходят до анального отверстия. Спинные
плавники разделены узким промежутком. D2 выше D( и анального, а также
длиннее анального. Анальный плавник начинается на некотором расстоянии
от анального отверстия.
Dj VI-VII; D2 13-15; А 13-14; Р 13-14 (15). Жаберных тычинок 5-7. Пило-
рические придатки отсутствуют, это единственный вид среди байкальских
Cottoidei, который не имеет пилорических придатков. Позвонков 34 (Сиделева,
1982). Подвидов нет (Аннотированный каталог..., 1998).
Распространение. Эндемик озера Байкал, обитает на подводном склоне в
широком диапазоне глубин от 150 до 1400 м.
Возраст и размеры. Достигает длины 78 мм, обычно 55-72 мм, масса
2,1-4,0 г.
Образ жизни. Донный вид, обитает на скальных, присыпанных илом грун-
тах. Особи впервые созревают в возрасте 3+, при длине 58 мм и массе 2,1 г.
Плодовитость при этом составляет 19 икринок. Обычные размеры производи-
телей 64—72 мм и 2,9—4,0 г, их плодовитость 20-33 икринки при среднем диамет-
ре 2,2 мм. Нерест проходит в мае-июле, половозрелые особи встречались на
глубинах 440-580 м. Глубоководная широколобка растет медленно, в возрас-
те 1+ она имеет среднюю длину 24,9 мм, в 2+ -47,8, в 3+ - 55, в 4+ - 61,4 ив воз-
расте 5+ - 69 мм. Пища состоит на 100% из мелких форм гаммарид:
Poekologammarus sp., Macropereiopus flori, Carinogammarus seidlitzi, Parapallasea
lagowskii, Carinurus sp. (Талиев, 1955).
Статус вида. Промыслового значения не имеет.
Автор-составитель: В.Г. Сиделева.
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Abyssocottidae
Род Asprocottus Berg, 1906
276. Asprocottus herzensteini Berg, 1906 -
шершавая широколобка Герценштейна
Описание и систематика. Маленькая рыбка с удлиненным телом и сравни-
тельно большой и широкой головой. Кожа, кроме головы, сплошь покрыта гу-
стыми мелкими костными шипиками. Хвостовой стебель довольно длинный.
Окраска тела серо-розовая, однотонная. Плавники светлые, темных попереч-
ных полос не имеют. Нижняя челюсть слегка выдается вперед и заходит за вер-
тикаль переднего края глаза. Этмоидальная кость образует .хорошо выражен-
188
ный бугорок, на предкрышке - 4 хоро-
шо развитых шипа. Края лобных кос-
тей приподняты над глазами, заверну-
ты внутрь и образуют валик. Подглаз-
ничные кости образуют неширокий
гребень. Зубы мелкие, простые, густо
сидят на челюстях и нёбных костях.
Глаза большие. Лоб узкий, меньше
диаметра глаза. Передние ноздри име-
ют вид длинных прямых кожистых
трубочек. Жаберная щель довольно
широкая, а межжаберный промежу-
ток узкий. Сейсмосенсорная система
представлена наружными невромаста-
ми, расположенными на пигментиро-
ванных папиллах. Латеральная линия
проходит посередине туловища и име-
ет 39 невромастов на теле и 9 на луче
хвостового плавника. Сверху лате-
ральной линии имеются одиночные
невромасты, также расположенные на
папиллах. Грудные плавники корот-
кие, оканчиваются на вертикали пос-
ледних лучей D1 и имеют усеченную
форму. Спинные плавники разделены
узким промежутком. Второй спинной
плавник (ОД длиннее Db он же длин-
нее и выше анального. Анальный
плавник начинается на значительном
расстоянии от анального отверстия. Брюшные плавники короткие, далеко не
доходят до анального отверстия.
Dj (V) VI-VIII; D2 14—16; А 12-15; Р (12) 15-16. Жаберных тычинок 7. Пило-
рических придатков 4, они имеют разную длину (1 длинный, превышает длину
желудка, 2 равные по длине желудку и 1 очень короткий). Позвонков 33-34 (Та-
лиев, 1955; Сиделева, 1982). Подвидов нет (Аннотированный каталог..., 1998).
Распространение. Эндемик озера Байкал, обитает на глубинах 200-800 м.
Особенно многочислен в Северном Байкале.
Возраст и размеры. Достигает длины 117 мм и массы 17,5 г (Талиев, 1955).
Образ жизни. Донный вид, обитает на песчано-илистых грунтах. Спектр
питания у особей из пролива Малое Море крайне узок. У рыб размерами
20-60 мм пища состояла из амфипод, копепод и остракод; у особей более
80 мм - только из амфипод (Базикалова, Вилисова, 1959). Половое созревание
начинается в возрасте 4 года при^лине 48 мм и массе тела 1,7 г (плодовитость
11 икринок). Обычные размеры производителей от 65 до 117 мм, масса от 3,5
до 17,5 г. Плодовитость составляет от 45 до 140 икринок при их диаметре
2,1-2,3 мм. Нерест проходит в мае-июле.
Статус вида. Промыслового значения не имеет.
Автор-составитель: В.Г. Сиделева.
189
Семейство Abyssocottidae
Род Asprocottus Berg, 1906
Класс Osteichthyes
Отряд Scorpaeniformes
277. Asprocottus korjakovi Sideleva, 2001 - широколобка Корякова
Описание и систематика. Новый вид назван в честь исследователя байкаль-
ских коттоидных рыб Е.А. Корякова. Небольшая рыбка с крупной головой и
укороченным туловищем. Кожа толстая. Окраска тела бледно-розовая с тем-
ными мелкими пятнами на спине и боках. Плавники светло-серые, иногда на
грудных и спинных плавниках имеются слабо выраженные полосы. Тело, кро-
ме брюшной части (включая область под грудными плавниками), покрыто гу-
стыми костными шипиками. На спине шипики сидят пучками на одном основа-
нии. На голове очень мелкие шипики имеются только в затылочной части.
Хвостовой стебель короткий. Голова
большая, ширина головы почти равна
ее длине. Нижняя челюсть слегка вы-
дается вперед. На этмоидальной кости
бугорок слабо выражен. На пред-
крышке 4 хорошо развитых шипа. Рот
маленький, угол рта достигает верти-
кали переднего края глаза. Зубы мел-
кие, многочисленные, густо сидят на
челюстях и нёбных костях. Внешние
края лобных костей приподняты над
глазами и образуют надбровные дуги.
Глаза довольно большие. Лоб узкий,
меньше продольного диаметра глаза.
Передние ноздри в виде длинных ров-
ных трубочек. Жаберная щель и меж-
жаберный промежуток узкие. Сейсмо-
сенсорная система представлена на-
ружными невромастами, расположен-
ными на маленьких пигментирован-
ных папиллах. Латеральная линия со-
держит 38 невромастов на туловище и
по 8 и 10 невромастов на 2 срединных
лучах хвостового плавника. Сверху,
параллельно латеральной линии име-
ется дополнительный ряд из 10 редко
сидящих невромастов. Лучи во всех
плавниках толстые. Грудные плавники
190
короткие, оканчиваются на вертикали начала D2. Брюшные плавники корот-
кие и широкие. Спинные плавники разделены промежутком. Анальный плав-
ник длинный, длиннее D2 и начинается сразу за анальным отверстием.
D] V-VII; D2 14—16; А 14—15; Р 15. Жаберных тычинок 5. Пилорических при-
датков 3, все длинные. Кишечник длинный, делает петлю. Всего позвон-
ков 33-34, из них 10 туловищных и 23-24 хвостовых. В районе дельты р. Селен-
ги описан подвид A. korjakovi minor Sideleva, 2001.
Распространение. Эндемик озера Байкал, несколько экземпляров было вы-
ловлено донным тралом в районе пролива Малое Море, на глубине 115-125 м.
Возраст и размеры. Достигает длины 13 см.
Образ жизни. Донный вид, обитает на песчаных грунтах. Типовые экзем-
пляры данного вида, выловленные в июне, представлены половозрелыми осо-
бями, готовыми к нересту. Плодовитость самки длиной 106 мм 220 икринок,
диаметр икринки 2,8 мм (Sideleva, 2001).
Статус вида. Редкий вид в Байкале. Хозяйственного значения не имеет.
Автор-составитель: В.Г. Сиделева.
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Abyssocottidae
Род Asprocottus Berg, 1906
278. Asprocottus parmiferus Taliev, 1955 - панцирная широколобка
Описание и систематика. Маленькая рыбка с коротким туловищем и боль-
шой головой (длина головы равна */3 длины тела). Ширина головы почти равна
ее длине. На этмоидальной кости бугорок слабо выражен. Окраска тела светло-
серая, с отдельными черными пятнами на спине и хвостовом стебле. Голова
сильно вооружена, на предкрышке имеются 4 хорошо развитых шипа. Подглаз-
ничные кости гребня не образуют. Нижняя челюсть слегка выдается вперед.
Зубы мелкие, простые, на челюстях и нёбных костях. Глаза маленькие, сверху
прикрыты толстыми кожистыми веками. Лоб широкий, больше продольного
диаметра глаза. Передние ноздри в виде широких, слегка зауживающихся квер-
ху пигментированных трубочек. ^Каберная щель и межжаберный промежуток
узкие. Кожа покрыта костными шипиками, одиночными на брюшной стороне,
а на спине, боках и верхней части головы образующими “щитки” (термин
Д.Н. Галиева), т.е. на одном основании сидят по 6-18 шипиков. Нижняя часть
головы без шипиков. Хвостовой стебель сравнительно длинный. Сейсмосенсор-
ная система представлена наружными невромастами, расположенными на низ-
ких, бугорковидных папилах. Латеральная линия проходит посредине туловища
191
и имеет 39 невромастов. Выше этой
линии имеется дополнительный ряд из
9 редко сидящих невромастов. Плавни-
ки имеют толстые лучи, пигментиро-
ваны, меланофоры сгруппированы в
отдельные пятна и не образуют попе-
речных полос. Грудные плавники ко-
роткие, оканчиваются на вертикали
первых лучей второго спинного плав-
ника. Брюшные плавники довольно
длинные, немного не доходят до аналь-
ного отверстия. Первый спинной плав-
ник ниже второго. Между спинными
плавниками имеется довольно широ-
кий промежуток, равный по величине
продольному диаметру глаза. Второй
спинной плавник длиннее анального.
Анальный плавник начинается на не-
котором расстоянии от ануса.
D, V-VI; D2 14-16; А 12-14;
Р 15-16; VI 3. Жаберных тычинок 5-6.
Пилорических придатков 3, все корот-
кие. Позвонков 29-31 (Талиев, 1955;
Сиделева, 1982). Подвидов нет (Анно-
тированный каталог..., 1998).
Распространение. Эндемик озера
Байкал, встречается редко на глуби-
нах 10-500 м.
Возраст и размеры. Достигает длины 78 мм (Талиев, 1955).
Образ жизни. Донный вид, обитает на песчаных и песчано-илистых грун-
тах. В уловах встречается единично. Скопления молоди длиной 20-34 мм и мас-
сой 0,1-0,4 г были обнаружены на глубинах 50-60 и 190-250 м.
Статус вида. Редкий вид. Промыслового значения не имеет.
Автор-составитель: В.Г. Сиделева.
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Abyssocottidae
Род Asprocottus Berg, 1906
279. Asprocottus platycephalus Taliev, 1948 - плоскоголовая широколобка
192
Описание и систематика. Неболь-
шая рыбка с уплощенной головой. Тело
удлиненное. Окраска тела однотонная
серо-розовая. Все плавники светлые, не-
пигментированные. Кожа на голове и
брюшной части голая; спина, бока и
хвостовой стебель густо покрыты ши-
пиками. Хвостовой стебель короткий.
Голова большая, ширина головы мень-
ше ее длины, она уплощена в дорсовен-
тральном направлении и по форме напо-
минает клюв утки. Среди видов рода
Asprocottus, только плоскоголовая ши-
роколобка имеет уплощенную голову.
Бугорок на этмоидальной кости слабо
выражен. Рот маленький, челюсти рав-
ной длины и почти достигают вертикали
переднего края глаза. Зубы мелкие,
простые, густо сидят на челюстях и от-
сутствуют на нёбных костях. Глаза ма-
ленькие, расположены в глубоких глаз-
ницах. Лоб широкий, слегка превышает
диаметр глаза. Передние ноздри в виде
коротких трубочек. Жаберная щель уз-
кая, межжаберный промежуток очень
узкий. Сейсмосенсорная система пред-
ставлена наружными невромастами: од-
ни из них расположены на папиллах,
другие сидят в углублениях эпидермиса.
Латеральная линия состоит из 32 невромастов на теле и 9 невромастов на луче хво-
стового плавника. Сверху этой линии имеется дополнительный ряд из 9 неврома-
стов. Грудные плавники короткие, оканчиваются на вертикали последних лу-
чей D, и имеют усеченную форму. Брюшные плавники узкие и короткие, далеко
не доходят до анального отверстия. Между спинными плавниками имеется неболь-
шое расстояние. Второй спинной плавник и анальный длинные и сходны по высо-
те. Анальный начинается на значительном расстоянии от ануса.
D, VI-VII; D2 (15) 16-17; А 14-15; Р 15, V I 3. Жаберных тычинок 6-7. Пи-
лорических придатков 3, все они одинаковой длины. Позвонков 31-32 (Берг,
19496; Талиев, 1955; Сиделева, 1982). Кариотип: 2п = 48, NF = 66 (Сиделева и
др., 1995). Подвидов нет (Аннотированный каталог..., 1998).
Распространение. Эндемик озера Байкал, встречается на глубинах SO-
SOO м.
Возраст и размеры. Достигает длины 11 см (Талиев, 1955).
Образ жизни. Донный вид, обйтает на песчаных и илисто-песчаных грунтах.
Начинает созревать в возрасте 3+ , самки длиной 85-90 мм и массой 7-8 г, самцы -
80 мм и 7 г. Средняя плодовитость 110 икринок, диаметр икринки 3,0 мм.
Статус вида. Промыслового значения не имеет.
Автор-составитель: В.Г. Сиделева.
13 Атлас..., том 2
193
Семейство Abyssocottidae
Род Asprocottus Berg, 1906
Класс Osteichthyes
Отряд Scorpaeniformes
280. Asprocottus pulcher (Taliev, 1948) - острорылая широколобка
Описание и систематика. Маленькая пестрая рыбка, внешне похожа на дру-
гие виды рода Asprocottus, но в отличие от них не имеет на теле шипиков. Тело
укороченное. Окраска тела и головы серо-коричневая с темно-коричневыми
пятнами неправильной формы. Брюшная сторона светлая. Спинные, грудные
и хвостовой плавники имеют темные поперечные полосы. Брюшные и аналь-
ный плавники - светлые, однотонные. Хвостовой стебель очень короткий. Го-
лова большая, ширина головы мало уступает ее длине; она сильно вооружена,
на предкрышке имеются 4 развитых шипа. Рот средних размеров, челюсти рав-
ной длины и доходят до вертикали пе-
редней трети глаза. Зубы мелкие, оди-
наковые по форме и размерам, густо
сидят на челюстях. Глаза большие.
Лоб узкий, меньше продольного диа-
метра глаза. Передние ноздри в виде
коротких трубочек, постепенно сужи-
вающихся кверху. Задние ноздри в ви-
де пор и не выделяются над поверхно-
стью кожи. Жаберная щель и межжа-
берный промежуток узкие. Сейсмо-
сенсорная система состоит из линий
наружных невромастов, расположен-
ных на коротких папиллах. Латераль-
ная линия проходит по середине тела и
содержит 32 невромаста на туловище
и 32 на 9-м луче хвостового плавника.
Выше этой линии, ближе к спине, про-
ходит длинный ряд из 17 наружных
невромастов, который заканчивается
на вертикали последних лучей D2. Ни-
же основной латеральной линии нахо-
дятся 2 коротких ряда, которые имеют
по 3 и 5 невромастов. Грудные плавни-
ки короткие, заканчиваются на верти-
кали последних лучей D,. Брюшные
плавники также короткие и далеко не
доходят до анального отверстия. Спин-
ные плавники расположены близко
194
друг к другу, промежуток между ними очень узкий. Второй спинной плавник
выше первого спинного и анального. Анальный плавник длинный, начинается
на некотором расстоянии от анального отверстия и заканчивается на одной
вертикали со вторым спинным плавником.
D, VI; D2 16-17; А 15-16; Р 14; VI 3. Жаберных тычинок 5, они бугорковид-
ные, покрыты сверху мелкими шипиками. Пилорических придатков 3, их дли-
на совпадает с длиной желудка. Позвонков 32, из них: 10 туловищных и 22 хво-
стовых (Берг, 19496; Талиев, 1955; Сиделева, 1982). Подвидов нет (Аннотиро-
ванный каталог..., 1998).
Распространение. Эндемик озера Байкал, обитает на глубинах 50-250 м,
наиболее часто присутствует в уловах в районе Селенгинского мелководья.
Возраст и размеры. Достигает длины 80 мм.
Образ жизни. Предпочитает илисто-песчаные грунты. Обычная длина
55-60 мм. Нерест происходит в мае-июле. Плодовитость составляет
115-128 икринок. Диаметр икринки 2,6-2,8 мм (Талиев, 1955).
Статус вида. Промыслового значения не имеет.
Автор-составитель: В.Г. Сиделева.
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Abyssocottidae
Род Cottinella Berg, 1907
281. Cottinella boulengeri (Berg, 1906) -
короткоголовая широколобка Буленже
Описание и систематика. Маленькая рыбка с коротким туловищем и боль-
шой круглой головой. Тело укороченное, кожа голая. Хвостовой стебель до-
вольно длинный. Окраска тела однотонная светло-розовая. Плавники светлые
однотонные. Межлучевые перегородки тонкие, прозрачные. Голова широкая,
ширина головы почти одинакова на всем ее протяжении и равна ее длине. На
предкрышке имеются 3 хорошо развитых шипа. Рот большой, челюсти равной
длины и достигают вертикали середины глаза. Зубы мелкие, простые, густо си-
дят на челюстях и нёбных костях. Глаза маленькие. Лоб очень широкий. Перед-
ние ноздри в виде длинных трубочек. Задние ноздри в виде широких и сравни-
тельно высоких трубочек. Жаберная щель широкая, а межжаберный промежу-
ток очень узкий. Сейсмосенсорная система представлена крупными (0,7 мм в
диаметре) наружными невромастами, которые расположены на слегка выдаю-
щихся над поверхностью кожных бугорках. Невромасты в линиях соединены ме-
195
жду собой. Латеральная линия фазу за
грудным плавником изогнута книзу,
имеет 35 невромастов на теле и 9 нев-
ромастов на срединном луче хвостово-
го плавника. Над латеральной линией
имеются 2 дополнительных ряда невро-
мастов, под ней - 1 ряд. Грудные плав-
ники короткие, усеченной формы
оканчиваются на вертикали последнего
луча DP Спинные плавники разделены
широким промежутком; первый спин-
ной плавник низкий, вдвое ниже второ-
го; второй спинной плавник по длине и
высоте сходен с анальным. Анальный
плавник начинается на небольшом
расстоянии от анального отверстия.
Брюшные плавники короткие, с узким
основанием и не доходят до ануса на
расстояние, почти равное их длине.
Di V-VT; D2 13-14; А 11-12; Р 16-
18, V I 3. Жаберных тычинок 6, ты-
чинки бугорковидные, сплошь покры-
ты мелкими шипиками. Пилориче-
ских придатков 3, они толстые и зна-
чительно превышают длину желудка.
Кишечник длинный (65% длины тела)
и имеет петлю (Талиев, 1955; Сиделе-
ва, 1982). Кариотип: 2п = 48, NF = 66
(Сиделева и др., 1995). Подвидов нет (Аннотированный каталог..., 1998).
Распространение. Обитает в глубоководной части Байкала на глубинах
400-1620 м.
Возраст и размеры. Максимальный возраст 10 лет, максимальные размеры
самок 130 мм и 40 г, самцов - 114 мм и 23 г.
Образ жизни. Донный вид, обитающий на илистых грунтах. Максимум чис-
ленности (7 экз./га) приходится на глубины 820-970 м. Особи этого вида актив-
но передвигаются по дну. С помощью подводных аппаратов “Пайсис” были по-
ставлены ловушки с приманкой на глубину 800 м. Особи этого вида посещали
места расположения ловушек и активно питались многочисленными гаммари-
дами, привлекаемыми приманкой. В ловушки, выставленные с корабля в цент-
ральной котловине озера на максимальную глубину 1620 м, из рыб попадались
только экземпляры этого вида. В желудках С. boulengeri найдены гаммариды,
коконы планарий и детрит. Гаммариды представлены мелкими формами
Parapallasea lagowskii и видами рода Ommatogammarus (Талиев, 1955). Средние
размеры половозрелых самок - 110 мм и 18 г, самцов - 107 мм и 17 г. Этот вид
созревает в возрасте 6 лет, при длине 91-94 мм и массе 10-11 г. Средняя плодо-
витость - 69 икринок диаметром 2,5 мм. Нерест проходит в июне-июле.
Статус вида. Довольно редкий глубоководный вид. Промыслового значе-
ния не имеет.
Автор-составитель: В.Г. Сиделева.
196
Семейство Abyssocottidae
Род Cyphocottus Sideleva, 2001
Класс Osteichthyes
Отряд Scorpaeniformes
282. Cyphocottus eurystomus (Taliev, 1955) - широкорылая широколобка
Описание и систематика. Небольшая рыбка с крупной головой и корот-
ким туловищем. Тело укороченное, кожа толстая, голая, под грудными плав-
никами имеются костные шипики. Хвостовой стебель сравнительно длин-
ный. Окраска тела, кроме светлой брюшной части, пестрая (леопардовая), на
светло-коричневом фоне имеются темные, почти черные пятна. У рыб, оби-
тающих в различных районах озера, степень контрастности пятен подверже-
на изменчивости (Талиев, 1955). Наиболее яркую окраску имеют особи из
Южного Байкала. Спинные, грудные и хвостовой плавники имеют темные
поперечные пятна, сгруппированные в полосы. На лучах анального плавника
имеются группы меланофоров. Голова очень большая и широкая. Этмои-
дальная кость образует хорошо выраженный крупный бугорок. Внутренние
края лобных костей приподняты и над глазами образуют валик. Подглазнич-
ные кости образуют гребень. Челюсти равной длины и достигают вертикали
середины глаза. Зубы мелкие, простые, густо сидят на челюстях, на нёбных
костях отсутствуют. Глаза большие, лоб узкий. Передние ноздри в виде тон-
ких, зауживающихся кверху трубочек, их длина в 2 раза больше основания.
Жаберная щель довольно широкая, а межжаберный промежуток узкий.
Сейсмосенсорная система представлена наружными невромастами, купулы
которых крупные и цилиндрические возвышаются над поверхностью кожи,
создается впечатление, что невромасты располагаются на бугорках. Лате-
ральная линия проходит по середине туловища и имеет 50 невромастов на те-
ле и 26 очень мелких невромастов на луче хвостового плавника. Над лате-
ральной линией имеются 2 дополнительных ряда, состоящих из 15 и 8 невро-
мастов, ниже нее - один ряд из 9 невромастов. Лучи в плавниках (кроме хво-
стового) толстые, неветвистые. Спинные плавники расположены близко друг
к другу, между ними имеется очень узкий промежуток. Второй спинной плав-
ник длиннее и выше анального. Анальный плавник начинается на некотором
расстоянии от ануса. Грудные плавники короткие, оканчиваются на вертика-
ли последнего луча Dt. Брюшные плавники короткие, не доходят до анально-
го отверстия на расстояние, равное их длине.
D] VI-VI I; D2 13-14; А 12-13; Р 16-17, VI 3. Жаберных тычинок 5-6. Пило-
рических придатков 3: два коротких и один длиннее остальных. Позвон-
ков 33-34 (Талиев, 1955; Сиделева, 1982). Подвидов нет (Аннотированный ка-
талог..., 1998).
Распространение. Эндемик озера Байкал, обитает на глубинах 50-600 м.
197
Возраст и размеры. Достигает дли-
ны 165 мм, обычно 100-110 мм (Тали-
ев, 1955).
Образ жизни. Донный вид, пред-
почитает илисто-песчаные грунты.
Наиболее многочислен в районах по-
вышенной биологической продук-
тивности: Селенгинское мелководье,
северный выход из пролива Малое
Море, створ Баргузинского залива.
Пища этого вида из района пролива
Малое Море состояла из гаммарид,
рыбы, ручейников и олигохет; в же-
лудках отмечались также раститель-
ность и грунт. У особей длиной до
130 мм рыба в желудках отсутствова-
ла. По мере увеличения размеров ши-
роколобки доля гаммарид в пищевом
комке постепенно уменьшалась (от
100 до 58%) и возрастала доля рыбы
(Базикалова, Вилисова, 1959). Из гам-
марид наиболее обычны виды родов
Pallasea, Poekilogammarus, Acantho-
gammarus и Crypturopus. Доля личи-
нок ручейников и олигохет не превы-
шала 7-8%. Средняя абсолютная пло-
довитость широколобки 189 икринок
диаметром 1,6 мм. Нерест проходит в
мае-июле.
Статус вида. Широко распространенный массовый вид. Хозяйственного
значения не имеет.
Автор-составитель: В.Г. Сиделева.
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Abyssocottidae
Род Cyphocottus Sideleva, 2001
283. Cyphocottus megalops (Gratzianow, 1902) - горбатая широколобка
198
Описание и систематика. Сравни-
тельно большая (для пресноводных
коттоидных рыб) рыбка с крупной го-
ловой и коротким туловищем. Спина
резко отделена впадиной от затылоч-
ной части головы, образуя горб. Анте-
анальное расстояние большое, почти
равно половине длины тела. Кожа го-
лая, только под верхней частью груд-
ных плавников имеются редкие кост-
ные шипики. Хвостовой стебель длин-
ный. Окраска тела однотонная, темно-
серая у половозрелых особей и светло-
коричневая у молодых экземпляров.
Плавники однотонные, темных попе-
речных полос не имеют. Спинные и
грудные плавники темно-серого цвета,
остальные светло-кремовые. У самцов
первый спинной плавник имеет по
краю темную кайму. На этмоидальной
кости имеется хорошо выраженный
крупный бугорок. Внутренние края
лобных костей приподняты. Подглаз-
ничные кости образуют широкий гре-
бень. Рот довольно маленький, челю-
сти сходной длины и достигают верти-
кали передней трети глаза. Зубы мел-
кие, простые, имеются на челюстях и
нёбных костях. Глаза большие. Лоб
меньше продольного диаметра глаза. Передние ноздри в виде тонких заужива-
ющихся кверху трубочек. Жаберная щель неширокая, а межжаберный проме-
жуток очень узкий. Сейсмосенсорная система представлена наружными невро-
мастами, которые в линиях расположены близко друг к другу. Латеральная ли-
ния проходит по середине туловища и имеет 58 невромастов на теле и 19 нев-
ромастов на 10-м луче хвостового плавника. Над этой линией имеются еще
2 коротких дополнительных ряда, состоящих из 15 и 8 невромастов. Ниже ла-
теральной линии также имеется 1 дополнительный ряд из 8 невромастов. Спин-
ные плавники расположены близко друг к другу. Лучи в плавниках, кроме хво-
стового, неветвистые. Первый спинной плавник немного ниже второго спинно-
го. Анальный плавник начинается на некотором расстоянии от ануса. Грудные
плавники короткие, заканчиваются на вертикали последних лучей D1; нижние
лучи грудного плавника имеют длинные свободные кончики. Брюшные плав-
ники не доходят до ануса и почти вдвое короче грудных.
Dj VI-VII; D2 16; А 12-14; Р 15—17; V I 3. Жаберных тычинок 7, они бугор-
ковидной формы, покрыты мелкими шипиками. Пилорических придатков 3,
они короткие, толстые и одинаковой длины. Позвонков 32-34 (Талиев, 1955;
Сиделева, 1982). Кариотип: 2п = 48, NF = 64 (Сиделева и др., 1995). Подвидов
нет (Аннотированный каталог..., 1998).
Распространение. Эндемик озера Байкал, обитает на глубинах от 25 до
600 м.
199
Возраст и размеры. Предельный возраст 8 лет, достигает длины 17 см и
массы 55 г.
Образ жизни. Донный вид, обитает на песчано-илистых и илистых грунтах.
Пища состоит из 4 компонентов: донные гаммариды, пелагический гаммарус
М. branickii, олигохеты и рыба (Базикалова, Вилисова, 1959). Половое созрева-
ние наступает в возрасте 4—5 лет при длине 100-105 мм и массе 8-10 г. Обычно
длина производителей колеблется в пределах 115-165 мм и масса 15-50 г.
Плодовитость составляет 250-375 икринок диаметром 2,2-2,6 мм. Нерест про-
ходит в июне-июле, половозрелые особи встречаются на глубинах 150-250 м.
Статус вида. Промыслового значения не имеет.
Автор-составитель: В.Г. Сиделева.
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Abyssocottidae
Род Limnocottus Berg, 1906
284. Limnocottus bergianus Taliev, 1935 - плоская широколобка
Описание и систематика. У всех представителей рода Limnocottus удли-
ненное тело с длинным хвостовым стеблем, кожа голая и тонкая, костные
шипики имеются только под грудными плавниками. Этмоидальная кость об-
разует хорошо заметный бугорок. Плоская широколобка - довольно боль-
шая для пресноводных коттоидных рыба, с крупной сильно уплощенной
головой и широко расставленными глазами. Окраска тела темная, серо-ко-
ричневая, с лиловым оттенком на спине и боках, и светло-кремовая на
брюшной части. Все плавники светлые, непигментированные. Голова очень
большая, но неширокая. На предкрышке один редуцирующийся шип. Рот
большой, верхняя челюсть слегка выдается вперед и оканчивается на верти-
кали середины глаза. Зубы мелкие, одинаковые по форме и размерам, загну-
ты назад и густо сидят на челюстях, образуя 6 неправильных рядов. На неб-
ных костях зубы отсутствуют. Глаза большие выпуклые, лоб широкий. Пе-
редние ноздри в виде узких, недлинных трубочек почти одинакового диамет-
ра на всем протяжении. Жаберная щель широкая, межжаберный промежу-
ток узкий. Сейсмосенсорная система представлена наружными невромаста-
ми, которые соединены между собой. Невромасты мелкие и имеют вид бе-
лых точек на поверхности кожи. Латеральная линия из 33 невромастов про-
ходит по середине тела и имеет 3 волнообразных изгиба. Над этой линией
имеются 2 дополнительных ряда, состоящих из отдельно сидящих неврома-
стов: внутренний ряд короткий, имеет 5 невромастов, наружный, располо-
200
женный ближе к спине, оканчивается
на вертикали последнего луча D2 и
имеет 26 невромастов. Под латераль-
ной линией имеется также короткий
ряд, представленный 10 невромаста-
ми. Грудные плавники короткие,
заканчиваются на вертикали первого
луча D2. Грудной плавник имеет усе-
ченную форму, длина его нижних лу-
чей втрое меньше, чем верхних.
Спинные плавники разделены широ-
ким промежутком. Первый спинной
плавник низкий, почти вдвое ниже
второго спинного и анального плав-
ников. Анальный плавник короткий.
Брюшные плавники довольно длин-
ные, они немного не доходят до
анального отверстия.
Dj V-VI; D212-14; А 9-11; Р 13-15;
V I 3. Жаберных тычинок 4. Пилори-
ческих придатков 4, они длинные и
толстые. Кишечник имеет петлю.
Позвонков 32-34 (Талиев, 1955; Сиде-
лева, 1982). Подвидов нет (Аннотиро-
ванный каталог..., 1998).
Распространение. Эндемик озера
Байкал, эврибатный вид, распростра-
нен в широком диапазоне глубин от
100 до 1000 м. Наиболее многочислен
в районах повышенной биологической продуктивности: Селенгинское мелко-
водье, северный выход из пролива Малое Море.
Возраст и размеры. Максимальный возраст 12 лет. Максимальные разме-
ры самок 226 мм, масса 170 г, самцов - 211 мм, масса ПО г. Д.Н. Талиев (1955)
указывал максимальную длину 250 мм.
Образ жизни. Донный вид, обитающий на илистых и илисто-песчаных
грунтах. Пища этого вида состоит из двух компонентов: гаммарид и рыбы.
Гаммариды составляют 94% по встречаемости и представлены 5 видами:
Paragarjaewia petersi, Plesiogammarus zienkowiczi, Garjaewia cabanisi,
Macropereiopus flori, Odontogammarus margaritaceus (Сиделева, Механикова,
1990). Средняя длина половозрелых самок 151 мм, масса 40 г, самцов -
150 мм и 37 г. Половое созревание начинается в возрасте 4 года при длине
116-119 мм и массе 10-13 г. Средняя плодовитость у самок длиной
133-208 мм составляет в среднем 133-834 икринки. Диаметр икринки варьи-
рует от 2,2 до 2,8 мм (Сиделева, Нагорный, 1985). Нерест L. bergianus прохо-
дит в июне-июле.
Статус вида. Обычный вид в глубоководной области Байкала. Промысло-
вого значения не имеет.
Автор-составитель: В.Г. Сиделева.
201
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Abyssocottidae
Род Limnocottus Berg, 1906
285. Limnocottus godlewskii (Dybowski, 1874) - крапчатая широколобка
Описание и систематика. Небольшая рыбка с довольно длинным узким
телом. Окраска тела, кроме светлой брюшной части, пятнистая, на пале-
вом фоне располагаются многочисленные коричневые пятна. Плавники,
кроме брюшных и анального, имеют темные поперечные полосы. Голова
большая, ее ширина больше высоты у затылка. Челюсти заканчиваются на
вертикали передней трети продольного диаметра глаза. Зубы мелкие, про-
стые, густо сидят на челюстях, присутствуют также на нёбных костях. Гла-
за большие, лоб вдвое меньше диа-
метра глаза, с возрастом ширина лба
увеличивается. Передние ноздри в
виде тонких трубочек. Жаберная
щель и межжаберный промежуток
узкие. Сейсмосенсорная система со-
стоит из мелких наружных неврома-
стов, которые имеют вид белых то-
чек. Латеральная линия проходит
строго по середине тела, содержит
58 невромастов на туловище и
19 невромастов на луче хвостового
плавника. Над ней имеются 2 допол-
нительных ряда, состоящих из 15 и
7 невромастов, ниже нее - 1 ряд из
8 невромастов. Спинные плавники
разделены широким промежутком.
Анальный плавник короткий, начи-
нается на некотором расстоянии от
ануса. Грудные плавники довольно
длинные, доходят до вертикали пер-
вых лучей D2. Брюшные плавники с
узким основанием, длинные, у самок
доходят, а у самцов заходят за аналь-
ное отверстие.
D, IV-V; D213-14; А 10-12; Р 12-14.
Жаберных тычинок 5. Пилорических
придатков 4, все они длинные, сходные
по размерам. Позвонков 32, из них ту-
202
ловищных 10, хвостовых 22 (Талиев, 1955; Сиделева, 1982). Подвидов нет (Анно-
тированный каталог..., 1998).
Распространение. Эндемик озера Байкал, встречается на глубинах
25-600 м.
Возраст и размеры. Максимальные размеры 130-135 мм, масса 21-23 г.
Образ жизни. Донный вид, обитающий на илисто-песчаных грунтах. Пища
особей из Южного Байкала состояла из мелких особей гаммарид (Талиев,
1955). Половое созревание начинается в возрасте 4+ при длине 77 мм и массе
3,4 г. Плодовитость 28-37 икринок диаметром 1,9-2,0 мм. Нерест проходит в
мае-июле. Молодь обитает на глубинах от 50 до 190 м. В возрасте 1 года име-
ет длину 20-23 мм и массу 0,06-0,08 г.
Статус вида. Обычный вид в Байкале, промыслового значения не
имеет.
Автор-составитель: В.Г. Сиделева.
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Abyssocottidae
Род Limnocottus Berg, 1906
286. Limnocottus griseus Taliev, 1948 - темная широколобка
Описание и систематика. Небольшая рыбка с узким прогонистым туло-
вищем. Окраска тела однотонная, темно-коричневая, брюшная часть свет-
лая. После нереста самцы приобретают интенсивную коричнево-шоколад-
ную окраску тела, грудных и спинных плавников. Плавники светлые, одно-
тонные, темных полос не имеют. Голова большая, уплощена в дорсовент-
ральном направлении, ее ширина почти вдвое больше ее высоты у затылка.
Челюсти равной длины и достигают вертикали переднего края глаза. Зубы
мелкие, простые, густо сидят на челюстях и отсутствуют на нёбных костях.
Глаза большие, лоб узкий. Передние ноздри в виде тонких трубочек. Жа-
берная щель и межжаберный промежуток узкие. Сейсмосенсорная система
представлена мелкими наружными невромастами, которые на коже имеют
вид белых точек, соединенных друг с другом. Латеральная линия проходит
по середине туловища и имеет 39 невромастов на теле и 7 невромастов на
срединном луче хвостового плавника. Два дополнительных ряда, располо-
женных выше латеральной линии, состоят из 10 и 6 невромастов, а допол-
нительный ряд ниже этой линии имеет 7 невромастов. Лучи в плавниках
(кроме хвостового плавника) неветвистые. Межлучевые перегородки тон-
кие, прозрачные. Между спинными плавниками имеется широкий промежу-
ток. Второй спинной плавник длиннее и ниже анального. Анальный плав-
203
ник начинается на небольшом рас-
стоянии от ануса. Грудные плавники
короткие, оканчиваются на вертика-
ли последних лучей DP Брюшные
плавники не доходят до анального
отверстия.
Dj V; D2 12; А 12; Р 14. Жаберных
тычинок 5. Пилорических придатков
3, все они одинаковой длины и тол-
стые. Позвонков 33, из них 10 туло-
вищных и 23 хвостовых (Талиев, 1955;
Сиделева,1982). Подвидов нет (Анно-
тированный каталог..., 1998).
Распространение. Эндемик озера
Байкал, обитает в широком диапазоне
глубин от 100 до 1600 м (Аннотиро-
ванный каталог..., 1998).
Возраст и размеры. Максималь-
ные размеры самок 165 мм, масса 45 г,
самцов - 147 мм и 40 г.
Образ жизни. Донный вид, обитает
на илистых грунтах. Половое созрева-
ние в возрасте 4+. Средняя длина по-
ловозрелых самок 150 мм, средняя
масса 32,2 г. Плодовитость составляет
280-512 (в среднем 370) икринок диа-
метром около 1,9 мм. Нерест бывает в
мае-июле (Талиев, 1955; Сиделева,
1982).
Статус вида. Довольно малочисленный глубоководный вид, промыслового
значения не имеет.
Автор-составитель: В.Г. Сиделева.
Класс Osteichthyes Семейство Abyssocottidae
Отряд Scorpaeniformes Род Limnocottus Berg, 1906
287. Limnocottus pallidus Taliev, 1948 - узкая широколобка
Описание и систематика. Небольшая рыбка с узким туловищем, уплощенной
головой и очень большими глазами. Окраска тела однотонная, на спине и боках
204
светло-коричневая, на брюшной части
светло-кремовая. Плавники однотон-
ные, темных полос не имеют. Голова
большая и значительно уплощенная,
ширина головы больше ее высоты у за-
тылка. Челюсти равной длины и почти
достигают вертикали середины глаза.
Зубы мелкие, простые, густо сидят на
челюстях. На нёбных костях зубы от-
сутствуют. Глаза очень большие, выпу-
клые. Лоб узкий. Передние ноздри в
виде коротких кожистых слегка заужи-
вающихся кверху трубочек, длина ко-
торых в 2 раза больше основания. Зад-
ние ноздри в виде широких коротких
трубочек. Жаберная щель узкая, меж-
жаберный промежуток очень узкий.
Сейсмосенсорная система представле-
на наружными невромастами в виде то-
чек, которые соединены друг с другом.
Латеральная линия имеет 63 неврома-
ста на теле и 7 невромастов на луче
хвостового плавника. Над латеральной
линией имеются 2 ряда из 15 и 10 нев-
ромастов, ниже этой линии - ряд из 11
невромастов. Лучи в плавниках тол-
стые и неветвистые, межлучевые пере-
городки тонкие и прозрачные. Между
спинными плавниками имеется широ-
кий промежуток, второй спинной плавник немного длиннее и выше анального.
Анальный плавник начинается на некотором расстоянии от ануса. Грудные
плавники короткие, не доходят до начала второго спинного плавника и имеют
усеченную форму. Брюшные плавники короткие, короче грудных и не доходят
до анального отверстия, они имеют узкое основание.
Dj IV-V; D2 11-12; А 8-11; Р 14-16. Жаберных тычинок 7, тычинки малень-
кие бугорковидные. Пилорических придатков 4, они длинные и сходные по
размерам (Талиев, 1955; Сиделева, 1982). Подвидов нет (Аннотированный ка-
талог..., 1998).
Распространение. Эндемик озера Байкал, эврибатный вид, распространен в
широком диапазоне глубин от 150 до 1200 м.
Возраст и размеры. Максимальные размеры самок 146 мм, масса 17 г, сам-
цов - 131 мм и 16 г. Максимальный возраст 12 лет.
Образ жизни. Донный вид, обитает на илистых и илисто-песчаных грунтах.
Пища L. pallidus длиной 31-90 Um в районе пролива Малое Море состоит из
4 компонентов: хирономиды, копеподы, гаммариды и рыба; у более крупных
рыб количество компонентов уменьшается до 2: гаммариды и рыба. Гаммари-
ды в пищевом комке рыб в районе пролива Малое Море были представлены
Crypturopus inflatus, Poekilogammarus (Базикалова, Вилисова, 1959), в районах
Селенгинского мелководья и Баргузинского залива - Carinogammarus seidlitzi,
Crypturopus inflatus, Acanthogammarus и Macrohectopus (Базикалова и др., 1937).
205
Самки становятся половозрелыми и впервые идут на нерест при длине 115—
116 мм и массе 9,2-12,0 г, при этом плодовитость составляет 65-95 икринок.
Самцы созревают при длине 84 мм и массе 3,4 г. Средние размеры половозре-
лых самок 129 мм, средняя плодовитость 105 икринок, диаметр икринки 2,4 мм.
Нерест проходит в июне-июле.
Статус вида. Промыслового значения не имеет. Иногда встречается в пище
взрослых особей видов Batrachocottus nikolskii и В. talievi.
Автор-составитель: В.Г. Сиделева.
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Abyssocottidae
Род Neocottus Sideleva, 1982
288. Neocottus thermalis Sideleva, 2002 - тепловодная широколобка
Описание и систематика. Представители нового рода Neocottus похожи на
рыб рода Limnocottus; у них сравнительно длинный хвостовой стебель. Этмои-
дальная кость имеет хороню выраженный бугорок, зубы мелкие и простые, рот
маленький и угол рта не доходит до вертикали переднего края глаза. Новый
вид, его название указывает на своеобразное его распространение в районе под-
водного гидротермального источника. Небольшая рыбка с крупной головой и
коротким туловищем. Кожа голая, толстая. Окраска тела однотонная, светло-
серая со спины и светло-кремовая с брюшной части. Спинные плавники темно-
серые, однотонные; все другие - светло-кремовые. Голова большая, ширина го-
ловы почти равна ее длине. Продольный диаметр глаза 5 раз содержится в дли-
не головы. Подглазничные кости на внешнем крае образуют выступающий гре-
бень, ширина гребня равна ширине лба. Внешние края лобных костей приподня-
ты и выступают над орбитами, поэтому глаза расположены в глубоких глазни-
цах. Лоб узкий. Передние и задние ноздри имеют вид коротких трубочек. Жабер-
ная щель узкая. Сейсмосенсорная система представлена наружными невромаста-
ми, которые расположены на плоских папиллах. Латеральная линия состоит из
31 невромаста на теле и 12 невромастов на луче хвостового плавника. Выше ла-
теральной линии имеется дополнительный ряд из 11 невромастов, ниже ее рас-
положен ряд всего из 4 невромастов. Грудные плавники короткие, оканчивают-
ся на вертикали первых лучей D2. Брюшные плавники короче грудных и не до-
ходят до анального отверстия. Спинные плавники следуют друг за другом без
промежутка. Первый спинной плавник очень низкий, почти в два раза ниже вто-
рого. Второй спинной выше и длиннее анального.
206
D, VI-VII; D2 (12) 14—15; A 10-11;
P (11) 13-16. Жаберных тычинок 5.
Позвонков 33-34. Подвидов нет (Си-
делева, 2002).
Распространение. Локальный эн-
демик, встречается в Северном Байка-
ле, в районе бухты Фролиха, в месте
расположения гидротермального поля
на глубинах 430—465 м.
Возраст и размеры. Предельный
возраст 8 лет, достигает длины 16 см.
Образ жизни. Обитает на илистых
грунтах с большой примесью расти-
тельного детрита. Большие концент-
рации тепловодной широколобки об-
наружены на периферии бактериаль-
ных матов гидротермальных полей. За
20 мин траления 4-метровым бимтра-
лом было поймано 70 экз. этого вида.
Численность донных Cottoidei, вылов-
ленных в 1990 и 1991 гг. в районе гид-
ровента, на 80% была представлена
этим видом; этот же вид был обнару-
жен во время исследования гидровента
из подводных аппаратов “Пайсис” в
1990 г. В результате многочисленных
тралений, проведенных в Северном
Байкале до этого времени, 21 июня
1978 г. Г.Н. Сиделевым был пойман всего 1 экз. данного вида вблизи от бухты
Фролиха на глубине 260-350 м (коллекция ЗИН РАН № 50885).
Статус вида. Массовый вид в районе бухты Фролиха, в месте расположения
гидровента. В других районах Байкала не встречается и хозяйственного значе-
ния не имеет.
Автор-составитель: В.Г. Сиделева.
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Abyssocottidae
Род Neocottus Sideleva, 1982
289. Neocottus werestschagini (Taliev, 1935) - рыхлая широколобка
207
Описание и систематика. Неболь-
шая рыбка светло-серого цвета. Тело
удлиненное, антеанальное расстояние
больше половины длины тела. Кожа го-
лая, тонкая. Окраска тела светло-серая
со спины и светло-кремовая с брюшной
стороны. Все плавники однотонные, не-
окрашенные. Голова большая, ее длина
больше ширины и почти в 2 раза боль-
ше высоты у затылка. Верхняя челюсть
слегка выдается вперед. Глаза малень-
кие. Подглазничные кости на внешнем
крае образуют выступающий гребень.
Внешние края лобных костей припод-
няты и слегка выступают над орбитами
таким образом, что глаза располагают-
ся в глубоких глазницах. Лоб узкий,
меньше диаметра глаза. Передние нозд-
ри в виде коротких широких трубочек.
Жаберная щель и межжаберный проме-
жуток узкие. Сейсмосенсорная система
представлена линиями наружных нев-
ромастов, которые расположены на
прозрачных уплощенных папиллах. У
N. werestschagini необычное строение
папилл: они в поперечном разрезе име-
ют форму восьмерки, т.е. похожи на ло-
пасть пропеллера. Посредине, в углуб-
лении сидит невромаст, внутри папиллы
хорошо виден подходящий к невромасту нерв. Латеральная линия имеет 4 волно-
образных изгиба и содержит 32 невромаста на туловище и 6 невромастов на 9-м
луче хвостового плавника. Лучи и межлучевые перегородки в плавниках тонкие.
Грудные плавники короткие, оканчиваются на вертикали первых лучей D2 и
длиннее брюшных. Лучи в грудных плавниках имеют длинные свободные кончи-
ки. Брюшные плавники далеко не доходят до анального отверстия. Спинные
плавники расположены близко друг к другу и значительно различаются по высо-
те: первый спинной плавник в 2 раза ниже второго. Анальный плавник короткий,
начинается на сравнительно большом расстоянии от анального отверстия и ко-
роче второго спинного.
D, VI-VII; D2 13-15 (16); А 11-13; Р 16-17. Жаберных тычинок 6, они бу-
горковидные, с мелкими шипиками. Пилорических придатков 3, все они прево-
сходят длину желудка. Кишечник делает петлю (Талиев, 1955; Сиделева, 1982).
Подвидов нет (Аннотированный каталог..., 1998).
Распространение. Вид очень редкий, встречается в Южной и Центральной,
наиболее глубоких, котловинах Байкала, на глубинах 900-1400 м.
Возраст и размеры. Предельный возраст 9 лет, достигает длины 130 см и
массы 30 г.
Образ жизни. Биология почти неизвестна. Распространена рыхлая широко-
лобка на илистых грунтах. В желудках найдены амфиподы, рыба и детрит. Ам-
фиподы представлены тремя видами одного рода: Macropereiopus flori, М. albu-
208
lus и Macropereiopus sp. (Сиделева, Механикова, 1990). Время нереста не выяс-
нено, в летние месяцы, добытые экземпляры были уже отнерестовавшими.
Статус вида. Крайне малочисленный глубоководный вид. Промыслового
значения не имеет.
Автор-составитель: В.Г. Сиделева.
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Abyssocottidae
Род Procottus Gratzianow, 1902
290. Procottus gotoi Sideleva, 2001 - широколобка Гото
Описание и систематика. У всех ви-
дов рода Procottus оба спинных плав-
ника слиты в один, состоящий из более
низкой передней части (Dt) и более
высокой задней (D2); кожа голая, хво-
стовой стебель короткий, голова без
шипов. Новый вид назван в честь япон-
ского ихтиолога Акира Гото - исследо-
вателя пресноводных коттоидных рыб.
Довольно большая рыба ярко-изумруд-
ного цвета с укороченным телом и бор-
дово-коричневыми пятнами, на тулови-
ще крупными, а на голове мелкими. Все
плавники хорошо пигментированы и
имеют темные поперечные полосы.
Отличительной особенностью данного
вида среди других видов рода Procottus
является наличие в грудном плавнике от
одного до трех ветвистых лучей. Голова
очень большая, ее ширина меньше дли-
ны. Рот небольшой, челюсти равной
длины и заканчиваются на одной вер-
тикали передней трети глаза. Зубы
мелкие, на челюстях образуют 7 непра-
вильных рядов. Глаза большие, выпук-
лые “тубулярные”, выступают над по-
верхностью головы. Лоб узкий, меньше.
209
14 Атлас.... том 2
диаметра глаза. Передние ноздри в виде коротких кожистых трубочек. Межглаз-
ничный промежуток широкий, а жаберная щель узкая. Сейсмосенсорная система
состоит из линий наружных невромастов. Латеральная линия расположена ближе
к спинной стороне тела и содержит 72 невромаста на туловище и 7 невромастов на
10-м луче хвостового плавника. Над этой линией имеется короткий ряд из 8 невро-
мастов. Грудные плавники короткие, оканчиваются на вертикали 8-го луча спинно-
го плавника. Брюшные плавники короткие, далеко не доходят до анального отвер-
стия, они короче грудных. Анальный плавник довольно короткий, начинается на
некотором расстоянии от анального отверстия, а заканчивается почти на одном
уровне со спинным плавником. Лучи в плавниках и межлучевые перегородки тол-
стые.
D( VII—IX; D218-19; А 13-15; Р 17-18. Жаберных тычинок 4. Пилорических
придатков 3, все длинные. Позвонков 33-35. Подвидов нет (Сиделева, 2002в).
Распространение. Эндемик озера Байкал, встречается в основном в южной час-
ти озера на глубинах от 3 до 150 м.
Возраст и размеры. Предельный возраст 5 лет, достигает длины 16 см и
массы 85 г.
Образ жизни. Приурочен к каменистым биотопам. В наших уловах присут-
ствовали половозрелые пятилетние особи длиной 14-16 см и массой
57-85 г. Нерест проходит в феврале-марте подо льдом. Икра крупная, диамет-
ром около 3 мм, зеленого цвета.
Статус вида. Промыслового значения не имеет.
Автор-составитель: В.Г. Сиделева.
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Abyssocottidae
Род Procottus Gratzianow, 1902
291. Procottus gurwici Taliev, 1946 - карликовая широколобка
Описание и систематика. Самая маленькая рыбка среди байкальских
коттоидных рыб. Тело удлиненное. Общий фон туловища темно-крас-
ный, с равномерно распределенными точечными меланофорами. Отдель-
ные меланофоры присутствуют также на голове, плавниках, ноздрях и
губах. Все плавники темные, так как сильно и равномерно пигментирова-
ны лучи и межлучевые перегородки. Первый спинной плавник имеет по
210
краю черную широкую кайму. Го-
лова большая, ее длина больше ши-
рины. Подглазничные кости не об-
разуют гребня. На этмоидальной
кости имеется маленький бугорок.
Рот маленький, челюсти слегка за-
ходят за вертикаль переднего края
глаза, верхняя челюсть слегка выда-
ется вперед. Зубы в большом коли-
честве присутствуют на челюстях и
нёбных костях. Глаза большие,
межглазничный промежуток очень
узкий, жаберная щель узкая, а меж-
жаберный промежуток довольно
широкий. Передние ноздри в виде
узких пигментированных трубочек.
Длина ноздри в 3 раза больше ее ос-
нования. Сейсмосенсорная система
состоит из наружных невромастов,
которые имеют вид очень мелких
светлых точек. Латеральная линия в
передней части туловища делает из-
гиб наверх, расположена близко к
спине и содержит 47 невромастов на
теле и 10 невромастов на луче хво-
стового плавника. Над латеральной
линией, почти у основания спинных
плавников имеются отдельные нев-
ромасты. Грудные плавники срав-
нительно длинные и имеют усеченную форму. Брюшной плавник длин-
ный, доходит до анального отверстия, имеет очень узкое основание. Лучи
в плавниках, за исключением спинных, имеют длинные свободные кончи-
ки.
D1 VII-VIII; D2 19-21; А 13-15 (16); Р 16-18; Жаберных тычинок 6. Пилори-
ческих придатков 4, из них 2 длинных, 1 короткий и один очень короткий. Под-
видов нет (Талиев, 1955; Сиделева, 1982; Аннотированный каталог..., 1998).
Распространение. Приурочен к прибрежной зоне южной части Байкала, к
глубинам от 5 до 250 м.
Возраст и размеры. Предельный возраст 7 лет, достигает длины 58 мм и
массы 2,5 г.
Образ жизни. Обитает на каменисто-песчаных и песчаных грунтах, часто
держится в колониях байкальских губок. Плодовитость 19-25 икринок диамет-
ром 1,7-1,9 мм (Талиев, 1955). (
Статус вида. Из-за малочисленности и маленьких размеров промыслового
значения не имеет.
Автор-составитель: В.Г. Сиделева.
211
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Abyssocottidae
Род Procottus Gratzianow, 1902
292. Procottus jeittelesii (Dybowski, 1874) - красная широколобка
Описание и систематика. Красная широкоболка - небольшая рыбка с валько-
ватым и укороченным телом. Окраска тела темно-бордовая, со светлыми непра-
вильной формы пятнами по бокам. Все плавники однотонные темно-серые, поч-
ти черные, поперечных полос не имеют. Голова большая, ее ширина в 2 раза
меньше длины. Подглазничная кость не выдается. Этмоидальная кость имеет бу-
горок. Рот маленький, заканчивается на вертикали переднего края глаза. Нижняя
челюсть слегка выдается перед. Зубы мелкие конические, на челюстях образуют
7 неправильных рядов и в большом количестве присутствуют на нёбных костях.
Глаза большие, выпуклые, выступают над поверхностью головы. Межглазнич-
ный промежуток и жаберная щель уз-
кие, а межжаберный промежуток ши-
рокий. Передние ноздри в виде пигмен-
тированных коротких трубочек. Сейс-
мосенсорная система состоит из линий
наружных невромастов. Невромасты
очень мелкие, имеют вид маленьких
светлых точек, расположенных близко
друг к другу. Латеральная линия смеще-
на ближе к спинной стороне туловища и
содержит 89 невромастов на теле, и еще
17 невромастов заходят на 10-й луч хво-
стового плавника. Над латеральной ли-
нией расположен дополнительный ряд
из 12 редко сидящих невромастов, кото-
рый оканчивается на вертикали послед-
них лучей хвостового плавника. В спин-
ном плавнике выше задняя часть.
Анальный плавник начинается сразу за
анальным отверстием и заканчивается
почти на одном уровне со спинным
плавником. Брюшные плавники длин-
ные, заходят за анальное отверстие, они
короче грудных.
D) VII-IX; D2 18-21; А 12-15;
Р 17-18. Жаберных тычинок 5. Пилори-
ческих придатков 4, из них 3 коротких и
один длинный. Позвонков 35-37 (Тали-
212
ев, 1955; Сиделева, 1982). Подвидов нет (Аннотированный каталог..., 1998). Внут-
ри вида Р. jeittelesii Д.Н. Талиев (1955) различал 2 формы, которые обитают в раз-
ном диапазоне глубин и имеют различную окраску: прибрежная форма - ярко-зе-
леную, глубоководная - коричнево-красную. Форма с зеленой окраской выделена
нами в самостоятельный вид Р. gotoi.
Распространение. Эндемик озера Байкал, обитает на глубинах от 15 до 200 м.
Возраст и размеры. Предельный возраст 8 лет, достигает длины 13 см, ча-
ще 10-12 см (Талиев,1955).
Образ жизни. Вид приурочен только к каменистым биотопам и многочислен в
прибрежной зоне Южного Байкала и истока р. Ангары. У красной широколобки
прослеживается возрастная смена состава пищевого комка: у молоди длиной
20-40 мм преобладают низшие ракообразные, которые исчезают у рыб длиной
80-100 мм. Состав пищи крупных особей - амфиподы, рыба, личинки ручейников
и редко олигохеты. Наибольшее значение в питании взрослых рыб имеют амфи-
поды, которые встречаются практически у всех особей (100%). Из амфипод встре-
чаются виды родов Poekilogammarus, Cripturopus, Acanthogammarus, Parapallasea и
Garjajewia (Базикалова, Вилисова, 1959).
Половая зрелость наступает при длине 95 мм и массе 12,5 г. Нерест бывает
с января до середины марта на глубинах 5-8 м, плодотовитость - 150-286 икри-
нок (Талиев, 1955).
Статус вида. Промыслового значения не имеет.
Автор-составитель: В.Г. Сиделева.
Класс Osteichthyes
Отряд Scorpaeniformes
Семейство Abyssocottidae
Род Procottus Gratzianow, 1902
293. Procottus major Taliev, 1944 - большая широколобка
Описание и систематика. Самая большая рыба среди байкальских коттоидных
рыб. Тело укороченное. Окраска тела темно-розовая, с темными коричнево-бор-
довыми пятнами неправильной формы. Спинные плавники имеют поперечные по-
лосы, грудные, анальный и хвостовой однотонные темно-серые, почти черные.
Голова большая, ее ширина почти равна длине. Рот маленький, заканчивается на
вертикали переднего края глаза, нижняя челюсть немного выдается вперед. Зубы
мелкие, конические, на челюстях образуют 6 неправильных рядов. Глаза большие,
выпуклые, выступают над поверхностью головы. Лоб узкий, в 2 раза меньше про-
дольного диаметра глаза. Межжаберный промежуток широкий, составляет поло-
вину длины жаберной щели. Сейсмосенсорная система состоит из линий мелких
213
наружных невромастов, которые соеди-
нены друг с другом. Латеральная линия
содержит более 100 невромастов на ту-
ловище и 18 на 10-м луче хвостового
плавника. Ряд невромастов над лате-
ральной линией доходит до середины
спинного плавника и содержит 22 редко
сидящих невромаста. Грудные плавники
короткие, заканчиваются на вертикали
9-го луча спинного плавника, они име-
ют хорошо выраженную усеченную
форму и широкое основание. Аналь-
ный плавник короткий, начинается пра-
ктически сразу за анальным отверсти-
ем. Брюшные плавники короткие, коро-
че грудных и далеко не доходят до
анального отверстия.
Dj VIII-IX; D2 19-21; А 13-15;
Р 16-18. Жаберных тычинок 4, как у
всех донных видов, они бугорковид-
ные, с мелкими шипиками на поверх-
ности. Пилорических придатков 3.
Позвонков 34—36 (Талиев, 1955; Сиде-
лева, 1982). Кариотип: 2п = 48, NF = 74
(Сиделева и др., 1995). Подвидов нет
(Аннотированный каталог..., 1998).
Распространение. Эндемик озера
Байкал. Распространен на глубинах от
50 до 900 м.
Возраст и размеры. Предельный возраст 12 лет, достигает длины 30 см и
массы 267 г.
Образ жизни. Предпочитает песчаные и песчано-илистые грунты. Как показа-
ли наблюдения из подводных аппаратов “Пайсис”, большая широколобка может
делать норы в мягком илистом грунте и использовать их в качестве убежища (Ко-
жова и др., 1979). В ее пище наибольшее значение имеют амфиподы, рыба, моллю-
ски, коконы планарий, единично олигохеты и личинки ручейников. Из амфипод
наиболее часто встречаются Acanthogammarus victorii maculosus, Garjajewia cabanisi,
Parapallasea puzulli и виды рода Poekilogammarus (Базикалова, Вилисова, 1959).
Обычные размеры рыб в уловах 18-22 см. Половозрелые самки длиной 25-26 см и
массой 245-267 г встречались в наших сборах на глубинах 240-500 м. Плодовитость
составляет 1600-2140 икринок, диаметр икринок 2,8-3,0 мм. Время нереста - конец
сентября-начало декабря. Кладка икры этого вида была поднята тралом с глуби-
ны 100-140 м, икринки перед выклевом имели диаметр 4,5 мм (Абросов, 1946).
Статус вида. В конце 1940-х-начале 1950-х годов во время нерестовых ско-
плений этот вид имел промысловое значение. В районе пролива Малое Море в
1943-1946 гг. добывалось 30-40 т нерестовых особей (Талиев, 1955). В настоя-
щее время из-за низкой численности промыслом не используется.
Автор-составитель: В.Г. Сиделева.
214
ЛИТЕРАТУРА
Абакумов В.А. Об образе жизни балтийской проходной миноги // Вопр. ихтиологии. 1956.
Вып. 6. С. 122-128.
Абакумов В.А. О систематике и экологии дальневосточной ручьевой миноги Lampetra reissneri
Dybowski из бассейна Амура // Там же. 1960. Вып. 15. С. 43-54.
Абакумов В.А. Систематика и экология европейской ручьевой миноги Lampetraplaneri (Bloch) //
Там же. 1964. Т. 4, вып. 3. С. 423-432.
Абакумов В.А. Систематика и экология украинской миноги (Lampetra mariae Berg) // Там же.
1966. Т. 6, вып. 4. С. 609-616.
Абдурахманов Ю.А. Рыбы пресных вод Азербайджана. Баку: Изд-во АН АзССР, 1962. 408 с.
Абдусамадов А.С. Биология белого амура, белого толстолобика и пестрого толстолобика, акк-
лиматизированных в Терском районе Каспийского бассейна И Вопр. ихтиологии. 1986. Т. 26,
вып. 3. С. 425-433.
Абросов В.Н. Осенний нерест бычка Procottus jeittelesii в Байкале // Природа. 1946. № 7. С. 66.
Александрова А.И. Сравнительная морфометрическая характеристика носаря Acerina acerina
(Giild.) из рек Северного Донца и Случь // Тр. Зоол. музея Ин-та зоологии АН УССР. 1976.
№36. С. 77-81.
Александрова Е.Н., Кузнецов В.В. Морфометрический анализ дифференциации муксуна р. Ле-
ны. 1. Морфометрическая характеристика четырех форм муксуна//Вести. МГУ. Биология,
почвоведение. 1972. № 4. С. 16-23.
Алексеев Г.А., Кудерский Л.А. Морская минога Petromyzon marinus (Petromyzontidae) в восточной
части Финского залива // Вопр. ихтиологии. 2001. Т. 41, вып. 1. С. 135-137.
Алексеев С.С. Морфо-экологическая характеристика ленков (Salmonidae, Brachymys-
tax) из бассейна Амура и из р. Уды // Зоол. журн. 1983. Т. 62, № 7. С. 1057-1067.
Алексеев С.С., Кириллов А.Ф. К вопросу о морфологии и распространении двух форм ленков
рода Brachymystax Gunther (Salmonidae) в бассейне Лены // Вопр. ихтиологии. 1985. Т. 25,
вып. 4. С. 597-602.
Алексеев С.С., Свириденко М.А. Микижа Salmo mykiss Walbaum (Salmonidae) Шантарских остро-
вов // Там же. 1985. Т. 25, вып. 1. С. 68-73.
Алиев Д.С., Суханова А.И., Шакирова Ф.М., Малахова Т.М. Растительноядные рыбы в Туркме-
нистане. Ашгабат: Ылым, 1994. 326 с.
Алтухов Ю.П., Салменкова Е.А., Омельченко В.Т. и др. Балансирующий отбор как возможный
фактор поддержания единообразия аллельных частот ферментных локусов в популяциях
тихоокеанского лосося - горбуши Oncorhynchus gorbuscha (Walb.) // Генетика. 1987. Т. 23,
№ 10. С. 1884-1896.
Андрияшев А.П. Рыбы северных морей. М.; Л.: Изд-во АН СССР, 1954. 566 с.
Андрияшев А.П., Панин К.И. О нахождении тихоокеанского осетра Acipenser medirostris Ayres в
Беринговом море Ц Зоол. журн. 1954. Т. 32, № 5. С. 932-936.
Аннотированный каталог круглоротых и рыб континентальных вод России / Под ред. Ю.С. Ре-
шетникова. М.: Наука, 1998. 218 с. '
Арефьев ВА. Поликариограммный анализ шипа Acipenser nudiventris Lov. (Acipenseridae,
Chondrostei) // Вопр. ихтиологии. 1983. T. 23, вып. 2. С. 209-218.
Артюхин Е.Н. Персидский осетр в реках Северного Каспия и перспективы его использования в
осетровом хозяйстве // Биологические основы развития осетрового хозяйства в водоемах
СССР. М.: Наука, 1979. С. 105-115.
Артюхин Е.Н. Дифференциация популяций персидского осетра и перспективы его заводского
разведения на Волге Ц Биологические основы осетроводства. М.: Наука. 1983. С. 54—61.
215
Артюхин Е.Н., Андронов А.Е. Морфобиологический очерк зеленого осетра Acipen-
ser medirostris (Chondrostei, Acipenseridae) из p. Тумнин (Датта) и некоторые ас-
пекты экологии и зоогеографии осетровых // Зоол. журн. 1990. Т. 69, № 12. С. 81-90.
Артюхин Е.Н., Заркуа З.Г. К вопросу о таксономическом ранге осетра р. Риони (бассейн Чер-
ного моря) // Вопр. ихтиологии. 1986. Т. 26, вып. 1. С. 61-67.
Асхаев М.Г. Новые породы рыб в водоемах бассейна Байкала // Рыбы и рыбное хозяйство в бас-
сейне озера Байкал. Иркутск: Кн. изд-во, 1958. С. 420-428.
Атлантический лосось / Под ред. Р.В. Казакова. СПб.: Наука, 1998. 575 с.
Багров А.М. Рост и развитие гонад растительноядных рыб в условиях тропического климата в
связи с их искусственным воспроизводством: (На примере Кубы): Автореф. дис. ... канд.
биол. наук. М., 1985. 26 с.
Багров А.М. Гаметогенез и половые циклы растительноядных рыб в разных климатических зонах
в связи с искусственным воспроизводством: Автореф. дис. ... д-ра биол. наук. М., 1993. 58 с.
Базикалова А.Я., Вилисова И.К. Питание бентосоядных рыб Малого моря // Тр. Байкал, лимнол.
станции АН СССР. 1959. Т. 17. С. 382-497.
Базикалова А.Я., Калинникова Т.В., Михин В.С., Талиев Д.Н. Материалы к познанию бычков
Байкала Ц Там же. 1937. Т. 7. С. 109-212.
Балушкин А.В., Черешнев И.А. Систематика рода Daltia (Umbridae, Esociformes) // Тр. Зоол.
ин-та АН СССР. 1982. Т. 114, С. 36-56.
Баранникова И.А. Биологическая дифференциация стада волго-каспийского осетра в связи с за- <
дачами промышленного осетроводства в дельте Волги // Учен. зал. Ленингр. ун-та. Сер. «
биол. наук. 1957. Вып. 4. № 228. С. 54—71. |
Баран Т.П. Рыбы пресных вод. Тбилиси, 1941. 287 с. (Фауна Грузии; Т. 1). s
Беляева В.Н., Власенко А.Д., Иванов В.П. Каспийское море: Ихтиофауна и промысловые ресур-
сы. М.: Наука, 1989. 235 с.
Берг Л.С. Рыбы пресных вод СССР и сопредельных стран. М.; Л.: Изд-во АН СССР, 1948. Т. 1.
468 с.
Берг Л.С. Рыбы пресных вод СССР и сопредельных стран. М.; Л.: Изд-во АН СССР, 1949а.
Т. 2. С. 469-929.
Берг Л.С. Рыбы пресных вод СССР и сопредельных стран. М.; Л.: Изд-во АН СССР, 19496. Т. 3.
С. 930-1381.
Билько В.П. Размножение черноморских бычков в Днепровско-Бугском лимане // Вопр. ихтио-
логии. 1968. Т. 8, вып. 4. С. 669-4578.
Биология и промысловое значение рыбцов (Vimba) Европы. Вильнюс: Минтис, 1970. 517 с.
Бирман И.Б. Некоторые вопросы биологии симы Oncorhynchus masu // Изв. ТИНРО. 1972. Т. 82.
С. 235-246.
Бирман И.Б. Морской период жизни и вопросы динамики стад тихоокеанских лососей. М.: Аг-
ропромиздат, 1985. 208 с.
Богданов Б.Э. Экология реофильных видов подкаменщиков (Cottidae) в водоемах
Байкальской рифтовой зоны: Автореф. дис. ... канд. биол. наук. Иркутск, 2000. 18 с.
Богданов В.Д. Морфологические особенности развития и определитель личинок сиговых рыб
р. Оби. Екатеринбург: ИЭРиЖ УрО РАН, 1998. 53 с.
Богданов ВД., Богданова Е.Н., Мельниченко ИЛ. Гидробионты // Мониторинг биоты п-ова
Ямал в связи с развитием добычи и транспортировки газа. Екатеринбург: УРЦ “Аэрокосмо-
экология”, 1997. С. 100-152.
Богданов ВД., Богданова Е.Н., Госькова О.А., Мельниченко И.П. Ретроспектива ихтиологиче-
ских и гидробиологических исследований на Ямале. Екатеринбург: ИЭРиЖ УрО РАН, 2000.
88 с.
Богданов В.Д., Кижеватов Я.А. Динамика ихтиофауны р. Собь//Научный вестник. Вып. 4. Ма-
териалы к познанию фауны и флоры Ямало-Ненецкого Автономного округа. Салехард,
2000. С. 3-15.
Богуцкая Н.Г. К вопросу о систематическом отношении видов родов Abramis, Blicca, Vimba
(Cyprinidae) И Вопр. ихтиологии. 1986. Т. 26, вып. 4. С. 576-584.
Богуцкая Н.Г. О таксономическом статусе ельца Данилевского (Cyprinidae) // Тр. Зоол. ин-та
АН СССР. 1987. Т. 162. С. 73-80.
Богуцкая Н.Г. Особенности изменчивости некоторых признаков алтайских османов в связи с ди- ।
агностикой рода Oreoleuciscus (Cyprinidae) // Там же. 1990. Т. 222. С. 110-131.
Богуцкая Н.Г, Насека А.М. Круглоротые и рыбы оз. Ханка (система р. Амур): Аннотированный
список видов с комментариями по их таксономии и зоогеографии региона // Науч, тетради
ГосНИОРХ. 1996. № 3. С. 1^15.
216
i
Богуцкая Н.Г., Позняк В.С. Переописание афипсского ельца Leuciscus aphipsi Aleksandrov
(Leuciscinae, Cyprinidae) // Вопр. ихтиологии. 1994. T. 34, вып. 3. С. 308—315.
Бодали Р.А., Вуоринен Д.А., Решетников Ю.С., Рист Д.Д. Генетические связи пяти видов сиго-
вых рыб Сибири // Там же. 1994. Т. 34, вып. 2. С. 195-203.
Бодареу Н.Н. Животный мир Молдавии: Рыбы, земноводные, пресмыкающиеся. Кишинев, 1981.
131 с.
Борзенко М.П. Каспийская севрюга: (Систематика, биология и промысел) // Изв. Азерб, науч,-
исслед. рыбохоз. станции. 1942. Вып. 7. С. 3-114.
Борисов П.Г. Новый вид гольца Salvelinus jacuticus n. sp. // Материалы к ихтиофауне бассейна
р. Лены: Сб. науч. тр. М.: Изд-во АН СССР, 1932. С. 1—42.
Борисова А.Т. Случайные вселенцы в водоемах Узбекистана // Вопр. ихтиологии. 1972. Т. 12,
вып. 1. С. 49-53.
Бэнэреску П., Налбант Т. К систематике и номенклатуре пескарей подсемейства Gobioninae
(Pisces, Cyprinidae) в бассейне Амура // Вопр. ихтиологии. 1968. Т. 8, вып. 4. С. 628-636.
Варпаховский Н. 1886. Очерк ихтиологической фауны Казанской губернии // Прилож. к 52 то-
му Записок Импер. Акад. наук. СПб., 1886. № 3. 70 с.
Васильев В.П. Эволюционная кариология рыб. М.: Наука, 1985. 300 с.
Васильев В.П., Соколов Л.И. Метод изучения кариотипов хрящевых ганоидов (Chondrostei) // Ци-
тология. 1980. Т. 22, № 9. С. 1106-1109.
Васильева ЕД. Озерный голец Salvelinus alpinus (L.) из бассейна р. Зарубиха: (К вопросу о голь-
цах и палиях Кольского полуострова) // Вопр. ихтиологии. 1981. Т. 21, вып. 2. С. 232-246.
Васильева ЕД. Систематическое положение некоторых видов рода Benthophilus (Gobiidae) Кас-
пийского и Азовского морей // Там же. 1983. Т. 23, вып. 4. С. 544-556.
Васильева Е.Д. О таксономическом статусе некоторых форм рода Oreoleuciscus (Cyprinidae) //
Там же. 1985. Т. 25, вып. 2. С. 196-211.
Васильева ЕД. Переописание, морфо-экологическая характеристика и распространение Cobitis
granoei (Teleostei, Cobitidae) // Зоол. журн. 1988. Т. 67, № 7. С. 1025-1036.
Васильева ЕД. Новые данные по распространению щиповки Лебедева С. lebedevi (Cobitidae) //
Вопр. ихтиологии. 1994. Т. 34, вып. 6. С. 839-840.
Васильева ЕД. Об отсутствии каспийской щиповки Sabanejewia caspia (Cobitidae) в бассейне Се-
верного Каспия // Там же. 1995. Т. 35, вып. 6. С. 822-824.
Васильева Е.Д., Васильев В.П. Новый вид щиповки - С. lebedevi sp. n. (Osteichthys, Cobitidae) из
бассейна Амура // Зоол. журн. 1985. Т. 64, № 3. С. 463—468.
Васильева Е.Д., Васильев В.П. Исследование внутривидовой структуры Sabanejewia aurata
(Cobitidae) с описанием нового подвида Sabanejewia aurata kubanica subsp. nova // Вопр. их-
тиологии. 1988. T. 28, вып. 2. С. 192-212.
Васильева Е.Д., Васильев В.П. Систематика кавказских речных бычков (Gobiidae) в свете совре-
менных данных с описанием нового вида Negobius rhodioni sp. nova // Там же. 1994. Т. 34,
вып. 6. С. 747-758.
Васильева ЕД., Васильев В.П. Виды-двойники в роде Cobitis (Cobitidae). 1. Южнорусская щипов-
ка Cobitis rossomeridionalis sp.n. // Там же. 1998. Т. 38, вып. 5. С. 604—614.
Васильева Е.Д., Козлова М.С. О таксономии востробрюшек рода Hemiculter (Cyprinidae) Совет-
ского Союза // Там же. 1988. Т. 28, вып. 6. С. 883-895.
Васильева Е.Д., Макеева А.П. Морфологические особенности и таксономия головешковых рыб
(Pisces: Eleotridae) фауны СССР//Зоол. журн. 1988. Т. 67, № 8. С. 1194-11204.
Васильева Е.Д., Мина М.В. Сравнительный анализ морфологических признаков ленков из разных
частей ареала рода Brachymystax (Sahnoniformes, Salmonidae) // Там же. 1980. Т. 59, № 1. С. 79-90.
Васильева ЕД., Позняк В.Г. Морфологическая характеристика предкавказской щиповки
Sabanejewia caucasica (Berg) (Cobitidae) // Вопр. ихтиологии. 1986. Т. 26, вып. 3. С. 402-409.
Введенская ТЛ. Возрастная структура, размеры и рост гольцов (р. Salvelinus) Кроноцкого озера
// Морфология, структура популяций и проблемы рационального использования лососевид-
ных рыб. Л.: Наука, 1983. С. 28-29.,
Величко А.М. Опыт получения икры сибирского чукучана на Северо-Западе СССР // Сб. науч,
тр. ГосНИОРХ. 1986. Вып. 247. С. 14-17.
Веригин Б.В. Проблема биологической мелиорации водоемов и охладителей ТЭС и их рыбохо-
зяйственного использования // Проблемы рыбохозяйственного использования растительно-
ядных рыб в водоемах СССР. Ашхабад. 1963. С. 62.
Веригин Б.В., Макеева А.П., Шубникова Н.Г. Случай естественной гибридизации толстолобиков
Hypophthalmichthys molitrix х Aristichthys nobilis (Pisces, Cyprinidae) // Зоол. журн. 1979. T. 58,
№ 2. С. 190-196.
217
Веригин Б.В., Негоновская И.Т. Растительноядные рыбы в естественных водоемах и водохра-
нилищах: (Результаты акклиматизации) // Сб. науч. тр. ГосНИОРХ. 1989. Вып. 301: Расти-
тельноядные рыбы в водоемах разного типа. С. 5-59.
Викторов Л.В. Краткая история изучения фауны, распространения и перспективы фаунистических
исследований водных позвоночных Тверского края. Тверь: Изд-во Твер. ун-та, 1992. С. 28-56.
Викторовский Р.М. Механизмы видообразования у гольцов. М.: Наука, 1978. 111 с.
Викторовский Р.М., Глубоковский М.К., Ермоленко Л.Н., Скопец М.Б. Гольцы рода Salvelinus
из оз. Эльгыгытгын (Центральная Чукотка) // Рыбы в экосистемах лососевых рек Дальне-
го Востока. Владивосток: ДВНЦ АН СССР, 1981. С. 67-78.
Викторовский Р.М., Макоедов А.Н., Шевчишин А.А. Хромосомные наборы ленка и сибирского
тайменя и дивергенция родов лососевых рыб // Цитология. 1985. Т. 27, № 6. С. 706-709.
Вилер А. Определитель рыб морских и пресных вод Североевропейского региона. М.: Лег. и
пищ. пром-сть, 1983. 432 с:
Виноградов В.К. Основные аспекты проблемы освоения представителей североамериканской
ихтиофауны в рыбном хозяйстве внутренних водоемов СНГ // Результаты работ по аккли-
матизации водных организмов. СПб., 1995. С. 85-89.
Виноградов В.К., Ерохина Л.В. Новые формы и новые объекты промышленного рыбоводства
// Тр. ВНИПРХ. 1973а. Т. 21. С. 3-6.
Виноградов В.К., Ерохина Л.В. Представители североамериканской ихтиофауны как объекты
рыбоводства и акклиматизации во внутренних водоемах СССР // Тр. ГосНИОРХ. 19736.
Т. 103. С. 220-225.
Вовк П. С. Биология дальневосточных растительноядных рыб и их хозяйственное использование
в водоемах Украины. Киев: Наук, думка, 1976. 245 с.
Волобуев В.В. Систематика и экология нейвы Salvelinus neiva Taranetz оз. Уегинского (бассейн
р. Охоты) // Вопр. ихтиологии. 1976. Т. 16, вып. 6. С. 989-999.
Волобуев ВВ. О карликовой форме нейвы Salvelinus neiva (Salmoniformes, Salmonidae) из
оз. Корраль (бассейн р. Охоты) // Зоол. журн. 1977. Т. 56, № 3. С. 405-411.
Волобуев В.В. О биологии кунджи Salvelinus leucomaenis (Pallas) материкового побережья Охот-
ского моря // Биология пресноводных рыб Дальнего Востока. Владивосток: ДВО АН СССР,
1987. С. 89-100.
Волобуев В.В., Максимов В.А., Рогатных А.Ю. Жилая кунджа Salvelinus leucomaenis (Pallas)
озерно-речной системы Чукча: (Материковое побережье Охотского моря) // Вопр. ихтиоло-
гии. 1985. Т. 25, вып. 4. С. 546-552.
Волобуев В.В., Никулин О.А. О биологии кунджи Salvelinus leucomaenis (Pallas) Мотыклейского
залива // Гидробиологические исследования внутренних водоемов Северо-Востока СССР.
Владивосток: ДВНЦ АН СССР, 1975. С. 354-375.
Вронский Б.Б. Материалы по биологии сомов р. Амур // Вопр. ихтиологии. 1960. Вып. 14.
С. 34-58.
Вронский Б.Б. Горчак Pseudoperilampus lighti amurensis subsp. nov. в бассейне Амура // Там же.
1967. Т. 7, вып. 1. С. 23-32.
Гавренков Ю.И., Иванков В.Н. Таксономический статус и биология дальневосточных краснопе-
рок рода Tribolodon Южного Приморья // Там же. 1979. Т. 19, вып. 6. С. 1014—1024.
Глубоковский М.К. Внутривидовая изменчивость и межвидовые отличия особенностей строения
черепа у гольцов {Salvelinus, Salmoniformes) // Фауна пресных вод Дальнего Востока. Влади-
восток: ДВНЦ АН СССР, 1980. С. 136-184.
Глубоковский М.К. Salvelinus albus sp. п. из бассейна р. Камчатки // Биология моря. 1977. № 4.
С. 48-56.
Глубоковский М.К. Эволюционная биология лососевых рыб: Автореф. дис. ... д-ра биол. наук.
Владивосток, 1990. 48 с.
Глубоковский М.К. Эволюционная биология лососевых рыб. М.: Наука, 1995. 345 с.
Глубоковский М.К., Фролов С.В., Ефремов В.В. и др. Филогенетические связи и систематика
гольцовых рыб оз. Эльгыгытгын // Природа впадины оз. Эльгыгытгын. Магадан, 1993.
С. 149-177.
Глубоковский М.К., Черешнев И.А. Спорные вопросы филогении гольцов рода Salvelinus Го-
ларктики. 1. Изучение проходных гольцов бассейна Восточно-Сибирского моря // Вопр. их-
тиологии. 1981. Т. 21, вып. 5. С. 771-786.
Глубоковский М.К, Шевчук С.А. Систематика гольцов оз. Кроноцкое (Камчатка) // Системати-
ка, биология и биотехника разведения лососевых рыб. СПб., 1994. С. 44-46.
Голубцов А.С. Эколого-генетический анализ популяций ротана Perccottus glenii Dyb. в естествен-
ном и приобретенном ареалах: Автореф. дис. ... канд. биол. наук. М., 1990. 24 с.
218
Гомелюк В.К. Репродуктивное поведение колюшковых: (Популяционный аспект). Автореф. дис.
... канд. биол. наук. М., 1978. 24 с.
Горбач Э.И. Биология белого амура в бассейне Амура. Автореф. дис. ... канд. биол. наук. М..
1973.24 с.
Горшков С.А., Дорофеева Е.А., Клюканов В.А., Куликова Н.И. Остеологические особенности
тихоокеанских лососей рода Oncorhynchus // Вопр. ихтиологии. 1979. Т. 19, вып. 6.
С. 963-982.
Гринюк И.Н. О миноге Petromyzon marinus L., выловленной у побережья Мурмана // Там же.
1970. Т. 10, вып. 1. С. 164-166.
Гриценко О.Ф. Рост, созревание и плодовитость гольца Salvelinus alpinus (L.) рек о-ва Сахалин //
Там же. 1971. Т. 11, вып. 4. С. 664—672.
Гриценко О.Ф. Систематика дальневосточных красноперок рода Tribolodon Sauvage, 1883
(Leuciscus brandti (Dybowski)) (Cyprinidae) Ц Там же. 1974. T. 14, вып. 5. С. 782-795.
Гриценко О.Ф. 1975. Систематика и происхождение сахалинских гольцов рода Salvelinus // Тр.
ВНИРО. 1975. Вып. 106. С. 141-160.
Гриценко О.Ф. Проходные рыбы острова Сахалин: Автореф. дис. ... д-ра биол. наук. М., 1990.
21 с.
Гриценко О.Ф., Иванков В.Н. Таксономический статус и биология дальневосточных краснопе-
рок рода Tribolodon Южного Приморья // Вопр. ихтиологии. 1979. Т. 19, вып. 6. С. 1014—1024.
Гриценко О.Ф., Костюнин Г.М. Амурский сиг Coregonus ussuriensis Berg и калуга Huso dauricus
(Georgi) в сахалинских водах // Там же. 1979. Т. 19, вып. 6. С. 1125-1128.
Гриценко О.Ф., Малкин Е.М., Чуриков А.А. Сахалинский таймень Huchoperryi р. Богатой (вос-
точное побережье Сахалина) // Изв. ТИНРО. 1974. Т. 93. С. 91-100.
Гриценко О.Ф., Пичугин М.Ю., Демьянов Т.В. Ихтиофауна пресных водоемов о-ва Шикотан
(южные Курильские о-ва) // Вопр. ихтиологии. 2002, Т. 42.
Гриценко О.Ф., Савваитова К.А., Груздева М.А., Кузищин К.В. О таксономическом положении
гольцов рода Salvelinus северных Курильских о-вов // Вопр. ихтиологии. 1998. Т. 38, вып. 2.
С. 189-198.
Гриценко О.Ф., Чуриков А.А. Биология гольцов рода Salvelinus и место их в ихтиоценозах зали-
вов северо-восточного Сахалина. I. Миграции, возраст, рост и созревание // Там же. 1976.
Т. 16. Вып. 6. С. 1012-1022.
Гриценко О.Ф., Чуриков А.А. Биология гольцов рода Salvelinus и место их в ихтиоценозах зали-
вов северо-восточного Сахалина. II. Питание // Там же. 1977. Т. 17, вып. 4. С. 668-676.
Гриценко О.Ф., Чуриков А.А. Систематика малоротых корюшек рода Hypomesus (Salmoniformes,
Osmeridae) азиатского побережья Тихого океана Ц Зоол. журн. 1983. Т. 62, № 4. С. 553-563.
Гриценко О.Ф., Чуриков А.А., Омельченко В.Т. и др. Генетическая дифференциация симпатрич-
ных популяций гольцов рода Salvelinus оз. Аччен (Чукотский полуостров) // Генетика. 1998.
Т. 34, № 1. С. 399-405.
Громов И.А. Новый вид косатки Mystus mica Gromov sp.n. (Pisces, Bagridae) в бассейне Амура //
Вопр. ихтиологии. 1970. Т. 10, вып. 3. С. 400-405.
Гудков П.К. О некоторых особенностях биологии гольца Таранца Salvelinus taranetzi из оз. Ач-
чен // Там же. 1994. Т. 34, вып. 1. С. 58-63.
Гудков П.К. Материалы по биологии кунджи Salvelinus leucomaenis бассейна Охотского моря //
Там же. 1991. Т. 31, вып. 6. С. 898-909.
Гудков П.К. Берингийская даллия Dallia pectoralis на Чукотке // Там же. 1998. Т. 38, вып. 2.
С. 252-256.
Гудков П.К., Скопец М.Б., Черешнев И.А. К биологии гольцов рода Salvelinus (Salmonidae) бас-
сейна Охотского моря. Характеристика биологических параметров симпатрических проход-
ных гольцов из рек залива Шелихова // Биология гольцов Дальнего Востока. Владивосток:
ДВО АН СССР, 1991. С. 21-36.
Гундризер А.Н. Рыбы Тувинской АССР: Автореф. дис. ... д-ра биол. наук. Томск, 1975. 34 с.
Гундризер А.Н., Иоганзен Б.Г., Кафанова^В.В., Кривощеков Г.М. Рыбы Телецкого озера. Ново-
сибирск: Наука, 1981. 159 с.
Гундризер А.Н., Иоганзен Б.Г., Кривощеков Г.М. Рыбы Западной Сибири. Томск: Изд-во Том.
ун-та, 1984. 120 с.
Гурова Л.А., Пастухов В.Д. Питание и пищевые взаимоотношения пелагических рыб и нерпы
Байкала. Новосибирск: Наука. 1974. 186 с
Дадикян М.Г. Рыбы Армении. Ереван: Изд-во АН АрмССР, 1968. 245 с.
Двинин П.А. Лососи Южного Сахалина // Изв. ТИНРО. 1952. Т. 37. С. 69-108.
Делямуре СЛ. Природа Крыма: Рыбы пресных водоемов. Симферополь: Крым, 1964. 69 с.
219
Дмитриева Е.Н. Морфо-биологический анализ бычка-песочника Neogobius fluviatilis (Pallas) в
течение эмбрионального периода развития // Морфо-биологические исследования развития
рыб. М.: Наука, 1968. С. 90-113.
Дмитриева Е.Н. Нерестилища судака Lucioperca lucioperca (L.) и берша L. volgensis в р. Урал //
Вопр. ихтиологии. 1973. Т. 13, вып. 5. С. 934-937.
Долгий В.Н. Ихтиофауна бассейнов Днестра и Прута. Кишинев: Штиинца, 1993. 319 с.
Дорофеева Е.А. Использование данных кариологии для решения вопросов систематики и фило-
гении лососевых рыб // Основы классификации и филогении лососевых рыб. Л.: Наука,
1977. С. 86-95.
Дорофеева Е.А. Таксономический статус, морфология и распространение атлантического лосо-
ся // Атлантический лосось. СПб.: Наука, 1998. С. 11-31.
Дорофеева Е.А., Горшков С.А., Романов Н.С. Остеологические особенности атлантических и
тихоокеанских лососей родов Salmo и Parasalmo // Тр. ГосНИОРХ. 1992. Вып. 304.
С. 194-205.
Дорошев С.И. Особенности икры, личинок и молоди полосатого окуня М. saxatilis (Walb.) в свя-
зи с проблемами его акклиматизации в СССР // Вопр. ихтиологии. 1970. Т. 10, вып. 2.
С. 341-359.
Дренски П. Рибите в България. С.: Изд. Бълг. АН, 1951. 270 с.
Дрягин П.А. О некоторых морфологических и биологических отличиях осетра, обитающего в
реках Якутии, от сибирского осетра Acipenser baeri Brandt // Зоол. журн. 1948. Т. 27, № 6.
С. 371-374.
Евланов И.А., Козловский С.В., Антонов П.И. Кадастр рыб Самарской области. Тольятти:
ИЭВБ РАН, 1998. 222 с.
Еловенко В.Н. Систематическое положение и географическое распространение рыб семейст-
ва Eleotridae (Gobioidei, Perciformes), интродуцированных в водоемы европейской части
СССР, Казахстана и Средней Азии// Зоол. журн. 1981. Т. 60, № 10. С. 1517-1522.
Еловенко В.Н. Морфо-экологическая характеристика ротана Perccottus glenii Dyb. в границах ес-
тественного ареала и за его пределами: Автореф. дис. ... канд. биол. наук. М., 1985. 24 с.
Емтыль М.Х. Рыбы Краснодарского края и Республики Адыгея. Краснодар: Кубан. гос. ун-т.
1997. 157 с.
Жерех A. aspius (L.): Характеристика вида в пределах ареала. Баку: Эльм, 1995. 249 с.
Жизнь животных. Т. 4; Рыбы. М.: Просвещение, 1983. 551 с.
Жуков П.И. Рыбы Белоруссии. Минск: Наука и техника, 1965. 412 с.
Зверева О.С., Кучина Е.С., Остроумов Н.А. Рыбы и рыбный промысел среднего и нижнего те-
чения Печоры. М.: Изд-во АН СССР, 1953. 230 с.
Зиновьев Е.А. Подкаменщик Средней Камы // Изв. Естеств.-науч. ин-та при Перм.
ун-те. 1963. Т. 14, вып. 6. С. 93-104.
Зубин А.А. Питание байкальских бентопелагических подкаменщик овых рыб Scorpaeniformes
(Cottoidei) //Вопр. ихтиологии, 1992. Т. 32, вып. 1. С. 147-151.
Зуянова О.В. Результаты пробной интродукции судака в оз. Воже // Сб. Науч. тр. ГосНИОРХ.
1989. Вып. 293. С. 80-83.
Зуянова О.В., Решетников Ю.С., Болотова НЛ„ Зуянов Е.А. Ряпушка озер Белое и Воже (Во-
логодская область) // Биология и биотехника разведения сиговых рыб: Матер. V Всерос. со-
вещания. СПб.: ГосНИОРХ, 1994. С. 62—64.
Зюганов В.В. Фауна СССР. Т. 5, вып. 1; Рыбы. Семейство колюшковых (Gasterosteidae) мировой
фауны. Л.: Наука, 1991. 261 с.
Иванова М.Н. Популяционная изменчивость пресноводных корюшек. Л., 1968. 145 с. (Тр. Ин-та
биологии внутр, вод; Вып. 15 (53)).
Илясова В.А. Гаметогенез и половые циклы веслоноса Polyodon spathula (Walbaum): Автореф.
дис. ... канд. биол. наук. М., 1989. 24 с.
Кагановский А.Г. Голец из бассейна Берингова моря //Вопр. ихтиологии. 1955. Вып. 3. С. 54-56.
Казакова И.И. Молодь бычков Северного Каспия//Тр. ВНИРО. 1951. Т. 18. С. 66-98.
Казанчеев Е.Н. Рыбы Каспийского моря. М.: Лег. и пищ. пром-сть, 1981. 167 с.
Калашников Ю.Е. Рыбы бассейна р. Витим. Новосибирск: Наука, 1978. 189 с.
Калинина Э.М. Размножение и развитие азово-черноморских бычков. Киев: Наук, думка, 1976.
118 с.
Карасев ГЛ. Рыбы Забайкалья. Новосибирск: Наука, 1987. 295 с.
Картавцев Ю.Ф., Глубоковский М.К., Черешнев И.А, Генетическая дифференциация и изменчи-
вость двух симпатричных видов гольцов (Salvelinus. Salmonidae) Чукотки // Генетика. 1983.
Т. 19. № 4. С. 584-592.
220
Кафанова В.В., Ширинский А.А. К биологии гольца Черского Salvelinus czerskii Drjagin озер бас-
сейна р. Алазеи // Лососевидные рыбы: (Морфология, систематика и экология). Л.: ЗИН
АН СССР, 1976. С. 45-46.
Кириллов Ф.Н. Рыбы р. Идигирки //Изв. ГосНИОРХ. 1955. Т. 35. С. 141-167.
Кириллов Ф.Н. Рыбы Якутии. М.: Наука, 1972. 360 с.
Кирпичников В.С. Генетика и селекция рыб. М.: Наука, 1987. 518 с.
Кифа М.И. Морфология двух форм ленка (род Brachymystax, семейство Salmonidae) из бассей-
на Амура и их систематическое положение // Зоогеография и систематика рыб. Л.: ЗИН АН
СССР, 1976. С. 142-156.
Клюканов В.А. Новые данные о распространении малоротых корюшек в водах СССР // Докл.
АН СССР. 1966. Т. 166, № 4. С. 990-991.
Клюканов В.А. Морфологические основы систематики корюшек рода Osmerus (Osmeridae) //
Зоол. журн. 1969. Т. 48, № 1. С. 99-109.
Клюканов В.А Систематика малоротых корюшек рода Hypomesus (Osmeridae) // Там же. 1970.
Т. 49, № 10. С. 1534-1542.
Клюканов В.А Систематика и родственные отношения родов Osmerus и Hypomesus и их рассе-
ление // Там же. 1975. Т. 54, № 4. С. 590-596.
Клюканов В.А Происхождение, расселение и эволюция корюшковых (Osmeridae) // Основы
классификации и филогении лососевидных рыб: Сб. науч. тр. Л.: ЗИН АН СССР, 1977.
С. 13-27.
Коблицкая А.Ф. Новые данные о биологии бычка-бубыря Pomatoschistus caucasicus (Kawrajsky
in Berg, 1916) из авандельты p. Волги //Вопр. ихтиологии. 1961. Т. 1, вып. 2. С. 253-261.
Коблицкая А.Ф. Определитель молоди пресноводных рыб. М.: Лег. и пищ. пром-сть, 1981. 167 с.
Ковтун И.Ф. Материалы по экологии Pomatoschistus microps leopardinus (Nordmann) Азовского
моря // Вопр. ихтиологии. 1976. Т. 16, вып. 1. С. 33-40.
Кожова О.М., Сиделев Г.Н., Резников Н.С. Наблюдения планктонных и нектобентических сооб-
ществ // Геолого-геоморфологические и подводные исследования оз. Байкал. М.: Наука,
1979. С. 87-91.
Колюшев А.И. Гольцы и палии (род Salvelinus) крупных озер Кольского полуострова: Автореф.
дис. ... канд. биол. наук. Л., 1969. 22 с.
Колюшев А.И. Некоторые остеологические признаки гольцов и палий (род Salvelinus) Северо-
Запада СССР в связи с вопросом их систематического положения // Вопр. ихтиологии. 1971.
Т. 11, вып. 4. С. 565-574.
Константинов К.Г. Нерест сома // Зоол. журн. 1941. Т. 20, № 4/5. С. 583-586.
Константинов К.Г. Сравнительный анализ морфологии и биологии окуня, судака и берша на
разных этапах развития // Работы по этапности развития костистых рыб. М., 1957.
С. 181-236. (Тр. ИМЖ АН СССР; Вып. 16).
Константинов К.Г. Возраст и темп роста амурского верхогляда Erythroculter erythropterus
(Basilewsky) // Тр. Амур, ихтиол, экспедиции, 1945-1949 гг. М.: Изд-во МГУ, 1958. Т. 4.
С. 103-114.
Коробочкина З.С. Основные этапы развития промысла осетровых в Каспийском море // Тр.
ВНИРО. 1964. Т. 52. С. 59-86.
Королев В.В. Влияние экстремальных погодных условий на биоту рек бассейна Верхней Печо-
ры //Экология. 1995, №2. С. 159-161.
Королев В.В. Изменчивость обыкновенного подкаменщика бассейна Печоры // Вопр. ихтиоло-
гии. 2002. Т. 42.
Коряков Е.А. Биология, ресурсы и хозяйственное значение голомянок // Тр. Лимнол. ин-та СО
АН СССР. 1964. Т. 2 (22). Ч. 3. С. 3-75.
Коряков Е.А. Пелагические бычковые Байкала. М.: Наука, 1972. 155 с.
Коряков Е.А., Сиделев Г.Н. Бычки-подкаменщики (Cottidae) из оз. Агата (плато Путорана) //
Вопр. ихтиологии. 1976. Т. 16, вып. 3. С. 553-555.
Котляр А.Н. Калинка Leuciscus borysthepicus в бассейне р. Снов. // Там же. 1991. Т. 31, вып. 5.
С. 862-864.
Кохненко С.В. Американский угорь в южных районах СССР // Рыбоводство и рыболовство.
1975. №2. С. 15.
Кохненко С.В.. Безденежных В .А., Горовая СЛ. Эколого-физиологическая пластичность у евро-
пейского угря Anguilla anguilla L. Минск: Наука и техника, 1977. 191 с.
Кошелев Б.В., Рябов И.Н., Зимин ВЛ. Ихтиологические исследования Кодорской губы Финско-
го залива в 1979-1980 гг. // Экологические аспекты исследований водоемов-охладителей
АЭС: Материалы совещ., март, 1983. М., 1986. С. 43-53.
221
Краев Н.П. Тиляпия в прудах-охладителях ГРЭС // Рыб. хоз-во. 1966. № 4. С. 23.
Красикова В.А. Тугун Coregonus tugun (Pallas) из норильских озер // Вопр. ихтиологии. 1967. Т. 7,
вып. 4. С. 601-608.
Красная книга Карелии. Петрозаводск: Карелия, 1995. 286 с.
Красная книга Российской Федерации: (Животные). М.: ACT, Астрель. 2001. 860 с.
Красная книга Ямало-Ненецкого автономного округа. Екатеринбург: Изд-во Урал, ун-та, 1997.
237 с.
Кривцов В.Ф., Калмыков Л.В., Калмыкова Б.В., Бурдонова Е.В. Основные морфо-экологиче-
ские признаки буффало // Сб. науч. тр. ВНИПРХ. 1983. Вып. 38. С. 11-23.
Крогиус Ф.В., Крохин Е.М. Об урожайности молоди красной // Изв. ТИНРО. 1948. Т. 28. С. 3-27.
Крохин Е.М. Оценка биомассы и численности трехиглой колюшки в оз. Дальнем на основании по-
требления корма рыбами-планктофагами Ц Вопр. ихтиологии. 1970. Т. 10, вып. 4. С. 637-642.
Крыжановский С.Г. Эколого-морфологические закономерности развития карповых, вьюновых
и сомовых рыб И Тр. Ин-та морфологии животных АН СССР. 1949. Вып. 1. С. 5-332.
Крыжановский С.Г. Особенности зрелых яиц костистых рыб // Вопр. ихтиологии. 1953. Вып. 1.
С. 37-62.
Крыжановский С.Г., Смирнов А.И., Соин С.Г. Материалы по развитию рыб р. Амура // Тр.
Амур, ихтиол, экспедиции, 1945-1949 гг. М.: Изд-во МГКУ, 1951. Т. 2. С. 5-222.
Крыхтин МЛ. Материалы о речном периоде жизни симы // Изв. ТИНРО. 1962. Т. 48. С. 84-132.
Крыхтин МЛ. Современное состояние и перспективы развития осетрового хозяйства в бассей-
не Амура И Биологические основы развития осетрового хозяйства в водоемах СССР. М.:
Наука. 1979. С. 68-74.
Кудерский Л.А. Условия существования и перспективы расселения судака водоемов Карелии //
Рыб. хоз-во Карелии. 1964. Вып. 8. С. 154—209.
Кудерский Л.А. Самоакклиматизация американского канального сомика в Черепетском водо-
хранилище // Сб. науч. тр. ГосНИОРХ. 1982. Вып. 187. С. 219-232.
КудерскийЛ.А. Осетровые в бассейне Онежского и Ладожского озер // Там же. 1983. Вып. 205.
С. 128-149.
Кудерский Л.А. Распространение сибирской миноги в водоемах Европейского Севера // Совре-
менные проблемы систематики рыб. СПб.: ЗИН РАН, 1998. С. 33-34.
Кудерский Л.А., Мельникова М.Н. Сибирская минога в бассейне Северной Двины // Рыбохоз.
изучение внутренних водоемов. 1970, № 4. С. 16-17.
Кузнецов В.В. Экологические взаимоотношения популяций промысловых рыб и проблемы их хозяй-
ственного использования: Автореф. дис.... д-ра биол. наук. М., 1998. 73 с.
Куликова Н.И., Фандеева В.Н. О порционности икрометания азовского бычка-кругляка (Gobius
melanostomus Pallas) // Тр. ВНИРО. 1975. Т. 196, С. 18-27.
Куренков С.И. Красная оз. Сарайного // Изв. ТИНРО. 1978. Т. 78. С. 49-60.
Лагунов И.И., Константинов КГ. Балтийский осетр в Белом море // Природа. 1954. № 3.
С. 113-114.
Лебедев В.Д. Материалы по биологии размножения амурского пескаря Pseudogobio rivularis
(Bas.) Pisces, Cyprinidae Ц Докл. АН СССР. 1948. Т. 59, № 3. С. 597.
Лебедев ВД. Пресноводная четвертичная ихтиофауна европейской части СССР. М.: Изд-во
МГУ, 1960. 401 с.
Лебедев В.Д., Спановская В.Д., Савваитова К.А. и др. Рыбы СССР. М.: Мысль, 1969. 447 с.
Левин Б.А. Находка украинской миноги Eudontomyzon mariae (Petromyzontidae) в Волжском бас-
сейне // Вопр. ихтиологии. 2001. Т. 41, вып. 6. С. 849-850.
Летичевский М.А. Опыт определения численности белорыбицы и эффективность ее заводско-
го разведения в условиях дельты и Нижней Волги // Там же. 1975. Т. 15, вып. 4, С. 632-634.
Линдберг Г.У., Красюкова З.В. Рыбы Японского моря и сопредельных частей Охотского и Жел-
того морей Л.: Наука, 1975. Ч. 4. 463 с.
Линдберг Г.У., Красюкова З.В. Рыбы Японского моря и сопредельных частей Охотского и Жел-
того морей. Л.: Наука, 1987. Ч. 5. 525 с.
Линдберг Г.У., Легеза М.И. Рыбы Японского моря и сопредельных частей Охотского и Желтого
морей. М.; Л.: Наука, 1965. Ч. 2. 391 с.
Лукин А. А. Интродукция радужной форели Parasalmo ту kiss в озеро Имандра (Кольский полу-
остров) Ц Вопр. ихтиологии. 1998. Т. 38, вып. 4. С. 485—491.
Лукьяненко В.И., Умеров Ж.Г., Каратаева Б.Б. Южнокаспийский осетр - самостоятельный вид
рода Acipenser // Изв. АН СССР. Сер. биол. 1974. № 5. С. 736-739.
Макеева А.П. Эмбрионально-личиночный период развития амурского плоскоголового жереха //
Вопр. ихтиологии. 1960. Вып. 16. С. 154—163.
222
Макеева А.П. Особенности развития некоторых сорных рыб, завезенных в Среднюю Азию //
Биологические основы рыбн. хоз-ва Средней Азии и Казахстана: Тез. докл. конф. Ташкент;
Фергана: ИЗИП АН УзбССР, 1972. С. 211-212.
Макеева АЛ. Особенности размножения, созревания и развития белого амура, белого и пестро-
го толстолобиков // Зоология позвоночных. М.: ВИНИТИ. 1974. Т. 5: Биология, разведение
и использование растительноядных рыб. С. 11-60.
Макеева А.П., Павлов Д.С. Ихтиопланктон пресных вод России: (Атлас). М.: Изд-во МГУ, 1998.
215 с.
Макеева А.П., Рябов И.Н. Размножение и развитие троегуба Opsariichthys uncirostris (Temminck
et Schlegel) и его положение в системе карповых рыб // Вопр. ихтиологии. 1973. Т. 13, вып. 3.
С. 471-478.
Макоедов А.Н. Кариология, биохимическая генетика и популяционная фенетика лососевых рыб
Сибири и Дальнего Востока. М.: УМК “Психология”, 1999. 291 с.
Максимов В.А., Долгов В.А. Вспышка численности трехиглой колюшки на Камчатке // Рыб.
хоз-во. 1983. № 1. С. 37-38.
Малинин Л.К. Поведение налима // Природа. 1971. № 8. С. 77-79.
Мамонтов А.Н., Яхненко В.М. Кариотипы шести видов байкальских бычков // Эколого-физио-
логические исследования рыб Байкала. Иркутск, 1981. С. 8-14.
Марти В.Ю. Систематика и биология русского осетра кавказского побережья Черного моря //
Зоол. журн. 1940. Т. 19, № 6. С. 865-872.
Мартино К.В. Естественное размножение белого амура в водоемах Нижней Волги // Гидроби-
ол. журн. 1974. Т. 10, вып. 1. С. 91-93.
Медников Б.М., Шубина Е.А., Мельникова М.Н. и др. Проблема родового статуса тихоокеанских
лососей и форелей: Геносистематический анализ // Вопр. ихтиологии. 1999. Т. 39, вып. 1.
С. 14-21.
Международный кодекс зоологической номенклатуры. Л.: Наука. 1988. 202 с.
Меньшиков М.И. К систематике сибирской стерляди Acipenser ruthenus marsiglii Brandt // Изв.
биол. НИИ при Перм. ун-те. 1937. Т. И, вып. 3/4. С. 55-75.
Меньшиков М.И. О географической изменчивости сибирского осетра Acipenser baeri Brandt //
Докл. АН СССР. 1947. Т. 55, № 4. С. 371-374.
Меньшиков М.И. Некоторые закономерности возрастной и географической изменчивости
рыб //Тр. Карело-Фин. отделения ВНИОРХ. 1951. Т. 3. С. 292-306.
Мина М.В. Микроэволюция рыб. М.: Наука, 1986. 207 с.
Миронова Н.В. О биологии Tilapia mossambica Peters в природных и лабораторных условиях //
Вопр. ихтиологии. 1969. Т. 9, вып. 4. С. 628-639.
Митрофанов В.П. К систематике ленка из оз. Марка-Куль // Сб. работ по ихтиологии и гидро-
биологии Ин-та зоологии АН КазССР. 1959. Вып. 2. С. 267-275.
Мишарин К.И. К биологии икры и молоди некоторых промысловых рыб оз. Байкал и р. Анга-
ры // Тр. Иркут, гос. ун-та, 1942. Т. 2, вып. 3. С. 89-118.
Михин В.С. Рыбы и рыбный промысел р. Хатанги и Хатангского залива // Тр. НИИ земледелия,
животноводства и промыслового хоз-ва. Сер. пром, хоз-во. Л., М.: Изд-во Главсевморпути.
1941. С. 37-41.
Михин В.С. Рыбы оз. Таймыр и Таймырской губы // Тр. ГосНИОРХ. 1955. Т. 35. С. 35—43.
Москаленко Б.К. Биологические основы эксплуатации и воспроизводства сиговых рыб Обского
бассейна. Тюмень: Кн. изд-во, 1958. 245 с.
Москалькова К.И. Некоторые особенности роста и размножения бычка Книпови-
ча в Таганрогском заливе // Тр. Азов. НИИ рыб. хоз-ва. 1960. Т. 1, вып. 1. С. 441-446.
Москалькова К.И. О питании и распределении бычка Книповича Knipowitschia longicaudata
(Kessler) в Таганрогском заливе //Вопр. ихтиологии. 1962. Т. 2, вып. 3 (24). С. 492-505.
Москалькова К.И. Развитие азовского бычка-кругляка Neogobius melanostomus (Pallas) в связи с
особенностями его существования в индивидуальном и историческом развитии // Эколого-
морфологические и эколого-физиолргические исследования развития рыб. М.: Наука, 1978.
С. 72-88.
Москалькова К.И. Экологические и морфо-физиологические предпосылки к расширению ареа-
ла у бычка-кругляка Neogobius melanostomus в условиях антропогенного загрязнения водо-
емов Ц Вопр. ихтиологии. 1996. Т. 36, вып. 5. С. 615-621.
Москул Г.А. Рыбохозяйственное освоение Краснодарского водохранилища. СПб.: ГосНИОРХ,
1994. 136 с.
Мухамедова А.Ф., Аксенов С.В., Шаповалов Н.П. Черный амур в Цимлянском водохранилище //
Сб. науч. тр. ГосНИОРХ. 1989. Вып. 200. С. 149456.
223
Мухачева ВЛ. К биологии амурского чебачка (Pseudorasbora parva Schlegel) // Труды Амур, их-
тиол. экспедиции, 1945-1949 гг. М.: МГУ, 1950. Т. 1. С. 365-374.
Мухомедияров Ф.Б. Биология ледовитоморской рогатки Myoxocephalus quadricornis labrado-
ricus (Girard) Онежского залива Белого моря // Вопр. ихтиологии. 1967. Т. 7, вып. 4.
С. 609-617.
Назаркин М.В. Пресноводные рыбы из позднечетвертичных отложений побережья Восточно-
Сибирского моря Ц Там же. 1992. Т. 32, вып. 5. С. 48-56.
Насека А.М., Богуцкая Н.Г. Новый вид пескаря Romanogobiopentatrichus (Gobioninae, Cyprinidae)
из бассейна р. Кубань // Там же. 1998. Т. 38, вып. 2. С. 173-181.
Негоновская И.Т. О результатах и перспективах вселения растительноядных рыб в естествен-
ные водоемы и водохранилища СССР // Там же. 1980. Т. 20, вып. 4. С. 702-712.
Неелов А.В. Сейсмосенсорная система и классификация керчаковых рыб (Cottidae:
Myoxocephalinae, Artediellinae). Л.: Наука, 1979. 208 с.
Неелов А.В. Природа Ленинградской области: Рыбы. Л.: Лениздат, 1987. 157 с.
Неелов А.В., Сиделева В.Г. О параллелизме в строении сейсмосенсорной системы у двух эколо-
гически различных представителей бычков (Cottoidei) // Журн. эволюц. биохимии и физио-
логии. 1979. Т. 15. С. 433-436.
Никольский Г.В. Рыбы Таджикистана. М., Л.: Изд-во АН СССР, 1938. 226 с.
Никольский Г.В. Рыбы бассейна Амура. М.: Изд-во АН СССР, 1956. 551 с.
Новоселов АЛ. Современное состояние рыбной части сообщества в водоемах европейского Се-
веро-Востока Росиии: Автореф. дис. ... д-ра биол. наук. М„ 2000. 50 с.
Новоселов А.П., Решетников Ю.С. Пелядь в новых местах обитания // Биология сиговых рыб.
Мл Наука, 1988. С. 78-114.
Озера Карелии. Петрозаводск: Карел, кн. изд-во, 1959. 618 с.
Олейник А.Г., Скурихина Л.А. Родственные взаимоотношения проходных гольцов рода
Salvelinus (Salmonidae, Salmoniformes) по данным рестриктазного анализа ядерной ДНК // Ге-
нетика. 1999. Т. 25, № 9. С. 1258-1268.
Олейник А.Г., Скурихина Л.А., Брыков В.А. и др. Дивергенция митохондриальной ДНК двух
подвидов мальмы Salvelinus malma (Salmonidae, Pisces) // Докл. РАН, 2002.
Омельченко ВТ., Политое Д.В., Салменкова Е.А. и др. Генетическая дифференциация симпат-
ричных гольцов рода Salvelinus р. Ямы // Генетика. 1996. Т. 32, № 11. С. 1562-1568.
Омельченко В.Т.. Салменкова Е.А. Генетические различия гольцов арктической группы
(Salvelinus alpinus L., Salvelinus taranetzi Kaganowsky) и тихоокеанской мальмы (Salvelinus
malma Walbaum) // Там же. 1998. T. 34, № 11. С. 1518-1522.
Орлова Э.А., Попова О.А. Особенности питания хищных рыб - сома и щуки - в дельте Волги по-
сле зарегулирования стока реки // Вопр. ихтиологии. 1976. Т. 16, вып. 1. С. 84—98.
Орлова Э.А., Попова О.А. Возрастные изменения в питании сома и щуки в авандельте Волги //
Там же. 1987. Т. 27, вып. 1. С. 140-148.
Осиное А.Г., Павлов С.Д. Аллозимная изменчивость и генетическая дивергенция популяций арк-
тического гольца и мальмы (Salvelinus alpinus - S. malma complex) // Там же. 1998. T. 38,
вып. 1. С. 47-61.
Остроумов В А. Этолого-физиологическая характеристика половых феромонов байкальских быч-
ков-подкаменщиков: Автореф. дис.... канд.биол. наук. Иркутск, 1987. 18 с.
Павлов Д.С., Савваитова К.А., Кузищин К.В. и др. Тихоокеанские благородные лососи и форе-
ли Азии. М.: Научный мир, 2001. 200 с.
Павлов Д.С., Савваитова К.А., Соколов Л.И., Алексеев С.С. Редкие и исчезающие животные:
Рыбы. М.: Высш, шк., 1994. 334 с.
Павловская Л.П. Аральский усач. Ташкент: ФАН, 1976. 148 с.
Парпура И.З. Биология сахалинского тайменя Parahucho perryi и гольцов рода Salvelinus в водах
Северного Приморья: Автореф. дис. ... канд. биол. наук. Владивосток, 1991. 22 с.
Пенязь В.С., Шевцова Т.М., Нехаева Т.П. Биология рыб водоемов Белорусского Полесья.
Минск: Наука и техника, 1973. 237 с.
Первозванский В.Я. Сибирский чукучан - новый компонент в ихтиофауне Ладожского озера //
Вопр. ихтиологии. 1999. Т. 39, вып. 4. С. 567-568.
Песериди Н.Е. Сезонная динамика хода осетровых в низовьях р. Урал //Тр. ЦНИОРХ. 1971. Т. 3.
С. 355-358.
Пинчук В.И. Новые данные о бычках Rhinogobiuspflaumi (Bleeker) и Rhinogobius similis similis Gill
в Приморском крае //Вопр. ихтиологии. 1974. Т. 14, вып. 3. С. 431435.
Пинчук В.И. Систематика бычков родов Gobius Linne (отечественные виды), Neogobius Illjin и
Mesogobius Bleeker // Там же. 1976. Т. 16, вып. 4. С. 600-609.
224
Пинчук В.И. Замечания и дополнения к семейству бычковых Gobiidae в книге Г.У. Линдберга,
З.В. Красюковой “Рыбы Японского моря и сопредельных частей Охотского и Желтого мо-
рей”, ч. 4, 1975 с описанием нового вида Chaenogobius taranetzi sp, nov. // Там же. 1978. Т. 18,
вып. 1. С. 3-18.
Пинчук В.И. Новый вид бычка Книповича Knipowitschia georghievi Pinchuk sp. n. (Pisces, Gobiidae)
из западной части Черного моря // Зоол. журн. 1978. Т. 57, № 5. С. 796-799.
Пинчук В.И. О фенетических и филетических отношениях малоротых бычков дальневосточных
родов Chaenogobius и Rhodoniichthys (Perciformes, Gobiidae) // Там же. 1981. Т. 60, № 10.
С. 1508-1516.
Пинчук В.И. Определительная таблица видов рода Chaenogobius Gill и двух близких монотипи-
ческих родов Rhodoniichthys Takagi и Paleatogobius Takagi (Gobiidae) // Вопр. ихтиологии.
1984. Т. 24, вып. 4. С. 545-551.
Пинчук В.И., Рагимов Д.Б. Новый вид пуголовки - рода Benthophilus svetovidovi Pinchuk et
Rahimov, sp. n. (Pisces, Gobiidae) из Каспийского моря и определительная таблица видов ро-
да Benthophilus Ц Зоол. журн. 1979. Т. 58, № 4. С. 515-519.
Пинчук В.И., Савчук М.Я. О видовом составе бычковых рыб рода Pomatoschistus (Gobiidae) мо-
рей СССР // Вопр. ихтиологии. 1982. Т. 22, вып. 1. С. 9-14.
Позняк В.Г. Животный мир Калмыкии. Рыбы. Элиста: Калм. кн. изд-во, 1987. 58 с.
Покровский В.В. Материалы по исследованию внутривидовой изменчивости окуня // Тр. Каре-
ло-Фин. отделения ВНИОРХ. 1951. Т. 3. С. 95-149.
Попова О.А. Некоторые особенности экологии щуки и окуня в дельте Волги // Вопр. ихтиоло-
гии. 1960. Вып. 15. С. 55-70.
Попова О А. Экология щуки и окуня в дельте Волги // Питание хищных рыб и их взаимоотно-
шения с кормовыми организмами. М.: Наука, 1965. С. 91-170.
Попова О.А. Биологические показатели щуки и окуня в водоемах с различным гидрологическим
режимом и кормностью // Закономерности роста и созревания рыб. М.: Наука, 1971.
С. 102-152.
Попова О.А. Особенности питания хищных рыб Псковско-Чудского водоема в период резкого
падения численности снетка // Основы биопродуктивности внутренних водоемов Прибалти-
ки. Вильнюс, 1975.С. 90-93.
Попова О.А. Роль хищных рыб в экосистемах // Изменчивость рыб пресноводных экосистем. М.:
Наука, 1979а. С. 13^17.
Попова О.А. Питание и пищевые взаимоотношения судака, окуня и ерша в водоемах разных ши-
рот Ц Изменчивость рыб пресноводных экосистем. М.: Наука, 19796. С. 93-112.
Попова О.А. Питание хищных рыб Сямозера после вселения корюшки // Изменение структуры
рыбного населения эвтрофируемого озера. М.: Наука, 1982. С. 106-160.
Попова О.А., Андреев ВЛ., Макарова Н.П., Решетников Ю.С. Изменчивость морфологических
показателей речного окуня Perea fluviatilis L. в пределах ареала // Биология речного окуня.
М.: Наука, 1993. С. 4-55.
Потапова О.И. Крупная ряпушка Coregonus albula L. Л.: Наука, 1978. 132 с.
Правдин И.Ф. Сиги водоемов Карело-Финской ССР. М.; Л.: Изд-во АН СССР, 1954. 324 с.
Пробатов А.Н. Хариус р. Кары // Изв. Биол. ин-та при Перм. гос. ун-те. 1936. Т. 10, вып. 9/10.
С. 393-401.
Промысловые рыбы СССР. М.: Пищепромиздат, 1949. С. 407-409.
Прохоров В.Г., Грачев Л.Е. О нахождении трехзубой миноги Entosphenus tridentatus (Gairdner) в
западной части Берингова моря Ц Вопр. ихтиологии. 1965. Т. 5, вып. 4. С. 723-726.
Раб П. Кариотип европейского налима Lota lota (L.) (Gadidae) // Там же. 1986. Т. 26, вып. 1.
С. 161-164.
Рагимов Д.Б. Видовой состав, биология и запасы бычковых западного побережья Среднего и
Южного Каспия: Автореф. дис.... канд. биол. наук. Баку, 1966. 19 с.
Рагимов Д.Б. Биология размножения бычков у западного побережья Среднего и Южного Кас-
пия // Изв. АН АзССР. Сер. биол. нау,к. 1968. Xs 2. С. 51-57.
Рагимов Д.Б. Benthophilus mahmudbejovi Rahimov sp.n. (Pisces, Gobiidae) из Каспийского моря //
Зоол. журн. 1976. Т. 60, № 8. С. 1196-1200.
Рагимов Д.Б. О систематическом положении некоторых видов рода Benthophilus (семейство
Gobiidae) Каспийского и Азовского морей // Вопр. ихтиологии. 1978. Т. 18, вып. 5. С. 791-798.
Рагимов Д.Б. Распространение и численность бычковых (Gobiidae) в Северном Каспии // Там же.
1981. Т. 21, вып. 2. С. 223-231.
Рагимов Д.Б. Новые подвиды каспийских пуголовок (Gobiidae, Benthophilus') // Зоол. журн. 1982.
Т. 61, № 1. С. 47-55.
15 Атлас.. , том 2
225
Рагимов Д.Б. Материалы по размножению некоторых каспийских видов пуголовок рода
Benthophilus Eichwald (Gobiidae) //Вопр. ихтиологии. 1985а. Т. 25, вып. 2. С. 242-247.
Рагимов Д.Б. О некоторых каспийских видах пуголовок рода Benthophilus (Gobiidae) // Там же.
19856. Т. 25, вып. 6. С. 917-924.
Рагимов Д.Б. Бычковые рыбы Каспийского моря: (Систематика, экология, значение): Автореф.
дис. ... д-ра биол. наук. Л., 1988. 32 с.
Разнообразие рыб Таймыра. М.: Наука, 1999. 207 с.
Редкие животные Монголии: (Позвоночные). М.: ИПЭЭ РАН, 1996. 184 с.
Решетников Ю.С. Об изменчивости сигов // Зоол. журн. 1963. Т. 42, № 8. С. 1187-
1199.
Решетников Ю. С. Питание разных внутривидовых форм сига из ряда озер Лапландского запо-
ведника // Вопр. ихтиологии, 1964. Т. 4, вып. 4. С. 679-694.
Решетников Ю.С. Особенности роста и созревания сигов в водоемах Севера // Закономерности
роста и динамики численности рыб Белого моря и его бассейна. М.: Наука, 1966. С. 93-155.
Решетников Ю.С. О периодичности размножения у сигов // Вопр. ихтиологии. 1967. Т. 7, вып. 6.
С. 1019-1031.
Решетников Ю.С. О систематическом положении сиговых рыб // Зоол. журн. 1975. Т. 54, №11.
С. 1656-1671.
Решетников Ю. С. Омуль р. Пенжина // Систематика и экология рыб континентальных водоемов
Дальнего Востока. Владивосток: ДВНЦ АН СССР, 1979а. С. 99-105.
Решетников Ю.С. О связи сиговых рыб Сибири и Северной Америки // Изменчивость рыб пре-
сноводных экосистем. М.: Наука, 19796.С. 48-73.
Решетников Ю.С. Экология и систематика сиговых рыб. М.: Наука, 1980. 301 с.
Решетников Ю.С. Биологическое разнообразие и изменение экосистем // Биоразнообразие:
Степень таксономической изученности. М.: Наука, 1994. С. 77-85.
Решетников Ю.С. Современные проблемы изучения сиговых рыб // Вопр. ихтиологии. 1995.
Т. 35, вып. 2. С. 156-174.
Решетников Ю.С. Состояние биоразнообразия и функционирование водных экосистем // Изуче-
ние и охрана разнообразия фауны, флоры и основных экосистем Евразии. М.: ИПЭЭ РАН,
2000. С. 264-270.
Решетников Ю.С., Богуцкая Н.Г., Васильева Е.Д. и др. Список рыбообразных и рыб пресных
вод России Ц Вопр. ихтиологии. 1997. Т. 37, вып. 6. С. 723-771.
Решетников Ю.С., Мухачев И.С., Болотова НЛ. и др. Пелядь Coregonus peled (Gmelin, 1788)
(Pisces: Coregonidae). M.: Наука, 1989. 302 c.
Решетников Ю.С., Новиков A.C., Слугин И.В. и др. Валек Prosopium cylindraceum (Pallas et
Pennant) Чукотки // Вопр. ихтиологии. 1975. Т. 15, вып. 5. С. 788-804.
Решетников Ю.С., Попова О.А., Стерлигова О.П. и др. Изменение структуры рыбного населе-
ния эвтрофируемого водоема. М.: Наука, 1982. 247 с.
Решетников Ю.С., Слугин И.В., Штундюк Ю.В. и др. Систематика и экология лососевидных
рыб рек Амгуэма, Анадырь и Пенжина // Экология и систематика лососевидных рыб. Л.:
ЗИН АН СССР, 1976. С. 82-87.
Решетников Ю.С., Шакирова Ф.М. Зоогеографический анализ ихтиофауны Средней Азии по
спискам пресноводных рыб // Вопр. ихтиологии. 1993. Т. 33, вып. 1. С. 37-45.
Романов В.И. Морфологические особенности сиговых рыб Хантайских озер и Хантайского во-
дохранилища в процессе его формирования: Автореф. дис. ... канд. биол. наук. Иркутск,
1985. 18 с.
Романов В.И. К вопросу о популяционной структуре гольцов (род Salvelinus) оз. Кета // Задачи и
проблемы развития рыбного хозяйства внутренних водоемов Сибири. Томск, 1996. С. 97-98.
Рубан Г.И. О структуре вида сибирского осетра Acipenser baerii Brandt // Вопр. ихтиологии. 1998.
Т. 38, вып. 3. С. 307-327.
Рубан Г.И. Сибирский осетр Acipenser baerii Brandt: (Структура вида и экология). М.: ГЕОС,
1999. 235 с.
Рубан Г.И., Акимова Н.В. Особенности экологии сибирского осетра Acipenser baerii Brandt р. Ин-
дигирки//Вопр. ихтиологии. 1991. Т. 31, вып. 4. С. 596-605.
Рыбы Казахстана. Карповые. Алма-Ата: Наука КазССР, 1987. Т. 2. 198 с.
Рыбы Казахстана. Карповые. Алма-Ата: Наука КазССР, 1988. Т. 3. 304 с.
Рыбы Казахстана. Вьюновые и др. Алма-Ата: Наука КазССР, 1989. Т. 4. 311 с.
Рыбы Казахстана. Акклиматизация, промысел. Алма-Ата: Гылым, 1992. Т. 5. 461 с.
Рыбы Монгольской Народной Республики. М.: Наука, 1983. 276 с.
Рыбы Подмосковья. М.: Наука, 1988. 141 с.
996
Сабанеев Л.П. Рыбы России. М., 1911. 452 с.
Сабанеев Л П. Жизнь и ловля пресноводных рыб. Киев: Госсельхозиздат УССР, 1959.
667 с.
Савваитова К.А. Кунджа - Salvelinus leucomaenis (Pallas) озер Южного Сахалина // Озера Южно-
го Сахалина. М.: Изд-во МГУ, 1964. С. 154—167.
Савваитова К.А. Кунджа - Salvelinus leucomaenis (Pallas) некоторых озер о-ва Кунашир из груп-
пы Курильских островов // Вести. МГУ. 1966. № 4. С. 35-42.
Савваитова К.А. Арктические гольцы: (Структура популяционных систем, перспективы хозяй-
ственного использования). М.: Агропромиздат. 1989. 224 с.
Савваитова К.А., Груздева М.А., Кузищин К.В., Стыгар В.М. Озерные гольцы рода Salvelinus
(Salmonidae) о-ва Парамушир, Курильские острова // Вопр. ихтиологии. 2001. Т. 41, вып. 1.
С. 5-18.
Савваитова К.А., Максимов В.А. О нересте тихоокеанских миног род Lampetra в связи с проб-
лемой таксономического статуса мелких форм // Вопр. ихтиологии. 1978. Т. 18, вып. 4.
С. 636-641.
Савваитова К.А., Максимов В.А. О симпатрических формах гольцов рода Salvelinus (Salmonidae)
из Пегтымельских озер Чукотки Ц Биология гольцов Дальнего Востока. Владивосток: ДВО
АН СССР. 1991. С. 37-56.
Савваитова К.А., Максимов В.А., Мина М.В. и др. Камчатские благородные лососи. Воронеж:
Изд-во Воронеж, ун-та, 1973. 118 с.
Савваитова К.А., Максимов В.А., Нестеров В.Д. К систематике и экологии гольцов рода
Salvelinus (сем. Salmonidae) водоемов п-ова Таймыр // Вопр. ихтиологии. 1980. Т. 20, вып. 2.
С. 195-210.
Салихов Т.В., Камилов Б.Г. Ихтиофауна бассейна среднего течения Сырдарьи // Там же. 1995.
Т. 35, вып. 2. С. 229-235.
Сальников В.Б. Возможные изменения в составе ихтиофауны после завершения строительства
Каракумского канала в Туркменистане //Там же. 1995. Т. 35, вып. 3. С. 365-373.
Сальников В.Б., Решетников Ю.С. Формирование рыбного населения искусственных водоемов
Туркменистана // Там же. 1991. Т. 31, вып. 4. С. 565-575.
Самуилов А.Е., Свирский В.Г. Список рыб оз. Ханка // Биология рыб Дальнего Востока. Влади-
восток: Дальневост, ун-т, 1976. С. 87-90.
Сафронов С.Н., Никифоров С.Н. Видовой состав и распределение ихтиофауны пресных и соло-
новатых вод Сахалина // Материалы XXX науч.-метод. конф, преподавателей ЮСГПИ (ап-
рель 1995 г.). Южно-Сахалинск, 1995. Ч. 2. С. 112-124.
Сафронов С.Н., Никифоров С.Н. Список рыбообразных и рыб пресных и солоноватых вод Са-
халина // Вопр. ихтиологии. 2002. Т. 42.
Световидов АН. Европейско-азиатские хариусы genus Thvmallus Cuvier )) Тр. Зоол. ин-та АН
СССР. 1936. Т. 3. С. 186-301.
Световидов А.Н. Фауна СССР. Рыбы: Трескообразные. Т. 9, вып. 4. М.; Л.: Изд-во АН СССР,
1948. 221 с.
Световидов А.Н. Фауна СССР. Рыбы: Сельдевые (Clupeidae). М.; Л.: Изд-во АН СССР, 1952.
331с.
Световидов А.Н. Рыбы Черного моря. М.; Л.: Наука, 1964. 550 с.
Свидерская А.К. Возраст и рост амурского жереха Pseudaspius leptocephalus (Pallas) // Труды
Амур, ихтиол, экспедиции, 1945-1949 гг. М.: МГУ, 1958. Т. 4. С. 83-101.
Сиделева В.Г. Сейсмосенсорная система и экология байкальских подкаменщиковых рыб
(Cottoidei). Новосибирск: Наука, 1982. 152 с.
Сиделева В.Г. Карликовые формы у Abyssocottus korotneffi Berg // Новое о фауне Байкала. Ново-
сибирск: Наука, 1985. Вып. 1. С. 83-85.
Сиделева В.Г. Новый вид эндемичного для Байкала рода Batrachocottus (Cottidae) 11 Вопр. ихтио-
логии. 1999. Т. 39, вып. 2. С. 149-154.
Сиделева В.Г. Новый вид тепловодной щироколобки из Байкала Ц Там же. 2002. Т. 42, № 2.
С. 274-278.
Сиделева В.Г., Карабанов Е.Б., Зубин А.А. и др. Экология нереста байкальской желтокрылки
майской генерации // Подводные ландшафты Байкала. Новосибирск: Наука, 1990.
С. 112-122.
Сиделева В.Г., Механикова И.В. Пищевая специализация и эволюция керчаковых рыб // Эколо-
гия и морфология рыб. Л.: Наука, 1990. С. 144—162.
Сиделева В.Г., Нагорный В.К. Особенности размножения и плодовитость байкальских подка-
менщиковых рыб (Cottoidei) // Зоол. журн. 1985. Т. 64, № 3. С. 384—392.
227
Сиделева В.Г., Природина В.П., Ханаев И.В. Хромосомные наборы донных байкальских котто-
идных рыб (Cottoidei) с замечаниями о кариологической продвинутости в связи с батиметри-
ческим распределением //Вопр. ихтиологии. 1995. Т. 35, вып. 6. С. 796-803.
Сидоров ГЛ. Рыбные ресурсы Большеземельской тундры. Л.: Наука, 1974. 163 с.
Скрябин А.Г. Рыбы Баунтовских озер Забайкалья. Новосибирск: Наука, 1977. 231 с.
Скрябин А.Г. Сиговые рыбы юга Сибири. Новосибирск: Наука, 1979. 229 с.
Слынько Ю.В., Кияшко В.И., Яковлев В.Н. Список видов рыбообразных и рыб бассейна р. Вол-
га // Каталог растений и животных водоемов бассейна Волги. Ярославль. 2000. С. 252-308.
Смирнов А.И. Ревизия таксонов рода подуст Chondrostoma Agassiz, 1835 (Pisces, Cyprinidae) Вос-
точной Европы в пределах бывшего СССР. Киев, 1992. 63 с.
Смирнов А.И., Зиновьев Е.А., Пушкин Ю.А. Уточнение таксономического статуса камской по-
пуляции подуста Chondrostoma Agassiz, (Pisces, Cyprinidae) //Сб. науч. тр. ГосНИОРХ. 1988.
Вып. 281. С. 121-126.
Смирнов В.В. Структура и продукционные возможности ихтиоценозов // Грамнинские озера в
зоне влияния БАМ. Новосибирск: Наука, 1980. С. 74—81.
Смирнов В.В., Шумилов ИЛ. Омули Байкала. Новосибирск: Наука, 1974. 160 с.
Смирнов Ю.А. Лосось Онежского озера. Л.: Наука, 1971. 142 с.
Смирнов Ю.А. Пресноводный лосось. Л.: Наука, 1979. 155 с.
Смольянов И.И. Развитие ленка Brachymystax lenok (Pallas) //Вопр. ихтиологии. 1961. Т. 1,вып. 1.
С. 136-148.
Соин С.Г. Размножение и развитие малоротой корюшки Hypomesus olidus (Pallas) // Изв. ТИНРО.
1947. Т. 25. С. 210-220.
Соин С.Г. О размножении и развитии черного байкальского хариуса (Thymallus arcticus baicalen-
sis Dyb.) // Зоол. журн. 1963. T. 42, № 12. С. 1817-1839.
Соин С.Г., Турдаков А.Ф. Развитие трех видов байкальских бычков-подкаменщиков (Cottoidei) //
Вопр. ихтиологии. 1966. Т. 6, вып. 4. С. 696-707.
Соколов Л.И. Питание китайского окуня в среднем течении Амура // Вести. МГУ. 1962. № 1.
С. 13-17.
Соколов Л.И. Сем. Acipenseridae - осетровые // Аннотированный каталог круглоротых и рыб
континентальных вод России. М.: Наука, 1998. С. 19-23.
Соколов Л.И. О зимовке ротана Perccottus glenii Dyb. в амурских водоемах // Вопр. ихтиологии.
2001. Т. 41, вып. 4. С. 572-573.
Соколов Л.И., Цепкий Е.А. Севрюга Acipenser stellatus Pallas в среднем и позднем голоцене // Там
же. 1969. Т. 9, вып. 4. С. 587-598.
Соколов Л.И., Цепкий Е.А. Стерлядь Acipenser ruthenus L. в среднем и позднем голоцене // Бюл.
МОИП. Отд. биол. 1971. Т. 75, вып. 3. С. 137-145.
Соколов Л.И., Цепкин Е.А., Барабанова Е.А. Экология размножения европейской ручьевой ми-
ноги Lampetra planeri (Petromyzontidae) И Вопр. ихтиологии. 1992. Т. 32, вып. 2. С. 181-185.
СоловкинаЛ.Н. Рыбы Коми АССР. Сыктывкар: Коми кн. изд-во, 1975. 166 с.
Сорокин ВЛ. Налим оз. Байкал. Новосибирск: Наука, 1976. 144 с.
Сорокин В.Н., Сорокина А.А. Биология молоди промысловых рыб Байкала. Новосибирск: Нау-
ка, 1988.216 с.
Спановская В.Д. О систематике амурских пескарей // Зоол. журн. 1953. Т. 32, № 2. С. 259-271.
Спановская В.Д. Возраст и рост пескарей подсемейства Gobioninae в бассейне Амура // Труды Амур,
ихтиол, экспедиции, 1945-1949 гг. М.: Из-во МГУ. 1958. Т. 4. С. 175-225.
Спановская В.Д., Макеева АЛ. О фаунистической принадлежности владиславии (Ladisla-
via taczanowskii Dyb.) бассейна Амура // Зоол. журн. 1962. Т. 41, № 7. С. 1051-1055.
Стариков ГВ. Голомянки Байкала. Новосибирск: Наука, 1977. 94 с.
Стребкова Т. Т. Рыбоводное освоение полосатого окуня // Результаты работ по акклиматизации
водных организмов. СПб.: ГосНИОРХ. 1995. С. 126-135.
Сулоев А.Т. Современное состояние ихтиофауны Брянской области // Рациональное природо-
пользование на территории Брянской области. М., 1983. С. 75-79.
Талиев Д.Н. Бычки-подкаменщики Байкала (Cottoidei). М.; Л.: Изд-во АН СССР, 1955.
602 с.
Таранец А.Я. О видах рода Hypomesus в бассейне Японского моря // Докл. АН СССР. 1934. Т. 3,
№ 8/9. С. 675-776.
Таранец А.Я. Пресноводные рыбы бассейна северо-западной части Японского моря // Тр. Зоол.
ин-та АН СССР. 1936. Т. 4. С. 483-537.
Тархова Ю.Н. Материалы по внутривидовой изменчивости песчаной широколобки // Крат, со-
общ. Бурят, комплекс, ин-та. 1962. Вып. 3. С. 101-115.
Трифонов ГЛ. Биология размножения азовских бычков // Тр. Карадаг. биол. станции АН СССР.
1955. Вып. 13. С. 5-46.
Троицкий С.К., Цуникова ЕЛ. Рыбы бассейнов Нижнего Дона и Кубани. Ростов н/Д: Рост. кн.
изд-во, 1988. 112 с.
Тугарина П.Я. Хариусы Байкала. Новосибирск: Наука, 1981. 281 с.
Фадеев Н.С. Промысловые рыбы северной части Тихого океана. Владивосток, 1984. 198 с.
Федоров А.В. Ихтиофауна бассейна Дона в Воронежской области Ц Рыбы и рыбное хозяйство
Воронежской области. Воронеж: Изд-во Воронеж, ун-та, 1960. С. 149-248.
Фортунатова К.Р., Попова О.А. Питание и пищевые взаимоотношения хищных рыб в дельте
Волги. М.: Наука, 1973. 298 с.
Фролов С.В. Кариотипы кунджи Salvelinus leucomaenis (Pallas) и белого гольца 5. albus
Glubokovsky из бассейна р. Камчатки // Биология гольцов Дальнего Востока. Владивосток:
ДВО АН СССР, 1991а. С. 103-Ш.
Фролов С.В. Механизмы дифференциации половых хромосом у лососевых рыб // Биология мо-
ря. 19916. №5. С. 3-17.
Фролов С.В. Дивергенция кариотипов чукотских гольцов рода Salvelinus // Докл. РАН. 1997.
Т. 357, № 5. С. 703-705.
Фролов С.В. Изменчивость и эволюция кариотипов лососевых рыб. Владивосток: Дальнаука,
2000. 229 с.
Фролов С.В., Скурихина Л.А. Анализ кариотипов и ядерной ДНК двух видов вальков (род
Prosopium) Ц Биология и биотехника разведения сиговых рыб: Материалы Всерос. совещ.
СПб.: ГосНИОРХ, 1994. С. 155-156.
Фролов С.В., Фролова ВЛ. Кариологическая дифференциация гольца Леванидова Salvelinus lev-
anidovi и мальмы 8. malma р. Ямы // Биология моря. 1997. Т. 23, № 6. С. 376-379.
Фролов С.В., Фролова В.Н., Молодиченко А.В. Кариотип мальмы Salvelinus malma р. Ямы
и таксономический статус северной и южной мальмы // Там же. 1997. Т. 23, № 5.
С. 309-313.
Ханаев И.В. Особенности строения кариотипа у вторично-пелагического вида рода Comephorus
(Comephoridae, Cottidae) оз. Байкал // Тез. докл. Первый конгресс ихтиологов России. М.:
ВНИРО, 1997. С. 27.
Ханаев И.В., Сиделева В.Г. Кариотипы двух придонно-пелагических видов рода Cottocomephorus:
С. grewingkii и С. inermis (Cottocomephorinae) // Тез. докл. II Верещагинской конф. Иркутск,
1995. С. 218-219.
Цепкин ЕЛ. Изменения промысловой фауны рыб континентальных водоемов Восточной Евро-
пы и Северной Азии в четвертичном периоде // Вопр. ихтиологии. 1995. Т. 35, вып. 1.
С. 3-17.
Цепкин Е.А., Соколов Л.И. Русский осетр Acipenser gilldenstadti Brandt в среднем и позднем голо-
цене // Там же. 1970а. Т. 10, вып. 1. С. 24-36.
Цепкин Е.А., Соколов Л.И. Белуга Huso huso (L.) в позднем голоцене // Биол. науки. 19706. № 5.
С. 11-16.
Цепкин Е.А., Соколов Л.И. О максимальных размерах и возрасте некоторых осетровых рыб //
Вопр. ихтиологии. 1971. Т. 11, вып. 3. С. 541-542.
Цыплаков Э.П. Расширение ареалов некоторых видов рыб в связи с гидростроительством на
Волге и акклиматизационными работами // Там же. 1974. Т. 14, вып. 3. С. 396-405.
Черешнев И.А. О систематическом положении бычка-подкамешцика рода Cottus (Cottidae,
Pisces) Чукотского полуострова // Пресноводная фауна Чукотского полуострова. Владиво-
сток: БПИ ДНЦ АН СССР, 1976. С. 123-128.
Черешнев И.А. Материалы по биологии проходных лососевых Восточной Чукотки // Рыбы в
экосистемах лососевых рек Дальнего Востока. Владивосток: ДВНЦ АН СССР, 1981.
С.116-148.
Черешнев И.А. К вопросу о таксономическом статусе симпатричных проходных гольцов
(Salvelinus, Salmonidae) Восточной Чукотки // Вопр. ихтиологии. 1982. Т. 22, вып. 6.
С. 922-936.
Черешнев И.А. Новый для фауны СССР вид сига Coregonus laurettae Bean (Salmonidae) - берин-
гийский омуль из бассейна р. Чегитунь (арктическое побережье Чукотского полуострова) //
Там же. 1984. Т. 24, вып. 6. С. 888-894.
Черешнев И.А. Состав ихтиофауны и особенности распространения пресноводных
рыб в водоемах Северо-Востока СССР //Там же. 1990. Т. 30, вып. 5. С. 836-844.
Черешнев И.А. Пресноводные рыбы Северо-Востока Азии: Фауна, систематика, история рассе-
ления: Автореф. дис. ... д-ра биол. наук. Владивосток, 1992. 75 с.
229
Черешнев И.А. Сравнительная краниология омулей Голарктики // Биология и биотехника разве-
дения сиговых рыб. Матер. 5-го Всерос. совещания. СПб.: Изд-во ГОСНИОРХ. 1994.
С. 157-161.
Черешнев И.А. Круглоротые и рыбы // Позвоночные животные Северо-Востока России. Влади-
восток: Дальнаука, 1996а. Раздел 1. С. 21-61.
Черешнев И.А. Биологическое разнообразие пресноводной ихтиофауны Северо-Востока Рос-
сии. Владивосток: Дальнаука, 19966. 197 с.
Черешнев ИЛ Биогеография пресноводных рыб Дальнего Востока России. Владивосток: Даль-
наука, 1998. 130 с.
Черешнев И.А. Голец нейва // Красная книга Севера Дальнего Востока России. М.: Пента, 1998.
С. 30-31.
Черешнев И.А., Балушкин А.В. Новый вид черной рыбы Dallia admirabilis Chereshnev, sp. п.
(Umbridae, Esociformes) из бассейна p. Амгуэма (Арктическая Чукотка) // Вопр. ихтиологии.
1980. Т. 20, вып. 6. С. 800-805.
Черешнев И.А., Гудков П.К., Скопец М.Б. К биологии гольцов рода Salvelinus (Salmonidae) бас-
сейна Охотского моря. Находка гольца Леванидова Salvelinus levanidovi Chereshnev, Skopetz
et Gudkov в p. Пенжина // Биология гольцов Дальнего Востока. Владивосток: ДВО АН
СССР, 1991. С. 57-69.
Черешнев И.А., Жариков С.И. О первой находке американской сельди шед Alosa sapidissima в
р. Анадырь // Вопр. ихтиологии. 1989. Т. 29, вып. 3. С. 501-503.
Черешнев И.А., Скопец М.Б. Salvethymus svetovidovi gen. et sp. nova - новая эндемичная рыба из
подсемейства лососевых (Salmoninae) из оз. Эльгыгытгын (Центральная Чукотка) //Там же.
1990. Т. 30, вып. 2. С. 201-213.
Черешнев И.А., Скопец М.Б. Новый для фауны России вид сига - карликовый валек Prosopium
coulteri (Eigenmann et Eigenmann ) из бассейна p. Амгуэма (Чукотский полуостров) // Там же.
1992. Т. 32, вып. 1. С. 21-28.
Черешнев ИЛ., Скопец М.Б. Биология гольцовых рыб оз. Эльгыгытгын // Природа впадины
озера Эльгыгытгын (проблемы изучения и охраны). Магадан: СВКНИИ ДВО РАН. 1993.
С. 105-127.
Черешнев ИЛ., Скопец М.Б., Гудков П.К. Новый вид гольца Salvelinus levanidovi sp. п. из бассей-
на Охотского моря // Там же. 1989. Т. 29, вып. 5. С. 691-704.
Черешнев И.А., Шестаков А.В., Скопец М.Б. Определитель пресноводных рыб Северо-Востока
России. Владивосток: Дальнаука, 2001. 197 с.
Черешнев И.А., Шестаков А.В., Юсупов Р.Р. и др. Биология нельмы Stenodus leucichthys nelma
(Coregonidae) бассейна p. Анадырь (Северо-Восток России) // Вопр. ихтиологии. 2000. Т. 40,
вып. 4. С. 537-550.
Черницкий А.Г., Матишов Г.Г., Ермолаев В.В. Полярная марикультура. Апатиты: Кол. фил.
АН СССР, 1987. 38 с.
Черняев ЖЛ. Эмбриональное развитие байкальского омуля. М.: Наука, 1968. 91 с.
Черняев ЖЛ. Размножение и развитие байкальского озерного сига в связи с вопросами его ис-
кусственного разведения // Вопр. ихтиологии. 1973а. Т. 139, вып. 2. С. 259-274.
Черняев ЖЛ. О генезисе фауны байкальских бычков-подкаменщиков (Cottoidei) // Зоол. журн.
19736. Т. 52, № 3. С. 459-464.
Черняев Ж.А. Морфо-экологические особенности размножения и развития песчаной широко-
лобки оз. Байкал Paracottus (Leocottus) kessleri (Dyb.) //Вопр. ихтиологии. 1977. Т. 29, вып. 3.
С. 501-503.
Черняев ЖЛ. Размножение и развитие большеголовой широколобки Batrachocottus baicalensis
(Dyb.) оз. Байкал // Там же. 1979. Т. 19, вып. 6. С. 1053-1067.
Черняев ЖЛ. Воспроизводство байкальского омуля. М.: Лег. и пищ. пром-сть, 1982. 127 с.
Черняев ЖЛ. Сравнительный эколого-морфологический анализ размножения и развития пела-
гических бычков-подкаменщиков рода Cottocomephorus оз. Байкал // Эколого-морфологи-
ческие исследования раннего онтогенеза позвоночных. М.: Наука, 1984. С. 149-167.
Черняев ЖЛ. Эколого-морфологические особенности размножения и развития черного бай-
кальского хариуса (Thymallus arcticus baicalensis Dyb.) популяции реки Фролихи (Северный
Байкал) // Тр. кафедры зоологии позвоночных Иркут, гос. ун-та. 2001. Т. 1. С. 136-147.
Черняев Ж.А., Пичугин М.Ю. Особенности раннего онтогенеза весенненерестующего баунтов-
ского сига // Вопр. ихтиологии. 1999. Т. 39, вып. 1. С. 78-88.
Чикова В.М. Состояние нерестовых стад и размножение рыб в Черемшанском и Сусканском за-
ливах Куйбышевского водохранилища // Биология рыб Волжских водохранилищ. М.; Л.:
Наука, 1996. С. 29 45. (Тр. ИБВВ АН СССР; Вып. 10 (13)).
230
Чугунов НЛ., Чугунова Н.И. Сравнительном промыслово-биологическая характеристика осет-
ровых рыб Азовского моря // Тр. ВНИРО. 1964. Т. 52. С. 87-182.
Шедько С. В. Экспедиция на НИС “Академик Опарин”, рейс № 2 // Пресс-релиз. Морские экспе-
диции ДВО РАН. Владивосток: Дальнаука, 1999. С. 13-16.
Шедько С.В. О видовом составе корюшек из Приморья // Вопр. ихтиологии, 2001. Т. 41, вып. 2,
С. 261-264.
Шестаков А.В. Биология молоди сиговых рыб бассейна р. Анадырь: Автореф. дис.... канд. биол.
наук. М., 1996. 24 с.
Шестаков А.В. Биология молоди сиговых рыб бассейна р. Анадырь. Владивосток: Дальнаука,
1998. 111 с.
Шмидт П.Ю. Рыбы Охотского моря. М.; Л.: Изд-во АН СССР, 1950. 370 с.
Шмидтов А.И. Стерлядь (Acipenser ruthenus L.) // Учен. зап. Казан, ун-та. 1939. Т. 99, кн. 4/5.
С. 3-279.
Щербуха А.Я. К систематике ельцов подрода Leuciscus из Сев. Донца и Днепра // Гидробиол.
журн. 1972. Т. 8, № 3. С. 9-75.
Экология и хозяйственное значение рыб МНР. М.: Наука, 1985. 276 с.
Эланидзе Р.Ф. Ихтиофауна рек и озер Грузии. Тбилиси: Мецниереба, 1983. 318 с.
Abe S. Note on the chromosomes of two species of freshwater cottid fishes // Chromosomes Inform. Serv.
1972. N 13. P. 25-27.
Airaksinen KJ. Preliminary notes on the winter-spawning vendace (Coregonus albula L.) in some Finnish
lakes //Ann. Zool. Fenn. 1968. Vol. 5, N 3, P. 312-314.
Alt K.T. Contribution to the biology of Bering cisco (Coregonus laurettae) in Alaska // J. Fish. Res. Board
Canada. 1973. Vol. 30, N 12. Pt 1. P. 1885-1888.
Banarescu P. Fauna Republicii Populate Romine. Pisces - Osteichthyes. Vol. 13. Buc.; Academiei, 1964.
959 p.
Banarescu P. Revision of the genus Hemiculter (Pisces, Cyprinidae) // Trav. Mus. Hist. Natur.
“Gr. Antipa”. 1968. Vol. 8. P. 523-529.
Banarescu P. A critical updated checklist of Gobioninae (Pisces, Cyprinidae) // Ibid. 1992. Vol. 32.
P. 303-330.
Banarescu P., Nalbant T.T. Revision of the genus Microphysogobio (Pisces, Cyprinidae) // Vestn. Os.
spolec. zool. 1966. Sv. 30, N 3. S. 149-209.
Behnke R.J. Native trout of Western North America I I Amer. Fish. Soc. Monogr. 1992. N 6. P. 1-
275.
Booke H.E. Cytotaxonomic studies of the Coregonine fishes of the Great Lakes, USA: DNA and karyotype
analysis // J. Fish. Res. Board Canada. 1968. Vol. 25, N 8. P. 1667-1687.
Booke H.E. Cytotaxonomy of the salmonid fish Stenodus leucichthys // Ibid. 1975. Vol. 32, N 2.
P. 295-296.
Brylinska M. Ryby slodkowodne Polski. Warszawa: Panstw. Wyda-wo Naukowe, 1986. 428 s.
Burgner R.L. Life history of sockeye salmon (Oncorhynchus nerka) // Pacific salmon life histories. UBC
Press. 1991. P. 3-117.
Chen T.R., Reisman H.M. A comparative study of the North American species of sticklebacks (Teleostei:
Gasterosteidae) H Cytogenetics. 1970. N 9. P. 321-332.
Choi Ki-Chul, Leon Sang-Rin, Kim Ik-Soo, Son Yeong-Mok. Coloured illustrations of Freshwater Fishes of
Korea. 1990. 277 p.
Crossman J.E., Rab P. Chromosome-banding study of the Alaska blackfish, Dallia pectoralis (Euteleostei:
Esocidae), with implications for karyotype evolution and relationship of esocoid fishes // Canad. J.
Zool. 1996. Vol. 74. P. 147-156.
Dgebuadze Yu.Yu. The land/inland-water ecotones and fish population of Lake Valley (West Mongolia) //
Hydrobiologia. 1995. Vol. 303. P. 235-245.
Dotu Y. The bionomics and life history of two Gobiid fishes Tridentiger undicivis and Tridentiger trigono-
cephalus (Gill) in innetmost part of Ari^ke Sound // Sci. Bull. Fac. Agr. Kyushu Univ. 1958. Vol. 16,
N 3. P. 343-358.
Dotu Y. The life history and bionomics of the Gobiid fish Aboma lactipes (Hilgendorf) // Bull. Fac. Fish.
Nagasaki Univ. 1959. Vol. 10. P. 133-139.
Dotu Y., Mito S. On the breeding-habitats, larvae and young of a Goby, Acanthogobius flavimanus (Temm.
et Schl.). // Jap. J. Ichthyol. 1955. Vol. 4, N 4/6. P. 153-161.
Freshwater Fisches in Japan. 1987. Tokyoi: Tokai Univ, press, 1987. 187 p.
Frolov S.V. Karyotype and chromosomal variability in small-mouth char, Salvelinus elgyticus, from lake
Elgygytgyn // J. Ichthyol. 1992. Vol. 32, N 1. P. 61-66.
231
Golubtsov A.S., Berendzen P.B., Annett CA. Morphological variation and taxonomic status of Altai osmans,
Oreoleuciscus, from the upper reaches of the Ob River system // J. Fish Biol. 1999. Vol. 54.
P. 878-899.
Goto A. Sexual dimorphism in a river sculpin Cottus hangiongensis //Jap. J. Ichthyol. 1984. Vol. 31, N 2.
P. 161-166.
Goto A. Movement and population size of the river sculpin Cottus hangiongensis in the Daitobetsu River
of Southern Hokkaido // Ibid. 1986. Vol. 32, N 4. P. 421-430.
Goto A. Life history variation in males of the river sculpin, Cottus hangiongensis, along the course of a river
//Environ. Biol. Fish. 1987. Vol. 19, N 2. P. 81-91.
Goto A., Nakano Sh. Distribution and ecology of freshwater fishes in Hokkaido // Biodiversity and ecolo-
gy in the Northern Japan. Sapporo: Hokkaido Univ, press, 1993. P. 114-126.
Gruchy C.G. Salmon nomenclature //Nature. 1971. Vol. 234. P. 360.
Hardisty M.W. Lampetra fluviatilis (Linnaeus, 1758) and other lamprey species // The Freshwater Fishes of
Europe. Wiesbaden: AULA-Veri., 1986. Vol. 1. Pt 1. P. 94-116, 249-278, 279-304.
Holcik J., Hensel K., NieslankJ., Skacel L. The Eurasian Huchen Hucho hucho. Dordrecht etc., 1988. 239 p.
Huggins RJ., Thompson A. Communal spawning of brook and river lampreys, L. fluviatilis and L. planeri //
J. Fish Biol. 1970. Vol. 2. P. 53-54.
Imamura T., Hashitani S. On the food habitat of four fishes in Marsh Hinuma // Bull. Fac. Lib. Arts. Ibaraki
Univ. Sci. 1957. Vol. 7. P. 45-56.
Koli L. Geographical variation of Cottus gobio L. (Pisces, Cottidae) in Northern Europe // Ann. Zool. Fenn.
1969. Vol. 6. P. 353-390.
Koli L. Suomen Kalat. Suomi: Werner Soderstrom Osakeyhtio, 1990. 342 p.
Lee D.S., Gilbert C.R., Hocutt C.H. et al. Atlas of North American Freshwater Fishes. USA: North Carol.
State Mus. Natur. Hist., 1980. 854 p.
McAllister D.E. A revision of the smelt family Osmeridae // Bull. Nat. Mus. Canada. 1963. N 191. P. 1-53.
McCart P. Evidence for the existence a sibling species of pygmy whitefish (Prosopium coulteri) in three
Alaskan lakes // Biology of coregonid fishes. Winnipeg: Univ. Manitoba press, 1970. P. 81-98.
McPhail JD. The Coregonus autumnalis in Alasca and northwestern Canada // J. Fish. Res. Board Canada.
1966. Vol. 23, NLP. 141-148.
McPhail J.D., Lindsey C.C. Freshwater fishes of Northwestern Canada and Alaska. Ottawa, 1970. 381 p.
(Fish. Res. Board Canada. Bull.; N 173).
Miller PJ. Gobiidae // Check-list of the fishes of the North-Eastern Atlantic And of the Mediterranean
(CLOFNAM). P.: UNESCO, 1986. P. 483-515.
Miyazaki J. Studies on the Japanese common Goby Acanthogobius flavimanus (Temm. et Schl.) //Bull. Jap.
Soc. Fish. 1940. Vol. 9, N 4. P. 159-180. На яп. яз.
Morrow J.E. The Freshwater Fishes of Alaska. Anchorage (Alaska): Alaska Northwest publ., 1986. 248 p.
Munehara H., Sideleva V.G. Intra- and interspecific competition between two Baikal sculpins for spawning
resources. Abstr. Intern. Symp. Lake Baikal. Yokogama, Japan. 1998. P. 74.
Muramoto J., Igarashi K., Itoh M., Makono S. A study of the chromosomes and enzymatic patterns of stick-
lebacks of Japan // Proc. Jap. Acad. 1969. Vol. 45, N 3. P. 803-807.
Nall H. The life of the sea trout. L., 1930. 335 p.
Naseka A.M. Comparative study on the vertebral column in the Gobioninae (Cyprinidae, Pisces) with spe-
cial reference to its systematics // Bull. Espec. Inst. Esp. Ocean. 1996. N 21. P. 149-167.
Nichols J.T. The Freshwater Fishes of China. N. Y.: Amer. Mus. Natur. Hist., 1943. Vol. 9. 322 p.
Nikolsky G.V., Reshetnikov Yu.S. Systematics of coregonid fishes in the USSR: Intraspecies variability and
difficulties in taxonomy // Biology of coregonid fishes. Winnipeg: Univ. Manitoba press. 1970.
P. 251-266.
Nordenskiold A.E. Vegas fard Kring Asien och Europe: Jenite eu historisk aterblick pa foregaende resor
langs galma verdens nordkust. Stockholm, 1881. 510 p.
Payne R.H., Child AJR., Forrest A. Geographical variation in the Atlantic salmon // Nature. 1971. Vol. 231.
P. 250-252.
Pacific salmon life histories. Vancouver: UBC press, 1991. 230 p.
Pethon P. Aschehougs store fiskebok. Stockholm: Aschehoug, 1989. 447 p.
Pflieger W.L. The fishes of Missouri. Missouri Dep. of Conserv., 1991. 343 p.
Pirogovsky M.L, Sokolov L.I., Vasil’ev V.P. Huso huso (Linnaneus, 1758) // The freshwater fishes of
Europe. Wiesbaden: AULA-Verl., 1989. Vol. 1. Pt 2. P. 156-200.
Popova О A., Reshetnikov Yu.S., Andreev V.L., Makarova N.P. An analysis of the variability in the perch,
Perea fluviatilis, with its range // Russ. J. Aquat. Ecol. 1997. Vol. 6, N 1/2. P. 75-84.
Popova О A., Reshetnikov Yu.S., Kiyashko V.I. et al. Ruffe from the former USSR: variability within the
largest part of its natural range //1. Great Lakes Res. 1998. Vol. 24, N 2. P. 263-284.
232
Rab P. Karyotype of European catfish Silurus glanis (Siluridae: Pisces) with remarks on cytogenetics of sil-
uroid fishes // Folia zool. 1981. Vol. 30, N 3. P. 271-286.
Rab P. The karyotype of the Cyprinid fish Pseudaspius leptocephalus // Jap. J. Ichthyol. 1991. Vol. 38, N 3.
P. 329-331.
Rab P., Karakousis У., Peios C. Karyotype of Silurus aristotelis with reference to the cytotaxonomy of the
genus Silurus (PISCES, Siluridae) // Folia Zoologica. 1994. Vol. 43, N 1. P. 75-81.
Rab P., Mayr B., Roth P. Chromosome banding of European catfish, Silurus glanis (PISCES, Siluridae) //
Genetica. 1991. Vol. 83- P. 153-157.
Rab P., Roth P., Arefjev VA. Chromosome studies of European Leuciscine fishes (Pisces, Cyprinidae):
Karyotype of Aspius aspius Ц Caryologia. 1990. Vol. 43, N 3/4. P. 249-255.
Rab P., Roth P., Mayr B. Karyotype study of eight species of European percid fishes (Pisces, Percidae) //
Ibid. 1987. Vol. 40, N 4. P. 307-318.
Robinson E.S., Potter I.C., Webb C.J. Homogeneity of Holarctic lamprey karyotypes // Ibid. 1974. Vol. 27.
p. 443-454.
Saruwatari T„ Lopez J A., Pietsch T.W. A revision of the osmerid genus Hypomesus Gill (Teleostei:
Salmoniformes), with the description of a new species from the Southern Kuril Island // Species
Diversity. 1997. Vol. 2. P. 59-82.
Scott W.B., Crossman EJ, Freshwater fishes of Canada. Ottawa, 1973. 966 p. (Fish. Res. Board Canada.
Bull.; N 184).
Sideleva V.G. List of fishes from Lake Baical with descriptions of new taxa of cottoid fishes // New con-
tributions of freshwater fish research. Proc. Zool. lust Russ. Acad. Sci. 2001. Vol. 287. P. 45-79.
Smith G.R., Stearley R.F. The classification and scientific names of rainbow and cutthroat trouts //
Fisheries. 1989. N 14 (1). P. 4-10.
Sokolov L.I., Vasil’ev VP. Acipenser nudiventris Lovetsky, 1928. Acipenser ruthenus Linnaeus, 1758.
Acipenser baeri Brandt, 1869 // The Freshwater Fishes of Europe. Wiesbaden: AULA-Verl., 1989.
Vol. 1. Pt 2. P. 201-284.
Suciu P., Rab P. A note on the kaiyotype of Percarina demidoffi (PISCES: Percidae) // Folia zool. Brno.
1991 (1992). Vol. 41, N 1. P. 93-95.
Svetovidov A.N. Clupeidae // Check list of the Fishes of the North-Eastern Atlantic and of the
Mediterranean (CLOFNAM). P.: UNESCO, 1973. Vol. 1. P. 99-109.
Takagi K. Taxonomic and nomenclatural status in chaos of the gobiid fish, Chaenogobius annularis Gill,
1858. II. Specific heterogeneity of G. annularis Gill sensu Tomiyama, with description of the genus
Rhodonichiihys, gen. nov. //1. Tokyo Univ. Fisch. 1966. Vol. 52, N 1. P. 55-70. На яп. яэ.. вступл.
англ.
The Freshwater Fishes of China in coloured illustrations. 1982. Pt 1. 182 p.
The Freshwater Fishes of China in coloured illustrations. 1988. Pt 2. 180 p.
The Freshwater Fisher of Europe / Ed. J. Holcik. Wiesbaden: AULA-Verl., 1986. Vol. 1. Pt 1. 313 p.
The Freshwater Fisher of Europe / Ed. J. Holcik. Wiesbaden: AULA-Verl., 1986. Vol. 2. 447 p.
Vasil’eva E., Sty gar V. Salvelinus gritzenkoi - a new species of char from the North Kuril Islands
(Salmonidae, Salmoniformes) // Folia Zool. 2000. Vol. 49, N 4. P. 317-320.
Vasil’eva E.D., Vasil’ev V-P. The description of a new species Neogobius iljini sp. n. within former N. kess-
leri (Gobiidae, Pisces) // Acta Univ. Carol. Biol. 1996. Vol. 39, N 3/4. P. 261-270.
Virbiikas J. Lietuvos zuvys. Vilnus: Mokslas, 1986. 149 p.
Vladykov V D. A review of salmonid genera and their broad geographical distribution // Trans. Roy. Soc.
Canada. Ser. 4, sect. 3. 1963. NLP. 459-504.
Vlasenko F.D., Pavlov A.V., Sokolov L.I., Vasil’ev V.P. Acipenser gueldenstaedti Brandt, 1833. Acipenser
persicus Borodin, 1897 // The Freshwater Fishes of Europe. Wiesbaden: AULA-Verl., 1989. Vol. 1.
Pt 2. P. 249-366.
Whitehead PJ.P. FAO species catalogue: Clupeoid fishes of the world. P. 1985. 303 p. (FAO Fish Synop.;
Vol. 7, N 125).
УКАЗАТЕЛЬ
РУССКИХ НАЗВАНИЙ РЫБ
Названия отрядов даются жирным шрифтом и прописными буквами, им соответ-
ствует номер римскими цифрами. Названия семейств представлено таким же образом,
но их номер дан арабскими цифрами. Название рода дается жирным шрифтом строч-
ными буквами, кроме первой - прописной. Названия видов даются только строчными
буквами, и они чаще расположены под родом. Цифры после отряда, семейства и рода
означают их соответствующий номер в “Систематическом указателе”, а цифра после
вида - соответствующий номер в основном тексте “Атласа”.
А
Абботтины 29
абботина китайская 86
Абомы 109
абома японская 220
Акаитогобиусы НО
Алозы 10
алоза европейская 22
Амуры белые 43
амур белый ИЗ
Амуры черные 58
амур черный 137
Аухи 100
ауха 209
Б
БАЛИТОРИЕВЫЕ 14
Барбатули 81
белоглазка 89
- южнокаспийская 89
белорыбица 72
Белуги 8
белуга 20
берш 216
бешенка 24
бобырец 129
Буффало 80
буффало большеротый 173
-малоротый 172
- черный 174
Быстрянки 32
быстрянка восточная 92
- обыкновенная 92
- русская 92
- южная 92
БЫЧКОВЫЕ 29
Бычки 113-122
Бычки дальневосточные 113
бычок большеротый 232
- каштановый дальневосточный 231
- кольчатый дальневосточный 230
- дальневосточный Таранца 233
Бычки Книповича 115
бычок-бубырь 235
- длиннохвостый бычок Книповича 236
Бычки-лысуны 118
- леопардовый лысун 245
Бычки-мар товики 116
бычок-кнут 237
- мартовик 237
Бычки носатые 120
- амурский бычок 247
Бычки-родоииихты 121
бычок-родониихт 248
- голый родониихт 248
Бычки трехзубые 122
бычок полосатый трехзубый 250
- темный трехзубый 249
Бычки тупоносые 119
бычок-цуцик 246
- мраморный тупоносый 246
Бычки черноморско-каспийские 117
бычок-гонец 239
- бычок-губан 242
- бычок-кругляк 241
234
- бычок-песочник 238 - бычок-ширман 244 - бычок-головач каспийский 240 - речной бычок 238 - речной бычок Родиона 243 - черноротый 241 бычок Берга 234 бычок японский речной 221 бычок четырехрогий 269 - белый 54 - восточносибирский 41 - длинноголовый 55 - Дрягина 44 - Леванидова 56 - таймырский 48 - Таранца 49 - Черского 43 - чукотский 41
в - Шмидта 57
Вальки 21 валек карликовый 70 - обыкновенный 71 Верховки 54 верховка обыкновенная 127 Верховки китайские 34 верховка китайская 94 Верхогляды 41 верхогляд 107 ВЕСЛОНОСОВЫЕ 3 Веслоносы 9 веслонос 21 владиславия 126 вобла 157 Востробрюшки 51 востробрюшка корейская 123 - уссурийская 124 вырезуб 156 ВЬЮНОВЫЕ 15 Вьюны 85 вьюн 186 - амурский 185 - якутский 45 Гольцы усатые, барбатули 81 голец Крыницкого 176 - сибирский голец-усач 177 - усатый 175 Гольцы восьмиусые 82 голец восьмиусый 178 Гольяны 63 гольян амурский 144 - Лаговского 144 - обыкновенный 146 - озерный 145 - суйфунский 143 - Чекановского 143 - Черского 143 горбуша 31 горбушка 106 Горчаки 67 горчак 151 - амурский 151 - европейский 151 - Лайта 150 - обыкновенный 151
Г Горчаки колючие 31 горчак колючий 90
Гамбузии 94 гамбузия обыкновенная 200 - хольбрукская 200 Гирканогобиусы 114 - бычок Берга 234 ГЛУБОКОВОДНЫЕ ШИРОКОЛОБКИ 33 голавль 130 Голавли усатые 74 голавль усатый 164 ГОЛОВЕШКОВЫЕ 28 Головешки 108 < головешка-ротан 219 ГОЛОМЯНКОВЫЕ 32 Голомянки 131 голомянка большая 270 - малая 271 Гольцы 18 голец арктический 41 - ханкинский 91 Густеры 38 густера 102 д даватчан 41 ДАЛЛИЕВЫЕ 10 Даллии 27 даллия 84 - амгуэмекая 82 - берингийская 84 - пильхыкайская 83 Е езо-угай 167 Ельцы 55 елец 133 - афипсский 128 - Данилевского 131
235
- сибирский 133 Ерши 102 - ерш донской 211 - обыкновенный 212 каспиосома 229 КЕРЧАКОВЫЕ 31 кета 32 кижуч 33
Ж Желтокрылки 125 желтокрылка северобайкальская 256 - длиннокрылая широколобка 258 - желтокрылая широколобка 257 Желтоперы 64 желтопер мелкочешуйный 147 Желтощеки 46 желтощек 116 Жерехи 36 жерех аральский 96 - красногубый 96 - обыкновенный 96 Жерехи амурские 65 жерех амурский плоскоголовый 148 - красноперый 148 КОЛЮШКООБРАЗНЫЕ X КОЛЮШКОВЫЕ 22 Колюшки трехиглые 96 колюшка трехиглая 202 Колюшки многоиглые 97 колюшка аральская 203 - девятииглая 204 - малая южная 203 - китайская, или амурская 205 - сахалинская 206 Кони 50 конь-губарь 121 - пятнистый 122 КОРЮШКОВЫЕ 8 Корюшки 25 корюшка азиатская зубатая 79 - европейская 78
з Корюшки малоротые 24
залом 24 зельдь 59 ЗМЕЕГОЛОВЫЕ 30 Змееголовы 123 змееголов амурский 251 - обыкновенный 251 золотая рыбка 103 зубатка 32 корюшка колымская 77 - малоротая 77 - тымская 76 - японская 76 КОСАТКОВЫЕ 17 косатка-крошка 195 Косатки 89 косатка Бражникова 192 - Герценштейна 193
И ИГЛООБРАЗНЫЕ XI ИГЛОВЫЕ 23 Иглы 98 игла каспийская 207 - черноморская 207 ИКТАЛУРОВЫЕ 18 -уссурийская 194 Косатки-скрипуны 91 косатка-скрипун 196 красная 35 краснопер 148 краснопер монгольский 108 Красноперки 72 красноперка 162
к Красноперки-угаи 76
калинка 129 калуга 19 Караси 39 карась золотой 104 - китайский 103 - серебряный 103 - якутский 104 КАРПООБРАЗНЫЕ VI КАРПОВЫЕ 12 Карпы 45 карп обыкновенный 115 КАРПОЗУБООБРАЗНЫЕ VIII Каспиосомы 112 красноперка-угай мелкочешуйная 166 - крупночешуйная 168 - сахалинская 167 красная 35 кумжа 40 - обыкновенная 40 - предкавказская 40 - черноморская 40 кунджа 52 кутум 156 Л Лавраки 99 лаврак полосатый 208
236
Ладиславии 53 ладиславия 126 Ленки 12 ленок 29 - острорылый 29 - тупорылый 29 Лептобоции 84 лептобоция маньчжурская 184 лефуа 178 Лещи 30 лещ 88 Лещи амурские черные 56 лещ амурский черный 135 Лещи амурские белые 61 лещ амурский белый 141 Лини 75 линь 165 листопадка 40 ЛОСОСЕОБРАЗНЫЕ IV ЛОСОСЕВЫЕ 5 Лососи 17 лосось американский 39 - атлантический 39 - европейский 39 - кавказский 40 - куринский 40 - стальноголовый 38 Лососи тихоокеанские 14 лысун леопардовый 245 - сибирская 7 - тихоокеанская 6 Миноги трехзубые 2 минога трехзубая 2 Мистусы 90 муксун 64 Н НАЛИМОВЫЕ 21 Налимы 95 налим обыкновенный 201 - тонкохвостый 201 нейва 47 Нельмы 22 нельма 72 нерка 35 О огуречник 79 ОКУНЕОБРАЗНЫЕ XII ОКУНЕВЫЕ 26 Окуни пресноводные 103 окунь желтый 213 - речной 213 - североамериканский 213 Окуни черные 101 окунь большеротый 210 - черный 210 окунь китайский 209
М мальма 53 - курильская 53 - северная 53 - южная 53 мартовик 237 медака 199 микижа 39 Микроперкопсы 107 микроперкопс 218 - китайский элеотрис 218 МИНОГООБРАЗНЫЕ I МИНОГОВЫЕ 1 Миноги зубатые 3 минога украинская 3 Миноги каспийские 1 минога каспийская 1 • Миноги морские 6 минога морская 9 Миноги обыкновенные 4 минога европейская ручьевая 5 - речная 4 Миноги тихоокеанские 5 минога дальневосточная ручьевая 8 окунь полосатый 208 омуль арктический 60 - байкальский 60 - беринговоморский 62 - ледовитоморский 60 ОРИЗИЕВЫЕ 19 Оризии 93 орфа 132 ОСЕТРООБРАЗНЫЕ II ОСЕТРОВЫЕ 2 Осетры 7 осетр амурский 16 - атлантический 18 - байкальский 10 - восточносибирский 10 - западносибирский 10 - зеленый 12 -колхидский 11, 14 - персидский 14 - русский 11 - сахалинский 12 - сибирский 10 -черноморско-азовский 11,14 - южнокаспийский 11, 14 - якутский 10
237
Османы алтайские 60
осман Потанина 140
-карликовый 139
П
палия 46
- боганидская 42
- есейская 50
- кряжевая 46
- лудожная 46
- малоротая 51
- сегозерская 46
Палии длинноперые 19
палия длиннойоперая Световидова 58
пелядь 66
Перкарины 104
перкарина 214
- азовская 214
- черноморская 214
ПЕРЦИХТОВЫЕ 24
Пескари амурские ящерные 71
пескарь ящерный 161
- длиннохвостый колючий 161
Пескари восьмиусые 49
пескарь восьмиусый 120
Пескари дальневосточные 47
пескарь маньчжурский 117
-чебаковидный 117
Пескари носатые 57
пескарь амурский носатый 136
Пескари обыкновенные 48
пескарь ленский 119
- обыкновенный 118
- сибирский 118
- Солдатова 119
- терский 118
- туркестанский 118
Пескари румынские 68
пескарь амурский белоперый 155
- белоперый 152
- северокавказский длинноусый 153
- кубанский длинноусый 154
Пескари-лени 70
пескарь-лень 159
- пескарь-губач Солдатова 160
- пескарь-губач Черского 158
пескарь ханкинский 163
пескоройка 1- 9
ПЕЦИЛИЕВЫЕ 20
Плотвы 69
плотва обыкновенная 157
-вырезуб 156
Подкаменщики 126
подкаменщик западноевропейский 262
- обыкновенный 262
- пестроногий 264
- русский 262
- сахалинский 259
- слизистый 260
- сибирский 265
- Черского 261
- японский 263
Подкаменщики-широколобки 128
- амурская широколобка 267
Подусты 42
подуст волжский 112
- колхидский 109
- обыкновенный ПО
- терский 111
Подусты-чернобрюшки 78
подуст-чернобрюшка 170
- амурский 170
Псевдоразборы 66
псевдоразбора 149
- амурский чебачок 149
Пуголовки 111
пуголовка Абдурахманова 226
- азовская 226
- Бэра 222
- звездчатая 228
- зернистая 224
- казахская 223
- каспийская 225
- ленкоранская 226
- магистра 226
- Махмудбекова 227
пузанок азовский 23
- астрабадский 23
- дунайский 23
- ильменский 23
- каспийско-черноморский 23
- палеостомский 23
- северо-восточный 23
- северокаспийский 23
- эйзелинский 23
Р
Рогатки 130
рогатка четырехрогая 269
родониихт голый 248
ротан 219
рыба черная 71
Рыбцы 77
рыбец 169
-каспийский 169
- малый 169
- обыкновенный 169
7TR
ряпушка баунтовская 67
- европейская 59
- переславская 59
- сибирская 67
С
сазан амурский 115
- европейский 115
сарделька 27
саурей 59
севрюга 17
- азовская 17
СЕЛЬДЕОБРАЗНЫЕ III
СЕЛЬДЕВЫЕ 4
сельдь астраханская 24
- волжская 24
- каспийская проходная 24
- кесслеровская 24
- черноморско-азовская проходная 25
семга 39
- обыкновенная 39
i - камчатская 38
I > СИГОВЫЕ 6
| Сиги 20
| сиг амурский 69
| - баунтовский 63
J - волховский 63
I — европейский 63
| - многотычинковый 63
| - обыкновенный 63
| - уссурийский 69
| - чудской 63
I сиг-востряк 63
s сиг-лудога 63
J сиг-пыжьян 63
? сиг-хадары 61
[ сима 34
> синец 87
f Сквалидусы 73
h снеток 78
СКОРПЕНООБРАЗНЫЕ XIII
СОМООБРАЗНЫЕ VII
СОМОВЫЕ 16
Сомики американские 92
сомик американский 197
- канальный 198
Сомы обыкновенные 88
сом европейский 190
- обыкновенный 190
- Солдатова 191
Сомы дальневосточные 87
сом амурский 189
стерлядь 15
- сибирская 15
Судаки 105
судак 215
- волжский 216
- обыкновенный 215
сырок 66
сырть 169
Т
Таймени 13
таймень дунайский 30
- обыкновенный 30
Таймени сахалинские 15
таймень сахалинский 37
Тилапии 106
тилапия мозамбикская 217
тинда 40
Толстолобики белые 52
толстолобик белый 125
Толстолобики пестрые 35
толстолобик пестрый 95
Трегубки 59
трегубка 138
- амурская 138
ТРЕСКООБРАЗНЫЕ IX
тугун 68
Тюльки 11
тюлька абрауская 27
- каспийская 28
- черноморско-азовская 28
- черноморско-каспийская 28
У
УГРЕОБРАЗНЫЕ V
УГРИ РЕЧНЫЕ 11
Угри речные 28
угорь американский 85
- речной 85
- японский 85
узо-угай 167
Уклеи 44
уклей 114
Уклейки 33
уклейка кавказская 93
- обыкновенная 93
Усачи 37
усач аральский 98
- днепровский 97
- булат-маи 99
- колхидский 101
- короткоголовый 98
- крымский 101
- кубанский 101
- обыкновенный 97
- терский 100
239
ф
Форели дальневосточные 16
Форель обыкновенная 40
- радужная 38
- эйзенамская 40
X
ХАРИУСОВЫЕ 8
Хариусы 23
хариус амурский 73
- аляскинский 73
- байкальский 73
- восточносибирский 73
- европейский 75
- западносибирский 73
- камчатский 73
- косогольский 73
- монгольский 74
— обыкновенный 75
- сибирский 73
- хубсугульский 73
ц
ЦЕНТРАРХОВЫЕ 25
ЦИХЛОВЫЕ 27
Ч
чавыча 36
чебак амурский 134
чебачок амурский 149
черная рыба 84
чернобрюшка 170
черноспинка 24
Черные окуни 101
- окунь большеротый 210
Чехони 62
чехонь 142
чир 65
ЧУКУЧАНОВЫЕ 13
Чукучаны 79
чукучан обыкновенный 171
- сибирский 171
Ш
Шемаи 40
шемая аральская 105
- дунайская 105
- каспийская 105
- черноморская 105
шип 13
Широколобки песчаные 127
широколобка амурская 267
- арахлейская 266
- гусиноозерская 266
- песчаная 266
Широколобки большеголовые 124
широколобка большеголовая 252
- жирная 254
- пестрокрылая 253
- Талиева 255
Широколобки каменные 129
широколобка каменная 268
- путоранская 268
ШИРОКОЛОБКИ ГЛУБОКОВОД-
НЫЕ 33
Широколобки глубинные 132
широколобка белая 273
- елохинская 272
- малоглазая 274
Широколобки горбатые 135
широколобка горбатая 283
- широкорылая 282
Широколобки короткоголовые 134
широколобка Буленже 281
Широколобки красные 138
широколобка большая 293
- Гото 290
- карликовая 291
- красная 292
Широколобки озерные 136
широколобка крапчатая 285
- плоская 284
- темная 286
- узкая 287
Широколобки рыхлые 137
широколобка рыхлая 289
- тепловодная 288
Широколобки шершавые 133
широколобка шершавая Герценштейна
276
- глубоководная 275
- Корякова 277
- острорылая 280
- панцирная 278
- плоскоголовая 279
- селенгинская 277
шэд американский 26
щ
Щиповки 83
щиповка каспийская 188
- байкало-селенгинская 181
- Лютера 180
- обыкновенная 183
- Сабанеева 187, 188
- сибирская 181
- Чоя 179
240
- южнорусская 182
Щиповки Сабанеева 86
щиповка золотистая 187
-переднеазиатская 187
- предкавказская 188
щокур 65
ЩУКОВЫЕ 9
Щуки 26
щука амурская 81
- обыкновенная 80
Э
элеотрис китайский 218
Я
Язь 132
- амурский 134
- аральский 132
- золотой 132
16 Атлас..., том 2
УКАЗАТЕЛЬ
ЛАТИНСКИХ НАЗВАНИЙ РЫБ
Названия отрядов даются жирным шрифтом и прописными буквами, им соответ-
ствует номер римскими цифрами. Названия семейств представлено таким же образом,
но их номер дан арабскими цифрами. Название рода дается строчными буквами, кро-
ме первой - прописной, название и номер рода выделены жирным шрифтом. Название
вида и цифра после него даются только строчными буквами, и они расположены под
родом, к ним добавлены подвидовые названия, если они имеются. Цифры после отря-
да, семейства и рода означают их соответствующий номер в “Систематическом указа-
теле”, а цифра после вида - соответствующий номер в основном тексте “Атласа”.
А - b. chatys 10
- b. stenorrhynchus 10
Abbottina 29 - gueldenstaedtii 11
- esocinus 86 - medirostris 12
- rivularis, 86 - mikadoi 12
Aboma 109 -nudiventris 13
- lactipes 220 - persicus 14
Abramis 30 - p. colchicus 14
- ballerus 87 - p. persicus 14
- brama 88 -ruthenus 15
- b. orientals 88 - r. marsiglii 15
- sapa 89 - schrenckii 16
- s. bergi 89 - stellatus 17
abrau, Clupeonella 27 - s. donensis 17
abyssalis, Asprocottus 275 - sturio 18
ABYSSOCOTTIDAE 33 aculeatus, Gasterosteus 202
Abyssocottus 132 adonis, Oncorhynchus 35
- elochini 272 admirabilis, Dallia 82
- gibbosus 273 affinis, Gambusia 200
- korotneffi 274 albipinnatus, Romanogobio 152
Acanthogobius 110 albula, Coregonus 59
- flavimanus 221 Alburnoides 32
Acanthorhodeus 31 - bipunctatus 92
- asmussii 90 - b. bipunctatus 92
- chankaensis 91 - b. eichwaldi 92
acerinus, Gymnocephalus 212 - b. fasciatus 92
ACIPENSERIFORMES II - b. rossicus 92
ACIPENSERIDAE 2 Alburnus 33
Acipenser 7 - alburnus 93
- baerii 10 - a. alburnus 93
- b. baerii 10 - a. hohenackeri 93
- b. baicalensis 10 alburnus, Alburnus 93
7.42
albus, Salvelinus 54
alexandrae, Cottocomephorus 256
Alosa 10
- alosa 22
- caspia 23
- c. caspica 23
- c. knipowitschi 23
- c. nordmanni 23
- c. palaeostomi 23
- c. persica 23
- c. salina 23
- c. tanaica 23
- kessleri 24
- k. kessleri 24
- k. pontica 24
- k. volgensis 24
- macedonica 22
- maeotica 25
- pontica 25
- p. borystenis 25
- p. issattschenkov 25
- sapidissima 26
alosa, Alosa 22
alpinus, Salvelinus 41
amblycephala, Megalobrama 135
amblystomopsis, Cottus 259
amurensis, Rostrogobio 136
amurensis, Saurogobio 136
ANGUILLIFORMES V
ANGUILLIDAE 11
Anguilla 28
- anguilla 85
- rostrata 85
anguilla, Anguilla 85
anguillicaudatus, Misgumus 185
annularis, Chaenogobius 230
aphipsi, Leuciscus 128
Aphyocypris 34
- chinensis 94
Aplocheilus latipes 199
arcticus, Thymallus 73
argentea, Xenocypris 170
argus, Channa 251
Aristichthys 35
- nobilis 95
asmussii, Acanthorhodeus 90
asotus, Parasilurus 189
Aspius 36
- aspius 96
- a. aspius 96
- a. iblioides 96
- a. taeniatus 96
aspius, Aspius 96
Asprocottus 133
- abyssalis 275
- herzensteini 276
- korjakovi 277
- k. minor 277
- parmiferus 278
- platycephalus 279
- pulcher 280
aurata, Sabanejewia 187
auratus, Carassius 103
autumnalis, Coregonus 60
В
baeri, Benthophilus 222
baerii, Acipenser 10
BAGRIDAE 17
baicalensis, Batrachocottus 252
baicalensis, Comephorus 270
BALITORIDAE 14
ballerus, Abramis 87
bambusa, Elopichthys 116
Barbatula 81
- barbatula 175
- merga 176
- toni 177
barbatula, Barbatula 175
Barbus 37
- barbus 97
- b. borysthenicus 97
- brachycephalus 98
- b, brachycephalus 98
- b. caspius 98
- capito 99
- c. capito 99
- c. conocephalus 99
- ciscaucasicus 100
- tauricus 101
- 1. escherichii 101
- 1. kubanicus 101
barbus, Barbus 97
batrachocephalus, Mesogobius 237
Batrachocottus 124
- baicalensis 252
- multiradiatus 253
- nikolskii 254
- talievi 255
Benthophilus 111
- baeri 222
- casachicus 223
- granulosus 224
- macrocephalus 225
- magistri 226
- m. abdurahmanovi 226
- m. lencoranicus 226
- m. magistri 226
- mahmudbejovi 227 - stellatus 228 bergi, Hyrcanogobius 234 bergianus, Limnocottus 284 bipunctatus, Albumoides 92 bjoerkna, Blicca 102 Blicca 38 - bjoerkna 102 - b. derjavini 102 - b. transcaucasica 102 boganidae, Salvelinus 42 borysthenicus, Leuciscus 129 boulengeri, Cottinella 281 brachycephalus, Barbus 98 Brachymystax 12 - lenok 29 - savinovi 29 brama, Abramis 87 brandtii, Tribolodon 166 brashnikowi, Leiocassis 192 brevicauda, Culter 114 brevirostris, Thymallus 62 brunneus, Rhinogobius 247 bubalus, Ictiobus 172 bulgarica, Sabanejewia 187 chadary, Coregonus 61 Chaenogobius 113 - annularis 230 - castaneus 231 - macrognathus 232 - taranetzi 233 - urotaenia 232 Chalcalburnus 40 - chalcoides 105 - c. aralensis 105 - c. chalcoides 105 - c. mento 105 chalcoides, Chalcalburnus 105 chankaensis, Acanthorhodeus 91 chankaensis, Acheilognathus 91 chankaensis, Gobio 163 chankaensis, Gnathopogon 163 CHANNIDAE 29 Channa 123 - argus 251 - a. argus 251 - a. warpachowskii 251 Chanodichthys 41 - dabryi 106 - eiythropterus 107
C capito, Barbus 99 Carassius 39 - auratus 103 - a. auratus 103 - a. gibelio 103 - carassius 104 - c. jacuticus 104 carassius, Carassius 104 carpio, Cyprinus 115 casachicus, Benthophilus 223 caspia, Alosa 23 caspia, Sabanejewia 188 Caspiomyzon 1 — wagneri 1 Caspiosoma 112 - caspium 229 caspium, Caspiosoma 229 castaneus, Chaenogobius 231 CATOSTOMIDAE 13 Catostomus 79 -catostomus 171 - c. rostratus 171 catostomus, Catostomus 171 caucasica, Knipowitschia 235 caucasica, Sabanejewia 188 cephalus, Leuciscus 130 cemuus, Gymnocephalus 212 - e. eiythropterus 107 - mongolicus 108 cherskii, Chilogobio 158 cherskii, Sarcocheilichthys 158 Chilogobio cherskii 158 Chilogobio soldatovi 160 chinensis, Aphyocypris 94 chishimaensis, Hypomesus 76 Chloea nakamurae 248 - senbae 248 choii, Cobitis 179 Chondrostoma 42 - colchicum 109 -nasus 110 - n. ohridanus 110 - n. nasus 110 - oxyrhynchum 111 -prespense 110 - vardarense 110 - variabile 112 chuatsi, Siniperca 209 cinctus, Micropercops 218 ciscaucasicus, Barbus 100 ciscaucasicus, Romanogobio 153 CLUPEIFORMES Ш CLUPEIDAE 4 Clupeonella 11 - abrau 27 - a. muhlisi 27
- cultriventris 28
- c. caspia 28
- c. cultriventris 28
COBITIDAE 15
Cobitis 83
- choii 179
- lebedevi 179
- lutheri 180
- melanoleuca 181
- m. olivai 181
- rossomeridionalis 182
- taenia 183
cognatus, Cottus 260
colchicum, Chondrostoma 109
COMEPHORIDAE 32
comephoroides, Cottocomephorus 258
Comephorus 131
- baicalensis 270
- dybowski 271
constructor, Neogobius 242
COREGONIDAE 6
Coregonus 20
- alascanus 60
- albula 59
- autumnalis 60
- a. autumnalis 60
- a. migratorius 60
- chadary 61
- clupeaformis 63
- laurettae 62
- lavaretus 63
- 1. baeri 63
- 1. baunti 63
- 1. lavaretus 63
- 1. maraenoides 63
- 1. pallasi 63
- 1. pidschian 63
- muksun 64
- nasus 65
- peled 66
- pollan 60
- sardinella 67
- subautumnalis 60
- tugun 68
- ussuriensis 69
costata, Lefua 178
COTTIDAE 31
Cottinella 134 ’
- boulengeri 281
Cottocomephorus 125
- alexandrae 256
- comephoroides 258
- grewingkii 257
- inermis 258
Cottus 126
- amblystomopsis 259
- cognatus 260
- c. cognatus 260
- c. gracilis 260
- czerskii 261
- gobio 262
- g. gobio 262
- g. koshewnikowi 262
- hangiongensis 263
- poecilopus 264
- p. volki 264
- sibiricus 265
coulteri, Prosopium 70
Ctenopharyngodon 43
- idella 113
Culter 44
- albumus 114
- brevicauda 114
cultratus, Pelecus 142
cultriventris Clupeonella 28
cylindraceum, Prosopium 71
Cyphocottus 135
- eurystomus 282
- megalops 283
cyprinellus, Ictiobus 173
CYPRINIFORMES VI
CYPRINIDAE 12
Cyprinus 45
- caipio 115
- c. carpio 115
- c. haematopterus 115
CYPRINODONTIFORMES VIII
cyrus, Neogobius 242
czekanowskii, Phoxinus 143
czerskii, Chilogobio 158
czerskii, Cottus 261
czerskii, Salvelinus 43
czerskii, Sarcocheilichthys 158
D
dabryi, Saurogobio 161
DALLIIDAE 10
Dallia 27
- admirabilis 82
- delicatissima 83
- pectoralis 84
danilewskii, Leuciscus 131
dauricus, Huso 19
delicatissima, Dallia 83
delineates, Leucaspius 127
demidoffi, Percarina 214
drjagini, Salvelinus 44
dybowski, Comephorus 271
245
igenmanni, Hemiculter 123 ^LEOTRIDIDAE 28 Cleotris 218 dgyticus, Salvelinus 51 :lochini. Abyssocottus 272 Elopichthys 46 - bambusa 116 Entosphenus 2 - tridentatus 2 - 1. ciliaris 2 - 1. tridentatus 2 eperlanus, Osmerus 78 Erythroculter - erythropterus 107 - mongolicus 108 - oxycephalus 106 erythropterus, Chanodichthys 107 erythrophthalmus, Scardinius 162 ESOCIDAE 9 Esox 26 - lucius 80 - reichertii 81 Eudontomyzon 3 - mariae 3 - m. mariae 3 Eupallasella perenurus 145 eurycephalus, Neogobius 242 eurystomus, Cyphocottus 282 ezoe, Tribolodon 167 Gnathopogon 47 -chankaensis 163 - strigatus 117 - ussuriensis 163 GOBIIDAE 28 Gobio 48 - chankaensis 163 - gobio 118 - g. cynocephalus 118 - g. gobio 118 - g. lepidolaemus 118 - soldatovi 119 - s. minulus 119 - s. soldatovi 119 - s. tungussicus 119 gobio, Cottus 262 gobio, Gobio 118 Gobiobotia 49 -pappenheimi 120 godlewskii, Limnocottus 285 gorbuscha, Oncorhynchus 31 gotoi, Procottus 291 granulosus, Benthophilus 224 grewingkii, Cottocomephorus 257 griseus, Limnocottus 286 gueldenstaedtii, Acipenser 11 gurwici, Procottus 291 Gymnocephalus 102 - acerinus 211 - cemuus 212 - schraetser 211
F flavescens, Perea 213 flavimanus, Acanthogobius 221 fluviatilis, Lampetra 4 fluviatilis, Neogobius 238 fluviatilis, Perea 213 fossilis, Misgumus 186 frisi, Rutilus 156 fulvidraco, Pelteobagrus 196 gymnotrachelus, Mesogobius 239 gymnotrachelus, Neogobius 239 H haitej, Mesocottus 267 hakuensis, Tribolodon 168 hangiongensis, Cottus 263 Hemibarbus 50 - labeo 121
G GADIFORMES IX Gambusia 94 - affinis 200 - holbrooki 200 GASTEROSTEIFORMES X GASTEROSTEIDAE 22 Gasterosteus 96 - aculeatus 202 georghievi, Knipowitschia 236 gibbosus, Abyssocottus 273 glanis, Silurus 190 glenii, Perccottus 219 - maculatus 122 Hemiculter 51 - eigenmanni 123 - leucisculus 123 - lucidus 124 herzensteini, Asprocottus 276 herzensteini, Leiocassis 193 holbrooki, Gambusia 200 Hucho 13 - hucho 30 - taimen 30 hucho, Hucho 30 humilis, Oreoleuciscus 139 Huso 8
246
- dauricus 19 - huso 20 - h. maeoticus 20 - h. ponticus 20 huso, Huso 20 Hypomesus 24 - chishimaensis 76 - nipponensis 76 - olidus 77 - o. bergi 77 - o. drjagini 77 - transpacificus 76 Hypophthalmichthys 52 - molitrix 125 Hypseleotris swinhonis 218 Hyrcanogobius 114 - bergi 234 L labeo, Hemibarbus 121 lactipes, Aboma 220 Ladislavia 53 - taczanowskii 126 laevis, Rhodoniichthys 248 lagowskii, Phoxinus 144 Lampetra 4 - fluviatilis 4 - planeri 5 latipes, Aplocheilus 199 latipes, Oryzias 199 laurettae, Coregonus 62 lavaretus, Coregonus 63 lebedevi, Cobitis 179 Lefua 82
I ICTALURIDAE 18 Ictalurus 92 - nebulosus 197 - n. nebulosus 197 -punctatus 198 Ictiobus 80 - bubalus 172 - cyprinellus 173 - niger 174 idella, Ctenopharyngodon 113 idus, Leuciscus 132 iljini, Neogobius 240 inermis, Cottocomephorus 258 - costata 178 Leiocassis 89 - brashnikowi 192 - herzensteini 193 -ussuriensis 194 lenok, Brachymystax 29 Leocottus 127 - kesslerii 266 - k. arachlensis 266 - k. gussinensis 266 lepechini, Salvelinus 46 Leptobotia 85 - mantschurica 184 leptocephalus, Pseudaspius 148 Lethenteron 5
J jacuticus, Salvelinus 45 japonicum, Lethenteron 6 jeittelesii, Procottus 292 - alaskanse 8 - japonicum 6 - kessleri 7 - reissneri 8 Leucaspius 54
К kennerlyi, Salmo 35 kessleri, Alosa 24 kessleri, Lethenteron 7 kessleri, Neogobius 240 kesslerii, Leocottus 266 keta, Oncorhynchus 32 kisutch, Oncorhynchus 33 knerii, Paracottus 268 Knipowitschia 115 - caucasica 235 - georghievi 236 - longecaudata 236 korjakovi, Asprocottus 277 korotneffi, Abyssocottus 274 kronocius, Salvelinus 55 - delineatus 127 leucichthys, Stenodus 72 leucisculus, Hemiculter 123 Leuciscus 55 - aphipsi 128 - borysthenicus 129 - cephalus 130 -danilewskii 131 - idus 132 f - i. idus 132 - i. oxianus 132 -leuciscus 133 - 1. baikalensis 133 - 1. leuciscus 133 - squaliusculus 129 - waleckii 134 - w. tumensis 134
247
leuciscus, Leuciscus 133
leucomaenis, Salvelinus 52
levanidovi, Salvelinus 56
lighti, Pseudoperilampus 150
lighti, Rhodeus 150
Limnocottus 136
- bergianus 284
- godlewskii 285
- griseus 286
- pallidus 287
Lindbergiana 233
longecaudata, Knipowitschia 236
LOTIDAE 21
Lota 95
- lota 201
- 1. leptura 201
- 1. lota 201
lota, Lota 201
lucidus, Hemiculter 124
lucioperca, Stizostedion 215
lucidus, Hemiculter 124
lucius, Esox 80
lutheri, Cobitis 180
M
macrocephalus, Benthophilus 225
macrolepis, Xenocypris 170
maculatus, Hemibarbus 122
magistri, Benthophilus 226
mahmudbejovi, Benthophilus 227
major, Procottus 293
malma, Salvelinus 53
mantschurica, Leptobotia 184
mariae, Eudontomyzon 3
marinus, Petromyzon 9
marmoratus, Pomatoschistus 245
marmoratus, Proterorhinus 246
masou, Oncorhynchus 34
medirostris, Acipenser 12
Megalobrama 56
- amblycephala 135
- terminalis 135
- 1. terminalis 135
megalops, Cyphocottus 283
melanoleuca, Cobitis 181
melanostomus, Neogobius 241
merga, Barbatula 176
Mesocottus 128
- haitej 267
Mesogobius 116
- batrachocephalus 237
mica, Mystus 195
microlepis, Plagiognatops 147
Microphysogobio 57
248
- tungtingensis 136
- 1. amurensis 136
Micropercops 107
- cinctus 218
- swinhonis 218
microps, Pomatoschistus 245
Micropterus 101
- salmoides 210
mikadoi, Acipenser 12
Misgurnus 85
- anguillicaudatus 185
- fossilis 186
molitrix, Hypophthalmichthys 125
mongolicus, Chanodichthys 108
mongolicus, Erythroculter 108
mordax, Osmerus 79
Morone 99
- saxatilis 208
mossambicus, Sarotherodon 217
muksun, Coregonus 64
multiradiatus, Batrachocottus 253
mykiss, Parasalmo 39
Mylopharyngodon 58
-piceus 137
Mystus 90
- mica 195
N
nakamurae, Chloea 248
nasus, Chondrostoma 110
nasus, Coregonus 65
nebulosus, Ictalurus 197
neiva, Salvelinus 47
Neocottus 137
- thermalis 288
- werestschagini 289
Neogobius 117
- constructor 242
- cyrius 242
- eurycephalus 242
- fluviatilis 238
- f. pallasi 238
- gymnotrachelus 239
- g. macrophthalmus 239
- iljini 240
- kessleri 240
- melanostomus 241
- m. affinis 241
- platyrostris 242
- rhodioni 243
- syrman 244
- s. eurystomus 244
nerka, Oncorhynchus 35
niger, Ictiobus 174
nigrolineatus, Syngnathus 207 nikolskii, Batrachocottus 254 nipponensis, Hypomesus 76 nobilis, Aristichthys 95 nudiceps, Pelteobagrus 196 nudiventris, Acipenser 13 - m. mykiss 38 - m. irideus 38 - penshinensis 38 Parasilurus 87 - asotus 189 parmiferus, Asprocottus 278
О obscurus, Tridentiger 249 olidus, Hypomesus 77 Oncorhynchus 14 - adonis 35 - gorbuscha 31 - kennerlyi 35 - keta 32 - kisutch 33 - masou 34 - m. masou 34 - m. macrostomus 34 - nerka 35 - tschawytscha 36 Opsariichthys 59 - uncirostris 138 - u. amurensis 138 Oreoleuciscus 60 - humilis 139 - potanini 140 ORYZIATIDAE 19 Oryzias 93 - latipes 199 OSMERIDAE 8 Osmerus 25 - eperlanus 78 - e. eperlanus 78 - mordax 79 - m. dentex 79 - m. mordax 79 - m. spectrum 79 oxycephalus, Erythroculter 106 oxyrhynchum, Chondrostoma 111 parva, Pseudorasbora 149 pectoralis, Dallia 84 pekinensis, Parabramis 141 Pelecus 62 - cultratus 142 peled, Coregonus 66 Pelteobagrus 92 - fulvidraco 196 -nudiceps 196 pentatrichus, Romanogobio 154 perryi, Parahucho 37 PERCIFORMES ХП PERCIDAE 26 Perea 103 - flavescens 213 - fluviatilis 213 Percarina 104 - demidoffi 214 - d. demidoffi 214 - d. maeotica 214 Perccottus 108 - glehni 219 - glenii 219 PERCICHTHYIDAE 24 perenurus, Eupallasella 145 perenurus, Phoxinus 145 perryi, Parachucho 37 persicus, Acipenser 14 PETROMYZONTIFORMES I PETROMYZONTIDAE 1 Petromyzon 6 - marinus 9 - m, dorsatus 9 Phoxinus 63
P pallidus, Limnocottus 287 pappenheimi, Gobiobotia 120 Parabramis 61 - pekinensis 141 Paracottus 129 - knerii 268 - k. putorania 268 Parahucho 15 - perryi 37 Paraleucogobio strigatus 117 Parasalmo 16 - mykiss 38 - czekanowskii 143 - c. czerskii 143 - c. suifunensis 143 - lagowskii 144 — perenurus 145 — phoxinus 146 ( phoxinus, Phoxinus 146 piceus, Mylopharyngodon 137 Plagiognathops 64 - microlepis 147 planeri, Lampetra 5 platycephalus, Asprocottus 279 platygaster, Pungitius 203 platyrostris, Neogobius 242
249
POECILIIDAE 20 - s. amarus 151
poecilopus, Cottus 264 - s. sericeus 151
POLYODONTIDAE 3 rhodioni, Neogobius 243
Polyodon 9 Rhodoniichthys 121
- spathula 21 - laevis 248
Pomatoschistus 118 Romanogobio 68
- marmoratus 245 - albipinnatus 152
pontica, Alosa 25 - a. albipinnatus 152
potanini, Oreoleuciscus 140 - a. belingi 152
prespense, Chondrostoma 110 - a. vladykovy 152
Procottus 138 -ciscaucasicus 153
- gotoi 290 - pentatrichus 154
- gurwici 291 - tenuicorpus 155
- jeittelesii 292 rostrata, Anguilla 85
- major 293 Rostrogobio amurensis 136
Prosopium 21 ruthenus, Acipenser 15
- coulteri 70 Rutilus 69
- cylindraceum 71 - frisii 156
Proterorhinus 119 - f. frisii 156
- marmoratus 246 - f. kutum 156
Pseudaspius 65 - rutilus 157
- leptocephalus 148 - r. caspicus 157
Pseudoperilampus - r. rutilus 157
- lighti 150 rutilus, Rutilus 157
Pseudorasbora 66 - parva 149 S
pulcher, Asprocottus 280 Sabanejewia 86
punctatus, Ictalurus 198 - aurata 187
Pungitius 97 - a. aralensis 187
- platygaster 203 - a. aurata 187
- p. aralensis 203 - a. balcanica 187
- p. platygaster 203 - a. baltica 187
- pungitius 204 - a. kubanica 187
- p. laevis 204 - a. radnensis 187
- p. pungitius 204 - a. vallachica 187
- sinensis 205 - bulgarica 187
- s. kaibarae 205 - caspia 188
- tymensis 206 - caucasica 188
pungitius, Pungitius 204 SALMONIFORMES IV
Q quadricomis, Triglopsis 269 SALMONIDAE 5 Salmo 17 - salar 40
R - s. salar 40
reichertii, Esox 81 - s. americanus 40
reissneri, Lethenteron 8 - trutta 41
rivularis, Abbottina 86 -1. caspius 41
rivularis, Pseudogobio 86 -1. ciscaucasicus 41
Rhinogobius 120 — t. ezenami 41
- brunneus 247 -1. labrax 41
- b, lindbergi 247 -1. trutta 41
- similis 247 salmoides, Micropterus 210
Rhodeus 67 Salvelinus 18
-lighti 150 - albus 45
- sericeus 151 ~ alpinus 41
250
- a. alpinus 41 sinensis, Pungitius 205
- a. andriashevi 41 sinensis, Sarcocheilichthys 159
- a. erythrinus 41 Siniperca 100
- a. orientalis 41 - chuatsi 209
- boganidae 42 - scherzeri 209
- czerskii 43 soldatovi, Gobio 119
- drjagini 44 soldatovi, Chilogobio 160
- elgyticus 45 soldatovi. Sarcocheilichthys 160
- jacuticus 46 soldatovi, Silurus 191
- kronocius 55 spathula, Polyodon 21
- lepechini 46 Squalidus 73
-1. melanostomus 46 - chankaensis 163
- leucomaenis 52 Squaliobarbus 74
- levanidovi 56 - curriculus 164
- malma 53 squaliusculus, Leuciscus 129
- m. krascheninnikovi 53 stellatus, Acipenser 17
- m. malma 53 stellatus, Benthophilus 228
- neiva 47 Stenodus 22
- schmidti 57 - leucichthys 72
- taimyricus 48 -1. leucichthys 72
- taranetzi 49 -1. nelma 72
- tolmachoffi 50 Stizostedion 105
Salvethymus 19 - lucioperca 215
- svetovidovi 58 - volgense 216
sapa, Abramis 89 strigatus, Gnathopogon 117
sapidissima, Alosa 26 strigatus, Paraleucogobio 117
Sarcocheilichthys 70 sturio, Acipenser 18
- czerskii 158 squaliusculus, Leucisculus 129
- sinensis 159 svetovidovi, Salvethymus 58
- s. lacustris 159 swinhonis, Eleotris 218
- soldatovi 160 swinhonis, Hypseleotris 218
sardinella, Coregonus 67 swinhonis, Micropercops 218
Sarotherodon 106 swinhonis, Perccottus 218
- mossambicus 217 SYNGNATHIFORMES XI
savinovi, Brachymystax 29 SYNGNATHIDAE 23
Saurogobio 71 Syngnathus 98
- amurensis 136 - nigrolineatus 207
- dabryi 161 - n. caspius 207
saxatilis, Morone 208 Scardinius 72 syrman, Neogobius 244
- erythrophthalmus 162 T
scherzeri, Siniperca 209 taczanowskii, Ladislavia 126
schmidti, Salvelinus 57 taenia, Cobitis 183
schrenckii, Acipenser 16 taimen, Hucho 30
SCORPAENIFORMES XIII taimyricus, Salvelinus 48
senbae, Chloea 248 talievi, Batrachocottus 255
sericeus, Rhodeus 151 ’ f taranetzi, Chaenogobius 233
sibiricus, Cottus 265 taranetzi, Salvelinus 49
SILURIFORMES VII tauricus, Barbus 101
SILURIDAE 16 tenuicorpus Romanogobio 155
Silurus 88 terminalis, Megalobrama 135
- glanis 190 thermalis, Neocottus 288
- soldatovi 191 THYMALLIDAE 7
similis, Rhinogobius 247 Thymallus 23
251
- arcticus 73
- a. arcticus 73
- a. baicalensis 73
- a. brevipinnis 73
- a. grubei 73
- a. mertens 73
- a. pallasi 73
- brevirostris 74
- thymallus 75
thymallus, Thymallus 75
Tinea 75
- tinea 165
tinea, Tinea 165
tolmachoffi, Salvelinus 50
toni, Barbatula 177
transpacificus, Hypomesus 76
Tribolodon 76
- brandtii 166
- ezoe 167
- hakuensis 168
tridentatus, Entosphenus 3
Tridentiger 122
- obscurus 249
- o. brevispinis 249
- trigonocephalus 250
Triglopsis 130
- quadric omis 269
trigonocephalus, Tridentiger 250
trutta, Salmo 41
tschawytscha, Oncorhynchus 36
tungtingensis, Mycrophysogobio 136
tugun, Coregonus 68
tymensis, Pungitius 206
U
uncirostris, Opsariichthys 138
urotaenia, Chaenogobius 232
ussuriensis, Coregonus 69
ussuriensis, Gnathopogon 163
ussuriensis, Gobio 163
ussuriensis, Leiocassis 194
V
vardarense, Chondrostoma 110
variabile, Chondrostoma 112
Vimba 77
- vimba 169
- v. persa 169
- v. tenella 169
- v. vimba 169
vimba, Vimba 169
volgense, Stizostedion 216
W
wagneri, Caspiomyzon 1
waleckii, Leuciscus 134
werestchagini, Neocottus 289
X
Xenocypris 78
- agrentea 170
- macrolepis 170
СОДЕРЖАНИЕ
Предисловие ........................................................... 5
Систематический указатель видов........................................ 6
Отряд VII. SILURIFORMES - СОМООБРАЗНЫЕ ............................ 11
Сем. 16. Siluridae - сомовые .................................... 11
Сем. 17. Bagridae - косатковые................................... 18
Сем. 18. Ictaluridae - икталуровые............................... 28
Отряд УШ. CYPRINODONTIFORMES - КАРПОЗУБООБРАЗНЫЕ................... 32
Сем. 19. Oryziatidae - оризиевые ................................ 32
Сем. 20. Poeciliidae - пецилиевые ............................... 34
Отряд IX. GADIFORMES - ТРЕСКООБРАЗНЫЕ .............................. 37
Сем. 21. Lotidae - налимовые .................................... 37
Отряд X. GASTEROSTEIFORMES - КОЛЮШКООБРАЗНЫЕ........................ 40
Сем. 22. Gasterosteidae — колюпгковые............................ 40
Отряд XI. SYNGNATHIFORMES - ИГЛООБРАЗНЫЕ ........................... 51
Сем. 23. Syngnathidae - игловые ................................. 51
Отряд XII. PERCIFORMES - ОКУНЕОБРАЗНЫЕ ............................. 53
Подотряд PERCOIDEI - окуневидные
Сем. 24. Percichthyidae - перцихтовые ........................... 53
Сем. 25. Centrarchidae - центрарховые............................ 57
Сем. 26. Percidae - окуневые .................................... 60
Сем. 27. Cichlidae - цихловые.................................... 74
Подотряд GOBIOIDEI - бычковидные
Сем. 28. Eleotrididae - головешковые ............................ 77
Сем. 29. Gobiidae - бычковые .................................... 82
Подотряд CHANNOIDEI - змееголововидные
Сем. 30. Channidae - змееголовые ............................... 141
Отряд ХШ. SCORPAENIFORMES - СКОРПЕНООБРАЗНЫЕ ...................... 144
Подотряд COTTOIDEI - рогатковидные
Сем. 31. Cottidae - керчаковые ................................. 144
Сем. 32. Comephoridae - голомянковые............................ 179
Сем. 33. Abyssocottidae - глубоководные широколобки............. 182
Литература........................................................... 215
Указатель русских названий рыб....................................... 234
Указатель латинских названий рыб .................................... 242
Научное издание
АТЛАС
ПРЕСНОВОДНЫХ РЫБ
РОССИИ
В двух томах
Том 2
Второе издание
Утверждено к печати
Ученым советом
Института проблем экологии и эволюции
им. А.Н. Северцова
Российской академии наук
Зав. редакцией
Н.А. Степанова
Редактор
А.М. Гидалевич
Художественный редактор
Е.А. Быкова
Технический редактор
ТА. Резникова
Корректоры
Р.В. Молоканова, ЕЛ. Сысоева,
Т.И. Шеповалова
Подписано к печати 05.02.2003
Формат 70 х lOO'/is
Гарнитура Таймс. Печать офсетная
Усл.печ.л. 20,8. Усл.кр.-отт. 21,3. Уч.-изд.л. 24,1
Тираж 1030 экз. Тип. зак. 4007
Издательство “Наука”
117997 ГСП-7, Москва В-485, Профсоюзная ул., 90
E-mail: secret@naukaran.ru
Internet: www.naukaran.ru
Санкт-Петербургская типография “Наука”
199034, Санкт-Петербург В-34, 9-я линия, 12