/



































































































































































































































Текст
Э. БЭКЕР и Г. УАРТОН
ВВЕДЕНИЕ
АКАРОЛОГИЮ
Э. БЭКЕР и Г. УАРТОН
ВВЕДЕНИЕ
АКАРОЛОГИЮ
Перевод с английского
А. А. ЗЕМСКОЙ
Под редакцией и с вступительной статьей
икну. 1'.. II. ПАВЛОВСКОГО
и * л
ИЗДАТЕЛЬСТВО
ИНОСТРАННОЙ ЛИТЕРАТУРЫ
Москва — 1 95S
AN INTRODUCTION TO ACAROLOGY
By
E. W. BAKER and G. W. WHARTON
1952
ВСТУПИТЕЛЬНАЯ СТАТЬЯ
Клещи (Acarina) представляют собой сборную группу, объединяю-
щую три отряда класса паукообразных (Arachnoidea). Среди клещей
имеются как свободноживущие формы, так и паразитические, пол-
ностью или временно связанные со своими хозяевами. Хозяевами клещей
служат многие наземные позвоночные, а среди беспозвоночных — насе-
комые, ракообразные и, редко, скорпионы. Известны клещи, питаю-
щиеся соком растений; другие виды питаются зерновыми продуктами
(мучные клещи); наконец некоторые клещи принимают участие в раз-
рушении трупов. Этот краткий перечень показывает, сколь велико
экологическое разнообразие, характеризующее эту группу пауко-
образных.
Многие растительноядные клещи являются важными вредителями
сельского хозяйства. Многие свободноживущие панцырные клещи (Ori-
batidae) играют роль промежуточных хозяев некоторых ленточных
червей, паразитирующих у домашних животных. Но особенно вредны
паразитические клещи, переносящие возбудителей трансмиссивных
болезней человека и домашних и промысловых животных. Установлено,
что в большинстве случаев клещи являются единственными посредни-
ками в передаче возбудителей самых разнообразных болезней чело-
века— вирусных, бактериальных, протозойных, риккетсиозов и др.
Это придает особое значение изучению биологии клещей, а также
условий нападения клещей на своих хозяев и заражения их возбуди-
телями передаваемых болезней, так как все эти знания совершенно
необходимы для борьбы с клещами в целях предупреждения этих
болезней.
Для изучения всех этих вопросов необходимо прежде всего уметь
правильно определить вид данного клеща, что в свою очередь требует
изучения систематики клещей. Между тем в коллекциях клещи до сих
пор представлены очень бедно; это объясняется тем, что многие из них
имеют крайне малые размеры, паразитические же виды требуют особых
4
Вступительная статья
методов сбора. Кроме того, различные группы клещей изучены крайне
неравномерно, так как клещи изучались главным образом в связи
с выявлением вреда, наносимого ими здоровью человека и сельскому
хозяйству. Поэтому одни группы изучаются уже давно и литература
по ним огромна, тогда как исследование других групп началось срав-
нительно недавно. Так, иксодовые клещи, переносящие возбудителей
протозойных болезней домашних животных (гемоспоридиозов, пиро-
плазмозов, тейлериозов и др.) давно уже представляли излюбленный
объект исследований в ветеринарии. Проявляемый к ним интерес
станет понятным, если вспомнить, что гемоспоридиозы домашних
животных в ряде местностей серьезно понижают продуктивность
животноводства. Хорошо изучены и многие чесоточные клещи, повреж-
дающие шерсть различных домашних животных, в частности овец.
Вместе с тем краснотелковые и гамазовые клещи привлекли внимание
сравнительно недавно в связи с изучением риккетсий и риккетсиозов, и
литература по систематике этих групп за последние годы быстро растет.
Необходимо также указать, что литература по клещам разбросана
по множеству различных изданий, порой трудно доступных или вовсе
недоступных для широкого использования. На основании всего сказан-
ного становится понятным, сколь велика потребность в сводках,
определителях и других подсобных изданиях по акарологии.
В связи с этим книга Э. Бэкера и Г. Уартона „Введение в акаро-
логию" представляет определенный интерес. Авторы ее поставили себе
задачу дать сводку материалов по мировой фауне клещей, касаясь
главным образом их систематики. Объем включенного в книгу мате-
риала огромен: 209 семейств и 1390 родов клещей. Это определяет
сжатость изложения и делает книгу более доступной для лиц, имеющих
некоторую зоологическую подготовку.
Основному содержанию книги авторы предпосылают введение,
в котором изложены общие сведения по методике изучения, морфологии
и жизненному циклу клещей. Далее описываются по группам все
семейства клещей. Даны диагнозы и определительные таблицы систе-
матических подразделений клещей до семейства включительно и списки
входящих в них родов и подродов (синонимика дается в минимальном
объеме). Как правило, для каждого семейства приведены краткие све-
дения по биологии, жизненному циклу и вредоносному значению
(обычно применительно к отдельным наиболее хорошо изученным
видам), а также основная литература.»
Вступительная статья
5
Сводка такого обширного материала полезна и необходима для
дальнейшей разработки акарологии, значение которой растет все более
и более по мере выяснения роли клещей в передаче возбудителей
многих трансмиссивных болезней и некоторых паразитических червей.
Это и сделало желательным выпуск книги Бэкера и Уартона в русском
переводе.
В СССР изучение клещей в последнее время бурно развивается.
Несмотря на это, в книге Бэкера и Уартона русская литература почти
не использована. Поэтому мы сочли необходимым дополнить книгу
списком важнейшей русской литературы по клещам.
В заключение следует отметить, что авторы книги придержи-
ваются устаревших взглядов на систему Acarina и'новые представления
о составе группы, развитые безвременно скончавшимся советским
акарологом А. А. Захваткиным, не отражены в книге.
Акад. Е. И. Павловский.
ОСНОВНАЯ РУССКАЯ ЛИТЕРАТУРА
ПО КЛЕЩАМ!
I. РАБОТЫ ОБЩЕГО ХАРАКТЕРА
За хват кин А. А., 1952. Разделение клещей (Acarina) на отряды и их
положение в системе Chelicerata, Паразит, сб. Зоол. инет. Акад, наук
СССР, т. 14, 5—46.
3 а х в а т к и н А. А., 1953. Сборник научных работ, Изд. Московского уни-
верситета. (Работы по морфологии, развитию и фауне различных групп
клещей.)
II. МЕТОДИЧЕСКИЕ ПОСОБИЯ
Галузо И. Г., 1946. Кровососущие клещи Казахстана, т. I, Алма-Ата.
Олсуфьев Н. Г., 1941. К методике лабораторного разведения иксодовых
клещей, Л1<?6. паразит., 10 (3—4), 436.
Павловский Е. Н., 1928. Наставление к собиранию и исследованию кле-
щей (1хобо1беа),Изд. Акад, наук СССР, Ленинград.
П о с п с л о в а-Ш т р о м М. В., 1941. К методике кормления клещей 1хо-
doidea в лаборатории, Мед. паразит., 10 (3—4), 433.
Померанцев Б. И., С с р д ю к о в а Г. В., 1940. Методика экологических
исследований в тайге над клещами сем. Ixodidae, переносчиками
весенне-летнего энцефалита и других болезней, Сб. изобрст. рациона-
лизат. предл. Воен.-мед. акад. им. С. М. Кирова, № 1.
III. РАБОТЫ ПО ОТДЕЛЬНЫМ ГРУППАМ КЛЕЩЕЙ
Gatnasoidea (гамазовые клещи)
Брегетов а Н. Г., 1949. Материалы по фауне клещей сем. Haemogamasi-
dac в СССР, Паразит, сб. Зоол. инет. Акад, наук СССР, т. И, 161—185.
Брсгетова Н. Г., 1949. О паразитических клещах рода Myonyssus (Ga-
masoidea, Liponyssidae), Докл. Акад. наук СССР, 67 (4), 751 753.
Брегетова Н. Г., 1950. Новые виды гамазовых клещей (Acarina, Gamasoi-
dea), Паразит, сб. Зоол. инет. Акад, наук СССР, т. 12, 13—16.
Брсгетова Н. Г., 1951. Клещи, паразитирующие в носовой полости птиц,
Паразит, сб. Зоол. инет. Акад, наук СССР, т. 13, 111—119.
Брсгетова Н. Г., 1952. Сбор и изучение гамазовых клещей, Изд. Акад,
наук СССР, М. — Л., 1—39.
Брсгетова Н. Г., 1952. Новые виды клещей рода Haemolaelaps (Gama-
soidea, Laelaptidae), паразитирующие на грызунах, Зоол. жури., 31 (6),
866—874.
Брсгетова Н. Г., 1953. К фауне гамазовых клещей Дальнего Востока,
Паразит, сб. Зоол. инет. Акад, наук СССР, т. 15, 302—338.
Брсгетова Н. Г., 1953. Клещи рода Bdellonyssus Fonseca, 1941 (Gama-
soidea, Liponyssidae), Труды. Зоол. инет. Акад, наук СССР, 13, 310—319.
f В список включены преимущественно работы по систематике, морфо-
логии и методике изучения клещей. Из работ по медицинскому и ветери-
нарному значению клещей приведены лишь наиболее важные.
8 Основная русская литература по клещам
Гроховская И. М., 1954. Изучение гамазовых клещей в приходном очаге
риккетсиоза и разработка системы мероприятий по ликвидации очага,
Тезисы докл. научи, сессии Мин. здрав. СССР, АМН СССР и Инет,
эпидемиол. и микробиол., Медгиз, 21—23.
За хват кин А. А., 1948. Систематика рода Laelaps (Acarlna, Parasitifor-
mes) и вопросы его эпидемиологического значения, Паразит, сб. Зоол.
инет. Акад, наук СССР, т. 10, 50—75.
Земская А. А„ 1951. Биология и развитие куриного клеща Dermanyssus
gallinae в связи с его эпидемиологическим значением, Зоол. журн.,
30 (1), 51-62.
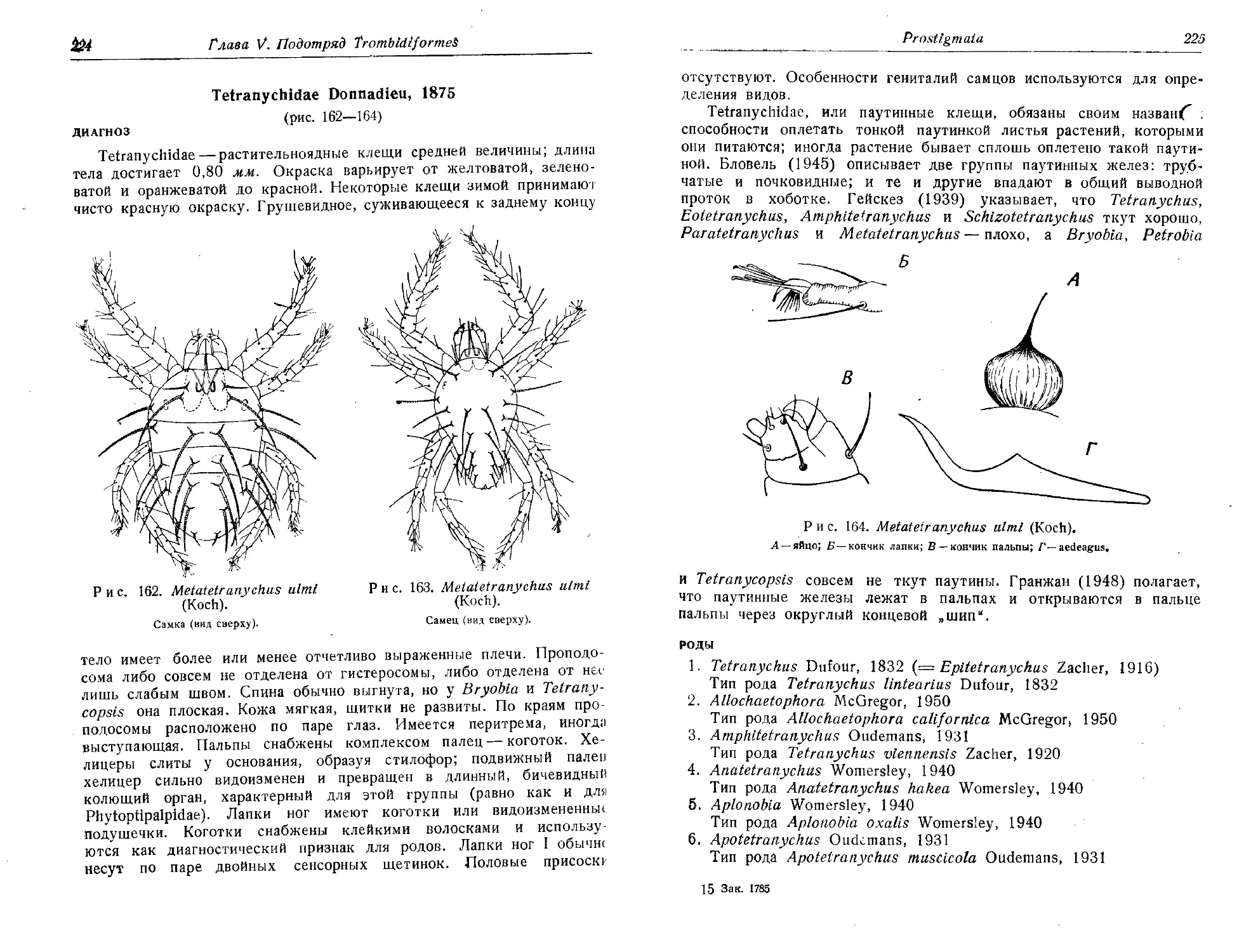
Земская А. А., 1951. Биология и развитие клещей сем. Dermanyssldae,
паразитирующих на рептилиях, в связи с проблемой возникновения
пастбищного паразитизма, Бюлл. Моск. общ. испыт. природы, 56 (3),
42—57.
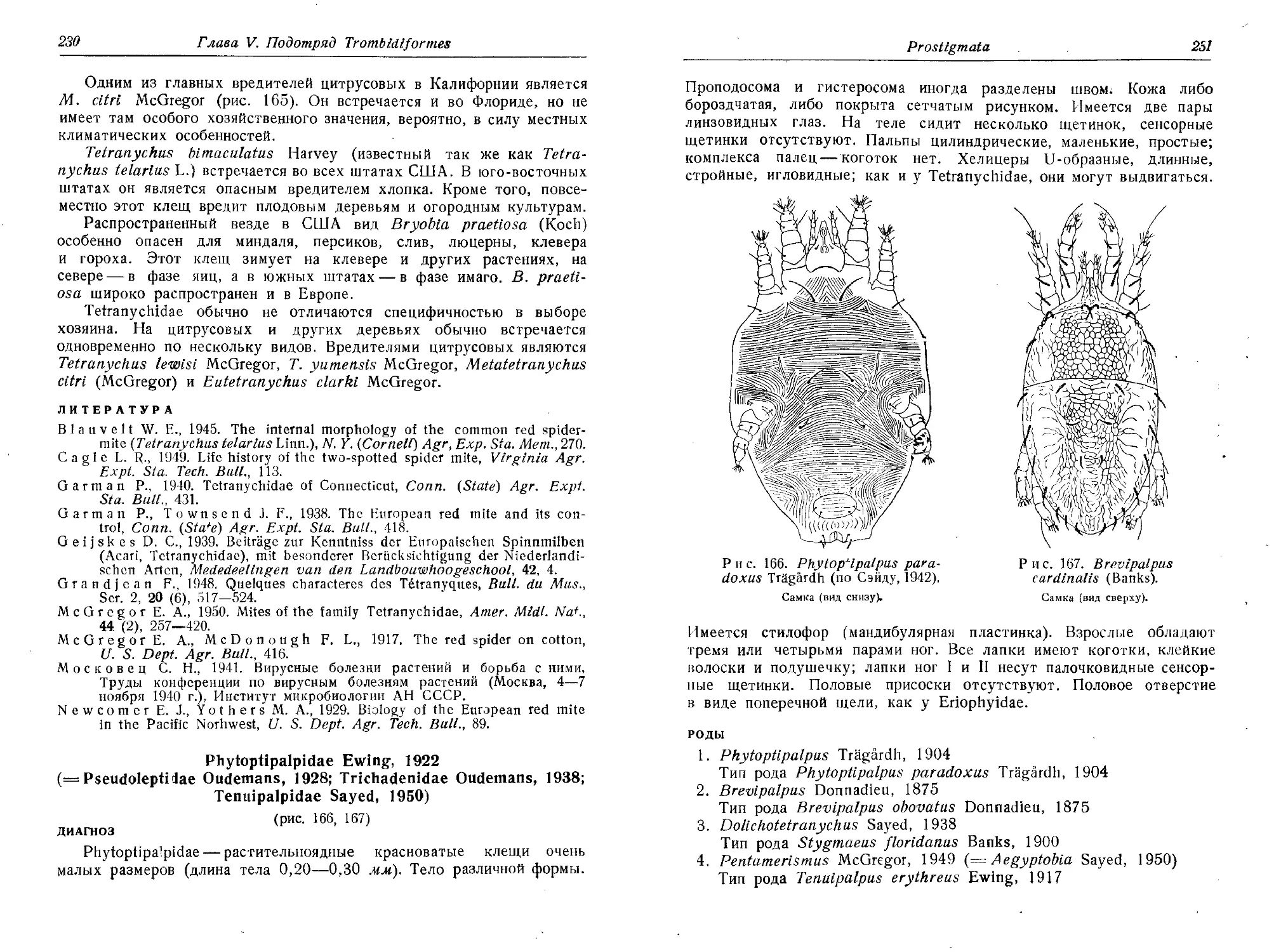
Кулагин С. М., Земская А. А., 1953. Гамазоидный клещ Alloderma-
nyssus sanguineus Hirst как переносчик везикулезного риккетсиоза,
Вопросы краевой, общей, эксперимент, паразит, и медиц. зоол., 8,
34—40.
Ланге А. Б., 1948. О некоторых Laelaps s. str. (Acarina, Parasitiformes),
паразитирующих на мышах подссм. Mtirinae, Паразит, сб. Зоол. инет.
Акад, наук СССР, т. 10, 77—86.
Нельзина Е. Н., 1951. Крысиный клещ, Изд. АМН СССР, 1—100.
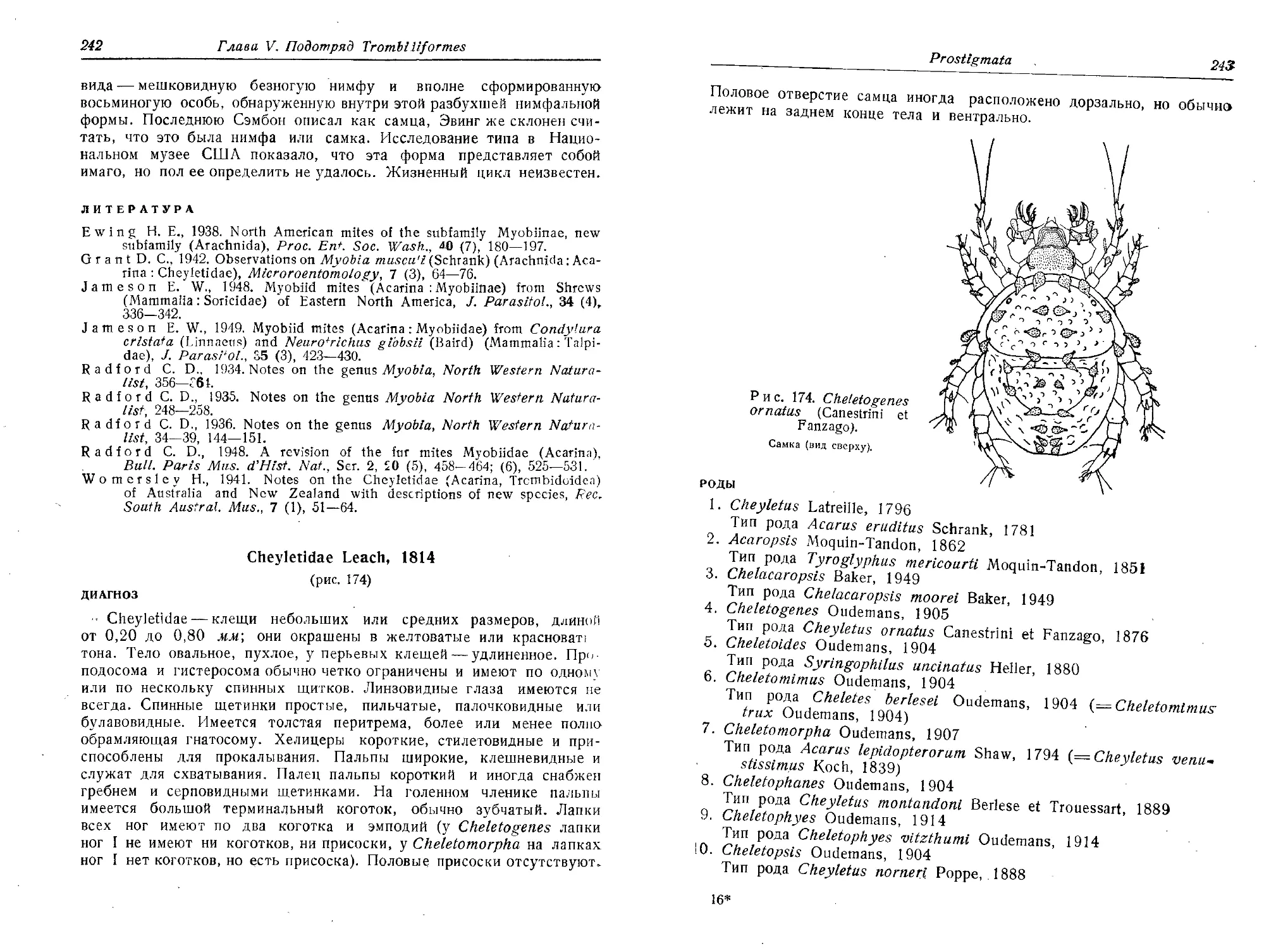
Нельзина Е. Н., Барков И. 11., 1951. Носительство туляремийного
микроба {В. iularense) некоторыми видами гамазовых клещей в есте-
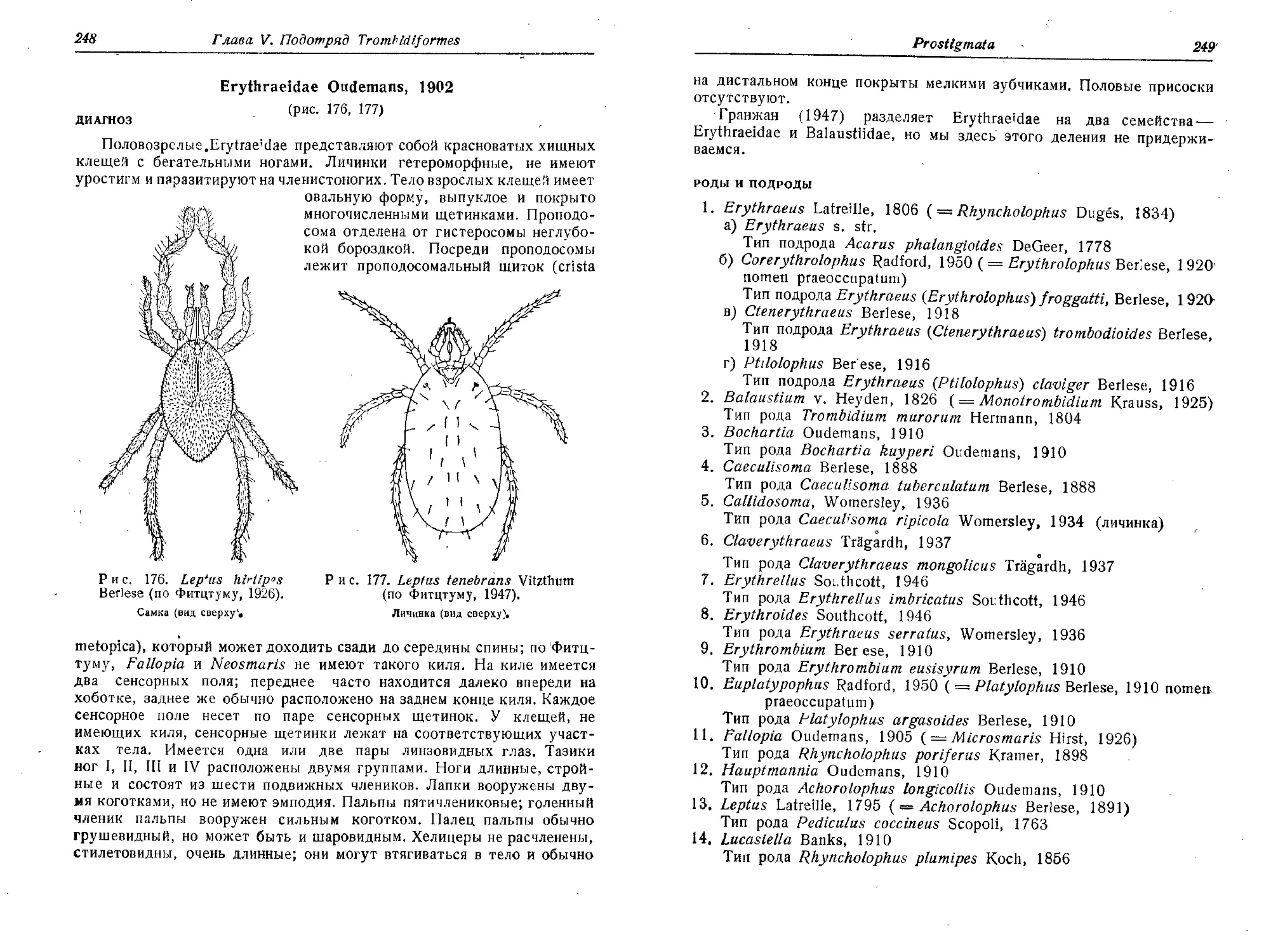
ственных условиях, Докл. Акад, наук СССР, 78 (4), 829—831.
Нельзина Е. Н., Романова В. П., 1951, Способ передачи туляремий-
ного микроба (В. iularense) гамазовыми клещами, Докл. Акад, наук
СССР, 78 (1), 179—180.
Павловский Е. Н., Штейн А. К., 1933. О действии Dermanyssus gallinae
Red! (Acarina) на кожные покровы человека, Энтомол. обозр., 25 (1—2),
104—105.
Разумова И. Н., 1953. Новые виды гамазовых клещей сем. Liponyssidae,
Паразит, сб. Зоол. инет. Акад, наук СССР, т. 15, 339—344.
AROAS1DAE
Галузо И. Г., 1952. Кровососущие клещи Казахстана, т. 5, Алма-Ата.
Павловский Е. Н., Лотоцкий Б. В., 1948. Наружная морфология
переносчика клещевого возвратного тифа в Средней Азии клеща
Orniihodorus paplllipes, Паразит, сб. Зоол. инет. Акад, наук СССР, т. 9.
Павловский Е. Н„ Скрынник А. Н., 1937. Наблюдения над биологией
клеща Ornithodorus paplllipes переносчика клещевого тифа в СССР,
Труды Воен. мед. акад., 8, 277.
Поспслова-Штром М. В., 1953. Клещи-орнитодорины и их эпидемио-
логическое значение, Москва, 1953.
Ixodidae1
Галузо И. Г., 1946—1950. Кровососущие клещи Казахстана, т. 1—4,
Алма-Ата.
Померанцев Б. И., 1950. Иксодовые клещи (Ixodidae). Фауна СССР,
Паукообразные, т. 4 (2), Изд. Акад, паук СССР, Ленинград.
Померанцев Б. И., Сердюкова Г. В., 1947. Экологические наблюдения
над клещами сем. Ixodidae, переносчиками весеппс-летпего энцефалита
на Дальнем Востоке, Паразит, сб. Зоол. инет. Акад, наук СССР, т. 9.
1 Русская литература по иксодовым клещам очень велика и здесь при-
водятся лишь наиболее важные работы.
Основная русская литература по клещам
9
1 Tetranychoidea (паутинные и плоские клещи)
Багдасарян А. Т., 1951. К фауне паутинных клещей (сем. Tetranychidae)
' Еревана и его окрестностей, Изв. Акад, наук Армянок. ССР, 4 (4).
Беккер Э. Г., 1935. Челюстной аппарат паутинного клещика и его отпра-
вления в связи с вопросом о химической борьбе с клещиком, Зоол.
журн., 14 (4).
Бондаренко Н. В., 1950. Влияние укороченного дня на годичный цикл
развития обыкновенного паутинного клещика, Докл. Акад, наук СССР,
70 (6).
Бондаренко Н. В., 1952. Паутинный клещик и борьба с ним в парниках
и теплицах, Изд. Акад, наук СССР, М. — Л., 1—53.
Никольский В. В., 1947. Видовой состав паутинных клещей семейства
Tetranychidae на хлопчатнике, Труды Азербайджанок, научно-исслед.
инет, земледелия, 55.
Р е к к Г. Ф., 1941. Клещи, вредящие культурным растениям, Изд. Акад,
наук Груз. ССР, Тбилиси, 1—96.
Рекк Г. Ф., 1948. Род ВгуоЫа по материалам из Грузии, Сообщ. Акад,
наук Груз. ССР, 8 (9—10).
Рекк Г. Ф., 1948. Род Schizo^efranychus по материалам из Грузии, Сообщ.
Акад, наук Груз. ССР, 9 (6).
Рекк Г. Ф., 1949. К установлению возрастных различий у паутинных кле-
щей, Сообщ. Акад, наук Груз. ССР, 10 (7).
Рекк Г. Ф., 1950. Материалы к фауне паутинных клещей Грузии, Труды
Инет, зоолог. Акад, наук Груз. ССР, 9.
Cheyletidae (хищные клещи)
Волгин В. И., 1949. Материалы по систематике хищных клещей рода
Cheyletus Latr. (Cheyletidae, Acarina), Докл. Акад, наук СССР, 64
(4), 583—586.
Захват кин А. А., 1935. Краткий определитель амбарных клещей, Изд.
Комит. заготов. с.-х. продуктов при СНК СССР, Москва, 1—31.
За хватким А. А., 1949. Новые виды амбарных клещей из Азербайджан-
ской ССР, Энтомол. обозр., 30 (3—4) 287—290.
Родендорф Б. Б., 1940. Определитель хищных и пузатых клещей (Chey-
letidae и Pediculoididae), Учен. зап. Моск. гос. унив,, Зоология, № 42,
69—98.
Demodicoidea (железницы)
Соколовский В. А., 1952. Железница (демодекоз) собак и ее лечение,
Автореф. докт. диссерт., Изд. Харьковск. ветерин. инет., Харьков, 1—26.
Trombidiidae (краснотелки)
Высоцкая С. О., Шлугер Е. Г.. 1953. Личинки краснотелок, пара-
зиты грызунов Ленинградской области, Паразит, сб. Зоол. инет. Акад,
наук СССР, т. 15, 345—352.
Соснина Е. Ф., 1950. Исследование фауны клещей краснотелок в Таджи-
кистане, Сообщ. Тадж. фил. Акад, наук СССР, 28, 7—9.
Шлугер Е. Г., 1948. Новые краснотелки (Acari, Trombidiidae) фауны СССР,
Энтомол. обозр., 30 (1—2), 157—164.
Шлугер Е. Г., 1949. Некоторые данные по биологии Trombicula pavlov-
skyl, Вопросы краевой, общей и экспериментальной паразитоло-
гии, 4, 65—66,
10
Основная русская литература по клещам
Ш л у г е р Е. Г., 1950. Паразитические клещи подсемейства Trombiculinae
фауны СССР, Автореф. канд. диссерт., Зоол. инет. Акад, наук СССР,
Ленинград, 1—17,
Hydracarina (водяные клещи)
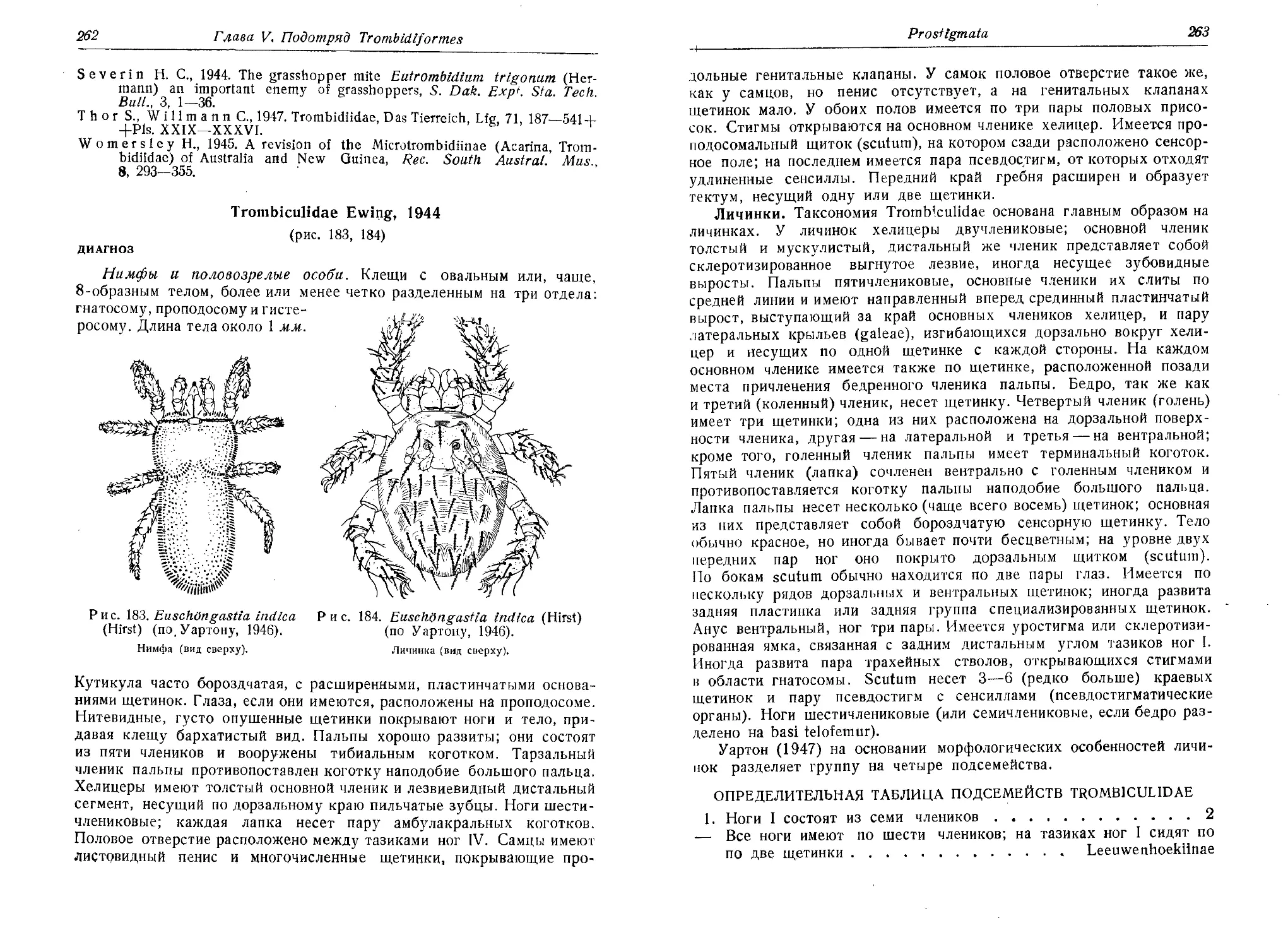
Соколов И. И., 1940. Hydracarina — водяные клещи (I, Hydrachnellae).
Фауна СССР, Паукообразные, т. 5 (2), 1—511, Изд. Акад, наук СССР,
Ленинград.
Соколов И. И., 1952. Водяные клещи (II, Halacarae). Фауна СССР, Пауко-
образные, т. 5 (5), 1—201, Изд. Акад, наук СССР, Ленинград.
Tyroglyphoidea (хлебные клещи)
Волгин В. И., 1940. Материалы по систематике клещей рода Tyrophagus
Ouds., 1923 (Tyroglyphidae, Acarina), Докл. Акад, наук СССР, 65 (3),
385 --388.
Волгин В. И., 1952. Новые виды клещей рода Rhizoglyphus Clap. (Aca-
rina, Tyroglyphidae), Зоол. журн., 31 (2), 249—252.
Захваткин А. А., 1936. О распрос гранении хлебных клещей в полевых
условиях, Зоол. журн., 15 (4), 697—719.
Захваткин А. А., 1941. Тироглифоидные клещи (Tyroglyphoidea), Фауна
СССР, Паукообразные, т. 6 (2), 1—475, Изд. Акад, наук СССР, Ленинград.
Захваткин А. А., 1953. Исследования по морфологии и постэмбриональ-
ному развитию тироглифид (Sarcoptiformes, Tyroglyphoidea (см.
А. А. Захваткин, Сб. научных работ, Изд. Московского универси-
тета, 19—120).
Каменский А. Ф., 1940. Хлебные клещи в целинных степях Казахстана.
Материалы о происхождении амбарного комплекса тироглифид, Зоол.
журн., 19 (4), 603—617.
Павловский Е. II., 111 г е й и А. К., 1930. Экспериментальное исследо-
вание над действием клеща Rhizoglyphus hyacinth!, вызывающего
у человека своеобразную форму дерматита, Паразит, сб. Зоол. ипст.
Акад, паук СССР, т. 1, 19—26.
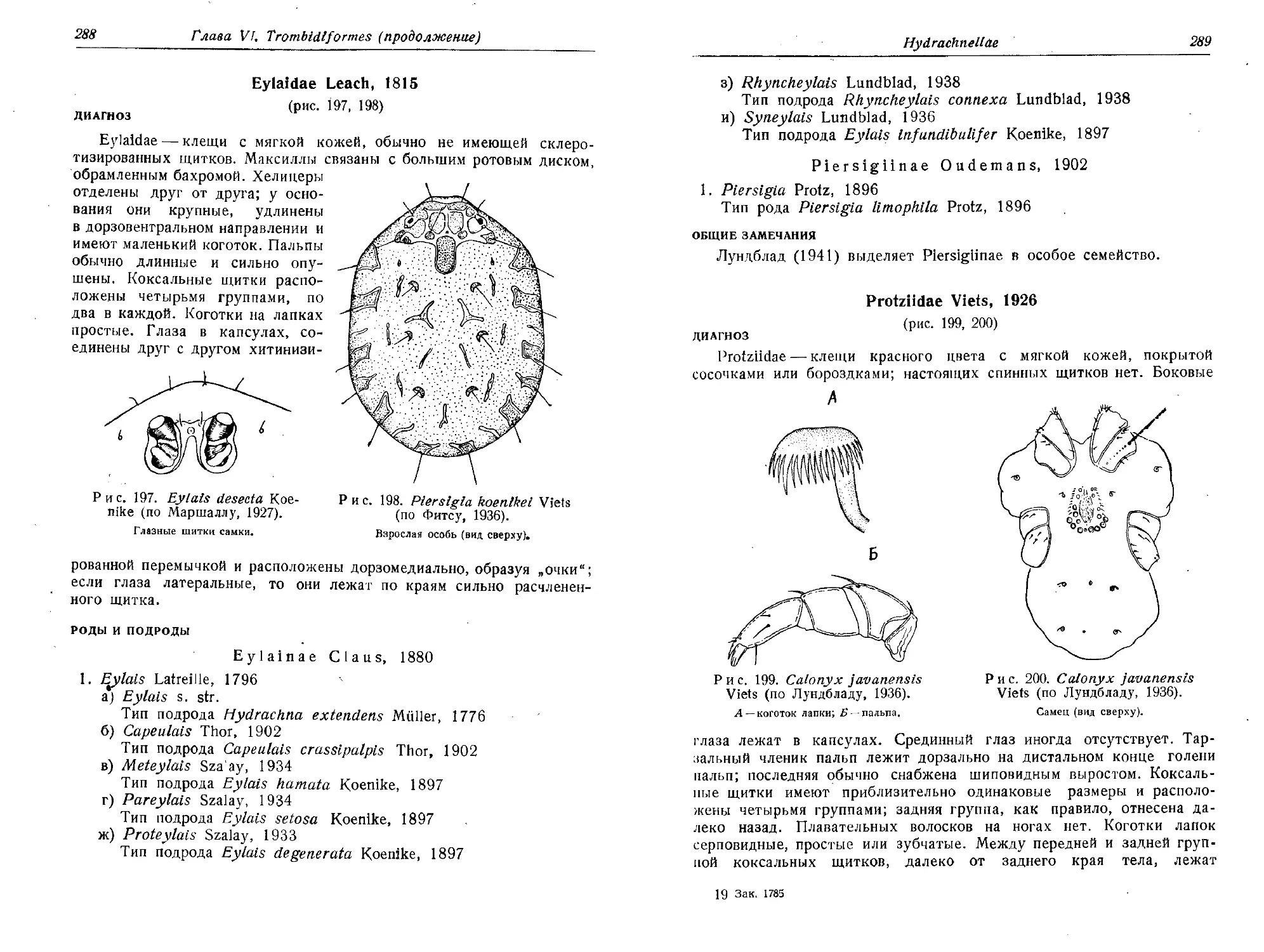
Родионов 3. С., 1937. Условия массового развития хлебных клещей,
Зоол. журн., 16 (3), 511—546.
Соколов И. И., 1952. Наблюдения над эмбриональным развитием амбар-
ных клещей, Тр. Ленингр. общ. естество аспытп., 71 (4), 243—260.
Сорокин С. В., 1949. Экология хлебных клещей в колхозных зернохра-
нилищах Вологодской области, Зоол. журн., 28 (4), 323—340.
Сорокин С. В., 1951. Причины образования гипопусов хлебных клещей,
Зоол. журн., 30 (6), 523—539.
Сорокин С. В., 1953. Особенности распространения хлебных клещей
в гнездах мышевидных грызунов на сельскохозяйственных землях,
Зоол. журн., 32 (1), 60—76.
У ш а т и нс кая Р. С., 1945. Влияние температуры и влажности на образование
гипопусов Qlycyphagus destructor Schr., Зоол. журн., 24 (3), 165—174.
ПРЕДИСЛОВИЕ АВТОРОВ
За последние несколько лет со стороны акарологов, а также
исследователей, работающих в смежных областях (энтомология, зоо-
логия, паразитология), отмечается рост интереса как к элементарным,
так и к достаточно подробным сводкам по клещам. До 1940 г. в США
клещами занимались лишь немногие исследователи. В настоящее время
в связи с выявлением роли клещей в переносе возбудителей заболе-
ваний человека и с проблемами, возникшими в сельском хозяйстве
в результате применения ДДТ, расчистившим почву для ряда неиз-
вестных и не описанных ранее видов, ставших серьезными вредите-
лями растений, число специалистов, заинтересованных в расширении
знаний о клещах, значительно возросло. Именно для таких лиц и
написана эта книга.
В 1942 г. немецкий акаролог Фитцтум закончил раздел по клещам
в сводке Бронна „Klassen und Ordnungen des Tierreichs“. Эта все-
объемлющая и исчерпывающая работа слишком трудна для того, чтобы
ее можно было рекомендовать начинающему исследователю. Для начи-
нающих исследователей одним из наиболее ощутимых недостатков
монографии Фитцтума является полное отсутствие определительных
таблиц и описаний семейств в последовательном порядке. При напи-
сании настоящего руководства мы широко использовали труд Фитцтума
и следуем здесь его классификации за исключением тех случаев,
когда наши собственные исследования указывали на необходимость
ее изменений.
В введении к настоящей книге содержатся основные сведения
о строении, развитии, и экологии клещей; в дальнейших главах после-
довательно рассматривается каждое из семейств клещей в целом.
В книге приведены разработанные нами определительные таблицы
всех основных групп клещей, пользуясь которыми читатель сможет
правильно определить семейство, к которому принадлежит любой
интересующий его клещ. Описания семейств построены по единому
плану и содержат диагноз, перечень родов и их типов1, общие заме-
чания и список основной литературы по группе. В разделе „Общие
замечания" приведены основные данные по биологии и хозяйственному
и медицинскому значению описываемой группы; особое внимание
1 См. S. A. Neave, Nqmenclator goologicus, 1939.
12
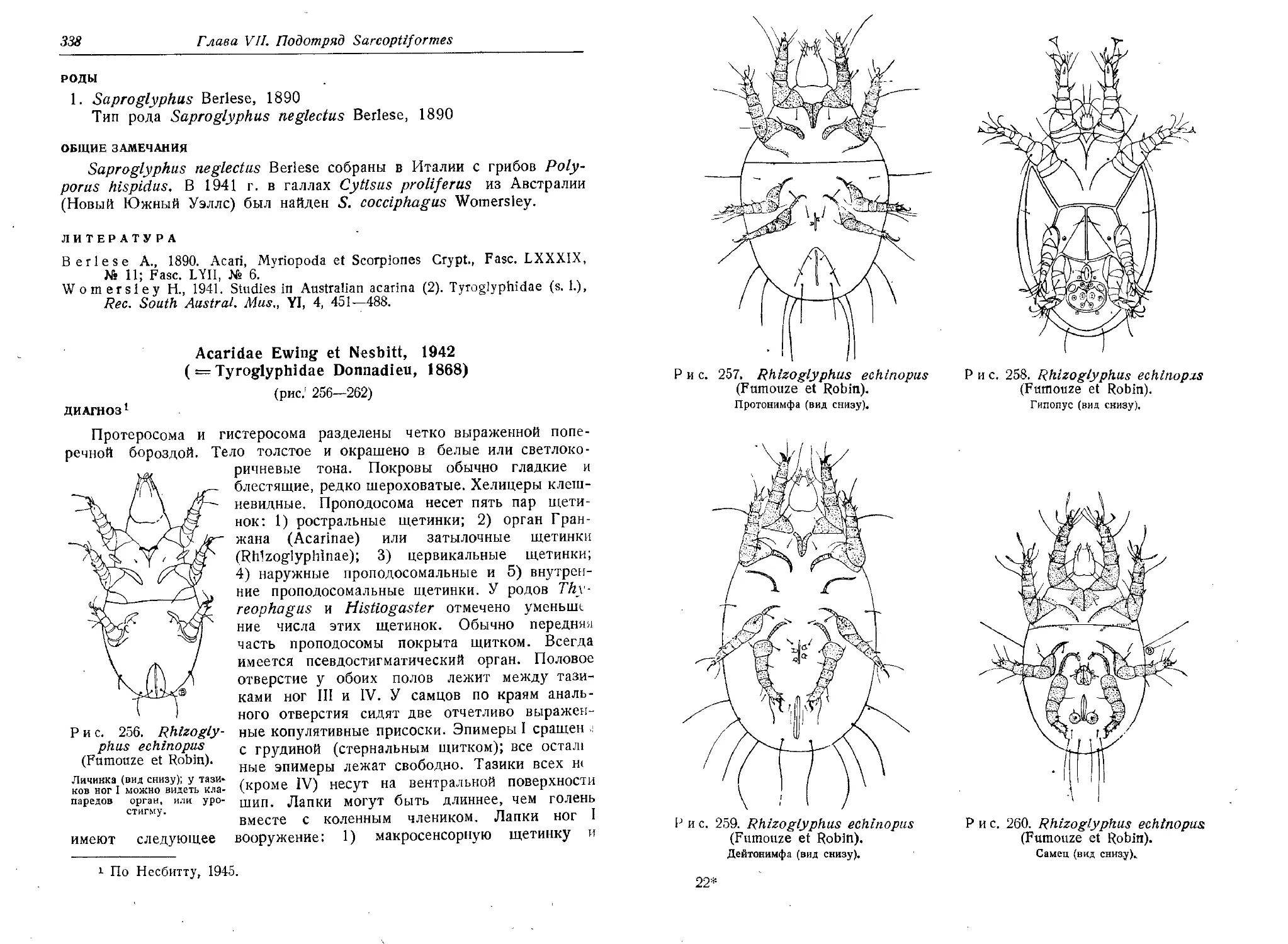
Предисловие авторов
уделено видам, имеющим большее практическое значение. Помимо всего
этого, в тексте помещены рисунки, изображающие типичных предста-
вителей каждого из описываемых семейств.
Клещи, как группа в целом, еще мало известны даже специали-
стам, работающим в этой области. Вероятно, время внесет еще много
изменений и дополнений, прежде чем работу по этой группе можно
будет считать близкой к концу. Во многих случаях очень мало
известно даже о целых семействах клещей. И когда всем станет ясно,
что клещи представляют нетронутую область для работ по система-
тике, экологии и прикладной зоологии, мы убедимся, что действи-
тельно находимся на пороге новой и очень интересной отрасли знания
Э. У. Бэкер и Г. У, Уортон.
Глава 1
ВВЕДЕНИЕ
Клещи встречаются повсюду, где имеется жизнь. В любой при-
горшне почвы при внимательном исследовании можно найти несколько
клещей (рис. 1), а в некоторых местах их можно найти тысячи. Перья
птиц так же, как и шерсть млекопитающих, часто буквально кишат
клещами. Многие люди являются носителями колоний чесоточных
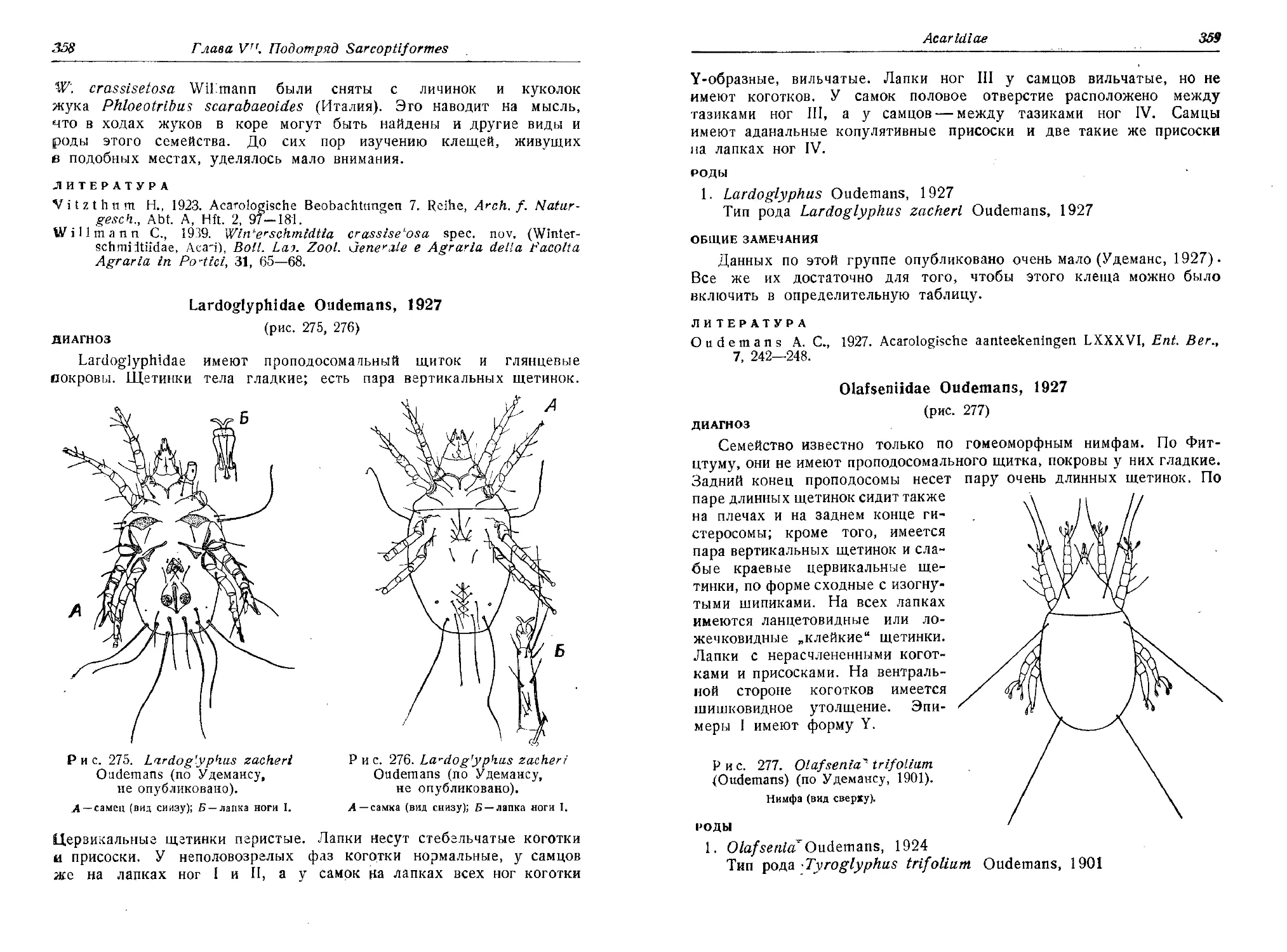
клещей, паразитирующих в коже. Вода ручьев, озер, прудов имеет
свою фауну клещей. Даже океан не свободен от них. Есть виды
клещей, которые приспособились к жизни в горячих источниках неко-
торых вулканических областей. Всевозможные насекомые являются
хозяевами многих видов клещей. Некоторые клещи проникают во
внутренние органы человека, позвоночных и беспозвоночных животных.
Убежища для клещей предоставляют и многие растения, и при обсле-
довании мусора в развилках стволов различных деревьев всегда можно
найти особей нескольких видов клещей. Однако мир клещей остается
неизвестным большинству людей. Некоторые зоологи и даже энтомологи
никогда не видели клещей в их природной обстановке. Очень немногие
из них представляют себе, какое разнообразие видов клещей встре-
чается повсюду.
Однако некоторые клещи известны даже и неспециалистам. Обычно
они привлекают внимание своей активностью. Так, иксодовые клещи
знакомы почти всем: они достаточно крупны, хорошо заметны невоору-
женному глазу и к тому же часто нападают на людей и на домашних
животных. На юге США большинство людей имели несчастье позна-
комиться с „chiggers'1, хотя очень немногие знают, что это личинки
клещей-краснотелок. Плодоводы хорошо знают паутинных клещей и
другую группу клещей — эриофиид. Многие владельцы собак своим
аппкомством с клещами обязаны собачьему клещу, нападающему на
их любимцев. Люди, работающие по хранению пищевых продуктов,
особенно таких, как зерно и копра, знают клещей, заражающих
и массе эти продукты, так как они вызывают у людей раздражающий
зуд, В периоды сильной скученности населения, как, например,
и Англии во время второй мировой войны, чесоточные клещи, вызы-
вающие у человека зуд, получают значительное распространение и
причиняют людям большие неприятности. Но, несмотря на то, что
реиультаты деятельности некоторых видов клещей хорошо известны
14
Глава I. Введение
людям, мало кто из пострадавших может правильно сказать, что
именно является причиной испытываемого вреда.
Р и с. 1. Почвенные клещи, извлеченные из небольшой пробы подстилки
и почвы.
Недостаточность знаний о клещах в значительной мере объясняется
их малой величиной. Хорошо изучен только один подотряд Acarina —
иксодовые клещи, видимые невооруженным глазом.
Специалисты по иксодовым клещам знают их так же хорошо, как
энтомолог знает крупных насекомых. Для изучения их не требуется
Глава I. Введений
13
:пециальной техники, так как необходимые диагностические признаки
ложно рассмотреть при помощи бинокуляра или даже сильной ручной
1упы. Со всеми остальными клещами дело обстоит иначе. Здесь тре-
гуется иная техника, чем та, которой пользуются энтомологи, и другие
летоды сбора материала. Из исследуемого экземпляра обычно приго-
говляют специальные препараты и изучают их под микроскопом
: увеличением до 1000 раз. Более того, методы изготовления микро-
скопических препаратов из клещей отличаются от методов, приме-
гяемых при изготовлении микроскопических препаратов в других
отраслях зоологии, хотя многие специальные методы исследования
гкарологи заимствовали у зоологов и энтомологов. Дело в том, что
слещи не настолько малы, чтобы с ними можно было обращаться
<ак с простейшими, не такие мягкие, чтобы их можно было обраба-
тывать так же, как и червей, и слишком малы, чтобы их можно было
изучать так же, как насекомых. Поэтому-то изучением клещей оди-
наково мало занимались как зоологи, так и энтомологи.
Бедность наших сведений о клещах иногда объясняют относи-
тельно малым практическим значением этой группы. Это совершенно
неверно. В настоящее время нам все еще мало знакомы многие клещи,
вредоносное значение которых несомненно. По мере накопления наших
знаний о клещах будет выявляться практическое значение многих
других видов, пока же акарология находится в таком же положении,
какое занимала энтомология пятьдесят или сто лет назад.
ИСТОРИЧЕСКИЙ ОБЗОР
Некоторые мелкие клещики и иксодовые клещи известны издавна.
Еще древние греки были знакомы с иксодовыми и некоторыми дру-
гими клещами. Само название Acari — латинизированная форма гре-
ческого слова. Ко времени Линнея в литературе было описано только
90 видов клещей, и в десятом издании „Systema Naturae" приведено
лишь 29 видов клещей рода Acarus. Однако к 1850 г. стали известны
уже многие виды клещей. Вся литература, опубликованная до 1850 г.
включительно, рассмотрена в трехтомной сводке Удеманса „Kritisch
Hlstorisch Overzicht der Acarologie" (1926—1937). Работа Удеманса
остается неоценимым источником сведений о всех ранних работах
но клещам.
Начиная с 1850 г. и по настоящее время, большинство работ по
Клещам общего характера выполняется европейскими исследователями.
Ценный вклад в акарологию сделали Налепа, Г и Р. Канестрини,
Верлене, Труссар, Хирст, Майкл, Удеманс, Фитцтум, Тор, Трегорд и
некоторые другие исследователи. Андре, Гранжан, Лундблад, Зельник,
Бильман и Фитс продолжают публиковать важные работы. В США
И продолжение большей части текущего столетия только двое ученых
иктинпо работали по общей акарологии, хотя изучению водяных клещей,
НИугннпых клещей, панцырных клещей и некоторых других групп
16
Глава А Введение
содействовали работы ряда исследователей. Первым американским
акарологом был Бэнкс, а его преемник по Национальному музею США
Эвинг сделал для акарологии больше, чем любой другой американский
ученый.
В настоящее время в США клещей активно изучает около
20—30 исследователей и число их быстро возрастает. Правитель-
ственные учреждения США, особенно министерство сельского хозяй-
ства, военное министерство, морское министерство и управление
здравоохранения, проявляют все возрастающий интерес к изучению
клещей, так что в ближайшее время следует ожидать значительного
развития акарологии в США.
МЕТОДИКА ИЗУЧЕНИЯ
Сбор материала. Собирать клещей одновременно и легко, и
трудно. Вследствие своего малого размера многие из клещей неза-
метны в естественных местообитаниях и для сбора их применяют
особые методы. Очень часто в поле собирают субстрат, заселенный
клещами, а клещей обнаруживают в нем лишь при лабораторном
исследовании. Во всех случаях при сборах материала следует при-
держиваться некоторых общих правил.
При сборах материала необходимо делать записи, включающие
дату, место, фамилию сборщика, описание местообитания1 и данные
о его температуре и влажности и об окружающих животных или
растениях, кажущихся важными по той или иной причине; без таких
записей собранный материал в значительной мере теряет свою ценность.
Записи в тетради должны быть возможно более подробными. Это
важно потому, что при обработке материала всегда .легко отбросить
излишнюю подробность, но часто совершенно невозможно восстановить
важные сведения, показавшиеся при сборе материала несущественными.
Все сборы должны сопровождаться этикеткой, номер которой дол-
жен соответствовать записи в тетради, что значительно облегчает
работу. '
Клещей, собранных в природе, можно неограниченно долгое время
сохранять в 85-процентном этиловом спирте. Обычно их помещают
в маленькие стандартные пробирки размером 5 X 28 мм. После того
как клещи и этикетка помещены в спирт, пробирку плотно затыкают
ватной пробкой и помещают в сосуд с 85-процентным спиртом.
В таком виде сборы могут храниться долгое время. Для хранения
клещей очень удобны также пробирки с резиновыми пробками из-под
новокаина; такие пробирки всегда можно достать у зубных врачей.
1 Под местообитанием следует понимать общий ландшафт местности и
конкретное указание на место обнаружения клеща. Например: „песчаная
пустыня Кара-Кумы в такой-то географической точке; нора толстохвостой
песчанки" или „смешанный широколиственный лес в таком-то районе; в толще
сухой подстилки из опавших листьев".—Прим. ред.
Глава I, Введение
17
Малые размеры клещей затрудняют работу исследователя. Для
сбора материала в природе пользуются влажными кистями для аква-
рельных красок из верблюжьего волоса. Лучше всего пользоваться
небольшими кистями (№ 0, 1 и 2); кистями больших размеров работать
труднее. Даже очень маленькие и хрупкие клещи почти не повреж-
даются, если брать их на кончик кисти. Глазной и анатомический
пинцеты применяют только при манипуляциях с крупными экземпля-
рами; ими пользуются также при манипуляциях с мелкими энтомоло-
гическими пробирками. Даже крупных клещей следует захватывать
не кончиками пинцета, а на расстоянии нескольких миллиметров от
кончиков; тогда кончики пинцета, смыкаясь, защищают клеща от
повреждений при сжатии.
Для сбора свободноживущих почвенных клещей обычно берут
пробы из различных органических остатков, таких, как верхние слои
почвы. Много почвенных клещей можно найти в подстилке или в ана-
логичных скоплениях в дуплах и в развилках деревьев и кустарников.
Гнилые бревна и почва под ними — излюбленное местообитание не-
которых клещей. Часто изобилует почвенными клещами и почва под
камнями. Много растительноядных клещей можно найти вблизи кор-
ней, листьев, бутонов растений или в грибах; обычно очень богаты
клещами и слои мха. При сборах следует особенно тщательно осмат-
ривать растения или органы растений, которые по внешнему виду
отличаются от нормальных здоровых растений или их органов. Боль-
шинство клещей нуждается во влажной среде. Поэтому влажная
почва и навоз обычно богаче клещами, чем сухие. В дождливую
погоду многие виды, обычно живущие в верхних слоях почвы, уходят
в подстилку.
При разборе материала в природе очень удобны белое или черное
эмалированное или фарфоровое блюдо или поднос. Исследуемый мате-
риал небольшими порциями помещают на поднос и просматривают
порцию за порцией. При этом многие клещи падают на поднос, на
гладкой блестящей поверхности которого они хорошо заметны. При
исследовании почвы следует соблюдать некоторые предосторожности.
Клещей, обнаруженных в исследуемом материале, сначала отодвигают
от материала на чистую часть подноса; затем их очищают от частиц
почвы и только после этого переносят кистью в пробирку. Все это
позволяет избежать переноса в спирт клеща, загрязненного частицами
почвы, приставшими к нему. Такого загрязненного клеща, попавшего
в спирт, в дальнейшем очистить практически невозможно. Поднос
может быть использован и для другой цели. Многие клещи, особенно
краснотелки, наползают на всякий новый объект, помещенный в их
окружение. Если поднос положить на землю и оставить на несколько
минут, то нередко на него наползают клещи, которых можно затем
собрать и поместить в пробирку.
Клещи часто наползают на фланель, протаскиваемую над ними-
Этот метод хорош для сбора неприсосавшихся иксодовых клещей-
2 Зак. 1785
Глава I. Введение
Клещевую волокушу (рис. 2) обычно делают из куска фланели вели-
чиной около 1 м'1 * 3, к одной из сторон которого прикрепляют палки
длиной 1 м. К концам палки привязывают веревку, за которую воло-
кушу тянут по обследуемой площадке. Через определенные интервалы
материю осматривают и снимают клещей пинцетом или руками.
Гораздо более эффективным методом сбора почвенных клещей
является разбор субстрата в лаборатории. Субстрат помещают в бу-
мажный пакет (№ 5), концы которого закручивают и перетягивают
Рис. 2. Волокуша для сбора клещей.
резиновым кольцом. Когда собирают влажный материал, например
влажный мох, используют клеенчатый или прорезиненный мешок К
В лаборатории собранный таким образом материал исследуют под
бинокуляром или пропускают его через воронки Берлезе.
Воронки Берлезе различных типов представляют собой наиболее
эффективное приспособление для отделения клещей и мелких насеко-
мых от субстрата, с которым они собраны. Существует много типов
воронок Берлезе. Такую воронку, вполне пригодную для разбора мате-
риала, легко сделать самому исследователю. Воронка Берлезе пред-
1 При переносе в лабораторию клеенчатых мешочков с влажным мате-
риалом их следует оберегать от перегрева солнечными лучами. В лаборато-
рии собранный материал следует без излишнего промедления переложить
з стеклянную посуду. — Прим. ред.
Глава I. Введение
19
, ншливт собой открытый с обеих сторон конус из листового железа
иысптой около 45 см. Большое верхнее отверстие должно иметь диа-
Mi'ip около 30 см, а маленькое нижнее—не более 2,5 см. Воронку
\ i । ||||||плинают в вертикальном положении на треножнике маленьким
iHiivpiTHCM вниз. Треножник должен быть достаточно высоким для
loiо. чтобы под воронку можно было подставить чашку. Желательно
ннжний конец воронки делать с нарезкой, чтобы чашечку для сбора
мигериала можно было привинчивать к воронке. Это уменьшает воз-
можность загрязнения сбора извне и не позволяет клещам расползаться
ил чашечки. Для верхнего отверстия делают крышку, снабженную
электрической лампой. Прекрасной крышкой может служить рефлек-
юр, пригнанный к патрону лампочки. Для некоторых специальных
целей удобнее воронки большего или меньшего размера.
11еред началом разбора материала субстрат заворачивают в марлю
И помещают в воронку. Воронку слегка встряхивают для того, чтобы
субстрат переместился книзу, после чего устанавливают на место
чашечку для сбора материала. Затем включают свет и ждут, пока
подсыхание субстрата не заставит клещей передвинуться ко дну
коронки. Иногда проходит не менее трех дней, прежде чем содержимое
коронки станет совсем сухим. В течение этих трех дней клещи будут
падать в чашку. Для разбора большого количества материала приме-
няют целую батарею воронок Берлезе (рис. 3). Сортировку по видам
клещей и других членистоногих, попавших в чашечку для материала,
при необходимости производят под бинокуляром.
Для сбора паразитических клещей необходимо в первую очередь
собирать их хозяев. Клещи паразитируют почти на всех животных,
превосходящих их по размеру, и не только на наземных, но также
и на водных животных. Большую часть паразитических клещей со-
с г.являют эктопаразиты, меньшую — эндопаразиты и эктопаразиты,
живущие в жилищах хозяина. Мы не’будем подробно останавливаться
на способах сбора всех типов хозяев и отсылаем читателя к руковод-
ству по сбору коллекций „Collectors Manual", изданному Смитсонов-
ским институтом. При сборе паразитических клещей следует помнить,
ч1о определение видовой принадлежности хозяина не менее важно,
чем определение самого клеща. Для определения хозяина необходимо
сохранить его и в дальнейшем передать для изучения специалисту.
1,ля более полного изучения паразитофауны какого-либо животного
лсс лательно добыть большое число экземпляров хозяина Ч Сбор пара-
зитических клещей облегчается тем, что клещи в отличие от неко-
торых насекомых, например блох, не покидают хозяина сразу после
его смерти. Поэтому для отлова грызунов и других мелких наземных
1 Для добывания клещей из нор грызунов применим способ сбора бес-
крылых паразитов из нор грызунов (см. Л. И. Ш и р а н о в и ч, Н. П. Миро-
нов и А. С. Фомичева, Методы сбора бескрылых паразитов из нор
। рызунов, серия „В помощь работающим на полезащитных лесных полосах
и па великих стройках коммунизма", Ха 6, изд. АН СССР, 1952).—Прим. ред.
о*
20
Глава I. Введение
позвоночных, являющихся хозяевами клещей, пользуются ловушками.
Для сбора птиц единственным удовлетворительным методом является
отстрел. Для этой цели желательно пользоваться двухствольным
ружьем, чтобы в распоряжении сборщика всегда имелась и мелкая
и крупная дробь. Животного-хозяина сразу же после отлова поме-
щают в индивидуальный непроницаемый для клещей мешок, что
не позволяет паразитам переползать с одного хозяина на другого.
Р и с. 3. Воронки Берлезе.
На переднем плане изображены небольшие воронки, употребляемые для сбора эктопа-
разитичсских клещей. Две из них находятся в работе. Животных-хозяев помещают
в небольшие*клетки и собирают клешей из сосудов с водой, стоящих под воронками.
Названием плане изображены большие воронки для сбора клещей из мусора. Над во-
ронками установлены лампы, используемые для сушки материала.
Обследование хозяина в лаборатории значительно облегчает при-
менение бинокуляра. Прежде всего тщательно осматривают наружную
поверхность тела. Осмотр кожи млекопитающих и птиц затрудняется
шерстью и перьями, но уже при небольшом навыке это легко удается
сделать, осторожно раздвигая мех и перья. При осмотре насекомых
необходимо раздвигать крылья, а у жуков — надкрылья, так как
многие эктопаразитические клещи селятся на участках, покрытых
крыльями. Прежде чем перейти к внутренним органам, осматривают
уши, глаза, анальное и половое отверстия. В пищеварительном тракте
животных паразитируют лишь немногие клещи; чаще клещи встре-
чаются в органах дыхания позвоночных и беспозвоночных животных.
Глава I. Введение
21
С живого животного много эктопаразитов удается собрать и без
осмотра покровов. Для этого живых животных помещают в клетки
С дном, затянутым металлической сеткой, а под клетку ставят поднос;
(Клещи, падающие с животного, не задерживаются сеткой и попадают
на поднос, с которого их и выбирают при осмотре. Правда, помет
•и моча хозяина затрудняют этот метод сбора, но при разумно подоб-
ранном рационе для животных и частом обследовании и чистке клеток
Это затруднение удается преодолеть.
Для сбора свободноживущих водных клещей используют два при-
способления: небольшой сачок с узким сетчатым мешочком, которым
можно вычерпывать различных клещей из воды, и сеть Бирге (рис. 4).
Р и с. 4. Сеть Бирге.
Водные клещи встречаются практически в любых водоемах, но наи-
более богатые сборы можно получить из небольших озер, ручьев и
прудов. При сборе клещей сетью Бирге нужно следить за тем, чтобы
мешок находился в воде в расправленном состоянии. Сеть Бирге
предназначена для использования в заросших местах, так как она
редко запутывается в водных растениях. Именно в таких заросших
Местах у берегов обитает большинство водных клещей.
Подготовка материала для изучения. Только иксодовых клещей
Можно успешно изучать не на препаратах. Для рассмотрения их
систематических признаков достаточно пользоваться бинокуляром
С большим увеличением.
Для изучения мелких клещей приготовляют препараты, которые
ятем рассматривают под микроскопом в проходящем свете. Для этой
Цели препараты следует предварительно просветлить и смонтировать
1111 предметном стекле. Прежде применяли много разных способов
ИИГотовления препаратов, описание которых можно найти в литературе.
Недавно разработали новый метод заключения в среду с поливинило-
НЫМ спиртом, дающий хорошие результаты при изготовлении препа-
ратов из многих клещей. При приготовлении соответствующей среды
и ипключении препарата следует придерживаться следующих указаний:
I, Растворить, помешивая, „Эльванол" 71-24 (поливиниловый
Спирт Дю Понта) в четырех объемах воды при температуре 90°,
Фильтровать раствор до просветления.
22
Глава I. Введение
3. Выпаривать светлый фильтрат на водяной бане до тех пор,
пока он не превратится в вязкий сироп. (Пена, образующаяся на
поверхности в процессе выпаривания, растворяется при помешивании.)
4. Добавить 22 части молочной кислоты к 56 частям концентрата
поливинилового спирта.
5. В полученную готовую среду клещей заключают обычным спо-
собом. Для приготовления препарата можно использовать как свежий
материал, так и материал, сохраняющийся в любом зодном растворе
или в спирте.
Значительная усушка поливинилового спирта вызывает сморщива-
ние клещей с мягким телом (Tetranychidae) и разрушение некоторых
крупных и хрупких (Oribatei). Это следует учитывать при примене-
нии описанной смеси.
В большинстве случаев применяют другие методы. Временные
препараты можно изготовлять в любом просветляющем веществе,
например в глицерине, минеральном масле, феноле, молочной кислоте
и т. п. Хорошие постоянные препараты могут быть изготовлены
в водном растворе гуммиарабика, хлоралгидрата и глицерина. Одним
из лучших является раствор Гойера (модифицированный раствор Бер-
лезе). В Национальном музее США образцы хранятся в этой жидкости
в течение 20 лет и не претерпевают никаких изменений. Эта среда
хороша для Tetranychidae, которые являются одной из самых трудных
групп для изготовления препаратов. Среда эта имеет следующий
состав:
Дестиллированпая вода......................... 50 г
Гуммиарабик (чистый, кристаллический) . . . 30 „
Хлоралгидрат................................. 200 „
Глицерин...................................... 20 „
Ингредиенты смешивают при комнатной температуре в указанной
выше последовательности. В более влажных областях, например
у побережья Мексиканского залива, покровное стекло следует окай-
млять полоской вещества, поглощающего влагу1. Для заключения
клещей применяют и другие модификации смеси Берлезе, но идеаль-
ной среды для заключения пока еще не разработано. 1 * * * * * * В
1 Обычно для окаймления покровного стекла используют какое-либо
водонепроницаемое вещество с подходящим коэффициентом преломления
(жидкий канадский бальзам-, менделеевская замазка, замазки специального
состава, препятствующие пересыханию среды, в которую заключен препарат
и пеподвергающиеся действию влажности внешней среды). В нормальных
условиях работы и хранения препаратов хорошие результаты даст и гумми-
арабиковая смесь, в особенности при изготовлении тотальных препаратов.
В гуммиарабиковой смеси препараты не достигают той степени прозрачности,
которая характерна для препаратов, заключенных в бальзам, однако в по-
следних скрадываются многие топкие, но важные детали строения клещей.
Поэтому на практике рекомендуется параллельное использование обеих сред.
Следует отметить, что в некоторых условиях препараты, заключенные в гум-
миарабиковую среду, не выдерживают длительного хранения. — Прим. ред.
Глава I. Введение
23
В Калифорнийском университете исследователи разработали среду
из метацеллюлозы, которая дает хорошие результаты при заключении
многих клещей. Эта среда имеет следующий состав:
Метацеллюлоза............................ 5 г
Карбовакс 4000 .......................... 2 »
Диэтиленгликоль......................... 1 мл
95-процентный этиловый спирт............ 25 „
Молочная кислота....................... 100 „
Дестиллированпая вода................... 25 „
Перед заключением рекомендуют просветлять материал в лакто-
феноле, но некоторые более нежные клещи не нуждаются в специаль-
ной обработке перед заключением, так как молочная кислота в среде
просветляет их вполне достаточно.
Ньюэлл в 1947 г. предложил новый метод изготовления постоян-
ных глицериновых препаратов. Необработанных клещей нельзя заклю-
чать в жирорастворимые смолы, так как коэффициенты преломления
этих веществ и ткани наружного скелета клещей крайне близки друг
к другу. Если клещей заключают в такие вещества, как даммарлак
или канадский бальзам, то необходимо предварительно просветлить
материал молочной кислотой или обработать его слабым раствором
щелочи для того, чтобы уничтожить мягкие ткани. Затем наружный
скелет окрашивают так, чтобы он контрастировал со средой, в ко-
торую заключают препарат. При фазовоконтрастной микроскопии
надобность в окраске отпадает и препараты можно непосредственно
заключать в такую среду.
Готовые препараты следует немедленно надписывать. Обычно
принято па правой стороне стекла записывать дату сбора, а на левой —
наименование клеща.
МОРФОЛОГИЯ и ЖИЗНЕННЫЙ цикл
Терминология
Акарология долгое время развивалась в недрах энтомологии и
hi,телилась из нее. Поэтому терминология, употребляемая в описа-
тельной анатомии клещей, заимствована из энтомологии. К сожалению,
энтомологические термины в приложении к клещам часто приобретают
Совершенно иное значение, ибо при переносе термина с одной группы
пн другую руководствовались главным образом аналогией, а не гомо-
логией. Можно было бы разработать совершенно новую терминологию
специально для клещей, но это, вероятно, привело бы лишь к увели-
чению путаницы. Поэтому в настоящей работе мы будем пользоваться
нрминами, получившими более или менее общее признание среди
,||<11рологов, определяя их только в приложении к клещам, но не учи-
ii.iiiiiii их первоначального значения в энтомологии. В большинстве
lyniicii мы следуем терминологии Фитцтума (1940).
24
Глава 1. Введение
Сегментация
Многие клещи кажутся состоящими из одного сегмента. У других
тело явственно разделено на головогрудь (цефалоторакс) и брюшко,
а у некоторых и брюшко кажется сегментированным. Следы первич-
ной сегментации можно обнаружить у примитивных Notosfigmafa.
У некоторых Tarsonemidae расчленена задняя часть тела, но эта
сегментация, вероятно, вторичного, а не первичного происхождения.
Фитцтум (1940) приводит следующую схему расчленения тела
клеща на отделы (рис, 5):
Рис. 5. Схема разде-
ления тела клеща на
отделы на примере
Schizo'etranychus
schizopus (Zacher),
1913 (по Фитцтуму,
1940).
Принимаемые
лагаемой схеме:
другими авторами подразделения показаны на при-
Область, состоящая из
ротового отверстия
и ротовых органов
Гнатосома
Протеросома
Область, включающая
ноги I и II
Область, включающая
ноги III и IV
Задний отде'л
Проподосома
1 Просома.
Подосома
Метаподосома
Опистосома
Идиосома
)
Гистеросома
Общий обзор воззрений о первичной сегментации клещей сделали
Андре и Лэми (1937). Эти авторы, следуя Генкингу (1882) и Уде-
Глава I. Введение
25
miiiicy (1909), приходят к заключению, что у большинства клещей
ггло состоит из 13 сегментов. Вентральная часть трех передних сег-
ментов сливается, образуя гнатосому. Первый, или преоральный,
сегмент не имеет придатков, второй несет хелицеры, а третий —
иедипальпы. Подосома образована дорзальными частями первых трех
сегментов и четырьмя полными сегментами (IV—VII), каждый из ко-
торых несет пару ног. Опистосома обычно состоит из 6 сегментов.
Установлено, что число опистосомальных сегментов может варьиро-
вать, чем и объясняются отмеченные выше различия в общем коли-
честве сегментов. Notostigmata могут иметь до 10—11 опистосомаль-
пых сегментов; Pachygnathus имеет 8 или 9, a Parky pocthonlus— 7.
Гнатосома обычно выдается вперед, по крайней мере со спинной
стороны тела, но в некоторых группах (Uropodina, Spelaeorhynchidae,
( nyptostigmatina и др.) сегменты проподосомы прикрывают гнатосому
и включают ее в особую полость — камеростом. В одном семействе
Prostigmata (Smaridiidae) гнатосома может выдвигаться на длинном,
узком стволовидном основании, которое может достигать длины тела
клеща. Гнатосома ни в коем случае не может быть приравнена
к голове других членистоногих. Ее часто называют капитулум, но
термин гнатосома предпочтительнее, так как эго образование несет
только рот и ротовые части.
Проподосома с первыми двумя парами пог часто бывает отделена
от метаподосомы глубокой бороздкой; у Tronibiculidae это разделение
выражено очень хорошо. Это позволяет некоторым авторам подразде-
лять тело клеща на два основных отдела: передний — протеросома,
образованная гнатосомой и проподосомой, и задняя — гистеросома,
образованная метаподосомой и опистосомой, обычно незаметно сливаю-
щимися друг с другом.
Просома соответствует головогруди других паукообразных, но
передние сегменты ни в одном случае не сливаются так, чтобы обра-
зовать отдельную часть тела, которую можно было бы назвать голово-
। рудью, противопоставляя ее задней брюшной части. Впрочем,
Speleorchestes отличается от остальных клещей тем, что у него гнато-
сома, проподосома, метаподосома и опистома внешне обособлены
руг от друга.
Подосочой называют часть тела, несущую ходильные ноги. Идио-
сомой называют все тело клеща позади гнатосомы. У многих мелких
клещей и у всех иксодовых тело внешне подразделено лишь на гнато-
сому и идиосому.
Покровы
Покровы тела у клещей так же, как и у других членистоногих,
состоят из одного слоя эпителиальных клеток — гиподермы — и из
склеротизированных слоев — кутикулы, являющейся ее секретом.
Кутикула (рис. 6) подразделяется на четыре различных слоя, которые
Фитцтум (1940) характеризует следующим образом:
26
Глава I. Введение
1. Текстостракум — поверхностный слой, всегда очень тонкий,
прозрачный и никогда не окрашивающийся.
2. Эпиостракум— верхний слой.
3. Эктостракум — средний слой, обычно хорошо окрашиваю-
щийся кислыми красками.
4. Гипоспгракум — внутренний слой, обычно хорошо окрашиваю-
щийся основными красками.
Эти четыре слоя выражены не у всех клещей. Один (а иногда и
больше) из хитинизированных слоев может отсутствовать или же так
комбинироваться с другим слоем, что оба они становятся неразличи-
мыми друг от друга. В некоторых случаях (например, у Pyemotes)
Р и с. б. Покровы Hyalomma marginatum brionicum
Schultze et Schlottke, 1929 (по Фитцтуму, 1940).
На срезе видны два внутренних, склеротизированпых слоя (эктостракум
и гипостракум) и подстилающая их гиподерма.
сама гиподерма так уплощается при набухании опистосомы, что ее
невозможно обнаружить.
Хотя структура покрова в основе сходна у всех видов, внешний
вид, строение и физические свойства склеротизированного слоя покро-
вов сильно варьируют и могут быть весьма различными у отдельных
групп. Так, Notostigmata обладают кожистой, тонкой гранулированной
кутикулой. Многие клещи полностью или частично покрыты мощным
панцыревидпым щитом. Все тело Holothyroidea, многих Mesostgmata
и большинства Oribatei покрыто твердыми защитными щитками.
Trombidiiformes и Acaridiae обладают эластичной прозрачной кутику-
лой, имеющей местами (например, у оснований щетинок) твердые
участки в виде щитков.
Красота рисунка, присущая клещам, во многом определяется орна-
ментом кутикулы (поры, гребни, складки) и пигментом, заложенным
в разных ее слоях (рис. 7). Орнамент кутикулы обычно постоянен
для вида и для некоторых групп используется в качестве диагно-
стического признака.
Многие структурные образования покровов являются производными
клеток гиподермы. Так, различные железы, щетинки и специальные
сенсорные органы образуются целиком или частично специальными
участками гиподермы и ее выделениями (рис. 8). За исклю-
Глава I. Введение
27
пением щетинок и гиподермальных желез различные производные
покровов рассматриваются ниже, в- системе тех органов, к кото-
рым они принадлежат.
Покровы многих клещей снабжены
железами. У иксодовых клещей железы
представлены увеличенными клетками
гиподермы, выделяющими свой секрет
непосредственно на поверхность тела.
Железы водных клещей открываются
па специальных участках по всему телу
(рис. 9). Эти железы сложнее желез
иксодовых клещей. Большинство Sar-
coptiformes имеет пару жировых же-
лез, которые выделяют маслянистую
жидкость, окрашенную в желтый или
темнокоричневый цвет. Аналогичные же-
лезы найдены у некоторых Mesostigmata,
как, например, у Eulaelaps stabularls
и Euhaemogamasus horridus.
Щетинки у клещей бывают самой
разнообразной формы и, как и у боль-
шинства членистоногих, выполняют раз-
личные функции. Многие щетинки слу-
жат органами осязания; в них чувствую-
щие окончания нервных клеток распо-
Р и с. 7. Стернальный щиток
нимфы ParasUus fucorum (De-
Geer), 1778 (по Фитцтуму, 1940)
Рис. 8. Поперечный срез через покровы Trom.bidiu.tn
holosericeum (Linnaeus), 1758 (по Фитцтуму, 1940).
ложены так, чтобы воспринимать давление, оказываемое на щетинку.
Некоторые щетинки имеют хеморецепторную функцию. Наконец,
28
Глава I. Введение
некоторые щетинки по-настоящему защищают своих обладателей от
нападения хищников подобно тому, как дикобраза защищают его иглы.
У большинства клещей различают два основных типа щетинок: обыч-
ные щетинки и немногочисленные сенсорные щетинки, которые резко
отличаются от обычных щетинок тела.
А Б
Рис. 9. Расположение кожных желез у Megaluracarus
globator (Muller), 1776 (но Фитцтуму, 1940).
А — на спине; Б —на брюшке; В—сбоку.
Собственно щетинки или обычные щетинки—„eigentliche Нааге“
Фитцтума (1940), „polls proprement dits“ Гранжана (1935) — чрезвы-
чайно разнообразны (рис. 10). Они бывают простые, опушенные,
пуховидные, булавовидные, ланцетовидные, сердцевидные, перепон-
чатые, обволошенные, перистые, содержащие камеру, зубчатые,
вильчатые или настолько неправильные по форме, что только подроб-
ное описание может дать представление о них. Трудно понять зна-
чение такого многообразия форм щетинок. У некоторых видов клещей
форма и количество щетинок постоянны, благодаря чему они исполь-
зуются как таксономические признаки.
Глава /. Введение
29
। Повидимому, примитивными являются опушенные щетинки, за-
остренные к вершине. Первично щетинки располагались соответ-
ственно первичной сегментации клещей, каждый сегмент по заднему
краю был снабжен рядом из 2—6 спинных щетинок. Такое прими-
iiiHiioe расположение щетинок удерживается с небольшими измене-
ниями у Trombldiformes и Sarcoptiformes. Во всех крупных группах *.
Рис. 10. Типы туловищных щетинок клещей.
Л простые (JJeocheyletiella rohweri Baker, 1949); Б — опушенные (KeQcheyletlella rohweri
Hnkcr, 1949); В — перистые (Euschdngastla peromyscl (Ewing), 1929); Г—булавовидные (Spathu-
fnthrombium southcotti (Womersley), 1935); Д — ланцетовидные {Spathalathrombium. maximum
Womersley, 1945); E — сердцевидные (Spathulathrombium maximum Womersley, 1945); JK—пере-
пончатые (Cheleta genes ornatus (Canestrini et Fanzago, 1876); 3 — обволошенные (Holcotrom-
bhlium securigerum (Canestrini), 1897); И — вильчатые {Hiotrombidium iubbi (Womersley), 1937;
А -содержащие камеру (Camerotrombidium opuJentum. (Womersley), 1945); JI — зубчатые
{Ac ar opsis docta (Berlese), 1886).
число щетинок неоднократно уменьшалось или увеличивалось, и
у многих видов первичное распределение щетинок затемнилось.
Наиболее распространенным типом сенсорных щетинок являются
бороздчатые сенсорные щетинки, служащие, вероятно, хеморецепто-
рами. Фитцтум называет их соленидиями, а Гранжан—соленидионами.
()пи полые, и в их просвет проникают живые отростки связанных
г ними клеток. Эти образования непосредственно связаны с перифе-
рической нервной системой и чаще всего встречаются на дистальных
члениках конечностей.
Помимо бороздчатых сенсорных щетинок, имеются микросенсор-
ные щетинки (фамулус по Гранжану (1935)), постоянно встречаю-
щиеся на колене, на голени и на лапках ног. Их функции неизвестны,
30
Глава I. Введениё
Рис. 11. Трито-
стернум Наето-
laelaps traubi
(Strandtmann),
1948 (по Страндт-
ману, 1948).
.ч
но маловероятно, чтобы столь мелкие структуры имели тактильную
функцию Бичевидные щетинки (акантоиды по Гранжану (1935))
также обычно встречаются на ногах. Они отличаются от обычных
щетинок тем, что лишены каких-либо придатков и своей большой
длиной. Эти щетинки, возможно, имеют тактильную функцию.
Некоторые щетинки часто бывают связаны с определенными струк-
турами. Сенсиллы, представляющие собой щетинковидные образова-
ния, которые поднимаются из специальных ямок
(псевдостигмы, или основания сенсилл), обычно
встречаются на проподосоме. Сенсиллы могут иметь
самую различную форму, но почти всегда они хо-
рошо отличимы от щетинок тела. У большинства
Mesost’gmata на тарзальном членике пальпы располо-
жена специализированная разветвленная щетинка.
Гранжан изучал структуру хитина щетинок раз-
личных групп клещей. Он нашел, что хитин
ще тинок Тrum bidiformes и Sarcoptiformes опти-
чески активен. Щетинки у этих двух групп легко
окрашиваются иодом. Щетинки у Mesostigmata и
Holothyroidea оптически неактивны и не окрашиваются
иодом. Гранжан называет оптически активный хитин
актинохитином и объединяет Trombidiformes и Sar-
coptiformes в одну группу — Actinochitinosi, a Meso-
stigmata и Holothyroidea — в группу Anactinochitinosi.
Для того чтобы определить значение этой системы,
основанной на оптической активности хитина щети-
нок, требуются еще дальнейшие исследования.
Щитки. Толщина покровов в различных частях тела
у большинства клещей варьирует. Некоторые участки
тела покрыты более толстым слоем хитина, и эти утол-
щенные участки образуют пластинки, или щитки.
Щитки клещей соответствуют склеритам насекомых,
но отличаются от них тем, что обычно покрывают
более одного сегмента, а в некоторых случаях и всю
идиосому. Кроме того, в большинстве случаев щитки у клещей
являются не видоизменением первичных склеритов, а развиваются
вторично.
У клещей не остается неизмененным ни один из первичных стер-
нитов. У Notostigmata и Mesostigmata сохраняется рудиментарный
остаток тритостернума в виде своеобразного щетинковидного обра-
зования перед стернальным щитком (рис. И). Тритостернум состоит
из основной части, обычно непарной, от которой берут начало парные
опушенные доли (lacinia); у Notostigmata основная часть тритостер-
1 Микрэсенсорные щетинки могут иметь значение проприорецепторэв,
раздражаемых при сгибании и разгибании суставов ноги.—Прим. ред.
Глава I. Введение
31
пума парная, а у Uropodina лацинии у основания слиты, но дистально
могут подразделяться на три доли. Все другие щитки на брюшке
клещей вторичного происхождения и не происходят от первичных
ггерпитов.
У немногих клещей все спинные щитки являются настоящими
видоизмененными тергитами. Их легко распознать у Tarsonemini
(рис. 12); гистеросомальные щитки у Protoplophoridae и Mesoplopho-
ildae также выводятся из первичных тергитов. У остальных клещей
Рис. 12. Resinacarus resinatus Vitzthum, 1927
(по Фитцтуму, 1910).
Видны тергиты.
только проподосомальный щиток представляет собой остаток первич-
ного тергита. Он образован слиянием тергитов сегментов первых двух
пар ног, т. е. сегментов IV и V.
Проподосомальный щиток в разных группах клещей носит разные
названия. У иксодовых клещей, у личинок Trombidiidae, Trombiculi-
dae и близких к ним групп клещей он известен под названием scutum.
У половозрелых клещей из группы Prostigmata его называют crista
metopica. У Oribatei, способных втягивать гнатосому и ноги в углуб-
ление гистеросомы, проподосомальный щиток прикрывает снаружи
ио углубление наподобие крышки люка. Среди Oribatei у Ptyctima
проподосомальный щиток называют aspis. Передняя часть спинных
щитков (или щита) других клещей, а в некоторых случаях и весь
i винной щит в действительности представляет собой проподосо-
мальный щиток.
Вторичные щитки формируются в разных группах по-разному.
У Pterygosomidae щитки образуются в результате расширения и
влияния оснований щетинок. Анальный и генитальный щитки форми-
руются в результате утолщения кутикулы в области полового и аналь-
ного отверстий. Иногда роговица глаза бывает укреплена по перифе-
рии склеротизированным ободком, который, расширяясь, образует
.• нктой щиток. Иногда покровы тела могут уплотняться без види-
мой причины. В местах выхода протоков желез или прикрепления
мышц кутикула часто превращена в щитки. В некоторых группах
г к'риальные щитки формируются за счет слияния тазиков, часть
которых может опускаться под покровы, образуя внутренние
32
Глава [. Введение
аподемы. Расположение вторичных щитков может быть весьма различным
у разных групп клещей, и поэтому оно будет рассматриваться для
каждой группы в отдельности.
Г натосома
Гнатосома несет ротовые органы — хелицеры и педипальпы, кото-
рые закрывают ротовое отверстие. Сама гнатосома редуцирована;
на первый взгляд создается впечатление, что она состоит только
Рис. 13. Тектум различных клещей (по Фитцтуму, 1940).
А — Pergamcsus olivaceus Vitzthum, 1927; Б — Discopoma regia Vitzthum, 1921;
3 —-Neopodocinum coprophllutn Vitzthum, 1926; Г—Trigonholaspis salti Vit-
zthum, 1930; Д— Cyrtolaelaps capreolus Berlese, 1904.
из роговых органов и их выростов. В разных группах клещей гнато-
сома формируется различными путями. Поскольку слияния, разраста-
ния и дегенерация составных частей гнатосомы шли в разных группа:
своими путями, то и отношение гнатосомы к остальному телу не мо
жет быть идентичным во всех группах. Гнатосома может быть заклю
чена в камеростом (специальная полость идиосомы) или же може'
образовывать выдающийся вперед носик, или рострум. Обычно гнато
сома хотя бы частично прикрыта сверху тектумом (Снодграсс
1948) или, как называют его другие авторы, эпистомом. Текту!
представляет собой направленный вперед вырост и состоит из тонко!
хитиновой пластинки, зазубренной на своем свободном конце (рис. 13)
Тектум может быть сильно редуцирован, а у некоторых клещей пол
ностью отсутствует.
Глава. I. Введение
33
Рис. 14. Гипостом
Ixodes rasas Neu-
mann, 1899.
Под тектумом отходят хелицеры, иногда прикрытые дорзальным
)асширением педипальп. Хелицеры имеют большое значение, и поэтому
йы их рассмотрим ниже в особом разделе.
Между хелицер и под ними находится ротовое отверстие. Оно
расположено вентрально по отношению к верхней губе (labrum) и
дорзально по отношению к гипостому. Как губа, так и гипостом
могут иметь очень сложное строение, но их
редко удается разглядеть, так как они при-
крыты педипальпами и хелицерами. С боков
ротовое отверстие защищено paralabrum, кото-
рые помогают закрывать рот, а также, ве-
роятно, направляют пищу в ротовое отверстие.
У Notostigma и Holothyroidea имеется зубчатый
теркообразный орган, развивающийся из верх-
ней губы, который также связан с ротовым
отверстием.
Педипальпы образуют латеро-вентральную
Поверхность гнатосомы. У некоторых групп
основания педипальп расширяются медиально
И дорзально; сливаясь по срединной линии,
они образуют трубку, в которой находятся
Хелицеры.
Гипостом образует вентро-медиальную
Стенку гнатосомы. У большинства групп он не-
заметно сливается с педипальпами, но у иксо-
довых клещей он имеет вид зубчатого обра-
зования, расположенного между педипальпами
(рис. 14). Так называемый „гипостом*1 иксодо-
пых клещей не гомологичен гипостому других
групп; у иксодовых клещей он образован глав-
ным образом из педипальп, хотя в образовании
его основания частично принимает участие и
Истинный гипостом.
Хелицеры. Хелицеры у клещей обычно кончаются клешней, кото-
рая состоит из неподвижного дорзального и подвижного вентрального
Пальцев. Такие клешневидные хелицеры имеются у всех подотрядов
Клещей. У Notostigmata, Holothyroidea и Tetrapodili имеются только
Хелицеры с клешней. У Tetrapodili хелицеры видоизменены, подвиж-
ный палец превращен в стилетовидное образование и является един-
ственным эффективным колющим элементом (рис. 15). Клешня обра-
зована из голени и лапки.
11ервичная сегментация хелицер не вполне ясна. У большинства
Паукообразных хелицеры состоят из трех члеников. На более ранних
Стадиях некоторые Mesostigmata имеют хелицеры, состоящие из шести
Члеников; однако у большинства других клещей хелицеры разделены
На два или три членика.
,| Зак. 1785
34
Глава f. Введение
Примитивные клешневидные хелицеры имеют поверхностное сход-
ство с клешнями большинства десятиногих раков. Каждый из пальцев
хелицер снабжен зубцами, которым иногда противопоставлены зубцы
Рис. 15. Хелицера Erlo-
phyes pint (N al ера), 1887.
P и с. 16. Хелицера Parasitus fucorum (De-
Gecr), 1778 (по Фитцтуму, 1940).
другого пальца. Такой тип клешни (рис. 16) служит для схватывания
добычи или раздавливания пищи. У самцов большинства Mesostigmata
подвижный палец превращается в дополнительный копулятивный орган
Рис. 17. Хелицера самца HeMerozercon audax Berlese, 1910
(по Фитцтуму, 1940).
(рис. 17) и используется для переноса сперматофоров из полового
отверстия самца в половое отверстие самки. Во всех случаях, как бы
ни изменялся подвижный палец, не-
подвижный палец остается неизме-
ненным.
Хелицеры исходного клешневид-
Р и с. 18. Хелицера Dermanyssus ного типа претерпевают у разных
galllnae (DeGeer), 1778. форм различные видоизменения. Так,
оба элемента могут уменьшаться
и делаются игловидными, образуя вместе колющий аппарат (напри-
мер, у Dermanyssus, рис. 18). Иногда неподвижный палец исчезает,
а подвижный занимает скорее концевое, чем боковое положение.
Подвижный палец может удлиняться в виде нити, как, например,
у паутинных клещей. У иксодовых клещей хелицеры клешневид-
ные, но подвижный палец располагается латерально, а не вентрально,
Глава I. Введение
33
зубцы же лежат на наружной поверхности клешни, а не на внутрен-
ней (рис. 19). Хелицеры иксодовых клещей служат для прикрепления
к телу хозяина, и их строение хорошо приспособлено для этой цели.
Специальные чувствующие (сенсорные) органы и щетинки имеются
па хелицерах у многих клещей. У основания подвижного пальца
некоторых Mesostigmata суставная перепонка удлиняется и образует
подушечку, напоминающую у одних клещей пучок щетинок, а у дру-
гих— венчик. Часто имеется щетинка на конце неподвижного пальца
хелицер, но на подвижном пальце она отсутствует. Щетинки и сен-
сорные поры встречаются также и на хелицерах других групп клещей.
Рис. 19. Хелицеры Ixodes reduvius (Linnaeus), 1758
(по Фитцтуму, 1940).
Д —самца (вид сверху); Б — самца (вид снизу); В —самки (вид сверху);
Г—самки (вид снизу).
Педипальпы. Педипальпы являются второй парой конечностей
паукообразных. У Acarina они отходят латерально от тазиков пальп,
которые, вероятно, гомологичны слившимся основным членикам пер-
вичных, разделенных на две ветви придатков. У современных Acarina
педипальпы никогда не разделены более чем на шесть члеников,
между тем как у ископаемого Protacarus crani Hirst из девона
пальпы состоят из семи члеников. Членики педипальп носят те же
названия, что и членики ног, а именно (начиная с проксимального
членика): тазик (соха), вертлуг (trochanter), основное бедро (basife-
mur), концевое бедро (telofemur), коленный членик (genu), голень
(tibia) и лапка (tarsus). Редукция числа члеников от первичных 7
то 6 — максимальное число члеников, отмеченное у современных кле-
щей,— достигается путем слияния basifemur и telofemur, которые
образуют единое нерасчлененное бедро.
Относительно неизмененные пальпы встречаются у Onychopalpida,
Mesostigmata, Glyptognathidae, Sarcoptiformes, Limnocharidae и
3*
36
Глава I. Введение
Eylaidae. У некоторых из названных выше групп клещей пальпы шести-
члениковые, причем их лапка и голень сходны с одноименными
Рис. 20. Тарзальные членики пальпы.
A — Holothyras austrclasiae Womersley, 1935; Б — Neocarus texanus
Chamberlain et Mulaik, 1942. Щетинки ие изображены, что позволяет
видеть коготки.
члениками ног. Лапка пальп Onychopalpidae, так же как и лапки
ног, снабжена коготком (рис. 20).
два членика педи-
пальпы EuschOnga-
stia indtca (Hirst),
1915.
Рис. 22. Педипальпа
клеща Cheyletidae.
Первично простые, похожие на ноги педипальпы во многих груп-
пах значительно видоизменяются. У некоторых паразитических групп
педипальпы редуцированы до зачаточных выступов, снабженных спе-
Глава I. Введение
37
анализированными шипами или угловыми выростами, что является
приспособлением к паразитическому образу жизни. У Trombidiformes
пальпы могут увеличиваться и функционируют как дополнительные
хелицеры.
У некоторых Trombidiformes соотношения между лапкой и
голенью пальп изменяются. Голень этих форм часто имеет концевой
крепкий коготь, который оттесняет лапку пальп книзу. Это смеще-
ние приводит к образованию клешневидных педипальп (рис. 21).
Такие клешневидйые педипальпы первично выполняют функцию рук.
Дорзальный коготь в них противопоставлен лапке пальп, которая
играет роль большого пальца; клещи, имеющие такие педипальпы,
способны схватывать и хорошо удерживать не только пищу, но и
другие предметы.
У Cheyletidae клешневидные педипальпы значительно усилены
(рис. 22). Голень снабжена крепким когтем, а медиально сдвинутая
лапка несет гребневидную или серповидную когтеобразную щетинку.
В этой группе правая и левая педипальпы противопоставлены одна
другой и образуют мощный хватательный аппарат, при помощи кото-
рого добыча прижимается к сравнительно небольшим хелицерам.
Педипальпы у Cunaxidae и Bdellidae имеет такую же функцию, но
„больши<^пальцы“ пальп у них отсутствуют.
Конечности
Большинство клещей обладает тремя парами ходильных ног в фазе
личинки и четырьмя парами — на всех последующих фазах. Eriophyi-
dae имеют только две пары ног на всех фазах, у некоторых Poda-
polipodidae и Phytoptipalpidae взрослые клещи имеют три пары ног,
а иногда всего лишь одну пару.
Ноги, так же как пальпы, разделяются на шесть члеников, рас-
полагающихся в следующем порядке: тазик, вертлуг, бедро, коленный
членик, голень и лапка. Часто бедро разделяется на две части basi-
feinur (основное бедро) и telofemur (концевое бедро); в этих случаях
насчитываются семь члеников. У некоторых форм происходит слияние
члеников, в результате чего число члеников ног может уменьшаться
до 5 или даже до 4, как, например, у Schizocarpus. У Chirodlscus
ноги первой и второй пары состоят каждая из одного членика. С дру-
гой стороны, у Tarsotomus лапка вторично подразделяется на боль-
шое число члеников (до 18).
Лапка ходильных ног в типичных случаях несет пару хорошо
развитых коготков. Кроме коготков, на каждой лапке может быть
По одному нежному прозрачному присосковидному образованию (ca-
runcle). Такие образования всегда развиты на лапках у клещей группы
Mesostigmata и у иксодовых клещей (рис. 23). У Trombidiformes
Между двумя коготками часто имеется эмподий, а в некоторых слу-
чаях срединный эмподий превращается в коготок, в то время как
Рис. 24. Нога II самца Parasitus
bombophilus Vitzthum, 1930
(по Фитцтуму, 1940).
Рис. 23. Лапка
ноги I Haemolaelaps
glasgowi (Ewing),
1925 (вид сверху).
Р и с. 25. Самец Analges nitzschii
Haller, 1878 (по Фитцтуму, 1940).
Обращает на себя внимание чрезмерное
развитие ног IIL
Рис. 26. Нога I Protomyobia cla-
paredel (Poppe), 1896 (вид снизу).
Видны шероховатые отростки, служащие
для обхватывания волосков хозяина.
Глава I. Введение
39
парные коготки редуцируются или видоизменяются, превращаясь
в клейкие волоски. Если ноги утрачивают локомоторную функцию,
то коготки становятся рудиментарными или вовсе исчезают. Так,
передняя пара ног клещей семейства Macrochelidae используется
скорее как тактильные органы, чем как органы передвижения; ко-
готки на них либо очень малы, либо совсем отсутствуют. Многие
из водяных клещей, особенно активно плавающие формы, также не
имеют коготков на лапках.
Ноги используются, в первую очередь, для хождения и бега, но
в некоторых группах они имеют и другие функции. Ходильные и
тактильные типы ног уже рассматривались выше. Часто у самцов
ноги используются для удерживания самки во время копуляции. Вто-
рая пара ног у самцов многих видов семейства Parasitidae сильно
видоизменена (рис. 24). Самцы Analgesidae имеют сильно развитые
задние ноги (рис. 25). Они так массивны, что возможность исполь-
зования их для передвижения исключена. Ноги вод .ных клещей во-
оружены многочисленными длинными густо сидящими волосами, кото-
рые увеличивают поверхность ног и делают их весьма совершенными
веслами. Иногда некоторые пары ног изменены для обхватывания
волоска (рис. 26). Задние ноги клещей рода Spelorchestes и Nanor-
chestes имеют специализированную мускулатуру, что позволяет этим
клещам довольно хорошо прыгать. Надо думать, что Eupodes, имею-
щие сильно удлиненные бедра четвертой пары ног, также хорошо
прыгают.
Тазики ходильных ног многих клещей неподвижны и, будучи по-
гружены в кожу, часто образуют эндоскелет.
Мускулатура
Так же как и другие членистоногие, клещи имеют поперечно-
полосатую мускулатуру. Мышцы конечностей проникают в них из
гела; каждая нога снабжена как разгибателями, так и сгибателями.
Мускулатура тела хорошо развита у многих групп; она приводит
в движение половой аппарат, анальные щитки и гнатосому. Мышцы
отходят и прикрепляются к щиткам, аподемам или эпимерам, или
к неспециализированной мягкой кутикуле между щитками. Места
отхождения и прикрепления мышц различимы с поверхности тела
в виде ясных рубцов на щитках и ямок на мягкой части покрова
гела. У клещей с мягким телом форма идиосомы может изменяться
под действием мышц.
Пищеварительная система
Пищеварительный тракт у клещей представляет собой простую
трубку. Ее передняя часть, или передняя кишка, имеет эктодер-
мальное происхождение (stomadaeum); она подразделена на муску-
40
Глава I. Введение
листую глотку и трубчатый пищевод. Средняя кишка, или желудок
(ventriculus), развивается из энтодермы и обладает широким просве-
том и хорошо развитым пищеварительным эпителием. Задняя кишка
развивается из эктодермы (proctodaeum) и часто подразделяется на
переднюю трубчатую тонкую кишку и заднюю мускулистую прямую
кишку, которая открывается анальным отверстием.
У клещей различают три основных типа пищеварительной системы:
1. Мезостигматический тип, характерный для Onychopalpida,
Mesostigmata и Ixodides.
2. Тромбидиоидный тип (краснотелковый), характерный для Trom-
bidiformes.
3. Саркоптоидный тип, характерный для Sarcoptiformes.
В мезостигматическом типе передняя кишка, состоящая из муску-
листой глотки и из длинного узкого пищевода, который переходит
Рис. 27. Продольный срез через тело Tetranychus
(по Бловслю, 1945).
в небольшую центральную часть средней кишки, вполне типична. Для
средней кишки характерно наличие относительно маленького желудка,
от которого отходят большие боковые слепые выросты (диверти-
кулы). Тонкая кишка у всех клещей, за исключением иксодовых,
длинная; впадает она в круглую прямую кишку, открывающуюся
анальным отверстием. У Notostigmata имеется слабо выраженная
толстая кишка (colon). Экскреторные трубочки (мальпигиевы со-
суды) впадают в заднюю кишку между тонкой и толстой кишкой.
Кишечник тромбидиоидного типа характеризуется отсутствием
толстой и прямой кишки. Передняя кишка типична и имеет хорошо
развитые глотку и пищевод. Желудок обширный; его дивертикулы
соединяются с ним широкими просветами. У Trombidiformes задняя
кишка преобразована в экскреторный орган. До недавнего времени
большинство авторов считало, что у Trombidiformes задняя кишка
не соединяется с желудком, но Влонель (1945) показал наличие
такого отверстия у Tetranychus (рис. 27).
Глава f. Введение
41
Кишечник саркоптоидного типа стоит ближе к мезостигматиче-
скому, чем к тромбидиоидному типу. Все его подразделения — глотка,
пищевод, желудок, тонкая кишка, толстая кишка, а также прямая
кишка — обычно хорошо различимы. Желудок обычно крупнее, чем
его задние слепые отростки. По этому признаку саркоптоидный тип
органов пищеварения легко отличается от мезостигматического.
На гнатосоме около ротового отверстия открывается несколько
слюнных желез. Эти железы, возможно, выделяют секрет, содержа-
щий пищеварительные ферменты. Личинки многих Trombidiformes
могут переваривать кутикулу своих хозяев. У паутинных клещей
некоторые железы выделяют шелковистую нить. Клетки эпителиаль-
ной стенки желудка и его слепых выростов — железистые. Они,
повидимому, вырабатывают пищеварительные ферменты, и весьма
вероятно, что пищеварение и всасывание происходят в просвете и
в клетках средней кишки. Фекалии формируются в тонких кишках
и обычно выделяются в виде мелких шариков, смешанных с экскре-
тами. У дейтонимф некоторых Uropodidae экскреты используются на
образование анальных ножек (стебельков), которыми они прикре-
пляются к крупным насекомым, чаще всего к жукам.
Система органов выделения
У Acarina имеется три типа органов выделения. Самый прими-
тивный из них это коксальные железы', наиболее распространенным
типом являются экск-реторные трубочки, открывающиеся в заднюю
кишку, но происходящие из энтодермы. Как упоминалось выше,
у Trombidiformes задняя кишка преобразуется в экскреторный орган.
В дополнение к этим органам выделения клетки средней кишки также
имеют экскреторную функцию. Во время пищеварения клетки напол-
няются экскреторными продуктами, которые выделяются в просвет
средней кишки (желудок), оттуда переходят в тонкую кишку.
Конечным продуктом белкового обмена клещей является гуанин. Noto-
stigmata имеют две экскреторные трубочки и пару коксальных желез,
которые открываются у тазиков первой или второй пары ног. Клещи
Ho’othyroidea тоже имеют две пары экскреторных трубочек и пару
коксальных желез, открывающихся у тазиков первой пары ног.
Клещи Mesostigmata имеют одну пару экскреторных трубочек и
от одной до четырех пар коксальных желез.
Trombidiformes не имеют экскреторных трубочек обычного типа,
по у многих из них задняя кишка превращена в экскреторный орган.
У некоторых Trombidiformes имеются коксальные железы.
Sarcoptiformes имеют пару маленьких экскреторных трубочек.
У Oribatei были обнаружены коксальные железы.
Кровеносная система
Кровеносная система клещей лакунарная, и у большинства форм
нет не только сесудор, но и рсрдца. Кровь клещей бесцветна и
42
Глава I. Введение
омывает все органы тела. Имеются амебовидные лейкоциты, и их лучше
всего наблюдать в периоды покоя, предшествующие линькам. У Holo-
thyroidea и у некоторых Mesostigmata имеется простое сердце.
Дыхательная система
Из всех анатомических особенностей клещей наибольшее таксо-
номическое значение имеют признаки, связанные с дыхательной си-
стемой. Подотряды клещей установлены главным образом на основа-
нии особенностей этой системы. У клещей, имеющих трахеи, число
и положение стигм имеет (в систематическом отношении) первосте-
пенное значение. Функционально дыхательная система клещей подобна
дыхательной системе большинства наземных членистоногих. Главные
трахеи подразделяются на трахеолы, которые пронизывают почти
все ткани и обеспечивают газообмен, необходимый для общего обмена
веществ в клетках и тканях. Некоторые клещи не имеют трахейной
системы. Это обычно мелкие формы, и дыхание у них происходит
через кутикулу кожных покровов; некоторые формы способны к анаэ-
робному дыханию.
Notostigmata имеют четыре пары стигм, расположенных дорзально
на первых четырех сегментах опистосомы. Они открываются в кожи-
стой кутикуле и не поддерживаются стигмальными щитками. Тра-
хеолы ответвляются от трахейных стволов, начинающихся от стигм,
не образуя анастомозов между собой.
Holothyroidea имеют две пары стигм. Передняя пара расположена
по бокам, над тазиками третьей пары ног. Они сообщаются с пред-
дверием, от которого многие трахеи проходят в ткани тела. Задняя
пара стигм сообщается с атриумами, от которых отходит несколько
тонкостенных отростков.
Mesostigmata и взрослые особи иксодовых клещей имеют одну
пару стигм, но личинки иксодовых клещей могут иметь по нескольку
пар стигм. Стигмы сообщаются с атриумом, от которого .отходит
трахейный ствол. Со стигмами и трахеями связана хитинизированная
трубочка — перитрема. У Mesostigmata стигмы лежат вентрально
сбоку от тазиков, на уровне или впереди тазиков третьей пары ног.
Перитремы обычно длинные и направлены вперед; они могут также
идти назад на некоторое расстояние. В то время как у одних форм
перитремы почти прямые, у других они сильно изогнуты. У иксодо-
вых клещей перитремы никогда не бывают трубчатые; они ограничены
полем, прилегающим к стигмам, которые лежат позади или сбоку
от тазиков четвертой пары ног.
У Trombidiformes стигмы обычно открываются на гнатосоме или
между гнатосомой и проподосомой. Самки Tarsonemini имеют
1—2 пары стигм на протеросоме. У самцов многих клещей дыха-
тельная система вовсе отсутствует. У Prostigmata стигм.ы открываются
на гнатосоме; часто имеются стигмальные рожки^ или перитремы
Глава I. Введение
43
(рис. 28). Перитремы простигматид дают начало трахейному стволу,
и они не похожи на перитремы Mesostigmata. У большинства Bdelli-
dae в дополнение к обычной трахейной
системе простигматического типа имеется
генитальная трахейная система. У неко-
торых мелких клещей-краснотелок стигмы
и трахеи редуцированы или отсутствуют.
У Sarcoptiformes трахеи и стигмы либо
совсем отсутствуют, либо представлены
лишь мельчайшими трахеями. У Acaridiae
специализированная дыхательная система
утрачена. Большинство Oribatei замеча-
тельны тем, что они имеют развитую
систему трахей, которая открывается че-
рез стигмы и пористые поля на многих
участках тела. У этой группы даже псевдо-
стигмы связаны с дыхательной системой.
Рис. 28. Перитремы
Alloihrombium. sp.
Гранжан (1934) дал подробное описание строения дыхательной си-
стемы этой группы. Некоторые орибатиды все же утратили и дыха-
тельные отверстия, и трахеи.
Нервная система
Ганглии нервной системы, первично располагавшиеся сегментально,
у Acarina настолько сближены друг с другом, что вся центральная
нервная система у них представляет собой
елиную цельную массу, окружающую пищевод.
В эмбриональной и личиночной фазах слияние
ганглиев еще не столь полное, как это наблю-
дается у нимф и у взрослых. Часть мозга,
располагающаяся под пищеводом, дает начало
нервам, которые иннервируют глотку, хелицеры
и глаза. Нервы, идущие к педипальпам, ногам
и внутренним органам задней части тела, бе-
рут начало от части мозга, расположенной
вентрально от пищевода (рис. 29). Нервы, отхо-
Р и с. 29. Мозг Tetra-
nychus (по Бловелю,
1945).
дящие к придаткам, не иннервируют другие части тела. Повидимому.
все нервы содержат как двигательные, так и чувствующие волокна,
Органы чувств
Как и у других членистоногих, у Acarina щетинки несут прежде
всего сенсорные функции. Различные типы сенсорных щетинок уже
описывались в разделе о наружных покровах. Остается упомянуть
еще о специальных щетинковидных псевдостигматических органах,
или сенсиллах, которые развиты у большинства Trombidiformes и
Sarcoptiformes, Сенсилль; обычно сидят в специальных углублениях
44
Глава I. Введение
на проподосомальном щитке или же на месте этого щитка, если он
отсутствует. Эти углубления называются псевдостигмами, и у Ori-
batei они связаны с воздушным мешком, который участвует в дыха-
нии. Функции псевдостигм и их сенсилл неизвестны. То, что они
являются сенсорными органами, вероятно, правильно, так как сен-
силлы непосредственно связаны с нервной системой. У наземных
клещей, обладающих сенсиллами, эти образования обычно хорошо
развиты, но у близкородственных групп водяных клещей они редуци-
рованы или вовсе
структуре так
смотря на это,
отсутствуют. Сенсиллы могут варьировать по
широко, как и щетинки тела (рис. 30), но, не-
легко распознаются у любых видов клещей, обла-
дающих ими. Они всегда сильно отли-
чаются от обыкновенных щетинок.
Пальпы и ноги всех клещей более
или менее часто покрыты щетинками
же
они
Сенсиллы.
Рис. 31. Срез через галлеров
орган Dermacentor venusius,
Banks, 1908.
Рис. 30.
A’-Trotnbicula (Eufrombiculc.)
alfreddugesi (Oudemans), 1910;
Б — СеЛатпа sp.
и в большинстве случаев снабжены бороздчатыми сенсорными ще-
тинками. Иногда пальпы или ноги (одна или несколько пар) пре-
вращены в такргильные органы. Лапки обычно богато оснащены
специальными сенсорными щетинками, а у иксодовых клещей и у
Rhagidiidae на лапках передних ног имеются еще специальные органы.
У всех иксодовых клещей на всех фазах развития (за исключением ним-
фальной и, возможно, также личиночной фазы Ceratixodes) имеется
галлеров орган. Галлеров орган связан с обонянием. Он расположен на
лапке первой пары ног и обычно состоит из углубления, вмещающего
специализированные щетинки (рис. 31). Клещи Notostigmata имеют похо-
жий на него орган, a Mesostigmata обладают скоплением щетинок
на лапке первой пары ног, напоминающих щетинки галлерова органа.
Клещи Rhagidiidae имеют хорошо развитые рагидиевы органы на лап-
ках первой и второй пары ног; родственные группы имеют похожий,
хотя и не столь сильно осложненный орган.
На покровах у большинства клещей есть специальные поля, со-
стоящие из пор, продольных щелей или углублений, которые непо-
Глава I. Введение
45
средственно или косвенно связаны с нервными волокнами. Функция
таких образований неизвестна, но они, вероятно, по своей природе
являются сенсорными образованиями. Клещи реагируют на разнообраз-
ные раздражители, но специализированных органов, воспринимающих
их в ряде случаев не обнаружено. Возможно, что ими служат именно
Эти непонятные образования1.
Глаза имеются у многих клещей, и многие клещи, не обладаю-
щие ими, вероятно, воспринимают изменения интенсивности света.
Глаза клещей всегда являются простыми; сложные глаза у клещей
неизвестны. Клещи, у которых отсутствуют глаза, обычно имеют
тонкие прозрачные поля на дорзальной поверхности, так что лучи
света проникают и в глубже лежащие ткани. Наиболее примитивные
глаза представляют собой скопления пигментных зерен, связанных
с глазным нервом. Пигмент обычно красного цвета, но встречается
также черный или синий пигмент. В ряде случаев скоплениями зерен
пигмента связана линзовидная роговица. Наиболее сложные глаза имеют
некоторые водяные клещи. Некоторые клещи имеют один срединный
глаз, другие — одну или две пары проподосомальных глаз и третьи,
как, например, большинство водяных клещей,—все пять глаз вместе.
Даже самые сложные глаза, вероятно, способны только различать
интенсивность света, и сомнительно, чтобы при их помощи клещи
могли различать форму предметов.
Половая система
Все клещи раздельнополы. У большинства групп самцов удается
отличать от самок, даже если они внешне похожи друг на друга.
Оплодотворение внутреннее, но способы введения сперматофоров
в половую систему самок сильно варьируют. Некоторые клещи спо-
собны размножаться партеногенетически, но нет таких, которые раз-
множались бы только этим путем. Самки бывают яйцекладущими
или яйцеживородящими. Бесполое размножение неизвестно.
У самцов половая система состоит из семенника или пары семен-
ников, семепроводов, придаточной железы или желез, семеизверга-
тельного канала и совокупительного органа. Последний отсут-
ствует у Mesostigmata, самцы которых вводят в самку сперматофоры
при помощи хелицер. У многих самцов одна или несколько пар ног
видоизменяются в органы, служащие для схватывания самок во время
копуляции.
Половая система самок состоит из яичника или пары яичников,
яйцевода, матки, семеприемника, придаточных желез, иногда вла-
галища. У некоторых групп (Oribatei) имеется специальный яйцеклад,
но обычно яйца откладываются через половое отверстие без помощи
яйцеклада.
1 Эти шелевидные образования были названы В. М. Шимкевичем лиро-
видными органами. — Прим. ред.
46
Глава J. Введение
Как у самца, так и у самки половое отверстие обычно прикрыто
специальными щитками, или клапанами, часто связанными с поло-
выми присосками, расположенными на самих щитках или рядом
с ними. Для большинства клещей исследование полового отверстия
является достаточным для определения пола; для других же единст-
венным надежным признаком служит наличие или отсутствие копу-
лятивного органа, а для определения пола некоторых форм прихо-
дится даже исследовать половые железы.
Сперматозоиды, вводимые в половые пути самки, часто бывают
незрелыми, и их созревание заканчивается только после копуляции.
У большинства клещей развивается единовременно одно яйцо или
немногим больше, но у иксодовых клещей и у некоторых Tarsone-
midae одновременно развивается очень большое количество яиц.
Большинство яиц имеет оболочку, которая предохраняет разви-
вающийся зародыш. Иногда яйца переносятся ветром на далекое рас-
стояние.
Жизненный цикл
Примитивный жизненный цикл характеризуется наличием следую-
щих последовательно развивающихся фаз1: а) яйцо, в котором раз-
вивается бластула, б) дейтовум, образуемый хорионом, являющимся
продуктом бластодермы, в) шестиногая личинка, г) протонимфа,
д) дейтонимфа, е) тритонимфа и ж) половозрелые самец и самка.
Однако во многих семействах клещей этот примитивный жизненный
цикл значительно изменяется и его следует рассматривать скорее как
исключение, чем как правило. Жизненные циклы многих клещей до
сих пор еще не раскрыты.
У большинства клещей личинка отличается от взрослого клеща
отсутствием полового отверстия и последней пары ног. У некоторых
простигматических клещей личинка совершенно отличается от взрос-
лого и превращается в нимфу в результате настоящего метаморфоза.
Нимфы обычно похожи на взрослых; однако у Oribatei, Uropodldae и
Acaridae некоторые из нимф заметно отличаются от взрослых..
КЛАССИФИКАЦИЯ
Большинство акарологов, энтомологов и зоологов считают Acarina
отрядом класса Arachnida. Если Arachnida подразделять непосред-
ственно на отряды, то при такой концепции группы клещей их
1 В жизненном цикле клещей, как и других членистоногих, следует раз-
личать фазы: яйцо, личинка, нимфа, имаго. В пределах фазы клещ может
проходить несколько стадий (например, нимфа I, II, III); в пределах одно-
членной фазы и стадии различают молодых и старых особей. Авторы книги
не придерживаются этой терминологии, которая ныне приобретает все боль-
шую популярность. При редактировании перевода нами была принята новая
терминология. — Прим. ред.
Глава t. Введение
41
родство с другими отрядами, так же как и родственные отношения
других групп класса, будет отражено более или менее правильно. Но
если отряды Arachnida группировать в подклассы, как это делал
Петрункевич (1949), то, пожалуй, правильнее рассматривать Acarina
как отдельный подкласс. Acarina легко отличимы от других пауко-
образных тем, что обладают особой гнатосомой и никогда не рас-
членены отчетливо на головогрудь и брюшко. Филогения Acarina
неясна, и большинство изучающих эту группу считают ее полифи-
летической по происхождению. В настоящей работе Acarina рассма-
тривается как отряд.
Систематическое положение Acarina явствует из следующей схемы:
Тип Arthopoda(членистоногие). Метамерные животные с наружным ске-
летом и членистыми придатками.
Подтип Chelicerata (хелицеровые). Членистоногие, лишенные антенн
или мандибул; ротовые части состоят из педипальп и хелицер.
Класс Arachnida (паукообразные). Хелицеровые, лишенные пластин-
чатых легких.
Отряд Acarina (клещи). Паукообразные с ротовыми органами, более
или менее отчетливо выделенными из тела в виде ложной головы
(capitulum, или гнатосома). Сегментация тела слабо выражена
или вовсе утрачена. Первичные склериты в значительной мере
замещены вторичными щитками различного происхождения. Личи-
ночная фаза, как правило, имеет три пары ног; нимфальные и
взрослые фазы обычно имеют четыре пары ног. Редкие исклю-
чения из этого отмечены для иксодовых клещей и немногих мелких
клещиков.
Схема подразделений клещей на подотряды приведена ниже.
Отряд Acarina.
Подотряд Onychopalpida (гл. II). Клещи с типичными амбулакраль-
ными коготками на педипальпах и более чем одной парой стигм
на идиосоме.
Подотряд Mesostigmata (гл. III). Клещи с одной парой стигм, откры-
вающихся снаружи от основания ног. Стигмы обычно связаны
с удлиненными перитремами; у немногих специализированных пара-
зитов дыхательных путей позвоночных они вторично утрачены.
Галлерова органа нет. Гипостом не приспособлен для прокалы-
вания.
Подотряд Ixodides (гл. IV). Клещи с парой стигм, расположенных
позади или кнаружи от тазиков; стигмы связаны со стигм аль-
ным щитком, а не с удлиненными перитремами. Галлеров орган
имеется. Гипостом превращен в орган для прокалывания покро-
вов хозяина и снабжен загнутыми назад зубцами.
Подотряд Trombidiformes (гл. V и VI). Клещи с парой стигм, распо-
ложенной на гнатосоме или вблизи ее; иногда стигмы совсем
48
Глава I. Введение
отсутствуют. Пальпы обычно свободны и хорошо развиты.
Хелицеры часто приспособлены для прокалывания. Анальные при-
соски всегда отсутствуют.
Подотряд Sarcoptiformes (гл. VII и VIII). Клещи без стигм или с тра-
хейной системой, открывающейся через стигмальные и пористые
поля на различных частях тела. Тазики образуют аподемы под
покровами брюшной поверхности тела. Ротовые органы обычно
жующие, хелицеры снабжены сильной клешней; у немногих па-
разитических формы хелицеры преобразованы для специальных
функций. Пальпы простые. У одних форм имеются псевдостигмы
или псевдостигмальные органы, у других они отсутствуют. Аналь-
ные присоски часто имеются.
ЛИТЕРАТУРА
Banks N., 1915. The Acarina or mites, U. S. Dept. Agric. Rpt., 108, 1—153
Berlese A., 1882—1903. Acari, Myriopoda et Scorpiones hucusque in Italia
reperta, Fasc. 1 -101, Padua.
Ewing H. E„ A manual of external parasites, 1929. 1—225+XIV, Baltimore.
О u d e m a n s A. C., 1926—1937. Kritisch Historisch Overzicht der Acarologie,
vol. 1 (850 V. C. —1758), 1—150 +VII, 1926, Tijdschr. Ent. Leiden, 69,
Suppl.; vol. 2(1759—1804), 1—1097+XVII, 1929, Tijdschr. Ent. Leiden,
69, Suppl.; vol. 3 (1805—1850), 1—3379 + CI, 1936—1937 (Bd. I—VI).
Petrunkevitch A., 1949. A study of Palaeozoic Arachnida, Trans. Conn.
Acad. Art. Sci., 37, 69—315.
Radford C. D., 1950. Systematic check list of mite genera and type species,
Union Internat. des Sci. Biol., Ser. C. (Sect. Ent.), 1, 1—252.
Reuter E., 1909. ZurMorphologic und Ontogenie der Acariden, Acta Soc. Sci.
Fenn., 36, № 4, 1—228.
Snodgrass R. E., 1948. The feeding organs of Arachnida including mites and
ticks, Smithsonian Misc. Coll., 110, № 10, 1—93.
Vitzthum H. G., 1929. Acari (Die Tierwelt Mitteleuropas, 3, № 7, 1—112).
Vitzthum H. G., 1931. Acari. Kiikenthals Handbuch der Zoologie, 3, 2 Half,
1—160.
Vitzthum H. G., 1940—1942. Acarina, Bronns, Klassen und Ordnungen des
Tierreiches, 5, Sect. 4, Buch 5,1—1011 + XI.
Глава II
ПОДОТРЯД ONYCHOPALPIDA WHARTON, 1947
У большинства клещей (Acarina) педипальпы специализированы
в том отношении, что они утратили коготки на тарзальном членике; у не-
которых форм имеются развивающиеся вторично коготки на голени
пальп. Группы Notostigmata, и Holothyroidea, объединяемые в подот-
ряд Onychopalpida, отличаются от всех остальных клещей тем, что
они сохраняют редуцированные амбулакральные коготки на педипаль-
пах и обладают теркообразным органом на верхней губе. Хотя еще
неясно, имеют ли эти две группы монофилетическое происхождение,
однако объединение их в один подотряд оправдано их примитивно-
стью и тем, что обе эти группы, возможно, представляют собой
живые остатки прежней обширной фауны примитивных клещей.
Морфология. О наличии коготков на пальпах уже упоминалось
выше. Клещи этого подотряда уникальны в том отношении, что их
представители имеют, по крайней мере, четыре расположенные лате-
рально стигмы. К сожалению, вследствие бедности Onychopalpida
видами общей сводки по морфологии этой группы нет и характер-
ные морфологические особенности группы еще не изучены. Для спра-
вок рекомендуется монографическая сводка Вита (1904) и работа
Гранжана (1938).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ГРУПП ONYCHOPALPIDA
1. Имеются тритостернум и, по крайней мере, четыре пары стигм,
открывающихся латерально. Глаз две пары. Кутикула покровов
кожистая.........................................Notostigmata
— Тритостернум отсутствует. Имеется только две пары стигм, откры-
вающихся латерально. Глаз нет. Кутикула покровов склеро-
тизирована......................................Holothyroidea
NOTOSTIGMATA WITH, 1903
В состав группы входит бедное видами семейство Opilioacaridae.
Чэмберлен и Малэйк (1942) установили семейство Neocaridae, вклю-
чив в него новые виды Notostigmata, обнаруженные этими авторами
в Техасе. Это в значите гьной мере оправдано с морфологической
точки зрения, но нам кажется нежелательным выделить новое
4 Зак. 1785
Ад
Глава II. Подотряд Onycbopalpida
семейство, так как и без того в состав группы в целом входит лишь
малое число видов. Поэтому мы в составе Notostigmata рассматриваем
лишь одно семейство.
Opilioacaridae With, 1902
(рис. 32)
Рис. 32. Opilioacarus segmentatus
With, 1902 (по Виту, 1903).
Самка (вид сверху); видны четыре пары
опистосомальных стигм.
ДИАГНОЗ
Клещи средней величины, длиной
около 1 мм\ тело овальной формы.
Гистересома на спинной стороне
имеет следы сегментации. Кутикула
исчерчена бороздками, образованными
многочисленными порами. Тритосте-
рнум расчлененный. На каждой сто-
роне проподосомы расположено по
два глаза. Щетинки простые или
перистые. Педипальпы имеют ре-
дуцированные амбулакральные ко-
готки; хелицеры не видоизменены.
Теркообразный орган снабжен за-
гнутыми назад зубцами и располо-
жен между хелицерами, которые
его прикрывают. Вертлуги ног
III и IV вторично расчленены.
Тазики свободно подвижны. На
каждой лапке имеется пара не-
измененных амбулакральных когот-
ков. Половое отверстие у обоих
полов лежит на брюшной стороне
между тазиками ног III; самки имеют
яйцеклад, а самцы — специальные
поля по бокам полового отвер-
стия. На спинной стороне по бокам
гистеросомы открывается четыре
пары стигм, тогда как у Neocarus
есть только одна стигма, расположенная по средней линии тела.
РОДЫ
1. Opilioacarus With, 1902 (=Eucarus With, 1903)
Тип рода Opilioacarus segmentatus With, 1902
2. Neocarus Chamberlain et Mulaik, 1942
Тип рода Neocarus texanus Chamberlain et Mulaik, 1942
3. Paracarus Chamberlain et Mulaik, 1942
Тип рода Opilioacarus hexopthalmus Redikorzev, 1937
Глава 11. Подотряд Onychopalpida
51
ОБЩИЕ ЗАМЕЧАНИЯ
Хотя Opilioacaridae и представляют собой примитивную группу,
они подобно многим другим примитивным животным специализиро-
ваны во многих отношениях. Теркообразный орган гнатосомы, вторич-
ная сегментация ног, расчленение тритостернума — все это следует
рассматривать как результаты специализации.
Opilioacaridae ведут скрытый образ жизни; встречаются они под
камнями и в почвенном гумусе. В кишечнике этих клещей попадаются
хитиновые остатки других членистоногих. Это указывает на то, что
они питаются, по крайней мере частично, мелкими членистоногими.
Представители этого семейства обнаружены в Средиземноморской
области и в Техасе. До сих пор добыто мало экземпляров этих кле-
щей, и поэтому лишь очень немногое известно об их жизненном
цикле. Нет сведений и об их хозяйственном и медицинском значении.
ЛИТЕРАТУРА
Chamberlain R., Mulaik S., 1942. On a new family in the Notostigmata,
Proc. Biol. Soc. Washington, 55, 125—131.
Grandjean F., 1936. Un acarien synthetiqne: Opilioacarus segmentatus
With., Bull. Soc. Hist. Nat. Afr. Nord., 27, 413—444.
With C. J., 1904. The Notostigmata a new suborder of Acari, Vidensk. Medd.
fra den Naturk. Foren. i. Kbhvn, 137—192-|-Pls. IV—VI.
HOLOTHYROIDEA REUTER, 1909
Группа Holothyroidea представлена только одним семейством Holo-
thyridae.
Holothyridae Thorell, 1882
(рис. 33)
ДИАГНОЗ
Holothyridae —крупные (длиной до 7 мм) клещи полусфериче-
ской формы; длина их заметно превышает ширину. Следы первичной
сегментации отсутствуют, тело подразделено на гнатосому и идиосому.
Темнокоричневая гладкая кутикула у взрослых сильно склеротизиро-
вана. Глаза отсутствуют. Щетинки по форме относительно не видо-
изменены. Педипальпы состоят из пяти подвижных члеников, лапки
пальп снабжены коготками. Хелицеры клешневидные. На конце ниж-
ней губы имеется теркообразный орган. Каждая нога имеет пару ам-
булакральных коготков. По средней линии между тазиками третьей
и четвертой пары ног расположено половое отверстие; у самца оно
прикрыто двумя щитками, а у самки — четырьмя.
РОДЫ
1. Holothyrus Gervais, 1842
Тип рода Holothyrus coccinella Gervais, 1842
4*
52
Глава II. Подотряд Onychopalpida
ОБЩИЕ ЗАМЕЧАНИЯ
Виды Holothyrus найдены в Новой Гвинее, на Цейлоне, в Австра-
лии и на островах Индийского океана. Жизненный цикл и биология
клещей Holothyridae неизвестны. Хирст (1922) указывал, что Holo-
thyrus довольно обычен на Маскаренских островах и что эти клещи
Рис. 33. Holothyrus
longlpes Thorell, 1882
(по Хирсту, 1922).
Самец (вид снизу).
выделяют раздражающий яд, который вызывает смерть уток и гусей,
если последние заглатывают их. Названный автор отмечает также,
что болезнетворному действию яда этих клещей подвержены дети.
ЛИТЕРАТУРА
Hirst S., 1922. Mites injurious to domestic animals, Econ. Ser. British Mas.
(Nat. Hist.), 13, 1—107.
Thon K., 1906. Die aiissere Morphologie und die Systematlk der Holothyriden,
Zool. Jahrb., Cyst., 23, 677—724 -f- Pls. 28—29.
Womersley H., 1935. A species of Acarina of the genus Holothyrus from
Australia and New Zealand, Ann. Mag. Nat. Hist., 16, 151—154.
Глава HI
ПОДОТРЯД MESOSTIGMATA G. CANESTRINI, 1819
Группы Mesostigmata и Ixodtdes обычно объединяют в один под-
отряд— Parasitiformes. Основанием для этого служит отчетливое
морфологическое сходство, особенно в положении стигм и в строе-
нии гнатосомы. Однако два других подотряда клещей—Trombidifor-
mes и Sarcoptiformes, — повидимому, более близки друг к другу, чем
Mesostigmata и Ixodides; тем не менее связь между этими подотря-
дами не отражена в классификации. Поэтому желательно не поль-
зоваться термином Parasitiformes для классификации клещей; исклю-
чение этого термина не означает отрицания связи между названными
подотрядами, но повышает практическую ценность классификации.
Mesostigmata легко распознать по строению гнатосомы, наличию
всего лишь одной пары стигм, расположенных латерально, наличию
тритостернума, строению спинных и брюшных щитков, а также по
строению полового аппарата и ног. Различные представители хотя и
отличаются друг от друга в деталях строения, но, безусловно, обра-
зуют единую группу.
В 1931 г. Фитцтум насчитывал в этой группе 1290 видов, но
с тех пор было описано много новых форм. Сейчас в каждой, даже
небольшой коллекции клещей обычно можно встретить несколько
новых видов Mesostigmata. Поскольку непрерывно открывают и опи-
сывают все новые и новые формы, классификация этой группы все
время изменяется.
Недавно Трегорд опубликовал ряд статей, посвященных сравни-
тельной анатомии склеротизированных образований, и ему на осно-
вании изучения главным образом брюшных щитков самок и структур,
связанных с половыми отверстиями, удалось внести некоторую ясность
в царивший в этой области хаос. Трегорд выделяет группы Meso-
stigmata на основании особенностей строения самок, но его последние
исследования показали, что родственные связи клещей группы Mes-
ostigmata могут быть установлены и на основании изучения строения
брюшных щитков самцов.
Даже после появления превосходных работ Трегорда для сколько-
нибудь удовлетворительного понимания Gamasides недостает еще
очень многих сведений. Gamasides являются наиболее много-
численной из групп Mesostigmata. Из 1290 видов, приведенных Фитц-
тумом (1939), 889 видов (т. е. более 75°/0) относятся к Gamasides,
54
Глава IJI. Подотряд Mesostigmata
Трегорд придерживается общепринятой системы клещей (Acarina)
и придает Mesostigmata ранг надкогорты. В настоящей работе над-
когорты Трегорда рассматриваются как подотряды. В основном мы
придерживаемся классификации Трегорда за исключением наимено-
ваний систематических категорий и некоторых его подразделений,
которые опущены.
Морфология. Тело клещей Mesostigmata обычно покрыто хорошо
развитыми коричневыми или более темными склеротизованными щит-
ками, или пластинками. Почти у всех Mesostigmata (за исключением
Rhodacarus) оно состоит из двух обособленных отделов: небольшого
переднего — гнатосомы и заднего — идиосомы. У Uropodina— одной
из групп Mesostigmata — гнатосома часто заключена в особую по-
лость идиосомы — камеростом.
Характерный облик клещей Mesostigmata позволяет даже неопыт-
ному исследователю установить принадлежность к Mesostigmata более
чем 90% входящих сюда видов. Все виды, имеющие даже один из
таких признаков, как наличие тритостернума или стигм, располо-
женных по бокам тела, могут быть с уверенностью отнесены к Me-
sostigmata. Особое значение в классификации подотряда имеет
расположение на брюшке щитков, щетинок и пор. Так как нами
используется классификация Трегорда, то для обозначения различных
структур Mesostigmata будет по возможности применяться его но-
менклатура.
Гнатосома клещей Mesostigmata устроена очень сложно. Хелицеры
и дистальные членики педипальп вполне типичны и обычно мало
видоизменены, однако базальные членики педипальп, которые, сли-
ваясь, образуют вентральную стенку футляра вокруг хелицер, вытя-
нуты вперед и дорзально наподобие выступов. Дорзально передний
выступ разрастается над гнатосомой. Снодграс (1948) называет его
тектумом, а другие авторы — эпистомом. На нижней стороне гнато-
сомы по средней линии тянется желобок с направленными назад зуб-
чиками. По бокам от желобка расположены парные главные отделы
нижней стенки гнатосомы. Впереди их медиальная часть дифференци-
руется в гипостом, а боковая — в рожки и лежащую более медиально
нижнюю губу (paralabrum).
Ноги обычно сильно склеротизированы. Как правило, они состоят
из шести или семи подвижных члеников. Лапки оканчиваются пред-
лапками (претарзусами), которые обычно несут присоскообразный со-
сочек и пару амбулакральных коготков. На вершине лапок ног I на
дорзальной стороне часто имеется пучок чувствительных щетинок, но
галлеров орган, который развит на лапках ног I у иксодовых клещей,
не дифференцирован (Харлов, 1943).
Идцосома защищена хорошо склеротизированными щитками. На
спинной стороне расположен обычно цельный щит, который, однако,
может быть полностью или частично разделен на два или большее
число щитков. По бокам тела могут располагаться щитки, соединяющие
Подотряд Mesostigmata
55
спинные и брюшные щитки; если боковые щитки отсутствуют,
то спинные и брюшные щитки соединяются мягкой исчерченной пере-
понкой.
Расположение щитков на брюшной поверхности тела служит
прекрасным признаком для определения подотрядов (рис. 34). Непо-
средственно позади гнатосомы по средней линии расположен трито-
стернум, который состоит из основной части и двух или трех щетинко-
образных дистальных отростков. Тритостернум представляет собой
---Тритостернум
— Престернальная щетинка
— Преэндоподальный щиток
— Югулярный щиток
— Перитремальный щиток
— Параподальный щиток
-- ЭнОоподальный щиток
— Стернальный щиток
- - Метастернальный щиток
— Латеральный щиток
— Эпигинальный щиток
—. Метаподалмый щиток
— Вентральный щиток
— Анальный щиток
Рис. 34. Схема' расположения щитков брюшной поверхности
тела у Mesostigmata.
Срединный щиток, если он развит, занимает такое же положение, как
и эпигинальный щиток.
видоизмененный остаток стерпитов третьего сегмента и является
единственным рудиментом брюшных склеритов. По бокам от
тритостернума расположена пара престернальных щетинок и (или)
преэндоподальных щитков, всегда лишенных щетинок. Эти щитки
Эвинг(1928) называет югулярными, однако Трегорд югулярными на-
зывает щитки, лежащие впереди стернального щитка, которые обычно
несут щетинки и иногда снабжены порами.
Позади тритостернума расположен так называемый стернальный
(грудной) щиток. У некоторых родов этот щиток несет четыре пары
щетинок и три пары пор. Можно предполагать, что стернальный щиток
образуется за счет слияния коксальных пластинок с брюшной стенкой
56
Глава 1П. Подотряд Mesostigmaia
тела. Наличие у некоторых форм четырех пар щетинок на стернальном
щитке подтверждает эту гипотезу. Как исключение стернальный
щиток у отдельных видов может нести только две пары щетинок.
В этом случае первая пара щетинок находится на югулярных щитках,
а четвертая — на метастернальных (промежуточных) щитках, чуть
позади и сбоку от стернального щитка, между тазиками III и IV
пары ног. Чаще всего на стернальном щитке имеется три пары ще-
тинок и две пары пор, а четвертая пара щетинок и третья пара
пор находятся на метастернальных щитках. У самок половое
отверстие обычно расположено позади стернального щитка, между
метастернальными щитками. Половое отверстие самки может при-
крываться либо срединным эпигинальным щитком, иногда снабженным
щетинками, либо боковыми щитками со щетинками; в ряде случаев
имеются и те и другие щитки одновременно, иногда же половое
отверстие вообще лишено щитков. Под эпигинальным щитком иногда
имеется срединный щиток, образованный склеротизацией дорзальной
стенки влагалища. У форм, вторично утративших эпигинальный
щиток, срединный щиток может лежать поверхностно, как это на-
блюдается у рода Fedrlzzia. Позади полового отверстия самки на-
ходится вентральный щиток. Вентральный щиток снабжен щетинками
и может сливаться с эпигинальным или же с анальным щитком. Эндо-
подальные (межкоксальные) щитки обычно расположены между тази-
ками и стернальными щитками. Параподиальные (прикоксальные) щитки
расположены кнаружи от тазиков ног. Позади тазиков четвертой пары
ног и кнаружи от вентрального щитка располагаются метаподальные
щитки. У самцов половое отверстие расположено или в середине
стернального щитка, или у его переднего края. Как правило, у самцов
наблюдается слияние щитков брюшной поверхности тела.
Личинки мезостигматических клещей имеют только три пары
ног и недоразвитые щитки. Нимфы имеют четыре пары ног и
легко распознаются по отсутствию полового отверстия. Дорзаль-
ный щит нимфы часто разделен на несколько щитков, а стер-
нальный и вентральный щитки обычно слиты в один вентральный
щиток.
Трегорд (1946) провел сравнительноморфологическое изучение
этих клещей с целью установления родственных отношений между
группами. У трех групп эпигинальный щиток отсутствует, и нет дан-
ных, свидетельствующих о том, что он утрачен вторично. Эти три
группы — Megisthanina, Liroaspina и Microgynina — составляют еди-
ную группировку. Другая группировка включает формы, у которых
у самок вблизи полового отверстия расположены эпигинальный или
боковые щитки или те и другие одновременно. Сюда входят восемь
групп: Gamasides, Celaenopsina, Fedrizziina, Zerconina, Thinozerconina,
Trachytina, Diarthrophallina и Uropodina.
Определительная групп таблица Mesostigmata с изменениями поТре-
горду (1946) приведена ниже.
Megisthanina
37
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ГРУПП MESOSTIGMATA
1. Эпигинальный щиток у самок хорошо развит; если же он недо-
развит или отсутствует, то его функцию выполняют срединный
или боковые щитки........................................2
— Первичный эпигинальный щиток отсутствует. Срединный и боко-
вые щитки не прикрывают половое отверстие..............8
2. Боковые щитки имеются...................................3
— Боковые щитки отсутствуют.................................4
3. Метастернальные щитки обособлены..............Celaenopsina
— Метастернальные щитки слиты с грудным щитком Fedrizziina
4. Эпигинальный щиток подвижно соединен с вентральным щитком
или слит с ним по поперечному шву. Перитремы расположены на
брюшной стороне (или один из этих признаков).................5
— Эпигинальный щиток не сочленяется основанием. Перитремы рас-
положены на спинной стороне.....................Thinozerconina
5. Эпигинальный щиток снабжен одной парой генитальных щети-
нок..........................................................6
— Эпигинальный щиток лишен щетинок...........................7
6. Половое отверстие самца расположено на стернальном щитке,
метастернальные щитки редуцированы, сохраняются только мета-
стернальные щетинки. Хелицеры самца не используются при ко-
пуляции и не видоизменены в связи с этим.............Zerconina
— Половое отверстие самца расположено впереди стернального
щитка. Имеются метастернальные щитки, различные по размерам и
форме. Хелицеры самца в той или иной степени видоизменены
и используются для переноса сперматофоров при копуляции
.................................................... Gamasides
7. Метастернальные щитки хорошо развиты.............Trachytina
— Метастернальные щитки сильно редуцированы или отсутствуют 10
8. Престернальные щетинки отсутствуют. Половое отверстие самки
имеет вид поперечной щели....................................9
— Престернальные щетинки расположены по бокам от тритостер-
нума. Половое отверстие самки в виде полулунной щели распо-
ложено между стернальным и вентральным щитками Megisthanina
9. Половое отверстие самки большое, расположено на границе
стернального и вентрального щитков..................Liroaspina
— Половое отверстие самки маленькое и не соприкасается со стер-
нальным или вентральным щитком....................Microgyniina
10. Престернальные щетинки расположены по бокам тритостернума
...............................................Diarthrophallina
— Престернальные щетинки отсутствуют.................Uropodina
MEGISTHANINA TRAGARDH, 1946
К Megisthanina относится только одно семейство клещей, выделе-
ние которого а самостоятельную группу не вызывает сомнения. Клещи,
58
Глава III. Подотряд Mesostigmata
принадлежащие к этой группе, уникальны по положению полового
отверстия, по отношению к которому грудной щиток выполняет
функцию эпигинального.
Megisthanidae Berlese, 1914
(рис. 35)
ДИАГНОЗ
Megisthanidae — крупные клещи (от 1 до 4 мм) с овальным телом,
разделенным на гнатосому и идиосому. Кутикула на спинной стороне
Рис. 35. Megisthanus flo-
ridanus Banks, 1904.
Самка (вид снизу).
поперечная щель. Стигмы
образует единственный дорзальный щит,
покрывающий всю спинную поверхность
тела. На брюшной стороне находится
удлиненный вентральный и стернальный
щиток, разделенный на две части, задняя
из которых имеет сходство с эпиги-
нальным щитком. Глаза отсутствуют. По
бокам тритостернума располагается пара
престернальных щетинок; пальпы не ви-
доизменены. Хелицеры мощные, с силь-
ными зубцами. Ноги крепкие, массивные,
на четвертой паре их имеются зубовид-
ные выросты; лапки ног I лишены ко-
готков.
Половое отверстие самки имеет вид
серповидной щели и лежит непосред-
ственно позади стернального щитка; по-
ловое отверстие самца расположено на
стернальном щитке и прикрыто двумя
пластинками, между которыми находится
расположены на уровне задней пары ног и
имеют удлиненные перитремы.
РОДЫ
1. Megisthanus Thorell, 1882
Тип рода Megisthanus caudatus Thorell, 1882
2. Celaenogamasus Berlese, 1901
Тип рода Celaenogamasus hlrtellus Berlese, 1901
3. Cyclothorax v. Frauenfeld, 1868
Тип рода Cyclothorax carcinicola v. Frauenfeld, 1868
4. Hoplomeglstus Berlese, 1903
Тип рода Megistanus1 armiger Berlese, 1888
5. Stenosternum Kramer, 1898
Тип рода Stenosternum blpilosum Kramer, 1898
i Megistanus — наименование, ошибочно употребляемое вместо Megisthanus.
Liroaspina
59
ОБЩИЕ ЗАМЕЧАНИЯ
Приведенный выше диагноз дан на основании рода Megisthanus.
Фитцтум (1942) высказал мысль, что не все роды соответствуют
этому диагнозу. Однако знания о других родах слишком ограничены,
чтобы можно было вносить в него какие-либо изменения. При под-
готовке этой работы исследовался только род Megisthanus.
Виды Megisthanus обычно находят на крупных жуках во влаж-
ных местообитаниях. Клещи Megisthanus в США чаще всего попа-
даются на жуках Popllius disjunctus liliger. На этом жуке найден
М. florldanus Banks. Насколько известно, эти клещи не имеют су-
щественного хозяйственного и медицинского значения.
ЛИТЕРАТУРА
Tragardh I., 1943. Further contributions towards the comparative morphology
of the Mesostigmata (Acarina) the Antennophoridae and the Megisthanidae, ‘ •
Arkiv. far Zoologi., 34A, № 20, 1—10.
Tragardh I., 1946. Outlines of a new classification of the Mesostigmata (Aca-
rina) based on comparative morphological data, Lunds U niversifets Ars-
skrift, N. F„ Avd. 2, 42, № 4, 1—37.
Tragardh 1., 1946. Contributions towards the comparative morphology of the
Mesostigmata (Acarina) VII. The praesternal hairs and the male genital
aperture, Sarlryk ur Entomologsk Tidskrift, 67, H. 3, 88—108.
LIROASPINA TRAGARDH, 1946
Группа Liroaspina содержит только два семейства мелких клещей,
которые подобно Megisthanina и Microgyniina лишены эпигинального
щитка. Трегорд (1946) на основании изучения особенностей полового
отверстия самцов предлагает оба семейства, входящие в Liroaspina,
рассматривать отдельно и поместить в разные группы. Однако, при-
держиваясь первоначальной классификации Трегорда, основанной на
различиях строения брюшных щитков самок, мы рассматриваем Ltro-
aspina как единую группу.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ LIROASPINA
1. Стернальные щитки Ш и IV слиты в узкий поперечный щит;
стернальные щитки I и II самостоятельны или сливаются друг
с другом у самок. Половое отверстие самца прикрыто круглым
диском, лишенным щетинок.......................Liroaspidae
— У самок стернальные щитки II, III и IV слиты, I — обособлен.
У самцов половое отверстие прикрыто двулопастным круглым
диском, снабженным только одной парой щетинок . . Epicriidae
60
Глава III. Подотряд Mesostigmata
Liroaspidae Tragardh, 1946
(рис. 36)
ДИАГНОЗ
Диагностическим признаком семейства служат особенности рас-
положения спинных щитков. Один крупный щиток покрывает боль-
шую часть спинной поверхности подосомы. На заднем конце тела
расположен цельный или расчлененный пигидиальный щиток, который
иногда имеет направленные назад выступы, несущие щетинки. Между
этими двумя щитками располагается целый набор мелких щитков,
или пластинок. Стернальный щиток самки расчленен или частично
слит. Эпигинальный Щиток отсутствует. Половое отверстие самца
прикрыто округлым, лишенным щетинок диском; оно расположено
у переднего края стернального щитка. Каждая из лапок снабжена
предлапкой с коготками.
РОДЫ И ПОДРОДЫ
1. Liroaspls Banks, 1902 (== Dwigubskyia Oudemans, 1936)
Тип рода Liroaspls americana Banks, 1902
2. Berlesiatia Turk, 1943
Тип рода Epicrtus cirratus Berlese, 1917
3. Epicroseius Berlese, 1904
a) Epicroseius s. str.
Тип подрода Epicroseius angelioides Berlese, 1904
Liroaspina
61
6) Iphtdinychus Berlese, 1913
Тип подрода Echinoselus1 (/phidinychus) manicatus Berlese, 1913
4. Wlllmannla Balogh, 1938
Тип рода Wlllmannia sejlformls Balogh, 1938
5. Zuluacartis Tragardh, 1906
Тип рода Zuluacartis termitophllus TrMgSrdh, 1906
ОБЩИЕ ЗАМЕЧАНИЯ
Liroaspidae распространены главным образом в тропических стра-
нах, но виды Liroaspls найдены в Северной Европе и в северной
части Северной Америки. Строение хелицер указывает на то, что
эти клещи — хищники. Отдельные экземпляры найдены на растениях;
Фокс (1947) нашел два экземпляра на двух крысах в Пуэрто-Рико.
Впрочем, находка Liroaspls armatus на крысах не доказывает того,
что эти клещи эктопаразиты. Они могли оказаться на крысе совер-
шенно случайно или в качестве добычи многочисленных эктопарази-
тов, обитающих на крысах. За исключением случаев нахождения
Liroaspidae на крысах, данные об их хозяйственном и медицинском
значении отсутствуют.
ЛИТЕРАТУРА
Balogh J„ 1938. Systematische Studien liber eine nene Milbengattung, Wil-
Imannia gen. nov. Zool. Anz., 123, 259—265.
Fox I., 1947. Seven new mites from rats in Puerto Rico, Ann. Ent. Soc. Ame-
rica, 40, 598—603.
Tragardh I., 1943. Further contributions towards the comparative morphology
of the Mesostigmata (Acarina), the Antennophoridae, and the Megistha-
nidae, Arkiv. fOr Zoologi, 34A, № 20, 1—10.
Tragardh I., 1946. Outlines of a new classification of the Mesostigmata (Aca-
rina) based on comparative morphological data, Lunds Universitets Ars-
skrift, N. F., Avd, 2, 42, № 4, 1—37.
Trag ardh I., 1946. Contributions towards the comparative morphology of the
Mesostigmata (Acarina) VII. The praesternal hairs and the male genital
aperture, Sdrtryk ur Entomologisk Tidskrift, 67, H. 3, 88—108.
Epicriidae Berlese, 1885
(рис. 37)
ДИАГНОЗ
Клещей Epicriidae легко распознать по наличию на спинной
поверхности небольших бугорков, которые создают впечатление
изящной, геометрически правильной сетки. Стернальные щитки самки
!) Echlnoseius, по всей вероятности, наименование, ошибочно употребляе-
мое вместо Epicroseius.
62
Глава lit. Подотряд Mesostigmata
разобщены; щиток I отделен от щитков II, III и IV. Половое отвер-
стие самца находится на стернальном щитке и прикрыто расчлененным
Рис. 37. Eplcrius mollis (Kramer), 1876 (по Трегорду, 1942).
А —щитки брюшной поверхности тела самки; Б—самка (вид сверху).
округлым диском, снабженным одной парой щетинок. Epicriidae—
клещи обычно малых размеров (0,5 мм), золотисто-коричневой
окраски.
РОДЫ И ПОДРОДЫ
1. Eplcrius G. Canestrini et Fanzago, 1877
a) Eplcrius s. str.
Тип подрода Oamasus mollis Kramer, 1876 (= Eplcrius geo-
metricus G. Canestrini et Fanzago, 1877)
6) Dlepicrius Berlese, 1916
Тип подрода Eplcrius {Dlepicrius) parisiensis Berlese, 1916
2. Cornubia Turk, 1943
Тип рода Cornubia ornata Turk, 1943
ОБЩИЕ ЗАМЕЧАНИЯ
Клещей Epicriidae обычно находят во мху или на корнях расте-
ний и ризоидах. На Гуаме найден Eplcrius sp. среди корней и
ризоидов эпифитного папоротника, растущего на вершинах деревьев
во влажных лесах. Хозяйственное и медицинское значение Epicriidae
неизвестно.
MicrogynitnH
63
ЛИТЕРАТУРА
Tragardh I., 1942. Zur Kenntnis def Gattung Eplcrius Berlese (Acarina),
Arkiv. fOr Zoologi, 34A, № 4, 1—10.
Tragardh I., 1943. Further contributions towards the comparative morphology
of the Mesostigmata (Acarina) the Antennophoridae and the Megisthani-
dae, Arkiv. for Zoologi, 34A, № 20, 1—10.
Tragardh I., 1946. Outline of a new classification of the Mesostigmata (Aca-
rina) based on comparative morphological data, Lunds Unlversitets Ars-
skrift, N. F., Avd. 2, 42, № 4, 1—37.
Tragardh I., 1946. Contributions towards the comparative morphology of the
Mesostigmata (Acarina) VII. The praesternal hairs and the male genital
aperture, Sdrtryk ur Entomologsk Tidskrift, 67, H. 3, 88—108.
MICROGYNIINA TRAGARDH, 1942.
Эта группа была выделена Трегордом в отдельную когорту,
включающую два новых вида, найденных Форслундом в Швеции;
Каждый вид рассматривается Трегордом как представитель нового
рода и семейства.
Трегорд характеризует эту группу следующим образом: половое
отверстие у самца расположено на стернальном щитке и никогда
не бывает округлым; щиток не расчленен и направлен вперед. Поло-
вое отверстие самки в виде небольшой поперечной щели; оно сильно
сдвинуто назад и не связано со стернальным или с вентральным щит-
ками. Имеется три или четыре спинных щитка. Перитремы короткие.
Лапки ног I лишены предлапки.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ MICROGYNIINA
1. На спинной стороне имеется 3 щитка. Вентральный щиток самки
разделен продольно.................................Microgyniidae
— На спинной стороне имеется 4 щитка. Вентральный щиток самки
цельный............................................. Microsejidae
Microgyniidae Tragardh, 1942
(рис. 38)
ДИАГНОЗ
Микрогинииды — мелкие (0,3—0,4 мм) клещи, варьирующие по
окраске от почти бесцветных до желтоватых или коричневых, с тремя
спинными щитками и расчлененным у самки вентральным щитком.
РОДЫ
1. Micro gynium TrSgtirdh, 1942
Тип рода Microgynium rectangulatum TrSgSrdh, 1942
64
Глава lit. Подотряд Mesostigmata
ОБЩИЕ ЗАМЕЧАНИЯ
М. rectangulatum найден в рыхлой коре старых пней. Собраны
личинки, нимфы, самцы и самки, но описана только нимфальная
Рис. 38. Microgynium rectan-
gulatum Tragardh, 1942
(по Трегорду, 1942).
Самка (вид снизу).
фаза. Хозяйственное и медицинское значение этих клещей не уста-
новлено.
ЛИТЕРАТУРА
Tragirdh I., 1942. Microgynlina, a new group of Mesostigmata, Enfomolo-
gisk Tidskrift, 63, H. 3—4, 120—133.
Tragardh I., 1943. Further contributions towards the comparative morphology
of the Mesostigmata (Acarina) the Antennophoridae and the Megisthani-
dae, Arkiv. for Zoologi, 34A, № 20, 1—10.
Tragardh I., 1946. Outlines of a new classification of the Mesostigmata(Aca-
rina) based on comparative morphological data, Lunds Untversite's Ars-
skrift, N. F., Avd. 2, 42, № 4, 1—37.
Tragardh I„ 1946. Contributions towards the comparative morphology of the
Mesostigmata (Acarina) VII. The praesternal hairs and the male genital
aperture, Sdrtryk ur Entomologisk Tidskrift, 67, H. 3, 88—108.
Microsejidae TrSgardh, 1942
(рис. 39)
ДИАГНОЗ
Клещи Microsejidae очень сходны с предыдущим семейством, но
отличаются тем, что имеют четыре спинных щитка, а вентральный
щиток у самок не расчленен.
Gam asides
65
РОДЫ
1. Microsejus TragSrdh, 1942
Тип рода Microsejus truncicola Tragardh, 1942
Рис. 39. Microsejus truncicoia
Tragardh, 1942
(no Трегорду, 1942).
Самка (вид снизу).
ОБЩИЕ ЗАМЕЧАНИЯ
Этих клещей находят в рыхлой коре пней. Известны только
нимфы и взрослые. Хозяйственное и медицинское значение не уста-
новлено.
ЛИТЕРАТУРА
Tragardh L, 1942. Microgyniina, a new group of Mesostigmata, Entomologisk
Tidskrift, 63, H, 3—4, 120—133.
Tragardh I., 1943. Further contributions towards the comparative morphology
of the Mesostigmata (Acarina) the Antennophoridae and the Megisthani-
dae, Arkiv. for Zoologi, 34A, № 20, 1—10.
Tragardh I., 1946, Outlines of a new classification of the Mesostigmata(Aca-
rina) based on comparative morphological data, Lunds Untversitets Ars-
skrift, N. F., Avd. 2, 42, № 4, 1—37.
Tragardh I., 1946. Contributions towards the comparative morphology of the
Mesostigmata (Acarina) VII. The praesternal hairs and the male genital
aperture, Slirtryk ur Entomologsck Tidskrift, 67, H. 3, 88—108.
GAM ASIDES LEACH, 1815
В число Gamasides входят наиболее обычные почвенные и экто-
паразитические (на позвоночных и беспозвоночных) клещи, встре-
чающиеся в большинстве сборов. Со времени монументальных работ
Берлезе никто из авторов не достиг действительной ясности в пони-
мании этого обширного собрания видов. В классификации этой группы
мы придерживаемся главным образом системы Фитцтума (1941).
5 Зак. 1785
66
Глава III. Подотряд Mesostigmata
Особое значение в классификации этих клещей имеет развет-
вленная щетинка у основания лапки пальп (рис. 40). Виды, у которых
эта щетинка имеет больше двух ветвей, могут быть легко отнесены
к соответствующим семействам, но виды, у которых эта щетинка
двураздельная, значительно труднее поддаются определению.
Gamasides легко определять по самцам, так как их хелицеры
разнообразно видоизменены и служат для переноса сперматофора
в половое отверстие самки при копуляции; соответственно этому
Рис. 40. Специализированная щетинка тарзального
членика пальпы.
Л —клеша из семейства Laclaptidstc; Б- клеша из другого се-
мейства Gamasides. Остальные щетинки тарзального членика
пальпы не изображены.
у самцов Gamasides половое отверстие расположено впереди стер-
нального щитка. Самки распознаются по характеру расположения
щитков на брюшной поверхности, что особо подчеркивалось Трегор-
дом (1946). Латеральные щитки отсутствуют, а метастернальные щитки
либо обособлены, либо слиты с соответствующими эндоподальными
щитками, или же очень малы и слиты со стернальным щитком.
Эпигинальный щиток снабжен парой щетинок и в той или иной мере
может сливаться со стернальным щитком.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ GAMASIDES
1. По бокам анального отверстия нет чашевидных присосков . . 2
— По бокам анального отверстия имеются две крупные чашевидные
присоски.....................................Discozerconidae
2. У самцов половое отверстие лежит на переднем крае стернального
щитка. Стернальный щиток нимф без темной поперечной полосы 3
Gamasides
67
— У самцов половое отверстие открывается на специальном пре-
стернальном щитке. У нимф на стернальном щитке имеется
темная поперечная полоса..........................Poecilochiridae
3. Самки не имеют югулярных щитков. У самцов половые при-
соски отсутствуют...............................................4
— У самок югулярные щитки имеются. У самцов половые при-
соски расположены на возвышающемся конусе вентрального
щитка............................................. Aceosejidae
4. Подосома и опистосома слиты. Тело между этими двумя отделами
не сужено.......................................................5
— Тело удлиненное. Опистосома отделена от подосомы глубокой
бороздой.................... . . .................. . Rhodacaridae
5. Боковые, вентральные и передние выросты тазиков педипальп не
имеют формы гарпуна. Хелицеры клешневидные или стилето-
видные .........................................................6
-— У самок боковые, вентральные и передние выросты тазиков •
педипальп имеют форму гарпуна. Хелицеры лишены неподвижного
пальца, их подвижный палец снабжен большими загнутыми
зубцами............................................Ixodorhynchidae '
6. Основание гнатосомы образует узкий футляр вокруг хелицер . 7
— Основание гнатосомы образует широкую полость или камеростом,
в которой помещаются хелицеры (но не пальпы) Spelaeorhynchidae
7. Тазики подвижные...........•................................8
— Тазики неподвижные и расположены радиально. Ноги мощные,
крепкие. Паразитируют на летучих мышах .... Spinturnicidae
8. Разветвленная щетинка на лапке пальп подразделена более чем
на две ветви...................................................10
— Разветвленная щетинка лапок пальп двураздельная ............. 9
9. Дорзальный щиток цельный или расчленен на больший передний
и меньший задний...............................................16
— Дорзальный щиток расчленен на два равных щитка, которые
покрывают большую часть спинной поверхности . . . Ascaidae
10. Разветвленная щетинка лапки пальп трехраздельная ......... 11
— Разветвленная щетинка лапки пальп четырехраздельная ....
.................................................Veigaiaidae
11. Эпигинальный щиток расположен между метастериальными щит-
ками, вершина его не заострена.................................12
— Эпигинальный щиток расположен между метастериальными щит-
ками, вершина его заострена........................Parasitidae
12. Ноги I не отличаются от остальных..........................13
— Ноги I тоньше других, лишены типичной предлапки, присоски
или коготков....................................... Macrochelidae
13. У самки параподальные, перитремальные и метаподальные щитки
обособлены друг от друга и никогда не разрастаются назад
кнаружи от вентрального щитка или же все боковые щитки
слиты с вентральным щитком................................... 14
5*
Глава HI. Подотряд Mesosiigmata
— Параподальные, перитремальные и метаподальные щитки слиты
и простираются за тазики ног IV по бокам вентрального щитка
PachylaeJaptidae
14. Дорзальный щиток цельный.........................15
— Дорзальный щиток разделен на две части . . Gamasolaelaptidae
15. Предлапка и коготки ног I сходны с таковыми других ног . .
................................................Neoparasitidae
— Предлапка и коготки ног I недоразвиты по сравнению с тако-
выми других ног..................................Pseudoparasitidae
16. Тритостернум имеется, у самок — с лациниями..................20
— Тритостернум отсутствует или редуцирован, у самок—лишен
лациний.................................................... 17
17. Хелицеры редуцированы или лишены клешни (за исключением
некоторых паразитов носовой полости птиц). У самцов ноги II
без апофизов....................................................18
— Хелицеры клешневидные. Паразиты наружного слухового прохода
рогатого скота и антилоп. 1/ самок основная часть тритостер-
нума редуцирована или вовсе утрачена, самцы с апофизами на
ногах II..............................................Raillietidae
18. Тело овальной формы. Генитальный щиток самки может быть
хорошо развит. Паразиты дыхательных путей птиц и змей . 19
— Тело удлиненно-овальное или червеобразное. Генитальный щиток
рудиментарный или утрачен. Паразитируют в легких млекопи-
тающих ...............................................Halarachnidae
19. Дорзальный щиток цельный и позади заходит за подосому, если
же дорзальный щиток ограничен подосомой, то его задний край
снабжен парой длинных щетинок. Паразитируют в легких змей
.................................................................. Entonyssidae
— Дорзальный щиток обычно разделен или размеры его ограничены
подосомой. Паразитируют в носовой полости птиц....................
.................................................Rhinonyssidae
20. Щетинки разбросаны по телу сравнительно редко. Тектум
не имеет формы языка, по бокам его развиты длинные зуб-.
чики.....................................................21
— Тело покрыто многочисленными щетинками, как меховым покро-
вом. Тектум имеет форму языка и снабжен длинными направлен-
ными вперед зубчиками.............................Haemogamasidae
21. Клешня хелицер редуцирована...................Dermanyssidae
— Клешня хелицер обычная . ................................22
22. Эпигинальный щиток „каплевидной" формы, иногда вырезан
сзади и прилегает к анальному щитку.......................23
— Эпигинальный щиток сзади усечен или вогнут . . . Phytsoeiidae
23. Перитремы очень короткие, короче суммы пяти диаметров стигмы
..............................................................Iphiopsidae
— Перитремы длинные, длиннее суммы пяти диаметров стигмы . .
..............................................................Laelaptidae
Gamasides
69
Discozerconidae Berlese, 1910
. (рис. 41, 42)
ДИАГНОЗ
Диагностический признак — наличие больших чашевидных присо-
сок по бокам анального отверстия — позволяет сразу отличить пред-
ставителей этого семейства от всех других форм. Даже при отсут-
ствии присосок эти клещи легко отличаются от других Gam asides
Рис. 41. Heterozercon oudemansl
Finnegan, 1931 (по Финнегану, 1931).
Самка (вид снизу).
Рис. 42. Dlscozercon mirabllis
Berlese, 1910 (но Берлезе, 1914)
Самка (вид снизу).
гем, что стернальные щитки разделены на пару передне-боковых
и пару задне-боковых, которые слиты с эндоподальными щитками.
Все лапки снабжены предлапками, каждая из которых несет присоску
и маленькие зачаточные коготки. Щетинки тела очень короткие и
крепкие.
РОДЫ
1. Dlscozercon Berlese, 1910
Тип рода Dlscozercon mirabllis Berlese, 1910
2. Allozercon Vitzthum, 1926
Тип рода Allozercon fecundissimus Vitzthum, 1926
3. Atacoselus Berlese, 1905
Тип рода Atacoseius pellucens Berlese, 1905
70
Глава Ш. Подотряд Mesostigmata
4. Discomeglstus Trag&rdh, 1911
Тип рода Discomeglstus pectinatus Tragardh, 1911
5. Heterozercon. Berlese, 1888.
Тип рода Heterozercon degeneratus Berlese, 1888.
ОБЩИЕ ЗАМЕЧАНИЯ
Положение Atacoselus в семействе несколько сомнительно, так
как тип известен только по неполовозрелым особям, которые не
обладают всеми характерными признаками семейства. Фитцтум
сообщает, что Берлезе (1892) предложил выделить эту группу
в особое семейство Heterozerconidae, но поиски, предпринятые для
подтверждения этого предположения, не увенчались успехом.
Клещи Discozerconidae— эктопаразиты различных крупных много-
ножек, термитов, змей. Имеются также свободноживущие формы
(по крайней мере, некоторые формы были обнаружены не на хозяине).
Все виды распространены в тропиках или субтропиках. Хозяйствен-
ное и медицинское значение не выяснено.
ЛИТЕРАТУРА
Berlese А., 1910. Brevi diagnosi di generi e specie nuovi di Acari, Redia, 6
(2), 346—388.
Finnegan, S., 1931. On a new species of mite of the family Heterozerconidae
parasitic on a snake, Proc. Zool. Soc. London, 1931, 1349—1357.
Vitzthum H. G., 1926. Malayische Acari, Treubia, 8, 106.
Poecilochiridae Willmann, 1940
(рис.
Рис. 43. Poecilochirus necrophori
Vitzthum, 1930 (по Курмэну, 1943).
Стернальный щиток дейтоцимфы.
43)
ДИАГНОЗ
Семейство охарактеризовано
Бильманом на основании осо-
бенностей строения самцов и дей-
тонимф следующим образом: по-
ловое отверстие расположено на
специальном престерпальном щит-
ке; по другим признакам пред-
ставители семейства близки к ро-
ду Parasitus (у самцов на но-
гах II имеются хорошо развитые
апофизы). Дейтонимфы четко ге-
тероморфны, с очень длинными
ногами и очень крупными жест-
кими щетинками. Клешни хели-
цер с передними выступающими
листовидными придатками. Стер-
нальный щиток дейтонимф обыч-
но имеет темную поперечную
Gamasides
71
полоску. Нойман отмечает, что самки очень сходны с самками семей-
ства Parasitidae.
РОДЫ
1. Poecilochlrus G. et R. Canestrini, 1882
Тип рода Poecilochiras carabi G. et R. Canestrini, 1882
ОБЩИЕ ЗАМЕЧАНИЯ
Клещей Poecilochiras находят на жуках, а также в пещерах
и норках, заселенных жуками. Описаны виды из Европы и Азии.
Их медицинское и хозяйственное значение не выяснено.
ЛИТЕРАТУРА
Neumann К. W., 1943. Die Lebensgeschichte der Kafermilbe Poecilochirus
necrophi Vitzthum, nebst Beschreibung aller Entwickiungstufen, Zool. Anz.,
142, 1—21.
Vitzthum H. G., 1930. Milben als Pesttrager? Zool. Jahrb. Syst., 60,381—428.
Willmann C., 1940. Neue Milben aus Hohlen der Balkanhabiinsel, gesam-
melt von Prof. Dr. K. Absolon, Briinn, Zool. Anz., 130, 209—218.
ДИАГНОЗ
Aceosejidae N. F.
(рис. 44)
Самок легко распознать по наличию югулярных щитков: перед-
ний стернальный щиток I у них обособлен от других стернальных
щитков и, кроме того, разделен
на две части, располагающиеся
по бокам тритостернума. Стер-
нальные щетинки I находятся на
этих щитках. Эпигинальный щиток
снабжен одной парой щетинок.
Позади эпигинального щитка рас-
положен большой вентроанальный
щиток. Самцы замечательны тем,
Рис. 44. Aceosejus murica^us (Koch),
1839 (по Зельиику, 1941).
что ИХ вентральный ЩИТОК имеет Самец (вид сбоку),
конусовидное возвышение, на ко-
тором расположена пара половых присосок. Все ноги снабжены пред-
лапками, имеющими присоску и коготки.
РОДЫ
1. Aceosejus Sellnick, 1941
Тип рода Sejus murlcatus Koch, 1839
ОБЩИЕ ЗАМЕЧАНИЯ
К этому семейству принадлежит только один вид. Расчленение
стернального щитка у самки и наличие присосок на возвышении
центрального щитка самца настолько ртдичают этот вид от
72
Глава III. Подотряд Mesostigmata
остальных форм, что оказалось необходимым выделить его в особое
семейство. Эти клещи живут в гниющей древесине. Их хозяйствен-
ное и медицинское значение неизвестно.
ЛИТЕРАТУРА
Sellnick М., 1941. Einige Milbenarten С. L. Kochs, Zooi. Anz., 133, 146—155.
ДИАГНОЗ
ной от 0,3
Rhodacaridae Oudemans, 1902
(рис. 45)
Приведенный диагноз заимствован из Фитцтума (1941). Rhoda-
caridae— бесцветные, золотистые или розовато-красные клещи дли-
до 0,5 мм. Тело удлиненное, его длина превышает ши-
рину обычно в 3 раза. Опистосома отделена от подо-
сомы глубокой бороздой, и эти отделы тела сохраняют
некоторую подвижность относительно друг друга. Раз-
деляющая линия имеется также между проподосомой и
метаподосомой. Стернальный щиток выдается вперед,
прикрывая у самцов половое отверстие. Лапки ног II,
III и IV снабжены предлапками, имеющими присоску
и коготки. Лапки ног I не имеют амбулакрального
аппарата, или предлапки, и коготки на них рудиментар-
ные. Передний край тектума трехзубчатый, средний
зубец обычно превышает длину боковых.
Рис. 45. Rhodacarus roseus Oudemans, 1910
(ио Бэнксу, 1915).
Самец (вид снизу).
РОДЫ
1. Rhodacarus Oudemans, 1902
Тип рода Rhodacarus roseus Oudemans, 1902
2. Rhodacaropsis Willmann, 1935
Тип рода Rhodacaropsis inexpectatus Willmann, 1935
3. Rhodacarellus Willmann, 1935
Тип рода Rhodacarellus subterraneus Willmann, 1935
ОБЩИЕ ЗАМЕЧАНИЯ
Клещей Rhodacaridae обычно находят в подстилке. Семейство еще
слабо изучено, повидимому, в дальнейшем будет описано много ви-
дов. Хозяйственное и медицинское значение не установлено.
ЛИТЕРАТУРА
Willman С., 1935. Ш. Ueber eine eigenartige Milbenfauna im Kiistengrund-
wasser der Kielen Bucht, Schr, naturw. Ver. Schl.-Holst., 20, 422—434,
Gamasides
73
Ixodorhynchidae Ewing, 1923
(рис. 46)
ДИАГНОЗ
Клещи Ixodorhynchidae легко распознаются по гарпуновидным
передне-боковым выростам на тазиках пальп самок, протонимф и
дейтонифм. Эти фазы имеют также своеобразные хелицеры, которые
лишены неподвижного пальца; на подвижном пальце хелицер имеются
Б
Р и с. 46. Ixodorhynchas bu.ta.nta.ne ns is (Fonseca), 1934
(по Фонзека, 1934).
А —самка (вид снизу); Б —гнатосома.
большие, загнутые назад зубцы. Личинка и тритонимфы не описаны;
возможно, они не существуют вообще. У самцов обе эти особенности,
столь характерные для других фаз, отсутствуют. Самцов все же
можно определить, так как у них отсутствуют лишь гарпуновидные
выросты на передних боковых отростках педипальп, а основная часть
этих отростков сходна с таковой других фаз. Хелицеры самцов
клешневидные и используются для переноса сперматофора во время
копуляции.
РОДЫ
1. Ixodorhynchas Ewing, 1923 ( = Ixobloldes Fonseca, 1934)
Тип рода Ixodorhynchas llponyssoldes Ewing, 1923
ОБЩИЕ ЗАМЕЧАНИЯ
Эти своеобразные клещи известны только как паразиты змей.
'Гидицнь^ вид был собран ид глаз з^ей в Мадриде (Айова), Другой
74
Глава III. Подотряд Mesostigmata
вид I. butantanensls (Fonseca), 1934, был найден на змее Optis
merreml Wagler, 1824, в Бразилии. Хозяйственное и медицинское
значение их не установлено.
ЛИТЕРАТУРА
Ewing Н. Е., 1923. The dermanyssid mites of North America, Proc. U. S. Nat.
Mas., 62, Art. 13, 1—26.
Fonseca F. da, 1934. Der Schlangenparasit Ixobioldes novi generis n. sp.
(Acarina, Ixodorhynchidae nov. fam.), Zeit. Parasitenk, 6, 508—527.
Spelaeorhytichidae Oudemans, 1902
(рис. 47)
ДИАГНОЗ
Spelaeorhynchidae легко определить по особенностям положения
тектума и прикрепления хелицер и пальп. Тектум окружает край
Рис. 47. Spelaeorhynchus sp. (вид снизу).
отверстия камеростома в идиосоме, в который хелицеры могут втя-
гиваться. Стигмы маленькие и расположены по бокам промежутка
между тазиками ног третьей и четвертой пар. Срединная часть ги-
постома имеет форму стилета. Хелицеры крупные и заканчиваются
двумя большими крючками. Дорзальный щиток покрывает только
подосому, задняя часть спинной поверхности тела не склеротизиро-
вана. Со спинной стороны край тектума широко соединяется с дор-
зальным щитком,
(Jamasides
75
РОДЫ
1. Spelaeorhynchus Neumann, 1902
Тип рода Spelaeorhynchus praecursor Neumann, 1902
ОБЩИЕ ЗАМЕЧАНИЯ
Spelaeorhynchidae паразитируют на летучих мышах Центральной
и Северной Америки. Они резко отличаются от остальных клещей и
не имеют сколько-нибудь тесных родственных связей с другими
Gamasides. Все же их принадлежность к Mesostigmata не вызывает
сомнений, хотя Бэнкс и Нойман придерживаются того мнения, что
Spelaeorhynchidae родственны иксодовым клещам. Spelaeorhynchidae
причисляются к Gamasides на основе работ Фитцтума (1941). Какие-
либо данные о медицинском или хозяйственном значении отсут-
ствуют.
ЛИТЕРАТУРА
Neumann G., 1902. Spelaeorhynchus praecursor n. g. n. sp., nouvel acarien
parasite, Arch, de Parasitol., 5, 31—37.
ДИАГНОЗ
Spinturnicidae Oudemans, 1901
(рис. 48, 49)
Spinturnicidae — коричневатые клещи средних размеров, длиной
от 0,5 до 1 мм. Тело обычно уплощено, а у Periglischrus (рис. 49)
Рис. 48. Splnturnix carloshoffmanni Hoffmann, 1944
(по Гофману, 1944).
А —самка (вид сверху); Б — самка (вид снизу).
опистосома имеет боковые отростки. Дорзальный щиток цельный
или разделен на две части. 4|оги относительно длинные, но очень
76
Глава III. Подотряд Mesostigmata
крепкие. Тазики всех ног неподвижны и направлены радиально.
Лапки снабжены короткими предлапками, каждая из которых имеет
большую присоску и сильные коготки. Тритостернум редуцирован до
маленькой пластинки или замещен относительно крупным преэндопо-
дальным щитком. Анальное отверстие расположено терминально на
Рис. 49. Perigllschrus vargasl Hoffmann, 1944
(по Гофману, 1944).
Л —самка (вид сверху); Б — самка (вид снизу).
небольшом выступе или же у самого конца тела. Перитремы и
стигмы расположены чаще на спинной стороне, чем на брюшной.
РОДЫ
1. Splnturnlx V. Heyden, 1826 ( = Pteroptus Dufour, 1832 et
Diplostaspis Kolenati, 1857)
Тип рода Splnturnlx niyotl Kolenati, 1856
2. Ancystropus Kolenati, 1856 ( = Leiostapsls Kolenati, 1857)
Тип рода Ancystropus zeleborii Kolenati, 1856
3. Meristaspls Kolenati, 1857
Тип рода Meristaspls lateralis Kolenati, 1857
4. Perigllschrus Kolenati, 1857
Тип рода Perigllschrus caligus Kolenati, 1857
5. Tympanospinctus Berlese, 1918
Тип рода Tympanospinctus paradoxus Berlese, 1918
ОБЩИЕ ЗАМЕЧАНИЯ
Клещей Spinturnicidae почти всегда находят только на летучих
мышах. Исключение составляет Tympanospinctus paradoxus, кото-
рый найден на Crassarchus fasciatus. Из описания Берлезе нельзя
понять, принадлежит ди Т. paradoxus к Spinturnicidae, Бердезе при-
Gamasides
11
числил T. paradoxus к этому семейству, и возможно, что для этого
есть достаточные основания. Виды семейства Spinturnicidae парази-
тируют на летучих мышах во всех районах земного шара. Клещи
рода Spinturnix живородящие. Не установлено, имеют ли клещи
этого семейства какое-либо медицинское или хозяйственное значение.
ЛИТЕРАТУРА
Hirst S., 1927. Note он Acari, mainly belonging to the genus Spinturnix von
Heyden, Proc. Zool. Soc. London, Pt. 11, 323—338.
Vitzthum H. G., 1931. Neue parasitische Flcdermatts-milben aits Venezuela,
Ziet. f. Parasitenk., 4, 1—47.
Ascaidae Oudemans, 1905
ДИАГНОЗ
(рис. 50)
Клещей Ascaidae отличают от других групп по наличию двуразделыюй
щетинки у основания лапки пальпы и по разделению дорзального щитка
па переднюю и заднюю часть приблизительно равной величины. Лапки
всех ног имеют предлапку с при-
соской и коготками. Генитальный
щиток самки усечен или закруглен
спереди и отделен от вентраль-
ного щитка прямой или слегка
изогнутой линией. Фитцтум (1941),
следуя Удемансу (1939), называет
это семейство Al'.olaelaptidae. Мы
не делаем этого, так как пред-
ложенное Удемансом наименование
грубо нарушает существующую
практику образования наименова-
ний семейств.
Рис. 50. Asca quinquesetosa Whar-
ton, 1941 (по Уартону, 1941).
Самка (вид снизу).
РОДЫ
1. Asca v. Heyden, 1826
Тип рода Acarus aphldioides Linnaeus, 1758.
2. Cyrtolaelaps Berlese, 1887 ( = Protolaelaps Traghardh, 1912, не
Cyrtolaelaps Berlese, 1892)
Тип рода Gamasus mucronatus G. et R. Canestrini, 1881
(= Gamasellus brevispinosus Tragardh, 1910
— Asca affinis Oudemans, 1902)
3. Dendrolaelaps Halbert, 1915
Тип рода Dendrolaelaps oudemansi Halbert, 1915
78
Глава III. Подотряд Mesostigmata
4. Gamasellus Berlese, 1892
Тип рода Qamasus falciger G. et R. Canestrini, 1882
5. Laelogamasus Berlese, 1904
Тип рода Qamasus {Laelogamasus) simplex Berlese, 1904
6. Lobocephalus Kramer, 1898
Тип рода Lobocephalus acuminatus Kramer, 1898 (плохо описан,
положение в системе неясно)
ОБЩИЕ ЗАМЕЧАНИЯ
Клещей Ascaldae обычно находят в подстилке и в других ско-
плениях органических остатков. Они довольно обычны, но данных
об их жизненном цикле и образе жизни пока мало. Уартон (1941)
нашел этих клещей в остатках гнезда глупыша, тогда как Фокс
(1947) сообщил, что он обнаружил одного клеща из этой группы
на крысе в Пуэрто-Рико. Однако находку Фокса нельзя считать
доказательством паразитизма Ascaidae, тем более что хелицеры
у этих клещей велики и сильно зазубрены, как у хищных форм.
Медицинское и хозяйственное значение этих клещей не установлено.
ЛИТЕРАТУРА
Oudemans А. С., 1939. Neue Funde auf dem Gebiete der Systematik und
der Nomenclatur der Acari. III., Zool. Anz., 126, 20—24.
Wharton G. W., 1941. Acarina collected on the presidential cruise of 1938,
Smithsonian Miscellaneous Collections, 99, № 12, 1—8.
Рис. 51. Veigaia transisa-
lae (Oudemans), 1901
(по Бильману, 1936).
Самка (вид снизу).
Veigaiaidae Oudemans, 1939
ДИАГНОЗ
(рис. 51)
Клещей Veigaiaidae можно распознать
не только по четырехраздельной ще-
тинке на лапке пальп, но и по своеоб-
разной форме тектума. Тектум имеет два
небольших боковых зазубренных отростка
и длинный тонкий срединный вырост, ко-
торый может нести зубец на своем перед-
нем небольшом расширении. Дорзальный
щиток полностью или частично разделен на
две части; при неполном разделении раз-
деляющая борозда не достигает средней
линии тела. Эпигинальный щиток самки
сильно склеротизирован только между
тазиками ног IV; сам щиток довольно
обширен, его передний край утончен. Скле-
ротизированные структуры имеются также
на стенках влагалища. Самцы имеют апо-
физы на передних ногах и хорошо разви-
тый придаток подвижного пальца хелицер,
который используется при копуляции.
Gamasides
7д
РОДЫ
1- . Veigaia Oudemans, 1905 ( = Cyrtolaelaps Berlese, 1892, не Ber-
lese, 1887)
Тип рода Gamasus nemorensis Koch, 1836
2. Cyrthydrolaelaps Berlese, 1904
Тип рода Cyrthydrolaelaps hirtus Berlese, 1904
ОБЩИЕ ЗАМЕЧАНИЯ
О биологии этих клещей известно очень мало. Вильман нахо-
дил их в разнообразнейших укрытиях, но клещей можно встретить
и в открытых местообитаниях, на поверхности. Насколько сейчас
известно, Veigaiaidae не имеют медицинского и хозяйственного зна-
чения.
ЛИТЕРАТУРА
Berlese А., 1904. Acari nuovi, Redia, 2, 10—32.
Willmann С., 1936. Mitteleuropaische Arten derGattung Veigaia (Parasitidae
Acarij, Zool. Anz., 116, 249—258.
ДИАГНОЗ
Parasitidae Oudemans, 1901
(рис. 52)
Самок клещей семейства Parasitidae легко рас-
познать по наличию больших метастернальных
щитков, расположенных по бокам эпигинального
щитка. У самцов имеются чрезвычайно большие
апофизы на ногах II, однако сходные образования
встречаются у самцов ряда других семейств. Лапки
всех ног снабжены предлапкой с коготками и при-
соской. Вильчатая щетинка на лапках пальп имеет
три ветви. У самцов подвижной палец хелицер
сильно видоизменен в связи с использованием его
при копуляции. Дорзальный щиток цельный или,
что встречается чаще, разделен на два щитка.
Тектум в своей основе трехзубчатый. Брюшные
щитки имеют разнообразное строение, но эпиги-
нальный щиток всегда трехугольный с основанием,
направленным назад, и вершиной, обращенной
вперед. На стенках влагалища имеются склероти-
зированные образования.
Рис. 52. Pergama-
sus probsti Oude-
mans, 1912 (по
Фитцтуму, 1940).
Самка (вид снизу).
РОДЫ
1. Parasitus Latreille, 1795 ( = Carpais Latreille, 1796
— Gamasus Latreille, 1802)
Тип рода Acarus fucorum DeGeer, 1778
80
Глава til. Подотряд Mesostigmata
2. Ambly gamasus Berlese, 1903
Тип рода Gamasus tiberlnus G. et R. Canestrini, 1882
3. Eugamasus Berlese, 1893
Тип рода Gamasus magnus Kramer, 1876
4. Holoparasitus Oudemans, 1936
Тип рода Gamasus calcaratus Koch, 1839
5. Nemtiichia Oudemans, 1936
Тип рода Zercon elegantulus Koch, 1839 (сведения неполные,
положение в системе неясно)
6. Oocarpais Berlese, 1916
Тип рода Oocarpais donlsthorpel Berlese, 1916
7. Parasitellus Willmann, 1939
Тип рода Eugamasus ferox Tragardh, 1910
8. Pergamasus Berlese, 1903
Тип рода Acarus crassipes Linnaeus, 1758
9. Trachy gamasus Berlese, 1906
Тип рода Gamasus pusillus Berlese, 1892
ОБЩИЕ ЗАМЕЧАНИЯ
Parasitidae обычно встречаются в скоплениях материала органи-
ческого происхождения, например в подстилке или гниющих брев-
нах. Медицинское и хозяйственное значение этих клещей не изу-
чено, но так как Parasitidae — хищники, то вероятно, что они могут
способствовать уничтожению многих мелких насекомых.
ЛИТЕРАТУРА
Berlese А., 1905. Monografia del genere Gamasus Latr., Redla, 3, 66—
304 + 18 pl.
Oudemans A. C., 1939. Neue Funde auf detn Gebiete der Systematik und
der Nomenklatur der Acari. IV, Zool. Anz., 126, 195 -201.
a
Macrochelidae Vitzthum, 1930
(рис. 53)
ДИАГНОЗ
Macrochelidae отличаются от других Gamasides удлиненной пер-
вой парой ног, обычно лишенной коготков. Строение брюшных щит-
ков самок — также важный диагностический признак. Стернальный
щиток несет три пары щетинок и две пары пор. Метастернальные
щитки мелкие и обособлены от эндоподальных. Каждый метаподаль-
ный щиток несет у переднего края щетинку и пору. Эпигинальный
щиток усечен спереди и сзади; он несет только одну пару щетинок.
Вентральный щиток слит с анальным и может быть в значительной
степени редуцирован. Тектум состоит из срединного узкого развет-
Gamasides
81
вленного или зазубренного элемента, по бокам которого могут рас-
полагаться небольшие боковые элементы. Дорзальный щиток обычно
цельный.
Рис. 53. Macrocheles
coprophila Womersley, 1942
(по Уомерсли, 1942).
Самка (вид снизу).
РОДЫ И ПОДРОДЫ
1. Macrocheles Latreille, 1829
a) Macrocheles s. str.
Тип подрода Acarus muscae Scopoli, 1772 ( = Acarus margina-
tus Hermann, 1804)
6) Coprholaspis Berlese, 1918
Тип подрода Holostaspis glabra Muller, 1859 (= Gamasus
stercorarlus Kramer, 1876)
в) Gamasholaspis Berlese, 1903
Тип подрода Gamasholaspis gamasoides Berlese, 1903
r) Geholaspis Berlese, 1918
Тип подрода Gamasus longispinosus Kramer, 1878
д) Holasptilus Berlese, 1903
Тип подрода Holostaspsis (Holaspulus) tenuipes Berlese, 1903
e) Nothroholaspis Berlese, 1903
Тип подрода Holostaspis tridentinus G. et R. Canestrini, 1882
2. Calholaspis Berlese, 1918
Тип рода Calholaspis super bus Berlese, 1918
6 Зак. 1785
82
Глава III. Подотряд Mesostigmata
3. Euepicrltts Womersley, 1942
Тип рода Euepicrlus fllamentosus Womersley, 1942
4. Holocelaeno Berlese, 1910
a) Holocelaeno s. str.
Тип подрода Holocelaeno mitts Berlese, 1910
6) Euholocelaeno Berlese, 1918
Тип подрода Holocelaeno bursiformes Berlese, 1910
5. Holostaspella Berlese, 1903
aj Holostaspella s. str.
Тип подрода Holostaspis {Holostaspella) sculpta Berlese, 1903
6) Prholaspina Berlese, 1918
Тип подрода Holostaspella micrarhena Berlese, 1916
в) Tricholocelaeno Berlese, 1918
Тип подрода Holocelaeno longicoma Berlese, 1910
6. Macrolaspis Oudemans, 1931
Тип рода Gamasus opacus Koch, 1839
7. Neopodocinum Oudemans, 1902
Тип рода Neopodocinum jaspersi Oudemans, 1902
8. Parholaspls Berlese, 1918
Тип рода Purholaspis desertus Berlese, 1918
9. Trlgonholaspis V’tzthum, 1930
Тип рода Trigonholaspis salti Vitzthum, 1930
ОБЩИЕ ЗАМЕЧАНИЯ
Клещи Macrochelidae довольно обычны; их можно встретить в почве,
а также на теле различных беспозвоночных и позвоночных. Хотя
все Macrochel’dae связаны с различными хозяевами, они, вероятно,
не являются паразитами в обычном смысле этого слова. Перере и
де Кастро в 1945 г. нашли причину такой связи, по крайней мере,
для одного случая. Оказалось, что Macrocheles muscae (Scopoli, 1772)
на всех фазах, кроме первой (которая вообще не питается), поедали
яйца комнатной мухи. Характер питания этого клеща указывает на
то, что Macrochel’dae могут иметь определенное хозяйственное и
медицинское значение, хотя до сих пор они в этом отношении мало
исследованы.
ЛИТЕРАТУРА
Cooreman J., 1943. Note sur la faune des Hautes-Fagnes en Belgique, Bull.
Mas. Roy. Hist. Na*ur. Belgique, 19, N? 63, 1—28.
Pereira C-, de Castro M. P., 1945. Contribuicoo para о conhecitnento da
especie tipo de „Macroche’es Latr.“ („Acarina"): „М. museaedomesticae
(Scopoli, 1772)“ emend, Arquivos Ins\ Biol., 16, 153—186.
Oudemans A. C., 1931. Acarologische Aanteekenigen. CVII., Ent. Ber., 8.
Vitzthum H. G., 1926. Malayische Acari, Treubia, 8, Linn. 1—2, 1—198.
(jamasides
83
Pachylaelaptidae Vitzthum, 1931
ДИАГНОЗ
(рис. 54)
Строение щитков брюшной поверхности тела самок позволяет
отличать Pachylaelaptidae от большинства других Gamasides. Парапо-
дальные, перитремальные и
метаподальные щитки слиты
в один и заходят за тазики
ног IV. Иногда они примы-
кают к вентральному щитку
и вместе с ним покрывают
большую часть нижней по-
верхности тела позади тази-
ков. Разветвленная щетинка
на лапке пальп имеет три
ветви. Все лапки снабжены
предлапками.
Рис. 54. Pachylaelaps roose-
velti (Wharton), 1941
(по Уартону, 1941).
Самка (вид снизу).
РОДЫ И ПОДРОДЫ
1. Pachylaelaps Berlese, 1886
a) Pachylaelaps s. str.
Тип подрода Gamasus pectinifer G. et R. Canestrini, 1881
6) Brachylaelaps, Berlese, 1910
Тип подрода Pachylaelaps (Brachylaelaps) rotundas Berlese, 1910
в) Elaphrolaelaps Berlese, 1910
Тип подрода Pachylaelaps {Elaphrolaelaps) fenestratus Berlese,
1910
r) Olopachys Berlese, 1910
Тип подрода Pachylaelaps (Olopachys) scutatus Berlese, 1910
д) Onchodellus Berlese, 1904
Тип подрода Pachylaelaps (Onchodellus) retieulatus Berlese, 1904
e) Paralaelaps TrSgdrdh, 1908
Тип подрода Paralaelaps kibonotensis Tragardh, 1908
ж) Platylaelaps Berlese, 1905
Тип подрода Pachylaelaps (Platylaelaps) latus Berlese, 1905
2. Coprolaelaps Berlese, 1908
Тип рода Qoprolaelaps caputmedusae Berlese, 1908
3. Megalolaelaps Berlese, 1892
Тип рода Pachylaelaps haeros Berlese, 1888.
6*
84 Глава Ilf. Подотряд Mesostigmata
4. Pachylaella Berlese, 1916
Тип рода Pachylaella robustlssima Berlese, 1896
5. Sphaerolaelaps Berlese, 1903
Тип рода Laelaps holothyroides Leonard!, 1896
ОБЩИЕ ЗАМЕЧАНИЯ
Клещей Pachylaelaptidae находят в гниющих остатках органических
веществ или на насекомых, особенно часто на жуках. Хозяйственное
и медицинское значение этих клещей не установлено.
ЛИТЕРАТУРА
Berlese А., 1950. Lista di ntiove specie e nuovi generi di Acari, Redia, 6,
242—271.
Vitzthum H. G., 1931. Resultats scientifiques du voyage aux Indes Orientales
Neerlandaises de LL. AA. Belgique, Acarinen, Mem. Mus. Hist. Nat.
Belgique, (Hors. Serie), 3, Fasc. 5, 1—55.
Gamasolaelaptidae Oudemans, 1939
(рис. 55)
ДИАГНОЗ
Представителей этого семейства можно рас-
познать по разделенному дорзальному щитку и
закругленному переднему краю сильно склеро-
тизированного эпигинального щитка. Все ноги
имеют коготки и присоски. Разветвленная ще-
тинка на лапках пальп имеет три ветви.
Тектум, по крайней мере у некоторых видов,
имеет направленный вперед длинный срединный
выступ.
Рис. 55. Digamaselltis frenzeli Willmann, 1936
(по Бильману, 1936).
Самка (вид снизу).
РОДЫ
1. Gamasolaelaps Berlese, 1903 (= Metaparasitus Oudemans et Voigts,
1904)
Тип рода Cyrtolaelaps aurantiacus Berlese, 1903
(= Metaparasitus soholes Oudemans et Vo'gts, 1904)
2. Digamasellits Berlese, 1905
Тип рода Gamasellus (Digamasellus) perpusillus Berlese, 1905
3. Euryparasitus Oudemans, 1901
Тип рода Gamasus emarginata Koch, 1839
4. Halolaelaps Berlese et Trouessart, 1889
Тип рода Gamasus marinas Brady, 1875 (—Halolaelaps glub-
rinsculus Berlese et Trouessart, 1889)
Gamasides
85
5. Gamasodes Oudemans, 1939
Тип рода Gamasodes spiniger Oudemans, 1936 (= Gamasus spi-
nipes Koch, 1841, не Gamasus spinipes Say, 1821)
6. Salntdidlerla Oudemans, 1939
Тип рода Parasitus sexclavatus Oudemans, 1902
ОБЩИЕ ЗАМЕЧАНИЯ
Клещи Gamasolaelaptidae живут во влажной почве или в болотах,
где они, 'вероятно, питаются другими мелкими членистоногими или их
яйцами. Saintdidieria sexclavatus обычно встречают совместно с чесо-
точными клещами лошадей. Хозяйственное и медицинское значение не
выяснено.
ЛИТЕРАТУРА
Oudemans А. С., 1939. Neue Funde auf dem Gebiete def Systematik und
der Nomenklatur der Acari. IV., Zool. Anz., 126, 195—201.
Womersley H., 1942. Additions to the Acarina — Parasitoidea of Australia,
Trans. Roy. Soc. South. Australia, 66, 142 -171.
Neoparasitidae Oudemans. 1939
(рис. 56)
ДИАГНОЗ
Клещи этого семейства в це-,
лом очень сходны с Gamasolaelap-
tidae, но все же легко отличаются
от них по дорзальному щитку,
который у Neoparasitidae никогда
не бывает явственно подразделен
на передний и задний. Эпигиналь-
пый щиток никогда не имеет тре-
угольной формы, его передний
край перепончатый и может соби-
раться в складки. Все ноги снаб-
жены коготками и присосками.
Специализированная щетинка на
лапке педипальп трехраздельная.
Рис. 56. Neoparasitus oudemansi
(Oudemans) 1901 (по Фитцтуму, 1926).
Самка (вид снизу).
РОДЫ И ПОДРОДЫ
1. Neoparasitus Oudemans, 1901 (= Pachyseius Berlese, 1910)
Тип рода Neoparasitus oudemansi Oudemans, 1901
(= Pachyseius quartus Vitzthum, 1926)
86
Глава III. Подотряд Mesostigmata
2. Austrogamasus Womersley, 1942
Тип рода Cyrtolaelaps graclllipes Banks, 1916
3. Beaurielna Oudemans, 1929
Тип рода Neopodocinum nederveeni Oudemans, 1903 (= Megalo-
laelaps spinirostris Berlese, 1910)
4. Donia Oudemans, 1930
Тип рода Hypoaspis gehennalis Oudemans, 1916
5. Epicriopsis Berlese, 1916
a) Epicriopsis s. str.
Тип подрода Gamasus horridus Kramer, 1876
6) Actinoseius Berlese, 1916
Тип подрода Actinoseius terrificus Berlese, 1916
6. Epiphis Berlese, 1916
Тип рода Gamasiphis (Epiphis) rarior Berlese, 1916
7. Gamasiphis Berlese, 1903
Тип рода Gamasus pulchellus Berlese, 1887
8. Hydrogamasus Berlese, 1892
Тип рода Gamasus salinus Labonlbene, 1851 (= Gamasus lifto-
ralls G. ct R. Canestrini, 1881)
9. Megaliphis Willmann, 1938
Тип рода Gamasiphis (Megaliphis) giganteus Willmann, 1938
10. Ologamasus Berlese, 1888 (=Ologamasellus Berlese, 1914)
a) Ologamasus s. str.
Тип подрода Gamasus aberrans Berlese, 1888
6) Micriphis Berlese, 1914
Тип подрода Gamasiphis gamasellus Berlese, 1913
11. Periphis Berlese, 1914
Тип рода Eumaeus hemisphacricus Koch, 1842
12. Sphaeroselas Berlese, 1904
Тип рода Laelaps ecitonis Wasmann, 1901
ОБЩИЕ ЗАМЕЧАНИЯ
Клещи Neoparaeitidae встречаются в почве, во мху и в разлагаю-
щихся органических остатках. Хозяйственное и медицинское значение
не выяснено.
ЛИТЕРАТУРА
Vitzthum Н. G., 1926. Malayschc Acari, Treubln, 8, 1—198.
Womersley H., 1942. Additions to the Acarina — Parasit ioidea of Australia,
Trans. Roy. Soc. South Australia, 66, 142—171.
ДИАГНОЗ
Pseudoparasitidae Vitzthum, 1941
(рис. 57)
Pseudoparasitidae имеют много общего с предыдущими двумя
семействами, но отличаются от них редукцйей предлапки и присоски
на ногах I и изменением коготков этих конечностей. Эпигинальный
Gamasides
87
щиток никогда не бывает треугольным, в некоторых случаях сливается
с вентральным щитком. Дорзальный щиток не разделен, хотя имеется
слабо выраженный поперечный шов. Щетинка на лапке пальпы трех-
раздельная.
Рис. 57. Luppgamasus
suecicus Tragardh, 1936
(по Трегорду, 1936).
Я— ланка ноги I самки с видо-
измененными коготками и при-
соской (у самца коготки па
ноге I не видоизменены);
Б —щитки брюшной поверх
ности тела самки.
РОДЫ И ПОДРОДЫ
1. Pseudoparasitus Oudemans, 1902 (= Hoplolaelaps Berlese, 1903,
но не 1910)
a) Pseudoparasitus s. str.
Тип подрода Laelaps meridional is G. et R. Canestrini, 1882
6) Alloparasitus Berlese, 1920
Тип подрода Pseudoparasitus angulatus Berlese, 1916
в) Praeparasitus Berlese, 1920
Тип подрода Pseudoparasltus {Praeparasitus) collaris Berlese,
1920
r) Pseudopachys Berlese, 1916
Тип подрода Pseudoparasitus {Pseudopachys) parasltlzans Ber-
lese, 1916
2. Leptogamasus TragSrdh, 1936
Тип рода Leptogamasus suecicus, TragSrdh, 1936
3. Sessiluncus G. Canestrini, 1898
Тип рода Gamasus heterotarsus G. Canestrini, 1897
ОБЩИЕ ЗАМЕЧАНИЯ
Gamasolaelaptidae, Neoparasitidae и Pseudoparasit’dae очень слабо
изучены, и количество неописанных видов в этих семействах очень
велико. Большинство Gamas’des, обитающих в почве, во мху или
в органических остатках, относится к одному из этих трех семейств.
88
Глава Iff. Подотряд Mesostigmata
Вероятно, в будущем, когда накопится больше сведений, окажется
необходимым пересмотреть и перегруппировать эти роды. Хозяйствен-
ное и медицинское значение клещей Pseudoparasitidae не изучено.
ЛИТЕРАТУРА
Tragardh I., 1936. Leptogamasus, a new genus of Acari from Sweden, Ent.
Tidsk., 1936, 228—234.
Vitzthum H. G,, 1926. Malayische Acari, Treabia, 8, 1—198.
Raillietidae Vitzthum, 1941
ДИАГНОЗ
(рис. 58)
Самок можно распознать по редукции или отсутствию основной
части тритостернума, довольно хорошо склеротизированному эпиги-
нальному щитку и по овальной
форме тела. У самцов на ногах II
имеются апофизы; кроме того,
они отличаются от самцов других
клещей тем, что имеют двураз-
дельную щетинку на лапках пальп.
Дорзальный щиток у обоих полов
не разделен. Все ноги имеют пред-
лапки, каждая из которых снаб-
жена присоской и коготками.
Railietidae паразитируют в наруж-
ном слуховом проходе крупного
рогатого скота и антилоп.
Рис. 58. Raillietia auris (Leidy),
1872 (по Хирсту, 1922).
Самка (вид снизу).
РОДЫ
1. Raillietia Trouessart, 1902
Тип рода Gamasus auris Leidy, 1872
ОБЩИЕ ЗАМЕЧАНИЯ
Из этого семейства описано только два вида: Raillietia auris Leidy
(1872) и R. hopklnsi (1938). Поданным Хирста' (1922) клещи R. auris
распространены в Северной Америке и Европе; в некоторых местах
они довольно обычны. R. auris питается ушной серой и отмирающими
эпидермальными клетками. Очевидно, этот вид не сосет крови.
R. hopklnsi был найден в ушах антилоп Cobus defassa ugandae Neu-
mann; вероятно, он ведет образ жизни, подобный образу жизни
R. auris из ушей рогатого скота. Зараженность Raillietidae не вызы-
вает патологических изменений. R. auris проходит личиночную фазу
Gamasides
89
в яйце, так что самка рождает нимф или откладывает яйца, в кото-
рых заключены уже сформировавшиеся нимфы.
ЛИТЕРАТУРА
Hirst S., 1922. Mites injurious to domestic animals, Brit. Mas. {Nat. Hist.),
Econ. Ser., 13, 1—107.
Radford G. D., 1938. Notes on some new species of parasitic mites, Parasl-
tol., 30, 427—440.
Halarachnidae Oudemans, 1906
ДИАГНОЗ
(рис. 59)
Клещи Halarachnidae — эндопаразиты дыхательных путей млеко-
питающих. Они, повидимому, являются крайне измененными потом-
ками Lelaptidae. По форме тела они варьируют от овальных {Рпеито-
nyssus) до червеобразных {Orthohalarachne attenuata). Дорзальный
Рис. 59. Pneumonyssas stmicola Banks, 1901 (по Фитцтуму, 1931).
А —самка (вид снизу); Б — самка (вид сверху); В —самец (вид снизу).
щиток редуцирован, но цельный, тритостернум отсутствует. Стер-
нальный щиток редуцирован и несет только три пары стернальных
щетинок. Генитальный щиток самки рудиментарный; половое отвер-
стие самца расположено на переднем крае стернального щитка. Аналь-
ное отверстие находится в задней части анального щитка, который
несет три или четыре щетинки. Стигмы с небольшими перитремами
расположены кнаружи от тазиков ног IV. Тектум простой, треугольный,
гипостом треугольный и опушенный. Подвижный палец хелицер раз-
вит сильнее неподвижного. Пальпы разделены на четыре сегмента.
90
Глаба III. Подотряд Mesostigmata
Разветвленная щетинка тарзального членика пальп имеет две ветви.
Все ноги имеют предлапку с присоской и коготками.
РОДЫ
1. Halarachne Allman, 1847
Тип рода Halarachne halichoerl Allman, 1847 {—Rhlnixodes
Nehring, 1884 et Rhlnacarus Nehring, 1884)
2. Pneumonyssus Banks, 1901 (= Pneumotuber Landois et Hoepke,
1914)
Тип рода Pneumonyssus slmlcola Banks, 1901
3. Orthohalarachne Newell, 1947
Тип рода Halarachne attenuata Banks, 1910
Общие замечания
Ньюэлл (1947) опубликовал превосходный обзор морфологии и
систематики этого семейства. Его работа положена в основу приво-
димого ниже обзора, хотя нами учитывались данные и других авторов.
Клещи Halarachnidae живут в дыхательных путях млекопитающих.
Жизненный цикл их полностью не изучен, и известны лишь немногие
факты относительно их биологии. Дойчмап (1944) и другие авторы
находили шестиногих личинок вместе со взрослыми самцами и сам-
ками. Нимфы этого семейства неизвестны. Следует иметь в виду,
что жизненный цикл Halarachnidae полностью еще не изучен, и пути
заражения новых хозяев нам неизвестны. Клещей рода Halarachne нахо-
дят исключительно в дыхательных путях тюленей (Phocidae). Orthoha-
larachne паразитирует у представителей других семейств ластоногих,
a Pneumonyssus найден у обезьян, собак и других млекопитающих.
Хотя заражение видами рода Pneumonyssus довольно обычно для
некоторых приматов, у человека эти клещи не встречались. Легочный
акароз человека является обычным заболеванием в некоторых районах,
например в Индии. Во время второй мировой войны синдром, вызван-
ный заражением дыхательных путей человека клещами, был описан
под названием тропической эозинофилии. Однако клещи, которые
были найдены в случаях этого заболевания, не относились к семей-
ству Halarachnidae, а оказались типичными свободноживущими видами.
ЛИТЕРАТУРА
Carter Н. F„ D’Abrera V. St. Е., 1946. Mites (Acarina) — a probable factor
in the aetiology of spasmodic bronchitis and asthma associated with high
eosinophilia, Trans. Roy. Soc. Trap. Med. a. Hyg., 39, 373—395.
Chandler W. L., Ruhc D. S., 1940. Pneumonyssus canlnum n. sp., a mite
from the frontal sinus of the dog, J. Parasltol., 26 (1), 59—70.
Doetschman W. H., 1944. A new species of endoparasitic mite of the family
Halarachnidae (Acarina), Trans. Amer. Mier. ~ioc., 63, 68—72.
Newell I. M., 1947. Studies on the morphology and systematics of the family
Halarachnidae Oudemans 1906 (Acari, Parasitoidea), Bull. Bingham. Ocea-
nogr. Coll.. 10 (4), 235-266.
Vitzthum H. O., 1931. Pneumonyssus simicola Banks, Z. Parastienk., 4,
48—74.
Gamasides
Si
Entonyssidae Ewing, 1923
(рис. 60)
ДИАГНОЗ
Entonyssidae паразитируют в легких змей. Их тело имеет оваль-
ную форму и может достигать 1 мм в длину. У самок тритостернум
рудиментарный; согласно Редфорду (1937), тритостернум отсутствует
у обоих полов Е. bedfordi Radford, 1937. Дорзальный щиток цель-
ный и обычно простирается до заднего края тела. Если дорзальный
щиток ограничен проподосомой, он несет на своем заднем конце две
крупные щетинки. Все ноги имеют предлапку, присоску и коготки.
А Б
Рис. 60. Entonyssus glasmacheri Vitzthum, 1935
(по Фитцтуму, 1935).
Л —самка (вид сверху); 5 —самка (вид снизу).
Хорошо развитых зубцов на хелицерах нет. Разветвленная щетинка
на лапках пальп двураздельная.
Ниже приводится заимствованная у Терка (1947) определительная
таблица подсемейств Entonyssidae. Терк придает этим подсемействам
ранг семейств.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ ENTONYSSIDAE
1. Клешня хелицер самки с подвижным пальцем серповидной формы
и гарпуновидным неподвижным пальцем, зубцы которого напра-
влены назад, или же оба пальца имеют серповидную форму и
обычно лишены щетинок...........................Entonyssinae
— Клешня хелицер самки со слабо склеротизированным длинным
подвижным пальцем, который сильно изогнут кнаружи и не имеет
ни зубцов, ни щетинок; неподвижный палец редуцирован . . .
...........•.......................... Pneumophionyssinae
92
Глава 1П. Подотряд Mesostigmata
РОДЫ
Entonyssinae Ewing, 1923
1. Entonyssas Ewing, 1923
Тип рода Entonyssas Haiti Ewing, 1923
2. Hammertonia Turk, 1947
Тип рода Entonyssas bedfordl Radford, 1937
3. Ophlopneumicola Hubbard, 1938
Тип рода Ophlopneumicola colubri Hubbard, 1938
Pneumophionyssinae Turk, 1947
4. Pneumophlonyssus Fonseca, 1940
Тип рода Pneumophlonyssus aristoterisl Fonseca, 1940
ОБЩИЕ ЗАМЕЧАНИЯ
Клещей Entonyssidae находили только в дыхательных путях змей.
Их жизненный цикл еще не совсем ясен, но он, вероятно, сильно
упрощен. В половых путях самок находили вполне развитых личинок,
поэтому предположение о том, что эти клещи яйцеживородящие, можно
считать обоснованным. Редфорд (1937)—-единственный автор, который
сообщил о находке нимф этого семейства. Терк (1947) предположил,
что наблюдения Редфорда ошибочны и что личинки линяют прямо во
взрослых самцов и самок. Этот вопрос остается открытым, так как
еще никто не проследил полного жизненного цикла ни у одного вида.
Entonyssidae найдены в Америке и Африке. Нет оснований пред-
полагать, что в будущем они не будут найдены и в других районах.
Насколько известно, эти клещи не причиняют большого беспокой-
ства своим хозяевам. Их хозяйственное и медицинское значение не
выяснено.
ЛИТЕРАТУРА
Radford С. D., 1937. A new species of mite of the genus Entonyssas Ewing,
The North Western Naturalist, Mar. 1937, 38—42.
Turk F. A., 1947. Studies of Acari IV. A review of the lung mites of snakes,
Parasitol., 38, 17—26.
Vitzthum H. O., 1935. Ueber die Gattung Entonyssas Ewing (Acari), Zeit.
Parasitenk., 7, 709—716.
Rhinonyssidae Trouessart, 1895
(рис. 61)
ДИАГНОЗ
Rhinonyssidae — довольно крупные клещи (длиной 0,5—1,6 мм).
Форма тела овальная, щитки слабо склеротизированы. Ноги хорошо
развиты, каждая лапка имеет предлапку с коготками и присоской.
Gamasides
03
Дорзальный щиток ограничен проподосомой, у некоторых форм
подразделен на два или три щитка или же совсем отсутствует.
Рис. 61. Larinyssus orbicularis Strandtmann, 1948
(по Стрэндтмэну, 1948).
А —общий вид самки сверху; 5 —пальпы и тектум (вид сверху); В —клешня хелицеры;
Г— гнатосома (вид снизу); Д— обший вид самки снизу.
Щетинки на теле и конечностях короткие. У некоторых форм стигмы
связаны с короткими дорзальными перитремами. Хелицеры обычно
имеют маленькую рудиментарную клешню. Тритостернум отсутствует.
94
Глава III. Подотряд Mesostigmata
РОДЫ И ПОДРОДЫ
1. Rhinonyssus Trouessart, 1894
a) Rhinonyssus s. str.
Тип подрода Rhinonyssus conlventris Trouessart, 1894
6) Cas (новое название для Rhinacarus Castro, 1948 {Arq. Inst.
S. Paulo, 18,257), но не для Rhinacarus Nehring, 1884)
Тип подрода Rhinonyssus {Rhinacarus) angrensis Castro, 1948
2. Larlnyssus Strandtmann, 1948
Тип рода Larlnyssus orbicularis Strandtmann, 1948
3. Neonyssus Hirst, 1921 (== Neonyssotdes Hirst, 1924)
a) Neonyssus s. str.
Тип подрода Neonyssus Inter medias Hirst, 1921
6) Paraneonyssus Castro, 1948
Тип подрода Neonyssus {Paraneonyssus) enriettii Castro, 1948
в) Rochanyssus Castro, 1948
Тип подрода Neonyssus {Rochanyssus) werneri Castro, 1948
r) Travanyssus Castro, 1948
Тип подрода Neonyssus {Travanyssus) paranensls Castro, 1948
д) Vltznyssus Castro, 1948
Тип подрода Dermanyssus nitzchi Giebel, 1871
4. Ptylonyssoides Vitzthum, 1935
Тип рода Ptylonyssoides triscutatus Vitzthum, 1935
5. Ptilonyssus Berlese et Trouessart, 1889
a) Ptilonyssus s. str.
Тип подрода Ptilonyssus echlnatus Berlese et Trouessart, 1889
6) Flavionyssus Castro, 1948
Тип подрода Ptilonyssus {Flavionyssus) rabellol Castro, 1948
в) Rhinonyssoides Hirst, 1921
Тип подрода Rhinonyssoides trouessarti Hirst, 1921
6. Rallinyssus Strandtmann, 1948
Тип рода Rallinyssus caudlstlgmus Strandtmann, 1948
7. Rhinoecius Cooreman, 1946
Тип рода Rhinoecius oti Cooreman, 1946
8. Sommatrlcola Tragardh, 1904
Тип рода Sommatrlcola levlnseni TragSrdh, 1904
9. Sternostoma Berlese et Trouessart, 1889
Тип рода Sternostoma cryptorhyneum Berlese et Trouessart, 1889
10. Sternostomum Trouessart, 1895
Тип рода Sternostomum rhinoletrum Trouessart, 1895
ОБЩИЕ ЗАМЕЧАНИЯ
Rhinonyssidae— живородящие клещи; на свет появляются личинки,
под покровами которых лежат уже сформированные нимфы. Rhino-
nyssidae паразитируют в носовой полости птиц. Распространены эти
клещи во всех частях земного шара. Вероятно, описана только часть
видов этого семейства, так как очень мало птиц исследовалось с этой
Gam asides
96
целью. Труссар указывает, что Rhinonyssidae могут наносить значи-
тельный вред цыплятам. Стрэндтмэн (1948) считает, что интересно было
бы исследовать возможную роль клещей в „орнитозе" (болезнь птиц).
ЛИТЕРАТУРА
БрегетоваН. Г., 1950. Новые виды эндопаразитических клещей из се-
мейства Rhinonyssidae (Gamasoidea, Acarina), Докл. Акад. наук СССР,
71 (5), 1005—1008.
Castro М. Р. de, 1948. Reestruturacao generica de familia ^Rhinonyssidae
Vitzthum, 1935“ (Acari Mesostigmata Gamasides) a descrifao de algumas
especies novas, Arq. Inst. Biol. San Paulo Brasil, 18, 253—284.
Strandtmann R. W., 1948. The mesostigmatic nasal mites of birds. 1. Two
new genera from shore and marsh birds, J. Parasitol., 34, 505—514.
Strandtmann R. W., 1950. The mesostigmatic nasal mites of birds. II. New
and poorly known species of Rhinonyssidae, J. Parasitol., 37 (2), 129—140.
Vitzthum H. G., 1935. Milben aus der Nasenhole von Vogeln., J. Ornith.,
83, 563—587.
ДИАГНОЗ
Haemogamasidae Oudemans, 1926
(рис. 62)
Haemogamasidae — овальные клещи средних размеров. Тело как
с брюшной, так и со спинной стороны покрыто густыми, как шерсть,
щетинками. У личинок, нимф и у взрослых
клещей дорзальный щиток цельный. Брюшные
щитки, несущие у представителей других
семейств лишь несколько щетинок, бывают
иногда покрыты щетинками. Лапки всех ног
имеют предлапку, коготки и присоску. Три-
тостернум имеется. Метаподальные щитки
мелкие; стигмы с длинными, вытянутыми
вперед перитремами. Тектум языковидный и
имеет многочисленные неправильные выросты.
Рис. 62. Haemogamasus mandschuricus Vitzthum
1930 (по Фитцтуму, 1930).
Самка (вид снизу).
РОДЫ
1. Haemogamasus Berlese, 1889
Тип рода Haemogamasus hirsutus Berlese, 1889
2. Acanthochela Ewing, 1933
Тип рода Acanthochela chiliensis Ewing, 1933
3. Euhaemogamasus Ewing, 1933
Тип рода Euhaemogamasus onychomydts Ewing, 1933
4. Ischyropoda Keegan, 1951
Тип рода Ischyropoda splniger Keegan, 1951
96
Г лав п f[f. Подотряд Mesostigmata
ОБЩИЕ ЗАМЕЧАНИЯ
Кигэн (1950) выделил Acanthochela из этого семейства, так как
не смог определить природы хелицер и тектума на материале типа
в Национальном музее США. Просмотр этого материала в фазово-
контрастном микроскопе показал, что у Acanthochela тектум и хели-
церы типичны для семейства Haemogamasidae.
Haemogamasidae паразитируют на мелких млекопитающих во всех
областях мира. Обычно большую часть жизни они проводят в гнездах
и переходят на хозяина только для сосания его крови. Жизненный
цикл у них типичен и состоит из фаз личинки, двух нимф и поло-
возрелого клеща. Тот факт, что эти клещи обычно паразитируют
на мелких млекопитающих, говорит за то, что они могут играть
определенную роль в переносе чумы, тифа, туляремии и других
болезней. Необходимо изучить возможность передачи этими клещами
возбудителей различных болезней.
ЛИТЕРАТУРА
Ewing Н. Е., 1933. New genera and species of parasitic mites of the super-
family Parasitoidea, Proc. U. S. Nat. Mas., 82, Art. 30, 1—14.
Keegan H. L., 1951. The mites of the subfamily Haemogamasinae (Acari;
Laelaptidae), Proc. U. S. Nat. Mas., 101 (3275), 203—268.
Vitzthum H., 1930. Milben als Pesttrager? Zool. Jahrb., 60, 381—428.
Dermanyssidae Kolenati, 1859
(рис. 63, 64)
ДИАГНОЗ
Dermanyssi dae — клещи средних размеров. Дорзальный щиток
обычно цельный или разделен у самок на большой передний щиток
Рис. 63. Allodermanyssus sanguineus (Hirst), 1914.
А — самка (вил сверху); Б — самка (вид снизу).
и очень маленький задний щиток. У самок щитки брюшной поверх-
ности тела типичны для подотряда, за исключением тех случаев,
Gamasides
97
когда они несколько редуцированы. Стернальный щиток имеет три
пары щетинок, метастернальные щитки редуцированы и лежат кнаружи
от генитального щитка. Генито-вентральный щиток в задней части
принимает каплевидную форму. Анальный щиток несет три характер-
ные щетинки. Хелицеры у Dermanyssidae иногда очень длинные
и заканчиваются маленькой ножницевидной клешней, так что они
А
Рис. 64. Dermanyssus gallinae (DeGeer) (по Хирсту, 1922).
А —самка (вид снизу); Б—самец (вид снизу).
кажутся игловидными. В других случаях хелицеры более обычные,
но с уменьшенной клешней. Тритостернум имеется. Каждая из ног
имеет предлапку с коготками и присоской.
РОДЫ
1. Dermanyssus Duges, 1834
Тип рода Acarus gallinae DeGeer, 1778
2. Allodermanyssus Ewing, 1923
Тип рода Dermanyssus sanguineus Hirst, 1914
3. Bdellonyssus Fonseca, 1941 (= Leiognathus Canestrini, 1885
гомоним = Liponyssus различных авторов = Fonsecaonyssus
Radford, 1950).
Тип рода Leiognathus bacoti H'rst, 1913
4. Brevisterna Keegan, 1949
Тип рода Euhaemogamasus utahensis Ewing, 1933
7 Зак. 1785
98
Глава HI. Подотряд Mesostigmata
5. Echinonyssus Hirst, 1925
Тип рода Echinonyssus nasutus Hirst, 1925
6. EllszDorthia Turk, 1945 (= Hetnilaelaps Ewing, 1923 гомоним)
Тип рода 'Hetnilaelaps americanus Ewing, 1923
7. Hirstesia Fonseca, 1948
Тип рода Liponyssus sternalis Hirst, 1921
8. Hirstonyssus Fonseca, 1948
Тип рода Dermanyssus arcuatus Koch, 1839
9. Ichoronyssus Kolenati, 1858 (= Chiroptonyssus Auguston,
1945 = Leptonyssus Kolenati, 1858 == Spinolaelaps Radford, 1940)
Тип рода Dermanyssus scutatus Kolenati, 1856
10. Kolenationyssus Fonseca, 1948
Тип рода Kolenationyssus athleticus Fonseca, 1947
11. Lepronyssoides Fonseca, 1943
Тип рода Liponissus pereirai Fonseca, 1935
12. Liponysella Hirst, 1925 (= Geneiadolaelaps Ewing, 1929)
Тип рода Liponyssus madagascarensis Hirst, 1921
13. Liponyssoides Hirst, 1913
Тип рода Dermanyssus (Liponyssoides') muris Hirst, 1913
14. Macronyssus Kolenati, 1858
Тип рода Caris longimana Kolenati, 1856
15. Manisicola Lawrence, 1939
Тип рода Manisicola africana Lawrence, 1939
16. Manitherlonyssus Vitzthum, 1925
Тип рода Liponyssus heterotarsus Vitzthum, 1925
17. Myonyssoides Hirst, 1925
Тип рода Myonyssoides capensis Hirst, 1925
18. Myonyssus Tiraboschi, 1904 ( -Tetragonyssus Ewing, 1923)
Тип рода Myonyssus decumani Tiraboschi, 1904
19. Neolchoronyssus Fonseca, 1943
Тип рода Liponissus wernecki Fonseca, 1935
20. Neoliponyssus Ewing, 1929 (= Oudemansiella Fonseca, 1948)
Тип рода Liponyssus gordonensis Hirst, 1923
21. Ophionyssus M6gnin, 1884 (= Serpenticola Ewing, 1923)
Тип рода Ophionyssus nt. tricis Megnin, 1884
22. Tur (новое название для Protonyssus Turk, 1947 (Ann. Mag.
Hist, 11, 14, 348), но не Protonyssus Trouessart, 1915)
Тип рода Protonyssus unlscutatus Turk, 1946
23. Radfordiella Fonseca, 1948
Тип рода Radfordiella oudemansi Fonseca, 1948
24. Steatonyssus Kolenati, 1858
Тип рода Amarus musculi Schrank, 1803
ОБЩИЕ ЗАМЕЧАНИЯ
Как правило, Dermanyssidae проходят одну личиночную и две
нимфальные фазы, после чего превращаются во взрослых самцов и
Gamasides 99
самок. Они паразитируют на позвоночных и имеют большое хозяй-
ственное и медицинское значение. Семейство обычно разделяют на
два подсемейства: Dermanyssinae с игловидными хелицерами и Lipo-
nyssinae, у которых хелицеры менее изменены. К подсемейству
Dermanyssinae относятся только три рода — Dermanyssus, Liponys-
soides и Allodermanyssus. Некоторые авторы возводят эти два
подсемейства в ранг семейства. Фонзека (1948) написал моногра-
фию по той группе родов, которая иногда включается в подсе-
мейство Liponyssinae или семейство Liponyssidae; сам он обозна-
чает эту группу как Macronyssidae. Принятая нами группировка
родов основана главным образом на монографии Фонзека (1948),
на работе Фитцтума (1941) и на опубликованных в печати заме-
чаниях Стрэндтмэна. Dermanyssidae еще плохо изучены, так что
приводимый здесь общий обзор носит лишь предварительный ха-
рактер.
Dermanyssus gallinae (DeGeer, 1778) во всех областях земного
шара является подлинным бичом домашних птиц, особенно кур и
голубей. Большая зараженность этими клещами может вызвать смерть
хозяев, в то время как слабая зараженность приводит к понижению
яйценоскости и плохому росту. Клещи обычно питаются ночью и
посещают своих хозяев именно в это время. Днем они сидят в щелях
насестов и курятников и в гнездах. Иногда клещей в гнезде бывает
настолько много, что наседки бросают насиживание яиц. D. gallinae
нападает и на человека, особенно на детей. В Северной Каролине
известен случай, когда в одной семье клещи вызвали у ребенка
сильный дерматит, а родители в том же помещении не были пора-
жены. При обследовании этого дома на электрических часах в жилой
комнате было обнаружено большое число клещей D. gallinae. Часы
были несколько нагреты по сравнению с комнатным воздухом;
именно это, вероятно, и привлекло клещей. На водосточном желобе
крыши дома любили сидеть голуби, и их помет скопился на крыше
над крыльцом; после того как убрали помет и разогнали голубей,
клещи исчезли и ребенок поправился. Недавно из куриных клещей
был выделен вирус энцефалита Сен-Луи, и это дает основание пред-
полагать, что D. gallinae является резервуаром названной инфекции
(Смит и др., 1948). Поскольку можно судить по результатам изу-
чения эпидемиологии болезней человека, вероятно, что многие клещи
играют в этом важную роль.
Allodermanyssus sanguineus (Hirst) обыкновенно паразитирует
на мелких грызунах. Иногда он нападает на человека; об этом
имеются сообщения из больших городов восточных штатов, как
Бостон, Нью-Йорк и Балтимора. Недавно было установлено, что
в Нью-Йорке этот клещ является переносчиком нового заболевания —
риккетсиозной оспы. Это заболевание первоначально смешивали
с ветряной оспой, но подробное исследование установило, что это —
новая болезнь (Хебнер, 1947).
7*
100
Глава Ш. Подотряд Mesostigmata
Bdellonyssus sylvlarum (Canestrini et Fanzago) — типичный птичий
клещ. Известен как резервуар нейротропных вирусов (Ривс и др.,
1949). Во время второй мировой войны было установлено, что
В. bacoti является промежуточным хозяином филарии Litomosoides
carinii, которая паразитирует на хлопковых крысах. Благодаря тому,
что клещей, крыс и филярий можно легко культивировать, они были
использованы в широком масштабе при изучении филяриозов (Уилльямс
и Браун, 1945).
ЛИТЕРАТУ PjA
Anderson С. R., 1944. Rat mite dermatitis. Acariasis caused by the tropical
rat mite, Liponyssus bacoti Hirst, 1914, Arch. Dermatol, and Syphiol.
50 (2), 90—95.
BishoppF. C., Wood H. P., 1939. Mites and lice on poultry, U. S. Dept'
Agr. Farmer’s Bull., 801, 1—25.
Fonseca F. da, 1948. A monograph of the genera and species of Macronyssi-
dac Oudemans, 1936 (synom. Liponyssidae Vitzthum, 1931) (Acari), Proc.
Zool. Soc. London, 118, 249—334.
HttebncrR. J., 1947. Rickettsialpox a new deseases, Amer. J. Clin. Path.,
17, 970—971.
R oueche B., 1950. The alerting of Mr. Pomerantz, MD., 5 (3), 73—82.
Smoth Margaret G, et al., 1948. Experiments on the role of the chicken mite
Dermanyssus gallinae and the mosquito in the epidemiology of St. Louis
encephalitis, J. Exp. Med., 87, 119—138.
Williams R. W., Brown H. W., 1945. The development of Litomosoides
carinii filariid parasite of the cotton rat in the tropical rat mite, Science,
102, 482—483.
Phytoseiidae Berlese, 1916
(рис. 65)
ДИАГНОЗ
Это семейство объединяет весьма разнородную группу родов,
которая характеризуется формой эпигинального щитка, усеченного
А Б
Рис. 65. Seiulus bakeri Garman, 1948 (по Джэрмэну, 1948).
Л —самка (вид снизу); Б — самка (вид сверху).
или вогнутого сзади. Разветвленная щетинка на лапке пальп дву-
раздельная.
Gamasides
101
Phytoseidae можно подразделить на два подсемейства: Phytosei-
inae,—недавно установленное и изученное Джармэном (1948), и
Podocininae — еще слабо изученную группу родов. Эти подсемейства
можно различать по признакам, указанным в приведенной ниже
определительной таблице.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ PHYTOSEI1DAE
1. На дорзальном щитке несколько щетинок. Предлапка, коготки
и присоска имеются на всех ногах. Дорзальный щит не разделен
......................................................Phytoseiinae
— Сочетание признаков иное.............................Podocininae
РОДЫ И ПОДРОДЫ
Phytoseiinae Berlese, 1916
1. Phytoseius Ribaga, 1902
Тип рода Gamasus plumifer Canestrini et Fanzago, 1876
2. Amblyselus Berlese, 1904 (= Borinquolaelaps Fox, 1946)
Тип рода Seius obtusus Berlese, 1889
3. Amblysiopsis Garman, 1948
Тип рода Amblyselus (Amblysiopsis} americanus Garman, 1948
4. Ameroseius Berlese, 1903
Тип рода Acarus corbicula Sowerby, 1806 (= Seius echinatus
Koch, 1839 = Seius muricatus Koch, 1839 == Se/zrx hirsutus
Berlese, 1887, но не Seius muricatus Berlese, 1887)
5. Iphidulus Ribaga, 1902
Тип рода Iphidulus communis Ribaga, 1902
6. Lasioseius Berlese, 1916
a) Lasioseius s. str.
Тип подрода Lasioseius aba, новое название для Seius muri-
catus Berlese, 1887
6) Cheiroseius Berlese, 1916
Тип подрода Seius unguiculatus Berlese, 1916
в) Leioseius Berlese, 1916
Тип подрода Ameroseius minusculus Berlese, 1905
r) Zercoseius Berlese, 1916
Тип подрода Seius spathultger Leonard!, 1899
д) Zygoseius Berlese, 1916
Тип подрода Zygoseius furciger Berlese, 1916
7. Melichares Hering, 1838
Тип рода Melichares agilis Hering, 1838
8. Seiopsis Berlese, 1923
Тип рода Amblyselus (Seiopsis) brevipilis Berlese, 1923
. 9. Seiulus Berlese, 1887 (= Echinoseius Ribaga, 1902)
Тиц рода Seiulus hirsutigenus Berlese, 1887 .
102
Глава III. Подотряд Mesostigmata
10. Seius Koch, 1836
Тип рода Seius togatus Koch, 1836
11. Tristomus Hughes, 1948
Тип рода Tristomus butlerl Hughes, 1948
12. Typhlodromus Scheuten, 1857
Тип рода Gamasus vepallidus Koch, 1839 (= Typhlodromus pyri
Scheuten, 1857)
Podocininae Berlese, 1916
1. Podocinum Berlese, 1882
Тип рода Laelaps sagax Berlese, 1882
2. Antennoseius Berlese, 1916
Тип рода Antennoseius delicatus Berlese, 1916
3. Asternolaelaps Berlese, 1923
Тип рода Asternolaelaps fecundus Berlese, 1923
4. Asternoseius Berlese, 1910
Тип рода Asternoseius ciliatus Berlese, 1910
5. Blattisoclus Keegan, 1944
Тип рода Typhlodromus tineivorus Oudemans, 1929
(= Blattisoclus triodons Keegan, 1944)
6. Episeiella Willmann, 1938
Тип рода Episeiella heteropoda Willmann, 1938
7. Hoploseius Berlese, 1914
Тип рода Zercon cometa Berlese, 1910
8. Platyseius Berlese, 1916 (= Episeius Hull, 1918)
Тип рода Platyseius copillatus Berlese, 1916
9. Thinoseius Halbert, 1920
Тип рода Thinoseius berlesei Halbert, 1920
10. Zerconopsts Hull, 1918
Тип рода Gamasus remiger Kramer, 1876
11. Iphiseius Berlese, 1916 (этот род установлен Фитцтумом в 1941 г.,
но тип рода не был обозначен; его первоописание неизвестно).
ОБЩИЕ ЗАМЕЧАНИЯ
Многих клещей семейства Phytoseiidae обычно находят на расте-
ниях; такие виды очень полезны, так как они, будучи хищниками,
уничтожают вредителей растений. Джэрмэн (1948) указывал, что
опыление растений инсектицидами, которые должны убивать насе-
комых, может принести больше вреда, чем пользы, так как эти
инсектициды ведут, в первую очередь, к гибели клещей, которые
уничтожают насекомых-вредителей. Джэрмэн обсуждает также клас-
сификацию этих клещей, но многим его положениям мы не следуем
в настоящей работе.
Gamasides
103
ЛИТЕРАТУРА
Garman P., 1948. Mite species from apple trees in Connecticut, Conn. Agr.
Expt. St a. Bull. 520, 1—27.
Hughes A. M., 1948. The mites associated with stored food products, Ministry
of Agriculture and Fisheries, London, p. 1—168.
Keegan H. L„ 1944. On a new genus and species of parasitid mite, J. Para-
sltol., 30, 181—183.
Lord F. T., 1949. The influence of spray programs on the fauna of apple
orchards in Nova Scotia. III. Mites and their predators, Canad. Ent., 81
(8), 202—214.
ДИАГНОЗ
Iphiopsidae Kramer, 1886
(рис. 66)
Самой характерной особенностью этого семейства является нали-
чие крошечных перитрем. Каждая из ног имеет предлапку с присоской
Щетинка лапок пальп дву- «
раздельная. Эпигинальный /Т
щиток или имеет капле- /У
видную форму, или выре- р/
зан сзади. \А к ЛЛ
Рис. 66. Jacobsonia
tertia Vitzthum, 1931
(по Фитцтуму, 1931).
Самка (вид снизу).
РОДЫ
1. Iphiopsis Berlese, 1882
Тип рода Iphis mlrabllls Berlese, 1882
2. Berlesla Canestrini, 1884
Тип рода Berlesla rapax Canestrini, 1884
3, Jacobsonia Berlese, 1910
Тип рода Iphiopsis (Greeniella) submollis Berlese, 1910 (Iphis
гомоним Iphis Me'gen, 180Q; Greeniella гомоним Greenlella
Cockerell, 1897)
104
Глава ш. Подотряд Mesostigmata
ОБЩИЕ ЗАМЕЧАНИЯ
Клещи этого семейства являются паразитами или комменсалами
насекомых и многоножек. Их хозяйственное и медицинское значение
не выяснено.
ЛИТЕРАТУРА
Vitzthum Н. О., 1931. Eine afrikanische Jacobsotiia (Acari), Zool. Anz., 96,
153—159.
ДИАГНОЗ
Laelaptidae Berlese, 1892
(рис. 67, 68)
У клещей Laelaptidae щетинка у основания лапки пальп двураз-
дельная. Эпигинальный щиток имеет каплевидную форму или же
Рис. 67. Мугто-
nyssus chapmani
Baker et Strandt-
mann, 1948 (по Бэ-
керу и Стрэндт-
мэну, 1948).
А —самка (вид сверху);
Б~ гнатосома (внд
сверху); В —клешня
хелицеры; Г—лапка
ноги I; Д—гнатосома
(вид снизу); Е—самка
(вяд снизу).
Gamasides
105
вырезан по заднему краю; перитремы удлиненные. Большинство видов
имеет на вс.ех ногах предлапку, присоску и коготки. Дорзальный
щиток цельный. Тазики ног II у переднего края имеют зубовидные
выросты.
Семейство Laelaptidae даже в том ограниченном объеме, в котором
оно рассматривается здесь, включает большое число разнообразных
Рис. 68. Echinolaelaps echidninus (Berlese), 1887
(по Хирсту, 1922).
Самка (вид снизу).
родов. Фитцтум (1941) включал в семейство Laelaptidae 13 подсе-
мейств, оправдывая такое усложнение системы неясностью, суще-
ствующей в отношении границ подсемейств. Эта неясность сохра-
няется до настоящего времени. Несмотря на то, что некоторые
последние работы проливают свет на этот вопрос, и девять из под-
семейств (Railliettidae — Laelaptidae) были возведены в ранг семейства,
а подсемейство Podocininae включено в семейство Phytoseiidae, раз-
личия между семействами во многих случаях остаются нечеткими. Мы
включаем в семейство Laelaptidae три подсемейства. Группа родов,
включенная в семейство Laelaptidae, требует дальнейшего пересмотра,
и приведенную ниже классификацию, основанную главным . образом
па данных Фитцтума (1941), нельзя считать удовлетворительной.
Потребуется длительная работа многих исследователей, прежде чем
/можно будет достичь удовлетворительной классификации этой группы
106
Глава III. Подотряд Mesostigmata
клещей. Ниже приводится определительная таблица, позволяющая
правильно определить подсемейство, к которому относится тот или
иной экземпляр.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ LAELAPT1DAE '
1. Тектум спереди выпуклый, с гладким или зазубренным краем 2
— Передний край тектума с ланцетовидным отростком.............
............................................Hyletastinae
2. Паразитируют на насекомых. Вентральные щетинки обычно неж-
ные ...........................................Hypoaspidinae
— Паразитируют на млекопитающих. Вентральные щетинки обычно
шиповидные.......................................Laelaptinae
РОДЫ И ПОДРОДЫ
Hyletastinae Vitzthum, 1941
L. Hyletastea Gistel, 1884 (—Iphis Koch, 1836)
Тип рода I phis globulus Koch, 1839
2. Copriphts Berlese, 1910
a) Copriphts s. str.
Тип подрода Iphis pterophilus Berlese, 1882
6) Peletiphls Berlese, 1911
Тип подрода Copriphts (Peletiphls) insignis Berlese, 1911
в) Alliphis Halbert, 1923
Тип подрода Gamasus halleri G. et R. Canestrini, 1881
3. Cosmlphis Vitzthum, 1926
Тип рода Etnaeus bosschai Oudemans, 1901
4. Eviphis Berlese, 1903
a) Eviphis s. str.
Тип подрода Eumaeus pyrobolus Koch, 1839
6) Oloiphis Berlese, 1916
Тип подрода Eviphis magnificus Berlese, 1916
5. Iphldozercon. Berlese, 1903
Тип рода Eviphis gibbus Berlese, 1903
6. Melitttphls Berlese, 1918
Тип рода Laelaps (Iphis) alvearius Berlese, 1895
7. Uroiphis Berlese, 1903
Тип рода Uroiphis scabratus Berlese, 1903
Hypoaspidinae Vitzthum, 1941
1. Hypoaspls G. Canestrini, 1885
a) Hypoaspls s. str.
Тип подрода Laelaps kramerli G. et R. Canestrini, 1881
6) Androlaelaps Berlese, 1903
Тип подрода Laelaps hermaphrodita Berlese, 1903
Gamasides
107
в) Cosmolaelaps Berlese, 1903
Тип подрода Laelaps claviger Berlese, 1883
r) Gymnolaelaps Berlese, 1920
Тип подрода Laelaps myrmecophilus Berlese, 1892
д) Holostaspis Kolenati, 1858
Тип подрода Holostaspis isotricha Kolenati, 1858
e) Laelaspts Berlese, 1903
Тип подрода Iphis astronomicus Berlese, 1889
ж) Leptolaelaps Berlese, 1918
Тип подрода Hypoaspls (Leptolaelaps) elegans Berlese, 1918
з) Pneumolaelaps Berlese, 1920
Тип подрода [phis bombicoletis G. Canestrini, 1885
и) Stratlolaelaps Berlese, 1916
Тип подрода Laelaps (Iphis) miles Berlese, 1882
2. Anystipalpus Berlese, 1911
Тип рода Anystipalpus percicola Berlese, 1911
3. Arctoseius Thor, 1930
Тип рода Arctoseius lateroinclsus Thor, 1930
4. Coleolaelaps Berlese, 1914
Тип рода Laelaps (Iphis) agrestls Berlese, 1887
5. Dlnogamasus Kramer, 1898 (= Greenia Oudemans, 1901
= Greeniella Banks, 1904 = Dolaea Oudemans, 1912)
Тип рода Dlnogamasus crasstpes Kramer, 1898
6. Iphldosoma Berlese, 1892 (Parasltldae?)
Тип рода Holostaspis fimetaria Muller, 185S)
7. JOrdensia Oudemans, 1937
Тип рода Gamasus cossi Duges, 1-834
8. Julolaelaps Berlese, 1916
Тип рода Julolaelaps dlspar Berlese, 1916
9. Laelantennus Berlese, 1916
Тип рода Laelantennus lagena Berlese, 1916
10. Ljunghia Oudemans, 1932
Тип рода Ljunghia selenocosmlae Oudemans, 1932
11. Mellponaspis Vitzthum, 1930
Тип рода Mellponaspis debllipes Vitzthum, 1930
12. Myrmeclphis Hull, 1923
Тип рода Myrmeclphis crawleianus Hull, 1923
13. Myrmolaelaps Tragardh, 1906
Тип рода Myrmolaelaps equitans TragSrdh, 1906
14. Myrmoleichus Berlese, 1903
Тип рода Myrmoleichus coronatus Berlese, 1903
15. Myrmonyssus Berlese, 1903
a) Myrmonyssus s. str.
Тип подрода Myrmonyssus diplogenlus Berlese, 1903
6) Laelaspulus Berlese, 1903
Тип подрода Myrmonyssus acuminatus Berlese, 1903
108
Глава III. Подотряд Mesostigmata
16. Myrmozercon Berlese, 1902
Тип рода Myrmozercon brevipes Berlese, 1902
17. Heoberlesia Berlese, 1892
Тип рода Neoberlesia equitans Berlese, 1892
18. Neocypholaelaps Vitzthum, 1941 (= Cypholaelaps Berlese гомо-
ним)
Тип рода Laelaps ampullula Berlese, 1892
19. Ololaelaps Berlese, 1903
a) Ololaelaps s. str.
Тип подрода Laelaps venetus Berlese, 1903
6) Cypholaelaps Berlese 1916 (но не — Cypholaelaps Berlese, 1918)
Тип подрода Ololaelaps {Cypholaelaps) haemisphaericus Ber-
lese, 1916)
20. Paradoxiphis Berlese, 1910
Тип рода Paradoxiphis tennibrachiatus Berlese, 1910
21. Phytojacobsonia Vitzthum, 1925
Тип рода Phy tojacobsonia irregularis Vitzthum, 1925
22. Podolaelaps Berlese, 1888
Тип рода Podolaelaps ambulacralis Berlese, 1888
23. Proctolaelaps Berlese, 1914
Тип рода Proctolaelaps productus Berlese, 1923
24. Stamfordia Tragardh, 1906
Тип рода Stamfordia carabicola TrMgardh, 1906
25. Stylochirus G. et R. Canestrini, 1882
Тип рода Stylochirus rovennensis G. et R. Canestrini, 1882
26. Urozercon Berlese, 1901 (= Termitacarus Tr3gSrdh, 1906)
Тип рода Urozercon paradoxus Berlese, 1901
27. Varroa Oudemans, 1904
Тип рода Varroa jacobsoni Oudemans, 1904
Laelaptinae Berlese, 1892
1. Laelups Koch, 1839
a) Laelaps s, str.
Тип подрода Acarus tnuris Ljungh, 1799 {—Laelaps agilis
Koch, 1839
6) Eugynolaelaps Berlese, 1918
Тип подрода Laelaps {Eugynolaelaps) coriaceus Berlese, 1918
в) Heterolaelaps Hirst, 1926
Тип подрода Heterolaelaps antipodianus Hirst, 1926
r) Macrolaelaps Ewing, 1929
Тип подрода Laelaps sanguisugus Vitzthum, 1924
д) Mesolaelaps Hirst, 1926
Тип подрода Mesolaelaps anomalus Hirst, 1926
e) Tricholaelaps Vitzthum, 1926
Тип подрода Laelaps {Tricholaelaps) comatus Vitzthum, 1926
2- Cavilaelaps Fonseca, 1935
Gamasides
160
Тип рода Cavllaelaps bresslaui Fonseca, 1935
3. Echinolaelaps Ewing, 1929
Тип рода Laelaps echidninus Berlese, 1887
4. Eubrachylaelaps Ewing, 1929 (= Cyclolaelaps Ewing, 1931)
Тип рода Laelaps hollisteri Ewing, 1925
5. Eulaelaps Berlese, 1903 (= Hemilaelaps Hull, 1918)
a) Fulaelaps s. str.
Тип подрода Laelaps stabularis Koch, 1839
6) Pseudolaelaps Berlese, 1916
Тип подрода Laelaps (Hoplolaelaps) doderoi Berlese, 1910
6. Glgantolaelaps Fonseca, 1939
Тип рода Glgantolaelaps vitzthumi Fonseca, 1939
7. Haemolaelaps Berlese, 1910 (= Atrlcholaelaps Ewing, 1929 =
= Ischnolaelaps Fonseca, 1935)
Тип рода Laelaps (Haemolaelaps) marsuplalls Berlese, 1910
8. Longolaelaps Vitzthum, 1926
Тип рода Longolaelaps longulus Vitzthum, 1926
9. Mysolaelaps Fonseca, 1935
Тип рода Mysolaelaps parvlsplnosus Fonseca, 1935
10. Neolaelaps Hirst, 1926
Тип рода Liponyssus magnlstigmatus Vitzthum, 1918
11. Neoparalaelaps Fonseca, 1935
Тип рода Neoparalaelaps blspinosus Fonseca, 1935
12. Ophidilaelaps Radford, 1947
Тип рода Ophidilaelaps imphalensls Radford, 1947
13. Scissuralaelaps Womersley, 1945
Тип рода Scissuralaelaps nova-guinea Womersley, 1945
14. Ugandolaelaps Radford, 1942
Тип рода Ugandolaelaps protoxera Radford, 1942
Роды неопределенного положения
По особенностям строения нимф и самцов было выделено три
рода, которые оказалось невозможным включить в какое-либо под-
семейство, так как классификация семейства первоначально основы-
валась на строении самок. Редфорд (1942) сообщил, что он нашел
самок из Ligilaelaps и Mungosicola, но его рисунок самки Mungo-
sicola ugandae, безусловно, изображает не самку, а нимфу.
РОДЫ
1. Rad (новое название для Banksia Radford, 1942 (Parasitol., 34,
299), не Banksia Voigts et Oudemans, 1905)
Тип рода Laelaps longiseta Banks, 1909
2. Ligilaelaps Radford, 1942
Тип рода Eulaelaps evuingl Pearse, 1930
3. Mungosicola Radford, 1942
Тип рода Mungosicola ugandae Radford, 1942
il(> Глава III. Подотряд Mesostigmata
ОБЩИЕ ЗАМЕЧАНИЯ
Клещи Laelaptidae широко распространены по всему свету. Они
паразитируют как на беспозвоночных, так и на позвоночных живот-
ных и являются самыми обычными энтопаразитами млекопитающих.
Жизненный цикл варьирует, и у некоторых форм личиночная и ним-
фальная фазы выпадают. Пока нет указаний на то, что Laelaptidae
могут переносить болезни от животного к человеку. Однако при
столь большом числе видов и столь больших возможностях для пере-
носа возбудителя вполне вероятно, что в будущем роль Laelaptidae
в передаче заболеваний будет установлена.
Echinolaelaps echidninus (Berlese) переносит Hepatozoan muris
(Balfour) от крысы к крысе. Это простейшее использует клещей
в качестве окончательного хозяина. Оно проникает в тело крысы при
проглатывании ею зараженных клещей. Haetnolaelaps arcuatus (Koch)
переносит другое сходное простейшее Hepatozoan crlceti (Danilewsky)
среди хомяков (Cricetus frumentarius Noller).
Как уже указывалось, систематика семейства Laelaptidae крайне
трудна и еще недостаточно тщательно изучена в связи с обилием
включаемых в него видов. Приведенный список литературы может
послужить лишь введением в огромную массу имеющейся литературы.
ЛИТЕРАТУРА
Berlese А., 1904. Acari tnirmccofili, Redia, 1, 300—4744~pls. VII—XX,
Ewing H. E., 1925. New parasitic mites of the genus Laelaps, Proc. Ent. Soc.
Wash., 27, 1-7.
Fonseca F. da, 1939. Notas de acarcologia XXV. Os Laelaptidae gigantes,
parasitas de roedores sul-amcricanos; genero e especies novas (Acari), Mem.
Inst. Butantan., 12, 7—101.
Radford C. D., 1943. Genera and species of parasitis mites (Acarina), Para-
sit о I., 35, 58—81.
Vitzthum H. G., 1930. Mllben als Pesttrager, Zool. Jahrb. Syst., 60, 381—428.
Womersley H., 1937. Studies on Australian Acarina. Laelaptidae, Parasitol.,
29, 530-538.
THINOZERCONINA TRAGARDH, 1946
Дорзальный щиток разделен на большой проподосомальный щиток
и маленький задний щиток. Перитремы у обоих полов на спинной
стороне. Стернальный щиток у самок отсутствует или слабо склеро-
тизирован; исключение составляют Thinozerconidae, у которых хорошо
развиты югулярные щитки. Эпигинальный щиток узкий, несет одну
пару щетинок и не сочленяется с вентральным щитком. Половое
отверстие у самца расположено на стернальном щитке.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ THINOZERCON1NA
1. Югулярные щитки у самки хорошо развиты. Половое отверстие
у самца расположено между тазиками третьей пары ног . . .
.............................................Thinozerconidae
Thinozerconina
111
— Склеротизированные югулярные щитки у самки отсутствуют.
Половое отверстие у самца расположено между тазиками второй
пары ног........................................Dasyponyssidae
Thinozerconidae Halbert, 1915
(рис. 69)
ДИАГНОЗ
Thinozerconidae — клещи средней величины, имеющие удлиненно-
овальное тело, несколько заостренное сзади. Дорзальный щиток раз-
делен на две части и слегка прикрывает сверху перитремальные пла-
стинки. Эпигинальный щиток узкий, языковидный, с одной парой
Рис. 69. Thinozercon michaeli Halbert, 1915 (по Трсгорду, 1946).
Я—тритостернум самки; Б — щитки брюшной поверхности тела самки.
щетинок, сидящих на его заднем крае. Половое отверстие самца
маленькое и прикрыто двумя очень маленькими полукруглыми щит-
ками, которые спереди несут пару крепких щетинок.
РОДЫ
1. Thinozercon Halbert, 1915
Тип рода Thinozercon michaeli Halbert, 1915
2. Dithinozercon Berlese, 1916
Тип рода Thinozercon (Dithinozercon) halberti Berlese, 1916
ОБЩИЕ ЗАМЕЧАНИЯ
Это семейство еще очень мало изучено. Два входящие в него рода
монотипичны. Thinozercon распространен в странах Старого Света,
a Dithinozercon — в Новом Свете. Вероятно, будет открыто еще
много новых форм из этой группы.
ЛИТЕРАТУРА
Berlese А., 1916. Centuria terza di acari nuovi, Redia, 12, 295.
Halbert J. H., 1915. Clare Island Survey, part 39, 11, Proc. Roy. Irish Acad.,
31, 82.
112
Глава II/. Подотряд Mesostigmata
Dasyponyssidae Fonseca, 1940
(рис. 70—73)
ДИАГНОЗ
У обоих полов дорзальный щиток разделен. У самок перитремы
направлены дорзально и поперечно, располагаясь между передним и
задним отделами дорзального щитка, в то время как у самцов
Рис. 70. Dasyponyssus neivai Fonseca, 1940
(по Фонзека, 1940).
Самец (вид сверху).
перитремы на некотором протяжении идут вдоль переднего отдела
дорзального щитка. Ноги I у обоих полов крайне увеличены; эти
ноги более чем вдвое толще ног II—IV. В щетинках, покрывающих
тело, проявляется половой диморфизм: у самок щетинки крупнее,
а у самцов они обычные. Вентральный щиток у самцов разделен. Пер-
вые и вторые стернальные щитки слиты в единый стернальный щи-
ток, на котором расположено половое отверстие. Третьи и четвертые
стернальные щитки слиты попарно с каждой стороны, но по средней
линии они остаются разделенными. Вентральный щиток редуцирован
Рис. 71. Dasyponyssus neival Fonseda, 1940 Рис. 72. Dasyponyssus nelvai Fonseca, 1940
(по Фонзека, 1940). (по Фонзека, 1940).
Самец (вид снизу). Самка (вид сверху).
а Зак. 1785
114
Глава. III. Подотряд Mesostigmata
и несет лишь одну пару щетинок. Анальный щиток снабжен семью
парными и одной срединной непарной щетинкой, расположенной
у заднего края щитка.
Рис. 73. Dasyponyssus neival Fonseca, 1940
(по Фонзека, 1940).
Самка (вид снизу).
РОДЫ
1. Dasyponyssus Fonseca, 1940
Тип рода Dasyponyssus neival Fonseca, 1940
ОБЩИЕ ЗАМЕЧАНИЯ
Клещи Dasyponyssus neival были обнаружены в Бразилии на
броненосце. Этот вид в настоящей работе отнесен к Thinozerconina,
так как он обладает определенным морфологическим сходством
с Thinozercon. Однако этот клещ обнаруживает некоторое сходство
и с Gamasides и, может быть, его правильнее выделять в особую
группу. Необходимы еще дополнительные исследования, прежде чем
место этого семейства в системе будет окончательно определено.
ЛИТЕРАТУР А
Fonseca F. da, 1940. Notas de Acarcologia. XXIV. Dasyponyssus neival
gen. n. sp. n., acariano parasita de Euphractus sexcinctus (L.) Acari,
Dasyponyssidac fam. n., Rev. Ent. Rio de Janeiro, 11, 104—119, 5 figs.
biarlhrophallina
115
DIARTHROPHALLINA TRAGARDH, 1946
Пенис двуколенчатый, направлен назад, расположен в желобке
между тазиками ног третьей пары. Имеются престернальные щетинки.
Эпигинальный щиток сливается с вентральным и стернальным щитками
и не отделяется от вентрального щитка даже швом. Известно только
одно семейство.
Diarthrophallidae TrSgardh, 1946
(рис. 74, 75)
ДИАГНОЗ
Diarthrophallidae в дополнение к основным, уже упомянутым в диа-
гнозе подотряда признакам характеризуются рядом других особен-
Р и с. 74. Diarthrophallus quercus (Pearse et Wharton), 1936
(по Трегорду, 1946).
Нимфа (вид снизу).
постей. Ноги первой пары лишены коготков и присоски и превра-
щены в осязательные органы, а все остальные ноги имеют хорошо
8*
116
Глава 111. Подотряд Mesostigmaia
развитый амбулакральный аппарат. Щетинки на спинной стороне длин-
ные и жесткие, у живого клеща они торчат вверх. Дорзальный щи-
ток цельный.
Рис. 75. Diarthrophallus quercus (Pearse et Wharton), 1936
(по Трегорду, 1946).
Щитки брюшной поверхности тела; Л —самца; Б — самки.
РОДЫ
1. Diarthrophallus TrSg&rdh, 1946
Тип рода Urosejus quercus Pearse et Wharton, 1936
2. Brachytremella Tragardh, 1946
Тип рода Brachytremella spinosa TragSrdh, 1946
3. Passalobia Lombardini, 1926
Тип рода Passalobia quadtricaudata Lombardini, 1926
ОБЩИЕ ЗАМЕЧАНИЯ
Клещи Diarthrophallidae по внешнему виду очень сходны с Uropodidae.
Однако Трегорд (1946) указал на ряд существенных отличий. Все
клещи этого семейства были найдены на жуках из семейства Passali-
dae. Они обнаружены в Южной и Северной Америке и в Новой
Гвинее.
D. quercus Pearse et Wharton, 1936 обычен для жуков-пассалид
в Северной Каролине (Даргэм).
ЛИТЕРАТУРА
Tragardh I., 1946, Diarthrophalllna, a new group of Mesostigmata, found on
passalid beetles, Eniom. Medd., 24, 370—394.
Zerconina
117
ZERCONINA TRAGARDH, 1944
Эта группа очень близка к Gamasides, отличаясь от них лишь
положением полового отверстия самца в центре стернального щитка.
К этой группе относится только одно семейство, в связи с чем мор-
фологическая характеристика этой группы дается при характеристике
единственного семейства.
ДИАГНОЗ
Zerconidae Berlese, 1892
(рис. 76)
Zerconidae — мелкие клещи с плоским телом, задний конец кото-
рого расширен, что придает телу более или менее треугольную форму
Дорзальный щиток может быть
разделенным. Лапки ног I лишены
присоски, часто на них редуциро-
ваны и предлапка, в то время как
лапки всех остальных ног имеют
предлапку, присоску и коготки.
Вентро-анальный щиток крупный,
широкий. У самца половое отвер-
стие расположено в середине стер-
нального щитка и прикрыто двой-
ным щитком, отходящим от его
переднего края. Эпигинальный щи-
ток несет только пару щетинок
и не сливается с эндоподальными
щитками. Метастернальные щитки
гак слабо склеротизированы, что
их местонахождение можно опре-
делить только по положению ме-
тастернальных щетинок. Стерналь-
ный щиток имеет три обычные
пиры стернальных щетинок.
Рис. 76. Prozercon kochi Willmann,
1943 (по Бильману, 1943).
Самка (вид снизу).
РОДЫ
1. Zercon Koch, 1836
Тип рода Zercon dimidatus Koch, 1841
2. Parazercon Tragardh, 1931
Тип рода Zercon sarakensls Willmann, 1939
( = Zercon ornatus Tragardh, 1910, ноне Zercon ornatus Berlese,
1904)
3. Prozercon Willmann, 1943
Тип рода Zercon fimbriatus Koch, 1839
118
Глава 111. Подотряд Mesostigmata
4. Seiodes Berlese, 1887-
Тип рода Seiodes ursinus Berlese, 1887
5. Triangulazercon Jacot, 1938
Тип рода Zercon peltatus Koch, 1836
( = Zercon triangularis Koch, 1836)
6. Trizerconoides Jacot, 1938
Тип рода Zercon radiatus Berlese, 1910
ОБЩИЕ ЗАМЕЧАНИЯ
Клещи Zerconidae составляют часть обширной фауны, заселяющей
верхние слои почвы, подстилку, слоевища мха. Их хозяйственное и
экономическое значение не выяснено.
ЛИТЕРАТУРА
Jacot А. Р., 1938. The Geenton mites of Florida, Florida Ent., 21, 49—57.
Tragardh I., 1946. Outlines of a new classification of the Mesostigmata
(Acarina) based on comparative morphological data, Lands Univ.
K. Fysiogr. Sallsk. Handl., N. F-, 57, No 4, 1—37.
TRACHYTINA TRAGARDH, 1937
Трегорд (1941) опубликовал обзор этой группы (когорты по Тре-
горду) и разделил ее на три семейства, явно родственных друг.другу.
Поэтому диагнозы этих семейств даются вместе с определительной
таблицей Трегорда для семейств, а отдельный диагноз для каждого
семейства опускается. Во избежание повторений общие замечания
и список литературы приводятся также для группы в целом.
ДИАГНОЗ
/ *
Эта группа представляет собой промежуточное звено между груп-
пами Gamasides и Uropodina. Сочленения тазиков первой пары ног
сходны с таковыми у Gamasides: тазики не прикрывают тритостер-
нума своей расширенной основной частью. Половое отверстие по
положению весьма схоже, но не идентично таковому у Uropodina.
У самок метастернальные щитки не сливаются со стернальным и
вентральным щитками, а расположены кнаружи от эпигинального
щитка. У взрослых особей спинная поверхность покрыта нимфальной
шкуркой. Сохранение нимфальной шкурки при линьке несвойственно
Mesostigmata; во всем подотряде это характерно лишь для Trachy-
tina. Подобную же своеобразную особенность имеют и некоторые
клещи из группы Oribatel,
Trachytina
119
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ TRACHYTINA
(ПО ТРЕГОРДУ, 1941)
1. Эпигинальный щиток подвижно сочленен с вентральным щит-
ком ......................................................2
— Эпигинальный щиток слит с вентральным щитком . . . Polyaspidae
2. Метастернальные щитки узкие, удлиненные и расположены по
бокам эпигинального щитка.......................Trachytidae
— Метастернальные щитки почти полностью утрачены, остаются
только их зачатки в задних углах полового отверстия ....
...........................................Polyaspinidae
Р и с. 77. Trachytes aegrota
(Koch), 1841
(по Трегорду, 1941).
Самка (вид сверху).
СЕМЕЙСТВА И РОДЫ
Т г а с h у t id ае Т rag ard h, 1938
(рис. 77)
1. Trachytes Michael, 1894
Тип рода Celaeno aegrota Koch, 1841
2. Neoseius Oudemans, 1903 (? известна только фаза дейтонимфы)
Тип рода Uroseius nevus Oudemans, 1902
3. Uroseius Berlese, 1888
Тип рода Uropoda acuminata Koch, 1847
120
Глава III. Подотряд Mesostigmata
Pol у as р i d а е В е rl е se, 1917
(рис. 78)
1. Polyaspis Berlese, 1881
Тип рода Polyaspis patavinus Berlese, 1881
2. Calotrachytes Berlese, 1916
Тип рода Trachynotus fimbriatus Michael, 1908
3. Dipolyaspis Berlese, 1916
Тип рода Polyaspis {Dipolyaspis') sansonei Berlese, 1916
Рис. 78. Dipolyaspis sansonei Ber-
lese, 1916 (по Трегорду, 1941).
Самка (вид снизу).
Рис. 79. Polyaspinus cylindricus
Berlese, 1916 (по Трегорду, 1941).
Самка (вид снизу).
Р о 1 у a s р i п i d а е Tragardh, 1941
(рис. 79)
1. Polyaspinus Berlese, 1916
Тип рода Polyaspinus cylindricus Berlese, 1916
ОБЩИЕ ЗАМЕЧАНИЯ
О жизненном цикле этой интересной группы клещей известно еще
очень немногое. Трегорд находил их под корой гниющих пней;
Uropodina
121
вероятно, эти клещи обитают в скоплениях гниющих органических
остатков, богатых целлюлозой. Хозяйственное и медицинское значе-
ние Trachytina не установлено.
ЛИТЕРАТУРА
Tragardh I., 1941. Further contributions towards the comparative morphology
of the Mesostigmata, III. On the Polyaspididae, Berl. Zool. Bidrag. F.
Uppsala, 20, 345—357.
UROPODINA KRAMER, 1881
Uropodina (за исключением промежуточной группы родов, объеди-
няемых в семейство Discourellidae) характеризуются строением гнато-
сомы. Брюшная часть тела спереди вырезана, так что гнатосома и
тритостернум заключены в полость, дорзальная стенка которой обра-
зована стенкой тела; с нижней стороны гнатосому могут прикрывать
тазики передней пары ног. Эту полость обычно называют камеро-
стомом. Метастернальные щитки у Uropodina или сильно редуциро-
ваны (Urodiaspis и Uroplifella'), или совсем отсутствуют. Половое
отверстие самца, так же как эпигинальный щиток самца, располо-
жены в центре стернального щитка. Тритостернум в передней части
часто разделен на три лацинии. Дорзальный щиток у взрослых цель-
ный. На нижней поверхности тела находятся желобки для ног, в ко-
торые могут укладываться короткие ноги, плотно прижимающиеся
при этом к телу. Стигмы расположены против ног второй и третьей
пары. Перитремы обычно изогнуты соответственно гребням, ограни-
чивающим желобки для вкладывания ног. Хелицеры обычно на неко-
тором протяжении втянуты в идиосому и в некоторых случаях почти
достигают заднего конца тела.
К этой группе с уверенностью можно отнести лишь 12 семейств.
Положение остальных семейств, включаемых в эту группу, несколько
сомнительно. По первоначальному описанию семейство Trematuridae
Berlese, 1917 характеризуется наличием четырех стигм. Трегорд (1942)
показал, что Берлезе ошибался в отношении числа стигм, но сам
Трегорд, исследуя экземпляры Берлезе, не смог установить система-
тическое положение Trematura patavina (Canestrini), 1885. Клещи
семейства Discourellidae лишены камеростома и занимают промежу-
точное положение между Uropodina и Trachytina. Ниже приводится
с некоторыми изменениями определительная таблица семейств, со-
ставленная Трегордом (1944).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ UROPODINA
1. Камеростом имеется .................................... 2
— Камеростом отсутствует ......................Discourellidae
2. Дорзальный щиток покрывает только спинную поверхность тела 3
122
Глава III. Подотряд Mesostigmata
— Дорзальный щиток переходит на боковые и брюшную поверх-
ности тела, так что только центральная часть брюшной поверх-
ности тела не покрыта дорзальным щитком . . Coxequesomidae
3. На брюшной стороне тела позади нет краевых щитков ... 4
— На брюшной стороне тела позади имеется пара краевых щит-
ков Planodtscidae
4. Спинной краевой щиток имеется................................5
— Спинной краевой щиток отсутствует..Circocyllibanidae
Рис. 80. Расположение щитков спинной поверх-
ности тела у различных родов Uropodina
(по Трегорду, 1944, и Фитцтуму, 1935).
A — Phaulodiaspis', Б — Urodlaspls', В — Oodinychas',
P—Phaulodinychiis-, Д—Trematurella', E-Phyllodi-
nychas; )K—Eutrachytes-, 3—Cllliba; U — Urodlnychus.
5. Стигмы открываются против тазиков ног II или между тазиками
ног II и III..............................................7
— Стигмы открываются против тазиков ног III...................6
6. Тритостернум прикрыт тазиками ног I.............Trematuridae
— Тритостернум не прикрыт тазиками ног I.............Cillibidae
7. Передние концы краевого щитка слиты впереди с дорзальным
щитком ...................................................... 8
— Передние концы краевого щитка не слиты с дорзальным щит-
ком .........................................Trematurellidae
8. Желобки для вкладывания ног имеются.......................10
—- Желобки для вкладывания ног отсутствуют....................9
9. Самостоятельный задний дорзальный щиток имеется Eatrachytidae
Uropodina
123
— Задний дорзальный щиток неразличим.................Prodinychidae
10. Задний дорзальный щиток отсутствует.........................11
— Задний дорзальный щиток имеется.....................Urodiaspidae
11. Краевой щиток слит с дорзальным щитком......................12
— Краевой щиток слит с вентральным щитком . . Trachyuropodidae
12. Краевой щиток полностью окружает дорзальный щиток . . 13
— Краевой щиток сзади редуцирован.................Phaulodinychidae
13. Краевой щиток по внутреннему краю с фестонами . . Urodinychidae
— Фестоны на краевом щитке отсутствуют..................Uropodidae
Для большей наглядности на рис. 80 приведено схематическое
изображение дорзальных щитков для наиболее характерных родов.
Discourellidae о
(новое название для Protouropodidae Tragardh, 1941)
(рис. 81, 82)
ДИАГНОЗ
К Discourellidae относятся те роды, которые лишены камеростома,
прикрытого первой парой тазиков, но обладают эпигинальным щитком,
Рис. 81. Discourella то-
desta Leonard!, 1900
(по Трегорду, 1941).
Самка (вид снизу).
Рис. 82. Apionoseius lagenae-
formis (Berlese), 1904
(по Трегорду, 1941).
Самка (вид снизу).
характерным для подотряда Uropodina. Тритостернум у представителей
этого семейства необычен тем, что его основание расширено,
124
Глава III. Подотряд Mesostigmata
РОДЫ
1. Discourella Berlese, 1910
Тип рода Discopoma venusta Berlese, 1884
( = Discourella dis copo moi des Berlese, 1910)
2. Apinoseius Berlese, 1904
Тип рода Trachytes lagenaeformis Berlese, 1904
3. Polyaspidella Berlese 1910
Тип рода Polyaspidella berenicea Berlese, 1910
ОБЩИЕ ЗАМЕЧАНИЯ
Трегорд (1941) предложил для этого семейства название Protou-
ropodidae. В настоящей работе для группы предлагается название
Discourellidae и род Discourella Berlese, 1910 принимается за тип
для семейства. Семейство Discourellidae, очевидно, является проме-
жуточным между Uropodina и Trachytina. Виды этого семейства были
найдены в Канаде, Австрии и на Яве; возможно, что клещи этого
семейства распространены очень широко. По образу жизни они сходны
с большинством клещей группы Uropodina. Хозяйственное и меди-
цинское значение Discourellidae не выяснено.
ЛИТЕРАТУРА
Tragardh I., 1941. Further contributions towards the comparative morphology
of the Mesostigmata, III. On the Polyaspididae, Berl. Zool. Bidrag. Uppsala,
20, 345—357.
ДИАГНОЗ
Coxequesomidae Sellnick, 1926
(рис. 83)
Тритостернум прикрыт первой парой ног. Желобки для вклады-
вания ног плоские. Половое и анальное отверстия окружены каждое
Рис. 83. Coxequesoma
colleglanorum Sellnick,
1926 (по Зельнику, 1926).
Самка (вид снизу).
Uropodina
125
отдельным щитком. Дорзальный щиток заходит на нижнюю сторону
тела и покрывает все животное, за исключением центральной части
нижней поверхности тела.
РОДЫ
1. Coxequesotna Sellnick, 1926
Тип рода Coxequesotna colieglanorum Sellnick, 1926
2. Antennequesoma Sellnick, 1926
Тип рода Antennequesoma reichenspergeri Sellnick, 1926
ОБЩИЕ ЗАМЕЧАНИЯ
Все Coxequesomidae являются мирмекофилами. Их хозяйственное
и медицинское значение не выяснено.
ЛИТЕРАТУРА
Sellnick М, 1926. Alguns novos Acaros (Uropodidae) myrmecophiles e
termitophilus, Arch. Mas. Rio de Janeiro, 26, 29—56, 27 figs.
ДИАГНОЗ
Planodiscidae Sellnick, 1926
(рис. 84)
Тритостернум прикрыт тазиками первой пары ног. Желобки для
вкладывания ног хорошо развиты. Анальное и половое отверстия
Р и с. 84. Pianodiscus squamaftim Sel-
lnick, 1926 (по Зельнику, 1926).
Самка (вид снизу).
f26 Глава III. Подотряд Mesostigmaia
расположены на неразделенном вентральном щитке. Имеются брюш-
ные краевые щитки, расположенные по бокам заднего конца тела.
Дорзальный щиток не окружен спинным краевым щитком.
РОДЫ
1. Planodiscus Sellnick, 1926
Тип рода Pianodiscus squamatum Sellnick, 1926
ОБЩИЕ ЗАМЕЧАНИЯ
Хозяйственное и медицинское значение клещей Planodiscidae не
выяснено. Представители этого семейства связаны с муравьями.
ЛИТЕРАТУРА
Sellnick М., 1926. Alguns novos Acaros (Uropodinae) myrmecophilos e ter-
mitophilus, Arch. Mas. Rio de Janeiro, 26, 29—56, 27 figs.
Circocyllibanldae Sellnkk, 1926
(рис. 85)
ДИАГНОЗ
Тритостернум прикрыт тазиками первой пары ног. Желобки для
вкладывания ног хорошо развиты. Половое и анальное отверстия
Рис. 85. Circocylliba earner ata
Sellnick, 1926 (по Зельнику, 1926)
Самка (вид снизу).
расположены на отдельных щитках. Дорзальный щиток не окружен
спинным краевым щитком.
РОДЫ
1. Circocylliba Sellnick, 1926
Тип рода Circocylliba camerata Sellnick, 1926
Uropodina
127
ОБЩИЕ ЗАМЕЧАНИЯ
С. camerata является мирмекофилом; его хозяйственное и меди-
цинское значение не выяснено.
литература
Sellnick М., 1926. Alguns novos Acaros (Uropodidae) myrmecophilos e ter-
mitophilus. Arch. Mus. Rio de Janeiro, 26, 29—56, 27 figs.
Trematuridae Berlese, 1917
(рис. 86)
ДИАГНОЗ
Тритостернум прикрыт тазиками первой пары ног. .Желобки для
вкладывания ног сильно развиты. Дорзальный щиток окружен
Рис. 86. Trematara patavina
(Canestrini), 1885
(по Трегорду, 1942).
Часть брюшка; видны стигйа
и перитрема.
краевым щитком, имеющим ровный внутренний край. Стигмы откры-
ваются в желобке для ног III. Щитки имеют хорошо выраженную
скульптуру.
РОДЫ
1. Trematara Berlese, 1917
Тип рода Uropoda patavina G. Canestrini, 1885
ОБЩИЕ ЗАМЕЧАНИЯ
Трегорд (1942) изучал экземпляры вида Т. patavina из коллекций
Берлезе. Он не смог достаточно хорошо описать их морфологию и
поэтому в опубликованном им в 1944 г. обзоре не включил это
семейство в группу Uropodina. Мы включаем здесь в группу Uropo-
dina семейство Trematuridae, а также три предыдущих семейства,
предложенных Зельником в 1926 г., потому что они включены в эту
группу в определительных таблицах Фитцтума за 1941 г. Хозяйст-
венное и медицинское значение Т. patavina не выяснено.
ЛИТЕРАТУРА
О
Triigardh I., 1942. Ueber die Trematurini Berlese (Acarina), Zool. Anz., 139,
109—112.
128
Глава III. Подотряд Mesostigmaia
ДИАГНОЗ
Дорзальный
Краевой щиток
Trematurellidae Tragardh, 1944
(рис. 80)
и вентральный щитки глубоко скульптурированы.
полностью отделен тонкой перепонкой от вентраль-
ного и дорзального щитков. Дорзальный щиток не разделен, но по
краевому щитку спереди проходит шов.
РОДЫ
1. Trematurella Tragardh, 1942
Тип рода Trematurella stylifera Tragardh, 1942
ОБЩИЕ ЗАМЕЧАНИЯ
Практическое значение Т. stylifera не выяснено.
ЛИТЕРАТУРА
Tragardh I., 1942. Ueber die Trematurini Berlese (Acarina), Zool. Anz.,
139, 109—112.
Cillibidae Tragardh, 1944
диагноз
(рис. 87)
Тритостернум не прикрыт тазиками ног I. Краевой щиток впереди
сплошной, но обособлен от спинного щитка. Задняя часть дорзаль-
Р и с. 87. Thrichocylllba
comata (Leonardi), 1895
(по Берлезе, 1904).
Самка (вид снизу).
ного щитка отделена от остальной части щитка швом. Стигмы рас-
положены против тазиков ног Ш. Желобки для вкладывания ног
хорошо развиты.
РОДЫ
1. Gilliba v. Heyden, 1827 ( = Discopoma G. et R. Canestrini, 1882)
Тип рода Notaspts cassideus Hermann, 1804
2. Thrichocylllba Berlese, 1904
Тип рода Discopoma pomata Leonardi, 1895
Uropodina
129
ОБЩИЕ ЗАМЕЧАНИЯ
Хозяйственное и медицинское значение этих клещей не выяснено.
ЛИТЕРАТУРА
Tragardh I.,1944. Zur Systematik der Uropodiden, Enft Tidsk.,G5, 173—186.
Eutrachytidae Trftgardh, 1944
(рис. 80)
ДИАГНОЗ
У клещей этого семейства краевой щиток слит впереди с дор-
зальным щитком. Сзади дорзальный щиток вполне самостоятелен.
Желобки для вкладывания ног не развиты.
РОДЫ
1. Eutrachytes Berlese, 1914
Тип рода Celaeno tru.nca.ta Berlese, 1888
ОБЩИЕ ЗАМЕЧАНИЯ
Хозяйственное и медицинское значение клещей этого семейства
не выяснено.
ЛИТЕРАТУРА
Tragardh 1., 1944. Zur Systematik der Uropodiden, Ent. Tidsk., 65, 173—186.
Prodinychidae Berlese, 1916
(рис, 88)
ДИАГНОЗ
Тритостернум прикрыт тазиками ног I. Желобки для вкладывания
ног или отсутствуют, или выражены очень слабо. Краевой щиток
сзади сплошной, а спереди слит с дорзальным щитком. Если же
дорзальный щиток обособлен, то краевой щиток присоединен к вен-
тральному щитку.
РОДЫ
1. Prodinychus Berlese, 1913
Тип рода Dlnychus fimicolus Berlese, 1903
2. Clausiadinychus Sellnick, 1930
Тип рода Clausiadinychus cristatus Sellnick, 1930
3. Dentidinychus Sellnick, 1926
Тип рода Dentidinychus zikani Sellnick, 1926
4. - Dlnychus Kramer, 1882
Тип рода Dinychus perforatus Kramer, 1882
6. Discotrachytes Berlese, 1916
Тип рода Discotrachytes splendidiformis Berlese, 1916
9 Зак. 1785
130
Глава III. Подотряд Mesostigmata
6. Metadlnychus Berlese, 1916
Тип рода Metadlnychus argaslform.es Ber'ese, 1916
7. Trlchodinychus Berlese, 1916
Тип рода Uropoda vulplna Ber'ese, 1888
8. Urolaelaps Berlese, 1916
Тип рода Urolaelaps macropl Berlese, 1916
Рис. 88. Prodinychas for-
micarius Vitzthum, 1925
(по Фитцтуму, 1940).
Самец (вид снизу).
ОБЩИЕ ЗАМЕЧАНИЯ
Фитцтум (1931) предложил для этого семейства название Diny-
chidae. Хозяйственное и медицинское значение этого семейства не
установлено-
ЛИТЕРАТУРА
Tragardh I., 1944. Zur Systematic der Uropodiden, Ent. Tidsk., 65, 173—186.
Urodiaspidae Tragardh, 1944
. . . (рис. 89)
ДИАГНОЗ
Краевой щиток впереди слит с дорзальным щитком; при препа-
ровке обнаруживается, что краевой щиток подвешен к дорзальному
щитку. Имеется крупный и вполне самостоятельный задний дорзаль-
ный щиток. Метастернальные щитки также имеются, но они очень
малы и располагаются у задних углов эпигинального щитка.
Uropodina
131
РОДЫ
1. Urodlaspis Berlese, 1916
Тип рода Uropoda tecta Kramer, 1876
2. Diurodinychus Berlese, 1916
Тип рода Urodlaspis {Diurodinychus') rectangulovatus Berlese, 1916
Рис. 89. Urodlaspis tecta (Kramer), 1876 (по Берлезе, 1887).
А — самка (вид сверху); Б — самка (вид снизу).
ОБЩИЕ ЗАМЕЧАНИЯ
Экономическое и медицинское значение этого семейства не вы-
яснено.
ЛИТЕРАТУРА
Tragardh I., 1944. Zur Systematik der Uropodiden, Ent. Tidsk., 65, 173—186.
Trachyuropodidae Berlese, 1917
ДИАГНОЗ
(рис. 90)
У этих клещей тритостернум скрыт тазиками ног I. Желобки
для вкладывания ног хорошо развиты. Все щитки явно скульпти-
ронаны. Краевой щиток слит с вентральным щитком. Задний дорзаль-
ный щиток отсутствует.
РОДЫ И ПОДРОДЫ
1. Trachyuropoda Berlese, 1888 (— Michaellella Berlese, 1904)
a) Trachyuropoda s. str.
Тип подрода Trachyuropoda festiva Berlese, 1888
6) Dinychura Berlese, 1913
Тип подрода Trachyuropoda (Urojanetia) rectangula Ber-
lese, 1913 • '
9*
Глава III. Подотряд Mesostigmata
в) Urojatietla Berlese, 1913 Janetiella Berlese, 1904 nomen
praeoccupatum)
Тип подрода Uropoda coccinea Michael, 1891
2. Cephalouropoda Berlese, 1903
Тип рода Uropoda berlesiana Berlese, 1887
3. Comyditiychus Berlese, 1917
Тип рода Uropoda capatmedusae Berlese, 1901
4. Deralophorus G. Canestrini, 1897
Тип рода Deralophorus chyzeri Canestrini, 1897
Рис. 90. Trachyuropoda crustosa Vitzthum, 1926
(по Фитцтуму, 1926).
А — самка (вид сбоку); fi —самка (вид снизу).
5. Dinychopsis Berlese, 1916
Тип рода Dinychopsis fractus Berlese, 1916 (не Dirtychus appen-
dlculatus Berlese, 1910)
6. Leonardiella Berlese, 1904
Тип рода Uropoda canestrtniana Berlese, 1891
7. Neodiscopotna Vitzthum, 1941 (== Discopoma G. et R. Canestrini
в понимании Berlese, 1904)
a) Neodiscopotna s. str.
Тип подрода Uropoda splendida Kramer, 1882
6) Capitodiscus Vitzthum, 1931 (=Cephalodiscus Berlese, 1916
nomen praeoccupatum)
Тип подрода Discopoma venusta Berlese, 1884
в) Crinitodiscus Sellnick, 1931
Тип подрода Discopoma (Crinitodiscus') beierl Sellnick, 1931
r) Olodiscus Berlese, 1917
Тип рода Discopoma Integra Berlese, 1910
д) Phymatodiscus Berlese, 1917
Тип рода Discopoma miranda Berlese, 1904
Uropodina .
133
8. Uropolyaspls Berlese, 1903
Тип рода Uropoda hamulifera Mihael, 1894
9. Urospina Sellnick, 1931
Тип рода Uropoda plana Sellnick, 1931
ОБЩИЕ ЗАМЕЧАНИЯ
Подобно представителям многих других семейств Uropodina,
клещей Trachyuropodidae часто находят вместе с муравьями. Их
хозяйственное и медицинское значение не выяснено.
ЛИТЕРАТРУА
Tragardh I., 1944. Zur Systematik der Uropodidcn, Erd. Tidsk.,t>5, 173—186.
Phaulodinychidae Berlese, 1917
(рис. 91)
ДИАГНОЗ
Тритостернум прикрыт тазиками ног I. Желобки для вкладывания
ног хорошо развиты. Вентральный щиток цельный, он не сливается
Рис. 91. Phaulodiaspis advena (TragArdh), 1912 (по Трегорду, 1912).
А —самка (вид сверху); Б —самка (вид снизу); В —тритостернум.
с краевым или с дорзальным щитком. Краевой щиток в задней
части редуцирован.
РОДЫ
I. Phauloclinychus Berlese, 1903 (= Halouropoda Halbert, 1915)
Тип рода Phaulodinychus repletus Berlese, 1903
2. DiphaulocylUba Vitzthum, 1925
Тип рода Phaulocylllba amplior Berlese, 1923
134
Глава Hi. Подотряд Mesostigmata
3. PhaulocyUiba Berlese, 1903
Тип рода PhaulocyUiba ventricosa Berese, 1903
4. Phaulodiaspis Vitzthum, 1925
Тип рода Urodiscella advena Tragardh, 1912
ОБЩИЕ ЗАМЕЧАНИЯ
Хозяйственное и медицинское значение этого семейства не вы-
яснено.
ЛИТЕР АТУ РА
Tragirdh I., 1944. Zur Systematik der Uropodiden, End. Tidsk., 65, 173—136.
Urodinychidae Berlese, 1917
ДИАГНОЗ
(рис. 92)
Тритостернум прикрыт тазиками ног I. Имеются хорошо разви-
тые желобки для вкладывания ног. Краевой щиток сплошной позади;
его внутренний край снабжен фестонами.
Рис. 92. Urodinychus polyphemus Vitzthum, 1935
(по Фитцтуму, 1935).
Л —самец (вид сверху); 5 — самец (вид снизу)»
РОДЫ и ПОДРОДЫ
1. Urodinychus Berlese, 1903
• a) Urodinychus s. str.
Тип подрода Uropoda carinata Berlese, 1888
6) Leiodinychus Berlese, 1917
Тип подрода Uropoda krameri G. et R. Canestrini, 1882
в) Macrodinychus Berlese, 1917
Тип подрода Urodinychus parallelepipedus Berlese, 1916
2. Calurodlscus Radford, 1950 (= Urodiscus Berlese, 1916, nomen
praeoccupatum)
Тип рода Urodiscus obesus Berlese, 1916
Uropodina
135
ОБЩИЕ ЗАМЕЧАНИЯ
Хозяйственное и медицинское значение клещей этого семейства
не выяснено.
ЛИТЕРАТУРА
Trag&rdh I., 1944. Zur Systematik der Uropodiden, Ent. Tidsk., 65, 173—186.
Uropodidae Berlese, 1917
(рис. 93)
ДИАГНОЗ
Тритостернум прикрыт тазиками ног I. Желобки для вкладывания
ног хорошо развиты. Краевой щиток впереди сливается с дорзаль-
ным. щитком; внутренняя поверхность краевого щитка не разделена
Рис. 93. Uropoda pearsei Wharton, 1938 (по Уартону, 1938).
Д —щитки брюшной поверхности тела самки; самец (вид снизу).
на фестоны. Щитки кажутся ровными как зеркало, даже когда они
покрыты более или менее тонким точечным рисунком.
РОДЫ И ПОДРОДЫ
1. Uropoda Latreille, 1806 (==Nummulus Berlese, 1884)
Тип рода Acarus orbicularis Miiller, 1776 (== Acarus 'vegetans
Latreille, 1806 = Discopoma romana G. et R. Canestrini, 1882
в понимании Berlese, 1884, но не = Acarus vegetans
DeGeer, 1768)
136 Глава Ш. Подотряд Mesostigmata
2. Centrouropoda Berlese, 1916
Тип рода Uropoda rhotnbogyna Berlese, 1910
3. Cyclacarus Ewing, 1933
Тип рода Cyclacarus aberrans Ewing, 1933
4. Cylllbula Berlese, 1916
Тип рода Cilllbatw {Cylllbula) infumata Berlese, 1916
5. Encylliba Berlese, 1917
Тип рода Cllliba bordagei Oudemans, 1912
6, Fuscuropoda Vitzthum, 1924
Тип рода Notaspls marglnatus C.. L. Koch, 1839
7. Marglnura Sellnick, 1926
Тип рода Marglnura adhaerens Sellnick, 1926
8. Metagynella Berlese, 1919
Тип рода Metagynella paradoxa Berlese, 1919
9. Olouropoda Berlese, 1916
Тип рода Uropoda (Olouropoda) nitidissima Berlese, 1916
10. Oplitls Berlese, 1884 (= Uroplitella Berlese, 1904)
Тип рода Uropoda paradoxa G. Canestrini et Berlese, 1884
11. Pseudouropoda Oudemans, 1936 {—Notaspls C. L. Koch, 1836,
но не Notaspls Hermann, 1804; — Uropoda Berlese, 1917)
a) Pseudouropoda s. str.
Тип подрода Acarus vegetans DeGeer, 1768
6) Calouropoda Berlese, 1916
Тип подрода Uropoda pergltta Berlese, 1904
в) Trichouropoda Berlese, 1916
Тип подрода Uropoda longiseta Berlese, 1888
12. Urodlscella Berlese, 1903
Тип рода Uropoda ricasoliana Berlese, 1889
13. Uroobovella Berlese, 1903
a) Uroobovella s. str.
Тип подрода Uropoda obovata G. Canestrini et Berlese, 1884
6) Urocyclella Berlese, 1913
Тип подрода Uroobovella (Urocyclella) parvula Berlese, 1913
14. Uroplltana Sellnick, 1926
Тип рода Uroplltana acinaca Sellnick, 1926
15. Urosternella Berlese, 1903
Тип рода Uropoda {Urosternella) foraminlfera Berlese, 1903
Сомнительные роды
1. Liponlssus Kolenati, 1858 (не Llponyssus различных авторов)
Тип рода Dermanissus setosus Kolenati, 1856
2. Neuterla Oudemans, 1905
Тип рода Uropoda tropica Oudemans, 1905
3. Paulitzia Oudemans, 1915
Тип рода Uropoda africana Oudemans, 1905
Celaenopsina 137
ОБЩИЕ ЗАМЕЧАНИЯ
Подобно другим семействам хозяйственное и медицинское зна-
чение Uropodidae не выяснено. Они распространены по всему свету
и обычно связаны с насекомыми особенно в нимфальных фазах.
Возможно, что эти клещи не приносят вреда своим хозяевам.
ЛИТЕРАТУРА
Tragardh I., 1944. Zur Systematik der Uropodiden,Ent. Tidsk.,&5, 173—186.
CELAENOPSINA TRAGARDH, 1938
Для клещей этой группы характерно наличие пары боковых щит-
ков, лежащих на брюшной поверхности тела и представляющих
собой как бы продолжение вперед вентрального щитка. Эти боковые
щитки принимают функцию эпигинального щитка, и последний реду-
цируется или полностью утрачивается. Кроме того, у Celaenopsina.
не развит и срединный щиток. Метастернальные щитки утрачивают
связь со стернальными щитками I—III, делаются самостоятельными,
в некоторых случаях сильно уменьшаются в размерах. Метастерналь-
ные щитки значительно варьируют внутри группы. У Celaenopsidae
они крупные, неприкрытые, самостоятельные и несут типичную ще-
тинку и пору; у Lobogynold.es и некоторых других родов метастер-
нальные щитки маленькие, скрытые, щетинки на них отсутствуют,
но имеются две поры, а у Lobogynium и некоторых других родов,
метастернальные щитки сливаются со стернальным щитком.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ CELAENOPSINA
1. Метастернальные щитки частично или полностью прикрыты бо-
ковыми щитками .......................................... 2
— Метастернальные щитки не прикрыты боковыми щитками . .
..........................................Celaenopsidae
2. Передние боковые доли вентрального щитка (боковые щитки).
не отделены от вентрального щитка..............................3
— Боковые щитки отделены от вентрального щитка швом; эпиги-
нальный щиток редуцирован и заметен только у заднего соеди-
нения боковых щитков...................................Diplogyniidae
3. Анальный щиток отделен от вентрального щитка. . Euzerconidae
— Анальный щиток слит с вентральным щитком . . Schizogyniidae
Diplogyniidae Trdgardh, 1941
(рис. 94)
ДИАГНОЗ
Клещей Diplogyniidae можно распознать по наличию пары тре-
угольных боковых щитков, которые по наружному краю подвижно
связаны с вентральным щитком. Боковые щитки впереди соприка-
саются друг с другом по средней линии тела, а сзади отделены друг
138
Глава III. Подотряд Mesostigmata
от друга остатком эпигинального щитка. Метастернальные щитки
редуцированы и частично прикрыты передними краями боковых щит-
ков. Кзади от последних вплоть до заднего конца тела простирается
вентральный щиток, с которым сливается анальный щиток. По бокам
от вентрального щитка расположена пара метаподальных щитков,
Рис. 94. Dlplogyniam tropica
(Oudemans), 1927 (по Удемансу, 1928).
А — самка (вид сверху); /> —самка (вид снизу).
Б
которые доходят до заднего конца тела. У самца половое отверстие
расположено впереди на стернальном щитке и защищено по бокам
парной престернальной щетинкой. Трегорд (1950) описал четырнад-
цать новых родов Diplogyniidae и разделил это семейство на пять
подсемейств. Четыре подсемейства являются монотипическими, тогда
как Diplogyniinae содержит все остальные роды. Ниже приводится
определительная таблица подсемейств Diplogyniidae, заимствованная
у Трегорда (1950).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ DIPLOGYNIIDAE
1. Анальный щиток отделен от вентрального щитка................
............................................Neodiplogyniinae
— Анальный щиток не отделен от вентрального щитка . . . . . 2
2. Вдоль краев тела тянется узкая полоска из мелких шипиков . .
.........................................Heterodiplogyniinae
— Краевая полоска из шипиков отсутствует ......... 3
Celaenopsma
139
3. Спинная сторона тела густо покрыта мелкими щетинками . . .
..........................................Trichdiplogyniinae
— Спинная сторона тела лишена густого покрова щетинок ... 4
4. Дорзальный щиток несет ряд крючковидных щетинок...............
..............................................Meinertulinae
— Дорзальный щиток не имеет таких щетинок . . . Diplogyniinae
РОДЫ
1. Diplogynium Canestrini, 1889 (= Anoplocelaeno Berlese, 1910)
Тип рода Diplogynium acuminatum Canestrini, 1889
2. Antennocelaeno Berlese, 1903
Тип рода Antennophorus braunsl Wasmann, 1902
3. Brachysternum Tragardh, l£50
Тип рода Brachysternum acuminatum Tr&gdrdh, 1950
4. Cryptometasternum Tragardh, 1950
Тип рода Cryptometasternum natalense Tragardh, 1950
5. Diplogynlella Tragardh, 1950
Тип рода Diplogyniella levinseni Tragdrdh, 1950
6. Diplogyniopsls Tragardh, 1950
Тип рода Diplogyniopsls multidentata TragSrdh, 1950
7. Heterodiplogynium Tragardh, 1950
Тип рода Heterodiplogynium vestitum Tragardh, 1950
8. Lobogynioides Tragardh, 1950
Тип рода Lobogynioides obtusum TragSrdh, 1950
9. . Lobogynium Tragardh, 1950
Тип рода Lobogynium rotundatum Tragardh, 1950
10. Megachaetochela Tragardh, 1950
Тип рода Megachaetochela warreni Tragardh, 1950
11. Meinertula Tragardh, 1950
Тип рода Meinertula hamifera TragSrdh, 1950
12. Microdiplogynium Tragardh, 1950
Тип рода Microdiplogynium retlculatum Tragardh, 1950
13. Neodiplogynium Tragardh, 1950
Тип рода Neodiplogynium schubarti Tragdrdh, 1950
14. Passalacarus Pearse et Wharton, 1936
Тип рода Passalacarus sylvestrls Pearse et Wharton, 1936
15. Schizodiplogynium Tragardh, 1950
Тип рода Schizodiplogynium capillatum Tragardh, 1950
16. Trichodiplogynium Tragardh, 1950.
Тип рода Trichodiplogynium hlrsutum Tragardh, 1950
17. TrIdipIogynium Tragardh, 1950
Тип рода Tridlplogynium Inexpectatum TragSrdh, 1950
ОБЩИЕ ЗАМЕЧАНИЯ
Эти крупные красивые клещи являются эктопаразитами или ком-
менсалами крупных жуков, а возможно, и других насекомых. Их
хозяйственное и медицинское значение не установлено.
140
Глава III. Подотряд Mesostigmata
ЛИТЕРАТУРА
Pearse A. S. et al., 1936. The ecology of Passalus cornutus Fabricius, a beetle
that lives in rotting logs, Ecol. Monogr., 6, 455—490.
Tragardh I., 1946. Outline of a new classification of the Mesostigmata (Aca-
rina), based on comparative morphological data, K. Fysiogr. Siillsk.
Handl., N. F„ 57, № 4, 1—37.
Tragardh I., 1950. Studies on the Celaenopsidae, Diplogyniidae and Schizo-
gyniidae, Arkiv. for Zoologi, Ser. 2,' 1, № 25, 361—451.
Euzerconidae Tragardh, 1938
(рис. 95)
ДИАГНОЗ
Клещи Euzerconidae близки к . Diplogyniidae, но отличаются от
последних тем, что в семействе Euzerconidae боковые щитки у самок
Рис. 95. Euzercon latus (Banks), 1909.
Самка (вид снизу).
не отделены швом от вентрального щитка, вследствие чего половое
отверстие имеет Т-образную форму. Анальный щиток отделен от
вентрального.
РОДЫ
1. Euzercon Berlese, 1888
Тип рода Euzercon balzanl Berlese, 1888
Celaenopstna
141
ОБЩИЕ ЗАМЕЧАНИЯ
Представители этого семейства связаны с жуками и широко рас-
пространены по всему свету. Их практическое значение неясно.
ЛИТЕРАТУРА
Tragardh I., 1946. Outline of a new classification of the Mesostigmata (Aca-
rina), based on a comparative morphological data, AT. Fysiogr. Sdllsk.
Handl., N. F., 57, № 4, 1—37.
Celaenopsidae Berlese, 1892
(рис. 96)
ДИАГНОЗ
Celaenopsidae напоминают клещей предыдущего семейства, но
эпигинальный щиток у них замещен боковыми щитками, которые по
средней линии сливаются
друг с другом, а сзади —
с вентральным щитком. Ме-
тастернальные щитки круп-
ные и обособлены; они имеют
форму прямоугольника и вы-
полняют роль клапанов поло-
вого отверстия. Последнее
расположено позади мета-
стернальных щитков.
Рис. 96. Celaenopsis cusptdata
(Kramer), 1876
(по Фитцтуму, 1940).
Самка (вид снизу).
РОДЫ
1. Celaenopsis Berlese, 1886 (== Antennocelaeno Berlese, 1903)
Тип рода Gamasus cuspidatus Kramer, 1876
"2. Ceratocelaenopsls Tragardh, 1950
Тип рода Ceratocelaenopsls 'womersleyl TrSgSrdh, 1950
3. Dinocelaeno Oudemans, 1936
Тип рода Gamasus gigas Duges, 1834
4. Neocelaeno Berlese, 1910
Тип рода Celaenopsis cryptodonata Berlese, 1901
5. Pleuronectocelaeno Vitzthum, 1926
Тип рода Celaenopsis {Pleuronectocelaeno) austrlaca Vitzthum, 1926
142
Глава III. Подотряд Mesostigmata
ОБЩИЕ ЗАМЕЧАНИЯ
Хозяйственное и медицинское значение этих клещей, так же как
и других представителей Celaenopsina не выяснено.
ЛИТЕРАТУРА
Tragardh I,, 1946. Outline of a new classification of the Mesostigmata (Aca-
rina), based on comparative morphological data, K, Fysiogr. Sllllsk. Handl.,
N. F., 57, № 4, 1—37.
Traghrdh I., 1950. Studies on the Celaenopsidae, Diplogyniidae and Schizo-
gyniidae, Arkiv. for Zoologi, Ser. 2, 1, № 25, 361—451.
Schizogyniidae Tragardh, 1950
ДИАГНОЗ
(рис. 97)
Клещи этого семейства в общем близки к семейству Diplogyniidae,
но отличаются от последних тем, что у Schizogyniidae с вентральным
щитком слиты и боковые, и аналь-
ный щитки. Боковые щитки не
сливаются друг с другом по сред-
ней линии, как это имеет место
у Celaenopsidae. Стернальный щи-
ток несет посередине гребень,
имеющий форму полумесяца. Сам-
цы не описаны.
Рис. 97. Schizogynium intermedium
Tragardh, 1950 (по Трегорду, 1950).
Самка (вид снизу).
РОДЫ
1. Schizogynium Tragardh, 1950
Тип рода Schizogynium intermedium TragSrdh, 1950
ОБЩИЕ ЗАМЕЧАНИЯ
Трегорд (1950) рассматривает это семейство как промежуточное
между Euzerconidae и Diplogyniidae. Подобно другим представителям
Fedrizziina
143
Celaenopsina виды этого семейства паразитируют на жуках. Оба вида,
объединяемые в это семейство, описаны из Африки.
ЛИТЕРАТУРА
Tragardh I., 1950, Studies on the Celaenopsidae, Diplogyniidae and Schizo-
gyniidae, Arkiv. fdr Zoologi, Ser. 2, 1, № 25, 301—451.
FEDRIZZIINA TRAGARDH, 1937
Самки этой группы легко распознаются по наличию у них сре-
динного щитка в дополнение к эпигинальному или взамен последнего.
Срединный щиток представляет собой склеротизированную часть стенки
влагалища. Если метастернальные щитки имеются, то они оказываются
слитыми со стернальным щитком. Боковые щитки имеются или же
утрачиваются вторично. У самца половое отверстие лежит в центре
стернального щитка; последний прикрыт щитком, прикрепленным к его
переднему краю.
Fedrizziina для удобства разделяют на семь семейств. Определи-
тельная таблица для этих семейств, представляющая собой несколько
видоизмененную таблицу Трегорда (1946), приводится ниже.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ FEDRIZZIINA
1. Стернальный и метастернальные щитки склеротизированы . . 2
— Стернальный и метастернальный щитки у самки не склеротизи-
рованы; их положение выдает только наличие стернальных и
метастернальных щетинок...........................Parantennulidae
2. Боковые щитки обособлены.............,,....................3
•— Боковые щитки слиты друг с другом по средней линии, средин-
ный щиток редуцирован ...........................Syngynapsidae
3. Срединный щиток не сливается со стернальным щитком ... 4
— Срединный щиток сливается со стернальным щитком .....
.............................................Cercomegistidae
4. Боковые щитки полностью склеротизированы..................5
— Боковые щитки склеротизированы только вдоль внутреннего края
.............................................Antennophoridae
5. Боковые щитки треугольные....................Paramegistidae
— Боковые щитки вытянуты в длину.............. Fedrizziidae
Parantennulidae Willmann, 1940
(рис. 98)
ДИАГНОЗ
Дорзальный щиток не разделен; у самки стернальный и метастер-
нальный щитки отсутствуют. На лапках ног I нет амбулакрального
аппарата, в то время как лапки ног II, III и IV имеют большие при-
соски и тонкие коготки. Хелицеры клешневидные..
144
Глава III. Подотряд Mesostigmata
РОДЫ
1. Parantennulus Berlese, 1904
Тип рода Antennophorus scolopendrarum Berlese, 1886
2. Diplopodophilus Willmann, 1940
Тип рода Dlplopodophilus antennophoroldes, Willmann, 1940
Рис. 98. Dlplodophilus antennophoroldes Willmann, 1940
(по Бильману, 1941).
А — самец (вид снизу); Б — самка (вид снизу).
ОБЩИЕ ЗАМЕЧАНИЯ
Этих клещей находят на многоножках и на жужелицах. Экономи-
ческое и медицинское значение их не выяснено.
ЛИТЕРАТУРА
Willmann С, 1940. Neue Milben aits HOhlen der Balkanhalbinsel, gesam-
melt von Prof. Dr. K. Absolum, Briinn, Zool. Anz., 130, 209—218.
Willmann C., 1941. Die Acari der Hohlen der Balkanhalbinsel. Studien aus
dem Gebiete der Allgemeinen Karstforschung, der Wissenschaftlichen
Hohlenkunde. der Eiszeitforschung und den Nachbargebieten, Biological
series, № 8, 1—80.
ДИАГНОЗ
Syngynaspidae Tragardh, 1938
(рис. 99)
Боковые щитки у самки сливаются по средней линии. Срединный
щиток редуцирован.
РОДЫ
1. Syngynaspis
Тип рода Syngynaspis trdgardhi n. sp.
ОБЩИЕ ЗАМЕЧАНИЯ
В 1937 г. Трегорд описал новый род Syngynaspis, но не да*л
специального названия его единственному виду, по которому им был
Fedrizzitna
145
выделен род. Этот вид, описанный
Zool., 29, № 11, 6, рис. 9), назван
Рис. 99. Syngynaspis tret-
О
gardhi, п. sp. (по Трегорду,
1946).
Щитки брюшной поверхности тела
самки.
Трегордом в 1937 г. {Ark.
нами Syngynaspis trdgardhi.
Семейство Syngynaspidae для названного вида установлено Трегордом
в 1938 г.
ЛИТЕРАТУРА
Tragardh I.,
1—8.
1937. Zur Systcmatik der Mesostigmata, Ark. Zool., 29, № 11,
Cercomegistidae Tragardh, 1938
(рис. 100)
со стернальным щитком. Боковые щитки
они
покрывают эпигинальный щиток.
щиток слит
ДИАГНОЗ
Срединный
удлиненные и узкие, сзади
Рис. 100. Cercomegistus sp.
(по Трегорду, 1937).,
Щитки брюшной поверхности
самки.
ю Зак. 1785
146
Глава Ill. Подотряд Mesostigmata
РОДЫ
1. Cercomegistus Berlese, 1914
Тип рода Cercomegistus bruckianus Berlese, 1914
2. Neo-Oudemansla
Тип рода Neo-Oudemansia trtigardhi n. sp.
ОБЩИЕ ЗАМЕЧАНИЯ
Род Neo-Oudemansia установлен Трегордом в 1938 г. для формы,
которой он не дал названия (Tragardh I., 1938, Ent. Tidsk., 3—4,
137, рис. 19). Здесь мы предлагаем для этого вида наименование
N. triigurdhi.
ЛИТЕРАТУРА
Т t a g й г d h I., 1938. Further contributions towards the comparative morphology
and classification of the Mesostigmata, Ent. Tidsk., 3—4, 123—158.
Antennophoridae Berlese, 1892
(рис. 101)
ДИАГНОЗ
Клещи Antennophoridae имеют широкий, овальный по форме, не-
расчлененный дорзальный щиток. Лапки ног 1 лишены амбулакрального
Рис. 101. Antennophorus
foreli Berlese, 1904
(по Берлезс, 1904).
Самка (вид снизу).
аппарата, в то время как каждая из лапок ног II—IV имеет пред-
лапку с присоской, а иногда и со слабыми коготками. Срединный
щиток представляет собой поперечную, сильно склеротизированную
ленту. Боковые щитки склеротизированы посредине, а эпигинальный
щиток сливается с вентральным щитком.
Pedrizzilna
147
ОДЫ И ПОДРОДЫ
1. Antennophorus Haller, 1877
Тип рода Antennophorus uhlmanni Haller, 1877 (не Jannet, 1897)
2. Antennurella Berlese, 1904 (— Eufedrlzzia Sellnick, 1938)
Тип рода Antetinurella trouessarti Berlese, 1904
3. Celaenopsoldes Gunther, 1942 (сомнительный)
Тип рода Celaenopsoldes huloloensis Gunther, 1942
4. Celaenosthanns Vitzthum, 1930
Тип рода Celaenosthanus trlgonophllus Vitzthum, 1930
5. Messoracarus Silvestri, 1912
a) Messoracarus s. str.
Тип подрода Messoracarus mirandus Silvestri, 1912
6) Leptantennus Berlese, 1916
Тип подрода Messoracarus {Leptantennus) pendulipes Berlese,
1916
6. Ophiomeglstus Banks, 1914
Тил рода Ophiomeglstus luzonensls Banks, 1914
7. Physalozercon Berlese, 1903
Тип рода Antennophorus raffray Wasmann, 1902
8. Ptoc hares Silvestri, 1910
Тип рода Ptochares daveyli Silvestri, 1910
ОБЩИЕ ЗАМЕЧАНИЯ
Эти клещи связаны с насекомыми и крупными многоножками.
Хозяйственное и медицинское значение Antennophoridae не выяснено.
ЛИТЕРАТУРА
Tragardh I., 1943. Further contributions towards the comparative morphology
of the Mesostigmata (Acarina) the Antennophoridae and the Megisthanidae,
Arkiv. fdr Zoologi, 34A, № 20, 1 — 10.
Paramegistidae Tragardh, 1946
(рис. 102)
ДИАГНОЗ
Боковые щитки хорошо развиты и подвижно сочленяются или
совершенно сливаются с вентральным щитком. Срединный щиток
у самок разделен на две части. У самца половое отверстие располо-
жено позади стернального щитка.
РОДЫ И ПОДРОДЫ
1. Parameglstus TragSrdh, 1906
Тип рода Parameglstus conf rater Triigardh, 1906
2. Echinomeglstus Berlese, 1903
a) Echinomeglstus s. str.
Тип подрода Antennophorus wheeleri Wasmann (cm. Berlese,
1903)
10*
148
Глава III. Подотряд Mesostigmata
б) Antennomegistus Berlese, 1904
Тип подрода Antennophorus caputcarabi Berlese, 1903
3. Mlcromegistus Tragardh, 1948
Тип рода Mlcromegistus bakeri Tragardh, 1948
4. Neomegistus Tragardh, 1906
Тип рода Neomegistus lulidicola Tragardh, 1906
Рис. 102. Mlcromegistus bakeri Tragardh, 1948 (по Трегорду, 1948).
А —самка (вид снизу); Б —самец (вис снизу).
ОБЩИЕ ЗАМЕЧАНИЯ
Клещей этого семейства находят на насекомых и многоножках.
Их хозяйственное и медицинское значение не выяснено.
ЛИТЕРАТУРА
Tragardh I., 1943. Description of Mlcromegistus, a new genus of the
Paramegistidae, with notes on Neomegistus, Paramegistas and Echino-
niegistus (Acarina), Ent. Tidsk., 69, 127—131.
Fedrizziidae TrMgardh, 1937
(рис. 103)
ДИАГНОЗ
Клещи Fedrizziidae имеют уплощенное широкое тело, овальное по
форме. Их дорзальный щиток нерасчленен. Ноги короткие; ноги I
явно тоньше остальных. Лапки ног I не имеют предлапок; лапки ног
П—IV снабжены предлапками, каждая из которых имеет присоску и
коготки. Хелицеры клешневидные, с придатком причудливой формы.
Тектум треугольный и имеет срединный вырост. У самок срединный
щиток очень велик, боковые щитки вытянуты в длину, а эпигинальный
щиток отсутствует. Половое отверстие самца лежит позади стерналь-
ного щитка.
Fedrizziina
149
РОДЫ
1. Fedrlzzla Canestrini, 1884 (= Toxopeusia Oudemans, 1927)
Тип рода Fedrlzzla grossipes Canestrini, 1884 (= Toxopeusia
strandl Oudemans, 1927)
2. Kltncko-wstroemia
Тип рода Klincko'wstroemiu trdgardhi n. sp.
Рис. 103. Fedrlzzla strandl
(Oudemans), 1927
(по Удемансу, 1928).
Самка (вид снизу).
ОБЩИЕ ЗАМЕЧАНИЯ
В 1938 г. Трегорд выделил род Klinckowstroemia, но не дал для
типа видового названия (Tragardh I., 1938, Ent. Tldsk., 59, 133—134,
рис. 14). Этот вид—-тип рода KHnckowstroemla — назван здесь К.
trdgardhi. Клещи семейства Fedrizziidae связаны с насекомыми и мно-
гоножками, Их хозяйственное значение не выяснено.
ЛИТЕРАТУРА
Tragardh I., 1946. Outlines of a new classification of the Mesostigmata (Aca-
rina), K- Fysiogr. Sdtlsk. Handl., N. F., 57, № 4, 1—37.
Глава IV
ПОДОТРЯД IXODIDES LEACH, 1815
Все иксодовые клещи — паразиты. Разные виды иксодовых клещей
имеют много общего не только в биологии, но и в морфологии; все
они имеют колющий гипостом с загнутыми назад зубцами, хелицеры
с боковыми зубцами на подвижном пальце, особый орган чувств,
расположенный в углублении на лапках ног I (галлеров орган). У всех
Ixodides стигмы, открывающиеся по бокам тела, не имеют извилистых
перитрем. Иксодовые принадлежат к наиболее крупным представите-
лям Acarina. Они близки к клещам подотряда Mesostigmata, но так
как переходных форм между этими двумя группами нет, то, вероятно,
выделение Ixodides в самостоятельный подотряд вполне оправдано.
Фитцтум (1941) разделил Ixodides на три подгруппы, каждая из
которых представлена только одним семейством. Мы не считаем это
целесгсбразным и в настоящей работе подразделяем иксодовых кле-
щей непосредственно на семейства. Останавливаться здесь на морфо-
логии иксодовых клещей мы не будем, так как определительная
таблица их семейств вполне понятна и без этого.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ IXODIDES
1. Проподосомальный щиток (scutum) имеется на всех фазах раз-
вития ......................................................2
— Проподосомальный щиток (scutum) отсутствует на всех фазах
развития.........................................Argasidae
2^ Тарзальный членик пальп занимает терминальное положение;
scutum по строению не отличается от несклеротизированных по-
кровов тела самки...............................Nuttalliellidae
— Тарзальный членик пальп погружен в углубление на верхушке
голени пальп; scutum по строению отличается от несклеротизи-
рованных покровов тела.................................Ixodidae
Argasidae G. Canestrini, 1890
(рис. 104, 105)
ДИАГНОЗ
Приводимый ниже диагноз составлен по работе Кули и Колса (1944).
Argasidae представляют собой иксодовых клещей, не имеющих scutum
и обладающих лишь слабо выраженным половым диморфизмом. Как
половозрелые особи, так и нимфы имеют кожистые, морщинистые и
зернистые покровы, усеянные или сосочками, или бугорками. Capitulum
Подотряд Ixodides
151
(гнатосома) у взрослых и у нимф расположен или субтерминально,
или на брюшной стороне на некотором расстоянии от переднего края
тела; у личинок гнатосома занимает или субтерминальное, или терми-
нальное положение, У голодных взрослых клещей и особенно у нимф
гнатосома помещается в более или менее заметном углублении идиосомы
(камеростом). Сочленения пальп на всех фазах превращения подвижны
(членики никогда не бывают слиты). Пористые поля отсутствуют
Рис. 104. Antrieola coprophilus
(McIntosh), 1935
(по Кули и Колсу, 1944).
Самка (вид снизу).
Рис. 105. Ar gas persicus
(Oken), 1918
(по Кули и Колсу, 1944).
Самка (вид сверху).
у обоих полов. Глаза, если они имеются, лежат на супракоксальной
складке. Дыхальца как у взрослых, так и у нимф обычно находятся
перед тазиками ног IV. Подушечки (присоски) на лапках у взрослых
клещей и нимф обычно рудиментарные или отсутствуют, но у личи-
нок иногда хорошо развиты. Нимфальных фаз несколько, и число их
может варьировать1.
1 „Варьирование" числа нимфальных стадий у ряда клещей рода Orni-
thodorus оказывается вполне закономерным явлением; так, по наблюдениям
Троицкого, Е. Павловского и А. Скрынник (см. Е. Н. Павловский, Руковод-
ство но паразитологии человека, т. II, 5-е изд., Изд. Академии Наук СССР,
1948), полный цикл превращения Ornithodorus papilltpes Biz. протекает
в следующей последовательности:
яйцо^личинка, нимфа I нимфа Ц -
нимфа Ш нимфа нимфа V
152
Глава IV. Подотряд Ixodides
РОДЫ
. 1. Argus Latreille,. 1795
Тип рода Acarus reflexus Fabricius, 1794
2. Antricola Cooley et Kohls, 1942
Тип рода Ornithodoros coprophilus McIntosh, 1935
3. Ornithodoros Koch, 1844
Тип рода Argas savignyi Audouin, 1826
4. Otobius Banks, 1912
Тип рода Argas megnini Duges, 1884
ОБЩИЕ ЗАМЕЧАНИЯ
Клещи Argasidae распространены во всех областях земного шара
и паразитируют на самых разнообразных позвоночных животных. Их
хозяевами могут быть змеи, черепахи, многие птицы и представители,
по крайней мере, восьми отрядов млекопитающих К Как правило,
Argasidae питаются периодически, причем периоды питания чере-
дуются с линькой подобно тому, как это имеет место у постельных
клопов.
Самки откладывают по нескольку яиц за один раз. Вылупившаяся
из яйца личинка питается на хозяине довольно продолжительное время,
после чего она открепляется от хозяина и при линьке превращается
в нимфу 1. Нимфа обычно питается лишь в течение одного-двух
часов; насосавшись крови, она покидает хозяина и снова линяет,
превращаясь в нимфу II. Затем следует еще один короткий период
питания, после чего проходит третье превращение; при этом воз-
можно появление самцов и самок'2. Во многих случаях самцы внешне
отличаются от самок только незначительными различиями в строении
полового отверстия: у самцов половое отверстие обычно несколько
меньше и сильнее изогнуто. У Ornithodoros moubata и О. savignyi
личинка не питается и в жизненном цикле первой активной фазой
является нимфа I. Наоборот, у Otobius megnini не питаются взрос-
лые клещи; соответственно они не имеют на гипостоме загнутых
назад зубцов.
Многие Argasidae являются обычными обитателями нор, гнезд и
других жилищ своих хозяев и периодически питаются на последних.
Так, например, Ornithodoros talaje живет в южной Мексике в щелях
домов, а О. moubata и Argas persicus являются домовыми парази-
тами в Африке и Иране. Недавно было описано несколько новых
Процесс метаморфоза может длиться очень долго (25 лет и более); по-
этому требуется время, чтобы полностью проследить его в лаборатории. Во
многих случаях, если наблюдается небольшое число нимф данной семьи
клеща, жизненный цикл может завершиться после линьки нимф III или IV;
этим и объясняются указания ряда авторов на „варьирование" числа ним-
фальиых стадий. —- Прим. ред.
t Argasidae встречаются также и па жабах. — Прим. ред.
'Л См. примечание на стр. 151. Прим. ред.
Подотряд Ixodides
153
видов клещей этого семейства с летучих мышей Северной и Централь-
ной Америки. Кули и Колс (1944) полагают, что, по всей вероятности,
имеются еще и другие представители Argasidae, которые паразити-
руют на летучих мышах в США, Мексике и в Центральной Америке.
Нахождение клещей Argasidae на летучих мышах неудивительно, так
как последние являются обычными обитателями жилищ человека.
Клещи Argasidae имеют большое хозяйственное и медицинское
значение. В Иране Argas persicus является неприятным домовым па-
разитом. Кроме того, этот клещ повсеместно нападает на домашних
птиц, причиняя последним много вреда своими укусами и распро-
страняя среди них спирохетоз. Крупный рогатый скот также страдает
от ушных клещей {Otobius megnini), и известны случаи гибели скота,
приписываемые этим клещам. Многие виды нападают на человека, и
их укусы бывают очень болезненными. Кули и Колс (1944) приводят
следующий список клещей, нападающих на человека в Северной Аме-
рике: Argas persicus, Otobius megnini, Ornithodoros coriaceus, O. her-
nisi, O. nicollei, 0. parkeri, O. rudis, O. stageri, O. talaje и О. tu-
ricata. По крайней мере 11 разных видов Ornithodoros передают
человеку спирохет возвратного тифа Borrella spp. Экспериментально
установлено, что О. parkeri может переносить туляремию. В экспе-
риментах было также показано, что различные Argasidae передают
также пятнистую лихорадку Скалистых гор и американскую Q-лихо-
радку.
ЛИТЕРАТУРА
(о о 1 е у R. А., К о h 1 s G. М., 1944. The Aigasidae of North America, Central
America and Cuba, Am. Midland Naturalist, Mongr., VI f- 152.
N и 11 a 11 G. H. F., W a r b u r t о n С., С о о p e r W. F., Robinson L. E.,
1908. Ticks. A monograph of the Ixodidae, Part I, The Argasidae, 1—104,
Cambridge Univ. Press.
Nuttallieilidae Schulze, 1935
(рис. 106)
ДИАГНОЗ
Это семейство занимает промежуточное положение между Argasi-
dae и Ixodidae. Scutum у Nuttallieilidae слабо склеротизирован и по
внешнему виду плохо отличим от кожистого покрова, покрывающего
все тело. Покровы тела несут сосочки, напоминающие сосочки, свой-
CI венные Argasidae. Членики пальп подвижны, но в отличие от Arga-
sidae на внутренней стороне второго членика имеется желобок. Гна-
юсома выступает вперед и, как у представителей Ixodidae, хорошо
заметна при рассмотрении клещей сверху.
РОДЫ
I. Nuttalliella Bedford, 1931
Тип рода Nuttalliella namaqua Bedford, 1931
154
Глава IV. Подотряд Ixodides
ОБЩИЕ ЗАМЕЧАНИЯ
Экземпляр, по которому был описан род, был найден под камнем в
Малом Намакваленде (Южно-Африканский Союз). Данные о каком-либо
хозяйственном или медицинском значении этого клеща отсутствуют.
Рис. 106. NMali tel la
namaqua Bedford, 1931
(по Бедфорду, 1931).
Самка (вид сверху).
ЛИТЕРАТУРА
Bedford G. А. Н„ 1931. Nuttalllella naniaqua, a new genus and species
of tick, Parasitol., 23, 230—232 Pl. X.
Ixodidae Murray, 1877
(рис. 107)
ДИАГНОЗ
Представители этого семейства имеют тело с выступающей вперед
гнатосомой. Scutum у самцов достигает заднего края тела, в то время
как у самок он ограничивается задним краем проподосомы; у личи-
нок и у нимф scutum развит примерно так же, как у самок. Одни
представители имеют глаза, у других они отсутствуют. Личинка имеет
две пары вентральных стигм, открывающихся кнаружи от ног. Нимфы
и половозрелые клещи имеют только одну пару стигмальпых щит-
ков. Терминальный членик пальп сильно уменьшен и лежит на вен-
тральной поверхности предпоследнего членика пальп. Членики пальп
соединены друг с другом малоподвижно. У самок у основания гнато-
сомы имеются пористые поля.
Подотряд Ixodides
7<55
Ixodidae можно разделить на три подсемейства в соответствии
с нижеследующей определительной таблицей.
Рис. 107. Dermacenfor varlabllis (Say), 1821 (по Кули, 1938).
Самка (вид сверху).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ IXODIDAE
1. Анальная бороздка не окружает анальное отверстие спереди.
У самцов брюшная поверхность тела никогда не бывает почти
полностью покрыта рядами щитков.............................2
— Анальная бороздка окружает анальное отверстие спереди. У сам-
цов брюшная поверхность тела почти полностью покрыта рядами
щитков.................................................Ixodinae
2. У самцов брюшная поверхность тела лишена щитков. Scutum
обычно орнаментирован............................Amblyomminae
— У самцов брюшная поверхность сзади покрыта четырьмя щитками.
Поверхность щитков обычно гладкая...............Rhlpicephaliпае
РОДЫ И ПОДРОДЫ
Ixodinae Vitzthum, 1941
1. Ixodes Latreille, 1795
a) Ixodes s. str.
Тип подрода Acarus ricinus Linnaeus, 1758
6) Ceratixodes Neumann, 1902
Тип подрода Hyalomma puta Picard, 1876
в) Endopalpiger Schulze, 1935
Тип подрода Endopalpiger luxuriosus Schulze, 1935
156
Глава IV. Подотряд Ixodides
г) . Eschatocephalus v. Frauenfeld, 1863
Тип подрода Ixodes vcspertilionls Koch, 1844
( = Eschatocephalus gracilipes v. Frauenfeld, 1853)
д) Exopalpiger Schulze, 1935
Тип подрода Ixodes priscicollaris Schulze, 1932
e) Lepidixodes Schulze, 1935
Тип подрода Eschatocephalus kopsteini Oudemans, 1925
ж) Sternalixodes Schulze, 1935
Тип подрода Ixodes cordifer Neumann, 1908
Amblyomminae Neumann, 1911
1. Amblyomma Koch, 1844 ( — Haemalastor Koch, 1844)
Тип рода Acarus cajennensis Fabricius, 1787
2. Alloceraea Schulze, 1918
Тип рода Haemaphysalis inertnls Birula, 1895
3. Amblycentor Schulze, 1932
a) Amblycentor s. str.
Тип подрода Ixodes rhinocerinus Denny, 1843
6) Punctlcentor Schulze, 1933
Тип подрода Dermacentor crlcumguttatus Neumann, 1897
4. Anocentor Schulze, 1937 ( = Otocentor Cooley, 1938)
Тип рода Dermacentor nttens Neumann, 1897
( = Anocentor columbianus Schulze, 1937)
5. Aponomma Neumann, 1899
Тип рода Ixodes gervaisi Lucas, 1847
6. Dermacentor Koch, 1844
a) Dermacentor s. str.
Тип подрода Acarus reticulatus Fabricius, 1794
6) Dermacentorites Olenev, 1931
Тип подрода Cynorhaestes plctus Hermann, 1804
7. Haemaphysalts Koch, 1844
Тип рода Haemaphysalts concinna Koch, 1844
8. Indocentor Schulze, 1933
Тип рода Dermacentor auratus Supino, 1897
Rhipicephalinae Vitzthum, 1941
1. Rhipicephalus Koch, 1844
a) Rhipicephalus s. str.
Тип подрода Ixodes sanguineus Latreille, 1806
6) Pterygodes Neumann, 1913
Тип подрода Rhipicephalus (Pterygodes) fulvus Neumann, 1913
2. Boophtlus Curtice, 1891
a) Boophilus s. str.
Тип подрода Ixodes annulatus Say, 1821 ( -x= Ixodes bovis Riley,
1867)
Подотряд Ixodides
157
б) Palpoboophilus Minning, 1934
Тип подрода Rhipicephalus decoloratus Koch, 1844
в) Uroboophilus Minning, 1934
Тип подрода Uroboophilus sharift Minning, 1934
3. Cosmiomma Schulze, 1919
Тип рода Ixodes hippopotamensls Denny, 1843
4. Hyalomma Koch, 1844
a) Hyalomma s. str.
Тип подрода Acarus aegyptius Linnaeus, 1758
6) Hyalommasta Schulze, 1930
Тип подрода Hyalomma syrtacum Koch, 1844
в) Hyalommlna Schulze, 1919
Тип подрода Hyalomma rhlplcephaloides Neumann, 1901
5. Margaropus Karsch, 1879
Тип рода Margaropus ‘Winthemt Karsch, 1879
6. Nosomma Schulze, 1919
Тип рода Hyalomma monstrosum Nuttall et Warburton, 1908
7. Rhipiceutor Nuttall et Warburton, 1908
Тип рода Rhipicentor bicornls Nuttall et Warburton, 1908
ОБЩИЕ ЗАМЕЧАНИЯ
Клещи подотряда Ixodides и особенно семейства Ixodidae поль-
зуются, вероятно, большей известностью, чем какие-либо другие
клещи. Это и неудивительно, если принять во внимание, что Ixodi-
des— самые крупные из клещей, паразитирующих на человеке и на
домашних и диких животных. Эти клещи являются к тому же пере-
носчиками многих серьезных заболеваний. Несмотря на широкую
известность иксодовых клещей, среди специалистов по систематике
этой группы существуют значительные расхождения в понимании
объема родов и их группировок. Мы здесь следуем главным обра-
зом работе Фитцтума (1941), который, в свою очередь, придержи-
вался системы Шульца. Другие же специалисты склонны рассматри-
вать многие выделенные Шульцем роды и подроды -как синонимы.
Жизненный цикл клещей Ixodidae довольно прост. Самки откла-
дывают яйца прямо на почву; число яиц в кладке очень велико. Из
яиц вылупляются шестиногие личинки, которые находят себе подхо-
дящего хозяина и питаются на нем. Насосавшаяся кровью личинка
линяет, превращаясь в нимфу. Нимфа имеет восемь ног и внешне
похожа на взрослую самку, отличаясь от последней значительно мень-
шими размерами и отсутствием полового отверстия. Нимфы насы-
щаются кровью и при линьке превращаются во взрослых самок и
самцов. Как самки, так и самцы сосут кровь. У самцов покровы
тела мало эластичны; поэтому они лишь незначительно раздуваются
при кровососании. Самки же, имеющие весьма эластичные покровы,
очень сильно раздуваются после насыщения. После копуляции насо-
савшиеся кровью самки откладывают яйца.
153
Глава IV. Подотряд Ixodides
Типичный жизненный цикл в отдельных деталях подвержен много-
численным видоизменениям. Большинство Ixodidae на половозрелых
и неполовозрелых фазах паразитирует на разных хозяевах. Например,
личинки и нимфы Dermacentor venustus Neumann (= D. andersoni
Stiles) паразитируют на мелких млекопитающих, в то время как
взрослые особи нападают на овец, коз, человека и т. д. D. venu-
stus для линьки и откладки яиц отваливаются от хозяина; таким
образом, для завершения жизненного цикла этот клещ должен пере-
менить, по крайней мере, трех хозяев. Между тем Boophilus аппи-
latus (Say) в личиночной фазе присасывается к хозяину, которым
служит крупный рогатый скот, и остается на нем до момента яйце-
кладки. Наттэлл (19П) указывает, что Ixodes ricinus (Linnaeus)
может пройти полный жизненный цикл в очень короткое время — за
170 дней, но что при неблагоприятных условиях продолжительность
жизненного цикла может достигнуть 2—3 лет.
Ixodidae имеют большое хозяйственное и медицинское значение.
Сильное заражение иксодовыми клещами домашних животных может
быть причиной анемии последних. Ixodidae передают домашним жи-
вотным некоторые опасные заболевания; так, Boophilus annulat'us
(Say) передает техасскую лихорадку крупного рогатого скота. У че-
ловека укусы Ixodidae могут вызывать клещевой паралич; считается
также, что они передают человеку различные заболевания, а именно:
пятнистую лихорадку Скалистых гор и другие виды пятнистых лихо-
радок Северной и Южной Америки, марсельскую лихорадку, тиф Кении,
южноамериканскую клещевую лихорадку, Q-лихорадку, туляремию, а
возможно, и некоторые другие (Макки, Хантер и Уорс, 1945). Под-
робные сведения о роли иксодовых клещей как переносчиков заболе-
ваний можно найти во многих книгах по паразитологии и тропиче-
ской медицине.
ЛИТЕРАТУРА
Cooley R. А., 1938. The genera Dermacentor and Oiocentor (Ixodidae) in
the United States, with studies in variation, Nat. Inst. Health Bull.,
№ 171, 1— 85-J-Pls.
Cooley R. A., 1946. The genera Boophilus, Rhipicephalus, and Haemaphysalis
(Ixodidae) of the New World, Nat. Inst. Health Bull., № 187, 54 J- III.
Cooley R. A., Kohls G. M., 1945. The genus Ixodes in North America,
Nat. Inst. Health Bull., № 184, 246 + 111.
Douglas J. R., 1943. The internal anatomy of Dermacentor andersoni Stiles,
Univ, of Calif. Publ. in Ent., 7 (10), 207—272+Pls.
Fielding J. W., 1926. Australasian Ticks, Dept, of Health Commonwealth
of Australia Service Publ. (Trap. Div.), № 9, 1—114.
Neumann L. G., 1911. Ixodidae. Das Tierreich. Acarina. 26 Lief., 1—169.
Nuttall G. H. F., Warburton C., 1911. Ticks. A monograph of the
Ixodoidea, pt. II, The Ixodidae, 105—348, Cambridge Univ. Press.
Nuttall G. H. F., W ar b ur t о n C., 1915. Ticks. A monograph of the Ixo-
doidea, pt. Ill, The genus Haemaphysalis, 349—550 + Pls., Cambridge
Univ. Press.
Robinson L. E., 1926. Ticks: A monograph of the Ixodoidea, pt. IV, The
Genus Amblyomma, 1—302 +Pls., Cambridge Univ. Press.
Глава V
ПОДОТРЯД TROMBIDIFORMES REUTER, 1909
Клещи этого подотряда обычно обладают одной парой стигм, от-
крывающихся на гнатосоме или около гнатосомы; лишь изредка
стигмы отсутствуют. Пальпы обычно подвижны и хорошо развиты;
у одних форм они построены по типу клешнеобразных хватательных
органов, у других функционируют как сенсорные органы. Хелицеры
обычно приспособлены для прокалывания и видоизменены в связи
с этим; лишь у представителей более примитивных семейств, веду-
щих хищный образ жизни, изредка можно встретить хелицеры
с клешнями, служащие, вероятно, для схатывания добычи. В неко-
торых случаях развиты половые присоски, но анальные присоски
всегда отсутствуют.
Trombidiformes разделены здесь на три группы: Tetrapodili, Tar-
sonemini и Prostigmata. Первая группа включает крошечных черве-
образных растительноядных клещей, имеющих на всех фазах только
две пары ног; дыхательная система у них не развита. Все эти клещи
объединяются в одно семейство Eriophyidae. В группу Tarsonemini
входят мелкие клещи, паразитирующие на растениях или членистоно-
гих; они обладают маленькими ротовыми органами, небольшими прижа-
тыми к роструму пальпами и очень маленькими игловидными хелицерами.
У самок Tarsonemini стигмы открываются позади педипальп, у самцов
же стигмы отсутствуют. Многие клещи этой группы имеют неполное
число пар ног. Повидимому, группа в целом сильно дегенерировала.
Группа Prostigmata включает клещей, у которых стигмы открываются
у основания хелицер. Это, как правило, сравнительно крупные клещи,
обладающие крупными и весьма подвижными ротовыми органами.
Во многих случаях пальпы хорошо развиты и служат как осязательные
или же как хватательные органы. Хелицеры несколько видоизменены,
но во всех случаях они хорошо развиты и служат для прокалывания
или как хватательные органы. Tarsonemini представляют собой единую,
более или менее однородную группу; в пределах же Prostigmata
мы находим весьма разнородные формы, в том числе формы, у ко-
торых лапка пальп противопоставляется коготку пальп, так что по-
следние превращены в хватательный орган (Trombidiidae), формы
с очень простыми пальпами (Rhagidiidae), с колющими игловидными
хелицерами (Tetranychidae) или с хватательными или дробящими
ротовыми органами (Rhagidiidae). По всей вероятности, перечисленные
160 Глава V, Подотряд Trombidiformes
семейства Prost'gmata могут быть объединены с некоторыми дру-
гими формами надсемейства; однако ввиду того, что классифика-
ция Prostigmata находится пока в стадии разработки, мы не пытаемся
здесь вводить какие-либо категории более высокого ранга, чем семейства.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ГРУПП TROMBIDIFORMES
1. Тело обычной для клещей формы, не червеобразное (исключе-
ние— Demodicidae). Имеются типичные четыре пары ног (за ред-
кими исключениями среди Podapolipodidae и Phytoptipalpidae).
Гнатосома у одних форм хорошо развита, у других же плохо
различима. Дыхательная система имеется ...................... 2
— Тело червеобразное, кольчатое. Имеются только две пары ног,
расположенные спереди. Дыхательная система не развита. Гнато-
сома сильно редуцирована. Пальпы очень маленькие и плотно
прилегают к роструму. Хелицеры стилетообразные. Группа вклю-
чает мелких растительноядных клещей . . Tetrapodili (Eriophyidae)
2. Хелицеры очень маленькие, стилетовидные. Пальпы маленькие,
прижатые к роструму. Гнатосома в целом сильно редуцирована и
мало заметна. Мелкие клещи с обычным или неполным числом
пар ног. У самок стигмы открываются за максиллами, самцы же
не имеют стигм и трахей.............................Tarsonemini
— Хелицеры гораздо крупнее и легко различимы. Пальпы крупные
и не прижаты к роструму (исключение составляют червеобразные
Demodicidae, имеющие четыре пары ног). Гнатосома обычно хо-
рошо различима. Как правило, крупные клещи с четырьмя парами
ног (исключение составляют Phytoptipalpidae, среди которых
один род обладает только тремя парами ног). Стигмы откры-
ваются у основания хелицер (Demodex не имеет стигм и трахей)
................................................... Prostigmata
TETRAPODILI BREMI, 1872
Группа включает мелких червеобразных растительноядных клещей
длиной до 0,20 мм. Tetrapodili имеют лишь две пары ног, располо-
женных впереди; коготков на лапках нет, зато развит „перистый “
эмподий. Тело кольчатое; половое отверстие расположено непосред-
ственно позади ног и покрыто поперечным щитком. Глаза, сердце,
выделительная система и дыхательная система не развиты. На теле
имеется всего несколько пар щетинок. В эту группу входит только
одно семейство — Eriophyidae.
Eriophyidae Nalepa, 1898
(рис. 108)
ДИАГНОЗ
Eriophyidae представляют собой очень мелких червеобразных кле-
щей с щитовидной проподосомой, покрытой отчетливым характерным
Tetrapodili
161
рисунком. Гистеросома удлиненная и кольчатая. Дыхательная система
отсутствует. На всех фазах метаморфоза имеются две передние пары
ног; лапки всех ног снабжены лучистыми коготками. Половое от-
верстие открывается на передней части брюшной поверхности тела
непосредственно позади ног; генитальный щиток по форме приближается
Рис. 108. Oxypleuri'es aesculifoliae Keifer
(по Кейферу, 1942).
А — яйцо; Б — нимфа I; В —нимфа II; Г—протогенная самка;
Д—дейтогенная самка.
к узкой поперечной полоске. Пальпы короткие, простые; хелицеры
короткие, стилетовидные, колющие. Анальное отверстие располо-
жено на заднем конце тела.
Все Eriophyidae живут на растениях. Одни из них — свободно-
живущие формы, другие — галлообразующие. Фитцтум подразделяет
эту группу на два семейства — Eriophyidae и Phyllocoptidae, но мы,
так же как и Кейфер, рассматриваем ее как единое семейство.
Несмотря на то, что Eriophyidae выделяют из простигматиче-
ских клещей и относят к особой группе Tetrapodili, их близость к
11 Зак. 1735
162
Глава V. Подотряд Trombidiformes
нормальным восьминогим клещам легко показать. Известно, что в неко-
торых группах восьминогих клещей (например, в семействе Podapo-
lipod’dae, представители которого паразитируют на насекомых)
отдельные роды утрачивают одну пару ног. И это неудиви-
тельно, так как при образе жизни, свойственном представителям
этого семейства, вырабатываются мешкообразные паразитические
формы. Среди растительноядных Phytoptipalpidae, находящихся в близ-
ком родстве с Tetranyclrdae, представители рода Phytoptipalpus
имеют только три пары ног. В том же семействе клещ Tetiuipalpus
eriophyoides Baker хотя и имеет четыре пары ног, однако на фазе
нимфы весьма напоминает Eriophyidae своим сильно вытянутым коль-
чатым телом. Поперечные генитальные щитки также указывают на
родственные связи Eriophyidae и Pliytoptipalpidae. Только у представи-
телей этих двух семейств половое отверстие имеет вид поперечной
щели, у всех же остальных форм оно имеет вид продольной щели.
Однако, в то время как у Eriophyidae половое отверстие расположено
непосредственно позади ног и, следовательно, отодвинуто далеко
вперед, у Phytoptipalidae оно находится на заднем конце тела. Это
различие, впрочем, не так существенно, как можно было бы думать;
повидимому, у клещей типа Pliytoptipalpidae произошло слияние сег-
ментов задней части тела, благодаря чему внешне кажется, что по-
ловое отверстие расположено на самом конце тела, а не в обыч-
ном положении. Лучистый тарзальный придаток Eriophyidae несколько
напоминает таковой у Dolichotetranychus, относящегося к семейству
Phytoptipalpidae. Стилетовидные хелицеры и особенности биологии
различных представителей этой группы также указывают на то, что
Tetrapodili имеют общее происхождение с растительноядными Tetra-
nychidae и Pliytoptipalpidae.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ ERIOPHYIDAE
(ПО КЕЙФЕРУ)
1. На спинном- щите имеется три или четыре бугорка, покры-
тых щетинками; на задней паре бугорков щетинки всегда
направлены вперед. На брюшке передняя пара субдорзальных
щетинок иногда отсутствует. Голень передних ног обычно имеет
расположенную латерально шпору. Задние щетинки присосок
часто расположены латерально. Генитальный клапан самки глад-
кий. Яичники эллиптические или продолговатые; от них начинаются
выступающие вперед протоки гонад, которые затем загибаются
в стороны.................................................. 2
— На спинном щите имеется лишь одна пара покрытых ще-
тинками бугорков (иногда эти бугорки отсутствуют). На брюшке
субдорзальных щетинок нет. Расположенная латерально шпора на
лапках передних ног не развита. Задние щетинки присосок всегда
отходят сверху. Генитальный клапан самки в различной степени
Tetrapodili
163
скульптурирован или гладкий. Яичники более или менее округлые
и сидят на коротких направленных кнаружи или диагонально
протоках, отходящих от заднего края полового отверстия . . 3
2. Спинной щит не имеет переднего выроста над рострумом.
Брюшко червеобразное. Кольца на спине и на брюшке одинако-
вые ................................................Phytoptinae
— Спинной щит имеет передний вырост, выступающий над
рострумом. Рострум иногда очень короткий. Брюшко разделено
сбоку на широкие тергиты и более многочисленные узкие стер-
ниты.......................................Sierraph у toptin ае
3. Спинной щит никогда не имеет выроста, нависающего над
рострумом. Брюшко червеобразное; кольца на спине и на брюшке
одинаковые, по крайней мере в передней части тела. Если рос-
трум крупный, то он не причленен к телу под прямым углом и
снабжен изогнутыми хелицерами.......................Eriophyinae
— Спинной щит обычно имеет передний вырост, нависающий
над рострумом. Брюшко чаще разделено по бокам на ши-
рокие тергиты и более узкие стерниты; реже имеется крупный
рострум, суживающийся к концу и причлененный к телу под
прямым углом. Хелицеры крупные, круто изогнуты книзу и
кнутри от основания рострума.....................Phyilocoptinae
РОДЫ
Phytoptinae Keifer, 1944
1. Phytoptus DuJardin, 1851
Тип рода Erlophyes avellanae Nalepa, 1889
2. Setoptus Keifer, 1944
Тип рода „Platyphytoptus“ jonesi Keifer, 1938
Sierraphytoptinae Keifer, 1944
(= M a c k i el 1 i n a e Keifer, 1946)
1. Slerraphytoptus Keifer, 1939
Тип рода Slerraphytoptus alnivagrans Keifer, 1939
2. Austracus Keifer, 1944
Тип рода Austracus havrylenkonis Keifer, 1944
3. Mackiella Keifer, 1939
Тип рода Mackiella phoenicis Keifer, 1939
4. Nalepella Keifer, 1944
Тип рода Phyllocoptes triceras Borner, 1906
Eryophyinae Nalepa, 1898
1. Erlophyes von Siebold, 1850
Тип рода Erlophyes vltls (Pgst.), 1857
2. Aceria Keifer, 1944
Тип рода Erlophyes tulipae Keifer, 1938
11*
164
Глава V. Подотряд Trombidiformes
3. Cecidodectes N al ера, 1917
Тип рода Cecidodectes euzonus Nalepa, 1917
4. Cecldophyes Nalepa, 1887
Тип рода Phytoptus galii Karpelles, 1884
5. Cymoptus Keifer, 1946
Тип рода Cymoptus spinlventris Keifer, 1946
6. Monochetus Nalepa, 1898 (= Monaulax Nalepa, 1892 nomen
praeoccupatum)
Тип рода Monaulax sulcatus Nalepa, 1892
7. Novophytoptus Roivainen, 1947
Тип рода Novophytoptus rostratae Roivainen, 1947
8. Paraphytophus Nalepa, 1896
Тип рода Paraphytophus paradoxus Nalepa, 1896
9. Phytoptochetus Nalepa, 1917
Тип рода Phytoptochetus tristichus Nalepa, 1917
10. Trichostigma Gerber, 1902
Тип рода Trichostigma crodii Gerber, 1902
P h у 11 о с о p t i n a e Nalepa, 1898
1. Phyllocoptes Nalepa, 1889
Тип рода Phyllocoptes carpini Nalepa, 1887
2. Abacarus Keifer, 1944
Тип рода Calepitrimerus acalyptus Keifer, 1939
3. Abacoptes Keifer, 1939
Тип рода Abacoptes platynus Keifer, 1939
4. Acamina Keifer, 1944
Тип рода Calepitrimerus nolinae Keifer, 1939
5. Acaphylla Keifer, 1943
Тип рода Acaphylla steimweideni Keifer, 1943
6. Acarelliptus Keifer, 1940
Тип рода Acarelliptus cocciformis Keifer, 1940
7. Acarlcalus Keifer, 1940
Тип рода Acarlcalus segundus Keifer, 1940
8. Anthocoptes Nalepa, 1892
Тип рода Phyllocoptes loricatus Nalepa, 1889
9. Calacarus Keifer, 1940
Тип рода Calacarus pulviferus Keifer, 1940
10. Calepitrimerus Keifer, 1938
Тип рода Calepitrimerus cariniferus Keifer, 1938
11. Caliphytoptus Keifer, 1938
Тип рода Caliphytoptus quercilobatae Keifer, 1938
12. Callynlrotus Nalepa, 1894
Тип рода Callyntrotus schlectendali Nalepa, 1894
13. Caroloptes Keifer, 1940
Тип рода Caroloptes faglvagrans Keifer, 1940
Tetrapodili
165
14. Coptophylla Keifer, 1944
Тип рода Phyllocoptes lamlmani Keifer, 1939
15. Cupacarus Keifer, 1943
Тип рода Cupacarus cuprlfester Keifer, 1943
16. Dlptilomiopus Nalepa, 1916
Тип рода Dlptilomiopus javanlcus Nalepa, 1916
17. Epltrimeras Nalepa, 1898
Тип рода Epitrlmerus getnmicola Nalepa, 1898
18. Gammaphytoptus Keifer, 1939
Тип рода Gammaphytoptus camphorae Keifer, 1939
19. Oxypleurites Nalepa, 1891
Тип рода Acanthonotus heptacanthus Nalepa, 1889 по Кейферу,
1938 (Tegonotus trouessartl Nalepa, 1890 по Фитцтуму, 1943)
20. Phyllocoptruta Keifer, 1938
Тип рода Typhlodromus olelvorus Ashmead, 1879
21. Phyllocoptyches Nalepa, 1922
Тип рода Phyllocoptyches galllcolus Nalepa, 1922
22. Platyphytoptus Keifer, 1938
Тип рода Platyphytoptus sabinianae Keifer, 1938
23. Quadracus Keifer, 1944
Тип рода Quadracus urtlcae Keifer, 1944
24. Rhyncaphytoptus Keifer, 1939
Тип рода Rhyncaphytoptus ficlfoliae Keifer, 1939
25. Tegonotus Nalepa, 1890
Тип рода Tegonotus fastigatus Nalepa, 1890
26. Tetra Keifer, 1944
Тип рода „Phyllocoptruta* concava Keifer, 1939
27. Thamnacus Keifer, 1944
Тип рода Phyllocoptes rhamnicola Keifer, 1938
28. Tumescoptes Keifer, 1939
Тип рода Tumescoptes trachycarpi Keifer, 1939
29. Vasates Shimer, 1869
Тип рода Vasates quadripedes Shimer, 1869
, Роды неопределенного положения
1. Flexlpalpus Scheuten, 1857
Тип рода Flexlpalpus tillae Scheuten, 1857
ОБЩИЕ ЗАМЕЧАНИЯ
Полное представление о жизненном цикле некоторых Eriophyidae
удалось получить только после появления работы Кейфера (1942)
о чередовании поколений у Oxypleurites aesculifoliae Keifer — возбу-
дителя ржавчины у конского каштана. Правда, ранее Йозерс с со-
трудниками (1930) изучали клещей, вызывающих ржавчину цитрусо-
вых, «к Бэкер (1939)—клеща Aceria [i.cus (Cotte), вредителя винных
166
Глава V. Подотряд Trombidiformes
ягод, но исследованные этими авторами виды имеют простой жизнен-
ный цикл без чередования поколений. Упомянутая выше работа Кей-
фера проливает свет также и на жизненный цикл клещей Epitrimerus
pirifollae Keifer, паразитирующих на листьях грушевого дерева.
Некоторые самки клеща Oxypleurites aesciilifoliae имеют соот-
ветствующих им самцов и называются протогинными. Другие самки
этого же вида морфологически отличаются от протогинного, или
первичного, типа и не имеют соответствующих им самцов; этих
самок Кейфер называет дейтогинными. Протогинные и дейтогинные
самки, отличающиеся друг от друга характером штриховки тела,
ранее описывались как разные виды разных родов — Phyllocoptes
aesciilifoliae (протогинные) и Oxypleurites neocarinatus (дейтогинные).
Оказалось, что самки Oxypleurites (протогинные) дают при выращи-
вании обе формы самок, дейтогинные же самки откладывают яйца,
из которых вылупляются только яйцекладущие протогинные самки.
Ниже приводится описание жизненного цикла Oxypleurites aescu-
llfoliae, заимствованное из работы Кейфера (1942).
„Дейтогинные самки становятся активными к концу зимы,
в феврале; они покидают зимние убежища на ветвях и, когда начи-
нают набухать почки, проникают под наружные почечные чешуйки
и питаются зеленой тканью внутренних чешуек. По мере развития
ранней весенней листвы дейтогинные самки откладывают яйца, из
которых вылупляются нимфы, превращающиеся в так называемых
первичных клещей, паразитирующих на листьях. Эти первичные
клещи вскоре начинают активно размножаться, воспроизводя допол-
нительных первичных клещей. Примерно к концу апреля или
в начале мая среди клещей первичного типа начинают появляться
новые — дейтогинные самки. Напитавшись полностью, дейтогинные
самки спускаются по стеблю примерно на 15 см или ниже и заби-
ваются в щели или иные убежища; там они остаются до листопада,
наступающего в середине июля. Первичные же клещи остаются на
листьях и в зеленых тканях растения и отмирают вместе с ними,
причем размножение их затухает уже к началу июля. Клещи пред-
почитают нижнюю сторону листьев, но обычно популяция их стано-
вится настолько многочисленной, что заселяются обе стороны листьев,
покрывающиеся „ржавчиной". Дейтогинные самки, забившись в щели,
впадают в состояние покоя1 и к концу лета частично высыхают,
но следующей весной с наступлением дождей и холодов вновь ста-
новятся активными. Смертность среди этих самок очень высока и
связана главным образом с недостатком убежищ.
Весеннее потомство дейтогинных самок Oxypleurites aesculifoliae
специально не исследовалось, но все же удалось установить, что
в первой же партии клещей первичного типа встречаются яйцекладу-
щие самки; это говорит либо об оплодотворении дейтогинных самок
1 Состояние анабиоза. — Прим. ред.
Tetrapodili
167
до начала зимовки, либо о том, что эти клещи обладают способ-
ностью к партеногенетическому воспроизведению как самцов, так и
самок".
Описание, которое приводит Патмэн (7th Ann. Rpt. Ent. Soc.
Ont., 1939, p. 33), работавший c Phyllocoptes fockeul в Канаде,
явно относится к дейтогинной самке. Автор констатирует, что из
яиц, откладываемых зимующими самками, развиваются и самцы, и
самки. Патмэн считает, что эти перезимовавшие самки (дейтогинные)
могли быть оплодотворены до зимовки, так как ему удалось дока-
зать, что обычные неоплодотворенные самки (протогинные) дают
только самцов. Далее Патмэн отмечает, что появление „зимующих
форм" связано, повидимому, с отвердением листвы; это предположе-
ние в основном согласуется с наблюдениями над Oxypleurites aescu-
lifoliae*.
Клещ Aceria ficus (Cotte), имеющий простой жизненный цикл,
в отличие от некоторых других Eriophyidae, непосредственно не при-
чиняет серьезного вреда растению-хозяину — фиговым деревьям Кали-
форнии. Клещи Aceria ficus (Cotte) живут под почечными чешуйками
на поверхности листьев и в зеленых плодах. Если популяция сильно
разрастается, то отдельные почечные чешуйки покрываются рубцами
и часть листьев в почке отмирает недоразвитыми. Подобные рубцы,
или „ржавчина", редко бывают заметны на развитых листьях, но
хорошо различимы на плодах, на которых обычно бывают повреж-
дены только чешуйки глазков; при очень большой численности кле-
щей поражается и внутренняя часть плода. Кроме того, некротические
участки, возникающие в результате поедания клещами тканей плода,
легко поражаются головней и плесневыми грибами (Aspergillus,
Hormodendrum, Caldosporlum, Rhizopus). Клещи зимуют в верху-
шечных почках, зараженность которых может достигать 10%, причем
в одной почке зимует от десятка до нескольких сот клещей. Размно-
жение происходит зимой. Весной, когда почки лопаются, клещи дер-
жатся на поверхности листьев. Численность популяции достигает
максимального уровня в июне и июле. Клещи откладывают яйца на
молодые нежные листья (не отдавая при этом предпочтения верхней
или нижней), но сами держатся преимущественно на нижней поверх-
ности развитых листьев, более сочной и лучше защищенной от
солнца; лишь при очень большой численности клещи заселяют и
верхнюю поверхность листьев. Винные ягоды первого урожая зара-
жаются в течение двух недель с момента превращения плодовой
почки в плод, причем бывает поражено около 90% урожая. Винные
ягоды второго сбора заражаются клещами уже через восемь дней
после их завязывания и к первому июля обычно бывает поражен
почти весь второй урожай. Но когда плод созревает и чешуйки
глазков расходятся, на фиговых клещей нападают хищные клещи и
трипсы, так что в зрелых винных ягодах паразитов почти не остается.
Клещи, проникшие в верхущечные почки в течение лета, зимуют и
168
Глава V. Подотряд Trombidiformes
весной воспроизводят новую популяцию. Осмотр почек, произведен-
ный летом, показал, что 50°/о их было заражено.
Ниже приводится выдержка из работы Кейфера (1946), в которой
он говорит об экономическом значении Eriophyidae.
„Эриофиидные клещи, известные под наименованиями галловых
клещей, пузырчаток и почечных клещей, являются самыми мелкими
из животных с наружным скелетом, с которыми приходится вести
борьбу в сельском хозяйстве. Eriophyidae вызывают деформацию и
ржавчину листьев и плодов, разрушают почки и, если не принимать
мер против них, часто приводят к гибели урожая.
Eriophyidae дают примеры чрезвычайно тесной связи между па-
разитом и хозяином и обладают значительной специфичностью при
выборе этого хозяина. Одним из выражений этой специфичности
является образование различного рода галлов; кроме того, Eriophyidae
в большинстве случаев используют естественные особенности строе-
ния хозяина и не наносят ему видимых повреждений. Клещи лока-
лизуются в местах питания и за немногими исключениями там же
и размножаются, когда наступают благоприятные температурные
условия. Исключение представляют специализированные формы, пе-
реживающие лето в состоянии покоя в сухих щелях древесной коры.
Различают два типа эриофиидных клещей: 1) червеобразные,
мягкие галловые клещи, которые питаются и размножаются в галлах,
и 2) ржавчинные клещи, более широкие и плотные, часто уплощен-
ные, с толстыми спинными щитками, защищающими от света и вы-
сыхания. Эти клещи питаются и размножаются на более или менее
открытых поверхностях листьев. За редкими исключениями ржав-
чинные клещи живут на листьях.
В Северной Америке насчитывается около 50 видов Eriophyidae,
имеющих большее или меньшее хозяйственное значение. Из них,
пожалуй, лучше всего известен Epitrimerus pirifoliae, вызывающий
пузырчатость листьев груши. Этот вид принадлежит к числу наи-
более опасных; на пораженном дереве может погибнуть весь урожай.
Eriophyidae, паразитирующие на виноградной лозе, приносят
меньше вреда главным образом потому, что виноградники постоянно
окуриваются против мильдью. Все же в США широкую известность
получили два вида виноградных клещей Erlophyes vitts (Pgst.) и
Calepitrlmerus •orfe (Сап.). При поражении растения первым клещом
на нижней поверхности листьев образуется налет (эринэум). Повиди-
мому, разные сорта винограда обладают разной устойчивостью
к этому. Так, однажды мы видели виноградные лозы, сплошь по-
крытые налетом, переплетенные с лозами другого сорта, на которых
налета почти не было. Другой вид-—ржавчинный клещ Calepitri-
merus vltls (Can.)—в США мало изучен, Известно, что он имеет
осеннее поколение дейтогинных самок.
Из вредителей цитрусовых в наиболее, жарких районах зоны
выращивания цитрусовых в США широко известен ржавчинный клещ
Tetrapodili
169
Phyllocoptruta oleivorus (Ashm.). Этот клещ, паразитирующий на
вечнозеленых рестениях, зимует на листьях и не имеет специальных
зимующих форм.
Из всех видов Eriophyidae, вредящих цитрусовым, в Калифор-
нии наиболее известен галловый клещ Aceria sheldoni (Ewing),
Рис. 109. Повреждения цветков, почек и плодов лимона, причиняемые
клещом Aceria sheldoni (Ewing) (по Бойсу, 1948).
распространенный во всех прибрежных районах зоны выращивания цит-
русовых. Зараженные им лимонные деревья дают чахлые и гроздевид-
ные побеги и уродливые листья и плоды (рис. 109). Этот клещ вы-
зывает характерное побурение пораженных почек; такие же бурые
пятна появляются и в завязях плодов, под которыми этот клещ
встречается иногда в очень большом количестве.
Aceria sheldoni (Ewing) поражает также апельсиновые деревья и
грейпфрут. Они, невидимому, распространены во всех районах зоны
170
Глава V. Подотряд Trombidiformes
выращивания цитрусовых, примыкающих к Тихоокеанскому побе-
режью. Весьма схожие, а возможно, и тождественные клещи парази-
тируют на цитрусовых на Гавайских островах, в Австралии и на Яве.
На персиковых деревьях паразитирует так называемый серебри-
стый клещ Vasates cornutus (Banks). Дейтогинные самки этого клеща
зимуют около боковых почек, на 8—25 см ниже верхушечных.
Ранней весной клещи выходят из зимних убежищ и поражают вер-
хушки нежных растущих побегов; если в это время у дерева-хо-
зяина не имеется листовых желез, выделяющих клейкий секрет, или
если эти железы еще мало развиты, то клещи вызывают продольное
скручивание листьев, которые иногда покрываются множеством мел-
ких желтых точек. Это заболевание не следует смешивать с по-
серебрением листьев, наблюдающимся после продолжительного пита-
ния клещей весной и летом. Персиковый клещ объедает листья
сверху и снизу. Дейтогинные самки появляются только при пониже-
нии температуры осенью.
Клещи Vasates fockeui (Nal. et Trt.), поражающие молодые сли-
вовые деревья, встречаются во всех северных штатах США и
в Канаде. Этот вид очень близок к серебристому персиковому клещу;
никаких морфологических различий между ними до сих пор не об-
наружено.
Орешник и смородина также страдают от галловых клещей. На
орешнике паразитирует Phytoptus avellanae Nal., а на смородине —
Cecidophyes rlbis Nal. У некоторых сортов орешника зараженные
верхушечные почки весной сильно увеличиваются и превращаются
в галлы, не давая побегов. Phytoptus avellanae Nal. уникален в том
отношении, что он имеет два типа нимф: нормальных, развивающихся
в пораженных увеличенных почках, и уплощенных, которые проводят
лето на нижней поверхности листьев.
Клещ Aceria essegi (Hassan), паразитирующий на ежевике, встре-
чается по всему Тихоокеанскому побережью США. Этот клещ сам
подготовляет себе место для размножения. На развивающихся ягодах,
которыми питается этот клещ, все или, по крайней мере, нижние
костянки не чернеют, оставаясь красными. Пораженные ягоды не со-
зревают и непригодны для еды.
Еще более своеобразным образом жизни отличается клещ Aceria
tulipae (К), поражающий луковичные растения — лук, чеснок, тюль-
пан и т. д. Клещи живут под землей, между чешуйками луковиц.
Эта особенность, насколько известно, не встречается ни у каких
других видов клещей. Aceria tulipae вызывает сморщивание и высы-
хание луковиц; вместе с пораженными луковицами клеща завозят на
склады, где он может перезимовывать".
ЛИТЕРАТУРА
Baker Е. W., 1939. The fig mite, Eriophyes ficus Cotte, and other mites
of the fig tree, Ficus carica Linn., Bull. Calif. Dept. Ayr., 28 (4),
266—275,
Tarsonemini
171
Boyce A. M., Korsmeier R. B., 1941. The citrus btid mite, Eriophyes,
sheldoni Ewing, J. Econ. Erd., 34 (6), 745—756.
Hodgkiss H. E., 1930. The Eriophyidae of New York: II. The maple mites,
N. Y. State Agr. Expt. Sfa. Tech. Bull., 163.
Ke if er H. H., 1938. Eriophyid studies, Bull. Calif. Dept. Agr., 27 (2),
181—206.
Ke if er H. H., 1942. Eriophyid studies XII, Bull. Calif. Dept. Agr., 31 (3),
117-129.
Keif er H. H., 1946. A review of North American economic eriophyid mites,
J. Econ. Ent., 39 (5), 563—570.
Parrott P. J., H о d g к i s s H. E-, S c h о e n e W. S., 1906. The apple and
pear mites, N. Y. State Agr. Expt. Sta. Bull,, 283.
Y о t h e r s W. W., Mason A. C„ 1930. The citrus rust mite and its control,
U. S. Dept. Agr. Tech. Bull., 176.
TARSONEMINI CANNESTRINI ET FANZAGO, 1877
Самки имеют систему трахей, расположенных за педипальпами;
у самцов иногда бывают рудиментарные стигмы. Самки обычно обла-
дают парой латеровентральных бочкообразных псевдостигматических
органов, лежащих между тазиками ног I и II. Ротовые органы реду-
цированы. Обычно выражен половой диморфизм.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ TARSONEMINI
>: 1. Самцы и самки имеют по четыре пары ног.....................2
— Самка имеет одну — три пар ног; у самцов три, редко четыре пары
ног...............................................Podapolipodidae
2. Передний дорзальный щиток тела не образует широкого козырька
над клещом. Ноги IV не кончаются многочисленными бичевидными
щетинками ................................................... 3
— Передний дорзальный щиток образует широкий козырек над кле-
щом. Ноги IV кончаются многочисленными бичевидными щетин-
ками; одна „щетинка" может нести пару коготков и является
сильно видоизмененной частью лапки...................Scutacaridae
3. У самок ноги IV снабжены амбулакральными образованиями (ко-
готки и чашечка)....................Pyemotidae (= Pedtculoididae)
— У самок ноги IV не имеют амбулакрального аппарата и кончаются
терминальными и субтерминальными бичевидными щетинками . .
................................................Tarsonemidae
Podapolipodidae Oudemans, 1939
(рис. ПО)
ДИАГНОЗ
Podapolipodidae паразитируют на насекомых. Самки не имеют круглых
псевдостигматических органов; число ног уменьшено, или же часть ног
дегенерирует; во взрослой фазе самки некоторых форм совсем не
имеют ног. Хелицеры игловидные, пальпы рудиментарные. Ноги имеют
коготки и присоску.
172
Глава V. Подотряд Trombidiformes
Представители этого семейства принадлежат к наиболее дегенери-
ровавшим или специализированным клещам. Podapolipodidae остана-
вливаются в своем развитии на стадии, близкой к личиночной. Первая
фаза представляет собой личинку, имеющую три пары ног; задняя
пара ног отделена от остальных и расположена почти на заднем конце
тела. Пол личинки недифференцирован. На заднем конце тела имеется
Рис. 110. Locustacarus trachealis Ewing.
Молодая самка.
небольшой бугорок, несущий
две срастающиеся друг с дру-
гом длинные бичевидные ще-
тинки; у взрослых эти щетинки
утрачены.
У Locustacarus trachealis
Ewing половозрелые самки со-
храняют внешний вид личинки;
в отличие от остальных предста-
вителей этого семейства взрос-
лые особи Locustacarus trachea-
lis сохраняют задние щетинки.
Из всех Podapolpodidae наименее
дегенерировавшие формы при-
надлежат к роду Tarsopolipus.
Самцы этого рода имеют четыре
пары нормальных ног, но по
внешнему виду походят на сам-
цов остальных родов, обладаю-
щих только тремя парами ног.
Самки Tarsopolipus имеют три
пары ног; третья пара распо-
ложена в задней части тела.
В период паразитической жизни
тело сильно раздувается. В роде Eutarsopolipus как самцы, так и самки
имеют три пары ног и этим походят на личинок. У самок в период
паразитической жизни гистеросома вытягивается так, что третья пара
ног отодвигается назад, придавая клещу колбообразную форму.
У Tetrapolipas самцы имеют три пары ног, самки же утрачивают
третью пару ног. Ноги I и II очень короткие, но сохраняют нор-
мальное расчленение. В период паразитической жизни тело Tetrapollpus
раздувается настолько, что ширина его в три раза превышает длину.
Самец Podapolipus имеет три пары ног, взрослые же самки пред-
ставляют собой совершенно дегенерировавшие формы, сохраняющие
после превращения только первую пару ног; ноги эти крайне корот-
кие, но разделены на 3—5 члеников. В том месте, где должны
были бы причленяться отсутствующие ноги II, имеются два выроста,
которые идут по сторонам гнатосомы и придают переднему концу
тела характерный вид. Тело Podapolipus может сильно раздуваться;
форма его зависит от различных внешних влияний,
Tarsonemlnt
173
РОДЫ
1. Podapolipus Rovelli et Grassi, 1888 (= Plmelofia Tragardh, 1902)
Тип рода Podapolipus reconditus Rovelli et Grassi, 1888
2. Eutarsopolipus Berlese, 1913
Тип рода Eutarsopolipus lagenaeformis Berlese, 1913
3. Locustacarus Ewing, 1924
Тип рода Locustacarus trachealis Ewing, 1924
4. Tarsopolipus Berlese, 1911
Тип рода Tarsopolipus corrugatus Berlese, 1911
5. Tetrapollpus Berlese, 1911
Тип рода Tetrapollpus batocerae Berlese, 1911
ОБЩИЕ ЗАМЕЧАНИЯ
Эти клещи паразитируют на различных членистоногих. Клещ Locus-
tacarus trachealis Ewing паразитирует в трахеях кузнечиков США
и Южной Африки, Tetrapolipus rhynchophori Ewing был найден в Пана-
ме под надкрыльями пальмового долгоносика Rhynchophorus palma-
rum, a Podapolipus reconditus Rovelli et Grassi — под надкрыльями
некоторых европейских жуков. Podapolipus grassi Berlese, P. bacileus
Berlese и P. diander Volkonsky являются наружными паразитами
Orthoptera.
Наиболее полное исследование этих клещей принадлежит Волкон-
скому (1940), изучавшему Р. diander, паразита саранчи Locusta mlg-
ratoria L. в Алжире. На саранче первых трех возрастов моло-
дые самки этих клещей держатся под задним выступом пронотума.
На саранче четвертого и пятого возрастов клещи переходят на
тело и прикрепляются под ориментами надкрылий и крыльев, где и
откладывают яйца. Когда саранча линяет, незрелые клещи, только
что вылупившиеся из яиц и еще не насосавшиеся, а также не отло-
жившие яиц самки переползают с линочной шкурки обратно на хозяина.
11а взрослой саранче клещи живут в складках между среднегрудыо
и заднегрудью, на частях груди, покрытых надкрыльями, на нижней
поверхности надкрылий — у основания последних и вдоль главных жилок
крыльев, обычно с нижней стороны. Взрослые самки Podapolipus dian-
der линяют из самок первой стадии. Эти взрослые самки созревают
и откладывают партеногенетически одно яйцо, из которого немедленно
вылупляется „маленький" самец. „Маленький" самец оплодотворяет
породившую его самку, после чего последняя откладывает яйца, из
которых через пять-шесть дней выходят самки первой стадии и
„большие" самцы. „Большие" самцы паразитируют на самках, пробо-
цая их брюшко, а иногда убивая их. Самки первой стадии развития
прикрепляются к межсегментным перепонкам саранчи, где они питаются,
прокалывая кожный покров хозяина. Постепенно самки первой стадии
мигрируют к заднему к!онцу тела саранчи и концентрируются у гени-
талий последней. Клещи переходят с одной особи саранчи на другую
при их спаривании. Если температура повышается до 27°, то многие
174
Глава V. Подотряд Trombidiformes
клещи ночью покидают хозяина и забираются в трещины в почве
или заползают на верхушки травинок; при этом они пытаются при-
крепиться к любому предмету, с которым приходят в соприкосновение.
Если они прикрепляются к саранче Locasta migratoria, то их жизнен-
ный цикл продолжается; в противном же случае клещи погибают.
Клещи могут развиваться непрерывно, в течение всего лета, и пере-
зимовывать на саранче в фазе яйца или самок первой стадии.
Locustacarus trachealis Ewing на всех фазах превращения парази-
тирует в главных трахеях и воздушных мешках саранчи. Клещи,
повидимому, питаются, прокалывая стенки трахей и воздушных меш-
ков и высасывая кровь хозяина. В период вынашивания потомства
тело самки увеличивается в несколько раз; самка иногда содержит
в половых путях от 6 до 8 вполне зрелых яиц и значительно большее
число их на разных стадиях развития. Locustacarus trachealis отклады-
вает яйца в трахеях хозяина.
ЛИТЕРАТУРА
Ewing Н. Е., 1924. New tarsonemid mites (Order Acarina, family Tarsonemi-
dae), Proc. Ent. Soc. Wash., 26 (3), 66—69.
Rovelli G., Grass i B., 1888. Di tin singolare acaride, Podapolipus recon-
ditus, nobis, Bol. Soc. Ent. Ital., 20, 59—63, Pl. XV.
Volkonsky M., 1940. Podapolipus diander, n. sp., acarien heterostigmate
parasite du criquet migratcur (Locasta migratoria L.), Archiv. Inst.
Pastear Algeril, 18 (3), 321—340.
Wohrle L. P., Welch P. S., 1925. The occurrence of mites in the tracheal
system of certain orthoptera, Ann. Ent. Soc. Amer., 28 (1), 35—44.
Scutacaridae Oudemans, 1916
(рис. Ill)
ДИАГНОЗ
Спинные щитки самки разделены на тергиты, передний щиток
покрывает гнатосому как козырек. Тело обычно круглое и плоское.
Рис. 111. Scutacarus medio-
critarsus Vitzthum
(по Фитцтуму, 1925).
Самка (вид снизу).
У самок ноги IV удлинены и иногда имеют коготки, кончающиеся
крепкими щетинками.
Tarsonemini
175
РОДЫ И ПОДРОДЫ
1. Scutacarus Gros, 1845 (= Disparipes Michael, 1884)
Тип рода Acarus acarorutn Goeze, 1780 (= Scutacarus femoris
Gros, 1845 = Disparipes bombi Michael, 1884)
2. Acarapts Hirst, 1921
Тип рода Tarsonemus woodi Rennie, 1921
3. Diversipes Berlese, 1903
a) Diversipes s. str.
Тип подрода Disparipes exhamulatus Michael, 1886
6) Microdispus Paoli, 1911
Тип подрода Microdispus obovatus Paoli, 1911
4. Imparipes Berlese, 1903
a) Imparipes s. str.
Тип подрода Imparipes hystricinus Berlese, 1903
6) Heterodispus Paoli, 1911
Тип подрода Imparipes elongatus Tragardh, 1904
5. Microdispodides Vitzthum, 1914
Тип рода Pedlculoides amaniensts Oudemans, 1914
(= Microdispodides wichmanni Vitzthum)
6. Pygmodispus Paoli, 1911
a) Pygmodispus s. str.
Тип подрода Pygmodispus equestris Paoli, 1911
6) Allodispus Paoli, 1911
Тип подрода Pygmodispus {Allodispus) latisternus Paoli, 1911
7. Variatipes Paoli, 1911
Тип рода Dlspaires nudus Berlese, 1886
ОБЩИЕ ЗАМЕЧАНИЯ
Scutacaridae — крошечные, странные существа, встречающиеся по
всему земному шару во мху, в почве и дерне, а также на разных насе-
комых, например на муравьях Formica и Lasius. Один из членов этого
семейства паразитирует на медоносной пчеле. Это распространенный
в Европе Acarapis woodi (Rennie), являющийся возбудителем „болезни
о-ва Уайт“. Acarapis woodi паразитирует в трахеях пчел и иногда
вызывает смерть хозяина. Все фазы превращения протекают внутри
трахей, но взрослые клещи встречаются и в наружном опушении заражен-
ных пчел; они могут переходить с одной пчелы на другую. Acarapis woodi
наносит пчеле тройной вред: 1) питается гемолимфой хозяина и тем
самым причиняет последнему активные повреждения, 2) вероятно,
выделяет токсический секрет и 3) механически закупоривает трахеи,
преграждая тем самым доступ воздуха к отдельным органам и
к нервным центрам, управляющим движениями пчел. Особенно серьез-
ные расстройства причиняет пчелам механическая закупорка тра-
хей. Пораженные пчелы иногда теряют способность к полету и пи-
танию.
176
Глава V. Подотряд trombidiformes
ЛИТЕРАТУРА
Hirst S., 1922. Mites injurious to domestic animals, Brit. Mas. (Nat. Hist.)
Econ. Ser., № 13.
Jacot A. P., 1936. Two unrecorded species of Scutacaridae from the southern
Appalachians, Canad. Ent., 68, 225—229, figs 1—3.
Paoli C., 1911. Monografia del „Tarsonemidi", Redia, 7, 215—281, pls VII—XI.
Rennie J. et al., 1921. Isles of Wight disease in hive bees, Trans. Roy. Soc.
Edinburgh, 52 (4), № 29, 737—779.
Pyemotidae Oudemans, 1937
(= Pediculoididae Berlese, 1907)
(рис. 112—114)
ДИАГНОЗ
Передний спинной щиток не выступает спереди в виде козырька.
Ноги IV у самок по строению подобны ногам II и III. Самки (за
Рис. 112. Pyemotes ( = Pediculoides) ventricosus (Newport).
Л —самка (вид сверху); К—самец (вид сверху).
исключением рода Acarophenax) имеют булавовидный орган между
тазиками ног I и II. Хелицеры стилетообразные; пальпы небольшие и
простые.
РОДЫ
1. Pyemotes Amerling, 1862 (= Heteropus Newport, 1850 nomen
praeoccupatum, = Pediculoides Targioni-Tozzetti, 1878)
Тип рода Pyemotes eccoptogasteri prunl Amerling, 1862
2. Acarophenax Newstead et Duvall, 1918
Тип рода Acarophenax tribolii Newstead et Duvall, 1918
3. Pavania Lombardini, 1949
Тип рода Pavania fusiformis Lombardini, 1949
Tarsonemini
177
4. Phthlroides Oudemans, 1937
, Тип рода Phthlroides megnini Oudemans, 1937
5. Pygmephorus Kramer, 1877
Тип рода Pygmephorus spinosus Kramer, 1877
6. Resinacarus Vitzthum, 1927
Тип рода Resinacarus resinatus Vitzthum, 1927
1’ и c. 113. Pyemotes ( = Pedicu-
loides) ventricosus (Newport).
Оплодотворенная самка, задняя часть
гола которой растянута развивающи-
мися эмбрионами.
Рис. 114. Siteroptes ( = Pediculopsis) gra-
minum (Reuter).
Дыхательная система беременной самки.
7. Siteroptes Amerling, 1861 (= Therlsmoptes Amerling, 1861 = Pedi-
culopsis Reuter, 1907)
Тип рода Acarus graminlsugus Hardy, 1851
8. Tarsonemoldes Tragardh, 1904
Тип рода Tarsonemoldes terniitophilus Tragardh, 1904
ОЫЦИЕ ЗАМЕЧАНИЯ
Жизненный цикл изучен лишь для немногих форм. Клещи Pyemotes
( -= Pediculoides) ventricosus (Newport)—живородящие. Самки рождают
половозрелых самцов и самок. Оплодотворение происходит немедленно
после рождения самки. После этого самка отыскивает подходящего хо-
шина и питается на нем, прокалывая его покровы стилетообразнымц
12 Зак. 1785
178 Глава У. Подотряд Trambidiformes
хелицерами. В период питания конец брюшка самки крайне разрастается,
превращаясь в мешок, в котором созревают яйца. Клещи проходят все фа-
зы развития в теле матери и рождаются уже вполне зрелыми. Первыми
на свет появляются самцы, число которых обычно невелико. Самцы до-
бывают себе пищу, прокалывая материнский „шар"; пищи им требуется
очень немного. Самцы не покидают тело матери, постепенно скапли-
ваются у ее полового отверстия и оплодотворяют новорожденных самок
по мере их появления. Беременная самка, достигшая полного роста,
рождает до 203—300 особей, сохраняя при этом жизнеспособность.
Клещ Siteroptes (= Pediculopsis) graminum (Reuter) отличается
еще более интересным жизненным циклом. За время питания гисте-
росома самки увеличивается в 100—500 раз; в образующемся мешке
развиваются яйца, из которых выходят половозрелые клещи. Вылуп-
ление всего потомства происходит одновременно и сопровождается
разрывом гистеросомы; лишь отдельные ранние личинки, нимфы и
особенно самцы могут появляться на свет и до момента массового
вылупления. Самцов рождается очень мало, но в каждом поколении
имеется не менее одного самца. Только что вылупившиеся клещи
спариваются как до рождения — внутри материнского мешка, так и
после рождения. Неоплодотворенные самки рождают самцов с гапло-
идным набором хромосом. У этого вида, так же как у Pyemotes,
имеются гетероморфные самцы.
Другие члены этого семейства мало известны и, повидимому,
хозяйственного значения не имеют. Бэнкс (1915) обнаружил мигрирую-
щие формы Pygmephorus на кротах, на мухах и на трипсах. Фитц-
тум указывает, что Pygmephorus— плотоядны. Pygmephorus ameri-
•canus Banks был описан как „псевдопаразит" Musca domeslica и Sto-
moxys calcitrans. Джекот (1936) описал несколько видов с восточного
побережья США и составил таблицу для определения наиболее рас-
пространенных родов — Pyemotes ( = Pediculoid.es), Siteroptes ( = Pedi-
culopsis), Pygmephorus и Resinacarus.
Pyemotes ( = Pediculoides) ventiicosus (Newpot) является возбу-
дителем сенной, или зерновой, чесотки. Нормально он известен как
эктопаразит личинок различных насекомых, например зерновой моли
Sitohoga cerealella (Oliv'er), шелковистой моли Stllpnotia salicis (L.),
фруктовой полосатой моли Anarsia lineatelia Zeller и многих других,
но при обмолоте, хранении или погрузке зерна, содержащего зара-
женных личинок зерновой моли, клещи переходят на человека, осо-
бенно в жаркую погоду, когда они испытывают потребность во влаге.
Клещи Р. ventricosus предпочитают руки и шею, но поражают и
другие части тела и вызывают сильный зуд. На пораженном человеке
находят иногда следы сотен укусов. Хотя эти клещи не проникают
под кожу и долго на человеке не удерживаются, они все же вызы-
вают значительное раздражение кожи. Сван (1934) описывает сенную
чесотку следующими словами: „Следы укусов варьируют по величине
и форме у разных лиц. Они более четко выражены после расчесыва-
Tarsonentini
179
ния и имеют вид беловатого прыща с небольшим пузырьком в сере-
дине, отмечающим место укуса. Побелевший участок окружен розо-
вато-красной каймой. Места укусов сильно чешутся, особенно в тепле
(например, в постели или при физических упражнениях). При расче-
сывании центральный пузырек обычно лопается, причем возникает
возможность вторичной инфекции. Зуд обычно утихает через два-три
дня, но следы укусов сохраняются дольше". Уэбстер (1910) указы-
вает, что в тяжелых случаях сенная чесотка может сопровождаться
генерализованными симптомами, например „повышением температуры
до 37—39°; известен случай, когда температура достигла 40°. Пульс
в тяжелых случаях учащается до 100—110, а в одном случае — даже
до 130 уд./мин. Появляются такие симптомы, как сильная головная
боль, потеря аппетита, тошнота, в некоторых случаях рвота и не-
сильный понос. В очень тяжелых случаях бывают жалобы на боли
в суставах и спине; при этом в моче удалось обнаружить небольшое
количество белка, но ни цилиндров, ни крови не было. С исчезнове-
нием острых симптомов белок в моче исчезал". Имеются сообщения также
о случае астмы, вызываемой этими клещами. Обычно считается, что
Р. ventricosus не имеет особого значения в борьбе с вредными насе-
комыми, но в лабораторных условиях этот клещ может полностью
уничтожить колонии личинок. Р. ventricosus имеет, невидимому, по-
всеместное распространение. До последнего времени был известен
только один вид этого рода — Pyemotes (= Pediculoides) ventricosus,
но в 1937 г. Удеманс описал еще несколько других форм.
Siteroptes { — Pediculopsis) graminum (Reuter) внешне напоминает
предыдущий вид, но является вредителем трав и зерновых культур,
причиняя большой вред пшенице, овсу, ячменю и ржи. Кроме того,
этот клещ в США служит активным переносчиком спор ржавчины
Sporotrichum роае. Пораженные зерновые малорослы, уродливы и
часто приобретают серебристый оттенок, чему и обязано название
Гюлезни—„белоколосость“.
ЛИТЕРАТУРА
Cooper К. W., 1937. Reproductive behaviour and haploid parthenogenesis
in the grass mite, Pediculopsis graminum (Rent.) (Acarina, Tarsonemidae),
Proc. Natl. Acad. Set., 23 (2), 41—44.
Cooper K. W., 1949. Relations of Pediculopsis graminum ani Fusarium
poae to central but rot of carnation, Phytopathology, 30 (10), 853—859.
II erf s A., 1926. Okologische Untersuchungen an Pediculoides ventricosus
(Newp.), Berl. Zoologica, H. 74, Stuttgart.
.I.i cot A. P., 1936. An undescribed mite from the southern Appalachians,
Canad. Ent., 68 (4), 82—85.
tHidemans A. C., 1937. Tijdschr. v. Ent., 80, Verslag IV—VIII.
Swan D. C., 1934. The hay itch mite, Pediculoides ventricosus (Newp.)
(Acarina, Pedicttloididae) in South Australia, J. Agr. South Austral., 37,
1289—1299.
Webster F. M., 1910. A predaceous and supposedly beneficial mite, Pedi-
cttloides, becomes obnoxious to man, Ann. Ent. Soc. Amer., 3 (1), 15—39,
pls. Ill—V.
12*
180
Глава V. Подотряд Trombidiformes
Tarsonemidae Kramer, 1877
(рис. 115—116)
ДИАГНОЗ
Tarsonemidae имеют сегментированное тело. Передний спинной
щиток этих клещей не выступает впереди в виде козырька. У самок
ноги IV отличаются от ног И и III тем, что несут на конце апикаль-
ные и субапикальные бичевидные щетинки. У Tarsonemella и Traso-
nemus typhae Oudemans, 1929, между тазиками ног I и II отсут-
ствует имеющийся у остальных видов булавовидный орган. Хелицеры
Рис. 115. Tarsonemus setifer Ewing.
А — самец (вид сверху); Б — самка (вид сверху).
стилетообразные, пальпы простые. В этом семействе имеются гетеро-
морфные самцы. Группа родов, объединяемая ныне в семейство Tar-
sonemidae, имеет, повидимому, разное происхождение ; и поэтому
требует пересмотра.
РОДЫ
1. Tarsonemus Canestrini et Fanzago, 1876
Тип рода Chironemus minusculus Canestrini et Fanzago, 1876
2. Avrosia Oudemans, 1928
Тип рода Acarus translucens Nietner, 1861
3. Hemitarsonemus Ewing, 1939
Тип рода Tarsonemus tepidariorum Warburton, 1904
Tarsonemint
181
4. Pseudotarsone moides Vitzthum, 1921
Тип рода Pseudotarsonemoides eccoptogasteris Vitzthum, 1921
5. Torsonetnella Hirst, 1923
Тип рода Tarsonemella africana Hirst, 1923
ОБЩИЕ ЗАМЕЧАНИЯ
Tarsonemidae обладают крайне упрощенным жизненным циклом.
Имеется всего четыре фазы: яйцо, личинка, нимфальная покоющаяся
фаза, в течение которой личинка превращается в половозрелую взрослую
особь (активных нимф нет), и половозрелая фаза. Самцы гетероморфны;
четвертая пара ног используется для схватывания
самки во время копуляции. Строение этой пары ног р х.
имеет очень большое значение в качестве диагности- \ \
ческого признака при определении вида (рис. 116). 'Ps—<—7
Самки откладывают одновременно *по одному яйцу. \ /
Гистеросома, имеющая ряд частично перекрывающих к
друг друга члеников, допускает растяжение туло- /
нища. Tarsonemidae, невидимому, предпочитают сы- /у-—. «У /
рые, укрытые места и часто исчезают с наступле- \\ >/
нием высокой температуры. / 'у
Эта группа клещей широко распространена; их рИ€ jjg т
можно встретить во всех теплых странах. Большин- nem'us ' phyllo-
ство форм паразитирует на растениях и имеет хозяй- pherus Ewing
iтвенное значение, но известны также хищные и са- (по Эвингу, 1939).
профитные формы. По приведенному ниже перечню нога iv самца,
видов, наносящих вред своему хозяину, можно судить
о разнообразии растений, поражаемых клещами Tarsonemidae, и о хо-
зяйственном значении этого семейства.
Tarsonemus pallidus Banks является опасным вредителем цикламенов
и США и земляники в Европе. Т. VDaitei Banks повреждает персико-
вые деревья во многих районах США; Т. spirifex Marchal наносит
большой вред овсу в Европе; в США этот вид известен в Аризоне
и Канзасе. Луковый клещ Т. laticeps Halbert поражает луковицы
нарцисса в Европе и США, уменьшая количество приносимых цветов.
Г. ananas Tryon служит переносчиком ржавчины ананаса в Австралии:
I lemitarsonemus latus (Banks) известен по всему миру как вредитель
самых разнообразных растений: чая — на Цейлоне, Cinchona sp. — на
Суматре, Hevea sp.—в Индонезии, томатов, цикламенов и дельфини-
умов— в США, хлопка—-в Бельгийском Конго.
Известны случаи нахождения Tarsonemus sp. в легких лиц, стра-
дающих астмой и другими заболеваниями дыхательных путей.
ЛИТЕРАТУРА
Carter Н., W е d d М., D ’A b г е г а V. St. Е., 1944. The occurrence of mites
(Acarina) in human sputum and their possible significance, India Med,
Gaz., 79 (4), 163—168.
182
Глава V. Подотряд Trombidiformes
Ewing Н. Е., 1939. A revision of the mites of the subfamily Tarsoneminae of
North America, the West Indies, and the Hawaiian Islands, U. S. Dept.
Agr. Tech. Bull., 653.
Hambleton E. J., 1938. A ocorrencia do dcaro tropical „Tarsonemus latus
Banks" (Acar. Tarsonemidae). Causador da rasgadura das folhas nos
algodoais de S. Paulo, Arch. Inst. Biol. (San Paulo), 9 (19), 201—209,
pls. 19—23.
Moznette G. F., 1917. The cyclamen mite, J. Agr. Res., 10 (8), 373—390.
S о у s a E., J a у awardena M., 1945. Pulmonary acariasis: a possible cause
of asthma, Brit. Med. J., № 4383, 1—6.
Vrydagh J. M., 1942. Etude de 1’acariose du cotonnier, causee par Hemi-
tarsonemus latus (Banks) au Congo Beige, Pubs Inst. Natl, pour I’Etude
Agron. du Congo Beige, Ser. Sci., № 28.
PROSTIGMATA KRAMER, 1877
У этих клещей стигмы открываются на основании или у основа-
ния хелицер.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ PROSTIGMATA
1. Тело обычной формы, не червеобразное и не кольчатое. ... 2
— Тело маленькое, червеобразное, кольчатое. Всегда имеют восемь
ног. Паразитируют в кожных железах................Demodicidae
2. Наземные формы...........................................3
— Водные формы.................................Hydrachnellae1
3. На пальпах нет комплекса палец — коготок.................4
— На пальцах имеется комплекс палец — коготок, состоящий из
коготка на голенном членике пальпы и расположенного вентрально
пальца (тарзального членика пальпы) ................... 21
4. Гнатосома вытянута в очень длинный конус ............... 5
— Гнатосома короткая и не имеет конической формы .......... 7
5. Гнатосома не заключена в футляр, образованный стенкой тела . (>
— Гнатосома заключена в футляр, образованный продолжением стенки
тела. Тело жесткое. Проподосома и гистеросома не разделены швом.
Хелицеры удлиненные и имеют небольшую дистальную клешню.
Половых присосок две пары.....................Cryptognathidae
6. Пальпы длинные, направленные внутрь; дистальный членик ког-
тевидный и приспособлен для схватывания. Тело с наружными
щитками. Половых присосок две пары.................Cunaxidae
— Пальпы длинные, коленчатые и превращены в сенсорный орган;
дистальный членик пальпы обычно снабжен двумя длинными сен-
сорными щетинками. Хелицеры удлиненные и имеют каждая не-
большую дистальную клешню. Тело мягкое, без поверхностных
щитков. Половых присосок три пары..................Bdellidae
1 Для простоты изложения Hydrachnellae („водяные клещи") рассматри-
ваются как самостоятельная группа. Их описанию посвящена гл. VI.
Prostigmata
183
7. Подвижный палец хелицер острый, небольшой, приспособлен
для прокалывания и не противопоставляется неподвижному . . 8
— Подвижный палец хелицер обычно снабжен зубцами, противо-
поставляется неподвижному и приспособлен для кусания и хвата-
ния .............................................................12
— Подвижный палец хелицер длинный, бичевидный и приспособлен
для прокалывания. Имеется стилофор. Мелкие клещи, близкие
к Tetranichidae, но не имеющие на пальпах комплекса палец—•
коготок.........................................Pliytoptipalpidae
8. Половых присосок две пары..................................9
— Половые присоски отсутствуют...........................10
9. На проподосоме и на гистеросоме имеется по паре длинных тон-
ких сенсорных щетинок. Перитремы отсутствуют . . Ereynatidae
— Сенсорные щетинки развиты только на проподосоме. Перитремы
имеются. На лапках небольшие когтеобразные подушечки . . .
..................................................Paratydeidae
10. Перитремы отсутствуют. Ноги I нормальные. Свободноживущие
формы..........................................................11
— Перитремы имеются. Хелицеры игловидные, выдвижные и приспо-
соблены для прокалывания. Пальпы крайне малы и различимы с
трудом. Ноги I во многих случаях хорошо приспособлены для
обхватывания волосков................................Myobiidae1
И. Ноги покрыты сетчатым орнаментом. Подвижный палец клешни
очень маленький и почти незаметен. Паразиты носовой полости
птиц............................................Speleognathidae
— Ноги не покрыты сетчатым орнаментом. Подвижный палец клешни
большой и вполне различим невооруженным глазом. Свободно-
живущие формы........................................ Tydeidae
12. На проподосоме нет хорошо развитых псевдостигм или псевдо-
стигмальных органов..........................................13
— На проподосоме имеются хорошо развитые псевдостигмы или
псевдостигмальные органы (Endeostigmata по Гранжану, 1939)
.................................................................16
13. На лапках ног I имеется „рагидиев орган". Тазики ног I—II и
III—IV расположены двумя отдельными группами...................14
— „Рагидиева органа" на лапках ног I нет. Тазики ног пластинча-
тые и соприкасаются друг с другом. Имеются линзовидные глаза и,
кроме того, позади глаз обычно имеются линзовидные органы.
Половых присосок две пары. Концевой членик пальп маленький,
заостренный и несет длинные терминальные щетинки................
...............................................Lab’dostommidae
14. Клешни хелицер маленькие, искривленные и не приспособлены
для хватания...................................................15
1 Повидимому, разнородная группа родов, близкая к CheyletMae, но ли-
шенная сильно развитых пальп. Комплекс палец — коготок имеется, но ввиду
малых размеров различим с трудом.
184
Глава V. Подотряд Trombidiformes
— Клешни хелицер крупные и приспособлены для схватывания.
Тело мягкое. Половых присосок две пары. Концевой членик пальп
большой, закругленный и несет многочисленные короткие щетинки
....................................................Rhagidiidae
15. Тело мягкое. Козырька над рострумом нет. Половых присосок
две пары..............................................Eupodidae
— Тело покрыто твердым панцырем. Имеется козырек, нависающий
над рострумом. Половых присосок две пары . . . Penthalodidae
16. Половых присосок три пары. Глаза иногда отсутствуют . . 17
— Половых присосок две пары. Глаза иногда отсутствуют ... 18
17. Проподосома несет лишь одну пару сенсорных щетинок ....
.....................................................Terpnacaridae
— Проподосома несет две пары сенсорных щетинок . Pachygnathidae
18. Проподосома несет две пары сенсорных щетинок................19
— Проподосома несет лишь одну пару сенсорных щетинок . Лапки
с одним коготком. Глаза не развиты..................Alicorhagiidae
19. Лапки имеют по два или по три коготка.......................20
— Лапки с одним коготком. Ноги IV приспособлены для прыгания.
Глаза имеются, повидимому, у всех родов . . . Nanorchestidae
20. Ноги I—IV имеют по три коготка. Глаза не развиты. Передняя
пара сенсорных щетинок сидит в специальной ямке. Неподвиж-
ный палец клешни хелицер почти полностью редуцирован . . .
................................................Lordalychidae
— Ноги I имеют по два коготка, ноги II—IV — по три коготка. Глаз
две пары. Специальные ямки для сенсорных щетинок не раз-
виты. Неподвижный палец клешни хелицер нормальный . . .
..............................................Sphaerolichidae
21. Подвижный палец клешни хелицер преобразован в прямой, сти-
летообразный, выдвижной колющий орган...........................22
— Подвижный палец клешни хелицер мощный, невыдвижной, по-
движно причленен к основной части хелицеры и приспособлен для
прокалывания .................................................. 26
22. Подвижный палец клешни хелицер прямой, жесткий, различной
длины. Стилофор (мандибулярная пластинка) отсутствует . . 23
— Подвижный палец клешни хелицер длинный, бичевидный и изги-
бается внутри тела. Основные членики хелицер слиты и образуют
стилофоры (мандибулярные пластинки). Гребень отсутствует. Раз-
виты линзовидные глаза. Половые присоски отсутствуют 1 . . .
................................................Tetranychidae
23. Хелицеры длинные, узкие, могут втягиваться глубоко в тело. Имеются
дорзальные сенсорные поля и сенсорные щетинки...................24
— Хелицеры короткие и не могут втягиваться глубоко в тело. Пери-
трема выступает на гнатосоме в виде буквы М. Дорзальные сенсорные
1 К этому семейству близки Phytoptipalpidae, но они не имеют комплек-
са палец — коготок на пальпах.
Prostigmata
185
поля отсутствуют. Крупные пальпы снабжены клешней и исполь-
зуются для хватания..............................Cheyletidae
24. Половые присоски не развиты. Имеются сенсорные поля и сре-
динный киль или только один киль.............................25
— Половые присоски имеются. Сенсорные поля отсутствуют. Пальпы
небольшие. Все ротовые органы могут втягиваться в тело . . .
............................................Calyptostomidae
25. Ротовые органы небольшие, могут втягиваться в тело. Проподо-
сома обычно вытянута в виде длинного носика . . Smaridiidae
Ротовые органы крупные и не могут втягиваться в тело. Пропо-
досома не вытянута в длинный носик ...... Erythraeidae
26. Тело не покрыто мощным панцырем. Щетинки на ногах нормаль-
ные ........................................................ 27
— Имеется несколько сильно склеротизированных спинных щитков.
Ноги I и II несут несколько больших шипов или бугорков, осо-
бенно крупных на ногах I; эти типы расположены наподобие
зубьев грабель.......................................Caeculidae
27. Палец пальп небольшой и иногда различим с трудом ... 28
- Палец пальп крупный и хорошо заметен......................29
28. Половых присосок три пары. Дорзальные сенсорные щетинки
иногда сидят в характерных ямках, или псевдостигмах. Коготки
на лапках крупные, зубчатые.......................Teneriffiidae
- - Половые присоски не развиты. Специализированных сенсорных
щетинок на спине нет. Коготки на лапках не зубчатые. Пропо-
досома и гистеросома не разделены швом. Как правило, формы,
паразитирующие на ящерицах под чешуйками . . Pterygosomidae
— Половые присоски не развиты. Проподосома несет две пары сен-
сорных щетинок. Лапки ног имеют присоски или коготки. Тело
разделено швом. Свободноживущие клещи . . . Pseudocheylidae
29. Тарзальный членик пальп вентральный.......................30
Тарзальный членик пальп дистальный. Крупные красные клещи
с радиально расположенными ногами. Перитремы свободные . . .
..................................................Anystidae
30. Большие клещи с сенсорными полями на проподосоме ... 32
- Небольшие клещи. Дорзальные сенсорные поля отсутствуют . 31
31. Половых присосок три пары. Глаза не развиты. Лапки вооружены
коготками, но не имеют подушечки. Лапка ноги I несет семь
палочкообразных сенсорных щетинок................Pomerantziidae
— Половые присоски не развиты. Имеется одна или две пары глаз.
Лапки имеют коготки и подушечку. Лапка ноги I несет только
одну палочкообразную сенсорную щетинку . . . Raphignathidae
32. Взрослые особи не имеют 8-образной формы. Тектум, если он
имеется, покрыт многочисленными щетинками. Личинки обычно
имеют более одного спинного щитка. Щетинки на тазиках педи-
пальп обычно расположены впереди бедренного членика пальп
................................................ Trombidiidae
186
Глава V. Подотряд Trombidiformes
— Взрослые особи обычно имеют 8-образную форму. Тектум несет
только одну или две щетинки. Личинки имеют один спинной
щиток (за исключением одного рода, обладающего пигидиальным
щитком). Щетинки на тазиках педипальп расположены позади
бедренного членика пальп...............................Trombiculidae
Eupodidae Koch, 1842
( = Penthaleidae Oudemans, 1931)
(рис. 117)
ДИАГНОЗ
Eupodidae—подвижные, крайне хрупкие клещи небольшой или
средней величины. Длина тела варьирует от 0,160 до 1,0 мм. Их
грушевидное, суживающееся кзади тело имеет желтоватую или зеле-
новатую, реже пятнистую окраску; ноги обычно красноватые, реже
Рис. 117. Penthaleus major (Duges).
А — самка (вид сбоку); экземпляр несколько сплющен при изготовлении
препарата. Видно расположенное на спине анальное отверстие;
Б — хелицера.
черные с красными пятнами (Penthaleus, Halotydeus). Проподосома
отделена от гистеросомы в различной степени выраженным швом.
Кожа этих клещей нежная, мелкобороздчатая и покрыта редкими
волосками. На проподосоме по краям имеется по глазку и пара
псевдостигм со слабо дифференцированными сенсорными щетинками.
У Linopodes ноги I почти в четыре раза длиннее тела. У Eupodes
ноги IV несколько толще остальных ног и часто имеют очень толстые
бедра. Лапки имеют по два коготка и опушенный эмподий. Пальпы
состоят из четырех подвижных члеников, хелицеры клешнеобразные,
НО их клешня невелика, несколько деформирована или видоизменена.
Prosilgmata
187
Половых присосок две пары. У Penthaleus анальное отверстие располо-
жено на спинке, у Halotydeus — терминально, у остальных родов — на
брюшке.
Европейские исследователи разделяют Eupodidae Koch, 1842, на
три семейства: Penthaleidae Oudemans, 1931, Penthalodidae Thor, 1933,
и Eupodidae. Признаки, использованные для выделения Penthalodidae
в особое семейство, достаточно убедительны, но выделение семейства
Penthaleidae нельзя считать достаточно обоснованным. Поэтому в на-
стоящей работе Penthaleidae {Penthaleus и Halotydeus) вновь отнесены
к Eupodidae.
РОДЫ
1. Eupodes Koch, 1836
Тип рода Eupodes hiemalls Koch, 1838
2. Cocceupodes Thor, 1934
Тип рода Eupodes clavifrons R. Canestrini, 1886
3. Halotydeus Berlese, 1891
Тип рода Notophallus hydrodromus Berlese et Trouessart, 1889
4. Linopodes Koch, 1836
Тип рода Acarus motatorius Linnaeus, 1758
5. Penthaleus Koch, 1836 ( = Notophallus R. Canestrini, 1886)
Тип рода Tetranychus major Duges, 1834 { — Penthaleus haetno-
topus Koch, 1836)
6. Protacarus Hirst, 1923 f
.Тип рода Protacarus crani Hirst, 1923
7. Prctereunetes Berlese, 1923
Тип рода Micrereunetes {Protereunetes) agilis Berlese, 1923
ОБЩИЕ ЗАМЕЧАНИЯ
Жизненный цикл этих клещей прост: яйцо, личинка, нимфа и
имаго. Подробно Eupodidae были изучены на примере Halotydeus
destructor Tucker в Австралии. Период засухи эти клещи переживают
в фазе покоящихся яиц. В октябре самки откладывают большое
число яиц на поверхности почвы, в дерн и т. д.; яйца крайне устой-
чивы к жаре, сухости, суховеям. Для развития этих яиц требуются
влага и солнечный свет. Взрослые Eupodidae имеют мягкое тело,
их передние ноги используются как сенсорные органы. Эти клещи
живут преимущественно в легкой, хорошо дренируемой почве. Они
разрывают поверхность листьев скребущими ротовыми органами и
высасывают сок. Взрослые особи живут 25—50 дней и погибают
летом от высокой температуры и высыхания.
Изучено лишь очень мало видов этого семейства. Linopodes
antennaepes Banks (? возможно, L. motatorius L.) является вреди-
телем грибов. Эти длинноногие клещи отгрызают корешки растущего
растения, в результате чего стебель гриба суживается у основания
и развивающееся плодовое тело повреждается или погибает. Клещи
188
Глава V. Подотряд Trombidiformes
L. antennaepes встречаются на (или у) поверхности грибных гряд-
а также под досками и листьями. Самки откладывают яйца в почву
группами; из яиц вскоре вылупляются личинки, похожие на взрослых
и отличающиеся от последних только числом ног, отсутствием поло-
вых органов и некоторыми другими мелкими подробностями. Нимфы
имеют восемь ног и похожи на взрослых особей. L. antennaepes
встречается в США и Европе.
Клещи Eapodes прекрасно прыгают при помощи специально уве-
личенных ног III. Клещи Eapodes обычно живут во мху и под
листьями.
Имеются сообщения, что Penthaleus major (Duges) повреждает
горох, клевер, овес, дикую горчицу и лупин в Калифорнии, ячмень —
в Аризоне и пшеницу — в Оклахоме и Техасе. Он вредит также
пастбищам и кормовым культурам в Австралии и Южной Африке
и латуку и гороху во Франции.
Halotydeus destructor (Tucker) является опасным вредителем
овощных и бобовых культур в Австралии и Южной Африке.
ЛИТЕРАТУРА
Baker Е. W., 1946. New species of North and Central America mites of the
family Penthaleidae (Acarina), J. Wash. Acad. Sci., 36 (12), 421—425.
Solomon M. E., 1937. Experiments on the effects of temperature and humidity
on the survival of Halotydeus destructor (Tucker), Acarina fam. Pcnthafei-
dae, Austral. J. Expt. Biol. Med. Sci., 15, 1—16.
Solomon M. E., 1937. Behaviour of the red-legged carthmite, Halotydeus
destructor, in relation to environmental conditions, J. Animal Ecol., 2,
340—361.
Thor S., 1934. Einzelne neue, besonders Norwegische Eapodes Arten mit
einigen alteren Arten vcrglichcn, Zool. Anz., 105, 201—215.
Womersley H., 1941. The red-legged earth-mitc (Acarina, Penthaleidae) of
Australia, Trans. Roy. Soc. South Australia, 65 (2), 292—294.
Penthalodidae Thor, 1933
(рис. 118)
ДИАГНОЗ
Тело этих клещей черное, обычно с красными пятнами; членики
ног красные. По величине Penthalodidae колеблются от мелких до
средних, длина тела варьирует у них в пределах 0,330—1,5 мм.
Penthalodidae имеют грушевидное, суживающееся кзади тело. Пропо-
досома и гистеросома или совсем не разделены швом, или последний
только намечен. Наружные покровы сильно склеротизированы и
покрыты сетчатым или точечным орнаментом. По краям проподосомы
имеется по одному глазу (по Трегорду, у Tectopenthalodes глаза
отсутствуют). Щетинки как на ногах, так и на теле короткие, число
их невелико. Имеется пара псевдостигм с сенсорными щетинками.
Ноги длинные, узкие и обычно состоят из шести подвижных члени-
ков. На всех лацках между коготками лежат опушенные эмподии.
Prostigmata
189
Пальпы узкие и состоят из четырех подвижных члеников; клешни
небольшие, ножницеобразные, несколько искривленные. Над рострумом
имеется выступ, напоминающий тектум. Половых присосок две пары.
РОДЫ
1. Penthalodes Murray, 1877 (= Penthaleus Berlese, 1891 = Chro-
motydeas, Berlese, 1903)
Тип рода Mega merits ovalis Duges, 1834
2. Stereotydeus Berlese, 1901
Тип рода Stereotydeus notophalloldes Leonardi, 1901
3. Tectopenthalodes Tr3gSrdh, 1907
Тип рода Penthaleus vlllosus Trouessart, 1903.
ОБЩИЕ ЗАМЕЧАНИЯ
Penthalodidae живут во мху, в гумусе и т. д. Они, повидимому,
хищники, но об их биологии и хозяйственном значении никаких
точных данных нет. Неполовозрелые формы походят на взрослых,
но менее склеротизированы. В 1946 г. Бэкер описал много новых
видов из Северной Америки. Бэкер отнес их тогда к семейству
Penthaleidae, которое в настоящей работе рассматривается как часть
Eupodidae.
ЛИТЕРАТУРА
Baker Е. W., 1946. New species of North and Central American mites of the
family Penthaleidae (Acarina), J. Wash. Acad. Sci., 36 (12), 421—425.
Womersley H., 1935. On some Australian and South African species of
Acarina of the genus Stereotydeus (Penthalodidae), Proc. Linn. Soc.
New South Wales, 60 (1—2), 79—82.
1до
Глава V. Подотряд Trombidiformes
ДИАГНОЗ
Bdellidae Duges, 1834
(рис. 119)
Bdellidae имеют красноватую окраску, но в зависимости от содер-
жимого кишечника могут казаться зеленоватыми, коричневатыми
или же покрываться фиолетовыми пятнами (реже черными, как
Trachymolgas). Это клещи среднего или крупного размера, их длина
Рис. 119. Bdella willisi Baker et
Balock.
Самка (вид сверху); с левой стороны пока-
зан бороздчатый узор покровов.
(без гнатосомы) колеблется между
0,45 и 3,50 мм. Bdellidae имеют удли-
ненное грушевидное тело с острым
рострумом (почему их и называют
„носатыми" клещами). Проподосома
отделена от гистеросомы швом. По-
кровы обычно тонкие, бороздчатые
{Trachymolgas покрыт панцирем, не-
сущим круглые бугорки); на пропо-
досоме часто развит подкожный щи-
ток. По краям проподосомы распо-
ложены четыре псевдостигмы с сен-
сорными щетинками и две пары
разделенных глаз {Cyta и Monotrt-
chobdella имеют непарный срединный
глаз, расположенный на переднем
конце проподосомы). Ноги состоят
из шести подвижных члеников;
каждая из лапок имеет короткую
предлапку с двумя короткими когот-
ками и подушечкой. Пальпы состоят
из пяти подвижных члеников. Колен-
ный членик и голень пальпы очень
короткие, бедро очень длинное, лапка либо цилиндрическая и длинная,
либо короткая, боченкообразная, с двумя концевыми щетинками;
исключение составляют Monotrlchobdella, имеющие только одну
щетинку. Нормально коленный членик и голень согнуты, что придает
пальпам коленчатый вид. Хелицеры удлиненные (у Cyta более ком-
пактные), с одной или несколькими дорзальными щетинками и очень
маленькими и обычно слабыми клешнями. Имеется три пары половых
присосок. Гранжан (1938) разделяет семейство Bdellidae на четыре
подсемейства.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ BDELLIDAE
1. Имеются хорошо развитые половые трахеи. Нижняя сторона
рострума несет всего лишь две пары жестких и две пары мягких
передних щетинок . . . ;.............................2
Prostigmata
191
— Хорошо развитые половые трахеи отсутствуют. Нижняя сторона
рострума несет шесть пар жестких и две пары мягких передних
щетинок...............'..........................................3
2. Перед половым отверстием имеется непарная щетинка . . Cytinae
— Перед половым отверстием имеется пара щетинок . Spinibdellinae
3. На голенях ног I, II и IV и на лапках ног III и IV имеются
трихоботии (длинные сенсорные щетинки) .... Odontoscirinae
— Трихоботии имеются лишь на голенях ног I и IV и на лапках
ног III и IV.............................................Bdellinae
РОДЫ
Cytinae Grandjean, 1938
1. Cyta v. Heyden, 1826
Тип рода Scirus latirostrts Hermann, 1804
2. Trachymolgas Berlese, 1923
Тип рода Bdella nigerrima G. Canestrini et Fanzago, 1876
Spinibdellinae Grandjean, 1938
1. Spinlbdella Thor, 1930
Тип рода Spinlbdella reducta Thor, 1930
2. Bisclrus Thor, 1913
Тип рода Bdella sllvatica Kramer, 1881
3. Monotrichobdella Baker et Balock, 1944
Тип рода Monotrichobdella max-osburni Baker et Balock, 1944
Bdellinae Grandjean, 1938
1. Bdella Latreille, 1795 (= Scirus Hermann, 1804, не Thor, 1931)
Тип рода Acarus longicornis Linnaeus, 1758 (= Scirus vulgaris
Hermann, 1804)
Odontoscirinae Grandjean, 1938
1. Odontoscirus Thor, 1913
Тип рода Bdella virgulata G. Canestrini et Fanzago, 1876
2. Bdellodes Oudemans, 1937 (— Scirus Thor, 1931, не Hermann,
1804).
Тип рода Scirus longlrostris Hermann, 1804.
3. Neomolgus Oudemans, 1937 (= Mol gas Thor, 1931 nomen praeoc-
cupatum)
Тип рода Acarus littoralis Linnaeus, 1758
4. Thoribdella Grandjean, 1938
Тип рода Biscirus meridionalis Thor, 1931.
192
Глава К Подотряд Trombidtformes
Добавление к системе
Перечисленные ниже роды не были правильно описаны и не могут
быть причислены ни к одному из указанных выше подсемейств.
1. Caenobdella Oudemans, 1937
Тип рода Bdella crassipes С. L. Koch, 1839
2. Hoplomolgus Berlese, 1923
Тип рода Bdella capillata Berlese, 1891 (не Kramer, 1881)
(— Molgus {Hoplomolgus) tuberculatus Berlese, 1923)
3. Hoploscirus Thor, 1937
Тип рода Scirus dubitatas Womersley, 1933
4. Troglobdella Oudemans, 1937
Тип рода Scirus obisium Gervais, 1841
ОБЩИЕ ЗАМЕЧАНИЯ
Самцы и самки очень сходны друг с другом; обычно они раз-
личаются лишь строением половых органов, хотя иногда имеются
небольшие различия и в строении пальп. Яйца, несколько овальные
по форме и покрытые многочисленными валикообразными выступами
или шипами, откладываются в почву, лиственный перегной или
в любое другое место, где встречаются эти клещи. Из яиц вылу-
пляются шестиногие личинки, отличающиеся от взрослых особей
только числом ног и отсутствием генитальных щитков. Прото- дейто-
и тритонимфы имеют генитальные щитки, две пары половых присосок
и меньшее число генитальных щетинок, чем взрослые клещи, от
которых они отличаются еще и меньшими размерами.
Bdellidae, повидимому, распространены по всему земному шару
и встречаются в совершенно различных климатических районах.
Bdellidae — хищники, питающиеся другими клещами и мелкими насе-
комыми. Приносят ли они пользу тем, что уничтожают вредителей,—
сказать трудно, хотя об одном виде — Biscirus lapldarius (Kramer)—-
известно, что он уничтожает вредителя люцерны Smtnthurus viridis L.
и может регулировать его численность в некоторых районах Западной
Австралии. Bdellidae обычно встречаются во мху, лишайниках, лист-
венном перегное, мусоре — всюду, где имеются скопления мелких
насекомых и других клещей.
Поль де Бах в неопубликованном сообщении упоминает о виде
Bdella sp., распространенном на апельсиновых деревьях в графстве
Орендж (Калифорния) и уничтожающем ногохвосток. Этот клещ
овладевает своей добычей, брызгая на нее жидким „ шелком “
(паутиной), и, когда жертва запутывается, клещ высасывает ее. При
уничтожении этого клеща при помощи ДДТ численность ногохвосток
крайне возрастала.
В Северной Америке обычен Bdella oblonga Say, встречающийся
по всему восточному побережью до Панамы на юге; на запад этот
вид идет до Техаса. В. oblonga — один из самых крупных предста-
Prostigmata
193
цителей семейства Bdellidae; он достигает 1300 р. в длину. Его легко
можно отличить по сетчатому проподосомальному щитку, имеющему
форму буквы ,,U“. Известный в Европе и Австралии Biscirus lapi-
darlus (Kramer) был найден также и в Мексике. Bdella virgata
Ewing встречается в США и Мексике.
ЛИТЕРАТУРА
Baker Е. W., Balock J., 1944. Mites of the family Bdellidae, Proc. Ent.
Soc. Wash., 46 (7), 176—184.
Currie G. A., 1934. The Bdellid mite Biscirus lapidaritis Kramer, predatory
on the lucerne flea Sminthurus viridis L. in Western Australia, J. Austral.
Council Sci. a. Ind. Res., 1 (1), 19—20.
Grand jean F., 1938. Observations sur les Bdelles (Acariens), Ann. Soc.
Ent. France, 107, 1—24.
Thor S., 1931. Bdellidae, Nicoletiellidae, Cryptognathidae, Das Tierreich, 56,
1—65.
Womersley H., 1933. A preliminary account of the Bdellidae (Snout mites)
of Australia, Trans. Roy. Soc. South Austral., 57, 97—107.
Womersley H., 1933. A possible biological control of the clover springtail
or lucerne flea (Sminthurus viridis L.) in Western Australia, J. Austral.
Council Sci. a. Ind. Res., 6 (2), 83—91.
Rhagidiidae Oudemans, 1922
(рис. 120—123)
ДИАГНОЗ
Rhagidiidae — беловатые, желтые или розовые клещи небольших
или средних размеров (длина тела 0,350—1,80 мм). Они чувстви-
тельны к свету и очень подвижны. Продолговатое, стройное тело этих
клещей не защищено панцирем. Проподосома и гистеросома разделены
швом. Щетинки простые и длинные. На проподосоме расположена
пара слабо развитых псевдостигм, связанных с мало измененными
сенсорными щетинками. Глазки обычно имеются. Ноги этих клещей
сравнительно длинные и тонкие и состоят из пяти подвижных члени-
ков. Бедра ног I и II разделены вентрально, а бедра ног III и IV
полностью разделены на две части. Каждая из лапок имеет по два
коготка и опушенный эмподий. Пальпы состоят из четырех подвиж-
ных члеников; комплекса палец — коготок на пальпах нет. Тарзальный
членик пальп цилиндрический, примерно такой же толстый, как их
голенный членик. Хелицеры крепкие, клешневидные, хватательные;
пальцы клешни противопоставлены друг другу. Имеется две пары
половых присосок.
РОДЫ
1. Rhagldla Thorell, 1871 (— Scyphius С. L. Koch, 1836 nomen
praeoccupatum; = Noerneria Canestrini, 1886)
Тип рода Rhagldla gelid a Thorell, 1871
2. Coccorhagldla Thor, 1934
Тип рода Noerneria clavifrons R. Canestrini, 1886.
13 Зак. 1785
194
Глава V. Подотряд Trombidiformes
ОБЩИЕ ЗАМЕЧАНИЯ
Об этих клещах имеется мало сведений. Они пока известны лишь
в Европе и Северной Америке, но, невидимому, дальнейшие сборы
обнаружат их и в других частях света. Rhagidiidae, повидимому,
Рис. 120. Rhagidia
gellida Thorell
(по Удемансу, 1928).
Самка (вид сверху).
Рис. 121. Rhagidia
gelida Thorell
(по Удемансу, 1928).
Самка (вид снизу'.
Рис. 122. Rhagidia gelida Thorell
(по Удемансу, 1926).
Хелицера и трахея.
хищники, и так как они очень многочисленны в некоторых стадиях,
то возможно, что роль их в общих взаимоотношениях между мелкими
насекомыми и клещами более велика, чем это принято думать. Rha-
gid’idae живут в почве, гумусе и во мху. Rhagidia pallida Banks
найден в Нью-Йорке, Нью-Гэмпшире и Вашингтоне; его находят
Prostigmata
19$
па земле, под сырыми досками или камнями. R. cavicola Banks был
найден в Мамонтовой пещере в Кентукки. Бэнкс указывает, что
R. cavicola имеет довольно широкое распространение и что его часто
можно встретить под камнями, на которых имеются кладки яиц пе-
щерного паука Anthrobia mammonthia Telkpf.
Рис. 123. Rhagidia sp.
Лапка I (вид сбоку); виден рагидиев орган.
О жизненном цикле клещей Rhagidiidae пока ничего неизвестно.
Мы знаем лишь, что все фазы внешне очень сходны, но личинки
отличаются от нимф и имаго тем, что имеют только шесть ног.
Cryptognathidae Oudemans, 1902
(рис. 124)
ДИАГНОЗ
Cryptognathidae — яркокрасные мелкие клещи, длиной от 0,30
ю 0,40 мм. Тело овальное, проподосома и гистеросома не разделены
швом. Кожа с точечным орнаментом, спинка покрыта сетчатым узо-
ром. С каждой стороны проподосомы имеется по паре линзовидных
глаз. Хитиновый покров тела разрастается вперед, образуя трубку,
открытую снизу; через эту трубку гнатосома может выдвигаться или
втягиваться в тело. Хелицеры клешневидные, клешни мелкие, почти
прямые, без зубцов; они заострены и приспособлены для прокалыва-
ния. Половые присоски отсутствуют.
годы
1. Cryptognathus Kramer, 1878
Тип рода Cryptognathus lagena Kramer, 1878
13*
196
Глава V. Подотряд Trombidiformes
ОБЩИЕ ЗАМЕЧАНИЯ
Эти клещи живут во мху и под камнями. Тор (1931) насчитывает
только два вида и один подвид, объединяемые в единственный род
Cryptognathus. Cryptognathus lagena Kramer известен в Германии,
Нидерландах, Швейцарии, Италии, Франции, Англии, Ирландии и
Рис. 124. Cryptognathus cucurbitae Berlese.
Самка (вид сверху).
Норвегии. С. cucurbita Berlese обнаружен в Италии; отдельные экзем-
пляры этого вида были собраны в гнезде белки Scittrus ttiger niger
во Флориде. Из Сомали (Африка) описан С. cucurbita var. subnitida
Berlese. Об образе жизни всех этих клещей ничего неизвестно, кроме
того, что они, вероятно, хищники.
ЛИТЕРАТУРА
Thor S., 1931. Bdellidae, Nicoletiellidae, Cryptognathidae. Das Tierreich, 56,
78—81.
Labidostommidae Oudemans, 1904
(рис. 125—126)
ДИАГНОЗ
Labidostommidae — свободноживущие хищные клещи, полностью
покрытые панцирем. Их яйцевидное тело имеет желтоватую окраску.
Длина тела колеблется от 0,57 до 1,5 мм. Проподосома разделена
Prostigmata
197
на два отдела; задняя часть постепенно переходит в гистеросому и
нечетко обособлена от последней. Имеется две пары псевдостигмаль-
ных органов, по одному на каждом проподосомальном щитке. По
краям проподосомы расположено по одному линзовидному глазу; иногда
посреди проподосомы по ее переднему краю лежит еще один непар-
ный глаз. Позади парных глаз, по краям проподосомы, иногда имеется
по одному крупному линзовидному органу. Пальпы
состоят из четырех члеников, хелицеры воору-
жены большой крепкой клешней с противо-
поставленными пальцами. Ноги имеют по шести
подвижных члеников; коксальные щитки сопри-
касаются друг с другом. Все лапки вооружены
двумя коготками, а на лапках ног II, III и IV
имеются когтевидные эмподии. Имеется две
пары половых присосок. У самки половое и
анальное отверстия сливаются и лежат на общем
Рис. 125. Labidostom-
та luteum Kramer
(по Тору, 1931).
Хелицера (вид сбоку).
круглом щитке; у самцов половое отверстие самостоятельное, но
занимает сходное положение.
РОДЫ
1. Labidostomma Kramer, 1879 [— Panoplia v. Heyden, 1826 (nomen
praeoccupatum) = Nicoletlella R. Canestrini, 1883 — Ceratoacarus
Ewing, 1913)
Гии рода Labidostomma luteum Kramer, 1879 (некоторые исследо-
ватели считают, что это синоним .Acarus denticulatus Schrank)
2. Eutilcolana Berlese, 1911
Тип рода Eunicolana tuberculata Berlese, 1911
ОБПХИЕ ЗАМЕЧАНИЯ
Гранжан (1942) довольно подробно изучил это семейство, и при-
веденные ниже данные о биологии Labidostommidae заимствованы из
его исследования. Взрослые особи охотно питаются ногохвостками и
мелкими, неполовозрелыми панцырными клещами. Самки откладывают
по одному довольно крупному яйцу; они не отыскивают для кладки
специальных укрытых местечек. Взрослый клещ способен переносить
яйца с места на место, подхватывая его первой парой ног и поддер-
живая концами хелицер. Из яйца через двенадцать дней вылупляется
личинка, которая не питается и бывает подвижной в течение всего
лишь нескольких часов, пока не найдет себе места для превращения
и протонимфу. Прото-, дейто- и тритонимфы по строению и способу
питания походят на взрослых клещей.
Labidostommidae представляют собой небольшое семейство мало
изученных клещей, встречающихся в Европе, Южной Африке, Север-
ной и Центральной Америке и в Бразилии. Они живут во мху, в гумусе
и почве и охотятся за другими клещами и мелкими насекомыми,
198
Глава V. Подотряд Trombidiformes
Многие виды имеют позади парных глаз выросты, функция которых
неизвестна. Labidostommidae представлены в США видом Labidostomma
Рис. 126. Labidostomma neotropica (Stoll.).
Л —самка (вид сверху); Б — самка (вид снизу).
paclfica (Ewing), а в южных штатах и Центральной Америке до
Панамы — видом L. neotropica (Stoll).
ЛИТЕРАТУРА
Grandjean F„ 1942. Observations stir les Labidostommidae, Bull. Paris
Mas. Nat. d’Hist. Nat., Ser. 2, 14 (2), 118—125; (3) 185—192; (5) 319—326;
(6) 414—418.
Thor S., 1931. Bdellidae, Nicoletiellidae, Cryptognathidae. Das Tierreich, 56,
66-77.
Prostigmata
199
Ereynetidae Oudemans, 1931
(рис. 127)
ДИАГНОЗ
Ereynetidae — красновато-желтые или беловатые мелкие клещи
размером от 0,220 до 0,500 мм. Проподосома и гистеросома разделены
швом. Покровы мягкие, исчерчен-
ные. У Ereynetes проподосомаль-
ный щиток покрыт очень сложным
рисунком; у Opsereynetes рисунок
гораздо проще. Щетинок на теле
немного; на проподосоме имеется
пара длинных сенсорных щетинок,
другая пара щетинок располо-
жена на гистеросоме. Opsereynetes
имеют только глазки, но у Ereyne-
tes развита пара настоящих линзо-
видных глаз. Пальпы простые и
лишены или пальца, или коготка;
они состоят из 3—5 члеников.
Подвижный палец хелицер за-
острен и служит для прокалыва-
ния; он явно длиннее неподвижного
пальца. Все лапки снабжены ко-
готками и опушенными подушеч-
ками. Имеется две пары половых
присосок.
Р и с. 127. Opsereynetes robustus
Baker.
Самка (вид сверху).
РОДЫ
1. Ereynetes Befese, 1883 (—Mlcrereunetes Berlese, 1923)
Тип рода Ereynetes berleset Oudemans, 1928
2. Opsereynetes Thor, 1932
Тип рода Opsereynetes norveglcus Thor, 1932
3. Riccardoella Berlese, 1923
Тип рода Acarus litnacum Schrank, 1781
ОБЩИЕ ЗАМЕЧАНИЯ
Эти клещи известны из Европы, Северной Америки, с Явы, Су-
матры и из Японии и, вероятно, могут быть обнаружены повсюду,
если произвести тщательные сборы. Они живут во мху и лишайниках,
а иногда и на листьях. Все они, вероятно, хищники. Вид Riccardoella
Цтасит (Schrank) встречается в Европе и Северной Америке, где
200
Глава К Подотряд Trombidiformes
держится на виноградной улитке Helix pomatia L. Иногда на покры-
тых слизью органах улитки находят большое число клещей, но они.
повидимому, не причиняют хозяину никакого вреда. Терк и Филлипс
(1946), изучавшие этого клеща, дают следующее описание его жизнен-
ного цикла. Самки откладывают яйца в слизь мантии улитки. Из яиц
выходят шестиногие личинки, не имеющие задней пары брюшных
сенсорных щетинок и половых присосок. Прото- и дейтонимфы похожи
на взрослых особей. Терк и Филлипс установили у этого вида неоте-
нию: дейтонимфы (nympha generans) содержат яйца, между тем как
у исследованных взрослых особей ни разу не были обнаружены зре-
лые яйца. Такие женские дейтонимфы живородящи или яйцеживоро-
дящи. Клещи не постоянно находятся на хозяине, но иногда бегают
по земле; улитку они находят по слизистому следу. Клещи питаются
слизью, но могут находить пищу и вне хозяина. Клещи и улитки,
повидимому, прекрасно уживаются друг с другом.
ЛИТЕРАТУРА
Baker Е. W., 1945. Five mites of the family Ereynetidae from Mexico, J. Wash.
Acad. Set, 35 (1), 16—19.
Grandjean F., 1939. Observations sur les Acariens (5 serie), Bull. Paris
Mas. d’Hist. Nat., Ser. 2, 11 (4), 394—401.
Thor S., 1933. Tydeidae, Ereynetidae. Das Tierreich, 60, 58—84.
Turk F. A., Phillips S. M., 1946. A monograph of the slug mite —Ric-
cardoella limacum (Schrank), Proc. Zool. Soc. London, 115 (3, 4).
448—472. ...
Paratydeidae Baker, 1949
(рис. 128)
ДИАГНОЗ
Paratydeidae представляют собой простигматических клещей, имею-
щих такие же перитремы, как и хищные Cheyletidae. Пальпы у них
состоят из четырех члеников, комплекс палец — коготок не развит,
а тарзальный членик пальпы занимает герминальное положение. Осно-
вания хелицер слиты, подвижный палец клешни короткий и невтяж-
ной и потому не приспособлен для прокалывания. Тело удлиненное;
как проподосома, так и гистеросома лишены щитков. Кожа бороздчатая.
Гистеросома разделена хорошо выраженным дорзальным швом, про-
ходящим позади ног III. На теле имеется несколько пар простых
щетинок. Проподосома несет две пары длинных сенсорных (?) щети-
нок, одну пару коротких щетинок, две пары линзовидных глаз и две
пары боковых палочковидных щетинок. Анальное отверстие расположено
на заднем конце брюшка. Половое отверстие обособлено от анального и
помещается позади тазиков ног IV; оно имеет две пары половых при-
сосок и от двух до четырех пар генитальных щетинок. Тазики ног
I—II и III—IV расположены двумя группами, далеко отстоящими друг
от друга. Тазики ног слиты с телом. Ноги покрыты редкими волос-
Prostigmata
201
ками. Все лапки имеют по два коготка и небольшую когтевидную
подушечку. Лапки ног I несут по две короткие палочкообразные
сенсорные щетинки. Длина этих клещей достигает 0,366 мм.
Рис. 128. Paratydeus
alexanderi Baker. ,
Самка (вид сверху).
РОДЫ
1. Paratydeus Baker, 1949
Тип рода Paratydeus alexanderi Baker, 1949
2. Neotydeus Baker, 1950
Тип рода Neotydeus ardisannae Baker, 1950
3. Scolotydaeus Berlese, 1910
Тип рода Scolotydaeus bacillus Berlese, 1910
ОБЩИЕ ЗАМЕЧАНИЯ
Эти клещи, повидимому, живут в почве и, вероятно, ведут хищный
образ жизни. Известно только три вида.
ЛИТЕРАТУРА
Baker Е. W., 1949. Paratydeidae, a new family of mites (Acarina), Proc.
Ent. Soc. Wash., 51 (3), 119—122.
Baker E. W., 1950. Further notes on the family Paratydeidae (Acarina) with
a description of another new genus and species, J. Wash, Acad. Set.,
4 (9), 289—291.
202
Глава V. Подотряд Trombidiformes
ДИАГНОЗ
Speleognathidae Womersley, 1933
(рис. 129)
Speleognathidae — светлые, желтовато-коричневые мелкие клещи,
длиной около 0,80 мм. Кожа мягкая, щитки не развиты. На теле
имеется несколько коротких щетинок, проподосома несет пару длинных
сенсорных щетинок. Пальпы
короткие, простые, комплекс
палец — коготок не развит.
Хелицеры редуцированы. Под-
вижный палец клешни хелицер
представляет собой очень ма-
ленькое острие. На проподосоме
иногда имеются линзовидные
глаза. Проподосома и гистеро-
сома не разделены швом. Ноги
короткие и покрыты сетчатым
склеротизированным узором.
Тазики ног разделены на две
группы. Все лапки имеют ко-
готки и опушенную подушечку.
Половые присоски отсутствуют.
Рис. 129. Speleognathus sturni
Boyd (по Бойду, 1948).
Самка (вид снизу).
РОДЫ
1. Speleognathus Womersley, 1936
Тип рода Speleognathus australis Womersley, 1936
ОБЩИЕ ЗАМЕЧАНИЯ
Speleognathus australis Womersley были собраны во мху и вокруг
водопоев рогатого скота в Австралии. В восточных штатах США был
обнаружен Speleognathus sturni Boyd, паразитирующий в носовой
полости скворцов. Бойд (1948) нашла, что у птиц, зараженных кле-
щами, в носовой полости выделяется больше слизи, чем у птиц,
не зараженных клещами. Как по строению, так и по биологии Speleo-
gnathidae, повидимому, очень близки к Ereynetidae; так, Ricardoella
limacum (Schrank) живет в слизи улиток, a S. sturni — в теплой слизи
носовой полости скворцов. Бойд считает возможным, что оба эти
клеща имеют общего предка. S. sturni, вероятно, широко распростра-
нен и встречается повсюду, где водятся скворцы или родственные им
птицы.
Prostigmata
203
Личинка S. sturni Boyd в основном похожа на взрослых клещей.
Однако лапка ноги I не несет коготка или подушечки, а голень этой же
ноги снабжена длинным раздвоенным когтеобразным отростком.
ЛИТЕРАТУРА
Boyd Е„ 1948. A new mite from the respiratory tract of the starling (Acarina,
Speleognathidae), Proc. Ent. Soc. Wash., 50 (1), 9—14.
WJomersley H., 1936. On a new family of Acarina, with description of
a new genus and species, Ann. Mag. Nat. Hist., 18 (104), 312—315.
Tydeidae Kramer, 1877
(рис. 130, 131)
ДИАГНОЗ
Tydeidae имеют блеклую желтую, коричневую, красную или зеле-
ную окраску. Очень мелкие клещи длиной от 0,100 до 0,300 мм
(обычно около 0,200 мм) и различ-
ной, чаще всего яйцевидной формы.
Кожа мягкая, с точечными борозд^
ками; у некоторых видов кожа по-
крыта точечным или сетчатым узо-
ром. На теле щетинок немного;
Б
A .
starri Baker.
Рис. 131. Tydeus starri Baker.
Самка (вид снизу).
Рис. 130. Tydeus
,1 —самка (вид сверху); Б— хелицера (вид сбоку).
проподосома несет одну пару сенсорных щетинок. Проподосома и
гистеросома обычно разделены хорошо заметным швом. Ноги состоят
из 5—6 члеников; лапки имеют короткую предлапку, два коготка и
204
Глава V. Подотряд Trombidiformes
опушенную подушечку (у Pronematus на ногах I коготки и поду-
шечка не развиты; у Proctotydaeus ноги I не имеют ни коготков,
ни подушечки, а ноги II и IV несут только коготки). Хелицера имеет
очень широкое основание, на котором сидит короткий, резко заостряю-
щийся неподвижный палец клешни; от неподвижного пальца отходит
почти прямой, кинжаловидный подвижный палец. Так измененная
клешня хелицеры теряет свою типичную форму. У некоторых видов
глазки отсутствуют, у других имеется по два или по три глазка.
Половые присоски отсутствуют.
РОДЫ и ПОДРОДЫ
I. Tydeus Koch, 1836 (== Microtydeus Thor, 1931 = Brachytydeus
Thor, 1931 = Melanotydeus Berlese, 1910 = Stylotydeus Thor,
1933 = Lasiotydeus Berlese, 1908 ? = Calotydeus Oudemans,
1937 = Tectotydeus Oudemans, 1937)
a) Tydeus s. str.
Тип подрода Tydeus kochi Oudemans, 1928 = {Tydeus croceus
Koch, 1836 (не Acarus croceus Linnaeus 1758)
6) Tydaeolus Berlese, 1910
Тип подрода Tydaeus atomus Berlese, 1908
в) Triophtydeus Thor, 1932 (= Trldilatydeus Baker, 1946)
Тип подрода Tydeus triophthalmus Oudemans, 1929
2. Coccotydeus Thor, 1931
Тип рода Coccotydeus globlfer Thor, 1931
3. Lorryla Oudemans, 1925 ( = Raphltydeus Thor, 1933)
Тип рода Lorryia superba Oudemans, 1925
4. Coleotydaeus Berlese, 1910
Тип рода Coleotydaeus rhombicus Berlese, 1910
5. Pronematus G. Canestrini, 1886
a) Pronematus s. str.
Тип подрода Pronematus bonatli Canestrini, 1886
6) Pronecupulatus Baker, 1944
Тип подрода Pronecupulatus anahuacensis Baker, 1944.
в) Proctotydaeus Berlese, 1911
Тип подрода Proctotydaeus viator Berlese, 1911
6. Retetydeus Thor, 1931
Тип рода Retetydeus catenulatus Thor, 1931
Прежние авторы подразделяли это семейство на большее число
родов. Делалось это на основании таких признаков, как размеры
тела, ширина и длина щетинок, тип щетинок, присутствие или от-
сутствие глазков. Авторы считают, что принятое ими деление более
обосновано и более просто.
ОБЩИЕ ЗАМЕЧАНИЯ
Tydeidae распространены по всему земному шару. Они, невиди-
мому, хищники, питающиеся мелкими насекомыми, клещами и их
Prostigmata
205
яйцами. Возможно, что некоторые виды могут питаться на растениях,
но для подтверждения этого требуются дальнейшие наблюдения. В од-
ном районе Бельгии известен клещ Tydeus molestus (Moniez), являю-
щийся маловажным паразитом человека и домашних животных; укусы
этого клеща вызывают раздражение кожи. В южной Калифорнии
Tydeus californicus (Banks) истребляет цитрусового галлового клеща
Aceria sheldoni (Ewing), a Pronematus ubiquitus (McGregor), распро-
страненный по всей Калифорнии, обычен на фиговых деревьях, где
он пожирает фигового клеща Aceria fucus (Cotte). Европейский вид
Lorryia superba Oudemans найден и в Калифорнии, а другой евро-
пейский вид — Lorryia mali (Oudemans) — был обнаружен на листьях
яблони в Новой Шотландии.
Большинство видов живет во мху, лишайниках или на листьях
растений вместе с другими клещами. Жизненный цикл простой: самка
откладывает по одному яйцу, из которого выходит личинка, превра-
щающаяся в нимфу и затем во взрослую особь. Все фазы превра-
щения очень похожи друг на друга. Взрослые особи отличаются тем,
что имеют половое отверстие; самцы отличаются от самок лишь зна-
чительно меньшими размерами полового отверстия. Ни один из видов
Tydeidae не имеет особого значения в борьбе с вредителями расте-
ний, хотя, возможно, Tydeidae приносят некоторую пользу в этом
отношении.
ЛИТЕРАТУРА
Thor S., 1933. Tydeidae, Ereynetidae, Das Tierreich, 60, 1—57.
Cunaxidae Thor, 1902
(рис. 132)
ДИАГНОЗ
Cunaxidae — мелкие, красные клещи. Длина тела (не считая длин-
ной, конусообразной гнатосомы) варьирует у них от 0,350 до 0,500 мм.
Проподосома отделена от гистеросомы швом. Покровы мягкие, по-
крыты тонким бороздчатым узором с отдельными точечными пятнами;
у многих форм имеются большие щитки. Проподосома несет четыре
хорошо выраженные сенсорные щетинки. Глаза у некоторых форм не
развиты. |4оги состоят из шести подвижных члеников, лапки всех
ног имеют коготки и подушечку. Пальпы крупные, клешневидные и
служат для схватывания; на их внутренней стороне обычно развиты
апофизы и крепкие шипы. Концевой членик пальпы когтевидный.
Хелицеры удлиненные; их подвижный палец, занимающий дистальное
положение, очень невелик и имеет серповидную форму. Имеется две
пары половых присосок.
РОДЫ
1. Сипаха v. Heyden, 1826 ( = Scirus seraszr Berlese = Dactyloscirus
Berlese, 1916)
Тип рода Scirus setirostris Hermann, 1804
206
Глава V. Подотряд Trombidiformes
2. Bonzia Oudemans, 1927
Тип рода Bomia halacaroldes Oudemans, 1927
3. Coleoscirus Berlese, 1916
Тип рода Coleoscirus halacaroides Berlese, 1916
4. Cunaxoides Baker et Hoffmann, 1948 ( — Eupaliis Koch, 1838
nomen praeoccupatum = Haleupalus Radford, 1950)
Тип рода Eupalus croceus Koch, 1838
5. Rosenhofia Oudemans, 1922
Тип рода Rosenhofia machairodus Oudemans, 1922
6. Scirula Berlese, 1887
Тип рода Scirula impressa Berlese, 1887
Рис. 132. Cunaxa
capreolus (Berlese).
Самец (вид сверху).
ОБЩИЕ ЗАМЕЧАНИЯ
Ни один из видов Cunaxidae не имеет большого хозяйственного
значения, хотя они питаются другими клещами и мелкими насеко-
мыми. Это семейство имеет широкое распространение; многие виды
его распространены более или менее космополитически. Европейский
вид Cunaxa capreolis Berlese встречается также в Африке, США и
Мексике; С. setirostris (Hermann) распространен по всему свету; то
же самое можно сказать и о С. inermis (Tragardh) и С. taurus (Kra-
mer). Последний отличается чрезвычайно длинными пальпами, несу-
щими по внутреннему краю длинные шипы и апофизы. В северо-
восточных штатах США этот вид обычно встречается на листьях и
в лиственном перегное. Cunaxoides parvus (Ewing) считается в Айове
Prositgmaid
207
серьезным врагом яблоневой запятовидной щитовки, хотя там этот
клещ сравнительно немногочислен. С. pectinatus (Ewing) распространен
от США (Иллинойс) до Панамы.
Личинки и нимфы похожи на имаго. Самцы отличаются от самок
только значительно меньшими размерами генитальных щитков и не-
которыми другими второстепенными признаками; так, у самцов Cu-
naxa capreolus (Berlese) апофиз на третьем членике пальп значи-
тельно меньше, чем у самок. Нимфы, повидимому, имеют три пары
генитальных щетинок, а не четыре, как взрослые особи. Нимфа
Cunaxoides pectinatus (Ewing) имеет три пары генитальных щетинок,
небольшое тело и длинные ноги; дорзальный щиток у нее отсутствует,
но все тело покрыто бороздчатым узором. В остальном у этого вида
нимфы сходны с имаго. Cunaxoides pectinatus (Ewing) обычно живут
во мху, гумусе и на листьях.
Cunaxidae близкородственны Halacaridae; было высказано мнение,
что Bonzia является одной из Halacaridae, перешедшей к наземному
образу жизни.
ЛИТЕРАТУРА
Baker Е. W., Hoffmann А., 1948. Acaros de la familia Cunaxidae, Ana-
les de la Escuela National de Cienctas Biologicas, 5 (3—4), 229—273.
Ewing H. E., 1917. New Acarina. Part II. Descriptions of new species and
varieties from Iowa, Missouri, Illinois, Indiana and Ohio, Bull. Amer.
Mas. Nat. Hist., 37 (2), 151.
Thor S., Willmann C., 1941. Acarina. Prostigmata 6—11 (Eupodidae,
Penthalodidae, Penthaleidae, Rhagidiidae, Pachygnathidae, Cunaxidae),
Das Tierreich, 71a.
Lordalychidae Grandjean, 1939
(рис. 133—136)
ДИАГНОЗ
Lordalychidae — клещи средней
с шарообразным телом, суживаю-
щимся между проподосомой и
гистеросомой. Проподосома по
сравнению с гистеросомой очень
величины (длина от 0,30 до 0,40 мм)
Рис. 133. Lordalychus peraltus
Grandjean (по Гранжану, 1939).
Хелицера (вид сбоку).
Рис. 134. Lordalychus peraltus
Grandjean (по Гранжану, 1939).
Самка (вид сбоку).
невелика. Кожа гранулированная и покрыта бороздчатым узором. Пропо-
досома несет две пары сенсорных щетинок, передняя пара их помещается
208
Глава V. Подотряд Trombidiformes
в особой ямке, образованной впячиванием покровов. Глаза отсут-
ствуют. Хелицеры примитивные, с дегенерировавшими неподвижными
пальцами. Пальпы простые и не имеют комплекса палец — коготок.
Ноги расположены в передней части тела. Лапки всех ног снабжены
Рис. 135. Lordalychus
peraltus Grandjean
(по Гранжану, 1939).
Проподосома (вид сверху).
Рис. 136. Lordalychus
peralius Grandjean
(по Гранжану, 1939).
Самка (вид снизу).
двумя коготками и эмподием (у Lordalycus коготки имеют вентраль-
ные лучи, подушечка с лучами). Бедро разделено на два членик;
(basifemur и telofemur). Имеются две пары половых присосок.
РОДЫ
1. Lordalycus Grandjean, 1939
Тип рода Lordalycus peraltus Grandjean, 1939
2. Hybalicus Berlese, 1913
Тип рода Hybalicus flabelliger Berlese, 1913
ОБЩИЕ ЗАМЕЧАНИЯ
Наличие трахейных систем простигматического типа, открываю
щихся у оснований хелицер, обнаруженное при помощи фазово-контраст
ного микроскопа у четырех семейств Endeostigmata Гранжана, указывав
на определенное родство этих семейств с Prostigmita. Endeostigmat
образуют группу весьма близких друг к другу семейств (Terpnacari
dae, Pachygnathidae, Alicorhagididae, Nanorchestidae, Lordalychidae
Sphaerolichidae) и в настоящее время их не выделяют из Prostigmatr
Endeostigmata мало изучены; до сих пор описано очень немного ро
дов и видов, относящихся к этим семействам.
ЛИТЕРАТУРА
Grandjean F., 1939. Quelques genres d’acariens appartenant an groupe de
Endeostigmata, Ann. des Set. Nat., Zool., Ser. 11, 2, 1—122.
Prosiigmata
209
ДИАГНОЗ
Sphaerolichidae Grandjean, 1937
(рис. 137—139)
Sphaerolichidae — клещи небольших и средних размеров, длиной
от 0,30 до 0,40 мм, с шарообразным толстым телом. В спирту они
Рис. 137. Sphaerolichus
barbarus Grandjean
(по Гранжану, 1939).
Самка (вид сверху).
имеют . беловатую или желтоватую
окраску. Кожа мягкая и покрыта бо-
роздками или точечными бороздками.
Проподосома имеет тектум, или перед-
ний вырост, с непигментированным вен-
тральным глазом; кроме того, имеется
две пары боковых глаз. Проподосома
несет две пары сенсорных щетинок и
не отделена швом от гистеросомы. Ноги
расположены в передней части тела
Рис. 138. Sphaerolichus barbarus
Grandjean (по Гранжану, 1939).
Хелицера (вид сбоку).
двумя группами, очень близко одна от другой. Бедра ног 1 разделены
на три членика, бедра ног II, III и IV — на два. Лапки ног I имеют
по два пальца, лапки ног II, III и IV — по
три пальца (коготки и когтевидный эмпо-
дий с короткими лучами). Ноги приспо-
соблены для прыгания. Клешня хелицеры
имеет противопоставленные друг другу
пальцы. Пальпы состоят из пяти члени-
ков, комплекс палец — коготок не развит.
Имеется две пары половых присосок.
Рис. 139. Sphaerolichus barbarus Grandjean
(по Гранжану, 1939).
Самка (вид снизу).
14 Зак. 1785.
210
Глава V. Подотряд Trombidiformes
РОДЫ
1. Sphaeroltchus Befese, 1904
Тип рода Sphaeroltchus artnlpes Berlese, 1904
ОБЩИЕ ЗАМЕЧАНИЯ
Sphaerolichidae мало изучены; они живут в сухих местах, в мерт-
вой лесной подстилке. У единственного имевшегося в нашем распо-
ряжении вида, принадлежащего к неописанному роду из Калифорнии,
была обнаружена трахейная система, открывающаяся у основания
хелицер. Гранжан (1939) нашел трахеи также у Sphaeroltchus.
ЛИТЕР АТУРА
Grandjean F., 1939. Quelques genres d’acariens appartenant an groupc
des bndeostigmata, Ann. des Sci. Nat., Zoo!., Ser. 11, 2, 1—122.
Рис. 140. Spe'.eorchestres
podwoides Hirst
(по Хирсту, 1917).
Самка (вид сверху).
Nanorchesfidae Grandjean, 1937
(рис. 140-141)
ДИАГНОЗ
Мелкие бесцветные или красноватые
клещи с коричневатой задней частью тела,
длиной от 0,165 до 0,350 Мм. Тело более
или менее сильно вытянуто в длину, гисте-
росома мешкообразная. Тело обычно раз-
делено резко выраженными перетяжками
(у Oehserchestes неясно) на гнатосому,
проподосому, метаподосому и опистосому.
Кожа мягкая, с тонкими бороздками. На
спине сидят 64 характерные щетинки.
Рис. 141. Nanorchestes puivinar Grandjean
(по Гракжану, 1942).
Первая пара проподосомальных сенсорных щетинок;
видно расположение длинной „активирующей" ще-
тинки и короткой, подлинно сенсорной щетинки.
Пальпы имеют от четырех до пяти члеников, комплекс палец — кого-
ток не развит. Хелицеры клешневидные. Глаза или совершенно от-
Pf'ostigmatd
211
сутствуют (Oehserchestes), или же с каждой стороны проподосомы
имеется по небольшому линзовидному глазу. Проподосома несет
четыре сенсорные щетинки. Тектум отсутствует. Ноги отодвинуты
далеко вперед; ноги IV специализированы для прыганья. Лапки
имеют только по одному эмподиальному коготку, который может
втягиваться в очень глубокую ямку. Имеются три пары половых при-
сосок.
РОДЫ
1. Nanorchestes Topsent et Trouessart, 1890 (— Monalichus Ber-
lese, 1904)
Тип рода Nanorchestes amphibius Topsent et Trouessart, 1890
2. Speleorchestes TrUgardh, 1909 ( = Leptalicus Berlese, 1910)
Тип рода Speleorchestes formlcorum Tragardh, 1909
3. Oehserchestes Jacot, 1939 (= Coccorchestes Jacot, 1938 nomen
praeoccupatum)
Тип рода Coccorchestes humicolus Jacot, 1938
4. Caenonychus Oudemans, 1903
Тип рода Caenonychus fallax Oudemans, 1903
ОБЩИЕ ЗАМЕЧАНИЯ
Описано всего несколько видов Nanorchestidae. Эти клещи, ве-
роятно, распространены повсеместно и недостаточно известны только
вследствие своих малых размеров. Обычно их можно найти во мху
и в почве. Известно также, что Nanorchestes amphibius Topsent et
Trouessart является по существу морской формой. Nanorchestidae,
вероятно, хищники.
Один из видов — Nanorchestes puivinar Grandjean — отличается
интересным строением передней пары проподосомальных сенсорных
щетинок: длинные „чувствующие щетинки" активируют короткую
щетинку, которая, повидимому, и является подлинным сенсорным
органом.
Гранжан (1942) приводит следующие фазы превращения этих кле-
щей: личинка, три нимфы и имаго. Самцы отличаются от самок ббль-
шим числом и расположением генитальных щетинок.
Speleorchestes sp. из Мексики имеет трахейную систему, откры-
вающуюся у основания хелицер.
ЛИТЕРАТУРА
(i г a n d j е а п F., 1942. Observations sur les Acariens (7 Ser.), Bull. Paris
Mas. d'Hist. Nat., Ser. 2, 14 (4), 264—267.
Womersley H., 1944. Australian Acarina, families Aiycidae and Nanorches-
tidae, Trans. Noy. Soc. South Austral., 68 (1), 133—143.
14*
№
Глава V. Подотряд TrombidiformeS
Pachygnafhidae Kramer, 1877
(рис. 142—144)
ДИАГНОЗ
Небольшие беловатые клещи, длиной от 0,30 до 0,40 мм. Кожа
мягкая и покрыта бороздчатым, сетчатым или каким-либо иным узо-
Рис. 142. Petralycus
unicornis Grandjean
(по Гранжану, 1943).
Самка (вид снизу).
Рис. 143. Petralycus
unicornis Grandjean
(по Гранжану, 1943).
Самка (вид сверху).
ром. Спереди имеется выступ, или тектум, форма которого варьи-
рует. Срединный глаз не развит, иногда отсутствуют и боковые
Рис. 144. Petralycus
unicornis Grandjean
(по Гранжану, 1943).
Хелицера (вид сбоку).
шечка с лучами); у
глаза. Проподосома отделена от гистеросомы
швом и несет две пары сенсорных щетинок
(у Petralycus передняя пара щетинок имеет
шарообразную форму, у Bimichaelia шаро-
образной является задняя пара). Гистеросома
у некоторых форм расчленена. Пальцы клешни
хелицер противопоставлены; пальцы состоят из
пяти члеников и не имеют комплекса палец —
коготок. Ноги расположены двумя отдельными
группами. Каждая из лапок имеет по два ко-
готка и подушечку (коготки простые, а поду-
Petralycus бедра ног I, II и III нерасчленены,
бедра же ног IV разделены на basifemur и te'ofemur. Имеется три
пары половых присосок.
Prosiigmata
213
РОДЫ
1. Pachygnathus Duges, 1834
Тип рода Pachygnathus villosus Duges, 1836
2. Bimichaelia Thor, 1902 (= Michaelia Berlese, 1884 nomen prae-
occupatum)
Тип рода Michaelia augustana Berlese, 1884
3. Petralycus Grandjean, 1943
Тип рода Petralycus unicornis Grandjean, 1943
ОБЩИЕ ЗАМЕЧАНИЯ
Эти клещи встречаются в гумусе, в мертвой лесной подстилке
и в почве. Они, вероятно, хищники, но образ жизни их мало изучен.
Bimichaelia diadema Grandjean имеет личиночную, две • активные
иимфальные и взрослую фазы, a Bimichaelia arbusculosa Grandjean
и Petraycus unicornis Grandjean — три нимфальные фазы. У иссле-
дованного вида Bimichaelia sp. из южных штатов США обнаружена
трахейная система.
ЛИТЕРАТУРА
Grand jean F., 1942. Quelques genres d’acariens apparienant au groups
des Endeostigmata (2 Ser.) Premiere partie, Ann. des Sci. Nal, Zool.,
Ser. 11, 4, 85—135.
Grandjean F., 1943. Quelpues genres d’acariens appatrenant au groupe des
Endeostigmata (2 ser.) Deuxieme partie, Ann. des Sci. Nal, Zool., Ser. 11,
5, 1—59.
Terpnacaridae Grand jean, 1939
(рис. 145—147)
ДИАГНОЗ
Terpnacaridae — мелкие красноватые клещи, длиной около 0,35 мм.
I1окровы гранулированные, тело мягкое, гистеросома имеет несколько
мешкообразную форму и сегментирована. Проподосома и гистеросома
разделены швом. На проподосоме имеются одна пара сенсорных
щетинок, одна пара латеральных линзовидных глаз и непарный сре-
динный глаз, расположенный на нижней стороне тектума, или на пе-
реднем выросте проподосомы. Пальпы состоят из пяти члеников,
комплекс палец — коготок не развит. Хелицеры имеют противопо-
ставленные пальцы. Ноги расположены двумя группами; они имеют
но пяти члеников и довольно длинные (особенно ноги I и IV).
Ведра ног I, III и IV разделены на две части. Все лапки имеют по
два коготка и подушечку. Половых присосок три пары.
РОДЫ
1. Terpnacarus Grandjean, 1939
Тип рода Terpnacarus bouvieri Grandjean, 1939 '
214
Глава V. Подотряд Trombidiformes
2. Alycosmesls Grandjean, 1939
Тип рода Sebaia palmata Oudemans, 1904
3. Sebaia Oudemans, 1903 (= Monallchus Berlese, 1904)
Тип рода Sebaia rosacea Oudemans, 1903
ОБЩИЕ ЗАМЕЧАНИЯ
Эти мелкие клещи, встречаются в более или менее сухих местах,
в мертвой лесной подстилке. Terpttacarus и Sebaia в отличие от
Рис. 145. Terpnacarus bouvieri
Grandjean (по Гранжану, 1939).
Самка (вид сверху).
Рис. 146. Terpnacarus
bouvieri Grandjean
(по Гранжану, 1939).
Область анального и полового
отверстий.
Рис. 147. Terpnacarus bouvieri
Grandjean (но Гранжану, 1939).
Пальпа.
Alycosmesis— прыгающие формы. У рода, обнаруженного в Гвате-
мале и, повидимому, не описанного, имеется простигматическая трахей-
ная система.
ЛИТЕРАТУРА
Grandjean F., 1939. Qttelques genres d’acariens appartenant an grotipe des
Endeostigmata, Ann. des Sei, Naf^Zoal., Ser. Ц, 2, 1—122
Prostigmata
215
Alicorhagiidae Grandjean, 1939
Диагноз
(рис. 148—150)
Alicorhagiidae-—-мелкие клещи, длиной от 0,22 до 0,30 мм. Они
имеют беловатую окраску, но задняя часть тела окрашена в корич-
неватые тона. Тело несколько вытянуто, проподосома отделена от
гистеросомы слабым швом. Кожа мягкая, с бугорчатыми бороздками.
Проподосома несет одну пару сенсорных щетинок, большой перед-
ний выступ проподосомы несет две щетинки. Глаза не развиты.
Рис. 148. Alicorhagia
fragilis Berlese
(по Гранжану, 1939).
Рис. 149. Allcorhagia
fragilis Ber’ese
(по Гранжану, 1939).
Самка (вид сверху).
Рис. 150. Alicor-
hagia fragilis Ber-
lese (по Гранжану,
1939).
Самка (вид снизу).
Л— хелицера (вид сбоку);
Б — часть хелицеры при
большом увеличении.
Клешня хелицеры с противопоставленными пальцами, пальпы не
имеют комплекса палец — коготок. Ноги разделены на пять члеников.
Бедро разделено на bas'femur и telofemur, вертлуги ног I и II также
разделены. Все лапки имеют один втяжной коготок (подушечку).
Половых присосок две пары.
РОДЫ
1. Alicorhagia Ber’ese, 1910 (= Willania Oudemans, 1931 =
— Epistomalycas Thor, 1931)
Тип рода Alicorhagia fragilis Berlese, 1910
ОБЩИЕ ЗАМЕЧАНИЯ
Гранжан (1939) указывает, что во Франции Alicorhagia fragilis
Berlese обычен в лесном гумусе.
ЛИТЕР АТУР А
prandjean F., 1939. Quelques genres d’acariens appartenant au groupe des
Endeostigmata, Ann. des Sd. Na*., Zool., Ser. fl, 2, 1—122.
216
Глава V. Подотряд Trombidiformes
Raphignathidae Kramer, 1877
(= Stigmaeidae Oudemans, 1931; Caligonellidae Grandjean, 1944)
(рис. 151—154)
ДИАГНОЗ
Мелкие, красные круглые клещи. Проподосома и гистеросома
слиты или разделены лишь слабо выраженным швом. У некоторых
форм имеется различное число щитков. Кожа между спинными щит-
ками покрыта тонкими бороздками
линзовидных глаз. Сенсорные ще-
тинки на проподосоме не дифферен-
цированы. Rhaphignathidae имеют
перитремы, которые иногда входят
в основание хелицер. Умеренно длин-
ный цилиндрический тарзальный чле-
ник пальп расположен вентрально
на тибиальном членике пальп, под
тибиальным коготком; размеры этого
Имеются одна или две пары
Р и с. 152. Mediolata mall
(Ewing).
Самка (вид сверху).
Рис. 151. Homocaligus sp.
(по Гранжану, 1946).
Пальпа; видны палец и коготок.
коготка варьируют. Подвижный палец клешни стилетовидный и при-
способлен для прокалывания. Все лапки имеют по два коготка 'и
лучистую подушечку. Лапки ног I и II имеют по одному сенсор-
ному органу каждая. Тазики ног соприкасаются или разделены на
две группы. Половые присоски не развиты, анальное отверстие зани-
мает терминальное положение или сдвинуто несколько на спинную
сторону клеща.
Прежние авторы разделяли это семейство на три. После нахож-
дения двух родов с перитремами в основаниях хелицер, но с круп-
ным когтем на пальпах и терминальным анусом эти семейства были
объединены, поскольку они связаны друг с другом переходами и не
обладают четко выраженными семейственными признаками.
Prostigmata
217
РОДЫ
I. Raphignathus Duges, 1834 {— Syncaligus Berlese, 1910)
Тип рода Raphignathus ruberrimus DugSs, 1834
2. Acheles Oudemans, 1903
Тип рода Acheles mirabilis Oudemans, 1903
3. Apostigmaeus Grandjean, 1944
Тип рода Apostigmaeus navicella Grandjean, 1944
Рис. 153. Raphig-
nathus sp.
Гнатосома (вид сверху).
I. Barbutla Oudemans, 1927
Тип рода Stlgmaeus anguineus Berlese, 1910
5. Caligonella Berlese, 1910
Тип рода Stlgmaeus humilis Koch, 1838
6. Caligonus Koch, 1836
Тип рода Caligonus piger Koch, 1838
7. Eupalopsellus Sellnick, 1949
Тип рода Eupalopsellus olandicus Sellnick, 1949
8. Eupalopsis Canestrini, 1886
Тип рода Eupalus maseriensis Canestrini et Fanzago, 1876
9. Eustigmaeus Berlese, 1910 (= Liostigmaeus Thor, 1930)
Тип рода Stlgmaeus kermeslnus Koch, 1841
10. Homocaligus Berlese, 1910
Тип рода Stlgmaeus scapularis Koch, 1838
218
Глава V. Подотряд Trombidiformes
И. Ledermulleria Oudemans, 1923
Тип рода Caligonus segnis Koch, 1836
12. Macrostigmaeus Berlese, 1910
Тип рода Stigmaeus (Macrostigmaeus) serpentinus Berlese, 1910
13. Mediolata R. Canestrini, 1890 (— Zetsellia Oudemans, 1927)
Тип рода Stigmaeus longirostris Berlese, 1887
14. Neophyllobius Berlese, 1886
Тип рода Neophyllobius elegans Berlese, 1886
15. Podaia Oudemans, 1923
Тип рода Acarus rubens Schrank, 1781
16. Stigmaeus Koch, 1836 (= Stigmaeodes Canestrini, 1890)
Тип рода Stigmaeus cruentus Koch, 1836
17. Storchia Oudemans, 1923
Тип рода Caligonus robustus Berlese, 1885
18. Villersia Oudemans, 1927
Тип рода Villersia vietsi Oudemans, 1927
ОБЩИЕ ЗАМЕЧАНИЯ
Несмотря на то, что Raphignathidae распространены по всему
свету, до сих пор известны лишь немногие виды этих клещей. Они
встречаются во мху, лишайниках, соломе, листьях. В Санта-Паула
(Калифорния) на лимонном дереве был найден клещ Mediolata termi-
nalus (Quayle), питающийся цитрусовым галловым клещом Aceria
sheldoni (Ewing). М. terminalus вонзает хелицеры в тело жертвы и
высасывает ее гемолимфу. Однако вследствие малочисленности и о i -
носительно малой подвижности, а также потому, что мелкие эрио-
фиидные клещи, скрывающиеся под почечными чешуйками и в пло-
довых завязях, почти недосягаемы для М. terminalus, этот клещ не
имеет особого.хозяйственного значения. Биология Rhaphignathidae мало
изучена; известно только, что яйца красноватого света откладываются
по одному под завязь плодов. Европейский вид Eupalopsis pinicola
Oudemans был найден на яблонях в восточной Канаде; вероятно, он
питается другими яблоневыми клещами. В Европе этот клещ унич-
тожает червецов. Mediolata nova-scotiae Nesb’tt был собран с яблонь
в Новой Шотландии. В южной Калифорнии обнаружен вид Neophyl-
lobius sp., охотящийся на личинок червецов.
ЛИТЕРАТУРА
Grandjean F., 1944. Observations sur les Acarines de la famille des Stig-
maeidae, Archives des Science physiques et naiurelles, Perlode 5, 26,
103—131.
Grand jean F., 1946. Au sujet de 1’organe de Claparede, Archives des Scien-
ces physiques et naturalles, Periode 5, 28, 82—87.
McGregor E. A., 1950. Mites of the genus Neophyllobius, Bull. South.
Calif. Acad. Sci., 49 (2), 55—70.
Vitzthum H„ 1929. Acari, Die Tierwelt Mitteleiropas, 3 (3), 50—52.
Prostigmata
219
Pomerantziidae Baker, 1949
(рис. 155—157)
ДИАГНОЗ
На пальпах Pomerantziidae развит комплекс палец—коготок.
Пальцы клешней подвижные, короткие, стилетовидные. Короткие
перитремы не доходят до внутреннего основания хелицер. Имеется
Рис. 155. Pomerantzia
chariest Baker.
Область полового отверстия
самки.
Рис. 156. Pomerantzia
chariest Baker.
Самка (вид сверху),
Рис. 157. Pomerantzia
chariest Baker.
Лапка ноги I; видны
сенсорные щетинки.
несколько спинных щитков. Глаза не развиты. Проподосома лишена
дифференцированных сенсорных щетинок. Анальное отверстие зани-
мает терминальное положение. Имеется три пары половых присосок.
На лапках ног I имеются семь палочкообразных и одна конусообраз-
ная сенсорные щетинки, а на лапках ног II — две палочкообразные
сенсорные щетинки. Все лапки имеют коготки, но подушечка на них
не развита.
РОДЫ
1. Pomerantzia Baker, 1949
Тип рода Pomerantzia charlesl Baker, 1949
ОБЩИЕ ЗАМЕЧАНИЯ
Pomerantziidae представляют собой мелких клещей, длиной 0,34 мм,
напоминающих клещей Rhaphignathidae. Их можно найти в. почве
220
Глава V. Подотряд Trombidiformes
персиковых садов в штате Джорджия, США. Морфология и характер
местообитания указывают на хищный образ жизни.
ЛИТЕРАТУРА
Baker F. W., 1949. Pomerantziidae, anew family of mites, J. Wash. Acad.
Sci., 30 (8), 269—271.
ДИАГНОЗ
Pterygosomidae Oudemans, 1910
(рис. 158—159)
Pterygosomidae — красные,
мелкие или средние по величине
1,30 мм. Паразитируют на ящерицах
клещи, длиной от 0,160 до
Рис. 158. Pimeliaphilus podapo-
lipophagus Tragardh.
Самка (вид сверху).
за исключением Pimeliaphilus ро-
dapolipophagus Tragardh, паразити-
рующего на тараканах, и Р. iso-
metri Cunliffe, найденного на скор-
пионе. Клещи, паразитирующие на
ящерицах, имеют плоское, мешко-
образное тело, ширина которого при-
мерно в два раза превосходит длину.
Щетинки на теле обычно палоч-
ковидные, листовидные или вееро-
Рис. 159. Pierygosoma neumanni
Berlese (по Хирсту, 1925).
Типичный представитель клещей,
паразитирующих на ящерицах.
образные; число щетинок сильно варьирует. Проподосома иногда
покрыта цельным или расчлененным щитком. У одних форм имеется
пара линзовидных глаз, у других они отсутствуют. Гнатосома терми-
нальная или же глубоко погружена в тело; хелицеры узкие; у форм,
паразитирующих на ящерицах, подвижный палец клешни хелицеры
искривлен, у форм же, паразитирующих на тараканах, он острый,
колющий. У основания гнатосомы расположены выступающие пери-
тремы. Пальпа с комплеском палец—-коготок, но палец не высту-
пает. Тазики ног у форм, паразитирующих на ящерицах, сдвинуты
вперед, а у форм, паразитирующих на тараканах, расположены более
Prostigmata
221
обычно. Лапки ног имеют по два коготка, каждый из которых несет
пару клейких волосков. Половые присоски не развиты. Анальное
отверстие расположено на заднем конце тела.
РОДЫ
1. Pterygosoma Peters, 1849
Тип рода Pterygosoma agamae Peters, 1849
2. Qeckobla M£gnin, 1878
Тип рода Oeckobia latasti Mdgnin, 1878
3. Geckoblella Hirst, 1917
Тип рода Geckobta texana Banks, 1915
4. Hirstlella Berlese, 1920
Тип рода Geckoblella {Hirstiella) trombidiformes Berlese, 1920
5. Ixodlderma Lawrence, 1935
Тип рода Ixodlderma inverta Lawrence, 1935
0. Pimeliaphilus Tragardh, 1904 (= Plmeliaphiloides Vitzthum, 1942)
Тип рода Pimeliaphilus podapolipophagus, Tragardh, 1904
7. Scaphothrix Lawrence, 1935
Тип рода Scaphothrix convexa Lawrence, 1935
8. Zonurobia Lawrence, 1935
Тип рода Zonurobia cordylensis Lawrence, 1935
ОБЩИЕ ЗАМЕЧАНИЯ
Большая часть Pterygosomidae паразитирует на ящерицах, селясь
под чешуями хозяина. Хирст (1925) пишет: „Если на одной ящерице
живет более одной формы Geckobia, то одна из них обычно дер-
жится под брюшными чешуйками; тело этой формы уплощено и
ширина его превосходит длину, а брюшко покрыто не волосками,
а щитками. Вторая же форма на том же хозяине обычно живет на
пальцах, между когтем и подушечкой, между пластинками поду-
шечки или между самими пальцами; у этих клещей тело, как пра-
вило, круглое, а брюшко лишено щитков и покрыто волосками11.
Лоуренс (1935) установил, что „...паразитические клещи не встре-
чаются: 1) на ящерицах, принадлежащих к семействам, совершенно
утратившим чешуи или имеющим неперекрывающуюся чешую;
2) у родов с редуцированными конечностями, встречающихся как
среди семейств, ведущих роющий образ жизни (Scincidae), так и
среди семейств, живущих на поверхности земли (Zonuridae, Gerrho-
sauridae), и 3) у отдельных видов, принадлежащих к родам, в кото-
рых большинство форм заражено паразитами11.
Биология большинства видов мало изучена. Жизненный цикл со-
стоит из пяти фаз (личинка, нимфохризалида, нимфа, телеиохризалида
и половозрелая особь) у самок и только из трех фаз (личинка, хри-
шлида и половозрелая особь) у самцов. Лоуренс (1935) описал
личиночную нимфальную и половозрелую фазы Zonurobia cordy-
lensis Lawrence. Неполовозрелые формы очень похожи на взрослых;
Глава V. Подотряд trombidiformes
у личинок и нимф имеется небольшой глазной щиток, несущий одну
щетинку; у взрослых глазной щиток отсутствует, но имеется два
глаза. У паразита таракана Pimeliaphilus podapolipophagus Tragardh
все фазы очень похожи друг на друга; различия между ними незна-
чительны. Некоторые исследователи относили этот вид к другим
семействам, но в действительности он весьма близок к другим пред-
ставителям Pterygosomidae, паразитирующим на ящерицах, и пред-
ставляет собой более примитивный, свободноживущий вид. Р. iso-
metri Cunliffe, распространенный на Филиппинских островах, парази-
тирует на скорпионах.
В США Pimeliaphilus podapolipophagus Tragardh оказался на-
столько большой помехой при выращивании тараканов в экспери-
ментальных садках, что иногда препятствовал работе. Geckobiellu
texana (Banks) был описан с Sceloporus florldanus в Техасе. Hir-
stiella trombidiformes Berlese был найден в Сан-Луисе (Потози,
Мексика), но хозяин его неизвестен.
ЛИТЕРАТУРА
Hirst S., 1925. On the parasitic mites of the suborder Prostigmata (Trombidi-
oidea) found on lizards, J. Linn. Soc. London, Zool., 36, 173—200.
Lawrence R. F., 1935. The prostigmatic mites of South African lizards, Pa-
rasitol., 27 (1), 1—45.
Lawrence R. F., 1936. The prostigmatic mites of South African lizards, Pa-
rasitol., 28 (1), 1—39.
Caeculidae Berlese, 1893
(рис. 169, 161)
ДИАГНОЗ
Caeculidae — коричневато-черные клещи средней величины; длина
тела колеблется от 1,0 до 2,60 мм. Толстое, короткое и широкое
тело имеет трапециевидную форму. Проподосома и гистеросома не
разделены швом. Тело покрыто хорошо развитым панцырем; обычно
имеется семь спинных щитков (три непарных и два парных), окру-
женных слегка бороздчатой или морщинистой кожей. На проподо-
соме по краям имеется по две пары линзовидных глаз. Характерно
наличие ряда бугорков на внутренней стороне ног I; бугорки эти
несут очень длинные крепкие шипы, вследствие чего Caeculidae по-
лучили наименование ,,грабленогих“ клещей; аналогичные бугорки
развиты и на ногах II. Все лапки лишены предлапок, но имеют по
два амбулакральных коготка, часто неодинаковой величины. Эмпо-
дий отсутствует. Короткие пальпы вооружены мощными тибиаль-
ными коготками, тарзальный членик пальпы (палец) занимает вен-
тральное положение. Хелицеры короткие, толстые, с крепким серпо-
видным подвижным пальцем и рудиментом неподвижного пальца.
Половые присоски отсутствуют.
Prostigmata
223
РОДЫ
1. Caeculus Dufour, 1832
Тип рода Caeculus echitilpes Dufour, 1832
2. Procaeculus Jacot, 1936
Тип рода Procaeculus bryanl Jacot, 1936
Рис. 160. Caeculus
calechius Mulaik
(по Малэйку, 1945).
Палина.
Рис. 161. Caeculus pettiti Nevin
(по Нэвину, 1943).
Самка (вид сверху).
ОБЩИЕ ЗАМЕЧАНИЯ
Андрэ (1935) характеризует Caeculus как хищных, вялых, мало
подвижных клещей; они встречаются в солнечных, сухих местах на
голой земле и камнях. В холодное время прячутся во мху. Будучи
потревожены, притворяются мертвыми. Восьминогие нимфы походят
на взрослых, но менее хитинизированы. Шестиногие личинки также
походят на более поздние фазы.
Большинство американских видов было описано Малэйком из
юго-западных штатов США.
ЛИТЕРАТУРА
Andre М., 1935. Notes sur le genre Caeculus Dufour (Acariens) aves descri-
ptions d’spfeces nouvelles africaines, Bull. Soc. d’Hist. Nat. de I’Afrique
du Nord, 26, 79—127.
Jacot A. P., Some rake-legged mites of the family Cheyletidae, J N. Y.
Ent. Soc., 44, 17—31.
Lawrence R. F„ 1939. New South African species of the genus Caeculus
(Acari), J. Linn. Soc. London, Zool., 40 (273), 537—545.
Mulaik S., 1945. New mites in the family Caeculidae, Bull. Univ. Utah, 35
(17), 1-23.
Nevin F. R., 1943. Caeculuc pettiti, a new species of mite from Virginia,
Ann. Ent. Soc. Amer., 36 (3), 389—393.
Глава V. Подотряд Trombidiformes
Tetranychidae Donnadieu, 1875
(рис. 162—164)
ДИАГНОЗ
Tetranychidae — растительноядные клещи средней величины; длина
тела достигает 0,80 мм. Окраска варьирует от желтоватой, зелено-
ватой и оранжеватой до красной. Некоторые клещи зимой принимают
чисто красную окраску. Грушевидное, суживающееся к заднему концу
Рис. 162. Metatetr any chus ulmi
(Koch).
Самка (вид сверху).
Рис. 163. Metatetranychas ulmi
(Koch).
Самец (вид сверху).
тело имеет более или менее отчетливо выраженные плечи. Проподо-
сома либо совсем не отделена от гистеросомы, либо отделена от нес
лишь слабым швом. Спина обычно выгнута, но у ВгуоЫа и Tetrany-
copsis она плоская. Кожа мягкая, щитки не развиты. По краям про-
подосомы расположено по паре глаз. Имеется перитрема, иногда
выступающая. Пальпы снабжены комплексом палец — коготок. Хе-
лицеры слиты у основания, образуя стилофор; подвижный палеи
хелицер сильно видоизменен и превращен в длинный, бичевидный
колющий орган, характерный для этой группы (равно как и для
Pliytoptipalpidae). Лапки ног имеют коготки или видоизмененные
подушечки. Коготки снабжены клейкими волосками и использу-
ются как диагностический признак для родов. Лапки ног I обычнс
несут по паре двойных сенсорных щетинок. Половые присоскг
Prostigmata
225
отсутствуют. Особенности гениталий самцов используются для опре-
деления видов.
Tetranychidae, или паутинные клещи, обязаны своим назван^ ;
способности оплетать тонкой паутинкой листья растений, которыми
они питаются; иногда растение бывает сплошь оплетено такой паути-
ной. Бловель (1945) описывает две группы паутинных желез: труб-
чатые и почковидные; и те и другие впадают в общий выводной
проток в хоботке. Гейскез (1939) указывает, что Tetranychus,
Eotetranychus, Amphitetranychus и Schizotetranychus ткут хорошо,
Paratetranychus и Metatetranychas—плохо, а ВгуоЫа, Petrobia
Рис. 164. Metatetranychas ulmi (Koch).
А —яйцо; Б — кончик лапки; В —кончик пальпы; Г—aedeagus.
и Tetranycopsis совсем
что паутинные железы
пальпы через округлый
не ткут паутины. Гранжан (1948) полагает,
лежат в пальпах и открываются в пальце
концевой „шип“.
роды
1. Tetranychus Dufour, 1832 (= Epitetranychus Zacher, 1916)
Тип рода Tetranychus lintearius Dufour, 1832
2. Allochaetophora McGregor, 1950
Тип рода Allochaetophora californica McGregor 1950
3. Amphitetranychus Oudemans, 1931
Тип рода Tetranychus viennensis Zacher, 1920
4. Anatetranychus Womersley, 1940
Тип рода Anatetranychus hakea Womersley, 1940
5. Aplonobia Womersley, 1940
Тип рода Aplonobia oxalis Womersley, 1940
6. Apotetranychus Oudemans, 1931
Тип рода Apotetranychus muscicola Oudemans, 1931
15 Зак. 1785
Глава И Подотряд TrombidtformeS
7. Bryobia Koch, 1836 (= Schmiedleinia Oudemans, 1928)
Тип рода Bryobia praetiosa Koch, 1836
8. Eotetranychus Oudemans, 1931
Тип рода Acarus telarlus Linnaeus, 1758
9. Eutetranychus Banks, 1917 Anychus McGregor, 1920)
Тип рода Tetranychus banksi McGregor, 1914
10. Eurytetranychus Oudemans, 1931
Тип рода Tetranychus latus Canestrini et Fanzago, 1876
11. Hystrichonychus McGregor, 1950
Тип рода Tetranychus gracilipes Banks, 1900
12. Linotetranus Berlese, 1910
Тип рода Linotetranus cylindricus Berlese, 1910
13. Metatetranychus Oudemans, 1931
Тип рода Tetranychus ultni Koch, 1836
14. Monoceronychus McGregor, 1945
Тип рода Monoceronychus californicus McGregor, 1945
15. Neotetranychus Tragardh, 1915
Тип рода Neotetranychus rubi Tragdrdh, 1915
16. Oligonychus Berlese, 1886 (по Удемансу (1938) = Rhodaxes
Kirchner, 1863)
Тип рода Tetranychus minimus Targ’oni-Tozzetti, 1878
(= Oligonychus brevlpodus Berlese, 1886
17. Paratetranychus Zacher, 1913
Тип рода Tetranychus ununguis Jacobi, 1905
18. Petrobia Murray, 1877 {—Tetranobia Banks, 1917)
Тип рода Acarus latens Muller, 1776 (= Trombldium lapidum
Hammer, 1804)
19. Platytetranychus Oudemans, 1931
Тип рода Tetranychus gibbosus Canestrini, 1890
20. Pseudobryobia McGregor, 1950
Тип рода Pseudobryobia bakeri McGregor, 1950
21. Septanychus McGregor, 1919
Тип рода Tetranychus tumtdus Banks, 1900
22. Schizotetranychus Tragardh, 1915 (= Sttgmaeopsis Banks, 1917 =
= Divarinychus McGregor, 1930)
Тип рода Tetranychus schlzopus Zacher, 1913
23. Simplinychus McGregor, 1950
Тип рода Neotetranychus buxi Garman, 1935
24. Tetranychina Banks, 1917 (,— Tenuicrus Womersley, 1940)
Тип рода Tetranychina apicalis Banks, 1917
25. Tetranycopsis Canestrini, 1890
Тип рода Tetranychus horrldus Canestrini et Fanzago, 1876
ОБЩИЕ ЗАМЕЧАНИЯ
Биология Metatetranychus ulmi (Koch), известного в США как
Paratetranychus pllosus (Canestrini et Fanzago), служит хорошим
Prostigmata
Ml
примером одного из типов биологии паутинных клещей. Metatetra-
nychus ulmi зимуют на фазе яйца. Самки откладывают яйца на
сучья и ветви иногда в таком огромном количестве, что ветки
кажутся красными вследствие яркокрасной окраски яиц. Яйца сте-
бельчатые, но лишены якорных крючков, которые развиты на яйцах
/И. citri (McGregor). В северо-западных штатах США яйца созревают
весной, с появлением новой листвы. Яйцо лопается вдоль экватора,
так что сохраняется только небольшая часть его окружности, служа,
щая шарниром для обеих половинок яйца. Вылупляющаяся шести-
ногая личинка приподнимает верхнюю часть яйца (крышку) и выпол-
зает наружу, после чего крышка опускается, возвращаясь в исходное
положение. Мелкие яркокрасные или оранжевые личинки скапли-
ваются на молодых листьях и сразу же начинают питаться; в период
питания личинки немного передвигаются с места на ме'сто. Напитав-
шись, личинки впадают в состояние покоя. Этот период они проводят
обычно на нижней стороне листьев, около жилок. Стадия покоя
длится примерно столько же, сколько период питания. Постепенно
покровы клеща становятся ровными, блестящими и, наконец, жемчужно
белыми; последнее указывает на то, что тело клеща отделилось от
личиночной кожи. Затем кожа лопается поперечно на спине, между
второй и третьей парой ног, и появляется восьминогая протонимфа;
линька длится несколько часов. Сброшенные покровы остаются на
листьях и придают им серебристый оттенок. Протонимфальная фаза
длится меньше, чем личиночная. Протонимфа также имеет периоды
питания и покоя. После второй линьки появляется дейтонимфа;
дейтонимфальная фаза несколько продолжительнее личиночной или
протонимфальной. На дейтонимфальной фазе обнаруживается половой
диморфизм: самки постепенно становятся более крупными и более
округлыми, чем самцы. Самцы проходят каждую фазу на несколько
часов быстрее, чем самки, и на северо-востоке США самцы проходят
превращение за 8, а самки — за 9 дней. Только что вылупившиеся
половозрелые самцы бегают по листьям в поисках покоящихся дей-
тонимфальных самок. Найдя такую самку, самец останавливается
и выжидает вылупления половозрелой самки. Как только на спине
самки лопнет нимфальная кожица, самец начинает сдирать заднюю
половину последней своими передними ногами и ротовыми органами.
Самка выгибает спину и через несколько минут высвобождает
задние ноги, после чего, пятясь, выходит из передней части линной
шкурки. Тут же немедленно происходит копуляция, иногда даже
раньше, чем самка полностью освободится от нимфальной кожицы.
При этом самец подползает под самку сзади, а последняя приподни-
мает конец брюшка. Затем самец обхватывает передними ногами
брюшко самки, а ногами II и III — задние ноги последней и выгибает
конец брюшка вверх и вперед, пока оно не соединится с концом
брюшка самки. Копуляция длится 10—15 минут. В жаркую погоду
яйца откладываются через 1—2 суток после вылупления самки,
15*
228
Глава V. Подотряд Trombidiformes
в холодную же погоду откладывание яиц задерживается, иногда на
несколько дней или на неделю. Самки откладывают яйца на обе
поверхности листьев, обычно по краям или вдоль жилок. Инкуба-
ционный период этих летних яиц длится в среднем 8—10 дней.
Число поколений, появляющихся за год, зависит от климата. Имеются
сведения, что в Швеции их бывает четыре, в Коннектикуте, невиди-
мому, шесть, в северо-западных штатах США — от шести до восьми.
Зимующие яйца откладываются в течение длительного периода,
начиная примерно с середины августа и до листопада или до насту-
пления холодов, убивающих клещей; этот период иногда затягивается
до ноября. Самые поздние яйца откладываются самками шестого,
седьмого и восьмого поколений.
Metatetranychas citri McGregor имеет примерно такой же жизнен-
ный цикл, что и М. ulmi, однако в Калифорнии взрослые особи
М. citri зимуют на растении-хозяине и за год появляется от 12
до 15 поколений клеща.
У Tetranychus bimaculatus Harvey зимуют преимущественно зрелые
самки. На юге США яйца откладываются в теплую погоду и зимой
и до весны успевают достичь довольно поздних стадий развития.
Клещи этого вида зимуют на диких растениях; весной по мере роста
популяции клеща условия питания на растении-хозяине ухудшаются
и клещи переселяются на другие растения, например на хлопок.
Летом, повидимому, преобладают самки, но по мере приближения
холодной погоды соотношение полов выравнивается. Продолжитель-
ность жизни самки варьирует от 17 дней (в середине лета) до несколь-
ких месяцев (зимой); срок жизни самцов гораздо короче. Самцы
мельче и уже, чем самки. Их брюшко может сильно вытягиваться,
например при копуляции. Отмечены случаи партеногенеза, причем
из неоплодотворенных яиц вылупляются только самки.
Возможность передачи заболеваний растений паутинными клещами
мало изучена. Московец (1940) сообщает, что Tetranychus telarlus (L.)
является переносчиком вирусного заболевания хлопка.
Tetranichidae имеют, пожалуй, наибольшее хозяйственное значение
из всех растительноядных клещей. Трудно обнаружить растение, на
котором не нашлось хотя бы одного вида тетранихид; практически
все виды этого семейства являются, повидимому, потенциальными
вредителями. Tetranichidae прокалывают листья своими стилетовид-
ными хелицерами и высасывают содержимое клеток вокруг ранки.
При сильном заражении растения этими клещами листья отмирают
и опадают. Популяции красных паутинных клещей могут достигать
чудовищных размеров, если с ними не бороться. Так, в Портервилле
(Калифорния) клещи летом размножились на магнолиях настолько,
что в течение нескольких дней все листья этих деревьев опали и
земля под ними и окружающие изгороди были сплошь покрыты
клещами. Применение ДДТ в борьбе с сельскохозяйственными вре-
дителями, очевидно, нарушило равновесие комплекса паутинный клещ—
Prostigmata
229
хищник, вследствие чего Tetranychidae представляют теперь серьезную
опасность в таких районах, где они раньше имели лишь второстепен-
ное значение или вообще не были известны. Неописанные и мало
известные виды не только превратились в опасных вредителей, но
начали приобретать устойчивость к новым акарицидным средствам.
Рис. 165. Повреждения плодов и листьев лимона, причиняемые клещом
Metatetranychas citri (McGregor) (по Бойсу, 1948).
А — поврежденный материал; Б— нормальный материал.
Все же во многих случаях причины разрастания популяции паутинных
клещей сейчас еще неясны.
В США многие виды Tetranichidae имеют хозяйственное значение;
здесь мы перечисляем только некоторые из них. Metatetranychas ulmi
(Koch) распространен по всей Европе от Италии до Швеции и Норвегии
и от Франции до Советского Союза. В США этот клещ встречается
по всему Атлантическому побережью, от Новой Шотландии до Север-
ной Каролины и на запад до Онтарио, Мичигана, Индианы, Теннеси,
Кентукки, Висконсина, Иллинойса и Миссури. На западном побережье
М. ulmi распространен от Британской Колумбии до центральной
Калифорнии, а на восток—до Айдахо и северной части Юты. Эти
клещи предпочитают растения с опадающими плодами, особенно
сливы, груши и яблони, но они встречаются и на многих других
растениях.
230
Глава V. Подотряд Trombidiformes
Одним из главных вредителей цитрусовых в Калифорнии является
М. citri McGregor (рис. 165). Он встречается и во Флориде, но не
имеет там особого хозяйственного значения, вероятно, в силу местных
климатических особенностей.
Tetranychus blmaculatus Harvey (известный так же как Tetra-
nychus telarius L.) встречается во всех штатах США. В юго-восточных
штатах он является опасным вредителем хлопка. Кроме того, повсе-
местно этот клещ вредит плодовым деревьям и огородным культурам.
Распространенный везде в США вид Bryobla praetiosa (Koch)
особенно опасен для миндаля, персиков, слив, люцерны, клевера
и гороха. Этот клещ зимует на клевере и других растениях, на
севере — в фазе яиц, а в южных штатах — в фазе имаго. В. praeti-
osa широко распространен и в Европе.
Tetranychidae обычно не отличаются специфичностью в выборе
хозяина. На цитрусовых и других деревьях обычно встречается
одновременно по нескольку видов. Вредителями цитрусовых являются
Tetranychus lewlsi McGregor, Т. yumensis McGregor, Metatetranychus
citri (McGregor) и Eutetranychus clarkl McGregor.
ЛИТЕРАТУРА
Blauvelt W. E., 1945. The internal morphology of the common red spider-
mite (Tetranychus telarius Linn.), N. Y. (Cornell) Agr, Exp. Sta. Mem., 270.
Cagle L. R., 1949. Life history of the two-spotted spider mite, Virginia Agr.
Expt. Sta. Tech. Bull., 113.
Garman P., 1940. Tetranychidae of Connecticut, Conn. (State) Agr. Expt.
Sta. Bull., 431.
Garman P., Townsend J. F., 1938. The European red mite and its con-
trol, Conn. (Sta^e) Agr. Expt. Sta. Bull., 418.
Geijskes D. C., 1939. Beitrage zur Kcnntniss der Europaischen Spinnmilben
(Acari, Tetranychidae), mit besondcrer Bcriicksiehtigung der Niederlandi-
schen Artcn, Mededeelingen van den Landbouwhoogeschool, 42, 4.
Grand jean F., 1948. Quelques characteres des Tetranyques, Bull, du Mas.,
Ser. 2, 20 (6), 517—524.
McGregor E. A., 1950. Mites of the family Tetranychidae, Amer. Midi. Nah,
44. (2), 257—420.
McGregor E. A., McDonough F. L., 1917. The red spider on cotton,
U. S. Dept. Agr. Bull., 416.
Московец С. H., 1941. Вирусные болезни растений и борьба с ними,
Труды конференции по вирусным болезням растений (Москва, 4—7
ноября 1940 г.), Институт микробиологии АН СССР.
Newcomer Е. J., Yothers М. А., 1929. Biology of the European red mite
in the Pacific Norhwest, U. S. Dept. Agr. Tech. Bull., 89.
Phytoptipalpidae Ewing, 1922
(= Pseudoleptidae Oudemans, 1928; Trichadenidae Oudemans, 1938;
Tenuipalpidae Sayed, 1950)
(рис. 166, 167)
ДИАГНОЗ
Phytoptipalpidae — растительноядные красноватые клещи очень
малых размеров (длина тела 0,20—0,30 мм). Тело различной формы.
Prostigmata
231
Проподосома и гистеросома иногда разделены швом; Кожа либо
бороздчатая, либо покрыта сетчатым рисунком. Имеется две пары
линзовидных глаз. На теле сидит несколько щетинок, сенсорные
щетинки отсутствуют. Пальпы цилиндрические, маленькие, простые;
комплекса палец — коготок нет. Хелицеры U-образные, длинные,
стройные, игловидные; как и у Tetranychidae, они могут выдвигаться.
Р и с. 166. Phytoptipalpus para-
doxus Tragardh (по Сэйду, 1942).
Самка (вид снизу).
Рис. 167. Brevipalpus
cardinalis (Banks).
Самка (вид сверху).
Имеется стилофор (мандибулярная пластинка). Взрослые обладают
тремя или четырьмя парами ног. Все лапки имеют коготки, клейкие
волоски и подушечку; лапки ног I и II несут палочковидные сенсор-
ные щетинки. Половые присоски отсутствуют. Половое отверстие
в виде поперечной щели, как у Eriophyidae.
РОДЫ
1. Phytoptipalpus Tragardh, 1904
Тип рода Phytoptipalpus paradoxus Tragardh, 1904
2. Brevipalpus Donnadieu, 1875
Тип рода Brevlpalpus obovatus Donnadieu, 1875
3. Dolichotetranychus Sayed, 1938
Тип рода Stygmaeus floridanus Banks, 1900
4. Pentamerismus McGregor, 1949 (= Aegyptobia Sayed, 1950)
Тип рода Tenuipalpus erythreus Ewing, 1917
232
Глава V. Подотряд Trombidiformes
5. Phyllotetranychus Sayed, 1938 (согласно Фитцтуму, возможно,
тождествен Raoiella Hirst)
Тип рода Phyllotetranychus aegyptium Sayed, 1938
6. Pseudoleptus Bruyant, 1911
Тип рода Pseudoleptus arechavalatae Bruyant, 1911
7. Raoiella Hirst, 1924 {—Rondaniacarus Oudemans, 1938)
Тип рода Raoiella indica Hirst, 1924
8. Tegopalpus Womersley, 1940
Тип рода Tegopalpus conicus Womersley, 1940
9. Tenuipalpus Donnadieu, 1875
Тип рода Tenuipalpus palmatus Donnadieu, 1875
10. Trichadenus Rondani, 1879
Тип рода Trichadenus sericariae Rondani, 1879
ОБЩИЕ ЗАМЕЧАНИЯ
Типичный пример жизненного цикла этого семейства дает Brevi-
palpus inornatus (Banks), биология которого была описана Мак-Грего-
ром (1916). Яйца утолщенные и имеют форму эллипса; размеры их
составляют примерно 96X67 р. Самка обычно откладывает около
20 яиц в щели или трещины, в старую сброшенную шкурку или
в бороздки вдоль средней жилки листьев, где их иногда накапли-
вается по нескольку сотен плотными кучками. На листьях яйца
откладываются так, что их продольная ось ориентирована перпенди-
кулярно листу. В Южной Каролине шестиногая личинка летом
вылупляется уже через 8 дней. Личиночная фаза длится в среднем
от 4 до 7 дней; после линьки появляется протонимфа, живущая
4—5 дней; она линяет на дейтонимфу, живущую примерно 4 дня.
Таким образом, весь цикл развития продолжается примерно 21 день.
В Южной Каролине эти клещи дают 6-—7 поколений в год и встре-
чаются в течение всего года. Все фазы более или менее похояги
друг на друга.
Род Phytoptipalpus Tragardh является типичным представителем
Phytoptipalpidae, однако эти клещи утратили четвертую пару ног.
По Сэйду, клещи Phytoptipalpus, прежде чем достигнуть половой
зрелости, проходят две „личиночные" фазы. Phytoptipalpus и нимфа
Tenuipalpus erlophyoides Baker напоминают удлиненных кольчатых
Eriophyidae, имеющих две пары ног, и типичных растительноядных
Tetranychidae, что свидетельствует о филогенетическом родстве между
этими группами.
Многие Phytoptipalpidae, невидимому, не наносят особого вреда
растениям, на которых они питаются, но некоторые виды все же
имеют хозяйственное значение. Мак-Грегор (1916) отмечает потерю
листвы у бирючины, вызываемую клещом Brevipalpus inornatus
(Banks) во Флориде, Южной Каролине, Алабаме, Миссисипи и Луи-
зиане. Грин (1900) описывает вид, наносящий сильный вред чайным
плантациям Цейлона; этот клещ уничтожает листья на ветвях и целых
Prosiigmata.
233
кустах, повреждает верхушечные почки и может привести к гибели
растения. Люис (1944) показал, что Brevipalpus lewisl McGregor
вызывает появление рубцов на лимонах в Портервилле (Калифорния).
На плантации, которая не подвергалась опрыскиванию в течение
десяти лет, более 25°/0 лимонов было покрыто рубцами, между тем
как на соседней плантации, регулярно опрыскиваемой, совсем не было
клещей. Джонс и другие (1941) нашли, что Brevipalpus (В. phoenlcls
Geijskes) наносит иногда сильный вред плодам дынного дерева на
Гавайских островах; на пораженных плодах появляются серые чешуй-
чатые участки, причем плод трескается, чаще всего в местах сопри-
косновения со стволом. Этот же клещ повреждает цитрусовые
в Испании. Бланшар (1940) сообщает, что В. pseudocuneatus (Blanchard)
вредит цитрусовым в Аргентине. Другой вид этого же рода повре-
ждает плоды растений из семейства Passiofloraceae b Австралии.
Tenuipalpus paclficus Baker и Brevipalpus australis (Tucker)
вредят плодовым садам. Сэйд (1942) нашел, что Tenuipalpus granati
Sayed вызывает побурение виноградных листьев в Египте. Этого же
клеща обнаруживали на листьях, ветвях, а иногда и на плодах гра-
натового дерева.
Pseudoleptus и Dolichotetranychus встречаются на различных
солончаковых травах по всему земному шару. Оба вида вызывают
искривление верхушек у пораженных ими трав.
ЛИТЕРАТУРА
Baker Е. W., 1945. Mites of the genus Tenuipalpus (Acarina: Trichadenidae)
Proc. Ent. Soc. Wash., 7 (2), 33—38.
Baker E. W., 1948. A new trichadenind mite which further indicates a phy-
logenetic relationship between the Tetranychidae and Eriophyidae, Proc.
Ent. Soc. Wash., 50 (3), 59, 60.
Baker E. W., 1949. The genus Brevipalpus (Acarina: Pseudoleptidae), Amer.
Midland Nat., 42 (2), 350—402.
Blanchard E. E., (1939) 1940. Tres acaros danino para los cultivos argenti-
nos, Rev. Рас. Agron. La Plata, 3, (24), 11—18.
Green F., 1900. Tea-mites and some suggcsted-experimental work against
them, Roy. Bot. Gard., Ceylon, Cir. 17 (Ser. 1), 202, 203.
Jones W. W., Storey W. B., Parr os G. K., Hol da way F. G., Papaya
production in the Hawaiian Islands, Bull. Hawaiian Agri. Expt. S'a.,
87, 46.
Lawrence R. F., 1943. New South African mites of the genus Tenuipalpus
Donnadieu (Tetranychidae), Trans. Roy. Soc. South Africa, 30 (1), 35—48.
Lewis H., 1944. Injury to citrus by Tenuipalpus mites, Calif. Citrograph,
29 (4), 87.
McGregor E. A., 1916. The privet mite in the South, J. Econ. Ent., 9,
556—560.
McGregor E. A., 1949. Nearctic mites of the family Pseudoleptidae, Mem.
South Calif. Acad. Sci., 3 (2), 1—45.
Planes S., (1944) 1945. La rona de los frutos citricos, Bol. Patal. Veg. у
Ent. Agr., 13, 47—54.
Sayed M. T., 1942. Contribution to the knowledge of the Acarina of Egypti:
II. The genus Tenuipalpus Donnadieu Bull. Soc. Found 1-er Ent., 24,
93—113.
234
Глава V. Подотряд Trombidiformes
Teneriffiidae Thor, 1911
ДИАГНОЗ
(рис. 168, 169)
Tenerilfiidae — клещи средней величины, длиной от 0,70 до 0,90 мм,
с удлиненным телом и бегательными ногами. Покровы бороздчатые,
срединного киля нет, но имеется проподосомальный щиток. Имеется
две пары широко расставленных глаз. Анальное отверстие располо-
Рис, 168. Austroteneriffia hirsti Wo-
mersley.
Самка (вид сверху).
жено вентрально у самого
конца тела. Удлиненное по-
ловое отверстие прикрыто
серповидными клапанами
с несколькими щетинками.
Имеется три пары слабо
развитых половых присосок.
Ноги опушены и имеют
простое строение. Если не
на всех, то, во всяком слу-
чае, на ногах I и II имеется
Рис. 169. Austrotene-
riffia hirsti Womersley.
Дистальная часть пальпы.
по два гребневидных амбулакральных коготка; ноги III и IV иногда
имеют когтевидный эмподий. Рострум короткий и широкий. Хелицеры
серповидные, с двумя небольшими щетинками на дорзальной поверх-
ности основного членика. Пальпы пятичлениковые, толстые, сильно
согнутые и имеют длинный тибиальный коготь и нежные щетинки.
Тарзальный членик пальпы почти рудиментарен и несет щетинки раз-
ной длины. Имеются перитремы, открывающиеся у основания хелицер.
РОДЫ
1. Tetieriffia Thor, 1911 (= Tenerifflola Strand, 1911)
Тип рода Tetieriffia quadrlpapillata Thor, 1911
2. Austroteneriffia Womersley, 1935
Тип рода Austroteneriffia hirsti Womersley, 1935
3. Heteroteneriffia Hirst, 1926
Тип рода Heteroteneriffia marina Hirst, 1926
Prostigmata
235
4. Neotenerlfiola Hirst, 1924
Тип рода Neotenerifiolla luxoriensis Hirst, 1924
5. Paratenerlffta Thor, 1911
Тип рода Paratenerlffta blpectlnata Thor, 1911
ОБЩИЕ ЗАМЕЧАНИЯ
На Малайских островах под камнями, на скалистой отмели,
ежедневно заливаемой при приливах на 1,2—1,8 м, был обнаружен
Heteroteneriffia marina Hirst, 1926; при отливах между банкой
и берегом остается постоянная лужа морской воды шириной при-
мерно 6 м. Клещи очень подвижны, и их трудно поймать. Описаны
и другие виды и роды из самых разнообразных местностей, напри-
мер из Исландии, Тенерифа, Парагвая, Египта и Австралии.
Судя по строению этих клещей, они ведут хищный образ жизни.
ЛИТЕРАТУРА
Womersley Н., 1935. On the occurrence in Australia of Acarina of the fa-
mily Teneriffiidae (Trombidoidea), Pec. South Austral. Mus., 5, 333—338.
Pseudocheylidae Oudemans, 1909
(рис. 170)
ДИАГНОЗ
Pseudocheylidae — бесцветные
или красноватые клещи неболь-
шой или средней величины. Раз-
меры тела варьируют от 0,23 до
1,1 мм. Они имеют более или
менее удлиненное, несколько ром-
бовидное тело. Проподосома и
гистеросома разделены глубокой
бороздкой. Проподосомальный
щиток выражен очень слабо, а чаще
отсутствует совсем. Перитремы
расположены у основания хели-
цер. Проподосома несет две пары
сенсорных щетинок, по краям ее
расположено или по две пары
линзовидных глаз (Pseudocheye-
lus), или по две пары глазков
с общей роговицей (Neocheylus).
Иногда глаза отсутствуют. Пальпы
короткие, обычно несколько более
толстые, чем ноги. Бедренный
членик пальпы длиннее осталь-
ных и утолщен с наружной сто-
Рис. 170. Pseudocheylus (Anoplo-
cheylus) eiiropaeiis Berlese
(по Берлезе, 1910).
Самка (вид сверху).
236
Глава V. Подотряд Trombidiformes
роны, благодаря чему пальпы становятся пинцетовидными. Голен-
ный членик пальпы вооружен сильно согнутым коготком; тарзальный
членик пальпы занимает вентральное положение и очень слаб (или
совсем отсутствует?). На лапках ног либо имеется длинная стебель-
чатая присоска, а коготки отсутствуют (Pseudocheylus, Neocheylus),
либо имеется по два коготка, между которыми расположена при-
соска (Tarsocheylus), либо, наконец, эмподий отсутствует и имеются
нормальные, но очень маленькие коготки на передней лапке и гребне-
видные коготки на остальных лапках (Stigmocheylus). Половые при-
соски отсутствуют. У рода Heterocheylus на проподосоме имеется
только одна пара сенсорных щетинок. У этого рода на лапке ноги I
амбулакральные образования отсутствуют, но на лапках ног II, III
и IV имеются только присоски. Трегорд (1950) разделяет этих клещей
На два подсемейства.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ PSEUDOCHEYLIDAE
(ПО ТРЕГОРДУ)
1. Хелицеры стилетовидные. Пальпы имеют по крупному, сильному,
заостренному зубчатому терминальному зубу и различное число
гребневидных или серповидных щетинок. Все ноги вооружены
коготками.....................................Pseudocheylinae
— Хелицеры вооружены сильно выгнутым, острым терминальным
зубом, сходным с зубом личинок Trombidiidae. Пальпы воору-
жены тупым терминальным зубом без зубцов; гребневидные или
серповидные щетинки на них отсутствуют. Ноги I шупальцеобразны;
ноги III и IV снабжены зубьевидной субтерминальной щетинкой
и большим дискообразным эмподием..............Heterocheylinae
РОДЫ И ПОДРОДЫ
Pseudocheylinae Tragardh, 1950
1. Pseudocheylus Berlese, 1888
a) Pseudocheylus s. str. (= Rhagina Womersley, 1935)
Тип подрода Pseudocheylus biscalatus Berlese, 1888
6) Anoplocheylus Berlese, 1910
Тип подрода Pseudocheylus (Anoplocheylus) europaeus, Ber-
lese, 1910
2. Neocheylus Tragardh, 1906
Тип рода Neocheylus natalensis Tragardh, 1906
3. Stigmocheylus Berlese, 1910
Тип рода Stygmocheylus brevisetus Berlese, 1910
4. Tarsocheylus Berlese, 1904
Тип рода Tarsocheylus paradoxus Berlese, 1904
Heterocheylinae Tragdrdh, 1950
1. Heterocheylus Lotnbardini 1926
Тип рода Heterocheylus fuslfortnis Lotnbardini, 1926
Prostlgmala
237
ОБЩИЕ ЗАМЕЧАНИЯ
Небольшое семейство, представленное всего лишь несколькими
видами. Из США описан только Pseudocheylus amerlcanus (Ewing),
обнаруженный под корой клена в Урбане (Иллинойс). Pseudocheylus
biscalatus Berlese был собран под древесной корой в Уругвае и
Бразилии. Neocheylus natalensls Tragardh обнаружен во влажном мху
в Натале (Южная Африка), a Pseudocheylus protea (Womersley) —
во мху в Австралии. Виды остальных родов были найдены в Италии —
в почве, мху, гумусе и гниющей подстилке. В целом клещи этого
семейства, повидимому, ведут хищный образ жизни, но Heterocheylus
является эктопаразитом членистоногих.
ЛИТЕРАТУРА
Tragardh I., 1950. Description of a new species of Heterocheylus Lombar-
dini from Africa, with notes on the classification of the Pseudocheyleti-
dae, Ent. Tidskrift, 71, H. 2, 104—410.
ДИАГНОЗ
Anystidae Oudemans, 1902
(рис. 171)
Anystidae — красные или красноватые клещи средней величины.
Длина тела варьирует от 0,55 до 1,30 мм. Покровы мягкие; пропо-
досомальный щиток имеется
не всегда, но и в тех случаях,
когда он развит, фактически
имеет место не склероти-
зация покровов, а развитие
бороздчатого узора на коже.
Проподосома и гистеросома
не разделены швом. Имеется
перитрема, иногда свободно
выступающая, расположен-
ная у основания хелицер.
Рострум короткий и конусо-
образный. На клешне хе-
лицер развит дистальный,
крючкообразный подвижный
палец. Голенный членик
пальпы несет от одного до
Рис. 171. Anystis baccarum (Linnaeus)
(по Удемансу, 1936).
Самка (вид сверху).
трех внутренних коготков; тарзальный членик пальпы причленен к голени
дистовентрально. Гладкий шишкообразный выступ передней части
проподосомы несет пару сенсорных щетинок; пара сенсорных щети-
нок имеется и на проподосоме. Ноги отходят от тела лучеобразно;
тазики расположены группой или двумя очень близкими, но раздель-
ными группами. Все лапки или цельные, или же многочлениковые;
238 Глава V. Подотряд Trom1' idiformes
они имеют по два коготка (гребневидные, зубчатые или опушенные)
и эмподий. Эмподий бывает когтевидный, щеткообразный или же
чашевидный. Половые присоски отсутствуют. Это семейство подраз-
деляется на два подсемейства.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ AHYSTIDAE
1. Тело короткое и широкое. Имеется две пары глаз. Голенный
членик пальпы вооружен двумя коготками................Anystinae
— Длина тела больше его ширины. Имеется одна или две пары
глаз. Голенный членик пальпы вооружен одним широким коготком
или двумя перистыми коготками..................Erythracarinae
РОДЫ
А и у s t i и а е О u d е m a n s, 1936
1. Anystis v. Heyden, 1826 (= Actineda Koch, 1836)
Тип рода Trombidium cornigerum Hermann, 1804
2. Autenriethla Oudemans, 1936
Тип рода Actineda velox Berlese, 1905
3. Barella Oudemans, 1936
Тип рода Anystis sinensis Berlese, 1923
4. Scharfenbergia Oudemans, 1936
Тип рода Actineda hllarls, Koch, 1836
5. Snartia Oudemans, 1936
Тип рода Snartia nepenthus Oudemans, 1936
6. Tencateia Oudemans, 1936
Тип рода Tencateia besselingi Oudemans, 1936
7. Walzia Oudemans, 1936
Тип рода Actinella antiguensis Stoll, 1886
Erythracarinae Oudemans, 1936
1. Erythracarus Berlese, 1903
Тип рода Trombidium parletlnum Hermann, 1804 (= Erythraeus
comes Berlese, 1886)
2. Absoloniana Willmann, 1940
Тип рода Absoloniana diversipes Willmann, 1940
3. Anandia Hirst, 1927
Тип рода Anandia alticola Hirst, 1927
4. Bechsteinia Oudemans, 1936
Тип рода Bechsteinia schneideri Oudemans, 1936
5. Chabrleria Oudemans, 1936
Тип рода Tarsotomus terminalts Banks, 1916
6. Chaussieria Oudemans, 1937 (== Schellenbergia Oudemans, 1936
nomen praeoccupatum)
Тип рода Erytrhaeus domesticus Koch, 1847
Prostigmata
23$
7. Erythrocheylus Berlese, 1903
Тип рода Pseudocheylus erythraeoldes Leonardi, 1901
8. Siblyia Oudemans, 1936
Тип рода Erythraeus ignipes Duges, 1834
9. Tarsolarkus Thor, 1912
Тип рода Tarsolarkus articulosus Thor, 1912
10. Tarsotomus Berlese, 1882
Тип рода Erythraeus hercules Berlese, 1882
ОБЩИЕ ЗАМЕЧАНИЯ
Длинноногие подвижные клещи, питающиеся другими клещами и
различными мелкими насекомыми. Живут на растениях — листьях и
ветках деревьев и на траве. Обычно немногочисленны, но порой встре-
чаются в очень больших количествах, Anystis agllis (Banks) описан
из США; Walzia antlguensis (Stoll), описанный из Венецуэлы, был
обнаружен и во Флориде.
Личиночные, нимфальные и взрослые фазы походят друг на друга.
Известны две нимфальные фазы. Anystidae обладают, повидимому,
способностью ткать паутину, и молодые стадии обволакиваются коко-
нами (нимфохризалида или телеиохризалида). Бэнкс находил экзем-
пляры, только что окончившие линьку, уже заключенными в малень-
кие шелковистые белые коконы.
ЛИТЕРАТУРА
Oudemans А. С., 1936. Neues fiber Anystidae (Acari), Archlv f. Natur-
gesch., Neue Folge, 51, 364—446.
Myobiidae Megnin. 1877
(рис. 172, 173)
ДИАГНОЗ
Myobiidae — клещи небольших или средних размеров. Панцыря
нет, кожа бороздчатая. Перитремы имеются. Хелицеры маленькие,
стилетовидные. Пальпы простые, очень маленькие и не приспособлены
для схватывания; коготок на пальпах имеется не всегда, а палец если
и имеется, то с трудом различим. Первая пара ног иногда видоизме-
нена в связи с приспособлением к захватыванию волосков; остальные
ноги вооружены одним или двумя коготками. У самца половое отвер-
стие может быть расположено на дорзальной поверхности. Myobiidae
чрезвычайно разнородная группа клещей и при дальнейшем изучении
из этого семейства, возможно, будет выделено несколько групп.
Повидимому, Harpirhynchus, Ophioptes, Picobia, Psorergates и
Syrtngophilus образуют особое семейство, занимающее промежуточное
положение между Myobiidae и Cheyletidae, хотя, возможно, и более
близкое к последнему.
240
Глава V. Подотряд Trombidiformes
РОДЫ
1. МуоЫа v. Heyden, 1826
Тип рода Pediculus musculi Schrank, 1781
2. Amorphacarus Ewing, 1938
Тип рода МуоЫа alongata Poppe, 1898
3. Calcarmyobia Radford, 1948
Тип рода МуоЫа rhinolophia Radford, 1940
Рис. 173. Syringophilus columbae Hirst
(по Хирсту, 1922).
Самка (вид сверху).
Рис, 172. МуоЫа muscullnus
(Schrank).
Самка (вид сверху).
4. Eadiea Jameson, 1949
Тип рода Eadiea condylurae Jameson, 1949
5. Eutalpacarus Jameson, 1949
Тип рода Eutalpacarus peltatus Jameson, 1949
6. Emngatui Radford, 1948
Тип рода E'Wingana bispinosa Radford, 1948
7. Foliomyobia Radford, 1948
Тип рода МуоЫа chiropteralis Michael, 1884
8. Harpirhynchus Megnin 1877 (= Harpicephalus Canestrini 1885 =
= Sarcopterlnus Railliet, 1893 = Sarcopterus Giebel, 1871 =
— Sarcoborus Oudemans, 1904)
Prostipmafa
241
Тип рода Sarcoptes nidulans Nitzsch, 1818
9. Neomyobia Radford, 1948
Тип рода МуоЫа rollinatl Poppe, 1908
10. Ophioptes Sambon, 1928
Тип рода Ophioptes parkeri, Sambon, 1928
11. Picobia Haller, 1878
Тип рода Picobia heeri Haller, 1878
12. Protomyobla Ewing, 1938
Тип рода Myobia claparedei Poppe, 1896
13. Psorergates Tyrrell, 1883
Тип рода Psorergates simplex Tyrrell, 1883
14. Radfordla Ewing, 1938
Тип рода МуоЫа ensifera Poppe, 1896
15. Syringophilus Hel er, 1880
Тип рода Syringophilus blpectinatus Heller, 1880
ОБЩИЕ ЗАМЕЧАНИЯ
Клещи рода МуоЫа паразитируют на грызунах, насекомоядных и
на летучих мышах. У этих клещей ноги I сильно видоизменены для
захватывания волосков. По Эвингу (1938), клещи Myobia Питаются
у основания волос хозяина секретом, выделяемым железами в волося-
ные сумки, и если и сосут крозь, то лишь очень редко. Грант
(1942), изучавший Myobia musculi (Schrank), пишет: „Они питаются,
повидимому, не только в волосяных сумках, но и в других местах;
по всяком случае, хоботок часто вводится в кожу в таких местах,
где даже под бинокуляром нельзя обнаружить никаких следов волос.
Следует упомянуть, что у одной особи содержимое ротовой трубки
состояло, повидимому, из эритроцитов". Джемсон (1948) находил
насосавшихся кровью клещей Myobia simplex Ewing. Скидмор (1934)
сказывает, что Radfordia ensifera (Poppe) (== МуоЫа ratti Skidmore)
паразитирует на белых крысах. „Зараженные крысы расчесывали
голову, нос и шею. У многих из них на голове, вокруг ушей и на
верхней части шеи были сухие струпья. У части крыс струпья крово-
точили от сильных расчесов".
Myobi'dae, повидимому, распространены, по всему свету. В Австра-
лии Psorergates ovis Womersley, 1941 паразитирует на овцах, вызы-
вая у них хроническое раздражение кожи на боках и бедрах. Северо-
американские и европейские виды Syringophilus паразитируют в стволах
птичьих перьев. Эти клещи предпочитают питаться на внутреннем
опахале пера. Harpirhynchus nidulans (Nitzsch) живет колониями
в перьевых фолликулах и вызывает образование в коже хозяина
опухолей или цист. Этот клещ встречается на очень многих птицах,
в том числе и на голубях. Виды Ophioptes паратизируют на змеях
в Южной Америке; они живут в маленьких ямках, создаваемых ими
в роговом слое чешуи змеи. Эвинг (1933) описал вид, образующий
ямки в чешуе хозяина; Эвинг и Сэмбон описали две формы этого
16 Зак. 1785.
242
Глава V. Подотряд Tromblliformes
вида — мешковидную безногую нимфу и вполне сформированную
восьминогую особь, обнаруженную внутри этой разбухшей нимфальной
формы. Последнюю Сэмбон описал как самца, Эвинг же склонен счи-
тать, что это была нимфа или самка. Исследование типа в Нацио-
нальном музее США показало, что эта форма представляет собой
имаго, но пол ее определить не удалось. Жизненный цикл неизвестен.
ЛИТЕРАТУРА
Ewing Н. Е., 1938. North American mites of the subfamily Myobiinae, new
subfamily (Arachnida), Proc. Ent Soc. Wash., 40 (7), 180—197.
G r a n t D. C., 1942. Observations on Myobia muscu'i (Schrank) (Arachnida: Aca-
rina : Cheyletidae), Microroentomology, 7 (3), 64—76.
Jameson E. W., 1948. Myobiid mites (Acarina : Myobiinae) from Shrews
(Mammaiia: Soricidae) of Eastern North America, J. Parasitol., 34 (4),
336—342.
Jameson E. W., 1949. Myobiid mites (Acarina: Myobiidae) from Condylura
cristata (Linnaeus) and Neuro^rlchus gibbsii (Baird) (Mammalia: Talpi-
dae), J. ParasBol., 35 (3), 423—430.
Radford C. D., 1934. Notes on the genus Myobia, North Western Natura-
list, 356—<61.
Radford C. D„ 1935. Notes on the genus Myobia North Western Natura-
list, 248—258.
Radford C. D., 1936. Notes on the genus Myobia, North Western Natura-
list, 34—39, 144—151.
Radford C. D., 1948. A revision of the fur mites Myobiidae (Acarina),
Bull. Paris Mus. d’Hist. Nat., Ser. 2, 20 (5), 458—464; (6), 525—531.
Womersley H., 1941. Notes on the Cheyletidae (Acarina, Trcmbidoidea)
of Australia and New Zealand with descriptions of new species, Rec.
South Austral. Mus., 7 (1), 51—64.
Cheyletidae Leach, 1814
(рис. 174)
ДИАГНОЗ
• Cheyletidae — клещи небольших или средних размеров, длиной
от 0,20 до 0,80 мм; они окрашены в желтоватые или красноват!
тона. Тело овальное, пухлое, у перьевых клещей—удлиненное. Про-
подосома и гистеросома обычно четко ограничены и имеют по одному
или по нескольку спинных щитков. Линзовидные глаза имеются не
всегда. Спинные щетинки простые, пильчатые, палочковидные или
булавовидные. Имеется толстая перитрема, более или менее полно
обрамляющая гнатосому. Хелицеры короткие, стилетовидные и при-
способлены для прокалывания. Пальпы широкие, клешневидные и
служат для схватывания. Палец пальпы короткий и иногда снабжен
гребнем и серповидными щетинками. На голенном членике пальпы
имеется большой терминальный коготок, обычно зубчатый. Лапки
всех ног имеют по два коготка и эмподий (у Cheletogenes лапки
ног I не имеют ни коготков, ни присоски, у Cheletomorpha на лапках
ног I нет коготков, но есть присоска). Половые присоски отсутствуют.
Prostigmata
243
Половое отверстие самца иногда расположено дорзально, но обычна
лежит на заднем конце тела и вентрально.
РОДЫ
Рис. 174. Cheletogenes
ornatus (Canestrini et
Fanzago).
Самка (вид сверху).
1. Cheyletus Latreille, 1796
Тип рода Acarus eruditus Schrank, 1781
2. Acaropsis Moquin-Tandon, 1862
Тип рода Tyroglyphus mericourti Moquin-Tandon, 1851
3. Chelacaropsis Baker, 1949
Тип рода Chelacaropsis moorei Baker, 1949
4. Cheletogenes Oudemans, 1905
Тип рода Cheyletus ornatus Canestrini et Fanzago, 1876
5. Cheletoldes Oudemans, 1904
Тип рода Syringophilus uncinatus Heller, 1880
6. Cheletomimus Oudemans, 1904
Тип рода Cheletes berlesei Oudemans, 1904 (= Cheletomimus:
trux Oudemans, 1904)
7. Cheletomorpha Oudemans, 1907
Тип рода Acarus lepidopterorum Shaw, 1794 [—Cheyletus vena-
sttssimus Koch, 1839)
8. Cheletophanes Oudemans, 1904
Тип рода Cheyletus montandonl Berlese et Trouessart, 1889
9. Cheletophyes Oudemans, 1914
Тип рода Cheletophyes vitzthumi Oudemans, 1914
10. Cheletopsis Oudemans, 1904
Тип рода Cheyletus nornerl Poppe, 1888
16*
244
Глава V. Подотряд Trombidiformes
11. Cheletosoma Oudemans, 1905
Тип рода Cheletosoma tyrannus Oudemans, 1905
12. Chelonohis Berlese, 1893
Тип рода Chelonotus selenirhynchus Berlese, 1893
13. Cheyletla Haller, 1884
Тип рода Cheyletla laureata Haller, 1884
14. Cheilettella Canestrini, 1886 (= Ewlngella Vai: et Augnstson, 1944)
Тип рода Cheyletus parasitivorax Megnin, 1878
15. Eucheyletla Baker, 1949
Тип рода Eucheyletla blshoppi Baker, 1949
16. Eutogenes Baker, 1949
Тип рода Eutogenes foxl Baker, 1949
17. Neoeucheyla Redford, 1950 {—Eucheyla Berlese, 1913 nomen
praeoccupatum)
Тип рода Cheyletla (Eucheyla) lorlcata Berlese, 1913
18. Neocheyletiella Baker, 1949
Тип рода Neocheyletiella rohvueri Baker, 1949
ОБЩИЕ ЗАМЕЧАНИЯ
Cheyletidae имеют обычный жизненный цикл, состоящий из сле-
дующих фаз; яйцо, личинка, нимфы, половозрелая особь. У нимф
иногда бывает больше спинных щитков, чем у взрослых. У видов
рода Cheyletus самец часто отличается от самки строением гнатосомы.
У этих клещей пальпы сильно удлинены и на коготке пальпы или
совсем нет базальных зубцов, или же имеется лишь небольшое число
последних. Aedeagus имеется. У самок половое отверстие невелико.
У самцов сенсорный орган на лапках ног I сильно увеличен. У Che-
yletiella parasltivorax (Megn’n) половое отверстие самца лежит дор-
зально. У видов других родов, у которых известны оба пола, самцы
и самки более или менее похожи и отличаются друг от друга только
упомянутыми выше признаками. Эзинг (1912) изучил жизненный цикл
Cheyletus. По его наблюдениям, яйца откладываются друг около
друга, но не кучками, а поодиночке. Самка некоторое время остается
возле откладываемых яиц. Одна из наблюдавшихся самок отложила
25 яиц в течение шести дней, другая —22 яйца за два дня, а третья —
только одно яйцо. При комнатной температуре инкубационный период
длился в среднем 4,3 дня, фаза личинки продолжалась 7,66 дня,
нимфы 1—4,5 днл, нимфы II — не менее 5 дней. Половозрелые клещи
жили 13 дней. Личинка походит на взрослого клеща, но имеет только
шесть ног; нимфы также походят на взрослых и имеют четыре пары
ног. Эвинг установил, что излюбленной пищей молодых клещей являются
яйца клещей других видов, например Acaridae, совместно с которыми
обычно находят и Cheyletidae. В пробах молотой пшеницы эти Che-
yletidae за короткое время уничтожили 95°/0 всех Acaridae. В Совет-
ском Союзе Родионов и Фурман (1940), изучавшие возможность борьбы
с амбарными клещами {Acarus siro L., Tyrophagus castellanii Hirst
Prostigmata
245
и Glycyphagus destructor (Schrank)) при помощи Cheyletus eruditus
(Schrank), нашли, что этот клещ в данном отношении особой пользы
не приносит и что он только засоряет зерно. Cheyletidae имеют более
длительный жизненный цикл, чем амбарные клещи, и хотя они и более
устойчивы к высыханию, но зато более чувствительны к синильной
кислоте. Хорошо защищенные панцырем гипопусы зерновых клещей
не подвергаются нападению Cheyletidae.
Cheyletidae в целом представляют собой свободноживущих хищ-
ников, но некоторые из них паразитируют в перьях птиц или в шерсти
белок, кроликов и кошек и иногда причиняют вред хозяину. Свободно-
живущие формы встречаются совместно с другими вредными клещами,
например Acaridae и Tetranychidae, а также с червецами, но, пови-
димому, не играют особой роли в борьбе с ними. Благодаря свобод-
ному образу жизни многие виды расселились по всему свету.
ЛИТЕРАТУРА
Baker Е. W., 1949. A review of the mites of the family Cheyletidae in the
United States National Museum, Proc. U. S. Mas., 99 (3238), 267—320.
Cooper K. W., 1946. The occurrence of the mite, Cheyle4e'laparasitivo'ax
(Megnin) in North America, with notes on its synonymy and „parasitic"
habits, J. ParasPol., 32 (5), 480—482.
Ewing H. E., 1912. The life history and habits of Cheyletus semlnivorus
Packard, J. Econ. Enf., 5 (5), 416—420.
McGregor E. A., 1944. A new potential enemy of the bud mite, Catif.
Citrograph., 30 (2), 53.
Olsen S. J., Roth H., 1947. Cn the mite СЧеу'е*1е"а parasEtvorax, occur-
ring on cats, as a facultative parasite of man, J. Parasifol., 33 (5),
444—445.
Oudemans A. C., 1906. Revision des Cheletines, Mem. Soc. Zool. de Erance,
19, 36—219.
Demodicidae Nicolet, 1855
(рис. 175)
ДИАГНОЗ
Demodicidae — бледноокрашенные, очень мелкие клещи, длиной
от 0,10 до 0,39 мм. Тело червеобразное, кольчатое; восемь коротких
ног, имеющих форму обрубков, отодвинуты к передней части тела.
Хелицеры, как и у Cheylet’dae, стилетовидные. Капитулум прикры-
вает пальпы и хелицеры. Пальпы трехчлениковые; дистальный членик
несет четыре или пять палочковидных щетинок. С нижней стороны
капитулума ближе к основанию последнего расположена пара крошеч-
ных отверстий. Половое отверстие самки расположено позади тазиков
ног IV, а половое отверстие самца расположено дорзально, в центре
подосомы. Половые присоски отсутствуют.
РОДЫ
1. Demodex Owen, 1843
Тип рода Acarus folllculorutn Simon, 1842
246
Глава V. Подотряд Trombidiformes
ОБЩИЕ ЗАМЕЧАНИЯ
Типичный пример жизненного цикла Demodic’dae дает Detnodex
muscardini Hirst, паразитирующий на соне Muscardlnus avellanarius.
Из яйца вылупляется личинка, внешне отличающаяся от молодой
нимфы лишь тем, что имеет не четыре, а три пары ног, состоящих
из хитиновой пластинки, или эпимера. Каждый эпимер несет пару
миниатюрных мало заметных зубчатых бугорков. Протонимфа имеет
четыре пары ног, или эпимер, вооруженных тремя или четырьмя крошеч-
ными, но отчетливыми зубцами. На брюшной поверхности тела, ближе
Рис. 175. Demodex muscardini Hirst (по Хирсту, 1919).
А —самец (вид сверху);. Б — самка (вид снизу).
к середине, находится четыре пары своеобразных полукруглых эпидер-
мальных образований, изогнутый задний край которых снабжен острыми
зубцами. Конечности дейтонимф похожи на конечности протонимф,
но все же они более напоминают ноги и состоят из нескольких чле-
ников, несущих две группы зубцов. У дейтонимф на брюшке также
имеется четыре пары полукруглых эпидермальных образований, но
последние у них гораздо меньших размеров. Половозрелые клещи
имеют половое отверстие и пятичлениковые ноги; полукруглые обра-
зования на брюшке у них не развиты.
Хирст (1919) дал прекрасный обзор этого семейства, из которого
мы заимствуем приводимые ниже данные. Demidex folllculorum Oxven,
паразитирующий в кожных железах человека (особенно вокруг носа
и век), считался раньше возбудителем различных кожных заболеваний,
но теперь установлено, что он не причиняет вреда и встречается
Prostigmata
247
у большинства людей и не только в больных, но и в нормальных
тканях. Распространен этот клещ почти повсеместно. D. canis Leyd’g,
встречающийся у собак, больных фолликулярной чесоткой, или желез-
пицей, также распространен по всему свету. В типичных случаях
этот клещ присутствует в большом количестве одновременно со ста-
филококками {Staphylococcus pyogenes albus или близкие формы),
являющимися подлинными возбудителями заболевания. Участие клеща
в этиологии заболевания сводится к тому, что он расширяет волосяные
сумки и потовые железы и, возможно, заносит инфекцию в поры кожи.
Различают две формы железницы — пустулезную, сопровождающуюся
образованием струпьев, и чешуйчатую. В типичных случаях чешуй-
чатой железницы образуются небольшие лишенные волос участки,
покрытые красноватыми прыщиками и гнойничками. Кожа на поражен-
ных местах воспаляется и уплотняется. Заболевание .сопровождается
зловонным запахом. От грибковой инфекции это заболевание позволяет
отличить присутствие клещей. Клещ D. equi Pailliet, встречающийся
и у здоровых лошадей, может вызывать патологическое состояние
кожи, характеризующееся зудом разной интенсивности или же выра-
женными повреждениями, сопровождающимися выпадением волос. За-
болевание обычно начинается с поясницы или загривка. В поздних
стадиях появляются гнойники, кожа уплотняется. Заболевание, пови-
димому, не заразно. D. phylloides Csokor вызывает гнойное заболевание
кожи у свиней. Гнойники могут быть размером от песчинки до грец-
кого ореха; иногда они сливаются, прорываются и образуют на коже
язвы. Гнойники образуются только на местах, покрытых нежной кожей,
и распространяются от морды по шее и нижней части груди и брюха,
вдоль боков и на внутренние поверхности ног. На общем состоянии
здоровья животного это заболевание не отражается. D. bovis Stiles
вызывает у рогатого скота появление узелков или гнойников на коже.
Гнойники по размерам варьируют от булавочной головки до куриного
яйца и локализуются обычно вокруг ушей, на боках, плечах и между
бедрами. В творожистом или жидком содержимом узелков встречаются
клещи на разных фазах развития. Заболевание, повидимому, носит
инфекционный характер, но на общем состоянии здоровья животного
не отражается. Все же ходы, прокладываемые клещами, снижают
качество шкуры. Различные виды Demodicidae были собраны с овец,
ежей, полевых мышей, кошек, кроликов, полевок, крыс. Возможно,
что со временем они будут обнаружены и на многих других хозяевах.
ЛИТЕРАТУРА
Hirst S., 1919. The genus Demodex Owen, Brit. Mus. (Nat. Hist.), Studies on
Acari № 1.
Hirst S., 1922. Mites injurious to domestic animals, Brit. Mus. {Nat. Hist.),
Econ. Series, № 13.
Lombardini G., 1942. Contribute alia conoscenza della morfologia dei
Demodicidae. Chieve analitica del genere Demodex Owen, Redia, 28,
89—102»
248
Глава V. Подотряд Trombidiformes
ДИАГНОЗ
Erythraeidae Oudemans, 1902
(рис. 176, 177)
Половозрелые.Erytrae’dae представляют собой красноватых хищных
клещей с бегательными ногами. Личинки гетероморфные, не имеют
уростигм и паразитируют на членистоногих. Тело взрослых клещей имеет
овальную форму, выпуклое и покрыто
Рис. 176. Lep'us hlHipos
Berlese (по Фитцтуму, 1926).
Самка (вид сверху#
многочисленными щетинками. Проподо-
сома отделена от гистеросомы неглубо-
кой бороздкой. Посреди проподосомы
лежит проподосомальный щиток (crista
Рис. 177. Leptus tenebrans Vitzthum
(по Фитцтуму, 1947).
Личинка (вид сверху).
metopica), который может доходить сзади до середины спины; по Фитц-
туму, Fallopia и Neosmaris не имеют такого киля. На киле имеется
два сенсорных поля; переднее часто находится далеко впереди на
хоботке, заднее же обычно расположено на заднем конце киля. Каждое
сенсорное поле несет по паре сенсорных щетинок. У клещей, не
имеющих киля, сенсорные щетинки лежат на соответствующих участ-
ках тела. Имеется одна или две пары линзовидных глаз. Тазики
ног I, II, III и IV расположены двумя группами. Ноги длинные, строй-
ные и состоят из шести подвижных члеников. Лапки вооружены дву-
мя коготками, но не имеют эмподия. Пальпы пятичлениковые; голенный
членик пальпы вооружен сильным коготком. Палец пальпы обычно
грушевидный, но может быть и шаровидным. Хелицеры не расчленены,
стилетовидны, очень длинные; они могут втягиваться в тело и обычно
Prostigmata
249
на дистальном конце покрыты мелкими зубчиками. Половые присоски
отсутствуют.
Гранжан (1947) разделяет Erythraeidae на два семейства —
Erythraeidae и Balaustiidae, но мы здесь этого деления не придержи-
ваемся.
РОДЫ И ПОДРОДЫ
1. Erythraeus Latreille, 1806 ( = Rhyncholophus Duges, 1834)
a) Erythraeus s. str.
Тип подрода Acarus phalangioldes DeGeer, 1778
6) Corerythrolophus Radford, 1950 ( = Erythrolophus Berlese, 1920“
nomen praeoccupatum)
Тип подрода Erythraeus {Erythrolophus) froggatti, Berlese, 1920-
в) Ctenerythraeus Berlese, 1918
Тип подрода Erythraeus (Ctenerythraeus) trombodioides Berlese,
1918
r) Ptilolophus Ber’ese, 1916
Тип подрода Erythraeus (Ptilolophus) claviger Berlese, 1916
2. Balaustium v. Heyden, 1826 ( = Monotrombidium Krauss, 1925)
Тип рода Trombidium murorum Hermann, 1804
3. Bochartia Oudemans, 1910
Тип рода Bochartia kuyperl Oudemans, 1910
4. Caeculisoma Berlese, 1888
Тип рода Caeculisoma tuberculatum Berlese, 1888
5. Callidosoma, Womersley, 1936
Тип рода Caeculisoma rlpicola Womersley, 1934 (личинка)
6. Claverythraeus Tragardh, 1937
Тип рода Claverythraeus mongolicus Tragardh, 1937
7. Erythrellus Southcott, 1946
Тип рода Erythrellus imbricatus Southcott, 1946
8. Erythroides Southcott, 1946
Тип рода Erythraeus serratus, Womersley, 1936
9. Erythrombium Ber ese, 1910
Тип рода Erythrombium eusisyrum Berlese, 1910
10. Euplatypophus Radford, 1950 ( — Platylophus Berlese, 1910 nometr
praeoccupatum)
Тип рода Platylophus argasoides Berlese, 1910
11. Fallopia Oudemans, 1905 (== Microsmaris Hirst, 1926)
Тип рода Rhyncholophus poriferus Kramer, 1898
12. Hauptmannia Oudemans, 1910
Тип рода Achorolophus longicollis Oudemans, 1910
13. Leptus Latreille, 1795 ( = Achorolophus Berlese, 1891)
Тип рода Pediculus coccineus Scopoli, 1763
14. Lucaslella Banks, 1910
Тип рода Rhyncholophus plumipes Koch, 1856
250
Глава V. Подотряд Trombidiformes
-15. Neosmaris Hirst, 1926
Тип рода Neosmaris npvaezealandiae Hirst, 1926
16. Paraveenhoekia Paoli, 1937
Тип рода Parawenhoekia dectlci Paoli, 1937
17. Parerythraeus Southcott, 1946
Тип рода Parerythraeus gregoryi Southcott, 1946
18. Sphaerolophus Berlese, 1910
a) Sphaerolophus s. str.
Тип подрода Rhyncholophus globiger Berlese, 1885
6) Cavannea Berlese, 1910
Тип подрода Rhyncholophus cavannae Berlese, 1885
в) Eupodolophus Berlese, 1914 ( — Fessoniella Hirst, 1924)
Тип подрода Sphaerolophus {Eupodolophus) chubbi Berlese, 1914
-ОБЩИЕ ЗАМЕЧАНИЯ
Личинки этих клещей паразитируют на насекомых подобно тому,
как личинки Trombiculidae паразитируют на позвоночных. В одном
•случае Лоуренс (1940) насчитал на одной особи саранчи до 400 личинок
Erythraeus s-wazianus Lawrence, но, как правило, их бывает гораздо
меньше. Лоуренс указывает, что личинки, повидимому, прикрепляются
ротовыми органами к саранче на любой из фаз развития. На хозяине
часто можно обнаружить остатки шкурок, сброшенных при линьке.
Большинство личинок прикрепляется к нижней поверхности крыльев
и даже к барабанной перепонке внутри органа слуха и под краем
слуховой щели; на ногах встречаются редцо. В Натале с ног паука
Sattis sp. (Attidae) был снят Leptus atticolus Lawrence. Клещ, опре-
деленный как Leptus sp., был выведен из личинки, снятой с ящерицы
на о-ве Бугенвиль (в южной части Тихого океана). Эго первый извест-
ный случай паразитирования клеща из семейства Erythraeidae на
позвоночном. Большинство видов Erythraeidae известно или только
по личиночной, или же только по имагинальной фазе; лишь в редких
случаях представлялась возможность сопоставить разные фазы пре-
вращения одного вида.
Половозрелые Erythraeidae ведут хищный образ жизни. Их можно
встретить на листьях деревьев, в гумусе и в песке на пляжах. Balaus-
tliim aonidophagus (Ebeling) уничтожает цитрусовую щитовку в Кали-
форнии. Эбелинг (1934) приводит следующие данные: „Эти клещи
•обычно немногочисленны, и автор встречал их лишь в некоторых са-
дах. На одном дереве было собрано до тридцати клещей, но, как
правило, их бывает по одному, по два, а может не оказаться совсем.
Они попадаются и на земле, отчасти, вероятно, потому, что им трудно
удержаться на листьях; на земле клещи бегают вокруг деревьев».
Erythraeidae — каннибалы; если поместить несколько клещей в закры-
тую банку, то через день или два в ней остается только один клещ.
„Автор наблюдал однажды, как клещ ввел мандибулы, а потом и
леею головогрудь под панцырь цитрусовой щитовки и выел все ее
Prostigmata
251
it.io. Эти клещи охотятся также на гусениц цитрусовой щитовки,
на других мелких насекомых и на клещей. Если бы они были много-
численнее, то могли бы стать опасным врагом щитовки".
Интересной особенностью отличается южноафриканский клещ Pti-
lophus namaquensis (Lawrence), у которого задняя пара ног длиннее
остальных, а голени несут большие султаны из длинных щетинок.
При беге клещи вытягивают задние ноги горизонтально или верти-
кально и не пользуются ими для передвижения. Р. namaquensis бегает
очень быстро и на бегу вибрирует задними ногами. Лоуренс (1937)
высказал предположение, что задние ноги могут выпогнять функции:
а) локомоторные, б) сенсорные, в) защитные и г) половые.
ЛИТЕРАТУРА
Ebeling W., 1934. A new predaceous mite from southern California, Pan-
Pactfic Ent., 10 (1), 33—34.
Gr and j can F., 1947. Etude sur les Smarididae et quelques autres Erythroi-
des (Acariens), Arch. de Zoo!. Exp! et Jen., 85 (1), 1 —126.
Lawrence R. F., 1937. A new species of p’umefioted mite from South
Africa, Ann. Soti'h African Mus., 32 (3), 269—279.
Lawrence R. F., 1940. New larval forms of South African mites from arth-
ropod hosts, Ann. Nada'. Mas., 9 (3), 401—408.
Southcott R. V., 1946. Studies on Australian Erythraeidae (Acarina), Proc.
Linn. Soc. New South Wales, 71 (1—2), 6—48.
Smaridiidae Kramer, 1878
(рис. 178, 179)
ДИАГНОЗ
Smaridiidae имеют удлиненное, овальное тело, заостренное спе-
реди; плечи выражены умеренно. Проподосома обычно вытянута в длин-
ный „хоботок" и отделена от гистеросомы неглубокой бороздкой.
Спинная поверхность тела несколько выпуклая и иногда имеет несколько
продольных и поперечных вдавленностей. На дорзальной стороне
проподосомы имеется два сенсорных поля, каждое с парой сенсорных
щетинок. Иногда имеется срединный киль. С каждой стороны пропо-
досомы лежит по одному или по два глаза. Щетинки отличаются
своеобразным строением; обычно они имеют форму листка и покрыты
сильно согнутыми и сложенными продольно шипиками или воюсками.
’Газики ног I и П и III и IV соединены попарно. Стройные, иногда
чрезвычайно длинные ноги состоят из шести подвижных члеников.
На лапках имеется по два коготка, но нет ни предлапки, ни эмподия.
Пальпы пятичлениковые, комплекс палец — коготок имеется. Хелицеры
длинные, стилетовидные. Гнатосома может выдвигаться далеко вперед
и втягиваться обратно в тело. Половые присоски отсутствуют. По Гран-
жану, фаза протонимфы отсутствует. Личинки гетероморфны, имеют
уростигмы. Первая пара ног снабжена трихоботиями, или сенсорными
щетинками. У личинок имеется также проподосомальный щиток
(scutum), несущий две пары сенсорных и две пары простых щетинок.
252
Глава V. Подотряд Trombidiformes
РОДЫ
1. Smarts Latrei’le, 1796 { — Smariidia Latreille, 1817 = Dugesia
Berese, 1883)
Тип рода Trombidium squamatum Hermann, 1804
2. Aecosmaris Grandjean, 1947
Тип рода Aecosmaris pallitricha Grandjean, 1947
Рис. 178. Smarts
la*relllel Grandjean
(по Гранжану, 1947).
Л 1чинка (вид сверху'.
Самка (вид сверху); гнатосома
слегка выдвинута.
3. Fessonia v. Heyden, 1826
Тип рода Trombidium papillosa Hermann, 1804
4. Hirstiosoma Womersley, 1934
Тип рода Hirstiosoma scalaris Womersley, 1934
5. Phanolophus Andr6, 1927
Тип рода Phanolophus nasica АпОгё, 1927
6. Sclerosmaris Grandjean, 1947
Тип рода Sclerosmaris philopempta Grandjean, 1947
7. Sphazrotarsus Womersley, 1936
Тип рода Sphaerotarsus allmani Womersley, 1936
Prostigmata
253
Зюдкотт (1946) пересмотрел это семейство и разделил его на че-
тыре подсемейства: Smaridiinae, Fessoniinae, Hlrstiosominae и Phano-
lophinae. Мы здесь этой системы не придерживаемся.
ОБЩИЕ ЗАМЕЧАНИЯ
Smaridiidae представляют собой мало изученную группу клещей.
Личинки Smaridiidae паразитируют на насекомых, а взрослые особи
ведут хищный образ жизни. Известно очень мало видов; лучше других
изучены клещи Австралии и Европы. В Северной Америке это семей-
ство мало известно. Эвинг описал Hirstiosoma longilinealis (Ewing)
из штатов среднего Запада, а Сэй — Smarts sericeutn Say — с восточного
побережья США.
Уомерсли и Зюдкотт изучали биологию некоторых Smaridiidae.
Они выводили нимф австралийского вида Smarts prom'lnens (Banks)
из личинок, снятых с Psocidae. Уомерсли и Зюдкотт пишут: „Насо-
савшиеся клещи покидали хозяина и, прежде чем впасть в состояние
покоя, в течение нескольких дней бродили по пробирке. По прошествии
нескольких дней кожица поперечно лопалась и появлялась куколка.
Через три-четыре недели из разрыва на заднем конце куколки выхо-
дила нимфа. Бывает ли у этих клещей вторая стадия покоя и вторая
нимфа — не установлено, однако это маловероятно". О наличии одной
лишь нимфальной фазы говорит и Гранжан, утверждающий, что
у Smaridiidae стадия протонимфы отсутствует. Уомерсли и Зюдкотту
на основании морфологических признаков удалось связать нимф
Hirstiosoma и Sphaerotarsus со взрослыми особями. Гранжан делал то же
самое для личинок и взрослых, находимых совместно.
ЛИТЕРАТУРА
Grad jean F„ 1947. Etude stir les Smaridiidae et quelques autres Erythroi-
des (Acariens), Arch. de Zoo’.. Exp. et Qin., 85 (1), 1 — 126.
Southcott R. V., 1946. On the family Smaridiidae (Acarina), Proc. Linn. Soc.
New South Wa'es, 7d (3-4), 173—178.
Womersley H., S r u t h с э 11 R. V., 1)41. .4 Лез эп the Smandiidae (Aca-
rina) of Australia and New Zealand, Trans. Roy. Soc. Sou‘h Austral.,
65 (1), 61—78.
Calyptostomidae Oudemans, 1923
(рис. 180)
ДИАГНОЗ
Calyptostomidae — наземные клещи, обычно живущие в очень влаж-
ных местах. Тело квадратное, пухлое, усеченное сзади, тупое спереди.
Проподосома и гистеросома не разделены швом. Спинная поверхность
тела плоская, с многочисленными, лишенными щетинок углублениями,
маркирующими места прикрепления мускулатуры. Срединный киль и
сенсорные поля отсутствуют, но на передней части проподосомы
254
Глава V. Подотряд Trombidiformes
расположена пара псевдостигм, напоминающих круглую чашу, со дна
которой поднимаются короткие сенсорные щетинки. По краям пропо-
досомы расположено по паре линзовидных глаз. Основания спинных
щетинок соединены склеротизиро-
ванными полосками, образующими
на покровах звездчатый узор;
обычно из центра каждой звезды
поднимается тонкая листовидная
щетинка. Ноги стройные, не-
сколько удлиненные, состоят из
шести подвижных члеников. Ноги
не имеют предлапок и эмпо-
диев, но снабжены каждая двумя
коготками. Пальпы состоят из
пяти подвижных члеников; голен-
ный членик пальп несет корот-
кий слабый коготок. Тарзальный
членик пальпы причленен дисталь-
но и направлен вперед. Хелицеры
не расчлененные, очень длинные,
игловидные. Гнатосома может
сильно выдвигаться вперед; в со-
стоянии покоя она втягивается
в тело. Имеется две пары половых
Рис. 180. Calyptostcma velutinus (Miiller).
Л —самка (вид сверху), ноги не изображены; Б — проподосомальные сенсорные щетинки.
присосок. Личинки гомоморфны, не имеют уростигм и, повидимому,
паразитируют на членистоногих.
РОДЫ
1. Calyptostoma Cambridge, 1875 (= Smarts Berlese, 1887)
Тип рода Acarus velutinus Miiller, 1778 ( = Calyptostoma hardyi
Cambridge, 1875)
ОБЩИЕ ЗАМЕЧАНИЯ
Малоизученная группа хищных клещей.
Prostigmata
255>
ДИАГНОЗ
Trombidiidae Leach, 1815
(рис. 181, 182)
Нимфальные и взрослые фазы имеют густой покров щетинок,,
придающий им бархатистый вид; этих клещей позволяет опознать-
также более или менее отчетливое расчленение тела на три отдела:
гнатосому, проподосому и гистеросому. Тело никогда не имеет
Р'ис. 181. Microtrotr.bidlum hys-
tricinum (Canestrini)
(по Фитцтуму, 1926).
Половозрелая особь (вид сверху^
8-образной формы. Хелицеры
двухчлениковые; основной чле-
ник снабжен мускулатурой, а ди-
стальный изогнут, имеет форму
лезвия и по дорзальному краю
несет пильчатые зубцы. Стигмы
открываются у основания хели-
цер, на медиодорзальной поверх-
ности последних, и обычно-
Рис. 182. Allothrombium
neapolitanum Oudemans
(по Удемансу, 1912).
Личинка (вид сверху).
лишены перитрем. Пальпы состоят из пяти члеников; тарзальному чле-
нику пальпы, подобно большому пальцу, противопоставлен терминаль-
ный тибиальный коготок. Ноги состоят из шести (как исключение — из-
пяти) члеников. Дорзальный проподосомальный щиток (scutum или crista
metopica) лежит в средней части проподосомы, на его боковых расши-
рениях сидит одна (как исключение — две) пара сенсилл. Спереди
scutum заканчивается расширенной пластинкой, которая может высту-
пать над гнатосомой в виде тектума; пластинка покрыта многочи-
сленными щетинками. Иногда эта часть проподосомального щитка’
отсутствует, и щиток тогда представляет собой лишь расширенное
основание сенсилл. Одно подсемейство (Stygothrombiinae) сильно-
255
Глава V. Подотряд Trombidiformes
дегенерировало; у его представителей отсутствуют многочисленные ще-
тинки и типичные ротовые органы, само же тело имеет удлиненную,
червеобразную форму.
У личинок имеется одна уростигма, связанная с ногами I; обычно
имеется более одного дорзального щитка; щетинка на тазике педи-
пальпы лежит обычно впереди от основания бедра пальпы. Личинки
Trombidiidae паразитируют на беспозвоночных.
Тор и Вильман (1947) разделили это семейство на пять подсемейств. •
Одно из них — Trombiculinae—ныне возводится в ранг семейства;
мы разберем его в следующем разделе. Ниже приводится определи-
тельная таблица подсемейств для взрослых форм, составленная с видо-
изменениями по Тору и Вильману.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ TROMBIDIIDAE
1. Проподосома с гребнем, включенным в щитки разной формы или
же свободно лежащим на покровах............................2
— Проподосома без гребня, scutum рудиментарный. Сенсиллы слабо
выражены и не опущены.................................Trombel inae
2. Сенсилл одна пара..........................................4
— Сенсилл две пары...........................................3
3. Имеется три гистеросомальных щитка...........Notothrombiinae
— Гистеросомальные щитки отсутствуют .... Johnstonianinae
4. Сенсиллы сидят на сенсорных полях..........................6
— Сенсиллы сидят не на сенсорных полях.........................5
5. Scutum расширенный..........................Calothrombiinae
— Scutum прямой...................................Tanaupodinae
€. Имеются перитремы, связанные со стигмами хелицер. Глаза
на подвижных стебельках......................................7
— Перитремы отсутствуют. Глаза, если имеются, не сидят на под-
вижных стебельках .......................................... 8
7. Имеется подушечка между амбулакральными коготками ....
...................................................Allothrombiinae
— Подушечка отсутствует...............................Trombidiinae
8. Сенсорные поля не смещены вперед. Ноги с двумя коготками 9
— Сенсорные поля лежат в переднем отделе scutum. Ноги с тремя
коготками ........................................Stygothrombiinae
9. Сенсорные поля отодвинуты к задней части scutum .... 10
— Сенсорные поля лежат в средней части scutum.................11
10. С каждой стороны гребня имеется по одному глазу...............
...............................................Tanaupodasterinae
— С каждой стороны гребня имеется по двойному глазу (как исклю-
чение, глаза отсутствуют)........................Microtrombidiinae
11. Щетинки неопушенные..................................12
• — Щетинки опушенные....................................13
12. Scutum со свободным передним выступом . . Rhinothrombiinae
Prostigmata
257
— Scutum без свободного переднего выступа.........Ocypetinae
13. Имеется дорзальный лигидиальный щиток . . . Eutrombidiinae
— Дорзальный пигидиальный щиток отсутствует . Holcotrombidiinae
РОДЫ
Trombidiinae Thor, 1935
1. Trombidium. Fabricius, 1775 ( = Sericothrombium Berlese, 1910)
Тип рода Acarus hotoserlceus Linnaeus, 1758
2. Austrothromblum Womersley, 1934
Тип рода Allothromblum (Mesothrombium) australien.se Hirst, 1929
3. Caenothromblum Oudemans, 1927
Тип рода Caenothrombium calorls Oudemans, 1927
4. Dinothrombium Oudemans, 1910
Тип рода Acarus tinctorlus Linnaeus, 1767
5. Pliyllotrombidium Cooreman, 1946
Тип рода Trombidium barbarum Lucas, 1846
6. Xenothrombium Oudemans, 1927
Тип рода Xenothrombium Insulare Oudemans, 1927
Allothrombiinae Thor, 1935
1. Allothrombium Berlese, 1903 ( = Mesothrombium Hirst, 1926)
Тип рода Trombidium fuliginosum Hermann, 1804
2. Corethroth.rombium Oudemans, 1929
Тип рода Corethrothrombium vandermeermohrl Oudemans, 1929
С a 1 о t h r om b iin a e Thor et Willmann, 1947
1. Calothrombium Berlese, 1918
Тип рода Parathrombium paolli Berlese, 1918
Eutrombidiinae Thor, 1935
1. Eutrombidlutn Verdun, 1909
Тип рода Pediculus rostratus Scopoli, 1763 {—Trombidium trigonum
Hermann, R04j
2. Leptothrombium Berlese, 1912 o
Тип рода Trombidium oblongum Tragardh, 1904
Johnstonianinae Thor, 1935
1. Johnstoniana George, 1909 ( = Rohaultia Oudemans, 1911)
Тип рода Rhyncholophus errans Johnston, 1852
2. Centrotrombidium Kramer, 1896
Тип рода Centrotrombidium schneideri Kramer, 1896
|7 Зак. 1735
258
Глава V. Подотряд Trombidiformes
3. Diplothrombium Berlese, 1910
Тип рода Trombidlum longipalpe Berlese, 1887
4. Hirstithrombium Oudemans, 1940
Тип рода Diplothrombium australiense Hirst, 1928
5. Myrmicotrombiuni Womersley, 1935
Тип рода Myrmicotrombium brevicrtstatum Womersley, 1935
Microtrombidiinae Thor, 1935
1. Microtrombidium Haller, 1882 ( = Manriqula Boshell et Kerr, 1942)
Тип рода Trombtdium purpureum Haller, 1882
2. Anomalothromblum Andr6, 1936
Тип рода Anomalothrombium nasigerum Andre, 1936
3. Camerotrombidium Thor, 1936
Тип рода Ottonia vesiculosa Thor, 1900
4. Campylothrombizm Krausse, 1916
Тип рода Microtrombidium langhofferi Krausse, 1916
5. Dendrotrombidium Thor, 1916
Тип рода Trombtdium perligerum Berlese, 1888
6. Dromeothrombium Berlese, 1912
Тип рода Trombidlum macropodum Berlese, 1903
7. Echinothrombium Womersley, 1937
Тип рода Ottonia spinosa Canestrini, 1877
8. Enemothrombium Berlese, 1910
Тип рода Trombidlum bifoliosum Canestrini, 1884
9. Eutrlchothromblum Womersley, 1937
Тип рода Trombidlum euthrichum Berlese, 1904
10. Georgia Hui, 1918
Тип рода Ottonia ramosa George, 1909
11. Foliotrombidlum Womersley, 1945
Тип рода Enemothrombium evansi Womersley, 1937
12. Hiotrombidium Womersley, 1945
Тип рода Calot rombium tubbi Womersley, 1937
13. Lamtnothrombium Womersley, 1937
Тип рода Microtrombidium myrmlcum Womersley, 1934
14. Pedotrombidium Womersley, 1945
Тип рода Pedotrombidium kohlsi Womersley, 1945
15. Neotrombidium Leonardi, 1901
Тип рода Neotrombidium turcigerum Leonardi, 1901
16. Platy seta Wharton, 1938
Тип рода Platyseta yucatanicus Wharton, 1938
17. Platytrombidium Thor, 1936
Тип рода Trombtdium vagabundum Berlese, 1903
18. Spelaeothromblum W'llmann, 1940
Тип рода Spelaeothromblum caecum Willmann, 1940
19. Valgothrombium Willmann, 1940
Тип рода Ottonia valga George, 1909
Prositgmaia $59
Notothrombiinae Thor et Willmann, 1947
1. Notothromblum Storkan, 1934
Тип рода Notothromblum regis-borlsi Storkan, 1934
Ocypetinae Vitzthum, 1941
1. Ocypete Leach, 1814 ( = Podothrombium Berlese, 1910)
Тип рода Ocypete rubra Leach, 1814
R h i no t h r om b ii n ae Thor et Willmann, 1947
1. Rhlnothromblum Berlese, 1910
Тип рода Trombidlum nemorlcola Berlese, 1886
Holcotrombidiinae
(новое название для Scalothrombiinae Thor et Willmann, 1937)
1. Holcotrombidlum Womersley, 1945 (= Scalothromblum Thor et
Willmann, 1947)
Тип рода Euthrombtum scalaris Womersley, 1936 (Euthrombtum—
— наименование, употребляемое ошибочно вместо Eutrombtdlum)
Stygothrombiinae Тдог, 1935
1. Stygothrombtum Viets, 1932
Тип рода Stygothrombtum karamani Viets, 1932
2. Cerberothrombium Viets, 1934
Тип рода Stygothrombtum {Cerberothrombium) armatutn Viets, 1934
Tanaupodasterinae Thor et Willmann, 1947
1. Tanaupodaster Vitzthum, 1933
Тип рода Tanaupodaster montezuma Vitzthum, 1933
TanaupodinaeThor, 1935
1. Tanaupodus Haller, 1882
Тип рода Tanaupodus steudeli Hal er, 1882
2. Eothromblum Berlese, 1910
Тип рода Eothromblum echlnatum Berlese, 1910
3. Neotanaupodus Garman, 1925
Тип рода Neotanaupodus tuberculatus Garman, 1925
4. Tignyia Oudemans, 1937
Тип рода Acarus sulcatus Muller, 1776
5. Typhlothrombium Berlese, 1910
Тип рода Trombidlum hlstricinum Leonardi, 1901
17*
260
Глава V. Подотряд 'trombidiformes
Trombellinae Thor, 1935
1. Trombella Berlese, 1887
Тип рода Trombella glandulosa Berlese, 1887
2. Chyzeria G. Canestrini, 1897
Тип рода Chyzeria ornata G, Canestrini, 1897
3. Parachyzeria Hirst, 1926 (= Thaumatotrombium Andrd, 1938)
Тип рода Parachyzeria Indlca Hirst, 1926
Добавление к системе
Роды, предлагаемые для личинок, положение которых в системе,
не установлено:
1. Astigma Bruyant, 1911
Тип рода Astigma oudemarisi Bruyant, 1911
2. Atomus Latreille, 1795
Тип рода Acarus parasiticus De Geer, 1778
3. Cercothrombium Methlagi, 1928
Тип рода Cercothrombium loricatam Methlagi, 1928
4. Ettmiilleria Oudemans, 1911
Тип рода Trombidium sucidum Tr3gardh, 1910
5. HeterothrombldiumV&xdun, 1910
Тип рода Thrombidium granulatum Oudemans, 1902
6. Hexathrombium Cooreman, 1944
Тип рода Hexathrombium spatullferum Cooreman, 1944
7. Hoplothrombium Ewing, 1925
Тип рода Hoplothrombium quinquescutatum Ewing, 1925
8. Monunguis Wharton, 1938
Тип рода Monunguis streblida Wharton, 1938
9. Neothrombium Oudemans, 1909
Тип рода Allotrombidium neglectum Bruyant, 1909 (Allotrombidlttm—
наименование, употребляемое ошибочно вместо Allothrombium)
10. Nothotrombicula Dumbleton, 1947
Тип рода Nothotrombicula deinacridae Dumbleton, 1947
11. Paratrombium Bruyant, 1910
Тип рода Paratrombium egregium Bruyant, 1910 4
12. Polydisca Methlagi, 1928
Тип рода Polydisca squamata Methlagi, 1928
13. Womersleyia Radford, 1946
Тип рода Womersleyia minuta Radford, 1946
ОБЩИЕ ЗАМЕЧАНИЯ
Trombidiidae — очень обширное семейство как по численности
популяций клещей, так и по числу видов. Тор и Вильман (1947)
насчитывают 304 описанных вида, но, повидимому, это лишь незна-
чительная доля огромного числа действительно существующих видов;
Prostigmata
261
почти в каждом сборе клещей из подстилки можно обнаружить
несколько новых видов Trombidiidae. Лучше изучены Trombicu’idae,
близкие к Trombidiidae. Тор и Бильман в своей работе о Trombidii-
dae рассматривают эту группу как подсемейство Trombiculinae,
насчитывающее 150 видов. Но и Trombicu’idae изучены далеко не
полно, и приведенный Тором и Бильманом список 150 видов не вклю-
чает множество новых видов, описанных за годы второй мировой
войны. В настоящее время известно около 300 видов Trombiculidae,
но их фауна во многих областях земного шара еще недостаточно
полно обследована. Надо полагать, что по самой умеренной оценке
имеется еще несколько тысяч неописанных видов, принадлежащих
к обоим семействам (TrombidiidaeTrombiculidae) С
Trombidiidae распространены чрезвычайно широко. Они встречаются
повсюду за исключении областей, покрытых в течение всего года
снегом и льдом. В тропиках они многочисленнее, чем в странах
умеренного пояса. Trombidiidae составляют значительную часть фауны
клещей океанических островов.
Жизненный цикл Trombidiidae состоит из следующих фаз: яйцо,
дейтовум, личинка, нимфохризалида, нимфа, имагохризалида, имаго.
Северин (1944) подробно описал жизненный цикл Eutrombidlum rost-
ratus. Майкнер (1946) установил, что Microtrombidiidae могут прохо-
дить несколько постимагинальных линек. Соотношение полов в исследо-
ванных случаях составляло 1:1. Самцов отличают от самок по
наличию пениса.
Личинки паразитируют на членистоногих, и потому они потен-
циально имеют значение не только как орудие борьбы с насекомыми,
по, возможно, и как переносчики микроорганизмов с одного члени-
стоногого на другое. Нимфальные и половозрелая фазы питаются
яйцами и молодыми личинками многих членистоногих и могут исполь-
зоваться для борьбы с ними. Eutrombidlum rostratus является серьез-
ным врагом саранчи. Личинки питаются на самой саранче, а взрослые
клещи поедают ее яйца.
ЛИТЕРАТУРА
Berlese А., 1912. Trombidiidae, Redia, 8, 1—291.
F е i d е г Z., 1950. Cercetari asupra aparatului respirator la Trombidiidae si pro-
stigmatele superioare si lista speciilor de Trombidiidae din Republica Popu-
lara Romana, Ann. Acad. Repub. Pop. Romans, 3, mem. 5, 1—21, 1—185,
pls. 1—21.
Michener C. D., 1946. The taxonomy and bionomics of some Panamanian
trombidiid mites (Acarina), Ann. Ent. Soc. Amer., 39, 349—380.
1 Изучение систематики Trombidiidae идет энергично. Каждый год описы-
ваются новые виды, главным образом из тропических и субтропических
стран. В СССР исследования фауны Trombidiidae начаты Е. Г. Шлугер. При
обработке первого имевшегося у нес материала с Дальнего Востока и из
Средней Азии из 50 выявленных видов 48 оказались новыми для науки
(Е. Г. Шлугер, Новые краснотелки (Acari, Trombiculidae) фауны СССР,
Энтомол, обозр., 30 (1—2), 157—164, 1949). — Прим, ред.
262
Глава И Подотряд Trombidiformes
Severin Н. С., 1944. The grasshopper mite Eutrombldium trigonum (Her-
mann) an important enemy of grasshoppers, S. Dak. Expt. sta. Tech.
Bull., 3, 1—36.
Thor S., Willmann C., 1947. Trombidiidae, Das Tierreich, Lfg, 71, 187—541 +
+Pls. XXIX—XXXVI.
Womersley H., 1945. A revision of the Microtrombidiinae (Acarina, Trom-
bidiidae) of Australia and New Guinea, Rec. South Austral. Mas.,
8, 293—355.
ДИАГНОЗ
Trombiculidae Ewing, 1944
(рис. 183, 184)
Нимфы и половозрелые особи. Клещи с овальным или, чаще
8-образным телом, более или менее четко разделенным на три отдела
гнатосому, проподосому и гисте-
росому. Длина тела около 1 мм.
Рис. 184. Euschdngastia indica (Hirst)
(по Уартону, 1946).
Личинка (вид сверху).
Рис. 183. Euschdngastia indica
(Hirst) (по.Уартону, 1946).
Нимфа (вид сверху).
Кутикула часто бороздчатая, с расширенными, пластинчатыми основа-
ниями щетинок. Глаза, если они имеются, расположены на проподосоме.
Нитевидные, густо опушенные щетинки покрывают ноги и тело, при-
давая клещу бархатистый вид. Пальпы хорошо развиты; они состоят
из пяти члеников и вооружены тибиальным коготком. Тарзальный
членик пальпы противопоставлен коготку наподобие большого пальца.
Хелицеры имеют толстый основной членик и лезвиевидный дистальный
сегмент, несущий по дорзальному краю пильчатые зубцы. Ноги шести-
члениковые; каждая лапка несет пару амбулакральных коготков.
Половое отверстие расположено между тазиками ног IV. Самцы имеют
листовидный пенис и многочисленные щетинки, покрывающие про-
Pros-tigmata
263
дольные генитальные клапаны. У самок половое отверстие такое же,
как у самцов, но пенис отсутствует, а на генитальных клапанах
щетинок мало. У обоих полов имеется по три пары половых присо-
сок. Стигмы открываются на основном членике хелицер. Имеется про-
подосомальный щиток (scutum), на котором сзади расположено сенсор-
ное поле; на последнем имеется пара псевдостигм, от которых отходят
удлиненные сенсиллы. Передний край гребня расширен и образует
тектум, несущий одну или две щетинки.
Личинки. Таксономия Tromb’culidae основана главным образом на
личинках. У личинок хелицеры двучлениковые; основной членик
толстый и мускулистый, дистальный же членик представляет собой
склеротизированное выгнутое лезвие, иногда несущее зубовидные
выросты. Пальпы пятичлениковые, основные членики их слиты по
средней линии и имеют направленный вперед срединный пластинчатый
вырост, выступающий за край основных члеников хелицер, и пару
латеральных крыльев (galeae), изгибающихся дорзально вокруг хели-
цер и несущих по одной щетинке с каждой стороны. На каждом
основном членике имеется также по щетинке, расположенной позади
места причленения бедренного членика пальпы. Бедро, так же как
и третий (коленный) членик, несет щетинку. Четвертый членик (голень)
имеет три щетинки; одна из них расположена на дорзальной поверх-
ности членика, другая — на латеральной и третья — на вентральной;
кроме того, голенный членик пальпы имеет терминальный коготок.
Пятый членик (лапка) сочленен вентрально с голенным члеником и
противопоставляется коготку пальпы наподобие большого пальца.
Лапка пальпы несет несколько (чаще всего восемь) щетинок; основная
из них представляет собой бороздчатую сенсорную щетинку. Тело
обычно красное, но иногда бывает почти бесцветным; на уровне двух
передних пар ног оно покрыто дорзальным щитком (scutum).
По бокам scutum обычно находится по две пары глаз. Имеется по
нескольку рядов дорзальных и вентральных щетинок; иногда развита
задняя пластинка или задняя группа специализированных щетинок.
Анус вентральный, ног три пары. Имеется уростигма или склеротизи-
рованная ямка, связанная с задним дистальным углом тазиков ног I.
Иногда развита пара трахейных стволов, открывающихся стигмами
в области гнатосомы. Scutum несет 3—6 (редко больше) краевых
щетинок и пару псевдостигм с сенсиллами (псевдостигматические
органы). Ноги шестичлениковые (или семичлениковые, если бедро раз-
делено на basi telofemur).
Уартон (1947) на основании морфологических особенностей личи-
нок разделяет группу на четыре подсемейства.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ TROMBJCULIDAE
1. Ноги I состоят из семи члеников..........................2
-— Все ноги имеют по шести члеников; на тазиках ног I сидят по
по две щетинки........................... Leeuwenhoekiinae
264
Глава К Подотряд Trombidiformes
2. Ноги II и III состоят из семи члеников...................3
— Ноги II и III состоят из шести члеников Walchiinae
3. Scutum несет впереди первые субмедиальные щетинки и (или)
передний срединный вырост.........................Apoloniinae
— Ни субмедиальных щетинок, ни переднего выроста scutum не
имеет...........................................Trombiculinae
РОДЫ И ПОДРОДЫ
Leeuwenhoekiinae Womersley, 1944
1. Leeu'wenhoekia Oudemans, 1911
a) Leeu-wenhoekia s. str.
Тип подрода Heterothrombidium verdant Oudemans, 1910
6) Comatacarus Ewing, 1942
Тип подрода Comatacarus americanus Ewing, 1942
2. Acomatacarus Ewing, 1942
a) Acomatacarus s. str.
Тип подрода Acomatacarus arlzonensls Ewing, 1942
6) Austracarus Lawrence, 1949
Тип подрода Austracarus procaviae Lawrence, 1949
в) Austrombicula Lawrence, 1949
Тип подрода Leeuwenhoekia womersleyt Lawrence, 1949
r) Hyracarus Lawrence, 1949
Тип подрода Hyracarus typlcus Lawrence, 1949
3. Hannemania Oudemans, 1911
Тип рода Heterothrombidium hylodeus Oudemans, 1910
4. Odontacarus Ewing, 1929
Тип рода Trombicula dentata Ewing, 1925
5. Whartonia Ewing, 1944
Тип рода Hannemania nudosetosa Wharton, 1938
Walchiinae Ewing, 1946
1. Walchia Ewing, 1931
Тип рода Trombtdium glabrum Walch, 1927
2. Gafirliepia Oudemans, 1912
Тип рода Typhlothrombium nanus Oudemans, 1910
3. Gateria Ewing, 1938
Тип рода Gahrliepia fletcheri Gater, 1932
4. Schongastlella Hirst, 1915
Тип рода Schdngastiella bengalensls Hirst, 1915
Apoloniinae Wharton, 1947
1. ApoIonia Torres et Braga, 1938
Тип рода Apolonla tigipioensis Torres et Braga, 1938
Prostigmaia
26S
2. Sauracarella Lawrence, 1949 (положение в системе неясно)
Тип рода Sauracarella whartoni Lawrence, 1949
3. Nomersia Wharton, 1947
Тип рода Womersia strandtmani Wharton, 1947
Trombiculinae Ewing, 1929
1. Tromblcula Berlese, 1905
a) Tromblcula s. str. (—Crotiscus Ewing, 1944)
Тип подрода Tromblcula minor Berlese, 1905 (сборная группа
форм, принадлежащих к роду Trombicula, с неясным поло-
жением в системе)
б) Blankaartla Oudemans, 1911 (== Tragardhula Berlese, 1912 =
= Pentagonella Thor, 1936 = Megatrombicula Michener, 1946)
Тип подрода Trombidlum nilotlcum Tragardh, 1905
в) Eutrombicula Ewing, 1938 (= Trombiculoides Jacot, 1938 и
Acariscus Ewing, 1943)
Тип подрода Mlcrothrombidlum alfreddugesi Oudemans, 1910
r) Fonsecia Radford, 1942
Тип подрода Tromblcula ewitigi Fonseca, 1932
д) Leptotrombidium Nagayao et aL, 1916
Тип подрода Trombidlum akamushi Brumpt, 1910
e) Neotrombicula Hirst, 1925
Тип подрода Acarus autumnalls Shaw, 1790
ж) Tromblcullndus Radford, 1948
Тип подрода Tromblcullndus squamosus Radford, 1948
2. Ascoschdngastia Ewing, 1946
Тип рода Neoschongastla malayensis Gater, 1932
3. Doloisia Oudemans, 1910
Тип рода Doloisia synoti Oudemans, 1910
4. Endotromblcula Ewing, 1931
a) Endotromblcula s. str.
Тип подрода Endotromblcula penetrans Ewing, 1931
6) Phrynacarus Lawrence, 1949
Тип подрода Phrynacarus fitzsimonsi Lawrense, 1949
5. Euschdgastia Ewing, 1938
Тип рода Euschongastia americana Ewing, 1938
6. Guntherana Womersley et Heaslip, 1943
Тип рода Neoschdngastia kallipygos Gunther, 1939
7. Heasllpla Ewing, 1944 (= Trombiculoides Womersley et Heaslip,
1943)
Тип рода Trombiculoides gaterl Womersley et Heaslip, 1943
8. Myotrombicula Womersley et Heaslip, 1943
Тип рода Myotrombicula vespertillonis Womersley et Heaslip, 1943
9. Neoschdngastia Ewing, 1929 (= Paraschongastia Womersley, 1939)
Тип рода Scliongastia americana Hirst, 1921
266
Глава V. Подотряд Trombidiformes
10. Novotrombicula Womersley et Kohls, 1947
Тип рода Novotrombicula owiensis Womersley et Koh's, 1947
11. OenoschOngastia Womersley et Kohls, 1947
Тип рода Oenoschdngastia сапа Womersley et Koh's, 1947
12. Riedlitila Oudemans, 1914 .
Тип рода Riedlinia coeca Oudemans, 1914
13. Saariscas Lawrence, 1949
Тип рода Sauriscus ewingi Lawrence, 1949
14. Scbongastia Oudemans, 1910
Тип рода Thronibidium vandersandei Oudemans, 1905
15, Tecomatlana Hoffmann, 1947
Тип рода Tecomatlana sandovalt Hoffmann, 1947
16. Trisetlca Traub et Evans, 1950
Тип рода Trisetlca melvinl Traub et Evans, 1950
Добавление к системе
1. Speotrombicula Ewing, 1946
Тип рода Trombicula trifurca Ewing, 1933 (установлен для поло-
возрелой фазы; положение в системе неясно)
ОБЩИЕ ЗАМЕЧАНИЯ
Trombiculidae по своему поведению весьма близки к Trombidiidae.
Нимфы и взрослые особи питаются яйцами и ранними личиночными
стадиями мелких членистоногих. Жизненный цикл включает те же
фазы, что и у Trombidiidae, но у Trombiculidae неизвестно ни одного
случая постимагинальной линьки. Главное различие в поведении кле-
щей этих двух семейств проявляется при выборе личинками хозяина.
Trombiculidae, за редкими исключениями (например, Acomatacarus
paradoxa Andre), паразитируют на наземных позвоночных, в том числе
на человеке и домашних животных.
Личинки (краснотелки), принадлежащие к родам Trotnbicalu и
Schongastia, являются во многих странах чрезвычайно тягостными для
человека . паразитами. В США Т. (Д.) alfreddugesi (Oudemans),
Т. (Д.) splendens Ewing и Т. (Д.) batatas (Linnaeus) вызывают раз-
дражающий дерматит. Некоторые люди обладают иммунитетом, к уку-
сам краснотелок, у других же в месте укуса этих клещей образуются
волдыри диаметром в несколько сантиметров.
На Востоке виды Trombicula (Leptotrombidlum'} являются пере-
носчиками риккетсиозного заболевания, известного как кустарнико-
вый тиф, или болезнь цуцугамуши1. Во время второй мировой войны
в армии США было отмечено более семи тысяч случаев этого забо-
левания. Трудно судить о том, насколько это заболевание распро-
странено в разных областях среди туземного населения.
1 Чаше известна под названием „японская речная лихорадка*.— Прим,
ред.
Prostigmata
267
Trombiculidae не только паразитируют на человеке и переносят
заболевания с одного животного на другого, но и причиняют актив-
ный вред домашним животным. Так, в Австралии Trombicula (Eut-
rombicula) sarcina является важным паразитом овец, a ApoIonia tigi-
ploensis вызывает выпадение перьев у кур. Обычные краснотелки и
виды Neoschongastia также причиняют вред курам.
Trombiculidae принадлежат к числу наиболее полно изученных
клещей. По настоящее время описано более 300 видов, однако этим,
вероятно, не исчерпывается и половина существующих видов.
ЛИТЕРАТУРА
Berlese А., 1912. Trombidiidae, Redia, 8, 1—291.
Ewing Н. Е., 1944. The trombicuiid mites (chigger mites) and their relations
to disease, J. Parasitol., 30, 339—365.
Lawrence R. F., 1949. The larval trombicuiid mites of South African verte-
brates, Ann. Natal Mas., 11, 405—486.
Philip С. B., 1948. Tsutsugamushi disease (scrub typhus) in World War II,
J. Par as it о I., 34, 169—191.
Thor S., Willmann C., 1947. Trombidiidae, Das Tierreich, Lfg 71b,
187—541 + Pls. XXIX—XXXVI.
Wharton G. W., 1947. Studies on North American chiggers. II. The subfa-
milies and Womersia strandtmani, n. gn., n. sp., J. Parasitol., 33,
380—384.
Wharton G. W., Jenkins D. W., Brennan J. M., Fuller H. S.,
Kohls I. M., Philip С. B., 1951. The terminology and classification
of trombicuiid mites (Acarina: Trombiculidae), J. Parasitol., 37 (1), 13-31.
Глава. VI
TROMBIDIFORMES (ПРОДОЛЖЕНИЕ)
HYDRACHNELLAE LATREILLE, 1802
(HYDRACARINA)
Водяные клещи образуют группу, выделяемую на основании эко-
логических признаков; морфологически они довольно разнородны. Среди
водяных клещей имеются как плавающие, так и неплавающие формы.
Все они, за исключением морских Pontarachnidae, — обитатели прес-
ных вод. По принципу местообитания к группе Hydrachnellae относят
и близких к Cunaxidae морских Halacaridae. Все Hydrachnellae (за
исключением Pontarachnidae) обладают кутикулярными железами,
стигмами и трахеями. У них имеется две пары глаз, которые могут
быть разделены или не сливаются; иногда имеется также не вполне
развитый срединный глаз. Лапка обычно снабжена двумя коготками,
но не имеет эмподия или предлапки. Пальпы обычно состоят из пяти
подвижных члеников. Хелицеры имеют только подвижный палец
обычно серповидной формы. Личинки за исключением Hydrovolziidae
и Halacaridae гетероморфны и имеют уростигмы, расположенные
между тазиками ног I и II.
Данные об этой группе заимствованы нами из работ Лундблада,
Фитса и Фитцтума. Европейские виды хорошо известны, но амери-
канские Hydrachnellae мало изучены; в США по этой группе рабо-
тали только Уолкотт и в более позднее время Маршалл. Но ни у того,
ни у другого автора нет больших сводных работ, которые давали бы
начинающему специалисту общее представление о водяных клещах.
Биология этой группы в целом мало известна, хотя некоторые рас-
пространенные виды хорошо изучены. Имеется несколько работ по
неполовозрелым фазам, но исследования в этой области (как и во
многих других группах) затрудняет мало разработанная методика вы-
ращивания клещей.
Водяные клещи ярко окрашены. Они встречаются в озерах, пру-
дах, вдоль берегов и в проточных водах. Некоторые виды паразити-
руют на жабрах пресноводных двустворчатых моллюсков, личинки
других форм паразитируют на водных насекомых, напоминая в этом
отношении краснотелок, паразитирующих на позвоночных. Взрослые
формы — хищники и питаются рачками, личинками насекомых
и другими достаточно мелкими водными животными. Некоторые
водяные клещи на всех фазах паразитируют в жаберных полостях
крабов и имеют специальные приспособления для захватывания жабер-
ных нитей. Водяные клещи встречаются в течение всего года, но
Hydrachnellae 249
у большинства видов половозрелые фазы появляются в конце лета и
осенью.
Halacaridae, близко родственные к Cunaxidae, отнесены здесь
к Hydrachnellae, так как сходство среды обитания этих клещей де-
лает методически удобным их совместное изучение.
Ниже приведены составленные по Фитсу (1936) определительные
таблицы основных семейств и подсемейств. Определительные таблицы
более подробных групп можно найти в упомянутой работе Фитса.
ЛИТЕРАТУРА*
Lundblad О., 1927. Die Hydracarinen Schwedens. I., Zool. Bidr. Uppsala,
11, 185—535'.
Lun db lad O., 1931. Siidamcrikanische Hydracarinen, Zool. Bidr. Uppsala,
13, 1—86.
Lundblad O., 1941. Eine Ubersicht des Hydrachnellensysterns und der bis
jetzt bekannten Verbreitung der Gattungen dieser Cruppe, Zool. Bidr.
Uppsala, 20, 359—379.
Lundblad O., 1941. Die Hydracarinenfauna Siidbrasiliens tmd Paraguays,
Kangl. Svenska Vetenskapsakademiens Handlinger, Ser. 3, 19 (7),
1—183.
Lundblad O., 1942. Kangl. Svenska Vetenskapsakademiens Handlinger,
20 (2), 1—175; 20 (8), 1—171.
Viets K., 1936. Wassermilben Oder Hydracarina (Hydrachnellae und Hala-
caridae), Die Tierwelt Deutschlands, 31, 1—288; 32, 289 -574.
Wolcott R. H., 1918. The Water Mites (Hydracarina). (Cm. Ward, Whipple,
Fresh Water Biology, ch. XXVI, 851—875, John Wiley and Sons, Inc.)
3. ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА НАДСЕМЕЙСТВ HYDRACHNELLAE
1. Половых присосок нет. Красные, не плавающие, обитают в вер-
ховьях ручьев........................................Hydrovolziae
— Половые присоски имеются (исключение — морские формы) . 2
2. Рострум узкий, шиловидный. Хелицеры кинжаловидные, отчет-
ливого разделения между коготком и основным члеником нет.
Пресноводные формы ....................................Hydrachnae
— Узкого шиловидного рострума нет. Хелицеры не кинжаловидные.
Как правило, коготки и основные членики отчетливо отделены
друг от друга (исключение—морские формы)....................3
3. Половые присоски никогда не лежат на щитках, или клапанах,
или под ними; у одних форм эти присоски лежат свободно в
коже (и тогда обычно очень малы), у других же они в той или
иной степени прикрыты склеротизированным клапаном и тогда
часто бывают стебельчатыми и желудевидными. Глаза лежат
в глазных капсулах....................................Limnocharae
1 Здесь приведено немного названий. Авторы не ставили себе цели
привести исчерпывающий список литературы, так как водяные клещи при-
надлежат к числу наиболее изученных, и составление полного списка лите-
ратуры вывело бы нас за рамки настоящей работы. Все же авторы надеются,
что в приведенных здесь работах читатель найдет сведения, необходимые
тля лучшего ознакомления с этой группой клещей. При изучении фауны
Hydrachnellae США необходимы также все работы Маршалла.
270 Глава VI. Trombidiformes (продолжение)
—- Половые присоски обычно хорошо заметны и лежат либо на
щитках (или под ними), либо в коже; иногда они соединяются
в поля присосок, лежащие на брюшных щитках (исключение пред-
ставляют некоторые виды Pion , у которых дискоидальные при-
соски, расположенные в толще кожи, рассеяны по брюшной по-
верхности животного).......................................4
4. Гениталии с подвижными клапанами, или щитками............5
— Гениталии не имеют подвижных клапанов, или щитков . • . 6
5. Половые присоски более или менее шишечковидные и лежат либо
на щитках, либо в связи с ними. Если присоски лежат свободно,
то они рассеяны по брюшной поверхности тела . . Hydryphantae
— Половые присоски лежат около полового отверстия либо под
внутренним краем щитков, либо рядом с ними; они не соеди-
няются с щитками склеротизированными образованиями. . Lebertiae
6. Пальпы не имеют апикального коготка, лежащего между более
или менее шиповидным пятым члеником пальп и дистальным кон-
цом расширенного четвертого членика.........................7
— Пальпы несут апикальный коготок, лежащий между четвертым чле-
ником пальпы (дистальный край которого более или менее расширен,
а дистальный угол на изогнутой стороне более или менее сильно
вытянут вперед) и когтеобразным пятым члеником . . Arrlienurae
7. Гениталии с более или менее отчетливыми присасывательными
щитками и с дисковидными или поровидными присосками, ле-
жащими на последних.........................................8
— Гениталии с выпуклостью, похожей на клапан, но сливающейся
с брюшным панцырем. Присоски немногочисленны и расположены
в области полового отверстия (исключение составляют Midea,
у которых имеются присасывательные щитки; у Midea четвертый
членик пальп шиловидный и редуцирован).............Mideopsae
8. Тело обычно покрыто мягкой кожей, редко на спинной поверх-
ности развиты более или менее обширные щитки и еще реже
развит полный панцырь. Четвертый членик пальп обычно несет
склеротизи'рованный отросток, расположенный на изогнутой сто-
роне членика дистального среднего угла; этот отросток часто
очень мал.............................................Pionae
— Тело всегда покрыто панцырем с узким спинным сводом.
Четвертый членик пальп не имеет отростка, сидящего на изо-
гнутой стороне у дистального среднего угла .... Axonopsae
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ HYDRACHNELLAE1
1. Тело покрыто мощным панцырем из отдельных щитков. Половые
присоски отсутствуют. Анальное отверстие расположено на щитке
среднего или большого размера..............................2
1 Семейство Stygomonidae Szalay, 1943 (тип Stygomonia talipes Szalay,
1943), описано после опубликования этой таблицы и потому не включено
в нее.
Hydrachnellae
%l
— Тело если и покрыто панцырем, то не таким мощным, как
у предыдущей группы. Половые присоски имеются (за исключе-
нием Pontarachnidae). Анальное отверстие почти никогда не лежит
на большом щитке. Ноги (за исключением Oxinae) расположены
более или менее радиально......................................3
— Тело покрыто мощным панцырем из отдельных щитков. Половых
присосок три пары. Половое и анальное отверстия лежат на
общем брюшном щитке. Ноги I и II направлены вперед, ноги III
и IV — назад. Плавательных щетинок нет. Морские формы . .
............................................... Halacaridae
2. Ноги III и IV прикреплены латерально, так что эпимеры высту-
пают за край тела и видны сверху. Эпимеры отделены хорошо
заметными швами. Анальное отверстие лежит на большом щитке.
Эпимеры ног I не образуют трубки..................Hydrovolziidae
— Ноги II и IV расположены так, что эпимеры сверху не видны.
Эпимеры не разделены швом. Анальное отверстие лежит на ма-
леньком щитке. Эпимеры ног I смыкаются спереди, образуя
трубку....................................Rhynchohydracaridae
3. Рострум не удлинен и не шиловидный, часто имеет форму ко-
нуса и всегда значительно короче пальпы. Хелицеры явно разделены
на два членика: серповидный коготок и основной членик ... 4
— Рострум длинный, узкий, шиловидный, направлен более или ме-
нее вентрально и достигает приблизительно конца пальп. Хели-
церы немного изогнуты, стилетовидные; коготок хелицер и их
основной членик не отделены друг от друга ясным швом; задний
конец хелицер заходит далеко внутрь тела . . . Hydrachnidae
4. Гениталии не имеют створчатых клапанов, или щитков, но с при-
поднимающимся уступом склеротизированным образованием, окру-
жающем половые присоски. Половые присоски заложены в толще
мягкой кожи, очень малы или более или менее стебельчаты и
сзади обычно напоминают жолудь.................................5
— Гениталии со створчатыми или щитовидными склеротизированными
частями, на которых (или под которыми) лежат более или ме-
нее обособленные половые присоски (исключение Piona spp., у
которых дисковидные половые присоски рассеяны по коже
брюшка)........................................................7
5. Двойные глаза расположены либо медиально на концах попереч-
ного склеротизированного щитка, образуя так называемые очки,
либо латерально — на щитках или свободно в коже тела. Пальпы
нормальной величины и гораздо длиннее рострума ............... 6
— Двойные глаза расположены латерально, на более или менее
палочковидном продольном склеротизированном щитке. Пальпы
очень малы и лишь немного длиннее рострума. Все ноги сильно
опушены, но не покрыты настоящими плавательными щетинками.
Тело настолько мягкое, что стенки его при извлечении клеща
из воды спадаются................................ Limnocharidae
272
Глава Vi. Trombidiformes (продолжение)
6. Двойные глаза расположены либо медиально и тогда соединены
отдельными поперечными щитками, либо латерально и тогда со-
единены многочленистым щитком. Настоящего рострума нет.
Сосущий ротовой аппарат велик и окружен „воротничком". Тре-
тий членик пальп несет на дистальном конце группу щетинок.
У клещей с глазами, соединенными поперечным щитком, половые
присоски мелкие и рассеяны по всей поверхности тела; у дру-
гих групп присоски крупные и сидят на щитках, лежащих по
обе стороны полового отверстия.........................Eylaidae
— Двойные глаза лежат латерально в коже тела; соединяющего их
склеротизированного щитка нет. Рострум имеется. Ротовой аппа-
рат не сосущего типа и не окружен „воротничком". Дистальный
конец третьего членика пальп не несет группы щетинок. Половые
присоски часто стебельчатые и имеют форму жолудя . . Protziidae
7. Гениталии снабжены более или менее выпуклыми и подвижными
щитками, которые при движении открывают половые губы . . 8
— Гениталии не имеют подвижных щитков........................19
8. Половые присоски многочисленные; они либо мелкие и поровид-
ные, либо более крупные шишковидные. Сидят присоски или на
генитальных щитках, или в связи с этими щитками. Между щит-
ками в области половых губ встречаются лежащие в коже изо-
лированные присоски. На дистальном конце четвертого членика
пальп иногда имеется когтевидный или пальцевидный отросток,
который совместно с пятым члеником пальпы образует короткую
клешню. Фронтальный орган обычно имеется. Коготки ног обычно
простые, серповидные, эм подия нет............................9
— Половые присоски лежат под внутренним краем генитальных щит-
ков ^или около него), но никогда не соединяются с щитками.
Число присосок варьирует. Обычно их по три с каждой стороны,
но может быть и по две, по четыре, по шести и даже более;
во всех случаях присоски расположены в ряд, одна за другой.
Четвертый членик пальп не имеет дистального отростка, обра-
зующего с пятым члеником пальп клешню. Фронтальный орган,
за редкими исключениями, отсутствует. Коготки на лапках часто
двузубчатые; эмподий имеется.................................12
9. На дистальном конце четвертого членика пальп обычно имеется
сильный шип или конусовидный придаток, который всегда коро-
че пятого членика пальп и не образует с последним клешни. Пар-
ные боковые глаза иногда располагаются близко друг к другу
на одном склеротизированном щитке............................10
— На дистальном конце четвертого членика пальп имеется узкий,
пальцевидный отросток, которому противопоставляется последний
членик пальпы, образующий совместно с ним клешню. Боковые
глаза лишены капсул и отделены один от другого. Фронтальный
орган отсутствует. Тело покрыто тонкой кожей, щитки не раз-
виты .............................................Hydrodromidae
Hydrachnellae
273
10. Ноги лишены настоящих плавательных щетинок................11
— Ноги с плавательными щетинками. Покровы тела мягкокожистые.
В передней части тела имеются склеротизированные щитки, в ред-
ких случаях щитков много......................Hydryphantidae
II. Тело покрыто склеротизированными щитками с красивым лучи-
стым рисунком. Половых присосок много . . Clathrosperchonidae
— Тело покрыто обычными щитками. Половых присосок много. Оби-
татели горячих источников..........................Thermacaridae
— Тело не покрыто щитками, но усеяно кожными сосочками. Имеется
три пары крупных половых присосок.................Ctenothyasidae
12. Четвертый и пятый членики пальп не уплощены дорзовентрально
и имеют лопатовидную форму....................................13
— Четвертый и пятый членики пальп расширены медиолатерально
и сильно уплощены дорзовентрально; на пятом -членике пальп
сидят четыре медиолатеральных концевых коготка, образующих
лопатовидный орган................................Riitripalpidae
13. Пальпы более или менее узкие; второй членик пальп, имеющий
иногда булавовидную форму, не превышает по длине четвертый
членик пальп и обычно короче последнего; с дорзальной стороны
второй членик пальп никогда не равен по длине и обычно ко-
роче третьего и четвертого члеников, вместе взятых ... 14
— Пальпы короткие и булавовидные, особенно толст и крепок
второй членик пальп; четвертый членик пальп короче второ-
го членика; с дорзальной стороны второй членик пальп обыч-
но равен или даже превышает длину третьего и четвертого
члеников, вместе взятых. Коготок хелицер сравнительно мал . .
...................................................Mamersopsidae
14. Четвертый членик пальп не имеет склеротизированного выроста
на внутренней стороне; второй членик пальп обычно с шипами
на внутренней стороне. Покровы тела лишены толстых склеро-
тизированных сосочков.........................................15
— Четвертый членик пальп удлинен дорзодистально; второй членик
пальп на внутренней стороне не имеет выростов или шипов.
Тело покрыто тесно сидящими склеротизированными сосочками,
напоминающими толстые стержни .... Pseudohydryphantidae
15. Эпимеры расположены четырьмя группами (по две в каждой); эти
группы (по крайней мере, вторая и третья) отделены одна от
другой участками кожи.........................................16
- - Эпимеры объединены в одну группу и часто связаны между со-
бой посередине. Эпимеры II и III латерально полностью сливают-
ся друг с другом, иногда между ними сохраняется неполный шов.
Генитальные щитки плотно прилегают к эпимерам IV, вытесняя
лежащие вентрально промежуточные участки кожи............18
16. На четвертых эпимерах железы отсутствуют.................17
— На переднем внутреннем углу четвертых эпимеров имеются боль-
шие железистые органы . . . ........................Teutoniidae
18 Зак. 1785
274
Глава Vf. Trombidiformes (продолжение)
17. Боковые глаза объединены глазной капсулой. На внутренней сто-
роне второго членика пальп имеется покрытый шипами выступ.
Ноги IV снабжены коготками..............................Sperchonidae
— Боковые глаза не объединены капсулой и отделены один от дру-
гого участком кожи. На внутренней стороне второго членика
пальп имеются только шипы (а не несущий их выступ). Ноги IV
лишены коготков......................................Anisitsiellidae
18. Область эпимеров не сливается с щитками брюшной поверхности
тела, и гениталии остаются обособленными от эпимеров. Брюшко
позади гениталии, не покрыто панцырем и перепончатое; спинка,
как правило, не склеротизирована; дорзовентрального шва нет.
Имеется шесть (редко четыре) больших половых присосок . .
........................................................ Lebertlidae
— Область эпимеров сливается с брюшными щитками и образует
совместно с ними единый брюшной панцырь, оканчивающийся за
гениталиями и включающий их; этот панцырь покрывает бока
животного и отделяется от спинного панцыря дорзальным швом.
Имеется двенадцать маленьких половых присосок, лежащих под
внутренними краями генитальных щитков .... Atractideidae
19. Половые присоски имеются. Пресноводные клещи..................20
— Половые присоски отсутствуют. Обитатели литорали морей . .
.................................................Pontarachnidae
20. Четвертые и пятые членики пальп не образуют клешни . . 21
— Когтевидный пятый членик пальп подвижно сочленен с дорзаль-
ной стороной дистальной части четвертого членика, образуя сон
местно с ним клешню. Внутренний край пятого членика пальп
несет пару тонких сенсорных или же хватательных щетинок; пя-
тый членик пальп обычно вооружен крепким шипом. Тело покры-
то панцырем.......................................................30
21. Боковые глаза объединены (Astacocrotonidae имеют пару глаз).
Ноги IV, как правило, с коготками.............................22
— Боковые глаза отделены друг от друга. Ноги IV обычно не имею i
коготков.. Второй членик пальп часто имеет на внутреннем крас
хитинизированный конус. Половых присосок три пары ....
....................................................Limnesiidac
— Как и Limnesiidae, но половых присосок более трех пар ....
.............................................Neotorrenticolidar
22. Покровы, как правило, перепончатые, иногда имеются более или
менее обширные спинные щитки; очень редко брюшко и спинка
покрыты вполне развитыми спинным и брюшным щитами. Эпи
меры расположены четырьмя группами; швы между ними обычно
имеются, но в отдельных случаях срединный шов между эпимс
рами I исчезает или же шов между третьей и четвертой парой
щитков не полностью развит. Гениталии кожистые или перепои
чатые, с отдельными щитками, несущими более или менее дне
коидальные, реже поровидные присоски. Второй, трет.ий и чет.
Hydrachnellae
вертый членики пальп часто имеют бугорки, сидящие по их
внутреннему краю....................................... 23
— Тело почти полностью покрыто панцырем, спинной и брюшной
щиты, как правило, отделяются лишь узким дорзальным швом;
иногда гениталии лежат на перепончатом поле, или панцырь развит
лишь на брюшной стороне. Тело обычно плоское, часто дискоидаль-
ное. Швов между эпимерами нет. Гениталии лежат между областью
эпимеров и задним концом тела. Половых присосок шесть, восемь
или больше; они лежат на более или менее обособленных створча-
тых или округлых щитках, часто объединенных с брюшным щи-
том. Если гениталии лежат близ области эпимеров, то половое
отверстие обычно расположено в отверстии брюшного щита, а по-
ловые присоски лежат либо на створчатых щитках, либо на
внутреннем крае (или под ним) половых „валиков", соединяю-
щихся с брюшным щитом. Внутренний край дистальной части
четвертого членика пальп не имеет склеротизированного вы-
роста ...................................................... 27
23. Эпимеры I отделены один от другого швом или участком кожи,
расположенными позади основания рострума. Рострум, как пра-
вило, лежит свободно и не соединяется с областью эпимеров.
Ноги обычно имеют плавательные волоски. На внутреннем крае
второго членика пальп нет шипа или конуса (исключение Pseudo-
feltria); четвертый членик пальп несет на внутреннем крае много-
численные зубцы или опушенные бугорки........................24
— Эпимеры I обычно соединены друг с другом и не разделены
срединным швом. Задний край рострума часто соединяется с эпи-
мерами I. Ноги не имеют плавательных волосков. Гениталии
обычно с шестью присосками, реже присосок больше, но число
их никогда не превышает 20 с каждой стороны. Половые при-
соски дискоидальной формы. На внутренней стороне второго чле-
ника пальп имеется шип или хитинизированный зуб; четвертый
членик пальп не несет зубцов на внутреннем крае, но обычно
имеет маленький бугорок, несущий щетинку . . . Hygrobatidae
24. Узкого рострума нет, в редких случаях имеется короткий рото-
вой конус....................................................25
— Имеется узкий рострум, по ширине равный примерно одной трети
органа. Генитальные щитки снабжены многочисленными маленькими
поровидными присосками. Ноги снабжены плавательными волос-
ками ......................................., . . Nautarachnidae
25. Обычно встречаются в стоячих, а также в медленно текущих
водах, лишь редко живут в холодных ручьях. Тело обычно мягкое,
кожистое, гладкое и редко заключено в сплошной панцырь. Чет-
вертый членик пальп несет на внутреннем крае конусовидное
возвышение. Половые присоски обычно дискоидальные, хорошо
заметны, редко маленькие и поровидные. Ноги обычно с плава-
тельными волосками. Ноги III у самца без выростов ... 26
18*
276
Глава Vi. Trombidiformes (продолжение)
— Мелкие клещики, живущие обычно в проточных водах. Тело
плоское. На спине развит более или менее обширный щиток,
покрывающий переднюю и среднюю часть спины; этот щиток
в бороздчатой области окружен мелкими парными щитками. Чет-
вертый членик пальп не имеет на внутренней стороне конусовид-
ного выроста. Эпимеры IV с большим продольным расширением
на боковых краях. Ноги не имеют плавательных волосков. У сам-
цов ноги III обычно имеют выросты. Гениталии самцов имеют
многочисленные мелкие поровидные присоски, лежащие на боль-
шом, не разделенном посредине щитке. Половое отверстие у самцов
небольшое, у самок же —большое и расположено между двумя
генитальными щитками; оно часто имеет щелевидную форму и
расположено в задней части тела.......................Feltriidae
26. Тело без щитков. Дистальный членик пальп несет толстые шипы;
некоторые из них крючковидные и приспособлены для захваты-
вания жабр крабов, на которых паразитируют эти клещи. На
тазиках ног IV имеются крупные железы. Тазики расположены
двумя группами, снаружи обе группы соприкасаются. Половых
присосок четыре пары, они лежат на щитках. Половое отверстие
лежит у заднего конца тела, далеко позади области эпимеров.
Глаза у самки отсутствуют, у неполовозрелых самцов имеется
пара глаз.......................................Astacocrotonidae
— Протоки желез часто открываются на коже в конусовидных возвы-
шениях. Основные членики хелицер посредине обычно срастаются
друг с другом. Третий членик пальп часто имеет очень длин-
ный боковой шип. Задняя часть эпимер I и II часто имеет общий
жезловидный длинный вырост, который обычно заходит под
кожей под эпимеры III или даже IV. Задний край эпимеров IV
обычно лежит поперечно и, как правило, не имеет совсем или
имеет лишь незначительный центральный отросток; поэтому между
тазиками или эпимерами нет вырезки, носящей наименование
генитальной. Гениталии обычно лежат позади области эпимеров.
У ног I и II проксимальные и средние членики обычно утолщены
и покрыты парными кинжаловидными щетинками. Коготки на
лапках, как правило, простые, серповидные и не имеют отрост-
ков. Ноги IV у самцов обычно не имеют половых отличий . .
...................................................Unionicolidae
— Протоки желез на коже открываются не в конусовидных возвы-
шениях. Хелицеры отделены одна от другой. Третий членик
пальп не имеет бокового шипа. Задний отросток передних эпи-
меров короткий, имеет крючковидную вершину и, как правило,
не заходит под задние эпимеры. Задний конец эпимеров IV имеет
центральный отросток, образующий генитальную вырезку. Гени-
талии обычно лежат близ эпимеров, далеко от заднего края тела.
Ноги I и II не имеют особо дифференцированных щетинок и
обычно никогда не утолщены. Коготки на лапках обычно раз-
Hydrachnellae
277
ветвлены и имеют отростки. Ноги III и IV у самцов часто имеют
половые отличия.....................................Pionidae
27. Половых присосок шесть, восемь или более; они лежат либо
в генитальной вырезке на более или менее серповидных щитках,
либо образуют „губчатые" поля (исключение — самки Momonia,
у которых присоски лежат на створчатых щитках частично скле-
ротизированной кожи брюшной поверхности тела); эти поля рас-
положены у внутреннего края половых валиков или под ними.
Половые валики соединены с брюшным щитом. Гениталии прикрыты
эпимерами IV и часто расположены в глубокой генитальной вы-
резке. Тело, если смотреть сбоку, круглое. Второй членик пальп
не имеет возвышений на внутреннем крае; четвертый членик пальп
часто несет вырост на внутреннем крае.........................28
— Половых присосок шесть, восемь или более; они лежат на более
или менее обособленных округлых или створчатых щитках. Эти
щитки расположены на заднем конце тела; у самцов они более
или менее свободные, у самок сливаются с брюшным щитом.
Имеется настоящая генитальная вырезка. Тело более или менее
плоское; как правило, длина его превышает ширину. Эпимеры
часто соединяются одна с другой посередине. Второй членик пальп
иногда имеет возвышение на внутреннем крае; четвертый членик
пальп несет на внутреннем крае тонкие щетинки . . Axonopsidae
28. Четвертый членик пальп короче, чем второй и третий членики
вместе. Генитальная вырезка не достигает эпимеров 1; она обра-
зована краями задних эпимерных щитков или же совсем отсут-
ствует. Половые присоски образуют „губчатые поля", лежащие
у внутреннего края створчатых, но сплошных неподвижных поло-
вых складок; эти складки ограничивают по бокам половое
отверстие в брюшном панцыре (исключение — самки Momonia,
у которых складки узкие и не соединяются с брюшным, панцырем).
Тело обычно дисковидное и плоское.............................29
— Четвертый членик пальп длинный, шиловидный и превосходит по
длине второй и третий членики вместе. Генитальная вырезка
глубокая и достигает заднего конца соединенных друг с другом
эпимеров I; гениталии почти полностью лежат в этой вырезке. По-
ловые присоски многочисленны и лежат на щитках. Тело сильно
выпуклое и покрыто панцырем, спинной и брюшной щиты раз-
делены дорзальным швом.................................Mideidae
29. Тело полностью заключено в панцырь, спинной щит отделен от
брюшного швом. Внутренний край четвертого членика пальп по
длине не превышает проксимального конца пятого членика пальп;
четвертый членик пальп не расширен и не утолщен дорзовен-
трально. Гениталии снабжены 6—10 присосками. Эпимеры, как
правило, разделены швом по средней линии . . . Mideopsidae
— Тело не покрыто панцырем на спинной стороне, дорзального шва
нет. Внутренняя сторона четвертого членика пальп длиннее прокси-
278
Глава VI. Trombidiformes (продолжение)
мального конца пятого членика н образует совместно с последним
клешневидный орган; дистальная часть четвертого членика пальп
утолщена дорзовентрально. Гениталии снабжены многочисленными
присосками. Эпимеры не разделены срединным швом; последняя
группа эпимеров медиально соединяется друг с другом под углом
...................................................Acalyptonotidae
30. Половых присосок четыре, шесть, восемь или более; они обычно
дисковидные и образуют „губчатые поля", лежащие в отверстии
брюшного щита у внутреннего края более или менее створчатых,
сводчатых, но неподвижных половых „валиков".....................31
— Половых присосок обычно много, они, как правило, мелкие, поро-
видные. В отверстии брюшного щита лежат только половые губы,
а половые присоски лежат рядом с отверстием на более или ме-
нее обособленных, обычно крыловидных пластинках брюшного щита.
У самцов на заднем конце тела часто имеется придаток, или
стебелек.............................................. Arrenuridae
31. Имеется много мелких половых присосок . . A-Thienemanniidae
— Половых присосок от трех до восьми пар; они крупные и обра-
зуют на каждой стороне продольный ряд . . . Krendowskiidae
НАДСЕМЕЙСТВА И СЕМЕЙСТВА
Hydrovolziae
Hydrovo’ziidae Thor, 1905
Hydrachnae
Hydraclin’dae Leach, 1815
Limnocharae
Limnocharldae Kramer, 1877
Eylaidae Leach, 1815
Protziidae Viets, 1926
Hydryphantae
Hydryphant’dae Thor, 1900
Clathrosperchonidae Lundbland,
1936
Ctenothyasidae Lundb'and, 1936
Eupatrellidae Viets, 1935
Hydrodromidae Viets, 1936
Rhynchohydracaridae Lundblad,
1931
Thermacaridae Sokolow, 1927
Lebertiae
Lebertiidae Thor, 1900
Anisits'ellidae Viets, 1929
Atractideidae Thor, 1902
Maniersopsidae Lundblad, 1930
Pseudohydryphantidae Viets, 1926
Rutripalpidae Sokolow, 1934
Sperchonidae Thor, 1900
Teutoniidae Lundblad, 1927
Arrhenurae Oudemans, 1902
Arrenur’dae Thor, 1900
Mideopsae
M’deopsidae Thor, 1928
Acalyptonot'dae Thor, 1929
A-Thienemanniidae Lundblad,
1930
Krendowskiidae Lundblad, 1930
Mideidae Viets, 1929
Pionae Viets, 1930
Pionidae Thor, 1900
Astacocrotonidae Thor, 1927
Feltriidae Thor, 1929
Hygrobatidae Koch, 1842
Limnesiidae Thor, 1900
Nautarachnidae Viets, 1935
Neotorrenticolidae Lundblad,
1936
Pontarachnidae Thor, 1929
Unionicolidae Oudemans, 1909
Axonopsae
Axonopsidae Viets, 1929
Hydrachnellae
279
ДИАГНОЗ
Halacaridae Murrey, 1776
(рис. 185, 186)
Halacaridae примыкают к типичным Trombidiformes. Их эллиптическое
i ело обычно сильно уплощено, но изредка имеет в поперечном сечении
форму круга. Ротовые органы выделены в ясно выраженный capitulum,
гие. 185. Halacarus (Tha-
lassarachna) subterraneus
(Schulz) (по Ньюэллу, 1947).
Самец (вид сверху).
который почти всегда выступает за
передний край тела; лишь у немно-
гих форм ротовые органы скрыты
и не заметны со спинной стороны.
Рис. 186. Ротовые органы
Halacaridae (по Ньюэллу, 1947).
Л —пальпа самки Copidogncttuis
(Arhodeoporas) sitbm-.rinus Newell;
Б —хелицера самки Hc.lacorus (Tha-
lassarachna) capuzinus Lohmann.
Пальпы состоят из трех или четырех члеников и прикрепляются
латерально, дорзолатерально или дорзально к основанию capitulum.
Максиллярные части пальп полностью слиты. Хелицеры, лежащие
в дорзальном желобке рострума, способны втягиваться; вентральный
палец хелицер крупный и подвижный, дорзальный палец редуцирован
и превращен в остроконечную перепонку. Трахейная система рудимен-
тарная и, повидимому, не играет роли при дыхании. Кутикулу стенок
тела укрепляют обычно четыре спинных щитка (предорзальный, постдор-
зальный и правый и левый окулярные) и четыре брюшных и боковых
щитка (передний эпимерный, правый и левый эпимерные и генитоаналь-
ный). У различных родов, разных видов одного и того же рода и
реже у двух полов одного вида эти щитки в различной степени
расчленены или слиты. Ноги прикрепляются по бокам: ноги I и II
причленяются к переднему эпимерному щитку (или щиткам) и направ-
лены вперед, ноги 111 и IV отходят от правого и левого задних
эпимерных щитков и направлены назад. У взрослых ноги шестичлени-
ковые. Личинки с тремя парами пятичлениковых ног. У протонимфы
280
Г лава VI. Trombidiformes (продолжение)
ноги IV также пятичлениковые, но ноги I, П и III шестичлениковые.
У дейтонимфы все ноги шестичлениковые. На лапках всегда имеется
по два латеральных коготка, срединный коготок иногда отсутствует.
Слюнные железы хорошо развиты. Средняя кишка не сообщается
с анусом. Анальное отверстие есть, но функционирует исключительно
как выводное отверстие единственной дорзомедиальной секреторной
трубки. Раздельнополы; половой деморфизм выражен в строении
генитоанального щитка и гениталий; остальные органы обычно (но не
всегда) сходны по форме у обоих полов. Три простых глазка имеются,
повидимому, всегда; настоящих глаз с роговицей может быть два,
четыре или шесть; иногда они не развиты. Срединный глаз не имеет
роговицы. Половых присосок три пары.
Эти клещи в основном морские, но имеется много и пресноводных
форм (Porohalacaridae по Фитсу). Они неспособны плавать. Как
правило, это хищные или травоядные формы, но имеется три пара-
зитических рода.
РОДЫ И ПОДРОДЫ
R h о m b og п a t h i n a e V1 e t s, 1927
1. Rhonibognathides Viets, 1927
Тип рода Aletes pascens Lohmann, 1889
2. Isobactrus Newell, 1947
Тип рода Aletes setosus Lohmann, 1889
3. Metarhombognathus Newell, 1947
Тип рода Rhombognathus armatus Lohmann, 1893
4. Rhombognathus Trouessart, 1888
Тип рода Pachygnathus notops Gosse 1855
A ct a c a ri п a e Viet s, 1939
1. Actacarus Schulz, 1936
Тип рода Acatacarus pygmaeus Schulz, 1936
Halacarinae Viets, 1927
1. Halacarus Gosse, 1855
a) Halacarus s. str.
Тип подрода Halacarus ctenopus Gosse, 1855
6) Thalassarachna Packard, 1871
Тип подрода Acarus basteri Johnston, 1836
2. Agaue Lohmann, 1889
Тип рода Halacarus parvus Chilton, 1883
3. Agauopsis Viets, 1927
Тип рода Agaue brevipalpus Trouessart, 1889
4. Coloboceras Trouessart, 1889
Тип рода Coloboceras longiusculus Trouessart, 1889
Hydrachnellae
281
5. Copidognathus Trouessart, 1888
a) Copidognathus s. str. (=CopidognathopsisVtets, \№7=Werthella
Lohmann, 1907)
Тип подрода Copidognathus glyptoderma Trouessart, 1888
6) Arhodeoporrus Newell, 1947
Тип подрода Copidognathus (Arhodeoporus) arenarius Newell,
1947
6. Pontacarus Lohmann, 1901
Тип рода Halacarus basidentatus Trouessart, 1900
Porohalacarinae Viets, 1933
1. Porohalacarus Thor, 1923
Тип рода Halacarus alpinus Thor, 1910
2. Caspihalacarus Viets, 1928
Тип рода Caspihalacarus hyrcanus Viets, 1928
3. Lohohalacarus Viets, 1939 (—Walterella Romijn, 1924 nomen-
praeoccupatum)
Тип рода Walterella weberi Romijn, 1924
4. Troglohalacarus Viets, 1937
Тип рода Troglohalacarus dentipes Viets, 1937
Lоh m a n n e 11 i n a e Viets, 1927
1. Lohmannella Trouessart, 1901
Тип рода Leptognathus falcatus Hodge, 1863
2. Porolohmannella Viets, 1933
Тип рода Leptognathus violaceus Kramer, 1879
3. Scaptognathus Trouessart, 1889
Тип рода Scaptognathus tridens Trouessart, 1889
Sim ogna t h i n ae Vie ts, 1927
1. Simognathus Trouessart, 1889
Тип рода Pachygnathus sculptus Brady, 1875
2. Acaromantis Trouessart et Neumann, 1893
Тип рода Acaromantis squllla Trouessart et Neumann, 1893
3. Atelopsalls Trouessart, 1896
Тип рода Atelopsalls tricuspis Trouessart, 1896
4. Ischyrognathus Trouessart, 1900
Тип рода Simognathus coutleri Trouessart, 1899
Limno halacarinae Viets, 1927
1. Limnohalacarus Walter, 1917
a) Limnohalacarus s. str.
Тип подрода Halacarus wackerl Walter, 1914
6) Stygohalacarus Viets, 1934
Тип подрода Stygohalacarus scuplensis Viets, 1934
282
Глава VI. Trombidiformes (продолжение)
2. Hamohalacarus Walter, 1931
Тип рода Hamofialacarus subterraneus Walter, 1931
3. Soldanellonyx Walter, 1917
a) Soldanellonyx s. str.
Тип подрода Soldanellonyx chappuisi Walter, 1917
6) Parasoldanellonyx Viets, 1929
Тип подрода Soldanellonyx purviscutatus Walter, 1917
Halixodinae Viets, 1927
1. Hallxodes Brucker et Trouessart, 1900
Тип рода Agaue chitonis Brucker, 1897
Enterohalacarinae Viets, 1938
1. Enterohalacarus Viets, 1938
Тип рода Enterohalacarus mlnutipalpus Viets, 1938
A s t а с о p s i p h a g i n a e Viets, 1931
1. Astacopsiphagus Viets, 1931
Тип рода Astacopsiphagus parasiticus Viets, 1931
ОБЩИЕ ЗАМЕЧАНИЯ
Ньюэлл (1947) дал экологическую характеристику Halacaridae по
подсемействам. Мы приводим здесь его систему.
Halacaridae — хищные или травоядные, питающиеся лишайниками
формы; в трех родах имеются паразиты.
1. Rhombognathinae Viets, 1927. Встречаются преимущественно
в приливной зоне (на литорали) и лишь очень редко — в пресных
водах (непроверенные данные). Никогда не живут в условиях, препят-
ствующих развитию водорослей. Космополиты.
2. Actacarinae Viets, 1939. Встречаются в приливной зоне (на ли-
торали), псаммофилы. Никогда не живут в условиях, препятствующих
развитию водорослей. Известны из Северного моря.
3. Lohmannellihae Viets, 1927. Хищники. Морские и пресноводные
формы. Возможно, космополиты.
4. Limnohalacarinae Viets, 1927. Встречаются в пресных водах
Европы, Африки и Северной и Южной Америки.
5. Simognathinae Viets, 1927. Морские формы, встречающиеся
в Адриатическом, Северном, Ирландском и Караибском морях и
в Тихом океане. Хищные виды описаны из Бискайского залива и
западной части Индийского океана.
6. Enterohalacarinae Viets, 1938. Паразитируют в кишечнике
иглокожих (по крайней мере, личиночные и пимфальпые фазы). Встре-
чаются в западной части Тихого океана.
7. Halixodinae Viets, 1929. Паразитируют (по крайней мере нимфы)
на жабрах боконервных моллюсков. Известны из Новой Зеландии.
Hydrachnellae
283
8. Astacopsiphaginae Viets, 1931. Паразитируют (по крайней мере,
нимфы) на жабрах пресноводных десятиногих раков в Австралии.
9. Halacarinae Viets, 1927. Хищники, преимущественно морские
и реже пресноводные. Космополиты.
10. Porohalacarinae Viets, 1933. Живут в пресных или солоноватых
ночах. Вероятно, распространены повсюду, исключая моря Арктики
и Антарктики.
ЛИТЕРАТУРА
Newell 1., 1947. A systematic and ecological study of the Halacaridae of eas-
tern North America, Bull, of the Bingham. Oceanographic Collection,
Peabody Museum of Natural History, Yale Univ., 10 (3), 1—232.
Hydrovolziidae Thor, 1905
(рис. 187, 188)
ДИАГНОЗ
Hydrovolziidae—красные водные клещи, длиной от 0,80 до
0,95 мм. Они имеют маленький, передний и крупный задний дор-
зальные щитки. У переднебокового края заднего дорзального щитка
Рис. 187. Hydrovolzia placophora
(Monti) (по Фитсу, 1936).
Взрослая особь (вид вверху).
Рис. 188. Hydrovolzia placophora
(Monti) (по Фитсу, 1936).
Взрослая особь (вид снизу).
лежит пара крупных боковых щитков; заднийдорзальный щиток окружен
набором мелких пластинок или щитков. Двойные глаза, окруженные
капсулой, помещаются в боковых углублениях переднего дорзального
щитка (у пещерных видов глаза отсутствуют). Тазики расположены
284
Глава VI. Trombidiformes (продолжение)
двумя ясно обособленными группами. Ноги причленены к телу так, что
тазики ног III и IV выступают по бокам тела. Плавательных волосков
на ногах нет; коготки лапок гладкие, серповидные. Половых присосок
нет. Пальпы пятичлениковые, простые; последний членик в виде
коготка и до некоторой степени напоминает последний членик пальп
у Cunaxidae.
РОДЫ и ПОДРОДЫ
1. Hydrovolzia Thor, 1905
a) Hydrovolzia s. str.
Тип подрода Polyxo placophora Monti, 1905
6) Hydrovolziella Viets, 1935
Тип подрода Hydrovolzia lata Walter, 1935
2. Acheroatacarus Viets, 1932
Тип рода Acherontacarus halacaroides Viets, 1932
ОБЩИЕ ЗАМЕЧАНИЯ
Эти клещи встречаются в родниках и ручьях Центральной Европы;
плавать не могут.
Rhynchohydracaridae Lundblad, 1936
ДИАГНОЗ
(рис. 189, 190)
Тело как со спинной, так и с брюшной стороны покрыто крепким
панцырем. Спинной панцырь состоит из большого центрального и
Рис. 189. Rhynchohy dr acarus testudo
Lundblad (ио Лупдбладу, 1941).
Взрослая особь (вид сверху}.
Рис. 190. Rhynchohydracarustestudo
Lundblad (ио Лундбладу, 1941).
Взрослая особь (вид снизу).
Hydrachnellae
285
многих мелких краевых щитков. Коксальные щитки I достигают
переднего конца тела и, смыкаясь впереди, образуют короткую склеро-
тизированную трубку; коксальные щитки I и II сливаются. Плаватель-
ных волосков на ногах нет. Половое отверстие окружают два широких
щитка; половых присосок нет. Пальпы не несут шипов или возвы-
шений. Хелицеры длинные и тонкие.
роды
1. Rhynchohydracarus Lundblad, 1936
Тип рода Rhynchohydracarus testudo Lundblad, 1936
Hydrachnidae Leach, 1815
(рис. 191, 192)
ДИ АГНОЗ
Hydrachnidae — обитатели стоячих вод. Это по большей части
красные, большие, круглые плавающие клещи. Кожа покрыта сосоч-
ками. Имеются парные глаза, лежащие в капсулах. Между, глазами
Рис. 192. Hydrachna kenyensis Ltind-
blad (по Лундбладу, 1942).
Коксальные щитки.
Рис. 191. Hydrach.no.
kenyensis Lundblad
(по Лундбладу, 1942).
Дорзальный щиток.
лежат щитки различной величины (исключение— Anohydrachna) и
формы. Коксальные щитки обычно объединяются в четыре группы,
по два в каждой. Плавательных волосков на ногах нет; коготки на лап-
ках гладкие, серповидные. Хелицеры нерасчлененные, стилетовидные.
Генитальные щитки, подвижно причлененные впереди, посередине
сливаются и несут много маленьких половых присосок.
РОДЫ И ПОДРОДЫ
1. Hydrachna Muller, 1776 (= Atax Fabriclus, 1805)
a) Hydrachna s. str. (== Diplohydrachna Thor, 1916)
Тип подрода Hydrachna cruenta Muller, 1776
6) Anohydrachna Thor, 1916
Тип подрода Hydrachna perniformis Koenike, 1895
286 Глава VI. Trombidiformes (продолжение}
в) Chitohydrachna Habeeb, 1950
Тип подрода Hydrachna (Chitohydrachna) ennishonenses Ha-
beeb, 1950
г) Monohydrachna Thor, 1916
Тип подрода Hydrachna cruenta Krendowsky, 1884
д) Rhabdohydrachna Viets, 1913
Тип подрода Hydrachna bisignifera Viets, 1913
e) Schlzohydrachna Thor, 1916
Тип подрода Hydrachna uniscutata Thor, 1897
ж) Scutohydrachna Viets, 1933
Тип подрода Hydrachna (Scutohydrachna) dorsoscutata Viets,
1933
з) Tetrahydrachna Lundblad, 1934
Тип подрода Hydrachna globoso var. miliaria Berlese, 1888
2. Bargena Koenike, 1893
,Тип рода Bargena mirlfica Koenike, 1893
ДИАГНОЗ
Limnocharidae Kramer, 1877
(рис. 193—196)
Limnocharidae — красные клещи с мягким телом, встречающиеся
в стоячих и медленно текущих водах; плавать не способны. Глаза,
Рис. 193. Limnochares (Cyclothrix) natans Рис. 194. Limnochares
Lavers (по Ловерсу, 1941). (Cyclothrix) natans La-
Нога IV. vers (по Ловерсу, 1941).
Глазной щиток.
окруженные капсулами, расположены латерально на срединном па-
лочковидном склеротизированном щитке. Рострум цилиндрический,
широкий, занимает переднее положение и снабжен ротовым диском,
Hydrachnellae
287
окаймленным бахромой. Хелицеры очень крупные, сливаются одна
с другой и прикрывают максиллярные органы. Половых присосок
много, они малы и лежат свободно на покровах.
Рис. 195. Limnochares
aquatica (Linnaeus)
(по Фитсу, 1936).
Личинка (вид снизу).
Рис. 196. Limnochares aquatica
(Linnaeus) (по Фитсу, 1936).
Личинка (вид сверху).
РОДЫ И ПОДРОДЫ
Limnocharinae Claus, 1880
1. Limnochares Latreille, 1796 ( = Hemitromblcula Ewing, 1938)
a) Limnochares s. str.
Тип подрода Acarus aquaticus Linnaeus, 1758
6) Cyclothrix Wolcott, 1905
Тип подрода Limnochares crinita Koenike, 1898
2. Neollmnochares Lundblad, 1937
Тип рода Neollmnochares placophora Lundblad, 1937
Rhyncholimnocharinae Lundblad, 1936
1. Rhyncholimnochares Lundblad, 1936
Тип рода Rhyncholimnochares latnellipalpls Lundblad, 1936
2. Paralimnochares Lundblad, 1937
Тип рода Paralimnochares sursumhians Lundblad, 1937
288
Глава V!. Trombidiformes (продолжение)
ДИАГНОЗ
Eylaidae Leach, 1815
(рис. 197, 198)
Eylaidae — клещи с мягкой кожей, обычно не имеющей склеро
тизированных щитков. Максиллы связаны с большим ротовым диском
обрамленным бахромой. Хелицеры
отделены друг от друга; у осно-
вания они крупные, удлинены
в дорзовентральном направлении и
имеют маленький коготок. Пальпы
обычно длинные и сильно опу-
шены. Коксальные щитки распо-
ложены четырьмя группами, по
два в каждой. Коготки на лапках
простые. Глаза в капсулах, со-
единены друг с другом хитинизи-
Рис. 198. Piersigia koenike i Viets
(по Фитсу, 1936).
Взрослая особь (вид сверху).
Рис. 197. Eylals desecta Кое-
nike (по Маршаллу, 1927).
Глазные шитки самки.
рованной перемычкой и расположены дорзомедиально, образуя „очки“;
если глаза латеральные, то они лежат по краям сильно расчленен-
ного щитка.
РОДЫ И ПОДРОДЫ
Eylainae Claus, 1880
1. Eclats Latreille, 1796
a) Eylals s. str.
Тип подрода Hydrachna extendens Muller, 1776
6) Capeulais Thor, 1902
Тип подрода Capeulais crasslpalpis Thor, 1902
в) Meteylais Sza'ay, 1934
Тип подрода Eylals hamata Koenike, 1897
r) Pareylals Szalay, 1934
Тип подрода Eylals setosa Koenike, 1897
ж) Proteylais Szalay, 1933
Тип подрода Eylals degenerata Koenike, 1897
Hydrachnellae
289
з) Rhyncheylals Lundblad, 1938
Тип подрода Rhyncheylals connexa Lundblad, 1938
и) Syneylals Lundblad, 1936
Тип подрода Eylals Infundibulifer Koenike, 1897
Piersigiinae Oudemans, 1902
1. Piersigia Protz, 1896
Тип рода Piersigia lltnophlla Protz, 1896
ОБЩИЕ ЗАМЕЧАНИЯ
Лундблад (1941) выделяет Piersigiinae в особое семейство.
Protziidae Viets, 1926
ДИАГНОЗ
(рис. 199, 200)
Protziidae — клещи красного цвета с мягкой кожей, покрытой
сосочками или бороздками; настоящих спинных щитков нет. Боковые
Л
Рис. 199. Calonyx javanensis
Viets (по Лундбладу, 1930).
А—коготок лапки; Б — пальпа.
Рис. 200. Calonyx javanensis
Viets (по Лундбладу, 1936).
Самец (вид сверху)-
глаза лежат в капсулах. Срединный глаз иногда отсутствует. Тар-
зальный членик пальп лежит дорзально на дистальном конце голени
пальп; последняя обычно снабжена шиповидным выростом. Коксаль-
пые щитки имеют приблизительно одинаковые размеры и располо-
жены четырьмя группами; задняя группа, как правило, отнесена да-
леко назад. Плавательных волосков на ногах нет. Коготки лапок
серповидные, простые или зубчатые. Между передней и задней груп-
пой коксальных щитков, далеко от заднего края тела, лежат
19 Зак. 1785
290
Глава Vl. Trombidiformes (продолжение)
гениталии. Длинные стебельчатые половые присоски обычно многочис-
ленны и довольно заметны; они лежат прямо на покровах и варьи-
руют по положению. Сзади присоски имеют шишковидную или
желудевидную форму. Генитальные щитки если имеются, то всегда
опушены.
РОДЫ
1. Protzla Piersig-, 1897
Тип рода Thyas exitnia Protz, 1896
2. Calonyx Walter, 1907
Тип рода Calonyx latus Walter, 1907
3. Neocalonyx Walter, 1919
Тип рода Neocalonyx godeti Wader, 1919
4. Partnunia Piersig, 1897
Тип рода Thyas angusta Koenike, 1893
5. Partnuniella Viets, 1938
Тип рода Partnuniella thermalls Viets, 1938
6. Protziella Lundblad, 1934
Тип рода Protziella hutchlnsoni Lundblad, 1934
7. Wandesla Schechtel, 1912
Тип рода Wandesla thori Schechtel, 1912
ДИАГНОЗ
Hydrodromidae Viets, 1936
(рис. 201, 202)
Hydrodromidae — красные клещи с мягкой кожей, покрытой со-
сочками. Спинные щитки не развиты. Боковые глаза отделены друг
Рис. 201. Hydrodroma despiciens (Midler)
(по Луидбладу, 1944).
Коксальные и генитальные щитки.
от друга и не окружены капсулой; срединный глаз отсутствует. Кок-
сальные щитки расположены четырьмя группами, по два в каждой.
Hydrachnellae
291
Ноги обычно с плавательными волосками. Коготки на лапках имеют
снаружи маленькие дополнительные зубцы. Голень пальп имеет на
Рис. 202. Hydrodroma despiciens (Muller).
Пальпа (по Лундбладу, 1944).
торзальной поверхности дистальный отросток, лежащий над тарзаль-
ным члеником пальп. Имеются генитальные щитки с
присосками.
РОДЫ
1. Hydrodroma Koch, 1837
Тип рода Hydrachna despiciens Muller 1776
umbrata Koch, 1837)
2. Oxopsis NordenskiOld, 1905
Тип рода Oxopsls diplodontoides Nordenskiold,
многочисленными
(— Hydrodrotnci
1905
Hydryphantidae Thor, 1900
(рис. 203, 204)
ДИАГНОЗ
Hydrophantidae — клещи красного цвета с кожей, покрытой со-
сочками. Иногда имеется фронтальный щиток, редко тело одето в пан-
цырь. Срединный глаз имеется только
щитка. Голень пальп имеет дисталь-
ный вырост на дорзальной стороне.
Коксальные щитки расположены че-
тырьмя группами. Все ноги с пла-
вательными волосками и простыми
серповидными коготками на лапках.
при наличии фронтального
Р и с. 204. Hydryphantes ruber
(DeOeer) (по Маршаллу, 1944).
Взрослая особь (вид снизу).
Рис. 203. Hydryphantes tenuabilis
Marshall, 1946 (по Маршаллу, 1946).
Передний отдел тела: показаны глаза и щиток.
19*
292
Глава VI. Trombidiformes (продолжение)
Гениталии снабжены двумя подвижными щитками и тремя парами
или большим числом половых присосок.
РОДЫ И ПОДРОДЫ
Ну dг ур h a ntin а е Piersig, 1896
1. Hydryphantes Koch, 1841
a) Hydryphantes s. str.
Тип подрода Acarus aquations ruber DeGeer, 1778
6) Octohydryphantes Lundblad, 1927
Тип подрода Hydryphantes octoporus Koenike, 1896
в) Polyhydryphantes Viets, 1926
Тип подрода Hydrodroma flexnosa Koenike, 1885
2. Georgella Koenike, 1907
Тип рода Hydrodroma helvetica Haller, 1882
3. Papilloporus Walter, 1935
Тип рода Hydryphantes incertus Koenike, 1893
D i p 1 о d on t i n a e Vitzthum, 1942
1. Diplodontus Dug£s, 1833 ( = Eupatra Koenike, 1896)
Тип рода Diplodontus scapularls Dug£s, 1834
Mamersinae Viets, 1931
1. Mamersa Koenike, 1898
Тип рода Mamersa testudinata Koenike, 1898
Thyasinae Viets, 1926
1. Thyas Koch, 1836
Тип рода Thyas venusta Koch, 1836
2. Ja-vathyas Viets, 1929 ( = Allothyas Viets, 1929)
Тип рода favathyas triumvirorum Viets, 1929
3. Kashmirothyas Lundblad, 1934
Тип рода Kashmirothyas hutchinsoni Lundblad, 1934
4. Lundbladia Viets, 1929
a) Lundbladia s. str. ,<
Тип подрода Lundbladia feuerborni Viets, 1929
6) Octolundbladia Vitzthum, 1942
Тип подрода Octolundbladia ladakiana Vitzthum, 1942
5. Octothyas Lundblad, 1945
Тип рода Octothyas he wit hi Lundblad, 1945
6. Panisellus Viets, 1925
Тип рода Panisus thienemanni Viets, 1920
7. Panisoides Lundblad, 1926
Тип рода "Thyas setipes Viets, 1911
Hydrachnellae
293
8. Panlsus Koenike, 1896
Тип рода Panlsus michaeli Koenike, 1896
9. Parathyas Lundblad, 1926
Тип рода Thyas thoracatus Piersig, 1896
10. Placothyas Lundblad, 1926
Тип рода Thyas octopora Viets, 1914
11. Pleslothyas Viets, 1935
Тип рода Trichothyas multlpora Walter, 1935
12. Thyasella Viets, 1926
Тип рода Thyas mandibularis Lundblad, 1924
13. Thyopsis Piersig, 1898
Тип рода Thyas cancellata Protz, 1896
14. Trichothyas Viets, 1926
Тип рода Thyas pennata Viets, 1913
15. Vietsia Lundblad, 1926
Тип рода Thyas scutata Protz, 1923
E u t h у a s i n ae A^ie t s, 1931
1. Euthyas Piersig, 1898 ( = Bradybates Neuman, 1874 nomen
praeoccupatum)
Тип рода Bradybates truncatus Neuman, 1874
2..Panisopsis Viets, 1926
Тип рода Thyas vigilans Piersig, 1896
3. Thyasides Lundblad, 1926
Тип рода Thyas dentata Thor, 1897
4. Zschokkea Koenike, 1892
Тип рода Zschokkea oblonga Koenike, 1892
T e r a t о t h у a s i n a e Viets, 1929
1. Teratothyas Viets, 1929
Тип рода Teratothyas reticulata Viets, 1929
Tartarothyasinae Viets, 1934
1. Tartarothyas Viets, 1934
Тип рода Tartarothyas mlcrommata Viets, 1934
Clathrosperchonidae Lundblad, 1936
(рис. 205, 206)
ДИАГНОЗ
Покровы склеротизированные, с тонкими шипами и радиальным сетча-
тым узором как на спинной, так и на брюшной поверхности. Кожные
железы открываются на слабо приподнятых участках кожи; эти уча-
стки сильно рдлеротцзированы и имеют форму кратера. Рострум
294
Глава VI. Trombidiformes (продолжение)
либо длинный, узкий и трубчатый на переднем конце (Clathrosper-
chori), либо сравнительно короткий и не трубчатый (Clathrospercho-
nella). Хелицеры тонкие у первого из названных родов и крупные
у второго. Пальпы никогда не бывают клещевидными. Плавательных
Р и с. 205. Clath.rosperch.on minor
Lundblad (по Лупдбладу, 1941).
Самец (вид сверху).
Самец (вид снизу).
волосков на ногах нет. Коготки лапок с дополнительными вентраль-
ными зубцами. Имеются два генитальных щитка с многочисленными
половыми присосками.
РОДЫ
1. Clathrosperchon Lundblad, 1936
Тип рода Clathrosperchon crasslpalpls Lundblad, 1936
2. Clathrosperchonella Lundblad, 1937
Тип рода Clathrosperchonella asterifera Lundblad, 1937 -
Hydrachnellae
295
ДИАГНОЗ
Ctenothyasidae Lundblad, 1936
(рис. 207)
Покровы тела плотные, кожистые, с сосочками; панцырь не раз-
вит. Повидимому, имеется рудиментарный срединный глаз, а боковые
глаза покрыты общей капсулой. Железистые поля заметны в виде
сильно приподнятых бородавчатых участков кожи. Рострум такого
же типа, как и у Thyas.
Рис. 207. Ctenothyas
verrucosa Lundblad
(по Лундбладу, 1936).
Самка (вид сверху).
Голени пальп не имеют дистального отростка- Коксальные щитки
расположены четырьмя группами, лапки с двумя простыми когот-
ками. Плавательных волосков на ногах нет. Наружный половой ап-
парат самок с двумя большими, сильно склеротизированными пори-
стыми щитками и с тремя крупными присосками на боковых краях
каждого из этих щитков.
РОДЫ
1. Ctenothyas Lundblad, 1936
Тип рода Ctenothyas verrucosa Lundblad, 1936
Thermacaridae Sokolow, 1927
(рис. 208, 209)
ДИАГНОЗ
Тело Thermacaridae целиком покрыто панцырем. Центральный
дорзальный щиток соединяется с вентральным щитком тонким, но креп-
ким швом. Впереди по бокам тела лежат по два глаза, окруженных
капсулами; имеется также и срединный глаз. Коксальные щитки I
сливаются друг с другом и не разделены швом; коксальные щитки
295
Глава. VJ. Trombidiformes (продолжение)
I, II, III и IV отделены друг от друга швами. Все коксальные щитки
соприкасаются друг с другом. У самок гениталии лежат между кок-
сальными щитками III и IV; у самцов коксальные щитки IV сопри-
касаются посередине и гениталии лежат позади них. Слившиеся кок-
сальные щитки I образуют камеростом, в который может втягиваться
гнатосома. Вертлуги ног I, II и III несут по сильной чашевидной
выемке, к которой причленяется основной членик бедра (basifemur);
коготки на лапках крепкие, серповидные и не несут дополнительных
коготков, однако имеют острые зубцы на вогнутой стороне. Имеется
Рис. 208. Thermacarus
nevadensis Marshall
(но Маршаллу, 1928).
Щитки брюшной поверхности
тела самки.
Рис. 209. Thermacarus
nevadensis Marshall
(по Маршаллу, 1928).
Самец (вид сверху).
два подвижных генитальных щитка, которые подразделены на перед-
нюю и заднюю части; последняя несет многочисленные маленькие
половые присоски. Thermacaridae встречаются в горячих источниках.
РОДЫ
1. Thermacarus Sokolow, 1927
Тип рода Thermacarus thermobius Sokolow, 1927
ОБЩИЕ ЗАМЕЧАНИЯ
Эти клещи встречаются в горячих источниках с температурой
42—46°.
ЛИТЕР АТУРА
Marshall R., 1928. A new species of water mite from thermal springs,
Psyche, 35 (2), 92—96.
Hydrachnellae
297
ДИАГНОЗ
Rutripalpidae Sokolow, 1934
(рис. 210)
Rutripalpidae имеют мягкие покровы со склонностью к образованию
склеротизированных пластиночек или сосочков. Коксальные щитки
расположены четырьмя группами,
по две в каждой. Плавательных
волосков на ногах нет. На лапках
передних трех пар ног имеются
хорошо развитые коготки, снаб-
женные многочисленными тонкими
дополнительными шипами (на но-
гах IV эти дополнительные шипы
почти всегда редуцированы). Сбоку
два последних членика пальп
кажутся лопатовидными и расши-
ренными. Неподвижный палец хе-
лицер редуцирован.
Рис. 210. Rutrlpalpus limlcola Soko-
low (по Соколову, 1934).
Взрослая особь (вид снизу).
РОДЫ
1. Rutrlpalpus Sokolow, 1934
Тип рода Rutrlpalpus limlcola Sokolow, 1934
Mamersopsidae Lundblad, 1930
(рис. 211, 212)
ДИАГНОЗ
Спинной и брюшной щиты, покрывающие тело Mamersopsidae,
отделены друг от друга дорзальным швом. Спинной щит образован
у одних форм единственным щитком, у других же он состоит из
нескольких слившихся щитков. Протоки желез открываются
в области дорзального шва на специальных щитках. Пальпы булаво-
видные, особенно их бедра, которые сильно утолщены дорзально и
несут на внутреннем крае шип или шип и вырост. Коленный членик
пальп короче бедра или голени. Голень пальп сильно укорочена; она
короче бедра пальп и на внутреннем крае несет вырост, покрытый
298
Глава Vr. Trombidiformes (продолжение)
щетинками. Тарзальный членик пальп на конце шиповидный. Кок-
сальные щитки образуют одну группу и соединяются с вентральным
щитком; все щитки отделены друг от друга слабо выраженными
швами. Ноги иногда с плавательными волосками. Лапки ног IV часто
редуцированы и либо совсем не несут коготков, либо имеют лишь
Рис. 211. Mamersopsis
thoracica NordenskiOld
(по Норденскьёлду, 1905).
Взрослая особь (вид сверху).
Рис. 212. Mamersopsis
thoracica NordenskiOld
(по Норденскьёлду, 1905).
Взрослая особь (вид снизу).
один простой коготок. Гениталии либо примыкают, либо более или
менее соединяются с тазиками ног IV и снабжены двумя подвижными
щитками и тремя парами половых присосок.
РОДЫ
1. Mamersopsis NordenskiOld, 1905
Тип рода Mamersopsis thoracica NordenskiOld, 1905
2. Bandakia Thor, 1913
Тип рода Bandakia concreta Thor, 1913
3. Platymamersopsis Viets, 1914
Тип рода Platymamersopsis nordenskioldi Viets, 1914
Pseudohydryphantidae Viets, 1926
(рис. 213)
ДИАГНОЗ
Кожа у Pseudohydrophantidae мягкая, без склеротизированных
щитков, хотя места прикрепления мышц слабо склеротизированы.
Спина покрыта короткими сильными многовершинными сосочками.
Боковые глаза лежат в капсулах; срединный глаз также окружен
маленьким склеротизированным кольцом. Коксальные щитки располо-
Hydrachnellae
299
жены четырьмя группами, по два в каждой. Ноги III и IV с плава-
тельными волосками. Коготки лапок простые, серповидные. Голенный
Рис. 213. Pseudohydryphantes parvulus Viets (по Фитсу, 1936).
Л —гениталии самки; Б — пальпа; В —кожные сосочки.
членик пальп на конце несет дорзальный отросток. Гениталии
с двумя подвижными щитками и тремя парами половых присосок.
РОДЫ
1. Pseudohydryphantes Viets, 1907
Тип рода Pseudohydryphantes parvulus Viets, 1907
Teutoniidae Lundblad, 1927
ДИАГНОЗ
(рис. 214, 215)
Teutonidae имеют мягкие кожистые покровы. Боковые глаза их
лежат близко друг к другу, но не окружены капсулой; срединного
Рис. 214. Teutonia cometes (Koch)
(по Фитсу, 1936).
Щитки брюшной поверхности тела самца.
глаза нет. Бедро пальп на внутрен-
нем крае несет конус. Коксаль-
ные щитки расположены четырьмя
группами, по два в каждой.
Рис. 215. Teutonia cometes (Koch)
(по Фитсу, 1936).
Пальпа*
300
Глава VI. Trombidiformes (продолжение)
Коксальные щитки III и IV расположены почти под прямым углом
друг к другу. На переднем внутреннем углу коксального щитка IV
имеется отверстие железы. Ноги III и IV с плавательными волосками.
Коготки лапок ног II и III имеют дополнительные зубцы. Лапки IV
на конце сужены и лишены коготков.
РОДЫ
1. Teutonia Koenike, 1889
Тип рода Teutonia primaria Koenike, 1889
ДИАГНОЗ
Sperchonidae Thor, 1900
(рис. 216 -218)
Sperchonidae имеют мягкие кожистые покровы, иногда несущие
щитки с бугорками или с тонко заостренными сосочками в борозд-
чатых полях. Боковые глаза лежат в капсулах; срединный глаз
Рис. 216. Sperchon
glandulosus Koenike
(по Маршаллу, 1943).
Генитальные щитки самки.
Рис. 217. Sperchonopsis
verrucosa (Protz)
(по Маршаллу, 1943).
Самец (вид снизу).
Рис. 218. Sperchon сог-
nutus Viets
(по Лундбладу, 1936).
Палыта самца.
имеется редко. Бедренный членик пальп на внутреннем крае несет шпе-
нек, покрытый шипами. Голенный членик пальп без концевого дорзаль-
ного выроста; его внутренняя поверхность несет два сенсорных шипа.
Коксальные щитки образуют четыре группы. Коксальные щитки IV
трех- или четырехугольные. Ноги без плавательных волосков.
Коготки лапок, как правило, с дополнительными зубцами. Гениталии
лежат между задними коксальными щитками и снабжены двумя бо-
ковыми подвижными щитками и тремя парами половых присосок.
Hydrachnellae 301
” ——•-----------—‘'"'r -------------------------т----------
РОДЫ И ПОДРОДЫ
1. Sperchon Kramer, 1877
a) Sperchon s. str.
Тип подрода Sperchon squamosa Kramer, 1879
6) Hlspidosperchon Thor, 1901
Тип подрода Sperchon hispldus Thor, 1898
в) Mixosperchon Viets, 1926
Тип подрода Sperchon papillosus Thor, 1901
r) Scutosperchon Viets, 1926
Тип подрода Sperchon thori Koenike, 1900
2. Sperchonopsis Piersig, 1896 (= Pseudosperchon Piersig, 1901)
Тип рода Sperchon verrucosus Protz, 1896
ДИАГНОЗ
Anisitsiellidae Viets, 1929
(рис. 219, 220)
Anisitsiellidae имеют мягкие кожистые покровы, иногда же тело
покрыто панцырем с дорзальным швом. Боковые глаза не окружены
Рис. 219. Mamersellides
ventriperforatus Lundblad
(по Лундбладу, 1941).
Самка (вид сверху).
Рис. 220. Mamersellides
ventriperforatus Lundblad
(по Лундбладу, 1941).
Самка (вид снизу).
капсулами и отделены один от другого. На внутренней стороне бед-
ренного членика пальп имеется отросток. На переднем внутреннем
углу коксальных щитков IV нет крупных отверстий желез. Лапки
ног IV на конце редуцированы и лишены коготков. Гениталии
с двумя щитками и тремя или четырьмя парами половых присосок.
302
Глава Vf. Trombidiformes (продолжение)
РОДЫ И ПОДРОДЫ
Anisit siel 1 inae Koenike, 1910
1. Anisitsiella Daday, 1905
Тип рода Anisitsiella aculeata Daday, 1905
2. Hydrobaumla Halik, 1930
Тип рода Hydrobaumla tnalacetisls Halik, 1930
3. Mamersella Viets, 1929
Тип рода Mamersella thlenemanni Viets, 1929
4. Mamersellides Lundblad, 1937
Тип рода Mamersellides ventriperforatus Lundblad, 1937
5. Matnersides Viets, 1935
Тип рода Mamersides saranganensls Viets, 1935
6. Mamersopides Viets, 1916
Тип рода Mamersopides sigthori Viets, 1916
7. Rutacarus Lundblad, 1937
Тип рода Rutacarus pyriformis Lundblad, 1937
8. Sigthoria Koenike, 1907 (== Amasis NordenskiOld, 1905 nomen
praeoccupatum)
Тип рода Amasis niloticus NordenskiOld, 1905
Nilotoniinae Viets, 1929
1. Nilotonla Thor, 1905
Тип рода Teutonia loricata NordenskiOld, 1905
2. Dartia Soar, 1917
a) Dartia s, str.
Тип рода Dartia harrlsl Soar, 1917
6) Dartlella Viets, 1929
Тип рода Dartia longtpora Walter, 1925
3. Dartonla Viets, 1929
Тип рода Dartonla caerulea Viets, 1929
4. Kawamuracarus Ucliida, 1937
Тип рода Kawamuracarus elongatus Uchida, 1937
5. Limnolegeria Motas, 1928
Тип рода Limnolegeria longiseta Motas, 1928
6. Manotonia Viets, 1935 (= Mania Walter, 1935 nomen praeoccupatum)
Тип рода Mania muscicola Walter, 1935
7. Ranautonia Viets, 1929
Тип рода Ranautonia dentipalpis Viets, 1929
Lebertiidae Thor, 1900
(рис. 221)
ДИАГНОЗ
Lebertiidae обладают мягкими кожистыми покровами; как правило,
щитков на спинной стороне нет, но брюшная поверхность часто
сильно склеротизирована. Боковые глаза двойные и заключены в неж-
Hydrachnellae
303
ную склеротизированную капсулу. Пальпы более или менее тонкие,
узкие. Все коксальные щитки объединены в одну группу и средин-
ный шов между ними часто отсутствует. Гениталии снабжены двумя
Рис. 221. Lebertia qulnquemaculosa
Marshall (по Маршаллу, 1932).
Брюшко взрослой особи.
щитками и тремя парами половых присосок; часто имеется только
аве пары присосок.'
роды и ПОДРОДЫ
Lebertiinae Wolcott, 1905
1. Lebertia Neumann, 1880
a) Lebertia s. str. (= Neolebertia Thor, 1905)
Тип подрода Pachygaster tau-lnslgnatus Lebert, 1879
6) Distolebertla Husiatinschi, 1937
Тип подрода Distolebertla bucovlnensis Husiatinschi, 1937
в) Hexalebertia Thor, 1906
Тип подрода Lebertia stigmatifera Thor, 1900
r) Mixolebertia Thor, 1906
Тип подрода Lebertia brevipora Thor, 1899
д) Pilolebertia Thor, 1900
Тип подрода Lebertia insignis Neumann, 1880
e) Pseudolebertia Thor, 1897
Тип подрода Pseudolebertia glabra Thor, 1897
Oxinae Viets, 1926
1. Oxus Kramer, 1877
a) Oxus s. str.
Тип подрода Oxas oblongus Kramer, 1879
6) Pseudoxus Thor, 1901
Тип подрода Pseudoxus integer Thor, 1901
304
Глава VI. Trombidiformes (продолжение)
2. Frontipoda Koenike, 1891
Тип рода Hydrachna muscalus Muller, 1776
3. Gnaphiscus Koenike, 1898
a) Gnaphiscus s. str.
Тип подрода Gnaphiscus setosus Koenike, 1898
6) Gnaphoxus Thor, 1913
Тип подрода Gnaphiscus (Gnaphoxus) ekmani Thor, 1913
Atractideidae Thor, 1902
(рис. 222, 223)
ДИАГНОЗ
Atractideidae имеют цельный брюшной щит, спинной же щит
подразделен у них швами на несколько щитков. Пальпы обычно
Рис. 222. Airactides sierrensis
Marshall (по Маршаллу, 1943).
Самец (вид снизу).
Рис. 223. Airactides sierrensis
Marshall (по Маршаллу, 1943).
Самец (вид сверху).
маленькие; их бедренный и голенный членики обычно несут на вну-
треннем крае шипы или конусы. Коксальные щитки I не разделены
срединным швом и сливаются друг с другом; коксальные щитки II
и III разделены неполным швом, а коксальные щитки IV сзади и
снаружи слабо отделены от брюшного щита. Ноги без плавательных
волосков; коготки на лапках с дополнительными зубцами. Гениталии
снабжены двумя подвижными щитками и тремя или шестью парами
половых присосок.
РОДЫ И ПОДРОДЫ
1. Atractldes Koch, 1837
a) Atractides s. str.
Тип подрода Atractides animalus Koch, 1837
I iydrachnellae
305
6) Monatractides Viets, 1926
Тип подрода Atractides aniscutatus Viets, 1925
в) Rusetria Thor, 1897
Тип подрода Rusetria spinirostris Thor, 1897
r) Rusetriella Viets, 1931
Тип подрода Atractides acutiscutatus Viets, 1913
2. Pseadotorrenticola Walter, 1906
Тип подрода Pseadotorrenticola rhynchota Walter, 1906
3. Testudacaras Walter, 1928
Тип подрода Testudacaras tripeltatus Walter, 1928
Pontarachnidae Thor, 1929
(рис. 224)
ДИАГНОЗ
Pontarachnidae — морские клещи, не имеющие трахейной системы.
Коксальные щитки расположены двумя группами, по четыре в каждой.
Рис. 224. Pontarachna
punctulitm Philippi
(по Уолтеру, 1925).
Самка (вид снизу).
Гениталии без подвижных щитков и половых присосок. Пальпы
простые и не имеют на внутреннем крае выростов.
РОДЫ
1. Pontarachna Philippi, 1840
Тип рода Pontarachna punctulum Philippi, 1840
2. Litarachna Walter, 1925
Тип рода Litarachna incerta Walter, 1925
20 Зак. 1785
396
Глава VI. Trombidiformes (продолжение)
Neotorrenticolidae Lundblad, 1936
(рис, 225, 226)
ДИАГНОЗ
Neotorrenticolidae — мягкокожие клещи с
друг от друга. У самцов спина покрыта
глазами, отделенными
сплошным панцырем,
Рис. 225. Neotorrenticola
violacea Lundblad
(по Лундбладу, 1942).
Брюшко молодой самки.
у самок же — многочисленными мел-
кими склеротизированными щитками.
Голенные членики пальп имеют опу-
шенные выросты, а бедренные чле-
ники— покрытые шипиками зубцы.
У самцов коксальные щитки слиты;
у половозрелых самок коксальные
щитки разделены на четыре группы,
а у неполовозрелых они сближены.
Ноги длинные и сильные, плаватель-
ных волосков на них нет, но имеются
Рис. 226. Neotorrenticola
violacea Lundblad
(по Лундбладу, 1942).
Палыта.
шипы различной толщины. Коготки на лапках крупные и на нижней
стороне имеют дополнительные зубцы. Гениталии окружены много-
численными половыми присосками.
РОДЫ
1. Neotorrenticola Lundblad, 1936
Тип рода Neotorrenticola violacea Lundblad, 1936
ЛИТЕРАТУРА
Lundblad О., 1936. Dritte Mitteilung fiber Wassermilben aus Santa Catha-
rina in Sfldbrasilien, Zool. Anz., 118, 200—211.
Lundblad O., 1942. Die Hydracarinenfauna Siidbrasiliens und Paraguays — II,
Af. Svenska Vetensk Akad. Handl., 20 (3), 3—175.
Hydrachnellae
307
Limnesiidae Thor, 1900
ДИАГНОЗ
(рис. 227, 228)
Limnaesiidae обычно имеют мягкие
кожистые гладкие покровы,
не несущие сосочков; редко их тело покрыто полным панцырем.
Рис. 227. Centrollmnesia bondl
Lundblad (по Лундбладу, 1935).
А —нога III; Б — нога IV.
В задней части спинной поверхности
имеется либо один маленький не-
парный щиток, либо два щитка. Бо-
ковые глаза отделены друг от друга
и лежат порознь. Бедренные чле-
ники пальп обычно несут на внутрен-
ней стороне шпенек, или маленький
Рис. 228. Centrollmnesia bondl
Lundblad (по Лундбладу, 1935).
Самец (вид снизу).
типик, а голенные членики — маленькую щетинку. Коксальные щитки
расположены четырьмя группами по два в каждой; изредка они со-
единяются в одну группу. Коксальные щитки IV крупные, треуголь-
ные с вертлужной впадиной, расположенной на задне-наружном
крае. Ноги обычно с плавательными волосками. Лапки ног IV, как
правило, редуцированы и лишены коготков. Гениталии прикрыты
коксальными щитками; генитальные щитки несут три, реже четыре
или больше присосок. У самцов генитальные щитки соединяются друг
с другом.
20*
308
Глава Vl. Trombidiformes (продолжение)
РОДЫ И ПОДРОДЫ
Limnesiinae Koenike, 1909
1. Limnesla Koch, 1836
a) Limnesla s. str.
Тип подрода Limnesla fulgida Koch, 1836
6) Allolimnesia Viets, 1936
Тип подрода Allolimnesia polypora Viets, 1936
в) Limnesiopsldes Viets, 1938
Тип подрода Limnesiopsldes pectungulatus Viets, 1938
r) Limnesiopsis Piersig, 1897
Тип подрода Limnesla anomala Koenike, 1895
д) Neolimnesia Lundblad, 1936
Тип подрода Limnesla (Neolimnesia) plaumanni Lundblad, 1936
e) Parallmnesia Lundblad, 1938
Тип подрода Limnesla (Paralimnesia) mlcrodon Lundblad, 1938
ж) Tetralimnesia Thor, 1923
Тип подрода Limnesla aspera Koenike, 1898
2. Centrollmnesia Lundblad, 1935
Тип рода Centrollmnesia bondi Lundblad, 1935
3. Daralimnesia Viets, 1923
Тип рода Daralimnesia tenuipalpis Viets, 1923
4. Limnesiella Daday, 1905
a) Limnesiella s. str.
Тип подрода Limnesiella pusilia Daday, 1905
6) Limnesiellula Viets, 1935
Тип подрода Limnesiellula brasiliana Viets, 1935
5. Llmnesicula Viets, 1937
Тип рода Limnesicula verrucosa Viets, 1937
6, Pllollmnesia Viets, 1938
Тип рода Pllollmnesia rostrata Viets, 1938
7. Tubophora Walter, 1935
Тип рода Tubophora limnesioides Walter, 1935
Protolimnesiinae Viets, 1940
1. Protolimnesia Lundblad, 1927
Тип рода Limnesla unguiculafa Walter, 1919
2. Crenolimnesla Lundblad, 1938
Тип рода Crenolimnesla placophora Lundblad, 1938
3. Limnesides Lundblad, 1936
Тип рода Limnesides epimerata Lundblad, 1936
Tyrelliinae Koenike, 1910
1. Tyreilia Koenike, 1895
Тип рода Tyreilia circularis Koenike, 1895
2. Neotyrellla Lundblad, 1938
Тип рода Neotyrellla petricola Lundblad, 1938
Hydrachnellae
309
Рис. 229. Hygrobates plebejus
Lundblad (по Лундбладу, 1930).
Самец (вид снизу).
Hygrobatidae Koch, 1842
(рис. 229)
ДИАГНОЗ
Hygrobatidae, как правило, имеют мягкие кожистые покровы.
Бедренные членики пальп часто несут вырост на внутренней сто-
роне, их передний внутренний край часто зубчатый. Голенные чле-
ники пальп на внутренней стороне несут
мелкие щетинки; часто имеется также
срединная кинжаловидная щетинка.
Коксальные щитки обычно расположены
тремя группами (коксальные щитки I
соединяются друг с другом позади мак-
силл); реже эти щитки образуют четыре
группы, по два в каждой, или же
все соединяются друг с другом. Коксаль-
ные щитки I, II и III обычно узкие и
вытянутые, коксальные щитки IV —
крупные. Шов между щитками III и IV
обычно не достигает их внутреннего
края. Плавательных волосков на ногах
обычно нет, реже ноги снабжены
отдельными длинными волосками. Гени-
талии снабжены щитками и, как пра-
вило, тремя парами присосок (иногда
присосок бывает меньше или больше).
Самцы имеют округлый щиток с более или менее щелевидным половым
отверстием, самки же имеют два (реже четыре) отдельных щитка.
РОДЫ И ПОДРОДЫ
Hygrobatinae Claus, 1880
1. Hygrobates Koch, 1837
a) Hygrobates s. str.
Тип подрода Hydrachna longipalpls Hermann, 1804
6) Capobates Thor, 1898
Тип подрода Capobates sarsl Thor, 1898
в) Dekabates Thor, 1923
Тип подрода Hygrobates decaporas Koenike, 1895
r) Diktyobates Thor, 1927
Тип подрода Acarus fluviatllis StrOm, 1768
д) Dodecabates Viets, 1926
Тип подрода Hygrobates dodecaporus Koenike, 1895
e) Monobates Walter, 1935
Тип подрода Hygrobates falclpalpls Koenike, 1907
ж) Neohygrobates Viets, 1935
Тип подрода Neohygrobates puberulus N'tA.'S, 1935
310
Глава Vl. Trombidiformes (продолжение)
з) Rhabdotobates Thor, 1927
Тип подрода Hygrobates calliger Piersig, 1896
и) Rlvobates Thor, 1897
Тип подрода Rivobates norveglcus Thor, 1897
к) Tetrabates Thor, 1923
Тип подрода Hygrobates octoporus Daday, 1913
2. Cortlcacarellus Lundblad, 1937
Тип рода Cortlcacarus labialis Lundblad, 1936
3. Cortlcacarus Lundblad, 1936
Тип рода Cortlcacarus dentlpalpls Lundblad, 1936
4. Crenohygrobates Lundblad, 1938
Тип рода Crenohygrobates multiporus Lundblad, 1938
5. Hygrobatella Viets, 1926 (= Placobates Lundblad, 1927)
Тип рода Hygrobates placophorus Walter, 1919
6. Hygrobatldes Lundblad, 1936
Тип рода Hygrobatldes pachydermis Lundblad, 1936
7. Hygrobatopsls Viets, 1924
Тип рода Hygrobatopsls levlpalpis Viets, 1924
8. Kyphohygrobatella Lundblad, 1936
Тип рода Kyphohygrobatella serratlpalpls Lundblad, 1936
9. Kyphohygrobates Viets, 1935
Тип рода Hygrobates verrucifer Daday, 1905
10. Megabates Viets, 1924
Тип рода Megabates rectlpes Viets, 1924
11. Paraschizobates Lundblad, 1937
Тип рода Paraschizobates megapoldes Lundblad, 1937
12. Schizobates Thor, 1927
Тип рода Hygrobates distunctus Walter, 1924
13. Schubartella Viets, 1937
Тип рода Schubartella longipes Viets, 1937
14. Stylohygrobates Viets, 1935
Тип рода Stylohygrobates longtpalpls Viets, 1935
15. Subcortlcacarus Lundblad, 1937
Тип рода Subcortlcacarus dlgitatus Lundblad, 1937
16. Thonla Hallk, 1941
Тип рода Thonla barbata Halik, 1941
17. Thoracohygrobates Lundblad, 1936
Тип рода Thoracohygrobates cancellatus Lundblad, 1936
M eg a p u sin a e Thor, 1927
1. Megapus Neumann, 1880
a) Megapus s. str.
Тип подрода Atractides splnipes Koch, 1837
6) Octomegapus Viets, 1926
Тип подрода Megapus octoporus Piersig, 19Q4
Hydrachnellae
311
в) Polymegapus Viets, 1926
Тип подрода Megapus polyporus, 1922
г) Tetramegapus Viets, 1926
Тип подрода Megapus gabretae Thon, 1901
д) Tympanomegapus Thor, 1923
Тип подрода Atractides pavesil Maglio, 1905
2. Hygrobatomegapus Lundblad, 1927
Тип рода Hygrobatomegapus spathullferus Lundblad, 1927
3. Megapella Lundblad, 1936
Тип рода Megapella longlmaxlllaris Lundblad, 1936
4. Megapoldes Lundblad, 1936
Тип рода Megapoldes porosus Lundblad, 1936
5. Mesobatella Viets, 1931
Тип рода Megapus serratlsetus Viets, 1916
6. Mesobates Thor, 1901
Тип рода Mesobates forcipatus Thor, 1901
7. Mlxobates Thor, 1905
Тип рода Mlxobates processlfer Thor, 1905
ДИАГНОЗ
Atractidellidae Lundblad, 1936
(рис. 230- - 233)
Atractidellidae имеют крепкие
в передней части спинной
поверхности.
Рис. 230. Atractidella hamata
Lundblad (по Лундбладу, 1942).
Самка (вид снизу).
кожистые покровы с щитками
Хелицеры длинные, тонкие.
Бедренный и коленный членики
пальп имеют каждый по вентраль-
ному шипу. Коксальные щитки
Рис. 231. Atractidella hatnaia
Lundblad (по Лундбладу, 1942).
Пальпа самки.
расположены тремя группами. Две передние пары щитков соединены
друг с другом позади максилл. Задний край коксальных щитков IV
312
Глава VI. Trombidiformes (продолжение)
склеротизирован и образует крючкообразный вырост. Передняя часть
коксальных щитков I заходит за край тела. Ноги длинные; плава-
Рис. 232. Atractidella hamaia .
Lundblad (по Лундбладу, 1942).
Самец (вид сверху).
Р и с. 233. Atractidella hamata
Lundblad (но Лундбладу, 1942).
Самка (вид сверху).
гельных волосков нет. Гениталии снабжены двумя
парами половых присосок.
щитками и тремя
РОДЫ
1. Atractidella Lundb’ad, 1936
Тип рода Atractidella hatnata Lundblad, 1936
2. Atractidopsls Lundblad, 1936
Тип рода Atractidopsls dlgitata Lundblad, 1936
ЛИТЕРАТУРА
Lundblad O., 1936. Neue Wassermilben aus Santa Catharina in Stidbrasilien,
Zool. Anz., 115, 29—51.
Nautarachnidae Viets, 1935
(рис. 234)
ДИАГНОЗ
Nautarachnidae имеют твердую кожу, покрытую сосочками. Бо-
ковые глаза лежат в капсулах. Хелицеры двухчлениковые. Пальпы
не имеют шиповидных отростков, и их голенные членики не несут
склеротизированного дистального шпенька. Коксальные щитки рас-
положены четырьмя группами, по два в каждой. На заднем крае
Hydrachnellae
313
коксальных щитков. IV посередине сидят направленные назад от-
ростки. Плавательные волоски на ногах имеются. Гениталии снабжены
Рис. 234. Naatarachna
crassa (Koenike)
(по Фитсу, 1936).
Самка (вид снизу).
двумя расположенными по бокам длинными щитками, несущими мно-
гочисленные присоски.
РОДЫ
1-. Nautarachna Moniez, 1888
Тип рода Nautarachna asperrltnum Mon’ez, 1888
Feltriidae Thor, 1929
ДИАГНОЗ
(рис. 235—237)
Feltriidae имеют более или менее бороздчатые покровы. У самцов
имеются некрупные склеротизированные щитки, у самок — мелкие
Рис. 235. Pelt ria
zschokkei Koenike
(по Фитсу, 1936).
Самка (вид снизу).
склеротизированные щитки. Коксальные щитки расположены четырьмя
группами, по два в каждой. У самцов коксальные щитки часто тесно
314
Глава VI. Trombidiformes (продолжение)
примыкают друг к другу. Коксальные щитки IV расширяются кнаружи
и часто не имеют настоящего
Рис. 236. Feltria minuta
Koenike (по Фитсу, 1936).
Самец (вид снизу).
внутреннего края; задний край их
лежит поперечно. Ноги без плава-
тельных волосков. Лапки ног III
обычно отличаются по строению
у разных полов. Гениталии круп-
ные, с многочисленными мелкими
поровидными присосками. У самцов
Рис. 237. Feltria minuta
Koenike (по Фитсу, 1936).
Самка (вид сверху).
щитки, несущие присоски, соединяются друг с другом и небольшое
половое отверстие открывается в этом сложном щитке.
РОДЫ И ПОДРОДЫ
1. Feltria Koenike, 1892
a) Feltria s. str.
Тип подрода Feltria minuta Koenike, 1892
6) Feltriella Viets, 1930
Тип подрода Feltria rubra Piersig, 1898
в) Mesofeltria Uchida, 1934
Тип подрода Feltria {Mesofeltria} torrenticola Uchida, 1934
2. Kongsber gia Thor, 1899
Тип рода Kongsbergia materna Thor, 1899
Unionicolidae Oudemans, 1909
(рис. 238)
ДИАГНОЗ
Unionicolidae обычно имеют мягкую кожу, но иногда покровы
у них твердые, а у некоторых родов тело покрыто сравнительно
Слабым панцырем. Кожа обычно гладкая и не несет округлых сосоч-
Hydrachnellae
315
ков. Кожные железы часто конусовидные. Бедренный и коленный
членики пальп, как правило, не имеют отростков на внутренней
стороне. Коленные членики пальп часто несут на наружной стороне
длинные шипы, тогда как их голенные членики несут по внутреннему
краю дистальный шип. Коксальные щитки расположены четырьмя
группами, по два в каждой; они со-
единены также друг с другом и с брюш-
ным щитом. Передние коксальные
щитки снабжены более или менее длин-
ными, направленными назад подкож-
ными выростами, которые часто дости-
гают задних коксальных щитков. Задние
коксальные щитки (особенно IV), как
правило, прямоугольные и большие.
Ноги I (или ноги I и II) часто утол-
щены и нередко несут парные кинжа-
ловидные щетинки, сидящие на выро-
стах. Ноги обычно с плавательными
волосками. Гениталии мягкие, кожистые
и снабжены двумя генитальными щит-
ками. У некоторых родов самки имеют
четыре щитка и не меньше пяти или
шести пар, а часто и больше, половых
присосок (редко присоски лежат прямо
Рис. 238. Neumania curvipes
Lundblad (по Лундбладу, 1930).
Взрослая особь (иид снизу).
на покровах). У панцирных форм гениталии снабжены двумя (или
большим числом) слабо дифференцированными щитками и множеством
присосок.
РОДЫ И ПОДРОДЫ
U n i о n i col i n a e Koenike, 1909
1. Unlonicola Haldeman, 1842
a) Unlonicola s. str.
Типы подрода Acarus ypsilophorus Bonz, 1783
6) Atax Koch, 1842 {—Hexatax Thor, 1926)
Тип подрода Hydrachna crassipes Muller, 1776
в) Pentatax Thor, 1926
Тип подрода Atax bonzi Claparede, 1868
r) Polyatax Viets, 1933
Тип подрода Polyatax japonicus Viets, 1933
д) Unionlcolides Lundblad, 1937
Тип подрода Unlonicola (Unionlcolides) sica Lundblad, 1937
2. Atacella Lundblad, 1937
a) Atacella s. str.
Тип подрода Atacella clathrata Lundblad, 1937
316 Глава VI. Trombidiformes (продолжение)
б) Atacellides Lundblad, 1941
Тип подрода Atax rugosus Koenike, 1890
3. Polyatacides Lundblad, 1941
Тип подрода Unionlcola prominens Koenike, 1914
Encentridophorinae Viets, 1935
1. Encentridophorus Piersig, 1897
Тип рода Atax spinifer, 1893
Huitfeldtiinae Viets, 1924
1. Huitfeldtia Thor, 1898
Тип рода Huitfeldtia rectipes Thor, 1898
Najadicolinae Viets, 1935
1. Najadlcola Piers'g, 1897
Тип рода Atax ingens Koenike, 1895
Neumaniinae Viets, 1931
1. Neu mania Lebert, 1879
a) Neumania s. str.
Тип подрода Hydrachna spinipes Muller, 1776
6) Lemienia Koenike, 1910
Тип подрода Atax multiporus Daday, 1901
в) Tetraneumania Lundblad, 1930
Тип подрода Neumania (Tetraneumania) curvlpes Lundblad,
1930
2, Amazonella Lundblad, 1930
Тип рода Amazonella ribagai Lundblad, 1930
3. Ecpolus Koenike, 1898
a) Ecpolus s. str.
Тип подрода Ecpolus tuberatus Koenike, 1898
6) Subneumania Viets, 1930
Тип подрода Subneumania dura Viets, 1930
4. Koenikea Wolcott, 1900
a) Koenikea s. str.
Тип подрода Koenikea concava Wo’cott, 1900
6) Diplokoenikea Lundblad, 1936
Тип подрода Koenikea (Diplokoenikea) pectlnata Lundblad,
1936
в) Ecpolopsis Piersig, 1906
Тип подрода Ecpolopsis multiscutata Piersig, 1906
r) Koenikella Lundblad, 1936
Тип подрода Koenikea (Koenikella) crassipalpis Lundblad, 1936
Hydrachnellae
317
д) Monokoenikea Lundblad, 1941
Тип подрода Koenikea mellni Lundblad, 1930
e) Neokoenikea Lundblad, 1936
Тип подрода Koenikea {Neokoenikea') artnipes Lundblad, 1936
ж) Nyangalla Viets, 1935
Тип подрода Koenikea acanthophora Viets, 1935
з) Parakoenikea Lundblad, 1936
Тип подрода Koenikea {Parakoenikea) curvipalpis Lundblad,
1936
и) Pseudokoenikea Lundblad, 1941
Тип подрода Koenikea rutae Lundblad, 1930
к) Recifella Viets, 1935
Тип подрода Recifella laminipes Viets, 1935 .
л) Tanaognathella Lundblad, 1941
Тип подрода Koenikea marshallae Viets, 1930
m) lanaognathus Wolcott, 1900
Тип подрода Tanaognathus spinipes Wolcott, 1900
5. Leptopterotrichophorns Viets, 1914
Тип рода Leptopterotrichophorns verrucosus Viets, 1914
Pionatacinae Viets, 1916
1. Pionatax Viets, 1916
Тип рода Pionatax uncipes Viets, 1916
Po 111 c i p a 1 p i n a e Viets, 1925
1. Pollicipalpns Viets, 1924
Тип рода Pollicipalpns scutatns Viets, 1924
Schadeellinae Lundblad, 1938
1. Schadeella Lundblad, 1938
Тип рода Schadeella crassipalpis Lundblad, 1938
Pionidae Thor, 1900
(рис. 239, 240)
ДИАГНОЗ
Pionidae обычно имеют мягкие кожистые покровы, не несущие
сосочков; щитки развиты редко. Хелицеры не соединены друг с дру-
гом. На внутреннем крае голенного членика пальпы сидит маленькая
щетинка, а часто и вырост; на дистальном конце членика, на вну-
треннем его крае, имеется склеротизированный шпенек. Коксальные
щитки расположены четырьмя группами, по два в каждой; у самцов
коксальные щитки прикрывают друг друга или в различной степени
318
Глава VI. Trombidiformes (продолжение)
соединяются друг с другом. На заднем крае коксальных щитков IV
имеется по более или менее длинному подкожному отростку. Задний
вогнутый край коксальных щитков IV
образует границу генитальной вырезки.
Ноги обычно с плавательными волосками.
Коготки лапок имеют дополнительные
зубцы. Гениталии снабжены щитками и
шестью или большим числом присосок.
Присоски лишь редко расположены в коже.
Рис. 239. Tiphys tor г is var americanus
Marshall (по Маршаллу, 1937).
Hora IV самца.
Рис. 240. Piona interrupta
Marshall (по Маршаллу, 1937).
Самка (вид снизу).
У самцов щитки, несущие присоски, обычно сливаются друг с дру-
гом. В строении ног самцы имеют ряд вторичнополовых отличий.
РОДЫ И ПОДРОДЫ
Pioninae Wolcott, 1905
1. Piona Koch, 1836
a) Piona s. str.
Тип подрода Hydrachna fuscata Hermann, 1804
6) Disperslpiona Viets, 1926
Тип подрода Hydrachna clavicornis Muller, 1776
в) Tetrapiona Viets, 1926
Тип подрода Nesaea variabllis Koch, 1836
Foreliinae Viets, 1926
1. Foreilia Haller, 1882
Тип рода Hydrachna lillacca Muller, 1776
2. Pseudofeltria Soar, 1904
Тип рода Pseudofeltria scourfieldi Soar, 1904
Pionellinae Viets, 1937
1. Pionella Viets, 1937
Тип рода Pionella karamani Viets, 1937
Tiphysinae Vitzthum, 1942
1. Tiphys Koch, 1836
a) Tiphys s. str. (—Acercus Koch, 1842)
Тип подрода Tiphys sagulatus Koch, 1837
Hydrachnellae
Bld
6) Acercopsis Viets, 1926
Тип подрода Acercus pistillifer Koenike, 1908
в) Pionides Thor, 1901
Тип подрода Piona ensifera Koenike, 1895
2. Hydrochoreutes Koch, 1837
Тип рода Spio ephippiata Koch, 1836
3. Pionacercus Piersig, 1894
a) Pionacercus s. str.
Тип подрода Pionacercus leuckarti Piersig, 1894
6) Pionacercopsis Viets, 1936
Тип подрода Tiphys vatrax Koch, 1837
4. Pionopsis Piersig, 1894
Тип рода Hydrachna lutescens Hermann, 1804
5. Wettina Piersig, 1892
Тип рода Tiphys podagricus Koch, 1837 (= Wettina macroplica
Piersig, 1892)
Astacocrotonidae Thor, 1927
(рис. 241)
ДИАГНОЗ
Astacocrotonidae паразитируют на жабрах десятиногих раков. Кожа
мягкая, спинных щитков нет. Кожных пор, стигм и трахей, по всей
вероятности, нет, У самцов глаза
имеются, тогда как у половозрелых
самок глаз нет. Гнатосома лежит
между широко расставленными пе-
редними коксальными щитками и не
сливается с последними. Все кок-
сальные щитки каждой стороны
слиты друг с другом. На каждом из
коксальных щитков IV открывается
по крупной железе (см. рис. 241).
У самки пальпы приспособлены для
захватывания жаберных лепестков
хозяина; пальпы очень короткие и
толстые (толще, чем ноги). Пла-
вательных волосков на ногах нет.
Коготки лапок трехзубчатые. У са-
мок половое отверстие отодвинуто
к заднему концу тела, далеко на-
зад от коксальных щитков; на
каждой стороне полового отверстия
Рис. 241. Astacocroton molle Has-
well (по Гасвеллу, 1922).
Самка (вид снизу).
имеется по генитальному щитку с продольным рядом из четырех
половых присосок. У самцов половое отверстие лежит между двумя
320
Глава VI. Trombidiformes (продолжение)
группами коксальных щитков. Клещей этого семейства находили
также на жабрах ильной рыбы (Amia).
РОДЫ
1. Astacocroton Haswell, 1922
Тип рода Astacocroton molle Haswell, 1922
ДИАГНОЗ
Axonopsidae Viets, 1929
(рис. 242, 243)
Тело Axonopsidae покрыто пористым панцырем, разделенным
дорзальным швом на спинной и брюшной. Коксальные щитки объе-
динены в одну группу; коксальные щитки I сливаются друг с другом
Рис. 242. Axonopsella
spinigera Lundblad
(по Лундбладу, 1930).
Самка (вид сверху).
Рис. 243. Axonopsella
spinigera Lundblad
(по Лундбладу, 1930).
Самка (вид снизу).
посередине. На внутренней стороне голени пальп иногда имеются
выросты. Половых присосок обычно три или четыре пары. Плаватель-
ные волоски иногда имеются.
РОДЫ И ПОДРОДЫ
Axonopsinae Viets, 1929
1. Axonopsis Piersig, 1893
a) Axonopsis s. str.
Тип подрода Hydrachna complanata Muller, 1781
6) Hexaxonopsis Viets, 1926
Тип подрода Axonopsis violacea Viets, 1911
Hydrachnellae '
321
2. Axonopsalbia Viets, 1913
a) Axonopsalbia s. str.
Тип подрода Axonopsalbia curvlsetifera Viets, 1913
6) Sumatralbia Viets, 1929
Тип подрода Sumatralbia rugosa Viets, 1929
3. Axonopsella Lundblad, 1930
a) Axonopsella s. str.
Тип подрода Axonopsella spinigera Lundblad, 1930
6) Neoaxonopsella Lundblad, 1937
Тип подрода Axonopsella {Neoaxonopsella) trifida Lundblad,
1937
в) Paraxonopsella Lundblad, 1937
Тип подрода Axonopsella {Paraxonopsella) filunguis Lundblad,
1937
I. Barbaxona Viets, 1924
Тип рода Barbaxona barbata Viets, 1924
5. Brachypoda Lebert, 1879
a) Brachypoda s. str.
Тип подрода Hydrachna versicolor Muller (= Brachypoda
paradoxa Lebert, 1879)
6) Hemibrachypoda Viets, 1937
Тип подрода Brachypodella mutila Walter, 1928
в) Parabrachypoda Viets, 1929 (= Brachypodella Viets, 1926)
Тип подрода Brachypoda montii Maglio, 1924
6. Kalobrachypoda Viets, 1929
Тип рода Kalobrachypoda rhopalopoda Viets, 1929
7. Lethaxona Viets, 1932
Тип рода Lethaxona pygmaea Viets, 1932
8. Ljania Thor, 1898
Тип рода Ljania bipapillata Thor, 1898
9. Miraxona Lundblad, 1936
a) Miraxona s. str.
Тип подрода Miraxona clavipes Lundblad, 1936
6) Miraxonella Lundblad, 1936
Тип подрода Miraxona {Miraxonella) compllcata Lundblad,
1936
10. Miraxonides Lundblad, 1938
Тип рода Miraxonides alata Lundblad, 1938
11. Neoalbia Lundblad, 1936
a) Neoalbia s. str.
Тип подрода Neoalbia violacea Lundblad, 1936
6) Pentalbia Lundblad, 1937
Тип подрода Neoalbia {Pentalbia) walteri Lundblad, 1936
12. Neoaxona Lundblad, 1936
a) Neoaxona s. str.
Тип подрода Neoaxona oblonga Lundblad, 1936
21 Зак. 1785
322 Глава VL Trombidiformes (продолжение)
6) Lamellaxona Lundblad, 1937
Тип подрода Neoaxona (Lamellaxona) abtiormipes Lundblad,
1937
13. Neoaxonopsis Lundblad, 1938
Тип рода Neoaxonopsis odontogaster Lundblad, 1938
14. Neobrachypoda Koenike, 1914
Тип рода Axonopsis (?) ekmani Walter, 1911
15. Subtniraxona Lundblad, 1937
Тип рода Submiraxona crasslpes Lundblad, 1937
Albiinae Viets, 1925
1. AIbia Thon, 1899
2. Тип рода Albia stationis Thon, 1899
Parasitalbia Viets, 1935
Тип рода Parasitalbia sumatrensis Viets, 1935
3. Subalbia Viets, 1913
Тип рода Subalbia proceripalpis Viets, 1913
Aturinae Wolcott, 1905
1. Aturus Kramer, 1875
a) Aturus s. str.
Тип подрода Aturus scaber Kramer, 1875
6) Crinaturus Thor, 1930
Тип подрода Aturus mirabilis Piersig, 1897
в) Subaturus Viets, 1916
Тип подрода Subaturus sulcatus Viets, 1916
2. Aturides Lundblad, 1937
Тип рода Subaturus dentatus Lundblad, 1936
3. Hjartdalia Thor, 1901
Тип рода Hjartdalia runcinata Thor, 1901
4. Pseudokongsbergia Walter, 1935
Тип рода Pseudokongsbergia longipalpis Walter, 1935
F г о n t ip о d о p s i n a e Viets, 1931
1. Frontipodopsis Walter, 1919
Тип рода Frontipodopsis staheli Walter, 1919
Mideidae Viets, 1929
(рис. 244, 245)
ДИАГНОЗ
Mideidae имеют сильно выпуклое тело, одетое в панцирь, под-
разделенный дорзальным швом. Бедра и коленные членики пальп
массивные, голени пальп тонкие, узкие и более длинные, чем бедро-
Hydrachnellae
и голень, взятые вместе. Голени пальп не несут на внутренней ста-<
роне выростов. Коксальные щитки образуют одну группу; из них
передние (I) соединены по средней линии, задние (IV) имеют тре-
угольную форму. Ноги И, III и IV с плавательными волосками.
У самцов лапки ног III превращены в хватательный орган. Гениталь-
ная вырезка большая и достигает заднего края тазика ног I. Гени-
талии расположены между коксальными щитками IV. У самок гениталии,
Р и с. 244. Midea expansa Marshall
(по Маршаллу, 1940).
Брюшные шитки взрослой особ' •
Рис. 245. Midea expansa Marshal?
(по Маршаллу, 1940).
Взрослая особь (вид сверху).
снабжены серповидными щитками: каждый из них несет пять или
шесть пар присосок. У самцов генитальные щитки имеют треугольные^
подвижные склеротизированные крылья. ...
РОДЫ
1. Midea Bruzel, 1854
Тип рода Hydrachna orbiculata Mill er, 1776
Mideopsidae Thor, 1928
(рис. 246)
ДИАГНОЗ
Mideopsidae имеют более или менее уплощенное тело, одетое
н панцирь, подразделенный дорзальным швом. Спинной щит обычно
сплошной. Голенный членик пальпы короче бедренного и коленного,
взятых вместе; обычно на их внутреннем крае имеются отростки
или же на их дистальном конце сидят шипы или маленькие волоски.
Коксальные щитки в той или иной степени объединены в одну группу.
21*
324
Глава VI. Trombidiformes (продолжение)
Генитальная вырезка простирается до заднего края коксальных щит-
ков I. На ногах иногда имеются плавательные волоски. Гениталии,
Рис. 246. Mideopsella
forficicalpis Lundblad
(по Лундбладу, 1943).
Самка (вид сверху).
снабженные 3—6 парами половых присосок, обычно лежат в вырезке
между коксальными щитками IV.
<*ОДЫ И ПОДРОДЫ
Mideopsinae Koenike, 1910-
l. Mideopsis Neumann, 1880
a) Mideopsis s. str.
Тип подрода Hydrachna orbicularis Miiller, 1776 (-- Mideopsis
depressa Neumann, 1880)
6) Neoxystonotus Lundblad, 1927
Тип подрода Mideopsis torrei Marshall, 1927
в) Octomideopsis Viets, 1931
Тип подрода Mideopsis minutus Soar 1910
r) Paraxystonotus Lundblad, 1936
Тип подрода Xystonotus 'Willmanni Viets, 1920
д) Xystonotus Wolcott, 1900
Тип подрода Xystonotus asper Wolcott, 1900
2. Notomideopsis Wolcott, 1905
Тип рода Mideopsis spinipes NordenskiOld, 1904
Djeboinae Viets, 1935
i. Djeboa Viets, 1911
a) Djeboa s. str.
Тип подрода Djeboa multidentata Viets, 1911
6) Djeboella Viets, 1935
Тип подрода Djeboella elongata Viets, 1935
Hydrachnellae 32г-
Momoniinae Viets, 1926
1. Momonia Halbert, 1906 (= Kondia Sokolow, 1926)
Тип рода Momonia falcipalpis Halbert, 1906
2. Momoniella Viets, 1929
Тип рода Momoniella sumatrensis Viets, 1929
Mideopsellinae Lundblad, 1937
1. Mideopsella Lundblad, 1937
Тип рода Mideopsella forcipalpis Lundblad, 1937
Acalyptonotidae Walter, 1911
(рис. 247)
ДИАГНОЗ
Тело Acalyptonotidae не защищено панцырем со спинной стороны,
дорзального шва нет. Дистальная часть голенного членика пальпы
Рис. 247. Acalyptonotas
violaceus Walter
(по Уолтеру, 1911).
Самка (вид снизу).
расширена дорзовентрально, его внутренний край удлинен и совместно
с тарзальным члеником образует клешню. Передние коксальные щитку
не разделены срединным швом. Тазики задних ног образуют вырезку,
в которой расположены гениталии, снабженные многочисленными
присосками.
РОДЫ
1. Acalyptonotas Walter, 1911
Тип рода Acalyptonotas violaceus Walter, 1911
-326
Глава VL Trombidiformes (продолжение)
ДИАГНОЗ
A-Thienemanniidae Lundblad, 1930
(рис. 248)
Тело A-Thienemanniidae покрыто панцырем; дорзальный шов
имеется. Челюсти обычно образуют короткий рострум. Дистальная
часть голени пальп выдается вентрально, образуя с лапкой пальп
небольшую клешню. Коксальные щитки в известной степени сра-
стаются. Гениталии обычно располагаются в отверстии брюшного
Р и с. 248. Plaumannia
crenophila Lundblad
(по Лундбладу, 1936).
Самка (вид снизу).
панцыря. Как правило, половые лопасти покрыты щетинками и несут
многочисленные половые присоски.
<»оды
А-Т h i е n е m a n n i i n а е Viets, 1923
1. A-Thienemannla Viets, 1923
Тип рода A-Thienemannia schermeri Viets, 1923
Harpagоp a 1 pinae Viets, 1924
1. Harpagopalpus Viets, 1924
Тип рода Harpagopalpus octoporus Viets, 1924
Mundamellinae Viets, 1931
1. Mundamella Viets, 1913
Тип рода Mundamella germanica Viets, 1913
2. Hungarohydracarus Motas et Tanaschi, 1947
Тип рода Hungarohydracarus subterraneus Motas et Tanaschi,
1947
3. Sty go hydracar us Viets, 1931
Тип рода Stygohydracarus troglobius Viets, 1931
Hydrachnellae
327
Plaumanniinae Lundblad, 1936
1. PlaumanMa Lundblad, 1936
Тип рода Plau.ma.nnta arrenurlpalpis Lundblad, 1936
Eupatrellidae Viets, 1935
(рис. 249, 250)
ДИАГНОЗ
Eupatrellidae описаны по нимфальным фазам и в настоящей работе
диагноз семейства не приводится.
Рис. 249. Eupatrella
reticulata Walter
(по Уолтеру, 1935).
Пальпа нимфы.
Рис. 250. Eupatrella
reticulata Walter
(по Уолтеру, 1935).
Брюшные щитки нимфы.
РОДЫ
1. Eupatrella Walter, 1935
Тип рода Eupatrella reticulata Walter, 1935 (возможно, Hydry-
phantes incertus Koenike, 1893)
Arrenuridae Thor, 1900
(рис. 251)
ДИАГНОЗ
Arrenuridae покрыты панцырем, обычно подразделенным дорзаль-
ным швом. Пальпы вооружены клешней, образованной голенным и
тарзальным члениками. Коксальные щитки соединяются с брюшным
щитом. Гениталии расположены в отверстии брюшного щита. Половые
328
Глава VI. Trombidiformes (продолжение)
присоски многочисленные, мелкие и поровидные и частью распола-
гаются не только на генитальных щитках, но и за их пределами.
Рис. 251. Arrenurus valencius
Marshall (по Лундбладу, 1935).
Самой (вид сверху).
Самцы часто имеют придаток на задней части тела; кроме того
половые отличия обнаруживаются в строении коленного членика
ног IV.
РОДЫ И ПОДРОДЫ
1. Arrenurus Dugds, 1833
a) Arrenurus s. str. (= Petlolurus Thon, 1900)
Тип подрода Arrenurus viridis Duges, 1834
6) Megaluracarus Viets, 1911
Тип подрода Hydrachna globator Miiller, 1776
в) Micruracarus Viets, 1911
Тип подрода Arrhenurus forplcatus Neumann, 1880
r)' Mlcruracaropsis Viets, 1939
Тип подрода Mlcruracaropsis phytotelmaticola Viets, 1939»
д) Truncaturus Thor, 1900
Тип подрода Arrenurus knauthel Koenike, 1895
2. Africasia Viets, 1931
Тип рода Mundamella arrhenuripalpls Viets, 1913
3. Dadayella Koenike, 1907
Тип рода Arrhenurella minima Daday, 1905
4. Rhinophoracarus Viets, 1916
Тип подрода Ehinophoracarus praeacutus Viets, 1916
5. Thoracophoracarus Viets, 1914
Тип рода Thoracophoracarus arrhenuroides Viets, 1914
Hydrach-tieilae <321>
6. Wuria Viets, 1916
Тип рода Wuria falclseta Viets, 1916
7. Wuriella Viets, 1935
Тип рода Wuriella sumatrensis Viets, 1935
Krendowskiidae Lundblad, 1930
(рис. 252, 253)
ДИАГНОЗ
Тело Krendowskiidae покрыто панцырем, дорзальный шов имеется-
Парные боковые глаза лежат в капсулах. Кожа покрыта сосочками-
Рис. 252. Krendowskia
megalopsis Lundblad
(по Лундбладу, 1930).
Рис. 253. Krendowskia
megalopsis Lundblad
(по Лундбладу, 1930).
Самка (вид снизу).
Самка (вид сверху).
Имеется короткий рострум. Тарзальный членик пальпы образует
клешню. Коксальные щитки в известной степени сливаются. Гени-
тальные щитки неподвижны; половые присоски внутренние, число их.
варьирует от трех до восьми пар. Гениталии расположены в вырезке,,
образованной тазиками ног IV.
РОДЫ И ПОДРОДЫ
1. Krendowskia Piersig, 1895
a) Krendo'wskia s. str. (= Arrhenurella Ribaga, 1902)
Тип подрода Krendowskia latissima Piersig, 1895
6) Krendowskiella Viets, 1931
Тип подрода Krendowskia megalopsis Lundblad, 1930
в) Neokrendowskia Lundblad, 1941
Тип подрода Krendowskia (Neokrendowskia) quadripustulatac.
Lundblad, 1941
ЛЗО Глава VI. Trombidiformes (продолжение)
2. Allokrendo'wskia Lundblad, 1941
Тип рода Allokrendowskia dentipes Lundblad, 1941
3. Geayia Thor, 1897
a) Geayia s. str.
Тип подрода Geayia venezuelae Thor, 1897
6) Geayella Lundblad, 1936
Тип подрода Geayia (Geayella) catharinensis Lundblad, 1936
в) Geayidea Lundblad, 1941
Тип подрода Geayia coeruleocruciata Lundblad, 1936
r) Pirapama Viets, 1935
Тип подрода Pirapama schubarti Viets, 1935
-4. Roqueella Lundblad, 1930
a) Roqueella s. str.
Тип подрода Rogueella flabellifera Lundblad, 1930
6) Neoroqueella Lundblad, 1937
Тип подрода Roqueella (Neoroqueella) maculata Lundblad, 1937
rr) Pararoqueella Lundblad, 1937
Тип подрода Roqueella (Pararoqueella) striata Lundblad, 1937
Глава VII
ПОДОТРЯД SARCOPTIFORMES REUTER, 1909
Стигм нет, но иногда развита трахейная система, открывающаяся
через пористые поля, расположенные в различных частях тела. Та-
зики образуют аподемы, лежащие под кожей на брюшной стороне
тела. Хелицеры грызущего типа обычно вооружены сйльной клешней,
имеющей форму ножниц; изредка встречаются специализированные,
или видоизмененные, хелицеры. Пальпы простые. Псевдостигматиче-
ские органы иногда отсутствуют. Часто у самцов имеются анальные
присоски.
Sarcoptiformes разделяются на Acaridiae и Oribatei. К первым от-
носятся сырные, чесоточные и перьевые клещи, а ко вторым — пан-
цирные клещи. Обе эти группы, хотя и легко отличаются друг от
друга, тесно связаны между собой и обладают многими общими при-
знаками (сходная форма тела многих родов, грызущие ротовые
органы, положение коксальных щитков, наличие у личинок обеих
групп клапаредова органа).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ГРУПП SARCOPTIFORMES
1. Покровы мягкие, панцыря нет. Псевдостигм и крупных булаво-
видных псевдостигматических органов нет (исключение — Pedicu-
lochelidae). Присосок на лапках нет. Половой диморфизм иногда
хорошо выражен. Самцы часто с копулятивными присосками,
расположенными на лапках или в анальной области . . Acaridiae
— Покровы перепончатые или сильно склеротизированные. За ред-
кими исключениями имеются крупные булавовидные псевдостиг-
матические органы. Присосок на лапках нет. Половой диморфизм
не выражен.............................................Oribatei
ACARIDIAE LATREILLE, 1802
Гнатосома обычно заметна сверху, реже она втянута в камеро-
стом. Трахей и стигм нет. Псевдостигматические органы не развиты.
Тело, как правило, мягкое, кожистое, но на спинной поверхности
обычно имеется один или несколько щитков. Тазики образуют ха-
рактерные щитки. На лапках обычно имеются стебельчатые или си-
дячие эмподиальные коготки и присоска; если же коготков нет, то
332
Глава Vil. Подотряд Sarcoptiformes
присоска имеет чашевидную форму. Пальпы простые, обычно с двумя
свободными, реже — с четырьмя подвижными члениками. Иногда вы-
ражен половой диморфизм. Половых присосок обычно две пары, а
у самцов в большинстве случаев имеются крупные аданальные присоски.
По некоторым морфологическим признакам, как, например, по
степени склеротизации и бороздчатости покровов, Acaridiae разде-
ляют на свободноживущих (Acaridae, или сырные клещи), паразити-
ческих клещей (Sarcoptidae, или чесоточные клещи) и перьевых и
шерстьевых клещей. Следует отметить, что различия между этими,
группами, так же как и между группами других подотрядов, нельзя
считать четкими.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ ACARIDIAE
1. Щитков или нет совсем, или же имеется лишь слабо склероти-
зированный проподосомальный щиток (изредка имеется гистеро-
сомальный щиток). Ноги не имеют приспособлений для захваты-
вания волосков. Как правило, свободноживущие клещи или кож-
ные паразиты ................................................. 2
— На теле обычно больше одного хорошо развитого щитка, все
они сильно склеротизированы. Ноги (или максиллы) приспособ-
лены для захватывания волосков. Перьевые и шерстьевые клещи.
................................................................27
2. Гистеросома не сегментирована .............................. 3
— Гистеросома разделена поперечными швами на четыре отдела.
Проподосома несет пару пузыревидных сенсорных щетинок.
Лапки с присосками, но коготков нет . . • . Pediculochelidae
3. Хелицеры с клешней; ее пальцы, хотя иногда и маленькие, про-
тивопоставлены друг другу.................................• .4
— Неподвижный палец клешни хелицер редуцирован, подвижный
палец развит нормально .................................... 17
4. Присоски с коготками, иногда очень мелкими (Enslinielliidae и
Canestriniidae :)............................................. 5
— Присоски без коготков.........................................19
5. Присоски сидячие, с коготками.................'..............6
-— Присоски более или менее стебельчатые, на конце их имеются
коготки, часто очень мелкие..................................3
6. У самцов на лапках ног IV или близ анального щитка развиты
копулятивные присоски (исключение — Caloglyphus anomalus
Nesbitt).................................................... 7
— У самцов ни на лапках, ни близ анальных щитков нет копуля-
тивных присосок. Личинки без уростигмы. Половое отверстие
расположено между тазиками ног IV...................Saproglyphidae
1 По Фитцтуму. Canestriniidae стоят очень близко к Sarcoptidae. У не-
которых их представителей коготки кажутся исчезнувшими; на лапках,
всегда развиты присоски различного типа.
Acaridiae
333
7. У взрослых тело покрыто редкими длинными, бичевидными
щетинками. У неполовозрелых особей щетинки часто жесткие,
стержневидные..........................................Acaridae
— Тело покрыто перистыми, гребенчатыми и булавовидными щетин-
ками ............................................Forcelliniidae
8. Покровы гладкие, тонкие, не склеротизированные.............9
— Покровы склеротизированные; они кожистые, чешуйчатые и не
гладкие......................................................15
9. На лапках всех ног присоски занимают концевое положение . 10
— На когтевидных лапках ног I и II, несущих концевые коготки,
присоски занимают латеральное положение; присоски длинные,
цилиндрические, гибкие. Лапки ног III и IV нормальные . . .
................................................Hyadesidae
10. Проподосома и гистеросома разделены швом.................11
— Шва между проподосомой и гистеросомой нет. Проподосомаль-
ного щитка нет. Щетинки спинной стороны тела простые, по-
зади на спинке сидят две пары длинных щетинок. Эпимеры ног
I и II слиты и у самок частично прикрывают половое отверстие,
лежащее между ногами II и III....................Carpoglyphidae
11. Цервикальных щетинок нет. У самцов присоски на лапках ног
IV или около анального отверстия не развиты..................12
— Цервикальные щетинки имеются. У самцов на лапках ног IV или
около анального отверстия развиты присоски .............. 14
12. На месте цервикальных щетинок линзовидные глаза не раз-
виты ....................................................... 13
— На месте каждой из цервикальных щетинок развито по линзо-
видному глазу. Проподосомальный щиток отсутствует. Тело несет
четыре пары длинных бичевидных щетинок; из них две пары
сидят на заднем конце тела, одна пара — на плечевых выступах
гистеросомы и одна пара — в заднем ряду проподосомальных
щетинок. Половое отверстие расположено между тазиками ног III
и IV. Самцы неизвестны..............................Oulenziidae
13. Половое отверстие у самок расположено между тазиками ног III,
у самцов — между тазиками, ног IV. Щетинки на спинной сто-
роне тела простые, умеренной длины. Проподосомальный щиток
имеется..........................................Ensliniellidae
— Половое отверстие у самок расположено между тазиками ног III
и IV. Проподосомальный щиток развит. Щетинки на спинной
стороне тела, как правило, короткие. Спинная поверхность тела
покрыта очень тонкими морщинами.................Czenspinskiidae
— Половое отверстие у самок расположено между тазиками ног IV.
Проподосомальный щиток развит. Щетинки на спинной стороне
тела простые, умеренной длины...........Winferschmidtiidae
14. Цервикальные щетинки длинные опушенные и занимают краевое
положение. Половое отверстие у самок расположено между тази-
ками ног III, у самцов — между тазиками ног IV. Коготки лапок
336
Глава Vlf. Подотряд Sarcoptiformes
— Максиллы или одна (или несколько) пара ног превращены
в органы для захватывания волос хозяина . . . Listrophoridae
28. Ноги I и II нормальные; их членики цилиндрические и не несут
шиповидных выростов...............................................29
— Членики ног I и II несут латеровентральные выросты . . . . .
...................................................Analgesidae
29. Задний край тела у обоих полов округлый или слабо двулопаст-
ной .................................................Dermoglyphidae
— Задний край тела у самок (а часто и у самцов) сужен и отчет-
ливо двулопастной.................•...............Proctophyllodidae
ДИАГНОЗ
Pediculochelidae Lavoipierre 1946
(рис. 254)
Pediculochelidae — мелкие беловатые клещи. Длина тела варьи-
рует у них от 0,184 до 0,24 мм. Покровы мягкие, бороздчатые;
щитков нет. Гистеросома, отделенная от проподосомы отчетливым
Рис. 254. Pediculochelus raulti Lavoipierre.
Самка (вид сверху'.
швом, разделяется тремя дорзальными поперечными швами. Глаз на
проподосоме нет, но имеется несколько длинных бичевидных и пара
дорзальных булавовидных сенсорных щетинок; проподосома несет
также пару вертикальных щетинок. Хелицеры крупные, пальцы их
клешни крепкие, противопоставлены друг другу и, как и у панцир-
ных клещей, приспособлены для разжевывания или раздавливания.
Пальпы четырехчлениковые, простые и несут немногочисленные ще-
тинки. Ноги относительно слабые. Коготков на лапках нет, но
имеются стебельчатые присоски, по одной на каждой лапке. Анальное
отверстие находится в задней части тела. Половых присосок две пары.
РОДЫ
1. Pediculochelus Lavoipierre, 1946
Тип рода Pediculochelus raulti Lavoipierre, 1946
Acaridiae
337
ОБЩИЕ ЗАМЕЧАНИЯ
Единственный известный вид — Pediculochelus raulti Lavoipierre—
был описан Лавуапьером (1946) с пчел в Дурбане (Натал, Южная
Африка). Позднее один экземпляр этого клеща был обнаружен на
крысе во Флориде при обследовании эктопаразитов крыс и несколько
экземпляров было собрано с банкивского петуха (Gallus gallus) на
острове Самар (Филиппины). Тело сегментировано и по общему виду
соответствует примитивному акаридному типу; возможно, что Pedi-
culochelidae представляют собой промежуточные формы между Аса-
ridae и Oribatei. В таком случае допустима мысль об общем сход-
стве образа жизни Pediculochelidae и Acaridae. Pediculochelidae,
повидимому, не так редки, но из-за малых размеров часто остаются
незамеченными при сборах.
ЛИТЕРАТУРА
Lavoipierre М., 1946. A new acarine parasite of bees, Nature, 158 (4004),
130, 131.
ДИАГНОЗ
Saproglyphidae Oudemans, 1924
(рис. 255)
Saproglyphidae, возможно, не имеют проподосомального щитка.
Покровы гладкие. Проподосома отделена от гистеросомы швом. На
задней части проподосомы рас-
положен поперечный ряд из
четырех щетинок; внутренняя
пара их значительно короче на-
ружной. Проподосома несет
также пару вертикальных ще-
H1HOK. Краевые щетинки очень
длинные и бичевидные. Лапки
всех ног несут коготки и при-
соску. Половое отверстие у са-
мок лежит между тазиками ног
111 и IV, у самцов же — между
газиками ног IV. У самцов нет
пи аданальных копулятивных
присосок, ни копулятивных при-
> осок на лапках ног IV.
I' и с. 255. Saprogiyphus neglectus
Berlese (по Берлезе, 1890).
Самка (вид'сверху).
22 Зак. 1785
338
Глава VII. Подотряд Sarcoptiformes
РОДЫ
1. Saproglyphas Berlese, 1890
Тип рода Saproglyphus neglectus Berlese, 1890
ОБЩИЕ ЗАМЕЧАНИЯ
Saproglyphus neglectus Berlese собраны в Италии с грибов Poly-
porus hispidus. В 1941 г. в галлах Cytisus proliferus из Австралии
(Новый Южный Уэллс) был найден S. cocciphagus Womersley.
ЛИТЕРАТУРА
Berlese А., 1890. Acari, Myriopoda et Scorpiones Crypt., Fasc. LXXXIX,
№ 11; Fasc. LYII, № 6.
Womersley H., 1941. Studies in Australian acarina (2). Tyroglyphidae (s. 1.),
Rec. South Austral. Mus., YI, 4, 451—488.
ДИАГНОЗ1
Acaridae Ewing et Nesbitt, 1942
(== Tyroglyphidae Donnadieu, 1868)
(рис.' 256—262)
гистеросома разделены четко выраженной попе-
Тело толстое и окрашено в белые или светлоко-
ричневые ~
блестящие, редко шероховатые. Хелицеры клеш-
невидные. Проподосома несет пять пар щети-
нок: 1) ростральные щетинки; 2) орган Гран-
жана (Acarinae) или затылочные щетинки
(Rhlzoglyphinae); 3) цервикальные щетинки;
4) наружные проподосомальные и 5) внутрен-
ние проподосомальные щетинки. У родов Thy-
reophagus и Histiogaster отмечено уменьше
ние числа этих щетинок. Обычно передняя
часть проподосомы покрыта щитком. Всегда
имеется псевдостигматический орган. Половое
отверстие у обоих полов лежит между тази-
ками ног III и IV. У самцов по краям аналь-
ного отверстия сидят две отчетливо выражен-
ные копулятивные присоски. Эпимеры I сращен
с грудиной (стернальным щитком); все остал!
ные эпимеры лежат свободно. Тазики всех н<
можно видеть кла- (кроме IV) несут на вентральной поверхности
орган, или уро- шип. Лапки могут быть длиннее, чем голень
стигму. J _ т
вместе с коленным члеником. Лапки ног 1
следующее вооружение: 1) макросенсорную щетинку и
Протеросома и
речной бороздой.
Рис. 256. Rhizogly-
phus echinopus
(Fumouze et Robin).
Личинка (вид снизу); у тази-
ков ног I
народов
имеют
тона. Покровы обычно гладкие и
1 По Несбитту, 1945.
Рис. 257. Rhizoglyphus echinopus
(Fumouze et Robin).
Протонимфа (вид снизу).
Рис. 258. Rhizoglyphus echinopus
(Fumouze et Robin).
Гипопус (вид снизу).
Рис. 260. Rhizoglyphus echtnopus
(Fumouze et Robin).
Самец (вид снизу)».
P и с. 259. Rhizoglyphus echinopus
(Fumouze et Robin).
Дейтонимфа (вид снизу).
22*
340
Глава VII. Подотряд Sarcoptiformes
расположенную впереди ее микросенсорную щетинку, сидящие у осно-
вания лапки; 2) суббазальную щетинку и присутствующую иногда
дополнительную парасуббазальную щетинку; 3) группу из четырех
медиальных щетинок; 4) вентротерминальную группу, состоящую из
трех-пяти шипиков; 5) дорзотерминальную группу, состоящую из четырех
длинных, иногда видоизмененных щетинок и 6) коготок, окруженный
присоской, которая никогда не бывает стебельчатой. Лапки ног II
несут одну макросенсорную ще-
тинку, но никогда не имеют ни
микросенсорной, ни парасуббазаль-
ной щетинки. Лапки ног III у
Рис. 261. Rhizoglyphus echinopus
(Fumouze et Robin).
Гетероморфный самец (вид снизу}.
Рис. 262. Rhizoglyphus echinopus
(Fumouze et Robin).
Беременная самка (вид снизу).
гетероморфных самцов превращены в крупный коготок. Лапки ног IV
у самок снабжены двумя боковыми шипами; у самцов шипы заме-
щены двумя присосками (исключение — род Thyreophagus, имеющий
только одну присоску). У более примитивных представителей Aca-
ridae щетинки на лапках имеют вид тонких, нежных волосков; у более
высокоорганизованных форм они превращаются в крупные шипы.
Личинки имеют хорошо развитый цилиндрический или кеглевидный
личиночный орган — уростигму („Bruststiele").
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ ACARIDAE
,1. Половой диморфизм не выражен. Внутренние проподосомальные
щетинки по длине равны или превышают наружные. Ноги тонкие,
щетинки на них волосовидные; как наружные, так и внутренние
Acaridiae 341
щетинки внутренней поверхности лапок не видоизменены. Микро- и
макросенсорная щетинки не имеют общего основания , . . Acarinae
— Половой диморфизм обычно хорошо выражен. Внутренние про-
подосомальные щетинки короче наружных. Ноги крепкие, ще-
тинки на них во многих случаях превращены в толстые шипы.
Микро- и макросенсорная щетинки сидят на общем основании
................................................ Rhizoglyphinae
Несбитт (1950) включил Pontoppidanidae в семейство Acaridae и
подразделяет группу на подсемейства на основании следующих при-
знаков:
I. Затылочные щетинки тонкие, разветвленные и плотно прижаты
к наружной стенке тела..............................Acarinae
— Затылочные щетинки крупные, роговидные и свободно торчат на
теле..........................................Rhizoglyphinae
— Затылочные щетинки крупные, гребенчатые, свободно торчат на
теле и легко заметны сверху.................Pontoppidaniinae
РОДЫ И ПОДРОДЫ
Acarinae Nesbitt, 1945
1. Acarus Linnaeus, 1758 ( = Tyroglyphus Latreihe, 1796)
Тип рода Acarus siro Linnaeus, 1758
2. Aleuroglyphus Zakhvatkin, 1940
Тип рода Tyroglyphus ovatus Troupeau, 1879
3. Ebertia Oudemans, 1924
Тип рода Tyroglyphus australis Oudemans, 1917
4. Podoglyphus Oudemans, 1937
Тип рода Glyciphagus buski Murray, 1877
5. Tyrolichus Oudemans, 1924
Тип рода Tyrolichus casei Oudemans, 1910
(>. Tyrophagus Oudemans, 1924 ( = Coelognathus v. Heffling, 1852)
a) Tyrophagus s. str.
Тип подрода Acarus putrescentiae Schrank, 1781
6) Tyroborus Oudemans, 1924
Тип подрода Tyroborus lini Oudemans, 1924
в) Povelsenia Oudemans, 1924
Тип подрода Tyroglyphus neotropicus Oudemans, 1917
Rhizoglyphinae Zakhvatkin, 1941
I. Rhizoglyphus Claparede, 1869
Тип рода Tyroglyphus echinopus Fumouse et Robin, 1868
Caloglyphus1 Berlese, 1923 (? = Achropodophorus Rosas Costa,
1927)
1 Деление рода Caloglyphus на подроды приводится по Фитцтуму.
342
Глава VII. Подотряд Sarcoptiformes
a) Caloglyphus s. str.
Тип подрода Tyroglyphus berlesei Michael, 1903 (= Tyrogly-
phns mycophagus Berlese, 1891, но не Mdgnin, 1874)
6) Pachyglyphus Berlese, 1923
Тип подрода Tyroglyphus pergandis Berlese, 1920
в) Lagenoglyphus Berlese, 1923
Тип подрода Tyroglyphus lamermannl Berlese, 1923
r) Isoglyphus Zakhvatkin, 1937
Тип подрода Isoglyphus sphaerogaster Zakhvatkin, 1937
3. Ceroglyphus Vitzthum, 1919 (=Petzschia Oudemans, 1923)
Тип рода Ceroglyphus monstruosus Vitzthum, 1919
4. Eberhardia Oudemans, 1924 (== Acotyledon Oudemans, 1903)
a) Eberhardia s. str.
Тип подрода Eberhardia mlchaell Oudemans, 1924 (= Rhizo-
glyphus agilis Michael, 1903)
6) Cosmoglyphus Oudemans, 1932
Тип подрода Tyroglyphus kramerii Berlese, 1881
5. Froriepia Vitzthum, 1919 (дейтонимфа)
Тип рода Froriepia vimariensis Vitzthum, 1919
6. Garsaultia Oudemans, 1916 (дейтонимфа)
Тип рода Garsaultia testudo Oudemans, 1916
7. Histiogaster Berlese, 1883
Тип рода Tyroglyphus carpio Kramer, 1882
8. Megninietta Jacot, 1936
Тип рода Megninietta ulmi Jacot, 1936
9. Mycetoglyphus Oudemans, 1932
Тип рода Mycetoglyphus fungivorus Oudemans, 1932
10. Myrmoglyphus Vitzthum, 1935 (дейтонимфа)
Тип рода Myrmoglyphus bipilis Vitzthum, 1935
11. Sancassania Oudemans, 1916 (дейтонимфа)
Тип рода Sancassania chelone Oudemans, 1916
12. Sch'wlebea Oudemans, 1916
Тип рода Sch'wlebea talpa Oudemans, 1916
13. Stereoglyphus Berlese, 1923
Тип рода Stereoglyphus haemisphaericus Berlese, 1923
14. Suidasla1 Oudemans, 1905 (= Aphelenia Oudemans, 1923)
Тип рода Suidasla pontifica Oudemans, 1905
15. Thyreophagus Rondani, 1874 (= Monieziella Berlese, 1897)
Тип рода Acarus entomophagus Laboulb£ne, 1852
16. Tyroglyphopsis Vitzthum, 1926 (дейтонимфа)
Тип рода Tyroglyphopsis ocellata Vitzthum, 1926
1 Несбитт не включает этот род в семейство Acarldae, а приводит его
как добавление к системе. Мы здесь следуем системе Фитцтума, относящего
Suidasla к Rhizoglyphinae.
Acaridiae
343
17. Valmontia Oudemans, 1923
Тип рода Valmontia mlra Oudemans, 1923
18. Vledebanttta Oudemans, 1929 (дейтонимфа)
Тип рода Viedebanttia schmitzi Oudemans, 1929
Pontoppidaniinae Oudemans, 1925
1. Pontoppidanla Oudemans, 1923
Тип рода Tyroglyphus littoralis Halbert, 1920
2. Calvolia Oudemans, 1911
Тип рода Calvolia hagensis Oudemans, 1911 {—Tyroglyphus
heterocomus Michael, 1913, гипопус)
3. Diphtheroglyphus Nesbitt, 1950
Тип рода Diphtheroglyphus maculata Nesbitt, 1950
ОБЩИЕ ЗАМЕЧАНИЯ
Представители семейства Acaridae известны своим широким рас-
пространением. Их можно встретить в самых разнообразных местах —
от арктических тундр до тропических лесов, — повсюду, куда человек
заносил клещей вместе с пищевыми продуктами и различными пред-
метами. Acaridae питаются всевозможными органическими веществами;
они заражают различные съестные припасы, свежие и соленые шкуры,
семена, мучные продукты и т. п. В зернохранилищах они причиняют
огромные убытки, определяющиеся не столько количеством съедаемого
ими зерна (хотя численность этих клещей в зернохранилищах дости-
гает порой астрономических цифр), сколько изменением содержания
влаги в среде, что благоприятствует развитию плесени. В естествен-
ных условиях Acaridae живут в гниющих листьях, лесной подстилке,
древесной коре, испорченных луковицах и клубнях, в свежих и гни-
лых грибах, а также в гнездах птиц и норах млекопитающих, где
они, повидимому, питаются органическими остатками и перьями или
шерстью хозяев. Интересно отметить, что это семейство, подразделяю-
щееся таксономически на два четко разграниченные подсемейства на
основании морфологических признаков, разделяется на те же две
группы и по признаку местообитания. Представители одной группы
почти без исключения предпочитают среду с низким содержанием
влаги (20—30%); их можно найти в пшенице, семенах, мучных про-
дуктах. Для второй группы непременным условием существования
является высокая влажность среды; многие из ее представителей пред-
почитают столь насыщенные влагой места, что они нормально бывают
покрыты пленкой воды. Автор в результате проведенных им исследо-
ваний убедился в том, что клещи первой группы могут питаться
плотными органическими веществами, например зародышем и эндо-
спермом семян, тогда как клещи второй группы питаются почти исключи-
тельно грибами и плесенью, растущими на влажном субстрате. Многие
Acaridae едят мертвых и разлагающихся насекомых; к числу их
344
Глава VI/. Подотряд Sarcoptiformes
относятся, прежде всего, Caloglyphus. Среди Acaridae нет ни хищников,
ни паразитов; исключение составляют только несколько видов рода
Thyreophagus, паразитирующих на червецах и других насекомых.
Сообщений о нахождении ископаемых Acaridae или их непосредствен-
ных предшественников не имеется, что и неудивительно, так как
Acaridae благодаря своим малым размерам с трудом поддаются фосили-
зации С
Жизненный цикл клещей этого семейства мсйкно описать на при-
мере Rhizoglyphus echinopus (Fumouze et Robin). Джэрмэн (1937)
установил, что при комнатной температуре (15—23°, в среднем 20°)
клещ проходит весь жизненный цикл за 17—27 дней, а при 21—26°
этот срок сокращается до 9—13 дней. При понижении температуры
до 12—10° и повышении ее до 35° клещ впадает в оцепенение.
Непременным условием существования R. echinopus является очень
высокая влажность. По Джэрмэну, жизненный цикл R. echinopus скла-
дывается из следующих фаз 1 2:
А. Цикл без фазы гипопуса (вторая нимфальная фаза)
^половозрелая самка
Яйцо -> личинка -> нимфа Г-> нимфа Ш-> нормальный самец
^гетероморфный самец
Б. Цикл с фазой гипопуса (вторая нимфальная фаза)
Яйцо -»• личинка -> нимфа I -> гипопус ->
^половозрелая самка
-> нимфа III ->нормальный самец
^гетероморфный сам#ец
Многие Acaridae имеют фазу гипопуса, или „блуждающей нимфы".
Гипопус — крохотное существо, снабженное присосками или хвататель-
ными приспособлениями для прикрепления к насекомым, служащим
средством расселения. Гипопусов неоднократно принимали за парази-
тов. В фазе гипопуса клещи, повидимому, наиболее стойки к высыханию.
Гипопус одного вида был обнаружен в жаберной полости моллюска,
а гипопус другого-—в гонадах многоножки. Фаза гипопуса длится
5—13 дней, тогда как все остальные фазы превращения занимают
3—8 дней или менее. Из гипопусов выходят нимфы, линяющие в самок,
а также в нормальных и в гетероморфных самцов.
Майкл (1901) нашел, что у разных Acaridae гипопусы появляются
при самых различных условиях, но не смог установить причин их
появления. Джэрмэн (1939) показал, что у Rhizoglyphus echinopus
(Fumouze et Robin) гипопусов появляется больше при жизни в усло-
виях влажной вязкой среды. То же наблюдается и у видов Histio-
stoma и Caloglyphus.
1 Приведенные выше сведения заимствованы из работы Несбитта (1945).
2 После всех этих фаз, кроме фазы яйца и имаго, клещ впадает в состоя-
ние покоя.
Acaridiae
315
Гетероморфные самцы известны у многих Acaridae; гетероморфный
самец Rhizoglyphus echinopus изображен на рис. 261. У некоторых
видов гетероморфные самцы редки, но у R. echinopus они могут
составлять до 2О°/о всех самцов; их легко можно отличить от нор-
мальных по увеличенным ногам III. Гетероморфные самцы спариваются
с самками и дают потомство, состоящее из самцов обоих типов и
самок. Такое же смешанное потомство дают и нормальные самцы.
Хорошо известный луковый клещ Rhizoglyphus echinopus (Fumouze
et Robin) распространен повсюду и встречается в луковицах крокусов,
лилий, гладиолусов, гиацинтов, нарциссов, тюльпанов и некоторых
других растений. Пищей ему служат луковицы самых различных
растений, особенно те, у которых неплотно прилегающие чешуйки
облегчают доступ клещей внутрь. Собранные луковицы обычно бывают
заражены этим клешем, уничтожающим при хранении порой до 15—20°/о.
урожая. Надо, однако, заметить, что порча луковиц вызывается не
только клещами, но и многими другими факторами. В поле R. echi*
nopus не причиняет особого вреда здоровым растениям и проникает
только в уже поврежденные загнивающие луковицы; на складах же
этот клещ, по всей вероятности, способствует порче луковиц. R. echi-
nopus обычно питается гниющими или разложившимися тканями, но
может заражать и здоровые ткани при соприкосновении с ними.
Поэтому, чтобы предотвратить распространение клеща, испорченные
луковицы следует удалять из хранилища. Повидимому, клещи R. echi-
nopus переносят также грибковые и бактериальные заболевания С
Широко распространенный клещ Tyrophagus Untneri (Osborne)9
является опасным вредителем пищевых продуктов на складах, а также
культивируемых грибов. Этот клещ поедает споры и выгрызает корешки
и шляпки грибов, и если с ним не вести борьбы, то он может уничто-
жить весь урожай (Девис, 1944). Т. lintneri встречается в буфетах
с продуктами, на загрязненных стойках с газированной водой и т. д.
Thyreophagus entomophagus Laboulbene часто находят в коллек-
циях насекомых, а в полевых условиях — вместе со щитовками. Имеются
отдельные сообщения о нападении этого клеща на щитовок, но факти-
чески он питается сброшенной кожицей и отмершими чешуйками,
если они слегка влажны. Вообще Т. entomophagus живет на любых
сухих животных или растительных остатках. Этот клещ имеет космо-
политическое распространение.
1 Луковый клещ Rhizoglyphus hyacinth! может быть причиной раз-
вития своеобразной формы дерматита у лиц, разводящих лук и держащих
его в жилье под потолком (Е. Н. Павловский и А. К. Штейн, Экспери-
ментальное исследование над действием клеща Rhizoglyphus hyaclnthi,
вызывающего у человека своеобразную форму дерматита, Паразитологиче-
ский сборник Зоологического музея Академии наук СССР, I, 1930, стр.
19—26). — Прим. ред.
2 В последние годы на грибах, культивируемых в Мерилэнде и Пенсиль-
вании, этот вид был вытеснен Caloglyphus sp.
346
Глава VI/. Подотряд Sarcoptiformes
Acarus stro Linneaus также является серьезным вредителем, имею-
щим космополитическое распространение. Встречается этот клещ
в зерне и муке, в сыре, сухих фруктах и овощах. Он выедает зерно,
оставляя только шелуху. Самцы легко опознаются по большому
зубовидному выросту на бедрах ног I.
Tyrophagus longlor (Gervais) — хорошо известный европейский
вид, встречающийся иногда и в Америке. Майкл (1903) сообщает,
что однажды в Ирландии целый стог сена был буквально уничтожен
этими клещами. Иногда они размножаются в сене и фураже миллио-
нами; они заражают также' пищевые продукты и встречаются в чело-
веческих фекалиях. При наличии этих клещей в пищеварительном
тракте человек испытывает боли, тошноту; у него наблюдается рвота
и понос. Когда клещи выводятся наружу, то симптомы эти исчезают,
если только не происходит повторного заражения при новом приеме
пищи, зараженной клещами. Откладывание яиц и вылупление личинок
может совершаться в кишечнике, но сомнительно, чтобы там мог
завершиться полный жизненный цикл клеща. Хинман и Кампмейер
(1934) сообщили об описанном в литературе случае заражения этим
клещом мочеполовой системы, вероятно, через инфицированный ка-
тетер.
Ванденбрюль (1940) сообщает о том, что Tyrophagus dimldiatus
(Hermann) уничтожил в Бельгии 8О°/о урожая шпината в неутеплен-
ных теплицах. Пластинки внутренних листьев были короткие, смор-
щенные, деформированные, черенки же оставались нетронутыми.
В некоторых случаях эпидерма была усеяна мелкими дырочками,
окруженными корковой тканью. Пораженные листья бурели. Заболе-
вание было вызвано клещом, занесенным с навозом и размножившимся
в большом количестве. Имеются сведения о подобном же заболевании
шпината в восточных штатах США на полях, где были найдены те же
клещи.
Tyrophagus castellan.il (Hirst) вызывает так называемый „копровый
зуд“ у лиц, работающих с копрой. Сигрянский (1940) указывает
также, что Tliyrophagus castellan.il питается спорами Tilletia tritici
и переносит эти споры на теле и на щетинках на здоровую пшеницу.
Клещи, питающиеся луком, зараженным Botrytis allii, переносили
споры на здоровые луковицы. Эти клещи оказались также переносчи-
ками вирусного заболевания картофеля.
Гипопус клеща Caloglyphus julidicolus Lawrence был найден
в семенниках южноафриканской многоножки. Лоуренс (1939) пишет,
что клещ, вероятно, проникает через „узкое отверстие между вер-
шинами гоноподиев и окружающей их оболочкой и, прежде чем
достигнуть места окончательного прикрепления у основания гонопо-
диев, проходит вперед и вверх примерно на 5 мм. Клещи, повидимому,
выискивают наиболее укрытое место в половых органах хозяина11.
Хальберт (1920) приводит следующие данные о Pontoppidania
littoralis (Halbert): „Две самки и самец были найдены во влажных
Acaridiae
347
гниющих морских водорослях среди гальки близ гавани Хоутс (граф*-
ство Дублин). Это место расположено чуть выше метки высокой
воды и, вероятно, заливается при особо высоких приливах (сентябрь
1918 г.)“.
Гипопус Calvo Ila hagensts Oudemans отличается той особенностью,
что имеет пару больших линзовидных глаз у переднего конца тела.
Diphtheroglyphus maculata Nesbitt был собран с соленых бычьих
кож в Буэнос-Айресе (Аргентина).
ЛИТЕРАТУРА
Andre М„ 1931. Acariens nuisables aux produits pharmaceutiques, Assoc.
Fran?, pour 1’Avanc. des Sci, p. 395—399.
Andre M., 1935. Acariens infestant les milieux de culture dans les laboratoires,
Assoc. Franc, pour FAvanc. des Sci., p. 469—471.
Davis A. C., 1944. The mushroom mite (Tyrophagus llntneri (Osborne)) as
a pest of cultivated mushrooms, U. S. Dept. Agr. Tech. Bull., 879.
Ewing H. E., Nesbitt H. H. J., 1942. Some notes on the taxonomy of
grain mites (Acarina: Acaridae, formerly Tyroglyphidae), Proc. Biol.
Soc. Wash., 55,121—124.
Grandjean F., 1937. Sur quelques caracteres des Acaridiae libras, Bull. Soc.
Zool. Prance, 62 (6), 388—398.
Halbert J. N., 1920. The Acarina of the seashore, Proc. Roy. Irish Acad.,
35, Sect. B, № 7, 106—152, pls. 21—23.
H i n m a n E. H„ К am p me i er R. H., 1934. Intestinal Acariasis due to Tyro-
glyphus longior Gervais, Amer. J. Trop. Med., 14 (4), 355—362.
Hughes A. M., 1948. The mites associated with stored food products, Mini-
stry of Agriculture and Fisheries, London, p. 1—168.
Nesbitt H. H. J., 1945. A revision of the family Acariadae (Tyroglyphidae),
order Acari, based on comparative morphological studies, Canad. J. Res.,
D, 23, 139—188.
Nesbitt H. H. J., 1950. On a new Argentinian mite. Diphtheroglyphus macu-
lata n. sp., n. gen., and the taxonomic position of the family Pontoppi-
daniae Oudms, 1925, Can. Ent., 82 (10), 211—216.
Snyder W. C„ Hansen H. N„ 1946. Control of culture mites by cigarette
paper barriers, Mycologia, 38 (4), 455—462.
Solomon M. E., 1943. Tyroglyphid mites in stored products. I. A survey of
published information, Dept, of Scientific and Industrial Research, London,
p. 1—36.
Solomon M. E., 1944. Tyroglyphid mites in stored products. 1. A survey of
published information. Supplement, 1944, Dept, of Scientific and Industrial
Research, London, p. 1—7.
Solomon M. F., 1945. Tyroglyphid mites in stored products. Methods for
the study of population density, Ann. Appl. Biol., 32 (1), 71—75.
Solomon M. E., 1946. Tyroglyphid mites in stored products. Nature and
amount of damage to wheat, Ann. Appl. Biol., 33 (3), 280—289.
Solomon M. E.. 1946. Tyroglyphid mites in stored products. Ecological Stu-
dies, Ann. Appl. Biol., 33 (1), 82—97.
Van den Bruel M. E., 1940. Un ravageur de 1’epinard d’hiver: Tyrogly-
phus dlmldlatus Herm. (longior Gerv.), Bull. Inst. Agron. Gembloux, 9,
(1—4), 81—99.
Захваткин A. A., 1940. Определитель клещей, вредящих запасам сель-
скохозяйственных продуктов в СССР, Ученые записки Моск, госуд.
университета, № 42, 7—68.
348
Глава VII. Подотряд Sarcoptiformes
ДИАГНОЗ
Forcelliniidae Oudemans, 192.7
(рис. 263, 264)
Проподосомальный щиток имеется. Покровы глянцевые, гладкие, (?).
На задней части проподосомы имеется поперечный ряд из четырех
Рис. 263. Forcellinia wasmanni (Moniez)
(по Майклу, 1903).
Самка (вид сверху).
щетинок; внутренняя пара их несколько короче наружной, щетинки
тела обычно палочковидные или узкие, булавовидные и перистые.
Цервикальные щетинки расположены на спинной стороне; они пери-
стые, гребенчатые и лежат на уровне вертлугов ног I. Имеется также
пара вертикальных щетинок. У самок половое отверстие лежит между
тазиками ног Ш и IV, у самцов — между тазиками ног IV. У самцов
развиты аданальные копулятивные присоски и две копулятивные при-
соски на лапках ног IV. Forcellinidae не были найдены ни разу после
первоначального описания, и поэтому подробных данных об их строе-
нии не имеется.
Acaridiae
349
РОДЫ
1. Forcellinla Oudemans, 1924
Тип рода Tyroglyphus wastnanni Moniez, 1892
2. Scatoglyphas Berlese, 1913
Тип рода Scatoglyphus polytrematus Berlese, 1913
ОБЩИЕ ЗАМЕЧАНИЯ
Европейский вид Forcellinla 'wasmannl (Moniez) описан из мура-
вейников, где были найдены все фазы превращения этого клеща.
Рис. 264. Forcellinla wasmannl (Moniez)
(по Майклу, 1903).
Нимфа (вид сверху)»
Майкл (1903) сообщил, что иногда число этих клещей в муравейни-
ках очень велико и тогда их гипопусы прикрепляются к муравьям
А таких количествах, что муравьи погибают. На одном муравье можно
насчитать от 50 до 1000 экземпляров клещей; особенно много гипо-
пусов на голове, брюшке и ногах муравья, лишь редко они держатся
ин грудном отделе. Гипопусы, повидимому, располагаются вдоль про-
дольной оси того членика, к которому они прикрепляются, причем
передний конец гипопуса ориентирован в направлении дистального
конца членика.
350
Глава VII, Подотряд Sarcoptiformes
Scatoglyphus polytrematus Berlese был обнаружен в экскрементах
домашних птиц вместе с другими клещами.
ЛИТЕРАТУРА
Berlese А., 1913. Acari Ntiovi, Redia, 9, 77—105.
Michael A. D., 1903. British Tyroglyphidae, 2, 131—136.
Hyadesidae Halbert, 1915
(рис. 265, 266)
ДИАГНОЗ
Hyadesidae имеют толстое тело, сильно выпуклое со спинной сто-
роны. Проподосома отделена от гистеросомы швом. На проподосоме
сидит пара вертикальных щетинок, а иногда имеется также нежный
Р и с. 265. Hyadesia algivorans
(Michael) (по Майклу, 1901).
Самка (вид. сверху).
Рис. 266. Hyadesia sp.
А —нога I Н. algivorans (Michael) (по
Майклу, 1901); Б — нога I Н. fusee Loh-
man (по Андрэ, 1931).
проподосомальный щиток. В остальных частях тела покровы мягкие
и покрыты очень тонкими бороздками. Хелицеры обычные. У обоих
полов лапки ног I и II имеют форму большого когтя, с дополнитель-
ным коготком на внутренней стороне. У основания дополнительного
коготка на чрезвычайно длинном стебельке сидит маленькая присоска.
Присоска несет на дистальном конце крошечный настоящий коготок.
Лапки ног III и IV у самок имеют по одному большому коготку,
сидящему на коротком стебельке; у самцов большой коготок сидит
на длинном стебельке. Половой диморфизм слабо выражен. Как у самок,
так и у самцов половое отверстие лежит значительно позади тазиков
Acaridiae
351
йог IV. Половые присоски отсутствуют, у самцов нет также и ада-
нальных копулятивных присосок.
РОДЫ
1. Hyadesia M6gnin 1889 (= Lent ungula Michael, 1893)
Тип рода Hyadesia uncinifer Megnin, 1889
ОБЩИЕ ЗАМЕЧАНИЯ
Hyadesidae встречаются в приливной зоне на морских водорослях.
Майкл (1901) сообщил о нахождении большого числа Hyadesia algi-
vorans (Michael) в Корнуэлле у „края земли" на скоплениях зеленых
водорослей, в месте, „где пресная вода небольшого ручейка стекает
тонкой струйкой по гранитному утесу, смешиваясь с долетающими
туда брызгами морской воды". Майкл считает, что эти клещи не
плавают, но „ползают по водорослям и камням в. мелкой воде или
даже на суше в приливной зоне. Живут они также в местах, где
пресная вода, стекающая по скалам, смешивается с солеными брыз-
гами; там обычно растут зеленые водоросли. Вероятно, эти клещи
чувствуют себя вполне хорошо и под водой".
"Хальберт (1920) обнаружил в июне 1916 г. у Малахайда большое
число Hyadesia fusca (Lohmann) на скалах, по краям расщелин,
в зонах роста Pelvetla и Spiralis. В этой же местности в июле и
сентябре клещи довольно часто встречались в зоне роста оранжевого
лишайника в литоральных ваннах, содержащих много Enteromorpha.
Н. algivorans (Michael) впервые был описан с острова Клэр Айленд
(Великобритания), где в литоральных ваннах на кораллиновых водо-
рослях нашли большое число этих клещей. Ломан указывает, что
Н. fusca распространен в Северном и Балтийском морях.
С Огненной Земли описан Hyadesia uncinifer Megnin — клещ,
ведущий полуводный образ жизни. Всего известно четыре вида Hya-
desidae— Н. uncinifer Megnin, Н. algivorans (Michael), Н. fusca
(Lohmann) и H. kerguelenesis Lohmann.
ЛИТЕРАТУРА
Andre M., 1931. Snr le genre Hyadesia Megnin, 1889 (Sarcoptides hydro-
philes), Bull. Paris, Mas. d'Hist. Nat., Ser. 2, 3 (6), 496—506.
Halbert J. N., 1915. Acarinida: II. Terrestrial and Marine Acarina, Clare Is-
land Survey, 31 (39), Section II, 45—136, pls. 4—8.
Halbert J. N., 1920. The Acarina of the sea-snore, Proc. Roy. Irish Acad.,
35, Sect. B, 106—152.
Michael A. D., 1901. British Tyroglyphidae, 1, 200.
Carpoglyphidae Oudemans, 1923
(рис. 267)
ДИАГНОЗ
Carpoglyphidae имеют гладкие, но не глянцевые покровы. Они
иногда имеют проподосомальный щиток (Carро glyph us), в других же
случаях они покрыты панцырем, а специальный проподосомальный
352
Глава VII. Подотряд Sarcoptiformes
щиток отсутствует (Fertninia). Имеется пара вертикальных щетинок.
Проподосома и гистересома не разделены швом. Все лапки имеют
стебельчатые коготки и при-
соску. У самок половое отвер-
стие достигает впереди аподем
тазиков ног II, слитых друг
с другом по средней линии.
У самца половое отверстие ле-
жит между тазиками ног IV
(Carpoglyphus') или позади них
(Fermlnid). Самцы не имеют
аданальных копулятивных при-
сосок и копулятивных присосок
на лапках ног IV.
Рис. 267. Carpoglyphus I act is
(Linnaeus).
Самка (вид снизу).
РОДЫ
1. Carpoglyphus Robin, 1869
Тип рода Acarus lactis Linnaeus, 1758 (= Carpoglyphus anonymus
Haller, 1882 = C. passulorum Robin, 1869)
2. Fermlnia Oudemans, 1928
Тип рода Glycyphagus fuscus Oudemans, 1902
ОБЩИЕ! ЗАМЕЧАНИЯ
Широко распространенный вид Carpoglyphus lactis (Linnaeus)
встречается в сухих фруктах, молочных продуктах, глюкозе, гнию-
щем картофеле, муке и многих других пищевых продуктах. По
сообщениям из Парижа, этот клещ иногда размножается в большом
количестве в бутылках с вином; там клещи держатся на плавающих
кусочках пробки и питаются вином. Carpoglyphus altenus Banks,
1904 был найден в моче человека, страдавшего болезнью почек.
При каждом мочеиспускании выделялись клещи, окруженные гной-
ными отделениями. Местонахождение типа этого клеща неизвестно,
и поэтому точно определить его таксономическое положение нельзя,
Acaridiae
333
однако приведенный рисунок свидетельствует о том, что это под-
линный Carpoglyphus. По местообитаниям этот клещ сходен с Der-
matophagoldes takeuchii Sasa.
ЛИТЕРАТУРА
Andre M., 1931. Presence d’Acariens dans les vins sucres, Bull. Soc. Zool.
de Prance, 56, 335—340.
Banks N., 1904. An alleged parasitic Tyroglyphid, Proc. Ent. Soc. Wash.,
8 (1), 40—42.
Michael A. D., 1903. British Tyroglyphidac, 2.
Oudemans A. C., 1926. Krifisch historisch overzicht der Acarologie, I,
Tijdschr. v. Entom., 69, Suppl., p. 218, 225.
ДИАГНОЗ
Oulenziidae Oudemans, 1928
(рис. 268)
У Oulenziidae проподосомальный щиток
зирован, как у представителей некоторых
морщинистые. На задней части
проподосомы сидят четыре ще-
тинки, образующие поперечный
ряд; внутренняя пара щетинок
короче наружной. По краям
проподосомы на месте церви-
кальных щетинок находится по
линзовидному глазу. Имеется
пара вертикальных щетинок.
У самок половое отверстие рас-
положено между тазиками ног
ill и IV. Все лапки тонкие, со
стебельчатыми коготками и при-
соской, а также с маленькими
шипиками. Самцы неизвестны.
Рис. 268. Oulenzia arborlcola
(Oudemans).
Самка (вид сверху); на краях проподосомы
можно видеть глаза.
РОДЫ
не так сильно склероти-
других групп. Покровы
1928 nomen^prae-
1. Oulenzia Radford 1950 (= Lenzla Oudemans,
occupatum)
Тип рода Lenzla arborlcola Oudemans, 1928
ОК1ЦИЕ ЗАМЕЧАНИЯ
Единственный известный вид был описан с листьев Hevea на
Суматре; кроме того, отдельные особи Lenzla arborlcola собраны
23 Зак. 1785
354
Глава VII. Подотряд Sarcoptiformes
с джута в Индии. L. arborlcola, так же как и Czenspinskia, ве-
роятно, питаются растениями.
ЛИТЕРАТУРА
Oudemans А. С., 1928. Acarologische aanteekeningen XCI. Lenziidae, n.
fam., Ent. Ber., 17 (161), 327, 328.
Radford C. D., 1950. Systematic check list of mite genera and type species,
Union Internal, des. Sci. Biol., Ser. G (Sec. Ent.), 1, 152.
Ensliniellidae Vitzthum, 1924
(рис. 269—271)
ДИАГНОЗ
Ensliniellidae имеют проподосомальный щиток и гладкие, глян-
цевые покровы. На заднем конце проподосомы сидят четыре щетинки,
образующие поперечный ряд; внутренняя пара их по длине либо
равна наружной {Ensliniella), либо
гораздо короче последней (Riemiad.
Цервикальные щетинки отсут-
ствуют, пара вертикальных
имеется. На каждой из лапок
имеются стебельчатые коготки и
присоски. У самок половое отвер-
стие лежит между тазиками ног
III и IV, у самцов же оно поме-
щается между тазиками ног IV.
Аданальных копулятивных присо-
сок и копулятивных присосок на
лапках ног IV у самцов нет.
Рис. 269. Ens’iniella parasitica
Vitzthum (по Фитцтуму, 1925).
Самка (вид снизу).
РОДЫ
1. Ensliniella Vitzthum, 1925
Тип рода Ensliniella parasitica Vitzthum, 1925
2. Horstia Oudemans, 1905
Тип рода Trichotarsus ornatus Oudemans, 1899
3. Mantldoglyphus V’tzthum, 1940
Тип рода Mantldoglyphus anastati Vitzthum, 1940
.4. Riemla Oudemans, 1925
L < Тип рода Riemla hesperidum Oudemans, 1925
Acaridiae
355
5. Tortonia Oudemans, 1911
Тип рода Trichotarsus intermedius Oudemans, 1901
6. Vidia Oudemans, 1905
Тип рода Vidia undulata Oudemans, 1905
ОГ.1ЦИЕ ЗАМЕЧАНИЯ
Ensliniellidae мало изучены. Клещи Ensliniella parasitica Vitztftum
были собраны в Германии с покоющихся личинок (Ruhelarve) осы
Рис. 270. Ensliniella parasitica
Viizthum (по Фитцтуму, 1925).
•{ Самец (вид снизу).-
Рис. 271. Ensliniella parasitica
Vitzthum (по Фитцтуму, 1925). |
Гипопус (вид снизу).
Odynerus (Lionotus} delphinalis Giraud (сем. Vespidae), а в Италии
Vidia undulata Oudemans нашли на Prosopis conformis Foerst (сем-
Colletidae, Hymenoptera).
ЛИТЕРАТУРА
Vitzthum H., 1925. Eine neue Milbengattung und-art ale Parasit von Odyne-
rus (Llonotus) delphinalis Giraud 1866, Deut. Ent. Ztschr., 4, 289—305-
ДИАГНОЗ
Czenspinskiidae Oudemans, 1927
(рис. 272)
Czenspinskiidae — клещи с проподосомальный щитком и . глянце-
вой кожей, покрытой тонкими бороздками. На заднем конце пропо-
досомы сидят четыре щетинки, образующие поперечный ряд; вну-
тренняя пара щетинок гораздо короче наружной. Цервикальных
23*
356
Глава VII. Подотряд Sarcoptiformes
щетинок нет, но имеется пара вертикальных щетинок. Лапки всех ног
покрыты маленькими шипами; каждая из лапок несет стебельчатые
коготки и длинную тонкую присоску. У самок половое отверстие
лежит между тазиками ног III и IV. Самцы неизвестны.
Рис. 272. Czenspinskia
lordi Nesbitt
(по Несбитту, 1946).
Самка (вид снизу)»
РОДЫ
1. Czenspinskia Oudemans, 1927 (—Donndorfla Oudemans, 1931)
Тип рода Tyroglyphus heterocomus Michael, 1903 (имаго)
ОБЩИЕ ЗАМЕЧАНИЯ
Czenspinskia heterocomus (Michael) были собраны с дубовых
деревьев в Гэмпшире (Англия); кроме того, в большом числе их со-
бирают во мху летних гнезд белок. Майкл (1903) разводил этих кле-
щей в большом количестве, выкармливая их кусочками грибов. Другой
вид — Czenspinskia lordi Nesbitt, 1946 — был обнаружен на яблонях
в Новой Шотландии. Колонии С. lordi держатся у срединной жилки
листьев. Питаются эти клещи растительными тенями. Зимуют они,
повидимому, под лишайниками и под сброшенными шкурками щито-
вок Lepidosaphes ulmi. Самцы не обнаружены; повидимому, С. lordi
размножается партеногенетически.
ЛИТЕРАТУРА
Michael A. D., 1903. British Tyroglyphidae, 2, 106—109, pl. 33.
Nesbitt H. H., 1946. Three new mites from Nova Scotian apple trees, Canad.
Ent., 78, 15—22.
Acaridiae
3SI
Winterschmidtiidae Oudemans, 1923
ДИАГНОЗ
(рис. 273, 274)
Winterschmidtiidae — клещи с проподосомальным щитком и глян-
цевыми покровами. Задний конец проподосомы несет поперечный
ряд из четырех щетинок одинаковой длины. Цервикальные щетинки
отсутствуют. Имеется пара вертикальных щетинок. Ноги короткие и
сильные.. Все лапки со стебельчатыми коготками и присосками.
Рис. 273. Wlnterschmidtia
hamadryas (Vitzthum)
(по Фитцтуму, 1923).
Самка (вид снизу).
Рис. 274. Wlnterschmidtia
hamad-yas (Vitzthum)
(по Фитцтуму, 1923).
Самка (вид сверху).
Лапки снабжены одним вентродистальным и двумя терминальными
шинами; терминальные шипы очень сильные, особенно на лапках ног
I и И. Как у самцов, так и у самок половое отверстие расположено
между тазиками ног IV. У самцов нет аданальных присосок и копу-
1ИГИВНЫХ присосок на лапках ног IV, но на лапках ног I имеется
но одной латеровентральной копулятивной присоске.
1'<»Д1>1
I. Wlnterschmidtia Oudemans, 1923
Гии рода Suidasia (?) hamadryas Vitzthum, 1923
n.iUHi; замечания
Wlntersi hmldlla hamadryas (Vitzthum) был обнаружен в ходах
/ Htoplugaster rugulosus Ratz. в терносливе Prunus Itisiiiila (Германия).
358
Глава V,r. Подотряд Sarcoptiformes
W'. crassisetosa WiEmann были сняты с личинок и куколок
жука Phloeotribas scarabaeoid.es (Италия). Эго наводит на мысль,
что в ходах жуков в коре могут быть найдены и другие виды и
роды этого семейства. До сих пор изучению клещей, живущих
в подобных местах, уделялось мало внимания.
ЛИТЕРАТУРА
Vitzthum Н., 1923. Acarologische Beobachtungen 7. Reihe, Arch. f. Natur-
gesch., Abt. A, Hft. 2, 97—181.
Willmann C., 1939. Wingrschmldtia crassise’osa spec. nov. (Winter-
schmHtiidae, Aca~i), Boll. Laj. Zool. generate e Agrarla della P'acolta
Agr ar la in Po-tici, 31, 65—68.
ДИАГНОЗ
Lardoglyphidae
покровы. Щетинки тела
Lardoglyphidae Oudemans, 1927
(рис. 275, 276)
имеют
проподосомальный
гладкие; есть
щиток и глянцевые
пара вертикальных щетинок.
Рис. 275. Lardog'yphtis zacheri
Oudemans (no Удемансу,
не опубликовано).
А —самец (вид снизу); Б —лапка ноги I.
Рис. 276. La-dog'yphas zacheri
Oudemans (по Удемансу,
не опубликовано).
А—самка (вид снизу); Б —лапка ноги I.
Цервикальные щетинки перистые. Лапки несут стебельчатые коготки
и присоски. У неполовозрелых фаз коготки нормальные, у самцов
же на лапках ног I и II, а у самок на лапках всех ног коготки
Acaridiae
359
Y-образные, вильчатые. Лапки ног III у самцов вильчатые, но не
имеют коготков. У самок половое отверстие расположено между
тазиками ног III, а у самцов — между тазиками ног IV. Самцы
имеют аданальные копулятивные присоски и две такие же присоски
на лапках ног IV.
РОДЫ
1. Lardoglyphus Oudemans, 1927
Тип рода Lardoglyphus zacheri Oudemans, 1927
ОБЩИЕ ЗАМЕЧАНИЯ
Данных по этой группе опубликовано очень мало (Удеманс, 1927).
Все же их достаточно для того, чтобы этого клеща можно было
включить в определительную таблицу.
ЛИТЕРАТУРА
Oudemans А. С., 1927. Acarologlsche aanteekeningen LXXXVI, Ent. Вег.,
7, 242—248.
Olafseniidae Oudemans, 1927
(рис. 277)
ДИАГНОЗ
Семейство известно только по гомеоморфным нимфам. По Фит-
цтуму, они не имеют проподосомального щитка, покровы у них гладкие.
Задний конец проподосомы несет пару очень длинных щетинок. По
паре длинных щетинок сидит также
на плечах и на заднем конце ги-
стеросомы; кроме того, имеется
пара вертикальных щетинок и сла-
бые краевые цервикальные ще-
тинки, по форме сходные с изогну-
тыми шипиками. На всех лапках
имеются ланцетовидные или ло-
жечковидные „клейкие" щетинки.
Лапки с нерасчлененными когот-
ками и присосками. На вентраль-
ной стороне коготков имеется
шишковидное утолщение. Эпи-
меры I имеют форму Y.
Рис. 277. Olafsenia'trifoliam
(Oudemans) (по Удемансу, 1901).
Нимфа (вид сверху).
РОДЫ
1. OlafsetiiaTOudemans, 1924
Тип рода -Tyroglyphus trifolium Oudemans, 1901
360
Глава VF. Пос отряд Sarcoptiformes
ОБЩИЕ ЗАМЕЧАНИЯ
Единственный вид Olafsenia trifolium (Oudemans) известен по
нимфам, собранным с Heliocopris bucephalus на Яве.
ЛИТЕРАТУРА
Oudemans А. С„ 1901. Notes on Acari. Third series, Tijd. Dierkundige
*Ver„ Ser. 2, 50—88.
Chortoglyphidae Berlese, 1897
(рис. 278—280)
ДИАГНОЗ
Тело овальное. Шва между проподосомой и гистеросомой нет.
Покровы жесткие, гладкие и глянцевые. Спинные щетинки гладкие
РОДЫ
и короткие (за исключением вну-
тренних лопаточных щетинок у
Fusacarus). Имеется пара верти-
кальных щетинок. Хелицеры нор-
мальные, ножницеобразные или
клешневидные. На лапках всех ног
имеются' присоски и коготки, по-
следние сидят в передней или
дистальной части присосок. У са-
мок половое отверстие лежит
между тазиками ног 111 и IV,
а у самцов — между тазиками
ног [ и II.
Рис. 278. Chortoglyphus arcuatus
(Troupeau) (по Захваткину, 1940).
Самка (вид сверху).
1. Chortoglyphus Berlese, 1884
Тип рода Tyroglyphus arcuatus Troupeau, 1879 (== Chortogly-
phus nudus Berlese, 1884)
2. Fusacarus Michael, 1903
Тип рода Fusacarus latninipes Michae', 1903
ОБЩИЕ ЗАМЕЧАНИЯ
По данным Майкла (1903), клещи Chortoglyphus arcuatus (Trou-
peau) были найдены: в Англии — на полу и в балках старого амбара
и в старой конюшне; в Италии — в конюшнях и сене; во Франции-—
Acaridiae
361
на экскрементах кролика, а также в муке; в США (Массачузетс)—
н мусоре на грузовых судах и в курятниках. Клещи Fusacarus:
Рис. 279. Chortoglyphus
arcuatus (Troupeau)
(по Захваткину, 1940).
Рис. 280. Chortoglyphus
arcuatus (Troupeau)
(по Захваткину, 1940).
Самец (вид снизу).
Самка (вид снизу).
Jaminipes Michael были собраны Майклом в Англии из гнезд кротов
(но не на самих кротах).
ЛИТЕРАТУРА
Michael A. D., 1903. British Tyroglyphidae, 2, 1—12, pls. XX, XXI.
Захваткин А. А., 1940. Определитель клещей, вредящих запасам сельско-
хозяйственных продуктов в СССР, Ученые записки Моск, сосуд, уни-
верситета, № 42, 7—68.
Glycyphagidae Berlese, 1887
(рис. 281—284)
ДИАГНОЗ
Glycyphagidae имеют довольно широкое тело, обычно неразделенное
швом между проподосомой и гистеросомой. Имеется пара вертикальных
щетинок. Покровы либо гладкие (но не глянцевые), либо в ероховатые,
тонко гранулированные или покрытые точечным орнаментом. Спинные
щетинки обычно опушенные, перистые или веерообразные. Хелицеры
нормальные, ножницеобразные. На всех ногах лапки постепенно сужи-
ваются в дистальном направлении и снабжены присосками. На перед-
ней или дистальной части присосок сидит тонкий коготок, а иногда;
очень большой эмподиальный коготь.
Я>ию. ”281. 'Glycyphagus genlculatus
Vitzthum (по Фитцтуму, 1931).
Самка (вид сверху).
iP и с. 283. Pusoherleia incedbHis
Vitzthum (по Фитцтуму, 1931).
Гипопус (вид снизу).
Рис. 282. Glycyphagus
domesticus (DeGeer) (по
Хьюджесу и Хьюджес,
1938).
Гипопус (вид сверху).
Рис. 284. Pusohericia incredibilb
Vitzthum (по Фитцтуму, 1931).
Самка (вид сверху).
Acarldiae
363
РОДЫ И ПОДРОДЫ
1. Glycyphagus Hering, 1838
a) Glycyphagus s. str.
Тип подрода Glycyphagus prunorum Hering, 1838
6) Oudemanslum Zakhvatkin, 1936
Тип подрода Acarus domesticus DeGeer, 1771
2. Blomia Oudemans, 1928
Тип рода Glycyphagus tjibodas Oudemans, 1910
3. Cerophagus Oudemans, 1902
Тип рода Cerophagus bomborum Oi demans, 1932
4. Chaetodactylus Rondani, 1866 (= Trichotarsus Canestrini, 1888)
Тип рода Trlchodactylus osrnlae Dufour, 1839
-5. Cometacarus Zakhvatkin, 1936
Тип рода Cometacarus smirnovi Zakhvatkin, 1936 (дейтонимфа)
6. Crastidoglyphus Oudemans, 1937
Тип рода Acarus hyalinus Koch, 1841
7. Ctenoglyphus Berlese, 1928
Тип рода Glycyphagus canestrinli Armanelli, 1887
( — Acarus plumiger Koch, 1835)
8. Dermacar us Haller, 1878
Тип рода Homopus sclurlnus Koch, 1842
9. Fusoherlcia Vitzthum, 1931
Тип рода Fusoherlcia tncredibilis Vitzthum, 1931
10. Gohiera Oudemans, 1938
Тип рода Glycyphagus fuscus Oudemans, 1903
11. Hericla Canestrini, 1888
Тип рода Glycyphagus hericius Robin, 1868
12. Labidophorus Kramer, 1877
Тип рода Labidophorus talpae Kramer, 1877
13. Lepidoglyphus Zakhvatkin, 1936
Тип рода Acarus destructor Schrank, 1781 (—A. cadaverum
Schrank, 1781)
14. Melisla Lombardini, 1944
Тип рода Melisla mellsll Lombardini, 1944
15. Sennertla Oudemans, 1905
Тип рода Pedlculus cerambycinus Scopoli, 1763
16. Stroemla Oudemans, 1923
Тип рода Dermacarus cantharoblus Oudemans, 1905
О1ИЦИЕ ЗАМЕЧАНИЯ
Род Glycyphagus известен лучше остальных. О. domesticus (DeGeer)
исгречается в сухих фруктах и в таких материалах органического
происхождения, как кожа и перья. Клещи этого вида часто размно-
жаются в огромном количестве в домах и в складских помещениях.
<1. domesticus является возбудителем „чесотки продавцов бакалейных
1онаров“, которой болеют лица, имеющие дело с зараженными товарами.
364
Глава VII. Подотряд Sarcoptlforn es
Lepidoglyphus destructor (Schrank), известный в литературе как Gly-
cyphagus destructor, или G. cadaverum, поражает семена в зерно-
хранилищах, понижая их всхожесть. Семена итальянского райграсса
и лугового мятлика весьма восприимчивы к поражению этим клещом,
между тем как у других растений клещ поражает только семена с уже
поврежденным ядром. Прескотт (1933) пишет, что основной пищей
клещей является битое зерно, полова и другой инертный материал, и
только в отдельных случаях клещи поражают хорошие, всхожие се-
мена и причиняют им значительный ущерб. Жуайе и Бэр (1945) счи-
тают Glycyphagus domesticus (DeGeer) промежуточным хозяином лен-
точного червя Catenotaenia pusilia (Goeze), паразитирующего у грызунов.
Эти данные весьма интересны и заставляют вспомнить о роли, которую
родственные панцирные клещи играют по отношению к ленточному
червю Moniezia expansa (Rudolphi), паразитирующему у овец.
Хьюджес и Хьюджес (1938) опубликовали работу по анатомии и
постэмбриональному развитию Glycyphagus domesticus (DeGeer). Жиз-
ненный цикл этого клеща включает обычные фазы — яйцо, личинку и
протонимфу. Протонимфа либо впадает в состояние покоя, превращаясь
затем в активную дейтонимфу, либо превращается в гипопуса, несущего
на себе линную шкурку протонимфы. Гипопус может жить в течение
шести месяцев. Гипопусы никогда не питаются. Майкл (1901) сооб-
щает, что гипопус G. spinipes (Koch) вполне сформирован и способен
двигать конечностями, хотя и не способен передвигаться. У Acaridae же
гипопусы активны и обычно прикрепляются к насекомым или к круп-
ным клещам, расселяясь с ними. Гипопус превращается в активную
дейтонимфу, очень похожую на взрослую особь. Дейтонимфа превра-
щается в имаго после короткого периода покоя.
ЛИТЕРАТУРА
Andre И., 1941. Invasions de Glycyphages (Acarines), Bull. Soc. Zool. de
France, 66, 142—148.
Hughes T. E., Hughes A. M., 1939 (1938). The internal anatomy and
post-ejnbryonic development of Glycyphagus domesticus (DeGeer), Proc.
Zool. Soc. London, Ser. B, 108 (4), 714—733.
Joyeux Ch., Baer G., 1945. Morphologie, evolution et position systemati-
qtie de Catenotaenia pusilia (Goeze, 1782), Cestode parasite de Ron-
geurs, Pev. Suisse de Zool., 52 (2), 13—51.
Presscott R. T. M., 1933. Mites in seeds, J. Victoria Dept. Agr. (10),
519—523.
Canestriniidae Berlese, 1S84
ДИАГНОЗ
(рис. 285)
Тело уплощенное, округлое или овальное, пятиугольное или ром-
бическое; его длина лишь слегка превышает ширину. Покровы либо
кожистые, либо чешуйчатые. Имеется пара вертикальных щетинок.
Хелицеры обычные ножницеобразные. Коготки лапок часто малы и
Acaridiae
365
полностью погружены в стебельчатые присоски. Обонятельных и па-
лочковидных сенсорных щетинок на лапках ног I или II нет. Половых
присосок две пары. У самцов иногда имеются аданальные копулятив-
ные присоски.
Рис. 285. Grandlella
escaudaia Lombardini
(по Ломбардини, 1938).
Самка (вид сверху).
РОДЫ
1. Canestrinia Berlese, 1881
Тип рода Canestrinia dorcicola Berlese, 1881
Acrotacarus Banks, 1915
Тип рода Acrotacarus mirabilis Banks, 1915
3. Amansia Oudemans, 1937
Тип рода Dermaleichus chrysomelinus Koch, 1841
4. Catiestriniella Berlese, 1910
Тип рода Canestriniella amplexans Berlese, 1910
5. Caraboecius Cooreman, 1950
Тип рода Caraboecius coriacei Cooreman, 1950
(i. Coleoglyphus Berlese, 1910
Тип рода Coleoglyphus fuscipes Berlese, 1910
7. Coleopterophagus Berlese, 1882
Тип рода Dermoglyphus megninii Berlese, 1881
.8, Dicanestrinia Berlese, 1911
Тип рода Dermaleichus cerambycis G. Canestrini, 1878
366
Глава VIT. Подотряд Sarcoptiformes
9. Grandlella Lombardini, 1938
Тип рода Grandlella tetracaudata Lombardini, 1938
10. Megacanestrinia Tragardh, 1906
Тип рода Megacanestrinia mucronata Tragardh, 1906
11. Paramansia Cooreman, 1950
Тип рода Paramansia menthastri Cooreman, 1950
12. Percanestrinia Berlese, 1911
Тип рода Alloptes blaptis Canestrini et Berlese, 1880
13. Photia Oudemans, 1904 ( = Eucanestrinia Berlese, 1911)
Тип рода Canestrinia pro rustidis Berlese, 1881
14. Pr >cericola Cooreman, 1950
Тип рода ProcerEola ichthyoides Cooreman, 1950
15. Pseudamansia Cooreman, 1950
Тип рода Dermaleichus chrysomelinus Koch, 1841
16. Rosensteinia Oudemans, 1923
Тип рода Rosensteinia sieversi Oudemans, 1923
ОБЩИЕ ЗАМЕЧАНИЯ
Клещи Canestrinia dorcicola Berlese собраны в Италии с жуков..
Dorcus parallelepipedus (L.) (сем. Lucanidae). Клещи Coleopterophagus
megninii (Berlese, собраны из-под накрылий различных видов Cetonia
в Италии. В Бразилии из-под накрылий жука Procrustes corlacei (L.)
(сем. Passalidae) были собраны клещи Photia procrustidis (Berlese),
а на жуках Phanaeus sp. (сем. Scarabaeidae) были обнаружены клещи,
Grandiella tetracaudata Lombardini.
ЛИТЕРАТУРА
Cooreman J., 1950. Etude de qttelques Canestriniidae (Acari) vivant sur
des Chrysomelidae et snr des Carabidae (Insecta Coleoptera), Bull. Inst.
Poy. Sci. Nat. de Belgique, 26, (33), 1—54.
Lombardini G., 1938. Acari Novi II, Mem. Soc. Ent. Ital., 17, 118—120.
Lombardini G., (1943) 1944. Acari, Un biuovo Genere della famiglia .Tyro-
glyphidae", Pedia, 30, 1—24.
Lombardini G., 1950. Canestriniidae dell’America del Sud (Acarina), Ar-
thropoda, 1 (2/4), 279—290.
Hemisarcoptidae Oudemans, 1908
(рис. 286)
ДИАГНОЗ
Hemisarcopt’dae имеют яйцевидное тело длиной от 0,23 до 0,34 мм..
Покровы гладкие, глянцевые. Имеются проподосомальный щиток и
вертикальные щетинки. Хелицеры обычные, ножницеобразные, зубцов
на них нет. Ноги короткие и сильные. Коготков на лапках нет, но на
концах их сидят по два крепких когтевидных шипа; между шипами
проходит длинный стебелек чашевидной присоски. Половой диморфизм
не выражен. Как у самцов, так и у самок половое отверстие лежит
далеко позади тазиков ног IV. Половых присосок две пары. Аданаль-
Acaridiae
367
пых копулятивных присосок у самцов нет. Лапки ног Ш и IV несу-г
по Одной длинной бичевидной щетинке.
Рис. 286. Hemisarcoptes
ma'.us (Shimer).
Самка (вид снизу).
РОДЫ
1. Hemisarcoptes Lignieres 1893
Тип рода Acarus malus Shimer, 1868 (= Hemisarcoptes coccisugu?
L’gnieres, 1893)
ОБЩИЕ ЗАМЕЧАНИЯ
Единственный вид этого семейства — Hemisarcoptes malus (Shi-
mer)— распространен широко; его находят обычно вместе с щитовками..
Н. malus питается яйцами щитовок или самими насекомыми. Иногда
пот клещ размножается в таком количестве, что получает заметную-
роль в уменьшении численности щитовок. Установлено, что он охо-
тится за многими видами щитовок, в том числе;
l.epidosaphes ficus (Sign.) — на инжире в Калифорнии;
/.. ulmi L.
becki (Newman)
t’arlatoria oleae
Aspidiotus latanae
Diaspis carueli Targ.
Aonidiella perniciosa
('.on stock
Chionaspis salicis L.
— во Франции, Германии, CHIA (Пенсиль-
вания) и восточной Канаде;
— на цитрусовых в Калифорнии;
— на маслинах в Калифорнии;
— в инсектариях в Калифорнии;
— на кипарисах на Бермудских островах'
— во Франции;
— в Германии.
Н. malus составляют, повидимому, яйца
Излюбленную пищу
l.epidosaphes. Тотхилл (1918), изучавший Н. malus, нашел, что уги
368
Глава VII. Подотряд Sarcoptiformes
клещи, пожалуй, приносят больше пользы, чем обычно думают. Тотхилл
установил, что в некоторых районах Канады Н. malus регулирует
численность яблоневой запятовидной щитовки {Lepidosaphes ulmi).
По этому поводу он пишет, что „в годы высокой численности L. ulmi
в восточной Канаде Н. malus является важнейшим фактором в борьбе
с ними. В тех районах, где щитовка-хозяин менее многочисленна, там
и клещ соответственно менее эффективен". Известен случай нападения
Hemlsarcoptes malus (Shimer) на щитовку Aspidiotus latanae, выра-
щиваемую в инсектарии Риверсайдской опытной цитрусовой станции
(Калифорния). При этом Н. malus так сильно размножился, что по
письменному сообщению Бартлетта и де Баха работу по выращиванию
этой щитовки пришлось прекратить.
ЛИТЕРАТУРА
An d гё М., 1942. Snr ГHemlsarcoptes malus Shimer ( = coccisugus Lignieres)
(Acariens), Bull. Paris Mus. d'Hist. Nat., Ser. 2, 14 (3), 173—180.
Ewing H. E., Webster R. L., 1912. Mites associated with the oyster-shell
scale {Lepidosaphes ulmi Linne), Psyche, 19, 121.
Lord F. T., 1947. The influence of spray programs on the fauna of apple
orchards in Nova Scotia: II. Oyster-shell scale, Canad. Ent., 79 (11, 12),
196—209.
Shimer H,, 1868. Notes on the apple bark-louse {Lepidosaphes conchifor-
mis, Gmelin sp.), with a description of a supposed new Acarus, Trans.
Amer. Ent. Soc., 1, 368.
Tot hill J. D., 1918. The predaceous mite, Hemlsarcoptes malus Shimer, and
its relation to the natural control o; the oyster-shell scale, Lepidosaphes
ulmi L., Agr. (Jaz. Canada, 5 (3), 234—239.
Ewingidae Pearse, 1929
(рис. 287)
ДИАГНОЗ
Ewingidae имеют яйцевидное тело, несущее несколько щетинок. Вер-
тикальные щетинки имеются. Задний конец проподосомы несет попе-
речный ряд из двух длинных щетинок. Проподосома и гистеросома
не разделены швом. На передней части гистеросомы сидит одна пара
длинных щетинок. Кроме того, тело несет несколько пар коротких
щетинок. Двучлениковые пальпы образуют неполную трубку вокруг
видоизмененных хелицер. Последние не имеют клешни и сохраняют
только подвижный палец. Лапки ног I и П имеют коготки, но присосок
на них нет. Ноги III и IV резко утолщены и превращены в хватательный
орган; тарзальных коготков на этих ногах нет, но сами лапки когте-
видные. Половое отверстие лежит между тазиками ног III и IV и снаб-
жено (?) двумя парами половых присосок.
РОДЫ
1. Ewingta Pearse, 1929
Тип рода Etpingia cenobitae Pearse, 1929
Acaridiae
369
ОБЩИЕ ЗАМЕЧАНИЯ
E-wingia cenobitae Pearse был обнаружен в жабрах наземного рака-
отшельника Cenobita diogenes (Latreille) близ Логгерхида, Драй-Тортугас
(Флорида). Клещи прикрепляются ближе к жаберным лепесткам при
помощи хватательных приспособлений на ногах III и IV; клещи дер-
жатся ближе к наружному краю жабер и принимают такое положение,
чтобы их тело лежало между жаберными лепестками. Во многих слу-
чаях в теле самок были обнаружены яйца, содержащие вполне развитых
личинок, повидимому, готовых к вылуплению1. Личинки шестиногие,
Рис. 287. Ewingia cenobitae
Pearse (по Пирсу, 1929).
Самка (вид снизу).
похожие на взрослых клещей. Пирс исследовал также крабов Ocypoda
albicans (Bose.), но не обнаружил у них этих клещей. Возможно, это
объясняется тем, что Ocypoda часто заходят в море и смачивают жабры,
между тем как основные хозяева этих клещей — Cenobita diogenes —
заходят в море только раз в год для размножения.
ЛИТЕРАТУРА
Pearse A. S., 1929. Two new mites from the gills of land crabs, Ca-negie
InSt. Wash. Publ., 391, 225—230.
Anoetidae Oudemans, 1904
(рис. 288, 289)
ДИАГНОЗ
На теле часто имеются большие выросты. Шов между проподо-
сомой и гистеросомой выражен не всегда. Проподосомальный щиток
также иногда отсутствует. Имеются как цервикальные, так и верти-
кальные щетинки. Покровы обычно гладкие, но иногда покрыты тонким
точечным рисунком. Хелицеры имеют только один пильчатый палец
I исключение — Cederhjehnia, у которых хелицеры имеют два пильчатых
пальца). Тарзальный членик пальпы с двумя крупными, выступаю-
1 Это же явление известно и для чесоточных клещей. —Прим. ред.
24 Зак. 1785
370
Глава VII. Подотряд Sarcoptiformes
щими по бокам выростами. На всех лапках имеются слегка изогнутые,
сидячие коготки, присосок же на лапках нет. У половозрелых особей
Рис. 289. Histiostoma humidiatus
(Vitzthum) (по Фитцтуму, 1927).
Самка (вид снизу).
в области тазиков обычно имеется четыре крупных округлых или
подковообразных органа. Половых присосок две пары, они развиты
только у дейтонимф.
РОДЫ
1. Anoetus Dujardin 1842 (= Nodipalpus Karpelles, 1893)
Тип рода Hypopus alicola Dujardin, 1849 Anoetus discrepans
Oudemans, 1903)
2. Anoetoglyphus Vitzthum, 1927
Тип рода Anoetoglyphus ateuchi Vitzthum, 1927
3. Cederhejlmia Oudemans, 1931
Тип рода Cederhjelmia quadriuncinata Oudemans, 1931
4. Chiropteranoetus Womersley, 1942
Тип рода Chiropteranoetus challnolobus Womersley, 1942
5. Creutzeria Oudemans, 1932
Тип рода Creutzeria tobaica Oudemans, 1932
6. Glyphanoetus Oudemans, 1929
Тип рода Glyphanoetus fulmeki Oudemans, 1929
7. Histiostoma Kramer, 1876 Zschachta Oudemans, 1929)
Тип рода Hypopus feroniarum Dufour, 1839 (= Tyroglyphus
rostroserratus M6gnin = Histiostoma pectineum Kramer, 1876)
8. Mauduytla Oudemans, 1929
Тип рода Anoetus tropicus Oudemans, 1911
9. Myianoetus Oudemans, 1929
Тип рода Acarus muscarum Linnaeus, 1758
Acaridiae
ЗН
10. Prowlchmannla Radford, 1950 (—Wichmannia Oudemans, 1929
nomen praeoccupatum)
Тип рода Histiostoma spitliferum Michael, 1901
11. Sellea Oudemans, 1929
Тип рода Histiostoma pulchrum Kramer, 1886
12. Zwickia Oudemans, 1924
Тип рода Anoetus guentheri Oudemans, 1915
ОБЩИЕ ЗАМЕЧАНИЯ
Anoetidae обычно предпочитают влажную среду; их находят
в соке деревьев, вытекающем из ранок на стволе, в гниющих гриб-
ках, культурах дрозофилы, испорченном картофеле и в аналогичных
местообитаниях. Дейтонимфы типа гипопуса (странствующие формы)
встречаются на насекомых. К числу наиболее ‘распространенных
в США форм принадлежит Histiostoma feronlarum (Dufour). О ме-
стообитании этого клеща Майкл (1901) сообщает следующее: „Менье
первоначально обнаружил И. feronlarum в покрывающей гниющие
грибы тонкой пленке жидкости, в которой эти клещи встречались
в очень большом количестве. Можно сказать без преувеличения, что
клещей Н. feronlarum можно найти в любых влажных разлагающихся
мягких материалах растительного происхождения, обладающих доста-
точной плотностью. Пожалуй, чаще всего эти клещи встречаются на
грибах и в корнях, но, вообще говоря, они широко распространены
и чрезвычайно многочисленны. Н. feronlarum был отмечен во Франции,
Германии, Италии и Швейцарии и встречается повсеместно в Англии.
Я думаю, что сам по себе этот клещ не вызывает гниения, а только
сопутствует ему“. Жизненный цикл Zwickia guentheri Oudemans
хорошо описан Фитцтумом (1931). В его работе приведены изобра-
жения: личинки, протонимфы, дейтонимфы (гипопус, или странствую-
щая фаза), тритонимфы и половозрелых самца и самки. Уомерсли
(1941) на основании изучения дейтонимф составил определительную
таблицу для родов Anoetidae.
ЛИТЕРАТУРА
Cooreman J., 1941. Etudes Biospeologiques. XXY11I (1). Note sur Myiano-
etus diadematus Willmann, 1937 (Acariens Sarcoptiformes) Pseudopara-
site des Helomyzidae (Dipteres) de Transylvanie, Bull. Musee Royal
d’Hist. Hat. Brussels, 17 (42), 1—16.
Cooreman J., 1947. Le stade adulte de Myianoetus muscarum (Linne),
Bull, et Ann. Soc. Ent. de Belg., 73, 141—149.
Hughes R. D., 1950. The genetics laboratory mite Histiostoma laboratorium,
n. sp. (Anoetidae), J. Wash. Acad. Sci., 40 (6), 177—182.
Michael A. D„ 1901. British Tyroglyphidae, 1.
Vitzthum H., 1931. Terrestrische Acarinen der Deutschen Limnologischen
Sunda-Expedition, Suppl., Bd. IX „Tropische Binnengewasser', Arch. f.
Hydrobiol., 2, 59—134.
Womersley H., 1941. Studies in Australian Acarina (2) Tyroglyphidae (s. 1.),
Rec. South Austral. Mus., 6 (4), 451—488.
24*
.572
Глава VII. Подотряд Sarcoptiformes
Linobiidae Oudemans, 1908
(рис. 290, 291)
ДИАГНОЗ
Linobiidae имеют широкое овальное или пятиугольное тело. Про-
подосомального щитка и вертикальных щетинок нет. Покровы гладкие.
Хелицеры имеют только подвижный пильчатый палец. Все лапки по
форме напоминают коготки, но самих коготков на них нет; на всех
Рис. 290. Linobia
coccinellae (Scopoli)
(по Берлезе, 1887).
Самка (вид сверху).
Р и с. 291. Linobia
coccinellae (Scopoli)
(по Берлезе, 1887).
Хелицера.
лапках имеется лишь по стебельчатой присоске. На лапках ног I и
Л нет палочковидных сенсорных щетинок. Половых присосок две
пары. Копулятивных присосок у самцов нет.
РОДЫ
1. Linobia Ber'.ese, 1884 (= Linocoptes Berlese, 1887)
Тип рода Acarus coccinellae Scopoli, 1763
ОБЩИЕ ЗАМЕЧАНИЯ
Единственный вид — Linobia coccinellae (Scopoli) — был обнаружен
под надкрыльями жука Melasoma populi в Италии.
ЛИТЕРАТУРА
Berlese А., 1887. Acari, Myriopoda et Scorpiones, Crypt. Fasc. 39, №7.
Acaridiae
373
Nanacaridae Oudemans, 1923
ДИАГНОЗ
(рис. 292, 293)
Nanacaridae — клещи с глянцевыми покровами. Проподосомального
щитка, повидимому, нет. На задней части проподосомы имеется по-
перечный ряд из четырех щетинок, внутренние щетинки гораздо
Р и с. 292. Nanacarus
minutus (Oudemans)
(по Удемансу, 1928).
А — самка (вид сверху);
Б —лапка иоги II.
Р и с. 293. Nanacarus
minutus (Oudemans)
(по Удемансу, 1928).
Самка (вид снизу).
короче наружных. Цервикальных щетинок нет, но имеется пара верти-
кальных щетинок. Как у самцов, так и у самок отверстие располо-
жено позади тазиков ног IV. Самцы имеют аданальные копулятивные
присоски, но копулятивных присосок на лапках ног IV у них нет.
11а всех лапках имеется по присоске, но коготков на лапках нет;
юлько у неполовозрелых фаз на лапках имеется по одному слабо
развитому коготку. Концы лапок заострены, напоминая по форме
коготок, и изогнуты. Длинных бичевидных щетинок на ногах нет.
роды
I. Nanacarus Oudemans, 1902 (= Frowelnia Oudemans, 1923)
Тип рода Hypopus minutus Oudemans, 1902
2. Giardius Perraud, 1896
Тип рода Giardius vitis Perraud, 1896
374
Глава VII. Подотряд Sarcoptiformes
ОБЩИЕ ЗАМЕЧАНИЯ
Клещи Nanacarus mlnutus (Oudemans) были собраны на островах
Буру с Megapodius buruensis (куриные), на Яве — с Koptorthosoma
tenuiscapa и в Германии с летучих мышей Vesperugo serotinus, Ves-
pertilio plpistrellus и с землеройки Sorex vulgaris. Помимо этого
вида, известна лишь одна форма — Giardius vitls Perraud.
Sarcoptidae Trouessart, 1892
(— Acaridae Oudemans, 1904 европейских исследователей)
(рис, 294, 295)
ДИАГНОЗ
Sarcoptidae — кожные паразиты теплокровных позвоночных. Они
имеют шаровидное тело; явственного шва между проподосомой и
гистеросомой у них нет. Проподосомальный щиток имеется не всегда.
Рис. 294. Notoedres
cedi (Hering)
(по Гранжану, 1938).
Лапка ноги I.
Рис. 295. Sarcoptes scabiei var. equi
(Gerlach) (no Хирсту, 1922).
Самка (вид сверху).
Проподосома несет пару вертикальных щетинок; в остальных отде-
лах кожа покрыта тонкими бороздками, которые часто прерываются
чешуйчатыми полями или участками, покрытыми мелким точечным
рисунком или шипиками. Ноги очень короткие, часто на них имеются
присоски или коготки. Половых присосок нет.
Acaridiae
375
РОДЫ
1. Sarcoptes Latrei le, 1802 (—Acarus Фитцтума)
Тип рода Acarus scabiei DeGeer, 1778
2. Knemldokoptes Fiirstenburg, 1870 (= Cnemidocoptes американских
исследователей)
Тип рода Sarcoptes mutans Robin et Lanquetin, 1859
3. Notoedres Railliet, 1893
Тип рода Sarcoptes cati Hering, 1838 (—Sarcoptes minor var.
cati Railliet, 1893)
4. Nyctertdocoptes Oudemans, 1897
Тип рода Nyctertdocoptes poppei Oudemans, 1897
5. Prosopodectes Canestrini, 1897
Тип рода Sarcoptes chlropteralis Trouessart, 1896
6. Teincoptes Rodheim, 1923
Тип рода Teinocoptes epomorphi Rodheim, 1923
7. Trlxacarus Sellnick, 1944
Тип рода Trlxacarus divers us Sei'nick, 1944
ОБЩИЕ ЗАМЕЧАНИЯ
Чесоточные зудни — Sarcoptes scabiei, — паразитирующие на че-
ловеке, собаках, крупном рогатом скоте, свиньях, овцах, козах, верб-
людах, кроликах и лошадях, морфологически не отличаются друг от
друга, но, вероятно, принадлежат к различным биологическим расам.
Гак, клещи, паразитирующие на лошади, могут переходить и на
человека, но, как правило, заражение в этом случае носит временный
характер.
Английский исследователь Мелланби (1943) дал прекрасный обзор
чесоточных зудней, из которого мы заимствуем большую часть при-
водимых ниже данных1. Жизненный цикл очень прост и включает
фазы яйца, личинки, нимфы и половозрелого самца и неполовозрелой
и зрелой самок. Превращение неполовозрелой самки в самку, содер-
жащую яйца, происходит, повидимому, после оплодотворения. Новый
хозяин, вероятнее всего, заражается только что оплодотворенными
самками, которые очень подвижны и быстро передвигаются по теплой
коже. Клещи внедряются в роговой слой кожи хозяина, затрачивая
па это около часа времени; глубже этого слоя клещи не проникают.
При внедрении клещ прикрепляется к коже присосками и вбуравли-
нается в нее при помощи ног и ротовых органов. Половозрелая
самка обычно остается в проделанном ею ходе всю жизнь, реже она
покидает его уже через три недели. Самка начинает откладывать
яйца через несколько часов после внедрения в кожу и продолжает
откладку с перерывами в 2—3 дня в течение примерно двух месяцев.
1 Автор выражает свою признательность Г. В. Эдди, просмотревшему
•>гот раздел и пополнившему его результатами собственных наблюдений
над „самозаряжением" беременными самками. ;
376
Глава VII. Подотряд Sarcoptiform.es
Все это время самка продолжает удлинять ход, оставляя за собой
отложенные яйца. Если клещ проделывает ходы в толстой оро-
говевшей коже, то вздутий на последней не образуется; на тонкой
же коже появляются небольшие вздутия. При температуре 35° ли-
чинки вылупляются через 3—8 дней, чаще всего через 5 дней.
Они покидают ход и ползают по коже хозяина в поисках убежища,
а возможно, и пищи. Личинок и нимф можно найти в кожных фол-
ликулах. Личинки превращаются в зрелых самцов и неполовозрелых
самок через 4—6 дней после вылупления. Немногочисленные самцы
также выгрызают короткие ходы, но остаются в них недолго и
большую часть времени проводят на поверхности кожи в поисках
неоплодотворенных самок. Неоплодотворенные самки также прогры-
зают небольшие ходы, но остаются в них всего один день или не-
много более. Спаривание, повидимому, происходит на коже хозяина.
Весь цикл — от яйца до половозрелой яйценосной самки — длится от
10 до 14 дней. Более 90°/о отложенных яиц гибнет до окончания
цикла. После откладывания яйца клещ обычно продолжает проделы-
вать ход в коже.
Клещи вызывают сильный зуд, не дающий спать ночью. Зуд не-
посредственно связан с пробуравливанием хода. Каждый раз, когда
клещи вызывали раздражение кожи или зуд, оказывалось, что они
передвигаются в коже. От укусов клещей и расчесов на коже по-
является характерная сыпь; подмышками, вокруг пояса, на запястьях,
между ног, на бедрах и щиколотках образуются эритематозные
пятна и участки, покрытые фолликулярными папулами. Большинство
симптомов со стороны кожи обусловлено вторичной инфекцией, со-
провождающей деятельность клещей. Считается, что первичное зара-
жение клещом зуда не вызывает. Зуд начинается примерно через
месяц, когда в области хода появляется сыпь и к этому времени
больной приобретает особую чувствительность к клещу. При вторич-
ной инфекции пораженный участок кожи воспаляется уже через не-
сколько часов. Сенсибилизированный больной начинает чесаться не-
медленно после заражения и при этом ему нередко удается удалить
клеща и тем самым механически прервать болезнь. У несенсибили-
зированных лиц заболевание нарастает примерно в течение месяца,
прежде чем они заметят, что заражены клещами.
Чесотка хорошо известна в истории человечества и, повидимому.
распространяется волнами. Сенсибилизация приводит к выработке так
называемого иммунитета; в таких случаях лиц, зараженных клещами,
легко удается выявить и уничтожить на них клещей. Но так как
поблизости всегда имеются незаряженные и несенсибилизированные
люди, то численность клещей и число зараженных ими лиц продол-
жают возрастать. Истинные причины вспышки и спада заболевания
до сих пор неясны. Чаще всего чесоткой болеют зимой, когда люди
нередко спят вместе, чтобы согреться, и реже моются. Чесотка не
передается при обычном общении людей друг с другом, заражение
Acaridiae
377
происходит лишь тогда, когда здоровый человек спит в одной по-
стели с зараженным.
Вторичные инфекции, возникающие в результате расчесов, часто
бывают более серьезными, чем сама чесотка. Кроме того, вредное
влияние на кожу оказывают различные серусодержащие соединения,
употребляемые для борьбы с клещами.
Возбудителем чесотки у собак является Sarcoptes scabiei var.
ranis. Заболевение обычно начинается с головы и быстро распростра-
няется по телу. Хирст (1922) описывает симптомы чесотки собак
следующим образом: „Сначала появляются красноватые пятнышки,
похожие на укус блохи, затем, как следствие расчесов, покраснение
распространяется на целые участки кожи и образуются папулы. По-
раженные места часто подсыхают и покрываются желтоватой коро-
стой. Шерсть выпадает, кожа утолщается и сморщивается. Появляется
сильный зуд. Главным диагностическим признаком заболевания слу-
жит нахождение паразита". Собачий клещ переходит и на человека.
Sarcoptes scabiei имеет космополитическое распространение. Чесотка
свиней, вызываемая клещом 5. scabiei var. suis обычна в США и=
многих других странах. Общие симптомы чесотки свиней те же, что
и при чесотке собак. Свиной клещ также может заражать человека.
Sarcoptes scabiei var. Megnin паразитирует на овцах; он поражает
главным образом область головы, но в тяжелых случаях заболева-
ние распространяется также и на ноги, а как исключение и на
туловище. Клещей находят только на участках кожи, не покрытых
шерстью. Овечий клещ передается и козам. Sarcoptes scabiei var.
caprae Fiirstenburg паразитирует на козах; он нередко вызывает
смерть хозяина. Этого клеща находили также на человеке, лошадях,
овцах, рогатом скоте и свиньях.
Род Notoedres включает виды, вызывающие чесотку у различных
животных. N. cati Hering является возбудителем чесотки у кошек.
Заболевание обычно начинается с головы. На пораженных местах
образуются струпья, кожа твердеет, утолщается и сморщивается. Как
правило, поражаются только голова и шея. Этот же клещ вызывает
чесотку и у собак. Эссиг (1929) отмечает его как паразита кали-
форнийской белки Sciurus fossor, у этих белок заболевание иногда
кончается даже смертью. Notoedres cati var. cuniculi Gerlach пара-
зитирует на кроликах; заболевание также начинается с головы и
в тяжелых случаях распространяется на ноги и область гениталий.
Notoedres marts M£gnin паразитирует на серых крысах, как диких,
так и домашних. Заболевание иногда кончается смертью.
Knemidokoptes mutans Robin et Lanquetin вызывает у кур и мел-
кой домашней птицы болезнь, известную под именем „чешуйчатые ноги".
Хирст дает следующее описание болезни: „Сначала поражаются круп-
ные чешуйки цевки, под краями которых образуются приподнимаю-
щие их скопления беловатой мучнистой массы, иногда смешан-
ной с серозным экссудатом, В тяжелых 94учаях ноги целиком
378
Глава VII. Подотряд Sarcoptiformes
покрываются беловатыми, ноздреватыми корками. Иногда поражаются
также гребень и шея“. Заболевание это чрезвычайно заразно. Кпе-
midokoptes laevis var. gallinae (Railliet) вызывает у птиц зуд и вы-
падение перьев. Клещи внедряются в ткани или в чешуйки у осно-
вания пера и вызывают опадание перьев с больших участков тела.
Заболевание распространено как в Северной Америке, так и в Европе.
Описано заражение клещами К. laevis Railliet и голубей. Во Фран-
ции этот клещ поражает и других птиц, например фазанов и гусей.
ЛИТЕРАТУРА
Baker D. W., 1946. Barn Itch. N. Y. State Vet. Col., Vet. Expt. Sta., Para-
sit. Lab., 36 p.
Friedman R., 1942. Biology of Acarus scabiei, 183 p., Froben Press, N. Y.
Grandjean F„ 1938. Observations sur les acaridiae (1 serie), Ball. Soc.
Zool. France, 63 (4—5), 214—224.
Hirst S., 1922. Mites injurious to domestic animals, Brit. Mus. (Nat. Hist.),
Econ. Ser., No 13.
'lines M„ 1918. Cattle Scab, U. S. Dept. Agr. Farmers' Ball., 1017 (revi-
sed 1935).
Imes M., 1924. Sheep Scab, U. S. Dept. Agr. Farmers' Bull., 713 (revised
1927, 1935).
Mell an by K., 1943. Scabies, Oxvord Univ, Press, London, 1—81, illus.
van Eyndhoven G. L., 1947 (1945). Beschrijving van een nieuwen vleer-
muisparasiet Notoedres vanschaiki v. Eyndh., 1946 (Acar.), Tijdschrift
v. Ent., 88, 132—154.
ДИАГНОЗ
Cytoditidae Oudemans, 1908
(рис. 296)
Cytoditidae— яйцевидные клещи средней величины. Длина тела
варьирует у них от 0,45 до 0,60 мм. Покровы на Спинной стороне
Рис. 296. Cytodites nttdas
(Vizioli) (по Хирсту, 1922).
Самка (вид снизу).
гладкие, на других частях тела — с тонкими бороздками. Вертикаль-
ных щетинок нет, имеется несколько тонких дорзальных щетинок.
Acaridiae
379
Гнатосома занимает терминальное положение. Хелицеры и педипальпы
сливаются, образуя сосущую трубку. На лапках всех ног имеются
присоски, сидящие на длинных стебельках; тарзальных коготков нет.
Половых присосок нет. У самцов нет копулятивных аданальных при-
сосок.
РОДЫ
1. Cytodites Megnin, 1877 ( = Cytolelchiis Megnin, 1880)
Тип рода Sarcoptes nudus Vizioli, 1870 (= Cytodites glaber
Megnin, 1877)
ОБЩИЕ ЗАМЕЧАНИЯ
Клещи Cytodites nudus (Vizioli) паразитируют у кур в воздушных
мешках, в дыхательной системе и в полости тела (на поверхности
долей печени и в аналогичных местах). Если клещей немного, то
они не причинают особого вреда хозяину, но в большом количестве
они становятся опасными. Считается, что С. nudus вызывают энте-
риты и перитониты у домашней птицы; при больших скоплениях
клещей в дыхательных путях и в воздушных мешках птицы иногда
погибают от удушья. Мейер (1923) установил, что С. nudus живо-
родящи; новорожденные клещи имеют три пары ног.
ЛИТЕРАТУРА
Myers J. G., 1923. The internal mite of the domestic fowl, New Zealand J.
Sci. and Tech., 6 (1), 59—61.
Latninosioptidae Vitzthum, 1931
(рис. 297)
ДИАГНОЗ
Laminos’optidae — мелкие клещики с удлиненным, слегка упло-
щенным телом. Их длина варьирует от 0,20 до 0,26 мм. Покровы
па спинной стороне гладкие, в остальных местах — с тонкими борозд-
ками. На проподосоме вертикальных щетинок нет. Тело несет не-
сколько длинных щетинок. Гнатосома нормальная и сверху незаметна.
Ноги короткие, особенно ноги I и II. Лапки ног I и II не имеют ни
коготков, ни присосок, лапки ног III и IV — с маленькими присос-
ками, сидящими на длинных стебельках, или ножках. Половых при-
сосок нет. У самцов аданальных копулятивных присосок нет.
РОДЫ
1. Laminosloptes M6gnin, 1880 ( = Symplectoptes Railliet, 1885)
Тип рода Sarcoptes cysticolq yigioli, 1870
380
Глава VII. Подотряд Sarcoptiformes
ОБЩИЕ ЗАМЕЧАНИЯ
Laminosloptes cysticola (Vizioli) — эндопаразиты домашней птицы.
В Европе этих клещей находят в огромных количествах у индеек.
Клещи проникают в подкожную клетчатку и разрушают волокна ткани.
Присутствие клещей можно выявить по наличию обызвествленных
цист в подкожной ткани, но следует учитывать, что цисты обра-
зуются только вокруг мертвых клещей; живые же паразитируют
Рис. 297. Laminosioptes cysticola (Vizioli)
(по Хирсту, 1922).
Самка (вид снизу).
в самой ткани. Известны случаи гибели птиц при сильном заражении
этим клещом. L. cysticola был .описан с домашней птицы и в США.
ЛИТЕРАТУРА
Hirst S., 1922. Mites injurious to domestic animals, Brit. Mus. (Nat. Hist.),
Econ. Ser., № 13.
Taylor T., 1884. Microscopic observations. Internal parasites in domestic
fowls, U. S. Dept. Agr., Dept. Rpt., 34, 1—5.
Heteropsoridae Oudemans, 1908
(рис. 298)
ДИАГНОЗ
Heteropsoridae — очень мелкие клещи, длиной около 0,22 мм.
Они имеют очень толстое, почти круглое тело. Вертикальных щети-
нок нет. Аподемы тазиков всех ног оканчиваются свободно. Все ноги
Acaridiae
381
очень толстые, одинаковой длины, расположены радиально. На каж-
дой из лапок имеется по одной громадной, широкой, листовидной
присоске. Лапки ног III и IV с вентральными когтевидными шипами,
направленными назад.
Рис. 298. Heteropsorus pteroptopus Trouessart
et Neumann (по Фитцтуму, 1929).
Самка (вид сверху).
РОДЫ
1. Heteropsorus Trouessart et Neumann, 1887'
Тип рода Heteropsorus pteroptorus Trouessart et Neumann, 1887
ОБЩИЕ ЗАМЕЧАНИЯ
Единственный описанный вид паразитирует на коже птиц. Он был
собран в Италии и во Франции на камышовке Acrocephalus arundl-
miceus, зарянке Erithracus cyanecula, и овсянках Emberiza clrlus и
E. schoeniclus. Повидимому, co времени первоначального описания
>тих клещей больше не находили.
литература
Berlese А., 1897. Acari, Myriopoda et Scorpiones. Ordo Crytpostigmata
(Sarcoptidae), Fasc. 78, № 4.
Myialgesidae Trouessart, 1907
(рис. 299, 300)
ДИАГНОЗ
Myialgesidae паразитируют на мухах-кровососках (Hippoboscidae)
и иногда на пухоедах (Mallophaga). Самки имеют широкое, овальное
тело; в период беременности оно расширяется и становится груше-
382
Глава V1I. Подотряд Sarcopilformei
видным. Самцы неизвестны. Проподосома и гистеросома не разде-
лены швом. Вертикальных щетинок нет, но проподосомальный щиток
имеется. В остальных частях тела покровы мягкие, с тонкими бо-
роздками. На внутренней стороне пальп имеется желобок и пальпы
совместно образуют трубку, почти полностью окружающую концы
искривленных хелицер. Лапки ног II, III и IV лишены коготков, но
имеют по одной стебельчатой
присоске каждая; лапки ног I
не имеют ни коготка, ни при-
соски и по внешнему виду
напоминают якорь. Ноги поме-
Р и с. 299. Myialges caulotoon
Spieser (по Феррису, 1928).
Самка (вид сверху).
. Рис. 300. Myialges sp.
(по Феррису, 1928).
А — лапка ноги I, М. anchora
Sergent et Trouessart; Б — лапка I
М. caulotoon Spieser.
щаются на передней части тела,
между тазиками ног IV, Половых
У самок половое отверстие лежит
присосок нет.
РОДЫ
1. Myialges Sergent et Trouessart, 1907
Тип рода Myialges anchora Sergent et Trouessart, 1907
2. Myialgopsis Cooreman, 1944
Тип рода Myialgopsis trinotoni Cooreman, 1944
ОБЩИЕ ЗАМЕЧАНИЯ
Известно всего три вида этого семейства: Myialges anchora Ser-
gent et Trouessart, M. caulotoon, Spieser и Myialgopsis trinotoni
Cooreman. Феррис (1928) указывает, что Af. anchora прикрепляется
к телу хозяев (мух) хоботком. Обычно они держатся на брюшке
мухи, но один экземпляр клеща был обнаружен на груди. Сержан и
Acaridiae
383
Труэссар находили М. anchora как на брюшке, так и на груди
мух. Эти клещи являются настоящими паразитами мух. На местах
их прикрепления вследствие отложения хитина образуются рубцы,
что свидетельствует о длительности периода прикрепления. Самцы
неизвестны; есть предположение, что их следует искать на птицах,
являющихся хозяевами мух-кровососок.
ЛИТЕРАТУРА
С. о о rem an J., 1944. Un nouveau cas d’hyperparasitism partni les Acaridiae:
Myialgopsis trinotoni n. gen. n. sp., parasite d’un Mallophage, Bull.
Mas. Roy. d'Hist. Nat. de Belgique, 20 (26), 1—12.
Ferris G. F., 1928. The genus Myialges (Acarina: Saicoptidac), Ent. News,
39 (5), 137—140.
Ou'demans A. C„ 1935. Description du Myialges anchora Sergent et
Trouessart, 1907 (Acarien), Ann. de Parasitol. Humaine et Compar., 13
(1), 5-11.
Psoroptidae Canestrini, 1892
ДИАГНОЗ
(рис. 301, 302)
Psoroptidae за немногими исключениями
паразитируют на млеко-
нет. На спинной поверхности тела
питающих. Вертикальных щетинок
Рис. 301. Psoroptes equi var. ovis
(Hering) (по Хирсту, 1922).
Самец (вид сверху).
имеются щитки. У самцов на
лапках всех ног, а у самок —
на лапках ног I, II и IV имеются
чашевидные присоски, сидящие
либо на длинных сегментиро-
ванных (Psoroptes), либо на
коротких несегментированных
стебельках; исключение со-
ставляют самцы Psoroptes и
самки Otodectes и Caparinae,
не имеющие присосок на лап-
ках ног III и IV. У самцов
ноги IV значительно короче
ног III. Лапки ног IV у самок
'Рис. 302, Otodectes cynotls
(Hering) (по Гранжану, 1937).
Дистальная часть ноги I.
384
Глава VII. Подотряд Sarcoptiform.es
оканчиваются длинной бичевидной щетинкой. Самцы с аданальными
копулятивными присосками. Задний край брюшка у самца слабо вы-
резан или подразделен на две доли.
РОДЫ
1. Psoroptes Gervais, 1841 ( = Dermatodectes Gerlach, 1857 = £>e/--
matokoptes Fiirstenburg, 1861)
Тип рода Sarcoptes equi Hering, 1838
2. Caparinia Canestrini, 1894
Тип рода Chorioptes setiferas M6gnm, 1880
3. Chorioptes Gervais, 1859 ( = Symbiotes Gerlach, 1857 nomen
praeoccupatum, = Dermatophagas Fiirstenburg, 1861)
Тип рода Sarcoptes caprae Delafond, 1854
4. Otodectes Canestrini, 1894
Тип рода Sarcoptes cynotls Hering, 1838
ОБЩИЕ ЗАМЕЧАНИЯ
Повидимому, род Psoroptes включает всего один вид, разновид-
ности которого живут на разных хозяевах. Эти разновидности мало
дифференцированы морфологически (Хирст, 1922), но переход кле-
щей с одного животного-хозяина на животное-хозяина другого вида
очень затруднен или даже невозможен.
Psoroptes equi var. ovis является возбудителем чесотки у овец.
Болезнь эта ведет к порче меха, а иногда и к смерти зараженных
овец. Чесотка, вызываемая этим клещом, отличается от чесотки, вы-
зываемой клещами Sarcoptes, тем, что поражает места, покрытые
шерстью. Клещ прокалывает кожу хелицерами, вызывая характерное
раздражение, сопровождающееся образованием струпьев и выделением
серозного экссудата. Шерсть сбивается в клочья и иногда выпадает.
Клещи живут под струпьями, образуемыми серозным экссудатом,
выделяющимся в месте прокола, и в результате их деятельности
струпья продолжают утолщаться. Ван Эс (1904) указывает, что
у слабых животных или при плохом уходе это заболевание ведет
к потере веса, анемии и слабости и может окончиться смертью. За-
ражение клещом происходит легко как при прямом контакте, так и
через инфицированные предметы. S. equi var. ovis распространен по
всему свету.
Psoroptes equi Nit. caprae паразитирует на козах, у которых
поражает только уши. Обычно клещ не причиняет вреда, но иногда
его присутствие вызывает глухоту и потерю аппетита; в тяжелых
случаях болезнь может кончиться смертью животного.
Psoroptes equi var. cuniculi паразитирует в ушах кроликов. Он
поражает, повидимому, только ушную раковину. Марин (1924) опи-
сывает симптомы болезни следующими словами: „Первое проявление
болезни заключается в гиперемии и образовании красновато-корич-
невых струпьев на дне ушной раковины. Эти явления распростра-
Acaridlae
385
пяются все шире и за'несколько месяцев могут охватить всю внут-
реннюю поверхность уха. Тяжелым осложнением заболевания является
гнойная инфекция среднего уха, которая может перейти на внутрен-
нее ухо и на мозговые оболочки. Клещи распространяются быстрее
н теплое время года. Заболевание это широко распространено, и нет
почти ни одной лаборатории, куда бы оно не проникало время от
времени".
Клещи Psoroptes вызывают чесотку также у лошадей и рогатого
скота. Чесотка лошадей обычна во Франции и встречается также и
в Англии. Возможно, она имеет очень широкое распространение.
Psoroptes equi var. bovis является возбудителем чесотки у рога-
того скота. Этот клещ, как и все остальные представители рода
Psoroptes, проходит весь жизненный цикл на хозяине. Имс (1918)
пишет: „Повреждения появляются, прежде всего, на загривке, на
шее, в области, лежащей непосредственно перед загривком, или во-
круг корня хвоста. Отсюда они распространяются по спине и бокам
животного, охватывая иногда все тело. Р. equi var. bovis, попав на
хозяина в поисках пищи, прокалывает кожу последнего. При этом
клещ, повидимому, вводит в ранку ядовитый секрет. В месте укола
появляется легкое воспаление, которое в ранней стадии по большей
части остается незамеченным. По мере того как численность клещей
на зараженном животном возрастает, в коже появляется все больше
мелких ранок; в результате начинается сильный зуд, образуются па-
пулы, развивается воспалительная реакция и выделяется серозный
экссудат. Серозные отделения, просачивающиеся на поверхность
кожи, смешиваются с частицами грязи и в большей или меньшей
степени инфицируются микроорганизмами. Серозные отделения быстро
затвердевают, образуя желтоватые или серые струпья, часто окра-
шенные кровью. На ранних стадиях заболевания струпья невелики
и обычно по величине не превышают горошину; в дальнейшем, по
мере того как клещи поражают здоровую кожу вокруг раны, струпья
постепенно увеличиваются".
Род Chorioptes содержит несколько видов, которые, возможно,
представляют собой такие же биологические расы, как и „виды"
Psoroptes. Различные виды Chorioptes паразитируют на лошадях,
крупном рогатом скоте, овцах, козах и некоторых других животных.
Chorioptes bovis (Gerlach), паразитирующий на крупном рогатом
скоте, обычно поражает только ноги или основание хвоста и аналь-
ную впадину. Лишь редко заболевание распространяется также и на
шею и спину или же на живот и промежность. Чесотка, вызываемая
этим клещом, обычно протекает легко. Она захватывает строго огра-
ниченные участки тела животного и распространяется медленно.
К этому роду принадлежат также С. equi (Hering), паразитирующий
па лошадях, С. caprae (Delafond), паразитирующий на козах, С.
ovis (Railliet), паразитирующий на овцах и С. cuniculi (Zurn), пара-
зитирующий на кроликах.
25 Зак. 17S5
381
Глава VlL Подотряд Sarcoptiformes
Представители рода Otodectes паразитируют в ушах собак, кошек,
хорьков и лис. Как и виды рода Chorioptes, они по строению не от-
личаются друг от друга и рассматриваются теперь как биологиче-
ские расы Otodectes cynotis Hering. В Англии эти клещи вызывают
у кошек появление язв в ушах. Собаки, зараженные клещами Oto-
dectes, испытывают, повидимому, сильный зуд и непрерывно трясут
головой.
ЛИТЕРАТУРА
Good Е. S., 1909. Sheep scab, Kentucky Agr. Expt. Sta. Bu'l., 1437,
219—246.
Gfjjndjean F., 1937. Otodectes cynotis (Hering) et les pretendues trachees
des Acaridiae, Bull. Soc. Zool. trance, (4), 280—299.
Hirst S., 1922. Mites injurious to domestic animals, Brit. Mus. (Nat. Hist.),
Econ. Ser., № 13.
I m e s M., 1918. Cattle scab and methods of control and eradication, U. S.
Dept. Agr. Farmers’ Bull., 1017, 1—29 (revised 1935).
Itnes M., 1924. Sheep scab, U. S. Dept. Agr. Farmers’ Bull., 7—13
(revised 1935).
Marine D., 1924. The cure and prevention of ear cancer in rabbits, Science,
60 (1546), 158.
Van Es L., 1904. Scabies in sheep and cattle and mange in horses, N. Dak.
Agr. Col. Expt. Sta Bull., 61, 399—435.
Epidermoptidae Trouessart, 1892
(-—Mealiidae Oudemans, 1923)
(рис. 303, 304)
ДИАГНОЗ
Epidermoptidae — очень мелкие клещи длиной от 0,17 до 0,39 мм.
Они имеют несколько уплощенное обычно очень короткое и более
или менее округлое тело. У самок на проподосоме развит щиток.
Вертикальных щетинок на проподосоме нет. Покровы нежные, борозд-
чатые. Задний край тела у самцов часто вырезан или двулопастной,
у самок он закруглен. Самцы обладают копулятивными присосками.
Лапки всех ног оканчиваются присосками.
РОДЫ
1. Epidermoptes Rivo’ta, 1876
Тип рода Epidermoptes bllobatus Rivolta, 1876
2. Dermatlon Trouessart et Neumann, 1887
Тип рода Epidermoptes bihamatus Trouessart et Neumann, 1887
3. Dermatophagoides Bogdanow, 1864 ( = Pachyllchus G. Canestrini,
1894 = ЛТеа/ш Berlese, 1897)
Тип рода Dermatophagoides scheremetewskyi Bogdanow, 1864
4. Microlichus Trouessart et Neumann, 1887
Тип рода Symbiotes avus Trouessart et Neumann, 1887
Acaridiae
337
5. Rivoltasia G. Canestrini, 1894
Тип рода Epidermoptes bifurcatus Rivolta, 1876
6. Turbinoptes Boyd, 1949
Тип рода Turbinoptes strandtmanni Boyd, 1949
ОБЩИЕ ЗАМЕЧАНИЯ
Epidermoptidae обычно паразитируют на коже птиц. Клещи Epi-
dermoptes bilobatus Rivolta паразитируют на коже кур. Иногда их
находят в огромном количестве при „питириазисе" (чешуйчатая кожа)
Рис. 303. Dermatophagoides
scheremetewskyi Bogdanow.
Самка (вид сверху).
Рис. 304. Dermatophagoides
scheremetewskyi Bogdanow.
Самка (вид снизу).
кур, и потому клещей Е. bilobatus считают причиной этого заболе-
вания. Dermatophagoides scheremetewskyi Bogdanow, 1864 был
описан как клещ, вызывающий дерматит у человека. Травер (1951)
сообщил о шестнадцатилетних наблюдениях над заражением кожи
волосистой части головы у человека этими клещами. Первыми симп-
। омами заражения было появление мелких зудящих красных папул,
сопровождающееся ощущением „ползания мурашек11 и царапания.
В дальнейшем эпидермис в пораженных местах утолщался, появля-
лась припухлость и болезненность. Клещи не оставались постоянно
па волосистой части головы и при применении для борьбы с ними
химических препаратов спускались на лицо и проникали в глаза,
они и ноздри. Глаза при этом сильно распухали, так что их дви-
жсиия становились невозможными. При проникновении клещей в ноздри
Глава Vll. Подотряд SarcoptiformeS
развивалось раздражение горла, трахей и бронхов. В ушах клеши,
повидимому, поражали только раковины, в которых при этом обра-
зовывались чесоточные папулы. Пораженные места часто покрывались
небольшими струпьями. Возможным источником заражения были до-
машние кошки и собаки, страдающие чесоткой. Фишер и др. (1951)
сообщили, что D. scheremetewskyi был обнаружен вместе с грибком
на ноге у пациента, страдавшего дерматитом. Основываясь на исто-
риях болезни, авторы выдвинули предположение, что развитие дер-
матита предшествует появлению клеща и что дерматит лишь спо-
собствует заражению клещом. Саза (1950) дал обзор японских видов
рода Dermatophagoid.es и описал два новых вида: D. takeuchil
Sasa — из мочи человека и D. saitoi Sasa — из мокроты больного
с синдромом Лефлера. Кроме того, он обнаружил клещей Dermato-
phagoides sp. в мокроте больного с хронической бронхиальной
астмой.
ЛИТЕРАТУРА
Boyd Е., 1949. A new genus and species of mite from the nasal cavity of
the ring-billed gull (Acarina, Epidermoptidae), J. Parasitol., 35 (3),
295—300.
Cooreman J., 1950. Snr un acaricn nouveau, prejudiciable aux matieres
alimentaires entreposecs: Mealia maynei n. sp., Bull, et Ann. Soc. Ent.
Belg., 86, 164—168.
Fisher A. A., Franks A. C., Wolf M., L eider M., 1951. Concurrent
infestation with a rare mite and infection with a common dermatophyte,
Arcliiv of Dermatology and Syphilology, 63 (3), 336—342.
Sasa M., 1950. Mites of the genus Dermatophagoides Bogdanoff, 1864 found
from three cases of human acariasis, Japanese J. Expt. Medicine, 20,
519—525.
Traver J. R., 1951. Unusual scalp dermatitis in humans caused by the mite
Dermatophagoides, Proc. Ent. Soc. Wash., 53 (1), 1—25.
Psoralgidae Oudemans, 1908
ДИАГНОЗ
Psoralgidae — уплощенные, короткотелые клещи. У самцов зад-
ний край тела вырезан посредине с брюшной стороны (двулопаст-
ной). Покровы тела мягкие, морщинистые. Срединный дорзальный
щиток всегда имеется, других щитков нет (?). Вертикальных щети-
нок нет. Хелицеры клешневидные; максиллы с двучлениковыми паль-
пами. Ноги почти одинаковой величины, но у самцов ноги III заметно
удлинены и ноги III и IV утолщены. У обоих полов имеются чаше-
видные присоски, сидящие на короткой несегментированной ножке.
Самцы с аданальными копулятивными присосками.
РОДЫ
1. Psoralges Trouessart, 1896
Типы рода Psoralges libertus Trouessart, 1896
Acaridiae
389
|>ЫЦИЕ ЗАМЕЧАНИЯ
Эти клещи были найдены в шерсти муравьеда Tamandua sp.
Личинки и нимфы живут колониями в подкожных тканях и вызы-
вают род чесотки. Взрослые клещи живут свободно в шерсти хо-
зяина. Изображение этого клеща в печати не опубликовано.
Listrophoridae Canestrini, 1892
(рис. 305)
ДИАГНОЗ
Listrophoridae имеют продолговатое тело (исключение—Myocoptes),
часто сильно сжатое с боков. Спинные щитки варьируют по числу
и по форме. Покровы имеют различный
вид на различных участках тела; они бы-
вают гладкие, морщинистые, покрытые
поперечными бороздками или чешуйчатые.
Имеется пара вертикальных щетинок.
Либо максиллы, либо часть ноги или вся
нога, либо стернальное образование пре-
вращены в хватательный орган, при по-
мощи которого клещ защемляет волосок
хозяина. Половые присоски за немногими
исключениями имеются. Самцы с аланиль-
ными копулятивными присосками; послед-
ние иногда мало заметны или даже вовсе
11 icy тствуют.
Рис. 305. Chirodiscoides caviae Hirst
(ио Хирсту, 1922).
Самец (вид снизу).
роды
1. Listrophorus Pagenstecher, 1861
Тип рода Listrophorus leucharti Pagenstecher, 1861
2. Alabidocarpus Ewing, 1929
Тип рода Labidocarpus megalonyx Trouessart, 1895
3. Atopomelus Trouessart, 1918
Тип рода Atopomelus locusta Trouessart, 1918
I. Austrochirus Womersley, 1943
Тип рода Austrochirus queenslandicus Womersley, 1943
5. Campylochirus Trouessart, 1893
Тип рода Campylochirus chelopus Trouessart, 1893
390
Глава VII. Подотряд Sarcoptiformes
6. Chirodiscoides Hirst, 1917 (Womersley, 1943 синоним Campylo-
chlrus Trouessart)
Тип рода Chirodiscoides caviae Hirst, 1917
7. Chirodiscus Trouessart et Neumann, 1890
Тип рода Chirodiscus amplexans Trouessart et Neumann, 1890
8. Eulabidocarpus Lawrence, 1948
Тип рода Labidocarpus compressus Ewing, 1910
9. Eurychiroides Womersley, 1943 (= Euryzonus Trouessart, 1918
nomen praeoccupatum)
Тип рода Euryzonus ventricosus Trouessart, 1918
10. Labidocarpus Trouessart, 1895
Тип рода Labidocarpus rollinati Trouessart, 1895
11. Llstrophoroides Hirst, 1923
Тип рода Llstrophoroides aethiopicus Hirts, 1923
12. Marquesania Womersley, 1943 ( = Llstrophoroides Ferris, 1932
nomen praeoccupatum)
13. Myocoptes Claparede, 1869 (= Criniscansor Poppe, 1887)
Тип рода Sarcoptes musculinus Koch, 1844
14. Neolabidocarpus Gunther, 1942
Тип рода Labidocarpus buloloensis Gunther, 1940
15. Olabidocarpus Lawrence, 1948
Тип рода Labidocarpus belsorum van Eyndhoven, 1940
16. Protabidocarpus Lawrence, 1948
Тип рода Protabidocarpus canadensis Lawrence, 1948
17. Schizocarpus Trouessart, 1896 (= Haptosoma Kramer, 1896)
Тип рода Schizocarpus mingaudi Trouessart, 1896
18. Schizocoptes Lawrence, 1944
Тип рода Schizocoptes conjugates Lawrence, 1944
19. Trichoecius Canestrini, 1899 ( = Trichoblus Canestrini, 1897
nomen praeoccupatum).
Тип рода Myocoptes brevipes Canestrini et Trouessart 1895
ОБЩИЕ ЗАМЕЧАНИЯ
Гюнтер (1942) разделил эту группу на четыре подсемейства,
основываясь на типе строения хватательного органа. Уомерсли (1943)
дал следующую определительную таблицу этих подсемейств.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ LISTROPHORIDAF
1. В хватательный орган превращены ноги I и II............
— В хватательный орган превращены ноги II и IV . . Myocoptlnar
— В хватательный орган превращены максиллы . . Listrophorinae
2. Ноги I и II с присосками и слабо видоизменены; иногда на них
развит дополнительный хватательный орган . . . Atopomelinar
— Ноги I и II без присосок и сильно видоизменены; дополнитель-
ного хватательного органа на них нет . , . , . Labidocarpinae
Acaridiae
391
ОБЩИЕ ЗАМЕЧАНИЯ
Listrophoridae живут в шерсти мелких млекопитающих и как
исключение в оперении птиц {Chirodiscus). Myocoptes musculinus
(Koch) вызывает у лабораторных белых мышей особую форму
чесотки, которую предложено называть „миокоптической". М. musculi-
nus распространен по всему свету. Часто встречается также Chiro-
discoides caviae Hirst, прикрепляющийся к волоскам на спине мор-
ских свинок; ноги этого клеща уплощены и служат для захватыва-
ния волосков шерсти хозяина. Labidocarpus nasicolus Lawrence был
собран с вибрисс на кожных выростах носа летучей мыши или близ них;
клещи прикрепляются к основанию одного из тактильных волосков
хозяина. Лоуренс (1938) пишет: „L. nasicolus, так же как и МуоЫа,
неспособен быстро передвигаться с места на место, так как его пе-
редние ноги крайне видоизменены. Поэтому можно предполагать,
что этот клещ лишь передвигается взад и вперед по волоску, к ко-
торому он прикреплен, и питается секретом сальных желез, распо-
ложенных у основания волосков. У летучих мышей Rhinolophus кожные
выросты носа снабжены крайне многочисленными сальными железами,
секрет которых поддерживает влажность поверхности кожных выро-
стов, что облегчает восприятие воздушных течений. Интересно отме-
тить, что эти клещи до сих пор были найдены только у листоносых
летучих мышей, принадлежащих к подотряду Mlcrochiroptera. В по-
лости тела яйценосных самок Labidocarpus rollinatl и L. nasicolus
находили шестиногих личинок".
ЛИТЕРАТУРА
Gunther С., 1942. Notes on the Listrophoridae (Acarina: Sarcoptoidea), Proc.
Linn. Soc. N. S. Wales, (Л (3, 4), 109—110.
Lawrence R. F., 1938. A new acarine parasite of bats, Parasitol., 30 (3),
309—313.
Lawrence R. F., 1948. Studies on some parasitic mites from Canada and
South Africa, J. Parasitol., 34 (5), 364—379.
Womersley H., 1943. Australian species of Listrophoridae Canest. (Acarina)
with notes on the new genera, Trans. Roy. Soc. South Australia, 67
(1), 10—19.
Analgesidae Trouessart, 1915
(рис. 306)
ДИАГНОЗ
Analgesidae паразитируют в оперении птиц. Как правило, у самок
щитков на гистеросоме нет (исключение Mesalges). Они имеют невыре-
занный, округлый задний край тела. Ноги I и II обычно снабжены сидя-
щими на латеровентральной стороне треугольными, манжетообразными
выростами. Ноги III и IV (или те и другие) у самцов крупнее осталь-
ных ног и часто очень сильно развиты.
392
Глава VII. Подотряд Sarcoptiformes
РОДЫ
1. Analges Nitzsch, 1818
Тип рода Analges passerinus Nitzsch, 1818
2. Analgopsis Trouessart, 1919
Тип рода Acarus passerinus Linnaeus, 1758 ( = DeGeer, 1778)
3. Berlesella Trouessart, 1919
Тип рода Berlesella alata Trouessart, 1919
4. Hemialges Trouessart, 1888
Тип рода Megninla pappus Trouessart et Neumann, 1888
Рис. 306. Megninla columbae
Buchholz (по Хирсту, 1922).
Самеи (вид снизу).
5. Hyperalges Trouessart, 1915
Тип рода Megninla magnlflca Trouessart, 1895
6. Ingrassla Oudemans, 1905
Тип рода Megninla veliger Oudemans, 1904
7. Megninla Berlese, 1881
Тип рода Dermaleichus cubitalis Megn’n, 1877
8. Mesalges Trouessart, 1888
Тип рода Dermaleichus abbreviates Buchholz, 1869
9. Metanalges Trouessart, 1919
Тип рода Megninla elongata Trouessart, 1886
10. Nealges Trouessart, 1886
Тип рода Nealges poppei Trouessart, 1886
11. Plesialges Trouessart, 1919
Тип рода Plesialges mimus Trouessart, 1919
12. Protalges Trouessart, 1885 ( = Hartingia Oudemans, 1897)
Тип рода Protalges robinl Trouessart, 1885
Acaridiae
393
13. Psoroptoid.es Trouessart, 1919
Тип рода Megninia psoroptopus Trouessart, 1885
14. Pteralloptes Trouessart, 1884 ( = Analloptes Trouessart, 1885)
Тип рода Dermaleichus stellarls Buchholz, 1869
15. Varchia Oudemans, 1905
Тип рода Pteralloptes gambettae Oudemans, 1904
16. Xolalges Trouessart, 1885
Тип рода Xolalges scaur us Trouessart, 1885
ЛИТЕРАТУРА
Gaud J., P e t i t о t M. L., 1948. Sarcoptides plumicoles des oiseaux du Ma-
roc, Ann. de Parasit., 23 (1—2), 35—46.
Gaud J., P e t i t о t M. L., 1948. Sarcoptides plumicoles des oiseaux d’Indo-
chine, Ann. de Parasit., 23 (5—6), 337—347.
Dermoglyphidae Megnin et Trouessart, 1883
( = Falculiferidae Oudemans, 1908;
Pterolichidae Trouessart, 1916;
Syringobiidae Berlese, 1907)
(рис. 307 -309)
ДИАГНОЗ
Dermoglyphidae паразитируют в оперении птиц. У обоих полов имеется
й щиток, а часто и другие спинные
по крайней мере проподосомальны
щитки. Задний край тела
или округлый, или слегка
вырезан. Проподосома несет
пару вертикальных щетинок.
11оги III и IV часто толще
остальных, а как исключе-
ние самыми толстыми бы-
вают ноги I или II или и те
и другие. Аподемы ног I и II
или сливаются друг с другом,
или концы их остаются сво-
бодными. Иногда встречают-
ся гетероморфные самцы,
очень сильно отклоняющиеся
от нормы строением ног IV
и хелицер (Bdellorh.ynch.us'),
или же строением ног I и II
и хелицер (Falculifer), или
строением пальп (Cheiloce-
nis), или же строением ног
III и IV (Protolichus).
Ранее считавшиеся само-
стоятельными семейства
Рис. 307. Pterolichus obtusus Robin,
(по Хирсту, 1922).
Самка (вид сверху),
394
Г лава VII. Подотряд Sarcopiifortnes
Falculiferidae, Pterolichidae и Syringobiidae объединены здесь в одно
семейство Dermoglyphidae. До настоящего времени признаков.
Рис. 309. Falciiltfer rostratus (Buchholz).
Самец (вид снизу).
Рис. 308. I'alcalifer
rostratus (Buchholz)
(по Хирсту, 1922).
Гипопус.
оправдывающих выделение этих семейств, не обнаружено, признаки же,
использованные Фитцтумом, трудно поддаются определению и недоста-
точно постоянны.
РОДЫ
1. Dermoglyphus Megnin, 1877
Тип рода Dermaleichus elongatus Megnin, 1877
2. Anasicudion Trouessart et Neumann, 1888
Тип рода Dermaleichus landoisii Buchholz, 1869
3. Anoplonotus Trouessart, 1915
Тип рода Pterolichus semaphorus Trouessart, 1886
4. Avenzoarta Oudemans, 1905
Тип рода Dermaleichus totani Canestrini, 1878
5. Bdellorhynchus Trouessart, 1885
Тип рода Bdellorhynchus polymorphus Trouessart, 1885
Acaridiae
395
6. Bonnetella Trouessart, 1934 ( = Buchholzia Trouessart, 1915
nomen praeoccupatum)
Тип рода Analges fuscus Nitzsch, 1818
7. Ceratothrix Trouessart, 1915
Тип рода Pterolichus corniger Trouessart et Neumann, 1888
8. Chauliacla Oudemans, 1905
Тип рода Pterolichus securlger Robin, 1877
9. Cheiloceras Trouessart, 1898
Тип рода Cheiloceras cervus Trouessart, 1898
10. Cheylabis Trouessart, 1885
Тип рода Cheylabis latus Trouessart, 1885
11. Columellala Oudemans, 1904
Тип рода Dermoglyphus various Trouessart, 1886
12. Eustathia Oudemans, 1905
Тип рода Pterolichus cultrifer Robin, 1868
13. Falculifer Railliet, 1896
Тип рода Dermaleichus rostratus Buchho’z, 1869
14. Freyana Haller, 1877
Тип рода Dermaleichus anatinus Koch, 1841
15. Gabucinia Oudemans, 1905
Тип рода Pterolichus dellbatus Tobin, 1877
16. Halleria Trouessart et Megnin, 1885
Тип рода Freyana {Halleria) hirsutirostris Trouessart et Megnin
17. Kramerella Trouessart, 1915 { = Grameria Haller, 1878 nomen
praeoccupatum)
Тип рода Crameria lunulata Haller, 1878
18. Mlchaeltchus Trouessart et Megnin ( = Michaelia Trouessart, 1885,
но не Michaelia Berlese, 1884)
Тип рода Dermaleichus heteropus Michael, 1881
19. Mlcrochelys Trouessart 1915
Тип рода Freyana {Microspalax) delicatula Trouessart, 1898
20. Microspalax Megnin et Trouessart, 1884
Тип рода Freyana {Microspalax) manicata Megnin et Trouessart,
1884
21. Neumanntella Trouessart, 1915 ( = Neutnannla Trouesart et Neu-
mann, 1888 nomen praeoccupatum)
Тип рода Neumannia chelifer Trouessart et Neumann, 1888
22. Oustaletia Trouessart, 1885
Тип рода Pterolichus {Oustaletia) pegasus Trouessart, 1885
23. Paralges Trouessart, 1885
Тип рода Paralges pachycnemls Trouessart, 1885
24. Plutarchla Oudemans, 1904
Тип рода Plutarchia chelopus Oudemans, 1904 (partim = Syrin-
gobla chelopus Trouessart et Neumann, 1888)
25. Protolichus Trouessart, 1884
Тип рода Pterolichus {Protolichus) brachiatus Trouessart, 1884
396
Глава VII. Подотряд Sarcoptiformes
26. Protonyssus Trouessart, 1915
Тип рода Protalges larva Trouessart, 1885
27. Pseudoglebelia Radford, 1950 ( = Giebelia Trouessart, 1915 nomen
praeoccupatum)
Тип рода Dermaleichus puffini Buchholz, 1869
28. Pterolichus Robin, 1868
Тип рода Pterolichus obtusus Robin, 1868
29. Pteronyssus Robin, 1868
Тип рода Pteronyssus striatus Robin, 1877
30, Pseudalloptes Trouessart et Megnin, 1884
Тип рода Pterolichus bisubulatus Robin, 1877
31. Sammonica Oudemans, 1904
Тип рода Syringobia ovalis Trouessart, 1898
32. Sphaerogastra Trouessart, 1897
Тип рода Sphaerogastra thylacodes Trouessart, 1897
33. Syringobia Trouessart et Neumann, 1888
Тип рода Syringobia chelopus Trouessart et Neumann, 1888
34. Thecarthra Trouessart, 1896
Тип рода Pterolichus theca Megnin et Trouessart, 1884
35. Xoloptes Canestrini, 1879
Тип рода Pterolichus claudicans Robin, 1877
ОБЩИЕ ЗАМЕЧАНИЯ
Жизненный цикл дсрмоглифид описал Удеманс (1922). По ею
Данным, имеется два типа яиц — с твердой и с мягкой скорлу-
пой. Последние уже в теле матери содержат сформированную ли-
чинку; таким образом, самки могут быть как яйцеживородящими,
так и живородящими. Из яйца вылупляется шестиногая личинка,
линяющая в восьминогую протонимфу, которая, в свою очередь, пре-
вращается в дейтонимфу. В этой фазе у самок уже появляется по-
ловое отверстие, и по этому признаку их можно отличить от самцов.
Дейтонимфы превращаются в восьминогих половозрелых особей.
Самцы копулируют с самками, находящимися в фазе дейтонимфы.
Спарлок и Имлен (1942) изучали нимф, собранных в Калифорнии
с дятла Colaptes cafer collaris, и отнесли их к новому виду Нуро-
dectes chapini. Эти клещи „лежали в неплотных цистах в соедини-
тельной ткани, окружающей трахею и (реже) пищевод. Особенно
много клещей было у гортанной щели птицы, но заражена ими была
вся ткань от легких до языка. Насколько можно было установить,
никакого движения клещей в цистах не было. Не удалось обнару-
жить и заметных патологических изменений в зараженных клещами
тканях хозяина". Изучив доступную им литературу, Спарлок и Имлен
обратили внимание на особенности жизненного цикла Falculifer re-
strains (Buchholz). Они пишут, что „при неблагоприятных условиях,
например при линьке птицы-хозяина и т. д., нормальные нимфы
Falculifer rostratus (Buchholz) превращаются в гипопусов. Гипопусы
Acaridlae
Зд7
через перьевые фолликулы или через дыхательные пути проникают
к ткани хозяина и залегают в подкожной ткани или в тканях трахеи;
там гипопусы живут и растут до восстановления нормальных усло-
вий, после чего снова выходят наружу и превращаются в прежнюю
форму. Робин и Менье рассматривают такое отклонение от нормаль-
ного жизненного цикла как защитное приспособление, спасающее
колонию от гибели при неблагоприятных условиях.
Pterolichus obtusus Robin паразитирует на разных птицах —ку-
рах, куропатках и т. д. В Южной Африке и Калифорнии со страу-
сов был собран Р. bicaudatus Gervais. Dermoglyphus elongatus
Megnin и D. minor Norner живут в стволах перьев домашней птицы.
ЛИТЕРАТУРА
Oudemans А. С., 1922. Ueber die metamorphose der vogetoewohnenden
Acaridiae, Tijdschr, v. Ent., 65, 184—191.
Spurlock G. M., Emlen I. T., 1942. Hypodectes chapini n. sp. (Acarina)
from the red-shafted flicker, J. Parasitol., 28 (4), 341—344.
Vitzthum H., 1933. Die endoparasitischc Deutonymphc von Pterolichus
nisi, Zeitschrift f. Parasitenkunde, 6 (2), 151—1G9.
Proctophyllodidae Megnin et Trouessart, 1883
(рис. 310, 311)
ДИАГНОЗ
Proctophyllodidae паразитируют в оперении птиц. У обоих полов име-
ются дорзальный и дорзолатеральный проподосомальные щитки, гисте-
росомальный щиток и несколько других мелких щитков. У самок зад-
ний край тела суживается и подразделяется на две лопасти; каждая из
лопастей снабжена мечевидным или щетинкообразным придатком.
У самцов задний край тела также часто бывает двулопастной и часто
несет листовидные придатки. Вертикальных щетинок на проподосоме
нет. На голенях ног I и II имеются палочковидные притупленные на
конце тактильные щетинки. Все лапки снабжены присосками.
РОДЫ
1. Proctophyllodes Robin, 1868
Тип рода Dermaleichus glandarinus Koch, 1840
2. Alloptes G. Canestrini, 1879
Тип рода Dermaleichus crassipes Canestrini, 1878
3. Favettea Trouessart, 1915
Тип рода Favettea heteroclyta Trouessart, 1915
4. Monojoubertia Radford, 1950 ( = Joabertia Oudemans, 1905
nomen praeoccupatum)
Тип рода Pterodectes microphyllus Robin et M6gnin, 1877
5. Montesauria Oudemans, 1905
Тип рода Pterodectes cylindricus Robin, 1868
№
Г лава Vll. Подотряд Sarcoptiformes
6. Pseudalges Trouessart, 1885 ( —Allaualges Trouessart, 1886)
Тип рода Pterocolus atialgoides Trouessart, 1885
7. Pterodectes Robin, 1868
Тип рода Pterodectes rutllus Robin, 1868
8. Pterophagus Megnin, 1877
Тип рода Pterophagus strictus Megnin, 1877
Рис. 310. Trouessartla Рис. 311. Trouessartia rosteri Berlese.
rosteri Berlese. Самка (вид сверху).
Самец (вид сверху).
9. Trouessartla Canestrini, 1899 ( = Pterocolus Haller, 1878 nomen
praeoccupatum)
Тип рода Dermaleichus corvitius Koch, 1840
ОБЩИЕ ЗАМЕЧАНИЯ
Proctophyllodidae — перьевые клещи, внешне очень похожие на
Analgesidae и Dermoglyphidae.
ЛИТЕРАТУРА
Vitzthum Н., 1929. Acari, Die Tierwelt Mitteleuropas, 3 (3), 1—112.
Глава VIII
SARCOPTIFORMES (ПРОДОЛЖЕНИЕ)
ORIBATEI DUGES, 1833
У Oribatei, или панцирных клещей, гнатосома обычно скрыта
в камеростоме. Имеющиеся иногда стигмы и трахеи открываются
через пористые поля. Обычно на проподосоме расположены псевдо-
стигматические органы. Тело обычно покрыто сильно склеротизиро-
нанным темноокрашенным панцырем, если же панцырь не развит, то
покровы тела кожистые. Аподемы тазиков погружены под кожу, но
все же заметны внешне, хотя и не так отчетливо, как у Acaridiae.
Каждая из лапок несет от одного до трех коготков, присосок на лап-
ках нет. Пальпы обычно пятичлениковые, все их сегменты подвижны.
11оловой диморфизм не выражен. Половых присосок три пары. У сам-
цов нет аданальных копулятивных присосок. У обоих полов половое
и анальное отверстия обычно прикрыты створчатыми щитками.
Европейские авторы разделяют Oribatei на много крупных групп,
однако эти клещи четко разделяются всего на две группы — Aptyctima
и Ptyctima. У Aptyctima проподосома неподвижно связана с гистеро-
сомой, а у Ptyctima — подвижно.
В пределах Aptyctima выделяют несколько естественных групп,
различающихся между собой по расположению генито-анальных щитков
и наличию или отсутствию птероморф (крыловидных придатков).
Ввиду сложности и недостаточной четкости этих различий мы не будем
здесь останавливаться на этих вопросах и рекомендуем читателю обра-
щаться за справками к работам Фитцтума, Бильмана и Зельника.
Возможность правильной оценки этих признаков может дать только
подробное изучение группы Aptyctima в целом.
Панцырные клещи ранее представляли только таксономический
интерес. Их хозяйственное значение выявилось после работ, показавших,
что Oribatei являются хозяевами различных ленточных глистов. В даль-
нейшем исследования почвенной фауны, значительную часть которой
составляют эти клещи, показали, что Oribatei, разлагая органическое
вещество в процессе пищеварения, способствуют повышению плодо-
родия почвы; в этом отношении они играют такую же роль, как дож-
итые черви. Оба эти обстоятельства достаточно убедительно показы-
вают целесообразность подробного изучения панцырных клещей. Пови-
цимому, особенно велико практическое значение панцырных клещей
как фактора, повышающего плодородие почвы. В США первые шаги
в изучении Oribatei сделал Джэкот. В Швеции некоторые данные
4М
Глава УШ. Sarcoptiformes (продолжение)
об этих клещах собрал Трегорд, изучавший лесные почвы. Ныш
в Дании Хаммер и Туксен ведут большую работу по изучению почвенной
фауны вообще и панцырных клещей в частности. При исследовании
почвенной фауны большую помощь оказывают воронки Берлезе, Туль
грена и различные их модификации; полученные результаты дают пред-
ставление о том огромном количестве материала, который можно
найти в почве.
Кэтс и Рункель (1948) составили обзор литературы, посвященной
выявлению роли панцырных клещей в качестве промежуточных хозяев
ленточных червей. Эти авторы показали, что многие виды ленточных
червей, в том числе паразитирующий у овец Moniezia expansa, исполь
зуют Oribatei в качестве промежуточных хозяев. Повидимому, клещи,
являющиеся промежуточными хозяевами ленточных червей, не обра
зуют единой систематической группы. Переносчиками могут служить
любые клещи, обычные на пастбищах и способные проглотить яйца
ленточного червя. Многие из клещей, встречающихся на пастбищах,
слишком малы для этого; другиЬ же, способные служить промежу-
точными хозяевами в лабораторных условиях, живут не на пастбищах,
а в лесном гумусе и в природе ленточными червями не заражаются.
В настоящее время в качестве промежуточных хозяев различных лен-
точных червей известны представители семейств Galumnidae, Oribatu-
lidae, Carabodidae, Notaspididae, Pelopidae, Liacaridae и Haplozetidae;
соответствующие виды перечисляются в разделах, посвященных этим
семействам. Этот список отнюдь не следует считать исчерпывающим;
он только дает представление о числе клещей, являющихся промежу-
точными хозяевами ленточных червей.
В качестве отправной точки для изучения Oribatei можно реко-
мендовать: работы Эвинга по систематике панцырных клещей штата
Иллинойс; отдельные работы Джэкота, перечисленные в посмертном
списке его трудов (см. Florida Entomologist, 24, № 2, 43—47, 1941);
прекрасно иллюстрированные статьи Гранжана; двухтомный труд
Майкла (1881—1883) по панцырным клещам Англии и два превос-
ходных немецких издания — Бильмана (1931) и Зельника (1929).
Имеется еще много других исследований, которые следует изучить,
прежде чем приступать к работе над панцырными клещами, но ука
занных выше работ достаточно для того, чтобы дать начинающем \
общее представление об этой группе.
В настоящее время имеется еще очень мало работ по фауне пан-
цырных клещей штатов США, расположенных на запад от Миссисипи
и практически еще совершенно необследованных в этом отношении.
Восточные и особенно северо-восточные штаты США были более или
менее подробно исследованы Джэкотом, который обнаружил там н<
сколько европейских видов. Один из таких видов — Scheloribates leav
gatus (Koch) — был найден Кэтсом и Рункелем (1948) в Северной Дакоп
5. laevigatas распространен в Германии и встречается также в Советской
Союзе. Таким образом, при изучении американских форм рекомеп
Oribatei
401
дуется соблюдать некоторую осторожность и сопоставлять находимые
виды с азиатскими и европейскими видами. Это правило распростра-
няется на всех клещей как свободноживущих, так и паразитических.
Образ жизни панцырных клещей мало изучен. Поэтому в даль-
нейшем изложении в разделах, посвященных отдельным семействам,
данные о биологии приводятся только в тех случаях, когда достоверность
их очевидна. В общем панцырные клещи, как правило, живут в почве
и в подстилке; они встречаются также на стволах деревьев, во мху,
лишайнике и других аналогичных местах. Исключение составляют
Hydrozetes, живущие в воде. Питаются панцырные клещи органиче-
скими веществами.
Мы не придерживаемся здесь сложных подразделений, введенных
европейскими авторами (когорты, фаланги и т. д.), и рассматриваем
панцырных клещей как единую большую группу, подразделяя ее только
па семейства. В некоторых случаях семейства охарактеризованы недо-
статочно четко и их, вероятно, в будущем придется пересмотреть.
В некоторых случаях от выделения семейств пришлось отказаться
совсем, но в общем мы придерживаемся здесь общепринятой системы
Oribatei.
ЛИТЕРАТУРА
Michael A. D., 1881—1883. British Oribatidae, 1, 1881; 2, 1883.
Scllnick М., 1929. Fortnenkreiss. Hormilben, Oribatei (см. Die Tierwelt
Mittelauropas, 3, 1—42).
Willmann C., 1931. Moosmilben oder Oribatiden (Oribatei) (cm. Tierwelt
Deutschland, 22, 79—200).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СВОЙСТВ ORIBATEI 1
1. Ротовые органы сверху незаметны. Тело мягкое или твердое
................................................................2
— Ротовые органы сверху заметны. Тело всегда мягкое ..............
...............................................Palaeacaridae
2. Просома и опистосома сочленяются друг с другом неподвижно
................................................................3
— Просома и опистосома сочленены друг с другом подвижно . . 32
3. Половое и анальное отверстия никогда не лежат на общем
брюшном щите; шов, разделяющий эти отверстия, проходит
непосредственно позади генитальных щитков. Анальное отверстие
лежит на заднем конце гистеросомы...............................4
— Половое и анальное отверстия лежат на общем брюшном
щите .......................................................... 5
4. Гистеросома цилиндрическая. Половое отверстие небольшое, ото-
двинуто далеко назад и лежит непосредственно впереди аналь-
ного отверстия . .............................. Eulohtnanniidae
1 Составлена по Зельнику и Бильману.
26 Зак. 1785
402
Глава Vf[[. Sarcoptiformes (продолжение)
— Гистеросома мешковидная. Половое отверстие отодвинуто далеко
вперед, расстояние между ним и анальным отверстием равно длине
анального отверстия..............................Nanhermanniidae
— Половое и анальное отверстия расположены каждое на отдельном
брюшном щитке...................................................Epilohmanniidae
5. Половое и анальное отверстия крупные и соприкасаются друг
с другом. Совместно они простираются на всю длину опистосомы
...........................................................6
— Половое и анальное отверстия средней величины; они имеют
более или менее трапециевидную или овальную форму и отделены
друг от друга значительным участком брюшного щита .... 13
6. Половое и анальное отверстия окружены общей узкой рамкой
...............................................................7
— Половое и анальное отверстия не заключены в узкую рамку, но
расположены на широком общем брюшном щите, достигающем
боковых краев гистеросомы.................................10
7. Гистеросома уплощена снизу, а сверху или уплощена, или слабо
выпуклая................................................................... 3
—- Гистеросома цилиндрическая.................'..... Lolnnanniidac
8. Гистеросома со спинной стороны слабо выпуклая.................................9
— Гистеросома со спинной стороны уплощена; обычно ее края
приподняты или же спинная поверхность впалая . . Camisiidae
9. Имеются псевлостигмы и псевдостигматические органы. Поперечные
швы на гистеросоме иногда выражены............. Hypochthoniidae
— Псевдостигм и псевдостигматических органов нет, на их месте
развиты только простые щетинки. Поперечных швов на гистеро-
соме никогда не бывает...........................Malaconothridae
10. Взрослые клещи сохраняют на спинной поверхности линочные
шкурки неполовозрелых фаз.....................................11
— Взрослые не несут на спинной поверхности гистеросомы линоч-
ных шкурок. Гистеросома сильно выпуклая . . . Hermanniidae
И. Трубчатых или других выростов на боковых краях гистеросомы
нет..............................................................................12
— На боковых краях гистеросомы имеются трубчатые или иные
выросты..........................................Hermanniellidae
12. Спинная поверхность гистеросомы плоская или слабо выпуклая.
Генитальные щитки разделены поперечным швом . . Neoliodidae
— Спинная поверхность гистеросомы плоская. Тело и ноги покрыты
толстым слоем секретов. Генитальные щитки не разделены попе-
речным швом.......................................Plateremaeidae
13. Гистеросома без птероморф, или крыловидных придатков . . 14
— Гистеросома с явственными, хотя иногда и небольшими, птеро-
формами, или крыловидными придатками.............................................21
14. Хелицеры обычного типа, клешневидные, жующие.................................15
— Хелицеры длинные, узкие, ножевидные; на дистальном конце они
покрыты зубчиками.................................. Gustaviidae
Orlbatei
403
15. Ноги IV, как и все остальные, не видоизменены в связи с при-
способлением к прыганию.........................................16
— Ноги IV прыгательные; у мертвых ноги направлены вперед
................................................Zetorchestidae
16. Боковые и задний края спинного щита не загибаются вентрально
................................................................17
Боковые и задний края спинного щита загибаются вентрально
так, что значительная часть спинного щита видна с брюшной
стороны тела *...............................Cymbaereniaeidae
17. Ноги Ш и IV расположены на боковых краях брюшка или неда-
леко от них ....................................................18
— Ноги III и IV расположены под брюшком на значительном рас-
стоянии от боковых краев............................Liacaridae
18. Ноги обычного строения; они всегда короче тела' и никогда не
бывают четковидными. Проподосома с пластинчатым, гребневид-
ным или щитовидным выростом.....................................19
— Ноги длинные, обычно длиннее тела; в противном случае
ноги в виде четок с члениками, вздутыми как луковицы . .
................•.............•.....................Belbidae
19. Шов между проподосомой и гистеросомой иногда имеется. Псев-
достигмы и псевдостигматические органы имеются*.................20
— Шва между проподосомой и гистеросомой нет. Псевдостигм и
псевдостигматических органов пет. Тектоподий II и III пет
.................................................. Ameronothridae
20. Спинная поверхность гистеросомы либо гладкая, либо покрыта
легким точечным орнаментом. Бедра ног I и II никогда не имеют
длинного стебелька.............................Eremaeidae
— Спинная поверхность гистеросомы обычно с хорошо развитой
скульптурой, реже она покрыта легким точечным орнаментом.
Бедра ног I и II часто несут тонкий стебелек и на дистальном
конце вздуты......................................Carabodidae
21. Птероморфы сочленены с телом неподвижно.................22
- Птероморфы сочленены с телом подвижно...................28
22. Птероморфы причленены только к гистеросоме и не прикрывают
сколько-нибудь значительной части проподосомы..............23
— Птероморфы причленены как к гистеросоме, так и к пропо-
досоме; они прикрывают псевдостигмы, псевдостигматические
органы и значительную часть проподосомы. Птероморфы неболь-
шие, не загибаются вентрально. Генитальных щетинок две пары
...................................................... Oripodidae
23. Птероморфы заходят далеко за передний край гистеро-
сомы ..........................................................24
— Птероморфы не заходят за передний край гистеросомы ... 25
24. Птероморфы от места прикрепления направлены вперед, но не за-
гибаются вентрально. Ламеллы иногда довольно крупные, но не
обращают на себя внимания.............................Tenuialidae
26*
Глава VIII. Sarcoptiformes (продолжение)
— Птероморфы заходят далеко вперед за передний край гистеро-
сомы и сильно загибаются вентрально. Ламеллы прикрываю,
проподосому...........................................Notaspididac
25. Птероморфы небольшие и не загибаются вентрально . ... 26
— Птероморфы крупные и загибаются вентрально. Ламеллы покры-
вают проподосому.....................................Oribatellidae
26. Ламеллы не прикрывают сколько-нибудь значительной части
проподосомы. Хелицеры обычные, без выростов.....................27
— Ламеллы прикрывают большую часть проподосомы. Неподвиж-
ный палец клешни хелицер снабжен направленными вперед рого-
видными выростами.....................................Microzetidac
27. Птероморфы лежат в одной плоскости и не загибаются вентрально.
У взрослых имеются четыре пары генитальных щетинок ....
.................................................Oribatu idae
— Птероморфы загибаются вентрально. У взрослых генитальных щети-
нок шесть пар........................................Ceratozetidae
28. Птероморфы не заходят вперед или назад за линию их сочлене-
ния с телом....................................................2!)
— Птероморфы заходят вперед или назад за линию их сочленения
с телом.........................................................30
29. Генитальные щитки несут пять пар щетинок (по Фитцтуму, иногда
только четыре пары). Хелицеры обычные, крепкие. Интерламел-
лярные щетинки нормальные........................... . Haplozetidae
— Генитальные щитки несут шесть пар щетинок. Хелицеры всегда
(за исключением Propelops) длинные, тонкие, с маленькой кле-
шней. Интерламеллярные щетинки нормальные или ложечковидные;
иногда они отсутствуют...............................Pelopidae
30. Ламеллы не покрывают сколько-нибудь значительной части про-
подосомы .......................................................31
— Ламеллы покрывают большую часть проподосомы. Генитальные
щитки с пятью парами щетинок. Передняя пара циркуманальных
щетинок сидит у переднего края анального отверстия ...............
................................................Epactozetjdae
31. Генитальные щитки с шестью парами щетинок. Передняя пара
циркуманальных щетинок сидит по бокам от анальных щитков .
. . ..............................................Galuninidae
— Генитальные щитки с пятью парами щетинок. Передняя пара
циркуманальных щетинок сидит впереди анального щитка . . .
...............................................Parakalummidae
32. Гистеросома не расчленена...................33
— Гистеросома расчленена.......................Protoplophoridae
33. Генитальное и анальное отверстия расположены на крупном
брюшном щите; они отделены друг от друга . . . Mesoplophoridae
— Генитальное и анальное отверстия не расположены на крупном
щите и соприкасаются друг с другом..................Phthiracaridac
Oribatei
403
Palaeacaridae Grandjean, 1932
(рис. 312—315)
ДИАГНОЗ
Palaeacaridae имеют очень тонкую кутикулу, не склеротизирован-
пую в сколько-нибудь значительной степени. Тело разделено на про-
нодосому и гистеросому; между ними имеется шеевидное сужение.
11роподосома несет пять пар щетинок (ростральные, ламеллярные,
Рис. 312. Grandjeanacarus
araneola Grandjean
• (по Граижапу, 1932).
Самка (вид сверху).
Р и с. 313. Acaronychus trd-
gardhl Grandjean
(no Гранжану, 1932).
Самка (вид сверху).
иитерламеллярные, экзостигматические и угловые). Псевдостигмати-
ческие органы нитевидные или ланцетовидные; они сидят в простых,
чашевидных псевдостигмах. Хелицеры крупные, неприкрытые и высту-
пают далеко вперед от рострального щитка. Клешня хелицер зуб-
чатая, приспособленная для разжевывания растительных тканей.
Пальпы пятичлениковые, простые. Максиллы с двумя парами при-
датков: (а) — сильно склеротизированные, двучлениковые, зубчатые
па конце наружные максиллярные лопасти и (б) — тонкие, снабженные
гремя парами щетинок, внутренние максиллярные щитки. Ноги пятн-
или шестичлениковые. На гистеросоме иногда имеются поперечные
швы и поперечные ряды щетинок. Дыхательная система состоит из
стигм, расположенных на наружной стороне основания хелицер, и
коротких трахей (Трегорд).
Гранжан (1932) разделил Palaeacaridae на три семейства Palae-
acaridae, Acaronychidae и Parhypochthonidae, но мы полагаем, что
406
Глава VIII. Sarcoptiformes (продолжение)
лучше пока объединять этих клещей в одно семейство—Palaeacaridae,
подразделяя его на подсемейства. Вопрос в целом остается неясным
Трегорд (1932) выделил этих клещей в особый подотряд—• Palae
acariformes. Гранжан (1932) дока-
зывает, что Palaeacaridae являются
сборной группой примитивных
панцырных клещей. Захваткин
(1945, 1946) без дальнейших
пояснений утверждает, что его
исследования позволяют принять
предложение Трегорда о выде-
лении Palaeacaridae в особый
подотряд. Таким образом, в на-
стоящее время систематики при-
держиваются более или менее
произвольных взглядов на система-
тическое положение наиболее при-
митивных панцырных клещей —
Рис. 315. Parkypochlhoniiis aphi-
dinus Berlese (по Всрлсзс, 1904).
v
! 1
Р и с. 314. Acaronychus trligardhi
Grandjean (по Гранжану, 1932).
Самка (вид сбоку).
Самка (вид сверху).
Pa'aeacaridae. Окончательное решение вопроса требует дальнейших
работ специалистов, интересующихся этими примитивными формами.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ PALAEACARIDAE
1. Ноги шестичлениковые, все сегменты их подвижны.............2
— Ноги пятичлениковые, все сегменты их подвижны. Гистеросома
с поперечными швами и отделена от проподосомы швом . . .
.........................................Parhypochthoniinae
2. Гистеросома с поперечным швом; проподосома и гистеросома
разделены швом. Половых присосок две или три пары ....
...................................................Palaeacarinae
— Гистеросома без поперечных швов; настоящего расчленения тела
на проподосому и гистеросому нет. Половых присосок три пары.
Гениталии снабжены шиповидными выростами . . Acaronychinae
Oribatei.
407
РОДЫ
Palaeacarinae Grandjean, 1932
I. Palaeacarus Tragardh, 1932
Тип рода Palaeacarus hystricinus Tragdrdh, 1932
2. Aphelacarus Grandjean, 1932
Тип рода Parhypochthoninus acarittus Berlese, 1910
3. Archeonothrus Tragardh, 1906
Тип рода Archeonothrus natalensis Tragdrdh, 1906
4. Beklentishevla Zakhvatkin, 1945
Тип рода Beklemlshevta galeodula Zakhvatkin, 1945
5. Grand]eanacarus Zakhvatkin, 1945
Тип рода Palaeacarus araneola Grandjean, 1932
6. Trdgardhacarus Zakhvatkin, 1945
Тип рода Trdgardhacarus lapshovl Zakhvatkin, 1945
Acaronychinae Grandjean, 1932
1. Acaronychus Grandjean, 1932
Тип рода Acaronychus trdgardhi Grandjean, 1932
P a r h у p о c h t h о n i i n a e Grandjean, 1932
1. Parhypochthonius Berlese, 1904
Тип рода Parhypochthonius aphidittus Berlese, 1904
2. Gehypochthonius Jacot, 1936
Тип рода Gehypochthonius rhadamantus Jacot, 1936
ОБЩИЕ ЗАМЕЧАНИЯ
Эти клещи мало изучены, и их знают лишь немногие акарологи.
Palaeacaridae живут в лесной подстилке и в верхних слоях почв;
в дальнейшем, когда будут сделаны более тщательные сборы,
вероятно, окажется, что они распространены гораздо шире, чем это
теперь известно.
ЛИТЕРАТУРА
G г а и d j с а п Р„ 1932. Au sujct des Palaeacariformes Tragardh, Bull. Paris
Mus. d’Hist. Nat., Ser. 2, 4 (4), 411—426.
Jacot A. P., 1936. Some primitive moss-mites of North Carolina, J. Elisha
Mitchell Sci. Soc., 52 (1), 20—26, pl. 1.
Jacot A. P., 1936. More primitive moss-mites of North Carolina, J. Elisha
Mitchell Sci. Soc., 52 (2), 247—353, pl. 20.
Jacot A. P., 1938. More primitive moss-mites of North Carolina — III, J. Eli-
sha Mitchell Sci. Soc., 54 (1), 127—137, pls. 12.
Tragardh I., 1932. Palaeacariformes, a new suborder of Acari, Arkiv. f Or
Zoologi, 24.B (2), 1—6.
Захваткин A. A., 1945. О нахождении Palaeacariformes (Acarina) в СССР,
Докл. Акад, наук СССР, 47 (9), 702—705.
Захваткин А. А., 1945. К морфологии Bek emishevia ga'eodula ri. g.,
n. sp. — нового представителя Palaeacariformes (Acarina), Бюлл. Моск-
общ. исп. прир., 50 (3—4), 60—71.
408
Глава VIII. Sarcoptiformes (продолжение)
ДИАГНОЗ
Eulohmanniidae Grandjean, 1931
(рис. 316)
Eulohmanniidae — стройные клещи с продолговатым телом, у кото-
рых проподосома и гистеросома соединены тонкой перепонкой.
Рис. 316. Eulohmannia riba-
gat Berlese (по Берлезе, 1910).
Самка (вид сверху).
Гистеросома цилиндрическая. Генитальные щитки отодвинуты далеко
назад и отделены от анальных щитков швом гистеросомального щитка.
РОДЫ
1. F.ulolimannla Berlese, 1910 (= Arthronothrus Tragardh, 1910)
Тип рода Lohmannia (Eulohmannia) ribagal Berlese, 1910
2. Collohmannia Sellnick, 1922
Тип рода Collohmannia gigantea Sellnick, 1922
3. Perlohmannia Berlese, 1916
Тип рода Lohmannia insignis Berlese, 1904
ОБЩИЕ ЗАМЕЧАНИЯ
Collohmannia и Perlohmannia помещены в сем. Eulohmanniidae
условно и требуют более подробного изучения.
ДИАГНОЗ
Nanhermanniidae Sellnick, 1924
(рис. 317, 318)
Nanhermanniidae — удлиненные, узкотелые клещи с мешковидной
гистеросомой. Анальные щитки лежат в задней части брюшка. Гени-
тальные щитки смещены вперед и отделены От анального отверстия
Orlbaiel
409
швом, проходящим между спинной и брюшной сторонами гистеросомы.
?)тот шов на брюшке смещен вперед так, что он не захватывает
Рис. 318. Nanhermannia е'.е-
gantula Berlese
(по Берлезе, 1913).
Самка (вид сверху).
Р и с. 317. Nanhermannia папа
(Nicolet).
Гистеросома взрослой особи (вид снизу);
показан шов, разделяющий генитальные
и анальные щитки.
анального отверстия, которое, таким образом, фактически расположено
в брюшной части спинного щита.
РОДЫ
1. Nanhermannia Berlese, 1913
Тип рода Nothrus nanus Nicolet, 1855
ЛИТЕРАТУРА
Hartman A. G., 1949. A new species of Nanhermannia with notes on (he
genus, Proc. Ent. Soc, Wash,, 51 (4)t 169—J71.
410
Глава VfIL Sarcoptiformes (продолжение)
Epilohmanniidae Oudemans, 1923
(рис. 319, 320)
ДИАГНОЗ
Epilohtnanniidae — мелкие клещи, обладающие двумя вырезанными
по заднему краю вентральными щитками, расположенными один за
другим. В вырезке одного из щитков расположены генитальные,
Рис. 319. Epilohmannia
cylindrica (Berlese)
(по Удемансу, 1915).
Самка (вид снизу); показаны
границы между щитками.
Рис. 320. Epilohmannia
cylindrica (Berlese)
(по Гранжану, 1946).
Личинка (пид снизу); между та-
зиками ног I и II можно видеть
клапарсдов орган (см. рис. 255).
а в вырезке другого — анальные щитки. Пластинки и тектоподии
отсутствуют. Последние четыре членика ног почти равны по длине
и снабжены одним коготком. Пальпы двучлениковые; ротовые органы
прикрыты сверху. Личиночные формы Epilohmannia cylindrica (Berlese)
обладают клапаредовым органом.
РОДЫ .
1. Epilohmannia Beriese, 1916 (== Lesseria Oudemans, 1916)
Тип рода Lohtnannia cylindrica Berlese, 1904
2. Archegozetes Grandjean, 1931
Тип рода Epilohmannia (?) magna Sellnick, 1925
Oribatei
411
ДИАГНОЗ
Lohmanniidae Grandjean, 1931
(рис. 321, 322)
Lohmanniidae — клещи с цилиндрической гистеросомой, широко
сочленяющейся с проподосомой. Генитальные и анальные щитки
непрерывные. Пальпы состоят из четырех подвижных члеников.
Рис. 321. Torpacwus
omittens Grandjean
(по Гранжану, 1950).
Самка (вид снизу).
Рис. 322. Torpacarus
omittens Grandjean
(по Гранжану, 1950).
Самка (вид сверху).
РОДЫ
1. Lohmannia Michael, 1898 (= Michaella Haller, 1884 nomen
praeoccupatum)
Тип рода Michaella par ado xa Haller, 1884
2. Annectacarus Grandjean, 1950
Тип рода Annectacarus mucronatus Grandjean, 1950
3. Cryptacarus Grandjean, 1950
Тип рода Cryptacarus promecus Grandjean, 1950
4. Merlstacarus Grandjean, 1934
Тип рода Merlstacarus porcula Grandjean, 1934
5. Thamnacarus Grandjean, 1950
Тип рода Lohtnannia desertlcola Grandjean, 1934
6. Torpacarus Grandjean, 1950
Тип рода Torpacarus omittens Grandjean, 1950
ОБЩИЕ ЗАМЕЧАНИЯ
Гранжан (1950) сохранил в этом семействе только перечисленные
выше роды, считая, что они являются обособленной однородной груп-
пой, характеризующейся примитивными признаками. Эти клещи не
412
Глава VIII. Sarcoptiformes (продолжение)
имеют трахей и боковых гистеросома г ических желез и отличаются
постепенным развитием без метаморфоза: личиночная, три нимфаль-
ные и взрослая фазы походят одна на другую.
ЛИТЕРАТУРА
Grandjean F., 1950. Etude snr les Lohnianniidae (Oribates, Acariens),
Archie. Zool. Exper. et Generate, 87 (2), 95—161.
Hypochthoniidae Berlese, 1910
ДИАГНОЗ
(рис. 323—326)
У Hypochthoniidae спинная
делена одним или несколькими
поверхность гистеросомы часто раз-
поперечными швами; эти поперечные
Рис. 324. Hypochthonius ги-
fulus var. paucipectinatus
Jacot.
Самка (вид сверху).
Рис. 323. Pterochthonlus
angelus (Berlese)
(ио Гранжану, 1950).
А —самка (вид сверху); Б — де-
таль проподосомальной щетинки;
В —пальпа; /'—когтевидная ще-
тинка яйцеклада; Д— хелицера.
швы могут и отсутствовать. Тело слабо склеротизировано. Гениталь-
ные щитки обычно шире анальных. Одновременно развивается только
одно яйцо (исключение составляют Trhypochthonlus и Trhypochtho-
nlellus, у которых одновременно могут созревать 4—6 яиц; Вильман
выделяет эти два рода в особое подсемейство — Trhypochthoniinae,
а Фитцтум — в особое семейство). Гранжан (1946) разделил Нуро-
рhthoniidae на семейства Hypochthoniidae, Brachychthoniidae, Cosmo-
Oribatei
41з
chthoniidae и Sphaerochthoniidae; основанием для этого послужило
число и положение поперечных швов. Позднее Гранжан (1950) выделил
Pterochthonlus angelus Berlese в особое семейство Pterochthoniidae;
основанием к этому послужило примитивное строение у этого клеща
хелицер, сохранение преанального сегмента, наличие когтевидных
щетинок яйцеклада и крупных красивых щетинок на теле. Р. ange-
lus— примитивный клещ, встречающийся в Западной Европе и в горах
Рис. 325. Cosmochthonius
plumatus Berlese.
Самка (вил сверху).
Рис. 326. Eniochthonius
pallidulus (Michael)
(по Гранжану, 1933).
Щитки брюшной поверхности
тела.
Мексики. Зельник объединяет все роды в одно семейство, и мы
следуем здесь его системе, хотя возможно, что последующие иссле-
дования подтвердят точку зрения Гранжана.
РОДЫ
1. Hypochthonius Koch, 1836 (= Hypochthoniella Berlese, 1910 =
= Arthrochthonius Ewing, 1917)
Тип рода Hypochthonius rufulus Koch, 1836
2. Amnemochthonius Grandjean, 1948
Тип рода Amnemochthonius taeniophorus Grandjean, 1948
3. Atopochthonius Grandjean, 1948
Тип рода Atopochthonius artiodactylus Grandjean, 1948
4Н Глава VIII. Sarcoptiformes (продолжение)
4. Brachychochthonius Jacot, 1938
Тип рода Brachychochthonius jugatus Jacot, 1938
5. Brachychthonius Berlese, 1910
Тип рода Hypochthonius brevis Michael, 1887
6. Cosmochthonius Berlese, 1910
Тип рода Hypochthonius lanatus Michael, 1887
7. Eniochthonius Grandjean, 1933
Тип рода Hypochthonius pallidulus Michael, 1887
8. Eobrachychthonius Jacot, 1936
Тип рода Eobrachychthonius sexnotatus Jacot, 1936
9. Eohypochthonius Jacot, 1938
Тип рода Hypochthonius gracilis Jacot, 1936
10. Epilohmannoides Jacot, 1936
Тип рода Epilohmannoides terrae Jacot, 1936
11. Haplochthonius Willmann, 1930
Тип рода Haplochthonius simplex Willmann, 1930
12. Heterochthonius Berlese, 1910
Тип рода Cosmochthonius (Heterochthonius) gibbus Berlese, 1910
13. Malacoangella Berlese, 1913
Тип рода Malacoangella remigera Berlese, 1910
14. Poecilochthonius Balogh, 1943
Тип рода Brachychthonius italicus Berlese, 1910
15. Pterochthonlus Berlese, 1913
Тип рода Cosmochthonius angelus Berlese, 1910
16. Sphaerochthonlus Berlese, 1910
Тип рода Hypochthonius splendldus Berlese, 1904
17. Trhypochthoniellus Willmann, 1928
Тип рода Trhypochthoniellus setosus Willmann, 1928
18. Trhypochthonius Berlese, 1904 (=Tumidalvus Ewing, 1908)
Тип рода Hypochthonius tectorum Berlese, 1896
ЛИТЕРАТУРА
Grandjean F., 1928. Sur un oribatidS pottrvu d’yeux, Bull. Soc. Zool.
France, 53 (4), 235—242.
Grandjean F., 1946. Les Enarthronota (Acariens) 1 serie, Annales Sciences
Naturelies, ser, Botanique et Zoologie, 11 Ser., 8, 213—248.
Grandjean F., 1948. Les Enarthronota (Acariens) 2 serie, Annales Sciences
Naturelles, ser. Botanique et Zoologie, 11 Ser., 10, 29—58.
Grandjean F., 1950. Les Enarthronora (Acaries) 3 serie, Annales Sciences
Naturelies, ser. Botanique et Zoologie, 11 Ser., 12, 85—107.
Jacot A. P., 1936. Some primitive moss-mites of North Carolina, J. Elisha
Mitchell Sci. Soc., 52 (1), 20—26, pl. 1.
Jacot A. P., 1936. More primitive moss-mites of North Carolina, J. Elisha
Mitchell Sci. Soc., 52 (2), 247—353, pl. 20.
Jacot A. P., 1938. More primitive moss-mites of North Carolina, J. Elisha
Mitchell Sci. Soc., 54 (1), 127—137, pl. 12.
Womersley H., 1945. Australian Acarina. The genera Brachychthonius
Berl. and Cosmochthonius Bert. (Hypochthonidae-Oribatoidea), Records
South Austral. Mus., 8 (2), 219—223.
Oribatei
413
Malaconothridae Berlese, 1916
(рис. 327)
ДИАГНОЗ
Malaconothridae обычно имеют серовато- или желтовато-коричневую
окраску. Спинная сторона тела слегка выпуклая. Настоящих псевдо-
стигм или псевдостигматических органов нет, и на их месте сидят
простые щетинки.
Рис. 327. Malaconothrus egregia
Berlese (по Берлезе, 1905).
Самка (вид сверху); обратить внимание на
отсутствие псевдостигматических органов.
РОДЫ
1. Malaconothrus Berlese, 1905
Тип рода Nothrus monodactylus Michael, 1888
2. Mucronothrus Tragardh, 1931
Тип рода Mucronothrus rostratus TragSrdh, 1931
3. Trimalaconothrus Berlese, 1916
Тип рода Malaconothrus {Trimalaconothrus') indusiatus Berlese,
1916
Camisiidae Sellnick, 1928
(рис. 328)
ЦИАГНОЗ
У Camisiidae половое и анальное отверстия соприкасаются друг
с другом. Рядом с половым и анальным отверстиями расположены
небольшие склеротизированные пластинки, или щитки. Анальное
отверстие прикрыто парой таких продольных склеротизированных
416
Глава VlII. Sarcopttformes (продолжение)
щитков, рсевдостигматические органы имеют форму воронки или
раковины. Спинная поверхность гистеросомы плоская, края ее обычно
Рис. 328. Camisia segnis (Hermann)
(по Гранжану, 1936).
Л —самка (вид сверху); /> —сильно увеличен-
ная дорзальная щетинка.
приподняты. У многих видов Camisia на заднем крае тела сидят
своеобразные удлиненные выросты.
РОДЫ
1. Camisia v. Heyden, 1826
Тип рода Notaspls segnis Hermann. 1804
2. Acronothrus Berlese, 1916
Тип рода Nothrus cophinarius Michael, 1907
3. Hemlnothrus Berlese, 1913
Тип рода Nothrus targionii Berlese, 1885
4. Nothrus Koch, 1836 (== Angelia Berlese, 1885 = Gymnonothrus
Ewing, 1917)
Тип рода Nothrus palustrls Koch, 1839
5. Platynothrus Berlese, 1913
Тип рода Nothrus peltifer Koch, 1839
6. Uronothrus Berlese, 1913
Тип рода Nothrus segnis Koch, 1839
Hermanniidae Sellnick, 1928
(рис. 329—331)
ДИАГНОЗ
Hermanniidae имеют сильно выпуклую гистеросому. Половое и
анальное отверстия лежат у них на общем брюшном щите и сопри-
касаются друг с другом.
Рис. 329. Hermannia
gibba (Koch).
Самка (вид сверху).
Рис. 330. Hermannia gibba
(Koch).
Взрослая особь (вид снизу); пока-
зано расположение щитков.
Рис. 331. Masthermannia
mammillaris (Berlese)
(по Берлезе, 1913).
Взрослая особь (вид сверху); обратите
внимание на причудливое строение
тела.
27 Зак. 1785
418 Глава VIII. Sarcoptiformes (продолжение)
РОДЫ И ПОДРОДЫ
1. Hermannia Nicolet, 1855
a) Hermannia s. str.
Тип подрода Nothrus gibbus Koch, 1839
6) Phyllhertnannia Berlese, 1916
Тип подрода Hermannia phyllophora Michael, 1908
2. Masthermannia Berlese, 1913
Тип рода Angelia mammillaris Berlese, 1904
ДИАГНОЗ
Neoliodidae Willmann, 1913
(рис. 332, 333)
У Neoliodidae боковые и задние края спинного щита загибаются
под углом вентрально так, что значительная его часть заходит на
Рис. 332. Teoliodes madininensts
Grandjean (по Гранжану, 1934).
Самка (вид сверху)»
Рис. 333. Neollodes sp.
Расположение щитков на брюшной
поверхности.
брюшко. Спинная сторона гистеросомы целиком покрыта линочными
шкурками молодых фаз; эти шкурки лежат одна на другой. Клапаны
полового отверстия разделены на две части поперечным швом.
РОДЫ
1. Neoliodes Berlese, 1888 (= Liodes Heyden, 1826 nomen praeoc-
cupatum = Udetaliodes Jacot, 1929)
Тип рода Notaspis theleproctus Hermann, 1804
Orlbatei
419
2. Embolacarus Sellnick, 1918f
Тип рода Embolacarus pergratus Sellnick, 1918 (согласно Фитц-
туму, систематическое положение этого рода точно не устано-
влено).
3. Platyliodes Berlese, 1916
Тип рода Nothrus doderleinii Berlese, 1883
4. Porollodes Grandjean, 1934
Тип рода Nothrus theleproctus Michael, 1888
5. Teleioliodes Grandjean, 1934
Тип рода Teleioliodes madininensis Grandjean, 1934
ДИАГНОЗ
Cymbaeremaeidae Willmann, 1931
(рис. 334, 335)
Птероморфы отсутствуют. Спинной щит переходит на Орюшную
сторону так, что снизу видна значительная его часть. Тело скуль-
птировано, но в различной степени.
Рис. 334. Cymbaeremaeus
(Scapheremaeus) marginalis
(Banks).
Самка (вид сверху).
Рис. 335. Cymbaeremaeus
{Scapheremaeus) marginalis
(Banks).
Самка (вид снизу); видно располо-
жение щитков.
РОДЫ[И ПОДРОДЫ
1. Cymbaeremaeus Berlese, 1896
a) Cymbaeremaeus s. str.
Тип подрода Eremaeus cymba
6) Scapheremaeus Berlese, 1910
Тип подрода Eremaeus cymba
Nicolet, 1855
(== Mulvius Sellnick,- 1918
Berlese, 1886
27*
420
Глава VIII. Sarcoptiformes (продолжение)
2. Eremella Berlese, 1913
Тип рода Eremella vestita Berlese, 1913
3. Micreremus Berlese, 1908
Тип рода Eremaetis brevipes Michael, 1888
4. Tectocymba Sellnick, 1918 f
Тип рода Tectocymba rar a Sellnick, 1918
ДИАГНОЗ
Belbidae Willmann, 1931
(рис. 336—338)
Птероморфы отсутствуют. Хелицеры обычные. Спинной щит обыч-
ный и не загибается на брюшную сторону. Ноги III и IV расположены
Рис. 336. Belba sp.
Hora I.
Рис. 337. Belba jacoti Wilson
(по Уилсону, 1936).
Самка (вид сбоку); на спине заметны ним-
фальные линочные шкурки.
Рис. 338. Belba jacoti Wilson
(по Уилсону, 1936).
Самка (вид сверху); линочных шкурок
на спине нет.
на краю или близко к краю тела. Как правило, крупные клещи, ноги
у которых длиннее тела. У мелких видов ноги четковидные. Иногда
на спинной поверхности взрослых клещей сохраняются линочные
шкурки неполовозрелых фаз.
Oribatei
42!
РОДЫ И ПОДРОДЫ
1. Belba v. Heyden, 1826
Тип рода Notaspls corynopus Hermann, 1804
2. Allodamaeus Banks, 1947
Тип рода Allodamaeas ewingi Banks, 1947
3. Amerus Berlese, 1896
a) Amerus s. str.
Тип подрода Belba troisli Berlese, 1883
6) Neamerus Willmann, 1939
Тип подрода Amerus {Neamerus) lundbladl Willmann, 1939
4. Damaeus Koch, 1836
Тип рода Damaeus auritus Koch, 1836
5. Dameobelba Sellnick, 1928
Тип рода Orlbata mitiutissitnus Sellnick, 1920
6. Gymnodamaeus Kulczynski, 1902
Тип рода Damaeus bicostatus Koch, 1836
7. Jacotella Banks, 1947
Тип рода Gymnodamaeus qaadrlcaudlcalus Jacot, 1937
8. Porobelba Grandjean, 1936
Тип рода Orlbata spinosus Sellnick, 1920
ДИАГНОЗ
Eremaeidae Willmann, 1931
(рис. 339, 340)
У Eremaeidae боковые и задние края гистеросомального щита не
загибаются вентрально. Ноги III и IV причленяются к боковому краю
брюшной поверхности тела или на близком расстоянии от него. Ноги
обычного строения и короче, чем тело. Бедра ног I и II не несут
заметных стебельков или выростов лопаточного угла. Проподосома
несет листовидные ламеллы. Спинная сторона гистеросомы гладкая
или покрыта легким точечным рисунком.
РОДЫ И ПОДРОДЫ
1. Eremaeus Koch, 1836
Тип рода Eremaeus hepaticus Koch, 1836
2. Amerobelba Berlese, 1908
Тип рода Amerobelba decedens Berlese, 1908
3. Autogneta Hull, 1916
Тип рода Notaspis longllamellata Michael, 1888
4. Caleretnaeas Berlese, 1910
Тип рода Notaspis monilipes Michael, 1882
5. Calvoppia Jacot, 1934
Тип рода Calvoppia perkinsl Jacot, 1934
6. Ceratoppia Berlese, 1908
Тип рода Notaspis bipilis Hermann, 1804
422
Глава VIH. Sarcoptiformes (продолжение)
7. Conoppia Berlese, 1908
Тип рода Oppla microptera Berlese, 1885
8. Damaeolus Paoli, 1908
Тип рода Damaeosoma aperatum Berlese, 1903
9. Datnpfiella Sellnick, 1931
Тип рода Datnpfiella procera Sellnick, 1931
10. Dolicheremaeus Jacot, 1938
Тип рода Dolicheremaeus rubripedes Jacot, 1938
Самка (вид сверху).
Рис. 339. Lucoppia sp.
Самка (вид снизу); виден
характер причленения ног.
11. Eremobelba Berlese, 1908
Тип рода Eremaeus leporosus Haller, 1884
12. Eremulus Berlese, 1908
a) Eremulus s. str.
Тип подрода Eremulus flagellifer Berlese, 1908
6) Epieremulus Berlese, 1916
Тип подрода Eremulus (Epieremulus) geometricus Berlese, 1916
13. Gradidorsum Sellnick, 1918f
Тип рода Gradidorsum asper Sellnick, 1918
14. Gymttodampia Jacot, 1937
Тип рода Amerobelba setata Berlese, 1916
15. Imparatoppla Jacot, 1934
Тип рода Imparatoppia imparata Jacot, 1934
Oribatei
423
16. Halozetes Berlese, 1916
Тип рода Notaspis marina Lohmann, 1907
17. Hydrozetes Berlese, 1902
Тип рода Notaspis lacustrls Michael, 1882
18. Lawrencoppia Jacot, 1934
Тип рода Lawrencoppia mauritius Jacot, 1934
19. Licneremaeus Paoli, 1908
Тип рода Notaspis licnophorus Michael, 1882
20. Llcnobelba Grandjean, 1931
Тип рода Llcnobelba alestensis Grandjean, 1931
21. Licnodamaeus Grandjean, 1931
Тип рода Licneremaeus undulatus Paoli, 1908
22. Licnollodes Grandjean, 1931
Тип рода Licnollodes andrei Grandjean, 1931
23. Lucoppia Berlese, 1908
a) Lucoppia s. str.
Тип подрода Zetes lucorutn Koch, 1840
6) Phauloppia Berlese, 1895
Тип подрода Oppla conformis Berlese, 1895
24. Metrioppia Grandjean, 1931
Тип рода Metrioppia Helvetica Grandjean, 1931
25. Nasozetes Sellnick, 1930
Тип рода Nasozetes sumatrensis Sellnick, 1930
26. Oppia Koch, 1836 (= Dameosoma Berlese, 1892 = Amolops Hull,
1916 = Dissorhina Hull, 1916 = Zetobelba Hull, 1916)
Тип рода Oppia nitens Koch, 1936
27. Oppiella Jacot, 1937
Тип рода Dameosoma corrugatum Berlese, 1904
28. Oribata Latreille, 1802
Тип рода Acarus geniculatus Linnaeus, 1758
29. Oribella Berlese, 1908
Тип рода Notaspis pectinata Michael, 1885
30. Paraliodes Hall, 1911
Тип рода Paraliodes incurvata Hall, 1911
31. Peloppia Sellnick, 1931
Тип рода Peloppia serrata Sellnick, 1931 ;
32. Phyllotegeus Berlese, 1913
Тип рода Leiosoma palmlcinctum Michael, 1883
33. Polypterozetes Berlese, 1916
Тип рода Polyterozetes cherubin Berlese, 1916
34. Rhynchorlbates Grandjean, 1929
Тип рода Rhynchorlbates rostratus Grandjean, 1929
35. Strieremaeus Sellnick, 1918f
Тип рода Strieremaeus illlbatus Sellnick, 1918
36. Suctobelba Paoli, 1908
Тип рода Notaspis trigona Michael, 1888
424
Глава VIII. Sarcoptiformes (продолжение)
37. Sactobelbella Jacot, 1937
Тип рода Suctobelbella serratlrostrum Jacot, 1937
38. Suctobelblla Jacot, 1937
Тип рода Suctobelblla punctillata Jacot, 1937
39. Tricheremaeus Berlese, 1908
Тип рода Notaspis serrqta Michael, 1885
40. Trizetes Berlese, 1904
Тип рода Trizetes pyramidalis Berlese, 1904
41. Tuberemaeus Sellnick, 1930
Тип рода Tuberemaeus slngularls Sellnick, 1930
ОБЩИЕ ЗАМЕЧАНИЯ
Peloppla и Metrloppla были выделены в отдельное семейство Pelop-
piidae Balogh, 1943. Основанием к этому послужило наличие длинных
тонких хелицер, аналогичных встречающимся у многих Pelopidae.
Здесь мы придерживаемся более распространенной системы.
ЛИТЕРАТУРА
Grandjean F., 1948. Sur les Hydrozetes (Acariens) de 1'Europe Occidental,
Bull. Paris Mas. d'Hist. Nat., Ser. 2, 20 (4), 328—335.
Newell I. M., 1945. Hydrozetes Berlese (Acari, Oribaroidea): The Occur-
rence of the genus in North America, and the phenomenon of levitation,
Trans. Conn. Acad. Arts and Sci., 36, 253—275.
S t r e n z k e K., 1950. Bestimmungstabelle der holsteinischen Suctobelba-M&n
(Acarina: Oribatei), Arch.lv. far Hydrobiologie, 44, 340—343.
ДИАГНОЗ
Carabodidae Willmann, 1931
(рис. 341)
У Carabodidae боковые и задние края гистеросомального щита не
загибаются вентрально. Спинная поверхность гистеросомы покрыта
Рис. 341. Cepheus mirabi-
loides Jacot.
Самка (вид сверху).
Oribatei
425
обычно грубой скульптурой или, по крайней мере, отчетливым точеч-
ным узором. Ноги III и IV причленены к телу у бокового края брюш-
ной поверхности или очень близко от него. Ноги обычного строения
и всегда короче тела. Бедра ног I и II часто имеют тонкие стебельки
и сильно раздуты дистально. Проподосома несет пластинчатые выросты
или небольшие листки.
РОДЫ И ПОДРОДЫ
1. Carabodes Koch, 1836
a) Carabodes s. str.
Тип подрода Carabodes coriaceus Koch, 1836
6) Carabocepheus Berlese, 1910
Тип подрода Carabodes (Carabocepheus) lounsburyl Berlese,
1910
в) Otocepheus Berlese, 1910
Тип подрода Car abodes (Otocepheus) longior Berlese, 1904
2. Carabodoides Jacot, 1937
Тип рода Carabodoides saccharomycetoides Jacot, 1937
3. Cepheus Koch, 1836 (= Tegeocranas Nicolet, 1855)
a) Cepheus s. str.
Тип подрода Cepheus latus Koch, 1836
6) Chaunoproctus Pearse, 1906
Тип подрода Chaunoproctus cancellatus Pearse, 1906 (плохо
описан; положение в системе неясно)
в) Microtegeus Berlese, 1916
Тип подрода Tegeocranas (Microtegeus) undalatus Berlese, 1916
г) Oribatodes Banks, 1895
Тип подрода Oribatodes mirabllis Banks, 1895
д) Protocepheus Jacot, 1928
Тип подрода Tegeocranas hericlus Michael, 1887'
4. Cerocepheus TrSgardh, 1931
Тип рода Cerocepheus mirabllis TrSgSrdh, 1931
5. Charassobates Grandjean, 1929
Тип рода Charrassobates cavernosas Grandjean, 1929
6. Eremaeozetes Berlese, 1913
Тип рода Eremaeozetes tuberculatus Berlese, 1913
7. Eremobodes Jacot, 1937
Тип рода Eremobodes pectinatus Jacot, 1937
8. Eupterotegaeus Berlese, 1916
Тип рода Tegeocranas ornatissimas Berlese, 1908
9. Eutegaeus Berlese, 1916
Тип рода Oribata bostocki Michael, 1908
10. Neocepheus Willmann, 1936
Тип рода Neocepheus hummelincki Willmann, 1936
11. Niphocepheus Balogh, 1943
Тип рода Cepheus nivalis Schweizer, 1922
426
Глава VIP. Sarcoptlformes (продолжение)
12. Ommatocepheus Berlese, 1913
Тип рода Cepheus ocellatus Michael, 1882
13. Passalozetes Grandjean, 1932
Тип рода Passalozetes afrlcanus Grandjean, 1932
14. Plategeocranus Sellnick, 1918 f
Тип рода Nothrus sulcatus Karsch, 1884
15. Pseudocepheus Jacot, 1928
Тип рода Cepheus vulgaris Nicolet, 1855
16. Scutorlbates Sellnick, 1918 f
Тип рода Scutorlbates perornatus Sell nick, 1918
17. Scutovertex Michael, 1879
Тип рода Scutovertex sculptus Michael, 1879
18. Tectocepheus Berlese, 1913
Тип рода Tegeocranus velatus Michael, 1880
19. Tegeocranellus Berlese, 1913
Тип рода Tegeocratius laevis Berlese, 1905
20. Tritegeus Berlese, 1913
Тип рода Cepheus blfidatus Nicolet, 1855
21. Xenillus Robineau-Desvoidy, 1839 (—Cepheus Nicolet, 1855 —
= Banksia Voigts et Oudemans, 1905)
Тип рода Notaspls tegeocratius Hermann, 1804 (= Xenillus cly-
peator Robineau-Desvoidy, 1839)
ОБЩИЕ ЗАМЕЧАНИЯ
Некоторые виды Carabodidae известны как промежуточные хозяева
ленточных червей. Так, Scutovertex minutus — промежуточный хозяин
Bertiella studeri, Clttotaenia ctenoides и C. denticulata в Германии;
Xenillus tegeocranus — промежуточный хозяин Cittotaenia ctenoides и
С. denticulata в Германии; Cepheus cephelformis — промежуточный
хозяин С. ctenoides и С. denticulata в Германии; Carabodidae sp. —
промежуточный хозяин Anoplocephala perfollata в СССР.
Ameronothridae Willmann, 1931
(рис. 342)
ДИАГНОЗ]
У Ameronothridae сплошной спинной панцирь покрывает проподо-
сому и гистеросому, и настоящего шва между ними нет. Покровы
тела кожистые. Псевдостигм, сенсорных щетинок и тектоподиев II и
III нет.
РОДЫ И ПОДРОДЫ
1. Ameronothrus Berlese, 1896
a) Ameronothrus s. str.
Тип подрода Eremaeus lineatus Thorell, 1871
6) Hygroribates Jacot, 1934
Тип подрода Nothrus (?) tnarinus Banks, 1896
Oribatei
427
ОБЩИЕ ЗАМЕЧАНИЯ
Клещи Ameronothrus живут у побережий морей, обычно в прилив-
ной зоне. Несколько видов их известно в Европе. Ameronothrus
(Hygroribates) marinus (Banks) был найден на скалах в приливной
зоне близ Си-Клиффа (Нью-Йорк). Позднее Джэкот (1934) нашел
его в небольшом районе вдоль берегов штата Нью-Йорк, а Гранжан
(1947) собрал этих клещей на берегах Франции. Джэкот пишет:
„А. marinus — явно морская форма. Встречается этот вид по большей
части в эстуариях и гаванях, т. е. в тех местах, где нет слишком
Рис. 342. Ameronothrus
schnetderi (Oudemans)
(по Удемансу, 1905).
Самка (вид сверху); обратить
внимание на отсутствие шва
между проподосомой
и гистеросомой.
больших наносов песка и где богато развиваются одноклеточные
водоросли. Поэтому нельзя рассчитывать найти этих клещей у откры-
тых мысов между заливами, где скалы, обросшие водорослями, встре-
чаются лишь редко. Полагаю также, что этих клещей лишь очень
редко можно встретить на скалах, в которых нет расщелин и трещин.
Неровные скалы, состоящие, например, из сланцев, наиболее подхо-
дящи для этого клеща. Другим фактором, ограничивающим распро-
странение A. marinus, является то, что самки его живородящи.
Поэтому яйца не разносятся течением, и молодь рождается на той же
скале, на которой живут родители".
Приведенный рисунок (рис. 342) заимствован у Удеманса. Он
недостаточно точен, но дает представление о признаках семейства.
Джэкот (1934) дает детальное описание североамериканских форм,
но не приводит рисунка.
ЛИТЕРАТУРА
Grandjean F„ 1947. Observations sur les Oribates (17® serie), Bull, d'Hist.
Nat., Ser. 2, 19 (2), 165—172.
Jacot A. P., 1934. An introduced moss mite in America, J. N. Y. Ent. Soc.,
42,. 329—337.
428
Глава VIII. Sarcoptiformes (продолжение)
Hermanniellidae Grandjean, 1934
(рис. 343—345)
ДИАГНОЗ
У Hermanniellidae по краям опистосомы сидят выступающие вперед
трубки, окружающие протоки сальных желез. Если трубок нет, то
Рис. 344. Plasmobates
pagoda Grandjean
(по Гранжану, 1929).
Самка (вид сверху); линочная
шкурка и секрет удалены.
Рис. 345. Plasmobates
pagoda Grandjean
(по Гранжану, 1929).
Хелицера,
Рис. 343. Plasmobates pagoda
Grandjean (по Гранжану, 1929).
Самка с линочной шкуркой на спине
и секретом на ногах (вид сверху).
на их месте развиты сильно склеротизированные листовидные или
бородавчатые выросты.
РОДЫ
1. Hermanniella Berlese, 1908
Тип рода Hermannla granulata Nicolet, 1855
2. Plasmobates Grandjean, 1929
Тип рода Plasmobates pagoda Grandjean, 1929
3. Solenozetes Grandjean, 1931
Тип рода Plasmobates cribratus Grandjean, 1929
Oribatei
429
ДИАГНОЗ
Plateremaeidae Tragardh, 1931
(рис. 346—348)
Гистеросома уплощенная; у взрослых особей она несет четыре
концентрически расположенные линочные шкурки (одну личиночную и
три нимфальных). Тело и ноги покрыты толстым слоем секрета.
Рис. 346. Plateremaeus ve-
st it us Tragardh
(по Трегорду, 1931).
A — сочленение между голенью и
лапкой IV; Б — сочленение между
бедром и коленным члеником III;
S—сочленение между коленным
члеником и голенью III.
Рис. 347. Plateremaeus
vestitus Tragirdh
(по Трегорду, 1931).
Самка (вид сверху).
Рис. 348. Plateremaeus
vestitus Tragardh
(по Трегорду, 1931).
Самка (вид снизу).
Дистальные концы бедра, коленного членика и голени ног узкие и
выпуклые; сочленяются они соответственно с впадинами коленного
членика, голени и лапки. Лапка несет маленький стебелек, на конце
которого сидят три коготка.
РОДЫ
1. Plateremaeus Berlese, 1908
Тип рода Damaeus ornatlsslmus Berlese, 1888
ЛИТЕРАТУРА
Trag&rdh I., 1931. Acarina from the Juan Fernandez Islands. The Natural
History of Juan Fernandez and Easter Islands, 3, 553—628.
Liacaridae Willmann, 1931
(рис, 349, 350)
ДИАГНОЗ
У Liacaridae боковые и задний края спинного щита гистеросомы
не загибаются вентрально. Ноги III и IV расположены на брюшке,
обычно на большом расстоянии от бокового края тела.
430
Глава VIII. Sarcoptiformes (продолжение)
РОДЫ
1. Liacarus Michael, 1898 (== Letosoma Nicolet, 1855 nomen praeoc-
cupatum)
Тип рода Orlbata nitens Gervais, 1844
2. Adoristes Hull, 1916
Тип рода Oribates ovatus Koch, 1840
Рис. 349. Caltroribula divergens Jacot.
Самка (вид сверху)^
Рис. 350. Liacarus cora-
cinus (Koch).
Самка (вид снизу); виден харак-
тер причленения4 тазиков ног.
3. Astegistes Hull, 1916 (= Cultrozetes Sellnick, 1922)
Тип рода Acarus muscorum Scopoli, 1763 (= Zetes pllosus Koch,
1840)
4. Caltroribula Berlese, 1908
Тип рода Notaspis juncta Michael, 1885
ОБЩИЕ&ЗАМЕЧАНИЯ
Из числа видов, относящихся к этому семейсТву, три известны
как промежуточные хозяева различных ленточных червей. Liacarus
coraclnus (Koch) является промежуточным хозяином Cittoiaenia ctenoides
и С. denticulata в Германии; Adoristes ovatas (Koch) — промежуточ-
ный хозяин Moniezia sp. в СССР; Liacarus sp. — промежуточный
хозяин Anoplocephala perfoliata.
Oribatei
431
Zetorchestidae Michael, 1898
ДИАГНОЗ
(рис. 351, 352)
У Zetorchestidae ноги IV приспособлены для прыганья; у мертвых
клещей эти ноги вытянуты вперед. Птероморфы не развиты.
Рис. 351. Zetorchestes micronychus Ber-
lese (по Берлезе, 1883).
Самка (вид сверху)»
Рис. 352. Zetorchestes
micronychus Berlese
(по Берлезе, 1883),
Самка (вид снизу); видно
положение ног IV.
РОДЫ
1. Zetorchestes Berlese, 1888
Тип рода Carabodes micronychus Berlese, 1883
2. Zetorchella Berrlese, 1916
Тип рода Zetorchella pedestris Berlese, 1916
Рис. 353. Qustavla microcephala
(Nicolet) (по Берлезе, 1884).
Самка (вид сверху).
Gustaviidae Willmann, 1931
(рис. 353, 354)
ДИАГНОЗ
Gustaviidae имеют длинные узкие
ножницевидные хелицеры, пильчатые
на дистальном конце. Птероморфы
отсутствуют.
Рис. 354. Gustavia microcephala
(Nicolet) (по Берлезе, 1884).
Хелицера.
432
Глава VIII, Sarcoptiformes (продолжение)
РОДЫ
1. Gustavla Kramer, 1879 (= Serrarius Michael, 1883)
Тип рода Leiosoma mlcrocephala Nicolet, 1855 (= Gustavla sol
Kramer, 1879)
Oripodidae Jacot, 1925
(рис. 355)
ДИАГНОЗ
Oripodidae принадлежат к группе Pterogasterina. Птероморфы
у них хорошо развиты и впереди соединяются друг с другом по
Рис. 355. Oripoda
elongata Banks.
Самка (вид сверху).
переднему краю опистосомы. Сросшаяся часть птероморф образует
над проподосомой козырек, прикрывающий основания псевдостигма-
тических и интерламелларных щетинок; иногда козырек целиком
покрывает эти щетинки. У некоторых форм козырек срастается с про-
подосомой. Генитальных щетинок две или три пары.
РОДЫ
1. Oripoda Banks, 1904
Тип рода Oripoda elongata Banks, 1904
2. Cryptoribatula Jacot, 1934
Тип рода Cryptoribatula taishanensls Jacot, 1934
Oribatei
433
3. Gytnnobates Banks, 1902
Тип рода Oymnobates glaber Banks, 1902
4. Jurabates Jacot, 1929
Тип рода Oribata pseudofusiger Schweizer, 1922
ЛИТЕРАТУРА
Banks N., 1904. A treatise on the Acarina, or mites. Proc. U. S. Nat. Mas.,
28 (1382), 71.
Tenuialidae Jacot, 1929
(рис, 356)
ДИАГНОЗ
Tenuialidae имеют широкие хелицеры с хорошо развитой клешней.
Ламеллы развиты умеренно. Птероморфы, продолжающиеся вперед
Рис. 356. Tenalala nuda
Ewing.
Самка (вид сверху); ноги не
изображеные
в виде пластинок, позади идут вдоль всей опистосомы и не отделены
от нее швами. По Джэкоту, это семейство вследствие наличия птеро-
морф должно быть отнесено к Pterogasterina.
РОДЫ
1. Tenalala Ewing, 1913
Тип рода Tenalala nada Ewing, 1913
2. Hafenrefferla Oudemans, 1906
Тип рода Orlbates gllvipes Koch, 1840
28 Зак. 1785
434
Глава VIII. Sarcoptiformes (продолжение)
Oribatulidae Jacot, 1929
( = Scheloribatidae Grandjean, 1933)
(рис. 357, 358)
ДИАГНОЗ
Oribatulidae принадлежат к группе Pterogasterina. Птероморфы
расположены в одной плоскости, не загибаясь вентрально для защиты
ног. Если смотреть сверху, то передний край птероморф кажется
загнутым назад. Каждый из генитальных щитков несет четыре щетинки
(исключение составляет один неописанный вид из Панамы, у которого
на каждом щитке имеются только по две передних щетинки).
Рис. 357. Protoschelobates seghettil
Runkel et Kates.
Передний конец тела самки (вид сверху); видны Р и С. 358. Oribatula minuta (Ewing),
птероморфы, псевдостигма, псевдостигмати- ' te/
ческие органы, ламеллы и щетинки. Самка (вид сверху).
Гранжан (1933) выделил клещей, неполовозрелые фазы которых
отклоняются по числу анальных щетинок, в особое семейство Schelo-
ribatidae. Эти клещи на фазе личинки вообще не имеют анальных
щетинок; на фазе прото-, дейто- и тритонимфы они имеют соот-
ветственно два, три и четыре щетинки и на фазе имаго — четыре
анальные щетинки. Личинки Oribatulidae имеют только одну анальную
щетинку.
Эти две группы трудно отделить друг от друга, поэтому они рас-
сматриваются здесь как единое целое.
РОДЫ И ПОДРОДЫ
1. Oribatula Berlese, 1896
a) Oribatula s. str.
Тип подрода Notaspls tibialis Nicolet, 1855
6) Zygoribatula Berlese, 1916 ( = Neoribatula Ewing, 1917)
Тип подрода Oribatula connexa Berlese, 1916
в) Henillelus Berlese, 1916
Тип подрода Protoribates (Scheloribates) initialis Berlese, 1908
Oribatei
435
2. Cardiorlbates Jacot, 1934
Тип рода Orlbata orlformls Pearse, 1910
3, Drymobates Grandjean, 1930
Тип рода Drymobates silvicola Grandjean, 1930
4. Drymobatoides Jacot, 1936
Тип рода Drymobatoides mauritius Jacot, 1936
5. Eporibatula Sellnick, 1928
Тип рода Eremaeus rauschensis Sellnick, 1928
6. Exoribatula Jacot, 1936
Тип рода Exoribatula biundatus Jacot, 1936
7. Liebstadia Oudemans, 1906
Тип рода Not aspis similis Michael, 1888
8, Protoschelobates Jacot, 1934
Тип рода Murcia Insularis Oudemans, 1917
9. Scheloribates Berlese, 1908
a) Scheloribates s. str.
Тип подрода Zetes latipes Koch, 1844
6) Paraschelobates Jacot, 1934
Тип подрода Scheloribates {Paraschelobates) mumfordi Jacot,
1934
10. Styloribates Jacot, 1934
Тип рода Styloribates pectinatus Jacot, 1934
11. Unguizetes Sellnick, 1925
Тип рода Oribates sphaerula Berlese, 1905
12. Zetomimus Hull, 1916
Тип рода Oribata furcata Warburton et Pearse, 1905
13. Zetomotrichus Grandjean, 1934
Тип рода Zetomotrichus lacrimans Grandjean, 1934
ОБЩИЕ ЗАМЕЧАНИЯ
Среди Oribatulidae основным промежуточным хозяином ленточных
червей является Scheloribates laevigatas (Koch). Помимо него, в насто-
ящее время известны четыре других вида, служащих переносчиками
ленточных червей. Scheloribates laevigatas служит промежуточным
хозяином Moniezia expansa в США и СССР, М. betiedeni в СССР,
Bertiella staderi, Cittotaenia ctenoides и C. denticulate в Германии и
Anoplocephala perfoliata, A. magna и Thysaniezia giardi в СССР;
Scheloribates latipes (Koch) служит промежуточным хозяином Anoplo-
cephala perfoliata и Thysaniezia giardi в СССР; Protoschelobates
seghettii Runkel et Kates и Oribatula minuta (Ewing) являются про-
межуточными хозяевами Moniezia expansa в США; Liebstadia similis
(Michael) — промежуточный хозяин Cittotaenia ctenoides в Германии.
Кэйтс и Рункель (1948) опубликовали превосходный обзор биологии
и распространения панцырных клещей— промежуточных хозяев ленточ-
ных червей. В этой работе можно найти более подробные сведения
об Oribatulidae.
28*
436
Глава VIII. Sarcoptiformes (продолжение)
ЛИТЕРАТУРА
Grandjean F., 1933. Etude stir le developpement des Oribates, Bull. Soc.
Zool. France, 58 (1), 30—61.
Kates К. C., Runkel С. E., 1948. Observations on oribatid mite vectors
of Moniezia expansa on pastures, with a report of several new vectors
from the United States, Proc. Helminth. Soc. Wash., 15 (1), 18—33.
ДИАГНОЗ
Ceratozetidae Jacot, 1925
(рис. 359)
Ceratozetidae принадлежат к Pterogasterina. Они имеют маленькие
птероморфы, не выступающие за переднюю часть опистосомы. Внутрен-
ние края ламелл прикрепляются к проподосоме; передняя часть ламелл
Рис. 359. Jugatala
tuberosa Ewing.
Самка (вид сверху).
выступает свободно. Трансламеллы имеются не всегда. Генитальные
щитки несут шесть пар щетинок.
РОДЫ И ПОДРОДЫ
1. Ceratozetes Berlese, 1908
a) Ceratozetes s. str.
Тип подрода Oribata gracilis Michael, 1884
6) Allozetes Berlese, 1913
Тип подрода Ceratozetes (Allozetes) pusillus Berlese, 1913
2. Allorlbates Banks, 1947
Тип рода Alloribates singularls Banks, 1947
3. Balzania Jacot, 1929
Тип рода Oribata microptera Canestrini, 1896
4. Banksinus Jacot, 1938
Тип рода Oribata arborea Banks, 1895
Oribatei
437
5. Calyptozetes Thor, 1929
Тип рода Oribata sarekettsls Tragardh, 1910
6. Chamobates Hull, 1916
Тип рода Oribata cuspidata Michael, 1884
7. Diapterobates Grandjean, 1936
Тип рода Sphaerozetes {Trichoribates) numerosas Sellnick,
1924
8. Edtvardzetes Berlese, 1914
Тип рода Oribata edwardsii Nicolet, 1855
9. Frischia Oudemans, 1915
Тип рода Frischia elongata Oudemans, 1915
10. Globozetes Sellnick, 1928
Тип рода Globozetes longipiltts Sellnick, 1928
11. Hamtneria Sellnick, 1944
Тип рода Hatntneria groenlandica Sellnick, 1944
12. Humerobates Sellnick, 1928
Тип рода Notaspis humeralis Hermann, 1804
13. lugoribates Sellnick, 1944
Тип рода lugoribates gracilis Sellnick, 1944
14. Jugatala Ewing, 1913
Тип рода Jugatala tuberosa Ewing, 1913
15. Ltmnozetes Hull, 1916
a) Ltmnozetes s. str.
Тип подрода Acarus ciliatus Schrank, 1803 {—Oribata sphagni
Michael, 1884)
6) My cobates Hull, 1916
Тип подрода Oribata parmeliae Michael, 1884
16. Melanozetes Hull, 1916
Тип рода Orlbates molllcomus Koch, 1840
17. Minunthozetes Hull, 1916
Тип рода Zetes simlrufus Koch, 1840 {—Oribata fusigera
Michael, 1884)
18. Mochlozetes Grandjean, 1930
Тип рода Mochlozetes penetrabills Grandjean, 1930
19. Nesiotizetes Jacot, 1934
Тип рода Nesiotizetes adamsoni Jacot, 1934
20. Oromurcla Thor, 1930
Тип рода Oromurcia bicuspidata Thor, 1930
21. Propeschelobates Jacot, 1936
Тип рода Oribata albida Ewing, 1907
22. Sphaerobates Sellnick, 1928
Тип рода Sphaerozetes (?) gratus Sellnick, 1921
23. Sphaerozetes Berlese, 1885 (= Euzetes Berlese, 1908)
Тип рода Oribates orbicularis Koch, 1836
24. Storkania Jacot, 1929
Тип рода Oribata simplex Storkan, 1925
438
Глава VIII. Sarcoptiformes (продолжение)
25. Svalbardla Thor, 1930
Тип рода Svalbardla paludlcola Thor, 1930
26. Tegeozetes Berlese, 1913
Тип рода Tegeozetes tunicatus Berlese, 1913
27. Terr azetes Jacot, 1936
Тип рода Orlbates sphaerula Berlese, 1905
28. Trachyorlbates Berlese, 1908
a) Trachyorlbates s. str.
Тип подрода Orlbates ampulla Berlese, 1904
6) Indorlbates Jacot, 1929
Тип подрода Protoribates punctulatus Sellnick, 1925
29. Trichorlbates Berlese, 1910
a) Trichoribates s. str.
Тип подрода Murcia trlmaculata Koch, 1836
6) Sphaerozetella Jacot, 1929
Тип подрода Orlbates orbicularis Berlese, 1883 (не C. L. Koch,
1886)
ОБЩИЕ ЗАМЕЧАНИЯ
Trichorlbates inclsellus (Kramer) известен как промежуточный
хозяин Clttotaenla ctenoides в Германии.
ЛИТЕР АТУР А
Hammer М., 1944. Studies on the oribatids and collemboles of Greenland,
Meddelelser от Grtfnland, 141 (3), 1—210.
Hammer M., 1946. The zoology of East Greenland, Meddelelser от QrHn-
land, 122 (1), 1—39.
Oribatellidae Jacot, 1925
(рис. 360)
ДИАГНОЗ
Oribatellidae также принадлежит к Pterogasterina. Птероморфы
у них не выступают вперед за пределы опистосомы. Ламеллы при-
крепляются к проподосоме только своим задним краем; как правило,
они крупные и покрывают значительную часть проподосомы. Многие
виды имеют чрезвычайно своеобразный облик.
РОДЫ И ПОДРОДЫ
1. Oribatella Banks, 1895
Тип рода Oribatella quadrldentata Banks, 1895
2. Anachipterla Grandjean, 1932
Тип рода Anachipterla deflclens Grandjean, 1932
3. Joelia Oudemans, 1906 (—Coggiella Berlese, 1916)
Тип рода Oribates florli Coggi, 1898
Oribatei
439
4. Tectoribates Berlese, 1910
a) Tectoribates s. str.
Тип подрода Orlbata tecta Michael, 1883
6) Unduloribates Balogh, 1943
Тип подрода Tectoribates undulatus Berlese, 1915
Рис. 360. Oribatella
magniseta Ewing.
Самка (вид сверху).
Microzetidae Grandjean, 1936
(рис. 361, 362)
ДИАГНОЗ
Microzetidae принадлежат к группе Pterogasterina. Хелицеры у них
снабжены направленным вперед рожком, сидящим на дорзальной сто-
роне неподвижного пальца. Тектоподий 1 имеет большую створку
или пластинку, которая является продолжением боковой стенки про-
подосомы. Свободный край тектоподия имеет сложную форму; он
состоит из больших лопастей, отделенных друг от друга глубокими
вырезками; эти лопасти изогнуты таким образом, что тектоподий
образует выпуклый свод по отношению к длинной оси тела. Ламеллы
всегда очень большие; они подвижны или неподвижно причленены
к телу своим основанием и не связаны друг с другом. Ламеллы
имеют различную форму; иногда они изогнуты вдоль своей длинной
оси. Когда клещ принимает оборонительную позу, его ноги прячутся
под ламеллы, располагаясь в их продольных складках. От бокового
края ламелл отходит перепончатая лопасть; у одних форм она
имеет простую форму, у других она гарпуновидная и иногда снабжена
440
Глава VIII. Sarcoptiformes (продолжение)
крупными зубцами. Подосома и опистосома на брюшной стороне
отделены друг от друга широким темным валиком, представляющим
собой утолщение аподем IV. Сверху можно различить тонкий попе-
речный шов, который соединяет передний боковой угол полового
Рис. 362. Mierozetes арра-
lachicola Jacot.
Самка (вид сверху,'»
Рис. 361. Phyla-
cozetes membrami-
lifer Grandjean
(по Гранжану,
1936).
Кончик хелицеры с рож*
ком на клешне.
отверстия со склеротизированными полями, окружающими ноги. Подо-
сома крупнее опистосомы.
РОДЫ
1. Mierozetes Berlese, 1913
Тип рода Sphaerozetes mirandus Berlese, 1908
2. Acaroceras Grandjean, 1936
Тип рода Acaroceras odontotus Grandjean, 1936
3. Nellacarus Grandjean, 1936
Тип рода Nellacarus petrocorlensls Grandjean, 1936
4. Phylacozetes Grandjean, 1936
Тип рода Phylacozetes membranulifer Grandjean, 1936
ЛИТЕРАТУРА
Grandjean F., 1936. Les Microzetidae n. fam. (Oribates), Bull. Soc. Zool.
de France, 61 (2), 60—93.
Notaspididae Oudemans, 1900
(рис. 363)
ДИАГНОЗ
Notaspididae принадлежит к Pterogasterina. Птероморфы у них
выступают вперед в виде конуса, достигающего почти до конца ро-
струма. Ламеллы, покрывающие почти всю проподосому сзади, под-
Orlbatei
441
вижно причленены к телу и соединяются друг с другом на небольшом
центральном поле.
РОДЫ
1. Notaspls Hermann, 1804 {= Achlpteria Berlese, 1885)
Тип рода Acarus coleoptratus Linneaus, 1758
2. Achlpterina Berlese, 1916
Тип рода Achlpteria {Achlpterina) orlbatelloid.es Berlese, 1916
Рис. 363. Notaspls magnus
Sellnick.
Самка (вид сверху).
3. Cerachlpterla Grandjean, 1935
Тип рода Cerachlpterla digltata Grandjean, 1935
4. Fuscozetes Sellnick, 1928
Тип рода Orlbata fusclpes Koch, 1844
ОБЩИЕ ЗАМЕЧАНИЯ
Notaspis coleoptratus (L) является промежуточным хозяином Ber-
tiella studeri и Cittotaenia ctenoides в Германии. Achlpteria {Notaspls)
sp. — промежуточный хозяин Anoplocephala perfoliata в СССР.
Haplozetidae Grandjean, 1936
(рис. 364)
ДИАГНОЗ
Haplozetidae имеют подвижно сочленяющиеся с телом птероморфы.
Тектоподии IV у них удлинены и на конце переходят в листовидное
образование. Генитальные щитки несут пять пар щетинок. Гранжан (1936)
указывает, что некоторые виды имеют всего четыре пары гениталь-
ных щетинок.
442
Глава VIII. Sarcoptiformes (продолжение)
РОДЫ
1. Haplozetes Willmann, 1935
Тип рода Pelorlbates vindobonensls Willmann, 1935
2. Neogymnobates Ewing, 1917
Тип рода Oribata multipilosa Ewing, 1907
3. Pelorlbates Berlese, 1908 ( = Parazetes Willmann, 1930 = £«ry-
parazetes Radford, 1950)
Тип рода Oribata peloptoides Berlese, 1888
4. Protorlbates Berlese, 1908
Тип рода Oribata monodactyla Haller, 1884
5. Rostrozetes Sellnick, 1925
Тип рода Rostrozetes foveolatus Sellnick, 1925
6, Xylobates Jacot, 1929
Тип рода Oribata lophothrichus Berlese, 1904
ОБЩИЕ ЗАМЕЧАНИЯ
Среди Haplozetidae один вид — Pelorlbates curtipilus Jacot — из-
вестен как промежуточный хозяин паразитирующего у овец ленточ-
ного червя Mordezla expansa.
ЛИТЕРАТУРА
Grandjean F., 1936. Observations sur les Orlbates (10 serie), Bull. Paris
Mus. d'Hist. Nat., Ser. 2, 8 (3), 246—249.
Oribatei
443
Pelopidae Ewing, 1917
(рис. 365, 366)
ДИАГНОЗ
Pelopidae принадлежит к группе Pterogasterina. Они имеют крупные
птероморфы, подвижно сочленяющиеся с телом. Впереди (но не сзади)
птероморфы заходят за линию своего сочленения с телом (исключение
составляют Peloptulus и Galumnella, у которых задний конец птеро-
морф вытянут, заострен и заходит за задний край сочленения с телом).
Рис. 365. Pelops sylvestris Jacot.
.4 —хелицера самки; Б— самка (вид сверху).
Рис. 366. Lepidozetes
singularls Berlese.
Самка (вид сверху)»
Средняя часть переднего края гистеросомы (между птероморфами)
заходит за границу между проподосомой и гистеросомой, а иногда —
даже за передний конец птероморф. Интерламеллярные щетинки либо
ложечковидные {Pelops, Eupelops, Tectopelops, Parapelops), либо обыч-
ные, волосовидные {Peloptulus, Pelopsis, Propelops), либо вообще
отсутствуют {Galumnella, Galumnopsis). Хелицеры, широкие у осно-
вания, быстро суживаются к концу; их концевой сегмент длинный и
тонкий и несет маленькую клешню (исключение—- Propelops, у кото-
рых хелицеры обычные). Генитальных щетинок шесть пар.
РОДЫ и ПОДРОДЫ
1. Pelops Koch, 1836
Тип рода Notaspis htrsutus Koch, 1836
2. Eupelops Ewing, 1917
Тип рода Pelops uraceus Koch, 1840
3. Galumnella Berlese, 1916
Тип рода Galumnella paradoxa Berlese, 1916
4. Galumnopsis Grandjean, 1931
Тип рода Galumnopsis holoscripta Grandjean, 1931
444 Глава, VIII. Sarcopf(formes (продолжение)
5. Lepidozetes Berlese, 1910
a) Lepidozetes s. str.
Тип подрода Lepidozetes slngularis Berlese, 1910
6) Tegoribates Ewing, 1917 ( = Lepidoribates Sellnick, 1920)
Тип подрода Tegoribates subnlger Ewing, 1917
6. Parapelops Jacot, 1938
Тип рода Pelops blfurcatus Ewing, 1909
7. Pelopsis Hall, 1911
Тип рода Pelopsis undiuscula Hall, 1911
8. Peloptulus Berlese, 1908
Тип рода Pelops phaeonotus Koch, 1844
9. Propelops Jacot, 1937
Тип рода Propelops plnicus Jacot, 1937
10. Tectopelops Jacot, 1929
Тип рода Pelops laevigatas Nicolet, 1855
ОБЩИЕ ЗАМЕЧАНИЯ
Два вида — Pelops tardus Koch и P. planicornis (Schrank) — из-
вестны как промежуточные хозяева ленточного червя Cittotaenia cte-
noides в Германии.
Род Lepidozetes не типичен для этого семейства и правильнее
выделить его в особое подсемейство или даже семейство. Клещи
этого рода отличаются главным образом наличием больших сросшихся
ламелл, покрывающих всю проподосому. Интерламеллярные щетинки
у них маленькие. Гистеросома частично покрывает проподосому. Ге-
нитальных щетинок шесть пар. Хелицеры нормальные.
Galumnidae Grandjean, 1936
(рис. 367, 368)
ДИАГНОЗ
Galumnidae также принадлежат к группе Pterogasterina. Они имеют
большие крыловидные птероморфы, подвижно сочленяющиеся с телом.
Закругленные передние и задние края птероморф заходят далеко за
линию соединения их с телом. Шов между проподосомой и гистеро-
сомой бывает не всегда. Ламеллы слабо развиты или отсутствуют.
Каждый генитальный щиток несет по шести щетинок. Передняя
(первая) пара циркуманальных щетинок расположена по бокам аналь-
ного отверстия, у переднего края последнего.
РОДЫ И ПОДРОДЫ
1. Qalumna v. Heyden, 1826 (= Zetes Koch, 1836 = Centroribates
Berlese, 1914)
a) Qalumna s. str.
Тип подрода Notaspis alatus Hermann, 1804
Oribatei
446
6) Pergalumna Grand jean, 1936
Тип подрода Orlbates nervosus Berlese, 1914
в) Stictozetes Berlese, 1916
Тип подрода Orlbates (Stictozetes) scaber Berlese, 1916
r) Vaghla Oudemans, 1917
Тип подрода Orlbates (Stictozetes) stupendus Berlese, 1916
2, Allogalumna Grandjean, 1936
Тип рода Galumna alamellae Jacot, 1935
Рис. 367. Galumna vlrginiensis
Jacot (no Джэкоту, 1934).
Самка (вид сверху).
Рис. 368. Galumna vlrginiensis
Jacot (по Джэкоту, 1934).
Самка (вид снизу).
3. Holokalumma Jacot, 1929
Тип рода Holokalumma coloradensls Jacot, 1929
4. Pilizetes Sellnick, 1837
Тип рода Pilizetes afrlcanus Sellnick, 1937
5. Psamnogalumna Balogh, 1943
Тип рода Stictozetes hungaricus Sellnick, 1925
ОБЩИЕ ЗАМЕЧАНИЯ
Galumnidae, или „большекрылые” панцырные клещи, представляют
собой весьма своеобразных клещей, обладающих большими подвиж-
ными птероморфами. Известно несколько видов, являющихся проме-
жуточными хозяевами ленточных червей. Galumna vlrginiensis Jacot
является хозяином Monlezia expansa в США; О. nervosus (Berlese) —
хозяином Cittotaenia ctenoides в США и Anoplocephala perfoliata —
446
Глава VIII. Sarcoptlformes (продолжение)
в СССР; G. obvius (Berlese) хозяин Moniezia expansa, М. benedenl,
Anoplocephala perfoliata и Paranoplocephala tnamillana в СССР и
для Cittotaenia ctenoides в Германии; О. emarginatum (Banks) и О. nig-
ra (Ewing) — промежуточные хозяева Moniezia expansa в США; Ga-
lanina sp. играют ту же роль по отношению к Moniezia expansa
в США и Bertiella studerl в Германии; Allogalumna longipluma (Ber-
lese) является промежуточным хозяином Paranoplocephala mamillana
в СССР.
ЛИТЕРАТУРА
Jacot А. Р„ 1929. American oribatid mites of the subfamily Galumninae,
Bull. Mas. Compar. Zool., 69 (1), 1—37.
Jacot A. P., 1933. The primitive Galumninae (Oribatoidea-Acarina) of the
Middle West, Amer. Midland Nat., 14 (6), 680—703.
Jacot A. P., 1934. The Galumnas (Oribatoidea-Acarina) of the northeastern
United States, J. N. У. Ent. Soc., 42, 87—125.
Jacot A. P,. 1935. The large-winged mites of Florida, Florida Ent., 19 (1),
1—15; 19 (2), 17—31; 19 (3), 43—47.
Parakalummidae Grandjean, 1936
(рис. 369, 370)
ДИАГНОЗ
Parakalummidae принадлежат к группе Pterogasterina. Они имеют
очень большие крыловидные птероморфы, подвижно сочленяющиеся
Рис. 369. Parakalumma lydia
(Jacot) (по Джэкоту, 1929).
Самка (вид сверху).
Рис. 370. Parakalumma lydia
(Jacot) (по Джэкоту, 1929).
Самка (вид снизу).
с телом. Как спереди, так и сзади птероморфы заходят за линию
своего сочленения с телом. Проподосома и гистеросома отделены друг
от друга швом. Ламеллы слабо развиты или вообще отсутствуют.
Каждый генитальный щиток несет по пяти щетинок. Первая пара цир-
куманальных щетинок сидит впереди анального отверстия.
Oribatei
447
РОДЫ
1. Parakalumma Jacot, 1929
Тип рода Neoribates lydia Jacot, 1923
2. Holozetes Jacot, 1929
Тип рода Galanina texana Banks, 1906
3. Kratzensteinia Oudemans, 1917
Тип рода Orlbata rugifrons Stoll, 1891
4. Neoribates Berlese, 1914
Тип рода Oribates roubali Berlese, 1900
5. Neorizetes Jacot, 1933
Тип рода Orlbata rugosala Ewing, 1909
6. Protokalumma Jacot, 1929
Тип рода Orlbata depressa Banks, 1905
7. Sandenia Oudemans, 1917
Тип рода Galumna georglae Oudemans, 1914
ЛИТЕРАТУР A
Grandjean F., 1936. Les Oribates de Jean Frederic Hermann et de son
Pere, Ann. Soc. Ent. de France, 105, 27—110.
Epactozetidae Grandjean, 1936
ДИАГНОЗ
(рис. 371)
Epactozetidae принадлежат к Pterogasterina и имеют большие кры-
ловидные птероморфы, подвижно сочленяющиеся с телом. Как спереди,
Рис. 371. Epactozetes
imitator Grandjean
(по Гранжану, 1936).
Самка (вид сверху).
так и сзади птероморфы заходят далеко за линию своего сочленения
с телом. Проподосома и гистеросома разделены швом. Имеются очень
448
Глава VIII. Sarcoptiformes (продолжение)
крупные ламеллы, покрывающие значительную часть проподосомы.
Генитальные щитки несут пять пар щетинок. Передняя пара цирку-
манальных щетинок расположена у переднего края анального отверстия.
РОДЫ
1. Epactozetes Grandjean, 1930
Тип рода Epactozetes imitator Grandjean, 1930
ЛИТЕРАТУРА
Grandjean F., 1936. Les Oribates de Jean Frederic Hermann et de son
Рёге, Ann. Soc. Ent. de France, 105, 27—110.
Protoplophoridae Jacot, 1923
(рис. 372)
ДИАГНОЗ
Protoplophoridae способны свертываться подобно броненосцам, чтобы
защитить ротовые органы и ноги. У свернувшегося клеща половое
Рис. 372. Aedoplophora glomerata Grandjean
(по Гранжану, 1932).
А —самка (вид сбоку); Б — свернувшаяся самка (вид сбоку);
В —свернувшаяся самка (вид спереди); Г—лапка ноги I.
отверстие, ноги и гнатосома прикрываются проподосомальный щитком
(aspis) и снаружи видно только анальное отверстие. Собственно тело
покрыто четырьмя щитками: передним дорзальным (pronotaspis), задним
дорзальным (metanotaspis, или пигидий) и двумя боковыми симметрично
расположенными щитками, покрывающими большую часть брюшной
поверхности гистеросомы (pleuraspis). Пигидий подвижный и может
полностью втягиваться под pronotaspis или же выдвигаться из-под
Oribatei
449
него. Генитальные щитки расположены непосредственно позади тази-
ков ног IV, а анальные щитки — непосредственно позади генитальных,
между pleuraspis.
РОДЫ
1. Protoplophora Berlese, 1910
Тип рода Protoplophora palpalis Berlese, 1910
2. Aedoplophora Grandjean, 1932
Тип рода Aedoplophora glomerata Grandjean, 1932
3. Arthroplophora Berlese, 1910
Тип рода Arthroplophora paradoxa Berlese, 1910
4. Cryptoplophora Grandjean, 1932
Тип рода Cryptoplophora abscondita Grandjean, 1932
5. Prototritta Berlese, 1916
Тип рода Arthroplophora (Prototritta) armadillo Berlese, 1916
ОБЩИЕ ЗАМЕЧАНИЯ
Protoplophoridae живут в гумусе и в гниющих корнях. Описано
очень мало видов, что, вероятно, в значительной мере обязано труд-
ности и малочисленности сборов.
ЛИТЕРАТУРА
Or and jean F., 1932. La Famille des Protoplophoridae (Acariens), Ball. Soc.
Zool. de Prance, 58 (1), 10—36.
Mesoplophoridae Jacot, 1923
ДИАГНОЗ
(рис. 373, 374)
У Mesoplophoridae гистеросома не расчленена на сегменты. По-
ловое и анальное отверстия лежат на боль-
шом брюшном щите, который соединяется
со спинным щитом.
Р ис. 373. Mesoplophora michae- Рис. 374. Mesoplophora pulchra Sellnick
liana Berlese (по Берлезе, 1904). (по Гранжану, 1938).
Самка (вид снизу). Самка (вид сбоку).
29 Зак. 1785
450
Глава VIII. Sarcoptiformes (продолжение)
РОДЫ
1. Mesoplophora Berlese, 1904
Тип рода Mesoplophora michaeliana Berlese, 1904
ЛИТЕРАТУРА
Grandjean F., 1933. Oribatcs de 1’Afrique du Nord, Bull. Soc. d'Hist,
de 1’Afrique du Nord, 24, 308—323.
Phthiracaridae Perty, 1941
ДИАГНОЗ
(рис. 375—377)
У Phthiracaridae гистеросома не разделяется поперечными швами.
Генитальные и анальные щитки лежат на брюшном щитке, который
не соединен со спинным щитом.
Рис. 375. Pseudotritia
ardua (Koch)
(по Джэкоту, 1930).
Самка (вид сбоку).
Р и с. 376. Pseudo-
tritia ardua (Koch)
(по Джэкоту, 1930).
Область полового и
анального отверстий.
Рис. 377. Phthiracarus setosellum
Jacot (по Джэкоту, 1930).
Взрослая особь (вид сбоку); проподосома
и ноги (обозначены пунктиром) втянуты
в полость в верхней части гистеросомы;
отверстие покрыто проподосомальный
щитком.
Джэкот (1930) указывает, что „замечательной особенностью Phthira-
caridae является способность втягивать головогрудь (цефалоторакс)
и ноги под своды нотогастера подобно тому, как втягивают голову
Oribatei
451
и ноги в панцирь черепахи, броненосцы и некоторые другие живот-
ные. Нередко приходится читать, что у Phthiracaridae aspis подвижно
прикреплен к брюшку и способен опускаться, как крышка, подвешен-
ная на петлях. Это совершенно неверно. У Phthiracaridae такого под-
вижного сочленения aspis с телом не имеется; у них головогрудь
просто втягивается между ногами в полость в верхней части гисте-
росомы. При этом хитиновый щиток, или aspis, находящийся на верх-
ней части головы, прикрывает отверстие как крышка. Aspis несет
на переднем конце козырек, и концы ног (лапки и коготки) скры-
ваются под этим козырьком".
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ PHTHIRACARIDAE
1. Генитальные и анальные щитки короткие и широкие. Брюшной
гистеросомальный щиток сзади широко окружает анальное отвер-
стие ..........................................Phthiracarinae
— Генитальные и анальные щитки длинные и узкие. Брюшной гис-
теросомальный щиток сзади тесно окружает: анальное отверстие
...............................................Euphthiracarinae
РОДЫ И ПОДРОДЫ
Phthiracarinae Pert у, 1841
1. Phthiracarus Perty, 1841
a) Phthiracarus s. str.
Тип подрода Acarus piger Scolpoli, 1763 ( = Phthiracarus
contractilis Perty, 1841)
6) Tropacarus Ewing, 1917 (= Calhoplophora Ber'ese, 1923)
Тип подрода Hoplophora carinata Koch, 1841
в) Trachyhoplophora Berlese, 1923
Тип подрода Hoplophora magna Nicolet, 1855
r) Hoplophorella Berlese, 1923
Тип подрода Hoplophora cucullatum Ewing, 1909
2. Steganacarus Ewing, 1917
Тип рода Hoplophora anomala Berlese, 1883
3. Hoplophthiracarus Jacot, 1933
Тип рода Hoploderma histricinutn Berlese, 1908
4. Atropacarus Ewing, 1917
Тип рода Hoplophora stricula Koch, 1836
5. Ginglytnacarus Ewing, 1917
Тип рода Hoplophora dasypus Duges, 1834
6. Hoploderma Michael, 1898 (= Hoplophora Koch, 1836 nomen
praeoccupatum)
Тип рода Hoplophora laevigata Koch, 1844
Euphthiracarinae Jacot, 1930
1. Oribotrltia Jacot, 1924(= Tritia Berlese, 1883 nomen praeoccupatum)
Тип рода Hoplophora decumana Koch, 1836
29* Зак. 1785.
452 Глава VIII. Sarcoptlformes (продолжение)
2. Indotritia Jacot, 1928
Тип рода Tritla krakatauensls Sellnick, 1924
3. Pseudotritia Willmann, 1920
a) Pseudotritia s. str.
Тип подрода Tritia monodactyla Willmann, 1919
6) Phthiracarulus Berlese, 1920
Тип подрода Phtiracarus (P Mir acar ulus) perexiguus Berlese,
1920
4. EupMhiracarus Ewing, 1917
Тип рода PMhiracarus flavus Ewing, 1908
5. Hummelia Oudemans, 1916
Тип рода Humtnella karpellesl Oudemans, 1916 (= Hoplophora
ardua Karpelles, 1893)
6. Perldromotritia Jacot, 1923
Тип рода PMhiracarus rotundus Ewing, 1908
7. Acrotritia Jacot, 1923
Тип рода PMhiracarus atnericanus Ewing, 1909
ЛИТЕРАТУРА
Jacot A. P., 1930. Oribatid mites of the subfamily Phthiracarinae of the north-
eastern United States, Proc. Boston Soc. Nat. Hist., 36 (6), 209—261.
УКАЗАТЕЛЬ
ЛАТИНСКИХ НАЗВАНИЙ
Abacarus 164
Abacoptes 164
Absoloniana 238
Acalyptonotidae 278, 325
Acalyptonotus 325
Acamina 164
Acanthochela 95, 96
Acanthonotus 165
Acaphylla 164
Acarapis 175
Acarelliptus 164
Acarlcalus 164
Acaridae 46, 244, 245, 333, 334, 338,
340, 342—345, 364 , 374
Acaridiae 26, 43, 331, 332, 337, 399
Acarina 3, 35, 43, 46—49, 54, 150
Acarinae 338, 341
Acaroceras 440
Acaromantis 281
Acaronychidae 405
Acaronychinae 406, 407
Acaronychus 405—407
Acarophenax 176
Acaropsis 29, 243
Acarus 77, 79—81, 97, 98, 101, 108,
135, 136, 152, 155—157, 175, 177,
180, 187, 191, 197, 199, 204, 218,
226, 243—245, 249, 254 , 257 259,
260, 265, 280, 287, 292, 309, 315,
341, 342, 346, 352, 363, 367, 370,
372, 375, 392, 423, 430, 437, 441,
451
Aceosejidae 67, 71
Aceosejus 71
Acer copsis 318
Acercus 318, 319
Aceria 163, 165, 167, 169, 170, 205,
218
Acheles 217
Acherontacarus 284
Achlpteria 441
Achipterina 441
Achorolophus 249
Achropodophorus 341
Acomatacarus 264, 266
Acotyledon 342
Acrocephalus 381
Acronothrus 416
Acrotacarus 365
Acrotritia 452
Actacarinae 280, 282
Actacarus 280
Actineda 238
Actinochitinosi 30
Actinoseius 86
Adoristes 430
Aecosmaris 252
Aedoplophora 448, 449
Aegyptobia 231
Ajricasla 328
Agaue 280, 282
Agauopsis 280
Alabidocarpus 389
Albia 322
Albiiпае 322
Aleuroglyphus 341
Alicorhagia 215
Alicorhagididae 184, 208, 215
Allanalges 398
Alliphis 106
Alloceraea 156
Allochaetophora 225
Allodamaeus 421
Allodermanyssus 96, 97, 99
Allodispus 175
Allogalumna 445, 446
Allokrendowskia 330
Allolaelaptidae 77
Allolimnesia 308
Alloparasitus 87
Alloptes 366, 397
Alloribates 436
Allothrombiinae 256, 257
Allothrombium 43, 255, 257, 260
Allotrombidium 260
Allothyas 292
Allozercon 69
Allozetes 436
Alycosmesis 214
Amansia 365
Amasis 302
Amazonella 316
Amblycentor 156
Amblygatnasus 80
454
Указатель латинских названий.
АтЫуотта 156
Amblyomminae 155, 156
Amblyseius 101
Amblysiopsis 101
Amerobelba 421, 422
Ameronothridae 403, 426
Ameronothrus 426, 427
Ameroseius 101
Amer us 421
Amnemochthonius 413
Amolops 423
Amorphacarus 240
Amphitetranychus 225
Anachipterla 438
Anactinochitinosi 30
Analges 38, 392, 395
Analgesidae 39, 336, 391, 398
Analgopsis 392
Analloptes 393
Anandia 238
Anarsia 238
Anasicudion 394
Anatetranychus 225
Ancystropus 76
Androlaelaps 106
Angelia 416, 418
Anisitsiella 302
Anisitsiellidae 274, 278, 301
Anisitsiellinae 302
Annectacarus 411
Anocentor 156
Anoetidae 334, 369, 371
Anoetoglyphus 370
Anoetus 370, 371
Anohydrachna 285
Anomalothrombium 258 .
Anoplocelaeno 139
Anoplocephala 426, 430, 435, 441, 445,
446
Anoplocheylus 236
Anoplonotus 394
Antennequesoma 125
Antennocelaeno 139, 141
Antennomegistus 148
Antennophoridae 143, 146, 147
Antennophorus 139, 144, 146—148
Antennoseius 102
Antennurella 147
Anfhocoptes 164
Anfhrobia 195
Antricola 151, 152
Anychus 226
Anystidae 237—239
Anystinae 238
Anystipalpus 107
Anystis 237—239
Aonidiella 367
Aphelacarus 407
Aphelenia 342
Apionseius 123, 124
Aplonobia 225
ApoIonia 264, 267
Apoloniinae 263, 264
Aponomma 156
Apostigmaeus 217
Apotetranychus 225
Aptyctima 399
Arachnida 46, 47
Archegozetes 410
Archeonothrus 407
Arctoseius 107
Argas 151—153
Argasidae 150, 152, 153
Arhodeoporus 281
Arrenuridae 278, 327
Arrenurus 328
Arrhenurae 270, 278
Arrhenurella 328, 329
Arrhenurus 328
Arthrochthonius 413
Arthronothrus 408
Arthroplophora 449
Arthropoda 47
A sea 77
Ascaidae 67, 77, 78
Ascoschongastia 265
Aspergillus 167
Aspidiotus 367, 368
Astacocroton 319, 320
Astacocrotonidae 274, 276, 278, 319
Astacopsiphaginae 282, 283
Astacopsiphagus 282
Astegistes 430
Asternolaelaps 102
Asternoseius 102
Astigma 260
Atacella 315
Atacellides 316
Atacoseius 69, 70
Atax 285, 315, 316
Atelopsalis 281
A-Thienemannia 326
A-Thienemannidae 278, 326
A-Thienemanniinae 326
A tomus 260
Atopochthonius 413
Atopomelinae 390
Atopomelus 389
Atractideidae 274, 278, 304
Atractidella 311, 312
Atractidellidae 311
Atractides 304, 305, 310
Atractidopsis 312
Atricholaelaps 109
Указатель латинских названий
455
т
Atropacarus 451
Attidae 250
Aturides 322
Aturinae 322
Aturus 322
Austracarus 264
Austracus 263
Austrochirus 389
Austrogamasus 86
Austrombicula 264
Aastroteneriffla 234
Austrothromblum 257
Autenriethia 238
Autogneta 421
Avenzoaria 394
Avrosia 180
Axonopsae 270, 278
Axonopsalbia 321
Axonopsella 320, 321
Axonopsidae 277, 278, 320
Axonopsinae 319, 320
Axonopsis 320, 322
Balaustiidae 249
Balaustium 249, 250
Balzania 436
Bandakia 298
Banksia 426
Banksinus 436
Barbaxona 321
Barbutia 217
Barella 238
Bargena 286
Bdella 190—193
Bdellidae 37, 43, 182, 190, 192, 193
BdelUnae 191
Bdellodes 191
Bdellonyssus 97, 100
Bdellorhynchus 393, 394
Beaurieina 86
Bechsieinia 238
Beklemishevia 407
Belba 420, 421
Belbidae 403, 420
Berlesella 392
Berlesia 103
Berlesiana 60
Bertiella 426, 435, 441, 446
Bimichaelia 212, 213
Biscirus 191—193
Blankaartia 265
Blattisocius 102
В lamia 363
Bochartia 249
Bonnetella 395
Bonzia 206
Boop hilus 156, 158
Borinquolaelaps 101
Borrelia 153
Botrytis 346
Brachychochthonius 414
Brachychthoniidae 412
Brachychthonius 414
Brachylaelaps 83
Brachypoda 321
Brachypodella 321
Brachysternum 139
Brachytremella 116
Brachytydeus 204
Bradibates 293
Brevipalpus 231—233
Brevisterna 97
ВгуоЫа 224—226, 230
Buchholzia 395
Caeculidae 185, 222
Caeculisoma 249
Caeculus 223
Caenobdella 192
Caenonychus 211
Caenothrombium 2B7
Calacarus 164
Calcarmyobia 240
Calepitrimerus 164, 168
Caleremaeus 421
Calholaspis 81
Calhoplophora 451
Caligonella 217
Caligonellidae 216
Caligonus 217, 218
Caliphytoptus 164
Callidosoma 249
Callyntrotus 164
Caloglyphus 332, 341, 342, 344—346
Calonyx 289, 290
Calothrombiinae 256, 257
Calothrombium 257, 258
Calotrachytes 120
Calotydeus 204
Calouropoda 136
Cal urodiscus 134
Calvolia 343, 347
Calvoppia 421
Calyptostoma 254
Calyptostomidae 185, 253
Calyptozetes 437
Camerotrombidium 29, 258
Camisia 416
Camisiidae 402, 415
Campylochirus 389, 390
Campylothrombium 258
Canestrinia 62, 71, 365, 366
Canestriniella 365
Canestriniidae 332, 334, 364
436
Указатель латинских названий
Caparinia 383, 384
Capeulais 288
Capitodiscus 132
Capobates 309
Carabocepheus 425
Car abodes 425, 431
Carabodidae 400, 403, 424, 426
Carabodoides 425
Caraboecius 365
Cardioribates 435
Car is 98
Caroloptes 164
Carpais 79
Carpoglyphidae 333, 351
Carpoglyphus 351—353
Cas 94
Caspihalacarus 281
Catenotaenia 364
Cavannea 250
Cavilaelaps 108, 109
Cecidodectes 164
Cecidophyes 164, 170
Cederhjelmia 369, 370
Celaeno 119, 129
Celaenogamasus 58
Celaenopsidae 137, 141
Celaenopsina 56, 57, 137, 142, 143
Celaenopsis 141
Celaenopsoides 147
Celaenosthanus 147
Cenobita 369
Cenlrolimnesia 307, 308
Centroribates 444
Cenirotrombidium 257
Centrouropoda 136
Cephalodiscus 132
Cephalouropoda 132
Cepheus 424—426
Cerachipteria 441
Ceratixodes 44, 155
Ceratoacarus 197
Ceratocelaenopsis 141
Ceratoppia 421, 422
Ceratothrix 395
Ceratozetes 436
Ceratozetidae 404, 436
Cerberothrombium 259
Cercomegistidae 143, 145
Cercomegistus 145, 146
Cercothrombium 260
Cerocepheus 425
Ceroglyphus 342
Cerophagus 363
Cetonia 366
Chabrieria 238
Chaetodactylus 363
Chamobates 437
Charassobates 425
Chauliacia 395
Chaunoproctus 425
Chaussieria 238
Cheiletiella 244
Cheiloceras 393, 395
Cheiroseius 101
Chelacaropsis 243
Cheletes 243
Cheletogenes 29, 242, 243
Cheletoides 243
Cheletomimus 243
Cheletomorpha 242, 243
Cheletophanes 243
Cheletophyes 243
Cheletopsis 243
Cheletosoma 244
Chelicerata 47
Chelonotus 244
Cheylabis 395
Cheyletia 244
Cheyletidae 36, 37, 183, 185, 200, 239,
242, 244, 245
Cheyletiella 244
Cheyletus 243—245
Chionas-pis 367
Chirodiscoides 390, 391
Chirodiscus 37, 390, 391
Chironemus 180
Chiropteranoetus 370
Chiroptonyssus 98
Chitohydrachna 286
Chorioptes 384, 385
Chortoglyphidae 334, 360, 361
Chorloglyphus 360
Chromotydeus 189
Chyzeria 260
Cilliba 122, 128, 136 ,
Cillibidae 122, 128
Cinchona 181
Circocylliba 126
Circocyllibanidae 122, 126
Cittotaenia 426, 430, 436, 438, 441,
444—448
Cladosporium 167
Clathrosperchon 294
Clathrosperchonella 294
Clathrosperchonidae 273, 278, 293
Clausiadinychus 129
Claverytheraeus 249
Cnemidocoptes 375
Cocceupodes 187
Coccorchestes 211
Coccorhagidia 193
Coccotydeus 204
Coelognathus 341
Coggiella 438
Указатель латинских названий
457
Colaptes 396
Coleoglyphus 365
Coleolaelaps 107
•Coleopterophagus 365, 366
Coleoscirus 206
Coleotydaeus 204
Colletidae 355
Collohmannia 408
Coloboceras 280
Columellaia 395
Comatacarus 264
Cometacarus 363
Comydinychus 132
Conoppia 422
Copiaognathopsis 281
Copidognathus 281
Coprholaspis 81
Copriphts 106
Coprolaelaps 83
Coptophijlla 165
Corerythrolophus 249
Corethrothrombium 257
Cornubia 62
Corticacarellus 310
Corticacarus 310
Cosmiomma 157
Cosmiphis 106
Cosmochthoniidae 412
Cosmochthonius 413, 414
Cosmoglyphus 342
Cosmolaelaps 107
Coxequesoma 124, 125
Coxequesomidae 122, 124, 125
Crameria 395
Crassarchus 76
Crastidoglyphus 363
Crenohygrobates 310
Crenolimnesia 308
Creuizeria 370
Cricetus 110
Crinaturus 322
Criniscansor 390
Crinitodiscus 132
Crotiscus 265
Cryptacarus 411
Cryptognathidae 182, 195, 196
Cryptognathus 195, 196
Cryptometasternum 139
Cryptoplophora 449
Cryptoribatula 432
Cryptostigmatina 25
Ctenerythraeus 249
Ctenoglyphus 363
Ctenothyas 295
Ctenothyasidae 273, 278, 295
Caltroribula 430
Caltrozetes 430
Cunaxa 205—207
Cunaxidae 37, 182, 205—207, 268, 269,
284
Cunaxoides 206, 207
Cupacarus 165
Cyclacarus 136
Cijclolaelaps 109
Cyclothorax 58
Cyclothrix 286, 287
Cyllibano 136
Cyllibula 136
Cymbaeremaeidae 403, 419
Cymbaeremaeus 419
Cymoptus 164
Cynorhaestes 156
Cypholaelaps 108,
Cyrthydrolaelaps 79
Cyrtolaelaps 32, 77, 79, 84, 86
Cyta 190, 191
Cytinae 191
Cytisus proliferus 398
Cytodites 378
Cytoditidae 335, 378, 379
Cytoleichus 379
Czenspinskia 354, 356
Czenspinskiidae 333, 355
Dactylosclrus 205
Dadayella 328
Damaeus 421, 429
Dameobelba 421
Damaeolus 422
Dameosoma 422, 423
Dampfiella 422
Dartia 302
Dartiella 302
Dartonia 302
Dasyponyssidae 111, 112
Dasyponyssus 112—114
Dekabates 309
Demodex 160, 245—247
Demodicidae 160, 182, 245—247
Dendrolaelaps 77
Dendrotrombidium 258
Dentidinychus 129
Deratophorus 132
Dermacarus 363
Dermacentor 44, 155, 156, 158
Dermacentorites 156
Dermaleichus 365, 366, 392—397
Dermanissus 136
Dermanyssidae 68, 96, 98, 99
Dermanyssinae 99
Dermanyssus 34, 94, 97—99
Dermation 386
Dermatodectes 384
Dermatokoptes 384
458
Указатель латинских названий
Dermatophagoides 353, 386—388
Dermatophagus 384
Dermoglyphidae 336, 393, 394, 398
Dermoglyphus 365, 394, 395, 397
Diapierobates 437
Diarthrophallidae 115, 116
Diarthrophallina 56, 57, 115
Dirthrophallus 115, 116
Diaspis 367
Dlcanestrinia 365
Diepicrius 62
Digamasellus 84
Diktyobates 309
Dinocelaeno 141
Dinogamasus 107
Dlnothrombium 257
Dinychidae 130
Dinychopsis 132
Dinychura 131
Dinychus 129, 132
Diphaulocylliba 133
Diphtheroglyphus 343, 347
Diplodontinae 292
Diplodontus 292
Diplogyniella 139
Diplogyniidae 137, 138, 140, 142
Diplogyniinae 139
Diplogyniopsis 139
Diplogynium 138, 139
Diplohydrachna 285
Diplokoenikea 316
Diplopodophilus 144
Diplostaspis 76
Diplothrombium 258
Dipolyaspis 120
Dlptilomiopus 165
Discomegistus 70
Discopoma 32, 124, 128, 132, 135
Discotrachytes 129
Discourella 123, 124
Discourellidae 121, 123, 124
Discozercon 69
Discozerconidae 66, 69, 70
Disparipes 175
Dispersipiona 318
Dissorhina 423
Distolebertla 303
Dithinozercon 111
Diurodinychus 131
Divarinychus 226
Diversipes 175
Djeboa 324
Djeboella 324
Djeboinae 324
Dodecabates 309
Dolaea 107
Dolicheremaeus 422
Dolichotetranychus 162, 231, 233
Doloisia 265
Donia 86
Donndorfia 356
Dorcus 366
Dromeothrombium 258
Drymobates 435
Drymobatoides 435
Dugesia 252
Duralimnesia 308
Dwigubskyia 60
Eadiea 240
Eberhardia 342
Ebertia 341
Eccoptogaster 357
Echinolaelaps 105, 109, 110
Echinomeglstus 147
Echinonyssus 98
Echinoseius 61, .'101
Echinoihrombium 258
Ecpolopsis 316
Ecpolus 316
Edwardzetes 437
Elaphrolaelaps 83
Ellsworthia 98
Etnaeus 106
Embolacarus 419
Emberiza 381
Encentridophorinae 316
Encentridophorus 316
Encylliba 136
Endeostigmata 208
Endopalpiger 155
Endotrombicula 265
Enemothrombium 258
Eniochthonius 413, 414
Ensliniella 354, 355
Ensliniellidae 332, 333, 354, 355
Enterohalacarinae 282
Enterohalacarus 282
Enteromorpha 351
Entonyssidae 68, 91, 92
Entonyssinae 91, 92
Entonyssus 91, 92
Eobrachychthonius 414
Eohypochthonius 414
Eotetranychus 225, 226
Eothronibium 259
Epactozetes 447, 448
Epactozetidae 404, 447
Epicriidae 59, 61, 62
Epicriopsis 86
Epicrius 60, 62
Epicroseius 60, 61
Epidermoptes 386, 387
Epidermoptidae 335, 386, 387
Указатель латинских названий
459
Epieremulus 422
Epilohmannia 410
Epilohmanniidae 402, 410
Epilohmannoides 414
Epiphis 86
Episeiella 102
Episeius 102
Epistomalycus 215
Epitetranychus 225
Epitrimerus 165, 166, 168
Eporibatula 435
Eremaeidae 403, 421
Eremaeozetes 425
Eremaeus 419—421, 426, 435
Eremella 420
Eremobelba 422
Eremobodes 425
Eremulus 422
Ereynetes 199
Ereynetidae 183, 199, 202
Eriophyes 34, 163, 168
Eriophyidae 37, 159—162, 165, 167—
169, 231, 232
Eriophyinae 163
Erithracus 381
Erythracarinae 238
Erythracarus 238
Erythraeidae 185, 248—250
Erythraeus 238, 239, 249, 250
Erythrellus 249
Erythrocheylus 239
Erythroides 249
Erythrolophus 249
Erythrombium 249
Eschatocephalus 156
Ettmillleria 260
Eubrachylaelaps 109
Eucanestrinia 366
Eucarus 50
Eucheyla 244
Eucheyletla 244
Euepicrius 82
Eufedrizzia 147
Eugamasus 80
Eugynolaelaps 108
Euhaemogamasus 27, 95, 97
E uholocelaeno 82
Eulabidocarpus 390
Eulaelaps 27, 109
Eulohmannia 408
Eulohmanniidae 401, 408
Eumaeus 86, 106
Eunicolana 197
Eupalopsellus 217
Eupalopsis 217, 218
Eupalus 206, 217
Eupatra 292
Eupatrella 327
Eupatrellidae 278, 327
Eupelops 443
Euphthiracarinae 451
Euphthiracarus 452
Euplatylophus 249
Eupodes 39, 186—188
Eupodidae 184, 186, 187, 189
Eupodolophus 250
Eupterotegaeus 425
Eurychiroides 390
Euryparasitus 84
Euryparazetes 442
Eurytetramjchus 225
Euryzonus 390
Euschongastia 29, 36, 262, 265
Eustathia 395
Eustigmaeus 217
Eutalpacarus 240
Eutarsopolipus 172, 173
Eutegaeus 425
Eutetranychus 226, 230
Euthrombium 259
Euthyas 293
Euthyasinae 293
Eutogenes 244
Eutrachytes 122, 129
Eutrachytidae 122, 129
Eutrichothrombium 258
Eutrombicula 265, 267
Eutrombidiinae 257
Eutrombidium 2Ы, 259, 261
Eutrombium 259
Euzercon 140
Euzerconidae 137, 140, 142
Euzetes 437
Eviphis 106
Ewingana 240
Ewingella 244
Ewingia 368, 369
Ewingidae 334, 368
Exopalpiger 156
Exoribatula 435
Eylaidae 35, 272, 278, 288
Eylainae 288
Eylais 288, 289
Falculifer 393—396
Falculiferidae 393, 394
Fallopio. 248, 249
Favettea 397
Fedrizzia 56, 149
Fedrizziidae 143, 148
Fedrizziina 56, 57, 143
Feltria 313, 314
Feltriella 314
Feltriidae 276, 278, 313
460
Указатель латинских названий
Ferminia 352
Fessonia 252
Fessoniella 250
Fessoniinae 253
Flaoionyssus 94
Flexipalpus 165
Foliomyobia 240
Foliotrombidium 258
Fonsecaonyssus 97
Fonsecia 265
Forcellinla 348, 349
Forcelliniidae 333, 348
Forelia 318
Foreliinae 318
Formica 175
Freyana 395
Frischia 437
Frontipoda 304
Frontipodopsinae 322
Frontipodopsis 322
Froriepia 342
Froweinia 373
Fusacarus 360, 361
Fuscozetes 441
Fuscuropoda 136
Fusohericia 362, 363
Gabucina 395
Gahrliepia 264
Gallus 337
Galumna 445—447
Galumnella 443
Galumnidae 400, 404, 444, 445
Galumnopsis 443
Gamasellus 77, 78, 84
Gamasholaspis 81
Gamasides 53, 56, 57, 65, 66, 69, 75,
80, 83, 87, 114, 117, 118
Gamasiphis 86
Gamasodes 85
Gamasolaelaps 84
Gamasolaelaptidae 68, 84, 85, 87
Gamasus 62, 77—88, 101, 102, 106,
107, 141
Gammaphytoptus 165
Garsaultia 342
Gateria 264
Geayella 330
Geayia 330
Geayidea 330
Geckobia 221
Geckobiella 221, 222
Geholaspis 81
Gehypochthonius 407
Geneiadolaelaps 98
Georgella 292
Georgia 258
Giardius 373, 374
Giebelia 396
Glgantolaelaps 109
Ginglymacarus 451
Globozetes 437
Glycyphagidae 334, 361
Glycyphagus 245, 341, 352, 362—364
Glyphanoetus 370
Gnaphiscus 304
Gnaphoxus 304
Gohiera 363
Gradidorsum 422
Grandiella 365, 366
Grandjeanacarus 405, 407
Greenia 107
Greeniella 103, 107
Guntherana 265
Gustavla 431, 432
Gustaviidae 402, 431
Gymnob ales 433
Gymnodamaeus 421
Gymnodampia 422
Gymnolaelaps 107
Gymnonothrus 416
Haetnalastor 156
Haetnaphysalis 156
Haemogamasidae 68, 95, 96
Haemogamasus 95
Haemolaelaps 30, 38, 109, 110
Hafenrefferia 433
Halacaridae 207, 268, 269, 271, 279,
282
Halacarinae 280, 283
Halacarus 279—281
Halarachne 90
Halarachnidae 68, 89, 90
Haleupalus 206
Halixodes 282
Halixodinae 282
Halleria 395
Halolaelaps 84
Halotydeus 186—188
Halouropoda 133
Halozetes 423, 442
Hammeria 437
Hammertonia 92
Hamohalacarus 282
Hannemania 264
Haplochthonius 414
Haplozetes 442
Haplozetidae 400, 404, 441, 442
Haptosoma 390
Harpagopalpinae 326
Harpagopalpus 326
Harpicephalus 240
Harpirhynchus 239—241
Указатель латинских названий.
461
Hartingia 392
Hauptmannia 249
Heaslipia 265
Heliocopris 360
Helix 200
Hernialges 392
Ilemibrachypoda 321
Hemilaelaps 98, 109
Hemileius 434
Heminothrus 416
Hemisarcoptes 367, 368
Hemisarcoptidae 335, 366
Hemitarsonemus 180, 181
Hemilromblcula 287
Hepatozoan 110
Hericia 363
Hermannia 417, 418, 428
Hermanniella 428
Hermanniellidae 402, 428
Hermanniidae 402, 416
Heterocheylinae 236
Heterocheylus 236, 237
H eterochthonius 414
Heterodiplogyniinae 138
Heterodiplogynium 139
Heterodis pus 175
Heterolaelaps 108
Heteropsoridae 335, 380
Heteropsorus 381
Heteropus 176
Heteroteneriffia 234, 235
Heterothrombidium 264
Heterothrombidium 260
Heterozercon 34, 69, 70
Heterozerconidae 70
Hevea 181, 353
Hexalebertla 303
Hexatax 315
H exathrombium 260
Hexaxonopsis 320
Hiotrombidium 29, 258
Hippoboscidae 381
Hirstesia 98
Hirstiella 221, 222
Hirstiosoma 252, 253
Hirstiosominae 253
Hirstithrombium 258
Hristonyssus 98
Hispidosperchon 301
Histiogaster 338, 342
Histiostoma 344, 370, 371
Hjartdalia 322
Holaspulus 81
Holcotrombidiinae 257, 259
Holcotrombidium 29, 259
Holocelaeno 82
Holokalumma 445
3Q Зак. 17S5
Holoparasitus 80
Holostaspella 82
Holostaspis 81, 82, 107
Holothyridae 41, 42, 49, 51
Holothyroidea 30, 33, 51
Ilolothyrus 36, 51, 52
Holozetes 447
Homocaligtts 216, 217
Homopus 363
Hormodendrum 167
Hoploderma 451
Hoplolaelaps 87
Hoplomegistus 58
Hoplomolgus 192
Hoplophora 451, 452
Hoplophorella 451
Hoplophthiracarus 451
Hoploscirus 192
Hoploseius 102
Hoplothrombium 260
Horstia 354
Huitfeldtia 316
Huitfeldtiinae 316
Humerobafes 437
Hummelia 452
Hungarohydracarus 326
Hyadesia 350
Hyadesidae 333, 350, 351
Hyalomma 26, 155, 157
Hyalommasta 157
Hyalommina 157
Hybalicus 208
Hydracarina 268
Hydrachna 285, 286, 288, 291, 304,
309, 315, 316, 319—321, 323, 324,
328
Hydrachnae 269, 278
Hydrachnellae 182, 268—270
Hydrachnidae 271, 278, 285
Hydrobaumia 302
Hydrochoreutes 319
Hydrodroma 290—292
Hydrodromidae 272, 278, 290
Hydrogamasus 86
Hydrovolzia 283, 284
Hydrovolziae 269, 278
Hydrovolziella 284
Hydrovolziidae 268, 271, 278, 283
Hydrozetes 401, 423
Hydryphantae 270, 278
Hydryphantes 291, 292, 397
Hydryphantidae 273, 278, 291
Hydryphantinae 292
Hygrobatella 310
Hygrobates 309, 310
Hygrobatidae 275, 278, 300
Hygrobatides 310
452
Указатель латинских названии
Hygrobatinae 309
Hygrobatomegapus 311
Hygrobatopsis 310
Hygroribates 426, 427
Hyletastea 106
Hyletastinae 106
Hymenoptera 355
Hyperalges 392
Hypoaspidinae 106
Hypoaspls 86, 106, 107
Hypochthoniella 413
Hypochthoniidae 402, 412
Hypochthonius 412—414
Hypodectes 396
Hypopus 370, 373
Hyracarus 264
Hystrichonychus 226
Ichoronyssus 98
Imparatoppia 422
Imparipes 175
Indocentor 156
Indoribates 438
Indotriiia 452
Ingrass la 392
Iphidinychus 61
Iphidosoma 107
Iphidozercon 106
Iphidulus 101
Iphiopsidae 68, 103
Iphiopsis 103
Iphis 103, 106, 107
Iphiseius 102
Ischnolaelaps 109
Ischyrognathus 281
Ischyropoda 95
Isobactrus 280
Isoglyphus 342
lugoribates 437
Ixobioides 73
Ixodes 33, 35, 155—158
Ixodidae 153—155, 157, 158
Ixodiderma 221
Ixodides 40, 47, 53, 150, 157
Ixodinae 155
Ixodorhynchidae 67, 73
Ixodorhynchus 73
Jacobsonia 103
Jacotella 421
Janetiella 132
Javathyas 292
Joelia 438
Johnstoniana 257
Johnstonianinae 256, 257
Jordensia 107
Joubertia 397
Jugatala 436, 437
Julolaelaps 107
Jurabates 433
Kalobrachypoda 321
Kashmirothyas 292
Kawamuracarus 302
Klinckowstroemia 149
Knemidokoptes 375, 377, 378
Kobus 88
Koenikea 316, 317
Koenikella 316
Kolenationyssus 98
Kondia 325
Kongsbergia 314
Koptorihosoma 374
Kramerella 395
Kratzensteinia 447
Krendowskia 329
Krendowskiella 329
Krendowskiidae 278, 329
Kyphohygrobatella .310
Kyphohygrobates 310
Labidocarpinae 390
Labidocarpus 389—391
Labidophorus 363
Labidostomma 196—198
Labodostommidae 183, 196—198
Laelantennus 107
Laelaps 84, 86, 87, 102, 106—109
Laelaptidae 66, 68, 89, 104—106, 110
Laelaptinae 106, 108
Laelaspis 107
Laelaspulus 107
Laelogamasus 78
Lagenoglyphus 342
Lamellaxona 322
Laminosioptes 379, 380
Laminosioptidae 335, 379
Laminothrombium 258
Lardoglyphidae 334, 358
Lardoglyphus 358, 359
Larlnyssus 93, 94
Lasioseius 101
Lasiotydeus 204
Lasius 175
Lawrencoppia 423
Lebertia 303
Lebertiae 270, 278
Lebertiidae 274, 278, 302
Lebertiinae 303
Lebermulleria 218
Leeuwenhoekia 264
Leeuwenhoekiinae 263, 264
Leiodinychus 134
Leiognathus 97
Указатель латинских названий
463
Leioseius 101
Leiosoma 423, 430, 432
Leiostaspis 76
Lemienia 316
Lentungula 350
Lenzia 353
Leonardiella 132
Lepidixodes 156
Lepidoglyphus 363, 364
Lepidoribates 444
Lepidospahes 356, 367, 368
Lepidozetes 443, 444
Lepronyssoides 98
Lepronyssus 98
Leptalicus 211
Leptantennus 147
Leptogamasus 87
Leptognathus 281
Leptolaelaps 107
Leptonyssoides 98
Leptonyssus 98
Leptopterotrichophorus 317
Leptothrombium 257
Leptotrombidium 265
Leptus 248—250
Lesseria 410
Lethaxona 321
Liacaridae 400, 403, 429
Liacarus 430
Licneremaeus 423
Licnobelba 423
Licnodamaeus 423
Licnoliodes 423
Lieb stadia 435
Ligilaelaps 109
Limnesia 308
Limnesicula 308
Limnesides 308
Limnesiella 308
Limnesiellula 308
Limnesiidae 274, 278, 307
Limnesiinae 308
Limnesiopsides 308
Limnesiopsis 308
Limnocharae 269, 278
Limnochares 286, 287
Limnocharidae 35, 271, 278, 286
Limnohalacarinae 281, 282
Limnohalacarus 281
Limnolegeria 302
Limnozetes 437
Linobia 372
Linobiidae 334, 372
Linocoptes 372
Linopodes 186—188
Linotetranus 226
Liodes 418
Lionotus 355
Liostigmaeus 217
Liponissidae 99
Liponissinae 99
Liponissus 98, 136
Liponysella 98
Liponyssoides 98, 99
Liponyssus 97, 98, 109, 136
Liroaspidae 59, 60
Liroaspina 59
Liroapis 60, 61
Listrophoridae 336, 389—391
Listrophorinae 390
Listrophoroides 390
Listrophorus 389
Litarachna 305
Litomosoides 100
Ljania 321
Ljunghia 107
Lobocephalus 78
Lobogynioides 137, 139
Lobogynium 139
Locusta 173, 174
Locustacarus 172—174
Lohmannella 281
Lohmannellinae 281, 282
Lohmannia 410, 411
Lohmanniidae 402, 411
Lohohalacarus 281
Loloisia 265
Longolaelaps 109
Lordalychidae 184, 207, 208
Lordalycus 207, 208
Loriaspina 56
Lorryia 204, 205
Lucanidae 366
Lucasiella 249
Lucoppia 422, 423
Lundbladia 292
Mackiella 163
Mackiellinae 163
Macrocheles 81, 82
Macrochelidae 39, 67, 80, 82
Macrodinychus 134
Macrolaelaps 108
Macrolaspis 82
Macronyssidae 99
Macronyssus 98
Macrostigmaeus 218
Malacoangelia 414
Malaconothridae 402, 415
Malaconothrus 415
Mallophaga 381
Mamersa 292
Mamersella 302
Mamersellides 301, 302
30*
464
Указатель латинских Названий
Mamersid.es 302
Mamersinae 292
Mamersopsidae 273, 278, 297
Mamersopides 302
Mamersopsis 298
Mania 302
Maniricola 98
Manisicola 98
Manitherionyssus 98
Manotonia 302
Manriquia 258
Mantldoglyphus 354
Margaropus 157
Marginura 136
Marquesania 390
Masthermannia 417, 418
Mauduytia 370
Mealia 386
Mealiidae 386
Mediolata 216, 218
Megabates 310
Megacanestrinia 366
Megachaetochela 139
Megaliphis 86
Megalolaelaps 83, 86
Megaluracarus 328
Megamerus 189
Megapella 311
Megapodius 374
Megapoides 311
Megapus 310, 311
Megapusinae 310
Megatrotnbicula 265
Megistanus 58
Megisthanidae 58, 59
Megisthanina 56, 57, 59
Megisthanus 58, 59
Megninia 392, 393
Megninietta 342
Meinertula 139
Meinertulinae 139
Melanotydeus 204
Melanozetes 437
Metasoma 372
Melichares 101
Meliponaspis 107
Melisia 363
Melittiphis 106
Meristacarus 411
Meristaspis 76
Mesalges 391, 392
Mesobatella 311
Mesobates 311
Mesofeltria 314
Mesolaelaps 108
Mesoplophora 449, 450
Mesoplophoridae 31, 404, 449
Mesostigmata 26, 27, 30, 33—35, 40—
45, 47. 53. 55. 56, 59, 75, 118,
150
Mesothrombium 257
Messoracarus 147
Metadinychus 130
Metagynella 136
Metanalges 392
Metaparasitus 84
Metarhombognathus 280
Metatetranychus 224—230
Meteylais 288
Mefrioppia 423, 424
Michaelia 213, 395, 411
Michaelichus 395
Michaeliella 131
Micreremus 420
Micrereunetes 187, 199
Micriphis 86
Microchelys 395
Microchiroptera 391
Microdiplogyhiutn 139
Microdispus 175
Microgyniidae 63
Microgyniina 56, 57, 59, 63
Microgynium 63, 64
Microlichus 386
Micromegistus 148
Microsejidae 63—65
Microsejus 65
Microsmaris 249
Microspalax 395
Microtegeus 425
Microtrombidiidae 261
Microtrombidiinae 256, 258
Microtrombidium 255, 258, 265
Microtydeus 204
Mierozetes 440
Microzetidae 404, 439
Micruracaropsis 328
Micruracarus 328
Midea 270, 323
Mideidae 277, 278, 322
Mideopsae 270, 278
Mideopsella 324, 325
Mideopsellinae 325
Mideopsidae 277, 278, 323
Mideopsinae 324
Mideopsis 324
Minunthozetes 437
Miraxona 321
Miraxonella 321
Miraxonides 321
Mixobates 311
Mixolebertia 303
Mixosperchon 301
Mochlozetes 437
Указатель латинских названий
465
Molgus 191, 192
Momonia 277, 325
Momoniella 325
Momoniinae 325
Monalichus 211, 214
Monatractidxs 305
Monaulax 163
Moniezia 364, 400, 430, 435, 442, 445,
446
Monieziella 342
Monobates 309
Monoceronychus 226
Monochetus 163
Monohydrachna 286
Monojouberiia 397
Monokoenikea 317
Monotrichob della 190, 191
Monotrombidium 249
Monlesauria 397
Monunguis 260
Mucronothrus 415
Mulvius 419
Mundamella 326, 328
Mundamellinae 326
Mungosicola 109
Murcia 435, 438
Musca 178
Muscardinus 246
Myceloglyphus 342
Mycobates 437
Myialges 382, 383
Myialgesidae 335, 381
Myialgopsis 382
Myianoetus 370
Myobia 240, 241, 391
Myobiidae 183, 239, 241
Myocoptes 390
Myocoptinae 390
Myonyssoides 98
Myonyssus 98
Myotrombicula 265
Myrmeciphis 107
Myrmicotrombium 258
Myrmoglyphus 342
Myrmolaelaps 107
Myrmoleichus 107
Myrmonyssus 104, 107
Myrmozercon 108
Mysolaelaps 109
Najadicola 316
Najadicolinae 316
Nalepella 163
Nanacaridae 335, 373
Nanacarus 373, 374
Nanhermannia 409
Nanhermanniidae 402, 408
Nanorchestes 39, 210, 211
Nanorchestidae 184, 210, 211
Nasozetes 423
Nautarachna 313
Nautarachnidae 275, 278, 312
Nealges 392
Neamerus 421
Nellacarus 440
Nemnichia 80
Neoalbia 321
Neoaxona 321, 322
Neoaxonopsella 321
Neoaxonopsis 322
Neoberlesia 108
Neobrachypoda 322
Neocalonyx 290
Neocaridae 49
Neocarus 36, 50
Neocelaeno 141
Neocepheus 425
Neocheyletiella 29, 244
Neocheylus 236, 237
Neocypholaelaps 108
Neodiplogyniinae 138
Neodiplogynium 139
Neodiscopoma 132
Neoeucheyla 244
Neogymnobates 442
Neohygrobates 309
Neoiochoronyssus 98
Neokoenikea 317
Neokrendowskia 329
Neolabidocarpus 390
Neolaelaps 109
Neolebertia 303
Neolimnesia 308
Neolimnochares 287
Neoliodes 418
Neoliodidae 402, 418
Neoliponyssus 98
Neomegistus 148
Neomolgus 191
Neomyobia 241
Neonyssoides 94
Neonyssus 94
Neo-Oudemansia 146
Neopar alaelaps 109
Neoparasitidae 68, 85—87
Neoparasitus 85
Neophyllobius 218
Neopodocinum 32, 82, 86
Neoribates 447
Neorib atula 434
Neorizetes 447
Neoroqueella 330
Neoschongastia 265, 267
Neoseius 119
466
Указатель латинских названий
Neosmaris 248, 250
Neotanaupodus 259
Neoteneriffiola 235
Neotetranychus 226
Neothrotnbium 260
Neotorrenticola 306
Neotorrenticolidae 274, 278, 306
Neotrombicula 265
Neotrombidium 258
Neotydeus 201
Neotyrellia 308
Neoxystonotus 324
Nesiotizetes 437
Neumania 315, 316
Neumaniinae 316
Neumannia 395
Ne.urnanru.ella 395
Neuteria 136
Nicoletiella 197
Nilotonia 302
Nilotoniinae 302
Niphocepheus 425 .
Nodipalpus 370
Noerneria 193
Nosomma 157
Notaspididae 400, 404, 440
Notaspis 128, 136, 418, 421, 423, 424,
426, 430, 434, 435, 437, 441, 444
Nothotrombicula 260
NothroholaspiS 81
Nothrus 409, 415, 416, 418, 419, 426
Notoedres 374, 375, 377
Notomideopsis 324
Notophallus 187
Notostigmata 24—26, 30, 33, 40—42,
44, 49, 51, 54
Notothrombiinae 256, 259
Notothrombium 259
Novophytoptus 164
Novotrombicula 266
Nummulus 135
Nuttalliella 153, 154
Nuttalliellidae 150, 153
Nya.nga.lla 317
Nycteridocoptes 375
Ocipoda 369
Octohydryphantes 292
Octolundbladia 292
Octomegapus 310
Octomideopsis 324
Octothyas 292
Ocypete 259
Ocypetinae 257, 259
Odontacarus 264
Odontoscirinae 191
Odontoscirus 191
Odynerus 355
Oehserchestes 210, 211
Oenoschongastia 266
Olabidocarpus 390
Olafsenia 359, 360
Olafseniidae 334, 359
Oligonychus 226
Olodiscus 132
Ologamasellus 86
Ologamasus 86
Oloiphis 106
Ololaelaps 108
Olopachys 83
Olouropoda 136
Ommatocepheus 426
Onchodeltus 83
Onychopalpida 35, 36, 40, 47, 49
Oocarpais 80
Ophidilaelaps 109
Ophiomegistus 147
Ophionyssus 98
Ophiopneumicola 92
Ophioptes 239, 241
Opiliocaridae 49—51
Opitioacarus 50, 51
Oplitis 136
Oppia 422, 423
Oppiella 423
Opsereynetes 199
Optis 73
Oribata 421, 423, 425, 430, 433, 435—
437, 439, 441—443
Oribatei 22, 26, 31, 41, 43—46, 118,
331, 337, 399—401
Oribatella 438, 439
Oribatellidae 404, 438
Oribates 430, 433, 435, 437, 438, 445,
447
Oribatodes 425
Oribatula 434, 435
Oribatulidae 400, 404, 434, 435
Oribella 423
Oribotritia 451
Oripoda 432
Oripodidae 403, 432
Ornithodoros 151—153
Oromurcia 437
Orthohalarachne. 89, 90
Orthoptera 173
Otobius 152, 153
Otocentor 156
Otocepheus 425
Otodectes 383, 384, 386
Ottonia 258
Oudemansiella 98
Oudemansium 363
Oulenzia 353, 354
Указатель латинских названий
467
Oulenziidae 333, 353
Oustaleiia 395
Oxinae 271, 303
О хор sis 291
Oxus 303
Oxypleurites 161, 165—167
Pachygaster 303
Pachyglyphus 342
Pachygnathidae 184, 208, 212
Pachygnathus 25, 213, 280, 281
Pachylaelaps 83
Pachylaelaptidae 68, 83, 84
Pachylaella 84
Pachylichus 386
Pachyseius 85
Palaeacaridae 401, 405—407
Palaeacarinae 406, 407
Palaeacarus 4G7
Palalaioria 367
Palpoboophilus 157
Panisellus 292
Panisoides 292
Panisopsis 293
Panisus 292, 293
Panoplia 197
Papilloporus 292
Parabrachypoda 321
Paracarus 50
Parachyzeria 260
Paradoxiphis 108
Parakalumma 446, 447
Parakalummidae 404, 446
Parakoenikea 317
Paralaelaps 83
Paralges 395
'Paralimnesia 308
Paralimnochares 287
Paraliodes 423
Paramansia 366
Paramegistidae 143, 147
Paramegistus 147
Paraneonyssus 94
Paranoplocephala 446
Parantennulidae 143
Parantennulus 144
Parapelops 443, 444
Paraphytoptus 164
Pararoqueella 330
Paraschelobates 435
Paraschizobates 310
Paraschongastia 265
Parasitalbia 322
Parasitellus 80
Parasitidae 39, 67, 79, 80, 107
Parasitiformes 53
Parasitus 27, 34, 38, 70, 79, 85
Parasoldanellonyx 282
Paratenerlffta 235
Paratetr any chus 225, 226
Parathyas 293
Parathrombium 257, 260
Paratydeidae 183, 200
Paratydeus 201
Parawenhoekia 250
Paraxonopsella 321
Paraxystonotus 324
Parazercon 117
Parazetes 442
Parerythraeus 250
Pareylais 288
Parholaspls 82
Parliypochthonidae 405
Parhypochthoninae 406, 407
Parhypochthoninus 407
Parhypochthonius 25, 406, 407
Partnunia 290
Partnuniella 290
Passalacarus 139
Passalidae 366
Passalobia 116
Passalozetes 426
Paulitzia 136
Pavania 176
Pediculochelidae 331, 332, 336, 337
Pediculochelus 336, 337
Pediculoides 175—179
Pediculoididae 171, 176
Pediculopsis 177—179
Pediculus 240, 249, 257, 363
Pedotrombidium 258
Peletiphls 106
Pelopidae 400, 404, 443
Peloppia 423, 424
Peloppiidae 424
Pelops 443, 444
Pelopsis 443, 444
Peloplulus 443, 444
Peloribates 442
Pelvetia 351
Pentagonella 265
Pentalbia 321
Pentamerismus 231
Penlalax 315
Penthaleidae 186, 187, 189
Penthaleus 186—189
Penthalodes 189
Penthalodidae 184, 187—189
Percanestrinia 366
Pergalumma 445
Pergamasus 32, 79, 80
Peridromotritia 452
Perigllschrus 75, 76
Periphis 86
468
Указатель латинских названий
Perlohmannia 408
Petiolurus 328
Petralycus 212, 213
Petrobia 225, 226
Petzschia 342
Phanaeus 366
Phanolophinae 253
Phanolophus 252
Phaulocylliba 133, 134
Phaulodiaspis 122, 133, 134
Phaulodinychidae 123, 133
Phaulodinychus 122, 133
Phauloppia 423
Phloeotribus 358
Phoci dae 90
Photia 366
Phrynacarus 265
Phthiracaridae 404, 450, 451
Phthiracarinae 451
Phthiracarulus 452
Phthiracarus 450—452
Phthiroides 177
Phylacozetes 440
Phyllhermannia 418
Phyllocoptes 163—167
Pliyllocoptidae 161
Phyllocoptinae 163, 164
Phyllocoptruta 165, 169
Phyllocoptyches 165
Phyllodlnychus 122
Phyllotegeus 423
Phyllotetranychus 232
Phyllotrombidium 257
Phymatodiscus 132
Physalozercon 147
Phytojacobsonia 108
Phytoptinae 163
Phytoptipalpidae 37, 160, 162,
184, 224, 230
Phytoptipalpus 162, 231, 232
Phytoptochetus 164
Phytoptus 163, 164, 170
Phytoseiidae 68, 100—102, 105
Phytoseiinae 101
Phytoseius 101
Picobla 241
Piersigia 289
Piersigiinae 289
Pilizetes 445
Pilolebertia 303
Pllollmnesia 308
Pimeliaphiloides 221
Pimeliaphilus 220—222
Pimelofia 173
Plena 217, 318, 319
Pionacercopsis 319
Pionacercus 319
Pionae 270, 278
Pionatacinae 317
Pionatax 317
Pionella 318
Pionellinae 318
Pionidae 277, 278, 317
Plonides 319
Pioninae 318
Plonopsis 319
Pirapapa 330
Placobates 310
Placothyas 293
Planodiscidae 122, 125, 126
Pianodiscus 125, 126
Plasmobates 428
Ptategeocranus 426
Plateremaeidae 402, 429
Plateremaeus 429
Platylaelaps 83
Platytiodes 419
Platylophus 249
Platymamersopsis 298
Platynothrus 416
Platyphytopius 163, 165
Platyseius 102
. Platyseta 258
Platytelranychus 226
Platytrombidium 258
Plaumannia 326, 327
Plaumanniinae 327
Plesialges 392
Plesiothyas 293
Pleuronectocelaeno 141
Plutarchia 395
Pneumolaelaps 107
Pneumonyssus 89, 90
Pneumophionyssinae 91, 92
183, Pneumophlonyssus 92
Pneumotuber 90
Podaia 218
Podapolipodidae 37, 160, 162, 171, 172
Podapolipus 172, 173
Podocininae 101, 102, 105
Podocitium 102
Podoglyphus 341
Podolaelaps 108
Podothrombium 259
Poecilochiridae 67, 70
Poecilochirus 70, 71
Poecilochthonius 414
Poilicipalpinae 317
Pollicipalpus 317
Polyaspidella 124
Polyaspidae 119, 120
Polyaspinidae 119, 120
Polyaspinus 120
Polyaspis 120
Указатель латинских названий
469
Polya.tacid.es 316
Polyatax 315
Polydisca 260
Polyhydryphantes 292
Polymegapus 311
Polyporus 338
Polypterozetes 423
Polyxo 284
Pomerantzia 219
Pomerantziidae 185, 219
Pontacarus 281
Pontarachna 305
Pontarachnidae 268, 271, 274, 278,
305
Pontoppidania 343, 346
Pontoppidaniinae 341, 342
Popilius 59
Porobelba 421
Porohylacarinae 281
Porohalacarus 281
Poroliodes 419
Porolohmannella 281
Povelsenia 341
Praeparasitus 87
Prholaspina 82
Procaecttlus 223
Procericola 366
Procrustes 366
Proctolaelaps 108
Proctophyllodes 397
Proctophyllodidae 336, 397, 398
Proctotydaeus 204
Prodinychidae 123, 129
Prodinychus 129, 130
Prolabidocarpus 390
Pronecupulatus 204
Pronematus 204, 205
Propelops 443, 444
Propeschelobates 437
Prosopis 355
Prosopodectes 375
Prostigmata 25, 31, 159, 160, 182, 208
Protacarus 35, 187
Protalges 392, 396
Protereunetes 187
Proteylais 288
Protocepheus 425
Protokalumma 447
Protolaelaps 77
Protolichus 395
Protolimnesia 308'
Protolimnesiinae 308
Protomyobia 38, 241
Protonyssus 98, 396
Protoplophora 449
Protoplophoridae 31, 404, 448, 449
Protoribates 434, 438, 442
Protoschelobates 434, 435
Prototritia 449
Protouropodidae 123, 124
Protzia 290
Protziella 290
Protziidae 272, 278, 289
Prowichmannia 371
Prozercon 117
Prunus 357
Psamnogalumna 445
Pseudalges 398
Pseudalloptes 396
Pseudamansia 366
Pseudobryobia 226
Pseudocepheus 426
Pseudocheylidae 185, 235, 236
Pseudocheylinae 236
Pseudocheylus 235—237, 239
Pseudofeltria 318
Pseudogiebelia 396
Psetidohydryphanles 299
Pseudohydryphantidae 273, 278, 298
Pseudokoenikea 317
Pseudokongsbergia 322
Pseudolaelaps 109
Pseudolebertia 303
Pseudoleplidae 230
Pseudoleptus 232, 233
Pseudopachys 87
Pseudoparasitidae 68, 86—88
Pseudoparasiius 87
Pseudosperchon 301
Pseudotarsonemoides 181
Pseudolorrenticola 305
Pseudotritia 450, 452
Pseudouropoda 136
Pseudoxus 303
Psoralges 388
Psoralgidae 335, 388
Psorergates 239, 241
Psoroptes 383—385
Psoroptidae 335, 383
Psoroptoides 393
Pteralloptes 393
Pterochthoniidae 413
Pterochthonius 412—414
Pterocolus 398
Pterodectes 397, 398
Pterogasterina 432—434, 436, 438—
440, 443, 444, 446, 447
Pterolichidae 393, 394
Pterolichus 393—397
Pteronyssus 396
Pterophagus 398
Pteroptus 76
Pterygodes 156
Plerygosoma 220, 221
470
Указатель латинских названий
Pterygosomidae 31, 185, 220—222
Ptilolophus 249, 251
Ptilonyssus 94
Ptoc hares 147
Ptyctima 31, 399
Ptylonyssoides 94
Puncticenior 156
Pyemotes 26, 176—179
Pyemotidae 171, 176
Pygmephorus 177, 178
Pygmodispus 175
Quadracus 165
Rad 109
Radfordia 241
Radfordiella 98
Raillietia 88
Raillietidae 68, 88, 105
Rallinyssus 94
Ranautonia 302
Raoiella 232
Raphignathidae 185, 216, 218
Raphignathus 217
Raphitydeus 204
Recifella 317
Resinacarus 31, 177, 178
Retetydeus 204
Rhabdohydrachna 286
Rhabdoiobates 310
Rhagidia 193—195
Rhagidiidae 159, 184, 193
Rhagina 236
Rhaphignathidae 219
Rhinacarus 90, 94
Rhinixodes 90
Rhinoecius 94
Rhinolophus 391
Rhinonyssidae 68, 92, 94, 95
Rhinonyssoides 94
Rhinonyssus 94
Rhinophor acarus 328
Rhinothrombiinae 256, 259
Rhinothrombium 259
Rhipicentor 157
Rhipicephalinae 155, 156
Rhipicephalus 156, 157
Rhizoglyphinae 338, 341, 342
Rhizoglyphus 338—342, 344, 345
Rhizopus 167
Rhodacarellus 72
Rhodacaridae 67, 72
Rhodacaropsis 72
Rhodacarus 54, 72
Rhodaxes 226
Rhombognathides 280
Rhombognathinae 280, 282
Rhombognathus 280
Rhyncaphytoptus 165
Rhyncheylais 289
Rhynchohydracaridae 271, 278, 284
Rhynchohydracarus 284
Rhyncholimnochares 287
Rhyncholimnocharinae 287
Rhyncholophus 249, 250, 257
Rhynchophorus 173
Rhynchoribates 423
Riccardoella 199
Riedlinia 266
Riemia 354
Rivobates 310
Rivoltasia 387
Rochanyssus 94
Rodaia 218
Rohaultia 257
Rondaniacarus 232
Roqueella 330
Rosenhofia 206
Rosensteinia 366
Rostrozetes 442
Rusetria 305
Ruselriella 305
Rutacarus 302
Rutripalpidae 273, 278, 297
Rutripalpus 297
Sainididieria 85
Saitis 250
Sammonica 396
Sancassania 342
Sandenia 447
Saproglyphidae 332, 337
Saproglyphus 337, 338
Sarcoborus 240
Sarcopterinus 240
Sarcopterus 240
Sarcoptes 241, 374—377, 379, 384
Sarcoptidae 332, 335, 374, 375
Sarcoptiformes 27, 29, 30, 35, 40, 41,
43, 48, 53, 331, 399
Sauracarella 265
Sauriscus 266
Scalothrombiinae 259
Scalothrombium 259
Scapheremaeus 419
Scaphothrix 221
Scaptognathus 281
Scarabacidae 366
Scatoglyphus 349, 350
Sceloporus 222
Schadeella 317
Schadeellinae 317
Scharfenbergia 238
Schelienbergia 238
Указатель латинских названий
471
Scheloribates 400, 434, 435
Scheloribatidae 434
Schizobates 310
Schizocarpus 37, 390
Schizocoptes 390
Schizodiplogynium 139
Schizogyniidae 137, 142
Schizogynium 142
Schizohydrachna 286
Schizotetrahychus 24, 225, 226
Schmiedleinia 226
Schongastia 265, 266
Schongastiella 264
Schubartella 310
Schwiebea 342
Scincidae 221
Scirula 206
Scirus 191, 192, 205
Scissuralaelaps 109
Sciurus 196
Sclerosmaris 252
Scololydaeus 201
Scutacaridae 171, 174, 175
Scutacarus 174, 175
Scutohydrachna 286
Scutoribates 426
Scutosperchon 301
Scutovertex 426
Scyphius 193
Sebaia 214
Seiodes 118
Seiopsis 101
Seiulus 100, 101
Seius 101, 102
Sejus 71
Sellea 371
Senneriia 363
Septanychus 226
Sericothrombium 257
Serpenticola 98
Serrarius 432
Sessiluncus 87
Setoplus 163
Siblyia 239
Sierraphytoptinae 163
Sierraphytopius 163
Sigthoria 302
Simognathinae 281, 282
Simognathus 281
Simplinychus 226
Siteroptes 177—179
Sitoiroga 178
Smaridiidae 25, 185, 251, 253
Smaridiinae 253
Smariidia 252
Smarts 252—254
Sminthurus 192
Snartia 238
Soldanellonyx 282
Sotenozetes 428
Sommatricola 94
Sorex 374
Spelaeorhynchidae 25, 67, 74, 75
Spelaeorhynchus 74, 75
Spelaeothrombium 258
Speleognathidae 183, 202
Speleognathus 202, 203
Speleorchestes 25, 39, 210, 211
Speotrombicula 266
Sperchon 300, 301
Sperchonidae 274, 278, 300
Sperchonopsis 300, 301
Sphaerobates 437
Spliaerochthoniidae 413
Sphaerochthonius 414
Sphaerogastra 396
Sphaerolaelaps 84
Sphaerolichidae 184, 208—210
Sphaerolichus 209, 210
Sphaerolophus 250
Sphaeroseius 86
Sphaerotarsus 252, 253
Sphaerozelella 438
Sphaerozeles 437, 440
Spinibdella 191
Spinibdellinae 191
Spinolaelaps 98
Spinturnicidae 67, 75—77
Spinturnix 75—77
Spio 319
Spiralis 351
Stamfordia 108
Staphylococcus 247
Sieatonyssus 98
Steganacarus 451
Slenoslernum 58
Slereoglyphus 342
Slereoiydeus 189
Slernalixodes 156
Sternostoma 94
Sternostomum 94
Siictozetes 445
Stigmaeidae 216
Stigmaeodes 218
Stigmaeopsis 226
Stigmaeus 217, 218
Stigmocheylus 236
Stilpnotina 178
Stomoxys 178
S tor chia 218
Storkania 437
Stratiolaelaps 107
Strieremaeus 423
Stroemia 363
472
Указатель латинских названий
Stygmaeus 231
Stygohatacarus 281
Stygohydracarus 326
Stygomonia 270
Stygomonidae 270
Stygothrombiinae 255, 256, 259
Stygothrombium 259
Stylochirus 108
Stylohygrobates 310
Styloribates 435
Stylotydeus 204
Subalbia 322
Subaturus 322
Subcortlcacarus 310
Submiraxona 322
Subneumania 316
Suctobelba 423
Suctobelbella 424
Suctobelbila 424
Suidasia 342, 357
Sumatralbia 321
Svalbardia 438
Symbiotes 384, 386
Symplectoples 379
Syncaligus 217
Syneylais 289
Syngynaspidae 143—145
Syngynaspis 144, 145
Syringobia 395
Syringobiidae 393, 394, 396
Syringophilus 239—241
Tamandua 389
Tanaognathella 317
Tanaognaihus 317
Tanaupodaster 259
Tanaupodasterinae 256, 259
Tanaupodinae 256, 259
Tanaupodus 259
Tarsocheylus 236
Tar solar kus 239
Tarsonemella 180, 181
Tarsonemidae 171, 180, 181
Tarsonemini 31, 42, 159, 160, 171
Tarsonemoides 177
Tarsonemus 175, 180, 181
Tarsopolipus 171, 173
Tarsotomus 37, 238, 239
Tartarothyas 293
Tartarothyasinae 293
Tecomatlana 266
Tectocepheus 426
Tectocymba 420
Tectopelops 443, 444
Tectopenthalodes 188, 189
Tectoribates 439
Tectotydeus 204
Tegeozetes 438
Tegeocraneltus 426
Tegeocranus 425, 426
Tegonotus 165
Tegopalpus 232
Tegoribates 444.
Teincoptes 375
Teleioliodes 418, 419
Tencateia 238
Teneriffia 234
Teneriffiidae 185, 234
Teneriffiola 234
Tenuiala 433
Tenuialidae 403, 433
Tenuicrus 226
Tenuipalpidae 230
Tenuipalpus 162, 231—233
Teratothyas 293
Teratothyasinae 293
Termitacarus 108
Terpnacaridae 184, 208, 213
Terpnacarus 213, 214
Terrazetes 438
Testudacarus 305
Tetra 165
Telrabates 310
Tetragonyssus 98
Tetrahydrachna 286
Tetralimnesia 308
Tetramegapus 311
Tetraneumania 316
Tetranichidae 228
Tetranobia 226
Tetranychidae 22, 159, 162, 184, 224,
228, 230, 231, 245
Tetr any china 226
Tetranychus 40, 43, 187, 225—227,
230
Tetranycopsis 224, 226
Tetrapiona 318
Tetrapodili 33, 159—162
Tetrapolipus 172, 173
Teutonia 299, 302
Teutoniidae 273, 278, 299
Thdlassarachna 279, 280
Thamnacarus 411
Thamnacus 165
Thaumatotrombium 260
Thecarthra 396
Therismoptes 177
Thermacaridae 273, 278, 295, 296
Thermacarus 296
Thinoseius 102
Thinozercon 111, 114
Thinozerconidae 110, 111
Thinozerconina 56, 57, 110, 114
Thonia 310
Указатель латинских названии
473
Thoracohygrobates 310
Thoracophoracarus 328
Thoribdella 191
Thrichocylliba 128
Thrombidium 266
Thyas 290, 292, 293, 295
Thyasella 293
Thyasides 293
Thyasinae 292
Thyopsis 293
Thyreophagus 338, 340, 342,
345
Thysaniezia 435
Tignyia 259
Tillett a 346
Tiphys 318, 319
Tiphysinae 318
Torpacarus 411
Torionia 355
Toxopeusia 149
Trachygamasus 80
Trachyhoplophora 451
Trachynetus 120
Trachymolgas 190, 191
Trachyoribates 438
Trachytes 119, 124
Trachytidae 119
Trachytina 56, 57, 118, 119,
124
Trachyuropoda 131, 132
Trachyuropodidae 123, 131, 133
Trdgdrdhacarus 407
Tragardhula 265
Trasonemus 180
Travanyssus 94
Trematura 121, 127
Tremaiurella 128
Trematurellidae 122, 128
Trematuridae 121, 122, 127
Trhypochthoniellus 412, 414
Trhypochthoniinae 412
Trhypochthonius 412, 414
T riangulazercon 118
Trichadenidae 230
Trichadenus 232
Tricheremaeus 424
Trichobius 390
Trichodactylus 363
Trichodinychus 130
Trichodiplogyniinae 138
Trichodiplogynium 139
Trichoecius 390
Tricholaelaps 108
Tricholocelaeno 82
Trichorlbates 437, 438
Trichostigma 164
Trichotarsus 354, 355, 363
Trichothyas 293
Trichouropoda 136
Tridilatydeus 204
Tridiplogynium 139
Trigonholaspis 32, 82
Trimalaconothrus 415
Triophtydeus 204
Trisetica 266
Tristomus 102
Tritegeus 426
344, Tritia 451, 452
Trlxacarus 375
T rizerconoides 118
Trizeles 424
Troglobdella 192
Troglohalacarus 281
Trombella 260
Trombellinae 256
Tromblcula 44, 264—267
Trombiculidae 25, 31, 186, 250, 261— ,
263, 266, 267
Trombiculinae 256, 261, 263, 265
Trombiculindus 265
Trombiculoides 265
Trombidiformes 26, 29, 30, 37, 40—43,
47, 53, 159, 268, 279
Trombidiidae 31, 159, 185, 255, 256,
121, 260, 261, 266
Trombidiinae 256, 257
Trombidlum 27, 226, 238, 249, 252,
257—260, 264, 265
Tropacarus 451
Trouessartia 281, 398
Truncaturus 328
Tuberemaeus 424
Tubophora 308
Tumescoptes 165
Tumidalvus 418
Tur 98
Turbinoptes 387
Tydaeolus 204
Ту doe us 204
Tydeidae 183, 203—205
Tydeus 203—205
Tympanomegapus 311
Tympanospinctus 76, 77
Typhlodromus 102, 165
Typhlothrombium 259, 264
Tyreilia 308
Tyrelliinae 308
Tyroborus 341
Tyroglyphidae 338
Tyroglyphopsis 342
Tyroglyphus 243, 341—343, 349, 356,
359, 360, 370
Tyrolichus 341
Tyrophagus 244, 341, 345, 346
474
Указатель латинских названий
Udetaliod.es 418
U gandolaelaps 109
Unduloribates 439
Unguizetes 435
Unionicola 315, 316
Unionicolidae 276, 278, 314
Unionicolides 315
Unionicolinae 315
Uroboophilus 157
Urocyclella 136
Urodiaspidae 123, 130
Urodiaspis 121, 131
Urodinychidae 123, 134
Urodinychus 122, 134
Urodiscella 134, 136
Urodiscus 134
Uroiphis 106
Urojanetia 131, 132
Urolaelaps 130
Uronothrus 416
Uroobovella 136
Uroplitana 136
Uroplitella 121, 136
Uropoda 119, 127, 130—136
Uropodidae 41, 116, 123, 135
Uropodina 25, 31, 46, 54, 56, 57, 118,
121, 123, 124, 127, 133
Uropolyaspis 133
Uroseius 119
Urosejus 116
Urospina 133
Urostemella 136
Urozercon 108
Vaghia 445
Valgothrombium 258
Valmontia 343
Varchia 393
Variatipes 175
Varroa 108
Vasates 165, 170
Veigaia 78, 79
Villersia 218
Veigaiaidae 67, 78, 79
Vespertugo 374
Vidia 355
Viedebanttia 343
Vietsia 293
Villersia 218
Vitznyssus 94
Walchia 264
Walchiinae 264
Г alter ella 281
Walzia 238, 239
Wandesia 290
Werthella 281
Wetiina 319
Whartonia 264
Wichmannia 371
Willanla 215
Willmannia 61
Winterschmidtia 357, 358
Winterschmidtiidae 333, 357
Womersia 265
Womersleyia 260
Wuria 329
Wuriella 329
Xenillus 426
Xenothrombium 257
Xolalges 393
Xoloptes 396
Xylobates 442
Xystonotus 324
Zaelaps 108
Zercon 80, 102, 117, 118
Zerconidae 117, 118
Zerconina 56, 57, 117
Zerconopsis 102
Zercoseius 101
Zetes 423, 430, 435, 437, 444
Zetobelba 423
Zelomimus 435
Zetomotrichus 435
Zetorchella 431
Zetorchestes 431
Zetorchestidae 403, 431
Zetsellia 218
Zonuridae 221
Zonurobia 221
Zschachia 370
Zschokkea 293
Zuluacarus 61
Zwickia 371
Zygoribatula 434
Zygoseius 101
ОГЛАВЛЕНИЕ
Вступительная статья . . .'........................................ 3
Основная русская литература по клещам.............................. 7
Предисловие автора................................................ 11
Глава /. Введение................................................. 13
Исторический обзор ......................................... 15
Методика изучения........................................... 16
Морфология и жизненный цикл................................. 23
Классификация............................................... 46
Глава II. Подотряд Onychopalpida Wharton, 1947 ................... 49
Глава III. Подотряд Mesostigmata G. Canestrini, 1819.............. 53
Megisthanina Tragardh, 1946 ................................ 57
Liroaspina Tragardh, 1946 .................................. 59
Microgyniina Tragardh, 1942 ................................ 63
Gamasides Leach, 1815....................................... 65
Thinozerconina Tragardh, 1946 ............................. 110
Diarthrophallina Tragardh, 1946 ........................... 115
Zerconina Tragatdh, 1944 .................................. 117
Trachytina Tragardh, 1937 ................................. 118
Uropodina Kramer, 1881......................................121
Celaenopsina Tragardh, 1938 ............................... 137
Fedrizziina Tragardh, 1937 ................................ 143
Глава IV. Подотряд Ixodides Leach, 1815...........................150
Глава V. Подотряд Trombidiformes Reuter, 1909 ................... 159
Tetrapodili Bremi, 1872 ................................... 160
Tarsonemini Cannestrini et Fanzago, 1877 .................. 171
Prostigmata Kramer, 1877 ................................ 182
Глава VI. Trombidiformes (продолжение)............................268
Hydrachnellae Latreille, 1802 (Hydracarina).................268
Глава VII. Подотряд Sarcoptiformes Ranter, 1909 ................. 330
Acaridiae Latreille, 1802 ................................. 330
Глава VIII. Sarcoptiformes (продолжение)..........................399
Oribatei Duges, 1833 ...................................... 399
Указатель латинских названий .....................................453
Э. Бэкер и Г. Уартон
ВВЕДЕНИЕ В АКАРОЛОГИЮ
Редактор Л. П. ТАТАРИНОВ
Технический редактор Е. С. Герасимова
Переплет художника Завьялова А. И.
Корректор А. А. Смирнова
Сдано в производство 6/XI 1954 г.
Подписано к печати 29/1 1955 г.
Т-00852. Бумага 60х9?Уи=15 бум. л.
29,8 печ. л.
Уч.-издат. л. 30. Изд. № 4/2132.
Цена 23 р. Зак. 1785.
Издательство иностранной литературы
Москва, Ново-Алексеевская, 52
Министерство культуры СССР.
Главное управление полиграфической
промышленности.
4-я тип. им. Евг. Соколовой.
Ленинград, Измайловский пр., 29.
ОПЕЧАТКИ
Страница Строка Напечатано Следует читать
194 5 снизу . стадиях стациях
212 8 снизу пальцы пальпы
/ 218 16 снизу света цвета
377 21 сверху var. Megnin var. ovls M6gnin
383 5 снизу Capartnae Caparinla
Зак. 1785