




































































































































































































































































































































Автор: Меттлер Л. Грегг Т.
Теги: общая генетика общая цитогенетика иммуногенетика эволюционное учение видообразование филогенез биология генетика
Год: 1972




ОСНОВЫ СОВРЕМЕННОЙ ГЕНЕТИКИ
Л. МЕ"1—msp=, Т“_ I—F=EI—Г
ГЕНЕТИКА ПОПУЛЯЦИЙ И ЭВОЛЮЦИЯ
POPULATION
GENETICS
AND EVOLUTION
LAWRENCE E. METTLER
North Carolina State University, Raleigh
THOMAS G. GREGG
Miami University, Oxford, Ohio
PRENTICE-HALL, INC.
ENGLEWOOD CLIFFS, NEW JERSEY 1969
Л. Меттлер, Т. Грегг
ГЕНЕТИКА ПОПУЛЯЦИЙ И ЭВОЛЮЦИЯ
Перевод с английского и предисловие канд. биол. наук Б. В. ШИЛЕНКО
ИЗДАТЕЛЬСТВО «МИР» МОСКВА 1972
УДК 575 + 577.1
Очередной выпуск из серии «Основы современной генетики», которую издательство «Мир» публикует начиная с 1966 г. В прошлые годы в этой серии вышли следующие книги: Дж. Джинкс, «Нехромосомная наследственность» (1966); Ф. Сталь, «Механизмы наследственности» (1966); В. Маккьюсик, «Генетика человека» и др.
Данный выпуск посвящен основным генетическим закономерностям эволюции. В нем рассмотрены современные представления о виде, подвиде и расе, внутривидовые генетические процессы, явление дрейфа генов, генетическая изменчивость и ее источники, мутационный процесс, проблема генетического разнообразия популяций, концепция «генетического груза». Очень хорошо изложены вопросы о роли малых популяций в эволюции, проблемы гомеостаза и канализации.
Предназначена для преподавателей биологии высших и средних учебных заведений, для студентов университетов, медицинских, педагогических и сельскохозяйственных институтов, а также для научных работников различных специальностей, интересующихся современным состоянием генетики в целом или отдельных ее разделов.
Редакция биологической литературы
ПРЕДИСЛОВИЕ
К РУССКОМУ ИЗДАНИЮ
Еще совсем недавно в русской биологической литературе дарвинизм рассматривали не как составную часть теории эволюции, а как собственно теорию эволюции. Иначе говоря, принималась не только дарвиновская интерпретация эволюционного становления целесообразности, но и дарвиновское толкование механизмов создания того разнообразия, которое под действием постулируемого естественного отбора должно постоянно элиминироваться. Ясно, что при таком подходе не оставалось места ни генетике, ни генетической теории эволюции, составляющей ядро современного эволюционного учения. Несмотря на то что ламаркистская концепция наследования благоприобретенных признаков, которую в свое время вынужден был принять Дарвин, поскольку истинные источники разнообразия тогда еще не были известны, давно уже обнаружила свою полную несостоятельность, влияние ее еще долго ощущалось. Уже после вторичного открытия законов наследственности, после того как была установлена ненаправленность наследственных новообразований, в течение долгих лет роль естественного отбора в становлении приспособленности сознательно или бессознательно принижали, выдвигая на первый план воображаемые возможности благоприобретенной приспособленности в адекватном изменении наследственности. Центр тяжести в эволюционном учении был перенесен с реально действующего, всеобъемлющего фактора на мнимый, кажущийся, нереальный. Не удивительно, что эта идеалистическая в своей сущности концепция изначальной целесообразности неоднократно давала поразительные рецидивы. «Главное
в биологии — это выяснить для чего и почему происходит тот или иной процесс в организме, а не как происходят химические превращения и физическое движение» 1 (курсив мой. —Б. Ш.). Так, в 1960 году —сто лет спустя после дарвиновских слов о том, что «в изменчивости живых существ и в действии отбора не больше преднамеренного плана, чем в том направлении, по которому дует ветер», — программной проблемой биологии вновь стал пресловутый, откровенно метафизический вопрос «для чего?»
...«Велика сила установившихся неверных взглядов, — писал Чарлз Дарвин, — но история науки показывает, что, к счастью, эта сила недолговечна». Современная научная теория эволюции совершенно свободна от надуманных постулатов о существовании сознательного начала, Высшей Воли. Она базируется на безусловном отрицании представления об эволюции как способе достижения заранее поставленной цели. Природа слепа в своем творчестве, ее действия абсолютно бесцельны и совершенно случайны. Только естественный отбор формирует порядок, называемый жизнью, и только он совершенствует этот порядок. В действиях ящерицы не больше изначальной целесообразности, чем в бездействии камня, за которым, она скрылась.
Естественно, что современная теория эволюции далеко ушла от представлений дарвиновских времен, но в главном, а именно в признании принципа становления приспособленности путем естественного отбора, она осталась верной этим представлениям. Конкретизация эволюционного процесса достигла максимума— современная теория эволюции базируется на генетических представлениях о связи поколений и оперирует понятиями, за которыми стоят не признаки или свойства, а их первопричины — гены и генотипы. Зная зависимость между генетическими параметрами популяции, можно с большой точностью описать все происходящие в ней микроэволюционные процессы.
1 Т. Д. Лысенко, в сб. «Дарвинизм живет и развивается», М., 1960.
И наоборот, по данным об изменениях структуры популяции за единицу времени — поколение — можно судить о количественных и качественных взаимосвязях этих процессов.
Предлагаемая читателю книга представляет собой доступное для широкого круга читателей и вместе с тем строго научное изложение генетических основ современного эволюционного учения. Существенно, что авторы легко и естественно переходят от таких крупномасштабных концепций, как эволюция, вид, борьба за существование, к таким в высшей степени конкретным понятиям, как инверсия, ген, генотип, селективная ценность отдельного локуса. Надо надеяться, что эта книга вслед за вышедшими в последние годы монографиями Н. П. Дубинина «Эволюция популяций и радиация» и Н. П. Дубинина и Я. Л. Глембоцкого «Генетика популяций и селекция» сыграет свою роль в дальнейшем формировании наших представлений о процессе органической эволюции.
Авторов, излагающих метод определения «платы за эволюцию» и рассуждающих о зависимости Скорости эволюционных изменений от частоты ненаправленных изменений наследственности и адаптационной ценности гомозиготных и гетерозиготных комбинаций, никак нельзя упрекнуть в приверженности к концепции самопостроения новых форм жизни через адекватное условиям среды изменение наследственности. И тем не менее они чаще (чем это необходимо, чтобы придать изложению большую образность) подчеркивают как бы некую активность популяции в самосохранении, своеобразную целенаправленность ее действий. Так, например, седьмая глава начинается со следующих слов: «Для того чтобы существовать, популяция должна...»
Не столько для того, чтобы подчеркнуть анахронизм такой фразы, сколько для того, чтобы еще раз напомнить читателям о втором авторе гениального открытия — А. Уоллесе (тем более, что авторы книги, увы! не нашли этому имени места в основном тексте, а ограничились лишь упоминанием о нем в сноске на стр. 13), приведем здесь его слова, блестяще схва
тывающие сущность этого открытия и замечательные не только в отношении научной аргументации, но и в отношении образности описания процесса эволюции. «Жирафа приобрела свою длинную шею не потому, что, желая достать листву с самых верхних веток, она постоянно вытягивала шею, но потому, что из всех разновидностей ее прототипа, та, у которой шея была длиннее, чем обычно, сразу расширила площадь своего пастбища на том же пространстве, где паслись ее товарищи с короткими шеями, и поэтому при первой же нехватке количества кормов могла легче выжить».
Б. В. Шиленко
ИЗ ПРЕДИСЛОВИЯ
АВТОРОВ
Уже при беглом знакомстве с историей генетики бросается в глаза определенная тенденция в выборе подопытных организмов — постепенный переход ко все более простым, более удобным для генетического и биохимического анализа формам.
Вполне естественно, что первые генетические наблюдения относились к таким привычным видам, как сам человек, домашние животные и культурные растения. Однако ранние попытки понять сущность явления наследственности, как правило, оказывались тщетными— сложность этих организмов сильно затрудняла разграничение «однофакторной» (менделевской) и «многофакторной» (количественной) наследственности. Огромная заслуга Менделя в том и состоит, что, работая с различными сортами гороха, он разработал такую систему экспериментов, которая позволила ему преодолеть эти трудности и вскрыть основные законы наследственности.
После вторичного открытия в 1900 году законов Менделя главным генетическим объектом, пришедшим на смену курам, кроликам, мышам, гороху и ослиннику, стала плодовая мушка (Drosophila), используемая в качестве подопытного организма начиная с 1910 года. Дальнейшими своими успехами экспериментальная генетика обязана красной плесени — нейроспоре, с которой стали работать с сороковых годов. В конце сороковых годов генетики начали работать на организмах предельной генетической простоты — бактериях, а затем и на бактериальных вирусах (бактериофагах). В результате мы теперь располагаем пусть неполными, но весьма глубокими представлениями о генетических механизмах и об их действии в индивидуальном организме.
В то же время постепенно становилось ясно, что в природе существуют более высокие уровни органи
зации, имеющие важнейшее значение. Это группы взаимодействующих индивидуумов, принадлежащих к одному виду, — популяции и группы взаимодействующих популяций, принадлежащих к различным видам,— биотические сообщества. Следовательно, для того чтобы понять сложность природы на уровне, лежащем выше уровня индивидуума, и для того чтобы понять процесс эволюции, мы должны начинать с проблем популяции.
Фундаментальная связь между частотой генов и частотой генотипов в популяции (гл. 3) была открыта в 1908 году английским математиком X. Харди. Поскольку зависимость между этими частотами довольно проста, он ограничился письмом к редактору журнала «Сайенс». В том же году эту зависимость обнаружил немецкий врач В. Вайнберг. Таким образом, параллельно с развитием генетики индивидуума стала развиваться и популяционная генетика Главная заслуга в разработке популяционной генетики, а особенно ее теоретического и математического аспектов, в этот ранний период (1920—1940), безусловно, принадлежит С. С. Четверикову, С. Райту, Р. Фишеру и Дж. Холдейну. Среди опубликованных к настоящему времени исследований по генетике популяций едва ли можно найти хотя бы одну, в которой не были бы так или иначе упомянуты эти первые работы.
Преобладающее внимание в современных исследованиях по генетике популяций уделяется экспериментальному направлению. Поэтому настоящая книга наряду с анализом некоторых теоретических построений содержит основы экспериментальной генетики популяций. Развитие популяционных исследований, особенно экспериментальных, нередко задерживалось в связи с отставанием в разработке основных представлений о генетике индивидуума. Последние достижения в познании генетических механизмов открыли новые возможности, и популяционная генетика, по-видимому, стоит на пороге новых больших открытий.
Л.Е.М. т Г Г.
Глава 1
ТЕОРИЯ ЭВОЛЮЦИИ
Концепция эволюции — процесса, породившего фантастическое многообразие ныне живущих организмов, — одна из основных концепций биологии. От до-клеточных агрегатов органических молекул, возникших в первичном океане более трех миллиардов лет назад, и до исключительно сложных многоклеточных организмов нашего времени эволюция, а точнее естественный отбор, всегда была формообразующей силой. Не удивительно поэтому, что понятие эволюция пронизывает все области биологии как основная объединяющая идея наших представлений о жизни и о функциях организмов.
В простейшей формулировке эволюция это то, что Чарлз Дарвин назвал «происхождением, сопровождаемым видоизменением» (descent with modification). Видоизменения возникают вследствие того, что особи, обладающие разными наследственными признаками, различаются по способности к воспроизводству. По определению Ф. Добжанского: «Эволюция есть изменение генетической структуры популяций». Однако, когда мы начинаем рассматривать все многочисленные и разнообразные доказательства существования эволюции и подробно исследуем различные механизмы, вызывающие модификации у организмов разных типов, а также участвующие в эволюции разнообразные факторы и взаимодействия между ними, проблема эта становится гораздо более обширной и сложной. В самом деле нам известны далеко не все
стороны процесса эволюции. Как подчеркнул сам Дарвин в книге «Происхождение видов»1: «Всякий, кто склонен придавать больше веса неразрешенным затруднениям, чем объяснению известного числа фактов, конечно, отвергнет мою теорию». Это замечание справедливо и сегодня, хотя число фактов, получивших объяснение, сейчас гораздо больше, а число трудностей, оставшихся необъясненными, значительно меньше, чем во времена Дарвина.
Доказательства происхождения, сопровождаемого водоизменением, носят косвенный характер, но в совокупности они чрезвычайно убедительны. Сегодня ученые, да и большинство других людей, единодушно считают, что эволюция происходила в прошлом и происходит до сих пор и что этот процесс полностью объясняет разнообразие ныне существующих организмов. Мы не намерены давать в этой книге обзор доказательств эволюции (это уместно в книгах более общего характера); не будем мы также излагать в хронологической последовательности историю происхождения живых форм. Мы сосредоточим все свое внимание на эволюции как процессе и на анализе движущих сил этого процесса, в частности на связи его с генетикой и генетическими механизмами.
Замечательная работа Дарвина «Происхождение видов», опубликованная в 1859 году, в корне изменила наши представления о природе. Она завершила длительные поиски естествоиспытателей и систематиков, которые пытались найти причинное объяснение многих черт сходства, наблюдаемых у организмов, явно принадлежащих к разным «сортам». Предложенное Дарвином объяснение кажется теперь банальным: сходные виды близкородственны по происхождению. В общем, чем больше родство между двумя группами, тем больше сходство между ними, и наоборот. Таким образом, главная идея эволюции заключается в том, что все существующие организмы произошли от одного или нескольких примитивных организмов, впер-
Все цитаты приводятся по изданию; Чарлз Дарвин, Сочинения, т. 3, Изд. АН СССР, М. — Л., 1938.
вне возникших несколько миллиардов лет назад. Некоторые авторы распространяют эту концепцию еще дальше в прошлое, до молекулярных агрегатов, еще не достигших клеточного уровня организации, но уже способных к самовоспроизведению (и к воспроизведению мутантных форм), а следовательно, и подверженных естественному отбору. А из этого следует, что такие агрегаты могли дать начало новым направлениям развития. В итоге, за исключением случаев конвергентной эволюции, на любом уровне филогенетического древа черты сходства, присущие разным группам, обусловлены единством их происхождения, т, е. общностью их предковых форм, тогда как различия возникли при последующей изоляции этих групп.
Дарвин отнюдь не был первым, выдвинувшим теорию эволюции, однако именно он впервые представил последовательную и убедительную картину эволюционного процессаЕго успеху благоприятствовало несколько обстоятельств. Тщательно и объективно подобранные им доказательства того, что виды не «неизменны», что они изменялись и изменяются, иными словами, что эволюция действительно существует, были так многочисленны, что опровергнуть их практически невозможно. Не менее важен тот факт, что Дарвин сумел дать логическое и убедительное объяснение возникновению этих изменений, т. е. естественному отбору. Дарвин опять-таки не был первым, предложившим концепцию естественного отбора, однако именно он впервые по-настоящему оценил связь между естественным отбором и наследственными изменениями популяции. В этом и заключается его успех; он не только сумел показать существование изменчивости, но и объяснил, каким образом она возникает.
1 В период подготовки Дарвином книги «Происхождение видов» Альфред Рассел Уоллес — натуралист, проводивший обширные исследования на Малайском архипелаге, — сформулировал теорию эволюции, по существу идентичную дарвиновской. Уоллес и Дарвин совместно доложили сущность теории Лондонскому королевскому обществу в 1857 году, после чего Дарвин подробно изложил ее в своей книге «Происхождение видов»; эта книга получила огромное и заслуженное признание, и поэтому Дарвина принято считать главным автором теории.
Основные положения дарвиновской теории эволюции можно сформулировать следующим образом.
/. В любой популяции численность особей стремится увеличиваться в геометрический прогрессии, если условия благоприятствуют выживанию всех потомков. «Не существует ни одного исключения из правила, по которому любое органическое существо естественно размножается столь быстро, что, не подвергайся оно истреблению, потомство одной пары очень скоро заняло бы всю Землю».
2. Потенциальная возможность быстрого увеличения численности реализуется редко. «На каждом виде, вероятно, отражается влияние самых разнородных препятствий, действующих в различные возрасты, в разные времена года или в различные годы, и хотя одно из них или небольшое число может оказаться более сильным, тем не менее средняя численность или даже существование вида будет зависеть от их совокупного действия».
3. Из этих фактов Дарвин сделал вывод о наличии конкуренции, или «борьбы» за выживание, в результате которой происходит элиминация большого числа особей. «Поэтому, так как производится больше особей, чем может выжить, в каждом случае должна возникать борьба за существование...»
4. Изменчивость в форме индивидуальных различий существует в каждом виде и в каждой популяции. «Никто, конечно, не станет утверждать, что все особи одного вида отлиты как бы в одну форму. Эти индивидуальные отличия крайне для нас важны, так как они часто наследственны, как всякому известно...» Хотя представления о механизме наследственности были весьма запутанны и Дарвин знал о них очень мало, он правильно оценил самый факт наследования и использовал его в качестве отправной точки для построения теории эволюции. «Ненаследственное изменение для нас несущественно».
5. Анализируя различия между индивидуумами и между разновидностями, Дарвин пришел к выводу, что процессу элиминации свойственна избирательность. «Можно ли, видя несомненное появление изме
нений, полезных для человека, считать невероятным, чтобы другие изменения, полезные в каком-нибудь отношении для существ в их великой и сложной жизненной битве, появлялись в длинном ряде последовательных поколений. Но если такие изменения появляются, то можем ли мы (помня, что родится гораздо больше особей, чем может выжить) сомневаться в том, что особи, обладающие хотя бы самым незначительным преимуществом перед остальными, будут иметь больше шансов на выживание и продолжение своего рода». Выжившие рассматриваются как наиболее приспособленные. Однако для Дарвина понятие «приспособленность» организма не ограничивается относительной способностью организма бороться с конкурентами за пищу, за пространство, за партнера в спаривании или шансами избежать гибели от хищников и болезней. По мнению Дарвина, самое лучшее — это определить приспособленность как относительную способность оставить потомство. «Я применяю этот термин в широком и метафорическом смысле, включая сюда зависимость одного существа от другого, а также включая (что еще важнее) не только жизнь одной особи, но и успех ее в оставлении после себя потомства».
6. Эволюция представляет собой постепенное изменение наследственной структуры вида. «Не подлежит сомнению, что до настоящего времени не удалось провести ясной пограничной черты между видами и подвидами, т. е. формами, которые, по мнению некоторых натуралистов, приближаются к видам, но не вполне достигают этой степени... Эти различия примыкают одни к другим, нечувствительно сливаясь в один непрерывный ряд, а всякий ряд производит на наш ум впечатление действительного перехода.
На основании этого я считаю индивидуальные различия, хотя они интересны для систематика, крайне важными для нас, так как они представляют собой первые шаги к образованию разновидностей, настолько незначительные, что о них, как обыкновенно полагают, не стоит даже упоминать в естественноисторических сочинениях. Разновидности, которые в некоторой
степени более различаются между собой и в некоторой степени постоянны, я рассматриваю как ступени к более резко выраженным и постоянным разновидностям, а эти последние — как ступени к подвидам, а затем к видам. Переход с одной степени различия на другую во многих случаях мог представлять собой простой результат особенностей самого организма и различных физических условий, которым он долго подвергался; но по отношению к важнейшим приспособительным признакам переход с одной ступени на другую можно с уверенностью приписать накопляющему действию естественного отбора... Ясно выраженная разновидность может быть вследствие этого названа «зарождающимся видом».
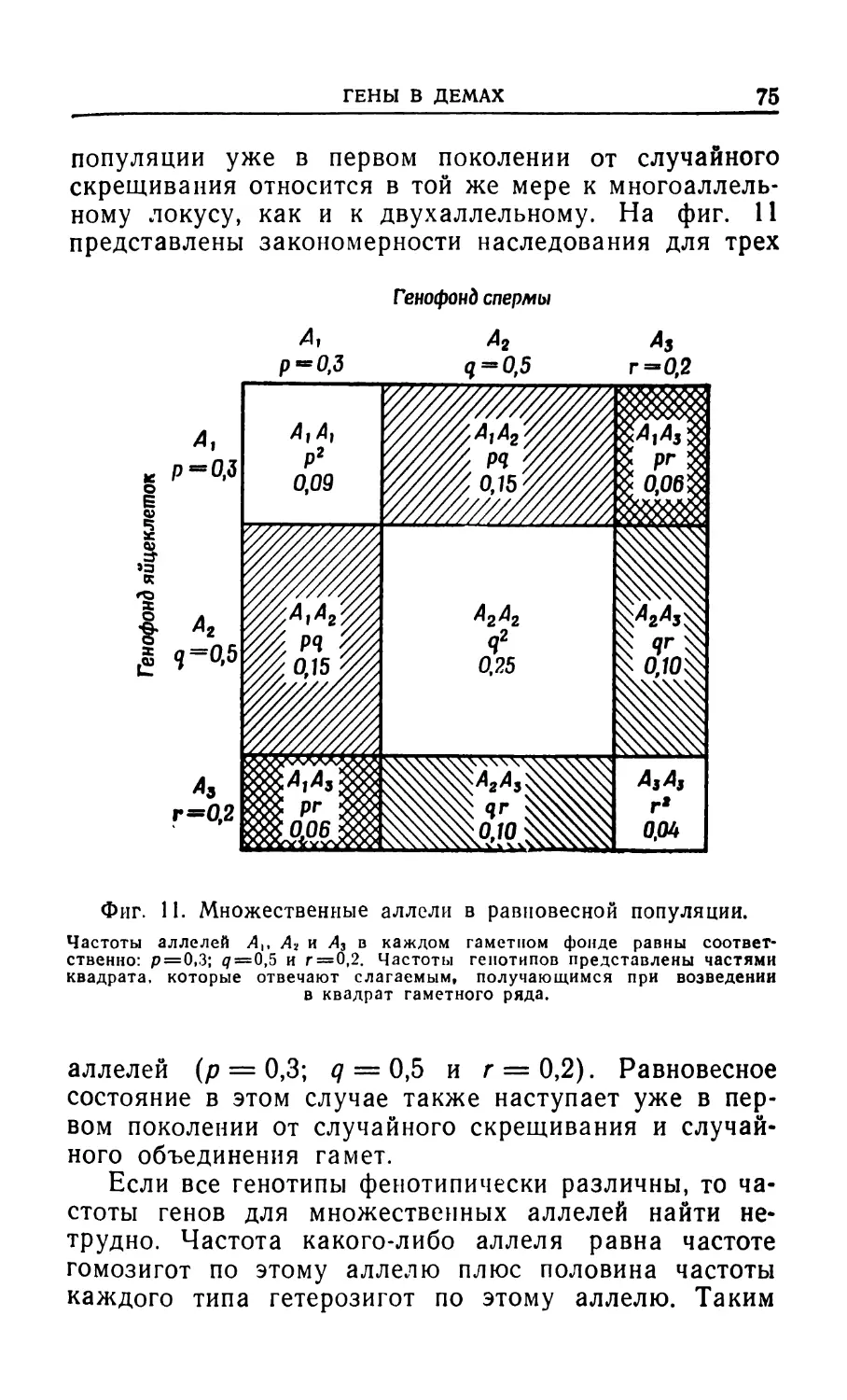
В связи с этим Дарвин указал, что существование в природе множества зарождающихся видов служит убедительным доказательством эволюции (фиг. 1).
В этом кратком очерке речь шла только о самых существенных чертах дарвинизма. Основные представления Дарвина о естественном отборе и эволюции или о «происхождении, сопровождаемом видоизменением», а следовательно, и «Происхождение видов» возникли в результате наблюдения фактов наследственности, высокой плодовитости и избирательной гибели.
Эволюционное учение нашего времени, которое называют «современным синтезом» или просто «неодарвинизмом», содержит ряд положений, которых не было в дарвиновской теории. В этом синтезе главной формообразующей силой по-прежнему считается естественный отбор, хотя наши представления о корпускулярной природе генов дают нам возможность более полно представить себе возникновение изменчивости в результате мутаций, сохранение изменчивости в скрытом состоянии в диплоидных организмах, перетасовку генов в процессе генетической рекомбинации, обеспечивающую постоянный источник новых генных сочетаний, на которые мог бы действовать естественный отбор. Все эти дополнения не изменили основных принципов, впервые постулированных Дарвином. Эволюция слагается из двух стадий: возникно
вения изменчивости и изменения направления этой изменчивости под действием естественного отбора.
Фиг. 1.
«Любопытно созерцать густо заросший берег, покрытый многочисленными, разнообразными растениями, птиц, поющих в кустах, насекомых порхающих вокруг, червей, ползающих в сырой земле, и думать, что все эти прекрасно построенные формы, столь отличающиеся одна от другой и так сложно одна от другой зависящие, были созданы благодаря законам, еще и теперь действующим вокруг нас. Эти законы —в самом широком смысле —Рост и Воспроизведение; Наследственность, почти необходимо вытекающая из вр-спроизведения; Изменчивость, зависящая от прямого или косвенного действия жизненных условий и от упражнения и неупражнения; Прогрессия Размножения, столь высокая, что она ведет к Борьбе за жизнь и ее последствию — Естественному Отбору, влекущему за собой Расхождение признаков и Вымирание менее совершенных форм. Таким образом, из войны природы, из голода и смерти непосредственно вытекает самый высокий результат, какой ум в состоянии себе представить, — образование высших животных. Есть величие в этом воззрении, по которому жизнь, с ее различными проявлениями, творец первоначально вдохнул в одну или ограниченное число форм; и между тем как наша планета продолжает вращаться согласно неизменным законам тяготения, из такого простого начала развилось и продолжает развираться бесконечное число самых прекрасных и самых изумительных форм» (Ч. Дарвин, «Происхождение видов»).
Следует также указать, что изменения, возникающие под действием естественного отбора, могут иметь разные последствия в зависимости от условий существования. Один процесс наблюдается в том случае, когда условия среды, определяющие естественный
отбор, весьма однородны на всем протяжении ареала вида или популяции. При этом приспособленность вида к своей среде неуклонно возрастает, а в случае изменения этой среды изменяется и вид в целом. За достаточно продолжительный период времени таким путем могут возникнуть весьма заметные изменения. Следовательно, генетическая структура отдельного ряда последовательных поколений постепенно и равномерно изменяется от поколения к поколению. Этот процесс называется филетической эволюцией. Если бы мы располагали машиной времени, позволяющей непосредственно сравнивать первоначальные признаки вида с его признаками спустя миллион лет, то различия между ними могли бы оказаться столь значительными, что эти формы по существу пришлось бы отнести к разным видам. Другой способ эволюционной дифференцировки наблюдается в том случае, когда разные популяции одного вида тем или иным образом изолируются друг от друга (см., гл. 8) и оказываются в разных условиях среды. Поскольку при этом естественный отбор действует на них по-разному, в разных популяциях возникают разные изменения. Таким образом, изолированные популяции будут все более дивергировать, пока, наконец, единый исходный вид не распадется на два или более новых вида. Описанный процесс известен под названием видообразования. Это единственный возможный способ увеличения числа видов.
Видообразование имеет очень важное значение для эволюции; оно ведет к дифференцировке, т. е. к дроблению единой генетической популяции на ряд подгрупп, каждая из которых представляет собой независимую эволюционную линию со своими особыми возможностями для дальнейших филетических изменений.
Таким образом, дивергенция может быть непрерывной; она не прекращается после того, как данная группа достигла ранга вида, а продолжается и дальше, приводя к возникновению более высоких таксономических категорий (фиг. 2),
Фиг. 2. Некоторые виды вьюрков, обитающие на Галапагосских островах. Эти птицы оказали заметное влияние на дарвиновскую аргументацию концепции естественного отбора.
«Я собрал двадцать шесть видов наземных птиц: все они совершенно особенные и ни один из них нигде больше не встречается, за исключением одной похожей на жаворонка певчей птички из Северной Америки . . . Прочие двадцать пять птиц состоят, во-первых, из хищника, представляющего по своему строению любопытный переход от коршунов к группе американских стервятников-каракар. . . Во-вторых, здесь имеются две совы . . . В-третьих, королек, три тиранна-мухоловки... и горлица —все виды, аналогичные американским, но отличающиеся от них .. . В-пятых, имеются три вида дроздов-пересмешников . . . Остальное наземные птицы составляют исключительно любопытную группу вьюрков, близких по строению клюва, короткому хвосту, форме тела и оперению: их тринадцать видов, которые м-р Гоулд разделил на четыре подгруппы. Все эти виды встречаются только на этом архипелаге . . . Рассматривая эту постепенность и разнообразие в строении, обнаруживающиеся в пределах одной небольшой и притом связанной тесными узами родства группы птиц, можно действительно представить себе, что в связи с первоначальной бедностью птиц на этом архипелаге был взят один вид и модифицирован в различных концах архипелаг».
Эволюция — явление, связанное с генетическими изменениями на видовом и популяционном уровнях организации. Популяции и виды представляют собой основные единицы эволюции, и для того, чтобы понять, какие силы вызывают изменения в этих группах, необходимо использовать наши знания по генетике популяций. Но, прежде чем обсуждать движущие силы эволюции, необходимо дать четкие определения понятиям «вид», «популяция» и пр. Именно об этом и пойдет речь в следующей главе.
ЛИТЕРАТУРА
Darwin Ch., The Origin of Species. (1859; paperback available, New York, Washington Square Press, 1963.) (Чарльз Дарвин, Происхождение видов, Сочинения, т. 3, Изд. АН СССР, М. — Л., 1938.)
Eiseley L., Darwin’s Century, New York, Doubleday and Co., 1958. (Paperback available, Anchor Books, 1961.)
Grant V., The Origin of Adaptations, New York, Columbia University Press, 1963. Part 1, Chapters 1, 2, 3 and 4. Philosophical aspects of science. Causal theories in biology. Abiogenesis and the course of evolution.
G reb stein S.. N., Monkey Trial, Boston, Houghton Mifflin, 1960. Houghton Mifflin Research Series, Number 4.
N о g a r F. J., The Wisdom of Evolution, Garden City, N. Y. Doubleday and Co., 1963.
Stebbins G., Processes of Organic Evolution, Englewood Cliffs, N. J., Prentice-Hall, Inc., 1966, p. 148.
Г лав a 2
ПОПЫТКИ ПРИВЕСТИ
В СИСТЕМУ
РАЗНООБРАЗИЕ ЖИЗНИ
«Что такое вид?» Мы попытаемся ответить иа этот интереснейший вопрос. Быть может, наш ответ не будет исчерпывающим и в процессе поисков перед нами возникнет множество других, не менее интересных вопросов.
ИСКУССТВО НАУЧНОЙ КЛАССИФИКАЦИИ ОРГАНИЗМОВ
На планете Земля жизнь появилась более трех миллиардов лет назад; за это время, непрерывно эволюционируя, органические формы достигли поразительного разнообразия. Трудами натуралистов и систематиков XIX и XX веков нам уже известно более миллиона форм животных и более четверти миллиона растений. К этому следует добавить множество вымерших форм, обнаруженных палеонтологами. По приблизительной оценке общее число видов организмов за все время существования жизни превышает миллиард. По-видимому, не менее 415 миллионов из них населяют Землю в настоящее время. Хотя по некоторым группам, таким, как птицы или млекопитающие, уже имеются достаточно полные перечни видов, несомненно, остается еще много форм, которые предстоит обнаружить и формально признать; особенно это касается насекомых — группы, занимающей первое место по числу описанных форм.
Большинство людей в общем представляют себе это разнообразие организмов и могут указать некото-
рые различия между особями, относящимися к разным группам. Так, например, пресмыкающихся легко отличить от птиц; группу животных, обозначаемую термином «кошки», никто не спутает с группой, обозначаемой как «собаки»; львы составляют группу, отличную от тигров. Мы замечаем также различия между членами одной группы. Совершенно очевидно, что каждый индивидуум уникален и что нет двух людей и, по-видимому, вообще двух организмов, которые были бы совершенно идентичны. Ни одна мать не спутает своего ребенка с другим и ни один ребенок не примет чужого щенка за своего любимца. Несмотря на различия между разными группами и разными особями в пределах одной группы, большинство из нас ясно видит и черты сходства, обусловленные родственным происхождением. Эта осведомленность о сходстве между особями отражена в таких, выдержавших испытание временем, поговорках как «от яблони — яблочко, а от елки — шишка», «яблоко от яблони недалеко падает», «одного поля ягода».
Существование сходства между особями и различий между группами особей дало возможность привести все многообразие форм органического мира в определенную систему. Были созданы и получили общеупотребительные названия категории живых организмов. Кроме того, каждая группа, формально признанная в системе классификации, получила научное название. Систематика, или таксономия, занимается изучением множества организмов, их отличительных признаков, их классификацией, основанной на изучении всех и каждой связей между разными организмами.
На заре систематики ее задачи ограничивались обнаружением группы, определением ее диагностических признаков и выбором названия.
Считалось, что каждый вид создан независимо от других, имеет вполне определенные морфологические признаки, неизменные во времени и пространстве, и легко отличим от других форм. Классификация заключалась в раскладывании морфологически различающихся между собой «подлинных» сущностей, позже
названных видами, в некую «естественную» систему. Считалось, что особи, составляющие вид, так же сходны между собой, как отливки, полученные из одной формы, а поэтому классификация форм считалась делом несложным. Индивидуумы, отклоняющиеся от основного типа, рассматривались как исключения, и их появление объясняли ошибками развития («спорты»). Черты сходства между разными видами, или родство, пытались относить за счет капризов природы или действия сверхъестественных сил в процессе независимого сотворения каждого типа.
Ранние систематики стремились не только дать названия группам организмов, но и выяснить естественную систему или «план творения». Наиболее удовлетворительного решения этой проблемы достиг Дарвин. Благодаря Дарвину эволюционная история популяций в конечном счете стала рациональной основой классификации, а филогенетическая интерпретация родства между различными организмами дала возможность объяснить естественную систему органического мира. Он обратил внимание на то, что принятая в таксономии иерархическая система классификации, т. е. объединение родственных популяций в один вид, родственных видов — в один род, родственных родов — в одно семейство, семейств — в отряд, отрядов — в класс и т. д., отражает существование в природе групп, подчиненных другим группам, а вовсе «не произвольна, как произвольна, например, группировка звезд в созвездия». Иерархическая классификация отражает реальную ситуацию, естественную систему, в основе которой лежат различия в уровнях филогенетической дивергенции, достигнутых в процессе эволюции. Согласно этой гипотезе, ныне существующие виды возникли от общих предковых форм в различные периоды эволюционной истории, причем признаки отдельных форм склонны дивергировать путем постепенного изменения, и степень дивергенции (степень различия между группами) отражает давность их отхождения от общей предковой формы. Однако использование филогенетической концепции в классификации организмов встречает ряд серьезных трудностей.
Трудности построения естественной системы связаны с необходимостью такого толкования имеющихся данных, которое подкрепляло бы филогенетическую классификацию групп. Основным критерием служит родственное происхождение. О степени родства обычно судят по сходству морфологических и анатомических признаков изучаемых групп. Прямые доказательства, т. е. данные о наличии репродуктивной связи в родословных, имеются лишь в очень редких случаях. В сущности очень многие виды организмов описываются и получают названия на основании изучения всего лишь нескольких экземпляров, хранящихся в музейных коллекциях. Лишь в относительно немногих случаях имеются подробные сведения о популяционной структуре, биологии размножения, экологических потребностях, цитологических и физиологических признаках и распространении группы, и совсем уже редки формы, по которым собраны адекватные выборки из популяции, в полной мере отражающие изменчивость. Однако сколько бы взаимосвязей ни удалось обнаружить, филогению можно лишь постулировать. Даже построение филогенетических рядов ископаемых форм основывается на допущении существовавших между ними репродуктивных связей, которые, конечно, никто не мог наблюдать непосредственно.
Естественная система классификации в современной таксономии базируется на филогении, которая в свою очередь основывается на эволюционном принципе дивергенции форм, имеющих общее происхождение. Поэтому в любой такой классификации категории, хотя они определяются в эволюционных терминах, устанавливаются почти исключительно на основании сходства между формами, т. е. выводы о происхождении носят чисто умозрительный характер. Это тонкое различие помогает понять Симпсон (1961, стр. 69) на примере однояйцовых (идентичных) близнецов: «Мы определяем таких близнецов как двух индивидуумов, развившихся из одной зиготы. Никто никогда не наблюдал у человека, как это происходит, по мы умеем различить те случаи, когда данные о сходстве достаточны, чтобы оправдать допущение об однояйцово-
сти. Такие индивидуумы являются близнецами не потому, что они идентичны, напротив — они идентичны потому, что они являются близнецами». Утверждение о том, что биологическая классификация самая «естественная», поскольку она базируется на эволюционных принципах, не означает, однако, что не может быть другой системы или что хоть какая-то категория объективно отражает некую реально существующую в природе единицу. Один систематик может назвать данную группу родом, тогда как другой считает ее семейством или видит в ней два рода. Но ведущие современные систематики утверждают, что основная категория классификации — вид — действительно соответствует обособленным единицам, так же как, например, особи, клетки или хромосомы представляют собой обособленные структурные единицы.
КАТЕГОРИЯ ВИДА
Перед систематиком стоит обширный ряд биологических форм, которые можно рассортировать в соответствии с разного рода системами классификации. Однако, хотя бы для одного того, чтобы стандартизировать номенклатуру, необходимо привести все это многообразие к какой-то одной официальной системе. В течение двух последних столетий иерархическая система Линнея (изложенная в его книге «Systema naturae», вышедшей в 1758 году) принята в зоологии, а принципы этой системы — и в ботанике. Основная единица линнеевской системы — вид. По традиции каждый организм относится к определенному виду и к надвидовым категориям более высокого ранга (роду, семейству, классу и т. д.). Каждый индивидуум имеет двойное название, состоящее из названия рода и названия вида, к которому он принадлежит, — так называемая биноминальная система номенклатуры. У систематиков вошло в традицию давать видам названия и определять их в соответствии с какой-либо концепцией категории вида. А это приводит к так называемой «проблеме» вида.
Как объективно определить категорию «вид»? Иными словами, какие признаки следует считать диагностическими и как велико должно быть различие между популяциями, чтобы отнести их к разным видам? Эта проблема постепенно проясняется (Майр, 1963; Симпсон, 1961), однако объективного определения вида, которое бы содержало критерий для универсального его применения, все еще не существует. Стало понятно, что критерии распознавания (практический уровень) не могут быть установлены до тех пор, пока определение вида базируется на эволюционных концепциях и включает в себя динамические системы (см. стр. 36). Тем не менее это не должно препятствовать определению (теоретический уровень) категории вида на основании какого-либо разумного и объективного критерия. В последущих разделах мы попытаемся отыскать такой критерий.
ЭВОЛЮЦИОННАЯ КОНЦЕПЦИЯ ВИДА
Симпсон (1961) определил эволюционирующий вид как «последовательный ряд популяций родителей — потомков, эволюционирующий изолированно от других и имеющий собственную эволюционную роль и собственную тенденцию». Легко видеть, что это скорее абстрактное, нежели практическое определение: его трудно или даже невозможно использовать на практике ввиду отсутствия правил, на основании которых можно дать бесспорное описание «эволюционной роли и тенденций». Вместе с тем это наиболее содержательное определение — некий теоретический стандарт, с которым можно сравнивать операциональные определения.
Вид мыслится при этом как дискретная, репродуктивно изолированная система популяций, имеющих общую эволюционную историю. Такие истории схематически изображают в виде «древес», что позволяет графически представить подлинную филогению, или родословную (фиг. 3). Концевые точки ветвей такого древа, достигающие на шкале времени современности,— это ныне существующие виды, а концевые точки, лежащие в прошлом, — вымершие популяции.
Фиг. 3. Филогенетическое древо лошади, построенное на основании палеонтологических данных.
Главный ствол ведет от Eohippus к современному Equus. Многие его боковые ветви вымерли. Эволюция главного ствола происходила преимущественно в Северной Америке. Hyracotherium (по-видимому, как и Eohippus) и его потомки обитали в Старом Свете, так же как Anchitherium, Hippar ion и Stylohippar ion. Эволюция от Pliohippus до Hippidiutn происходила в Южной Америке. Только те формы Equus, которые распространились в период плейстоцена по Евразии и Африке, дали современную лошадь, осла и зебру (по Мэттью, Лумису и Симпсону).
О взаимосвязях родословных во времени обычно судят на основании родственных отношений между ныне существующими группами; ископаемые остатки позволяют повысить вероятность правильности описания филогении.
Если бы мы располагали полной палеонтологической летописью, мы могли бы проследить филети-ческую историю отдельного эволюционного ряда (называемого филумом) как ряда популяций родители — потомки. Но это не значит, например, что все члены ряда, прослеженного в обратном направлении от Equus через Pliohippus, Merychippus, Miohippus, Mesohippus и Eohippus, принадлежат к одному виду. Согласно теории филетической эволюции, с течением времени могут происходить изменения, достаточные для превращения одного вида в другой в пределах одной и той же родословной. По всей вероятности, в филум от Eohippus до Equus входило большое число различных видов. На основании принципа последовательного видообразования путем филетической эволюции мы делим любую такую родословную на ряд последовательных видов, или палеовидов (Кэйн, 1954). Но здесь мы сталкиваемся с одним из аспектов проблемы вида: как велико должно быть различие между популяцией родителей и популяцией потомков (т. е. между двумя ископаемыми формами), чтобы, исходя из эволюционной концепции вида, считать их разными видами? Наше решение было бы совершенно субъективным, если бы мы располагали полным палеонтологическим материалом и могли бы наблюдать постепенное превращение одного вида в другой. Однако в действительности опознание палеовидов облегчается благодаря наличию в палеонтологической летописи разрывов, по обе стороны которых наблюдаются четко выраженные виды, а также точек ветвления родословных.
Согласно эволюционной концепции, вид рассматривается как последовательность популяций «родители— потомки», образующая один из отрезков единой родословной. Предлагали и другие концепции вида, акцентирующие внимание на одной или нескольких
связях между существующими группами организмов, но не придающие особого значения эволюционной истории видов. Майр (1963) рассматривает три концепции: 1) типологическая концепция вида; 2) концепция безмерного вида и 3) концепция многомерного вида.
ТИПОЛОГИЧЕСКАЯ КОНЦЕПЦИЯ (морфологический вид)
В основе представлений о виде лежат две противоположные гипотезы: гипотеза о неизменности организмов и гипотеза об их превращениях; в процессе развития науки предпочтение неоднократно отдавалось то одной, то другой гипотезе. Как уже говорилось, в настоящее время эволюция рассматривается как процесс, ведущий к образованию видов и обеспечивающий их постоянство. Эта последняя стадия в развитии систематики возникла под влиянием идей Ламарка, но окончательно установилась лишь после появления книги Дарвина «Происхождение видов». В течение же предшествующих 100 лет в систематике господствующими были идеи Линнея.
В линнеевской иерархии вид был низшим рангом над уровнем особи. Считалось, что особи одного вида относительно однородны по анатомическому строению и четко отличаются от особей других видов. Весьма существенным для этих представлений была вера в то, что каждый вид создан независимо от других и что морфологические признаки каждого вида остаются неизменными, варьируя лишь в известных пределах, установленных Творцом.
Считалось, что, поскольку вид состоит из особей, построенных по единому плану, для изучения основных свойств какой-либо группы достаточно изучить одного типичного представителя этой группы. Таким образом, типологическая концепция базируется на идее неизменности видов. Согласно этой концепции, главным критерием для описания видов служит степень морфологических различий. Описанные по такому принципу виды называются морфологическими видами, или морфовидами (Кэйн, 1954).
Ранние систематики приняли статический морфо-вид по двум причинам: во-первых, под влиянием общепринятого буквального толкования библейской легенды о божественном творении, записанной в Книге бытия; во-вторых, вследствие того что описание видов обычно производилось на основании изучения всего лишь нескольких особей, собранных в одной местности, причем локальной и географической изменчивости особей, составляющих группу, уделялось очень мало внимания.
КОНЦЕПЦИЯ БЕЗМЕРНОГО (NONDIMENSIONAL) ВИДА
Эта концепция наиболее близка к представлениям о виде местных натуралистов, которые изучают сосуществующие во времени и пространстве популяции различных организмов. Например, орнитологи-любители за сравнительно короткий промежуток времени могут встретить на своих кормовых столиках несколько разных видов птиц (кардиналы, голубые сойки, юнко, скворцы, зонотрихии белогорлые и американские певчие воробьи). Каждый из этих видов хорошо отличается от других, и большинство, если не всех, особей можно без труда отнести к тому или иному виду. Каждая отличающаяся популяция представляет здесь отдельный вид.
Чтобы яснее представить себе эту концепцию, следует пополнить нашу терминологию. Конспецифичные особи (члены одного вида) у перекрестнооплодотво-ряющихся форм объединяются в популяции. В любой момент времени каждый вид занимает в пространстве некий ограниченный ареал, слагающийся из суммы всех территорий, занятых всеми популяциями этого вида. Величина ареала, а также численность и пространственное распределение в нем размножающихся колоний зависят: 1) от системы скрещивания; 2) от подвижности, или способности к расселению, проявляющейся в активной (индивидуальная подвижность) или пассивной форме (расселение с помощью таких факторов, как ветер, вода или же с помощью других
организмов); 3) от распределения благоприятных для данного вида экологических условий (климатические и биотические преграды, препятствующие расширению ареала) и 4) от природы географических преград, препятствующих расселению (таких, как океаны, горы, реки, озера, ледники и пустыни). Ареал вида изменяется во времени. Его границы изменчивы, подобно границам водоема. В одних условиях виды «затопляют» огромные территории, тогда как в других область их распространения сокращается, приобретая четкие границы. Широко распространенные виды, приспособленные к широкому диапазону условий среды, обычно равномерно распределены по всему ареалу. Но популяции, обитающие в пограничных частях ареала, нередко представляют собой полу-изолированные «группы», которые по мере колебаний границ ареала то объединяются с остальными популяциями, то отделяются от них. Менее широко распространенные формы, приспособленные к более специфическим условиям, распределены по ареалу в виде небольших скрещивающихся колоний.
Ареалы разных видов могут перекрываться или не перекрываться. Особи всех сосуществующих в одной и той же географической области видов называют симпатрическими. Особи, популяции или виды, занимающие различные географические области, называют аллопатрическими. Аллопатрические популяции бывают смежными (соприкасающиеся популяции) или разобщенными (популяции, разделенные широкой разделяющей зоной или географическими преградами). Смежные популяции в большинстве случаев характеризуются специфической приспособленностью к примыкающим друг к другу местообитаниям — это географические соседи. Эти популяции считаются географически аллопатрическими, однако, если в пограничной области особи или половые клетки этих популяций вступают в контакт, так что между ними становится возможным скрещивание, их рассматривают как генетически симпатрические популяции. Вслед за Грантом (1963) мы называем такие соприкасающиеся популяции территориально симпатрическими (или
соседними) популяциями, а популяции, занимающие различные экологические ниши в пределах одного и того же местообитания, — биотически симпатрически-ми популяциями. Если между территориально или биотически симпатрическими популяциями не происходит обмена генами в результате скрещивания, то их относят к разным видам; если же обмен генами, о котором судят по наличию промежуточных форм, имеет место, то их относят к одному виду.
Каждый вид, существующий симпатрически (имеющий неразорванный ареал) в ограниченной области, представлен одной популяцией (см. гл. 3), в которой не обнаруживается ни географической изменчивости, ни дифференцировки на расы. Локальные популяции различных видов представляют собой по определению независимые скрещивающиеся единицы; чаще всего такие популяции легко отличить от других благодаря морфологической прерывистости или «видовым разрывам». Так, например, в горных районах Юго-Западной Африки от округа Каоковелд в нагорье Дамараленд до реки Оранжевой зебру Гартмана легко идентифицировать по некоторым простым морфологическим признакам. В районе озера Этоша на севере Дамараленда стада этих зебр заходят в западную часть ареала распространения столь же хорошо отличимой формы, называемой зеброй Чэпмена (фиг. 4). Эти два вида различаются по ряду морфологических признаков —по общим размерам, окраске, характеру полосатости, экстерьеру и по голосу. Между ними существует четкий разрыв. Различие между популяциями этих двух видов подчеркивается тем, что в зоне перекрывания они объединяются в видоспецифичные стада. Подобный же «межвидовой разрыв» существует между зеброй Греви и зеброй Гранта. Их ареалы сливаются на равнине между озером Рудольфа и горой Кения. Зебра Греви в два раза крупнее, а полосы у нее более узкие и многочисленные. Подобным же образом две различные формы — почти вымершая капская горная зебра и квагга (вымершая в 1870 году)—сто лет назад иногда встречались совместно в небольшом районе южнее реки Оранжевой в Капской провинции. Все 6
упомянутых форм получили названия еще до начала нашего века, когда данных об их распространении не было и они рассматривались как отдельные морфови-
Фиг. 4. Распространение зебр и типы полосатости.
Распространение зебры в областях, расположенных ниже пунктирной линии, основано на старых (до 19С0 года) данных о формах, теперь уже вымерших или близких к вымиранию. Зебра Гранта, зебра Чэпмена и бурчеллева зебра (а возможно, и квагга) обнаруживают кливальную изменчивость полосатости. Эти формы представляют собой расы одного многомерного вида Equus bur-chelli, отличного от распространенной севернее зебры Греви (Equus grevyi) и распространенной южнее горной зебры (Equus zebra).
/ — зебра Греви; 2 —зебра Гранта; 3 — зебра Чэпмена; 4 — бурчеллева зебра; 5 —квагга; 5 —зебра Гартмана; 7 —капская горная зебра.
ды. Другая вымершая форма — бурчеллева зебра,— распространенная в прошлом севернее реки Оранжевой, по-видимому, существовала аллопатрически
вместе с кваггой и была также выделена в самостоятельный вид. В таких случаях «виды» характеризуются нескрещиваемостъю двух сосуществующих локальных популяций. В этом и состоит концепция безмерного вида.
КОНЦЕПЦИЯ МНОГОМЕРНОГО ВИДА
После того как натуралисты получили возможность часто и много путешествовать, обнаружилось, что между многими локальными популяциями, которые в своих специфических местностях казались совершенно обособленными, существуют промежуточные группы. Следовательно, отдельную популяцию уже нельзя было рассматривать непременно как отдельный вид. Стало ясно, что «вид» — групповое явление (система, состоящая из множества популяций, рассредоточенных по географическому ареалу). При описании таких систем нельзя игнорировать межпопуляционную изменчивость.
Вернемся к примеру с зебрами. Семь разновидностей, получивших названия как морфовиды, не давали промежуточных фенотипов в тех трех районах, где их популяции симпатричны. Каждая разновидность размножается в чистоте (гибридизация отсутствует). Однако после описания дополнительных популяций, обитающих в отдаленных районах, стало совершенно ясно, что популяции зебры Гранта, зебры Чэпмена и бурчеллевой зебры интерградируют морфологически и географически замещают одна другую. Они образуют одну широко распространенную сеть популяций, в которой не удается обнаружить морфологических «видовых разрывов». У этих популяций можно отметить слабо выраженный градиент в изменении характера полосатости, идущий с севера на юг. Зебре Гранта свойственна резко выраженная волосатость— широкие отчетливые полосы, идущие до самых копыт. Начиная с популяций, распространенных южнее реки Замбези, полосатость становится все менее четкой; между некоторыми полосами образуются дополнительные, слабо выраженные полосы
(зебра Чэпмена). Вместе с тем уменьшается количество пигментированных полос на ногах. У обитавшей еще южнее вымершей бурчеллевой зебры брюхо и конечности были почти белыми. Такие географические преграды, как реки (Замбези, Оранжевая и Вааль) и пустынные районы (Ботсвана), препятствовали свободному обмену генами между этими популяциями. Именно поэтому градиент этого признака не совсем плавный, а напоминает изменчивость, характерную для географически изолированных популяций. Такие градиенты признаков, называемые кланами, служат морфологическим доказательством генетического родства сопряженных популяций, образующих надпопуляционную систему — единый вид. На этом основании три рассмотренных локально ограниченных «вида» зебры были объединены в одну мультипопуля-ционную систему под названием Equus burchelli.
Квагга остается загадкой. Очень сильная редукция полосатости и светлое брюхо, казалось бы, заставляют поместить ее на самый конец клины, образуемой Е. burchelli по окраске. И действительно, некоторые таксономисты включают кваггу в этот вид. Однако, по мнению других, квагга слишком своеобразна, чтобы объединять ее с формами, имеющими резко выраженную полосатость. Кроме того, имеются старые данные о том, что севернее реки Оранжевой квагга и бурчеллева зебра паслись вместе, но оставались репродуктивно изолированными. Если это действительно так, их следует относить к разным видам. Однако, судя по другим данным, квагга могла быть аллопатрической; в таком случае решение вопроса о ее видойой принадлежности становится субъективным.
Географическая изменчивость £. burchelli послужила причиной создания нескольких безмерных мор-фовидов. Это относится и к Е. zebra (зебра Гартмана и капская горная зебра). Подобные виды, обнаруживающие географическую изменчивость и состоящие из идентифицируемых' групп более низкого ранга (или рас), называются политипическими. Вид Е. grevyi имеет более ограниченное распространение и его таксономически значимые признаки не обнаруживают
географической изменчивости. Это монотипический вид. С тех пор как во всех основных группах растений и животных были обнаружены многочисленные примеры политипических видов, интересы исследователей резко сместились, и вместо характеристики особей или локальных популяций они уделяют главное внимание описанию популяционных систем. Попытаемся теперь определить видовую категорию с позиций многомерной концепции.
ОПРЕДЕЛЕНИЕ ПОНЯТИЯ «ВИД» С ПОЗИЦИЙ МНОГОМЕРНОЙ КОНЦЕПЦИИ
Первые концепции вида создавались под давлением чисто практических потребностей — необходимости давать названия различным формам жизни и определять их. Концепция вида должна была быть универсальной, т. е. приемлемой для всех встречающихся форм. Морфологическая концепция вполне соответствовала этой цели. Систематики-практики и по сей день занимаются определением особей и именованием видов, однако на совершенно иной концептуальной основе. Установление морфологических различий между индивидуумами остается необходимым и даже главным аспектом процедуры в систематике, однако теперь вид понимают как совокупность изменчивых популяций, динамически эволюционирующих, как некую собирательную единицу, репродуктивно изолированную от остального биотического мира. Теперь, когда ясно, что единое репродуктивное сообщество может состоять из морфологически различающихся субъединиц, главным критерием определения вида служит не морфологическое различие между индивидуумами, а репродуктивная связь между популяциями. Именно на репродуктивной связи основано так называемое современное биологическое определение вида. Это определение включает в себя следующие четыре существенных положения.
1. Биологический вид возможен только у форм, размножающихся половым путем.
2. Вид состоит из одной или нескольких популяций родственных особей (в соответствии с многомерной концепцией).
3. Взаимоотношения между членами любой популяции вида обусловлены их зависимостью друг от друга в репродуктивном отношении (разобщенные популяции одного вида считаются потенциально способными к обмену генами).
4. Главный критерий биологического вида — репродуктивная изоляция входящих в него популяций от всех других (это как раз то, что имеется в виду при формулировании понятий «видовые разрывы» в безмерной концепции и «собственная эволюционная роль и тенденции» в эволюционной концепции вида). Короче говоря; биологический вид есть «группа действительно или потенциально скрещивающихся естественных популяций, репродуктивно изолированная от других таких же групп» (Майр, 1940).
Ясно, что симпатрические, морфологически отличные популяции, принадлежащие к одному виду, не могут долгое время оставаться обособленными, так как в результате скрещивания и обмена генами происходит интеграция их отличительных признаков. Отсюда следует, что различающиеся симпатрические популяции относятся к разным видам и что они различны потому, что гибридизация между ними невозможна. Человек, а также некоторые домашние животные и культурные растения составляют исключение. Искусственное ограничение свободного скрещивания приводит к тому, что несколько отдельных пород (рас) даже при совместном существовании сохраняют свои специфические особенности. Например, если собаковод содержит в питомнике собак нескольких разных пород, то, ввиду того что они потенциально способны к скрещиванию, их, по крайней мере в период течки, следует изолировать, чтобы они не испортили породу.
Что касается человека, то разные расы, будучи симпатрическими, могут оставаться генетически изолированными ввиду многочисленных социально-бытовых преград, препятствующих смешанным бракам. Однако эти преграды далеко не всегда действуют, и
смешанные браки довольно обычны. Межрасовые гибриды способны к воспроизводству, а, следовательно, все люди независимо от расы представляют собой (в соответствии с биологическим определением) один вид.
Наиболее существенная особенность биологического вида — репродуктивная изоляция. По той или иной причине какие-то формы оказываются неспособными скрещиваться друг с другом или же производить жизнеспособное и плодовитое потомство, которое обеспечило бы передачу генетического материала последующим поколениям. Самая очевидная причина этого — пространственная изоляция видов. Однако географическая изоляция — сугубо внешняя причина репродуктивной изоляции, и при этом разобщенные формы вовсе не обязательно утрачивают потенциальную способность к скрещиванию. С другой стороны, даже занимая соседние территории или будучи биотически симпатрическими, популяции могут оставаться изолированными. Обмену генами между популяциями препятствует одна или несколько внутренних преград, называемых изолирующими механизмами (см. гл. 8). К изолирующим механизмам относятся такие факторы, как утрата способности к спариванию или перекрестному опылению, резкое снижение эффективности оплодотворения, а также нежизнеспособность или бесплодие гибридов. Согласно биологическому определению, популяции, принадлежащие к одному виду, скрещиваются между собой и дают жизнеспособное плодовитое потомство; если такие популяции разобщены, мы подразумеваем, что они потенциально способны к эффективному скрещиванию. Популяции же, принадлежащие к разным видам, репродуктивно изолированы благодаря существованию одного или нескольких изолирующих механизмов.
ОГРАНИЧЕННОСТЬ БИОЛОГИЧЕСКОЙ КОНЦЕПЦИИ
Биологическое определение вида применимо к эволюционной концепции на любом отрезке времени в пределах родословной интересующих нас форм. Фило
генетические ряды остаются обособленными до тех пор, пока их популяции не смешиваются генетически. Подчеркнем еще раз, что эти понятия объективно применимы для определения категории вида, но что данные, свидетельствующие о его применимости, обычно носят косвенный характер и скорее всего совершенно субъективны. Подобно тому как наши представления о репродуктивной связи между членами последовательного ряда видов (интерградирующие ископаемые остатки) выводятся на основании логических рассуждений, так и о репродуктивных взаимоотношениях современных форм чаще всего судят по косвенным данным. При решении вопроса о том, принадлежат ли две формы (популяции) к разным видам, используют три главных критерия.
1. Формы настолько различны морфологически, что их трудно принять за один вид. Делается вывод, что степень репродуктивной изоляции коррелирует с величиной различий. Совершенно очевидно, что в таких случаях решение систематика субъективно.
2. Репрезентативные «представители двух форм, помещенные в экспериментальные условия, либо не скрещиваются, либо дают нежизнеспособных или стерильных гибридов.
3. Формы сохраняют обособленность, даже если их ареалы соприкасаются или если они биотически сим-патричны. Данные о скрещивании этих форм или образовании гибридов между ними отсутствуют (т. е. существует видовой разрыв).
Только третий из этих критериев в какой-то степени приемлем. Однако, если даже применять этот строгий критерий, при выделении видов часто возникают трудности. Эти трудности можно разбить на три главных типа.
I. Данные недостаточны, неубедительны или противоречивы. Мы уже знаем, например, что кваггу можно рассматривать как крайнюю форму зебры Е. burchelli или как отдельный вид. В этом случае критерий 1 не позволяет решить вопрос, критерий 2 неприменим, а возможность использования критерия 3 остается неясной.
Особенно трудно установить истину в тех случаях,
Фиг. 5. Географическое распространение двух видов-двойников Drosophila pseudoobscura и D. persimilis.
Эти два вида остаются симпатрическими на большом пространстве от Британской Колумбии до горных районов Калифорнии.
когда критерии 1 и 2 приводят к противоречивым выводам. Степень репродуктивной изоляции между популяциями не всегда соответствует степени морфологических различий между ними. Например, среди тысяч особей, взятых из районов симпатрического сосуществования двух близкородственных видов Drosophila pseudoobscura и D. persimilis не было обнаружено ни одного гибрида (фиг. 5). По-видимому, между этими двумя формами не происходит обмена генетическим
материалом (критерий 3). Ясно, что эволюционные пути этих форм разошлись, и, согласно данному выше определению, их следует рассматривать как два разных вида. Вместе с тем они морфологически так сходны (критерий 1), что в течение многих лет считались идентичными, и только в лабораторных испытаниях на скрещиваемость (критерий 2) их удалось разделить. Такие морфологически очень сходные виды называют видами-двойниками.
Теперь известно, что между видами-двойниками все же существуют некоторые весьма тонкие различия, по которым специалистам удается их распознавать. К ним относятся различия в половых органах самцов, в половых гребешках (толстые щетинки на передних лапках), в форме крыльев и в хромосомах. Кроме того, эти виды различаются по экологическим требованиям и ареалам.
Виды-двойники обнаружены во всех группах высших животных и растений. Они очень часто встречаются среди насекомых, особенно в таких хорошо изученных родах, как Drosophila. Поскольку различия между видами-двойниками, как правило, очень невелики и весьма трудоемкие методы, необходимые для их выявления, могут быть применены только к ограниченному числу популяций, естественно предположить, что таких видов существует значительно больше, чем мы в состоянии обнаружить. Полиплоидия (кратное увеличение хромосомного набора) — весьма обычный способ видообразования у растений — привела к возникновению криптических «видов во многих семействах высших растений. В некоторых местностях наряду с двумя или несколькими диплоидными видами встречаются также полиплоидные формы, возникшие в результате гибридизации от этих диплоидных форм, причем все эти формы морфологически неразличимы. Эта группа представляет собой полиплоидный комплекс видов-двойников, которые тем не менее репродуктивно изолированы. К такому комплексу морфологическая концепция вида совершенно неприменима.
Следует помнить, что морфологические различия между группами — это внешние проявления более
глубоких генетических различий, действующих на процесс развития и подверженных влиянию внешних условий. Когда таксономист определяет популяции морфологически и присваивает им ранг вида, он стремится выделить репродуктивно независимую эволюционирующую единицу. Во многих случаях степень и характер генетических различий, необходимые для возникновения репродуктивной изоляции, одновременно вызывают внешне заметные изменения в строении тела. Опытный таксономист нередко может точно предсказать генетическое различие на основании анализа коррелированной с ним изменчивости морфологических признаков. Многие из выделенных таким образом морфовидов представляют собой настоящие биологические виды. Однако эту корреляцию можно использовать не всегда, так как степень морфологического различия между некоторыми видами так велика, что один вид можно отличить от другого даже на расстоянии (например, зебра Гранта и зебра Греви). В других же случаях (виды-двойники) для этого приходится прибегать к специальным тестам.
Особенно трудно решать вопрос о видовой принадлежности в случае родственных, но дифференцированных аллопатрических’популяций, поскольку в этом случае прямое определение репродуктивной изоляции (критерий 3) невозможно. Обратимся снова к двум видам дрозофилы — D. pseudoobscura и D. persimilis. Было показано, что в лабораторных условиях эти виды способны к гибридизации и дают высокожизнеспособное потомство Fi. Однако гибридные самцы бесплодны. Хотя гибридные самки обладают нормальной плодовитостью, жизнеспособность их потомства при возвратных скрещиваниях мала. Таким образом, генный обмен между этими видами сильно ограничен (критерий 2). Эти факты подтверждают имеющие более важное значение данные об отсутствии гибридов в зоне симпатрического сосуществования этих популяций (критерий 3). Таким образом, несмотря на чрезвычайно большое сходство между этими формами, можно считать доказанным, что это разные виды.
Гораздо сложнее обстоит дело с двумя близкородственными пустынными формами D. tnojavensis и D. arizonensis. D. tnojavensis образует три аллопатрические группы популяций: на юге Калифорнии, в Нижней Калифорнии и в Мексике (штат Сонора). Родственная форма arizonensis встречается главным образом в штатах Аризона и Сонора (фиг. 6). В штате Сонора эти две формы биотически симпатричны, но репродуктивно изолированы (критерий 3), что свидетельствует об их принадлежности к разным видам. Однако вопрос о том, объединять ли разобщенные калифорнийскую популяцию и популяцию Нижней Калифорнии в один вид tnojavensis, решается в значительной степени субъективно. В лабораторных условиях члены этих трех аллопатрических популяций tnojavensis легко скрещиваются, давая многочисленное жизнеспособное и плодовитое потомство (критерий 2). Эти данные можно принять как косвенные доказательства принадлежности трех разобщенных популяций к одному виду tnojavensis. Дальнейшие исследования показали, что в лабораторных условиях члены разобщенных популяций tnojavensis скрещиваются с особями arizonensis и дают немногочисленное, но жизнеспособное и плодовитое потомство. Больше того, F2 и потомки от возвратного скрещивания не только жизнеспособны, но адаптивная ценность некоторых из них даже выше, чем у исходных родительских форм. Конечно, вопрос о возможности эффективной гибридизации между arizonensis и аллопатрическими популяциями tnojavensis, если бы они оказались в условиях симпатрии, остается спорным. По-видимому, в штате Сонора такой гибридизации не происходит. Между прочим следует указать, что калифорнийская популяция tnojavensis морфологически отлична от двух других, которые выглядят как двойники arizonensis (критерий 1). Все это вместе заставляет предположить, что эти три формы представляют собой очень близкородственные или пограничные формы, которые, возможно, находятся в процессе видообразования. Трудность здесь состоит в том, ч4ю эти разобщенные популяции систематик может отнести
к одному, двум или трем разным видам. В данном случае выделили два вида. Терминология Майра помогает
Фиг. 6. Распространение близкородственных видов Drosophila mojavensis и D. arizonensis.
Вид mojavensis представлен тремя большими разобщенными популяциями; популяция, обитающая на юге Калифорнии, представляет собой морфологически отличную геэг >афическую расу (полувид). Хотя в лабораторных условиям особи эти' Д1ух видов способны к гибридизации, в природе (штат Сонора в Мексике) они биотически симпатричны и репродуктивно иэоли-г рованы.
застраховаться от подобных неопределенностей и путаницы в анализе видообразования: аллопатрические «пограничные» формы он называет полувидами, а
группы родственных полувидов — надвидами. Применение такой терминологии ие решает проблемы наименования, но по крайней мере при этом признается существование самой проблемы.
II. Другая трудность выделения биологических видов связана с тем, что исследуемые группы находятся в состоянии эволюционной промежуточности. Процесс эволюции непрерывен, и поэтому в некоторых случаях невозможно провести строгую границу между тем, что называется расой, и тем, что называется видом. Экологические, морфологические и воспроизводительные различия между современными популяциями могут быть развиты лишь частично. Например, виды-двойники достигли репродуктивной изоляции, но почти не дифференцировались морфологически. Другие же группы свободно скрещиваются и дают потомство в лабораторных условиях или на опытных участках, хотя морфологическая и экологическая дифференцировка этих групп достигла такого уровня, что их вполне можно считать отдельными морфовидами; это относится, например, к евразиатскому платану (Platanus orientalis) и американскому платану (Р. occidentalism которые на протяжении миллионов лет были изолированы друг от друга географически. Есть также формы, состоящие из ряда географических рас, между которыми возникла частичная репродуктивная изоляция.
Ситуация еще более усложняется, когда отличительные видовые признаки варьируют по степени проявления у разных популяций данного вида. Интересен хорошо проанализированный случай с красноглазым тауи (Pipilo erythrophthalmus). Этот вид распространен по всей территории США и на юге заходит в Мексику, встречаясь преимущественно в дубравах и кустарниковом подлеске. На юге Центрального плато отдельные популяции этого вида разбросаны среди популяций другого вида тауи — Р. ocai\ это более южная форма, обитающая в хвойных лесах. В этом районе существует несколько популяций, у которых перемешаны признаки обоих видов, причем интерградация этих признаков достигает такой степени, которая
оправдывает отнесение двух «чистых» форм к одному виду (фиг. 7). Но по крайней мере в одной местности (штат Оахака в Мексике) эти виды не смешиваются, хотя и сосуществуют симпатрически. Было высказано предположение, что эти виды до самого недавнего времени сохраняли свою обособленность благодаря репродуктивной изоляции экологического характера. Вследствие изменений в условиях среды, вызванных прежде всего в результате сведения лесов местными жителями, возникли новые и промежуточные местообитания, пригодные для обоих видов, для их гибридов и потомков этих гибридов. Напротив, в Оахака зоны растительности все еще остаются нетронутыми и виды здесь сохранили обособленность.
Известно много других примеров межвидовой гибридизации, особенно среди растений. То обстоятельство, что некоторые виды в известной мере гибридизируют друг с другом, не обязательно противоречит биологической концепции. Вряд ли можно сомневаться, что смежные популяции, свободно обменивающиеся генетическим материалом в зонах контакта, принадлежат к одному виду, а репродуктивно изолированные симпатрические формы принадлежат к разным видам. Однако критерий, основанный на репродуктивной изоляции, не всегда можно применять так объективно. Ведь репродуктивная изоляция — характеристика популяции, и создается она путем постепенных эволюционных изменений. В некоторых случаях межпопуляционные преграды могут быть еще неполными, т. е. время от времени между этими популяциями может происходить скрещивание с образованием гибридов, правда, менее жизнеспособных и менее плодовитых, чем потомки от скрещивания между конспецифичными особями. Кроме того, даже хорошо развитые изолирующие механизмы могут разрушаться, допуская некоторое скрещивание. В результате то, что считалось двумя хорошими видами (полная изоляция), может оказаться пограничным случаем (частичная изоляция). Межвидовая гибридизация чаще всего дает несколько маложизнеспособных потомков F! и потомков от некоторых возвратных скрещиваний. При
этом лишь относительно небольшая часть генов одного вида соединяется с генами другого, так что «эволюционная роль» каждого вида остается относительно независимой. Этот процесс взаимного проникновения генов называется интрогрессивной гибридизацией, или интрогрессией. Чаще всего интрогрессия наблюдается между видами, изолированными экологически, как в случае с тауи. Изоляция, основанная на экологической специализации, представляет собой очень слабый механизм сохранения обособленности популяций и может быстро разрушаться при самых незначительных колебаниях условий среды. Если различия между ин-трогрессирующими формами сохраняются, то из этого следует, что они все еще остаются различными видами.
Популяции, принадлежащие к одному виду, характеризуются фактической или потенциальной скрещиваемостью, тогда как между видами существует репродуктивная изоляция. Смежные популяции со средним уровнем гибридизационного обмена генами могут представлять собой расы с частичной репродуктивной изоляцией или виды с частично разрушенными преградами. Во всяком случае их нельзя с полным правом возвести ни в ранг расы, ни в ранг вида. Для этих пограничных видов, как и для аллопатрических форм, удобно использовать термин полувиды.
Полувиды создают много неудобств таксономисту, но зато они не противоречат биологической концепции; напротив, это понятие подчеркивает динамическую природу эволюционирующих единиц.
Если бы мы могли найти действительно объективный, всеобъемлющий и практический критерий для определения вида, то эволюционная теория оказалась бы в опасности. Репродуктивному критерию придается особое значение потому, что он отражает основную биологическую характеристику организмов, размножающихся половым путем (их генетическую независимость в процессе эволюции), и, таким образом, применительно к таким единицам его следует считать объективным.
III. И наконец, главный критерий биологического определения вида неприменим к бесполым формам,
о
50 100 150 км
Фиг. 7. Распространение и интерградация популяций тауи в Мексике.
Числа указывают средние показатели признаков, характерные для гибридных популяций, начиная с «чистой» Pipilo erytnrophthalrnus (24) до «чистой» Р. осей (0). Обратите внимание, что на юго-востоке близ Оахака формы разобщены, оставаясь симпатрическими.
Два предшествующих недостатка репродуктивного критерия для классификации и номенклатуры имеют практическое значение. Существование же бесполых форм порождает теоретическую проблему, поскольку критерий «репродуктивная изоляция видов» применим только к формам, размножающимся половым путем. Половым размножением называют любой способ размножения, обеспечивающий комбинацию и рекомбинацию генов, происходящих из различных родословных. Это чаще всего достигается за счет двух независимых процессов: 1) процесса рекомбинации генов при мейозе и гаметогенезе; 2) процесса сингамии или слияния двух гаплоидных гамет для образования одной диплоидной зиготы. В результате этих процессов происходит объединение генетической информации отдельных особей, принадлежащих к одному поколению, или объединение генетической информации разных поколений. Таким образом создается организационная единица более высокого уровня — популяция. Но по определению среди строго бесполых организмов не существует ни генетической рекомбинации, ни генетически определенной популяционной структуры, а следовательно, не может быть и видообразования в биологическом смысле. Одна из неотложных проблем, стоящих перед систематиками, состоит в том, чтобы включить в рамки эволюционной теории многочисленные «парасек-суальные» механизмы рекомбинации генов, обнаруженные у низших организмов, прежде считавшихся бесполыми. Работ, посвященных анализу популяционных систем у таких организмов, пока еще мало.
Таким образом, мы установили, что термин вид используется двояко: 1) в общем смысле — как таксономическое обозначение различных организмов, которые экологически объединены, а морфологически различаются (в том числе и бесполых) и 2) в специфическом смысле — для обозначения репродуктивных изолятов, каждый из которых состоит из популяций фактически или потенциально скрещивающихся организмов. Использование термина в его первом значении просто облегчает процедуру классификации форм жизни. При этом во многих случаях происходит объ
единение или дробление эволюционных родословных, но тем не менее этот термин повсеместно применяется для описания (в первом приближении) наблюдаемой в природе упорядоченности. Второе значение термина с концептуальной «точки зрения вполне приемлемо; в этом смысле он служит некой рабочей абстракцией, с помощью которой можно обобщать популяционные явления, однако в большинстве случаев применение термина затрудняется отсутствием фактических данных, необходимых для принятия совершенно однозначного решения.
ВНУТРИВИДОВЫЕ КАТЕГОРИИ
(расы и подвиды)
Согласно многомерной и эволюционной концепциям, виды состоят из популяций, распределенных в пространстве и во времени. Каждая локальная популяция какого-либо вида может рассматриваться как колония скрещивающихся между собой особей, приспосабливающихся к специфическому комплексу условий в их местообитании. Родственные колонии, обитающие в различных частях одной общей области обитания, по-видимому, будут занимать сравнительно сходные экологические ниши. Члены данной локальной популяции спариваются преимущественно между собой просто в силу своей территориальной близости, но в пограничных районах происходит некоторое взаимопроникновение особей и половых клеток. Поэтому для смежных популяций характерно большое число общих генов и признаков. Для более удаленных друг от друга и разобщенных популяций одного вида степень обмена генами ниже. Следует также ожидать, что популяции, обитающие в удаленных частях ареала, будут более своеобразны в результате приспособленности к несходным условиям среды: они обычно обладают различными генными комплексами и разными признаками. Хотя каждая популяция вида представляет собой продукт единственной в своем роде генетико-физиологической реакции на условия некой локальной среды, группа родственных популяций,
обитающих в одном районе, будет иметь много общих признаков, но может по некоторым признакам или частотам генетических вариантов отличаться от других групп, обитающих в других районах. Вследствие сходства и различия между популяциями иногда удобно делить их на внутривидовые группы, называемые расами или подвидами. Мы видели, например, что вид Е. burchelli состоит по крайней мере из трех довольно различных (но интерградирующих) рас: зебра Гранта, зебра Чэпмена и бурчеллева зебра. Расами можно считать группы локальных популяций, распространенных в различных частях видового ареала и отличающихся по одному или нескольким признакам. Подвиды — это те же расы, только удостоенные формального таксономического наименования. Как уже отмечалось, виды, формально разделенные на расы, называются поли-типическими, а виды, не имеющие рас, — монотипическими. Человек, несомненно, принадлежит к числу по-литипических видов. Чаще всего политипия встречается у форм, представленных широко расселенными и разобщенными популяциями, которые приспособлены к различным условиям. Когда разные расы встречаются друг с другом в природе, то наблюдается слияние их отличительных признаков в результате гибридизации. Степень интерградации зависит прежде всего от степени пограничных контактов между расами.
Деление на расы — сугубо субъективная процедура, основанная на существовании двух или большего числа групп популяций, различия между которыми достигли такой величины, которая оправдывает формальное признание этого факта. Хотя различия между расами — объективно установленные факты, число рас, которые мы решили различать, определяется соображениями удобства и необходимости привести в систему наблюдаемое внутривидовое разнообразие, с тем чтобы облегчить общение между теми, кто занимается изучением видов и эволюции.
Всякое эволюционное событие очень сильно зависит от величины и природы изменчивости среди членов локальной популяции и от различий между популяциями или расами, составляющими вид. Поэтому
полное описание и классификация органического разнообразия — необходимая предпосылка для понимания эволюционного процесса. Кроме того, попытки определить элементарные категории биологической классификации имеют большую эвристическую ^ценность.
Мы получили некоторое представление о проблемах, стоящих перед нами. Для того чтобы понять раздел эволюции, который называется видообразованием, нужно изучить факторы, участвующие в возникновении изолирующих механизмов. Эти факторы действуют в локальной скрещивающейся популяции, или деме/ анализ которой с генетических позиций будет произведен в следующей главе.
ЛИТЕРАТУРА
Cain A. J., Animal Species and Their Evolution, New York, Harper and Row, Publishers, Incorporated, 1954. (А. Кэйн, Вид и его эволюция, ИЛ, М., 1958.)
Ehrlich Р. R., Holm R. W., The Process of Evolution, New York McGraw-Hill Book Company, 1963. (П. Эрлих, P. Холм, Процесс эволюции, изд-во «Мир», М., 1966.)
Grant V., The Origin of Adaptations, New York, Columbia University Press, 1963.
Heslop-Harrison J., New Concepts in Flowering-Plant Taxonomy, London, William Heinemann, Ltd., 1953.
Ma yr E., Speciation Phenomena in Brids, Amer. Naturalist, 74 (1940), 249-78.
Ma yr E., Animal Species and Evolution, Cambridge, Mass., Harvard University Press, 1963. (Э. Май p, Зоологический вид и эволюция, изд-во «Мир», М., 1968.)
Simpson G. G., Principles of Animal Taxonomy, New York, Columbia University Press, 1961.
Глава 3
ГЕНЫ В ДЕМАХ
Эволюция — это по существу процесс изменения генетической структуры популяций. Поэтому, для того чтобы действительно понять этот процесс, необходимо знать генетику популяций.
Зная тип скрещивания, можно предсказать генотипический состав потомства, ожидаемого от этого скрещивания, но для точного предсказания необходимо знать целый ряд факторов: встречающиеся в популяции локусы и аллели, генотипы родительских особей, относительную жизнеспособность гамет, относительную жизнеспособность генотипов. Частоты фенотипов могут совпадать с частотами генотипов или отличаться от них, что зависит от таких дополнительных факторов, как доминирование, эпистаз, пенетрантность. Менделевская генетика изучает эти факторы преимущественно для того, чтобы выяснить основные механизмы передачи генетического материала от родителей потомкам. Популяционная генетика также имеет дело с предсказанием частот генотипов и фенотипов в ряде последовательных поколений, но при этом мен-делевские принципы распространяются не на какое-либо одно скрещивание, а на все скрещивания, возможные в данной популяции. И в этом случае для того, чтобы делать предсказания, необходимо знать множество факторов.
В этой главе мы изложим основные представления, необходимые для применения менделевской генетики на популяционном уровне. Читатель убедится, что
предсказание частот генотипов и фенотипов не представляет особого труда. В процессе изложения указывается, какими данными следует располагать для такого предсказания, а также рассматриваются возможные затруднения и недостатки метода.
ПОПУЛЯЦИИ И ДЕМЫ
В природе очень немногие особи живут поодиночке. Обычно живые существа вступают в пространственно-временные взаимоотношения; в частности, они склонны к образованию скоплений. Причина образования скоплений тривиальна — организмы приспосабливаются к определенному комплексу условий среды, и естественно, что одинаково приспособленные особи стремятся скопиться в одной и той же экологической нише.
В широком смысле термин популяция означает любую совокупность каких-либо объектов, но биологи используют этот термин специально для обозначения группы одинаково приспособленных особей. Специалисты по генетике популяций еще более сужают этот термин, понимая под популяцией группу раздельнополых форм, которых объединяет способность скрещиваться между собой и сходство экологических потребностей. В генетическом смысле популяция (или, более точно, генетическая популяция) —это пространственно-временная группа скрещивающихся между собой особей одного вида. Вследствие репродуктивной связи между поколениями генетическая популяция непрерывна во времени, а внутрипопуляционные скрещивания обеспечивают ей пространственное единство. Численность популяции может возрастать или уменьшаться в результате миграции особей в популяцию и из нее, а также изменений рождаемости и смертности. Популяция может объединиться с другими популяциями или исчезнуть вследствие эмиграции всех ее членов или полного вымирания.
Главным предметом обсуждения в этой главе будет обмен генами между членами популяции. Насколько
это важно, видно из того, что многие фундаментальные открытия в популяционной генетике были сделаны на основе анализа популяций генов, а не популяций особей. С этой точки зрения вся генетическая информация, распределенная среди особей свободно скрещивающейся группы, в совокупности образует некий генофонд, который в данное время представлен набором отдельных генотипов и сохраняется в них. Зиготы (генотипы), развивающиеся в особей одного поколения, возникают вследствие объединения гамет, образованных предшествующим поколением. Аллели сортируются, перетасовываются и комбинируются в гаметы в процессе мейоза. Гаметы объединяются в процессе оплодотворения, образуя новый набор генотипов. В результате всех этих процессов в каждом поколении происходит реконструкция генофонда.
Концепцию генофонда можно особенно наглядно иллюстрировать на примере маленьких изолированных или полуизолированных колоний, скрещивание в которых приближается к панмиксии, т. е. скрещивающиеся пары объединяются случайно. Для обозначения таких популяций предлагались разные термины: гамодем (Гилмур и Грегор), панмиктическая единица (Райт), локальная менделевская популяция (Добжан-ский), однако чаще всего для них используется термин генетическая популяция или просто дем.
ЧАСТОТЫ ГЕНОТИПОВ в популяциях
Совершенно очевидно, что изучение любой популяции начинается с изучения фенотипов особей. Обычно среди членов популяций наблюдается некоторая изменчивость. Это легко обнаружить при беглом внешнем осмотре членов какого-либо дема. В определенной степени наблюдаемая фенотипическая изменчивость базируется на генетическом разнообразии. Поэтому при описании популяции в генетических терминах нашим следующим шагом будет определение генетической природы интересующих нас признаков путем анализа потомков от известных скрещиваний. Когда тип наследования какого-либо признака из
вестей, популяцию можно охарактеризовать в терминах относительного числа генотипов среди взрослых особей (диплоидная фаза) и относительного числа аллелей в гаметической (гаплоидной) фазе.
Рассмотрим простейший случай наследования двух аллелей Айа одного аутосомного локуса. В отсутствие доминирования, т. е. когда все три фенотипа отличаются один от другого, число особей в каждом из этих фенотипических классов равно соответственно числу особей каждого из трех генотипов АА, Аа, аа. Пусть Пь п2, п3 — число особей каждого из трех генотипов АА, Аа и аа соответственно, и пусть п\ + +м2 + п3 = Л^. В этом гипотетическом случае N обозначает общее число особей в популяции. Если анализируется реально существующая большая популяция, то определить генотип каждой особи невозможно; в таком случае У есть число особей в выборке из такой популяции. Репрезентативность популяционной выборки, конечно, существенно зависит от квалификации исследователя.
Обозначим процент или долю генотипов АА, Аа и аа соответственно через х, у и z или через частоты генотипов:
= х (частота АА)
= у (частота Аа)
-jy- = z (частота аа)
Сумма х + у + х равна единице, или 100%.
ЧАСТОТЫ ГЕНОВ в популяциях
Общее число диплоидных особей в популяции (Af) развивается из 2N гамет, образованных предшествующим поколением. Некоторые из этих гамет содержат аллель А, а остальные — аллель а. Гаметы, содержащие аллели А и а, свободно комбинируясь в процессе оплодотворения, дают популяцию диплоидных потомков. Число особей с генотипом АА,
обозначаемое п{, образовалось в результате объединения вдвое большего числа гамет (2/ti), каждая из которых содержит по одному аллелю А. Гетерозиготы образовались из 2/12 гамет, из которых п2 содержат аллель А и п2 — аллель а. Общее число Д-гамет (т. е. гамет, содержащих аллель Д), принявших участие в образовании диплоидных особей, равно сумме 2/ii + + п2. Относительное число этих гамет (р) среди всех гамет популяции равно
2hj + и2 __ «! + '/2п2 Р 2N
Относительное число гамет с аллелем a(q) можно определить на основании числа а-гамет, принявших участие в образовании особей Аа и аа. Следовательно,
___ /^2 "h 2/1з _ Уд^2 + п3 Ч 2N ~ N
Такие пропорции называются частотами генов. Поскольку речь идет о диаллельном локусе, р + р = 1. Таким образом,
I __ п1 + Уд^2 I Уд^г + п3 _ п\ + п2 + Из _ |
РТУ— N т N — м —1
И
p=l-q.
Частоты генов можно найти из частот генотипов, так как
р = = ^ = x + i/2y (3.1)
и
q = ^^=^+^- = '/2y + z. (3.2)
Проще говоря, чтобы найти частоту аллеля Д, нужно к частоте гомозигот АА прибавить половину частоты гетерозигот Да; для нахождения частоты аллеля а нужно к частоте гомозигот аа прибавить половину частоты гетерозигот Аа.
Для иллюстрации метода определения частот генов рассмотрим распространение групп крови системы М—N у человека. Данные семейного анализа показывают, что М—N группы крови контролируются парой аллелей Ам и LN одного аутосомного локуса. Все три группы крови М, MN и N четко идентифицируются и соответствуют трем генотипам LMLM, LMLN и LNLN. В табл. 1 приведены данные о численности и пропорции каждого из этих генотипов в выборках из трех популяций.
Таблица 1
Число лиц с разными группами крови М — N и частота этих групп в трех группах людей
Группа людей Число обследованных М MN N Частота аллелей
число % число % число % LM lN
Белые (США) 6129 1787 29,16 3039 49,58 1303 21,26 0,5395 0,4605
Индейцы племени Навахо (США) 361 305 84,49 52 14,40 4 1.11 0,917 0,083
Коренные жители Австралии 730 22 3,01 216 29,59 492 67,40 0,178 0,822
Частоты генов LM и LN в каждой из этих популяций можно найти либо с помощью данных о числе людей в выборках, либо с помощью частот генотипов (уравнения 3.1 и 3.2). Так, частоты генов LM и LN среди 730 коренных жителей Австралии равны
/ ,м\ 22 + !/2 (216)
р (частота L ) =-------—- =
= 0,0301 + ’/2 (0,2959) = 0,178;
/ ’/г (216)+ 492
q (частота L ) = ------=
/ Ом
= >/2 (0,2959) 4- 0,6740 = 0,822.
РАВНОВЕСНАЯ ПОПУЛЯЦИЯ
До сих пор мы описывали одно определенное поколение некой популяции. А каковы частоты генов и генотипов в последовательном ряду поколений? Генотипы каждого следующего поколения, или генотипы потомков, образуются из тех гамет, которые дает предшествующее поколение. Аллельный состав этих гамет определяется несколькими факторами, оказывающими влияние на процесс передачи генов из поколения в поколение. Не зная всех этих факторов, невозможно получить ответ на поставленный нами вопрос. Некоторые из этих факторов будут подробно обсуждаться позже, однако нам придется уже сейчас представить их читателю, чтобы было ясно, что выводы, сделанные в этой главе, основаны на нескольких важных допущениях.
Тип наследования
В дальнейшем речь пойдет преимущественно о раздельнополых диплоидных организмах, образующихся в результате объединения двух гаплоидных гамет. Как уже говорилось, прежде чем рассчитывать фенотипический состав будущего потомства, нужно знать местоположение локуса (аутосомный или сцепленный с полом), число аллелей, число локусов, закономерности фенотипического проявления аллелей — доминантность или эпистаз.
Величина популяции
Изменчивость выборки, обусловленная только случаем, может оказать существенное влияние на то, какие гены содержатся в гаметах, образующих поколение потомков. Численность любой популяции ограничена; вследствие ошибки выборки с уменьшением величины популяции увеличивается вероятность того, что частоты генов в гаметах будут отличаться от частот соответствующих генов в родительской популяции. Пока мы будем говорить только о «больших» популя
циях, т. е. о популяциях, в которых случайные изменения частот генов настолько незначительны, что ими можно пренебречь.
Системы скрещивания
Для облигатно раздельнополой популяции наиболее обычная система скрещивания — это система, приближающаяся к случайному размножению (панмиктическая единица), при которой данная особь имеет одинаковую вероятность спариться с любой особью противоположного пола независимо от генотипа последней. Но в большой популяции, занимающей большую территорию, дело обстоит несколько иначе, так как обычно спаривание происходит между особями, находящимися вблизи друг от друга. Такое ограничение панмиксии не имеет существенного значения, если популяция не склонна образовывать преимущественно родственные пары (инбридинг). Инбридингу способствует деление популяции на маленькие подгруппы, сопровождающееся частичной изоляцией этих групп, или — как у многих растений — самоопыление. Важно подчеркнуть, что фенотипические выражения рассматриваемых аллелей не относятся к числу тех, которые служат критерием при выборе партнера (неслучайное скрещивание).
Мутационный процесс и миграция
В результате мутации может произойти переход гена из одного аллельного состояния в другое. При мутировании частоты генов среди гамет не соответствуют частотам тех же генов среди родителей, за исключением тех случаев, когда число прямых мутаций (аллель А мутирует в аллель а) равно числу обратных мутаций (аллель а мутирует в аллель Л).
Изменение частоты генов в популяции может произойти также в результате введения в популяцию или выведения из нее генотипов в количестве, непропорциональном частотам соответствующих генотипов в популяции, т. е. изменения генотипической структуры
популяции. Такие процессы, как иммиграция «чужих» особей в популяцию или эмиграция членов популяции, изменяют частоты продуцируемых гамет (изменяя состав родительского поколения), если частоты генов и мигрантов отличаются от частот соответствующих генов в исходной популяции.
Отбор
Нередко в число фенотипических проявлений различных генотипов входят различия в плодовитости и жизнеспособности. Различная способность родительских генотипов участвовать в создании фонда функциональных гамет, а также различная жизнеспособность гамет и/или зигот, безусловно, изменяет частоту генов и генотипов в поколении потомков по сравнению с родительским поколением. В неодинаковой передаче генов последующим поколениям разными генотипами и заключается действие отбора, которому посвящена гл. 5. Сравнивая частоты генов в разных поколениях какой-либо популяции, нельзя игнорировать силы отбора. Однако для того, чтобы максимально упростить наши рассуждения, рассмотрим сначала гипотетический случай, полностью пренебрегая действием этих сил.
Попытаемся ответить на поставленный выше вопрос: каковы частоты генов и генотипов в поколении потомков? Рассмотрим сначала гипотетический случай наследования пары аллелей одного аутосомного локуса при условии, что генотипы образуют одну большую панмиктическую и замкнутую (без миграции) популяцию, в которой отсутствуют мутации и отбор. Короче говоря, предположим, что на популяцию не действуют никакие факторы, способные изменить частоту генов в популяции потомков по сравнению с родительской популяцией.
Очевидный, хотя и трудоемкий, подход к решению проблемы состоит в том, чтобы определить: 1) типы скрещивания в популяции; 2) частоту каждого типа скрещивания; 3) ожидаемое соотношение генотипов в потомстве от каждого типа скрещивания; 4) частоты
каждого из возможных генотипов среди всех потомков от всех вступающих в скрещивание особей родительского поколения.
Возможные типы скрещивания находят как сочетания из всех генотипов по два (самец—самка). При трех генотипах АА, Аа и аа возможны девять таких попарных сочетаний.
При случайном скрещивании частота каждого типа скрещивания зависит только от частот генотипов. Когда индивидуум совершенно случайно выбирает партнера для спаривания, то вероятность того, что партнер будет иметь некоторый определенный генотип, равна частоте этого генотипа в популяции. Таким образом, частота каждого типа скрещивания равна произведению частот участвующих в этом скрещивании генотипов. Например, среди всех возможных в популяции скрещиваний скрещивание АА X АА встречаете?! с частотой, равной х2. При этом следует иметь в виду, что частоты появления одного и того же генотипа среди самцов и среди самок одинаковы (xd'=x$). В табл. 2 представлены 9 возможных при этом типов скрещиваний и их частоты.
Таблица 2
Частота различных типов скрещивания в популяции
Самки Самцы
АА X Аа У аа г
АА ААХ АА АА X Аа АА X аа
X X2 ху XZ
Аа АаХАА Аа X Аа Аа X аа
У ХУ У2 Уг
аа аа Х АА аа X Аа аа X аа
Z XZ Уг Z2
Эти скрещивания дают потомство, состоящее из трех генотипов АА, Аа и аа, однако соотношение этих
генотипов у потомков может отличаться от их соотношения в поколении родителей. Ранее мы условились игнорировать действие отбора, т. е. мы исходим из того, что каждое скрещивание независимо от входящих в него генотипов дает одинаковое число потомков. Таким образом, доля потомков, производимых каждым из девяти типов скрещивания, соответствует частоте каждого типа скрещивания. Например, частота скрещивания АА X АА равна х2. Доля потомков, получаемых от этого отдельного скрещивания, во всем производимом популяцией потомстве составит х2. Каждый потомок от этого скрещивания будет иметь генотип АА. Частота скрещивания Аа X Аа равна у2. Среди потомков от этого скрещивания будут все три генотипа АА, Аа и аа с частотами соответственно V4p2, Ч2У2 и Ч аУ2 • Продолжая подобный расчет для всех возможных скрещиваний, можно найти общую частоту каждого генотипа среди потомков. В табл. 3 представлены: девять типов скрещивания, их частоты и доля потомков, произведенных скрещиваниями каждого типа.
Генотипы АА ожидаются в потомстве от следующих четырех типов скрещивания: ААхАА, ААХ^а, АаХ^А и АаХАа с общей частотой х24-хр-|- ЧьУ2 = = (х4-1/2#)2. Но х+72г/ равно р (уравнение 3.1); следовательно, частота генотипа АА среди всех потомков равна р2, т. е. квадрату частоты аллеля Д.
Гетерозиготы Аа возможны среди потомков 7 типов скрещивания, а общая частота генотипа Аа равна ху + 2xz + Ч2У2 + yz. Эту сумму можно представить как 2(х+ Ч2У) (Ч2У + 2) и, наконец, в виде 2pq. Следовательно, частота генотипа Аа в поколении потомков равна удвоенному произведению генных частот. Наконец, суммируя частоты потомков с генотипами аа, возникающих от скрещиваний АахАа, Аахаа и ааХаа, находим, что общая частота этого генотипа равна (Ч2У + *)2 = У2- Итак, после случайного скрещивания в одном поколении частоты генотипов, равные в исходной родительской популяции х, у, z, в популяции потомков составляют р2, 2pq, q2 соответственно.
Таблица 3
Типы скрещивания и потомки в свободно скрещивающейся популяции
Тип скрещивания Частота скрещивания Потомки
АА Аа aa
ЛЛ X аа X2 X2
АА X Аа 2ху 9 ху ху
Аа X аа 2xz !) 2ху
АА X аа аа X АА 2xz 9 2xz
Аа X Аа У2 хКу2 Ч2У2 Х1ьУ2
Аа X аа аа X Аа 2yz') У* yz
аа X аа z2 z2
Итого (x + y + z)2 = = 1 (х + Ч2у)2 = = р2 2 (х + */гУ)Х X (*/2.V + *)= = ->7 ('/2у + ^ = = <72
г) Реципрокные скрещивания объединены в один класс суммированы.
Зная частоты генотипов, можно определить частоты генов среди потомков. Как было установлено ранее, частота аллеля А равна сумме частоты гомозигот АА и половины частоты гетерозигот Аа. Пусть pi — частота гена А в поколении потомков, тогда
Pi = Р2 + Уг (2р?) = р2 + pq = р(р + <?)•
Но р А~ Я = 1, следовательно, р[ = р. Таким образом, частота гена А в популяции потомков равна частоте этого гена в популяции родителей! Естественно, что это относится и к частоте гена а. В этом нет ничего удивительного; ведь мы преднамеренно исключили из нашей теоретической модели все факторы, способные изменить частоту гена.
С другой стороны, если частоты генотипов АА, Аа и аа среди родителей равны к, у, г, то частоты тех же генотипов среди потомков от случайного скрещивания
3 Зак. 88
будут равны р2, 2pq, q2. Популяция р2: 2pq : q2 называется равновесной, поскольку в такой популяции частоты генов и частоты генотипов остаются неизменными во всех последующих поколениях. Читатель может убедиться в этом, используя описанный выше метод расчета и подставляя вместо х, у и z соответственно р2, 2pq и q2.
Рассмотрим другой, значительно более простой метод определения частот генотипов в последовательном ряду поколений. Если, как постулировалось выше, оба процесса — образование родительских пар и соединение гамет — представляют собой совершенно случайные процессы, то генотипический состав потомства можно определять более простым способом, а именно исходя только из случайного объединения всех гамет, производимых родительским поколением, и отбросив типы скрещивания. Приближение к такому общему случайному «фонду» и случайному соединению гамет представляет собой наружное оплодотворение, характерное для ряда водных форм. Под таким же углом зрения можно рассматривать и популяции других организмов, считая, что при случайном скрещивании они обладают неким общим генофондом.
Состав поколения потомков установить нетрудно — достаточно определить типы и частоты гамет, производимых предшествующим поколением, и, постулируя случайное оплодотворение, найти частоты разных сочетаний этих гамет в генотипе. Допустим, что все самцы вносят одинаковый вклад в фонд спермы, а все самки — одинаковый вклад в фонд яйцеклеток. Можно считать, что каждый фонд содержит р гамет, несущих аллель А, и q гамет, несущих аллель а. Частоты зигот АА, Аа и аа, образующихся при слиянии сперматозоидов и яйцеклеток, поступающих случайно из двух гаметных фондов, будут равны соответственно р2, 2pq, q2. Эти значения находят, умножая частоты разных гамет из фонда самок на частоты гамет из фонда самцов + q^(p# + q#)- Частоты генотипов среди потомства равны квадрату частот генов среди гамет, если =pd и q<>=q<r
Таким образом,
(р + <7)2 = Р2 + 2р/7-!-<72. (3.3)
Приведенное на фиг. 8 графическое изображение связи между частотами генов в гаметах р = 0,6 и q = 0,4
Фиг. 8. Равновесная популяция.
Диаграмма изображает равновесную’ популяцию, образующуюся при с л у* чайном объединении фондов гамет. Длина каждой из четырех линий, обра* зующих квадрат, принимается за единицу. Нижняя и левая стороны квадрата разделены каждая на две части, представляющие собой частоты генов в фонде гамет самцов и самок (р=0,6 и <7=0,4). Вертикальная и горизонтальная линии, проведенные из точек деления нижней и боковой сторон соответственно, делят квадрат на четыре части. Площади этих частей равны относительным частотам генотипов в потомстве, возникшем в результате случайного объединения мужских и женских гамет. Общая площадь квадрата равна единице, и, таким образом, сумма сгенотипов» также равна единице. Вертикальные и горизонтальные линии пересекаются в точке, лежащей на диагонали, идущей из левого верхнего в правый нижний угол. Точка пересечения рассматривается как популяционная точка, делящая диагональ на два отрезка, пропорциональные частотам генов в фонде гамет.
и частотами образующихся генотипов помогает лучше понять эти рассуждения. В этом примере потомство от случайного объединения гамет будет состоять из 0,36ДД, 0,48Ла и 0,16аа. Такое соотношение генотипов
называют равновесным, так как оно сохраняется в последующих поколениях. Если первоначальные условия остаются неизменными, то при любом значении q (и р=\—q) популяция в конечном счете приходит в некое равновесное состояние. Например, при q = 1,0 частоты генотипов составляют соответственно р2 = 0; 2pq = 0; g2=l,0. Это пример тривиального равновесия, так как в этом случае популяция будет состоять только из гомозигот аа. Если частоты генов одинаковы (р = д = 0,5), то квадрат, изображенный на фиг. 8, будет разделен на четыре равные части, каждая из которых составляет 25% всей площади. Такая популяция обладает одним замечательным свойством, отличающим ее от всех возможных равновесных популяций: она содержит максимально возможный процент гетерозигот — 50% (фиг. 9).
Популяции, имеющие одинаковые частоты генов, вовсе не обязательно идентичны по частотам генотипов. Например, при частотах генов 0,6Д и 0,4а возможны следующие четыре популяции (табл. 4).
Таблица 4
АА Аа аа Р Q
/ 0,20 0,80 0 0,6 0,4
II 0,36 0,48 0,16 0,6 0,4
III 0,50 0,20 0,30 0,6 0,4
IV 0,60 0 0,40 0,6 0,4
Хотя они отличаются по частотам генотипов, равновесное состояние всех популяций при изложенных выше условиях, наступающее в первом поколении после случайного скрещивания (а популяция // уже находится в этом состоянии), совершенно одинаково — 0,86ДД : 0,48Ла : 0,16аа (фиг. 9).
Из поколения в поколение в равновесных популяциях будет воспроизводиться один и тот же генотипический состав. Эта концепция постоянства генотипиче-
A
2pq=O,2
(p)
(q) Z
X
Фиг. 9. Частоты генотипов, представленные одной точкой в равностороннем треугольнике.
А. Высота треугольника принята за единицу, а перпендикуляры, опущенные из точки Р (называемой популяционной точкой) на все три стороны треугольника, соответствуют соотношению х, у и z (или р2, 2pq, q2). Проекция точки Р делит основание треугольника на отрезки XY и YZ, пропорциональные генным частотам р и q соответственно. Б. Четыре популяции (/, //, /// и IV) имеют одну и ту же частоту генов (47 = 0,4), однако только популяция // находится в состоянии равновесия Харди — Вайнберга. Ее популяционная точка лежит на параболе, представляющей собой локус всех равновесных популяций. Обратите внимание, что вершина параболы (обозначена светлым кружком) соответствует той равновесной популяции, в которой р-=^а=0,5 и в которой частота гетерознгот максимальна (2р<7»0,5).
70 ского состава в случайно скрещивающихся популяциях при постоянстве частоты генов известна как закон Харди—Вайнберга, названный так в честь двух исследователей, сформулировавших его независимо друг от друга в 1908 году. Этот закон является краеугольным камнем всей популяционной генетики.
АНАЛИЗ ПОПУЛЯЦИОННОЙ ВЫБОРКИ
Вернемся к примеру с М—N группами крови человека (табл. 1) и определим, совпадают ли частоты, наблюдаемые в случайной выборке коренных жителей Австралии, с частотами, ожидаемыми в случайно скрещивающейся равновесной популяции, согласно закону'Харди—Вайнберга. Если группу в 730 индивидуумов можно считать представительной выборкой и если популяция действительно находится в равновесии, то частоты, полученные для этой выборки (р = 0,178, q = 0,822), будут представлять собой истинные частоты генов в популяционном генофонде. Больше того, ожидаемые частоты генотипов, определявшиеся в соответствии с законом Харди—Вайнберга как квадрат генных частот, равны
(о, 178 LM + 0,822LN)2 = 0,0317LMLM + 0,2926LMLN +
+ 0,6757LnLn.
В табл. 5 представлено фактическое распределение генотипов среди 730 индивидуумов и ожидаемые частоты каждого генотипа при условии случайного скрещивания.
Генные частоты оценены по случайной выборке, поэтому они не абсолютно точно совпадают с фактическими частотами генов во всей популяции. Эти оценки использовались для определения доли каждого генотипа в популяции в предположении равновесия Харди—Вайнберга. Обычно для установления степени соответствия между теоретическим ожиданием и фактическими частотами, наблюдаемыми в выборке, используется критерий х2. В данном случае сравнение наблюдаемых в выборке и ожидаемых частот геноти
пов дает х2 = 0,083, что при одной степени свободы соответствует очень хорошему согласию (вероятность совпадения равна 0,76). Следовательно, мы вправе считать, что скрещивание членов популяции происходит совершенно случайно (панмиктически) по отношению к группам крови М—N. Это представляется правдоподобным еще и потому, что очень немногие люди знают, к какой они принадлежат группе по системе М—N, а если и знают, то едва ли принимают во внимание этот признак при выборе будущего супруга.
Таблица 5
Наблюдаемое и ожидаемое распределение генотипов по группам крови
Генотип Наблюдаемая частота Ожидаемая ч
22 23,1
lmln 216 213,6
£nLn 492 493,3
Всего . 730 730,0
Достоверное несовпадение с теоретически ожидаемым значением свидетельствует либо о том, что популяция не панмиктична, либо о том, что в ней действует какой-то другой из упомянутых выше «возмущающих» факторов.
Может показаться, что ожидаемые величины все-гда должны точно отражать наблюдаемые значения, поскольку первые вычисляются по генным частотам, найденным из вторых. Однако это не всегда так, в чем можно убедиться, изучив гипотетическую «популяцию», о которой априори известно, что она не является панмиктической. Примем условно, что суммарные данные, полученные для популяции коренных жителей Австралии и индейцев племени Навахо, на самом деле относятся к одной гипотетической популяции (табл. 6).
Таблица 6
Частота групп крови в гипотетической популяции
Популяция м M-N N Всего
Австралийцы 22 216 492 730
Индейцы племени 305 52 4 361
Навахо
Гипотетическая по- 327 268 496 1091
пуляция
Что представляют собой частоты генов в этой искусственной популяции? Что представляют собой частоты и ожидаемые численности генотипов при условии случайного скрещивания и других допущениях Харди—Вайнберга? Проверка с помощью критерия х2 свидетельствует о том, что наблюдаемые отклонения вряд ли могут быть чисто случайными. Однако нужно помнить, что этот критерий не дает ответа на вопрос о том, какой именно фактор (неслучайность скрещивания, отбор, миграция и т. п.) повинен в отсутствии совпадения.
Помимо хорошего совпадения с моделью Харди— Вайнберга, распределение по группам крови М—N удобно использовать для демонстрации равновесной популяции еще и потому, что все три генотипа фенотипически различны и, следовательно, для повышения точности оценки генных частот можно использовать данные о частоте гетерозигот М—N. Если же рассматриваемый признак контролируется рецессивным геном, то гетерозиготы неотличимы от гомозигот по доминантному гену и составляют с ними один фенотипический класс; в этом случае для оценки генных частот используют данные о частоте гомозигот по рецессивному признаку и допускают наличие генетического равновесия (z = q2). Например, такой рецессивный признак, как фенилкетонурия, встречается у одного человека из 40 000. Предполагая равновесие популя
ции по этому признаку, находим, что частота пораженных индивидуумов равна q2, т. е. что q2 = 1/40 000. Частота рецессивного аллеля равна квадратному корню из частоты генотипа: q = 1/200. Следовательно,
Генофонд спермы
А р=0,95
А
р = 0,95
q = 0,05
а
q = 0,05
Фиг. 10. Пример равновесной популяции, в которой один аллель (а) является редким и находится преимущественно в гетерозиготном состоянии.
В этом случае д=0,05 и частота гетерозигот в 34)8 раз больше частоты гомозигот по редкому аллелю (аа).
частота доминантного аллеля равна (1—q) = = (1 — 1/200) = 199/200. Пропорции генотипов при равновесии находят путем разложения бинома (Р + <7)2: 2_/199 \2_ 39 601 .
Р ~ \ 200 ) ~ 40 000 ’
2 =9(199V 4= 398 • 2 = ( 1 V= 1
\ 200 / \ 200 J 40000 ’ * \ 200 ) 40 000 *
В подобных случаях, когда прямое сравнение теоретических и наблюдаемых значений провести нельзя и невозможно установить закономерности образования пар, исходным моментом в определении генетических параметров служит допущение о наличии генотипического равновесия.
На этом примере можно продемонстрировать еще одну особенность: если аллель редкий, то большинство носителей этого аллеля — гетерозиготы (фиг. 10). Так, в случае фенилкетонурии частота носителей соответствующего гена (гетерозигот) в 398 раз больше частоты больных индивидуумов (гомозигот). Гены, контролирующие еще более редкий рецессивный признак, еще чаще оказываются в гетерозиготном состоянии. Например, такой рецессивный признак, как алькап-тонурия — нарушение обмена веществ, связанное с отсутствием определенного фермента, — встречается примерно у одного человека на миллион. Отсюда частота рецессивного гена, контролирующего это заболевание, равна примерно 0,0001, а отношение гетерозигот к гомозиготам по этому гену равно 1998:1. Это ясно выражает тот факт, что редкие аллели существуют преимущественно в гетерозиготном состоянии. Отсюда следует, что у большинства людей, имеющих редкий рецессивный признак, родители фенотипически нормальны, хотя и гетерозиготны.
МНОЖЕСТВЕННЫЕ АЛЛЕЛИ
Законы Харди—Вайнберга остаются верными и для множественных аллелей, если все рассмотренные выше условия сохраняются. Рассмотрим локус с тремя аллелями Alt Л2, А3 с частотами соответственно р, q и г, где p + q + r=\. Генотипическая структура равновесной популяции имеет вид
p’GMO + 2р<?(Л1Л2) + 2рг(Л,Д3) +
4- q2 (А2А2) + 2qr (Д2Д3) + г2 (43Д3) где частоты генотипов представляют собой члены разложения квадрата суммы частот генов (р + q + г)2. Принцип установления равновесия в неравновесной
популяции уже в первом поколении от случайного скрещивания относится в той же мере к многоаллельному локусу, как и к двухаллельному. На фиг. 11 представлены закономерности наследования для трех
Генофонд спермы
Фиг. И. Множественные аллели в равновесной популяции.
Частоты аллелей А,, А2 и А3 в каждом гаметном фонде равны соответственно: р=0,3; <7=0,5 и г = 0,2. Частоты генотипов представлены частями квадрата, которые отвечают слагаемым, получающимся при возведении в квадрат гаметного ряда.
аллелей (р = 0,3; q = 0,5 и г = 0,2). Равновесное состояние в этом случае также наступает уже в первом поколении от случайного скрещивания и случайного объединения гамет.
Если все генотипы фенотипически различны, то частоты генов для множественных аллелей найти нетрудно. Частота какого-либо аллеля равна частоте гомозигот по этому аллелю плюс половина частоты каждого типа гетерозигот по этому аллелю. Таким
ГЛ АР A 3
образом, в случае трех аллелей р = р2 + pq + рг, q = q2 4- pq 4- qr, г = г2 + pr + qr.
Иногда нас интересует не вся серия аллелей, а только один ее член. В этом случае нужно рассматривать множественную группу как пару аллелей. Если частота одного аллеля, скажем аллеля А2, равна 9, тогда частота всех остальных аллелей равна 1 — q; отсюда по биному Харди—Вайнберга получаем
[q + (1 - q)]2 = q2 + 2q (1 - q) + (1 - qf.
РАВНОВЕСНЫЕ ПОПУЛЯЦИИ И ПОЛ
Закон Харди—Вайнберга исходит из предположения о равенстве частот генов среди самок и среди самцов. Если генные частоты самцов и самок различны, то равновесие наступает через два поколения — в первом поколении происходит уравнивание пропорций аллелей в обоих полах, а уже в следующем наступает генотипическое равновесие. Графически это представлено на фиг. 12. В исходных поколениях сперма содержит 80% гамет с аллелем Л, а среди яйцеклеток таких гамет только 40%. Точка пересечения вертикальной и горизонтальной линий, которые делят квадрат на части, соответствующие потомству первого поколения, не лежит на диагонали квадрата. Популяция не достигла равновесия. Равновесие наступает в следующем поколении при р = 0,6. На графике эта величина определяется как середина отрезка диагонали, заключенного между точками пересечения ее с горизонтальной и вертикальной линиями. Проще говоря, равновесная частота данного аллеля равна полусумме частот этого аллеля среди мужских и женских гамет.
Локус, аллели которого располагаются в половых хромосомах, называется сцеплением с полом. Рассмотрим случай, когда самки гомогаметны (XX), а самцы гетерогаметны (XY). Х-хромосомы распределяются от-
носйтельно пола непропорционально — две трети среди самок и одна треть среди самцов; поэтому и соответствующие аллели в популяции распределяются
Генофонд спермы
Фиг. 12. Генотипический состав популяции в случае, когда самцы и самки различаются по частоте генов.
Если провести диагональ из левого верхнего угла, то окажется, что точка пересечения горизонтальной й вертикальной линий не лежит на этой диагонали, поскольку частоты генов среди самцов и среди самок неодинаковы (q^^q^). Популяционная точка будет лежать на диагонали только при достижении равновесия; ее точное местоположение —середина между точками пересечения диагонали с вертикальной и горизонтальной линиями. Точка, расположенная на 1/3 этого расстояния, используется для случая локуса, сцепленного с полом (белый кружок на диагонали).
относительно неравномерно. Проследим за изменением генотипического состава популяции, имеющей следующие генотипы:
Самки Самцы
Генотипы А А Аа аа А а
Частоты х у z т п
При определении генотипических и генных частот сцепленных с полом локусов каждый пол
рассматривается как отдельная субпопуляция. Частота гена среди самок определяется так же, как в случае аутосомных генов, т. е. = х + ЧъУ, а среди самцов частоты генов равны частотам генотипов pd = tn и q& = п. Если q$ = q&, значит, популяция находится в состоянии равновесия или же достигнет равновесного состояния в следующем панмиктическом поколении. Если в популяции численность самцов равна численности самок, то частота аллеля а во всей популяции
ЧТ = М + '/з^-
Популяция не является равновесной, если q^ =И= qd Такая популяция не достигает равновесия ни в первом, ни во втором поколении, но тем не менее на протяжении ряда поколений она быстро приближается к этому состоянию. Как и в случае с аутосомным локусом, общая величина qT в популяции, приближающейся к равновесию, остается неизменной. Однако частота отдельных генов у каждого пола (q$ и q^ колеблется от поколения к поколению. Это связано с тем, что qd для каждого поколения равно q$ для предшествующего поколения, a q$ для любого поколения равно V2?$ + ЧъЯ# для предшествующего.
Для иллюстрации этих закономерностей можно вновь использовать фиг. 12, но с учетом особенностей локуса, сцепленного с полом. Генофонд самцов: /^ = 0,8 и q& = 0,2, а генофонд самок: р?=0,4 и <7? = О,6.
Исходя из этого, находим
?г = 2/з(0,6) + 7з(0,2) = 0,467, отсюда
рт = 0,533.
График демонстрирует случайное объединение мужских и женских гамет. Отдельные части квадрата в данном случае соответствуют частотам генотипов
только у самок(ЛД? =0,32, Да9=0,56 и аа?=0,12). Частота аллелей а среди потомков женского пола <7' = = 0,12 + V2 (0,56) = 0,4. Частоты генотипов у самцов (т и п) можно представить как длины двух отрезков, составляющих правую сторону квадрата. Две прилегающие к этим отрезкам части квадрата также соответствуют частотам генов у потомков мужского пола (Pd и ?*)• Очевидно, частоты генотипов среди потомков мужского пола (р^и^) равны частотам генов среди их матерей и 7$), что видно из равенства соответствующих отрезков на сторонах квадрата.
К чему приведет случайное скрещивание в одном поколении в этом случае? Мы рассмотрим здесь только частоту q, поскольку р равно 1—q.
Поколение родителей
<7? = 0,6 ^=0.2 qT = 0,467
Поколение потомков /s = 0,4 q'g = 0,6 q'T = 0,467
Общая частота qr гена остается неизменной, так что qT = q'T- Равновесное значение q («ку с крышкой») частоты аллеля а будет равно исходной величине q-r-На фиг. 12 равновесная частота находится в точке (показана белым кружком), расположенной на отрезке диагонали, заключенном между пересекающимися горизонтальной и вертикальной линиями; от горизонтальной линии эта точка лежит на расстоянии, равном 7з расстояния между точками пересечения диагонали с горизонтальной и вертикальной линиями.
В поколении родителей q^ меньше равновесного значения (0,467), a q'^ — больше. Для самок справедливо обратное > 0,467 > </'). В последующих поколениях генные частоты у обоих полов колеблются, становясь то выще, то ниже общей частоты данного
гена, остающейся неизменной. В этом и состоит колебательный характер упоминавшегося выше изменения генных частот.
Разность между значениями q'& и q? у потомков равна половине разности между qd и q? в поколении родителей, т е. за каждое поколение разница в частоте гена у двух полов уменьшается вдвое, приближаясь в конце концов к нулю. Популяция достигает равновесия.
МНОЖЕСТВЕННЫЕ ЛОКУСЫ И ГЕНЕТИЧЕСКОЕ РАВНОВЕСИЕ
Все аутосомные локусы, рассматриваемые независимо, достигают равновесия в отношении частоты генотипов уже в первом поколении от случайного скрещивания. Но если два локуса или больше сцеплены, то полифакториальные генотипы, такие, как ААВВ, AABb, ААЬЬ, АаВВ, достигают генетического равновесия лишь постепенно, как правило, в течение не одного, а нескольких поколений. Это достаточно пояснить на простом примере. Рассмотрим альтернативные аллели, относящиеся к двум независимо расщепляющимся локусам (Д, а и В,Ь), где частоты соответствующих аллелей равны р(Д), q(a), r(B), s(b). Частоты монофакториальных генотипов при равновесии равны р2(АА) :2pq(Aa) :q2(aa) для локуса А и r2(BB) :2rs(Bb) :s2(bb) для локуса В. Рассматриваемые совместно пропорции бифакториальных генотипов задаются выражением (p2 + %pq + q2)(r2-]-4-2rs-f-s2), которое получается возведением в квадрат частот гамет АВУ АЬУ аВ и ab. Если преобладает равновесие, то частота гамет каждого типа является функцией частот содержащихся в гаметах аллелей. Тогда, если частоты гамет обозначить следующим образом
Тип гамет АВ Ab аВ ab
Частота h i j k,
то при равновесии h = pr; i = ps\ j = qr\ k = qs. Кроме того, находим, что при равновесии hk = ip
В качестве численного примера рассмотрим популяцию, имеющую следующие частоты гамет*.
Тип гамет АВ Ab аВ ab
Частота h = 0,40 i = 0,30 / = 0,20 k = 0,1
Генные частоты в этом случае имеют следующие значения: р = 0,7; q = 0,3; г = 0,6; s = 0,4. Популяция не находится в равновесии относительно рассматриваемых нами аллелей. Это следует из того, что, во-первых, частоты гамет не равны теоретически ожидаемым, рассчитанным из генных частот, и, во-вторых, равенство hk = ij не выполняется. Разность hk— ij в случае двух несцепленных локусов уменьшается за каждое поколение вдвое. Это продолжается до достижения точки равновесия, в которой hk — ij = 0. Когда популяция достигнет равновесия, частоты гамет будут иметь следующие значения:
Тип гамет АВ Ab аВ ab
Частота h=pr = 0,42 Z = ps = 0,28 / = qr = 0,18 k = qs = 0,l2
Чтобы пояснить идею достижения равновесного состояния в случае двух локусов, рассмотрим результаты последовательного ряда скрещиваний в двух упрощенных ситуациях.
Рассмотрим сначала популяцию, содержащую в исходном поколении только гетерозиготы АаВЬ. Эти особи будут давать четыре типа гамет, частоты которых, допуская несцепленность локусов, одинаковы (VMB, '/^АЬ, xhaB, 1/4аЬ). В результате случайного объединения гамет возникнет 9 генотипов, возможных при двух локусах по два аллеля в каждом. Частоты этих генотипов будут соответствовать частотам, ожидаемым в F2 для обычного менделевского расщепления при дигибридном скрещивании. В следующем поколении четыре типа гамет будут вновь воспроизводиться с одинаковой частотой, и случайное их объединение даст те же генотипы и в том же соотношении, что в предыдущем поколении. Следовательно, в этом случае равновесие достигается уже за одно поколение случайного скрещивания,
Рассмотрим второй простейший пример. Допустим, что исходная популяция содержит в равном числе особей ААВВ и aabb. В такой популяции образуется только два типа гамет ('/2АВ и x/zab). При случайном объединении они дадут только три генотипа ААВВ,
Фиг. 13. Приближение двух сцепленных лэкусов к равновесию.
Графическое изображение зависимости разности (d) между частотами гамет в «фазе отталкивания» и в «фазе сцепления» от числа поколений. Исходные популяции содержат в равном числе индивидуумы ААВВ и aabb. Пять кривых отражают различную скорость рекомбинации (R) между двумя локусами. di/dQ — доля сохраняющегося исходного различия.
АаВЬ и aabb в соотношении 1:2:1. В следующем поколении фонд гамет будет содержать 3/зАВ, х1зАЬ, х1%аВ и 3/e ab. В третьем поколении соотношение гамет составит 5/|бЛВ; 3/16Л&; 31\заВ\ 5/^ab. Таким образом, из поколения в поколение увеличивается частота гамет, находящихся в фазе отталкивания (т. е. АЬ и аВ), и пропорционально снижается частота гамет в фазе сцепления (т. е. ДД и ab), пока частоту всех гд-
мет не станут одинаковыми, т. е. пока не наступит равновесие.
Поскольку в первом поколении образуются только гаметы в фазе сцепления, величины hk и ij не равны и d = V4; во втором поколении d = 7в, т. е. (3/в • 3/в) — — (7e* 7s); в третьем 716» в четвертом 7зг и т. д., уменьшаясь вдвое в каждом последующем поколении вплоть до наступления равновесия.
Равновесие достигается значительно быстрее, если локусы не сцеплены (/? = 7г). Если же оба локуса расположены в одной хромосоме, скорость (число поколений) наступления равновесия обратно пропорциональна силе сцепления (фиг 13).
Из этого вытекает важное следствие: при равновесии независимо от силы сцепления частота гамет, находящихся в фазе сцепления и в фазе отталкивания, будет одинакова. Это значит, что в равновесной популяции не наблюдается специфической связи между признаками, которые контролируются аллелями сцепленных локусов. Таким образом, совместное появление в популяции каких-либо признаков может быть обусловлено аллелями, принадлежащими к разным локусам и находящимися в разбалансированном состоянии в результате недавно просочившихся в популяцию генетических примесей. Кроме того, это явление может быть обусловлено плейотропным действием аллелей, относящихся к одному локусу. Подобно доминированию, сцепление нельзя обнаружить непосредственно из популяционных данных; для этого необходимы специальные эксперименты с применением инбридинга.
ДРЕЙФ ГЕНОВ
Абсолютное равновесие может быть достигнуто только в бесконечно большой популяции случайного скрещивания. Это неоднократно подчеркивалось в предыдущих разделах. Однако в действительности популяция в лучшем случае только приближается к равновесию, поскольку любая природная популяция, как бы велика она ни была, все же ограничена по величине. Вспомним, что особи, составляющие данное
поколение, всегда образуются из 2W гамет предшествующего поколения. Гаметы абсолютно точно воспроизводят генофонд родительского поколения только в том случае, если JV— величина бесконечно большая. Если же W — величина конечная, то гаметы несут лишь выборку из генофонда родительского поколения, которая, как всякая выборка, подвержена случайным колебаниям. Чем меньше численность популяции и соответственно меньше выборка гамет, тем больше размах возможных колебаний состава выборки.
Частота гена (q) может случайно изменяться в каждом последующем поколении, оказываясь то больше, то меньше соответствующего значения в предшествующем поколении. Хотя направление и точная величина этих изменений не детерминированы, мы можем оценить вероятность того, что это изменение будет иметь данную величину или будет заключено в данных пределах. Случайное изменение частоты гена обозначают греческой буквой «дельта» (6). Изменение частоты гена q можно обозначить как bq, где bq = q\ — qQ (числовые индексы означают поколения). Для всех значений bq, возможных для данного признака в одном поколении некоторой ограниченной популяции, можно построить теоретический график распределения частот. Число точек на любом таком графике равно 2W+ 1» т. е. ограничено только величиной популяции (значением AQ. Изменение частоты гена, обусловленное ошибкой выборки, выражается изменением формы графика. График можно описать с помощью таких статистических понятий, как среднее значение изменения генной частоты (bq) и дисперсия (cf|7). Это среднее равно нулю, поскольку положительные и отрицательные отклонения равновероятны. Дисперсия за одно поколение
равна квадрату стандартного отклонения
а ___ 1/ РоЯо
у 2N •
С уменьшением численности популяции (2V) дисперсия возрастает. Чем меньше популяция, тем больше вероятность того, что частота гена будет подвержена ошибке выборки.
Пусть частоты аллелей Айа аутосомного локуса в панмиктической популяции р0 = qQ = 0,50, Положим далее, что в следующем поколении численность популяции ограничена 50 особями. Поскольку &q = 0, средняя величина возможных значений qi равна 0,5, а стандартное отклонение
0,5-0,5
V 2-50
= 0,05.
Распределение вероятностей всех возможных значений q (частота в поколении потомков) приближается к нормальному распределению со средним значением 0,5. Вероятности различных отклонений в нормальном распределении можно найти в соответствующих статистических таблицах. Так, например, вероятность того, что отклонение вправо или влево от среднего значения будет больше одного стандартного отклонения (6), приблизительно равна 0,32. Это значит, что с вероятностью 32% q\ будет больше 0,55 или меньше 0,45. Подобные случайные колебания частоты генов продолжаются и в последующих поколениях. Эти частоты как бы смещаются к некой определенной точке, хотя явно выраженной траектории при этом не обнаруживается. Конечная судьба аллеля— его закрепление (q = \) или потеря (q = 0). Как только частота аллеля станет равной нулю или единице, дальнейшие изменения в данном локусе прекращаются; все члены популяции становятся гомозиготными по этому локусу (гомоаллельная популяция), если, конечно, утраченный аллель не появится вновь в результате мутаций или иммиграции. Таким образом, будет ли данный аллель потерян или же он закрепится в популяции, зависит от случая. В очень маленькой популяции все локусы стремятся к гомоал-лелизму, что ведет к снижению гетерозиготности и утрате изменчивости.
4
Величину можно рассматривать также как дисперсию частоты генов или в первом поколении после случайного скрещивания среди многих популяций, равных по численности и одинаковых по исходной частоте генов. Эта величина имеет совершенно тот же смысл, что и рассмотренная выше дисперсия возможных изменений частот генов в одной популяции; разница состоит лишь в том, что теперь мы рассматриваем дисперсию частот генов в нескольких популяциях. Пусть имеется множество лабораторных популяций, одинаковых по численности и исходной частоте генов. В следующем поколении частота генов в каждой из этих популяций изменяется и приобретает значение q\. Эти новые значения будут различными, и, таким образом, произойдет случайное распределение частот среди популяций. Средняя частота гена во всех популяциях будет равна исходной величине В этом случае дисперсия служит количественной мерой дивергенции популяций по частоте генов (фиг. 14).
С каждым поколением дисперсия увеличивается и в некотором поколении t ее величина равна
< = ад.[1 -(| (3-5>
В конце концов, когда t становится очень большим, все популяции становятся гомоаллельными, а дисперсия
Фиг. 14. Разброс генных частот среди 400 гипотетических популяций (моделирование на быстродействующей вычислительной машине методом Монте-Карло).
Исходные допущения модели: 1) каждая популяция в каждом поколении содержит только 8 диплоидных особей —4 случайно образованные родительские пары; 2) каждый индивидуум спаривается один раз, а число потомков, образующихся от каждого спаривания, распределяется по закону Пуассона со средней равной двум; 3) отбор не действует и мутаций не происходит; 4) каждая популяция имеет следующий исходный состав: 2АА, 4Аа, 2аа, так что исходные частоты каждого аллеля для одного аутосомного локуса каждой популяции равны 0,5. Было смоделировано поведение 400 таких популяций на протяжении 32 поколений, на диаграмме показано распределение популяций по частоте генов в 1-, 2-, 4-, 8-, 16-, 32-м поколениях. Вследствие случайной изменчивости в таких небольших популяциях для бодьщинства из них характерна повышенная скорость фиксации того иди иного алледд.
принимает максимальное значение o2q = pQqQ. Иначе говоря, в одних популяциях происходит закрепление аллеля a(q = \), а в других — полная элиминация (q = 0) этого аллеля. Отношение числа первых к числу вторых прямо пропорционально отношению ро: q0. Как видно из уравнения (3.5) дисперсия в данном поколении зависит от величины популяции, числа прошедших поколений и исходной частоты генов. В табл. 7 показана эта зависимость для популяций с q = 0,5.
Таблица 7
Дисперсия в популяциях различной величины на протяжении ряда поколений при ро = qQ = 0,5 1
Число поколений Дисперсия в популяциях разной величины
6 10 50 100 500 1000
1 0,02 0,01
2 0,04 0,02 0,01
3 0,06 0,04 0,01 0,01
4 0,07 0,05 0,01 0,01
5 0,09 0,06 0,01 0,01
10 0,15 0,10 0,02 0,01
50 0,25 0,23 0,10 0,06 0,01
100 0,25 0,25 0,16 0,10 0,05 0,01
*) Предполагается, что численности особей постоянны, а поколения дискретны.
Усреднение частот генов для нескольких субпопуляций показывает, что средняя частота остается неизменной в ряду поколений, а дисперсия частот увеличивается. Если же рассматривать отдельную популяцию из группы, то здесь генная частота q колеблется от поколения к поколению, пока не произойдет либо закрепление аллеля, либо полная его элиминация. Это явление случайных колебаний, обусловленных
ошибкой выборки, называют дрейфом генов (значение дрейфа генов для структуры природных популяций рассматривается в гл. 6).
ИНБРИДИНГ
Одно из главных допущений модели Харди—Вайнберга — случайность скрещивания в больших популяциях — в реальных популяциях встречается очень редко. Обычно скрещивание происходит между особями, находящимися поблизости друг от друга. В результате большая популяция оказывается более или менее подразделенной на небольшие инбредные субпопуляции; степень этого подразделения зависит от размеров области, занимаемой популяцией, и от подвижности индивидуумов. В предыдущем разделе мы видели, что в малых популяциях генные частоты подвержены дрейфу, обусловленному ошибкой выборки гамет. В результате этого наблюдается тенденция к закреплению в каждом локусе какого-либо одного аллеля. Следовательно: 1) в небольших популяциях наблюдается снижение генетической изменчивости (увеличивается генотипическое сходство индивидуумов); 2) поскольку доля гетерозигот при средних значениях частот генов больше, чем при крайних значениях частот, характерных для случайных изменений, в малых популяциях наблюдается снижение общей гетерозиготности; 3) поскольку дрейф генов — процесс случайный, происходит дифференциация родственных субпопуляций; одни будут содержать только широко распространенный аллель, другие — только редкий.
Мы уже излагали идею о случайных изменениях частот генов в терминах ошибки выборки. Эту идею можно изложить в понятиях инбридинга. Ясно, что ограничение возможностей выбора партнера в малых субпопуляциях увеличивает родство особей, вступающих в скрещивания. Хотя скрещивания могут быть случайными (фенотипически не ассортативными), в небольших популяциях сходство между скрещивающимися особями выше, чем в больших. Генетическое
значение инбридинга состоит в постепенном увеличении гомозиготности. В связи с этим важно указать, что в инбредной популяции редкие рецессивные гены оказываются в гомозиготном состоянии чаще, чем в большой панмиктической популяции. Именно поэтому в животноводстве и растениеводстве широко применяется систематический инбридинг с целью получения гомозиготных особей для последующего разведения их в чистоте (см. Д. Брюбейкер, «Сельскохозяйственная генетика»1).
Популяция, подвергнутая постоянному инбридингу (многократное самооплодотворение, скрещивание сибсов и т. п.), оказывается разделенной на ряд репродуктивно независимых линий (субпопуляций). В каждой отдельной линии увеличение гомозиготпости сопровождается изменениями частоты генов и тенденцией к закреплению в каждом локусе какого-либо одного аллеля. Таким образом, происходит дивергенция линий, имеющих общее происхождение, по частоте генов. Но если все без исключения инбредные субпопуляции (считая, что не происходит ни потерь, ни отделения) объединить и рассматривать как одну большую «популяцию», то окажется, что влияние инбридинга не зависит от средней частоты гена во всех линиях. Следовательно, степень инбридинга лишь определяет, каким образом аллели комбинируются в генотипы — в инбредных линиях доля гомозигот увеличивается.
Это можно продемонстрировать на примере изменений генотипического состава локуса (аллели А и а) на протяжении ряда поколений в «популяции» со строгим самооплодотворением. Каждая из гомозигот АА или аа производит в отношении анализируемого
1 Автор постоянно ссылается на другие книги этой серии, выпущенные на русском языке издательством «Мир»: Дж. Джинкс, «Нехромосомная наследственность» (1966 г.); В. Маккьюсик, «Генетика человека» (1967 г.); Ф. Сталь, «Механизмы наследственности» (1966 г.); К. Суонсон, Т. Мерц и В. Янг, «Цитогенетика» (1969 г.); Ф. Хартман и 3. Саскайнд, «Действие гена» (1966 г.). Книга Д. Брюбейкера «Сельскохозяйственная генетика» вышла в издательстве «Колос» в 1966 г. — Прим. ред.
локуса только один тип гамет. Объединение двух гамет, производимых одним и тем же гомозиготным индивидуумом, если не считать редкие случаи мутаций, дает тот же самый (родительский) генотип.
Гетерозиготы дают гаметы двух типов, А и а, и, таким образом, производят три класса потомков АА, Аа, аа, появляющихся со средними частотами соответственно 74: 7г : 74. Поскольку перекрестное оплодотворение невозможно, гомозиготы не могут давать гетерозиготное потомство; что же касается потомков гетерозигот, то из них лишь половина также гетерозиготна. Иными словами, частота гетерозигот среди потомков вдвое ниже, чем среди родителей. Одновременно происходит соответствующее увеличение часто* ты гомозигот, в равных долях для каждого типа. При постоянном самооплодотворении снижение гетерозиготности продолжается до тех пор, пока все генотипы не станут гомозиготными.
Проследим за изменением генотипического состава панмиктической популяции, находящейся в равновесии при р = q = 7г, которая внезапно и полностью переходит на такой способ размножения, как самооплодотворение. Частоты генотипов в исходной популяции равны 7|ЛЛ х^Аа: х^аа. Половина потомков в следующем поколении будет получена от гомозигот двух классов (7^^ и xhaa), а другую половину потомков дадут гетерозиготы; потомство гетерозигот распределится в каждой семье в отношении 1:2:1 или Х1%АА : х^Аа: х!%аа. Таким образом, суммарные частоты генотипов в первом поколении будут равны 3!%АА, 21ъАа, 3/8аа. В табл. 8 представлены частоты генотипов, образуемых при самооплодотворении в ряду последовательных поколений. Видно, что частота гетерозигот за каждое поколение уменьшается вдвое.
При наиболее интенсивной форме инбридинга — самооплодотворении — полная гомозиготность достигается очень быстро. Как упоминалось выше, инбридинг может иметь место и в раздельнополых популяциях, если скрещивающиеся особи находятся в родстве (скрещивания между сибсами, полусибсами, двоюродными сибсами). Скрещивания между родствен-
ними особями также снижают гетерозиготность, но это происходит значительно медленнее, чем при самооплодотворении (фиг. 15).
Таблица 8
Снижение гетерозиготности при систематическом самооплодотворении ’)
(исходная популяция находится в равновесии: р = q = ’/2)
Поколения Частоты генотипов F Q
АА Аа аа
0 '/< Чг ч< 0 Чг
1 7» 7. Чг Чг
2 7/16 7» 7и Ч< Чг
3 *6/з2 7,в 7з2 Ч, Чг
4 3'/«. 732 3761 ,5Л« Чг
п 1 ~ (7г)п (72)п 1 - Шп 1 - шп Чг
2 2
оо 7г 0 Чг 1 Чг
’) Инбридинг не влияет на частоты генов: генный состав всех потомков от самооплодотворения остается неизменным. Коэффициент инбридинга F растет из поколения в поколение до тех пор, пока все индивидуумы не станут гомозиготными. Поскольку исходная частота каждого гена равна */а, то п( и F=1 доли двух типов гомозигот будут равны. Обратите внимание, что с увеличением числа поколений инбридинга приближение F к предельной величине (F = l) становится все медленнее.
Степень инбридинга в популяции измеряется обычно коэффициентом инбридинга Райта F, который выражает потерю гетерозиготности. В табл. 9 представлены соотношения генотипов в популяциях с различной степенью инбридинга (см. также значения F в ряду поколений при самооплодотворении в табл. 8). Общая формула Райта для определения отношения равновесных частот АА:Аа:аа имеет вид
р2 + Fpq : 2pq — F2pq : q2 + Fpq.
Видно, что для данной степени инбридинга частота гетерозигот уменьшается на величину F2pq, а частота каждого типа гомозигот увеличивается на поло-
вину этой величины. В панмиктической популяции коэффициент инбридинга F равен нулю, и поэтому формула Райта, выражающая частоты генотипов, сводится к формуле Харди—Вайнберга.
Коэффициент инбридинга не только выражает среднюю степень инбридинга всех членов популяции
Фиг. 15. Процент гомозиготных потомков при систематических скрещиваниях с разной степенью родства.
I — самоопдодртворение; // — скрещивание между сибсамц’» /// — скрещивание между двойными (и по отцу и по матери) двоюродными сибсами; IV — скрещивание между двоюродными сибсами; V — скрещивание мейсду троюродными сибсами.
в определенном поколении, но и служит мерой вероятности идентичности аллелей у отдельной инбредной особи.
Под идентичностью аллелей мы понимаем идентичность двух аллелей одного локуса диплоидного организма по происхождению, т. е. тот случай, когда гаметы, образующие зиготу, получены от родственных по происхождению родительских особей и содержат аллели, возникшие в результате удвоения одного гена общего предка. Поясним это подробнее. В диплоидном организме каждый аутосомный локус
представлен дважды. Соответствующие два гена могут быть представлены одинаковыми или разными аллелями — гомозиготное состояние в первом и гетерозиготное во втором случае. Хотя два аллеля гомозиготной особи сходны по структуре и функции («одинаковое состояние локуса»), они не обязательно должны происходить от одного предкового аллеля: если эти аллели происходят из разных источников (от одинаковых аллелей неродственных особей), их называют независимыми; если же они представляют собой реплики одного и того же аллеля, существовавшего в одном из предшествующих поколений, их называют идентичными. Гомозиготы могут быть независимыми или идентичными. Величина F есть ожидаемая доля идентичных гомозигот. Она показывает, насколько уровень гомозиготности превышает тот, который был бы достигнут, если бы все скрещивающиеся особи были неродственными и содержали независимые аллели.
Таблица 9
Частоты генотипов в случайно скрещивающейся популяции в отсутствие инбридинга, при частичном инбридинге и при полном закреплении
Поколения инбридинга F Частоты генотипов
АА Аа аа
Ни одного (формула Харди—Вайнберга) F = 0 р2 ^РЯ я2
Одно или больше (формула равновесия Райта) 1 >F>0 р2 + Fpq %ря — 2Fpq Я2 + Рря
Бесконечное множество поколений (полная гомозиготность) F = 1 р2 + р<1 0 я2+ ря
Концепция идентичности аллелей придает ясность понятию инбридинга. Среднюю величину инбридинга в популяции (приведенную выше) можно выразить в
понятиях идентичности аллелей, представив соотношения Райта в ином виде:
Независимые Идентичные
АА р2 (1 — F) + Fp
Аа 2pq (1 — F)
аа q2 (1 — F) + Fq
Как мы видим, та часть популяции, которая обозначена F, состоит из идентичных гомозигот, а остальная часть (1 —F) — из гетерозигот и независимых гомозигот. Здесь уместно подчеркнуть, что все особи некоторой популяции в какой-то степени родственны между собой. Поэтому, для того чтобы коэффициент инбридинга имел смысл, нужно рассматривать родословную (особи или популяции) лишь до какой-то определенной точки в прошлом, за которой идентичность аллелей уже не прослеживается. Та точка родословной, за которой все аллели считаются независимыми, называется основанием. В большинстве исследований основанием служит или большая панмиктическая популяция или одна или несколько особей, совершенно случайно выбранных из этой популяции.
Можно вычислить коэффициент инбридинга ограниченной панмиктической популяции при тех или иных определенных условиях. Формула для определения степени инбридинга в данном поколении и связанного с инбридингом снижения гетерозиготности вполне сравнима с уравнением (3.5) (подробности см. Штрик-бергер, 1968, гл. 32). Здесь достаточно указать, что степень инбридинга в популяции, подобно дисперсии генных частот среди субпопуляций, можно найти из эффективной величины популяции N. Можно найти коэффициент инбридинга для некоторого определенного поколения данной популяции, но на практике это делается редко. Обычно определяют степень инбридинга для отдельных особей, используя анализ родословных.
Величина F для отдельной особи зависит от степени генетического родства ее родителей и может быть определена непосредственно путем анализа родослов-
ной этой особи. Определим, например, вероятность того, что данный потомок гомозиготен по идентичным генам, если его родители сибсы, а дед и бабка неродственны. Здесь мы примем, что дед и бабка есть основание; они неродственны и несут независимые аллели, которые можно обозначить А{ и А2 у деда и Л3, и А4 — у бабки. Ниже приведены родословная и процедура определения вероятности того, что гены идентичны.
Не связанные родством дед и бабка Связанные родством родители (сибсы) Инбредное потомство
Дед Бабка
Сын
1. Отец получает половину генов от деда. Вероятность того, что он получит ген Ah равна 1/2. Если отец получил ген Ah то вероятность передачи этого гена сыну равна также 1/2. Таким образом, вероятность того, что сын получит от деда через отца ген
Дь равна 72-72 = 74
2. Подобным же образом вероятность того, что ген А\ получит мать, равна 7г, а вероятность того, что этот ген будет передан от деда через мать сыну, равна 74-
3. Вероятность того, что сын получит два гена А\ (т. е. что он будет гомозиготен по идентичным генам Л i Л i), равна произведению двух найденных выше вероятностей 74 • 74 = 716-
4. Точно так же вероятность того, что инбредный сын имеет по два гена Л2, Л3 или Л4, равна 716-Так как существует четыре возможных типа идентичности аллелей — образование генотипов Л1ЛЬ Л2Л2, Л3Л3, Л4Л4 с вероятностью, равной 716 для каждого, — общая вероятность идентичности аллелей по данному локусу (величина F для инбредного сына) равна 4’716=74. Это значит, что в потомстве от скрещивания сибсов в среднем 25% всех локусов будут го
мозиготны по идентичным аллелям. Коэффициенты инбридинга потомства, полученного от объединения: 1) полусибсов (особей, имеющих одного общего родителя) и 2) двоюродных сибсов, равны соответственно 7в и 716- Пользуясь изложенным выше приемом и данными родословных, можно самостоятельно проверить правильность этих цифр (см. В. Маккьюсик, «Генетика человека»).
РАВНОВЕСНАЯ ПОПУЛЯЦИЯ И ФАКТОРЫ ЭВОЛЮЦИИ
Согласно модели Харди—Вайнберга, бесконечно большая «идеальная» популяция достигает равновесия, или устойчивого состояния; подобное состояние — полная антитеза эволюции. Органическая эволюция — это изменение наследственной структуры популяций, проявляющееся в виде различий между предками и их потомками. Мы уже говорили, что популяция эволюционирует как путем прогрессивных изменений внутри родословных, так и в результате разделения единой популяции на множество отдельных популяционных единиц, имеющих общее происхождение, и последующего обособления этих единиц. Это изменения разного порядка. Простые изменения генных частот, происходящие за относительно небольшие периоды времени, называют ступенями микроэволюции. Эти изменения обратимы и могут возникать многократно. Микроэволюция протекает в столь короткие сроки, что ее можно наблюдать непосредственно в природе или в лабораторном эксперименте.
Изменения, в результате которых происходит дивергенция отдельных линий, т. е. изменения, приводящие к возникновению высших систематических категорий, таких, как роды, семейства, отряды и классы, называют макроэволюцией. Эта классификация изменений по порядку величины сугубо описательна и относительна. Все имеющиеся у нас филогенетические и экспериментальные данные относительно причинных
факторов эволюции подтверждают представление о том, что все изменения более или менее постепенны и что высокодивергентные популяции (макроэволю ция) образуются путем накопления изменений, каждое из которых само по себе невелико (микроэволюция). Исходя из этого органическую эволюцию можно определить в более общей форме — как всякое прогрессивное изменение в частотах генов (слово прогрессивное включено в это определение для того, чтобы изъять из него обратимые изменения, приводящие к небольшим колебаниям вокруг некой стабильной точки равновесия).
По определению всякий фактор, способный вызывать изменения частоты генов, является потенциальным источником эволюции. Как было показано, случайная ошибка при передаче гена из одного поколения в другое в ограниченной по величине популяции нарушает установившееся равновесие. Таким образом, к факторам, вызывающим эволюционные изменения популяций, можно отнести случайный дрейф генов. В этом случае изменения оказываются
ненаправленными. Существуют иные__факторы, вызывающие направленные изменения частоты, ко-
торые систематически неуклонно смещают популяцию к некоторому определенному состоянию равновесия.
Направленные изменения предсказуемы как по величине, так и по направлению, тогда как для генетического дрейфа можно предсказать только величину. Направленные изменения вызываются тремя факторами: повторными (reccurent) мутациями, повторными волнами иммиграции и массовым отбором. Эти силы могут привести к «устойчивому дрейфу». К случайным процессам относятся любые колебания интенсивности или действия направляющих факторов плюс ошибка выборки (которая существенна лишь при малом N). Эти ненаправленные процессы вызывают «случайный дрейф».
Итак, суммарное изменение генной частоты за одно поколение, обозначаемое Д<7, можно разделить па два компонента: изменения, вызванные случайны
ми событиями, и изменения, вызванные направленным процессом, где kq + bq=\q.
При построении модели равновесия Харди — Вайнберга вышеупомянутые факторы, способные нарушить это равновесное состояние, сознательно игнорировались. Поскольку лишь очень немногие из природных популяций не испытывают воздействия этих возмущающих факторов, само существование состояний равновесия имеет лишь чисто теоретический смысл. Однако в действительности популяции могут находиться в состоянии равновесия иного рода, нежели равновесие Харди — Вайнберга. Эти другие устойчивые состояния могут возникнуть, когда силы действия одного или нескольких эволюционных факторов направлены в противоположные стороны. Например, равновесие может сохраняться в том случае, если мутации увеличивают частоту данного аллеля, а естественный отбор уменьшает ее (см. гл. 7). Для любого неизменного комплекса условий среды (например, в некой необычной, но вместе с тем постоянной экологической обстановке) частоты аллелей по всем локусам стремятся к генетическому равновесию. Конечно, закрепление какого-либо одного аллеля в данном локусе рассматривается здесь как тривиальный случай равновесия.
Итак, можно сформулировать два главных принципа, на которых основано распространение законов Менделя на популяционный уровень.
1. В отсутствие сил, вызывающих изменение генных частот, большая популяция со случайным скрещиванием будет находиться в состоянии генетического равновесия.
2. Эволюция представляет собой постепенное изменение генных частот. К известным эволюционным факторам, ответственным за изменения генных частот, относятся отбор, мутационный процесс, миграция и дрейф генов. Эти силы действуют совместно таким образом, что популяция приходит в состояние равновесия и остается в этом относительно устойчивом состоянии, пока условия среды остаются постоянными. При изменении условий или в некоторых
случаях, когда существуют случайные отклонения от начальной точки равновесия, популяция стремится к новому равновесию.
ЛИТЕРАТУРА
Falconer D. S., Introduction to Quantitative Genetics, New York, The Ronald Press Company, 1960.
L i С. C., Population Genetics, Chicago, University of Chicago Press, 1955.
Rasmuson M., Genetic on the Population Level, Stockholm, Sweden, Scandinavian University Books, 1961.
Strickberger M. W., Genetics, New York, The Macmillan Company, 1968.
Глава 4
ГЕНЕТИЧЕСКАЯ ИЗМЕНЧИВОСТЬ И ЕЕ ИСТОЧНИКИ
Значение биологической изменчивости, как это молчаливо подразумевалось в предшествующих главах, состоит в следующем: в результате изменения наследственной структуры видов становится возможной эволюция (гл. 1). О необходимости изменчивости свидетельствует поразительное разнообразие жизни как на популяционном, так и на видовом уровне (гл. 2). Если эволюционные силы бездействуют или находятся в равновесии, изменчивость отсутствует (гл. 3). Сейчас твердо установлено, что в популяциях раздельнополых перекрестнооплодотворяющихся организмов существует множество генотипов, т. е. изменчивость— это правило, а не исключение. Генетическая изменчивость — результат мутационного процесса, обмена генетическим материалом между популяциями, а также рекомбинации генетических факторов; все это в сочетании с давлением отбора определяет характер изменчивости. В дальнейших главах мы будем рассматривать главным образом возникновение, сохранение и формирование специфических типов изменчивости. Но прежде мы познакомимся с некоторыми общими аспектами изменчивости и ее источниками.
ТИПЫ ИЗМЕНЧИВОСТИ
Когда мы говорим об изменчивости, мы по сути дела лишь используем другие слова, чтобы констатировать существование различий между индивидуумами.
Можно абстрактно рассуждать об «общей» изменчивости, но на практике обычно рассматривают только один или несколько признаков. Изменчивость обычно характеризуется степенью идентичности особей в популяции. Члены одного дема проявляют вну-трипопуляционную (индивидуальную) изменчивость, тогда как различия между индивидуумами, принадлежащими к разным популяциям, составляют межпопуляционную (географическую) изменчивость. Для оценки последней сначала находят среднюю индивидуальной изменчивости в каждой группе, а затем сравнивают эти средние. Займемся пока изменчивостью в пределах дема, т. е. индивидуальной изменчивостью.
Естественно, что при описании внутрипопуляцион-ной изменчивости мы стремимся охватить все существенные различия между особями исследуемой популяции. Но как бы ни была велика и представительна выборка, многообразие популяционных вариантов и несовершенство методов анализа ограничивают наши возможности. Поэтому биолог вынужден довольствоваться анализом тех свойств и признаков, которые он считает наиболее существенными для понимания эволюции изучаемой группы. По существу же выбор признаков определяется в конечном счете тем, какими методами анализа может воспользоваться исследователь, а чаще всего — его личными интересами.
По традиции для исследования различий между индивидуумами использовались хорошо заметные морфологические признаки. С течением времени характер изменчивости многих популяций стали изучать более тщательно, привлекая для этого цитологические, гистологические, серологические, физиологические и поведенческие признаки. В последнее время для описания прежде неизвестных признаков стали с большим успехом использовать биохимические методы (анализ белков, ферментов, ДНК). Тем не менее в большинстве случаев основой для сравнения все еще остаются морфологические признаки. Морфологическая изменчивость затрагивает такие структурные признаки, которые легко охарактеризовать, изме
рить и по которым популяцию легко разбить на классы (например, вес, высота или длина тела, узоры окраски, ее оттенки, число каких-либо структурных образований, типы белковых структур).
Изменчивость того или иного признака обычно носит либо количественный, либо полиморфный характер. Изменчивость называется количественной, если различные морфологические типы постепенно переходят один в другой, образуя непрерывный ряд. Подобная изменчивость свойственна таким признакам, как, например, рост, вес, цвет кожи у человека. Изменчивость называется полиморфной, если внутри популяции существует два, три или несколько ясно различающихся классов «морф». Примерами полиморфизма служат половой диморфизм, группы крови у человека или структура хромосом. Зачастую бывает нелегко отличить количественный признак от полиморфного. Например, цвет глаз у человека считается полиморфным признаком — полиморфизм по окраске (голубая, коричневая, черная),—но благодаря существованию множества промежуточных оттенков между этими тремя типами наблюдается известная интерградация. Следует иметь в виду, что деление признаков на количественные и полиморфные до некоторой степени произвольно, так как те и другие имеют, в сущности, одну и ту же генетическую основу. Главное различие между ними — это число генетических факторов, определяющих данный признак, и способ их действия. Количественные признаки контролируются множеством генов, действующих кумулятивно. Полиморфные признаки, по-видимому, контролируются генами-переключателями, т. е. генами, действие которых вызывает переключение эпигенотипа на тот или иной путь развития. Возможно также, что появление полиморфного признака непосредственно обусловлено первичным продуктом действия отдельного гена (или системы сцепленных генов, действующих как одна функциональная единица). Оба типа изменчивости достаточно подробно рассматриваются в большинстве работ, посвященных популяционному анализу (более подробно о наследовании
количественных признаков см. Д. Брюбейкер, «Сельскохозяйственная генетика»). Как правило, полиморфные варианты легко дифференцировать, классифицировать и анализировать. Поэтому для иллюстрации внутрипопуляционной изменчивости обычно приводят примеры полиморфизма, и притом чаще всего — морфологически различающиеся формы у термитов и перепончатокрылых (фиг. 16).
Взрослые члены колонии термитов образуют несколько различных каст, морфологически специализированных для выполнения специфических функций. У большинства видов колонии состоят из особей первичной половой касты, вторичной половой касты, рабочих особей и солдат. В каждой касте есть и самцы, и самки, однако у рабочих особей половые органы недоразвиты, а у солдат они не функционируют. Особи первичной половой касты—это полностью развитые в половом отношении крылатые самцы и самки. В конце весны они вылетают из гнезда, сбрасывают крылья и спариваются. Некоторые пары, найдя подходящее место, строят гнездо, закладывают колонию и становятся ее царем и царицей. Постепенно брюшко царицы увеличивается за счет увеличения в размерах половых органов и образования яиц. Если один из членов этой пары основателей колонии гибнет, на смену ему приходит бескрылая особь вторичной половой касты, вполне развитая в половом отношении, но морфологически напоминающая нимфу. Почти всю работу в колонии выполняют маленькие бескрылые рабочие, слепые и бесплодные. Главная роль солдат, которых легко отличить по очень сильно развитой голове и мощным жвалам, — защита колонии от врагов. У некоторых родов термитов вместо касты таких солдат есть каста так называемых носатых солдат, имеющих маленькую головку с характерным длинным хоботовидным отростком; из этого отростка выделяется клейкая жидкость, используемая для защиты от врагов и как материал для постройки гнезда. Незрелые нимфы всех каст проходят частичный метаморфоз и переходят в разряд взрослых особей, находясь на разных стадиях развития.
Фиг. 16. Специализация (деление на касты) у муравьев.
/ — солдат; 2 — рабочий; 3 —крылатый самец; 4 —самка, сбросившая крылья все четыре особи (/—4) —представители вида Pheidole instabilus\ 5 —«медовая бочка» — Myrtnecocystus hortideorutn; 6 — солдат-привратник — Colobop-sis etiolalir, 7 —вид его головы спереди.
У общественных видов насекомых в одной колонии представлено несколько классов особей, морфологически различающихся между собой. Пример с термитами был выбран потому, что он иллюстрирует все четыре основных, или «популяционноспецифичных», типа изменчивости, непосредственно связанных со структурой, функцией и системой размножения популяции. В популяции термитов можно наблюдать:
1) возрастную изменчивость (нимфы и взрослые особи);
2) половую изменчивость (плодовитые и бесплодные самцы и самки);
3) функциональную или социальную изменчивость (производители, рабочие, солдаты);
4) сезонную изменчивость (крылатые и бескрылые особи первичной половой касты).
Конечно, термиты как общественные насекомые с делением на касты представляют собой исключительный пример крайне выраженной индивидуальной изменчивости. Однако можно утверждать, что демы всех раздельнополых видов в той или иной степени подвержены индивидуальной изменчивости. В большинстве случаев это изменчивость «общего» типа, наблюдаемая среди особей одной касты, одного пола, одного возраста и (или) одной сезонной группы. Под «общей» мы имеем в виду изменчивость любого признака (например, белокурый, рыжий или темный цвет волос у человека), непосредственную связь которого со специфическими морфологическими и физиологическими свойствами, необходимыми для существования популяции, установить не удается. Это не означает, что такие признаки непременно неадаптивны. Вполне возможно, что гены, направляющие развитие этих признаков, прямо или косвенно влияют на приспособленность, и притом в различной степени. Мы просто считаем, что эти признаки не играли большой роли в эволюции вида.
ГЕНОТИП — ФЕНОТИП
Для понимания движущих сил эволюции важно выяснить значение изменчивости, а вовсе не заниматься
ее классификацией. В этом нам может помочь краткое обсуждение терминов генотип и фенотип (введенных В. Иогансеном). Изменчивость признаков внутри популяции проявляется в виде фенотипических различий.
Фенотип — это общая сумма морфологических, физиологических, экологических и поведенческих признаков индивидуума, проявляющихся в течение всех и каждого периодов его существования. Короче говоря, фенотип организма — это его внешний вид, то, что можно наблюдать. Фенотипическое проявление организма, постоянно изменяющееся в течение всей жизни — от стадии зиготы до смерти — представляет собой результат взаимодействия факторов внешней среды с направляющим действием генотипа. Генотип, или сумма полученного зиготой наследственного материала предков, определяет ход развития особи в тех или иных определенных условиях.
В зависимости от условий существования развитие одного и того же генотипа может идти разными путями и давать самые разные фенотипы (фиг. 17). Например, у большинства животных половой диморфизм генетически детерминирован. Однако не всегда пол определяется типом и числом хромосом. Так, у морского червя Boneilia viridis (эхиуриды) пол зависит от условий, в которых развивается личинка. Личинки, подверженные действию химического вещества, имеющегося на поверхности хобота взрослой самки, развиваются в самцов, а свободно плавающие личинки превращаются в самок. Морфологические различия между двумя полами чрезвычайно резкие. Самцы не имеют органов пищеварения и так малы по размерам, что могут обитать по нескольку одновременно в не-фридиях самки, где они ведут паразитический образ жизни.
С другой стороны, феминизация стерильных фримартинов у крупного рогатого скота есть прямое влияние хромосомного набора этих особей (генетический контроль), а маскулинизация — следствие внутриутробного действия гормонов, выделяемых братом-близнецом (фактор среды).
Все возможные варианты фенотипического проявления генотипа в условиях, при которых организм может существовать, называются диапазоном реакций или нормой реакции. В некоторых случаях «норма» ограничена очень узкими пределами, т. е. во всем
Фиг. 17. Морфологические изменения стрелолиста Sagittaria sagittifolia при росте на суше (Л), в полупогруженном состоянии (Б) или под водой (В).
Эти различия не обусловлены генетически, а возникли в результате взаимодействия генотипа и среды. Тонкие листья, развивающиеся под водой, лишены кутикулы и поглощают питательные вещества непосредственно из воды; они не имеют структурной прочности и будучи вынуты из воды поникают. Надводные листья или листья растений, произрастающих на суше, напротив, сохраняют вертикальное положение, а под действием потока воды они, вероятно, отрывались бы. У растения, произрастающего на суше, корневая система развита сильнее, чем у водных форм; оно поглощает питательные вещества из почвы. Эта изменчивость стрелолиста имеет генетическую основу. Многие наземные растения гибнут при затоплении водой, а многие водные растения не могут жить на суше. Таким образом, в известных условиях адаптивные реакции, подобные таковым у стрелолиста, могут давать преимущество при отборе.
диапазоне условий, при которых генотип может существовать, его фенотипическое проявление остается неизменным (канализация развития). Если же диапазон реакций широк, то развитие оказывается более гибким, и в различных условиях среды возникают
разные фенотипы. Кроме того, если диапазоны реакций нескольких генотипов перекрываются, то, несмотря на генотипические различия, индивидуумы могут быть фенотипически схожи. Изучению процесса развития и их значению для теории эволюции стали уделять должное внимание лишь в последнее время. Например, в современных разногласиях между ботаниками и зоологами о способах видообразования у растений и у животных легче разобраться, если ясно представлять себе, что животные подвижны — они имеют возможность выбрать себе местообитания (поведенческая гибкость) и поэтому более специализированы и менее изменчивы, чем растения. Большинство же растений неразрывно связано с условиями существования, и поэтому их развитие должно быть достаточно гибким, чтобы обеспечить им возможность существования в тех условиях, в каких они оказались (широкая «норма реакции»). На популяционном уровне генетическая изменчивость многих видов растений, создающая дополнительные возможности для выживания в новых условиях, имеет чрезвычайно большое значение. Таким образом, при оценке эволюционной роли тех или иных факторов следует не только сравнивать их между собой, но и учитывать специфику изучаемых видов и характерные для них пути приспособления к внешним условиям.
Всю нормальную фенотипическую изменчивость популяции (обозначаемую как VP) теоретически можно разделить на генетическую изменчивость (VG) и изменчивость, определяемую условиями среды (V#). Для экспериментального разделения фенотипической изменчивости на ее составляющие необходимо знать степень родства (двоюродные сибсы, сибсы, родители — потомки) исследуемых индивидуумов. Степень генетического родства отражается в степени фенотипического сходства между родственными особями. При различных упрощающих допущениях, но с непременным учетом взаимодействия между генами и взаимодействия генов с внешней средой увеличение фенотипического сходства пропорционально отражает степень генетической изменчивости. Такие расчеты очень
сложны и их совершенствование составляет одну из перманентных проблем количественной генетики.
Генетическая изменчивость связана с различиями в диапазонах реакций, а изменчивость, определяемая условиями среды, — с фенотипическими различиями внутри этих диапазонов. Негенетическая изменчивость, которую часто называют фенотипической пластичностью, дает возможность особи лучше приспосабливаться в процессе развития. Поэтому генотипы с широким диапазоном реакций считаются в общем лучше приспособленными, поскольку они могут развиваться в широком диапазоне условий среды; такие генотипы более гибки. В гл. 7 будет показано, что большее генетическое разнообразие придает аналогичную пластичность популяции.
Изменчивость большинства популяций —хорошо установленный факт, однако следует иметь в виду, что нормы реакций всех особей популяции во многом сходны (перекрываются). Канализация — способность процесса развития сглаживать генетические и экологические различия — стремится к тому, чтобы все генотипы в данной популяции развивались в направлении единого фенотипа, максимально приспособленного к существующим условиям среды. В природных популяциях большая часть генетической изменчивости скрыта за неким пластичным фенотипом (такой фенотип часто называют диким типом) \ образующимся в результате сходства норм реакции и действия канализации.
ФАКТОРЫ, ВЛИЯЮЩИЕ НА ГЕНЕТИЧЕСКУЮ ИЗМЕНЧИВОСТЬ
Итак, совершенно ясно, что для понимания механизмов эволюции как процесса, необходимо иметь
1 Как было показано в гл. 2, подчеркивание сходства между близкородственными формами обычно ведет к типологическим представлениям; однако данная здесь интерпретация сходства допускает широкую генетическую изменчивость и представляет отклоняющиеся формы как естественную и отнюдь не неожиданную принадлежность изменчивой популяции.
представление об изменчивости популяций и в частности об их генетической изменчивости. Во времена Дарвина и ранних эволюционистов исходным тезисом всех эволюционных построений был взгляд на наследственную изменчивость как на необходимое условие филогенетического развития. В то же время считалось, что естественный отбор стремится к созданию единственной формы («тип», который особенно хорошо приспособлен). Считалось, что почти вся наследственная изменчивость постоянно утрачивается. Кроме того, в то время господствовала «теория слитной наследственности». Подумав немного, легко понять, что в случае слитной наследственности у пере-крестнооплодотворяющихся раздельнополых форм генетическая изменчивость должна снижаться вдвое за одно поколение, если только нет высокой корреляции между наследственной конституцией скрещивающихся особей (Фишер, 1930). Если наследственная изменчивость теряется как в результате слияния, так и в результате отбора, направленного против неприспособленных форм, то единственный разумный вывод состоит в том, что, во-первых, генетическая изменчивость, необходимая для эволюционного процесса, возникла относительно недавно и, во-вторых, что независимо от ее источника частота отклоняющихся особей очень велика. Кроме того, недавно возникшие варианты должны быть приспособлены к существующим условиям, так как в противном случае они будут немедленно элиминированы отбором — в этом заключалась главная предпосылка теорий эволюции Бюф-фона и Ламарка, согласно которым новые формы возникают как прямой ответ на условия среды (наследование приобретенных признаков). Эти же идеи легли в основу мутационной теории эволюции, согласно которой эволюция происходит скачкообразно, путем генных и хромосомных изменений, ведущих к появлению новых функций и новых структур.
Современные представления об источниках, сохранении и потере генетической изменчивости сильно модифицировались под влиянием мснделевской генетики. В гл. 3 мы видели, что в свете корпускулярной
теории наследственности тенденции к утере генетической изменчивости не наблюдается (теория Харди — Вайнберга). Истощение генетического разнообразия популяции может быть вызвано только направленным изменением частот аллелей под действием отбора или же ненаправленным действием дрейфа генов. Теперь предполагается, что популяциям нетрудно сохранять генетические варианты и что они могут порождать множество генотипов с различной приспособленностью. В каждом поколении формы, возникающие на неадаптивном конце шкалы приспособленности, устраняются отбором, однако их постоянное возникновение обеспечивает некий оперативный резерв, который послужит источником предковых особей для будущих поколений в случае внезапного изменения условий. Это «преадаптированные» генотипы для тех новых обстоятельств, с которыми может столкнуться популяция. Такой вывод отнюдь не значит, что для любого генетического варианта существует соответствующая среда. Некоторые из этих новых вариантов не могли бы существовать ни при каком мыслимом комплексе условий. Однако популяция, оказавшаяся в новых необычных условиях и не имеющая генотипов, преадаптированных к новым условиям, постепенно вымирает.
Итак, всякая популяция должна обладать достаточным запасом генетической изменчивости (преадаптированные типы); в противном случае она не застрахована от гибели в будущем. Однако чрезмерное количество отклоняющихся вариантов в каком-то одном поколении также очень неблагоприятно, так как при этом резко нарушается приспособленность. Таким образом, приспособленность к данным условиям должна совмещаться с достаточной генетической гибкостью, которая обеспечила бы в случае необходимости соответствующую перестройку генофонда. Популяция должна каким-то образом поддерживать равновесие между этими диаметрально противоположными тенденциями.
Прежде чем перейти к анализу различных форм равновесия, рассмотрим основные факторы, опреде
ляющие степень генетического разнообразия популяции. Начнем с источников генетической изменчивости.
МУТАЦИИ КАК ИСТОЧНИК ПЕРВИЧНОЙ ГЕНЕТИЧЕСКОЙ ИЗМЕНЧИВОСТИ
Наследственность в противоположность эволюции консервативна. Наследственность обусловливает сходство между особями, родственными по происхождению, тогда как эволюция порождает различие. Изменчивость в пределах родословной проявляется лишь при возникновении отклонений от общей наследственности, т. е. при появлении потомков, не являющихся полными копиями своих предков.
В настоящее время общепризнано, что генетическая информация («генетический код») записана в виде специфической последовательности пар азотистых оснований вдоль цепи (или цепей) генетического материала ДНК. У большинства высших организмов ДНК входит в состав нуклеопротеидов, которые в свою очередь входят составной частью в еще более высокоорганизованную функциональную единицу — хромосому. Одним из самых больших достижений в биологии было открытие уникальной способности одного из двух компонентов хромосомы—теперь мы знаем, что это молекула ДНК, — к точному само-удвоению в процессе клеточных делений. Этот процесс и лежит в основе явления наследственности. Возникновение генетической изменчивости возможно только в тех случаях, когда точность копирования нарушается. При этом в структуре молекулы ДНК возникают изменения, иногда приводящие к появлению в цепи ДНК новых пар оснований, метаболический эффект которых будет отличен от метаболического эффекта исходной структуры. Как правило, новая молекула почти так же устойчива и почти так же точно воспроизводится в последовательном ряду клеточных поколений, как и исходная. Такие изменения называют генными мутациями, а участок молекулы ДНК,
затронутый таким изменением, называют мутоном ’. Однако мутацией можно считать «любое дискретное изменение, обладающее генетическим эффектом» (Майр, 1963). Хотя это определение несколько туманно, оно удобно тем, что учитывает не только основные химические изменения, происходящие на молекулярном уровне, но и перестройки на уровне хромосом.
Мутируют не только отдельные локусы, но и сами хромосомы подвергаются значительным изменениям. В процессе деления может произойти утрата отдельных частей хромосомы и даже целых хромосом или же, напротив, умножение их числа; возможны также перестройки некоторых участков хромосом. Все виды хромосомных аберраций удобно делить на два класса: изменения числа хромосом и структурные изменения.
СТРУКТУРНЫЕ ИЗМЕНЕНИЯ ХРОМОСОМ И СУПЕРГЕНЫ
Степень генетической изменчивости популяции определяется содержащейся в ней генетической информацией, которая зависит от аллельного состава всех локусов, частоты аллелей среди членов популяции и частоты всех отдельных аллельных комбинаций, т. е. генотипов. Несомненно, что источником первичной изменчивости служит генная мутация, ответственная за различные аллельные состояния. Существенным дополнительным источником разнообразия служит сочетание аллелей в хромосомах. Объяснение этому нужно искать в строении генетического материала (см. Ф. Сталь, «Механизм наследственности») и в его
1 Новые определения гена и новая терминология (мутон, цистрон и рекон) были предложены Бензером в результате исследований, проведенных главным образом на микроорганизмах (см. Ф. Хартман и 3. Саскайнд, «Действие гена»). Однако в генетике высших организмов и в генетике популяций эти термины пока еще не нашли широкого применения. На популяционном уровне удобно пользоваться классическими терминами гены (локусы) и аллели. Прекрасный обзор на эту тему сделал Грант (1964),
функции (см. Ф. Хартман и 3. Саскайнд, «Действие гена»).
Согласно классическим представлениям, закодированная генетическая информация рассматривалась как интегрированная система отдельных генов, линейно расположенных в хромосомах и определяемых как участки хромосом, отделяющиеся от других при кроссинговерс. Помимо такого определения гена как структурной единицы, существует и другое — определение гена как функциональной единицы, которая направляет предписанную последовательность развития таким образом, чтобы это привело к некоторой внешне различимой особенности фенотипа — признаку. Эта концепция гена во многом сохраняется до сих пор. Гены действительно расположены линейно в хромосомах, данным кроссинговера продолжают придавать важное значение и отдельные участки хромосом считаются, как и прежде, ответственными за первичную функцию (определение аминокислотной последовательности ферментов — этих специфических регуляторов химических реакций, протекающих в строго установленной последовательности).
Однако эта концепция оказывается непригодной, если рассматривать функциональную и структурную единицу как одно целое. После обнаружения у высших форм и у микроорганизмов псевдоаллелизма стало ясно, что внутри гена как функциональной единицы может иногда происходить кроссинговер, т. е. ген оказался делимым. С помощью цис-транс-теста на комплементацию было обнаружено, что функциональный ген состоит из субгенов.
Кроме того, биохимические исследования показали, что гены объединяются в системы генов более высокого порядка. Действие отдельных генов — это своеобразные ступени в последовательной цепи реакций биосинтеза. Примером может служить образование аминокислоты аргинина у нейроспоры. Хотя каждый ген действует независимо в определенном участке метаболической цепи, все гены любой последовательности метаболических реакций, по-видимому, кооперируются, действуя сообща для получения
конечного продукта. Функционально связанные совокупности генов взаимодействуют, образуя «линейную систему генов» (Грант, 1964). Гены, образующие такую систему, могут быть разбросаны по всему геному или же тесно сцеплены в определенном участке одной хромосомы. Как было точно установлено у салмонел-лы и довольно убедительно — у некоторых других видов, ряд генов, контролирующих последовательные этапы одного метаболического пути, склонны «скапливаться». Такое расположение подобно расположению множественных субгенов одного псевдоаллель-ного локуса, поскольку смежные субгены, по-ви-димому, также функционально связаны. Однако комплементация — это свойство отдельных членов сцепленной генной системы, тогда как субгены, относящиеся к одному цистрону, более едины в функциональном отношении.
Хотя эти многочисленные определения гена могут внести некоторую путаницу, они вместе с тем многое проясняют. Взаимодействие генов и непрерывность распределения генетического материала вдоль хромосомы приобретает новое значение. Хромосомы уже не рассматриваются лишь как футляр для независимых локусов. Хромосомы состоят из генов и субгенов, структурно объединенных в линейную последовательность и в различной степени взаимосвязанных функционально. Существует определенная иерархия функциональных единиц: азотистые основания ДНК, субгены, гены, генные системы, весь материал генома. Изменения, возникающие на любом из этих структурно-функциональных уровней и завершающиеся генетическим эффектом, и представляют собой по определению мутации.
Хромосомные мутации заключаются в разрыве и структурной перестройке хромосом. При этом расположение генов вдоль хромосомы изменяется. Сами локусы при мутации остаются неизменными, хотя фенотип может измениться. Любое подобное фенотипическое проявление, зависящее только от пространственного расположения генов и нередко напоминающее реоулыат i синей мутации, но не связанное
A
BCD E F в H
t f
Нехватка A 8 c F F G H &
» б П -_-J o
Ацентрический фрагмент утрачивается
А В С D Е F G н с_ рс~~
t
Дупликация
А В С В С D Е EG Н
Дуплицированный участок
А В С D Е Г 6 Н
t \t
Пара центрическая^ инверсия
А Е F G Н
D. £ _ Г - — Перицентрическая ABC G н
f | инверсия
А В С D Е F G Н
М N О Р Q R
Транслокация
D С В
Центрическое слияние г
ОФ
Центрический фрагмент\ утрачивается
Фиг. 18. Основные типы хромосомных перестроек (а — g).
Слева показаны стандартные (или нормальные) хромосомы, а справа — хромосомы с перестройками. Маленькие стрелки указывают места разрывов нормальных хромосом, необходимые для получения соответствующих перестроек. Причиной возникновения дуплицированного участка может быть необычный кроссинговер (как показанная здесь тандемная дупликация) или разрыв гомологичной хромосомы (смещенная дупликация).
с такой мутацией, называется эффектом положения. Многие, по не все хромосомные перестройки обладают эффектом положения; большая часть таких перестроек обычно сопровождается снижением приспособленности их носителей.
Хромосомные мутации, или аберрации, представляют собой перестройки генных последовательностей хромосом. Образование хромосомных перестроек начинается с появления разрывов, послр чего происходит воссоединение частей хромосом в новых комбинациях. Основные типы хромосомных аберраций — нехватки, дупликации, инверсии и транслокации (фиг. 18).
Нехватки и дупликации
Гаплоидный набор хромосом содержит один полный набор генов, или один геном, выступающий в процессе развития организма как единый интегрированный комплекс. Целостность генома является необходимым условием жизнеспособности гаметы и ее нормального функционирования. В норме два таких набора генов служат главной предпосылкой развития диплоидной фазы. При этом некоторые вредные в гомозиготном состоянии аберрации могут существовать в гетерозиготном состоянии, скрываясь от действия отбора, так же как рецессивные генные мутации скрываются за проявлением доминантных аллелей. Недостаток (нехватка) или избыток (удвоение) какого-либо из эухроматиновых участков хромосомы означает изменение дозы генов. Если развитие организма контролируется непосредственно генетической информацией, закодированной в молекуле ДНК хромосом, то нетрудно видеть, что разрушение или утрата даже небольшого участка «программирующей информации» приводит к нарушению процесса развития. И в самом деле, генетическое действие многих нехваток аналогично действию рецессивных леталей. Дупликации, напротив, только увеличивают дозу некоторых генов, и их действие на развитие носит в основном количественный характер. Они менее вредны.
Для эволюционистов эти типы аберраций интересны по двум причинам. Во-первых, гаметы с дупликациями и нехватками могут образовываться после кроссинговера в хромосомах особей, гетерозиготных по инверсиям и транслокациям (см. следующий раздел). Это значит, что эти гетерозиготы будут обладать пониженной плодовитостью, если только у них нет компенсаторных механизмов, помогающих предотвратить развитие аберрантных гамет. Во-вторых, дупликации и увеличения плоидности (кратное увеличение числа хромосом) представляют собой единственные известные нам способы увеличения генетического материала. В свете наблюдаемой эволюции живых существ от простых форм к более сложным естественно ожидать добавления новых локусов и дивергенции их функций.
Инверсии и транслокации
При возникновении единичных разрывов в двух негомологичных хромосомах дистальные концы хромосом могут, поменявшись местами, соединиться с центральными частями «чужих» хромосом. Этот тип обмена называется транслокацией. При таком взаимном обмене локусы, бывшие прежде разобщенными, соединяются, т. е. образуются новые группы сцепления. Для обнаружения транслокации используют специальные генетические тесты. Если обменивающиеся участки неодинаковой длины, то транслокацию можно также обнаружить цитологически при митозе. Кроме того, транслокации обнаруживаются по наблюдаемым на пахитенной стадии мейоза комплексам, состоящим из четырех хромосом.
Транслокации плеча возникают в тех случаях, когда разрывы произошли вблизи центромер. Особый вид транслокации плеча — это центрическое слияние, возникающее в результате воссоединения длинных плеч акроцентрических хромосом (хромосомы, у которых центромера располагается очень близко к одному концу); их называют также палочковидными
хромосомами или барабанными палочками. Такая транслокация может возникнуть, если один разрыв возник близ центромеры в длинном плече одной хромосомы, а другой — в коротком плече негомологичной хромосомы. При этом ацентрический участок одной хромосомы соединяется с содержащим центромеру участком другой, в результате чего образуется одна огромная хромосома. Положение центромеры в новой хромосоме зависит от относительных размеров исходных акроцентрических хромосом. Хотя один из двух маленьких фрагментов, остающихся от исходных хромосом, содержит центромеру, тем не менее оба они обычно теряются в процессе деления. Чаще всего это происходит, если фрагменты содержат только гетерохроматин.
Поскольку два акроцентрических участка «сливаются», образуя одну хромосому, а один центрический фрагмент исключается из хромосомного набора, самым значительным следствием слияния является снижение числа хромосом. Известно, что подобный процесс имеет место у дрозофилы. Хотя у примитивных форм рода Drosophila гаплоидное число хромосом равно шести, основное число хромосом у ряда видов равно пяти, четырем и даже трем. У многих видов удается определить, какие именно хромосомы образовались в результате транслокаций.
При одновременном разрыве хромосомы в двух местах образуются три участка. Только один из них содержит центромеру. Если средний участок хромосомы повернется на сто восемьдесят градусов и его концы вновь соединятся с дистальными участками, то последовательность генов в среднем участке окажется обратной по отношению к исходной. Такое изменение в строении хромосомы называется инверсией. Если в этот средний участок входит центромера, то инверсия называется перицентрической\ если же оба разрыва произошли в одном плече хромосомы, то инверсию называют парацентрической. В любом случае сцепление генов изменяется Именно при изучении групп сцепления и были впервые обнаружены инверсии.
Парацентрические инверсии не затрагивают центромеру. Их можно обнаружить только у некоторых организмов, у которых в пахинеме легко наблюдать конфигурации, образующиеся при конъюгации у гетерозигот. Кроме того, о существовании инверсий свидетельствуют аномалии процесса расхождения хромосом и образование хроматидных мостов на стадии
Фиг. 19. Апериодическая структура и конъюгация политенных хромосом (третья хромосома Drosophila tnojavensis).
На вид кажется, что здесь одна хромосома, но на самом деле это две гомологичные хромосомы, тесно спаренные, диск к диску, по всей своей длине (ср. с фиг. 20, на которой можно различить отдельные хромосомы).
анафазы мейоза. Особенно ценные данные по парацентрическим инверсиям были получены на дрозофиле. Насекомые из отряда двукрылых, в частности виды, принадлежащие к роду Drosophila, замечательны тем, что у них в клетках слюнных желез имеются гигантские политенные хромосомы. Эти хромосомы обладают не только очень большими размерами, но, что гораздо важнее, и чрезвычайно своеобразным строением: наличием ясно выраженных поперечных дисков, вздутий (пуффы) и других хорошо заметных «ориентиров» (фиг. 19). Опытный цитолог может использовать эти специфические особенности строения хромосом в качестве маркеров для идентификации не только отдельных хромосом, но и самых мелких участков внутри хромосом, а также любых изменений в расположении дисков, возникающих при
хромосомных перестройках. Для гигантских хромосом можно составить карту практически всех хромосомных аберраций, за исключением самых незначительных. Гигантские хромосомы имеют еще одну очень ценную особенность: гомологичные хромосомы конъюгируют с исключительной точностью и постоянством, каждый диск одной хромосомы плотно конъюгирует со своим гомологом. Изменение структуры одной
Фиг. 20. Конъюгация двух гомологичных хромосом, различающихся по двум перекрывающимся инверсиям.
Это хромосомы (III) гибридной особи, полученной от скрещивания Drosop-Jiila mojavensis и D. arizonensis. Для каждого вида характерно особое строение третьей хромосомы.
хромосомы сопровождается изменением последовательности дисков, поэтому у гетерозигот при конъюгации измененной хромосомы с нормальной хромосомам приходится изгибаться, образовывать петли или каким-то иным способом компенсировать различие, с тем чтобы каждый участок одной хромосомы оказался расположенным точно против гомологичного участка другой. Для каждого типа перестроек характерна специфическая, свойственная ему одному фигура конъюгации (фиг. 20). При нехватках и дупликациях образуется выступ, поскольку некоторый участок одной хромосомы не имеет гомолога в другой.
Гетерозиготы по инверсиям характеризуются петлями, а гетерозиготы по транслокациям — крестами.
Такие сложные фигуры появляются и при конъюгации в клетках зародышевого пути. Представим себе, что происходит при кроссинговере и расхождении перепутанных хроматид у гетерозигот. По-видимому, различные типы аберраций по-разному влияют на рекомбинацию хромосом и образование гамет. Рассмотрим случай с гетерозиготной транслокацией. Конъюгация гомологичных участков четырех хромосом (две пары, участвующие в транслокации) приводит к образованию ассоциации, имеющей форму креста (фиг. 21). При случайном расхождении хромосом к разным полюсам в первой анафазе мейоза две трети гамет получат хромосомы с дупликациями или нехватками. Такие гаметы нежизнеспособны. Сбалансированные геномы образуются только в том случае, если ядро получает «родительскую» комбинацию хромосом, т. е. когда чередующиеся хромосомы (хромосомы, расположенные в фигуре креста одна против другой) отходят в одно ядро.
Гетерозиготные хромосомы можно отличить и цитологически — по особым комбинациям аллелей. Посредством кроссинговера родительские комбинации генов рекомбинируют в гетерозиготах по транслокациям так же, как и в обычных цитологических гомозиготах (гомокариотипах), с той разницей, что гены вблизи мест разрывов оказываются сцепленными теснее.
У гетерозигот по инверсиям положение иное. Если одна из двух гомологичных хромосом имеет «стандартную» последовательность генов ABCDEFGHI, а вторая содержит парацентрическую инверсию ABCDGFEHI, то во время мейоза эти хромосомы образуют характерную петлю. Кроссинговер, происходящий вне петли, приводит к рекомбинации любых различающихся аллелей, хромосомы расходятся нормально и включаются в дочерние ядра. Однако если внутри инверсионной петли произойдет один обмен, то две центромеры оказываются соединенными хроматидным мостом. На стадии анафазы этот мост
растягивается и обычно разрывается. Второй продукт этого обмена — ацентрический фрагмент (фиг. 22), который элиминируется из веретена деления. Затем
Фиг. 21. Конъюгация и расхождение хромосом у гетерозиготы по транслокации.
а. Строение двух гомологичных пар хромосом, вовлеченных в транслокацию, б. Ожидаемая картина конъюгации этих хромосом (хроматиды не показаны). В течение анафазы первого мейотического деления эти четыре хромосомы могут разойтись тремя способами (в — д). Если соседние хромосомы дви жутся к одному полюсу (I и II; III и IV —как в случае в, или I и III; II и IV —как в случае г), то гаметы будут содержать дупликации и нехватки и, следовательно, не будут нормально функционировать. Если к одному полюсу отходят измененные, а к другому — неизмененные хромосомы (I и IV; II и III —как в случае д), то каждая гамета получит нормальный геном и будет нормально функционировать. Это можно показать, пометив анафазные хромосомы, как это сделано на а и б.
образуются гаметы с дупликацией и нехваткой (здесь следует заметить, что в отличие от транслокаций, где причиной образования таких гамет является нерве-хождение, в данном случае их образование обусловлено кроссинговерохм в инверсии). Кроссинговер
гетерозигот по псрицентрическим инверсиям также ызывает частичную стерильность.
Хотя образование аномальных (с нехватками и упликациями) гамет у гетерозигот по транслока-
дв С Р Е F^
ОЗС_____ Г-J
А В С D Е F
В С D Е Г
Фиг. 22. Три возможных генотипа (а — в) в популяции, содержащей стандартную хромосому и хромосому с парацентрической инверсией.
Конъюгация и кроссинговер, как у гомозигот по нормальной, так и у гомозигот по инвертированной хромосоме, идут совершенно нормально. Гетерозиготы при конъюгации образуют петлю, компенсируя структурные различия между гомологами (г). Кроссинговер между двумя хроматидами из четырех в участке C — D приводит в анафазе / (е) к образованию хроматидного моста и ацентрического фрагмента. Этот фрагмент обычно теряется в цитоплазме, а хроматидный мост может разорваться или не разорваться. Только те ядра, которые во втором делении получили интактную стандартную хромосому ABCDEF или инвертированную хромосому AEDCBF, дают жизнеспособные гаметы. Кроссоверные гаметы содержат дупликации и нехватки и нежизнеспособны.
циям и инверсиям и вызывает снижение плодовитости, в природных популяциях широко распространен полиморфизм по этим перестройкам. Например, некоторые расы растений рода Oenothera состоят исключительно из гетерозигот по транслокациям. У этих форм обнаружен механизм, обеспечивающий чередующееся расхождение хромосом в мейозе, что
позволяет нейтрализовать цитологические трудности, связанные с транслокациями. Еще более обычны в природных популяциях гетерозиготы по инверсиям. Они широко распространены у дрозофилы, где их легко обнаружить с помощью цитологического анализа гигантских хромосом слюнных желез. У самцов дрозофилы кроссинговер вообще отсутствует, а у самок овогенез осуществляется таким образом, что большая часть кроссоверных хроматид не включается в функциональные ядра яиц. Поэтому плодовитость инверсионных гетерозигот почти не отличается от нормы. Для чего же виду могли понадобиться эти цитологические осложнения? Какие преимущества дает инверсия? По-видимому, она обеспечивает образование сцепленных блоков адаптивных аллелей.
Супергены
Поскольку рекомбинационные гаметы нежизнеспособны и частота кроссинговера в инвертированных участках гетерозигот понижена, все гены, входящие в этот участок, наследуются совместно, как одна группа сцепления. Гены инвертированного участка не обязательно связаны функционально как сцепленные линейные системы генов, однако они взаимозависимы в адаптивном отношении и отбор действует на них, как на единую группу. Такая группа генов представляет собой в эволюционном смысле отдельную физическую и функциональную единицу. Этот комплекс сцепленных локусов можно считать супергеном или отдельным гигантским локусом. Аллели инвертированного участка не рекомбинируют с аллелями неин-вертированного гомологичного «локуса». Эти различающиеся последовательности генов можно считать «аллельными», и их распространенность в популяции можно выразить как частоты аллелей.
В результате рекомбинации у раздельнополых видов непрерывно возникает огромное множество аллельных комбинаций. Некоторые из этих комбинаций дают высокоадаптированные фенотипы, однако рекомбинация может и разрушить эти гармоничные группы. В любой популяции постоянно возникают
также структурные мутации, но только те из них, которые благоприятствуют объединению в одну группу генов, обладающих более высокой адаптационной ценностью, закрепляются в популяции, а это случается довольно редко. Но если такому «аллелю» удалось закрепиться, он воспроизводится и накапливается. Фактически многие популяции полиморфны по двум и большему числу «супераллелей». Например, среди отличающихся наибольшим полиморфизмом популяций Drosophila willistoni уже обнаружено более 50 инверсионных «морф».
Если превосходство инверсий проявляется в гомозиготном состоянии, то инвертированная последовательность может совершенно вытеснить из популяции «стандартную» последовательность генов. Так, например, при сравнении многих близких видов (а также рас или популяций) было обнаружено, что они структурно гомозиготны, но различаются по расположению пуффов и дисков гигантских хромосом. Другими словами, в разных популяциях закрепились различные супергены. Возможно, что в некоторых случаях возникновение этих различий и замещение нормальных аллелей инверсионными сверхаллелями произошло уже после дивергенции видов. Согласно другой модели, структурно гомозиготные виды могут происходить из одной полиморфной предковой популяции. По-видимому, видоспецифичность генной последовательности есть следствие закрепления супергенов в отдельных изолятах полиморфной популяции, которые позже становятся отдельными видами.
Итак, первопричина большинства изменений — генная мутация. Хромосомные мутации, хотя они иногда и сопровождаются эффектом положения, играют главным образом механическую роль, влияя на рекомбинацию и сцепление.
СУДЬБА ОТДЕЛЬНОГО МУТАНТА
Между генными мутациями и перестройками имеется еще одно существенное отличие. Химические изменения, связанные с превращением одного аллеля
в другой, время от времени повторяются, т. е. каждый тип изменений характеризуется определенной частотой мутирования. Между тем любая хромосомная перестройка представляет собой редчайшее, а может быть, даже уникальное явление. Мы уже видели, что хромосомы слюнных желез имеют весьма сложное строение. Например, Х-хромосома Drosophila melano-gaster содержит более 1000 ясно выраженных дисков. Экстраполируя эту величину на весь геном, находим, что в хромосомах этого вида имеется около 5000 микроскопически отличающихся участков. Поскольку большинство спонтанных перестроек — результат двух разрывов хромосомы, причем место возникновения разрывов по длине хромосомы зависит только от случая, вероятность того, что данная перестройка произойдет еще раз (т. е. что обе точки разрыва совпадут с точками разрыва какой-либо из уже существующих перестроек), исключительно мала. Хромосомные мутации не имеют измеримо?! частоты — каждую мутацию можно рассматривать как событие, происходящее лишь один раз.
Какова судьба отдельного мутанта? Фишер (1930) разработал следующую модель: пусть большая ограниченная по численности популяция состоит исключительно из гомозигот АА. Допустим далее, что в этой популяции произошла мутация (Л->а) и появилась единственная гетерозигота (Да) по новому аллелю. Будет ли частота мутантного гена увеличиваться от поколения к поколению? Судьба нового мутантного гена зависит от числа потомков, оставляемых гетерозиготой, и их ценности для популяции. На этой стадии анализа удобно считать новый аллель эволюционно нейтральным, т. е. считать, что естественный отбор не действует и судьбы гена зависят только от численности потомства.
Очевидно, что если гетерозигота Аа не оставит потомства, то новый ген будет немедленно потерян. Даже в том случае, если гетерозигота оставит потомство, только половина се потомков будет иметь мутантный аллель (партнером гетерозиюты Аа всегда будет гомозигота АА). Это значит, что вероят
ность потери гена а равна (’/2)где п — число потомков в каждой семье.
Число потомков (п) 0 12 3 п
Вероятность потерн 1 l/2 Vi Ve С/г)*
гена a(Ln)
Конечно, точно предвидеть число потомков, оставляемых гетерозиготой, невозможно, оно может варьировать от 0 до п. Однако мы знаем, что при неизменной величине популяции среднее число потомков равно двум. Фишер показал, что по некоторым данным о числе потомков распределение различных классов по частоте очень близко к распределению Пуассона (где и среднее и дисперсия равны 2).
Число потомков (и) 0 1 2 3 п
Частота (Fn) е~2 2^“2
Умножая вероятность исчезновения гена (Ln) на частоту соответствующего числа потомков (Fn) и суммируя все полученные значения вероятностей, находим математически точную вероятность потери гена а за одно поколение. Эта величина равна е~\ или 0,3679. Следовательно, вероятность того, что мутант сохранится, т. е. что он перейдет во второе поколение, равна 1 — 0,3679, или 0,6321. Иными словами, из ста появившихся отдельных мутаций около 2/3 перейдет в следующее поколение. Этот процесс потери генов повторится и во втором поколении, но теперь в нем примут участие уже не 100, а только 63 мутации. В результате третье поколение будет содержать только 53 мутации. Процесс потери генов продолжается и дальше, пока не исчезнет последний мутантный ген. Даже мутации, обладающие некоторым селективным преимуществом по сравнению с нормальными аллелями, не могут избежать этой участи — случай медленно вытесняет их из популяции. Следовательно, почти единственная возможность для новой перестройки укрепиться в популяции состоит
5 Зак. 88
в том, чтобы иметь с самого начала большое селективное преимущество (именно ту «ценность», которую мы выше игнорировали).
Генные мутации многократно возникают в популяции. Поток мутаций действует как эволюционный фактор («давление мутаций»), непрерывно поставляющий в генофонд популяции идентичные мутантные гены, даже если эти гены вредны.
ДИНАМИКА ГЕННЫХ ЧАСТОТ ПРИ ПОСТОЯННОМ ДАВЛЕНИИ МУТАЦИЙ
Как мы показали в гл. 3, эволюционное состояние популяции с генетической точки зрения характеризуется относительными частотами аллелей (частоты генов) в отдельных локусах в данный момент времени. Мутационный процесс как фактор эволюции направляет изменение частоты гена, переводя его из одного аллельного состояния в другое с определенной скоростью. Эти изменения генных частот проявляются в характере и степени генетической изменчивости контролируемого этими генами признака. Некоторые генетические варианты существуют скрыто в популяции, как, например, рецессивные мутантные аллели, частота которых так мала, что они почти всегда находятся в гетерозиготном состоянии. Эта потенциальная изменчивость (резервная) становится свободной изменчивостью (проявляющейся), когда под действием мутационного давления частота новых аллелей достигает такой величины, что становится возможным переход их в гомозиготное состояние, т. е. новый ген проявится фенотипически и станет сырьем для естественного отбора.
Хотя в реальном эволюционном процессе мутирование и естественный отбор неразделимы, для упрощения расчетов мы вначале исключим действие отбора и найдем количественную связь между частотой мутаций и изменением частоты гена. Частота мутаций обычно измеряется вероятностью мутирования одного аллеля в течение одного поколения. Иными словами, если в среднем за одно поколение мутирует
один аллель из 100 тысяч таких аллелей, значит, частота мутаций равна 1/100 000 (или 10-5). Частота мутирования зависит от типа аллеля, а также от внутренних и внешних условий. В естественных условиях частота мутирования обычно равна 10-5—10~6. Но существуют и более мутабильные локусы (например, частота появления у человека гена нейрофиброматоза и некоторых изоферментов у дрозофилы равна 10-4). Как следует из низкой частоты мутирования, значение давления мутаций в эволюции тем выше, чем больше величина популяции (см. гл. 6). Именно поэтому мы будем рассматривать бесконечно большие популяции, существующие лишь теоретически.
Мутирование в одном направлении (необратимые мутации)
Пусть ро — частота аллеля А в исходной популяции, и— скорость мутирования А к другому аллелю a, a pi — частота аллеля А в первом поколении потомков. Согласно определению скорости мутирования, частота гена А уменьшится за одно поколение на величину Др = иро. Таким образом,
Pi = Ро — Ар = Ро — иро = Ро (1 — «)• (4.1)
Подставляя в это выражение вместо pi и р0 соответственно р2 и pi, находим частоту аллеля А в следующем поколении:
P2 = Pl(l — «),
и так как pi — ро(1 — и), то
Рг = Ро(1 - и)2-
Повторяя этот процесс t раз, найдем, что частота гена спустя t поколений будет равна
Р/ = Ро(1—и)'- (4.2)
Это значит, что аллель А постепенно исчезает из популяции. Величина и значительно меньше единицы, поэтому выражение (1—и)* можно приближенно заменить величиной e~ut (е — основание натуральных
логарифмов, равное 2,718...). Таким образом, формулу (4.2) можно записать в виде
Pt = Ptf~ut (4-3)
и qt = 1 — (4.4)
где q — частота гена а. Нетрудно заметить удивительное сходство этих выражений с уравнениями, описывающими ход каталитической реакции в химии и S-образную кривую роста в биологии. Различие состоит только в том, что в одном случае речь идет о частоте аллеля, в другом — о концентрации химических веществ, а в третьем — о высоте растения. Все эти переменные описываются одной и той же величиной q, которая в начальный момент времени (/ = 0) равна qQ = 1 — pQ и которая с течением времени экспоненциально стремится к предельному значению, равному 1. Скорость превращения одного состояния в другое постоянна (и) для каждого аллеля, для каждой молекулы или каждого клеточного деления.
Чтобы представить себе скорость эволюционного сдвига, обусловленного только давлением мутаций, рассчитаем число поколений, необходимых для снижения частоты гена А в два раза. Подставив вместо р* в выражение (4.3) V2po и найдя натуральные логарифмы обоих частей равенства, получим, что t = = 0,69/w. Таким образом, при и= 10~5 для снижения частоты гена А в два раза потребуется 69 000 поколений. Рассмотрим случай, когда р0 = 0,96. За 69 000 поколений частота гена А снизится до 0,48; пройдет еще 69 000 поколений, и частота снизится еще в два раза — станет равной 0,24, и т. д. Таким образом, пройдет около 5-69 000 поколений, т. е. примерно треть миллиона, прежде чем частота гена в результате мутационного процесса снизится с 0,96 до 0,03, т. е. для мутационной замены одного аллеля другим требуется очень большое число поколений. Этот расчет приводит к выводу, что, по-видимому, наблюдае
мую в природе скорость эволюционных изменений нельзя объяснить одним лишь мутационным процессом.
Мутирование в двух направлениях (обратимые мутации)
Если аллель А мутирует к аллелю а со скоростью и и аллель а вновь мутирует к аллелю А со скоростью v, то за одно поколение частота гена Л, равная pt, уменьшается на —ptu и увеличивается на (1—Pt)v. Следовательно, чистое приращение этого гена hpt равно —ptu + (\—Pt}v> з частота гена А в следующем поколении равна
Pi+\=Pt — Др< = » + (1 — и — v)pt. (4.5)
Главное различие между мутационным процессом, происходящим в одном и в двух направлениях, состоит в том, что прямые и обратные мутации образуют систему противодействующих сил. Равновесие между двумя направленными в противоположные стороны процессами мутирования можно уподобить равновесию в системе, состоящей из пружины и подвешенного к ней груза. В точке равновесия частота гена не изменяется (Др< = 0). Таким образом, приравнивая в уравнении (4.5). величины pt и pt+\ равновесной частоте р, находим
р=—^— • (4.6)
Например, частота гена А в системе, в которой происходят мутации в обоих направлениях, где и = — 2-Ю"5, a v = 10-5 (т. е. скорость процесса А— ->а вдвое выше скорости обратного процесса а— ->Л), стремится к равновесной величине р= ’/з-
Согласно (4.3), частота гена спустя t поколений равна
P< = ^ = (po + j6)e-(“+o)'. (4.7)
где р0 обозначает частоту аллеля А в исходном поколении. При увеличении t величина e-<u+”)f уменьшается
до нуля. При ро разность между исходной частотой гена и равновесной частотой отрицательна. Таким образом, как следует из (4.7), при Ро<р величина pt, стремясь к р, монотонно возрастает. Соответственно если ро > р, то pt монотонно уменьшается.
Проведенные выше математические выкладки показывают, что мутационный процесс — эволюционный фактор, способный изменять частоты генов. В большой, не подверженной действию отбора популяции давление мутаций приводит к закреплению (замене аллеля) или к сбалансированному равновесию. Однако частота мутаций обычно очень мала и поэтому давление мутаций может изменять частоты генов лишь в слабой степени по сравнению с отбором или с дрейфом генов. Итак, главная роль мутаций в эволюции состоит в создании изменчивости, а не в определении ее характера.
ПОТОК ГЕНОВ
Новые аллели, как мы видели выше, могут поступать в генофонд популяции в результате мутаций; но, кроме того, они могут поступать в генофонд в виде гамет, привнесенных иммигрантами из других популяций. Этот процесс поступления «чужих» генов мы назовем обменом генами или, если этот процесс происходит все время, потоком генов. Мутационный процесс и генный поток сходны в том отношении, что оба Эти процесса поставляют популяции новые или дополнительные аллели, т. е. оба служат источником изменчивости.
Демы, географические изоляты, расы и даже виды очень редко представляют собой замкнутые системы. Обычно между ними всегда происходит некоторый обмен генами, причем чем теснее пространственная и генетическая близость популяций, тем больше поток генов. Особенно велик поток генов между смежными демами одного вида. Поскольку степень миграции (частота скрещивания между популяциями) в таких случаях велика, следует ожидать, что соседние демы
имеют сходные генофонды. В таком случае поток генов не может привести ни к изменению генных частот, ни к дополнительной изменчивости популяций — ведь частоты аллелей у мигрантов по существу мало отличаются от частот аллелей в тех популяциях, в которые эти мигранты поступают.
Географические изоляты (популяции одного вида, разделенные географическими преградами) более обособленны, обмен генами между ними ограничен и отбор может привести к локальной адаптации. В таких популяциях закрепляются либо разные аллели, либо одни и те же аллели, но со значительными различиями по частоте. По определению поток генов между изолятами — явление относительно редкое, но если уж обмен генами произошел, то он приводит к серьезным последствиям — адаптированные генные комплексы разрушаются, частоты генов изменяются и все генетические различия между популяциями уменьшаются. Этот процесс генетического сближения особенно обостряется при вторичных контактах популяций и усилении потока генов.
Эффективность обмена генами зависит от структуры популяции или, точнее, от степени миграции (частота миграции равна Л4) и от степени генетической дивергенции участвующих в нем популяций (от величины различий между популяциями по частотам генов).
Для того чтобы эти общие соображения выразить в строгой математической форме, построим еще одну теоретическую модель. Для облегчения расчетов примем, что миграция происходит лишь в одном направлении, например из большой материковой популяции в маленькую полуизолированную островную популяцию. Примем также, что все допущения, используемые в модели Харди — Вайнберга, справедливы и для этого случая с той разницей, что миграция (поток генов) происходит с постоянной скоростью М из материковой популяции в островную. Частоты аллеля а в материковой и островной популяциях равны соответственно Q и q, Величина Q считается постоянной
(снижение частоты гена а незначительно, так как величина М — представительная выборка из материковой популяции — ничтожно мала по сравнению с величиной материковой популяции). Однако частота
До миграции
Популяция 2
Популяция 1
р 0,6 ч 0,4
Чо=О,4О
После миграции
qt=0,2(0,7) + 0,8(0,4)= 0,46
Фиг. 23. Влияние миграции на частоты генов.
Предполагается, что частоты генов в популяциях 1 и 2 различны и что происходит миграция из популяции 2 в популяцию /. После миграции популяция 1 будет содержать 1 — М эндемиков, у которых частота генов сходна с частотой их в этой популяции до миграции, и М мигрантов, у которых частота генов соответствует таковой популяции 2.
гена в островной популяции (q) будет с каждым поколением значительно изменяться, так как вклад мигрантов и их потомков в генофонд этой популяции будет неуклонно возрастать.
В каждом отдельном поколении часть островной популяции состоит из мигрантов Му а остальная часть — из эндемичных особей (1—М), Величина q для каждого поколения — просто средняя из генных частот Q и q, умноженных соответственно на долю
мигрантов и долю эндемичных особей в островной популяции (фиг. 23). Таким образом,
Частота гена а в исходном (нулевом) поколении = ?0 Частота гена а в первом поколении = q\
<71=<7o(l-Af) + QM =
= q0 — ЯоМ + QM —
= q0 — M(q0 — Q). (4.8)
Изменение в частоте гена на одно поколение равно Д</ = q\ — qo. Подставляя значение qx из уравнения (4.8), получим
kq = qx—qQ = qQ — M(qa — Q) — q0 = — M (q0 — Q).
(4.9)
Таким образом, как говорилось выше, влияние миграции, измеряемое приращением генной частоты Д<7, зависит от скорости миграции М и генетических различий между популяциями q — Q.
Долгое время на роль потока генов в развитии изменчивости природных и экспериментальных популяций не обращали должного внимания. Фактически большая часть экспериментальных популяций —это замкнутые системы, которые тщательно оберегаются от генетического загрязнения. Выводы из таких работ нельзя непосредственно переносить на природные популяции, поскольку последние представляют собой системы открытые. Однако в последние годы, главным образом благодаря проведенной Майром (1963) блестящей переоценке роли географической изоляции в видообразовании (см. гл. 8), миграции и обмену генами стали придавать большое значение. Суть этих представлений состоит в том, что поток генов задерживает видообразование. Это, безусловно, верно, если М велико, — при этом дивергенция популяций сводится к минимуму, расы объединяются в результате скрещивания между ними, и самое большее, к чему это может привести, — это образование клины.
Но миграцию чужих особей в популяцию можно рассматривать и как источник дополнительной
генетической изменчивости. Эта концепция применима к тем случаям, когда разница в частоте генов (q—Q) велика, как это можно ожидать в случае хорошо дифференцированных рас или отдельных видов. Так, например, Кемин и Эрлих (1958) нашли, что популяции водяных ужей Natrix sipedon, обитающие на островах в западной части озера Эри, изменчивы по окраске особей. Буквально все популяции sipedon, населяющие большие пространства в Огайо и Онтарио, представлены полосатыми формами, тогда как частично изолированные островные популяции содержат обе формы — полосатую и неполосатую, с явным преобладанием последней. Среди потомков, полученных в неволе от выловленных в природе самок, полосатые особи встречаются чаще, чем среди взрослых особей островных популяций. Известно, что характер окраски остается неизменным на протяжении онтогенеза; поэтому был сделан вывод, что эти две формы отличаются по выживаемости, причем неполосатые формы более жизнеспособны. На этих островах нет внутренних водоемов, и популяции sipedon обитают только в береговой зоне, где этих змей можно видеть отдыхающими на плитах известняка. Здесь полосатые особи более заметны, чем неполосатые (с точки зрения человека). Поэтому было высказано предположение, что отбор происходит в результате дифференциального уничтожения змей хищниками. Судя по наблюдениям, чайки и цапли уничтожают преимущественно молодых особей. Возникает вопрос, почему, несмотря на такой отбор, островные расы остаются полиморфными. Кемин и Эрлих считают, что полиморфизм сохраняется благодаря иммиграции. На острова постоянно мигрируют полосатые особи с материка и вливаются в островную популяцию, хотя здесь их ожидают менее благоприятные условия. Подводя итог, можно сказать, что изменчивость этих островных популяций возникла и поддерживается благодаря периодическим миграциям.
В большинстве случаев гибридизация между близкими видами не ведет к обмену генами, поскольку гибриды обычно обладают слабой приспособлен
ностью. Несмотря на это, плохо приспособленные гибриды иногда оказываются достаточно плодовитыми, чтобы дать F2 и потомство от возвратного скрещивания. Когда исходные виды репродуктивно обособлены, мы также ожидаем, что некоторое количество рекомбинантного потомства окажется приспособленным. Но в некоторых условиях наблюдаются возвратные скрещивания гибридов с той или другой из родительских форм на протяжении ряда поколений. Рекомбинантные генотипы от многократных возвратных скрещиваний все более и более приближаются к тому родительскому типу, с которым происходили эти скрещивания. Чем больше рекомбинантные генотипы приближаются к генотипам «чистых» видов, тем больше вероятность того,, что они будут адаптивны и вольются в генофонд родительской линии, с которой происходили многократные возвратные скрещивания. В процессе такой интрогрессивной гибридизации некоторое количество отдельных генов или блоков генов может перейти из одного вида в другой. Это, так сказать, «генные ручейки», соединяющие близкие виды. В результате этого процесса интрогрессии в генетическую архитектуру контактирующих форм проникают новые гены и генные комбинации, однако они, по-видимому, не нарушают целостность вида (см. пример с видом Aquilegia, приведенный в гл. 8).
Естественно, что интрогрессивный обмен возможен только тогда, когда есть хотя бы несколько гибридов. Однако в действительности гибриды между хорошо дифференцированными видами, особенно у животных, очень редки. Но можно привести множество случаев по крайней мере экспериментального получения плодовитых межвидовых гибридов у близких видов растений. Когда такие плодовитые при скрещиваниях формы сосуществуют в природе, то часто оказывается, что они экологически изолированы, будучи приспособлены к специфическим условиям соседних местообитаний, что весьма неблагоприятно для существования гибридов. Хотя гибриды и могут возникнуть, они редко могут выполнять роль инициаторов обмена генами между соседними видами. Тем
не менее потенциальные возможности для такого обмена существуют, если изменение внешних условий окажется в каких-либо отношениях благоприятным именно для выживания гибридных форм и их производных.
Вопрос о том, сколь велико значение интрогрессии в эволюции, остается открытым. Видимо, она играет достаточно серьезную роль, поскольку имеется множество примеров интрогрессии среди высших растений. Тем не менее можно утверждать, что в некоторых из этих примеров речь идет на самом деле о «скоплениях гибридов», образующихся между географическими (экологическими) расами, которые наивно принимают за морфовиды. У высших животных происходит выбор партнера для спаривания, и поэтому гибридизация между близкими видами—явление редкое. Кроме того, гибриды эти обычно стерильны и нежизнеспособны. Это еще одно доказательство того, что зоологические морфовиды больше соответствуют «настоящим» биологическим видам, чем выделенные на основании морфологических признаков формы среди растений. Тем не менее, известно несколько случаев интрогрессии (см. пример с тауи в гл. 2), и несомненно, что существует много других необнаруженных случаев.
РЕКОМБИНАЦИЯ КАК ИСТОЧНИК ИЗМЕНЧИВОСТИ
До сих пор мы говорили о происхождении генной и хромосомной изменчивости, т. е. изменчивости, обусловленной в конечном счете мутациями и их перераспределением среди популяций под действием потока генов и интрогрессии. Но, действуя на фенотипы, отбор тем самым действует на первопричины этих фенотипов — генотипы. Непосредственная причина изменчивости на генотипическом уровне — это не мутация, а перераспределение (рекомбинация) мутантных аллелей, накопленных ранее и сохраняющихся в виде внутрипопуляционных и межпопуляционных вариантов. Содержащиеся в родительских
гаметах комбинации генов объединяются в процессе оплодотворения. Эти комбинации нарушаются при мейозе, когда негомологичные хромосомы распределены независимо (второй закон Менделя), и между сцепленными локусами происходит кроссинговер.
Даже при наличии лишь небольшого числа локусов, содержащих по нескольку разных аллелей, при рекомбинации вследствие перемешивания генных комплексов может возникнуть колоссальное множество генотипов. Если п — число отдельных локусов, а г— число аллелей в каждом локусе, то возможное при рекомбинации число генотипов составит
При 10 локусах, содержащих по 4 возможных аллеля каждый, теоретическое число диплоидных генотипов равно 10 миллиардам! Многочисленность генотипических вариантов может быть обусловлена либо многочисленностью расщепляющихся локусов (/?), либо многочисленностью аллелей в каждом из большого числа локусов (г), либо, наконец, многочисленностью тех и других.
Чтобы лучше представить себе значение рекомби* нации как источника изменчивости, возьмем пример, в котором рекомбинация отсутствует, а именно бесполый организм. Изменчивость бесполых форм целиком зависит от мутаций. При бесполом размножении обмен генами между клонами невозможен, поэтому, для того чтобы мог сложиться благоприятный генный комплекс, в одной линии в результате независимого мутирования должны возникнуть все аллели, составляющие такой комплекс. Именно в связи с этим мы можем оценить всю важность полового размножения. Оно делает возможным обмен генами между отдельными родословными; мутации могут накапливаться, а затем перетасовываются посредством рекомбинирования, образуя множество различных генотипов.
У бесполых микроорганизмов большая скорость размножения и высокая плодовитость компенсируют низкую частоту мутаций и, таким образом, обеспечивают
приспособляемость этих форм к постепенным изменениям внешней среды. За счет одних только мутаций в каждом поколении возникает несколько пре-адаптированных типов. Низкая численность популяций, большая продолжительность жизни и сложность развития высших организмов наряду с их эволюционной гибкостью возможны лишь благодаря наличию у них полового размножения, обеспечивающего рекомбинацию.
Рекомбинация представляет собой, по-видимому, одно из основных свойств ДНК и поэтому вполне естественно, что она обнаружена в той или иной форме у большинства организмов (например, при половом или парасексуальном размножении). Те высшие формы, для которых характерно бесполое размножение, по-видимому, в большинстве случаев произошли от раздельнополых форм (Стеббинс, 1960). Многие из них не являются облигатно бесполыми. А те, которые, по-видимому, способны размножаться только бесполым путем, отошли от главного пути эволюционного прогресса вследствие потери гибкости.
ЛИТЕРАТУРА
С a m i n J. H., E h г 1 i c h P. R., Natural Selection in Water Snakes (Natrix sipedon L.) on Islands in Lake Erie, Evolution, 12 (1958), 504—11.
Carlson E. A., The Gene, A Critical History, Philadelphia, W. B. Saunders Company, 1966.
Fisher R. R., The Genetical Theory of Natural Selection, London, Oxford University Press (Clarendon), 1930.
Grant V., The Architecture of the Germplasm, New York, John Wiley and Sons, 1964.
M а у r E., Animal Species and Evolution, Cambridge, Mass., Harvard University Press, 1963. (Э. Май p, Зоологический вид и эволюция, изд-во «Мир», М, 1968.)
Pontecorvo G., Trends in Genetic Analysis, New York, Columbia University Press, 1958.
Stebbins G. L., The Role of Hybridization in Evolution, Proc. Am. Phil. Soc., 103 (1959), 231—51.
Stebbins G. L., The Comparative Evolution of Genetic Systems, in Evolution After Darwin, Sol Tax, ed. Vol. 1, Chicago, University of Chicago Press, 1960, pp. 197—226.
Глава 5
ЕСТЕСТВЕННЫЙ ОТБОР
Источниками генетической изменчивости служат мутации и поток генов. В результате размножения популяция пополняется множеством генотипов, образующихся путем рекомбинации аллелей. Из бесчисленного множества возможных генотипов лишь сравнительно немногие действительно возникают в популяции в каком-либо поколении. Поскольку численность любой популяции ограничена, а число возможных генотипов невообразимо велико, преобладающее большинство теоретически возможных сочетаний не реализуется чисто случайно; более того, даже некоторые из уже возникших генотипов впоследствии исчезают по случайным причинам. Конечно, чем меньше популяция, тем меньше генотипов в ней создается. Мы уже отмечали в гл. 3, что в очень маленьких популяциях закрепление аллелей может произойти в результате ненаправленного процесса — дрейфа генов. Это один из путей потери изменчивости популяции.
Хотя и в большой популяции число генотипов не бесконечно, все же здесь вероятность того, что частоты аллелей будут изменяться только под действием случая, меньше (модель Харди—Вайнберга). Однако если между особями популяции происходит конкуренция, то некоторые из них элиминируются избирательно, а не случайно. Эта избирательная элиминация, или отбор, направленный против некоторых генотипов, может привести к направленному изменению
частоты гена. Оба фактора — и дрейф генов, и естественный отбор — стремятся снизить генотипическое разнообразие популяции, но, как будет показано ниже, отбор нельзя считать случайным фактором; он создает определенную упорядоченность, увеличивая приспособленность популяции; он формирует новую картину изменчивости.
Рассмотрим теперь более детально факторы, снижающие генетическое разнообразие популяций, и подчеркнем еще раз, что постулированный Дарвином естественный отбор до сих пор считается главной движущей силой эволюции.
ЕСТЕСТВЕННЫЙ ОТБОР
Между особями, составляющими вид, существует множество различий. Те члены популяции, которые обладают признаками, способствующими выживанию особи и оставлению потомства, считаются лучше приспособленными. Естественно, что если в популяции происходит конкуренция ввиду ограниченности таких необходимых для существования факторов, как пища, партнер для спаривания, жизненное пространство и т. п., то победителями оказываются эти лучше приспособленные особи. Благоприятные признаки могут выражаться просто в физической силе, помогающей спастись от нападения хищника или победить в драке; а иногда это могут быть менее явно выраженные признаки, как, например, те, которые повышают устойчивость к заболеваниям. В некоторых случаях решающую роль могут играть признаки, кажущиеся вовсе несущественными, как, например, небольшое улучшение аэродинамических характеристик крылат* ки кленового семени. Главная особенность приспособительного признака состоит в том, что он тем или иным способом повышает, пусть даже очень незначительно, шансы носителя этого признака участвовать в создании популяции следующего поколения, т. е. оставить потомство. Конечно, способность выжить — существенный элемент приспособленности, поскольку, прежде чем дать какое-то потомство, организм дол
жен дожить до половозрелого состояния. Однако это лишь один из множества факторов, определяющих успех размножения. Например, альтруистическое поведение (повышенная забота о других особях в ущерб себе) может стать адаптивным признаком, если оно направлено на родственных особей и приводит к увеличению числа выживших потомков. Это справедливо даже в том случае, если альтруизм снижает вероятность выживания самой проявляющей его особи. В качестве примера можно привести поведение родителей, голодающих в период острой нехватки пищи, но обеспечивающих пищей своих потомков, или случаи самопожертвования, когда родители, играя роль приманки, уводят хищников от убежища, где укрываются их детеныши. В основе «выживания наиболее приспособленных» не всегда лежит «эгоизм».
Признаки, повышающие приспособленность, если они наследуются, накапливаются из поколения в поколение, и, таким образом, генетический состав популяции постепенно меняется. Такие постепенные изменения и составляют содержание филетической эволюции, цель которой — максимизация общей приспособленности популяции, а движущая сила этого процесса — естественный отбор. Естественный отбор состоит в неслучайном избирательном сохранении изменчивых генотипов. Этот процесс может иметь два исхода: 1) создание новой или сохранение прежней картины изменчивости популяции путем дифференциального сохранения некоторых генотипов; 2) потеря изменчивости, обусловленная преимущественной элиминацией других генотипов. В конечном счете это два аспекта одного и того же процесса.
Все популяции, в том числе популяции человека, домашних или лабораторных животных, а также сельскохозяйственных растений, подвержены естественному отбору. В дарвиновском смысле варьирующим признаком является приспособленность. Но в большинстве популяций домашних животных отбор самцов и самок, оставляемых на племя, определяется интересами животновода или экспериментатора. Это искусственный отбор, цели и задачи которого
определяются человеком. При искусственном отборе приспособленность определяется так же, как и при естественном отборе, но ситуация усложняется тем, что дарвиновская приспособленность накладывается на хозяйственную ценность того или иного признака. Эта разница весьма существенна; при искусственном отборе приспособленность имеет лишь второстепенное значение (см. Д. Брюбейкер, «Сельскохозяйственная генетика»).
В природе существуют репродуктивные единицы различных уровней: гены, комплексы генов, хромосомы, особи, клоны, популяции и виды. В классическом понимании (дарвинизм) отбор действует на особей, составляющих популяцию. Однако действие отбора может проявляться и при конкуренции между репродуктивными единицами на других уровнях сложности. Если предположить, что на ранней стадии эволюционного развития существовали гены в виде обособленных, ничем не защищенных единиц, между ними могла происходить конкуренция. Среди известных форм к такому состоянию близки только неклеточные нуклеопротеидные частицы — вирусы. У клеточных организмов отбор генов или генных комплексов осуществляется уже не прямо, а опосредованно, через фенотипическое проявление всего генотипа. Отбор действует на фенотипы. В той мере, в какой фенотип отражает генотипическую изменчивость, вторичное действие отбора проявляется в отборе генотипов. В этом же смысле можно говорить, что аллели, способствующие росту приспособленности, также подвержены отбору, направленному на их сохранение. В самом деле, силу отбора мы обычно выражаем в терминах превосходства аллелей и изменений частот генов, хотя в действительности конкурируют не гены, а особи.
ДЕМОНСТРАЦИЯ ЕСТЕСТВЕННОГО ОТБОРА В ЛАБОРАТОРИИ
В описанном ниже эксперименте действие естественного отбора в замкнутой популяции Drosophila проявлялось в виде изменений частоты генов во вре
мени. При постановке этого эксперимента соблюдались все условия модели Харди — Вайнберга, за исключением того, что популяция испытывала влияние естественного отбора. Хотя популяция находилась в контролируемых условиях лаборатории, состав особей, служивших родителями для каждого поколения потомков, контролировался не искусственным, а естественным отбором.
В пластмассовый популяционный ящик (фиг. 24) поместили две линии Drosophila mulleri. Одна линия имела нормальные хромосомы, а вторая была гомозиготна по аутосомной парацентрической инверсии (индуцированной рентгеновскими лучами). Инверсия была выбрана в качестве изучаемого фенотипического признака, потому что она позволяет, во-первых, исключить из рассмотрения обратные мутации и, во-вторых, идентифицировать хромосомные комбинации (генотипы).
Исходная популяция содержала 2000 мух, из которых 15% были гомозиготны по нормальной последовательности генов, а 85% — гомозиготны по инверсии. В клетку периодически добавляли питательную среду. В силу ограниченности пищи и пространства величина популяции вскоре стабилизировалась на уровне 5000—6000 особей. Через каждые 30 дней из популяции брали пробы яиц. Для определения числа гомо- и гетерокариотипов среди личинок, развившихся из этих яиц, проводили анализ гигантских хромосом слюнных желез. Затем, исходя из частот генотипов, находили частоты нормальных и мутантных хромосом. На фиг. 24 представлен процент мутантных хромосом в каждой пробе. Результаты очевидны. Примерно за 20 поколений (610 дней) частота мутантных хромосом упала с 0,85 до 0,0075.
Исследуемая популяция была очень велика (слабый Дрейф генов) и замкнута (отсутствие миграции). Давление мутаций совершенно отсутствовало. Кроме гого, по результатам предварительных тестов было известно, что все три генотипа: 1) норма/норма; 2) норма/мутант, 3) мутант/мутант — спариваются почти случайно. Таким образом, мы логически вправе
отнести изменение частоты за счет действия естественного отбора, поскольку все остальные параметры соответствовали модели Харди — Вайнберга. Сни-
Фиг. 24. Естественный отбор в замкнутой популяции Drosophila mulleri, оцениваемый по изменению частоты мутантной инверсии в сравнении с частотой нормальной хромосомы (см. текст).
Плавная кривая, проведенная через точки, обозначающие наблюдаемые частоты мутантных хромосом, соответствует теоретически ожидаемым изменениям частот, выведенным на основании относительных жизнеспособностей генотипов нормальных гомозигот, гетерозигот и гомозигот по инверсии, равных соответственно 1,0; 0,74; 0,63. Вверху фотография обычного популяционного ящика для дрозофил. Популяция мух может поддерживаться в таком ящике на протяжении многих поколений. Корм находится в стаканчиках, закрепляющихся в отверстиях в дне ящика с помощью резиновых прокладок. Стаканчики эти периодически меняют. Ящик сделан из плексигласа и на обоих концах имеет отверстия, затянутые сеткой. Пробы яиц берут из чашек со свежим кормом и помещают в стаканчики с питательной средой. Развивающихся из яиц личинок используют для приготовления цитологических препаратов.
жению частоты мутантной формы дается тавтологическое объяснение: особи, содержащие хромосомы с нормальной последовательностью, обладают большей приспособленностью.
адаптационная ценность и коэффициент ОТБОРА
В популяции, размножающейся половым путем, не может существовать двух совершенно идентичных генотипов (особей). Большая часть этой генетической изменчивости проявляется как фенотипическая изменчивость, которая подвержена действию естественного отбора. Отбор—это проявление различий между особями по приспособленности, которая, как было сказано, оценивается на основании их относительной способности передать свои гены будущим поколениям. В каждом отдельном случае адаптационная ценность в идеале должна выражать как непосредственную приспособленность, или число реальных потомков, производимых каждым генотипом, так и «долгосрочную» приспособленность, или вклад этих генотипов в последующие генофонды. Хотя мы сейчас говорим о генотипе в целом, относительную адаптационную ценность отдельных генов, генных блоков или даже популяций также можно выразить с учетом их приспособленности к существующим условиям и их преадаптированности к будущим ситуациям.
Это дополняет идею о том, что степень непосредственной приспособленности зависит от условий среды, в которых она складывалась, поскольку выживание особи и ее способность к размножению коррелируют с физиологическими условиями (вспомним описание нормы реакции). Степень приспособленности строго относительна и в другом смысле. Степень приспособленности отдельной особи вне сравнения ее с приспособленностью других членов данного дема лишена смысла. Таким образом, говоря об отборе, следует иметь в виду именно относительную приспособленность, а не просто приспособленность.
Рассмотрение адаптационной ценности и отбора на уровне целых генотипов позволяет нам приблизиться к истинной природе процесса отбора. Но это сугубо теоретический подход, неизбежно приводящий к ряду сложных концепций плейотропии, полигении, канализации развития, гомеостаза и взаимодействия
аллелей и генов. Такой подход лежит в основе многих современных представлений об отборе, видообразовании и адаптивной норме (Майр, 1963), тем не менее количественная оценка при этом почти невозможна, поскольку каждый случай по существу уникален. Преобладающее большинство наших современных идей построено на упрощенных моделях особь — ген, связанных с краевым генотипом (определяемым как генетический надел одной или большего числа особей или гамет в рассматриваемом локусе) и с изменением частоты аллелей в одном локусе. Кроме того, приспособленность рассматривается лишь в ее непосредственном аспекте, поскольку оценить «долговременную» приспособленность практически невозможно.
Несколько особей могут быть идентичны по одним компонентам генотипа и различаться по другим; они обладают специфической комбинацией определенных аллелей. Пусть, например, локус А расщепляется по аллелям а\ и а2. При этом популяция оказывается разделенной на три группы (генотипа): aia2,
а2а2. Обозначим адаптационную ценность этих генотипов соответственно №0, и W2 (что представляет собой средние адаптационной ценности отдельных особей по каждой группе). Поскольку приспособленность— понятие относительное, одну из этих величин (обычно характеризующую наиболее приспособленный генотип) принимают за единицу, выражая приспособленность остальных генотипов в долях этого стандарта.
Генотип ахах аха2 а2а2
Степень приспособ- W2
ленности Го W] w
НЛП Го — I Го — 1 S1 «70 — 1
Коэффициенты отбора (s) показывают интенсивность отбора против менее приспособленных форм. Каждая величина ($i и s2) означает пропорциональное снижение воспроизводства этого генотипа по сравнению со стандартом (в данном случае за стан
дартный генотип произвольно принят генотип а^ц). Приспособленность имеет диапазон от 1 (s = 0; такую оценку получает генотип, вносящий наибольший вклад в будущие поколения) до 0 ($ = 1, летальный или совершенно стерильный генотип).
Как было показано в гл. 3, отбор среди генотипов может привести к изменению частоты генов. Если частота одного аллеля, скажем аллеля а\, увеличивается, то можно считать, что отбор благоприятствует этому аллелю и действует против аллеля «2» хотя в действительности процесс отбора идет на уровне особей (генотипов), а не на уровне аллелей. Предпочтение, оказываемое отбором тому или иному аллелю, редко подчиняется принципу «все или ничего». Одни особи, имеющие аллель а\, могут оказаться хорошо приспособленными фенотипами, другие, имея тот же аллель, — приспособленными плохо. То же самое касается и аллеля а2. Это объясняется тем, что приспособленность особи определяется не одним только этим аллелем: адаптационная ценность каждого генотипа зависит от действия и взаимодействия всех генов, имеющихся у данной особи. Как видно, действие отбора на генотипы и на взаимно исключающие аллели имеет статистическую природу. Если в целом чистая селективная ценность всех генотипов, содержащих аллель а^, больше селективной ценности всех генотипов с аллелем а2, то отбор будет направлен на сохранение аллеля ait и поэтому частота этого аллеля среди прошедшей отбор группы будет выше, чем в популяции, не подвергавшейся отбору. Увеличение частоты аллеля а\ будет продолжаться из поколения в поколение до тех пор, пока отбор будет благоприятствовать этому аллелю. В конце концов аллель закрепится.
Большинство высших организмов проходят в своем жизненном цикле две стадии — гаплоидную и диплоидную. Можно представить себе, что существует один набор критериев приспособленности для гамет, а другой—для диплоидных генотипов. Приспособленность гаплоидной фазы зависит от таких факторов, как жизнеспособность гамет или их конкуренте-
способность при оплодотворении, тогда как действие отбора на диплоидные генотипы может происходить благодаря существующим у них различиям по жизнеспособности, относительной плодовитости или половой активности. Рассмотрим вначале последствия отбора на стадии гамет.
ОТБОР ГАМЕТ
Свободно скрещивающаяся популяция с соотношением генотипов 0,16ai<2j; 0,48«ia2; 0,36a2tz2 должна производить 0,4 гамет с аллелем ai и 0,6 гамет с аллелем а2 (см. гл. 3). Объединяясь попарно, эти гаметы образуют зиготы, из которых развиваются особи следующего поколения. Однако возможно, что вмешательство отбора произойдет в период между стадией образования гамет и стадией образования зигот. Может оказаться, что гаметы разных классов обладают дифференцированной способностью к выживанию или же какими-то различиями, влияющими на оплодотворение. Допустим, например, что средняя жизнеспособность гамет, содержащих аллель а2, составляет только 2/з жизнеспособности гамет, несущих аллель а\. Это значит, что доли тех гамет а\ и а2, которые действительно образуют зиготы, равны, т. е. что после отбора частота гамет а\ равна 0,4, а частота гамет а2 равна 2/3-0,6 = 0,4; или, приняв весь фонд гамет за 1, находим, что ах = 0,4/0,8=0,5 и а2= = 0,4/0,8=0,5. Следовательно, частота аллеля а2 уменьшилась с 0,6 до 0,5, а частота аллеля а\ увеличилась с 0,4 до 0,5. Такие изменения отразятся на генотипическом составе потомства, который будет следующим: 0,25ai«i 0,50«ia2: 0,25а2а2.
В общем виде отбор гамет можно представить следующим образом. Популяция, состоящая из ^-генотипов 2pq генотипов aifl2 и q2 генотипов a2a2t образует р гамет с аллелем а\ и q гамет с аллелем а2. Если относительную приспособленность более приспособленных гамет, скажем гамет сц, принять за единицу, то относительная приспособленность гамет а2 будет равна 1 — s, где s — коэффициент отбора или
степень отбора, направленного против этих гамет. Умножая частоту гамет каждого класса на их относительную приспособленность, находим, что эффективный фонд гамет (после отбора) определяется частотами р для а\ и б?(1 —s) для «2, а сумма этих частот равна p + q(\—s) = l—sq. Относительные доли гамет а\ и а2 после отбора равны соответственно р/(1—sq) и б?(1—s)/(l — sq). Здесь важно отметить, что сумма частот гамет после отбора равна уже не единице, а 1 — sq. Деление на эту величину производят для того, чтобы сумма относительных частот аллелей была равна единице.
Вызванное отбором изменение частоты генов обозначается обычно как Aq. Здесь \q есть разность между qi (частота аллеля а2 после отбора) и qo (частота аллеля а2 до отбора). Поэтому
\л = п — л = ~~s) — п =
У У® 1 — sq 1 — sq-
Здесь мы видим, что Aq величина отрицательная. Иными словами, под влиянием отбора гамет частота аллеля а2 уменьшилась. Если такой отбор будет продолжаться на протяжении ряда поколений, то частота аллеля а2 будет становиться все меньше, и в конце концов этот аллель совершенно исчезнет из популяции.
Среди высших организмов отбор гамет, видимо, значительно чаще встречается у растений, чем у животных. Это связано с тем, что в более дифференцированных гаплоидных гаметофитах растений аллели проявляются в большей степени. Но и у животных известны примеры отбора гамет. Нарушения соотношения гамет наблюдались у дрозофилы и у мыши. Особенно большое значение отбор гамет имеет для микроорганизмов и таких низших форм, как грибы и водоросли, у которых основное поколение — гаплоидное. У высших растений и животных, напротив, отбор проявляется главным образом на диплоидной стадии жизненного цикла — в процессе развития из зиготы взрослого организма.
ОБЩАЯ МОДЕЛЬ ОТБОРА
Проследим теперь за изменением частот генов, приписав различным генотипам различную адаптационную ценность и считая, что отбор происходит только на уровне генотипов. Хотя при определении адаптационной ценности каждого генотипа следует учитывать и способность к спариванию, мы для простоты будем считать, что скрещивание носит случайный характер и что действие отбора сводится к дифференциальному выживанию. При этом не следует забывать, однако, что в уравнении должны быть отражены все процессы отбора, действующие на всех стадиях жизненного цикла от одного поколения до другого. Результаты одного цикла отбора можно представить так, как это сделано в табл. 10.
Таблица 10
Результаты одного цикла отбора
Генотип Всего
Частота в нулевом поколении (до отбора) р2 2pq q- 1
Приспособленность w. wt —
Пропорциональный вклад P2W0 2pqW, q*W2 w
Частота в первом поколении (после отбора) p'Wo w 2pqW, w q2W2 W 1
Обратите внимание, что пропорциональный вклад находят как произведение частоты каждого генотипа на его приспособленность. Поскольку происходит отбор, лишь один или два генотипа могут иметь адаптационную ценность, равную единице. А так как по крайней мере одна из величин W меньше единицы, математически невозможно, чтобы общий вклад всех генотипов был равен единице. Эта суммарная величина, которую можно обозначить W, называется средней приспособленностью группы, подверженной от-
бору. Она равна просто сумме частот генотипов после отбора:
i^_=p2^o + 2p^i + ^2. (5.1)
Величину W можно представить также как разность между единицей и элиминируемой из популяции частью каждого из менее приспособленных (относительно стандартного) генотипов. Это вовсе не означает, что под действием отбора популяция непременно уменьшается; вполне может быть, что элиминируемые отбором особи относятся к тем формам, которые обычно гибнут в результате перенаселения. Увеличение или уменьшение популяции зависит от многих факторов, и интенсивность отбора — только один из них. Это значит, что воспроизводительная способность популяции ниже той, которую она имела бы, если бы все генотипы давали столько же потомков, сколько дает в среднем каждая особь, принадлежащая к классу наиболее приспособленных. К этому важному аспекту отбора мы еще вернемся при обсуждении межпопуляционного отбора (см. гл. 7).
Как следует из уравнений (3.1) и (3.2), частота каждого из аллелей в первом поколении, р' или q', слагается из частоты гомозигот по данному аллелю и половины частоты гетерозигот Так, частота аллеля «2 в популяции, подверженной отбору, равна
, = q^ + pqWx = q (pUZ2J-р^) 2)
Отсюда изменение частоты гена за один цикл отбора равно \q=q' — q, или, подставляя значение q' из уравнения (5.2), находим
д = q(qW2 + PWx) = q(qW2 + pWi)-qW
4 W 4 W
и далее, используя значение W в уравнении (5.1), получаем
д.в д (дУг + pu^i) - д (р^о + ZpgW, + ^ir2) = w
. (5.3)
Это выражение представляет собой комбинацию следующих трех членов:
Первый член: pq Третий член: q (UZ2 — U^i) + р (W — WQ) Второй член: W
Первый член означает, что приращение \q пропорционально величине pq при условии постоянства остальных членов. Произведение pq принимает максимальное значение при ^=’/2, а минимальное — при q=0 или при q=l (если нет генетической изменчивости, то нет и отбора).
Второй член означает, что_приращение Aq обратно пропорционально величине W при условии постоянства остальных членов. Это значит, что при некоторых определенных значениях 1Г0, W2 с увеличением средней приспособленности уменьшается приращение, и наоборот. Иначе говоря, скорость изменения частоты генов снижается, когда средняя приспособленность увеличивается, приближаясь к максимуму, и увеличивается, когда популяция далека от «потенциальной» репродуктивной способности. Это сходно с установленным ранее положением, что отбор стремится максимизировать приспособленность популяции, т. е. частота___генов продолжает изменяться до
тех пор, пока W не достигнет максимально возможного значения.
Самая существенная часть выражения для Aq — третий член. Этот член называется аддитивным или «усредненным» эффектом замещения гена. Он состоит из двух слагаемых, содержащих разности
—1^1) и p(Wi — 1FO), которые выражают влияние на приспособленность замещения одного аллеля, т. е. а2а2-+ сцс^. Аддитивный эффект есть средняя этих двух разностей, взятых с весами q и р. Таким образом, мы выразили приспособительную ценность генов, усредненную по тем генотипам, в которые входят эти гены. Любое изменение частоты гена пропорционально «среднему» эффекту. Если замещение слабых аллелей (аллели, которые дают в среднем генотипы с меньшей адаптационной ценностью)
более благоприятными приводит к увеличению средней приспособленности популяции, то аддитивный эффект— величина положительная. Когда \q равно нулю, аддитивный эффект также равен нулю. Лучший пример этого — случаи, когда пррисходит закрепление более благоприятного аллеля или когда величины Wif W2 и Wo одинаковы; при этом никакого отбора не происходит и частоты генов постоянны. Однако, поскольку «средний» эффект является также функцией частот р и q, возможна такая ситуация, когда адаптационная ценность разных генотипов различна, а средний эффект равен нулю (см. ниже пример повышенной приспособленности гетерозигот).
Величины Wi в числителе и знаменателе уравне^ ния (5.3) представляют собой линейные функции. Поэтому можно разделить все величины W на самую большую из них и, таким образом, выразить приспособленность через коэффициент отбора, как это было сделано в предшествующем разделе. Значения Aq можно найти, выражая адаптационную ценность любым из этих двух способов.
Подобным же образом можно выразить изменения частот множественных аллелей в одном локусе. Изменение частоты какого-либо одного аллеля из системы т аллелей
bql=^-w)
4 W
(5.4)
где qi — частота f-го аллеля среди аллелей а2, аз, . • •, • • •, Средняя приспособленность всех
генотипов равна Wf a Wt — средняя приспособленность всех генотипов, содержащих i-й аллель (т. е. генотипов a\ait a2aiy •••» атаг).
Так как приспособленность есть проявление фенотипа, следует ожидать, что гены, частоты которых мы анализируем, будут обладать некоторой доминантностью по признаку, определяющему приспособленность. В этом смысле можно говорить о нулевом, частичном, полном доминировании или о сверхдоминировании. Кроме того, отбор может благоприятствовать либо одному, либо другому аллелю; по своей
интенсивности отбор может быть частичным (вредные генотипы) или полным (летальные или стерильные генотипы). Располагая точными данными о степени доминирования и интенсивности отбора, можно упростить общую формулу изменения частот для какого-либо отдельного случая. Для этого в формулу подставляют значения W, выраженные через «коэффициент отбора», а р заменяют разностью 1 — q. Ниже мы рассмотрим лишь некоторые из упомянутых здесь сочетаний условий.
Полное доминирование
Если отбор направлен против вредных рецессивных гомозигот, то $ больше 0, но меньше 1 и, следовательно, №1 = 1Го=1, а 1^2 = 1—5 (фиг. 25,/). Изменение частоты рецессивного аллеля
Д<7 =
(1-<7)<7[?(1-s-D + (1-<?)(!-D1 _
1 — sq2
_ -sq2(l-q)
1 — sq2
(5.5)
Поэтому Aq всегда отрицательна и частота рецессивного аллеля q? убывает, пока не обратится в нуль. Полный отбор наблюдается в тех случаях, когда рецессивный аллель в гомозиготном состоянии детален или приводит к стерильности; в этом случае s=l. Выражение (5.5) при этом упрощается и принимает вид
д„=-^(l-?) = ~«г (56)
4 I — q2 (1 +<?)(!-<Z) (1+<7) V j
Если коэффициент отбора $ или частота гена q очень малы, то знаменатель в уравнениях (5.5) и (5.6), а также_в других моделях отбора почти равен единице, т. е. W можно приближенно считать равной единице. Величина &q выражается при этом только через числители; иначе говоря, в выражении (5.6) \q = —q2, Здесь интересно обратить внимание на то, что снижение частоты редкого рецессивного летального гена равно частоте образующихся летальных зи-
гот. Это значит, что с уменьшением частоты гена скорость его элиминации резко падает. Например,
Шкала Элиминируемая После отбора
приспособленности часть
-здг( 1-д) 1-sq2
- '/itq(l-q) l-sq
I - s.O-qf-Saq1
Фиг. 25. Схема действия отбора при разной степени доминирования в отношении приспособленности (Z — IV).
/ — полное доминирование —отбор против а2; // —полное доминирование — отбор против ar. /II — отсутствие доминирования — отбор против а2; /V —сверхдоминирование —отбор против и а2а2. Относительная приспособленность трех генотипов дана на шкале приспособленности (столбец Б). Интенсивность отбора против данного генотипа показана в графической форме в столбце В (ср. с квадратами, изображенными в гл. 3) и в алгебраической форме —в столбце Г. Приспособленность популяции W до отбора равна единице, а после отбора уменьшается на долю генотипов, элиминированных отбором. В столбце Г приведены ожидаемые изменения ча стоты генов для каждой модели.
число поколений, необходимых для снижения q с 0,01 до еще более малых величин, составляет:
Частота 0,01000
Число поколений 0
0,00250 300
Можно показать, что в общем случае частота летального гена уменьшается вдвое за 1/^ поколений. При неполном отборе процесс выведения вредного гена происходит еще медленнее. Эти расчеты позволяют судить, сколь низка эффективность евгенической программы, основанной на принудительной стерилизации (s = l) людей с тяжелыми врожденными заболеваниями, обусловленными рецессивными генами.
Отбор против доминантов при тех же частотах генов и той же интенсивности оказывается значительно более эффективным, чем против рецессивов (фиг. 25,//), поскольку в этом случае отбор действует не только против гомозигот, но и против гетерозигот по доминантному вредному гену (Wq= = U71 = 1—s; №2=1). При этом изменение частоты генов
Ал7 —— ^7) ^{^7 — ^)1 + (1 — ^7) [(1 — s) — (1 — s)]} _ _
q 1 —2(1 — q)qs — (1 -q)2s
= q\. (5.7)
1 — s (1 — q2) v '
Если доминантный ген летален, то это выражение принимает форму
1 — 1 — q2 q2 4
Это значит, что если выживают и размножаются только рецессивы, то частота гена уже за одно поколение возрастает от q до I, т. е. аллель а2 фиксируется уже в первом поколении.
(5.8)
Сверхдоминирование
Если в каком-либо локусе проявляются полное или частичное доминирование или рецессивность, то отбор действует до полного исчезновения из популяции менее приспособленных аллелей. Приращение Ад будет либо положительным, либо отрицательным в зависимости от благоприятного или неблагоприятного действия отбора на аллель а2. Если, однако, локус проявляет сверхдоминирование (гетерозиготы превос
ходят гомозигот по селективной ценности), то в популяции остаются оба аллеля и частота генов стремится к некоторому равновесному значению, заключенному в интервале между 0 и 1; это равновесное значение определяется относительной степенью «непригодности» гомозигот. Можно выразить относительную приспособленность генотипов по сверхдоминантному локусу, как Ц70=1— 51, U^i = l и WZ2=1—$2 (рис. 25./V); из уравнения (5.3) находим, что изменение частоты гена
Л_ (1 — <7) <7 {? 1(1 — s2) — 1] Ч- (1 — ?) [1 — (1 - sj]) _ 1 - (1 - Q)* - S2^
= (j —q)q(—s.q + si -stf) = 1 - «I (1 - q)2 - s2q2
_ -q)q [$i — (s2 + sj] - 1- S1 (1 _ q}2 __ S2q2 •
Приращение Aq имеет тот же знак, что и аддитивный эффект Si — #(52 4- 5i). Частота гена, достигнув равновесного значения, обозначаемого q, больше не изменяется, т. е. Д^ = 0. Равновесную частоту q можно найти, полагая аддитивный эффект равным нулю и решая относительно q уравнение
0 = $1 — <7(«2 + «1).
Получаем
(5.Ю)
S| -f- $2
Знак и величина \q определяются разностью между q и q. Если q меньше равновесного значения, kq положительно и частота гена возрастает, приближаясь к 5i/(5j Н-52); если q больше q, то частота гена снижается до точки равновесия. Частный случай такого изменения частоты продемонстрирован на фиг. 26. Когда q достигает значения, равного q, наступает устойчивое равновесие. Равновесие считается устойчивым, если после отклонения от q в ту или другую сторону частота гена снова стремится к точке q, как показано на фиг. 26 для случая, когда отбор благоприятствует гетерозиготам. Равновесие считается
6 Зак. 88
Фиг. 26. Модели сверхдоминировапия (11^0=1—5); 1Т1 = 1; Г2 = 1 - s2).
Кривые Л, В, С и D отражают приращение частоты генов (\q) при разных коэффициентах отбора. График построен на основе уравнения (5.8). Коэффициенты отбора и s2 равны соответственно: Л) 0,4; 0,267; В) 0,6; 0,4j_ С) 0,2; 0,133; D) 0,15; 0,35. Кривые а, Ь, с показывают изменение величины W для первых трех случаев. Равновесная частота в случаях Л, В и С одинакова (^ = 0,6), поскольку во всех этих случаях одинаково отношение (уравнение 5.9). В случае D это отношение меньше и равновесная частота также меньше (^ = 0,3). Приращение b.q во всех случаях положительно, если q меньше, чем и отрицательно, если q больше, чем <?. Заметьте, что W имеет максимальную величину при равновесии. Кроме того, приближение к равновесию происходит тем быстрее, чем больше интенсивность отбора (ср. кривые Л, В и С).
неустойчивым, если некоторое отклонение частоты генов от равновесного состояния по крайней мере в одном направлении все более и более отклоняется от точки q. Отбор, не благоприятствующий гетерозиготам, приводит к неустойчивому равновесию ($i и s2 в уравнении (5.9) будут при этом отрицательны). Это можно проверить, анализируя изменение частоты генов для случая, когда ITi меньше 1Г0 и меньше 1Г2.
Равновесие генных частот, обусловленное превосходством гетерозигот, означает, что оба вида аллелей могут сохранять в популяции высокую частоту. Это происходит даже в тех случаях, когда селективная ценность гомозигот одного типа, скажем а2а2, очень мала, поскольку гетерозиготы, расщепляясь, будут непрерывно пополнять фонд аллеля а2. Сохранение в популяции обоих аллелей при сверхдоминировании служит одним из способов поддержания полиморфизма на фенотипическом уровне (см. гл. 7). Когда большинство генов или супергенов и их аллеломорфов селективно сбалансированы, то достижение равновесия или приближение к этому состоянию создаст сбалансированный полиморфизм. Крайний пример такого полиморфизма — система сбалансированных леталей, т. е. такая ситуация, когда Si и s2 равны единице и, следовательно, потомство дают только гетерозиготы. (Какими будут частоты генов в линии, сбалансированной по деталям? Ответить на этот вопрос будет легче, если вспомнить, что q зависит только от коэффициентов отбора.)
ОПРЕДЕЛЕНИЕ АДАПТАЦИОННОЙ ЦЕННОСТИ
Математическая модель, описанная в предыдущем разделе, основана на определенных допущениях, в частности на ряде допущений об относительной адаптационной ценности генотипов. Мы проследили зависимость изменения частоты генов от степени доминирования признака, определяющего приспособленность. Эти данные помогают понять процессы, происходящие в реальных популяциях. Однако исследование живых популяций представляет собой полную
противоположность исследованию теоретических построений: в этом случае не известен коэффициент отбора и для нахождения величины \q удобнее использовать собранные данные. Коэффициент отбора и соответствующие величины адаптационной ценности определяются на основании данных о соотношении генотипов до и после отбора. Такой анализ гораздо легче проводить для какого-либо одного, чем одновременно для всех компонентов приспособленности.
К компонентам приспособленности относятся такие особенности особи данного генотипа, которые тем или иным способом влияют на конечную адаптационную ценность (такие свойства, как жизнеспособность, способность к оплодотворению, плодовитость, половая активность). Каждый компонент представляет собой приспособительную «черту». Определение этих компонентов у форм, которые можно скрещивать в условиях лаборатории или опытного поля, ведут специальными стандартными методами. Примером может служить определение относительной жизнеспособности упоминавшихся выше хромосомных типов у D. inulleri.
Эти три кариотипа (или хромосомных типа, или генотипа) мы обозначим здесь + 4-, -Н, и, где i означает мутантную инвертированную последовательность, а Н---нормальную последовательность генов. Жизне-
способность на всех стадиях развития от яйца до взрослого состояния определяется по дифференциальному выживанию каждого из трех кариотипов в изучаемой партии в условиях конкуренции. Большая партия гетерозиготных особей, полученных от скрещивания двух линий, была помещена в популяционный ящик, содержащий свежий корм. Поскольку эта исходная популяция содержала только один генотип, характер образования пар и плодовитость не имели значения. Поскольку исходная генерация состояла из одних гетерозигот, среди отложенных на поверхность корма яиц должно быть 25% гомозигот + +» 50% гетерозигот + 1 и 25% гомозигот Н Взрослые особи,
Именно такое соотношение было обнаружено среди хорошо развитых личинок, выращивавшихся при оптимальных условиях.
появившиеся в следующем поколении, были изъяты из ящика и спарены каждая по отдельности с известными гомозиготами. Генотип каждой мухи был определен затем методом испытания потомков — путем анализа хромосом слюнных желез нескольких личинок, развившихся в индивидуальных тест-культурах. Соотношение генотипов в популяции потомков (после отбора) составило 0,35 ++, 0,48 + i и 0,17 Н. Отсюда следует, что в период развития от яйца до имаго разные генотипы обладают разной выживаемостью (т. е. происходит отбор).
Теперь, зная соотношение генотипов до и после отбора, нетрудно найти относительную выживаемость каждого из трех генотипов (табл. 11).
Таблица 11
Определение относительной выживаемости генотипов
Генотип + + + 1
Частота до отбора 0,25 0,50 0,25
Частота после отбора 0,35 0,48 0,17
Относительная выживае- 0,35 , л °’48 о оя 0,17
= 1.4 = 0.96 = 0.68
мость 0,25 0,50 ’ 0,25 ’
1,4 0.96 0,68 п ,
или 77 ='-° 77” = 0,7 1,4
Коэффициент отбора 0 0,3 0,6
Как видно из табл. 11, признаки, определяющие выживание, не являются доминантными: коэффициент отбора для гомозигот (0,6) вдвое выше, чем для гетерозигот (0,3).
Жизнеспособность — один из главных компонентов приспособленности, однако она не эквивалентна адаптационной ценности генотипа. Теоретически адаптационная ценность — интегральная мера всех признаков, от которых зависит приспособленность. В ряде работ была проведена оценка нескольких адаптивных признаков для данного набора генотипов, но до сих
пор не создано метода, который бы позволял выразить эти отдельные оценки в виде единой величины общей селективной ценности. Кроме того, оценка этих компонентов приспособленности обычно производится в условиях, отличающихся от условий жизни популяции, поэтому полученные величины далеко не всегда сравнимы с истинным значением признака в популяции.
Единственный известный способ определения общей адаптационной ценности — использование данных об изменении частот генов на протяжении ряда последовательных поколений. На фиг. 24 представлены такие данные, полученные в эксперименте с D. mul-leri. Каждая точка графика представляет собой частоту мутантной последовательности генов в данный момент исследования. Для нахождения этой частоты из популяции брали случайную выборку особей, определяли в ней соотношение особей различных хромосомных типов и по данным о частотах генотипов находили частоты нормальных и инвертированных последовательностей генов.
Нередко ошибочно полагают, что относительную адаптационную ценность генотипов можно определить на основании данных лишь по одной выборке, сравнивая наблюдаемые частоты генотипов с частотами, ожидаемыми по формуле Харди — Вайнберга. Ожидаемыми мы считаем частоты генотипов р2, 2pq, q2, где р и q — частоты генов, обнаруженные в одной выборке. Этот метод расчета мы уже применяли в гл. 3 для определения частот групп крови М—N у коренных жителей Австралии и нашли, что наблюдаемые частоты не отличаются достоверно от тех, которых следует ожидать на основе вычисленных значений р и q. Однако Уоллес (1958) показал, что если бы даже и существовало различие между теоретически ожидаемой и наблюдаемой величиной (хотя при этом, конечно, равновесие Харди — Вайнберга не имеет места), это различие нельзя использовать в качестве основы для сравнения адаптационной ценности исследуемых генотипов. Дело в том, что адаптационная ценность зависит от двух переменных:
частоты генов до отбора и относительной приспособленности генотипов, причем ни та, ни другая нам не известны. Если использовать этот метод для сбора данных об адаптационной ценности, то мы должны допустить, что «частоты гамет остаются постоянными с момента образования поколения, из которого взята выборка». Это — необоснованное допущение, если только не известно заранее (благодаря повторным выборкам), что аллели изучаемого локуса находятся в состоянии равновесия.
Если равновесия нет и частоты генов изменяются от поколения к поколению, что наблюдалось, в частности, в описанном эксперименте (фиг. 24), то степень приспособленности можно найти по кривой, проведенной через все точки, соответствующие отдельным выборкам. Частоты, отложенные на графике, не лежат на плавной кривой, поскольку на каждую точку влияет ошибка выборки; кроме того, наклон кривой непостоянен, поскольку с уменьшением значений q различие между точками (А#) уменьшается. Ввиду всех этих осложнений для анализа изменений частот генов и определения коэффициентов отбора необходимы специальные методы. Методы, подобные, например, тем, которые разработаны С. Райтом и Г Лейви-ном (см. Спиз, 1962), сводятся к выбору (путем итерации) такой адаптационной ценности, которая, будучи введена в уравнение (5.3), дает теоретическую кривую изменения частоты гена, лучше всего совпадающую с кривой, построенной на основе экспериментальных частот (т. е. такая кривая, сумма квадратов отклонений каждой точки которой от соответствующих точек опытной кривой минимальна). Найденная таким образом величина характеризует «общую» приспособленность генотипов за данный период. Плавная кривая на фиг. 24 соответствует изменениям, ожидаемым в популяции, если допустить, что адаптационная ценность трех генотипов + +, + i и и равна соответственно 1,00; 0,74 и 0,63 и что эти величины оставались неизменными на протяжении всего эксперимента. Как показывают эти данные, инвертированная последовательность генов частично доминирует над
нормальной. Хотя значения общей адаптационной ценности довольно близки к тем, которые найдены для одного компонента приспособленности, а именно для жизнеспособности, надо думать, что и другие компоненты общей приспособленности внесли определенный вклад в репродуктивный успех одних генотипов и неудачу других. И в самом деле, дополнительные исследования показали, что разные хромосомные типы различаются по способности к откладке яиц и по скорости развития.
ТРИ ТИПА ОТБОРА
Отбор внутри популяции делят на три типа, стабилизирующий, направленный и дизруптивный (разрывающий). Эти термины характеризуют отбор по его внешнему проявлению — сохранению или отбрасыванию фенотипов, которые могут стать родителями последующих поколений. Кроме того, эти термины дают представление о типах изменений, которые могут возникать в генетической изменчивости, лежащей в основе фенотипических различий. Ответ популяции на действие селективных сил зависит не только от их характера и мощности, но и от действия и взаимодействия генов в подвергающихся отбору фенотипах, а также от изменчивости и генетической системы популяции (Мазер, 1955). Последняя зависит от системы скрещивания, численности популяции и состояния ее приспособленности. Что же касается взаимодействия генов, то следует иметь в виду, что каждый тип отбора может действовать на признаки, генетически детерминированные главным образом отдельными генами (мы их рассматривали при описании математической модели отбора), или же на количественную изменчивость, зависящую от действия и взаимодействия множества генов.
Основные типы отбора представлены на фиг. 27 в виде диаграмм, отражающих направление действия отбора на существующую группу фенотипов, варьирующих по гипотетическому метрическому признаку; у большинства особей в каждой группе этот признак
близок к средней величине, а у меньшинства, располагающегося на концах кривой распределения, значение признака близко к максимуму или к минимуму. Такое резкое дифференцирование популяции на отбираемую и элиминируемую части, как это показано на фиг. 27, конечно, условно. Только искусственному
Фиг. 27. Три основных типа отбора и вызываемые ими изменения в генетической изменчивости.
А. Стабилизирующий отбор. Б. Направленный отборлВ. Дизруптивный отбор.
отбору свойственна четкая отбраковка по типу «все или ничего». Для естественного отбора характерна дифференциальная способность к воспроизводству всей массы особей и определенная вероятность выживания для каждого класса фенотипов. Однако такие упрощенные схемы позволяют легче понять разные типы отбора. Кроме того, легче представить себе отбор как дифференциальное выживание в период, непосредственно предшествующий случайному спариванию (за исключением примера на фиг. 27, В, где скрещивание может быть до некоторой степени избирательным).
Стабилизирующий отбор
Популяция всегда находится в зависимости от условий среды. Для того чтобы существовать, популяция вынуждена постоянно приспосабливаться к условиям своего местообитания. На протяжении всей истории жизни популяции естественный отбор стремится сохранить те генотипы, которые приводят к образованию фенотипов, наиболее хорошо соответствующих комплексу внешних условий, преобладающих в данное время. Если в течение нескольких поколений условия существования остаются неизменными, то популяция имеет возможность достичь высокого уровня приспособленности (т. е. та составляющая величины Ж, на которой сказывается долговременное действие отбора, стремится к максимуму). Если это произойдет, то может наступить стабилизация генетической изменчивости и частоты генов в большинстве локусов приблизятся к равновесным значениям (0; 1 или q)—популяция достигает «селективного пика». Но отбор не прекращается. В этих условиях его роль заключается в стабилизации создавшегося положения, в сохранении статус-кво. Это значит, что некоторая группа хорошо приспособленных генотипов (адаптивная норма) сохраняется из поколения в поколение, тогда как формы, отклоняющиеся от нормы, элиминируются в каждом цикле отбора. Отбор не прекращается потому, что эти слабо приспособленные формы постоянно возникают за счет одного или нескольких источников изменчивости — мутационного процесса, миграции, рекомбинации и выщепления. Такую элиминацию резко выраженных фенотипических отклонений неоднократно наблюдали в природных популяциях. Эти наблюдения показали, что нередко именно эти формы, соответствующие крайним значениям метрического признака, наиболее слабо приспособлены.
В качестве классического примера можно привести работу Бампуса, более 60 лет назад изучавшего дифференциальное выживание в группе воробьев (Passer domestica), которых он подобрал в жестокую пургу
и принес в лабораторию. Из всех этих беспомощных и полузамерзших птиц выжила лишь половина. Проведя измерения некоторых морфологических признаков у живых и мертвых птиц, Бампус пришел к выводу, что у выживших воробьев признаки близки к средним для вида значениям (адаптивная норма), тогда как среди погибших птиц эти признаки сильно варьировали. Позднее Керн и Пенроуз установили связь между смертностью и весом новорожденных. Оказалось, что наибольшая смертность в первые 28 дней жизни наблюдалась среди детей, вес которых при рождении был значительно больше или значительно меньше среднего (3,6 кг).
Другой пример отбора такого типа, касающийся признака, непосредственно связанного с размножением, — это работа Лэка, изучавшего величину кладки у птиц. Лэк исходил из предположения, что для каждого вида существует некая оптимальная величина кладки, отрегулированная естественным отбором и обеспечивающая максимальное Зисло потомков. Хотя число яиц в кладке, казалось бы, является одним из компонентов приспособленности, тем не менее оказалось, что вследствие отрицательной корреляции между числом птенцов и способностью родителей обеспечить их пищей слишком большие кладки невыгодны. Птенцы при этом получают меньше корма и даже в более позднем возрасте гибнут чаще. Он показал, например, что у скворца в Швейцарии оптимальное число яиц в кладке равно 5. Хотя число оперившихся птенцов для кладок, содержащих более 5 яиц, было выше, однако даже после вылета из гнезда такие плохо выкормленные птенцы гибли чаще.
Здесь мы видим, как давление отбора на один признак (выживание) уравновешивается другим (откладка яиц), устанавливая оптимум (величина кладки). Это служит иллюстрацией сформулированного ранее положения о том, что приспособленность слагается из всех тех особенностей фенотипа, которые участвуют в оставлении жизнеспособного и
плодовитого потомства. Особь с ее генотипом и фенотипом, популяция и ее генофонд — все это в высшей степени сложно организованные системы, каждая часть которых подгоняется к другой с помощью естественного отбора. Естественный отбор ведет к такому равновесию компонентов, которое обеспечивает высокую приспособленность особей и популяций к преобладающим условиям существования. Вследствие такого равновесия отбор нередко благоприятствует не максимальному проявлению отдельного признака, а какому-либо промежуточному уровню.
Отбор, благоприятствующий сохранению особей, количественные признаки которых близки к среднему значению или равны ему, называется стабилизирующим отбором (фиг. 27, Л). Стабилизирующий отбор иногда приводит к некоторому снижению изменчивости признака, причем это снижение не сопровождается изменением среднего значения признака (обратите внимание на заштрихованную площадь). Размах изменчивости сужается, когда отбор благоприятствует «среднему» значению, но популяция еще не достигла селективного пика. Сужение изменчивости может-наблюдаться и в том случае, если популяция, уже достигшая сбалансированного состояния, внезапно подвергается более интенсивному стабилизирующему отбору. Однако если элиминация отклоняющихся особей полностью уравновешивается их поступлением в популяцию в результате размножения, то структура популяции может длительное время оставаться неизменной. Во всяком случае, если отбор все время благоприятствует «ординарным» значениям, то среднее значение признака остается неизменным.
Стабилизирующий отбор по признакам, контролируемым одним локусом, характеризуется силами, удерживающими частоты генов вблизи их равновесных значений, т. е. 1 или q. Поддержание частот аллелей на промежуточном уровне (q) иногда связано со сбалансированным полиморфизмом, характеризуемым адаптивным преимуществом гетерозигот (примеры такого полиморфизма см. гл. 7).
Направленный отбор
Отбор способствует сохранению признаков, благоприятных при существующих условиях, и создаваемая таким образом приспособленность отнюдь не гарантирует каких-то преимуществ ее носителям в будущем. Чем дольше популяция находится в условиях стабильного местообитания, тем выше ее шансы достигнуть приспособленности и сохранить ее впредь. Если популяция оказывается в новой среде, то следует ожидать, что вместо стабилизации изменчивости будет наблюдаться прямо противоположная картина — резкое ускорение эволюционного процесса. При этом может оказаться, что наиболее редкие варианты, прежде сравнительно плохо приспособленные, будут сохраняться отбором и, следовательно, фенотипический оптимум последующих поколений будет отличаться от фенотипической нормы, характерной для предков. Поскольку IV под влиянием отбора стремится достигнуть максимума, гены и генотипы, ответственные за такие варианты, постепенно изменяются, пока не будет достигнута новая точка равновесия. Отбор, при котором оптимальной приспособленностью обладают фенотипы, расположенные не в середине, а ближе к одному из концов кривой распределения частот фенотипов, называется направленным (фиг. 27, Б). На такой отбор популяция отвечает сдвигом среднего значения признака для популяции в направлении к его среднему значению для отбираемой группы. В терминах модели для одного гена это можно сравнить со случаями, когда отбор благоприятствует одному аллелю, а величина W не достигла максимального значения. Концепция о том, что всякое направленное изменение в наследственной структуре популяции составляет отдельную ступень микроэволюции, представляет собой самую сущность дарвиновской теории механизма эволюции.
Примеров направленного отбора так много, что даже просто перечислить их нет возможности. Некоторые из них рассмотрены в следующем разделе книги; есть такие примеры и в других книгах (см., например, Добжанский, 1955).
Дизруптивный (разрывающий) отбор
Третий тип отбора можно рассматривать как обратный по отношению к стабилизирующему отбору в том смысле, что при этом множество различных сил действует на различные формы одной популяции. Отбор, благоприятствующий более чем одному фенотипическому оптимуму, т. е. совершенно различным генотипам, и действующий против промежуточных форм, называется дизруптивным, или разрывающим. В этом случае, как показано на фиг. 27, В, оптимумы образованы крайними типами, располагающимися на концах кривой распределения частот фенотипов.
Ожидаемый ответ популяции на дизруптивный отбор—установление прерывистости в изменчивости. В итоге дизруптивный отбор может привести: 1) к полиморфизму — состоянию, при котором каждый оптимум представлен различной морфой; 2) к дивергенции и изоляции — состояниям, при которых оптимумы представлены особями, образующими локально адаптированные «субпопуляции». Первое условие, необходимое для установления полиморфизма, — некоторая функциональная взаимозависимость отбираемых генотипов, тогда как репродуктивная изоляция ожидается (в теории) при дизруптивном отборе независимых групп, которые в гетерогенной среде становятся пространственно ориентированными в силу своих различных экологических потребностей (Мазер, 1955). В любом случае конечный результат сводится к направляющему действию, т. е. каждому классу отбираемых вариантов свойственно свое направление изменений, свой оптимум, к которому и стремится структура группы (фиг. 27, В)1. Распределение фенотипов по количественному признаку, зависящее от интенсивности отбора, различий между оптимальными фенотипами и продолжительности дизруптивного отбора, с возрастанием дисперсии под действием дизруптив-
1 Случаи дивергенции между «субпопуляциями» также можно анализировать с точки зрения направленного отбора в отдельных демах, учитывая при этом и величину потока генов между ними (см. гл. 4).
ного отбора превращается в мультимодальное. Именно к такому результату привели опыты на дрозофиле; в условиях случайного скрещивания отбор одновременно благоприятствует увеличению и уменьшению числа щетинок (Гибсон и Тудэй, 1964).
По-видимому, в природе некоторые виды полиморфизма обусловлены именно дизруптивным отбором. В качестве примера можно привести множество мимикрирующих форм бабочек у многих географических рас африканского парусника Papilio dardanus. Каждая из нескольких обычных для данной местности форм сходна с каким-либо обитающим в той же местности несъедобным видом бабочек. Это сходство сравнительно съедобных форм с несъедобными и предохраняет первых от уничтожения птицами (фиг. 28). Как показывают многочисленные данные, такой тип мимикрии особенно эффективен в том случае, когда численность подражающих особей значительно меньше численности особей, служащих моделью. Это объясняется тем, что хищникам необходимо многократно убеждаться в несъедобности модели, для того чтобы научиться избегать ее. Если же несъедобная модель встречается редко, то мимикрирующая форма не сможет сохраниться в течение сколько-нибудь долгого времени, поскольку отвращение у хищников к внешнему виду модели будет выражено слабо. Отсюда был сделан вывод, что значительно благоприятнее для таких видов, как Р. dardanus, подражать не одной, а нескольким несъедобным*формам. По-видимому, полиморфизм у этого вида возник в результате дизруптив-ного отбора — различные морфы (оптимумы) функционально связаны, поскольку успех каждой из них зависит от наличия других морф.
Исследования последних лет, собранные в обзоре Шеппарда (1961), позволяют считать, что главные аллели, контролирующие подражательную окраску, вначале малоэффективны и что совершенствование этой окраски происходит постепенно, путем отбора модификаторов, специфических для каждой расы, содержащей мимикрирующие формы. Более того, известно, что среди серий множественных аллелей,
Фиг. 28. Мимикрирующие и немимикрирующие формы Papilio dardanus и их модели.
Самка (/) и самец (5) Р. dardanus с Мадагаскара — немимикрирующая раса. Неприятная на вкус модель Danaus chrysippus (2), обитающая на востоке и на юге Африки, и подражающая ей самка Р. dardanus (3). (4). Гибрид между мимикрирующей и немимикрирующей расами, не имеющий защитной окраску.
появляющихся в каждой популяции, имеется доминирование. Однако при скрещивании рас и сочетании в гетерозиготах главных генов из разных генофондов возникают особи с промежуточным фенотипом (которые, естественно, обладают менее совершенной мимикрией). Это отсутствие доминантности между аллопатрическими формами заставляет предполагать, что доминантность, наблюдающаяся между принадлежащими к одному дему типами, обусловлена спецификой генетического фона (модификаторы), сложившегося в каждой расе.
Рис. 28. (Продолжение)
Модель, принадлежащая к другому роду Amauris hiavlus dominlcanus (5)’ ее самка, которой подражает Р. dardanus (7), и особь с не вполне выраженной подражательной окраской, выщепившаяся при гибридизации с немимикрирующей расой (5). Неполная мимикрия возникает под действием модифицирующих комплексов, появляющихся в районах, где модели редки.
Обсуждая модели приспособленности и отбора, мы исходили из упрощающего допущения об однородности среды и постоянстве адаптационной ценности на протяжении данного периода отбора. Такое положение редко встречается в природе. Условия среды варьируют во времени и пространстве и соответственно изменяются силы отбора. Разные виды по-разному приспосабливаются к изменениям среды.
1) Одни становятся высокоспециализированными; обычно это редкие формы, занимающие ареал с довольно однородными и постоянными условиями.
2) Другие, напротив, состоят из генотипов с широкой нормой реакции: благодаря фенотипической гибкости они морфогенетически и физиологически приспосабливаются к возникающим в разное время изменениям условий среды.
3) Третьи приспосабливаются к гетерогенным условиям благодаря генотипической гибкости (полиморфизму) вплоть до возникновения множества морф, каждая из которых приспособлена к определенной субнише в пределах одной области.
4) Ведущие сидячий образ жизни или малоподвижные виды животных и большинство растений обычно приспособлены к непосредственно окружающей их среде. Обычно, помимо генотипической и фенотипической гибкости, у многих из этих видов наблюдается подразделение на группы (экотипы), каждая из которых приспособлена к специфическим экологическим условиям в пределах одной или нескольких субобластей видового ареала. Такие «субпопуляции» обычно соприкасаются, но, несмотря на поток генов из других соседних «экологических» рас, они сохраняют свою самостоятельность благодаря экологически специфичным давлениям отбора.
5) Виды, отличающиеся большой подвижностью, как, например, многие высшие животные, обычно занимают более обширные ареалы с широким спектром физических и биологических условий. Некоторые из них отвечают на изменения экологической обстановки образованием географических рас, каждая из которых занимает сравнительно большую часть ареала вида (политипические виды). Таким образом, различие между группами 4 и 5 носит главным образом количественный характер (экотипы можно считать микро-географическими единицами).
Ответы популяции, относящиеся к типам 3 и 4, т. е. экологический полиморфизм и дивергенция, очевидно, осуществляются путем дизруптивного отбора, поскольку в каждом из этих случаев многочисленность оптимумов совмещается со скрещиванием между группами, выделенными отбором. Однако среди генетиков-популяционистов нет единого мнения
о том, может ли дивергенция среди «субпопуляций» вести к генетической изоляции (симпатрическое видообразование) при наличии обмена генами между группами. Мазер (1955) предположил, что «там, где изменения условий существования приводят к тому, что в различных группах популяций отбор благоприятствует различным оптимумам, возникнет тенденция... к генетической изоляции этих групп друг от друга». Однако для реализации такой тенденции «.. оптимальные фенотипы должны быть независимы друг от друга... группы должны быть достаточно отличными... (и) разделение популяций должно продолжаться во времени».
Теоретически очень мощные разобщающие силы могут привести к изоляции генотипов в пределах одной популяции, и это было даже подтверждено в лабораторных опытах с дрозофилой (Тудэй и Гибсон, 1962). Однако Майр (1963) считает, что необходимая для такого разделения интенсивность отбора значительно больше той, которая возможна в природе. Хотя все признают, что дизруптивный отбор может привести к фенотипическому полиморфизму (пункт 3), включая генетическую дивергенцию, связанную с гетерогенностью условий в пространстве (пункт 4), Майр считает, что это никогда не может привести к симпатрическому видообразованию, учитывая изменяющиеся давления отбора и поток генов из соседних демов. Эти разногласия, по-видимому, сводятся к вопросу о том, какова должна быть величина потока генов, чтобы предотвратить генетическое смешение экологически различных субгрупп популяции. Сейчас мы еще не готовы ответить на этот вопрос.
ОТБОР В ПРИРОДНЫХ ПОПУЛЯЦИЯХ
Все примеры прогрессивного отбора можно разбить на четыре категории.
1) Непрерывное повышение урожайности; улучшение химического состава; повышение устойчивости к болезням; повышение молочной и мясной продуктивности, увеличение яйценоскости, достигаемые
с помощью искусственного отбора среди сельскохозяйственных растений и животных. Успехи искусственного отбора лучше всяких других доказательств свидетельствуют о колоссальных возможностях отбора в смысле изменения признаков вида.
2) Результаты многочисленных лабораторных опытов по искусственному и естественному отбору. Один из примеров — описанный выше опыт с D. mulleri. Главная цель экспериментов такого типа—проверка теоретических моделей отбора и получение дополнительных данных для построения более совершенных моделей (о наследовании количественных признаков см. Фалконер, 1960).
3) Бесчисленное множество самых изощренных способов специфического приспособления к условиям существования, наблюдаемое у растений и животных.
4) Принято считать, что хотя адаптивные свойства сохраняются благодаря действию стабилизирующих факторов, первоначально они включаются в адаптивную «норму» вида под действием направленного отбора, т. е. примерно таким же образом, как происходит постепенное улучшение хозяйственно-важных признаков у домашних животных и культурных растений. Хотя эти данные косвенные, поскольку об отборе судят a posteriori по его результатам, однако они хорошо дополняются последней группой примеров — прямым наблюдением длительных изменений в природных популяциях. Среди этих последних почти все исследования касаются полиморфизма в популяциях, недавно оказавшихся в новых условиях.
Наиболее детально проанализированные и наиболее наглядные примеры такого полиморфизма — увеличение меланистических форм бабочек (обзор Форда, 1964). Это явление известно под названием «индустриального меланизма», так как изменения частоты генов связаны во времени и пространстве с интенсивным развитием промышленности в последние несколько десятков лет. Оно распространено вокруг промышленных центров на севере Европы, США и особенно Англии, где известно более 70 видов бабочек, содержащих меланистические формы. Наиболее детально
английские исследователи изучили бабочку Biston be-tularia (L). Все экземпляры этого вида, собранные до середины XIX века, имели серовато-белую окраску с темными пятнышками на теле и на крыльях. Эта типичная окраска обеспечивала насекомым надежную маскировку во время дневного отдыха на покрытых лишайником стволах деревьев. Однако многие из современных популяций beiularia полиморфны, т. е. содержат и совершенно черные меланистические морфы (carbonaria). Этот полиморфизм контролируется аллелями одного локуса, причем меланистические формы доминируют над типичными. Первый меланистический экземпляр был выловлен в Манчестере в 1848 году. Согласно расчетам, в то время меланисты составляли только 1 % всей популяции. Однако частота этих форм в Манчестере и в других больших городах постепенно увеличивалась до тех пор, пока в конце века они не стали преобладающими (свыше 90% в отдельных популяциях). Чрезвычайно интересно то обстоятельство, что распределение этих популяций заметно коррелирует с промышленными районами Англии. В сущности, популяции с преобладанием типичной формы обитают в настоящее время только в Ирландии, на севере Шотландии и на юго-западе Англии.
Кеттлуэлл (см. Форд, 1964) исследовал факторы, ответственные за это явное преобладание меланисти-ческой формы в промышленных районах. Он нашел, что фантастическая скорость замены светлой морфы есть следствие сильного направленного отбора, главным фактором которого служит избирательное уничтожение птицами. Кеттлуэлл исходил из того, что в задымленных лесах вокруг промышленных городов на стволах деревьев нет лишайников, а деревья почернели от копоти. В таких местах покровительственной окраской является черная, тогда как на покрытых лишайниками деревьях в незагрязненных сельских районах менее заметной оказывается светлая пятнистая окраска (фиг. 29).
Для исследования были выбраны два района — загрязненные леса близ Бирмингема, где популяция на 87% состояла из меланистов, и незагрязненный
район в Дорсете, где популяция была мономорфной и состояла из одних только светлых особей. В обоих районах провели ряд экспериментов; бабочек вылавливали, метили, выпускали на волю, а затем вновь вылавливали, регистрируя соотношение темных и светлых особей. Изменение этого соотношения среди
Фиг. 29. Типичная и меланистическая формы Biston betularia.
А. Бабочки на покрытом лишайником древесном стволе в сельской местности с незагрязненным воздухом. Б. Бабочки, сидящие на закопченном стволе дерева близ Бирмингема (Англия). Jia фото А типичная форма сидит ниже и правее меланистической.
бабочек, выловленных вторично, подтвердило гипотезу об избирательной элиминации разных морф. Среди вторично выловленных бабочек число особей, окраска которых контрастировала с цветом древесных стволов, было пропорционально меньше, чем особей с защитной окраской. Так, в одном эксперименте были выпущены в загрязненный лес Бирмингема 151 мела-нистические и 64 типичные светлые бабочки. Среди вторично выловленных меченых особей оказалось 82 темные и 16 светлых, т. е. 52% выпущенных тем
ных и 25% выпущенных светлых. В незагрязненном районе Дорсета были выпущены 406 темных и 393 светлые особи. Выловить вторично удалось 19 темных и 54 светлые бабочки, что составляет 4,7 и 13,7% соответственно. Совокупность всех этих результатов показывает, что отбор действительно происходит и что он благоприятствует сохранению особей, обладающих защитной окраской.
Один из факторов отбора, действующего против хорошо заметных форм, был обнаружен путем непосредственного наблюдения (Кеттлуэлл и Тинберген). На стволы деревьев поместили равные количества темных и светлых бабочек, а затем наблюдали за ними из укрытия и фотографировали в течение всего дня. Вскоре были замечены птицы, разыскивавшие бабочек, находившие и поедавшие их. В Бирмингеме горихвостки уничтожили 43 хорошо заметных светлых особей и только 15 меланистов. А в Дорсете были получены противоположные результаты: пять видов птиц уничтожили 164 меланистов и только 26 светлых бабочек, причем светлых особей птицы поедали только после того, как были съедены все меланисты.
Такое простое толкование несколько усложняется некоторыми обстоятельствами. Во-первых, известна третья форма пятнистых бабочек — островная (insu-laria). По внешнему виду она занимает промежуточное положение между светлой и меланистической. Так же как меланистическая, эта окраска доминирует над светлой, хотя контролируется аллелем, находящимся в другом локусе, не сцепленном с локусом, который контролирует две другие окраски. Частота in-sularia (которую можно отдифференцировать только от типичной светлой формы, поскольку ген меланистической окраски обладает эпистатическим действием) редко бывает высокой. Эта форма чаще всего встречается в пограничных по степени загрязнения районах. Такое распространение служит еще одним подтверждением гипотезы о том, что покровительственная окраска коррелирует со степенью загрязненности среды — промежуточным формам благоприятствуют промежуточные условия.
Во-вторых, аллели, контролирующие меланистиче-скую окраску, по-видимому, обладают плейотропным действием. Они определяют не только характер окраски бабочки, но ее поведение и относительную жизнеспособность. Кеттлуэлл нашел, что особи каждой морфы предпочитают отдыхать на неконтрастирующей поверхности. Кроме того, судя по некоторым данным, полученным при смешанном разведении в лаборатории, меланистические особи обладают некоторым физиологическим преимуществом на личиночных стадиях, особенно в случае голодания личинок. Однако эти данные вызывают некоторое сомнение, поскольку возвратные скрещивания, проведенные в 1900 году, не обнаружили достоверного различия между разными типами окраски по жизнеспособности. Возможно, что превосходство меланистических особей по жизнеспособности вполне реально, но возникло сравнительно недавно. Возможно, что по мере увеличения численности меланистических особей весь генный комплекс вида перестроился с образованием новой генетической среды (гены-модификаторы), изменившей поведение и жизнеспособность экологически более совершенных темных особей. Это объяснение приемлемо и для толкования следующего обстоятельства, касающегося этого вида.
В-третьих, доминантность гена, определяющего меланистическую окраску, очевидно, сформировалась за последние сто лет. Первые из обнаруженных меланистических особей были темнее, чем insularia, но далеко не такие темные, как современные гетерозиготы по гену меланистической окраски, которые неотличимы от гомозигот по этому гену. Можно утверждать, что первые меланистические особи были гетерозиготны, поскольку частота мутантного гена была очень мала. Если, как предполагается, ранние гомозиготы были и в самом деле темнее гетерозигот, то отсюда следует, что доминирование возникло между взаимоисключающими аллелями.
Индустриальный меланизм — пример направленного отбора. Ген, однажды возникший как редкий мутант, внезапно оказался благоприятным, быстро рас
пространился в популяции и низвел своего предшественника до уровня мутанта. В течение переходного периода от редкости до преобладания и «мутантный», и «нормальный» аллели имели довольно высокую частоту, т. е. вид стал полиморфным. По определению Форда, генетический полиморфизм — это одновременное существование в популяции двух или нескольких различных форм в таких соотношениях, что для сохранения самой редкой из них недостаточно только обратных мутаций. Полиморфизм, подобный индустриальному, называется переходным полиморфизмом в силу временной природы замещения гена (сравните с описанным ранее в этой главе сбалансированным полиморфизмом).
ВЗАИМОДЕЙСТВИЕ ЕСТЕСТВЕННОГО ОТБОРА С ДРУГИМИ ФАКТОРАМИ ЭВОЛЮЦИИ
Действие каждой из основных эволюционных сил— мутационного процесса, миграции, отбора и дрейфа генов, известных как элементарные факторы, ответственные за изменение частот генов, — мы охарактеризовали количественно с помощью простых моделей. Мы выделили также два аспекта эволюционного процесса: создание сырья для эволюции (разнообразие аллелей и генотипов) и преобразование создаваемой таким образом изменчивости. Мутационный процесс и миграция наряду с рекомбинацией, присущей раздельнополым видам, служат источниками генетической изменчивости. Однако только часть возникающих генотипов представлена в последовательном ряду генетически преемственных популяций — остальные теряются. Мы только что видели, что подобная потеря генетических вариантов на второй стадии процесса приводит к дифференцированности, когда непрерывность (чередование гаметных и зиготных фаз) проходит через сито естественного отбора, а несколько ранее мы показали, что в любой популяции, численность которой ограничена, причиной элиминации может быть также случайная ошибка выборки или дрейф генов.
В разное время «направляющим» фактором или «причинным» агентом эволюционного процесса считали то один, то другой из этих основных факторов. Были периоды, когда в эволюционной теории преобладало влияние мутационистов, селекционистов и даже сторонников дрейфа. Современная синтетическая теория, получившая почти всеобщее признание, учитывает глубокую связь этих двух аспектов процесса и несомненное взаимодействие различных факторов в создании приспособленности.
Защитники современного неодарвинизма все еще утверждают, что «ведущим» фактором эволюции служит естественный отбор, поскольку это единственная сила, направленная на достижение некого эталона приспособленности. Когда и какой тип изменчивости возникнет в популяции или будет потерян в силу действия дрейфа генов, определяется главным образом случайно. Например, мутации и рекомбинации в отсутствие какого-либо контролирующего влияния будут создавать множество аберрантных вариантов и аномалий, поскольку большинство новообразованных и повторно возникших мутантов вредны и поскольку процесс рекомбинации (половое и парасексуальное размножение) просто пересортировывает существующие сочетания генов, причем делает это без учета потребностей организма. Крайне маловероятно, чтобы высокоинтегрированные генотипы, ответственные за совершенную структуру главных приспособлений, таких, как глаз позвоночных или шишка хвойных, могли возникнуть в результате случайных комбинаций случайных изменений.
Теория естественного отбора пришла на смену старым автогенетическим и финалистическим представлениям о силах, направляющих процесс эволюции, наблюдаемый a posteriori. Как правильно заметил Фишер (1954), естественный отбор — это тот «процесс, благодаря которому вероятность некоего случайного события, a priori крайне малая, так увеличивается со временем, что крайне маловероятным становится не наличие, а отсутствие этого события». Приспособления возникают посредством естественного отбора. От
бор снижает разнообразие организмов, но действует при этом по-разному, сохраняя одни и уничтожая другие генотипы и накапливая с течением времени адаптивные аллели и генные комбинации. В результате можно сказать, что действие отбора «порождает» органические формы и функции. В классической метафоре действие отбора сравнивается с творчеством скульптора, который извлекает из глыбы мрамора скрытую в ней статую, откалывая ненужные куски.
ЛИТЕР АТУ РА
Dobzhansky Th., Evolution, Genetic and Man, New York, John Wiley and Sons, 1955.
Falconer D. S., Introduction to Quantitative Genetics, New York, The Ronald Press Company, 1960.
Fisher R. A., Retrospect of the Criticisms of the Theory of Natural Selection, in Evolution as a Process, Julian Huxley, A. C. Hardy and E. B. Ford, eds. (London, George Allen and Unwin, Ltd., 1954), pp. 84—98.
Fisher R. A., The Genetical Theory of Selection, London, Oxford University Press (Clarendon), 1930. (New York, Dover Publications, Inc., 1958.)
Ford E. B., Ecological Genetics, New York, John Wiley and Sons, 1964.
Lerner I. M., The Genetic Basis of Selection, New York, John Wiley and Sons, 1958.
Mather K-, Polymorphism as an Outcome of Disruptive Selection, Evolution, 9 (1955), 52—61.
M а у r E., Animal Species and Evolution, Cambridge, Mass., Harvard University Press, 1963. (Э. Май p, Зоологический вид и эволюция, изд-во «Мир», М., 1968.)
Sheppard Р. М., Some Contributions to Population Genetics Resulting from the Study of the Lepidoptera, in Advances in Genetics, vol. 10, E. W. Caspari and J. M. Thoday, eds. (New York, Academic Press, Inc., 1961), pp. 165—216.
S p i e s s E. B., Papers on Animal Population Genetics, Boston, Little, Brown and Company, 1962.
Thoday J. M., Gibson J. B., Isolation by Disruptive Selection, Nature, 193 (1962), 1164—1166.
W a 11 а се B., The Comparison of Observed and Calculated Zygotic Distributions, Evolution, 12 (1958), 113—115.
Глава 6
ДРЕЙФ ГЕНОВ
Из всех факторов, ответственных за существенные эволюционные изменения, главным является естественный отбор. Как мы уже говорили некоторое уменьшение генетической изменчивости — характерный признак, сопровождающий увеличение приспособленности генотипов. Другой фактор, снижающий наследственную изменчивость популяций, — это дрейф генов (см. гл. 3). Дрейф генов связан со случайными колебаниями частот аллелей, вызванными случайностью выборки; при этом наблюдается тенденция к фиксации того или иного аллеля, особенно ясно выраженная в очень маленьких популяциях. Поскольку случайность выборки математически вытекает из мен-делевского механизма наследования, во всякой ограниченной по численности, а следовательно в любой, природной популяции происходит дрейф генов. Тем не менее эволюционное значение таких случайных изменений все еще остается предметом дискуссий.
ВЗАИМОДЕЙСТВИЕ СЛУЧАЙНЫХ И НЕСЛУЧАЙНЫХ ФАКТОРОВ
Теоретические представления о случайных процессах в популяциях, разработанные главным образом Райтом, были подвергнуты экспериментальной проверке как на реально существующих, так и на теоретических популяциях.
Результаты исследований Шаффера с помощью вычислительной машины, особенно наглядно проде-
монстрировавшие процесс дисперсии, были целиком обусловлены случайными процессами, поскольку отбор, мутационный процесс и миграция в модели отсутствовали (см. фиг. 14).
В серии опытов, поставленных на популяциях Drosophila melanogaster, миграцией и мутациями можно было пренебречь, но отбор оставался в силе. Используя в качестве родоначальников каждого следующего поколения по 8 случайно взятых особей (4 самки и 4 самца), Керр и Райт создали 96 небольших популяций, которые содержались по отдельности. Исходно каждая «линия» содержала с одинаковой частотой (по 50%) два аллеломорфных гена — ген forked (раздвоенные щетинки) и нормальный аллель этого гена (нормальные щетинки). Через 16 поколений оба эти гена имелись только в 26 линиях, в 29 линиях произошло закрепление гена forked, а в 41 линии — закрепление гена нормальных щетинок. Таким образом удалось показать, что дрейф генов стремится сделать популяцию гомозиготной, причем закрепление того или иного аллеля в каждой данной популяции определяется главным образом случайно. Однако следует заметить, что в этих экспериментах отбор несколько повлиял на результаты, так как он в некоторой степени благоприятствовал сохранению нормального аллеля. Действие отбора было еще более заметным в других опытах, где использовались аллели Ваг (полосковидные глаза) и дикий тип (нормальные глаза). Закрепление нормального аллеля наблюдалось в 95 из 108 линий, а аллель Ваг достиг 100% только в 3 линиях. Самый существенный вывод из этих опытов состоит в том, что гомозиготным становится большинство линий, и при этом иногда происходит потеря того аллеля, который, казалось бы, обладает преимуществом. Подобный результат (случайное закрепление слабого аллеля) вряд ли возможен в больших популяциях, где преобладает влияние отбора. Керр и Райт показали также, что возможность случайного закрепления резко уменьшается, если отбор сильно благоприятствует гетерозиготам. Так, в третьей серии опытов, где малые популяции содержали
аллели, дающие гетерозис (селективное превосходство гетерозигот над гомозиготами обоих типов), также наблюдался разброс по частоте аллелей, но в этом случае за 10 поколений стали гомозиготными только 8 линий из 113.
Таким образом, эти опыты подтвердили математический результат, согласно которому в малых популяциях частоты генов подвержены случайным флуктуациям. Справедливость этого математического результата не вызывает сомнения. Вопрос состоит в том, имеет ли генетический дрейф какое-то эволюционное значение. Мы уже показали, что мутационный процесс, миграция и рекомбинация играют важную роль в возникновении генетической изменчивости, а следовательно, через естественный отбор и в создании адаптации органического мира, но роль в эволюции дрейфа генов нам не совсем ясна. Дрейф генов, подобно мутированию или рекомбинации, — процесс биологически ненаправленный, но в отличие от последних он не создает разнообразия, а напротив, уничтожает его. Дрейф генов — непременный спутник отбора в его преобразующем действии на популяцию, но в противоположность отбору он не детерминирован и лишь нарушает упорядоченность природы. Если дрейф генов играет какую-либо положительную роль, то она неизбежно должна быть связана с действием факторов, вызывающих систематические генетические изменения в популяции; в частности, он может способствовать фиксации благоприятных аллелей или сочетаний генов или же противодействовать консерватизму стабилизирующего отбора. Однако даже самые горячие защитники значения дрейфа генов как важного фактора эволюции неизменно подчеркивают, что это лишь фактор, дополняющий естественный отбор.
Проблема эволюционной роли случайного блуждания генных частот, естественно, не ограничивается изложенными выше взаимоотношениями дрейфа с отбором. Она значительно сложнее, и для ее решения необходимо рассмотреть взаимодействие всех факторов эволюции в количественном аспекте. Дрейф генов как таковой зависит прежде всего от эффективной репро
дуктивной величины популяции (AQ, а его эффект определяется уровнем и направлением действия таких систематических факторов, как давление мутаций (и), поток генов (Л4) и в особенности отбор (s). Расчеты Райта показывают, что вероятность случайного закрепления увеличивается с уменьшением всех этих параметров (N, М, s и и), т. е. в случае небольших изолированных популяций, которые содержат селективно нейтральные аллели и в которых частота мутаций мала, что препятствует замещению утраченных аллелей. Но понятие «малая частота» относительно. Его нужно охарактеризовать количественно определенными значениями параметров s, М и и, а также N.
Именно это и сделал Райт. Математическая сторона расчетов сложна и выходит за рамки нашего изложения, но главный вывод Райта совершенно очевиден: чем меньше эффективная величина популяции, тем больше случайные флуктуации генных частот; и в то же время чем сильнее систематические факторы, тем менее действенным становится этот недетерминированный процесс. Рассматривая по отдельности каждый из имеющих известное направление факторов в его совместном действии с дрейфом, можно придать определению размеров популяции количественный оттенок, считая, что популяция «относительно» мала (т. е. преобладает дрейф), если 4Ns, 4Nu или 4NM меньше единицы, или «велика» (генетические изменения преимущественно детерминированы), если одно из этих произведений больше единицы. Таким образом, замкнутая популяция численностью в четверть миллиона мала, если частота мутирования нейтрального аллеля равна 10-5, а популяция в 250 особей очень велика, если коэффициент отбора больше 1%.
Хотя элиминируемые в результате дрейфа аллели могут пополняться за счет мутаций, тем не менее в малых популяциях повторные мутации нельзя считать силой, способной противостоять дрейфу, поскольку частота мутаций обычно очень мала. Однако может оказаться, что это не относится к многочисленным изоаллелям, недавно обнаруженным с помощью
электрофореза. Имеются некоторые данные, свидетельствующие о том, что частота мутаций в случае изоаллелей значительно выше, чем известная нам частота мутаций, характерная для так называемых «главных» генов. Тем не менее мы будем считать, что мутационное давление едва ли может предотвратить снижение генетической изменчивости в малых популяциях, обусловленное дрейфом. Этого нельзя сказать об отборе или потоке генов, так как они действуют гораздо более интенсивно. Для того чтобы роль дрейфа в изменении генных частот стала второстепенной, достаточно самой низкой скорости миграции (т. е. М = 1/4W, или один иммигрант за четыре поколения независимо от величины Л/). Отбора интенсивностью 0,01 достаточно для предотвращения случайного закрепления аллелей во всех природных популяциях, за исключением самых маленьких (содержащих менее 50 особей).
ИЗУЧЕНИЕ ПРИРОДНЫХ ПОПУЛЯЦИЙ
Естественно встает вопрос, существуют ли в природе явления, возникающие главным образом в результате генетического дрейфа? Подобное объяснение предлагалось для ряда явлений, однако во всех случаях вопрос оставался спорным, ибо любое эволюционное изменение, возникновение которого можно пытаться приписать генетическому дрейфу, почти всегда можно объяснить также различиями давления отбора, изменяющегося либо в пространстве, либо во времени. Чтобы быть уверенным, что причинным фактором явления действительно служит дрейф, необходимо сначала полностью исключить вероятность влияния отбора, а это едва ли возможно. Каким бы нейтральным в адаптивном отношении ни казался признак, мы никогда не гарантированы, что связанные с ним гены не оказывают глубокого влияния на какой-либо еще неизвестный компонент приспособленности. Однако в тех случаях, когда доказательства такого влияния полностью отсутствуют и различные другие условия кажутся подходящими, мы, по-види
мому, вправе привлечь в качестве объяснения дрейф генов.
Дрейф генов — процесс случайного изменения, проявляющийся в виде: 1) различий между последовательными поколениями одной и той же популяции, наблюдаемых во времени; 2) различий между одновременно существующими родственными популяциями, наблюдаемых в пространстве. По чисто практическим причинам большинство исследований посвящено различиям второго типа, хотя все получаемые при этом данные можно считать лишь косвенными, поскольку они получены a posteriori. Поскольку дисперсия частоты генов накапливается и относительно постоянна, действие дрейфа ощутимо не только в популяциях, численность которых всегда невелика, но и в популяциях, численность которых периодически сокращается (эффект «горлышка бутылки»), или в тех, которые невелики при своем образовании, как, например, популяции, образованные несколькими эмигрантами, или основателями, и несущие лишь небольшую долю генетической изменчивости исходной большой популяции («принцип основателя»).
Некоторые из наиболее убедительных примеров случайной генетической изменчивости касаются человека. Человечество на самых ранних стадиях своей истории разделилось на множество различных по величине популяций. Эти популяции разделены в большей или меньшей степени географическими, политическими, религиозными и социальными барьерами. Хотя рост человечества можно охарактеризовать как «популяционный взрыв» и человек обитает сейчас почти во всех частях земного шара, тем не менее все еще существуют некоторые племена и изолированные религиозные секты с очень небольшой эффективной репродуктивной величиной. Считается, что даже некоторые из больших популяций человека возникли сравнительно недавно от небольшого числа основателей. Почти все популяции человека детально исследованы в отношении полиморфизмов по различным системам групп крови; оказалось, что часто небольшие изолированные популяции значительно отличаются от
смежных с ними или окружающих их популяций по некоторым наследственным признакам.
Большинство популяций человека полиморфно по группам крови АВО, т. е. содержат все три аллеля /А, /в, /°. На основе относительных частот этих аллелей жителей различных областей земного шара можно характеризовать как разные «серологические расы». Большинство рас содержат особей со всеми четырьмя группами крови О, А, В, АВ и различаются только по распространенности отдельных групп. Однако у некоторых рас те или иные группы крови вовсе отсутствуют. Например, южноамериканские индейские племена были до прихода европейцев или остаются до сих пор изогенными по группе О. Аллели /А и /в довольно редки также среди индейцев Северной Америки. Индейцы племен блад и черноногих, обитающие на юге Канады и в Монтане, представляют исключение; в отличие от южноамериканских племен они содержат аллель /А, причем частота этого аллеля очень высока, достигая 80%, чего не наблюдается ни в одной другой популяции человека. Американские индейцы произошли, по-видимому, от азиатской (монголоидной) расы, небольшая группа представителей которой в свое время пересекла Берингов пролив. Хотя у азиатской расы встречаются все четыре группы крови, частота группы О относительно мала; поэтому среди этих основателей, пришедших из Азии, или в небольших сформировавшихся позже изолятах могли сложиться условия, благоприятствовавшие дрейфу генов, и либо просто случай, либо случай в сочетании с естественным отбором мог привести к такому необычному распределению.
В большинстве популяций человека частота аллеля /А колеблется между 15 и 30%, а аллеля /в — между 5 и 20% (фиг. 30). Например, частоты этих аллелей в больших популяциях эскимосов, распространенных по всему континенту от Аляски до Гренландии и Лабрадора, равны соответственно 30 и 6%. Но в небольшом племени северных эскимосов в Туле (Гренландия), которое насчитывает менее 300 человек и почти лишено притока генов из другцх популяций, частота
генов /А составляет всего 9%. Кроме того, в некоторых изолятах Лабрадора и Баффиновой Земли ген /в совершенно отсутствует. По-видимому, все эти случаи можно отнести за счет дрейфа генов.
Фиг. 30. Распределение частот аллелей, определяющих группы крови АВО в 215 характерных популяциях человека со всего земного шара.
Треугольник в целом соответствует всему диапазону возможных частот аллелей /л, /в, А в которых процент аллелей 1® равен единице минус процент /А +/В. Можно видеть, что все популяции группируются лишь в пределах небольшого участка всей площади треугольника.
Интересные данные получили Гласс с сотрудниками (1952) по общине дункеров в Пенсильвании. Дункеры — сектанты, эмигрировавшие в начале XVIII века из Германии и с тех пор живущие небольшими общинами, генетически почти изолированными
от остального населения. Следовало ожидать, что частоты аллелей в этой группе будут промежуточными между частотами, наблюдаемыми на родине дункеров в Рейнской области, и частотами среди живущих вокруг них восточноамериканских популяций. Однако эти ожидания не подтвердились. Среди американцев, не принадлежащих к дункерам, так же как среди немцев (по данным 1939 года), люди с группой крови А составляют 40—45%, а среди дункеров — почти 60% (/А =0,38); что касается частоты аллеля /в, то она снижена почти до нуля (/в = 0,025). Частота аллеля LM (система MN) в Западной Германии и в США почти одинакова (LM = 0,54), но среди дункеров частота этого аллеля выше (LM = 0,65). Эти данные вместе с данными о распространении ряда других признаков свидетельствуют о том, что дрейф действительно имел место среди 27 семей, заложивших основу этих общин, или же происходил в этих общинах в течение 200 лет добровольной изоляции (см. Гласс, 1954). Позже Гласс получил доказательства в пользу второго объяснения, т. е. что с момента возникновения общины в ней произошли некоторые генетические изменения. Все члены популяции были сгруппированы в три возрастные группы: от 3 до 27 лет, от 28 до 55 лет, от 55 лет и старше. Различия между этими «поколениями» по группам крови системы АВО и Rh оказались недостоверными, однако по частотам аллелей системы MN наблюдается некоторая зависимость от возраста: в старшем поколении обнаружено соотношение 0,55 Iм и 0,45 LN, что соответствует частотам этих аллелей среди населения Рейнской области и США; в среднем поколении соотношение равно 0,66 LM и 0,34 LN, в младшем же различия между частотами еще больше: 0,735 и 0,265 LN. Как было показано ранее, данные такого типа служат прямым доказательством существования изменений. Однако они ничего не говорят о причине этих изменений.
Есть много других примеров резких различий по частоте генов между небольшими родственными племенами или между религиозными изолятами и соседними популяциями с более высокой численностью.
О том, что изоляция играет большую роль в такой дифференциации, свидетельствует совершенно иное распределение по частоте групп крови в больших популяциях человека. В больших смежных популяциях с практически неограниченной миграцией частоты генов обычно сходны. В тех случаях, когда различия между популяциями все же существуют, частота изменяется постепенно, от одной популяции к другой, т. е. изменчивость носит клинальный характер. Так, частота гена /в, особенно высокая среди жителей Северной Индии и Центральной Азии, постепенно снижается в западном направлении к Европе (см. А. Маккьюсик, «Генетика человека»), в северо-восточном направлении — через Китай и Аляску к Америке и в юго-восточном направлении — к Австралии. Аллель /А имеет несколько центров высокой частоты (например, северо-западная Европа, Австралия и северные районы Северной Америки — вспомним индейцев из племени черноногих), от которых его частота изменяется клинально по многим направлениям. Таким образом, обмен генами между большими популяциями, по-видимому, перекрывает любую тенденцию к четкой дифференциации, наблюдаемой в небольших изолятах.
Все это наводит на мысль, что причиной генетического разнообразия небольших изолятов человека служит случайность выборки. В течение многих лет распространение групп крови в популяциях человека по всему земному шару приводили в качестве лучшего примера неадаптивной изменчивости, поскольку считалось, что гены, ответственные за этот признак, селективно нейтральны. Теперь это представление пересматривается. Некоторое время назад Фишер на основании теоретического анализа пришел к выводу, что лишь немногие гены могут достигнуть значительной частоты и сохранять ее, не обладая каким-либо селективным преимуществом. Особенно это касается генов, связанных с давно существующим в популяции полиморфизмом. Доказательством того, что АВО-по-лиморфизм возник давно, служит распространенность некоторых антигенов во всех популяциях человека,
обитающих на земле. А поскольку у некоторых других приматов были обнаружены те же самые или сходные антигены, можно считать, что полиморфизм по группам крови АВО входит в наследство, полученное человеком от весьма далеких предков.
Форд уже давно, опираясь на многочисленные данные, выдвинул концепцию, что полиморфизм контролируется главными генами (или генами-переключателями) , проявляющими заметное плейотропное действие, что его появление и сохранение в популяциях обусловлено равновесием сил отбора, действующих на все развивающиеся при этом морфологические и физиологические признаки. «Поэтому существование полиморфизма обычно свидетельствует о важности затрагиваемых им признаков...» (Форд, 1964). В начале сороковых годов он предсказал, что полиморфизм в отношении групп крови сбалансирован и что селективное значение участвующих в этом генов обусловлено их влиянием на плодовитость, жизнеспособность или устойчивость к болезням.
За прошедшее с тех пор время была досконально изучена зависимость такой хорошо известной гемолитической болезни новорожденных как эритробластоз плода от Rh-несовместимости матери и плода (подробный обзор см. Стерн, 1960). Вкратце, эритробластоз возникает у Rh-положительного плода, развивающегося во чреве Rh-отрицательной матери, сенсибилизированной чужеродными для нее Rh-антигенами плода во время данной или чаще предыдущих беременностей. Клетки крови плода содержат Rh-антиген. Если этот антиген попадет в кровоток матери (через плаценту), то он индуцирует образование в плазме матери Rh-антител (изоиммунизация). Это ведет к разрушению эритроцитов плода (гемолиз) и анемии (см. Маккьюсик, «Генетика человека», фиг. 50).
Несовместимость матери и плода по антигенам системы АВО также ведет иногда к эритробластозу новорожденных или чаще к выкидышам на ранних сроках беременности. Одна из АВО-несовместимых комбинаций такова: наличие у матери анти-А- или анти-В-антител против одноименных антигенов, имеющихся
у плода. У женщин группы О, в крови которых содержатся оба вида антител, вероятность гибели плода максимальна, а у женщин группы АВ — минимальна. Ознакомившись с основными генетическими и серологическими механизмами, характерными для систем групп крови, читатель может самостоятельно рассчитать потенциально неблагоприятные браки и показать, что внутриутробный отбор неизменно выступает во всех случаях против гетерозигот. Отсюда следует, что одни лишь силы отбора не могут обеспечить сохранение полиморфизма, поскольку отбор против гетерозигот теоретически приводит к состоянию неустойчивости, благоприятствующей закреплению наиболее распространенного аллеля.
Была установлена связь с АВО-группами крови и ряда других заболеваний. Представляется весьма вероятным, что люди с группой крови А более склонны к раку желудка, раку женских половых органов, к пернициозной анемии и сахарному диабету. С другой стороны, больные с язвой желудка или язвой двенадцатиперстной кишки чаще относятся к группе О. Но ни в одном случае не удалось показать, что гетерозиготы обладают каким-либо селективным преимуществом или же установить расовые различия в частоте заболеваний, связанных с группами крови системы АВО.
Тем не менее становится все более и более очевидной справедливость предположения Форда о том, что дивергенцию по частоте групп крови нельзя объяснить одними лишь случайными флуктуациями. Если бы в этом был замешан только дрейф генов, то вряд ли можно было бы ожидать, что частоты, обнаруженные во всех (а их было немало) изученных популяциях, займут лишь одну пятую всего возможного диапазона частот (фиг. 30). Кроме того, Моурант (1959) показал, что различия между популяциями по частоте аллелей системы АВО значительно больше, чем различия по аллелям систем MN и Rh; это показывает, что аллели систем АВО изменяются под влиянием внешних условий быстрее, чем другие аллели.
Дрейф генов выдвигали в качестве причины во многих случаях дифференцировки, наблюдаемых среди различных растительных и животных популяций, в особенности когда популяции малы и сильно разобщены (что зависит от подвижности рассматриваемых форм). Но только в нескольких случаях была предпринята попытка описать, какие именно силы отбора принимают в этом участие. Пожалуй, наиболее ярким примером предполагаемого дрейфа генов, который был действительно изучен, служит полиморфизм наземной улитки Сераеа nemoralis по окраске раковины, изучавшийся М. Ламоттом и А. Кейном и П. Шеппардом. Эти улитки имеют прерывистое распространение; они образуют популяции, или «колонии», в различных местообитаниях — лесах, живых изгородях, высокотравье и на открытых лужайках, причем эти местообитания более или менее отделены друг от друга лежащими между ними неподходящими участками. Почти каждая популяция характеризуется чрезвычайно своеобразной окраской и ее узором. Раковины бывают коричневые, желтые или розовые; одни окрашены равномерно, а другие полосатые с одной-пятью темными продольными полосами. Различия окраски контролируются серией множественных аллелей, причем ген желтой окраски (у) рецессивен по отношению к гену розовой (У), а ген коричневой окраски (Yb) доминирует и над У, и над у. Равномерная окраска (В) доминирует над любой формой по-лосатости (&); другие аллели контролируют число полос и их расположение. Кейн и Шеппард показали, что локусы Y и В тесно сцеплены и наследуются как един «суперген». Установлено, что полиморфизм по цвету и узорам окраски существует в популяциях наземных улиток очень давно. У ископаемых раковин из плейстоценовых отложений Англии и Франции были обнаружены различные типы полосатости, причем соотношение этих типов сходно с тем, которое наблюдается в популяциях улиток, обитающих в этих районах и в настоящее время.
Широкое исследование распространения Сераеа во Франции провел Ламотт. Он нашел, что средняя
частота двух главных признаков — цвета раковины и типа окраски (характера полос) — широко варьируют в разных областях. Обнаружены также различия между колониями, населяющими одну область. Помимо главного фактора, эти различия между популяциями усиливаются вследствие ограниченности потока генов, связанного со столь характерной для улиток малой подвижностью. Возможно, что различия между отдельными колониями Сераеа обусловлены спецификой сил отбора, действующих в разных местообитаниях. Вместе с тем различия между колониями можно объяснить случайными флуктуациями частоты генов. При этом, если принять сделанное Ла-моттом допущение о независимости естественного отбора от величины популяции, с уменьшением величины колонии случайные изменения должны увеличиваться. Анализируя распространение генов полосато-сти и отсутствия полос в нескольких сотнях колоний различной величины, Ламотт нашел, что небольшие популяции варьируют сильнее, чем большие, и чаще бывают гомоаллельными. Более того, частоты генов в соседних небольших популяциях совершенно различны (фиг. 31), в то время как их частоты в больших смежных популяциях довольно сходны. Эти данные ясно показывают, что степень дифференциации зависит от величины популяции. Не отрицая действия селективных сил, определяемых условиями местообитания, Ламотт (1959) объясняет увеличение разнообразия малых популяций дрейфом генов.
Кейн и Шеппард считают наше исходное допущение о том, что действие отбора не зависит от величины популяции, неприемлемым, по крайней мере для колоний улиток, поскольку условия среды в их колониях сильно варьируют, несмотря на незначительность расстояний между ними. По их мнению, большие популяции, занимающие более обширные территории, располагают большим числом местообитаний, чем малые популяции. Следовательно, большие колонии более сходны, чем малые, так как они приспособлены к каким-то усредненным условиям (средние обычно обнаруживают стремление к сходству), тогда
к * » ill-1 I 1 i-J и 5 10 км
Допитале
Фиг. 31. Относительные частоты неполосатых (белые части кружков) и полосатых (черные части кружков) улиток в ряде соприкасающихся колоний, обитающих вдоль реки Лрьеж во французских Пиренеях.
Можно видеть, что изменение частот от колонии к колонии в значительной степени случайно.
как малые популяции специфически приспособлены к частным, локально однородным местообитаниям.
Данные, подтверждающие этот тезис Кейна и Шеппарда, были получены при анализе популяций Сераеа близ Оксфорда (Англия). Помимо типичной изменчивости от одной колонии к другой, здесь была обнаружена связь между фенотипическим составом популяций и типом местообитания (фиг. 32). В каждом местообитании наиболее часто встречаются те варианты, окраска которых близка к окраске субстрата. Так, в зелени живых изгородей и в траве чаще встречаются особи с желтой раковиной, а в лесу, где гниющие листья и участки голой земли имеют темную окраску, преобладают коричневые раковины. В местах, где преобладают цветовые контрасты (живые изгороди и неровности почвы), преобладают полосатые раковины, тогда как сплошная окраска характерна для местообитаний с однотипной поверхностью. Эти корреляции убедительно свидетельствуют о происходящем здесь естественном отборе, направленном на соответствие окраски раковины окраске субстрата. Нетрудно догадаться, что это еще один пример покровительственной окраски, предохраняющей от уничтожения хищниками. Улиток поедает певчий дрозд; он разбивает раковину, используя подходящие камни («наковальню»), и поедает ее содержимое. Избирательный характер уничтожения улиток был установлен при сравнении частоты различных типов окраски среди разбитых раковин вокруг «наковален» и в популяции улиток, обитающих в данной местности. Среди раскрытых раковин частота окраски, контрастирующей с преобладающим для данной местности субстратом, была неизменно выше, чем среди живых особей. Доказательства в пользу избирательного уничтожения хищниками были пополнены данными, полученными при выпуске и повторном отлове улиток. Однако, хотя относительная частота различных форм в разных местообитаниях регулируется визуальным отбором со стороны хищников, здесь, как и в случае полиморфизма человека по группам крови, факторы, определяющие сбалансированность
О Короткоподстриженныи дерн
Q Грубая трава
ф Зеленые изгороди
ф Буковый пес
Смешанный лиственный лес ф Дубравы
Раковины без полос, %
Фиг. 32. Корреляция между процентом желтых и процентом неполосатых раковин в шести различных местообитаниях.
Цвет и узор раковины хорошо коррелируют с субстратом каждого местообитания. Даже исключения подтверждают справедливость тезиса о защитной роли окраски. Например, необычная для смешанного лиственного леса популяция была найдена в атипичном лесу, поросшем короткой зеленой травой, усыпанной опавшими листьями. В таких условиях и следовало ожидать встретить преимущественно желтые формы.
полиморфизма, обнаружены нс были; селективное преимущество гетерозигот не было доказано, хотя несть основания считать, что гетерозис связан с физиологическими признаками, одновременно контролируемыми этими генами. Возможны и другие способы достижения сбалансированного полиморфизма (см. гл. 3), как, например, обмен генами между популяциями, приспособленными к разным условиям, однако ни один из них, по-видимому, не может служить достаточным объяснением в этом случае. И наконец, следует признать, что, поскольку описанные корреляции выражены недостаточно четко, надо думать, что дрейф, возможно в форме упоминавшегося ранее «принципа основателя», играет определенную роль в дивергенции малых колоний.
Обсуждение приведенных выше примеров могло создать впечатление, что следует исходить из дилеммы «или — или», т. е. что дрейф и отбор — две альтернативы, из которых в каждом случае за наблюдаемую генетическую изменчивость ответственна только одна. Повторим еще раз, что кажущееся отсутствие адаптивного значения отнюдь не свидетельствует об исключительной роли дрейфа генов, равно как не следует сбрасывать дрейф со счетов, если удалось установить, что гены оказывают какое-то влияние на приспособленность. Дрейф генов—математическая, а отбор — биологическая реальность. В процессе эволюции они дополняют друг друга; в теории мы разделяем эти два фактора, выражая их в величинах $ и W, но на практике разделить их не так легко.
РОЛЬ В ЭВОЛЮЦИИ МАЛЫХ ПОПУЛЯЦИЙ
Теперь, когда существование дрейфа генов в малых популяциях доказано теоретически и экспериментально, вопрос о его роли в эволюции легче всего решить, рассмотрев эволюционную роль малых популяций.
Большинство исследователей согласны с мнением Райта, что дрейф генов в его классической форме (т. е. случайные колебания частот генов в изолиро
ванных популяциях, которые все время остаются небольшими), по-видимому, не играет большой роли. Такие популяции скорее всего будут мало приспособлены, поскольку в них наблюдается гомозиготность по большинству локусов, причем в некоторых из них в результате дрейфа происходит закрепление худшего в селективном отношении аллеля. Это снижение приспособленности, связанное с гомозиготностью, обсуждается в гл. 7 с позиций инбредной депрессии — обычного явления, сопровождающего самооплодотворение или другие типы тесного инбридинга в линиях животных и растений, для которых нормой служит перекрестное оплодотворение. Здесь можно говорить об инбредной депрессии, поскольку с уменьшением размножающейся группы независимо от типа скрещивания в популяции увеличивается доля близкородственных спариваний (по существу дрейф генов можно представить как процесс инбридинга и выразить через коэффициент инбридинга; см. гл. 3). Длительный инбридинг сопровождается увеличением числа закрепленных аллелей и сведением к минимуму изменчивости; поэтому возможности естественного отбора повысить уровень приспособленности генотипов быстро истощаются, что приводит к «вырождению и вымиранию».
Совершенно очевидно, что вымирание — наиболее вероятная судьба популяции, величина которой упала ниже некоторого критического уровня; такая же участь ждет вновь образованную популяцию, основанную несколькими мигрантами за пределами обычного ареала вида. Однако следует заметить, что неизбежность такой судьбы несколько преувеличена. Известны высокоинбредные линии, сохраняющиеся на протяжении многих поколений. Известны также небольшие природные изоляты, специфически приспособившиеся к какому-либо определенному комплексу условий среды и успешно поддерживающие свое существование в течение продолжительного времени. И все-таки существованию таких популяций постоянно грозит опасность: в случае резкого изменения условий среды, к которым они не могут приспособиться,
они также вымирают. Вместе с тем может случиться, что изменение среды приведет к расширению экологической ниши данной колонии и тем самым будет благоприятствовать распространению популяции. При этом у популяции возникают огромные потенциальные возможности для эволюции (см. ниже).
Для небольшого изолята существует и третья возможность—слияние с другими популяциями. Однако в данном контексте подобное развитие событий не представляет интереса, поскольку любая дифференциация, развивающаяся в колонии за время ее изоляции, быстро исчезает, как только восстанавливается свободный обмен генами. В своем превосходном труде Майр (1963) постоянно возвращается к этой теме: «Поток генов и его последствия оказывают тормозящее действие на процесс эволюции».
Нужно подчеркнуть также, что инбридинг сам по себе не всегда наносит вред виду. Существует немало хорошо приспособленных растительных форм, для которых обычным способом размножения служит самоопыление. Кроме того, совершенно ошибочно полагать, что инбридинг переводит все локусы генома в гомозиготное состояние. Даже в отсутствие отбора и при самом тесном инбридинге закрепление аллелей идет очень медленно. В действительности же отбор действует практически всегда и везде и, как говорилось выше, отбор, благоприятствующий гетерозиготам, сильно снижает эффективность дрейфа. Например, в нашей лаборатории были специально выведены линии дрозофилы, гомозиготные по нескольким инверсиям, найденным в одной лабораторной популяции. Множество линий поддерживали путем спаривания сибсов в течение более чем 20 поколений, однако произошло закрепление только одного из нескольких возможных гомозиготных сочетаний; большинство же линий оставалось полиморфным. Экспериментальная популяция, полученная из гибридов между двумя близкородственными видами, перешла в состояние «облигатной хромосомной гетерозиготпости» вследствие сильного отбора, благоприятствовавшего гетерозиготам, и сохраняла сбалансированный хромосомный
полиморфизм более 10 лет. Есть много данных, свидетельствующих о сохранении генетической изменчивости в небольших изолированных природных популяциях.
Количество данных, доказывающих существование географического видообразования, поистине огромно (см. Майр, 1963). Принято считать, что происходящие от нескольких эмигрантов изолированные периферические популяции особенно подвержены резким генетическим преобразованиям и быстрому эволюционному изменению (подробное объяснение см. гл. 8). В такой ситуации наиболее благоприятное для дрейфа время — лишь самое начало процесса (т. е. он должен действовать в ограниченной группе основателей). Поскольку численность большинства выживших колоний впоследствии возрастает, влияние дрейфа носит одноразовый характер; это обусловлено случайностью образования зигот в маленьких группах мигрантов, несущих лишь небольшой запас генетической изменчивости. Этот аспект дрейфа получил название принципа основателя. Главная особенность такой системы состоит в том, что разные колонии, возникающие от нескольких основателей (или популяции, численность которых резко сократилась), будут иметь различные (и притом довольно истощенные) генофонды. Как только такая группа основателей образовалась, естественный отбор начнет накапливать наиболее благоприятные из возникающих генотипов. Таким образом, хотя эволюция такой линии направляется отбором, она сильно зависит от первоначальных генетических ресурсов колонии основателей. Различия в исходном генетическом багаже обусловят разные пути эволюции.
Добжанский и Павловская (см. Спиесс, 1962, в библиографии к гл. 5) продемонстрировали взаимодействие отбора и дрейфа генов в экспериментальных популяциях Drosophila melanogaster. Они показали, что создаваемая в условиях отбора изменчивость тем больше, чем меньше популяция основателей (фиг. 33). Сначала была создана исходная популяция, состоявшая из мух F2, полученных от гибридов между техас
ской и калифорнийской популяциями. Мухи из Техаса были гомозиготны по последовательности генов в третьей хромосоме, обозначенной AR, а мухи из Калифорнии— гомозиготны по другой последовательности генов, обозначенной РР. Таким образом, в исходной линии частоты третьих хромосом типа AR и РР
Фиг. 33. Частота в процентах (йо вертикали) хромосом РР в 20 экспериментальных популяциях Drosophila pseudoobscura смешанного географического происхождения (Техас и Калифорния).
В группе из 10 популяций, основанных небольшим числом мух-родоначальников, спустя 21 поколение наблюдается гораздо более сильная дивергенция, чем среди популяций, которые все время имеют высокую численность.
были равны. Из этой исходной линии были случайно отобраны особи, послужившие основателями для 20 лабораторных популяций. Для 10 популяций было взято по 20 основателей, а для 10 других — по 4000 основателей. Спустя два поколения «небольшие» популяции достигли размеров «больших», после чего все 20 популяций оказались в сходных условиях в отношении отбора. Популяции анализировались дважды: спустя 3 и спустя 17 месяцев (примерно после 21 поколения отбора). Как показывает фиг. 33, частоты хромосом РР в популяциях, происходящих от небольшого числа основателей, ко второму анализу распределялись в диапазоне от 16 до 47%. Частоты
хромосом РР в популяциях, происходящих от 4000 особей, варьируют меньше — от 20 до 35%. Известно, что во всех популяциях частоты хромосом РР и AR контролируются жестким отбором, благоприятствующим гетерозиготам РР/AR. Однако большая гетерогенность среди «малых» популяций по сравнению с «большими» свидетельствует о том, что конечные результаты отбора в них до некоторой степени обусловлены дрейфом генов, который, очевидно, начался еще в популяции основателей. Эти данные подтверждают тезис о том, что взаимодействие отбора и дрейфа может привести к эволюционным изменениям даже в том случае, когда снижение численности популяции — явление временное.
Приведенные выше факты не раз привлекали для объяснения дивергенции некоторых природных популяций, которые, по-видимому, прошли через период резкого уменьшения численности, как, например, островные популяции, основанные несколькими иммигрантами. Интересные примеры межпопуляционной изменчивости бабочки Maniola jurtina, обитающей на островах Силли у юго-западного побережья Англии, приводят Даусвелл и Форд (см. Форд, 1964). У особей этого вида на нижней поверхности задних крыльев имеется несколько маленьких пятнышек (от 0 до 5), по распределению которых можно охарактеризовать ту или иную популяцию. У популяций больших островов распределение по числу пятен довольно сходно, а популяции малых островов отличаются по этому параметру как одна от другой, так и от больших популяций (фиг. 34). За некоторыми исключениями, характер распределения по числу пятен среди островных популяций не изменяется из года в год. Сходство популяций больших островов нельзя объяснить обменом генами, поскольку море служит хорошей изолирующей преградой; кроме того, между большими островами разбросаны мелкие острова с их весьма своеобразными популяциями.
Добжанский и Павловская предположили, что популяции мелких островов, хотя они теперь и велики, происходят от небольшого числа иммигрантов, т. е.
популяция каждого из мелких островов имела изначально своеобразный генетический состав, что при-
Фиг. 34. Северовосточная часть островов Силли.
Три нижних графика показывают распределение по числу точек на крыльях для самок Mandola jurtina на трех больших островах. На остальных графиках представлено распределение по числу точек для популяций пяти маленьких островов. Обратите внимание на сходство кривых для больших и различие их для малых островов. По оси абсцисс — число точек на крыльях; по оси ординат — процент бабочек.
вело к образованию относительно стабильных, но различных генофондов. Форд не согласен с таким объяснением. Он привел два факта, которые, по-видимому,
прямо опровергают участие в этой ситуации дрейфа генов. На одном из маленьких островов (Теан) в большой популяции недавно возникло новое, но опять-таки стабильное распределение по числу пятен, причем численность популяции не уменьшилась. Изменение это было связано с резким изменением растительного покрова острова, что в свою очередь произошло в результате удаления с острова стада коров. Напротив, полностью изолированная популяция одного из больших островов в один из засушливых годов сократилась чуть ли не до полного исчезновения, однако затем после восстановления растительности вновь достигла нормальной величины и, что особенно интересно, сохранила прежнее распределение по пятнистости! Форд предполагает, что каждая популяция приспособлена к конкретным условиям своего острова; морфологическое сходство популяций бабочек больших островов объясняется тем, что в них происходил отбор на соответствие неким усредненным условиям, тогда как популяции малых островов подвергались отбору на приспособленность к различным, но довольно однородным в пределах каждого острова экологическим условиям. Он утверждает, что наблюдаемую у Maniola изменчивость легко объяснить сильным давлением отбора, происходящим в природе (и проявляющимся в тестах на скрещиваемость).
Итак, математическая теория дрейфа генов хорошо подтверждается экспериментальными данными. Споры о том, какова эволюционная роль дрейфа генов, будут, вероятно, продолжаться вечно; во всяком случае, их не удастся разрешить до тех пор, пока мы не откажемся от чисто спекулятивных построений и не займемся вместо этого подлинными научными исследованиями.
ЛИТЕРАТУРА
Ford Е. В., Ecological Genetics, New York, John Wiley and Sons, 1964.
Glass B., Genetic Changes in Human Populations, Especially Those Due to Gene Flow and Genetic Drift, Advan. Genet., 6 (1954), 95-139.
G 1 a s s В., S а с к s M. S., J a h n E. F., Hess C., Genetic Drift in a Religious Isolate: An Analysis of the Causes of Variation in Blood Group and Other Gene Frequencies in a Small Population, Amer. Naturalist, 86 (1952), 145—160.
Lamotte M., Polymorphism of Natural Populations of Cepaea nemoralis, Cold Spring Harbor Symp. Quant. BioL, 24 (1959), 65—86.
Ma yr E., Animal Species and Evolution, Cambridge, Mass., Harvard University Press, 1963. (Э. Май p, Зоологический вид и эволюция, изд-во «Мир», М., 1968.)
Moura nt А. Е., Human Blood Groups and Natural Selection, Cold Spring Harbor Symp. Quant. Biol., 24 (1959), 57—63.
Stern C., Principles of Human Genetics, 2nd ed., San Francisco, W. H. Freeman and Company, 1960. (К. Штерн, Основы генетики человека, Медгиз, М., 1965.)
Wright S., On the Roles of Directed and Random Changes in Gene Frequency, in the Genetics of Populations, Evolution, 2 (1948), 279-295.
Глава 7
ГЕНЕТИЧЕСКАЯ ИЗМЕНЧИВОСТЬ ПОПУЛЯЦИЙ
Для того чтобы существовать, популяция должна быть приспособлена к непосредственно окружающей ее среде. Это значит, что большая часть особей в популяции должна иметь такие генотипы и фенотипы, которые обеспечивают ей существование в данных условиях. Чтобы популяция могла существовать и в дальнейшем, она должна иметь возможность сохранять свою приспособленность к среде, изменяя свою генетическую структуру при неизбежных изменениях среды. Это значит, что в популяции должны возникать и воспроизводиться новые генотипы и фенотипы, приспособленные к изменившимся условиям.
Генетическая гибкость или пластичность, дающая возможность популяции совершить адаптивный сдвиг, достигается у разных организмов различными способами. У микроорганизмов с их многочисленными популяциями и огромной скоростью размножения в каждый данный момент времени имеется вполне достаточное число новых мутантов, причем среди них обычно находятся такие, которые приспособлены к новым условиям; быстрое размножение этих мутантов обеспечивает сохранение популяции. Величина популяции и скорость размножения у многоклеточных раздельнополых организмов обычно несравненно меньше. Поэтому вновь возникшие мутации редко могут служить основой для адаптивного ответа популяции на изменение условий. У этих организмов генетическая гибкость обеспечивается за счет рекомбинации запасен-
ной или скрытой генетической изменчивости, имеющейся в популяции, но, как правило, не выражающейся фенотипически.
Эта глава посвящена главным образом обзору некоторых из наиболее важных механизмов сохранения и использования генетической изменчивости у раздельнополых видов.
СОХРАНЕНИЕ ГЕНЕТИЧЕСКОЙ ИЗМЕНЧИВОСТИ
Несмотря на то что большинство популяций довольно хорошо стабилизировалось каждая в своей среде, для непрерывной эволюции необходимо наличие в периоды изменений среды генетических вариантов, преадаптированных к этим возможным изменениям.
В популяциях диплоидных раздельнополых организмов может храниться в гетерозиготном состоянии, совершенно не проявляясь, огромный запас генетической изменчивости. Это возможно благодаря тому, что во многих случаях для нормального фенотипического проявления признака вполне достаточно нормальной активности одного аллеля данного локуса.
У полиплоидов количество запасенной генетической изменчивости может быть еще больше, потому что за фенотипическим проявлением одного нормального аллеля может скрываться не один, а несколько мутантных аллелей. Вместе с тем, хотя запас генетической изменчивости у полиплоидов и больше, он менее полезен популяции, так как создавать новые фенотипы в этом случае сложнее. Например, для проявления рецессивного признака у тетраплоидов необходимо, чтобы соответствующий рецессивный аллель был представлен в генотипе четырехкратно. Таким образом, потенциальные возможности приспосабливаться к резким изменениям условий среды, имеющиеся у диплоидов, по-видимому, являются наилучшим компромиссом между очень небольшим запасом изменчивости у гаплоидов и чрезвычайно глубоко скрытым избытком ее у полиплоидов,
Поиски источников изменчивости и способов ее сохранения занимают значительное место в истории эволюционной теории и до сих пор крайне важны для дальнейшего развития наших представлений об эволюции. Хотя эволюция и зависит от постоянного наличия генетической изменчивости, одно из самых значительных ее последствий для популяции — это неизбежное появление слабо приспособленных особей. Таким образом, приспособленность популяции всегда ниже той, которая характерна для оптимально приспособленных индивидуумов. Это снижение средней приспособленности популяции за счет особей, приспособленность которых ниже оптимальной, называют генетическим грузом. Известный английский генетик Холдейн так охарактеризовал генетический груз: «Это та цена, которую вынуждена платить популяция за право эволюционировать». Займемся теперь вопросом об истинной величине генетической изменчивости природных популяций.
ОПРЕДЕЛЕНИЕ СКРЫТОЙ ГЕНЕТИЧЕСКОЙ ИЗМЕНЧИВОСТИ
Обнаружение и количественная оценка скрытой генетической изменчивости — непростая задача, все еще остающаяся одной из главных проблем популяционной генетики.
Большинство методов определения изменчивости основано на использовании систематического инбридинга. Инбридинг приводит к повышению гомозиготности, что в свою очередь благоприятствует проявлению рецессивных аллелей, скрывавшихся прежде в гетерозиготах. Многие аллели в гомозиготном состоянии приводят к аномалиям или к гибели, поэтому инбредные линии обычно имеют пониженную приспособленности или, иначе, среднюю адаптационную ценность (РК). Это особенно касается видов, для которых аутбридинг — правило, а не исключение. Несколько примеров инбредной депрессии в популяциях домашних животных приводит Фалконер (1960,
стр. 249). Обычно чем больше скрытая генетическая изменчивость популяции, тем больше инбредная депрессия. У видов, для которых инбридинг является нормой, т. е. которые подвергались инбридингу и отбору в течение длительных периодов своей эволюционной истории, скрытая изменчивость невелика и снижение адаптационной ценности (если оно вообще происходит) при дальнейшем инбридинге незначительно. Но в аутбредных популяциях скрытая изменчивость может быть очень велика, и в этих случаях наблюдается очень сильная инбредная депрессия. В принципе уровень скрытой генетической изменчивости, проявляющийся при инбридинге, соответствует достигнутой степени инбридинга. Например, поколение F2 от скрещивания сибсов будет иметь коэффициент инбридинга (величина F), равный V4. Следовательно, в среднем четвертая часть скрытой генетической изменчивости родительских особей перейдет в гомозиготное состояние и проявится фенотипически. В этом случае изменчивость популяции на самом деле в четыре раза выше того уровня, который был выявлен при инбридинге.
Для обнаружения скрытой генетической изменчивости важно знать, какие параметры следует оценивать. Иногда о количестве изменчивости судят по частоте видимых мутаций, но мы знаем, что видимые мутации составляют лишь небольшую часть новых мутаций и скрытых рецессивов, также сохраняющихся в популяции (превосходный обзор этого аспекта скрытой изменчивости сделан Спенсером, 1947). Частота рецессивных деталей — более объективная мера скрытой изменчивости, но рецессивные летали — весьма ограниченная группа вариантов, которые не могут дать адекватного представления относительно общего уровня генетической изменчивости. Поскольку большинство генов оказывает некоторое воздействие на приспособленность организма, более точную оценку общей изменчивости можно получить, определяя снижение средней адаптационной ценности (эквивалент средней приспособленности) популяции при инбридинге.
При определении изменения средней адаптационной ценности популяции нам хотелось бы измерить общую приспособленность, но практически это сделать трудно. Поэтому инбредную депрессию обычно оценивают как снижение средней величины таких измеримых компонентов общей приспособленности, как число яиц в кладке, процент вылупления, процент особей, достигших взрослого состояния, и т. д. Для иллюстрации рассмотрим процедуру определения ин-бредной депрессии у дрозофилы, измеряемую жизнеспособностью, начиная со стадии яйца и до взрослой особи. Выборку наловленных в природе оплодотворенных самок помещают по одной в пробирки с питательной средой. Потомство этих самок систематически скрещивают для получения групп особей с различными коэффициентами инбридинга. Например, при спаривании сибсов, т. е. развившихся в одной индивидуальной культуре потомков одной дикой самки, получается потомство с F = !/4. Спаривание неродственных самцов и самок, т. е. потомков мух из разных индивидуальных культур, дает аутбредное потомство с F = 0. Если потомство из одной культуры Fi скрестить в отдельном спаривании с потомством двух других культур Fi, то потомки от этих скрещиваний будут двоюродными сибсами, а коэффициент инбридинга потомства от скрещивания их между собой будет равен Vie- С помощью соответствующих скрещиваний можно получить группы особей с другими коэффициентами инбридинга. Как получить двойных двоюродных сибсов, потомство которых имело бы коэффициент инбридинга, равный Vs? Наконец, определяется средняя жизнеспособность (или смертность) для аутбредных особей (F = 0) и для особей каждой из инбредных групп, для которых величина F известна. В табл. 12 представлены данные о жизнеспособности разных инбредных групп, полученных от нескольких различных природных популяций Drosophila.
Данные, приведенные в табл. 12, которые типичны для множества аналогичных работ, свидетельствуют о снижении выживаемости с увеличением F.
Таблица 12
Зависимость выживания (от стадии яйца до взрослой стадии) от степени инбридинга у Drosophila (числа, выделенные жирным шрифтом, — средняя по нескольким линиям из разных местностей)
Вид F = 0 F = 7io F = ’A
р. willistoni 84,3 — 76,0 63,5
D. pseudoobscura 90,2 83,2 — 80,5
D. pseudoobscura 88,7 — 71,4 73,6
D. pseudoobscura 88,0 — 81,9 78,2
D. arizonensis 77,0 73,3 69,1 66,2
D. tnojavensis 62,4 46,1 48,4 44,1
D. ananassae 90,8 75,0 — 65,5
D. ananassae 85,2 79,3 — 63,4
D. ananassae 70,9 71,0 — 67,2
D. novaniexicana 91,7 — — 86,7
Сходные данные получены для человека. В этом случае коэффициент инбридинга определяется a posteriori, по родословным. Типичным примером служит анализ церковных записей о бракосочетаниях и смертях в одном из сельских районов Франции за прошлое поколение. Все браки разделили на две группы— родственные и неродственные и подсчитали для каждой группы детскую смертность и число мертворожденных. При этом были получены следующие результаты:
Неродственные родители (F = 0): 210 из 1628, т. е. 0,129
Браки между двоюродными братьями 115 из 461, т. е. 0,249
и сестрами (F = ’/1б):
Мортон, Крау и Миллер (1956) разработали метод использования инбридинга для количественной оценки летального действия скрытых в популяции мутаций. Рассмотрим упрощенный вариант этого метода, используя приведенные выше данные о смертности. Смертность среди потомков от браков двоюродных
сибсов почти на 12% выше, чем среди потомков от неродственных браков (0,249 и 0,129). Мы знаем, что у потомков от браков двоюродных сибсов Vie генов, в норме гетерозиготных, переходят в гомозиготное со* стояние. Поскольку переход Vie генов в гомозиготное состояние привел к увеличению смертности на 12%, то, если бы все гены оказались в гомозиготном состоянии, т. е. если бы индивидуумы могли возникнуть вследствие удвоения каждой гаметы, смертность должна была бы увеличиться в 16 раз. Это значит, что увеличение смертности составило бы 200% (0,12-16 ж 2). Отсюда был сделан вывод, что в среднем каждая гамета исследуемой популяции несет два летальных эквивалента. «Летальным эквивалентом» может быть отдельный летальный ген или группа частично летальных мутантных генов. Это просто означает, что если данная особь гомозиготна по нескольким генам, каждый из которых до некоторой степени снижает жизнеспособность, то их кумулятивное действие приводит к гибели особи. Напротив, если частично летальные гены, в сумме равные одному летальному эквиваленту, распределены по разным особям и переходят в гомозиготное состояние, то их действие сведется в среднем к гибели одной особи или к соответствующему снижению плодовитости.
Каждая зигота образуется из двух гамет; следова тельно, из приведенных выше данных можно сделать вывод, что каждый человек содержал по 4 скрытых в гетерозиготном состоянии летальных эквивалента. Эта величина близка к полученным для дрозофилы, у которой число летальных эквивалентов равно в среднем 0,4 ... 4,0.
Точность такой оценки, а следовательно, и ее биологическое значение зависят от того, насколько обоснованы исходные допущения модели. Одно из самых существенных допущений — это допущение о незави симом действии генов на жизнеспособность, или, как обычно говорят, об отсутствии синергизма в действии генов. Если такое допущение верно, то действие алле- । лей, относящихся к отдельным локусам, должно быть j мультипликативным (на логарифмической шкале ве- j
роятность выживания s есть линейная функция от F).
В последних работах показано, что это условие выполняется далеко не всегда, особенно при высоких степенях инбридинга, как в опытах с дрозофилой (табл. 12). Особи с коэффициентом инбридинга F=1/a нередко обладают почти такой же жизнеспособностью, как и особи, у которых коэффициент инбридинга в четыре раза меньше, т. е. F = 716- Эти и другие несоответствия, порождаемые допущением о независимом действии локусов, — одна из самых сложных проблем, стоящих сейчас перед популяционной генетикой (Доб-жанский, 1963). Однако, несмотря на эти и другие недостатки модели Мортона — Крау — Меллера, ее тем не менее можно использовать для приближенной оценки общего генетического груза, содержащегося в популяциях по отношению к измеряемому компоненту приспособленности.
Для дрозофилы некоторые типы скрытой генетической изменчивости можно определить более прямым способом, основанным на переводе той или иной хромосомы целиком в гомозиготное состояние; для этого используют специальные аналитические линии. К сожалению, такие линии имеются только в нескольких видах Drosophila, а среди других родов их пока нет вообще. Аналитическая линия обычно гетерозиготна по двум различным инверсиям (см. гл. 4), причем каждая хромосома данной пары гомологичных хромосом содержит по одной инверсии. Каждая такая инверсия включает в себя доминантные гены, фенотипически проявляющиеся в гомозиготном состоянии как летали. У D. melanogaster такими маркерами служат гены Curly (Су) и Plum (Рт). Инверсии, содержащие эти гены, образуют сбалансированную по деталям систему (фиг. 35). Поскольку гомозиготы CytCy и Рт/Рт нежизнеспособны, в каждом поколении потомков выживают только гетерозиготы Су/Рт. Для получения гомозигот по интересующей нас второй хромосоме проводится скрещивание по схеме, показанной на фиг. 36. Самок аналитической линии скрещивают с самцами дикого типа. Все потомство от этого скрещивания оказывается гетерозиготным по
одной из инверсий матери и по одной из нормальных ( + ) хромосом отца. При возвратном скрещивании таких особей с аналитической линией, скажем при скрещивании Cyl-}- X Су/Рт, 3 из 4 вторых хромосом у обеих родительских особей содержат одну инверсию и один доминантный маркер. Потомство от этого
Dth_+ __+
= X -
4- 4- О212
D|lf + Dtlt + + P2l2
DjJ + + ~2 4- D2l2
Гибнцт _ ~ Гибнут
(l гомозиготы) Выживают только (^гомозиготы) гетерозиготы
Фиг. 35. Система сбалансированных деталей.
Каждая маркированная мутантом хромосома гетерозиготы DJDz содержит рецессивный летальный фактор, проявление которого подавляется доми нантным аллелем дикого типа в соответствующем локусе гомологичной хромосомы. В простейшем случае, изображенном здесь, участвуют по крайней мере два локуса. Летальные гены сцеплены в состоянии комплементарное™ («трано-положение), а наличие инверсии препятствует кроссинговеру. Вследствие проявления рецессивных леталей гомозиготы обоих типов гибнут, поэтому линия Dx/D2 сохраняется в чистоте.
скрещивания будет относиться к трем хромосомным типам: CylPm, Cyl+ и Рт! + . Основная особенность такого скрещивания состоит в том, что все (+)-хромосомы потомков — продукт репликации одной-единственной ( 4-)-хромосомы одной из родительских особей. Таким образом, среди потомков от скрещивания Q//+ X Су!+ У Чь зигот эта хромосома будет гомозиготна (+/+). Если ( + )-хромосома содержит деталь (или несколько частичных леталей, в сумме соответствующих одному летальному эквиваленту), то +/+ зиготы погибнут и все потомство будет состоять только из гетерозигот Су/4-. Если (+)-хромосома нормальна, то соотношение фенотипов потомства составит 1 дикий тип +/+ : 2Q//+ (мухи Су!+ отличаются от мух +/+ загнутыми
Крыльями — фенотипическое проявление доминант-ного гена Су).
Таким образом, этот метод позволяет за три поколения выделить из природной популяции отдельную
ММ Z\ м
Гибнут 2 х I
Фиг. 36. Обобщенная схема скрещиваний, позволяющих перевести отдельные хромосомы в гомозиготное состояние с целью обнаружения рецессивных мутантов, обычно скрытых в природных популяциях Drosophila.
I — скрещивание; // — второе (возвратное) скрещивание; /// — третье скрещивание (между сибсами).
хромосому, реплицировать ее, сохранить интактной (поскольку инверсии препятствуют кроссинговеру) и перевести в гомозиготное состояние. Каждый ряд последовательных скрещиваний, начиная со скрещивания одного дикого самца, представляет собой тест на одну и ту же хромосому. Частичные летали (так
называемые субвитальные факторы, полулетали, дет-риментали и т. п.) приводят к снижению численности диких особей в разной степени. Если среди потомства нет ни одной мухи дикого типа, испытуемая хромосома считается летальной. Испытуемая хромосома считается полулегальной, если число диких особей в два или более раз меньше ожидаемого. Культуры, содержащие более половины ожидаемой одной трети (или 16,7%) особей +/+, называются квазинормаль-ными, субвитальными или детриментальными. Следует заметить, что при наличии мутаций, обладающих видимым эффектом, они могут проявиться у особей +/+•
У некоторых видов подобным же образом можно испытать другие хромосомы и получить представление о генетической структуре больших хромосом. Такие исследования дают лучший результат, чем простое близкородственное скрещивание.
В качестве примера на фиг. 37 приведены данные о 113 гомозиготных и 90 гетерозиготных комбинациях вторых хромосом в выборке D. pseudoobscura, взятой из природной популяции на юго-востоке Аризоны. Как видно, средняя жизнеспособность гетерозигот равна 32,24%, т. е. довольно близка к теоретически ожидаемым 33,33%. Заметьте, что большинство испытанных комбинаций было нормально в отношении жизнеспособности или очень близко к норме. Однако некоторые были отчетливо ниже нормы. Причины этого, видимо, могут быть разными: переход в гомозиготное состояние субвитальных рецессивных аллелей, наличие субвитальных генов, которые обладают частичной доминантностью, и др. Жизнеспособность некоторых из испытанных гетерозиготных комбинаций была выше нормальной. Это так называемые «супер-витали».
То обстоятельство, что средняя жизнеспособность гомозигот равна 22,36%, т. е. значительно ниже нормальной жизнеспособности, равной 33,33%, свидетельствует о существовании в популяции в скрытом состоянии значительного числа вредных генов. С другой стороны, почти 24% (27 из 113) испытанных вто-
Фиг. 37. Влияние гомозиготности по вторым хромосомам на жизнеспособность Drosophila pseudoobscura из аризонской популяции.
По оси ординат — число хромосомных комбинаций, соответственно гомозиготных (Б) и гетерозиготных (Д), характеризующихся той или иной жизнеспособностью. Гомозиготы делятся на три класса: летали (черные столбцы), полулетали (заштрихованные столбцы) и квазииормали (белые столбцы). Пунктирными линиями показаны средняя жизнеспособность гетерозигот (4) и средняя жизнеспособность гомозигот (Б). Сплошная линия па диаграмме Б— средняя жизнеспособность квазинормальных гомозигот.
8 Зак. 88
рых хромосом оказалось летальными или полулегальными в гомозиготном состоянии. В панмиктической популяции эти хромосомы распределяются случайно. Каждая муха несет по две вторых хромосомы. Поскольку частота хромосом, содержащих летальные или полулегальные факторы, равна 24%, вероятность того, что обе хромосомы отдельной особи легальны или полулегальны, равна (0,24)2, или около 6%. Однако такие особи обычно не гибнут, поскольку чаще всего летальные или полулегальные факторы неаллельны и их действие маскируется действием нормальных доминантных аллелей соответствующих локусов. Соответственно (0,76)2, или 58% особей популяции, будут совершенно свободны от этих очень вредных мутаций вторых хромосом, а 42% особей будет содержать по крайней мере одну такую хромосому. Подобный же расчет можно провести и для двух других пар больших аутосом. Было показано, что в упомянутой аризонской популяции 25% третьих и 32% четвертых хромосом также содержат летали и полулетали. Соответственно вероятность того, что особи не будут нести таких генов ни в одной из трех больших аутосом, равна (0,76)2-(0,75)2-(0,68)2= 15%. Для Х-хромосомы подобных данных нет.
Таким образом, пока мы рассмотрели только мутанты, присутствие которых можно определить по их сильному влиянию на жизнеспособность. Но большая часть скрытой генетической изменчивости представлена хромосомами, содержащими субвитальные мутанты с незначительным вредным действием. Как отмечено выше, субвитали относятся к так называемому квазинормальному классу. Присутствие субвиталей проявляется в том, что средняя приспособленность при гомозиготности по квазинормальным хромосомам, равная 29,12%, существенно ниже жизнеспособности контрольных гетерозигот (32,24%) (фиг. 37). Распределение по приспособленности носит непрерывный характер — между различными классами жизнеспособности нет резких разрывов. Поэтому вопрос о том, какие хромосомы считать субвитальными, решается произвольно. Желательно, однако, чтобы в по
нятие «субвитальный» вкладывался какой-то определенный смысл. Нормой обычно считают те классы жизнеспособности, которые лежат в пределах двух стандартных отклонений (2а) генетической изменчивости гетерозиготных контролей, т. е. двух стандартных отклонений вправо и влево от средней. Особи, жизнеспособность которых более чем на 2а выше средней жизнеспособности, относят к классу суперви-талей. В большинстве экспериментов главную часть квазинормальных хромосом составляют субвитали (жизнеспособность более чем на 2а ниже средней) (табл. 13). Общий генетический груз популяции, отражающийся в снижении такого компонента общей приспособленности, как жизнеспособность, слагается из леталей, полулеталей и сублеталей. Однако приспособленность включает и ряд других компонентов, которые мы не рассматривали, как, например, плодовитость (что также показано в табл. 13), фертиль-
Таблица 13
Частота в природной популяции Drosophila хромосом, которые в гомозиготном состоянии снижают плодовитость особей ’)
Вид Хромосома Леталь или полу-леталь Субвиталь Стерильность самок Стерильность самцов
D. prosaltans | Вторая 32,6 33,4 9,2 11,0
Третья 9,5 14,5 6,6 4,2
Z). willistonl | Вторая 28,4—41,2 57,5 40,5 64,8
Третья 25,6—32,8 47,1 40,5 66,7
Вторая 18,3-32,1 56,0—93,5 10,6 8,3
D. pseudoobscura Третья 16,4—25,0 29,5—86,7 13,6 10,5
Четвертая 32,2 95,4 4,3 11,8
D. Вторая 25,5 84,4 18,3 13,2
persimilis Третья 22,7 74,2 14,3 15,7
Четвертая 28,1 98,3 18,3 8,4
*) Границы изменчивости приведены в тех случаях, когда были изучены популяции из разных местообитаний. Частота субвиталей выражена как процент от квазинормалей.
ность, скорость развития и др., которые также вносят свой вклад в скрытую генетическую изменчивость.
Следовательно, можно сделать вывод, что в среднем каждая особь вида Drosophila гетерозиготна по одной видимой мутации, двум летальным эквивалентным и четырем или большему числу субвиталей. Конечно, в разных популяциях эти средние величины различны в зависимости от численности популяции, степени инбридинга, силы отбора, изменчивости экологических и географических факторов и т. д. Но в общем можно видеть, что степень генетической изменчивости и, следовательно, потенциальные возможности для эволюционных изменений огромны.
АДАПТИВНАЯ НОРМА И КОНЦЕПЦИЯ ГЕНЕТИЧЕСКОГО ГРУЗА
Определим теперь, какой ценой удается популяции сохранять эту изменчивость. В идеале каждый член популяции должен был бы иметь генотип и фенотип, наилучшим образом соответствующие конкретным условиям существования. Однако мы приходим к заключению, что в силу разных обстоятельств (см. раздел «Гомеостаз и канализация», стр. 266) огромное разнообразие генотипов будет давать довольно ограниченное число весьма сходных фенотипов. Это неразрывно связано с концепцией скрытой генетической изменчивости. Отсюда следует, что разные генотипы могут быть одинаково хорошо приспособлены (именно это препятствует точной оценке степени генетической изменчивости популяции, поскольку наличие этого типа изменчивости установить очень трудно). Следовательно, большинство генотипов, имеющихся в данной популяции, дает хорошо приспособленных и «нормальных» особей. Такие особи составляют группу, называемую адаптивной нормой популяции. Концепция адаптивной нормы графически представлена на фиг. 38. На участке IJ пунктирная кривая образует плато, которое соответствует группе одинаково или почти одинаково приспособленных генотипов и фенотипов. Концепция адаптивной нормы
отвергает типологическую концепцию вида (см. гл. 2), согласно которой существует некий «наилучший генотип» и «наилучший фенотип», так что популяция состоит в значительной мере из идентичных «типичных» особей.
Фиг. 38. Различные концепции вида.
Сплошная кривая иллюстрирует типологическую концепцию вида, согласно которой существует некий «наилучший» генотип (и «наилучший» фенотип), соответствующий точке At. Точки /[ и /? представляют собой определяемые средой границы выживания при нормализующем отборе. Пунктирная кривая иллюстрирует концепцию адаптивной нормы, согласно которой существует множество генотипов, дающих сходные фенотипы с одинаковой степенью приспособленности.
Уоллес и Мадден (1953) дали количественное определение концепции адаптивной нормы как группы особей, расположенных в пределах двух стандартных отклонений от средней приспособленности репрезентативной выборки гетерозиготных особей. Хотя «наилучшего генотипа» и «наилучшего фенотипа» не существует, каждой особи выгодно быть хорошо приспособленной к данным определенным условиям своей среды. Однако, как мы уже говорили, изменения среды неизбежны, так что некоторые генотипы, менее приспособленные в данный момент, могут оказаться
преадаптированными к условиям, которые, возможно, возникнут в будущем, и поэтому их роль в «долгосрочном» выживании популяции весьма велика. Благодаря наличию таких генотипов популяция может легко приспосабливаться в случае изменения условий, т. е. сохранить приспособленность и в новых условиях. Как уже упоминалось, изменчивость, поставляющая плохо приспособленные фенотипы, называется генетическим грузом популяции.
Холдейн был первым, кто привлек внимание исследователей к существованию генетического груза, а сам термин был введен в конце сороковых годов Меллером. За прошедшее с тех пор время концепция генетического груза получила широкое распространение и развитие. Нетрудно видеть, что высокая степень генетической изменчивости необходима популяции для того, чтобы иметь возможность непрерывно приспосабливаться к изменениям условий существования, и что это обстоятельство неизбежно снижает общую приспособленность популяции, порождая слабо приспособленных особей. Генетический груз в его широком смысле — это просто всякое снижение (действительное или потенциальное) приспособленности популяции, возникающее в силу наличия генетической изменчивости. Однако существует много факторов, усложняющих конкретное представление о генетическом грузе; к ним относятся, в частности, различного рода мутантные гены (летали, полулетали и др.), способствующие увеличению груза, и различные типы отбора, уточняющие связанные с этим «расходы» популяции.
Концепцию генетического груза как таковую понять нетрудно, однако дать количественную оценку этого груза, установить его подлинное влияние на популяцию или же хотя бы точно определить это влияние оказалось гораздо сложнее. До сих пор нет единого мнения даже о том, как подступиться к этой проблеме. Крау (1958) предложил определять генетический груз как «относительное снижение жизнеспособности популяции (или какого-либо другого рассматриваемого признака) по сравнению с оптималь-
Н1
HbiM генотипом». Это определение было подвергнуто критике на том основании, что идентифицировать оптимальные генотипы трудно, что оптимум выбирается совершенно произвольно и что теоретически величина груза может резко измениться просто вследствие включения в популяцию лишь одной новой благоприятной мутации. Насколько обоснованна эта критика, сказать трудно. Так или иначе, приведенное определение служит отправным пунктом для всех недавних попыток подойти к генетическому грузу с количественной стороны.
Добжанский (1965) определил генетический груз в понятиях адаптивной нормы популяции. Его определение основано на средней приспособленности гетерозиготных особей популяции. Носителями генетического груза считаются индивидуумы, приспособленность которых более чем на два стандартных отклонения ниже средней приспособленности гетерозигот.
Перейдем теперь к более детальному анализу генетического груза.
МУТАЦИИ И МУТАЦИОННЫЙ ГРУЗ
По существу имеется три вида генетического груза: мутационный, сбалансированный и субституцией-ный (или переходный). Из этих трех видов груза слагается общий генетический груз популяции — одни локусы участвуют в создании мутационного груза, другие — сбалансированного, третьи — субституцион-ного. Мутационный груз — это та доля общего генетического груза, которая возникает за счет мутантных аллелей; поскольку естественный отбор направлен против этих аллелей (нормализующий отбор), их частота невелика и они поддерживаются в популяции только благодаря повторным мутациям. Такое положение вещей характерно для того, что принято считать типичным локусом. Рассмотрим более подробно влияние на эти локусы мутационного процесса и отбора, одновременно действующих на популяцию.
В гл. 4 (стр. 132) мы показали, что влияние одного лишь давления мутаций на частоту гена
задается формулой
\р = — ptu + (\ —pt).
Соответственно приращение частоты другого гена равно
= ptu — qtv.
Рассматривая действие отбора, направленного против рецессивов (уравнение 5.5), мы показали, что
sq4 \ - д)
1 — sq2
Таким образом, давление мутаций стремится повысить частоту гена, а естественный отбор, напротив, снижает ее. Когда эти противодействующие силы приходят в равновесие, то
. sq2 (1 — q) pu-qv — -)- .
Здесь обычно допускают, что во всякой реальной популяции частота аллеля, создающего неблагоприятный фенотип (в данном случае q), относительно мала. Следовательно, величину qv как приближающуюся к нулю можно исключить из формулы, а величину 1 — sq2 практически приравнять единице. Таким образом,
pu = sq2(\—q),
(1 — q)u = sq2(\ —q),
и — sq2 (приблизительно); (7.1)
«2=т._
= (приблизительно), (7.2)
Произведение доли (q2) пораженных особей такой популяции на величину вызванного этим геном снижения приспособленности (з) и представляет собой численное выражение мутационного груза (sq2 в уравнении 7.1).
Заметим, что для любого данного s это равно частоте мутаций и. Уравнение (7.2) в действительности применимо только в том случае, когда аллель А полностью доминирует над аллелем а, а отбор направлен против гомозигот аа. Если отбор выступает против доминантного аллеля, то
s?2(i -д)
1-5(1 -^2)
и при равновесии
sq2 (1 — q) pu-vq= (5.7)
На этот раз пренебречь можно величиной ри. Знаменатель 1— s(l — q2) опять считается равным единице. Решая уравнение относительно 2pq, находим
qv = s<72(1 — q), д = s q2( \ —q) v '
2 v 2pq=— (приблизительно). (7.3)
В отсутствие доминирования и при отборе, направленном против аллеля а,
1 — sq *
а при равновесии
2м
Я = — (приблизительно) (7.4)
Для вычисления мутационного груза при разной степени доминирования (Крау, 1958) рассмотрим локус, в котором генотипы АА, АА' и А'А' имеют относительную частоту 1, 1—hs и 1—s соответственно. Здесь h — степень доминирования аллеля А' Поскольку частота гомозигот А'А' очень мала, их вкладом в мутационный груз можно пренебречь, т. е.
считать, что отбор против аллеля А' идет только в гетерозиготах. Следовательно, в состоянии равновесия за каждое поколение частота гетерозигот будет снижаться на величину hs, но эта потеря будет восполняться благодаря мутационному процессу (Д—►Д'), в результате чего часть гомозигот ДД будет переходить в группу ДД' Поскольку каждая гомозиготная особь ДД содержит пр два аллеля Д, скорость превращения генотипов ДД в ДЛ? вдвое выше скорости превращения Д->Д', т. е. равна 2и. Таким образом,
(2pq)
Мы видим вновь, что мутационный груз зависит (если исключить случай, когда h близко к нулю) только от частоты мутаций и равен удвоенной частоте мутаций. В тех случаях, когда й = б, можно использовать уравнение (7.1). Общий генетический груз определяется как сумма генетических грузов отдельных локусов.
До сих пор еще не удалось определить ни для одной реальной популяции, какую часть общего генетического груза составляет мутационный груз, хотя это одна из важнейших задач, стоящих перед нами в настоящее время. По мнению одних авторов, мутационный груз составляет главную часть общего генетического груза популяции; другие отводят ему второстепенную роль, считая, что генетический груз создается в основном за счет сбалансированного груза (см. следующий раздел). Доля участия того или другого в создании общего генетического груза зависит как от доли локусов, сохраняющихся в популяции благодаря уравновешивающему, а не нормализующему отбору (т. е. от того, в какой степени описанная нами ситуация действительно типична), так и от величины генетического груза, приходящегося на один локус, для каждого типа отбора. Следует также подчеркнуть, что изменение условий среды может вызвать сдвиг типа отбора таким образом, что какой-либо локус, участвовавший в создании сбалансированного груза, будет участвовать в создании мутационного груза, или наоборот.
Сдвиг такого типа можно хорошо проиллюстрировать на примере серповидноклеточной анемии. В малярийных районах Африки аллель серповидноклеточ-ности сохраняет высокую частоту благодаря сильному уравновешивающему отбору. Это связано с тем, что гетерозиготы невосприимчивы к малярии, а анемия у них бывает выражена лишь в слабой степени. Гомозиготы (Sk/Sk) отличаются высокой восприимчивостью к малярии и вовсе не подвержены анемии, а гомозиготы другого типа sk/sk страдают тяжелой анемией. В другой части света — в Северной Америке, — где проблемы малярии не существует, аллель sk встречается очень редко. В этих областях происходит жесткий отбор против генотипа skjsk, так как эти гомозиготы по существу нежизнеспособны, но этот отбор носит нормализующий характер, поскольку гомозиготы Sk/Sk обладают преимуществом по сравнению с гетерозиготами, у которых имеется слабо выраженная анемия. Частота гена sk в этих областях значительно ниже, так что данный локус участвует в создании мутационного груза.
УРАВНОВЕШИВАЮЩИЙ ОТБОР И СБАЛАНСИРОВАННЫЙ ГРУЗ
При изучении природных популяций внимание привлекает множество таких ситуаций, как при серповидноклеточное™, в системе групп крови АВО или во многих ферментных системах (фиг. 39), когда два или больше аллелей какого-либо локуса сохраняют относительно высокую частоту. Естественно, что в этом случае частоты разных генотипов и соответствующих им фенотипов также сохраняются на высоком уровне. Такие полиморфные локусы отличаются от нашего «типичного» локуса не по числу возможных фенотипов, а по их частотам. Эти локусы сохраняются благодаря какой-то форме уравновешивающего отбора, в котором участвуют селективные силы, действующие в противоположных направлениях. Мы уже видели, например, что сбалансированное равновесие может возникнуть, если аллель, против которого
направлен отбор, когда он находится в гомозиготном состоянии, сохраняется вследствие превосходства гетерозигот (сверхдоминирование). Состояние равновесия может быть также достигнуто в следующих случаях:
1) отбор благоприятствует данному аллелю на одной стадии развития и направлен против него на дру-
Фиг. 39. Электрофореграмма D. ananassae, выявляющая полиморфизм эстеразы-6, контролируемой одним из локусов третьей хромосомы.
Во всех популяциях обнаружена «быстрая» и «медленная» формы эстеразы, а у гетерозигот — оба варианта. Места локализации ферментов на крахмальном геле обнаруживаются по цветной реакции взаимодействия субстрата и красителя. Самые темные полосы —это полосы эстеразы-6; светлые полосы —следы активности других эстераз. Особи А, Е и F гетерозиготны (FS), В и D — медленные варианты (SS), а С —быстрый вариант (FF).
1 — быстрая полоса; 2 — медленная полоса.
гой стадии (частный случай представляет собой «мейотический дрейф» — нарушение мейоза, в результате которого преобладают гаметы, содержащие один из аллелей);
2) отбор благоприятствует сохранению аллеля у особей одного пола, но действует против него в отношении особей другого пола;
3) в одни периоды (например, в определенное время года) отбор направлен на сохранение аллеля, а в другое время — против него;
4) разные генотипы используют различные экологические ниши в пределах одного ареала, что приводит к снижению конкуренции и, следовательно, к ослаблению элиминирующего действия отбора—так называемый эффект Людвига;
5) в разных субпопуляциях, занимающих местообитания с различными условиями среды, отбор благоприятствует разным аллелям;
6) отбор благоприятствует сохранению аллеля, когда он редок, и направлен против него, когда он встречается часто.
Каждый из этих механизмов сохранения полиморфизма, за исключением, возможно, четвертого, создает генетический груз, называемый сбалансированным грузом.
Рассмотрим более подробно сохранение полиморфизма благодаря сверхдоминированию. Можно привести множество примеров, когда гетерозиготы превосходят по приспособленности гомозиготы обоих типов. Но в большинстве случаев причины такого сверхдоминирования остаются неясными (см. Д. Брю-бейкер, «.Сельскохозяйственная генетика»). Однако одну из возможных причин сверхдоминирования можно понять, обратившись к механизму действия гена. Ген совместно с рибосомами и такими молекулами, как молекулы информационной РНК, транспортной РНК и различных связанных с ними ферментов, контролирует последовательность аминокислот в таких специфических функциональных белках, как ферменты, антитела и т. п. Если в данном локусе может содержаться два разных аллеля, то гетерозигота будет потенциально более гибкой, чем любая из гомозигот, поскольку она может обеспечить синтез не одного, а двух разных полипептидов. Это может давать преимущество в разных случаях. Если оба аллеля обеспечивают синтез белков, полезных в любое время, то особь, несущая оба аллеля, будет обладать селективным преимуществом. Или, быть может, в разные периоды жизни, при разных условиях внешней или внутриклеточной среды организм испытывает потребность в разных белках, и опять-таки наличие двух аллелей окажется благоприятным.
В связи с этим интересно напомнить полученные в последнее время многими исследователями данные о том, что многие соединения, считавшиеся одним чистым ферментом, на самом деле представляют собой
группу близких ферментов. В отдельном организме в такую группу входят ферменты, катализирующие одну и ту же реакцию, но различающиеся по строению или по условиям, в которых они проявляют оптимальную активность. Такие ферменты были названы изоферментами или изозимами', в каждую группу может входить до десятка и даже более изоферментов. Известно, что на активность ферментов сильно влияют даже незначительные изменения таких факторов, как температура, pH и концентрация различных ионов. Считают, что наличие в клетке или в организме двух или нескольких изоферментов лучше удовлетворяет потребности организма и притом в более широком диапазоне условий, чем один фермент. По-видимому, это можно считать примером сбалансированного полиморфизма. Исследованные системы изоферментов подтверждают предположенье о том, что одновременное присутствие двух или большего числа сходных, но не идентичных ферментов обеспечивает сохранение стабильности особи в более широком диапазоне внешних условий, чем это может обеспечить один фермент. Кроме того, становится очевидным большое значение взаимодействия между генами разных локусов, а у гетерозигот возможности для такого взаимодействия гораздо больше, чем у гомозигот. Однако остается фактом, что наши знания относительно однолокусного гетерозиса очень ограниченны, а объяснения спекулятивны. Эта ограниченность знаний отягощается тем, что очень часто гетерозис связан с аллелями, которые считаются рецессивными и которые в гомозиготном состоянии детальны. Мы принимаем, хотя, возможно, и ошибочно, что большинство этих аллелей неактивно и что их рецессивность обусловлена именно тем, что они не активны. Спрашивается, как же в таком случае эти гены могут привести к гетерозису?
У Drosophila наблюдается более сложный тип гетерозиса, особенно подробно изученный в большой серии работ Добжанского с сотрудниками. Здесь гетерозис связан не с отдельными генами, а с ком
плексом генов, включенных в инверсию и благодаря этому сохраняющихся как единое целое.
Как правило, большинство особей, выловленных в природных популяциях, гетерозиготны по различным инверсиям. Так, в хорошо изученных случаях (например, у D. pseudoobscura) было найдено, что распространение в популяциях тех или иных инверсий характеризуется определенной закономерностью. Имеется ряд инверсий, широко распространенных во многих популяциях и встречающихся с высокой частотой по всему ареалу вида. Некоторые из этих наиболее распространенных инверсий получили названия по месту, где они были впервые обнаружены («Стандарт», «Эрроухед», «Чирикахуа», «Санта-Крус», «Три Лайн», «Олимпик», «Истс Парк» и «Пайке Пик»). Анализ особей, выловленных в ряде районов северной Мексики и на юго-западе США, показывает, что хотя большинство изученных популяций содержит несколько инверсий, преобладает обычно одна или две. Больше того, инверсия «Стандарт», преобладающая на побережье Калифорнии (фиг. 40), оказывается обычной и во внутренних районах, тогда как в других частях ареала вида эта инверсия встречается редко. Инверсия «Чирикахуа» преобладает на нагорьях Мексики, обычна в Калифорнии и редка в остальных районах юго-запада США. Инверсия «Эрроухед» преобладает в Юте, Аризоне, Колорадо и Нью-Мехико, а инверсия «Пайке Пик» — на восточном склоне Скалистых гор и в Западном Техасе.
Показано также, что в горных районах частота инверсий изменяется в зависимости от высоты над уровнем моря (фиг. 41). Например, в Калифорнии — вю-рах Сьерра-Невада — частота трех больших инверсий изменяемся следующим образом: при подъеме от 250 до 3000 м частота инверсии «Стандарт» (St) постепенно уменьшается от 46 до 10%, частота инверсии «Эрроухед» (Аг) увеличивается от 25 до 50%, а частота инверсии «Чирикахуа» (Ch) колеблется, увеличиваясь от 16 до 20%.
В другой серии экспериментов была показана сезонная изменчивость частоты некоторых инверсий
Фиг. 40. Частота (в процентах) различных инверсии в третьей хромосоме в популяциях D. pseudoobscura, распространенных от юга Калифорнии до севера Техаса.
в отдельных популяциях. Так, в горах Сан-Джасинто (Калифорния) частота инверсии «Стандарт» максимальна ранней весной (53%), затем падает до 27% в июне, после чего постепенно вновь увеличивается, достигая к октябрю 50%. Частота инверсии «Чирикахуа» меняется в обратном направлении. В марте,
g St 46 41 32 26 14 II 1010
g Аг 25 35 38 44 45 55 5050
О
Фиг. 41. Зависимость частоты инверсий «Стандарт» (S/) и «Эрроухед» (Лг) в популяциях D. pseudoobscura от высоты над уровнем моря (Йосемитская долина, Сьерра-Невада).
Места взятия проб указаны на схематическом профиле местности. Цифры над схемой — процент инверсий St и Аг в каждой местности.
когда сократившаяся за зиму популяция начинает увеличиваться, частота этой инверсии равна 23%, к июню она достигает 40%, а затем постепенно вновь уменьшается, достигая к октябрю 20% (фиг. 42).
Все эти данные показывают, что в разных условиях разные последовательности генов имеют разную адаптационную ценность. Это неоднократно подтверждалось лабораторными экспериментами, в которых Разные хромосомные комбинации помещали в разные Условия. В большинстве случаев в опытах варьировали относительную влажность или температуру. Но
в некоторых опытах изменяли другие факторы, например вводили ограничение диеты. При относительной влажности 100% инверсия «Чирикахуа» более жизнеспособна (91%), чем «Эрроухед» (74%) или «Стандарт» (84%). При относительной влажности 92%
Фиг. 42. Изменение частоты инверсий «Стандарт» (/) и «Чирикахуа» (//) в природных популяциях D. pseudoobscura в горах Сан-Джасинто в период с марта по октябрь.
жизнеспособность «Стандарт» (94%) выше, чем «Чирикахуа» (81%) и «Эрроухед» (85%). При нулевой относительной влажности наиболее жизнеспособна инверсия «Эрроухед» (74, 54 и 59% соответственно) При температурах 28.. .30° С наиболее жизнеспособен кариотип St/St, а при 0. .4° С — кариотип St/Ch.
Хотя по многим тестам гетерозиготы оказываются более жизнеспособными, чем гомозиготы обоих типов, но, как показывают приведенные цифры, это не всегда так. Наблюдения хорошо соответствуют данным множества экспериментов в популяционных клетках, проведенных на D. pseudoobscura и на многих других видах. В этих условиях на каждой стадии жизненного цикла происходит конкуренция между геноти
пами и можно определить относительную ценность разных инверсий для выживания (включая и воспроизводство). Если в популяционный ящик, в котором поддерживаются определенные условия, вводят несколько инверсий, то в нем устанавливается некоторое равновесие с характерной для каждой инверсии частотой. Чрезвычайно важно, что соотношение инверсий в такой равновесной популяции не зависит от их первоначальных частот — во всех случаях точки равновесия почти совпадают. Однако, изменяя условия в популяционном ящике, можно изменить и точку равновесия. Так, для изменения относительной жизнеспособности двух инверсий бывает достаточно ввести в исходную популяцию третью инверсию.
В гл. 4 мы уже говорили о том, что гены, входящие в инверсию, наследуются совместно, в виде единого блока, и что если большинство особей в популяции гетерозиготно по данной инверсии, то отбор действует на всю эту группу генов, а не на отдельные гены. Как уже отмечалось, существует множество данных, подтверждающих, что адаптационная ценность инверсий у D. pseudoobscura зависит от условий существования. Так, адаптационная ценность отдельной инверсии различна в разных географических областях. На фиг. 43 показано изменение четырех популяций, исходный состав которых был одинаков (20% Si и 80% Ch). Все четыре популяции с одинаковой скоростью приближались к состоянию равновесия, причем во всех четырех равновесная частота инверсии St равна 80%. Это равновесие говорит о том, что гетерозиготы St/Ch имеют преимущество по сравнению с гомозиготами обоих типов. Все эти популяции содержат инверсии, взятые из одного и того же географического района (Пиньон-Флэт, Калифорния). Эти результаты резко отличаются от тех, которые представлены на фиг. 44. В этом случае исходным материалом для лабораторных популяций служили инверсии, взятые из разных мест: инверсия
—из Пиньон-Флэт, а инверсия Ch — из шести Различных мексиканских популяций. Здесь не наблюдается того сходства в динамике популяции, которая
характерна для инверсий, взятых из одного района. К концу эксперимента в четырех популяциях происходит, хотя и с разной скоростью, закрепление инверсии St, а в двух устанавливается равновесие, но па различном уровне. Очевидно, Cft-хромосомы из разных географических районов генетически различны.
Фиг. 43. Изменение относительной частоты инверсий в четырех популяционных ящиках D. pseudoobscura.
Исходное соотношение: 20% St и 80% Ch, взятых из одного и того же гео графического района (нагорная терраса Пиньон-Флэт, Калифорния). Во вес х четырех ящиках изменение соотношения происходит одинаково вплоть до наступления равновесия.
Эти данные привели Добжанского (1951) к выводу о том, что инверсии предохраняют генные комплексы от кроссинговера, благодаря чему в процессе эволюции образуются интегрированные группы renoi, которые подвергаются отбору как единое целое (как супергены). Создается впечатление, что, по крайней мере у D. pseudoobscura, разные комплексы становятся приспособленными к различным нишам той или иной местности. При этом особи, гетерозиготные по двум таким комплексам, по-видимому, могут лучше
противостоять колебаниям условий среды, поскольку они несут по два интегрированных и внутренне сбалансированных генных комплекса, каждый из которых обеспечивает устойчивость к иным или по меньшей мере частично отличающимся условиям. Это
Фиг. 44. Результаты эксперимента, аналогичного приведенному на фиг. 43, с той разницей, что исходные популяции были взяты из разных географических районов.
В этом случае изменение относительной частоты инверсий в разных ящиках происходит далеко неравномерно, а кроме того, наблюдается элиминация хромосомы с инверсией Ch. По-видимому, генетический состав одних и тех же инвертированных участков в разных местностях различен.
порождает инверсионный полиморфизм, носящий приспособительный характер. Иными словами, полиморфизм по самой своей природе адаптивен, поскольку он дает возможность популяции лучше выдержать все испытания, каким она может подвергнуться при изменениях условий среды, чем если бы она содержала всего один генотип или один адаптивный генный комплекс.
ОГРАНИЧЕННОСТЬ ЧИСЛА СВЕРХДОМИНАНТНЫХ ЛОКУСОВ
Сохранение полиморфизма благодаря явлению сверхдоминирования представляет собой один из типов уравновешивающего отбора, позволяющего поддерживать в популяции довольно высокую частоту двух и более аллелей. Плата (выраженная в форме генетического груза) за поддержание полиморфизма с помощью сверхдоминирования может быть очень высокой. При равновесии
5) “Г *2
При этом вследствие расщепления потомков гетерозигот в популяцию постоянно поступает множество слабо приспособленных гомозиготных особей. Эти особи £ создают часть сбалансированного груза, и притом, по-видимому, большую его часть. Крау (1958) назвал его сегрегационным грузом, поскольку он возникает в результате расщепления гетерозигот. Если относительные приспособленности генотипов АА, АА' и А'А' равны соответственно 1 —sb 1 и 1 —s2, то этот груз равен Sip2 + s2^2, т. е. суммарному снижению приспособленности, вызванному гомозиготами обоих типов. Используя формулу (5.10) для q и зная, что
л _ $2
Р “ 5, + s2 ’
находим значения р2 и q2 и получаем, что
(s2)2 , (Si)2
сегрегационный груз = s, ^| + ^- + s2 (s, + s2~ =
___ S| (s2y + s2 (s,)2
(Si + s2)2
_ s,s2 (S| + s2) __ (S) + s2)2
5i + 52
Очень большой сегрегационный груз несет локус, в котором гомозиготы обоих типов летальны: в этом
случае 50% потомков гибнет. Это крайний случай, редко встречающийся в природе; однако он заставляет поставить весьма важный вопрос о том, сколько сверхдоминантных локусов может содержать популяция, не рискуя оказаться под бременем непосильного для нее генетического груза. Например, если выживают только гетерозиготы, то в каждом поколении доля выживших особей снижается на 50% за счет лишь одного такого локуса. Если допустить, что популяция содержит 10 таких сбалансированных по деталям локусов, то вероятность того, что какой-либо индивидуум окажется гетерозиготным по всем этим 10 локусам, а следовательно, будет жизнеспособным, равна (7г)10 или V1024. Это означает, что только одна из 1024 зигот избежит генетической гибели. Вряд ли существует много видов, репродуктивный потенциал которых мог бы обеспечить выживание при таком сильном давлении отбора. Конечно, допущение о том, что все сверхдоминантные локусы наследуются независимо, необоснованно. Часть этих локусов или даже все они могут оказаться сцепленными. В этом случае генетический груз популяции снижается, поскольку вероятность того, что один элиминируемый индивидуум унесет с собой не одну, а несколько пар летальных комбинаций, возрастает и соответственно возрастает вероятность для каждой данной зиготы быть свободной от летальных комбинаций. Но и в этом случае число сверхдоминантных локусов, которые популяция в состоянии выдержать, несомненно, ограничено.
Генетический груз переносится значительно легче, если гомозиготы нелетальны. Допустим, что гомозиготы не гибнут, но эффективность их размножения (способность потомков достигнуть половозрелого состояния) составляет 98% эффективности размножения гетерозигот (si = s2 = 0,02) . Тогда вероятность того, что зигота избежит генетической гибели, равна 396/400 (если равновесная частота # = 0,5). В начале каждого поколения соотношение зигот составит 7ИД Ч2Аа Ч^аа, что в круглых числах удобно записать как 100 :200 :100. Генотипический состав
половозрелых особей будет уже иным — 98:200:98. Отсюда вероятность для данного индивидуума достигнуть половозрелости равна 0,99. Если популяция содержит 10 таких независимых локусов, то вероятность выживания для данной особи будет равна (0,99) 10 = 0,904, при 100 локусах (0,99) 100=0,363, а при 1000 локусах (0,99)1000=0,000043. Исходя из этих расчетов, можно сделать вывод, что среди большинства видов общее число локусов, сохраняющих свой полиморфизм благодаря сверхдоминированию, не может сильно превышать 100, даже если наша оценка летальности несколько завышена.
Большинство генетиков считают, что даже такой относительно простой организм, как дрозофила, содержит не менее 10 000 локусов. Если это действительно так, то даже в том случае, когда гомозиготы лишь незначительно менее приспособлены, чем гетерозиготы, сверхдоминантными могут быть одновременно менее 10% локусов. Логичность такого вывода кажется безупречной. Тем не менее проблема сверхдоминирования стала в последние годы одной из наиболее острых проблем популяционной генетики. Дискуссия началась с выдвинутого Уоллесом (1958) предположения о том, что 50% (или даже больше) всех локусов дрозофилы находятся в гетерозиготном состоянии. Так как эти результаты были получены в радиационном эксперименте, их широкое толкование было воспринято с некоторым сомнением. Однако совсем недавно Джонсон с сотр. (1966) и Хэбби п Левонтин (1966) предприняли первые прямые попытки определить долю полиморфных локусов в природных популяциях. Используя недавно разработанный метод электрофореза ферментов на крахмальном геле, группа Джонсона исследовала D. ananassae, а Левонтин и Хэбби — D. pseudoobscura. Левонтин и Хэбби обнаружили в природных популяциях изменчивость по 21 ферменту. Многократно показано, что синтез ферментов находится под прямым генетическим контролем; основываясь на широко принятой н подтверждаемой многими данными теории одни ген — один фермент (или вернее, один цистрон
один полипептид), можно считать, что синтез этих ферментов контролируется 21 локусом. Хэбби и Левонтин обнаружили, что по всему ареалу вида 9 локусов из 21 (или 39%) полиморфны. В среднем каждая локальная популяция содержит 30% полиморфных локусов.
Основываясь на этих данных, которые в силу ряда причин, таких, как небольшая величина выборки, преуменьшают число полиморфных локусов, число полиморфных локусов у дрозофилы по скромной оценке можно считать равным 2000. Эти данные полностью совпадают с данными, полученными на D. ananassae. Если при этом принять во внимание предыдущие рассуждения о том, что популяция может содержать одновременно немногим более 100 сверхдоминантных локусов, то, по-видимому, придется признать, что большая часть этих полиморфных локусов сохраняется с помощью каких-то других механизмов. Именно такое предположение высказали Хэбби и Левонтин. Однако другие авторы (Свед, Рид и Бодмер, 1967; Кинг, 1967) усомнились в правильности исходного допущения о независимом действии каждого локуса на процесс снижения приспособленности. Это значит, что в приведенном выше примере величина (0,99)п не соответствует вероятности выживания. Хотя доводы этих авторов опираются на более сложную модель, читателю, достаточно знакомому с математикой, их работы будут интересны. Предположим, что два локуса независимы и что приспособленность гомозигот по каждому из этих локусов равна 0,99; тогда для гомозигот по обоим локусам вероятность гибели будет вдвое выше, чем для гомозиготы по одному локусу (точнее, вероятность гибели будет равна 0,0199, а не 0,02, поскольку вероятность того, что особь будет гомозиготна по деталям обоих локусов, равна 0,0001). Если исходить из более реального допущения о функциональном взаимодействии между локусами, то можно представить себе ситуацию, когда с увеличением числа гомозиготных локусов будет прогрессивно уменьшаться доля генетического груза, вносимого каждым дополнительным локусом. При этом вскоре
будет достигнут такой уровень, при котором дальнейшее введение в генотип гомозиготных локусов уже не будет сопровождаться снижением приспособленности.
Мукаи (1968) предложил другой подход к проблеме генетического груза, помогающий объяснить некоторые кажущиеся противоречия между степенью гетерозиготности и величиной генетического груза. Он показал, что, по крайней мере для некоторого класса генов, называемых полигенами, существует некоторое оптимальное число гетерозиготных локусов. Полигенами называются гены, которые в отличие от леталей и полулеталей оказывают слабое влияние на приспособленность. Сущность метода состоит в следующем. Из природной популяции D. melanogaster выделяли одну вторую хромосому, переводили ее в гомозиготное состояние и создавали ряд линий по схеме скрещиваний, приведенной на фиг. 45. Скорости накопления полигенных мутаций в каждой линии регистрировали, периодически определяя жизнеспособность вторых хромосом. Для этого, как показано в нижней половине фиг. 45, потомков Q//+, полученных от скрещивания Pml+ X CylPm, скрещивали между собой. Для определения жизнеспособности гетерозигот особей Q//+ скрещивали с особями +о/+о, взятыми из анализирующей линии, у которой хромосомы +о имеют нормальную жизнеспособность.
В течение 25 поколений жизнеспособность гомозигот неуклонно снижалась вследствие накопления полигенных мутаций — в среднем 0,1411 на одну вторую хромосому за поколение. (Эта скорость гораздо выше скорости появления летальных мутаций, равной 0,006.) Что же касается гетерозигот, то их жизнеспособность варьирует. Если тест проводили таким образом, что остальные хромосомы (генетический фон) были гомозиготны, то жизнеспособность гетерозигот по испытуемой хромосоме была выше нормальной; это показывает, что мутантные локусы были сверхдоминантны. Если же генетический фон уже был гетерозиготным, то жизнеспособность гетерозигот по испытуемой хромосоме в лучшем случае оказывалась нормальной, хотя обычно она ниже нормы. Таким об-
разом, влияние новых мутаций на приспособленность зависит от степени уже имеющейся гетерозиготности.
St
Рт
I
(5?)^ X
I-------------1
Су Су
с» +, +,
1 2 1
Гибнут Фиг. 45. Схема скрещиваний для выделения и поддержания независимых линий D. melanogaster, гомозиготных по второй хромосоме.
®се линии содержат копии одной и той же исходной второй хромосомы, выделенной из природной популяции.
На фиг. 46 показана зависимость между жизнеспособностью гетерозигот и гомозигот в случае гомозиготного генетического фона. На фиг. 46, Л все мутантные гены гетерозигот находятся в фазе «сцепле-
Фиг. 46. Эффект «сцепления — отталкивания» и оптимальная гетерозиготность полигенов, контролирующих жизнеспособность
D. melanogaster.
На графиках (Л и Б) показано относительное влияние на жизнеспособноегь особи гетерозиготности или гомозиготности но различному числу спонтанных полигенных мутаций в копиях одной исходной хромосомы. У гетеро-зигот в «фазе сцепления» (Д) одна хромосома свободна от мутантов, т. е. имеет «нормальную жизнеспособность» ( + //+о)- Гетерозиготы в «фаiC отталкивания» (Б) содержат две различные мутантные хромосомы ( + //+/)• Жизнеспособность гетерозигот в «фазе сцепления» во всех случаях выai<j жизнеспособности нормальных гомозигот (+о/+о)« принятой за 1, тогда 1 1К жизнеспособность гетерознгот в «фазе отталкивания» всегда ниже. О’! 11 мальный уровень сверхдоминирования проявляют гетерозиготы в «Ф 1 ,с сцепления», которые содержат хромосомы, дающие в гомозиготном сос!"я нии жизнеспособность 0,3... 0,5 (по приблизительной оценке такие хромосома содержат от 8 до 11 мутантных полигенов). Оценка жизнеспособности зигот и гетерозигот (оси X и У) для каждой из двух кривых (Д и Б) относительно жизнеспособности стандартных гомозигот (+о/+о). При э1'’. коэффициент регрессии 0,39 (кривая Б) служит мерой средней степе1 доминирования, h, вновь возникших мутантных полигенов.
ния (т. е. все лежат в одной и той же хромосоме). С увеличением числа мутантных генов, приводящим к снижению жизнеспособности гомозигот, жизнеспособность гетерозигот возрастает, а затем, достигнув максимума, падает. Точка оптимальной гетерозиготности соответствует примерно 9 локусам. Совершенно неожиданные результаты получены в том случае, когда полигенные мутации гетерозигот находятся в фазе «отталкивания» (мутантные гены лежат в обеих хромосомах; фиг. 46, Б); при этом снижение жизнеспособности гомозигот сопровождается снижением жизнеспособности гетерозигот. На фиг. 46, Б +//+/ обозначает гетерозиготу по хромосомам из двух разных экспериментальных линий. В этом случае по оси ординат откладывают жизнеспособность гетерозигот -р7+/, а по оси абсцисс — нормированную среднюю жизнеспособностей двух классов гомозигот -Н7+* и +}/+}• Какими бы ни были причины этого странного явления, оно позволяет объяснить несоответствие между результатами, полученными Уоллесом (1958), и результатами, полученными Гринбергом и Крау (1960). Уоллес нашел, что облученные хромосомы в гомозиготном состоянии обычно менее жизнеспособны, а в гетерозиготном — более жизнеспособны, чем нормальные хромосомы, т. е. индуцированные мутации сверхдоминантны. Но Гринберг и Крау, изучая хромосомы природных популяций, не обнаружили у них этого интересного явления. В природных популяциях хромосомы, жизнеспособность которых в гомозиготном состоянии была ниже средней жизнеспособности гомозигот, оказывались менее жизнеспособными и в гетерозиготном состоянии. Таким образом, в этом случае новые мутации проявляли некоторую доминантность, но не сверхдоминантность. Но, оказывается, эти два эксперимента проводились по-разному; Уоллес изучал жизнеспособность гетерозигот, находившихся в «фазе сцепления», а Гринберг и Крау работали с гетерозиготами в «фазе отталкивания», видимо, это и вызвало расхождение в результатах.
Другие исследователи указывали на то, что, Даже если гены различных локусов функционально
независимы, гомозиготность по более чем одному локусу увеличивает вероятность гибели для данной особи. Таким образом, гибель одной особи сопровождается элиминацией многих гомозиготных локусов (или большой доли груза).
Возможна и другая причина сохранения полиморфизма в природных популяциях: специфическая приспособленность каждого генотипа к своей микронише, обеспечивающая ему безусловное преимущество в этой нише. В течение нескольких последних десятилетий главные проблемы, касающиеся генетической структуры природных популяций, и, в сущности, главные проблемы всей эволюционно-генетической теории, сводились к следующим:
1) Какова степень генетической изменчивости панмиктической популяции диплоидных организмов?
2) Какова в среднем доля гетерозиготных локусов у одной особи?
3) Каков генетический груз, сопряженный с генетической изменчивостью?
4) Каким образом, несмотря на этот груз, сохраняется генетическая изменчивость?
Наши знания по всем этим вопросам далеко не достаточны, тем не менее уже сейчас ясно, что ответы на первые три вопроса будут заключать в себе гораздо более высокие оценки, чем мы ожидали. Дальнейшие исследования в этой области могут привести к некоторым неожиданным изменениям в наших представлениях о генетической структуре популяций.
ЭФФЕКТ ЛЮДВИГА, ДИЗРУПТИВНЫЙ ОТБОР И ВОЗНИКНОВЕНИЕ РАЗНООБРАЗИЯ
МЕЖДУ ДЕМАМИ
Другой важный источник так называемого сбалансированного груза — сбалансированный полиморфизм, сохраняющийся благодаря дизруптивному отбору, т. е. отбору, направленному на сохранение одновременно двух или большего числа генотипов. Если отбор благоприятствует обеим гомозиготам, то даже при селективной слабости гетерозигот частоты обоих
аллелей в популяции могут оставаться на довольно высоком уровне. Это явление повышенного разнообразия особенно свойственно давно сложившимся популяциям, размещенным в центре ареала вида. Его часто называют «эффектом Людвига». Людвиг (1950) утверждал, что генотип, способный использовать незанятую субнишу, может выжить даже в том случае, если его приспособленность ниже средней приспособленности генотипов, занимающих другие субниши. В пользу этой концепции свидетельствует то обстоятельство, что у многих видов полиморфизм, как морфологический, так и хромосомный, гораздо сильнее выражен в центральных частях ареала, чем на периферии (Карсон, 1958). По всей вероятности, центральные районы ареала по некоторым основным условиям более благоприятны для вида. Поэтому естественно, что вид укрепляется в этих наиболее подходящих для него частях ареала до тех пор, пока он не сможет постепенно, развивая ряд полиморфизмов (путем дизруптивного отбора), начать проникать в новые ниши, получая, таким образом, возможность еще более эффективно использовать свою среду. В периферических районах основные условия гораздо менее благоприятны; поэтому популяции, сумевшие здесь обосноваться, специфически приспособлены к одной или нескольким субнишам, тогда как в остальном эти области для них непригодны. Дизруптивный отбор, распространяясь на популяции различных географических районов, приводит к возникновению разнообразия между демами и к образованию рас (гл. 8). Со временем возрастает разнообразие и внутри периферических демов, если они не вымирают.
Что касается полиморфизмов вообще, то здесь следует остерегаться слишком жестких определений и не рекомендуется классифицировать их на основании вызывающих их механизмов. Вполне возможно, цто в некоторых случаях полиморфизм поддерживается благодаря сочетанию ряда описанных выше механизмов. Сверхдоминирование и «эффект Людви-Га>> вовсе не исключают один другого; возможно, Что во многих случаях полиморфизм обусловлен
взаимодействием этих факторов. Некоторые доказательства этого получены Хейтсом (1948) на инверсиях «Стандарт» и «Чирикахуа» вида D. pseudoobscura. При температурах 28 .. 30° С средняя жизнеспособность гомозигот St/St выше жизнеспособности гомозигот Ch/Ch и гетерозигот Ch/St. Но при 0... 4° С жизнеспособность гетерозигот была выше, чем у обеих гомозигот. Таким образом, можно сделать вывод, что и в природных популяциях при некоторых определенных условиях среды (субниши) гомозиготы могут оказаться более приспособленными, хотя в других частях ниши или при некоторых других условиях та же самая инверсия сохраняется благодаря сверхдоминированию. Следовательно, источником полиморфизма может быть действие дизруптивного отбора в сочетании со сверхдоминированием.
Другой интересный способ сохранения сбалансированного полиморфизма — отбор, зависящий от частоты признака. Это значит, что когда ген редок, то отбор направлен на его сохранение, а когда этот ген становится обычным для популяции, отбор действует против него. Иначе говоря, селективная ценность гена зависит от его частоты. Примером такой переменчивости направления отбора могут служить уже упоминавшиеся случаи мимикрии (см. фиг. 28 и 29). Так, при наличии у какого-либо полиморфного вида мимикрирующей формы, которая подражает крайне несъедобной модели, отбор благоприятствует аллелю, контролирующему мимикрию, когда частота этих форм невелика. Такое действие отбора обусловлено тем, что в силу самой редкости мимикрирующих особей хищники научаются «распознавать» их как несъедобную модель. Впоследствии, с увеличением частоты этого аллеля и числа мимикрирующих особей, хищники уже не могут так эффективно обучаться избегать их, и отбор может начать действовать против аллеля, обеспечивающего подражание.
В работе Койима с сотр. (1967) приводится более подробно документированный пример зависимости отбора от частоты. Эти авторы изучали локус эстераза-6 у D. melanogaster, контролирующий синтез опрс-1
деленного фермента, а именно эстеразы-6. В этом локусе имеется два аллеля F и S, которые определяют синтез молекул фермента, отличающихся по электрофоретической подвижности — быстрая (F) и медленная (5) формы (фиг. 39). Исходным материалом для этих опытов послужила большая лабораторная популяция, в которой за 15 поколений частоты аллелей F и S стабилизировались на уровнях 0,3 и 0,7. Затем из этой популяции были выделены путем индивидуальных скрещиваний линии, гомозиготные по аллелю F или S. Скрещивая эти линии между собой, они получили гетерозиготы FS; оказалось, что самки FS на двух различных питательных средах откладывают больше яиц, чем гомозиготные самки FF или SS, причем гомозиготы FF превосходят в этом отношении гомозиготы SS. Так, среднее число яиц, отложенных самками FF, FS и SS на кукурузной среде за 4 дня, равно соответственно 49,83 ±1,65; 52,97 ± ± 1,62; 46,71 ± 1,65. Видимо, существуют какие-то приспособительные признаки, компенсирующие пониженную плодовитость генотипов SS (ведь равновесная частота гена S равна 0,7).
Для определения жизнеспособности генотипов от стадии яйца до взрослого состояния проводили скрещивания между гомозиготными линиями FF и SS, и оплодотворенных самок помещали в популяционные ящики, где они могли откладывать яйца. Таким образом было создано несколько популяций с различными частотами генов, находящимися в состоянии равновесия Харди — Вайнберга. Ожидаемая доля каждого генотипа была определена из уравнения Харди — Вайнберга с учетом обнаруженных различий в плодовитости между самками FF, FS, SS. Эти теоретические расчеты для всех четырех -клеток представлены в табл. 14.
Однако анализ потомства показал, что фактические частоты не всегда совпадают с теоретическим ожиданием. В табл. 15 приведены оценки относительных жизнеспособностей генотипов (отношение наблюдаемой доли к ожидаемой). Заметим, что когда наблюдаемые величины равны ожидаемым, относитель-
Таблица 14
Ожидаемые частоты генотипов FF, FS, SS
Частота гена F FF FS SS Bcci
0,7 0,511 0,393 0,096 1,000
0,5 0,260 0,497 0,243 1,000
0,3 *) 0,105 0,403 0,492 1,000
0,15 0,047 0,250 0,703 1,000
*) Та же, что и в основной популяции.
Таблица 15
Оценки жизнеспособности разных генотипов и соответствующие стандартные ошибки
Частота гена F FF FS SS
0,7 0,759 ± 0,048 1,140 ± 0,061 1,713 ± 0,151
0,5 0,842 ± 0,078 1,034 ± 0,049 1,099 ± 0,084
о,з 1,148 ± 0,151 1,016 ± 0,062 0,954 ± 0,052
0,15 1,677 ±0,195 1,726 ± 0,075 0,697 ± 0,028
ная жизнеспособность равна 1. Интересно, что когда частота аллеля F выше равновесной частоты, наблюдаемой в исходной популяции, то жизнеспособность гомозигот FF значительно ниже, а гомозигот SS значительно выше ожидаемой. Это особенно четко проявляется в ящике, где частота аллеля F=0,7. Если же частота аллеля F ниже равновесной, то жизнеспособность гомозигот FF выше, а гомозигот SS ниже ожидаемой. Эти данные ясно показывают, что относительная жизнеспособность генотипа зависит от частоты гена. В данном случае в ящике, где частота аллеля F меньше равновесной, отбор направлен на его сохранение, а в ящике, где его частота больше равновесной, отбор действует против этого аллеля.
Фиг. 47. Изменение частоты аллелей в локусе эстераза-6 у D. те-lanogasier на банановой питательной среде.
Фиг. 48. Изменение частоты аллелей в локусе эстераза-6 у D. те-lanogaster на кукурузной среде.
Относительная приспособленность (наблюдаемая/ ожидаемая)
Фиг. 49. Относительная приспособленность популяции (по данным фиг. 47 и 48) D. nielanogaster.
Когда популяция еще далека от конечного состояния равновесия (поколения 1..10), адаптационная ценность аллелей F иЗ зависит от того, выше или ниже равновесной их исходные частоты. При равновесии адаптационные ценности обоих аллелей F и S одинаковы и равны единице, т. е. эти аллели селективно нейтральны. Это один из возможных способов сохранить в по пуляции большое число полиморфных локусов, не обременяя ее непосильным генетическим грузом. Кружками обозначены генотипы FFt треугольниками — генотипы FSt квадратами — генотипы 55,-
Относительная приспособленность ( наблюбаемая/ожидаемая)
Частота аллеля S изменяется в противоположном направлении. В популяции, находящейся в состоянии равновесия, жизнеспособности всех трех генотипов очень близки к ожидаемым. Это можно видеть и на Фиг. 47 и 48. Если исходная частота аллеля F в
популяционном ящике выше или ниже равновесной, то эта частота уменьшается или увеличивается и через 30 поколений становится равной примерно 0,3. К тому же, когда частота аллеля F выше равновесной, отбор направлен против него, и наоборот. На кривых, представленных на фиг. 49, можно видеть, что в ранних поколениях оценки жизнеспособностей (приравниваемые к оценкам приспособленности) выше или ниже ожидаемых, но в более поздних поколениях они приближаются к единице (ожидаемой). Кривые на фиг. 49 построены на основании данных об изменении структуры популяций, приведенных на фиг. 47 и 48. Из этих данных следует, что при равновесии все три генотипа селективно нейтральны в отношении жизнеспособности. По определению этот локус нс несет генетического груза, поскольку все генотипы равноценны в отношении приспособленности. Поэтому Койима и сотр. пришли к выводу, что обнаруженный ими механизм можно считать неким общим механизмом, ответственным за большую степень генетического полиморфизма в природных популяциях.
Другой пример отбора, зависящего от частоты, связан с брачным поведением дрозофилы (Эрман, 1966). В нескольких экспериментах по определению половой активности особей различных генотипов было показано, что особи одного и того же генотипа в популяции, где этот генотип редок, значительно более активны, чем в популяции, где они составляют большинство.
Итак, мы видим, что сверхдоминирование — не единственный способ сохранить полиморфизм. Следовательно, сбалансированный генетический груз популяции создается не только за счет сверхдоминантных локусов.
НАПРАВЛЕННЫЙ ОТБОР, ПЕРЕХОДНЫЙ ГРУЗ И ПЛАТА ЗА ЭВОЛЮЦИЮ
Существует еще один тип генетического груза, возникающий при динамическом изменении частот генов в популяции в процессе замены одного аллеля другим.
Этот процесс, называемый «замещением генов»,— одна из основных черт эволюции.
Подобное замещение аллелей обычно происходит в ответ на какое-либо изменение в условиях среды, которое приводит к сдвигу направления отбора, так что неблагоприятные аллели становятся благоприятными, и наоборот. В отличие от уравновешивающего и нормализующего отбора этот тип отбора называется линейным или направленным. При этом частота одного аллеля уменьшается по мере увеличения частоты другого. В какой-то промежуток времени в течение этого переходного периода частоты обоих аллелей довольно велики. Соответственно при этом возникает период переходного полиморфизма (по генам, оказывающим большое влияние на фенотип). Конечно, при таких сдвигах давления отбора, в результате которых самый распространенный аллель начинает подвергаться отрицательному отбору, большая часть популяции становится плохо приспособленной и возникает переходный груз, называемый также субституционным грузом.
Холдейн (1957) произвел очень интересные расчеты, показывающие, во что обходится популяции замещение аллелей. Согласно его расчетам, число смертей (или их эквивалентов в виде снижения плодовитости), сопровождающих изменение направления отбора, не зависит от интенсивности отбора, а зависит только от исходной частоты первоначально неблагоприятного аллеля. Однако, как и следовало ожидать, число поколений, на которое распределяется эта гибель, определяется непосредственно интенсивностью отбора. Для среднего локуса (q = 0,005) зависимость между интенсивностью отбора и числом поколений, на протяжении которых особи гибнут, имеет вид / = 30/гг, где / — интенсивность отбора, п — число поколений, а 30—величина, показывающая, во сколько раз общее число смертей во всех поколениях выше числа особей, имеющихся в любом данном поколении. Если / = 0,1 (так большинство исследователей оценивают среднюю интенсивность отбора в процессе замещения аллелей), то а? — 300 Таким образом, для замещения одного
гена необходимо в среднем 300 поколений и в 30 раз больше смертей, чем имеется особей в каждом поколении. Величина 30 найдена из уравнения
n 1 fi , A In (1 — А) 1
D = —[-1пРо+----J,
где D — число смертей при следующих значениях приспособленности и частоты генотипов:
Генотип
Частота
Приспособленность
АА Р2 1
Аа 2pq 1 -k
аа
q2
1 -К
При этом k = ХК. Когда А = 0, Л полностью доминирует и D становится равным просто — In pQ (подробное объяснение этой формулы и ее вывод см. Холдейн, 1957).
Сколько же генов могут заменяться одновременно без такого резкого снижения приспособленности популяции, которое угрожали бы ее существованию? Холдейн, рассуждая в общем таким же образом, как при оценке максимально возможного числа сверхдоминантных локусов, пришел к выводу, что та цена, которой популяции приходится расплачиваться за замещение аллеля, очень высока. К подобному же выводу пришел Кимура (1960). Так, например, численность бабочек Biston betularia, искусственно введенных в промышленный район, ежедневно уменьшалась в два раза вследствие уничтожения птицами (см. стр. 183). Таким образом, если принять интенсивность отбора по локусу, контролирующему покровительственную окраску, за 0,5, то при 10 таких локусах вероятность выживания снизится до 71024, или (’/г)10- Конечно, некоторые виды, обладающие достаточно большой плодовитостью, могут выдержать такое давление отбора, но для большинства видов это привело бы к вымиранию. Таким образом, снова встает вопрос о том, какое число аллелей должно быть заменено, чтобы популяция могла выжить в случае изменения условий или могла лучше приспособиться к новой нише? По-видимому, для этого необходимо замещение аллелей не в 10, а в значительно большем числе локусов. Однако
Холдейн считает, что замещение аллелей — процесс очень медленный и что увеличение числа одновременно эволюционирующих локусов за счет снижения интенсивности отбора (т. е. за счет менее резкого снижения приспособленности) по каждому из них не поможет делу. Просто в этом случае увеличивается /г, т, е. число поколений, на протяжении которых происходит замещение. Среднее число поколений, необходимых для замены одного аллеля, равно 300.
Как отметил Брюс (1964), как бы ни была велика плата за эволюцию, за то, чтобы «не эволюционировать», приходится платить еще дороже. В изменившихся условиях отбор, направленный на сохранение редких особей, обладающих более высокой приспособленностью, приведет к повышению приспособленности данной популяции, хотя за это приходится расплачиваться дорогой ценой — гибелью множества слабо приспособленных генотипов.
Скорость эволюции, основанная на расчетах Холдейна, хорошо согласуется с палеонтологическими данными о скорости процесса видообразования. Если мы примем вслед за Холдейном, что для образования нового вида достаточно 1000 замещений, то на это потребуется не менее чем 300 тысяч поколений. Эта величина очень хорошо совпадает с оценками скорости эволюции для многих видов 'млекопитающих, сделанных на основании палеонтологических данных.
Вместе с тем, по-видимому, есть много примеров, когда скорость эволюции выше, чем можно ожидать из расчета 1 замещение за 300 поколений. Так современного человека и его далекого предка Homo erectus разделяет миллион лет. Если принять среднее время между поколениями равным 20 годам (что, быть может, несколько завышено), то оказывается, что всего за 50 000 поколений объем черепа и ряд морфологических и физиологических признаков человека претерпели огромные изменения. Приняв, что для замещения одного гена требуется 300 поколений, легко подсчитать, что 50 000 поколений могло хватить для замещения только 166 генов. А различие по столь небольшому числу генов представляется слишком незначительным
по сравнению с масштабами и сложностью происшедших изменений. Можно привести и другие примеры большой скорости эволюции.
Кажущемуся несоответствию между низкой скоростью замещения генов и большой скоростью эволюции давали и другие, может быть более правдоподобные, объяснения. Майр (1963) высказал теоретическое предположение о синергическом снижении приспособленности по различным локусам, понимая под этим, что наличие в генотипе одновременно двух слабых аллелей по разным локусам оказывает на приспособленность этого генотипа более сильное влияние, чем сумма тех влияний, которые оказывал бы каждый из этих аллелей по отдельности. Это значит, что вероятность гибели особей, содержащих несколько нежелательных генов, значительно выше суммы вероятностей гибели каждого гена в отдельности. Иначе говоря, потенциальные генетические смерти концентрируются в меньшем числе особей. Другой возможный фактор снижения приспособленности — это заметное снижение внутривидовой конкуренции, сопровождающее падение численности.
Независимо от того, какое объяснение окажется верным, в ближайшие годы нам представится интересная возможность следить за развитием исследований, которые помогут проникнуть в суть этих проблем.
ГОМЕОСТАЗ И КАНАЛИЗАЦИЯ
Принимая во внимание большую генетическую изменчивость, свойственную большинству менделевских популяций, и огромное число возможных генотипов, кажется несколько неожиданным, что нормальный фенотип (или, иначе, «дикий тип») популяции так однообразен. Конечно, общая изменчивость популяций значительно выше той, которую можно обнаружить визуально, тем не менее остается фактом, что существует множество генотипов, дающих фенотипы, которые неотличимы один от другого даже при различных условиях среды. Это явление понято еще не до конца, но частично его можно объяснить на основе
концепций гомеостаза и канализации развития. Эти понятия частично перекрываются. Гомеостаз развития— широкий термин, относящийся к способности данного генотипа создавать один определенный фенотип в широком диапазоне условий. Канализация — несколько более ограниченный термин, относящийся к генетическим взаимодействиям, которые постепенно, по мере развития организма, сужают возможный диапазон фенотипического проявления генотипа.
Гомеостаз развития не следует путать с генетическим гомеостазом, который означает устойчивость популяции к изменению ее генетического состава под влиянием отбора. Эта устойчивость к изменению или, во всяком случае, к резкому изменению в значительной степени обусловлена тем, что организм представляет собой высокоинтегрированную, тонко уравновешенную систему; поэтому, изменяя какую-то одну часть, можно нарушить равновесие интегрированного генотипа, что приведет, к появлению слабо приспособленных генотипов. Хотя генетический гомеостаз, несомненно, связан с интеграцией процессов индивидуального развития, его нельзя отождествлять с гомеостазом развития, поскольку первое явление свойственно группе особей, а второе — отдельной особи. Детальное обсуждение генетического гомеостаза можно найти в книге Лернера (1954).
Концепция канализации была развита в работах К. Уоддингтона. Он утверждал, что стабилизирующий отбор имеет два более или менее обособленных аспекта: это, во-первых, нормализующий отбор, т. е. отбор, приводящий к элиминации аномальных фенотипов, и, во-вторых, канализирующий отбор, направленный против особей, в развитии которых наблюдаются отклонения и которые плохо защищены от воздействия меняющихся условий среды. В канализации развития участвуют главные гены, называемые иногда генами-переключателями; они стабилизируют фенотип, придавая ему устойчивость как к генетической изменчивости других локусов, так и к изменениям условий среды. Например, у высших животных в определенный момент индивидуального развития вступают в дей
ствие некоторые главные гены, определяющие пол данной особи. Это каким-то образом ограничивает деятельность многих других генов, участвующих в определении пола. Иными словами, фенотипическое выражение вторичных половых признаков зависит от множества генов, но, несмотря на свойственную этим генам потенциальную изменчивость, конечный результат их действия довольно однороден. Необходим кумулятивный эффект очень многих локусов, чтобы перейти границы, установленные «генами-переключателями», или «канализирующими генами».
Многие авторы считают, что если какой-либо фенотип выгоден популяции, то отбор будет стремиться сохранить гены, канализирующие развитие этого фенотипа, так что пути развития в сущности всех организмов в значительной степени канализированы. В пользу такой точки зрения свидетельствуют главным образом две группы данных: 1) у многих организмов известно большое число видов-двойников, которые практически неотличимы один от другого по морфологическим признакам. По-видимому, в этих случаях высокая канализация развития создает возможность весьма обширных изменений и перестроек генотипа, не сопровождающихся изменением фенотипа; 2) у большинства организмов дикого типа многие морфологические признаки очень устойчивы к искусственному отбору. Например, из дикой линии дрозофилы трудно выделить путем отбора линию, которая отличалась бы от исходной по числу скутеллярных щетинок или глазных фасеток. По-видимому, это связано с тем, что пути развития, определяющие число щетинок и число фасеток, канализированы. Однако в мутантных линиях, характеризующихся измененным числом щетинок или фасеток, как, например, scute или Ваг, отбор на отклонения от нормального числа щетинок и глазных фасеток идет вполне успешно. Scute — рецессивный мутантный ген, вызывающий полное исчезновение четырех скутеллярных щетинок; этот ген до некоторой степени действует также на другие шетинки и волоски. Следует заметить, что и среди мух scute наблюдается значительная вариабельность по числу щетинок. Эф-
фективпость отбора на увеличение числа щетинок показывает (в добавление к тому, что мутанты scute варьируют по степени изменения числа щетинок), что аллель дикого типа играет канализирующую роль в образовании щетинок; при замене же его мутантным аллелем scute этот механизм канализации нарушается. Такое же нарушение канализации наблюдается и при введении в генотип гена Ваг. Этот мутант уменьшает число фасеток в глазу, причем его действие зависит от температуры, при которой происходит развитие. При 25 °C среднее число фасеток равно 55, а при 18 °C— 156 (нормальный глаз содержит около 750 фасеток). По-видимому, ген Ваг нарушил канализацию и образование фасеток стало чувствительно к температуре. Однако вместе с тем нарушение канализации создает возможность для эффективного отбора — различие по числу фасеток между мухами, развивающимися при 18 и 25°С, снижается у мутантов с 300% до всего лишь 10%. Другой пример, сходный с геном scute, касается мутантного гена Tabby, контролирующего число вибрисс у мышей. В диких линиях число вибрисс довольно постоянно и устойчиво к отбору. В линии Tabby возможен отбор на увеличение или уменьшение числа вибрисс. Очевидно, канализация развития существует не только у дрозофилы. Описанные здесь эксперименты провели Уоддингтон (1950), Рендел (1959) и Дан и Фрезер (1958).
Что касается участия отдельных локусов в гомеостазе развития, то следует вновь указать, что сама ге-терозиготпость, хотя она и создает во многих случаях изменчивость, должна приводить к большей фенотипической стабильности, поскольку она допускает синтез Двух различных форм полипептидов, каждая из которых лучше удовлетворяет потребности организма при разных условиях.
РЕКОМБИНАЦИЯ и генетические системы
Один из наиболее важных аспектов генетического Строения популяции — генетическая система вида и роль рекомбинации в создании новых генотипов. Под диетической системой мы понимаем способ организа
ции генов у отдельной особи и способ передачи их потомству. Генетическая система слагается из ряда факторов, таких, как число генов, число хромосом, типы строения хромосом, плоидность, средняя частота мутаций, частота кроссинговера, особенности мейоза, способ размножения, способ определения пола, пути регулирования генной активности и т. д. Как показал Грант (1958), генетические системы сильно варьируют от вида к виду и в меньшей степени — внутри одного вида. Поскольку компоненты генетической системы контролируются генотипом, они подвержены естественному отбору. В результате генетическая система не более постоянна, чем морфология или физиология организма, и под действием естественного отбора адаптируется к условиям среды, в которых организм должен существовать. Другими словами, генетическая система, как и всякий другой признак, подвержена дивергенции, конвергенции и параллелизму. Дарлингтон и Мазер показали это в конце 30-х годов, но до последнего времени этому вопросу не уделяли должного внимания. Многие свойства генетических систем универсальны, и эта универсальность, несомненно, есть следствие общности происхождения от исходного типа. Например, все имеющиеся данные говорят о том, что триплеты РНК (кодоны; см. Ф. Хартман и С. Са-скайнд, «Действие гена»), кодирующие отдельные аминокислоты, одинаковы для всех организмов. Компоненты генетического контроля белкового синтеза взаимозаменяемы в бесклеточных экстрактах таких различных организмов, как бактерия Е. coli, садовый горох и кролик. Хотя эти компоненты взаимозаменяемы, поскольку здесь используется один и тот же код РНК, однако они не идентичны, как доказал Спигелман (1966) в своих изящных экспериментах по комплементации нуклеиновых кислот. Другие особенности, например присутствие хромосом и наличие мейоза, широко распространены, ио не универсальны и, по всей вероятности, возникали в процессе эволюции неоднократно.
Рассматривая генетические системы как относительно постоянные и неизменные, мы по мере изучения
рее большего числа видов уже не раз сталкивались с неожиданностями. Определение пола у дрозофилы связано с равновесием генов, детерминирующих мужской и женский пол. Эти гены распределены между Х-хромосомами и аутосомами; участие У-хромосомы очень ограниченно — опа содержит несколько генов, контролирующих подвижность сперматозоидов. Зигота, лишенная одной Х-хромосомы (XY или ХО, где О означает полное отсутствие второй половой хромосомы), развивается в самца независимо от того, есть в ней Y-хромосома или пет. Зигота с двумя Х-хромосомами (XX или XXY) развивается в самок, опять-таки независимо от того, есть в пей Y-хромосома или нет. Очевидно, именно отсутствие одной Х-хромосомы (и, конечно, находящихся в ней генов, детерминирующих развитие самки), а не присутствие У-хромосомы детерминирует мужской пол. Таким образом, нормальные самки дрозофилы несут хромосомы XX, а нормальные самцы — ХУ. Такую же хромосомную конституцию имеют самцы и самки у всех млекопитающих, в том числе и у человека. Однако в генетической системе млекопитающих мужской пол определяет У-хро-мосома. У особей с аномальным составом половых хромосом XXY преобладают тенденции к развитию мужского пола, а особи ХО — самки (см. В. Мак-кыосик, «Генетика человека»). Другая неожиданность связана с толерантностью в отношении анеуплоидии (состояние, характеризующееся избытком или недостатком хромосом) по Х-хромосоме. У самок дрозофилы две Х-хромосомы — норма, но особи XXX резко аномальны и очень редко выживают. Такие самки всегда бесплодны и обычно гибнут на стадии куколок. Их необоснованно называют «сверхсамками» ввиду избыточной Х-хромосомы, которая, помимо того, что °на увеличивает дозу генов, детерминирующих развитие женского пола, вызывает нарушение генетического равновесия. Поэтому первое сообщение о «сверхсамках» у человека было встречено с большим скептицизмом. Однако оказалось, что генетические системы человека и дрозофилы различны и в этом отношении. У человека и других млекопитающих все Х-хромосо-
мы, кроме одной, в значительной степени инактивируются на ранних стадиях эмбрионального развития.
Установив, что генетические системы подвержены естественному отбору, а следовательно, неоднородны, перейдем к рассмотрению одного из универсальных свойств генетических систем — рекомбинации. Процесс рекомбинации присущ всем живым существам — от вирусов до высокоорганизованных многоклеточных животных; в рекомбинации заключается весь смысл полового размножения. Вопрос о существовании неких единых для всех организмов механизмов рекомбинации, интересный сам по себе, здесь не рассматривается. Нас интересует эволюционная роль рекомбинации, т. е. не столько механизмы этого явления, сколько его распространенность. Ясно, что свободная рекомбинация всех генов в каждом поколении порождала бы полный хаос, поскольку это касается естественного отбора. Это, а также необходимость регулировать распределение множества элементов в процессе клеточного деления — два существенных фактора, обусловливающих необходимость объединения генов в более крупные единицы — хромосомы. Вместе с тем, для того чтобы рекомбинация могла сохраняться, должен существовать какой-то способ перетасовки генов, расположенных в одной и той же хромосоме. Этот процесс осуществляется у всех организмов путем так называемого кроссинговера, суть которого состоит в разрыве и воссоединении хромосом, обычно гомологичных (см. К. Суонсон, Т. Мерц и В. Янг, «Цитогенетика»). У вирусов рекомбинация, по-видимому, происходит редко, поскольку необходимое для этого одновременное присутствие в одной клетке двух разных вирусов— явление крайне редкое. Среди бактерий изредка происходит конъюгация между клеткой-донором и клеткой-реципиентом. Этот процесс сводится к переносу целых хромосом или их отдельных частей, и поэтому рекомбинация возможна только в клетке реципиента. Однако в природе конъюгация у бактерии происходит довольно редко. У высших организмов кроссинговер, или перекрест, между гомологичными хромосомами при мейозе является правилом.
Число кроссинговеров на одну пару хромосом зависит от ряда факторов, но главным образом от величины хромосом. В очень маленьких хромосомах почти не происходит обменов, тогда как у больших хромосом в среднем бывает по одному и более кроссинговеров на каждое мейотическое деление. Конечно, поскольку расщепление пар хромосом в процессе мейоза происходит независимо, гены, лежащие в разных парах хромосом, тем не менее свободно рекомбинируют. Судя по ее универсальности, рекомбинация даже сцепленных генов (гены одной и той же пары хромосом) создает большое селективное преимущество. Это нетрудно понять, принимая во внимание существование интегрированных генных комплексов и взаимодействие между генами. Если бы новая мута-щия представляла собой единственный возможный ^способ возникновения нового сочетания генов, то процесс создания и испытания новых генотипов происходил бы недопустимо медленно (если только не предполагать, что скорость мутирования чрезмерно высока) и интегрированные комплексы никогда не могли бы образовываться. Однако эффективность создания новых генетических комбинаций безмерно увеличивается, если мутации, возникшие у различных особей, Смогут включиться в одну и ту же хромосому путем рекомбинации в гетерозиготах. Вместе с тем существует много способов ограничения кроссинговера, а следовательно, и рекомбинации. По-видимому, причина состоит в том, что, несмотря на важнейшую роль рекомбинации в построении интегрированного генотипа, слишком частая рекомбинация губительна для сохранения интегрированных комплексов — она приводит к их разрушению.
В качестве примера ограничения рекомбинации можно привести гетерозиготы по инверсиям у дрозофилы и других насекомых, ограничение кроссинговера полом индивидуума, изменение количества гетерохроматина в хромосоме или физиологическое снижение частоты кроссинговера при партеногенезе и инбридинг. Кроме того, по-видимому, степень рекомбинации, создающая преимущество, в разных случаях различна.
Так, у D. melanogaster и у D. hydei Х-хромосомы имеют практически одинаковую физическую длину. Однако генетическая карта Х-хромосомы у D. hydei вдвое длиннее, чем у D. melanogaster. Поскольку строение генетической карты зависит от кроссинговера, его частота у D. hydei должна быть вдвое выше. Как показал Карсон (1958), полиморфизм по инверсиям гораздо более резко выражен в центральных районах ареала данного вида, чем в периферических областях. Таким образом, разрушение интегрированных комплексов, бывших высокоадаптированными на протяжении длительных периодов времени, в условиях центральных областей ареала, по-видимому, неблагоприятно. В относительно недавно освоенных периферических частях ареала высокоадаптированные интегрированные комплексы еще не успели сложиться и здесь более благоприятна большая частота рекомбинации, создающая возможность для образования таких комплексов и для возникновения большого числа генотипов, которые подвергнутся испытанию в этих относительно новых условиях.
Итак, мы видели, что для большинства популяций характерна значительная генетическая изменчивость, скрытая за фасадом кажущейся нормальности. Мы обсудили также некоторые способы сохранения этой изменчивости, ту цену, которой приходится расплачиваться за нее популяции, и некоторые пути использования изменчивости. Наконец, мы показали, что допущение о «независимом влиянии разных локусов на приспособленность», невзирая на его полезность в других областях, быть может, придется модифицировать при определении границ возможной изменчивости популяции.
ЛИТЕРАТУРА
В г u е s А. М., The Cost of Evolution Versus the Cost of Not Evolving, Evolution, 18 (1964), 379—383.
Carson H. L., Response to Selection Under Different Conditions of Recombination in Drosophila, Cold Spring Harbor Symposia on Quantitative Biology, 23 (1958), 291—306.
Crow J. F., Some Possibilities for Measuring Selection Intensities in Man, Human Biology, 30 (1958), 1—13,
pobzhansky Th., Genetics and the Origin of Species, New York, Columbia University Press, 1951.
pobzhansky Th., Evolution as a Creative Process, Proceedings of the 9th International Congress of Genetics (1954), 435—449; reprinted in Papers on Animal Population Genetics, Eliot B. Spiess, ed. (Boston, Little, Brown and Co., 1954).
pobzhansky Th., How Do the Genetic Loads Affect the Fitness of Their Carriers in Drosophila Populations'? American Naturalist, 98 (1964), 154—166.
pobzhansky Th., Spassky B., Genetics of Natural Populations XXXIV, Adaptive Norm, Genetic Load, and Genetic Elite in Drosophila pseudoobscura, Genetics, 48 (1963), 1467—1485.
Pobzhansky Th., Spassky B., Tidwell T., Genetics of Natural Populations XXXII. Inbreeding and the Mutational and Balanced Genetic Loads in Natural Populations of Drosophila pseudoobscura, Genetics, 48 (1963), 361—373.
Dun R. B., F г a s e r A. S., Selection for an Invariant Character — «Vibrissa Number» — in the House Mouse, Nature, 181 (1958), 1018—1019.
E h г m a n L., Mating Success and Genotype Frequency in Drosophila, Animal Behavior, 14 (1966), 332—339.
Falconer D. S., Introduction to Quantitative Genetics, New York, The Ronald Press Company, 1960.
Grant V., The Regulation of Recombination in Plants, Cold Spring Harbor Symposia on Quantitative Biology, 23 (1958), 337—363.
Greenberg R., Crow J. F., A Comparison of Lethal and Detrimental Chromosomes from Drosophila Populations, Gene tics, 45 (1960), 1153—1168.
H a 1 d a ne J. B. S., The Cost of Natural Selection, Journal of Ge* netics, 55 (1957), 511—524; reprinted in Papers on Animal Population Genetics, E. B. Spiess, ed. (Boston, Little, Brown and Co., 1962).
Hartman P. E., S u s к i n d S. R., Gene Action. Englewood Cliffs, N. J., Prentice-Hall, Inc., 1964 (2nd ed., 1969). (Ф. Хартман, 3. С а с к а й н д, Действие гена, изд-во «Мир», М., 1966.)
Н е u t s М. J., Adaptive Properties of Carriers of Certain Gene Arrangements in Drosophila pseudoobscura, Heredity, 2 (1948), 63—74.
Hubby J. L., Le won tin R. C., A Molecular Approach to the Study of Genic Heterozygosity in Natural Populations. I. The Number of Alleles at Different Loci in Drosophila pseudoobscura, Genetics, 54 (1966), 595—609.
Johnson F. M., К a n a p i C. G., Richardson R. H., Wheeler M. R., Stone W. S., An Analysis of Polymorphisms among Isozyme Loci in Dark and Light Drosophila ananassae Strains from American and Western Samoa, Proceedings of the National Academy of Sciences U. S., 56 (1966), 119—125.
К i in и г а М., Optimum Mutation Rate and Degree of Dominance as Determined by the Principle of Minimum Genetic Load, Journal of Genetics, 57 (1960), 21—35.
King J. L., Continuously Distributed Factors Affecting Fitness, Genetics, 55 (1967), 483—492.
К о j i m a K., Yarbrough K. N., Frequency Dependent Selection at the Esterase-6 Locus in Drosophila melanogaster, Proceedings of the National Academy of Sciences U. S., 57 (1967), 645—649.
Lerner I. M., Genetic Homeostasis, New York, John Wiley and Sons, 1954.
L i С. C., Population Genetics, Chicago, University of Chicago Press, 1955.
Ludwig W., Zur Theorie der Konkurrenz, Die Annidation (Einni-schung) als fiinfter Evolutionsfaktor, Neue Ergeb. Problemc Zoologie, Klatt-Festschrift (1950), 516—537.
McKusick V., Human Genetics, Englewood Cliffs, N. J., Prentice-Hall, Inc. 1964 (2nd ed., 1969). (В. Маккьюсик, Генетика человека, изд-во «Мир», М., 1967.)
Malogolowkin-Cohen Ch., Levene Н., Dobzhan-sky N. P., Simmons A. S., Inbreeding and Mutational and Balanced Loads in Natural Populations of Drosophila willi-stoni, Genetics, 50 (1964), 1299—1311.
Ma yr E., Animal Species and Evolution, Cambridge, Mass., Harvard University Press, 1963. (Э. Май p, Зоологический вид и эволюция, изд-во «Мир», М., 1968.)
Milkman R. D., Heterosis as a Major Cause of Heterozygosity in Nature, Genetics, 55 (1967), 493—495.
Morton N. E., Crow J. F., Muller H. J., An Estimate of the Mutational Damage in Man from Data on Consanguineous Marriages, Proceedings of the National Academy of Science U. S., 42 (1956), 855—863.
Mukai T., The Coupling-Repulsion, Optimum Heterozygosity Theory of Polygenic Variation, Proceedings of the 12th International Congress of Genetics, vol. II, (1968), 159—160.
Mukai T., Yamazaki T., The Genetic Structure of Natural Populations of Drosophila melanogaster, V. Coupling-Repulsion Effect of Spontaneous Mutant Polygenes Controlling Viability, Genetics, 59 (1968), 513—535.
Mukai T., Sado Chigusa and Isao Yoshikawa, The Genetic Structure of Natural Populations of Drosophila melanogaster, III, Dominance Effect of Spontaneous Mutant Polygenes Controlling Viability in Heterozygous Genetic Backgrounds, Genetics, 52 (1965), 493—501.
R e n d e 1 J. M., Canalization of the scute Phenotype of Drosophila, Evolution, 13 (1959), 425—439.
Spencer W. P., Mutations in Wild Populations of Drosophila, in Advances in Genetics, M. Demerec, ed., New York, Academic Press, Inc., 1947.
Spiegelman S., Hybrid Nucleic Acids, Scientific American, 211 K (1964), 48. (См. Молекулы и клетки», вып. I, стр. 49, изд-во «Мир». М., 1966.)
Stone W. S., Wilson F. D., Gerstenberg V. L., Genetic Studies of Natural Populations of Drosophila-. Drosophila pseudoobscura, a Large Dominant Population, Genetics, 48 (1963), 1089—1106.
Sved J. A., Reed T. E., Bodmer W. F., The Number of Balanced Polymorphisms That Can Be Maintained in a Natural Population, Genetics, 55 (1967), 469—481.
Swanson С. P., Merz T., Young W. J., Cytogenetics, Englewood Cliffs, N. Y., Prentice-Hall, Inc., 1967. (К. Суонсон, T. Мерц, У. Янг, Цитогенетика, изд-во «Мир», М., 1969.)
Thoday J. М., Effects of Disruptive Selection, 1. Genetic Flexibility, Heredity, 13 (1959), 187—203.
Waddington С. H., Experiments on Canalizing Selection, Genetical Research, 1 (1953), 140—150.
Waddington С. H., New Patterns in Genetics and Development, New York, Columbia University Press, 1962.
Wallace B., The Average Effect of Radiation-Induced Mutations on Viability in Drosophila melanogaster, Evolution, 12 (1958), 532—525.
Wallace B., Madden C., The Frequencies of Sub- and Supervitals in Experimental Populations of Drosophila melanogaster, Genetics, 38 (1953), 456-470.
Глава 8
СПОСОБЫ ВИДООБРАЗОВАНИЯ
В предшествующей главе мы рассмотрели те черты структуры популяции, которые, с одной стороны, приводят к стабилизации каждой популяции в ее особой среде, а с другой — обеспечивают поддержание в ней разнообразия, достаточного для адаптивного ответа на изменения средыг При этом мы исходили из того, что эволюционные силы, способные вызвать из менения, т. е. мутационный процесс, естественный отбор, дрейф генов, миграция, находятся в почти полном равновесии с частотами генов, и поэтому изменений почти не происходит. В этой главе мы рассмотрим некоторые условия, сдвигающие это равновесие и приводящие к изменению структуры популяции.
Для иллюстрации сказанного мы приводим здесь изображение предложенного Райтом «адаптивного поля» (фиг. 50). В этой модели группа пиков и впадин соответствует приспособленности различных ге нотипов, определяемой природными условиями. Каждая точка на карте изображает генотип, приспособленность которого равна высоте точки. Генотипы, имеющие одинаковую приспособленность, соединены пунктирными изолиниями. При помощи этой модели легко видеть, каким образом с помощью детерминирующего воздействия одного лишь естественного отбора вид займет устойчивое положение на вершине одного из адаптивных пиков. Но как популяция перехо дит с одного пика на другой, т. е. как возникают новые виды? Как установил Райт (1960), «Генетическая
гистема может перейти от менее высокого к более высокому адаптивному пику только с помощью какого-либо неселективного процесса. Это может произойти в результате новой мутации, которая создаст новый пик, однако подобные события очень редки. Единственная
Фиг. 50. Адаптивное поле.
Гипотетическое двумерное изображение относительной приспособленности различных генотипов как функции условий существования. Пики соответствуют хорошо приспособленным, а впадины — плохо приспособленным генотипам. Изолинии обозначают генотипы с одинаковой пригодностью. Обратите внимание, что пики имеют разную высоту.
альтернатива — случайное отклонение от строго детерминирующего действия разных процессов. Такое случайное отклонение может быть следствием какого-то чрезвычайного события в истории популяции или же может произойти в результате накопления мелких случайных отклонений... Случайный дрейф сам по себе может быть только вредным именно потому, что он случаен. Но, с другой стороны, совместное действие направленного и ненаправленного процессов оказывается более прогрессивным, чем действие одного только
направленного процесса. ...Допустим, что некто в густой туман идет по холмистой местности, намереваясь все время подниматься вверх по крутому склону. Почти наверняка через некоторое время он окажется на вершине небольшого холма, откуда уже некуда идти, если не изменить своему намерению идти все время в гору. Если он будет бродить, совершенно не выбирая пути, т. е. не ограничивая себя никакими условиями, то почти наверняка большую часть времени будет оставаться на сравнительно небольшой высоте, поскольку холмов относительно мало. Если же он в основном идет в гору, но, кроме того, время от времени допускает значительные отклонения в совершенно случайных направлениях, то он не дойдет ни до какой высокой вершины, но зато и не будет вынужден остановиться на вершине какого-нибудь низкого холма. В конце концов он при этом окажется на значительно большей высоте, чем в том случае, когда он все время неуклонно идет по самому крутому склону». Несомненно, однако, что наибольшее значение имеют изменения во внешних условиях. Эти изменения вызывают сдвиг давления отбора (направленный отбор), что приводит к перераспределению самого адаптивного поля. С изменением приспособленности генотипов меняется и рельеф адаптивного поля.
Если изменение условий захватило весь ареал, то вид в целом подвергнется более или менее однородному изменению. Этот процесс называется филетической эволюцией. Поскольку он может привести к образованию новых видов, его нередко считают одним из способов видообразования. Однако при этом, как мы уже отмечали, не происходит увеличения числа существующих видов.
Когда разные популяции одного вида подвергаются воздействию разных условий, то возникающие в них изменения также различны. Если эти изменения достаточно велики, то один исходный вид может разделиться на два или большее число одновременно существующих видов. Мы рассматриваем этот процесс как видообразование и считаем его другим способом возникновения новых видов,
Существует еще одна возможность для образования новых видов — слияние двух существующих видов в один новый вид. Этот процесс иногда называют вТОричным видообразованием. На фиг. 51 схематически представлены все три способа видообразования, распространенность вторичного видообразования еще остается предметом споров, но несомненно, что этот способ имеет большее значение для растений, чем для
Время
Фиг. 51. Способы видообразования.
А. Филетическая эволюция. Б. Первичное видообразование. В. Вторичное видообразование.
животных, поскольку у растений скрещивание между разными видами происходит значительно легче.
К любому из этих способов видообразования применимо неоднократно повторявшееся здесь положение о том, что естественный отбор, используя существующую изменчивость, производит эволюционные изменения. Рассмотрим теперь явление видообразования в более узком смысле, а именно в смысле увеличения числа видов.
ОБРАЗОВАНИЕ РАС
На первой стадии видообразования необходимо, чтобы разные популяции, относящиеся к одному виду, находились в разных условиях среды. В процессе приспособления популяции к этим различным условиям Увеличивается и генетическое различие между популяциями. Это дивергентное филетическое изменение, лежащее в основе превращения различных популяций
в расы, и составляет самую сущность видообразования. В результате накопления генетических различий популяции приобретают четкие морфологические и физиологические различия. Популяции, различия между которыми достигли достаточного уровня, чтобы их можно было ясно отличать одну от другой, обычно считаются расами и им можно давать подвидовые названия. Это первая различимая ступень видообразования. Популяции становятся генетически различными в процессе приспособления к различным условиям среды. По определению расы сохраняют потенциальные способности к скрещиванию, иначе говоря, между ними еще не возникла репродуктивная изоляция. Судьба рас переменчива и зависит от множества факторов.
Если различия в условиях обитания сохраняются или усугубляются, то дивергенция рас усиливается, достигая, наконец, такого уровня, когда расы столо различны генетически, что теряют способность к свободному скрещиванию. На этой стадии дивергенции расы уже считаются отдельными видами. Возможен другой вариант, когда расы сохраняются на одном и том же уровне дивергенции в течение неопределенно долгого времени, или же, в случае восстановления между ними потока генов, вновь сливаются в одну скрещивающуюся популяцию. На фиг. 52 схематически изображены разные взаимоотношения между популяциями в процессе образования рас и видообразования.
Важно иметь в виду, что вследствие постепенности и непрерывности образования рас и видообразования невозможно четко разграничить эти процессы и дать им точные определения (см. гл. 2). В какой момент гомогенный вид становится разделенным на расы? Кик определить тот момент, когда расы становятся обособленными новыми видами? В истории биологии непрерывно происходили и до сих пор продолжаются споры о правильной классификации различных групп. Попытки классифицировать различные уровни дивергенции привели к появлению множества терминов — «раса», «географическая раса», «экологическая раса», «разно-1 видность», «местная разновидность», «подвид» и т. i j
1
Локальная скрещивающаяся популяция
2
3
[§] § а.
Щ Локальные популяции, постепенно переходящие одна в другую (клина первичной интерградации)
П
Географические изоляты "расы”
Подвиды „расы”
А
== Интрогрессия
-----(просачивание гибридов)
Виды
«НИТ! [Я1 Под^иды
1111,11111,1111] ] М [ =^1 ’’виды”
Фиг. 52. Различные типы взаимоотношений между популяциями, наблюдаемые при образовании рас или видов (/) или после гибридизации групп, которые до того были обособлены (//).
+ Частичная репродуктивная изоляция; + полная репродуктивная изоляция.
Возможные пути первичного видообразования (столбец /): 1 — 3—4—5—7; 1 —2—4—5—7; 1-3-6. Возможные пути вторичного видообразования 7-8-D-C-B-4.
Возможные типы гибридизации (столбец II) после устранения географических преград (начиная с наиболее вероятного): ЗД; 4С; 5D (или С, или В, пли Е)\ 8Е (или D, или С, или В).
Некоторые авторы используют понятия «разновидность», «раса» и «подвид» для обозначения постепенно повышающихся уровней дивергенции. Однако в силу отсутствия каких-либо подлинных различий между этими категориями они используются более или менее как синонимы; одни авторы предпочитают один, другие — другой термин (термин «подвид» — формальная таксономическая категория). Теперь мы знаем, что большинство трудностей классификации связано с попыткой разбить непрерывный процесс на дискретные категории. Конечно, у этого процесса есть стадии, идентификация которых не вызывает разногласий. Например, известны виды, явно состоящие из отдельных рас, или же виды, столь хорошо различающиеся между собой, как тигр (Fells tigris) и пума (Felts cougar), видовая самостоятельность которых не вызывает сомнений. Но эти четко отличающиеся стадии достигаются в процессе постепенного накопления менее четких различий.
ИЗОЛИРУЮЩИЕ МЕХАНИЗМЫ
И ВИДООБРАЗОВАНИЕ
Изолирующие механизмы играют первостепенную роль в процессе видообразования, и поэтому их интен сивно изучают. Для более полного знакомства с предметом следует обратиться к работам Добжанского (1951), Паттерсона и Стоуна (1952), Гранта (1963) и Майра (1963).
Как мы уже видели, необходимым условием видообразования является репродуктивная изоляция между популяциями. Но прежде чем сможет установиться репродуктивная изоляция, необходимо появление какого-либо физического механизма, ограничивающего обмен генами. По-видимому, наиболее обычный тип физического разделения — географическое разобщение, т. е. тот случай, когда особи обитают в разный географических районах.
Иногда понятие «географическая изоляция» означает не более чем простое пространственное разобщение. Лягушки Северной Каролины не могут спарп-
ваться с лягушками из штата Огайо, а малиновки штата Мэн не могут спариваться с малиновками Мичигана. Когда лягушки Северной Каролины размножаются, они подвергаются отбору в соответствии с местными условиями среды, которые во многом отличны от условий, характерных для Огайо. В результате нередко возникают виды, в которых фенотипы, характерные для одной местности, постепенно и незаметно переходят в фенотипы, характерные для другой местности, т. е. образуется так называемая клина. Это связано с тем, что геологические, климатические и другие условия в пределах ареала вида также изменяются постепенно. Кроме того, если по некоторым факторам среды наблюдаются резкие изменения, то обмен генами между соседними популяциями нередко затемняет картину. Такой тип географической изоляции характерен для леопардовой лягушки (Rana pipiens) — вида, широко распространенного по всей восточной половине США. Если брать выборки из популяций с небольшими интервалами, начиная с любого места в пределах ареала этого вида и двигаясь в любом направлении, мы не обнаружим между этими выборками сколько-нибудь значительной прерывистости. Морфологические различия становятся заметны только в том случае, если выборки взяты на значительном расстоянии одна от другой. Наконец, в испытаниях на плодовитость при скрещивании было показано, что лягушки из Техаса и из Вермонта в значительной степени изолированы, хотя по всей клине соседние популяции дают плодовитых гибридов (Моор, 1949). В настоящее время создается впечатление, что в природных условиях изолирующим механизмом между расами служит различие в брачных песнях. В результате может оказаться, что эти популяции придется возвести в ранг видов. Тем не менее пространственное разобщение может привести к образованию рас даже в том случае, когда между группами нет четкого разграничения (фиг. 52,2). Такие расы следует считать алло-иатрическими.
Во многих других случаях в пределах ареала вида существуют более или менее четкие экологические
разрывы и при этом наблюдается соответствующее резкое разграничение между различными географическими расами (фиг. 52,1,4). Так, например, в районе Сан-Франциско имеются две расы певчего воробья — горная и низменная, встречающаяся только на соленых болотах. Подобное же деление характерно для нескольких рас гоферов в штате Вашингтон, каждая из которых обитает лишь на почве одного определенного типа. Эти расы также считают аллопатрическими. Важно понять, однако, что между такими расами могут существовать полные или почти полные разрывы, если важные для них экологические факторы встречаются лишь в некоторых частях ареала, подобных изолированным островкам. Если же экологические факторы изменяются постепенно, то и расы также постепенно переходят одна в другую.
Многие другие факторы могут привести к географической изоляции. Например, причиной разобщении наземных и водных животных может быть опускание и соответственно поднятие суши. Примером возникших таким образом эффективных географических преград служит Берингов пролив для наземных и Панамский перешеек — для морских животных. Симпсон (1950) показал, сколь большую роль в формировании южноамериканской фауны сыграло погружение перешейка, соединявшего Северную и Южную Америку в конце третичного периода. К географической изоляции могут также привести оледенения или изменения климата. Имеются весьма убедительные доказательства того, что когда-то ареалы аллигаторов и магнолий, а также некоторых других видов занимали огромные территории от юго-восточного Китая до юго-востока США. Эти ареалы были разделены и сильно сократились в результате развития плейстоценового оледенения. Кроме того, у клинально распределенных видов, как, например, упомянутая выше лягушка Rana pi* piens, периферические расы, находившиеся на пролы воположных концах ареала, могли достигнуть рапы1 несомненных видов просто в силу элиминации промежуточных популяций. К географической изоляц!11] могут привести такие события, как образование рсы|
ных долин, горных массивов, пустынь или, например, большие лесные пожары. Однако, хотя эти крупные события иногда играют значительную роль, особенно в создании длительной изоляции, они случаются не так часто, чтобы можно было отнести за их счет огромное множество видов, разделенных на расы. Значительно большее значение имеет прерывистость в распределении экологических факторов.
Как мы говорили в гл. 5, Майр (1963) убедительно доказывает, что преобладающим способом образования новых видов служит географическое видообразование, происходящее путем дальнейшей генетической дивергенции аллопатрических рас; он считает также, что если не все, то большинство известных примеров симпатрического видообразования в действительности представляют собой случаи вторичной симпатрично-сти, возникшей после аллопатрического видообразования.
Однако многие исследователи все еще утверждают, что симпатрические расы существуют и что они могут в конечном счете привести к симпатрическому видообразованию (см. Грант, 1963). Предполагается, что этот процесс чаще всего начинается с распределения особей, имеющих разные генотипы, по разным экологическим субнишам в пределах одного географического ареала. Благодаря тому что каждый генотип ограничивает свою активность пределами предпочитаемой субниши, возникают субпопуляции, физически отделенные от остальной части популяции. В этом случае физическое разделение имеет генетическую основу. Такой тип изоляции обычно называют экологической изоляцией, однако иногда его определяют как изоляцию по местообитанию или биотопическую изоляцию. Противники концепции симпатрического видообразования возражают, что экологическая изоляция сама по себе (т. е. без географической изоляции) не может продолжаться так долго, как это необходимо для процесса видообразования, хотя они и допускают, что ее продолжительность достаточна для того, чтобы до Разрушения экологических преград успели образоваться симпатрические расы.
Конечно, существует несколько очень интересных примеров экологической изоляции симпатрических видов, но можно ли считать, что эти виды произошли в результате дальнейшей дивергенции симпатрических рас? Этот вопрос все еще не решен. Яркий пример экологической изоляции одного из видов дрозофилы, Drosophila pachea, обнаружили Хид и Кирхер (1965). Этот вид дрозофилы известен только в пустыне Сонора на юго-западе США и на севере Мексики; он размножается только на стволах кактуса Lophocereus schottii и совершенно не способен размножаться на лабораторной среде, если не добавить в пробирку с кормом кусочек этого кактуса. Не менее своеобразная черта этого вида состоит в том, что если в пробирку, содержащую личинки других видов дрозофилы из этой же пустыни, положить кусочек того же кактуса, то личинки гибнут или почти перестают развиваться. Таким образом, один и тот же кактус, абсолютно необходимый для одного вида, ядовит для другого. Когда это необходимое для вида D. pachea вещество было выделено, оказалось, что это стерин — А7-стигмастенол-Зр. Из этого кактуса выделили, кроме того, несколько алкалоидов, летальных для всех видов, обитающих в пустыне Сонора, но не для D. pachea. Этот пример показывает, как очень мелкие генетические изменения могут привести к полной экологической изоляции Быть может, изменений лишь в нескольких локуса \ оказалось достаточно, чтобы начался синтез фермен тов, дающих возможность D. pachea превращать это! новый стерин в форму, в которой он может быть использован в метаболизме стеринов. Одновременно насекомое утратило способность синтезировать ферменты, необходимые для метаболизма более обычных стери-нов, и приобрело способность синтезировать фермент, расщепляющий ядовитый алкалоид. Случаи, подобные этому, когда потенциально простая генети ческая основа приводит к развитию полной изоляции, позволяют заключить, что это могло бы произойти и в условиях симпатричности, быть может, на основе ОДНОГО ИЗ ВОЗМОЖНЫХ генотипов полиморфной ПО' пуляции.
‘А9 Зак.
Я
Фиг. 53. Часть карты южной Австралии, показывающая современные границы распространения членов «прибрежной» формы саранчи группы viatica.
Во времена плейстоценового оледенения морские заливы между полуостровами Эйр, Йорк и Флорьо были сушей; остров Кенгуру также был частью материка. Идеограммы соответствуют гаплоидным кариотипам различных форм. Хромосомы вида viatica обозначены (слева направо) А, В, С, D, 1, 2. 3, 4, 5, 6, X. Кариотип расы ХО вида Р45Ь не показан; он практически не отличается от кариотипа расы ХО вида Р25. Эти виды еще не получили видовых названий и пока условно обозначаются как Р24, Р25 и т. д. Парапатрическое распространение таких карио-типически обособленных видов не удается объяснить ни с позиций симпатрического, ни с позиций аллопатрического видообразования. Такой тип видообразования назвали стаси-патрическим (Уайт. 1968).
Недавно Уайт (1968) предложил новую альтернативу видообразованию через аллопатрические или симпатрические расы. Речь идет о так называемом стасипатрическом видообразовании. Работая с кузнечиками Moraba scurra (сем. Morabinae), Уайт с сотрудниками обнаружили несколько видов, распределенных парапатрически (фиг. 53). Некоторые из этих видов. различаются кариотипически, однако, судя по их географическому распространению, они вряд ли возникли из аллопатрических рас; в то же время они не являются и симпатрическими. Наиболее приемлемое объяснение этих фактов состоит в том, что в пределах ареала старого вида возникает новый вид, стремящийся захватить как можно больше пространства путем элиминации старого вида. Это совсем иная ситуация, чем все описанные ранее. Ее бы рассматривали как случай аллопатрического видообразования, если бы не столь необычное распределение хромосомных типов.
Так или иначе, рассмотренных здесь типов изоляции (географическая и экологическая изоляция) вполне достаточно, чтобы прервать поток генов и положить начало образованию рас. Но, для того чтобы расы могли превратиться в самостоятельные виды, обычно необходимо развитие дополнительных изолирующих механизмов, обеспечивающих эффективную репродуктивную изоляцию.
РЕПРОДУКТИВНАЯ ИЗОЛЯЦИЯ
Рассмотрим некоторые механизмы, создающие репродуктивную изоляцию. Их можно представить следующим образом:
А. Презиготические механизмы — предотвращение оплодотворения и образования зиготы.
1. Биотопическая изоляция. Популяции сосуществуют в одной местности, но занимают разные биотопы.
2. Сезонная изоляция. Популяции сосуществуют в одном географическом районе, но половые продукты созревают в разное время.
3. Этологическая изоляция (только у животных). Изоляция популяций создается различиями и несовместимостью брачного поведения, предшествующего спариванию.
4. Механическая изоляция. Перекрестному оплодотворению препятствуют (пли ограничивают его) различия в строении органов размножения (половые органы у животных и цветки у растений).
5. Несовместимость гамет. Гаметы не развиваются в половых путях чужого вида.
5, Постзиготические механизмы — оплодотворение происходит и гибридные зиготы образуются, но они нежизнеспособны; если гибриды все же развиваются, то они слабые или бесплодные.
1. Слабость или нежизнеспособность гибридов.
2. Гонадная или конъюгационная стерильность гибридов. Стерильность гибридов вызвана аномальным развитием гонад или аномалиями мейоза.
3. Сегрегационная стерильность гибридов. Стерильность гибридов обусловлена аномальным распределением по гаметам целых хромосом, их отдельных участков или сочетаний генов.
4. Гибель F2. Гибриды Fi нормальны, жизнеспособны и плодовиты, но среди потомков F2 много слабых или стерильных особей.
Разные авторы несколько по-разному классифицируют эти механизмы, но сущность самих механизмов остается неизменной. Презиготические механизмы сводятся к тому, что особи изолированных видов либо не могут встретиться, либо, встречаясь, не могут спариваться друг с другом. Невозможность встречи чаще всего обусловлена сезонной или биотопической изоляцией (изоляция по местообитанию). Сезонная изоляция распространена как среди животных, так и среди растений. Так, сезон размножения жабы Bufo ameri-canus не совпадает с сезоном размножения жабы В. fowleri — первый вид размножается раньше (Блэйр, 1928). Хотя эти виды размножаются в одном водоеме и хотя, регулируя овуляцию, их можно скрещивать в лаборатории, в природе они остаются изолированными. Для многих растений решающие факторы размножения — созревание яйцеклеток и Пыльцы — очень строго зависят от ряда климатических Условий. Если сроки созревания пыльцы сдвинуты всего на неделю, это может привести к полной изоляции. Известно множество других примеров изоляции, Основанной «а несовпадении сезонов размножения.
Важно подчеркнуть, что изоляция между видами не ограничивается лишь одним изолирующим механизмом. Так, между Bufo americanus и В. fowleri существует не только сезонная, но и биотопическая изоляция. В. fowleri не только позже размножается, но и проявляет явное предпочтение таким местам обитания, как пруды, большие лужи и реки со спокойным течением. В. americanus, напротив, предпочитает размножаться в небольших мелких лужах и небольших водоемах в долинах пересыхающих ручьев. Тем не ме нее сезоны размножения этих видов несколько перекрываются, и иногда оба вида размножаются в одной луже, так что становится возможным образование межвидовых гибридов. Однако в целом эти два вида дают пример одновременного существования двух изолирующих механизмов. Кроме того, показано (Блэйр, 1958), что препятствие к скрещиванию между этими видами создает различие в брачных песнях. Ко всему этому следует добавить, что механизмы, изолирующие вид А от вида В, могут отличаться от механизмов, изолирующих тот же вид А от вида С.
Главным препятствием к спариванию между особями разных видов, способных встретиться в период размножения, служит этологическая изоляция. Этология— это наука о сравнительном поведении животных в природных условиях. Следовательно, этологическая изоляция — это изоляция, основанная на различиях в поведении. Иногда это явление называют «психологической» или «половой изоляцией». Естественно, что среди растений этого типа изоляции не существует. Брачное поведение большинства животных отличается сложностью и множеством тесно скоординированных действий, усиливающих одно другое и достигающих кульминации в момент спаривания. Если на одной из стадий развития этой тонко скоординированной цепи событий специфический сигнал, посланный одним из партнеров, не вызовет должною ответа со стороны другого партнера, то брачный ритуал нарушается и скрещивания не происходит. Обычно, хотя и не всегда, выбор партнера осуществляет самка. Самцы обычно менее разборчивы в выборе
партнера для спаривания, что иногда приводит к нелепым попыткам спариться с самцом или же с самками довольно далекого вида. Половые стимулы, из совокупности которых слагается процедура ухаживания, бывают зрительными, обонятельными, слуховыми и осязательными. Яркое оперение птиц, брачные песни птиц и амфибий, привлекающие запахи самцов у бабочек, обоняние или его эквиваленты—органы химического чувства у дрозофилы и других насекомых, распускание перьев у птиц — все это компоненты видового опознавания и ухаживания. Были поставлены специальные опыты, чтобы выяснить, какие органы чувств играют наибольшую роль в брачном поведении ряда организмов. При этом удалось установить некоторые компоненты этого поведения. Так, например, оказалось, что если у самок D. pseudoobscura, ищущих партнера для спаривания, удалить антенны, то они теряют способность отличить самца своего вида от самца вида-двойника D. persimilis. Самцы дрозофилы, у которых удалены крылья, теряют способность стимулировать самок к спариванию. Зрительные стимулы также имеют существенное значение, о чем свидетельствует неспособность некоторых видов спариваться в темноте. У светляка решающий фактор привлечения самцов своего вида — частота световых вспышек. В общем эти эксперименты показывают, что брачное поведение основано на множестве различных строго координированных стимулов, каждый из которых необходим для достижения конечного результата — оплодотворения.
Этологическая изоляция и другие презиготические механизмы значительно более эффективны (в смысле экономии энергии), чем постзиготические механизмы, которые не всегда предотвращают появление потомства. Другая особенность этологической изоляции состоит в том, что этологические преграды, хорошо Действующие в естественной среде, нередко разрушаются при изменении условий, например в лаборатории. Это означает, во-первых, что этологическая изоляция, по крайней мере у близких видов, отнюдь не
безукоризненна и, во-вторых, что она зависит от условий существования.
Другой тип презиготической изоляции — механическая изоляция, по-видимому играющая большую роль у многих членистоногих, в том числе у насекомых. Механическая изоляция может быть обусловлена различиями в размерах тела, но чаще она связана с различиями в размерах и морфологии наружных половых органов. У насекомых эти различия столь характерны, что нередко используются в качестве таксономических признаков.
Последний из презиготических механизмов, которые мы здесь упоминаем, — это невозможность оплодотворения и неспособность спермы сохранять жизнеспособность в половых путях самок чужого вида. Один из наиболее хорошо документированных примеров такой изоляции у насекомых — гибель спермы у дрозофилы вследствие так называемой «реакции на осеменение» (Паттерсон и Стоун, 1952). В норме сперма поступает во влагалище самок, а затем переносится в семяприемники, где она хранится. При межвидовых скрещиваниях секрет, выделяемый стенками влагалища, вызывает как бы коагуляцию спермы, сгустки которой могут на несколько дней блокировать влагалище, так что спаривание оказывается бесплодным. Интенсивность и продолжительность реакции на осеменение варьирует в зависимости от степени родства скрещивающихся видов или линий. Сходные механизмы, по-видимому, существуют и у других животных, но они пока мало изучены. У растений наблюдается аналогичная реакция: пыльца некоторых видов (или даже некоторых генотипов, как, например, в случае с самостерильными аллелями клевера) не способна прорастать на рыльцах других видов. Это явление иногда называют физиологической изоляцией.
К числу постзиготических механизмов относятся такие, как неспособность зигот к развитию, пониженная жизнеспособность гибридов (если они вообще жизнеспособны), пониженная оплодотворяемость, пониженная плодовитость или же разрушение у гибридов коадаптированных генных комплексов в результате
рекомбинации, приводящей к возникновению плохо приспособленных особей. Постзиготические механизмы более расточительны в отношении расходования энергетических ресурсов вида, чем презиготиче-ские. Особенно дорого обходится виду производство гибридных потомков, в той или иной степени нежизнеспособных, бесплодных или в свою очередь производящих слабо приспособленных потомков. Существует несколько причин нежизнеспособности гибридов, по главная из них — просто генетическая несбалансированность, приводящая к нарушениям развития или к физиологическим аномалиям. Естественно, что нежизнеспособные гибриды одновременно попадают и в категорию бесплодных. Однако известно немало примеров, когда жизнеспособность гибридов не ниже, а иногда даже и выше, чем у родительских форм, но при этом они частично или полностью стерильны. Лучшим примером такого рода служит мул. Причиной стерильности гибридов являются обычно, если не всегда, вызываемые хромосомными различиями нарушения. Нередко это обусловлено отсутствием между хромосомами гомологии, которая обеспечила бы их правильное расхождение в мейозе.
В условиях, благоприятствующих генетической дифференцировке популяций, которая дает им возможность приспосабливаться к различным условиям, дальнейшее развитие механизмов репродуктивной изоляции между членами разных рас создает несомненные преимущества. Часто оказывается, что гибриды не приспособлены ни к тем, ни к другим условиям, в которых обитали родительские особи (нежизнеспособность или слабость гибридов). В таких случаях отбор благоприятствует любым различиям в физиологии, поведении и т. п., которые предотвращают скрещивание между двумя популяциями (половая изоляция). Отбор будет действовать против родительских особей, дающих этих слабых гибридов, поскольку гены таких особей попадают к слабо приспособленным гибридам; по существу, генетический вклад этих особей в следующее поколение пропадает впустую. Это, несомненно, представляет собой один из способов
IQ*
упрочения презиготической репродуктивной изоляции. Справедливость этого утверждения была продемонстрирована Купменом (1950) в опыте, в котором он вручную удалял гибридов между D. preudoobscura и
Фиг. 54. Эволюция хромосом у дрозофил группы virilis.
Прописными буквами обозначены инверсии, фиксированные у каждого вида; строчными буквами обозначены инверсии, присутствующие не всегда. Стрелками показана последовательность образования кариотипов.
D. persimiUs из популяционных ящиков. Число межвидовых скрещиваний снижалось при этом от поколения к поколению. Однако вполне возможно также, что в некоторых случаях географическая изоляция продолжается достаточно долго и популяции успевают накопить такое множество генетических различий, т. е. дивергировать так сильно, что необходимость в репродуктивной изоляции отпадает, В этом случае гиб
риды генетически несбалансированы и нежизнеспособны.
Не следует забывать также о роли хромосомных аберраций в качестве изолирующего механизма. Паттерсон и Стоун (1952) представили обширные данные о хромосомных различиях между близкородственными видами Drosophila. Эти различия в основном сводятся к различиям по хромосомным инверсиям (фиг. 54), но иногда также и по транслокациям. Транслокации, если они есть, почти всегда представляют собой центрические слияния, что приводит к превращению двух акроцентрических хромосом в одну метацентрическую, которая буквально вдвое больше (см. фиг. 16). Мы уже говорили об инверсиях как о супергенах, но здесь не лишне повторить, что особи, гетерозиготные по транслокациям и инверсиям, могут оказаться частично стерильными, поскольку они дают несбалансированные по хромосомам (анеуплоидные) гаметы (гл. 4).
Хромосомная изоляция имеет одну особенность, отличающую ее от других рассмотренных нами изолирующих механизмов. Появление хромосомной аберрации — довольно редкое событие, происходящее в отдельном организме. Для того чтобы выжить, такая особь должна обладать каким-то селективным преимуществом, однако в остальном носители аберрантных хромосом генетически мало отличаются от других особей. Эта ситуация прямо противоположна описанным выше случаям, в которых репродуктивная изоляция выступает не как следствие, а как причина генетических различий. Однако возможно, что вследствие генетического сходства двух хромосомных типов роль хромосомных различий в видообразовании ограничена. В этом случае они будут просто конкурировать друг с другом, что приведет к элиминации того или другого типа.
Широкое распространение хромосомных различий У некоторых видов, например таких, как Drosophila, иногда считают указанием на то, что эти различия представляют собой непременные факторы видообразования. Ошибочность подобного мнения была ясно
показана на примере гавайских видов Drosophila (Карсон и др., 1967). На Гавайских островах обитает очень много сильно дивергировавших видов дрозофилы, однако межвидовые хромосомные различия практически отсутствуют.
Хотя хромосомная изоляция не имеет существенного значения для эволюции, она может играть важную роль в стасипатрическом видообразовании. Новый хромосомный тип, возникший в какой-либо одной точке ареала вида, может распространяться на обширные области до тех пор, пока, наконец, не встретится с популяциями, достаточно отличающимися от него генетически и лучше приспособленными к своим собственным местообитаниям; такие популяции могут приостановить дальнейшее распространение нового типа. При этом возникают парапатрические виды.
Таким образом, мы приходим к выводу, что существует множество путей установления репродуктивной изоляции; главная их особенность состоит в том, что они приводят к возникновению генетически независимых линий развития.
ПЕРИФЕРИЧЕСКИЕ ИЗОЛЯТЫ И ВИДООБРАЗОВАНИЕ
Ареал любого вида непрерывно сдвигается и перемещается, его границы то расширяются, то сужаются в зависимости от условий, причем все эти изменения сказываются прежде всего на популяциях, обитающих в периферических частях ареала. Считается, что именно в этих периферических популяциях создаются особенно благоприятные условия для видообразования (Карсон, 1959; см. также Майр, 1963). Здесь постоянно образуются новые географически изолированные популяции, называемые периферическими изолятами или популяциями основателей (см. фиг. 52, /3); в этом случае географической преградой обычно служит расстояние. Периферические изоляты обычно малы и их приспособленность минимальна: поскольку данный вид прежде не обитал в этих районах, он не мог подвергнуться отбору на приспособленность к их специфическим условиям. Такие популяции обычно испы
тывают жесткий отбор на приспособленность к экологическим нишам, имеющимся в новых областях. В принципе этот процесс не отличается от рассмотренного выше в связи с географической изоляцией, когда вид делится на несколько больших групп в процессе образования рас. Различие состоит только в том, что периферические изоляты малы и поэтому подвержены дрейфу генов (гл. 6); кроме того, гомозиготность в таких популяциях выше вследствие инбридинга (гл. 3) и возникают они с очень высокой частотой. Периферические изоляты, несомненно, возникают все время, тогда как основная масса вида, занимающая центральные части ареала, остается хорошо приспособленной к условиям, на которые она подвергалась отбору. Таким образом, бесконечные попытки вида проникнуть в новые экологические ниши не угрожают его сохранению.
Это создает также возможность для «генетической революции» (Майр, 1963) — процесса очень быстрых и резких изменений генетического состава популяции, которые иногда бывают столь значительны, что могут повлечь за собой серьезные эволюционные сдвиги и привести к появлению новых родов и более крупных таксономических групп. Такой революции может способствовать ряд факторов. Прежде всего, генетическая структура колонии основателей, как правило, несколько отличается от генетической структуры родительской популяции. Это может быть вызвано либо случайными причинами, либо тем, что лишь некоторые генотипы исходной популяции способны существовать в новой области. Уже по одному тому, что численность изолята невелика, степень генетической изменчивости в нем будет ниже, чем в родительской популяции. Кроме того, под действием инбридинга эта изменчивость может стать еще меньше. Главный аргумент Майра состоит в том, что адаптационная ценность гена находится под непосредственным влиянием не только внешних условий, но и генетической среды. Следовательно, вся генетическая система изолята Должна подвергнуться перестройке, прежде чем в пей вновь установится генетическое равновесие. Кроме
того, тенденция к гомозиготизации будет способство-вать отбору гомозиготных генотипов, узко приспособленных к ограниченному и потому более специфическому комплексу внешних условий. В родительской популяции, напротив, преобладает гетерозиготность, обеспечивающая каждой особи широкую приспособленность к различным нишам центрального района.
Для описания процесса, идущего в периферических изолятах, Карсон ввел термин гомоселекция, а отбор на повышенную забуференность, характерную для популяции центральных районов, назвал гетероселекцией. Большинство исследователей полагают, что высокогетерозиготная, сильно забуференная родительская популяция слишком стабильна, чтобы в ней могло происходить видообразование. Гомоселекция в небольших изолированных популяциях способствует видообразованию, тогда как гетероселекция замедляет этот процесс (фиг. 55).
Кроме того, высказывалось предположение, что генетическая революция — процесс, который может происходить в природных популяциях, вызывая в них быстрые и крупные изменения. Некоторых палеонтологов смущает внезапность появления высших таксономических групп, т. е. отсутствие среди ископаемых остатков общих предковых форм, которые могли бы в результате дивергенции дать начало этим группам. Теория генетической революции помогает объяснить эти факты, поскольку, согласно этой теории, значительные преобразования происходят быстро и притом в небольших изолированных популяциях, т. е. сильно ограничены во времени и в пространстве, что делает крайне маловероятным обнаружение следов этого процесса в палеонтологической летописи. Однажды воз никнув, периферический изолят может начать расширять свою территорию и экспериментировать с буферными системами и полиморфизмом, что характерно для вполне сложившихся, высокоприспособленных видов На основе изучения ряда видов дрозофилы (в частности D. robusta) Карсон высказал мнение, что гомоселекция в периферических изолятах распространяется и па инверсионные комплексы. Этот процесс восстанавли-
Малая популяция ±
А
Б
Фиг. 55. Два противоположных типа генетического регулирования.
А. Гомоселекция, приводящая к генетическому закреплению приспособительных признаков в небольших (например, периферических) популяциях. Это обычно инбредные, полуизолированные популяции. Отбор благоприятствует увеличению гомозиготности и ведет к закреплению различных приспособительных признаков, что способствует видообразованию. Б. Гетероселекция, приводящая к генетическому равновесию, в условиях, характерных для центральных популяций географически дифференцированных видов, у которых равновесие обычно сдвинуто в сторону гетерозиготности по отдельному локусу; гетерозиготность обеспечивает повышенную устойчивость во всех экологических нишах ареала (гетеротическое забуфферивание). Благодаря большой численности и аутбри-дингу такая популяция может позволить себе роскошь постоянно производить гомозиготы, которые менее приспособлены, чем гетерозиготы.
вает рекомбинацию, нарушенную в родительских популяциях. Характерный для родительских популяций сбалансированный полиморфизм, основанный на ко-адаптированных генных комплексах и сохраняемый благодаря гетерозиготности по инверсиям, оказывается неприемлемым для новых условий. Гомоселекция дает начало популяциям, которые можно считать рекомбинационно «открытыми», тогда как гетероселекция создает рекомбинационно «замкнутые» популяции. Эти понятия отражают относительную частоту возможной рекомбинации. «Открытость» периферических изолятов увеличивает рекомбинационные возможности, что повышает эффективность образования новых генотипов, необходимых для приспособления к новым условиям.
АДАПТИВНЫЕ ПОВЕРХНОСТИ И МЕЖДЕМОВЫЙ ОТБОР
До сих пор мы ограничивали свои рассуждения отбором внутри популяций. Именно этому аспекту популяционной генетики уделялось до сих пор наибольшее внимание. Стабилизирующий отбор, уравновешивающий отбор, направленный отбор, нормализующий отбор, канализирующий отбор и, наконец, дизруптив-пый отбор — все эти термины относятся к дифференциальному выживанию особей, относящихся к одной популяции. Что же касается дифференциального выживания самих популяций, то этот аспект эволюции до сих пор почти игнорировался. Небольшая локальная свободно скрещивающаяся популяция — дем — особенно важна в этом отношении, а ее генетическая структура — существенный фактор ее выживания. Это особенно относится к периферическим изолятам. Наиболее детально исследовал это явление Левонтии (1965), давший ему название междемовый отбор в отличие от внутридемового отбора. Концепция межде-мового отбора помогает до известной степени понять некоторые явления, кажущиеся парадоксальными, если рассматривать их только с позиций внутрипояу-ляционного отбора. Например, существует ряд при
знаков, имеющих высокую приспособительную ценность для особи, за сохранение которых выступает внутрипопуляционный отбор, но которые тем не менее угрожают существованию популяции. Один из таких признаков — размеры тела. Обычно отбор внутри популяции направлен на увеличение размеров тела. Но такой отбор не может продолжаться безгранично, иначе наступит истощение пищевых ресурсов популяции и она начнет вымирать. В таких ситуациях, вероятно, действует междемовый отбор, благоприятствующий сохранению тех демов, в которых величина особи не вступает в резкое противоречие с пищевыми ресурсами. Таким образом, внутридемовая тенденция к увеличению размеров может оказаться не реализованной.
Для того чтобы междемовый отбор мог действовать, необходимо существование различий между демами по селективным оптимумам для ряда признаков. Возможность таких различий была подтверждена на построенной Левонтином математической модели. В качестве простейшего случая рассмотрим полиморфную систему, состоящую из двух локусов с двумя аллелями в каждом. В потомстве двойных гетерозигот возможны девять генотипов: AABb, ААЬЬ и т. д. Каждый генотип характеризуется известной приспособленностью, которую мы обозначим где I просто указывает, что мы рассматриваем некий конкретный генотип. Пусть частота генотипа равна Zj. Эту величину можно выразить через частоты pi и q\ и р% и г/о соответственно в одном и другом локусе. Таким образом, зная приспособленность каждого из 9 генотипов^ можно найти_величину средней приспособленности W по формуле = 2 Средняя приспособленность популяции определяется относительными частотами различных генотипов, которые в свою очередь определяются частотами генов в двух локусах. Найдя (лучше с помощью вычислительной машины) значения W аля большого числа разных генпых частот, можно построить «адаптивную поверхность», подобную изображенной на фиг. 56. Адаптивная поверхность и
Фиг. 56. Топографическая карта двухлокусной адаптивной поверхности.
Топографические контуры (см. фиг. 50) получены с помощью вычислительной машины. По осям отложены частоты генов в каждом из двух локусов. Цифры обозначают приспособленность популяции для каждого сочетания генных частот.
адаптивное поле, представленное па фиг. 50, — не одно и то же. Каждая точка адаптивной поверхности соответствует не генотипу, а популяции. Каждая точка этой поверхности_представляет собой среднюю приспособленность (Ж) для определенной совокупности генных частот, отложенных на оси абсцисс и на оси ординат. Следует указать, что приспособленности различных генотипов, необходимые для построения такого поля, не обязательно выбирать произвольно; )для некоторых популяций их можно определить экспериментально. Так, адаптивная поверхность, изображенная на фиг. 56, построена на основе данных Левонтина и Райта (1960) по двум инверсиям (которые в нашей модели можно рассматривать как гены), распространенным в популяциях саранчи Moraba scurra.
Приспособленности популяций определялись на вычислительной машине на основании данных о частоте генотипов и об относительной приспособленности каждого из возможных генотипов. Для данной реальной популяции, в которой приспособленность генотипа aabb была условно принята за единицу, приспособленности генотипов были определены эмпирически еще до того как стала известна относительная приспособленность других генотипов.
bb ВЬ ВВ
аа 0,79 1,0 0,834
Аа 1,006 0,901
АА 0,657 0,657 1,067
Популяция с наименьшей средней приспособленностью целиком состоит из особей A Abb {р\ = 1, р2 = 0) и имеет приспособленность 0,657. Наибольшую среднюю приспособленность 1,067 имеет популяция, состоящая только из особей ААВВ (р, = 1, р2=1). Диапазон приспособленности (1,067—0,657) был разделен на десять равных частей. Таким образом, цифра / на карте обозначает величины приспособленности в интервале от 0,657 до 0,657 + 0,041; Цифра 2—величины в интервале от 0,698 до 0,739 и т. д. Четные цифры не напечатаны для большей наглядности. Самый низкий участок данной адаптивной
поверхности лежит в нижнем левом углу. Поверхность поднимается по диагонали вправо до высоты 7, которая служит как бы водоразделом. В направлении к верхнему правому углу поверхность вновь снижается, а к нижнему правому углу — повышается, образуя здесь самый высокий участок. (В действительности при более мелкой градации видно, что гребень из семерок имеет форму седла с некоторым повышением посредине вверху.) Эту карту можно использовать также для демонстрации изменений в частотах генов в популяции под действием отбора. Это представлено дорожкой из звездочек. Исходная популяция (pi = = 0,95 и р2 = 0,05) имеет низкую среднюю приспособленность. Под влиянием отбора частоты генов меняются так, что средняя приспособленность популяции возрастает. Увеличение приспособленности от единицы через тройки, пятерки и семерки сопровождается уменьшением pi и увеличением рг. Но когда Pi начинает увеличиваться, средняя приспособленность популяции возрастает до максимума при pi = 1 и Р2= 1. (Однако она могла бы достигнуть и другого максимума — вверху в центре.) Таким образом, изменения частот генов под влиянием отбора всегда сопровождаются увеличением приспособленности. Однако это движение обычно не прямолинейно и частоты генов не всегда изменяются равномерно или в одном направлении; как мы уже упоминали, это может привести к тому, что популяция стабилизируется на одном из субмаксимальных пиков.
Самая существенная особенность адаптивной поверхности заключается в том, что она состоит из ряда пиков и впадин, различающихся соответственно по высоте и глубине. Популяция, представленная на этой поверхности точкой, стремится двигаться «в гору». Это отражает действие отбора, увеличивающего среднюю приспособленность популяции. Двигаясь вверх, популяция достигнет какого-то пика, но не обязательно самого высокого. Поэтому какой-либо дем может оказаться на вершине одного из субмаксимальных пиков, откуда он не может быть сдвинут силами вну-тридемового отбора. Это обусловлено тем, что для
достижения наивысшего пика нужно сначала спуститься с достигнутого пика, а это сопряжено со снижением приспособленности, которому противодействует стабилизирующий отбор. Следовательно, по размерам тела или по любому другому признаку, который мы изберем для анализа, многие демы могут оказаться на одном из субмаксимальных пиков и дальнейшее движение вверх станет невозможным без того, чтобы дем не прошел через селективную долину. Стабилизация разных популяций на субмаксимальных пиках по разным признакам и создает те различия между демами, на которые может действовать меж-демовый отбор.
ПОЛИПЛОИДИЯ КАК СПОСОБ ВИДООБРАЗОВАНИЯ
Полиплоидией называется увеличение числа хромосомных наборов. У животных, по крайней мере высших, это явление встречается редко, однако у растений полиплоидия встречается довольно часто и служит одним из важных способов видообразования (подробности см. Стеббинс, 1950). Для удобства полиплоидию обычно делят на автополиплоидию и аллополиплои-дию. Автополиплоиды образуются путем умножения числа хромосомных наборов у одного вида. Так, у автотетраплоидов каждая хромосома представлена четырьмя гомологами, и в мейозе при спаривании они образуют квадриваленты. Такая мультивалентность нередко затрудняет мейоз, приводя к частичной стерильности, однако обсуждение механизмов, вызывающих эти трудности, не входит в наши задачи (см. К. Суонсон, Т. Мерц и В. Янг, «Цитогенетика»). Наиболее распространенный способ образования полиплоидов — соматическое удвоение числа хромосом в отдельных побегах и цветках или же случайное образование и объединение гамет с нередуцированным числом хромосом. Для образования полиплоидов у самоопыляющихся видов вполне достаточно соматического удвоения в цветке. При этом гаметы будут содержать 2п хромосом, а зиготы — 4м. У перекрестно-опыляющихся видов образование полиплоидов — про
цесс более сложный. Сначала должно произойти соматическое удвоение в цветках двух разных растений, и если затем произойдет случайное объединение гамет 2п от каждого из этих растений, то образуется зигота 4п. Если гамета 2п объединится с гаплоидной гаметой, то образуется зигота Зп. Существует и другой, еще менее вероятный путь образования тетраплои-дов — это объединение двух гамет с нередуцированным числом хромосом (гамет 2п, образовавшихся в процессе аберрантного мейотического деления).
Чаще всего встречаются тетраплоиды, однако нередко основное число хромосом увеличивается не только вдвое, но и в большее число раз. Обычно автополиплоиды очень сходны с диплоидными видами, отличаясь от них более крупными размерами. Автополиплоиды довольно часто оказываются репродуктивно изолированными; эта их черта имеет, возможно, определенное эволюционное значение. Например, гибрид между тетраплоидом и диплоидом будет триплоидом, а триплоиды отличаются стерильностью, поскольку каждая хромосома представлена тремя гомологами и в процессе мейоза образуются анеуплоидные гаметы (гаметы с необычным числом хромосом). В такой клетке гомологичные хромосомы встречаются в виде тривалентов, а не парами, как в норме. В лучшем случае при расщеплении триплоидов в одну дочернюю клетку отходит один набор хромосом, а в другую — два набора. А поскольку различные триваленты распределяются в процессе деления клетки независимо друг от друга, во вторичных гаметоцитах одни хромосомы будут представлены унивалентами, а другие — бивалентами, и лишь изредка [2(72)п, где л—число хромосомных пар] будут появляться сбалансированные гаплоидные (и) или диплоидные (2и) гаметы. Таким образом, тетраплоиды — это другой пример возникновения репродуктивной изоляции до начала генетической дивергенции. Кроме того, образование тетраплои-дов можно рассматривать как способ увеличения числа локусов.
Аллополиплоиды образуются при гибридизации различных вполне сложившихся видов. Эти гибриды
I
Фиг. 57. Образование амфидиплоидов.
Поскольку при мейозе хромосомы не конъюгируют, они направляются либо к одному, либо к другому полюсу по воле случая, так что иногда все хромо-Гомы попадают в одну гамету. Образовавшийся в результате слияния двух таких гамет амфидиплоид будет вести себя как диплоидный организм, и у него будет происходить нормальный мейоз.
обычно стерильны, так как хромосомы уже в значительной мере утратили свою гомологичность, неспособны нормально конъюгировать и мейоз нарушен. Однако если такие гибриды сохраняются, размножаясь бесполым путем, до тех пор пока, как в случае с автополиплоидами, не произойдет соматического удвоения или не образуются гаметы, содержащие все хромосомы, то могут возникнуть сбалансированные полиплоидные комплексы. В природе существует немало таких аллополиплоидов, а ряд аллополиплоидов создан человеком, например табак Nicotiana digluta (N. tabacum X N. glutinosa), хлопчатник Gossypium hirsutum (G. arbor earn X G. thurberi) и Raphanobras-sica — межродовой гибрид, полученный в результате скрещивания капусты (Brassica) с редисом (Rapha-nus). Если скрещиваемые виды настолько далеки, что гомология между хромосомами практически отсутствует, то цитологически гибрид между этими видами выглядит просто как диплоид (фиг. 57). Такие алло-полиплоиды называют амфиплоидами или амфиди-плоидами. Эволюционное значение аллополиплоидов состоит в том, что они по многим признакам отличаются от обеих родительских форм и, будучи репродуктивно изолированы от этих исходных видов, образуют новый вид, сочетающий в себе многие признаки обоих исходных видов. Таким образом, всего лишь за два поколения полового размножения может возникнуть совершенно новый вид.
РОЛЬ ГИБРИДИЗАЦИИ В ЭВОЛЮЦИИ
Гибриды — это потомки отчетливо различающихся родителей. Термин гибридизация используется весьма широко и в разных значениях, но в эволюционном контексте под гибридизацией обычно понимают скрещивание между разными популяциями, расами или видами.
Подобно полиплоидии межвидовая гибридизация свойственна почти исключительно растениям. Среди животных гибридов дают только очень близкие виды,т. е. виды, между которыми еще не установилась
полная репродуктивная изоляция. Мы не знаем, почему межвидовая гибридизация более обычна среди растений. Частично это можно объяснить тем, что у животных большую роль играет этологическая изоляция, устанавливающаяся у них быстро и довольно легко, тогда как у растений этот тип изоляции совершенно отсутствует.
Мы уже подчеркивали значение гибридизации как первой ступени в образовании амфидиплоидов, но гибридизация важна и в других отношениях. Основное эволюционное значение гибридизации состоит в том, что она способствует объединению разных генотипов, а это приводит в ряде случаев к резким рекомбинационным изменениям, затрагивающим не только отдельные гены, но и целые коадаптировапные генные комплексы. Достигнуть таких резких изменений обычными способами, т. е. с помощью мутационного процесса и замещения генов, было бы гораздо сложнее и дольше. Даже в тех случаях, когда межвидовые гибриды лишь частично плодовиты при скрещивании с родительскими формами, гибридизация служит механизмом переноса генных комплексов (и даже целых хромосом) от одного вида к другому.
В большинстве случаев гибриды хуже приспособлены к определенной среде, чем любая из родительских форм. Следовательно, даже если гибриды плодовиты, отбор действует против них, препятствуя росту их численности; поэтому гибриды обычно редки. Если же появление гибридов совпадает с резкими изменениями условий среды (даже если это лишь локальные изменения), то гибриды могут стать обычными. Более того, поскольку вначале гибриды относительно редки, они будут скрещиваться преимущественно не между собой, а с родительскими формами. У потомков от таких скрещиваний в среднем 3/4 хромосомного набора будет представлено хромосомами одного, а 1/4 — хромосомами другого родительского вида. Последующие возвратные скрещивания гибридов с той или другой родительской формой или с обеими приводит к изменению этой формы (или форм) в направлении другого вида. Этот процесс включения отдель
ных генов в юном какого-либо вида, который при этом сохраняет свою самостоятельность, называется интрогрессивной гибридизацией (Андерсон, 1949). Схематически этот процесс изображен на фиг. 52, //, Е.
Фиг. 58 Цветки Aquilegia pubescens (Л) и A. formosa (иаг.) truncata (Б).
Замечательным примером интрогрессивной гибридизации служат два вида водосбора Aquilegia pubescens и A. formosa, растущие высоко в горах Сьерра-Невада. Эти виды различаются по ряду признаков: некоторые из них перечислены в табл. 16. На фиг. 58 изображены цветки этих видов: у A. pubescens цветок обращен вверх, а у Л. formosa он обращен вниз. Ареалы распространения этих видов перекрываются, но незначительно; Л. formosa обычно встречается в сравнительно низких и сырых местах, тогда как Л. pubescens распространен в более сухих и холодных возвышенных местообитаниях. Однако в некоторых местах, например в районе Саддлбег-Лэйк, произрастают оба вида, и здесь они иногда образуют гибриды. Интересно, как при этом происходит опыление. Обращенные
вверх цветки A. pubescens опыляют бабочки бражники: с наступлением сумерек они подлетают к цветкам и, погружая длинный хоботок в столь же длинный шпорец, сосут нектар. У A. formosa пыльцу переносят колибри. Благодаря своей способности висеть в воздухе в перевернутом положении эти птицы высасывают нектар из обращенных вниз цветков A. formosa, вводя клюв в их короткие шпорцы. Кроме того, цветки formosa окрашены в яркие привлекающие птиц красные и желтые тона, хорошо заметные в дневное время. А бледножелтые и белые цветки pubescens, напротив, во всяком случае для человека, более заметны при слабом вечернем освещении, в период активности бражников. Таким образом, бражники редко посе-
Таблица 16
Сравнение Aquilegla formosa и A. pubescens по 5 признакам цветка и их гибридные индексы
№ пп. Признак A. formosa Промежуточные формы A pubescens
1 Положение цветка Обращен вниз Горизонтальное Обращен вверх
2 Окраска шпорца и чашелистиков Красная Оранжевая, бледно-оранжевая, розовая или бледно-розовая Светло-желтая или белая
3 Окраска лепестков Темно-желтая Умеренножелтая Светло-желтая или белая
4 Длина шпорца от 10 до 17 мм от 18 до 28 мм от 29 до 37 мм
5 Длина лепе- от 2 до от 5 до от 9 до
стка 4 мм 8 мм 12 мм
Гибридный индекс для каждого признака 0 1 2
Общий гибридный индекс 0 5 10
щают цветки formosa, а колибри редко посещают цветки pubescens', при этом опыления все равно не происходит—бабочкам почти невозможно добраться
Гибридный индекс
Фиг. 59. Диаграмма частоты различных гибридных индексов в трех популяциях Aquilegia в высокогорной зоне Сьерра-Невады. Можно сделать вывод, что между видами A. formosa и A. pubescens происходит интрогрессивный обмен генами, причем гены A. formosa перешли в этой области к популяции A. pubescens.
до нектара в обращенных книзу цветках, а колибри — высосать его из глубины длинных шпорцев цветков, обращенных вверх. Поэтому гибриды встречаются редко, а если и возникают, то ни бабочки, ни колибри их не посещают. В результате обмен генами между этими видами очень незначителен, причем чаще гены поступают от formosa к pubescens, чем наоборот.
Характеризуя соседние популяции величиной гибридного индекса, Грант нашел (табл. 16), что боль
шинство растений вида formosa из Слэйт-Крик относятся к крайнему типу с гибридным индексом, равным нулю (фиг. 59). В популяции Уайт-Маунт очень немногие растения являются ярко выраженными pubescens. У большинства из них цветки розоватые, а там и сям в популяции встречаются экземпляры, у которых цветки довольно отчетливо розовые. В местах, удаленных от зоны контакта, среди растений pubescens преобладают экземпляры с белыми цветками. Таким образом, мы видим, что ограниченный обмен генами может оказывать определенное влияние на видовые признаки, хотя вид при этом не утрачивает своих отличительных особенностей. На фиг. 59 показан также состав гибридной популяции в районе Саддлбег-Лэйк.
Иногда гибридизация может происходить на всем протяжении ареалов двух видов, если эти ареалы перекрываются, приводя к довольно полной интерградации между ними и к образованию одного нового вида. Этот процесс слияния двух видов с образованием нового вида Грант (1963) назвал вторичной интерградацией (фиг. §2JIyD). В процессе такого слияния исходные виды теряют свою самостоятельность.
Позднее Грант (1966) показал, что гибридизация имеет важное значение как необходимая предпосылка того, что он называет «хромосомным видообразованием». В некоторых отношениях эта гибридизация сходна с амфидиплоидией, но в отличие от последней при этом не происходит удвоения числа хромосом. Единственный известный случай «хромосомного видообразования» произошел в экспериментальных условиях, а в природе оно не обнаружено. Для получения хромосомного вида были скрещены виды Gilia malior и Gilia modocensis. Потомки Fi были почти стерильны, а по фенотипу промежуточны между двумя родительскими формами. По-видимому, стерильность обусловлена межвидовыми различиями в строении хромосом, делающими невозможной конъюгацию в мейозе. После отбора на плодовитость в течение нескольких поколений плодовитость стала повышаться и вернулась к норме. Была обнаружена высокая корреляция между плодовитостью и числом конъюгирующих хромо
сом. Таким образом, через несколько поколений геном стал структурно гомозиготным по хромосомам того или другого из исходных видов и стабилизировался. После восстановления плодовитости эта линия была испытана в скрещиваниях с обеими родительскими формами. При этом была обнаружена почти полная стерильность, т. е. между гибридной линией и родительскими формами возникла репродуктивная изоляция. Трудно сказать, возможен ли подобный процесс в природе; ведь плодовитость гибридов первых поколений, как правило, очень понижена.
У животных различия между родительскими формами при гибридизации обычно очень незначительны, и поэтому здесь трудно ожидать таких эволюционных последствий, как у растений. Однако у них гибридизация может способствовать устранению генетических различий между популяциями, которые прежде были географически разобщены.
ВЫВОДЫ
В этой главе мы показали, как происходит дифференциация генофондов, за которой следует постепенная изоляция и обособление филетических линий. Она происходит в результате прекращения обмена генами между популяциями, что ведет к образованию рас. Путем дальнейшей генетической дивергенции, сопровождающейся усилением репродуктивной изоляции, расы превращаются в отдельные виды.
Тем, кто сомневается в возможности возникновения таким путем всего разнообразия органического мира, окружающего нас сегодня, следует напомнить, что этот процесс происходил на протяжении огромных периодов времени и что событие, вероятность которого в каждый данный момент очень мала, при наличии достаточного времени непременно когда-нибудь произойдет.
ЛИ ТЕРАТУРА
Anderson R., Introgressive Hybridization, New York, Jopn Wiley and Sons, 1949.
В 1 a i r W. F., Mating call in the speciation of anuran amphibians, American Naturalist, 92, (1958), 27—51.
Blair А. Р., Variation, Isolation Mechanisms, and Hybridization in Certain Toads, Genetics, 26 (1941), 398—417.
Carson H. L., Clayton F. E., Stalker H. D., Karyotypic stability and speciation in Hawaiian Drosophila, Proceedings of the National Academy of Sciences U. S., 57 (1967), 1280—1285.
Carson H. L., Genetic Conditions Which Promote or Retard the Formation of Species, Cold Spring Harbor Symposia on Quantitative Biology, 24 (1959), 87—105.
D о b z h a n s к у T., Genetics and the Origin of Species, New York, Columbia University Press, 1951.
Grant V., The Origin of Adaptations, New York, Columbia University Press, 1963.
Grant V., The origin of a new species of Gilia in a hybridization experiment, Genetics, 54 (1966), 1189—1199.
Heed W. B., Kircher H. W., Unique Sterol in the Ecology and Nutrition of Drosophila pachea, Science, 149 (1965), 758—761.
Koopman K. F., Natural Selection for Reproductive Isolation Between Drosophila pseudoobscura and Drosophila persitnilis, Evolution, 4 (1950), 135—148.
Le won tin R. X., Selection in and of Populations, in Ideas in Modern biology, J. A. Moore ed., Proceedings of the 16th International Congress of Zoology, Vol. 6. Garden City, N. Y., Natural History Press, 1965.
L e w о n t i n R. C., W h i t e M. J. D., Interaction between Inversion Polymorphisms of Two Chromosome Pairs in the Grasshopper Moraba scurra, Evolution, 14 (1960), 116—129.
M а у r E., Animal Species and Evolution, Cambridge, Mass., Harvard University Press, 1963. (Э. Майр, Зоологический вид и эволюция, изд-во «Мир», М., 1968.)
Moore J. A., Patterns of evolution in the genus Rana, in Genetics, Paleontology and Evolution, G. L. Jepsen, E. Mayr and G. G. Simpson, eds. (Princeton, N. J., Princeton University Press, 1949).
Patterson J. T., Stone W. S., Evolution in the Genus Drosophila, New York, Macmillan Company, 1952.
Simpson G. G., History of the Fauna of Latin America, American Scientist, 38 (1950), 361—389.
Stebbins G. L., Variation and Evolution in Plants, New York, Columbia University Press, 1950.
White M. J. D., Models of speciation, Science, 159 (1968), 1065-1070.
Wright S., The Roles of Mutation, Inbreeding, Crossbreeding and Selection, Proceedings of the 6th International Congress of Genetics, Vol. 1 (1932), 356—366.
Wright S., in Evolution after Darwin, S. Tax, ed. Vol. I, Chicago, University of Chicago Press, 1960.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Адаптационная ценность 149, 150, 155
Адаптация 187
Адаптивная норма 171, 228 — поверхность 302, 303 -----топографическая карта 304
Адаптивное поле 278—280 Адаптивный пик 278, 279 Алкалоиды 288
Аллели 74, 75
— идентичные 94
— множественные 74, 75
— независимые 94
Аллопатрический 31 Алькаптонурия 74 Американские индейцы 194 Африканский парусник (Papi-lio dardanus) 175, 176
Березовая пяденица (Biston betularia) 181, 182
Бесполый организм 141 Бражник 313
Величина кладки у птиц 171
Вес при рождении 171
Взаимопревращения видов 12 Видовой разрыв 32, 37 Видообразование 18, 279—316 — вторичное 281 — географическое 135 — симпатрическое 287 — способы 278—316 — стасипатрическое 288 — хромосомное 316 — через полиплоидию 307
Виды 12, 13, 16, 23, 25, 26, 29
— безмерные 30
— бесполые 47, 50
— биологические 38
Виды-двойники 41, 42
— зарождающиеся 16
— критерий ' определения 26, 39
— многомерные 34
— множественные популяционные системы 34
— монотипические 36 — морфологические 29, 39 — палеонтологические 28
Виды-двойники политипические 35
— полувиды 29, 44
— число 21
— эволюционная концепция 26 Водосбор (Aquilegia) 108, 139 Водяной уж (Natrix sipedon) 138
Воробей 170
Вымершие формы 21
Вьюрки 19
Гаметы с нехватками и дупликациями 124
Гамодем 56
Ген 85, 88, 128
— закрепление 85, 88, 189, 191
— потеря 128, 129, 191
Генетическая изменчивость
113, 214
— революция 299
— система 254, 269, 270
Генетический груз 216, 228, 230
----мутационный 231, 233
----сбалансированный 235
----сегрегационный 246
— код 113
Генный поток 134, 207
Генотип интегрированный 273
Генофонд 66, 67
Гены-переключатели 103
Географическая изменчивость 34, 210
— изоляция 137, 286 Географические преграды 31 Гетерозис 242
Гетероселекция 300, 301
Гибридизация 32, 34, 37, 39, 123, 310, 311
— вторичная интерградация 315
— интрогрессивная 47, 138, 312
Гомеостаз 266
— развития 266, 267
Гомоселекция 300, 301
Группы крови 59, 70—73, 194, 195, 198, 199, 235
----система АВО 194, 195
-------М — N 59, 70—72
Дем 56
Дивергенция 18, 174, 284
Дисперсия 84, 85, 87
ДНК 102, ИЗ, 116
Доклеточные агрегаты 11
Дрейф генов 83, 84, 144, 185, 186, 279
----- в природных популяциях 190, 299
Дункеры 195, 196
Дупликации 122, 124
Естественная система 23
Закон Харди — Вайнберга 10, 69, 70, 97, 135, 143, 147, 166, 258
Зебра 32—35
— бурчеллева 33—35
— Гартмана 32, 33, 35
— Гранта 32—34
— Греви 32, 33
— капская горная 32—35
— квагга 32, 33, 35
— Чэпмена 32, 33, 35
Идентичные аллели 94
Изменчивость 14, 17, 101, 214 — внутрипопуляционная 102 — вызванная внешними условиями 109
— генетическая 107, 109, 185
— географическая 102
----- вероятность 63
----- определение 215
----- сохранение 215
— индивидуальная 101, 102
— количественная 103, 168
— морфологическая 43
— наследственная 15, 24
— резервная 130, 214
— скрытая 74
— фенотипическая 104
Изолирующие механизмы 284— 298
Изоляты периферические 38, 208, 298, 299
Изоляция 37, 38, 284, 285 — географическая 38, 284,
285
— половая 295
Изоляция репродуктивная 37 — сезонная 292
Изоферменты 238
Инбредная депрессия 217
Инбридинг 89, 91, 92, 206, 207. 217
— коэффициент (F) 92, 217 Инверсии 119, 125, 239—242, 250, 251, 272, 296, 297
— гетерозиготность 123
— изменение частоты 239— 242
— относительная ценность 243
— парацентрические 120
— перицентрические 125
Индустриальный меланизм 180
Канализация 110
— и отбор 267
Клина 35, 137
Колибри 313
Кроссинговер 123, 272
Летальные эквиваленты 220
Локальная менделевская популяция 56
Макроэволюция 97
Междемовое разнообразие 254
Мейоз 270, 295
Мертворожденные 219
Механизмы изоляции 38
-----географические 284, 285
----- постзиготические 291
----- презиготические 290
----- «психологические» 292
----- физиологические 294
-----хромосомные 289, 297
-----экологические 287
----- этологические 290, 292
Миграция 134—137
Микроэволюция 97
Мимикрия 175
Множественные аллели 74, 75
Модификация доминирования 159
Мул 295
Муравьи 105
Мутации 61, 113, 186, 191, 192, 273
Мутации генные 113
— давление 130
— детриментальные 224
— летальные 222, 225, 227
— обратимые 133
— полулетальные 225—227
— субвитальные 224, 227
— супервитальные 224, 227
— частота 131 —133
Мутон 114
Мышь 153
Надвиды 45
Неодарвинизм 16, 186
Нередуцированные гаметы 308
Нехватки 118
Норма реакции 108, 178
Определение пола 271
Оптимальная гетерозиготность 252
Основатели популяции 193, 208, 298
Отбор 98, 99, 142, 152, 154, 157, 228
— гаметический 152, 153
— гетеросел екционный 300,
301
— гомоселекционный 300, 301
— давление 178
— дизруптивный 168, 174
— естественный 5, 6, 13, 17, 62, 130, 142, 144, 146, 170, 185, 186, 270
— искусственный 145, 146
— коэффициент 150, 158
— модель 169
— общая модель 154
— междемовый 302, 303
— направленный 168, 173,
280
— нормализующий 229, 267
— против доминантного аллеля 154, 155
----- рецессивного аллеля 158
— стабилизирующий 168, 170, 172
Палеовиды 28
Панмиктическая единица 56
Периферические изоляты 38, 208, 298, 299
Плата за эволюцию 264 Плейотропия 83, 184 Подвиды 15, 52, 283, 284 Подвижность 30, 178
Покровительственная окраска 203
Пол, определение 271
Полигены 250
Полиморфизм 103, 163, 178, 179, 193, 197, 154, 255 — адаптивный 245 — генный 184, 185 — групп крови 194, 195, 203, 235
— инверсионный 185, 245, 274 — переходный 185
— при балансирующем отборе 205, 236
----- дизруптивном отборе 174, 175
-----сверхдоминировании 163, 256
-----частотнозависимом отборе 258, 259
— улиток 201—204
— хромосомный 239, 240, 241 — экологический 178
Популяции 18, 20, 30, 55, 80, 81 — биотически симпатрические 32 — величина 60 — в равновесии 60, 66—69, 72—76, 99 — замкнутые 302 — открытые 302
Преадаптация 112, 149
Приспособленность 14, 15,
111, 112, 142, 149, 151, 170, 186, 217, 274 — компоненты 168 — общая 145, 167 — средняя 154, 157 — хромосомы 242
Происхождение 11
— родственное 22
Псевдоаллелизм 115
Разновидности 16, 284
Равновесие 66, 68
— в периферических изолитах 298, 301
— — популяциях 60, 73, 74, 100
— множественного локуса 75, 80
— нарушение 84
— определение 68, 71, 72
*— при сверхдоминировании 161, 163
— — стабилизирующем отборе 170
---сцеплении с полом 76— 78
Распределение Пуассона 129
Расы 37, 51, 281—283, 316
— аллопатрические 285—287
— географические 286
— образование 281—284
— парапатрические 288
— симпатрические 287, 288
— экологические 286
Рекомбинация 126, 140—142, 185, 272, 273
Рецессивные летали 118
Соматическое удвоение 308
Степень доминирования (Л)
159
Сцепление с полом 76, 77
Таксономия 22
Термиты 104, 105
Транслокации 119, 124
— гетерозиготность 124
— целого плеча 119
— центрическое слияние 119
Улитка Сераеа nemoralis 200— 204
Фенилкетонурия 72
Филетическая эволюция 18, 280
Хи-квадрат 70
Хромосомные перестройки 114, 117, 118
Хромосомы 113
— слюнных желез 121
Самооплодотворение 91—93
Саранча Moraba scurra 288, 305
Сверхдоминирование 160, 162, 236
— предельное число локусов 248, 249
— цена 247
Селективный пик 170
Серповидноклеточная анемия 235
Симпатрия 31, 32
Сингамия 50
Систематика 22
Системы классификации 22— 24
Скопления гибридов 140
Скрещивание возвратное 222
Частота генов 57, 58
----- в равновесной популяции 65
----- изменения при инбридинге 94, 95
---------- отборе 155
Частоты скрещиваний 63, 64
Эволюция 18, 98, 280
Экологическая ниша 51
Экотипы 178
Энотера 125
Эритробластоз плода 198
Эстераза-6 236—259
Эффект Людвига 254, 255 — «сцепления — отталкивания» 252
ОГЛАВЛЕНИЕ
Предисловие к русскому изданию 5
Из предисловия авторов 9
Глава 1. Теория эволюции 11
Литература 20
Глава 2. Попытки привести в систему разнообразие жизни 21
Искусство научной классификации организмов 21
Категория вида 25
Эволюционная концепция вида . ... . 26
Типологическая концепция (морфологический вид) 29
Концепция безмерного (nondimensional) вида 30
Концепция многомерного вида . 34
Определение понятия «вид» с позиций многомерной концепции 36
Ограниченность биологической концепции 38
Внутривидовые категории (расы и подвиды) 51
Литература 53
Глава 3. Гены в демах 54
Популяции и демы 55
Частоты генотипов в популяциях 56
Частоты генов в популяциях 57
Равновесная популяция . 60
Анализ популяционной выборки 70
Множественные аллели 74
Равновесные популяции и пол 76
Множественные локусы и генетическое равновесие 80
Дрейф генов 83
Инбридинг . 89
Равновесная популяция и факторы эволюции 97
Литература 100
Глава 4. Генетическая изменчивость и ее источники 101
Типы изменчивости 101
Генотип — фенотип . 106
Факторы, влияющие на генетическую изменчивость ПО
Мутации как источник первичной генетической изменчивости 113
Структурные изменения хромосом и супергены 114
Судьба отдельного мутанта 127
Динамика генных частот при постоянном давлении мутаций 130
Поток генов . 134
Рекомбинация как источник изменчивости 140
Литература 142
Глава 5. Естественный отбор 143
Естественный отбор .... 144
Демонстрация естественного отбора в лаборатории 146
Адаптационная ценность и коэффициент отбора 149
Отбор гамет 152
Общая модель отбора . 154
Определение адаптационной ценности 163
Три типа отбора 168
Отбор в природных популяциях . . . 179
Взаимодействие естественного отбора с другими факторами эволюции 185
Литература 187
Глава 6. Дрейф генов 188
Взаимодействие случайных и неслучайных факторов 188
Изучение природных популяций 192
Роль в эволюции малых популяций 205
Литература 212
Глава 7. Генетическая изменчивость популяций 214
Сохранение генетической изменчивости 215
Определение скрытой генетической изменчивости 216
Адаптивная норма и концепция генетического груза 228
Мутации и мутационный груз 231
Уравновешивающий отбор и сбалансированный груз 235
Ограниченность числа сверхдоминантных локусов . . 246 Эффект Людвига, дизруптивный отбор и возникновение разнообразия между демами 254
Направленный отбор, переходный груз и плата за эволюцию 262
Гомеостаз и канализация 266
Рекомбинация и генетические системы 269
Литература 274
Г лава 8. Способы видообразования , 278
Образование рас 281
Изолирующие механизмы и видообразование 284
Репродуктивная изоляция . . 290
Периферические изоляты и видообразование 298
Адаптивные поверхности и междемовый отбор 302
Полиплоидия как способ видообразования 307
Роль гибридизации в эволюции 310
Выводы 316
Литература 316
Предметный указатель 318
УВАЖАЕМЫЙ ЧИТАТЕЛЬ!
Ваши замечания о содержании книги, ее оформлении, качестве перевода и другие просим присылать по адресу: 129820, Москва, И-110, ГСП, l-й Рижский пер., д. 2, издательство «Мир».
Л. МЕТТЛЕР, Т. ГРЕГГ
«Генетика популяции и эволюция»
Редактор Н. О. Фомина
Художник А. В. Шипов Художественный редактор Ю. Л. Максимов Технический редактор В. П. Сизова Корректор О. Ф. Иванова
Сдано в набор 17/11 1972 г. Подписано к печати 10/VII 1972 г. Бума-
га Ха 2 84X1081/jjb"5,06 бум. л. 17,01 печ. л. Уч.-иэд л. 15,92. Изд. Хе 4/6055 Цена 1р. 11 к. Зак. 88
ИЗДАТЕЛЬСТВО <МИР> Москва, 1-й Рижский пер., 2
Ордена Трудового Красного Знамени Ленинградская типография Ха 2 имени Ев1ении Соколовой Главполиграфпрома Комитета по печати при Сонете Министров СССР.
Измайловский проспект, 29
1 р. 11 к.
дж. джинке
НЕХРОМОСОМНАЯ НАСЛЕДСТВЕННОСТЬ
2.
В. МАККЬЮСИК
ГЕНЕТИКА ЧЕЛОВЕКА
К. МАРКЕРТ
ГЕНЕТИКА РАЗВИТИЯ
Ф. СТАЛЬ
МЕХАНИЗМЫ НАСЛЕДСТВЕННОСТИ
К. СУОНСОН, Т. МЕРЦ и В ЯНГ ЦИТОГЕНЕТИКА
Ф. ХАРТМАН, 3. САСКАЙНД
ДЕЙСТВИЕ ГЕНА