/































































































































































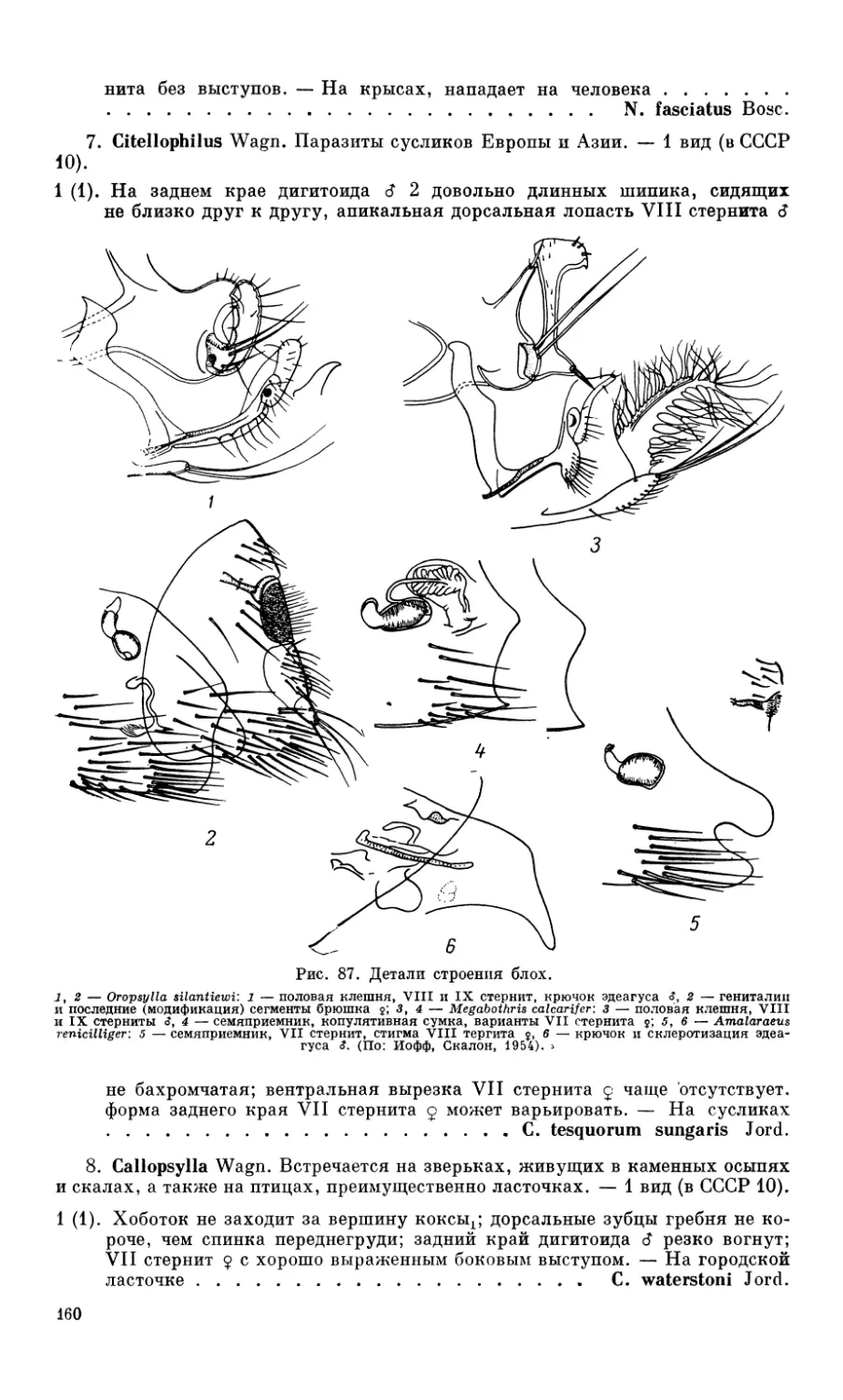























































































































































Похожие




Текст
АКАДЕМИЯ Н AV К
СССР
НАСЕКОМЫЕ
И КЛЕЩИ
ДАЛЬНЕГО ВОСТОКА
ИМЕЮЩИЕ
МЕДИКО-ВЕТЕРИНАРНОЕ
ЗНАЧЕНИЕ
АКАДЕМИЯ НАУК СССР
ДАЛЬНЕВОСТОЧНЫЙ НАУЧНЫЙ ЦЕНТР
БИОЛОГО-ПОЧВЕННЫЙ ИНСТИТУТ
НАСЕКОМЫЕ И КЛЕЩИ
ДАЛЬНЕГО ВОСТОКА,
ИМЕЮЩИЕ
МЕДИКО - ВЕТЕРИНАРНОЕ
ЗНАЧЕНИЕ
В
ЛЕНИНГРАД
ИЗДАТЕЛЬСТВО «НАУКА?
ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ
1987
УДК (595.79+595.77) : 61 (571.6)
Насекомые и клещи Дальнего Востока, имеющие медико-ветеринар-
ное значение.—Л.: Наука, 1987. 309 с.
Книга является первой фундаментальной сводкой о насекомых и клещах
Дальнего Востока, приносящих вред здоровью человека и животных.
В ней приводятся сведения о внешнем строении, биологии и вредоносном
значении 2 классов членистоногих, в которые входят 7 отрядов, 61 се-
мейство, 227 родов, 631 вида. Представлены следующие насекомые: крово-
сосущие двукрылые, или «гнус» (комары, мошки, мокрецы, слепни),
оводы, синантропные и зоофильные мухи, вши, блохи, клопы, пухоеды,
тараканы, муравьи; из клещей — иксодовые, гамазовые, краснотелковые,
перьевые, пылевые, чесоточные и возбудители демодекоза животных.
По указанным группам насекомых и клещей приводятся определительные
таблицы семейств, родов, видов. Библиогр. — 141 назв. Ил. — 146.
Составители:
С. Д. Артамонов, Ю. Д. Бодрова, В. М. Глухова, Е. В. Дубинина,
Т. Н. Досжанов, В. Н. Зарубина, Г. В. Колонии, И. М. Кержнер,
А. Н. Купянская, Н. И. Кудряшова, С. В. Миронов, Н. А. Никулина,
Б. К. Петрова, Т. П. Ромашова, Р. Г. Соболева, С. Ю. Стороженко
Ответственный редактор
Р. Г. СОБОЛЕВА
Рецензенты
Э. П. НАРЧУК, В. Н. КУЗНЕЦОВ
„ 2005000000-597
Н 042(02)-87 27<)-87-//
ф Издательство «Наука», 1987 г.
ПРЕДИСЛОВИЕ
На огромной территории Дальнего Востока, охватывающей все ландшафтно-
географические зоны и характеризующейся в связи с этим очень разнообраз-
ными климатическими условиями и своеобразной растительностью, обитают де-
лтки тысяч насекомых и клещей, многие из которых являются вредителями
гельско хозяйственных культур, охотничьего и лесного хозяйства, животно-
водства, а также переносчиками возбудителей некоторых инфекционных, про-
тозойных и инвазионных болезней человека и животных.
В современных условиях в связи с интенсификацией и специализацией раз-
личных областей сельского хозяйства (в том числе и животноводства) и необ-
ходимостью окончательной ликвидации болезней большое значение приобре-
тают глубокие знания систематики, географического распространения
ж биологии вредных насекомых и клещей, без чего невозможно правильное,
научно обоснованное планирование и осуществление противоэпидемических,
эпизоотологических и защитно-истребительных мероприятий.
Как известно, особенно опасны кровососущие насекомые и клещи, являю-
щиеся потенциальными переносчиками возбудителей трансмиссивных и неко-
тэрых природно-очаговых болезней. Плодотворное систематическое изучение
zx в СССР начато лишь в годы Советской власти работами академика Е. Н. Пав-
ловского и продолжено его учениками. За эти годы проделана огромная работа,
однако Дальний Восток до сих пор остается слабоизученным в этом направле-
нии, несмотря на то что первые экспедиции Зоологического института АН СССР
под руководством Е. Н. Павловского начали исследование фауны и экологии
насекомых и клещей, а также выяснение их роли в переносе клещевого энце-
фалита еще в 30-е годы. Уже тогда было установлено, что переносчиком возбу-
дителя этого опасного заболевания людей являются иксодовые клещи, а также
доложено начало изучению фауны и экологии кровососущих двукрылых на-
секомых.
Наиболее интенсивные и разноплановые исследования вредных насекомых
л клещей Дальнего Востока проведены учеными в основном в последние 20 —
25 лет, но сведения, полученные по отдельным группам этих членистоногих,
неравноценны. Причиной тому их большое видовое разнообразие, недостаток
специалистов, огромная территория, которую в такой короткий срок невоз-
можно было охватить. Тем не менее накоплен достаточно большой материал,
который до сих пор не обобщен, что крайне затрудняет его использование.
А между тем практические работники санитарно-эпидемической службы,
зооветеринарные специалисты и биологи давно нуждаются в доступном пособии,
1*
3
которое поможет им разобраться в сложном многообразии отдельных отрядов
и семейств насекомых и клещей Дальнего Востока, имеющих медико-ветеринар-
ное значение.
Предлагаемая читателям сводка не претендует на исчерпывающую полноту,
так как в нее не вошли некоторые редкие и плохо изученные виды, например,
известные с Дальнего востока 2 вида аргасовых клещей из гнезд диких птиц,
медико-ветеринарное значение которых пока не ясно, и др. Из-за ограничен-
ности объема невозможно было включить все семейства (их более 20) синантроп-
ных и зоофильных мух; пришлось ограничиться лишь самыми важными. Не
включены также многие насекомые (жуки, стрекозы, двукрылые и др.), являю-
щиеся промежуточными хозяевами некоторых гельминтов (это большой само-
стоятельный раздел гельминтологии). По этой же причине не представлены
определительные таблицы самцов кровососущих двукрылых насекомых, кото-
рые к тому же не приносят непосредственного вреда людям и животным, так
как не являются кровососами. Вполне возможно, что при дальнейшем изуче-
нии материковой и островной фаун (особенно в пограничных районах) она
пополнится новыми находками.
Для каждой группы насекомых и клещей приводятся сведения по морфо-
логии (внешнее строение), биологии (образ жизни и размножение), вредонос-
ному значению, а также — определительные таблицы (в основном, по взрослым
насекомым, но там, где это необходимо, и по личинкам) родов и видов. Для
каждого рода указывается число видов, обитающих на Дальнем Востоке
и в СССР, в отдельных случаях — и число видов, имеющих медико-ветеринар-
ное, значение. Для некоторых групп членистоногих приводится географическое
распространение, которое дано очень кратко с указанием крупных регионов,
областей Дальнего Востока или их частей. Размеры тела приводятся в конце
тезы или антитезы без указания «мм».
В работе над монографией принялй участие 16 специалистов из разных
учреждений и городов. Сотрудники Зоологического института АН СССР (Ле-
нинград) подготовили следующие разделы: В. М. Глухова — по кровососущим
мокрецам, И. М. Кержнер — по клопам, Е. В. Дубинина — по «аллергенным»,
чесоточным клещам, железницам, общую часть к разделу «Клещи» и к Акари-
формным клещам, С. В. Миронов — по перьевым клещам; Н. И. Кудряшова
(Зоомузей МГУ, Москва) — по клещам-краснотелкам; сотрудники Биолого-
почвенного института ДВНЦ АН СССР подготовили несколько разделов: Р. Г. Со-
болева — по слепням и оводам, а также общую часть в разделе «Насекомые»,
К). Д. Бодрова — по кровососущим комарам и мошкам, С. Ю. Стороженко —
по пухоедам и тараканам, А. Н. Купянская — по муравьям; Б. К. Петровой
(НИИ птицеводства Министерства сельского хозяйства РСФСР, г. Ломоносов
Ленинградской области) написана общая часть к разделу «Синантропные мухи»,
а также семейства Muscidae и Calliphoridae; С. Д. Артамоновым сем. Sarco-
phagidae (Уссурийский государственный педагогический институт); Г. В. Ко-
лониным — раздел по иксодовым клещам (Тихоокеанский институт географии),
В. Н. Зарубиной по вшам, Т. П. Ромашовой по блохам (Иркутский научно-
исследовательский противочумный институт); Н. А. Никулиной по гамазовым
клещам (Иркутский государственный университет); Т. Н. Досжановым по му-
хам-кровососкам (Институт зоологии АН КазССР, Алма-Ата).
Большую работу по оформлению иллюстраций проделали О. В. Звягенцева
и Т. В. Кучина, выполнившие тушевые рисунки по некоторым группам члени-
стоногих. Техническую помощь в работе над рукописью оказали Ю. Д. Бод-
4
рова и А. А. Мещеряков . Пользуясь случаем, выражаю им сердечную призна-
тельность.
Хочется надеяться, что эта монографическая сводка будет способствовать
дальнейшему, еще более углубленному изучению вредных насекомых и клещей
Дальнего Востока, имеющих эпидемиологическое и эпизоотологическое зна-
чение.
Р, Г. Соболева
СПИСОК СОКРАЩЕНИЙ И УСЛОВНЫХ ОБОЗНАЧЕНИЙ
В ОПРЕДЕЛИТЕЛЬНЫХ ТАБЛИЦАХ
Аз. ч. — Азиатская часть СЗ — северо-запад, северо-за-
Алтайск. — Алтайский край падный
Амур. — Амурская обл. Сиб. — Сибирь
Бурят. — Бурятия Хаб. — Хабаровский край
В — восток, восточный ДВ — Дальний Восток Ц- — центральный
Чук. — Чукотка
евр. ч. — европейская часть Ю — юг, южный
3 — запад, западный Заб. — Забайкалье ЮВ — юго-восток, юго-вос-
точный
Иркут. — Иркутская обл. ЮЗ — юго-запад, юго-западный
Камч. — Камчатка Якут. — Якутия
Коряк. — Корякский националь- сх (сх1ч
ный округ сх21 сх3) — тазики ног (передние,
Кур. — Курильские о-ва средние, задние)
Маг. — Магаданская обл. f (fl, fz,
Приам. — Приамурье /з) — бедра (передние сред-
Приб. — Прибайкалье Прим. — Приморье * (*1, *2> ние, задние) — гблени (передние, сред-
С — север, северный *з)
Сах. — Сахалин ние, задние), или го-
СВ — северо-восток, северо- лениJ * 2t з — самец
восточный
— самка
Римскими цифрами в тексте и на рисунках обозначены номера сегментов, арабскими —
номера члеников усиков и других придатков тела. Пояснения к условным обозначениям
жилок и ячеек крыла, щетинок и шипиков на голове, груди, ногах, теле приведены в под-
рисуночных подписях, диагнозах семейств и родов, в отдельных случаях — в определитель-
ных таблицах.
I. НАСЕКОМЫЕ (Класс INSECTA)
Насекомые — это мелкие животные (от 1 до 50 мм), относящиеся к типу
членистоногих (Arthropoda), тело которых покрыто в разной степени хитини-
зированной кутикулой (оболочкой), расчленено на 3 отдела (голову, грудь
и брюшко) и снабжено членистыми ногами. На голове имеются глаза и простые
глазки, ротовые оргдны (у некоторых они редуцированы), пара усиков и щупи-
ков.
Класс насекомых очень обширный. Мировая фауна насчитывает более
1 млн, в СССР — не менее 100 тыс. видов, на Дальнем Востоке — более 40 тыс.,
но фактически их известно гораздо меньше, так как многие группы очень слабо
изучены. В целом число видов насекомых составляет около 85 % от всех живот-
ных и растений вместе взятых.
ОБЩИЕ СВЕДЕНИЯ О СТРОЕНИИ ВЗРОСЛОГО НАСЕКОМОГО
(Морфология)
Окраска тела насекомых различна, что зависит от пигмента кутикулы или
покрывающих его волосков и налета. Нередко на отдельных частях тела и на
крыльях хорошо выражены разной ширины полосы (перевязи) и пятна, имею-
щие большое диагностическое значение. Верхняя часть тела называется спин-
ной (дорсальной), а нижняя — брюшной (вентральной). Основанием того или
иного органа является та его часть, которая прикрепляется к телу или распо-
ложена ближе к его центру, а вершиной — та часть, которая находится дальше
от тела (апикальный конец). Длина тела (мм) определяется от головы до конца
брюшка, не считая усиков.
Голова нечленистая; твердой скелетной основой ее является черепная
коробка, или капсула. На передней поверхности головы находится лоб, по бо-
кам которого — орбиты (имеются не у всех насекомых), выше лба — темя,
а за ним затылок. По бокам лба — сложные фасеточные глаза, а в верхней
части — глазковый треугольник с 1—3 простыми глазками. Ниже лобной по-
лосы (или лба) находится лицо, на котором различают наличник, скулы и щеки,
резких границ между которыми нет (они условны). На голове имеются усики.
Они очень разнообразны: трех- или многочлениковые, с щетинкой (аристой)
или без нее, перистые, жгутиковые, шарообразные и др. Эти признаки, а также
окраска усиков имеют большое таксономическое значение.
Грудь состоит из трех частей: передне-, средне- и заднегруди, которые
у многих слиты между собой. Верхняя часть груди называется спинкой, а ниж-
няя — грудкой. Боковые стороны называются бочками, или плейритами.
Они состоят из нескольких пластинок (склеритов), разделенных швами. Позади
груди расположен округло-треугольный щиток (у некоторых насекомых име-
ется и подщиток).
Ноги состоят из тазика, или коксы (членик, соединяющий ногу с грудью),
малозаметного вертлуга, бедра, голени и 2—5-члениковой лапки (считать от го-
лени!). У некоторых насекомых лапка не членистая; у многих на последнем
членике лапки имеются подушечки, коготки, пульвиллы (боковые присоски),
а иногда еще и эмподий (срединная присоска).
7
г
6
Рис. 1. Схема строения насекомого.
1 — общий вид мухи: б — брюшко, г — голова, гл — глаз, гр — грудной отдел, к — крыло, т — тергит,
I, II, III — передняя, средняя и задняя пара ног (по: Штакельберг, 1956); 2 — схема строения ноги мухи:
б (/) — бедро, в — вертлуг, г (О — голень, л — лапка, т (сх) — тазик, или кокса, 1—5 — членики лапки;
3 — крыло двукрылых насекомых (слепня) с полным жилкованием (по: Соболева, 1974); 4 — крыло мухи-
кровососки с неполным жилкованием (по: Досжанов, 1980); 5—11 — схема строения усиков некоторых
насекомых: 5,6 — перистые усики 8 и комара, 7 — усик подкожного овода с концевой оголенной ари-
стой, 8 — усик с кольчатым 3-м члеником (слепня), 9 — то же, муравья, 10 — усик мухи со спинной
голой аристой, 11 —то же, с перистой аристой (по: Штакельберг, 1956).
Представители разных групп насекомых имеют 1 или 2 пары крыльев, кото-
состоят из тонкой (часто прозрачной) перепонки (мембраны), укрепленной
: крепких продольных и поперечных жилках, ограничивающих крыловые
тйкл. Жилкование крыльев неодинаково и имеет большое диагностическое
-чсние (рис. 1).
Б разных группах насекомых строение крыльев имеет свои особенности,
некоторых насекомых летательную функцию осуществляют задние крылья,
передние превращаются в надкрылья (элитры), являющиеся покрышками
is задних крыльев. В процессе эволюции в ряде случаев задние крылья соеди-
Lziicb друг с другом и работают как один орган, а у наиболее прогрессивных
тм задняя пара крыльев постепенно исчезает и остаются только передние
:ылья (у двукрылых насекомых).
Брюшко у разных насекомых имеет неодинаковое число члеников — от 5—6
! 1. не считая хвостовых придатков (у некоторых высших форм число члеников
нет быть меньше пяти). В типичных случаях верх брюшка состоит из тер-
т:в (верхние полукольца) и стернитов (нижние полукольца), которые соеди-
:э:тся по бокам мягкой межсегментарной перепонкой (мембраной). Последние
ннлкп брюшка обычно сильно видоизменены и представляют собой половой
пгграт (гениталии, терминалии). У самок они образуют яйцеклад, а у самцов
пулятивный аппарат. Строение гениталий в ряде групп имеет существенное
п^же ведущее значение в систематике, но у многих насекомых они еще очень
изучены, и потому не представляется возможным использовать их при
згностике разных таксонов (особенно на видовом уровне). Брюшко у насе-
*сых — разной формы, неодинаковой окраски, со своеобразным рисунком
голос и пятен; оно может быть матовым или блестящим. У ряда насекомых
z признаки имеют ведущее или вспомогательное значение при определении
г: в.
Особенности строения тела и его частей у разных групп интересующих нас
. :екомых будут рассмотрены в соответствующих разделах.
*
МЕТОДИКА ХРАНЕНИЯ И ОПРЕДЕЛЕНИЯ НАСЕКОМЫХ
Большинство крупных насекомых с твердыми покровами хранятся в сухом виде, нако-
zo-rz на энтомологические булавки или на ватных матрасиках (тонкий слой ваты, вложен-
z 1 иесклеенный продолговатый конверт с загнутыми со всех сторон краями) в плотно
гьгтых коробках. Насекомые с мягкими покровами и мелкие, а также личинки многих
-г- :мых помещаются в 70—96°-ный спирт, формалин или другую консервирующую жид-
:ть. При хранении насекомых в спирту этикетки пишутся на плотной бумаге (лучше «пер-
четной») тушью или мягким простым карандашом; при хранении в других жидкостях —
карандашом.
г. ели собранные насекомые успели подсохнуть, то перед накалыванием их следует но-
гтя г ь во влажную камеру (стеклянную шпрокогорлую банку с влажным песком на дне-
zziTHon крышкой) и выдержать там до состояния гибкости (необходимо следить, чтобы
I не заплесневели). Накалывают насекомое на энтомологическую булавку примерно на
: 2/з ее длины и помещают в специальные коробки с пенопластом на дне и хорошо при-
гнБсй крышкой, чтобы не съели кожееды. В коробки с ватными матрасиками и с наколо-
izz насекомыми помещается нафталин и тимол (последний — от плесени). Методы сбора
izzrx групп насекомых достаточно подробно описаны в имеющихся руководствах.
-Для определения многих мелких насекомых необходимо делать временные или постоян-
- zit-параты. В первом случае насекомое или его часть помещается на время рассматри-
вая в каплю жидкости (фиксирующая жидкость, глицерин, дистиллированная вода) и
zzp^ae необходимости покрывается покровным стеклом.
_1ля изготовления постоянного препарата объект вначале помещается в 70°-ный спирт
:—30 мин, затем обезвоживается в 95°-ном спирту также 5—30 мин, после чего просвет-
<:я в течение 3—15 мин в гвоздичном или кедровом масле или в карболксилоле (1 часть
хгталлпческой карболовой кислоты и 3 части ксилола). Затем переносится на предметное
г~:. покрывается каплей жпдкого канадского бальзама (загустевший бальзам необхо-
* _ разбавить ксилолом) и осторожно прикрывается покровным стеклом.
--гп определении некоторых групп насекомых приходится готовить препараты гени-
zzz половых придатков) или других склеротизованных структур. С этой целью они пред-
ттольно подвергаются медленному кипячению (в специальных маленьких тигельках)
:.-ном растворе едкого калия в течение 3—5 мин. По мере испарения раствора в него
ппле добавляется дистиллированная или кипяченая вода. После этого объект тщательно
. кывается водой или даже кипятится в небольшой ее порции в течение 1—2 мин, после
. готовится временный пли постоянный препарат. Гениталии удобно хранить подколо-
iz приклеенными на кусочки картона или в мелких пробирках с глицерином) вместе
ггекомымп, от которых они взяты.
9
Уложенные на вату и наколотые насекомые, а также препараты снабжаются подробной
этикеткой с указанием даты, места сбора (иногда метода сбора), фамилии сборщика. Видо-
вое определение насекомых и изучение препаратов производится с помощью микроскопа.
Определить вид насекомого (его латинское, а если есть, то и русское название) можно
с помощью приводимых в этой книге определительных таблиц, которые состоят из тез (цифра
без скобок) и антитез (цифра в скобках), т. е. из серии противопоставлений. Тщательно рас-
сматривая насекомое, необходимо выбрать совокупность наиболее подходящих признаков.
Начинают определение с тезы № 1. Если она подходит, то переходят к следующей тезе,
а, если нет, то обращаются к антитезе, указанной в скобках. Необходимо продвигаться только
вперед, пока не будет достигнуто латинское название.
В определительных таблицах латинские названия родов снабжены порядковыми номе-
рами, по которым их в дальнейшем легко отыскать в тексте. Для упрощения определения
в таблицы по некоторым группам насекомых не включены более высокие систематические
категории, такие как надсемейство, подсемейство, триба.
ДВУКРЫЛЫЕ (Отряд DIPTERA)
КРОВОСОСУЩИЕ КОМАРЫ (Сем. CULICIDAE)
Средней величины (до 10 мм) насекомые отряда двукрылых (Diptera) под-
отряда длинноусых (Nematocera) имеют стройное, вытянутое тело, маленькую
голову и обычно длинные ноги. Крылья по жилкам, тело и ноги покрыты уз-
кими прилегающими чешуйками.
Семейство подразделяется на три подсемейства: Anophelinae, Taxorhynchi-
tinae) с одним родом каждое (Anopheles и Тaxorhynchites) и Culicinae с 6 родами:
Uranotaenia, Orthopodomyia, Manzonia, Culiseta, Aedes, Culex. На территории
Дальнего Востока отсутствуют представители первых трех родов подсем.
Culicinae, а виды рода Тaxorhynchites не нападают для кровососания.
Комары встречаются по всей территории СССР, за исключением участков
безводных пустынь. Наиболее широко во всех климатических зонах распро-
странены комары рода Aedes. На территории tCCP зарегистрировано 87 видов
комаров, на Дальнем Востоке — 47 [Гуцевич и др., 1970; Данилов, Филиппова,
1978].
ВНЕШНЕЕ СТРОЕНИЕ
Тело комаров, как и у всех насекомых, подразделяется на голову, грудь
и брюшко (рис. 2, 7).
Голова (рис. 2, 1) шарообразная. Глаза почковидные, фасеточные, сбли-
женные или соприкасаются в верхней и нижней частях головы. На лбу за-
частую имеется хохолок из направленных вперед волосков и чешуек. Налич-
ник без волосков и чешуек.
На границе между лбом и наличником располагаются усики. Усики сбли-
жены у основания и состоят из 15 члеников. 1-й членик усика (скапус) имеет
вид кольца, 2-й (торус) — шаровидный и несет сложно устроенный орган слуха
(джонстонов орган), открывающийся узкой щелью. Остальные 13 члеников
образуют жгутик; у самок они почти одинаковой формы. Усики покрыты
волосками, собранными в розетки, или роговидными придатками (рис. 2, 7).
Ротовые органы имеют вид хоботка. Длина его в несколько раз превышает
диаметр головы. Состоит из верхней губы, двух передних (мандибул) и двух
нижних (максилл) челюстей, подглоточника и нижней губы.
Щупики причленяются к нижней челюсти. Состоят из 5 члеников, но иногда
5-й и даже 4-й членики редуцированы. Щупики часто покрыты чешуйками
(рис. 2, 7).
Грудь обычного для двукрылых строения с некоторыми особенностями,
характерными для группы (рис. 2, 2).
Переднегрудь представлена пластинкой переднеспинки (пронотумом); на
бочках — эпистернами и эпимерами. Среднегрудь подразделяется на средне'
спинку (пластинку значительной величины), за которой располагается щиток —
трехлопастный или цельнокрайний. Заднегрудь представлена только боковыми
склеритами — метэпистерном и метэпимером.
На отделах груди различают следующие щетинки и пятна. По средней линии
среднеспинки проходит ряд акростихальных, а кнаружи от него (на половине
10
расстояния от средней линии тела до боковых краев) — дорсоцентральных
щетинок (рис. 2, 3, 4). На пронотуме расположена группа пронотальных щети-
нок. на заднем краю (обращенном к стигме) — проэпимеральные; перед перед-
ним дыхальцем — дыхальцевые (спиракулярные); позади него (в верхнем отделе
мезэпистерна) — задыхальцевые (постспиракулярные); на метаэпистерне задне-
груди — проэпистернальные щетинки.
Пятна образованы сгруппированными чешуйками, щетинками и волосками.
Перед передним дыхальцем, на верхнем мезэпистерне лежит гипостигмальное
пятно (рис. 2, 5), ниже (на границе с вентральными мезэпистернами) — переднее
и заднее парастигмальные; сзади, на границе со среднеспинкой — метастиг-
мальное, на вентральных мезэпистернах — вентральное мезэпистермальное;
между передними тазиками, проэпистерном и передним краем вентральных
мезэпистерн — посткоксальное пятно.
Крылья удлиненно-овальные (рис. 2, 6). Среди жилок крыла различают:
костальную (С), ограничивающую крыло со всех сторон, лежащую за ней суб-
костальную (Sc) и далее 4 радиальные жилки. Последние составляют радиаль-
ный сектор крыла. Жилки г2+гз, расходясь недалеко от вершины крыла, обра-
зуют переднюю, или радиальную, вилку. Отрезок г4+5 начинается либо от об-
щего ствола со второй и третьей радиальной жилки (подсем. Culicinae), либо
самостоятельно, в центре крыла перед пересечением с поперечной радио-меди-
альной жилкой (подсем. Anophelinae).
За радиальными следуют медиальные жилки. При впадении в верхнюю часть
крыла они образуют среднюю, или медиальную, вилку. Кубитальных жилок
также одна-две — си± и си2. При впадении в задний край крыла они образуют
заднюю вилку крыла. Анальная жилка (одна) впадает около середины заднего
края крыла.
Поперечных жилок три: плечевая (Л), соединяющая вблизи основания крыла
костальную и субкостальную. Вблизи середины крыла имеется радио-медиаль-
ная жилка (гт). При этом она может состоять из двух (2—3 и 2—4) либо из од-
ного отрезка. Медиальные и кубитальные жилки соединяет медио-кубитальная
। тси или шт).
Крылья покрыты чешуйками, которые по заднему краю образуют бахрому.
Чешуйки сгруппированы в темные и светлые пятна, из которых слагается
рисунок крыла. Пластинка крыла, кроме того, покрыта очень мелкими волос-
ками — микротрихиями.
Ноги обычного для насекомых строения: 5-члениковая лапка заканчивается
коготками. На внутренней поверхности задних голеней вблизи вершины (части
противоположной месту прикрепления) располагается ряд плоских шипиков..
Коготки разных пар лапок имеют обычно одинаковую форму, но размеры их.
могут уменьшаться от передней к задней паре. На нижней вогнутой поверхности
гни могут быть вооружены зубцами (род Anopheles) или быть без них (род Си-
1ех). Под коготком располагаются присоски (пульвиллы), хорошо развитые
род Culex) или рудиментарные (род Aedes), а между ними непарное образова-
ние — эмподий (рис. 2, 7, а, б, в).
Брюшко удлиненно-цилиндрическое. Состоит из 10 члеников; IX и X сег-
менты приспособлены к функции размножения и носят название гениталий
рпс. 2, 1).
В качестве таксономических признаков у комаров фигурируют разнообраз-
ные придатки кожных покровов: чешуйки (удлиненные, широкие, продольно-
поперечные), шипы, щетинки, волоски, размеры различных частей тела (хо-
ботка), строение щитка, жилкование крыла, окраска тела, наличие на теле
пли крыльях пятен, полос {Виноградская, 1974].
Яйца продолговато-овальные, почти цилиндрические, с расширенным перед-
ним концом и суженным притупленным задним (подсем. Culicinae), с сетчатой
поверхностью, с более мелкими (род Culiseta) или более крупными (род Manzo-
nia) ячейками сетки или в виде лодочки (подсем. Anophelinae) с вогнутой верх-
ней поверхностью и выпуклой нижней. В этом случае на них имеется узкий
плавательный поясок, который у середины прерывается более крупными пла-
вательными камерами, с различным числом составляющих их секций (рис. 3,
М. а, б). Поверхность яиц с рисунком.
11
Рис. 2. Строение взрослых комаров.
1 — тело комара: I — голова и ее придатки, II — грудь, III — брюшко, б — бедро, г — голень, гл —
глаза, ж — жужжальца, л — лапка, у — усик, ц — церки, х — хоботок, щп — щупик. 2 — грудь ко-
мара, сбоку: шс — шейный склерит; переднегрудь: псп — переднеспинка, пэм — проэпимеры, пэс — про-
эпистерны; среднегрудь: 1д — переднее дыхальце, к — место прикрепления крыла, мэп — мезэпимеры,
мэс — мезэпистерны (верхний отдел), пск — постскутеллум, сг — среднегрудка, сп — мезэпистерны (ниж-
ний отдел — «стерноплевры»), срсп — среднеспинка, щ — щиток; заднегрудь: 2д — заднее дыхальце,
мтэп — метаэпимеры, мтэс — метаэпистерны; ноги: 1к, Пк, Шк — коксы; брюшко: 1с — стернит,
1т — тергит. 3, 4 — расположение щетинок на бочках груди: 3 — Culiseta, 4 — Aedes: вмэп — верхние
мезэпимеральные, нмэп — нижние мезэпимеральные, пк — предкрыловые, пн — протонотальные, пспр —
Таксономическое значение имеют форма яйца, наличие или отсутствие пла-
вательного пояска с плавательными камерами, рисунок верхней поверхности
яиц [Шипицина, 1974].
Тело личинки подразделяется на голову, грудь и брюшко.1
Голова плоская снизу и выпуклая сверху. Форма колеблется от яйцевидной
(род Anopheles) до поперечно-овальной (роды Manzonia, Culex и др.) или ближе
к прямоугольной (Тaxorhynchites) (рис. 3, Л, Б).
Окраска головной капсулы от светлой, полупрозрачной (род Culex) до жел-
товато-коричневой, темно-коричневой (многие Aedes) и черной (некоторые
из Culiseta). На щечных и лобных щетинках (в заднем отделе) зачастую имеются
закономерно расположенные темные пятна (следы прикрепления мускулатуры).
На спинной стороне головы различают лобный (фронтальный) щиток
(рис. 3, В). У заднего края щитка личинок I стадии располагается яйцевой зуб.
Впереди лба лежит суженный посредине и расширяющийся по бокам налич-
ник (клипеус), с боков к лобному щитку примыкают щечные щитки, которые
в заднем отделе средней линии разделяются коротким (теменным) коронарным
швом.
На щечных щитках, в местах наибольшего расширения головы, располага-
ются глаза: у личинок I стадии простые глазки в виде маленьких округлых
пигментных пятен, а с III стадии — еще и пара сложных глаз. В передней части
щечного щитка находятся усики (антенны). Они одночлениковые, палочковид-
ные, иногда слегка изогнутые, суживающиеся к вершине, вооруженной чувстви-
тельными (от 1 до 3) волосками. Более толстый (базальный) отдел усиков отде-
ляется от концевого (более тонкого) волоском, состоящим из нескольких вееро-
образно расположенных ветвей (рис. 3, Г, 1—3).
На голове личинки комара около 20 пар волосков и щетинок, которые имеют
диагностическое значение. У переднего края, на боковых выступах налич-
ника, находится пара приклипеальных щетинок различной длины и толщины,
изогнутых внутрь и заостренных (рис. 3, В). За ними идет пара задних волосков
наличника, развитых слабее передних (Anopheles) или находящихся между
основаниями следующего ряда, более сильно развитых волосков. Следующим
рядом волосков являются лобные (фронтальные или постнотальные). Их 3 пары
(наружные, средние, внутренние), которые располагаются в один изогнутый
дугообразно ряд (Anopheles. Culiseta) или в виде двух треугольников, когда
средние волоски лежат впереди задних. Лобные волоски могут быть перистыми
(Anopheles), веерообразными (у многих Aedes. Culex) или простыми. По бокам
заднего отдела лобного щитка теменные волоски.
На щечных щитках различают глазные волоски: у наружного края глаз —
транссутуральные, лежащие между лобным швом и внутренним краем глаз,
и волоски у основания усика.
Ротовые органы личинок состоят из верхней губы с надглоточником, пары
верхних (мандибул) и пары нижних (максилл) челюстей, нижней губы и под-
глоточника. Верхняя губа имеет 3 лопасти: 2 боковых и 1 (непарную) — сре-
динную.
По способу питания личинок делят на фильтратов, обскребывателей и хищ-
ников. Способ питания отражается на строении головы. Так, у Anopheles, яв-
ляющихся фильтраторами, удлиненная форма головы и большое количество
длинных щетинок в боковых и срединных лопастях верхней губы. У обскребы-
вателей (Culex, некоторые Aedes) сильно развиты щечные склериты, особенно
в заднем отделе, вследствие чего голова становится широкой; внутренние во-
1 Описание личинок дается более подробно, так как достоверное определение многих
видов возможно только при наличии этой фазы.
задыхальцевые (постспиракулярные), пэс — проэпистернальные, сп — стерноплевральные, спр — дых а ль-
де в ые (спиракулярные). 5 — пятна чешуек на бочках груди Aedes: г — гипостигмальное, .метл — метастиг-
мальное, мэп — мезэпимеральное, мэс — мезэпистернальное (стерноплевральное), пар — парастигмаль-
ное, пк — посткоксальное, пэп — проэпимеральное. 6 — крыло Culex: жилки: с — костальная (краевая),
— субкостальная, гп г2, г3, г4+б — радиальные, т., тп2, тп3+4 — медиальные, си. — кубитальная, ап —
анальная, rm — передняя поперечная (радиомедиальная), тити — задняя поперечная, h — плечевая. 7 —
концевые придатки лапки комара: а — последний членик лапки CuZex, б — коготки самки Aedes, в — ко-
готки самки Culex, п — присоски (пульвиллы). 1—6 — по: Гуцевич и др., 1970; 7 — из Е. Н. Павлов-
ского, по: Гуцевич и др., 1970.
13
Рис. 3. Строение личинок комаров.
А, Б — личинки: А — Anopheles, Б — Cnlicidae: б — брюшко, г — голова, гл — глаза, гп — гребень,.
гр — грудь, д — дыхальце, ж — жабры, п — пучок волосков, пв — пальмовидные волоски, пл — пле-
чевые лопасти, с — сифон, у — усик, щ — щетка, I—X — сегменты брюшка. В — голова личинки Ano-
pheles (VI стадия) со спинной стороны: зш — затылочный шов, зщ — затылочный щиток (воротничок),
к — клипеус, лш — лобный шов, лщ — лобный щиток, тш — теменной шов, щщ — щечные щитки, 1,2 —
внутренний и наружный волоски наличника, з — задний волосок наличника, 4—6 — внутренний, средний,
наружный лобные волоски, 7 — теменной волосок, 8 — транссутуральный волосок, .9 — волосок у осно-
вания усика, 10 — глазной волосок. Г — разные типы усиков личинок: 1 — Culcx morsitans, 2 — Aedes,.
3 — Culex pipiens. Д — VI членик брюшка личинки Anopheles: а — спинная, б — брюшная стороны, птП
и эт — передняя и задняя тергальные пластинки, 1—13 — волоски членика (1 — звездчатый, 6 — бо-
ковой). Е — звездчатый волосок при большом увеличении.
Рис. 3 (продолжение).
Ж — волоски груди личинки Anopheles’. I — брюшная сторона: 8—12 — боковые волоски, 15, 14 — прочие
волоски; II— спинная сторона; пг— переднегрудь: 1—3—срединные волоски (внутренний, средний,
наружный), 4—7 — плечевые волоски; срг — среднегрудь: 1 — внутренний, срединный волосок; зг — задне-
грудь: 1 — срединный волосок (часто звездчатый). 3 — сифон личинки Culicinae: 1 — стигмальная плас-
тинка, 2 — размер сифона. И — задний конец личинки: VII—VIII — членики брюшка: 1 — зубец гребня,
2 — рулевые волоски, 3 — хвостовые волоски спинной стороны, 4—8 — волоски позади щетки, 9 — че-
шуйка щетки. К — задний конец брюшка личинки Anopheles: а — аппарат, поддерживающий стигмальную
пластинку, ж — жабры, с — седло, о—13 — волоски. Л — стигмальная пластинка: бл, зл, пл — боковая,
задняя, передняя лопасти, «л, нп — верхняя и нижняя (основание) поверхности задней лопасти, зо, по,
со — задний, передний, средний отделы центральной пластинки, ст — стигмы, сто — стигмальные от-
ростки, цп — центральная пластинка, a, b, с, d, f, 1—5 — волоски стигмальной пластинки. М — яйцо:
а — Anophelinae, б — Culicinae. А, В, 3, И, Л, Г — по: Гуцевич и др., 1970; В, Ц, Е, Ж — по Пури:
К — по Мартини, из: Гуцевич и др., 1970; М — по: Шипицина, 1974.
лоски лопастей на верхней губе уплощенные с зубчиками на концах. У актив-
ных хищников (Lutzia, Тaxorhynchitehs) боковые лопасти верхней губы, служа-
щие для схватывания добычи, сильно раздвинуты в стороны, средняя лопасть
редуцирована; количество волосков уменьшается, а некоторые из них превра-
щаются в изогнутые щетинки, с зубчиками или зубцами.
Грудь состоит из 3 члеников (передне-, средне- и заднегрудного), слитых
вместе. Это — наиболее широкий отдел тела. Форма ее колеблется от округло-
6-угольной до округло-овальной. У личинок рода Anopheles по бокам переднего
края спинной стороны груди расположены спинные лопасти (плечевые кла-
паны) — прозрачные выросты, состоящие из 2 лопастей, направленных диаго-
нально вперед и вбок.
На груди различают следующие, имеющие диагностическое значение во-
лоски. У личинок Anopheles на спинной стороне переднегруди — срединные
волоски (внутренний, срединный, наружный) и плечевые; на среднегруди —
внутренний волосок, на заднегруди — волоски звездчатого типа, а на брюшной
стороне — боковые волоски (рис. 3, Ж). У личинок Culicinae при определении
видов могут иметь значение срединные (медиальные) волоски переднегруди.
Брюшко состоит из 9 сегментов. У первых 7 члеников учитывают строение
и развитие волосков, особенно звездчатых (рис. 3, Д’, Е). На спинной стороне
VIII членика у личинок рода Anopheles располагается стигмальная пластинка
(рис. 3, Л). На ней различают стигмы со стигмальными отростками, окружен-
ные 5 лопастями: непарной передней лопастью, парой боковых и парой задних
лопастей, и центральную пластинку. По бокам ее основания 3 пары коротких
волосков. На верхней поверхности боковых лопастей лежит темная удли-
ненной формы хитиновая опорная пластинка, с чувствительным волоском на су-
женной вершине. Задние лопасти своими верхними поверхностями слиты
с центральной пластинкой в одно целое. В последней выделяют 3 отдела: пе-
редний, сужающийся в лежащий между стигмами передний отросток, средний
и задний. По характеру скульптуры (результат прикрепления мышц) разли-
чают срединную и боковые части пластинки.
У личинок Culicinae от спинной поверхности VIII сегмента отходит дыхаль-
цевая трубка — сифон (рис. 3, 3, И, К). На вершине сифона располагаются ды-
хальцевые отверстия — стигмы, окруженные стигмальной пластинкой. На зад-
ней поверхности сифона по бокам — два продольных ряда зубцов или гребней.
Кроме них на сифоне имеется от одной до 5—6 пар различно расположенных
волосков. Над основанием сифона размещается состоящая из чешуек щетка.
Спинная сторона последнего сегмента брюшка несет пластинку — седло, кото-
рое у некоторых видов охватывает весь сегмент в виде кольца. Позади этой
пластинки помещаются две пары хвостовых волосков. На брюшной стороне
последнего сегмента расположены рулевые волоски — плавник. Вокруг аналь-
ного отверстия лежат две пары жабер [Шипицина, 1974].
Куколка состоит из широкой нерасчлененной головогруди и узкого брюшка,
обычно подогнутого под головогрудь. Сквозь покровы головогруди просвечи-
вают детали строения имаго. На спинной стороне головогруди имеются 2 ды-
хательные трубки: у Anophelinae — конической формы, у Culicinae — цилин-
дрической. На I членике брюшка расположены 2 древовидных волоска, на зад-
нем крае VIII сегмента — 2 листовидные пластинки (плавники). Короткий
IX сегмент несет зачатки наружных половых органов имаго.
БИОЛОГИЯ
Созревание яиц у самок комаров начинается после принятия крови. В зави-
симости от температуры окружающей среды созревание яиц продолжается
от 1.5—2 до 10—12 сут. За сезон самка может сосать кровь и откладывать
яйца от 3—5 до 17—18 раз.
Количество откладываемых самками яиц различно: от 120 до 350—450 (у ви-
дов урода Anopheles) и от единичных до 260 и более у видов родов Aedes м. Cu-
lex. Весенние комары обычна более крупные и плодовитые. В течение лета
размеры комаров уменьшаются, соответственно уменьшается и количество'
откладываемых яиц.
16
Самки родов Anopheles и Culex откладывают яйца на поверхность воды
постоянных или длительно существующих временных стоячих водоемов, по-
росших растительностью, в крупных заболоченных и поросших камышами
плавнях рек, в дуплах деревьев.
Самки большинства видов рода Aedes откладывают яйца не на поверхность
воды, а на влажную почву, дно различных углублений в земле, которые могут
быть залиты водой в результате дождей, снеготаяния, разлива рек.
После откладки яиц в них происходит процесс созревания личинок. У кома-
ров рода Aedes сформировавшиеся личинки могут длительное время (до не-
скольких лет) оставаться в яйце (покоящиеся яйца). Продолжительность раз-
вития яиц зависит от температуры окружающей среды и составляет от 1 до
22 сут (Anopheles, Culex). Летом массовый выплод личинок из яиц можно наблю-
дать через 1.5—2 ч после заполнения водоема водой (например, у Aedes vexans).
Развитие личинки и куколки в зависимости от природно-климатических
условий может продолжаться от 5—7 до 11—26 дней или затягиваться до 22—
47 дней.
Большинство видов рода Aedes зимует в фазе яйца, Anopheles^ Culex — в фазе
имаго: в погребах, подпольях, овощехранилищах и т. д., а в природе — в пе-
щерах, трещинах скал, норах животных, растительном сухостое, дерне.
Виды родов Anopheles и Culex имеют в основном несколько поколений за се-
зон (полициклические виды); виды рода Aedes могут быть как поли-, так и моно-
циклические (с одним поколением за сезон).
Комары появляются ранней весной, еще до окончания таяния снега и ле-
тают до самой осени. Наиболее короткий период лёта комаров отмечен в зоне
тундры — 1.5—2 мес.; в таежно-лесной зоне — 3—4 мес. При этом в северных
и умеренной зонах наблюдается один, реже два подъема численности этих на-
секомых, а в южных — два или три.
Летом комары наиболее активны перед заходом (20—22 ч) и восходом (4—
б ч) солнца; осенью — днем (16—18 ч). Весной, когда температура воздуха еще
не высокая, комары нападают и днем. В жаркие солнечные дни комары не
активны и находятся в укрытии: в нижнем ярусе растительности, вблизи водо-
емов, и нападают лишь иногда в густой лесу, кустарнике, зарослях тростника.
При достаточно высокой температуре воздуха комары активны в светлые ночи.
Комары наиболее активно нападают при температуре воздуха 9—30 °C. Более
высокая или низкая температура воздуха действует на них угнетающе. Высо-
кая влажность и моросящий дождь не снижают их активность. Нападают они
и при слабом ветре — 2—3 м/с, а при более сильном (4—5 м/с) лёт их прекра-
щается.
ВРЕД
В отдельных районах кровососущие комары являются преобладающими
среди кровососущих двукрылых насекомых. Массовое нападение комаров
снижает производительность труда людей и рабочего скота (лошадей, волов,
верблюдов). Снижаются также надои молока у коров и привесы у молодняка.
Многочисленные укусы комаров, кроме значительной потери крови, вызывают
из-за интоксикации слюной специфическое заболевание, характеризующееся
подъемом температуры, одышкой, вялостью, слабостью скелетной мускулатуры.
Ко всему вышеизложенному необходимо добавить, что комары являются
механическими и специфическими переносчиками возбудителей многих инфек-
ционных и инвазионных заболеваний людей и животных: малярии, японского
энцефалита, инфекционной анемии лошадей, туляремии, лихорадки долины
Рифт, омской геморрагической лихорадки, инфекционной катаральной го-
рячки («синего языка») овец, африканской чумы и экзантемы свиней, анаплаз-
моза крупного рогатого скота, лошадиного энцефаломиелита, филяриотозов
и др.
Диагноз сем. Culicidae
Ротовые органы в виде хоботка, длина которого значительно превышает
диаметр головы. Усики 15-члениковые, покрыты волосками. Крылья узкие:
костальная жилка обходит все крыло; г2+г3, и ^.з+4+ш1 с развилком.
2 Зак. № 990
17
По ходу жилок обязательны чешуйки. Средней величины насекомые, самки ко-
торых, как правило, нападают для кровососания. Личинки и куколки разви-
ваются преимущественно в стоячих водоемах.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
Самки
1 (2). Щупики по длине равны хоботку. Брюшко полностью или только стер-
ниты на значительном протяжении без
Рис. 4. Детали строения взрослых комаров.
1— щитки: а—Anophelinae, б — Culicinae; 2 — брюшко: а —
Aedes, б —Culex. 1 — по О. Н. Сазоновой, из: Шипицина, 1974;
— по Сое, Freeman, Mattinlgy, из: Гуцевич и др., 1970.
чешуек. Щиток цельнокрайний
j (рис. 4, Id) ............
... 1. Anopheles Mg.
2 (1). Щупики короче хо-
ботка. Брюшко пол-
ностью покрыто чешуй-
ками. Щиток трехло-
пастный (рис. 4, 16).
3 (4). Дыхальцевые щетинки
имеются (рис. 2, 3) . .
.... 2. Culiseta Felt.
4 (3). Дыхальцевые щетинки
отсутствуют (рис. 2, 4).
5 (6). Задыхальцевые ще-
тинки имеются. Церки
выступают (рис. 4, 26).
Коготки с зубчиком,
пульвиллы отсутствуют
.... 3. Aedes Mg.
6 (5). Задыхальцевые ще-
тинки отсутствуют.
* Церки не выступают.
Коготки без зубчика, пульвиллы имеются (рис. 2, 7а, в; 4, 2d). Чешуйки
крыла узкие...........................................4. Culex L.
Личинки IV стадии
1 (2). Стигмы помещаются на спинной стороне VIII членика брюшка; окружены
стигмальной пластинкой.............................1. Anopheles Mg.
2 (1). Стигмы, окруженные стигмальной пластинкой, помещаются на конце
дыхательной трубки — сифона, отходящей от спинной стороны VIII чле-
ника брюшка.
3 (6). Сифон с 1 парой пучков на задней поверхности.
4 (5). Пучки на сифоне помещаются у его основания ... 2. Culiseta Felt.
5 (4). Пучки на сифоне помещаются у его середины или ближе к вершине . . .
......................................................3. Aedes Mg.
6 (3). Сифон с несколькими парами пучков на задней и боковых его поверхно-
стях ...................................................4. Culex L.
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
1. Anopheles Mg. Щупики равны хоботку. Щиток цельнокрайний. Ствол
медиальной жилки прямой. Брюшко, кроме гениталий, без чешуек
Личинки с сильно развитыми перистыми волосками на члениках тела. Го-
лова вытянутая. Усики короткие, палочковидные, с 1 волоском. Грудь с пле-
чевыми клапанами. Брюшко со звездчатыми волосками. Стигмы окружены
стигмальной пластинкой. — 3 вида (в СССР 7).
18
Самки
1 (2). Крылья с пятнами, образованными скоплениями чешуек темного цвета;
передние бедра тонкие, цилиндрические . . An. (An.) maculipennis Mg.
2 (1). Крылья с пятнами, образованными скоплением чешуек белого цвета;
передние бедра при основании утолщенные.
3 (4). Костальная жилка с 1 белым пятном вблизи вершины. Задние бедра
с явственным предвершинным белым кольцом. Передние бедра при осно-
вании слабо утолщены ........................An (An.) lindesayi Giles
а (б). Белые чешуйки нижней поверхности задних бедер развиты при-
мерно на V3 длины бедра от его основания ........................
.............................. An. (An.) lindesayi lindesayi Giles
б (а). Белые чешуйки нижней поверхности задних бедер развиты только
на V6 длины бедра от его основания..............................
....................... An. (An.) lindesayi japonicus Jam.
4 (3). Костальная жилка с 2 белыми пятнами. Задние бедра без белого пред-
вершинного кольца. Передние бедра при основании явственно утолщены
.............................................An. (An.) hyrcanus Pall.
Личинки IV стадии
1 (2). Наружные волоски наличника простые или слабо разветвленные на кон-
цах или начиная с середины...................An. (An.) lindesayi Giles
2 (1). Наружные волоски наличника древовидно разветвлены (рис. 5, 7).
3 (4). Волосок на усике длинный, около V2 длины последнего; отходит от его
середины или лишь немного смещен к основанию (рис. 5, 2а)............
...................................... An. (An.) hyrcanus Pall.
4 (3). Волосок на усике очень короткий, не превышает ширину последнего
у места его отхождения, около основания (рис. 5, 26).................
................................... An. (An.) maculipennis Mg.
2. Culiseta Felt. (=Theobaldia NeveVLemaire). Средней величины или круп-
ные комары. Имеются только дыхальцевые щетинки. Коготки ног простые,
пульвиллы отсутствуют. Церки не выступают. Ствол радиальной жилки крыла
в основной части с волосками.
Личинки крупные или очень крупные. Щетка на VIII членике брюшка
состоит из большого числа чешуек, не имеющих главного шипа. Сифон различ-
ной длины. Пучок всегда у основания сифона. Стигмальная пластинка с зад-
ним отростком рычага в виде более или менее массивной нерасщепленной пла-
стинки. Плавник сильно развит, часть пучков лежит впереди общего основания
и прободает кольцо седла. — 4 вида (в СССР 9).
Самки
1 (4). rm(ta) и mm (tp) расположены обычно на одной прямой. Крылья часто с тем-
ными пятнами.
2 (3). Лапки одноцветно темные. Крылья с более или менее выраженными не-
резкими темными пятнами ..................С. (Culiseta) bergrothi Edw\
3 (2). Лапки с белыми кольцами, только 1-е членики задних лапок посредине
без светлого кольца и бедра без предвершинного светлого кольца. Крылья
с явственно выраженными темными пятнами . . С. (С.) alaskaensis Ludl.
а (б). Рисунок на теле и ногах контрастный, черно-белый. Светлые че-
шуйки на крыльях отсутствуют или их немного; темные пятна выра-
жены резко. Светлые чешуйки в задней половине тергитов отсут-
ствуют ........................... С. (С.) alaskaensis alaskaensis Ludl.
б (а). Рисунок на теле и ногах расплывчатый, буро-охристый. На крыльях
значительное количество светлых чешуек; темные пятна выражены
не резко. Светлые чешуйки в небольшом количестве вкраплены по
всей поверхности тергитов ... С. (С.) alaskaensis indica Edw.
4 (1). mm (tp) ближе к основанию крыла, чем rm (ta). Крылья без темных пятен.
19
2*
ч
Рис. 5. Детали строения личинок IV стадии.
1 — передний край лобного щитка Anopheles maculipennis; 2 — усики личинок: а — A. maculipennis, б —
A. hyrcanus\ 3 — сифоны: а — Culiseta alascaensis, б — С. ochroptera\ 4 — голова сверху: а — С. bergro-
thi,6 — С. ochroptera (I, 2, За, 4 — по: Гуцевич и др., 1970; 36 — по Пеусу из: Гуцевич и др., 1970).
5 (6). Узкие перевязи светлых чешуек расположены только при основании
тергитов. Стерниты брюшка без рисунка из темных чешуек в виде буквы V.
Чешуйки на крыле не образуют пятна.....................................
................................. С. (Culicella) morsitans Theob.
6 (5). Узкие перевязи светлых чешуек расположены при основании, у вершины
тергитов или отсутствуют вовсе; иногда тергиты сплошь покрыты свет-
лыми чешуйками. Стерниты брюшка также без рисунка из темных чешуек
в виде буквы V. Чешуйки в средней части крыла, при основании г4+5 могут
образовывать неясное темное пятно................С. (С.) ochroptera Peus.
Личинки IV стадии
1 (4). Сифон относительно короткий и толстый (индекс не более 4) (рис. 5, За).
Главные трахейные стволы широкие, лентовидные. Усики короче головы,
со слаборазвитым пучком.
2 (3). Усики в длину достигают половины длины головы (рис. 5, 4а). Средние
лобные волосики из 4—8 ветвей. Щетка в среднем более чем из 60 чешуек
.......................................... С. (Culiseta) bergrothi Edw.
3 (2). Усики менее половины длины головы. Средние лобные волоски не более
чем из 3 ветвей. Щетка в среднем менее чем из 50 чешуек................
..................................... С. (С.) alaskaensis Ludl.
4 (1). Сифон относительно длинный и тонкий (индекс выше 5) (рис. 5, 36). Глав-
ные трахейные стволы тонкие, округлой формы. Усики длиннее головы,
с хорошо развитым пучком.
5 (6). Внутренние лобные волоски из 5—9 ветвей. Чешуйки у заднего края
щетки с темной продольной полосой. Волосок на задних клапанах стиг-
мальной пластинки крючкообразно изогнут на самом конце. Жабры длин-
нее седла ................................С. (Culicella) ochroptera Peus.
6 (5). Внутренние лобные волоски из 2—3 ветвей. Чешуйки у заднего края
щетки без темной продольной полосы. Волосок на задних клапанах стиг-
мальной пластинки крючкообразно изогнут по всей длине. Жабры короче
седла ......................................4. С. (С.) morsitans Theob.
3. Aedes Mg. Отсутствуют только дыхальцевые щетинки. Церки самки
более или менее выступают. Коготки с зубчиком на всех ногах, реже на 2 или
на 1 паре ног; пульвиллы отсутствуют. Личинки с палочковидными короткими
усиками, с пучком у середины или перед нею. Средние и боковые волоски
переднегруди умеренно развиты. Сифон короткий (индекс не более 4), всегда
с гребнем и парой пучков на заднебоковой поверхности, иногда с дополни-
тельными передними волосками на передней стороне и боках трубки. Стигмаль-
ная пластинка с двуветвистым задним отростком рычага. — 27 видов (в СССР 51).
Самки
1 (28). Лапки со светлыми кольцами.
2 (7). Светлое кольцо опоясывает вершину предыдущего и основание после-
дующего члеников (рис. 6, 4J.
3 (4). Брюшко с продольной полосой светлых чешуек. Крылья в светлых и
темных чешуйках. Хоботок и щупики в светлых чешуйках..................
....................................A. (Ochlerotatus) caspius Pall,
а (б). Среднеспинка в золотистых или ржаво-желтых чешуйках с 2 про-
дольными светлыми полосками . . А. (О.) caspius caspius Pall.
б (а). Среднеспинка с 1 коричневой продольной полосой ............
..................................А. (О.) caspius dorsalis Mg.
4 (3). Брюшко без продольной полосы светлых чешуек. Крылья в темных че-
шуйках. Хоботок и щупики в темных чешуйках, иногда с белой вершиной.
5 (6). Светлые кольца лапок узкие, развиты в основном на задних ногах.
Щупики в одноцветных темных чешуйках . .A. (Finlaya) alektorovi Stack.
6 (5). Светлые кольца широкие, развиты на всех лапках. Последний членик
лапки темный целиком или со светлым кольцом при основании. Щупики
21
в черных чешуйках с белой вершиной (рис. 7).....................
...............................................A. (F.) togoi Theob
7 (2). Светлые кольца расположены при основании члеников лапки (рис. 6, А2)
8 (23). Хоботок длиннее передних бедер. Чешуйки щитка желтоватые ил1
беловатые, узкие, изогнутые.
9 (12). Церки короткие, едва выступающие. Светлые пятна по бокам брюшк;
с серебристым оттенком.
10 (11). Белые кольца имеются на первых 3 члениках задней лапки . . .
...........................................A. (F.) japonicus Theob
Рис. 6. Детали строения ног комаров рода Aedes.
А — лапки: 1 — A. caspius, 2 — A. cantans, 3 — A. vexans’, Б — коготки лапок: 1 — A. pullatus, 2 -
A. cinereus, 3 — A. intrudens, 4 — A. nigripes, 5 — A. cantans, 6 — A. cataphylla, 1 — A. excrucians, 8 -
A. beklemishevi, 9 — A. riparius, 10 — A. impiger. (А — по: Гуцевич и др., 1970; Б — по О. Н. Сазоно
вой из: Гуцевич и др., 1970).
11 (10). Белые кольца имеются на члениках задней лапки................
............................................. A. (F.) koreicus Edw
12 (9). Церки длинные, явственно выступающие. Светлые пятна по бокат
брюшка без серебристого оттенка.
13 (14). Белые кольца на лапках узкие, не превышающие V4 длины соответ
ствующего членика (рис. 6, А3)................A. (Aedimorphus) vexans Mg
а (б). Вершинная половина каждого тергита брюшка в одноцветных тем
ных чешуйках . , . . ,...................A. (A.) vexans vexans Mg
б (а). Вершинная часть тергитов со светлыми пятнами, иногда образуй:
щими прерывистую продольную полоску..............................
.................................A. (A.) vexans nipponii Theol
14 (13). Белые кольца на лапках широкие, занимающие на средних членика
задней лапки V3 длины членика.
15 (16). Окраска тела охряно-желтая. Среднеспинка в золотисто-желтых ч(
шуйках. Бочки груди в кремовых чешуйках. Нижние мезэпимеральны
щетинки имеются. Брюшко сверху в одноцветных светлых чешуйка
с единичными темными ......................A. (Ochlerotatus) cyprius Lud.
16 (15). Окраска тела желтовато-серая. Среднеспинка в ржаво-коричневы
чешуйках. Бочки груди в серовато-белых чешуйках. Нижние мезэпимс
ральные щетинки отсутствуют. Брюшко сверху также в одноцветных ceei
лых чешуйках.....................................А. (О.) flavescens Mill
22
17 (20). Общая окраска тела более темная. Среднеспинка большей частью
в коричневых, боковая поверхность груди в белых чешуйках.
18 (19). Среднеспинка без продольной или с намечающейся продольной темной
полосой, в шоколадно-кремовых чешуйках с небольшими неясными пят-
Рис. 7. Aedes togoi Theob. (По: Гуцевич и др., 1970).
нами светлых чешуек. Коготок лапки круто изогнут (рис. 6, Б5) . . . •
................................................А. (О.) cantans Mg.
19 (18). Среднеспинка с продольной темной полосой; боковые отделы средне-
спинки в золотистых чешуйках. Коготок лапки полого вытянут (рис. 6, Б9)
....................................................А. (О.) riparius D. К.
а (б). Крылья, хоботок, щупики с темными и светлыми чешуйками . . .
..................................А. (О.) riparius riparius D. К.
б (а). Крылья, хоботок, щупики в одноцветных темных чешуйках . . .
......................................... А. (О.) riparius ater Guts.
23
20 (17). Общая окраска тела более светлая. Среднеспинка большей частью
в золотистых или кремовых чешуйках, иногда с продольной полосой тем-
ных чешуек.
21 (22). Среднеспинка в золотистых чешуйках, без продольной темной полосы.
Коготок лапки круто изогнутый, крупный; зубчик направлен почти
параллельно вершинной части коготка (рис. 6, Б7)...........................
...........................................А. (О.) excrucians Walk.
22 (21). Среднеспинка также в золотистых чешуйках, без продольной темной
полосы. Коготок лапки менее изогнутый, с большим резко отставленным
зубчиком (рис. 6, Б8)...........................А. (О.) beklemishevi Den.
23 (8). Хоботок не длиннее передних бедер. Чешуйки щитка широкие, прямые,
серебристо-белые.
24 (27). Боковые светлые полоски среднеспинки развиты по всей ее длине.
25 (26). 5-й членик задней лапки темный; 3-й членик задних и 2-й средних
светлые сверху........................A. (Stegomyia) sibiricus Dan. et Fil.
26 (25). 5-й членик задней лапки светлый; 3-й членик задних и 2-й средних
темные в вершинной половине...........................A. (S.) galloisi Jam.
27 (24). Боковые светлые полоски выражены только в задней половине средне-
спинки .........................................A. (S.) flavopictus Jam.
28 (1). Лапки без светлых колец.
29 (46). Хоботок явственно длиннее передних бедер.
30 (31). Церки короткие, едва выступающие. Светлые пятна на брюшке бле-
стящие, серебристые. Среднеспинка с 2 большими серебристо-белыми
пятнами, иногда сливающимися в 1 пятно....................................
.............................A. (Finlaya) nipponicus La Casse et Jam.
31 (30). Церки длинные, явственно выступающие. Светлые пятна на брюшке
не имеют блестяще-серебристого оттенка. Среднеспинка без серебристых
пятен.
32 (35). Среднеспинка и голова сверху в длинных и густых черных щетинках.
Щетинки разбросаны по всей задней половине проэпимер.
33 (34). Пятно светлых чешуек на стерноплеврах доходит до переднего их угла
(у основания передних ног). Коготок лапки слабо изогнут (рис. 6, Z>4)
....................................*. . A. (Ochlerotatus) nigripes Zett.
34 (33). Пятно светлых чешуек на стерноплеврах не доходит до переднего
их угла. Коготок лапки резко изогнут (рис. 6, £>10)......................
.............................................А. (О.) impiger Walk.
35 (32). Среднеспинка и голова в менее длинных и густых коричневых или золо-
тистых волосках.
36 (37). Передние бедра спереди пестрые; светлые чешуйки перемешаны с тем-
ными. Светлые чешуйки имеются не только при основании крыла, но
и в различных его частях. Коготок как на рис. 6, Б6......................
...........................................А. (О.) cataphylla Dyar.
37 (36). Передние бедра спереди не пестрые, преимущественно в темных чешуй-
ках или с небольшой примесью светлых. Светлые чешуйки на крыле
имеются только при их основании или их вовсе нет.
38 (39). Гипостигмальное пятно (пятно светлых чешуек книзу от переднего
дыхальца) имеется. Коготок как на рис. 6, Б3 . . А. (О.) intrudens Dyar.
39 (38). Гипостигмальное пятно отсутствует.
40 (45). Пятно белых чешуек на мезэпимерах доходит до их нижнего края.
Нижние мезэпимеральные щетинки имеются. Коготок как на рис. 6, Бг
......................................................А. (О.) pullatus Cog.
41 (42). Посткоксальное пятно отсутствует (т. е. нет чешуек на перепончатом
участке между передними тазиками и стерноплеврами) ......................
.............................................А. (О.) communis Deg.
42 (41). Посткоксальное пятно светлых чешуек имеется.
43 (44). Основание костальной жилки в темных чешуйках ...................
.............................................А. (О.) punctor Kirby
44 (43). Основание костальной жилки в светлых чешуйках..................
............................................. А. (О.) pionips Dyar.
45 (40). Пятно белых чешуек на мезэпимерах не доходит до их нижнего края.
24
Нижние мезэпимеральные щетинки отсутствуют . .А. (О.) sticticus Mg.
46 (29). Хоботок не длиннее передних бедер, или длина хоботка лишь немного
превышает длину передних бедер.
47 (48). Среднеспинка с продольной темной полосой. Крылья со светлыми
чешуйками. Затылок с пятном золотистых чешуек ...........................
.............................................A. (sgen.?) aureus Guts.
48 (47). Среднеспинка без продольной темной полосы. Крылья в темных че-
шуйках. Затылок без пятна золотистых чешуек.
49 (50). Тергиты брюшка с белыми пятнами по бокам, без светлых перевязей
................................................ A. (Aedes) nobuconis Jam.
50 (49). Тергиты брюшка без светлых пятен по бокам, одноцветно темные,
иногда со светлыми перевязями. Коготок как на рис. 6, Б2.................
.....................................................A. cinereus Mg.
а (б). Брюшко сверху без светлых перевязей . . A. cinereus cinereus Mg.
б (а). Брюшко со светлыми перевязями при основании тергитов . . .
................................... A. cinereus esoensis Jam.
Личинки IV стадии
1 (8). Лобные волоски расположены в 1 дугообразный ряд (рис. 8, 7).
2 (3). Лобные волоски находятся в средней части лобного щитка. Щетка на
VIII членике брюшка из 20 чешуек с остроконечным главным шипом.
Дистальные зубцы гребня широко расставлены, заходят за середину дыха-
тельной трубки; пучок ближе к вершине, из 3—5 коротких ветвей. (Под-
род Aedes Mg.).
а (б). Типичные личинки: с признаками подрода ..................
............................. A. (A.) cinereus cinereus Mg.
б (а). Типичные личинки: на боковых поверхностях дыхательной трубки
у ее вершины имеются 4 пары дополнительных, очень коротких и
тонких волосков ....................A. (A.) cinereus esoensis Jam.
3 (2). Лобные волоски смещены к переднему краю лобного щитка. Щетка на
VIII членике брюшка из 40—50 чешуек, без главного шипа.
4 (5). Дыхательная трубка короткая (индекс около 2), одной длины с послед-
ним члеником; пучок около вершины трубки. Жабры короткие, тупокони-
ческие ......................................A. (Finiaya) togoi Theob.
[5 (4). Дыхательная трубка более длинная (индекс около 3), длиннее послед-
него членика; пучок ближе к ее середине. Жабры длинные, ланцетовидной
формы.
6 (7). Зубцы гребня расположены на приблизительно одинаковом расстоянии
друг от друга; из них не более 2 дистальных заходят за основание пучка
................................................A. (F.) koreicus Edw.
7 (6). Зубцы гребня, наиболее дистальные, в числе 1—4 более широко рас-
ставлены в виде крупных шипов, лежащих под более острым углом к про-
дольной оси трубки .............................A. (F.) japonicus Theob.
8 (1). Лобные волоски с каждой стороны расположены треугольником; сред-
ние впереди внутренних.
9 (10). Средний и внутренний лобные волоски смещены к переднему краю
головы; задние волоски наличника одной длины с ними, лежат между
основаниями средних; все они сильно ветвятся.........................
............................A. (F.) nipponicus La Casse et Jam.
10 (9). Средний и внутренний лобные волоски не смещены к переднему краю
головы; задние волоски наличника короче их; если не короче, то лобные
волоски слабо ветвятся.
11 (16). Ушки по бокам основания дыхательной трубки, ближе к ее заднему
краю, отсутствуют (рис. 8, 2). Усики с небольшим простым волоском,
короткие, не покрыты шипиками, или как исключение шипики очень
слабо развиты. (Подрод Stegomyia Theob.).
12 (13). Жабры длиннее дыхательной трубки. Задние волоски наличника короче
лобных, из 9—10 ветвей. Боковой волосок на последнем членике не длин-
нее седла.........................................A. (S.) galloisi Jam.
25
Рис. 8. Детали строения личинок IV стадии комаров рода Aedes.
1 — голова сверху A. cinereus cinereus; 2—8 — задний конец личинки сбоку: 2 — A. galloisi, 3 — A. alek-
iorovi, 4 — A. nigripes, 5 — A. sibiricus, 6 — A. vexans, 7 — A. excrucians, 8 — A. communis (1—3, 7,8 —
по: Гуцевич и др., 1970; 4 — по: Carpenter, La Casse; 5 — по: Данилов, Филиппова, 1978; 6 — по: Mar-
shall из: Гуцевич и др., 1970).
13 (12). Жабры короче дыхательной трубки. Задние волоски наличника короче
лобных, из 2—5 ветвей. Боковой волосок или его нижняя ветвь короче
седла.
14 (15). Чешуйки щетки VIII сегмента брюшка крупные, крупнее дистального
зубца гребня сифона...............................A. (S.) flavopictus Jam.
15 (14). Чешуйки щетки VIII сегмента брюшка мелкие, лишь немного крупнее
дистального зубца гребня сифона (рис. 8, 5)..............................
.......................................A. (S.) sibiricus Dan. et Fil.
16 (11). Ушки по бокам основания дыхательной трубки, ближе к ее заднему
краю, хорошо развиты (рис. 8, 5). Усики различного типа, обычно с хорошо
заметными шипиками, реже гладкие, с простым или чаще ветвящимся
волоском.
17 (18). Средние лобные волоски длинные, вдвое длиннее разветвленных вну-
тренних. Жабры длинные, заостренные на концах, более короткая нижняя
пара одной длины с дыхательной трубкой...................................
.................................... A. (Finlaya) alektorovi Stack.
18 (17). Средние лобные волоски примерно одной длины с внутренними.
Жабры самой различной длины.
19 (22). Дистальные зубцы гребня широко расставлены и заходят за основание
пучка дыхательной трубки (рис. 8, 4),
20 (21). Седло кольцом опоясывает последний членик брюшка................
...............................................А. (О.) nigripes Zett.
21 (20). Седло последнего членика брюшка заходит только за середину его
боковых сторон....................................А. (О.) cataphylla Dyar.
22 (19). Дистальные зубцы гребня не заходят за основание пучка дыхательной
трубки.
23 (36). Впереди плавника отходят 4—6 более коротких пучков, не объединен-
ных общим основанием. Пучок у середины дыхательной трубки в длину
не меньше диаметра трубки у места его отхождения (рис. 8, 7).
24 (25). Поверхность тела покрыта мелкими темными шипиками, лежащими
густыми рядами......................................А. (О.) cyprius Ludl.
25 (24). Поверхность тела без мелких шипиков.
26 (27). Щетка на VIII членике брюшка из 6—12 чешуек.....................
....................................А. (О.) riparius Dyar. et Knab.
27 (26). Щетка на VIII членике брюшка из 15—45 чешуек.
28 (29). Волоски на вершине задних клапанов стигмальной пластинки крючко-
образно изогнуты и утолщены.......................А. (О.) excrucians Walk.
29 (28). Волоски на вершине задних клапанов стигмальной пластинки только
слегка изогнуты и не утолщены.
30 (33). Дистальные зубцы гребня расставлены.
31 (32). Наиболее дистальные 1—3-й зубцы гребня лежат за серединой дыха-
тельной трубки, ближе к вершине. Пучок из 2—4 ветвей.....................
.......................................15. А. (О.) beklemishevi Den.
32 (31). Наиболее дистальные зубцы гребня заметно не доходят до середины
дыхательной трубки. Пучок из 4—7 (чаще 4—6) ветвей.......................
....................................А. (О.) flavescens Mull, (частично)
33 (30). Дистальные зубцы гребня не расставлены; промежуток между 2 по-
следними зубцами в среднем равен таковому между 2 предыдущими.
34 (35). Впереди плавника имеется 6—7 более коротких пучков, не окруженных
общим основанием.........................А. (О.) flavescens Mull, (частично)
35 (34). Впереди плавника 4—5 более коротких пучков, не окруженных общим
основанием ..........................................А. (О.) cantans Mg.
36 (23). Впереди плавника не более 3 пучков, не объединенных общим основа-
нием. Если их 4, то пучок на дыхательной трубке короткий и лежит в на-
чале вершинной трети.
37 (38). Жабры, как правило, короче седла, реже не более чем на х/3 длиннее
его, не пигментированы..............................А. (О.) caspius Pall,
а (б). Пучок на дыхательной трубке лежит за ее серединой, из 5—10 вет-
вей. Внутренние хвостовые волоски из 12—15 ветвей. Средний во-
лосок.позади щетки из 7—14 ветвей. . А. (О.) caspius caspius Pall.
27
б (а). Пучок на дыхательной трубке лежит у ее середины, из 3—5 (редко
больше) ветвей. Внутренние хвостовые волоски из 4—12 ветвей.
Средний волосок позади щетки из 5—8 ветвей.........................
..................................А. (О.) с as pi us dorsalis Mg.
38 (37). Жабры не менее чем в 1.5 раза длиннее седла, чаще пигментированные.
39 (44). Число чешуек в щетке более 40 (рис. 8, 8).
40 (41). Средние и внутренние лобные волоски простые, как исключение 1 илп
2 из них 2—3-ветвистые...................А. (О.) communis De Geer
41 (40). Средние и внутренние волоски из 3—7 ветвей.
42 (43). Усики тонкие, длинные, до 2/3 длины головы или немного длиннее.
Все чешуйки щетки без главного шипа............А. (О.) pionips Dyar.
43 (42). Усики толстые, короткие, около V2 длины головы. Некоторые из чешуек
щетки имеют слабо выраженный главный шип . . А. (О.) pullatus Cog.
44 (39). Число чешуек в щетке меньше 30.
45 (46). Седло опоясывает кольцом последний членик брюшка, или на его
брюшной стороне между боковыми краями седла имеется узкая щель
....................................................А. (О.) punctor Kirby
46 (45). Седло не опоясывает кольцом последний членик брюшка, может до-
ходить только до нижнего края его боковых сторон.
47 (48). Внутренние лобные волоски простые. Дистальные зубцы гребня не
расставлены.........................................А. (О.) impiger Walk.
48 (47). Внутренние лобные волоски из 2—5 ветвей.
49 (52). Дистальные зубцы гребня (1—3) всегда широко расставлены, заметно
крупнее остальных.
50 (51). Пучок на дыхательной трубке за серединой, ближе к вершине, всегда
впереди наиболее дистального из зубцов гребня; из 3—8 (чаще 4—6)
тонких ветвей, более коротких, чем ширина трубки у места отхождения
пучка (рис. 8, 6)........................24. A. (Aedimorphus) vexans Mg.
51 (50). Пучок на дыхательной трубке у середины или немного за ней из 3—8
(чаще 4—6) ветвей, равных ширине трубки у* места его отхождения или
длиннее. При числе расставленных зубцов гребня больше 2 пучок лежит
между их основаниями ...................A. (Ochlerotatus) intrudens Dyar.
52 (49). Дистальные зубцы гребня расположены на равном расстоянии друг
от друга; как исключение один наиболее дистальный может быть немного
удален от остальных. В щетке 16—27 (чаще 22—24) чешуек..................
.............................................. А. (О.) sticticus Mg.
4. Culex L. Щупики короткие. Дыхальцевые щетинки отсутствуют; мез-
эпимеральных щетинок 1, реже они отсутствуют вовсе, еще реже их от 2 до 4.
Большая часть поверхности груди лишена чешуек. 1-й членик задних лапок
за редким исключением по длине равен или длиннее задних голеней. Коготки
лапок простые, с присосками (пульвиллами). Крылья с длинной радиальной
вилкой.
Личинки с большой широкой головой и длинными усиками, несущими пучок
в виде веера вторичноперистых ветвей и длинные предвершинные щетинки,
часто стоящие на некотором расстоянии от вершины. Волоски груди обычно
длинные и разветвленные. Щетка по бокам VIII членика брюшка обычно из боль-
шого числа чешуек. Сифон за редким исключением длинный или очень длин-
ный. — 13 видов (в СССР 20).
Самки
1 (2). Брюшко сверху темное. Голени3 длиннее 1-го членика задней лапки
..........................................С. (Barraudius) modestus Fic.
2 (1). Брюшко сверху, как правило, с поперечными перевязями светлых че-
шуек. Голени3 не длиннее 1-го членика задней лапки.
3 (6). Нижние мезэпимеральные щетинки многочисленны, их не менее 4,
обычно 6—10. Комары крупных размеров. (Подрод Lutzia Theob.).
4 (5). Все тергиты брюшка по заднему краю со светлыми перевязями, б. м.
одинаковой ширины. Основная половина передней поверхности задних
бедер с многочисленными темными чешуйками . .С. (Lutzia) vorax Edw.
28
5 (4). II—IV тергиты брюшка одноцветно темные или с очень узкими свет-
лыми перевязями по заднему краю; V—VIII тергиты на всем протяжении
покрыты светлыми чешуйками или же имеют очень широкие перевязи
светлых чешуек по заднему краю. Основная половина передней поверх-
ности задних бедер в белых чешуйках..............С. (L.) fuscanus Wied.
6 (3). Нижние мезэпимеральные щетинки немногочисленны: чаще 1, реже
вовсе отсутствуют, еще реже в числе 2—3. Комары средней величины
или мелкие. (Подроды Culex L., Neoculex Dyar.).
7 (16). Хоботок и лапки с белыми колечками, иногда очень узкими.
8 (И). Крылья с большими светлыми пятнами.
9 (10). Вершинная часть жилки сиг в светлых чешуйках...................
.........................................С. (Culex) orientalis Edw.
10 (9). Вершинная часть жилки сиг в темных чешуйках....................
............................................С. (С.) jacksoni Edw.
*1 (8). Крылья без пятен, с вкраплениями белых чешуек..................
.................................... С. (С.) bitaeniorhynchus Giles
12 (15). Среднеспинка резко двухцветная: передние 2/3 среднеспинки покрыты
светлыми, задняя треть темно-бурыми чешуйками.
13 (14). Перевязи светлых чешуек расположены по заднему краю тергитов
брюшка. Передняя часть среднеспинки в светло-желтых чешуйках.
Крылья в узких чешуйках..........................С. (С.) sinensis Theob.
14 (13). Перевязи светлых чешуек расположены по переднему краю тергитов
брюшка. Передняя часть среднеспинки в белых чешуйках. Чешуйки
крыльев более широкие............................С. (С.) whilmorei Giles
15 (12). Среднеспинка на всем протяжении покрыта одноцветными темными
чешуйками ...............................С. (С.) tritaeniorhynchus Giles
16 (7). Хоботок и лапки без светлых колец.
17 (18). Брюшко сверху одноцветно бурое, без поперечных перевязей или боко-
вых пятен, образованных светлыми чешуйками............................
.......................................«С. (Neoculex) hayasbii Jam.
18 (17). Брюшко с б. м. развитыми поперечными перевязями, образованными
скоплениями белых или желтоватых чешуек.
19 (20). Передние и средние бедра и голени спереди темные, со светлой про-
дольной полосой; реже лишь передние бедра имеют такую полосу . . .
....................................................С. (С.) vagans Wied.
20 (19). Передние и средние бедра и голени спереди без светлой полосы.
21 (22). Светлые поперечные перевязи расположены по заднему краю тергитов
брюшка. Проэпистерны покрыты широкими светлыми чешуйками . . .
............................................. С. (N.) territans Walk.
22 (21). Светлые поперечные перевязи или пятна расположены по переднему
краю тергитов брюшка. Проэпистерны покрыты узкими светлыми че-
шуйками ..............................................С. (С.) pipiens L.
Личинки IV стадии
1 (2). Боковые лопасти верхней губы из 30—40 крепких, S-образно изогнутых
щетинок, несущих на дистальной половине гребень из ряда коротких
шипиков. Сифон короткий, не длиннее последнего членика брюшка, резко
сужен на конце, с гребнем, занимающим всю его длину до сужения (рис. 9,
7, 2). (Подрод Lutzid)..............................С. (L.) vorax Edw.
С. (L.) fuscanus Wied.
2 (1). Боковые лопасти верхней губы обычного строения. Сифон длиннее
последнего членика брюшка, не резко сужен на конце, с гребнем, занима-
ющим не более V2 его длины.
3 (8). Щетка по бокам VIII членика брюшка из 4—8 крупных чешуек с острым
главным шипом (рис. 9, 5).
4 (5). Нижнегубная пластинка по краю с многочисленными мелкими зубчи-
ками одинаковой величины...................С. (С.) bitaeniorhynchus Giles
5 (4). Нижнегубная пластинка из небольшого числа увеличивающихся к сере-
дине зубцов.
29-
6 (7). Нижнегубная пластинка с каждой стороны состоит из 10—15 мелких
зубчиков у основания и 6—8 увеличивающихся к середине зубцов. Гре-
бень из 2—4 маленьких зубцов у основания сифона.........................
............................................С. (С.) sinensis Theob.
Рис. 9. Детали строения личинок IV стадии комаров рода Culex.
1 — щетинки верхней губы С. fuscanus; 2—4 — дыхательная трубка сбоку: 2 — Culex fuscanus, 3 — С. Ы-
taeniorhynchus, 4 — С. tritaeniorhynchus (1—3 — по: Гуцевич и др., 1970; 4 — по: Kirkpatrick из: Гуцевич
и др., 1970).
7 (6). Нижнегубная пластинка только с более крупными зубцами. Гребень из
8—14 зубцов занимает не менее V4 длины сифона . . С. (С.) whitmorei Giles
8 (3). Щетка по бокам VIII членика брюшка не менее чем из 20 чешуек, у боль-
шинства видов не имеющих главного шипа, а несущих по краю ряд в раз-
личной степени развитых зубчиков или шипиков.
30
/ ЦО). Сифон на задней стороне в дистальной четверти имеет дополнительные-
шипы в числе от 2 до 9............................С. (С.) jacksoni Edw.
(9) . Сифон в дистальной четверти не имеет дополнительных шипов.
11 (16). Сифональные пучки (кроме 1 или 2 у вершины) расположены на задней
поверхности сифона зигзагообразно или сильно сближенными по средней
линии парами.
*....................................................2 (13). Внутренний и средний лобные волоски не менее чем вдвое короче на-
ружных. Зубцы гребня в виде длинных тонких шипов. Лопасти жирового
тела темно пигментированы и придают личинке поперечно-полосатый вид.
Стигмальная пластинка типа «territans».............С. (N.) hayashii Jam.
13 (12). Внутренний и средний лобные волоски длинные, одной длины с наруж-
ными волосками или длиннее их. Лопасти жирового тела не содержат
темного пигмента, и личинки не имеют поперечно-полосатого вида. Стиг-
мальная пластинка также типа «territans».
14 (15). Ближайшие к основанию 1—3 пучка волосков на сифоне лежат между
широко расставленными дистальными зубцами гребня. Стигмальная пла-
стинка типа «pipiens» ............. С. (В.) modestus Fic.
15 (14). Ближайший к основанию пучок волосков на сифоне расположен ди-
стальнее последнего из зубцов гребня. Стигмальная пластинка также типа
«pipiens».................................. . С. (С.) orientalis Edw.
Id (11). Сифональные пучки волосков расположены парами по краям задней
поверхности сифона; часть их может быть на боковой его поверхности
(рис. 9, 4).
17 (18). Сифон наиболее широк в средней трети, к вершине сужен сильнее,
чем к основанию.
18 (17). Сифон наиболее широк у основания и явственно расширен у вершины
.................................................С. (N.) territans Walk.
19 (20). Все пучки на сифоне одной длины...............................
................................ . С. (С.) tritaeniorhynchus Giles
20 (19). По меньшей мере первые от основания пары пучков на сифоне длиннее
остальных и больше его диаметра у мест их отхождения.
21 (22). Сифональных пучков 4 пары. Ближайшая к основанию сифона пара
лежит на заметном расстоянии ближе к вершине от наиболее дистального-
зубца гребня. Боковой волосок на последнем членике, как правило,
простой...............................................С. (С.) pipiens L.
22 (21). Сифональных пучков 5 пар. Ближайшая к основанию сифона пара
лежит на одном уровне или между 1 и 2 из наиболее дистальных зубцов-
гребня. Боковой волосок на последнем членике, как правило, двухвет-
вистый ............................................С. (С.) vagans Wied.
ОСНОВНАЯ ЛИТЕРАТУРА I
Виноградская О. Н. Взрослые комары. — В кн.: Руководство по медицинской энто-
мологии. М., 1974, с. 59—78
Гуцевич А. В., Мончадский А. С., Штакельберг А. А. Комары (сем. Culicidae). Л.,.
1970. 384 с. (Фауна СССР. Насекомые двукрылые; Т. 3; Вып. 4).
Данилов В. Н., Филиппова В. В. Новый вид комара Aedes (Stegomyia) sibiricus sp. п.
Culicidae). — Паразитология, 1978, т. 12, вып. 2, с. 170—176.
Шипицина Н. К. Комары (сем. Culicidae). Преимагинальные фазы. — В кн.: Руко-
водство по медицинской энтомологии. М., 1974, с. 48—58.
КРОВОСОСУЩИЕ МОКРЕЦЫ (Сем. CERATOPOGONIDAE)
Кровососущие мокрецы — самый мелкий компонент гнуса (длина тела
от 1—2.5 мм) (подотряд Nematocera). Они широко распространены во всех
ландшафтных зонах. Особенно многочисленны они в необжитой тайге, где до-
стигают огромной численности.
Основная масса кровососущих мокрецов (свыше 90 %) относится к роду
Culicoides. На Дальнем Востоке — 37 видов (в СССР — 132). Мировая фауна
насчитывает около 1000 видов.
31
ВНЕШНЕЕ СТРОЕНИЕ
Голова почти шаровидная, несет усики, щупики, хоботок; по бокам
фасеточные глаза, которые соприкасаются на некотором расстоянии или
разделены так называемой лобной полоской. В этом случае сверху над лобной
щетинкой имеется верхний поперечный шов, иногда бывает и нижний. Между
фасетками глаза могут располагаться короткие шипики или же глаза голые.
У разных видов мокрецов хоботок бывает неодинаковой длины. Для опреде-
ления видов используется хоботковый индекс ХИ — отношение высоты головы
от лобной щетинки до склеротизованных поперечно лежащих образований
(торм) (рис. 10, 2) к длине хоботка, которая измеряется от торм до его вершины.
3-й членик нижнечелюстного щупика может быть тонким и удлиненным, более
коротким и умеренно утолщенным или сильно утолщенным. При диагностике
видов учитывается отношение длины щупика к его наибольшей ширине (щупи-
ковый индекс — ЩИ). Чувствительный орган на щупиках разбросанный,
в виде отдельных маленьких ямок, или компактный (одна ямка). Он может
быть поверхностным, когда ямка неглубокая, или погруженным, когда глубина
ямки не менее или более ее диаметра. Усики состоят из 15 члеников, из них
13 составляют нить усика. Первые 8 члеников нити (проксимальные) (1-й чле-
ник является 3-м) более короткие, последние 5 (дистальные) более удлиненные,
иногда значительно. Отношение длины 5 дистальных члеников к 8 проксималь-
ным — усиковый индекс (УИ). На всех или нескольких члениках усика распо-
ложены чувствительные образования в виде округлых ямок — сенсиллы, окру-
женные выростами. Сенсиллы всегда имеются на 3-м членике. Различное
расположение сенсилл на усиках — важный диагностический признак.
Большую часть спинной поверхности у мокрецов занимает среднеспинка.
Ее окраска важна при определении видов. Среднеспинка бывает одноцветной
или в мелких округлых пятнышках (крапчатая). У некоторых видов на светлом
фоне среднеспинки есть крупные. пятна, часто неправильной формы.
Крылья в покое сложены одно над другим; длица крыла 0.8—2.2 мм. Имеются
следующие продольные жилки (рис. 10, /): костальная, радиальные, медиаль-
ные, кубитальная и поперечная радио-медиальная (г-тп). Жилки ограничи-
вают ячейки: 1-ю и 2-ю радиальные, базальную, 1-ю и 2-ю медиальные, куби-
тальную и анальную. Вся поверхность крыла покрыта очень мелкими волос-
ками — микротрихиями — и более крупными, заметными под бинокуляром—
макротрихиями. Расположение последних имеет значение при определении ви-
дов. У большинства мокрецов крылья пятнистые, реже — бесцветные. Обычно
на светлом (молочном, сероватом или кремоватом) фоне расположены темные
пятна или же на сером фоне —светлые пятна. Рисунок крыла — важнейший
диагностический признак.
Семяприемники, или сперматеки, мокрецов различной формы, в числе от 1
до 3, также используются при определении видов.
Яйца мокрецов рода Culicoides очень мелкие, сигаровидной формы; сразу
после откладки они грязновато-белые, но через несколько часов темнеют до
светло- или темно-коричневого цвета. Длина яиц у разных видов мокрецов
неодинаковая: у самых крупных — 440—460 мкм, ширина — 55—80 мкм,
у мелких мокрецов — 278—300 мкм и 48—50 мкм соответственно.
Личинки этого рода червеобразные (рис. 10, 4), лишенные каких-либо при-
датков, независимо от того, где они выплаживаются — в водоемах, влажной
почве или подстилке. Тело их состоит из хорошо выраженной головной кап-
сулы, окрашенной в различные оттенки желтого или коричневого цвета, 3 груд-
ных и 9 брюшных цилиндрических, мало отличающихся по длине и толщине
сегментов. Последний, анальный сегмент, несет на заднем конце несколько
коротких тонких волосков (кроме дуплового вида С. dendrophilus, у которого
эти волоски длинные).
Куколка — обычной для длинноусых двукрылых формы; ее длина от 2
до 4 мм (рис. 10, 5, 6). Куколка свободна от личиночной оболочки; состоит
из неподвижной головогруди и подвижного брюшка из 9 сегментов. Дыхатель-
ные органы в виде трубочки с дыхательными отверстиями. Окраска ее чаще
всего коричневая, желтоватая или буровато-коричневатая.
32
Рис. 10. Детали строения мокрецов.
1 — самка мокреца рода Culicoides: б — бедро, в — вертлуг, г — голень , ж — жужжальце, л — лапка,
т — тазик, ус — усик, х — хоботок, щ — щупик; жилки крыла: с — костальная, г — радиальная, си —
кубитальная, т — медиальная; ячейки: — 1-я радиальная, R2 — 2-я радиальная, — 5-я радиаль-
ная, М — медиальная, гт — радио-медиальная, Си — кубитальная, Ап — анальная, В — базальная;
2
2 — схема строения головы: г — глаза, лп — лобная полоска, лщ — лобная щетинка, т —тормы, —----уси-
ковый индекс (УИ), — щупиковый индекс (ЩИ), у — хоботковый индекс (ХИ); 3 — яйца мокрецов:
с — яйцо С. pulicaris, б — оболочка яйца С. pulicaris с выходящей из нее личинкой, в, г — оболочка яйца
С. obsoletus; 4 — личинка; 5 — куколка, вид сбоку; 6 — то же с брюшной стороны. (По: Глухова, 1962;
Гуцевич, Глухова, 1970).
3 Зак. ^я 990
БИОЛОГИЯ
Мокрецы летают с мая до октября. В разных районах численность их в от-
дельные периоды сезона может меняться, но наибольшая обычно наблюдается
в первой половине лета: конец мая—июнь. Нападение мокрецов наиболее силь-
ное в утренние и вечерние часы, в моменты восхода и захода солнца. Разви-
ваются мокрецы в разных мелких водоемах и по берегам крупных, во влажной
почве и лесной подстилке.
Созревшие яйца откладываются самками через 5—7 дней после кровососания
поодиночке или небольшими группами в виде причудливых линий и фигур,
никогда не прикрепляются к субстрату. Длительность развития личинки в яйце
в зависимости от температуры и вида мокреца колеблется от 3 до 14 дней (в сред-
нем — 3—7 дней), а после вылупления из яйца до окукления — от 2 до 10 не-
дель. Личинка проходит в своем развитии 4 стадии. Вылет насекомого из ку-
колки через 2—5 дней. В связи с тем, что зимовка проходит в фазе личинки,
весь цикл развития в природе растягивается на несколько месяцев. Большин-
ство видов имеют одну генерацию в году, редко — две.
ВРЕД
Мокрецы — это массовые кровососы человека и животных. Нападая на
человека в огромных количествах — до 5000 экз. за 5 мин (под учетным поло-
гом), они делают невозможным пребывание человека на открытом воздухе.
Мокрецы, кроме того, являются переносчиками возбудителей некоторых вирус-
ных, бактериальных, протозойных, гельминтозных заболеваний человека,
сельскохозяйственных животных и домашних птиц.
Диагноз рода Culicoides
Усики длинные, 15-члениковые, у самки 5, у самца 3 дистальных членика
длиннее проксимальных. Щупики 5-члениковые. На переднем крае среднеспинки
2 маленьких плечевых ямки. Крылья чаще пятнистые или одноцветные. Ко-
стальная жилка заканчивается у середины крыла. 2 радиальные ячейки, обычно
равные, Поверхность крыла покрыта эдикротрихиями (вся) и макротрихиями
(вся или частично). Коготки лапок маленькие, без зубчиков. Эмподий слабо
развит. Сперматек 1—3.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА САМОК
1 (4). Среднеспинка полностью или в задней половине ярко-желтая. 2-я ради-
альная ячейка длиннее и шире 1-й. 3 сперматеки.
2 (3). Среднеспинка полностью ярко-желтая (рис. 11, 9) •
................................... С. flavidorsum Schev. et Lis.
3 (2). Задняя половина среднеспинки ярко-желтая, в передней половине
большое коричневое пятно............................С. humeralis Okada
4 (1). Окраска среднеспинки иная. 2-я радиальная ячейка равна 1-й. 1 или
2 сперматеки.
5 (22). Сперматека одна. Среднеспинка чаще крапчатая.
6 (9). Крылья с темными пятнами неправильной формы. Кубитальная ячейка
с темным пятном. Сенсиллы на III и VIII—XI члениках усика. Крапинки
на среднеспинке отчетливые, крупные.
7 (8). Сперматека в форме реторты с изогнутой шейкой. Крыло более контраст-
ное, пятно в ячейке Иъ заметно выделяется (рис. 11, 15)..............
............................................С. nubeculosus (Mg.)
8 (7). Сперматека яйцевидной или вытянутой формы. Рисунок крыла менее от-
четливый; оно сероватое, пятно в /?5 слабо выделяется (рис. И, 2) . . .
.................................................... С. riethi Kieff.
9 (6). Крылья с многочисленными светлыми округлыми пятнами. Темного
пятна в кубитальной ячейке нет. Сенсиллы на всех члениках либо отсут-
ствуют на некоторых проксимальных члениках. Крапинки имеются или
очень слабо выражены. Сперматека обычно правильной овальной или
округлой формы.
34
15
Рис. 11. Детали строения мокрецов.
2 — крыло: 1 — Culicoides salinarius, 2 — С. riethv, 3—7 — С. koreensis: 3 — крыло, 4 — щупик, 5 —
лоб, 6 — усик, 7 — сперматека; 8 — С. toyamaruae, конец брюшка; 9 — С. flavidorsum, крыло; 10—14 —
€. arakawae: 10 — крыло из Японии, 11 — крыло из Приморского края, 12 — сперматека, 13 — усик,
14 — щупик; 15 — С. nubeculosus, крыло. (По: Гуцевич, 1973).
10 (И). 7-й членик брюшка с заметно склеротизованным неправильной формы
кольцом (рис. 11,5)...................................С. toyamaruae Агп.
11 (10). 7-й членик брюшка без склеротизованного кольца.
12 (15). Среднеспинка с отчетливо выраженными крапинками.
13 (14). Темное пятнышко в центре светлого пятна на поперечной радио-меди-
альной жилке крупное, отчетливое. Задняя ветвь кубитальной жилки чаще
9
Ю
Рис. 12. Детали строения мокрецов.
1,2 — Culicoides manchuriensis: 1 — крыло, 2 — усик; «3, 4 — С. variifrons: 3 — крыло, 4 — варианты
строения лба; 5—7 — С. actoni: 5 — крыло, 6, 7 — вариации крыла; 8 — С. chiopteius, лоб; 9, 10 — крыло:
9 — С. sphagnum crisis, 10 — С. circumscripius (3, 4 — по: Иванов, Глухова, 1967; 7—10 — по: Гуце-
вич, 1973; остальное — ориг.).
светлая. Рисунок крыла контрастный (рис. 12, 10)...........................
................................................ С. circumscriptus Kieff.
14 (13). Темное пятнышко в центре светлого пятна на поперечной радио-меди-
альной жилке отсутствует или слабо выражено. Задняя ветвь кубиталь-
ной жилки чаще темная (рис. 11, 1)..........................С. salinarius Kieff.
15 (12). Крапинки на среднеспинке отсутствуют или слабо выражены.
16 (19). УИ высокий: 1.7—2.1.
17 (18). Вершина крыла с крупными светлыми пятнами (рис. 11, 3—7) . . . .
............................................................С. koreensis Агп.
18 (17). Светлые пятна у вершины крыла отсутствуют или очень маленькие
.......................................................С. sphagnumensis Will.
19 (16). УИ менее 1.6, редко 1.7.
36
20 (21). Крыло всегда с многочисленными отчетливыми светлыми пятнами на
темном фоне. Базальная ячейка без макротрихий. Сперматека удлиненно-
грушевидная (рис. 11, 10—14).......................С. arakawae (Агак.)
21 (20). Рисунок крыла неотчетливый, размеры пятен варьируют. Базальная
ячейка с макротрихиями. Сперматека овальная (рис. 12, 1)..............
..........................................С. manchuriensis Tok.
22 (5). Две сперматеки. Среднеспинка без крапинок, одноцветная или с круп-
ными пятнами.
23 (66). Крылья с пятнами.
24 (47). Глаза соприкасаются на некотором расстоянии, если же разделены, то
имеется нижний поперечный шов. Сенсиллы обычно на III и XI—XV чле-
никах.
25 (38). Рисунок крыла довольно слабо развит: на сероватом фоне немного
светлых пятен.
26 (27). Макротрихии покрывают большую часть крыла. Глаза соприкасаются
или разделены и имеется нижний поперечный шов (рис. 12, 5, 4) . . . .
.........................................С. variifrons Glukh. et Ivan.
27 (26). Макротрихии покрывают вершинную треть крыла. Глаза всегда сопри-
касаются.
28 (31). Большая часть межфасеточного пространства глаза покрыта заметными
шипиками.
29 (30). Крыло с заметными светлыми пятнами, В5 со светлым пятном. Сперма-
теки грушевидные, неравные, с длинными шейками (рис. 12, 5)...........
...................................................С. actoni Smith
30 (29). Пятна на крыле мало заметны, их только 2 у переднего края или еще
небольшие светлые пятна расположены в проксимальной части медиаль-
ной ячейки, в кубитальной и анальной ячейках, в Я5 пятна нет (рис. 12,
6, 7). Сперматеки иной формы.......................С. chiopterus (Mg.)
31 (28). Глаза голые или же шипики расположены только в области соприкосно-
вения глаз.
32 (35). Вершина крыла темная. В /?5 светлых пятен нет. Макротрихии более
развиты. В анальной ячейке их более 50.
33 (34). Сперматеки крупные, овальные*, их длина 70—80 мкм. Светлые пятна
у переднего края крыла относительно небольшие, округлые или слегка
треугольные, 1-е не достигает основания медиальных ячеек. 1-я и 2-я
медиальные ячейки имеют продолговатые светлые пятна (рис. 13, 1—3)
................................................. С. sinanoensis Tok.
34 (33). Сперматеки округлые, мельче, их длина не более 50 мкм. Светлые пятна
у переднего края крыла крупнее, с параллельными краями; 1-е из них
простирается в основание 1-й медиальной ячейки (рис. 13, 4)...........
...............................................С. abchazicus Dzhaf.
35 (32). Вершина крыла со светлым пятном. Макротрихии слабее развиты.
В анальной ячейке их не более 20 (рис. 13, 5) ... . С. filicinus Gorn.
36 (25). Рисунок крыла отчетливый: из темных пятен на светлом фоне или
с многочисленными светлыми пятнами на темном фоне.
37 (40). Кубитальная ячейка с темным пятном.
38 (39). Вершины медиальных жилок светлые............С. punctatus (Mg.)
39 (38). Вершины медиальных жилок темные ..............С. pulicaris (L.)
40 (37). Кубитальная ячейка со светлым пятном.
41 (42). Крыло темное, с небольшими отчетливыми светлыми пятнами. 2-я ра-
диальная ячейка с темной основной половиной. Пятно в /?5 двойное, узкое,
поперечно вытянутое (рис. 13, 6—9).................С. nipponensis Tok.
42 (41). Крыло с крупными светлыми пятнами. 2-я радиальная ячейка цели-
ком светлая. Пятно в Т?5 овальное, продольно вытянутое.
43 (44). Светлые пятна на крыле очень сильно развиты. 1-е и 2-е пятна у перед-
него края почти сливаются. Пятно в Т?5 очень большое, достигает вершин-
ного края крыла (рис. 13, 10)...........................С. dubius Агп.
44 (43). Светлые пятна на крыле умеренно развиты. 1-е и 2-е пятна у переднего
края отчетливо разделены широким темным пространством. Пятно Иъ
37
небольшое, отделено от вершинного края крыла (рис. 13, 11)...........
................................................. С. aterinervis Tok.
45 (24). Глаза разделены лобной полоской. Сенсиллы имеются на всех или
нескольких проксимальных и дистальных члениках.
Рис. 13. Детали строения мокрецов.
1—з — Culicoides sinanoensis: 1 — крыло, 2 — лоб, 3 — щупик; 4, 5 — крыло: 4 — С. abchazicus, 5 —
С. filicinus', 6—9 — С. nipponensis: 6 — крыло, 7 — голова, 8 — щупик, 9 — усик; 10, 11 — крыло: 10 —
С. dubius, 11 — С. aterinervis. (4, 5, 10, 11 — ориг.; остальное — по: Гуцевич, 1973).
46 (53). Рисунок крыла отчетливый, из светлых пятен на темном фоне. В осно-
вании 1-й медиальной ячейки имеется светлое пятно.
47 (52). Крыло с многочисленными светлыми пятнами во всех ячейках.
48 (49). В 1-й медиальной ячейке 3 светлых пятна. Чувствительный орган
щупика из многих чувствительных ямок, сконцентрированных на одной
площадке........................................С. erairai Kono et Так.
49 (48). В медиальной ячейке 2 светлых пятна. Чувствительный орган иного
строения.
50 (51). На передней ветви медиальной жилки имеется светлое пятно. Чувстви-
тельный орган щупика в виде одной неглубокой ямки. Среднеспинка
со сложным рисунком из темных пятен на сером фоне......................
. » . > ......................................... С. odibilis Aust.
38
51 (50). Светлого пятна на передней ветви медиальной жилки пет. Хоботок
короткий, ротовые части со слаборазвитыми зубчиками......................
...............................................С. dendrophilus Amos.
Рис. 14. Детали строения мокрецов.
1,2 — Culicoides fascipennis; 3, 4 — С. pallidicornis\ 5, 6 — С. arboreus', 7, 8 — С. subfascipennis’, 9—11 —
С. amossovae', 12 — С. cubitalis. 1, 3, 5, 7 9, 12 — крыло; 2, 4, 6, 8, 10 — лоб; 11 — щупик. (По: Гуце-
вич, 1973).
52 (47). Крыло с немногими крупными светлыми пятнами: в области радио-
медиальной жилки и 4 сливающихся пятна за 2-й радиальной ячейкой.
Чувствительный орган разбросанный ...................С. epactius Агп.
53 (46). Рисунок крыла из немногочисленных пятен или из большего числа
неотчетливых светлых пятен. В основании 1-й медиальной ячейки нет
светлого пятна.
3&
54 (59). Сенсиллы имеются только на III и дистальных (XI—XV) члениках
усика.
55 (56). Макротрихии в базальной ячейке есть. 1-е светлое пятно в виде преры-
вистой поперечной полосы (рис. 14, 1,2) .... С. fascipennis (Staeg.)
56 (55). Макротрихий в базальной ячейке нет.
57 (58). Крыло только с 2 светлыми пятнами у переднего края (рис. 14, 5, 4)
............................................ С. pallidicornis Kieff.
58 (57). Кроме 2 пятен у переднего края крыла имеются светлые пятна в куби-
тальной и анальной ячейках (рис. 14, 7—8) ... С. subfascipennis Kieff.
59 (54). Сенсиллы имеются на всех или некоторых проксимальных члениках.
60 (61). Крыло только с 2 неясными пятнами у переднего края и иногда с не-
ясными посветлениями в кубитальной ячейке. Щупик утолщен, чувстви-
тельная ямка большая, но неглубокая. Сенсиллы на всех члениках усика
(рис. 14, 12).........................................С. cubitalis Edw.
61 (60). На крыле несколько светлых пятен.
62 (63). Дистальные членики усика очень длинные; У И — 1.5—1.6. Чувстви-
тельная ямка на щупике большая, но неглубокая (рис. 14, 9, 11) ....
....................................................С. amossovae Remm
63 (62). Дистальные членики усика более короткие; УИ — 1.03—1.3. Чувстви-
тельная ямка довольно глубокая.
64 (65). со светлым пятном. Хоботок умеренно удлинен. ХИ — 1.36—1.55
............................................С. subsylvarum Remm
65 (64). без светлого пятна. Хоботок короткий. ХИ — 1.72—1.94 (рис. 14,
5, 6) ..........................................С. arboreus Guts.
66 (23). Крылья без пятен.
67 (68). 3-й членик щупика сильно утолщен. Чувствительный орган глубоко
погруженный, в виде длинного цилиндра. Глаза без шипиков...............
.................................... С. reconditus Camp, et P.-CL
68 (67). 3-й членик щупика умеренно утолщен. Чувствительный орган в виде
неглубокой ямки. Глаза с волосками.
69 (70). Глаза разделены лобной полоской.........С. comosioculatus Tok.
70 (69). Глаза соприкасаются . . .....................С. tokunagai Агп.
ОСНОВНАЯ ЛИТЕРАТУРА
Глухова В. М. Кровососущие мокрецы (Diptera, Heleidae) Карелии. — Тр. Зоол. ин-та
АН СССР, 1962, т. 31, с. 197—249.
Гуцевич А. В. Кровососущие мокрецы (сем. Ceratopogonidae). Л., 1973. 296 с. (Фауна
СССР. Насекомые двукрылые; Т. 3; Вып. 5).
Гуцевич А. В., Глухова В. М. Методы сбора и изучения кровососущих мокрецов. —
В кн.: Методы паразитологических исследований. М.; Л., 1970. 103 с.
Иванов К. С., Глухова В. М. Новые данные по фауне кровососущих мокрецов (Diptera,
Ceratopogonidae) прибрежной части Приморского края. — Энтомол. обозрение, 1967, т. 46,
вып. 4, с. 808—813.
КРОВОСОСУЩИЕ МОШКИ (Сем. SIMULIIDAE)
Мошки мелкие (от 2—2.5 до 5—5.5 мм), темноокрашенные, иногда со свет-
лыми пятнами на спинке и лапках, двукрылые, длинноусые (подотряд Nema-
tocera) насекомые.
ВНЕШНЕЕ СТРОЕНИЕ
Тело взрослых насекомых короткое, плотное, подразделяется на голову,
грудь и брюшко (рис. 15, 1).
Голова (рис. 15, 2, 3) подогнута под грудь, по ширине несколько уже послед-
ней. Лоб расширяется вверху перед теменем и внизу перед усиками. Внизу
ко лбу примыкает почти квадратный наличник (лицо). Между лбом и налични-
ком располагаются 9—11-члениковые усики. В нижней части головы находятся
ротовые придатки, состоящие из щупиков и хоботка. Щупики 4-члениковые;
2-й членик наиболее крупный, имеет круглый или удлиненный чувствительный
40
лаутерборнов) орган; 4-й — тонкий ^длинный. Верхние и нижние челюсти
вооружены зубцами.
Диагностическое значение имеют отношение высоты и ширины лба, окраска,
?пушение и налет (или отсутствие) их на лбу и лице, количество члеников уси-
Рис. 15. Строение взрослых мошек.
1 — взрослое насекомое; 2 — голова самки; 3 — верхняя и нижняя челюсти при большом увеличении;
4 — грудь сбоку, бр — брюшко, вг — верхняя губа, г — голова, гл — глаза, гр — грудь, 1д, 2д — перед-
нее и заднее дыхальца, ж — жужжальце, к — крыло, кс^з — кокситы, кт — катэпистернум, лб — лоб,
ло — чувствительный (лаутерборнов) орган, м — мембрана, мд — мандибула, мк — максилла, мпк — место
прикрепления крыла, н — нога, на — наличник, нг — нижняя губа, пг — переднегрудь, пп — плевраль-
ные полосы, сп — спинка, стж — стволик жужжалец, у — усик, щ — щупик, щт — щиток, эпс2 — эпи-
стерн, эпм2— эпимер. (I —по: Рубцов, 1956; 2—4 — по: Stone, Jamnback, 1955).
ков, форма и окраска первых 2 из них, строение и соотношение величины члени-
ков щупиков, величина и характер строения лаутерборнова органа, количество
п расположение зубцов на верхних и нижних челюстях.
Грудь (рис. 15, 4) состоит из 3 сросшихся склеритов. Наиболее развита
среднегрудь, передне- и заднегрудь сильно редуцированы. На спинной стороне
груди различают выпуклую, хорошо развитую среднеспинку, позади которой
41
/4
Рис. 16. Детали строения мошек.
1, 2 — крылья: 1 — подсем. Prosimuliinae, 2 — подсем. Simuliinae; з, 4 — участки крыла: 3 — подсем.
Prosimuliinae, 4 — подсем. Simuliinae; 5 — голень и лапка задней ноги самки; 6 — конец задней ноги
самки; 7—12 — коготки самок разных родов: 7, 8 — Simulium, 9 — Wilhelmia, 10 — Gnus, 11 — Cnetha,
12 — Prosimulium; 13 — брюшко самки кровососущих видов; 14 — задний конец брюшка самки, апл —
анальная пластинка, бс — базистернум, бя — базальная ячейка, в — волоски, вв — ветви вилочки, вкв —
внутренний край вилочки, вув — внутренний угол вилочки, го — голень, гп — генитальные пластинки,
кп — кальципала, л — лапка, мчс — межчлениковый склерит лапок, нкв — наружный край вилочки,
пс — педисулькус, св — стволик вилочки, скл — складка крыла, су — склеротизованные участки, VIIIc —
стернит, 1Хт — тергит, ш — шипы, ц — церки. Жилки крыла: с — костальная, sc — субкостальная,
rt, г4, гл — радиальные, mlt тг — медиальные, cult си2 — кубитальные, ап — анальная, ах — ансилляр-
ная. 1,3 — по: Stone, Jamnback, 1955; 2, 4 — ориг.; 5—14 — по: Рубцов, 1956, 15 — ориг.
располагается щиток. На бочках груди отграничены швами вторичные скле-
риты: эпистерны и располагающиеся позади них эпимеры (рис. 15, 4). Между
:редним эпистерном и передним эпимером находится неправильной формы ко-
ричневая мембрана (рис. 15, 4). К мембране вблизи передней части средне-
хинки примыкает небольшое переднее дыхальце, заднее дыхальце располо-
т-но на заднеспинке у основания стебелька жужжалец (рис. 15, 4). Между 1-м
и 2-м кокситами располагается катэпистернум.
При диагностике учитывается опушение среднеспинки, наличие на ней пя-
тен и полос, цвет и опушение мембраны, катэпистернума, а также окраска
расположенных вблизи основания крыльев жужжалец.
Крылья (рис. 16, 7, 2) прозрачные, короткие и широкие, округло-овальные»
Поперечные жилки крыла почти полностью редуцированы; продольные сдви-
нуты к переднему краю. Костальная жилка не доходит до вершины крыла.
Радиальных жилок 2 или 3. Медиальные жилки и 1-я кубитальная без стебелька.
В основании медиальных жилок иногда имеется базальная ячейка. Костальная,
рубкостальная и 1-я радиальная жилки могут быть в волосках или (2 первые)
в волосках и шипиках (рис. 16, 5, 4).
Таксономическое значение имеют количество радиальных жилок, характер
: пушения первых 3 (костальной, субкостальной, 1-й радиальной), отсутствие
или наличие базальной ячейки.
Ноги (рис. 16, 5, 6) короткие, толстые, обычного для насекомых строения.
Первые членики лапок (базитарзусы) всех пар ног обычно крупные, пятые —
с коготками. Коготки простые или с зубцами у основания (рис. 16, 7—12).
На 1-х члениках (базитарзусах3) задних ног иногда имеется выступ (пятка
или кальципала), на 2-х бороздка (педисулькус). Ноги могут быть одноцветно
темными или со светлыми пятнами.
При диагностике имеет значение величина и форма первых члеников (бази-
тарзуса]) передних пар ног (отношение его наибольшей ширины к длине),
наличие или отсутствие пятки на первых и бороздки на вторых члениках лапок
задних пар ног, окраска ног и строение коготка.
Брюшко (рис. 16, 13) продолгорато-овальное, постепенно суживающееся
к заднему концу. Образовано 11 сегментами. Для кровососущих видов харак-
терно отсутствие на нижней стороне брюшка (со II по VII сегмент) хитиновых
пластинок (стернитов). Последние сегменты (с VIII по XI) преобразованы
з половые придатки: VIII сегмент представлен пластинкой базистернума
и 2 генитальными пластинками. IX сегмент преобразован в вилочку, X —
в анальные пластинки, XI — в церки (рис. 16, 14, 15). В вилочке различают
стебелек, ветви, лопасти ветвей — наружный и внутренний края и углы,
хитиновые бугорки и склеротизованные участки.
При диагностике учитывается строение базистернума, форма и расположе-
ние генитальных пластинок, стебелька и лопастей вилочки, расположение
л форма склеротизованных участков на ветвях, характер углов.
Яйцо неправильно треугольной, приближающейся к овальной формы, с за-
кругленными краями (рис. 17, 1). Размеры яйца в зависимости от вида колеб-
лются от 0.1 до 0.4 мм, ширина — от 0.1 до 0.2 мм. Оболочка яиц гладкая, без
каких-либо структур. Свежеотложенные яйца белого или светло-желтого цвета;
: развитием эмбриона становятся ярко-желтыми, желтоватыми или коричне-
выми.
Личинка червеобразная, с уплощенной брюшной и выпуклой спинной сторо-
нами, с утолщенными передними и задними отделами тела (рис. 17, 2). Окраска
тела различная: желтоватая, зеленовато-желтая, сероватая, коричневатая. Раз-
меры личинки — от 0.3—0.5 до 0.8—1.5 мм.
Голова цилиндрическая, темная, иногда черная, слегка подогнута под грудь.
На ней различают отграниченный фронтальными швами лоб и щечные скле-
риты; на границе щечного и лобного склеритов 3-члениковые усики, под ними —
боковые выросты нижней губы — веера, состоящие из целой системы щетинок.
На вентральной стороне головной капсулы расположен вентральный вырез
1рис. 17, 2).
Ротовые органы представлены сложно устроенными верхней и нижней гу-
бами, верхней и нижней челюстями, подглоточником и надглоточником.
43
Тело личинки подразделяется на 11 сегментов. Грудной отдел тела снизу
снабжен непарной «ногой», на конце с венцом крючков. У зрелых личинок
по бокам под покровами тела просвечивают свернутые в клубок дыхательные
нити куколки.
Рис. 17. Преимагинальные фазы мошек.
J — яйцо; 2 — личинка; 3 — куколка; 4 — куколка в коконе, вид сверху; 5—12 — разные формы коко-
нов: 5 — простой, 6 — с воротничком, 7 — башмаковидный, 8 — сапожковидный, 9 — бесформенный,
10 — с отверстиями по бокам, 11 — с двумя роговидными выростами, 12 — с одним роговидным выростом.
в — веера, дн — дыхательные нити, зпо — задний прикрепительный орган, лб — лоб, н — нога, см — суб-
ментум, у — усик. (По: Рубцов, 1956).
Брюшко на заднем конце имеет прикрепительный орган, состоящий из мно-
жества расположенных в ряд крючков и хитиновой рамы, а также ректальные
придатки.
Куколка полностью или частично покрыта сплетенным личинкой коконом.
Форма и характер плетения кокона разнообразны (рис. 17, 3—12). На теле ку-
44
колки имеются бляшки, кожные шипы, волоски и щетинки. Внутри кокона
находится формирующееся насекомое, которое в целом отражает строение тела
имаго, за исключением расположенного на плечевых полях дыхательного органа —
дыхательных нитей (Рубцов, 1956; Усова, 1963].
БИОЛОГИЯ
Самки большинства видов мошек имеют ротовые придатки кровососущего
типа, но не всегда нападают для кровососания на человека и животных. На
Дальнем Востоке в качестве кровососов зарегистрировано пока 27 видов мо-
шек из 3 подсемейств и 13 родов. При этом количество кровососущих видов,
число нападающих особей и сроки лёта мошек неодинаковы в различных ланд-
шафтно-географических зонах. Так, на юге советского Дальнего Востока,
в Приморском крае, в качестве кровососов зарегистрировано 10 видов и 1 под-
вид мошек. В долинах численность нападающих на человека и животных мо-
шек колеблется от 40 до 500 особей в одном 5-минутном учете. Наименьшая чис-
ленность (около 40 особей за учет) наблюдалась вблизи морских побере-
жий.
В горах нападают для кровососания в основном 2 вида: Prosimulium irritans
Rubz. и Gnus rostratum Rubz. Численность нападающих особей увеличивается
по мере подъема в горы: в лесах переходного растительного пояса она колеб-
лется от 100 до 600 особей за 5 мин, а в предгольцовой растительности и гипсо-
хтонной тундре на вершинах гор (900—1000 м над ур. м. и более) — от 900
до 3000 особей за учет. В долинах мошки нападают с мая по июль; в горах —
с конца июля до сентября.
На севере Амурской обл. (Тындинский р-н) кровососами являются 4 вида
мошек, а массовыми — только виды рода Gnus. Численность нападающих мо-
шек колеблется от 1—2 до 350 особей за 5 мин. Лёт их продолжается с конца
июня до конца августа—начала сентября {Бодрова, 1978а].
В Хабаровском крае в качестве кровососов отмечены 5 видов. Наиболее
массовым среди них был Byssodon rftaculata Mg. Максимальная численность
нападающих мошек составляла 1300—1500 особей в учете. Лёт их наблюдается
с июня по конец августа—начало сентября [Боброва, 1980].
На севере Дальнего Востока (Магаданская обл.) кровососами являются
9 видов мошек, среди которых преобладали Simulium aff. truncatum, Gnus
decimatum Dor. et Rubz. и G. rostratum Rubz. Максимальная численность
за 5 мин — в среднем около 3000 особей. Мошки активны с июня до сентября
[Боброва, 1973].
На одном из самых южных островов Курильской гряды — о-в Кунашир —
в качестве кровососов зарегистрированы 5 видов мошек. Среди них преобла-
дают по численности Simulium japonicum Mats., S. venustum Say. Численность
нападающих мошек колеблется от 20 до 170 особей за 5 мин [Бодрова, 19786].
Нападению самок мошек предшествует спаривание (копуляция). Сразу же
после кровососания у самок начинается быстрое развитие яичников и форми-
рование яиц. В зависимости от температуры окружающей среды оно продолжа-
ется от 5—8 до 10—20 дней.
Созревшие яйца самки разных видов мошек откладывают следующими
способами: 1) сбрасывают их в воду на лету (Prosimulium irritans Rubz.); 2) под
водой на камнях и палках (Eusimulium Ысогпе Radz.); 3) на урезе воды (Odag-
mia ornata Mg., Gnus rostratum Rubz., G. cholodovskii Rubz.).
Число яиц в яйцекладке одной самки колеблется от 10—50 до 500—600
и зависит от вида мошек и количества принятой самками порции крови. У боль-
шинства самок, принявших кровь, созревание яиц и их откладка происходит
один раз — один гонотрофический цикл. Реже наблюдаются два гонотрофи-
ческих цикла [Прокофьева, 1957; Рубцов, 1958; Конурбаев, 1983].
Развитие яиц, личинок и куколок (преимагинальных фаз) происходит только
в проточных водотоках, где они обычно концентрируются на участках наиболее
быстрого течения воды: на камнях перекатов, на застрявших между камнями
ветвях деревьев, на водной растительности или спускающейся с берега в воду.
45
Длительность развития отдельных фаз неодинакова у различных видов
и в различных природно-климатических районах. На юге Дальнего Востока
(Приморский край) у зимующих в фазе личинки моновольтинных (имеющих
одно поколение в году) видов (Twinnia sedecimfistulatus Rubz.) наиболее
продолжительной является эта фаза. Она составляет 7—7.5 мес., а длительность
развития яйца — около 1.5—2 мес. У моновольтинных видов, зимующих в фазе
яйца (Prosimulium irritans Rubz., Gnus rostratum Rubz.), наиболее продолжи-
тельной является эта фаза — 8.5—9 мес. и короткой фаза личинки — 1 —
1.5 мес.
Сходным образом изменяется продолжительность отдельных фаз у би-
вольтинных (имеющих два поколения в году) видов. У зимующих в фазе ли-
чинки бивольтинных видов (Simuliumiaponicum Mats.) наиболее продолжитель-
ной является фаза личинки 1-го поколения (7—7.5 мес.), а у зимующих в фазе
яйца (Gnus malyschevi clbipes Rubz., Simulium venustum Say.), напротив, фаза
яйца 1-й генерации — 6.5—7 мес. Фаза личинки 1-й генерации этих видов
составляла 1—1.5 мес. Наиболее короткой была фаза яйца (0.5—1 мес.)
2-го поколения всех бивольтинных видов.
ВРЕД
Вред, наносимый кровососущими мошками, складывается из механического
воздействия их на человека и животных и последствий укуса. Зачастую мошки,
скапливаясь на границе между одеждой и открытыми участками тела, могут
постепенно проникать под одежду, в результате чего множественные укусы
регистрируют на запястьях, голенях и других местах. Постоянное ползание мо-
шек около глаз, носа, рта чрезвычайно возбуждает людей и животных, а порой
доводит их до исступления. С попаданием мошек в дыхательные пути при вдохе
возникает долгий, мучительный кашель. У животных известны случаи гибели
в результате закупорки мошками верхних дыхательных путей. Не случайно
поэтому во время лёта мошек резко падают гфоизводительность труда людей,
занятых работой в лесу и в поле, надои и привесы у животных, яйценоскость
кур.
При укусах мошек в ранку попадает слюна, содержащая гистамин, вызы-
вающий антикоагуляцию крови и агглютинацию тканей птиц и млекопитающих.
На месте укуса мошек возникают небольшие наполненные кровью пузырьки,
кожа вокруг которых краснеет, появляется отечность, а затем сильный зуд.
При расчесах в ранку попадает микрофлора и вызывает нагноение.
Многочисленные укусы мошек являются причиной специфического заболе-
вания — симулидотоксикоза (мелузинотоксикоза). У человека при этом забо-
левании отмечают головную боль, одышку, сердцебиение, боль за грудиной,
иногда потерю сознания. Смертных случаев, как правило, не наблюдается.
У животных после массового нападения мошек появляются сильные отеки,
повышение температуры тела, серозно-пенистое истечение из носа, затрудненное
дыхание, сердцебиение. Через 6—13 ч после массового нападения мошек может
наступить гибель животного.
Однако этим не ограничивается вредоносность мошек. Доказано, что неко-
торые виды мошек являются специфическими или механическими переносчи-
ками возбудителей некоторых инфекционных и инвазионных болезней сельско-
хозяйственных животных и человека: онхоцеркоза, кровепаразитарных болез-
ней домашней птицы, сибирской язвы, сапа, туляремии, проказы, чумы и др.
Вредоносное значение кровососущих мошек возрастает с освоением новых,
ранее малонаселенных земель.
Диагноз сем. Simuliidae
Небольшие двукрылые насекомые с коренастым темным, часто со светлыми
пятнами, телом, сильно выпуклой среднеспинкой и короткими ногами. Усики
9—11-члениковые. Крылья короткие и широкие, без поперечных жилок и пя-
тен. Вилка ЛГ3_4и Сщ без стебелька. Преимагинальные фазы развиваются только
в проточной воде. Личинки фильтраторы, с крупной головой и специфическими
46
выростами нижней губы — веерами. Куколка прикреплена к субстрату, пол-
ностью (род Prosimulium) или частично (все остальные роды) покрыта коконом.
На Дальнем Востоке — 37 видов.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (4). R2 с развилком. 3 передние жилки в волосках. Кальципала на 1-м чле-
нике задней лапки отсутствует или слабо развита (рис. 16, 1, 3).
2(3). Усики 9-члениковые................... 1. Twinnia Stone et Jamnb.
3 (2). Усики 9—11-члениковые ....................2. Prosimulium Roub.
4 (1). Z?2 без развилка. 3 передние жилки в волосках или в волосках и шипи-
ках. Кальципала на 1-м членике задней лапки имеется (рис. 16, 2, 4, 6).
5 (6). Педисулькус отсутствует.....................3. Stegopterna End.
6 (5). Педисулькус имеется.
7 (16). Зубец у основания коготка крупный, иногда раздвоенный (рис. 16,77).
8 (9). Спинка с 3 продольными темными полосами .... 4. Byssodon End.
9 (8). Спинка без продольных темных полос.
10 (11). 1-й членик передней лапки уплощен и расширен; его длина в 4—6 раз
превосходит ширину................................5. Parabyssodon Rubz.
11 (10). 1-й членик передней лапки чаще цилиндрический; его длина в 7—8 раз
превосходит ширину.
12 (13). Задние края генитальных пластинок не вытянуты языковидно. Ветви
вилочки сближены, с широкими лопастями..................6. Cnetha End.
13 (12). Задние края генитальных пластинок вытянуты языковидно. Ветви
вилочки чаще широко расставлены, без широких лопастей.
14 (15). Задняя голень с темными кольцами в основании и на вершине ....
...................................................7. Chelocnetha End.
15 (14). Задняя голень с одним темным кольцом на вершине..............
.......................................... 8. Eusimulium Roub.
16 (7). Коготок с небольшим зубцом у основания или без него (рис. 16, 7—9).
17 (20). Зубец у основания коготка небольшой.
18(19). Мембрана голая. Генитальные пластинки не вытянуты языковидвс;
их медиальные края без 5-образного изгиба...............9. Gnus Rubz.
19 (18). Мембрана в волосках. Генитальные пластинки языковидно вытянуты;
их медиальные края с 5-образным изгибом.............10. Odagmia End.
20 (17). Коготок без зубца у основания.
21 (24). 1-я радиальная жилка крыла в волосках на всем протяжении.
22 (23). Спинка без серебристых пятен. Ветви вилочки и анальные пластинки
широкие..........................................11. Schoenbaueria End.
23 (22). Спинка с неявственными серебристыми пятнами. Ветви вилочки и
анальные пластинки узкие...........................12. Boophthora End.
24 (21). 1-я радиальная жилка крыла на V3—1/2 от основания без волосков
................................................. . 13. Simulium Latr.
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
1. Twinnia Stone et Jamnb. Усики 9-члениковые» щупики 4-члениковые,
короткие, толстые. Ротовые придатки по длине короче наличника. Я2 с развил-
ком. Спинка темная, в длинных золотистых волосках. Ноги сплошь черные.
Коготки на лапках простые.
1 (1). Усики 9-члениковые. Ротовые придатки кровососущего типа: мандибулы
и максиллы со слабыми рвуще-режущими зубцами. Генитальные пла-
стинки округлые. Внутренние лопасти вилочки не выражены (рис. 18,
1—4) . • • • .........................Т. sedecimfistulatus Rubz.
W 2. Prosimulium Roub. Тело темное. Z?2 с развилком, С и Sc только в волосках.
1-й членик задней лапки без пятки, 2-й — без бороздки. Лоб широкий: высота
и ширина его примерно равны. Коготок простой или с небольшим зубцом у осно-
вания. Генитальные пластинки удлиненные, со склеротизованными внутрен-
ними краями.
47
1 (4). Усики 11-члениковые. Анальные пластинки состоят из одного склерита.
2 (3). Переднебоковые края спинки красноватые. Генитальные пластинки
языковидно вытянуты, без четкого 6-образного изгиба. Наружные углы
вилочки без зазубрин (рис. 18, 5—8)................Р. erythronotum Rubz.
1—4 — Tioinnia sedecimfistulatus; 5—8 — Prosimulium erythronotum; 9 — P. irritans; 10, 11 — P. isos; 12r
13 — Stegopterna asema; 14 — S. mutata. an — анальные пластинки, бс — базистернум, в — вилочка,
гп — генитальные пластинки, гр — грудь, к — коготок, л — лапка, мд — мандибула, мк — максилла,
н — нога, у — усик, ц — церки. {1—6, 8—14 — Рубцов, 1956; 7 — ориг.).
3 (2). Спинка черная. Генитальные пластинки языковидно вытянуты, с четким
5-образным изгибом. Наружные углы вилочки с зазубринами (рис. 18, 9}
....................................................... Р. irritans Rubz.
4 (1). Усики 9-члениковые. Анальные пластинки состоят из 2 склеритов
(рис. 18, 10, 11)...........................................Р. isos Rubz.
3. Stegopterna End. Усики 11-члениковые. Тело и ноги сплошь темные.
R2 без развилка, только в волосках. Лоб расширяется кверху. 1-й членик задней
48
лапки с пяткой; 2-й — без бороздки (рис. 18, 12), Коготки с очень маленьким
ппшиком у основания. Генитальные пластинки почти прямоугольные. Вилочка
с глубокой арковидной выемкой и массивными лопастями.
1 (2). Спинка в прилегающих серебристых волосках. Ноги в серебристом
опушении. Жужжальца охряно-желтые. Анальные пластинки непра-
вильно-полулунной формы, с выступами по переднему и заднему краям
(рис. 18, 13)..........................................S. asema Rubz.
2 (1). Спинка в прилегающих золотистых волосках. Ноги в золотистом опу-
шении. Жужжальца коричневато-охряные. Анальные пластинки без
выступа по переднему краю (рис. 18, 14)................S. mutata Mall.
4. Byssodon End. Лоб узкий, в волосках, с серебристым налетом. Средне-
спинка темно-серая, без пятен, с 3 узкими, продольными темными полосками.
R2 без развилка. Ноги в основном черные. Кальципала и педисулькус имеются.
1-й членик задней лапки с параллельными краями. Генитальные пластинки
прямоугольные. Ветви вилочки узкие, короткие, расставлены широко, без вну-
тренних выростов на лопастях. На ветвях обычно склеротизованные шипы.
1 (2). Лицо и лоб серебристо-серые. Ноги сплошь черные . . В. maculata Mg.
2 (1). Лицо и лоб черные, в темно-серебристом опушении. Сочленения ног
желтоватые .........В. pseudonearctica Rubz.
5. Parabyssodon Rubz. Ноги co светлыми пятнами. 1-й членик передних
лапок уплощен: отношение его длины к ширине 3—4 : 1.
1 (1). Спинка черная, блестящая. Усики сплошь черные. Генитальные пла-
стинки прямоугольные, в редких коротких волосках. Стебелек вилочки
короткий, лопасти почти прямоугольной формы, с крепкими склероти-
зованными шипами (рис. 19, 7, 2)................. . . . Р. transiens Rubz.
6. Cnetha End. Лоб узкий, в волосках. Длина 4-го членика щупиков равна
(или несколько меньше) сумме длин двух предыдущих. R2 без развилка, в во-
лосках и шипиках. Ноги темные на всем протяжении, пятка и бороздка имеются.
Коготок сужается к концу, не удлинен, с крупным зубцом у основания. Гени-
тальные пластинки прямоугольные. Ветви вилочки широкие, лопастевидные.
1 (2). Внутренние края вилочки закруглены. Хитиновые глыбки на ветвях
в виде небольшого бугорка (рис. 19, 3) ... С. pugetense Dyar et Shan.
2 (1). Внутренние края вилочки вытянуты. Хитиновые глыбки на ветвях пальце-
видные (рис. 19, 4)..........................С. elburnum Rubz. et Carls.
3 (6). Хитиновые участки расположены в средней части лопастей ветвей ви-
лочки.
4 (5). Хитиновые участки расположены на границе с внутренними выростами
лопастей ветвей вилочки (рис. 19, 5) .... С. latipes shutovae Rubz.
5 (4). Хитиновые участки расположены на некотором расстоянии от внутрен-
них выростов лопастей ветвей вилочки (рис. 19, 6)......................
....................................... С. bicorne Dor. et Rubz.
6 (3). Хитиновые участки расположены вблизи основания рукоятки вилочки
на переднебоковом краю лопастей.
7 (8). Хитиновые выросты направлены от основания рукоятки вилочки
(рис. 19, 7)..................................С. curvans Rubz. et Carls.
8 (7). Хитиновые выросты направлены к основанию рукоятки вилочки
(рис. 19, 8)..........................................С. amurense Rubz.
7. Chelocnetha End. Спинка без полос. R2 без разветвления, в волосках и
шипиках. Лоб расширяется кверху. Спинка серовато-черная, иногда с вытяну-
тыми темными пятнами. Ноги коричневато-черные, причем темные участки че-
редуются с более светлыми. Кальципала и педисулькус имеются. Коготок
с крупным зубцом у основания. Генитальные пластинки языковидно вытянуты
и округлены. Анальные пластинки серповидные. Ветви вилочки узкие.
1 (1). Ноги коричневые; у основания голеней темное кольцо. Ветви вилочки
узкие (рис. 19, 9).....................................С. brunnea Jank.
4 Зак. Кв 990
49
Рис. 19. Детали строения мошек.
1, 2 — Parabyssodon transient 3 — Cnetha pugetense\ 4 — C. elburnum', 5 — C. latipes schutovae; в — С. bi-
come; 7 — C. curvans', 8 — C. amurense', 9 — Chelocnetha brunnea\ 10, 11 — Eusimulium aureum. (1—7, 10,
11—по: Рубцов, 1956; 8— ориг.; 9— по: Янковский, 1977). Остальные обозначения как на рис. 18.
5. Eusimulium Roub. Спинка темная, без полос. Максиллы зазубрены
зной стороны. 4-й членик щупиков длиннее 2-го и 3-го вместе взятых. Ноги
I значительном протяжении желтые; голени задних ног затемнены только
з з?ршпне. 1-й членик задней лапки удлинен и сужен; его длина превосходит
з’тнну в 6—8 раз (рис. 19, 11).
- — Gnus malyschevi albipes] 3 — G. subvariegatum', 4 — G. decimatum', 5, 6— G. cholodkovskii; 7, 8 — G.
z-2-lovskii; 9, 10 — G. rostratum. (По: Рубцов, 1956). Остальные обозначения как на рис. 18.
!.». Спинка в светло-золотистых волосках. 2 первых членика усика коричне-
вые. Задние концы генитальных пластинок языковидно вытянуты; ло-
пасти ветвей вилочки не выражены (рис. 19, 10) . . . . Е. aureum Fries
?. Gnus Rubz. Спинка с неявственными, не соприкасающимися серебри-
—уи пятнами. Мембрана голая. 7?х на 3Д с волосками, в вершинной половине
—кликами. Ноги всегда со светлыми пятнами. 1-й членик передней лапки
"<^генно расширен и уплощен: его длина относится к ширине как 5:1. Зубец
- хнования коготка небольшой, приостренный. Генитальные пластинки по-
-1з,тены косо к базистернуму или прямоугольные. Анальные пластинки мо-
т быть с узкой выемкой по нижнему краю. Ветви вилочки широко расстав-
со склеротизованными шипами.
4*
51
1 (6). Генитальные пластинки расположены косо по отношению к базистер-
нуму.
2 (3). Голень3 затемнена только на вершине. Хитиновые глыбки на ветвях
вилочки направлены вниз (рис. 20, 1,2).. G. malyschevi albipes Rubz.
3 (2). Голень3 темная почти на всем протяжении. Косо срезанная светлая
полоска проходит лишь по заднему краю. В основании ветвей вилочки
бесформенные глыбки хитина (рис. 20, 3) . . . G. subvariegatum Rubz.
4 (5). Голень3 светлая на х/3 от основания. Наружные углы ветвей вилочки без
зазубрин (рис. 20, 5,6)............................G. cholodkovskii Rubz.
5 (4). Голень3 светлая менее чем на V3 от основания. Наружные углы ветвей
вилочки с зазубринами (рис. 20, 4)............G. decimatum Dor. et Rubz.
6 (1). Генитальные пластинки расположены почти под прямым углом по отно-
шению к базистернуму. Спинка в редких золотистых волосках.
7 (8). Спинка в редких золотистых волосках. Голень3 светлая более чем на
V2 от основания. Наружные углы ветвей вилочки с частыми мелкими за-
зубринами. Анальные пластинки почти квадратные (рис. 20, 9, 10) . . .
................................................... G. rostratum Rubz.
8 (7). Спинка в редких серебристых волосках. Голень3 светлая на 3/4 от осно-
вания. Наружные углы вилочки с более редкими крупными зазубринами.
Анальные пластинки неправильно прямоугольные (рис. 20, 7,8) . . .
................................................... G. pavlovskii Rubz.
10. Odagmia End. Лоб широкий. Спинка с хорошо выраженными, почти
соприкасающимися светлыми пятнами. Мембрана в волосках. Ноги со светлыми
пятнами. Пятка и бороздка хорошо выражены. Коготок с более крупным (чем
у видов предыдущего рода), дальше отставленным зубцом. Генитальные пла-
стинки языковидно вытянуты, внутренние края их S-образно изогнуты. Аналь-
ные пластинки вытянуты, с намечающимся делением на две доли. Ветви вилочки
узкие, широко расставленные, со склеротизованными шипами.
1 (1). Зубец у основания коготка более крупный и дальше отставленный.
Мембрана в волосках. Генитальные пластинки с выемкой по внутреннему
краю. Серебристые пятна на спинке почти соприкасаются . О. о гпа t a Mg.
11. Schoenbaueria End. Среднеспинка без полос и пятен. Мембрана голая.
Я/на всем протяжении в волосках. Лоб в волосках. Ноги темные. Коготок ко-
роткий, без зубца у основания. 1-й членик передней лапки слегка уплощен
и расширен: отношение длины к наибольшей ширине 5—6 : 1. Генитальные
пластинки прямоугольные, в редких волосках. Ветви вилочки сближены,
с массивным хитиновым бугорком.
1 (2). Генитальные пластинки длинные: почти достигают середины арковидной
выемки вилочки. Анальные пластинки с сильно вытянутым заднелатераль-
ным краем (рис. 21, 7)..................................Sch. pusilia Fries
2 (1). Генитальные пластинки короткие: не достигают середины арковидной
выемки вилочки. Анальные пластинки с менее вытянутым заднелатераль-
ным краем (рис. 21, 2).............................Sch. subpusilia Rubz.
12. Boophthora End. Лоб широкий, блестящий, по высоте превосходит
ширину в 1.2—1.5 раза. Спинка с неявственными серебристыми пятнами и по-
лосками. Мембрана голая. 7?х на всем протяжении в волосках. Коготок без зуб-
чика. Генитальные пластинки прямоугольные; анальные — узкие, серповид-
ные. Ветви вилочки тонкие, широко расставлены, с хитиновыми выростами.
1 (1). Спинка черная, блестящая. Ноги в большей части темные. Ветви вилочки
и анальные пластинки узкие (рис. 21, 5) . .В. erythrocephala De Geer
13. Simulium Latr. Спинка с пятнами на плечевых полях или с неясными
полосами на темном или сером фоне. Мембрана голая. 7?х голая наполовину
от основания. Ноги со светлыми пятнами. Коготки чаще простые, реже с зуб-
чиком или утолщением у основания. Генитальные пластинки довольно короткие,
52
широко расставлены у основания, ближе к прямоугольным, с закругленным
внутренним краем. Анальные пластинки округлые или вытянутые. Ветви ви-
лочки узкие, широко расставленные, с хитиновыми выростами.
Рис. 21. Детали строения мошек.
1 — Schoenbaueria pusilia} 2 — Sch. subpusilia', 3 — Boophthora erythrocephala', 4 — Simulium tuberosum',
3 — S. vulgare; 6—8 — S. japonicum; 9, 10 — S. honakovi; 11, 12 — S. kurilense. (По: Рубцов, 1956).
Остальные обозначения как на рис. 18.
1 (4). Лоб и лицо черные.
2 (3). Светлые пятна на передних и задних ногах. Хитиновые выросты ветвей
вилочки направлены вовнутрь (рис. 21, 4).............S. tuberosum Lund.
3 (2). Светлые пятна на средних и задних ногах. Хитиновые выросты ветвей
вилочки направлены наружу (рис. 21, 5)................S. vulgare Rubz.
4 (1). Лоб черный, блестящий, лицо чаще в сероватом налете.
53
5 (10). Спинка с продольными полосами (рис. 21, 6).
6 (9). Анальные пластинки с отростком (рис. 21, 7). 2 первых членика усиков
красноватые.
7 (8). Бедра передних и средних пар ног светло-желтые, задних — желтые
(рис. 21, 8).........................................S. japonicum Mats.
8 (7). Бедра передних и средних пар ног темные; задних — желтоватые у вер-
шины (рис. 21, 9, 10)................................S. konakovi Rubz.
9 (6). Анальные пластинки без отростков. 2 первых членика усиков охряно-
желтые. Бедра, голени, базитарзусы2 3 со светлыми пятнами (рис. 21,
11, 12)..............................................S. kurilense Rubz.
Рис. 22. Детали строения мошек.
1 — Simulium verecundum’, 2, 3 — S. venustum; 4 — S. truncatum; 5 — S. morsitans; 6 — S. curvitarse; 1 —
S. longipalpe. (По: Рубцов, 1956). Остальные обозначения как на рис. 18.
10 (5). Спинка без продольных полос; на плечевых полях спинки неправиль-
ной формы пятна.
11 (18). 4-й членик щупиков короче 2-го и 3-го члеников вместе взятых.
12 (13). Анальные пластинки с добавочной долей (рис. 22, 1)............
....................................S. verecundum Stone et Jamnb.
13 (12). Анальные пластинки без добавочной доли.
14 (17). Генитальные пластинки прямоугольные, в крупных редких волосках.
Внутренние края без или со слабым 5-образным изгибом (рис. 22, 2).
15 (16). Голень3 светлая более чем на х/2. Граница между темным и светлым
участками четкая (рис. 22, 3).........................S. venustum Say.
15 (15). Голень3 светлая менее чем на х/2. Граница между темным и светлым
участками не четкая (рис. 22, 4) ......... S. truncatum Lund.
17 (14). Генитальные пластинки прямоугольные, в многочисленных и слабых
волосках. Внутренние края с более выраженным S-образным изгибом
(рис. 22, 5)..........................................S. morsitans Edw.
18 (11). 4-й членик щупиков длиннее 2-го и 3-го члеников вместе взятых.
19 (20). Анальная пластинка с вырезом по переднему краю (рис. 22, 6) . . .
.................................,............. S. curvitarse Rubz.
20 (19). Анальная пластинка без выреза по переднему краю; край неровный
(рис. 22, 7)..........................................S. longipalpe Belt.
54
ОСНОВНАЯ ЛИТЕРАТУРА
Боброва С. И. Фауна и экология мошек (Diptera, Simuliidae) бассейна р. Анадырь Ма-
гаданской области. — Паразитология, 1973, т. 7, вып. 5, с. 457—464.
Боброва С. И. Мошки (Diptera, Simuliidae). — В кн.: Паразитические насекомые и
клещи Сибири. Новосибирск, 1980, с. 28—47.
Бодрова Ю. Д. Особенности фауны мошек (Diptera, Simuliidae) юга острова Кунашир. —
В кн.: Новые данные о насекомых Сахалина и Курильских островов. Владивосток, 1978а,
с. 102—124. (Тр. Биол.-почв. ин-та. Нов. серия, т. 50 (153)).
Бодрова Ю. Д. Видовой состав и особенности экологии мошек (Simuliidae) в районе
строительства БАМа. — Паразитология, 19786, т. 12, вып. 3, с. 233—239.
Конурбаев Э. О. Особенности биологии и экологии мошек (Diptera, Simuliidae) Сред-
ней Азии. — Энтомол. обозрение, 1983, т. 62, вып. 4, с. 702—707.
Прокофьева К. К. О гонотрофическом цикле кровососущих мошек Восточной Сибири.—
Зоол. журн., 1957, т. 36, № 12, с. 1831—1840.
Рубцов И. А. Мошки (сем. Simuliidae). М.; Л. 1956. 859 с. (Фауна СССР. Насекомые
двукрылые; Т. 6; Вып. 6).
Рубцов И. А. Гонотрофический цикл у кровососущих мошек. — В кн.: Паразитол. сб.
Зоол. ин-та АН СССР, 1958, т. 18, с. 255—282.
Усова 3. В. Фауна мошек Карелии и Мурманской области (Diptera, Simuliidae). М.;
Л., 1961. 286 с.
Янковский А. Б. Новые виды мошек (Diptera, Simuliidae) из Хабаровского края. —
В кн.: Систематика и фаунистика насекомых. Л., 1977, с. 96—102.
Stone A., Jamnback Н. The blackflies of New York State (Diptera, Simuliidae). — N. Y.
State Mus. Bull., 1955, N. 349. 144 p.
СЛЕПНИ (Сем. TABANIDAE)
Слепни относятся к подотряду короткоусых двукрылых (Brachycera) к группе
прямошовных (Orthorapha). Самки являются факультативными (периодиче-
скими) кровососами, нападающими на людей и животных; самцы — раститель-
ноядны. Слепней нередко путают с оводами, от которых они легко отличаются
яркоокрашенными глазами и хорошо развитым колюще-сосущим ротовым аппа-
ратом, который у оводов редуцирован.
Слепни распространены повсеместно, за исключением Антарктиды, Грен-
ландии и Исландии. Наиболее пышного расцвета они достигают во влажных
тропиках. В мировой фауне известно 3119 видов, в СССР — 189 видов из 13 ро-
дов, а на Дальнем Востоке — 60 видов и подвидов, относящихся к 2 подсемей-
ствам и 5 родам. Наиболее богатый видовой состав слепней на юге Дальнего
Востока. По мере продвижения к северу и с поднятием в горы он сильно обед-
няется и не только за счет исчезновения теплолюбивых видов, но и целых родов.
Так, если в Приморском крае отмечено 48 видов и 1 подвид из 5 родов, то
в Амурской обл. — 39, в Хабаровском крае — 36, в Магаданской обл. (в южной
половине) — 20, в разных местах Чукотской тундры и лесотундры 1—8 из 2 ро-
дов, на Камчатке — 13 видов из 3 родов. Островная фауна слепней также бед-
нее материковой: на о-ве Сахалин обитает 22 вида, а на Курильских островах
(в основн м на южном Шикотане) — 15 [Виолович, 1968; Соболева, 1971, 1974,
1978; Соболева, Петрова, 1973; Олсуфьев, 1981; Соболева, Попов, 1983].
ВНЕШНЕЕ СТРОЕНИЕ
В настоящее время видовое определение слепней ведется преимущественно
по взрослым насекомым, так как личинки и куколки пока слабо изучены.
Слепни — это средних или крупных размеров мухи (от 6—7 до 28—30 мм),
тело которых окрашено в черно-серые, черно-желтые, коричнево-черные или
желтовато-палевые (род Atylotus) тона.
Голова довольно крупная, по ширине равна груди или шире ее (рис. 23).
Большую часть головы занимают сложные фасеточные глаза. У слепней родов
Chrysops и Haematopota на глазах имеются различной конфигурации переливча-
тые волнистые полосы и пятна, у остальных трех родов (Hybomitra, Atylotus,
Tabanus) от 1 до 4 темных поперечных полос, но у большинства видов рода Ta-
banus они отсутствуют. Самок легко отличают от самцов по более или менее
широкой лобной полоске, разделяющей глаза; у самцов глаза соприкасаются.
Важное диагностическое значение имеют форма, цвет и характер поверхности
55
Рис. 23. Детали строения слепней.
1 — общий вид слепня Hybomitra. Обозначения на крыле по Н. Г. Олсуфьеву (1937). Жилки: с — ко-
стальная, sc — субкостальная, г — радиальные, т — медиальные, си — кубитальные, ап — анальная,
ар — придаток т4. Ячейки: С — костальная, S — субкостальная, R — радиальные, Р — заднекрайние,
D — дискоидальная, В — базальные (Bt — передняя и В2 — задняя базальные ячейки), А — анальная;
ш — шпоры; 2 — последние членики лапки Chn/sops: к — коготок, п — пульвиллы, э — эмподий (по:
Олсуфьев, 1937); 3 — голова Hybomitra lundbechi (усики удалены): гб — глазковый бугорок, гл — глаза,
лб — лобная полоса, лт — лобный треугольник, н — наличник, нлм — нижняя лобная мозоль, слм —
срединная лобная мозоль, щ — щупальца, щк — щеки, х — хоботок; 4—11 — усики: 4 — Chrysops diss&c-
tus, 5 — Ch. vanderwulpi, 6 — Tabanus buddha, 7 — Hybomitra brevis, 8 — Haematopota tamerlani, 9 —
H. desertorum, 10 — H. koryoensis, 11 — Haeptatoma (виды этого рода на ДВ отсутствуют); д — дорсальный
угол 3-го членика усиков. (По: Соболева, 1974).
нижней и срединной лобных мозолей, а также ширина и высота лобной полосы.
Последняя покрыта налетом или волосками, которые могут частично или пол-
ностью закрывать лобные мозоли.
Ниже лобной полоски располагается лобный треугольник, в нижней части
которого имеются трехчлениковые усики; 3-й членик их самый крупный, с ши-
рокой площадкой (центральная часть) и дорсальным углом. Заканчивается
Рпс. 24. Личинка, куколка и яйцекладка слепней.
I — общий вид личинки Haematopota tamerlani с дорсальной стороны; 2 — последний сегмент тела ли-
чинки сбоку; 3 — общий вид куколки того же вида сбоку; 4 — задний конец куколки («розетка») сбоку;
5 — яйцекладка Hybomitra tarandina, (По: Соболева, 1977).
он палочкой, состоящей из 3—7 колец (у слепней Дальнего Востока палочка
имеет только 3—4 кольца). Форма усиков и соотношение длины их члеников
неодинаковы у слепней разных родов. У большинства слепней лицо и щеки по-
крыты матовым налетом и волосками различного цвета, но у представителей
родов Chrysops и Pangonia (на Дальнем Востоке последний род отсутствует)
на лице и щеках имеются оголенные пятна хитина, носящие название лицевых
мозолей. У отдельных видов (например, Ch, japonicus) лицо целиком лаково-
блестящее, без налета.
По бокам хоботка располагаются парные двучлениковые нижнечелюстные
щупики. Форма, окраска и опушенность волосками конечного членика щупиков
имеют большое диагностическое значение.
Грудь обычного для насекомых строения. Большое значение для видового
определения слепней имеют цвет нотоплевр, а также наличие волосков, их
57
окраска и характер расположения на боковых сторонах (плеврах) грудиг
которые состоят из отдельных склеритов.
Крылья широкие, прозрачные или слегка затененные (роды Hybomitra,
Atylotus, Tabanus) с темными перевязями (род Chrysops) или сетчатым «мрамор-
ным» рисунком (род Haematopota), Жилкование у представителей всех родов
однотипное (рис. 23, 7). У основания крыла снизу имеется небольшое прозрач-
ное крылышко и парные закрыловые чешуйки. Сзади основания крыльев
на груди прикрепляются жужжальца, являющиеся рудиментами задней пары
крыльев.
Ноги обычного для насекомых строения. На лапках по 2 коготка и три рав-
новеликие присоски (пульвиллы и между ними — эмподий). При видовом опре-
делении слепней имеет значение окраска ног.
Брюшко широкое, сжатое в дорсовентральном направлении, состоит из 7 ви-
димых члеников. Окраска его у разных видов неодинакова, что имеет диагно-
стическое значение.
Яйцо. Яйца слепней мелкие, удлиненные, с зауженными концами. У разных
видов длина яиц от 1.23 до 2.65 мм, толщина — 0.19—0.55 мм. Яйца обычно
откладываются самками в виде компактных 2—5-слойных пирамидок или 1—2-
слойных бляшек белого, желтоватого, сероватого, коричневого или черного
цвета. В зависимости от вида слепня в одной яйцекладке может быть от 50—
100 до 600—1200 яиц (рис. 24, 5).
Личинка. Тело тонкое, веретеновидное, с зауженными концами, состоит
из 12 сегментов и головы, которая может втягиваться внутрь (рис. 24, 1, 2).
Длина личинок разного возраста и вида — от 0.5 до 10—40 мм. Внешние по-
кровы у разных видов белые, кремовые, зеленоватые, розовато-коричневатые.
На заднем конце тела имеются узкая дыхательная трубка и анальное отверстие.
На брюшной стороне расположены валикообразные утолщения и бородавчатые
выступы кутикулы, служащие для передвижения.
Куколка слепня напоминает куколку бабочки (рис. 24, 3, 4). Тело ее состоит
из неподвижной головогруди и подвижного брюшка из 8 сегментов. У молодой
куколки покровы тела светлые и сравнительно мягкие, у зрелой хорошо хитини-
зированные, от светло-коричневатого до бурого и темно-коричневого цвета.
Голова у дальневосточных видов без шипов и выступов. На ней имеются неболь-
шие теменные бугорки, покрышки усиков и швы. На грудном отделе видны
грудные дыхальца, зачатки крыльев и ног. Дыхальца есть и на каждом членике
брюшка. Конец брюшка с 6 выростами, образующими своеобразную розетку.
Куколки разных видов слепней трудно различимы, систематика их не разра-
ботана.
БИОЛОГИЯ
Слепни — свето- и теплолюбивые насекомые и поэтому наиболее активны
в ясную, солнечную погоду, с 7—8 до 20—21 ч. С наступлением темноты лёт их,
как правило, прекращается. Выплаживаются слепни во временных и постоян-
ных водоемах и на переувлажненных пойменных лугах. На хорошо освоенных
человеком территориях места выплода слепней сосредоточены в основном
вокруг крупных животноводческих хозяйств, так как сельскохозяйственные
животные (главным образом крупный рогатый скот) являются основными про-
кормителями слепней. В неосвоенных лесах места выплода рассеяны диффузно
[Соболева, 1977].
Наши исследования показали, что в разных ландшафтно-географических
зонах Дальнего Востока активность, численность и сроки лёта слепней неодина-
ковы [Соболева, 1974]. Наиболее многочисленны слепни в хорошо освоенных
человеком районах лесной и таежной зон, на юге этого региона (особенно
в Приморском крае), где благодаря мягкому климату численность слепней
в долинных широколиственных лесах достигает колоссальных показателей:
на одно животное в стаде нападает от 150 до 700 слепней за 15 мин. В нетронутой
человеком или слабоосвоенной тайге, где нет животноводческих хозяйств, чис-
ленность слепней в 3—20 раз меньше. По мере продвижения к северу и с подня-
тием в горы постепенно сокращается их численность. Здесь массовое размноже-
ние слепней сдерживается суровыми климатическими условиями. На гольцах
58
и в горных тундрах слепни единичны. Сравнительно мало их также в зонах
тундры и лесотундры (Чукотский автономный округ), где вдали от оленьих
стад, в разгар лёта в одном учете регистрируется 10—40 экз. этих кровососов.
В тундре и лесотундре слепни концентрируются около оленьих стад, но даже
и на оленях их во много раз меньше, чем в лесной и таежной зонах юга Дальнего
Востока.
Лёт слепней на разных географических широтах Дальнего Востока продол-
жается от 1 до 4 мес.; наиболее продолжительный он в южных регионах (Амур-
ская обл., Приморский и Хабаровский края).
Слепни относятся к насекомым с полным превращением. Они проходят
3 фазы развития: яйцо, личинка, куколка. После полного насыщения кровью
у самок происходит синхронное переваривание крови и созревание яйцевых
фолликулов (гонотрофическая гормония), которое в зависимости от температуры
продолжается от 5—10 до 15—32 дней [Соболева, 1977].
Самка откладывает яйца на растительность во влажных местах (берега
рек, озер, прудов, больших луж, канав, ям, на болотах и т. п.). Из яиц через 5—
18 дней вылупляются личинки, которые падают в воду или во влажную почву,
где развиваются и растут.
Личинки большинства видов слепней — хищники, питающиеся мелкими
личинками насекомых, дождевыми червями и другой доступной им живностью.
В течение жизни личинки разных видов слепней линяют 5—12 раз, имеют
VI—XIII возрастов, зимуют 2—4 раза [Соболева, 1977]. Ранней весной перед
окуклением они перебираются в более сухие места, где и превращаются в ку-
колку, развитие которой продолжается от 6 до 30 дней. Отрождение слепней
из куколок происходит в жаркие, солнечные дни. Продолжительность жизни
слепней — в среднем 20—30 дней. Весь жизненный цикл от яйца до взрослого
насекомого (онтогенез) у разных видов слепней продолжается 2—5 лет, реже —
1 год.
ВРЕД
Слепни причиняют значительные экономические убытки животноводству и
сильно досаждают людям. Уколы слепней очень болезненны, поэтому их массо-
вое нападение вызывает перевозбуждение нервной системы. Животные испыты-
вают значительные кровопотери вследствие высасывания самками крови,
а также из-за вытекания ее из ранок (в слюне слепней содержатся антикоагули-
рующие вещества, препятствующие свертыванию крови).
Слюна слепней, вводимая в ранку в момент укола, весьма токсична и вызы-
вает неприятные болезненные ощущения, местный воспалительный процесс и
общую интоксикацию [Павловский и др., 1932; Соболева, 1974].
Слепни препятствуют нормальному выпасу скота, вызывая снижение удоев
(на 20—40 %) и привесов молодняка, снижается также работоспособность ло-
шадей.
В тундре и лесотундре слепни в комплексе с другими компонентами гнуса
(комарами, мошками, мокрецами) причиняют значительные экономические
убытки оленеводству. В лесу, где нет поблизости основных прокормителей
этих кровососов — стад крупного рогатого скота, — они не менее активно ата-
куют бригады лесозаготовителей и другие коллективы людей, работающие
в тайге. Производительность труда при этом заметно снижается, появляется
нервозность, учащаются травмы. Мешают они и нормальному отдыху.
Особую опасность слепни представляют как потенциальные механические
(реже — биологические) переносчики возбудителей болезней человека и живот-
ных: туляремии, сибирской язвы, эмфизематозного карбункула крупного рога-
того скота, инфекционной анемии лошадей, трипанозомозов, гемоспорадиозов,
филяриотозов и др.
Диагноз сем. Tabanidae
Тело коренастое, покрыто мелкими волосками, без грубых щетинок. Мухи
средних или крупных размеров (10—30 мм), реже мельче (6—7 мм).
Голова крупная, полушаровидная. Глаза у живых насекомых ярко окра-
шены. Усики прямые, трехчлениковые; палочка 3-го членика из 3—7 колец,
59
ариста отсутствует. Щиток по заднему краю без шипов. Брюшко из 7 сегментов,
реже из 6. Крылья большие, костальная жилка обходит не все крыло; радиаль-
ные жилки не сдвинуты к переднему краю крыла. 4-я заднекрайняя ячейка
преимущественно открытая, анальная ячейка обычно замкнутая. Жужжальца
и закрыловые чешуйки хорошо развиты. Лапки с хорошо развитой срединной
присоской (эмподием). Средние и задние голени не всегда со шпорами. Самки —
кровососы, самцы растительноядны, личинки развиваются в воде и почве.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ 1 (рпс. 25)
1 (2). Задние голени на вершине с концевыми шпорами (шипами). На темени,
как правило, имеются 3 глазка. Лицевые мозоли хорошо развиты. Крылья
в большинстве случаев с коричнево-черным рисунком из поперечных
перевязей и пятен.....................................1. Chrysops Mg.
2 (1). Задние голени на вершине без шпор. Темя без глазков, но иногда имеется
небольшой глазковый бугорок. Лицевые мозоли отсутствуют. Крылья
имеют иной рисунок.
3 (4). Крылья с сетчатым рисунком («мраморные»). Мелкие виды с узким телом:
6—12.5.............................................5. Haematopota Mg.
4 (3). Крылья прозрачные или слегка затемнены, но без сетчатого рисунка.
Чаще всего крупные коренастые мухи: 12—28, очень редко мельче.
5 (6). Глаза бледные охряно-желтые или с салатным оттенком, с тонкой, не
достигающей внешнего края полоской или без нее; у сухих экземпляров
окраска глаз обычно сохраняется или приобретает коричневатый оттенок.
У $ мозоли на лбу лишь в виде рудиментарных округлых точек или отсут-
ствуют совершенно. Светло-серые или охряно-желтые небольшие мухи
(9—16) ..................................3. Atylotus Ost.-Sack.
6 (5). Глаза яркие, изумрудно-зеленые, синие, коричневые, с полосками или
без них; у сухих экземпляров глаза становятся черными, у д иногда
двухцветные: верхняя часть с крупными фасетками остается светлой,
нижняя мелкофасеточная часть темнеет.^ У $ мозоли на лбу, как правило,
хорошо развиты. Черно-серые или черно-коричнево-желтые виды различ-
ных размеров (10—20).
7 (8). Темя имеет б. м. блестящий глазковый бугорок (или его рудимент);
глаза покрыты волосками (в редких исключениях у $ волоски плохо
заметны) и, как правило, с 3 полосками.............2. Hybomitra End.
8 (7). Глазковый бугорок на темени отсутствует; иногда он имеется у д, но
целиком покрыт налетом; глаза голые или в волосках, число полосок на
глазах варьирует от 1 до 4 или последние отсутствуют совершенно
(у дальневосточных видов отсутствуют)..................4. Tabanus L.
Подсем. CHRYSOPSINAE
Шпоры на задних голенях хорошо развиты. Как правило, имеются 3 темен-
ных глазка. 3-й членик усиков имеет 5 сегментов. Базикоста крыла голая,
без щетинок.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ САМОК СЛЕПНЕЙ
1. Chrysops Mg. — златоглазики или пестряки. Средних размеров мухи
(7—12.5), тело сравнительно узкое, стройное; брюшко обычно с ярким черно-
желтым рисунком, реже целиком черное. Дорсальный (угловидный выступ)
угол 3-го членика усиков почти не выражен. Нижняя часть головы с хорошо
развитыми лицевыми мозолями (блестящие участки хитина). Крылья пестрые
за счет темных перевязей и пятен на бесцветном фоне. — 12 видов (в СССР —
27), из которых 11 относится к подроду Chrysops и 1 вид — к Heterochrysops.
1 Помимо собственных материалов при составлении определительных таблиц нами
использовались таблицы и описания Н. Г. Олсуфьева [1937, частично 1977], X. Такахаси
[Takahasi, 1962], В. Мурдоча и X. Такахаси [Mardoch, Takahasi, 1969].
60
Рис. 25. Представители разных родов слепней и детали их строения.
1 — Hybomitra nigella; 2 — Chrysops suavis; 3 — Tabanus buddha; 4 — Aiylotus miser; 5 — Haematopota ta-
merlani; 6 — голова Chrysops suavis. гл — глаза, г — глазки, лм — лицевые мозоли, нлм — нижняя лоб-
ная мозоль, им — наличниковая мозоль, щм — щечные мозоли, щ — щупалыта, х — хоботок. (По: Со-
болева, 1974).
1 (22). Дискоидальная ячейка одноцветно темная (изредка слегка просвет-
лена).
2 (3). Лицо блестяще-черное, лицевые и щечные мозоли слиты вместе. Грудь,
брюшко и ноги целиком черные. Вершинное пятно крыла соединено
с перевязью очень узкой перемычкой, не превосходящей 1/2 ширины
ячейки Z?r — Лесной вид. Немногочислен (рис. 26, 1)......................
..................................... Ch. (s. str.) japonicus Wied.
Рис. 26. Детали строения слепней рода Chrysops.
1 — Ch. japonicus; 2,3 — Ch. anthrax; 4, 5 — Ch. divaricatus; 6,7 — Ch. nigripes; 8, 9 — Ch. maherovi.
1, 2, 4,7,9 — крыло; 3, 5,6,8 — брюшко. (2—5 — по: Олсуфьев, 1977; остальное — по: Соболев;),
197 4).
3 (2). Лицевые и щечные мозоли явственно обособлены между собой окружаю-
щим их желтым или серым налетом. По крайней мере II тергит брюшка
по бокам желтоватый. Остальные признаки иные.
4 (5). Вершинное пятно крыла не достигает г4 и очень узкое, не шире половины
ячейки (рис. 26, 2, 3)......................Ch. (s. str.) anthrax Ols.
5 (4). Вершинное пятно достигает г4 и обычно не уже ширины ячейки Rv
6 (15). Голени черные, лишь иногда средние голени в основании коричневые.
7 (10). Вершинное пятно крыла узкое, его вершина налегает на г4 не более
чем на V2 ее длины. Внешний край темной перевязи крыла с резким вы-
ступом в ячейке Z?2.
8 (9). II тергит брюшка преимущественно желтый с черным пятном, похожим
на латинскую букву V. III стернит брюшка желтый, с черным поперечным
пятном посредине. Крупнее: 9—12. — Таежный вид. Редок (рис. 26, 4, 5)
..........................................Ch. (s. str.) divaricatus Lw.
9 (8). II тергит брюшка преимущественно черный с неявными желтыми пят-
‘62
нами по бокам и небольшим серым треугольником посредине. III стернит
брюшка черный. Мельче: 7—11. — Таежный вид. Обычен (рис. 26, 6, 7)
...................................... Ch. (s. str.) nigripes Ztt.
10 (7). Вершинное пятно крыла широкое, налегает на г4 явственно больше, чем
на г/2 ее длины. Внешний край темной перевязи крыла ровный или лишь
с небольшим уголком в ячейке /?2.
Рис. 27. Детали строения слепней рода^Chrysops.
1 — Ch. caecuiiens caecutiens’, 2,3 — Ch. validus', 4,5 — Ch. suavis suavis; 6,7 — Ch. angaricus; 8, 9 — Ch.
dissecius', 10, 11 — Ch. (H.) vanderumlpi. 1, 2, 4, 6, 8, 10 — брюшко; 3, 5, 7, 9, 11 — крыло. (J, 6,7 —
по: Олсуфьев, 1977; остальное — по: Соболева, 1974).
11 (14). Вершинное пятно крыла соединено с перевязью перемычкой по всей
вершине ячейки Rv
12 (13). Брюшко (начиная с III тергита) с резкими светлыми каемками по зад-
нему краю тергитов. II тергит брюшка желтый с крупным раздвоенным
черным пятном, широко примыкающим к I тергиту. 10—12.5. — Таежный
вид. Немногочислен (рис. 26, S, 9)............Ch. (s. str.) makerovi Pl.
13 (12). Тергиты брюшка не имеют светлых каемок. II тергит брюшка желтый,
посредине с черным пятном, похожим на перевернутую букву V. III тер-
63-
гит целиком черный. 7.5—11. — Лесной подвид (рис. 27, 7)..........
. .......................... Ch. (s. str.) caecutiens caecutiens L.
14 (11). Вершинное пятно крыла явственно обособлено от перевязи просветом
в ячейке Rv II тергит брюшка желтый с крупным раздвоенным черным
пятном посредине. 10—11. — Восточносибирский таежный вид (рис. 27,
2, 3)...........................................Ch. (s. str.) validus Lw.
15 (6). По крайней мере голени средней пары ног коричневые или желтые;
II тергит брюшка желтый.
16 (21). Голени задних ног черные. Вершинное пятно крыла узкое, не шире
ячейки Z?4. Своей вершиной оно лишь незначительно налегает на жилку г4.
Черные пятна на II тергите в виде продольных полос или остроконечных
треугольников.
17 (20). II тергит брюшка посредине с двумя продольными пятнами, продол-
жающимися на последующих тергитах в виде двух параллельных полос,
разделенных желтой полосой.
18 (19). Черные продольные пятна II тергита не достигают заднего края I тер-
гита. III и IV тергиты обычно по бокам с желтыми пятнами. 7.9—9.6. —
Дальневосточный лесной , вид. Обычен (рис. 27, 4, 5)...................
...................................Ch. (s. str.) suavis suavis Lw.
19 (18). Черные продольные пятна II тергита доходят до I тергита и сливаются
с его черным пятном. III и последующие тергиты по бокам черные. 9—
10. — Лесной островной подвид. Обычен .................................
............................ Ch. (s. str.) suavis sakhalinensis Pl.
20 (17). II тергит брюшка с черными узкими остроугольными пятнами, соеди-
ненными в пару в передней части тергита. Последующие тергиты черные
с узкими желтыми каемками по заднему краю. Уже и мельче. 7—9. —
Обитатель открытых степных пространств (рис. 27, 6, 7).................
. . . • ............................... Ch. (s. str.) angaricus OIs.
21 (16). Голени задних ног коричневые или желтые. Вершинное пятно крыла,
как правило, налегает на жилку г4 не более чем на V4 ее длины. II тергит
брюшка обычно с двумя маленькими круглыми пятнами, расположенными
отступя от края тергита (иногда они соединены в пару, а у варьетета
Ch. dissectus apuncta они отсутствуют). 8—10. 4. — Дальневосточный
лесной и луго-лесной вид (рис. 27, S, 9) . . . Ch. (s. str.) dissectus Lw.
22 (1). Дискоидальная ячейка в средней части явственно просветлена (с «око-
шечком»). 8—9. —Луго-лесной вид. Немногочислен (рис. 27, 70, 77) . . .
............................ - Ch. (Heterochrysops) vanderwulpi Krob.
Подсем. TABANINAE
Глазки на темени, как правило, неразвитые; вместо них имеется глазковый
бугорок (иногда отсутствует). 3-й членик усиков поделен на 5—4 сегмента
(реже на меньшее число).
Базикоста крыла в коротких волосках, реже без них.
2. Hybomitra End. Средних размеров (11—23) или крупные коренастые
мухи, окрашенные в черно-серые и черно-коричневые тона; брюшко нередко
с коричнево-желтыми пятнами по бокам (чаще у ?). Глаза, как правило, с 3 яр-
кими поперечными полосками (хорошо видны только у свежих экземпляров;
для их восстановления насекомое необходимо поместить во влажную камеру).
Глазковый бугорок хорошо выражен. Лобные мозоли у ? обычно хорошо раз-
виты. Палочка 3-го членика усиков чаще всего 3-члениковая. — 30 видов и
подвидов (в СССР 50).
1 (56). Жужжальца коричневые или черные (иногда вершина головки желто-
ватая).
2 (35). Концевой членик шупалец узкий, его длина приблизительно в 3.5—
4 раза превосходит толщину. Щупальца коричневые, оранжевые или чер-
ные, в последнем случае иногда в основании утолщены.
3 (6). Крылья с хорошо выраженными темными пятнами на поперечных жил-
ках, в особенности в основании жилки г4.
64
4 (5). Лобный треугольник покрыт серым налетом. 13—16. — Восточносибир-
ский лесной вид (рис. 28, 1—3)..................Н. (s. str.) astur Erichs.
5 (4). Лобный треугольник частично оголен (по бокам), блестяще-черный.
13—17. — Дальневосточный лесной вид (рис. 28, 4—6) .............
.......................................Н. (s. str.) stigmoptera Ols.
6 (3). Крылья без темных пятен на поперечных жилках.
7 (12). Черные тергиты брюшка с яркими золотисто-желтыми каемками.
8 (9). Лобный треугольник и лобные мозоли блестяще-черные. 15—19. —
Тундровый вид (рис. 28, 7—9)................И. (s. str.) aequetincta Beck.
9 (8). Лобный треугольник частично оголен, блестяще-желтый. VIII стернит
иной конфигурации.
10 (И). Бочки груди в желтых волосках. Срединная лобная мозоль черная,
в виде узкой длинной полоски. 17.5—22.3. — Таежный вид (рис. 28,
10—12) ................................И. (s. str.) tarandina L.
11 (10). Бочки груди в черных волосках. Срединная лобная мозоль сравни-
тельно короткая, коричневая, на конце вильчато расщеплена. VIII стер-
нит более узкий. 17.5—21.3. — Лесной вид (рис. 29, 1—3).................
.................................. Н. (s. str.) tarandinoides Ols.
12 (7). Тергиты брюшка без золотисто-желтых каемок по заднему краю. Лоб-
ный треугольник целиком покрыт налетом.
13 (24). Брюшко сверху с небольшими темно-коричневыми пятнами по бокам II
(реже I—III) тергита или целиком черное. Снизу оно одноцветно черное,
иногда по бокам II стернита небольшие коричневые пятна или I—III стер-
ниты по бокам коричневые.
14 (17). Лобная полоска широкая, ее высота превосходит ширину основания
в 2—3 раза.
15 (16). Брюшко по бокам с небольшими коричневыми пятнами. Щупальца
темно-коричневые. 11—15. — Тундрово-таежный вид (рис. 29, 4—6)
............................................Н. (s. str.) sexfasciata Hine
16 (15). Брюшко одноцветно черное с серым рисунком из 3 продольных рядов
пятен. Щупальца бледно-коричневые. 14.3. — Тундровый вид (рис. 29,
7—9) .......................*..............Н. (s. str.) astuta O.-S.
17 (14). Лобная полоска заметно уже, ее высота превосходит ширину основания
в 3.5—4 раза.
18 (21). Брюшко черно-серое, без оливкового налета; светлые волоски на тер-
гитах беловато-серые.
19 (20). Нижняя лобная мозоль крупная, б. м. прямоугольная, достигает или
почти достигает края глаз. II тергит брюшка черный с небольшими буро-
ватыми пятнами по бокам. Брюшко с неясными и узкими каемками по
заднему краю тергитов. 11.5—14. — Таежный вид (рис. 29, 10—12)
.............................................Н. (s. str.) lapponica Wahlb.
20 (19). Нижняя лобная мозоль небольшая, чаще округло-треугольная, часто
коричневатая, не достигает края глаз. II тергит брюшка с хорошо
выраженными небольшими каштаново-коричневыми пятнами по бокам.
Брюшко с четкими и более широкими беловатыми каемками по заднему
краю тергитов. 13.5—17.5. — Дальневосточный лесной вид (рис. 30, 1—3)
.............................................Н. (s. str.) stenopselapha Ols.
21 (18). Брюшко черное, в оливковом налете; светлые волоски на тергитах
золотисто-желтые.
22 (23). Брюшко сверху в интенсивном оливковом налете и в коротких густых
золотисто-желтых волосках. Концевой членик щупалец оранжево-жел-
тый. покрыт такого же цвета волосками. 16. — Редок (рис. 30, 4—6)
.............................................. Н. (s. str.) bagheera Viol.
23 (22). Оливковый налет на брюшке более слабый. Тергиты по заднему краю
в желтых волосках, а в передней части — в черных. Концевой членик
щупалец коричневатый, в черных волосках. 14—15. — Островной лесной
вид. Редок (рис. 30, 7—9)....................И. (s. str.) obscurinervis Viol.
24 (13). Брюшко сверху с хорошо развитыми желто-коричневыми или золотисто-
оранжевыми пятнами хотя бы по бокам I — III тергитов. Снизу оно обычно
5 Зак. № 990
65
в основании желто-коричневое, лишь изредка (темные Н.'brevis) черное
или черно-коричневатое с желтыми каемками по заднему краю стернитов.
25 (26). Брюшко резко двухцветное: основная часть его ярко-желтая, в золо-
1—3 — Н. astur\ 4 — 6 — Н. stigmoptera', 7 — 9 — Н. aequetincta', 10—12 — Н. tarandina. 1, 4, 7, 10 — лоб-
ная полоска; 2, 5, 8, 11 — щупик; з, 6, 9, 12 — усик. (По: Соболева, 1974).
тисто-желтых волосках; конец брюшка (от 4-го кольца) одноцветно чер-
ный, в черных волосках. 15.8—19. — Лесной вид (рис. 30, 10—12) . . .
......................................... Н. (s. str.) nigella Szil.
26 (25). Брюшко иной окраски; 4-й и последующие кольца брюшка всегда
с б. м. выраженными светлыми каемками или в светлых волосках.
27 (32). Усики оранжевые или желто-коричневые, палочка одноцветная с пло-
щадкой.
66
28 (31). Светлые волоски брюшка рыжие или золотистые.
29 (30). Щупальца желто-оранжевые; 3-й членик усиков с резким дорсальным
выступом. 16—21. — Лесной вид (рис. 31, 1—3)..............................
......................................... Н. (s. str.) pavlovskii Ols.
30 (29). Щупальца черные; 3-й членик усиков с б. м. прямым дорсальным углом.
Мельче: 13—16. — Лесной вид (рис. 31, 4—6) . . Н. (s. str.) brevis Lw.
Рис. 29. Детали строения слепней рода Hybomitra.
1—3 — Н. tarandinoides\ 4—6 — Н. sexfasciatcr, 7 — 9 — Н. astuta\ 10—12 — Н. lapponica. (По: Соболева,
1974). Обозначения как на рис. 28.
31 (28). Светлые волоски брюшка серебристо-серые. 16.6—18.5. — Местами
многочислен (рис. 31, 10—12}.....................Н. (s. str.) ussuriensis Ols.
32 (27). Усики коричневые с черной палочкой.
33 (34). 3-й членик усиков с резким дорсальным выступом. Нижняя лобная
мозоль не достигает края глаз, слегка морщинистая. 13—18.5. — Таеж-
ный вид (рис. 31, 7—9).............................Н. (s. str.) arpadi Szil.
34 (33). 3-й членик усиков с б. м. прямым дорсальным углом. Нижняя лобная
мозоль гладкая, выпуклая, блестящая, достигает края глаз. Мельче:
12—16.5. — Горно-таежпый вид (рис. 32, 1—3)..............................
........................................... Н. (s. str.) olsoi Takah.
35 (2). Концевой членик щупалец утолщен, его длина превосходит толщину
не более чем в 3 раза. Щупальца светлые, желтовато-белые, бледно-
5*
67
желтовато-серые пли сероватые, но никогда не черные пли темно-корич-
невые.
36 (39). Лобный треугольник на б. м. значительном протяжении блестяще-чер-
ный или черно-коричневатый.
12
Рис. 30. Детали строения слепней рода Hybomitra.
1—3 — Н. stenopselapha', 4 — 6 — Н. bagheera', 7— 9— Н. obscurinervis; 10—12 — Н. nigella. — 9 — по:
Олсуфьев, 1977; остальное — по: Соболева, 1974). Обозначения как на рис. 28.
37 (38). Сильно выпуклый лобный треугольник целиком блестяще-черный.
Нижняя лобная мозоль, как правило, черная. 15—16.2. — Таежный вид
(рис. 32, 4—6).....................Н. (s. str.) lurida Flln., var. sordida Ols.
38 (37). Умеренно выпуклый лобный треугольник лишь в центральной части
блестящий, коричнево-черный, по краям в сером налете. Нижняя лобная
мозоль чаще коричневая, тонкоморщинистая. 14—17.2 (рис. 32, 7—9)
.................................. Н. (s. str.) nitidifrons nitidifrons Szil.
39 (36). Лобный треугольник целиком покрыт налетом, матово-серый.
68
40 (45). Брюшко сверху одноцветно черное или черно-серое, без следов корич-
нево-желтых пятен по бокам.
41 (44). Брюшко сверху с тремя рядами серых пятен, треугольных в середине
и б. м. ромбических по бокам.
42 (43). Лоб сильно сужен к основанию. Его высота превосходит ширину осно-
вания приблизительно в 4.5—5 раз. 3-й членик усиков с хорошо выражен-
ным дорсальным углом, коричнево-красный, палочка черная. — Таежно-
лесной вид...................Н. (s. str.) bimaculata, var. bisignata Jaenn.
43 (42). Лоб лишь слегка сужен к основанию, относительно широкий: его вы-
сота превосходит ширину приблизительно в 3—3.5 раза. 3-й членик уси-
ков черного цвета, очень узкий, с прямым или слегка изогнутым под углом
69
нижним краем; дорсальный угол усиков слабо выражен. — Таежный вид
(рис. 33, 1—3)................................Н. (s. str.) nigricornis Ztt.
44 (41). Сверху на брюшке имеются 2 продольные сплошные широкие яркие
голубовато-серые полосы, разделенные черной срединной полоской.
14.3—16. — Таежный вид (рис. 33, 4—6) . Н. (s. str.) adachii Takah.
1—3 — H. olsoi', 4—6 — H. luridcr, 7— 9 — H. nitidifrons: 10—12 « H. bimaculata. (По: Соболева, 1974).
Обозначения как на рис. 28.
45 (40). Брюшко с коричнево-желтыми пятнами по бокам.
46 (51). Лобная полоса сильно сужена к основанию; лоб узкий, его высота
превосходит основание в 4.5—6 раз. Нижняя лобная мозоль б. м. прямо-
угольная, плоская, мелкоморщипистая и неблестящая. Концевой членик
щупалец в основании сильно утолщен (иногда умеренно утолщен).
47 (50). II тергит брюшка по бокам покрыт преимущественно черными воло-
сками или со значительной их примесью.
48 (49). II тергит брюшка со значительной примесью черных волосков. Отноше-
ние основания лба к его высоте 1 : 4.5—5. Вершинная половина задних
70
голеней черная. Концевой членик щупалец сильно утолщен (вздут).
14—17...................................И. (s. str.) bimaculata Macq.
49 (48). II тергит брюшка покрыт преимущественно черными волосками с не-
большой примесью желтых. Отношение основания лба к его высоте 1 : 5—6.
1—3 — Н. nigricornis\ 4—6 — Н. adachii; 7 — 9 — Н. muehefeldi; 10—12 — Н. distinguenda contigua. (По
Соболева, 1974). Обозначения как на рис. 28.
Голени целиком желто-коричневые. Концевой членик щупалец умеренно
утолщен. 11 —16.5. — Таежно-лесной вид (рис. 33, 7—9)...............
.............................................. Н. (s. str.) miihlfeldi (Br.)
50 (47). II тергит брюшка в золотисто-желтых волосках с небольшой примесью
черных. Голени коричнево-желтые. Концевой членик щупалец сильно
утолщен. 12.5—16. — Восточносибирский лесной вид (рис. 33, 10—12)
............................................... Н. (s. str.) distinguenda contigua Ols.
71
51 (46). Лобная полоса лишь незначительно сужена к основанию; лоб более
широкий, его высота превосходит ширину основания приблизительно
в 3—4 раза. Нижняя лобная мозоль округлая, треугольная или прямо-
угольная, в последнем случае сильно блестящая. Концевой членик щупа-
лец нормально утолщен.
Рис. 34. Детали строения слепнет! рода Hybomitra.
1—3 — Н. lundbecki sibiriensis; 4—6 — Н. montana montana; 7 — 9 — Н. expollicata orientalis: 10—12 —
H. montana morgani. (По; Соболева, 197 4). Обозначения как на рис. 28.
52 (55). Нотоплевры, как правило, черные (редко коричневатые).
53 (54). 3-й членик усиков наполовину затемнен, с черной палочкой. 12.5—
16.5. — Таежно-лесной подвид .............................................
................................Н. (s. str.) lundbecki lundbecki Lyneb.
54 (53). 3-й членик усиков целиком красновато-желтый, лишь иногда самый
кончик палочки слегка затемнен. 12.3—16 (рис. 34, 1—3) ..........
............................. К. (s. str.) lundbecki sibiriensis Ols.
55 (52). Нотоплевры желто-коричневые (иногда красно-бурые), очень редко
темные. 3-й членик усиков относительно узкий, целиком черный или же
в основании красновато-коричневый. 12.8—16.8. — Таежно-лесной подвид
(рис. 34, 4—6)...........................Н. (s. str.) montana montana Mg.
56 (1). Жужжальца (хотя бы головка) белые или желтоватые (у потертых экзем-
пляров принимают грязно-желтую пли коричневатую окраску).
57 (58). Брюшко сверху посредине с черной блестящей полосой, вдоль которой
тянется узкая линейная полоска из желтовато-серых волосков. Снизу
72
брюшко повторяет рисунок верха (посредине — резкая блестящая черная
полоса). 13.8—16. — Степной подвид (рис. 34, 7—9) ..............
............................ Н. (s. str.) expollicata orientalis Ols.
58 (57). Брюшко сверху с черной полосой, посредине которой имеется ряд серо-
вато-белых треугольников. Черная сплошная полоса на нижней стороне
брюшка отсутствует. Она прерывистая с неровными краями, зачастую
в центральной части совсем стертая. В этом случае на I—II стернитах
имеется черное продолговатое пятно; конец брюшка затемнен. 13.9—
16.7. — Степной подвид (рис. 34, 10—12) . . . ,..........................
. . ............................И. (s. str.) montana morgani Sure.
3. Atylotus Ost.-Sack. Светло-серые или охряно-желтые, небольшие (8—16)
слепни, обычно с сильно выпуклой головой. У живых слепней глаза бледные:
зеленоватые, желтые или сероватые, с одной тонкой, не достигающей внешнего
края полоской или без нее. У сухих экземпляров глаза темнеют до красно-
коричневых тонов. Глаза в волосках или голые. У самок глазковый бугорок
отсутствует. Лобные мозоли у самки небольшие, округлые, плоские, далеко
не достигают края глаз и обычно обособлены; иногда мозоли прикрыты налетом.
Крылья прозрачные; г4 у большинства видов с придатком. — 4 вида и 1 подвид,
входящие в 2 подрода Atylotus и Surcoufiella (в СССР 18 видов).
1 (4). Глаза явственно покрыты волосками.
2 (3). Затылок с рядом заметно выступающих над поверхностью глаз черных
волосков. Брюшко одноцветное, черно-серое. 8—10. — По-видимому,
не кровосос (рис. 35, 1—3)....................A. (S.) sublunaticornis Ztt.
3 (2). Затылок в коротких желтоватых не выступающих волосках. I (или I—III)
тергит с небольшими грязно-желтыми пятнами по бокам. Светлые волоски
брюшка желтоватые. — Редок. Не кровосос (рис. 35, 4, 5)..................
....................................A. (S.) plebejus sibiricus Ols.
4 (1). Глаза голые (или в едва заметных волосках).
5 (6). Желто-бурые виды. Светлые волоски брюшка охряно-желтые. Брюшко
сверху и снизу с темной срединной полосой, занимающей около 1/3 ширины
брюшка (иногда полоса еще уже, особенно снизу). Желтые пятна по бокам
этой полосы крупные*(на I — IV тергитах). Лобные мозоли небольшие, лоб
более широкий. 13—16.4. — Дальневосточный лесной вид (рис. 35, 6—8)
.............................................. A. (s. str.) horvathi Szil.
6 (5). Голубовато-серые или черно-серые виды. Светлые волоски брюшка бело-
вато-серые. Остальные признаки в ином сочетании.
7 (8). Бедра ног, как правило, затемнены почти целиком, исключая самые кон-
чики. Средние и задние лапки желтые, с коричневыми кончиками. Уже
и мельче (сравни с A. (s. str.) horvathi). Желтые пятна по бокам брюшка
маленькие (на II, реже на III тергитах), иногда они отсутствуют. 12—16. —
Восточносибирский степной (или луголесной) вид (рис. 35, 9—11) . . .
.................................................A. (s. str.) miser Szil.
8 (7). Бедра ног почти целиком желтые, лишь иногда в основании слегка серо-
ватые. Брюшко сверху с широкой темной продольной полосой из черных
волосков; II тергит (реже I—III) по бокам с желтовато-коричневыми пят-
нами. Снизу брюшко темно-серое. 10—13. — Степной вид. Малочислен
(рис. 35, 12—14).................................A. (s. str.) pallitarsis Ols.
4. Род Tabanus L. Слепни различных размеров (10—28), главным образом
темноокрашенные. Глазковый бугорок у самок отсутствует (иногда на темени
бывает небольшое блестящее пятно — мозоль). Глаза чаще всего голые, реже
покрыты волосками. Лобные мозоли у самок обычно хорошо развиты; ниж-
няя — прямоугольная пли овальная, верхняя — веретеновидная или в виде
обособленного пятна. Крылья прозрачные, реже затемнены; г4 иногда с хорошо
развитым придатком. — 8 видов и 1 подвид (в СССР 65).
1 (16). Лобный треугольник покрыт налетом.
2 (9). 1-я заднекрайняя ячейка крыла сильно сужена к основанию, почти
замкнутая или замкнутая, иногда со стебельком, упирающимся в край
крыла.
73
Рис. 35. Детали строения слепней рода Atylotus.
1—3 —A. (S.) sublunaticornis; 4, 5 — A. (S.) plebejus sibiricus; 6 — 8 — A. (s. str.) horvathi; 9—11 —'A.
(s. str.) miser; 12—14 — A. (s. str.) pallitarsis. 1, 4, 6, 9, 12 — лобная полоска; 2, 5, 7, 10, 13 — щупик;
3, 8, 11, 14—усик. (1—5— по: Олсуфьев, 1977; остальное — по: Соболева, 1974).
3 (8). Брюшко имеет черно-серую окраску. Дорсальный угол 3-го членика
усиков б. м. прямой, слегка заостренный или в виде маленького зубчика.
4 (5). Брюшко сверху с одним срединным рядом серых треугольников и с се-
рыми пятнами по бокам II (реже III) тергитов. 1-я заднекрайняя ячейка
Рис. 36. Детали строения слепней рода Tabanus.
1—3 — Т. trigeminus', 4—6 — Т. amaenus griscinus', 7 — 9 — Т. signatipennis\ 10—12 — Т. sapporoenus (1—
9 — по: Соболева, 1974; 10—12 — по; Олсуфьев, 1977). Обозначения как на рис. 28.
крыла сильно сужена, иногда почти замкнутая или замкнутая, но без
стебелька. 16.5—20. — Лесной вид (рис. 36,1—3) . . . Т. trigeminus Coq.
5 (4). Брюшко сверху с 3 продольными рядами серых пятен, треугольных по
центру и косо направленных по бокам.
6 (7). Продольные ряды серых пятен на брюшке неясные, покрыты желтова-
тыми волосками. 1-я заднекрайняя ячейка крыла всегда замкнутая и на
конце с коротким (иногда с более длинным) стебельком. Усики оранжевые
с черной палочкой. Дорсальный угол 3-го членика усиков в виде очень
75
маленького зубчика. 13.2—17. — Дальневосточный лесной вид (рис. 36,
4—6)......................................Т. amaenus griseinus Phil.
7 (6). Продольные ряды серых пятен на брюшке ярко выражены (особенно
по центру), покрыты серыми и белыми волосками. 1-я заднекрайняя ячейка
крыла почти замкнутая или замкнутая (иногда со стебельком). Усики
темно-коричневые с черной палочкой; дорсальный угол 3-го членика
б. м. прямой, слабо выражен. 15—18. — Лесной вид (рис. 36, 7—9) . . .
...............................................Т. signatipennis Portsch.
8 (3). Брюшко сверху коричнево-желтое, посредине с неясной темной про-
дольной полосой, а снизу темно-бурое с желтыми каемками по заднему
краю стернитов. ДорЬальный угол 3-го членика усиков в виде крупного,
хорошо выраженного зубца (немного меньше, чем у Т. buddha и Т. chrysu-
rus). 1-я заднекрайняя ячейка крыла заметно сужена к основанию, но не
замкнутая. 16—20. — Редок (рис. 36, 10—12) . . . Т. sapporoenus Shir.
9 (2). 1-я заднекрайняя ячейка крыла открытая.
10 (11). Брюшко черно-серое, без следов коричневой или желтой окраски.
Нижняя лобная мозоль расположена несколько выше основания лобной
полоски. Небольшой голубовато-серый вид. 10—12, реже 13—14. — Лес-
ной вид (рис. 37, 1—3)..................................Т. geminus Szil.
11 (10). Брюшко с коричневыми пятнами по бокам I—III тергитов или резкими
золотистыми каемками по заднему краю тергитов. Нижняя лобная мозоль
расположена на уровне основания лба и даже примыкает к нему. Более
крупные (20—28), яркоокрашенные виды.
12 (15). Черные тергиты брюшка с резкими золотисто-желтыми каемками по
заднему краю. Дорсальный выступ 3-го членика оранжево-желтых уси-
ков в виде резкого зубца.
13 (14). Передние лапки желто-коричневые. Бедра ног коричневые. 19—23. —
Таежно-лесной вид (рис. 37, 4—6).....................Т. buddha Portsch.
14 (13). Передние лапки черные. Бедра ног в основной части черные. 25—28. —
Таежно-лесной вид (рис. 37, 7—9)........................Т. chrysurus Lw.
15 (12). Брюшко без бросающихся в глаза резких золотистых каемок по зад-
нему краю тергитов. Оно с коричневыми пятнами по бокам I—III терги-
тов и срединным рядом серовато^белых треугольников. Снизу коричнево-
черное, с резкой черной полоской посредине. Дорсальный выступ 3-го
членика усиков более умеренный. 20—26.8. — Лесной вид (рис. 37,10—12)
.....................................................Т. pleskei Krob.
16 (1). Лобный треугольник блестяще-черный, без следов налета. 14—16. —
Редок (рис. 37, 13—15) . . 9 . • . . ♦ . . . Т. kinoshitai Kono et Так.
5. Haeinatopota Mg. Небольшие серые мухи с узким телом, крылья серые,
с сетчатым («мраморным») рисунком; в покое они складываются кровлеобразно
вдоль тела; г4 с придатком. Глаза в волнистых поперечных полосках и б. м.
заметных волосках. Усики длиннее головы: 1-й членик довольно крупный,
цилиндрический или яйцевидный, часто сильно блестящий; 2-й членик малень-
кий, бокаловидный, 3-й б. м. длинный, с трехчлениковой палочкой; площадка
узкая, дорсальный угол слабо выражен. Лоб широкий, в основании с поперечно
вытянутой лобной мозолью, посредине лба 3 бархатистых пятна, расположен-
ных по треугольнику, — два более крупных боковых и одно маленькое вершин-
ное (иногда отсутствует). Глазки или глазковый бугорок на темени отсут-
ствуют. На щеках черные крапинки. Хоботок короткий, не длиннее высоты
головы. Задние голени без концевых шпор. — 6 видов (в СССР 18).
1 (2). Крылья с интенсивным коричневым рисунком. Лобная мозоль темно-
коричневая. Усики почти целиком желтые; 1-й членик длинный, б. м. ци*
линдрический; его длина превосходит ширину более чем в 3 раза. Мелкий
вид: 6—7. — Луго-леспой вид (рис. 38, 1—3) . . . . Н. koryoensis Shir.
2 (1). Крылья с серым или буровато-серым рисунком. Лобная мозоль черная.
1-й членик усиков более короткий, б. м. утолщен, его длина превосходит
свою собственную толщину не более чем в 3 раза, иногда с резкой перетяж-
кой у вершины. Обычно более крупные виды, с черными, серыми, реже
красновато-желтыми усиками.
76
Рис. 37. Детали строения слепней рода Tabanus,
I—3’— Т. geminus\ 4—6 — Т. buddhcr, 7 — 9 — Т. chrysurus’, 10—12 — Т. pleskei’, 13—15 — Т. kinoshitai.
Z, 4,V- 10, 13— лобная полоска; 5, 8, 11, 14—щупик; 3, б, 9, 12, 15 — усик. (1—12 — по: Собо-
лева, 1974; 13—15 — по: Олсуфьев, 1977).
Рис. 38. Детали строения слепней рода Haematopota.
1—3 — Н. koryocnsis', 4— 7 — II. dcscrtorum', 8—11 — II. iamci lani: 12 —16 — Н. slachelbergi: 12 — лоб,
13 — усик, 14, 15 — спинка и б) юшко с дорсальной стороны, 7 6 — щупик; 17, 18 — II. tamerlani, спинка
и брюшко с дорсальной стороны. 1,4, 8 — лобная полоска; 2,5. 9 — крыло; 3, 7 — усик; 6 — щупик.
(7—13, 16— по. Соболева, 1974; 14, 15, 17, 18 — по; Олсуфьев, 1 977).
3 (6). 3-й членик усиков по крайней мере в основании коричневый, желто-
коричневый или красно-желтый. Крылья с беловатой каемкой по заднему
краю.
4 (5). 1-й членик усиков блестяще-черный, вздутый (овальный), без перетяжки
у вершины. Крылья буро-серые с мелким сетчатым рисунком. Белая
кайма крыла на его вершине в виде широкой одинарной полоски. 7.2—
8. — Степной вид (рис. 38, 4—7)............................Н. desertorum Szil.
5 (4). 1-й членик усиков черный или изнутри коричнево-желтый, более тонкий
и в средней части муфтообразно утолщен, целиком покрыт серым налетом,
с резкой перетяжкой перед вершиной. Крылья светло-серые. Белая
каемка по краю крыла очень узкая (иногда отсутствует), а на вершине
крыла она в виде двух полос, из которых внешняя более широкая. 8.5—
11. — Крайне редок ..........................Н. turkestanica orientalis Ols.
6 (3). 3-й членик усиков целиком черный. Задний край крыла не имеет белова-
той каймы. Белые пятна на крыльях сгруппированы в четкие розетки.
7 (8). 1-й членик усиков с резкой перетяжкой у вершины. Во внешней поло-
вине крыла ближе к вершине имеются 2 широких белых кольца, в середине
которых нет белых пятен. 7—12. — Дальневосточный лесной островной
подвид...............................................Н. pluvialis tristis Big.
8 (7). 1-й членик усиков без перетяжки у вершины. Во внешней половине
крыла беловатые розетки с густым рисунком, образованным несколькими
извилистыми полосками и пятнами.
9 (10). На спинке имеются 3 хорошо выраженных светлых продольных по-
лосы. На IV—VI тергитах брюшка — парные округлые светлые пятна,
между которыми тянется светлая срединная полоса или хотя бы имеется
серый треугольник на II тергите. 9—10.5. — Восточносибирский лесной
вид (рис. 38, 8—11, 17, 18)................................Н. tamerlani Szil.
10 (9). Вместо сплошных светлых полос на спинке имеются разобщенные между
собой продолговато-овальные или клиновидные серые островки (не у всех);
срединная серая полоса на брюшке отсутствует или выражена слабо, округ-
лые серые пятна имеются лишь на VI (иногда и на VII) тергите. 10—12.5. —
Лесной вид. Малочислен (рис. 38, 12—16)....................Н. stackelbergi Ols.
ОСНОВНАЯ ЛИТЕРАТУРА
—
Виолович Н. А. Слепни Сибири. Новосибирск, 1968. 281 с.
Олсуфьев Н. Г. Слепни (Tabanidae). Л., 1937. 433 с. (Фауна СССР. Насекомые дву-
крылые; Т. VII; Вып. 2).
Олсуфьев Н. Г. Слепни (Tabanidae). Л., 1977. 434 с. (Фауна СССР, Новая сер. № ИЗ.
Насекомые двукрылые; Т. VII; Вып. 2).
Олсуфьев Н. Г. Итоги изучения фауны слепней (Diptera, Tabanidae) советского Даль-
него Востока. — Энтомол. обозрение, 1981, т. 60, вып. 1, с. 110—116.
Павловский Е. Н., Штейн А. Н., Олсуфьев Н. Г. Экспериментальные исследования
над действием слюны слепней на кожные покровы человека. — В кн.: Паразиты, перенос-
чики и ядовитые животные. М., 1932, с. 426—447.
Соболева Р. Г. Токсичность слюны слепней для организма животных. — Зоол. журн.,
1965, т. 44, вып. 3, с. 396—402.
Соболева Р. Г. К изучению фауны слепней (Diptera, Tabanidae) Магаданской области. —
В кн.: Биологические ресурсы суши севера Дальнего Востока. Владивосток, 1971, т. 2,
с. 266—270.
Соболева Р. Г. Слепни юга Приморского края. Новосибирск, 1974. 262 с.
Соболева Р. Г. Биология слепней Приморского края. М., 1977. 197 с.
Соболева Р. Г. К изучению фауны слепней (Diptera, Tabanidae) Курильских остро-
вов# — в кн.: Новые данные о насекомых Сахалина и Курильских островов. Владивосток,
1978, с. 137—141. (Тр. Биол.-почв, ин-та ДВНЦ АН СССР. Новая сер. Т. 50 (153)).
Соболева Р. Г., Петрова Б. К. К изучению фауны и экологии слепней (Diptera, Taba-
nidae) севера Чукотки. — Изв. Сиб. отдел. АН СССР, Сер. биол. наук. Новосибирск, 1973,
вып. 3, с. 99—105.
Соболева Р. Г., Попов В. Д. К изучению фауны слепней (Diptera, Tabanidae) Хабаров-
ского края и Еврейской автономной области. — В кн.: Систематика и эколого-фаунисти-
ческий обзор отдельных отрядов насекомых Дальнего Востока. Владивосток, 1983, с. 69—77.
Murdoch W., Takahasi Н. The female Tabanidae of Japan, Korea and Manchuria. —
Mem. Entomol. Soc. Washington, 1969, 230 p.
Takahasi H. Fauna Japonica (Tabanidae). Tokyo, 1962. 143 pp.
79
СИНАНТРОПНЫЕ И ЗООФИЛЬНЫЕ МУХИ
К синантропным и зоофильным мухам относятся двукрылые, связанные с че-
ловеком и его окружением, с домашними и разводимыми «дикими» животными,
встречающиеся в населенных пунктах или вне их, но в силу эколого-биологиче-
ских особенностей имеющие санитарно-эпидемиологическое или эпизоотологи-
ческое значение.
Подавляющее большинство синантропных и зоофильных мух развивается
в навозе домашних животных, экскрементах человека. Однако их личинки мо-
гут жить в различных разлагающихся субстратах (в помойках и на свалках,
в гнилых фруктах и овощах и т. п.).
В фауне синантропных и зоофильных мух Дальнего Востока насчитывается
более 22 семейств. Причем основное ядро этого комплекса составляют высшие
двукрылые семейств Muscidae, Calliphoridae и Sarcophagidae, поэтому ниже
приводятся определительные таблицы по этим трем семействам, а также по кро-
вососущим мухам сем. Hippoboscidae.
ВНЕШНЕЕ СТРОЕНИЕ
Строение тела обычное для двукрылых насекомых, но имеет свои особен-
ности.
Рис. 39. Схема строения головы мухи.
1 — спереди, 2 — сбоку, з — сзади, ар — ариста, вк — вибриссальный киль, гл — фасеточные глаза,
л — лунка, лп — лобная полоса, лш — лицевой шов, м — медиана, ор — орбиты, ск — скулы, т — те-
менной (глазковый) треугольник, ус — усики, фк — фронтоклипсус (лицо), щ — щеки, щуп — щупик.
Щетинки: вибр — вибриссы, втщ — внутренние теменные, глгц — глазковые, згл — заглазничные, зглк—
заглазковые, зтщ — затемненные, 'лщ — лобные, или фронтоорбитальные, птщ — наружные теменные,
ощ — орбитальные, сти. — стомальныс, пли ротовые. (По: Петрова, 1974а).
Голова мускоидного типа мух несет щетинки, расположение которых имеет
большое диагностическое значение (рис. 39).
Верхняя сторона груди, или среднеспинка, подразделяется на прескутум,
скутум и скутеллум, или щиток, отделенные друг от друга поперечными швами;
80
Рис. 40. Детали строения мух.
2 — грудь сверху, 2 — грудь сбоку. Части груди: впт — вентральная часть плевротергита, гпл — гипо-
плевра, дц д2 — передние и задние грудные дыхальца, ж — жужжальца, .зб — закрыловой бугорок, мпл—
мезоплевра, мф — мезофрагма, нпл — нотоплевра, окр — основание крыла, пб — плечевой бугорок,
ппл — птероплевра, прпл — проплевра, пек — постскутеллум, скр — супрасквамальное ребро, срсп —
среднеспинка, стпл — стерноплевра, т1)2, з — тазики 1-й, 2-й, 3-й пары ног, ч1г 2 — чешуйки, ш — по-
перечный шов среднеспинки, щ — щиток. Щетинки: акр (ас) — акростихальные, б — боковые, в — вер-
шинные, диск — дискальные, дц (de) — дорсоцентральные, зкр — закрыловые, зпл — заплечевые,
иа (га) — интраалярные, нкр — надкрыловые, нпл — нотоплевральные, осн — основные, пл — плечевая,
про — предосновные, прк — предкрыловые, прш — предшовные (по: Петрова, 1974а); 3 — расположение
супрасквамального ребра: — переднее грудное дыхальце, осн — основание крыла, скр — супрасквамаль-
ное ребро и одноименные волоски: пв — передние, зв — задние; чешуйки: ч^ — крыловая, ч2 — грудная,
щ — щиток (по: Зимин, 1951); 4 — схема строения лапок мух: к — коготки, п — пульвиллы, э — гмподий;
5 — схема строения крыла: Вс — базикоста, кр — крылышко, с — костальная жилка, sc — субкосталь-
ная, т — медиальные, г — радиальные, th — плечевая поперечная жилка, ап — анальная, тси — медио-
кубитальная, erv — изгиб медиальной жилки, тт — радиомедиальная, или средняя поперечная, tp — зад-
няя поперечная жилка. Ячейки: — радиальные, Basx_2 — базальные, М2 (=Д) — медиально-дисно-
идальная, Pi_2 — первая и вторая заднекрайние ячейки, Си — кубитальная (по: Петрова, 1974а). 6—9 —
яйцо: 6 — Musca autumnalis, 7 — Muscina stabulans, 8 — Fannia canicularis, 9 — Calliphora erytrocephala,
10 — личинка мускоидного типа III стадии (по: Зимин, 1948).
6 Зак. № 990
средний поперечный шов отделяет прескутум от скутума (рис. 40). В боковых
передних углах прескутума расположено по плечевому бугорку.
Бока груди, или плевры, состоят из следующих склеритов: проплевр
(над основанием передней пары ног), мезоплевр (несколько кзади), нотоплевр
(над мезоплеврами), птероплевр (под основанием крыльев), стерноплевр (между
основаниями передних и средних ног). От основания щитка протягивается
к основанию крыла супрасквамальное ребро (рис. 40, 3).
Среднеспинка у мускоидных двукрылых несет щетинки, расположенные
продольными рядами. Количество и расположение щетинок на груди имеют
3 4
Рис. 41. Схема строения конца брюшка самцов мух.
1 — сем. Muscidae (по: Зимин, 1951); 2 — Scoliocentra engeli\ 3 — Sargus rufifrons; 4 — Parasarcophaga al-
biceps. ao — анальное отверстие, апф — апикальная часть парафалла, гк — гонококсит, гп — гипандрий,
гс — гоностиль, дб_7 — дыхальца V—VII сегмента брюшка, зп — задние парамеры, к-9 (кс) — кокситы
(эдиты) IX тергостернита, мо — мембрана и мембранальные отростки, п (эд) — пенис (эдеагус), пм —
парамеры (гониты), пп — передние парамеры, пф — парафалт, 4с—7с — стерниты IV—VII, ср — сур-
стпли, т — тека, 5т—Ют — тергиты V—X, 9 тс — IX тергостернит, ф — фаллосома, г; — церки, эп —
эпандрий (по: Петрова, 1974а).
значение при определении видов (рис. 40, 7). Щетинки на боках груди назы-
ваются так, как склерит, к которому они прикреплены: нотоплевральные,
проплевральные, стерноплевральные и т. п.
Ноги обычного для насекомого строения. К вершине лапки, которая обычно
состоит из 5 члеников, прпчленяются коготки и две присоски — пульвиллы,
между которыми имеется щетинка или образование в виде короткого узкого
язычка (рис. 40, 4). На ногах большинства двукрылых присутствуют щетинки,
которым присваивается название соответственно месту их прикрепления: перед-
ние, задние, дорсальные, вентральные, передне- и заднедорсальные и передне-
и задневентральные. Иногда бедра и голени мух несут зубцы (самцы Hydrotaed)
пли шипы (Sepsidae), бугорки (самцы родов Fannia и Morellid):, у некоторых они
бывают искривлены (Mesembrina, Morellia aenescens R.-D. и др.).
Крылья обычного строения, но количество крыловых жилок различно у раз-
ных групп. Жилки ограничивают собой отдельные участки крыла — ячейки
(рис. 40, 5).
82
Брюшко состоит из 11 сегментов, но часть их редуцируется или сливается
друг с другом, поэтому количество видимых сегментов бывает неодинаково.
Концевые сегменты брюшка модифицированы у самок в яйцеклад, у самцов —
в генитальный аппарат, строение которого не сходно у разных групп мух
(рис. 41), что успешно используется в диагностических целях. Наиболее широко
в настоящее время этот признак используется при диагностике серых падальных
мух (сем. Sarcophagidae).
Личинки мух, как и у других двукрылых, безногие и безголовые. Тело со-
стоит из 11 —12 сегментов (рис. 40, 10). У большинства групп они удлиненные,
червеобразные, с зауженным передним концом. Но у личинок некоторых родов
и видов на теле могут быть выросты и бугорки разной конфигурации. На зад-
нем конце тела располагаются дыхальца, строение которых используется в диаг-
ностике видов.
Куколка представляет собой пупарий продолговато-овальной формы, с твер-
дыми покровами от светло- до темно-коричневого цвета.
БИОЛОГИЯ
Мухи относятся к насекомым с полным превращением. Большинство мух
яйцекладущие, но есть и живородящие, у которых отрождаются готовые личинки.
Если среди настоящих (сем. Muscidae) и синих падальных мух (сем. Calliphori-
dae) это явление не частое, то серые падальные (мясные) мухи (сем. Sarcophagi-
dae) исключительно живородящие. Мухи могут откладывать как личинок I ста-
дии (например, дальневосточная пастбищная муха Musca saishuensis Ouch.),
так и II стадии, т. е. уже взрослых личинок (род Dasyphora).
Яйца и личинки откладываются мухами в различные субстраты: в навоз,
фекалии, пищевые отбросы, гниющие органические и растительные остатки,
в трупы и т. д. [Петрова, 1968, 1974а, 19746; Артамонов, 1977].
Быстрота созревания яиц и развития личинок зависят от гидротермических
условий. При благоприятных условиях весь цикл развития у большинства мух
завершается за 5—10 дней, а при низких температурах затягивается до 20—
30 дней. Перед окукливанием личинки из переувлажненного субстрата мигри-
руют в более сухие места. Окукливание большинства видов происходит в почве,
реже в периферических частях питавшего субстрата, где они развивались.
Большинство синантропных и зоофильных мух зимует в стадии взрослой ли-
чинки, реже — в стадии куколки, отдельные виды — в стадии имаго.
В открытой природе большинство мух, как правило, размножается в теплый
период года, а в закрытых отопляемых помещениях и скотных дворах — кругло-
годично. На юге Дальнего Востока (Приморский край) активных мух нередко
можно видеть зимой во время оттепелей прямо на снегу.
Мухи очень плодовиты и за всю жизнь дают несколько поколений.
Жизнь мух-кровососок тесно связана с жизнедеятельностью животных-
хозяев. Они отрождаются в мае—июне и погибают с наступлением осенних
холодов. Наиболее многочисленны мухи в июле—августе.
У большинства мух наших широт наблюдается одно поколение в году с очень
длинной осенне-зимне-весенней куколочной диапаузой. За всю жизнь одна
самка откладывает от 10 до 17 личинок (по 1 личинке через каждые 7—9 дней).
Личинки, отложенные с мая по октябрь, перезимовывают и дают новое поколе-
ние летом следующего года. Некоторые виды мух-кровососок (Craterina hirundi-
nis) в течение года дают 2 генерации. На более южных широтах (в Средней Азии)
возможны 3 генерации в году.
У представителей рода Lipoptcna появление окрыленных мух наблюдается
с середины июля до октября. Паразитирующие особи на оленях встречаются
с августа до середины марта.
Гиппобосциды рода Melophagus характеризуются круглогодичным безотрыв-
ным паразитизмом; полный жизненный цикл у них завершается вне зависимости
от времени года за 33—36 дней. Откладка куколок и появление имаго происхо-
дят в течение всего года.
6*
83
ВРЕД
Вредоносное значение мух разнообразно. Перелетая с навоза, фекалий,
с разлагающихся трупов, из помоек и со свалок в жилые дома, они загрязняют
продукты питания и могут быть переносчиками большого числа возбудителей
бактериальных и глистных болезней. Известно более 63 микроорганизмов, кото-
рые переносятся мухами, велика их роль в переносе кишечных инфекций.
Некоторые виды мух принимают участие в биологическом цикле развития
ряда гельминтов. Например, промежуточным хозяином паразита глаз человека
и животных на Дальнем Востоке (Thelazia callipaeda Pall, et Henry) является
мушка Phortica variegata Fll. из сем. Drosophilidae. Пастбищная муха Musca
saishuensis — один из наиболее многочисленных видов Приморского края, яв-
ляющихся промежуточным хозяином гельминта глаз крупного рогатого скота
{Thelazia rhodesi Dest.).
Помимо того, некоторые мухи могут вызывать миазы — заболевания, свя-
занные с паразитированием личинок в мягких тканях человека и животных.
В этом случае личинки мух могут поражать у человека носовую и гайморовы
полости, лобные пазухи, десны, глаза, подкожную клетчатку, уши (слуховой
проход), что приносит людям страшные мучения (Павловский, 1948). Иногда
такие миазы кончаются даже смертью. Но чаще всего мухи вызывают миазы
у животных.
Кровососущие мухи (роды Haematobia, Lyperosia) своими укусами сильно
досаждают сельскохозяйственным животным на выпасах.
В связи с тем, что численность многих пастбищных мух {Musca saishuensis,
М. tempestiva, М. amita и др.) очень велика, они нередко сплошь покрывают
морду и другие части тела коров, причиняя сильное беспокойство и затрудняя
пастьбу.
Как и многие другие кровососущие двукрылые насекомые, мухи-кровососки
сильно беспокоят животных своими укусами и ползанием. На месте укуса
возникает воспаление (прппухание, краснота и сильный зуд).
При высокой численности мух (особенно у птенцов и молодняка млекопитаю-
щих) наблюдается резкое истощение животных, отставание в росте и в конеч-
ном счете — гибель.
Большой вред наносит оленеводству (лосям, благородным и пятнистым оле-
ням) Lipoptena cervi — массовый назойливый кровосос оленьих. Частота встре-
чаемости этих паразитов доходит до 100 %, а численность от 200—300 до 1000—
1200 особей на одном животном.
Существенный вред животноводству наносит возбудитель малафагоза
овец — овечий рунец {Malophagus ovinus), резко снижающий мясную и шерст-
ную продуктивность овец. На одной овце могут одновременно паразитировать
от 250—300 до 3700 кровососок этого вида. Пораженность овец достигает 60—
100 %.
Но этим не ограничивается вред кровососок. Они могут быть механическими
переносчиками возбудителей некоторых кровепаразитарных, гельминтозных и
бактериальных болезней.
НАСТОЯЩИЕ МУХИ (Сем. MUSCIDAE)
Как правило, средней величины или крупные мухи. Тело в большинстве
случаев серое, бурое или черное, реже желтое или металлически-синее или ме-
таллически-зеленое. Тело и ноги усажены многочисленными волосками и ще-
тинками. Глаза <5 в большинстве случаев сильно сближены, /г имеются в виде «ал-
лейки» щетинок, ограничивающих с боков лобную полосу и идущих от усиков
к темени. Лоб иногда с перекрещивающимися щетинками. Жилка крыла М1+2
в вершинной части прямая, реже изогнута дугой или под углом, не доходит
до заднего края крыла. Закрыловые чешуйки сильно развиты.
Мухи попадаются на цветках, траве, листьях кустарников, стволах деревьев,
залетают в дома и животноводческие постройки и т. п. Питаются соком расте-
ний, разлагающимися веществами: некоторые — хищники; небольшое число
видов — кровососы.
84
Личинки развиваются в разлагающихся веществах растительного и живот-
ного происхождения или в навозе. — 52 вида.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). Жилка крыла Cj короткая и заметно С- или S-образно изогнутая, резко
оканчивающаяся. У <П2 хотя бы с вентральной стороны в вершинной поло-
вине в густых торчащих волосках. У $ орбиты очень широкие с направ-
ленными наружу or. Ариста в коротком опушении ... 1. Fannia R.-D.
2 (1). Жилка крыла а1, как правило, длинная; если короткая, то прямая,
утончающаяся к концу. У J t2 иного строения; орбиты $ умеренно широ-
кие, ариста, как правило, в длинных волосках, реже в коротких волосках
у основания.
3 (14). £3 без длинной развитой pd.
4 (И). Место разветвления /?4+5 и /?2+3 сверху и снизу усажено щетинками.
5 (8). М\+2 в вершинном отделе изогнута вперед.
6(7). Передние stpl имеются..........................25. Myospila Rd.
7 (6). Передние stpl отсутствуют...................6. Graphomyia R.-D.
8 (5). /?4+5 и М1+2 в вершинном отрезке б. м. параллельны друг другу или
расходящиеся.
9 (10). Край рта сильно выступает вперед. Ширина лба 3 равна V2 ширине
глаза..............................................4. Subphaonia Rind.
10(9). Край рта не выступающий. Лоб в уже..............2. Mydaea R.-D.
11 (4). Место разветвления жилок RM и Д2+3 лишь с верхней поверхности крыла
усажено щетинками, но часто голое и сверху и снизу.
12 (13). У 3 голова сверху явно сплющенная. У ? яйцеклад относительно
короткий; конец брюшка начиная от VI сегмента с явно заметными тер-
гитами и стернитами. Церки большие, их ширина превышает длину
...................................................3. Hebecnema Schnabl.
13 (12). У 3 голова сверху не сплющенная. У ? яйцеклад длинный; конец
брюшка без явно заметных тергитов и стернитов. Церки небольшие, их
длина превышает ширину *...............................5. Helina R.-D.
14 (3). t3 с 1 явственно развитой pd.
15 (20). Хоботок сильно хитинизированный, блестящий, выступающий из
ротового отверстия, с узкими сосательными лопастями. Птероплевры
с мелкими щетинками или волосками.
16 (19). Ариста в длинных волосках только по верхней стороне.
17 (18). Щупики тонкие, в 3.5—5 раз короче подбородка. Проплевры и гипо-
плевры перед дыхальцами в тонких волосках. Глаза узкие, затылок внизу
слабовыпуклый.......................................22. Stomoxys Geoff.
18 (17). Щупики сплющены с боков, расширены к вершине, не более чем вдвое
короче подбородка. Проплевры и гипоплевры перед дыхальцами без воло-
сков. Глаза широкие. Затылок в нижней половине сильно выпуклый
....................................................24. Lyperosia Rd.
19 (16). Ариста в волосках по верхней стороне и с волосками в дистальном
отделе на нижней стороне. Щупики в дистальной половине сильно рас-
ширены, длинные....................................23. Haematobia R.-D.
20 (15). Хоботок б. м. мясистый, втяжной, как правило, с широкими сосатель-
ными лопастями. Если хоботок стройный, с небольшими сосательными
лопастями, то птероплевры голые.
21 (52). 66: глаза на лбу, как правило, б. м. сближены.
22 (23). /х снизу перед вершиной с вырезкой и зубцами; t± близ основания
с длинной плоской вырезкой..........................11. Hydrotaea R.-D.
23 (22). Д снизу перед вершиной без вырезки и зубцов; t± близ основания без
вырезки.
24 (39). Птероплевры голые. Ариста перистая или опушенная.
25 (28). сх3 с задней стороны с группой небольших щетинок.
26 (27). t2 без pv ....................................7. Thricops Rd.
27 (26). t2 с 1 pv.................................8. Alloeostylus Schn.
28 (25). Задний край cx3 без щетинок.
85
29 (30). Вершинная часть Л/1+2 слегка изогнута к Т?4+5. Вершина щитка всегда
красноватая........................»....................13. Muscina R.-D.
30 (29). ТИ1+2 на вершине не изогнутая.
31 (34). t3 с несколькими одинаково длинными и крепкими pd.
32 (33). Глаза в густых длинных волосках. Хоботок короткий, с широкими
сосательными лопастями .......................9. Trichopticoides Ringd.
33 (32). Глаза без явного волосяного покрова, если с таковым, то хоботок не
удлиненный и с небольшими сосательными лопастями иного строения.
34 (31). t3 лишь с 1 длинной и крепкой pd', если pd более 1, то остальные явно
короче и слабее.
35 (36). Грудь и брюшко темно-синие, металлически блестящие. Скулы и щеки
очень узкие. Глаза в вентральном направлении сильно удлиненные . . .
..................................................... 10. Ophyra R.-D.
36 (35). Грудь и брюшко не темно-синие, металлически блестящие. Скулы и
щеки более широкие.
37 (38). or длиннее и крепче, чем jr..........12. Dendrophaonia Malloch
38 (37). or не длиннее и крепче, чем /г..............14. Phaonia R.-D.
39 (24). Птероплевры с мелкими щетинками или волосками. Ариста перистая.
40 (41). М1+2 прямая, не изогнутая по направлению к переднему краю крыла.
Грудная чешуйка узкая, языковидная......................15. Polietes Rd.
41 (40). Af1+2 в вершинном отрезке б. м. сильно изогнута (дуговидно или под
углом) по направлению к переднему краю крыла. Грудная чешуйка ши-
рокая, в задней части прилегающая к щитку.
42 (43). Щиток с многочисленными (10—16) краевыми щетинками. Базальная
часть крыльев и закрыловые чешуйки ярко-желтые. Очень крупные мухи,
часто в густых волосках..............................21. Mesembrina Mg.
43 (42). Щиток с обычным количеством краевых щетинок. Базальная часть
крыла не ярко-желтая.
44 (47). t2 без ри.
45 (46). Л71+2 изогнута под углом .........................17. Musca L.
46 (45). ТИ1+2 изогнута дугой........................20. Morellia R.-D.
47 (44). t2 с pv.
48 (49). Супрасквамальное ребро усажено волосками . . 16. Orthellia R.-D.
49 (48). Супрасквамальное ребро без волосков.
50 (51). 7?! в основной части усажена короткими щетинками. Глаза, как пра-
вило, в густых длинных волосках......................19. Dasyphora R.-D.
51 (50). Н1 голая. Глаза лишь в коротких волосках . . . 18. Pyreilia R.-D.
52 (21). глаза на лбу широко расставлены.
53 (54). Птероплевры с мелкими щетинками или волосками (определяются
по тезе 40 (41)).
54 (53). Птероплевры голые.
55 (56). Задний край сх3 с группой небольших щетинок . . 7. Thricops Rd.
56 (55). Задний край сх3 без щетинок.
57 (64). Направленные вперед or сильно развиты.
58 (59). t3 с несколькими почти одинаковыми и крепкими pd (необходимо
исключить род Pogonomyia, у которого край рта в профиль явно высту-
пает вперед).
59 (58). t3 лишь с 1 длинной и крепкой pd, если pd более одной, то остальные
явно короче и слабее.
60 (61). Ариста перистая; длина ее волосков по меньшей мере равна V2 ширины
3-го членика усиков. Нотоплевры голые, stpl1+2..........................
.........................................12. Dendrophaonia Malloch.
61 (60). Ариста лишь опушенная. Нотоплевры с несколькими волосками;
если их нет, то тело блестящее, темно-синее, stplM.
62 (63). Грудь и брюшко блестящие, темно-синие, почти без светлого налета.
Среднеспинка без продольных полос. Глазковый треугольник заходит
на переднюю половину лба................................10. Ophyra R.-D.
63 (62). Грудь и брюшко в явственном сером налете. Среднеспинка с темными
продольными полосами. Глазковый треугольник не заходит за среднюю
линию лба............................................11. Hydrotaea R.-D.
86
64 (57). Как правило, имеется лишь верхняя пара направленных назад хорошо
развитых or.
65 (66). Вершинная часть М слегка изогнута к RM. Вершина щитка всегда
Красноватая........................................13. Muscina R.-D.
66 (65). Вершинная часть Мне изогнута к Т?4+5......14. Phaonia R.-D.
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ (рис. 42-44)
1. Fannia R.-D. — 5 синантропных видов.
1 (2). Тергиты средней части брюшка с удлиненными срединными и округ-
лыми черными боковыми пятнами. 4.3—3.5. — Центр, Ю евр. ч. СССР,
Кавказ, Ср. Азия, Приам., Прим., Ю Хаб.; ср. полоса и юг 3 Европы,
Тайвань (Китай), Ява............................F. leucosticta Mg.
2 (1). Брюшко без рисунка, если с рисунком, то без округлых черных пятен.
3 (4). Боковые отделы основной части брюшка желтые, просвечивающие.
Среднеспинка в коричневато-сером налете с тремя продольными темными
полосами. Ноги черные, колени желтоватые, средние голени без вентраль-
ных выростов. Ариста голая. 6—7 — Космополит..................
............................F. canicularis L. — Малая комнатная муха
4 (3). Брюшко бурое, в темном или светлом налете, без желтых просвечиваю-
щих пятен.
5 (10). 33: лоб много уже 3-го членика усиков или равен ему по ширине.
6 (7). Средние тазики сх2 без шипов. Брюшко в светло-сером налете с темной
продольной полосой. t2 с вентральной стороны в очень коротких торчащих
волосках, /2 с рядом крепких щетинок посредине и у основания в тонких,
слабых щетинистых волосках. 6—8. — С и Ю евр. ч. СССР, Ср. Азия,
Прим., Хаб.; 3 Европа, С и Ю Америка...............F. incisurata Ztt.
7 (6). сх2 с 1—3 крепкими, на вершине резко изогнутыми шипами.
8 (9). сх2 с 3 изогнутыми назад шипами. /3 снизу посредине с пучком крепких
щетинок, t2 с вентральной стороны в вершинной трети с резко выражен-
ным бугорком; tr сплошь черные. 6—7. — Космополит . . F. scalaris F.
9 (8) сх2 с 1 изогнутым назад шипом. /3 снизу без пучка крепких щетинок,
t2 в вершинной половине с вздутием; красно-желтые, лишь в вершинной
трети черные. 6—7.5. — С и Ю евр. ч. СССР, Якут., Чук., Камч.; 3 Ев-
ропа, С Африка.....................................F. manicata Mg.
10 (5). лоб значительно шире 3-го членика усиков.
11 (14). tr с 1 тонкой переднедорсальной щетинкой ad.
12 (13). /2 снизу при основании с 1 длинным щетинистым волоском....
............................................ 1. F. scalaris F.
13 (12). /2 снизу при основании без щетинистого волоска . . F. incisurata Ztt.
14 (11). Ц без ad .................... F. manicata Mg.
2. Mydaea R.-D. — 1 вид.
1 (1). Щиток темно-серый. Передние дыхальца с темно-бурыми волосками
перитремы. Голени и бедра в основной части желтые. Задние бедра не-
сколько искривлены. 9.2—10. — СЗ евр. ч. СССР; Прим.; С и ср. полоса
3 Европы ...........................................М. detrita Ztt.
3. Hebecnema Schnabl. — 1 вид.
1 (1). Глаза в явственно волосистые. Усики и щупальцы черные. Ноги на всем
протяжении черные. Среднеспинка и брюшко черные, слабо блестящие.
Брюшко с темной срединной полоской. 3—5. — Вся Палеарктика;
С Америка.....................................Н. umbratica Mg.
4. Subphaonia Ringd. — 1 вид.
1 (1). Близок к Phaonia. Черный буроопыленный. /2 снизу с 2 рядами длинных
щетинистых волосков. 5. — Чук..................S. nitidiventris Ringd.
87
5. Helina R.-D. — 1 вид.
1 (1). Брюшко в желтовато-серой пыльце, в основной части не просвечивающее,
с парными темными пятнами на II и III тергитах. По меньшей мере перед-
ние. 42. Мухи ср.м. Muscidae.
1 — Faunia canicular is, о; 2 — Hydrotca dentipes, <?. (По: Штакельберг, 1936).
ние бедра при основании затемненные. Арпста длинноперистая (ширина
оперения в 2—3 раза превышает ширину 3-го членика усиков). Тело
в желто-серой пыльце. Среднеспинка с 4 темными продольными полосами.
Ноги желтые, бедра при основании и лапки черные. 7—9. — СЗ евр. ч.
СССР, Урал, Таджикистан, Камч., Кур.; 3 Европа . . . Н. depuncta F.
88
6. Graphomyia R.-D. — 1 вид.
1 (1). Среднеспинка <3 черная, с серыми продольными полосками; 9 серая
с черными волосками; брюшко серое, с черными пятнами. 9—10. — С и Ю
Сиб., лесная зона ДВ; 3 Европа...........................G. maculata Scop.
7. Thricops Rd. — 1 вид.
1 (1). Позади шва 4 пары de. Ариста коротко опушенная. /2 сзади близ сере-
дины с крепкими щетинками. 4.5—5. — С, СЗ евр. ч. СССР, Урал, Кавказ,
3 Сиб., Чук., Камч.; С и ср. полоса 3 Европы, Япония.......................
.................................................. Th. hirsutula Ztt.
8. Alloeostylus Schn. — 1 вид.
1 (1). Основание T?4+5 без щетинок. Щиток снизу голый. Позади шва 4 пары de.
t желтые или красноватые. 6—7. — Камч.; С и 3 Европы.......................
. .............................................. A. albibasalis Ztt.
9. Trichopticoides Ringd. — 1 вид.
1 (1). Среднеспинка черная, в слабом буроватом налете, с 4 темными продоль-
ными полосами. Брюшко черное, в сером налете, у <3 с темной продольной
полосой. Ноги черные. 6—7.5. — СЗ евр. ч. СССР, Урал, Маг.; 3 Европа,
Гренландия. Редок............................................Т. decalor F1L
10. Ophyra R.-D. — 3 вида.
1(2). Закрыловые чешуйки буро-коричневые. Дорсоцентральные щетинкн на
прескутуме крепкие, явственно выделяющиеся среди волосков. Тело
черное с синеватым отливом, жужжальца коричневатые; t3 <3 сильно
изогнутые, на внутренней стороне в длинных волосках. Ариста почти
голая. 6—8. — По всей Голарктике, за исключением Крайнего Севера
.........................................................О. leucostoma Wd.
2 (1). Закрыловые чешуйки желто-белые.*
3 (4). Задний край глаз с выемкой. Ширина лба <3 равна ширине 3-го членика
усиков или чуть меньше; de перед швом среднеспинки волосовидные, едва
различимы среди длинных черных волосков груди. Лапки сплошь черные.
Тело оливково-черное. 4.5—7. — Прим.; Япония, Китай, п-ов Корея,
Шри-Ланка, Австралия.....................О. spinigera Stein. (=nigra Wd.).
4 (3). Задний край глаз без выемки. Глаза <3 сближены на V4 ширины 3-го
членика усиков; de хорошо развиты. Тело черное, брюшко более узкое,
тергиты со II по IV в сером налете и с черной продольной полосой по-
средине. Дистальные концы члеников лапок светло-желтые. 5—7. —
Ю Прим.; Япония, Китай. Обычен в Эфиопской, Ориентальной и Австра-
лийской областях, юг С Америки...........................О. chalcogaster Mg.
11. Hydrotaea R.-D. — 8 видов.
1 (16). 3d: глаза на лбу сильно сближены; снизу с зубцом.
2 (5). /3 у основания с 2 близко стоящими друг к другу, крючковидно изогну-
тыми на конце щетинками.
3 (4). Глаза в довольно густых светлых волосках. t2 близ вершины без av.
4—5. — СЗ евр. ч. СССР, Ср. Азия, Хаб., Приам., Прим.; 3 Европа,
С Америка...................................................Н. occulta Mg.
4 (3). Глаза голые. t2 близ вершины с 1 очень длинной av. Крупнее: 7—7.5. —
Якут., Чук., Камч.; Скандинавия, С Америка . . . Н. ringdahli Stein.
5 (2). /3 снизу у основания без крючковидных щетинок.
6 (7). t2 несут кроме задних щетинок еще по 2—3 pv; t3 посредине с внутрен-
ней стороны с крепкой, крючковидно изогнутой на конце щетинкой.
5.5—7.5. — Приам.; Скандинавия, С Америка . . . Н. scambus Ztt.
7 (6). t2 с 1 pv или без нее.
8 (9). /3 снизу посредине с 1 толстой шиповидной щетинкой. /2 с 4 крепкими
шипами, t3 с клиновидным пучком щетинистых волосков посредине вну-
тренней поверхности, а к вершине от него — с несколькими длинными
89
прямыми щетинками и изогнутыми волосками. 4—6. — СЗ евр. ч. СССРГ
Кавказ, Урал, Ср. Азия, Приб., Приам., Прим.; вся 3 Европа, С Африка,
С Америка.....................Н. armipes F11. — Конская зубоножка
9 (8). /3 снизу без шиповидных щетинок.
10 (И). Жужжальца желтые. 1-й членик средних лапок снизу в густых щетини-
стых волосках. Глаза почти соприкасаются. 5—6. — СЗ евр. ч. СССР,
Кавказ, Урал, Прим.; 3 Европа...........................Н. irritans F1L
11 (10). Жужжальца черные. 1-й членик средних лапок снизу без щетинистых
волосков.
12 (13). Глаза явственно разделены лбом. Лобные щетинки достигают глазко-
вого треугольника; Д снизу близ основания с несколькими крепкими шипи-
Рис. 43. Строение ног мух сем. Muscidae.
1 — Hydrotaea vilutina; 2 — Н. armipes', 3 — Н. dcntipes; 4 — Н. scambus; 5 — Н. meteorica; 6 — Н. irri-
tans. (По: Рингдал, из: Зимин и Эльберг, 1970).
ками, с крупным зубцом у вершины. 6.5—8. — СЗ, ср. полоса и Ю евр. ч.
СССР, Ср. Азия, Сиб., повсюду на ДВ; 3 Европа, С Америка...........
.................................Н. dentipes F. — Обыкновенная зубоножка
13 (12). Глаза почти соприкасаются. Ряды лобных щетинок доходят лишь до
половины высоты лба.
14 (15). Д с очень характерными шипами, нижний из которых в виде птичьей
головы, /3 снизу почти голые. Среднеспинка черная, слабо блестящая.
3—5. — СЗ евр. ч. СССР, Украина, Урал, Амур., Прим.; 3 Европа,
Пер. Азия, С Америка.....................................Н. meteorica L.
15 (14). Д с обычным зубцом, без шипа в форме птичьей головы. /3 снизу с зад-
ней поверхности покрыты длинными нежными волосками. Среднеспинка
черная, блестящая, без налета. 5—8. — СЗ евр. ч. СССР, Прибайкалье^
Амур., Прим.; 3 Европа, кроме Крайнего Севера, Мал. Азия................
................................................. Н. velutina R.-D.
16 (1). глаза разделены широким лбом. Передние бедра простые.
17 (18). t2 с 1 ри.......................................Н. scambus Ztt.
18 (17). t2 без pv.
19 (22). Головки жужжалец желтые.
20 (21). Верхний край гипоплевр с 1 или несколькими короткими щетинками,
Д с 1 ad...............................................Н. ringdahli Stein.
21 (20). Верхний край гипоплевр голый, Д без ad . . . . Н. irritans F]L
22 (19). Головки жужжалец черные.
90
23 (24). t2 с крепкой ad...................................Н. dentipes F.
24 (23). t2 без ad.
25 (26). Теменной треугольник и скулы у основания усиков блестяще-черные.
Тело черно-серое............................................Н. occulta Mg.
26 (25). Теменной треугольник и скулы у основания усиков покрыты пыльцой.
27 (28). Ариста голая. Щетинка на спинной поверхности задних голеней почти
всегда отсутствует ......................................Н. armipes Flln.
28 (27). Ариста в коротком пушке. Щетинка на спинной поверхности задних
голеней всегда имеется.
29 (30). Среднеспинка в едва заметном налете, черная, блестящая...........
....................................................Н. velutina R.-D.
30 (29). Среднеспинка пепельно-серая ....................Н. meteorica L.
12. Dendrophaonia Malloch. — 1 вид.
1 (1). Длина 3-го членика усиков вдвое больше ширины. Среднеспинка в сером
налете, с 3 темными продольными полосками; брюшко в сером налете
с узкой темной срединной продольной полосой. Ноги черные. 6—7.5. —
СЗ евр. ч. СССР, Ср. Азия, Амур.; 3 Европа, на север до Ю Швеции . . .
.................................................... D. querceti Bouche
13. Muscina R.-D. — 6 видов.
1 (4). t2 и t3 темно-бурые или красновато-желтые. Тело бурое в желтовато-
сером налете.
2 (3). Базикоста и щупальца желтые. Задняя треть / желтовато-коричневая.
Лоб 3 шире 3-го членика усиков. 6—9.5 (рис. 44, 7). — Космополит
........................<..........................М. stabulans F11.
3 (2). Базикоста черная, щупальца темно-коричневые. / сплошь черные.
Лоб <3 уже 3-го членика усиков. 7—9. — Приам., юг Хаб., Сах., Кур.
(Кунашир), Прим.; Япония.................................М. august if rons L.
4 (1). Ноги на всем протяжении черные. Посредине в сером налете. Средне-
спинка с 4 темными продольными полосами, брюшко с темной срединной
полосой.
5 (6). Щупики черные. М у вершины крыла слабо изогнута. Брюшко со слабым
шашечным рисунком. 7—8 — Голарктика. Обычен...............................
................................. М. assimilis F11. — Муха черноногая
6 (5). Щупики желто-оранжевые. М сильно изогнута у вершины крыла.
7 (8). Закрыловые чешуйки затемненные, спинка перед поперечным швом без
ясно различимых удлиненных полос. 9—10. — Ю Прим.; Япония . . .
................................. М. lapponica Sehin. ( = nigra Wied.)
8 (7). Закрыловые чешуйки светлые, желтовато-белые.
9 (10). Грудная чешуйка относительно широкая, ее внутренний край приле-
гает к боковому краю щитка. Брюшко ? без отливающих серебристо-
серых пятен, с темной продольной полосой. 7—9. — Голарктика, кроме
севера...................................................М. pasquorum Mg.
10 (9). Грудная чешуйка уже, ее внутренний край расположен под углом к бо-
ковому краю щитка. Брюшко ? с отливающими серебристо-серыми пят-
нами. 7—9. — Голарктика, кроме севера. Редок . . М. pabulorum F11.
14. Phaonia R.-D. — 2 вида.
1 (2). ас перед швом 1—2 пары. Гипоплевры голые; желтые. Щиток сплошь
желтый. Около 8.5. — СЗ евр. ч. СССР, Кур. (Кунашир); Англия . . .
................................................. Ph. umbraticola Fonseca.
2 (1). ас перед швом отсутствуют, гипоплевры с несколькими мелкими воло-
сками. 7—9.5 — СЗ евр. ч. СССР, Урал, Ср. Азия, Маг., Камч.; 3 Европа,
С Америка...................................................Ph. errans Mg.
15. Polietes Rd. — 3 вида.
1 (2). Чешуйки темно-бурые, особенно крыловая. 9—12. — СЗ, Ю Урал,
Камч., Сах., Кур. (Кунашир); СЗ, 3 Европы, СВ Китай, Япония . . .
.................................................. Р. nigrolimbata Bonsd.
91
2 (1). Чешуйки светлые.
3 (4). Жужжальца желтые. Вершинный отрезок жилки М явственно длиннее
предвершинного ее отрезка. Среднеспинка блестяще-черная, посредине
и с боков в беловатом налете. Брюшко в сером налете с темной продоль-
ной полосой, темными задними краями тергитов и шашечным рисунком.
Рис. 44. Мухи сем. Muscidae.
1 — Muscina stabulans, 6,2 — Pyreilia cadaveriva, 3 — Stomoxys calciira'ns, ?. (По: Штакелъберг, 1956/.
7—8. — СЗ евр. ч. СССР, ДВ, кроме крайнего Севера; С и ср. полоса
3 Европы ............................................Р. albolineata F11.
4 (3). Жужжальца черные. Длина вершинного отрезка М равна длине ее пред-
вершинного отрезка или короче. Среднеспинка серая, брюшко с темной
срединной продольной полосой и шашечным рисунком. 10—13. — СЗ.
ср. полоса евр. ч. СССР, Прим.; 3 Европа, Япония, Мал. Азия . . . .
.............................................................Р. lardaria F.
16. Orthellia R.-D. — 3 вида.
1 (2). ас 1 пара впереди шва; de перед швом 2, позади шва 3 пары. 4.5—8. —
С евр. ч. СССР, юг, Кавказ, Ср. Азия, Сиб., ДВ — повсеместно, кроме
92
севера; 3 Европа, С Африка, Монголия, Китай, Япония, С и Ю Америка
............................О. caesarion Mg. — Большая зеленая коровница
2 (1). ас перед швом нет.
3 (4). de перед поперечным швом нет; если есть, то они едва отличимы от во-
лосков; позади шва 2 пары de. 6—9.2. — Прим.; Япония...................
................................................ О. pacifica Zim.
4 (3). Перед швом 2, позади 4 пары крепких длинных de. 6.5—9. — С и Ю
евр. ч. СССР (Кавказ), Ср. Азия, Сиб., Хаб., Сах., Приам., Прим.;
3 Европа, С Монголия.................О. cornicina F. — Зеленая коровница
17. Musca L. — 5 видов.
1 (14). 33: глаза сближены; лоб в наиболее узком месте много уже глаза.
2 (5). I стерпит по краям без волосков.
3 (4). Брюшко бурое в желтовато-сером налете, с бурыми задними краями
тергитов и узкой срединной продольной полосой. Глаза голые. Передние
дыхальца груди коричневые или бурые. Лоб в 2—3 раза уже ширины
3-го членика усиков. 2.5—4.2. — Ю евр. ч. СССР, Кавказ, Ср. Азия,
Сиб., Амур., Прим.; 3 Европа, С Африка, Пер. Азия, Индия, Центр.
Китай................................................М. tempestiva F11.
4 (3). Брюшко с отчетливыми желтыми пятнами на боках. Глаза в волосках.
Передние дыхальца груди белые и желтые. Лоб равен ширине 3-го чле-
ника усиков или немного уже его. 3—5.5. — Ср. полоса и юг, на В до
Уральского хр., Кавказ, Закавказье, Ср. Азия, Ю Прим.; 3 Европы,
Африка, Пер. Азия, С Индия...............................М. osiris Wd.
5 (2). I стернит по краям в волосках.
6 (7). Передняя группа супрасквамальных волосков (под основанием крыльев)
отсутствует; проплевры в волосках. Орбиты в 6—8 раз уже лобной по-
лосы. Среднеспинка в сером или серо-желтом налете, с 4 черными про-
дольными полосами. Брюшко черное, с б. м. развитыми желтыми боковыми
пятнами и переливчатыми пятнами светлого налета. 4—9. — Космополит
............................................М. domestica L.
а (б). Лоб примерно вдвое уж*е глаза. 7—9. — На юг до Главного Кав-
казского хребта, Сиб. — повсюду, кроме мест обитания приведен-
ного ниже вида .............................М. domestica domestica L.
б (а). Лоб в 3—5 раз уже глаза. 4.5—8. — Кавказ (местами), Закав-
казье, Ср. Азия, Амур, Ю Прим., о-в Кунашир. Тропическая и суб-
тропическая зона земного шара . . . М. domestica vicina Mcq.
7 (6). Передняя группа супрасквамальных волосков имеется; проплевры по-
средине без волосков. Орбиты шире лобной полосы.
8 (11). Задние супрасквамальные волоски отсутствуют.
9 (10). Ширина скулы на уровне 2-го членика усиков меньше ширины 3-го
членика усиков. V тергит с резко выраженными темными переливчатыми
продольными полосами налета. Щеки в переднем отделе в длинных во-
лосках, которые заходят полукругом значительно далее нижнего края
глаза. 4—8. — СЗ, Ю евр. ч. СССР, Кавказ, Ср. Азия, 3 Сиб., Амур.;
3 Европа, С и С-В Африка...........................М. autumnalis Deg.
10 (9). Ширина скулы на уровне 2-го членика усиков явственно превосходит
ширину 3-го членика усиков. V тергит равномерно покрыт серовато-
желтым налетом, не образующим резких темных продольных полос. Щеки
в коротких волосках, едва заходящих за линию, соединяющую нижний
край глаз с угловой вибриссой. 4.5—6.7. — Ю Сибг, и ДБ; Монголия,
С Китай..................................М. amita Henn. (— arnica Zim.)
11 (8). Задние супрасквамальные волоски имеются так же, как и передние
супрасквамальные щетинки.
12 (13). Жилка г4+5 снизу в основной части со щетинками, нередко заходящими
за rm. V тергит с 3 резкими узкими темными продольными полосками,
развитыми по всей длине тергита, чередующимися с сильно блестящими
промежутками налета между ними. 7—9. — Амур., Прим.; Япония,
п-ов Корея, Китай ................................. М. saishuensis Ouchi
93
13 (12). Жилка г4+5 снизу, как правило, со щетинками, развитыми лишь у самого
ее основания. V тергит без резких продольных темных полос, развитых
по всей длине тергита. Мельче: 4.5—8.3. — СЗ и юг евр. ч. СССР, Ср.
Азия; ср. полоса и юг 3 Европы, С Африка, Монголия......................
............................................ М. larvipara Portsch.
14 (1). глаза на лбу широко расставлены.
15 (18). 1 стернит без волосков.
16 (17). Передние дыхальца груди темно-бурые или рыжие. Глаза голые.
Лобная полоса в 2—2.5 раза шире орбиты; скулы равны или чуть шире
3-го членика усиков....................................М. tempestiva F11.
17 (16). Передние дыхальца груди светлые: белые или палево-желтые. Глаза
в редких светлых волосках, иногда плохо заметных. Лобная полоса равна
по ширине орбите или в 1 и 1.5 раза шире ее; скулы почти в 2 раза шире
3-го членика усиков.......................................М. osiris Wd.
18 (15). I стернит в черных волосках по краю или по краю и посредине.
19 (20). Проплевры впереди дыхалец посредине в волосках. Супрасквамальное
ребро без волосков........................................М. domestica L.
а (б). III и IV тергиты с маленькими красно-желтыми боковыми пятнами,
или брюшко сверху сплошь бурое, в переливчатом налете . . . .
..........................................М. domestica domestica L.
б (а). Ill и IV тергиты с крупными красно-желтыми пятнами, или брюшко
красно-желтое, с узкой срединной продольной бурой полосой . . .
....................................... М. domestica vicina Mcq.
20 (19). Проплевры посредине без волосков. Супрасквамальное ребро по край-
ней мере с передней группой волосков (под основанием крыла).
21 (24). Супрасквамальные щетинки имеются лишь в передней части ребра, под
основанием крыла; задняя часть супрасквамального ребра без волосков.
22 (23). Лобные щетинки в задней или в средней и задней частях орбиты рас-
положены в 2—3 ряда, из которых наружные неправильные...................
........................................... . М. autumnalis Deg.
23 (22). Лобные щетинки расположены в 1 ряд...............М. amita Henn.
24 (21). Супрасквамальное ребро и в передней и в задней частях с черными щети-
нистыми волосками.
25 (26). Жилка г4+5 снизу в основной части, как правило, с многочисленными
щетинками, часто заходящими за гтп. Темные переливчатые пятна по
бокам III и IV тергитов в виде полоски заходят на нижнюю сторону
брюшка. 7—9 ..............................М. saishuensis Ouchi
26 (25). Жилка г4+5 снизу со щетинками лишь у самого основания. Темные
пятна по бокам III и IV тергитов не заходят на нижнюю сторону брюшка.
Мельче: 4.8—8.3 ...........................М. larvipara Portsch.
18. Pyreilia R.-D. — 2 вида.
1 (2). Переднее дыхальце груди бурое или буровато-рыжее. Тело лишено
налета. 4—7. — СЗ и юг евр. ч. СССР, Кавказ, Казахстан, Ср. Азия, Сиб.,
Ю Прим.; 3 Европа, С Африка, 3 Азия, Япония.............................
. . Р. cadaverina L. — Зеленая навозница (малая зеленая коровница)
2 (1). Переднее дыхальце груди белое или светло-палевое. Тело с полоской
белого налета в переднем отделе среднеспинки. 4.5—6.5. — В зоне тайги
и лиственных лесов от Ленинграда до 10 Прим., в Сиб. — местами, горы
Кавказа, вся 3 Европа, С Монголия . . Р. ignita R.-D. (—aenea Ztt.)
19. Dasyphora R^-D. — 1 вид.
1 (1). Жилка R± сверху у основания с несколькими мелкими щетинками. Грудь
и брюшко темно-синие, металлически-блестящие. Среднеспинка в перед-
ней половине с широкой продольной полосой сероватого налета. 6—9. —
С и юг евр. ч. СССР, С Кавказ, Киргизия, Сиб., Ю Прим., Камч.; С, ср.
полоса 3 Европы, Япония, С Америка.....................D. cyanicolor Ztt.
20. Morellia R.-D. — 5 видов.
1 (12). Переднегрудь без волосков.
94
2 (7). dd: ширина лба приблизительно равна ширине 3-го членика усиков.
3 (4). tx изогнуты спереди, с рядом мелких коротких волосков, сзади вблизи
середины с 1 крепкой щетинкой; /2 в основной трети с 1 крепкой толстой
щетинкой снизу; первые 2—3 членика задних лапок снизу с длинными
толстыми щетинками, резко выделяющимися своей длиной от прочих ще-
тинок на лапках. Тело черное. Среднеспинка с переливчатыми пятнами
белого налета. 4.5—6.5. — С и юг евр. ч. СССР, Крым, Кавказ, Казах-
стан, Сиб., Прим.; вся 3 Европа.......................................
.............М. aenescens R.-D. — Конская или кривоногая садовница
4 (3). tx сзади с несколькими крепкими щетинками или без них. t2 прямые;
/2 без крепкой щетинки у основания; все членики задних лапок снизу
в мелких почти одинаковых щетинках.
5 (6). t2 у основания на дорсальной стороне с резко выступающим бугорком,
покрытым многочисленными толстыми шипами. 7—9.5. — СЗ и юг евр. ч.
СССР (Дагестан); Сиб., Ю Прим.; 3 Европа, С. Америка..................
.................................................М. podagrica Lw.
6 (5). t2 без бугорка у основания. 6.5—9. — СЗ и юг евр. ч. СССР (Украина),
Казахстан, Сиб., до Якут., Ю Прим.; 3 Европа, Монголия................
.........3. М. hortorum F11. — Обыкновенная или коровья садовница
7 (2). ширина лба равна примерно V3 ширины головы.
8 (9). tx с одной хорошо развитой задней щетинкой . . . М. aenescens R.-D.
9 (8). Задних щетинок на tr более одной или они неявственные (тонкие, корот-
кие).
10 (11). снизу с короткими согнутыми щетинистыми волосками; III и IV тер-
гиты брюшка с черными задними краями и слабо заметными переливча-
тыми пятнами..........................................М. podagrica Lw.
11 (10). Д снизу, помимо обычных мелких волосков в щетинистых волосках,
почти такой же длины, как задневентральные щетинки или немного короче
их; темные пятна на боках III и IV тергитов ясные; края тех же тергитов
не темнее их поверхности..............................М. hortorum Fall.
12 (1). Переднегрудь в тонких волосках по краям.
13 (14). de впереди поперечного шва сцинки и 2—3 пары позади него в виде
тонких волосков. t3 в вершинной половине с одним рядом волосков по
задневентральной стороне. 6—8. — Ю Прим.; Индия, Шри-Ланка, Индо-
китай, Китай (Тайвань), Ява, Япония, Австралия . . М. hortensia Wd.
14 (13). de шва крепкие, длинные; t3 в вершинной половине с неполными 2 ря-
дами редко стоящих волосков. 6.2—8.3. — Сиб., от Челябинска на В до
Прим., С Казахстан; Польша............................................
........................М. saishuensis Ouchi { = simpUcissima Zim.)
21. Mesembrina Mg. Личинки обитают в помете крупного рогатого скота,
взрослые мухи встречаются на помете, экскрементах в лесу, на деревьях и
кустарниках. — 4 вида.
1 (2). Вся среднеспинка и дистальная половина брюшка в светлых (желтых или
серовато-желтых) длинных и густых волосках. Щупики красно-желтые;
t2 и f2 3 длинные, изогнутые. Вершинные членики лапок часто рыжеватые.
12—16. — Хвойные леса Сиб., ДВ; СЗ Европы, Монголия, п-ов Корея
......................................................М. decipiens Lw.
2 (1). Среднеспинка и брюшко в черных волосках. Щупики бурые, t2 и /2 d
обычного строения, передние лапки черные; 5-й, реже 4-й и 3-й членики
красно-желтые.
3 (4). 1-я заднекрайняя ячейка крыла сильно сужена к вершине, поэтому отре-
зок костальной жилки между Т?4+5 и М в 2—4 раза короче предыдущего
отрезка этой жилки. Скулы в верхнем отделе с жесткими волосками,
в золотисто-желтом палете. Тело блестяще-черное. 11—12. — С и Ю
евр. ч. СССР, Кавказ, Сиб., лесная зона ДВ.............М. meridiana L.
4 (3). 1-я заднекрайняя ячейка слабо сужена к вершине, поэтому отрезок
костальной жилки между RM и М равен или едва короче предыдущего
отрезка.
95
5 (6). Продольная срединная полоса светло-серого налета на среднеспинке
широкая; ширина ее равна расстоянию между наружным ее краем и вну-
тренней границей плечевого бугорка. 11—13. — Лесная и лесостепная
зоны от Ленинградской обл. до Ю Прим.; 3 Европа (от Швеции до Венг-
рии) ..............................................М. resplendens Wahl.
6 (5). Полоса светлого налета на среднеспинке узкая, ширина ее в 1V2—2 раза
меньше расстояния между наружной ее границей и внутренним краем
плечевого бугорка. 7—11. — Лесная зона Сиб., Прим., Камч., Сах. . . .
..................................................... М. gracilior Zimin
22. Stomoxys Geoffr. — В СССР и на ДВ — 1 вид.
1 (1). М]+2 изогнута дуговидно. Лоб у обоих полов широкий. Ариста с длин-
ными волосками только по верхнему краю. Щупики тонкие, цилиндриче-
ские, в 3—5 раз короче направленного вперед тонкого хоботка. Грудь и
брюшко в густом желтовато-сером налете: среднеспинка с 4 продольными
полосами; III и IV тергиты посредине с треугольным, а по бокам заднего
края — с округлым бурым пятном. 5.5—7. — С и Ю евр. ч. СССР, За-
кавказье, Ср. Азия, В Сиб. и центр, часть ДВ — местами, Амур., Прим.
Космополит..............................S. calcitrans L. —Осенняя жигалка
23. Haematobia R.-D. — 2 вида.
1 (2). Проплевры впереди дыхалец и центральная часть гипоплевр без волосков.
Жилка с волосками в основной половине. Щупальца красно-желтые.
Грудь и брюшко темно-серые. Брюшко с 3 рядами крупных бурых пятен.
Ноги чернйе, колени желтые. 5—7. — Лесная зона евр. ч. СССР, Крым,
Алтай, Прим.; вся 3 Европа, С Монголия..................................
.................................Н. stimulans Mg. — Коровья жигалка
2 (1). Проплевры и гипоплевры в тонких волосках. Жилка без волосков.
Щупальца бурые или буровато-рыжие. В остальном сходен с предыду-
щими видами. 4—5.5. — Украина (включая Крым), Кавказ, Казахстан,
Ср. Азия, Прим.; юг 3 Европы .........................Н. atripalpis Bezzi
24. Lyperosia R. — 2 вида.
1 (2). Ариста в длинных волосках только по верхней стороне. Основные чле-
ники задних лапок d сплющенные с боков и расширены, по верхнему краю
с длинными щетинистыми волосками, но не превосходящими поперечник
члеников; крылья слегка дымчатые; закрыловые чешуйки коричневатые;
анальная жилка, постепенно утончаясь к вершине, немного не доходит
до заднего края крыла. 2.7—4.5. — СЗ и юг евр. ч. СССР; Казахстан,
Ср. Азия, Ю Сиб.; вся 3 Европа, Африка, С и Ю Америка, Гавайские о-ва,
о-в Гаити..................................................L. irritans L.
2 (1). Членики задних лапок 3 не расширены, цилиндрические; щетинистые
волоски почти вдвое длиннее поперечника соответствующих члеников
лапок. В остальном похож на L. irritans L. — Ю Прим.; Япония, Китай,
Австралия ...........................................L. exigua de Meij.
25. Myospila Rd. — В СССР и на ДВ — 1 вид.
1 (1). Крыловые чешуйки узкие, языковидные. Брюшко коническое, не шире
груди, с большими квадратными у в и мелкими округлыми у $ парными
пятнами на III и IV тергитах. Грудь серая с 4 продольными полосами,
узкими у о и широкими у <J. 5—9. — Вся Палеарктика, С и Ю Америка
.......................М. meditabunda F. — Обыкновенная личинкоедка
СИНИЕ И ЗЕЛЕНЫЕ (МЯСНЫЕ) МУХИ (Сем. CALLIPHORIDAE)
Мухи крупные или средней величины, редко мельче (4—5 мм), часто метал-
лически-синие или металлически-зеленые, hpl развиты; подщиток отсутствует
или слабо развит. Вершина М, как правило, изогнута вперед под углом, реже
дугообразно. У синантропных видов голова обычной формы, без угловидного
выступа вперед. Распространение всесветное, большинство представителей
в тропических областях, но есть и высокоширотные виды. — 34 вида.
96
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). Лоб широкий у обоих полов. Щеки очень широкие, их высота больше
х/2 высоты глаза. Л/1+2 изогнута дугообразно.........16. Booponus Aldr.
2 (1). Лоб d значительно уже лба $. М1+2 изогнута под углом.
3 (22). Ремигий (ствол радиальных жилок) голый.
4 (5). Вибриссальные углы расположены много выше края рта. Грудь в до-
полнение к макрохетам в длинных светлых волнистых волосках. Брюшко
без металлической окраски, проплевры голые ... 9. Pollenia R.-D.
5 (4). Вибриссальные углы расположены близко к краю рта. Грудь без свет-
лых длинных волнистых волосков. Брюшко обычно металлически-синее
или зеленое. Проплевры со щетинками.
6 (7). Задняя и передняя половины супрасквамального канта с группами щети-
нистых волосков. Грудная чешуйка голая. Скулы без волосков. Грудь
и брюшко металлически-зеленые, синие или медного цвета................
.................................................8. Lucilia R.-D.
7 (6). Задняя половина супрасквамального ребра (над основанием грудной
чешуйки) без групп щетинистых волосков. Грудная чешуйка обычно
с волосками. Скулы по меньшей мере в верхних частях с волосками,
иногда слабо заметными.
8 (9). Ариста с короткими волосками, по длине лишь немного превосходящими
ширину ее основной сильно утолщенной части .... 4. Abonesia Vill.
9 (8). Ариста с длинными волосками.
10 (11). Верхняя поверхность грудной чешуйки голая, брюшко без металличе-
ской окраски...................................... 7. Paradichosia S.-W.
11 (10). Верхняя поверхность грудной чешуйки в волосках, если голая, то
брюшко металлически темно-синее или зеленое, часто в сером налете.
12 (15). Крыловая и грудная чешуйки сильно затемненные. Крупные коре-
настые мухи с широким металлически-синим, со светлым налетом брюш-
ком.
13 (14). Предшовные ia имеются ...1. Calliphora R.-D.
14(13). Предшовные ia отсутствуют...................2. Aldrichina Aldr.
15 (12). Крыловая и грудная чешуйки светлые, желтоватые или слегка коричне-
ватые.
16 (17). Щеки двухцветные: спереди желтые или серебристые, сзади черные.
Кант крыловой чешуйки темный, ia 0-]-2. Брюшко синее, без светлого
налета. У d очень крупный гипопигий, у $ 5-й тергит с очень толстыми
щетинками.............................................6. Cynomya R.-D.
17 (16). Щеки одноцветные. Кант обеих чешуек светлый.
18 (19). Предшовная ia отсутствует................3. Triceratopyga Rohd.
19 (18). Предшовная ia имеется.
20 (21). У d V стернит сильно выдается над поверхностью брюшка, у ? вер-
шина V стернита с гребнем толстых щетинок (ктенидий) . 5. Acrophaga В. В.
21 (20). У d V стернит не выдается или незначительно выдается над поверх-
ностью брюшка, у ? ктенидий отсутствует..................Melinda R.-D.*
BellardiaR.-D. *
Onesia R.-D.*
22 (3). Ремигий co щетинистыми волосками, иногда светлыми и нежными.
23 (24). Грудная чешуйка в волосках, очень широкая, ее задний край прямой,
наружный — с неглубокой вырезкой, предкрыловый бугорок на груди
в длинных волосках..................................10. Chrysomya R.-D.
24 (23). Грудная чешуйка голая, округлая. Предкрыловый бугорок на груди
без волосков.
25 (28). Среднеспинка позади поперечного шва выпуклая. Базикоста желтая.
Лоб d самое большее немного шире глазкового треугольника, обычно
уже. Переднее дыхальце желтое или коричневое.
26 (27). На среднеспинке ас и de довольно слабые. Переднее дыхальце желтое.
Закрыловый склон с волосками. Крыловая и грудная чешуйки белые, со
* Виды этих родов не синантропы, но могут встречаться в населенных пунктах.
7 Зак. № 990 97
светлым кантом. У $ fr имеются. Окраска среднеспинки и брюшка у d
и $ одинаковая....................................13. Phormia R.-D.
27 (26). На среднеспинке ас и de крепкие. Переднее дыхальце коричневое.
Закрыловый склон голый. Крыловая и грудная чешуйки желтые, с жел-
тым кантом. У $ /г отсутствуют. В окраске среднеспинки и брюшка явст-
венный половой диморфизм................... . . 15. Tripocalliphora Pens.
28 (25). Среднеспинка позади поперечного шва плоская или несколько вы-
гнутая. Базикоста темная. Лоб d значительно шире глазкового треуголь-
ника. Переднее дыхальце черное или темно-бурое.
29 (30). Крыловая и грудная чешуйки белые (иногда грудная чешуйка корич-
неватая), голые и со светлым налетом. На среднеспинке ас по меньшей
мере 3—рЗ.......................... . . ♦ # е 14. Protocalliphora Hough.
30 (29). Крыловая и грудная чешуйки очень темные с бурым кантом, первая
с темными волосками. На среднеспинке ас перед поперечным швом отсут-
ствуют.
31 (32). Ариста с очень короткими волосками. На среднеспинке ia 0-|-2. Крылья
с почти замкнутой Рг. Передние дыхальца очень крупные, бархатисто-
черные ..................................11. Boreellus Oldr. et Schan.
32 (31). Ариста с длинными волосками. На среднеспинке ia 1—(-2. Крылья
с открытой Рх. Передние дыхальца обычные • .^12.JProtophormia Hough.
j ОПРЕДЕЛИТЕЛЬНЫЕЦТАБЛИЦЫЦВИДОВЦ
1. Calliphora R.-D. — 5 видов.
1 (2). Базикоста, как правило, желтая или желтовато-оранжевая; лицо"и щеки
в большей части оранжевато-желтые; ширина лба d составляет 1/1—V8
длины глаза, кокситы гипопигия на вершине притупленные. 6—12. —
Повсеместно...........................С. vicina Mg. — Синяя мясная муха
2 (1). Базикоста черная; лоб d значительно уже.
3 (4). Передние дыхальца груди яркие оранжево-желтые; щеки сплошь по-
крыты черными волоскамй; ширина лба d составляет —V22 длины
глаза, кокситы широкие. 8—16. — Ю ДВ; Япония, Китай . . С. lata Coq.
4 (3). Передние дыхальца груди темно-бурые.
5 (6). Щеки черные, покрыты черными и частично, особенно в задней части,
желто-оранжевыми волосками. Снизу голова окаймлена густыми длин-
ными желтыми или желто-оранжевыми волосками, ширина лба d соста-
вляет 1/14 длины глаза. Кокситы длинные и узкие. 11—14. — Вся
территория СССР; 3 Европа, С Америка, Китай, Япония, Индия, Филип-
пинские о-ва, частично в Африке, Гавайские о-ва ... С. vomitoria L.
6 (5). Щеки и лицо в большей части оранжево-желтые, покрытые черными
волосками; если щеки лишь частично с красноватым оттенком или пол-
ностью черные, то лобная полоса и медиана красновато-коричневые.
7 (8). Щеки и лицо в большей части оранжево-желтые; ширина лба d соста-
вляет х/10 длины глаза; кокситы гипопигия при взгляде в профиль с носи-
ком вниз, а концы церок чуть загнуты вверх. 8—14. — Вся террито-
рия СССР; 3 Европа, Гренландия. Обычен (рис. 45) . . С. uralensis Vill.
8 (7). Щеки черные или передняя часть их красновато-коричневая, медиана
также красновато-коричневая; у ? лобная полоса с красным пятном почти
во всю ширину нижней трети лобной полосы; у d лобная полоса почти
прерванная; церки гипопигия в отличие от таковых предыдущего вида на
концах прямые, а кокситы без загнутого вниз выроста, в виде носика.
9—12. — ДВ; С Америка................................С. terraenovae Mcq.
2. Aldrichina Aldr. — 1 вид.
1 (1). Среднеспинка с 3 широкими темными полосами, отчетливо заметными
в верхней части; ширина лба $ занимает около 2/3 длины глаза. V стернит
треугольный и много длиннее других стернитов. У d гипопигий с вен-
тральной стороны с 2 длинными палочковидными придатками. 6.6—19. —
98
Прим., Приам.; Япония, Китай, п-ов Корея, занесен в Калифорнию . . .
• . ...............................................A. grahami Aldr.
3. Triceratopyga Rohd. — 1 вид.
1 (1). Среднеспинка с 4 узкими продольными полосами; ширина лба ? занимает
около 4/б длины глаза, V стернит брюшка удлиненно-овальный; гипопигий
с вентральной стороны с коротким перовидным отростком. 7—8. — Сиб.
(местами), ДВ; Китай, Япония...................Т. calliphoroides Rohd.
4. Abonesia Vill. — 1 вид.
1 (1). Голова черная; орбиты и скулы матовые с серебряным отливом, щеки
блестящие. Среднеспинка черная с серым налетом, ас 2—|—3. Передние
дыхальца черные. Крыловые и грудные чешуйки белые, с белым кантом,
последняя — с темными волосками. Брюшко металлически сине-зеленое
со светлым налетом. 7—9. — С, включая высокие широты, 3. Саяны, ДВ
.......................................................... A. genarum Ztt.
99
5. Acrophaga В. В. — 1 вид.
1 (1). На среднеспинке перед швом расстояние между ас и между ас и de обычно
одинаковое. По бокам щитка с каждой стороны по 4 макрохеты. Лоб 3
равен 0.09 ширины головы, на орбитах нет or, 11—12. — Евр. ч. СССР,
повсеместно, Сиб., ДВ; 3 Европа, Япония..............A. subalpina Ringd.
6. Cynomya R.-D. — 2 вида.
1 (2). Голова спереди желто-оранжевая с золотистым отливом; волоски скул
короткие и нежные. 8—16. — В СССР повсеместно. На ДВ в центр, ч.
и на С обычен, в Прим. — редок; Гренландия, Аляска. Личинки в трупах
позвоночных ...........................................С. mortuorum L.
2 (1). Голова спереди серебристо-белая; волоски скул едва различимы. —
Чук. Крайне редок....................................С. саdaverina R.-D.
7. Paradichosia S.-W. — 3 вида.
1 (2). Грудные дыхальца светло-коричневые. Бедра и голени одноцветные,
коричневые. Основание и боковые поверхности верхних тергитов брюшка
желтые. 6—9. — Ю Прим.; Япония (Хоккайдо, Хонсю), Китай (Тайвань).
Имеется предположение, что личинки паразитируют на мелких животных
....................................... Р. pusilla Vill. (=japonica Hori)
2 (1). Грудные дыхальца темно-бурые, только передние голени светло-коричне-
вые. Бедра черные, если коричневые, то с темно-коричневым пятном
снизу в вершинной части. Брюшко темное, с серыми переливчатыми
пятнами.
3 (4). Передние дыхальца светло-коричневые, бедра черные. Паразит улиток.
7—8. — Прим.; Япония (центр, ч.).......................Р. itoi Капа
4 (3). Передние и задние дыхальца темно-бурые, бедра коричневые с темно-
коричневым пятном снизу в вершинной части. 6—7. — Прим.; Япония
(Хонсю, Кюсю)..........................................Р. okazaki Капо*
8. Lucilia R.-D. Мухи средних размеров яркой металлической окраски от
сине-зеленого до бронзового цвета*, блестящие, очень редко в светло-сером
опылении в верхней части среднеспинки и у основания брюшка. — 8 видов.
1 (6). Субкостальный склерит в коротких волосках.
2 (3). Базикоста темно-коричневая до черного цвета; срединные краевые-
щетинки на III тергите сильные. 5—8. — Ю Прим.; Китай . . L. chini Fan.
3 (2). Базикоста слегка коричневая до оранжевого цвета; срединные краевые-
щетинки III тергита отсутствуют.
4 (5). Лоб равен 0.11—0.18 ширины головы, V стернит со щетинистыми во-
лосками такой же длины, как и на прочих стернитах. 5—10. — Всесветно
................................................. . L. sericata Meig.
5 (4). Лоб шире, чем у L, sericata, V стернит с очень длинными тонкими неж-
ными волосками, превосходящими длину щетинистых волосков прочих
стернитов и даже краевых щетинок V тергита. 5—9. — Прим. (окр.
оз. Ханка, Хасанский р-н).............................L. hirsutula Grun.
6 (1). Субкостальный склерит в щетинках.
7 (8). Задние края тергитов с темной полосой, у $ посредине III тергита по-
лоса узкая. 6—12. — Ю Прим.; Япония, Китай................L. bazini Seg.
8 (7). Задние края тергитов по краю затемнены.
9 (10). На среднеспинке позади поперечного шва 3 ас. Лоб d в несколько раз
шире поперечника глазка, составляет 0.13—0.16 ширины головы, у $ —
около 0.4. V стернит сильно выступает. 3-й членик усиков $ длинный и
очень широкий. 5—9. — Циркумполярно в высоких широтах . . . .
................................................. L. fuscipalpis Ztt.
10 (9). На среднеспинке позади поперечного шва 2 ас,
11 (16). dd: лоб очень узкий, лишь иногда в 2 раза превосходит поперечник
глазка.
12 (15). Лоб около 0.03 ширины головы. III тергит без крепких щетинок.
100
13 (14). Анальный (последний видимый) тергит очень крупный. 5—11. — Вей
Палеарктика, кроме Крайнего Севера. Весьма обычен . . . L. caesar L.
14 (13). Анальный тергит маленький. 5—11. — В евр. ч. СССР редок, в В ч.
СССР — обычен; 3 Европа, Япония, Австралия . . . L. ampulacea VilL
15 (12). Лоб равен 0.07—0.11 ширины головы. 5—10. — В СССР от Заполярья
до южной границы; 3 Европа, Китай, Япония, С Америка .. L. illustris Mg.
16 (11). $$: лоб широкий, около 0.3 ширины головы.
17 (18). I тергит на спинной стороне (см. в профиль) выпуклый. Длина III
стернита почти в 1.5 раза превышает длину III тергита . . L. caesar L;
18 (17). I тергит на спинной стороне без выпуклостей. Длина III стерший
равна длине III тергита; если больше, то совсем незначительно.
19 (20). I стернит напоминает прямоугольник, не грушевидный, ширина его
составляет 4/5 длины....................................L. ampulacea VilL
20 (19). I стернит грушевидный, ширина его в самом широком месте (в вер-
шинной половине) составляет примерно 2/3 длины . . L. illustris Mg.
9. Pollenia R.-D. Мухи серого или черного цвета, грудь в длинных светлых
волосках, брюшко с шашечным рисунком из светлого налета. — 2 вида
(в СССР 13).
1 (2). Ширина лба <J равна поперечнику простого глазка, лицевой киль едва
намечается, широкий и плоский. Щупики коричневатые. Базикоста чер-
ная. Задние дыхальца бурые. Членики задних лапок d и ? снизу с 2 греб-
нями из толстых тупых зубцов, длина которых почти равна толщине.
Церки с перетяжкой в середине, расщеплены на конце на десятую часть
их длины. 7—10. — От Урала до Прим, и Камч.; Монголия....................
................................................ Р. pectinata Grunin
2 (1). Ширина лба <J значительно превосходит поперечник простого глазка.
Лицевой киль хорошо развит, узкий, острый. Щупики темные. Базикоста
коричневая. Задние дыхальца желтые. Членики задних лапок простые.
Церки без перетяжек в середине, расщеплены на вершине на V8 длины.
4—12. — Широко распространен в Голарктике......................Р. rudis F.
10. Chrysomya R.-D. — 2 вида.
1 (2). Скулы, лицо и щеки покрыты серебристо-белым налетом, в нежных белых
волосках. Усики черные, с бурыми основными члениками. 4—12. — Крым,
Азербайджан, Грузия, Ср. Азия, Ю ДВ; Ю Европа, Азия, Африка, Австра-
лия .....................................................Ch. albiceps Wd.
2 (1). Скулы, лицо и щеки желтые, в золотисто-желтых волосках. Усики жел-
тые. Крупнее. 8—13. — Очень редко на Ю Прим.; Китай, Индия . . . .
.......................................................Ch. megacephala F.
11. Boreellus Oldr. et Schan. Близок к Protophormia R.-D., но несколько
мельче. Грудь и брюшко блестящие черно-зеленые, без налета. Известен
1 циркумполярный вид, личинки которого развиваются в трупах млекопитаю-
щих, в частности в норах леммингов.
1 (1). Голова черная. Лоб в широкий, равен 0.23 ширины головы, крыловая
и грудная чешуйки сильно затемненные, с темным кантом; грудная че-
шуйка, кроме того, с темными волосками. Переднее дыхальце довольно
крупное. 6—9. — Новая Земля; Гыданский и Таймырский п-ова, Анабар-
ская губа, р. Оленек, Усть-Янск, о-в Новая Сибирь, в тундровой зоне ДВ
и на о-ве Врангеля .......................................В. atriceps Ztt.
12. Protophormia Haugh. Среднеспинка позади плоская, даже слегка вогну-
тая. Грудь и брюшко блестящие темно-синие, иногда с зеленоватым оттенком.
На крайнем С ДВ встречаются мухи с более узким брюшком, покрытым не-
ясными волосками. Личинки в трупах позвоночных, бытовых отходах. — 1 вид.
1 (1). Лоб в широкий: 0.13—0.14, у $ — 1/3 ширины головы; ас перед швом
отсутствуют. 8—12. — Повсеместно, часто...............Р. terraenovae R.-D.
101
13. Phormia R.-D. Среднеспинка позади шва слегка выпуклая, на передней
части — с едва различным налетом, окраска тела темно-зеленая. Личинки
в трупах позвоночных животных. — 1 вид.
1 (1). Лоб d узкий: 0.13—0.14, у $ — около 1/5 ширины головы; ас перед швом
слабые. 7—9. — Повсеместно, кроме севера. В ц. ч. ДВ не часто, на юге
обычен ................................................Ph. regina Mg.
14. Protocalliphora Hough. — 4 вида.
1 (4). Поперечные морщинки на скулах едва выражены.
2 (3). Лобная полоса d в 3—5 раз шире одной орбиты. 8—10. — Ленинград-
ская обл., Украина, Челябинская обл., Киргизская ССР, Прим............
..................................................Р. proxima Gr.
3 (2). Лобная полоса d приблизительно равна ширине одной орбиты. — Иркут.,
Прим. ...»...........................................Р. amblyogma Pens.
4 (1). Поперечные морщинки на скулах хорошо развиты.
5 (6). Тело темно-синее, средние голени с 1 щетинкой на вентральной поверх-
ности; вентральные поверхности лапок темно-коричневые до черных, /2
d обычные. 8—12. — Широко распространен в виде нескольких подвидов
по всей Палеарктике, Япония (о-ва Хоккайдо и Хонсю) . . Р. azurea F11.
6 (5). Тело темно-зеленое, средние голени без вентральной щетинки, вентраль-
ные поверхности лапок слегка коричневатые или оранжевые, у d на вну-
тренней стороне /2 ктенидий из толстых щетинок. 8—9. — О-ва Кунашир,
Сах.; Япония (Хоккайдо, Хонсю, Сикоку) ...............................
............................... Р. maruyamensis Kano et Sehin.
15. Tripocalliphora Pens. Личинки развиваются под кожей птиц, главным
образом на птенцах. — В Палеарктике 2 вида, в СССР 1.
1 (1). d с грязно-зеленой среднеспинкой и синим брюшком, ? с желтовато-
зелеными и среднеспинкой; и брюшком; у d среднеспинка и брюшко почти
без налета. — СЗ, центр евр. ч. СССР, Прим. (Владивосток) . . . .
.....................................................Т. lindneri Pens.
16. Booponus Aldr.
1 (1). Преимущественно светло-желтая муха с затемненной грудью и средне-
спинкой, покрытой светло-серым налетом, образующим рисунок в виде
продольных полос (5 — перед швом и 3 — за швом), с темными задне-
крайними боковыми полосами на брюшке у $>. — Ю Прим. Паразитирует
на кабарге и в пантах пятнистых оленей................................
........................... В. inexpectata Gr. — Кабарожья муха
СЕРЫЕ ПАДАЛЬНЫЕ (МЯСНЫЕ) МУХИ
(Сем. SARCOPHAGIDAE)
Тело удлиненное, преимущественно серого цвета, с массивным грудным от-
делом и относительно узкой головой (рис. 46, 1). На голове имеются два пра-
вильных ряда лобных щетинок. Скулы умеренно широкие, обычно покрыты
волосками или щетинками. На краю рта имеются вибриссальные щетинки,
одна пара наиболее длинная. Глаза средних размеров, без волосков. Грудь
с правильно расположенными в продольном направлении рядами щетинок:
эти ряды прерваны поперечным швом. Наибольшее таксономическое значение
имеет количество дорсоцентральных и акростихальных щетинок впереди и по*
зади шва. Различия в жилковании крыла у видов, приведенных в определителе,
незначительны и не используются при их идентификации. Брюшко с контраст-
ными или переливчатыми черными пятнами; у самца оно конусовидное,
у самки — яйцевидное, с хорошо заметными щетинками на краях тергитов.
Определение саркофагид строится в основном на различиях в строении постаб-
домена $ и его придатков — церок, сурстилей и фаллосомы (рис. 46, 47). Иден-
102
Рис. 46. Общий вид мухи, яйца и личинки.
1 — Angiometopa mihalyii, J; 2, 3 — Boettcherisca septentrionalis: 2 — личинка, 3 — пупарий. (Ориг.).
Рис. 47. Детали^строения гениталий самцов.
7 — Wohlfahrtia sp., постабдомен в профиль: ат — анальный терг., гт — генитальный терг., с — сур*
стили, ф — фаллосома, ц — церки; 2 — Parasarcophaga sp., в профиль, фаллосома (слева) и церки (справа):
апф — апикальная часть парафалла, боапф — боковые отростки апикальной части парафалла, вл — вен-
тральные лопасти, м — мембрана, мо — мембральные отростки, пф — парафалл, соапф^— срединный
отросток апикальной части парафалла, ст — стилеты. (Ориг.).
тификация самок, как правило, затруднена, поскольку для этого необходимо
детальное изучение концевых стернитов и тергитов, скрытых под основными
сегментами брюшка.
Диагноз сем. Sarcophagidae
Тело покрыто серым налетом, без синего или зеленого металлического
блеска, с продольными полосами на груди и черным рисунком в виде переливча-
тых или контрастных пятен на брюшке. 3-й членик антенн всегда больше 2-го.
Стерниты брюшка свободные, не прикрыты краями тергитов.
На Дальнем Востоке обитает 119 видов 41 рода 4 подсемейств. Из них меди-
цинское и ветеринарное значение имеет 47 видов, относящихся к 21 роду 2 под-
семейств (в СССР 375 видов).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (12). Задние коксы (тазики — сх3) сзади без волосков. VI тергит d слит
с генитальным тергитом, в месте слияния имеется ряд крепких щетинок
(рис. 47, 7).
2 (7). Ариста с очень короткими прилегающими волосками.
3 (4). Скулы голые; 2-й сегмент аристы очень вытянутый, 3-й членик усиков
в 2.5—3 раза длиннее 2-го (рис. 48, 7)............1. Goniophyto Town.
4 (3). Скулы покрыты щетинками; 2-й сегмент аристы нормальный, 3-й членик
усиков более короткий.
5 (6). Брюшко с переливчатым блестящим рисунком ... 4. Brachicoma Rd.
6 (5). Брюшко с крупными округлыми черными пятнами на сером фоне . . .
............................................. 6. Wohlfahrtia В. В.
7 (2). Ариста перистая, с длинными волосками.
8 (11). Тело в густом светло-сером налете. Брюшко с черными округлыми
пятнами.
9 (10). Орбитальные щетинки (or) у d отсутствуют. Щупики оранжевые.
Церки d длинные и тонкие (рис. 48, 4).........3. Angiometopa В. В.
10 (9). Имеются 2 пары крепких qf. Щупики черные. Церки d короткие
(рис. 49, 2).....................................5. Saгсophila Rd.
11 (8). Тело темное, иногда почти черное. Пятна на брюшке образуют нечеткий
переливчатый шашечный рисунок...........................2. Agria R.-D.
12 (1). сх3 на задней поверхности с небольшими тонкими волосками. VI тергит
d редуцирован.
13 (14). На внешней поверхности средних бедер (/2) обоих полов имеется оваль-
ное пятно из очень коротких оранжевых, иногда желто-серых щетинок
............................................ 11. Sarcotachinella Town.
14 (13). /2 без каких-либо пятен.
15 (16). Ряды лобных щетинок (/г) в нижней части лба не расходятся в стороны,
параллельны..........................................7. Ravinia R.-D.
16 (15). Ряды (/г) книзу расходятся в стороны.
17 (24). За швом на спинке груди 3 пары дорсоцентральных щетинок (de)
примерно равной длины.
18 (19). Проплевры покрыты черными волосками. Мембранальные лопасти
фаллосомы в виде крупного широкого выроста (рис. 51, 3) ........
............................................ 12. Phallantha Rohd.
19 (18). Проплевры голые. Мембранальные лопасти иные.
20 (21). Фаллосома удлиненная, без расширений, несет короткие крючко-
видные выросты (рис. 50, 7).......................8. Helicophagella End.
21 (20). Фаллосома обычно значительно расширена в дистальной (задней)
части; короткие крючковидные выросты отсутствуют.
22 (23). Апикальная часть (верхушка) парафалла в профиль в виде ковша.
Мембранальные лопасти длинные, своими концами сближаются с апи-
кальной частью парафалла (рис. 51, 7)............10. Bellieriomima Rohd.
23 (22). Апикальная часть парафалла не ковшевидная. Мембранальные лопасти
более короткие (рис. 50, 5)..........................9. Pierretia R.-D.
104
24 (17). За швом на спинке груди 4—5 пар de, передние пары часто короче
задних.
25 (32). Проплевры в верхней части покрыты черными волосками.
26 (27). Акростихальные щетинки (ас) перед швом на груди хорошо заметны
............................................20. Sarcorohdendorfia Ваг,
27 (26). ас перед швом отсутствуют.
28 (29). Фаллосома в срединной части прозрачная, с четко обособленной апи-
кальной частью (рис. 52, 1)....................14. Boettcherisca Rohd.
29 (28). Фаллосома значительно склеротизирована, в том числе и в срединной
части.
30(31). Церки в профиль короткие и широкие, топоровидные (рис. 53, 3).
III тергит брюшка на середине заднего края без крепких щетинок . . .
..................................................15. Kramerea Rohd,
31 (30). Церки длинные, сравнительно узкие (рис. 57, 4). III тергит с парой
крепких срединных краевых щетинок.........17. Phallosphaera Rohd.
32 (25). Проплевры в верхней части без волосков.
33 (34). ас на груди отсутствуют..................13. Вегсаеа R.-D.
34 (33). Имеются зашовные ас.
35 (36). Апикальная часть парафалла довольно узкая, перепончатая (рис. 59, 3)
........................ 22. Sarcophaga Mg.
36 (35). Апикальная часть парафалла хорошо склеротизирована, обычно несет
боковые отростки.
37 (38). Апикальная часть парафалла крупная, разделенная на два больших
раздвоенных на концах отростка. Срединный отдел апикальной части
парафалла не выражен (рис. 58,1)................18. Robineauella End,
38 (37). Апикальная часть парафалла с хорошо выраженным срединным отде-
лом.
39 (40). Апикальная часть парафалла очень длинная, несет почти прямые
боковые отростки (рис. 58, 3)..................19. Sinonipponia Rohd.
40 (39). Апикальная часть парафалла более короткая.
41 (42). Апикальная часть парафалла без отростков (рис. 59, 2) .....
................................?.................21. Takanoa Rohd.
42 (41). Апикальная часть парафалла с боковыми отростками (рис. 56, 1)
..............♦................. 16. Parasarcophaga Johns, et Tiegs
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
1. Goniophyto Town. Средней величины мухи серого цвета. Основание
антенн в профиль расположено выше уровня середины глаз, 3-й членик усиков
в 2.5—3 раза длиннее 2-го, ариста голая. Угловые вибриссальные щетинки (vi)
расположены на краю рта, очень длинные; ас отсутствуют; de за швом в коли-
честве 3 пар; сх3 сзади без волосков. Личинки некрофаги. — В СССР 1 дальне-
восточный вид.
1 (1). Щеки высокие, около 0.2 высоты глаза (рис. 48, 1). Задний край церок
почти прямой. 6—8. — Ю Прим.; Япония, Китай, п-ов Корея. Личинки
в трупах морских животных........................G. honshuensis Rohd.
2. Agria R.-D. Средней величины темноокрашенные (иногда почти серые)
мухи. 3-й членик усиков в 2 раза длиннее 2-го, ариста с волосками. Скулы
узкие, с 1 рядом щетинок. За швом на груди 3 пары de; сх3 голые. Постабдомен d
крупный, черный. Личинки некрофаги и хищники бабочек. — В СССР и на ДВ
3 вида (медико-ветеринарное значение имеют 2).
1 (2). Церки отогнуты назад (рис. 48, 2). Тело почти черное. Шашечный рису-
нок на брюшке плохо развит. Крылья затемнены. 6—9. — Амур., Ю Хаб.,
Прим., Сибирь; Европа, Монголия, Япония. В природе и населенных
пунктах лесной зоны. Личинки в трупах животных и в куколках некото-
рых бабочек.......................................A. monachae Kram.
2 (1). Церки не отогнуты назад (рис. 48, 3). На генитальном и анальном терги-
тах d хорошо выражены парные выросты в виде больших бугров. Тело
105
более светлое. Брюшко с типичным шашечным рисунком. Крылья не
затемнены. 6—9. — Амур., Ю Хаб., Ю и 3 Прим., Ю Сиб., евр. ч. СССР,
Закавказье, Ср. Азия. В окрестностях населенных пунктов; личинки —
хищники гусениц и куколок бабочек......................................
A. mamillata Pand. (=pactfica Karazeeva)
Рис» 48. Детали строения самцов»
1 — Coniphyto honshuensis, голова; 2—4 — постабдомен: 2 —
Agria monachae, 3 — A. mamillata, 4 — Angiometopa mihalyil*
(2, 3 —jno: Артамонов, 1985; остальное — оригЛ
3. Angiometopa В. В. Сред-
них размеров стройные свет-
лоокрашенные мухи. 3-й чле-
ник усиков в 1.5—2 раза
длиннее 2-го, ариста с длин-
ными волосками. Скулы уз-
кие, с длинными щетинками;
or у $ отсутствуют. За швом
3 пары dc\ сх3 голые. Тело
покрыто очень густым светло-
серым налетом. Брюшко с чер-
ными пятнами и срединной
полосой на тергитах. Постаб-
домен 3 средних размеров.
Личинки — возбудители миа-
зов человека и копытных,
некрофаги, хищники и пара-
зиты насекомых., — 1 вид
(в СССР 4).
1 (1). Церки длинные и
узкие, саблевидные
(рис. 48, 4). Брюшко
с 8 черными, хорошо
отграниченными округ-
лыми боковыми пят-
нами и срединной по-
лоской из слитых уд-
линенных пятен. 7—
12. —Ю Прим., В Сиб.,
В Европа; Монголия,
С п-ова Корея. В посел-
ках лесной зоны; ли-
чинки в трупах грызу-
нов ......................
A. mihalyii Rohd. et Verv*
Brachicoma Rd. Крупные или средней величины темноокрашенные мухи,
тело в крепких щетинках. 3-й членик усиков в 2 раза длиннее 2-го, ариста голая.
Скулы с многочис тонными щетинками, vi очень крепкие. За швом 3 пары cfc;
сх3 без волосков. Постабдомен 5 средней величины. Личинки — хищники личи-
нок и куколок шмелей. Мухи на продуктах питания, слизывают потовые вы-
деления. — 3 вида (в СССР 5) (медико-ветеринарное значение имеет 1).
IJl). Цер<и короткие, слегка крючковидно изогнуты, сурстили короткие
(рис. 49, Z). Гениталии черные. 8—12. — Маг., Хаб., Прим.; Голарктика.
Долины равнинных рек и агроценозы . » • ...................В. devia jFll.
5. Sarcophila Rd. Мелкие, реже средней величины светлоокрашенные мухи
с черными пятнами на брюшке. 3-й членик усиков в 1.5—2.5 раза длиннее 2-го,
ариста с длинными волосками. За швом 3 пары dc\ сх3 голые. Постабдомен
3 крупный. Личинки — некрофаги. — 1 вид (в СССР 6).
1 (1). Скулы в 2 раза уже щек, с двойным рядом тонких волосков. Церки по
заднему краю слегка вогнутые, короче сурстилей (рис. 49, 2). Брюшко
с черными пятнами на III— V тергитах. 4—8. — Ю Прим.; Япония,
106
п-ов Корея. Мухи близ берега моря и равнинных рек, на пастбищах
...............................................S. japonica Rohd.
6. Wohlfahrtia В. В. Крупные мухи с блестящим черным рисунком в виде
пятен и полос на груди и брюшке. Ариста неперистая, с очень короткими воло-
сками; сх3 сзади без волосков. Постабдомен S крупный. Личинки — некрофаги,
вызывают миазы позвоночных. — 2 вида (вредоносное значение имеет 1 вид),
в СССР 15.
1 (1). 3-й членик усиков по длине равен 2-му. 1-й и 2-й членики усиков и щупи-
ков красно-желтые. Церки короткие, сурстили прилегают к анальному
тергиту (рис. 49, 3). Тело очень темное, блестящее. III—V тергиты с оваль-
ными черными пятнами. 8—15. — Ю Хаб., Прим.; Голарктика. Открытые
Рис. 49. Постабдомен самцов.
1 — Brachicoma devia; 2 — Sarcophila japonica; 3 — Wohlfahrtia meigeni; 4 — Ravinia striata: a — фалло-
сома, 6 — церки. (Ориг.).
биотопы на окраинах населенных пунктов, агроценозы. Личинки вызы-
вают миазы у человека, млекопитающих и земноводных (лягушки) . . .
................................. ..............W. meigeni Sehin.
7. Ravinia R.-D. Мелкие или средней величины мухи серой окраски. Ряды
/г книзу не расходятся в стороны. Ариста перистая, сх3 с волосками. Про-
плевры голые. Брюшко с типичным шашечным рисунком. Постабдомен обоих
полов красного цвета. Личинки — копро- или некрофаги. — В СССР 1 вид»
1 (1). Лоб и скулы серебристо-желтые. Постабдомен d крупный. Церки к концу
равномерно сужающиеся (рис. 49, 4). 4—9. — На ДВ встречается повсе-
местно (на север до 64° с. ш.); Палеарктика и Ориентальная зоогеогра-
фическая область. Полусинантроп; обычен. Личинки в фекалиях, экскре-
ментах млекопитающих и птиц, гниющем мясе; миазы человека и домаш-
них животных...............................................R. striata F.
8. Helicophagella End. (=Bellieria End.). Крупные или средней величины
мухи серого цвета, иногда с коричневатым налетом. Ариста перистая. 3 пары
зашовных de примерно равной длины, ас перед швом отсутствуют, за швом —
1 пара; сх3 с волосками. Проплевры голые. Фаллосома длинная, в профиль
без прозрачной зоны. Постабдомен d очень крупный, черный. Личинки
копро-некрофаги. — 4 вида (вредоносное значение имеют 2), в СССР 14.
1 (2). Фаллосома узкая, длинная. Мембранальные лопасти незначительно вы-
ступающие; боковые отростки парафалла короткие (рис. 50, 7). 6—14. —
На ДВ — повсеместно, на север до 65° с. ш.; Голарктика, Ориентальная
область. Массовый вид, полусинантроп. Личинки в экскрементах, трупах,,
мясных отбросах; вызывает миазы у позвоночных . . . Н. melanura Mg.
107
2 (1). Фаллосома массивная, более короткая. Мембранальные лопасти заметно
выступающие вперед; все боковые отростки парафалла крупные (рис. 50, 2).
8—12. — Амур., Ю Прим., С Казахстан. Лесостепные биотопы . . . .
..................................................Н. inopinata Rohd.
9. Pierretia R.-D. Средней величины или мелкие мухи с темноокрашенным
телом. Ариста перистая. 3-й членик усиков в 1.5—2.5 раза длиннее 2-го. Скулы
с длинными щетинками. За швом на груди 3 пары de; сх3 с волосками. Про-
плевры голые. Тергиты по заднему краю с длинными щетинками. Брюшко
Рис. 50. Фаллосома (а) и церки (б) самцов.
1 — Helicophagella melanura’, 2 — Н. inopinata’, 3 — Pierretia villeneuvei’, 4 — P. ugamskii. (Ориг.).
с резким шашечным рисунком. Личинки — некрофаги, паразиты наземных мол-
люсков или хищники в яйцевых коконах пауков. — 7 видов (вредоносное зна-
чение имеют 2), в СССР 15.
1 (2). Щетинки на скулах очень крепкие. Апикальная часть парафалла нечетко
обособленная, длинная и тонкая, несет острые выросты (рис. 50, 3).
4—8. — Ю и В Прим., 3 и Ю Сиб.; Европа, СВ Китай. Влажные луга,
пастбища. Мухи на пищевых продуктах в природных условиях ....
................................................... Р. villeneuvei Boett.
2 (1). Щетинки на скулах менее крепкие. Апикальная часть парафалла хорошо
обособленная, лишена острых выростов. Мембранальные лопасти в виде
острых светлых углов (рис. 50, 4). 5—9. — Амур., Ю Хаб., Прим ;
СВ Китай, п-ов Корея, Япония. Лесостепные и лесные биотопы. Полу-
синантроп. Личинки — нектрофаги на трупах грызунов, птиц, на мясных
отбросах ...............................................Р. ugamskii Rohd.
10. Bellieriomima Rohd. Средней величины или крупные серые мухи.
3-й членик усиков в 2—3.5 раза длиннее 2-го, ариста перистая. Скулы с длин-
ными щетинками. За швом на груди 3 пары de; ех3 с волосками. Проплевры го-
108
1 (1). Фаллосома с многочисленными длинными выростами; апикальная часть
парафалла с резко выдающимся назад светлым срединным отделом
(рис. 51, 5). 5—9. — Ю Хаб., Прим.; Япония, п-ов Корея. Многочислен.
В лесной зоне. Личинки в трупах насекомых . . Р. sichotealini Rohd.
13. Вегсаеа R.-D. ( = Coprosarcophaga Rohd.). Крупные серые мухи с крас-
ными гениталиями. 3-й членик усиков в 2 раза длиннее 2-го; ас отсутствуют;
сх3 с волосками. Проплевры голые. Фаллосома очень крупная и узкая; мембра-
нальные лопасти в виде одной пары крупных отростков. VI тергит g разделен
на две ярко-красные пластинки. Личинки — копро-некрофаги. — В СССР 1 вид.
Рис. 52. Фаллосома (а) и церки (б) самцов.
1 — Boettcherisca peregrina; 2 — В. septentrionalis\ 3 — Kramerea schuetzei’, 4 — Parasarcophaga argyrostoma.
(Ориг.).
1 (1). Фаллосома по заднему краю почти прямая; мембранальные лопасти
серповидные, заостренные. Церки тонкие, в базальной части с глубокой
выемкой (рис. 51, 4). 7—16. — Ю Хаб., Ю и 3 Прим.; космополит. Полу-
синанТроп, в Ю Прим. — синантроп (залетает в помещения). Личинки
в трупах животных, мясных отбросах, экскрементах; миазы человека и
млекопитающих......................В. cruentata Mg. (=haemorrhoidalis Fll.)
14. Boettcherisca Rohd. Крупные, реже средних размеров серые мухи.
3-й членик усиков в 2—4 раза длиннее 2-го. Скулы с крепкими щетинками;
ас перед швом на груди отсутствуют; сх3 с волосками. Проплевры с хорошо за-
метными волосками; IV тергит с парой крепких щетинок на середине заднего
края. Личинки — некрофаги. — В СССР и на ДВ — 2 вида.
1 (2). Церки в профиль по заднему краю почти прямые, на конце с зубцом
(рис. 52, 1). Анальный тергит красно-коричневый. 7—14. — Ю Прим.;
В и Ю Китай, п-ов Корея, Япония, Ориентальная и Австралийская зоо-
географические области. Синантроп. Личинки в мясных и рыбных отбро-
сах, трупах позвоночных..................................В. peregrina R.-D.
2 (1). Церки изогнуты, с равномерно сужающейся к концу апикальной частью
(рис. 52, 2). Анальный тергит черный. 7—15. — К) Прим., Ю Кур.;
110
Китай, Япония. Полусинантроп. Многочислен. Личинки в трупах мелких
млекопитающих и птиц, пищевых отбросах . .В. septentrionalis Rohd.
15. Kramerea Rohd. Крупные, реже средней величины мухи серой окраски.
3-й членик усиков в 2.5 раза длиннее 2-го. Скулы с многочисленными щетин-
ками. За швом на груди 4 пары de, две задние сх3 пары наиболее длинные. Про-
плевры в густых черных волосках, волосистые. III тергит без срединных задне-
крайних щетинок. Личинки — некрофаги и хищники куколок и гусениц чешуе-
крылых. — В роде 1 вид.
1 (1). Фаллосома сильно склеротизирована, в профиль округлая. Церки корот-
кие и широкие, топоровидные (рис. 52, 5). Гениталии черные. 7—16. —
Рис. 53. Фаллосома (а) и церки (б) самцов.
1 — Parasarcophaga crassipalpis\ 2 — Р. macroauriculata; 3 — Р. unguitigris\ 4 — Р. albiceps. (Ориг.).
Амур., Ю Хаб., Прим., Сах., Ю Кур.; Палеарктика. Лесные и лесостеп-
ные биотопы; довольно обычен в поселках. Массовый полусинантроп.
Личинки в трупах грызунов, птиц, пресноводных рыб, в гусеницах и
куколках бабочек...............................К. schuetzei Kram.
16. Parasarcophaga Johns, et Tiegs. Крупные или средней величины серые
мухи. 3-й членик усиков в 2—4 раза длиннее 2-го. Скулы с щетинками средней
длины, ас перед швом на груди отсутствуют; de за швом в числе 4—6 пар, две
задние пары наиболее длинные; сх3 волосистые. Проплевры без волосков.
Церки (5 длинные. Анальный тергит черный, коричневый или красный. Брюшко
с типичным шашечным рисунком. Личинки могут быть некрофагами, копро-
фагами, хищниками, паразитами. — В СССР 32 вида, на ДВ 22 (вредоносное
значение имеет 21).
1 (4). Анальный тергит 3 и гениталии $ всегда красные. Генитальный тергит d
по заднему краю с крепкими щетинками.
2 (3). Мембранальные лопасти крупные, пластинчатые, на конце заостренные.
Боковые отростки апикальной части парафалла короткие (рис. 52, 4).
111
Анальный тергит и церки красные. 7—16. — Амур., Ю Хаб., С Прим.
Космополит, синантроп. Личинки — некро- и копрофаги; миазы позво-
ночных ......................................Р. argyrostoma R.-D.
3 (2). Мембранальные лопасти не пластинчатые, не заострены. Боковые от-
ростки апикальной части парафалла длинные (рис. 53, 7). Генитальный и
анальный тергит красные. 9—17. — Ю Прим. Космополит, синантроп.
Личинки — некрофаги; в мясных и рыбных отбросах, трупах; миазо-
образователи.......................................Р. crassipalpis Mcq.
4 (1). Анальный тергит d, как правило, черный, у некоторых видов коричне-
вый. Генитальный тергит d по заднему краю без щетинок.
5 (12). Мембранальные лопасти в виде непарного выроста, сидящего на узком
стебельке.
6 (7). Щупики у обоих полов светло-коричневые, иногда желтые. 8—15. —
Ю Прим.; Китай, п-ов Корея, Ориентальная и Австралийская области.
Полусинантроп, приуроченный к морскому побережью. Личинки —
копро-некрофаги...................... . Р. sericea Walk. (=knabi Park.)
7 (6). Щупики всегда темные.
8 (9). Церки короткие и заостренные, в срединной части очень широкие, на
переднем крае несут крупные вздутия, усаженные шипами (рис. 53, 2).
7—17. — ЮВ Прим.; Китай, Непал, Япония, ЮВ Азия. Прирусловые*
леса и окраины таежных сел. Личинки в фекалиях или трупах ....
........................................... Р. macroauriculata Но
9 (8). Церки сравнительно длинные, в срединной части расширены незначи-
тельно, с небольшими вздутиями или без них.
10 (11). Церки очень тонкие, слегка изогнуты вентрально в апикальной части,
их базальная часть несет шипы (рис. 53, 3). 10—16. — Ю Хаб., Прим.;
Китай, Япония, п-ов Корея. Долины рек. Личинки в экскрементах, реже
в трупах..........................................Р. unguitigris Rohd.
11 (10). Церки более широкие, почти прямые (рис. 53, 4), 10—17. — Амур.г
Ю Хаб., Прим., Сах., Кур.; Палеарктика, Ориентальная и Австралий-
ская области. Полусинантроп. Личинки — полифаги; в экскрементах
позвоночных, фекалиях, трупах, мясных и рыбных отбросах, куколках
бабочек ..............................................Р. albiceps Mg-
12 (5). Мембранальные лопасти без стебелька, в виде бугра или пластинчатых
парных отростков.
13 (16). Апикальная часть парафалла с плохо выраженным срединным отделом
или последний отсутствует.
14 (15). Боковые отростки апикальной части парафалла широкие, к концу почти
не сужаются. Срединный отдел апикальной части парафалла отсутствует.
Мембранальные лопасти примерно одной длины, нижняя пара клешне-
видная (рис. 54, 7). 6—14. — Амур., Хаб., Маг., Камч.; Голарктика.
Лесной вид, па севере ДВ — обычный полусинантроп. Личинки — хищ-
ники бабочек..........................................Р. aratrix Pand.
15 (14). Боковые отростки апикальной части парафалла к концу сужаются.
Срединный отдел апикальной части парафалла в виде тонкого прозрач-
ного выроста. Верхние мембранальные лопасти склеротизированные,
крючковидные, нижние — прозрачные, не расщеплены (рис. 54, 2).
9—17. — Амур., Ю Хаб., Прим., Сах.; Голарктика. Лесные биотопы и
зеленая зона населенных пунктов. Личинки — хищники некоторых бабо-
чек. Мухи посещают продукты питания.................Р. uliginosa Kram.
16 (13). Апикальная часть парафалла с хорошо развитым срединным отделом.
17 (24). Мембранальные лопасти в числе двух пар, обычно тонкие и острые.
18 (23). Обе пары мембранальных лопастей узкие, примерно одинаковой ве-
личины.
19 (22). Обе пары мембранальных лопастей равномерно сужаются к переднему'
концу. Апикальная часть парафалла снизу без зубцов. Передние пара-
меры к концу сужающиеся.
20 (21). Апикальная часть парафалла длинная и заостренная. Мембранальные*
лопасти прямые или слегка загнуты вверх (рис. 54, 3). На внешней стороне*
апикальной части церок имеется небольшое вздутие. 8—15. — Ю Хаб.г
112
Ю и В Прим., Ю Сиб.; С Китай, п-ов Корея, Япония. Травянистые берега
озер и рек, агроценозы, окраины населенных пунктов. Полусинантроп.
Личинки — некрофаги ..............................Р. polystylata На
21 (20). Апикальная часть парафалла более короткая. Мембранальные лопасти
загнуты вниз. Церки равномерно сужены к концу, без вздутия (рис. 54, 4).
7—17. — Амур., Ю Хаб., Прим., Сах., Кур. (кроме севера); Палеарктика
и север Ориентальной области. Массовый полусинантроп. Личинки —
Рис. 54. Фаллосома (а) и церки (б) самцов.
1 — Parasarcophaga aratrix\ 2 — Р, uliginosa', 3 — Р, polystylata', 4 — Р. similis. (Ориг.).
копро-некрофаги, хищничают на наземных моллюсках и вызывают миазы
у человека......................................Р. similis Meade*
22 (19). Верхняя пара мембранальных лопастей на конце резко расширена.
Апикальная часть парафалла снизу с многочисленными зубцами (рис. 55,.
7). 6—10. — Ю Кур.; Япония. Приречные низкотравные луга и паст-
бища. Личинки — копро-некрофаги..............Р. kawayuensis Капо
23 (18). Нижняя пара мембранальных лопастей широкая, в виде вытянутого
треугольника (рис. 55, 2). Апикальная часть парафалла красно-коричне-
вая. 6—17. — Амур., Ю Хаб., Прим., Ю Сиб.; С Китай, п-ов Корея, Япо-
ния. В лесных районах — типичный полусинантроп. Личинки — некро-
фаги .............................................Р. kanoi Park
24 (17). Мембранальные лопасти в числе одной пары пластинчатых выростов.
25 (26). Боковые отростки апикальной части парафалла на конце невильчатые,
без зубцов. Мембранальные лопасти длинные, узкие и сильно склероти-
8 Зак № 990 113
зированы (рис. 55, 5). 9—12. — Ю Прим. Только в населенных пунктах
..................................... Р. ussuriensis Kolom. et Art.
26 (25). Боковые отростки апикальной части парафалла на конце вильчатые
или с зубцом. Мембранальные лопасти склеротизированы в меньшей
степени.
27 (28). Конец церок в профиль значительно вытянут и утончен; апикальная
часть церок волнообразно изогнута (рис. 55, 4). 6.5—16. — Ю Хаб.,
Прим., Ю Сах.; Голарктика. Обитает в диких и окультуренных биотопах
Рис. 55. Фаллосома (а) и церки (б) самцов.
1 — Parasarcophaga kawayuensis; 2 — Р. kanoi; 3 — Р. ussuriensis; 4 — Р. tuberosa. (Ориг.).
лесостепи, дубовых и кедрово-широколиственных лесов. Личинки в тру-
пах животных и куколках некоторых бабочек . . . * Р. tuberosa Pand.
28 (27). Конец церок в профиль вытянут в меньшей степени, иногда с резким
зубцом; волнообразный изгиб апикальной части церок не выражен.
29 (30). Конец церок не сужен, б. м. притупленный, с коротким зубцом
(рис. 56, 7). 8—17. — Амур., Ю Хаб., Прим., Сах.; Голарктика, Ориен-
тальная область. В Ю Прим. — обычный полусинантроп. Личинки в тру-
пах позвоночных; хищники куколок бабочек; миазы у животных и чело-
века ..................................................Р. harpax Pand.
30 (29). Церки постепенно утончаются к концу.
31 (32). Анальный тергит коричнево-красный, иногда на заднем крае желтый.
Мембранальные лопасти очень длинные, сильно склеротизированные,
к концу резко сужающиеся (рис. 56, 2). 7—14. — Ю Прим. (оз. Ханка),
Закавказье, Ср. Азия, Казахстан; Европа, Монголия, С Китай, п-ов Ко-
рея. Личинки в трупах позвоночных и фекалиях . . . Р. jacobsoni Rohd.
32 (31). Анальный тергит черный.
33 (34). Срединный отдел апикальной части парафалла короткий неширокий,
в профиль треугольный, с коротким выростом на вершине. Боковые от-
ростки апикальной части парафалла прямые, их зубцы, образующие
«вилку», равны по величине (рис. 56, 3). 9—14. — Ю и В Прим.; Китай,
п-ов Корея, Япония, Индо-Малайская и Австралийская области. В местах
отдыха людей близ берега моря. Полусинантроп; личинки в трупах мор-
ских животных......................................Р. brevicornis Но
Рис. 56, Фаллосома (а) и церки (б) самцов,
1 — Parasarcophaga harpax; 2 — Р, jacobsoni', 3 — Р, brevicornis', 4 — Р, kobayashii» (Ориг.).
34 (33). Срединный отдел апикальной части парафалла узкий, вытянутый,,
с длинным заостренным выростом на вершине. Боковые отростки апикаль-
ной части парафалла изогнуты.
35 (36). Мембранальные лопасти очень большие, широкие, прозрачные. Вен-
тральный зубец на боковых отростках апикальной части парафалла зна-
чительно больше дорсального (рис. 56, 4). 13—18. — Ю Прим., Забай-
калье; Япония, Китай. Долины рек; личинки — некрофаги......................
........................................ . . . . Р. kobayashii Hori
36 (35). Мембранальные лопасти сравнительно небольшие, узкие. Вентраль-
ный зубец на боковых отростках апикальной части парафалла меньше
дорсального.
37 (38). Передние парамеры резко изогнуты. Мембранальные лопасти очень
острые (рис. 57, 7). 7—16. — Ю Прим. (оз. Ханка), евр. ч. СССР, Закав-
казье, Ср. Азия, Ю Сибирь; Китай, Монголия. Полусинантроп. Личинки
в фекалиях и трупах позвоночных.......................Р. portschinskyi Rohd.
38 (37). Передние парамеры без резких изгибов.
39 (40). Срединный отдел апикальной части парафалла с ясным выступом на
внутренней стороне. Мембранальные лопасти на конце с крючком, на-
правленным вверх (рис. 57, 2). 8—15. — Маг. (на сев. до 62° с. ш.), Камч.,
Амур., Хаб., Прим, (кроме Ю), горы Ср. Азии, Сибирь; С Монголия.
На юге ДВ — приречные луга, на севере — сельские поселки. Полусинан-
троп. Личинки — некрофаги, возможна копрофагия . . Р. pleskei Rohd.
40 (39). Срединный отдел апикальной части парафалла на внутренней стороне
без выступа. Мембранальные лопасти без крючка на конце, слегка ото-
Рис. 57. Фаллосома (а) и церки (б) самцов.
1 — Parasarcophaga portschinshyi; 2 — Р. pleskei', 3 — Р. jaroschevskyi’, 4 — Phallosphaera konakovi. (Ориг.).
гнуты вниз (рис. 57, 3). 9—14. — Ю и В Прим.; Монголия, СВ Китай.
Лесной вид. Личинки — некрофаги..............Р. jaroschevskyi Rohd.
17. Phallosphaera Rohd. Крупные мухи серого цвета. 3-й членик усиков
в 3—4 раза длиннее 2-го. Скулы с многочисленными щетинками, нижние наи-
более длинные, оформлены в ряды; ас перед швом отсутствуют; за швом 4 пары
dc\ сх3 с волосками. Проплевры с несколькими короткими волосками. Церки
длинные. V стернит d с резким срединным выступом, хорошо заметным в про-
филь. Фаллосома округлая, без длинных отростков и лопастей. Личинки —
некрофаги и хищники куколок бабочек. — В СССР 2 дальневосточных вида
(вредоносное значение имеет 1).
1 (1). Церки с пучком щетинок в апикальной части. Задние парамеры на конце
расщеплены. Гениталии см. рис. 57, 4. 10—17. — Ю Прим., Ю Кур.
(Кунашир); СВ Китай, п-ов Корея, Япония. Приурочен ко всем лесным
формациям, встречается в зеленой зоне населенных пунктов. Личинки
в трупах животных; хищники в куколках бабочек . . Р. konakovi Rohd.
116
18. Robineauella End. Крупные темно-серые, иногда почти черные, мухи.
3-й членик усиков в 2—3 раза длиннее 2-го. Скулы умеренно волосистые. Про-
плевры голые; сх3 с волосками, ас перед швом отсутствуют; за швом 4—6 пар de.
Гениталии 3 крупные. Апикальная часть парафалла с двумя большими боко-
выми отростками, вильчатыми на конце. Личинки — хищники бабочек и некро-
фаги. — В СССР 7 видов, на ДВ 4 (вредоносное значение имеют 2).
1 (2). Генитальный тергит по заднему краю без крепких щетинок. Церки почти
параллельнокрайние, с очень коротким крючком на конце (рис. 58, 1).
Рис. 58. Фаллосома (а) и церки (б) самцов.
I _Robineauellapseudoscoparia\ 2 — R. scoparia' 3 — Sinonipponia musashinensis; 4 — S. henebazini. (Ориг.).
тл™ тАллтл-селый. 8—17. — Маг. (на С до 65° с. ш.), Амур.,
Хаб., Прим., евр. ч. СССР, Сибирь; СВ Китай, Монголия, п-ов Корея,
Япония. Массовый вид лесной зоны. Обычный полусинантроп. Личинки
в трупах животных; хищники гусениц и куколок бабочек.................
..............................................R. pseudoscoparia Кгаш.
2 (1). Генитальный тергит по заднему краю с рядом щетинок. Церки саблевид-
ные, к концу утончаются (рис. 58, 2). Общий фон тела почти черный.
9—19. — Чук. (на сев. до 68° с. ш.), Маг., Коряк., Камч., Амур., Хаб.,
Прим., Сах., Кур.; Голарктика. Лесной вид. Личинки в трупах позвоноч-
ных ....................................................R. scoparia Pand.
19. Sinonipponia Rohd. Мухи средней величины серого цвета. 3-й членик
усиков в 2—3 раза длиннее 2-го. Щетинки на скулах слабые, расположены
в один ряд. Проплевры голые; сх3 с волосками; ас перед швом отсутствуют;
за швом 3—4 пары крепких de. Гениталии средних размеров. Апикальная
часть парафалла очень узкая, вытянутая, несет прямые боковые отростки.
117
Церки на конце с зубцом. Личинки — копрофаги или некрофаги. — В СССР
2 дальневосточных вида.
1 (2). Боковые отростки апикальной части парафалла узкие, шиловидные;
апикальная часть парафалла без шипов (рис. 58, 5). 6—10. — Ю Прим.;
п-ов Корея, Япония. Долины рек ЮЗ Сихотэ-Алиня, таежные села.
Личинки — в трупах пресноводных животных ..............................
.................................. S. musashinensis Kano et Okaz
2 (1). Боковые отростки апикальной части парафалла широкие, листовидные;
апикальная часть парафалла с двумя длинными шипами, направленными
вперед (рис. 58, 4), 6—10. — Ю Прим.; В Китай, п-ов Корея, Япония.
Травянистые биотопы, в местах отдыха людей; личинки в фекалиях . . .
...................................... S. hervebazini Seg. (=erecta Но)>
Рис. 59. Фаллосома и церки (б) самцов.
1 — Sarcorohdendorfia antilope', 2 — Takanoa holomyietzi', 3 — Sarcophaga camaria, (Ориг.)>
20. Sarcorohdendorfia Bar. (=Tricholioproctia Ваг.). Крупные серые мухи
с хорошо выделяющимися черными полосами на груди. 3-й членик усиков
в 3—4 раза длиннее 2-го. Скулы умеренно волосистые; ас перед швом на груди
имеются, за швом 4 пары dc\ сх3 с волосками. Проплевры полностью в густых
длинных волосках. Церки с резким изгибом на середине апикальной части. Пе-
редние парамеры широкие, вильчатые. Личинки — некрофаги и хищники ба-
бочек. — В СССР 1 вид.
1 (1). Апикальная часть церок после изгиба почти параллельнокрайняя.
Апикальная часть парафалла вытянута вперед (рис. 59, 1), Церки черные;
анальный и генитальный тергиты, парамеры и большая часть фаллосомы
коричнево-красные. 12—17. — Ю Хаб., Прим.; В Китай, п-ов Корея,
Япония, Океания. Массовый лесной вид в окультуренных биотопах;
личинки в трупах животных, хищники куколок бабочек......................
.................................................. S. antilope Boett.
21. Takanoa Rohd. Крупные серые мухи. 3-й членик усиков в 1.7—3 раза
длиннее 2-го; ас перед швом на груди развиты или отсутствуют; за швом 4 пары
de. Проплевры без волосков; сх3 с волосками. Генитальный тергит удлиненный.
Апикальная часть парафалла крупная и широкая. Личинки — некрофаги. —
В СССР 2 дальневосточных вида (вредоносное значение имеет 1).
1 (1). Церки саблевидно изогнутые, к концу сужающиеся. Мембранальные
лопасти изогнуты почти под прямым углом. Апикальная часть парафалла
к концу заужена (рис. 59, 2), 12—15. — Ю Прим. Берега пресных водо-
емов, на окраинах сельских поселков. Личинки в трупах рыб и водных
моллюсков ..............................................Т. kolomyietzi Art.
118
22. Sarcophaga Mg. Крупные мухи серого цвета. 3-й членик усиков в 1.5—
3 раза длиннее 2-го. Скулы волосистые. Проплевры без волосков; сх3 с нежными
волосками; ас перед швом на груди отсутствуют; за швом — 4 пары de. Фалло-
сома в профиль в середине прозрачная. Апикальная часть парафалла перепон-
чатая. Личинки — некрофаги или паразиты дождевых червей, хищничают
на бабочках. — 1 вид (в СССР И).
1 (1). Мембранальные лопасти узкие и длинные. Стилеты крупные, на конце
расширенные (рис. 59, 3). 6—18. — Амур., В и С Прим., Ю Сах.; лесная
зона Палеарктики, в селах и зверосовхозах. Полусинантроп. Личинки
в останках животных, дождевых червях, гусеницах и куколках бабочек
................................................ S. earn a ria L.
МУХИ-КРОВОСОСКИ (Сем. HIPPOBOSCIDAE)
Кровососки входят в подотряд Brachycera (короткоусые), группу Cyclor-
rhapha (круглошовные). Семейство Hippoboscidae состоит из 3 подсемейств:
Ornithomyinae, Hippoboscinae и Lipopteninae.
ВНЕШНЕЕ СТРОЕНИЕ
Мухи-кровососки — мелких или средних размеров (длина тела от 1.2 до
10—12 мм) кровососущие двукрылые насекомые с плоским телом (рис. 60).
Голова уплощенная, с выдвинутым вперед ротовым аппаратом. Сложные
глаза у большинства видов хорошо развиты, со множеством мелких омма-
тидиев и лишь у некоторых строго специфичных видов глаза небольшие, но
всегда разделены у обоих полов широким лицом (медиовертексом). Ротовой
аппарат приспособлен к прокалыванию кожи и сосанию крови теплокровных
животных. Усики состоят из 3 сегментов и находятся в антеннальном углуб-
лении по краям лба.
Грудь вдавленная и одинаково ейлеротизована со спинной и брюшной
сторон. Сильные ноги отходят от боковых краев груди и расположены более
или менее горизонтально; они оканчиваются двумя мощными коготками.
Крылья разные по размеру и форме. У большинства видов они хорошо
развиты и функционируют, довольно длинные и широкие по отношению к раз-
мерам тела. У некоторых видов они атрофированы или редуцированы и не
функционируют. Иногда они полностью сформированы, но отпадают вблизи
от основания, как только насекомое достигает хозяина.
Брюшко на большей части его поверхности мембранозное, сверху и снизу
со склеротизованными пластинками. Тергиты у большинства видов редуци-
рованы или атрофированы, стерниты сильно расширены. У всех видов крово-
сосок имеется 7 брюшных сегментов и 7 пар брюшных дыхалец.
Личинка плоская, грязно-желтого или белого цвета, за исключением по-
липнейстических долей, которые окрашены в коричневый или черный цвет.
Она неподвижна и не питается.
Постепенно наружный покров личинки делается каштаново-коричневым,
а затем черным; кутикула твердеет.
Куколка представляет собой пупарий овальной формы, коричневого или
черного цвета, внутри которого образуется настоящая куколка, как у всех
круглошовных двукрылых. Швы передней части чехлика, где разрывается
оболочка при выходе взрослого насекомого, мало заметны. Размеры пупария
от 1.7 X 1.5 до 3.5x3 мм.
Диагноз сем. Hippoboscidae
Голова плоская, клинообразная, ротовой аппарат направлен вперед. Грудь
сильно уплощена дорсовентрально. Покровы головы и груди склеротизованы.
Сложные глаза у большинства групп развиты или у некоторых родов редуци-
рованы (Crataerina, Lipoptena, Melophagus). Крылья обычно развиты; у пред-
ставителей некоторых родов они в той или иной мере редуцированы (Сга-
119
taerina), отламываются у основания после перехода на хозяина (Lipoptend)*
реже — совсем отсутствуют (Melophagus). Характерно аденотрофическое жи-
ворождение, когда все три стадии личинки развиваются в организме самки,
в маткообразном расширении яйцевода. Временные или постоянные пара-
зиты — кровососы теплокровных животных. — В СССР — 11 родов, 41 вид.
На Дальнем Востоке — 15 видов из 9 родов.
3 4
Рис. 60. Общий вид и детали строения мух-кровососок.
1 — самка Omithomya avicularia: бр — брюшко, гл — глаза, гр — грудь, кр — крыло, Hit Ht, Н9 — пе-
редняя средняя и задняя ноги, нп — нотоплевры, пб — плечевые бугорки, у — усики, щ — щупики, щк —
щиток (по: Досжанов, 1980); 2 — схема головы сверху: вщ — вертикальные щетинки, л — лунка, лр —
лобные рожки, лш — лобные швы, мв — медеовертекс, пг — простые глазки, пф — парафронталии, щт —
щетинки; 3 — голова снизу: глс — горловая мембрана, ющ — югальные щетинки; 4 — усик: ар — ариста,
2сг — второй сегмент, Зег — третий сегмент; 5 — простой коготок; двойной коготок: бп — базальная
пятка, бтис — дистиотарзус, к — коготки, п — пульвилла, э — эмподий (по: Theodor, Oldroyd, 1964).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (4). Голова слегка удлиненная, эллиптическая; глаза небольшие, узкие.
Крылья и жужжальца обычно не развиты; если развиты, то с 3 продоль-
ными жилками; при переходе на хозяина у основания обламываются.
2 (3). Крылья и жужжальца отсутствуют. Глаз уже усиков. Длина глаз пре-
вышает ширину в 3—4 раза (рис. 74, 1)............9. Melophagus Latr.
3 (2). Крылья и жужжальца имеются. Крылья с 3 продольными жилками
или обламываются близ основания. Глаз шире усиков. Длина глаз пре-
вышает ширину в 2 раза (рис. 72, 1)..............8. Lipoptena Nitsch.
120
4 (1). Голова круглая; глаза большие. Крылья и жужжальца хорошо раз-
виты, имеют 6—7 продольных жилок; только у одного рода (Crataerina)
они уменьшены и не функционируют.
5 (14). Крылья с 3 поперечными жилками; Ап замкнута, простые глазки
развиты.
6 (7). Коготки простые, но выглядят двухвершинными; плечевые бугорки
ровно закруглены (рис. 60, 5)....................1. Ornithoica Rond.
6
Рис. 61. Детали строения Ornithoica unicolor.
1 — голова; 2 — грудь; з — брюшко ?; 4 — гениталии <У; 5 — брюшко $ (снизу, задняя часть); 6 —
крыло.
7 (6). Коготки двойные, но выглядят трехвершинными; плечевые бугорки
конусообразные (рис. 60, 6).
8 (9). Крылья рудиментарные (их вершина острая или далеко не достигает
конца брюшка), не функционируют. Глаза маленькие (рис. 67, 2) . . .
................................................... 4. Crataerina Olf.
9 (8). Крылья нормальные (их вершина значительно заходит за конец брюшка),
функционируют. Глаза большие (рис. 63, 7).
10 (И). г2+3 прилегает к С; перерыв на С между впадением Sc и гх. Мембрана
крыла без микротрихий (рис. 63, 2) ....... 2. Ornithophila Rond.
11 (10). г2х3 не прилегает к С; перерыв на С перед впадением Sc. Мембрана
крыла с микротрихиями (рис. 64, 4).
12 (13). Длина усиков составляет V3 длины головы, они направлены вбок и
вниз, без канта с наружной стороны. Кант грудной чешуйки с редкой
бахромой светлых волосков............................3. Ornithomya Latr.
13 (12). Длина усиков составляет 2/3 длины головы, они направлены вперед,
с кантом с наружной стороны (рис. 68, 4). Кант грудной чешуйки с густой
бахромкой темных щетинистых волосков . . , 5. Ornithoctona Speis.
121
14 (5). Крылья с 1 или 2 поперечными жилками; Ап открыта, простые глазки
отсутствуют.
15 (16). Поперечная жилка на крыле одна (ta)\ tp отсутствует. Щиток в виде
длинного поперечного прямоугольника с короткими пальцевидными
отростками по бокам заднего края (рис. 71, 3) . .7. Pseudolynchia Beq.
16 (15). Поперечных жилок две (ta и tp)\ щиток округлен, без коротких пальце-
видных отростков по бокам заднего края (рис. 70) ... 6. Icosta Speis.
Рис. 62. Детали строения Ornithoica stipituri.
1 — голова; 2 — грудь; 3 — брюшко <?; 4 — брюшко $; 5 — брюшко $ (снизу, задняя часть); 6 — крыло,.
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
1. Ornithoica Rond. Плечевые бугорки ровно закруглены. Вершинная
треть прилегает к С, Мембрана крыла с микротрихиями. На птицах. —
2 вида Yb СССР 3).
1 (2). Длина ячейки ап в 3 раза больше ширины. На щитке 8—10 длинных чер-
ных щетинок (рис. 61). Длина тела 1.8—2, крыла 3.9—4.4. — Ю Прим.;
Япония, Бирма, Таиланд, Малайзия..................О. unicolor Speis.
2 (1). Длина ячейки ап в 2 раза больше ширины. На щитке 4 длинные, черныа
щетинки (рис. 62). Длина тела 1.3—1.5, крыла 2—2.8. — Ю Прим.;
Новая Гвинея, Соломоновы о-ова, Австралия ... О. stipituri Sehin.
2. Ornithophila Rond. Плечевые бугорки сферические, сильно выступают.
На птицах. — 1 вид (в СССР 2).
1 (1). На щитке только светлая поперечная полоска у основания (рис. 63).
Длина тела 1.8—2.6, крыла 3.2—4.4. — Прим., Украина, Крым, Кавказ,
Ср. Азия; 3 Европа, Африка, Индия, Китай, Япония. Преимущественно
на воробьиных.....................................О. metallica Sehin.
122
3. Ornithomya Latr. Плечевые бугорки большие, в виде широких долей.
г2+3 и 5 оканчиваются острым углом у косты. Мембрана крыла с микротри-
хиями. На птицах. — 3 вида (в СССР 5).
1 (2). Крыло 5.7—7 мм. На щитке 7—10, часто 8 предвершинных щетинок.
На нижней стороне головы нет коричневых пятен (рис. 64). 2.7 3.4.
В лесной и таежной зонах. На воробьиных, хищных и голубиных . . . .
.....................................................О. avicularia L.
5
Рис. 63. Детали строения Ornithophila metallica.
1 — голова; 2 — крыло; 3 — грудь; 4 — задняя часть брюшка $ снизу; 5 — брюшко 3; 6 — гениталии <?.
(По: Theodor, Oldroyd, 1964; Досжанов, 1980).
2 (1). Крыло 3.5—5.5 мм. На щитке 3—6 предвершинных щетинок, на нижней
стороне головы имеются коричневые пятна.
3 (4). Крыло 3.5—4.5 мм. На щитке 4 предвершинные щетинки. Окраска светло-
коричневая. На нижней стороне головы 2 коричневых треугольника, не
достигающих югулярной щетинки (рис. 65). 2—2.5. — Ю Прим., Молда-
вия, Украина, Крым, Кавказ, Ср. Азия, 3 и СЗ РСФСР; 3 Европа,
С Америка. Преимущественно на воробьиных ... О. fringillina Curt.
4 (3). Крыло 4.5—5.5 мм. На щитке 6 предвершинных щетинок. Окраска темно-
коричневая. Треугольные темные пятна на нижней стороне головы дости-
гают югулярной щетинки (рис. 66). 2.1—2.6. — Сах., Амур., Прим., С,
СЗ, ср. полоса и ЮЗ РСФСР, Ср. Азия, Алтай, Сиб. На воробьиных,
хищных, куриных ....................................О. chloropus Bergr.
4. Crataerina Olf. На птицах. — 1 вид (в СССР 4).
1 (1). Крылья узкие, лентообразные; длина их в 7—8 раз превышает ширину.
Глаза маленькие, длина глаз составляет х/2 длины головы, простые глазки
123
имеются (рис. 67). — Прим., СЗ и 3 РСФСР, Украина, Поволжье 3 Сиб.г
Казахстан, Алтай. На ласточках . ...................С. hi run din is L.
5. Ornithoctona Speis. — 1 вид (в СССР 2).
1 (1). Усики ложкообразные, широкие и тупые у вершины. Ячейки крыла 7?3
с узкой полосой микротрихий на вершине; длина крыла более 10 мм
(рис. 68). 4.9. — Ю Прим.; Ю и ЮВ Азия, Австралия, о-ов Мадагаскар.
На хищных, голубиных, кукушковых..........................О. plicata Speis.
6. Icosta Speis. На птицах. — 2 вида (в СССР 6).
1 (2). Темнее. Щупики короче длины медиовертекса. Мембрана крыла пол-
ностью и равномерно покрыта микротрихиями, достигающими Cl^A
(рис. 69). — Ю Прим., Кур., Украина, Кавказ, дельта Волги, Казахстан;
тропики, субтропики и умеренные зоны Старого Света. На цаплевых
....................................................1. а г de а е Macq.
2 (1). Светлее. Щупики равны длине медиовертекса. В мембране крыла микро-
трихиями покрыта полностью 7?3, 3/5 части между M3+i и СиАп (рис. 70). —
Ю Прим. На глухой кукушке.................................I. maai Dosz.
7. Pseudolynchia Beq. На предщите 10—30 длинных латероцентральных
волосков. По бокам заднего края щитка 2—5 пальцевидных отростков. На пти-
цах. — 1 вид (в СССР 2).
1 (1). На предщите 20—30 длинных тонких желтых волосков. Щиток прямо-
угольный, ширина его в 4 раза больше длины, по бокам 4—5 пальцевидных
отростков (рис. 71). 2.6—3.2. — Ю Прим., Молдавия, Украина, Кавказ,
Ср. Азия; Африка, Средиземноморье, К) Азия, Ю Америка. На голубиных
.....................................................Р. canariensis Macq.
8. Lipoptena Nitsch. Крылья и жужжальца имеются. Крылья отпадают,
как только мухи достигают хозяина. Простые глазки имеются. На млекопитаю-
щих. — 2 вида (в СССР 4).
124
1 (2). Крыло 5.5—6 мм. Парафронталии х/3 шириш глаза. Ширина медиовер-
текса больше длины; щупики по длине равны лобной части головы; 15—
18 латероцентральных щетинок (рис. 72). — Амур., Прим., Ю Сах., 3,
СЗ и центр РСФСР, Белоруссия, Украина, СЗ Сиб., С Казахстан, Алтай;
Европа, СЗ Индии, С Америка. На оленьих, полорогих . . . L. cervi L.
2 (1). Крыло 3—3.5 мм. Парафронталии х/2 ширины глаза. Ширина медиовер-
текса равна или несколько меньше длины. Щупики в 2 раза короче лоб-
Рис. 65. Детали строения Ornithomya fringillina.
1 — голова; 2 — брюшко $; 3 — крыло; 4 — гениталии <J; 5 — грудь. (По: Theodor, Oldroyd, 1964; Дос-
жанов, 1980).
ной части головы; 2—5 латероцентральных щетинок (рис. 73). — Амур.,
Прим., Ю Сах., Кавказ, В Тянь-Шань, 3 и В Сиб., Алтай; Япония.
На оленьих ....................................L. fortisetosa Маа
9. Melophagus Latr. Крылья и жужжальца отсутствуют. Глаза редуци-
рованы до узких полосок. Простых глазков нет. На млекопитающих. — 2 вида
(в СССР 8).
1 (2). Переднегрудное дыхальце заметно шире самого узкого места между
коксальными углублениями 1 и 2; на уровне дыхальца 6 имеется 18—
20 щетинок. Щетинки груди и волоски брюшка короткие, крепкие
(рис. 74, S, 9). — Амур., Прим., повсеместно в умеренных зонах. На овцах
.............................................. . . . . М. ^ovinus L.
125
Рис. 66. Детали строения Ornithomya chloropus.
1 — голова; 2 — брюшко $; 3 — крыло; 4 — гениталии 5 — грудь. (По: Theodor, Oldroyd, 1964; Дос-
жанов, 1980).
Рис, 67, Детали строения Crataerina hirundinis.
1 голова; 2 ** грудь; 3 — брюшко $; 4 — крыло; 5 — гениталии 3. (По: Досжанов, 1980).
4
Рис. 69. Детали строения Icosta
~ S — гениталии Л; « — 1енитальвый склерит (По::
2 - грудь; з - брюшко £ *О-«Р*^654; д^нов, 1980).
ardeae.
j’— голова;
2 (1). Переднегрудные дыхальца уже самого узкого места между коксальными
углублениями 1 и 2; на уровне дыхальца 6 имеется 9—10 щетинок. Ще-
тинки груди и волоски брюшка темные, длинные, тонкие (рис. 74,1—7). —
Камч. На овцах ............................... М. kamtschaticus Dosz.
Рис. 70. Детали строения Icosta maai9
1 — голова; 2 — грудь; 3 — крыло; 4 — брюшко $ с вентральной сторовы; 5 — то же с дорсальной
стороны. (Ориг.).
ОСНОВНАЯ ЛИТЕРАТУРА
Артамонов С. Д. К изучению мух семейства Sarcophagidae центральной части Мага-
данской области. — В кн.: Животный мир Дальнего Востока. Благовещенск, 1977, вып. 2,
с. 168—173.
Зервес Ю. Г. Новые и малоизвестные виды Sarcophagidae (Diptera) Сибири и Дальнего
Востока. — В кн.: Таксономия и экология членистоногих Сибири. Новосибирск, 1978,
с. 184—187.
Грунин К. Я. Определительная таблица родов сем. Hippoboscidae. — В кн.: Опреде-
литель насекомых европейской части СССР. М.; Л., 1970, т. 5, ч. 2, с. 596—601.
Дербенева-Ухова В. П. Высшие двукрылые, или мухи (кроме семейств Sarcophagidae
и Calliphoridae). — В кн.: Определитель членистоногих, вредящих здоровью человека.
М., 1958, с. 126—144.
Досжанов Т. Н. Мухи-кровососки Казахстана. Алма-Ата, 1980. 208 с.
128
1 — голова; 2 — верхняя часть груди; 3 — генитальная пластинка (а) и склерит (б) $; 4 — брюшко ?;
5 — щиток; 6 — крыло; 7 — гениталии <?. (По: Досжанов, 1980).
Рис. 72. Детали строения Lipoptena cervi.
1 — голова; 2 — грудь; з — брюшко ?; 4 — крыло; 5 — гениталии <?. (По: Theodor, Oldroyd, 1964; Дос*
жанов, 1980).
9 Зак. № 990
Рис. 73. Детали строения Lipoptena forttsetosa.
1 — голова; 2 — грудь; 3— брюшко $; 4 — I стернит 5 — гениталии о. (По: Маа, 1965; Дос-капов.
1980).
9
Рис. 74. Мухи рода Melophagus.
1—7 —'М. kamtschaticus: 1 — голова, 2 — грудь, 3 — брюшко $ с дорсальной стороны, 4 — брюшко о
с вентральной стороны, 5 — I стернит, 6 — генитальный склерит j, 7 — гениталии о (ориг.); 6. —
ovinus: 8 — голова, 9 — гениталии <? (по: Theodor, Oldroyd, 1964).
Зимин Л. С. Определитель личинок синантропных мух Таджикистана (по III стадии).
Л., 1948. 116 с. (Определители по фауне СССР, изд. Зоол. ин-том АН СССР, № 28).
Зимин Л. С. Сем. Muscidae. Настоящие мухи (трибы Muscini, Stomoxidini). М.; Л.,
1951. 286 с. (Фауна СССР. Насекомые двукрылые; Т. 18. Вып. 4).
Лобанов А. М. Определитель самок сем. Calliphoridae (Diptera) фауны СССР по яйце-
кладам. Л., 1976. 47 с. (Определители по фауне СССР, изд. Зоол. ин-том АН СССР, № 109).
Павловский Е. Н. Мухи. — В кн.: Руководство по паразитологии человека. М.; Л.,
1948, т. 2, с. 860—904.
Петрова Б. К. Фаунистико-экологический обзор синантропных двукрылых (Diptera)
юга Приморского края. — Энтомол. обозрение, 1968, т. 47, вып. 1, с. 95—104.
Петрова Б. К. Определитель синантропных двукрылых Приморского края. Новоси-
бирск, 1974а. 103 с.
Петрова Б. К. Фауна синантропных каллпфорпд (Diptera, Calliphoridae) Дальнего
Востока. — Изв. Сиб. отдел. АН СССР. Серия биол. наук, Новосибирск. 19746, вып. 15,
с. 51—55.
Родендорф Б. Б. Семейство Sarcophagidae. М.; Л., 1937. 501 с. (Фауна СССР. Насе-
комые двукрылые; Т. 29. Вып. 1).
Родендорф Б. Б. Определитель членистоногих, вредящих здоровью человека. М., 1958,
с. 132-143.
Сычевская В. И. О синантропных мухах Сахалина, Камчатки и Курильских островов.—
В кн.: Фауна и экология двукрылых насекомых Дальнего Востока, 1973, с. 82—86. (Тр.
Бпол.-почв. ин-та. Нов. сер., т. 19 (122)).
Штакельберг А. А. Определитель мух европейской части СССР. М., 1932. 742 с.
Штакельберг А. А. Синантропные двукрылые фауны СССР. М.; Л., 1956. 163 с.
Маа Т. A synopsis of the Lipopteninae. — J. Med. Ent., 1936, N 6.
Theodor O., Oldrooyd H. Hippoboscidae. — In: E. Lindner. Die Fliegen der Palaearkti-
scken Region. 1964, p. 65.
ОВОДЫ
Оводы — это короткоусые двукрылые насекомые (мухи), в стадии личинки
паразитирующие на сельскохозяйственных и диких животных, а также на
человеке. В зависимости от локализации личинок в теле хозяина различают
желудочных, носоглоточных и подкожных оводов, которые внешне в общих
чертах похожи друг на друга, но имеют свои морфологические и биологические
особенности.
На Дальнем Востоке обитает 15 видов оводов, относящихся к 3 семействам
и 8 родам, из которых медико-ветеринарное значение имеют 12 видов из 7 ро-
дов. Для некоторых видов подкожных оводов оно пока неясно (род Portschtn-
skla Sem.).
ВНЕШНЕЕ СТРОЕНИЕ
Мухи средних или крупных размеров (8—22 мм), тело которых голое или
в разной степени покрыто волосками или мельчайшими щетинками — микро-
хетами (рис. 75, 7, 2).
Голова обычного для двукрылых строения, но с характерными особенно-
стями (рис. 75, 3). У самок м самцов крупные фасеточные глаза разделены
лбом, который у первых шире. Расстояние между простыми глазками не оди-
наковое у разных видов; оно определяется по отношению к поперечнику одного
глаза. Ниже усиков находится клипеус, ограниченный по бокам возвышенными
валиками — вибрисионными килями; верхняя часть последних сливается
с усиковыми лунками. Ротовые органы редуцированы (отсутствуют) или пред-
ставлены только двумя шарообразными щупиками. Усики у представителей
всех трех семейств 3-члениковые, с голой аристой (без волосков и щетинок).
Особенности их строения используются при видовом определении оводов.
В ряде случаев в видовой диагностике имеет значение также отношение ширины
орбит и лобной полосы (например, в сем. Hypodermatidae).
Грудь обычного для двукрылых строения; наиболее развита среднегрудь
(среднегрудка), на которой имеются плечевые и закрыловые бугорки, а также
продольные полосы, конфигурация и окраска которых имеют диагностическое
значение. У желудочных оводов среднеспинка делится на 2 приблизительно
равные части глубоким поперечным швом, широко прерванным в середине,
что является хорошим таксономическим признаком этого семейства. У носо-
глоточных оводов всех видов на среднеспинке различаются 4 продольных
полосы, которые поперечным швом делятся на 2 отрезка. Степень выражен-
9*
131
ности этих полос не одинакова у разных видов (от очень узких полос до широ-
ких выпуклых валиков-мозолей), что является надежным родовым, а иногда
и видовым признаком. У подкожных оводов на среднеспинке имеются 4 отчет-
5
Рис. 75. Общий вид и детали строения оводов.
1 — самец желудочного овода Gastrophilus intesiinalis (из: Грунин, 1955); 2 — самец подножного овода
Hypoderma diana\ 3 — схема строения головы носоглоточного овода: вк — вибрисионный киль, г— про-
стые глазки, гл — фасеточный глаз, гт — глазковый треугольник, л — лунка, лб — лобная полоса, at —
медиана, н — наличник (клипеус), ор — орбиты, оро — остатки ротовых органов, ск — скулы, у — усик
с аристой на 3-м членике, щ — щеки; 4 — схема жилкования крыла носоглоточного овода: жилки: с — ко-
стальная, sc — субкостальная, rt, г2+3, г4+ь — радиальные, тг, т2, т3+4 — медиальные, ап — анальная,
сщащ — кубитально-анальная, h — плечевая, гт — радио-медиальная поперечная, тт — медиальная
поперечная; ячейки: R — передняя основная, ВБ — первая заднекрайняя, М — задняя основная, М2 —
дискоидальная, Си — кубитальная; 5 — яйцо подкожного овода Oedemagena tarandi: а — яйцо с развер-
нутым прикрепительным придатком после мацерации в едком кали, б — прикрепительный придаток (по:
Грунин, 1955, 1957, 1962).
ливые голые (без волосков) продольные, прерванные посредине полоски.
У некоторых видов подкожных оводов щиток разделен по заднему краю выем-
кой или с небольшим вдавлением посредине, что учитывается при видовом
определении. Подщиток имеется.
132
Крылья (рис. 75, 4). Общее строение крыльев у представителей всех трех
семейств сходное, но имеются некоторые различия в жилковании и окраске.
Для желудочных оводов характерны следующие особенности в жилковании
крыла: 1) почти прямая, чуть изогнутая кзади жилка т, не имеющая в вер-
шинной части резкого изгиба к переднему краю крыла (сравни с семействами
Oestridae и Hypodermatidae); 2) разная степень выраженности поперечной
жилки tp до полного ее исчезновения (у разных видов).
У носоглоточных оводов крылья в покое складываются крышеобразно.
Особое значение в видовом определении имеют расположение жилок г2+3,
г4+5тп, гт, а также конфигурация первой заднекрайней ячейки, которая может
быть открытой (роды Cephenomyia и Pharingomyia) или замкнутой, со стебель-
ком (род Oestrus). У подкожных оводов жилка гт чаще всего располагается
между вершинами sc и гх. Ячейка /?5 открытая, реже замкнутая; вершинный
отрезок жилки т]+2 всегда плавно (не резко) изогнут к переднему краю крыла;
в месте изгиба отросток (т2) отсутствует (т2 слита с т2). Ремигиум (основание
радиального ствола жилок) голый, без волосков.
Ноги обычного для насекомых строения. В диагностике разных родов и ви-
дов оводов имеют значение соотношение длины члеников лапок (желудочные
оводы), соотношение длины лапок и голеней (носоглоточные и подкожные
оводы), а также длина коготков (подкожные оводы).
Брюшко состоит из 5—6 основных члеников и нескольких сильно изме-
ненных концевых члеников, склериты которых образуют наружный половой
аппарат. Оно может быть голым или в разной степени покрыто волосками,
гладкое или с крупными мозолями и бородавками. Окраска брюшка у разных
родов и видов даже в пределах одного семейства сильно варьирует в зависимости
от цвета и густоты покрывающих его волосков (черно-желтое, ярко-рыжее
с примесью черного, желто-белое с черным, серебристое и т. д.).
У разных семейств оводов строение брюшка имеет свои особенности. У же-
лудочных оводов I и II тергиты слиты вместе, но граница слияния хорошо
видна. Задний край I тергита закругленным концом вдается в середину II тер-
гита. Стерниты очень тонкие, чуть плотнее широкой боковой перепонки (мем-
браны) брюшка. Они прозрачные или слабо пигментированы. У некоторых
подкожных оводов брюшко может быть со сложным серебристым рисунком
из пятен разной конфигурации {Pharyngomyia picta, Oestrus ovis).
Строение яйцеклада неодинаковое у разных родов и видов оводов. Он мо-
жет быть вытянуто-телескопическим, выступающим наружу и составляющим
х/3 длины брюшка (например, у Gastrophilus intestinalis) либо очень коротким,
почти неподвижным и невыступающим за конец брюшка (у многих видов).
Яйцо (рис. 75, 5). Яйца у желудочных и подкожных оводов очень мелкие,
продолговатые, зауженные на концах. Верхний конец яйца закруглен, а на
нижнем имеется прикрепительный придаток, служащий для фиксирования яиц
на волосяном покрове животных. Цвет яиц чаще белый, желтоватый, серова-
тый, реже черный. Особенностью яиц желудочных оводов является отпадаю-
щая при выходе личинки крышечка. У подкожных оводов личинка выходит
через образующуюся дуговидную щель хориона (оболочки), крышечки у них
нет. У самок с разным строением яйцеклада (с длинным и коротким) яйца
не одинаковые. У некоторых видов этого семейства яйца не имеют прикрепи-
тельного придатка.
Личинки (рис. 76 и др.) оводов безголовые и безногие. Тело зрелых личинок
состоит из 12—13 члеников: псевдоцефала (ложноголова), 3 грудных и 8—
9 брюшных члеников. На кутикуле имеются мелкие и более крупные шиппки,
форма, величина, степень развития и расположение которых имеют значение
при видовом определении. На переднем и заднем концах тела располагается
по 2 дыхальца. Ротовой аппарат как и у личинок других двукрылых. При
определении видов имеют значение величина и форма ротовых крючьев. Их
размер определяется по прямой линии, соединяющей переднюю и заднюю
вершины ротового крючка (так называемый диаметр крючка).
Куколка. У пупария те же внешние признаки, что и у личинок III стадии,
тело которых при окуклении сокращается и затвердевает, приобретая корич-
невую окраску.
133
БИОЛОГИЯ
Желудочные и подкожные оводы — яйцекладущие; яйца прикрепляются
ими к шерстному покрову животных. Носоглоточные оводы — живородящи;
самка выпрыскивает в носовые ходы животных готовых личинок [Грунин,
1955, 1957, 1962]. У представителей всех семейств оводов личинки линяют
два раза и имеют 3 стадии; окукление их происходит во внешней среде (в земле).
Длительность фазы личинки исчисляется месяцами (от 1 до 9 мес.), фаза ку-
колки — 2—4 недели. Взрослые оводы не питаются, поэтому ротовые органы
у них редуцированы; самки после спаривания и оставления потомства вскоре
погибают.
Личинки желудочных оводов паразитируют в пищеварительном тракте
животных. Плодовитость разных видов — от 200—500 до 1000—2500 яиц.
Вылупившиеся личинки попадают в ротовую полость, а затем в пищевари-
тельный тракт, где продолжают свое развитие до III стадии. Зимовка проис-
ходит в теле хозяина. На следующий год личинки выпадают во внешнюю
среду с экскрементами и окукливаются. В течение года развивается одно по-
коление.
Личинки носоглоточных оводов паразитируют в полостях головы животных
(парнокопытных и непарнокопытных). Самка носоглоточного овода впрыски-
вает в носовые ходы готовых личинок. Достигнув III стадии развития, они
весной выпадают через ноздри во внешнюю среду. На северных широтах СССР
у носоглоточных оводов развивается одно поколение в году, а на южных —
два.
Личинки подкожных оводов обитают под кожей мелких и крупных млеко-
питающих, но прежде чем выйти под кожу они проделывают в организме слож-
ные миграции. Свое развитие личинки заканчивают в желваках под кожей,
питаясь окружающими тканями; достигнув III стадии, они через образовав-
шийся свищ выпадают наружу. У одной самки созревает от 300 до 800 яиц.
Средняя продолжительность развития у пищеводника и строки — от 2 до
3 мес.
ВРЕД
Единичные личинки оводов не вызывают заметных изменений в общем
состоянии животных-хозяев. Массовое паразитирование приводит к резкому
снижению мясо-молочной продуктивности, малокровию, ухудшению общего
состояния, резкому истощению и гибели, поэтому наше животноводство в ряде
областей терпит значительные убытки от массового паразитирования оводов.
Зараженность животных личинками оводов бывает очень большой. Напри-
мер, в желудке одной лошади могут одновременно паразитировать от 1000 до
1800 личинок желудочного овода, что приводит к нарушению функции желу-
дочно-кишечного тракта, ороговевающему раку желудка, злокачественному
малокровию и смертельному исходу.
Люди (взрослые и дети) нередко заражаются личинками I стадии при кон-
такте с лошадьми. При этом личинки могут мигрировать под кожей в течение
5—10 мес., вызывая болезнь «волосатик», «ползучая болезнь» («порикожа»),
сопровождающуюся мучительным зудом. В таких случаях личинок удаляют
из окончания ходов (место покраснения кожи). Известны также случаи пара-
зитирования личинок III стадии в желудке человека, что причиняет ему не-
стерпимые мучения.
Сильное заражение овец и северных оленей личинками носоглоточных
оводов вызывает ухудшение общего состояния или даже тяжелую болезнь —
ложную вертячку, которая приводит к смерти; у лошадей возникает болезнь
ринэстроз. В период лёта оводов неполноценно используются пастбища, так
как оводы в комплексе со слепнями причиняют сильное беспокойство скоту.
Происходит резкое понижение привесов и удоев коров, обесцениваются шкуры
и шерсть. Большие убытки от оводов терпит и охотничье хозяйство, поскольку
носоглоточные оводы паразитируют у диких животных.
Нередки случаи паразитизма носоглоточных оводов в глазах человека (муха
выбрызгивает личинок в глаз), что вызывает резкую боль и воспаление. Иногда
134
личинки выбрызгиваются самкой в нос человека, что приводит к тяжелому
заболеванию (высокая температура, головная боль, кровотечение из носа).
Прежде чем вымыть личинок из глаза струей воды (из спринцовки) их нужно
обездвижить 3%-ным раствором кокаина или 0.25%-ным раствором дикаина.
Прекратить движение личинок в глазу можно также народным средством —
закапыванием 2—3 капель сока табака.
Подкожные оводы также причиняют большой вред сельскохозяйственным
и некоторым диким животным, вызывая заболевание гиподерматоз. Шкуры
зараженного крупного рогатого скота, оленей и других животных теряют
товарную ценность или становятся непригодными. К косвенным убыткам от-
носятся и средства, затрачиваемые на борьбу с подкожными оводами.
Личинки подкожных оводов строки и пищеводника могут быть случай-
ными паразитами человека, вызывающими серьезное заболевание и даже
смерть. Известно 145 случаев паразитизма личинок подкожных оводов у че-
ловека (64 % найденных личинок локализовались в области головы и 36 % —
на других частях тела). Поражаются взрослые и дети. Очень часто личинки
заканчивают миграцию выходом в глаз [Грунин, 1962].
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ
1 (2). Жилка крыла Л/1+2 прямая.......................1. Gastropbilidae
2 (1). Жилка ЛГ1+2 изогнута под углом.
3 (4). Наличник в середине не сужен, яйцеклад длинный, телескопический
.....................................................3. Hypodermatidae
4 (3). Наличник посредине резко сужен, яйцеклад короткий . . 2. Oestridae
ЖЕЛУДОЧНЫЕ ОВОДЫ (Сем. GASTROPHILIDAE)
Среднего размера мухи, тело которых покрыто волосками. Ротовые органы
отсутствуют, иногда имеются 2 рудиментарных шаровидных щупика. Ариста
голая. Жилка т на крыле одна. Яйцеклад склеротизованный, длинный, по-
догнутый под брюшко; он может быть и коротким, но никогда не втяжной.
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
1. Gastrophilus Leach. Личинки паразитируют на лошадиных и носорогах,
развиваясь в пищеварительном тракте животных-хозяев. — 3 вида (в СССР 7).
Мухи
1 (4). Крылья бесцветные, прозрачные, без темных пятен.
2 (3). Задняя поперечная жилка крыла (tp) очень бледная; она расположена
гораздо дальше от основания крыла, чем поперечная жилка ta. Коготки
всех лапок $ одинаковой длины. 9—10. — Евр. ч. СССР, Ср. Азия, Сиб.,
ДВ; 3 Европа, С Африка, Турция, Монголия, Китай, Япония, С н
Ю Америка, Австралия, Нов. Зеландия. На лошади ........
................................ G. haemorrhoidalis L. — Овод усоклей
3 (2). tp явственная, расположена против ta, иногда отклоняется в обе стороны
менее чем па длину ta. Коготки передних лапок $ вдвое короче, чем на
остальных лапках. 12—13. — Евр. ч. СССР, 3 и В Сиб., Прим., Кур.,
Ср. Азия; 3 Европа, Турция, Ирак, Монголия, Китай, Индия, Япония,
С и Ю Америка, Австралия, Нов. Зеландия. На лошади...................
..................... G. veterinus Cl. — Овод двенадцатиперстник
4 (1). Крылья с темной перевязью посредине, 2 небольшими темными пятнами
вблизи вершины крыла и черной точкой на (в месте ответвления ради-
альных жилок). Коготки передних лапок у $ вдвое короче, чем на осталь-
ных лапках. 12—15. — Всесветно. На лошади и осле, иногда на собаке
(рис. 76, 1—3).......................G. intestinalis De Geer — Крючок
135
Личинки III стадии
1 (2). Шипы на члениках располагаются в один ряд. I грудной членик кониче-
ский, расширяющийся по направлению к II членику, в связи с чем ширина
заднего края I членика и нередко края II членика примерно одинаковая.
II грудной членик, по крайней мере на спинной стороне, вооружен рядом
шипов. На спинной стороне зоны шипов без перерыва в середине (рис. 76, 6)
..................................................G. veterinus Cl.
2 (1). Шипы на члениках располагаются в 2 ряда. Остальные признаки в ином
сочетании.
3 (4). Ротовые крючки с явственной выемкой перед коленообразным изгибом.
Шипы на члениках тупые, как бы обрубленные. На спинной стороне во-
оружение обычно заканчивается на VII членике (рис. 76, 4)..........
...................................... G. intestinalis De Geer
4 (3). Ротовые крючки без выемки перед коленообразным изгибом. Шипы на
члениках острые. На спинной стороне вооружение не простирается далее
VI брюшного членика (рис. 76, 5)...............G. haemorrhoidalis L.
НОСОГЛОТОЧНЫЕ ОВОДЫ (Сем. OESTRIDAE)
Крупные или средней величины мухи, густо опушенные или почти лишен-
ные волосков. Ротовые части сильно редуцированы (сохраняются рудименты
хоботка и шаровидные щупики). Лоб очень широкий. Усики очень короткие,
располагаются в усиковой ямке, чаще разделенной лицевым килем. Ариста
голая. На крыльях вершинная часть тп1 резким углом изогнута к переднему
краю крыла. Яйцеклад неподвижный, не выдается за вершину брюшка. Ноги
короткие, задние короче длины брюшка.
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ПОДСЕМЕЙСТВ И РОДОВ
Мухи
1 (4). Крылья с открытой ячейкой Л5; в месте изгиба всегда с отростком
(пг2). Подщиток едва различимый, линейный (подсем. Cephenemyinae).
2 (3). Отросток ш1 (т2) длинный, постепенно утончающийся к вершине.
Длина VI тергита брюшка приблизительно равна длине V тергита. Тело
в густых длинных волосках........................1. Cephenemyia Latr.
3 (2). Отросток ml (m2) короткий, резко обрывающийся на вершине. VI тергит
брюшка в 2 раза короче V тергита. Тело в редких коротких волосках,
кажется голым..................................2. Pharingomyia Shin.
4 (1). Крылья с замкнутой стебельчатой ячейкой Т?5; в месте изгиба чаще
всего без отростка. Подщиток веретеновидный, длина его лишь в 5—8 раз
меньше ширины (подсем. Oestrinae).
5 (6). Орбиты в углублениях с черными бляшками на дне. Брюшко гладкое,
с переливчатым рисунком................................3. Oestrus L.
6 (5). Орбиты с крупными коричневыми бородавками. Брюшко с крупными
черными бородавками...............................4. Rhinoestrus Вт.
Виды этого рода известны на сопредельных территориях (В Сиб.,
Монголия, Китай) и могут быть найдены на ДВ.
Личинки III стадии
1 (2). На задних дыхальцах рубец замкнутый в центре дыхальцевой пластинки;
замыкающий шов идет к середине ее внутреннего края. Шипы в зонах
на брюшной стороне члеников расположены беспорядочно. Среднебоковые
шипы на члениках и зона шипов у заднего края спинной стороны VII
брюшного членика отсутствуют (рис. 79, 3, 4)...........3. Oestrus L.
136
4 5 6
76. Желудочные оводы.
1—4 — Gastrophilus intestinalis:
кубитальная жилка, ip — задпл,ж
роны; 5, 6 — личинка III стадии: 5
Рис.
1 — самка, 2 — голова, з — схема жилкования крыла: тси — медио-
задняя поперечная, си — кубитальная, 4 — личинка III стадии с брюшной сто-
стадии: 5 — G. haemorrhoidalis, 6 — G. veterinus. (По: Грунин, 1955). Остальные
обозначения как на рис. 75.
2 (1). На задних дыхальцах рубец находится в выемке внутреннего края
дыхальцевой пластинки.
3 (4). Расположение шипов спинной стороны (если они имеются) резко отли-
чается от расположения шипов брюшной стороны. Шипы на брюшной
стороне члеников образуют правильные ряды .... 4. Rhinoestrus Вг.
4 (3). Расположение шипов на спинной и брюшной сторонах члеников одина-
ковое.
Рис. 77. Носоглоточные оводы.
1—4 — Cephenomyia trompe: 1 — самка, 2 — голова, 3 — личинка III стадии, брюшная сторона, 4 —
VIII брюшной членик сзади; 5—7 — С. ulrichi: 5 — передняя часть тела личинки (спинная сторона), 6 —
то же с вентральной стороны (брюшная сторона), 7 — VIII брюшной членик сзади. (По: Грунин, 1957);
5 (6). Нижние лопасти внутреннего края обеих дыхальцевых пластинок обычно
находятся друг от друга па таком же расстоянии, как и верхние, реже
расстояние между ними превышает расстояние между верхними лопа-
стями, но не более чем в 2 раза. У зрелых личинок черные точки над зад-
ними дыхальцами сливаются, образуя коропу (рис. 77, 4).................
.............................................1. Cephenomyia Latr.
6 (5). Нижние лопасти внутреннего края обеих дыхальцевых пластинок далеко
отстоят друг от друга — не менее чем в 3 раза по сравнению с верхними
лопастями. У зрелых личинок черные точки над задними дыхальцами
никогда не образуют короны........................2. Pharyngomyia Shin.
138
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
1. Cephenomyia Latr. Среднего размера мухи (13—18 мм), тело коренастое,
покрытое густыми пушистыми волосками, усики относительно крупные, крылья
слегка затемненные. Личинки паразитируют на олене, косуле, лосе. — 3 вида
(в СССР 5).
Мухи
1 (2). Лицевой киль на дне усиковой ямки отсутствует. Узкая полоска черных
волосков вдоль средней линии III—V тергитов брюшка отсутствует.
Брюшко сверху покрыто ярко-рыжими волосками, реже волоски желтые,
но тогда по бокам брюшка на II и III тергитах расположено по пятну
ярко-рыжих волосков. 14—16. — Евр. ч. СССР, Алтай, Бурятия, ДВ
(Амур.); Бельгия, ГДР, ФРГ, Швейцария, Австрия, Венгрия. На косуле
....................................................С. stimulator Cl.
2 (1). Лицевой киль на дне усиковой ямки развит. Узкая полоска черных
волосков вдоль средней линии III—V тергитов брюшка имеется.
3 (4). Перепонка крыла по сторонам жилки гт сильно затемнена, в связи с чем
эта часть крыла резко выделяется. Все бедра в черных волосках. 17—18. —
От Ленинградской обл. и Белоруссии до Урала, ДВ (Сихотэ-Алинь).
На лосе................................................С. ulrichi Вг.
4 (3). Перепонка крыла по сторонам жилки гт с едва заметным затемнением,
в связи с чем это место крыла не выделяется. Все бедра в основной части
с длинными желтоватыми волосками. Мельче: 14—16. — С евр. ч. СССР
и Сиб., Якут., Маг., Сах.; С Скандинавского п-ова, Аляска, С Канады.
На северном олене (рис. 77, 1—4)...................................
. ...........С. trompe Modeer — Сяну, или северный носоглоточник
Личинки III стадии
1 (2). На заднем крае верхней подушки подталкивателя расстояние от каждого
бокового сосочка до ближайшего срединного в 3—4 раза превышает
расстояние между срединными сосочками. Шипы в зонах на передних
краях члеников расположены в беспорядке. Дыхательная трубка перед-
них дыхалец в 6—7 раз тоньше атриума. Задние дыхальца достигают
в высоту 1.15 мм. На спинной стороне VII брюшного членика в середине
переднего края шипы отсутствуют (рис. 77, 5—7) .... С. ulrichi Вг.
2 (1). На заднем крае верхней подушки подталкивателя расстояние от каждого
бокового сосочка до ближайшего срединного лишь в 2 раза превышает
расстояние между срединными сосочками. Шипы в зонах на передних
краях члеников, особенно на брюшной стороне, местами сгруппированы
в короткие поперечные ряды. Дыхательная трубка передних дыхалец
в 3—4 раза тоньше атриума. Задние дыхальца высотой 0.85 мм.
3 (4). Посредине переднего края спинной стороны VII брюшного членика шипы
всегда имеются. Шипы спинной стороны у зрелой личинки в большинстве
сплошь черные ......................................С. stimulator Cl.
4 (3). Посредине переднего края спинной стороны VII брюшного членика шипы
обычно отсутствуют. Шипы спинной стороны черные на вершине и с чер-
ными точками на основной части (рис. 77, 5, 4) . . . .С. trompe Modeer
2. Pharyngomyia Shin. Личинки паразитируют на оленях, изюбре, марале,
лосе, косуле. Нередко приводит к гибели молодняка. — 1 вид (в СССР 2).
Мухи
1 (1). Коренастые мухи с почти голым телом. Брюшко округлое, немного короче
и шире груди. Оно буровато-черное с переливчатым рисунком. Средне-
спинка беловато-серая или серовато-желтая с черно-коричневым рисунком.
Экзотического вида мухи. 13—14. — Крымские горы, Таджикистан,
Алтай, ДВ (Сихотэ-Алинь). На обыкновенном олене, изюбре (рис. 78, 7, 2)
.......................................................Ph. picta Mg.
139
Личинки III стадии
1 (1). На переднем крае VIII брюшного членика на брюшной стороне имеются
шипы. У зрелой личинки на спинной стороне шипы окрашены только
3
Рис. 78. Посоглоточиый овод Pharyngomyia picta.
1 — самка; 2 — се голова; 3 — передняя часть личинки JII стадии, брюшная сторона; 4 — VII брюшной
членик сзади. (По: Грунин, 1957).
на вершине; яркие боковые группы черных точек на нескольких последних
брюшных члениках отсутствуют (рис. 78, 3,4).................Ph. picta Mg.
3. Oestrus L. Среднего размера мухи с очень маленькими усиками. Ячейка
/?5 всегда замкнутая, со стебельком; отросток т2 отсутствует. Личинки парази-
тируют у домашней овцы в носовой полости, хоанах, лобных пазухах и в поло-
стях у основания рогов. — 1 вид (в СССР 2).
140
Мухи
1 (1). Коренастые мухи, тело выглядит голым. Голова сильно вздута. Брюшко
овальное, чуть короче груди, бархатистое, от коричневого до черно-
коричневого, с переливчатым серебристым рисунком. Жилки крыла светло-
коричневые. 10—12. — Повсеместно, за исключением севера (рис. 79, 2, 2)
О. ovis L. — Кручак, или овечий овод
1 — самка; 2 — ее голова; з — личинка
III стадии, спинная сторона; 4 — VIII
брюшной членик сзади. (По: Грунин,
1957).
Личинки III стадии
1 (1). Тело стройное, зауженное кзади. Вооружение из шиников на спинной
стороне отсутствует; редко немногочисленные шипы имеются по перед-
нему краю грудных и I брюшного члеников (рис. 79, 3,4) . . О/ovis L.
4 Rhinoestrus Вг. Коренастые мухи среднего размера, тело голое. Орбиты
с темными крупными бородавками, на скулах более мелкие блестящие черные
мозольки. Брюшко сверху с бородавками, крылья с 3 черными пятнами у осно-
вания. — (На советском ДВ пока не найден. В СССР 4 вида).
141
Мухи
1 (2). Четыре продольные полосы среднеспинки блестяще-черные. Бородавки
на орбитах не слиты в плотную группу, между ними видны белые участки
поверхности орбит. Наличник в самой узкой части равен V6—V3 его ши-
рины на уровне усиков. 9—11. — Евр. ч. СССР, Кавказ, Казахстан,
Узбекистан, Ю Сиб., Заб.; Ю Европа, Африка, Монголия, Китай, Индия.
В полостях головы лошади и осла.................................
............................ Rh. purpureus Вг. — Овод малошип
2 (1). Продольные полосы среднеспинки красно-коричневые, матовые, только
передний край двух внутренних полос блестящий. На орбитах бородавки
слиты в плотную группу, занимающую верхнюю половину орбит. Налич-
ник в самой узкой части равен х/3—х/2 его ширины на уровне усиков.
11—13. — Евр. ч. СССР, Казахстан, Узбекистан; Монголия, Китай.
Личинки в полостях головы лошади ...............................
............................. Rh. latifrons Gan — овод-коротыш
Личинки III стадии
1 (2). Тело личинок стройное, заужено кзади; его длина превышает ширину
приблизительно в 3.5 раза. Группы среднебоковых шипов без мелких
шипиков с внутренней стороны. Внутренние края задних дыхалец парал-
лельные ...»....................................Rh. purpureus Вг.
2 (1). Тело личинки широкое, концы его лишь немного закруглены, длина пре-
вышает ширину не более чем в 2.5 раза. Группы среднебоковых шипов
с многочисленными мелкими шипиками с внутренней стороны. Внутрен-
ние края задних дыхалец расходятся книзу .... Rh. latifrons Gan
ПОДКОЖНЫЕ ОВОДЫ (Сем. HYPODERMATIDAE)
*
Крупные или средней величины мухи, тело которых покрыто густыми
длинными волосками или почти голое. Ротовые органы сильно редуцированы
или вообще отсутствуют (иногда сохраняются рудименты хоботка и шаровид-
ные щупики). Короткие усики спрятаны в усиковой ямке, разделенной лице-
вым килем. Ариста голая. Наличник похож на округленный щит. Вершинная
часть 7п]+2 всегда плавно изогнута к переднему краю крыла. Ноги довольно
длинные, задние не короче длины тела. Яйцеклад длинный, сильно выдается
за вершину брюшка, втяжной.
Паразиты зайцеобразных, парнокопытных и грызунов. Медико-ветеринар-
ное значение имеют представители 2 родов.
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ РОДОВ
Мухи
1 (2). Тело в густых длинных волосках. Щиток по заднему краю не раздвоен.
Имеются шарообразные щупики. Вершинный отрезок жилки т1+2 не
пигментирован. Голени тонкие, не расширены в середине...........
..........................................1. Oedemagena Latr.
2 (1). Тело в густых длинных или коротких волосках. Щиток по заднему краю
двухвершинный или только с неглубокой продольной бороздкой. Вер-
шинный отрезок 77?1+2 коричневатый, редко бесцветный. Голени в середине
заметно утолщены................................2. Hypoderma Latr.
i
Личинки III стадии
1 (2). Над псевдоцефалом имеется 1 ряд крупных шипов. Спинная и брюшная
стороны по степени вооружения шипами почти равны. Мелкие шипы
вокруг задних дыхалец расположены узкой зоной . . 1. Oedemagena Latr.
142
2 (1). Над псевдоцефалом шипы мелкие, образуют не менее 2 рядов, реже —
отсутствуют. Спинная сторона вооружена шипами гораздо слабее брюш-
ной. Мелкие шипы вокруг задних дыхалец образуют широкую зону или
отсутствуют..................................2. Hypoderma Latr.
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
t. Oedemagena Latr. В СССР —- 1 вид.
Мухи
1 (I). II тергит брюшка в соломенно-желтых, остальные тергиты — в ярко-
рыжих волосках. 16. — Распространение циркумполярное: С евр. ч.
СССР и Сиб., на юг до Алтая, С и Ю ДВ; Скандинавский п-ов, Аляска,
Канада. На северном олене (рис. 80, 7, 2) . . * . О. tarandi L. — Пилю
Личинки III стадии
1 (1). Пара сенсорных органов псевдоцефала расставлена. 1-й грудной членик
вооружен только на переднем крае спинной стороны одним рядом круп-
ных шипов (над псевдоцефалом). Шипы переднего и заднего краев чле-
ников одинаковы по форме и характерно окрашены: основание и при-
остренная часть шипов красно-бурые, с широким черным пятном в месте
перехода к приостренной части. На брюшной стороне шипы мельче,
поэтому зоны шипов уже, чем на спинной стороне. Отношение длины тела
к ширине от 1.5 до 1.9, длина личинки до 30 мм (рис. 80, 3—6). О. tarandi L.
2. Hypoderma Latr. Среднего размера мухи, тело в густых длинных и корот-
ких волосках. Рудиментов хоботка и щупиков нет. Ячейка Z?5 открытая, очень
редко замкнутая у края крыла. Личинки паразитируют на полорогих или оле-
нях. — 3 вида (в СССР 10).
Мужи
1 (4). Щиток по заднему краю с неглубокой продольной бороздкой. Тело
в длинных волосках.
2 (3). На среднеспинке впереди и позади поперечного шва окраска волосков
различная: перед поперечным швом они желтоватые, а позади шва черно-
бурые. Длина видимой части 3-го членика усиков значительно короче
2-го членика. 13—15. — От Карелии до Якут, и Маг.; Монголия. На круп-
ном рогатом скоте, буйволах, зебу, редко на лошади, овце, человеке
(рис. 81,1) . . Н. bovis De Geer — Строка, или обыкновенный подкожник
3 (2). На среднеспинке впереди и позади поперечного шва окраска волосков
одинаковая (черно-бурая). Длина видимой части 3-го членика усиков
у обоих полов приблизительно равна длине 2-го членика. Мельче: 11 —
13. — Ср. полоса евр. ч. СССР, Урал, 3 и В Сиб., Якут., побережье Охот-
ского моря, на юг до Ростова-на-Дону и Байкала; Монголия, СВ Китай,
Пакистан, Индия, Япония, С Африка, Америка, Гавайские о-ва, о-в Таити.
На крупном рогатом скоте, яке, зебу, редко на лошади, кошке, человеке
(рис. 80, 7, 8)..............................................
.............Н. lineatum De Vill. — Пищеводник, или южный подкожник
4(1'. Щиток двухвершинный. Тело в густых коротких волосках. В нижней
трети орбиты светло-коричневые, остальная часть (2/3) темные. Коготки
коричневые, на вершине черные. 11—12. — От Калининградской обл.
и Украины до Прим., Ср. Азии; Англия, ГДР, Австрия, Болгария, Фран-
ция, Бельгия. На косуле, оленях, изюбре (рис. 75, 2) . . Н. diana Вт.
Личинки III стадии
1 (4). Мелкие шипы вокруг задних дыхалец образуют широкую зону.
2 (ЗУ. Шипы заднего края заканчиваются на спинной стороне на IV—V, на
брюшной — на VI брюшном членике. Задние дыхальца воронкообразные
(рис. 81, 2—5) . »••••••••••••••. ,,H. bovis De Geer
143
Рис. 80. Подкожные оводы.
1—6 — Oedemagena tarandi: 1 — самка (по: Бреев, Савельев, 1958, из: Грунин, 1962), 2 — ее голова,.
3 — личинка III стадии с брюшной стороны, 4 — передняя часть тела этой личинки спереди, 5 — VIII брюш-
ной членик сзади, 6 — одно из задних дыхалец; 7, 8 — Hypoderma lineaium: 7 — личинка III стадии со
спинной стороны, 8 — задние дыхальца (по: Грунин, 1962).
Рис. 81. Подкожный овод Hypoderma bovis.
1 — самка (по: Штакельберг, 1933, из: Грунин, 1962); 2 — личинка III стадии со спинной стороны; 3 —
передняя часть личинки спереди; 4 — VIII брюшной членик сзади; 5 — одно из задних дыхалец (по: Гру-
нин, 1962).
Ю Зак. № 990
3 (2). Шипы заднего края заканчиваются на спинной стороне на VI—VII,
на брюшной — на VII брюшном членике. Задние дыхальца плоские
(рис. 80, 7, 8).....................................Н. lineatum De Vill.
4 (1). Мелкие шипы вокруг задних дыхалец либо отсутствуют, либо не обра-
зуют широкой зоны.
5 (6). На спинной стороне шипы заднего края члеников обычно отсутствуют;
изредка они имеются до III—V члеников включительно . . Н. diana Вг.
6 (5). На спинной стороне шипы заднего края члеников всегда имеются и за-
канчиваются на VI—VII (реже на V) брюшных члениках.......................
............................................... Н. capreola Rubtz.1
ОСНОВНАЯ ЛИТЕРАТУРА
Грунин К. Я. Личинки оводов домашних животных СССР. М.; Л., 1953. 124 с. (Опре-
делители по фауне СССР, изд. Зоол. ин-том, № 51).
Грунин К. Я. Желудочные оводы (Gastrophilidae). М.; Л., 1955, 95 с. (Фауна СССР.
Двукрылые; Т. XVII. Вып. 1).
Грунин К. Я. Носоглоточные овода (Oestridae). М.; Л., 1957. 145 с. (Фауна СССР.
Двукрылые; Т. XIX. Вып. 3).
Грунин К. Я. Подкожные овода (Hypodermatidae). М.; Л., 1962. 237 с. (Фауна СССР.
Двукрылые; Т. XIX. Вып. 4).
Грунин К. Я. Желудочные овода (Gastrophilidae). — В кн.: Определитель насекомых
европейской части СССР. Л., 1970, т. 5, ч. 2, с. 595—596.
Грунин К. Я. Носоглоточные овода (Oestridae). Подкожные овода (Hypodermatidae).—
В кн.: Определитель насекомых европейской части СССР. Л., 1970, т. 5, ч. 2, с. 674—678.
БЛОХИ
(Отряд SIPHONAPTERA = APHANIPTERA, SUCTORIA)
Мелкие (0.75—5 мм) кровососущие насекомые, паразитируют на млекопитаю-
щих и птицах. Крылья отсутствуют. Тело сплюснуто с боков, покрыто направ-
ленными назад щетинками, волосками, гребнями (ктенидиями), хорошо при-
способлено к передвижению в шерсти и в гнезде хозяина. Цвет покровов от
соломенно-желтого до темно-бурого и смоляно-черного. 60 видов.
ВНЕШНЕЕ СТРОЕНИЕ
Голова разделена усиковой ямкой на две части: переднюю и заднюю. На
переднем (лобном) крае головы бывает более или менее развитый зубчик или
бугорок. В нижнем отделе передней части головы различают ротовой, щечный
края и щечную лопасть, расположенную позади и ниже глаза. В задней части
головы находятся темя и задний край головы. В усиковой ямке с каждой сто-
роны помещается усик (рис. 82), состоящий из 3 члеников; 3-й членик (була-
ва) делится еще на ряд члеников. У самцов, а также у самок некоторых родов
теменные ямки или бороздки, идущие от усиков вверх (межусиковая бороздка),
соединяются на темени. Глаза помещаются перед ямкой усика и развиты раз-
лично у представителей разных родов или совсем отсутствуют. На поверх-
ности головы имеются волоски, щетинки и другие кутикулярные образования.
Кроме щетинок на голове могут быть на лбу толстые темные шипы и более или
менее развитые ряды плоских зубцов (головные ктенидии), носящие название
по месту расположения. Строение ротовых органов показано на рис. 83, 4.
Грудь состоит из 3 сегментов. В каждом различают верхнее полукольцо —
переднеспинка (пронотум), среднеспинка (мезонотум), заднеспинка (метано-
тум) — и груднобоковые пластинки (стерночлевры). Для определения исполь-
зуют строение, степень развития стеноплевр средне- и заднегруди. Плевра
среднегруди (мезоплевра) может быть с внутренним вертикальным склероти-
зованным тяжом или без него. На заднегруди различают еще эпистерн (мет-
1 Мухи этого вида пока не найдены, но их обнаружение на Дальнем Востоке вполне
возможно.
146
эпистерн) и эпимер (метэпимер) — большую крыловидную пластинку, форма
переднего края которой имеет значение при определении. На члениках груди
имеются ряды щетинок; у многих блох по краю сегментов — гребни (чаще на
пронотуме) и апикальные зубчики. Число зубцов в гребнях, а также на тер-
гитах груди и брюшка подсчитывается всегда с обеих сторон вместе.
К членикам груди прикрепляются 3 пары ног, из которых наиболее раз-
вита задняя — прыгательная пара. Нога состоит из следующих члеников:
тазик (кокса, ляжка), вертлуг, бедро, голень, пятичлениковая лапка. При
1 — схема строения блохи: аз — апикальные зубчики тергитов, анс — анальный стернит, ант — аналь-
ный тергит, зсп — заднеспинка, кт — гребень (кненидист), лз — лобный зубчик, мк — максиллы, мкп —
максиллярные пальпы, пиг — пигидий (впереди от него — стигма VIII тергита), прсп — переднеспинка,
I—VIИс — стерниты брюшка, сем — семяприемник, срсп — среднеспинка, I—VI 11т — тергиты брюшка,
у — усик, х — хоботок, ц — церки; членики задней ноги: б — бедро, в — вертлуг, гол — голень, л —
лапка (I—V — членики лапки), сх3 — тазик (кокса) III пары ног; 2,3 — V членик лапки блохи: 2 —
рода Ceratophyllus, 3 — рода Dasypsryllus (по: Иофф, Тифлов, 1954); 4 — голова и грудь Pulex irritans; 5,
6 — Archaeopsylla sinensis: 5 — голова и переднеспинка, 6 — половая клешня. (По: Иофф, Скалой, 1954).
определении смотрят наличие или отсутствие на тазиках2 внутреннего склеро-
тизованного тяжа; на тазиках3 у некоторых видов имеется группа или жосой
ряд шипиков. Используется также число и расположение щетинок на внутрен-
ней и наружной поверхности бедра3, на наружной поверхности голени3, шпор
на заднем крае голени, длина апикальных щетинок на 2-м членике и вооруже-
ние 5-го членика лапкп3. На нем различают, кроме коготков, боковые и подош-
венные щетинки, волоски, прикогтевые шипики.
Брюшко состоит из 10 сегментов; I членик брюшка не имеет стернита, его
место занято метэпимером. На тергитах и стернитах имеются различные ряды
щетинок и одиночные щетинки. Подсчет щетинок на тергитах и стернитах
10*
147
брюшка ведется с каждой стороны тела блохи. У блох многих родов на заднем
крае тергитов имеются апикальные зубчики, а иногда гребни из плоских зуб-
цов.
Очень характерные признаки при видовом определении дает строение по-
следних (модифицированных) сегментов брюшка. У самки очень характерно
Рпс. 83. Схема строения последних сегментов брюшка, гениталии и наружных ротовых
органов блохи.
1 — последние сегменты брюшка и гениталии ?: анс — анальный стернит, ант — анальный тергит, в —
влагалище, кп — копулятивный проток, кс — копулятивная сумка, пиг — пигидий, сем — семяприемник,
слп — слепой проток, сп — семенной проток, стиг — стигма I тергита брюшка, ц — церки; 2 — послед-
ние сегменты брюшка <?: бр — брюшко IX стернита, вв — вертикальная ветвь, гв — горизонтальная ветвь,
к — тело половой клешни, м — рукоятка половой клешни, р — неподвижный палец, яз — язычок IX стер-
нита, F — дигитоид; з — схема строения фалласома <?: ат — анодемальный тяж, аэ — аподема эдеагуса,
вп — вентральный парамер, вт — внутристенные тяжи, дп — дорсальный парамср, кэ — крючок эдеа-
гуса, са — суппорп аподемы, тп — тяжи пениса, тэ — внутренняя трубка эдеагуса, ша — шейка апо-
демы, эд — эдеагус, энд — эндосфаллус; 4 — наружные ротовые органы блохи: гб — нижняя губа, гп —
губные пальпы, л — лацпнии, мк — максиллы, мкп — макспллярные пальпы, н — надглоточнпк-
строение VII стернита. Его апикальный край может быть вертикальный или
закругленный (рис. 83, 7), с дорсальными и боковыми вырезками и выступами.
Форма его очень характерна для вида, хотя может и варьировать в определен-
ных пределах. VIII тергит у самок занимает большую часть заднего конца
брюшка; VII и IX стерниты развиты слабо, между ними находится влагалище.
Последний сегмент — анальный. У самок к анальным тергитам прикрепдя-
148
ются церки (анальные стилеты), по одному с каждой стороны. Они представляют
собой цилиндрические или конические выросты с сильными щетинками на
конце. Внутри брюшка на просветленных препаратах видны склеротизован-
ные части полового аппарата. Это — семяприемник (чаще непарный). Форма
его является хорошим родовым, а иногда видовым признаком. В нем различают
резервуар и придаток, последние могут быть резко отграничены друг от друга
или постепенно переходить один в другой. Для определения используются
форма и степень склеротизации некоторых других частей полового аппарата
самки: копулятивная сумка, слепой и семенной протоки,
У самца для видовой дифференциации используются форма и вооружение
так называемой половой клешни. Она служит для удержания самки при ко-
пуляции. Ее принято относить к IX сегменту. В половой клешне (рис. 82, 5,
83) различают тело, рукоятку, неподвижный палец, подвижный палец (диги-
тоид), дорсальную пластинку. На заднем крае половой клешни в месте сочле-
нения с дигитоидом (сочленовная ямка) у многих видов блох находятся длин-
ные ацетабулярные щетинки. Очень характерным видовым признаком является
форма и вооружение дигитоида. Кроме щетинок на нем бывают развиты раз-
личные шипы, шпоры, выросты. У некоторых родов блох половая клешня
устроена несколько иначе — может быть 2 дигитоида и пластинчатый вырост.
IX стернит самца (субгенитальный) примыкает снизу к половым органам, часто
бывает слит с ними. Форма его напоминает изогнутую под углом ленту. Раз-
личают две основные части: вертикальную (дорсальную или проксимальную)
ветвь, горизонтальную (вентральную или дистальную) ветвь. Форма и воору-
жение ее используются как диагностические признаки. Анальный сегмент
самца покрыт щетинками. При определении видов учитывается соотношение
длины анального тергита и стернита. Развитие VIII тергита самца резко варьи-
рует у разных родов. У некоторых видов с внутренней стороны его дорсальной
части имеется участок с мелкими шиппками — дорсальное поле шипиков.
VIII стернит может быть широкий, узкий, с апикальными щетинками и раз-
личными лопастями. У самцов в диагностических целях все чаще использу-
ется строение сложно устроенного половрго аппарата — фаллосома (прово-
дящий, интромиттирующий орган), состоящего из апикальной наружной
части — эдеагуса и внутренней — эндофаллуса. От основания эдеагуса внутрь
брюшка над эндофаллусом идет широкая склеротизованная аподема эдеагуса.
Следует учитывать, что все модифицированные сегменты и элементы поло-
вого аппарата самцов и самок являются подвижными органами. Их взаимо-
расположение не является постоянным. Поэтому на рисунках важно не распо-
ложение, а форма отдельных органов.
БИОЛОГ И Я
Блохи — насекомые с полным превращением. Паразитический образ жизни
ведут взрослые насекомые — самцы и самки. Питаются кровью млекопитаю-
щих и птиц. Самки откладывают яйца, разбрасывая их свободно в гнездах
своих хозяев. Синантропные виды используют скопление мусора и пыли
в углах и щелях полов различных построек. Яйца овальные, светлые, разме-
ром до 0.5 мм. Личинки червеобразные, безногие, питаются мелкими твердыми
органическими частицами субстрата. Личинка линяет 2 раза, вырастает до 5 —
8 мм, крупные виды — до 1 см. Зрелые личинки III стадии перед окукливанием
перестают питаться и передвигаться, складываются подковой и окружают себя
шелковистым коконом, к которому прилипают снаружи соринки. Внутри
кокона личинка линяет в куколку, а последняя — во взрослую блоху, которая
выходит из кокона, разрывая его оболочку с помощью лобного зубчика.
ВРЕД
Массовые укусы блох сильно досаждают людям, при расчесах могут возни-
кать раздражение кожи и нагноение. Но наибольший вред приносят блохи
как переносчики возбудителей некоторых опасных болезней и в первую оче-
149
редь чумы. В настоящее время насчитывается около 50 государств в Азии,
Европе, Африке, Америке, на территории которых обнаружены или предпо-
лагаются природные очаги чумы. На территории Дальнего Востока таких
очагов нет. Но существует реальная возможность заноса больных грызунов,
зараженных блох и укоренение последних.
Кроме того, в природе обнаружены блохи, зараженные микробами туля-
ремии. Выделили из блох возбудителей псевдотуберкулеза, кишечного иер-
синиоза, листериоза, эризипелоида, эндемического сыпного тифа. Находят
в природе блох, зараженных вирусами лимфоцитарного хориоменингита,
геморрагического нефрозонефрита, клещевого энцефалита.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ
1 (2). Передневерхний край эпимера заднегруди идет от плевры б. м. верти-
кально вверх. Стигма I тергита брюшка расположена высоко над уровнем
эпистерна заднегруди (рис. 82, 4). Метанотум и тергиты брюшка без апи-
кальных зубчиков или гребней. Наружная стенка кокс2 без внутреннего
склеротизованного тяжа .....................................1. Pulicidae
2 (1). Передневерхний край эпимера заднегруди идет от плевры косо кзади и
кверху под углом примерно 45°. Стигма I тергита брюшка лежит не выше
или немного выше эпистерна заднегруди. Наружная стенка кокс2 с вну-
тренним склеротизованным тяжом, идущим от сочленения с грудью вниз.
3 (4). Гребней или отдельных апикальных зубчиков на тергитах груди и
брюшка нет. На голове нет ктенидия. Предсенсиллиальные щетинки
отсутствуют. У $ нет церок..............................2. Vermipsyllidae
4 (3). На пронотуме имеется ктенидий. Гребни или отдельные апикальные
зубчики есть еще на некоторых тергитах брюшка. Предсенсиллиальные
щетинки и у $ церки имеются.
5 (10). На голове нет ктенидия, если имеется, то расположен под глазом, на
щечной лопасти или вдоль всего ротового края.
6 (7). Метанотум без апикальных зубчиков, но псевдосеты могут быть. IX стер-
нит d без аподемального тяжа. СешТиллий б. м. выпуклый. Церки обычно
с 1 длинной апикальной и 2 мелкими тонкими субапикальными щетинками
...................................................3. Hystrichopsyllidae
7 (6). Метанотум с апикальными зубчиками. IX стернит d с аподемальным
тяжом, направленным вперед от места сочленения вертикальной и гори-
зонтальной ветвей. Дорсальная сторона сенсиллия прямая. Церки с 1
или 2 довольно длинными субапикальными щетинками в добавление
к апикальной.
8 (9). Щечный гребень отсутствует. В глазном ряду 3 щетинки, верхняя сидит
впереди глаза. Передний тяж тентория, как правило, не развит. VIII стер-
нит d узкий, иногда маленький или рудиментарный. Глаз круглый . . .
....................................................... 4. Ceratophyllidae
9 (8). Со щечным гребнем или без него. Верхняя щетинка глазного ряда сидит
близ края усиковой ямки. Передний тяж тентория имеется, хотя иногда
скрыт щечным гребнем. Глаз часто выемчатый или рудиментарный . . .
....................................................... 5. Leptopsyllidae
10 (5). На голове имеется своеобразный предротовой ктенидий из двух лопато-
образных зубцов. Блохи летучих мышей....................6. Ischnopsyllidae 1
1. Сем. PULICIDAE
На внутренней поверхности кокс2 имеется ряд шипиков. Метанотум и тер-
гиты брюшка без апикальных зубчиков. Эпимер заднегруди простирается
далеко вверх. Стигма I тергита брюшка расположена высоко над уровнем эпи-
стерна заднегруди. Брюшко сравнительно короткое и высокое. Стигмы округ-
лые.
1 Медико-ветеринарное значение нс изучено, поэтому определительная таблица не
приводится.
150
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). Плевра среднегруди без внутренней склеротизованной перекладины, иду-
щей от места прикрепления кокс2 к верхнему краю плевры . .1. Pulex L.
2 (1). Склеротизованная перекладина плевры среднегруди имеется.
3 (4). Ктенидии на голове и пронотуме отсутствуют ... 2. Xenopsylla Glink.
4 (3). На щечном крае головы имеются ктенидии, развитые или рудиментарные.
5 (6). Щечный гребень из 2 (реже из 1—3) зубцов. В гребне на пронотуме не
более 8 зубцов с обеих сторон вместе.............3. Archaeopsylla Dampf
6 (5). Щечный гребень из 7—8 зубцов. В гребне на пронотуме около 16 зубцов
...................................... 4. Ctenocephalides Stil. et Coll.
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
1. Pulex L. Паразитирует на человеке, домашних животных и диких хищ-
ных млекопитающих. — В СССР — 1 вид.
1 (1). Глаза хорошо развиты; имеются 2 глазные щетинки, расположенные под
глазом (рис. 82, 4); цвет тела темно-бурый. — Космополит..............
.............................Р. irritans L. — Блоха человеческая
2. Xenopsylla Glink. Паразитирует на крысах, песчанках и некоторых
других млекопитающих. — 1 вид (в СССР — 11).
1 (1). От 1-го и 2-го теменных рядов сохранились хотя бы по 1 щетинке в каж-
дом; у <3 подвижные пальцы половой клешни перекрещиваются, пластинча-
тый вырост недоразвит; у $ VIII тергит с многочисленными щетинками на
боковой поверхности, придаток семяприемника длинный, его основание
лишь слегка шире резервуара. На крысах.................X. cheopis Roth.
3. Archaeopsylla Dampf. Паразиты ежей рода Erinaceus. — 1 вид (в СССР 2).
1 (1). Большая волосатая пластинка пЪловой клешни 3 короткая, тупая . . .
...............• ......................A. sinensis Jord, et Roth.
4. Ctenocephalides Stil. et Coll. Паразитируют на домашних кошках и соба-
ках, нападают на человека, иногда на диких животных; 1 вид — на козах. —
2 вида (в СССР 3).
1 (2). Лоб пологий, особенно у $ (рис. 84, 2)\ на заднем крае голени3 между
средней и апикальной группами шпор имеется обычно лишь 1 толстая
щетинка; па метэпистерне не более 2 щетинок; стигмы брюшных тергитов
сравнительно небольшие; у 3 рукоятка половой клешни слабо расширяется
к переднему концу. — Космополит. Блоха кошки. Нападает на человека,
встречается в подвалах жилых домов....................................
............................... С. felis Bouche — Блоха кошачья
2 (1). Лоб крутой у обоих полов (рис. 84, 7); на заднем крае голени3 между сред-
ней и апикальной группами шпор имеются обычно не менее 2 толстых
щетинок; на метэпистерне не менее 3 щетинок; стигмы брюшных тергитов
крупнее; у 3 рукоятка половой клешни сильно расширяется к переднему
концу. Распространение всесветное ... С. canis Curtis — Блоха собачья
2. Сем. VERMIPSYLLIDAE
На голове, тергитах груди и брюшка нет гребней, одиночных или апикаль-
ных зубчиков. На 5-м членике лапок3 4 пары боковых щетинок. Предсенсил-
лиальные щетинки у 9 и (?, а также церки у 9 отсутствуют. Семяприемник 1.
На хищных и копытных в Палеарктике и Неарктике. На Дальнем Востоке —
1 род.
1. Chaetopsylla Koch. — Распространены в Палеарктике во всех субрегио-
нах, некоторые виды встречаются и в Неарктике на Аляске. Паразитируют
на хищных. Отдельные виды проявляют большую специфичность к своим
151
хозяевам, например, Ch. tuberculaticeps — блоха медведя, Ch. zibellina —
блоха соболя. Блохи этого рода недостаточно изучены, медико-ветеринарное
значение не установлено. — На ДВ — 7 видов (в СССР 15).
5
Рис. 84. Детали строения блох.
1,2 — голова и грудь: 1 — собачья блоха Ctenocephalides canis, 2 — кошачья блоха С. felis', 3,4 — Hysi-
richopsylla microti: 3 — половая клешня, VIII и IX стерниты с. 4 — VII стернит и семяприемники $; 5,
6 — Stenoponia sidimi: 5 — VII стернит и семяприемник 6 — половая клешня, IX — стернит, парамер
эдеагуса <?. (По: Иофф, Скалой, 1954).
3. Сем. HYSTRICHOPSYLLIDAE
На голове имеется ктенидий (исключая род Catallagia). Сенсиллий более
или менее выпуклый. IX стернит (J без аподемального тяжа. Церки £ с 1 длин-
ной апикальной и 2 мелкими субапикальными щетинками. Паразитируют на
многих видах грызунов и зайцеобразных.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (18). На голове имеется ктенидий.
2 (17). В головном ктенидии более 2 зубцов.
3 (6). Зубцы головного ктенидия узкие, их не менее 4; на некоторых тергитах
брюшка имеются настоящие или хотя бы зачаточные гребни в виде ряда
152
редко расположенных апикальных зубчиков на боках, отступя от спины;
блохи очень крупные.
4 (5). На 5-м членике лапок3 5 пар боковых щетинок, подошвенная пара от-
сутствует; ? с 2 семяприемниками .... 1. Hystrichopsylla Tasch.
5 (4). На 5-м членике лапок3 4 пары боковых и 1 пара подошвенных щетинок,
расположенных между боковыми 1-й пары; на I тергите брюшка имеется
гребень из многочисленных зубцов; $> с 1 семяприемником................
.....................................2. Stenoponia Jord, et Roth.
6 (3). Зубцы головного ктенидия широкие; гребни на тергитах брюшка отсут-
ствуют, апикальные зубчики расположены близко к спине.
7 (10). Головной ктенидий, расположенный па ротовом или щечном крае,
состоит из 4 тупых зубцов.
8 (9). Лоб выпуклый с лобным зубчиком; головной ктенидий расположен на
щечной лопасти и прикрывает ее; апикальный край VII тергита d без
выроста над предсенсиллиальными щетинками .............................
.................................... Doratopsylla Jord, et Roth.*
9 (8). Лоб довольно покатый, без ясного зубчика; головной ктенидий начи-
нается от переднего конца нижнего края головы, последний его зубец
находится впереди от края щечной лопасти; апикальный край VII тер-
гита 3 с резким выростом над предсенсиллиальными щетинками . . .
................................................. Corrodopsylla Wagn.*
40 (7). Головной ктенидий, расположенный на ротовом или щечном крае,
состоит из заостренных зубцов различной формы.
11 (14). На 5-м членике лапок3 3—4 пары боковых щетинок и 1 пара подошвен-
ных.
12 (13). Головной ктенидий состоит из 3 зубцов, направленных косо книзу
.............................................. 3. Ctenophthalmus Koi.
13 (12). Головной ктенидий состоит из 4 зубцов, из которых 2-й сверху длин-
нее других, вытянут в острие........................Palaeopsylla Wagn.*
14 (11). На 5-м членике лапок3 4—5 пар боковых щетинок, основная подошвен-
ная пара отсутствует.
15 (16). Головной ктенидий обычно из 5—6 (редко из 4—8) зубцов примерно
одинаковой формы 4. Rhadinopsylla Jord, et Roth.
16 (15). Головной ктенидий из 5 зубцов, из которых 2-й снизу самый длинный
.................................................. Nearctopsylla Roth.*
17 (2). Головной ктенидий из 2 зубцов, из которых один более короткий, на-
правлен вниз, другой более длинный перекрещивает его, направляясь
кзади ..............................................5. Neopsylla Wagn.
18 (1). На голове не имеется ктенидия; глаз рудиментарный, но заметный;
на внутренней поверхности коксы3 имеется группа шипиков................
...............................................6. Catallagia Roth.
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
1. Hystrichopsylla Tasch. Очень крупные блохи. Обитают в гнездах лесных
грызунов и насекомоядных Европы, Азии, С Африки, Америки. — 1 вид
<в СССР 4).
1 (1). Щечный гребень состоит из 5—6 зубцов; в гребне пронотума 26—31 зу-
бец; на II — IV тергитах брюшка на боках имеются лишь зачаточные гребни
из редких зубчиков; у обоих полов по 3 предсенсиллиальных щетинки;
вершина горизонтальной ветви IX стернита d конусовидно суживается
(рис. 84, 5); VII стернит $ как на рис. 84, 4. — В гнездах мелких млеко-
питающих, на серой крысе ...............................Н. microti Seal.
2. Stenoponia Jord, et Roth. Крупные блохи, живущие преимущественно
в гнездах мышевидных, цокоров, песчанок на юге Европы, в Азии, С Африке,
Америке. — 2 вида (в СССР 9).
* Роды, отмеченные звездочками, не рассматриваются, так как медико-ветеринарное
-значение их на Дальнем Востоке слабо изучено.
153
1 (2). Вершина горизонтальной ветви IX стернита d усеченная с густым пучком
толстых шпорообразных щетин, парамер эдеагуса шлемовидный; VII стер-
нит $ чаще без выступающего вентрального угла, резервуар семяприем-
ника грушевидный (рис. 84, 5, 6). — На мышевидных грызунах . . . .
........................................................ S. sidimi Marik.
2 (1). Вершина горизонтальной ветви IX стернита d округленная, пучок щетин
на ней менее развит, парамер эдеагуса крючковидный; VII стернит
обычно с заметно выступающим вентральным углом, резервуар семя-
приемника более округлый (85, 7, 2). — На лесных полевках ......
........................................................ S. mon tana Desk.
3. Ctenophthalmus Koi. Паразиты грызунов, распространенные преимуще-
ственно в Европе и Африке. — 3 вида (в СССР 43).
1 (4). Стигма VIII тергита широкая, выпуклая.
2 (3). Дигитоид 3 топорообразной формы с хорошо выраженным задневерхним
углом; задний край VII стернита д с развитой вырезкой, разделяющей его*
на 2 выступа, из которых верхний лишь слегка выдается над нижним.—
На домовых мышах, крысах, в гнездах мелких грызунов, нападает на
человека ..........................................С. congeneroides Wagn.
3 (2). Дигитоид 3 с усеченным задневерхним углом, боковая вырезка VII стер-
нита $ менее выражена, верхний боковой выступ уже и значительна
выдается над нижним. Вид, близкий к предыдущему............................
............................................. С. truncus loff et Seal.
4 (1). Стигма VIII тергита узкая, Y-образная; наиболее длинные щетинки
2-го членика лапки3 далеко не достигают вершины 4-го членика; генита-
лии S и j как па рис. 85, 3, 4. — На лесных грызунах.....................
.................................. С. pisticus pacificus loff et Seal.
4. Rhadinopsylla Jord, et Roth. Паразитирует на грызунах и зайцеобразных
в Азии и Европе. —- 4 вида (в СССР 19).
1 (2). Хоботок из 6—7 члеников, достигает вершины коксь^ или заходит за нее;
основная половина горизонтальной ветви IX стернита d шире апикальной;
VII стернит ? с раздвоенным боковым выступом. — На хомячках, поле-
вых мышах и др., живущих в степях и на лугах .... Rii. insolita Jord.
2 (1). Хоботок ИЗ 5 члеников, большей частью далеко ие достигает вершины
коксыг
3 (4). На всех или некоторых II—VII тергитах брюшка ниже стигм имеются
2 щетинки (у ? как правило, yd — иногда); VIII стернит d с многочис-
ленными щетинками (часто более 10), образующими более одного ряда;
IX стернит d с характерным апикальным расширением горизонтальной
ветви; VII стернит ? с усеченным боковым выступом. — В гнездах поле-
вок, иногда других мелких грызунов и хищников, часто на крысе . . .
..................................................... Rh. rothschildi loff
4 (3). На III—VI тергитах брюшка ниже стигм лишь 1 щетинка; на VIII стер-
пите d в большинстве случаев не более 7 щетинок, расположенных в 1 ряд.
5 (6). Задненижний угол VIII стернита d вырезан сильно, так что ряд из 6
7 щетинок расположен близко к краю; апикальная часть горизонтальной
ветки IX стернита d расширена, вершина округлена (рис. 85, 5); гени-
талии $ — рис. 85, 6; $ отличается от $ Rh. integella лишь слегка менее
вертикальным щечным гребнем. — На различных грызунах .....
........................................... Rh. pseudodahurica Seal.
6 (5). Задняя часть VIII стернита d срезана не сильно, ряд из 3—5 щетинок
расположен далеко от заднего края; апикальная часть горизонтальной
ветви IX стернита d менее расширена, с более выраженным апикально-
дорсальным углом. — В гнездах мелких млекопитающих ....................»
.................................... Rh. integella Jord, et Roth.
(На ДВ — два подвида: Rh. integella casta Jord, и Rh. integella con-
cava loff et Tifle).
154
5. Neopsylla Wagn. Паразитируют на мышевидных и некоторых других
грызунах. Распространены преимущественно в Азии, немногие виды проникли
в Европу, несколько известно из С Америки. — 3 вида (в СССР — 12).
1 (4). На наружной поверхности голени3 2—3 продольных ряда щетинок, не
считая передних маргинальных или субмаргинальных.
2 (3). Задний край дигитоида 3 с ясным выступом; VIII стернит d с гривой
длинных извилистых щетин, IX стернит с 2 группами толстых темноватых
Рис. 85. Детали строения блох.
2, 2’— Stenoponia montana: 1 —половая клешня, IX стернит, парамер эдеагуса <?, 2 —семяприемник,
варианты конфигурации VII стернита $; 3, 4 — Ctenophthalmus pisticus pacificus: 3 — VII стернит и семя-
приемник $, 4 — половая клешня и IX стернит <J; 5, 6 — Rhadinopsylla pseudodahurica: 5 — половая
клешня, VIII и IX стерниты <?, 6 —VII стернит и семяприемник %. (По: Иофф, Скалой, 1954). ।
тупых шипов; VII стернит $ с резким и довольно узким выступом. —
На грызунах (мышах, крысах, в жилье человека), зайцеобразных, земле-
ройках .........................................N. bidentatiformis Wagn.
3 (2). Дигитоид 3 узкий, его задний край без выступа, VIII стернит d без гривы
из длинных извилистых щетин, шипы IX стернита d не толстые, светлые;
боковой выступ VII стернита $ шире и короче, чем у $ N. bidentatiformis. —
Степной подвид ...........................N. pleskei orientals loff et Argy.
4 (1). На наружной поверхности голени2 1 продольный ряд щетинок, впереди
от которого нет или имеются единичные щетинки (не считая маргинальных
155
или субмаргинальных); в нижней части дигитоида d характерная склере*
тизация в виде темного узкого язычка, оставляющего у заднего края свет-
лую полоску; на VIII стерните $ гривы нет, а бледные апикальные ло-
Рис. 86. Детали строения блох.
1,2 — Catalagia ioffi'. 1 — половая клешня, VIII и IX стерниты <?, 2 — семяприемник, VII — стернит
и стигма VII стернита 3, 4 — С. docenkoi: з — половая клешня, VIII и IX стерниты <?, 4 — семяприем-
ник, VII стернит и стигма VIII тергита $; 5, 6 — Oropsylla alaskensis: 5 — половая клешня, VIII и IX стер-
ниты <J, 6 —VII стернит, семяприемник (По: Иофф, Скалой, 1954).
пасти слабо развиты; IX стернит d с многочисленными бледноокрашен-
ными шипами и резким горбом в апикальной трети горизонтальной ветви;
$ отличаются от 9 N. р. orientalis вооружением голени и более крупной
стигмой VIII тергита. — Сибирский вид, в лесных и лесостепных районах.
Может паразитировать на крысах . . . N. acanthina Jord, et Roth.
156
6. Catallagia Roth. Преимущественно на лесных полевках рода Cleihrio-
nomys, но встречаются и на многих других видах мелких лесных млекопитаю-
щих. — 3 вида (в СССР — 4—5).
1 (2). Стигмы II—VII брюшных тергитов расширенные, яйцевидной формы;
гениталии d и $ как на рис. 86, 5, 4. — На лесных полевках............
.............................................. С. dacenkoi loff
2 (1). Стигмы II—VII брюшных тергитов не расширенные, соскообразно вытя-
нутые назад.
3 (4). Дигитоид d с закругленным задненижним краем и слегка превышает
неподвижный палец (рис. 86, 7, 2); стигма VIII тергита $ расширена, гени-
талии ее как на рис. 86, 2. — На лесных полевках .... С. ioffi Seal.
4 (3). На задненижнем крае дигитоида d 2 глубокие выемки, он значительно
короче неподвижного пальца; стигма VIII тергита $ маленькая ....
..................................................... С. striata Seal.
4. Сем. CERATOPHYLLIDAE
На переднеспинке имеется гребень. Метанотум и несколько тергитов
брюшка, как правило, несут апикальные зубчики, но гребни на тергитах
брюшка отсутствуют. Сенсиллий у $ и d невыпуклый. У d IX стернит с аподе-
мальным тяжом, идущим кпереди под эндофаллусом. У $ один семяприемник.Х
На грызунах и других млекопитающих, а также на птицах. Многие виды имеют
эпидемиологическое и эпизоотологическое значение. — На ДВ — 13 родов.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). 1-й членик лапок3 длиннее 2—4-го вместе взятых................
............................................ Tarsopsylla Wagn.*
2 (1). 1-й членик лапок3 не длиннее 2—4-го вместе взятых.
3 (8). 1 или более члеников губных пальп заходят за вершину бедра!.
4 (5). Значительная часть наружной поверхности бедра! свободна от щетинок.
Отсутствует неподвижный палец половой клешни..........................
............................................1. Aenigmopsylla Sych.
5 (4). Наружная поверхность бедра! довольно равномерно усеяна щетинками,
\ Неподвижный палец половой клешни имеется.
6 (7). Половая клешня d с большим широким дигитоидом. Семяприемник $
с узким резервуаром...........................Paraceras Wagn.*
7 (6). Половая клешня d с дигитоидом удлиненно-овальной формы. Семя-
приемник $ с грушевидным резервуаром. Форма VII стернита $> как
на рис. 87, 2............................2. Oropsylla Wagn. et loff
8 (3). Губные пальпы короткие или за вершину бедрах заходит менее одного
членика.
9 (10). 3-я пара боковых щетинок 5-го членика лапки3 сильно сдвинута на по-
дошву, а ее место занято тонкими волосками .... 3. Dasypsyllus Bak.
10 (9). 3-я пара боковых щетинок 5-го членика лапки3 не сдвинута на подошву.
11 (12). Ктенидий пронотума состоит из коротких зубцов................
............................................4. Mioctenopsylla Roth.
12 (11). Ктенидий пронотума состоит из довольно длинных зубцов.
13 (14). Стигма VIII тергита большая, особенно у $. Дигитоид d всегда с силь-
ными шпорами.......................................5. Megabothris Jord.
14 (13). Стигма VIII тергита небольшая. На дигитопде d короткие шипы или
щетинки.
15 (16). VIII стернит d рудиментарен, без щетинок. Копулятивная сумка $
спирально изогнута.................................6. Nosopsyllus Jord.
16 (15). VIII стернит d удлиненный, со щетинками и лопастями различной
формы. Копулятивная сумка, семяприемник $ иной формы.
* Диагноз этих родов не приводится, так как медико-ветеринарное значение их на
Дальнем Востоке не выяснено.
157
17(20). На внутренней поверхности кокс2 и 3 щетинки имеются не только
на апикальной половине, но и на базальной (не считая сидящих по перед-
нему краю). У в анальный стернит намного длиннее тергита. Копулятив-
ная сумка $ мешкообразно расширена.
18 (19). Дигитоид d широкий, примерно треугольной формы, с 2 шипами, уда-
ленными друг от друга. У $ фронтальные щетинки отсутствуют, имеются
2 предсенсиллиальные..............................7. Citellophilus Wagn.
19 (18). Вершина дигитоида в оттянута назад, на задневерхнем выступе его
2 близкосидящих шипа. У $ фронтальные щетинки имеются, предсенсил-
лиальных 3..........................................8. Callopsylla Wagn.
20 (17). На внутренней поверхности кокс2 и 3 щетинки имеются лишь на апи-
кальной половине. У 8 анальный стернит не намного длиннее тергита.
Копулятивная сумка $ иной формы.
21 (24). В гребне пронотума менее 24 зубцов. Поле шипиков VIII тергита d
отсутствует.
22 (23). Во 2-м теменном ряду более 2 щетинок. Продольный диаметр глаза
меньше, чем расстояние от глаза до вершины щечной лопасти . . . .
.................;....................................9. Amalaraeus loff
23 (22). Во 2-м теменном ряду 2 щетинки. Продольный диаметр глаза больше,
чем расстояние от глаза до вершины щечной лопасти......................
...............................................10. Monopsyllus Koi.
24 (21). В гребне пронотума более 24 зубцов. Поле шипиков VIII тергита d
развито. Большинство видов — паразиты птиц.............................
............................... И. Ceratophyllus Curt.
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
1. Aenigmopsylla Sych. — В СССР — 1 вид.
1 (1). Неподвижный палец половой клешни d отсутствует; край VII стер-
нита ? округлый, резервуар семяприемника бочкообразный. — На бел-
ках и других лесных грызунах . . . *..................A. grodekovi Sych.
2. Oropsylla Wagn. et loff. Паразиты сурков и сусликов С. Америки, Азии
и Европы. — 2 вида (в СССР 3).
1 (2). Хоботок длинный, не менее 1-го членика, а чаще и часть следующего
заходят за вершину коксых; глаз небольшой; лобный зубчик ясно высту-
пает; зубцы пронотума расположены не тесно друг к другу, они короче
его спинки по дорсальной линии; пронотум удлинен. Гениталии d и $
как на рис. 87, 7, 2, — На сурках.....................О. silantievi Wagn.
2 (1). Хоботок короче, чем у предыдущего вида, за вершину коксыт захщит
меньше целого членика; глаз крупнее и сильнее пигментирован; лобный
зубчик очень слабый; зубцы пронотума расположены густо и несколько
длиннее его по дорсальной линии. Гениталии d и $ как на рис. 86, 5, 6. —
На длиннохвостом суслике..........................................
......................... О. alaskensis Bak. ( = 0. asiatica Wagn.)
3. Dasypsyllus Bak. Паразиты различных птиц в Европе, Ц и ЮВ Азии,
С Америке. — 1 вид (в СССР 2).
1 (1). На передней части головы 2 ряда щетинок; дигитоид d конической формы
с 2 шпоровидными щетинками у задненижнего угла и 1 — около середины
заднего края; у J 3 предсенсиллиальные щетинки; край VII стернита
с боковой вырезкой, разделяющей небольшой верхний и широкий ниж-
ний выступы; семяприемник с широким округлым резервуаром. — Пара-
зит птиц-норников . . ................................D. stejnegeri Jord.
4. Mioctenopsylla Roth. Паразиты морских птиц. — На ДВ и в СССР —
2 вида.
1 (2). Неподвижный палец половой клешни d узкий, его высота превышает
ширину почти в 2 раза; основание нижней шпоры дигитоида расположено
158
на небольшом выступе; вершина VIII стернита d без изгиба кверху,
с апикальной мембранозной лопастью, щетинки на нем расположены
в ряд по вентральному краю; у ? VII стернит с глубокой вырезкой. —
На моевке, трехпалой чайке.........................М. arctica Roth.
2 (1). Неподвижный палец половой клешни d широкий, его высота не превы-
шает ширину; у основания нижней шпоры дигитоида нет выступа; вершина
VIII стернита d загнута кверху, мембранозная лопасть развита слабо,
щетинки расположены группой на боковой поверхности; у $ вырезка
на VII стерпите неглубокая. — На бакланах, моевке, топорке . . . .
...................................М. traubi kurilensis Violovich
5. Megabothris Jord. Паразиты лесных грызунов в лесных и влажных мест-
ностях Европы, Азии, С Америки. — 6 видов (в СССР 9).
1 (2). На пронотуме впереди от главного ряда имеется еще ряд мелких щетинок;
характерна форма дигитоида d: верхний и нижний выступы с шипами,
узкая средняя часть (рис. 87, 3); у $ широкая складчатая копулятивная
сумка без склеротизации, форма VII стернита как на рис. 87, 4. — На по-
левках, леммингах и других грызунах; могуть пить кровь человека . . .
................................................... М. calcarifer Wagn.
2 (1). На пронотуме 1 ряд щетинок; признаки d и 9 иные.
3 (4). Кроме обычных рядов щетинок на голове имеется префронтальный,
а впереди него уд — еще добавочные щетинки; анальный стернит d очень
длинный; дигитоид узкий, пальцеобразный, без выступов, с характерным
вооружением; у д боковой выступ VII стернита острый, под ним хорошо
выраженная вырезка. — На северной пищухе. Редкий вид....................
....................................................М. taiganus Seal.
4 (3). На голове не имеется префронтального ряда.
5 (6). Дигитоид d с оттянутым задненижним углом, на котором расположена
сильная шпора; боковой выступ VII стернита д заострен, вырезка под ним
и вентральный выступ сглажены. — На лесных полевках.....................
..................................................М. sokolovi Gersh.
6 (5). Задненижний угол дигитоида* d не оттянут; боковой выступ VII стер-
нита д закруглен.
7 (8). Задневерхний угол дигитоида d образует узкий выступ, вершина кото-
рого вооружена 2 шипами, в середине заднего края дигитоида 2 шиловид-
ные щетинки; апикальный край VII стернита д с выступающим закруглен-
ным боковым углом, без выемки под ним. — На красно-серой полевке
.....................................................М. bispinosus Sych.
8 (7). Задневерхний угол дигитоида d не образует узкого выступа, закруглен,
по заднему краю дигитоида расположены не шиповидные щетинки, а на-
стоящие шипы; апикальный край VII стернита д со сглаженными боковым
и субвентральным выступами и выемкой (хотя бы небольшой) между ними.
9 (10). Тело половой клешни d вытянутое, анальный стернит довольно широ-
кий, по заднему краю дигитоида сидят 2 довольно длинных шипа и кривая
шпора; край VII стернита д с выемкой под сглаженным боковым углом,
который выступает обычно меньше субвентрального. — На лесных гры-
зунах .............................................М. advenarius Wagn.
10 (9). Тело половой клешни d не вытянутое, анальный стернит удлиненный,
шипы на задневерхнем крае дигитоида сближены, на задненижнем нахо-
дится сильный прямой шип с острой вершиной; д внешне очень похожа
на д М. advenarius, отличается более сглаженными (но не всегда) боко-
вым и субвентральным выступами VII стернита и несколько меньшими
стигмами брюшка; более точное определение д по сопутствующим d. —
На высокогорной лемминговидной полевке, большеухой пищухе . . . .
.....................................................М. beljaevi Yemel.
6. Nosopsyllus Jord. Паразиты крыс, мышей, полевок в Европе, Азии и
С Америке. — 1 вид (в СССР 14).
1 (1). Дигитоид d с угловатым передним краем; вершина IX стернита d асим-
метричная; у д фронтальных щетинок не больше 2, задний край VII стер-
159
нита без выступов. — На крысах, нападает на человека........................
........................................................ N. fasciatus Возе.
7. Citellophilus Wagn. Паразиты сусликов Европы и Азии. — 1 вид (в СССР
10).
1 (1). На заднем крае дигитоида <J 2 довольно длинных шипика, сидящих
не близко друг к другу, апикальная дорсальная лопасть VIII стернита d
Рис. 87. Детали строения блох.
J, 2 — Oropsylla silantiewi: 1 — половая клешня, VIII и IX стернит, крючок эдеагуса <?, 2 — гениталии
и последние (модификация) сегменты брюшка 3, 4 — Megabothris calcarifer: 3 — половая клешня, VIII
и IX стерниты <?, 4 — семяприемник, копулятивная сумка, варианты VII стернита $; 5, 6 — Amalaraeus
renicilliger: 5 — семяприемник, VII стернит, стигма VIII тергита $, 6 — крючок и склеротизация эдеа-
гуса <?. (По: Иофф, Скалой, 1954). j
не бахромчатая; вентральная вырезка VII стернита 5 чаще отсутствует,
форма заднего края VII стернита 9 может варьировать. — На сусликах
.........................................С. tesquorum sungaris Jord.
8. Callopsylla Wagn. Встречается на зверьках, живущих в каменных осыпях
и скалах, а также на птицах, преимущественно ласточках. — 1 вид (в СССР 10).
1 (1). Хоботок не заходит за вершину коксых; дорсальные зубцы гребня не ко-
роче, чем спинка переднегруди; задний край дигитоида d резко вогнут;
VII стернит 9 с хорошо выраженным боковым выступом. — На городской
ласточке............................................ С. waterstoni Jord.
160
9. Amalaraeus loff. Паразиты преимущественно полевок лесной, луговой
: горной фауны Европы, Азии и С Америки. — В СССР и на ДВ — 3 вида.
(2). Вершина дигитоида 3 шире, чем его середина, у задневерхнего угла ди-
гитоида сидит тупой шип, а около середины заднего края — удлиненная
шпора с 2 толстыми шпорами под ней. — На лесных полевках . . . .
......................................................A. ioffi Darsk.
(1). Дигитоид 3 расширен близ середины и вооружен 2 тупыми шипами.
(4). Крючок эдеагуса 3 с массивным широким основанием и относительно
коротким «клювом», темнопигментированная горизонтальная часть скле-
ротизации эдеагуса без выступа (рис. 87, 6); VII стернит $ как на рис. 87,
5. От A. dissinilis надежнее отличать по сопутствующим 3. — Лесной вид
................................................A. penicilliger Grube
(3). Крючок эдеагуса 3 с небольшим основанием и вытянутым «клювом»,
темнопигментированная склеротизация эдеагуса с вертикальным высту-
пом (рис. 88, 7, 2). От A. penicilliger надежнее отличать по сопутствую-
щим 3. — Лесной вид..................................A. dissimilis Jord.
10. Monopsyllus Koi. Паразиты различных видов грызунов, зайцеобразных,
:елких хищных Европы, Азии, С Америки. — 4 вида (в СССР 7).
(2). Стернит анального сегмента d не заходит или мало заходит за тергит;
на заднем крае дигитоида 3 имеются шипы или толстые шпоры; апикаль-
ный край VII стернита $ с широкой и глубокой боковой вырезкой. —
Северный таежный вид..................................М. tolli Wagn.
(1). Стернит анального сегмента 3 далеко заходит за тергит; на заднем крае
дигитоида нет развитых шипов и шпор; апикальный край VII стернита $
без боковой вырезки.
(4). Задневерхний угол тела половой клешни d без развитого неподвижного
пальца; апикальная часть горизонтальной ветви IX стернита d расширена;
задний край VII стернита $ с глубоким вентральным вырезом и характер-
ным выступом, имеющим выраженный апикальный дорсальный угол. —
На бурундуке и других лесных грызунах, в гнездах птиц.................
............................... М. indages Roth. ( = tamias Wagn.)
(3). Неподвижный палец половой клешни 3 развит; апикальная часть гори-
зонтальной ветви IX стернита 3 не расширена; задний край VII стернита $
без выраженного апикального дорсального угла.
(6). Лобный зубчик развит; неподвижный палец половой клешни d узкий,
конусовидный; дигитоид без коротких шипиков по заднему краю; апикаль-
ная лопасть VIII стернита d почти не развита (рис. 88, 7); слепой проток $
без склеротизации, форма ее VII стернита и семяприемника как на рис. 88,
3. — На крысах.......................................М. anisus Roth.
(7). Лобный зубчик рудиментарен; неподвижный палец половой клешни 3
хорошо развит, высокий; по заднему краю дигитоида d 2 коротких шипа;
апикальная лопасть VIII стернита d хорошо развита; исключительно
сильно развиты тяжи пениса; слепой проток $ с частичной склеротиза-
цпей. — На соболе ...................................М. paradoxus Seal.
11. Ceratophyllus Curt. Все виды, кроме одного, — паразиты птиц, иногда
ереходят на других животных, имеющих контакт с птичьими гнездами. Род
аспространен в Евразии и С Америке. — 11 видов (в СССР около 30).
(6). Задняя часть метанотума (cillare) и находящийся на ней зубчик заметно
редуцированы.
(3). Задний край тела половой клешни 3 образует резкий выступ, на котором
сидят ацетабулярные щетинки; край VII стернита $ без боковой вы-
резки. — В гнездах ласточки.......................С. maculatus Wagn.
(2). Половая клешня 3 и боковой выступ VII стернита $ иной формы.
(5). Дигитоид 3 резко суживается к вершине; апикальный дорсальный
отросток VIII стернита 3 узкий, без бахромчатой лопасти; край VII стер-
нита $ с боковой и вентральной вырезками и хорошо выраженными 2 бо-
11 Зак. № 990
161
новыми выступами, копулятивная сумка склеротизована. — На город-
ской ласточке......................................С. farreni Roth.
5 (4). Дигитоид с? к вершине не суживается; апикальный дорсальный отросток
VIII стернита 3 слабо развит и с небольшой бахромой по заднему краю;
вырезки заднего края VII стернита $ менее глубокие, хорошо выражен
только нижний боковой выступ, копулятивная сумка без склеротиза-
ции. — В гнездах городской ласточки .... С. olsufjevi Seal, et Viol.
6 (1). Задняя часть метанотума и находящийся на ней зубчик развиты нор-
мально.
Рис. 88. Детали строения блох.
1,2 — Amalaraeus dissimilis-. 1 — часть половой клешни, VIII и IX стерниты <?, 2 — крючок и склероти-
зация эдеагуса <?; 3, 4 — Monopsyllus anisus: 3 — гениталии и VII стернит $, 4 — половая клешня, VIII
и IX стерниты 3. (По: Иофф, Скалой, 1954).
7 (8). Стигмы брюшных тергитов большие; в гребне пронотума от 23 до 28 зуб-
цов; на наружной поверхности задней голени много щетинок; тело поло-
вой клешни 3 узкое; резервуар семяприемника $ удлиненный, но более
широкий, чем у птичьих блох. — На горностае и лесных полевках . . .
..............................................С. lunatus Jord, et Roth.
8 (7). Стигмы брюшных тергитов небольшие; признаки 3 и $ иные.
9 (10). Зубцы гребня короче, чем пронотум по дорсальной линии. — На мор-
ских птицах...................................С. hagoromo Jam. et Sak.
10 (9). Зубцы гребня не короче, чем пронотум по дорсальной линии.
11 (20). В гребне пронотума 22—27 зубцов.
12 (13). На конце VIII стернита 3 среди обычных щетин имеются утолщенные
и укороченные; мембранозная лопасть этого стернита большая, округлой
формы с характерным передним отростком; резервуар семяприемника $
суживается к концам (лимонообразный), склеротизация слепого протока
не развита. — В птичьих гнездах на поверхности земли и в кустарниках
........................................................С. garei Roth.
162
13 (12). На конце VIII стернита 3 сидят обычные щетинки, его лопасть иной
формы; резервуар семяприемника иной формы.
14 (19). На внутренней поверхности бедра3 ряд из 4—6 щетинок; дорсальный
отросток VIII стернита в узкий без бахромчатой лопасти; склеротизован-
ный участок слепого протока ? короче булавы усика.
15 (16). Горизонтальная ветвь IX стернита d к вершине суживается; дорсаль-
ный отросток VIII стернита с? узкий, расположен отступая от вершины;
стигмы на тергитах брюшка почкообразные. — На воронах..................
.............................................С. rossitensis Dampf.
16 (15). Горизонтальная ветвь IX стернита d с б. м. закругленной вершиной;
дорсальный отросток VIII стернита d расположен на его вершине; стигмы
на тергитах брюшка $ округлые.
17 (18). Дигитоид d выступает над вершиной неподвижного пальца примерно
наполовину, его задний край ниже середины слегка выгнут, а 2 (редко)—
3 щетинки на нем обычно толще, чем апикальная маленькая щетинка,
расположенная ниже апикальной, чаще находится приблизительно посре-
дине между нею и верхней заднекрайней; край VII стернита $ довольно
округлен. — В гнездах воробьев, скворцов и других птиц.................
.................................................С. gallinae Schr.
18 (17). Дигитоид d выступает над неподвижным пальцем менее чем наполо-
вину, его задний край без выгнутости ниже середины или слегка выпук-
лый; щетинки на его заднем крае обычно тоньше, чем апикальная малень-
кая щетинка, расположенная между апикальной и верхней из заднекрай-
них, сидит близко к апикальной; край VII стернита $ имеет слабые вы-
ступы и выемки........................................С. tribulis Jord.
19 (14). На внутренней поверхности бедра3 ряд состоит не менее чем из 7 щети-
нок; дорсальный отросток VIII стернита d с бахромчатой лопастью; скле-
ротизованный участок слепого протока ? длиннее булавы усика. —
Паразиты птиц, гнездящихся в скалах и норах ... С. vagabundus Boh.
20 (11). В гребне пронотума не мене§ 30 зубцов. — На береговой ласточке
...............................................С. riparius Jord, et Roth.
5. Сем. LEPTOPSYLLIDAE
Верхняя щетинка глазного ряда сидит у края усиковой ямки, выше уровня
глаза (а при отсутствии развитого глаза — выше обычного места его расположе-
ния). Впереди и выше глаза просвечивает развитый тяж тентория, иногда он
скрыт гребнем. Сенсиллий невыпуклый. Паразиты многих видов грызунов
и зайцеобразных. Наибольшее число видов в Палеарктике и Неарктике.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (4). Щечный гребень имеется.
2 (3). Щечный гребень состоит из 3—4 или многочисленных зубцов . . . .
..........................................1. Leptopsylla Jord, et Roth.
3 (2). Щечный гребень состоит из 2 зубцов .... 2. Peromiscopsylla I. Fox
4 (1). Щечный гребень отсутствует.
5 (6). Глаз уменьшенный или рудиментарный; 5-й членик лапок3 с 4 парами
боковых щетинок, между щетинками 1-й пары вдвинута подошвенная
пара..............................................3. Amphipsylla Wagn.
6 (5). Глаз нормально развит; 5-й членик лапок3 с 5 парами боковых щетинок,
причем 1-я пара может быть несколько сдвинута на подошвенную поверх-
ность, но не вдвинута между щетинками 1-й пары.
7 (8). Теменные ряды щетинок недоразвиты, в каждом от 0 до 2 щетинок . . .
...................................... 4. Paradoxopsyllus Miy. et Koid.
8 (7). Имеются 2 ряда развитых теменных щетинок.
9 (10). По заднему краю голени3 многочисленные пучки (не менее 10 групп)
очень длинных щетин; щетинки, сидящие вдоль лобного края, превра-
щены в шипы.........................................Ctenophyllus Wagn.1
1 Медико-ветеринарное значение этого рода не выяснено.
10 (9). По заднему краю голени3 обычные шпоры; вдоль лобного края нет шипов
• .................................. в . . . 5. Frontopsylla Wagn. et loff.
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
1. Leptopsylla Jord, et Roth. Паразиты мелких грызунов, широко распро-
страненные в Азии и С Америке. — 2 вида (в СССР 7).
1 (2). В щечном гребне 4 зубца; у переднего угла лба 2 толстых темных шипа. —
На домовых мышах.......................................L. segnis Schor.
2 (1). В щечном гребне 13—17 зубцов; у переднего угла лба нет толстых темных
шипов; на заднем крае дигитоида в нет шипика; на VIII стерпите в рас-
положены многочисленные щетинки; VII стернит $ без добавочных щети-
нок впереди от главного ряда или с единичными. На лесных мышах . . .
.................. L. pectiniceps Wagn. (=Pectonoctenus pectiniceps Wagn.)
2. Peromiscopsylla I. Fox. — 2 вида (в СССР 4).
1 (2). У переднего угла лба имеются 2 толстых шипа, а остальные щетинки
хотя и утолщены, но, как правило, все же отличаются от этих шипов;
у $ VII стернит с узким длинным боковым выступом и глубокой вырезкой
под ним. — На мелких лесных зверьках.................Р. ostsibirica Seal.
2 (1). Морфологически очень близок к Р. ostsibirica. Отличается деталями
строения эдеагуса; $ отличают по сопутствующим в. — На мелких лес-
ных грызунах............................Р. hamifer takahasii Ono et Has.
3. Amphipsylla Wagn. Паразиты мелких грызунов (полевок, хомячков, др.)
в Азии. Немногие виды проникли в Европу и С Америку. — 3 вида (в СССР 22).
1 (2). Неподвижный палец половой клешни d высокий, значительно подни-
мается над ее телом, задний край дигитоида закруглен, его шипы нахо-
дятся в верхней половине; край VII стернита $ с небольшой боковой
выемкой. — На хомячках и других мелких грызунах. Степной вид . . .
.....................................................A. vinogradovi loff
2 (1). Неподвижный палец половой клешни d широкий, мало поднимается
над ее телом, на заднем крае дигитоида ясно обозначены верхний и ниж-
ний углы, на которых расположены шипы; край VII стернита ? без боко-
вой выемки.
3 (4). Задневерхний угол дигитоида d на одном уровне с передним, оба угла
почти прямоугольные; VII стернит $ с небольшим боковым выступом. —
На мышевидных грызунах.........................A. marikovskii loff et Tif.
4 (3). Задневерхний угол дигитоида d выше переднего; край VII стернита $>
с большим боковым выступом и вентральной вырезкой под ним. —
На маньчжурском цокоре.....................*...........A. aspalacis Jord.
4. Paradoxopsyllus Miy. et Koid. Паразитируют на различных грызунах
и зайцеобразных в Азии. — 1 вид (в СССР 14).
1 (1). На VII тергите d группа многочисленных щетинок, подвижный и непо-
движный пальцы половой клешни d короткие и широкие; апикальная
доля горизонтальной ветви IX стернита d треугольной формы. — На кры-
сах .......................................Р. curvispinus Miy. et Koid.
5. Frontopsylla Wagn. et loff. Паразиты различных грызунов. Большинство
видов неразборчивы в выборе хозяина. Подрод Orfrontia паразитирует на
птицах. Распространены преимущественно в Азии, немногие — в Европе. —4
вида (в СССР 16).
1 (4). В гребне пронотума не более 24 зубцов; метанотум не укорочен, его зуб-
чики развиты не сильнее, чем зубчики тергитов брюшка (подрод Fronto-
psylla).
2 (3). Дигитоид d мало заходит за вершину неподвижного пальца; щетинка
на верхнем его крае сидит близко к шипу, ряд щетин под шипом располо-
жен поперечно; апикальная часть горизонтальной ветви IX стернита d
164
(«брюшко») отделена вырезкой от базальной (рис. 89, 4); край VII стер-
нита $ с боковой вырезкой; расположенный под ней боковой угол острый
и выступает несколько меньше, чем вентральный (рис. 89, 3). — На мно-
гих грызунах в степных и лесостепных местностях....................
.... *..............................F. (F.) luculenta Jord, et Roth.
3 (2). Дигитоид 3 значительно заходит за вершину неподвижного пальца;
щетинка на его верхнем крае заметно отодвинута от шипа кпереди, ряд
щетин под шипом идет косо вниз; апикальная часть горизонтальной
ветви IX стернита 3 («брюшко») не отделена вырезкой от базальной и
2 Ч
Рис. 89. Детали строения блох.
1,2 — Frontopsylla lapponica prior: 1 — часть половой клешни, IX и часть VIII стернитов <?, 2 — гени-
талии и VII стернит j; 3, 4 — F. luculenta: 3 — семяприемник, VII стернит $, 4 — часть половой клешни»
IX стернит <?. (По: Иофф, Скалой, 1954).
слегка «свисает»; боковая вырезка VII стернита § шире, чем у F. lucu-
lentar, расположенный над ней боковой угол более тупой, выступает
не меньше, чем вентральный.............F. (F.) elata botis Jord.
4 (1). В гребне пронотума более 24 зубцов (34—38); метанотум укорочен; па-
разиты ласточек (подрод Orfrontid).
5 (6). Апикальный шип дигитоида J небольшой (89, 7); задний край VII стер-
нита $ с боковой вырезкой (рис. 89, 2) ...............
....................................... F. (О.) lapponica prior Seal.
6 (5). Апикальный шип дигитоида J в виде длинной толстой щетинки; задний
край VII стернита $ без боковой вырезки. — На городской ласточке
.................................................F. setigera Smit.
ОСНОВНАЯ ЛИТЕРАТУРА
Дарская Н. Ф. Определитель птичьих блох рода Ceratophyllus. — В кн.: Эктопара-
зиты. М., 1950, вып. 2, с. 85—105.
Дудолкина Л. А. О паразите кур Ceratophyllus gallinae и близких видах блох. — В кн.:
Эктопаразиты. М., 1950, вып. 2, с. 106—111.
165
Иофф И. Г., Микулин М. А., Скалой О. И. Определитель блох Средней Азии и Казах-
стана. М., 1956. 370 с.
Иофф И. Г., Скалой О. И. Определитель блох Восточной Сибири, Дальнего Востока
и прилегающих районов. М., 1954. 275 с.
Скалой О. И. Блохи Сибири, Дальнего Востока и Монгольской Народной Республики.—
Доклад, представленный на соиск. уч. степени канд. биол. наук по совокупности опублик.
работ. Ставрополь-на-Кавказе, 1966. 58 с.
Скалой О. И. Отряд Siphonaptera (Aphaniptera, Suctoria) — блохи. — В кн.: Опреде-
литель насекомых европейской части СССР. Л., 1970, т. 5, ч. 2, с. 799—844.
Тифлов В. Е., Скалой О. И., Ростигаев Б. А. Определитель блох Кавказа. Ставрополь,
1977. 278 с.
ВШИ (Отряд ANOPLURA)
Мелкие бескрылые насекомые со сжатым сверху вниз телом, 3—5-членико-
выми нитевидными усиками, колюще-сосущим ротовым аппаратом, слитной,
ясно не расчлененной на сегменты грудью и хватательными ногами; облигатные
кровососущие паразиты млекопитающих с неполным превращением. На Даль-
нем Востоке — 4 семейства, 10 родов, 21 вид.
ВНЕШНЕЕ СТРОЕНИЕ
Тело (рис. 90, 7, 2) с малоподвижными основными отделами (головой,
грудью, брюшком), дорсовентрально уплощенное, узкопродолговатое или
широкояйцевидное, с рядом переходов. Покровы часто со склеротизированными
участками в виде пластинок, с волосками и разнообразными по форме и вели-
чине щетинками и шипами. Длина тела от 0.5 до 6 мм.
Голова (рис. 90, 4) небольших размеров, варьирует по форме от узкооваль-
ной до шестиугольной. Ротовой аппарат колюще-сосущего типа. На голове
имеются 3—4- или 5-члениковые усики. У самцов 1-й членик усиков заметно
крупнее, чем у самок; 3-й с выступом, несущим шип или щетинку. Глаза реду-
цированы или отсутствуют, слабо пигментированы, находятся позади усиков.
Грудь (рис. 90, 7, 2) состоит из трех слитых сегментов, несущих три пары
ног. На груди имеются среднегрудные дыхальца, срединная спинная ямка,
стернальная пластинка и стернальные апофизальные ямки.
Ноги (рис. 90, 3) хватательного типа, такого же строения, как у других
насекомых. Лапка одно- или двучлениковая с коготком. Передние ноги меньше
средних и задних, с тонким коготком. Голень и лапка бывают слитыми или
неясно разделенными. Голени дистально расширены, с шипами или щетин-
ками, внутренним пальцевидным выступом, иногда с подушкообразным претар-
зальным придатком.
Брюшко (рис. 90, 7, 2) узкопродолговатое с рядом переходов или широко-
яйцевидное, состоит из 9 более или менее явственных сегментов: I сегмент
сильно редуцирован, IX у самок несколько раздвоенный, у самцов — округлен-
ный и конусообразно суженный. На брюшке имеются тергальные, стернальные,
среди них генитальные, относящиеся к половой области, и плейральные пла-
стинки. Число дыхалец, открывающихся латерально на III—VIII сегментах,
колеблется от одной до шести пар. У самок по бокам половой створки, на
VIII стерпите, часто имеются половые придатки — гоноподы, или гонапофизы.
Придатки кожи (хетотаксия) в виде волосков, разнообразных по форме и вели-
чине щетинок и шипов, расположенных на теле разбросанно, пучками или
рядами, нередко многочисленны.
Личинки внешне похожи на взрослых насекомых, отличаются меньшими
размерами, типом хетотаксии, рисунком брюшных пластинок, отсутствием
полового отверстия, гонапофиз или функционально заменяющих их щетинок
(у самок) и копулятивного аппарата (у самцов). У личинок I стадии размеры
тела наименьшие, голова относительно крупная, брюшко короткое, покровы
мягкие и светлые, с неполной хетотаксией. Во II и III стадиях размеры личинок
увеличиваются, отделы тела становятся более соразмерными, склеротизация и
пигментация его усиливаются, число щетинок возрастает.
166
БИОЛОГИЯ
Вши являются постоянными эктопаразитами, связанными со своими хозяе-
вами в течение всей жизни. Они живут в волосяном покрове или на одежде
хозяев близ кожи, а при массовом размножении — по всему покрову.
Рис. 90. Детали строения вшей.
1 — тело вши (схема), дорсально; 2 — то же вентрально: бр — брюшко, брд — брюшные дыхальца, гл —
голова, гон — гонапофизы, гр — грудь, грд — грудное дыхальце, пп — плейральные пластинки, стр —
стернальная пластинка груди, стбр — стернальные пластинки брюшка, тпбр — тергальные пластинки-
брюшка, у — усик; з — нога: б — бедро, в — вертлуг, гол — голень, л — лапка, пв — пальцевидный
выступ, т — тазик; 4 — Linognathus stenopsis, голова; 5 — род Haematopinus, голова с постантеннальными
углами (пауУ, 6 — Phthirus pubis, кокса ноги; 7 — Hoplopleura acanthopus, плейральные пластинки; 8 —
род Ender leinellus, парные пластинки II сегмента брюшка; 9 — род Hoplopleura, стернальная пластинка
II сегмента брюшка. (По: Благовещенский, 1960; Сергиенко, 1974).
Локализация вшей на хозяине может изменяться в зависимости от физиче-
ских факторов среды, физиологического состояния хозяина и паразита. Вошь
Haematopinus eurysternus встречается у коров на многих частях тела, тогда
как летом она попадается преимущественно на голове и хвосте. Платяная вошь
переходит для питания с одежды на кожу человека [Благовещенский, 1960].
Вши — это насекомые с неполным превращением. Они размножаются бес-
прерывно. Самки откладывают за сутки от 2 до 14 яиц, за всю жизнь — до 330.
16Т
Яйца прикрепляются задним концом к волосу или одежде секретом клеевых
желез. Личинки свиной вши отрождаются из яиц на хозяине через 12—20 дней.
Постэмбриональное развитие их, сопровождающееся трехкратной линькой,
занимает 7—14 дней. Яйцо платяной вши на человеке созревает за 7—10 дней.
Личиночная фаза длится 2—3 дня, а полный жизненный цикл от яйца до имаго
проходит за 15—45 дней. При относительно коротком цикле развития и беспре-
рывности размножения вши дают в год несколько поколений. Вылупившиеся
из яиц и слинявшие личинки, как и только что появившиеся взрослые насеко-
мые, способны к передвижению и сосанию крови через 20—45 мин. Через
1—7 дней по достижении половой зрелости самка приступает к откладке яиц
независимо от спаривания, неоплодотворенные яйца не развиваются. Макси-
мальный срок жизни самок вшей составляет 46 дней, самцов — 32 дня [Пав-
ловский, 1948; Благовещенский, 1960].
Характер завшивленности подвержен сезонным колебаниям, зависящим
от изменений физиологического состояния хозяев и паразитов, а также от усло-
вий среды обитания хозяина и паразита. Заражение крупного рогатого скота
вшами начинает увеличиваться осенью, достигая максимума зимой и ранней
весной, и резко уменьшается к концу весны и летом, по мере выпаса и нагула.
На погибших хозяевах вши поднимаются на поверхность волосяного по-
крова или одежды и частично отпадают и расползаются. Вне хозяина они
погибают в течение одного-нескольких дней. Вши расселяются путем переполза-
ния при контакте хозяев.
Вши характеризуются по отношению к своим хозяевам строгой или относи-
тельной специфичностью. На млекопитающих паразитируют по 1—2 и более
видов вшей. Несмотря на приуроченность этих паразитов к одному виду или
к группе видов хозяев, имеются случаи находок их на случайном хозяине.
Более того, у некоторых видов прослежен полный жизненный цикл при пита-
нии на неспецифическом хозяине.
ВРЕД
Как постоянные кровососущие эктопаразиты вши вызывают у хозяина
механическое раздражение и зуд кожи* во время передвижения и уколов при
питании, которое сопровождается введением слюны, обладающей раздражаю-
щими свойствами.
Сильная завшивленность вызывает беспокойное состояние у животных,
нарушение правильного режима их питания. Животные становятся вялыми,
худеют и могут доходить до полного истощения, ведущего к гибели. Работо-
способность рабочих животных понижается, у молочного скота уменьшается
продукция молока. На пораженных вшами местах кожи возникают разной
формы дерматиты, происходит выпадение или сваливание шерсти, что у коз
и овец ухудшает ее товарный выход. При работе с завшивленными животными,
свежими их трупами или шкурами возможно случайное попадание вшей на чело-
века, поскольку для некоторых видов установлена способность сосания крови
человека.
Эпидемиологическое и эпизоотологическое значение вшей обусловливается
их биологическими особенностями: быстрым размножением, многократным
питанием кровью в течение суток и частой дефекацией.
Вши являются переносчиками возбудителей некоторых болезней человека
и животных: сыпного и возвратного тифа, чумы свиней, свиной лихорадки,
инфекционной анемии лошадей, туляремии, бруцеллеза, салмонеллеза, эризи-
пелоида и др.
К настоящему времени накоплены обширные сведения о естественной зара-
женности вшей диких млекопитающих возбудителями ряда инфекционных
заболеваний.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ
1 (6). Брюшко с б. м. развитыми плейральными пластинками.
2 (5). Плейральные пластинки брюшка колпачкообразно (рис. 91, 2) покры-
вают боковые сегментальные выступы, не свободные или частью очень
слабо апикально свободные.
168
3 (4). Глаза не пигментированы. Грудь с хорошо ограниченной спинной ямкой.
Ноги почти одинаковой величины. На парно- и непарнокопытных . . .
....................................................... 1. На emat opin idae
Рис. 91. Детали строения вшей.
1,2 — плейральные пластинки брюшка: 1 — род Polyplax, 2 — Haematopinus asini\ 3 — Solenopotes capil-
latus, брюшные дыхальца; 4 — род Phthirus, бугорковидные выступы брюшка; 5—13 — стернальные груд-
ные пластинки: 5 — Haematopinus suis, 6 — Н. asini, 1 — Н. eurysternus, 8 — Enderleinellus dolichocephalus*
9 — E. disparilis, 10 — Polyplax serrata, 11 — P. spinulosa, 12 — Haemodipsus ventricosus, 13 — Neohaema-
iopinus laeviusculus’, 14, 15 — 6—7-я пары плейральных пластинок: 14 — Hoplopleura captiosa, 15 — H. af-
finis', 16—20 —терминальная часть брюшка самки снизу: 16 — Solenopotes capillatus, 17 — Linognathus
vituli, 18 — L. stenopsis, 19 — L. setosus, 20 — L. pedalis', 21—22 — голова: 21 — L. ovillus, 22 — L. seto-
sus. (По: Благовещенский, 1960; Сергиенко, 1974).
4 (3). Глаза с пигментным пятном. Грудь с неявственной спинной ямкой.
Ноги почти одинаковой величины или передние — заметно меньше дру-
гих. На приматах (обезьянах, людях).......................2. Pediculidae
5 (2). Плейральные пластинки брюшка не колпачкообразно (рис. 90, 7) по-
крывают боковые сегментальные выступы, частично апикально свобод-
ные. На грызунах и насекомоядных......................3. Hoplopleuridae
6 (1). Брюшко без плейральных пластинок. На парнокопытных...............
...................................................4. Linognathidae
169
1. Сем. HAEMATOPINIDAE
Усики 5-члениковые. Глаза обычно без пигмента. Грудь с явственной средин-
ной спинной ямкой. Передние, средние и задние ноги почти одинаковой вели-
чины, со сходными по размерам и форме коготками. Плейральные пластинки
брюшка колпачкообразно покрывают боковые сегментальные выступы, без
свободных полей. На парно- и непарнокопытных. — 1 род.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1. Haematopinus Leach. Тело довольно крупное. Голова с постантенналь-
ными углами (рис. 90, 5). Грудь трапециевидная или прямоугольная, обычно
имеет стернальную пластинку. Плейральные пластинки на III—VIII сег-
ментах. — 3 вида (в СССР 5).
1 (4). Голова длинная и узкая.
2 (3). Стернальная пластинка груди неправильно четырехугольная, поперечно
вытянута (рис. 91, 5). Брюшко без конусообразных боковых выступов.
5 — 4.2—5.0, 3 — 3.6—4.2. — Населенные пункты. На домашней свинье
....................................................Н. suis L.
3 (2). Стернальная пластинка груди неправильно четырехугольная, продольно
удлинена, кувшинообразная (рис. 91, 6). Брюшко с конусообразными
боковыми выступами (рис. 91, 2), $ — 2.7—3.5, 3 — 2.6—3.2. — Насе-
ленные пункты. На осле, лошади...........................Н. asini L.
4 (1). Голова короткая и широкая. Стернальная пластинка груди неправильно
четырехугольная, слегка стиснута с боков, со срединным переднекрайним
выступом (рис. 91, 7). ? — 2.6—3.4, 3 — 2.1—2.5. — Населенные пункты.
На домашнем крупном рогатом скоте.................Н. eurysternus N.
2. Сем. PEDICULIDAE
Усики 5-члениковые. Глаза с пигментным пятном. Ноги приблизительно
одинаковой величины и формы или передние маленькие и тонкие, а средние
и задние — большие и массивные. Брюшко чаще мембранозное, у ? обычно без
тергал ьных и стернальных пластинок (за исключением генитальных), ус? —
с узкими тергальными пластинками. Плейральные пластинки некоторых сег-
ментов брюшка колпачкообразно покрывают боковые лопасти, несвободные
или частично слабо апикально свободные. — 2 рода.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). Тело удлиненное, ноги почти одинаковой величины и формы. Брюшко
без боковых бугорковидных выступов..................1. Pediculus L.
2 (1). Тело короткое и широкое. Передние ноги заметно меньше других.
Брюшко с сегментальными боковыми бугорковидными выступами (рис. 91,
4)................................................2. Phthirus Leach.
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
1. Pediculus L. Грудь со спинной ямкой и стернальной пластинкой. Ноги
примерно одинаковой величины и формы, с тонким коготком. Сегменты брюшка
не имеют боковых выступов — лопастей, межсегментальные вырезки более или
менее развиты. Плейральные пластинки темнопигментированные, развиты
на большинстве сегментов. Дыхальца в количестве 6 пар, открываются на плей-
ральных пластинках. Брюшко у $ дорсально мембранозное или со слабо раз-
витыми пластинками, ус? — обычно с небольшими тергальными пластинками. —
1 вид.
1 (1). Плейральные пластинки брюшка без боковых лопастей, хорошо развиты
на III—VIII сегментах, у — 2.2—4.7, с? —2—3.7. — Космополит. На че-
ловеке ..............................................Р. humanus L.
2. Phthirus Leach. Грудь очень широкая, без спинной ямки и стернальной
пластинки. Передние ноги значительно меньше других, тонкие, с тонким ко-
готком; средние и задние — большие, массивные, с массивным коготком.
V—VIII сегменты брюшка с боковыми бугорковидными выступами. Дыхальца
III—V сегментов тесно сближены. — 1 вид.
1 (1). Голова вентрально, с каждой стороны, близ переднего края со склеро-
тизованным бугорком и лопастевидным выступом. Усики относительно
большие и тонкие. Тазики всех ног с лопастевидным выступом (рис. 90,
6). Тело короткое и широкое. ? — 1.5—2, 8 — 1—1.3. — Космополит.
На человеке..............................................Р. pubis L.
3. Сем. HOPLOPLEURIDAE
Усики 5-члениковые, иногда с более или менее слитными последними двумя
или тремя члениками, нередко с половым диморфизмом. Глаза отсутствуют.
Стернальная пластинка груди хорошо развита, со свободным задним концом.
Брюшко обычно с тергальными и стернальными пластинками и поперечными
рядами щетинок. Плейральные пластинки хорошо развиты и не покрывают
колпачкообразно боковые лопасти сегментов. Паразиты грызунов, зайце-
образных, насекомоядных. — 5 родов (в СССР — 8).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). Передние и средние ноги маленькие и тонкие, с тонким коготком. II стер-
нит брюшка обычно с парой небольших склеротизованных пластинок,
из которых каждая с отростком (рис. 90, 8) . . .1. Enderleinellus Fahr.
2 (1). Передние ноги меньше, чем средние и задние. II стернит брюшка без пары
склеротизованных пластинок.
3 (8). Стернальная пластинка II сегмента брюшка не удлиненная, не сочленя-
ется с соответствующими плейральными пластинками.
4 (7). Плейральные пластинки в количестве не менее 7 пар развиты на II —
VIII сегментах брюшка.
5 (6). Плейральные пластинки II сегмента брюшка без продольного разделения
............................................2. Neohaematopinus Mjob.
6 (5). Плейральные пластинки II сегмента брюшка с признаком продольного
разделения (рис. 91, 1).............................3. Polyplax End.
7 (4). Плейральные пластинки в количестве не менее 4 пар имеются на III —
VI сегментах брюшка (очень маленькие, с выступающим зубчиком) или
отсутствуют....................................4. Haemodipsus End.
8 (3). Стернальная пластинка II сегмента брюшка удлиненная, сочленяется
с соответствующими плейральными пластинками (рис. 90, 9)............
........................................... 5. Hoplopleura End.
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
1. Enderleinellus Fahr. Усики без зубцевидных придатков. Передняя и сред-
няя пары ног одинаковой величины — маленькие, со слабым коготком.
На II сегменте брюшка обычна пара маленьких склеротизованных вентральных
пластинок, из которых каждая с направленным назад отростком. Каждый
сегмент брюшка дорсально и вентрально с одним поперечным рядом шипов
и щетинок. Тергиты и стерниты брюшка чаще без хитинизированных пласти-
нок. — 2 вида (в СССР 6).
1 (2). Стернальная пластинка груди (рис. 91, 8) четырехсторонняя, несколько
удлиненная, с округленными углами и х-образным пигментным рисун-
ком. $ — 0.8—1.1, 8 — 0.8—0.9. — Аз. ч. СССР. На камчатском сурке
............................................ Е. dolichocephalus Blag.
2 (1). Стернальная пластинка груди (рис. 91, 9) почти квадратная, несколько
суженая впереди, с округленными углами и «двугнездным» пигментным ри-
сунком. —0.7—0.9. —Аз. ч. СССР. На длиннохвостом суслике . . .
............................................Е. disparilis Blag.
171
2. Neohaematopinus Mjob. Голова с явственными среднебоковыми углами.
Усики 5-члениковые, часто с половым диморфизмом. Передние ноги маленькие,
с тонким коготком, средние и задние — более крупные, с массивным коготком,
нередко почти одинаковой величины. Плейральные пластинки развиты на II —
VIII сегментах брюшка, пластинки II сегмента продольно неразделенные.
Брюшко $ с двумя или тремя рядами щетинок на тергитах и стернитах. Тер-
гальные и стернальные пластинки совсем или частично отсутствуют, d —
с двумя рядами щетинок на II тергите и стерпите брюшка. — 1 вид (в СССР 6).
1 (1). Первый членик усика без заднекрайнего шипа. Стернальная пластинка
груди грубоовальная или многоугольная с закругленными углами
(рис. 90, 2). Сегменты брюшка ? в большинстве без тергальных и стерналь-
ных пластинок, со сплошными рядами (до 30) щетинок. Брюшко d с груп-
пами срединных и боковых щетинок, с тергальными и стернальными
пластинками. $ — 1.6—1.7, d — 1.1—1.3. — Евр. и аз. ч. СССР. На длин-
нохвостом суслике..................................N. laeviusculus Gr.
3. Polyplax End. Голова с явственными среднебоковыми углами. Усики
5-члениковые, часто с половым диморфизмом. Передние ноги маленькие, с тон-
ким коготком, задние — массивные, с массивным коготком. Брюшко узкое,
вытянутое. Плейральные пластинки всегда развиты на II—VIII сегментах, пла-
стинки II сегмента брюшка продольно разделенные. Задний край плейритов
с двумя короткими или длинными щетинками. Тергиты и стерниты, за некото-
рым исключением, у ? обычно с двумя пластинками, yd — с одной. По заднему
краю тергальных и стернальных пластинок расположены ряды щетинок. —
2 вида (в СССР 11).
1 (2). Стернальная пластинка груди б. м. сердцевидная (рис. 91, 10). Плейраль-
ные пластинки 2-й, 4-й и 5-й пар по заднему краю с двумя короткими ще-
тинками, пластинки 3-й пары с короткой и длинной щетинками. $ —
1—1.2, d — 0.6—0.8. — Евр. и аз. ч. СССР. На домовой, лесной, полевой
мышах и полевках...................*..................Р. serrata Burm.
2 (1). Стернальная пластинка груди неправильно шестиугольная, с почти па-
раллельными боковыми краями (рис. 91, 11). Плейральные пластинки
2—5-й пар по заднему краю, с дорсальным зубцом и 2 короткими щетин-
ками. $ —0.9—1.7, d — 0.8—0.9. — Евр. и аз. ч. СССР. На крысах
...................................................Р. spinulosa Burm.
4. Haemodipsus End. Усики 5-члениковые, без полового диморфизма. Перед-
ние ноги наименьшие, средние и задние — более крупные, почти одинаковой
величины. Брюшко большое и широкое, большая часть сегментов без тергаль-
ных и стернальных пластинок. Тергиты и стерниты с одним рядом щетинок.
Плейральных пластинок нет или они маленькие, слабохитинизированные, зуб-
цевидные на II—VI сегментах. Брюшные дыхальца маленькие, расположены
на III—VIII сегментах. — 1 вид (в СССР 3).
1 (1). Стернальная пластинка груди поперечно расширена, почти прямоуголь-
ная (рис. 91, 12). Плейральные пластинки в виде светлых зубцевидных
пластинок на III—VI сегментах. § — 1.4—1.5, d — 1—1.2. — Евр.
и аз. ч. СССР. На кроликах.........................Н. ventricosus Denny
5. Hoplopleura End. Голова co слабо выраженными постантеннальными
углами. Усики 5-члениковые. Грудь немного шире головы, с хорошо развитой
стернальной пластинкой. Передние ноги маленькие, средние и задние — боль-
шие, с более мощным коготком. Брюшко яйцевидное или узкоовальное. Плей-
ральные пластинки развиты на I—VIII сегментах: 1-я пара маленькая, распо-
ложена под дорсумом. Брюшко с тергальными и стернальными пластинками
и рядами щетинок. Стернальная пластинка II сегмента и обычно первая стер-
нальная пластинка III сегмента удлиненные, сочленяются с соответствующими
плейральными пластинками. Плейральные пластинки без продольного разделе-
ния. — 3 вида (в СССР 8).
1 (4). Плейральные пластинки 6-й пары с двумя одинаковыми лопастями: кли-
новидными или прямоугольными.
2 (3). Плейральные пластинки 6-й пары с клиновидными лопастями, пластинки
7-й пары без лопастей. Плейральные пластинки 3-й пары с узкой, зао-
стренной дорсальной и прямоугольной вентральной лопастями (рис. 90,
7). У $ IV—VII сегменты брюшка с 3—4 боковыми тергальными и 2—
3 стернальными щетинками; у с? IV—VII сегменты брюшка с 1—3 боко-
выми тергальными и 1—2 боковыми стернальными щетинками. $ — 0.9—
1.2, б — 0.7—0.9. — Евр. и аз. ч. СССР. На полевках и других мелких
грызунах, на зайцеобразных, насекомоядных . . . Н. acanthopus Burm.
3 (2). Плейральные пластинки 6-й пары с прямоугольными лопастями (рис. 91,
14), пластинки 7-й пары у $ с клиновидными лопастями, у d с заостренной
дорсальной частью и небольшим вентральным зубцом; пластинки 8-й пары
у $ с дорсальной лопастью, у с? без лопастей. На IV—VIII сегментах
брюшка у $ 1—2 боковые стернальные щетинки, у с? 1 боковая стерналь-
ная щетинка. $ — 1—1.3, d — 0.7—0.9. — Населенные пункты. На до-
мовых мышах........................................Н. captiosa John.
4 (1). Плейральные пластинки 6-й пары с двумя неодинаковыми лопастями:
прямоугольной дорсальной и клиновидной вентральной (рис. 91, 15).
Плейральные пластинки 3-й пары с узкими лопастями, разделенными ши-
рокой выемкой; пластинки 7-й пары с клиновидной дорсальной лопастью.
Брюшко $ с одной боковой щетинкой между 1-й тергальной пластинкой
и плейральными пластинками V—VII сегмента и несколькими боковыми
стернальными щетинками. Брюшко с? с 4 боковыми тергальными щетин-
ками на IV—VII сегментах и несколькими боковыми стернальными ще-
тинками. $ — 0.9—1.4, с? — 0.8—1. — Евр. и аз. ч. СССР. На лесных
и полевых мышах, хомякообразных, насекомоядных . . Н. affinis Burm.
4. Сем. LINOGNATHIDAE
Усики 5-члениковые. Глаза отсутствуют. Грудь чаще без стернальной пла-
стинки. Передняя пара ног наименьшая, средняя и задняя — примерно одина-
ковой величины. Плейральные пластинки не развиты. Тергальные и стерналь-
ные пластинки отсутствуют, за исключением IX сегмента. На парнокопытных
и хищных. — 2 рода.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). Сегменты брюшка с одним рядом щетинок. Брюшные дыхальца откры-
ваются на б. м. выдающихся бугорках ..............1. Solenopotes End.
2 (1). Сегменты брюшка более чем с одним рядом щетинок. Брюшные дыхальца
не расположены на бугорках .......................2. Linognathus End.
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
1. Solenopotes End. Грудь с несвободной стернальной пластинкой. Перед-
ние ноги значительно меньше, чем средние и задние. Брюшко широкое и округ-
лое. Брюшные дыхальца открываются на боковых выступах-бугорках. Сег-
менты брюшка дорсально и вентрально имеют по 1 ряду щетинок. — 2 вида
(в СССР 4).
1 (2). Брюшко дорсально без боковых пятен. Брюшные дыхальца открываются
на заметно выдающихся бугорках. Гонапофизы небольшие, апикально
округленные, окаймленные тонкими щетинками (рис. 91, 16). $ — 14—
1.6, d — 1.2—1.4. — Населенные пункты. На домашнем крупном рогатом
скоте.............................................S. capillatus End.
2 (1). Брюшко дорсально с боковыми рядами неправильных пятен. $ — 2.5,
d — 2. — Аз. ч. СССР. На северном олене ...... S. tarandi Mjob.
2. Linognathus End. Стернальная пластинка груди отсутствует или развита
слабо. Брюшко неясно сегментированное. Тергиты и стерниты брюшка более
чем с одним поперечным рядом щетинок. Дыхальца не расположены на бугор-
ках. — 5 видов (в СССР 8).
173
1 (6). Голова удлиненная.
2 (5). Брюшные дыхальца б. м. крупные.
3 (4). Передняя часть головы коротко- и ширококонусообразная (рис. 91, 21).
Гонапофизы широкоокругленные. $ — 2.8, в — 2.1. — Населенные
пункты. На домашней овце.........................L. ovillus Neum.
4 (3). Передняя часть головы удлиненно- и узкоконусообразная. Гонапофизы
с вогнутым задним краем (рис. 91, 17). ? — 1.8—2.6, d — 1.5—1.9. —
Населенные пункты. На домашнем крупном рогатом скоте.................
................................................... L. vituli L.
5 (2). Брюшные дыхальца мелкие. Задние боковые края головы ровно выгну-
тые. Гонапофизы усеченные, с заднекрайним внутренним зубцевидным
выступом (рис. 91, 18). $ — 2.2—3, d — 1.7—2.1. — Населенные пункты.
На домашней козе...................................L. stenopsis Burm.
6 (1). Голова короткая, ее длина немного превышает ширину.
7 (8). Передняя часть головы почти трапециевидная (рис. 91, 22). Усики при-
членены к средней части головы. Брюшные дыхальца крупные. Гонапо-
физы широкоокругленные (рис. 91, 19). $ — 1.8—2.4, d — 1.6—1.7. —
Населенные пункты. На домашней собаке...................L. setosus Olf.
8 (7). Передняя часть головы ширококонусообразная. Усики причленены
ближе кпереди. Брюшные дыхальца умеренной величины. Гонапофизы
с угловато изогнутым внутренним краем (рис. 91, 20). $ — 1.8—2.4,
d — 1.7. — Населенные пункты. На домашней овце . . . L. pedalis Osb.
ОСНОВНАЯ ЛИТЕРАТУРА
Благовещенский Д. И. Mallophaga и Anoplura с животных о. Врангеля. — Энтомол.
обозрение, 1958, т. 37, вып. 2, с. 374—379.
Благовещенский Д. И. Вши (Siphunculata) домашних млекопитающих. М.; Л., 1960. 86 с.
Благовещенский Д. И. Вши (Anaplura). — В кн.: Определитель насекомых европей-
ской части СССР. М.; Л., 1964, т. 1, с. 324—334.
Благовещенский Д. И. Новые виды вшей (Siphunculata), паразитирующих на грызунах.—
Энтомол. обозрение, 1965, т. 44, вып. 1, с. 151—165.
Павловский Е. Н. Руководство по паразитологии человека. М.; Л., 1948, т. 2. 1016 с.
Серпенко Г. Д. Фауна Украшп (Воши). Кшв, 1974, т. 22, вып. 3, 108 с.
*
клопы, ИЛИ ПОЛУЖЕСТКОКРЫЛЫЕ
(Отряд HETEROPTERA, или HEMIPTERA,
Сем. CIMICIDAE)
Наземные или водные насекомые. Тело чаще овальное и умеренно уплощен-
ное, ротовые органы колюще-сосущие, в виде прикрепленного к передней
части головы хоботка, состоящего из 3 или 4 члеников. Усики обычно 4-члени-
ковые, тонкие. Передние крылья (надкрылья) сложены на спинной стороне,
обычно состоят из кожистой основной и перепончатой вершинной частей,
иногда укорочены. Ноги чаще ходильные или бегательные. Щиток (открытая
сверху часть среднегруди) обычно небольшой, треугольный. У большинства
видов на заднегруди есть пахучие железы.
Отряд включает около 40 тыс. видов, распространен всесветно.
Большинство представителей — растительноядные или хищники. Кровью
теплокровных животных и людей питаются виды семейств Cimicidae (см. ниже),
Polyctenidae (паразиты летучих мышей в тропиках) и подсем. Triatominae
из сем. Reduviidae (обитают в тропических районах Центральной и Южной
Америки, переносят болезнь Шагаса, напоминающую сонную болезнь). При
случайных обстоятельствах хищные клопы могут прокалывать кожу человека.
Вводимая при этом слюна вызывает неприятные ощущения и воспаление на-
подобие ужаления пчелы. Представители сем. Cimicidae по облику более или
менее похожи на постельного клопа. Медико-ветеринарное значение в СССР
имеет 1 вид, постельный клоп, который встречается практически на всей тер-
ритории СССР, в том числе и на Дальнем Востоке.
174
ВНЕШНЕЕ СТРОЕНИЕ ПОСТЕЛЬНОГО КЛОПА
Тело овальное или округлое, сильно уплощенное, длиной 4.2—6.5 мм
(самцы в среднем меньше самок). Окраска варьирует от желтоватой до буро-
вато-красной. Все тело покрыто прилегающими щетинковидными волосками,
которые невооруженным глазом почти не различимы. Голова слегка вытянута
вперед, на вершине обрублена, простых глазков нет. Хоботок 3-члениковый.
Усики 4-члениковые; 1-й членик короткий, толстый, 3-й и 4-й членики тоньше
2-го (рис. 92). Переднеспинка с сильно выемчатым передним краем, сужается
к основанию; ее уплощенные бока в самом широком месте почти вдвое шире
глаза. Надкрылья сильно укорочены, кожистые, не прикрывают брюшко.
Нижних крыльев нет. Ноги ходильные, лапки 3-члениковые.
БИОЛОГИЯ ПОСТЕЛЬНОГО КЛОПА
Постельные клопы держатся обычно в жилище человека и питаются его
кровью, но могут также паразитировать на курах, домашних животных (кошках
и собаках), голубях, летучих мышах и мелких грызунах (в заброшенных по-
стройках и в дуплах деревьев).
ь Взрослые и личинки днем прячутся в щелях и других укрытиях, предпочи-
тая сухие шероховатые поверхности, особенно деревянные и бумажные. Они
образуют постоянные скопления
в кроватях, под обоями, плинту-
сами, выключателями, картинами
и коврами, иногда — в трещинах
стен и потолка, в чемоданах и
книгах. В этих местах обычно
много яиц, линочных шкурок и
экскрементов; клопы, как пра-
вило, возвращаются сюда после
каждого акта кровососания.
Наибольшая активность напа-
дения клопов отмечается около*
3 ч ночи, но голодные особи мо-
гут нападать даже при свете. Рас-
стояние от мест скопления до объ-
екта нападения может достигать
20 м. При прокалывании кожи
клопы вводят в ранку слюну, пре-
пятствующую свертыванию крови. рпс 92. Общий вид постельного клопа Cimex
Среднее количество высасываемой lecturius.
крови варьирует от 0.34 мг у ли-
чинок 1-го возраста до 7.81 мг у взрослых самок (максимум — 13.9 мг). Без
пищи при комнатной температуре личинки младших возрастов могут жить
в среднем около 80, а взрослые — около 130—140 дней; при низкой темпе-
ратуре отдельные особи могут голодать до года и более.
Клопы развиваются с неполным превращением. Копуляция (спаривание) —
травматическая: самец прокалывает парамером покровы самки в области вторич-
ных генитальных структур на брюшке. Сперматозоиды при этом поступают
в особый внутренний орган и из него сложным путем мигрируют в основание
яичников.
Как правило, самка откладывает яйца после очередного приема крови.
В течение жизни самка пьет кровь несколько раз, обычно с недельными пере-
рывами. Общее число отложенных одной самкой яиц может превышать 500 штук.
Из яиц через 4—20 дней (в зависимости от температуры) вылупляются личинки,
в целом напоминающие взрослых клопов, но с 2-члениковыми лапками, без
зачатков крыльев и с некоторыми другими отличиями. Личинки тоже питаются
кровью, линяют 4 раза и имеют 5 возрастов. Продолжительность развития
от яйца до взрослого насекомого составляет около 4 мес. при 18 °C, около
2 мес. — при 22 °C и около месяца — при 27—33 °C.
175
ВРЕД
Клопы сильно беспокоят людей и животных своими болезненными укусами
и ползанием. В местах укусов появляется припухлость, покраснение и сильный
зуд. В расчесанные ранки может быть внесена вторичная инфекция. Но около
20 % людей мало чувствительны к их укусам. Хотя потери крови от паразити-
рования клопов невелики, при длительном и частом массовом их нападении
может снижаться процент гемоглобина в крови (развивается малокровие).
Иногда клопы в массе размножаются в курятниках, нанося серьезный ущерб
птицеводству.
В постельных клопах могут длительное время сохраняться возбудители
некоторых болезней, но переносчиками их клопы не являются.
Диагноз сем. Cimicidae
Мелкие насекомые с округлым плоским телом, окраска желтая или буро-
вато-красная, голова на вершине обрублена, хоботок 3—4-члениковый, усики
4-члениковые, переднеспинка с выемчатым передним краем, надкрылья укоро-
чены.
В семействе 22 рода и более 70 видов. В СССР — 2 рода и 5—7 видов, на
Дальнем Востоке — 1 род и 2 вида, но медико-ветеринарное значение имеет
1 вид. Предполагается, что он перешел на человека с летучих мышей в период
обитания древних людей в пещерах.
1. Cimex L. Переднеспинка не менее чем в 1.5 раза шире головы. Длина
2-го членика усиков почти равна расстоянию между глазами.
1 (1). У ? задний край 5-го (4-го видимого) сегмента брюшка снизу справа с уз-
кой выемкой, вокруг которой такие же щетинки, как на других участках
брюшка. 4.2—6.5. — Распространен всесветно .... С. lectularius L.
ОСНОВНАЯ ЛИТЕРАТУРА
Usinger R. L. Monograph of Cimicida’e (Hemiptera — Heteroptera). Baltimore, 1966.
585 p.
ТАРАКАНЫ (Отряд BLATTOPTERA)
ВНЕШНЕЕ СТРОЕНИЕ
Тело (рис. 93, 7) более или менее овальное, плотное. Размеры от крупных
до мелких. Голова треугольная или сердцевидная, плоская, почти целиком
прикрыта сверху переднеспинкой. Наличник или слабообособленный простой,
либо (сем. Polyphagidae) резко отделен ото лба сильной бороздкой и подраз-
делен на более светлую переднюю и более темную заднюю части (рис. 93, 2, 6).
Глаза большие, обычно почковидные; глазков 2, расположены они у внутрен-
него края глаз над основанием усиков; у бескрылых и короткокрылых видов
глазков нет. Ротовые органы грызущего типа, направлены вниз и назад; челюст-
ные щупики 5-члениковые, губные щупики 3-члениковые. Усики длинные,
щетинковидные. Переднеспинка большая, более или менее поперечная, плоская,
часто с прозрачными боковыми краями. Надкрылья лежат на спине плоско,
более плотные, чем крылья. Крылья нежные, прозрачные, в покое скрыты под
надкрыльями. Иногда надкрылья и крылья укорочены или отсутствуют. Ноги
бегательные. Бедра уплощенные, снизу вооружены шипами, либо без них.
Голени с сильными шипами. Лапки 5-члениковые, 2 коготка на последнем
членике одинаковой длины, симметричные или неравные, асимметричные; при-
соска между коготками имеется или отсутствует. Брюшко плоское, состоит
из 8—10 тергитов и у самца из 8—9, у самки 7 стернитов, но I стернит очень
маленький. Последний тергит называется анальной пластинкой, его форма
часто используется при определении видов (рис. 93, 4, 9). Под анальной пла-
стинкой расположены остатки X—XI стернитов — парапрокты. Последний
176
стернит называется генитальной пластинкой и обычно несет 1—2 грифелька;
у самки последний стернит крупный, без грифельков, иногда со створками.
На конце брюшка имеется пара членистоногих веретеновидных церков, реже-
(некоторые бескрылые виды) церки нечленистые. — В СССР более 50, из них
на Дальнем Востоке — 6.
Рис. 93. Детали строения тараканов.
1—5 — Blatta orienialis: 1 — ? сверху, 2 — голова спереди, 3 — лапка сверху, 4 — вершина брюшка в
сверху, 5 — то же $, снизу; 6 — Polyphaga sp., голова спереди; 7 — Periplaneta americana, лапки сверху;
8 — Р. australasiae, 3 сверху; 9, 10 — Blatiella germanica'. 9 — вершина брюшка <? сверху, 10 — в сверху.
ап — анальная пластинка, гл — голова, гп •— генитальная пластинка, гр — грифельки, зг — задняя го-
лень, зл — задняя лапка, ког — коготки, л — лоб, нал — наличник, надкр — надкрылья, пр — претар-
зус, прсп — переднеспинка, стпв — створки, у — усик, ц — церки. (По: Бей-Биенко, 1950).
Б И OXO ГИЯ
Насекомые с неполным превращением, личинки похожи на взрослых,
отличаясь меньшими размерами, окраской тела, меньшим числом члеников
в усиках, строением вершины брюшка и отсутствием крыльев (за исключением
бескрылых форм). Яйца откладывают в особую капсулу — оотеку, строение
которой характерно для отдельных родов. Количество яиц и размеры оотеки
у вида колеблются в зависимости от условий существования. У пруссака (Blat-
tella germanicd) длина оотеки изменяется от 6.5 до 9.1 мм, а число яиц в них —
от 17 до 41, причем одна самка за жизнь откладывает 3—4 оотеки. У пруссака
развитие при 20 °C занимает 115—130 дней, а при 30 СС сокращается до 75 дней.
У черного таракана (Blatta orientalis) биологический цикл длится более четырех
12 Зак. № 990 1 77
лет, причем откладка оотек происходит только на пятый год. Зимуют личинки,
взрослые насекомые или оотеки. Тараканы — преимущественно ночные на-
секомые, днем они скрываются в щелях, под камнями, под опавшей листвой
и в других укромных местах.
ВРЕД
Медико-санитарное значение тараканов заслуживает внимания, так как
некоторые тараканы (Blattella germanica, Blatta orientalis) являются весьма
обычными и широко распространенными синантропными видами и обитают
в жилищах человека даже за полярным кругом, другие (Periplaneta americana,
Р. japonica, Р, australasiae) характерны для широт с умеренным климатом,
где встречаются главным образом в портовых городах. Синантропные тараканы,
питаясь продуктами, отбросами и даже фекалиями, могут переносить возбуди-
телей болезней на ногах, а также рассеивать их вместе со своими испражне-
ниями. Так, американский таракан (Periplaneta americana) разносит на ногах
стафилококки и другие патогенные микробы. У пруссака и черного таракана
в кишечнике обнаружены яйца некоторых паразитических червей — лентеца
широкого, острицы и власоглава, поэтому тараканы могут служить разносчи-
ками инвазий. Кроме того, ряд синантропных видов тараканов являются
промежуточными хозяевами некоторых нематод — паразитов домашних птиц
и млекопитающих.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ
1 (4). Тело голое. Наличник неясно отделен от лба, без резкого разделения
на переднюю и заднюю части, плоский (рис. 93, 2).
2 (3). Генитальная пластинка d симметричная, с парой одинаковых грифель-
ков (рис. 93, 4). Последний стернит брюшка ? на вершине с парой створок,
отделенных друг от друга поперечной бороздкой (рис. 93, 5).........
................................................... 1. Blattidae
3 (2). Генитальная пластинка d асимметричная, с 1 или 2 неодинаковыми гри-
фельками (рис. 93, 9). Последний стернит брюшка $ простой ....
................................................. 2. Blattellidae
4 (1). Тело или хотя бы края переднеспинки и передний край надкрылий
в волосках и ресничках. Наличник нерезко ограничен от лба, явственно
подразделен на переднюю плоскую и более вздутую заднюю части (рис. 93,
6)............................................* . . . 3. Polyphagidae
1. Сем. BLATTIDAE
Усики не короче тела. Надкрылья вполне развиты или укорочены, задние
крылья не короче надкрылий или отсутствуют. Лапки с одинаковыми когот-
ками (подсем. Blattinae). Церки у бескрылых и короткокрылых видов члени-
стые. — В СССР 3 рода, 6 видов, на Дальнем Востоке — 2 рода, 4 вида.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). Присоска между коготками лапки хорошо развита (рис. 93, 7). Над-
крылья у d длиннее брюшка, у ? не боковые ... 1. Periplaneta Burm.
2 (1). Присоска между коготками лапок маленькая, плохо заметная (рис. 93,
5). Надкрылья у d короче брюшка, у $ боковые, не соприкасаются на
спине (рис. 93, 7)......................................2. Blatta L.
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
1. Periplaneta Burm. Известно около 50 видов, главным образом из тропиков
и субтропиков Старого Света. В СССР и на ДВ 3 вида, завезенные преимуще-
ственно в портовые города, где живут в отапливаемых помещениях.
178
1 (2). Переднеспинка одноцветная, черно-бурая. Надкрылья у $ укороченные,
не достигают вершины брюшка, в 2 раза длиннее переднеспинки. <J —
22—25, $ — 24—25. — Ю Прим.; Япония...................Р. japonica Кагпу
2 (1). Переднеспинка рыжая, с крупными черными или каштаново-бурыми
пятнами. Надкрылья у ? достигают вершины брюшка или заходят за него,
в 3—4 раза длиннее переднеспинки.
3 (4). Надкрылья одноцветные. Задний край переднеспинки и 2 крупных пятна
на ней каштаново-бурые. 6 — 27—34, $ — 28—35. — Космополит, си-
нантроп ......................Р. americana (L.) — Американский таракан
4 (3). Надкрылья со светлой полосой у основания переднего края. Задний
край переднеспинки и 2 крупных пятна на ней черные (рис. 93, 8). 8 —
23.5—25.5, $ — 24—29. — Космополит, синантроп...........................
..............................................Р. australasiae (Fabr.)
2. Blatta L. В роде 1 синантропный вид, развезенный по всему свету.
1 (1). Черно-бурый, одноцветный. Надкрылья у $ боковые (рис. 93, 1). Аналь-
ная пластинка у 3 поперечная, у $ с выемкой по заднему краю, в — 20—
26, $ — 18—30. — Космополит, синантроп. В домах и др. отапливаемых
помещениях......................В. orientalis L. — Черный таракан
2. Сем. BLATTELLIDAE
Усики длиннее тела. Средней величины или маленькие. Церки всегда чле-
нистые. — 1 род (в СССР 6).
1. Blattella Caud. В роде несколько десятков видов, преимущественно
в тропиках и субтропиках Африки и Азии. В СССР и на ДВ — 1 вид.
1 (1). Надкрылья вполне развитые, заходят за вершину брюшка (рис. 93, 10).
Промежуток между темными полосками на переднеспинке не уже каждой
отдельно взятой полосы, d — 10—13, $ — 11—13.5. — Космополит, си-
нантроп ..................................В. germanica (L.) — Пруссак
3. Сем. POLYPHAGIDAE
Усики у самца не длиннее, у самки короче тела. Бедра снизу без шипов.
Надкрылья и крылья часто отсутствуют, особенно у самок. Церки у бескрылых
самок нечленистые, короткие. — 1 род (в СССР 5).
1. Eupolyphaga Chop. Известно 5 видов, преимущественно из Китая и Юго-
Восточной Азии. В СССР и на ДВ — 1 вид.
1 (1). Тело у d светло-бурое, в густых волосках, у $ черноватое, коренастое,
выпуклое. Надкрылья у d заходят за вершину брюшка, $ полностью бес-
крылая. d — 19—22, $ — 25—26. — Ю Прим.; Китай, Монголия. Про-
никает в дома.......................................Е. sinensis (Walk.)
ОСНОВНАЯ ЛИТЕРАТУРА
Бей-Биенко Г. Я. Насекомые таракановые. М.; Л., 1950. 343 с. (Фауна СССР. Насеко-
мые прямокрылые; Нов. сер.; Т. 40).
ПУХОЕДЫ (Отряд MALLOPHAGA)
ВНЕШНЕЕ СТРОЕНИЕ
Тело 1 — 11 мм длиной, от узкопродолговатого до широкояйцевидного
(рис. 94, 1). Голова относительно большая, разнообразной формы, иногда
с зубцевидными или крючковидными отростками и с полосками, обозначае-
мыми по месту их положения (рис. 94, 2—4). Переднекрайняя полоска окайм-
ляет переднюю половину головы и, если прерывается на наличниковом шве,
12*
179-
то разделяется на полоски лба и наличника. Глазничные полоски находятся
перед глазами. Теменные (верхние задние продольные) полоски отграничивают
виски от темени, а горловые (нижние) — скулы от горла. Глотка обычно снаб-
жена глоточным склеритом. Усики головчатые, булавовидные или нитевидные,
короткие, 3—5-члениковые, в покое часто скрыты в боковых усиковых ямках.
Рис. 94. Общий вид и детали строения пухоедов.
1 — Anatoecus dentatus, общий вид $ сверху; 2 — Eomenacanthus stramineus, голова $ снизу; 3 — Oulocre-
pis dissimilis, голова 3 сверху; 4 — Columbicola columbae, передняя часть головы <? сверху; 5 — Bovicola
bovis, вершина брюшка $ снизу; 6 — Gyropus ovalis, гениталии <?. бп — базальная пластинка, вис — ви-
сок, ву — височный угол, г — глаз, гон — гонапофизы, гс — глоточный склерит, зу — затылочный угол,
лп — лобные полоски, лщ — ланцетовидные щетинки, мез — мезосома, мш — препуциальный мешок,
нал — наличник, нал. п — наличниковая пластинка, нал. пол — наличниковые полоски, отр — пост-
пальпальный отросток, пар — парамера, пк. пол — переднекрайняя полоска, пп — плейральные пластинки,
пр — придаток 3-го членика усика, птп — птероторакс, ств — половая створка, тп — тергальные пла-
стинки, /и. пол — теменные полоски, уз — предусиковый узел, у — усики, уя — боковая усиковая ямка,
ш — наличниковый шов, щ — челюстной щупик. (По: Благовещенский, 1964).
Глаза расположены по бокам головы, редуцированы, 1—2-линзовые. Грудь
с ясно отграниченной переднегрудью, часто средне- и заднегрудь слиты в единый
птероторакс, обычно с 1 парой среднегрудных дыхалец. Крыльев нет. Ноги бега-
тельные или хватательные; лапки 1—2-члениковые с 1—2 коготками.
Брюшко состоит из 8—10 видимых сегментов, обычно отличается у самца
и самки по строению последнего сегмента, с 5—6 парами дыхалец на II—VII
или III—VIII сегментах. Тергиты и стерниты с боковыми, срединными или
180
сплошными пластинками (пятнами). Плейриты с пластинками, пятнами или
аподемами, иногда с вентральными клиновидными выступами. На последних
стернитах часто бывают непарные (срединные) или парные (боковые) гениталь-
ные пластинки или пятна. Половая створка самки нередко с боковыми половыми
придатками — гонапофизами (рис. 94, 5). Гениталии самца, как правило,
состоят из базальной пластинки, парамер, мезозомы и препуциального мешочка
(рис. 94, 6).
Тело слабо или густо покрыто щетинками, иногда сидящими на светлых
округлых участках — пустулах. Задние бедра и брюшко нередко со скопле-
ниями или гребнями щетинок — псевдоктенидиями.
Известно около 2200 видов пухоедов с птиц и примерно 300 с млекопитаю-
щих. В определительные таблицы включено 42 вида, паразитирующих на до-
машних животных на Дальнем Востоке и широко распространенных по всему
свету.
БИОЛОГИЯ
Насекомые с неполным превращением. Яйца откладывают на бороздки пера
или между ними, на ствол, в полость очина или на волос поодиночке, рядами
или кучками. Развитие от яйца до половозрелой особи занимает 3—4 недели
и сопровождается 3 личиночными стадиями. Некоторые виды размножаются
партеногенетически. Одна самка откладывает за жизнь 20—35 яиц, приблизи-
тельно 1—2 яйца за 2—3 дня.
Пухоеды обитают на птицах и млекопитающих, причем одни виды живут
на перьях или на волосах, другие — на коже, внутри стержня пера или даже
в ротовой полости хозяев. Питаются кожными дериватами — частицами пера
или эпидермиса, выделениями кожи и кровью.
ВРЕД
Пухоеды встречаются почти на всех группах домашних животных. При
численном обилии пухоеды причиняют своим хозяевам сильное беспокойство.
У птиц наблюдается потеря в массе, снижение яйценоскости, нарушение нор-
мального роста цыплят. Зуд, причиняемый пухоедами домашним животным,
заставляет их чесать пораженные места, где шерсть обычно выпадает и об-
разуются проплешины. Кожные повреждения способствуют проникновению
патогенных микроорганизмов. Борьба с пухоедами сводится к ликвидации их
на самих животных — хозяевах, дезинсекцией помещений и профилактическим
мероприятиям.
К методам борьбы с пухоедами относятся купание, опрыскивание и приме-
нение мазей и порошковидных инсектицидов [Благовещенский, 1940].
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДОТРЯДОВ
1 (2). Глаз 2 пары. Нижнечелюстные щупики есть (рис. 95, 3)......
................................................1. АшЫусега
2 (1). Глаз 1 пара. Нижнечелюстных щупиков нет........2. Ischnocera
Подотряд AMBLYCERA
Нижнечелюстные щупики 2—4-члениковые. Усики головчатые или булаво-
видные, 4—5-члениковые, в покое скрыты в боковых усиковых ямках. Средне-
грудь и заднегрудь обычно не слиты. На Дальнем Востоке около 40 родов,
в определительную таблицу включены 16 видов 15 родов.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (22). Все лапки с 2 коготками. (Надсем. Menoponoidea).
2 (5). Крупные пухоеды, длина тела более 5 мм. (Сем. Trinotonidae, род Tri-
noton N.).
181
3 (4). Длина тела более 5.5 мм. Группа щетинок на глоточной пластинке широ-
кая (рис. 95, 7). 3 — 6—6.1, ? — 6—6.8. — На гусях .......
...................................................... Т. anserinum (F.)
4 (3). Длина тела 5—5.6 мм. Группа щетинок на глоточной пластинке узкая
(рис. 95, 2). 8 — 4.8—5.5, $ — 5.2—5.6. — На утках........................
................................................. Т. querquedulae (L.)
10
Рис. 95. Общий вид и детали строения пухоедов.
1,2 — хетотаксия глоточной области <?: 1 — Trinoton anserinum, 2 — Т. querquedulae', 3 — Uchida numi-
dae, голова j снизу; 4 — Neocolpocephalum turbinatum, голова снизу; 5 — Heterodoxus spiniger, голова
сверху; 6 — Cuclotogaster heterographus, голова и грудь <? сверху; 7 — Numidilipeurus lawrensis tropicalis,
j; 8 — Oxylipeurus deniaius, j; 9, 10 — усик <?: 9 — Holorstiella lata, 10 — Gallacanthus cornutus. гп —
глоточная пластинка, гс — глоточный склерит. (По: Злоторжицкая и др., 1974).
5 (2). Мелкие пухоеды, длина тела менее 5 мм.
6 (9). Тело одноцветное, без значительной склеротизации.
7 (8). Переднегрудь значительно уже головы. Брюшко покрыто волосками и
щетинками различной толщины. (Сем. Anistronidae, род Holomenopon
Eich.). 8 — 1.6—1.8, $ — 2—2.3. — На утках . . . Н. maxbeieri Eich.
8 (7). Переднегрудь такой же ширины, как голова. Брюшко покрыто очень
тонкими волосками. (Сем. Somaphantidae, род Bonomiella Con.). 8 —
1—1.1, $ — 1.9—2. — На голубях.............................В. columbae Em.
9 (6). Тело не одноцветное, со значительной склеротизацией.
10 (13). Голова с парными глазничными и затылочными пятнами (рис. 95, 4).
Задние бедра и III стернит брюшка с псевдоктенидиями. (Сем. Colpocepha-
lidae).
182
11 (12). Глазничные выемки узкие, остроконечные. Глазничные и затылочные
пятна развиты слабо. (Род Anseriphilus Eich.). 6 — 1.9, $ — 2.1—2.2. —
На гусях..........................................A. pectiniventris (Наг.)
12 (И). Глазничные выемки широкие, закругленные. Глазничные и затылоч-
ные пятна хорошо развитые (рис. 95, 4). (Род Neocolpocephalum Ew.).
d — 1.4, $ — 1.7. — На голубях.................... N. turbinatum (Denny)
13 (10). Голова с иной склеротизацией. Псевдоктенидии отсутствуют. (Сем. Ме-
noponidae).
14 (15). Голова без постпальпальных отростков. Плейриты брюшка с темными
пятнами. (Род Мепороп N.) d — 1.8, $ — 1.9. — На курах...................
................................................... М. gallinae (L.)
15 (14). Голова с парными постпальпальными отростками (рис. 95, 3). Плей-
риты брюшка большей частью без темных пятен.
16 (17). Глоточный склерит коричневый, колбовидный (рис. 95, 3). Плейриты
брюшка с темными пятнами. (Род Uchida Ew.). d — 1.3—1.6, $ — 1.5—
1.8. — На курах......................................U. pallidulus (N.)
17 (16). Глоточный склерит светлый, иной формы. Плейриты брюшка без тем-
ных пятен.
18 (19). Постпальпальные отростки слиты со склеротизованной нижней по-
верхностью головы (рис. 94, 2). Длина тела более 2.5 мм. (Род Eomena-
canthus Uch.), d, ? — 2.9—3.1. — На курах .... Ео. stramineus (N.)
19 (18). Постпальпальные отростки свободные. Длина тела менее 2.5 мм.
20 (21). 5-й членик усика больше 2-го членика (рис. 95, 10). (Род Gallacanthus
Eich.). d — 1.4—1.6, $ — 1.7—1.9. — На курах.......................
..............................................G. cornutus (Schom.)
21 (20). 5-й членик усика меньше 2-го членика (рис. 95, 9). (Род Hohorstiella
Eich.). d — 1.5, $ — 2.2. — На голубях......................Н. lata (Piag.)
22 (1). Лапки средних и задних ног с 1 коготком или совсем без коготков.
(Надсем. Gyropoidea).
23 (26). Брюшко с 6 парами дыхалец.
24 (25). Голова сверху с 2 крючками (рис. 95, 5). (Сем. Boopidae, род Hetero-
doxus Le Sou6f et Bui.), d — 1—1.1, j — 1.2—1.3. — На собаках . . .
..................................*................H. spiniger (End.)
25 (24). Голова сверху без крючков. (Сем. Gyropidae, род Gyropus N.). d —
1—1.1, ? — 1.3—1.4. — На морских свинках..................G. ovalis Burm.
26 (23). Брюшко с 5 парами дыхалец.
27 (28). Грудь со слитыми сегментами. (Сем. Trimenoponidae, род Trime-
пороп Cum.), d — 1.6—2.2, $ — 1.7—2.3. — На морских свинках . . .
....................................................Т. hispid urn (Burm.)
28 (27). Грудь с ясно отграниченными друг от друга сегментами. (Сем. Gliri-
colidae).
29 (30). Длина головы больше ее ширины. Брюшко лентовидное. (Род GU-
ricola Mjob.). d — 1.1—1.2, $ — 1.3—1.4. — На морских свинках . . .
..............*................................... G. porcelli (Schr.)
30 (29). Длина головы меньше ее ширины. Брюшко овальное. (Род Pitruf-
quenia Mar.), d — 1.2, $ — 1.4. — На нутриях . . . . Р. coypus Mar.
2. Подотряд ISCHNOGERA
Нижнечелюстные щупики отсутствуют. Усики обычно нитевидные, 3- или
5-члениковые, нередко сильно отличаются у самцов и самок, открыто высту-
пают по бокам головы. Среднегрудь и заднегрудь обычно слиты. На Дальнем
Востоке около 60 родов, в определительную таблицу включены 26 видов
19 родов.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (36). Все лапки с 2 коготками.
2 (13). Ширина головы больше ее длины. Виски б. м. угловидные. (Надсем. Go-
niodoidea, сем. Goniodidae).
3 (10). Длина тела меньше 3 мм. Голова и усики у обоих полов одинаковые.
4 (7). Боковые края висков тупоугольные. (Род Goniocotes Bum.).
5 (6). Переднекрайняя полоска на наличнике узкая, спереди не расширенная.
d, $ — 1.2—1.3. — На курах......................G. maculatus Tasch.
6 (5). Переднекрайняя полоска на наличнике широкая, особенно спереди. в —
1—1.1, $ — 1.5—1.6. — На курах...........................»G. gallinae (Deg.)
7 (4). Боковые края висков остроугольные.
8 (9). Мелкие пухоеды, длина тела менее 2 мм. Длина головы равна ее ширине.
(Род Campanulotes Кё1.). в — 0.9—1.2, $ — 1.3—1.6. — На голубях
................................................С. bidentatus compar (N.)
9 (8). Пухоеды средней величины, длина тела больше 2 мм. Ширина головы
больше ее длины. (Род Coloceras Tasch.). d — 2.5—2.8, $ — 2.8—3. —
На голубях.................................С. damicornis fahrenholzi (Eich.)
10 (3). Длина тела более 3 мм. Форма головы и иногда усиков различается
У d И ?.
11 (12). Усики у обоих полов одинаковые, нитевидные; длина 2-го членика
усика не меньше длины 3—5-го члеников, взятых вместе. (Род Stenocro-
taphus Кё1.). в — 3—3.1, ? — 4—4.1. — На курах . . S. gigas (Tasch.)
12 (11). Усики только у $ нитевидные, у d форма члеников усиков различная;
длина 2-го членика усика меньше длины 3—5-го члеников, взятых вместе
(рис. 94, 3). (Род Oulocrepis KeL). d — 2.2—2.5, $ — 2.8—3. — На ку-
рах .............................................О. dissimilis (Denny)
13 (2). Ширина головы меньше ее длины. Виски закругленные. (Надсем. Phi-
lopteroidea).
14 (23). Наличник с темной переднекрайней полоской.
15 (16). Усики у обоих полов одинаковые, нитевидные. Средней величины ко-
ренастые пухоеды. (Сем. Degeeriellidae, род Lagopoecus Wat.), d. — 1.7,
$ — 1.9. — На курах...................................L. sinensis (Sugimoto)
16 (15). Усики у $ нитевидные, у d членики усиков разнообразной формы.
Большие, б. м. стройные пухоеды. (Сем. Lipeuridae).
17 (18). Наличник спереди округленный, его ширина сзади не меньше ширины
висков. (Род Lipeurus N.). d — 2—2.3*, ? — 2.2—2.5. — На курах . . .
.........................................................L. caponis (L.)
18 (17). Наличник спереди б. м. заостренный, его ширина сзади меньше ширины
висков.
19 (20). Задний край птероторакса с 4 пучками длинных щетинок (рис. 95, 6').
(Род Cuclotogaster Саг.), d — 2.4, $ — 2.5. — На курах ....................
.................................................С. heterographus (N.)
20 (19). Задний край птероторакса с 2 пучками длинных щетинок (рис. 95,
7, S).
21 (22). Брюшко со слитыми тергоплейральными пластинками (рис. 95, $).
(Род Oxylipeurus Mjob.). d — 3—3.2, $ — 3.2—3.5. — На курах . . .
......................................................О. dentatus (Sugimoto)
22 (21). Брюшко с раздельными тергальными и плейральными пластинками
(рис. 95, 7). (Род N umidilipeurus Ten.), d, ? — 3.2—3.5. На курах . . .
............................................ N. lawrensis tropicalis (Pet.)
23 (14). Наличник без темной переднекрайней полоски. (Сем. Esthiopteridae).
24 (31). Стройные пухоеды. Крупнее: длина тела более 2 мм.
25 (28). Передний край наличника с 2 ланцетовидными щетинками (рис. 94, 4).
Форма усиков у d и $ сильно различается. (Род Columbicola Ew.).
26 (27). Голова сильно удлиненная (рис. 94, 4). Гениталии d с луковицеобраз-
ными парамерами (рис. 96, 7). d — 2—2.3, $ 2.4—2.8........................
....................................................С. columbae (L.)
27 (26). Голова менее удлиненная (рис. 97, 7). Гениталии d с шиловидными
парамерами (рис. 96, 2). в — 2.1—2.2, $ — 2.3—2.6. — На голубях . .
......................................................С. montschadskyi Blag.
28 (25). Передний край наличника без ланцетовидных выростов. Усики у обоих
полов различаются слабо. (Род Anaticola Cl.).
184
29 (30). Щетиночки посредине переднего края наличника явно толще соседних
щетиночек. Мельче. 6 — 2.5—2.9, ? — 3.2—3.4. — На утках.....................
................................................ A. crassicornis (Scop.)
30 (29). Щетиночки посредине переднего края наличника едва толще соседних
щетиночек. Крупнее, 'в — 3.5, $ — 4. — На гусях ... A. anseris (L.)
31 (24). Коренастые пухоеды (рис. 94, 7). Мельче: длина тела менее 2 мм.
(Род Anatoecus Cum.).
32 (33). Тело желто-коричневое. Длина тела обычно свыше 1.5 мм. Гениталии d
с зубцами (рис. 96, 3). S, $ — 1.6—1.7. —На утках ..........................
........................................A. (Benatoecus) dentatus (Scop).
33 (32). Тело желтоватое. Длина тела обычно менее 1.5 мм. Гениталии d без зуб-
цов (рис. 96, 4). (Подрод Anatoecus Cum.).
Рис. 96. Гениталии £ пухоедов.
1 — Columbicola columbae; 2 — С. montschadskyi", 3 — Anatoecus dentatus", 4 — A. ictrodes. (По: Злоторжиц-
кая и др., 1974).
34 (35). Наличниковая пластинка слабоудлиненная (рис. 97, 2). в — 1.3, $ —
1.5. — На гусях............................A. (A.) a Justus (N. et Gieb.)
35 (34). Наличниковая пластинка явно удлиненная (рис. 97, 5). d, $> — 1.5. —
На утках........................................A. (A.) ictrodes (N.)
36 (1). Все лапки с 1 коготком.
37 (48). Ноги, особенно бедра, длинные и стройные. Вершина брюшка $ пара-
болптическая (рис. 94, 5). (Сем. Bovicolidae).
38 (39). Вершина голеней с прозрачными пластиночками на ясном отростке.
(Род W erneckiella Eich.). d — 1.6, $ — 1.8. — На лошадях.................
.......................................................W. equi (Denny)
39 (38). Вершина голеней только с прозрачными пластиночками, без отростка.
На козах, овцах и крупном рогатом скоте.
40 (41). Тергиты брюшка, за исключением генитальной области, с длинными
щетинками. (Род Holakartices Kel.). d — 1.3—1.4, $ — 1.5—1.6. —
На козах..............................................Н. crassipes (Rudow)
41 (40). Тергиты брюшка, за исключением генитальной области, с очень корот-
кими щетинками.
42 (43). Средние и задние голени на вершине сильно расширены: наибольшая
ширина голени достигает длины лапки с коготком. (Род Lepikentron Kel.).
d — 1.4, j — 1.6. — На овцах................................L. ovis (Schr.)
43 (42). Средние и задние голени на вершине слабо расширены: наибольшая
ширина голени явно меньше длины лапки с коготком. (Род Bovicola Kel.).
44 (45). Наличник спереди параболический; ширина головы равна ее длине
(рис. 97, 7). d — 1.2, $ — 1.4. — На крупном рогатом скоте ....
.............................................................В. bovis (L.)
45 (44). Наличник спереди прямой или вогнутый; ширина головы больше ее
длины (рис. 97, 5, 6).
185
46 (47). Наличник спереди плоский, с короткими щетиночками одинаковой
длины (рис. 97, 5). d — 1.4—1.5, $ — 1.8—2. — На козах......................
.................................................. В. саргае (Gurlt.)
47 (46). Наличник спереди вогнутый, со сравнительно длинными щетиночками
различной длины (рис. 97, 6), d — 1.2, $ — 1.5—1.6. — На козах . . .
........................................................В. limbatus (Gerv.)
48 (37). Ноги, особенно бедра, сравнительно короткие и толстые. Вершина
брюшка $ полукруглая. (Сем. Trichodectidae).
5
Рис. 97. Детали строения пухоедов.
1 — Columbicola montschadskyi, голова <5 сверху; 2 — Anatoecus adustus, наличниковая пластинка j; 3 —
A. ictrodes, то же <?,; 4 — Bovicola bovis, голова о сверху; 5—7 — голова у сверху: 5 — В. саргае, 6 — В.
limbatus, 7 — Felicola subrostratus. (По: Злоторжицкая и др., 1974).
49 (50). Брюшко с 6 парами дыхалец. Наличник закругленный. (Род Trlcho-
dectes N.). d — 1.6—1.7, $ — 1.6—1.9. — На собаках............................
........................................................Т. canis (Deg.)
50 (49). Брюшко с 5 парами дыхалец. Наличник треугольный (рис. 97, 7).
(Род Felicola Ew.). d — 1.2, $ — 1.3. — На кошках.............................
.....................................................F. subrostratus (N.)
ОСНОВНАЯ ЛИТЕРАТУРА
Благовещенский Д. И. Определитель пухоедов (Mallophaga) домашних животных. М.: Л.,
1940. 89 с. (Фауна СССР. Нов. сер.; Т. 27).
Благовещенский Д. И. Пухоеды. М.; Л., 1959. 202 с. (Фауна СССР. Насекомые пухоеды.
Нов. сер. 72; Т. 1. Вып. 1).
Благовещенский Д. И. Отряд Mallophaga — пухоеды. — В кн.: Определитель насекомых
европейской части СССР. М.; Л., 1964, т. 1, с. 309—323.
Zlotorzycka J., Eichler W., Ludwig H. W. Taxonomie und Biologie der Mallophagen
und Lause mitteleuropaischer Hous- und Natztiere. — Parasitol. SchrReihe, 1974, Bd 22, S. 1 —
160.
186
МУРАВЬИ
(Отряд HYMENOPTERA,
сем. FOPMICIDAE)
Муравьи — широко известные перепончатокрылые насекомые. Они живут
в особых гнездах «обществами» — семьями. Как правило, семья состоит из одной
или нескольких бескрылых плодущих самок и большого количества (у не-
которых видов до 1.5 млн) рабочих особей.
ВНЕШНЕЕ СТРОЕНИЕ
Муравьи, обитающие в умеренных широтах, — мелкие или средние насеко-
мые: размеры рабочих — 2—14 мм, самцов и самок — 3—18 мм (рис. 98, 1—3).
'Окраска тела муравьев варьирует от светло-желтой до темно-бурой, коричневой
1 2
Рис. 98. Рыжий лесной муравей.
1 — самка; 2 — самец; 3 — рабочий; 4 — яйцо; 5 — личинка; 6 — куколка.
и черной. Тело обычно покрыто тонким серым или желтоватым опушением
разной плотности, кроме того, на теле могут быть более длинные и жесткие
отстоящие волоски, или хеты.
Голова продолговатая, квадратная или несколько суживающаяся к перед-
нему краю. Спереди головы располагаются наличник, ротовые органы (челюсти
и щупики) и коленчато согнутые усики, состоящие из 12 или 13 члеников.
Кроме округлых сложных (фасеточных) глаз, располагающихся по сторонам
головы, имеются небольшие простые глазки на темени.
Грудь состоит из 3 члеников (переднегруди, среднегруди и заднегруди),
но у рабочих муравьев швы между ними очень слабо заметны. У целого ряда
видов заднегрудь вооружена 2 зубцами, или шипами. Ноги обычного для на-
секомых строения. На нижнем конце голени зачастую имеется шпора. Крылья
у самцов и самок нежные, прозрачные или слабо окрашенные, с неполным
жилкованием.
Брюшко обычно продолговатое или сердцевидное и состоит из хорошо раз-
личимых 5—6 члеников. Один или 2 передних членика всегда отличаются по
форме от остальных и образуют стебелек, соединяющийся с грудью.
Если стебелек одночлениковый, то он несет вертикальную или наклонную
чешуйку, двухчлениковый стебелек имеет форму округлых или угловатых
узелков. У ряда видов на конце брюшка имеется жало.
187
Яйца мелкие, обычно овальные, белые или светло-желтые, с очень тонкой
нежной оболочкой. Личинки червеобразные, малоподвижные, светлоокрашен-
ные, белые или желтоватые, с тонкими нежными покровами. Как правило,
они безглазые и имеют зачатки усиков и слабо хитинизированные ротовые
органы. Куколки у одних видов свободные, у других — в коконах, имеющих
овальную форму (рис. 98, 4—6). Личинки и куколки самцов и самок обычно
более крупные, чем рабочих.
БИОЛОГИЯ
Муравьи обитают в почве, под камнями, в старых пнях, под корой гнилых
бревен. Многие виды устраивают холмики из выброшенной почвы, некоторые —
большие куполы из растительных частиц — хвоинок, тонких веточек и листьев.
Самки откладывают яйца, рабочие ухаживают за развивающимся потомством
(личинками и куколками), снабжают семью пищей, занимаются строительством
и поддерживают порядок в гнездах. В летний период выводятся крылатые
самцы и самки, которые вылетают из гнезд и спариваются. Самцы, как пра-
вило, погибают, а самки сбрасывают крылья и основывают новые гнезда.
С ранней весны и до поздней осени муравьи питаются сладким соком расте-
ний, сахаристыми выделениями (падью) тлей. В период кормления личинок
они приносят в гнезда огромные количества животной пищи, главным образом
разных насекомых, их личинок, гусениц, куколок. Многие лесные муравьи,
уничтожающие массу вредителей деревьев и кустарников и участвующие в поч-
вообразовании, несомненно полезны и заслуживают строгой охраны.
Некоторые муравьи приспособились к жизни в помещениях. Одним из са-
мых известных домовых насекомых является фараонов, или домовый муравей
(Monomorium pharaonts). Он имеет тропическое происхождение, сейчас рас-
пространился по всему свету, встречается и в городах Дальнего Востока.
Поселяется исключительно в жилых отапливаемых помещениях: квартирах,
больницах, столовых, ресторанах, банях, на кораблях, в теплицах. Устраивает
гнезда в трещинах и щелях стен, под штукатуркой, вдоль труб водяного отопле-
ния. Образует относительно крупные ^семьи, состоящие из нескольких самок
и большого количества рабочих (более 1000), лишь незначительная часть их вы-
ходит на поверхность, остальные, как правило, остаются в гнездах.
Обыкновенный, или черный садовый муравей {Lasius niger), широко рас-
пространенный и самый обычный вид в умеренных широтах. Обычно живет
в лесах, садах, на полянах, на обочинах дорог. Устраивает разнообразные
гнезда в почве, под камнями, в пнях, гнилой древесине, старых усохших де-
ревьях. В городах и поселках живет на клумбах, газонах, возле домов, может
поселиться и в старых деревянных домах, под полом, в гнилых балках, в ще-
лях, под плинтусами. Основные, самые крупные, его гнезда всегда находятся
снаружи, в них живут очень большие семьи, состоящие из одной бескрылой
самки и нескольких тысяч рабочих.
ВРЕД
Фараоновы муравьи питаются углеводной и мясной пищей. Проникая в по-
мещение (чаще всего ночью), они нападают на продовольственные запасы,
загрязняют и портят их. Могут быть механическими переносчиками болезне-
творных бактерий. В литературе описываются случаи нападения домовых
муравьев на новорожденных и недоношенных детей, разъедания кожи и об-
разования ранок [Малышева, 1981]. Бороться с домовыми муравьями исключи-
тельно трудно, так как они могут быстро восстанавливать численность своих
семей за счет тех половых особей, которые прячутся в укромных местах. Не-
обходимо терпеливое многократное применение сладких и мясных ядовитых
приманок, рассчитанное на то, что муравьи, которые переносят в своем зобу
пищу, постепенно накормят ядом всех членов семьи, находящихся в гнездах.
Черные садовые муравьи в природе питаются сладкими выделениями расте-
ний, падью тлей, мелкими насекомыми. В помещения проникают через щели
в поисках сладкой пищи, а обнаружив ее, собираются сплошной колонной,
загрязняя пищевые запасы. Вероятно, участвуют и в механическом переносе
188
возбудителей заболеваний, хотя точных сведений об этом не имеется. Чтобы
предотвратить проникновение черных садовых муравьев с улицы, достаточно
плотно заделать отверстия в стенах и полу. Если они поселились в доме, нужно
найти гнезда и заменить заселенные муравьями гнилые деревянные детали.
Диагноз сем. Formicidae
Усики коленчато согнутые, брюшко прикрепляется к груди стебельком,
состоящим из 1 или 2 члеников. Муравьи живут в гнездах и ведут общественный
образ жизни.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ И РОДОВ
1 (2). Стебелек состоит из 2 члеников, жало хорошо развито (рис. 99). Куколки
без коконов (подсем. Myrmicinae). Заднегрудь округлая, наличник впе-
зг ст
Рис. 99. Детали тела муравьев.
1 — фараонов муравей Monomorium pharaonis, рабочий; 2 — его голова; 3 — черный садовый муравей
Lasius niger, рабочий; 4 — его голова, г — голени, ж — жало, зг — заднегрудь, н — наличник, р — ру-
коять усика, ст — стебелек.
реди с небольшой выемкой и 2 продольными валиками, челюстные щупики
одно- или двухчлениковые. Тело стройное .... 1. Monomorium Мауг
2 (1). Стебелек состоит из 1 членика, имеющего форму чешуйки, жало не раз-
вито (рис. 99). Куколки с коконами (подсем. Formicinae). Заднегрудь
угловатая, наличник впереди округлый без выемки и продольных вали-
ков, челюстные щупики шестичлениковые. Тело более широкое . . . .
.......................................................2. Lasius L.
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
1. Monomorium Мауг. Очень мелкие муравьи. Гнездятся в разнообразных
полостях и почве. Широко распространены в тропиках и субтропиках. Больше
130 видов, несколько космополитов, расселившихся при помощи человека
по всему свету. — 1 вид (в СССР 2).
1 (1). Рабочие и $ желтые, голова и грудь матовые, густо пунктированные.
6 темнее. Рабочие — 1.7—2.3, $ — 3.5—4, 3 — 2.8—3..............
...........«... М. pharaonis L. — Фараонов, или домовый муравей
189
2. Lasius L. Насекомые средних размеров. Гнездятся в почве, под предме-
тами, в гниющей древесине. Связаны с тлями. Колонии от мелких до круп-
ных. — 1 вид (в СССР около 20).
1 (1). Тело одноцветное черное или буровато-черное. Рукоять усиков и голени
с многочисленными отстоящими волосками. Рабочие и d — 3—5, $ —
7—10..................................L. niger L. — Черный садовый муравей
ОСНОВНАЯ ЛИТЕРАТУРА
Арнольди К. В., Длусский Г. М. Сем. Formicidae — муравьи. — В кн.: Определитель
насекомых европейской части СССР. Л., 1978, т. 3, ч. 1, с. 519—556.
Дмитриенко В. К., Петренко Е. С. Муравьи таежных биоценозов Сибири. Новосибирск,
1976. 218 с.
Малышева М. С. Муравьи — Formicidae. — В кн.: Насекомые и клещи — вредители
сельскохозяйственных культур. Перепончатокрылые и двукрылые. Л., 1981, т. 4, с. 46—50.
II. КЛЕЩИ —ACARINA (Класс ARACHNIDA)
Клещи — мелкие паукообразные, различные по строению и образу жизни.
Это — одна из древнейших групп наземных беспозвоночных. Общей их особен-
ностью являются малые размеры тела (0.2—5.0 мм, сытые иксодовые увеличи-
вают размеры до 25.0 мм) и наличие в цикле развития трехногой личинки (первой
послезародышевой фазы). Все другие фазы развития (нимфы и взрослые), как
и все прочие паукообразные, имеют 4 пары ног. Кроме того, клещам свойственна
тенденция к слиянию различных отделов тела в одно нерасчлененное целое,
к исчезновению внешних признаков членистости. Указанные особенности
имеют массу отклонений, поэтому в современной таксономии нет единого взгляда
на эту группировку. Одни считают клещей единым отрядом Acarina класса
паукообразных Arachnida в подтипе хелицеровых Chelicerata, другие обосновы-
вают самостоятельность трех отрядов, объединяемых в отдельный подкласс
(или класс) Acaromorpha, или размещают их в различных подотрядах сборной
группы хелицеровых. Среди последних крупных исследований, проведенных
советскими учеными в области акарологии, первая точка зрения нашла свое
отражение в монографии Ю. С. Балашова [1982], второй придерживаются
авторы трехтомного «Определителя обитающих в почве клещей» (1975—1978).
Вслед за большинством исследователей, изучающих клещей, мы признаем
наличие двух больших группировок клещеобразных животных — Parasiti-
formes и Acariformes с подразделением последней на Sarcoptiformes и Trombidi-
formes.
Поскольку в настоящее время большинством исследователей признается
(для удобства), что клещи — это единая группа паукообразных животных,
а упомянутым выше группировкам Sarcoptiformes и Trombidiformes придают
ранг подотрядов, то по аналогии с этим группировкам Acariformes и Parasiti-
formes следует придать ранг отрядов. Тогда объединяющая их группировка
Acarina принимает вид надотрядного уровня — надотряда или подкласса.
Такой системы придерживаются Ланге [1984] (за исключением последней
группировки — Acarina) и Крантц [Krantz, 1978] в единственном в мире учеб-
нике акарологии, придавая ранг подотряда иксодовым (подотряд Ixodida
с единственным надсемейством Ixodoidea) и гамазовым (подотряд Gamasida
с 5 когортами, лишь одна из которых, Gamasina, представляет предмет на-
шего интереса) клещам.
В связи с тем, что в настоящее время не существует единого мнения специа-
листов относительно ранга этих таксонов, мы решили оставить их такими,
какими они даны в упомянутом выше учебнике. Подобная система представ-
ляется формальной, однако она позволяет сопоставить группы более или менее
одинаковой таксономической значимости.
Мировая фауна клещей насчитывает на сегодняшний день более 25 тыс.
(в СССР — около 12 тыс., на Дальнем Востоке — не менее 3 тыс.), но факти-
чески это, по мнению различных авторов, составляет от г/3 до х/10 всех действи-
тельно существующих видов.
191
ВНЕШНЕЕ СТРОЕНИЕ ВЗРОСЛЫХ КЛЕЩЕЙ
Следует оговориться, что содержащиеся в данном разделе сведения общие
и касаются только тех групп, которые могут представлять интерес для меди-
цины и ветеринарии. Более конкретная характеристика и уточнение деталей
строения, необходимых для определения, даны в диагнозах каждой конкретной
группы.
Как уже говорилось, тело большинства клещей характеризуется слит-
ностью его отделов в единое целое; выделяют лишь два отдела: головной, или
гнатосома (несущий ротовой аппарат), и туловищный, или идиосома (несущий
4 пары ног). Однако у большей части клещей (во всяком случае у большинства
Sarcoptiformes) тело явственно подразделено хорошо выраженной перетяжкой
или поперечной (сеюгальной) бороздой на передний (проподосома) и задний
(гистеросома) отделы, каждый из которых несет по две пары ног (рис. 119).
В некоторых группах наблюдается вторичное подразделение заднего отдела
на тагмы поперечными складками.
Кожные покровы двух типов: тонкие, эластичные, неокрашенные, гладкие
или с поперечными параллельными складками, сеткой и уплотненные в виде
щитов, которые могут быть или мягкими, отличающимися от окружающей
кожи тем, что на них никогда нет складок, или очень плотными, окрашенными
в буроватый цвет. У некоторых клещей покровы уплотнены так, что все туло-
вище оказывается как бы одетым в панцирь (у почвенных клещей Oribatei,
например, некоторых Gamasida).
Все тело и ноги клещей обычно несут щетинки, различающиеся по величине
(очень длинные, в несколько раз превышающие длину тела, и очень короткие,
еле заметные, называемые микрохетами), форме, положению на теле и числу.
Число и положение щетинок или их комплексов — один из наиболее стойких
диагностических признаков для таксонов как высокого, так и низкого ранга.
Головной отдел относительно невелик; в подавляющем большинстве случаев
свободен и обладает подвижностью. Занимает на теле терминальное или суб-
терминальное положение и может быть полностью скрыт передним краем тела.
В состав его входят 2 пары ротовых придцтков (хелицеры и пальпы), сохранив-
ших частичную членистость. Хелицеры чаще всего оканчиваются клешней,
образованной подвижным (соответствующим лапке ходильной конечности) и
неподвижным (соответствующим голени, сросшейся с последующим члеником)
пальцами. Строение гнатосомы крайне разнообразно, что зависит от типа пита-
ния. Иногда при почти полном слиянии члеников головной отдел представляет
собой монолитное образование. В некоторых случаях те или иные придатки
сильно выделяются, например органы заякоревания — зубчатый гипостом
иксодовых клещей (рис. 100). Только у гамазид позади гипостома на брюшной
поверхности тела медиально располагается особый вильчатый осязательный
придаток — тритостернум (рис. 115, г).
Туловищный отдел со спинной стороны лишен каких-либо органов. Он моно-
литный или разделен бороздами на отделы; полностью покрыт щитом или щиты
вовсе отсутствуют. Спина, как правило (исключение клещи рода Notoedres —
Sarcoptidae), не несет каких-либо отверстий и органов. Она более или менее
равномерно покрыта щетинками или выделяют отдельные их комплексы, ука-
занные при описании каждой группы клещей.
Туловищный отдел с брюшной стороны несет 4 пары ног (за исключением
самок рода Otodectes). Этим в препаратах она легко отличается от спинной
стороны. Ноги или прикрепляются непосредственно к брюшной стороне (как
у Parasitiformes — рис. 115, 7), или базальные их членики полностью срас-
таются с брюшной поверхностью (все Acariformes), что в одних случаях выгля-
дит как щиты — коксальные щиты (рис. 121, 6), в других — как сетка из более
темных полос или отдельных темных ребер — коксо-стернальный скелет
(рис. 124).
Ноги состоят из 6 члеников: тазик (соха), вертлуг (trochenter), бедро (femur),
колено (genu), голень (tibia), лапка (tarsus). Лапки всех ног заканчиваются
предлапкой (pretartus), которая имеет самое различное строение (присоска,
коготок или коготки, сочетание того и другого), и служит одним из значимых
192
признаков при диагностике высших таксонов. В строении ног часто проявля-
ются дополнительные специальные функции, которые они могут нести, а также
вторично-половые различия.
Индивидуальное развитие клещей включает несколько неполовозрелых
фаз: личинку, отличающуюся, как уже говорилось, наличием всего лишь
3 пар ног, 2 или 3 нимфы, имеющих 4 пары ног, но без сформированного поло-
вого аппарата, и половозрелых самцов и самок, похожих или не похожих
друг на друга (отличающиеся от нимф наличием половых отверстий). Различия
самцов и самок указаны специально при описании отдельных групп.
С брюшной стороны открываются половое и анальное отверстия, положение
и строение которых очень сильно варьируют у разных групп клещей. У самок
чаще всего имеются 2 половых отверстия: яйцевыводное и копулятивное;
у самцов — одно.
Половой аппарат самца значительно меньше по размерам, темнее окрашен
и более склеротизован, чем яйцевыводное отверстие самки. Он представлен
комплексом склеритов, окружающих половой орган — пенис (эдеагус). У гама-
зид половым органом служат видоизмененные хелицеры. У представителей
многих групп имеются дополнительные органы для скрепления копулирую-
щих пар: видоизмененные хелицеры, гипертрофированные ноги I или III пар,
тарзальные и анальные присоски и т. д.
Яйцевыводное отверстие $ представлено продольной или поперечной щелью,
часто прикрытой клапанами. В ряде случаев снабжено окаймляющими склери-
тами.
Анальное отверстие обычно занимает положение у заднего края брюшной
поверхности, иногда терминальное, в исключительных случаях смещено на
спинную сторону. Обычно оно прикрыто клапанами, иногда окружено аналь-
ным щитком или щитками.
Дыхание примитивных Acariformes кожное, у более продвинутых форм
имеются дыхательные трубочки — трахеи. Последние хорошо развиты у всех
Parasitiformes и открываются по бокам тела стигмами, расположенными на щит-
ках (перитремах) либо округлых, довольно крупных, либо лентовидных, тяну-
щихся вперед от стигм (дыхалец) и пронизанных порами (рис. 100, Б, 8;
115, А, Л).
МЕТОДИКА ХРАНЕНИЯ И ОПРЕДЕЛЕНИЯ
Методы сбора даже только клещей, имеющих медицинское и ветеринарное значение,
очень разнообразны в зависимости от того, какая группа представляет интерес. Однако опи-
сание их не входит в цель данного издания, так как предполагается, что те, кто будет пользо-
ваться данной книгой, уже будут иметь материал в руках.
Для консервирования всех клещей обычно применяют 70 %-ный спирт. Иногда на 95
частей разбавленного спирта добавляют 5 частей глицерина, что несколько облегчает при-
готовление препаратов. Материал только тогда представляет научную ценность и годится
для дальнейших исследований, если он подробно этикетирован с указанием хозяина, части
тела, с которой собран клещ, места (географического пункта) и даты сбора, фамилии сбор-
щика.
Клещей, собранных из одного места, одного хозяина или одной части его тела, помещают
в отдельный стеклянный пузырек (баночку из-под пенициллина пли других подобных ле-
карств) пли мелкую пробирку с 70 %-ным спиртом. Туда же помещается этикетка, которая
пишется на плотной бумаге или кальке мягким простым карандашом или тушью. Сосуд с соб-
ранным материалом доливается доверху спиртом и плотно закупоривается. Удобнее всего
пользоваться мелкими пробирками, которые можно затыкать ватной пробкой, смоченной
в спирте, и складывать в общую банку со спиртом.
Более крупных клещей (иксодовые, аргасовые) определяют с помощью бинокулярного
стереоскопического микроскопа (бинокуляра), прямо вынимая из спирта и слегка обсушив
фильтровальной бумагой. Полоску фильтровальной бумаги, на которой лежит клещ, на би-
нокулярном столике время от времени смачивают спиртом, чтобы избежать высыхания.
После определения клеща снова кладут в спирт.
Личинок этих клещей определяют как под бинокуляром, так и под микроскопом, изго-
товляя из них постоянные препараты.
Определить до вида клещей большинства групп (гамазовые, аллергенные, перьевые,
чесоточные, краснотелки и другие) можно только в микроскопических препаратах, используя
микроскоп. Для приготовления временных препаратов можно использовать 90 %-ную мо-
лочную кислоту, хорошо просветляющую клещей, хуже — глицерин. Затем препараты
могут быть размонтированы, и клещи снова помещены в спирт. Однако во временных препа-
ратах далеко не все детали можно рассмотреть, и поэтому лучше изготовлять постоянные
13 Зак. № 990
193
препараты, заключая клещей в специальную монтировочную гуммиарабиковую смесь по
рецепту Фора—Берлезе:
Дистиллированной воды — 50 мл,
Гуммиарабика сухого или вишневой камеди — 30 г,
Хлоралгидрата — 200 г,
Чистого глицерина — 20 мл.
Смесь приготовляется следующим образом. В хорошо закрытую банку с дистиллирован-
ной водой всыпают гуммиарабик и на водяной бане или в термостате (температура 40—50 сС)
доводят до полного растворения. Затем добавляют хлоралгидрат и глицерин и держат там же
до полного растворения хлоралгидрата. При приготовлении смеси ее нельзя мешать палоч-
кой, чтобы избежать попадания пузырьков воздуха, от которых позже трудно избавиться.
Полученную жидкость фильтруют через несколько слоев стиранной (чтобы не попали волоски)
марли. Фильтровать надо также в термостате, иначе жидкость густеет и фактически не фильт-
руется. Если химические компоненты, из которых приготовлена смесь, чистые, фильтрацию
можно не производить. Хранить жидкость следует в плотно закрытой банке и в темноте, так
как хлоралгидрат летуч, гидрофилен, легко разлагается на свету.
Изготовление препаратов требует внимания и аккуратности. На предметное стекло иго-
лочкой пли стеклянной палочкой наносится капля молочной кислоты или гуммпарабиковой
смеси. Клещи прямо из пробы или из спирта переносятся в эту каплю, которая затем накры-
вается покровным стеклом. Опускать стекло надо очень плавно, чтобы в препарат не попали
пузырьки воздуха. Желательно при изготовлении препаратов пользоваться лупой пли би-
нокуляром: прежде чем накрыть каплю стеклом, следует расположить в ней клещей рядами,
положить часть экземпляров спинной стороной вверх, другую часть вверх брюшной стороной.
Препараты с молочной кислотой оставляют просветляться на 1—2 сут. Препараты с гумми-
арабиковой смесью для лучшего и более быстрого просветления следует слегка погреть на
спичке, над пламенем спиртовки или держать в термостате (температура 50—60° С) в течение
1—3 сут.Чем лучше сделан препарат, чем лучше просветлен клещ, тем легче и быстрее удастся
его определить.
Препараты обязательно этикетируют. Надписывают или непосредственно на стекле
тушью, или на отдельной бумажной этикетке, которую затем приклеивают к стеклу клеем
БФ-2 или БФ-4. На этикетку переносят данные с этикетки, приложенной к материалу, а после
того, как клещ будет определен, его научное название.
Хранят препараты на картонных или фанерных лотках обязательно в горизонтальном
положении, чтобы объект не сполз с предметного стекла (смесь легко разжижается во влаж-
ной среде).
Правила пользования определительными таблицами описаны в разделе «Насекомые»
(с. 10).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ОТРЯДОВ И ПОДОТРЯДОВ
1 (4). Базальные членики ног (тазики) свободные или неподвижно соединены
с покровами тела, однако они хорошо различимы и полностью не сраста-
ются с брюшной поверхностью. По бокам тела располагаются отчетливо
видимые стигмы, число и положение которых на теле варьируют . . .
......................................................Отряд Parasitiformes
2 (3). Стигмы находятся в промежутке между коксами III и IV пар ног; от них
по направлению к переднему концу тела тянется длинная и узкая пери-
трема (рис. 115, д). Ротовой аппарат располагается субтерминально и
всегда виден при рассмотрении клеща сверху. Пальпы 5-члениковые,
имеют вид пары маленьких ног. Гипостом довольно сложного строения,
с вентральной стороны к нему примыкает тритостернум (рис. 115, а)
........................................................ Когорта Gamasina
3 (2). Стигмы находятся у кокс IV пары ног и окружены стигмальной пластин-
кой; длинные узкие перитремы отсутствуют. Ротовой аппарат распола-
гается субтермииально или вентрально (в углублении — камеростоме)
и может быть не виден при рассмотрении клеща сверху. Пальпы 3—4-
члениковые, массивные. Гипостом с брюшной стороны вооружен многочис-
ленными направленными назад зубцами (рис. 100, Б, 2, Г, 6). Тритостер-
нум отсутствует.........................................Надсем. Ixodoidea
4 (1). Базальные членики ног (тазики) полностью срастаются с брюшной по-
верхностью, образуя коксальные щиты или коксо-стернальный скелет.
Стигмы отсутствуют или незаметны........................Отряд Acariformes
5 (6). На 2-м от свободного конца членике ног I и II (голени) имеются длинные
бичевидные щетинки. Ротовые органы грызущие. Все 4 пары ног или
по крайней мере 3 передние пары заканчиваются 1 коготком или стебель-
194
чатой присоской, либо присоской с 1 коготком; J обычно имеют анальные
копулятивные присоски................1. Подотряд Sarcoptiformes
6 (5). На 2-м от свободного конца членике ног I и II (голени) длинная бичевид-
ная щетинка отсутствует. Ротовые органы чаще всего колюще-сосущие.
Все 4 пары ног или по крайней мере II и III заканчиваются 2 коготками
с перепончатой присоской между ними; в всегда без анальных копулятив-
ных присосок..............................2. Подотряд Trombidiformes
ОСНОВНАЯ ЛИТЕРАТУРА
Балашов Ю. С. Паразито-хозяинные отношения членистоногих с наземными позвоноч-
ными. Л., 1982. 320 с. (Труды Зоол. ин-та АН СССР; Т. 87).
Ланге А. Б. Подтип Хелицеровые (Chelicerata). — В кн.: Жизнь животных. М., 1984,
т. 3, с. 70—108.
Определитель обитающих в почве клещей. М., 1975, т. 1, 491 с.; 1977, т. 2, 718 с.; 1978,
т. 3, 271 с.
Krantz G. W. A manual of Acarology. 2nd ed. Corvallis, 1978. 509 p.
ПАРАЗИТИФОРМНЫЕ КЛЕЩИ
(Отряд PARASIT1FORMES)
ИКСОДОИДНЫЕ КЛЕЩИ (Надсем. I X О D О I D E A)
Занимают довольно обособленное положение в отряде Parasitiformes. Клещи
характеризуются наличием вытянутого гипостома, снабженного рядами зуб-
чиков; пальцами хелицер с зубчатой вершиной; парой перитрем; органом Гал-
лера на I лапке. Облигатные кровососы, паразиты млекопитающих, птиц,
рептилий и амфибий. Объединяет 3 семейства: Argasidae, Ixodidae и монотип-
ное Nuttalliellidae (вид Nuttalliella namaqua Bedf. встречается в Южной
Африке).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ
1 (2). Скутум имеется, он покрывает всю верхнюю поверхность тела (3) или
ее переднюю часть (?, нимфы, личинки). Гнатосома располагается на пе-
реднем конце идиосомы и направлена вперед (терминально). $ с поровыми
полями. Перитремы крупные, располагаются позади IV пары кокс. Глаза
отсутствуют или имеются по бокам скутума................1. Ixodidae
2(1). Скутум отсутствует. Гнатосома располагается на вентральной стороне
передней части идиосомы (имаго, нимфы) или терминально (личинки).
? без поровых полей. Перитремы мелкие, располагаются впереди или
на уровне IV пары кокс. Глаза отсутствуют (у видов фауны СССР) . . .
.................................................... 2. Argasidae 1
ИКСОДОВЫЕ КЛЕЩИ (Сем. IXODIDAE)
Диагноз семейства
Размеры до 10 мм (в голодном состоянии). Гнатосома расположена на перед-
нем конце тела и направлена вперед. IV членик пальп сильно редуцирован
и помещается на вентральной стороне III членика. Глаза имеются или отсут-
ствуют. Перитремы имеются (у личинки отсутствуют), они расположены по-
зади IV кокс. Спинной щиток (скутум) имеется. Он покрывает всю верхнюю
поверхность тела (самец) или только переднюю ее часть (самка, нимфа, ли-
чинка). Покровы гладкие, не морщинистые. Половой диморфизм резко вы-
ражен.
1 На ДВ представлено только одним видом А г gas vulgaris Fil., найденным лишь однажды
в Приморском крае в гнездах рыжепоясничной ласточки [Гордеева, Белослюдов, 1963;
Филиппова, 1966].
13*
195
Иксодовые клещи — временные наружные паразиты наземных позвоноч-
ных животных, облигатные кровососы. Они распространены по всему земному
шару, но наибольшего видового разнообразия достигают в тропическом поясе.
Большинство видов клещей паразитирует на млекопитающих, значительно
меньше — на птицах и пресмыкающихся и только два южноамериканских
вида — на земноводных (жабах).
ВНЕШНЕЕ СТРОЕНИЕ
Морфология иксодовых клещей подробно описана в ряде отечественных
специальных руководств [Померанцев, 1950; Сердюкова, 1956; Филиппова,
1977, и др.], которые и послужили основой данного раздела.
Взрослые клещи. Размеры в голодном состоянии от 1 до 10 мм
(у дальневосточных видов — от 2 до 5 мм). Тело уплощенное, округлой, оваль-
ной или дисковидной формы. После насыщения кровью тело самок сильно
увеличивается и приобретает яйцевидную или шаровидную формы, тело самца
становится выпуклым только с брюшной стороны. Хитиновый покров крепкий,
раздавить голодного клеща нелегко.
Тело клеща разделяется на идиосому и гнатосому, расположенную в перед-
ней части тела, основанием она входит внутрь идиосомы и имеет ограничен-
ную подвижность (может подгибаться вниз).
Гнатосома состоит из основания, пары пальп, пары хелицер и гипостома
(рис. 100, Г). На спинной (дорсальной) стороне основания ближе к заднему
краю лежат поровые поля. Внешне они представляют две неглубокие выемки
округлой или овальной формы с густой сетью маленьких ямочек. У многих
видов клещей задние углы основания гнатосомы оттянуты назад — их называют
корнуа. Снизу основание гнатосомы может иметь боковые выросты — аурикулы,
форма и расположение которых часто используются в диагностике.
Пальпы 4-члениковые (рис. 100, Г): I членик короткий, у некоторых видов
невидим со спинной стороны. Форма и строение II и III члеников разнообразны,
они определяют общую форму не только пальп, но в известной степени и гнато-
сомы в целом. В сложенном виде пальпы всегда имеют конусовидную форму,
облегчающую клещу продвижение в шерсти или перьях хозяина. IV членик
очень мал и расположен на нижней стороне III членика, поэтому сверху он
не виден. Вершина IV членика покрыта щетинками, а сам членик является
осязательным органом. Внутренняя сторона пальп имеет продольную выемку,
в которую при сложенных пальпах помещаются хелицеры и гипостом. Хели-
церы состоят из основания и двух пальцев, один из которых подвижен. Основа-
ния хелицер погружены в футляры, видимые со спинной стороны. Хелицеры
свободно двигаются в футлярах вперед-назад. Неподвижный палец на вершине
зазубрен. Хелицеры предназначены для разрезания кожи хозяина, куда вво-
дится гипостом. По средней линии верхней стороны гипостома проходит канал,
по которому движется кровь из тела хозяина в ротовое отверстие; нижняя
сторона покрыта рядами зубчиков, которыми клещ удерживается в теле хо-
зяина. У некоторых непитающихся самцов видов рода Ixodes наблюдается
разная степень редукции гипостома. Форма гипостома и число зубчиков варьи-
руют даже у близких видов и используются в диагностике.
Идиосома имеет как мягкие, растяжимые в процессе кровососания
покровы, так и твердые, нерастяжимые щитки. Имеется спинной щиток (ску-
тум), который у самцов занимает почти всю спинную часть идиосомы, а у са-
мок — только переднюю ее часть (рис. 100). Этот признак является основным
для различения полов у иксодовых клещей.
По бокам скутума у клещей большинства родов {Dermacentor и др.) располо-
жены глаза; их нет у представителей родов Ixodes, Haemaphysalts и Аропотта.
На скутуме имеются цервикальные и боковые бороздки (рис. 100, А, 4, 5),
щетинки и разное число маленьких ямок и точек, называемых обычно пункти-
ровкой. Выступы, обрамляющие с двух сторон переднюю выемку скутума, —
скапулы. У самцов Haemaphysalts и Dermacentor на задней стороне скутума
имеются насечки, образующие 11 фестонов. Растяжимый хитиновый покров
самки, лежащий за спинным щитком, называют аллоскутумом, где различают
196
краевую борозду, фовеа и фестоны (рис. 100, Л, 6—9). Фовеа и фестоны от-
сутствуют у клещей рода Ixodes.
Рис. 100. Схема строения иксодового клеща (по: Сердюкова, 1956; Филиппова, 1977).
А — идиосома самки сверху: 1 — скутум, 2 — скапулы, з — глаза, 4 — боковая борозда, 5 — цервикаль-
ная борозда, 6 — аллоскутум, 7 — фовеа, 8 — краевая борозда, 9 — фестоны. Б — самец Ixodes снизу:
1 — основание гнатосомы, 2 — гипостом; з—6 — членики пальп: з — I членик, 4 — II членик, 5 —
III членик, 6 — IV членик; 7 — генитальное отверстие, 8 — перитрема, 9 — анус; 10—14 — вентральные
щитки: 10 — анальный, 11 — аданальный, 12 — эпимеральный, 13 — срединный, 14 —стернальный;
15 — кокса, 16 — медиальный зубец, 17 — латеральный зубец, 18 — вертлуг, 19 — бедро, 20 — колен-
ный членик, 21 — голень, 22 — лапка, 23 — присоска, 24 — коготки, 25 — орган Галлера. В — аналь-
ные клапаны: 1 — Ixodes persulcatus’, 2 — Haemaphysails сопсгппа\ 3 — Dermacentor silvarum. Г — гнато-
сома самки Ixodes сверху (слева) и снизу (справа): 1 — основание гнатосомы, 2 — корнуа, 3 — поровые
поля, 4 — аурикулы, 5 — постгипостомальные щетинки, 6 — гипостом, 7 — футляры хелицер, 8 — пальпы
хелицер; 9—12 — членики пальп: 9 — I членик, 10 — II членик, 11 — III членик, 12 — IV членик.
На брюшной (вентральной) поверхности идиосомы расположены гениталь-
ное отверстие, анус и перитремы (по бокам). У самцов рода Ixodes вентральная
поверхность покрыта 7 твердыми щитками (рис. 100, Б); их разделяют бо-
розды — анальная, генитальная и преанальная.
197
Половое отверстие находится на уровне I —IV кокс (в зависимости от пола
и вида клеща) и представляет собой щель различной формы — прямой, изогну-
той, дуговидной. Анус помещается в задней части тела в виде продольной щели
и снабжен клапаном. На створках анального клапана имеется от 1 до 5 пар
щетинок (рис. 100, В). По бокам тела позади IV кокс располагаются перитремы
(одна пара). Они выполняют функции дыхания и регуляции водного баланса.
Ноги 6-члениковые (рис. 100, Б). Коксы уплощены, почти неподвижны и
могут нести разное число зубцов (шипов), имеющих важное диагностическое
значение. Коксы у самцов Dermacentor сильно увеличены, однако их размер
коррелирует с размером тела клеща — у мелких экземпляров клещей IV кокса
по своим размерам мало отличается от III коксы. Все последующие членики
подвижные, имеют цилиндрическую форму. Лапка заканчивается двумя когот-
ками и присоской. Коготками клещ цепляется за хозяина, а присоска помогает
ему передвигаться по гладкой поверхности (растений, тела хозяина или другого
субстрата).
Нимфа. Скутум покрывает только переднюю часть идиосомы, перитремы
имеются, ног 4 пары. Генитальное отверстие, твердые щитки на вентральной
поверхности тела и поровые поля на гнатосоме отсутствуют. У некоторых
видов Ixodes первый членик пальп с крупными выростами. У клещей рода
Dermacentor основание гнатосомы шестиугольное с острыми боковыми углами,
скутум без эмалевого пигмента. Нимфы Haemaphysalts больше похожи на самок,
чем у клещей родов Ixodes и Dermacentor.
Личинка. Ног 3 пары, перитремы отсутствуют. Пальпы обычно 4-члени-
ковые, но граница между I и II, II и III члениками неясная или отсутствует
совсем. На анальном клапане 1 пара щетинок. У некоторых видов Ixodes первый
членик пальп с крупными выростами. У клещей рода Dermacentor основание
гнатосомы шестиугольное с боковыми углами, скутум без эмалевого пигмента.
В диагностике личинок рода Ixodes важное значение имеет характер расположе-
ния щетинок на теле и их размер.
БИОЛОГИЯ
В процессе индивидуального развития иксодовые клещи проходят 4 морфо-
логические фазы: яйцо, личинка, нимфа, взрослый клещ (имаго).
Иксодовые клещи в каждой активной фазе (личинка, нимфа, имаго) пи-
таются только один раз, после чего, как правило, покидают хозяина и линяют
(или откладывают яйца) во внешней среде (в гнезде или норе хозяина, в верх-
нем слое почвы, под камнями, в трещинах грунта и т. п.). Дальневосточные
виды клещей в течение жизненного цикла сменяют трех хозяев (треххозяинный
тип развития): клещи лишь во время питания находятся на теле хозяина, а вся
остальная жизнь проходит в природной среде, как у свободноживущих организ-
мов. Поэтому факторы внешней среды (в особенности климатические) оказывают
существенное влияние на их жизнедеятельность.
По характеру местообитаний клеща во внешней среде и условиям встречи
с хозяином у иксодовых клещей различают два типа паразитизма — гнездово-
норный (убежищный) и подстерегающий пастбищный. В первом случае вся
жизнедеятельность клеща вне хозяина (линька, яйцекладка, зимовка, период
покоя и т. д.) проходит в гнезде хозяина, норе или в непосредственной близости
от него; здесь клещ нападает на хозяина, и здесь же происходит отпадение
напившихся крови клещей.
Клещи с пастбищным типом паразитизма не имеют постоянных убежищ, они
нападают на хозяина в открытой природе, там же отпадают сытые клещи, а их
линька, яйцекладка и период покоя проходят в лесной подстилке, в дерновине,
верхнем слое почвы и т. п.
На протяжении каждой активной фазы клещ проходит следующие стадии:
послелиночное доразвитие, активность, питание, линька или яйцекладка
(Балашов, 1967]. В период активности пастбищные клещи подстерегают хозяев
на поверхности земли, на траве и кустах, где они сидят «в позе ожидания», вы-
ставив в стороны переднюю пару ног, снабженных коготками. Личинки рас-
полагаются в приземном ярусе на высоте до 10 см, нимфы — до 50—70 см,
198
а имаго — до 1.5 м. Клещи могут совершать незначительные горизонтальные
перемещения, особенно их привлекают звериные тропы, к которым они ползут
с расстояния до 10 м.
Средняя продолжительность питания личинок составляет 3—5 сут, нимф —
3—8 сут, самок — 6—12 сут. Самцы клещей Haemaphysalts и Dermacentor,
как и самки, пьют кровь хозяина — это является необходимым условием для
завершения сперматогенеза и оплодотворения самок. Самцы клещей рода Ixodes
кровь не пьют. Самцы гнездово-норных видов иксодин редко встречаются нг1
хозяевах, а некоторые неизвестны совсем (самец I. pomerantzevt).
Во время питания личинка, нимфа и самка поглощают громадные порции
крови. Масса полностью напившихся клещей в десятки и сотни раз превосходит
их массу в голодном состоянии. У D. silvarum, например, масса сытых самок
достигает 600 мг, а у I. persulcatus — 790 мг [Беликова, 1975].
Всем иксодовым клещам свойственна жесткая гонотрофическая гармония —
однократное полноценное питание обеспечивает откладку единственной порции
яиц, после чего самка погибает. Оплодотворение самок — обязательное усло-
вие успешного завершения гонотрофического цикла. Некоторые популяции
клеща Н. longicornis могут развиваться партеногенетически. Полностью напи-
тавшиеся самки клещей откладывают до 2000—4000 яиц, а некоторые тропиче-
ские виды — до 10 000 яиц.
Продолжительность жизненного цикла известна только для части видов.
За один год полностью завершают свое развитие I. lividus и D. silvarum. Боль-
шинство других видов клещей завершает жизненный цикл на юге Дальнего
Востока за 2—3 года, на севере — за 3—5 лет.
ВРЕД
Иксодовые клещи являются хранителями и переносчиками возбудителей
ряда опасных болезней человека и животных. В патологии населения При-
морья, Амурской обл. и юга Хабаровского коая большое значение имеет кле-
щевой энцефалит — тяжелое вирусное заболевание, передающееся человеку
иксодовыми клещами. Основной хранитель и переносчик вируса — таежный
клещ Ixodes persulcatus. Дополнительными переносчиками являются и другие
пастбищные виды клещей — Н. japonica, Н. concinna, D. silvarum. Указанные
виды клещей сохраняют вирус в процессе метаморфоза и могут передавать его
человеку посредством укуса, но частота выделения вируса клещевого энцефа-
лита из этих видов ниже, чем из таежного клеща. Длительность эпидемического
сезона клещевого энцефалита в значительной степени обусловлена неодновре-
менностью сезона активности разных видов клещей. Эти же виды клещей могут
быть переносчиками возбудителей болезней человека и животных: клещевого
сыпного тифа, энцефалита Повассан, туляремии, тейляриоза, клещевого рик-
кетсиоза.
Эффективным методом борьбы с иксодовыми клещами является ранневесен-
няя обработка территорий акарицидами с самолета. Это ведет к резкому
(почти полному) сокращению численности клещей в природе на длительное
время. Из индивидуальных средств защиты от клещей самое надежное — пра-
вильное ношение одежды и частый самоосмотр. Верхняя одежда (рубашка или
куртка) всегда должны быть заправлены в брюки (а последние в носки) во избе-
жание заползания клещей на тело под одежду. Необходимо периодически
(через 2—3 часа) осматривать себя во время движения по лесу. Присосавшегося
клеща надо плотно захватить пинцетом или пальцами как можно ближе к коже
и резким рывком выдернуть. Если есть возможность, необходимо обратиться
в ближайший медицинский пункт.
В семействе выделяют 2 подсемейства — Ixodinae (с одним родом Ixodes)
и Amblyomminae, куда входят все остальные роды, включая Haemaphysails
и Dermacentor, представленные на Дальнем Востоке. Мировая фауна насчиты-
вает около 650 видов клещей 14 родов; в СССР обитают примерно 75 видов
7 родов, на Дальнем Востоке — 20 видов, относящихся к 3 родам.
199
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). Анальная борозда огибает анус спереди или, если не замыкается спереди,
то имеет вид двух продольных отрезков или складок, расположенных
по бокам или позади ануса. (Подсем. Ixodinae)........1. Ixodes
Рис. 101. Самки клещей рода Ixodes (по: Филиппова, 1977; Померанцев, 1950).
J, 2, 4 - - гнатосомы сверху: 1 — I. maslovi, 2 — I. lividus, 4 — I. arboricola\ 3 — гнатосома (сверху) и ску-
тум I. crenulatus', 5 — анус и анальная борозда 1. uriae.
2 (1). Анальная борозда огибает анус сзади или отсутствует. (Подсем. Amblyom-
minae).
3 (4). Глаз нет..........................................2. Haemaphysalis
4 (3). Глаза есть.........................................3. Dermacentor
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦЫ ВИДОВ
1. Ixodes Latr.1 Большинство видов мелкие (до 4 мм в голодном состоянии).
Анальная бороздка в форме полной дуги, огибающей анус спереди, или в виде
двух продольных незамыкающихся отрезков. Глаз нет. Перитремы округлые
1 Для видов рода Ixodes определительные таблицы составлены в основном по Н. А. Фи-
липповой [1977].
200
или овальные, без спинного отростка. Брюшная поверхность J покрыта 7 щит-
ками. Постгипостомальных щетинок обычно 2.пары,.реже — 4 пара. — 13 видов
(в СССР 34).
Самки
1 (2). I членики пальп смещены дорсально и слились в единую «платформу!
(рис. 101, 7). (Подрод Monoindex Em. et KozL). Очень редок . . . .
...........................................I. (М.) maslovi Em. et Kozl.
2 (i). I членики пальп раздельные и занимают нормальное положение.
Рис. 102. Самки клещей рода Ixodes (по: Филиппова, 1977; Yainaguli et al., 1971).
j, 2 — гнатосома сиерху: 1 — I. signaius, 2 — I. caledonicus\ 3 — гнатосома и скутум I. angustus; 4 — гна-
тосома и скутум 7. pomerantzeii', 5 — гнатосома (снизу) и анальная борозда I. turdus. _б — боковая^бо-
розда, бв — боковой валик.
3 (4). Анальная борозда состоит из двух продольных отрезков, не достигающих
ануса (рис. 101, 5). — Гнездово-порный паразит, в колониях морских
птиц. Многочислен. (Подрод Ceratixodes Neum.).............................
.............................I. (С.) uriae White (putus Pick.-Camb.)
4 (3). Анальная борозда дугой огибает анус спереди.
5 (10). Все коксы без зубцов. (Подрод Pholeoixodes Sch.).
6 (7). Дуговидные валики огибают поровые поля медиально. Боковые борозды
на скутуме отчетливые и глубокие (рис. 101, 5). — Гнездово-норный пара-
зит, основные хозяева — грызуны п хищные. Редок...........................
............................ I. (Ph.) crenulatus Koch (canisuga John.)
7 (6). Дуговидные валики между поровыми полями отсутствуют. Боковые бо-
розды на скутуме отсутствуют или выражены нечетко, поверхностные.
8 (9). Вершины заднебоковых углов основания гнатосомы смещены несколько
вперед по отношению к заднему краю основания. Поровые поля крупные,
близко сдвинуты друг с другом и достигают или почти достигают заднего
201
края основания (рис. 101, 2). — В гнездах береговых ласточек. Редок
I. (Ph.) lividus Koch (plumbeus Leach)
9 (8). Вершины заднебоковых углов основания гнатосомы лежат на уровне
заднего края основания или позади него. Поровые поля небольшие,
широко расставлены друг от друга и отделены от заднего края основания
Рис. 103. Самки клещей рода Ixodes (по: Филиппова, 1977).
1 — идиосома (сверху) и коксы I. pavlovskyi; 2 — гнатосома (снизу) и коксы I. nipponensis; 3 — гнатосома
(снизу) и коксы I. persulcatus; 4 — гнатосома I. sachalinensis ( = 1. persulcatus) сверху.
кромкой (рис. 101, 4). — В гнездах птиц (в дуплах или земляных норах).
Редок ... I. (Ph.) arboricola Sch. et Schl. (lagodechiensis Djapar.)
10 (5). Все или некоторые коксы с хорошо выраженными зубцами.
11 (16). На I — III вертлугах или хотя бы на одной из этих пар имеются вент-
ральные зубчики.
12 (13). Анальная борозда не достигает заднего края тела. Пальпы длинные.
Гипостом острый (рис. 102, 5). — Паразитирует на птицах. (Подрод Tri-
chotoixodes Reznik).......................................I. (T.) turdus Nak.
13 (12). Анальная борозда достигает заднего края тела. Пальпы короткие.
Вершина гипостома закруглена. (Подрод Scaphixodes Sch.).
202
14 (15). Корнуа в виде торчащих кверху бугорков. II членик пальп дорсальнб
с поперечивши бороздками (рис. 102, У). — В гнездах бакланов. Обычен
.................................................. I. (S.) signatus Bir.
15 , (14). Корнуа в виде зубцов, лежащих в одной плоскости с основанием.
II членик пальп дорсально без поперечных бороздок (рис. 102, 2). —
В гнездах белопоясничных стрижей. Редок . . I. (S.) caledonicus Nutt.
16 (И). Вертлуги без вентральных зубцов.
17 (20). Аурикулы отсутствуют. (Подрод Ixodiopsis FiL).
18 (19). Присоски I лапок достигают вершин коготков и заметно длиннее при-
сосок III и IV лапок. Боковые борозды и боковые валики не достигают
заднебоковых краев скутума (рис. 102, 5). — Гнездово-норный паразит;
на мышевидных грызунах. Немногочислен . . . I. (Ix.) angustus Neum.
19 (18). Присоски I лапок не достигают вершин коготков и лишь незначительно
длиннее присосок последующих лапок. Боковые борозды и боковые валики
достигают заднебоковых краев скутума (рис. 102, 4). — Гнездово-норный
паразит; на мышевидных грызунах. Немногочислен.........................
.......................................I. (Ix.) pomerantzevi Serd.
20 (17). Аурикулы имеются. (Подрод Ixodes s. str.).
21 (22). Боковые борозды отчетливые, латерально они ограничены гребневид-
ным или пологим валиком. Пунктировка скутума в задней части значи-
тельно крупнее, чем в передней. Корнуа хорошо выражены. Аурикулы
в виде крупных дуговидных зубцов (рис. 103, 7). — Пастбищный паразит;
имаго на птицах и ежах, нимфы и личинки на птицах и мелких млекопи-
тающих. Немногочислен. К человеку мало агрессивен......................
............................................I. (I.) pavlovskyi Pom.
22 (21). Боковые борозды отсутствуют, реже нечеткие, валик латеральнее их
тоже отсутствует. Пунктировка скутума почти однородная. Корнуа отсут-
ствуют или слабо выражены. Аурикулы в виде треугольных выростов.
23 (24). Скутум удлиненно-овальный. Аурикулы смещены немного назад, более
крупные и острые. Медиальный зубец на коксе I короткий (рис. 103,
2). — Пастбищный паразит; имаго на крупных и средних млекопитающих,
нимфы и личинки на мелки\ млекопитающих и птицах. Присасывается
к человеку. — Немногочислен .... I. (I.) nipponensis Kit. et Saito 2
24 (23). Скутум обычно округлый или широкоовальный, реже — удлиненно-
овальный. Аурикулы смещены кпереди, меньше и тупее. Медиальный
зубец на коксе I длиннее. Более крупные клещи (рис. 103, 5, 4). — Паст-
бищный паразит, имаго на средних и крупных млекопитающих, нимфы
и личинки на мелких и средних млекопитающих и птицах. К человеку
очень агрессивен. Многочислен..........................................
........I. (I.) persulcatus Sch. (sachalinensis FiL). — Таежный клещ
Самцы3
1 (2). I членики пальп смещены дорсально и слились в единую «платформу»
(рис. 104, 7).............................I. (М.) maslovi Em. et Kozl.
2 (1). I членики пальп раздельные и занимают нормальное положение.
3 (4). III членик пальп конусовидный (рис. 104, 2). По заднему краю тела име-
ется щетка из уплотненных щетинок...................I. (С.) uriae White
4 (3). Ill членик пальп округлый или притуплен на переднем конце. По заднему
краю тела щетинки не образуют щетки.
5 (6). На лапках I—III имеются крепкие апикальные зубчики (рис. 104, 7)
.................................................. I.(S.) signatus Bir.
6 (5). На лапках I — III апикальных зубчиков нет.
7 (16). Кокса I с медиальным и латеральным зубцами.
8 (9). Ветви анальной борозды по направлению назад сходятся (рис. 104, 8)
...............................................I. (S.) caledonicus Nutt.
2 Диагностика самок I. nipponensis и I. persulcatus представляет определенные труд-
ности, особенно в местах совместного обитания.
3 Самцы I. turdus и I. pomerantzevi неизвестны.
203
9 (8). Ветви анальной борозды по направлению назад параллельны или расхо-
дятся.
10 (11). Зубчики занимают меньше половины длины гипостома, они все при-
мерно одинаковые, сильно редуцированы (рис. 104, 3)....................
............................................ I. (I.) angustus Neum.
11J10). Зубчики занимают значительно больше половины длины гипостома.
Наружные зубчики значительно крупнее внутренних.
Рис. 104. Самцы клещей рода Ixodes (по: Филиппова, 1977; Arthur, 1963).
1, 2 — гнатосомасверху: 1 — I. maslovi, 2 — I. иггае’, 3 — гнатосома (снизу) и коксы I. angustus’ 4 — аналь-
ная борозда I. crenulatus; 5 — анальная борозда и неподвижный палец хелицер I. arboricola; 6 — неподвиж-
ный палец хелицер I. lividus; 7 — I, II, III лапки I. signatus; 8 — анальная борозда I. caledonicus.
12 (13). Корнуа имеются, если их нет, то задний край основания гнатосомы
со спинной стороны прямой или чуть вогнутый, но не выпуклый (рис. 110,
5)................................................1.(1.) pavlovskyi Pom.
13 (12). Корнуа отсутствуют. Задний край основания гнатосомы со спинной
стороны чуть выпуклый (рис. 110, 4).
14 (15). Перитрема удлиненно-овальная. Медиальный зубец коксы I короткий,
притуплен (рис. 110, 6)..................I. (I.) nipponensis Kit. et Saito
15 (14). Перитрема широкоовальная. Медиальный зубец коксы I средней длины,
заострен (рис. 110, 4)............................I. (I.) persulcatus Sch.
16 (7). Кокса I без зубцов.
17 (18). Анальная борозда впереди ануса дуговидная (рис. 104, 4).........
........................................... I. (Ph.) crenulatus Koch
18 (17). Анальная борозда впереди ануса срезана (рис. 104, 5).
19 (20). Неподвижный палец хелицер с 4 апикальными зубчиками (рис. 104, 5)
. . . . .................................I. (Ph.) arboricola Sch. et Schl.
204
20 (19). Неподвижный палец хелицер с 3 апикальными зубчиками (рис. 104, 6)
...................................... .... I. (Ph.) lividus Koch
Нимфы4
1 (4). I членик пальп неправильной формы, с выростами, отчленен от основания
гнатосомы (рис. 105, 2, 3).
Рис. 105. Нимфы клещей рода Ixodes (по: Филиппова, 1977).
1 — гнатосома I. crenulatus сверху и снизу; 2 — I. angustus, идиосома сверху, гнатосома снизу и I лапка;
3 — I. pomerantzevi, идиосома сверху, гнатосома снизу и I лапка; 4 — гнатосома I. arboricola снизу; 5 —
гнатосома I. lividus сверху, вк — вершинный конус, ду — дорсальный уступ, пк — предкраевые щетинки,
ср — срединные щетинки.
2 (3). Предкраевые щетинки аллоскутума длиннее срединных щетинок скутума
в среднем в 1.2 раза. На всех лапках в профиль переход дорсальной по-
верхности в вершинный конус постепенный, без уступа (рис. 105, 2)
................................................I. (Ix.) angustus Neum.
3 (2). Предкраевые щетинки аллоскутума длиннее срединных щетинок скутума
не менее чем в 1.8 раза. На лапке I в профиль при переходе дорсальной
4 Препмагпнальные фазы I. maslovi неизвестны.
205
поверхности в вершинный конус имеется уступ (рис. 105, 3)...........
..........................................I. (Ix.) pomerantzevi Serd.
4 (1). I членик пальп цилиндрический, без выростов, отчленён или слился
с основанием гнатосомы.
5 (6). Створки анального клапана с 1 парой щетинок. Скутум сильно удлинен
(рис. 106, 4).........................................I. (С.) uriae White
6 (5). Створки анального клапана с 2—3 парами щетинок. Скутум овальный
или слабо удлинен.
Рис. 106. Нимфы клещей рода Ixodes (по: Филиппова, 1977; Yamaguti et al., 1971k
1,2 — гнатосома (сверху и снизу) и коксы: 1 — 1. caledonicus; 2 — I. signatus; 3 — гнатосома I. turdus
снизу; 4 — идиосома сверху и анальный клапан I. uriae.
7 (12). Аурикулы отсутствуют. Все коксы без медиальных и латеральных зуб-
цов, небольшие латеральные бугорки могут быть па III и IV коксах.
8 (9). Корнуа имеются (рис. 105, 5)..................I. (Ph.) lividus Koch
9 (8). Корнуа отсутствуют.
10 (11). Гипостом наиболее широкий в передней трети («булавовидный»). Пальпы
дорсально несколько изогнуты с наружной стороны (рис. 105, 1) . . .
.......................................... I. (Ph.) crenulatus Koch
11 (10). Гипостом наиболее широкий посредине (рис. 105, 4). Пальпы дорсально
с ровными или слегка выпуклыми сторонами...................................
......................................I. (Ph.) arboricola Sch. et Schl.
12 (7). Аурикулы имеются. Коксы с зубцами.
13 (14). I членик пальп слит с основанием гнатосомы. Впереди аурикул имеется
зубцевидный выступ, несущий щетинку (рис. 106, 5)..........................
.................................................I. (Т.) turdus Nak.
206
(13). I членик пальп отделен от основания гнатосомы. Впереди аурикул
зубцевидного выступа нет.
15 (22), Пальпы длинные: длина II—III члеников превосходит их ширину бо-
лее чем в 3 раза. Граница между II и III члениками пальп отчетливая.
Вертлуги без вентральных зубчиков.
Рис. 107. Нимфы клещей рода Ixodes (по: Филиппова, 1977).
1 — I. nipponensis, идиосома сверху, коксы и гнатосома снизу; 2,3 — идиосома сверху, гнатосома снизу
и коксы: 2 — I. pavlovshyi, 3 — I. persulcatus. пк — предкраевые щетинки, ср — срединные щетинки.
16 (19). Щетинки скутума и аллоскутума резко отличаются по размеру: на ску-
туме короткие, на переднебоковых участках аллоскутума они в 2—3 раза
длиннее (рис. 107, 7, 2).
207
17 (18). Гипостом копьевидный: наиболее широкий в задней части, вершина
острая. Латеральный зубец на IV коксе в виде слабо выраженного бугорка
(рис. 107, 1)...........................I. (I.) nipponensis Kit. et Saito
18 (17). Гипостом наиболее широкий в средней части, к вершине сужен, но сама
вершина несколько округлена. Латеральный зубец на IV коксе отчетли-
вый, острый (рис. 107, 2)......................I. (I.) pavlovskyi Pom.
19 (16). Щетинки скутума и аллоскутума короткие и мало отличаются по раз-
меру: на переднебоковых участках аллоскутума они менее чем в 1.5 раза
длиннее срединных щетинбк скутума (рис. 107, 5).
Рис. 108. Личинки клещей рода Ixodes (по: Филиппова, 1977; Yamaguti et al., 1971).
I, 2 — гнатосома снизу и I лапка: 1 — I. angustus, 2 — I. pomerantzevi', 3 — идиосома с гнатосомой сверху
и гипостом I. turdus', 4,5 — гнатосома снизу и коксы: 4 — I. signatus, 5 — I. caledonicus.
20 (21). Латеральный зубец на IV коксе отчетливый, острый (рис. 107, 2)
...............................................1.(1.) pavlovskyi Pom.
21 (20). Латеральный зубец на IV коксе в виде слабо выраженного бугорка
(рис. 107, 3)...................I. (I.) persulcatus Sch. — Таежный клещ
22 (15). Пальпы короткие: длина II — III члеников превосходит их ширину
менее чем в 3 раза. Граница между II и III члениками пальп слабо выра-
жена или отсутствует. Вертлуги (хотя бы один) с вентральными зубчиками
(рис. 106, 7, 2).
23 (24). Латеральные зубцы на коксах I — IV мощные, острые. Кокса I с хорошо
выраженным медиальным зубцом. Вентральные зубчики имеются на I
и II вертлугах, а иногда на III. Корнуа крупные (рис. 106, 7) . . .
..............................................I. (S.) caledonicus Nutt.
24 (23). Латеральные зубцы на коксах I — IV в виде тупых бугорков. Кокса I
без медиального зубца. Вентральный зубчик имеется только на I вертлуге.
Корнуа почти не выражены (рис. 106, 2).............I. (S.) signatus Bir.
208
Личинки
1 (4). I членик пальп неправильной формы, с выростами, отчленен от основания
гнатосомы (рис. 108, 7, 2).
2 (3). Две передние пары краевых щетинок аллоскутума в среднем в 1.5 раза
длиннее срединных щетинок скутума. Длина задней дорсолатеральной
Рис. 109. Личинки клещей рода Ixodes (по: Филиппова, 1977).
1 — идиосома сверху и гнатосома снизу I. nipponensis’, 2,3 — гнатосома снизу и коксы: 2 — I. parlor&yi,
3 — I. persulcatus', 4 — идиосома сверху I. uriae.
щетинки II членика пальп меньше минимальной ширины членика более
чем в 2 раза. Лапка I в профиль при переходе дорсальной поверхности
в вершинный конус без уступа (рис. 108, 1) . . .1. (Ix.) angustus Neum.
3 (2). Две передние пары краевых щетинок аллоскутума в среднем длиннее
срединных щетинок скутума не менее чем в 2 раза. Длина задней дорсола-
теральной щетинки II членика пальп меньше минимальной ширины чле-
ника в среднем в 1.7 раза. Лапка I в профиль при переходе дорсальной
поверхности в вершинный конус имеет уступ (рис. 108, 2)..................
......................................... I. (Ix.) pomerantzevi Serd.
4 (1). I членик пальп цилиндрический, без выростов, отчленен или частично
слился с основанием.
5 (6). Скутум сильно удлинен, заднебоковые стороны скутума длиннее передне-
боковых примерно в 5 раз (рис. 109, 4)................I. (С.) uriae White
14 Зак. № 990 209
6 (5). Скутум округлой формы или немного удлинен, заднебоковые стороны
скутума длиннее переднебоковых не более чем в 2 раза.
7 (18). Все, или хотя бы I и II коксы, с латеральными зубцами.
8 (11). Пальпы короткие: длина II и III члеников превосходит их ширину
менее 3 раз (рис. 108, 4, 5).
9 (10). Латеральные зубцы на коксах крупные, острые. Аурикулы отчетливые,
зубцевидные (рис. 109, 5)......................I. (S.) caledonicus Nutt.
10 (9). Латеральные зубцы на коксах в виде небольших бугорков. Аурикулы
отсутствуют (рис. 108, 4)..........................I. (S.) signatus Bir.
Рис. НО. Личинки и самцы клещей рода Ixodes (по: Филиппова, 1977).
1—3 — гнатосома личинки сверху: 1 — I. crenulatus, 2 — I. arboricola, 3 — I. Hindus', 4 — самец I. per-
suicatus, гнатосома сверху коксы и перитрема; 5 — самец 1. pavlovshyi, гнатосома сверху и вариации ее
заднего края; 6 — самец I. nipponensis, перитрема и коксы.
11 (8). Пальпы длинные: длина II—III члеников превосходит их ширину
в 3 раза и более.
12 (13). На аллоскутуме срединных щетинок 5 пар, промежуточных 4 пары.
Вершина гипостома заострена (рис. 108, 3)...........I. (Т.) turdus Nak.
13 (12). На аллоскутуме срединных щетинок 3—4 пары, промежуточных 1 пара.
Вершина гипостома закруглена (рис. 109, 7).
14 (15). Аурикулы прямоугольные и направлены в стороны (рис. 109, 7) . . .
.....................................I. (I.) nipponensis Kit. et Saito
15 (14). Аурикулы остроугольные, направлены назад и в стороны (рис. 109,
2, 3).
16 (17). Медиальный зубец I кокс длинный и острый (рис. 109, 2)........
...........................................I. (I.) pavlovskyi Pom.
17 (16). Медиальный зубец I кокс короткий и тупой (рис. 109, 3)........
.......................... I. (I.) persulcatus Sch. — Таежный клещ
18 (7). Все коксы без латеральных зубцов.
19 (20). Корнуа имеются (рис. 110, 3)..............I. (Ph.) lividus Koch
20 (19). Корнуа отсутствуют.
210
21 (22). Вершины заднебоковых углов основания гнатосомы смещены вперед
по отношению к ее заднему краю. Пальпы длинные (рис. 110, 1) . . .
................................................ I. (Ph.) crenulatus Koch
22 (21). Вершины заднебоковых углов основания гнатосомы лежат на уровне
ее заднего края. Пальпы короткие (рис. 110, 2)...........................
. ...................................I. (Ph.) arboricola Sch. et Schl.
2. Haemaphysalis Koch. Мелкие клещи (до 4 мм в голодном состоянии).
Анальная борозда огибает анус сзади. Анальных щетинок обычно 5/5. Глаз нет.
6
Рис. 111. Самки клещей рода Haemaphysalis,
1—3 — гнатосома сверху и коксы: 1 — Н. эаропгса, 2 — Н. Цата, 3 — Н. сопсгппа\ 4—6 — гнатосома
сверху: 4 — Н. longicornis\ 5 — Н. (О.) phasiana, 6 — Н. filippovae. (По: Болотин, 1979).
Брюшная поверхность d без щитков. Задний край тела с фестонами, обычно
их 11. Боковая борозда на скутуме есть только у d. Вертлуг I с крупным зуб-
цом. Пальпы обычно широкие, выдающиеся за основание гнатосомы. Аурикулы
отсутствуют. Окраска однотонная коричневая, реже — желтоватая. — 6 видов
(в СССР 15).
Самки
1 (2). III членик пальп дорсально с крупным направленным назад дорсальным
зубцом (рис. 111, 4). — Пастбищный паразит, основной хозяин всех фаз —
одомашненный пятнистый олень. Многочислен. (Подрод Kaiseriana Santos
Dias).......................Н. (К.) longicornis Neum. (=neumanni Donitz)
2 (1). Ill членик пальп дорсально без зубца.
3 (4). Пальпы сильно расширены к основанию: II членик образует резкий
и длинный боковой выступ (рис. 111, 5). — Пастбищный паразит, хозяева
всех фаз — птицы. (Подрод Ornithophysalis Hoog. et Wassef)..............
..............................H. (О.) phasiana Saito, Hoog. et Wassef
14*
211
4 (3). Пальпы умеренно расширены к основанию: II членик образует небольшой
боковой выступ. (Подрод Haemaphysalts s. str.).
5 (6). Основание гнатосомы без корнуа (рис. 111, 6)........................
..............................................Н. (Н.) filippovae Bol.
6 (5). Основание гнатосомы с корнуа.
7 (8).^Зубец на коксе IV больше зубца на коксе I (рис. 111, 2). Окраска тела
голодных клещей светло-желтая. — Пастбищный паразит, имаго на круп-
ных и средних млекопитающих, нимфы и личинки на млекопитающих
и птицах. Редок........................................Н. (Н.) flava Neum.
Рис. 112. Самцы и личинки клещей рода Haemaphysalts»
I — коксы самца Н. flava’, 2—5 — гнатосома (сверху) самца: 2 — Н. concinna, 3 — Н. эаропгса, 4 — Н.
phasiana (по: Saito et al., 1971), 5 — Н. longicornis’, 6 — гнатосома (сверху) личинки Н. flava (по: Yama-
guti et al., 1971); 7, 8 — гнатосома сверху и коксы личинки: 7 — Н. эаропгса, 8 — Н. longicornis.
8 (7). Зубец на коксе IV меньше зубца на коксе I или равен ему. Окраска тела
голодных клещей коричневая.
9 (10). Края полового отверстия с двумя хитиновыми валиками. Зубец на коксе I
заметно крупнее зубца на коксе IV. Дорсальнобоковые края II и III чле-
ников пальп почти равны (рис. 111, 3). — Пастбищный паразит; имаго
на крупных и средних млекопитающих, нимфы и личинки на мелких
и средних млекопитающих и птицах. Многочислен........................
...............................................Н. (Н.) concinna Koch
10 (9). Края полового отверстия без хитиновых валиков. Зубец па коксе I
примерно равен зубцу на коксе IV или чуть больше него. Дорсальнобо-
ковой край II членика пальп заметно короче бокового края III членика
(рис. 111, 7). — Пастбищный паразит; имаго на крупных и средних млеко-
питающих, нимфы и личинки на птицах и средних млекопитающих. Обы-
чен ...........................................Н. (Н.) japonica Warb.
212
Самцы5
1 (2). Ill членик пальп дорсально с крупным направленным назад дорсальным
зубцом (рис. 112, 5)............................Н. (К.) longicornis Neum.
2 (1). Ill членик пальп без дорсального зубца.
3 (4). Пальпы сильно расширены к основанию; II членик образует резкийьи
длинный боковой выступ (рис. 112, 4)....................................
............................Н. (О.) phasiana Saito, Hoog. et Wassef
Рис. 113. Нимфы и личинки клещей рода Haemaphy salts,
1 ,2 — гнатосома сверху и коксы нимфы: 1 — Н. concinna, 2 — Н. phasiana (по: Saito et al., 1974); 3—5 —
гнатосома (сверху) нимфы: 3 — Н. longicornis, 4 — Н. flava (по: Yamaguti et al., 1971), 5 — Н. japonica',
6,7 — гнатосома (сверху) личинки: 6 — Н. concinna, 7 — Н. phasiana (по: Saito et al., 1974).
4 (3). Пальпы умеренно расширены к основанию; II членик образует небольшой
боковой выступ или таковой отсутствует.
5 (6). Зубец на коксе IV длинный, не менее половины ее ширины (рис. 112, 7).
Окраска тела голодных особей светло-желтая . . . Н. (Н.) flava Neum.
6 (5). Зубец на коксе IV короткий, значительно меньше половины ее ширины.
Окраска тела голодных особей коричневая.
7 (8). Заостренные вершины III члеников пальп загнуты медиально и при сло-
женных пальпах заходят одна под другую (рис. 112, 2).......................
...............................................Н. (Н.) concinna Koch
8 (7). Вершины III члеников пальп не загнуты (прямые) и при сложенных
пальпах не заходят одна под другую (рис. 112, 3)...........................
...............................................Н. (Н.) japonica Warb.
5 Самец Н. filippovae неизвестен.
213
Нимфы6
1 (4). Пальпы сильно расширены к основанию: II членик шире III членика
более чем в 1.5 раза (рис. ИЗ, 7, 2).
2 (3). II членик пальп шире III членика более чем в 2 раза. Зубец на коксе IV
почти не выражен. Гипостом с каждой стороны несет по 2 (2/2) полных
ряда зубчиков (рис. ИЗ, 1).......................Н. (Н.) concinna Koch
3 (2). II членик пальп шире III менее чем в 2 раза. Зубец на коксе IV хорошо
выражен, острый. Гипостом с каждой стороны несет по 3 (3/3) полных
ряда зубчиков (рис. ИЗ, 7)..............................................
............................Н. (О.) phasiana Saito, Hoog. et Wassef
4 (1). Пальпы умеренно расширены к основанию: II членик шире III членика
менее чем в 1.5 раза.
5 (6). Гипостом с 3/3 рядами зубчиков. Основание III членика пальп дорсально
в виде выгнутой назад линии (рис. ИЗ, 3) . . Н. (К.) longicornis Neum.
6 (5). Гипостом с 2/2 рядами зубчиков. Основание III членика пальп дорсально
в виде почти прямой линии.
7 (8). Вершина III членика пальп заострена (рис. ИЗ, 5).................
.............................................Н. (Н.) japonica Warb.
8 (7). Вершина III членика пальп закруглена (рис. ИЗ, 4)................
.............................................Н. (Н.) flava Neum.
Личинки
1 (4). Корнуа имеются.
2 (3). Вершина III членика пальп закруглена (рис. 112, 6)...............
.............................................Н. (Н.) flava Neum.
3 (2). Вершина III членика пальп заострена (рис. 113, 7)................
.................................Н. (О.) phasiana Saito, Hoog. et Wassef
4 (1). Корнуа отсутствуют.
5 (6). Пальпы резко расширяются к основанию: II членик образует отчетливый
боковой выступ (рис. ИЗ, 6) . . . . ..............Н. (Н.) concinna Koch
6 (5). Пальпы постепенно расширяются к*основанию: II членик не образует
резкого бокового выступа (рис. 112, 7, 8).
7 (8). II членик пальп значительно шире III членика, вершина III членика
заострена. Кокса I со слабо выраженным зубцом (рис. 112, 7) . . . .
.................................................Н. (Н.) japonica Warb.
8 (7). II членик пальп лишь немного шире III членика, вершина III членика
закруглена. Кокса I с отчетливым зубцом (рис. 112, 8)...................
........................................ Н. (К.) longicornis Neum.
3. Dermacentor Koch. Сравнительно крупные клещи (до 8 мм в голодном
состоянии). Анальная борозда огибает анус сзади. Анальных щетинок 5 пар.
Глаза краевые, плоские или слегка выпуклые. Вентральная поверхность d без
щитков. Задний край тела с 11 фестонами. Скутум имаго всегда с белым эмале-
вым пигментом разной интенсивности, пигмент часто покрывает также ноги и
гнатосому. Кокса IV у <5 сильно увеличена. Все коксы с латеральными зубцами.
Пальпы короткие. Аурикулы у <5 отсутствуют, но имеются обычно у нимф и
личинок и редко — у £. — 1 вид (в (ССР 10).
Самки
1 (1). Голени и предлапки II — IV без крупных зубцов. II членик пальп без дор-
сального зубца. Перитрема с выраженным спинным отростком, ее хитино-
вая каемка без спинного утолщения. Корнуа тупые. Зубец па коксе IV
длинный и острый, выступает за задний край коксы (рис. 114, 7). —
Пастбищный паразит; имаго — на крупных млекопитающих, нимфы и ли-
6 Преимагинальные фазы Я. filippovae неизвестны.
214
чинки — на мелких млекопитающих. Многочислен. (Подрод Dermacen-
tor s. str.)...............................................D. (D.) sil-
varum 01. ( = asiaticus Em. et Kozl.; =coreus Itagaki, Noda et Yam.)
Самцы
1 (1). Голени и предлапки с небольшими зубчиками. II членик пальп без дор-
сального зубца. Хитиновая каемка перитремы без спинного утолщения,
длинный и широкий спинной отросток перитремы доходит до края ску-
тума. Вертлуг I с острым дорсальным зубцом (рис. 114, 2)......
.........................................D. (D.) silvarum 01.
Рис. 114. Самка, самец, нимфа и личинка Dermacentor silvarum.
1 — самка снизу и ес гнатосома (сверху); 2 — самец снизу и его гнатосома (сверху); 3 — гнатосома сверху
и скутум нимфы; 4 — гнатосома сверху и скутум личинки.
Нимфы
1 (1). Основание гнатосомы с острыми, сильно оттянутыми боковыми высту-
пами (рис. 114, 3). Латеральный зубец на коксе I немного длиннее медиаль-
ного зубца. Зубец на коксе IV небольшой ......D. (D.) silvarum 01.
Личинки
1 (1). Основание гнатосомы с острыми, сильно оттянутыми боковыми высту-
пами (рис. 114, 4). Коксы с выраженными зубцами. Зубец на коксе III
острый, значительно выдается за край коксы . . . D. (D.) silvarum 01.
ОСНОВНАЯ ЛИТЕРАТУРА
Балашов Ю. С. Кровососущие клещи (Ixodoidea) — переносчики болезней человека
и животных. Л., 1967. 317 с.
Беликова Н. П. Иксодовые клещи. — В кн.: Природноочаговые болезни в Приморском
крае. Владивосток, 1975, с. 162—180.
215
Болотин Е. И. Новый вид Haemaphysalis (Parasitiformes, Ixodidae) с юга Дальнего Во-
стока. — Зоол. журн., 1979, т. 58, № 2, с. 267—269.
Гордеева В. П., Белослюдов Г. А. О первых находках аргасовых клещей из группы Argas
reflexus F. в Приморском крае. — Докл. Иркут, противочумного ин-та, 1963, т. 5, с. 194.
Померанцев Б. И. Иксодовые клещи (Ixodidae). М.; Л. 1950. 223 с. (Фауна СССР. Пауко-
образные; Т. 4. Вып. 2).
Сердюкова Г. В. Иксодовые клещи фауны СССР. М.; Л., 1956. 121 с.
Филиппова Н. А. Аргасовые клещи. М.; Л., 1966. 255 с. (Фауна СССР. Паукообразные;
Т. 4. Вып. 3).
Филиппова Н. А. Иксодовые клещи подсем. Ixodidae. Л., 1977. 396 с. (Фауна СССР.
Паукообразные; Т. 4. Вып. 4).
Arthur D. В. British ticks. London, 1963. 348 р.
Saito Y., Hoogstraal H., Wassef H. Y. The Haemaphysalis ticks (Ixodoidea, Ixodidae)
of birds. 4. H. (Ornithophysalis) phasiana sp. n. from Japan. — J. Parasitol., 1974, vol. 60,
N 1, p. 198—208.
Yamaguti N., Tipton V. J., Keegan H., Toshioka S. Ticks of Japan, Korea and the Ryukyu
Islands. — Brigham Young Univ. Sci. Bull. Biol, ser., 1971, vol. 15, N 1, p. 1—226.
ГАМАЗОВЫЕ КЛЕЩИ (Когорта GA MA SI NA)
Большинство гамазовых клещей — наружные паразиты многих теплокров-
ных животных: домашних и диких птиц, млекопитающих; способны нападать
на человека. По типу питания делятся на гематофагов, схизофагов и энтомо-
фагов.
В СССР когорта включает более 20 семейств, около 100 родов и более 500 ви-
дов, число которых пополняется. Однако медико-ветеринарное значение имеют
6 семейств и около 100 видов; на Дальнем Востоке — 6 семейств, 9 родов, более
30 видов.
ВНЕШНЕЕ СТРОЕНИЕ
Взрослые клещи. Тело чаще всего овальной или овально-округЛой
формы, разделено на идиосому и гнатосому (рис. 115). Покровы от желтого до
красновато-коричневого цвета. Они имеют определенную морщинистость,
исчерченность, а на щитках чешуйчатую структуру или сетчатый рисунок.
Гнатосома причленена к переднему концу тела с брюшной стороны. Основ-
ные ее части: базальное кольцо, несколько передних выступов и 2 пары конеч-
ностей. Та часть гнатосомы, которая непосредственно соединяется с телом,
называется гнатобазой. Она имеет вид трубки или кольца, на спинной стороне
средняя часть выдается вперед и образует тектум. К переднебоковым углам
гнатобазы (к неподвижным гнатококсам) подвижно причленены 5-члениковые
педипальпы, покрытые многочисленными щетинками.
С брюшной стороны на гнатобазе расположена пара гнатосомальных щети-
нок, кпереди от которых находится гипостомальная область (гипостом) с 3 парами
гипостомальных щетинок. От передних боковых углов со спинной стороны гипо-
стома выдаются вперед подвижно причлененные массивные склеротизованные
корникулы. Впереди гипостом раздваивается с образованием двух узких лопа-
стей. С брюшной стороны он разделен на правую и левую половины желобком
с поперечными рядами зубчиков, направленных вперед. В желобок вклады-
ваются ветви тритостернума.
Хелицеры 3-члениковые, оканчивающиеся клешней с пальцами: неподвиж-
ным (сверху) и подвижным (снизу). По внутреннему краю пальцев часто рас-
полагаются зубцы. На неподвижном пальце, ближе к вершине, находится
прозрачный придаток, форма и размеры которого имеют диагностический
признак. У основания неподвижного пальца сидит одна щетинка, а у основа-
ния подвижного пальца — пучок щетинок.
На перепончатой ткани, окружающей гнатосому позади ее основания, с вен-
тральной стороны прикрепляется тритостернум (в виде цельного щитка с цен-
тральным придатком-стволиком с ветвями-лациниями или в виде отдельных
склеритов; иногда имеется лишь стволик с ветвями), ветви которого в покое
вкладываются в желобок гипостома.
Спинная сторона тела с 1 или 2 щитами различной формы и размеров, покры-
вающих всю спину или оставляя неприкрытой значительную ее часть. В числе
216
и положении щитов брюшной стороны значительно сказываются возрастные и
половые отличия и это один из признаков различия пола и фаз развития. Самой
характерной особенностью является расположение щитов на брюшной стороне,
наличие одного пли нескольких крупных щитов: стернального (между II и
Рис. 115. Строение гамазовых клещей.
1 — схема строения самки с брюшной сто-
роны: а — хелицера, б — щупальце, педи-
пальпа, в — гнатосома, г — тритостернум, д —
перитерма, с — предгрудной щиток, ж —
грудной щиток, з — межкоксальный щиток,
и — промежуточный щиток, к — перитремаль-
ный щиток, л — дыхальце, м — прикокса ts-
ный щиток, н — генитальный щит, о — бо-
ковой брюшной щиток, п — брюшной, или
вентральный, щит, р — анальный щит, с —
тазик, тп — вертлуг, у — бедро, ф — колено,
х — голень, ц — лапка, ч — предлапка; 2 —
спинная сторона самки Laelaps hilaris: ще-
тинки: Fi — F3 — лобные, V — теменные, ETit
ЕТ2, 7\, Т2 — наружные и внутренние ви-
сочные, Sc — лопаточные, Dt—D3 — дорсаль-
ные, Si — Se — предкраевые, Mi — Ми — крае-
вые, Ii—I3— вставочные; на брюшной стороне:
Sti—St3 — грудные, MSt — промежуточные,
Vli—Vl4 — брюшные, Ad — аданальные, Pa —
постанальная, Пл — плевральные; з—брюш-
ная сторона самки Androlaelaps glasgowi', 4 —
хелицеры самца A. glasgowi.
Ill ногами), генитального, вентрального и анального. Кроме того, имеются
мелкие парные щитки: предгрудные, промежуточные, боковые брюшные, меж-
коксальные, прикоксальные, перитремальные. Иногда наблюдается слияние ге-
нитального с вентральным в генито-вентральный щит или могут быть другие
варианты (рис. 115). У самцов брюшные щиты либо слиты в один общий щит,
либо щит на середине тела разделен на два; иногда от брюшного щита отделен
анальный щит.
Половое отверстие самца обычно открывается у переднего края брюшного
щита позади основания тритостернума и имеет округлую форму.
217
Половое отверстие самки располагается позади грудного щита и прикрыто
генитальным щитом. Такие различия в положении и строении половых отвер-
стий — второй основной признак для различения полов у взрослых клещей.
По бокам тела в области III и IV кокс имеются стигмы, от которых отходят
узкие трубчатые перитремы.
Тело и конечности клещей покрыты многочисленными щетинками различной
формы и размеров. Характерной чертой имаго является то, что постанальная
щетинка никогда не достигает длины тела, иногда очень короткая (рис. 115).
Постэмбриональное развитие гамазовых клещей помимо имаго включает
фазы личинки, прото- и дейтонимфы.
Дейтонимфа. Отличается прежде всего тем, что между грудным и
анальным щитами нет крупных щитов; имеются несколько мелких, едва замет-
ных, округлых или почти линейных склеритов; иногда щиты вообще отсут-
ствуют. Отсутствует перитремальный щит. На спинной стороне щитов 2, реже 1.
Перитремы длинные. Грудной щит с 4, реже 5 парами щетинок (у имаго на
грудном щите, как правило, 3, реже 2 или 4 пары щетинок). По внешнему
строению дейтонимфы можно определить пол клеща: дейтонимфа самок круп-
нее дейтонимфы самцов.
Протонимфа. Покровы тела мягкие, беловатые (у ряда паразитиче-
ских клещей) или склеротизованные, желтоватые (у хищных клещей и у ряда
факультативных паразитов). На спинной стороне 2 более крупных щита (голово-
грудной и пигидиальный) и между ними несколько мелких склеритов. Пери-
тремы короткие. Грудной щит (если он имеется) с 3 парами щетинок.
Личинка. Дыхальца и перитремы отсутствуют. Постанальная щетинка
у некоторых видов достигает длины тела или превышает ее; иногда на заднем
конце тела располагается несколько пар длинных щетинок. Щиты имеются
(у хищных клещей) или отсутствуют (у паразитических клещей).
Гамазовые клещи яйцекладущие и живородящие.
Яйцо имеет овальную или округлую форму, очень крупное по отношению
к телу клеща, окраска молочно-белая, грязновато-белая, желтоватая.
БИОЛОГИЯ
По характеру паразито-хозяинных отношений среди гамазовых клещей
можно выделить несколько типов, отражающих способ питания, образ жизни
и тип развития. Клещи семейств Dermanyssidae и Macronyssidae являются
гнездово-норовыми облигатными кровососами; сем. Hirstionyssidae — обли-
гатные кровососы. В семействах Haemogamasidae, Laelaptidae и других встре-
чаются факультативные и облигатные кровососы, у которых наблюдается сме-
шанный тип питания, включающий гематофагию, схизофагию и энтомофагию.
Среди гамазовых клещей имеются и типичные нидиколы (род Eulaelaps).
Наиболее полно изучен жизненный цикл у куриного клеща Dermanyssus
gallinae. Обычно в теле самки одновременно развивается только одно яйцо;
она откладывает яйца с интервалом 8—10 ч в щели стен и пола птичников.
При температуре 16—24 °C эмбриональное развитие длится от 50—70 до 110—
120 ч. Отродившаяся личинка малоподвижна, питается за счет остатков желтка.
Через 20—30 ч линяет в протонимфу, которая способна прокалывать кожу
хозяина и сосать кровь. После кровососания протонимфа некоторое время
находится в состоянии покоя, а затем линяет в дейтонимфу (вторая активно
питающаяся фаза). При температуре 24 °C за 36—50 ч, а при 16 °C — за 96 ч
происходит переваривание крови и дейтонимфа превращается в самку. Если
дейтонимфа слабо напиталась, но способна быстро переваривать кровь, то она
превращается в самца.
При оптимальных условиях развития весь цикл куриного клеща от яйца
до имаго продолжается 7—8 дней. Эти клещи могут сосать также кровь человека
и животных.
В сем. Macronyssidae наиболее детально изучено питание некоторых пред-
ставителей из рода Ornithonyssus, которые помимо животных способны нападать
и на человека. У О. bacoti и у О. silviarum некровососущими фазами являются
личинка и дейтонимфа, и они кратковременны. Самки способны поглощать
218
большое количество крови, значительно превышающей собственную массу.
Откладка яиц тесно связана с процессом пищеварения. У О. bacott при опти-
мальных условиях (температура 20—25 °C и влажность 75—90 %) откладка яиц
протекает в течение 3 сут, а эмбриональное развитие проходит за 166 ч, у О. sil-
viarum — за 48 ч. Личинка линяет в протонимфу через 12—13 ч при темпера-
туре 30—35 °C; более низкие температуры (15—20 °C) замедляют этот процесс
до 40 ч. Протонимфа сосет кровь один, чаще — два раза и через 50 ч после
второго кровососания линяет в дейтонимфу. Последняя — непитающаяся фаза,
продолжительность которой от нескольких часов до суток. Дейтонимфы обоего
пола держатся вместе, так как сразу после превращения в имаго происходит
спаривание.
Весь цикл развития О. silviarum занимает максимально 5—7 дней.
Самки Haemogamasus nidi (сем. Haemogamasidae) при питании кровью от-
кладывают в среднем за месяц 1—4 яйца; при питании мелкими членистоногими
откладка яиц прекращается через 5 дней. Питание смешанной пищей повышает
яйцепродукцшо самок до 5—7 яиц в месяц. Размножение происходит кругло-
годично. Высокая численность наблюдается в июне и в ноябре.
Биология пчелиного клеща Varroa jacobsoni (сем. Varroidea) очень сложна и
тесно согласуется со сложной структурой и жизнью пчелиной семьи. Самка
V. jacobsoni откладывает от 2 до 5 яиц, прикрепляя их к стенке ячейки перед
запечатыванием ячейки сота. Развитие эмбриона в яйце продолжается несколько
дней. Самки оплодотворяются в ячейке перед выходом пчелы. Самки легко
переносят понижение давления и минусовую температуру на протяжении
одного-двух дней и сохраняют активность при голодании 2—3 дня. В летний
период времени самки живут 2—3, а осенняя генерация — 6—8 мес. При тем-
пературе 28 °C и влажности 85 % самки способны голодать до 9 сут, при тем-
пературе 35 °C и 50 % влажности — 3 дня, при температуре 35 °C и 10—20 %
влажности — сутки. Питаются V. jacobsoni гемолимфой пчел, поражая рабочие
и трутневые особи, причем чаще всего трутневый расплод. Наибольшая пора-
женность расплода в условиях Приморского края отмечена с марта по май и
в сентябре, минимальная — в июне—июле. С появлением в пчелиной семье
трутневого расплода пораженность увеличивается и достигает максимума
в августе (при этом снижается пораженность рабочих пчел), а в сентябре—
октябре происходит увеличение числа пораженных рабочих пчел.
Представители сем. Laelaptidae — обитатели гнезд и нор, типичные гнез-
дово-норовые факультативные кровососы. Все развитие этих клещей, наиболее
характерными представителями которых являются Eulaelaps tabularis и Andro-
laelaps glasgowi, происходит в гнезде хозяина, где встречаются все фазы разви-
тия. Экологически очень пластичные. Питаются мелкими членистоногими и
кровью теплокровных животных. Большинство представителей семейства живо-
родящие, но некоторые могут откладывать также и яйца (например, A. casalis).
В сем. Laelaptidae представители рода Laelaps осуществили переход к обитанию
на хозяине. Они характеризуются большим морфологическим сходством, еди-
ным типом развития: эмбрионизацией ранних фаз развития и живорождением
личинки. L. muris, питаясь кровью и лимфой полевок, способен прокалывать
кожу человека и приступать к кровососанию.
Клещи сем. Hirstionyssidae аналогично роду Laelaps обладают большим мор-
фологическим сходством и единым типом развития, для которого характерна
эмбрионизация ранних фаз развития (личинки и протонимфы), а первой крово-
сосущей фазой является дейтонимфа. Активные фазы клещей рода Hirstionyssus
пьют кровь неоднократно и небольшими порциями, в связи с чем у них отсут-
ствует гармония процессов пищеварения и линек и гонотрофическая гармония
у самок. В период развития зародыша в половых путях самка многократно
питается. Всем видам присуще яйцеживорождение, иногда живорождение.
ВРЕД
Паразитические виды гамазовых клещей являются переносчиками возбу-
дителей инфекционных болезней человека и животных.
Большой вред птицеводству приносит куриный клещ Dermanyssus gallinae,
слюна которого токсична и вызывает местное воспаление и зуд. Массовое пара-
219
зитирование клещей сильно ослабляет кур, снижает их яйценоскость и даже?
может вызвать падеж цыплят. Кроме того, клещ способен питаться кровью
людей, вызывая неприятные ощущения.
Крысиный клещ — Omlthonyssus bacoti — заселяет гнезда синантропных
птиц (голубей), мигрирует в помещения, нападает на людей, вызывая зудящую
сыпь и дерматиты, при этом особенно страдают дети.
Не менее опасен и клещ О. sylviarum, наносящий вред птицеводству; напа-
дает на человека, вызывая дерматиты.
Заметный урон нашему пчеловодству приносит пчелиный клещ V. jacobsoni
(питается гемолимфой пчел), паразитирование которого не только понижает
активность пчел и приводит к ослаблению медосбора, но практически может
сводиться к уничтожению не одной пчелиной семьи.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ
1 (2). Хелицеры со слабо развитой клешней: длинные, тонкие, игловидные или
стилетовидные........................................1. Dermanyssidae
2 (1). Хелицеры с хорошо развитой клешней.
3 (6). Спинной щит и все тело густо покрыты щетинками различного строения.
4 (5). Щетинки волосовидные, большей частью гладкие, реже перистые; мо-
гут быть шиповидноутолщенные, преимущественно на конечностях. Хе-
лицеры j с клешней, оба пальца с зубцами; у 8 хелицеры несут 2 прозрач-
ных придатка...................................3. Haemogamasidae
5 (4). Щетинки, снабженные мелкими бороздками: винтообразные, игольчатые,
уплощенно-кинжаловидные или уплощенные. Хелицеры $ имеют под-
вижный кинжаловидный палец с острым изогнутым кончиком и 2 зубцами
по верхнему краю; у 3 подвижный палец хелицер превращен в желобча-
тый придаток с ложковидным концом........................4. Varroidae
6 (3). Спинной щит и все тело не густо покрыты щетинками, число и форма
которых варьируют.
7 (8). На спинном щите 37—39 пар щетинок; могут быть непарные, несимме-
тричные щетинки в центральной части задней половины щита ....
................................*................. 5. Laelaptidae
8 (7). На спинном щите до 30 пар щетинок; непарные щетинки отсутствуют.
9 (10). На вентральной поверхности кокс имеются склеротированные шипы
.................................................6. Hirstionyssidae
10 (9). На вентральной поверхности кокс шипы отсутствуют..............
.................................................2. Macronyssidae
1. Сем. DERMANYSSIDAE
Покровы тела кожистые, с характерной морщинистостью, способны к силь-
ному растяжению. На дорсальной поверхности взрослых клещей и дейтонпмф
либо 1 щит с 10—20 парами щетинок, либо у самки позади щита есть еще не-
большой округлый щиток с 1 парой щетинок или неправильно очерченные скле-
риты, лишенные щетинок. Стернальный щит самки небольшой, короткий,
с 1—2 или 3 парами щетинок и обычно с 2 парами щелевидных органов. Генито-
вентральный щит значительно вытянут назад за уровень IV кокс, с закруглен-
ным или суженным задним концом с 1 парой щетинок. Анальный щит с 3 около-
анальными щетинками. Метаподальные щитки маленькие, иногда незаметные.
Перитремы от коротких до длинных. У самца на брюшной стороне 1 (голо-
вентральный) пли 2 (генито-стернальный и вентро-анальный или анальный)
щита. Тритостернум у обоих полов с 2 опущенными или гладкими ветвями.
Корникулы лопастевидные, прозрачные, никогда не имеют форму рога. Тектум
удлиненный, языковидный или заметно сужается к вершине. Хелицеры у самок
и нимф в виде длинных тонких иголок или гибких стилетов с крошечной клеш-
ней; у самцов хелицеры много короче; сперматодактиль целиком слит с подвиж-
ным пальцем, неподвижный палец узкий, может быть очень коротким, реже
равен подвижному. Педипальпы 5-члениковые; вильчатая щетинка 2-раздель-
220
пая. Все ноги с предлапками. Переднедорсальный край II кокс обычно без
шипа, реже с шипом. На II ногах у самца нет выростов или массивных шипов;
на III или IV лапках одна из щетинок модифицирована в короткий зубовидный
вырост. Яйцекладущие. Личинка не питается. Обитатели гнезд птиц, на которых
они паразитируют; реже в гнездах грызунов. Способны нападать на человека.
Переносчики различных природноочаговых болезней. — Семейство включает
2 рода. На Дальнем Востоке — 1.
1. Dermanyssus Duges. У $ и в по одному спинному щиту. Генито-вентраль-
ный щит $ с закругленным задним краем; анальный расширяется впереди
переднего края. У d подвижный палец хелицер длинный, неподвижный —
в виде короткого придатка у его основания. Паразиты птиц; некоторые виды
паразитируют на грызунах. — 1 вид 1 (в СССР 3).
1 (1). На брюшной стороне тела у $ все щетинки тонкие. Анальный щит не-
сколько вытянут в длину, закругленно-треугольный или почти овальный.
Брюшной щит 6 разделен на два поперечной линией, проходящей позади
IV кокс. Перитрема простирается вперед до середины или переднего
края II кокс. Спинной щит $ сзади как бы обрублен, с 8 парами спинных
щетинок. $ — 0.75—0.84, в — 0.6—0.63. — Паразит домашних и диких
птиц. Нападает на человека. Переносит различные природноочаговые за-
болевания. — Космополит.............................D. gallinae Redi
2. Сем. MACRONYSSIDAE
Покровы тела способны к сильному растяжению и увеличению объема.
На дорсальной поверхности самки 1 или 2 щита, прикрывающих почти все
тело сверху или оставляющих не покрытой значительную часть спины. У сам-
цов дорсальные покровы редко сформированы как у самок, обычно имеется
1 дорсальный щит; на вентральной поверхности 1 или 2 щита. Обычно на щите
самок 29 пар большей частью гладких щетинок; у самцов — 22 пары щетинок.
Стернальный щит самок короткий с 3 парами щетинок (длина его меньше ши-
рины) или почти квадратный с 2 парами щелевидных органов. Метастерналь-
ные щитки отсутствуют и щетинкй Mst расположены на мягкой кутикуле.
Генито-вентральный щит различной формы: языковидный с закругленным кон-
цом, клиновидный, резко суженный к концу или с перетяжкой в средней части
и расширенный сзади с 1 парой генитальных щетинок, расположенных у его
середины; иногда щит налегает на стернальный. Анальный щит при-
близительно треугольной формы, длина его обычно больше ширины с 3 около-
анальными щетинками. Перитремы короткие или длинные, иногда переходят
на дорсальную сторону. Тритостернум хорошо развит, лацинии длинные.
Тектум пленчатый, удлиненный, с гладкой пли слабо рассеченной вершиной.
Корникулы слабо развиты. Хелицеры протонимфы и взрослых клещей с хорошо
развитой клешней, пальцы которой лишены зубцов, но иногда с гиалиновыми
крючками или лопастями; сперматодактиль не превышает длину подвижного
пальца более чем в 1.5—2 раза, часто равен длине клешни, иногда с прозрач-
ными расширениями у его основания. На вертлуге педипальп у самок часто,
у самцов и протонимф реже имеется вентральный вырост; вильчатая щетинка
на лапке двураздельная (редко цельная). Все ноги с предлапками, с хорошо
развитыми коготками и присосками. Яйцекладущие. Личинка и дейтонимфа —
непитающиеся фазы. Паразитируют на мелких млекопитающих, птицах и реп-
тилиях. Семейство включает 7 родов. На Дальнем Востоке — I.1 2
1. Ornithonyssus Sambon. На спинной стороне 1 щит с длинными игольча-
тыми щетинками. Грудной щит 2 обычно с 3 парами, реже с 2 парами щетинок.
Генито-вентральный щит £' узкий, не заходит далеко за IV коксы. Строение
хелицер у $ и d одинаково: ножницевидные, с хорошо развитым подвижным и
1 В таблицу не включены D. grochovskae Zems. п D. quintus Vitzthum, медико-ветеринар-
ное значение которых слабо изучено.
2 Включен только род Ornithonyssus, имеющий медико-ветеринарное значение.
221
неподвижным пальцами без сильно склеротизованных зубцов по внутренней
стороне. Паразиты мелких млекопитающих и птиц. — 2 вида (в СССР 3).
1 (2). Грудной щит $ с 3 парами щетинок. На непокрытых щитами частях тела
многочисленные щетинки (приблизительно 100 пар). У 3 единый брюшной
щит, узкий, с 9 парами игольчатых щетинок. Клешня хелицеры 3 с более
короткими пальцами, чем у $; от основания подвижного пальца отходит
сперматодактиль. $ — 0.61—0.76, d — 0.4—0.52. — На крысах (пасюк,
черная, туркестанская), мышах; может встречаться на других грызунах.
Нападает на человека. Космополит....................О. bacoti Hirst.
2 (1). Грудной щит $ с 2 или 3 парами щетинок. На непокрытых щитами частях
тела щетинки малочисленны (с брюшной стороны 10—12 пар и со спинной
25—30 пар). У 3 единый брюшной щит, на котором отчетливо видно резкое
сужение перед анальной частью, вооружен только 6 парами щетинок.
$ — 0.62, в — 0.45. — Паразит кур, голубей и многих видов диких птиц.
Нападает на человека. Возможны случайные находки на грызунах.
Голаркт.............................О. silviarum Canestrini et Fanzago
3. Сем. HAEMOGAMASIDAE
Тело овальное, густо покрыто щетинками (неотрихия). Дорсальный щит
у обоих полов и дейтонимфы цельный, обычно с широко закругленным задним
краем; иногда щит сзади сужен или укорочен и тогда оставляет тело не при-
крытым с боков и сзади. На переднем конце спинного щита выделяется по
величине пара щетинок, которые у большинства видов более или менее пери-
стые. Стернальный щит самки крупный, с почти прямым или со слабо вогну-
тым, реже с глубоко выемчатым задним краем (щит в виде арки); на щите либо
обычные 3 пары щетинок (St±—St3), либо, кроме того, еще от 1 до 40—50 доба-
вочных щетинок; 2 пары щелевидных органов расположены всегда на щите,
третья пара иногда вне пределов щита, за его задним краем. Генито-вентраль-
ный щит сужен с боков и расширен сзади, иногда почти колбовидной формы,
всегда с многочисленными щетинками. Анальный щит обратногрушевидной
формы, с 2—12 добавочными щетинками, помимо обычных 3 околоанальных. Ме-
тастернальные щитки нередко сливаются с межкоксальными. У самца брюшная
поверхность покрыта одним общим головентральным щитом (род Haemogamasus)
или анальный щит отделен (род Ischyropodd). Постоянная (ортотрихическая)
система щетинок нарушена: среди густого «шерстевидного покрова», образован-
ного тонкими мелкими (гладкими или опушенными) щетинками, на спинном
щите выделяются 1 или 2—3 пары более крупных (обычно F± и Л/и), на щитах
брюшной стороны — парные St±—St3, Mst, Vl± и 3 околоанальные. Тектум
треугольный или с закругленным передним краем, который глубоко надрезан;
его отдельные лопасти в свою очередь имеют ответвления. Корникулы от хорошо
склеротизованных треугольных с острой вершиной до перепончатых с закруглен-
ной вершиной. Хелицеры самки с клешней, оба пальца с зубцами, реже без
зубцов. У самца клешни хелицер и сперматодактиль часто имеют сложное
строение; их форма служит хорошим диагностическим признаком. Педипальпы
с двураздельной вильчатой щетинкой. Ноги довольно длинные (I и IV длиннее
тела, II и III равны длине тела или слегка короче) и тонкие; у 3 ноги толще,
на II ногах нередко сильные шипы. Предлапки удлиненные, со слабыми когот-
ками. Яйцекладущие. Личинка и протонимфа не питаются. Обитатели нор и
гнезд грызунов, насекомоядных. Среди семейства встречаются хищники, факуль-
тативные и облигатные кровососы. Семейство включает 3 рода; в СССР и на
Дальнем Востоке — 1.
1 . Haemogamasus Berl. Грудной щит j либо с 3 парами щетинок (St1 — St3),
либо, помимо этих щетинок, имеет добавочные, часто в большом количестве.
У 3 все щиты на брюшной поверхности слиты в единый. II ноги 3 либо, помимо
обычных щетинок, с несколькими шиповидно утолщенными щетинками (на
бедре, колене, голени и лапке), либо только с обычными тонкими щетинками
(гладкими и перистыми); у $ II ноги без шиповидно утолщенных щетинок.
222
Хелицеры несут 2 прозрачных придатка: I расположен на неподвижном пальце
хелицер, II находится у основания клешни. Паразиты млекопитающих, встре-
чаются в жилых помещениях. — 8 видов (в СССР 16).
Рис. 116. Хелицеры клещей рода Haemogamasus.
1—10 — самки: 1 — Н. pontiger, 2 — Н. kusumotoi, 3 — Н. nidi, 4 — Н. nidiformes, 5 — Н. dauricus, 6 —
Н. serdjuhovae, 7 — Н. timofejeri, 8 — Н. ambulans, 9 — Н. hitanoi, 10 — Н. mandschuricus; 11—20 — самцы:
11 — Н. pontiger, 12 — Н. ambulans, 13 — Н. hitanoi, 14 — Н. timofejevi, 15 — Н. mandschuricus, 16 —
Н. serdjuhovae, 17 — Н. nidi, 18 — Н. nidiformes, 19 — Н. husumotoi, 20 — Н. dauricus.
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
Самки
1 (8). На грудном щите имеются только 3 пары щетинок.
2 (5). Прозрачный придаток неподвижного пальца хелицер (I) лентовидной
или листовидной формы.
3 (4). Прозрачный придаток (I) имеет листовидную форму, часто с загнутой
тонкой вершиной; прозрачный придаток II узкий, прямой, вершина его
значительно не доходит до основания придатка I (рис. 116, 5). 0.89—
1.03. — В гнездах грызунов и насекомоядных и на самих зверьках . . .
................................................ Н. nidi Michael
223
4 (3). Прозрачный придаток I чаще лентовидной формы; прозрачный прида-
ток II широкий, с заостренной, слегка изогнутой вершиной, которой он
почти достигает основания придатка I (рис. 116, 4), 0.89—1.03. — В гнез-
дах различных грызунов и на самих зверьках . . . . Н. nidiformes Breg.
5 (2). Строение прозрачного придатка I иное.
6 (7). Подвижный палец в основании шире неподвижного, постепенное сужение
к вершине начинается сразу от основания. 2 зубца на нем и клювовидно
загнутая вершина более массивны, чем зубчики неподвижного пальца
(рис. 116, 6). 0.9—1.1. — На мелких млекопитающих.......................
..............................................Н. serdjukovae Breg.
7 (6). Подвижный палец имеет вид «ложки». 1.0—1.4. — На мелких млекопи-
тающих. Голаркт..................................Н. liponyssoides Ewing
8 (1). Грудной щит, помимо обычных 3 пар щетинок, несет добавочные: от одной
до 50.
9 (12). На грудном щите добавочных щетинок от одной до шести, но в отдельных
случаях они могут отсутствовать.
10 (11). На заднем конце спинного щита несколько выделяются по величине
две слабоперистые щетинки. Прозрачный придаток I листовидной формы
(рис. 116, 5). 1.02—1.05. — В гнездах грызунов и на самих грызунах
..................................................... Н. dauricus Breg.
11 (10). На заднем конце тела несколько выделяются по величине две пары
слабоперистых щетинок. Прозрачный придаток I пиковидной формы
(рис. 116, 7). 0.93—1.14. — Обитает в подпольях жилых домов, возможны
находки на домовых мышах. Палеаркт......................................
......................................Н. timofejevi Bujk. et Gonch.
12 (9). На грудном щите добавочных щетинок более шести.
13 (14). Добавочные щетинки имеются на передней и задней частях грудного
щита, исключая передний конец. На переднем конце грудного щита доба-
вочные щетинки отсутствуют (имеются только StJ, Добавочных щетинок
от 8 до 27. Прозрачный придаток I листовидной формы (рис. 116, 10),
1.0—1.23. — На грызунах, насекомоядных и в их гнездах. Палеаркт
.............................*...............Н. mandschuricus Vitzthum
14 (13). Добавочные щетинки могут находиться на переднем конце грудного
щита. На переднем конце грудного щита, помимо щетинок St13 имеется
пара добавочных щетинок. В целом на грудном щите от 10 до 33 щетинок.
Прозрачный придаток I узкий (рис. 116, S), вытянутый. 0.9—1.1. —
На грызунах, в их гнездах. Космополит ...... Н. ambulans Thorell
Самцы
1 (2). Все щетинки тела и конечностей гладкие. Передняя часть брюшного щита
лишена добавочных щетинок до уровня заднего края IV кокс. На II ногах
имеются утолщенные щетинки: по одной на бедре, колене, голени и лапке.
0.91—1.05..........................................Н. liponyssoides Ewing
2 (1). Многие щетинки тела перистые.
3 М). На переднем крае брюшного щита, помимо St14 имеются две добавочные
щетинки, которые располагаются с двух сторон полового отверстия. Пе-
редняя часть брюшного щита до Mst редко покрыта добавочными щетин-
ками. Неподвижный палец хелицер (рис. 116, 12) без зубцов на внутрен-
ней стороне, 'с коротким и тонким прозрачным придатком у вершины.
0.70—0.77 ..........................................Н. ambulans Thorell
4 (3). На переднем конце брюшного щита на уровне Str нет добавочных щетинок.
5 (6). II нога имеет по одной или по две массивные вильчатые щетинки на лапке
и голени.
6 (5). II нога имеет двураздельную вильчатую щетинку лишь на лапке. 0.73—
0.78.......................................Н. timofejevi Bujk. et Gonch.
7 (10). Сперматодактиль с зубовидным выступом в месте его изгиба в сторону
неподвижного пальца.
224
8 (9). Добавочные щетинки густо покрывают брюшной щит начиная от поло-
вины расстояния между Str—St2, Неподвижный палец хелицер (рис. 116,
15) на середине внутренней стороны имеет небольшой зубец, второй зубец
образован крючковидно загнутой вершиной пальца; между зубцами рас-
полагается малозаметный короткий прозрачный придаток I. Спермато-
дактиль толстый, одинаковой длины с подвижным пальцем. На II ноге
щетинки большей частью игольчатые, некоторые из них перистые; иногда
перистые щетинки лапки могут быть дву- и трехраздельные. 0.77—0.9
.............................................Н. mandschuricus Vitzthum
9 (8). Добавочные щетинки на брюшном щите начинаются вблизи основа-
ния Stv На части щита между St2—St3 они редкие, а от основания St3
густо покрывают всю поверхность щита. Неподвижный палец хелицер
(рис. 116, 16) иного строения. Сперматодактиль тонкий и короткий, остав-
ляющий неприкрытым около половины большого зубца подвижного
пальца хелицер. На II ноге, кроме игольчатых и перистых щетинок,
имеются сильно утолщенные гладкие щетинки. 0.77—0.84 .................
...............................................Н. serdjukovae Breg.
10 Сперматодактиль без зубовидного выступа в месте его изгиба в сторону
неподвижного пальца.
11 42). Добавочные щетинки брюшного щита начинаются от уровня Mst, выше
иногда заходят лишь 1 —2 щетинки. Сперматодактиль близ своего изгиба
над подвижным пальцем имеет длинный узкий лентовидный прозрачный
придаток, как и неподвижный палец хелицер (рис. 116, 18). Сперматодак-
тиль значительно длиннее подвижного пальца хелицер. 0.67—0.8 . . .
................................................... Н. nidiformes Breg.
12 (11). Добавочные щетинки брюшного щита начинаются ниже уровня St^
Сперматодактиль и неподвижный палец хелицер не имеют длинного листо-
видного прозрачного придатка. Сперматодактиль почти одинаковой длины
с подвижным пальцем хелицер или короче его.
13 (14). Вершина подвижного пальца хелицер (рис. 116, 17) загнута внутрь
под прямым углом. Неподвижный палец хелицер без зубцов. Прозрачный
придаток неподвижного пальца хелицер широкий. Добавочные щетинки
брюшного щита начинаются от уровня St3. 0.7—0.85......................
................................................ Н. nidi Michael
14 (13). Вершина подвижного пальца хелицер слегка загнута по направлению
к неподвижному пальцу. Неподвижный палец хелицер (рис. 116, 20)
на внутренней стороне имеет маленький зубец. Сперматодактиль слабо
возвышается над подвижным пальцем хелицер, как бы налегая на него.
Прозрачный придаток неподвижного пальца хелицер маленький, тонкий,
заостряющийся к вершине. Добавочные щетинки брюшного щита начи-
наются на половине расстояния Str—St2. 0.76—0.85 .....................
.............................................. Н. dauricus Breg.
4. Сем. VARROIDAE
Форма тела поперечно-овальная (у самок) или округло-овальная (у самцов).
Дорсальный щит покрывает всю поверхность тела, у самок заходит на брюшную
сторону. Щетинки, покрывающие тело, имеют различное строение: игольчатые,
винтообразные, уплощенно-кинжаловидные. У самок по краям спинного щита
в один ряд расположено около 20 кинжаловидных утолщенных щетинок; у сам-
цов утолщенные щетинки на заднебоковой поверхности ряда не образуют.
На брюшной стороне самок имеются грудной щит полулунной формы, боковые
брюшные, околотазиковые, средний брюшной и анальный щиты. Все щиты
крупные, между ними располагаются только узкие полоски эластичной кути-
кулы. У самцов на брюшной стороне грудной щит слит с брюшным и неправильно
изрезан в задней части. Щетинки многочисленны только в преданальной об-
ласти. Перитремы самок в виде петлеобразной замкнутой трубки; у самцов —
маленькие с короткой перитремальной трубочкой, сросшейся с покровами.
Тритостернум подвижный, двуветвистый. Хелицеры самок имеют подвижный
кинжаловидной палец с очень острым изогнутым кончиком и 2 зубцами по
15 Зак. № 990
225
верхнему краю. По нижней и верхней кромкам имеются острые и тонкие скле-
ритные лезвия. Неподвижный верхнебоковой палец меньше подвижного п
имеет крепкий острый кончик. У самца подвижный палец хелицер превращен
в желобчатый придаток с ложковидным концом. Педппальпы 5-члениковые,
их лапки вооружены щетинками и предлапкой в виде двураздельной острой
вилочки. Ноги сильные, короткие. Лапки заканчиваются хорошо развитой
присоской. Яйцекладущие. Личинка не питается. Паразиты пчел. Семейство
включает 1 род.
1. Varroa Oudms. Грудной щит $ полулунной формы, густо покрыт щетин-
ками; у его заднего края расположена поперечная дугообразная половая щель.
Грудной щит d слит с брюшным и изрезан в задней его части. У переднего края
щита открывается воронковидное половое отверстие. Лапки имеют предлапкп,
на концах которых крупные присоски. На лапках передних ног различные сен-
силлы. — 1 (в СССР 1).
1 (1). Спинной щит $ сплошной, заходит на брюшную сторону. Щиты на брюш-
ной стороне крупные, практически закрывают всю брюшную поверхность,
между ними располагаются только узкие полоски эластичной кутикулы.
У d спинной щит не заходит на брюшную сторону. На брюшной стороне
один щит, изрезанный в задней части. Хелицеры $ имеют подвижный кин-
жаловидный палец с очень острым изогнутым кончиком и 2 зубцами
по верхнему краю. Неподвижный палец меньше подвижного и имеет креп-
кий острый кончик. Хелицеры d: подвижный палец хелицер превращен
в желобчатый придаток с ложковидным концом. $ —1.2x1.7, d —
0.85x0.8. —Паразиты пчел. Всесветно..............V. jacobsoni Oudms.
5. Сем. LAELAPTIDAE
Дорсальная поверхность взрослых клещей покрыта одним щитом, обычно
с цельными краями. Количество и форма щетинок на щите разнообразны: чаще
бывает 37—39 пар, но гипертрихия не редка. Особенно часто добавочные ще-
тинки (непарные, несимметрично расположенные) на центральной части задней
половины щита. Щетинки волосовидные, игольчатые, клиновидные, листовид-
ные и т. д., могут быть плоские или ребристые, гладкие или оперенные. Строение
щитов на брюшной стороне у самок сформировано различно и характерно для
различных родов. Стернальный щит самки обычно с 3 парами, редко с 2 или
4 парами щетинок и с 2 или 3 парами щелевидных органов. Генито-вентральный
щит вытянут позади IV кокс, иногда значительно, и достигает до анального
щита; на щите обычно 1—6 пар щетинок, реже щетинки многочисленны; иногда
щит слит с анальным (род Ololaelaps). Ширина щита также разнообразна — от
узкого, с почти параллельными краями, до широкого, покрывающего всю вен-
тральную поверхность. Анальный щит с обычными 3 околоанальными щетин-
ками, редко на его переднем крае появляются 1—2 добавочные щетинки (не-
которые виды рода Myonyssus). Метастернальные щитки имеются или отсут-
ствуют. Метаподальные щитки от небольших, овальных или линейных, до
крупных, округлых или треугольных; иногда они слиты с генито-вентральным
щитом. У самца на брюшной стороне щиты либо все слиты в единый, либо сво-
бодным остается анальный (роды Haemolaelaps, Hyperlaelaps). Перитремы от
коротких до длинных. Тритостернум у обоих полов с 2 опушенными или глад-
кими ветвями. Корникулы чаще хорошо склеротизованы, имеют форму лопасти
разнообразного строения — от простых гладких до разветвленных, бахромча-
тых. Тектум обычно с выпуклым цельным гладким или зубчатым краем, про-
зрачный или сильно склеротизованный. Хелицеры самок и обычно самцов
с хорошо развитой клешней, оба пальца которой примерно одинаковой длины;
иногда у самца неподвижный палец редуцирован; сперматодактиль свободный,
короткий или значительно длиннее подвижного пальца, иногда целиком слит
с последним. Педипальпы 5-члениковые, вильчатая щетинка лапки педипальп
дву-, реже трехраздельная.
Все ноги с предлапками, обычно с коготками и присосками, реже коготки
отсутствуют. На переднем крае II кокс часто имеется переднедорсальный шип.
226
На II ногах самцов никогда не бывает крупных выростов (апофизы), но иногда
имеются утолщенные щетинки в виде шипов или шпор; у самок они встречаются
реже. Яйцекладущие и живородящие. Обитатели нор и гнезд мелких млеко-
питающих, птиц, в муравейниках, ульях, гниющих органических веществах.
В СССР — 10 родов, на Дальнем Востоке — 10, медико-ветеринарное
значение имеют 4.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). Перитремальные щитки широкие. Генито-вентральный щит $ сзади рас-
ширен. В передней его части одна пара Vlr щетинок, в расширенной задней
части наблюдается значительное количество щетинок. К бокам генито-
вентрального щита примыкают крупные боковые брюшные щиты, имею-
щие различную форму. У в на брюшной стороне все щиты слиты в один
.......................................... 2. Eulaelaps Berl.
2 (1). Перитремальные щитки узкие или отсутствуют.
3(4). Хелицеры у обоих полов одинакового строения: пальцы хелицер с узкой
клешней, без зубчиков по внутреннему краю. Анальный щит у $ крупный,
широкий, с вогнутым передним краем. У 3 один брюшной щит с многочис-
ленными щетинками............................4. Myonyssus Tiraboschi
4 (3). Хелицеры у $ и d различного строения.
5 ’(6). Генито-вентральный щит $ с 4 парами щетинок. Брюшной щит 3 цельный
или разделен. Хелицеры $ с короткой клешней, несущей тупые зубцы
по внутренней стороне пальцев. Хелицеры 3 нередко представлены лишь
одним сперматодактилем и совершенно утрачивается их клешневидный
характер.......................................3. Laelaps С. L. Koch
6-’(5). Генито-вентральный щит $ с одной парой щетинок. Брюшной щит J
цельный. Пальцы хелицер $ хорошо развиты. Подвижный палец вооружен
зубчиками по внутреннему краю или без них. У d пальцы хелицер слабо
склеротизованы, без зубцов по внутреннему краю, неподвижный палец
нередко значительно короче подвижного. Сперматодактиль в виде желобка,
иногда изогнут..................................1. Androlaelaps Berl.
1. Androlaelaps Berl. Дорсальный щит с 38—39 парами тонких гладких
или слегка опушенных щетинок, иногда 40—41 парой или появляются непарные
добавочные щетинки. У $ стернальный щит обычно в ширину больше, чем
в длину (реже ширина равна длине), с вогнутым или почти прямым задним
краем; на щите 3 пары щетинок и 2 пары пор. Генито-вентральный щит узкий
пли расширенный с 1 парой щетинок. Анальный щит грушевидный или тре-
угольный с 3 околоанальными щетинками. Метаподальные щитки обычно не-
большие, неправильно овальные или удлиненные. Перитремальные щитки
узкие, свободные, иногда заметны лишь вокруг стигм и позади них. Пальцы
хелицер $> хорошо развиты. Подвижные пальцы обычно с 2 зубцами, прозрач-
ный придаток часто расширен у основания, реже — узкий. У <5 на брюшной
стороне обычно генито-вентральный щит с 8—11 парами щетинок, реже аналь-
ный щит отделен. Пальцы хелицер лишены зубцов; неподвижный палец часто
значительно короче подвижного. Сперматодактиль желобчатый, превышающий
длину пальцев или целиком слит с подвижным пальцем. — 2 вида (в СССР 10).
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
Самки
1 (2). На спинном щите имеется 38 пар щетинок; II пара внутренних височных
щетинок (Г2) отсутствует. Грудной щит довольно широкий: его ширина
превышает длину в 1.5—1.6 раза. Длина щетинок Str—St3 превышает
половину длины грудного щита. Генито-вентральный щит (рис. 115, 3)
с небольшой перетяжкой на уровне Vl19 сзади равномерно закруглен и
удален от анального щита. Паразит грызунов, насекомоядных, рукокры-
лых. реже птиц. Космополит........................A. glasgowi Ewing
15*
227
2 (1). На спинном щите имеется 39 пар щетинок или больше; II пара внутрен-
них височных щетинок (Т2) имеется. Грудной щит с несколько вогнутым
задним краем. Длина щетинок St± равна половине щита, St2 несколько
длиннее. Генито-вентральный щит довольно равномерной ширины, очень
длинный, почти вплотную приближается к анальному. — Паразит гнезд
птиц и грызунов и на самих грызунах. Космополит ... A. casalis Бег1.
1 — хелицеры самца Androlaelaps casalis', 2—8 — щиты брюшной стороны самок родов Laelaps и Нуро-
laelaps: 2 — L. echidninus, 3 — L. nuttalli, 4 — L. lemmi, 5 — L. hilaris, 6 — L. chethr гопоту dis, 1 — 1,. agr
Us, 8 — L. pavlovskyi.
Самцы
1 (2). На спинном щите отсутствуют щетинки Т2. Брюшной щит узкий, с неров-
ными, как бы выщербленными краями ниже IV кокс, с 9 парными и 1 не-
парной щетинками. Боковые брюшные щитки имеются. Сперматодактиль
(рис. 115, 4) крючковидно изогнут. 0.5—0.6......A. glasgowi Ewing
2 (1). На спинном щите присутствуют щетинки Т2. Брюшной щит с 11 парными
и 1 непарной щетинками, сильно расширен ниже IV кокс. Боковые брюш-
ные щитки отсутствуют. Сперматодактиль в виде желобка, почти прямой
(рис. 117, 7). 0.4—0.5.............................A. casalis Berl.
228
2. Eulaelaps Berl. Спинной щит у обоих полов один. У $ на брюшной стороне
грудной щит с полным набором щетинок Str—St3; генито-вентральный щит
с сильно расширенной задней частью; анальный щит треугольный. Ниже
IV кокс крупные треугольные или округлые боковые брюшные щитки, лишен-
ные щетинок. Перитремальные щитки расширены. Клешни хелицер крупные
с хорошо развитыми зубцами и с небольшим прозрачным придатком на непо-
движном пальце. У в на брюшной стороне все щиты слиты в единый. Клешни
хелицер крупные, неподвижный палец без зубцов, лишь с прозрачным придат-
ком, на подвижном пальце близ вершины имеется массивный зубец. Спермато-
дактиль трубчатый, слегка возвышается над подвижным пальцем. Тело густо
покрыто щетинками. — 1 вид (в СССР 3).
1 (1). На лапках II ног у обоих полов все щетинки тонкие. Задняя часть генито-
вентрального щита $ очень широкая, покрыта многочисленными щетин-
ками. Генито-вентральный щит достигает анального щита; последний
овально-треугольной формы. Боковые брюшные щитки треугольной
формы, крупные. У d цельный брюшной щит, сильно расширенный ниже
уровня IV кокс. 0.7—1.1. —Типичный нидикол и факультативный кро-
восос. Голаркт..............................Eu. stabularis С. L. Koch
3. Laelaps С. L. Koch. Форма широкоовальная или почти круглая. Спинной
щит крупный; в средней части щита у ряда видов имеется темно окрашенное
утолщение якореобразной или крестообразной формы. На щите 39 пар щетинок.
Грудной щит 5 большой, с почти прямым или вогнутым задним краем. Генито-
вентральный щит с 4 парами щетинок (VZX—VZ4). Анальный щит треугольный
или почти овальный. У d сплошной щит на брюшной стороне. Щетинки тела
и конечностей нередко шиповидно утолщены: на коксах некоторые щетинки
шиповидные (чаще передняя на I коксах и задняя на II и III коксах). Хелицеры
2 с короткой клешней, несущей тупые зубцы по внутренней стороне пальцев.
Хелицеры d нередко представлены лишь одним сперматодактилем и совершенно
утрачивают клешневидный характер. — 9 видов (в СССР 14).
i
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
Самки
1 (4). Щетинки спинного щита разнородные: краевые длинные, игольчатые
(за исключением которые короткие, шиповидные), в передней и сред-
ней частях щита щетинки короткие, шиповидно утолщенные.
2 (3). Анальный щит обратногрушевидный, спереди значительно шире, чем
сзади. Аданальные щетинки в 1.5 раза длиннее анального отверстия.
Постанальная щетинка длинная и толстая, превышающая длину аналь-
ного отверстия. Спинной щит широкояйцевидный, в,средней части с окра-
шенным якоревидным или крестообразным рисунком; длина его 0.55 —
0.70. На щите 39 пар щетинок. — На водяной полевке по всему ее ареалу
L. muris Ljungh (arvicola Oudms.)
3 (2). Анальный щит удлиненно-овальный, ширина его, спереди немногим
больше, чем сзади. Аданальные щетинки очень маленькие, короче аналь-
ного отверстия. Постанальная щетинка прямая, короткая, значительно
короче анального щита. Спинной щит без темного рисунка. На нем отсут-
ствуют щетинки и, Т2, D?, D8. На ногах I у основания наружной бедренной
щетинки имеется сильно склеротизованный выступ. Спийной щит 0.57 —
0.7. —На ондатре по всему ареалу.L. multispinosus Banks
4 (1). В передней и средней частях спинного щита нет шиповидно утолщенных
щетинок; в основном игольчатые.
5 (6). Задняя часть генито-вентрального щита сильно расширена; задний край
его вогнут и почти соприкасается с передним краем анального щита.
Постанальная щетинка утолщена, длинная. Длина грудного щита равна
его ширине. Щетинки на коксах толстые; на I — передняя щетинка шипо-
видно укорочена. Все щетинки спинного щита длинные, игольчатые
229
(рис. 117, 2). — На пасюке, на дальневосточном пасюке. Космополит
................................................. L. echidninus Berl.
6 (5). Задняя часть генито-вентрального щита не широкая, никогда не сопри-
касается с анальным щитом.
7 (16). Адайальные щетинки равны или лишь немногим превышают анальное
отверстие, расположены ниже уровня анального отверстия или на уровне
его заднего края.
5 6 7 8 9
Рис. 118. Детали строения самок гамазовых клещей.
1—4 — щиты на брюшной стороне: 1 — Laelaps micromydis, 2 — Hirstionyssus amphibius, 3 — H. arvalis.
4 — Myonyssus dubinini', 5—8 — II, III, IV коксы самок рода Hirstionyssus: 5 — H. sciurinus, 6 — H. isa-
bcllinus, 7 — H. musculi, 8 — H. criceti\ 9 — II лапка самки H. eusoricis.
8 (15). Постанальная щетинка равна длине анального отверстия.
9 (10). Аданальные и постанальная щетинки равны длине анального отвер-
стия. — На леммингах. Голаркт..................L. semitectus С. L. Koch
10 (9). Аданальные щетинки в 11/4 раза длиннее анального отверстия, располо-
жены на уровне его заднего края. Постанальная щетинка равна длине
анального отверстия. Генито-вентральный щит узкий; щетинки Vl4 рас-
ставлены так же широко, как Vl4. Боковые брюшные щитки удлиненно-
овальные (рис. 117, 3). —На дальневосточном пасюке, реже на черной
крысе, пасюке. Космополит...............................L. nuttalli Hirst
11 (12). Грудной щит без уступа по заднему краю. Генито-вентральный щит
равномерно широкий.
12 (11). Грудной щит с уступом по заднему краю, длинный, как бы сжат с бо-
ков; генито-вентральный щит (рис. 118, 1) впереди узкий, ширина его
230
переднего края в 2 раза меньше ширины на уровне щетинок Г/2: УЦ
сильно сближены между собой. — На мыше-малютке. Голаркт . . . .
..............................................L. inicromydis Zachv.
13 (14). Грудной щит почти квадратный, его длина равна ширине; без уступов
по заднему краю. Генито-вентральный щит узкий.
14 (13). Грудной щит более вытянут в ширину; углы уступа по заднему краю
не оттянуты. Генито-вентральный щит (рис. 117, 8) впереди широкий,
ширина его переднего края в полтора раза меньше ширины на уровне ще-
тинок VZ2; VZ4 сближены между собой мало. — На полевой и азиатской
мышах и других грызунах. Палеаркт..................L. pavlovskyi Zachv.
15 (8). Постанальная щетинка короткая, значительно короче анального отвер-
стия (рис. 117, 4). —На леммингах. Голаркт.............L. lemmi Grube
16 (7). Аданальные щетинки длинные, значительно превышают длину анального
отверстия. Щетинки брюшной поверхности, расположенные вне щитов,
неодинаковы: наряду с длинными игольчатыми имеются короткие, часто
притупленные. Генито-вентральный щит резко расширяется от Vlr с очень
выпуклыми краями; У1г расставлены широко, VZ4 сближены (рис. 117,
6). — На полевках. Полеаркт...................L. clethrionomydis Lange
Самцы
1 (4). Щетинки спинного щита разнородные: краевые —длинные, игольчатые,
прямые или искривленные (только Мг короткие, шиповидные), в передней
и средней частях щита короткие, несколько шиповидно утолщенные,
2 (3). На IV лапках с внутренней стороны 2 шиповидные щетинки (1 апикаль-
ная и 1 субапикальная), расположенные одна над одной. Постанальная
щетинка длинная, почти равна по длине щетинкам брюшного щита; ада-
нальные щетинки короткие (немногим длиннее анального отверстия). Бед-
ренные щетинки длинные, часто волнисто искривленные; наружная раза
в полтора длиннее внутренней. Перитрема более длинная —доходит
до заднего края I кокс.................................L. muris Ljungh
3 (2). На IV лапках с внутренней стороны 3 шиповидные щетинки (2 апикаль-
ные и над ними 1 субапикальная). Постанальная щетинка короткая, много
короче прочих щетинок брюшного щита; аданальные щетинки очень ко-
роткие (примерно в 2 раза короче анального отверстия). Бедренные при-
мерно в 1х/4 раза длиннее внутренней; над ними еще одна толстая короткая
щетинка. Перитрема короткая — доходит до заднего края или середины
II кокс..........................................L. inultispinosus Banks
4 (1). В передней и средней частях спинного щита нет шиповидно утолщенных
щетинок; большинство или все щетинки игольчатые.
5 (10). На лапках IV ног либо только игольчатые щетш/ки, либо наряду с ними
имеются утолщенные короткие шиповидные.
6 (7). На IV лапках щетинки только игольчатые. Брюшной щит сильно расши-
рен позади IV кокс ................................ L. echidninus BerL
7 (6). На IV лапках щетинки игольчатые и шиповидные.
8 (9). На IV лапках 2 или более утолщенные щетинки: 2 апикальные и 1 суб-
апикальная. На II лапках аналогичное вооружение, на III лапках —
2 апикальные щетинки (одна из них несколько длиннее другой). Бедрен-
ные щетинки I ног: наружная длинная, волнисто искривленная, внутрен-
няя в 2—3 раза короче наружной ........................L. lemmi Grube
9 (8). На II лапках 4 укороченные и шиповидно утолщенные щетинки (2 апи-
кальные, 1 субапикальная и 1 медиальная). Бедренные щетинки умерен-
ной длины, почти одинаковые..................... . . . L. nuttalli Hirst
10 (5). На IV лапках имеются длинные кинжаловидные щетинки.
11 (14). На IV лапке с наружной стороны одна длинная субмедиальная кин-
жаловидная щетинка.
12 (13). Спинной щит правильно овальный; сперматодактили короче, слабо
изогнуты...........................................L. inicromydis Zachv.
13 (12). Спинной щит овальный, почти круглый; сперматодактили длиннее,
с загнутыми концами................................L. pavlovskyi Zachv.
231
14 (11). На IV лапке с наружной стороны 2 или 4 длинные кинжаловидные
щетинки.
15 (16). Аданальные щетинки длиннее анального отверстия. Перитремы длин-
ные, впереди достигают переднего края II кокс............................
..........................................L. clethrionomydis Lange
16 (15). Аданальные щетинки значительно короче анального отверстия . . .
..........................................L. semitectus С. L. Koch
4. Myonyssus Tiraboschi. Довольно крупные клещи яйцевидной или оваль-
ной формы. Спинной щит большой, покрывает все тело. На щите 38 пар щетинок
(внутренних височных одна пара). Грудной щит j крупный, длина его лишь
немного меньше ширины или равна ей. Генито-вентральный щит либо недалеко
выдается за IV коксы, либо почти доходит до анального щита, с 6—7 щетин-
ками. Анальный щит большой, вытянут в поперечном направлении; помимо
3 околоанальных щетинок, иногда с 1—3 добавочными щетинками у переднего
края. У в на брюшной стороне один щит с многочисленными щетинками ниже
IV тазиков. Ноги коренастые: I и II могут быть толще, чем III и IV. Хелицеры
у обоих полов длинные, с тонкими пальцами без зубчиков по внутренней сто-
роне. Паразиты насекомоядных, грызунов. — 1 вид (в СССР 5).
1 (1). Скульптировка спинного щита ? нерезкая, неясная. Ширина анального
щита не превышает ширины генито-вентрального щита, на котором 24 —
29 щетинок. Между генито-вентральным и анальным щитами щетинки
отсутствуют. Постанальная щетинка равна длине аданальных. Боковые
брюшные щитки (рис. 118, 4) вытянутые, продолговатые, расположены
на уровне середины расширенной части генито-вентрального щита. У d
на IV лапке все щетинки игольчатые; одна наружная медиальная слегка
толще прочих, но не шиповидная. На II лапке одна субмедиальная ще-
тинка сильно утолщена в основной части и с острой вершиной. Брюшной
щит единый и несколько расширен в задней части. $ —1.05—1.1, J —
0.97—1.04. — Палеаркт...................................М. dubinini Breg.
6. Сем. HIRSTIONYSSIDAE
Форма тела овальная. У взрослых клещей и дейтонимф 1 дорсальный щит,
не прикрывающий тело с боков и сзади. На щите не свыше 27—30 пар обычно
коротких гладких щетинок. Непарных щетинок нет. Стернальный щит самки
небольшой, короткий, занимает пространство между II коксами; с 3 парами
щетинок п 2 парами щелевидных органов. Mst на мягкой кутикуле. Генито-
вентральный щит языковидный, широкозакругленный сзади, значительно вы-
ступает за уровень IV кокс, на щите 1 пара щетинок; передняя часть щита полу-
прозрачная с продольной штриховкой. Анальный щит небольшой с выпуклым
передним краем, кзади сужен, несет 3 околоанальные щетинки. У самца на
брюшной стороне 1 щит не слитый с перитремальными щитами; генитальное
отверстие у переднего края щита. Метаподальные щитки обычно не развиты или
небольшие, слабо склеротизованные. Перитремы обычно достигают I кокс,
иногда впереди переходят на дорсальную сторону, реже укороченные. Стволик
трито(;тернума постепенно сужается от основания к вершине, где переходит
в 2 слабо опушенные лацинии. Корникулы пленчатые, не имеют форму рога.
Тектум удлиненный, языковидный, спереди зазубренный или бахромчатый.
Хелицеры с хорошо развитой клешней, пальцы которой лишены зубцов, как
у взрослых, так и нимф; у <5 сперматодактиль целиком слит с подвижным паль-
цем, желобчатый. Педипальны 5-члениковые; на вертлуге 2 щетинки, на бедре —
5, на колене — 6, на голени — 12—14; вильчатая щетинка на лапке двураздель-
ная. Все ноги с предлапками, с хорошо развитыми1 коготками. На вентральной
поверхности кокс имеются склеротизованные треугольные шипы. На II коксах
всегда имеется переднедорсальный шип. У вершины II лапок у самцов всегда,
у самок часто имеются 2 толстые короткие когтевидные или шиповидные ще-
тинки. Личинка и протонимфа эмбрионизированы. Дейтонимфа и взрослые
клещи паразитируют на мелких млекопитающих, в основном на грызунах и
232
насекомоядных, способны нападать на человека. Космополиты. — В семействе
1 род.
1. Hirstionyssus Fonseca. Стернальный щит $ различной формы с 3 парами
щетинок и 2 парами щелевидных органов. Генито-вентральный щит закруглен-
ный сзади, умеренно выступает за уровень IV кокс. Анальный щит небольшой,
овальной формы с 3 околоанальными щетинками. У d на брюшной стороне один
щит не слитый с перитремальными щитами. На вентральной поверхности кокс II,
III и IV ног обоих полов имеются шипы, количество, форма, размеры и распо-
ложение которых служат диагностическим признаком видов. У вершины
II лапок у J всегда, а у j часто имеются 2 толстые короткие когтевидные или
шиповидные щетинки. Все ноги с предлапками и хорошо развитыми коготками.
Паразиты млекопитающих, способны нападать на человека. — 8 видов
(в СССР 18).
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
Самки
1 (10). На IV коксах имеется один шип.
2 (3). На II коксах передняя щетинка в виде короткого толстого шипика.
На спинном щите 6 или 5 пар предкраевых щетинок. Анальный щит
широкоовальный. У вершины II лапок с вентральной стороны имеется
пара утолщенных щетинок (рис. 118, 9). 0.53—0.57. —Паразит земле-
роек .................................................Н. eusoricis Breg.
3 (2). На II коксах передняя щетинка иного вида.
4 (5). На II коксах передняя щетинка длинная, крепкая. На спинном щите
предкраевых щетинок 7 пар. — На бурундуках............................
..............................................Н. pavlovskyi Zemsk.
5 (4). На II коксах передняя щетинка короткая, тонкая, часто с трудом раз-
личимая.
6 (9). На спинном щите предкраевцх щетинок 8 пар. Анальный щит узкий,
удлиненный.
7 (8). Шипы на коксах, особенно на II, крупные (рис. 118, 8), Спинной щит
сзади (начиная от 55) резко сужается. Щетинки, расположенные в средней
части щита 1±—/2), заметно мельче щетинок, расположенных
по краю щита (5Х—58, Z)8, Л7П). —На хомяках и сусликах. Сопутствует
своим хозяевам...................................Н. criceti Sulzer
8 (7). Шипы на коксах менее крупные (рис. 118, 7). Очертания спинного щита
не столь угловаты. Щетинки центральной части и щетинки, расположен-
ные по краю щита, мало отличаются по размеру. — На грызунах. Па-
леаркт ............................................Н. musculi Johnst.
9 (6). На спинном щите предкраевых щетинок 7 пар. Анальный щит относи-
тельно широкий, удлиненно-овальный. Стернальный щит короткий,
арковидный, изогнутый, длина его значительно короче ширины. —
На полевках..........................................Н. apodemi Zuevski
10 (1). На IV коксах шип отсутствует.
11 (12). Формула коксальных шипов 0—3—2—0 (рис. 118, 5). На II коксах
3 шипа: переднедорсальный и 2 задних — внутренний и наружный; пе-
редняя коксальная щетинка изогнута и сильно шиповидно утолщена.
0.7—0.81. —На обыкновенной белке. Сопутствует ее ареалу...............
.............................................. Н. sciurinus Hirst
12 (11). Формула коксальных шипов 0—2—2—0 (рис. 118, 6).
13(14). На II лапках имеется пара утолщенных щетинок. 0.87—0.92. —
На даурском ш северокитайском цокорах . . . . Н. confucianus Hirst
14(13). На II лапках пара утолщенных щетинок отсутствует. 0.6—0.76. —
На полевках и землеройках-бурозубках. Голаркт..........................
..............................................Н. isabellinus Oudms.
233
Самцы
1 (12). На IV коксах шип имеется. Формула коксальных шипов 0—2—2—1.
2 (9). На спинном щите предкраевых щетинок 8 пар.
3 (4). Передняя щетинка на II коксах короткая, шиповидная; шип на IV коксе
маленький, острый. 0.49—0.61...............................Н. eusoricis Breg.
4 (3). Передняя щетинка на II коксе иного строения.
5 (6). Спинной щит сзади сильно вытянутый; щетинки на щите резко разли-
чаются по длине.........................................Н. confucianus Hirst
6 (5). Спинной щит сзади широко закруглен; щетинки на щите незначительно
различаются по длине.
7 (8). Спинные и вставочные щетинки D±—D7, Ц—12 примерно одинакового раз-
мера с прочими щетинками................................ Н. musculi Johnst.
8(7), Спинные и вставочные щетинки Dr—D7, Д—/2 мельче прочих щетинок
........................................................... Н. criceti Sulzer
9 (2). На спинном щите предкраевых щетинок 7 пар; S3 отсутствуют.
10 (И). Вентральный шип на II коксе крошечный . . . . Н. apodemi Zuevski
11 (10). Шипы на II коксах крупные......................Н. pavlovskyi Zemsk.
12 (1). На IV коксах шип отсутствует. Формула коксальных шипов иная.
13(14). Формула коксальных шипов 0—3—2—0...................Н. sciurinus Hirst
14(13). Формула коксальных шипов 0—1—2 —0 . . . . Н. isabellinus Oudms.
ОСНОВНАЯ ЛИТЕРАТУРА
Беляев В. Г. Мелкие млекопитающие и их эктопаразиты на севере Дальнего Востока:
Автореф. дис. . . . канд. бпол. наук. Владивосток, 1973. 21 с.
Брегетова Н. Г. К фауне гамазовых клещей Дальнего Востока. — Паразптол. сб.
Зоол. ин-та АН СССР, 1953, № 15, с. 304—338.
Брегетова Н. Г. Гамазовые клещи (Gamasoidea). — В кн.: Краткий определитель. М.;
Л., 1956. 246 с.
Виолович Н. А. Материалы по фауне гамазовых клещей острова Сахалина и Куриль-
ских островов. — В кн.: Биол. сб. Иркутск, 1961, с. 251—265.
Виолович Н. А. Материалы по фауне гамазовых клещей (Gamasoidea) полуострова Кам-
чатка. — Тр. БИН СО АН СССР, 1963, № J0, с. 26—30.
Гончарова А. А., Буякова Т. Г. Клещи семейства Haemogamasidae (Parasitiformes,
Gamasoidea) в Восточной Сибири. — Изв. ИНИЧИ Сибири и ДВ. Иркутск, 1962, вып. 24,
с. 354—367.
Земская А. А. Паразитические гамазовые клещи и их медицинское значение. М., 1973.
167 с.
Ланге А. Б., Пацкий К. В., Таций В. М. Клещи Varroa. — Пчеловодство, 1976, № 3,
с. 16—20.
Ланге А. Б., Нацкий К. В., Таций В. М. О некоторых особенностях биологии клещей
Varroa Якобсони — паразитов пчел. — В кн.: Варроатоз пчел. М., 1977, с. 13—16.
Федоров В. Г. Случаи массового нападения клещей Dermanyssus gallinae (Redi, 1674)
на человека. — Мед. паразитол. и паразитар. болезни, 1961, № 1, с. 104.
Худяков И. С. Об изучении клещей надсемейства Gamasoidea в Южном Приморье. —
Зоол. журн., 1965, т. 44, N 4, с. 521—526.
Черных П. А. Гамазовые клещи мышевидных грызунов Хабаровского Приамурья:
Автореф. дис. . . . канд. биол. наук. Иркутск, 1969. 20 с.
Шапиро М. И. Клещевой сыпной тиф на островах Южного Приморья: Автореф. дис.
. . . канд. биол. наук. М., 1959. 23 с.
АКАРИФОРМНЫЕ КЛЕЩИ (Отряд ACARIFORMES)
САРКОПТИФОРМНЫЕ КЛЕЩИ
(Подотряд SARCOPTIFORMES)
ВНЕШНЕЕ СТРОЕНИЕ
Покровы тела большинства видов тонкие, растяжимые. Характерно расчле-
нение на 2 отдела — протеросому и гистеросому (рис. 119). У паразитов и кле-
щей с твердыми покровами наблюдается тенденция к слиянию этих отделов.
Ротовой аппарат грызущего типа, претерпевающий иногда изменения в связи
234
с паразитическим образом жизни. Глаза отсутствуют, однако клещи способны
различать свет и темноту; у некоторых имеются специальные светочувствитель-
ные образования. Пальпы простые, не видоизмененные в хватательные органы,
чаще всего выполняют чувствительную функцию.
При наличии даже тонких растяжимых покровов, гладких или складчатых,
отдельные участки тела (щиты) с утолщенной, склеротизованной и темноокра-
шенной кутикулой. Со спинной стороны всегда находится проподосомальный
щит, занимающий различную по величине площадь проподосомы, а у самцов —
и гистеросомальный щит, соответственно располагающийся на гистеросоме
(рис. 122, 3, 4). Набор щитов варьирует не только в пределах группы, но и
в пределах отдельных родов. Иногда все щиты сливаются воедино с образова-
нием панциря.
Все тело покрыто щетинками, набор которых не превышает 27 пар и постоя-
нен для рода или даже для таксонов более высокого ранга (рис. 119, 124).
Почти всегда имеются внутренние (vi) и наружные (ие) теменные щетинки перед-
него края тела, иногда значительно отодвинутые от него; внутренние (sei) и
наружные (see) лопаточные — щетинки в центре проподосомы. Они распола-
гаются в 1 поперечный ряд, приблизительно на уровне оснований ног II; ще-
тинки бокового ряда (Z2~^)? наружные (h) и внутренние (Zx) плечевые, сидящие
впереди уровня основания ног III, щетинки (dY—d5) спинного ряда, распола-
гающиеся по обеим сторонам от средней линии тела (рис. 119, <2; 124, 7).
Между основаниями ног I и II со спинной стороны часто помещается чувст-
вительный щетинковидный латерококсальный орган (рис. 119, 2; 120, 16—20).
Не путать с трихоботриями Oribatei — булавовидными или перистыми чувст-
вительными щетинками, основание которых погружено в глубокую воронко-
видную чашечку — ботрию, и которые располагаются по бокам протеросомы,
почти на границе с гистеросомой (рис. 119, 6).
Ноги состоят из 5 свободных члеников: вертлуга, бедра, колена, голени и
лапки. Тазики ног (шестой членик) полностью сливаются с покровами тела и
представлены обычно 2 дугообразными склеритами (коксо-стернальный скелет):
эпимеры — передние и эпимериты — задние склериты кокс, которые могут,
сливаясь, образовывать замкнутые коксальные поля, а иногда и сплошную сеть
(рис. 124, 2). Лапки чаще всего всех пар ног заканчиваются предлапкой, строе-
ние которой имеет большое значение в диагностике таксонов более высокого,
чем род, ранга. Она состоит либо из 1 коготка (иногда 3 — у Oribatei), либо из
разнообразной по форме присоски, несущей на конце более или менее развитый
непарный эмподиальный коготок (рис. 119, 4, 5, эк), либо только из присоски
(амбулакральный аппарат), состоящей из диска и стебля (рис. 123, 77; 124, 7;
127).
Нередко в связи с паразитическим образом жизни ноги претерпевают зна-
чительные изменения: слияние отдельных члеников друг с другом, редукцию их,
изменение формы члеников, связанное с изменением функции. На ногах распола-
гаются многочисленные щетинки и хеморецепторные чувствительные щетинко-
видные органы — соленидии, число и расположение которых является стойким
признаком, широко использующимся в систематике в пределах таксонов высо-
кого ранга (рис. 125, 3).
Оформленной дыхательной системы как таковой нет; стигмы отсутствуют,
трахейная же система, если имеется, открывается через поры и поровые поля.
Иногда по бокам гистеросомы располагаются отверстия жировых желез (рис. 123,
•ЯГиЛгЬ J •
Наружный половой аппарат построен различно в разных группах и обычно
различается у самцов и самок. У самок чаще всего присутствуют два половых
отверстия: яйцевыводное (рис. 119, яо) и копулятивное. Первое помещается
в промежутке между ногами II —III и обычно прикрыто складками покровов —
клапанами. У некоторых групп клещей впереди отверстия лежит полукруглая
более темно окрашенная пластинка (склерит) — эпигиний (рис. 123, а). Через
яйцевыводное отверстие происходит выведение яиц у яйцекладущих клещей и
личинок — у живородящих. Копулятивное отверстие находится либо на заднем
конце тела, терминально и тогда его трудно увидеть, или на вершине копулятив-
ной трубки (рис. 121, кт).
235
Однако в некоторых группах через прозрачные покровы тела можно раз-
глядеть трубочку (проток), ведущую в семяприемник, стенки которого бывают
частично склеротизованы и потому также видимы — копулятивная, половая
бурса (рис. 123, 6—10). В этих случаях форма и положение бурсы имеют важ-
ное систематическое значение.
У самцов половое отверстие одно, чаще всего оно сдвинуто несколько назад
по сравнению с положением яйцевыводного отверстия самки и располагается
между III и IV парами ног. Половой аппарат самцов значительно меньше по
размерам, чем яйцевыводное отверстие самок, значительно более уплотнен,
склеротизован и темнее окрашен (рис. 119, го).
Анальное отверстие у обоих полов находится у заднего конца тела в виде про-
дольной щели, положение которой несколько варьирует у разных групп: чаще
всего на брюшной стороне или терминально; у некоторых паразитов смещается
на спинную сторону. Анальное отверстие самцов в большинстве семейств несет
по бокам пару округлых анальных копулятивных присосок (рис. 119, акп).
У большинства видов в той или иной степени выражен половой диморфизм,
который проявляется не только в различном строении полового аппарата сам-
ца и самки, но и в строении одной или нескольких пар ног (I—III пары ног
самца значительно отличаются от таковых самки большими размерами, при этом
часто наблюдается редукция ног IV самца), наличием у самцов пары половых
(тарзальных) присосок на лапках IV (рис. 120,10,11) или особых лопастевидных
выростов, часто с мембранами (опистосомальные лопасти) на заднем конце тела
(рис. 126).
Индивидуальное развитие включает: трехногую личинку, 2 или 3 нимфы
(часто нимфа 2-я либо превращается в особую непитающуюся расселительную
или покоящуюся фазу — гипопус (рис. 120, 21—23), либо совсем выпадает) и
половозрелых самца и самку, которые легко отличаются от нимф по наличию
половых отверстий. В таблицах приводятся признаки, пригодные для различе-
ния лишь взрослых половозрелых клещей, по которым только и возможна диаг-
ностика видов большинства приводимых ниже групп клещей; неполовозрелые
фазы можно определять лишь до семейства, реже рода.
Иерархические отношения среди отдельных групп саркоптиформных кле-
щей в целом слабо изучены [Ланге, 1969]. Система их очень сложна и разные
исследователи (чаще всего специалисты по отдельным группам) придают им
различный ранг. Мы придерживаемся системы, изложенной О’Коннором [O’Con-
nor, 1982] в разделе по клещам в книге «Классификация живых организмов».
Саркоптиформные клещи — группа очень разнообразная по «жизненным
формам» и включает виды как свободноживущие, так и паразитические, причем
как «накожников» (эктопаразитов), так и «вкожников» (эндопаразитов).
Все эти клещи характеризуются различной степенью связи с их хозяевами,
чем и определяется степень их вредоносности. В связи с большим разнообразием
этих отношений даже в пределах одной группы клещей медико-ветеринарное
значение их будет рассмотрено в соответствующих разделах.
Помимо групп клещей приводимых ниже, следует обратить внимание на
панцирных клещей, или орибатид (Oribatei), которые являются промежуточ-
ными хозяевами ленточных червей сем. Anoplocephalidae. Единичные экзем-
пляры этих клещей встречаются в пыли домов, но практического значения там
не имеют.
Большинство специалистов выделяют Oribatei в самостоятельный подотряд
[Krantz, 1978 и др.], на что, по-видимому, отчасти сказалась довольно высокая
степень изученности этой группы по сравнению с другими саркоптиформными
клещами. Однако, учитывая значительную морфологическую общность с кле-
щами подотряда Sarcoptiformes, мы склонны относить их к этому подотряду,
выделяя в самостоятельную группировку более высокого, чем надсемейство,
ранга (по аналогии с Parasitiformes, — по-видимому, когорту или надкогорту).
Панцирные клещи из-за ограниченности объема этого издания нами не рас-
сматриваются, о чем говорилось в предисловии.
236
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА
КРУПНЫХ ТАКСОНОМИЧЕСКИХ ГРУПП КЛЕЩЕЙ
ПОДОТРЯДА SARCOPTIFORMES
1 (2). Покровы сильно склеротизованы, образуют сплошной панцирь. Трихо-
ботрии имеются. Лапка заканчивается 1—3 коготками (рис. 119, 6) . . .
..................................................... Группа Oribatei
2 (1). Покровы полностью или частично мягкие, не образуют сплошного пан-
циря. Трихоботрии отсутствуют. Лапка заканчивается присоской без ко-
готка или с одним коготком, окруженным в основании хотя бы небольшой
присоской (пульвилой).
3 (4). Покровы, как правило, совершенно гладкие, без параллельной штри-
ховки, иногда уплотненные. Голень I с 2 щетинками на брюшной стороне
членика. Предлапка, по крайней мере ног I и II, представлена присоской
? эмподиальным когтем (рис. 119, 4, 5). Обитатели кормовых запасов
и домашней пыли..................................Надсем. Acaroidea
4(3). Покровы всегда с хорошо выраженной параллельной штриховкой. Го-
тень I с 1 щетинкой на брюшной стороне членика. Предлапка по крайней
мере ног I и II представлена присоской (амбулакральным аппаратом);
коготь отсутствует (рис. 123, 77).
5 (6). Соленидии о»! и со3 расположены на вершине лапки I (рис. 122, Р, 10). —
Обитатели домашней пыли........................Надсем. Pyroglyphoidea
6 (5). Соленидий располагается в базальной половине лапки I.
7 (8). Паразиты млекопитающих, включая человека . . Надсем. Psoroptoidea
8 (7). Паразиты птиц............Группа надсем. Sarcoptiformes plumicoles
9 (10k Лапки всех ног с двузубчатыми на вершине щетинками р и q. Кондило-
форный направитель предлапки Г-образный (рис. 125, 3)...............
.......................................... Надсем. Pterolichoidea
10 (9). Лапки всех ног без щетинок р и q. Кондилофорный направитель пред-
лапки иного строения (рис. 127).
11 (12). Лапка IV с 6 щетинками (у в 2 из них могут быть представлены при-
сосками, не считая предлапки). Кондилофоры предлапки имеют вид ши-
роких пластинок (рис. 127, 3). Кондилофорный направитель отсутствует
.................................................Надсем. Freyanoidea
12 (И». Лапка IV с 5 щетинками (у в 2 из них могут быть представлены при-
сосками, не считая предлапки). Кондилофоры предлапки узкие, прямые
пли зигзагообразные. Кондилофорный направитель обычно хорошо за-
метен (рис. 127, 2)..............................Надсем. Analgoidea
КЛЕЩИ АМБАРНО-ЗЕРНОВОГО КОМПЛЕКСА
(Надсем. ACAROIDEA)
Гнатосома почти полностью открыта при рассмотрении клеща со спинной
стороны. Пальпы без пластинчатых разращений и бичевидных вершинных щети-
нок.
Покровы гладкие, зернистые или очень редко с еле заметной тонкой штри-
ховкой, совершенно иного вида, чем у паразитических Sarcoptiformes. В край-
нем случае покровы уплотнены, образуя сплошной жесткий панцирь; чаще име-
ются очень небольшие щиты: проподосомальный у обоих полов, реже и только
у самца — гистеросомальный. Латерококсальный орган очень изменчивый по форме.
Амбулакральный аппарат состоит из более или менее хорошо развитого пере-
пончатого претарзуса и непарного крючковидного (эмподиального) когтя
(рис. 119, 4, 5). Кондилофоры обычно очень маленькие, прямые, утонченные.
Кондилоформный направитель отсутствует. Соленидий расположен в сере-
дине или в базальной части лапки I (рис. 121, Р, 10).
Половой диморфизм иногда выражен. Яйцевыводное отверстие самки в виде
продольной щели, совершенно обособленное от анальной щели. Оба отверстия
находятся на брюшной стороне. Копулятивное отверстие располагается терми-
нально и открывается порой или на конце копулятивной трубки (рис. 121, 4).
237
Половой аппарат самца состоит из пениса, расположенного в генитальной
щели. В отдельных семействах (Acaridae) функционально с ним связаны аналь-
ные копулятивные присоски, сидящие по бокам анальной щели, и тарзальные
присоски.
В цикле развития большинства представителей помимо двух нимфальных фаз
имеется расселительная фаза — гипопус.
Надсемейство включает 4 семейства, из которых^медицинское и ветеринар-
ное значение имеют представители только двух.
Рис. 119. Общий вид и детали строения акароидных клещей.
I, 2 — Acarus siro: 1 — самка с брюшной стороны, 2 — со спинной стороны; I — гнатосома, II — про-
подосома, III — гистеросома; 3 — тело самца с брюшной стороны; 4, 5 — амбулакральный аппарат: 4 —
Aleuroglyphus ovatus, 5 — Glycyphagus destructor; 6 — Schiloribates laerigatus (Oribatei), co спинной стороны:
б — поперечная борозда: акп — анальные копулятивные присоски, ао — анальное отверстие, го — гени-
тальное отверстие самца, ло — латерококсальный орган, п — претарзус, т — трихоботрия, эк — эмпо-
диальный коготь, яо — яйцевыводное отверстие. Щетинки тела: vi — внутренние и ve — наружные те-
менные; sci — внутренние и see — наружные лопаточные, lt — внутренние и h — наружные плечевые,
12—13 — боковые, —d3 — спинные.
Большинство клещей — обитатели зерна, плодоовощей и других сельско-
хозяйственных продуктов. Они могут встречаться в почве, подстилке, на раз-
личных субстратах растительного и животного происхождения, в сене, соломе,
остатках обмолота, в гнездах грызунов, насекомоядных и птиц. Переносчиками
их расселительной фазы — гипопуса — служат самые различные насекомые
(амбарные вредители, шмели, жуки, блохи и т. д.), мелкие млекопитающие
и птицы.
В последние годы выявлено, что эти клещи обладают аллергенными свой-
ствами. Попадая в дома вместе с зараженными ими продуктами, кормом для
домашних животных, они живут и размножаются, являясь причиной аллерги-
ческих заболеваний человека (см. ниже надсем. Pyroglyphoidea). Кроме того,
совсем недавно выяснилось, что многие заболевания сельскохозяйственных
животных и птиц, этиология которых оставалась невыясненной (некоторые
238
легочные заболевания, облысение птиц и другие), носят аллергенный характер.
Причиной развития этих заболеваний являются сильно засоренные клещами
корма и фураж.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ
1 (2), Тело всегда разделено на 2 отдела; поперечная борозда, отделяющая их,
хорошо выражена на спинной стороне. Покровы полупрозрачные, глад-
кие. Проподосомальный щит хорошо развит. Лапка типичной для всех
саркоптиформных клещей формы. Присоска претарзуса сидячая, различ-
ной степени развития; коготок средних размеров, чаще крупный (рис. 119,
4).....................................................1. Acaridae
2 (Г). Тело цельное, не разделено на два отдела. Покровы слегка уплотненные,
как правило, с различной скульптуровкой. Проподосомальный щит либо
отсутствует, либо имеет вид узкого продольного склерита — киля. Лапка
сильно удлиненная, очень узкая. Присоска претарзуса длинная, пере-
пончатая, стебельчатая, коготок очень маленький, иногда плохо видимый,
располагается на ее конце (рис. 119, 5).............2. Glycyphagidae
1. Сем. ACARIDAE
Поперечная борозда, разделяющая тело на проподосому и гистеросому, хо-
рошо выражена (рис. 119, 7; 120, 7). Покровы тонкие, мягкие, беловатые, полу-
прозрачные, почти всегда гладкие. Проподосомальный щит хорошо развит,
занимает большую часть спинной стороны проподосомы от vi спереди до sci
и see сзади, слабо пигментированный или почти бесцветный. Отверстия жиро-
вых желез отсутствуют. Хетом спинной и боковых сторон тела представлен
7 — 14 парами чаще всего гладких волосовидных или реже коротко- и редко-
перистых щетинок. Длина и пропорции ног изменчивы, но их строение довольно
однообразно. Щетинки ног могут быть в виде мощных шипов, но никогда не бы-
вают густоперистыми или гребенчатыми. Коготки лапок средних размеров или
крупные, сидячие, причленяющиеся^к вершине с помощью небольших склери-
тов, окруженных у основания более или менее хорошо развитым мясистым
претарзусом.
У самца анальные и тарзальные копулятивные присоски хорошо развиты
(рис. 119, 5, акп; 120, 70); у самки яйцевыводное отверстие всегда расположено
между эпимерами III и IV ног, небольшое, без эпигиния. Копулятивной трубки
и половой бурсы нет.
Гипопус известен у большинства видов родов за исключенивхМ Aleurogly-
phys, Tyrolichus. Гипопус энтомохорного типа, с присасывательным диском
на вентральной стороне опистосомы (рис. 120, 21—23). — 6 родов (в СССР 15).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (4). Обе пары плечевых щетинок (Z1? h) крупные, приблизительно равной
длины.
2^(3). Передние боковые щетинки (Z2) много короче плечевых. Конец лапки
с 3 шипами (рис. 120, 5,6).......................4. Tyrophagus Ouds.
3 (2). Передние боковые щетинки не короче плечевых щетинок. Конец лапки
с 1 довольно крупным коническим шипом и 5 более мелкими хорошо раз-
витыми шипами (рис. 120, 7,8)....................3. Tyrolichus Ouds.
4(1). Внутренние плечевые щетинки значительно короче наружных или отсут-
ствуют.
5 (6) Две пары вершинных щетинок лапок I и II увеличены в размерах, дуго-
образно искривлены и дистально сильно расширены в виде полулунной
прозрачной пластинки (рис. 120, 9). Очень крупные формы: d — более
0.6; ? — более 0.8...............................6. Caloglyphus Berl.
6 (5). Все щетинки лапок волосовидные, не увеличены и не расширены ди-
стально. Формы не очень крупные: 8 — не более 0.5; $ — не более 0.7.
239
Рис. 120. Общий вид и детали строения клещей сем. Acaridae.
1—4 — самец со спинной стороны: 1 — Aleuroglyphus ovatus, 2 — Tyrolichus casei, 3 — Tyrophagus puircs-
centiae, 4 — Caloglyphus rodionovi; 5,6 — лапки I Tyrophagus tenuiclavus co спинной и брюшной сторон;
7, 8 — то же Tyrolichus casei; 9 — лапки I С. rodionovi со спинной стороны; 10, 11 — лапки IV самца:
10 — Tyrophagus putrescentiae, 11 — Т. humerosus; 12—15 — пенис в профиль самца: 12 — Т. putrescentiae,
13 — Т. perniciosus, 14 — Т. humerosus, 15 — Т. tenuiclavus, 16—20 — латерококсальный орган: 16 — Т.
putrescentiae, 17 — Т. perniciosus, 18, 19 — Т. humerosus, 20 — Т. tenuiclavus; 21, 22 — Acotyledon рата-
doxa: 21 — гипопус с брюшной стороны, 22 — то же, со спинной стороны; 23 — гипопус A. sokolovi, с брюш-
ной стороны.
7 (8). Наружные теменные щетинки (уе) расположены латерально, почти на
одной поперечной линии с внутренними (vi), короче последних в 1.5 —
2 раза........................................2. Aleuroglyphus Zachv.
8 (7). Наружные теменные щетинки расположены значительно позади внутрен-
них и представлены чаще всего микрохетами.
9 (10). Проподосомальный щит занимает всю поверхность проподосомы. На-
ружные теменные щетинки расположены латерально, на самом краю тела
.......................................................1. Acarus L.
10 (9). Проподосомальный щит небольшой, квадратный, занимает лишь сред-
нюю часть проподосомы. Наружные теменные щетинки расположены
на его боковых краях и значительно удалены от краев тела............
............................................ 5. Acotyledon Ouds.
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
1. Acarus L. Хелицеры нормальные, не превышают 3/5 длины проподосомы.
Наружные теменные щетинки расположены значительно позади внутренних
(yi) и в несколько раз короче их. Внутренние плечевые щетинки (Zx) много ко-
роче наружных (К). Ноги длинные и стройные. Все щетинки тела и ног волосо-
видные. Половой диморфизм резко выражен.
У 3 генитальное отверстие небольшое, располагается в пространстве между
ногами IV. Анальные копулятивные присоски небольшие, круглые, с плотно
склеротизованным периферическим кольцом. Тарзальные копулятивные при-
соски расположены в проксимальной половине лапки IV (рис. 119, 3). Ноги
I б. м. сильно утолщены и снабжены крупным зубцом на вентральной поверх-
ности бедер; у $ ноги I и II одинакового строения. Анальное отверстие распо-
лагается несколько кпереди от заднего края идиосомы. — 1 вид (в СССР 4).
1 (1). Внутренние и наружные лопаточные щетинки приблизительно одинаковой
длины. Большинство щетинок спинной поверхности тела короткие; отно-
сительно длинные — лопаточные и только 2 пары постанальных щетинок
у заднего края тела. Гипопусы расселительный и покоящийся. Перено-
сятся как насекомыми, так и мелкими млекопитающими и птицами.
3 —0.32—0.54; $ —0.34— 0?65. —Космополит. Повсеместно. В пищевых
продуктах растительного и животного происхождения; вредитель зерна
и продуктов его переработки; в гнездах мышевидных грызунов. В При-
морье найден в пыли жилых домов (рис. 119, 1—3)......................
........................................A. siro L. — Мучной клещ
2. Aleuroglyphus Zachv. Хелицеры очень крупные и массивные, особенно
у 3 (длина их составляет У4 длины тела). Обе пары теменных щетинок распола-
гаются почти на одном уровне вдоль переднего края проподосомального щита:
наружные короче внутренних не более чем в 2 раза. Внутренние лопаточные
щетинки очень короткие, в несколько раз короче наружных. Наружные пле-
чевые щетинки представлены почти микрохетами. Большинство щетинок спин-
ной поверхности тела короткие: крупные, кроме наружных лопаточных, 2 пары
крестцовых и 2 пары постанальных. Ноги относительно короткие, утолщен-
ные, более темно окрашены (насыщенного винно-красного цвета), чем тело.
Половой диморфизм выражен слабо.
У 3 ноги I несколько утолщены по сравнению с ногами II, но не вздуты
и без зубцов. Анальные копулятивные присоски яйцевидные; анальные щетинки
расположены на них. Тарзальные копулятивные присоски помещаются на при-
близительно одинаковом расстоянии от основания и вершины лапки IV; у j
анальная щель придвинута к заднему краю тела. Гипопус отсутствует. —
1 вид.
1 (1). 3 —0.48—0.55; j —0.58—0.67. — Голарктика. Евр. ч. СССР, ДВ; Тур-
ция, АРЕ, КНР, Япония, США. В муке, отрубях, в норах полевок.
Очень часто встречается в пыли домов (рис. 120, 7)...................
.............................A. ovatus Troup. — Темноногий клещ
3. Tyrolichus Ouds. Хелицеры нормального размера, не более 3/5 длины
проподосомы. Наружные теменные щетинки густо- и тонкоперистые, относи-
16 Зак. № 990
241
гельно крупные, расположены латерально, незначительно позади внутренних
теменных. Внутренние лопаточные равны или слегка длиннее наружных. Обе
пары плечевых щетинок почти равной длины. Большинство щетинок спинной
поверхности длинные, в том числе и передние боковые (1а). Ноги длинные,
стройные. Половой диморфизм очень слабо выражен.
(5: ноги I не видоизменены. Эдеагус крупный, клиновидный. Анальные копу-
лятивные присоски округлые, окружены склеротизованным кольцом. Тарзаль-
ные копулятивные присоски помещаются на одинаковом расстоянии от основа-
ния и вершины лапки IV. 5: анальная щель приближена к яйцевыводному
отверстию. Анальные щетинки длинные, волосовидные. Гипопус отсутствует. —
1 вид.
1 (1). з —0.36—0.48; $ — 0.42—0.73. — Палеарктика. Евр', ч. СССР, Сиб.,
ДВ; Европа. Синантропный вид, живущий в сыре, зерне, семенах льна.
Встречен в домашней пыли. На ДВ (Приморье) найден в пыли жилища
человека (рис. 120, 2)...................Т. casei Ouds. — Сырный клещ
4. Tyrophagus Ouds. Удлиненные клещи. Хелицеры нормального размера,
не более 3/5 длины проподосомы. Наружные теменные щетинки длинные, тонко-
перистые, расположены латерально почти на одной поперечной линии с вну-
тренними теменными. Внутренние лопаточные всегда заметно длиннее на-
ружных. Обе пары плечевых щетинок почти равной длины. Все щетинки спин-
ной поверхности длинные: микрохетами представлены лишь d1? 12, иногда d2.
Щетинки по большей части редко- и тонкоперистые, образуют сзади длинный
пучок. Ноги длинные и стройные. Половой диморфизм выражен очень незна-
чительно.
У 8 анальные копулятивные присоски овальные, без периферического скле-
ротизованного кольца. Положение тарзальных копулятивных присосок сильно
варьирует (рис. 120, 10, 11)\ у ? анальная щель подходит почти к заднему
краю тела. Часть анальных щетинок представлена микрохетами. Гипопус от-
сутствует. — 5 видов (в СССР 10).
1 (8). Срединные дорсальные щетинки d2 заметно длиннее очень коротких ще-
тинок dr (рис. 120, 3).
2 (5). d2 в 2.2—4 раза длиннее d±. Ноги относительно короткие, лапки I не длин-
нее голени и колена, взятых вместе.
3 (4). Латерококсальный орган ланцетовидный, с длинными часто расположен-
ными отростками, отходящими только от его сильно расширенной прокси-
мальной половины. У 8 пенис очень маленький, резко S-образно искрив-
ленный (рис. 120,12,16). в — 0.28—0.35; 9 — 0.32—0.42. — Космополит.
Повсеместно. На ДВ — в Прим. Самый распространенный и обычный
амбарный вид; в зерне и пищевых продуктах. Часто встречается в пыли
жилых домов, представляет серьезную опасность как источник аллергена
...................................................Т. putrescentiae Schr.
4 (3). Латерококсальный орган с узким стволом и редко посаженными длин-
ными отростками, располагающимися почти по всей его длине. У 8 пенис
очень крупный, дугообразно изогнутый (рис. 120, 13, 17). в — 0.45 —
0.54, $ — 0.55—0.70. — Голарктика. В СССР повсеместно; Европа, Ав-
стралия. Широко распространен в полевых условиях и в хранилищах
зерна (преимущественно овса, ячменя и различных продуктов их перера-
ботки). Встречен в Приморье в пыли домов . . . . Т. perniciosus Zachv.
5 (2). d2 не более чем в 1.6 раза длиннее dr. Ноги длинные и стройные, лапки I
чаще всего (в 1.1—1.5 ра?за) длиннее голени и колена, взятых вместе.
6 (7). Лапки I почти в 1.5, лапки IV 1.5—1.6 раза длиннее соответствующих
голени и колена, взятых вместе. Идиосома относительно узкая, почти
в 2 раза длиннее своей ширины. 8 —0.17—0.40, $ —0.20—0.75. —
Голарктика. Широко распространенный синантропный вид. Встречен
в Приморье в пыли домов.................................Т. longior Gerv.
7 (6). Лапки I не более чем в 1.2, лапки IV в 1.1 —1.3 раза длиннее соответствую-
щих голени и колена, взятых вместе. Идиосома более широкая. 8 — 0.53 —
0.67, $ —0.33—0.54. У 8 пенис тонкий, цилиндрический, постепенно
242
суживающийся к концу, по форме напоминающий носик кофейника
(рис. 120, 15, 20). — Голарктика. Евр. ч. СССР, Кавказ, Камч. Преиму-
щественно на гниющих овощах и фруктах . . . . Т. tenuiclavus Zachv.
8 (1). Срединные дорсальные щетинки d2 такие же мелкие, как dx. J — 0.28 —
0.47; $ — 0.35—0.77. Латерококсальный орган с узким стволом и корот-
кими отростками, редко расположенными по всей длине. У 3 пенис дважды
коленчато изогнутый, б. м. одинаковой ширины по всей длине (рис. 120,
14, 18, 19). — Палеарктика. Евр. ч. СССР, Кавказ, 3 Сиб., Прим.; Ев-
ропа. Широко распространен в полевых условиях; встречается в норах
грызунов и зернохранилищах........................Т. humerosus Ouds.
5. Acotyledon Ouds. Хелицеры небольшие. Наружные теменные щетинки
располагаются примерно посредине боковых краев проподосомального щита.
Наружные лопаточные и наружные плечевые щетинки значительно длиннее
соответствующих внутренних. Большинство щетинок спинной поверхности
короткие, в том числе анальные и аданальные. Ноги средних размеров.
У <5 анальные копулятивные присоски как у видов рода Tyrophagus. Тар-
зальные копулятивные присоски располагаются приблизительно на одинако-
вом расстоянии от основания и вершины лапки IV; у 9 анальная щель значи-
тельно отстоит от заднего края тела. Все анальные щетинки представлены
микрохетами. Гипопус всегда имеется. — 2 вида (в СССР — 10, 4 из которых
известны только по гипопусам). В пыли домов встречен лишь в фазе гипопуса.
Гипопусы
1 (2). Щетинки спинной поверхности тонкие, волосовидные, довольно короткие.
Присасывательный диск вытянутый, хорошо развит, с 3 рядами присосок:
2, 2 и 4. 0.28—0.35x0.19—0.25. —Палеарктика. Евр. ч. СССР, Прим.
В разнообразных растительных остатках и в норах грызунов. В Приморье
отмечен в домашней пыли пола (рис. 120, 23) .... A. sokolovi Zachv.
2 (1). Щетинки спинной поверхности очень тонкие, волосовидные, длинные.
Присасывательный диск представлен мягкой перепончатой подушкой,
хорошо очерченной лишь сзади и с боков, с 2 парами сильно редуцирован-
ных присосок. 0.24—0.29x0.16—0.20. — Палеарктика. Евр. ч. СССР,
Иркут., Прим. В амбарах в пшенице. Встречается в Приморье в пыли
домов чаще, чем предыдущий вид (рис. 120, <27, 22)..................
.............................................A. paradoxa Ouds.
6. Caloglyphus Berl. Очень крупные формы. Хелицеры небольшие, узкие.
Наружные теменные щетинки расположены вблизи середины боковых краев
проподосомального щита и представлены микрохетами. Наружные лопаточные
и наружные плечевые щетинки значительно длиннее соответствующих вну-
тренних. Многие щетинки спинной поверхности длинные.
(5 — диморфны. У гетероморфных 3 III пара ног резко утолщена с 2 мощ-
ными крючками на вершине; щетинки тела значительно удлинены (рис. 120, 4).
Анальные копулятивные присоски как у видов рода Tyrophagus. Тарзальные
копулятивные присоски располагаются ближе к вершине лапки IV; у 9 аналь-
ное отверстие удалено от заднего конца тела на расстояние равное или превы-
шающее его собственную длину. Гипопус имеется у всех видов. — 2 вида
(в СССР 10). Медико-ветеринарное значение может иметь один вид, встречаю-
щийся в СССР в пыли домов.
1 (1). Идиосома удлиненная, яйцевидная. Латерококсальный орган хорошо
развит, щетинковидный, значительно крупнее наружных теменных щети-
нок. Задний край небольшого проподосомального щита прямой. 3 —
0.6—1.0, $ —0.70—1.24. — Палеарктика. Евр. ч. СССР, Алтайск.,
Иркут.; Европа. Преимущественно синантропный амбарный вид (во влаж-
ном зерне, в гниющих корнеплодах). В полевых условиях чаще всего —
в гниющей соломе, в непосредственной близости от жилья (рис. 120, 4, 9)
............................................... С. rodionovi Zachv.
16*
243
Рис. 121. Общий вид и детали строения клещей сем. Glycyphagidae.
1—3 — Chortoglyphus arcuatus: 1 — самка с брюшной стороны, 2 — то же со спинной стороны, 3 — самец
с брюшной стороны; 4 — Glycyphagus domesticus, самка со спинной стороны; 5—7 — Gohieria fusca: 5 — самка
со спинной стороны, 6 — то же, с брюшной стороны, 7 — самец с брюшной стороны; 8 — нога III самки
G. michaeli; 9 — нога II самца G. fustifer, сбоку; 10 — нога II самца G. destructor, сбоку; 11—13 — темен-
ной киль: 11 — G. domesticus, 12 — G. ornatus, 13 — G. cadaverum. кт — копулятивная трубка, <Dt — со-
ленидий.
2. Сем. GLYCIPHAGIDAE
Идиосома цельная (рис. 121, 2, 4). Покровы уплотненные, как правило,
с различными скульптурными образованиями: бугорками, мелкими шипиками,
гранулами, штрихами, складками (но не равномерно складчатые); если гладкие,
то более или менее склеротизованы и окрашены в буроватый тон. Проподо-
сомальный щит или отсутствует, или имеет вид узкого продольного склерита —
теменной киль (рис. 121, 11—13). Отверстия жировых желез отсутствуют.
Хетом спинной и боковых сторон тела колеблется: от 15 до 18 пар щетинок.
Наиболее обычны в этом семействе простые короткоперистые щетинки, однако
нередко встречаются и длинноперистые, гребневидные, ланцетовидные и другие
модификации.
Ноги очень однотипные, стройные с сильно удлиненной лапкой. Щетинки
обычно волосовидные или перистые, однако встречаются крупные перистые
или гребневидные щетинки, чаще всего на коленях и голенях. Коготок чаще
всего маленький, всегда стебельчатый, т. е. находится на конце более или менее
длинного перепончатого претарзуса и не связан соединительными склеритами
с вершиной лапки (рис. 119, 5).
У самцов анальные и тарзальные копулятивные присоски отсутствуют у пред-
ставителей всех родов, кроме Chortoglyphus, у которого они имеют оригиналь-
ное строение, резко отличное от копулятивных присосок Acaridae (рис. 121, 5);
у самок положение и строение яйцевыводного отверстия изменчиво. Эпигиний
-обычно имеется (исключение род Chortoglyphus). Большинство видов с более
или менее развитой копулятивной трубкой (рис. 121, 4). Половая бурса отсут-
ствует. Гипопус известен у большинства видов семейства. Представлен всеми
основными типами, включая покоящийся (Glycyphaginae). — 3 рода (в СССР
12).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2k Покровы розовато-коричневые, сильно склеротизованы, тонкопористые.
Щетинки спинной поверхности короткие, игольчатые, гладкие, приблизи-
тельно одинаковой длины .............................3. Gohieria Ouds.
2 (1). Покровы желтоватые, слабо окрашены, эластичные, гладкие, почти
прозрачные; если они уплотнены, то без пор. Щетинки спинной поверх-
ности длинные и густоперистые или же тонкие, трудно заметные микро-
хеты.
3 (4). Все щетинки спинной поверхности тела в виде тонких, подчас трудно
заметных микрохет; кожа гладкая, довольно плотно склеротизована
.................................................1. Chortoglyphus Berl.
4 (3). Все щетинки спинной поверхности тела длинные, густоперистые. Покровы
мягкие, очень тонко точечно-бугорчатые...........2. Glycyphagus Hering
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
1. Chortoglyphus Berl. Покровы довольно сильно склеротизованы, плот-
ные, совершенно гладкие. Проподосомальный щит отсутствует. Все щетинки
представлены очень тонкими, едва различимыми микрохетами.
У $ генитальное отверстие располагается в пространстве между основани-
ями I и II ног. Анальная щель находится сразу же позади основания ног IV,
на значительном расстоянии от заднего конца тела. По бокам помещается пара
воронковидных перепончатых копулятивных присосок; у j генитальное отвер-
стие очень крупное, располагается на уровне ног III. Генитальные клапаны
дугообразно сходятся спереди под тупым углом, сильно утолщены и склеро-
тизованы. Анальное отверстие находится на значительном расстоянии кзади от
генитального. — На ДВ и в СССР — 1 вид.
1 (1). При жизни полупрозрачный, очень блестящий. Тело правильной яйце-
видной формы; длина в 1.5 раза больше максимальной ширины. Внутрен-
няя щетинка колен I и II изогнутая, крупногребенчатая, наружная пря-
мая, гладкая. 8 —0.27—0.33, $ —0.35—0.45. —Космополит. Евр. ч.
245
СССР, Кавказ, Прим.; Европа, Япония. Преимущественно амбарный вид
(в муке, зерне, семенах кормовых культур), обнаружен в сене и старой
соломе, в пыли домов (рис. 121, 1—3)...............................
..............................Ch. arcuatus Troup. — Гладкий клещ
2. Glycyphagus Hering. Волосатые клещи. Покровы мягкие, иногда заметно
тонко точечно-бугорчатые. Перистые щетинки тела цилиндрические, опушенные
по всей окружности, очень длинные, в 1.5—2 раза длиннее тела. Внутренние
теменные щетинки сильно сближены между собой, задние широко расставлены.
Латерококсальный орган с 1 опушенным стволом. Теменной киль имеется или
отсутствует. Лапки очень длинные, тонкие, вооружены зазубренными снизу
и с боков щетинками средней длины или одеты пушистой чехловидной чешуй-
кой (которая в препаратах может легко отделяться).
У <5 генитальное отверстие очень маленькое, расположено между основа-
ниями ног III; у $ генитальное отверстие удлиненное, покрытое парой хорошо
развитых клапанов, которые смыкаются друг с другом на большом протяжении
по средней линии тела; эпигиний небольшой, полулунный или полукруглый.
Гипопус покоящийся; стенки гипопальной капсулы покрыты сетчатым ри-
сунком. — 5 видов (в СССР 12).
1 (6). Теменной киль отсутствует.
2 (3). Базальная щетинка колена III превращена в крупную яйцевидную пу-
шистую чешую (рис. 121, 8). Очень крупная форма: 8 — более 0.45 (0.45—
0.56), $ — 0.6—0.81. — Палеаркт. Евр. ч. СССР, Иркут. В сене, соломе,
старых стогах, на сеновале, в амбарах; широко распространен в зерно-
хранилищах, обычный амбарный вредитель.................G. michaeli Ouds.
3 (2). Базальная щетинка колена III не расширена, такого же строения, как и
другие перистые щетинки ног. Более мелкие формы: 3 — менее 0.43,
$ — менее 0.55.
4 (5). Перистые щетинки голеней и колен I и II узкие (рис. 121, 10); длина ге-
нитального отверстия $ равна расстоянию между его задним краем и
анальной щелью. 8 — 0.34—0.43, $ — 0.38—0.55. — Голаркт. Евр. ч.
СССР, Кавказ, ДВ, Прим.; Европа, США. В запасах зерна, круп, сене,
соломе, в скоплениях растительных остатков вблизи жилья. Наиболее
широко распространенный и многочисленный обитатель домашней пыли.
В Приморье составляет 15 % фауны пыли жилищ; представляет наиболь-
шую опасность как источник аллергена.................G. destructor (Schr.)
5 (4). Перистые щетинки голени и колен I и II широкие (рис. 121, 9); длина
генитального отверстия $ значительно превосходит расстояние между его
задним краем и анальной щелью, 8 — 0.3—0.4; $ — 0.41—0.48. — Па-
леаркт. Ивановская обл., Прим. Синантропный вид. В амбарах (семенная
рожь) и в пыли жилых домов.............................G. fustifer Ouds.
6 (1). Теменной киль имеется.
7 (8). Внутренняя теменная щетинка (ri) расположена почти у самого переднего
края тела, на переднем конце линейного теменного киля, тянущегося
кзади до внутренней лопаточной щетинки sci (рис. 121, 13). в — 0.28—
0.36; ? — 0.37—0.45. — Палеаркт. Чувашская АССР, ДВ, Прим. В до-
мах и складах, преимущественно в семенах, реже в зерне; в Приморском
крае найден в пыли домов (в постели, мягкой мебели, дорожках, на полу)
.................................................... G. cadaverum (Schr.)
8 (7). Внутренняя теменная щетинка (rf) несколько удалена от переднего края
тела и расположена посредине теменного киля, тянущегося назад до на-
ружной теменной щетинки (ие); передняя половина теменного киля ши-
рокая, слабо склеротизована (рис. 121, 4, 11). 8 — 0.27—0.35, ? — 0.35—
0.68. — Палеаркт. Арктика. Евр. ч. СССР, Кавказ, ДВ, Прим.; Европа.
Синантропный, преимущественно домовый вид; на сухих субстратах
растительного и животного происхождения; имеет серьезное хозяйствен-
ное значение. На ДВ приурочен исключительно к ульям медоносных
пчел, где отмечен в 90 % случаев........................................
..............................G. domesticus (Deg.) — Домашний клещ
246
3. Gohieria Ouds. Гнатосома среднего размера, квадратная, почти целиком
прикрыта со спинной стороны ростральным выступом (рис. 121, 5). Покровы
сильно хптинизированы, гладкие, окрашены в темный буроватый цвет; на кок-
сальных полях они толще и покрыты более грубой пористой скульптуровкой.
Проподосомальный щит отсутствует. Все щетинки спинной поверхности корот-
кие, гладкие. Ноги длинные, массивные; бедра, колени и голени всех ног с мно-
гочисленными продольными ребрами; щетинки ног густоперистые, гребенчатые.
У $ коксальные поля очень сильно хптинизированы и образуют сплошной
стернальный панцирь. Генитальное отверстие очень маленькое, расположено
позади основания ног IV; эпимеры I слиты с эпигинием, образуя массивную
поперечную пластинку впереди генитального отверстия, окруженного оваль-
ным хитиновым кольцом. Генитальное отверстие очень велико и занимает все
пространство между основаниями ног II —IV. Гипопуса нет. — 3 вида (в СССР
4). Медико-ветеринарное значение имеет 1 вид, распространенный в пыли
жилищ и зернохранилищах.
1 (1). Розово-коричневый клещ. Все дорсальные щетинки одинаковой длины,
составляющей не более 15 % длины тела, в — 0.3—0.34, $ — 0.38—
0.41. — Транспалеаркт. Евр. ч. СССР, Кавказ, Ср. Азия, 3 Сиб.; Европа.
Синантропный вид; в муке, зерне и кормах. Часто встречается в пыли
жилых домов (рис. 121, 5—7)....................................
........................G. fusca Ouds. — Бурый хлебный клещ
«АЛЛЕРГЕННЫЕ» КЛЕЩИ (Надсем. PYROGLYPHOIDEA)
Включает единственное сем. Pyroglyphidae, одни представители которого
ведут свободноживущий образ жизни, другие — обитатели гнезд грызунов,
чаще птиц, случайно попадают на их покровы, третьи — постоянные эктопара-
зиты птиц.
ВНЕШНЕЕ СТРОЕНИЕ
Гнатосома частично прикрыта со спинной стороны выступом проподосомаль-
ного щита — эпистомом. Пальпы без пластинчатых разращений. Покровы
мягкие, чаще всего тонко параллельно складчатые. У обоих полов всегда имеется
проподосомальный щит, у самцов (у самок некоторых родов также) — гистеро-
сомальный щит. Латерококсальный орган отсутствует. Амбулакральный аппа-
рат без эмподиального когтя; состоит из диска и стебля. Всегда имеется кондило-
форный направитель (рис. 123, 11). Соленидии и <о3 расположены на вер-
шине лапки I (рис. 122, /). Половой диморфизм выражен. Яйцевыводное от-
верстие самки очень большое, занимает центральную часть брюшной поверх-
ности тела, отделено, полностью от анальной щели. Копулятивное отверстие
располагается субтерминально и ведет в просвечивающую через покровы поло-
вую бурсу; самцы имеют хорошо развитый аданальный щит с парой анальных ко-
пулятивных присосок. Резко выражено различие в величине, хетоме и по-
ложении на теле ног III и IV (рис. 122, III, IV). Задний конец тела нередко
с опистосомальными лопастями.
Цикл развития двунимфальный, гипопус отсутствует.
ВРЕД
Особый интерес к этой группе появился после того, как в 1964—1967 гг.
было выяснено, что аллергенная активность бытовой пыли, являющейся при-
чиной атопических форм бронхиальной астмы и других аллергических забо-
леваний (ринита, конъюнктивита, нейродермита и других), имеет прямую
связь с жизнедеятельностью обитающих в ней клещей. В связи с этим в меди-
цинской литературе их иногда условно называют «аллергенными» клещами.
В разных странах мира при исследовании жилых помещений обнаружено
много различных видов клещей (об акароидных клещах уже было сказано выше),
из которых наибольшее практическое значение имеет всесветно распростра-
247
00
пенный так называемый постельный клещ Dermatophagoides pteronyssinus (Trt.).
Аллергенное действие на дыхательные пути людей оказывают не столько сами
живые клещи (размеры взрослых особей достаточно велики — 0.2—0.6 мм),
сколько разрушенные мертвые особи, фрагменты линочных шкурок и про-
дукты жизнедеятельности. Поэтому постельные принадлежности даже с мерт-
выми сухими клещами служат источником аллергена. Пищей для этих клещей
служат чешуйки рогового слоя кожи человека, которые в огромных коли-
чествах скапливаются в постельных принадлежностях, а также плесневые
грибки, развивающиеся на этом субстрате. В меньшем количестве чешуйки
кожи и грибки скапливаются в мягкой мебели, однако п она также является
местом обитания клещей. Размножаясь в постелях, особенно сильно во влаж-
ное и теплое время года (весной и осенью), клещи расселяются по всей квартире
и обнаруживаются в значительном количестве на нательном белье, одежде
и обуви. Так из зараженных квартир клещи заносятся в другие дома, где при
наличии благоприятных условий также могут размножаться и служить источ-
ником мощного аллергезирующего воздействия на человека [Дубинина, 1985].
Исследование ведущих видов фауны домашней пыли является основой для
профилактики аллергических заболеваний и методом выявления этиологии
некоторых из них.
Сем. PYROGLYPHIDAE
Поперечная борозда, отделяющая проподосому от гистеросомы, имеется,
иногда плохо различима. Покровы различной степени склеротизации, чаще
мягкие с правильно параллельной исчерченностью (рис. 123, 7, 2). Форма
проподосомального щита очень варьирует: чаще он небольшой, занимает ме-
диальную часть проподосомы. Гистеросомальный щит всегда присутствует
у самцов, у самок — иногда. Отверстия жировых желез открываются между
щетинками 12 и Z3 (рис. 123, жж).
Хетом спинной и боковых сторон тела представлен 14 парами тонких волосо-
видных щетинок. Длина и пропорции ног изменчивы. Щетинки ног почти всегда
тонкие, волосовидные, редко шиповидные. Лапка с крупным округлым пре-
тарзусом, на ножке с небольшим коготком (рис. 122, 7, II).
У самца ноги IV всегда меньше ног III и располагаются на теле значительно
дальше от края, чем ноги III, приближаясь к основаниям последних своими
основаниями. Аданальный щит хорошо развит с парой анальных копулятивных
присосок (рис. 122, ащ). Тарзальные присоски имеются, иногда сильно реду-
цированные.
У самки яйцевыводное отверстие большое, начинается от задних концов эпи-
мер II и оканчивается на уровне вертлугов ног IV. Эпигиний полулунный,
иногда плохо различимый (рис. 123, 3—5, э). У заднего края тела через покровы
просвечивает копулятивная бурса (трубка, ведущая в семяприемник) различной
формы и степени склеротизации (рис. 123, 6—10). — На Дальнем Востоке
и в СССР — 3 рода.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). Внутренние и наружные лопаточные щетинки (sci, see) небольшие,
по длине равные между собой. Покровы тела значительно склеротизованы,
с неправильной волнистой исчерченностью (рис. 123, 1) . . . . ♦ . .
................................................1. Euroglyphus Fain.
2 (1). Наружные лопаточные щетинки крупные, в несколько раз длиннее вну-
тренних лопаточных. Покровы почти мягкие с правильной параллельной
исчерченностью (рис. 123, 2).
Рис. 122. Общий вид и детали строения самцов сем. Pyroglyphidae.
1, 2 — Euroglyphus longior: 1 — со спинной стороны, 2 — с брюшной стороны; 3,4 — гистеросомальный
щит: 3 — Dermatophagoides farinae, 4 — D. pteronyssinus’, 5—8 — Hirstia chelidonis’, 9—12 — D. farinae}
13—16 — D. evansi; 17—20 — D. pteronyssinus: 5, 6, 9, 10, 13, 14, 17, 18 — концевые части лапок I
и II; 7, 11, 15, 19 — половой аппарат; 8, 12, 16, 20 — ноги III и IV и аданальный щит. ащ — аданаль-
ный щит, па — половой аппарат, d2 — дорсальные щетинки, ш,, <о3 — соленидий, I—IV — ноги.
249
3 (4). На вершине лапок I отсутствуют какие-либо коготки (рис. 122, 5, I)
........................................................ 2. Hirstia Hull.
4 (3). На вершине лапок I имеется хотя бы 1 небольшой коготок (рис. 122,
6—8, 1)........................................3. Dermatophagoides Bogd.
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
1. Euroglyphus Fain. Покровы склеротизованы, с брюшной стороны силь-
нее, чем со спинной; исчерчены неравномерно расположенными, слегка волни-
стыми складками. Гистеросомальный щит имеется у обоих полов. Все^ще-
Рис. 123. Общий вид и детали строения самок сем. Pyroglyphidae.
J, 2 — Euroglyphus longior: 1 — общий вид со спинной стороны, 2 — яйцевыводное отверстие; 5, 4 —
Dermatophagoides pteronyssinus: 3 — общий вид со спинной стороны, 4 — яйцевыводное отверстие; 5 —
D. farinae, яйцевыводное отверстие; 6—10 — половая бурса: 6 — Е. longior, 1 — Hirstia chelidonis, 8 —
D. farinae, 9 — D. evansi, 10 — D. pteronyssinus', 11 — схема строения амбулакрального аппарата, жж —
отверстия жировых желез, к — кондилофоры, кн — кондилоформный направитель, э — эпигиний, see —
наружные лопаточные щетинки, sci — внутренние лопаточные щетинки.
тинки спинной поверхности тела, включая и обе пары лопаточных (sci, see), ко-
роткие. Лапки I без коготков; колено I с одним соленидием.
У 3 опистосомальные лопасти имеются, хотя бы слегка намеченные. Все
щетинки лапок III волосовидные; у j ноги III слегка короче ног IV. Копулятив-
ная бурса небольшая, слабо склеротизована. —- 1 вид (в СССР 2).
1 (1). Исчерченность покровов, где она имеется, слегка волнистая, неравномер-
ная, довольно частая. Гистеросомальный щит $ маленький, треугольный,
помещается вдоль заднего края тела (рис. 123, 7). У d этот щит занимает
почти всю гистеросому. Имеются 2 небольшие, хорошо выраженные
опистосомальные лопасти, на которых расположены щетинки заднего
конца тела (рис. 122, 7). Яйцевыводное отверстие $ полностью прикрыто
складкой, направленной от заднего края вперед и доходящей почти
250
до эпигиния (рис. 123, 2). Последний узкий, очень маленький, едва раз-
личимый. Копулятивная бурса с короткой склеротизованной трубкой и
круглым семяприемником (рис. 123, 6). 3— 0.24—0.26x0.13—0.14,
$ — 0.28—0.32x0.16—0.2. — Транспалеаркт. Евр. ч. СССР, ДВ, Прим.,
Хаб.; Канада, США, Европа, Азия. Обычный полевой вид (в остатках
зерна, в пыли амбаров). Найден также в волосяных остатках парикмахер-
ских и в пыли жилых домов. На ДВ распространен очень широко в местах
обработки зерна и в зернохранилищах (ячмене, горохе, просе и т. п.),
в пищевых складах, в гнездах воробьев ..........Е. longior (Trt.)
2. Hirstia Hull. Покровы мягкие с хорошо выраженной прямой, правиль-
ной параллельной исчерченностыо. Брюшная сторона склеротизована слабо.
Гистеросомальный щит имеется только у 8. Щетинки спинной поверхности
тела различной длины: всегда короткие — внутренние лопаточные (sei), всегда
длинные — наружные лопаточные (see), наружные плечевые (h) и щетинки
заднего конца тела. Лапки I со сглаженными вершинами без коготков, колено I
с двумя соленидпями (рис. 122, 5, /).
У 8 опистосомальные лопасти отсутствуют. Лапка III с 2 сильными, хо-
рошо выраженными коническими щетинками (рис. 122, 5, III); у 2 ноги III
длиннее ног IV. В копулятивной бурсе склеротизована только трубка, ведущая
в семяприемник, стенки которого не видны. — На ДВ и в СССР — 1 вид.
1 (1). У 8 гистеросомальный щит короткий, начинается позади щетинок d2.
Ноги IV в 2 раза меньше ног III. Пенис очень маленький, почти круглый,
как и весь половой аппарат; половые щетинки у его заднего края тесно
сближены. Аданальный щит линзообразный (рис. 122, 5, ащ). У 2 эпиги-
ний очень короткий. Копулятивная бурса представлена довольно длинной
петлеобразно изогнутой трубкой, оканчивающейся слепо. Снаружи она
открывается на небольшом половом сосочке, расположенном субтерми-
нально (рис. 123, 7). 8 — 0.24—0.26x0.15—0.16, 2 — 0.3—0.36x0.19—
0.25. — Всесветно. Евр. ч. СССР, ДВ, Прим.; США, Африка, Европа,
Япония. В Европе — обитатель гнезд синантропных птиц (воробьев, ла-
сточек, стрижей); встречен также на самих птицах. Обнаружен в домаш-
ней пыли; очень высокой численности достигает в пыли из подушек.
Один из основных аллергенов домашней пыли . . . . Н. chelidonis Hull.
3. Dermatophagoides Bogd. Покровы, степень их склеротизации и число
спинных щитов как у видов рода Hirstia. Соотношение длины щетинок спинной
поверхности тела такое же: sei — короткие, see — длинные (рис. 123, 3). Лапки
I ног обычно с 2 коготками на вершине, реже с 1; колено I с 2 соленидиями
(рис. 122, 6—8, I).
У 8 опистосомальные лопасти отсутствуют. Все щетинки лапок III волосо-
видные; у $ ноги III крупнее, иногда значительно ног IV. В копулятивной
бурсе склеротизована как трубка, так и стенки семяприемника. — 3 вида
(в СССР 5).
Самцы
1 (2). Гистеросомальный щит короткий, квадратный, начинается позади щети-
нок d2 (рис. 122, 3). Ноги I варьируют в своих размерах: часто сильно
расширены, с наружным шипом на бедре. Лапка I с одним хорошо выра-
женным сильным когтем. Ноги IV короче ног III не более чем на половину
длины лапки III. Половой аппарат большой, почти квадратный; пенис
очень маленький, заостренный на конце, ширина его составляет не более
х/5 ширины самого аппарата. Половые щетинки располагаются вдоль зад-
него края полового аппарата и расставлены на ширину пениса. 8 —
0.29—0.33x0.22—0.23, 2 — 0.4—0.44x0.29—0.31. — Космополит. Все-
светно. Евр. ч. СССР, Кавказ, Киргизия, Казахстан, ДВ. На токах в остат-
ках обмолота и муке; в гнездах грызунов, в пыли жилых домов; иногда
встречается на коже больных дерматитом. В Приморье встречен в пыли
4 % обследованных домов. Один из главных аллергенов домашней пили
(рис. 122, 3, 9—12)...............................D. farinae Hughes
251
2 (1). Гистеросомальный щит длинный, продолговатый; щетинки располагаются
на щите (рис. 122, 4). Ноги I не расширены, без наружного шипа на бедре.
3 (4). Лапка I с 2 коготками, лапка II — с 1. Ноги IV доходят вершиной лапки
до основания лапки III. Пенис грушевидный, в 1.5 раза длиннее широкого*
и короткого основания (генитальной арки); половые щетинки располага-
ются вблизи заднего края и расставлены на всю ширину полового аппарата,
d — 0.29—0.32x0.19—0.2, $ — 0.36—0.44x0.24—0.3. — Транспалеаркт.
Евр. ч. СССР, ДВ, Прим.; Канада, США, Европа. В гнездах ласточек.
Синантропный вид, часто встречающийся в пыли жилых домов (рис. 122,
13—16)................................D. evansi Fain, Hughes et Johnston
4 (3). Лапка I с 1 коготком, лапка II — без коготка. Ноги IV вершиной лапки
не достигают основания лапки III. Пенис по длине равен длине основания
(генитальной арки) узкого, длинного, с дугообразно изогнутыми аподе-
мами; половые щетинки значительно отстоят от заднего края полового
аппарата и расставлены на расстояние равное 2/3 его ширины, d — 0.24—
0.31x0.17—0.21, $ — 0.32—0.4x0.21—0.28. — Космополит. Всесветно.
В СССР повсеместно. Синантропный вид. Место обитания и размноже-
ния — постель человека. Широко распространен в пыли домов. Встречен
также у различных животных и человека на участках, пораженных чесот-
кой. Главный источник аллергена в домашней пыли (рис. 122, 17—20)
.................................................. D. pteronyssinus (Trt.)
Самки
1 (2). Эпигиний очень короткий, слабо изогнутый. Трубка копулятивной бурсы
одинаковой ширины па всем своем протяжении. Семяприемник воронко-
видно расширен, со слегка отогнутыми краями (рис. 123, 5, 5) ... .
......................................................D. farinae Hughes
2 (1). Эпигиний средних размеров, дугообразно изогнут. Трубка копулятивной
бурсы расширяется у наружного конца; семяприемник иной формы.
3 (4). Семяприемник имеет вид ухвата. Расширение трубки у наружного конца
равно V3—V2 длины всей трубкй (рис. 123, 9)................................
.................................D. evansi Fain, Hughes et Johnston
4 (3). Семяприемник имеет вид цветка или полулуния (в зависимости от поло-
жения). Трубка расширена на очень коротком участке у самого наруж-
ного конца — места ее выхода на поверхность тела (рис. 123, 5, 10) . . .
•................................................. D. pteronyssinus (Trt.)
ОСНОВНАЯ ЛИТЕРАТУРА
Дубинина Е. В. Эколого-фаунистические исследования клещей пыли в связи с пробле-
мой аллергии. — В кн.: Паразитол. сб. Л., 1985, т. 33, с. 209—229.
Дубинина Е. В., Плетнев Б. Д. Методы обнаружения и определения аллергенных клещей
домашней пыли. Л., 1977. 52 с.
Захваткин А. А. Определитель клещей, вредящих запасам сельскохозяйственных про-
дуктов в СССР. — Уч. записки МГУ. Зоология, 1940, вып. 42, с. 7—68.
Захваткин А. А. Тироглифоидные клещи (Tyroglyphoidea). М.; Л., 1941. 475 с. (Фауна
СССР. Паукообразные; Т. 6. Вып. 1).
Захваткин А. А. Некоторые итоги изучения фауны хлебных клещей СССР. — В кн.:
А. А. Захваткин. Сборник научных работ. М., 1953, с. 169—176.
Тареев В. Н. К вопросу о значении клещей Tyrophagus noxius для человека. — В кн.:
5-я науч. конф. Приморского с.-х. ин-та, Уссурийск, 1966а, с. 76—79.
Тареев В. Н. Особенности фауны амбарных клещей Дальнего Востока. — В кн.: 1-е
акарологич. совещ. М.; Л., 19666, с. 211—212.
Тареев В. Н., Дубинина Е. В. К фауне пылеобитающих клещей Приморья. — Парази-
тология, 1985, т. 19, вып. 1, с. 27—31.
252
ПЕРЬЕВЫЕ КЛЕЩИ
(Группа надсемейств SARCOPTIFORMES PLUMICOLES)
Обширная и многообразная группа постоянных эктопаразитов птиц, ранее
рассматривавшаяся в качестве единого таксона — надсемейства Analgoidea
(Дубинин, 1951, 1953). По современным представлениям этих клещей относят
к 3 самостоятельным надсемействам Analgoidea, Freyanoidea, Pterolichoidea,
по-видимому, независимо сформировавшимся на птицах (Gaud, Atyeo, 1977).
В акарологической литературе перьевых клещей часто традиционно именуют
«Sarcoptiformes plumicoles», понимая под этим термином акаридиевых клещей,
постоянно паразитирующих на птицах. Мировая фауна этих клещей насчиты-
вает около 2000 видов, в СССР известно свыше 370 видов. На домашних пти-
цах на Дальнем Востоке — 17 видов, принадлежащих к 14 родам 11 семейств.
ВНЕШНЕЕ СТРОЕНИЕ
Гнатосома открытая, со спинной стороны клеща полностью видна. Особен-
ностью строения гипостома (гипостомальной пластинки) является наличие
псевдорутеллярных отростков — мембраноподобных прозрачных образований,
обычно выступающих в стороны и вперед из-под пальп.
Покровы не занятые щитами, тонкоскладчатые. Боковые края тела могут
нести мембраноподобные прозрачные выросты. Проподосома обычно несет
3 щита: проподосомальный в центральной ее части и лопаточные по бокам.
Латерококсальный орган щетинковидный или отсутствует. Набор щитов ги-
стеросомы более разнообразен: в передних ее углах и на боковых краях имеется
пара плечевых щитов; большая часть дорсальной поверхности занята гистеро-
сомальным щитом, который иногда разделен поперечной бороздой на два —
метаподосомальный и опистосомальный — или разделен продольной бороздой
на пару продольных щитов (рис. 124, 7, 2; 125, 4\ 137, 7).
Претарзус лишен эмподиального когтя и преобразован в амбулакральный
аппарат, состоящий из диска и стебля (рис. 124, 7). Соленидий расположен
в середине или в базальной ч^сти лапки I. Анальная область лишь с 1—2 па-
рами щетинок.
Половой диморфизм резко выражен, особенно в строении опистосомы.
Яйцевыводное отверстие самки обычно с хорошо склеротизованными краями
(рис. 126, 5; 128, 8, 9). Копулятивное отверстие расположено терминально или
на конце копулятивной трубки.
Половой аппарат самца состоит из генитальной арки и стилетовидного эде-
агуса; спереди он может быть с полукруглым склеритом — эпиандрием, с бо-
ков — с продольными генитальными аподемами (рис. 130, 4; 132, 2). Позади
может располагаться генитальный щит (рис. 129, 4). Впереди и по бокам аналь-
ного отверстия могут находиться один или несколько анальных щитов (рис. 128,
2, 4). Анальные присоски обычно имеются, реже редуцированы (рис. 132, 70;
135, 5). Задняя часть опистосомы обычно двулопастная, реже закругленная,
как у самки. Боковые края лопастей могут нести плоские прозрачные выросты —
мембраны (рис. 130, 3, 4; 134, 7, 2). Терминальная выемка (выемка между ло-
пастями) может быть вся затянута мембраной (рис. 128, 5, 6).
БИОЛОГИЯ И ВРЕД
Цикл развития двунимфальный, гипопус отсутствует. Дейтонимфа, как
показали недавние экспериментальные исследования, полностью утрачена.
Приписываемые ранее перьевым клещам подкожные дейтонимфы (гипопусы)
в действительности принадлежат совершенно другим акаридиевым клещам
сем. Hypoderidae Murrey, 1877 [Fain, 1968].
Перьевые клещи — постоянные эктопаразиты, весь жизненный цикл кото-
рых проходит на одной особи хозяина, за исключением некоторых представи-
телей сем. Epidermoptidae (надсем. Analgoidea) (см. ниже). Большинство видов
характеризуется узким кругом хозяев, принадлежащих к одному или несколь-
253
Рис. 124. Общая схема строения перьевых клещей.
I — общий вид $ со спинной стороны; 2 — общий вид ? с брюшной стороны, гщ — гистеросомальный щит,
— лопаточный щит, плщ — плечевой щит, пщ — проподосомальный щит, эг — эпигиний, эм I—
эд IV — эпимеры соответствующих ног, эт 1—эт IV — эпимериты соответствующих ног. Щетинки:
vi — внутренние теменные, ve — наружные теменные, sci — внутренние лопаточные, see — наружные
лопаточные, h — плечевые, sh — субплечевые, dt—db — дорсальные, lt—16 — латеральные, рае — наруж-
ные постанальные, pai — внутренние постанальные, ап — анальные, cxt, сх3, сх4 — коксальные, сп сг —
генитальные; з — ноги I; 4 — ноги II; 5 — ноги III; 6 — ноги IV; 7 — амбулакральный аппарат лапки.
ад — амбулакральный диск, ас — амбулакральный стебель, бл — базилатеральный склерит, кн — конди-
лофорный направитель, кс — когтевидный склерит, кф — кондилофоры, лс — латеральный склерит, пл —
паралатеральный склерит, цс — центральный склерит (з—б и номенклатура щетинок по: Atyeo, Gaud,
1966: 7 — по: Gaud, Atyeo, 1977).
ким близким родам птиц. Как высокоспециализированные паразиты они при-
способлены к обитанию на покровах определенного типа или даже к отдельным
участкам этих покровов со строго определенными микроклиматическими усло-
виями и структурой.
Большинство клещей расселяется только при непосредственном контакте
хозяев: при насиживании птенцов и при спаривании. Они не способны покидать
погибшего хозяина, осуществлять самостоятельный поиск новой особи хозя-
ина. Вне хозяина они живут от 1 до 10 дней. При скученном содержании птиц
и значительной концентрации клещей их передача может осуществляться через
окружающую среду: через пылевые ванны, подстилку гнезд, выпавшие перья
и т. д.
Клещи сем. Epidermoptidae обитают на поверхности кожи (подсем. Derma-
tioninae), частично внедряются в нее и даже проделывают короткие ходы (под-
сем. Epidermoptinae). Они питаются слущивающимися чешуйками эпидермиса,
а также неороговевшими слоями кожи и, по-видимому, лимфой. Представители
подсем. Epidermoptinae, внедряющиеся в кожу, наиболее патогенны для птиц,
поскольку вызывают чесоткоподобное заболевание кожи. В местах поселения
клещей кожа сильно воспаляется, шелушится и покрывается корками, обра-
зующимися из засыхающей лимфы. Иногда наблюдается выпадение перьев из
пораженных участков кожи. При массовом размножении клещей птицы быстро
истощаются и погибают [Дубинин, 1951].
Большинство представителей подсем. Epidermoptinae расселяется с хозяина
на хозяина не только при их непосредственном контакте, но и используют
для расселения мух-кровососок сем. Hippoboscidae. Поскольку круг хозяев
для мух-кровососок обычно достаточно широк, большинство видов клещей,
расселяющихся с их помощью, также приспособлены к широкому кругу хозяев,
который может включать даже птиц из различных отрядов. В связи с этим
они представляют собой одну из наиболее опасных групп перьевых клещей.
Клещи подсем. Dermationinae, обитающие на поверхности кожи, менее пато-
генны для хозяев и расселяются только при непосредственном контакте хозяев.
Наиболее патогенными для домашних птиц являются клещи сем. Knemico-
ptidae. Вызываемое ими заболевание, кнемидокоптоз, проявляется в ножной
или кожной форме, в зависимости от специализации вида клеща. В случае
ножного кнемидокоптоза клещи внедряются под роговые чешуи на неоперенных
частях ног. Клещи проникают в кожу вплоть до неороговевающего слоя эпи-
дермиса. На первых этапах развития кнемидокоптоза наблюдается резкое воспа-
ление кожных покровов. На более поздних стадиях заболевания, которое мо-
жет тянуться в течение нескольких лет, воспаление исчезает, а роговой слой
эпидермиса разрастается в виде губчатой рыхлой корки, которая пронизана
многочисленными ячейками и ходами, где находятся все фазы клещей. Больные
участки обычно имеют трещины и сильно кровоточат. Перья и роговые чешуи
на пораженных участках выпадают; на поздних стадиях заболевания, сопровож-
дающихся некрозом, могут отпадать и фаланги пальцев. Заболевшие кнемидо-
коптозом птицы гибнут от истощения или от бактериальных и грибных инфек-
ций, проникающих через пораженные участки покровов [Дубинин, 1953].
Клещи сем. Laminosioptidae обитают в соединительной ткани кожи, про-
никая в нее через перьевые фолликулы. Процесс питания не изучен. Вокруг
погибших клещей в соединительной ткани образуются обызвествленные кап-
сулы, которые проступают в виде небольших бугорков на поверхности кожи.
По наличию капсул легко установить наличие заражения этими клещами.
При массовом заражении клещи вызывают гибель птицы.
Большинство перьевых клещей поселяются на маховых, кроющих, контур-
ных или пуховых перьях, питаясь обычно жировой смазкой. Представители
сем. Freyanidae и других могут иногда проникать в перьевые фолликулы, а виды
сем. Analgidae при массовом размножении внедряются в кожу птицы или про-
никают в носовую полость и легкие. В последних случаях клещи, вероятно,
питаются лимфой и слизистыми выделениями эпителия.
Клещи сем. Dermoglyphidae поселяются в полости очинов крупных перьев,
обычно маховых, рулевых и больших кроющих крыла. Они питаются роговыми
частями пера и дужкой. В одном пере клещи живут в течение 2—3 поколений.
255
Клещи покидают перо только в период линьки хозяина, в гнездовой период
для заражения птенцов, а также при заполнении очина линочными шкурками.
Выход клещей из старого пера и проникновение в новое происходит через верх-
нее отверстие в очине. На поверхность перьев выходят только самки и самцы
или телеонимфы [Дубинин, 1956].
Наименее патогенными для хозяев являются перьевые клещи, обитающие
на оперении. Однако, питаясь жировой смазкой, клещи ухудшают свойства
пера, увеличивают его смачиваемость. У птиц, ослабленных инфекционными
заболеваниями, клещи, в норме обитающие на оперении, размножаясь в массе,
способны проникать в носовую полость и в легкие птицы. Таким образом,
и эта экологическая группа перьевых клещей представляет собой потенциально
опасную паразитофауну птицы.
Надсем. PTEROLICHOIDEA
Идиосома уплощена дорсовентрально или выпуклая, веретеновидная.
Щетинки р, q двузубчатые на вершине, имеются на всех лапках; щетинки и
и v отсутствуют. Лапки IV с 3 вентральными щетинками (s, w, г). Эмподиальный
коготь отсутствует, на вершине лапки имеется амбулакральный аппарат, состоя-
щий из диска и стебля. Центральный и латеральные склериты диска всегда
крупные. Кондилофоры Г-образные, кондилофорный направитель отсутствует
(рис. 125, 5).
Представители надсемейства обитают на поверхности оперения птиц или
в полости очинов. — В СССР — 8 семейств. На домашних птицах встречаются
представители только одного семейства.
Сем. PTEROLICHIDAE
Идиосома сильно уплощена дорсовентрально. Спинная поверхность тела
занята крупными щитами. Гистеросомальный щит имеется у обоих полов.
Присутствует пара теменных щетинок vi. Ноги III и IV расположены на боко-
вых краях идиосомы. Вертлуги I—III советниками. Щетинка кТ на голенях
IV имеется. Лапки III с 3 вентральными щетинками. У самцов опистосо-
мальные лопасти хорошо развиты или едва намечены. Анальные присоски
имеются. Ноги IV гипертрофированы или сходного строения с ногами III.
У самок гистеросомальный щит цельный, доходит до заднего края тела или раз-
делен на метаподосомальный и опистосомальный щиты. Эпигиний имеется.
Щетинки с2 расположены впереди генитальных присосок. Копулятивная трубка
отсутствует или представлена небольшим коническим бугорком.
Клещи обитают на вентральной поверхности маховых и кроющих перьев
крыла. Представители семейства распространены на птицах многих отрядов,
но наиболее широко представлены на куриных. В СССР — 9 родов, на Даль-
нем Востоке — 1 род. На домашних птицах обитают представители только
одного рода.
1. Pterolichus Rob. Проподосомальный щит с глубокими боковыми выем-
ками вокруг оснований щетинок set и see. Щетинки sh волосовидные, короткие.
Эпимеры I свободные. Коксальные поля I и II не склеротизованы. У 3 опи-
стосомальные лопасти прямоугольные, разделены широкой выемкой или едва
намечены. Щетинки с2 расположены впереди уровня основания генитальной
арки. Ноги IV немного толще ног III, но одинаковой с ними длины. Лапки IV
цилиндрические, с шиповидным отростком, направленным медиально. Уд дор-
сальная поверхность гистеросомы несет крупный метаподосомальный щит
(рис. 125, 4). На заднем конце тела обычно имеется небольшой опистосомальный
щит. Эпигиний полукруглый или подкововидный. Обитают только на кури-
ных. — В СССР 7 видов, на домашних птицах известен только 1 вид.
1 (1). У в опистосома короткая и широкая, опистосомальные лопасти не выра-
жены. Все щетинки на заднем конце тела волосовидные. Анальные при-
соски мелкие, слегка овальные (рис. 125, 7, 2). 0.25—0.27. У $метаподосо-
256
Рис. 125. Клещ Pterolichus obtusus.
I — тело 3 co спинной стороны; 2 — общий вид 3 с брюшной стороны, з — лапка II со спинной стороны;
4 — гистеросома $ со спинной стороны; 5 — яйцевыводное отверстие $. кф — кондилофоры.
мальный щит трапециевидный, расширенный спереди, длина его 0.18—
0.19, ширина по переднему краю 0.16—0.17 (рис. 125, 4, 5). 0.35—0.42. —
Транспалеаркт. Евр. ч. СССР, Закавказье, Ср. Азия,3 Сиб., Якут. На до-
машних курах и многих видах фазановых........Р. obtusus Rob.
Надсем. FREYANOIDEA
Идиосома уплощена дорсовентрально. Щетинки р, q, zz, v отсутствуют на
всех лапках. Лапки IV с 3 вентральными щетинками (s, w, г). Эмподиальный ко-
готь отсутствует, на вершине лапок имеется амбулакральный аппарат, состоя-
щий из диска и стебля. Центральный и латеральные склериты диска крупные,
широкие. Кондилофоры широкие и короткие, имеют вид пластинок неправиль-
ной формы. Кондилофорный направитель отсутствует (рис. 127, 3).
Все представители надсемейства обитают на поверхности маховых и крою-
щих перьев крыла, иногда проникают в перьевые фолликулы. — В СССР —
1 семейство.
Сем. FREYANIDAE
Идиосома широкая, сильно уплощенная дорсовентрально. Почти вся дор-
сальная поверхность гистеросомы занята гистеросомальным щитом. Имеется
пара теменных щетинок vi. Плечевые щетинки h и sh волосовидные. Ноги III
и IV удалены от боковых краев тела и прикреплены вентрально. Лапки I и II
с 4 вентральными щетинками (wa, га, la, s), III — с 3 вентральными щетинками
(s, w, г). Щетинка кТ на голенях IV и соленидий о4 на коленах I — III присут-
ствуют. У самцов опистосомальные лопасти хорошо развитые, треугольные
или короткие прямоугольные. Анальные присоски имеются. Ноги III или IV
гипертрофированы или такого же строения, как у самок. У £ эпигиний имеется
(отсутствует только у рода Mlchaelid). Копулятивная трубка отсутствует.
На птицах, обитающих в водных биотопах. Наиболее широко семейство пред-
ставлено на пластинчатоклювых, в том числе на домашних утках. — В СССР —
6 родов. На домашних птицах встречаются клещи только одного рода.
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
1. Freyana Hal. Идиосома у обоих полов почти овальная. Эпимеры I слиты
У-образно. Боковые края гистеросомы обычно с широкой прозрачной мембра-
ной. Щетинки на заднем конце тела с листовидными выростами или полностью
листовидные. У $ опистосомальные лопасти короткие, имеют вид небольших
прямоугольных выступов. На лопастях и в терминальной выемке мембрана
отсутствует. Щетинки Z4, pai, d5 всегда с листовидными расширениями. Го-
лени I и II с дорсальным гребневидным выростом. У гетероморфных $ весь
коксостернальный скелет слит в единую сеть, опистосомальные лопасти четко
выражены. У гомеоморфных $ элементы коксостернального скелета проподосомы
всегда свободные, коксальные поля III замкнутые, коксальные поля IV —
открытые, вентрум отсутствует, опистосомальные лопасти выражены слабо.
У 2 задняя часть тела притупленная, широко закругленная. Все коксальные
поля не замкнуты. Эпигиний небольшой, дуговидный. На заднем краю тела
1—3 пары щетинок ланцетовидные или с листовидными расширениями. На ути-
ных. — В СССР — 14 видов. На домашних птицах на ДВ 3.
Самки
1 (4). Эпимеры II прямые или слабо изогнутые (рис. 126, 7). Щетинки pat
одновершинные, ланцетовидные или листовидные.
2 (3). Боковые мембраны гистеросомы отсутствуют. Щетинки Z4 и pai узкие,
ланцетовидные; щетинки cZ5 без ланцетовидного расширения в основании
(рис. 126, 5). 0.5—0.62. — Космополит. Евр. ч. СССР, Закавказье, Якут.,
3 Сиб., ДВ; С Америка. На сером гусе и других гусиных..........
..................................... F. anserina Meg. et Trt.
258
3 (2). Боковые мембраны гистеросомы имеются; простираются от уровня осно-
ваний ног III до щетинок Z4. Щетинки Z4 широкие, ланцетовидные, щетинки
pai имеют вид неправильного ромба; щетинки d5 с ланцетовидным расши-
Рис. 126. Клещи рода Freyana.
1 ,2 — передняя часть тела $, с брюшной стороны: 1 — F. anatina, 2 — F. largifolia; 3 — общий вид ?
F. anscrina с брюшной стороны; 4 — гетероморфный <? F. largifolia с брюшной стороны, 5 — гетероморфный
<? F. anatina с брюшной стороны; 6 — гомеоморфный <? того же вида; 7 — гомеоморфный о F. anscrina; 8, 9 —
задний конец тела $> снизу: 8— F. anatina, 9— F. largifolia; 10—13 —щетинки pai; 10 — гомеоморфный <J
F. anatina, 11 — гетероморфный о F. anatina, 12 — гетероморфный: F. largifolia, 13 — гомеоморфный <J
F. anscrina (по: Дубинин, 1953, с изменениями). — латеральная щетинка, pai — внутренняя постаналь-
ная щетинка.
рением в основании (рис. 126, 7, 8). 0.5—0.55. — Космополит. ДВ. На до-
машней утке и других видах утиных..........F. anatina Meg. et Trt.
4 (1). Эпимеры II резко изогнуты, почти под прямым углом (рис. 126, 2). Ще-
тинки pai двувершинные, ствол щетинки входит во внутреннюю вершину
(рис. 126, 9). 0.48—0.52. — Космополит. ДВ. На домашней утке и других
видах утиных...................................F. largifolia Meg. et Trt.
17*
259
Самцы
1 (4). Щетинки Z4 узкие, ланцетовидные или ножевидные. У гетероморфных 3
боковые мембраны гистеросомы начинаются от уровня оснований ног III
и плавно расширяются к заднему концу тела (рис. 126, 5). У гомеоморф-
ных d эпимеры II прямые или слабо изогнутые (рис. 126, 7).
2 (3). Внутренний мембранозный вырост щетинок pai широкий, обычно с хо-
рошо выраженным острым углом (рис. 126, 5, 6, 10, 11). Гомеоморфный
d — 0.45—0.53, гетероморфный d — 0.5—0.6.................................
.............................................F. anatina Meg. et Trt.
3 (2). Внутренний мембранозный вырост щетинок pai узкий, угол щетинки
закругленный (рис. 126, 7, 13). Гомеоморфный d — 0.55—0.60, гетеро-
морфные d неизвестны..............................F. anserina Meg. et Trt.
4 (1). Щетинки Z4 широкие, овальные. У гетероморфных d боковые мембраны
гистеросомы начинаются от плечевых щитов и имеют равную ширину
на всем протяжении до щетинок Z4 (рис. 126, 4, 12). У гомеоморфных d
эпимеры II изогнуты почти под прямым углом (рис. 126, 2). Гомеоморф-
ный d — 0.45—0.5, гетероморфный d — 0.5—0.58.............................
.................................... . . . F. largifolia Meg. et Trt.
Надсем. ANALGOIDEA
Идиосома сильно уплощенная дорсовентрально или яйцевидная, или ша-
ровидная. Щетинки р, q, и, v на лапках всех ног отсутствуют. Лапки IV с 2 вен-
тральными щетинками (w, г). Эмподиальный коготь отсутствует, на вершине
лапки имеется амбулакральный аппарат, состоящий из стебля и диска. Склериты
амбулакрального диска обычно узкие. Кондилофоры тонкие, стержневидные,
прямые или слабо изогнутые (рис. 127, 7, 2, 4, 5). Кондилофорный направитель
имеется или вторично редуцирован. В последнем случае амбулакральный диск
очень маленький, значительно меньше по диаметру, чем длина амбулакрального
стебля, или диск совершенно отсутствуем (сем. Knemidocoptidae) (рис. 135, 3,
4\ 136, 4). На поверхности оперения птиц, в полости очинов, на поверхности или
в толще кожи хозяев. — В СССР 12 семейств, на Дальнем Востоке на домашних
птицах отмечено 8 семейств клещей.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ
1 (2). Амбулакральный диск с 3—5 отростками на краю (рис. 137, 5, 4). Верт-
луги всех ног без щетинок . . ......................8. Laminosioptidae
2 (1). Амбулакральный диск округлый или сердцевидный. Вертлуги по крайней
мере ног I и II со щетинками.
3 (4). Идиосома шаровидная или яйцевидная, ноги I и II короткие, конические.
Проподосомальный щит с широкими, сильно склеротизованными боко-
выми гребнями (рис. 135, 7; 136, 7).................7. Knemidocoptidae
4 (3). Идиосома вытянутая в длину, сильно уплощенная дорсовентрально или
веретеновидная. Если идиосома яйцевидная, то ноги I и II относительно
длинные, не конические. Проподосомальный щит иного строения.
5 (8). Щетинка кТ голеней IV отсутствует. Бедро и колено всех ног слиты
в единый членик.
6 (7). Щетинки sh расположены впереди щетинок h. Щетинки vi всегда отсут-
ствуют. У $ гистеросомальный щит не доходит до заднего края идиосомы;
задний конец тела закругленный...........................2. Xolalgidae
7 (6). Щетинки sh расположены позади щетинок h. Щетинки vi обычно имеются.
У ? гистеросомальный щит всегда доходит до заднего края идиосомы;
задний конец тела раздвоен или не раздвоен .........4. Alloptidae
8 (5). Щетинка кТ голеней IV имеется (рис. 134, 4).
9 (10). Соленидий колен III отсутствует.............6. Epidermoptidae
260
10 (9). Соленидий колен III имеется.
11 (12). Склериты амбулакрального диска хорошо развиты; латеральные скле-
риты обычно с овальными лакунами; центральный склерит широкий,
имеет вид многоугольника (рис. 127, 5). Амбулакральный стебель I и
II пар ног тонкий, в 3—4 раза меньше диаметра амбулакрального диска
....................................................3. Avenzoariidae
Рис. 127. Лапки II и амбулакральный аппарат клещей.
1 — Paralges alwori (Dermoglyphidae); 2 — Megninia cubilalis (Analgidae); 3 — Frey ana anaiina (Freyani-
dae); 4 — Ingrassia velaia (Ingrasiinae); 5 — Bdellorhynchus polymorphus (Avenzoariidae). (3 — no: Gaud,
Atyeo, 1977). — кондплофоры, лс — латеральный склерит, г/с — центральный склерит.
12(11). Склериты амбулакрального диска мелкие; латеральные склериты уз-
кие, палочковидные, крючковидные или совсем редуцированы; центральный
склерит диска имеет вид узкой полоски или Т-образный (рис. 127, 7, 2, 4).
Амбулакральный стебель пог I и II утолщенный, лишь в 1.5—2 раза
меньше диаметра амбулакрального диска.
13 (14). Ноги 111 (у $ и ноги IV) расположены на краях тела. Проподосомальный
щит с медиальными продольными гребнями или без них. У 3 гистеросо-
мальный щит имеется. У $ эпигиний имеется (рис. 128, 1—4) . . . .
.......................................................1. Analgidae
14 (13). Ноги III и IV у обоих полов расположены на вентральной стороне тела,
удалены от его боковых краев. Проподосомальный щит с продольными бо-
ковыми гребнями. У 8 гистеросомальный щит, а у $ эпигиний отсутствуют
(рис. 132, 1—10)...................................5. Dermoglyphidae
261
1. Сем. ANALGIDAE
Идиосома умеренно уплощенная дорсовентрально. Проподосомальный щит
с медиальными продольными гребнями или без них. Щетинки sh расположены
впереди щетинок h. Членики всех ног соединены подвижно. Голени I и II с вен-
тральными шиповидными отростками. Вертлуги I —III со щетинкой. кТ голе-
ней IV имеется. Соленидий имеется на коленях I—III. Амбулакральный диск
небольшой, округлый, центральный и латеральные склериты узкие. Амбула-
кральный стебель длинный, утолщенный, иногда сильно вздутый в основании,
по длине значительно превышает диаметр диска (рис. 127, 2). У <5 гистеросо-
мальный щит имеется. Опистосомальные лопасти хорошо развиты или отсутст-
вуют. Анальные присоски имеются. Ноги III или IV гипертрофированы. У g
гистеросомальный щит отсутствует. Эпигиний имеется. Задний конец тела
с 2 парами макрохет.
Клещи обитают на контурных и пуховых перьях тела птиц, питаются обычно
жировой смазкой. При массовом размножении могут внедряться в кожу птицы
или проникать в носовую полость и легкие. В последних случаях клещи, ве-
роятно, питаются лимфой и слизистыми выделениями эпителия. Это наиболее
широко распространенное у птиц семейство перьевых клещей, его представители
встречаются на птицах 16 отрядов. — В СССР — 8 родов, на домашних птицах
на Дальнем Востоке представители только одного рода.
I. Megninia Berl. Проподосомальный щит трапециевидный, с продольными
гребнями. Вентральные отростки голеней I и II тонкие, ножевидные. Лапки
1 и II с короткими шиповидными вентральными отростками или без них. Бедро I
без наружного сильно склеротизованного крючковидного отростка.
У о эпимеры I обычно слиты, реже незначительно расставлены. Опистосо-
мальные лопасти треугольные, выемка между ними треугольная или прямо-
угольная. Ноги III гипертрофированы, значительно длиннее и толще ног IV.
Лапка III прямая, сильно вытянута, несколько суживается к вершине, две ее
вентральные щетинки (w, г) короткие и утолщенные, имеют ланцетовидную или
шиповидную форму. Щетинка кТ голеней III длинная, в 3—4 раза превышает
длину голени. У j эпимеры I свободные. Обитают только на куриных. —
В СССР — 6 видов, на домашних птицах на ДВ — 2.
1 (2). У в эпимеры I слиты У-образно (рис. 128, 2, 2). У $ эпигиний изогнут
слабо, почти прямой, расположен позади концов эпимер II (рис. 128, 8).
в — 0.32—0.35, $ — 0.72—0.34. — Космополит. На домашних курах
....................................................М. cubitalis (Meg.)
2 (1). У 8 эпимеры I свободные, сближены задними концами, по не слиты
(рис. 128, 4). У эпигиний дуговидно изогнут, расположен между кон-
цами эпимер II (рис. 128, 3). 8 — 0.3—0.33, $ — 0.33—0.35. — Космопо-
лит. На домашних курах и других куриных . . . М. gynglymura (Meg.)
2. Сем. XOLALGIDAE
Идиосома умеренно уплощенная дорсовентрально. Проподосомальный щит
без продольных гребней. Гистеросомальный щит имеется у обоих полов. Ще-
тинки sh расположены впереди щетинок h. Бедро и колено всех ног слиты в еди-
ный членик. Голени I и II с вентральными шиповидными отростками. Вертлуги
I —III или I —II со щетинками. Щетинка кТ голеней IV отсутствует. Соленидий
имеется на коленах I — III. Амбулакральный диск небольшой, округлый или
сердцевидный; центральный и латеральные склериты узкие. Амбулакральный
стебель тонкий и короткий, не превышает по длине диаметра амбулакрального
диска (рис. 127, 4).
У S опистосомальные лопасти хорошо развиты, обычно треугольные. Аналь-
ные присоски имеются. Ноги III и (или) IV гипертрофированы. У £ гисте-
росомальный щит не доходит до заднего края тела. Эпигиний имеется (за исклю-
чением подсем. Xo]alginae). Задний конец тела с 2 парами макрохет.
262
Рис. 128. Клещи родов Megninia и Ingrassta.
1,2 — М. cubitalis: 1 — общий вид в, 2 — идиосома о с брюшной стороны; 3,4 — М. gynglymura: 3 —
общий вид ? с брюшной стороны, 4 — идиосома <? с брюшной стороны; 5—7— 1. velata: 5 — общий вид<$
с брюшной стороны, 6 — тело <? со спинной стороны, 7 — тело ? со спинной стороны; 8 — передняя часть 9
М. cubitalis с брюшной стороны; 9 — тело 9 I. velata с брюшной стороны.
Обитают на контурных и пуховых перьях тела птиц многих отрядов, пре-
имущественно связанных с водными биотопами. — В СССР — 5 родов.
На домашних птицах на Дальнем Востоке 1 род.
Подсем. INGRASSIINAE
1. Ingrassia Oud. Проподосомальный щит трапециевидный или овальный,
отделен от лопаточных щитов; щетинки see расположены вне этого щита. Эпи-
меры I слиты V- или У-образно. Бедро II без крючковидного, направленного
наружу отростка. У 3 эпимеры IV сильно вытянуты, на их передних концах
расположены щетинки сг. Эпиандрий отсутствует. Опистосомальные лопасти
длинные, треугольные, окружены непрерывной мембраной, проходящей по
их наружному краю, по вершине и по краю терминальной выемки. Ноги III
гипертрофированы, в 1.5—2 раза длиннее и толще ног IV. У 9 гистеросомаль-
ный щит не соединен с плечевыми щитами. — В СССР — 5 видов, на домашних
птицах на ДВ — 1.
1 (1). Проподосомальный щит трапециевидный. Эпимеры I слиты V-образно.
У d опистосомальные лопасти широко расставлены, все пространство
между ними занято мембраной; на вершинах лопастей мембрана широко
закругленная, вырезка в мембране короткая, не доходит до уровня вер-
шин лопастей (рис. 128, 5, 6). У $ гистеросомальный щит имеет грибовид-
ную форму (рис. 128, 7, Р). в — 0.31—0.35, $ — 0.34—0.36. — Транспа-
леаркт. Евр. ч. СССР, Якут.; Европа. На домашней утке и других утиных
........... ? ... I. velata (Meg.)
3. Сем. AVENZOARIIDAE
Идиосома сильно уплощена дорсовентрально. Проподосомальный щит без
продольных гребней. Щетинки sh расположены впереди или на одном уровне со
щетинками h. Гистеросомальный щит имеется у обоих полов. Голени I и II без
вентральных шиповидных отросткоЪ. Все членики ног соединены подвижно.
Вертлуги I —III со щетинками. Щетинка кТ голеней IV имеется. Соленидий
имеется на коленах I—III. Амбулакральный диск крупный, сердцевидный,
центральный склерит диска широкий, имеет вид многоугольника, латеральные
склериты также крупные, обычно с лакунами. Амбулакральный стебель корот-
кий и тонкий, по длине значительно короче диаметра диска (рис. 127, 5).
|У самцов опистосомальные лопасти развиты или сильно укороченные. Аналь-
ные присоски имеются. Ноги III, а иногда и I—II гипертрофированы. У самок
гистеросомальный щит доходит или не доходит до заднего края тела, иногда
разделен на несколько отдельных щитов. Эпигиний имеется. Задний конец
тела с 1—2 парами макрохет.
Клещи обитают на маховых и кроющих перьях крыла; они приурочены пре-
имущественно к отрядам птиц, связанных с водными биотопами. — В СССР —
15 родов. На домашних птицах на Дальнем Востоке встречаются только клещи
рода Bdellorhynchus.
1. Bdellorhynchus TH. У в эпимеры I слиты У-образно. Опистосомальные
лопасти сильно укороченные, едва выражены. Генитальная арка окаймлена
с боков узкими серповидными генитальными аподемами. Ноги III гипертрофи-
рованы, всегда значительно толще ног IV. Самцы полиморфны. У гетероморф-
ных самцов проподосома сильно утолщенная с острым ростральным выступом;
теменные щетинки ив отсутствуют; ноги I—II и хелицеры гипертрофированы,
сильно удлинены. Последние могут составлять до х/2 длины тела (рис. 129, 7, 2)
У гомеоморфных в проподосома не утолщенная, ноги I и II такого же строения,
как и у самок. Пара теменных щетинок присутствует у обоих полов. У 9 эпи-
меры I слиты V- или У-образно. Гистеросомальный щит узкий, прямоугольный,
сильно вытянут в длину, но не доходит до заднего края тела. Эпигиний подко-
вовидный. Задний конец тела с 1 парой макрохет (рис. 130, 7, 2). В СССР —
3 вида. На ДВ на домашних птицах — 1 вид.
264
1 (1). У 2 эпимеры I слиты V-образно; мелкопунктированная часть гистеро-
сомального щита имеет форму узкого треугольника (рис. 130, 7, 2). У d
эпиандрий отсутствует (рис. 129, 1—4). Гомеоморфный 3 — 0.34—0.37,
гетероморфный 3 — 0.48—0.55, $ — 0.40—0.45. — Транспалеаркт.
Евр. ч. СССР, Закавказье, Ср. Азия, ЗСиб., Якут. На домашней утке
и других утиных....................................В. polymorphic Trt.
Рис. 129. Клещ Bdellorhynchus poly-
morphus.
1 — гетероморфный <J, идиосома со спин-
ной стороны; 2 — то же, общий вид с брюш-
ной стороны; 3 — гемеоморфный <J, об-
щий вид с брюшной стороны; 4 — то же,
генитальный аппарат. (По: Дубинин,
1956 с изменениями).
4. Сем. ALLOPTIDAE
Идиосома сильно уплощена дорсовентрально. Проподосомальный щит без
гребней. Щетинки sh расположены позади щетинок h. Гистеросомальный щит
имеется у обоих полов. Бедро и колено всех ног слиты в единый членик. Го-
лени I и II без вентральных шиповидных отростков. Вертлуги I —III со ще-
тинками. Щетинка кТ голеней IV отсутствует. Соленидий имеется на коленах
I —III. Амбулакральный диск небольшой, сердцевидный; центральный его
склерит Т-образный, латеральные склериты развиты слабо. Амбулакральный
стебель тонкий и короткий, по длине не превышает диаметр диска (рис. 131, 5).
У $ опистосомальные лопасти хорошо развиты, иногда вторично сливаются
внутренними краями. Анальные присоски имеются. Ноги III или IV гипертрофи-
рованы или развиты одинаково. У g гистеросомальный щит доходит до заднего
края тела. Эпигиний имеется. Задний конец тела с 2 парами макрохет (рис. 130,
5, 4- 131, 7, 2).
18 Зак. № 990
265
Клещи обитают на маховых и кроющих перьях крыльев, только на отрядах
птиц, связанных с водными биотопами (утиные, ржанкообразные, журавлеобраз-
ные). — В СССР — 3 рода, на домашних птицах на Дальнем Востоке — 1.
1. Brephosceles Hull. Эпимеры I слиты У-образно. Имеется 1 теменная
щетинка vi. У <5 опистосомальные лопасти вытянутые, прямоугольные, верши-
Рис. 130. Клещи родов Bdellorhynchus и Brephosceles.
1'2 — Bd. poli/morphus: 1 — тело $ <о спинной стороны, 2 — общий вид о с брюшной (тороны; 3, 4 —
Br. anatina'. 3 — гистеросома о (о < пигиой (тороны, 4 — гистеросома о ( брюшной (тороны. (2, 2 — по:
Дубинин, 1936 с изменениями; остальное — ориг.).
нами загнуты внутрь, разделены широкой терминальной выемкой. Лопасти
имеют хорошо выраженную мембрану на внутреннем крае и узкие мембраны
вдоль наружного края и на вершине. Гистеросомальный щит цельный или раз-
делен поперечной бороздой позади уровня ног IV. Ноги III и IV одинаковой
величины, не гипертрофированы. У < эпигиний полукруглый, отделен от эпи-
мер IV и эпимеритов III. Задний край тела закругленный или с небольшой
выемкой. На ржанкообразных, пластинчатоклювых и трубконосых. —
В СССР — 6 видов, на домашних птицах на ДВ — 1.
266
1 (1). У 8 генитальные аподемы длинные и узкие, слиты с эпимеритами III.
Передние концы аподем сильно сближены с эпимерами III. Опистомаль-
ные лопасти относительно короткие, длина их не более чем в 2.5 раза пре-
вышает ширину в основании. Длина терминальной выемки 0.047—0.062,
ширина 0.032—0.043. Длина тела 0.28—0.34 (рис. 130, 5, 4). У j гистеро-
сомальный щит на заднем крае с треугольной выемкой, длина которой
Рис. 131. Клещ Brephosceles anatina.
1 — тело $ со спинной стороны; 2 — общий вид $ с брюшной стороны; 3 — лапка’ II.
0.015—0.025 (рис. 131, 1—3). Длина тела 0.34—0.39, ширина 0.115—
0.135. — Транспалеаркт. Евр. ч. СССР, 3 Сиб., Якут. На домашней утке
и других видах утиных...............................В. anatina Dub.
5. Сем. DERMOGLYPHIDAE
Идиосома веретеновидная. Проподосомальный щит с боковыми склеротизо-
ванными гребнями. Гистеросомальный щит отсутствует у обоих полов. Щетинки
sh расположены впереди щетинок к. Все членики ног соединены подвижно. Го-
лени I и II без вентральных шиповидных отростков. Вертлуги I —III со щетин-
ками. Щетинка кТ голеней IV имеется. Соленидий имеется на коленах I—
III. Амбулакральный диск небольшой, округлый; центральный и латеральные
склериты узкие. Амбулакральный стебель короткий и толстый, по длине не-
много превышает диаметр диска (рис. 127, 1). У 5 опистосомальные лопасти
отсутствуют. Анальные присоски отсутствуют (за исключением рода Gaudo-
18*
267
glyphus). Ноги III гипертрофированы или имеют сходное строение с ногами IV.
У 9 эпигиний отсутствует. Задний конец тела с 2 парами макрохет.
Обитают в полости очинов крупных перьев (маховых, рулевых и больших
кроющих) на птицах самых разнообразных отрядов. — В СССР — 3 рода.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (4). Боковые гребни проподосомального щита параллельные, расстояние
между ними не превышает V3 ширины тела. Анальные присоски у 8 отсут-
ствуют (рис. 132, 6, 9).
2 (3). Эпимеры I слиты задними концами с эпимерами II. У в ноги III и IV при-
близительно равной длины и толщины (рис. 132, 7)....................
............................................2. Dermoglyphus Meg.
3 (2). Эпимеры I отделены от эпимер II. У d ноги III значительно длиннее
и толще ног IV (рис. 132, 8).........................3. Paralges Trt.
4 (1). Боковые гребни проподосомального щита расходятся в задней части щита,
расстояние между их задними концами более V2 ширины тела. Анальные
присоски у <J имеются (рис. 132, 1).......1. Gaudoglyphus Вг. et John.
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
1. Gaudoglyphus Вг. et John. Проподосомальный щит сильно расширен
в задней части, с выпуклым задним краем. Боковые гребни щита сзади расхо-
дятся, расстояние между их задними концами более 1/2 ширины тела. Эпимеры I
не слиты задними концами с эпимерами II. Щетинки с2 имеются. У $ анальные
присоски имеются. 3 полиморфны. У гетероморфных i ноги III более длинные
и толстые, чем ноги IV, с шипами на медиальной поверхности бедра. У гомео-
морфных 8 ноги III лишь немного длиннее, чем ноги IV, но одинаковой с ними
толщины и без шипов. — В роде 1 вид.
1 (1). Коксальные поля I полностью склеротизованы у обоих полов. У гомео-
морфных 8 бедро III цилиндрическое, у гетероморфных — сильно утол-
щенное, выпуклое на наружном крае; с 2 шипами на медиальном крае
(рис. 132, 1—4). Гомеоморфный 8 — 0.30—0.36, гетероморфный 8 —
0.35—0.38, j — 0.33—0.39. — Космополит. Евр. ч. СССР; Европа, США.
На домашних курах...................................G. minor (Nor.)
2. Dermoglyphus Meg. Проподосомальный щит почти прямоугольный, с пря-
мым задним краем. Боковые гребни щита параллельные, расстояние между
ними не более 1/3 ширины тела. Эпимеры I задними концами слиты с эпиме-
рами II. Щетинки с2 имеются. У 8 анальные присоски отсутствуют. Ноги III
и IV одинаковой длины и толщины или первые немного толще; шипы на бедрах
и вертлугах отсутствуют. — В СССР и на ДВ — 2 вида; медико-ветеринарное
значение имеет 1 вид.
1 (1). Эпимеры II дуговидные. Длина колена, бедра, вертлуга ног III и IV пре-
вышает ширину этих члеников (рис. 132, 5—7). 8 — 0.63—0.67, $ —
0.65—0.68. — Евр. ч. СССР; Европа. На домашних курах................
.............................................D. elongatus (Meg.)
3. Paralges Trt. Проподосомальный щит почти прямоугольный, с прямым
или вогнутым задним краем. Боковые гребни щита параллельные, расстояние
между ними не более 1/3 ширины тела. Эпимеры I не слиты с эпимерами II. Ще-
типки с2 отсутствуют. У 8 анальные присоски отсутствуют. Ноги III гипертро-
фированы, значительно длиннее и толще ног IV: бедро или вертлуг несут шип
на медиальном крае. — В СССР и на ДВ — 1 вид.
1 (1). Задние концы эпимер II резко загнуты наружу. У 8 концы эпимер III,
направленные внутрь, отделены от эпиандрия; бедро III с тупым шипом
на медиальной поверхности (рис. 132, 8—10). 8 — 0.45—0.50, $ — 0.49—
0.60. — Евр. ч. СССР; Индия. На домашней утке и других видах утиных
. . ................................................Р. alwari (Gaud)
268
Рис. 132. Клещи родов Gaudoglyphus^ Dermoglyphus и Paralges.
1—4 — Gaudoglyphus minor: 1 — тело гетероморфного о со спинной стороны, 2 — общий вид гетероморф-
ного 8 с брюшной стороны. 3 — гистеросома гомеоморфного о с брюшной стороны, 4 — проподосома с
с брюшной стороны; 5— 7 — Dermoglyphus elougalus: 5 — генитальный аппарат 3, 6 — общий вид $ со спин-
ной стороны, 7 — $ с брюшной стороны; 8—10 — Paralges alwari: 8 — общий вид j с брюшной стороны,
у — проподосома о со спинной стороны, 10 — гистеросома 8 с брюшной стороны.
6. Сем. EPIDERMOPTIDAE
Идиосома сильно уплощена дорсовентрально (за исключением рода Му lod-
ges, у которого идиосома грушевидная). Проподосомальный щит без продоль-
ных гребней. Щетинки sh расположены впереди или на одном уровне со щетин-
ками h. Все членики ног соединены подвижно. Голени I и II без вентральных
шиповидных отростков. Вертлуги I—III всегда со щетинками, вертлуги IV
со щетинками или без них. Щетинка кТ на голенях IV и соленидий на коле-
нах I—II присутствуют. Амбулакральный диск небольшой, его склериты сильно
редуцированы или полностью отсутствуют. Амбулакральный стебель тонкий и
длинный, по длине обычно в 1.5—2 раза превышает диаметр диска (рис. 133,
5). У $ опистосомальные лопасти хорошо развиты (рис. 134, 7, 2) или сильно
укорочены (рис. 133, 7, 2). Гистеросомальный щит имеется. Анальные присоски
имеются. Ноги III или IV гипертрофированы или обе пары одинакового строе-
ния. У 2 гистеросомальный щит имеется и не доходит до заднего края тела
или полностью отсутствует. Эпигиний имеется. Задний конец тела с 1 парой
макрохет.
Обитают на поверхности кожи (подсем. Dermationinae), частично внедряются
в нее и даже проделывают короткие ходы (подсем. Epidermoptinae).
В СССР — 8 родов, на домашних птицах на Дальнем Востоке 2.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВА И РОДОВ
1 (10). Щетинки Z4 и отверстия жировых желез имеются. Лапки I и II кониче-
ские, с крупными крючковидными или шиповидными отростками. Наруж-
ная щетинка колена II (mG) имеется (рис. 133, 2). У $ эпигиний располо-
жен позади концов эпимер I и не сливается с ними. (Подсем. Epider-
moptinae Trt).
2 (3). Вершина лапок I и II с шиповидным отростком. Концы эпимер I широко
расставлены...................................... 1. Epidermoptes Riv.
3 (2). Вершина лапок I и II с крючковидным или Т-образным отростком. Задние
концы эпимер I тесно сближены или*слиты.
4 (5). Дорсальные щетинки пальп dp2 веретеновидные...................
.......................................Microlichus Trt. et Neum.*
5 (4). Дорсальные щетинки пальп dp2 волосовидные.
6 (7). Крючковидный отросток лапок I и II одинаковой величины........
...........................................Metamicrolichus Fain ♦
7 (()). Крючковидный отросток лапки I значительно крупнее, чем на лапке II.
8 (9). Отросток лапки I когтевидный или серповидный. Гистеросомальный щит
имеется.......................................Promyialges Fain ♦
9 (8). Отросток лапки I Г- или Т-образный. Гистеросомальный щит отсутствует
..................................................Myialges Trt.*
10 (I). Щетинки Z4 и отверстия жировых желез отсутствуют. Лапки I и II ци-
линдрические, со слабым шиповидным отростком на вершине или без него.
Наружная щетинка колена II (rnG) отсутствует. У $ эпигиний слит с эпи-
мерами I таким образом, что они образуют U-образную фигуру (рис. 134, 4)
(подсем. Dermationinae). Соленидий на лапках I и II одинаковой ве-
личины, щетинки Z3 имеются.......................• . 2. Rivoltasia Сап»
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
1. Epidermoptes Riv. Проподосомальный щит с острыми передними углами.
Щетинки see расположены снаружи от set. Гистеросомальный щит имеется. Эпи-
меры I не слиты. Щетинки 12 и Z3 имеются. Все щетинки пальп волосовидные.
Крючковидные отростки на вершине лапок I и II короткие, имеют сходное строе-
ние (рис. 133, 2, 3, 5). У $ задний конец тела сильно сужен. Опистосомальные
* Представители отмеченных родов встречаются на домашних птицах, но в силу слабой
изученности на Дальнем В остоке пока не найдены, поэтому их диагнозы ниже не приведены.
270
лопасти короткие, сильно сближены с закругленными мембранами на вершинах.
Генитальный аппарат окаймлен с боков узкими генитальными аподемами,
сзади — генитальным щитом. Ноги IV немного толще ног III, но приблизительно
одинаковой с ними длины. Лапка III с апикальным шиповидным отростком.
У j гистеросомальный щит непарный, прямоугольный. Эпигиний короткий,
Рис. 133. Клещ Epidermoptes bilobatus.
1 — тело о со спинной стороны; 2 — общий вид <? с брюшной стороны; з — нога I; 4 — тело $ со спин-
ной ст эроны; 5 — общий вид $ с брюшной стороны. mG — наружная щетинка колена II, lt——лате-
ральные щетинки. (По: Fain, 1965).
дуговидный, расположен между концами эпимер I. Анальная щель расположена
терминально, аданальные щиты отсутствуют. Лапки III и IV с шиповидными
апикальными отростками. На куриных. — В СССР и на ДВ — 1 вид.
1 (1). У d задний конец тела постепенно суживается позади оснований ног III.
Терминальная выемка щелевидная. Мембраны на вершинах лопастей ко-
роткие, длина их меньше ширины в основании. Длина щетинок </5 меньше
расстояния между основаниями щетинок Z5. Отверстия жировых желез
расположены ближе к щетинкам Z3, чем к Z2 (рис. 133, 7, 2). 0.16—0.18.
У $ гистеросомальный щит имеет наибольшую ширину в передней части;
271
длина щита составляет менее 1/2 длины тела (рис. 133, 4, 5). 0.23—0.27. —
Голарктика. Евр. ч. СССР, Закавказье, Ср. Азия; Европа, Сев. Америка.
На домашних курах................................Е. bilobatus Riv.
2. Rivoltasia Сап. Проподосомальный щит со сглаженными боковыми
краями, передние углы не выражены. Щетинки see расположены медиальнее
Рис. 134. Клещ Rivoltasia bijurcata.
1 — тело <? со спинной стороны; 2 — тело <5 с брюшной стороны; 3 — тело $ со спинной стороны; 4 — об-
щий вид $ с брюшной стороны. кТ — вентральная щетинка голени IV. (По: Fain, 1963).
щетинок set. Гистеросомальный щит имеется. Эпимеры I не слиты. Щетинки 12
и 13 имеются. Все щетинки пальп волосовидные. Лапки I и II без апикальных
шиповидных отростков. Соленидий развит на лапках I и II одинаково.
Лапки III и IV с шиповидным отростком в базальной части. У 6 опистосомаль-
ные лопасти прямые, сильно вытянутые, широко расставлены. Внутренние края
и вершина лопастей с хорошо развитыми мембранами. Генитальный аппарат
без генитальных аподем и генитального щита. Ноги IV толще ног III, но при-
близительно одинаковой с ними длины. У 9 гистеросомальный щит непарный,
прямоугольный. Эпигиний слит с задними концами эпимер I в £7-образную
фигуру. Анальная щель расположена вентрально, по бокам имеется пара ада-
нальных щитов. На куриных. — В СССР и на ДВ — 1 вид.
272
1 (1). Бедра всех йог с вентральными крючковидными отростками (рис. 134,
1—4). У d между генитальным аппаратом и анальными присосками скле-
риты отсутствуют, d — 0.16—0.22, $ — 0.20—0.25. — Евр. ч. СССР;
Европа. На домашних курах........................R. bifurcata (Riv.)
5
Рис. 135. Клещ Knemidocoptes mutans.
1 — тело $ со спинной стороны; 2 — общий вид $ с брюшной стороны; з — лапка I $ с брюшной стороны;
4 — нога I $ со спинной стороны; 5 — общий вид 3 с брюшной стороны, h — плечевая щетинка, sh —
субплечевая щетинка. (По: Fain, Eisen, 1967).
7. Сем. KNEMIDOCOPTIDAE
Идиосома сильно выпуклая, шаровидная или яйцевидная. Проподосомаль-
ный щит с сильно склеротизованными латеральными гребнями. Щетинки sh
расположены впереди щетинок h. Все членики ног соединены подвижно или
лапка и голень слиты в единый членик. Вертлуги I — III со щетинками, вертлуги
IV — со щетинками или без них. Голени I и II без вентральных шиповидных
отростков. Щетинка кТ голеней IV имеется или отсутствует. Амбулакральный
диск, если имеется, маленький, округлый, склериты диска и кондилофорный
273
направитель сильно редуцированы, едва заметны; амбулакральный стебель
длинный и тонкий, в 3—4 раза превышает диаметр (рис. 135, 5; 136, 5).
У <5 задний конец тела закругленный, опистосомальные лопасти совершенно
не выражены. Гистеросомальный щит имеется. Анальные присоски имеются
или отсутствуют. Ноги III и IV развиты одинаково. Амбулакральный аппарат
развит на всех ногах. У £ гистеросомальный щит отсутствует. Идиосома всегда
шарообразная, ноги короткие, конические. Эпигиний отсутствует. Задний конец
тела с 1 парой макрохет. Амбулакральный аппарат всех ног может быть пол-
ностью или частично редуцирован (рис. 135, 4; 136, 4). — В СССР и на ДВ —
2 рода.
Рис. 136. Клещ Neocnemidocoptes gallinae.
1 — тело $ со спинной стороны; 2 — то же, с брюшной стороны; 3 — гистеросома ° с брюшной стороны;
4 — нога I ? со спинной стороны; 5 — нога IV ? с брюшной стороны. Ру — рудиментарный амбулак-
ральный стебель. (По: Fain, Eisen, 1967).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). Щетинка кТ голеней IV имеется. Вокруг анального отверстия располо-
жены 4 пары щетинок. У $ дорсальная поверхность гистеросомы с преры-
вистыми дуговидными чешуевидными или овальными складками; лапка
и голень всех ног слиты в единый членик. У d анальные присоски отсут-
ствуют (рис. 135, 1—5).........................1. Knemidocoptes Fur.
2 (1). Щетинка кТ голеней IV отсутствует. Анальное отверстие окружено 5 па-
рами щетинок. У самок дорсальная поверхность гистеросомы исчерчена
поперечными непрерывными складками, иногда несущими мелкие зубцы;
лапки и голени не слиты в единый членик. У самцов анальные присоски
имеются (рис. 136, 1—5)....................2. Neocnemidocoptes Fain
274
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
1 . Knemidocoptes Fiir. По бокам анального отверстия имеется только 4 пары
щетинок, щетинки рае отсутствуют. Вентральные щетинки кТ голеней IV
имеются. У 3 анальные присоски отсутствуют; у g дорсальная поверхность
гистеросомы в центральной части в прерывистых складках, имеющих вид ко-
ротких дуг, чешуек или овальных бугорков. Лапки и голени всех ног слиты
в единый членик с 1—3 зубцами. Амбулакральный аппарат полностью отсут-
ствует на всех ногах. — В СССР и на ДВ — 2 вида, медико-ветеринарное зна-
чение имеет 1.
Рис. 137. Клещ Laminosioptes cysticola.
1 — тело j со спинной стороны; 2 — общий вид ? с брюшной стороны; 3 — голень и лапки II со спинной
стороны; 4 — голень и лапка IV со спинной стороны. МПЩ — метаподосомальный щит, ЛТЩ — латераль-
ный щит, ОПЩ — опистосомальный щит. (По: Fain, 1981).
1 (1). У $ лопаточные щиты позади ног II отсутствуют; дорсальная поверх-
ность гистеросомы с чешуевидными складками, занимающими небольшой
округлый или овальный участок в ее центре и не превышающий х/3 ширины
тела. У d плечевые щетинки h значительно длиннее субплечевых sh
(рис. 135, 1—5); щетинки see в 1.5—2 раза превышают длину гребней
проподосомального щита, d — 0.22—0.24, $ — 0.37—0.48. — Космопо-
лит. Евр. ч. СССР, 3 Сиб. На домашних курах и диких куриных . . .
...............................................К. mutans Rob. et Lanq.
2 . Neocnemidocoptes Fain. По бокам анального отверстия имеется 5 пар
щетинок. Вентральные щетинки кТ голеней IV отсутствуют. У 3 анальные при-
соски имеются. У g дорсальная поверхность гистеросомы исчерчена частыми
275
непрерывными поперечными складками, иногда несущими мелкие склеротизи-
рованные зубцы. Лапки и голени всех ног не слиты. Имеется рудиментарный
амбулакральный стебель, амбулакральный диск отсутствует (рис. 136, 4, 5). —
В СССР и на ДВ — 1 вид.
1 (1). Вентральная поверхность тела в центральной части, у <3 позади эпимер 11.
у самок позади яйцевыводного отверстия, несет поперечные складки с мел-
кими зубцами (рис. 136, 1—5), такими же, как и на дорсальной стороне.
<3 — 0.20—0.22, $ — 0.34—0.44. — Космополит. Евр. ч. СССР. На до
машних курах и диких куриных.............................N. gallinae (Rail.)
8. Сем. LAMINOSIOPTIDAE
Идиосома сильно вытянутая в длину, уплощенная дорсовентрально. Про-
подосомальный щит без продольных гребней. Гистеросомальный щит имеется
у обоих полов. Щетинки sh расположены впереди h. Все членики ног соединены
подвижно. Голени I и II без вентральных шиповидных отростков. Вертлуги
всех ног без щетинок. Щетинка кТ голеней IV имеется. Соленидий имеется на
коленах I — II. Амбулакральный диск маленький, округлый, с 3—5 шиповид-
ными отростками по краю. Центральный и латеральные склериты диска и кон-
дилофорный направитель сильно редуцированы. Амбулакральный стебель тол-
стый и длинный, в 3—4 раза превышает по длине диаметр диска (рис. 137, 5, 4).
У $ опистосомальные лопасти отсутствуют. Анальные присоски отсутствуют.
Ноги III и IV имеют одинаковое строение. У £ гистеросомальный щит доходит
до заднего края тела; иногда он представлен несколькими отдельными щитами.
Эпигиний имеется. Задний конец тела с 1 парой макрохет. Обитают в соеди-
нительной ткани кожи, проникая в нее через перьевые фолликулы. — В СССР
на домашних птицах весьма вероятно нахождение представителей рода La-
minosioptes.
1. Laminosioptes Meg. Проподосомальный щит цельный, не разделен по-
перечной бороздой. Щетинки Z3, Z4 и имеются. Латеральные щетинки 1г—12
длинные, приблизительно такой же длины, как и see. Все щетинки ног короче
длины члеников, на которых расположены. У 9 гистеросома с 4 щитами: мета-
подосомальным, опистосомальным и парой латеральных (рис. 137, 7). Эпимеры I
слиты У-образно, конец стернума не соединен с эпимерами II. Эпигиний не
слит с эпимерами III. — В роде 1 вид.
1 (1). У j коксальные поля I склеротизованы частично, II — IV — полностью
склеротизованы. Пальцы хелицер с 3 зубцами на вершинах. Лапки I
и II с 2 короткими шиповидными отростками, III и IV — с 2 длинными
шиповидными отростками (рис. 137, 1—4). $ — 0.25—0.27. — Космопо-
лит. Европа, Африка. На домашних курах и диких куриных ....
........................................... . . . . L. cysticola (Viz.)
ОСНОВНАЯ ЛИТЕРАТУРА
Дубинин В. Б. Перьевые клещи (Analgesoidea). Ч. 1. М.; Л., 1951. 363 с. (Фауна СССР.
Паукообразные; Т. 6. Вып. 5). Ч. 2. М.; Л., 1953. 412 с. (Фауна СССР. Паукообразные; Т. 6.
Вып. 6). Ч. 3. М.; Л., 1956. 814 с. (Фауна СССР. Паукообразные; Т. 6. Вып. 7).
Atyeo W., Gaud J. The chaetotaxy of Sarcoptiform feather mites (Acarina: Analgoidea). —
J. Kansas Entomol. Soc., 1966, vol. 39, N 2, p. 337—346.
Fain A. A review of the family Epidermoptidae Trouessart parasitic on the Skin of birds
(Acarina: Sarcoptiformes). — Konink. 6, Acad. Wetensch Let. schone Hunst. Belgie. 84, 1965,
part. 1, 176 p.; part. 2, 144 p.
(Fain А.) Фэн А. О гетероморфных дейтонпмфах (гипопусы) Acaridiae (Acarina: Sarco-
ptiformes). — Паразитология, 1968, т. 11, вып. 5, с. 395—406.
Fain A. Note on the genus Laminosioptes Megnin, 1880 (Acari, Astigmata) with description
of three new species. — Systematic parasitology, 1981, vol. 2, p. 123—132.
Fain A., Eisen P. Les acariens de la famille Knemidocoptidae producteurs de gale chez les
oiseaux. — Acta Zool. et pathol. Antverpiensia, 1967, vol. 45. 142 p.
Gaud J., Atyeo W. W. Nouvelles superfamilies pour les Acariens Astigmates parasites
d’oiseoux. — Acarologia, 1977, vol. 19, N 4, p. 678—685.
276
ЧЕСОТОЧНЫЕ КЛЕЩИ (Надсем. PSOROPTOIDEA)
Гнатосома со спинной стороны почти полностью или хотя бы в основании
прикрыта эпистомом.
У представителей некоторых семейств пальпы с пластинчатыми разраще-
ниями — прозрачной мембраной.
Покровы мягкие, чаще всего тонко параллельно складчатые, иногда с ши-
пами (хетоидами) вдоль складок на небольших участках. Проподосомальный
щит обоих полов и гистеросомальный щит самцов чаще всего имеются (хотя оба
могут отсутствовать). Латерококсальный орган отсутствует.
Амбулакральный аппарат без эмподиального когтя; состоит из небольшого
диска и чаще всего очень длинного стержня. Кондилоформный направитель
имеется в базальной части диска. Соленидий оц у большинства семейств зани-
мает базальное положение на лапке I. Чаще всего хотя бы на одной паре ног
амбулакральный аппарат отсутствует, а лапка несет очень длинную вершинную
щетинку. Половой диморфизм резко выражен. Яйцевыводное отверстие с хо-
рошо развитым эпигинием или без него; полностью отделено от анального от-
верстия, которое у представителей некоторых родов смещено на спинную сто-
рону. Копулятивное отверстие часто располагается на половом сосочке и поло-
вая бурса может быть видна через покровы.
Самцы с сильно склеротизованным небольшим половым аппаратом; имеется
маленький аданальный щит. Тарзальные копулятивные присоски, как правило,
имеются. У большинства представителей по бокам анального отверстия рас-
полагаются анальные копулятивные присоски, а на заднем конце тела хотя бы
небольшие опистосомальные лопасти.
Цикл развития двунимфальный, гипопус отсутствует. У большинства
представителей неполовозрелая фаза (телеонимфа) имеет специальные органы
скрепления с самцом для копуляции (парные выступы на заднем конце тела).
У представителей некоторых родов неполовозрелые фазы развития на самца и
на самку значительно различаются морфологически.
Разными авторами в надсемейство включается от 8 до 14 семейств. Наиболь-
шее медицинское и ветеринарное значение имеют представители двух приведен-
ных ниже. Большинство современных исследователей [Krantz, 1978; Балашов,
1982; и др.] относят их к двум отдельным самостоятельным надсемействам:
Psoroptoidea и Sarcoptoidea. Вслед за В. Б. Дубининым (1954) и О’Коннором
(O’Connor, 1982) мы считаем необходимым объединить их, несмотря на значи-
тельные морфологические различия, носящие адаптивный характер в связи с раз-
личием мест обитания.
Накожники (сем. Psoroptidae) паразитируют на поверхности кожи, под че-
шуйками эпидермиса большинства домашних и многих диких животных. Пи-
таются эпидермисом кожи или лимфой и серозной жидкостью (не кровью!)
через проколы. Являются наиболее патогенными паразитами домашних жи-
вотных. В месте их поселения развивается воспаление, обильное выделение
экссудата, зуд. Последний влечет за собой расчесывание этих мест животными,
повреждение покровов, образование корок, облысение, гиперкератоз кожи.
«Вкожные» паразиты (сем. Sarcoptidae) человека и животных прогрызают
ходы в толще рогового слоя кожи на границе с мальпигиевым слоем, где и раз-
множаются. В ходах встречаются одновременно все фазы развития, хотя зара-
жение новых хозяев осуществляется лишь оплодотворенными самками, един-
ственной фазой, способной прогрызать ходы внутри кожи. Развитие чесоточного
процесса связано с зудом, сопровождающимся расчесыванием кожи, нарушением
целостности рогового слоя ее, воспалительными явлениями в более глубоких
частях кожи с обильной экссудацией.
Оба типа чесотки усугубляются токсическим и аллергическим воздействием
клещей на организм хозяина в целом.
Самым сложным и до сих пор неясным остается вопрос о видовой самостоя-
тельности форм чесоточных клещей. Существуют три точки зрения: 1) каждому
виду животного специфичен свой особый вид чесоточного клеща; 2) существует
лишь небольшое число полиморфных видов, представленных некоторым коли-
чеством разновидностей или вариаций на отдельных видах животных; 3) в каж-
277
дом роде существует лишь один вид, дающий некоторые морфологические вариа-
ции, носящие черты внутривидовой изменчивости. Последнего мнения придер-
живался В. Б. Дубинин (1954) на основании специальных опытов по пересадке
клещей с одних животных на другие и наблюдениями над их развитием,
а также — Фэн [Fain, 1968] в своей монографии по вариабельности Sarcoptes
scabiei.
Тем не менее этот вопрос требует очень тщательной экспериментальной
проверки в связи с появлением все новых данных о явлении «псевдосаркоптоза».
Чесоточные клещи одного хозяина (например, свиньи) вызывают на неспецифич-
ном хозяине (например, человеке — у женщин, ухаживающих за свиньями)
явления мало отличимые по своим проявлениям от настоящей чесотки. Однако
при отсутствии дополнительного заражения в течение 10 дней происходит само-
излечивание [Ланге, Клочко, 1977].
В связи со всем изложенным в определителе приведены лишь таблицы ро-
дов.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ
1 (2). Тело широкоовальное, уплощенное. На заднем конце тела d всегда име-
ются опистосомальные лопасти. Ноги длинные, развиты нормально; у d
ноги III в несколько раз длиннее и толще редуцированных ног IV. Эпи-
меры ног I свободные, не сливаются вместе (рис. 138, 4). Анальные копу-
лятивные присоски у d хорошо развиты....................1. Psoroptidae
2 (1). Тело почти шаровидное. Задний конец тела у обоих полов закруглен
без опистосомальных лопастей. Ноги очень короткие, толстые, кониче-
ские; у d ноги III и IV развиты почти одинаково. Эпимеры ног I сливаются
вместе и образуют стернум (рис. 140, 2, 3). Анальные копулятивные
присоски d отсутствуют............................* . . 2. Sarcoptidae
1. Сем. PSOROPTIDAE
(Чесоточные клещи-накожники и кожееды)
Гнатосома крупная, выдается из-под эпистома на 2/3 длины. У клещей од-
ного рода (Psoroptes^ хелицеры сильно вытянуты, образуя колющий хоботок
(рис. 138, 7).
Тело уплощенное, широкоовальное. Поперечная борозда плохо выражена.
Покровы поперечно-складчатые. Щиты — проподосомальный у обоих полов и
гистеросомальный у $ хорошо развиты. Все щетинки тела волосовидные, не-
которые достигают значительной длины. Анальное отверстие чаще всего тер-
минальное, редко смещено на брюшную сторону.
Ноги длинные, толстые. Эпимеры ног I свободные. Амбулакральный диск
различного строения у представителей отдельных родов, а длина стебля варьи-
рует (рис. 138, 9—11).
Самка. Яйцевыводное отверстие большое, складчатое. Эпигиний и гениталь-
ные аподемы обычно имеются, хорошо развиты, редко отсутствуют. Амбула-
кральный аппарат отсутствует на ногах III, где имеются 2 длинные волосовид-
ные щетинки — терминальная и субтерминальная.
Самец. Опистосомальные лопасти обычно хорошо развиты. Ноги III во много
раз толще и длиннее ног IV. Лапка III с очень длинной волосовидной щетинкой.
Анальные копулятивные присоски большие, хорошо развиты. Тарзальные ко-
пулятивные присоски имеются.
Паразитируют на коже под чешуйками эпидермиса большинства домашних
и некоторых диких млекопитающих.
На Дальнем Востоке — 3 рода (в СССР 4).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). Гнатосома узкая, вытянутая в виде заостренного конуса. Предлапка
с очень длинным 3-членистым стебельком, коротким лишь па ногах IV d
(рис. 138, 9).......................................1. Psoroptes Gerv.
278
2 (1). Гнатосома широкая, короткая, почти квадратная. Предлапка коротко-
стебельчатая.
3 (4). Опистосомальные лопасти d крупные, почти квадратные, сближенные,
разделены треугольной щелью; некоторые щетинки, расположенные
на опистосомальных лопастях, плоские, листовидно расширенные. Ноги IV
$ крупнее ног III, значительно выступают за задний край тела; пред-
лапка имеется на I, II и IV ногах (рис. 139, 2) .... 2. Chorioptes Gerv.
Рис. 138. Общий вид и детали строения клещей падсем. Psoroptoidea.
1—4 — Psoroptes equi\ 1 — гнатосома, 2 — отдельно хелицера, 3 — самец со спинной стороны, 4 — самец
с брюшной стороны; 5,6 — Chorioptes equi: 5 — самец со спинной стороны, 6 — самец с брюшной стороны:
7,8 — Otodectes cynotis: 7 — самец со спинной стороны, 8 — самец с брюшной стороны; 9—11 — амбу-
лакральный аппарат клещей: 9 — рода Psoroptes, 10 — рода Chorioptes, 11 — рода Sarcoptes.
4 (3). Опистосомальные лопасти d едва выраженные, широко расставленные
бугорки; все щетинки заднего края тела волосовидные. Ноги IV $ сильно
редуцированы, едва достигают заднего края тела; предлапка имеется
только на ногах I и II (рис. 139, 7)................3. Otodectes Сап.
1. Psoroptes Gerv. Гнатосома сильно вытянута, узкая; хелицеры тонкие,
длинные, с 2 зубцами на вершине, далеко выступают за передний край пальп,
образуя колющий хоботок (рис. 138, 7, 2). Проподосомальный щит трапецие-
видный, занимает все пространство спинной поверхности между ногами I.
279
Остальная половина проподосомы поперечно-складчатая. Лопаточные щетинки
помещаются у наружных углов заднего края проподосомального щита; set —
короткие, see — длинные. Амбулакральный диск в форме небольшого колоколь-
чика на очень длинном 3-членистом стебельке (рис. 138, 9).
Самка. Яйцевыводное отверстие помещается между эпимерами ног I и II и
имеет вид арки; его клапаны сильно склеротизованы. Ноги III короче ног IV
и оканчиваются на уровне середины лапки IV (рис. 139, 5).
Самец. Гистеросомальный щит довольно сильно склеротизован; начинается
на уровне плохо выраженной поперечной борозды и доходит до заднего края
тела, переходя на опистосомальные лопасти. Последние треугольные, расстав-
лены на ширину, равную ширине лопастей в основании, и несут 5 пар щетинок,
2 из которых очень длинные (рис. 138, 3). Половой аппарат расположен на
уровне кокс III, очень маленький. Анальные копулятивные присоски помеща-
ются впереди основания опистосомальных лопастей. Ноги IV в 2.5 раза меньше
ног III. Стебелек предлапки IV сильно укорочен.
На ДВ — 5 «форм» этого рода, известных под названием Р. equi или Р. сот-
munis (в СССР — 9). 8' — 0.58-0.68x0.37—0.45; q - 0.76—0.86x0.45—0.58.
Всесветно.
Как уже было сказано, морфологическая диагностика форм затруднена;
они различаются лишь по хозяевам, встречаясь в Приморье на лошадях, круп-
ном рогатом скоте, овцах, козах и особенно кроликах. Заболевание начинается
с ушей, поражая наружные ушные раковины, и поэтому называется также «уш-
ной чесоткой». Больные кролики принимают характерную позу: склоняют го-
лову набок и трясут ушами. В острых случаях заболевание приводит к гибели
животных. Клещи длительное время могут существовать во внешней среде (до
2 хмесяцев), что затрудняет борьбу с заболеванием.
2. Chorioptes Gerv. Гнатосома широкая, короткая, почти квадратная, ти-
пичного грызущего типа. Хелицеры не выступают за вершину пальп. Пропо-
досомальный щит очень узкий, слегка трапециевидно расширяющийся на зад-
нем конце; занимает почти всю срединную часть проподосомы (рис. 138, 5).
Длина и положение лопаточных щетинок такие же, как у клещей рода Psorop-
tes. Амбулакральный диск широкий, сиди? на коротком стебельке (рис. 138,
10).
Самка. Яйцевыводное отверстие очень большое, помещается в основании эпи-
мер II. его передний край укреплен хорошо развитым эпигинием (рис. 138,
2).
Самец. Гистеросомальный щит хорошо очерчен, начинается на уровне
ног III и достигает заднего края тела. Половой аппарат компактный, окружен
хорошо выраженным генитальным щитом. Анальные копулятивные присоски
очень крупные, помещаются посредине брюшной стороны опистосомы. Описто-
сомальные лопасти почти квадратной формы, разделенные треугольной щелью.
Каждая из них несет 4 пары длинных щетинок, 2 из которых плоские, расширен-
ные. Ноги IV в 3 раза мельче ног III. Длина волосовидных щетинок лапок III
превышает половину длины тела (рис. 138, 6).
На ДВ — 5 видов (в СССР — 6), диагностируемых лишь по хозяевам: в При-
морье Ch. equi — у лошадей, Ch. ovis — у овец, Ch. саргае — у коз, Ch. bovis —
у крупного рогатого скота и Ch. cuniculi — у кроликов. $ — 0.26—0.33; £ —
0.35—0.39. Всесветно.
Поражаются обычно у домашних копытных сначала путовые суставы зад-
них конечностей, почему иногда это заболевание называется «ножной чесоткой»
(лишь у коз заболевание затрагивает только кожу тела). У кроликов паразиты
поселяются сначала в ушных раковинах, а потом распространяются на другие
участки тела. Заболевание развивается медленно, протекает с зудом, образова-
нием корок и выпадением волос. Нижняя поверхность корок часто сильно мок-
нет. Однако заболевание протекает «доброкачественно» и легко поддается лече-
нию.
3. Otodectes Сап. Гнатосома широкая, короткая, коническая на вершине.
Хелицеры не выступают за вершину пальп.
280
Проподосомальный щит ромбовидный, занимает всю переднюю часть про-
подосомы, простираясь назад на 2/3 ее длины. По бокам от задней закругленной
вершины щита располагаются два маленьких плечевых щита, окружающих
только основания плечевых щетинок; sci в несколько раз короче see (рис. 138,
7).
Амбулакральный аппарат сходен с таковым клещей рода Chorioptes.
Самка. Яйцевыводное отверстие очень большое, занимает все пространство
между основаниями эпимер I и уровнем ног III. Его края укреплены мощным
полукруглым эпигинием. Ноги IV очень маленькие, сильно редуцированные,
едва достигают заднего края тела. Обе задние пары ног без предлапки с двумя
длинными щетинками (рис. 138, 7).
Самец. Гистеросомальный щит занимает всю среднюю часть гистеросомы,
квадратный с сильно склеротизованными боковыми краями. Опистосомальные
лопасти едва намечены в виде двух широко расставленных бугорков на заднем
крае тела. Из щетинок, сидящих на этих бугорках, две довольно длинные:
длина одной составляет х/з длины тела, другая — короче (рис. 138, 7). Половой
аппарат крупный, располагается на уровне ног III. Анальные копулятивные
присоски очень крупные, сидят на анальном щите, ограниченном спереди хо-
рошо выраженной складкой. Ноги IV не более чем в 1.5 раза короче ног III,
очень узкие. Лапки обеих пар с предлапками, кроме того лапка III с двумя
очень длинными срединными щетинками (рис. 138, S).
Род включает 1 вид.
1 (1). $ __ 0.19-0.31Х0.12-0.21; ? — 0.28-0.41x0.19-0.36. — Голарктика.
Евр. ч. СССР, Казахстан, Алтай, Сибирь, Приморье, Командорские о-ва;
Европа, Канада, США. Паразитируют в наружном слуховом проходе пес-
цов, лисиц и енотовидных собак в зверопитомниках, у домашних кошек
и собак. Заболевание сопровождается воспалением, зудом; больные жи-
вотные постоянно трясут головой, расчесывают лапами уши, часто сидят
с опущенной склоненной на бок головой. В острцх случаях процесс пере-
ходит на среднее и внутреннее ухо, приводит к прободению барабанной
перепонки; в запущенных случаях заканчивается гибелью животных.
В сочетании с другим заболеванием (унценариозом — случай, зареги-
стрированный на песцах Alopex lagopus о-в Медный Командорские о-ва)
клещевое поражение распространяется по всей поверхности тела, которое
сплошь покрывается коростой, и приводит к гибели уже молодняк [Овсян-
ников, 1981] . < > г < . •«•••• • О. cynotis Сап.
[2. Сем. SARCOPTIDAE
(Чесоточные зудни)
Гнатосома короткая и очень широкая, окружена прозрачной мембраной,
иногда полностью скрыта под эпистомом (Notoedres).
Тело почти округлое, иногда незначительно цилиндрическое. Поперечная
борозда плохо выражена. Покровы тонкие, складчатые на большей части по-
верхности. В роде Sarcoptes на спинной стороне тела имеется область с шипиками
(хетоидами), которая у S выражена слабее, чем у j. Щиты слабо склеротизо-
ваны, плохо очерчены, иногда полностью отсутствуют. Задний конец тела за-
круглен у обоих полов.
Щетинки спинной и боковых сторон тела представлены 9—15 парами, чаще
всего короткие волосовидные, ve всегда отсутствует. Некоторые щетинки спин-
ного и бокового рядов утолщены и имеют вид палочковидных шипов (рис. 140,
Z). Анальное отверстие терминальное или смещено на спинную сторону.
Ноги укорочены, конические с короткими и широкими члениками. Эпи-
меры ног I сливаются в стернум. Часто имеет место слияние голени и лапки ног III
и (или) IV в один членик. Амбулакральный диск небольшой на очень длин-
ном (в "несколько раз превышающем величину диска) стебле (рис. 138, II);
амбулакральный аппарат отсутствует на лапках III и IV g и лапке III 5, где вме-
сто него помещается по одной длинной волосовидной щетинке.
19 Зак № 990
281
Половой диморфизм выражен резко и прежде всего в величине полов:
2 вдвое крупнее о.
Самка. Яйцевыводное отверстие в виде поперечной щели на уровне середины
расстояния между II и III ногами, без эпигиния. Копулятивное отверстие от-
крывается на небольшом половом сосочке, расположенном со спинной стороны
у заднего края тела; иногда видна половая бурса.
Рис. 139. Общий вид самок клещей надсем. Psoroptoidea.
1—3 — с брюшной стороны: 1 — Otodectes cynotis, 2 — Chorioptes equi, 3 — Psoroptes equi\ 4 — Notoedre»
muris, co спинной стороны; 5,6 — Notoedres cati: 5 — co спинной стороны, 6 — с брюшной стороны, ао —
анальное отверстие, ко — копулятивное отверстие, ст — стер ну м, яо — яйцевыводное»отверстие.
Самец. Опистосомальные лопасти и анальные копулятивные присоски от-
сутствуют, тарзальные копулятивные присоски имеются.
Паразитируют в коже, проделывая ходы в толще эпидермиса, у многих
млекопитающих, включая человека.
На ДВ — 2 рода (в СССР 3).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). Анальное отверстие терминальное, иногда слегка сдвинуто на брюшную
сторону. Покровы спинной стороны с хетоидами, некоторые щетинки
в виде палочковидных шипов..................... • 1. Sarcoptes Lair.
282
ложены последовательно, а апикальный (лапка) противопоставлен им. Лапка
вооружена определенным числом разных щетинок (от 3 до 7), иногда гладкой
ST и соленидием. Последний присутствует постоянно, а число щетинок варьи-
рует, но постоянно для рода. Голень пальп несет коготь, разделенный на не-
сколько зубцов (от 2 до 7) и 3 щетинки: дорсальную, латеральную и вентраль-
ную. На колене и бедре пальп всегда по 1 щетинке. Последний членик пальп
(вертлуг) щетинок не имеет. На боковых частях гнатококсы, образованной
слиянием коксальных члеников пальп, немного ниже вертлуга располагается
по 1 коксальной щетинке. Характер опушения щетинок пальп используется
как таксономический признак. Хелицера состоит из неподвижного основания
и подвижного когтя с трехвершинной головкой и иногда с рядами зубчиков по
дорсальному или вентральному краю. Число и расположение последних харак-
терно для родов и подсемейств.
Идиосома. В апикальной части спинной поверхности расположен щит
(scutum). Он несет пару сенсорных органов — сенсилл (трихоботрий) и от 3
до 7 и более простых щетинок. Строение, расположение, число всех щетинок
щита являются важными таксономическими признаками родов, триб, подсе-
мейств. Поверхность щита часто покрыта точечной пунктировкой, а иногда
частично и линейной штриховкой. Форма щита очень разнообразна и характерна
для рода. По бокам щита, ближе к заднему краю, расположены простые глаза.
Обычно их 2 пары, иногда 1 пара или они отсутствуют. Ниже щита идиосома
несет спинные щетинки, число и расположение которых специфично для вида.
Первый неполный ряд щетинок по бокам одиосомы называют плечевыми (Я).
Их может быть 1 пара и более. Они хорошо обособлены или, наоборот, плохо
отграничены от первого ряда собственно спинных щетинок. Последние могут
располагаться правильными рядами или беспорядочно. Иногда выделяют ще-
тинки, сидящие по заднему краю идиосомы, называя их пигидиальными.
Брюшная сторона идиосомы несет стернальные щетинки (1-я пара между кок-
сами I пары ног и 2-я пара — между коксами III пары), подплечевые (Ни),
расположенные между коксами II и III пары ног, брюшные щетинки — ниже
уровня задних кокс и анус. Постанальные щетинки обычно сходны по строению
со спинными, а преданальные несколько тоньше и реже опушены. Границу
между ними не всегда можно четко провести. Трудно отграничить также зад-
ние спинные и брюшные щетинки и подсчет их числа носит часто элемент субъек-
тивности. Более объективно общее число щетинок тела (NDV).
Ноги. У всех личинок 3 пары ног. Число члеников ног различно, но по-
стоянно для подсемейств. Членистость ног выражается в виде формулы (напри-
мер, fsp—1—7—7; 6—6—6; 7—6—6). Все членики ног вооружены тактильными
опушенными щетинками, имеющими постоянное число и расположение в каждом
роде или подсемействе, и различными специализированными щетинками, ха-
рактерными для рода и вида. Среди последних различают соленидий, или spur
(S), гладкие относительно короткие щетинки, называемые по месту расположе-
ния — tarsala, tibiala, genuala, femorala (очень короткие микрощетинки —
microtibiala, microgenuala), гладкие длинные тонкие щетинки на члениках
III пары ног, называемые — mastitarsala, mastitibiala и т. д. В апикальной
части лапки I и II пары ног имеется по 1 (редко 2) pretarsala (PT), по 1 spur и
famulus (/) и только на I лапке subterminala (ST) и parasubterminala (pST).
Лапки сужены в апикальной части и на конце несут по 2 коготка и
эмподий.
В современной литературе при описании и определении видов личинок крас-
нотел ок пользуются набором формул, в которые включены признаки, обозначен-
ные латинскими буквами. Наиболее употребительны следующие: SIF — синте-
тическая определительная формула, в которую входят следующие таксономи-
ческие признаки: fT=lBS — формула, отражающая число и характер (В —
опушенная, S — гладкая) опушения щетинок на лапке пальп; Ga — галеальная
щетинка, опушенная (В) или гладкая (TV); Gr — число зубцов когтя пальп;
число различных щетинок на ногах: ga — genualae на колене I, gm — genualae
на колене II, gp — genualae на колене III, tp — tibialae на голени III, МТ —
mastitarsalae III пары ног, Mt — mastitibialae III пары, MG — mastigenualae
III пары, MF — mastifemoralae III пары. Все эти признаки входят в синтетиче-
288
Самец. На спинной стороне имеется 6 небольших склеротизованных щитов:
2 медиальных и 4 латеральных. Передние ноги значительно сильнее развиты,
чем задние. В коксо-стернальном скелете ног III и IV имеет место лишь слияние
эпимер IV друг с другом и с аподемами полового аппарата. Последний неболь-
4
Рис. 140. Общий вид Sarcoptes scabiei.
1 — самка со спинной стороны; 2 — то же, с брюшной стороны; 3 — самец, с брюшной стороны; 4 — то же,
со спинной стороны, ст — стернум, яо — яйцевыводное отверстие, vi — внутренние теменные щетинки,
sci — внутренние лопаточные щетинки, see — наружные лопаточные щетинки.
шой, занимает пространство между основаниями ног IV и отделен от заднего
края тела значительным участком складчатых покровов.
Паразитируют в роговом слое кожи, более поверхностно, чем клещи рода
Sarcoptes.
На ДВ — 3 вида, два из которых паразитируют на грызунах (в СССР 5).
Медицинское и ветеринарное значение имеют 2 вида.
/
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Чешуйки впереди анального отверстия довольно хорошо выражены и
занимают центральную часть спинной поверхности; анальное отверстие
284
помещается в задней половине тела и значительно отстоит от заднего края
(рис. 139, 5). Многие спинные щетинки утолщены, 5 — 0.14—0.15x0.12—
0.13; $ — 0.21—0.24x0.16—0.2. — Голоаркт. Евр. ч. СССР, Казахстан^
Приморье; Европа, США, Центр, и Ю Африка. Паразитирует на домаш-
них кошках (реже собаках?) и кроликах, поражает прежде всего уши и
морду. Отнесение паразитов с кошек и кроликов к одному виду [Fain,
1965] требует проверки. Зарегистрированные случаи перехода клещей
этого вида на человека [Дубинин, 1954] следует, по-видимому, рассматри-
вать как проявление «псевдочесотки» (см. с. 278)............................
. . . . N. cati (Hering) — Кошачий зудень (сип.? N. cuniculi Gerlach)
2 (1). Чешуйки впереди анального отверстия отсутствуют; анальное отверстие
помещается на спинной стороне вблизи его заднего края (рис. 139, 4).
Спинные щетинки очень короткие. <J — 0.16—0.17; ? — 0.29—0.35. —
Голоаркт. Евр. ч. СССР, Сибирь, ДВ; Европа, США, Ю Африка. Парази-
тирует на лабораторных белых крысах и мышах, на домовых крысах.
Заболевание первоначально поражает голову, откуда распространяется
на тело, преимущественно конечности и хвост. Воспаление кожи сопро-
вождается зудом, образованием корок, а затем обширных складчатых на-
ростов, выпадением волос. Животные сильно худеют и гибнут. В вивариях
заболевание принимает массовый характер. Нередко сопровождается раз-
личными дополнительными инфекциями: грибковыми, бактериальными,
конъюктивитами. Зарегистрированные случаи перехода на человека [Ду-
бинин, 1954] следует, по-видимому, рассматривать как проявление «псевдо-
чесотки» (см. с. 278)........................................N. muris (Megnin)
ОСНОВНАЯ ЛИТЕРАТУРА
Балашов Ю. С. Паразито-хозяинные отношения членистоногих с наземными позвоноч-
ными. Л., 1982. 320 с. (Труды Зоол. ин-та АН СССР; Т. 87).
Дубинин В. Б. Чесоточные клещи. Их биология, вред в сельском хозяйстве, меры про-
филактики и борьбы с ними. М., 1954. 172 с.
Ланге А. Б., Клочко Р. Т. К этиологии саркоптоза свиней. — В кн.: Проблемы ветери-
нарной санитарии. М., 1976, с. 138—149.
Ланге А. Б., Клочко Р. Т. К вопросу о роли возбудителя саркоптоза свиней в патологии
человека. — Вести, дерматол. и венерол., 1977, № 1, с. 19—24.
Овсянников И.Г. Песец острова Медного может исчезнуть. — Природа, 1981, № 3, с. 59—
62.
Fain A. Notes sur le genre Notoedres Railliet, 1893 (Sarcoptidae: Sarcoptiformes). — Aca-
rologia, 1965, t. 7, fasc. 2, p. 321—342.
Fain A. Etude de la variabilite de Sarcoptes scabiei avec une revision desSarcoptides. —
Acta Zool. et Pathol. Antverpiensia, 1968, N 47, 196 p.
Krantz G. W. A manual of Acarology. 2nd ed. Corvallis, 1978. 509 p.
O'Connor В. M. Astigmata. — In: Synopsis and classification of living organisms. New
York, 1982, vol. 2, p. 146—169.
ТРОМБИДИФОРМНЫЕ КЛЕЩИ
(Подотряд TROMBIDIFORMES)
Самая разнообразная группа акариформных клещей. Представлена очень
большим числом «жизненных форм», резко различающихся как образом жизни
(свободноживущие: почвообитающие, водные, растениеобитающие; паразиты
растений, рептилий, птиц и млекопитающих), так и соответственно степенью
тех морфологических изменений, которые они приобрели в результате при-
способления к данному образу жизни. Существенные различия в строении от-
дельных представителей обусловили сложную и весьма не устоявшуюся си-
стематическую структуру этого таксона. В данном разделе принимается система,
разработанная Б. А. Вайнштейном [Определитель обитающих в почве клещей.
Trombidiformes. М., 1978, т. 3. 271 с.].
Медико-ветеринарное значение имеют лишь 3 сравнительно небольшие
группы, рассматриваемые ниже, поэтому представляется нецелесообразным
285
давать общий очерк внешнего строения тромбидиформных клещей, и описание
морфологических особенностей дается по каждому отдельному таксону.
Единственное, на что следует указать, это то, что у всех фаз и у обоих полов
тромбидиформных клещей всегда отсутствуют длинные бичевидные щетинки на
голенях I—II, а у самцов — тарзальные и анальные копулятивные присоски,
характерные для большинства представителей саркоптиформных клещей.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА
। НЕКОТОРЫХ ТАКСОНОМИЧЕСКИХ ГРУПП [КЛЕЩЕЙ
ПОДОТРЯДА TROMBIDIFORMES
1 (2). Паразитируют только на фазе личинки. Проподосомальный щит с парой
волосовидных или булавовидных трихоботрий в центральной части. Мно-
гочисленные щетинки тела опушенные. — Ярко-красные или оранжевые,
реже белые клещи («краснотелки»), встречающиеся на позвоночных (осо-
бенно много в ушных раковинах) и сильно увеличивающиеся в размерах
во время питания. (Надсем. Trombidioidea, Когорта Parasitengona) . . .
..............................................1. Trombiculidae
2 (1). Паразитируют на всех фазах жизненного цикла. Проподосомальный щит
без трихоботрий. Немногочисленные щетинки тела неопушенные. (Когорта
Prostigmata).
3 (4). Покровы поперечно исчерченные. Тело лишь с несколькими микроще-
тинками. Ноги I—IV тесно сближены. Лапки с 2 видоизмененными мощ-
ными коготками. — Очень мелкие червеобразные клещи, обитающие
под кожей и в волосяных фолликулах млекопитающих, включая человека.
(Надсем. Demodicoidea).............................2. Demodicidae
4 (3). Покровы гладкие. Тело покрыто длинными волосовидными щетинками.
Ноги III—IV значительно удалены от ног I—II. Лапки с 2 маленькими
коготками и гребенчатой предлапкой между ними. — Мелкие вытянутой
формы клещи, обитающие в очинах перьев птиц. (Надсем. Cheyletoidea)
........................................... 3. Syringophilidae
КЛЕЩИ-КРАСНОТЕЛКИ (Сем. TROMBICULIDAE)
Очень мелкие клещи. Личинки — паразиты позвоночных животных нимфы
и имаго — почвенные обитатели, хищники. Распространены всесветно.
Личинки паразитируют на позвоночных животных во все сезоны года и часто
в большом числе, а потому легко доступны для сбора и изучения. Систематика,
морфология, анатомия, экология и другие вопросы биологии краснотелок ос-
нованы на изучении личиночной стадии. Сбор в природе нимф и имаго трудо-
емок и мало эффективен, а лабораторное разведение не всегда приводит к ус-
пешным результатам. Поэтому в мировой литературе описание видов красно-
телок принято делать по личинке, а последующих стадий — при получении их
в лабораторной культуре от точно определенного вида личинки. Не рекомен-
дуется делать описания по взрослым особям, так как это приводит к серьезным
ошибкам и путанице. Одновременный сбор взрослых особей и личинок в одном
местообитании не является основанием для объединения их в один вид. Ниже
пойдет речь только о личинках этих клещей.
( ВНЕШНЕЕ СТРОЕНИЕ
Тело личинки делится на гнатосому и идиосому (рис. 141). Величина по-
следней в зависимости от степени насыщенности личинки кровью колеблется от
50—70 до 500 мкм, а в отдельных случаях может достигать 4.5 мм (род Vataca-
rus). Кутикула, покрывающая тело клеща, мягкая, складчатая, легко растяжи-
мая. Твердые склеротизованные покровы тела представлены в виде щита и
коксальных члеников ног, величина которых не изменяется в процессе питания
личинки. Относительный размер клеща определяется индексом pedipus (Ip),
который слагается из суммы длины всех трех ног (из I, II, III пары ног). Окраска
тела зависит от пигмента кожных покровов и чаще представлена всеми оттен-
286
ками красного цвета, но может быть также оранжевой, желтой, белой, прозрач-
ной. На спинной поверхности идиосомы располагаются щит, глаза и спинные
щетинки. К апикальной части брюшной поверхности прикрепляется гнатосома,
по бокам — коксы трех пар ног, в нижей части располагается анальное отвер-
стие, а по всему брюшку — разные типы щетинок.
Рис. 141. Детали строения клещей-краснотелок.
1 — общий вид идиосомы с брюшной и спинной сторон: 1—3 — коксы I—III и щетинки на них, 4 — I и
II пара стернальных щетинок, 5 — брюшные щетинки, 6 — анальное отверстие, 7 — щит, 8 — глаза, 9 —
плечевые щетинки (H), 10 — спинные щетинки; 2 — общий вид гнатосомы со спинной и брюшной сторон:
1 — основание хелицеры, 2 — коготь хелицеры, з — геальная щетинка (Ga), 4 — щетинка на бедре пальп,
5 — щетинка на колене пальп, 6 — дорсальная и 7 — латеральная щетинки на голени пальп, 8 — коготь
пальп, 9 — гнатококса, 10 — щетинка гнатококсы, 11 — вертлуг пальп, 12 — бедро пальп, 13 — колено
пальп, 14 — голень пальп, 15 — вентральная щетинка на голени пальп, 16 — лапка пальп, 17 — опушен-
ная щетинка лапки пальп (В), 18 — соленидий или tarsala (So), 19 — субтерминальная щетинка лапки
пальп; 3 — ноги I—III, их специализированные щетинки и порядок измерения длины ног: 1—з — лапки
I—III, 4—6 — голени I—III, 7—9 — колени I—III; щетинки: PT1, РТ11 — pretarsaiae лапок I и II, ST—
subterminala, pST — parasubterminala, S2 — соленидий лапок I и II, Д, f2 — microtarsalae, или
famulus лапок I и II, ta, tm, ip — tibialae голеней I—III, mia — microtibiala голени I, ga, gm, gp —
genualae коленей I—III, mga — microgenuala колена I; длина ноги pp=aH-6-|-B-j-r. »
Гнатосома (ротовые органы) состоят из пары пальп, пары двухчле-
никовых хелицер, непарной гнатококсы, кпереди переходящей в гипостом. Бо-
ковые края последнего изгибаются дорсально вокруг хелицер и несут по 1 га-
леальной щетинке, характер опушения которой используется как таксономиче-
ский признак. Пальпа состоит из 5 члеников, из которых 4 дистальных распо-
287
скую формулу в виде цифр и букв, например: SIF—1BS—N—3—2—1—1 —
1-1-2-0-0.
Кроме основной употребляются еще и другие формулы: fD=2H—6—6—4—
4—2=24 — формула спинных щетинок, где последовательно указывается число
плечевых (Я) и число спинных по рядам. Если щетинки располагаются беспо-
рядочно, указывают их общее число: 2)5=80. /7 и VS — соответственно фор-
мула или общее число брюшных щетинок. NDV=DS-\-VS: jPp=N/N/NBB —
формула щетинок пальп, указывающая характер опушения их (5 — опушенная,
N — гладкая) последовательно на бедре, колене и голени (дорсальная, латераль-
ная, вентральная) пальп. fsp=7—7—7 число сегментов I—III пар ног, fSt=
=2—2 — число I и II пары стернальных щетинок, fCx—1—1 = 1 число щетинок
на коксах I—III пар ног.
Кроме таксономических признаков приводятся и их количественные пока-
затели. Классические стандартные промеры выражаются в мкм (рис. 142, /).
БИОЛОГИЯ
Личинки краснотелок ведут паразитический образ жизни. Они питаются
тканевой жидкостью и фрагментами эпителиальных и других клеток хозяев —
различных видов позвоночных животных. Сроки пребывания на хозяине колеб-
лются у разных видов личинок клещей в среднем от 1—2 до 30 дней. Насытив-
шись, личинки покидают хозяина и уходят в почву. Здесь они дважды линяют,
в результате чего появляется вначале нимфа, а потом взрослая особь (самец
или самка). Таким образом, цикл развития краснотелок складывается из яйца,
трех активных морфологических фаз (личинка, дейтонимфа, имаго) и между
активными — 2 покоящиеся фазы — протонимфа (нимфохризалис) и трито-
нимфа (имагохризалис). Длительность цикла различна и в зависимости от вида
клеща и условий среды колеблется от 60 до 90 и более дней. Оплодотворение
наружно-внутреннее, без сближения полов. Самец откладывает сперматофоры
на субстрат, а самка захватывает головку сперматофора со спермой, помещая
ее внутрь через половое отверстие. Яйца откладываются на субстрат пооди-
ночке или кучками. Каждая саамка откладывает несколько десятков яиц. В за-
висимости от вида клеща длительность откладки яиц колеблется от 30 дней до
нескольких месяцев. Данные о цикле развития и его продолжительности полу-
чены для некоторых видов при лабораторном культивировании (наблюдения
в природе отсутствуют).
Личинки, как правило, не обнаруживают специфичности в выборе хозяев,
однако некоторые роды и виды паразитируют только на определенных группах
позвоночных животных. Таковыми являются краснотелки летучих мышей, нор-
ных птиц, амфибий. Голодные личинки, поджидая хозяина, скапливаются в за-
тененных влажных местах на почве или в подстилке, заползают иногда на более
возвышенные предметы (камни, поверхность листьев травянистых растений
и т. п.). В почве личинки находятся чаще всего на глубине от 3, реже — до
18 см. В зависимости от вида личинки могут паразитировать на хозяевах круг-
лый год или только в определенные сезоны — летом, весной и осенью. Для
большинства краснотелок характерна приуроченность к определенным место-
обитаниям в лесной, таежной, степной и пустынной зонах. Хозяйственная
деятельность человека приводит обычно, за редким исключением, к уменьшению
численности и исчезновению краснотелок.
Нимфы и взрослые особи краснотелок менее изучены. Это — хищники, жи-
вущие в почве и питающиеся яйцами мелких беспозвоночных. Некоторым ви-
дам свойственны сезонные вертикальные миграции, связанные с их циклом
развития. Оставаясь большую часть года на глубине до 50—60 см, весной самки
поднимаются в менее глубокие слои (10—30 см) для откладки яиц. У других
видов сезонные миграции не отмечаются. Вся жизнедеятельность их проходит
в поверхностных слоях подстилки, где они обнаруживаются даже в зимний
период. Различные фазы развития клещей способны длительное время выдер-
живать низкие температуры и при наступлении благоприятных условий про-
должать нормальное развитие.
289
ВРЕД
Клещи-краснотелки воздействуют на здоровье человека двумя путями: не-
посредственно, вызывая укусами тяжелые кожные дерматиты или тромбидиоз,
и как переносчики возбудителей некоторых природноочаговых инфекций.
Тромбидиоз — широко распространенное заболевание, вызываемое укусами
некоторых видов клещей из родов Neotrombicula, Eutrombicula, Euschoengastia
и др. Случаи заболевания среди людей и сельскохозяйственных животных от-
мечены в Европе, Америке, Африке, Австралии. В месте укуса на теле жертвы
появляется покраснение, припухлость, ощущается сильный зуд. Длительность
заболевания 1—3 недели, если оно не осложняется вторичной инфекцией, про-
никающей через расчесы кожи. Заболеваемость часто носит сезонный характер,
проявляясь чаще летом или осенью, когда наиболее высока активность и чис-
ленность определенного вида клеща.
Естественное носительство риккетсий цуцугамуши установлено у многих
видов клещей, но доказанными переносчиками возбудителя лихорадки цуцу-
гамуши являются лишь некоторые виды.
Диагноз сем. Trombiculidae
Личинки мелкого размера (менее 1 мм). Голодные особи круглые, овальные,
удлиненные. Хелицеры двухчлениковые. Дистальный членик имеет форму вы-
гнутого лезвия с тремя вершинами в апикальной части, иногда с зубчиками по
дорсальному и вентральному краям. Пальпы пятичлениковые. Коксы их слиты
по средней линии и образуют гнатококсу с 2 щетинками позади бедренного чле-
ника пальп. Кпереди гнатококсы переходят в гипостом, боковые края которого
изгибаются дорсально вокруг хелицер и несут по 1 галеальной гипостомальной
щетинке с каждой стороны. На члениках пальп имеется постоянное число ще-
тинок разного строения. Лапка противопоставлена голени пальп и несет от 3
до 7 опушенных щетинок, соленидий (постоянно) и иногда гладкую субтерми-
нальную щетинку (ST). Щит несет от 3 и более обычных щетинок и пару ботри-
дий (псевдостигм) с сенсиллами (трихоботрцями). Анус вентральный. Ног
3 пары. Тазики I ног имеют уростигмы. Коксы I и II пар ног сближены,
а III пара лежит значительно отступя от них. Ноги могут состоять из 7 или
6 члеников. Лапки ног несут 2 коготка и эмподий. Личинки паразитируют на
разных группах позвоночных животных. Распространены всесветно. На Даль-
нем Востоке — около 30 видов (в СССР около 200).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ, ТРИБ И РОДОВ
1 (2). fsp=Q—6—6, AM=2, на коксе I всегда 2 щетинки, I пара стернальных
щетинок отсутствует............................Подсем. Leeuwenhoekiinae
2(1). fsp = l —7—7 или 7—6—6, AM~i или отсутствует, I пара стернальных
щетинок имеется.
3(18). /$/>=7—7—7, AM=i ..........................Подсем. Trombiculinae
4 (11). Сенсиллы бичевидные..........................Триба Trombiculini
5 (8). Щит почти прямоугольный, задний край его прямой или слегка выпуклый
или вогнутый посредине.
6 (7). fT=lBS, переднебоковые плечи щита выражены, на telofemur III пары
ног 4 щетинки...............................................Chiroptella 1
7 (6). Щит без переднебоковых плечей, на telofemur III пары ног 3 щетинки
............................1. Leptotrombidium Nag., Miyag., Mit., Imam.
8 (5). Щит трапециевидный с выпуклым задним краем.
9 (10). Щит пятиугольный, AL и PL сближены (рис. 145, 4)................
............................3. Miyatrombicula Sasa, Kawash., Egash.
10 (9). Щит трапециевидный с закругленным задним краем, если пятиугольный,
то AL и PL не сближены............................2. Neotrombicula Hirst
1 На ДВ известен 1 вид Ch. (Oudemansidium) subakamushi Schl., систематическое поло-
жение которого требует уточнений.
290
11 (4). Сенсиллы вздутые (шаровидные, булавовидные и т. п.).......
..........................................Триба Schoengastiini
12 (15). Щит трапециевидный с переднебоковыми плечами, коготь пальп раз-
делен на 3 зубца, mastitarsala III часто имеется.
13(14). ]Т=65, щит с точечной пунктировкой .... 4. Ascoschoengastia Ew.
14 (13). fT=7B. IBS. щит с точечной пунктировкой и часто с линейной штри-
ховкой .....................................7. Neoschoengastia Ew.
15 (12). Щит трапециевидный без переднебоковых плечей, коготь пальп разде-
лен на 3 и более зубцов, mastitarsala всегда III отсутствует.
16 (17). fT=4J3. SB обычно выше уровня PL. всегда 2 пары плечевых щетинок,
tibiala III отсутствует..............6. Cheladonta Lip., Cros., Loom.
17 (16). /Т=7В, IBS. SB обычно ниже уровня PL ... 5. Euschoengastia Ew.
18 (3). fsp=7— 6 — 6, AM=0......................Подсем. Gahrliepiinae
Подсем. TROMBICULINAE
Триба TROMBIGULINI
ОПРЕДЕЛИТЕЛЬНЫЕ ТАБЛИЦЫ ВИДОВ
1. Leptotrombidium Nag., Miyag., Mit. et Imam. Личинки среднего раз-
мера и крупные: Ip от 700 и свыше 1000. 5/5=75—5—3—2—1—1—1—0—0—
0—0. Щит близок прямоугольному, задний край'его прямой, слегка закруглен
или вогнут посредине. AL расположены всегда в углах щита, AM — ниже
уровня AL. PL — в заднебоковых углах или на боковых краях щита. Галеаль-
ная щетинка всегда опушенная. На колене I пары ног всегда 2 genualae и micro-
genuala. Гладкие щетинки на члениках III пары ног отсутствуют. Объем рода
и его разделение на подроды пока не имеет четких границ. В СССР отмечены
представители двух подродов — номинативного и Trombiculindus. Последний
характеризуется наличием листовидных щетинок на идиосоме и щите (чаще РЕ),
Распространение всесветное. Паразитируют на всех группах позвоночных жи-
вотных, чаще на грызунах. — 5 видов (в СССР более 20).
1 (4). fPp=N/N/BNB. щетинка на коксе III расположена на переднем крае
членика (рис. ,142, 5).
2 (3). PL лежат на боковых сторонах щита близко к AL. SB значительно
ниже PL. число щетинок в 3 первых рядах — 25—8—6—6, . . ., NDV ме-
нее 100. — Прим.; сев. п-ова Корея, Япония.........L. orientale Schl.
3 (2). PL расположены в закругленных заднебоковых углах щита, SB почти
на уровне PL или чуть ниже, число щетинок в 3 первых рядах —25—
12—14—12 . . NDV более 100. — Прим.; сев. п-ова Корея, Япония
(рис. 142, 2. 3)...........................L. palpale Mag., Mit., Tam.
4 (1). fPp=N/N/BNN. щетинка на коксе III расположена заметно ниже перед-
него края членика (рис. 142, 5).
5 (6). PL расположены на боковых сторонах щита близко к AL. SB значительно
ниже PL (рис. 142, 4). —Прим.; сев. п-ова Корея, Япония, Китай
(рис. 142, 4. 5).........................L. pallidum Mag., Mit., Таш.
6 (5). PL расположены в заднебоковых углах, SB почти на уровне PL (чуть ниже
или выше).
7 (8). SB на уровне или выше PL. заднебоковые углы щита не закруглены и
хорошо обозначены, АР более 30 мкм, число щетинок в 3 первых рядах _
25—10—14—14 . . ., длина спинных щетинок менее 50. — Япония,
Сев.-Вост. Китай, Таиланд, Зап. Малайзия......................
......................L. scutellare Nag., Miyag., Mit., Tam., Ten.
8 (7). SB на уровне или чуть ниже PL. заднебоковые углы щита закруглены,
АР менее 30 мкм, число щетинок в 3 первых рядах — 25—10—8—8 . . .,
длина спинных щетинок более 50 (рис. 143, 1. 2). — Прим., Хаб., Амур.,
Бурят., Кур., Монголия............................L. pavlovskyi Schl.
2. Neotrombicula Hirst. Личинки среднего размера и крупные, 1р более 800.
5/5=755-5 (А)-З-З (2)—1—1—1—1 (2-3)-0 (1)-0-0-(1). Щит трапе-
291
циевидныи с выпуклым закругленным или заостренным посредине задним краем.
Форма щита, число и характер специализированных щетинок на ногах, харак-
тер опушения пальп очень различны, что говорит о неоднородности рода и не-
Рис. 142. Детали строения клещей-краснотелок.
X — строение и промеры деталей щита (в мкм): AW — расстояние между основаниями переднебоковых
щетинок, PW — расстояние между основаниями заднебоковых щетинок (ширина щита), SB — расстояние
между основаниями сенсилл, ASB — расстояние от самой выступающей части переднего края щита до
центра ботридий, PSB — расстояние от самой выступающей части заднего края щита до центра ботридий,
SD=ASB-t-PSB — длина щита, AM — срединная щетинка щита, AL — переднебоковая щетинка щита,
PL — заднебоковая щетинка щита, АР — расстояние между основаниями AL и PL, Sens — сенсилла;
2, 3 — Leptotrombidium palpale: 2 — личинка со спинной стороны, 3 — расположение щетинки на коксе
III; 4, 5 — L. pallidum: 4 —личинка со спинной стороны, 5 — расположение щетинки на коксе III.
обходимости его ревизии. Распространение всесветное. Паразитируют на поз-
воночных животных, чаще на грызунах. Некоторые виды нападают на человека
и сельскохозяйственных животных, вызывая у них кожное заболевание — тром-
бидиоз. — 10 видов (в СССР более 30).
292
3 (8). Имеется только 1 mastitarsala III.
j (5). Галеальная щетинка опушенная, число спинных щетинок в 3 первых
рядах, как правило, 2Н—6—6—6.
3 (4). Плечевых щетинок 4 (по 2 с каждой стороны). —Евр. ч. СССР, Заб.,
Прим., Сах.; 3 Европа, Япония, сев. п-ова Корея (рис. 143, 3,4). . .
.........................................N. japonica Tan., Kaiw., Тег., Kag.
Рис. 143. Детали строения клещей-краснотелок.
1,2 — Leptotrombidium pavlovshyi’. 1 — личинка со спинной стороны, 2 — с брюшной стороны; 3,4 —
Neotrombicula japonica: 3 — личинка со спинной стороны, 4 — с брюшной стороны.
4 (3). Плечевых щетинок 2 (по 1 с каждой стороны). — Евр. ч. СССР, Ср. Азия,
Алтай, Амур., Хаб., Прим.; 3 Европа, Иран, п-ов Корея (рис. 144, 7, 2)
.......................................................N. talmiensis Schl.
5 (2). Галеальная щетинка гладкая, число спинных щетинок в 3 первых рядах
разное.
6 (7). Общее число спинных щетинок не более 30, расположение их в 3 первых
рядах — 2Н — 6— 6— 6. — Прим., Азербайджан, Армения; п-ов Корея
.......................................................N. gardellai Kard.
7 (6). Общее число спинных щетинок более 40, расположение их в 3 первых
рядах —2Н—8—10—10. —Алтай, Хаб., Прим., Сах., Камч., Кур.;
293
Япония, п-ов Корея (рис. 144, 3,4)....................................
.........................N. nagayoi Sasa, Hayas., Sato, Miura, Asah.
8 (1). Имеются 2 и более mastitarsala III.
9 (14). 3 mastitarsalae III и 1 mastitibiala III.
10(11). Галеальная щетинка гладкая. —Амур., Прим. (рис. 145, 1, 2) . . .
...................................................N. zhmajevae Schl.
Рис. 144. Детали строения клещей-краснотелок.
I, 2 — Neotrombicula ialmiensis, 1 — личинка со спинной стороны, 2 — с брюшной стороны; 3, 4 — Neo-
trombicula nagayoi: 3 — личинка со спинной стороны, 4 — с брюшной стороны.
11 (10). Галеальная щетинка опушенная.
12(13). PL > AL > AM. Кокса III имеет почти круглую форму, АР =
=26 мкм. —Прим.; Япония..................................................
...............................N. mitamurai Sasa, Hayas., Kum., Тег.
13 (12). PL > AM > AL. Кокса III имеет удлиненную форму, ЯР=19 мкм. —
Амур., Хаб., Прим.; Китай, п-ов Корея, Япония, Гавайские о-ва . . .
.................................................N. tamiyai Phill. et FulL
14 (9). 2 mastitarsalae и 1 mastitibiala III.
294
15 (18). Mastifemorala III имеется.
16 (17). Галеальная щетинка перистая, fPp=N/N/BNB. —Таджикистан, Ал-
тай, Саяны, Амур., Хаб., Прим., Сах., Кур.; Япония, Сев.-Вост. Китай,
С Америка............................................N. microti Ew.
" 5
дне. 145. Детали строения клещей-краснотелок.
1—3 — Neotrombicula zhmajerae: 1 — личинка со спинной стороны, 2 — с брюшной стороны, 3 — лапка;
4,5 — Miyatrombicula esoensis: 4 — личинка со спинной стороны, 5 — с брюшной стороны.
17[(16). Галеальная щетинка гладкая, fPp=BIBIBNB. —Татарская АССР,
Амур., Хаб., Прим., Сах., Кур.; Сев.-Вост. Китай, Япония, США . . .
..............................................N. pomeranzevi Schl.
18 (15). Mastifemorala III отсутствует, галеальная щетинка гладкая, fPp~
=BIBIBNB. - 3 Сибирь.................................N. radfordi Schl.
3. Miyatrombicula Sasa, Kawash. et Egash. Личинки среднего размера,
Ip от 600 до 800. Щит пятиугольный с точечной пунктировкой. AL и PL рас-
положены в углах щита, которые несколько сближены. SB лежит примерно на
одном уровне с PL. Сенсиллы бичевидные, опушены по всей длине короткими и
295
длинными бородками. На коксах III пары ног от 1 до 9 щетинок. Mastitarsala
III часто присутствует. Паразитирует в основном на грызунах. — 1 вид (в СССР
6).
1 (1). Все щетинки на пальпах опушенные, jT=lBS, на коксе III от 4 до 6 ще-
тинок, mastitarsala III гладкая. —Заб., Хаб., Прим., Сах., Азербайд-
жан; Япония, С Америка (рис. 145, 3, 4)...................................
............................................М. esoensis Sasa et Ogata
4. Ascoschoengastia Ew. Личинки мелкие и среднего размера, Ip около 600.
Щит трапециевидный с хорошо выраженными переднебоковыми плечами. Сен-
силлы булавовидные, покрыты равномерно мелкими шипиками. Основания
сенсилл расположены почти в центре щита значительно выше уровня PL.
Mastitarsala III часто присутствует. Паразитируют на мелких млекопитающих
и птицах. — В СССР обнаружено 2 вида, оба отмечены на ДВ.
1 (2). Число спинных щетинок более 40, расположены беспорядочно, общее
число щетинок тела около 100. — Евр. ч. СССР, Ср. Азия, Заб., Прим.,
Сах.; Монголия (?), Зап. Европа..........................A. latyshevi Schl.
2 (1). Число спинных щетинок менее 40, расположены правильными рядами —
2Н—8—6—6 . . ., общее число щетинок тела около 70. —Хаб. . . .
............................................... A. danrela Kudr. et Shat.
5. Euschoengastia Ew. Личинки мелкие и среднего размера, Ip от 700 до 900.
Щит сильно вытянут 6 ширину. AL и PL сидят довольно близко, SB обычно
значительно ниже уровня PL. Почти всегда PL по длине больше AL.
Коготь пальп разделен на 3 и более (до 7) зубцов. Гладкие щетинки
на члениках III пары ног всегда отсутствуют. Паразитируют преимуще-
ственно на грызунах. —2 вида (в СССР 6).
1 (2). Щит в 2 —2.5 раза шире своей длины, PL в 2 раза длиннее других щетинок
щита, спинных щетинок более 60, NDV около 150, щетинка на коксе III
расположена на переднем крае членика почти посредине. — Прим. . . .
.......................................................... Е. armata Schl.
2 J1). Щит в 4—5 раз шире своей длины, PL намного длиннее других щетинок
на щите, спинных щетинок менее 60, NDV около 100, щетинка на коксе III
лежит в центре членика значительно ниже его переднего края. — Евр. ч.
СССР, Ср. Азия, Заб., Прим.; Монголия, сев. п-ова Корея...................
...................................................Е. rotundata Scbl.
6. Cheladonta Lip., Cros. et Loom. Личинки мелкие и среднего размера,
Ip менее 900. Щит прямоугольный, задний край часто двулопастной. Сенсиллы
булавовидные или шаровидные с мелкими шипиками. Всегда имеются 2 пары
плечевых щетинок, masti —щетинки и tibiala III отсутствуют. Паразитируют
на грызунах. — 1 вид (в СССР 3).
1 (1). Галеальная щетинка гладкая, все щетинки пальп опушенные, на лапке
пальп 4 опушенных щетинки, спинных щетинок около 40 или чуть более,
щетинка на коксе III расположена почти на переднем крае. — Евр. ч.
СССР, Прим.; 3 Европа, Япония, п-ов Корея.................................
......................Ch. ikaoensis Sasa, Saw., Kano, Hayas., Kum.
7. Neoschoengastia Ew. Личинки среднего и крупного размера, Ip много
больше 1000. Щит трапециевидный с переднебоковыми плечами, с точечной и
линейной штриховкой в нижней половине. Сенсиллы шаровидные, реже булаво-
видные. Коготь пальп разделен на 3 зубца. Специализированные щетинки
на ногах варьируют в числе. Паразиты птиц. — 1 вид (в СССР более 10).
1 (1). Щит трапециевидный с закругленными углами, с параллельной штри-
ховкой по заднему краю и вдоль средней линии, галеальная щетинка
с 2—3 бородками, дорсальная щетинка пальп гладкая, щетинка на
коксе III расположена на переднем крае членика, спинные щетинки опу-
шены короткими бородками, расположены рядами, общее число их бо-
лее 40. — Евр. ч. СССР, Казахстан, Прим.; Малайзия, Вьетнам, Китай
(о-в Тайвань)..........................................N. gallinarum Hat.
296
ОСНОВНАЯ ЛИТЕРАТУРА
Гопаченко И. М. Клещи краснотелки. — В кн.: Природноочаговые болезни в Примор-
ском крае. Владивосток, 1975, с. 202—212.
Кудряшова Н. И. Ревизия клещей рода Miyatrombicula (Trombiculidae) фауны СССР. —
Паразитология, 1978, т. 12, вып. 2, с. 154—166.
Кудряшова Н. И. Современное состояние изученности клещей краснотелок (Acarifor-
mes, Trombiculidae) фауны СССР. — В кн.: Итоги науки и техн. ВИНИТИ АН СССР. Сер.
зоопаразитол., 1979, вып. 5, с. 5—112.
Шлугер Е. Г. V. Надсемейство Trombeae Turk. I. Семейство Trombiculidae Ewing. I.
Подсемейство Trombiculinae Ewing — краснотелки. — В кн.: Клещи грызунов фауны
СССР. М.; Л., 1955, с. 188—217.
Шлугер Е. Г. Два новых вида клещей краснотелок (Acariformes, Trombiculidae) При-
морского края. — Зоол. журя., 1960, т. 39, вып. 8, с. 1258—1261.
Шлугер Е. Г., Кудряшова Н. И. Клещи краснотелки (Trombiculidae) и Rickettsia tsu-
tsugamushi. — В кн.: Биология взаимоотношений кровососущих членистоногих с возбуди-
телями болезней человека. М., 1967, с. 143—150.
Надсем. DEMODICOIDEA
Эндопаразитические клещи с телом червеобразной (сем. Demodicidae) или
округлой (сем. Psorergatidae) формы.
Гнатосома хорошо выражена, крупная, резко выдается впереди тела.
Пальпы толстые, 3-члениковые, концевой членик сплющен и на вершине несет
несколько маленьких папилл или зубцов. Между пальпами располагаются
стилетовидные хелицеры, заключенные в перепончатый футляр. Заметных
дыхательных отверстий и перитрем на гнатосоме нет.
Ноги обычно короткие и тонкие, имеют вид обрубков или конусов, с 5 под-
вижными члениками. Лапки с 2 коготками, предлапка отсутствует. Коксо-
стернальный скелет развит различно: либо в виде слабо развитых тонких эпи-
мер и эпимеритов, не соединяющихся между собой, либо в виде сильно склеро-
тизованных пластинок коксальных полей.
Половой диморфизм выражен слабо. Половой аппарат самца расположен
со спинной стороны в центре спинного щита; половое отверстие самки —
с брюшной стороны непосредственно позади эпимер ног IV.
Цикл развития двунимфальный.
Включает 2 семейства; медико-ветеринарное значение имеет лишь одно из
них.
ЖЕЛЕЗНИЦЫ (Сем. DEMODICIDAE)
[ВНЕШНЕЕ СТРОЕНИЕ
Очень мелкие (0.16—0.30) эндопаразитические клещи с червеобразно вы-
тянутым телом. Гнатосома крупная, широкая, почти квадратная. Пальпы тол-
стые: второй членик со спинной стороны несет утолщенную щетинку; концевой
членик имеет на наружной поверхности крючковидный вырост, а на уплощен-
ной вершине несколько маленьких конусовидных сосочков-папилл. Хелицеры
тонкие, стилетовидные, окруженные футляром (рис. 146, 2, 5).
Имеется небольшой проподосомальный щит, вся остальная поверхность тела
с поперечными кольцевыми складками. Опистосома обычно очень длинная:
длина ее не меньше длины передней части тела, несущей плотно сидящие ноги.
Хетом тела редуцирован до нескольких пар спинных микрохет.
Ноги очень короткие, 5-члениковые, лапка с 2 видоизмененными мощными
коготками. Коксо-стернальный скелет представлен прямоугольными пластин-
ками — коксальными полями, тесно сближенными в два продольных ряда
вдоль средней линии тела.
Самец. Половой аппарат располагается на спинной стороне тела — на уровне,
соответствующем положению ног II и III, и представлен крупным пенисом
с сильно склеротизованным искривленным стилетом.
Самка. Половое отверстие в виде продольной щели находится с брюшной
стороны между коксальными полями ног IV (рис. 146, 7).
20 Зак. № 990 297
Неполовозрелые фазы, кроме отсутствия половых отверстий, отличаются гем,
что их ноги имеют вид сосочков с коготками; членистыми они становятся лишь
на фазе телеонимфы.
БИОЛОГИЯ
Клещи паразитируют в сальных железах кожи или в волосяных фолликулах
различных млекопитающих, включая человека. Вследствие очень быстрого раз-
вития в одной пораженной волосяной сумке может находиться до нескольких
сот особей всех фаз развития.
Рис. 146. Представители подотряда Trombidiformes.
I—3 — Demodex folliculorum: 1 — самка с брюшной стороны, 2 — гна-
тосома со спинной стороны, з — гнатосома с брюшной стороны (по:
Дубинин, 1955); 4 — Syringophilus bipectinatus, самка со спинной сто-
роны (по: Fritsch, 1958). н — ноги; кп — коксальные поля; яо — яйце-
выводное отверстие: х — хелицеры; членики пальп: кчп — концевой,
вчп — второй, очп — основной; щп — щетинка пальп; ро — ротовое
отверстие; до — дыхательное отверстие.
ВРЕД
Клещи вызывают атрофию эпителиальных желез, выпадение волос, гипере-
мию покровов. Заболевание протекает в «чешуйчатой» и «пастулезной» формах.
При первой пораженная поверхность покрывается чешуйками, принимает си-
невато-серую окраску с красными пятнами. При пастулезной форме на поверх-
ности развиваются узелки-пастулы, наполненные гноем с примесью крови или
сальной густой массы. Заболевание часто сопровождается вторичной стафило-
кокковой инфекцией, приводящей к изъязвлению покровов, сильному исто-
щению животных и даже гибели их от интоксикации или общей септической ин-
фекции [Дубинин, 1955].
Включает 1 род.
Demodex Owen. Характеризуется всеми признаками семейства. Половой
диморфизм выражен слабо. Самки отличаются от самцов положением полового
аппарата и обычно несколько более крупными размерами, за счет более длин-
ной и толстой опистосомы.
Основными диагностическими признаками являются форма и расположение
щетинок пальп, форма и размеры тела, форма склеритов копулятивного ор-
298
гана самцов. В силу очень мелких размеров тела диагностика видов крайне
затруднена и может вестись лишь с помощью иммерсионных увеличений ми-
кроскопа.
На Дальнем Востоке — 9 видов (в СССР более 20).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ1
1 (4). Общая длина тела d превышает таковую 9.
2 (3). Коксальные шипы с раздвоенной вершиной. $ —0.22, d —0.26. —По-
всеместно. Паразит домашнего крупного рогатого скота.....................
.....................................................D. bovis Stiles
3 (2). Коксальные шипы без раздвоенной вершины. $ — 0.24, d — 0.29. —
Повсеместно. Паразит лошади..............................................
........... D. caballi Desch, Nutting (folliculorum var. equi (Railliet))
4 (1). Общая длина тела $ превышает таковую d.
5 (6). Эпимеры d не сливаются на средней линии тела. $ —0.21, d —0.17. —
Повсеместно. Паразитирует в волосяных фолликулах человека . . . .
...................................................D. brevis Akbulatova
6 (5). Эпимеры d сливаются на средней линии тела.
7 (8). Ноги неполовозрелых фаз представлены в виде плоских бугорков и не вы-
даются за боковые края тела; коготки располагаются на этих бугорках.
9 — 0.23, d — 0.22. — Повсеместно. Паразит домашней козы.................
.................................................D. саргае (Railliet)
8 (7). Ноги неполовозрелых фаз выдаются за боковые края тела; коготки рас-
полагаются дистально.
9(10). Задний конец тела нимф загнут. $ — 0.22, d —0.17. — Повсеместно.
Паразит домашней свиньи................................D. phylloides Csokor
10 (9). Задний конец тела нимфы прямой.
11 (12). Длина опистосомы составляет менее половины общей длины тела.
9 —0.26, d —0.19. —Повсеместно. Паразит домашней лошади . . . .
....................................................D. equi (Railliet)
12 (11). Длина опистосомы превышает половину общей длины тела.
13 (14). Видоизмененная спинная щетинка основного членика пальп распола-
гается впереди ротового отверстия. 9 —0.21, d —0.15. —Повсеместно.
Паразит домашней овцы....................................D. ovis (Railliet)
14 (13). Видоизмененная спинная щетинка основного членика пальп распола-
гается по бокам ротового отверстия.
15 (16). Задняя кишка у d видна в виде трубки, направленной вперед. 9 —
0.25, d —0.21. — Повсеместно. Паразит домашней кошки.....................
..................................................... D. cati Megn.
16 (15). Задняя кишка у d не видна.
17 (18). Дорсальная проподосомальная щетинка d гантелевидная или в виде
восьмерки. Яйцо веретеновидной формы. 9 —0.22, d —0.17. —По-
всеместно. Паразит домашней собаки...................... D. canis Leydig
18 (17). Дорсальная проподосомальная щетинка d закругленная. Яйцо округ-
лой формы. 9 — 0.30, d — 0.28. — Повсеместно. Паразитирует в сальных
железах кожи человека (рис. 146, 1—3) . . . . D. folliculorum (Simon)
ОСНОВНАЯ ЛИТЕРАТУРА
Дубинин В. Б. Семейство Demodicidae Meg. — Железницы. — В кн.: Клещи грызунов
фауны СССР. М.; Л., 1955, с. 143—152. (Определители по фауне СССР, изд. Зоол. ин-том
АН СССР, № 59).
Desch С. Е., Nutting W. В. Demodicids (Trombidiformes; Demodicidae) of medical and
veterinary importance. — In: Proc, of the 3rd Intern. Congress of Acarology, Prague, 1971.
Prague, 1973, p. 499-505.
Desch С. E., Nutting W. B. Demodex folliculorum (Simon) and D. brevis Akbulatova of
man: redescription and reevaluation. — J. ParasitoL, 1972, vol. 58, N 1, p. 169—177.
1 Определительная таблица дается полностью по Дешу и Наттингу (Desch, Nutting,
1973).
20*
299
Надсем. CHEYLETOIDEA
Мелкие и средней величины клещи. Гнатосома сильно варьирует по форме
в разных семействах, значительно короче идиосомы, состоит из среднего отдела
(рострума) и пары лежащих по сторонам от него пальп. Средний отдел конусо-
видный, склеротизованный, с маленьким ротовым отверстием на конце, через
которое выдвигается пара стилетовидных хелицер. Со спинной стороны по
бокам хелицер располагается пара вытянутых перитрем с отходящими от них
трахеями. Пальпы крупные, различного строения в каждом семействе, как и
вооружение пальп и лапки.
Покровы тела обычно тонко штрихованные, иногда гладкие.
Ноги 5-члениковые, развиты весьма различно в зависимости от образа
жизни клещей разных семейств.
Половой диморфизм выражен слабо. Половой аппарат самца занимает суб-
терминальное положение или смещен на спинную сторону опистосомы; половое
отверстие самки располагается в задней трети брюшной стороны и, как правило,
вплотную приближено к анальному отверстию.
Цикл развития 2—3-нимфальный.
Включает 4 семейства; медико-ветеринарное значение имеет лишь одно из
них.
ОЧИННЫЕ КЛЕЩИ (Сем. SYRINGOPHILIDAE)
ВНЕШНЕЕ СТРОЕНИЕ
Относительно мелкие клещи с вытянутой веретеновидной формой тела, уз-
кие, с прямыми, почти параллельными друг другу боками; ширина тела в 3—
3.5 раза меньше длины. Гнатосома маленькая, треугольная, вытянутая в длину.
Пальпы 3—4-члениковые, концевой их членик закруглен и не выдается за
передний край гнатосомы. На вершине пальп располагаются щетинки и малень-
кие коготки или только щетинки. Стилетовидные хелицеры заключены в эла-
стичный футляр.
Покровы мягкие; на спинной стороне имеются прямоугольный проподо-
сомальный щит с 4—5 парами волосовидных щетинок и опистосомальный щит,
сильно варьирующий по форме.
Передние две пары ног причленяются по бокам переднего конца тела, а ноги
III и IV удалены от передних и располагаются в средней части тела или ближе
к заднему концу. Ноги всех пар развиты одинаково, либо ноги I—II утолщены
и видоизменены. Все ноги сравнительно короткие, конусовидные; лапки окан-
чиваются 2 маленькими крючковидными коготками и крупным У-образным пе-
ристым эмподием между ними.
Самец. Половое отверстие располагается терминально или субтерминально,
стилет пениса серповидно изогнут.
Самка. Копулятивное отверстие располагается терминально на заднем конце
тела, непосредственно примыкая к заднему краю анального отверстия.
Развитие протекает по двунимфальному типу.
БИОЛОГИЯ
Все представители живут в полости очинов маховых и крупных контурных
перьев тела птиц, откуда они могут проникать в подкожную клетчатку [Дуби-
нин, 1957]. Заселение новых здоровых очинов осуществляют лишь самки. В слу-
чае отсутствия оплодотворения имеет место факультативный компенсаторый
партеногенез [Гриценко, 1969].
ВРЕД
Клещи прокалывают стенки очинов длинными хелицерами, повреждают
сосочек пера и высасывают лимфу; это приводит к развитию воспалительных
процессов и преждевременному выпадению перьев. Кроме того, при подобном
300
раздражении кожи птицы часто сами начинают выщипывать перья, что приводит
к различного рода дерматитам.
Семейство включает 2 рода. Ветеринарное значение имеет лишь один из них.
Syringophilus Heller. Перитремы в виде одного ряда зерен различной вели-
чины. Ноги всех пар развиты примерно одинаково, толстые, конусовидные.
Лапки всех ног с одинаково развитыми 2 маленькими коготками и гребенчатым
эмподием; каждый выступ гребенки эмподия заканчивается маленьким треуголь-
ным расширением. Некоторые щетинки спинной стороны лапок расширены на
концах, опушены или зазубрены. Концевой членик пальп с 2 коготками на
вершине. Обитают в полости очинов различных птиц.
Род насчитывает около десяти видов, ветеринарное значение имеет лишь
один — паразит домашних кур.
1 (1). Перитремы состоят из ряда крупных зерен. Пальпы вздутые со спинной
стороны. Проподосомальный щит с 6 парами щетинок. Опистосомальный
щит имеет вид очень маленькой пластинки, лежащей субтерминально.
$ — 0.825—1.050, J —0.525—0.800. — Повсеместно. Паразитирует в очи-
нах маховых и крупных контурных перьев домашних кур (рис. 146, 4)
.......................................................S. bipectinatus Heller
ОСНОВНАЯ ЛИТЕРАТУРА
Гриценко Е. Ф. Биология и экология очинного клеща Syringophilus bipectinatus Heller,
1880: Автореф. дис. . . . канд. биол. наук. Ставрополь, 1969. 17 с.
Дубинин В. Б. Новая классификация клещей надсемейств Cheyletoidea W. Dub. и De-
modicoidea W. Dub. (Acariformes, Trombidiformes). — В кн.: Паразитол. сб., Л., 1957, т. 17,
с. 71—136.
Fritsch W. Die Milbengattung Syringophilus Heller, 1880 (Subordo Trombidiformes, fam.
Myobiidae Megnin, 1877). — Zool. Jahrbiicher, 1958, Bd 86, H. 3, S. 229—243.
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ НАСЕКОМЫХ 1
abchazicus, Culicoides 37, 38*
Abonesia 97, 99
acanthina, Neopsylla 156
acanthopus, Hoplopleura 167*, 173
actoni, Culicoides 36*, 37
adachii, Hybomitra 70, 71*
adustus, Anatoecus 185, 186*
advenarius, Megabothris 159
Aedes 10, 11, 12*, 13, 16—18
aenescens, Morelli a 82, 98
Aenigmopsylla 157, 158
aequetincta, Hybomitra 65, 66*
affinis, Hoplopleura 169, 173*
Agria 104, 105
alaskaensis, Culiseta 19, 20*, 21
alaskaensis, Culiseta alaskaensis 19
alaskensis, Oropsylla 158
albibasalis, Alloeostylus 89
albiceps, Chrysomya 101
albiceps, Parasarcophaga 111*, 112
albipes, Gnus malyschevi 46, 51*, 52
albolineata, Polietes 92
Aldrichina 97, 98
alektorovi, Aedes 21
Alloeostyllus 85, 89
Amalaraeus 158, 161
Amblycera 181
amblyogma, Protocalliphora 102
americana, Periplaneta 177*, 178, 179
arnica, Musca 93
amita, Musca 84, 93, 94
amossovae, Culicoides 39*, 40
Amphipsylla 163, 164
ampulacea, Lucilia 101
amurense, Cnetha 49, 50*
Anaticola 184
Anatoecus 185
angaricus, Chrysops 63*, 64
Angiometopa 104, 106
angustifrons, Muscina 91
Anistronidae 182
anisus, Monopsyllus 161, 162*
Anopheles 10, 11, 13, 14*, 15*, 16, 18*
Anophelinae 10, 11, 15*, 16
Anoplura 166
anserinum, Trinoton 182*
Anseriphilus 183
anseris, Anaticola 185
anthrax, Chrysops 62*
antilope, Sarcorohdendorfia 118*
Aphaniptera 146
arakawae, Culicoides 35*, 37
aratrix, Parasarcophaga 112, 113*
arboreus, Culicoides 39*, 40
Archaeopsylla 151
arctica, Mioctenopsylla 159
ardeae, Icosta 124, 127*
argyrostoma, Parasarcophaga 112, 113*
armipes, Hydrotaea 90*
arpadi, Hybomitra 67, 69*
asema, Stegopterna 48, 49*
asini, Haematopinus 169*, 170
aspalacis, Amphipsylla 164
assimilis, Muscina 91
astur, Hybomitra 65, 66*
astuta, Hybomitra 65, 67*
ater, Aedes riparius 23
aterinervis, Culicoides 38
atriceps, Boreellus 101
atripalpis, Haematobia 96
Atylotus 58, 60, 73
» aureum, Eusimulium 50*, 51
aureus, Aedes 25
australasiae, Periplaneta 177*, 178,7179
autumnalis, Musca 93, 94
avicularia, Ornithoica 123, 124*
azurea, Protocalliphora 102
bagheera, Hybomitra 65, 68*
bazini, Lucilia 100
beklemishevi, Aedes 22
beljaevi, Megabothris 159
Bellardia 97
Bellieriomia 104, 108
Bercaea 105, 110
bergrothi, Culiseta 19, 20*, 21
bicorne, Cnetha 49, 50*
bicorne, Eusimulium 45
bidentatiformis, Neopsylla 155
bimaculata, Hybomitra 69, 70*, 71
bisignata, Hybomitra bimaculata 69
bispinosus, Megabothris 159
bitaeniorhynchuSj Culex 29, 30*
Blatta 177*, 178, 179
Blattella 179
Blattellidae 178, 179
Blattidae 178
Blattinae 178
Blattoptera 176
Boettcherisca 105, 110
Bonomiella 182
1 Здесь и далее в указателях жирным шрифтом даны таксоны выше родового
ранга, курсивом — синонимы; звездочкой отмечены страницы с рисунками.
302
Boophinus 47, 52
Boophthora 47, 52
Boopidae 183
Booponus 97, 102
Boreellus 98, 101
botis, Frontopsylla lata 165
Bovicola 185
Bovicolidae 185
bovis, Boyicola 185, 186*
bovis, Hypoderma 143, 145*
Brachicoma 104, 106
Brachycera 55, 119
brevicornis, Parasarcophaga 115*
brevis, Hybomitra 56*, 66, 67,' 69*
brunnea, Chelocnetha 49, 50*
buddha, Tabanus 56*, 61, 76, 77*
Byssodon 47, 49
cadaverina, Cynomya 100
cadaverina, Pyreilia 94
caecutiens, Chrysops 63*, 64
caesar, Lucilia 101 1
caesarion, Orthellia 93
calcarifer, Megabothris 159, 160*
calcitrans, Stomoxys 92*, 96, 99*
Calliphora 97, 98
Calliphoridae 80, 83, 96
calliphoroides, Triceratopiga 99
Callopsylla 158, 160
Campanulotes 184
canariensis, Pseuodolynchia 124. 129*
canicularis, Fannia 87, 88*
canis, Ctenocephalides 151, 152*
canis, Trichodectes 186
cantans, Aedes 22*, 23, 27
capillatus, Solenopotes 169*, 173
caponis, Lipeurus 184
caprae, Bovicola 186*
capriola, Hypoderma 146
captiosa, Hoplopleura 169*, 173
carnaria, Sacrophaga 118*, 119
caspius, Aedes 21, 22*
caspius, Aedes caspius 21
casta, Rhadinopsylla integella 154
Catallagia 152, 153, 157
cataphylla, Aedes 22*, 24, 27
Cephenomyia 136, 138, 139
Ceratophyllidae 150, 157
Ceratophyllus 158, 161
Ceratopogonidae 31, 34
cervi, Lipoptena 84, 125, 129
Chaetopsylla 151
chalcogaster, Ophira 89
Chelocnetha 47, 49
cheopis, Xenopsylla 151
china, Lucilia 100
chiopterus, Culicoides 36*, 37
chloropus, Ornithoica 123, 126
cholodkovskii, Gnus 45, 51*, 52
Chrysomya 97, 101
Chrysops 55, 56, 57, 60
Chrysopsinae 60
chrysurus, Tabanus 76, 77*
Cimex 176
Cimicidae 174
cinereus, Aedes 25
cinereus, Aedes cinereus 25, 26*
circumscriptus, Culicoides 36*
Citellophilus 158, 160
Cnetha 42, 47, 49
Coloceras 184
Colpocephalidae 182
columbae, Bonomiella 182
columbae, Columbicola 180*, 184
Columbicola 184
communis, Aedes 24, 26*, 28
comosioculatus, Culicoides 40
compar, Campanulotus bidentatus 184
concava, Rhadinopsylla integella 154
congeneroides, Ctenophthalmus 153
contigua, Hybomitra distinguenda 71*
cornicina, Orthellia 93
cornutus, Gallacanthus 182*, 183
Corrodopsylla 153
coypus, Pitrufquenia 183
crassicornis, Anaticola 185
crassipalpis, Parasarcophaga 111*, 112
crassipes, Holokartikos 185
Crataerina 119—121, 123
cruentata, Bercaea 109*, 110
Ctenocephalidae 151
Ctenophthalmus 153
Ctenophyllus 163
cubitalis, Culicoides 39*, 40
Cuclotogaster 184
Culex 10, 11, 12*, 13, 16, 18*, 28
Culicidae 10, 14*, 15*, 17
Culicinae 11, 15*, 16, 18*
Culicoides 31, 32, 33, 34*
Culiseta 10, 11, 12*, 13, 18
curvans, Cnetha 49, 50*
curvispinus, Paradoxopsylla 164
curvitarse, Simulium 54, 54*
cyanicolor, Dasyphora 94
Cyclorrhapha 119
Cynomya 97, 100
cyprius, Aedes 22
dacenkoi, Catalagia 156*, 157
Dasyphora 83, 86, 94
Dasypsyllus 157, 158
decimatum, Gnus 45, 51*, 52
decipiens, Mesembrina 95
decolor, Trichopticoides 89
Degeeriel lidae 184
Dendrophaonia 86, 91
dendrophilus, Culicoides 32, 39
dentatus, Anatoecus 185*
dentatus, Oxylipeurus 182*, 184
dentipes, Hydrotaea 88*, 90*, 91
depuncta, Helina 88
desertorum, Haematopota 56*, 78, 79*
detrita, Mydaea 87
devia, Brachicoma 106, 107*
diana, Hypoderma 132*, 143, 146
Diptera 10
disparilis, Enderleinellus 169*, 171
dissectus, Chrysops 56*, 63*, 64
dissimilis, Amalaraeus 161, 162*
dissimilis, Oulocrepis 180*, 184
divaricatus, Chrysops 62*
dolichocephalus, Enderleinellus 169*, 171
domestica, Musca 93, 94, 99*
Doratopsylla 153
dorsalis, Aedes caspius 21
Drosophilidae 84
dubius, Culicoides 37, 38*
elburnum, Cnetha 49, 50*
Enderleinellus 167*, 171
Eomenacanthus 183
epactius, Culicoides 39
equi, Wernekiella 185
eraorai, Culicoides 38
erecta, Sononipponia 118
errans, Phaonia 91
erythrocephala, Boophthora 52, 53*
303
erythronofum, Prosimulium 48
esoensis, Aedes cinereus 25
Eupolyphaga 179
eurysternus, Haematopinus 167, 169
Eusimulium 47, 51
excrucians, Aedes 22*, 24, 27
exigua, Lyperosia 96
Heteroptera 174
Hippoboscidae 119, 255
Hippoboscinae 119
♦, 170 hirsutula, Lucilia 100
hirsutula, Tricops 89
hirundinis, Crataerina 83, 124
hispidum, Trimenopon 183
Hohorstiella 183
Holokartices 185
Holomenopon 182
honshuensis, Goniophyto 105
Hoplopleura 167*, 171, 172
Hoplopleuridae 169, 171
hortensia, Morellia 95
hortorum, Morellia 95
horvathi, Atylotus 73, 74*
humanus, Pediculus 170
humeralis, Culicoides 34
Hybomitra 55, 56*, 58, 60, 64
Hydrotaea 82, 85, 86, 89
Hymenoptera 187
Hypoderma 142,143
Hypodermatidae 131, 133, 135, 142
hyrcanus, Anophelles 19, 20*
Hystrichopsylla 153
Hystrichopsyllidae 150, 152
fahrenholzi, Coloceras damicornis 184
Fannia 82, 85, 87
farreni, Ceratophyllus 162
fasciatus, Nosopsylla 160
fascipennis, Culicoides 39*, 40
Felicola 186
felis, Ctenocephalides 151, 152*
filicinus, Culicoides 37, 38*
flavescens, Aedes 22, 27
flavidorsum, Culicoides 34, 35*
flavopictus, Aedes 24, 27
Formicidae 187, 189
Formicinae 189
fortisetosa, Lipoptena 125, 130*
fringillina, Ornithoica 123, 125*
Frontopsylla 164
fuscanus, Cullex 29, 30*
fuscipalpis, Lucilia 100
Gallacanthus 183
gallinae, Ceratophyllus 163
gallinae, Goniocotes 184
galloisi, Aedes 24, 25, 26*
Gastrophilidae 135
Gastr ophilus 135
geminus, Tabanus 76, 77*
genarum, Abonesia 99
germanica, Blattella 177*, 178, 179
gigas, Stenocrotaphus 184
Gliricola 183
Gliricolidae 183
Gnus 42, 45, 47, 51
Goniocotes 184
Goniodidae 183
Goniodoidea 183
Goniophyto 104, 105
gorei, Ceratophyllus 162
gracilior, Mesembrina 96
grahami, Adri china 98
Graphomyia 85, 89
griseinus, Tabanus amaenus 75
grodekovi, Aenigmopsylla 158
Gyropidae 183
Gyropoidea 183
Gyropus 183
Haematobia 84, 85, 96
Haematopinidae 169
Haematopinus 167*, 170
Haematopota 55, 58, 60, 76
Haemodipsus 171, 172
haemorrhoidalis, Bercaea 110
haemorrhoidalis, GastrOphilus 135, 136, 137*
Haeterochrysops 60, 64
hagoromo, Ceratophyllus 162
harpax, Parasarcophaga 114, 115*
hayashii, Culex 29, 31
Hebecnema 85, 87
Helicophagella 104, 107
Helina 85, 88
Hemiptera 174
hervebazini, Sinonipponia 117*, 118
Heterodoxus 183
heterographus, Cuclotogaster 184
304
Icosta 122, 124
ictrodes, Anatoecus 185, 186
ignita, Pyreilia 94
illustris, Lucilia 101
impiger, Aedes 22*, 24, 28
incisurata, Fannia 87
indages, Monopsyllus 161
indica, Culiseta alaskaensis 19
inexpectata, Booponus 102
inopinata, Helicophagella 108*
insolita, Rhadinopsylla 154
integella, Rhadinopsylla 154
intestinalis, Gastrophilus 122, 133, 135
intrudens, Aedes 22*, 24, 28
ioffi, Amalareus 161
ioffi, Catalagia 156*, 157
irritans, Prosimulium 45, 46, 48
irritans, Hydrotaea 90
irritans, Lyperosia 96, 99*
irritans, Pulex 147*, 151
Ischnocera 181, 183
isos, Prosimulium 48*
itoi, Paradichosia 100
jacksoni, Culex 29, 31
jacobsoni, Parasarcophaga 114, 115*
japonica, Paradichosia 100
japonica, Periplaneta 178, 179
japonica, Sareophila 107*
japonicum, Simulium 45, 46, 53*, 54
japonicus, Aedes 22, 25
japonicus, Anopheles lindesayi 19
japonicus, Chrysops 57, 62*
jaroschevskyi, Parasarcophaga 116*
kamtschaticus, Melophagus 128
kanoi, Parasarcophaga 113, 114*
kawayuensis, Parasarcophaga 113, 114*
kinoshi tai , Tabanus 76, 77*
kobayashii, Parasarcophaga 115*
kolomyietzi, Takanoa’ 118*
konakovi, Phallosphaera 116*
konakovi, Simulium 53*, 54
koreenvis, Culicoides 35*, 36
koreicus, Aedes 22
fcoryoensis, Haematopota 56*, 76, 78*
Kramerea 105, 111
kurilense, Simulium 53*, 54
kurilensis, Mioctenopsylla traubi 159
laeviusculus, Neohaematopinus 172
Lagopoecus 184
lapponica, Hybomitra 65, 67*
lapponica, Muscina 91
lardaria, Polietes 92
larvipara, Musca 94
Lasius 189, 190
lata, Calliphora 98
lata, Hohorstiella 182*, 183
latifrons, Rhinoestrus 142
lectularis, Cimex 175*, 176
Lepicentron 185
Leptopsylla 163, 164
Leptopsyllidae 150, 163
leucosticta, Fannia 87
leucostoma, Ophtra 89
limbatus, Bovicola 186*
lindesayi, Anopheles 19
lindneri, Tripocalliphora 102
lineatum, Hypoderma 146
Linognathid^e 169, 173
Linognathus 173
Lipeuridae 184
Lipeurus 184
Lipoptena 119, 120, 124
Lipopteninae 119
longipalpe, Simulium 54*
Lucilia 97, 100
luculenta, Frontopsylla 165*
lunatus, Ceratophyllus 162
lundbecki, Hybomitra 56*
lurida, Hybomitra 70
Lyperosia 84—86
maculata, Byssodon 45, 49
maculata, Graphomya 89
maculatus, Ceratophyllus 161
maculatus, Goniocotes 184
maculipennis, Anopheles 19, 20*
makerovi, Chrysops 62*, 63
Mallophaga 179
mamillata, Agria 106
manchuriensis, Culicoides 36*, 37
manicata, Fannia 87
Manzonia 10, 13
marikovckii, Amphipsylla 164
maruyamensis, Protocalliphora 102
maxbeieri, Holomenopon 182
meditabunda, Myospila 96
Megabothris 157, 159
megacephala, Chrysomyia 101
meigeni, Wohlfahrtia 107*
melanura, Helicophagella 107, 108*
Melinda 97
Melophagus 83, 119, 120, 125, 130*
Menopon 183
Menoponidae 183
Menoponoidea 181
meridiana, Mesembrina 95
Mesembrina 82, 86, 95
metallica, Ornithoica 122, 123*
meteorica, Hydrotaea 90, 91
microti, Hystrichopsylla 152, 153
mihalyii, Angiometopa 103*, 106*
Mioctenopsylla 150, 158
miser, Atylotus 61*, 73, 74*
modestus, Culex 28, 31
monachae, Agria 105, 106*
Monomorium 189
Monopsyllus 158
montana, Hybomitra 72
mon tana, Stenoponia 154, 155*
montschadskyi, Columbicola 184, 185*, 186*
Morellia 82, 86, 94
morgani, Hybomitra montana 72*, 73
morsitans, Culex 14*, 21
morsitans, Simulium 54*
mortuorum, Cynomia 100
muhlfeldi, Hybomitra 71*
musachinensis, Sinonipponia 117*, 118
Musca 86, 93
Muscidae 80, 83, 84
Muscina 86, 87, 91
mutata, Stegopterna 48*, 49
Mydaea 85, 87
Myospila 85, 96
Myrmicinae 189
Nearctopsylla 153
Neocolpocephalum 183
Neohaematopinus 171, 172
Neopsylla 153
nigella, Hybomitra 61*, 66, 68*
niger, Lasius 188, 189*, 190
nigra, Muscina 91
nigra, Ophyra 89
nigricornis, Hybomitra 70, 71*
nigripes, Aedes 22*, 24, 26*, 27
nigripes, Chrysops 62*, 63
nigrolimbata, Polietes 91
nipponensis, Culicoides 37, 38*
nipponicus, Aedes 24, 25
nipponii, Aedes vexans 22
nitidifrons, Hybomitra 68, 70*
nitidiventris, Subfaonia 87
nobuconis, Aedes 25
Nosopsyllus 157, 159
nubeculosus, Culicoides 34, 35*
*Numidilipeurus 184
obscurunervis, Hybomitra 65, 68*
occulta, Hydrotaea 89, 91
ochroptera, Culiseta 20*, 21
Odagmia 47, 52
odibilis, Culicoides 38
Oedemagena 142, 143
Oestridae 133, 135, 136
Oestrinae 136
Oestrua 136, 140
okazaki, Paradichosia 100
olsoi, Hybomitra 67, 70*
olsufjevi, Ceratophyllus 162
Onesia 97
Ophyra 86, 89
Orfrontia 164, 165
orientalis, Blatta 177*, 178, 179
orientalis, Culex 29, 31
orientalis, Haematopota turkestanica 79
orientalis, Hybomitra expollicata 72*, 73
orientalis, Neopsylla pleskei 155, 156
ornata, Odagmia 45, 52
Ornithoctona 121, 124
Ornithoica 121, 122
Ornithomya 121, 123
О rnithomyinae 119
Ornithophila 121, 122
Oropsylla 157, 158
Orthellia 86, 92
Orthopodomyia 10
Orthorapha 55
305
osiris, Musca 93, 94
ostsibirica, Peromisopsylla 164
Oulocrepis 184
ovalis, Gyropus 183
Oxylipeurus 189
pabulorum, Muscina 91
pacifica, Agria 106
pacifica, Orthellia 93
pacificus, Ctenophthalmus picticus 154, 155*
Palaeopsylla 153
pallidicornis, Culicoides 39*, 40
pallidulus, Uchida 183
pallitarsis, Atylotus 73, 74*
Pangonia 57
Parabissodon 47, 49
Paraceras 157
Paradichosia 97, 100
Paradoxopsyllus 163, 164
paradoxus, Monopsyllus 161
pasquorum, Muscina 91
pavlovskii, Gnus 51*, 52
pavlovskii, Hybomitra 67, 69*
pectinata, Pollenia 101
pectiniceps, Leptopsylla 164
pectiniceps, Pectonoctenus 164
pectiniventris, Anseriphilus 183
pedalis, Linognathus 169*, 174
Pediculidae 169, 170
Pediculus 170
penicilliger, Amalaraens 160*, 161
peregrina, Boettcherisca 110*
Periplaneta 177*, 178
Peromyscopsylla 163, 164
Phallantha 104, 109
Phallosphaera 105, 106
Phaonia 86, 87, 91
pharaonus, Monomorium 188, 189*
Pharingomyia 136, 138, 139
Philopteroidea 184
Phormia 98, 102
Phthirus 169*, 170, 171
picta, Pharingomyia 139, 140*
picticus, Ctenophthalmus pacificus 154, 155*
Pierretia 104, 108
pionips, Aedes 24, 28
pipiens, Culex 14*, 29, 31
Pitrufquenia 183
pleskei, Parasarcophaga 116*
pleskei, Tabanus 76, 77*
plicata, Ornithoctona 124, 127*
podagrica, Morellia 95
Polietes 86, 91
Poliphaga 177*
Pollenia 97, 101
Polyphagidae 178, 173
Polyplax 169*, 171, 172
polystylata, Parasarcophaga 113*
porcelli, Gliricola 183
Portschinskia 131
portschinskyi, Parasarcophaga 115, 116*
Prosimulium 42, 47
Protocalliphora 98, 102
Protophormia 98, 101
proxima, Protocalliphora 102
pseudodahurica, Rhadinopsylla 154, 155*
Pseudolynchia 122, 124
pseudonearetica, Byssodon 49
pseudoscoparia, Robineauella 117*
pubis, Phthirus 167*, 171
pugetense, Cnetha 49, 50*
Pulex 151
pulicaris, Culicoides 37
Pulicidae 150
pullatus, Aedes 22*, 24, 28
punctatus, Culicoides 37
punctor, Aedes 24, 28
purpureus, Rhinoestrus 142
pusilia, Paradichosia 100
pusilia, Schoenbaueria 52, 53*
Pyreilia 86, 94
querceti, Dendrophaonia 91
querquedulae, Trinoton 182*
Ravinia 104, 107
reconditus, Culicoides 40
regina, Phormia 102
resplendes, Mesembrina 96
Rhadnopsylla 153, 154
Rhinoestrus 138, 141
riethi, Culicoides 34, 35*
ringdahlei, Hydrotaea 89, 90*
Rinoestrus 136, 138, 141
riparius, Aedes 22*, 23, 27
riparius, Aedes riparius 23
riparius, Ceratophyllus 163
Robineauella 105, 117
rossitensis, Ceratophyllus 163
rostratum, Gnus 45, 46
rothschildi, Rhadinopsylla 154
rudis, Pollenia 101
saishuensis, Morellia 95
saishuensis, Musca 83, 84
sakhalinensis, Chrysops suavis 64
salinarius, Culicoides 35*, 36
sapporoenus, Tabanus 75, 76*
Sarcophaga 105, 119
Sarcophagidae 80, 83, 102, 104
Sarcophija 104, 106
Sarcorohdendorfia 105, 118
Sarcotachinella 104, 109
scalaris, Fannia 87
scambus, Hydrotaea 89, 90*
Schoenbaueria 47, 52
schuetzei, Kramerea 110*, 111
scoparia, Robineauella 117*
sedecemfistulatus, Twinnia 46, 47, 48*
segnis, Leptopsylla 164
Sepsidae 82
septentrionalis, Boettcherisca 102*, 110*, 111
sericata, Lucilia 100
sericea, Parasarcophaga 112
serrata, Polyplax 169*, 172
setigera, Frontopsylla 165
setosus, Linognathus 169*, 174
sexfasciata, Hybomitra 65, 67*
shutovae, Cnetha latipes 49, 50*
sibiricus, Aedes 24, 26*, 27
sibiricus, Atylotus plebejus 73, 74*
sibiriensis, Hybomitra lundbecki 72*
sichotealini, Phallantha 109*, 110
sidimi, Stenoponia 152*, 154
signatipennis, Tabanus 75*, 76
silantiewi, Oropsylla 158, 160*
similis, Parasarcophaga 113*
simplicissima, Morellia saishuensis 95^
Simuliidae 40, 46
Simuliinae 42
Simulium 42*, 47, 52
sinanoensis, Culicoides 37, 38*
sinensis, Archaeopsylla 147*, 151
sinensis, Culex 29, 30
sinensis, Eupolyphaga 179
sinensis, Lagopoecus 184
306
Sinonipponia 105, 117
sinuata, Sarcotachinella 109*
Siphonaptera 146
sipituri, Ornithoica 122*
sokolovi, Megabothris 159
Solenopotes 173
Somaphantidae 182
sordida, Hybomitra lurida 68
sphagnumensis, Culicoides 36*
spiniger, Heterodoxus 182*, 183
spinigera, Ophyra 89
spinulosa, Polyplax 169*, 172
stabulans, Muscina 91, 92*
stackelbergi, Bellieriomima 109*
stackelbergi, Haematopota 78*, 79
Stegopterna 47, 48
stejnegeri, Dasypsylla 158
Stenocrotaphus 184
Stenoponia 153
stenopselapha, Hybomitra 65, 68*
stenopsis, Linognathus 174
sticticus, Aedes 25, 28
stigmoptera, Hybomitra 65, 66*
stimulans, Haematobia 96, 99*
stimulator, Cephenomyia 139
Stomoxys 85, 96
stramineus, Eomenacanthus 180*, 183
striata, Catalagia 157
striata, Ravinia 107
suavis, Chrysops 61*, 63*, 64
subalpina, Acrophaga 100
subfascipennis, Culicoides 39*, 40
sublunaticornis, Atylotus 73, 74*
Subphaonia 85, 87
subpusilia, Sohoenbaueria 52, 53*
subrostratus, Felicola 186*
subsylvarum, Culicoides 40
subvariegatum, Gnus 51*, 52
Suctoria 146
suis, Haematopinus 169*, 170
sungaris, Citellophilus tesquorum 160
Tabanidae 55, 59
Tabaninae 60, 64
Tabanus 55, 58, 60, 73
taiganus, Megabothris 159
takahasii, Peromiskopsylla hamifer 164
Takanoa 105, 118
tamerlani, Haematopota 56*, 57*, 60, 78*, 79
tamias, Monopsyllus indages 161
tarandi, Oedemagena 132*, 143, 144*
tarandi, Solenopotes 173
tarandina, Hybomitra 57*, 65, 66*
tarandinoides, Hybomitra 65, 67*
Tarsopsylla 157
Taxorhynchites 10, 13, 16
Taxorhynchitinae 10
tempestiva, Musca 84, 93, 94
terraenovae, Calliphora 98
terraenovae, Protophormia 101
territans, Culex 29, 31
Thricops 85, 86, 89
togoi, Aedes 22, 23*
tokunagai, Culicoides 40
tolli, Monopsylla 161
toyamaruae, Culicoides 35*, 36
transiens, Parabyssodon 49, 50*
tribulis, Ceratophyllus 163
Triceratopyga 97, 99
Trichodectes 186
Trichodectidae 186
Tricholioproctia 118
Trichopticoides 86, 89
Tricops 86, 89
trigeminus, Tabanus 75*
Trinoton 181
Trinotonidae 181
Tripocalliphora 98, 102
Tristis, Haematopota pluvialis 79
tritaeniorhynchus, Culex 29, 30*, 31
Trimenopon 183
Trimenoponidae 183
trompe, Cephenomyia 139
tropicalis, Numidilipeurus la wrensis 182*, 184
truncatum, Simulium 45, 54*
truncus, Ctenophthalmus 154
tuberculaticeps, Chaetopsylla 152
tuberosa, Parasariophaga 114*
tuberosum, Simulium 52, 53*
turbinatum, Neocolpocephalum 183
Twinnia 47
Uchida 183
ugamskii, Pierretia 108*
uliginosa, Parasarcophaga 112, 113*
ulrichi, Cephenomyia 138*, 139
unguitigris, Parasarcophaga 111*, 112
unicolor, Ornithoica 121*, 122
umbratica, Hebecnema 87
umbraticola, Phaonia 91
uralensis, Calliphora 98, 99*
Uranotaenia 10
ussurionsis, Hybomitra 67, 69*
ussuri ensis, Parasarcophaga 114*
vagabundus, Ceratophyllus 163
vagans, Culex 29, 31
validus, Chrysops 63, 64*
vanderwulpi, Chrysops 56*, 63*, 64
variegata, Phortica 84
variifrons, Culicoides 36*, 37
velutina, Hydrotaea 90*, 91
ventricosus, Haemodipsus 169*, 172
venustum, Simulium 45, 46, 54*
verecundum, Simulium 54*
Vermipsyllidae 150, 151
veterinus, Gastrophilus 136, 137*
vexans, Aedes 22*, 26*, 28
vicina, Calliphora 98
vicina, Musca domestica 93, 94
villeneuvei, Pierretia 108*, 108
vinogradovi, Amphipsylla 164
vituli, Linognathus 169*, 174
vomitoria, Pierretia 98
vorax, Culex 28, 29
vulgare, Simulium 52, 53*
waterstoni, Callopsylla 160
Werneckiella 185
whitmorei, Culex 29, 30
Wilhelmia 42
Wohlfartia 103*, 104, 151
zibellina, Chaetopsylla 152
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЯ—КЛК
Acaridae 238—240, 245
Acariformes 191—194, 234
Acaroidea 237
Acaromorpha 191
Acarus 241
Acotyledon 241, 243
agilis, Laelaps 228*
Aleuroglyphus 239, 241
Alloptidae 260, 265
alwari, Paralges 261*, 268, 268*
Amblyomminae 199, 200
ambulans, Haemogamasus 223*, 224
amphibius, Hirstionyssus 230*
Analgidae 255, 261, 262
Analgoidea 237, 253, 260
anatina, Brephosceles 266*, 267*, 267
anatina, Freyana 259*, 259, 260, 261*
Androlaelaps 227
angustus, Ixodes 201*, 203—205,204*, 205*,
208*, 209
anserina, Freyana 258, 259*, 260
apodemi, Hirstionyssus 233, 234
Aponomma 196
Arachnida 191
arboricola, Ixodes 200*, 202, 204*,
204, 205*, 206, 210*, 211
arcuatus, Chortoglyphus 244*, 246
Argasidae 195
armata, Euschoengastia 296
arvalis, Hirstionyssus 230*
Ascoschoengastia 291, 296
asiaticus, Dermacentor 215
Avenzoariidae 261, 264
bacoti, Ornithonyssus 218—220, 222
Bdellorhynchus 264, 266
bifurcata, Rivoltasia 272*, 273
bilobatus, Epidermoptes 271*, 272
bipectinatus, Syringophilus 298*, 301
bovis, Chorioptes 280
bovis, Demodex 299
Brephosceles 266
brevis, Demodex 299
cadaverum, Glycyphagus 244*, 246
caledonicus, Ixodes 201*, 203, 204*, 206*,
208*, 208, 210
Caloglyphus 239, 243
canis, Demodex 299
canisuga, Ixodes 201
саргае, Chorioptes 280
саргае, Demodex 299
casalis, Androlaelaps 219, 228, 228*
casei, Tyrolichus 240*, 242
cati, Demodex 299
cati, Notoedres 282*, 285
Ceratixodes 201
Cheladonta 291, 296
Chelicerata 191
chelidonis, Hirstia 249*, 250*, 251
Cheyletoidea 300
Chorioptes 279*, 279—281
Chortoglyphus 245
clethrionomydis, Laelaps 228*, 2 31, 232
communis, Psoroptes 280
concinna, Haemaphysalis 197*, 199, 211 —
213*, 212-214
confucianus, Hirstionyssus 233, 234
corpus, Dermacentor 215
crenulatus, Ixodes 200*, 201, 204*, 204, 205*
206, 210*, 211
criceti, Hirstionyssus 230*, 233, 234
cubitalis, Megninia 261*, 262, 263*
cuniculi, Chorioptes 280
cuniculi, Notoedres 285
cynotis, Otodectes 279*, 281, 281*
cysticola,* Laminosioptes 275*, 276
danrela, Ascoschoengastia 296
dauricus, Haemogamasus 223*, 224, 225
Demodex 298
Demodicidae 286, 297
Demodicoidea 297
Dermacentor 196, 198—200, 214, 215
Dermanyssidae 218, 220
Dermanyssus 221
Dermationinae 255, 270
Dermatophagoides 250, 251
Dermoglyphidae 255, 261, 267
Dermoglyphus 268, 269
destructor, Glycyphagus 238*, 244*, 246
domesticus, Glycyphagus 244*, 246
dubinini, Myonyssus 230*, 232
echidninus, Laelaps 228*, 230, 231
elongatus, Dermoglyphus 268, 269*
Epidermoptes 270
Epidermoptidae 253, 255, 260, 270
Epidermoptinae 255, 270
equi, Chorioptes 279*, 280, 282*
equi, Demodex 299
equi, Psoroptes 279*, 280, 282*
esoensis, Miyatrombicula 295*, 296
Eulaelaps 218, 227, 229
Euroglyphus 249, 250
Euschoengastia 290, 291, 296
eusoricis, Hirstionyssus 230*, 233, 234
Eutrombicula 290
evansi, Dermatophagoides 249*, 250*, 252
308
f
arinae, Dermatophagoides 249*, 250*, 251,
252
fillippovae, Haemaphysalis 211*, 212—214
flava, Haemaphysalis 211—213*, 212—214
folliculorum, Demodex 298*, 299
Freyana 258, 259
Freyanidae 255, 258, 261
Freyanoidea 237, 253, 258
fusca, Gohieria 244*, 247
fustifer, Glycyphagus 244*, 246
Leeuwenhoekiinae 290
lemmi, Laelaps 228*, 231
Leptotrombidium 290, 291
liponyssoides, Haemogamasus 205, 205*
lividus, Ixodes 199, 200, 202, 204*, 205, 205*:
206, 210*, 210
longicornis, Haemaphysalis 199, 211—213*r
211, 213, 214
longior, Euroglyphus 249*, 250*, 251
longior, Tyrophagus 242
Gahrliepiinae 291
gallinae, Dermanyssus 218, 219, 221
gallinae, Neocnemidocoptes 274*, 276
gallinarum, Neoschoengastia 296
Gamasida 191, 192
Gamasina 191, 194, 216
gardellai, Neotrombicula 293
Gaudoglyphus 267—269.
glasgovi, Androlaelaps 217*, 219, 227, 228
Glycyphagidae 239, 244, 245
Glycyphaginae 245
Glycyphagus 245, 246
Gohieria 245, 247
grochovskae, Dermanyssus 221
gynglymura, Megninia 262, 263*
Haemogamasidae 218—220, 222
Haemogamasus 222, 223*
Haemalaelaps 226
Haemophysalis 196, 198—212, 211—213*
hilaris, Laelaps 217*, 228*
Hirstia 250, 251
Hirstionyssidae 218—220, 232
Hirstionyssus 219, 233
humerosus, Tyrophagus 240*, 243
Hyperlaelaps 226, 228*
Hypoderidae 253
ikaoensis, Cheladonta 296
Ingrassia 263, 264
isabellinus, Hirstionyssus 230*, 233, 234
Ischyropoda 222
Ixodes 196—210
Ixodida 191
Ixodidae 195
Ixodinae 199, 200
Ixodiopsis 203
Ixodoidea 191, 194, 195
jacobsoni, Varroa 219, 220, 226
japonica, Haemaphysalis 199, 211—213*,
212-214
japonica, Neotrombicula 293, 293*
Kaiseriana 211
kitanoi, Haemogamasus 223*
Knemidocoptes 274, 275
Knemidocoptidae 255, 260, 273
kusumotoi, Haemogamasus 223*
Laelaps 219, 227, 228*, 229
Laelaptidae 218—220, 226
laevigatus, Schiloribatus 238*
lagodechiensis, Ixodes 202
Laminosioptes 276
Laminosioptidae 255, 260, 276
largifolia, Freyana 259*, 259, 260
latyshevi, Ascoschoengastia 296
Macronyssidae 218, 220, 221
mandschuricus, Haemogamasus 223*, 224, 225
maslovi, Ixodes 200*, 201, 203, 204*, 205
Megninia 262, 263
Metamicrolichus 270
michaeli, Glycyphagus 244*, 246
Michaelia 258
Microlichus 270
micromydis, Laelaps 230*, 231
microti, Neotrombicula 289
minor, Gaudoglyphus 268, 269*
mitamurai, Neotrombicula 294
Miyatrombicula 290, 295
Monoindex 201
multispinosus, Laelaps 229, 231
muris, Laelaps 219, 229, 231
muris, Notoedres 282*, 285
musculi, Hirstionyssus 230*, 233, 234
mutans, Knemidocoptes 273*
Myialges 270
Myonyssus 226, 227, 232
nagayoi, Neotrombicula 294*, 294
namaqua, Nuttalliella 195
Neocnemidocoptes 274, 275
Neoschoengastia 291, 296
Neotrombicula 290, 291
neumanni, Haemaphysalis 211
nidi, Haemogamasus 219, 223*, 223, 225
nidiformes, Haemogamasus 223*, 225
nipponensis, Ixodes 202*, 203, 204, 207*,
208, 209*, 210*, 210
Notoedres 192, 281, 283
nuttalli, Laelaps 228*, 230, 231
Nuttalliellidae 195
obtusus, Pterolichus 257*, 258
Ololaelaps 226
Oribatei 192, 235—237
orientale, Leptotrombidium 291
ornatus, Glycyphagus 244*
Ornithonyssus 218, 221
Ornithophysalis 211
Otodectes 192, 279, 280
ovatus, Aleuroglyphus 238*, 240*, 241
ovis, Chorioptes 280
ovis, Demodex 299
pallidum, Leptotrombidium 291, 292*
palpale, Leptotrombidium 291, 292*
paradoxa, Acotyledon 240*, 243
Paralges 268, 269
Parasitiformes 191—195, 236
pavlovskyi, Hirstionyssus 233
pavlovskyi, Ixodes 202*, 203, 204, 207*, 20$
209*, 210*, 210
pavlovskyi, Laelaps 228*, 231
pavlovskyi, Leptotrombidium 291,^293*
perniciosus, Tyrophagus 240*, 242
30»
persulcatus, Ixodes 197*, 199, 202*, 203, 204,
207*, 208, 209*, 210*
phasiana, Haemaphysalis 211—213*, 211,
213, 214
Pholeoixodes 201
phylloides, Demodex 299
plumbeus, Ixodes 202
polymorph us, Bdellorhynchus 261*, 265*,
265, 266*
pomerantzevi, Ixodes 199, 201*, 203, 205*,
206, 208*, 209
pomeranzevi, Neotrombicula 295
pontiger, Haemogamasus 223*
Promyialges 270
Psoroptes 278—280, 279*
Psoroptidae 277, 278
Psoroptoidea 237, 277, 279, 282
Pterolichidae 256
Pterolichoidea 237, 253, 256
Pterolichus 256
pteronyssinus, Dermatophagoides 249, 249*,
250*, 252
putrescentiae, Tyrophagus 240*, 242
putuc, Ixodes 201
Pyroglyphidae 247, 249, 250
Pyroglyphoidea 237, 238, 247
quintus, Dermanyssus 221
radfordi, Neotrombicula 295
Rivoltasia 270, 272
rodionovi, Caloglyphus 240*, 243
rotundata, Euschoengastia 296
sachalinensis, Ixodes 202*, 203
Sarcoptes 279*, 281—284
Sarcoptidae 192, 277, 278, 281
Sarcoptiformes 191, 192, 195, 234, 236, 237
Sarcoptiformes plumicoles 237, 253
Sarcoptoidea 277
scabiei, Sarcoptes 278, 283, 284*
Scaphixodes 202
Schoengastiini 291
sciurinus, Hirstionyssus 230*, 233, 234
scutellare, Leptotrombidium 285
semitectus, Laelaps 230, 232
serdjukovae, Haemogamasus 223*, 224, 225
signatus, Ixodes 201*, 203, 204*, 206*, 208*,
208, 210
silvarum, Dermacentor 197*, 199, 215, 215*
silvarum, Ornithonyssus 218—220, 222
siro, Acarus 238*, 241
sokolovi, Acotyledon 240*, 243
stabularis, Eulaelaps 229
subakamushi, Chiroptella 290
Syringophilidae 286, 300
Syringophilus 301
tabularis, Eulaelaps 219
talmiensis, Neotrombicula 293, 294*
tamiyai, Neotrombicula 294
tenuiclavus, Tyrophagus 240*, 243
timofejevi, Haemogamasus 223*, 224
Trichotoixodes 202
Trombiculidae 286, 290
Trombiculinae 290, 291
Trombiculindus 291
Trombiculini 290, 291
Trombidiformes 191, 195, 285
turdus, Ixodes 201*, 202, 203, 206*, 206,
208*, 210
Tyrolichus 239, 241
Tyrophagus 239, 242, 243
Varroa 226
Varroidea 219, 220, 225
Vatacarus 286
velata, Ingrassia 261*, 263*, 264
vulgaris, Argas 195
uriae, Ixodes 200*, 201, 203, 204*, 206, 206*,
207*, 209
Xolalgidae 260, 262
Xolalginae 262
zhmajevae, Neotrombicula 294, 295*
ОГЛАВЛЕНИЕ
Стр.
Предисловие ................................................................ 3
Список сокращений и условных обозначений в определительных таблицах .... 6
I. Насекомые (Класс lusecta) ................................................. 7
Общие сведения о строении взрослого насекомого (Морфология)................... 7
Методика хранения и определения насекомых................................... 9
Двукрылые (Отряд Diptera).................................................... 10
Кровососущие комары (Сем. Culicidae) — Ю. Д. Бодрова..................... 10
Кровососущие мокрецы (Сем. Ceratopogonidae) — В, М. Глухова.............. 31
Кровососущие мошки (Сем. Simuliidae) — Ю. Д. Бодрова..................... 40
Слепни (Сем. Tabanidae) — Р. Г, Соболева................................. 55
Синантропные и зоофильные мухи........................................... 80
Настоящие мухи (Сем. Muscidae) — Б. К. Петрова....................... 84
Синие и зеленые (мясные) мухи (Сем. Calliphoridae) — Б. К. Петрова ... 96
Серые падальные (мясные) мухи (Сем. Sarcophagidae) — С. Д. Артамонов 102
Мухи-кровососки (Сем. Hippoboscidae) — Т. Н. Досжанов............... 119
Оводы — Р. Г. Соболева.................................................. 131
Желудочные оводы (Сем. Gastrophilidae).............................. 135
Носоглоточные оводы (Сем. Oestridae)................................ 136
Подкожные оводы (Сем. Hypodermatidae) .............................. 142
Блохи (Отряд Siphonaptera=Aphaniptera, Suctoria) — Т. П. Ромашова........... 146
Вши (Отряд Anoplura) — В. Н. Зарубина....................................... 166
Клопы, или полужесткокрылые (Отряд Heteroptera, Hemiptera, сем. Cimicidae) —
И. М. Кержнер .......................................................... 174
Тараканы (Отряд Blattoptera) — С. Ю. Стороженко............................. 176
Пухоеды (Отряд Mallophaga) — С. Ю. Стороженко............................... 179
Муравьи (Отряд Hymenoptera, сем. Formicidae) — А.Н. Купянская............... 187
II. Клещи —Acarina (Класс Arachnida) ....................................... 191
Внешнее строение взрослых клещей............................................ 192
Методика хранения и определения............................................. 193
Паразитиформные клещи (Отряд Parasitiformes)................................ 195
Иксодоидные клещи (Надсем. Ixodoidea) — Г. В. Колонии ......... 195
Иксодовые клещи (Сем. Ixodidae) — Г. В. Колонии ............ 195
Гамазовые клещи (Когорта Gamasina) — Н. А. Никулина .......... 216
Акариформные клещи (Отряд Acariformes) . . • . ............................. 234
Саркоптиформные клещи (Подотряд Sarcoptiformes) — Е. В. Дубинина .... 234
Клещи амбарно-зернового комплекса (Надсем. Acaroidea) — Е. В. Дубинина 237
«Аллергенные» клещи (Надсем. Pyroglyphoidea) — Е. В. Дубинина . . . 247
Перьевые клещи (Группа надсемейств Sarcoptiformes plumicoles) — С. В. Ми-
ронов ........................................................ 253
Чесоточные клещи (Надсем. Psoroptoidea) — Е. В. Дубинина ........... 277
Тромбидиформные клещи (Подотряд Trombidiformes) — Е. В. Дубинина . . . 285
Клещи-краснотелки (Сем. Trombiculidae) — Н. И. Кудряшова............ 286
Железницы (Сем. Demodicidae) — Е. В. Дубинина....................... 297
Очинные клещи (Сем. Syringophilidae) — Е. В. Дубинина ........ 300
Указатель латинских названий насекомых ................................. 302
Указатель латинских названий клещей ....................................... 308
НАСЕКОМЫЕ И КЛЕЩИ ДАЛЬНЕГО ВОСТОКА,
ИМЕЮЩИЕ МЕДИКО-ВЕТЕРИНАРНОЕ ЗНАЧЕНИЕ
Утверждено к печати
Биолого-почвенным институтом
Дальневосточного научного центра
Академии наук СССР
Редактор издательства А, А. Ройтман
Художник О, М. Разулевич
Технический редактор Л. М. Семенова
Корректоры Э. Г. Рабинович и Т. Г. Эдельман
ИВ № 21500
Сдано в набор 24.11.86. Подписано к печати 09.04.87.
М-17131. Формат 70X1081/ie- Бумага книжно-журнальная.
Гарнитура обыкновенная. Печать высокая. Усл,
печ. л. 27.30. Усл. кр.-от. 27.55. Уч.-изд. л. 29.87.
Тираж 1750. Тип. зак. № 990. Цена 3 р. 50 к.
Ордена Трудового Красного Знамени
издательство «Наука». Ленинградское отделение.
199034, Ленинград, В-34, Менделеевская лин., 1.
Ордена Трудового Красного Знамени
Первая типография издательства «Наука».
199034, Ленинград, В-34, 9 линия, 12.