/




























































































































































































































































































Текст
РОССИЙСКАЯ АКАДЕМИЯ НАУК
С И Б Л Р С К О Е О Т Д Е Л Е Н И Е
ИНСТИТУТ ЦИТОЛОГИИ И ГЕНЕТИКИ
Р. Л. БЕРГ
ГЕНЕТИКА
И ЭВОЛЮЦИЯ
ИЗБРАННЫЕ ТРУДЫ
Ответственные редакторы
доктор биологических наук С. И . Малецкий
кандидат биологических наук Л . Д Колосова
во «НАУКА»
НОВОСИБИРСК
1993
УДК 575 + 575.8
Генетика и эволюция. Избранные труды /Р . Л. Берг.—
Новосибирск: ВО «Наука». Сибирская издательская фирма,
1993.— 284 с.
ISBN 5 — 0 2 — 030591— X.
В книгу вошли статьи по вопросам генетических закономерностей
эволюции, опубликованные с 1937 по 1972 г., и не публиковавшаяся ра
нее статья «Случайна или закономерна эволюция?». Все статьи прониза
ны идеей закономерного строения живого компонента биосферы. С пози
ций генетика автор рисует приспособительную эволюцию живых систем
как восхождение по степеням прогресса, как высвобождение из-под дик
тата среды, как путь обретения свободы.
Книга предназначена для генетиков, цитологов, эволюционистов.
Р е ц е нзснты
доктора биологических наук В. В. Бабков, Э. X. Гинзбург
Утверждено к печати
Институтом цитологии и генетики СО РАН
Б
1903020000 - 024
К Б -5 0 -1 2 1 -1 9 9 2
042(02) - 93
© Р. Л. Берг, 1993
ISBN 5 — 02— 030591— X
(с) Российская Акачемия наук,
1993
ПРЕДИСЛОВИЕ
Решение ученого совета Института цитологии и генети
ки Сибирского отделения Российской Академии наук из
дать мои избранные труды преисполняет меня чувством
глубокой благодарности. Выход в свет тома «Генетика и
эволюция» приурочен к шестидесятилетию моих исследо
ваний в 1993 г.
Мне довелось сотрудничать с наиболее выдающимися
биологами двадцатого века. Это были: Герман Мёллер —
основатель радиационной генетики, создатель количествен
ных методов учета вновь возникающих мутаций; облучив
дрозофил лучами Рентгена, он повысил частоту возникно
вения мутаций в сто раз; Иван Иванович Шмальгаузен —
один из тех биологов-энциклопедистов, кому удалось наи
более глубоко проникнуть в закономерности эволюции,—
сформулировал теорию стабилизирующего отбора; теория
эта венчает новую эру в биологии, эру синтеза генетики и
теории эволюции, основанной на дарвиновском принципе
отбора; и Николай Владимирович Тимофеев-Ресовский —
генетик-натуралист, умевший блестяще сочетать сравни
тельный метод познания — главное орудие науки прошло
го века — с экспериментом — нынешним средством испы
тания природы.
В этой книге собраны теоретические статьи, посвящен
ные генетическим аспектам эволюции. Том открывается
статьей «О путях эволюции генотипа», написанной
Н. В. Тимофеевым-Ресовским в соавторстве со мной. Мне
принадлежит только маленький раздел «Эволюция мутабильности», но Тимофеев-Ресовский в перечне авторов по
ставил мое имя первым. С первых строк звучит голос этого
великого человека, умевшего как бы с высоты птичьего по
лета озирать подвластную его уму отрасль науки и загля
3
дывать в будущее. В сборник включена работа, сделанная
под руководством Мёллера еще в тридцатые годы. Весь бо
танический цикл исследований, вошедший в книгу, осно
вывается на работах И. И. Шмальгаузена, и одна из моих
статей по истории науки, посвященная И. И. Шмальгаузену, «Проблема целостности живых систем в трудах
И. И. Шмальгаузена» помещена в ней. Все статьи объеди
нены идеей эволюции самих путей, способов, механизмов
эволюции. Усовершенствованию подлежит все — сама из
менчивость ставится под контроль генотипа, эволюциони
рует сам аппарат наследственности, вырабатываются меха
низмы, обеспечивающие константное соотношение форм в
популяции, совершенствуется расположение генов по хро
мосомам, число наборов хромосом регулируется в соответ
ствии с нуждами не только жизни, но и эволюции. От
бор — жесткий и щедрый на смертные приговоры законо
датель и судья — вводится в определенное заранее струк
турированное русло, создаются устройства, умеряющие его
пыл, и рядом с ними, там, где надо, возникают механиз
мы, обеспечивающие его беспрепятственное буйство.
В строго дарвиновском — шмальгаузеновском духе я
трактую органическую эволюцию как явление, протекающее
на популяционно-видовом уровне, и вид — совокупность по
пуляций — я рассматриваю как единственную систему, чей
способ преобразования заслуживает названия эволюции.
В первом из трех разделов сборника речь идет об эво
люции аппарата наследственности, генома, в частности о
выработке механизмов, обеспечивающих нужное популя
ции соотношение форм.
Второй раздел — эволюция изменчивости. Здесь это не
мутабильность, а способность реагировать на воздействия
окружающей среды, норма реакций. В основе серии работ,
представленных в книге, лежит обнаруженное мной у рас
тений различие признаков одного и того же организма по
способности меняться под воздействием среды. Признаки,
сохраняющие стандарт в разнообразных условиях обита
ния, не реагируют, само собой разумеется, на воздействия,
исходящие от других признаков того же организма. Неспо
собность эта выражается в низких или нулевых коэффици
ентах корелляции между размерами стабильных частей и
размерами частей лабильных. Обнаружив у лягушки груп
пы признаков, коррелированных друг с другом и не корре
лированных с признаками других групп, П. В. Терентьев
назвал эти группы корреляционными плеядами. Гетеро
номный рост органов эмбриона цыпленка, описанный
4
И. И. Шмальгаузеном,— онтогенетическая манифестация
корреляционных плеяд.
Мне на растениях удалось вскрыть механизм образова
ния независимости признаков в эволюции. Несовпадение
формирующих и отбирающих факторов среды — общий
принцип возникновения жестких наследственных про
грамм. Категорический императив генного контроля, раз
возникнув, делает признак независимым от воздействия
внешней среды и вырывает его количественную характери
стику из корреляционных зависимостей организма. Насе
комые — переносчики пыльцы растений специфических
энтомофилов оказались калибровщиками опыляемых ими
цветков. Возникновение в эволюции жестких генетических
программ, наследственное программирование количествен
ных признаков выявлены сравнительным методом с по
мощью миллиметровой портняжной линейки. Теория ста
билизирующего отбора Шмальгаузена получила веское,
хотя и косвенное доказательство.
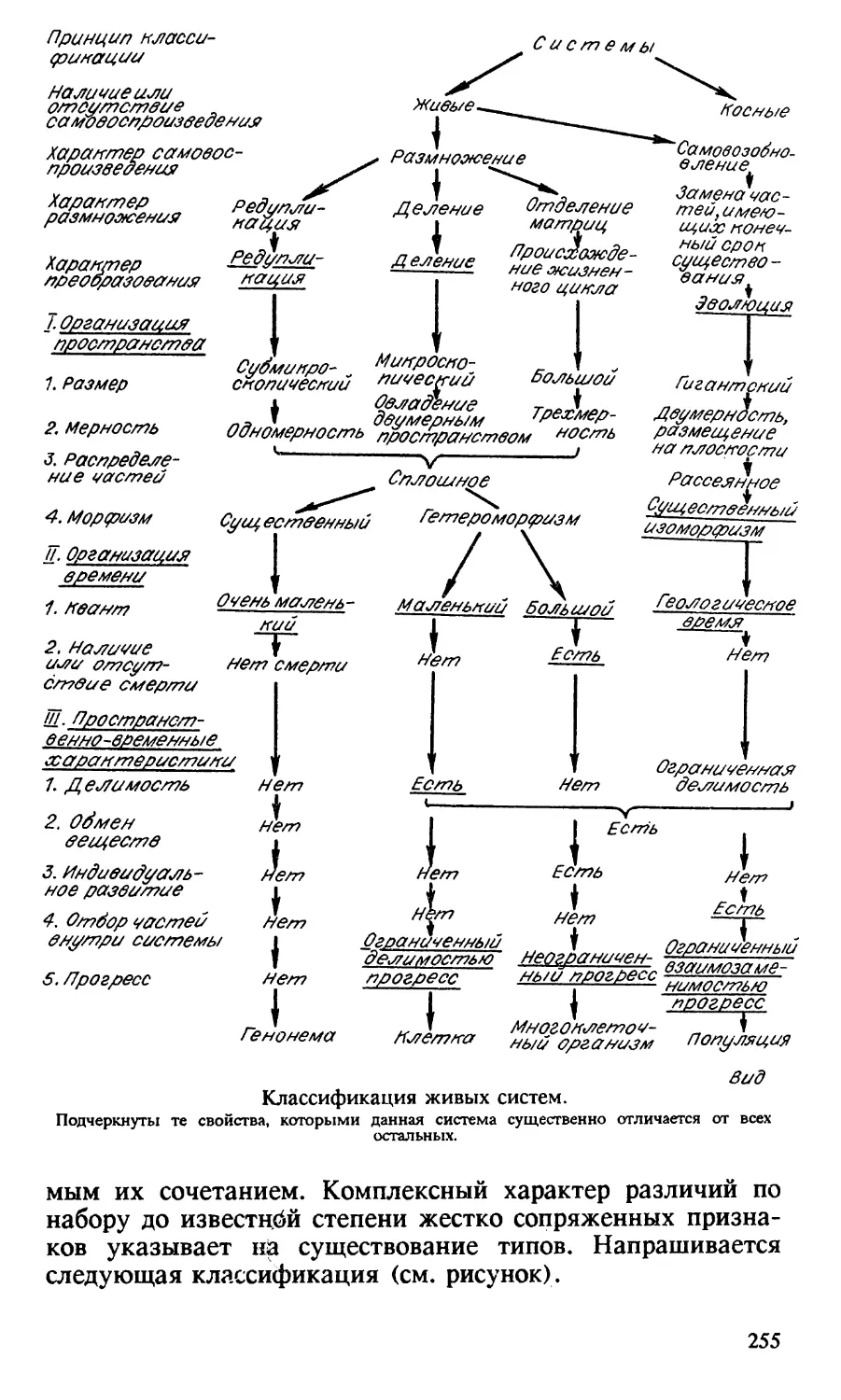
Третий раздел сборника посвящен классификации жи
вых систем. Основная мысль раздела — иерархии живых
систем соответствует иерархия отборов. Высшие ступени
иерархии — групповой, межпопуляционный, межвидовой
отборы — контролируют манипуляции отбора организмов.
Значению соподчинения форм отбора, осуществлющихся на разных уровнях организации живого компонента
биосферы, в приспособлении к среде и в обретении свобо
ды на путях прогрессивной эволюции будет уделено вни
мание в следующей книге, где собраны мои работы по ге
нетике популяций дрозофилы и человека.
Р. Л . Берг
Раздел
I
ЭВОЛЮЦИОННАЯ ГЕНЕТИКА
О ПУТЯХ ЭВОЛЮЦИИ ГЕНОТИПА*
Р. Л . Берг , # . В. Тимофеев-Ресовский
1. ВВЕДЕНИЕ
Так как генотип является той управляющей системой,
которая определяет онтогенез живых организмов и вместе
с тем представляет собой код наследственной информации,
передающейся из поколения в поколение, то можно счи
тать тривиальным утверждение, что происходящая на на
шей планете эволюция живых организмов связана с эволю
цией генотипа. В нашу задачу входит показать, что вся
сумма знаний, добытая современной генетикой, так же как
и результаты специальных наблюдений и опытов, в част
ности, изучение различий в структуре и изменчивости ге
нотипа у различных групп организмов заставляет думать,
что эволюции путем естественного отбора могут подлежать
и некоторые свойства самого генотипа.
Действительно, хотя генотип и является основной уп
равляющей системой и кодом наследственной информации,
передающейся из поколения в поколение, он в то же время,
несомненно, является определенным «признаком» организ
ма. При этом мы можем различать два свойства генотипа
как признака организма: его структуру и его изменчивость.
Общей особенностью структуры генотипа любого организ
ма является известная дискретность, связанная с одним из
определений понятия гена, как в значительной мере авто
номной структурной единицы в пределах набора хромосом.
Изменениями же генотипа являются различные мутации,
т. е. элементарные единицы наследственной изменчивости.
Таким образом, вполне обоснована постановка специ
ального вопроса об эволюции генотипа, в частности, как
его структуры, так и его изменчивости. Несомненно, что в
процессе эволюции естественным отбором направляется и
* Проблемы кибернетики.— 1961.— Вып. 5.
6
формируется не только морфологическая организация осо
бей; естественный отбор должен в какой-то мере изменять,
направлять и поддерживать в оптимальном состоянии так
же и вышеупомянутые основные свойства генотипа. Долж
но меняться число, феногенетические свойства и взаимные
связи генов, а также частота их изменчивости (степень
стабильности) и направления последней.
Несмотря на то, что даже грубыми методами кариосистематики устанавливаются резкие различия, а более тон
кими методами сравнительной цитогенетики и весьма зна
чительная разница в структуре генотипа у организмов, в
той или иной степени отдаленных друг от друга,— та же
сравнительная цитогенетика показывает, что у близких
форм значительные участки генотипа являются структурно
сходными и генетически гомологичными [Frolova, Astaurov,
1929; Розанова, 1930; Левитский, 1937; Синская, 1948; и
др. ]. При этом то обстоятельство, что степень структурной
схожести и генетической гомологичности различных участ
ков генотипа в общем соответствует степени систематикофилогенетической близости соответствующих форм, явля
ется наиболее ярким доказательством эволюции структуры
генотипа; особенно подробно этот вопрос изучен у различ
ных видов Drosophila [Dobzhansky, 1937— 1955; Liiers,
1939; Stone, 1955]. Точно также, наряду со все большими
различиями в наследственной изменчивости разных групп
живых организмов по мере уменьшения филогенетического
родства, у близких видов наблюдается ярко выраженный
параллелизм наследственной изменчивости, сохраняющий
ся в убывающей степени у близких родов, семейств и даже
более высоких систематических категорий, как это было
подмечено еще Дарвином [Darwin, 1859]. Это явление па
раллелизма наследственной изменчивости было изучено на
огромном материале культурных растений Н. И. Вавило
вым [1920, 1931 ] и сформулировано в его известном зако
не гомологичных рядов в наследственной изменчивости.
Аналогично тому как параллельное общефилогенетическое
уменьшение сходства и гомологии в структуре генотипов
является внешним показателем их эволюции, так и умень
шающийся с повышением степени филогенетических раз
личий параллелизм наследственной изменчивости является
внешним выражением эволюции второго основного свойст
ва генотипа — его изменчивости. При этом необходимо
подчеркнуть, что закон гомологичных рядов является,
строго говоря, эмпирическим обобщением, гениально сфор
мулированным Н. И. Вавиловым на основе сравнительного
изучения грандиозного по объему материала; он имеет
7
большое практическое значение, позволяя разумно плани
ровать селекцию новых растений или отбор старых форм в
новых направлениях, а также предсказывать возможность
и вероятность находок при поисках нужных форм среди
родственников хорошо изученного вида. Но причины (гене
тические и феногенетические механизмы), лежащие в ос
нове параллельных наследственных вариаций, как это бу
дет показано в конце работы, в отдельных случаях могут
быть различными.
Работы Н. И. Вавилова о параллелизме в наследствен
ной изменчивости, его работы о генетической структуре
видов культурных растений и их наследственной изменчи
вости, сформулированное им представление о линнеевском
виде как системе, а также его классические исследования о
центрах происхождения сельскохозяйственных культур яв
ляются важными вехами на самых трудных, ранних этапах
развития эволюционной генетики и общей связи генетики
с эволюционным учением.
Последующее изложение разделяется на две части, из
которых первая посвящена' преимущественно проблеме
эволюции структуры генотипа и самих генов, а вторая —
эволюции мутабильности. В настоящей работе используют
ся, однако, лишь генетические методы и не привлекаются
цитологические. Цель работы состоит в изучении эволю
ции генотипа и мутационного процесса как его важнейшей
составной части.
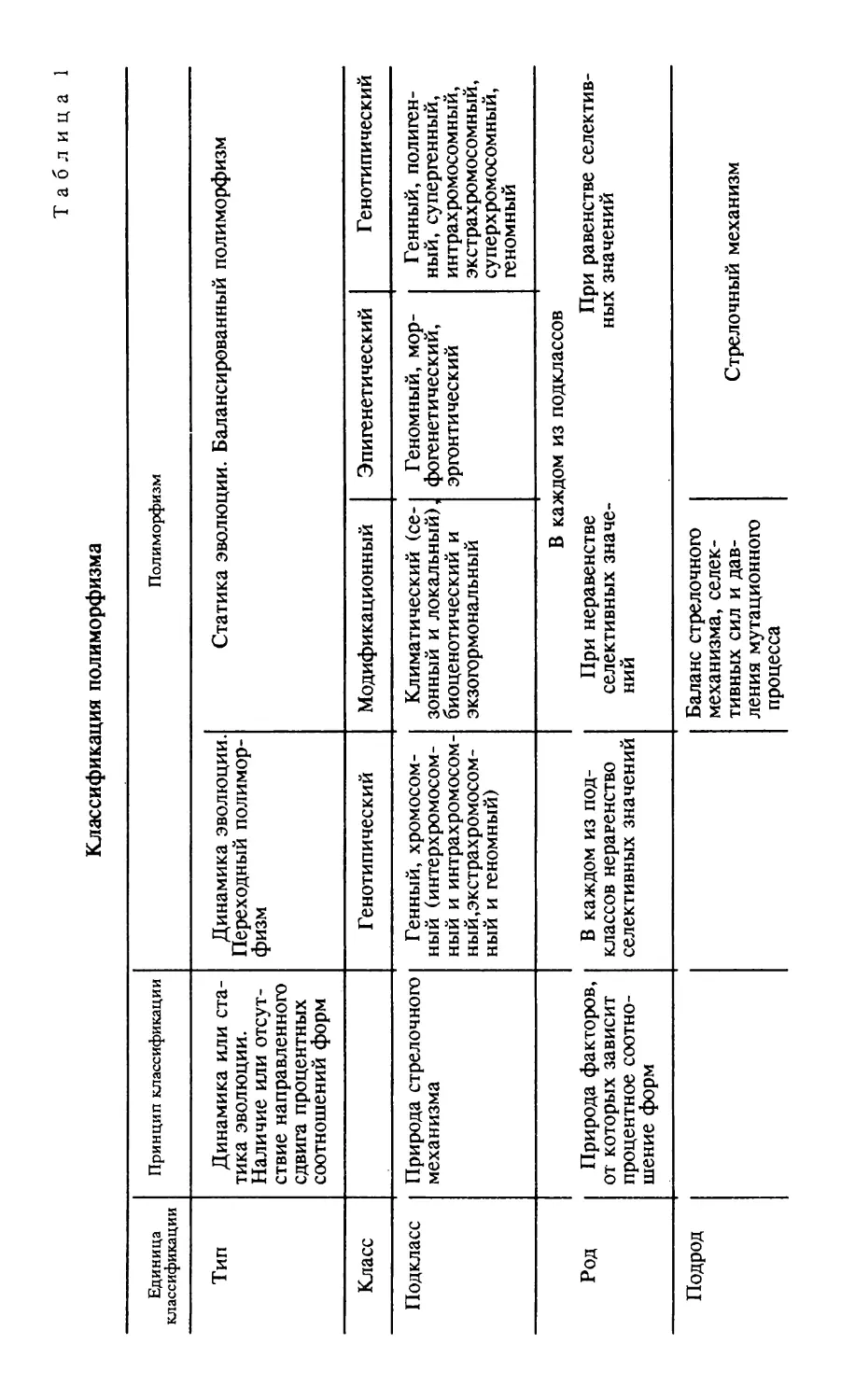
2. ВОЗМ О Ж Н Ы Е М ЕХАНИЗМ Ы ЭВОЛЮ ЦИИ ГЕНОТИПА
Теория эволюции доминантности Фишера и горячая
полемика, которая развернулась вокруг нее [Fischer, 1930,
1932; Haldane, 1932— 1941; Muller, 1935; и др], ознамено
вали рождение новой отрасли генетики — эволюционной
генетики. Впервые явления наследственности стали рас
сматриваться не только как фактор эволюции, но и как
объект эволюционных преобразований. Встал вопрос об
эволюции самих явлений наследственности и процессов из
менчивости. Приспособительный смысл таких фундамен
тальных генетических и феногенетических особенностей,
как линейное расположение генов, «чистота гамет», огра
ниченность поля действия единичной мутации (мера плейотропного действия гена), был раскрыт Малиновским
[1939, 1941].
Представление об эволюционном преобразовании самой
амплитуды изменчивости признака легло в основу теории
8
косвенного закрепления в процессе отбора первоначально
ненаследственных изменений [Кирпичников, 1935; Лукин,
1942; и др. ], а затем в основу теории стабилизирующего
отбора [Шмальгаузен, 1938, 1946; Камшилов, 1939, 1946;
Lerner, 1954, 1958; Берг, 1958; Berg, 1959].
Дарлингтон [Darlington, 1937, 1939, 1940], Гекели
[Huxley, 1942], Уайт [White, 1949] и Стеббинс [Stebbins,
1950], введя понятие генетической системы (genetic
system ), впервые указали на приспособительное значение
того или иного распределения наследственного материала
по хромосомам и открыли возможность трактовать эволю
цию кариотипа с помощью теории естественного отбора.
Генетическая система — это совокупность внутренних
факторов эволюции. Она охватывает не только строение
наследственного аппарата, в частности число и размеры
хромосом, но и воспроизводящую структуру популяции.
Свойства генетической системы выявляются в процессе
смены поколений. Они являются характеристикой не особи
и даже не данного поколения популяции, а группы особей,
будь то популяции или вид, отграниченной от подобных ей
групп в пространстве и длительно существующей во време
ни. Идея генетической системы, дополненная представле
ниями о групповом отборе [Берг, 1939, 1941а, б, 1942а—в,
1943, 1945а, 1957; Берг и др., 1941; Wright, 1955] и об он
тогенетических закономерностях эволюции [Шмальгаузен,
1938, 1946], представляет собой систему понятий, доста
точную для решения эволюционно-генетических задач,
В будущем учение об эволюции свяжет в единую систему
представлений не только цитологический и популяционный
уровень организации живого, как это делали Дарлингтон,
Гекели и Стеббинс, охватив оба уровня понятием генети
ческой системы вида, но и все уровни организации вида —
молекулярный, цитологический, онтогенетический и, нако
нец, популяционный.
Генетические системы эволюционируют. Главный фак
тор их эволюции — межгрупповой отбор. Межвидовой от
бор, или соревнование видов по путям и способам эволю
ции,— соревнование, в процессе которого менее успешно
эволюционирующие виды гибнут, а их более успешные
конкуренты вытесняют их с жизненной арены,— ведет к
усовершенствованию самих путей и способов эволюции.
Генетическая система вида меняется в процессе эволюции
как целое. Изменение охватывает не только количество
наследственного материала, характер его распределения по
хромосомам, условия и частоту перехода мутаций в гомо
зиготное состояние (у диплоидов и полиплоидов), условия
9
накопления гетерозиготных мутаций, но и тонкие особен
ности наследственного материала, в частности, его мутабильность.
„
В настоящее время привлекают внимание не только
способы преобразования популяций и видов, но и во все
возрастающей степени механизмы, обеспечивающие под
держание устойчивых состояний на всех уровнях организа
ции живого. Проблема биологического гомеостаза непос
редственно соприкасается с проблемами эволюционной ге
нетики [Lerner, 1954].
Теоретически мыслимы разные пути исследования воп
роса об эволюции генотипа: детальное сравнительно-цито
логическое изучение, точный сравнительный химический
анализ и различные косвенные генетические методы иссле
дования. Первые два метода пока имеют очень ограничен
ные возможности применения в рассматриваемой пробле
ме. Действительно, цитологический анализ, даже более
тонкий, позволяет уловить лишь относительно грубые ко
личественные изменения вещества хромосом; значительно
более точный сравнительный цитогенетический метод ис
следования различных хромосомных аберраций позволяет,
кроме того, устанавливать наличие весьма мелких струк
турных изменений хромосом и гомологию или негомологию
их частей. Но последнее возможно лишь у некоторых, осо
бенно благоприятных объектов, таких, как двукрылые с их
гигантскими хромосомами в ядрах слюнных желез. Значе
ние физико-химического анализа хромосом для понимания
природы самого гена, механизма его мутационных преоб
разований и механизма его действия в индивидуальном
развитии ясно понимал Кольцов [1936]. Однако значи
тельные успехи в этом направлении совместными усилия
ми биологов, физиков, химиков и математиков во всем ми
ре достигнуты лишь в последние пять лет. Возможности
физического и химического анализа отдельных генов так
же еще крайне ограничены и сводятся к установлению
лишь самых общих черт их химической природы и струк
туры. Глубже всего в тончайшие свойства генов и их отли
чий друг от друга до настоящего времени может проник
нуть лишь генетический анализ, в распоряжении которого
имеются три основных типа критериев различия: детальное
сравнение фенотипических эффектов и степени доминант
ности, детальное изучение явлений аллелизма, включая
псевдоаллелизм, и количественное и качественное изуче
ние мутабильности генов. Этими генетическими путями и
возможно в настоящее время относительно глубоко про
никнуть в природу гена и общий характер его возможных
Ю
изменений, а также экспериментально изучить пути его
эволюции. Поэтому во всем дальнейшем изложении мы бу
дем основываться преимущественно на данных, получен
ных этими генетическими путями.
Первые же попытки вскрыть генотипическое разнооб
разие в популяциях домашних и диких животных и расте
ний показали, что все исследованные популяции насыщены
огромным количеством рецессивных, частично доминант
ных и доминантных мутаций, в том числе летальных.
Дальнейший анализ показал, что хромосомные перестрой
ки, главным образом инверсии, играют заметную роль в
образовании генетического разнообразия популяций. Наи
более важным результатом генетико-популяционных исс
ледований в плане эволюции генотипа как такового явля
ется установление различий между популяциями в самом
характере генотипической изменчивости. Именно эти раз
личия позволили Вавилову [1926, 1927] сформулировать
представление о центрах происхождения видов, а Серебровскому [1929а] — понятие геногеографии. Дубинин
| Дубинин и др., 1934] отразил эти различия в понятии
экогенотипа. Мало-помалу раскрылись факторы преобразо
вания генотипического состава популяционно-генетических
исследований для систематики [Четвериков, 1926, 1927;
Е. Timofeeff-Ressovsky, N. Timofeeff-Ressovsky, 1927; Роза
нова, 1930; Wright, 1931, 1949, 1955; Дубинин и др., 1934;
Dobzhansky et al., 1938— 1955; Оленов и др., 1939; Muller,
1940; Timofeeff-Ressovsky, 1940а; Берг и др., 1941; Муретов, 1941; Bauer, Timofeeff-Ressovsky, 1943; Синская,
1948; Тимофеев-Ресовский, 1957; и др. ].
Из сравнительной кариологии и цитогенетики известны
несколько механизмов увеличения числа генов [Розанова,
1930; Карпеченко, 1935; Darlington, 1937; Левитский,
1937; Синская, 1948; Bauer, 1941; Stebbins, 1950; и др. ].
При весьма распространенной у растений полиплоидии и
гстероплоидии удваивается число всех генов или генов,
расположенных в определенных хромосомах. При этом на
блюдается весьма замечательное явление: сбалансирован
ное увеличение числа всех генов в 2 раза или более не
вносит почти никаких существенных изменений в деятель
ность генотипа, за ислючением некоторого изменения от
носительной жизнеспособности, а также размеров. Правда,
как показывает сравнительное изучение полиплоидных се
рий в пределах рода у растений, первоначально гомологич
ные гены с течением времени дифференцируются на пары,
которые в результате потери полной первичной гомологич
ности могут в дальнейшем изменять общий ход эволюции
п
генотипа, йапример, путем повышения его пластичности. |
Несбалансированные удвоения числа генов путем гетеро-/
плоидии ведут в большинстве случаев к более или менее
серьезным нарушениям гармонической системы онтогенеза
и часто являются летальными; в случае выживания и со
хранения в !историческом пути развития вида их значение,
по-видимом|у, сходно со значением полиплоидии. Увеличе
ние числа ^енов повышает число дискретных автономных
подразделений генотипа и вместе с увеличением числа
хромосом может повлиять на некоторые механизмы изме
нения генетических систем в популяционно-динамических
процессах. Как известно, полиплоидия, будучи распростра
нена у растений, относительно очень редко встречается у
животных [Muller, 1925], что весьма просто объясняется
широким распространением самоопыления у растений.
Аналогичных
механизмов
простого,
скачкообразного
уменьшения числа генов гаплоидного набора, по-видимо
му, не существует, так как внезапное исчезновение из ге
нотипа даже немногих генов влечет за собой летальный
эффект. Возможно лишь вторичное сокращение количества
генов из чи|сла первоначально удвоенных путем дуплика
ций, полиплоидии или гетерополиплоидии; эти явления
встречаются у гетеро- и полиплоидных форм растений, а
также в форме гомозиготно жизнеспособных делеций у лю
бых организмов, содержащих в исходной для этих делеций
форме дупликации соответствующих участков генома. На
конец, сокращение числа генов возможно в случаях обли
гатной гете!розиготности определенного комплекса генов
(например, ;Y-хромосомы). В этих случаях соответствую
щий блок генов, части хромосомы или целая хромосома
могут постепенно лишиться большинства или даже всех
первоначально расположенных в ней генов [Darlington,
1937, 1939, 1940; Берг, 1942а—в].
Весьма важным механизмом, ведущим к эволюцион
ным изменениям генотипа, является образование самых
различных типов хромосомных перестроек. Как уже упо
миналось, дупликации разной величины участков хромосо
мы ведут к увеличению числа генов и в некоторых случаях
к выгодной для вида дополнительной стабилизации тех или
иных онтогенетических процессов. На последнее указывает
то обстоятельство, что детальные сравнительные цитогене
тические исрюдования в роде дрозофила показали доволь
но широкое! распространение дупликации у разных видов.
Несомненно), очень интересным и важным, но далеко не
достаточно еще изученным является значение инверсий во
внутривидовой и межвидовой эволюции организмов
12
[Sturtevant, Dobzhansky, 1936; Дубинин и др., 1937;
Dobzhansky et al., 1938— 1955; Дубинин, Соколов, 1940;
Малиновский, 1940; Carson, 1955; Stone, 1955; и др. ]. Повидимому, в некоторых случаях инверсии создают условия
длительной гетерозиготности определенных участков хро
мосом, вероятной благодаря положительному отборному
значению самой частичной гетерозиготное™; может быть,
на этом пути благодаря отсутствию перекреста в пределах
гетерозиготйой инверсии [Mather, 1954] возможно созда
ние своеобразных «блоков» более тесно сцепленных генов.
При длительной гетерозиготное™ инверсии возможно про
текание в инвертированном участке хромосомы принципи
ально таких же «независимых» дальнейших эволюционных
изменений, какие особенно ярко проявляются в условиях
постоянной облигатной гетерозиготное™ Y-хромосомы, а
частично и в постоянно гомозиготной у гетерогаметного
пола Y-хромосомы. К своеобразному образованию «блоков»
ведут и «соседствующие» дупликации, т. е. те случаи, в
которых оба одинаковых участка хромосомы расположены
рядом. При всех условиях образования «блоков» возможно
селективное использование их для накопления мутацион
ных изменений, связанных с «укреплением» определенного
онтогенетического процесса; ясно, что в общей форме лю
бые состояния длительной гетерозиготное™ связаны с яв
лением балансированного генотипического полиморфизма
со всеми его эволюционными последствиями.
Жизнеспособные транслокации и сложные перестройки,
основанные на одновременном или последовательном обра
зовании хромосомных разрывов, могут привести, при всей
их редкости, к существенным изменениям структуры гено
типа, в результате которых возможно возникновение ярко
выраженной или даже абсолютной биологической изоляции
от исходногр типа со всеми вытекающими отсюда последст
виями [Кожевников, 1936]. Наконец, необходимо учиты
вать одно весьма важное и интересное явление, связанное
с хромосомными перестройками,— так называемый «эф
фект положения». Со времени классической работы Стертеванта [Stiirtevant, 1928], посвященной выяснению приро
ды изменений, происходящих в локусе bar у дрозофилы,
накопился уже довольно большой материал, показываю
щий, что на нормальное функционирование определенных
генов може+ влиять присутствие «новых» соседей в хромо
соме близ разрыва и перестройки. В первых опытах Стертеванта с bar, а также в опытах Дубинина и Сидорова
[1934],
Паншина
[1935]
и
Тимофеева-Ресовского
[Timofeeff-Ressovsky, 1940в] было показано (путем введе
13
ния соответствующих генов в непосредственную близость к
месту разрыва транслоцированной хромосомы и обратного
их выведения в нормальную хромосому), что эффект поло
жения связан не с мутационным структурным изменением
соответствующих генов, а с изменением, в большинстве
случаев ослаблением, их нормальных действий под влия
нием «необычных» соседей. Чаще всего понижается сте
пень доминантности нормального аллеля, что, разумеется,
должно (при длительном закреплении соответствующих
хромосомных перестроек) повести к дальнейшей эволюции
этих генов (хотя бы в смысле нового селекционного пути
накопления мутаций, вновь повышающих степень доми
нантности). Помимо этого, эффект положения может
иметь самостоятельное положительное или отрицательное
селекционное значение. В частности, в результате изме
нившегося положения могут меняться частота и характер
изменения генов. Эффект положения может выразиться,
таким образом, не только в изменении самого признака, но
и в изменении его мутационной изменчивости [Бельговский, 1938].
В предыдущих абзацах говорилось о тех механизмах, с
помощью которых происходит увеличение числа генов или
образование морфологических перестроек генома, в ре
зультате которых генотип может меняться тривиальным
путем мутационной замены одних аллелей другими. Но,
помимо этого, должны существовать какие-то пути измене
ния природы самих генотипов. Хромосомные и геномные
мутации изменяют число генов в пределах генома, но не
создают новых генов и не изменяют существенно природу
старых. Очень мало вероятно, что в процессе всей органи
ческой эволюции на нашей планете происходили в преде
лах геномов лишь перекомбинации и изменения числа ка
кого-то определенного количества исходных генов. Вряд ли
эволюция шла таким лотсианским, в широком смысле это
го слова, путем. Ниже будет показано, что имеются уже
экспериментальные данные, указывающие достаточно ясно
на наличие процессов внутригенной дифференцировки, ве
дущей к «рождению» новых генов и к изменению природы
генов, как специфических катализаторов и организаторов
клеточных и онтогенетических процессов.
Еще в конце двадцатых годов Серебровским с сотруд
никами [19296, 1938] была получена целая серия аллелей
гена scute у дрозофилы. Подробное изучение различий в
их фенотипическом проявлении и в проявлении соответст
вующих признаков у компаундов (гетерозигота по двум
мутантным аллелям одного и того же гена) позволило рас
14
положить эти аллели в совершенно определенный ряд: этот
ряд указывал на возможность существования в пределах
данного гена линейно (или в определенной последователь
ности) расположенных частей или субгенов. Одна мутация
всегда затрагивала либо один из этих субгенов, либо груп
пу из двух или более соседних друг с другом. Такой свое
образный ряд множественных аллелей был назван ступен
чатой аллелией. Мы здесь имеем дело, несомненно, с од
ним геном, в котором можно, однако, подозревать наличие
отдельных, в известном смысле автономных частей. Осо
бенно замечательным явилось открытие таких мутаций,
которые затрагивали одновременно «крайние» субгены гена
scute и соседний, считавшийся до того времени совершенно
независимым ген achaeta, а затем и таких, которые одно
временно вызывали признаки achaeta и соседнего с ним ге
на yellow. Все эти три гена — scute, achaeta и yellow —
расположены столь близко друг от друга в дистальном кон
це Х-хромосомы в слабо дифференцированном широком
диске на цитологической карте гигантских хромосом слюн
ных желез и перекресты между ними настолько редки, что
даже подвергались сомнению. На основании всех данных
по ступенчатой аллелии Н. П. Дубинин [1935, 1956] в
свое время развил представление о существовании непре
рывности и прерывистости в строении хромосом. Весьма
вероятно, что в данном случае мы имеем дело с внутригенной дифференцировкой, зашедшей так далеко, что перво
начально единая генетическая структура подразделялась
на ясно обособленный ген yellow, обособляющиеся гены
achaeta и scute и намечающуюся, но еще не осуществлен
ную генную дифференцировку внутри гена scute. Мы име
ем здесь, таким образом, новообразование независимых ге
нов in statu nascendi. К сожалению, работы по анализу
ступенчатой аллелии были у нас прерваны, и в этой обла
сти остается много неясного и незавершенного. В последнее
время множатся, однако, описания случаев у дрозофилы и
у других объектов так называемой псевдоаллелии, несом
ненно, имеющих ряд общих черт с явлением ступенчатой
аллелии [Lewis, 1954]. В некоторых случаях псевдоаллели
могут оказаться продуктами дифференцировки первона
чально единого гена, но в целом ряде случаев это, .несом
ненно, «соседствующие» дупликации, проявляющие или не
проявляющие эффект положения, в котором до известной
степени на наших глазах происходит процесс дифференци
ровки двух соседних первоначально одинаковых генов.
Возможно, что в данном случае мы имеем дело с тем же
процессом постепенной потери полной гомологии в резуль
15
тате нарушения истинной аллелии,который произошел и у
филогенетически закрепившихся случаев аутополиплоидии
у растений* Во всяком случае, явления ступенчатой алле
лии и псевдоаллелии заслуживают самого широкого анали
за и пристального внимания с точки зрения эволюционной
генетики, так как они в то же время являются одним из
путей глубокого исследования формирования и значения
генных «блоков» (поскольку они связаны с дупликациями)
и феногенетических действий определенных участков хро
мосом.
Все вышеизложенные механизмы изменения числа ге
нов и внутригенной дифференцировки изучались с по
мощью генетического, цитогенетического и феногенетиче
ского анализа возникавших мутаций; мутации являлись
материалом исследования. Но можно взять за основу коли
чественные и качественные исследования самого процесса
мутирования определенных генов с целью нахождения раз
личий между аллелями одного и того же гена в отношении
изменчивости, как второго из упоминавшихся в начале
свойств генотипа. Не следует забывать, что об участии оп
ределенного гена в тех или иных онтогенетических процес
сах мы узнаем в основном по фенотипическим эффектам
его изменений — мутаций; ведь доступны анализу, путем
скрещиваний, лишь те гены, у которых нам известна хотя
бы одна фенотипически проявляющаяся мутация. Отсюда
ясно, что качественный и количественный характер мути
рования может явиться хорошей характеристикой природы
самого гена.
На очень большом материале количественно изучалось
появление обратных и разнонаправленных мутаций под
влиянием рентгеновских лучей в пределах серии локуса
white у дрозофилы [Timofeeff-Ressovsky, 1932а, Ь, 1937,
1940а, с]. Исходным материалом здесь служили нормаль
ные культуры различного географического происхождения.
При обработке обширного материала в этом локусе оказа
лось, что как по общему проценту мутаций, так и, в осо
бенности, по относительному проценту крайней формы се
рии аллелей white систематически наблюдается реальная
разница между культурами из России и Северной Амери
ки. Путем специальных скрещиваний нормальные аллели
серии white из «американской» культуры были введены в
«русскую», а из «русской»— в «американскую». Проведен
ные затем на очень большом материале облучения рентге
новскими лучами показали, что обнаруженная разница в
мутабильности и направлении мутирования нормальных
аллелей white из «американской» и «русской» культур со
16
хранилась и что они связаны с самими нормальными алле
лями этой серии, а не с различиями в остальном генотипе.
Таким образом, была обнаружена фенотипически неулови
мая разница между так называемыми нормальными алле
лями определенного локуса из двух культур различного
географического происхождения; эта разница касается установимой стабильности и направления мутирования соот
ветствующих аллелей. Вскоре было установлено некоторое,
весьма незначительное различие в степени доминантности
двух этих аллелей [Muller, 1935].
В данном случае была найдена хотя и совершенно оп
ределенная, но относительно незначительная разница в на
правлении мутационной изменчивости двух нормальных
аллелей, указывающая на изменение возможной феногене
тической и эволюционной потенции данного гена. В связи
с этим были предприняты поиски других аналогичных слу
чаев. Положительные результаты получены в локусе
vestigial (II хромосома) и spineless (III хромосома) у дрозо
филы (Тимофеев-Ресовский, неопубликованные данные).
В локусе vestigial в культуре из Средней Германии было
получено значительно больше мутаций, сильно редуциру
ющих крылья и относительно мало влияющих на щетинки;
а в одной из «итальянских» культур большинство мутаций
в этом локусе заметно влияло на щетинки и относительно
слабо — на редукцию крыльев. Локус spineless у дрозофи
лы интересен тем, что часть мутантных аллелей сильно ре
дуцируют щетинки, а некоторые превращают усики в ноги
(так называемый аристопедиа) [Balkaschina, 1929]. В о д
ной из «русских» культур наряду со щетиночными были
получены и мутации аристопедиа; в одной же из «итальян
ских» культур на очень большом материале и при наличии
мутаций, редуцирующих щетинки, ни разу не были полу
чены аллели типа аристопедиа. В обоих случаях различия
статистически реальны: но не были закончены опыты с со
ответствующими нормальными аллелями, введенными с
помощью специальных скрещиваний в «чужой» генотип.
Упомянутые выше три серии опытов с достаточной яс
ностью указывают на возможность возникновения (среди
«малых» мутаций) таких структурных изменений нормаль
ных аллелей, которые не сказываясь достаточно резко в
изменении какого-либо морфологического признака (и по
тому будучи внешне неотличимыми друг от друга), вызы
вают заметную разницу в общем характере и направлении
своей дальнейшей мутационной изменчивости. Изменения
же в мутабильности определенного гена неизбежно должны
поставить его в иные условия естественного отбора и тем
17
самым явиться стимулом дальнейшей дифференциации.
Весьма возможно, что с изменением характера изменчиво
сти соответствующий ген может несколько изменить и свое
участие в определенных онтогенетических процессах; ведь
именно мутации гена, как уже упоминалось, показывают
нам те онтогенетические процессы, в которых данный ген
принимает специфическое участие.
Таким образом, мы видим, что к настоящему времени
уже накоплен громадный материал, показывающий раз
личные пути и возможности изменений структуры геноти
па, а в связи с этим создающий интересные предпосылки
для его эволюции. Необходимо, однако, стремиться к даль
нейшему увеличению материала, освещающего проблему
эволюции генотипа. Вместе с детальными сравнительно ге
нетическими анализами близких видов это позволит су
дить об относительном значении разных механизмов изме
нения числа и природы генов в общем ходе эволюции гено
типа.
3. ЭВОЛЮ ЦИЯ МУТАБИЛЬНОСТИ
Различие между видами и разновидностями одного ви
да по мутабильности [Timofeeff-Ressovsky, 1936; Stubbe,
1937; и др. ], а также генотипический контроль мутабиль
ности [Demerec, 1928; Шапиро, 1938; Rhoades, 1941;
Stebbins, 1950; Dobzhansky, 1954; Вагнер, Митчел, 1958; и
др. ] широко известны. Следовательно, наследственная из
менчивость по мутабильности, или материал для эволюци
онных преобразований, имеется. Сопоставляя разные спо
собы генотипического контроля мутабильности, ТимофеевРесовский [Timofeeff-Ressovsky, 1936] еще в 1936 г. ука
зывал, что мутабильность может претерпевать как кажу
щиеся, так и истинные изменения и что подлинные разли
чия между генами по мутабильности могут быть как внеш
ними, т. е. зависеть от других генов того же генотипа, так
и внутренними, т. е. зависеть от различий в структуре са
мих генов. Однако изменчивость еще не эволюция. Эволю
ционирует ли сама мутабильность, и каковы методы позна
ния эволюции мутабильности?
Сравнительный метод и моделирование естественного
отбора с помощью искусственного отбора применимы к
изучению факторов и путей эволюции мутабильности, как
и всех эволюционных проблем.
С помощью искусственного отбора можно пролить свет
на способ действия естественного отбора и частично моде
18
лировать его действие. Эффективность искусственного от
бора по мутабильности доказана всеми теми авторами, ко
торым удалось выделить высокомугабильные линии
[Demerec, 1937; Тиняков, 1939; Neel, 1942; Берг, 19446;
Mampell, 1946; Ives, 1950; и др. ]. Показано также, что ис
кусственный отбор мутантов приводит к повышению мута
бильности других локусов. По-видимому, отбор мутантов
сопровождается отбором генотипических причин, способст
вующих повышению мутабильности [Берг, 1948].
Сравнение мутабильности генов половых хромосом с
мутабильностью генов аутосом позволяет выяснить связь
между мутабильностью и интенсивностью естественного
отбора. У тех форм, где половая хромосома пребывает у
одного из полов в гемизиготном состоянии, отбор сцеплен
ных с полом мутаций более суров, чем отбор аутосомных
наследственных ' изменений. У D. melanogaster летальные
мутации возникают в половой хромосоме примерно в 2 ра
за реже на единицу длины эухроматинового района, чем в
аутосомах [Timofeeff-Ressovsky, Delbriick, 1936; Berg,
1937а, Ь]. Та же закономерность установлена и у тутового
шелкопряда [Эфроимсон, 1940]. Отсюда следует, что отбор
является условием стабилизации генотипа, по крайней ме
ре в отношении частоты появления летальных мутаций.
Сравнительное действие половых хромосом и аутосом и их
фрагментов на жизнеспособность, плодовитость и признаки
пола £>. melanogaster убеждают в том, что у этого вида ста
билизация генов половых хромосом сопровождалась час
тичной утратой их влияния на жизнеспособность [Берг,
1942в; Robertson, Reeve, 1953].
Сравнение диплоидных и полиплоидных форм одного и
того же вида по частоте фенотипического выявления мута
ций проливает свет на генотипические причины различий
по мутабильности [Сахаров, 1958]. Изучение влияния гиб
ридизации на мутабильность позволяет в некоторых случа
ях истолковать полученные результаты в эволюционном
аспекте [Sturtevant, 1939; Берг, 19416]. Наиболее важным
методом являются сравнение мутабильности популяций и
близких видов и сопоставление различий с различиями в
экологической ситуации. Именно этим способом выявляет
ся роль изоляции в эволюции мутабильности. Изоляция,
по-видимому, препятствует повышению мутабильности, ог
раничивая процесс замещения менее пластичных групп бо
лее пластичными. Изолированные лишь частично, равнин
ные популяции D. melanogaster более мутабильны, чем
изолированные в горных долинах и в лабораторных усло
виях популяции и линии того же вида [Берг, 1939, 19426;
19
19446, 1945а, б; а также неопубликованные данные]. Со
хранение популяциями различий по мутабильности в оди
наковых (лабораторных) условиях указывает на независи
мость мутабильности от колебаний в климатических фак
торах среды [Берг, 1942а], а автономность количественно
го показателя наводит на мысль о существовании биотиче
ских условий отбора [Берг, 1958]. Конкуренция групп по
пластичности и есть, по-видимому, условие повышения
мутабильности и вместе с тем причина ее независимости
от случайных колебаний в абиотических факторах среды.
К этой же категории исследований относится сравнение у
близких видов растений частоты возникновения мутаций в
стареющих семенах, отличающихся наличием или отсутст
вием периода покоя [Навашин и др., 1940], и у близких
видов дрозофил [Dobzhansky et al., 1954], обитающих в
разных температурных условиях и обладающих разной
структурой популяций. Вся совокупность этих исследова
ний, так же как теоретические соображения Sturtevant
[1937,
1938], Шапиро
[1938] и Буццати-Траверсо
[Buzzati-Traverso, 1954], с достаточной убедительностью
показывают, что мутабильность эволюционирует, что, яв
ляясь приспособительным признаком вида, она досягаема
для естественного отбора как в его индивидуальной, так и
групповой форме. Мутабильность входит как необходимый
компонент в генетическую систему вида и как таковой
участвует в целостных преобразованиях генетических сис
тем в процессе эволюции. Установление оптимальных со
стояний, закономерное сочетание с другими свойствами, в
частности с другими факторами эволюции,— итог эволю
ции мутабильности, как и всякого иного свойства вида.
Условия для быстрого изменения мутабильности в сто
рону как понижения, так и повышения имеются. Геноти
пический контроль мутабильности одних локусов со сторо
ны других локусов открывает широкие возможности повы
шения мутабильности. Всякая новая мутация, меняя в ка
кой-то степени химизм клетки, представляет собой хими
ческий агент, вызывающий мутации других локусов, и
притом независимый от проницаемости каких бы то ни бы
ло клеточных и тканевых барьеров. Положительный отбор
мутантов создает условие повышения мутабильности
[Берг, 1948]. Это не значит, конечно, что мутабильность
неуклонно эволюционирует в сторону повышения. По-ви
димому, главной чертой эволюции мутабильности являет
ся, как это ясно подчеркнул Шапиро [1938], согласование
частоты возникновения мутаций с другими свойствами ге
нетической системы.
20
4. ЗАКЛЮЧЕНИЕ
Обычный путь эволюции, вернее накопление элемен
тарного эволюционного материала, заключается в мутаци
онном изменении генов — появлении новых аллелей. Из
этого материала под влиянием элементарных эволюцион
ных факторов (популяционных волн, изоляции и естест
венного отбора) формируются элементарные эволюцион
ные явления — изменение генотипического состава попу
ляций. Но, как мы видели выше, этим не исчерпываются
возможности эволюционного процесса. Общетеоретические
соображения заставляют предполагать наличие более глу
боких изменений самого генотипа, его структуры и измен
чивости. Задачей нашего небольшого сообщения было по
казать, что в обширном материале современной генетики
уже имеются указания на целый ряд механизмов, с по
мощью которых могут меняться признаки самого гена.
Изменения генотипа, известные в настоящее время,
сводятся к следующим основным формам. Путем образова
ния различных геномных и хромосомных мутаций могут
изменяться число и морфология хромосом, сопровождаясь
или не сопровождаясь изменением общего числа генов.
С помощью полиплоидий, гетероплоидий, дупликаций,
вторичных делеций и некоторых небалансированных хро
мосомных аберраций может меняться число генов. Процес
сы формирования хромосомов и стабилизации некоторых
хромосомных перестроек могут вести, и в ряде случаев не
сомненно ведут, к образованию специализированных бло
ков генов, участвующих преимущественно в управлении
какими-либо специальными онтогенетическими процесса
ми. Явления ступенчатой аллелии и псевдоаллелии указы
вают на возможности внутренней дифференцировки генов
(а в некоторых случаях, возможно, их объединения). Точ
ное количественное и качественное изучение изменчивости
определенных генов в разных направлениях указывает на
возможности изменения как направления изменчивости,
так и онтогенетических связей отдельных генов. Наконец,
изучение общей средней мутабильности генов в разных по
пуляциях и генетических системах позволяет наметить
возможности изучения действия отбора на стабильность и
некоторые феногенетические особенности генов.
Конечно, все вышеперечисленные явления только на
мечаются. Специальной проблеме эволюции генотипа по
священо относительно небольшое количество работ. Необ
ходимо интенсивно и планомерно накапливать дальнейший
материал в точных опытах на удобных объектах. В настоя21
щее время такие работы приобретают особый интерес в
связи с представлениями о физико-химической природе ге
нов, основанными на впервые развиваемых теориях струк
туры, редупликации и ферментативно каталитических дей
ствий нуклеиновых кислот и нуклеопротеидов, как матери
альных основ наследственности; физико-химическая конк
ретизация вопроса о природе генов позволит конкретизиро
вать и проблему их эволюции. Необходимо еще указать на
значение всей области изучения возможных путей эволю
ции генотипа в связи с развитием идей кибернетики. Как
указывалось во введении, на языке кибернетики генотип
является кодом наследственной информации, передаваемой
от поколения к поколению, и управляющей системой онто
генеза. Поэтому изучение путей и механизмов эволюции
такой управляющей системы представляет особый интерес.
СПИСОК ЛИТЕРАТУРЫ*
Бельговский М. Л. Влияние инертных районов хромосом на частоту и ха
рактер изменений в соседних с ними активных участках / / Изв.
АН СССР. Отд. матем. и естеств. наук.— 1938.
Берг Р. Л. Различие природных и лабораторных линий Drosophila
melanogaster (гипотеза генетических корреляций): Дис.
канд.
биол. наук.— Ленинград, 1939.
Берг Р. Л. Роль малых мутаций в эволюции мутабильности / / Докл. АН
СССР.— 1941а.— Т. 32.— С. 71.
Берг Р. Л. Пониженная частота возникновения мутаций при внутривидо
вой гибридизации / / Докл. АН СССР.— 19416.—Т. 32.— С. 214.
Берг Р. Л. Сохранение различной мутабильности популяций Drosophila
melanogaster при переносе их в одинаковые условия / / Докл. АН
СССР.— 1942а.— Т. 3 4 .— С. 221.
Берг Р. Л. Зависимость между мутабильностью и степенью изоляции по
пуляций Drosophila melanogaster / / Докл. АН СССР.— 19426.—
Т. 3 6 .— С. 79.
Берг Р. Л. Мутабильность популяций Drosophila melanogaster, обитающих
на границе ареала распространения вида / / Докл. АН СССР —
1942в.— Т. 3 6 .— С. 171.
Берг Р. Л, Вид как эволюционирующая система / / Реферат работ учр.
Отд. биол. АН СССР 1941 — 1943.— 1943.— С. 185.
Берг Р. Л. Генетический анализ популяций Drosophila melanogaster Дилижана (Армянская ССР) / / Изв. АН СССР. Сер. биол.—
1944а.— Т. 1.— С. 51.
Берг Р. Л. Зависимое варьирование мутабильности и доминантности в
пределах одной естественной популяции Drosophila melanogaster / /
Изв. АН СССР. Сер. б и о л .- 19446.— Т. 5 . - С. 300.
Берг Р. Л. Корреляция между мутабильностью и регуляторной способно
стью организма и ее эволюционное значение / / Изв. АН СССР.
Сер. биол.— 1945а.— Т. 3 .— С. 367.
Берг Р. Л. Различная частота возникновения мутаций «yellow» в разных
популяциях Drosophila m elanogaster// Йзв. АН СССР. Сер. би
ол.— 19456.— Т. 3 .— С. 377.
* За недостатком места приведены лишь немногие работы экспери
ментального и обзорного характера, посвященные затронутой проблеме и
соде р ж а щ и е б ибл и о гра ф и ю .
22
Берг Р. Л. О взаимоотношении между мутабильностью и отбором в при
родных популяциях Drosophila melanogaster / / Журн. ошц. биол.—
1948.— Т. 9 .— С. 299.
Берг Р. Л. Типы п ол и м ор ф и зм а// Вести. ЛГУ.— 1957.— Т. 21.—
С. 115— 139.
Берг Р. Л. Дальнейшие исследования по стабилизирующему отбору / /
Бот. ж урн.— 1958.— Т. 43.— С. 12.
Берг Р. Л., Бриссенден Э., Александрийская В., Галковская К. Генетиче
ский анализ двух природных популяций Drosophila melanogaster / /
Журн. общ. биол.— 1941.— Т. 2 .— С. 143.
Вавилов Н. И. Закон гомологичных рядов.— Саратов, 1920.
Вавилов Н. И. Центры происхождения культурных растений / / Тр.
прикл. бот., генет., селекции.— 1926.— Т. 16.
Вавилов Н. И. Географические закономерности в распределении генов у
культурных растений / / Тр. прикл. бот., генет., селекции.—
1927.— Т. 17.
Вавилов Н. И. Линеевский вид как система / / Тр. прикл. бот., генет., се
лекции.— 1931.— Т. 26.
Вагнер Р., Митчел Г. Генетика и обмен веществ.— М.: ИЛ, 1958.
Дубинин Н. П. Дискретность и непрерывность в строении наследственно
го вещества / / Тр. по динамике развития.— 1935.— Т. 10.
Дубинин Н. П. Проблема физических и химических основ наследственно
сти / / Биофизика.— 1956.— Т. 1.— С. 8.
Дубинин Н. П. и др. Экспериментальный анализ экогенотипов Drosophila
melanogaster / / Биол. журн.— 1934.— Т. 3, № 1.
Дубинин Н. П., Сидоров Б. Н. Зависимость действия гена от его положе
ния в системе / / Биол. журн.— 1934.— Т. 3 .— С. 307.
Дубинин Н. П., Соколов Н. Н., Тиняков Г. Г. Внутривидовая хромосом
ная изменчивость / / Биол. журн.— 1937.— Т. 6 .— С. 1007.
Дубинин Н. П м Соколов Н. Н. Хромосомные мутации и система вида / /
Журн. общ. биол.— 1940.— Т. 1.— С. 543.
Камшилов М. М. Отбор как фактор, меняющий зависимость признака от
применения внешних условий / / Докл. АН СССР.— 1939.—
Т. 23, № 4.
Камшилов М. М. Первичная дивергенция нормы реагирования в зависи
мости от условий развития (наследственное и ненаследственное в
эволюции): Автореф. дис.
докт. биол. наук.— М., 1946.
Карпеченко Г. Д. Роль ненаследственной изменчивости в процессе естест
венного отбора / / Биол. ж урн.— 1935.— Т. 4.
Кирпичников В. С. Роль наследственной изменчивости в процессе естест
венного
о т б о р а //
Биол.
ж урн.—
1935.—
Т. 4,
№ 5 С. 343—353.
Кожевников Б. Ф . Экспериментальное получение расовой кариотипиче
ской изоляции / / Биол. ж урн.— 1936.— Т. 5.
Кольцов Н. К. Организация клетки.— М.; Л., 1936.
Левитский Г. А. Кариотип в систематике / / Тр. прикл. бот., ген., селек
ции.— 1937.— Т. 27.— С. 187.
Лукин Е. И. Приспособительные ненаследственные изменения организмов
и их эволюционная судьба / / Журн. общ. биол.— 1942.— № 4 .—
С. 235— 266.
Малиновский А. А. Роль генетических и феногенетических явлений в
эволюции вида. Ч. 1. Плейотропия / / Изв. АН СССР. Сер. биол.—
1939 — Т. 4 — С. 575.
Малиновский А. А. Роль хромосомных инверсий в эволюции вида / /
Журн. общ. биол.— 1940.— С. 565.
Малиновский А. А. Закономерности наследственности в свете дарвинов
ского учения об отборе / / Усп. соврем, биол.— 1941.— Т. 14.—
С. 171.
Муретов Г. Д. Возникновение физиологических мутаций и их распрост
ранение в популяциях / / Журн. общ. биол.— 1941.— Т. 5.
Навашин М. С., Герасимова Е. С., Беляева Г. М. О ходе мутационного
процесса в клетках зародыша покоящихся семян / / Докл. АН
СССР.— 1940.— Т. 26.
23
Оленов Ю. М. и др. Факторы, обеспечивающие состав природных попу
ляций Drosophila melanogaster / / Докл. АН СССР.— 1939.—
Т. 24.— С. 476.
Паншин И. Б. Новые доказательства справедливости гипотезы эффекта
положения / / Докл. АН СССР.— 1935.— № 4 .— С. 79.
Розанова М. А. Современные методы систематики растений / / Тр. прикл.
бот., ген., селекции.— Прил. 41.— 1930.
Сахаров В. В. Полиплоидия и радиация / / Совещание по полиплоидии у
растений.— М.: Моек, о-во испытателей природы при МГУ,
1958.— С. 16.
Серебровский А. С, Проблемы и методы геногеографии / / Тр. съезда ге
нетиков.— Л., 1929а.— Вып. 2 .— С. 71.
Серебровский А, С. Исследование ступенчатого аллеломорфизма / /
Журн. эксперим. биол.— 19296.— Т. 4, № 2.
Серебровский А. С. Гены scute и achaeta у дрозофилы и гипотеза дивер
генции гена / / Докл. АН СССР.— 1938.— Т. 29.
Синская Е. Н. Динамика вида.— Л., 1948.
Тимофеев-Ресовский Н. В. Микроэволюция / / Бот. журн.— 1957.—
Т. 43.
Тиняков Г. Г. Высокомутабильная линия из природной популяции / /
Докл. АН СССР.— 1939.— Т. 2 2 . - С. 609.
Четвериков С. С. О некоторых чертах эволюционного процесса с точки
зрения современной генетики / / Журн. эксперим. биол.— 1926.—
Т. 2 .— С. 3.
Четвериков С. С. Об одной эволюционной проблеме и ее эксперименталь
ном решении / / Тр. съезда зоол., анат. и гист.— Л., 1927.
Ш апиро Н. И. Мутационный процесс как адаптивный признак вида / /
Зоол. ж урн.— 1938.— Т. 17.— С. 592.
Ш мальгаузен И. И. Организм как целое и индивидуальное в историче
ском развитии.— М., 1938.
Ш мальгаузен И. И. Факторы и эволюция.— М.; Л., 1946.
Эфросимсон В. П. Изменение скорости мутационного процесса у тутового
шелкопряда / / Изв. АН СССР. Сер. биол.— 1940.— С. 6»8.
Balkaschina Е. G. Ein Fall der Erbhomoosis (die Genvariotion «aristopedia»)
Drosophila melanogaster / / Arch. Entw. Mech. Org.— 1929.— Bd 115.
Bauer H. Chromosomenforschung / / Fortschr. d. Zool., N. F .— 1941.— Bd 5.
Darlington C. D. Taxonomic species and genetic systems / / The New
Systematics.— Oxford, 1940.
Darwin Ch. The Origin of Species.— L., 1959.
D em erec M. The behaviour of mutable genes / / Verhandlungen des V Int.
Kongress
Zeitschrift
fur
Induktive
Abstammungs
und
Uererbungslehre.— Suppl. 1 — 1928.
Dem erec M. Frequency of spontaneous mutations in certain of Drosophila
melanogaster / / Genetics.— 1937.— Vol. 22.— P. 469—478.
Dobzhansky Th. Genetic nature in species differences / / Amer. Nat.—
1937.— Vol. 71.
Dobzhansky Th. Genetics and the Origin of Species.— 1937— 1954.
Dobzhansky Th. et al. Genetics of natural populations / / Genetics.—
1938— 1955.— Vol. 23—40.
Dobzhansky ThM Spassky B.. Spassky N. Rares of spontaneous mutation in
the second chromosome of the sibling species Drosophila pseudoobscura
and Drosophila persimilis / / Genetics.— 1954.— Vol. 39.
F ischer R. A. The Genetical Theory of Natural Selection.— Oxford, 1930.
F ischer R. A. The evolutionary modification of genetic phenomena / / Proc. 6.
Int. Congr. Genet. I.— 1932.
Frolova S. L., Astaurov B. L. Die Chromosomengarnitur als systematisches
Merkmal / / Z. Zellforsh. Mikr. Anat.— 1929.— Bd 10.
Haldane J. B. The Causes of Evolution.— L., 1932.
Haldane J* B. The theory of the evolution of dominance / / J. Genet.—
1939.— Vol. 37.
Haldane J. B. New Paths in Genetics.— L., 1941.
H uxley J. S. Evolution: the Modern Synthesis.— N. Y., 1942.
Ives P. T. The importance of mutation rate genes in evolution / / Evolution.—
1950.— Vol. 4.
24
Lerner J. M. Genetic homeostasis.— Edinburgh, 1954.
Lerner J. M. The Genetic Basis of Selection.— N. Y.; L., 1958.
Lewis E. B. Pseudoallelism and the gene concept / / Proc. 9. Int. Congr.
Genet. I.— 1954.
Liiers H. Zur vergleichenden Genetik der Drosophila Arten / / Verh. 7 Int.
Kongr. Entom.— Berlin, 1939.
Mampell R. Genic and nongenic transmission of mutator activity / /
Genetics.— 1946.— Vol. 31.— P. 589—597.
Mather K. The genetical units of continuous variation / / Proc. 9. Int. Gongr.
Genet. I.— 1954.
M uller H. J. Why polyploidy is rarer in animals than in plants? / / Amer.
Nat.— 1925.— Vol. 5$.
M uller H. J. On the incomplete dominance of the normal allelomorphs of
«white» in Drosophila / / J. G en.— 1935.— Vol. 30.
M uller H. J. Bearinge of the Drosophila work on systematics / / The New
Systematics.— Oxford, 1940.
N eel J. V. A study of a case of high mutation rate in Drosophila
melanogaster / / Genetics.— 1942.— Vol. 27.— P. 519—536.
Rhoades M. M. The genetic control of mutability in maize / / Cold. Spr.
Harbor. Symp. Quant. Biol.— 1941.
Robertson R. W. Reeve E. C. R. Studies in Quantitative Inheritance / / J.
G en.— 1953.— Vol. 51.
Stebbins G. L. Variation and Evolution in Plants.— N. Y., 1950.
Stone W. S. Genetic and chromosomal variability in Drosophila / / Cold Spr.
Harbor Symp. Quant. Biol.— 1955.— Vol. 20.— P. 256.
Stubbe H. Spontane und Strahleninduzierte Mutabiliat.— Leipzig, 1937.
Sturtevant A. H. A furher study of the so-colled mutation at the Bar locus in
Drosophila / / Genetics.— 1928.— Vol. 13.— P. 401.
Sturtevant A. H. Essays on evolution, I—III / / Quart. Rev. Biol.—
1937— 1939.— Vol. 12— 13.
Sturtevant A. H. Hing mutation frequency induced by hybridsation / / Proc.
Nat. Acad. Sci., Wash.— 1939.— Vol. 7.
Sturtevant A. H., Dobzhansky Th. Inversions in the third chromosome of wild
races of Drosophila pseudoobscura and their use in the study of the
history of the species / / Proc. Nat. Acad. Sci. (USA).— 1936.
Tim ofeeff-R essovsky E. A., Tim ofeeff-Ressovsky N. W. Genetische Analyse
einer freilebenden Drosophila melanogaster Population / / Roux Arch.
Entwmech.— 1927.— Bd 109.
Tim ofeeff-R essovsky N. W. Mutations of the gene in different directions / /
Proc. 6. Int. Congr. Genet. I.— 1932a.
Tim ofeeff-R essovsky N. W. The experimental production of mutations / /
Biol. Rev.— 1932b.— Vol. 9.
T im ofeeff-R essovsky N. W. Qualitativer Vergleich der Mutabilitat von
Drosophila funebris und Drosophila melanogaster / Z. ind. Abst.
Vererb.— 1936.— Bd 71.
Tim ofeeff-R essovsky N. W. Experimentelle Mutationsforschung in der
Vererbungslehre / / Verl. Steinkopf.— Dresden; Leipzig, 1937.
T im ofeeff-R essovsky N. W. Mutations and geographical variation / / The New
Systematics.— Oxford, 1940a.
T im ofeeff-R essovsky N. W. Positionseffect / / Handb. Erbbiol. Menschen.,
Berlin.— 1940b.— Bd 1.
T im ofeeff-R essovsky N. W. Spontane und strahleninduzierte Mutabilitat in
geographisch verschiedenen Stammen von Drosophila melanogaster / /
Biol. Zbl.— 1940c.— Bd 60.
Tim ofeeff-R essovsky N. W., Delbriick M. Strahlengenetische Versuche tiber
sichtbare Mutationen und die Mutabillitat einzelner Gene dei Drosophila
melanogaster / / Z. ind. Abst. Vererb.— 1936.— Bd 1.
White M. J. D. Animal cytology and evolution.— Cambridge, 1948.
Wright S. Evolution in Mendelian populations / / Genetics.— 1931.— Vol. 16.
Wright S. Adaptation and selection, Genetics, Paleontology and Evolution.—
Princeton, 1949.
Wright S Classification of the factors of evolution / / Cold Spr. Harbor
Symp. Quant. Biol.— 1955.— Vol. 20, N 16.
25
ОТНОСИТЕЛЬНАЯ РОЛЬ СТАБИЛИЗАЦИИ
И ПЕРЕДИФФЕРЕНЦИРОВКИ ГЕНА В ЭВОЛЮЦИИ
НАСЛЕДСТВЕННОЙ СУБСТАНЦИИ*
Р. Л . Берг
THE RELATIVE ROLES OF STABILIZATION
AND REDIFFERENTIATION
OF THE GENE IN THE EVOLUTION OF HEREDITY
SUBSTANCE
R. L . Berg
Как показали предыдущие исследования [Berg, 1937],
частота возникновения мутаций, понижающих жизнеспо
собность мух (суммарно деталей и семилеталей) в Х-хромосоме при пересчете на ген в 3 раза ниже, чем во второй
хромосоме. Показано также, что различие быть может за
висит не от пониженной частоты возникновения мутаций в
генах Х-хромосомы, а от утери в процессе эволюции неко
торым количеством генов Х-хромосомы способности влиять
на жизнеспособность особи и соответственно мутировать,
давая аллели, снижающие жизнеспособность. Эти гены
могли превратиться в гены, участвующие в развитии при
знаков пола. На такую возможность указывает относитель
но высокая, по сравнению со второй хромосомой, частота
возникновения мутаций стерильности в Х-хромосоме. Про
цесс передифференцировки не исключает, однако, стаби
лизации гена по отношению к мутагенному воздействию.
Настоящая статья представляет собой попытку пролить
свет на относительную роль стабилизации и перидифференцировки в уменьшении частоты возникновения мута
ций, снижающих жизнеспособность, в Х-хромосоме по
сравнению со второй хромосомой.
Можно количественно оценить роль стабилизации и пе
редифференцировки в снижении мутабильности генов
Х-хромосомы.
Если стабилизации нет и наблюдаемая разница в час
тоте возникновения мутаций целиком объясняется передифференцировкой генов, нехватка возникающих деталей
и семилеталей окажется восполненной мутациями стериль
* G enetics.— 1937.— Vol. 22.— Р. 402. В статье сохраняется терми
нология времен первой публикации работы.
26
ности, возникающими с большей, частотой в Х-хромосомс,
чем во второй хромосоме. Частота возникновения леталей
и семилеталей в Х-хромосоме в 3 раза меньше. Ожидалось,
что частота возникновения мутаций стерильности будет в
2 раза превышать частоту возникновения леталей и семи
леталей. Недостача двух третей мутаций будет ими ком
пенсирована и суммарная частота возникновения мутаций
на ген уравнивается. Если частота возникновения мутаций
стерильности будет меньше, чем в 2 раза превосходить ча
стоту возникновения леталей и семилеталей, мы вправе
допустить существование, помимо передифференцировки,
еще и стабилизацию тем большую, чем больше будет отли
чаться от соотношения 2:1 полученное нами соотношение.
В задачу входило также изучение соотношения между
степенью понижения жизнеспособности и наличием или
отсутствием стерильности у особей с пониженной жизне
способностью. Некоторые гены, не полностью утратившие
влияние на жизнеспособность, могли стать на путь превра
щения в гены, участвующие в реализации признаков пола.
ЭКСПЕРИМЕНТ
Учету подлежали летальные, семилетальные мутации и
мутации стерильности, возникающие под воздействием х-лучей в половой хромосоме самцов Drosophila melanogaster.
Чтобы выявить все степени понижения жизнеспособности гемизиготных по облученной Х-хромосоме самцов, в культу
рах F2 подсчитано соотношение самцов и самок. Число мух
в культурах было значительным (90—200).
Самцы wa были облучены дозой х-лучей равной 8000 г
и скрещены с C IB /v самками. В потомстве этого скрещи
вания девственные С 1 В /wa самки скрещивались индивиду
ально с несколькими самцами.
Всего заложено 850 культур. Каждые 3—4 дня мух
каждой культуры помещали на новый корм. Было получено
три реплики. Если самка погибала раньше, чем создава
лась реплика, культура исключалась из опыта. 827 куль
тур содержали плодовитых самок, в 23 культурах самки
были стерильные. Частота доминантных мутаций стериль
ности не превышает 2,7 %. В культурах F2 подсчитано
число самок и самцов. Сигнальные гены v и scsl позволяли
выявлять всех самцов, возникающих в результате нерасхождения хромосом у самки. C lB /w a и у самца scsl. Хромо
сома scsl содержит две инверсии, полностью исключающие
перекрест по всей длине хромосомы.
27
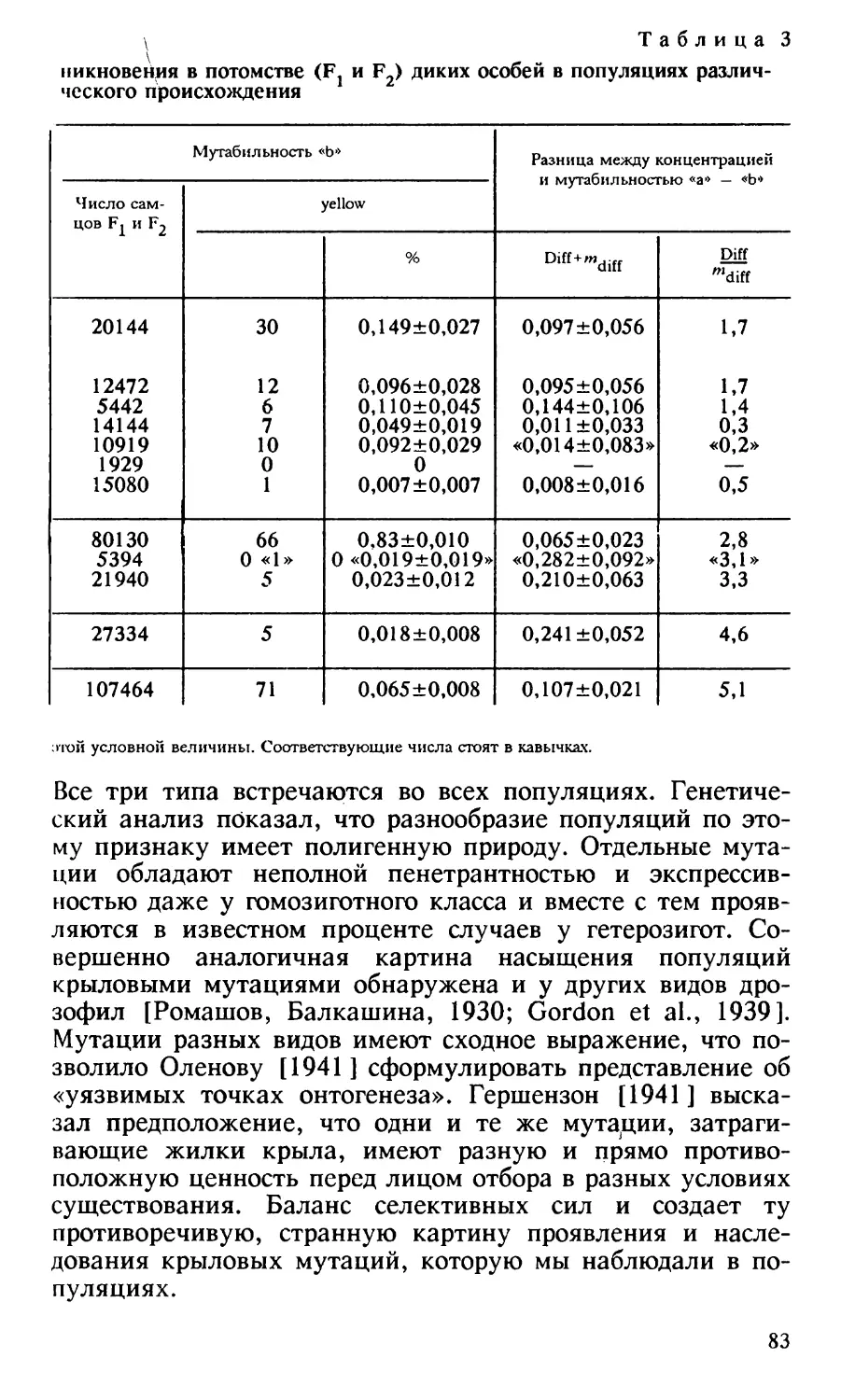
Таблица 1
Распределение мутаций стерильности среди культур с различной сте
пенью понижения жизнеспособности
Параметры
популяции
Культуры с указанным процентом самцов
80 80- 70- 6070 60 50
5040
40- 3030 20
2
4
146
100
0,4
0,9
0
0
0
0
0
0
20- 1010 0
0
Все
го
66
475
Число культур
р2
Процент культур
с указанной
жизнеспособ
ностью самцов
Число мутаций
стерильности
среди культур
соответствующего
класса
Процент мута
ций стерильнос
ти среди культур
соответствующего
класса
29
19
22
4,4 14,3 30,7 21,1 6,1
4,2
4,8 13,9
20
67
8
17
11,9 11,6
17
11
8
17 37,9 12,1
6
0
27,2
0
67
Исследовано 475 культур F2 и обнаружено 66 леталей
(13,6 %). .Деталями считались только те культуры, в кото
рых не было ни одного самца ни в культуре F2, ни при
проверке в F3. 408 культур, не содержащих летали, послу
жили для обнаружения мутаций стерильности. Испытание
самцов на фертильность проводилось путем массовых скре
щиваний самцов F2 со своими сестрами. Обнаружено
52 случая полной стерильности (12,7 %) и 15 случаев
сильно пониженной плодовитости.
Все мутации — летали, семилетали и мутации сте
рильности подвергались проверке в следующем поколении.
Число мутаций стерильности среди культур F2 с раз
личной степенью понижения жизнеспособности самцов по
казано в табл. 1.
ОБСУЖ ДЕНИЕ
Число мутаций стерильности, обнаруженных в опыте
(12,7 %), не превосходит число летальных мутаций
(13,9 %). Отношение тех и других приближается к 1:1.
Оно меньше соотношения 2:1, которое указывало бы на передифференцировку как на единственную причину мень
шего числа мутаций на ген, снижающих жизнеспособность
28
в Х-хромосоме по сравнению с аутосомой. Значит, передифференцировка не является единственной причиной ма
лого числа леталей в Х-хромосоме. Процессы стабилизации
должны были играть роль в эволюции части генов Х-хромосомы и роль эта в утере генами Х-хромосомы способно
сти мутировать к летальным аллелям не менее значитель
на, чем роль передифференцировки.
Это заключение имеет силу только в случае, если чис
ло мутаций стерильности соответствует числу генов, участ
вующих в реализации пола. Возможно, однако, что часть
генов, не влияющих на жизнеспособность, влияют на при
знаки пола так мало, что их мутации не приводят к пол
ной стерильности.
Эти гены не могут быть многочисленными, так как
отбор элиминирует их с той же суровостью, с ка
кой бы элиминировал гены Y-хромосомы [Muller, Pain
ter, 1932], так как эти гены, мутируя, не меняли бы
(или меняли бы в незначительной степени) фенотип
мутантов,
Таблица 1 показывает, что среди культур, содержащих
мутации, понижающие жизнеспособность, частота мутаций
стерильности выше, чем среди культур, таких мутаций не
содержащих. По-видимому, понижение плодовитости и по
нижение жизнеспособности являются плейотропными эф
фектами одних и тех же мутантных генов.
Отсюда следует, что среди культур с наиболее сильно
пониженной жизнеспособностью наблюдается много мень
ше мутаций стерильности, чем в классе культур с менее
сильным понижением жизнеспособности.
Плейотропия, быть может, является переходным состо
янием в процессе передифференцировки генов. Гены, став
шие на путь смены функции и начавшие принимать уча
стие в реализации признаков пола, вряд ли детерминиро
вали, в своем исходном состоянии очень важные жизненные
функции.
Фенотипическое проявление мутаций таких генов вы
разится в незначительном ослаблении жизнеспособности и
в сильном понижении фертильности вплоть до полной сте
рильности.
Гены, играющие важную роль в морфогенезе, подвер
жены смене функций в меньшей мере. На признаки пола
такие гены влияют мало. Их мутации, по всей вероятно
сти, выражаются в сильном понижении жизнеспособности,
но выживающие в небольшом количестве окажутся фертильны. В 16 культурах с наибольшим понижением жиз
неспособности самцов немногие выжившие оказались
29
фертильны, и только в 6 культурах этого класса низкая
жизнеспособность сочеталась со стерильностью. Было бы
очень важно подтвердить это наблюдение на большом
материале.
выводы
1. Выдвинута гипотеза передифференцировки генов,
влияющих на жизнеспособность, в гены, вовлеченные в ре
ализацию признаков пола.
2. Частота возникновения мутаций стерильности в
Х-хромосоме, будучи более высокой, чем частота возник
новения тех же мутаций в аутосомах, все же недостаточна,
чтобы объяснить более редкое возникновение летальных
мутаций в пересчете на ген в Х-хромосоме по сравнению с
аутосомами.
3. Помимо утери генами в процессе передифференци
ровки влияния на жизнеспособность происходит стабилиза
ция генов Х-хромосомы.
4. Процент мутаций стерильности больше среди куль
тур с пониженной жизнеспособностью самцов, чем в классе
культур, гд. жизнеспособность не отклоняется от нормы.
Это указывает на плейотропное действие генов Х-хромосо
мы, многие из которых затрагивают такие относительно
независимые стороны жизнедеятельности как выживае
мость и фертильность.
Автор выражает благодарность за руководство этой ра
ботой и помощь в подготовке к печати рукописи профессо
ру Г. Дж. Мёллеру и за интерес к работе и помощь в ос
мыслении ее результатов А. П. Владимирскому.
СПИСОК ЛИТЕРАТУРЫ
Berg R. L. The relative frequency of mutations in different chromosomes of
Drosophila melanogaster. 1. Lethal m u ta tio n s// Genetics.— 1937.—
Vol 2 2 .— P. 225— 240.
M uller H. J., Painther T. S. The differentiation of the sex-chromosomes of
Drosophila into genetically active and inert regions / / Z. Ind.
Abst.— 1932.— Bd 6 2 .— S. 316—365.
30
ИСТОРИЧЕСКОЕ РАЗВИТИЕ МЕХАНИЗМА
РЕАЛИЗАЦИИ ПОЛА*
Р . Л. Берг
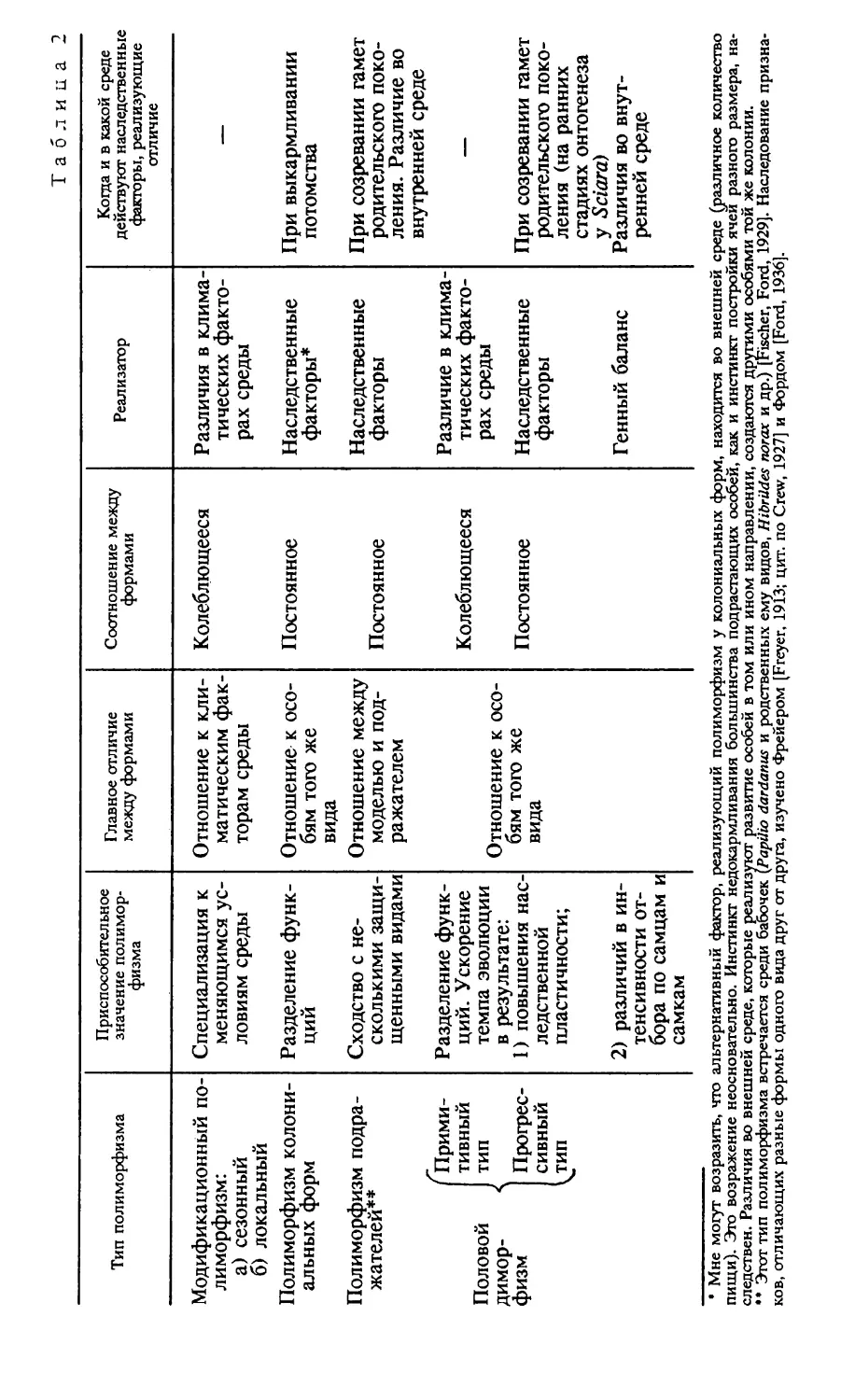
1. ПРИСПОСОБИТЕЛЬНОЕ ЗНАЧЕНИЕ ПОЛИМОРФИЗМ А
Половой диморфизм — это частный случай полимор
физма. Существует несколько типов полиморфизма — по
лиморфизм колониальных форм (муравьи, пчелы, терми
ты), полиморфизм подражателей, когда один и тот же вид
подражает разным моделям (бабочки, например, некото
рые виды Papilio , Hibrides и д р .), модификационный (се
зонный и локальный) полиморфизм и, наконец, половой
поли- или чаще диморфизм. Каждый тип полиморфизма
является приспособительным признаком вида.
Приспособительный признак вида — это признак, на
личие которого повышает уровень приспособительности ви
да в целом, обеспечивает победу в межгрупповом соревно
вании. Такой признак может не давать никаких преиму
ществ отдельной особи в процессе внутригруппового сорев
нования, или эти преимущества могут быть относительно
малы. Плате [Plate, 1913] называет такие признаки
Arterhaltende Merkmale [см. также: Северцов, 1940]. По
лиморфизм повышает темп эволюции. Он действует при
этом косвенно — благодаря увеличению численности вида.
Исключение в этом отношении представляет полиморфизм
подражателей, где численность подражателя находится в
строгой зависимости от численности модели. Модификаци
онный пстиморфизм обеспечивает сохранение вида при
резких сдвигах в условиях существования [Шмальгаузен,
1940]. Приспособительное значение полиморфизма может
заключаться в разделении функций. При этом достигается
более совершенная специализация. Это наиболее ясно для
колониальных форм. В случае полового диморфизма разде
ление функций, связанное с резкой морфологической диф
ференциацией полов, обеспечивает диморфизм яйцеклеток
и спермиев, приспособительное значение которого огромно.
Разделение функций при заботе о потомстве также повы
шает приспособленность вида. Достаточно указать на роль,
которую сыграло обеспечение зародышей большим количе
ством питательного материала, и развитие зародышевых
оболочек и скорлупы в эволюции рептилий [Северцов,
* Журнал общей биологии.— 1942.— Т. 3, № 3.
31
1939]. Эти приспособления могли выработаться на основе
специализации полов, на основе их морфологической диф
ференциации. Переход к внутриутробному развитию и вы
кармливанию детенышей молоком у млекопитающих по
требовал еще большей специализации. Во всех случаях
специализация шла не только по линии морфологической,
но и по линии специализации инстинктов. Например, если
у птиц самец обеспечивает пищей самку в то время, когда
она насиживает яйца, самка может не покидать гнезда в
течение всего насиживания. Это предохраняет яйца от пе
риодического охлаждения. Специализация инстинкта в
этом направлении позволяет виду расширить среду обита
ния, селиться в широтах с более холодным климатом, а
также удлинить время размножения — насиживать яйца в
более холодное время года. Разделение функций способст
вует тому, что вид активно изменяет среду, в которой жи
вет,— пчелы поддерживают относительно постоянную тем
пературу улья, птицы — гнезда, потомство обеспечивается
пищей и т. п. В других случаях приспособительное значение
полиморфизма заключается в специализации к меняющим
ся условиям среды (сезонный и локальный полиморфизм).
Приспособительное значение полиморфизма подража
телей заключается в возможности использовать несколько
видов в качестве моделей. Это особенно важно, если моде
лями служат малочисленные виды. Приспособительное
значение мимикрии может появиться только в том случае,
если модель значительно более многочисленна, чем подра
жатель [Bates, 1862; цит. по Darwin, 1871; Vallace, 1865,
1891; см. также Fischer, 1930; Карпентер, Форд, 1935].
Как и любой тип полиморфизма, половой димор
физм — это приспособительный признак вида. В этом его
сходство с другими типами полиморфизма. Но есть и ряд
особенностей, которые отличают половой диморфизм от
всех других типов полиморфизма. Половой диморфизм
обеспечивает ускорение темпа эволюции несколько иными
путями, чем другие типы Полиморфизма. Благодаря скре
щиванию создается возможность сочетания разных геноти
пов, перекомбинирования возникающих мутаций [Fischer,
1930; Muller, 1932], наследственная пластичность вида
возрастает. Однако ускорение темпа эволюции, основанное
на существовании полового диморфизма, зависит не только
от увеличения наследственной пластичности вида. Половой
диморфизм создает предпосылки возникновения различий
в интенсивности отбора внутри вида. Увеличение наследст
венной пластичности связано с увеличением абсолютного
числа вредных мутаций и их комбинаций, приспособление
32
все время нарушается. Становится необходимым оставле
ние потомства не всеми особями, а лишь наиболее приспо
собленной частью вида, даже если эта часть мала. Отбор
становится необходимым условием существования вида
[Берг и др., 1941 ].
Сильное сокращение воспроизводящей части популя
ции, неизбежное при повышении интенсивности отбора,
может повести к снижению численности вида со всеми ги
бельными последствиями такого снижения. Сохранение
большой численности вида и одновременно усиление отбо
ра может быть достигнуто при условии, если повышение
интенсивности отбора ограничено лишь частью вида. Поло
вой диморфизм и создает предпосылки возникновения раз
личий в интенсивности отбора внутри вида. Сокращение
воспроизводящей части популяции идет, главным образом,
за счет самцов. Отбор самцов более интенсивен, чем отбор
самок. Можно назвать ряд приспособлений, допускающих
к копуляции только безупречных самцов.
1. Более интенсивное уничтожение или устранение
от размножения самцов особями того же вида. Этому
способствуют: а) бой самцов; б) уничтожение самцов сам
ками того же вида (пчелы, пауки); в) предпочтение со сто
роны самок, оказываемое некоторым самцам; г) брачный
полет пчел и муравьев [Plate, 1913; Snell, 1932].
2. Более интенсивное уничтожение самцов хищника
ми. Этому способствуют: а) яркое оперение, бросающиеся
в глаза движения, пение самцов [Jager, 1874]; б) отсутст
вие подражательной окраски у самцов многих видов, где
самки являются подражательницами (бабочки); в) отсутст
вие паразитизма у самцов многих видов, где самки парази
тируют (паразитические раки); г) более раннее появление
самцов по сравнению с самками (истинная и ложная протерандрия) [Demoll, 1908]; д) избыток самцов.
3. Более интенсивное уничтожение самцов со сторо
ны абиотических факторов среды. Этому способствуют:
а) более раннее появление самцов; б) избыток самцов.
4. Более интенсивное уничтожение самцов под влия
нием возникающих неблагоприятных мутаций. Этому
способствуют:
а) гаплоидия
самцов
(Hymenoptera,
Anthotrips) [Schull, 1917]; б) гетерогаметия самцов, кото
рая встречается гораздо чаще, чем гетерогаметия самок
[Schrader, 1928; Correns, 1928]. Приспособления, ведущие
к дифференциальной интенсивности отбора, могут основы
ваться на морфологических различиях самцов и самок (яр
кая окраска самцов, отсутствие подражательной окраски,
наличие орудий борьбы и т. п.), на различии в инстинктах
2 Р. Л. Берг
33
(ложная протерандрия, уничтожение самцов особями того
же вида и т. п.), или на различиях в строении кариотипа
(гаплоидия, гетерогаметия).
Более интенсивное уничтожение мужского элемента по
сравнению с женским не ограничивается диплофазой. Оно
распространяется и на гаплофазу. Избыток спермиев, по
мимо обеспечения встречи спермия и яйцеклетки, может
играть роль и как фактор, повышающий интенсивность гаметического отбора [Лобашов, 1939]. Этому должно спо
собствовать сохранение спермы в семеприемниках самок
многих видов насекомых j прорастание большого количест
ва пыльцевых зерен на рыльце пестика растений. У тех ра
кообразных, спермин которых имеют мало шансов встре
титься с яйцеклеткой, спермин передвигаются в воде, сое
динившись попарно. Этим обеспечена конкуренция между
ними при встрече с яйцеклеткой.
В некоторых случаях повышение интенсивности отбора
может идти и за счет самок. Этому способствует бой маток
у пчел. У видов, размножающихся с помощью кладок, объ
ектом отбора становится целая кладка. Интенсивность от
бора возрастает, так как в случае уничтожения кладки
сразу погибает все потомство одной особи или, если кладок
несколько, значительная его часть. Это ведет к сокраще
нию воспроизводящей части популяции. Отбор действует
избирательно только в отношении самок. Производитель
ницами следующего поколения оказываются самки, кладки
которых лучше защищены.
В других случаях повышение интенсивности отбора мо
жет идти как за счет самцов, так и за счет самок в одина
ковой степени. В качестве примеров можно указать на сни
жение доминирования нормального фенотипа [Берг и др.,
1941] и инстинкт уничтожения слабой молоди со стороны
особей того же вида. Но гораздо чаще именно самцы уни
чтожаются наиболее интенсивно. Повышенная интенсив
ность отбора самцов по сравнению с самками обеспечивает
полноценность следующего поколения. Стоя на этой точке
зрения, мы можем дать единое объяснение множеству слу
чаев полового диморфизма, начиная с гаплоидности самцов
и кончая ярким оперением их. Егер [Jager, 1874] и
ШтоЛьцман [Stolzmann, 1885] отмечают, что уничтожение
большого количества самцов выгодно для вида. Поэтому
они считают образования, облегчающие уничтожение сам
цов хищниками или особями того же вида, видовыми при
способлениями, выработавшимися под влиянием естествен
ного отбора. Штольцман приводит в качестве примера гре
бень петуха, длинные перья хвоста, затрудняющие полет
34
многих птиц. Самцы, привлекая яркостью своего оперения
внимание врага, отвлекают его от самок [Jager, 1874]. По
сле уничтожения самцов количество пищи, которое прихо
дится в среднем на одну самку, возрастает [Stolzmann,
1885]. Уничтожением слабых со стороны хищников и со
братий по виду можно объяснить и непроявлением подра
жательной окраски у самцов многих видов бабочек, где
самки являются подражательницами.
Возникновение гаплоидности самцов, так же как при
чины преобладания гетерогаметии среди самцов, необъяс
нимо с этой точки зрения. Я думаю, что приспособитель
ное значение повышенной интенсивности отбора самцов
сказывается в следующем поколении. Победителями в
межвидовой борьбе оказываются те виды, у которых наи
более развиты приспособления, обеспечивающие оставле
ние потомства только немногими преодолевшими все пре
пятствия самцами.
Приспособления, повышающие интенсивность отбора,
могут выработаться в процессе межгруппового соревнова
ния. Победителями в нем оказывались те виды или относи
тельно изолированные группы особей внутри вида, в кото
рых приспособления, обеспечивающие оставление потомст
ва лишь наиболее приспособленными особями, лучше всего
развиты. Распространение на весь вид признака, способст
вующего уничтожению самцов, может совершаться в ре
зультате внутривидовой межгрупповой борьбы. Как извест
но, Дарвин [1939;. Darwin, 1871] указывал на возможность
сохранения признака, несущего гибель или бесплодие ин
дивидууму, если этот признак полезен для общины. Разви
тие такого признака совершается в результате конкурен
ции общин друг с другом. В качестве примеров Дарвин
приводит бесплодие рабочих пчел, смерть пчелы, жалящей
врага, истребление пчелиной маткой молодых маток, ее
дочерей, и, наконец, уничтожение рабочими пчелами их
братьев — трутней.
Резкий половой диморфизм возникает в результате
различий в интенсивности и формах отбора самцов и самок
[Darwin, 1871; Jager, 1874; Fischer, 1932]. Этим создаются
предпосылки для возникновения дальнейших различий в
интенсивности и формах отбора у разных полов. Часто по
вышение интенсивности отбора самцов связано с тем, что
один самец способен оплодотворить многих самок. Тогда
соотношение полов 1:1 уже обеспечивает избыток самцов и
дифференциальную интенсивность отбора самцов и самок.
У пчел трутень может оплодотворить лишь одну самку,
так как он погибает сейчас же после оплодотворения. В со35
Таблица
1
Корреляция по ряду приспособительных признаков
(по: [Dem oll, 1908])
Признак
Протерандрия
Eucera, Systropa
Apis mellifica
Самцы вылетают на 3 — 4
недели раньше самок
Большой
Выражен слабо или от
сутствует
Рудиментация ро Нет
товых частей
Избыток самцов
Паразитизм
Melecta
Самцы вылетают
на 2—3 дня раньше са
мок
Нет
Выражен сильно
Сильная у
иу ?
ответствии с этим в улье имеется громадный избыток сам
цов. Их количество доходит до 5000. Это избыток самцов
и обеспечивает вместе с протерандрией, брачным полетом
и гаплоидностью самцов повышенную интенсивность отбо
ра самцов. Повышенная интенсивность отбора имеет осо
бое значение для пчел, где инбридинг, а также инстинкты
заботы о потомстве, инстинкты совместной защиты улья —
все приспособления, понижающие интенсивность внутри
группового соревнования, могли бы повести вид по пути
дегенерации в результате накопления неблагоприятных
мутаций (табл. 1). Повышение интенсивности отбора при
одновременном сохранении большой численности вида —
это мощный фактор ускорения темпа эволюции. Оно по
зволяет беспредельно повышать наследственную пластич
ность вида, не ставя вид под угрозу дегенерации. Половой
диформизм и создает предпосылки как для повышения на
следственной пластичности вида, так и для повышения ин
тенсивности отбора. Степень повышения интенсивности от
бора зависит от избытка самцов, не принимающих участия
в оплодотворении. Эти преимущества полового диморфиз
ма основаны на относительно постоянном числовом соотно
шении между полами.
2. ПОСТОЯНСТВО ЧИСЛОВОГО СООТНОШЕНИЯ
М ЕЖ ДУ Ф ОРМ АМ И И СПОСОБ РЕАЛИЗАЦ ИИ
П ОЛИМ ОРФ ИЗМ А
Половой диморфизм и полиморфизм колониальных
форм основаны на различном отношении разных форм к
особям того же вида. Полиморфизм подражателей — на
взаимоотношении между хищником и жертвой, с одной
36
стороны, между моделью и подражателем — с другой. Во
всех этих случаях приспособительное значение полимор
физма основано на постоянном числовом соотношении
между различными организмами: между самцами и самка
ми, между маткой и рабочими пчелами, между моделью и
подражателем и т. п. Биотические факторы среды играют в
процессе отбора главную роль в регулировании числового
соотношения между формами. В случае сезонного поли
морфизма особи не находятся в каком-либо взаимоотноше
нии друг с другом. Числовое соотношение между формами
регулируется, главным образом, абиотическими (климати
ческими) факторами среды.
В основе полиморфизма лежит способность давать раз
личную морфологическую реакцию на разные воздействия.
Нужно различать способность реагировать различно на ре
ализующие воздействия и сами эти воздействия, реализую
щие развитие в том или другом направлении. На необхо
димость такого разграничения в случае полового димор
физма указывает Корренс [Correns, 1938], в случае поли
морфизма подражателей — Фишер [Fischer, 1930, гл. VII,
раздел «The theory of saltations» ]. Если способность давать
ту или иную реакцию и исход этой реакции всегда наслед
ственно обусловлены, то различия в среде, служащие реа
лизатором того или другого пути развития, могут нахо
диться вне организма, а могут образовываться и в процессе
дифференциации тканей у особей родительского поколе
ния. Приспособительное значение полиморфизма может
наилучшим образом проявиться в том случае, если комп
лексы признаков, отличающие различные формы друг от
друга, целиком возникают либо у одной, либо у другой
особи. Это возможно в том случае, если альтернативные
факторы, реализующие различия, находятся вне организма
(количество и качество пищи, температура и т. п.), или,
если альтернативные факторы расщепляются в каждом по
колении. В результате расщепления создается альтернатив
ная внутренняя среда, которая обусловливает различное
проявление всех признаков. В этом случае можно говорить
о генотипической реализации полиморфизма. Какой способ
реализации полиморфизма окажется наиболее приспособ
ленным и сможет закрепиться в процессе отбора, зависит
or того, какое числовое соотношение между формами —
постоянное или колеблющееся — характеризует тот или
другой тип полиморфизма. Имеется тесная взаимообуслов
ленность между приспособительным значением соотноше
ния между формами и способом реализации полиморфиз
ма.
37
Если полиморфизм приобретен видом как приспособле
ние, позволяющее лучше использовать разнообразные ус
ловия среды, в которых живет вид (сезонный и локальный
полиморфизм — Шмальгаузен [1940] называет этот тип
полиморфизма модификационным полиморфизмом), то
наиболее приспособительным способом реализации разли
чий и будет первый способ, когда роль реализатора при
надлежит различию в тех самых факторах внешней среды,
к которым приспособлены различные формы. Преимущест
ва этого типа полиморфизма связаны со специализацией к
тем или иным меняющимся условиям среды. Они не осно
ваны на постоянстве соотношения между разными форма
ми. Все особи, живущие в одних условиях, образуют одну
форму, живущие в других условиях — другую.
Все другие виды полиморфизма основаны на относи
тельно постоянном соотношении между формами. Постоян
ное соотношение между формами может осуществиться,
когда реализация признаков, на которых основан полимор
физм, относительно независима от колебания внешних ус
ловий. Поэтому наиболее приспособительный способ реа
лизации этих типов полиморфизма — реализация с по
мощью наследственных факторов или генного баланса.
В табл. 2 схематично представлены различия разных типов
полиморфизма.
Роль различий во внешних условиях в качестве реали
заторов отступает в процессе эволюции, на задний план.
Их заменяют различия внутренней среды. Отбор, который
приводит к этой замене, это стабилизирующий отбор
[Шмальгаузен, 1939]. Только здесь отбор стабилизирует
числовое отношение между формами, а не конечный ре
зультат развития отдельной особи по отношению к колеба
нию факторов внешней среды.
Числовое соотношение между формами — признак, ха
рактеризующий группу особей. Отбор такого признака со
вершается в результате межгрупповой борьбы. В соответст
вии с этим и усовершенствование механизмов реализации
полиморфизма также совершается в результате межгруп
повой борьбы.
Дарвин [Darwin, 1871] подчеркивает, что то или иное
числовое соотношение между полами в потомстве особи не
дает никакого преимущества этой особи в борьбе за суще
ствование. Он пишет: «Я раньше думал, что если тенден
ция производить оба пола в равных количествах приспосо
бительна для вида, она может повиноваться естественному
отбору, но теперь я вижу, что вся проблема настолько
сложна, что разрешение ее лучше оставить на будущее».
38
* Мне могут возразить, что альтернативный фактор, реализующий полиморфизм у колониальных форм, находится во внешней среде (различное количество
пищи). Эго возражение неосновательно. Инстинкт недокармливания большинства подрастающих особей, как и инстинкт постройки ячеи разного размера, на
следствен. Различия во внешней среде, которые реализуют развитие особей в том или ином направлении, создаются другими особями той же колонии.
** Этот тип полиморфизма встречается среди бабочек (Papilio dardanus и родственных ему видов, Hibrildes погах и др.) [Fischer, Ford, 1929]. Наследование призна
ков, отличающих разные формы одного вида друг от друга, изучено Фрейером (Freyer, 1913; цит. по Crew, 1927] и Фордом [Ford, 1936].
3, ПРИСПОСОБИТЕЛЬНОЕ ЗНАЧЕНИЕ ВНУТРЕННЕГО
(ХРОМОСОМАЛЬНОГО)
М ЕХАНИЗМ А РЕАЛИЗАЦ ИИ ПОЛА
У гермафродитов дифференцировка тканей в онтогене
зе особи создает альтернативные условия, которые приво
дят к тому, что одни ткани принимают участие в образова
нии органов одного пола, а другие ткани — в образовании
органов другого пола.
При генотипической реализации раздельнополости по
ложение обстоит точно так же, только дифференцировка
тканей, которая затем создает при оплодотворении альтер
нативные условия, происходит при образовании гамет в
родительском поколении у гетерогаметичного пола. Она со
вершается, правда, несколько необычно — путем неравно
наследственных делений. При этом образуются клетки, ко
торые содержат разные хромосомы (X или Y в сочетании с
набором аутосом) или разное число хромосом (X или 0 у
тех форм, где Y-хромосома отсутствует). У Sciara неравно
наследственные деления, в результате которых создаются
альтернативные условия, реализующие пол следующего
поколения, совершаются на ранних стадиях онтогенеза ро
дителей. Неравнонаследственные деления происходят раз
лично у разных особей Sciara и приводят к тому, что обра
зуются два сорта самок, из которых одни производят толь
ко самцов, другие — только самок [Bridges, 1939].
Наследственные факторы, обусловливающие различие
во внутренней среде, могут быть либо парой аллеломор
фов, члены которой отличаются друг от друга, либо двумя
различными комплексами генов, между которыми пере
крест отсутствует, или, по крайней мере, затруднен. Гены
или комплексы генов, от которых зависит различие во
внутренней среде, реализующее пол особи, могут быть ус
ловно названы генами-реализаторами [Correns, 1928]. Раз
личия во внутренней среде могут зависеть также от разли
чий в количестве наборов хромосом или в соотношении
между количеством разных хромосом. В этом случае фак
тором, реализующим различия, является генный баланс.
Роль отдельных генов отступает на задний план, весь гено
тип в целом участвует в реализации отличий. Во всех этих
случаях, за исключением тех, когда различия во внутрен
ней среде зависят от различий в количестве хромосом или
наборов хромосом (например, пчелы), реализация пола
связана с гетерозиготностью по отдельным генам, по участ
кам хромосом или по целым хромосомам.
40
Исходным способом реализации пола был, по всей ве
роятности, способ реализации под влиянием различий в
факторах внешней среды (его принято называть очень неу
дачным термином «латентный гермафродитизм»). Такой
способ реализации половых отличий ставит вид под угрозу
разобщения полов, так как в одном месте могут существо
вать условия, благоприятные для реализации одного пола,
и другом — другого. Резкая дифференциация полов может
оказаться поэтому неприспособительной, так как оба пола
будут требовать для своей реализаций резко различных ус
ловий, а это увеличит угрозу разобщения полов. Но важ
нее всего то, что этот способ реализации не обеспечивает
постоянства числовою отношения полов. Поэтому возник
новение внутреннего механизма, реализующего пол особи,
должно было представить больше преимуществ и могло
быть подхвачено естественным отбором. Этот механизм с
самого начала должен был удовлетворять некоторым усло
виям. 1. Факторы, реализующие пол, должны обладать до
статочной силой, чтобы преодолевать реализующие дейст
вие внешних условий и других вновь возникающих мута
ций. 2. Факторы, реализующие мужской и женский пол,
не должны при образовании гамет попадать в одну и ту же
гамету. Если факторы, реализующие мужской и женский
пол, локализованы в цитоплазме, точное расхождение их в
разйые гаметы не обеспечено в силу особенностей воспро
изведения цитоплазмы. Поэтому участие цитоплазматиче
ских элементов клетки в реализации половых отличий
должно характеризовать примитивные типы определения
иола. В процессе эволюции роль цитоплазмы в реализации
пола должна отступать на задний план по тем же причи
нам, как и роль колебаний внешних факторов. Это, конеч
но, относится только к реализации того или другого пола.
Осуществление признаков пола, так же как осуществление
любого признака организма, зависит и от ядерных и от
плазматических элементов клетки и возможно только в оп
ределенных условиях среды, обеспечивающих жизнедея
тельность организма.
Расхождение реализаторов пола в разные гаметы обес
печено в том случае, если они расположены в различных
гомологах одной пары хромосом и перекрест между ними
затруднен. Если они являются аллеломорфами, расхожде
ние их всегда в разные гаметы обеспечено, но вероятность
1Юзникновения такой системы очень мала, хотя и не иск
лючена [Jones, 1931, 1932, 1934; Emerson, 1932]. Более
вероятно возникновение обоих факторов поблизости друг
от друга. Наиболее благоприятным условием для отбора
41
таких факторов окажется возникновение их близ нити ве
ретена, где перекрест меньше, чем в других частях хромо
сом [Dobzhansky, 1930; Offermann, Muller, 1932; Mather,
1939].
После того как факторы, из которых один реализует
мужской пол, а другой — женский, распространились на
вид, возникновение под влиянием естественного отбора ба
лансированной системы, при которой один пол гомогаметичен, а другой гетерогаметичен, неизбежно. Хромосома, но
сительница фактора гомогаметичного пола, это Х-хромосома, ее гомолог — Y-хромосома. Исходные этапы эволюции
половых хромосом можно наблюдать у высших растений,
ее конечные этапы — у тех животных, у которых У-хромосома утеряна. Процесс эволюции половых хромосом мо
жет быть обрисован следующим образом.
4. ДАЛЬНЕЙШ АЯ ЭВОЛЮ ЦИЯ ПОЛОВОГО ДИ М О РФ ИЗМ А
Не только качество признака, но и характер его насле
дования являются предметом отбора. Характер наследова
ния и проявлений мутаций зависит от их положения в сис
теме генотипа. При этом могут играть роль: эффект поло
жения, величина перекреста с другими генами, та или
иная близость к месту прикрепления нити веретена, поло
жение в районе дупликации или вне его, локализация в
половых хромосомах и аутосомах.
Y-хромосома реализует своим присутствием гетерогаметичный пол [Kosswig, 1930]. Она присутствует только у
одного пола. Все мутации, возникающие в ней около гена,
реализатора пола, ограничены в своем проявлении гетерогаметичным полом. Теперь, когда угроза разобщения полов
исчезла, исчезли и препятствия, мешавшие резкой диффе
ренциации полов. Эволюция признаков, на которых осно
ван половой диморфизм, ускоряется, и количество призна
ков, отличающих самца от самки, возрастает. Признаком,
который отличает один пол от другого, может быть лишь
такой признак, который полезен одному полу и вреден
другому [Fischer, 1930а, Ь, 1932]. В противном случае, ес
ли он проявляется одинаково у обеих форм, он либо закре
пится у обеих, если он полезен, либо ни у одной, если он
вреден. До образования половых хромосом новый признак,
отличающий один пол от другого, мог закрепиться отбо
ром, если польза, приносимая им одному полу, велика, а
вред, приносимый им другому полу, мал. Тогда отбор,
включая возникшую мутацию в комплекс признаков поло
42
вого диморфизма, одновременно понижает ее проявление у
особей того пола, которому вреден возникший признак.
Среди особей этого пола выживают те, у которых мутация
проявляется наиболее слабо. Это, конечно, очень замедля
ет процесс включения. То же справедливо и теперь для
мутаций, возникающих в аутосомах. Совсем другое дело,
если мутация, полезная гетерогаметичному полу, возникла
в Y-хромосоме, около гена, реализатора пола. Она никогда
не попадает в тот пол, где ее действие приносит вред. Оче
видно, включение в комплекс признаков полового димор
физма такой мутации пойдет быстрее, чем включение
аутосомальной мутации. Это приведет к преобладанию ге
нов, осуществляющих свое действие у гетерогаметического
пола в Y-хромосоме. Гены, осуществляющие свое полезное
действие у гомогаметичного пола, имеют больше шансов
быть включенными в комплекс под действием отбора, если
они возникли в Х-хромосоме, чем если они возникли в
аутосомах, так как локализация в Х-хромосоме препятст
вует выщеплению их в гомозиготе у гетерогаметичного по
ла. Половые хромосомы эволюционируют в сторону все
большего и большего накопления в них мутаций, влияю
щих на признаки полового диморфизма. Их роль в процес
се дифференциации полов по сравнению с аутосомами воз
растает. Это подтверждают наиболее хорошо изученные
случаи наследования признаков полового диморфизма.
Винге [Winge, 1927], Кирпичников [1935] показали, что
признаки, обусловливающие яркую окраску самцов Lebistes
reticulatus, где гетерогаметичен мужской пол, преобладают
в половых хромосомах по сравнению с аутосомами. Они
преобладают в Y-хромосоме по сравнению с Х-хромосомой.
Показано, что большое количество генов, связанных с
развитием признаков пола у женского гаметофита, мха
Sphaerocarpus, локализовано в Х-хромосоме, которая при
сутствует только у женского гаметофита.
Накопление в половых хромосомах мутаций, из кото
рых одни осуществляют свое действие в самце, а другие в
самке, возможно только при подавлении перекреста между
X- и Y-хромосомами [Darlington, 1932]. Подавление пере
креста может быть достигнуто в результате отбора генов,
понижающих перекрест, или в разультате отбора хромо
сомных перестроек в X- или Y-хромосоме. Процесс возник
новения хромосомальных аббераций более быстр для
Х-хромосомы, чем для Y-хромосомы, так как перекрест у
гомогаметичного пола служит источником образования
вторичных перестроек, после того как возникла первичная
перестройка. Возникновение дупликации в Х-хромосоме
43
будет не только понижать перекрест между X- и Y-хромосомой, но и сразу поведет к различию доз генов, включен
ных в дупликацию у самца и у самки. Это различие доз
поведет к различию в их проявлении, и они могут принять
участие в образовании признаков полового диморфизма.
Сначала районы пониженного перекреста, а затем и райо
ны дупликаций явятся очагами наиболее быстрой диффе
ренциации Х-хромосомы.
Наиболее совершенное подавление перекреста во всей
хромосоме достигается при локализации хиазмы в той хро
момере, к которой прикреплена нить веретена [Darlington,
1932, 1934]. По-видимому, у Drosophila подавление пере
креста между X- и Y-хромосомами как раз и зависит от
локализации хиазмы близ этой хромомеры. Поэтому-то пе
рекрест между X- и Y-хромосомами совершается только в
этой области [Stern, 1929; Stern, Ogura, 1931; Kaufmann,
1933; Darlington, 1934; Philips, 1934]. Однако локализация
хиазмы и ограничение перекреста этой областью есть, по
всей вероятности, вторичное явление, возникшее тогда,
когда гомология между X- и Y-хромосомами была в значи
тельной степени утеряна.
5. ОБРАЗО ВАН ИЕ РАЗЛ И ЧИЙ В ГЕННОМ БАЛАНСЕ
М ЕЖ ДУ НАБОРАМ И ХРОМОСОМ САМЦА И САМКИ
Понижение перекреста между X- и Y-хромосомами
приводит к постоянной, вынужденной гетерозиготности Yхромосомы. Интенсивность отбора по отношению к мута
циям, возникающим в Y-хромосоме, понижена по сравне
нию с интенсивностью отбора по отношению к мутациям,
возникающим в других хромосомах. Это ведет к «опусте
нию» Y-хромосомы [Muller, 1918; Darlington, 1932; Muller,
Painter, 1932; Haldane, 1933]. У растений, где гаплофаза
значительно сильнее развита, чем у животных, отбор в
гаплофазе мутаций, возникающих в Y-хромосоме, препят
ствует возникновению резких различий между X- и Y-хро
мосомами. Гетерозиштность Y-хромосомы ограничена дип
лофазой. В соответствии с этим эволюция хромосомного
механизма реализации пола находится у растений на более
примитивной стадии, чем у животных. Однако и здесь най
дены случаи резких различий между X- и Y-хромосомами
у мха Sphaerocarpus [Allen, 1917, 1932], где X- и Y-хромосома находятся в диплофазе в состоянии постоянной гете
розиготности. Это показывает, какие глубокие последствия
может иметь даже ограниченное диплофазой понижение
44
интенсивности отбора. Опустение начнется с тех районов,
где перекрест наиболее подавлен — это районы, прилежа
щие к нити веретена, а также районы, гомологичные райо
нам дупликаций в Х-хромосоме. Дистальные части Х-хромосомы £>. melanogaster несут активный материал, в то
время как проксимальные части инертны. Сперва некото
рые районы Х-хромосомы, а затем и вся Х-хромосома ока
жутся в одиночной (или в двойной для районов дуплика
ций) дозе у самца и в двойной дозе (или четвертной для
районов дупликаций) у самки. Это различие доз генов
Х-хромосомы представляет очень совершенный механизм
реализации половых отличий. Как отмечает Мёллер
[Muller, 1932а], само различие доз генов у самца и у сам
ки поведет к различию в их проявлении, и они могут при
нять участие в образовании полового диморфизма.
Степень обратимости пола в течение онтогенеза может
служить критерием того, насколько далеко зашла эволю
ция пола. Но таким же критерием может служить и сте
пень подавления перекреста между X- и Y-хромосомами.
Бросается в глаза корреляция между рядом признаков. Од
ни из них характеризуют примитивный тип определения
пола, другие прогрессивный [Darlington, 1932], схема от
личий примитивного и прогрессивного типов, составленная
Дарлингтоном, приведена у Рыжкова [1936].
Примитивный тип определения пола характеризуется
легкой обратимостью пола в течение онтогенеза как под
влиянием колебания факторов среды, так и в результате
мутаций. Половой диморфизм развит слабо. Гомология
между X- и Y-хромосомами еще не утеряна, и они конъю
гируют по всей длине, или гомологичные части их сравни
тельно велики. Различное проявление мутаций, возникаю
щих в Х-хромосоме, у форм XX и XY зависит не от осо
бенностей Y-хромосомы. Перекрест между X- и Y-хромосомами совершается свободно или несколько понижен. Фор
мы строения YY жизнеспособны и являются самками у тех
форм, где гетерогаметичен женский пол, самцами — у тех
форм, где гетерогаметичен мужской пол [Winge, 1934].
Родственные формы имеют различный тип определения
пола. Этот тип встречается у некоторых рыб и лягушек.
Прогрессивный тип определения пола характеризуется
сильным развитием полового диморфизма. Пол реализует
ся в зависимости от различий в балансе половых хромосом
и аутосом. Обратимость пола в течение онтогенеза под
влиянием обычных условий среды невозможна. Мутации,
влияющие на реализацию пола сильнее, чем обычный реа
лизатор, встречаются очень редко. В роде Drosophila опи
45
саны 3 случая — один у D . melanogaster возник под влия
нием х-лучей [Muller, 1928], другие два спонтанно у
D. simulans [Sturtevant, 1921] и у D. virilis [Lebedeff,
1938 ]. Гомология между X- и Y-хромосомами в значитель
ной степени утеряна. Они конъюгируют не по всей длине,
а лишь на небольшом отрезке, обычно прилежащем к нити
веретена. Перекрест ограничен этим участком. Различное
проявление мутаций, возникающих в Х-хромосоме, у форм
XX и XY зависит от особенностей Y-хромосомы (отсутст
вие нормального аллеломорфа в Y-хромосоме). Формы YY
нежизнеспособны. Родственные формы имеют сходный тип
определения пола. Этот тип встречается у Drosophila.
Lebistes занимает промежуточное положение. У нее по
ловой диморфизм выражен очень сильно, перекрест между
X- и Y-хромосомами происходит сравнительно редко. Не
давно обнаружен случай, когда формы YY оказались не
жизнеспособными [Winge, Ditlevsen, 1938]. Эволюция по
ловых хромосом — блестящий пример, направленности эво
люционного процесса. Дарлингтон [1932] отмечает, что в
случае генотипического определения пола сходство конеч
ных результатов указывает не на общность происхожде
ния, а на сходство механических условий возникновения.
Один шаг эволюции с неизбежностью ведет за собой следу
ющий. Образование генов, реализаторов пола, ведет к
дифференциации половых хромосом — систем генов, пере
крест между которыми затруднен. Подавление перекреста
ведет к опустению и, наконец, к полному исчезновению
одного из гомологов и к образованию различий в генном
балансе между наборами хромосом самца и самки. Роль
отдельных генов в реализации половых отличий отступает
на задний план. Будет ли реализован тот или другой пол,
зависит теперь от соотношения между количеством поло
вых хромосом и количеством аутосом. Когда Y-хромосома
исчезла, нет генов, которые присутствовали бы только у
одного пола и отсутствовали бы у другого. Реакция ткани
на действие реализатора зависит теперь не от наличия спе
цифических генов мужского и генов женского пола, а от
различий доз генов половой хромосомы у самца и у самки.
По-видимому, одни и те же гены могут выступать как
мужские в одной дозе и как женские в другой. Опыты Добжанского и Шульца [Dobzhansky, Schultz, 1934], Паттер
сона и сотрудников [Patterson et al., 1935, 1937], Варшавер
[1940] и С. Бедичек-Пипкин [Bedichek-Pipkin, 1940] пока
зали, что прибавление фрагментов Х-хромосомы к набору
самца или интерсекса Drosophila сдвигает пол в сторону
самки, причем степень сдвига пропорциональна длине
46
фрагмента. Вслед за Бриджесом [Bridges, 1921, 1922, 1925,
1935, 1939], который изучал действие прибавления целых
хромосом, они объясняют это локализацией женских моди
фикаторов в половой хромосоме самки. Эти опыты, однако,
могут быть истолкованы в том смысле, что гены, действо
вавшие как мужские в редуцированной дозе начинают дей
ствовать как женские, когда доза их уравнивается с дозой
генов аутосом.
Прибавление участков Х-хромосомы выявляет способ
ность генов Х-хромосомы дать качественно другой эффект
в изменившихся условиях генотипа, в измененной дозе.
Оно совсем не доказывает превалирования специфических
женских модификаторов в половых хромосомах. Изучение
влияния гипо- и гиперплоидии по фрагментам аутосом на
самцов и самок не позволило выявить преобладания муж
ских модификаторов в аутосомах [Patterson et al., 1940].
Сравнение действия гипо- и гиперплоидии по участкам
Х-хромосомы с действием гипо- и гиперплоидии по участ
кам аутосом [Bedichek-Pipkin, 1940; Burdette, 1940;
Patterson et al., 1940] показывает, что изменение количе
ства материала половых хромосом сказывается в большей
мере на сдвиге полового типа и в меньшей мере на жизне
способности особей, чем изменение количества материала
аутосом. Это указывает нам на то, в каком направлении
действовал отбор. Он подавлял действие изменения дозы
генов Х-хромосомы на жизнеспособность (компенсация до
зы генов Х-хромосомы) [Muller, 1932b, 1935] и использо
вал, фиксировал и усиливал действие изменения дозы ге
нов Х-хромосомы на пол (передифференцировка генов).
Доза генов аутосом оставалась без изменения, и отбор не
мог действовать так же на них, как на гены Х-хромосомы.
Внутрихромосомный баланс в Х-хромосоме [Bridges, 1925;
Muller,
1930 ]> по-видимому, более развит, чем в
аутосомах.
Точно те же выводы можно сделать, изучая распреде
ление леталей и мутаций стерильности по хромосомам
Z). melanogaster [Берг, 1934, 1938; Berg, 1937, а—с]. Под
влиянием х-лучей в аутосомах чаще возникают летальные
мутации, чем мутации стерильности, в Х-хромосоме нао
борот. Частота возникновения мутаций стерильности в
Х-хромосоме больше, чем в одной из больших аутосом (во
второй хромосоме), а быть может, чем во всех хромосомах,
взятых вместе [Berg, 1937b, с; Prabhu, 1939]. Это также
указывает, что роль Х-хромосомы в процессах детермина
ции пола больше, чем роль аутосом. Эти данные ставят
под сомнение преобладание «мужских модификаторов» в
47
аутосомах. Мною была сделана попытка определить, пре
обладают ли в аутосомах мутации стерильности, ограни
ченные мужским полом, над мутациями стерильности, ог
раниченной женским полом [Берг, 1938]. Изучено 23 ре
цессивных мутаций стерильности, возникшие во 2-й хро
мосоме под влиянием Х-лучей. 21 затрагивала оба пола,
одна — только самок и одна — только самцов. Кроме того,
из природной популяции Никитского сада выделена 21 ли
ния, содержавшая рецессивную мутацию в 3-й хромосоме
(11,2 % от общего числа хромосом). Изучение идентично
сти мутаций показало, что они представляют собой изме
нения 11 различных локусов. Из 11 разных мутаций четы
ре затрагивают только самок, две — и самцов и самок, од
на — самок в сильной степени, самцов с слабой, и четыре
затрагивают больше самцов, чем самок. Из 11 возможных
случаев самки затронуты 11 раз; 7 раз сильно (полная сте
рильность) и 4 раза слабо (понижение плодовитости); сам
цы 7 раз — 6 раз сильно и один раз слабо. Никакого пре
обладания «мужских модификаторов» обнаружить не уда
лось. Прэбью [Prabhu, 1939] изучал частоту возникнове
ния доминантных, ограниченных одним полом, мутаций
под влиянием х-лучей у Z). melanogaster. Он пришел к вы
воду, что доминантные мутации стерильности, ограничен
ные в своем проявлении одним полом, встречаются в ауто
сомах очень редко.
Мутации стерильности, ограниченные одним полом
(женским), найдены мною в Х-хромосоме. Частота их воз
никновения под влиянием х-лучей 7,2 ± 1 , 9 % [Берг,
1938]. Частота возникновения леталей в том же опыте
19,2 %, а мутаций стерильности, затрагивающих самца —
18,8 + 2,6 % (затрагивают ли эти мутации также самку —
неизвестно). В Х-хромосоме установлено большое количе
ство мутаций стерильности, проявляющих свое действие в
самце. Затрагивают ли эти мутации также и самок — не
известно, так как получить гомозиготных самок по мута
ции, которая стерилизует самца, невозможно без примене
ния специальной методики. О соотношении мутаций, влия
ющих различно на самцов и самок в Х-хромосоме, нам по
ка ничего не известно. Но зато нам известно, что «муж
ские модификаторы» не преобладают в аутосомах. Этого
одного достаточно, чтобы усомниться в том, что «женские
модификаторы» преобладают в половой хромосоме. Но тог
да нет необходимости и принимать существование специ
фических «мужских» и «женских» модификаторов у форм,
где выработались различия в генном балансе между набо
рами хромосом самца и самки. У этих форм реализация
48
пола зависит от различий в концентрации реализующих
пол веществ, а не в количественном преобладании «муж
ских» генов над «женскими» или наоборот [Aida, 1936;
1>ерг, 1938]. Те же самые гены, которые в зависимости от
дозы выступают как реализаторы мужского и женского по
ла, определяют и исход реализации пола,— они действуют
как женские в одной генотипической среде, как муж
ские — в другой. Реализация пола — это начальный этап
развития мужского или женского пола. Мутации, действу
ющие сильнее на реализацию полов, чем различия в ген
ном балансе, конечно, могут возникать. У Drosophila опи
саны такие мутации [Sturtevant, 1921а, b; Muller, 1928;
Lebedeff, 1938]. Однако нет оснований предполагать, что в
норме реализация пола зависит преимущественно от нор
мальных аллеломорфов этих мутаций.
6. РАЗЛ И ЧНА Я РОЛЬ РАЗН Ы Х УЧАСТКОВ Х-ХРОМОСОМЫ
В РЕА Л И ЗА Ц И И ПОЛОВЫХ ОТЛИЧИЙ
Невозможность обнаружить преобладание «мужских
модификаторов» над «женскими модификаторами» в аутосомах Drosophila заставила нас отказаться от представле
ния, что реализация пола у форм с различиями в генном
балансе зависит от количественного преобладания женских
модификаторов у самки, мужских — у самца. Реализация
пола зависит от различий доз генов половой хромосомы по
отношению к дозам генов аутосом у самца и у самки. Дан
ные по влиянию гиперплоидии по участкам Х-хромосомы
на пол (сдвиг в женскую сторону) можно рассматривать
как доказательство того, что одни и те же гены, которые
действовали как мужские в одной дозе, начинают действо
вать как женские, когда их доза изменилась по отношению
к дозе генов аутосом. Отсюда следует, что нет надобности
различать специфические мужские и женские модификато
ры. Точно так же нет надобности и различать у этих форм
гены-реализаторы и гены сексуальности. Одни и те же ге
ны реализуют различие в зависимости от различий доз, и
от них же зависит и исход реализации — развитие всего
комплекса признаков, отличающих самца от самки. Это,
однако, не означает, что все гены принимают одинаковое
участие в реализации пола. Недавно Варшавер [1940] де
тально изучала влияние гиперплоидии по левому концу
Х-хромосомы на половые признаки самца D. simulans. Она
показала, что дупликации, не включающие район бульбуса, оказывают сильное действие на внутренние половые
органы самца, но не затрагивают наружные гениталии.
49
Дупликации, включающие районы бульбуса, изменяют на
ружные гениталии. Степень изменения внутренних половых
органов остается при этом без изменения. Варшавер объяс
няет это различие специфическим влиянием разных участ
ков хромосомы на разные признаки пола.
Если различие в балансе генов половых хромосом и
аутосом устанавливалось постепенно, не все участки хро
мосом одновременно оказывались представленными в раз
ных дозах у разных полов. Наиболее быстро возникнове
ние различий в генном балансе должно было совершаться в
тех районах, где перекрест подавлен — в районах инвер
сий, у нити веретена и в дуплицированных участках. Те
районы, которые раньше других стали на путь передифференцировки, должны быть наиболее активны в процессах
реализации пола и осуществления половых отличий. Поэ
тому есть основания предполагать, что и внутри Х-хромосомы между разными ее участками существует то же раз
личие, что и между половой хромосомой в целом и аутосомами. В районах дуплицированных участков Х.-хромосомы
должны были быть особенно благоприятные условия для передифференцировки генов в сторону их большого участия
в процессах реализации пола и осуществления половых от
личий.
Дупликации являются источником образования «сво
бодных генов». Передифференцировка генов, которая ле
жит в основе возникновения генов, связанных с определе
нием пола, совершается, по всей вероятности, за счет воз
никновения качественно новых функций генов, т. е. требу
ет образования «неоморфных» или «антиморфных» мута
ций (по терминологии Мёллера [ 1932b ]), а в этом случае
наличие нескольких нормальных аллеломорфов не может
препятствовать выявлению мутаций. Вместе с тем, нару
шение старых функций генов при возникновении мутаций
компенсировано наличием сходных генов в дуплицирован
ном участке. Кроме того, при возникновении дупликации в
половых хромосомах сразу возникает целая группа генов,
между которыми перекрест даже в гомозиготе понижен в
силу конфликта сил притяжения, и которые присутствуют
в различной дозе у самца и у самки. После того, как пере
крест полностью подавлен, и Y-хромосома опустела, райо
ны вновь возникающих дупликаций уже не представляют
столь существенных преимуществ для отбора генов, участ
вующих в дифференциации полов.
Наши эксперименты подтверждают предположение от
носительно различного участия разных участков Х-хромосомы в осуществлении половых отличий [Берг, 1941]. Ло
50
кализация мутаций стерильности, возникших под влияни
ем х-лучей, показала, что они распределены неравномерно
по длине хромосомы. Они превалируют в проксимальной
части хромосомы. Из 45 мутаций стерильности левее локу
са vermilion (середина хромосомы) расположено 6 (или
13,3 %), правее — 39 (86,7 %). В районе, прилежащем к
нити веретена, мутации стерильности особенно многочис
ленны. Из 20 мутаций стерильности, положение которых
определено с помощью линии sc v f car 8 расположены
между геном саг и местом прикрепления нити веретена.
Эти 8 мутаций не связаны с разломом в правой части
Х-хромосомы («малые перестройки» не могли быть учте
ны). По-видимому, в районе, прилежащем к месту при
крепления нити веретена, имеется скопление генов, спо
собных давать мутации стерильности. Другое скопление
мутаций стерильности расположено, по-видимому, в сере
дине хромосомы между генами vermilion и forked. Пробью
[Prabhu, 1939] также обнаружил скопление мутаций сте
рильности в середине Х-хромосомы.
Есть основания предполагать, что большое количество
мутаций стерильности, локализованных в середине Х-хро
мосомы, приурочено к району дупликации. Мёллер и
Пайнтер [Muller, Painter, 1932] нашли, что летальные му
тации в середине хромосомы возникают реже, чем в дру
гих районах Х-хромосомы. Косиков [1936] цитологически
доказал существование дупликации в этом же районе. Ха
рактер конъюгации этой дупликации односторонне ветви
стый, и конъюгация происходит очень редко. По-видимо
му, дупликация возникла очень давно, и вовлеченные в
нее гены почти полностью утеряли свою гомологию. В те
чение дальнейшей эволюции половых хромосом различие
между целыми участками Х-хромосомы должно сгладиться.
Те различия по длине Х-хромосомы, которые мы находим
сейчас,— это отражение ее предшествующей истории, того
состояния, когда она еще мало отличалась от своего гомо
лога — Y-хромосомы.
выводы
1.
Приспособительное значение полового диморфизма
основано на разделении функций в процессе размножения,
которое создает возможность расширить среду обитания.
Более совершенные способы заботы о потомстве понижают
интенсивность отбора. Это, однако, не сопровождается за
медлением темпа эволюции, т. е. половой диморфизм од
51
новременно создает предпосылки для ускорения темпа эво
люции. Это ускорение зависит: 1) от повышения наследст
венной пластичности вида в результате сочетания разных
генотипов и перекомбинации мутаций и 2) от возникнове
ния различий в интенсивности отбора самцов и самок. Со
кращение воспроизводящей части популяции идет, глав
ным образом, за счет самцов. Этому способствуют бой сам
цов, гаплоидность самцов, протерандрия, непроявление
подражательной окраски у самцов многих видов, где самки
являются подражательницами, избыток самцов и т. п.
Большая интенсивность отбора самцов по сравнению с сам
ками ведет к тому, что производителями следующего поко
ления оказываются наиболее приспособленные особи, без
того, чтобы от этого снизилась общая численность вида.
В практике сельского хозяйства эта особенность поло
вого диморфизма широко используется.
2. Приспособительное значение полового диморфизма
связано с определенным соотношением между полами. По
стоянное соотношение между формами может осуществить
ся, когда реализация признаков, на которых основан поли
морфизм, независима от колебания внешних условий. Реа
лизация с помощью наследственных факторов, расщепляю
щих в каждом поколении и создающих альтернативную
внутреннюю среду, или с помощью различий в генном ба
лансе является поэтому тем способом реализации, при ко
тором преимущества полового диморфизма могут про
явиться наилучшим образом.
3. Можно наметить три основных этапа эволюции пола:
1) роль реализатора пола принадлежит внешним условиям
или плазматическим элементам клетки; 2) возникновение
генов-реализаторов мужского и женского пола в гомологи
ческой паре хромосом; подавление перекреста между ни
ми; образование двух систем генов, перекрест между кото
рыми подавлен; 3) подавление перекреста ведет к опусте
нию одного из гомологов и к образованию различий в ген
ном балансе между наборами хромосом самца и самки. Это
различие и становится реализатором пола. Реакция ткани
на действие реализатора зависит не от различного соотно
шения «женских» и «мужских» модификаторов, а от раз
личного действия генов половой хромосомы у самца и у
самки в разной дозе.
4. Сравнение действия гипо- и гиперплоидии по участ
кам Х-хромосомы с действием гипо- и гиперплоидии по
участкам аутосом показывает, что изменение количества
материала половых хромосом сказывается в большой мере
на сдвиге полового типа и в меньшей мере на жизнеспо
52
собности особей, чем изменение количества материала
аутосом [Bedichek-Pipkin, Burdette, 1940; Patterson et al.,
1940]. Это указывает нам на то, в каком направлении дей
ствовал отбор. Он подавлял действие изменения дозы генов
Х-хромосомы на жизнеспособность (компенсация дозы ге
нов Х-хромосомы) [Muller, 1932b, 1935] и с такой же не
обходимостью использовал, фиксировал и усиливал дейст
вие изменения дозы генов Х-хромосомы на пол (передифференцировка генов).
Точно те же выводы можно сделать, изучая распреде
ление по хромосомам летал ей и мутаций стерильности,
возникших под влиянием х-лучей [Берг, 1934, 1938; Berg,
1937а, Ь, с]. Попытка выявить преобладание мутаций сте
рильности, ограниченных мужским полом в аутосомах
[Берг, 1938], не увенчалась успехом. Никакого преоблада
ния мужских модификаторов в аутосомах нет. Это вынуж
дает меня отказаться от представления, что пол определя
ется соотношением специфических мужских и женских мо
дификаторов.
У форм, где установились стойкие различия в балансе
между половыми хромосомами и аутосомами, нет необхо
димости принимать и различие между генами сексуально
сти и генами-реализаторами, как это делает Аида [Aida,
1936]. Те же самые гены, которые выступают в зависимо
сти от дозы как реализаторы мужского и женского пола,
определяют и исход реализации,— они действуют как
женские в одной дозе, как мужские — в другой. Доза ге
нов Х-хромосомы измеряется, конечно, по отношению к
дозам генов аутосом — это относительная, а не абсолют
ная величина.
5.
Различие между дозами генов у самца и у самки
возникло в течение эволюции, по всей вероятности, не од
новременно для разных участков Х-хромосомы. Наиболее
быстро возникновение различий в генном балансе должно
совершаться в тех районах, где перекрест подавлен,— в
районах инверсий, у нити веретена и в районах дуплика
ций. Эти районы являются очагами наиболее быстрой диф
ференциации Х-хромосомы. При возникновении дуплика
ций по участкам Х-хромосомы сразу возникает целая груп
па генов, между которыми перекрест понижен, и которые
присутствуют в различной дозе у самца и у самки.
Локализация мутаций стерильности, возникших под
влиянием х-лучей в половой хромосоме D . melanogaster
[Берг, 1941] показала, что мутации стерильности распре
делены неравномерно по длине Х-хромосомы. Они преоб
ладают в середине хромосомы и у нити веретена. Есть ос
53
нования предполагать, что большое количество мутаций
стерильности, локализованных в середине Х-хромосомы,
приурочено к району дупликации, обнаруженной Косиковым [1936].
Неравномерное распределение мутаций стерильности в
половой хромосоме есть результат действия естественного
отбора в процессе эволюции половых хромосом.
6.
Эволюция механизма реализации пола совершается
сходно в самых различных группах органического мира.
Она представляет собой блестящий пример направленности
эволюционного процесса. Каждый шаг эволюции пола кон
тролируется естественным отбором, который протекает на
основе межгруппового соревнования.
СПИСОК ЛИТЕРАТУРЫ
Берг Р. Л. / / Докл. АН СССР. — 1934.— Т. 4, № 4 . - С. 234— 236.
Берг Р. Л. / / Тр. Ленингр. О-ва естествоисп.— 1938.— Т. 67.—
С. 96— 108.
Берг Р. Л., Бриссенден Э. Б., Александрийская В. Т., Галков
ская К. Ф . / /
Журн. общ. биол.—
1941.— Т. 2, № 1.—
С. 143— 158.
Варшавер Н. Материалы по генетике пола. Дис.
канд. биол. наук.—
1940.
Дарвин Ч. Происхождение видов путем ёстественного отбора.— М.: Издво АН СССР, 1939.— Т. 3 .— 831 с.
Карпентер Г. Д. X., Ф орд Э. Б. Мимикрия.— М.; Л., 1935.
Кирпичников В. С. / / Биол. журн.— 1935.— Т. 4 .— С. 343— 353.
Косиков К. В. / / Докл. АН СССР.— 1936.— Т. 3 (1 2 ).— С. 297— 300.
Рыжков В. Л. Генетика пола.— Киев: Госмедиздат УССР, 1936.
Северцов А. Н. Морфологические закономерности эволюции.— М.; Л.,
1939.
Северцов С, А, / / Рефераты работ учрежд. Отд. биол. наук АН СССР.—
М.: Изд-во АН СССР, 1940.— С. 218.
Ш мальгаузен И. И. Пути и закономерности эволюционного процесса.—
М.; Л., 1939.
Ш мальгаузен И. И. / / Журн. общ. биол.— 1940.— Т. 1, № 4 .—
С. 5 09—528.
Aida Т. / / Genetics.— 1936.— Vol. 2 .— Р. 136— 153.
Allen Ch. Е. / / Science.— 1917.— Vol. 46.— P. 466.
Allen Ch. E. / / Amer. Nat.— 1932.— Vol. 66.— P. 97— 117.
Bedicher-Pipkin S. / / Univ. Texas Publ.— 1940.— N 4032.— P. 126— 156.
Berg R. L. / / Genetics.— 1937a.— Vol. 2 2 . - P. 225— 240.
Berg R. L. / / Genetics.— 1937b.— Vol. 22.— P. 241— 248.
Berg R. L. / / Genetics.— 1937c.— Vol. 22.— P. 402—405.
Bridges С. B. / / Science.— 1921.— Vol. 5 4 .— P. 252— 254.
Bridges С. B. / / Amer. Nat.— 1922.— Vol. 5 6 .— P. 5 1 — 63.
Bridges С. B. / / Amer. Nat.— 1925.— Vol. 5 9 .— P. 127— 137.
Bridges С. B. / / J. Hered.— 1935.— Vol. 26.— P. 60—64.
Bridges С. B. Sex. and Internal Secretion.— 2d ed. — 1939.— P. 15— 63.
Burdette N. J. / / Univ. Texas Publ.— 1940.— N 4032.— P. 157— 163.
Correns C. Bestimmung, Vererbung und Verteilung des Geschlechtes bei den
hoheren Pflanzen / / Hand. d. Vererbungswissenchaft.— 1928.—
Bd 11.
Grew F. A. E. The genetics of sexuality in animals.— Cambridge, 1927.
Darlington C. D. / /R e c e n t Advances in Cytology.— L., 1932.
54
Darlington C. D. / / Amer. Nat.— 1934.— Vol. 68.— P. 1— 4.
Darwin C. The Descent of Man, and Selection in Relation to Sex.— L., 1871.
Demoll R. / / Zool. Jahrd. (Syst.).— 1908.— Vol. 26.— P. 621— 628.
Dobzhansky Th. / / Biol. Zbl.— 1930.— Vol. 50.— P. 671—685.
Dobzhansky Th., Schaltz J. / / J. Gen.— 1934.— Vol. 28.— P. 349— 386.
Emerson R. A . / / Proc. VI Int. Congr. Gen.— 1932.— Vol. 1.—
P. 141 — 152.
Fischer R. A. / / Amer. Nat.— 1930a.— Vol. 64.— P 385—406.
Fischer R. A. / / The Genetical Theory of Natural Selection.— Oxford,
1930b.
Fischer R. A . / / Proc.
VI Int. Congr. Gen.— 1932.— Vol. 1.—
P. 165— 173.
Fischer R. A., Ford E. B. / / Trans. Ent. Soc. London.— 1929.— Vol. 76.—
P. 367— 384. (With an Appendix by E. B. Poulton).
Ford E. B. / / Trans. Royal Ent. Soc. London.— 1936.— Vol. 85.—
P. 335— 366.
Freyer J. P. / / Linn. Phil.— 1913.— N 204.— P. 227— 254.
Jager G. In Sachen D am n 's.— Stuttgart, 1874.
Jones D. E. / / Science.— 1931.— Vol. 2 .— P. 104— 107.
Kaufmann E. P. / / Proc. Nat. Acad. Sci.— 1933.— Vol. 19.— P. 830—838.
Lebedeff G. A. / / Proc. Nat. Acad. Sci— 1938.— Vol. 20.— P. 613— 616.
Mather K. / / GeneticS.— 1939.— Vol. 24, N 3.— P. 413—435.
M uller H. J. / / Genetics.— 1918.— Vol. 3.— P. 422—489.
M uller H. J. / / Verhandlungen des V Int. Kongress Zeitschrift fur Induktive
Abstammungs und Vererbungslehre.— Suppl. 1.— 1928.— S. 234—260.
M uller H. J. / / J. G en.— 1930.— Vol. 22.— P. 299— 334.
M uller H. J. / / Amer. Nat.— 1932a.— Vol. 66.— P. 118— 138.
M uller H. J. / / Proc. VI Int. Congr. G en.— 1932b.— N 1.— P. 213— 255.
M uller H. J. / / J. G en.— 1935.— Vol. 30, N 3 .— P. 407—414.
M uller H. J., G erschenson S. / / Proc. Nat. Acad. Sci. Wash.— 1935.—
Vol. 2 1 .— P .— 69— 75.
M uller H. J., Painter T. S. / / Verhandlungen des V Int. Kongress Zeitschrift
fur Induktive Abstammungs und Vererbungslehre.— 1930.— Bd 62.—
S. 316— 365.
Offermann C, A., M uller H. J. / / Proc. VI Int. Congr. Gen.— 1932.—
Vol. 2 .— P. 143— 145.
Patterson J. T., Stone W., Bedichek S. / / Genetics.— 1935.— Vol. 20.—
P. 259— 279.
Patterson J. T., Stone W., Bedichek S. / / Genetics.— 1937.— Vol. 22.—
P. 407— 426.
Patterson J. T., Suche M., Brown, Stone W. / / Univ. Texos Publ.— 1940.—
N 4032.— P. 167— 189.
Philips U. / / Nature.— 1934.— N 3367.
Plate L. Selectionsprinzip und Probleme der Artbildung.— 4 Aufl.— Leipzig;
Berlin, 1913.
Prabhu S. S. / / J. G en.— 1939.— Vol. 28, N 1 /2 .— P. 177— 191.
Schrader
F. / /
Zellen
und
Befruchtungslehre,
I.
Die
Geschlechtschromosomen.— Berlin, 1928.
Schull A, F. / / Genetics.— 1917.— Vol. 2 .— P. 480—488.
Snell G. D. / / Amer. Nat.— 1932.— Vol. 66, N 705.— P. 381— 384.
Stern C. / / Verhandlungen des V Int. Kongress Zeitschrift fur Induktive
Abstammungs und vererbungslehre.— 1929.— Bd 5 1 .— S. 253— 353.
Stern C., Ogura S. / / Verhandlungen des V Int. Kongress Zeitschrift fur Induk
tive Abstammung und Vererbungslehre.— 1931.— Bd 58.— S. 81 — 121.
Stolzmann J. / / Proc. Zool. London.— 1885.— N 3 .— P. 421— 432.
Sturtevant A. H. / / Science.— 1921a.— Vol. 5 1 .— P. 325—327.
Sturtevant A. H. / / Genetics.— 1921b.— Vol. 6 .— P. 179— 207.
Wallace A. R, / / Trans. Linn. Soc. London.— 1985.— Vol. 25, N 1.—
P. 1— 71.
Wallace A. R. Der Darwinismus.— Braunschweig, 1891.
W inge b . / / J. G enet.— 1927.— Vol. 18.— P. 1—43.
W inge b . / / C. R. Lab. Carlsb.— 1934.— Vol. 21,— P. 1—49.
W inge b ., D itlevsen E. / / C. R. Lab. Carlsb.— 1938.— Vol. 22.— P.
203— 210.
55
ГЕТЕРОЦИКЛИЧНОСТЬ ХРОМОСОМ
И ИХ КОМПЛЕКСОВ КАК МЕХАНИЗМ,
ОБЕСПЕЧИВАЮЩИЙ ПОСЛЕДОВАТЕЛЬНОСТЬ
ВСТУПЛЕНИЯ В ДЕЙСТВИЕ
ОТДЕЛЬНЫХ ЭЛЕМЕНТОВ
КОДА НАСЛЕДСТВЕННОЙ ИНФОРМАЦИИ*
Р. Л . Берг
Клетки разных тканей многоклеточного организма, как
бы различны они ни были, имеют одинаковые наборы хро
мосом, а следовательно, и одинаковые наследственные за
датки. К числу редких исключений относятся мошки
Sciara , у которых пара так называемых L-хромосом элими
нируется из соматических клеток. (Обзор соответствующих
исследований см.: White [1948].) Очевидно, существуют
механизмы, обеспечивающие последовательность вступле
ния в действие разных задатков в разных тканях.
Ж. А. Медведев [1965] в докладе на Симпозиуме по меди
цинской генетике выдвинул остроумные гипотезы относи
тельно биохимических механизмов, выключающих из дей
ствия отдельные участки генонемы.
Мне хотелось бы осветить тот же вопрос с цитологиче
ской точки зрения.
Хромосомы редуплицируются в интеркинезе, т. е. стро
ят рядом с собой своих гомологов, затем в фазе деления
клетки хромосомы спирализуются, одеваются хроматиновым футляром, проделывают ряд маневров, обеспечиваю
щих расхождение материнской и дочерней хромосомы каж
дой пары в разные клетки.
Начиная свое существование в новой клетке, хромосо
мы сбрасывают хроматиновый футляр и деспирализуются.
Деспирализованные хромосомы осуществляют свою органи
зующую функцию и редуплицируются.
Цитологические наблюдения показывают, что разные
участки хромосом в разное время вступают в цикл клеточ
ного деления. По данным Риса [I960], полностью развер
нутыми по всей длине хромосомные нити не бывают ни
когда.
Разновременное вступление в стадию митоза или диф
ференциальная спирализация разных участков хромосом и
представляет собой тот цитологический механизм, который
обеспечивает последовательность выключения из действия
отдельных элементов кода наследственной информации.
* Проблемы медицинской генетики.— М.: Медицина, 1965.
56
Мысль эта была впервые высказана Шульцем [Schultz,
1952] в 1952 г. Шульц считает, что разные участки хромо
сом в разное время вступают в действие, претерпевая
местную деспирализацию и полиплоидизацию. Спирализация хромосом переводит их в гетерохроматическое состоя
ние, в котором они теряют свою специфическую функцию.
Гетерохроматизация может быть как обратимой, так и не
обратимой. Гольдшмидт [Goldschmidt, 1955] подверг эту
гипотезу жесточайшей критике. Согласно его мнению, экс
периментальные данные, которые нуждались бы в соответ
ствующей интерпретации, отсутствуют. Однако точка зре
ния Шульца получила, как мы увидим, новое подтвержде
ние. Многочисленные наблюдения над половым хромати
ном, над структурами «ламповых щеток» в овогенезе птиц
и амфибий и над вздутиями на политенных хромосомах
двукрылых находят единое объяснение в рамках гипотезы
Шульца относительно разновременного включения в дейст
вие и выключения из действия разных районов хромосом
или целых хромосом с помощью дифференциальной деспирализации и эухроматизации.
Существование этого механизма, само собой разумеет
ся, не исключает других, чисто биохимических механизмов
и само требует биохимической интерпретации.
Разновременное вступление в митоз разных участков
хромосомного набора названо А. А. Прокофьевой-Бельговской [1948] гетероцикличностью системы клеточного ядра.
Гетероцикличность можно подразделить на внутрихромосомную, межхромосомную и межгеномную.
Внутри хромосомная
гетероциклич
ность
ярко выявляет себя в политенных хромосомах
двукрылых (Diptera). Хромосомы эти находятся в пикнотическом состоянии. Они как бы фиксированы на стадии ран
ней профазы. Сравнение структуры этих хромосом в раз
ных тканях и на разных стадиях жизненного цикла пока
зало, что отдельные участки хромосом деспирализуются,
увеличиваются в объеме как за счет удлинения, так и за
счет дальнейшего увеличения числа нитей в пучке, обра
зуя вздутия. Процессы эти носят обратимый характер и
строго локализованы по времени и в пространстве
[Beermann, 1952; Mechelke, 1953, 1958; Pavan, 1958].
Другой пример внутрихромосомной гетероциклично
сти — последовательное образование боковых петель в
овогенезе амфибий и птиц [Кольцов, 1938; Callan, Gall,
1962; Gall, Callan, 1962]. Изучение этих вздутий и петель
цитологическими методами и с помощью введения мече
ных предшественников ДНК, РНК и белков и посдедую57
щей авторадиографии показало, что в районе вздутия и пе
тель хромосома включает метку. Подводя итог работам,
касающимся различий во времени и в пространстве актив
ности разных участков хромосом, Дарлингтон [Darlington,
1958] пишет: «Структура хромомер меняется в течение
развития. Эти изменения обратимы и происходят независи
мо в разных хромомерах. Другими словами, каждая хромо
мера имеет свой генетически обусловленный тип развития,
или свою серию реакций. Эти реакции осуществляются
между всеми активными в данный момент генами и между
этими генами и цитоплазмой» (с. 69).
М ежхромосомная
гетероциклич
н о с т ь выявляет себя в наличии полового хроматина у
самок млекопитающих. Одна из двух Х-хромосом находит
ся у самки в метаболических (интеркинетических) ядрах
соматических клеток в спирализованном состоянии и обла
дает способностью окрашиваться, в то время как другие
хромосомы деспирализованы [Зыбина, 1959; Barr, Carr,
1962]. У многих организмов, в частности у большинства
грызунов, у змей, амфибий, некоторых моллюсков половой
хроматин маскируется наличием окрашенных участков
аутосом [Moore, 1962].
М е ж г е н о м н а я г е т е р о ц и к л и ч н о с т ь , или раз
новременное прохождение митотического цикла хромосо
мами материнского и отцовского комплексов, вскрыта
А. А. Прокофьевой-Бельговской
[1948].
Великолепные
примеры различного поведения отцовского и материнского
наборов хромосом можно наблюдать в сперматогенезе сам
цов Sciara coprophila и у некоторых видов коккцид.
В сперматогенезе самцов Sciara наблюдается элимина
ция одного из наборов хромосом [Metz, 1938]. У коккцид в
семействе Diapsidae наблюдается элиминация отцовского
набора хромосом на ранних стадиях эмбриогенеза у сам
цов. В группе Lecaaoidae также только у самцов один из
наборов хромосом инактивируется в эмбриогенезе, претер
певая гетерохроматизацию, а затем элиминируются в спер
матогенезе. Как генетические данные [Dickson, 1941], так
и наблюдения над фрагментированными хромосомами в
потомстве скрещиваний облученных самцов с необлученными самками и у потомков облученных самок, скрещен
ных с необлученными самцами, показывает, что элимина
ции подлежит именно отцовский комплекс хромосом
[Brown, Bennett, 1957; Brown, 1958, 1959].
Внутрихромосомная гетероцикличность обеспечивает
действие должных генов на должных местах и в надлежа
щее время. Межхромосомная гетероцикличность, касающа
58
яся половых хромосом, поддерживает, кроме того, соотно
шение половых хромосом и аутосом и нивелирует излиш
ние различия между самцом и самкой. Межгеномная гете
роцикличность, или более позднее вступление в действие
отцовского комплекса хромосом, привносимого спермием в
метафатическом состоянии, по сравнению с материнским
комплексом, который находится в момент кариогамии на
стадии ранней профазы, имеет, на наш взгляд, несколько
иное и притом многообразное значение.
Межгеномная гетероцикличность может служить регу
лирующим механизмом на популяционном и на онтогене
тическом уровне организации живого. Мыслимы следую
щие регулирующие функции разновременного вступления
в действие отцовского и материнского геномов:
1.
Межгеномная гетероциклич юсть выступает как ре
гулятор генотипического состава популяций, позволяя со
четать преимущества диплоидного состояния с преимуще
ствами гаплоидного состояния, делая организм как бы про
межуточным по валентности ядра между диплоидным и
гаплоидным. До вступления в действие отцовского набора
функционально активен один материнский набор. Вредные
мутации, привнесенные матерью, могут отметаться отбо
ром. Эта гибель генотипически неполноценных особей на
возможно более ранней стадии эмбриогенеза имеет значе
ние у тех видов животных, где забота о потомстве связана
с большой затратой сил или, в силу каких-либо обстоя
тельств, жестко лимитирована (пчелы, птицы, млекопита
ющие). Всякое понижение интенсивности отбора, создавая
угрозу накопления генотипически неполноценных особей,
сочетается с механизмами, повышающими интенсивность
отбора или переносящими элиминацию на возможно более
раннюю стадию онтогенеза. Предотвратить массовое появ
ление обреченных на гибель гомозигот можно с помощью
принудительной гемизиготизации, которая и обеспечивает
ся на некоторый срок гетероцикличностью. Благоденствие
вида покупается дорогой ценой: сохраниться в межвидовом
соревновании могут лишь виды, активно регулирующие
свои взаимоотношения со средой и предотвращающие на
копление генотипически неполноценных особей с помощью
специальных механизмов. В поколениях пчел таким биоевгеническим приспособлением служит принудительная гомозиготизация всех самцов; трутни развиваются из неоплодотворенных яиц. Диплоидный набор хромосом восста
навливается затем с помощью эндомитоза [Risler, 1954].
Гетероцикличность отцовского и материнского наборов
хромосом — временный аналог гаплоидного партеногенеза.
59
Ранние стадии эмбриогенеза находятся под контролем
гаплоидного набора хромосом матери. Питающие клетки
в яичниках насекомых с их гигантскими политенными яд
рами изливают свое содержимое в созревшую яйцеклетку.
Они усиливают контроль над развитием зародыша со сто
роны материнского ядра, умеряя значение межгеномной
гетероцикличности в качестве биоевгенического фактора.
2.
Межгеномная гетероцикличность позволяет сочетать
преимущества партеногенетического размножения с пре
имуществами двуполого размножения. При двуполом раз
множении создается угроза, что белки семян, развиваю
щихся на материнском растении, или белки плода, созре
вающего в утробе матери, окажутся несовместимыми с
белками материнского организма. При партеногенетическом размножении такой опасности нет. Более позднее
вступление в действие отцовских генов уменьшает эту
опасность.
В обоих случаях значение межгенной гетероциклично
сти оказывается в системах, включающих организм, как
подчиненный элемент системы (система мать — плод,
семья, колония). Помимо этого, межгеномная гетероцик
личность имеет значение и на онтогенетическом уровне.
а. Межгеномная гетероцикличность позволяет сочетать
высокий индекс рекомбинаций в половых клетках с подав
лением соматического перекреста. Самостоятельное и, до
известной степени, разновременное прохождение стадий
митотического цикла отцовским и материнским комплекса
ми хромосом уменьшает вероятность сегментного обмена
между хромосомами обоих комплексов.
б. В клетках двукрылых (Diptera), содержащих гигант
ские политенные хромосомы, межгеномная гетероциклич
ность, быть может обеспечивает сохранение порядка в рас
положении хромомер. Благодаря тому, что две половины
пучка, образованные отцовской и материнской хромосома
ми, вступают в процесс образования вздутия неодновре
менно, гигантские хромосомы не располагаются в местах
образования вздутий. Неодновременное участие двух пуч
ков в создании вздутия можно видеть на рисунках тех исс
ледователей, которые работали с гигантскими хромосомами
[Beermann 1952; Pariitz 1960; и др. ]. Сами эти картины
могут служить доказательством межгеномной гетероцик
личности.
в. Межгеномная гетероцикличность способствует умень
шению валентности полиплоидных ядер, способствуя обо
соблению целых геномов.
60
Уместно поставить вопрос, откуда исходят сигналы к
включению в действие или к выключению разных, участ
ков хромосом, целых хромосом и их комплексов?
Образ современных цитологических данных показыва
ет, что сигналы, управляющие поведением отдельных уча
стков хромосом, могут исходить из цитоплазмы, а могут
носить генетический и, наконец, эпигенетический харак
тер. Сигналы эпигенетической природы исходят из внеш
них по отношению к данному участку хромосомы участков
хромосомного набора в зависимости от того, находятся эти
участки в спирализованном или в деспирализованном со
стоянии.
Примером цитоплазматической сигнализации может
служить изменение мистического цикла хромосом нижне
го полярного ядра зародышевого мешка у некоторых пред
ставителей рода Gagea [Романов, 1961].
Образование полового хроматина у человека может
служить примером эпигенетической сигнализации. Поло
вой хроматин присутствует в интеркинетическом ядре, со
матических клеток некоторых тканей женщины и отсутст
вует в тех же тканях у мужчины.
*
Мужчина и женщина отличаются друг от друга: 1)
комплексом физиологических и биохимических признаков;
2) наличием Y-хромосомы только в геноме мужчины; 3)
наличием отцовской Х-хромосомы только в геноме женщи
ны. Ни одно из этих отличий не является причиной отли
чий полов по половому хроматину. Женщина, страдающая
синдромом Тернера— Шерешевского, имеющая одну Х-хромосому (ХО) не имеет полового хроматина, независимо от
того, получила ли она свою Х-хромосому заведомо от отца
(если нерасхождение половых хромосом произошло в онто
генезе матери) или от матери (если нерасхождение про
изошло в сперматогенезе отца). Мужчина с синдромом
Клейнфельтера, имеющий две Х-хромосомы и Y-хромосому
(XXY), имеет половой хроматин также вне зависимости от
того, получил ли он две Х-хромосомы от матери и Y-хро
мосому от отца или унаследовал от матери одну Х-хромо
сому, а от отца две другие хромосомы — X и Y [Barr,
Carr, 1962]. Отсюда следует, что наличие деспирализованной Х-хромосомы служит препятствием к деспирализации
второго партнера. Вступление активной Х-хромосомы в ми
тотический цикл, т. е. спирализация, служит сигналом к
деспирализации гомологичной Х-хромосомы, которая полу
чает возможность редупликации. Двойной половой хрома
тин возникает тогда, когда одна хромосома переходит из
61
активного в пикнотическое состояние, а другая еще не ус
певает полностью сбросить хроматин.
Анализ действия сцепленных с полом генов у человека
[Beutler et ah, 1962] и у мышей [Lyon, 1962; Searl, 1962]
на клеточном уровне показал, что в одних клетках прояв
ляет активность материнская, в других — отцовская хро
мосома. Как показывают данные Лайон по мозаичности ок
раски и структуре шерстного покрова у мышей, гетерози
готных по двум сцепленным с полом признакам, если одна
из Х-хромосом активна в данной клетке, она имеет шанс
проявить свою активность и в дочерних клетках, т. е. опе
режает своего партнера в прохождении цикла спирализации в течение ряда клеточных поколений. Узор шерстного
покрова слагается из крупных пятен.
Генотипический контроль над прохождением хромосо
мами цикла спирализации и деспирализации в мейозе ясно
виден на примере гена «sex-ratio», найденного у несколь
ких видов дрозофил [Stertevant, Dobzhansky, 1936]. В при
сутствии этого гена Y-хромосома дегенерирует, а Х-хромосома претерпевает добавочное деление. В результате спер
мин самца, содержащего ген «sex-ratio», за немногими ис
ключениями, содержат Х-хромосому, и от него родятся
преимущественно самки. Известны и другие не менее за
мечательные случаи генного контроля поведения хромосом
в мейозе.
Цитоплазматический, эпигенетический и генетический
механизмы сигнализации действуют, по-видимому, строго
согласованно и обусловливают действие друг друга. Один и
тот же участок хромосомы может, по-видимому, находить
ся в разное время и в разных клетках под воздействием
различных сигналов. Мы видели, что половой хроматин,
имеющийся в клетках некоторых тканей женщины, по
является в результате воздействия Х-хромосомы, находя
щейся в активном состоянии. Однако и в клетках некото
рых тканей мужчины имеется половой хроматин [Moore,
1962]. Здесь его появление имеет, очевидно другую приро
ду, чем в клетках женщины. По данным Парка [Park,
1957], у зародыша человека половой хроматин в разных
тканях появляется в разное время и с разной частотой.
У эмбрионов крысы наблюдается различие по половому
хроматину между полами. Однако у взрослых это различие
исчезает.
Гетероцикличность клеточного ядра выступает перед
нами как регулятор с разносторонними функциями и, вме
сте с тем, как регулируемый процесс.
62
СПИСОК ЛИТЕРАТУРЫ
Зыбина Е. В. / / Цитология.— 1959.— Т. 1, № 4 ,— С. 357.
Кольцов Н. К. / / Биол. ж урн.— 1938.— Т. 7, № 1.— С. 3.
Медведев Ж . А* / / Проблемы медицинской генетики.— М.: Медицина,
1965.— С. 4 5 — 96.
Прокофьева-Бельговская А. А. / / Журн. общ. биол.— 1948.— Т. 8,
№ 4 .— С. 247.
Рис Г. Химические основы наследственности: Пер. с англ.— М., 1960.—
С. 2 5 - 5 5 .
Романов И. Д. / / Докл. АН СССР.— 1961.— Т. 141, № 4 .— С. 984.
Barr М. L., Carr D. Н. / / Acta Cytologica.— 1962.— Vol. 6, N 1.— Р. 34.
Beermann W. / / Chromosoma.— 1952.— N 5 .— P. 139.
Benett F. D., Brown A. W. / / Canad. Entomol.— 1958.— Vol. 90, N 6 .—
P. 317.
Beutler EM Seh N M Fairbanks V. F. / / Proc. Nat. Acad. Sci. USA.— 1962.—
Vol. 48, N 1.— P. 9.
Brown S. W. / / Evolution.— 1958.— Vol. 12, N 1.— P. 115.
Brown S. W. / / Chromosoma.— 1959.— N 10.— P. 278.
Brown S. WM Benett F. D. / / Genetics.— 1957.— Vol. 49, N 4 .— P. 510.
Callan H. C., Gall J. G. Abstracts of papers on the 138 meeting of the
genetical society.— Univ. of Edinburgh, 1962.
Darlington C. D. Evolution of Genetic Systems.— 2d ed .— Edinburgh;
London, 1958.
Dickson R. S. / / Hilgardia.— 1941.— N 1 3 . - P. 515.
Gall J. G., Callan H. G. / / Proc. Natl. Acad. Sci. USA.— 1962.— Vol. 48, N
4 .— P. 562.
G oldschm idt R. Theoretical Genetics.— Los Angeles, 1955.
Lyon M. F. / / Abstracts of papers on the 138 meeting of the genetical
society.— Univ. of Edinburgh, 1962.
M echelke F. / / Chromosoma.— 1953.— N 5 .— P. 511.
M echelke F. / / Proc. X Int. Congr. G en.— 1958.— N 2 .— P. 185.
Metz C. W. / / Amer. Nat.— 1938.— N 7 2 .— P. 485.
Moore K. L. / / Acta Cytologica.— 1962.— Vol. 6, N 1.— P. 1.
Panitz R. / / Naturwissenschaften.— I960.— Bd 47, N 15.— S. 359
Park W. W. / / J. Anat.— 1957.— Vol. 91, N 3 .— P. 369.
Pavan P. / / Proc. X Int. Congr. G en.— 1958.— N 1 — P. 321.
Risler H. / / Zschr. Zellforsch.— 1954.— N 41.— S. 1.
Schultz J. / / Exp. Gell. Res.— 1952,— Suppl. 2 .— P. 17.
Seari A. G .— Abstracts of Papers on the 138 Meeting of the Genetical
Society.— Univ. of Edinburgh, 1962.
Sm ith-Stocking H. / / Genetics.— 1936.— Vol. 21.— P. 42.
Sturtevant A. H M Dobzhansky Th. / / Genetics.— 1936 — Vol. 21.—
P. 473.
White M. J. D. / / Animal Cytology and Evolution.— 2d ed.— Cambridge.
Univ. Press, 1948.
ТИПЫ ПОЛИМОРФИЗМА*
P. Л. Берг
Полиморфизмом называют разнообразие живых су
ществ. Различия между видами одного рода, прерывистое и
непрерывное разнообразие представителей одного вида с
равным правом могут быть отнесены к полиморфизму в
* Вестник Ленинградского университета.— 1957.— Вып. 4, № 21.
63
этом широком смысле слова* Полиморфизм, в узком смыс
ле слова,— это прерывистое разнообразие форм внутри ви
да, не связанное ни с какими барьерами для скрещивания
между представителями различных форм. Полиморфизм в
этом узком смысле слова и является предметом настоящей
статьи.
Внутривидовое разнообразие предстает перед нами в
двух формах. Первая из них, связанная с возникновением
барьеров для смешения и скрещивания,— разнообразие
групп. Вторая, не связанная ни с какими барьерами для
смешения и скрещивания,— разнообразие особей в преде
лах группы.
Первая форма таит в себе потенциальную возможность
дивергенции. Когда единая совокупность распадается на
две независимые совокупности, отделенные друг от друга
географическими, экологическими или генетическими
барьерами, препятствующими перемешиванию масс инди
видов у агамных и апомиктических видов и скрещиванию
у амфимиктических видов, сразу же возникают две тен
денции — к усилению различий между совокупностями и
к усилению их изолированности друг от друга. Обе сово
купности обретают независимые пути эволюции, и углуб
ление различий между ними и усиление их изолированно
сти ведет к возникновению двух видов из первоначально
единого вида. Внутривидовая дифференциация, связанная
с изоляцией, взрывает систему вида изнутри, ведет к рас
падению первоначально единого целого на части, не свя
занные друг с другом, друг другу не необходимые, а, на
против, находящиеся в антагонистическом отношении друг
к другу. В природе нет более антагонистических отноше
ний, чем отношения видов одного рода друг к другу. Имен
но к видам одного рода применим постулат Дарвина, что
конкуренция является наиболее ожесточенной между наи
более близкими друг к другу формами.
Разнообразие популяций в сочетании с изоляцией —
хотя бы потенциально деструктивный момент в жизни ви
да. Это во всяком случае момент динамический.
Вторая форма дифференциации, не связанная ни с ка
кими барьерами для скрещивания, и есть полиморфизм.
Полиморфизм — это прерывистое разнообразие форм
внутри вида, не связанное ни с какими барьерами для
скрещивания и совместного существования и осуществлен
ное на единой генотипической основе [Ford, 1940; Mather,
1955]. Хотя бы потенциально все формы полиморфного ви
да являются братьями и сестрами или, в случае чередова
ния поколений, родителями и детьми.
64
Полиморфизм — это сочетание коренного сходства
групп друг с другом, с разнообразием особей в пределах
группы. Вместе с тем, полиморфизм,— это выражение
соподчинения особи и вида. Различные формы полиморф
ного вида представляют собой необходимые части единого
целого.
Полиморфизм распространен в природе чрезвычайно
широко. Достаточно указать на возрастной полиморфизм и
на смену фаз жизненного цикла, чтобы сразу стало ясно,
что полиморфными являются все виды, у которых индиви
дуальное развитие характеризуется прерывистым разнооб
разием возрастных форм, и все виды, обладающие сменами
фаз в жизненном цикле.
Прерывистое разнообразие возрастных форм, чередова
ние поколений, смена гапло- и диплофазы относятся к по
лиморфизму. Половой диморфизм, гетеростилия, наличие
твердых и мягких семян у растений, разновременное созре
вание мужских и женских половых продуктов у гермафро
дитов, разнообразие особей в колониях перепончатокры
лых, наличие сезонных форм у многих беспозвоночных и
подражание разным моделям со стороны одного и того же
вида — таковы хорошо известные случаи полиморфизма.
Разнообразие типов полиморфизма, таким образом, огром
но. Полиморфизм может касаться морфологических, физи
ологических, биохимических признаков. Отличие форм
всегда носит комплексный характер, как это ясно на при
мере сезонного полиморфизма у божьей коровки (Adalia
bipunctata). Как показал Тимофеев-Ресовский [TimofeeffRessovsky, 1940], черные жуки более жизнеспособны ле
том, красные — зимой. Комплексные отличия между фор
мами существуют и у бабочки Colias [Hovanitz, 1953], и у
кукушек [Southern, 1954], и у дрозофил, гетерозиготных
по разным инверсиям [Dobzhansky, 1939; Дубинин, 1940].
Отличия между формами могут касаться немногих при
знаков, но могут носить чрезвычайно разносторонний
характер.
Разные типы полиморфизма встречаются в самых при
чудливых сочетаниях. Фенотипическое разнообразие мо
жет строиться на единой генотипической основе, как это
наблюдается в колониях пчел. Генотипические различия
могут строго соответствовать фенотипическому разнообра
зию, как это имеет место в случае гетеростилии и в случае
полового диморфизма высших форм. Слабое фенотипиче
ское разнообразие может сопутствовать глубоким геноти
пическим различиям, как это имеет место в случае поли
морфизма по инверсиям в популяциях дрозофил.
3 Р. Л. Берг
65
На возрастной полиморфизм может налагаться совсем
другой тип полиморфизма, затрагивающий только одну
возрастную группу. Половой диморфизм может сочетаться
с каким-либо другим полиморфизмом, который касается
одних самок или одних самцов. У всех видов бабочки
Colias самки диморфны, а в некоторых случаях триморфны
по окраске. Самцы мономорфны. Мономорфизм самцов ка
сается только конечного выражения признака. Генотипиче
ски, а следовательно и морфогенетически, самцы разнооб
разны; они распадаются на то же число групп, что и самки
[Hovanitz, 1953]. У бабочки Maniola jurtina [Dowdeswell,
Ford, 1953; Dowdeswell, 1956] количество пятен на нижней
стороне крыльев варьирует у обоих полов, образуя у сам
цов унимодальную, у самок бимодальную кривую.
Сочетание возрастного полиморфизма с половым ди
морфизмом встречается у всех раздельнополых многокле
точных животных. На фоне этих двух типов могут осуще
ствляться еще два или три типа полиморфизма, как это
имеет место у бабочек-подражательниц и у пчел.
Бывают случаи, когда полиморфны только зародыши.
Так обстоит дело у немертины Lineus ruber. В каж
дой кладке этого червя, как показал Г. А. Шмидт
[1936— 1941], часть зародышей гибнет, не завершив эмб
рионального развития. Гибнущие зародыши служат пищей
своим собратьям. Зародыши образуют две формы: гибну
щие в определенном проценте случаев и их выживающие
собратья — некрофаги.
Культуры Drosophila melanogaster дают возможность
моделировать этот случай. Экспериментальной моделью
полиморфизма зародышей при мономорфизме взрослых
могут служить двойниковые гибриды, полученные Мёлле
ром [Muller, 1918]. Генетической основой полиморфизма
двойниковых гибридов являются два доминантных гена,
летальных в гомозиготном состоянии и расположенных в
разных представителях одной пары гомологичных хромо
сом, и две инверсии, подавляющие перекрест между гомо
логичными хромосомами этой пары. Расщепление на три
типа в соотношении 1:2:1 осуществляется в каждом поко
лении. Представители обоих гомозиготных классов гибнут.
Гибриды, в точности повторяющие генотип и фенотип ро
дителей, сохраняются. Зародыши образуют две группы —
50 % гибнет, 50 % выживает. Взрослые мономорфны.
Полиморфизм зародышей может сочетаться с полимор
физмом взрослых форм. С таким случаем мы встречаемся
на примере тех видов бабочек-подражательниц, у которых
взрослые особи подражают разным моделям [Fisher, 1927].
66
Здесь два совершенно различных механизма сочетаются
друг с другом и служат общей цели. Именно их сочетание
позволяет виду-подражателю извлечь пользу из миметиче
ского сходства. Часть зародышей гибнет в силу перехода
леталей в гомозиготное состояние. Зародыши образуют две
формы — форму гибнущих и форму выживающих. Нали
чие леталей служит двоякой цели. Во-первых, летали под
держивают численность вида на низком уровне, что имеет
для подражателя первостепенное значение, так как малая
численность подражателя по сравнению с численностью
модели является необходимым условием использования ви
дом преимуществ подражательного сходства. Во-вторых,
летали поддерживают гетерозиготность по доминантным
генам, обусловливающим подражательное сходство и рас
щепляющимися в каждом поколении. Взрослые особи так
же полиморфны. Самки подражают разным моделям. В по
томстве каждой самки осуществляется расщепление как до
доминантным генам, обусловливающим различие в окраске
крыльев, так и по леталям. При этом полиморфны только
самки. Самцы мономорфны и никому не подражают. Летали
проявляются у самцов. Гены, влияющие на окраску самок,
не проявляются у самцов совсем, хотя и наследуются через
аутосомы [Freier, 1913; Fisher, Ford, 1929; Ford, 1936].
Сочетание полиморфизма, затрагивающего только за
родышей и обусловленного летальным геном, с полимор
физмом по морфологическим признакам, касающимся
только взрослых особей, и притом только самок, моделиро
вано в культуре Drosophila melanogaster, содержащей хро
мосому С1В [Muller, 1927]. Скрещивая самок С1В с нор
мальными самцами, мы получаем культуру, в которой за
родыши полиморфны, так как четверть из них гибнет, а
три четверти выживают. При этом зародыши самок моно
морфны — среди них выживает 100 %. Полиморфны толь
ко зародыши самцов — среди них выживает 50 %. Отно
шение полов в потомстве самки С1В — 2:1. Взрослые особи
тоже полиморфны — одна треть из них имеет полосковид
ные глаза Ваг, две трети имеют нормальные круглые глаза.
По этому признаку полиморфны только самки — 50 % из
них имеют полосковидные глаза, 50 % — нормальные
круглые глаза. Баланс форм и правильность числовых со
отношений зависят от наличия в генотипе самок С1В ин
версии (С), запирающей перекрест между леталью и геном
Ваг. Инверсия создает полиморфизм хромосомных струк
тур уже на клеточном уровне.
Многие случаи полиморфизма регулируются летальны
ми генами. Природные популяции всех изученных видов
67
дрозофил насыщены огромным количеством гетерозигот
ных мутаций [Дубинин и др., 1934; Sturtevant, 1937;
Dobzhansky, Queal, 1938b; Dobzhansky, 1939, 1952; Оленов
и др., 1939; Берг и др., 1941; Берг, 19426, 1943а, 1944а, б;
Дубинин, 1946; и др. ]. Каждая особь гетерозиготна по не
скольким мутациям. Некоторый процент зародышей регу
лярно гибнет в каждом поколении в результате гомозиготизации леталей. Их выживающие гетерозиготные со
братья в некоторых случаях обнаруживают пониженную
жизнеспособность [Берг и др., 1941; Dobzhansky, Wright,
1941; Берг, 1943а, 19446; Dobzhansky et al., 1955].
Известны парадоксальные на первый взгляд случаи,
когда летальный в гомозиготном состоянии ген обусловли
вает повышенную жизнеспособность гетерозигот [Мазинг,
1938, 1939; Берг, 1941, 19426, 1943а; Teissier, 1942;
Gustafsson, 1947; Stern et al., 1952; и др. ]. Более высокое
селективное значение гетерозигот по сравнению с обеими
гомозиготами, — предпосылка для образования балансиро
ванного полиморфизма. Повышенной жизнеспособностью
гетерозигот не исчерпывается приспособительный смысл
насыщенности популяций летальными и другими мутация
ми, снижающими приспособленность гомозигот. Особи, ге
терозиготные по разным генам, обнаруживают в разных
условиях различия норм реагирования [Dobzhansky et al.,
1955]. В этом разнообразии, в повышенной пластичности
популяции в целом и состоит биологическое значение
вредных мутаций. Благодаря гетерозиготным мутациям
комбинативная изменчивость в популяциях имеет такой
широкий размах и такое разностороннее проявление, что
сами понятия «нормального» генотипа и «нормального»
фенотипа приобретают условный характер [Шмальгаузен,
1946; Cordeiro, Dobzhansky, 1954]. «Норма» всегда поли
морфна.
Полиморфизм популяции строится не только на основе
генных мутаций, но и на основе перестроек хромосом (ин
версий). Популяции всех изученных видов дрозофил насы
щены огромным количеством инверсий. Инверсии относят
ся к ограниченному числу типов [Дубинин и др., 1937;
Dobzhansky, Quleai, 1938а; Carson, Stalker, 1947; Меладзе
и др. 1943; Brunetto, 1954; и др. ]. Обнаружены сезонные и
годичные циклы в концентрации различных инверсий
[Dobzhansky, 1947, 1951, 1956а, Ь]. Разные генные поряд
ки в различной степени и различным образом изменяют
жизнеспособность
гетерозигот
[Dobzhansky,
Spassky,
1954]. Инверсии обнаружены в популяциях и других пред
ставителей двукрылых [Acton, 1956; и др. ].
68
Приспособительный смысл полиморфизма по генным
порядкам заключается в использовании гетерозиса и в ог
раничении перекомбинаций генов. Гены инвертированного
участка наследуются как единый комплекс, или суперген.
Инвертированные участки и создают благоприятные гено
типические условия для совместного наследования тех ге
нов, которым, с точки зрения приспособления, надлежит
наследоваться совместно. Следует иметь в виду, что суще
ствование супергенов имеет смысл только в полиморфной
популяции и само служит обеспечению полиморфизма. Го
мозиготная популяция не нуждается ни в каких суперге
нах, инверсиях, ни в каком ограничении сцепления. Рас
пространение супергенов в природе очень велико. Доста
точно сказать, что они играют огромную роль в качестве
механизмов, реализующих пол. Половые хромосомы у
большинства форм это не просто хромосомы, несущие ге
ны — реализаторы пола. Вся половая хромосома выступает
у гетерогаметичного пола как суперген. У самцов дрозофи
лы перекрест отсутствует во всех хромосомах. Каждая хро
мосома самца — это суперген. Широкое распространение
супергенов свидетельствует в пользу первостепенного зна
чения полиморфизма в жизни вида. Сказанного достаточ
но, чтобы показать как разнообразно явление полиморфиз
ма в живой природе. Разные категории полиморфизма от
личаются друг от друга фундаментальными чертами: меха
низмом реализации различий, приспособительным смыслом
разнообразия, генетическими и экологическими механиз
мами, регулирующими процентное соотношение форм, и
множеством других черт. Каковы же черты, присущие всем
категориям полиморфизма?
1. ЧЕРТЫ, П РИ СУЩ И Е ВСЕМ КАТЕГОРИЯМ ПОЛИМ ОРФИЗМ А
Прерывистый характер различий между формами, воз
можность скрещивания, рождение всех форм в потомстве
каждой пары показывает, что полиморфизм осуществляет
ся на единой генотипической основе. Генотипически все
особи полиморфного вида сходны друг с другом по всем на
следственным факторам, за исключением тех, которые об
разуют стрелочный механизм, реализующий различия с
помощью расщепления. Рождение всех форм полиморфного
вида в потомстве одной особи или друг от друга — решаю
щий момент для определения, является ли разнообразие
форм полиморфизмом или экологическим обособлением
части вида. Отсутствие барьеров для совместного существо
69
вания и пространственного перемешивания особей — обя
зательный критерий полиморфизма.
Полиморфизм в узком смысле слова наблюдается у ви
дов, размножающихся амфимиктически. Половой процесс,
поддерживая единство генотипического состава популяции,
препятствует образованию на основе полиморфизма груп
пового разнообразия. Разнообразие клонов у клонального
вида или разнообразие, связанное с географической, эколо
гической или генетической изоляцией у видов, размножа
ющихся путем скрещивания, не входит, таким образом, в
понятие полиморфизма. Переход к апомиксису или агамному размножению представляет собою утерю одного из
интегрирующих механизмов вида и сопровождается распа
дом первоначально полиморфного вида на изолированные
друг от друга формы. Прерывистое межвидовое разнообра
зие возникает сразу, имитируя скачкообразную эволюцию
[Mather, 1955].
Существование полиморфизма возможно только в соче
тании с интегрирующими механизмами. Полиморфизм —
показатель наличия этих интегрирующих механизмов, по
казатель преобладания центростремительных сил над цент
робежными. Полиморфизм — это выражение целостности
вида.
В некоторых случаях решить, имеем ли мы дело с по
лиморфизмом или с намечающейся дифференциацией, не
так-то легко. Как известно, кукушки несут яйца, похожие
на яйца тех птиц, в гнездах которых осуществляется гнез
довой паразитизм [Промптов, 1941; Southern, 1954]. Меж
ду кукушками, обитающими в одной местности, никаких
других морфологических отличий, кроме отличий по окра
ске яиц, не наблюдается. Если бы дело касалось только ок
раски яиц, можно было бы объяснить разнообразие рас
щеплением генетических факторов, т. е. отнести данный
случай к полиморфизму. Однако дело осложняется тем,
что определенное яйцо должно быть отложено в опреде
ленное гнездо. Если птица привыкает к гнезду своих при
емных родителей, или к микроэкологическим условиям,
окружающим это гнездо, и затем кладет свое яйцо в такое
же гнездо, эта привязанность к гнезду будет передаваться
от матери к дочери. С самцами возникают затруднения.
«Высокое развитие мимикрии яиц у некоторых биологиче
ских рас кукушек, очевидно, может быть объяснено лишь
ограниченностью видов хозяев в данных случаях ареала
кукушек, т. е. значительным преобладанием в местности
самцов “общего происхождения“ с самками (выкормлен
ных тем же видом хозяев), — пишет А. Н. Промптов.—
70
Несомненно, что видовая специфичность (привязанность к
определенному виду-хозяину) у самок кукушек поддержи
вается из поколения в поколение их птенцовыми навыка^ми». Как установил Чапек [Сарек, 1896, цит. по: Southern,
1954] каждая кукушка несет яйца только одного типа.
Значит, разнообразие окрасок яиц у кукушек нельзя отне
сти к полиморфизму, а следует квалифицировать как на
мечающуюся эколого-генетическую дифференциацию вида.
Кукушки, несущие разные яйца, отличаются друг от друга
целым комплексом экологических признаков и, в частно
сти, привязанностью к определенному биотопу, причем
биологические различия касаются как самцов, так и самок.
Ограничение миграций, привязанность и самцов и самок к
биотопу, окружающему гнездо приемных родителей, при
водит к возникновению барьеров для скрещивания между
кукушками, выведенными разными видами хозяев. А су
ществование барьеров для скрещивания заставляет исклю
чить разнообразие окрасок яиц кукушек из категории по
лиморфизма.
Возникновение барьера для скрещивания между форма
ми сдвигает баланс центробежных и центростремительных
сил в сторону преобладания центробежных сил. И все же
барьеры для скрещивания существуют и в полиморфных
популяциях. Это совершенно своеобразные барьеры. Они
осуществляют свое действие только в пределах формы,
способствуя тем самым скрещиванию форм друг с другом.
Как известно, представители одного пола друг с другом не
скрещиваются. Скрещивания невозможны в пределах форм
и у тех растений, где перекрестное опыление обеспечено
гетеростилией, как у примулы Р. vulgaris [Ford, 1956].
У бабочки Panaxia dominula нормальные самки оказы
вают предпочтение самцам Р. bimaculata, гомозиготным по
меланистической мутации, и самцам Р. medionigrai гетеро
зиготным по той же мутации, перед нормальными самцами
[Fisher, Ford, 1947; Sheppard, 1953, 1956]. То же явление
наблюдал и Кэтлуэлл [Kettlewell, 1956а, b } у бабочки
Bistort betularia L. Нормальные самцы этого вида предпочи
тают темных самок нормальным, руководствуясь запахом.
Такое предпочтение, оказываемое какой-либо форме, со
стороны особей противоположного пола может способство
вать включению предпочитаемой формы в популяцию. Да
же в том случае, если ее селективное значение ниже, чем
у другой, может установиться баланс селективных сил, в
результате которого адаптивное значение обеих форм срав
няется или даже окажется выше у той из форм, селектив
ное значение которой ниже.
71
В некоторых случаях барьеры для скрещивания сущест
вуют и между формами. Если одна или некоторое количе
ство форм бесплодны (как это имеет место у колониальных
перепончатокрылых), или нежизнеспособны (наподобие
гибнущих зародышей немертины Lineus), или разобщены
во времени (сезонные формы, чередование поколений и т.
п .,), то между формами вида существует барьер, не грозя
щий, однако, разобщением групп именно в силу того, что
все формы рождаются в потомстве одной пары или друг от
друга.
Полиморфизм — это разнообразие особей в пределах
группы. Формы носят служебный, как бы подсобный ха
рактер. Полиморфизм — это групповое приспособление,
обеспечивающее жизненные и эволюционные возможности
вида как целого. Отношения между формами полиморфно
го вида конгруэнтны.
Форма полиморфного вида порождает наряду с форма
ми, подобными ей самой, формы, от нее отличающиеся,
или рождает только отличающиеся от нее самой формы (в
случае чередования поколений); она скрещивается преиму
щественно, или исключительно с другими формами того
же вида. Различие между формами носит прерывистый ха
рактер. Среда обитания разных форм часто тождественна,
за исключением того обстоятельства, что для формы «А»
компонентом среды является отличающаяся от нее форма
«В», а для формы «В» — форма «А». В сходных условиях
среды различия форм сохраняются. Они носят, таким обра
зом, ярко выраженный авторегуляторный, а чаще всего ав
тономный характер [Шмальгаузен, 1940].
2. КЛАССИФ И КАЦИ Я ПОЛИМ ОРФИЗМ А
Мы попытались охватить различные категории поли
морфизма и показать то общее, что присуще им всем. Раз
личия между категориями полиморфизма систематизирова
ны и представлены в табл. 1.
Наличие или отсутствие направленного сдвига в про
центном отношении форм — вот та демаркационная ли
ния, по обе стороны которой лежат два типа полиморфиз
ма, две главные его категории, отличающиеся друг от дру
га как по длительности существования, так и по эволюци
онным последствиям.
Первый тип — переходный — характеризуется направ
ленным сдвигом в процентном соотношении форм. Он
представляет собой переход от одного единообразного со72
со
S
н
СЗ
я
Стрелочный механизм
*и
<
ч
VO
сЗ
н
он >я
$
a5и3i $5*
gS
°*S
S«
Классификация полиморфизма
З хJQ
U
X
8 . »О
5х
S
э*8ч § §
§ а *1
8.я 4 сЗ
Ь | 5 >>$
сЗ сЗ Я Я о
3S вS <оЯ О,
с
И S НЧ с
| 2£ Ок
и;
ЭЬ
GЯ 2£
22»
Ях
1§ *8-31
* со S
2 о н
*уо
m g ч
38
о.
а§
с
стояния к другому единообразному состоянию. Переходный
тип полиморфизма — это динамика эволюции.
Второй тип — балансированный — характеризуется от
сутствием направленного сдвига числовых соотношений
форм во времени. Процентное соотношение форм либо ос
тается из поколения в поколение одним и тем же, либо ко
леблется вокруг какой-то постоянной величины. Размах
этих колебаний может быть сколь угодно широк. Отсутст
вие направленности, а не постоянство процентных соотно
шений форм, характеризует собой тип. Балансированный
полиморфизм — это, в противоположность переходному
типу, статика эволюции. Термины «динамика» и «статика
эволюции» принадлежат Холдэну [Haldane, 1954]. Со ста
тикой эволюции мы встречаемся и там, где имеется баланс
различных динамических сил. Будучи направленными в
противоположные стороны, динамические силы приводят к
равновесному состоянию. Примером именно такого равно
весного состояния и является балансированный полимор
физм. Термины «переходный» (transient) и «балансирован
ный» (balanced) полиморфизм принадлежат Форду [Ford,
1940]. Шмальгаузен [1940] называет балансированный по
лиморфизм равновесным гетероморфизмом, а Гекели
[Huxley, 1956] — морфизмом.
Примером переходного полиморфизма является так на
зываемый «индустриальный меланизм» бабочек. Из 780
видов Macrolepidoptera , которые населяют британские ост
рова, у 70 видов совершается процесс распространения в
популяциях темных особей. Сто лет назад «индустриаль
ный меланизм» бабочек не был известен. В запыленных,
лишенных лишайников лесах, прилежащих к индустриаль
ным районам Англии, темные особи менее заметны для
птиц, чем светлые представители тех же видов [Kettlewell,
1956а, Ь]. Гершензон [1940— 1946] описал распростране
ние меланистических форм у хомяка Cricetus cricetus L.
Меланистические формы вытесняют нормальные в самых
различных систематических группах [Huxley, 1956]. Один
из наиболее эффектных случаев направленного изменения
в соотношении форм — возрастание концентрации инвер
сии «РР» в популяциях Drosophila pseudoobscura. Возра
стание совершилось независимо в двух районах. Концент
рация генного порядка была в 1945 г. ничтожной. В 1954 г.
она возросла до 11 %. Ни мутационный процесс, ни миг
рации не могли быть причиной этого направленного про
цесса [Dobzhansky, 1956; Epling, Levine, 1956]. Гекели
[Huxley, 1956] предлагает назвать самую изменчивость в
соотношении форм полиморфного вида градиентом, или
74
клином соотношения (ratiocline). Градиент соотношения
форм подводит нас к проблеме видообразования.
Примеров балансированного полиморфизма множество:
половой диморфизм, возрастной полиморфизм, гетеростилия. Сезонный диморфизм некоторых видов бабочек, раз
нообразие окрасок у Mantis religiosa [Беляев, 1940], у
Adalia bipunctata [Timofeeff-Ressovsky, 1940], у Parattetix
[Fisher, 1927] и у Colias , полиморфизм по генным поряд
кам у дрозофил, москитов, хирономуса, разнообразие заро
дышей у Lineus ruber входят в категорию балансированного
полиморфизма.
Разные категории ею отличаются друг от друга приро
дой стрелочного механизма, реализующего различия меж
ду формами. Роль этого механизма в одних случаях вы
полняют условия внешней среды. В других случаях разли
чия между формами реализуются в процессе индивидуаль
ного развития в результате взаимодействия частей разви
вающегося организма. Стрелочным механизмом в этом
случае служит последовательность, с которой вступают в
действие наследственные факторы. И, наконец, наследст
венные факторы могут играть роль стрелочного механизма,
расщепляясь в каждом поколении. Балансированный поли
морфизм по разнообразию механизмов, реализующих раз
личия, делится на три класса: модификационный, эпигене
тический и генотипический.
К модификационному классу относится сезонный и ло
кальный полиморфизм. Различающиеся формы, хотя и
разобщены во времени или пространстве, развиваются на
единой генотипической основе и стоят друг к другу в отно
шении родительского и дочерного поколения или являются
братьями и сестрами. Этим сезонный и локальный поли
морфизм отличается от географического и экологического
пути расообразования. Процентное соотношение форм ус
танавливается как результат действия самого стрелочного
механизма. Оно варьирует в широких пределах. К модифи
кационному классу полиморфизма относятся и различия
между плодовитыми самками и их бесплодными сестрами в
колониях медоносной пчелы. Условия кормления определя
ют, разовьется та или другая форма. Среда служит пуско
вым механизмом реализации различия и в случае полового
диморфизма у червя ВопеШа.
Черви, прикрепляющиеся к субстрату, становятся сам
ками. Особи, поселяющиеся в половых путях самок, пре
вращаются в самцов.
Модификационный класс можно подразделить на под
классы в зависимости от характера того внешнего фактора,
75
который реализует различия. Таким фактором можеу быть
температура, влажность, пищевой режим или воздействия,
исходящие от представителей противоположного пола.
В качестве первого приближения к такой классификации
можно наметить два подкласса модификационного поли
морфизма: климатический и биоценотический. Климатиче
ский подкласс охватывает сезонный и локальный полимор
физм, биоценотический подкласс — различие между пло
довитыми и бесплодными самками в колониях перепонча
тых (трофический полиморфизм) и различие самцов и са
мок ВопеШа (экзогормональный полиморфизм).
К эпигенетическому классу балансированного полимор
физма относится возрастной полиморфизм, смена гапло- и
диплофазы, функционирование гермафродитных особей
сперва в качестве представителей одного, а затем в качест
ве представителей другого пола. К этому классу относятся
также различия между рабочими пчелами, выполняющими
различные работы в улье. Разделение труда между ни
ми — результат возрастной дифференциации. Каждая пче
ла проделывает все виды работы в определенной последо
вательности.
Эпигенетический класс можно подразделить на под
классы в зависимости от характера взаимозависимостей ча
стей в эмбриогенезе, реализующих прерывистое разнообра
зие возрастных форм, и в зависимости от времени осуще
ствления реализующего действия. В основном эта класси
фикация совпадает с классификацией корреляций, предло
женной Шмальгаузеном [1942]. Эпигенетический класс
разделяется на геномный, морфогенетический и эргонтический подклассы.
Генотипический класс балансированного полиморфизма
охватывает все случаи прерывистого разнообразия форм,
связанного с расщеплением. Постоянная гетерозиготность
является, за редким исключением, необходимой предпо
сылкой существования этого класса.
Известны случаи, когда различие между формами по
лиморфного вида зависит от одной пары факторов. Так об
стоит дело у Mantis religiosa [Беляев, 1940], у Colias
[Hovanitz, 1953].
Гораздо чаще балансированный генотипический поли
морфизм имеет полигенную природу. Расщепление по не
скольким наследственным факторам обусловливает разно
образие форм у Parattetix [Fisher, 1927] и у Papilio polytes
[Fisher, Ford, 1929]. Передаваясь из поколения в поколе
ние, реализаторы полиморфизма у Parattetix и у Р. polytes
перекомбинируются свободно по отношению друг к другу.
76
Летальный эффект гомозиготных комбинаций у Р. polytes,
малая жизнеспособность особей гетерозиготных по двум ге
нам, каждый из которых гам по себе повышает жизнеспо
собности гетерозигот у Parattetix поддерживают комбинативную изменчивость, препятствуя распадению всей систе
мы. Число форм, реализуемых с помощью комбинативной
изменчивости, больше двух. Диморфизм не мог бы сущест
вовать на такой основе. Различия между длиннопестичной
и короткопестичной формами у Primula sinensis [Ernst,
1936] и между мужскими и женскими растениями у
Melandrium [Westergaard, 1948] зависят от нескольких ге
нов. Все гены, от которых зависит реализация диморфизма
у обеих форм, образуют сцепленные комплексы — супер
гены [Sheppard, 1953b; Mather, 1955; Darlington, 1956].
Значение супергенов в качестве реализаторов половых раз
личий в природе очень велико. Половые хромосомы
Lebistes reticulatus — это в сущности супергены. У дрозо
филы, как и у всех видов, где один из полов гетерогаметичен, эволюция пола прошла через стадию супергена и до
стигла высшей ступени [Darlington, 1932, 1939, 1956; Берг,
1942а]. Реализатором пола является хромосомный баланс.
Расщепление в каждом поколении создает различия в соот
ношении половых хромосом и аутосом. Реализация пола с
помощью хромосомного баланса — своеобразная категория
генотипического балансированного полиморфизма.
Хромосомные перестройки — инверсии выступают в
качестве реализаторов различий в популяциях дрозофил.
Межхромосомные перестройки в сочетании с летальными
генами поддерживают постоянную гетерозиготность у эно
тер. Структурная гибридность существует у Campanula
percisifolia [Darlington, 1956].
Своеобразный полиморфизм обнаружен Валентейном
[Valentine, 1956; Darlington, 1956] у фиалок. У некоторых
видов особи имеющие В-хромосомы и лишенные их нахо
дятся в отношениях, характерных для балансированного
полиморфизма, и отличаются друг от друга рядом призна
ков.
Различия в числе целых наборов хромосом выступают
в качестве реализатора различий у перепончатокрылых,
где самцы развиваются из неоплодотворенных яиц. Самцы
начинают развитие с гаплоидным набором хромосом.
В процессе развития число хромосом восстанавливается до
диплоидного, числа хромосом самцов и самок выравнива
ются, но только после того, как гаплоидное состояние сам
ца оказало свое реализующее действие.
77
Таблица
2
Деление генотипического класса балансированного полиморфизма
Подкласс
Генный
Полигенный
Суперген
ный
Суперхро
мосомный
Интрахромосомный
Интерхро
мосомный
Экстрахромосомный
Геномный
Характер различий
Примеры форм, у ко
торых обнаружен соот
ветствующий подкласс
Mantis religiosa
Одна пара аллеColias
морфных генов
Несколько несцепParattetix
ленных друг с
Papilo polytes
другом генов
Комплекс сцепленLebistes
ных генов» пере
Melandrium
дающихся как це Primala
лое
Хромосомный баланс Drosophila
Межхромосомные пе Oenothera
рестройки
Campanula
Различия в порядке
генов в одной из
хромосом гомоло
гичной пары (ин
версии)
Наличие дополни
тельных В-хромосом
Различие в числе
целых наборов хро
мосом
Drosophila
j
Автор
Беляев
Hovanitz
Fisher,
Fisher, Ford
Winge
Westergaard
Ernst
Bridges
Берг
Renner
Darling
ton
Dobzhansky
Дубинин
Viola
Valentine
Hymenoptera
Valentine
Генотипический класс балансированного полиморфизма
делится, таким образом, на восемь подклассов, в соответст
вии с характером генотипических различий, выступающих
в роли реализаторов. Эти 8 подклассов представлены в сле
дующей схеме (табл. 2).
Полигенный подкласс встречается там, где число форм
больше двух, где формы отличаются друг от друга неболь
шим числом признаков. Супергенный подкласс приурочен
к диморфизму, будь то различие полов, или гетеростилия в
том случае, когда различия форм косят комплексный ха
рактер. Хромосомные перестройки в качестве реализаторов
полиморфизма обеспечивают постоянную гетерозиготность
по целому комплексу генов и создают предпосылки для ис
пользования видом гетерозиса. Различие в числе наборов
хромосом и хромосомный баланс выступают в роли реализа
торов полового диморфизма на высших этапах эволюции по
лового диморфизма [Берг, 1942а]. Среди всего разнообразия
стрелочных механизмов в каждом отдельном случае осуще
ствлен именно тот, который обеспечивает реализацию разли
78
чий с\наибольшей точностью, не допуская образования про
межуточных форм. Триморфизм и диморфизм требуют для
своей реализации разных механизмов, и различия механиз
мов реализации у полиморфных и диморфных видов нали
цо. Видовое приспособление несет черты целесообразности.
Каждый из подклассов балансированного полиморфиз
ма подразделяется в свою очередь на роды в зависимости
от природы фактора, определяющего процентное соотноше
ние форм. Процентное соотношение форм связано с их от
носительным селективным значением. Селективное значе
ние признаков, отличающих формы, может быть равным,
и тогда процентное соотношение форм определяется стре
лочным механизмом. Селективное значение форм может
отличаться, и тогда стрелочный механизм в каждом поко
лении возвращает соотношение форм к исходному значе
нию. Равенство селективных значений форм наблюдается в
случае полового диморфизма у моногамов. Соотношение
форм 1:1 определяется механизмом реализации пола. При
полигамии или полиандрии селективные значения форм
отличаются друг от друга. Представители одного пола
уничтожаются или отстраняются от размножения в боль
шей степени, чем представители другого пола. Процентное
соотношение форм при полигамии и полиандрии — это ре
зультат взаимодействия стрелочного механизма и селек
тивных сил. Встречаются и более сложные взаимодействия.
Селективное значение каждой из форм может варьировать
в зависимости от условий. Признак, отличающий одну
форму от другой, может подвергаться отрицательному от
бору в одних условиях, но оказаться полезным в других
условиях. Баланс селективных сил поддерживает соотно
шение форм на одном и том же уровне. Процентное соот
ношение форм складывается как результат взаимодействия
механизма, реализующего различия, и нескольких селек
тивных сил, действующих в прямо противоположных на
правлениях. Эти наиболее сложные случаи широко распро
странены в природе.
Когда селективное значение мутации очень мало, а по
стоянная гетерозиготность по ней отсутствует, ее концент
рация в популяции поддерживается балансом между часто
той возникновения мутации и частотой ее элиминации.
Балансированный полиморфизм в этом случае основан на
взаимодействии стрелочного механизма, давления мутаци
онного процесса и давления отбора.
В основе переходного полиморфизма, в противополож
ность балансированному полиморфизму, всегда лежит один
механизм реализации различий — генотипический. На79
правленный сдвиг в процентном соотношении форм Осуще
ствим только на основе генотипического разнообразия по
пуляций. Переходному типу полиморфизма соответствует
один класс — генотипический.
/
Деление на подклассы переходного полиморфизма
иное, чем балансированного. Не все подклассы балансиро
ванного полиморфизма возможны для переходного. Факто
ры, реализующие различия в случае балансированного поли
морфизма, могут носить как элементарный, так и комплекс
ный характер. Чаще всего стрелочный механизм, обусловли
вающий различия между формами полиморфного вида, но
сит комплексный характер — полигенный, супергенный или
суперхромосомный. Переходный полиморфизм, по смыслу
вещей, основан на элементарных стрелочных механизмах.
Основные его подклассы — генный и хромосомный (интер- и
интрахромосомный). Комплексные механизмы могут играть
роли реализаторов различий при переходном полиморфизме
только в том случае, если они ранее возникли в качестве реа
лизаторов балансированного полиморфизма. Там они приоб
рели свой комплексный характер, а когда баланс динамиче
ских сил нарушился, стали обусловливать различия между
формами переходного полиморфизма [Mather, 1955].
Суперхромосомный подкласс в случае переходного по
лиморфизма невозможен. Геномный подкласс можно лишь
условно отнести к полиморфизму. Различие между форма
ми по числу целых наборов хромосом само является изоли
рующим механизмом.
Пример генного подкласса — «индустриальный меланизм» бабочек. Пример интерхромосомного подкласса —
повышение концентрации инверсии «РР» в популяциях
Drosophila pseudoobscura. Каждому подклассу переходного
полиморфизма, в противоположность балансированному, со
ответствует один род: селективное значение форм не может
быть, по смыслу вещей, равно. Выравнивание селективных
значений переводит переходный полиморфизм в балансиро
ванный. Вместе с тем, балансированный полиморфизм не
связан, как мы видели, с равенством селективных значений
форм. Он основан на балансе сил. Уровень, на котором
произойдет взаимное погашение динамических сил эволю
ции, совершенно безразличен. Безразлично, будут ли пога
шать друг друга две селективные силы, или в игру вступит
еще и механизм расщепления, или давление мутационного
процесса. Какими способами достигнуто равновесие — не
имеет никакого значения. Равновесие достигнуто — перед
нами балансированный полиморфизм, равновесия нет и мы
имеем дело с переходным типом полиморфизма.
80
3. ПОЛИМОРФИЗМ В ПОПУЛЯЦИЯХ
Здесы уместно остановиться на двух случаях балансиро
ванного полиморфизма, которые наблюдались мной в по
пуляциях ^плодовой мушки D. melanogaster, Первый из них
касается разнообразия мух по окраске, второй — разнооб
разия в строении жилок крыла.
Во всех изученных мною популяциях D. melanogaster (за
исключением популяции г. Серпухова, где исследовано ма
лое количество особей) наряду с серыми мухами встречают
ся мухи ярко-желтой окраски. Генетический анализ пока
зал, что различия между серыми и желтыми мухами зави
сят от рецессивной сцепленной с полом мутации (yellow).
Концентрация мутации различна в разных популяциях.
Различна и частота ее возникновения [Берг и др., 1941;
Берг, 19486].
Если мутанты в каждом поколении доживают до стадии
imago, но не оставляют потомства, что равносильно их
полной элиминации, концентрация сцепленной с полом
мутации в популяции все же будет в 4 раза превосходить
мутабильность [Эфроимсон, 1940].
Сопоставление концентрации мутации yellow с часто
той ее возникновения показывает, что концентрация, как
правило, не превосходит мутабильность более, чем в 2 ра
за (табл. 3).
В популяциях, населяющих горные области и границу
распространения вида (Дилижан, Кашира), концентрация
мутации yellow в популяции меньше, чем частота ее воз
никновения. Это показывает, что селективное значение
мутации низкое. Мутанты не только не оставляют потом
ства, но и гибнут в большем проценте случаев, чем нор
мальные серые мухи. Вскрыты и условия этой элиминации.
Оказалось, что миграции являются одним из факторов от
рицательного отбора мутации yellow (табл. 4).
В 1946 г. в популяциях Тирасполя и Умани было впер
вые обнаружено резкое различие между концентрацией и
частотой возникновения мутации yellow (см. табл. 3).
В популяции Тирасполя концентрация в 10 раз превосхо
дила мутабильность. В Умани в 1937 г. концентрация му
тации yellow превосходила частоту ее возникновения ме
нее, чем в 2 раза. В 1946 г. мутабильность упала. Концен
трация осталась на прежнем уровне. Сохранение концент
рации на прежнем уровне, в то время как частота возник
новения соответствующей мутации снизилась, показывает,
что мутация стала подвергаться менее жесткой элимина
ции. Ее селективное значение возросло. Баланс мутацион81
Концентрация мутации yellow в естественных условиях и частота ее воз
ногр географи
Концентрация «а*/
Популяция
/
Годы
yellow
Число пой
манных сам
цов
число
%
Умань
Никитский сад
(Крым, близ
Ялты)
Ереван
Дилижан
Кашира
Серпухов
Кутаиси
1937
10159
25
0 ,2 4 6 ± 0 ,0 4 9
1937— 1938
1939
1939
1940— 1941
1940
1945
8375
2759
5238
1282
190
6564
16
7
2
0 «1»*
0
1
0,191 ± 0,048
0 ,2 5 4 ± 0 ,0 9 6
0,038 ± 0 ,0 2 7
0 «0,078±0,078»
0
0 ,0 1 5 ± 0 ,0 1 5
Итого...
Умань
Тирасполь
1937— 1945
1946
1946
34567
3652
6016
51
11
14
0,148±0,021
0,301 ± 0 ,0 9 0
0,2 3 3 ± 0 ,0 6 2
9668
25
0 ,2 5 9 ± 0 ,0 5 2
44235
76
0 ,1 7 2 ± 0 ,0 2 0
Итого...
Всего...
1946
1937— 1946
* Вместо полученного нуля условно принята единица, и все расчеты сделаны,
ного процесса и отбора сохранился на прежнем уровне.
Чем объясняется возрастание селективного значения мута
ции,— сказать трудно. Быть может, сыграла роль меньшая
активность самцов yellow по сравнению с нормальными
самцами. В условиях засухи 1946 г. меньшая активность и
связанная с ней большая оседлость желтых мух могли со
ставить преимущество, так как вынуждали мутантов дер
жаться в очагах размножения мух, куда везут фрукты дЛя
переработки. Селективное значение мутации yellow может,
по-видимому, варьировать в разные годы и в разные сезо
ны одного и того же года [Берг, 1948а].
Другой пример полиморфизма популяций D. melanogaster касается разнообразия в строении жилок крыла. Все
изученные популяции плодовых мух содержат в опреде
ленной концентрации наследственные изменения жилок
крыла, прямо противоположные по своему фенотипическо
му выражению. По строению жилок особи делятся на три
типа: один тип имеет нормальные жилки, другой редуци
рованные жилки, у третьего имеются добавочные жилки.
82
\
Таблица
3
никновения в потомстве (Fj и F2) диких особей в популяциях различческого происхождения
Мутабильность «Ь»
Разница между концентрацией
и мутабильностью «а» —«Ь*>
yellow
Число сам
цов Fj и F2
%
D,ff+mdifr
Diff
wdiff
и
20144
30
0,1 4 9 ± 0 ,0 2 7
0 ,0 9 7 ± 0 ,0 5 6
12472
5442
14144
10919
1929
15080
12
6
7
10
0
1
0 ,0 9 6 ± 0 ,0 2 8
0 ,1 10± 0,045
0 ,0 4 9 ± 0 ,0 1 9
0 ,0 9 2 ± 0 ,0 2 9
0
0,007 ± 0,007
0,0 9 5 ± 0 ,0 5 6
0,1 4 4 ± 0 ,1 0 6
0,011 ±0,033
«0,014 ± 0 ,083»
80130
5394
21940
66
0 «1»
5
27334
5
107464
71
—
1,7
1,4
0,3
«0,2»
—
0,00 8 ± 0 ,0 1 6
0,5
0 ,065± 0,023
«0,282±0,092»
0 ,210± 0,063
2,8
«3,1»
3,3
0 ,0 1 8 ± 0 ,008
0,241 ± 0 ,0 5 2
4,6
0,0 6 5 ± 0 ,0 0 8
0,107±0,021
5,1
0 ,8 3 ± 0 ,0 1 0
0 «0,019±0,019»
0 ,0 2 3 ± 0 ,0 1 2
:>той условной величины. Соответствующие числа стоят в кавычках.
Все три типа встречаются во всех популяциях. Генетиче
ский анализ показал, что разнообразие популяций по это
му признаку имеет полигенную природу. Отдельные мута
ции обладают неполной пенетрантностью и экспрессив
ностью даже у гомозиготного класса и вместе с тем прояв
ляются в известном проценте случаев у гетерозигот. Со
вершенно аналогичная картина насыщения популяций
крыловыми мутациями обнаружена и у других видов дро
зофил [Ромашов, Балкашина, 1930; Gordon et al., 1939].
Мутации разных видов имеют сходное выражение, что по
зволило Оленову [1941 ] сформулировать представление об
«уязвимых точках онтогенеза». Гершензон [1941 ] выска
зал предположение, что одни и те же мутации, затраги
вающие жилки крыла, имеют разную и прямо противо
положную ценность перед лицом отбора в разных условиях
существования. Баланс селективных сил и создает ту
противоречивую, странную картину проявления и насле
дования крыловых мутаций, которую мы наблюдали в по
пуляциях.
83
Связь между миграциями и концентрацией мутаций yellow
Место сбора
Завод
им. 1-го Мая
Завод
им. Т кр емко,
заготови
тельная
площадка
Завод
им. Ткаченко,
спиртовый
цех
yellow
Объект
сбора
Расстояние от
места лова до
места вылупления
Число
самцов
imagines
Далекое
2102
0
0
imagines
Близкое
Лов в самом
очаге раз
множения
Сбор в оча
ге размно
жения
1075
2
0 ,1 8 6 ± 0 ,131
1192
5
0 ,419± 0,187
1647
7
0,4 2 5 ± 0 ,1 6 2
Всего...
6016
14
0,2 3 3 ± 0 ,0 6 2
imagines
pupae
число
%
Приме чай и е. Вероятность случайного различия по Стивенсу <0,005.
Мною были предприняты детальные [Берг и др., 1941;
Берг, 1948а, б, 1957] исследования концентрации и насле
дования мутаций, затрагивающих жилки крыла, в изоли
рованных и неизолированных популяциях среди мух, толь
ко что вылупившихся из куколок, пойманных взрослыми и
мигрировавших на разные расстояния от очагов их выплода. Эти условия позволили пролить свет на природу селек
тивных сил, управляющих концентрациями крыловых му
таций. Большинство этих материалов не опубликовано.
Изучение зависимости концентрации обоих типов мутаций
от отбора в процессе миграций приводит к заключению об
относительной пользе обоих типов мутаций. В условиях,
где оседлые формы приобретают преимущество перед миг
рирующими, всякая порча крыла, связанная с отягчением
его жилками, приобретает положительное значение, под
хватывается отбором, и носители ее включаются в состав
популяции. Отбор мигрирующих форм включает в популя
ции некоторые мутации, связанные с редукцией жилок.
Отбор меняет не только процентное соотношение форм, но
и характер их наследования. В изолированных популяци
ях, где идет отбор на оседлость, мутации, отягчающие
крыло, обнаруживают при скрещивании мутантов с нор
мальными особями большую степень доминирования, чем в
неизолированных популяциях.
84
Таблица
4
к популяции D. melanogaster Тирасполя
Отношение
к мигра
циям
Число
самцов
Мигриро1 вавшие
3177
yellow
число
%
2
0 ,0 6 3 ± 0 ,0 4 5
Diff±mdiff
Diff
wdiff
0,3 6 0 ± 0 ,1 2 5
2,9
—
—
1
Не мигри
ровавшие
—
2839
12
0 ,4 2 3 ± 0 ,1 2 9
—
—
—
>
Концентрация мутации yellow в популяции Умани от
носится к генному подклассу генотипического класса
балансированного полиморфизма. Благодаря неравенству
селективных значений желтых и нормальных форм баланс
является результатом взаимодействия мутационного про
цесса, механизма расщепления и селективных сил.
Концентрации крыловых мутаций представляют собой
пример полигенного подкласса генотипического класса ба
лансированного полиморфизма. Баланс устанавливается в
результате взаимодействия механизма, реализующего раз
личия, и селективных сил, действующих в разных услови
ях в противоположных направлениях. Феногенетические
процессы (степень доминирования) также играют роль,
включаясь в это взаимодействие.
4. ПОЛИМОРФИЗМ и э в о л ю ц и я
Значение переходного полиморфизма для эволюции
очевидно. Переходный полиморфизм — главный путь эво
люции вида как целого, протекающий на основе интегри
рующих механизмов вида, без какого-то ни было участия
изоляции. Единственные барьеры, ограничивающие поле
деятельности отбора,— это барьеры, отделяющие виды од
ного рода друг от друга. Эволюция видов одного рода про
текает независимо. Однако эта независимость не абсолют
на. Идея зависимой эволюции близких видов, живущих в
сходной экологической обстановке, выдвинута в недавнее
85
время Шеппардом [Sheppard, 1956]. Генетическое измене
ние, увеличивающее защищенность жертвы одного вида,
повышает интенсивность атаки хищника на представителей
других видов жертв и тем самым повышает интенсивность
отбора защитных средств у этих видов жертв. Процесс
принимает характер «аутокатализа». В качестве иллюстра
ции Шеппард приводит мимикрию и индустриальный меланизм у бабочек и полиморфизм в окраске у улиток
Сереае . Результатом такого «аутокаталитического» харак
тера эволюции является параллелизм в образовании за
щитных приспособлений у многих видов, живущих в той
же области при сходных экологических условиях. Встав на
определенный путь эволюции, вид не только весь как це
лое вовлечен в процесс преобразования, но вовлекает на
этот путь и другие сходные с ним виды.
Гомологическая изменчивость [Вавилов, 1922, 1935] и
параллелизм процессов отбора выражаются в параллелизме
эволюционных преобразований.
Эволюционное значение балансированного полимор
физма также многообразно. Нарушение баланса динамиче
ских сил превращает балансированный полиморфизм в пе
реходный. По мнению Кэтлуэлла [Kettlewell, 1956], имен
но этим путем произошел «индустриальный меланизм» ба
бочек. На севере Шотландии в старых темных сосновых
лесах у нескольких видов бабочек меланические и нор
мальные формы образуют балансированный полиморфизм.
Направленное изменение условий существования оказалось
благоприятным для меланистической формы, сдвинуло ба
ланс селективных сил в ее пользу и превратило балансиро
ванный полиморфизм в переходный. Холдэн [Haldane,
1956а] считает, что так обстояло дело только первые деся
тилетия. Доминантная мутация распространялась беспре
пятственно, пока не достигла такой концентрации, когда
началось массовое появление гомозиготных мутантов. Бо
лее высокая жизнеспособность гетерозигот, по сравнению с
черными гомозиготами, и меньшее истребление гетерози
гот, по сравнению с нормальными особями, если не при
остановило, то замедлило этот процесс, снова придав ему
черты балансированного полиморфизма. Баланс динамиче
ских сил установился на новом, отличающемся от исходно
го, уровне.
Модификационный полиморфизм может стать источни
ком эволюционных преобразований, если редко наступав
шие или редко встречавшиеся условия приобретут господ
ствующий характер [Шмальгаузен, 1940]. «Несомненно,
что известный гетероморфизм, постоянно наблюдаемый в
86
популяциях вида, живущего в разнообразных условиях
внешней среды, значительно повышает его эволюционную
пластичность,— пишет И. И. Шмальгаузен.— Полимор
физм создает известную степень “преадаптации44 вида к
условиям существования, которые встречались уже и рань
ше в жизни вида, но которые не были господствующими.»
Модификационный полиморфизм, будучи сам выраже
нием авторегуляторного типа развития, создает предпосыл
ки для дальнейшей деятельности стабилизирующего отбо
ра. Форма, ставшая господствующей, стала вместе с тем
той «нормой», которую берет под свое покровительство
стабилизирующий отбор.
Могучим источником эволюционных преобразований
является возрастной полиморфизм. Прерывистый характер
возрастного полиморфизма через неотению и педоморфоз
создает предпосылку для возникновения различий высокого
таксономического ранга [Garstang, 1922; De Beer, 1951,
1954; Hardy, 1954; Тахтаджян, 1954; Alvarado, 1956; и
ДР. I
А. Л. Тахтаджян [1954, с. 102] выдвигает принцип воз
растающей прерывистой эволюции. Прерывистость эволю
ции возрастает по мере возрастания прерывистости онтоге
неза. Неотения и педоморфоз становятся преобладающими
путями эволюции.
До сих пор мы говорили о значении полиморфизма для
эволюционных преобразований вида, но и сам полимор
физм эволюирует. Это касается прежде всего стрелочных
механизмов, направляющих развитие по одному из немно
гих возможных русел. Их совершенствование в процессе
эволюции состоит в том, чтобы выполнять свою роль с воз
можно большей точностью, не допуская развития проме
жуточных форм. Внешние стрелочные механизмы заменя
ются генотипическими, основанными на расщеплении. Автономизация — столбовая дорога эволюции не только
форм, но и механизмов их реализации.
Блестящий пример прогрессирующей автономизации
стрелочного механизма представляет собой эволюция поло
вого диморфизма. Здесь можно проследить, как эпигенети
ческий способ реализации приходит на смену модификационному, как возрастает роль генетических факторов в реа
лизации половых различий. Генный способ реализации
сменяется полигенным, который при участии интерхромо
сомных перестроек превращается в супергенный. Суперген
ный способ реализации пола влечет за собой вынужденную
гетерозиготность целой хромосомы и ее постепенную де
градацию. С абсолютной неизбежностью из супергенного
87
способа реализации пола возникает суперхромосомный
способ реализации, когда уже не отдельные гены или бло
ки генов, а генный баланс выступает в роли реализатора
различий [Darlington, 1932, 1939, 1956; Берг, 1942а].
Прерывистый характер полиморфизма возрастает. Ус
ложняется стрелочный механизм, различия между форма
ми приобретают все более комплексный характер. Вместе с
тем, возрастает и соподчинение особи и вида. Чем больше
различие между формами, тем более необходимы все фор
мы для воспроизведения и эволюции вида. Дифференциа
ция вида идет бок о бок с развитием интегрирующих меха
низмов. Значение полиморфизма в качестве группового
приспособления возрастает.
Концепция групповых приспособлений все более овла
девает умами биологов. Она была ясна еще Дарвину. Пе
чать глубокой целесообразности лежит на явлениях измен
чивости и наследственности. Эта целесообразность сказы
вается в процессе смены поколений: группа существует не
только в пространстве, но и во времени. С особенной силой
подчеркнул приспособительное значение генетических и
феногенетических явлений А. А. Малиновский [1939—
1947]. Приспособление касается группы, а отнюдь не от
дельной особи. Тудей [Thoday, 1953] говорит о приспособ
ленности «эволюирующей единицы» (unit of evolution),
подразумевая под таковой совокупность факторов, обеспе
чивающих не только жизнь ближайших генераций, но и
отдаленных потомков данной популяции.
В сущности та же мысль заключена и в понятии «гене
тической системы» (genetic system) Дарлингтона [Darling
ton, 1939, 1956; Stebbins, 1951 ] и в концепции группы
Миллера [Miller, 1953].
Но какой же фактор несет ответственность за образова
ние групповых приспособлений? Индивидуальный отбор не
может быть этим фактором.
Дифференциация и интеграция группы как целого, со
подчинение особи и группы — это результат деятельности
группового отбора. На арену межгрупповой борьбы выхо
дят виды одного рода. Межвидовая групповая борьба явля
ется могущественным фактором эволюции. Но и внутри
вида отдельные, относительно изолированные группы, кон
курируют друг с другом в развитии групповых приспособ
лений. Полиморфизм является прекрасным косвенным до
казательством существования группового отбора. Неспособ
ность отдельных особей жить или оставить потомство
поставлена групповым отбором на службу виду. Гибель
зародышей дрозофил, бабочек-подражательниц, немертин
88
Lineus в результате перехода в гомозиготное состояние ле
гальных мутаций, бесплодие рабочих пчел — это выраже
ние соподчинения особи и вида.
Групповой отбор, так же как индивидуальный, высту
пает не только в движущей, или преобразующей форме, но
и в стабилизирующей форме.
Эволюция стрелочного механизма — результат дея
тельности стабилизирующего группового отбора. Признак
группы, который в данном случае берет под свое покрови
тельство стабилизирующий отбор,— это числовое соотно
шение форм. Замена внешних факторов реализации поли
морфизма внутренними обеспечивает независимость число
вого отношения форм от случайных колебаний в факторах
внешней среды. Приспособительный смысл такой замены
совершенно ясен, когда речь идет о половом диморфизме
или о гетеростилии.
В результате отбора групп сама эволюция предстает
перед нами как процесс, меняющийся в течение времени.
Отбор наиболее успешно, наиболее перспективно эволюирующих групп — это способ совершенствования самой эво
люции. Эволюция эволюирует. Наиболее могущественным
фактором этой эволюции является межгрупповой, и в
частности межвидовой отбор.
Идея группового отбора не нова [Шмальгаузен, 19466;
huzzati-Traverso, 1952; Fisher, 1954; Darlington, 1956]. Одна
ко эволюционные последствия группового отбора еще неясны
Концепция группового отбора открывает возможность
формулировать понятия приспособлений, обеспечивающих
не жизнь особи, а эволюцию группы. На вопрос: почему
существует то или иное устройство? — например, линей
ное расположение генов в хромосомах — мы можем дать
ответ: потому что оно обеспечивает наиболее успешную
эволюцию, ускоряет ее темп, ведет форму по пути про
гресса*. Без концепции группового отбора такая степень
целесообразности кажется неосуществимой.
* Приведенный пример — ошибка. Линейное расположение генов в
хромосомах не результат сохранения линейной структуры в процессе
группового отбора, сократившего разнообразие хромосомных структур раз
ных видов в угоду наиболее успешной эволюции, а неотъемлемый атрибут
субстрата жизни — конвариантно редуплицирующихся макромолекул.
Возникновение жизни — это возникновение линейной структуры, допу
скающей запись информации и ее считывание. В начале было Слово. От
бор не в его преобразующей, а в сохраняющей форме осуществлялся, ко
нечно, и на предоиологической стадии эволюции в мире макромолекул,
но это не был групповой отбор. Примером признака, возникшего под вли
янием группового отбора и обеспечивающего поставку материала эволю
ционных преобразований, может служить рекомбинация генов в процессе
образования половых клеток (примечание Р. Л. Берг, 1991 г.).
89
Полиморфизм, как мы видели, эволюирует сам и вме
сте с тем определяет дальнейший ход эволюции, делая ее
все более прерывистой.
Полиморфизм является выражением целостности вида.
Целостность организма и клетки не вызывает ни у кого со
мнений. Целостность всего органического мира — разно
родного живого вещества биосферы — обоснована трудами
Вернадского [1926, 1940]. Представление о целостности
вида [Берг, 19436; Малиновский, 1945; Северцов, 1951;
Завадский, 1954] и о целостности биоценоза все еще дис
кутируется. Пространственная разобщенность частей, от
сутствие «цельности» вида препятствуют признанию его
целостности.
Идея целостности вида, сравнимой с целостностью ор
ганизма, вытекает из представления о разнообразии строе
ния систем, которые остаются системами, будучи сплош
ными, как организм, клетка или кристалл, или рассеянны
ми, как Солнечная система и Галактика. Целостность не
есть цельность. Вид целостен. Однако его дискретность,
связанная с пространственной разобщенностью его конст
руктивных элементов, подавляет представление о наличии
взаимосвязи между отдельными конструктивными элемен
тами системы и препятствует признанию его целостности.
Дифференциация — необходимое условие существова
ния системы. Дифференциация основана на дискретности.
Всякая система дискретна, т. е. состоит из частей. Части
могут быть однородными. Систему, состоящую из однород
ных частей, мы назовем изоморфной. Наряду с изоморф
ными системами существуют системы гетероморфные —
системы, состоящие из разнородных частей. Изоморфной
системой является кристалл, обладающий атомной или мо
лекулярной решеткой, гетероморфной системой — кри
сталл, имеющий ионную кристаллическую решетку. Среди
живых систем, т. е. среди систем, обладающих свойством
самовоспроизведения, каковыми являются внутриклеточ
ные структуры — хромосомы, митохондрии и т. п., сами
клетки, индивиды и виды, есть только одна изоморфная
система — вид.
Вид относится к своеобразному типу систем. Среди
живых систем он система единственная в своем роде. По
некоторым признакам (изоморфизм, пространственная
разобщенность частей) он сходен с неживыми системами,
но от всех них отличается именно тем, что он живая сис
тема. Таким образом, вид — это система особого рода сре
ди всех систем вообще. Вид — это живая, изоморфная эволюирующая система, части которой разобщены в простран
90
стве. Ни одной другой живой, изоморфной, эволюирующей
системы, части которой разобщены в пространстве, кроме
вида, нет.
Вид изоморфен не только в том смысле, что он состав
лен из особей, сходных друг с другом в основных чертах.
На этот фундаментальный изоморфизм особей налагается
изоморфизм сходных групп. Состав вида из особей и засе
ление ареала распространения вида отдельными, относи
тельно изолированными группами особей является всеоб
щим законом его бытия. Изоморфизм вида представляет
его основную характеристику. Именно из сходства индиви
дов и групп индивидов в основных чертах своего строения
и строя вытекает морфофизиологическое единство вида,
единство его геохимической функции в биосфере, его гео
графическая и экологическая определенность.
На фундаментальный изоморфизм налагается диффе
ренциация, которая носит второстепенный характер. Диф
ференциация, или гетероморфизм вида, выражается в раз
нообразии особей и групп особей, слагающих вид.
Сходство групп может сопровождаться глубокими раз
личиями внутригруппового характера. Полиморфизм как
раз и является примером такого сочетания принципиальною изоморфизма групп с гетероморфизмом особей в преде
лах групп.
СПИСОК ЛИТЕРАТУРЫ
Ьеляев М. М. Гомохромия окраски у Mantis religiosa D., ее защитное зна
чение и отборная ц е н н о с т ь // Журн. оощ. биол.— 1940.— Т. 1,
№ 4 .— С. 529 —542.
Псрг Р. Л. Роль малых мутаций в эволюции мутабильности / / Докл. АН
СССР.— 1941.— Т. 32, № 1.— С. 71—74.
Псрг Р. Л. Историческое развитие механизма реализации пола / / Журн.
общ. биол.— 1942а.— № 3 . — С. 171 — 189.
Псрг Р. Л. Значение изоляции для эволюции доминантности в естествен
ных популяциях Drosophila melanogaster / / Докл. АН СССР.—
19426.— Т. 36, № 9 .— С. 304— 308.
Иерг Р. Л. Происходит ли элиминация особей гетерозиготных по леталь
ным
мутациям
в
естественных
популяциях
Drosophila
melanogaster? / / Изв. АН СССР. Отд. биол. наук.— 1943.—
№ 4 .— С. 243— 248.
Псрг Р. Л. Вид как эволюционирующая система / / Рефераты работ учрежд. Отд. биол. наук АН СССР за 1941 — 1943 гг.— 19436.—
С. 185.
Псрг Р. Л. Генетический анализ популяций Drosophila melanogaster Дилижана (Армянская ССР) / / Изв..АН СССР. Отд. биол. наук.—
1944а.— № 1.— С. 5 1 —58.
Псрг Р. Л. Зависимость между степенью проявления мутаций в гетерози
готном состоянии и их концентрацией в генотипе естественных по
пуляций Drosophila melanogaster / / Изв. АН СССР. Отд. биол. на
ук .— 19446.— № 2 .— С. 121 — 128.
91
Берг Р. Л. Различная частота возникновения мутации «yellow» в разных
популяциях Drosophila melanogaster / / Изв. АН СССР. Отд. биол.
наук.— 1945.— № 3 .— С. 374—382.
Берг Р. Л. Об условиях стабилизации и лабилизации фенотипа в естест
венных популяциях Drosophila melanogaster / / Тез. докл. конф. по
проблемам дарвинизма 3 —8 февраля 1948 г.— 1948а.— С. 61—62.
Берг Р. Л. О взаимоотношении между мутабильностью и отбсюом в при
родных популяциях Drosophila melanogaster / / Журн. общ. биол.—
19486.— Т. 9, № 4 .— С. 299—312.
Берг Р. Л* Коллоквиум на биологической станции «Миассово» (лето
1956 г.) / / Вести. ЛГУ Сер. биол.— 1957.— Т. 3, вып. 1.—
С. 144— 149.
Берг Р. Л., Бриссенден Э. Б., Александрийская В. Тм Галковская К. Ф.
Генетический анализ двух природных популяций Drosophila
m elan ogaster//
Журн.
общ.
биол.—
1941.—
Т. 2,
№ 1.— С. 143— 158.
Вавилов Н. И. Закон гомологичных рядов в наследственной изменчиво
сти / /
Генетические основы селекции.— М.; Л.,
1935.—
С. 75— 128.
Вернадский В. И. Биосфера.— Л., 1926.
Вернадский В. И. Биогеохимические очерки.— М.; Л., 1940.
Герш ензон С, М. «Мобилизационный резерв» внутривидовой изменчиво
сти / / Журн. общ. биол.— 1941.— Т. 2, № 1.— С. 85— 108.
Герш ензон С. М. Роль естественного отбора в распространении и динами
ке меланизма у хомяков ( Cricetus cricetus L.) / / Журн. общ. би
ол.— 1946.— Т. 7, № 2 .— С. 97— 130.
Герш ензон С. М., Полевой В. В. Наследование черной окраски у хомяка
( Cricetus cricetus L.) / / Докл. АН СССР.— 1940.— Т. 29.
Герш ензон С. М., Полевой В. В. Система скрещивания в природных по
пуляциях хомяка ( Cricetus cricetus L.) / / Докл. АН СССР —
1941.— Т. 30.
Дубинин Н. П. Мутабильность и проявление леталей в популяциях / / Зоол. ж урн.— 1940.— Т. 24, N9 6.
Дубинин Н. Пм Гептнер М. А., Бессмертная С. Я. и др. Эксперименталь
ный анализ экогенотипов Drosophila melanogaster, ч. 1 / / Биол.
журн. 1934.— Т. 3, № 1. С. 207—216.
Дубинин Н. П., Соколов Н. Н. Хромосомные мутации и система вида / /
Журн. общ. биол.— 1940.— Т. 1, № 4.— С. 543—554.
Дубинин Н. Пм Соколов Н. Н., Тиняков Г. Г. Внутривидовая хромосом
ная изменчивость / / Биол. журн.— 1937.— Т. 6, N9 5 / 6 .—
С. 1007— 1054.
Завадский К. М. Проблема вида и видообразования / / Вести. ЛГУ.—
1954.— № 10.— С. 3 — 15.
Мазинг Р. А. Повышенная жизнеспособность гетерозиготных по летали
мух Drosophila melanogaster / / Докл. АН СССР.— 1938.— Т. 20,
№ 2 — 3.
Мазинг Р. А. Анализ жизнеспособности мух Drosophila melanogaster, гете
розиготных
по леталям,
возникшим
в природе / /
Докл.
АН СССР.— 1939.— Т. 25, № 1.— С. 64—67.
Малиновский А. А. Роль генетических и феногенетических явлений в
эволюции вида, ч. I. Плейотропия / / Изв. АН СССР. Сер. биол.—
1939.— № 4 .— С. 575—614.
Малиновский А. А. Роль хромосомных инверсий в эволюции вида / /
Журн. общ. биол.— 1940.— № 4 .— С. 565—596.
Малиновский А. А. Закономерности наследственности в свете дарвинов
ского учения об отборе / / Усп. соврем, биологии.— 1941.— Т. 14,
вып. 1.— С. 171 — 176.
Малиновский А. А. Типы взаимодействия и их значение в развитии / /
Рефераты
работ учрежд. отд.
биол. наук АН СССР за
1941 — 1943 гг.— 1945.— С. 292—293.
92
Малиновский А. А. Вид как эволюционное целое / / Рефераты работ учрежд. биол. наук АН СССР за 1945 г.— 1947.— С. 286— 287.
М еладзе Д. Д м Джорджикия Г. Гм Болквадзе П. К. Некоторые результа
ты генетического анализа популяции Drosophila melanogaster / /
Сообщ. АН Груз. ССР.— 1943.— Т. 4, № 7 .— С. 701—710.
Оленов Ю. М. О влиянии предшествующей истории вида на его дальней
шее развитие / / Докл. АН СССР.— 1941.— Т. 31.
Оленов Ю. Мм Хармац И. С., Галковская К. С., Муретов Г. Д. Факторы,
обусловливающие генный состав природных популяций Drosophila
melanogaster / / Докл. АН СССР.— 1939.— Т. 24.— С. 475.
Промптов А. Н. Современное состояние изучения гнездового паразитизма
п т и ц / / Усп. соврем, биологии.— 1941.— Т. 14, вып. 1.—
С. 30—51.
Ромашов Д. Д., Балкашина Е. И. Материалы по генетике Drosophila
funebris / / Журн. эксперим. биол.— 1930.— Т. 4, вып. 3 .—
С. 201— 219.
Северцов С. А. О конгруэнциях и понятии целостности вида / / Пробле
мы экологии животных.— М., 1951.— Т. 1.— С. 30—57.
Тахтаджян А. Л. Вопросы эволюционной морфологии растений.— Л.,
1954.
Шмальгаузен И, И, Изменчивость и смена адаптивных норм в процессе
эволюции / / Журн. общ. биол.— 1940.— № 4 .— С. 510—528.
Шмальгаузен И* И. Организм, как целое в индивидуальном и историче
ском развитии.— М.; Л.: Изд-во АН СССР, 1942.
Шмальгаузен И. И. Факторы эволюции.— М.; Л.: Изд-во АН СССР,
1946а.
Шмальгаузен И. И. Проблемы дарвинизма.— М.: Сов. наука, 19466.
Шмидт Г. А. Закономерности смены типов эмбриональных приспособле
ний / / Биол. ж урн.— 1936.— Т. 5, вып. 4.
Шмидт Г. А. Биология размножения Linens gessere/zs/sSubsp. ruber из Роскова и Кольского залива / / Биол. журн.— 1940.— № 6.
Шмидт Г. А* Эмбриональный диморфизм у немертины Linens gesserensis
Subsp. ruber / / Tp. ин-та/И н-т цитол. гист. и эмбр. АН СССР.—
1941а.— Т. 1.
Шмидт Г. А. Коррелятивные соотношения между количеством потомства
и приспособлениями к его охране / / Усп. соврем, биологии.—
19416.— Т. 14, вып. 1.— С. 160— 170.
Шмидт Г. А., Янковская А. А. Биология размножения немертин из Роскова и Кольского залива / / Биол. журн.— 1937.— № 2.
Эфроимсон В. П. Измерение скорости мутационного процесса у тутового
шелкопряда / / Изв. АН СССР. Отд. биол. наук.— 1940.—
С. 688— 705.
Acton А. В. Chromosomal polymorphism in Chironomns / / Proc. Roy. Soc.—
1956.— Vol. 145, N 920.— P. 347—350.
Alvarado R. Poligenesis у cenogenesis en las formas larvarias de
invertebrados / / Ciencias.— 2956.— Vol. 21, N 3 .— P. 413—436.
Brunetto A. I cromosomi politenici delle ghiandole salivadi Drosophila
ambigna Pomini / / II inversione eterozigoti Scientia genet.— 1954.—
N 4 .— P. 214— 222.
Buzzati-Traverso A. A. Heterosis in population Genetics / / Heterosis/Ed. by
J. W. Gowen.— Jowa, 1952.— P. 149— 160.
Carson H. L. Marginal homozygosity for gene attangement in Drosophila
robnsta / / Science.— 1956.— Vol. 123, N 3198.— P. 630—631.
Carson H. LM Stalker H, D. Gene arrangements in natural populations of
Drosophila robnsta St / / Evolution — 1947.— Vol. 1.— P. 113— 133.
Cordiero A. R., Dobzansky Th. Combining ability of certain chromosomes in
Drosophila willistoni and invalidation of the «wild-tupe» concept / /
Amer. Nat.— 1954.— Vol. 99, N 839.— P. 75—86.
Crosby Y. L. Selection of an unfavorable genecomplex / / Evolution.—
1949.— Vol. 3 .— P. 212— 230.
Darlington C. D. Recent Advances in Cytology.— L., 1932.
Darlington C. D. The Evolution of Genetic Systems.— Cambr. Univ. Press,
1939.
93
Darlington C. D. Natural populations and the breakdown of classical
genetics / / Proc. Roy. Soc.— 1956.— B. 145, N 920.— P. 350—364.
D e Beer G. K. Embryos and ancestors.— Oxford, 1951.
De Beer G. K. The Evolution of Metazoa / / Evolution as a Process/Ed. by
J. Huxley, A. C. Hardy, E. B. Ford.— L., 1956.— P. 24— 33.
D obzhansky Th. Genetics of natural populations. IV Mexican and Guatemalan
populations of Drosophila pseudoobscura / / Genetics.— 1939.—
Vol. 2 4 .— P. 391— 412.
Dobzhansky Th. A directional change in the genetic constitution of a natural
population / / Heredity.— 1947.— Vol. 1.— P. 53—62.
Dobzhansky Th. Genetics and the Origin of Species.— Columbia: Univ. Press,
1951.
Dobzhansky Th. Nature and Origin of Heterosis / / Heterosis/Ed. by
J. W. Gomen.— Iowa, 1952.— P. 218— 223.
D obzhansky Th. Genetics of natural populations. XXV Genetic changes in
populations of Drosophila pseudoobscura and Drosophila persimilis in
some localities in California / / Evolution.— 1956a.— Vol. 10, N 1.—
P. 82— 92.
Dobzhahsky Th. What is an adaptive trait / / Amer. Nat.— 1956b.— Vol. 40,
N 855.— P. 337— 347.
Dobzhansky Th., Pavlovsky O., Spassky B., Spassky N. Genetics of natural
populations. XXIII Biological role of deleterious recessives in
populations of Drosophila pseudoobscura / / Genetics.— 1955.— Vol.
40.
Dobzhansky Th., Queal M. L. Genetics of natural populations variations in
populations of Drosophila pseudoobscura inhabiting mountain ranges / /
Genetics.— 1938a.— Vol. 23.— P. 239— 251.
Dobzhansky Th., Queal M. L. Genetics of natural populations of Drosophila
pseudoobscura inhabiting isolated mountain ranges / / Genetics.—
1938b.— Vol. 23.— P. 463— 484.
Dobzhansky Th., Spassky N. Genetics of natural populations. XXII. A
Comparison or the concealed variability in Drosophila pseudoobscura
and Drosophila persimilis / / Genetics.— 1954.— Vol. 39, N 6 .—
P. 899— 907.
Dobzhansky Th., Wright S. Genetics of natural populations. V. Relations
between mutation rate and assumulation of lethals in populations of
Drosophila pseudoobscura / /
Genetics.—
1941.—
Vol. 26.—
P. 23—51.
Dowdeswell W. H.
Isolation
and
Adaption
in populations of the
Lepidoptera / /
Proc. Roy. Soc.—
1956.—
B. 145,
N 920.—
P. 322— 329.
Dowdeswell W. H., Ford E. B. The Influence of Isolation on Variability in
Butterly Maniola jurtina L. / / Evolution Symp. Soc. Exp. Biol.
Cambridge.— 1953.— Vol. 7 .— P. 254— 273.
Fascher J. Evolution and Bird Sociality / / Evolution asa Process/Ed. by
J. Hyxlev, A. C. Hardy, E. B. Ford.— L., 1954.— P. 71—83.
Fischer R. A. On some oblections to mimicry theory statistical and genetic / /
Trans.— Oxford, 1927.
F ischer R. A., Ford E. B. The variability of species in the Lepidoptera with
reference to abundance and sex / / Trans. Ent. Soc. London.— 1929.—
Vol. 7 6 .— P. 367— 384.
Ford E. B. Polymorphism and Taxonomy / / The New Sistematics.— Oxford,
1940.— P. 493—513.
Ford E. B. The study of organic evolution by observation and experiment / /
Endeavour.— 1956.— Vol. 15, N 5 9 .— P. 149— 152.
Garstang W. The theory of recapitulation. A critical restatement of biogenetic
law / / J. Linn. Soc. London Zool.— 1922.— Vol. 35.—P. 81 — 101.
Gordon C., Spurway H., Street P. A. K. An analysis of three wild populations
of Drosophila subobscura / / J. G en.— 1939.— Vol. 38.— P. 37—90.
G ustafsson A. The advantageous effect of deleterious mutations / /
Hereditas.— 1947.— Vol. 33.— P. 573—575.
Haldane J. B. S. Animal populations and their regulation / / New Biology.—
1953.— Vol. 15.— P. 9 — 24.
94
Haldane J. В. S. The Statics of Evolution / / Evolution as a Process/Ed. by
J. H yxley, A. C. Hardy, E. B. Ford.— L., 1954.— P. 109— 121.
Haldane J. B. S. The theory of selection for melanism in Lepidoptera / /
Proc. Roy. Soc.— 1956a.— B. 145, N 920.— P. 303—306.
Haldane J. B. S. The relation between density regulation and natural
selection / /
Proc. Roy. Soc.—
1956b.—
B. 145,
N 920.—
P. 306— 308.
Hardy A. C. Escape from Specialization / / Evolution as a Process/Ed. by
J. Huxley, A. C. Hardy, E. B. Ford.— L., 1954.— P. 122— 142.
Hivanitz W. Polymorphism and Evolution / / Evolution Symp. Soc. Exp. Biol.
Cambridge.— 1953.— Vol. 7 .— P. 238— 253.
H uxley J. S. Morphism as a clue in the study of population dynamics / /
Proc. Roy Soc.— 1956.— B. 145, N 920.— P. 319—322.
Kettlewell H. B. D. A resume of investigations on the evolution of melanism in
the Lepidoptera / / Proc. Roy. Soc.— 1956a.— B. 145, N 920.—
P. 297— 305.
Kettlewell H. B. D. Further selection experiments on industrial melanism in the
Lepidoptera / / Heredity.— 1956b.— Vol. 10, pt 3 .— P. 387— 301.
Levine R. P.
Crossingover and
inversions in coadapted systems / /
Amer. Nat.— 1956.— Vol. 90, N 850.— P. 41—45.
Mather K. Polymorphism as an outcome of disruptive selection / / Evolution.—
1955. — Vol. 9, N 1.— P. 5 2 — 61.
Mather K. Polygenic mutation and variation in populations / / Proc. Soc.—
1956. — B. 145, N 920.— P. 293—297.
Miller N. A new concept in genetics the group / / Actes XI Congr. Internat
philos. Bruxelles.— 1953.— Vol. 6 .— P. 242— 246.
M uller H. J. Genetic variability, twin hybrids and constant hybrids, in a case
of balanced lethal factots / / Genetics.— 1918.— N 3.
M uller H. J. The problem of genic modification / / Verh. V. Int. Congr.
Vererbungswiss. Suppl.— 1927.— Bd 1.— S. 234—260.
Sheppard P. M. Polymorphism and population Studies / / Evolution Symp.
Soc. Exp. Biol. Cambridge VII.— 1953a.—Vol. 7 .— P. 274—289.
Sheppard P. M. Polymorphism, linkage and blood groops / / Amer. Nat.—
1953b.— Vol. 8 7 .— P. 283— 294.
Sheppard P. M. Evolution in bisexually reproducing organisms / / Evolution as
a Process/Ed. by J. Huxley ^ L. A. C. Hardy, E. B. Ford.— L.,
1954.— P. 201— 218.
Sheppard P. M. Ecology and bearing on population genetics / / Proc. Roy.
Soc.— 1956.— B. 145, N 920.— P. 308— 315.
Southern H. N. Mimicry in Cuckoos Eggs / / Evolution as a Process/Ed. by
J. H yxley, A. C. Hardy, E. B. Ford.— L., 1954.— P. 219—232.
Stebbins G. L. Variation and Evolution in Plants.— 1951.
Stern C., Carson G., Rinst M„ Novitski E., Uphoff D. The viability of
heterozygotes
for
leth als/ /
Genetics.—
1952.—
Vol. 37.—
P. 413—449.
Autosomal lethals in wild populations of Drosophila
pseudoobscura / / Biol. Bull.— 1937.— Vof 73.— P. 542—551.
Sturtevant A. H.
T iessier G. Vitalite et fecondite relative de diverses combinatins genetiques
comportant un gene lethal chez chez la Drosophila / / Compt. Rend.
Acad. Sci., Paris.— 1942.— Vol. 214.— P. 241— 244.
T im ofeeff-R essovsky N. W. Zur Analyze des Polimorphismus bei Adalia
bipunctata L / / Biol. Zbl.— 1940.— Bd 60.— S. 130.
Thoday J. M. Components of Fitness / / Evolution Symp. Soc. Exp. Biol.
Cambridge.— 1953.— Vol. 7 .— P. 96— 113.
Valentine D. H. Variation and polymorphism in Viola / / Proc. Roy. Soc.—
1956.— B. 145, N 920.— P. 315—319.
Vavilov N. I. The law of homologous series in variation / / J. G en.— 1922.—
Vol. 12, N 1.— P. 47— 89.
Westergaard M. The relation between chromosome constitution and sex in the
offspring of triploid Melandrium / / Hereditas.— 1948.— Vol. 34.—
P. 257— 279.
95
ЗНАЧЕНИЕ ВАЛЕНТНОСТИ ЯДРА
В Ж И ЗН И И ЭВОЛЮЦИИ ВИДА
(О НЕКОТОРЫХ МЕХАНИЗМАХ,
ОБЕСПЕЧИВАЮЩИХ ОПТИМАЛЬНОЕ
ЧИСЛО НАБОРОВ ХРОМОСОМ)*
Р. Л . Берг
ВВЕДЕНИЕ
Оптимальным называется некоторое количественное
выражение признака, обеспечивающее максимальную ус
тойчивость системы для систем существенно устойчивых,
или наилучшие условия преобразования систем преобразу
ющихся. Преобразование — программированное измене
ние. Живые системы сочетают в себе устойчивость в пре
делах данной фазы преобразования со способностью пере
хода из одной фазы преобразования в другую.
Органический мир нашей планеты представляет собой
иерархию
соподчиненных
систем
[Needham,
1937;
Bertalanffy, 1949, 1951; Ляпунов, 1962; Тимофеев-Ресов
ский, 1962]. Совокупность живых существ биосферы [Вер
надский, 1926], биоценозов [Сукачев, 1948], вид [Завад
ский, 1961], организм, клетка и макромолекулярный ком
плекс, обладающий способностью к самовоспроизведению,
представляют собой последовательные ступени этой иерар
хии. Каждая ступень представляет собой целостное образо
вание,
обладающее
всеми признаками
целостности
[Шмальгаузен, 1961 ].
Живые системы отличаются друг от друга типом преоб
разования [Берг, 1962]. Преобразование макромолекулярного комплекса выражается в саморепродукции путем по
строения гомолога-копии, клетка — система делящаяся,
организм — система циклическая, осуществляющая про
цесс онтогенетических преобразований и воспроизводящая
себя с помощью отделения маленькой, а подчас и особо ус
троенной, части, вид — система эволюционирующая. Пре
образования вида происходят по программам наименее же
стко детерминированным по сравнению с программами
преобразования организма, клетки и способного к аутореп
родукции макромолекуляркого комплекса. Эволюция из
* Доклад на II Конференции по полиплоидии 17.1.63 г. и на заседа
нии Московского Общества Испытателей Природы в секции генетики и
селекции.
96
всех преобразований в наибольшей степени совершается с
помощью вероятностного программирования и является в
наибольшей степени новообразованием.
Валентность ядра, или число наборов хромосом — важ
ный признак клетки и организма, состоящего из клеток с
соответствующей валентностью ядра. Валентность ядра —
один из диагностических признаков вида. Для биоценоза
как целого совсем не безразлично каков уровень валентно
сти ядра слагающих его видов. Понятие оптимума валент
ности ядра имеет смысл лишь применительно к определен
ному уровню организации живого.
Оптимальной для каждого уровня является такая ва
лентность ядра, которая обеспечивает устойчивость в пре
делах фазы преобразования и не налагает запрет на пере
ход из одной фазы преобразования в другую.
Как всякий признак, определенная валентность ядра
устанавливается под действием естественного отбора у ди
ких видов и в результате совместного действия искусствен
ного и естественного отбора у культурных видов.
Естественный отбор выступает во всем многообразии
форм. Если возникло изменение числа наборов хромосом,
оно прежде всего будет подвергнуто испытанию со стороны
клеточного отбора.
Множество условий должно быть соблюдено: соотноше
ние между объемом ядра и объемом клетки, между объе
мом клетки и ее поверхностью, качественно-количествен
ные соотношения между ядром и цитоплазмой, все то, от
чего зависят обмен веществ и делимость клетки. Рыжков
[1962] на Первом совещании по полиплоидии, Навашин
[1963] на Втором совещании частично осветили эти усло
вия. Сохранятся лишь те клетки, для которых изменение
числа наборов хромосом не влечет за собой гибели.
В результате индивидуального отбора, или отбора осо
бей, будет устранено каждое изменение, которое снижает
жизнеспособность организма и понижает его шансы оста
вить потомство.
Если валентность ядра превратилась в признак, прису
щий группе особей — популяции, а затем виду, она под
вергается испытанию со стороны группового отбора, как
внутривидового, совершающегося между изолированными
группами внутри вида, так и межвидового, оперирующего
целыми видами в пределах рода.
Пройдя через горнило всех этих форм отбора, новая
валентность ядра входит в гармоническое сочетание с дру
гими свойствами клетки, обеспечивая ее жизнедеятель
ность и делимость, она сочетается с признаками организ
4 Р. Л. Берг
97
ма, повышая его шансы выжйть и оставить потомство, она
входит в закономерный комплекс свойств, характеризую
щих вид, как целое, обеспечивая виду жизнь и возмож
ность эволюционных преобразований*
Оптимальным является число хромосом, которое удов
летворяет условиям существования и преобразования всех
уровней организации живого.
Оптимум валентности ядра различен для разных видов.
Жуковский [1963] указал, что биологический прогресс до
стигается разными видами на разном уровне валентности
ядра. Искусственный отбор дает наилучшие результаты
при разных уровнях плоидности у разных сельскохозяйст
венных культур. Наиболее продуктивны гексаплоидные ви
ды пшениц [см. также Вавилов, 1935, с. 11 — 12], наилуч
шие культурные виды земляники — октоплоиды, сахарная
свекла и арбузы дают максимальный выход сельскохозяй
ственной продукции на триплоидном уровне, хлопчат
ник — на тетраплоидном. Оптимальным уровнем валент
ности Жуковский называет тот уровень, который дает наи
больший хозяйственный эффект.
Интересы человека могут столкнуться с интересами са
мого растения. Естественный отбор пресечет тогда деятель
ность искусственного отбора. Среди отобранных лучших
будут выживать тогда одни лишь худшие. Хохлов [1963]
предложил различать экономический и биологический оптимумы валентности. Однако и биологический оптимум, т.
е. тот уровень валентности, который регулируется естест
венным отбором, различен по отношению к разным функ
циям живого организма. Астауров [1963] с особенной си
лой подчеркнул тот факт, что жизнь и воспроизведение
особи осуществляется наилучшим образом на разном уров
не валентности ядра.
Начальные этапы удвоения наследственной субстанции
в эволюции носили универсальный и прогрессивный харак
тер. Самовоспроизведение осуществляется на основе струк
тур ДНК, имеющих строение двойных нитей. Модель самовоспроизводящейся структуры, построенная Пенроузом
[Penrose, 1958, 1959], состоит из двух цепей. Такая струк
тура обеспечивает не только наращивание материала пу
тем притяжения из окружающей среды веществ, необходи
мых для построения копии, но и последующую регуляцию
размеров.
Именно программированная регуляция размеров отли
чает самовоспроизведение от кристаллизации, а она воз
можна только в спаренных гомологичных структурах
98
[Pontecorvo, 1958]. Возникновение жизни рисуется как
возникновение этих двойных структур.
Двойные молекулярные структуры лежат в основе мно
гоступенчатой иерархии микрофибрилл (молекул нуклеопротеидов), хроматид, политенных хромосом и повторения
целых наборов хромосом.
Прокофьева-Бельговская [1960], подводя итог исследо
ваниям по субмикроскопической организации хромосом,
склоняется к мнению, что единичные молекулы дезоксинуклеопротеида диаметром 20— 30 А представляют собой
низшую ступень этой иерархии.
Установлено, что в ядрах ранних сперматид содержатся
хромосомные нити толщиной 100—250 А. В процессе пре
образования ядра сперматиды в ядро спермия эти нити за
мещаются нитями с диаметром 28 А. Происходит это заме
щение путем деполиплоидизации или в результате отбра
сывания белкового компонента хромосомы, сказать сейчас
невозможно.
Робертис, Новинский и Саэс [1962] на основе данных
литературы приходят в тому же выводу о хромосоме, как о
иерархической системе подчиненных элементов, основной
единицей которой является нитевидный макромолекулярный компонент, или микрофибрилла. Толщина этого ком
понента совпадает, по-видимому, с толщиной одной моле
кулы нуклеопротеида и составляет 20— 30 А. Число микро
фибрилл в политенной хромосоме, возникшей в результате
эндомитоза, может достигать 1000. Иерархии структурных
элементов соответствует, вероятно, целая шкала размеров
волокон.
Гольдшмидт [Goldschmidt, 1961] сближает политению
и полиплоидию. «Возможно,— пишет он,— что полиплои
дия и политения в одном отношении совпадают: как та,
так и другая служит увеличению поверхности того шабло
на, который способствует синтезу протеинов».
«Различные наблюдения показывают, что число эле
ментарных нитей в хромосомах варьирует не только в раз
ных клетках вследствие эндомитоза, создающего политенные хромосомы, но и в сходных клетках у разных ви
дов»,— пишет Прокофьева-Бельговская [1960].
Загадочно соотношение между сложным характером
строения хромосом из повторяющихся структур и явления
ми перекреста, коньюгации, расхождения хромосом и в
особенности мутагенеза и расщепления, т. е. всеми теми
явлениями, в которых хромосома ведет себя, как целое.
99
ПРИСПОСОБИТЕЛЬНОЕ ЗНАЧЕНИЕ
РА ЗН Ы Х СТЕПЕНЕЙ ВАЛЕНТНОСТИ ЯДРА
Низшие организмы — бактерии, вирусы существуют на
гаплоидном уровне организации их наследственной суб
станции. Однако уже здесь при слиянии отдельных особей
друг с другом на короткий период возникает диплоидия и
создается возможность использовать те преимущества, ко
торые она дает: осуществляется процесс рекомбинации на
следственных задатков.
Переход к диплоидному состоянию знаменовал собой
прогрессивное развитие органических форм, обеспечивая
повышение энергии жизнедеятельности и оптимальные ус
ловия использования мутационного процесса. На высших
этапах эволюции гаплоидны только гаметы, а весь цикл
онтогенетических преобразований протекает на диплоид
ном уровне. Диплоидность — универсальное явление среди
высокоорганизованных представителей органического мира.
Дальнейшее повышение валентности ядра наталкивает
ся на преграды. Высшие степени плоидности могут давать
преимущества лишь в определенных сочетаниях призна
ков. Ряд прогрессивных признаков налагает на увеличение
валентности ядра запрет. Подвижность относится к тем
свойствам, которые не сочетаются, или сочетаются с тру
дом, с высокими степенями валентности. Гетеротрофный
способ питания требует подвижности. Как активные, так и
пассивные способы передвижения предъявляют жесткие ус
ловия к сохранению пропорций подвижного существа. По
вышение валентности ядра меняет не только размеры орга
низма в сторону увеличения, но, главным образом, его
пропорции [Навашин, 1951, 1963].
Перед гетеротрофными, подвижными существами от
крылись новые перспективы прогресса в виде разделения
полов. Разделение полов создало богатейшие возможности
использования перекрестного оплодотворения, а тем самым
гетерозиса и комбинативной изменчивости. В эволюции
возник генотипический механизм реализации пола. Вы
сшие ступени развития этого механизма завершились реа
лизацией пола с помощью генного баланса между половы
ми хромосомами и аутосомами.
Генотипический механизм реализации пола, как ука
зал Мёллер еще в 1925 г. [Muller, 1925], является серьез
нейшим барьером для увеличения валентности ядра. Жи
вотные с их гетеротрофным способом питания, подвижно
стью, разделением полов и механизмами определения пола
с помощью баланса половых хромосом и аутосом вынужде
100
ны были отказаться от увеличения в процессе эволюции
числа наборов хромосом [White, 1945; Darlington, 1953;
Баранов, Астауров, 1956; Дарлингтон, 1956; Уайт, 1956;
Астауров, 1962, 1963].
Другой барьер для повышения уровня валентности ядра
это гаметогенез, сопровождающийся редукцией числа хро
мосом. Нечетные степени валентности — как например,
три- и пентаплоидия, полностью исключают нормальное
течение гаметогенеза, четные степени, такие как тетра- и
гексаплоидия, затрудняют его, приводя к образованию га
мет, неспособных при оплодотворении дать зиготы с урав
новешенным числом хромосом.
Аутотрофный способ питания, оседлый образ жизни и
способы размножения исключающие редукцию хромосом,
т. е. вегетативное размножение и апомиксис представляют
собой тот комплекс условий, который благоприятствует
включению высших степеней валентности ядра в число
приспособительных признаков вида, и мы действительно
видим, как широко использовали растения полиплоидию
на путях своего биологического прогресса.
Полиплоидия у животных обнаружена в соответствии с
предсказанием Мёллера [Muller, 1925], лишь в сочетании с
гермафродитизмом у земляных червей [Мюльдаль, 1956] и
в сочетании с партеногенезом у жуков долгоносиков [Суомалайнен, 1956], у некоторых ракообразных, чешуекры
лых и прямокрылых. Сакс [1956] указывает на возмож
ность образования полиплоидов у раздельнополых форм,
размножающихся половым путем, в частности у хомяков,
а может и у песчанок. Включение полиплоидии в комплекс
приспособительных признаков вида должно было сопро
вождаться здесь перестройкой механизма определения пола.
Однако и у растений на пути повышения валентности
ядра то тут, то там встают преграды. Потеря способности к
вегетативному размножению и апомиксису, расселение се
мян с помощью ветра, наличие специфического переносчи
ка пыльцы затрудняют повышение валентности ядра.
Барьером для включения высоких степеней валентно
сти в комплекс приспособительных признаков вида являет
ся энтомофилия, при наличии специфического переносчика
пыльцы. Повышение валентности ядра таит в этом случае
опасность нарушения биоценотических связей.
Транспортировка пыльцы с помощью насекомого, отно
сящегося к определенному виду, возможна только в случае
строжайшего соблюдения стандарта размеров и пропорций
цветков данного вида растения [Берг, 1956, 1958, 1959,
1961]. Цветки растений, имеющих специфических пере
101
носчиков пыльцы, локализуют пыльцу всегда на одном и
том же маленьком участке тела насекомого, недосягаемом
для очистительных аппаратов транспортировщика пыльцы.
Конфигурация частей цветка такова, что насекомое дотра
гивается именно этим местом сперва до рыльца, а затем до
пыльников посещаемого им цветка. Явление локализации
пыльцы обнаружено Дарвином [1928] для орхидных и про
демонстрировано Шванвичем [1952, 1955, 1956] для мо
тыльковых. Явление это имеет первостепенное значение
для использования полиплоидии в качестве средства повы
шения продуктивности растений-медоносов, резко ограни
чивая возможности селекции полиплоидных форм. Нару
шение размеров и пропорций цветка может сделать расте
ние практически стерильным, так как нарушит механизм
локализации пыльцы. Одновременный отбор по способно
сти к самоопылению, или подбор новых переносчиков
пыльцы может спасти положение.
Как отмечает Стеббинс [Stebbins, 1950], «увеличение
размеров в результате полиплоидии чаще всего наблюдает
ся и сильнее всего проявляется в органах с конечным ти
пом роста, в таких, как чашелистики, лепестки, пыльники,
малосеменные плоды и семена», т. е. как раз в тех орга
нах, на изменение размеров которых природа в наиболь
шей степени наложила запрет, подвергнув их жесткому
стабилизирующему отбору со стороны биотических компо
нентов среды и специфических функций (энтомохория се
мян) [Берг, 1956, 1958, 1959, 1961].
Рамануджам и Партасарати [1956] пишут: «Автопо
липлоидия играла в видообразовании и эволюции растений
сравнительно небольшую роль. Главной причиной этого,
по-видимому, является пониженная способность автопо
липлоидов приспосабливаться к биотическим факторам».
Преграды на пути использования высоких степеней ва
лентности у растений связаны, как мы видим, с преодоле
нием оседлости. Транспортировка семян с помощью ветра
(анемохория) и транспортировка пыльцы с помощью насе
комых (энтомофилия) — эти средства выхода за пределы
места произрастания, предъявляя строгие требования к
соблюдению пропорций и размеров семян и цветков,
затрудняют использование полиплоидии в эволюции рас
тений. Распространение полиплоидии в растительном
царстве связано с характером их размножения [Muller,
1925; Stebbins, 1950; Баранов, Астауров, 1956; Липаева,
1963; Хохлов, 1963] и способами транспортировки семян и
пыльцы.
102
ВАЛЕНТНОСТЬ ЯДРА КАК УСЛОВИЕ
ИСПОЛЬЗОВАНИЯ МУТАЦИОННОГО ПРОЦЕССА
Валентность ядра является не только условием, обеспе
чивающим, в комплексе с другими условиями, жизнь и
воспроизведение вида* Как каждый признак, определенная
валентность ядра открывает, или облегчает одни эволюци
онные преобразования и закрывает, или затрудняет, дру
гие. Каждый признак организации клетки, организма и ви
да, как целого, представляет собою внутреннюю предпо
сылку дальнейшей эволюции вида, условие, которое опре
деляет деятельность естественного отбора, элемент про
граммы эволюционных преобразований.
Валентность ядра представляет собой один из важней
ших компонентов, генетической системы. Понятие генети
ческой системы введено в науку Дарлингтоном [Darlington,
1937, 1953]; Гекели [Huxley, 1942]; Стеббинсом [Stebbins,
1950] и Уайтом [White, 1945]. Генетическая система
включает строение наследственного аппарата и способ раз
множения популяции, т. е. представляет собой совокуп
ность структур, обеспечивающих функциональное единство
событий, разыгрывающихся на клеточном и популяцион
ном уровнях организации вида. Строгое согласование ва
лентности ядра со способом воспроизведения вида — одна
из главных характеристик генетической системы.
Использование мутационного процесса видом находится
в теснейшей зависимости от валентности ядра. Использова
ние комбинативной изменчивости в результате перекомбинации и сегментного обмена хромосом отцовского и мате
ринского наборов становится возможным лишь начиная с
диплоидного уровня. Только начиная с диплоидного уровня
возникает предпосылка для использования гетерозиса, для
создания различного рода балансированных систем, осно
ванных на гетерозиготности. Вместе с тем, условия исполь
зования вновь возникающих мутаций, равно как и условия
отсева всего вредоносного, наиболее благоприятны у гапло
идных организмов. Высшие степени валентности создают
свои преимущества и свои недостатки для использования
мутационного процесса видом.
С возрастанием валентности ядра возрастает число ло
кусов, способных мутировать и входить в новые комбина
ции с другими генами. Относительно большая часть мута
ций переводится в разряд малых, а тем самым полезных
мутаций [Берг, 1941]. Этому повышению наследственной
пластичности противостоит более могучее погашение про
явления вновь возникающих мутаций [Stadler, 1929; Ман
103
сурова и др., 1958; Эфроимсон, 19616; Астауров, 1962; Са
харов 1962], падение вероятности перехода мутаций в го
мозиготное состояние [Stebbins, 1950; Дубинин, Щерба
ков, 1963], и падение, по крайней мере у некоторых форм,
самой частоты возникновения мутаций. Сахаров [1963] и
Платонова [1963] продемонстрировали это удивительное
явление для хромосомных перестроек у гречихи.
Образование скрытого резерва мутаций, в частности
накопление гетерозиготных вредоносных для гомозигот му
таций, также стоит в тесной зависимости от валентности
ядра. Гаплоидность исключает его, диплоидность и высшие
степени валентности создают для него благоприятную
предпосылку.
Высокая плоидность ядра имеет тенденцию снижаться в
течение эволюции в промежутках между кратными увели
чениями валентности. Гомологичные хромосомы теряют
свою гомологию с каждой вновь возникшей мутацией. Вы
сокие степени плоидности создают богатейший материал
для дивергенции геномов.
ЭВОЛЮ ЦИЯ ПЛОИДНОСТИ КАК КОМПРОМИСС
Из всего сказанного выше следует, что оптимум вален
тности ядра различен для разных уровней организации жи
вого. Более того, жизнь и воспроизведение особи обеспечи
ваются наилучшим образом разными степенями валентно
сти. Но и этого мало.
Валентность ядра, обеспечивающая наилучшим обра
зом жизнь и воспроизведение вида на коротком отрезке
времени, может, в конце концов, привести вид к гибели.
Но и это еще не все.
Жизнь и воспроизведение вида могут обеспечиваться
одним уровнем валентности, а оптимальный темп эволю
ции, дающий виду победу в межвидовом соревновании,—
другим уровнем валентности.
Действительно, высокая валентность ядра всех кле
ток тела, помимо генеративных, дает преимущества по
сравнению с низкой валентностью. Она погашает мута
генез, и может играть таким образом канцеростатическую
роль в той мере, в какой соматический мутагенез является
причиной злокачественного роста [Эфроимсон, 1961а].
Высокая валентность повышает энергию жизнедеятель
ности соответствующих клеток и тканей. Вместе с тем,
генеративные клетки дают наибольшее количество дее
способных гамет при диплоидном уровне. Здесь интересы
104
жизни и воспроизведения приходят в противоречие друг с
другом.
Конфликт возникает и между жизнью и воспроизведе
нием вида на коротком отрезке времени и жизнью и восп
роизведением вида на длительном отрезке времени. Дипло
идный уровень валентности ядра позволяет накопиться ог
ромному количеству летальных и субвйтальных мутаций в
генотипе популяций вида. Гибель части особей в каждом
поколении в результате перехода этих мутаций в гомози
готное состояние с лихвой окупается наличием мобилиза
ционного резерва наследственной изменчивости и гетерози
сом, возникающим на основе гетерозиготности. Но есть ви
ды, которые не могут накапливать в своем генотипе ле
тальные мутации и жертвовать частью особей в каждом
поколении. Это те виды, у которых воспроизведение сопря
жено с большой затратой сил со стороны родительского по
коления для выращивания каждой отдельной особи. Выход
из положения может быть найден в гаплоидности части
особей вида в каждом поколении. Гаплоидность выступает
в данном случае, как внутренний регулятор интенсивности
естественного отбора и приобретает биоевгеническое значе
ние, способствуя очищению вида от генотипически непол
ноценных особей.
Установление оптимальной валентности ядра осуществ
ляется, как некий компромисс. Компромисс этот может
быть достигнут двумя путями.
В результате деятельности разнонаправленных процес
сов отбора валентность ядра устанавливается на опреде
ленном уровне, представляющем некую равнодействую
щую, которая удовлетворяет до известной степени требова
ниям жизнедеятельности и делимости клетки, соответству
ющим условиям циклических превращений и воспроизве
дения особи и эволюционным преобразованиям вида.
Другой выход из положения может быть найден по ли
нии сочетания различных уровней валентности в разных
тканях одного и того же организма, на разных стадиях
жизненного цикла, у разных особей одного и того же вида
у многоклеточных организмов и по линии сочетания раз
личных уровней валентности макро- и микронуклеуса у
высших представителей одноклеточных или, если угодно,
бесклеточных животных — инфузорий.
Оптимум валентности быть может не установился ни
где и никогда. Создается некоторая приемлемая величина,
удовлетворяющая требованиям сегодняшнего дня и не уст
раняющая возможностей дальнейших поисков оптимума
системой. Эшби [1962] подчеркнул общие причины, за
105
трудняющие приближение к оптимуму живых систем. Ос
новная преграда — длительное время, необходимое для до
стижения оптимального состояния.
М ЕХАН И ЗМ Ы ОБРАЗОВАНИЯ РАЗЛ И ЧИЙ
М ЕЖ ДУ КЛЕТКАМ И ОРГАНИЗМА
И М ЕЖ ДУ ОСОБЯМИ ОДНОГО ВИДА
ПО УРОВНЮ ВАЛЕНТНОСТИ ЯДРА
Четыре механизма лежат в основе образования внутри
видовых и внутриорганизменных различий по уровню ва
лентности ядра.
Их можно подразделить на два типа: I — онтогенети
ческую и II — видовую миксоплоидию. Каждый тип де
лится на два класса. Онтогенетическая миксоплоидия пред
ставлена хорологическим и хронологическим классами. Ви
довая миксоплоидия подразделяется на: 1 — смену жиз
ненного цикла и 2 — сочетание в пределах фазы жизненно
го цикла особей с разной валентностью ядра. Таблица 1
(см. вкладку) дает представление о классификации спосо
бов обеспечения оптимума валентности ядра.
I.
Хорологическая онтогенетическая миксоплоидия.
Сочетание высоких степеней валентности соматических
тканей с диплоидностью генеративных тканей у многокле
точных организмов позволяет извлечь максимальную поль
зу из полиплоидии и вместе с тем благополучно миновать
барьер, который ставит редукционное деление и определе
ние пола с помощью хромосомного баланса на пути дости
жения высших степеней валентности.
Сочетание в одном существе клеток с разной валентно
стью Астауров [1963] предложил назвать миксоплоидией.
Миксоплоидия распространена, по-видимому, очень широ
ко как у растений, так и у животных. Аналогом ее у про
стейших служит сочетание высокой степени валентности
макронуклеуса с диплоидностью микронуклеуса, характер
ное для высших ступеней эволюции инфузорий [Hartmann,
1952; Grell, 1953; Полянский, 1960; Полянский, Райков,
1960; Райков, 1963].
Можно указать 4 способа происхождения миксоплоидии.
1.
Механизм, позволяющий сочетать в пределах одного
и того же организма клетки с разной валентностью ядра,
это двойное оплодотворение у покрытосеменных растений.
Зародыши всех покрытосеменных — миксоплоиды.
106
2. Эндомитотическая полиплоидизация и политения
хромосом соматических тканей при сохранении диплоидности генеративными тканями. Впервые политенные гигант
ские хромосомы были обнаружены в ядрах клеток слюн
ных желез у двукрылых. Соматическая конъюгация хромо
сом облегчает исследование политении у этой группы жи
вотных. Уайт [White, 1945] уделил хромосомам слюнных
желез двукрылых специальную главу в сводке «Цитология
животных и эволюция». Раскрыто биологическое значение
полиплоидизации питающих клеток в яичнике насекомых
[Painter, Reindorp, 1939; Хвостова, 1947; Эфроимсон,
19616]. Полиплоидизация клеток в результате эндомитоза
обнаружена Соколовым [1963] в сперматогенезе многих
видов пауков. Клетки с гигантскими ядрами играют и здесь
трофическую роль, аналогичную питающим клеткам в он
тогенезе насекомых. В настоящее время эндомитотическая
полиплоидизация и политения известны у многих растений
и животных, в том числе у млекопитающих и у человека
[Geitler, 1954; Баранов, Астауров, 1956; Жинкин, 1962;
Щербаков, 1962а, б]. Явления эти привлекают все возра
стающее внимание цитологов всего мира. На Первом сове
щании по полиплоидии и эндомитозу у растений посвятили
свои доклады Липаева [1962] и Захарьева [1962]. Баранов
и Матвеева [1962] глубоко затронули этот вопрос. На Вто
ром совещании по полиплоидии явления эндомитоза и по
литении нашли глубокое и разностороннее освещение [Ас
тауров, 1963; Бродский, 1963; Горощенко, Чуксанова,
1963; Захарьева, 1963; Лагучев, 1963; Прокофьева-Бельговская, 1963; Рябинина, 1963; Соколов, 1963; Щербаков,
1963].
3. Третий способ создать различия в валентности ядер
в клетках одного и того же организма — это слияние на
правительных телец с ядром одного из бластомеров. Такую
ситуацию наблюдали у одного из видов червецов, а именно
у Pseudoanlacapsis pentagona (Targ.) Беннет и Броун
[Bennet, Brown, 1958]. Ядра спермия и яйцеклетки, несу
щие по 8 хромосом, сливаются. Начинается дробление. Яд
ро одного из бластомеров сливается с обоими направитель
ными тельцами, несущими соответственно 16 и 8 хромо
сом. Образовавшаяся пентаплоидная клетка, имеющая
16 + 16 + 8 = 8 х 5 = 40 хромосом, дает начало особой
ткани, служащей питательным материалом для однокле
точных симбионтов.
4. Соматическая редукция хромосом — процесс обрат
ный эндомитозу — обнаружена Сахаровым [1946] у тетраплоидной гречихи в качестве патологического явления.
107
В качестве элемента нормального морфогенеза соматиче
ская редукция наблюдалась у комаров в родах Culex, Aedes
и Anopheles рядом авторов [Berger, 1938; S. Grell, 1946а,
b, Risler, 1959, цит. по: White, 1945; Робертис, Новинский,
Саэс, 1962; Рыжков, 1962; Щербаков, 1962а], а также у
радиолярий [К. Grell, 1952, 1953а]. Согласно Холту [Holt,
1917], полиплоидные клетки многих тканей комаров, в
противоположность политенным хромосомам, вступают в
процесс деления. Бергер [Berger, 1938] утверждает, что в
течение метаморфоза такие, не утратившие способности
делиться, клетки претерпевают соматическую редукцию,
причем восстанавливается диплоидное число хромосом.
Уайт [White, 1945] допускает возможность другого процес
са — дегенерации клеток с высокой валентностью ядра. На
Втором совещании по полиплоидии вопрос о редукции чис
ла наборов хромосом получил освещение в докладах Ата
бековой [1963]; Рябининой [1963] и Бродского [1963]. Со
матическая редукция, описанная перечисленными автора
ми, по-видимому, явление вторичное, переводящее перво
начально увеличенное число наборов хромосом снова на
диплоидный уровень.
И. Хронологическая онтогенетическая миксоплоидия
(временная гетероцикличность). Выключение одного из
наборов хромосом во всех клетках организма, если оно на
ступает на определенной стадии онтогенеза, создает осо
бый тип миксоплоидии. Ранние стадии развития протекают
на диплоидном уровне, поздние — на гаплоидном. Этот
тип миксоплоидии можно назвать хронологическим в отли
чие от предыдущего — хорологического, где разные уровни
валентности разобщены пространственно, но сосуществуют
во времени. Наблюдается хронологическая миксоплоидия у
коккцид (Coccoidae) в семействе Lecanoidae. У самцов ком
плекс хромосом, привнесенный спермием, переходит на оп
ределенной стадии развития в пикнотическое состояние во
всех исследованных клетках тела. В мейозе он элиминиру
ется [White, 1945; Brown, 1958, 1959].
Различие в поведении отцовского и материнского
комплекса хромосом у коккцид наводит на мысль о су
ществовании еще одного механизма, обеспечивающего
оптимум валентности ядра. Механизм этот — гетероцик
личность отцовского и материнского наборов хромо
сом. Прокофьева-Бельговская [1948] привлекла внима
ние к этому явлению и указала на его биологическое
значение.
Межгеномная гетероцикличность, или разновременное
прохождение стадий митотического цикла отцовским и ма
108
теринским хромосомными наборами, функционально сни
жает валентность ядра зародыша. Хромосомный набор отца
вступает в действие несколько позже, чем материнский на
бор хромосом, в дальнейшем он может быть весь временно
выключен из действия, как мы это видели на примере
коккцид из семейства Lecanoidae или полностью элими
нирован, как у коккцид из семейства Diapsidae. Гетеро
цикличность хромосомных наборов делает организм проме
жуточным по валентности ядра между диплоидным и
гаплоидным.
III. Различие уровней валентности на разных стадиях
жизненного цикла. Смена дипло- и гаплофазы характерна
для всех видов, размножающихся половым путем, и не ут
ративших вторично процесс оплодотворения. Прогрессив
ная эволюция сопровождается сокращением гаплофазы и
удлинением диплофазы. Преимущества диплоидного состо
яния над гаплоидным носят, по-видимому, универсальный
характер. Однако в той мере, в какой гаплоидная фаза со
хранена, она может служить средством отсева вредоносных
мутаций. Следует ожидать, что популяции растений со
держат меньше гетерозиготных летал ей, чем популяции
животных.
IV. Различие уровней валентности ядра у разных осо
бей одного и того же вида в пределах фазы жизненного
цикла. Однако существуют виды животных, у которых
смена гапло- и диплофазы не обеспечивает оптимальную
для вида интенсивность отбора. Колониальный образ жиз
ни, коллективная защита потомства, выкармливание дете
нышей и личинок, все те ситуации, когда представители
следующего поколения ведут паразитический образ жизни
за счет предыдущего поколения, приводят к возникнове
нию внутренних регуляторов интенсивности естественного
отбора. Интенсивность индивидуального отбора, достаточ
ная для своевременного отсева всего неполноценного, т. е.
всего того, что легло бы тяжким бременем на стадо, коло
нию или популяцию, обеспечивается целым комплексом
закономерно сочетающихся свойств. Гаплоидия входит как
необходимый компонент в число этих биоевгенических
приспособлений, У медоносной пчелы интенсивность отбо
ра ослаблена, личинки избавлены от каких бы то ни было
опасностей бытия. Это ослабление отбора в одном отноше
нии компенсировано повышением отбора в другом отноше
нии. Лишь биологически полноценные могут оставить по
томство. Компенсаторами служат брачный полет, огром
ный избыток трутней, бои маток и, наконец, гаплоидия
самцов. Самки и рабочие пчелы медоносной пчелы дипло
109
идны и развиваются из оплодотворенных яиц. Самцы раз
виваются из неоплодотворенных яиц в результате гаплоид
ного партеногенеза. Гаплоидия самцов в сочетании с жест
кими условиями испытания их летных качеств и их органов
чувств с помощью брачного полета препятствует накоп
лению вредных мутаций в генотипе вида. Эндомитоз
восстанавливает диплоидное число хромосом в индиви
дуальном развитии трутня [Risler, 1954]. Самец оказыва
ется полностью гомозиготным по всем генам хромосомного
набора.
Гаплоидия присуща самцам многих перепончаток
рылых, в том числе самцам наездников. Гершензон [1946]
у одного из видов наездника (Mormoniella vitripennis)
проанализировал сотни хромосом самок с целью обнаружить
летальные и субвитальные мутации, скрытые в гетерози
готном состоянии в их генотипе, и не обнаружил ни одной
мутации. Подобный анализ у дрозофилы или тутового
шелкопряда выявил бы десятки мутаций среди равного ко
личества хромосом [Четвериков, 1926; Дубинин и др.,
1934; Эфроимсон, 1940; Dobzhansky, Wright, 1941; Берг,
1943].
У перепончатокрылых гаплоидия самцов является ре
зультатом
гаплоидного
партеногенеза.
У коккцид
(Coccoidae) дело обстоит иначе. У некоторых видов самцы
гаплоидны. Развиваются они из оплодотворенных яиц. Ге
нотипические механизмы определения пола отсутствуют.
У тех видов, которые подверглись тщательному изучению,
в частности у упоминавшегося уже Pseudoaulacapsis
pentagona (Targ.) обнаружен эпигенетический механизм
определения пола. Молодые самки откладывают яйца, из
которых развиваются самки, старые самки порождают сам
цов. И самки, и самцы диплоидны и имеют 16 хромосом во
всех клетках тела за исключением мицетоцитов, которые
пентаплоидны и имеют 40 хромосом. На ранних стадиях
эмбриогенеза самцов один из наборов хромосом во всех
клетках, имеющих по 16 хромосом, переходит в пикнотическое состояние и элиминируется. Самцы становятся гап
лоидными. Элиминации подвергается отцовский комплекс
хромосом. Так обстоит дело у коккцид из семейства
Diapsidae. У коккцид из семейства Lecanoidae (у которых
отцовский комплекс хромосом пребывает в пикнотическом
состоянии во всех клетках тела самца в течение всей жиз
ни и элиминируется лишь в мейозе) самцы также функци
онально гаплоидны.
по
ВАЛЕНТНОСТЬ ЯДРА КАК РЕГУЛЯТОР
Валентность ядра представляет собой внутренний регу
лирующий механизм* Скорость течения биохимических
процессов стоит в непосредственной связи с числом набо
ров хромосом, т. е. с числом матриц, способствующих син
тезу активных веществ в клетке. Многие биохимические,
физиологические, морфогенетические процессы находятся
под более прямым контролем валентности ядра. Так обсто
ит дело на клеточном и онтогенетическом уровнях органи
зации живого. На видовом уровне валентность ядра конт
ролирует вероятность перехода каждой возникшей мутации
в досягаемое для отбора (гомозиготное, или гемизиготное)
состояние, характер выявления мутаций и, до известной
степени, саму мутабильность. Валентность ядра выступает,
таким образом, как один из регуляторов эволюционного
процесса.
Изменение валентности ядра служит, быть может, од
ним из механизмов, регулирующих взаимоотношения меж
ду представителями разных поколений, в том случае, когда
они вступают в тесный контакт друг с другом и между тка
нями одного и того же организма, когда генотипы этих
тканей могут оказаться различными.
Триплоидный эндосперм лучше диплоидного не только
потому, что энергия жизнедеятельности триплоидных тканей
выше, чем тканей диплоидных. Прибавление мужского ядра
к ядрам зародышевого мешка при образовании эндосперма
уравнивает до известной степени генотипы клеток эндоспер
ма и всех остальных тканей, предотвращая опасность несов
местимости белков эндосперма и других тканей зародыша.
Угроза несовместимости белков матери и плода в ре
зультате различия их генотипов могла бы быть уменьшена
межгеномной гетероцикличностью, которая выражается в
выключении из действия отцовского набора хромосом, в
результате чего на определенной стадии развития генотипы
матери и плода функционально уравниваются. Существует
ли это явление в природе в настоящее время — неизвестно.
Различия в валентности ядра выступают в качестве
стрелочных механизмов, определяющих полиморфизм вида
[Берг, 1957]. У самцов перепончатокрылых гаплоидия яв
ляется стрелочным механизмом, определяющим возникно
вение самцов, диплоидия — самок. Это геномный род ге
нотипического класса механизма определения полимор
физма. Возрастные отличия по степени валентности ядра
у самцов коккцид служат причиной возникновения
возрастного полиморфизма по строению ядра. В семействах
ш
Классификация
Единица
класси
фикации
Тип
Принцип
классификации
Полиморфизм
Динамика эволюции
Динамика или ста
Переходный полиморфизм
тика эволюции
Направленный сдвиг процентных
Наличие или отсутст
вие направленного сдви соотношений форм имеется
га процентных соотноше
ний форм
Класс
Природа стрелочного
механизма
Генотипический
Подкласс
Механизм регуляции
полиморфизма
Генный
Хромосомный
(интерхромосомный и интрахромосомный)
Полигенный
Супергенный
Экстрахромосомный
Геномный
Механизм, регулиру
ющий соотношение
форм в случае геномно
го подкласса полимор
физма, и арена его дей
ствия
Селективное
преимущество
вновь
возникшей
формы
с
измененной валентностью над
исходной формой. Вытеснение
форм с низшими степенями
валентности ядра формами с
высшими степенями на грани
це ареала и в крайних услови
ях обитания в пределах аре
ала у растений и при переходе к
партеногенезу,
самоопылению
и апомиксиду у животных и
у растений
Lecanoidae и Diapsidae в индивидуальном развитии самцов
происходит гаплоидизация ядер (структурная у Diapsidae,
функциональная у Lecanoidae). Этот род полиморфизма мо
жет быть отнесен к эпигенетическому классу механизмов,
определяющих полиморфизм вида, представляя геномный или
генотипический подкласс генотипического класса. Классифи
кацию типрв полиморфизма, приведенную в работе 1957 г.,
следует дополнить. Таблица 2 представляет эту классифика
цию в расширенном виде.
112
Таблица
2
полиморфизма
Полиморфизм
Статика эволюции
Балансированный полиморфизм
Направленный сдвиг отсутствует
Модификационный Эпигенетический
Генотипический
Климатический
(сезонный и ло
кальный)
Биоценотический (трофиче
ский и экзогормональный)
Морфогенетический
Эргонтический
Геномный
Генный
Полигенный
Супергенный
Интрахромосомный
Интерхромосомный
Экстрахромосомный
Суперхромосомный
Геномный
Изменение числа набо
ров хромосом в отдельных
тканях и во всем организме
в онтогенезе: эндомитоз,
соматическая редукция,
восстановление диплоидности трутней, образова
ние полиплоидных ядер у
радиолярий, полиплоид
ного макронуклеуса у
инфузорий, гаплоидизация тканей у самцов в не
которых семействах коккцид, переход от дипло
фазы к гаплофазе
Возникновение
разли
чий по числу наборов
хромосом между особя
ми одной популяции.
Гаплоидный
партено
генез у пчел, гаплоидизация тканей у сам
цов в некоторых се
мействах коккцид, пере
ход от гаплофазы к
диплофазе
ВАЛЕНТНОСТЬ ЯДРА КАК РЕГУЛИРУЕМОЕ ЯВЛЕНИЕ
В нашу задачу входило вскрыть механизмы, которые
обеспечивают оптимальный уровень валентности ядра в
тех случаях, когда этот уровень различен для разных про
явлений жизнедеятельности, воспроизведения и эволюции.
Выяснилось, что изменение валентности ядра может совер
шаться как в процессе эволюционных преобразований в
результате деятельности естественного отбора, так и в каиз
честве реакции на изменение ситуации в процессе диффе
ренциации тканей. Возраст матери у коккцид определяет
подвергнется ли у ее потомков один из наборов хромосом
элиминации на ранней стадии эмбриогенеза, или нет. От
различий в состоянии самих наборов хромосом зависит
элиминация именно отцовского комплекса хромосом. Ос
новной набор хромосом — гаплоидное число хромосом —
является строго фиксированным с помощью отбора наслед
ственным признаком*. Валентность ядра находится, по-ви
димому, в несколько ином положении. Мы не знаем, явля
ется эндомитотическая полиплоидизация некоторых тканей
многоклеточного организма причиной или следствием дифференцировки. Эпигенетический контроль гаплоидизации
тканей у самцов. Р. pentagona показывает, что изменение
валентности ядра может быть следствием дифференцировки, а если это так, то валентность ядра рисуется, как до
известной степени регулируемый процесс. Астауров обра
тил внимание на различие размеров клеток, органов и це
лых организмов у ди-, три- и тетраплоидных особей туто
вого шелкопряда. В течение развития первоначальные раз
личия в размерах выравниваются. Бабочки начинают раз
витие, имея разные уровни валентности ядра, заканчивают
его, имея, по-видимому, сходный уровень полиплоидности
ядра, образовавшийся эндомитотически. Если это так, ва
лентность ядра также предстает перед нами как процесс,
подлежащий регуляции в онтогенезе.
Мы совсем не касались вопросов гетероплоидии, или
различий в валентности разных хромосом хромосомного
набора, а также различий в валентности разных участков
одной и той же хромосомы.
Пайнтер и Рейндорп [Painter, Reindorp, 1939] обнару
жили, что в питающих клетках в яичнике дрозофилы раз
ные хромосомы и разные участки хромосом имеют разную
степень валентности. В то время как валентность активных
участков хромосом достигала 512, валентность гетерохро
матических районов всех хромосом и Y-хромосомы равня
лась четырем.
Образование вздутий у политенных хромосом в сомати
ческих тканях двукрылых сопровождается, по-видимому,
полиплоидизацией соответствующих участков хромосом.
Дифференциальные включения меченных предшественни
ков ДНК, РНК и белков в районы вздутий [Pavan, 1958]
* Это положение имеет исключения. Изменение числа хромосом в
разных тканях, как константный элемент морфогенеза, наблюдается у
Sciara. Пара больших хромосом — так называемые L-хромосомы, выбра
сывается из хромосомного набора соматических клеток [Metz, 1938].
114
делает полиплоидизацию этих районов весьма вероятной.
Наблюдения над половыми хромосомами в генеративных и
соматических делящихся клетках и в интеркинезе в норме
и паталогии позволяют пролить свет на природу сигналов,
регулирующих функциональную валентность Х-хромосом.
Сигналы эти носят, по-видимому, эпигенетический харак
тер и связаны с функциональным состоянием половых хро
мосом [Берг, 1963]. Наличие деспирализованной Х-хромосомы в клетках женщины служит препятствием к деспирализации второго партнера. Он остается в спирализованном
состоянии, образуя так называемый половой хроматин.
Эволюция половых хромосом совершается по линии
превращения у гетерогаметного пола бивалента, несущего
гены мужской и женской стерильности (делающие гермаф
родитное существо представителем женского или мужского
пола) в унивалент [Darlington, 1937, 1958; Берг, 1942]. Из
генного механизм определения пола в течение эволюции
становится хромосомным, а хромосомный механизм, по ме
ре деградации обреченного на вечную гетерозиготность
партнера, превращается в механизм определения пола с
помощью баланса половых хромосом и аутосом. По мере
превращения половой хромосомы у гетерогаметного пола в
унивалент, развивается механизм компенсации дозы генов
половой хромосомы [Muller, 1932]. Этот механизм в свою
очередь компенсируется выключением одной половой хро
мосомы в процессе дифференциации тканей у гомогаметного пола (у самок млекопитающих) [Barr, Carr, 1962]. Xхромосома у самок млекопитающих так же как у самцов
пребывает в состоянии унивалента, у самцов это структур
ная унивалентность, у самок — функциональная. Как по
казали новейшие исследования [Beutler et al., 1962; Lyon,
1962; Searl, 1962], в одних клетках женщины и самок мы
шей фyнкциoниpveт отцовская, в других материнская Xхромосома. Анализ механизмов, обеспечивающих включе
ние в действие той или иной хромосомы выходит за рамки
поставленной здесь задачи.
Функциональная гетероплоидия распространена, по-ви
димому, чрезвычайно широко. По мере надобности одни
гены выключаются из действия, другие включаются.
Гетерохроматизация отдельных участков хромосом
(дифференциальная спирализация и гетеропикноз) быть
может представляет собой механизм выключения соответ
ствующих участков из действия. Это предположение было
впервые сделано Шульцем [Schultz, 1952] и подверглось
жестокой критике со стороны Гольдшмидта [Goldschmidt,
1961]. Гольдшмидт считал, что нет фактов, которые нуж
115
дались бы в объяснении с помощью гипотезы Шульца. На
блюдения над половым хроматином в норме и в патологии
и представляют собой те факты, которые нуждаются в до
пущении дифференциального включения и выключения из
действия отдельных унивалентов с помощью дифференци
альной спирализации и гетеропикноза. Выключение из
действия это сведение валентности локуса к нулю. Вклю
чение в действие сопровождается повышением валентности
соответствующего локуса. В соматических клетках двукры
лых это так. Образование ламповых щеток в овогенезе ам
фибий также сопровождается повышением валентности деспирализованных участков. Рис [Ris, 1957] с помощью
электронного микроскопа показал, что петли состоят из
пучков субмикроскопических фибрилл. Хромонемы, точно
так же как политенные хромосомы в ядрах двукрылых,
увеличиваются при этом не только в толщину, но и в
длину. Уменьшение размера хромосом типа ламповых ще
ток на более поздних стадиях профазы происходит, вероят
но, за счет удаления материала на субмикроскопическом
уровне.
Эпигенетический контроль над валентностью разных
локусов хромосом имеет первостепенное значение для эво
люции валентности ядра. Возможность дифференциально
регулировать валентность локусов хромосом кладет предел
изменению в эволюции валентности целого ядра.
Оптимум дифференциальной валентности ядра устанав
ливается с помощью эпигенетического механизма и делает
все изменения целых наборов хромосом излишними. Конт
роль над валентностью ядра создает предпосылку для су
ществования хромосом с разным числом структурных гомо
логичных элементов в пределах хромосом.
Политенная хромосома, состоящая из множества нитей,
ведет себй так, как если бы она состояла из двух нитей.
По-видимому, существуют силы, действующие в пределах
пучка и не действующие за его пределами. Наличие этих
сил превращает хроматиды в функциональное целое.
Деполитенизация хромосом, т. е. сведение числа нитей,
образующих политенную хромосому до одной, и межге
номная гетероцикличность могут объяснить противоречие
между сложным строением хромосомы и ее поведением в
качестве элементарной единицы в митозе, мейозе и в про
цессах менделевского расщепления и мутагенеза.
Причина независимости хроматид друг от друга заклю
чается, как мне кажется, в межгеномной гетероциклично
сти ядра. Хроматиды, привнесенные яйцом и спермием, со
храняют на протяжении всей жизни свою автономию. Если
116
это так, становится понятным, что хроматида выступает
как единое целое в процессах деления клетки, перекреста
и спирализации. Единственность хромосомы в процессе му
тагенеза остается необъяснимым. Но если число нитей в
пучке может меняться, доходя до одной нити, то становит
ся понятной и единственность хромосомы в процессе мута
генеза. Сочетание представлений об изменчивости числа
нитей в хромосоме с гипотезой межгеномной гетероцик
личности позволит, мне кажется, понять как разрешается
конфликт между сложностью строения и элементарностью
поведения хромосом в процессе самовоспроизведения, при
перекресте, при делении клетки и в процессе мутирования
и расщепления.
выводы
Оптимальной является валентность ядра, наилучшим
образом обеспечивающая жизнедеятельность и делимость
клетки, жизнеспособность и воспроизведение организма,
жизнь и эволюцию вида.
Оптимальная валентность различна для разных уров
ней организации живого. Жизнедеятельность организма
обеспечивается некоторым не слишком высоким, но превы
шающим диплоидный, уровнем валентности. Процессы со
зревания гамет, сопровождающиеся редукцией хромосом,
протекают наиболее успешно в диплоидном ядре. Диплоидия обеспечивает комбинативную изменчивость, которая
исключена у гаплоидов. Гаплоидность способствует отсеву
вредных мутаций и может иметь у видов с пониженной
интенсивностью отбора компенсаторное значение, повышая
интенсивность отбора и выступая в качестве биоевгенического фактора.
Эволюция плоидности рисуется как компромисс, удов
летворяющий противоречивым требованиям разных уров
ней организации живого. Миксоплоидия, или сочетание
клеток с разной валентностью ядра в пределах одного орга
низма — одно из таких компромиссных решений задачи.
Двойное оплодотворение, ведущее к образованию триплоидного эндосперма, эндомитотическая полиплоидизация
некоторых соматических тканей, слияние ядра одного из
бластомеров с направительными тельцами и соматическая
редукция приводят к миксоплоидии. Некоторое функцио
нальное снижение плоидности может достигаться с по
мощью гетероцикличности отцовского и материнского на
боров хромосом. В эволюции возникают различия между
117
валентностью ядра на разных стадиях жизненного цикла
(смена дипло- и гаплофазы) и различия по уровню валент
ности ядра между представителями одного и того же вида в
пределах фазы жизненного цикла (диплоидия самок и гаплоидия самцов).
СПИСОК ЛИТЕРАТУРЫ
Астауров Б. Л. Диплоидный и полиплоидный термический партеногенез у
двух видов шелковичного червя и их гибридов / / Вопросы эволю
ции, биогеографии, генетики и селекции: Сб., поев. 70-летию
Н. И. Вавилова.— М.; Л.: Изд-во АН СССР, I960.— С. 32— 42.
Астауров Б. Л. Ионизирующее излучение и наследственность. Генетиче
ская теория лучевого поражения / / Природа.— 1962.— № 4 .—
С. 5 5 —67.
Астауров Б. Л. Экспериментальная полиплоидия у животных / / Тез.
докл. Второго совещания по полиплоидии, 14— 18 января 1963 г.—
Л., 1963.— С. 4 —5.
Атабекова А. И. О механизме возникновения полиплоидных клеток у рас
тений / / Тез. докл. Второго совещания по полиплоидии, 14— 18
января 1963 г.— Л., 1963.— С. 5 —6.
Баранов П. Ам Астауров Б. Л. Вступительная статья к сборнику «Поли
плоидия».— М.: ИЛ, 1956.— С. 5 —22.
Баранов П. А., Матвеева Т. С. Значение полиплоидии в эксперименталь
ной ботанике / / Полиплоидия у растений. Тр. Совещания по по
липлоидии у растений, 25— 28 июля 1958 г.— M.: Изд-во АН
СССР, 1962.— С. 11— 20.
Берг Р. Л. Роль малых мутаций в эволюции мутабильности / / Докл. АН
СССР.— 1941.— Т. 32, № 1.— С. 71—74.
Берг Р. Л. Историческое развитие механизма реализации пола / / Журн.
общ. биол.— 1942.— Т. 3, № 3 .— С. 171 — 189.
Берг Р. Л. Происходит ли элиминация особей гетерозиготных по леталь
ным
мутациям
в
естественных
популяциях
Drosophila
melanogaster / / Изв. АН СССР. Отд. биол. наук.— 1943.— Т. 4 .—
С. 243— 248.
Берг Р. Л. Стандартизирующий отбор в эволюции цветка / / Бот. журн.—
1956.— Т. 41, № 3 .— С. 3 1 8 - 3 3 4 .
Берг Р. Л. Типы полиморфизма / / Вести. ЛГУ. Сер. биол.— 1957.—
Т. 21, № 4 .— С. 115— 139.
Берг Р. Л. Дальнейшие исследования по стабилизирующему отбору в эво
люции цветка / / Бот. ж урн.— 1958.— Т. 43, № 1.— С. 1l — 27.
Берг Р. Л. Экологическая интерпретация корреляционных плеяд / /
Вести. ЛГУ. Сер. биол.— 1959.— Т. 9, № 2 .— С. 142— 152.
Берг Р. Л. Корреляционные плеяды у растений / / Тез. докл. на Третьем
совещании по примен. мат. методов в биологии.— Л.: Изд-во ЛГУ,
1961.— С. 10— 12.
Берг Р. Л. Структура и развитие / / Тез. докл. на науч. сес :ии Биологи
ческого института.— Л.: Изд-во ЛГУ, 1962.— С. 8 — 10.
Бродский В. Я. Полиплоидия соматических клеток животных тканей / /
Тез. докл. Второго совещания по полиплоидии, 14— 18 января
1963 г.— Л., 1963.— С. 9.
Вернадский В. И. Биосфера.— М., 1926.
Герш ензон С. М.
Генетическое
строение
природных
популяций
Mormoniella vitripennis Wlk. (Chaloididae, Hymenoptera) / / Журн.
общ. биол.— 1946.— Т. 7, № 3 .— С, 165— 173.
Горощенко Ю. Л., Чуксанова Н. А. Эндоплоидия в клубнях картофеля / /
Тез. докл. Второго совещания по полиплоидии, 14— 18 января
1963 г.— Л., 1963.— С. 1 1 - 1 2 .
Дарвин Ч. Присг особление орхидных к оплодотворению насекомыми / /
Поли. собр. соч.— 1928.— Т. 4.
118
Дарлингтон К. Полиплоидия у животных. Происхождение полиплои
дии / / Полиплоидия.— М.: ИЛ, 1956.— С. 287—292.
Дубинин Н. П. и др. Экспериментальный анализ экогенотипов Drosophila
melanogaster / / Биол. ж урн.— Т. 3, № 1.— С. 166— 206.
Дубинин Н. П., Щербаков В. К. Новое в использовании полиплоидии в
селекции растений / / Тез. докл. Второго совещания по полиплои
дии, 14— 18 января 1963 г.— Л., 1963.— С. 13.
Жинкин Л, Н. Эндомитоз и соматическая полиплоидия млекопитаю
щ и х / / Архив, анат., гист. и эмбриол.— 1962.— Т. 42, № 1.—
С. 3 - 2 1 .
Жуковский П. М. Эволюция культурных растений на основе полиплои
дии / / Бюл. МОИП. Сер. биол — 1963.
Завадский К. М. Учение о виде.— Л.: Изд-во ЛГУ, 1961.
Захарьева О. И, Полиплоидия в онтогенезе растений / / Полиплоидия у
растений. Тр. Совещания по полиплоидии у растений, 25—28
июня 1958 г.— М.: Изд-во АН СССР, 1962.— fc. 98— 109.
Захарьева О. И. Новые данные по эндополиплоидии клеток периблемы
корня / / Тез. докл. Второго совещания по полиплоидии 14— 18 ян
варя 1963 г.— Л., 1963.— С. 23—24.
Лагучев С. С. Значение полиплоидии и двуядерности в процессах физио
логической регенерации / / Тез. докл. Второго совещания по поли
плоидии, 14— 18 января 1963 г.— Л., 1963.— С. 26— 27.
Липаева Л. И, Полиплоидизация тканей в онтогенезе растений / / Поли
плоидия растений. Тр. Совещания по полиплоидии у растений,
25— 28 июня 1958 г.— М.: Изд-во АН СССР, 1962.— С. 90— 97.
Липаева Л. И. Об апомиксисе и перспективах его использования в селек
ции полиплоидов / / Тез. докл. Второго совещания по полиплои
дии, 14— 18 января 1963 г.— Л., 1963.— С. 29—30.
Ляпунов А, А. Об управляющих системах живой природы и общем пони
мании жизненных процессов.— М.: Изд-во АН СССР, 1962.
Мансурова В. В., Сахаров В, В., Хвостова В. В. Чувствительность дипло
идных и аутотетраплоидных растений к гамма-излучению / / Бот.
ж урн.— 1958.— Т. 43, № 7 .— С. 989— 997.
Мюльдаль С. Хромосомы земляных червей. I. Эволюция полиплоидии / /
Полиплоидия.— М.: ИЛ, 1956.— С. 307—321.
Навашин М. С. О значении размера меристематических клеток для роста
и развития / / Тр. Ин-та/Бот. Ин-т.— 1952.— Т. 7, № 2 .—
С. 268— 293.
Навашин М. С. О некоторых малоизученных цитологических вопросах,
связанных с полиплоидией / / Докл. на Втором совещании по по
липлоидии, 14— 18 января 1963 г.— Л., 1963.
Платонова Р. Н. Отбор диплоидных и аутотетраплоидных форм гречихи
посевной на радиоустойчивость и устойчивость к химическим му
тагенам / / Тез. докл. Второго совещания по полиплоидии, 14— 18
января 1963 г.— Л., 1963.— С. 44—45.
Полянский Ю. И. О хромосомном аппарате у простейших / / Вопросы
эволюции, биогеографии, генетики и селекции: Сб., поев. 70-ле
тию Н. И. Вавилова.— М.; Л.: Изд-во АН СССР.— 1960.—
С. 204— 209.
Полянский Ю. И., Райков И. Б. Роль полиплоидии в эволюции простей
ших / / Цитология.— 1960.— Т. 2, № 5 .— С. 509—518.
Прокофьева-Бельговская А. А. Гетероцикличность системы клеточного
ядра / / Журн. общ. биол.— 1948.— Т. 8, № 4 .— С. 247—280.
Прокофьева-Бельговская А. А. Строение хромосомы. Серия «Итоги нау
ки» / / Ионизирующее излучение и наследственность.— М.: Изд-во
АН СССР, I960.— № 3 .— С. 7 —60.
Прокофьева-Бельговская А. А. Поведение хромосом в эндомитотическом
ядре / / Тез. докл. Второго совещания по полиплоидии, 14— 18 ян
варя 1963 г.— Л., 1963.— С. 45—46.
Райков И. Б. Новые данные о полиплоидных ядрах у простейших / / Тез.
докл. Второго совещания по полиплоидии, 14— 18 января 1963 г.—
Л., 1963.— С. 46—48.
Рамануджам С., Партасарати Н. Автополиплоидия / / Полиплоидия.— М.:
ИЛ, 1956.— С. 9 5 — 129.
119
Робертис Е. де, Новинский В., Саэс Ф. Общая цитология.— М.: ИЛ,
1962.
Рыжков В. Л. Полиплоидия и качественно-количественные отношения в
генетике / / Полиплоидия у растений. Тр. Совещания по полипло
идии у растений, 25— 28 июня 1958 г.— М.: Изд-во АН СССР,
1962.— С. 33— 38.
Рябинина 3. А. Полиплоидия в неоперированной и регенерировавшей пе
чени крыс / / Тез. докл. Второго совещания по полиплоидии,
14— 18 января 1963 г.— Л., 1963.— С. 49—50.
Сакс Л. Роль полиплоидии в эволюции и хромосомные комплексы млеко
питающих / / Полиплоидия.— М.: ИЛ, 1956.— С. 299— 306.
Сахаров В. В. Соматическая редукция, как причина своеобразной мозаич
ности у тетраплоидной гречихи / / Докл. АН СССР.— 1946.—
Т. 52, № 4 . - С. 3 4 9 - 3 5 2 .
Сахаров В. В. Полиплоидия и радиация / / Полиплоидия у растений. Тр.
Совещания по полиплоидии у растений, 25—28 июня 1958 г.— М.:
Изд-во АН СССР.— 1962.— С. 52—64.
Сахаров В. В. Особенности полиплоидных растений, выявляемые действи
ем проникающего облучения / / Тез. докл. Второго совещания по
полиплоидии, 14— 18 января 1963 г.— Л., 1963.— С. 5 1 —53.
Соколов И. И. Эндомитотическая полиплоидизация клеток семенника па
уков / / Тез. докл. Второго совещания по полиплоидии, 14— 18 ян
варя 1963 г.— Л., 1963.— С. 5 5 —56.
Сукачев В. Н. Фитоценология, биогеоценология и география / / Тр. Вто
рого Всесоюз. Географ, съезда.— 1948.— Вып. 1.— С. 186—201.
Суомалайнен Э. Полиплоидия у партеногенетических долгоносиков / /
Полиплоидия.— М.: ИЛ., 1956.— С. 322— 330.
Тимофеев-Ресовский Н, В. Некоторые проблемы радиационной биогеоценологии.— Свердловск, 1962.
Уайт М. Данные, говорящие против существования полиплоидии у живо
тных, размножающихся половым путем / / Полиплоидия.— М.:
ИЛ, 1956.— С. 293— 298.
Хвостова В. В. Развитие хромосом питающих клеток яичника мухи-тахины Ernestia consobrina Mg / / Докл. АН.— 1947.— Т. 57, № 6 .—
С. 6 2 1 - 6 2 4 .
Хохлов С. С. Полиплоидия и апомиксис у покрытосеменных растений / /
Тез. докл. Второго совещания по полиплоидии, 14— 18 января
1963 г.— Л., 1963.— С. 64—66.
Четвериков С. С. О некоторых моментах эволюционной теории с точки
зрения современной генетики / / Жури, эксперим. биол.— 1926.—
Т. 2, № 1.
Шванвич Б. Н, О механизме опыления красного клевера медоносной пче
лой / / Докл. Всесоюз. акад. с.-х. наук им. В. И. Ленина.—
1952.— Т. 8 .— С. 33— 36.
Шванвич Б. Н. О механизме опыления мотыльковых культур медоносной
пчелой / / Тез. докл. науч. сессии биолого-почв. фак. ЛГУ.— Л.:
ЛГУ, 1955.— С. 4 2 - 4 3 .
Шванвич Б. Н. О работе медоносной пчелы на мохнатой вике и на лядв е н ц е/ / Вести. ЛГУ
Сер. биол.— 1956.— Т. 11, № 9 С. 5 5 - 6 1 .
Шмальгаузен И. И. Интеграция биологических систем и саморегуля
ция / / Бюл. МОИП. Отд. биол.— 1961.— Т. 66.— С. 104— 134.
Щ ербаков В. К. Типы полиплоидизаци и редукции наборов хромосом / /
Цитология.— 1962а.— Т. 4, № 5 .— С. 477—489.
Щ ербаков В. К. Полиплоидизация и редукция наборов хромосом у расте
ний под влиянием различных факторов и роль ядра и цитоплазмы
в этих процессах / / Усп. соврем, биологии.— 1962.— Т. 54,
№ 2 .— С. 146— 157.
Щ ербаков В. К. Соотношение кариокинеза и цитокинеза в явлениях полиплоидизации и редукции наборов хромосом в связи с проблемой
120
митоза / / Тез. докл. Второго совещания по полиплоидии, 14— 18
января 1963 г.— Л., 1963.— С. 70— 71.
Эфроимсон В. П. Измерение скорости мутационного процесса у тутового
шелкопряда / / Изв. АН СССР. Отд. Биол. наук.— 1940.—
С. 6 88— 705.
Эфроимсон В. П. Анализ управляющих механизмов канцерогенеза / /
Пробл. кибернетики.— 1961а.— Вып. 5 .— С. 217— 243.
Эфроимсон В. П. Управляющие механизмы лучевой болезни / / Пробл.
кибернетики.— 19616.— Вып. 6 .— С. 183—205.
Эшби У. Р. Конструкция мозга.— М.: ИЛ, 1962.
Barr М. L., Carr D. Н. Correlation between sex chromatin and sex chromoso
mes / / Acta Cytologia.— 1962.— Vol. 6, N 1.— P. 34— 45.
Bennett F. D., Brown S. W. Life History and Sex Determination in the Diaspine Scale, Pseudoaulacaspis pentagona (Targ). (Coccoidea) / / Canad. Entomolog.— 1958.— Vol. 90, N 6 .— P. 317—325.
Berger C. A. Multiplication and Reduction of Somatic Chromocome Groups as
a Regular Developmental Process in the Mosquito Culex pipiens / /
Publ. Carn, Inst. Wash.— 1938.— N 476.— P. 209— 232.
Bertalanffy L. von. Das biologische Weltbild / / Die Stellung des Lebers in
Natur und Wissenschaft.— B., 1949.— Bd 1.
Bertalanffy L. von. Problems of General System Theory / / Symp. on General
System Theory: a New Aproach to Unity of Science. Human Biol.—
1951.— Vol. 25, № 4 .— P. 302—311.
Beutler E. et al. The normal human female as a mosaic of X-chromosome acti
vity. Studies using the gene for G-6-PD-deficiency as a marker / /
Proc. Nat. Acad. Sci USA.— 1962.— Vol. 1. № 48.— C. 9 — 16.
Brown S. W. Haplodiploidy in the Diapsidae-Confirmation of an Evolutionary
Hupothesis / / Evolution.— 1958.— Vol. 12, № 1.— P. 115— 116.
Brown S. W. Lecanoid Chromosome Behavior in Three more Families of the
Coccoides
(Homoptera) / /
Chromosoma.—
1959.— Vol. 10.—
P. 278— 300.
Darlington C. D. Recent Advances in Cytology.— L., 1937.
Dobzhansky Th., Wright S. Genetics of Natural Populations. V. Relations bet
ween Mutation Rate and Accumulation of Lethals in Populations of Dro
sophila pseudoobscura / / Genetics.— 1941.— Vol. 26.— P. 23—51.
Ceitler L. Endomitose und endomitotisce Polyploidisierung / / Protoplasmatologia.— 1954.— № 6 .— P. 1— 89.
G oldschm idt R. Theoretische Genetik.— B., 1961.
Grell K. Der Stand unserer Kenntnisse iiber der Bau der Protistenkerne / /
Verh. Deutsch. Zool. Ges. in Freiburg.— 1952.
Grell K. Die Chromosomes von Aulacantna scolymantha Haeckel / / Arch.
Protistenk.— 1953a.— Vol. 99.
Grell K. Die Koniugation von Ephelota gemmipara / / Arch. Protistenk.—
1953b.— Vol. 99.
Grell S. M. Cytological Studies in Culex. 1. Somatie Reduction Divisions / /
Genetics.— 1946a.— Vol. 31, № 1.— P. 60— 76.
Grell S. M. Cytological Studies in Culex. II. Diploid and Meiotic Divisions / /
Genetics.— 1946b.— Vol. 31, № 1.— P. 77—94.
Harmann M. Polyploide (polyenergide) Kerne bei Protozoen / / Arch. Proti
stenk.— 1952.— Vol. 98, № 1.— P. 1—54.
Hyxley J. S. Evolution. The Modern Synthesis.— N. Y.; L., 1942.
Lyon M. F. Attempts to test the inactive — X-theory of dosage Compensation
in Mammals / / Abstracts of Papers on the 138 Meeting of the genetical
society.— Univ. of Edinburgh, 1960.
Metz C. W. Chromosome behavior, inheritance and sex determination in Sciara / / Amer. Nat.— 1938.— Vol. 72.— P. 485—520.
M uller H. J. Why polyploidy is rarer in animals than plants / / Amer. Nat.—
1925.— Vol. 5 9 .— P. 346— 353.
M uller H. J. Further studies on the nature and causes of gene mutations / /
Proc. VI Int. Congr. G en.— 1932.— Vol. 1.— P. 213— 256.
Needham J. Integrative Levels.— Oxford, 1937.
Painter T. SM Reindorp E. C. Endomitosis in the nurse cells of the ovary of
Drosophila melanogaster/ /
Chromosoma.—
1939.— Vol. 1.—
P. 276— 283.
121
Pavan C. Morpological and Phisiological Aspects of Chromocomal Activities / /
Proc. X Congr. G en.— 1958.— Vol. 1.— P. 321— 336.
P enrose L. S. Mechanics of Seif-reproduction / / Ann. Hum. G en.— 1958.—
Vol. 23, № 1.— P. 5 2 —72.
P enrose L. S. Self-reproducing m a c h in e s// Sci. Amer.— 1959.— Vol. 200,
№ 1.— P. 9 5 — 114.
Pontecorvo G. The versatility of heredity / / J. Linnean Soc., London (Bot.,
Zool.) — 1958.— Vol. 5 6 /4 4 , № 3 6 5 /2 9 5 .— P. 116— 122.
Ris H. Chromosome structure / / Chemical basis of heredity.— Baltimore,
1957.
Risler H. Die somatische Polyploidie in der Entwicklung der Honigbiene (Apis
mellifera L.) und die Wiederherstellung der Diploidie bei den
Drohnen / / Zeitsch. Zellforsch.— 1954.— Bd 41.— S. 1— 78.
Risler H. Polyploidie und Reduction in der Larvenepidermis von Aedes
aegypty L. (Culicidae) / / Chromosoma.— 1959.— Vol. 10, № 2 .—
P. 184— 209.
Schultz J. Interrelations between nucleus and cytoplasm: problems at the biolo
gical level / / Exp. Cell R es.— 1952.— P. 17—43.
Searl A. G. Is sex-linked tabby reallv recessive in the M ouse?// Abstracts of
papers on the 138 meeting of the genetical society.— Univ. of Edin
burgh, 1962.
Stadler L. S. Chromosome number and mutative rate in Avena and Triticum / / Proc. Nat. Acad. Sci.— 1929.— № 12.
Stebbins G. L. Variation and Evolution in Plants.— N.Y., 1950.
White M. I. D. Animal Cytology and Evolution.— Cambr. Univ. Press, 1945.
Раздел
II
КОРРЕЛЯЦИОННЫЕ ПЛЕЯДЫ
ЭКОЛОГИЧЕСКАЯ ИНТЕРПРЕТАЦИЯ
КОРРЕЛЯЦИОННЫХ ПЛЕЯД*
Р. Л. Берг
ВВЕДЕНИЕ
Понятие корреляционных плеяд введено в науку
П. В. Терентьевым в 1931 г. Наличие плеяд основано на
существовании корреляций между размерами одних частей
(а также между промерами в пределах части) и на отсут
ствии корреляций между размерами других частей. Суще
ствование корреляционных плеяд ведет к важным теорети
ческим выводам.
Отсутствие корреляций между размерами одних частей,
наличие корреляций между размерами других частей у од
ной и той же совокупности организмов указывает на опре
деленную автономность одних показателей по отношению
к другим показателям в пределах организма. Плеяды —
это выражение диференциации.
По отношению к данной части организма другие части
того же организма являются внешней средой. Возникнове
ние корреляционных плеяд в эволюции — показатель автономизации жизненных явлений по отношению к внеш
ним воздействиям, включая и взаимодействие частей в
пределах организма. Изучение корреляционных плеяд по
зволяет устанавливать степень автономности. Введение ко
личественной характеристики создает основу для внедре
ния сравнительного метода в изучение процесса автономизации.
Исследование корреляционных плеяд имеет первосте
пенное практическое значение и в области научного ана
лиза, и в сельском хозяйстве, и в медицине.
Применение статистических методов в некоторых обла
стях систематики неразрывно связано с выбором некорре
лированных признаков (Гейнке, Heincke; Фишер, Fischer;
А. А. Любищев; П. В. Терентьев). Общая теория плеяд,
* Вестник Ленинградского университета.— 1959 .— № 9.
123
частью которой является их интерпретация, как раз и по
служит руководящей нитью для выбора некоррелирован
ных друг с другом признаков или, как назвал их П. В. Те
рентьев [1959] признаков-индикаторов.
В сельскохозяйственной практике, в особенности в свя
зи с промышленной обработкой продуктов животноводства,
все возрастающее значение приобретает стандартизация
сельскохозяйственного сырья. Стандарт — это выражение
автономности, независимости от колебаний в условиях
формирования. Принцип корреляционных плеяд подводит
нас вплотную к овладению процессом стандартизации. По
знать, как возникают плеяды — значит понять механизм
возникновения независимости от колебаний в факторах
среды.
Общая теория плеяд должна, очевидно, ответить на
вопрос: почему существуют корреляционные плеяды и по
чему именно данная часть, а не какая-либо другая, выры
вается из корреляционных зависимостей, сохраняя посто
янство размеров, или варьируя по своим собственным зако
нам, независимо от изменчивости других частей организ
ма? Сформулировать общую теорию плеяд можно, устано
вив некоторый общий принцип, позволяющий рассматри
вать каждый случай наличия или отсутствия плеяд как ча
стное его проявление. Принцип этот, будучи установлен,
позволит предсказывать корреляционные плеяды.
Очевидно, однако, что решение задачи в каждом конк
ретном случае ограничено рамками специальной отрасли
науки. Каждая отрасль биологии может дать ответ на лю
бой вопрос, касающийся живых объектов или их совокуп
ностей. Но она даст его в рамках тех и только тех сторон
объекта, которые она сама изучает. Сформулированный в
терминах строго ограниченной отрасли знания, принцип
этот тем не менее будет общим, так как позволит вскрыть
то общее, что присуще всему многообразию явления, если
рассмотреть его в определенном аспекте.
Наша задача — дать экологическую интерпретацию
корреляционных плеяд. А это значит, что в центр внима
ния ставится принцип приспособленности, или принцип
целесообразности.
Задача разделяется на две части. Прежде всего надле
жит ответить на вопрос: каково приспособительное значе
ние корреляционных плеяд? Для этого нужно, во-первых,
установить изменчивость в самом явлении корреляционных
плеяд, или, говоря иначе, показать, что для одних и тех
же признаков в одном случае корреляционные плеяды
есть, а в другом их нет. Во-вторых, нужно установить, в
124
какую систему закономерно сочетающихся экологических
свойств входит наличие или отсутствие данной плеяды. На
этом пути выявится приспособительное значение наличия
или отсутствия корреляционной плеяды и фактор отбора,
ответственный за возникновение плеяды.
Только после того, как такая операция произведена хо
тя бы для одного случая, следует ставить вопрос об общем
принципе возникновения корреляционных плеяд. Ответ на
него следует искать, сравнивая способы возникновения са
мых разнообразных корреляционных плеяд, с тем, чтобы
выявить то общее, что присуще им всем.
М АТЕРИАЛ И МЕТОДИКА
С целью выявления изменчивости характера зависимо
сти размеров различных частей организма друг от друга
было изучено 19 видов травянистых растений. В табл. 1
приведен список растений, число изученных особей каждо
го вида и характеристика растений по интересующим нас
признакам. Один из изученных видов — ячмень —
Hordeum vulgare представлен в нашем материале двумя
разновидностями (Паллидум, сорт Пушкинский озимый 1
и голозерный). Пшеница — Triticum aestivum (сорт Диа
мант) — изучена в 1956 г. С. С. Красновидовой и в 1957 г.
автором; в настоящей статье приведены результаты второй
серии 1957 г. Ниже приведен список промеров. Каждый
промер обозначен буквой. Не все виды перечисленных рас
тений были изучены по всем признакам.
Наименьшее число изученных признаков равно трем,
наибольшее — десяти.
Вегетативные
части
Стебель
) Лист
, Соцветие
г Венчик
Околоцветник
Генеративные
части
<
Чашечка
Цветок
Зерно
Высота
Длина
Ширина
Число
Длина
Число цветков
(колосков)
Диаметр цветка
Длина лепестка (длина
цветковой чешуи)
Длина трубчатых
частей
Длина чашелистика
(длина колосковой
чешуи)
Длина чашечки
Тычинка — длина
Пестик — длина
Длина
А
В
С
D
F
G
I
J
К
м
N
О
Р
L
125
оЗ
X
(D
О «и
Эколого-морфологическая характеристика изученных растений
г>
о
яА
5
О.
<ц
о
0Q
0)
оЗ
о
X
2
5§
й в
Й О
5 *
<ц о <о
ООО
о
S 21
§ й§ о?
й
gg*
5а
X
а
В
cd
cd cd S
Ч я
**
о
К
05
=г
к
оЗ
X
к
А
ёs*
н
Л
45
sк
А
Q
5
S .S
О.
<и
н
оЗ
5
§
оЗ
Он
W
х£
•s Ч
>, о ч
sо ss gс
rff§
яu §S
<
1S
с-е-
81
csd £X
U<
оо
S
cd
и
s я
S- sх
Н
§ gС оя
S
•§*<■> О
2<sd flJ
I
оS
н ин аа>
>? Э
S «О г;5?
£я .2С S o ;
■fCE
ш
£ -Ь 1о.1 =
£ 3 гиаз
К
X
A
К
cd
§
3
I§
N so.
* § *g
Ь
s £Й
О . и ; 5* е
Н cd
3
и
о.'вчо
«
cd
X
А
5
В
cd
О.
из
Wя
уs I«
85
80)
I г
.
С
03 я
Я S
J3 05
35я охя
►
U V D
*0©<0
N0*0Tfr^ x^ oСОo too o ot o
it!
S2
126
А
§■Л1оЗ
£С я
У
0©с0»0©
to CO to r-to
Характеристика C. bipinnata касается не цветка, а соцветия.
УPh. divaricata билатеральная симметрия касается расположения тычинок, но не частей околоцветника.
УL. viscaria. L. chalcedonica и М. silvestre ноготки лепестков, налегая друг на друга, образуют ложную трубочку. Чашечка трубчатая.
S
В число генеративных частей мы включили те органы,
от формы и размеров которых зависит успех или неуспех
переноса пыльцы. Соцветия космеи, аналогичные по своей
функции радиально-симметричным цветкам, мы включили
в число генеративных частей. Все остальные соцветия
включены в число вегетативных частей. Мы обозначали
одной и той же буквой наружные части околоцветника,
точно так же одной и той же буквой мы обозначали внут
ренние части околоцветника любого растения. Нас интере
совали аналогичные части, и мы нимало не заботились о
гомологиях. Принципиальное сходство функции наружных
частей околоцветника разных растений оправдывало наши
поиски наличия или отсутствия плеяд именно для их раз
меров; то же справедливо и для внутренних частей. Второ
степенные различия в функционировании оправдывали на
шу надежду понять, почему присутствует или отсутствует
корреляционная плеяда, и дать на этом пути экологиче
скую интерпретацию плеяд.
Размеры разных частей изучались во всех случаях у
одной и той же совокупности особей. Были высчитаны ко
эффициенты корреляции между размерами всех изученных
частей в пределах каждого вида.
Наш материал, как видно из табл. 1, давал нам воз
можность сравнивать растения, резко различающиеся как
по характеру переноса пыльцы, т. е. по экологическим
признакам, так и по строению, расположению, типу сим
метрии цветков, т. е. по морфологическим признакам.
Среди наших растений имеются виды, размножающие
ся самоопылением (Т. aestivumy Я. vulgare), растения-анемофилы (Я. arenarius), растения-энтомофилы, не имеющие
специфических переносчиков пыльцы (A nemorosa, F.
collina, Papaver sp., С. bipinnata), растения-энтомофилы,
чья пыльца переносится или переносилась на родине ба
бочками (L. viscaria, L. chalcedonica, М. silvestre, N. alata,
Ph. divaricata), и растения, имеющие в лице пчел и шме
лей специфических переносчиков пыльцы. Среди них име
ются растения, чьи цветки снабжены трубчатыми частями
{L. vulgaris у D. datum , Т. та jus), растения с цветками, це
ликом преобразованными в трубочку (Digitalis ambigua,
D. purpurea), и растения с цветками, лишенными трубча
тых частей (G. pratense, Ch. angustifolium). Избранные на
ми виды отличаются друг от друга и по степени олигоме
ризации и фиксации числа частей цветка и околоцветника.
127
РЕЗУЛЬТАТЫ
Выяснилось, что корреляционными плеядами по изу
ченным признакам обладает 12 видов растений: L. viscaria,
L. chalcedonica, М. silvestre, N. alata, PA. divaricata, G. pratense, Ch. angustifolium, L. vulgaris, £>. elatum, £>. ambigua,
D. purpurea и 71. та jus. Корреляционными плеядами по
тем же признакам не обладают 7 видов растений: 7\ aestfvwm, обе разновидности Я. vulgare, Я. arenarius, Л. петогоsa, F. collina, Papaver sp. и С. bipinnaia.
У первых 12 видов растений изученные признаки обра
зуют две группы: в пределах каждой из них все размеры
положительно коррелированы друг с другом, размеры при
знаков, относящихся к разным группам, не коррелированы
друг с другом или, обнаруживают небольшую степень кор
реляции. Такое распадение на группы мы, вслед за
П. В. Терентьевым, называем наличием корреляционных
плеяд.
У остальных 7 видов растений наблюдается отсутствие
корреляционных плеяд; размеры связаны друг с другом вы
сокими положительными коэффициентами корреляции, об
разуя как бы одну единственную корреляционную плеяду.
Такую ситуацию мы рассматриваем как отсутствие корре
ляционных плеяд, так как только отсутствие корреляций
между каким-либо размерами очерчивает плеяды и создает
разницу между наличием корреляций вообще и наличием
корреляционных плеяд.
В настоящем сообщении не приводятся все полученные
результаты. В табл. 2—5 представлены коэффициенты
корреляции между размерами всех частей, изученных у
данного вида, для двух видов растений, обладающих кор
реляционными плеядами (Chamaenerium angustifolium —
Иван-чай, табл. 2 и Delphinium elatum — живокость,
табл. 3), и для двух видов растений, не обладающих кор
реляционными плеядами ( Triticum aestivum — пшеница,
табл. 4 и Hordeum vulgare — ячмень, табл. 5). В рамке
стоят коэффициенты корреляции между размерами разных
вегетативных и генеративных органов друг с другом, над
рамкой — коэффициенты корреляции между размерами
вегетативных органов друг с другом, справа от рамки —
коэффициенты корреляции между размерами генератив
ных органов друг с другом. Как видно из табл. 2 и З, раз
меры вегетативных частей Иван-чая и живокости корре
лированы друг с другом, точно так же как размеры генера-
128
Корреляция (r ± m r ) между размерами вегетативных и генеративных частей у C h lam aen eriu m a n g u stifo liu m Scop.
сЗ
а-
1—
1
©
+
1
Tj-
О
©
+
_ _H
—<—
I
©
©
+
1
+
1
ON
2
©
г©
©
+1
©
©
+
fl C
O
1—
1 ча- T
—
1—
'
о © © ©
+
1
+
1
+
1
+
1
Г" оо (N<
© © 1—
о
©" © ©
+ + 1 +
f1 C
O
тг
—
4 4t 1T
—
1—
'
о © © ©
+
1
+
1
+
1
+
1
N
O
csN
—
■
« со CO O
о © © ©
+ + 1 +
Tf
CO
о © © ©
O
со
о
о
+1
00
оо
о
+
о о
о о
+1
о
+1
NO
•о
о о
+
+
(N NO
—Н о
г-
о
о о о
г\
+1
о
+1
Tf
г©
+
00
+
+
NO
СО
W—I
rt
Tf
СQ
+
<
о
+
о
+
+1
со
©
1
+1 +1 +1
<4 00
©
CS ©
©" ©
©
1
1 +
, (N
©
+1
тГ
TJ©
+
,
CO
— < —*
©
©
©
+1 +1 +1
O n
СО
©
+
CN
©
+
со
©'
+1
©
(N
©
+
n
©
+1
т*
CN
©"
+
©
+
-
©
©
+1 +1
oo 00
©^
©
©
+
+
+
+
+
•о
Tf
СО
тг
''t Tf
00
©"
+1
«о
©
О
+
©
©
©
+1 +1 +1
ON O n CN
CO
©
©
©
©
©
+
1
о о о о о
о о о о о
+1 +1 +1 +1 +1
CN о 00 •о O n
оо
жн
-еис!ц
+1
ON
NO
о о о о
о о о о
+1 +1 +1 +1
ТГ VO г* о
оо NO Г'' ON
о о о о
о
5 Р. Л. Берг
©
+ +
—
■
'O
©N
© ©
+1 r+1
©
•o •o
©
©
+ +
©
г-
00
о о о
+
+
+
CQ и Q Цн о
_н s
О
CO
+
Oh
129
Таблица
3
Приз
нак
Корреляция ( r ± m r) между размерами вегетативных и генеративных часг
тей у D elph in iu m elatu m L.
А
В + 0 ,5 3 ± 0 ,1 0
G + 0,67 ± 0 ,0 8
Н + 0,4 5 ± 0 ,1 1
К + 0 ,2 2 ± 0 ,1 3
м
+ 0 ,1 6 ± 0 ,1 3
в
G
н
к
+ 0 ,5 8 ± 0 ,0 9
+ 0 ,5 7 ± 0 ,0 9 + 0 ,4 9 ± 0 ,1 0
—0 ,1 0 ± 0 ,1 4 + 0 ,1 2 ± 0 ,1 4 —0 ,0 3 ± 0 Д 4
—0 Л 0 ± 0 ,1 4 —0 ,1 3 ± 0 ,1 4 - 0 ,0 8 ± 0 Л 4
+ 0,40± 0Л 2
тивных частей. Между размерами генеративных и веге
тативных частей корреляция мала или в подавляющем
большинстве отсутствует. Как показывают табл. 4 и 5, у
пшеницы и ячменя корреляции существуют между всеми
изученными признаками. Коэффициенты корреляции меж
ду размерами зерна (L) и размерами других частей расте
ния несколько меньше других коэффициентов. Этого сле
довало ожидать, ибо зерно — это организм следующего по
коления со своим генотипом, а не часть родительского ор
ганизма. Корреляционные плеяды по признакам цветка у
пшеницы и ячменя отсутствуют.
Таблица 6 показывает, что у луговбй герани (Geranium
pratense) размер венчика не коррелирован с размерами
стебля и листа, а у ветреницы (Anemone nemorosa) корре
лирован.
Мы установили, таким образом, изменчивость в самом
явлении корреляционных плеяд. Во всех случаях наличия
плеяд частью, вырвавшейся из корреляционных зависимо
стей, оказался цветок.
130
О
О
Я
Корреляция ( r± m r) между размерами вегетативных и генеративных частей у Triticum aestivum (сорт Диамамант)
+1
00
•о
о
+
•о £
СЗ
я S
S Э
ч
VO И
сз £
Н §
в
NO
о
о
+1
00
»о
о
+
*
g
оо
оо
+1 +1
г—
<
О О N©
об
0
0
О
в
S3
О.
г-гоо
о"о
+1 +1
t-N
•о «о
о б
+ +
*
#1
-Н
Й
.§>
++
S
«О «О 00
2
g
£
о
о
+1
<N
«о
о
+
2
ООО
Г" Г"г - о
О О О о
б б б б
+1 +1+1+1
-ч т* СЛЧО
о " о 'о " o '
+ + + +
тг
о*
+1
On
о
Ь
О
+
<0*0^00
ooSo
*•
Г.
0 0©0
+1+1+1+1
O
vOo^Uo
NOt^^Tf
ббоб
++++
>в^
lii
tr **
>в
* 2
£ S
* со
В
я
Е°
13»s
о- S
£В оа
9 я
. *5
X9
зс
Я Ui
в Ь
5 о
9
й
н
2
я
в
о
Lb
d
q
NO
г-о
Яо
о -н
+1об
N0*0
По
О+
+
+
<
NOONON
ООО
ООО
+1 +1 +1
■ОООГNOГОГО
боб
+++
оо2
S
? OnON
Я§ оо
б
°о о
+1® +1 +1
noa! 00
'Пго (ЧГО
°о о'о
++ ++
со^о
'*SU
0
>
35
C
U
VO
NO Г-ООО
оо
+1 +1
°® б°.
оо . о
+1 +1 ,, +1
го—
< О U-i
г - +1 го
г - Т|f-NO NO
бб^б
++
+ +° +
о б
S-Й
С X
U.O
зл и
U-)NO NONO
оо оо
о б об
+1 +1 +1 +1
0OQ NOON
N0>0
nO*0
o'O' о б
++ ++
<
-н
о
о
+1
ГО
On
о
§CU
S3
NONO NONO
оо оо
об о 'о
+1 +1 +1 +1
NO00 NOC4
N0*0 vqvo
о б о б
+ + + +
ризнак
боб
+1+1+1
n
OГ-rf
боб
+++
nO0О
оо
об
+1 +1
N00
N0*0
о б
+ +
Г*00
оо
об
+1 +1
iro
«TO
TJ-
00 00
оо
о"о
+1 +1
NtО
^ rf
бб об
++ ++
<
и
>
35
S
+1
о
с
в
Я'
к
м
8.
си
о
и*
С
Измерена длина верхнего междоузлия.
Ij —длина наружной цветковой чешуи; ^ —длина внутренней цветковой чешуи.
сЗ
131
Таблица
6
Корреляция (r ± m r ) между размерами вегетативных и генеративных час
тей у A n em o n e n em orosa и G eran iu m pra ten se
Geranium pratense
Anemone nemorosa
Признак
А
В
Признак
А
в
В
+0,75±0,05
в
+0,71±0,06
J
+0,79±0,05 +0,77±0,05
I
+0,01±0,13 —0,05±0,13
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Попытаемся теперь показать, в какой комплекс законо
мерно сочетающихся признаков входит наличие или отсут
ствие корреляционных плеяд. Как показывает табл. 1, рас
тения, обладающие корреляционными плеядами, и расте
ния, лишенные их, могут обладать цветками, характеризу
ющимися как радиальной, так и билатеральной симмет
рией, цветки тех и других растений могут иметь как лате
ральное, так и концевое расположение. Тип симметрии и
характер расположения цветков не связаны на первый
взгляд закономерным образом с наличием или отсутствием
корреляционных плеяд. Наличие или отсутствие плеяд не
разрывно связано с характером переноса пыльцы. Все рас
тения, обладающие специфическими переносчиками пыль
цы, будь то бабочки или пчелы, обладают корреляционны
ми плеядами. Все растения, лишенные специфических пе
реносчиков пыльцы, независимо от способа опыления —
самоопылители, анемофилы и энтомофилы, лишенные спе
цифических опылителей, корреляционными плеядами не
обладают. Наличие плеяд по изученным признакам озна
чает известную автономность размеров цветка.
Приспособительный смысл этой автономии совершенно
ясен. Размеры цветков энтомофильных растений, имеющих
специфических переносчиков пыльцы, не только не зави
симы от размеров растения в целом, но, как показали на
ши исследования [Берг, 1956, 1958], более стабильны, чем
размеры стеблей, листьев и соцветий. Эта стабильность и
независимость размеров цветков являются непременными
условиями переноса пыльцы специфическими переносчика
ми. Дело в том, что пыльца определенного вида энтомо
фильных растений, имеющих ограниченный круг пере
носчиков, локализуется на строго определенном участке
132
или на немногих (двух, трех) участках тела насекомого.
Это явление строгой локализации пыльцы обнаружено еще
Дарвином у орхидных [Дарвин, 1928]. В недавнее время
Шванвич [1952, 1956], обнаружил его и подробно исследо
вал у мотыльковых. Место локализации пыльцы жестко за
дано размером и формой цветка. Но точно так же жестко
задано размером и формой цветка и место, куда должна
быть транспортирована пыльца. Размеры цветков расте
ний, имеющих специфических переносчиков пыльцы оче
видно приспособлены не к условиям местообитания самого
растения, а к размерам цветков других растений того же
вида, обитающих, быть может, в иных условиях, обладаю
щих другими размерами вегетативных частей, и к разме
рам насекомых — переносчиков пыльцы. Независимость
размеров цветков этих растений от размеров других частей
растений и постоянство размеров цветков — непременное
условие выполнения их функции.
Все это прежде всего относится не к размерам лепест
ков и отгибов венчика, служащих дистантными сигнализа
торами и ориентирами для привлечения насекомых и для
ориентации их в отношении цветка, а к тем частям расте
ния, которые обеспечивают контакт сперва между пыльни
ками цветка и строго определенным местом на теле насе
комого, а затем между этим местом и рыльцем другого
цветка. Важнейшую роль в обеспечении контакта играют
тычиночные нити и трубчатые части цветков. Именно их
размеры и обнаружили в нашем материале наибольшую
стабильность и полное отсутствие корреляций с размерами
вегетативных частей того же растения.
Наличие корреляционных плеяд по изученным призна
кам закономерно сочетается с наличием специфических
переносчиков пыльцы, а тем самым с явлением строгой ло
кализации пыльцы на теле переносчика. В это закономер
ное сочетание признаков входят все приспособления, обес
печивающие надежность транспортировки пыльцы и ее
экономное использование, а все это как раз и достигается
путем локализации комочка пыльцы на строго определен
ном маленьком участке тела насекомого. Эффект локали
зации достигается: с помощью билатеральной симметрии,
сокращающей до минимума число возможных поз билате
рально симметричного существа, каким является насеко
мое; наличием трубчатых частей, т. е. срастанием частей
околоцветника друг с другом, в результате чего строго ог
раничивается пространство, где разыгрывается вся драма;
уменьшением числа (олигомеризацией) и строгой фикса
цией числа гомологов и, наконец, латеральным положени
133
ем цветков, которые, даже будучи радиально-симметрич
ными по строению, функционируют как билатерально-сим
метричные. Каждый из 12 видов растений, имеющих спе
цифических переносчиков пыльцы, обладает по крайней
мере
одним
из
перечисленных
приспособлений
(см. табл. 1).
Таким образом, имеются два главные комплекса зако
номерно сочетающихся свойств, один из которых включает
наличие корреляционных плеяд, а другой — их отсутст
вие. Наличие корреляционных плеяд сочетается с наличи
ем специфических переносчиков пыльцы, с эффектом ло
кализации пыльцы и с одним или несколькими структур
ными признаками, обеспечивающими локализацию.
Отсутствие корреляционных плеяд сочетается с отсут
ствием специфических переносчиков пыльцы, с отсутстви
ем эффекта локализации и всех приспособлений, обеспечи
вающих локализацию.
Мы показали, таким образом, в какой комплекс зако
номерно сочетающихся свойств входит как наличие корре
ляционных плеяд, так и их отсутствие.
выводы
Каков же механизм отбора, ответственный за возник
новение корреляционных плеяд по генеративным и вегета
тивным органам у энтомофильных растений, имеющих ог
раниченный круг переносчиков пыльцы?
Механизм отбора, ведущий к возникновению корреля
ционных плеяд, в данном случае совершенно ясен. Пре
имущества независимости размеров одних частей от дру
гих, точно так же как фактор, устраняющий всех облада
телей зависимым типом развития, очевидны. Зависимые
цветки просто выбрасывались с жизненной арены, так как
их пыльца не достигала рыльца цветка. Оставляли потом
ство их независимые собратья. Только они одни экономно
локализовали комочек своей пыльцы на том самом месте
тельца насекомого, которое при следующем посещении
цветка дотрагивалось до рыльца цветка. Насекомые и были
тем фактором отбора, который калибровал цветок, вырвал
его из корреляционных зависимостей и превратил его раз
меры в самостоятельную корреляционную плеяду. Вместе с
тем, только калиброванное существо может осуществить
стандартизующий отбор размеров.
Насекомые осуществляют отбор стандартных цветков.
В формировании цветка в процессе онтогенеза они, само
134
собой разумеется, никакого участия не принимают. Функ
ция в данном случае выступает лишь как испытание при
годности. Несовпадение формирующих и отбирающих фак
торов и есть, очевидно, общий экологический принцип автономизации размеров одних частей от размеров других
частей, одних свойств от других свойств организма, а сле
довательно, и общий принцип возникновения корреляци
онных плеяд.
В нашем случае выполнение определенной функции
требует строгого соответствия размеров существ, не спо
собных влиять друг на друга в процессе индивидуально
го развития. Такая ситуация отнюдь не уникальна. Био
тические компоненты среды, не принимающие ни ма
лейшего участия в формировании особей какого-либо вида,
зачастую осуществляют отбор. Хищники и жертвы, самцы
и самки, животные, разносящие семена растений, и сами
эти растения стоят именно в таких отношениях друг к
другу.
Органы, выполняющие соответствующие функции об
щения с биотическими компонентами среды, вырываются
из корреляционных зависимостей и образуют корреляцион
ные плеяды. У растений — цветки, оболочки плодов и се
мян, листья, вовлеченные в процесс воспроизведения, у
животных — нервная система, половой аппарат, органы
чувств, пищеварительная система, поведение, связанное с
воспроизведением, социальные инстинкты представляют
собой то огромное поле, где разыгрываются процессы автономизации и где следует искать корреляционные плеяды.
Во всех случаях общим принципом их возникновения яв
ляется несовпадение средств существования и назначения
части. П. В. Терентьев [Terentjev, 1931 ] показал, что у ля
гушки размеры органов чувств и органов, играющих ту
или иную роль в воспроизведении обнаруживают наимень
шую зависимость от размеров других частей организма.
Берталанфи и Пирозинский [Bertalanffy, Pyrozinsky, 1952]
обнаружили непропорциональность роста для печени и
мозга у крыс. С наших позиций аллометрический рост
именно этих органов вполне понятен.
Автономность жизненных процессов покоится на гоме
остатических аппаратах цитологического, онтогенетическо
го и популяционного ранга. Значение онтогенетических
(developmental homeostasis) и популяционных (genetic
homeostasis) гомеостатических аппаратов в эволюции не
давно подчеркнул Лернер [Lerner, 1954]. Совокупность на
следственных задатков представляет собой гомеостатиче
ский аппарат, осуществляющий свое действие на клеточ
135
ном, онтогенетическом и популяционном уровнях органи
зации живого. Наследственные задатки играют роль стаби
лизаторов, находясь внутри клеток развивающегося орга
низма.
Возникновение автономности в эволюции указывает на
повышение относительной роли внутренних факторов раз
вития по сравнению с внешними [Шмальгаузен, 1938,
1939, 1946; Камшилов, 1939]. Совершенно очевидно, что
размер автономного цветка контролируется факторами, ле
жащими внутри самой закладки, скорее всего с помощью
внутриклеточного механизма.
В нашем случае повышение роли внутренних факторов
в развитии является результатом несовпадения формирую
щих и отбирающих устройств. Эти устройства, в сущности,
не совпадают никогда. Достаточно указать на смену поко
лений, как высказанное положение становится очевидным.
Формирование организма проходит в одних условиях,
жизнь его протекает в других. Родители формируют детей,
предназначая их к условиям, которые не могут быть иден
тичными тем условиям, в которых протекала жизнь роди
телей, а тем более условиям формирования. У форм, обла
дающих индивидуальным развитием, родители порождают
существо, отличное от них самих, которое только в про
цессе онтогенеза приобретает сходство с родителями. Орга
низм наследует от родителей не признаки свои и свойства,
а некоторое устройство, содержащее в кодированном виде
программу развития. Совокупность наследственных задат
ков, представляя собой внутренние предпосылки развития,
обеспечивает известную степень автономности жизненных
процессов по отношению к факторам внешней среды. Само
программирование развития имеет значение только у
форм, обладающих чередованием поколений или индиви
дуальным развитием, или тем и другим. Весь смысл на
следственного кода заключается в несовпадении формиру
ющих и отбирающих факторов. Несовпадение формирую
щих и отбирающих факторов ответственно, таким образом,
за возникновение всех форм автономности и всех уст
ройств, обеспечивающих автономность, в том числе за воз
никновение самого наследственного программирования раз
вития.
Что же дает нам в практическом отношении общий
эколого-эволюционный принцип корреляционных плеяд?
Он дает нам уверенность, что отбор на стандартное прояв
ление в меняющихся условиях среды, отбор устойчивых
состояний каждого отдельного признака, тот отбор, кото
рый насекомые осуществили в отношении цветков, позво
136
лит в процессе селекции новых пород и сортов и нам рвать
по желанию зависимости, создавая стандартное, независи
мое от колебаний в факторах внешней среды состояние
признака.
СПИСОК ЛИТЕРАТУРЫ
Берг Р. Л. Стандартизирующий отбор в эволюции цветка / / Бот.
ж урн.— 1956.— № ' 3 . - С. 318—334.
Берг Р. Л. Дальнейшие исследования по стабилизирующему отбору в эво
люции цветка/ / Бот. журн.— 1958.— № 1.— С. 12— 27.
Дарвин Ч. Приспособления орхидных к оплодотворению насекомыми / /
Поли. собр. соч.— 1928.— Т. 14.
Камшилов М. М. Отбор как фактор, меняющий зависимость признака от
изменения внешних условий / / Докл. АН СССР.— 1939.— Т. 23,
№ 4 .— С. 361— 364.
Терентьев П. В. Метод корреляционных плеяд / / Вести. ЛГУ.— 1952.—
№ 9 .— С. 137— 141.
Шванвич Б. Н. О механизме опыления красного клевера медоносной пче
лой / / Докл. Всесоюз. акад. с.-х. наук им. В. И. Ленина.—
1952.— Т. 8 .— С. 33— 36.
Шванвич Б. Н. О работе медоносной пчелы на мохнатой вике и лядвенце / / Вести. ЛГУ.— 1956.— № 9.— С. 55— 61.
Ш мальгаузен И. И. Организм как целое в индивидуальном и историче
ском развитии.— M.; Л.: Изд-во АН СССР, 1938.
Шмальгаузен И, И. Пути и закономерности эволюционного процесса.—
М.; Л.: Изд-во АН СССР, 1939.
Ш мальгаузен И. И. Факторы эволюции (теория стабилизирующего отбо
р а).— М.; Л.: Изд-во АН СССР, 1946.
Bertalanffy L., Pyrozinsky W. J. Ontogenetic and evolutionary allometrv / /
Evolution.— 1952.— Vol. 7, № 4 .— P. 387—393.
Lerner M. Genetic Homeostasis.— Edinburgh; London, 1954.
Terentjev P. V. Biometrische Untersuchuneen tiber die morphologischer
Merkmale von Rana ridibunda Pall / / Biometrica.— 1931.— Ba 23.—
S. 23—51.
КОРРЕЛЯЦИОННЫЕ ПЛЕЯДЫ
И СТАБИЛИЗИРУЮЩИЙ ОТБОР*
Р . Л. Берг
ВВЕДЕНИЕ
Отсутствие корреляции размеров разных частей в пре
делах одной и той же совокупности организмов одного вида
открыто В. А. Алпатовым у медоносной пчелы [Алпатов,
Тюнин, 1925], а затем независимо от него — П. В. Терен
* Применение математических методов в биологии.— Л.: Изд-во
ЛГУ, 1964.— Т. 3.
137
тьевым [Terentjev, 1931 ] у озерной лягушки. Алпатов по
казал, что размер хоботка медоносной пчелы не коррели
рован с размерами всех остальных частей тела насекомого.
Наличие корреляции лишь в пределах комплексов призна
ков при отсутствии корреляции между признаками разных
комплексов Терентьев назвал корреляционными плеядами
[Terentjev, 1931; Терентьев, 1959, 1960]. Корреляционные
плеяды описаны теперь как у растений [Шмидт, 1963], так
и у животных [Петрушевская, 1961 ].
Начиная с 1955 г. мною исследованы корреляционные
плеяды у растений [Берг, 1956, 1958, 1959, 1961а, б; Berg,
1959, 1960]. Настоящая работа посвящена итогам этих ис
следований и рассмотрению их результатов в связи с тео
рией живых систем.
Отправной точкой для работы служила теория стабили
зирующего отбора И. И. Шмальгаузена [1946]. В мою за
дачу входило не только вскрыть экологические причины
образования корреляционных плеяд в процессе естествен
ного отбора, но и сформулировать общий принцип возник
новения независимости отдельных признаков, который по
зволил бы предвидеть характер связей.
Можно дать качественную характеристику комплекса,
указав на признаки, закономерно сочетающиеся друг с
другом, такие как моносимметрия, сростнолепестность,
олигомерия частей, наличие специфического переносчика
пыльцы у растений или наличие хитинового панциря, ма
лые размеры, мозаичный тип эмбрионального развития у
насекомых. Можно охарактеризовать комплексы признаков
с количественной стороны, показав, например, что разме
ры каких-либо частей согласованы с размерами одних час
тей в пределах системы и в то же время независимы от
размеров других частей той же системы. То, что сказано о
размерах, приложимо к любым количественным показате
лям.
Независимость части от других частей является выра
жением ее стабильности. Выяснение причин возникновения
независимости — это в то же время выяснение причин
стабилизации. Экологический анализ корреляционных пле
яд, таким образом, представляет собой анализ стабилизи
рующего отбора. Но можно сделать и еще один шаг. Воз
никновение корреляционных плеяд неразрывно связано со
стабилизацией. Стабилизация же означает повышение от
носительной роли внутренних факторов развития по срав
нению с внешними. Наличие внутренних факторов разви
тия, осуществляющих свое действие в меняющихся услови
ях среды, способных преодолевать помехи, придавая тече
138
нию процессов специфичность в соответствии со своими
особенностями, характеризует собой всякий процесс, осу
ществляющийся на основе программы. Создание внутрен
них предпосылок событий — существенная черта програм
мирования. Экологический анализ корреляционных плеяд
подводит нас, таким образом, к механизму наследственно
го программирования.
Взаимозависимость признаков организма является не
отъемлемым условием надежности существования организ
ма и сама возникает в результате отбора наиболее устой
чивых особей. Однако она имеет меру. Независимость од
них процессов по отношению к другим процессам и одних
признаков по отношению к другим признакам столь же не
обходима для организма. Будучи выражением специфично
сти и устойчивости признака, независимость частей в отно
шении друг друга является непременным условием дифференцировки.
Оптимум независимости одних частей по отношению к
другим частям той же системы представляет собой адапта
цию, направленную на сохранение устойчивости системы в
целом.
У.
Р. Эшби [1962], анализируя возможности моделиро
вать адаптивное поведение, говорит о мере взаимозависи
мости частей как об оптимальном условии функционирова
ния системы. Относительно независимые друг от друга час
ти приспосабливаются друг к другу по тем же принципам,
по каким организм в целом приспосабливается к среде.
Обилие связей имеет преимущество, но несет с собой и не
выгоды. Разные органы реагируют на разные компоненты
среды. Если эти компоненты среды независимы друг от
друга, то независимыми друг от друга должны быть и ре
акции организма. Если семена данного растения разносятся
ветром, а пыльца переносится пчелами, нет никакой нуж
ды, чтобы строение семян стояло в какой бы то ни было
зависимости от окраски венчика цветка.
Живой организм рисуется как система, состоящая из
отдельных подсистем, обладающих своими показателями
устойчивости (нормой реагирования). Каждая из подсистем
взаимодействует с определенным комплексом внешних ус
ловий.
Влияние среды на изменение подсистемы следует рас
членить. Можно выделить здесь функциональное, морфоге
нетическое и филогенетическое влияние среды. Последнее
осуществляется через отбор.
Все признаки организма образуются в процессе разви
тия в результате взаимодействия наследственных факторов
139
со средой. Роль внешних и внутренних факторов в разви
тии разных признаков различна и может меняться в эво
люции. Одни признаки ставятся под жесткий генный конт
роль, в отношении других внешние воздействия сохраняют
формирующее значение. Все эти события осуществляются
под контролем отбора. Отбор, в процессе которого конеч
ное выражение признака не меняется, а меняется характер
его развития, причем относительная роль внутренних фак
торов развития, по сравнению с внешними, возрастает,
И. И. Шмальгаузен [1946] назвал стабилизирующим отбо
ром. Стабилизирующий отбор, т. е. отбор более жестких
программ индивидуального развития особи, происходит
только в тех случаях, когда более жесткое программирова
ние развития дает организму преимущество перед менее
строго детерминированным развитием признака.
Стабилизация отдельного признака имеет две стороны.
Во-первых, развитие признака в результате повышения отт
носительной роли внутренних факторов, по сравнению с
внешними, становится независимым по отношению к коле
баниям среды. Во-вторых, ослабевают взаимные влияния
развивающихся частей друг на друга. В результате стаби
лизирующего отбора развитие становится мозаичным. Ста
билизирующий отбор создает меру взаимозависимости час
тей. Так возникают корреляционные плеяды.
Независимости частей в отношении друг друга и неза
висимости по отношению к формообразующему действию
среды — два взаимообусловленных явления, вырабатываю
щиеся в эволюции совместно. С замечательной ясностью
это понято и продемонстрировано В. А. Драгавцевым
[1962]. Отбор по независимости частей друг от друга име
ет своим следствием независимость по отношению к среде,
а избирательное высвобождение отдельных частей организ
ма из-под контролирующего действия среды приводит к ав
тономии частей.
Целенаправленность развития, характеризующая жи
вые существа, рисуется на современном этапе развития на
уки, как осуществление программы, а эволюция — как не
прерывный процесс самопрограммирования. Испытание все
новых и новых программ развития и преимущественное
размножение более совершенных программ и составляют
сущность естественного отбора.
Наше исследование преследовало частную цель — по
нять приспособительное значение независимости размеров
частей у травянистых растений и вскрыть механизм отбо
ра, ведущего к возникновению независимости размеров ча
стей друг от друга.
140
Мы прибегли к сравнительному методу. Следовало: 1)
найти в пределах растения части, отличающиеся друг от
друга по степени стабильности; 2) найти растения, отлича
ющиеся друг от друга по степени стабильности аналогич
ных частей, выполняющих одну и ту же функцию; 3) выя
вить адаптивное значение стабильности, проанализировав
комплекс свойств, включающих стабильность как непре
менное условие жизни и 4) найти фактор отбора, ответст
венный за возникновение высших степеней стабильности.
МАТЕРИАЛ И МЕТОДИКА
В качестве объекта исследования были избраны травя
нистые растения. Внимание было сосредоточено на разме
рах цветков, семян и односеменных плодов. Семена и од
носеменные плоды интересовали нас в качестве той части
растений, на которой лежит функция расселения. По ана
логии с семенами и односемянными плодами, т. е. с «еди
ницей расселения» мы рассматривали цветки как «единицу
опыления».
Строение цветков стоит в строгом соответствии с ха
рактером транспортировки пыльцы, строение семян и пло
дов — с характером расселения [Sernander, 1906; Koller,
1959]. Сходные способы переноса пыльцы и семян наблю
даются у растений далеких систематических категорий.
Здесь открываются широчайшие возможности для конвер
гентного и параллельного развития. Здесь и следует искать
взаимообусловленные комплексы свойств, независимых от
всех других признаков организма. Для того, чтобы выявить
роль среды в формировании этих комплексов, нужно свя
зать характер изменчивости и степень независимости при
знака с характером его взаимоотношений со средой. Сами
эти взаимоотношения следует разделить на типы и пока
зать, что каждому типу взаимоотношений соответствует
свой оптимум изменчивости и независимости.
Исследовано 20 видов растений. У 19 из них изучена
изменчивость и соотносительная изменчивость «единицы
опыления» цветка, у шести изменчивость «единицы рассе
ления».
По характеру переноса семян растения подразделены
на три группы. Семена I группы расселяются осыпанием;
во II группе наблюдается зоохория — семена разносятся
птицами или млекопитающими, которые поедают плоды; в
III группе царит анемохория — семена разносятся ветром
(табл. 1).
141
Морфоэкологическая характеристика плодов
Вид
Группа
1
T riticu m a e stiv u m L. — пшеница
Способ переноса плодов
и семян
Ветрорасселение у ди
ких предков, приоб
ретенное в культуре
осыпание
H ordeu m . vulgare L. var. p a l
lid u m — ячмень
И. vu lgare L. var. n u du m —
ячмень голозерный
II
T ropaeolu m m a ju s L. — настурция
То же
Осыпание
F ragaria m o scliata Duch. — клуб
Связан с поеданием
ника
III
C h am aen erion a n g u stifo liu m Scop.—
иван-чай
Ветрорасселение
T araxacum o ffic in a le Web. — оду
ванчик
»
По характеру переноса пыльцы изученные растения
подразделяются на четыре группы: I — не имеет и не име
ла в историческом прошлом живых переносчиков пыльцы,
она состоит из анемофилов и растений, размножающихся с
помощью самоопыления; II — состоит из энтомофилов, не
имеющих специфических переносчиков пыльцы; пыльца
III группы растений переносится бабочками, а IV — пче
лами и шмелями (табл. 2).
Мерилом изменчивости и соответственно стабильности
служили коэффициенты вариации (С%) и отношение наи
большего значения признака к наименьшему (Vmax/ V min).
Мерилом независимости служил коэффициент корреляции
(г). Исследован 151 признак и соответственно высчитано
столько же значений С% и Vmax/ V min. Число признаков,
изученных у каждого вида растений, варьировало от трех
до десяти. Между размерами всех изученных признаков
каждой совокупности растений одного вида подсчитаны ко
эффициенты корреляции. Общее их число 356.
Все количественные показатели разбиты в соответствии
с частями, к которым они относятся, и каждому типу при
дано буквенное обозначение:
Р — размеры частей «единицы расселения» (семян
иван-чая, зерновок злаков, семянок одуванчика, плодиков
клубники и летучек одуванчика, включая высоту носика
семянки, число и длину волосков);
142
Таблица
1
и семян изученных растений
Наличие
летучек
Съедобность плодов и семян
Размер плодов и семян
У ДИКИХпредков
есть, в
культуре
нет
Несъедобные оболочки
плодов
Крупные
То же
То же
»
Нет
—
»
Съедобные плоды
Мелкие
Есть
Несъедобные плоды и семена
»
То же
»
ЦП — размеры пластинчатых частей цветка и отгиба
венчика, лепестков и чашелистиков, цветковых и колоско
вых чешуй злаков и нитевидных частей (столбиков и ты
чиночных нитей);
ЦТ — размеры трубчатых частей венчика и чашечки;
В — размеры вегетативных органов (стеблей и листь
ев);
С — количественные показатели плодовитости: разме
ры соцветий, число цветков, колосков, число и размеры
многосемянных плодов, число плодиков и семянок.
Коэффициенты вариации и отношения максимальной
варианты к минимальной разделены на пять типов в соот
ветствии с частями, изменчивость которых они характери
зуют.
Коэффициенты корреляции подразделены на шесть ти
пов:
Р — коэффициенты корреляции, связывающие разме
ры «единицы расселения» со всеми другими показателями;
ЦТ — то же для трубчатых частей цветка;
ЦП — то же для пластинчатых и нитевидных;
ЦЦ — коэффициенты корреляции между размерами
различных частей цветка;
В — между размерами стеблей и листьев между собой;
С — между показателями плодовитости друг с другом
и с размерами стеблей и листьев.
143
144
Морфоэкологическая характеристика цветков изученных растений
I
Б>
о>
аЯ
G
сЗ
ая
«I
нu
s i s a.
<
W>
*> 2
s
J
«
e
e
t
e
2
S >Я
Я 5«
i iX оОЯ
а м 3 1S a g§
g5о Я
| g * i
•3 2 2 v§ I * о
С *
S
5 а
2 ЕГ
я
С
я
о,
о
S
о
*
Н
*
0Q
>3
L. цветок моносимметрический благодаря положению тычинок.
S : U »S «
\ох divaricata
mо
Я о
а, н
О,
>»
145
Значения коэффициентов корреляции, коэффициентов
вариации и отношений максимальной варианты к мини
мальной послужили для построения вариационных рядов.
Вариационные ряды построены для всех соответствующих
показателей отдельно для каждой группы растений и для
каждого типа показателей. Таким образом, открывается
возможность сравнить изменчивость одного и того же орга
на у разных групп растений и разных органов в пределах
группы.
РЕЗУЛЬТАТЫ
Сравним сперва коэффициенты корреляций, связываю
щие размеры цветка с размерами листьев, стеблей и соцве
тий у четырех групп растений, отличающихся способом
переноса пыльцы. Данные, суммированные по группам,
представлены в табл. 3. Из коэффициентов корреляции
данного типа подсчитана средняя величина, характеризую
щая связи в пределах вида, а из этих средних высчитана
средняя величина, характеризующая связь в пределах
группы.
Суммированы не абсолютные значения коэффициентов
корреляции, а значения, включающие знак. ОтрицательТаблица
3
Коэффициенты корреляций, связывающие размеры цветка с размерами
листьев, стеблей и соцветий у четырех групп растений, отличающихся
способом переноса пыльцы*
Группа
Способ переноса пыльцы
Тип коэффициентов
цц
В ис
ЦТ и ЦП
суммарно суммарно
п
I
11
III
IV
Самоопыление, ветроопыление
Энтомофилия, отсутст
вие специфического
переносчика пыльцы
Энтомофилия, наличие
специфического пере
носчика пыльцы
То же
п
ммг
п
ммг
15 0,69 33 0,53
9 0,57
12 0,81 21 0,78
3 0,61
ЦТ
п
ммг
11 0,60 34 0,21 11 0,35 21 0,09
25 0,67 50 0,06 11 0,27 12 0,02
* Втабл. 3 и 4 приняты следующие обозначения: п —число коэффициентов корреляции
соответствующею типа, М —среднее значение для вида коэффициентов корреляции соот
ветствующего типа, Мм —"среднее значение для групп из средних значений для вида.
146
ные коэффициенты в нашем материале, по понятным при
чинам, имеют значение, близкое к нулю. Погашение близ
ких к нулю коэффициентов корреляции с противополож
ными знаками резче выявляет отсутствие корреляции.
В I и II группах растений, не имеющих специфических
переносчиков пыльцы, коэффициенты корреляции, связы
вающие размеры цветка с размерами всего растения в це
лом и с показателями плодовитости (типы ЦТ и ЦП),
лишь немногим меньше коэффициентов корреляции, свя
зывающих размеры вегетативных частей — листьев и стеб
лей друг с другом (тип В) и показатели плодовитости друг
с другом и с размерами листьев и стеблей (тип С). Соот
ветствующие средние значения равны 0,53 и 0,78 для раз
меров цветков и 0,69 и 0,81 для размеров стеблей, листьев
и показателей плодовитости. Размеры отдельных частей
цветка также связаны друг с другом высокими коэффици
ентами корреляции (тип ЦЦ). Соответствующие значе
ния — 0,57 и 0,61. Корреляционных плеяд по размерам
цветка здесь нет.
Совершенно другая картина наблюдается в III и IV
группах растений, имеющих специфических переносчиков
пыльцы. Коэффициенты корреляции типов ЦТ и ЦП, свя
зывающие размеры цветков с размерами стеблей, листьев
и с показателями плодовитости, низки, в противополож
ность коэффициентам корреляции типов В и С, связываю
щим размеры листьев, стеблей и соцветий друг с другом.
Соответствующие значения равны 0,21 и 0,06 для разме
ров цветков и 0,60 и 0,67 — для стеблей, листьев и пока
зателей плодовитости. Коэффициенты корреляции типа
ЦЦ, связывающие размеры различных частей цветка друг
с другом, выше и достигают величин 0,35 и 0,27.
Размеры цветков образуют в этих группах растений
корреляционные плеяды. Наиболее четко они выражены в
IV группе^ т. е. там, где переносчиками пыльцы служат
пчелы и шмели — насекомые с жестко детерминированны
ми размерами.
Наибольшую независимость обнаруживают трубчатые
части цветков. Коэффициенты корреляции типа ЦТ, свя
зывающие их размеры со всеми другими количественными
показателями за пределами цветка, практически» равны
нулю.
Стандарт размеров трубчатых, а также нитевидных ча
стей цветка в наибольшей степени обеспечивает правиль
ность локализации пыльцы на тельце насекомого и прида
ет всему процессу транспортировки пыльцы надежность ис
тинного приспособления.
L47
Таблица
4
Коэффициенты корреляции, связывающие размеры «единицы расселе
ния» с размерами других частей растений в трех группах растений, от
личающихся способом расселения
Группа
Тип коэффициентов
Способ расселения
Ви С
суммарно
Р
Ммт
Ммт
I
II
III
Осыпание
Зоохория
Анемохория
21
13
10
0,46
0,09
0,37
13
36
9
0,67
0,68
0,72
Выявилось таким образом, что цветки энтомофилов,
имеющих специфических переносчиков пыльцы, независи
мы по размерам от размеров растения в целом. Цветки
всех прочих растений, лишенных специфических перенос
чиков пыльцы, зависимы по размерам. Только у растений,
связанных с определенными видами насекомых, размеры
цветков образуют корреляционные плеяды.
Теперь сравним коэффициенты корреляции, связываю
щие размеры «единицы расселения» с размерами других
частей растения, в грех группах растений, отличающихся
друг от друга способом расселения семян (табл. 4). Разме
ры семян независимы у клубники (II группа), которая по
способу расселения относится к зоохорам. Слабая зависи
мость наблюдается у анемохоров: иван-чая и одуванчика.
Наибольшая зависимость размеров семян наблюдается у
тех растений, которые,расселяются осыпанием. У анемохо
ров и зоохоров размеры семян образуют корреляционные
плеяды. У других растений плеяд по размерам семян нет.
Сравнительно низкий коэффициент корреляции, связыва
ющий размеры семян с размерами других органов в I груп
пе растений, зависит от выбора объектов. У культурных
злаков анемохория утеряна под влиянием искусственного
отбора, и сравнительно низкие коэффициенты корреляции,
связывающие размеры зерновок с размерами других частей
растения, являются ее пережитком. Помимо размеров
цветков энтомофильных растений, имеющих специфиче
ского переносчика пыльцы, и семян анемо- и зоохоров, не
зависимыми оказались размеры соцветий у смолки и высо
та стебля у наперстянки желтой.
У клубники сравнительно низкую степень зависимости
от размеров растений обнаруживает число крупных плоди
ков. Каждый плод клубники содержит наряду с крупными
148
Рис. 1. Изменчивость коэффици-
<%
ентов
ipynn
qq.
корреляции у четырех
растений, отличающихся
(CM.
qq.
табл. 1), и у одуванчика Taraxa
cum officinale Web.
54-
ПО
СПОСОбу
ОПЫЛенИЯ
I—IV —группы растений; V —Taraxacum
officinale ; 2 —суммарные данные.
----------------- :---------------------------------
_
42~
плодиками некоторое чис- 56~
ло мелких. Крупные и joмелкие плодики подсчиты
вались отдельно. Опыт по г*~
проращиванию семян с 1в~
протравливанием и без 12_
протравливания
серной
кислотой
показал,
что 6~]
прорастают только семена
I
крупных плодиков. Семена
мелких плодиков не прора
стают совсем. Мелкие пло
дики представляют собой,
по-видимому, неопыленные завязи. Число больших плоди
ков, или реальная плодовитость, обнаруживает более низ
кую степень зависимости от размеров растения, чем общее
число плодиков, или потенциальная плодовитость.
Рис. 1 представляет изменчивость всех коэффициентов
корреляции сумарно в четырех группах, подразделенных в
соответствии со способом переноса пыльцы, и отдельно у
одуванчика.
Кривая, изображающая распределение коэффициентов
корреляции в I группе растений, одновершинна и распола
гается в области больших значений г. Ни по размерам
цветков, ни по размерам семян здесь нет корреляционных
плеяд. Кривая, изображающая распределение значений г
во II группе растений,— двувершинна. Подъем на малых
величинах образован коэффициентами корреляции типа Р,
связывающими размеры «единицы расселения» у клубники
с размерами других частей растения. Все остальные коэф
фициенты образуют одновершинную кривую с подъемом
на больших значениях г. Корреляционные плеяды наблю
даются по размерам семян, но не по размерам цветков.
В III группе растений кривая имеет подъем на малых зна
чениях. Он образован независимыми размерами цветков.
В IV группе наблюдается та же картина: обе кривые рас
пределения коэффициентов корреляции носят двувершин
149
ный характер с одним подъемом на малых и одним — на
больших значениях.
Двувершинной является и кривая распределения всех
коэффициентов корреляции. Большой интерес представля
ют значения коэффициентов корреляции, относящиеся к
средним классам вариационного ряда. Коэффициенты кор
реляции принимают средние значения, когда: 1) стабили
зирующий отбор имеет слабую интенсивность; 2) искусст
венный и естественный отбор идут в противоположных на
правлениях и 3) естественный отбор осуществляется мно
гими разнообразными функциями, каждая из которых тре
бует своего размера и своей меры стабильности.
Несколько функций выполняют многие части растений.
Чашечки или наружные чешуи цветка участвуют затем в
образовании защитных оболочек плода. Листья растений
иногда участвуют в привлечении насекомых, экранируют и
способствуют вытеснению претендентов на занимаемое
пространство, защищают цветок от дождя. Отгибы венчи
ков цветков служат одновременно сигнализаторами и вы
нуждают насекомое принять определенную позу, при кото
рой пыльца попадает на нужное место на теле насекомого.
В нашем материале промежуточные классы коэффици
ентов корреляции включают: 1) цветки злаков, части кото
рых входят не только в состав «единицы опыления», но и в
состав «единицы расселения», 2) пластинчатые части цвет
ков; 3) длину пестика иван-чая; 4) чашечку гвоздичных,
служащую опорным образованием и защитным футляром
цветка и защитной оболочкой плода; 5) зерновки злаков;
6) летучки и семянки одуванчика и семена иван-чая, для
которых достижение критического размера исключило, повидимому, необходимость автономизации; 7) размеры со
цветий смолки; 8) размеры листьев наперстянки желтой.
В случае, когда многие функции предъявляют требова
ние к размерам части, вырабатывается некий компромисс,
до известной степени удовлетворяющий этим разнообраз
ным, а в некоторых случаях противоположным требовани
ям. Средние значения г и выражают это компромиссное
решение.
Рис. 2 представляет изменчивость коэффициентов кор
реляции, подразделенных на типы в соответствии с частя
ми растений, относительную изменчивость которых они
характеризуют.
Наименьшие значения коэффициентов г характерны
для размера трубчатых частей цветка, для семян зоохоров
и для пластинчатых частей цветка растений энтомофилов,
имеющих специфических переносчиков пыльцы. Кривые
150
Рис. 2. Изменчивость коэффицит
ентов корреляции, относящихся к
различным частям растений.
Корреляции: Р —размеров семян и одно
семенных плодов с другими частями рас
тений; ЦТ — размеров трубчатых частей
цветка; ЦП — размеров пластинчатых и
нитевидных частей с другими частями;
ЦЦ —размеров частей цветка друг с дру
гом; В — размеров листьев и стеблей
друг с другом; С —показателей плодови
тости с размерами листьев и стеблей
и друг с другом.
для коэффициентов корре
ляции типов В и С, харак
теризующих связи между
размерами
вегетативных
частей друг с другом и с
показателями плодовито
сти, располагаются в области больших значений.
Рис. 3 представляет изменчивость коэффициентов ва
риации различных частей растений и всех коэффициентов
вариации, подсчитанных для всех признаков.
Сопоставление показателей изменчивости размеров
(коэффициентов вариации и отношений максимального
значения к минимальному) разных органов у растений
разных группы с экологическими и морфологическими осо
бенностями групп выявило
точно ту же картину, что
и сопоставление показате
лей независимости. Наи
более стабильными оказа
лись размеры частей, об
ладающих наибольшей не
зависимостью.
Стабильность и неза
висимость — проявление
Рис. 3. Изменчивость всех коэф
фициентов вариации и коэффи
циентов вариации размеров раз
личных частей.
Р — семена и односеменные плоды;
ЦТ — трубчатые части цвегков; ЦП —
пластинчатые и нитевидные части цвет
ков; В —стебли и листья; С показате
ли плодовитости; 2 —все коэффициенты
вариации.
151
одной и той же стабилизирующей деятельности отбора.
Наименьшие коэффициенты вариации и наименьшие вели
чины отношения максимального значения к минимальному
присущи семенам, а также трубчатым, нитевидным и пла
стинчатым частям цветков растений, имеющих специфиче
ских переносчиков пыльцы.
Исключение составляют большие плодики клубни
ки. Коэффициенты корреляции, связывающие их число
с размерами растения, ниже, чем коэффициенты корре
ляции, связывающие общее число плодиков с размера
ми растения. Соответствующие средние значения из пя
ти показателей (Мг): 0,55 и 0,77. Коэффициент вариа
ции числа больших плодиков много выше (79,4 %),
чём коэффициент
вариации
общего
числа
плоди
ков (28,1 %). Почему число больших плодиков оказалось
в таком исключительном положении, будет разъяснено в
дальнейшем.
ОБСУЖ ДЕНИЕ РЕЗУЛЬТАТОВ
Сопоставление степени стабильности и независимости
размеров различных частей растений с их морфологически
ми и экологическими особенностями позволяет выявить
фактор, ответственный за возникновение высших степеней
стабильности и независимости.
В отношении размеров цветка стабилизаторами яв
ляются насекомые — переносчики пыльцы, в отноше
нии семян — специфические функции, например ха
рактер разноса семян. Анемохория и в особенности зоохория кладут запрет на укрупнение семян и вынуждают рас
тение поставить размер семян под жесткий генотипический
контроль.
Жесткость генотипического контроля размеров цветков
растений, пыльца которых транспортируется пчелами и ба
бочками, беспрецедентна. Независимость трубчатых частей
этих цветков не имеет себе равной среди других показате
лей независимости. Цветки сохраняют стандартный размер
как в пределах вида, так и в пределах одного растения в
разных его ярусах. Стандартизация размеров всех цветков
имеет громадное значение в жизни вида, позволяя эконом
но расходовать пыльцу. Связь здесь не двойная, а тройная:
приспособлены друг к другу не только цветки и насеко
мые, но и цветки данного вида растения друг к другу.
Стандарт и независимость размеров цветков — выражение
этой приспособленности. Стабильность цветка обеспечива
152
ет экономию и надежность доставки пыльцы. Цветки рас
тений, имеющих специфических переносчиков пыльцы, ло
кализуют пыльцу на строго определенном месте на теле
насекомого-переносчика. Каждый вид растения имеет свой
участок. Явление локализации пыльцы открыто Дарвином
[1928] у орхидных, а затем Шванвичем [1952, 1955, 1956]
у мотыльковых. Растение локализует пыльцу там, где она
недосягаема для очистительных аппаратов насекомого:
между глазами бабочки, на мембране, поддерживающей
хоботок, или на стебельке, соединяющем грудь и брюшко
пчелы. Принцип локализации пыльцы имеет смысл только
в том случае, если размеры всех цветков данного вида оди
наковы и строго согласованы с размерами насекомых. Кон
фигурация частей цветка заставляет насекомое принять
позу, при которой пыльца, несомая насекомым, попадает
на рыльце цветка, а новая порция пыльцы будет локализо
вана пыльниками в точности на том же месте, куда поло
жил свою пыльцу предыдущий цветок того же вида. Моносимметрия цветков, латеральное положение их на рас
тении, строгая фиксация частей, достигаемая на пути
срастания и стандартизации размеров частей, и олигоме
ризация числа частей цветка обеспечивают локализацию
комочка пыльцы. Наибольшая роль принадлежит трубча
тым и нитевидным частям цветка. От длины трубочки
цветка табака или флокса зависит, упрется ли бабочка го
ловой в пыльники, торчащие из трубочки цветка, и будет
ли пыльца локализована между глазами бабочки. Отступ
ление от стандартных размеров делает растение практиче
ски стерильным. Мы имели случай убедиться, что наибо
лее стабильны и независимы размеры трубчатых частей
цветков. Они стандартнее и независимее размеров пла
стинчатых частей, которые служат главным образом дис
тантными сигнализаторами и посадочными площадками
для насекомых.
Насекомые и выступают в роли отбирающего фактора.
Мозаичный характер развития насекомых, их жесткие про
граммы индивидуального развития делают насекомых пре
красными контролерами размеров цветков, чью пыльцу
они транспортируют. Полет налагает запрет на изменение
пропорций тела летающих форм. Среди них в процессе от
бора сохраняются лишь те, кго имеет стандартные пропор
ции. Пчелы и шмели в особенности пригодны в качестве
браковщиков, так как, развиваясь в ячейках стандартных
размеров и получая в течение развития строго ограничен
ное количество пищи, сами имеют стандартные размеры.
Гены рабочих пчел, контролирующие строительные инс
153
тинкты и инстинкты выкармливания молоди, контролируют
в конечном счете размеры тех цветков, с которыми пчелы
собирают пыльцу и нектар. Контроль размеров цветков
осуществляется, само собой разумеется, с помощью отбора.
Причины возникновения независимости в случае цвет
ков и в случае семян различны. Отличаются друг от друга
и факторы, ответственные за стабилизацию размеров пло
диков клубники и факторы стабилизации семян иван-чая.
В первом случае это — условия безопасного прохождения
через пищеварительный тракт птицы, во втором — аэроди
намические условия полета. Как бы различны ни были
факторы, ответственные за возникновение независимости,
все они имеют нечто общее. В качестве стабилизаторов вы
ступают функции, лишенные морфогенетического эффек
та, не принимающие ни малейшего участия в развитии тех
структур, размеры которых они контролируют. Насекомые,
переносящие пыльцу, птицы, расселяющие семена клубни
ки, условия полета иван-чая не участвуют в развитии
цветков и семян и не влияют на их размеры. Между тем
благополучный исход событий в сильной степени зависит
от размеров цветков и семян.
Очевидно, что здесь мы имеем дело не с единичными
событиями и не с разрозненными явлениями. Перед нами
определенная категория явлений или определенный класс
подсистем. Выявляя общие черты, присущие классу явле
ний, мы можем дать ответ на интересующий нас вопрос
относительно механизма возникновения в эволюции устой
чивости и независимости целенаправленных явлений. Ста
бильность и независимость размеров цветков специализи
рованных энтомофилов и семян зоо- и анемохоров достига
ются в эволюции, т. е. представляют собой примеры фило
генетического воздействия среды. Изменения, вызванные
этим воздействием, касаются сужения нормы реакции или,
иными словами, погашения функциональных и морфогене
тических воздействий среды. Наш материал позволяет
сформулировать общий принцип возникновения онтогене
тического гомеостазиса (developmental homeostasis).
Независимость данной части организма от форматив
ного действия внешней среды и других частей того же
организма достигается в процессе отбора в том случае,
если формирующие и отбирающие компоненты среды
не совпадают и функция выступает исключительно в ро
ли испытателя пригодности. В процессе отбора сохраня
ются особи, не ответившие формообразовательной ре
акцией на воздействие тех компонентов среды, от кото
рых не зависит затем исход событий. Это и есть стаби
154
лизирующий отбор, или отбор более жестких программ
развития, осуществляемый с помощью функции, лишенной
формативного действия.
Независимость и стандарт размеров цветков входят в
закономерно построенный комплекс свойств.
Во всех случаях, когда организм, независимо от своего
систематического положения, вступает во взаимодействие с
определенными компонентами среды, используя их для вы
полнения одной и той же специфической функции, возни
кает один и тот же комплекс свойств. В нашем случае спе
цифической функцией является воспроизведение с по
мощью перекрестного оплодотворения. Среда, с которой
растение вступает во взаимодействие,— это насекомое,
транспортирующее его пыльцу и относящееся к определен
ной систематической категории. Обретая специфического
переносчика пыльцы, суживая круг своих визитеров, рас
тение теряет в надежности переноса. Компенсаторно раз
вивается механизм локализации пыльцы на маленьком
участке тельца насекомого. Мы уже знакомы с тем комп
лексом свойств, который обеспечивает этот механизм. Вза
имодействие всех признаков, входящих в этот комплекс,
носит компенсаторный характер. Локализация пыльцы
обеспечивает экономию этого драгоценного продукта. На
секомое получает голодный паек. Растению грозит отказ со
стороны насекомого посещать цветки. Компенсаторно вы
рабатывается замена — нектар. Но и нектар нужно эконо
мить и уровень нектара в резервуарах, хранящих его, ото
двигается как можно дальше от пыльников, трубочки цвет
ков удлиняются. Насекомые вынуждаются сосать из этих
нелепых сосудов, плотно прижавшись головой к краям со
суда, вытянув хоботок на полную длину. Насекомое дотраги
вается головой сперва до рыльца, потом до пыльников, торча
щих из трубочки у самого входа в нее. Затрата материала на
трубочку и на тычиночные нити компенсируется экономией
пыльцы и нектара и надежностью локализации пыльцы на
очень ограниченном участке тельца насекомого.
Размеры цветков образуют корреляционную плеяду, и то
же справедливо и для семян анемо- и зоохоров. Сами плеяды
могут комбинироваться друг с другом в любых сочетаниях.
Растение, пыльца которого переносится специфически
ми переносчиками пыльцы, а семена ветром или животны
ми, должно обладать двумя корреляционными плеядами.
Таков в нашем материале иван-чай. Размеры его цветков
и семян связаны низкими коэффициентами корреляции с
размерами стеблей, листьев, соцветий, плодов. Растение —
специализированный энтомофил, расселяющееся осыпани
155
ем, дожно иметь одну плеяду, включающую размеры цвет
ков. Такова настурция. Растение, не вступающее во взаи
модействие с живыми компонентами среды ни в процессе
переноса пыльцы, ни в процессе расселения и не имеющее
способа переноса семян, требующего стандартного размера
семян, лишено корреляционных плеяд. Такова, например,
пшеница.
Знание корреляционных плеяд имеет большое значение
как в практической жизни, так и в практике научного исс
ледования. Медицинская генетика нуждается в показате
лях, характеризующих относительную роль внешних и
внутренних факторов в этиологии заболевания. Системати
ка нуждается в общей теории плеяд для выбора диагности
ческих признаков. П. В. Терентьев [1960] осветил значе
ние плеяд в биоценологии и практическое значение плеяд
в промысловом деле. В селекционной практике практиче
ское значение корреляционных плеяд заключается в воз
можности составить прогноз эффективности искусственного
отбора, зная характер и количественную характеристику
корреляций в пределах организма. Чем шире размах онто
генетической изменчивости признака и чем большими ко
эффициентами корреляции связаны размеры признака с
размерами организма в целом, тем менее эффективен от
бор по данному признаку, т. е. чем больше величина Мг
тем менее эффективен отбор [Берг, 19616]. Сейчас суще
ствует несколько способов определения наследуемости при
знака или относительной роли внешних и внутренних фак
торов в развитии признака. Все эти методы основаны на
сопоставлении степени сходства со степенью родства
[Dickerson, 1955; Lerner, 1958; Плохинский, 1963; и др. ].
Однако прогноз можно сделать с помощью изучения из
менчивости в популяции и корреляций, не прибегая к со
поставлению степени сходства со степенью родства. Отно
сительная роль внешних и внутренних факторов развития
выявляется при этом очень четко. Но следует иметь в ви
ду, что низкий коэффициент корреляции данного количе
ственного показателя с общими размерами тела только в
том случае является показателем высокой наследуемости
признака и, следовательно, высокой эффективности отбо
ра, если он указывает на независимость процессов разви
тия данного признака от процессов развития других при
знаков организма. Если количественный показатель нахо
дится под контролем внешних факторов среды, действую
щих на него формативно и притом избирательно и непос
редственно, а не опосредованно через процессы развития,
он связан низкими коэффициентами корреляции с другими
156
количественными показателями той же совокупности, и
это ни в коей мере не указывает на наличие жесткого ге
нотипического контроля, а означает как раз обратное. Раз
личить обе ситуации можно по коэффициенту вариации;
если оба показателя (г и С %) низкие, перед нами случай
жесткой программы; если г мал, а С % высок, то мы име
ем дело с избирательным формативным действием среды.
Мы встретились с подобным случаем на примере числа
больших плодиков в плодах клубники. Сколько завязей
будет оплодотворено, зависит от случайного наличия или
отсутствия и от числа насекомых переносчиков пыльцы,
посетивших цветок. Число больших плодиков и представ
ляет собой признак, на количественное выражение которо
го внешний агент действует непосредственно и избиратель
но. Высокий коэффициент вариации (С%), характеризую
щий изменчивость признака, указывает на отсутствие
жесткого генетического контроля и позволяет избежать
ошибки при составлении прогноза эффективности отбора.
Независимость и стабильность возникают в процессе
эволюции и сами налагают печать на дальнейшее течение
отбора. Наличие сильного генетического контроля не толь
ко не служит преградой для дальнейших эволюционных
преобразований, а, напротив, способствует изменению со
ответствующих структур в историческом развитии. Цветки
энтомофильных растений, плоды и семена с эндозойным
способом расселения претерпели наиболее глубокие преоб
разования в процессе эволюции [Тахтаджян, 1954а,
6;Tachtajan, 1959]. Это и понятно. Отбор по признаку, об
ладающему зависимым типом развития, постоянно подхва
тывает то, что в следующем поколении при изменении ус
ловий не повторится вновь. Отбор по независимому при
знаку избавлен от этих «ошибок». Его преобразующая дея
тельность не наталкивается на помехи в виде модифика
ций, случайных реакций на случайные воздействия среды
и т. п. Онтогенетическая устойчивость — предпосылка эво
люционных преобразований.
У покрытосемянных, имеющих специфических пере
носчиков пыльцы, выработался механизм, облегчающий
образование новых видов. Принципиальная возможность
образования гибридогенных симпатрических видов энтомо
фильных растений, имеющих специфических переносчиков
пыльцы, продемонстрирована Грантом [Grant, 1949, 1952]
для видов рода Aquilegia и Стро [Strow, 1955] для видов
рода Penstemon . Здесь видообразование осуществляется без
географической изоляции по типу экобиотической дивер
генции [Huxley, 1942].
157
Возникновению гибридных комбинаций под влиянием
насекомых сопутствует испытание новых географических и
экологических местообитаний в результате разноса семян
птицами [Anderson, Stebbins, 1954]. Зоохория, способствуя
стабилизации размеров семян, является фактором эволю
ции растений.
Стабилизированные структуры, находящиеся под конт
ролем живых компонентов биоценоза, эволюционируют,
таким образом, особенно быстро. Ускорение эволюции
[Шмальгаузен, 1943; Simpson, 1944; Рубцов, 1945] совер
шается под действием тех самых живых компонентов сре
ды, которые способствуют стабилизации онтогенеза.
Значение корреляционных плеяд в систематике очень
велико. Возможность предсказывать одни признаки орга
низма по другим признакам составляет самую суть систе
матики. Наличие плеяд вносит известные ограничения в
построение прогноза. Признаки, входящие в одну плеяду,
не могут быть предсказаны по признакам другой плеяды.
Прогноз возможен только в пределах плеяды.
Как известно, идея согласованных различий между
представителями разных систематических категорий была
использована Кювье для предсказания неизвестных при
знаков по признакам известным. Реставрируя скелеты по
звоночных по немногим ископаемым остаткам, Кювье дей
ствовал так, как если бы все признаки организма входили
в одну корреляционную плеяду. Естественно возникает
вопрос, почему на пути успешного прогноза в работах
Кювье не встали корреляционные плеяды. Чтобы ответить
на этот вопрос, попытаемся вскрыть условия, способствую
щие возникновению плеяд в эволюции.
Разные функции организма: питание, расселение, раз
множение — осуществляются с помощью структур, функ
ционирующих до известной степени независимо друг от
друга. Специализация функций создает предпосылку для
создания корреляционных плеяд. Если разные стадии жиз
ненного циж*ла специализируются в выполнении разных
функций, индивидуальное развитие приобретает резко вы
раженный прерывистый характер: вырабатываются корре
ляционные плеяды признаков, присущие каждой стадии
жизненного цикла. Предсказать свойство одной стадии по
свойствам другой становится невозможным. Такого рода
независимые комплексы признаков, присущие разным ста
диям жизненного цикла, можно назвать
хроно
логическими
плеядами. Независимость цветков
энтомофильных растений от размеров стеблей и листьев
представляет собой хронологическую плеяду.
158
Онтогенез насекомых, обладающих полным превраще
нием, изобилует хронологическими плеядами. Возникнове
нию в процессе эволюции плеяд у насекомых способствует
наличие у них хитинового скелета. Хитин ограничивает
рост, исключает возможность формативного действия фун
кции на все те части организма, которые состоят из него.
Компенсаторно развиваются линьки, создается предпосыл
ка для специализации разных стадий жизненного цикла.
Питание и рост становятся функциями одной — личиноч
ной стадии жизненного цикла, расселение и размноже
ние — функциями взрослого насекомого. Создается преры
вистость онтогенеза и возникают хронологические корреля
ционные плеяды. Сходным образом возникновению хроно
логических плеяд способствует прикрепленный образ жиз
ни в половозрелой стадии. Функция расселения ложится на
другую стадию. Происходит специализация разных стадий
жизненного цикла в отношении выполнения разных функ
ций. Все растения, за исключением планктонных, находят
ся в этом положении.
Признаки семян, связанные с особенностями расселе
ния растения, образуют корреляционную плеяду. Мы обна
ружили такого рода плеяду по размерам «единицы расселе
ния» у растений, расселяющихся с помощью ветра и с по
мощью птиц и млекопитающих. Независимость признаков
зародышей и семян от признаков материнского организма
образует особый класс плеяд. Плеяды эти можно назвать
р е п р о д у к ц и о н н ы м и . Их существование связано
с заботой о потомстве. Семя, или односеменной плод, при
надлежит уже следующему поколению, а в случае полово
го размножения обладает, кроме того, собственным геноти
пом, отличающимся от генотипа материнского растения.
Специализация функций отдельных органов и систем
органов в пределах фазы жизненного цикла создает плея
ды, которые целесообразно назвать
хорологиче
с к и м и . Хорологические плеяды включают одновремен
но функционирующие органы в пределах одной стадии
развития и выражают связь разных систем органов орга
низма с независимыми друг от друга компонентами среды.
Примером этого рода может служить независимость разме
ров цветков от показателей плодовитости у энтомофильных растений, имеющих специфических переносчиков
пыльцы.
Хорологические, хронологические и репродукционные
плеяды являются результатом специализации функций.
Если взять группу, имеющую единообразный способ
размножения и расселения со сходной специализацией фаз
159
жизненного цикла и разделяющуюся на подгруппы по спо
собу питания, открывается широчайшая возможность про
гнозов. Именно с такой группой и имел дело Кювье, когда
он сравнивал сочетания признаков подгрупп — хищных и
травоядных млекопитающих. Выявив закономерные соче
тания признаков, Кювье дал, в сущности, качественную
характеристику одной корреляционной плеяды.
Определение вида заключается в указании необходимо
го и достаточного числа таких признаков, на основании ко
торых все остальные признаки могут быть предсказаны.
Успешность таких прогнозов требует сочетания принципа
Кювье с принципом корреляционных плеяд, или принци
пом Терентьева, как я предлагаю его назвать.
Теперь несколько слов об эволюции самих плеяд. За
конно поставить вопрос, сопровождается ли морфофизиоло
гический прогресс возрастанием числа и объема независи
мых комплексов закономерно сочетающихся признаков,
вырабатывается ли в этом отношении некий оптимум или
прогрессивная морфофизиологическая эволюция сопровож
дается повышением взаимозависимости частей организма.
Само возникновение онтогенеза является, несомненно,
предпосылкой обособления и специализации разных стадий
и возникновения независимых комплексов признаков, при
сущих каждой стадии. Мозаичный тип эмбриональною
развития, смена фаз жизненного цикла, метаморфоз насе
комых, амфибий, наличие специализированной стадии,
осуществляющей расселение у всех прикрепленных форм,
включая все растения, кроме планктонных, сопровождает
ся образованием независимых комплексов свойств. Возник
новение независимости одних частей по отношению к дру
гим — элемент прогрессивной эволюции. В особенности
это касается растений. Наличие семян, расселяемых специ
фическим способом,— показатель прогрессивной эволюции
растений, и отказ от полового размножения не сопровож
дается утерей этой стадии жизненного цикла, обладающей
способностью пассивного передвижения [Хохлов, 1959].
Образование всех классов балансированного полимор
физма [Берг, 1957] сопровождается возникновением комп
лексов взаимообусловленных признаков, различных у раз
ных форм одного и того же вида. Возникновение полимор
физма сопровождается возникновением плеяд.
Однако наивысшие ступени эволюции характеризуют
ся, по-видимому, оптимумом стабилизации и оптимумом
независимости частей в отношении друг друга. Отказ от
метаморфоза, отсутствие резкой специализации стадий по
образу жизни, отсутствие афагии какой бы то ни было ста
160
дии, регуляторный тип эмбрионального развития — все
это отличает наиболее прогрессивное направление фи
логенеза.
Тем не менее и на высших этапах эволюции мы встре
чаемся с плеядами. Это касается главным образом процес
са воспроизведения у животных и у растений. Именно эта
область изобилует экологическими параллелями [Берг,
1935; Беклемишев, 1951, 1954; Левина, 1959] и конверген
циями. И у человека воспроизведение сохранило, по-види
мому, наибольшую независимость.
Но дело' отнюдь не ограничивается существованием
корреляционных плеяд в пределах организма, к какой бы
систематической категории он ни относился. Организм —
лишь одна из ступеней в иерархии живых систем. Комп
лексы взаимосвязанных признаков, независимые по отно
шению друг к другу, характеризуют все живые системы, к
какому бы уровню организации живого они ни относились.
Наш материал дает возможность рассмотреть корреля
ционные плеяды в плане теории систем. Системой мы на
зываем некоторую устойчивую совокупность явлений, от
граниченную от других совокупностей во времени и в про
странстве, состоящую из частей, подчиненных целому,
взаимодействующих друг с другом и несущих на себе чер
ты соподчинения. Система как целое взаимодействует со
средой; и ее реакции на воздействия, как внешние, так и
внутренние, носят регуляторный характер. Входя в систему
высшего порядка, система составляет ее часть, сохраняя
свою автономию.
Все перечисленные живые системы обладают этими
свойствами. Однако все они, в отличие от систем неживых,
обладают только им присущими чертами. Неживые систе
мы — это системы существенно устойчивые, не включаю
щие преобразование в качестве условия своего бытия. Жи
вые системы — это системы не только устойчивые, но и
преобразующиеся, несущие в себе существенную информа
цию своего преобразования. Преобразование — это изме
нение, осуществляющееся на основе программы. Рост, са
мовоспроизведение, деление, индивидуальное развитие и
эволюция — это изменения, в той или иной мере програм
мированные. Живыми являются системы открытые для ве
щества и энергии и сочетающие в себе способность к пре
образованиям с устойчивостью в пределах фазы преобразо
вания.
Устойчивость — неотъемлемая черта живой системы.
Живому присуща способность не только сохранять устой
чивость в пределах фазы преобразования. Сам переход из
6 Р. Л. Берг
161
одного состояния в другое, т. е. течение процессов, носит
устойчивый характер. Эту устойчивость жизненных про
цессов Уоддингтон [Waddington, 1957] предложил назвать
гомеорезисом, в отличие от устойчивости состояний, или
гомеостазиса.
Независимость одних частей от других частей той же
системы — только выражение этой устойчивости. И в той
же мере, в какой устойчивость характеризует систему, ха
рактеризуют ее и корреляционные плеяды. Корреляцион
ные плеяды присущи, таким образом, всем системам, как
живым, так и неживым. Сама возможность измерения вре
мени основана на существовании независимости одних яв
лений от других явлений, происходящих в той же системе,
в данном случае в Солнечной системе. Скорость вращения
Земли вокруг своей оси устойчива и независима, в частно
сти, от ее положения на орбите. Тот же принцип независи
мости лежит в основе исчисления времени по периоду рас
пада радиоактивных элементов. Зависимый от внешних
воздействий процесс не мог бы служить мерилом времени.
Другое доказательство независимости одних свойств систе
мы от других ее свойств заключается в самой возможности
моделирования. Тип организованности при моделировании
сохраняется вне зависимости от различий в материале, из
которого построены образец и его модель. Живая природа
сама создает модели на путях конвергентного развития.
Жизнь на нашей планете представлена иерархией жи
вых систем. Идея целостности всех уровней организации
живого все больше овладевает умами ученых. Вернадский
[1926, 1940], Нидгем [Needgam,1937], Сукачев [1947],
Мазер [Mather, 1955, 1961], Завадский [1961], Берталанфи [Bertalanffy, 1949, 1951], Шмальгаузен [1946, 1961],
Ляпунов [1962], Тимофеев-Ресовский [1962], Ушаков
[1963] являются яркими выразителями этой идеи.
В.
И. Вернадский [1926] назвал область распростране
ния жизни на планете биосферой. Совокупность живых ор
ганизмов или разнородное живое вещество находится в
теснейшей взаимосвязи с неживым, или косным, вещест
вом биосферы.
По идее Вернадского, неразрывная связь живого веще
ства со средой выражается прежде всего в том, что живое
вещество биосферы управляет геохимическими процесса
ми, протекающими в биосфере, само контролирует среду
своего обитания. Живое вещество биосферы представляет
собой высшую ступень иерархии живых систем. Системы
низшего порядка — сообщества живых систем, или биоце
нозы,— составляют следующую ступень иерархии. Они це
162
лостны и сами подчинены тому целому, частями которого
являются. Идея целостности биоценоза развита Сукачевым
[1947] и А. Д. Александровой [1963]. Биоценозы состоят
из видов. Вид и представляет собой следующую ступень со
подчинения. Целостность вида менее очевидна, чем цело
стность всякой другой системы. Части, из которых состоит
вид,— организмы, рассеяны в пространстве. Тем не менее
виду присущи все признаки системы [Берг, 1944]. Идею
целостности вида наиболее полно выразил К. М. Завадский
[1961 ]. И виды состоят из систем. Мы дошли до четвертой
ступени иерархии — до организма. Организм — самая
«системная» из всех систем. Он не только целостен, он це
лен. Взаимодействие его частей, разделение между ними
функций, подчинение каждой части целому достигает в ор
ганизме наибольшего выражения. Но и организм состоит
из систем. Это клетки. Каждая клетка представляет собой
до известной степени обособленное образование, подчинен
ное организму, но в какой-то ограниченной степени сохра
няющее свою автономию. Низшую ступень иерархии обра
зует макромолекулярный комплекс, способный воспроизво
дить себя и мутировать.
Мы называем именно этот ряд — живое вещество био
сферы, биоценоз, вид, организм, клетки и способный к са
мовоспроизведению молекулярный комплекс — потому,
что каждый из членов этого ряда обладает хотя бы ограни
ченной способностью к самовоспроизведению.
Самовоспроизведение можно подразделить на две боль
шие категории: связанное с порождением системы и не
связанное с таковым. Первая категория представлена тре
мя способами: построением копии, делением и отделением
части. Живые макромолекулы, клетки (и одноклеточные
организмы) и организмы обладают соответственно этими
тремя способами самовоспроизведения. Самовоспроизведе
ние второй категории, не связанное с порождением систе
мы, основано на способности к самовоспроизведению каж
дой части, входящей в состав системы. Части размножают
ся и гибнут, а целое обновляется и в конечном счете восп
роизводит себя заново. Вид, биоценоз и все живое вещест
во биосферы в целом обладают способностью самовоспро
изведения именно в этом последнем смысле.
Системы низшего порядка, строя систему высшего по
рядка, объединяются в комплексы, и комплексы эти обла
дают известной автономией и признаками целостности.
Клетки слагаются в ткани, ткани в органы, органы образу
ют системы органов в пределах организма. Организмы вхо
дят как подчиненные элементы в состав колоний, стад, се
163
мей. Они группируются в подчиненные виду низшие систе
матические категории, образуя группы внутри вида — ра
сы, подвиды и т. п. Отдельные группы видов в пределах
биоценоза вступают в более тесные взаимоотношения друг
с другом, чем другие группы вида, образуя цепи питания
или заселяя один и тот же ярус сообщества. Главными
членами иерархии все же остаются перечисленные шесть
ступеней, так как только они обладают хотя бы потенци
альной возможностью воспроизвести себя как целое. Ни
ткань, ни орган способностью этой не обладают. Группы
внутри вида представляют собой исключение. Популяция,
или совокупность особей определенного вида, обладающая
границами, совпадающими с границами биоценоза, частью
которого она является, воспроизводит себя как целое. Поэ
тому целесообразно, как это и делают Мазер [Mather,
1955], Шмальгаузен [1961] и Ушаков [1963], выделять
популяционный уровень организации.
На нашем материале мы имели дело с признаками, от
носящимися к разным уровням организации живого. Неза
висимость размеров цветков живокости, или татарского
мыла, от размеров листьев и стеблей того же растения яв
ляется выражением строгой согласованности размеров
цветков друг с другом в пределах популяции. Размеры
цветков всех представителей данного вида входят в одну
плеяду друг с другом и с наличием специфического пере
носчика пыльцы. Это уже плеяда более высокого, популя
ционно-видового уровня. Но в одну плеяду входят также
размер хоботка медоносной пчелы и разхмер трубочки опы
ляемых ею цветков. Это уже плеяда еще более высокого
уровня организации — уровня биоценотического. Она ох
ватывает многие стороны жизни обоих видов — строитель
ный инстинкт рабочих пчел и размер ячей, инстинкт вы
кармливания молоди и количество и качество корма, кото
рый получает личинка, размеры хоботка и положение ко
мочка пыльцы на тельце насекомого, позу, которую при
нимает насекомое, посещая цветок данного вида растения,
и согласованность размеров хоботков у всех представите
лей данного вида пчел. И далее, она включает размеры и
конфигурацию частей цветка и согласованность размеров
цветков у всех представителей данного вида растений и
много других свойств.
Любопытно отметить изменение знака зависимости при
переходе от одного уровня организации к другому. Так,
независимость размеров цветков от размеров листьев и
стеблей у энтомофильных растений, имеющих специфиче
ских переносчиков пыльцы, является условием образова
164
ния стандарта размеров цветков всех растений данного ви
да, условием согласованного функционирования всех рас
тений, а также условием согласованности размеров цветков
и насекомых. Независимость признаков одного уровня яв
ляется условием жесткой зависимости признаков вышеле
жащих уровней организации. Подобные отношения суще
ствуют и на других уровнях организации. Так, независи
мость элементов кода наследственной информации на макромолекулярном уровне сочетается с их согласованным
функционированием на клеточном и индивидуальном
уровнях организации.
Корреляционные плеяды имеют и еще одно важное
значение в плане теории живых систем. Именно существо
вание закономерных сочетаний свойств придает эволюции
канализованный характер. Эволюция представляет собой
процесс создания закономерных сочетаний взаимозависи
мых свойств, каждое из которых накладывает известные
ограничения на дальнейший ход эволюции. Каждому уров
ню организации присущ свой тип преобразования [Берг,
1962]. Есть четыре типа преобразования: конвариантная
редупликация, деление, индивидуальное развитие, включа
ющее цикл изменений от зачатия до смерти, и эволюция.
Макромолекулярный комплекс, несущий в себе информа
цию самовоспроизведения, обладает одним только типом
преобразования. Он воспроизводит рядом с собой свою ко
пию и отталкивается от нее. На этом, помимо мутационно
го процесса, заканчиваются возможности его преобразова
ния. Он пребывает на молекулярном уровне организации
материи и неделим.
Клетка — система делящаяся. Понятие индивидуально
го развития неприменимо к клетке. Клетка, проделывая
цикл своих преобразований, не приобретает индивидуаль
ности. Она остается делимой, дивидуальной. Когда клетки
многоклеточного существа теряют делимость, в них разви
ваются структуры, не подлежащие делению. Это уже собы
тия, связанные с дифференциацией организма как целого.
Делимость клетки наложила запрет на увеличение ее раз
меров и на ее морфофизиологический прогресс (в понима
нии А. Н. Северцова). Микроскопические размеры деля
щихся клеток и единство строения клеток во всем органи
ческом мире являются выражением этого запрета.
Индивидуальность, неделимость — удел другой систе
мы — организма. Организм — система циклическая. Восп
роизводит себя организм путем отделения маленькой, а
подчас и специфически устроенной части. Преобразование
организма — это индивидуальное развитие, т. е. цикл не
165
обратимых превращений, связанных с усложнением орга
низации и неуклонно ведущих к смерти. Неограниченная
возможность усложнения организации, увеличение разме
ров, смерть — признаки этой системы, сочетающиеся зако
номерным образом друг с другом и друг от друга неотъем
лемые. Из всех способов воспроизведения только отделение
маленькой частицы не кладет запрет на усложнение орга
низации. Обладая этим способом воспроизведения, орга
низмы достигли в процессе эволюции наибольшей высоты
организации из всех систем.
Системой, претерпевающей эволюционные преобразо
вания, является вид. Структура вида как системы обеспе
чивает ему возможность эволюционировать. Все виды со
стоят из разобщенных в пространстве, до известной степе
ни сходных взаимозаменимых частей — организмов, спо
собных воспроизвести себя с избытком и обладающих ко
нечным сроком существования. Части вида способны изме
няться и стойко сохранять возникшие изменения. Именно
эти черты строения делают виды ареной деятельности есте
ственного отбора, а тем самым обеспечивают ему возмож
ность эволюционировать. Воспроизводящие себя молекулы,
клетки, особи, популяции, биоценозы и весь живой компо
нент биосферы изменяются в течение эволюции, но только
вид является тем субстратом, структура которого обеспечи
вает испытание на устойчивость отдельных частей и заме
нимость одних частей другими. Взаимозаменяемость частей
вида обеспечивает эволюцию всех вышестоящих и всех
низших по отношению к виду уровней организации. Сам
вид взаимозаменимость частей обрекает на низкий уровень
организации. Изоморфизм (или гомологичность) частей
внутри системы максимален в пределах вида по сравнению
с другими системами. Взаимозаменимость предельно огра
ничивает дифференциацию. Части организма не обладают
способностью заменять друг друга или обладают этой спо
собностью в строго ограниченной степени. Дифференциа
ция в пределах организма сочетается с интеграцией; в пре
делах вида дифференциация частей, если она не принима
ет форму балансированного полиморфизма, завершается
дезинтеграцией, обособлением, ведущим к разделению ви
да на два вида, антагонистичных друг другу.
Мы говорили уже, что все типы преобразований: авто
репродукция, деление, индивидуальное развитие и эволю
ция — осуществляются на основе определенной програм
мы, от которой зависит конечный результат. Развитие ор
ганизма представляет собой ясный пример цикла преобра
зований, осуществляющихся на основе программы. Про
166
граммы развития закодированы в виде определенной по
следовательности нуклеотидов и авторепродуцирующейся
субстанции и передаются от родителей к потомкам в про
цессе смены поколений.
Наличие программы преобразования в случае эволю
ции далеко не столь очевидно. Сами программы онтогене
тического развития и эволюции в принципе различны.
Программирование может осуществляться с помощью
жестко заданных команд, но команды могут носить и веро
ятностный характер. Относительная роль жестких и веро
ятностных команд различна для программ разных типов
преобразований. Наименьшее значение вероятностные ко
манды имеют в программах самовоспроизведения, наиболь
шее — в программах эволюции. Программирование эволю
ции осуществляется с помощью вероятностных алгоритмов.
Алгоритм эволюции подобен платежной матрице любой иг
ровой ситуации. Тахтаджян [1959] определил эволюцию
как основанную на случайных ходах антиэнтропийную иг
ру живого со средой.
Вероятностный характер программирования имеет два
разных аспекта. В каждом данном случае вероятность пре
образования зависит от наличия наследственных измене
ний и от их характера, т. е. от той пользы, которую эти
изменения приносят своим обладателям. Кроме того, тече
ние процесса задано строением системы лишь с известной
жесткостью, допускающей различные варианты. Исход
процесса преобразования зависит от конкретного использо
вания платежной матрицы участниками игры. Задана лишь
некоторая стратегия, выбор тактики зависит от обстоя
тельств конкретной обстановки. Однако само наличие стра
тегии ограничивает роль случая в антиэнтропийной игре
живого со средой. Строение системы живого выступает как
фактор ее эволюции, как ее внутренняя предпосылка. Это
касается прежде всего строения вида, с присущей ему раз
общенностью его частей в пространстве, ограниченностью
срока существования частей по сравнению со сроком суще
ствования вида, способностью частей воспроизвести себя с
избытком, т. е. черт, которые присущи всем видам живых
существ как таковым. Ко взаимообусловленность структу
ры и преобразования выражается не только в строгой при
уроченности процессов отбора к системам, построенным
подобно виду и только к ним одним, но и в деталях эволю
ционного процесса. Каждая черта организации открывает
одни возможности перед системой и неизбежно закрывает
другие. Каждый признак отдельного организма и всей со
вокупности организмов данного вида является элементом
167
программы будущей эволюции вида. Действительно, каж
дый уже закрепленный отбором признак представляет ус
ловие дальнейшей деятельности отбора. Совершенно оче
видно, что химическое строение структурных основ наслед
ственности предопределяет в значительной степени воз
можности их химических изменений. Но и совершенно, ка
залось бы, второстепенные свойства веществ, используе
мых в качестве скелетных образований, налагают на эво
люцию глубочайший отпечаток. Неспособность хитина рас
тягиваться и упругость построенных из него пластин пред
определяют множество черт организации всех тех групп
организмов, которые использовали хитин в качестве на
ружного скелета. Являясь прекрасной защитой против ме
ханических повреждений и против высыхания, хитин обес
печил биологический прогресс насекомых [Четвериков,
1915] и создал предпосылки для выхода целых групп орга
низмов из воды на сушу [Гиляров, 1959]. Вместе с тем хи
тиновый наружный скелет, создавая преграду для выделе
ния продуктов обмена, ограничил морфофизиологический
прогресс и, в частности, возможности увеличения размеров
особи. В качестве строительного материала машущего кры
ла насекомых хитин наложил дальнейший запрет на про
грессивное увеличение размера насекомых [Берг, 1960].
Но физические свойства хитинового наружного скелета
имели и несколько неожиданные последствия.
Ограничивая рост, хитин требовал компенсации в виде
линек. Линьки, способствуя разделению функций между
стадиями, создали предпосылку метаморфоза насекомых и
способствовали разделению функции роста и питания, с
одной стороны, и функции размножения и расселения, с
другой стороны, по отдельным стадиям жизненного цикла.
Афагия, или ограничение питания взрослой формы, созда
ют благоприятную предпосылку для развития полета, стан
дартизуя вес насекомого. Хитиновые покровы не растяги
ваются и не состоят из элементов, способных к пролифера
ции. Части, покрытые хитином или состоящие из хитина,
не могут менять свои габариты в процессе функционирова
ния. Крыло птицы активно приспосабливается к полету,
меняясь под воздействием функции. По отношению к хи
тиновым частям крыла насекомого функция лишена какого
бы то ни было формообразующего значения. Все процессы
роста заканчиваются до начала функционирования. Фор
мирование органа и его функционирование разобщены во
времени. Функция выступает только в роли браковщика
или испытателя пригодности. Относительная роль внутрен
них факторов развития возрастает по сравнению с внешни
168
ми. Жесткий генный контроль индивидуального развития
или независимость по отношению ко всем тем факторам,
которые не решают исход дела, приобретает важное при
способительное значение и возникает под действием отбо
ра. Погашаются как воздействие внешних факторов на
развитие, так и формативные действия одних частей на
другие. Развитие приобретает мозаичный характер.
Мозаичный характер эмбрионального развития присущ
всем группам животных, у которых части наружного ске
лета состоят из неспособных к растяжению веществ и об
разуют сплошной покров, элементы которого располагают
ся параллельно защищаемым поверхностям. Таковы черви,
членистоногие и моллюски.
Все те организмы, у которых защитные образования
располагаются перпендикулярно защищаемым поверхно
стям, как шерсть, перья, чешуя позвоночных животных,
обладают регуляторным типом развития.
Таким образом, физические свойства хитина, использо
вание его в качестве наружного покрова, относительно не
большие размеры тела, наличие линек и метаморфоз, раз
деление функций по фазам жизненного цикла и мозаич
ный тип развития насекомых представляют собой взаимо
обусловленный комплекс свойств. Причина образования
каждого из свойств коренится в другом свойстве, наличие
которого предопределяет течение процессов отбора. Каж
дый признак входит в комплекс свойств, сочетающихся с
ним строго закономерным образом.
Логика вещей такова, что по одному признаку можно с
большой достоверностью предсказать множество других
признаков. Взаимозависимы не только части системы, но и
ее свойства. Мы остановились на эволюционных последст
виях, с неизбежностью вытекающих из использования хи
тина в качестве наружного покрова. Но в таком положении
находятся все признаки живого, присущие любому уровню
организации.
Эволюционная судьба различных видов различна и сто
ит в зависимости от таких важных свойств, как автотрофность или гетеротрофность, положение в цепях питания,
оседлость или подвижность, наличие или отсутствие актив
ных способов передвижения. Но и каждый ничтожный, ка
залось бы, признак обрастает множеством связанных с ним
признаков и тем самым становится элементом программы
будущей эволюции. Портативность корма, которым родите
ли обеспечивают или выкармливают личинок или детены
шей имеет грандиозные эволюционные последствия. Оди
наковый способ выкармливания потомства конвергентно
169
ведет к образованию у птиц и у млекопитающих сходных
комплексов свойств. Если корм портативен и родители но
сят его в гнездо или в нору, образуется комплекс призна
ков, взаимно компенсирующих друг друга. Наблюдается
моногамия, борьба за охотничью территорию, ритуал вза
имного ухаживания, слабое развитие вторичных половых
различий. Участие самца в выкармливании потомства рен
табельно. Детеныши моногамов рождаются беспомощными,
в поведении взрослых условно-рефлекторный компонент
играет относительно большую роль, по сравнению с безус
ловно-рефлекторным. Если корм не портативен и не может
быть принесен детенышам, развивается другой комплекс
свойств. Вид полигамен, самец не принимает участия в вы
кармливании потомков, в лучшем случае на нем лежит ох
рана самок и молоди. Мать водит молодь к корму. Нор или
гнезд нет, как нет и боев за охотничью территорию. Ра
венство численности самок и самцов при рождении превра
щается в этих условиях в избыток самцов. Самцы поли
гамных видов ведут за самку бои друг с другом, вторичные
половые различия развиты у этих видов сильно, они не
имеют защитных окрасок, взаимного ухаживания самцов и
самок нет, ритуал ухаживания носит односторонний харак
тер стимуляции самки со стороны самца. Детеныши полигамов рождаются относительно самостоятельными, способ
ными вскоре после рождения следовать за матерью, безус
ловно-рефлекторный компонент поведения относительно
преобладает над условно-рефлекторным.
Между г.тицами и млекопитающими наблюдаются эко
логические параллели по характеру семейного строя.
Сходные комплексы свойств характеризуют моногамов сре
ди птиц и среди мелкопитающих (ласточка подобна вол
ку)» другой комплекс свойств характерен для полигамов
(фазан подобен оленю).
Способ выкармливания молоди служит как бы стрелоч
ным механизмом, переключающим эволюцию на опреде
ленный путь. Развитие всего комплекса свойств — процесс
до известной степени канализованный.
Пример выбран наудачу. Можно было бы избрать не
эволюцию семейного строя вида, а эволюцию половых хро
мосом и показать, как и здесь каждый шаг эволюции пред
определен предыдущим. Сходные комплексы признаков
развиваются конвергентно у насекомых, рыб и растений.
Различия между механизмами определения пола у разных
объектов связаны не с положением этих объектов в систе
ме, а с той или иной высотой организации самого механиз
ма определения пола [Darlington 1937, 1958:, Берг, 1942].
170
Насекомые ушли дальше по пути прогрессивного развития
половых хромосом, чем позвоночные, включая человека.
Шли они одинаковым путем, хотя и совершенно независи
мо. Энто- и анемофилия, зоо- и анемохория, образование
стелющегося или лазающего облика, эфемерность и спо
собность засорять посевы определенной сельскохозяйствен
ной культуры и множество других признаков растений
могли бы с равным основанием послужить для иллюстра
ции той же мысли.
Комплексы признаков закономерно построены. Законо
мерность их строения заключается во взаимной компенса
ции признаков, которая создается с помощью отбора.
Уже Кювье показал, что весь взаимозависимый комп
лекс свойств, характеризующий внутреннюю организацию
животного, связан с условиями существования, образуя с
ними гармоничное и чрезвычайно устойчивое целое. Кювье
переоценил устойчивость. В. О. Ковалевский [1873] пока
зал, как сочетаются принцип корреляции и принцип усло
вий существования Кювье с идеей эволюции. Бергсон
[1914] с особенной силой подчеркнул канализованный ха
рактер эволюции. Фабр [1898], Бергсон [1914], Беклеми
шев [1951, 1954], Северцов [1941, 1951], Васнецов [1938,
1953], Крыжановский [1949], Гербильский [1956, 1957,
1962], Ушаков [1959а-в], Ушаков и Даревский [1959], Ги
ляров [1945, 1959], Беляев и Журавлева [1961], Воронцов
[1962] и многие другие описали корреляции между морфо
логическими, эмбриологическими, экологическими, психо
физиологическими, биогеографическими признаками у са
мых разнообразных групп живых существ.
Параллелизмы и конвергенции представляют собой
наиболее яркое доказательство канализованного характера
эволюции. Здесь важно, что сходства касаются именно
комплексов признаков. При всем разнообразии органиче
ского мира отдельные черты организации вида отнюдь не
образуют любых комбинаций, а лишь строго ограниченные
предсказуемые сочетания. Именно благодаря наличию этих
сочетаний органический мир поддается систематизации и
образует прерывистое разнообразие форм. В основе разви
тия комплекса закономерным образом сочетающихся
свойств лежит способ взаимодействия со средой — техно
логия жизненных процессов в самом широком смысле слова.
Коррелированы не только качественные характеристи
ки, но и количественные показатели.
Образование в процессе эволюции комплексов взаимо
обусловленных свойств указывает на программированный
характер эволюции и ставит в этом отношении вид с при
171
сущим ему типом преобразования в один ряд с другими си
стемами.
Каждая живая система характеризуется комплексом
взаимообусловленных свойств или корреляционными плея
дами. Можно предложить следующую классификацию кор
реляционных плеяд.
Плеяды по уровню организации
Ма кромол е кул ярн ые
Клеточные
Онтогенетические
Видовые
Биоценотические
Живого вещества
биосферы (включая
биогеохимическис
и биогеофизические)
Плеяды в пределах уровня
Хорологические
Хорологические
Xopi логические
Хронологические
Репродуктивные
Хорологические
Филогенетические
Хорологические
Хронологические
Филогенетические
Хорологические
филогенетические
Существенные черты макромолекулярного комплекса
образуют плеяду. Линейная организация, химический уро
вень строения, электронно-микроскопические размеры,
стационарное состояние — черты, неразрывно связанные с
воспроизведением путем построения копии и с мутабильностыо. Корреляционную плеяду образуют микроскопиче
ские размеры клетки, отсутствие у нее онтогенеза, ее по
тенциальное бессмертие.
Плеяды онтогенетического уровня связывают фунда
ментальные черты строения каждой особи друг с другом.
Все существенные черты организма вытекают из циклич
ности преобразований: неделимость, воспроизведение с по
мощью отделения маленькой части, начинающей весь цикл
преобразований сначала, большие размеры, сложность
строения, смертность.
Выше приведен комплекс признаков, основанный на
физических свойствах хитина. Комплекс этот образует он
тогенетическую плеяду. Онтогенетическую плеяду образу
ют и комплексы признаков, описанные Кювье.
Популяционно-видовые плеяды описаны Васнецовым
[1938, 1953 J под названием экологических корреляций.
Взаимные приспособления особей одного и того же вида
друг к другу, или конгруэнции [Северцов, 1951], являясь
показателями целостности вида, представляют собой плея
ды признаков популяционно-видового уровня. Закономер
ные сочетания свойств популяции как целого [Берг, 1945 ]
относятся к этой же категории. Свойства, обусловленные
172
способом выкармливания молоди, также образуют плеяды
видового уровня.
Биоценотические плеяды описаны П. В. Терентьевым
[1960]. Примеров биоценотических плеяд множество: вза
имные адаптации хищника и жертвы [Lotka, 1925; Северцов, 1941 ], связь между числом линек, характером полово
го созревания, образом жизни кровососущих насекомых и
особенностями тех животных, чьей кровью они питаются
[Беклемишев, 1951, 1954], согласованность длины хоботка
пчел с размерами цветков тех растений, которые они посе
щают [Алпатов, 1927; и др. ]. Терентьев указал на сущест
вование хорологических биоценотических плеяд. Законы
сукцессии создают хронологические биоценотические плея
ды. Плеяды каждого уровня организации обладают своим
характером распределения во времени и в пространстве.
Хорологические плеяды присущи всем уровням органи
зации живого. Хронологические плеяды встречаются только
у тех систем, которые обладают циклом превращений или
индивидуальным развитием, т. е. у организмов и биоценозов
в целом. Филогенетические плеяды присущи тем системам,
которые обладают соответствующей структурой, обеспечива
ющей возможность эволюционных преобразований, т. е.
виду и системам, включающим вид как подчиненный эле
мент системы,— биоценозу и живому веществу биосферы.
Репродукционные плеяды характерны только для одного
уровня организации. Они присущи только организму —
единственной системе, достигшей высшей интеграции час
тей, полностью утратившей делимость и воспроизводящей
себя с помощью отделения маленькой части существа, та
кой маленькой, что она потребовала заботы о потомстве.
Каждая черта организации данного уровня согласована
с другими чертами этого уровня и подчинена вышележа
щему уровню иерархии.
Изучение взаимоотношений адаптаций разных уровней
организации живого — одна из важнейших глав современ
ной теоретической биологии [Берг, 1945; Александров,
1956; Гербильский, 1956, 1957, 1962; Ушаков, 1959а — в;
Ушаков, Даревский, 1959; и др. ].
Особого внимания заслуживает только что опублико
ванная работа Б. П. Ушакова [1963], посвященная класси
фикации приспособлений в соответствии с уровнями орга
низации живсго. У нас на глазах возникает новая наука —
цитоэкология, в задачу которой, по мысли ее создателей,
входит изучение роли клеточных реакций в приспособле
нии организма к условиям обитания [см.: Роль клеточных
реакций..., 1963; Ушаков, 1963].
173
выводы
1. Анализ корреляционных плеяд позволяет внести ко
личественную меру в изучение стабилизирующего отбора.
Мера независимости размеров данной части от размеров
других частей того же организма является показателем ее
устойчивости. Мерилом стабильности части служили коэф
фициент вариации (С %) ее размеров и отношение макси
мальной варианты к минимальной Утах/ У т^ Мерилом не
зависимости служил коэффициент корреляции г, связыва
ющий размеры части с размерами всего организма в целом.
Между показателями устойчивости и независимости на
блюдается прямая корреляция.
2. Выяснилось, что размеры цветков устойчивы и неза
висимы по отношению к размерам всего растения в целом
у растений, имеющих специфических переносчиков пыль
цы, и что размеры «единицы расселения» (семян и односе
мянных плодов) устойчивы и независимы у растений, име
ющих специфические способы расселения (анемо- и зоохория).
Введение количественного показателя относительной
роли внешних и внутренних факторов в развитии и сопо
ставление меры независимости с экологической ситуацией
позволяют вскрыть эволюционный механизм образования
независимости одних частей по отношению к другим час
тям того же организма. Независимость признака возника
ет там, где формирующие факторы и факторы контроля
(отбирающие факторы) не совпадают, т. е. там, где выра
батывается приспособление к таким условиям среды, ко
торые не способны изменить признак ни в процессе его
формирования, ни в процессе функционирования соот
ветствующей части. Контроль со стороны биотических
компонентов среды и со стороны специфических функций
в том случае, когда те и другие не принимают участия в
формировании признака и не способны преобразовать
его, и представляет собой условие наследственного про
граммирования,
3. Каждая система характеризуется комплексом взаи
мообусловленных свойств. Если таких комплексов несколь
ко, перед нами — корреляционные плеяды признаков.
Мыслимы плеяды всех уровней организации живого. На
нашем материале мы встретились с комплексами согласо
ванных признаков трех уровней организации живого. Со
гласованность размеров листьев, стеблей и соцветий, с од
ной стороны, и размеров различных частей цветка, с дру
гой стороны, образует плеяды онтогенетического уровня.
174
Согласованность размеров насекомых и цветков образует
биоценотическую плеяду.
Выявляется теснейшая взаимообусловленность процес
сов и структур онтогенетического, популяционно-видового
и биоценотического уровней организации. Независимость
размеров цветков энтомофильных специализированных
растений от размеров стеблей и листьев — условие тесней
шей взаимозависимости между размерами всех цветков
данного вида друг с другом и с размерами насекомых —
переносчиков пыльцы.
СПИСОК ЛИТЕРАТУРЫ
Александров В. Я. Цитофизиологический анализ теплоустойчивости рас
тительных клеток и некоторые задачи цитоэкологии / / Бот.
ж урн.— 1956.— Т. 41, № 7 .— С. 939—961.
Александрова А. Д. Опыт анализа явлений саморегуляции в фитоценозе
с точки зрения некоторых идей кибернетики / / Применение мате
матических методов в биологии.— Л.: Изд-во ЛГУ, 1963.—
Вып. 2 .— С. 37— 45.
Алпатов В. В. Биометрическая характеристика среднерусской и украин
ской пчелы / / Русск. зоол. журн.— 1927.— Т. 7, № 4 .—
С. 31— 74.
Алпатов В. В., Тюнин Ф. А. К познанию изменчивости хоботка рабочей
пчелы / / Русск. зоол. ж урн.— 1925 — Т. 5, № 4 .— С. 79— 108.
Беклемишев В. Н. Паразитизм членистоногих на наземных позвоночных
и пути его возникновения / / Мед. паразитология и паразитар. бо
лезни.— 1951.— Вып. 2 .— С. 151 — 160; Вып. 3.— С. 233— 141.
Беклемишев В. Н. Паразитизм членистоногих на наземных позвоночных.
Основные направления его развития / / Мед. паразитология и пара
зитар. болезни.— 1959.— Вып. 1.— С. 3—20.
Беляев Д. К., Журавлева К. В. Связь жирномолочности с некоторыми се
зонно изменяющимися признаками крупного рогатого скота и ее
значение для селекции / / Тез. докл. Межвузовской конф. по эксперим. генетике.— Л.: Изд-во ЛГУ, 1961.— Вып. 1.— С. 19— 20.
Берг Л. С. Экологические параллели между миногами и лососевыми / /
Докл. АН СССР.— 1935.— Т. 3, № 2 .— С. 91—93.
Берг Р. Л. Историческое развитие механизма реализации пола / / Журн.
общ. биол.— 1942.— Т. 3. № 3 — С. 171 — 189.
Берг Р. Л. Вид как эволюционирующая система / / Рефераты работ учр.
Отд. биол. наук АН СССР за 1941 — 1943 гг.— М.: Изд-во АН
СССР.— 1944.— С. 184.
Берг Р. Л. Корреляция между мутабильносгью и регуляторной способно
стью организма и ее эволюционное значение / / Изв. АН СССР.
Отд. биол. наук.— 1945.— № 3 .— С. 367—376.
Берг Р. Л. Стандартизирующий отбор в эволюции цветка / / Бот. журн.—
1956.— Т. 41, № 3 . - С. 318—334.
Берг Р. Л. Типы полиморфизма / / Вести. ЛГУ.— 1957.— С. 115— 139.
Берг Р. Л. Дальнейшие исследования по стабилизирующему отбору в эво
люции цветка / / Бот. ж урн.— 1958.— Т. 43, № 1.— С. 12— 27.
Берг Р. Л. Экологическая интерпретация корреляционных плеяд / /
Весгн. ЛГУ.— 1959.— Вып. 2 .— С. 142— 152.
Берг Р. Л. Межвидовая и внутривидовая изменчивость жилкования крыла
в семействе дрозофилид (Drodophilidae) / / Применение мат. мето
дов в биологии.— Л.: Изд-во ЛГУ, I960.— С. 47—64.
Берг Р. Л. Корреляционные плеяды у растений / / Тез. докл. на третьем
совещании по примен. мат. методов в биологии.— Л.: Изд-во ЛГУ,
1961а.— С. 10— 12.
175
Берг Р. Л. Селекционно-генетическое значение корреляционных плеяд / /
Тез. докл. Межвузовской конф. по эксперим. генетике.— Л.: Издво ЛГУ, 19616.— Вып. 1.— С. 24.
Берг Р. Л. Структура и развитие / / Тез. докл. на науч. сессии Биол. инта ЛГУ.— Л.: Изд-во ЛГУ, 1962.— С. 8— 10.
Бергсон А. Творческая эволюция.— М; Спб, 1914.
Васнецов В. В. Экологические корреляции / / Зоол. журн.— 1938.—
Т. 17, № 4 .— С. 5 71—581.
Васнецов В. В. Целостность экологии вида у рыб (экологические корреля
ции) / / Очерки по общим вопросам ихтиологии.— М.: Изд-во АН
СССР, 1953.
Вернадский В. И. Биосфера.— Л.: Науч. хим.-техн. изд-во, 1926.
Вернадский В. И. Биогеохимические очерки.— М.: Изд-во АН СССР, 1940.
Воронцов Н. Н. Пути пищевой специализации и эволюции пищевари
тельной систем у некоторых мышеобразных (Muroidea) грызунов:
Автореф. дис. ... канд. биол. наук.— М., 1962.
Гербильский Н. Л. Специфика и задачи экологической гистофизиологии
как одного из направлений гистологических исследований / / Арх.
анат., гистол., эмбриол.— 1956.— Т. 33, № 2 .— С. 14—21.
Гербильский Н. Л. Внутривидовая биологическая дифференциация и ее
значение для evma в мире рыб / / Вести. ЛГУ.— 1957.— Вып. 4 .—
С. 82— 92.
Гербильский Н. Л. Теория биологического прогресса вида / / Тез. докл.
на науч. сессии Биол. Ин-та ЛГУ.— Л.: Изд-во ЛГУ, 1962.—
С. 3 - 4 .
Гиляров М. С, Влияние характера расселения на ход онтогенеза насеко
мых / / Журн. общ. биол.— 1945.— Т. 6, № 1.— С. 26—36.
Гиляров М. С. Закономерности изменения морфологических и физиологи
ческих особенностей членистоногих при переходе к наземному об
разу жизни / / Тр. ин-та/И н-т эвол. морф, животных им.
А. Н. Северцова.— М; Л.: Изд-во АН СССР, 1959.— Т. 27.—
С. 118— 133.
Дарвин Ч. Приспособление орхидных к оплодотворению насекомыми / /
Поли. собр. соч.— 1928.— Т. 4.
Драгавцев В. А. Методы анализа формообразования древесных растений
на основе изучения изменчивости черного саксаула: Автореф.
дис. ... канд. биол. наук.— Алма-Ата, 1962.
Завадский К. М. Учение о виде.— Л.: Изд-во ЛГУ, 1961.
Ковалевский В. О. Остеология Anchiterium aurelianense Cuv. как формы,
выясняющей генеалогию типа лошади (Equua).— М.: Изд-во АН
СССР, 1948.
Крыжановский С. Г. Экологическо-морфологические закономерности раз
вития карповых, вьюновых и сомовых рыб / / Тр. ин-та/Ин-т эвол.
морф, животных им. А. Н. Северцова.— М.; Л.: Изд-во АН СССР,
1949.— С. 237— 331.
Левина Р. Е. Типы плодов и их классификация.— Ульяновск, 1959.
Ляпунов А. А. Об управляющих системах живой природы и общем пони
мании жизненных процессов.— М.: Изд-во АН СССР, 1962.
Петрушевская М. Г. Сравнение групп Azagia по морфологическим при
знакам / / Тез. докл. Третьего совещания по примен. мат. методов
в биологии.— Л.: Изд-во ЛГУ, 1961.— С. 61— 62.
Плохинский Н. А. Индекс производителя и его ошибка репрезентативно
сти / / Применение мат. методов в биологии.— Л.: Изд-во ЛГУ,
1963.— Вып. 2 .— С. 230— 233.
Роль клеточных реакций в приспособлении многоклеточных организмов к
температуре среды / / Тез. докл. Между нар. симпозиума по гистоэ
кологии.— М.; Л.: Изд-во АН СССР, 1963.
Рубцов И. А. О неравномерности темпа эволюции / / Журн. общ. биол.—
1945.— Т. 6, № 6 .— С. 411—441.
Северцов С. А. Динамика населения и приспособительная эволюция ж и
вотных.— М.; Л.: Изд-во АН СССР, 1941.
Северцов С А. О конгруэнциях и понятии целостности вида / / Пробле
мы экологии животных.— М.: Изд-во АН СССР, 1951.— Вып. 1.—
С. 3 0 - 5 7 .
176
Сукачев В. Н. Основы теории биогеоценологии / / 30-летие Вел. Окт.
Соц. Рев.: Юбил. сб.— М.: Изд-во АН СССР, 1947.
Тахтаджян А. Л. Вопросы эволюционной морфологии растений.— Л.:
Изд-во ЛГУ, 1954а.
Тахтаджян А. Л. Происхождение покрытосемянных растений.— М., 19546.
Тахтаджян А. Л. Эволюция в терминах кибернетики и общей теории
игр / / Тез. докл. Второго совещания по примен. мат. методов в
биологии.— Л.: Изд-во ЛГУ, 1959.— С. 4J—49.
Терентьев П. В. Метод корреляционных п л е я д // Вести. ЛГУ.— 1959.—
№ 9 .— С. 137— 141.
Терентьев П. В. Дальнейшее развитие метода корреляционных плеяд / /
Применение мат. методов в биологии.— Л.: Изд-во ЛГУ, 1960.—
С. 27— 36.
Тимофеев-Ресовский Н. В. Некоторые проблемы радиационной биогеоце
нологии / / Докл. по опублик. работам, представл. для защиты уч.
степ, д-ра биол. наук.— Свердловск, 1962.
Ушаков Б. П. О механизме адаптации клеток животных / / Цитология.—
1959а.— Т. 1, № 1.— С. 35— 47.
Ушаков Б. П. Теплоустойчивость тканей — видовой признак пойкилотермных животных / / Зоол. ж урн.— 19596.— Т. 38, № 9 .—
С. 1292— 1302.
Ушаков Б. П. Физиология клетки и проблемы вида в зоологии / / Цито
логия.— 1959в.— Т. 1, № 5 .— С. 541—565.
Ушаков Б. П. О классификации приспособлений животных и растений и
о роли цитоэкологии в разработке проблем адаптации / / Проблемы
цитоэкологии животных.— М.; Л.: Изд-во АН СССР, 1963.—
С. 5 — 20.
Ушаков Б. П., Даревский И. С. Сравнение теплоустойчивости мышечных
волокон и отношения к температуре у двух симпатрических видов
полупустынных я щ е р и ц // Докл. АН СССР.— 1959.— Т. 128,
№ 4 .— С. 833— 839.
Ф абр Ж .-А . Инстинкт и нравы насекомых. Из энтомологических воспо
минаний Фабра (сокращенный перевод под ред. И. Ш евырева).—
Спб., 1898.
Хохлов С. С. О количестве видов растений, размножающихся бесполосе
менным путем / / Уч. Зап. Саратов. Ун-та. Вып. биолого-почвен
ный.— 1959.— № 64.— С. 117— 123.
Четвериков С. С. Основной фактор эволюции насекомых / / Изв. Моек,
энтомол. о-ва.— 1915.— Т. 1.
Шванвич Б. Н. О механизме опыления красного клевера медоносной пче
лой / / Докл. ВАСХНИЛ.— 1952.— № 8 .— С. 33— 36.
Шванвич Б. Н. О механизме опыления мотыльковых культур медоносной
пчелой / / Тез. докл. науч. сессии биолого-почв. фак. ЛГУ.—
1955.— С. 42— 43.
Шванвич Б. Н. О работе медоносной пчелы на мохнатой вике и на лядвенце / / Вести. ЛГУ.— 1956.— С. 5 5 — 61.
Шмальгаузен И. И. Темпы эволюции и факторы, его определяющие / /
Журн. общ. биол.— 1943.— № 4 / 5 .— С. 253— 185.
Шмальгаузен И. И. Факторы эволюции (теория стабилизирующего отбо
р а ).— М.; Л.: Изд-во АН СССР, 1946.
Шмальгаузен И. И. Интеграция биологических систем и саморегуля
ция/ /
Бюл. МОИП. Отд. биол.— 1961.— Т. 66, № 2 . —
С. 104— 134.
Шмидт В. М. Корреляционная структура признаков некоторых видов и
форм зубчатки Odontites Zinn. / / Применение мат. методов в био
логии.— Л.: Изд-во ЛГУ, 1963.— С. 81— 89.
Эшби У. Р. Конструкция мозга. Происхождение адаптивного поведе
ния.— М.: ИЛ.— 1962.
Anderson Е., Stebbins G. L. Jr. Hydridi/ation as an evolutionary stimulus / /
Evolution.— 1954.— Vol. 8, № 4 .— P. 378— 388.
Berg R. L, A. general evolutionary principle underlying the origin of develop
mental homeostasis / / Amer. Nat.— 1959.— Vol. 93.— P. 103 — 105.
Berg R. L. The ecological signiqicance of correlation pleiades / / Evolution.—
I9 6 0 .— Vol. 14, № 2 ,— P. 171 — 180.
177
Bertalanffy L. von. Das biologische Weltbild. Die Stellung des Lebens in Natur
und Wissenschaft.— Berlin, 1949.— Bd 1.
Bertalanffy L. von. Problems of general system theory Symp. on general
system theory: a new approach to unity of science / / Human Biol.—
1951.— Vol. 23, № 4 .— P. 302—311.
Darlington C. D. Recent Advances in Cytology.— Philadelphia, 1937.
Darlington C. D. Evolution of Genetic Systems.— 2d ed .— Cambridge Univ.
Press, 1958.
D ickerson G. E. Genetic slippage in response to selection for multiple
objectives / / Cold Spring Harb. Symp. on Quant. Biol.— 1953.—
Vol. 2 0 .— P. 213— 224.
Grant V. Pollination systems as isolating mechanism / / Evolution.— 1949.—
Vol. 3 .— P. 82— 97.
Grant V. The protection of the ovules the flowering plants / / Evolution.—
1950.— Vol. 4 .— P. 179— 201.
Grant V. Isolation and hybridization between Aquilegia formosa and A.
pubescetis / / El Aliso.— 1952.— № 2 .— P. 341—360.
H yxley J. S. Evolution. The Modern Synthesis.— N. Y.; L., 1942.
K oller D. Germination / / Scint. Amer.— 1959.— № 4 .— P. 75— 85.
Lerner M. I. Genetic Homeostasis.— N. Y., 1954.
Lerner M. I. The Genetic Basis of Selection.— N. Y:, L., 1958.
Lotka A. I. Element of Physical Biology.— Baltimore, 1925.
Mather K. Biological organisation / / * Adv. of Science.— 1955.— Vol. 22,
№ 4 6 . - P. 2 5 0 - 2 5 7 .
Mather K. Competition and co-operation / / Mechanisms in Biological
Competition. Symp. Soc. Exp. Biol.— Cambridge Univ. Press, 1961.—
Vol. 15.— P. 264— 281.
Needham , J. Integrative Levels.— Oxford, 1937.
Sernander R.
Entwurf
einer
Monographic
der
eupropaischen
Myrmecochoren / / Kungl. Svenska Vetenskapsacad. Handl.— 1906.—
Bd 41, № 7 .— S. 1— 410.
Sim pson G. G. Tempo and Mode in Evolution.— N. Y., 1944.
Strow R. M.
Hybridization,
homogamy
and
sympatric
speciation / /
Evolution.— 1955.— Vol. 9, № 4 .— P. 441—444.
Tachtajan A. L. Die Evolution der Angiospermen.— Jena, 1959.
Terentjev P. V. Biometrische Untersuchunger uber die morphologischen
Merkmale Ratia ridibunda P a ll.// Biometrica.— 1931.— Bd 23.—
§ 23_51.
Waddington С. H. The Strategy of the G ene.— L., 1957.
О ЗАКОНОМЕРНОСТЯХ эволюции
НА ПРИМЕРЕ КОМПЛЕКСНЫХ ПРЕОБРАЗОВАНИЙ
ПРИЗНАКОВ ПРИ ДИВЕРГЕНЦИИ ВИДОВ ВЕРОНИК
(Veronica L., Scrophulariaceae)*
Р. Л . Берг , Л . Д. Колосова
Закономерности сопряженной эволюции. Сопряжен
ная эволюция цветков и насекомых — пример закономер
ного хода истории, когда каждый предыдущий шаг предоп
ределяет направление последующего. Перенос пыльцы с
цветка на цветок с помощью насекомых диктовался необ
* Проблемы эволюции.— Новосибирск, 1975.
178
ходимостью более экономного использования пыльцы.
Дальнейшая экономия пыльцы вызвала к жизни ее лока
лизацию на теле насекомого-опылителя. Принцип локали
зации пыльцы был открыт Дарвином [1928] у орхидных и
долго не привлекал внимания исследователей, пока через
сто лет профессор Ленинградского университета энтомолог
Б. Н. Шванвич [1955] не переоткрыл его для бобовых.
Шванвич показал, что клевер локализует пыльцу на ниж
ней стороне головы шмеля, вика мохнатая — на мембране,
поддерживающей снизу хоботок, а лядвенец — на стебель
ке, соединяющем грудь и брюшко пчелы.
Локализация пыльцы на маленьком участке тела насе
комого, да еще там, куда не достигают очистительные ап
параты транспортера,— весьма эффективный способ эко
номить пыльцу, но смысл он имеет только в сочетании с
наличием узкого круга переносчиков пыльцы. По мере
уменьшения количества пыльцы, израсходованной для од
ного акта опыления, уменьшалось и число возможных ее
переносчиков. Стандарт размеров насекомого, осуществля
ющего перекрестное опыление, стал необходимым услови
ем эффективности действия. Стандарт до известной степе
ни обеспечен, если пыльца доступна насекомым только од
ного вида. Возникла специфическая энтомофилия и одно
временно с ней множество приспособлений, обеспечиваю
щих привлечение своего транспортера и защиту от нежела
тельных визитеров. Задача заключалась в том, чтобы по
местить пыльники и рыльце в резервуар, предохраняющий
их от воды, ветра и чужих животных и указать «своему»
тот единственный путь, который обеспечит контакт внача
ле с рыльцем, а затем с пыльниками цветка. Цветковые
растения «изобрели» средства привлечения и ориентации
насекомого (подачу сигналов на расстоянии путем воздей
ствия на его зрение и обоняние) и целый комплекс приспо
соблений, чтобы заставить насекомое принять позу, при
которой пыльца, локализованная на должном месте «свое
го» вида насекомого, попадает на рыльце. После этого в
ход должны быть пущены аппараты, заставляющие насеко
мое прижаться к пыльникам и притом тем участком тела,
которым оно только что касалось рыльца. Множество видов
насекомых и различия в деталях конструирования у расте
ний создает превеликое разнообразие мира растений —
специфических энтомофилов.
Билатеральная симметрия, малое число гомологичных
частей, строгая фиксация их положения в пространстве,
срастание частей друг с другом,— все это необходимые ус
тройства, обеспечивающие локализацию пыльцы на теле
179
насекомого-переносчика. Совершенно сходный комплекс
признаков специфического энтомофила возник на разной
морфологической основе конвергентно в различных семей
ствах — бобовых, норичниковых, губоцветных, лютико
вых и др.
Настурция (Tropaeolum та jus L.) локализует пыльцу
на нижней стороне шмеля. Шмель должен заметить цве
ток, сесть на посадочную площадку из трех нижних лепе
стков, привалиться грудью к пыльникам, засунув голову в
свод, образованный сросшимися верхними лепестками и
чашелистиками, вытянуть хоботок на полную длину и со
сать нектар, наполняющий шпорец цветка. Но стоит шме
лю нацелиться и сесть, как лепестки, снабженные тонкими
ноготками, разъезжаются в стороны: шмель сам включил в
действие аппарат, специально предназначенный для разво
да его ног. Насекомое вынуждено привалиться нижней сто
роной груди к пыльникам цветка. Возвышающийся над
входом в шпорец узорный свод мешает подойти к цветку с
другой стороны, помимо предательской посадочной пло
щадки. Но дотронуться до пыльников еще мало, устройство
цветка настурции заставляет шмеля плотно прижаться к
пыльникам. Нектар заполняет длинный шпорец настурции
ровно настолько, чтобы добраться до него можно было бы
только кончиком хоботка, распластавшись на цветке.
Билатеральная симметрия цветка целиком порождена
специфической энтомофилией. Билатеральная симмет
рия — свойство подвижных и присуща она целым организ
мам только в мире животных за исключением тех, кто
прикрепляется к грунту. У растений билатерально симмет
ричны те части, по которым идет ток питающих жидко
стей: листья, целые ветви, лепестки и чашелистики. Ис
ходная организация цветка — радиальная симметрия. Нет
иных факторов, помимо насекомых, которые сделали бы
цветок билатерально симметричным — он неподвижен и
питающие его токи могут сделать симметричной каждую
его часть, но не цветок как целое. Билатеральная симмет
рия цветков — приспособление для локализации пыльцы
на теле насекомого-переносчика пыльцы. Цветки разделе
ны плоскостью симметрии на две равные части, потому что
их опылители активно перемещаются в пространстве.
Уменьшение числа элементов симметрии — показатель
прогрессивной эволюции. Возникает оно конвергентно и
бесчисленное число раз.
В. А. Догель [1954] выдвинул принцип уменьшения
числа гомологичных частей как ярчайший образец законо
мерного хода эволюции животных, иллюстрируя это мно
180
жеством примеров. Он назвал его принципом олигомериза
ции. Олигомеризация элементов симметрии не попала в
поле его зрения. Для растений принцип олигомеризации
выдвинул К. С. Мережковский [1910], который так же,
как Догель, говорит об уменьшении числа гомологичных
частей. Олигомеризация частей цветка в процессе эволю
ции вырабатывалась в целях экономии пыльцы по мере то
го, как сужалось пространство на теле насекомого, на ко
торое пыльце надлежало попасть. Параллельно с уменьше
нием числа гомологов шла строгая фиксация их положения
в пространстве. Сростнолепестность, образование резервуа
ров, шпорцев, трубочек из лепестков и чашелистиков или
из налегающих пластинчатых частей цветка — все это ре
зультат приспособления цветка к насекомому.
Закономерное сочетание признаков, характерное для
растения специфического энтомофила, касается не только
расцветок, форм, положения и числа частей цветка. В ком
плексе свойств, отличающих цветки специфических энтомофилов от цветков, на опыление которых со стороны на
секомых не наложено никаких ограничений, входит и ха
рактер изменчивости цветка [Берг, 1958, 1959, 1964; Берг,
Колосова, 1971; Берг и др., 1973]. Цветки специфических
энтомофилов стандартны и независимы по размерам от ус
ловий произрастания растения и от размеров других частей
того же растения. Размеры цветков не следуют за измене
нием условий произрастания так же, как не следуют они
за изменением высоты стебля, размером листьев и соцве
тий. Они независимы как от внешней, так и от онтогенети
ческой среды, если позволительно назвать так взаимовлия
ние частей организма в процессе развития.
Итак, билатеральная симметрия цветка, малое число
гомологичных частей, строгая фиксация их положения в
иространсте, наличие трубочек, резервуаров, сводов, всех
дериватов сростнолепестности и всех ее аналогов, стандарт
и независимость от среды размеров цветка составляют ком
плекс свойств, сопряженный с наличием узкого круга пере
носчиков пыльцы. Каждое свойство комплементарно по от
ношению к другому, без другого не существует. Противо
положный комплекс свойств характерен для неспецифиче
ских энтомофилов. Мак или водяная лилия могут быть
примером таких растений. Радиальная симметрия, большое
число гомологичных частей, отсутствие строгой фиксации
их положения в пространстве, отсутствие срастания частей
околоцветника, широчайший размах изменчивости и зави
симость размеров цветка от размеров растения в целом со
ставляют этот комплекс. Он построен столь же закономер
181
ным образом, как и противоположный ему, и каждый его
компонент так же предопределен наличием других призна
ков комплекса.
Совокупность свойств образует единое целое в процессе
функционирования. Каждое Свойство комплекса служит
элементом программы эволюционных преобразований, на
правляет эволюцию в определенное русло, служит той
внутренней средой, по отношению к которой под действи
ем отбора создается приспособление. В случае специфиче
ской энтомофилии в основе комплекса лежит локализация
пыльцы на теле насекомого-опылителя. Экономия пыльцы
выдвинута на первый план. В случае неспецифической эн
томофилии ставка делается на повышение вероятности пе
реноса пыльцы.
Специфическая энтомофилия — алломорфоз. Специ
фическая энтомофилия, вызвав к жизни множество про
грессивных черт, касающихся предохранения пыльцы от
рассеяния ветром, намокания, иссушения и поедания насе
комыми, создав защитные камеры, обеспечила своим обла
дателям возможность экспансии за пределы ареала.
Стандарт и независимость размеров создали особо бла
гоприятные условия для ускорения эволюции. Узкий раз
мах онтогенетической изменчивости цветка, коль скоро он
создался в процессе стабилизирующего отбора, сам способ
ствовал повышению интенсивности отбора. Возможности
филогенетической изменчивости возрастают по мере того,
как возрастает доля генотипической изменчивости в попу
ляционном разнообразии. Отбор не наталкивается на моди
фикации, имитирующие в силу параллелизма наследствен
ной и ненаследственной изменчивости фенотипическое вы
ражение мутаций, и беспрепятственно творит новые фор
мы [Берг и др., 1973].
Но сама быстрота эволюционных преобразований на
пути совершенствования специфической энтомофилии таит
в себе угрозу вымирания. Сужение круга переносчиков
пыльцы понижает вероятность опыления. И дальнейший
прогресс лежит уже на путях отказа от одного единствен
ного способа перекрестного опыления. Разрушение всей си
стемы тонко и точно подогнанных друг к другу признаков
шло строго закономерно — стоило измениться одному при
знаку, как с неизбежностью менялся и другой. Виды раз
ных семейств конвергентно переходили от специфической
энтомофилии к неспецифической, к самоопылению и ветроопылению. Это расширение связей со средой, достигну
тое аппаратами с высокой степенью расчленения отдель
ных структур в пространстве и отдельных актов во време
182
ни, несомненно, было ароморфозом. К некоторым зако
номерностям в эволюции вероник, связанным с отказом от
специфической энтомофилии, мы и хотим привлечь вни
мание.
Эволюционные парадоксы. Узкий размах онтогенети
ческой изменчивости, выработанный в условиях специфи
ческой энтомофилии, обеспечил легкость эволюционных
преобразований на путях отказа от специфической энтомо
филии. Увеличение числа возможных способов переноса
пыльцы могло идти по линии совмещения неспецифиче
ской энтомофилии с самоопылением и ветроопылением.
Что выиграли новоявленные неспецифические энтомофилы
от своей былой узкой специализации? Множество различ
ных приспособлений — резервуары влаги, укрытия для са
мых уязвимых частей цветка, разнообразные изобретения
для экономного расходования пыльцы и нектара, для луч
шего использования пространства и времени. Вид растения,
превратившего шпорец с нектаром в хранилище влаги, по
добен народу, перед которым сосуд — бурдюк или амфо
ра — открыл возможности расселения. Но шпорец был со
здан как застольная утварь — рог или чаша, предназна
ченная для строго ограниченного круга визитеров. Без спе
цифической энтомофилии он не был бы выработан никог
да. Специфическая энтомофилия как предпосылка к рас
ширению связей со средой — парадокс в эволюционной
практике Природы и притом парадокс не единственный.
Переход насекомых к жизни в воде — не менее яркий
пример подобного эволюционного парадокса. Дж. Бимент
[1964] убедительно показал, что насекомые выработали
множество черт, связанных с водным образом жизни, поль
зуясь приспособлениями для полета. Слои воздушных пу
зырьков и воздушные мешки водяных жуков, подкрыловые
воздушные камеры плавунцов, строение дыхалец и способ
ность подвешиваться под поверхностью воды у личинок ко
маров, способность к скольжению как по поверхности во
ды, так и под водой, не намокая, свойственная многим на
секомым,— все это различные способы поставить себе на
службу ту несмачиваемость, которая как необходимое ус
ловие входит в план строения летающих организмов. По
гружаются под воду они и живут там, потому что их пре
дки или они сами летают.
Вероники. Не возникает ни малейшего сомнения, что
предок современных вероник был специфическим энтомофилом. Цветки всех представителей рода обладают всеми
признаками связи с узким в таксономическом отношении
кругом переносчиков пыльцы [Берг, Колосова, 1971]. Чис
183
ло гомологичных частей мало — рыльце одно, тычинок
две, лепестков четыре, чашелистников — четыре, пять.
Цветки зигоморфны. Нижние части лепестков и чашелист
ников образуют трубочки. Но все вероники являются не
специфическими энтомофилами.
Мы произвели измерения размеров многих признаков у
13 видов вероник и изучили корреляции между всеми из
меренными признаками в пределах каждого вида. Сборы
проведены в окрестностях г. Новосибирска ( V. chamaedrys L.,
F. krylovii Schischk., F. serpyllifolia L.), Владивостока
(F. sibirica L., F. tubiflora Fisch. et Mey), на Южном
Сахалине
(F.
sachalinensis
Boriss.,
F.
americana
(Rafin) Schweinitz), на Камчатке на горе Колдун
(F. grandiflora Gaertn), на Саянах (F. sajanensis Printz), в
степных районах юга Красноярского края (F. spicata L.,
F. incana L.).
В поле нашего зрения попали виды вероник, отлича
ющиеся друг от друга по множеству признаков. Есть
обитатели высокотравья (F. sibirica, F. sachalinensis),
пойменных лугов (F. longifolia ), высокогорной тундры
(F. grandiflora ) , субальпийских лугов (F. sajanensis ),
сильно увлажненных местообитаний (F. americana) и сте
пей (F. incana ). Не менее различаются изученные виды и
по общему облику, по размерам растения в целом и по
строению
цветков.
Здесь
и такие
гиганты,
как
F. sachalinensis, F. sibirica, и крошечные высокогорные F.
densiflora , F. grandiflora . Измерялись низкорослые виды с
короткотрубчатыми яркими цветками, собранными в рых
лые малоцветковые
соцветия
(F.
chamaedrys,
F. grandiflora) и гиганты с крошечными цветками, со
бранными в колосовидные соцветия тоже гигантских
размеров. Число цветков в последних достигает многих
сотен.
Все изученные нами виды вероник отказались от спе
цифической энтомофилии, и средства, которые они исполь
зовали, переходя к неспецифической энтомофилии, были
ограниченными. Наметилось два основных направления в
эволюции, причем каждое из них привело к строго законо
мерному сочетанию целого комплекса признаков. Некото
рое представление об этих сочетаниях дает таблица, в ко
торую включены шесть видов с длиннотрубчатым венчи
ком (F. sachalinensis у F. sibirica, V. tubiflora, F. longifolia,
F. incanat F. spicata) и три вида с короткотрубчатым
(F. chamaedrySy F. krylovii, F. serpyllifolia). Мы подобрали
этот контрастирующий признак, чтобы посмотреть, не по
ложит ли он грани между видами и по другим признакам,
184
Характеристика видов вероник по ряду признаков
Вид
У.
У.
У.
У
У
У.
V.
У.
У.
Высота
стебля
Длина Отношение
листа длины тру
бочки венчика кдли
не большего
лепестка
Отношение
количества
цветков
в соцветии
кдлине
соцветия
Единица
опыления
1
2
3
4
5
6
sachalinensis
sibirica
tubiflora
longifolia
spi catа
/пеана
krylovii
charnaedrys
serpyllifolia
1382л
837,8
578,8
694,0
221,5
161,4
191,9
132,8
114,4
149,5
131,1
85,3
90,3
45,4
33,4
29,7
21,1
12,3
2,10
2,04
2,40
0,63
0,43
0,68
0,20
0,17
0,25
3,29
2,63
2,17
Соцветие
1,71
1,79
1,52
0,24
0,49
0,26
Цветок
помимо длины трубочки венчика. И грань наметилась.
Длиннотрубчатые виды обладают более длинными листья
ми (см. таблицу, графа 3) и более высоким стеблем (гра
ф а ! ) , чем виды короткотрубчатых, хотя изменчивость по
последнему признаку и носит трансгрессивный характер.
Отношение длины трубочки венчика к длине самого боль
шого лепестка у них больше, чем у вероник с короткой
трубочкой венчика (графа 4). Число цветков на единицу
длины соцветия, или плотность соцветия у них во много
раз превышает тот же индекс у видов короткотрубчатых
(графа 5). Все эти признаки вовлечены в процесс диверген
ции.
Сочетания признаков отнюдь не равновероятны. Боль
шие размеры растения в целом сочетаются с наличием
плотных колосовидных соцветий, насчитывающих сотни
цветков, снабженных длинной трубкой и коротким отгибом
венчика. Более мелкие представители рода имеют рыхлые
соцветия, их цветки имеют крупные лепестки, отходящие
от короткой трубочки венчика.
В предыдущих работах [Берг, Колосова, 1971; Колосо
ва, Берг, 1972; Берг и др., 1973; Колосова, 1973] о Верони
ках получены данные о связи между степенью стабиль
ности признака внутри вида и его эволюционными воз
можностями в процессе дивергенции. Мы обнаружили, что
различия между видами носят комплексный характер,
причем комплекс закономерно сочетающихся друг с дру
гом признаков включает и размах изменчивости, и ха185
рактер корреляционных связей. Комплексные различия
стимулируют поиск того основного признака, который
сыграл роль переключателя, своего рода стрелочного ме
ханизма. Признак этот, будучи взят на вооружение,
направил дальнейшее течение событий по определенно
му руслу* Представляется вероятным, что такой при
знак у вероник — характер единицы опыления (графа 6
в таблице).
У всех длиннотрубчатых видов с плотными колосовид
ными соцветиями (первые шесть видов в таблице) едини
цей опыления служит не отдельный цветок, а целое соцве
тие. Плотное колосовидное соцветие — единая система,
отвечающая за оставление потомства. Архитектоника со
цветий у разных видов различна и зависит от действия ес
тественного отбора в различных экологических условиях
[Колосова, 1973]. Подробнее этот вопрос изложен в нашем
предыдущем сообщении в этом же сборнике. Характер вза
имозависимостей признаков у рассматриваемых видов
сложный. Структура единицы опыления — соцветия —
обусловлена степенью изменчивости и согласованности ча
стей цветка. В то же время каждая определенная структу
ра соцветия создается отбором только в определенной эко
логической нише.
Пример изучаемых нами вероник относится к эволю
ционным парадоксам, а именно — специфическая энто
мофилия создала у них все предпосылки для эволюци
онных преобразований в сторону неспецифической энто
мофилии. Дивергенция видов шла по целым комплек
сам признаков. Причиной большого комплекса следст
вий иногда бывает совсем неожиданное обстоятельство.
Таким обстоятельством, например, у рассматриваемых ве
роник стал характер единицы опыления — соцветие или
цветок.
Мы изучали корреляционные плеяды у вероник. Корре
ляционная плеяда — группа признаков, коррелированных
между собой и некоррелированных с признаками других
групп. По структуре корреляционных плеяд отличаются
вероники, обитающие в разных экологических условиях и
стоящие на разных ступенях филогенетического древа рода
Veronica. Дивергенция вероник также идет по целой плея
де признаков. Комплексирование признаков и причин —
образование корреляционных плеяд — неотъемлемая при
надлежность процесса эволюции. Эволюция — это процесс
согласования каждого мельчайшего свойства с бесчислен
ным числом других.
186
СПИСОК ЛИТЕРАТУРЫ
Берг Р. Л. Дальнейшие исследования по стабилизирующему отбору в эво
люции цветка / / Бот. ж урн.— 1958.— Т. 43, № 1.— С. 12— 27.
Берг Р. Л. Экологическая интерпретация корреляционных плеяд / /
Вести. ЛГУ. Серия биол.— 1959.— № 9, вып. 2 .— С. 142— 152.
Берг Р. Л. Корреляционные плеяды и стабилизирующий отбор / / Приме
нение математ. методов в биологии.— Л.: Изд-во ЛГУ, 1964.—
С. 2 3 - 6 0 .
Берг Р. Л., Калинин О. Мм Колосова Л. Д. Сопоставление внутривидо
вой и межвидовой изменчивости у вероник (род Veronica L.) / /
Журн. общ. биол.— 1973.— Т. 34, № 2.— С. 216—226.
Берг Р. Л., Колосова Л. Д., Корреляционные плеяды признаков у
V. serpylhfolia L., V. cfiamaearys L., V. krylovii Schischk. / / Бот.
ж урн.— 1971.— T. 56, № 8 .— С. 1 0 8 3 -1 0 9 4 .
Бимент Дж. Роль физиологии в адаптации и в конкуренции среди ж и
вотных / / Механизмы биологии. Конкуренция.— М., 1964.—
С. 82— 93.
Дарвин Ч. Приспособление орхидных к плодотворению насекомыми / /
Ноли. собр. соч.— М., 1928.— Т. 4.
Догель В. А. Олигомеризация гомологичных органов.— Л., 1954.
Колосова Л. Д. К вопросу о дивергенции корреляционных плеяд (на при
мере 12 видов вероник) / / Журн. общ. биол.— 1973.— Т. 34,
№ 1.— С. 5 8 — 65.
Мережковский К. С. Конспективный курс общей ботаники.— Казань,
1910.
Шванвич Б. Н. О механизме опыления мотыльковых культур медоносной
пчелой / / Тез. докл. иауч. сессии биолого-почвен. фак. ЛГУ.— Л.,
1 9 5 5 .- С. 42— 43.
Раздел
III
ЗАКОНОМЕРНОСТИ ЭВОЛЮЦИИ
МЕЖВИДОВАЯ И ВНУТРИВИДОВАЯ
ИЗМЕНЧИВОСТЬ ЖИЛКОВАНИЯ КРЫЛА
В СЕМЕЙСТВЕ ДРОЗОФИЛ ИД (Drosophilidae)*
Р. Л . Берг
Цель настоящего сообщения — вскрыть сравнительным
методом на одном конкретном примере факторы, лежащие
в основе направленной параллельной эволюции. Возмож
ность решения подобного рода задачи была впервые проде
монстрирована В. О. Ковалевским на примере эволюции
конечности копытных. В. О. Ковалевский [1873 ] показал,
что усиление функции мышц в онтогенезе ведет к утолще
нию костей. Усиление функции в филогенезе сопровожда
ется как абсолютным, так и относительным удлинением
костей. Отсюда Ковалевский делает вывод, что не прямое
приспособление, а отбор случайных уклонений приводит к
изменению соотношения длины среднего и бокового паль
цев в филогенезе копытных. Расчленив преобразующее и
отбирающее значение функции, Ковалевский выявил, та
ким образом, роль отбора в филогенетическом преобразо
вании органа.
Нашим объектом было крыло насекомого. Насекомые с
их хитиновым наружным скелетом, хитиновыми, изгибаю
щимися, эластичными, но не способными к растяжению
крыльями, с разобщенностью периодов роста и функциони
рования хитиновых структур являются весьма удобным
объектом для выяснения роли отбора в филогенетическом
преобразовании органа. Полет требует точного согласова
ния отдельных свойств крыла, в частности, его размеров и
структуры. Отбирающая роль функции полета в основных
чертах ясна. Преобразующие значение функции принци
пиально исключено характером материала, из которого по
строено крыло насекомого. Полет не может ни изменить
* Применение математических методов в биологии.— Л.: Изд-во
ЛГУ, 1960; Berg R. L. Inter- und intraspezifische Variabelitat der
Fliigeladerung in der Familie der Drosophilidae / / Biologische Beitrage.—
1962.— Bd 1, N 4.
188
летные качества крыла насекомого, ни увеличить его раз
меры, ни изменить его структуру, ни тем более вызвать
согласованные изменения размеров и структуры.
Эволюция крыла насекомого — строго закономерный
процесс, совершающийся сходным образом в различных от
рядах насекомых. Во всех случаях, когда условия жизни
ставят вид перед необходимостью выработать быстрый по
ступательный, хорошо управляемый полет, задача решает
ся сходным образом в самых различных, хотя и не во всех,
отрядах насекомых. Главным направлением эволюции ос
нащающего аппарата крыла — его жилок — является сме
щение сети жилок к переднему краю крыла. Процесс этот,
называемый костализацией, ведет к возникновению разли
чий в оснащенности жилками переднего (костального и за
днего (анального) края крыла [Гуссаковский, 1935;
Needham, 1935; Bodenstein, 1940; Залесский, 1943; Родендорф, 1946, 1949, 1951; и др. ]. Олигомеризация гомологич
ных образований [Догель, 1954] — основной, хотя и не
единственный, путь эволюции крыла.
Дифференциация по оснащенности передней и задней
части крыльев шла у разных видов по-разному. Она осу
ществлялась по ряду направлений: 1) сдвиг жилок к пере
днему краю крыла; 2) возникновение различий между
жилками переднего и заднего края крыла по толщине и по
структуре; 3) исчезновение жилок на заднем крае крыла и
на его вершине; 4) слияние жилок на переднем крае и у
основания крыла и 5) увеличение анальной лопасти крыла.
Все эти пути вели к усовершенствованию полета в ре
зультате разделения функций: передний край крыла, ук
репленный могучим скелетным остовом, режет воздух, за
дний — лишенный жилок совсем или снабженный тонкими
эластичными жилками, волнообразно изгибаясь, придает
полету пропеллирующий характер. Значение эластичности
заднего края крыла для усовершенствования летных ка
честв машин и организмов подчеркивает, в частности,
М. К. Тихонравов [1949]. Роль волнообразных движений в
динамике водных организмов выявил В. В. Шулейкин
[1953]. Б. Б. Родендорф [1946, 1949, 1951 ] осветил значе
ние костализации в усовершенствовании летных качеств
крыльев насекомых.
Однако польза костализации сказывается лишь при
строго ограниченных условиях. Увеличение размеров тела
насекомого кладет ей предел. Самые крупные представите
ли отряда двукрылых, являясь наилучшими летунами, та
кие, как оводы и сирфиды, имеют менее костализованные
крылья, чем более мелкие представители этого отряда [Ро189
дендорф, 1951 ]. Сплошная нерастягивающаяся пластина
крыла должна быть укреплена скелетными образованиями
во избежание разрыва. Противоречие между требованиями
прочности и эластичности разрешается образованием опти
мальных размеров тела насекомых и различными обходны
ми маневрами. Стрекозы, бабочки, а среди двукрылых —
оводы и сирфиды, сохранили жилкование, равномерно рас
положенное по всей поверхности крыла или даже сконцен
трированное на вершине крыла (Nemestrinidae), именно в
силу того, что усовершенствование летных качеств крыла
пошло у них в обход костализации. У бабочек жилки сами
приобрели эластичность, и волна, пробегая по крылу, вов
лекает их в свою кинематику [Залесский, 1954].
Эволюция прогрессивных форм насекомых сопровожда
лась образованием оптимальных размеров тела. Измельча
ние прогрессивных ветвей у насекомых осуществлялось не
только потому, что хитин «позволял» им мельчать без
ущерба защищенности [Четвериков, 1915], но и потому,
что хитин «запрещал» им увеличиваться, затрудняя диссимиляторную фазу обмена [Гиляров, 1959] и создавая пре
граду для увеличения их сплошных, неспособных к растя
жению крыльев. Насекомые малы не только потому , что
хитин хорош в качестве механической защиты, но и пото
му, что он плох в качестве фильтра для жизненных шла
ков и в качестве строительного материала большого крыла.
Хитиновый покров насекомых, увеличивая возможности
дифференциации группы, способствовал ее биологическому
прогрессу (в смысле А. Н. Северцова) и в то же время ог
раничивал ее морфопсихофизиологический прогресс [Ви
нер, 1958].
Измельчение давало насекомым преимущества для усо
вершенствования летных качеств крыла, конечно,, только в
том случае, если биология группы одновременно требовала
усовершенствования полета, если уменьшение размеров не
сопровождалось крайним угнетением группы в борьбе за
жизнь, переходом к скрытому образу жизни и к пассивным
способам пер^двигкения.
Мы ограничивали задачу изучением положения жилок
на
крыльях
различных
представителей
семейства
Drosophilidae. Крылья дрозофилид представляют собой ви
доизменение одного единственного типа. По Родендорфу
[1951], все они относятся к одному дрозофилоидному под
типу подъемного маложилкового (мускоидного) типа
крыльев двукрылых. Различия, которые существуют между
крыльями разных видов во взаимном расположении жилок,
особенно в положении поперечных жилок, настолько стой
190
кие, что служат диагностическим признаком при определе
нии видов [Sturtevant, 1921; Duda, 1935].
Прослеживая изменение положения жилок на крыле
при изменении размеров крыла в филогенетически единой
группе, мы исследуем согласованную изменчивость призна
ков, каждый из которых служит условием эволюции друго
го, но не способен оказать на него какое бы то ни было со
гласующее воздействие ни в предфункциональный, ни в
функциональный период жизни насекомого. Связь разме
ров крыла с положением жилок на нем — пример коорди
нации в историческом развитии при отсутствии корреля
ции в индивидуальном развитии. На различие между коор
динациями и корреляциями указал А. Н. Северцов [1914].
Для того чтобы выяснить, существует ли связь между раз
мерами крыла и положением жилок на крыле, были сдела
ны следующие промеры: а — от места слияния костальной
жилки и жилки, вертикально идущей к косте, и ограничи
вающей костальную ячею (гуморальная поперечная жил
ка) до дистального конца костальной ячеи; b — от первой
поперечной жилки до второй поперечной жилки вдоль по
IV продольной жилке; с — от второй поперечной жилки до
края крыла вдоль той же IV жилки; е — от дистального
конца костальной ячеи до места слияния костальной и вто
рой жилок (см. рисунок). Сумма b + с характеризует длину
свободной крыловой пластины и дает удовлетворительное
представление о размерах крыла. Индекс Ь /с характеризу
ет положение второй поперечной жилки крыла: чем более
близко к основанию крыла лежит вторая поперечная жил
ка, тем меньше 5, тем соответственно больше с и тем
меньше индекс b/ с . Индекс a / b + с характеризует относи
тельную длину костальной ячеи; сокращение костальной
ячеи выражается в уменьшении этого индекса. Индекс
e / b + с характеризует положение конца второй продольной
жилки; чем ближе к основанию крыла расположен конец
второй продольной жилки и чем короче краевая ячея, тем
меньше индекс. Очевидно, более костализованное крыло
будет обладать меньшими величинами индексов b/ с и
e / b + с . Связь между положением жилок на крыле и дли
ной крыловой пластины определялась путем вычисления
коэффициентов корреляции: 1) между b/ с и 5 + с; 2) меж
ду а /Ъ + с и Ь + с и 3) между е / b + с и 5 / с . Первый коэф
фициент корреляции даст нам представление о том, суще
ствует ли связь между положением второй поперечной
жилки крыла и длиной свободной крыловой пластины. Вто
рой коэффициент позволяет вскрыть непропорциональное
изменение длины костальной ячеи по мере уменьшения
191
е
Схема промеров крыла.
Пояснения в тексте.
крыла, и третий ко
эффициент раскрыва
ет судьбу конца вто
рой продольной жил
ки при сдвиге второй
поперечной жилки.
Семейство Drosophilidae делится на два подсемейства
[Duda, 1935] — Steganinae и Drosophilinae. Мною изучено
36 видов. Один вид, а именно Parascaptomyza disticha ,
представлен в нашем материале тремя формами. Изучен
ные виды относятся к 4 родам подсемейства Drosophilinae:
Leucophenga (L. muculata), Cacoxenus (C. indagator),
Gitona (G. vlasovi, C. beckeri, G. distigma) и к 7 подродам
рода Drosophila; Chymomyza (Ch. distincta, Ch. costata,
Ch, nigromana), Acrodrosophila (A. testacea), Ircisurifrons
(L nigriventris), Scaptomyza ( S. flaveola, 5. graminum.
5. unipunctum), Parascaptomyza (P . disticha, P, tfzs#cha v. flat a y P. disticha v. gracilis), Mycodrosophila
(M . poecilogastra) Drosophila s. str. CD. kuntzei, D. funebris,
D. miki, D. vibrissina, D. phalerata, £>. longictosa,
D . littoralis, D. repleta, D. fenestrarum, D. transversa,
Z). travittata, D. tristis, D. obscura, Z). deflexa, D. trivittata v.
us sirica, D. busckii, D. pokornyi, D.
D . rufifrons, D . ananassae, D. montium).
melanogaster,
Следует отметить, что Стертевант [Sturtevant, 1921]
считает подроды Drosophila самостоятельными родами. Мы
в дальнейшем следуем в этом ему, говоря о роде
Drosophila, имеем в виду то, что Дуда обозначает как под
род Drosophila s. str.
Коэффициент корреляции между 5 / с и 5 + с высчитан
для подсемейства Drosophilinae, для рода Drosophila и для
9 видов: 6 видов Drosophila, 1 вид Gitona, 1 вид
Scaptomyza , 1 вид (2 формы) из рода Parascaptomyza . Кро
ме собственных данных использованы индексы и промеры
крыльев, приведенные Стертевантом для 54 североамери
канских видов рода Drosophila, и промеры, сделанные
мною по фотографиям крыльев 45 видов, относящихся к
16 родам подсемейства Drosophilinae, приведенным у Д у
ды. Кроме того, коэффициент корреляции между 5 / с и
5 + с высчитан для четырех линий Z>. melanogaster, разво
димых в лаборатории и ведущих свое происхождение от
диких мух из популяций этого вида Каширы и Кутаиси.
192
Обе каширские линии К 74 и К 54 и одна кутаисская ли
ния Ку 26 выделены из третьего поколения опыта по изу
чению концентрации гетерозиготных мутаций в генотипе
популяций. Все они гомозиготны по второй хромосоме ди
ких мух. Линии К 74 и Ку 26 нормальны. Линия К 54 го
мозиготна по мутации, уменьшающей размеры тела и
крыльев. Вторая кутаисская линия Ку + нормальна и пред
ставляет собой потомков диких мух.
При вычислении коэффициента корреляции между Ъ /с
и 5 + с для какого-либо вида в качестве варианты в корре
ляционной решетке стоит особь. При вычислении коэффи
циента для рода в качестве варианты взят вид, который
характеризуется средними величинами для Ь /с и Ь + с,
или, если использован материал Дуды,— типичный пред
ставитель вида с его величинами Ь /с и b + с, полученными
при измерении крыльев по фотографиям.
При вычислении коэффициента корреляции для под
семейства в качестве варианты снова взяты величины,
средние для видов или характерные для типичного пред
ставителя вида. Любопытно было получить коэффициенты
корреляции для подсемейства, подставив в качестве вари
анты среднюю для рода, и сравнив результат с тем, кото
рый вычислен на основании видовой средней величины.
Это сделано как по собственным данным, так и на основа
нии промеров фотографий крыльев из Дуды. Числа полу
чились почти тождественные. Поэтому к данному методу
мы больше не возвращались. Правда, каждый род, за иск
лючением рода Drosophila, включал незначительное коли
чество видов.
Корреляция между индексами а /Ь + с и b/ с для подсе
мейства Drosophilinae и рода Drosophila установлена по фо
тографиям крыльев Дуды. Внутривидовая корреляция между
обоими индексами изучена для четырех лабораторных ли
ний Z). melanogaster, происходящих из Кутаиси и Каширы.
Корреляция между индексом a / b + с и длиной свобод
ной крыловой пластины b + с изучена на материале кол
лекции Зоологического музея АН СССР. Корреляция от
сутствует во всех изученных систематических категориях.
Костальная ячея, располагающаяся в основании крыла у
его переднего края, не вовлечена в процесс костализации.
Соответствующие данные в таблице не приведены.
Таблица 1 дает представление о соотносительном пове
дении длины крыла, второй поперечной жилки и конца
второй продольной жилки в различных систематических
категориях. Раскрывается любопытная и многозначитель
ная картина. В пределах подсемейства Drosophilinae на
7 I». Л.
Г>ерг
193
блюдается положительная корреляция между индексом
Ъ /с и длиной свободной крыловой пластины £ + с; чем
меньше крыло, тем ближе к основанию располагается вто
рая поперечная жилка. Она захвачена процессом костализации, который, сдвигая поперечную жилку вверх и к ос
нованию крыла, увлекает ее, по мере уменьшения крыла,
от заднего края, где ее присутствие в качестве опорного
элемента утрачивает свой смысл. Однако коэффициент
корреляции невелик ( + 0,40 ± 0,14).
В пределах рода Drosophila коэффициент корреляции
вдвое выше ( + 0,80 ± 0,12). Чем объясняется это разли
чие? По-видимому, в пределах подсемейства костализация
достигается разными путями, которые представляют собой
частные решения одной и той же аэродинамической зада
чи. Раз одно из решений дано, все остальные становятся
ненужными. Действительно, у некоторых видов рода
Cacoxenus дифференциация переднего и заднего края кры
ла достигнута в результате резких различий в толщине I,
II и III продольных жилок, с одной стороны, и IV и V про
дольных жилок, с другой; у Cacoxenus argyreator, кроме то
го, V продольная жилка не доходит до заднего края крыла.
Все это решение одной и той же задачи увеличения эла
стичности заднего края крыла по сравнению с его пере
дним краем. Исключением из общего правила является род
Gitona, в особенности вид G. distigma . Являясь самой круп
ной из всех изученных форм, G. distigma характеризуется
самым маленьким индексом Ъ /с . Можно предположить,
что изменение в положении жилки обусловлено здесь осо
бенностями материала, из которого построена крыловая
пластина, или, быть может, особенностями управления
крыла при полете.
В пределах рода Drosophila сдвиг жилки к основанию
крыла является главным способом, которым осуществляет
ся костализация. Наблюдается почти полная положитель
ная корреляция между длиной свободной крыловой пласти
ны и величиной индекса Ъ /с . Различие между подсемейст
вом и родом по соотносительному поведению длины крыла
и положением второй поперечной жилки сводится, по-ви
димому, к различному варьированию структуры крыла в
процессе эволюции в пределах разных систематических ка
тегорий. В пределах рода крыло от вида к виду варьирует
относительно меньше, и решение одной и той же аэродина
мической задачи осуществляется более сходными путями,
чем в пределах семейства, где у разных родов различия в
структуре крыла больше, они разностороннее, и решение
задачи осуществляется разными путями.
194
Таблица
1
Соотношение между размерами крыла и положение жилок у Drosophiliade
Корреляция
Систематическая
категория
1
b /c И Ь + С
2
Варианта
Пол
п
т
'г±~
—г
По
дан
ным
3
4
5
6
7
Особь
иу
и
и
Виды:
D. deflexa*
D. obscura*
D. rufifrons*
D . fenestraruniA
D.melanogastef
Линии Ky +
Ky 26
К 74
К 54
D.funebris*
Gitона vlasovi*
Scaptomyza
graminum*
Parascaptomyza
distisha*
P. distisha v.
gracilis*
Ь/с И Ь + С
Род (по Дуда
подрод)
Drosophila
b/ с И
длиной крыла
b/ с и Ь + с
Подсем.
Drosophilinae
14
17
20
21
28
U
25
п
г / 25
о о
4 * 22
6*6 25
ОО 25
►•
/б- 25
о о
♦ V 34
oV
50
о *
О 28
о *
О 21
ОУ
4 d 30
—0 ,1 7 ± 0 ,2 7 Берг
»
+ 0 ,3 2 ± 0 ,2 2
»
—0,21 ± 0 ,2 2
»
—0 ,0 6 ± 0 ,2 2
»
+ 0 ,4 0 ± 0 ,1 6
+ 0 ,2 9 ± 0 ,1 8
»
+ 0 ,2 2 ± 0 ,1 9
»
+ 0 ,2 9 ± 0 ,2 0
»
+ 0,31 ± 0 ,1 8
»
—0 ,0 4 ± 0 ,2 0
+ 0 ,1 6 ± 0 ,2 0
+ 0 ,2 2 ± 0 ,1 6
+ 0 ,1 8 ± 0 ,1 4
- 0 ,1 4 ± 0 ,1 9
—0 ,1 8 ± 0 ,2 2
- 0 ,0 5 ± 0 Л 9
48 + 0 ,3 4 ± 0 ,1 3
о *
4- о
Среднее для
вида
Типичный
представи
тель вида
Среднее для
вида
Среднее для
вида
Типичное
для вида
Среднее
для рода
То же
19
+
0,31 ± 0 ,2 2
21
+
0 8 0 ± 0 ,1 2
22
+
0 ,6 5 ± 0 ,1 3 Дуда,
1935
54
+
0 ,5 2 ± 0 ,1 0 Стертевант,
1921
38
+
0 ,4 0 ± 0 ,1 4 Берг
45
+
10
+
0,41 ± 0,13 Дуда,
1935
0 ,4 6 ± 0 ,2 5 Берг
16
+
Берг
0 ,4 4 ± 0 ,2 0 Дуда,
1935
195
Окончание
2
1
Ь/с и е/ b + с
4
5
»
}._
Ку 26
»
oV
О0
Ку 74
»
//
оо
К 54
»
о9
25
25
22
25
25
25
34
50
22
3
6
табл.
1
7
Вид
D. melanogas- Особь
ter
Линии К у+
Уо’
oV
Род
Drosophila
Подсем.
Drosophilinae
Типичный
представи
тель вида
То же
+ 0 ,0 5 ± 0 ,2 0 Берг
»
—0.01 ± 0 ,2 0
»
—0 ,3 8 ± 0 ,1 8
»
—0 ,0 2 ± 0 ,2 0
+ 0 ,0 6 ± 0 ,2 0
»
—0,01 ± 0 ,2 0
»
—0 ,1 2 ± 0 ,1 7
»
—0 ,0 0 2 ± 0 ,1 4
+ 0 ,8 7 ± 0 ,5 Дуда,
1935
45 + 0 ,4 6 ± 0 ,1 2 Дуда,
1935
* Измерения крыльев сделаны на материале коллекции Зоологического музея АН СССР.
Казалось бы, что чем более сходны условия, в которых
происходит изменчивость, тем больше должно быть сходст
во в достижении одного и того же результата и что в пре
делах систематической категории ниже рода, корреляция
между размерами крыла и положением второй поперечной
жилки будет еще выше, чем в пределах рода. Однако дело
не только в сходстве эколюционирующей структуры, но и
в способах согласования различных признаков. А. Н. Северцов [1914] делит способы согласования на корреляции,
основанные на взаимозависимости структур в индивиду
альном развитии, и на координации, осуществляющиеся в
филогенезе при отсутствии онтогенетической взаимообус
ловленности. Примером онтогенетической корреляции мо
жет служить соотносительное изменение положения зонтичка в соцветии Conundrum sativum и расположения лепест
ков в цветках [Смирнов, 1937]. В случае координации со
гласование независимо варьирующих признаков соверша
ется через отбор и достигается только в процессе смены
поколений на основе комбинативной изменчивости. Для
филогенетически координированных признаков нет ника
ких оснований ожидать наличия согласованной внутриви
довой изменчивости.
Как видно из табл. 1, коэффициенты корреляции при
внутривидовой изменчивости много ниже, чем для межви
довой изменчивости в пределах рода. Можно сказать, что
196
корреляция между длиной свободной крыловой пластины и
положением второй поперечной жилки в пределах вида от
сутствует. У 6 видов из 10 форм коэффициент корреляции
мал и имеет отрицательное значение, у 4 форм ( D.
obscura, D . melanogaster, Parascapiomysci disticha — 2 фор
мы) он имеет положительную величину, максимальное его
значение + 0 ,4 0 + 0,16 (Z>. melanogaster).
Можно считать доказанным, что «физиологическая»
корреляция между длиной свободной крыловой пластины и
положением второй поперечной жилки отсутствует или, по
крайней мере, что она не обнаруживается при обычных ус
ловиях изменчивости признака. Та согласованность изме
нений двух признаков, которую мы наблюдаем при пере
ходе от вида к виду в пределах рода, есть результат дея
тельности отбора, который целесообразно сочетает два не
зависимо варьирующих признака.
Установление корреляции между размерами крыла и
положением жилок на крыле для такого подсемейства, где
сами размеры насекомого лишь незначительно варьирует,
показывают, что отбор способен оперировать с очень тон
кими, едва уловимыми отличиями. По-видимому, даже
мелкие отличия в жилковании крыла имеют приспособи
тельное значение.
Костализация не ограничивается сдвигом поперечных
жилок. По мере уменьшения крыла сдвигается к основа
нию крыла и конец второй продольной жилки. Корреляция
между индексами Ь /с и е /Ь + с дает представление о соот
носительном поведении обеих жилок (см. табл. 1). В пре
делах подсемейства наблюдается положительная корреля
ция. Она не велика, но вполне реальна ( + 0,45 + 0,12).
В пределах рода она почти вдвое выше ( + 0,87 + 0,05).
В пределах вида корреляция отсутствует. Бросается в глаза
удивительное совпадение величин коэффициентов корреля
ции между Ь /с и 5 + с, с одной стороны, и между Ъ /с и
e / b + с, с другой, как в пределах подсемейства, так и в
пределах рода. Сходство это наводит на мысль, что сдвиг
второй поперечной жилки и сдвиг конца второй продоль
ной жилки к основанию крыла являются выражением од
ного эволюционного процесса, который захватывает все
крыло, равномерно протекая во всех точках, одинаково
удаленных от основания крыла. По-видимому, дистальный
отрезок радиальной жилки и вторая поперечная жилка
крыла являются теми участками крыла, где отбор ведет к
значительным эволюционным преобразованиям. Как пока
зали наблюдения над полиморфизмом популяций разных
видов мух, всевозможные наследственные уклонения от
197
нормального жилкования (отсутствие отрезков жилок, до
бавочные жилки) локализуются наиболее часто в районе
дистального конца радиальной жилки, а также близ первой
и второй поперечных жилок [Ромашов, Балкашина, 1930;
Gordon et aL, 1939; Bodenstein, 1940; Берг, 1941, 1957;
Оленов, 1941; Дубинин, 1948; и др. ]. Вопрос этот подробно
освещен В. Н. Беляевой [1944]. Районы эти являются не
только филогенетически, но и онтогенетически наиболее
лабильными.
Для изучения соотносительной изменчивости размеров
крыла и положения жилок на нем при мутационной из
менчивости было проделано два опыта. В первом сравнива
лись шесть линий мух Drosophila melanogaster, выведенных
при одинаковых условиях, две из них были нормальные
К 74 и К у + (см. выше), четыре несли мутации, затрагива
ющие размер крыловой пластины. Одна из мутантных ли
ний содержит мутацию miniature, которая выражается в
резком уменьшении размеров крыла за счет уменьшения
размеров клеток, строящих крыловую пластину. Другая
мутантная линия К2 3— 11 происходит из популяции
г. Каширы. Мутация повышает жизнеспособность как го
мозигот, так и гетерозигот и характеризуется укороченным
и расширенным телом и укороченными крыльями. Осталь
ные две мутантные линии выделены из популяции Кутаи
си в опыте по изучению концентрации гетерозиготных му
таций в генотипе популяции. Одна из них К у 26 отличает
ся маленькими размерами крыльев и тела, другая —
Ку 163 — удлиненными крыльями.
Во втором опыте линия К 74 сравнивалась с линией
К 54 (см. выше). Для того, чтобы уравнять условия, в пер
вой серии опыта одинаковое количество производителей
каждой линии рассаживалось по пробиркам, во второй се
рии — в пробирки раскладывалось по 20 яиц.
Результаты опытов представлены в табл. 2. Если бы
уменьшение размеров крыла, возникшее мутационным пу
тем, сопровождалось сдвигом жилок к основанию и к пере
днему краю крыла, мы наблюдали бы в первом опыте са
мые маленькие величины индексов Ь /с и е /Ь + с в линиях
miniature, К2 3— 11 и Ку 26, а самые большие в нормаль
ных линиях и в линии Ку 163. В действительности этого
не наблюдается. Самыми маленькими крыльями обладают
мухи линии miniature. Они отличаются самыми большими
индексами. Во втором опыте меньшие размеры крыла в ли
нии К 54 связаны с большими величинами индекса Ь /с и с
меньшими величинами индекса e / b + с . Очевидно, мутаци
онная изменчивость не вызывает такого согласованного из198
Таблица
2
Соотношение между размерами свободной крыловой пластины и поло
жением жилок при мутационной изменчивости у D. melanogaster
№
опы
та
1
2
Линия
miniature
К2 3 — 11
Ку 26
Ку 163
к 2 3 — 11
Ку 26
Ку 163
К74
К у+
К 74
Ку +
К
К
К
К
54
74
54
74
п
25
26
25
25
24
22
25
25
25
25
25
50
45
34
50
М ±т
Пол
о'/
с'о'
??
??
??
Ь+ с
Ь/с
е/Ъ + с
3,16 ± 0 ,0 4
4 ,6 0 ± 0 ,0 3
4 ,7 0 ± 0 ,0 4
5 ,4 9 ± 0 ,0 4
5 ,2 8 ± 0 ,0 3
5 ,4 0 ± 0 ,0 7
6 ,4 0 ± 0 ,0 3
5,11 ± 0 ,0 3
5 ,1 3 ± 0 ,0 4
5 ,8 8 ± 0 ,0 4
5 ,7 0 ± 0 ,0 5
0,445± 0,005
0,413±<\005
0,416 ± 0 ,003
0,453±0,007
0,420± 0,003
0 ,420± 0,002
0,440± 0,005
0,330± 0,003
0,426± 0,005
0,382± 0,004
0,422± 0,005
0 ,912± 0,005
0 ,865± 0,004
0,819± 0,003
0 ,857± 0,006
0,901 ±0,005
0,840± 0,003
0 ,880± 0,005
0 ,889± 0,004
0 ,856± 0,004
0,907± 0,007
0 ,878± 0,005
5 ,6 5 ± 0 ,0 2
5 ,9 0 ± 0 ,0 2
6 ,3 0 ± 0 ,0 3
6 ,6 4 ± 0 ,0 2
0,403± 0,002
0,381 ±0,003
0,402± 0,003
0,382± 0,002
0,887 ±0,003
0 ,905± 0,004
0 ,906± 0,003
0 ,924± 0,002
менения размеров крыла и расположения жилок на нем,
которое наблюдается при сравнении разных видов одного
рода друг с другом. Те различия, которые существуют
между изученными линиями, скорее производят впечатле
ние географической изменчивости по величине индекса
Ь/ с. Так все линии каширского происхождения имеют в
среднем меньший индекс Ь/ с, чем линии из Кутаиси.
Для изучения соотносительного изменения размеров
крыла и положения жилок на крыле при модификационной
изменчивости проделано два опыта. В первом опыте срав
нивались дикие мухи £>. melanogaster, пойманные в естест
венных условиях их обитания в г. Умани и в г. Тирасполе,
с их потомками первого поколения, выведенными в лабо
ратории в условиях сильного перенаселения. Как показы
вают данные (табл. 3), резкое измельчание животного со
провождалось незначительным сдвигом жилок в том же на
правлении, как это наблюдается при межвидовой изменчи
вости. У уманских мух разница между индексами Ь /с диких
мух и их измельчавших потомков недостоверна. У тирас
польских мух она близка к достоверной: Diff / m diff = 2,9. Ана
логичный результат получен Е. С. Смирновым и А. Н. Желоховцевым у £>. funebris [Smirnov, Zhelochovtsev, 1929].
199
Т а б л и ц а
3
Соотносительное изменение размеров крыла и положение второй попе
речной жилки на нем при модификационной изменчивости у самцов
D. melanogaster
Популяция
Л/±т
Условия
Ь±с
Ыс
Умань
Естественные
Перенаселенные
20
32
5 ,7 7 + 0 ,0 8
4 ,2 4 ± 0 ,0 5
0,418 ± 0 ,005
0,415 ± 0 ,007
Тирасполь
Естественные
Перенаселенные
2.8
28
5 ,1 4 ± 0 ,0 8
4 ,4 9 ± 0 ,0 7
0 ,413± 0,004
0,392 ± 0 ,0 0 6
Материалом для второго опыта послужила мутантная
каширская линия К 54, изогенная по второй хромосоме,
которая содержала мутацию, уменьшавшую размеры тела
и крыльев. Были изучены размеры крыльев и положение
жилок в исходной линии в условиях слабого и более плот
ного заселения. Для получения слабо и сильно заселенных
культур в пробирки с кормом помещались соответственно
по 20 и по 200 яиц. Затем из исходной линии были сдела
ны две отводки. Одна из них велась в условиях слабого за
селения. Для получения слабого заселения на корм поме
щалось небольшое количество производителей. Производи
телями каждого следующего поколения служили мухи из
наименее заселенных культур. Другая отводка велась в усло
виях сильного перенаселения. Большое количество произ
водителей помещалось в каждом поколении в пробирки
Соотносительное изменение размеров крыла и положение жилок на нем
с
'—
с
g
1
2
3
4
5
6
200
Условия
Исходная линия К 54
слабое заселение
Умеренное перенасе
ление
Слабое заселение
Резкое перенаселение
Слабое заселение
Резкое перенаселение
Л/:t m
Число по
колений,
проведен
ных ли
нией в
этих усло
виях
п
О
1
34
50
6 ,30± 0,03
5 ,6 5 ± 0 ,0 2
1
22
20
2
25
50
25
25
26
31
50
19
25
25
25
6 ,06± 0,03
6,2 2 ± 0 ,0 4
4 ,4 6 ± 0 ,0 6
6,5 2 ± 0 ,0 3
5 ,0 8 ± 0 ,0 7
5 ,4 9 ± 0 ,0 2
5 ,4 2 ± 0 ,0 3
4 ,1 4 ± 0 ,0 5
5 ,8 2 ± 0 ,0 4
4 ,6 9 ± 0 ,0 5
пр
Ь+с
dV
с кормом. Опыт был доведен до 22-го поколения в непере
населенной отводке и до 20-го поколения в перенаселен
ной, когда было препринято измерение крыльев в обеих
отводках.
Из 23-го поколения перенаселенной серии снова бы
ли следаны две отводки: одна из них в течение двух поко
лений разводилась в условиях слабого заселения, другая —
в условиях перенаселения. Мы преследовали цель полу
чить мух, как можно более отличающихся друг от друга
по размеру, поэтому неперенаселенная серия содержа
лась при температуре 13— 15 °С, а перенаселенная —
при температуре 22— 24 °С. В 25-м поколении были про
изведены измерения крыльев как в перенаселенной серии,
так и в ее слабо заселенной отводке. Результаты опытов
представлены в табл. 4.
В исходной линии небольшое уменьшение крыла, свя
занное с более плотным заселением, вообще не привело к
сдвигу второй поперечной жилки крыла (2-я строка).
В 22-м поколении неперенаселенной серии наблюдалось
некоторое уменьшение индекса b/ с (3-я строка). Этот
сдвиг жилки был вызван, по-видимому, случайными при
чинами или совершился коррелятивно в связи с отбором
на слабую заселяемость культуры. В 20-м и 25-м поколе
ниях перенаселенной серии сильное измельчание сопро
вождалось сильным уменьшением индексов Ъ /с и е /Ь + с
(4-я и 6-я строки). Сдвиг жилок к основанию крыла бро
сался в глаза и без измерения крыльев.
Т абл и ц а 4
при модификациопной изменчивости в мутантной каширской линии К 54
\1 ± т
bjc
ji>+г
??
п
<*V
0 ,4 0 2 ± 0 ,0 0 3
0 ,4 0 3 ± 0 ,0 0 2
0,906± 0,003
0,887± 0,003
0 ,4 0 7 ± 0 ,0 0 3
0 ,3 8 9 ± 0 ,0 0 4
0 ,3 6 9 ± 0 ,0 0 3
0 ,3 8 3 ± 0 ,0 0 4
0 ,3 7 3 ± 0 ,0 0 5
0,407 ± 0,003
0 ,3 7 9 ± 0 ,0 0 5
0 ,3 5 2 ± 0 ,0 0 5
0 ,399± 0;006
0 ,3 6 0 ± 0 ,0 0 4
0,887± 0,003
0,901 ± 0,003
0 ,832± 0,006
0 ,888± 0,004
0 ,839± 0,005
0 ,872± 0,003
0 ,894± 0,005
0 ,819± 0,004
0 ,866± 0,004
0 ,829± 0,005
201
Возвращение мух в неперенаселенные условия привело
к полному возврату прежних размеров тела и к почти пол
ному возврату в прежнее положение второй поперечной
жилки у самок и конца второй продольной жилки у
самцов и у самок (5-я строка). У самцов 25-го поколения,
возвращенных в условия слабого заселения, индекс
(0,399 + 0,006)
даже
превосходил
тот
же
индекс
(0,379 + 0,005) у самцов 22-го поколения из линии, кото
рая в течение всего этого срока разводилась в условиях
слабого заселения (3-я строка). Результаты этого опыта
показывают, таким образом, что модификационная измен
чивость идет в том же направлении, как и межвидовая, от
личаясь от нее в первую очередь обратимостью возникаю
щих изменений. Закрепления полученных сдвигов, как мы
видим, не произошло, хотя отбор на экономичность строе
ния, несомненно, должен был идти в течение всех 23 поко
лений. Напротив, создается впечатление, что пластич
ность линии в отношении сдвига жилок возросла.
Для изучения географической изменчивости мы
сравнили мух из популяций Кутаиси, Каширы, Умани,
Тирасполя и Одессы. До начала опыта кутаисская линия
просуществовала в лаборатории 1 год 2 месяца, кашир
ская — 3 месяца, тираспольская, одесская и у майская —
не более одного месяца. Мухи для опытов во всех пяти ли
ниях выведены одновременно при одинаковых условиях.
Заметных на глаз отличий в заселенности тех культур, из
которых брались мухи для измерения крыльев, не наблю
далось. Как видно из табл. 5, наличие какой-либо связи
между размерами крыла и положением второй поперечной
жилки сомнительно, хотя крылья, имеющие самую ма
ленькую среднюю величину b + с, обладают самым малень
ким значением индекса b/ с . Однако наибольшая средняя
величина Ъ+ с не сочетается с наибольшим значением ин
декса Ъ/ с (Тирасполь). Максимальное значение индекса
Ь / с сочетается со средними размерами крыла (Кутаиси).
Различия можно обнаружить, как это уже указывалось
раньше, в величине индекса b / с . Наибольший индекс Ь/ с
наблюдается в кутаисской популяции, наименьший — в
каширской. Популяции Умани, Одессы и Тирасполя мало
отличаются друг от друга и стоят ближе к каширской по
пуляции, чем к кутаисской. Сравнение крыльев мух ка
ширской и кутаисской линий дано в табл. 6. Различия
в индексе b/ с не сопровождаются различиями в индек
се е / Ъ + су как это имеет место при межвидовой измен
чивости.
202
Таблица
5
Сравнение размеров крыла и положение второй поперечной жилки на
нем у самцов D. meianogaster различного географического происхождения
М±т
Линия
25
25
25
25
25
Кашира
Умань
Одесса
Тирасполь
Кутаиси
Ь+с
Ь/с
4,91 ±0,03
5,30±0,05
5,05±0,06
5,48±0,03
5,22+0,03
0,389+0,005
0,397 ±0,004
0,410±0,005
0 ,412±0,005
0,444±0,005
Если положение второй поперечной жилки крыла игра
ет какую-либо роль при полете, то из этого следует, что
кутаисские мухи обладают худшими летными качествами,
чем мухи обширных, пространственно неизолированных
популяций степных и лесостепных районов СССР. Закав
казская популяция мух Кутаиси, быть может, действитель
но живет в условиях большей оседлости, чем четыре дру
гие изученные нами популяции. Очаги размножения мух в
Кутаиси характеризуются, в противоположность кашир
ской популяции, чрезвычайным изобилием пищи, что само
должно ограничивать необходимость миграций. В сезон ви
ноделия в Кутаиси дуют сильные ветры, которые ставят,
по нашим наблюдениям, мигрирующих мух в чрезвычайно
невыгодные условия, и превращают винные заводы — оча
ги размножения дрозофил в подобие океанических остро
вов. Кутаиси расположен в горной долине р. Риони, хотя
популяция, населяющая его, и занимает обширное про
странство между горами, она обладает все же более огра
ниченным ареалом возможных миграций, чем популяции
Умани, Тирасполя, Каширы и Одессы. По-видимому, в по
пуляции дрозофил Кутаиси идет отбор на оседлость. Его
Таблица 6
Сравнение размеров крыла и положение жилок на нем в каширской и
кутаисской линиях D. meianogaster
Линия
А/ ±т
Пол
Ь +с
bje
Кашира
26
25
5,61 ± 0 ,0 5
4,91 ± 0,03
0,385
0,389
Кутаиси
26
26
6 ,0 4 ± 0 ,0 4
5 ,2 2 ± 0 ,0 3
0,435: 0,004
0,444: 0,005
0,006
0,005
е/Ь + с
0,861
0,848
0,005
0,005
0,863 0,005
0,852: 0,003
203
последствие — ухудшение летных качеств крыла. Этот вы
вод подтверждается также данными по концентрации раз
личных уклонений от нормального жилкования крыла в
разных популяциях Drosophila melanogaster. Из всех пяти
популяций, уклонения от нормального жилкования крыла,
уменьшающего эластичность заднего края крыла и утяже
ляющие его, встречаются наиболее часто и выражаются
наиболее резко в кутаисской популяции.
Географические отличия по количественным показате
лям, характеризующим, в частности, размер крыла и поло
жение жилок на нем, были обнаружены Р. А. Мазинг
[1927] для различных видов рода Drosophila.
Изучение соотносительной изменчивости размеров кры
ла и степени его костализации в процессе мутационной, модификационной и географической изменчивости у D. mela
nogaster показало, что согласованное изменение размеров
крыла и положение жилок на нем наблюдается при модификационной изменчивости, намечается при географической
изменчивости и полностью отсутствует при мутационной
изменчивости. Согласованные изменения, наблюдаемые
при модификационной изменчивости, полностью обратимы.
Отбор является, таким образом, единственным факто
ром, ответственным за согласованное изменение размеров
крыла насекомого и положения жилок на нем в процессе
видообразования. Согласованная эволюция целого комп
лекса признаков, протекающая параллельно и независимо
в разных отрядах насекомых, совершается под его направ
ляющим влиянием.
СПИСОК ЛИТЕРАТУРЫ
Беляева В. Н. Изменчивость жилкования крыла в природных популяциях
Drosophila melanogaster: Дис.
канд. биол. наук.— М., 1944.
Берг Р. Л. Корреляция между различными формами изменчивости / / Ре
фераты работ учрежд. Отд. биол. наук АН СССР за 1940 г.— 1941.
Берг Р. Л. Типы п ол и м ор ф и зм а// Вести. ЛГУ Сер. биол.— 1957.—
№ 21, вып. 4.
Винер Н. Кибернетика и общество.— M.: ИЛ, 1958.
Гиляров М. С. Закономерности изменения морфологических и физиоло
гических особенностей членистоногих при переходе к наземному
образу
жизни / /
Тр.
ин-та/И н-т эвол.
морф,
животных
им. А. Н. Северцова.— 1959.— Вып. 27.
Гуссаковский В. В. Рогохвосты и пилильщики / / Фауна СССР. 4 . I. На
секомые. Перепончатокрылые.— M.; Л., 1935.— Т. 2, вып. I.
Догель В. А. Олигомеризация гомологичных органов.— Л.: Изд-во ЛГУ,
1954.
Дубинин Н. П. Экспериментальное исследование интеграции наследст
венных систем в процессах эволюции популяций / / Журн. общ.
биол.— 1948.— Т. 9, № 3.
Залесский Ю. М. Краткое изложение оснований к изменению терминоло
гии жилкования крыльев н а сек о м ы х // Зоол. журн.— 1943.—
Т. 22, вып. 3.
204
Залесский Ю. М. Сверхскоростная киносъемка полета бабочек / / Приро
д а.— 1954 — № 10.
Ковалевский В. О. Остеология Anchiteriun aurelianense Cuv. как формы,
выясняющей генеалогию типа лошади (Equus).— М.: Изд-во АН
СССР, 1948.
Мазинг Р. А. К вопросу о количественных различиях между расами и ви
дами мухи Drosophila / / Тр. Ленингр. о-ва естествоисп.— 1927.—
Т. 26, вып. 2.
Оленов Ю. М. О влиянии предшествующей истории вида на его даль
нейшее развитие / / Докл. АН СССР.— 1941.— Т. 31, № 2.
Родендорф Б, Б. Эволюция крыла и филогенез длинноусых двукрылых
Oligoneura (Diptera, Nematocera) / / Тр. ин-та/Палеонт. ин-т АН
СССР.— 1946.— Т. 13, вып. 2.
Родендорф Б. Б. Эволюция и классификация летательного аппарата на
секомых / / Тр. ин-та/Палеонт. ин-т АН СССР.— 1949.— Т. 16.
Родендорф Б. Б. Органы движения двукрылых насекомых и их проис
хождение / / Тр. ин-та/Палеонт. ин-т АН СССР.— 1951.— Т. 35.
Ромашов Д. Д., Балкашина Е. И. Материалы по генетике Drosophila funehris / / Журн. эксмерим. биол.— 1930.— Т. 4, вып. 3.
Северцов А. Н. Современные задачи эволюционной теории / / Собр.
соч.— 1914.— Т. 3.
Северцов А. Н. Морфологические закономерности эволюции.— М.; Л.:
Изд-во АН СССР, 1939.
Смирнов Е, С. Регуляции формы соцветия Coriandrum sativun L. (К воп
росу о теории поля) / / Уч. зап. МГУ.— 1937.— Вып. 13.
Тихонравов М. К. Полет птиц и машины с машущими крыльями.— М.:
Оборониздат, 1949.
Четвериков С. С. Основной фактор эволюции насекомых.— М.: Моек, энтомол. о-во, 1915.— Т. 1.
Шулейкин В. В. Физика моря.— 3-е изд.— М.: Изд-во АН СССР, 1953.
Bodenstein С* Die Auslosund von Modificationen und Mutationen dei Musca
domestica / / Roux' Arch. Entwmech.— 1940.— Bd 140, H 4.
Gordon С. H. Spurway. Street P. A. R. An analysis of three wild populations
of Drosophila subobscura / / J. G en.— 1939. — Vol. 38, N 1/2.
Duda O. Periscelidae, Astiidae, Aulacogastridae, Drosophilidae / / In Linder.
Die Fliegen der palearktischen Region.— Stuttgart, 1935.— Lief. 58.
Needham J. G. Some Basic Principles of Insect Wing Venation / / J.N.Y. Ent.
Soc.— 1935.— Vol. 43.
Smirnov E., Zhelochovtsev A.N. Uber den Einfluss verkiirzter latvaler Ernahrungszeit auf die morphologische Charaktere von Drosophila funebris
F. / / Zsch. Wiss. Zook— 1929.,— Bd 135, H 1/2.
Sturtevant A. H. Tne North American Species of Drosophila / / Publ. Carn.
Inst. Wash.— 1921.— N 301.
ПРОБЛЕМА ЦЕЛОСТНОСТИ ЖИВЫХ СИСТЕМ
В ТРУДАХ И. И. ШМАЛЬГАУЗЕНА*
Р. Л . Берг
Последние два столетия стали ареной столкновения
двух одинаково великих идей в области познания живой
природы. Бой между ними начался на рубеже прошлого и
позапрошлого веков и то одна, то другая брала верх.
* Проблемы кибернетики.— 1966.— Вып. 16.
205
К счастью для науки борьба была турнирной. Побежден
ная идея возрождалась вновь. Соперничающие концеп
ции — это эволюционная теория и идея гармонии црироды, которая в начале XIX века вылилась в теорию^типов
[Канаев, 1963; Беклемишев, 1964]. Одним из творцов и
ярчайшим выразителем теории типов был Кювье [1937].
Именно он на сравнительно-анатомическом и палеонтологи
ческом материале обосновал представление о типе организа
ции животного как о закономерно построенной целостной
системе. Его принцип корреляций и принцип условий суще
ствования дают углубленную трактовку одной из наиболее
существенных черт организаций живой природы — соот
ветствия структуры и функции, будь то часть организма —
отдельный орган или организм как целое. К. Бэр обосновал
теорию типов на материале сравнительной эмбриологии
[Бэр, 1950]. Подход к объекту исследования как к целост
ной системе, вскрытие законов, по которым сочетаются
друг с другом признаки организма, и возможность предска
зывать неизвестные признаки системы на основании того,
что уже познано, составляют действенную сторону теории
типов, которая не исчерпана и сейчас. Абсолютизация ус
тойчивости живых систем, отождествление устойчивости с
неизменностью составляют слабую сторону теории.
Теория эволюции — антипод теории типов. Раскрытие
механизма эволюционных преобразований Дарвином было,
как известно, решающим аргументом в пользу идеи эволю
ции. Победа эволюционной теории означала огромный
сдвиг в понимании живой природы. Только в рамках тео
рии эволюции могли быть познаны интимные стороны про
цессов изменчивости и наследственности. Раскрылись сами
механизмы поддержания устойчивости. Таинственная эквифинальность виталистов — последнее прибежище вита
лизма — получила ясное и исчерпывающее объяснение в
свете данных генетики на путях познания цитологических
основ наследственности. Эквифинальность, или способность
разных частей (разных как по размерам, так и по уже реа
лизованным потенциям) воссоздать целое, оказалась след
ствием равнонаследственных распределений хромосом в
процессе деления клеток. Точно так же как познание
структуры атома раскрыло причины его устойчивости, так
и познание механизма наследственности в широком смысле
слова, включая осуществление действия гена в онтогенезе,
позволило понять устойчивость и эквифинальность органи
ческих форм. Но те преимущества, которыми обладала те
ория типов, были, если не забыты окончательно, отодвину
ты на задний план.
206
\р некотором смысле теория типов была менее созерца
тельной, чем теория эволюции. Она прямо отвечала на
вопросы об устройстве организмов и о назначении их орга
нов, не вдаваясь в отвлеченный анализ способа их возник
новения. Рассматривая целесообразность как изначальное
свойство живого, она смело ставила вопрос: «для чего», не
опасаясь правомочности самого вопроса. Сфера ее прогноза
была ограничена. Предсказывать на ее основе связующие
звенья между разошедшимися формами невозможно, но
предсказывать одни признаки организма по другим можно.
Теория типов натолкнулась на преграду в лице изменя
емости органических форм, в частности той изменяемости,
в процессе которой возникали связующие звенья. Но и те
ория эволюции в своем поступательном развитии натолк
нулась на объективные факты устойчивости органических
форм, на ограниченный характер эволюционных преобра
зований, на канализованный характер эволюции. Оказа
лось, что кривая эфективности отбора имеет плато.
В настоящее время мы вступили в новую эру развития
биологии, которая ознаменовалась синтезом теории типов
и теории эволюции. Первой преградой на пути эволюцион
ной теории, в том виде, в каком ее сформулировал Ла
марк, встал факт реальности вида. Теория эволюции была
признана только тогда, когда возник синтез идеи реально
сти вида с идеей его изменяемости. Этот синтез был осу
ществлен Дарвином. Не случайно он озаглавил свой труд
«Происхождение видов». Признав, в отличие от Ламарка,
реальность вида, Дарвин первый встал на путь синтеза те
ории типов и эволюционной идеи.
Однако эра синтеза идеи типа и идеи эволюции наступи
ла лишь тогда, когда были раскрыты эволюционные механиз
мы возникновения самой устойчивости органических форм.
Впервые в истории науки вопрос о возникновении в
процессе эволюции самой устойчивости поставлен Иваном
Ивановичем Шмальгаузеном [Шмальгаузен, 1934а, б,
1935а-в, 1937а, б, 1938, 1939а, б, 1940].
Жизнь полна взаимоисключающих явлений. Живое
только в меру своей устойчивости является живым. Но са
ма устойчивость живого покоится на процессах самовосп
роизведения и самообновления. Без этих процессов жизнь
невозможна. Живой субстрат остается равным самому се
бе — он устойчив, только вовлекая вещество и энергию в
свой круговорот и используя их для конструктивных пре
образований. Элементарным жизненным актом является
построение своей копии и отторжение этой копии от себя,
как это делает самовоспройзводящая часть хромосомы. Д е
207
ление клетки, размножение многоклеточного организма
невозможны без этого процесса редупликации наследствен
ной субстанции. Способность к преобразованиям — т^кая
же неотъемлемая часть живого, как и устойчивость. Устой
чивость первых существ, впервые в истории планеты осу
ществивших синтез себе подобных существ, сама была ре
зультатом длительной эволюции материи, которая предше
ствовала их возникновению. Живые организмы в отличие
от неживых предметов являются устойчивопреобразующимися. Это касается не только непрерывности процесса во
времени, но и самого характера преобразования. Само пре
образование в сфере живого несет на себе черты устойчи
вости. Все процессы изменения живого до известной степе
ни предопределены характером живых структур. Измене
ние, осуществленное на основе предсуществующей струк
туры, я и называю преобразованием. Преобразованиями в
указанном смысле являются все типы воспроизведения.
Это относится прежде всего к элементарному жизненному
акту. Как построение копии по матричному принципу, так
и отторжение копии от шаблона возможно лишь на основе
определенной структуры. Химический состав, пространст
венная организация, включая взаимное расположение час
тей и размеры копии, заданы предсуществующей структу
рой шаблона. В этом смысле можно говорить о самокопировании как о программированном процессе. Разнообраз
ные способы воспроизведения живых систем подразделяют
ся на четыре главные типа. Это — редупликация по прин
ципу построения копии, свойственная макромолекулярному комплексу — носителю элементарного свойства жизни;
деление, присущее клетке; отделение маленькой части, за
которым следует рост и развитие особи; и самообновление
или замена отмирающих организмов вновь рождающимися,
свойственная всем надорганизменным уровням в иерархии
живых систем. В основе всех типов самовоспроизведения
лежит построение копий элементарных жизненных единиц.
Этот процесс с большой степенью жесткости задан струк
турой самовоспроизводящего материала. Деление клетки,
размножение многоклеточного организма и самообновление
популяций, биоценозов и живого компонента биосферы
также осуществляются в соответствии со структурой этих
систем. Развитие организма жестко контролируется геноти
пом. Но и эволюция популяций введена в определенные
рамки ее генотипом. Жесткость контроля со стороны гено
типа популяции над эволюционным процессом менее четко
выражена, чем жесткость контроля со стороны генотипа
особи над ее онтогенезом. Вероятностные процессы играют
208
в эволюции вида относительно большую роль, чем в инди
видуальном развитии особи, но и тут и там задана лишь
стратегия, а не тактика, и тут и там устойчивым является
не процесс и не состояние сами по себе, а определенная
смена устойчивых состояний. В этом смысле мы и назовем
живую |систему устойчиво преобразующейся. Самооргани
зация живых систем в процессе эволюции включает повы
шение недежности преобразований. Возрастание устойчи
вости органических форм в процессе эволюции осуществ
ляется путем повышения в онтогенезе структурных пред
посылок развития по сравнению с вероятностными процес
сами. Роль внутренних факторов по сравнению с внешни
ми возрастает. Механизм возникновения устойчивости рас
крыт И. И. Шмальгаузеном в процессе создания им его
знаменитой теории стабилизирующего отбора [Шмальгаузен, 1941, 1943, 1945а-в, 1946а, б].
В этой динамической форме, в форме вопроса — как
достигается неизменность организма в меняющихся усло
виях среды, частично возродилась теория типов. Проблемы
совершенствования регуляции, возникновения механизмов,
обеспечивающих сохранение неизменного состояния в меня
ющихся условиях среды, а также механизмов, поддерживаю
щих целостность живой системы в ее взаимодействии со сре
дой, с новой силой поставлены на новом кибернетическом
этапе развития биологии [Эшби, 1959]. Теория стабилизи
рующего отбора И. И. Шмальгаузена раскрыла способ воз
никновения в эволюции генетических, эмбриологических,
анатомических, физиологических механизмов обеспечения
целостности, а вместе с тем и устойчивости организма.
Устойчивость организма в трудах Ивана Ивановича
предстает перед нами не как статистическая изначально
существующая данность, а как приспособление, возникаю
щее как всякое другое приспособление при участии естест
венного отбора. Организм целостен и потому устойчив. Его
целостность покоится на взаимных влияниях одних частей
на другие в процессе развития и в процессе функциониро
вания. Но и в живом мы встречаемся с особым типом взаи
модействия — с управлением. Устойчивость возрастает в
эволюции в том случае, если устойчивое состояние призна
ка увеличивает шансы оставления потомства. В сложных
взаимоотношениях организма со средой его обитания в це
лом независимость от отдельных компонентов среды подчас
становится приспособлением. Среда расчленена, как рас
членен на системы органов сам организм. Разные системы
органов взаимодействуют с разными компонентами среды.
Независимость формирования органов от тех факторов сре
209
ды, с которыми эти органы не вступают в контакт в /Про
цессе функционирования, является необходимым условием
обеспечения надежности развития. Есть целые комплексы
признаков, вступающие в контакт с одним единственным
компонентом среды, да еще таким, который сам ни малей
шего участия в формировании этих признаков не принимает.
Таковы покровительственные— защитные, привлекающие,
распознавательные — окраски. Их достоинства в качестве
приспособлений контролируются биотическими компонен
тами среды — глазом хищника, симбионта, представителя
противоположного пола или просто собрата по виду.
По отношению к исходу событий — к тому, спасется
бабочка благодаря наличию приспособительной окраски
или будет уничтожена,— колебания всех, даже самых не
обходимых, самых обычных факторов среды случайны. Ни
количество корма, ни его состав, ни температурный ре
жим, ни влажность, освещение, никакие иные компоненты
среды, окружающей гусеницу, не влияют на этот исход со
бытий — не от них зависит, будет ли бабочка замечена
птицей или покровительственная окраска спасет ее.
Всякий формообразовательный ответ на воздействие
изменчивых условий существования при этих обстоятель
ствах ошибочен и ведет к гибели. Независимость формиро
вания окраски от воздействия среды способствует в этом
случае преимущественному сохранению наиболее устойчи
вых особей. Стабилизирующий отбор действует в пользу
нормы, отсекая все отклонения от нее — изменчивость со
кращается, жесткость сигналов управления возрастает.
Среднее выражение признака остается без изменения. Не
изменен лишь конечный результат развития. Способ осу
ществления признака в онтогенезе меняется. Онтогенез пе
рестраивается в направлении повышения порога реагирова
ния. Буферные системы обеспечивают неизменность конеч
ного результата развития. Само существование этих бу
ферных устройств, амортизирующих вредные воздействия
и возвращающих организм к норме при изменении внеш
ней и внутренней среды, служит косвенным доказательст
вом стабилизирующего отбора. Примерами неизменности
конечного результата при изменившейся ситуации могут
служить доминирование нормального аллеломорфа над му
тантным, физиологический принцип «все или ничего» в
его фазе «ничего», принцип множественного обеспечения
отдельных формообразовательных реакций индивидуально
го развития. Независимость конечного результата от воз
действий извне имеет место не только в морфогенезе, но и
в процессе жизнедеятельности и касается осуществления
210
функций, и в частности поведения. Мы сталкиваемся с ней
на примере инстинкта, или вернее безусловно-рефлектор
ного компонента поведения.
Но независимость формообразования и функционирова
ния встречается только там, где она нужна. Там, где нуж
но тонко и точно реагировать на все градации данного
фактора среды, вырабатывается зависимый тип развития.
Иван Иванович пишет: «Движущая форма естественного
отбора, т. е. регуляция с положительной обратной связью,
характерна для саморазвивающейся системы в условиях
адаптации к изменению в соотношениях с внешней средой
(биогеоценозом). Стабилизирующая форма естественного
отбора, т. е. регуляция с отрицательной обратной связью
характерна для системы, достигающей стационарного со
стояния при данных условиях существования» [Шмальгаузен, 1961, с. 126]. И. И. Шмальгаузен выделяет три типа
индивидуального развития, отличающиеся друг от друга по
относительной роли внешних и внутренних условий разви
тия в формировании признаков: зависимый, авторегуляторный и автономный типы развития. Внешние сигналы игра
ют наибольшую роль при зависимом типе развития. Веге
тативные части растений, вынужденных приспосабливаться
к условиям местообитания, обладают этим типом развития.
Специфична и своеобразна роль внешних воздействий при
авторегуляторном типе развития. Воздействие служит
лишь пусковым механизмом формообразовательной реак
ции, которая создает признаки, вступающие в контакт со
всем не с теми компонентами среды, которые послужили
сигналом к их развитию. Так, световой режим является
пусковым механизмом многих процессов у растений и жи
вотных, которые вовсе не являются приспособлениями к
свету и не связаны с использованием света. К таким про
цессам относятся половые циклы птиц и млекопитающих.
При автономном типе развития признака влияние любых
внешних воздействий на формообразование ничтожно или
полностью отсутствует. Функция в этом случае теряет
формообразующее значение и выступает в роли браковщи
ка. Развитие жестко детерминировано со стороны генотипа.
Формирование крыла насекомого как летательного аппара
та — прекрасный пример такого рода автономного типа
развития [Berg, 1960; Берг, 1964]. Стабилизирующий от
бор действует очень тонко, делая независимыми одни орга
ны и оставляя зависимыми другие органы одного и того же
вида. Примером могут служить разные органы растения,
имеющего специфического переносчика пыльцы: так, раз
меры листьев и стеблей наперстянки Digitalis purpurea L.
211
обладают зависимым типом развития, а размеры /цвет
ков — независимым, и гигантские растения наперстянки,
достигающие 2,5 м вышины, обладают ничуть не более
крупными цветками, чем их крошечные собратья, не пре
вышающие 0,3 м в высоту. Автономность формообразова
ния касается не только взаимоотношения организма как
целого со средой обитания, но и разных органов в пределах
организма. Целостный характер взаимоотношений организ
ма со средой обитания обеспечивается как зависимостью
одних систем органов друг от друга в процессе развития,
так и независимостью некоторых систем органов друг от
друга. Стабилизирующий отбор способствует возникнове
нию формообразования одних органов по отношению к
другим, делая эмбриональное развитие мозаичным. Сама
мозаичность эмбрионального развития — только способ,
обеспечивающий целостность в процессе формирования и
эволюции [Берг, 1964].
Иван Иванович выступил со своей теорией стабилизи
рующего отбора в 1939 г. В 1946 г. он подытожил свои со
ображения о роли стабилизирующего отбора в эволюции в
книге «Факторы эволюции», дав ей подзаголовок «Теория
стабилизирующего отбора» [Шмальгаузен, 1946а]. Однако
еще много раньше, чем была сформулирована эта теория,
он заложил ее основы. Сравнивая рост разных органов в
эмбриональном развитии у кур, И. И. Шмальгаузен
вскрыл автономию частей развивающегося зародыша. Вы
яснилось, что закладки разных органов отличаются друг от
друга по количеству исходного клеточного материала, по
темпу роста и по продолжительности периода роста.
И. И. Шмальгаузен сформулировал принцип гетерономно
го роста, который затем был независимо разработан
Дж. Гекели под названием принципа аллометрического ро
ста.
Исследования
по
феногенетике
кур
убедили
И. И. Шмальгаузена, что степень зависимости разных сис
тем друг от друга в процессе эмбрионального развития раз
лична. Так, эктодермальные системы органов, а именно —
покровы, нервная система, органы чувств, обладают боль
шей степенью независимости (и на высших ступенях эво
люции развиваются преимущественно под прямым воздей
ствием генотипа) по сравнению с мезодермальными и энтодермальными системами органов, такими, как кости и мус
кулатура, в развитии которых взаимные влияния развива
ющихся частей и у наиболее высокоорганизованных форм
продолжают играть большую роль.
Изменчивость степени автономных частей, показанная
И. И. Шмальгаузеном у кур, и возрастание автономности в
212
восходящем ряду форм, продемонстрированное А. А. Маш
ковцевым [1936] и Д. П. Филатовым [1941 ] у амфибий, по
служили затем косвенными доказательствами теории стаби
лизирующего отбора. Другие косвенные доказательства ста
билизирующего отбор были найдены позже учениками и со
трудниками И. И. Шмальгаузена. Так, А. Л. Зеликман
[1946] создал остроумную модель естественного стабилизи
рующего отбора в лабораторных условиях. Он проследил за
изменением плодовитости рачков-циклопов в процессе сме
ны поколений при изменении количества пищи и показал,
как сузилась норма реагирования в результате отбора на эко
номичность организации. М. М. Камшилов [1939, 1946,
1948] моделировал естественный стабилизирующий отбор с
помощью искусственного отбора и добился превращения
признака, зависимого от колебаний внешних условий, в
признак независимый. Р. Л. Берг [1956, 1958, 1964; Berg,
1959, 1960] указала на различие стабильности размеров
разных органов растений и вскрыла условия стабилизации.
Стабилизирующий отбор и является механизмом воз
никновения устойчивости и независимости в эволюции.
Способность остаться самим собой в меняющихся условиях
среды, регуляторный, направленный, на самосохранение
характер всех процессов, протекающих в живом, пока си
лы жизни одерживают верх над разрушением, сами возника
ют и совершенствуются в эволюции. Вместе с тем, стабилиза
ция и сопровождающее ее возрастание независимости отнюдь
не ограничивают возможность дальнейших эволюционных
преобразований, а наоборот, способствуют ей. Это и понят
но. Стабильность, выработанная в результате отбора, по
вышает шансы на выживание и способствует увеличению
численности вида. Одновременно с этим независимость по
отношению к случайным колебаниям факторов среды по
вышает эффективность отбора, так как доля генотипиче
ской изменчивости в общей изменчивости популяции у
стабилизированных признаков возрастает и темп их эво
люции соответственно также возрастает. Наиболее быстро
эволюционируют наиболее стабилизированные структуры.
Уже в работах по росту и по феногенетике кур, опублико
ванных в конце двадцатых и в начале тридцатых годов,
Иван Иванович подчеркнул, что гетерономный рост, т. е.
автономность в процессе эмбрионального развития, обнару
живают те структуры, которые отличаются наиболее быст
рым темпом эволюции, а именно — нервная система, органы
чувств, покровы, защитные образования. Вместе с тем, эти
же структуры в наименьшей степени вовлечены в рекапи
туляции, т. е. обладают наменьшим консерватизмом.
213
Не только онтогенетические изменения носят целост
ный характер, но и в процессе эволюции организм перест
раивается как целое.
Много лет своей жизни И. И. Шмальгаузен отдал
исследованию происхождения наземных позвоночных
[Шмальгаузен 1947а, б, 1957]. Он показал, что смена сре
ды обитания потребовала перестройки всего организма,
причем изменения разных органов носили характер взаим
ных компенсаций и регуляций. Орган, служащий у рыб
для использования кислорода воздуха,— плавательный пу
зырь — при переходе к наземному существованию теряет
свое назначение в качестве дыхательного аппарата в силу
отсутствия устройства, нагнетающего воздух. В воде цир
куляция воздуха обеспечивалась изменением давления при
всплывании и погружении рыбы. При выходе на сушу на
ружные покровы и слизистая оболочка ротовой полости бе
рут на себя функцию дыхания. Но кожные покровы одева
ются панцирями, защищающими от потери влаги. Компен
саторно происходит увеличение поверхности слизистой
оболочки ротовой полости. Череп непомерно разрастается в
ширину и становится все более плоским. На путях этой
инадаптивной эволюции многие филогенетические ветви
амфибий вымерли окончательно. Такая судьба постигла
стегоцефалов. Современные амфибии влачат жалкое суще
ствование именно в силу несовершенства дыхательных ап
паратов. Развитие механизмов, обеспечивающих дыхатель
ные движения, в первую очередь диафрагмы и межребер
ной мускулатуры, было условием прогрессивной эволюции
рептилий, и, в частности, условием центральной нервной
системы — мозга. Переход из воды на сушу сопровождался
перестройкой всех систем органов. Глубочайшие изменения
произошли в органах чувств, в органах движения, в кож
ных покровах и в нервной системе. Все они носили взаиморегуляторный характер и совершались под влиянием есте
ственного отбора. Загадочное расширение и уплощение че
репа амфибий получило в рамках дарвиновского понима
ния эволюции рациональное объяснение. Так пала послед
няя цитадель ортогенеза.
Изучение преобразований организма как целого в про
цессе эволюции показало, что каждый шаг по пути при
способления позвоночных к наземному существованию со
вершался в процессе взаимного согласования разных сис
тем органов друг с другом, что приспособление каждого ор
гана осуществлялось не только по отношению к среде, но и
к другим органам того же организма. Строясь на случай
ных по отношению к приспособлению наследственных из
214
менениях, эволюция приобретает закономерное течение
именно в силу компенсаторного характера преобразования
каждой части. Направленность эволюционного процесса
ничего общего с о]ртогенезом не имеет.
Таким образом, регуляторный характер развития не
только обеспечивает выживание организма в борьбе за су
ществование, но и налагает печать на эволюцию.
Норма защищена высоким порогом реагирования от
воздействий, идущих как извне, так и изнутри. Регулиру
ются и погашаются не только случайные внешние воздей
ствия, но и феногенетический эффект возникающих мута
ций. И. И. Шмальгаузен формулирует парадоксальный вы
вод относительно меньшей устойчивости генотипа по срав
нению с фенотипом. «Мы пришли к заключениям, каза
лось бы, парадоксальным,— пишет И. И. Шмальгаузен в
статье “Возникновение и преобразование системы морфоге
нетических корреляций в процессе эволюции“— механизм
индивидуального развития обеспечивает у высших живо
тных через сложную систему корреляций известную стой
кость организации, а аппарат наследственности (с его му
тациями), т. е. структура генома, гарантирует достаточную
ее пластичность в процессе эволюции» [1940, с. 365]. Но де
ло не только в ослаблении вредного действия мутаций. Про
явление мутаций вводится в определенное русло, что также
придает эволюции до известной степени канализованный ха
рактер. Феногенетические явления начинают играть в про
цессе возникновения новых форм все возрастающую роль.
В 1961 г. Иван Иванович писал: «Чем выше дифферен
циация организма, тем сложнее и выражение каждой от
дельной мутации. В особенности это касается высших орга
низмов. Преломляясь через эпигенетические процессы, на
следственные изменения приобретают свое специфическое
выражение в каждой обособленной части, в каждой функ
ции организма. Вместе с тем и в этих выражениях сказы
вается взаимозависимость частей в их развитии (элемен
тарные корреляции) и конечный эффект мутаций является
хотя и множественным, но вместе с тем и интегральным»
[1961, с. 109].
Сама система регулирования онтогенеза усложняется и
совершенствуется в процессе эволюции. Регулируются не
только уклонения от нормального течения, но и само тече
ние поддерживается в требуемом состоянии за счет все ус
ложняющейся системы защитных механизмов. В эволюции
происходит раздвигание нижнего и верхнего порога нор
мальной реактивности тканей и возрастание запаса реаги
рующего материала. Надежность развития возрастает за
215
счет увеличения кодирующего материала — образования
целых серий генов со сходным действием. У высших бес
позвоночных (насекомых) и у высших позвоночных (птиц
и млекопитающих) высшего развития достигает нервная
регуляция. Самонастройка и самоорганизация живых сис
тем, лежащие в основе поддержания устойчивости, в тече
ние эволюции совершенствуются. Это касается живых сис
тем, образующих все уровни организации живого [Шмальгаузен, 1958а, б, 1959, 1960а, б, 1961, 1964а, б].
Среди живых систем многоклеточный организм как си
стема претерпевает в процесе эволюции наиболее глубокие
преобразования. Действительно, стоит только вспомнить
поразительное единообразие строения структурных основ
наследственности у всех живых существ и не менее пора
зительное единообразие строения способных к делению
клеток всех многоклеточных организмов, как станет ясно,
что разнообразие органических форм осуществлено на организменном уровне. И тем не менее организм не является
субстратом эволюционных преобразований. Организм жи
вет, и для него жить означает претерпевать полный пре
вратностей жизненный цикл, начиная с одноклеточного су
щества — зиготы и кончая смертью. Развитие организма
во всем, что касается его физических свойств, представля
ет собой осуществление программы, закодированной с по
мощью определенной последовательности нуклеотидов в
молекуле ДНК в его генотипе. Изменение самой програм
мы и возникновение новых черт организации осуществля
ется в процессе эволюции под воздействием отбора, а от
бор протекает на уровне множеств организмов, т. е. на
уровне популяций. Впервые в истории науки это обстоя
тельство, а именно приуроченность эволюции к популяци
онно-видовому уровню организации живого, ясно понял, в
отличие от Ламарка, Дарвин и положил во главу угла
своей теории естественного отбора.
«Хотя эволюция живых существ находит свое наиболее
яркое выражение в повышении интегрированности особи,
она протекает, однако, не в особях, а в надиндивидуальных биологических системах — популяциях, видах и био
ценозах»,— писал И. И. Шмальгаузен в статье «Интегра
ция биологических систем и их саморегуляция» [1961, с.
113]. Эти системы, будучи полем деятельности естествен
ного отбора, в процессе которого меняются организмы, са
ми имеют организацию и сами эволюционируют.
Основным механизмом, поддерживающим единообразие
вида и его целостность, является скрещивание между пред
ставителями вида. Значение законов расщепления для по
216
нимания элементарных механизмов эволюции в популяци
ях диплоидных организмов, размножающихся половым пу
тем, было впервые раскрыто С. С. Четвериковым. Значе
ние комбинативной изменчивости в полной мере раскрыто
И. И. Шмальгаузеном.
И. И. Шмальгаузеи суммировал
данные филогенетики и популяционной генетики и пока
зал, что отбор никогда не имеет дела с отдельными мута
циями, а лишь с выражением комбинаций мутаций в фе
нотипе особей. Особенно важным в этом плане являются
наблюдения над возникновением качественно нового эф
фекта в результате комбинирования мутаций. Мутация
становится объектом естественного отбора сначала в гете
розиготном состоянии. Уже комбинация мутантного гена с
его нормальным аллеломорфом не всегда обладает ослаб
ленным выражением мутантного признака.
Как впервые показала Р. А. Мазинг, [1938, 1939], а за
тем многие исследователи, некоторые летальные в гомози
готном состоянии мутации повышают жизнеспособность ге
терозигот. Вскрыта большая устойчивость гетерозигот по
сравнению с гомозиготами по отношению к изменению
факторов внешней среды. В процессе отбора, идущего не
по отдельным мутациям, а по их комбинациям, соверша
ются глубокие изменения как в строении отдельных пред
ставителей популяции, так и в строении самой популяции.
Эволюционируют не только признаки организма, но и спо
соб наследования мутаций. И. И. Шмальгаузеи поднял те
орию эволюции доминантности Фишера на новый уровень.
Согласно теории эволюции доминантности Шмальгаузена или — как предпочитает говорить Иван Иванович —
эволюции рецессивности, погашение действия мутантного
аллеломорфа у гетерозигот осуществляется за счет отбора
комбинаций полезных и безвредных мутаций. Повышение
верхнего порога реагирования тканей в результате стаби
лизирующего отбора имеет побочным результатом повыше
ние доминирования полезных проявлений мутантных генов
и понижение доминирования или рецессивность вредных
эффектов. Отбор и скрещивание в результате испытания
все новых и новых комбинаций в меняющихся во времени
и в пространстве условиях среды ведут к интеграции гено
типа популяции и одновременно к возникновению геноти
пического разнообразия. В особенности большую роль иг
рает в поддержании генотипического разнообразия популя
ции положительный отбор гетерозигот с повышенной жиз
неспособностью в том случае, когда гомозигота обладает
пониженной жизнеспособностью. Образуется мобилизаци
онный резерв наследственной изменчивости, который мо
217
жет быть в любой момент использован популяцией в слу
чае изменения условий и в то же время служит гомеоста
том индивидуального развития отдельных особей. Устойчи
вость онтогенеза, а подчас и однообразие фенотипов в по
пуляции обеспечивается генотипической гетерогенностью
популяции. «Стабильность особи сочетается с ее приспосо
бительной реактивностью, а стабильность структуры попу
ляции — с ее исключительной эволюционной мобильно
стью»,— пишет И. И. Шмальгаузен [1961, с. 118].
Показав, как велико разнообразие генотипов в преде
лах популяции, И. И. Шмальгаузен ставит вопрос об ус
ловности самого понятия «нормы». Нормальными при этих
обстоятельствах следует считать всех особей, фактически
принимающих участие в воспроизведении следующего по
коления. Стабилизирующий отбор, отсекая все уклоняю
щееся от этой «нормы», ведет к сокращению размаха из
менчивости, но отнюдь не приводит к генотипическому
единообразию популяции. Условность «нормы» предполага
ет условность «пользы» и «вреда» отдельных мутаций. При
изменившихся условиях и в новых комбинациях мутации,
ранее понижавшие жизнеспособность гомозигот, могут
стать нейтральными или полезными.
И. И. Шмальгаузен с особенной силой подчеркнул це
лостный характер популяции. Но целостными, согласно
представлению И. И. Шмальгаузена, являются не только
организм и популяция. И. И. Шмальгаузен выделяет пять
главнейших уровней организации живого: молекулярный,
клеточный, организменный, популяционный, биоценотический. Само возникновение жизни связано с интеграцией на
молекулярном уровне. Основное свойство живого — само
воспроизведение — осуществляется в сравнительно устой
чивой системе, состоящей из нуклеиновых кислот и бел
ков, с помощью обратимых химических реакций, скорость
которых определяется относительной концентрацией реаги
рующих субстанций. Молекулярному уровню организации
жизни соответствует регуляторный механизм, включаю
щий поддержание устойчивости химических реакций. Ин
теграция на этом уровне выражается в объединении эле
ментов, только совместно способных выполнить функцию.
Простейшей биологической системой является клетка.
Ее целостность покоится на сложной системе взаимозави
симостей, в которые вовлекаются ее элементы в процессе
биосинтеза белков. Регуляция клеточного метаболизма
осуществляется под контролем унаследованных структур
ядра и органоидов клетки.
218
Наибольшей степени дифференциация и интеграция
частей системы достигают в многоклеточном организме.
Сложные регуляторные механизмы, построенные по типу
замкнутого цикла с обратной связью, встречаются в физи
ологических регуляциях, включая поведение, и в морфоло
гических процессах. Отбор особей в его движущей и в ста
билизирующей форме ведет к усложнению не только
форм, и тех регуляторных механизмов, которые поддержи
вают устойчивость форм.
Устойчивость целостных систем надорганизменного
уровня — популяций — также поддерживается отбором.
Но помимо отбора особей и в качестве механизма, ответст
венного за интеграцию популяций, выступает отбор груп
пы. «Именно межгрупповое состязание и групповой отбор
ведут к установлению оптимальных в данных условиях со
отношений между основными факторами эволюции — мутабильностью, формами и темпом размножения, а также
формами полиморфизма и генетическим механизмом его
поддержания
на
оптимальном уровне»,—
пишет
И. И. Шмальгаузен [1962, с. 117]. Именно в результате
межгруппового соревнования создается возможность опти
мизации самого процесса эволюции, так как в процессе
вытеснения менее успешно эволюционирующих групп
группами с более перспективными темпами и характером
эволюции, эволюция становится все более гармоническим
процессом и ее течение приобретает черты устойчивости,
превращаясь в гомеорезис. И. И. Шмальгаузен ставит воп
рос об эволюции самого механизма эволюции. Иван Ива
нович пишет: «Основным объектом исторических измене
ний является вид в целом. Эволюция вида определяется,
однако, не суммативно эволюцией его популяций. Она
представляет интегральный результат соревнования попу
ляций в качестве и скорости эволюции и сопровождается
распространением одних и вытеснением других популяций.
Таким образом, и отбор выражается здесь в новой форме
группового отбора, который также может иметь стабилизи
рующий характер» [1961, с. 129].
Целостной системой надпопуляционного ранга является
биоценоз. Основной управляющей системой биоценоза явля
ется система взаимоотношений между популяциями видов
животных, растений и микроорганизмов, входящих в его со
став. Взаимоотношения между организмами находят свое вы
ражение в поддержании видового состава биоценоза и в под
держании численности популяций разных видов, а видовой
состав и численность популяций выступают как качествен
ный и количественный регулятор тех потоков веществ и
219
энергии, которыми биоценоз как целое связан со средой свое
го обитания. И. И. Шмальгаузен подчеркивает подвижный
характер равновесия всех процессов, и в частности, соотно
шений численности популяций разных видов в биоценозе.
В процессе эволюции все уровни организации живого
взаимодействуют друг с другом, и эти взаимодействия но
сят регуляторный характер. В этой идее И. И. Шмальгаузена эволюционный подход к проблеме целостности живого
нашел свое высшее выражение.
Эволюция популяций протекает только в биогеоценозах.
И. И. Шмальгаузен был первым в мире, кто переложил
дарвинское понимание эволюции на язык кибернетики.
Регуляция эволюционного процесса осуществляется по
средством циклического механизма с обратной связью на
основе сопоставления полученных результатов (фенотипов)
в биогеоценозе. Сравнительная оценка особей осуществля
ется по их способности участвовать в воспроизведении сле
дующего поколения, т. е. с помощью естественного отбора.
Отбор, таким образом, является регулирующим механиз
мом эволюции. И. И. Шмальгаузен выделил ту целостную
систему, в которой осуществляется полный цикл регулиру
ющих потоков информации. Общая схема регулирующего
механизма популяций представлена на рисунке [Шмальга
узен, 1959, с. 110].
Общая схема регулирующего механизма эволюции популяций
о
Он
V
аO
Ео
Помехи абиотические
4
^
Передача наследственной информации
через зиготу и клеточные деления (хромосомы) |
■£ а
1
5 £
Преобразование информации Преооразование информации
~ %*1?в популяции (естественный
в индивидуальном развитии
G с. отбор'и половой процесс)
(реализация фенотипов)
£ °
Передача обратной информации
через фенотипы (активность особей)
Помехи абиотические и биотические
Контроль в биогеоценозе
220
Фенотипы особей
О)-S
| Б.
2
й~
cS'G*
Объектом управляемых изменений является популяция
данного вида организма. В качестве регулирующего меха
низма выступает, очевидно, биогеоценоз, включающий
данную популяцию. Отобранные особи данной популяции
приступают к размножению. Состав размножающейся час
ти популяции является итогом тех регулирующих воздей
ствий; которые испытала популяция со стороны биогеоце
ноза и через которые она прошла на завершающем этапе
предыдущего цикла. Каждый новый цикл начинается с пе
редачи наследственной информации от родительской попу
ляции к дочерней по каналу прямой связи. Образуется до
черняя популяция. Она представлена зиготами.
Реализация фенотипов представителей дочерней попу
ляции сопровождается переводом полученной от родителей
информации с языка генов на язык признаков. Обратная
информация передается от популяции к биогеоценозу че
рез посредство фенотипов на организменном уровне орга
низации. Прямая информация передается на клеточном
уровне через посредство гамет. Воздействие среды на все
стадии гаметогенеза и на сами гаметы приводит к возник
новению мутаций. Количество информации, передаваемой
по каналу прямой связи, в результате мутационного про
цесса повышается. Однако в канале обратной связи проис
ходит отбор, или испытание особей по их фенотипам. Ко
личество особей сокращается, сокращается и количество
передаваемой информации. Одним из самых существенных
последствий этого изменения количества информации в ка
налах прямой и обратной связи является перестройка гено
типа популяции в сторону обеспечения большой приспо
собляемости организмов к среде обитания, включая и жи
вые компоненты среды. Меняется не только количество пе
редаваемой информации, но и ее характер. Замыкание
цикла потоков информации сопровождается эволюционны
ми преобразованиями. Сама устойчивость организма, поко
ящаяся на целой системе сигналов управления, эволюцио
нирует. И в то же время, в результате состязания групп,
состязания, которое идет не только по признакам, но и по
процессам, включая сам эволюционный процесс, осуществ
ляется эволюция самой эволюции.
СПИСОК ЛИТЕРАТУРЫ
Беклемишев В. Н. Основы сравнительной анатомии беспозвоночных / /
Проморфология.— 3-е изд.— М.: Наука, 1964.— Т. 1.
Берг Р. Л. Стандартизирующий отбор в эволюции цветка / / Бот.
журн. — 1956.— Т. 41, № 3 ,— С. 318— 334.
221
Берг Р« Л. Дальнейшие исследования по стабилизирующему отбору в эво
люции цветка / / Бот. ж урн,— 1958.— Т. 43, № 1.— С. 12— 27.
Берг Р. Л. Корреляционные плеяды и стабилизирующий отбор / / Приме
нение математических методов в биологии.— Л.: Изд-во ЛГУ,
1964.— Т. 3. — С. 23— 60.
Бэр К. М. История развития животных.— М.; Л.: Изд-во АН СССР,
1 9 5 0 . - Т .1 .
Дриш Г. Витализм. Его история и система.— М., 1915.
Зеликман А. Л. Экспериментальное изучение стабилизирующего отбо
ра / / Журн. общ. биол.— 1946.— Т. 7, № 4 .— С. 239— 252.
Камшилов М. М. Отбор как фактор, меняющий зависимость признака от
изменений внешних условий / / Докл. АН СССР.— 1939.— Т. 23,
№ 4 .— С. 361— 364.
Камшилов М. М, Первичная дивергенция нормы реагирования в зависи
мости от условий развитий (наследственное и ненаследственное в
эволюции): Дис.
д-ра биол. наук.— М., 1946.
Камшилов М. М. Отбор как фактор усложнения организации / / Изв. АН
СССР. Сер. Биол.— 1948.— № 3. — С. 349— 356.
Канаев И. И. Очерки из истории сравнительной анатомии до Дарвина.
Развитие проблемы морфологического типа в зоологии.— М., Л.:
Изд-во АН СССР, 1963.
Кювье Ж . Рассуждение о переворотах на поверхности земного шара.—
М.: Биомедгиз, 1937.
Мазинг Р. А. Повышенная жизнеспособность гетерозиготных по летал ям
мух Drosophila melanogaster / / Докл. АН СССР.— 1938.—
Т. 20.— С. 2 — 3.
Мазинг Р. А. Анализ жизнеспособности мух Drosophila melanogaster, ге
терозиготных по леталям, возникшим в природе / / Докл.
АН СССР.— 1939.— Т. 25, № 1.— С. 64—67.
Машковцев А. А. Смена эндогенных и экзогенных факторов эмбриональ
ного развития в онтогенезе и филогенезе. Эволюция онтогенеза / /
Изв. АН СССР. Отд. матем. и естеств. наук.— 1936.—
С. 9 4 5 - 9 9 7 .
Филатов Д. П. Об историческом подходе к явлениям механики развития
и его значение / / Журн. общ. биол.— 1941.— С. 3— 18.
Шмальгаузен И. И. Корреляция в развитии некоторых расовых призна
ков у кур / / Тр. ин-та / Ин-т зоол. и биол. АН СССР.— 1934а.—
С. 3 — 23.
Шмальгаузен И. И. К феногенетике некоторых морфологических призна
ков у домашних кур / / Докл. АН СССР.— 19346.— Т. 2, № 5 .—
С. 331—336.
Шмальгаузен И. И. Определение основных понятий и методика исследо
вания роста / / Рост животных.— М.; Л.: Биомедгиз, 1935а. —
С. 8 — 60.
Шмальгаузен И. И. Рост и общие размеры тела в связи с их биологиче
ским значением / / Рост животных.— М.; Л.: Биомедгиз, 19356.—
С. 61—73.
Ш мальгаузен И, И. Рост и дифференцировка / / Рост животных.— М.;
Л.: Биомедгиз, 1935в. — С. 74—84.
Ш мальгаузен И. И. Развитие различных форм гребня у кур / / Тр. ин-та /
Ин-т зоол. и биол. АН УССР.— 1937а.— Вып. 17.— С. 61— 69.
Шмальгаузен
И. И.
Современные
задачи
феногенетики / /
Изв.
АН СССР. Отд. матемаг. и естеств. наук.— 19376.— С. 895—906.
Ш мальгаузен И. И. Организм как целое в индивидуальном и историче
ском развитии.— М.; Л.: Изд-во АН СССР, 1938.
Шмальгаузен И. И. Пути и закономерности эволюционного процесса.—
М.: Изд-во АН СССР, 1939а.
Шмальгаузен И. И. Значение корреляций в эволюции животных / / Сб.
памяти акад. А. Н. Северцова Ин-та эвол. морфологии животных
АН СССР.— 19396.— Вып. 1.
Ш мальгаузен И. И. Возникновение и преобразование системы морфоге
нетических корреляций в процессе эволюции / / Журн. общ. би
ол.— 1 9 4 0 .- Т. 1, № 3.— С. 3 4 9 - 3 7 0 .
222
Шмальгаузен И. И. Стабилизирующий отбор и его место среди факторов
э в о л ю ц и и //
Жури.
общ.
биол.—
1941.— Т. 2,
№ 3 С. 3 0 7 - 3 5 4 .
Ш мальгаузен И. И. Темп эволюции и факторы его определяющие / /
Журн. общ. биол.— 1943.— Т. 4, N9 5 .— С. 253—285.
Ш мальгаузен И. И. Стабилизирующий отбор и проблема передачи поло
вых признаков с одного пола на другой / / Журн. общ. биол.—
1945а — Т. 6, № 6 .— С. 363—380.
Ш мальгаузен И. И. Закономерности в эволюции формообразовательных
процессов и законы классической физиологии / / Природа.—
19456.— № 4 .— С. 34—46.
Ш мальгаузен И. И. Проблема устойчивости органических форм (онтоге
незов) в их историческом развитии / / Журн. общ. биол.—
1945в.— Т. 6, № 1.— С. 3 —25.
Ш мальгаузен И. И. Факторы эволюции (теория стабилизирующего отбо
р а ).— М.; Л.: Изд-во АН СССР, 1946а.
Ш мальгаузен И. И. Проблемы дарвинизма.— М.: Сов. наука.— 19466.
Шмальгаузен И. И. Внешние факторы, межвидовая борьба и внутривидо
вое соревнование и их взаимодействие / / Внутривидовая борьба у
животных и растений.— М.: Изд-во МГУ, 1947а.— С. 3 — 13.
Шмальгаузен И. И. Основы сравнительной анатомии позвоночных ж и
вотных.— М.: Сов. наука, 19476.
Шмальгаузен И. И. Биологические основы возникновения наземных по
звоночных / / Изв. АН СССР. Сер. Биол.— 1957.— С. 3 —30.
Ш мальгаузен И. И. Контроль и регуляция в эволюции / / Бюл. МОИП.
Отдел Биол.— 1958а.— Т. 63, № 5 .— С. 93— 121.
Шмальгаузен И. И. Регулирующие механизмы эволюции / / Зоол.
ж урн.— 19586.— Т. 37, № 9 .— С. 1291 — 1305.
Ш мальгаузен И. И. Перспективы применения точных методов для изуче
ния факторов эволюции / / Вести. ЛГУ. Сер. Биол.— 1959.—
С. 108— 118.
Шмальгаузен И. И, Естественный отбор и информация / /
Изв.
АН СССР. Сер. Биол.— 1960а.— С. 19—38.
Шмальгаузен И. И. Естественный отбор и его формы / / Вопросы эволю
ции, биогеографии, генетики и селекции: Сб. поев. 70-летию со
дня рождения Н. И. Вавилова.— М.: Изд-во АН СССР, 19606.—
С. 311— 319.
Шмальгаузен И. И. Интеграция биологических систем и их саморегуля
ц и я / / Бюл. МОИП. Отд. Биол.— 1961.— Т. 66, № 2 . —
С. 104— 134.
Ш мальгаузен И. И, Регуляция формообразования в индивидуальном раз
витии.— М.: Наука, 1964а.
Шмальгаузен И. И. Происхождение наземных позвоночных.— М.: Нау
ка, 19646.
Эшби Р. У, Введение в кибернетику.— М.: ИЛ, 1959.
Berg R. L. A general evolutiknary principle underlying the origin of
developmental homeostasis / / Amer. Nat.— 1959.— N 93.— P.
103— 105.
Berg R. L. The ecological significance of correlation pleiades / / Evolution.—
I9 6 0 .— Vol. 14, N 2 .— P. 171 — 180.
223
МЕСТО ТЕОРИИ СТАБИЛИЗИРУЮЩЕГО ОТБОРА
И. И. ШМАЛЬГАУЗЕНА
В СОВРЕМЕННЫХ ЭВОЛЮЦИОННЫХ ВОЗЗРЕНИЯХ*
Р. Л . Берг
Книга И. И. Шмальгаузена «Факторы эволюции (тео
рия стабилизирующего отбора)» опубликована в 1946 г.,
т. е.
за
22 года до ее теперешнего переиздания.
И. И. Шмальгаузен писал ее в годы войны, находясь в эва
куации в отрыве не только от мировой и отечественной ли
тературы, но и от своей библиотеки.
В 1964 г. вышла в свет книга И. И. Шмальгаузена «Ре
гуляция формообразования в индивидуальном развитии», в
которой он рассмотрел всю проблему органических регуля
ций в свете общей теории регулирующих устройств
[Шмальгаузен, 1964].
Со времени выхода в свет «Кибернетики» Н. Винера
наука об управлении завладела умами биологов. Первый,
кто переложил теорию Дарвина на язык кибернетики, был
И. И. Шмальгаузен. Он выделил тот целостный объект —
биогеоценоз,— в котором по каналам прямой и обратной
связи передаются от материнской популяции к дочерней и
от популяции к биогеоценозу сигналы управления. Эволю
ция вида в свете кибернетики обрисована Шмальгаузеном
как управляемый процесс, а главный фактор эволюции —
отбор — как ее регулятор. Статьи и неопубликованные
главы книги, посвященные вопросам биокибернетики, вош
ли в сборник, изданный Сибирским отделением АН СССР
в серии «Кибернетика в монографиях» [Шмальгаузен,
1968].
Вполне уместен вопрос, не устарела ли издаваемая
книга И. И. Шмальгаузена, созданная при обстоятельст
вах, когда подвести итог мировой литературе по условиям
военного времени автор не мог.
Книга Шмальгаузена — единственный труд, специаль
но посвященный вопросам эволюции изменчивости, круп
нейший вклад не только в теорию эволюции, но и в новую
отрасль знания — эволюционную генетику. В свете эволю
ции нормы реакции Шмальгаузен рассматривает возникно
вение всевозможнейших пороговых эффектов, защищаю
* Послесловие к книге И. И. Шмальгаузена «Факторы эволюции. Те
ория стабилизирующего отбора».— 2-е изд. — M.: Наука, 1968. Печата
ется с сокращениями.
224
щих организм от воздействий,— как внешних, исходящих
от среды, так и внутренних, связанных с мутационным
процессом и с комбинативной изменчивостью, начиная с
доминирования одного аллеломорфа над другим и кончая
отсутствием морфогенетической реакции при изменении
условий развития. Как прямое следствие стабилизирующе
го отбора, как частное проявление единого принципа вы
свобождения из-под контроля среды (понимаемой в самом
широком смысле слова), предстают перед нами самые раз
нообразные явления — полигенная обусловленность при
знака и мозаичный тип эмбрионального развития многих
беспозвоночных, неспецифичность индуктора в морфогене
зе позвоночных и гетерономный рост частей организма, ав
торегул яторный характер сезонных явлений в жизни расте
ний и высших животных и безусловно-рефлекторный ком
понент поведения и автономность функции внутренних ор
ганов животного, принцип «все или ничего» физиологиче
ских реакций и поддержание постоянства внутренней сре
ды, перенос в течении эволюции признаков одного пола на
другой пол и высвобождение морфогенеза вторичных поло
вых признаков из-под гормонального воздействия, и многое
другое. Закрепление модификаций с помощью отбора му
таций, совпадающих по своему проявлению с реакцией на
среду IWaddington, 1937], в свете теории стабилизирующе
го отбора рисуется не как прямое следствие отбора по дан
ному закрепляемому признаку, а как побочный эффект об
щей стабилизации признаков, получивших значение широ
ких адаптаций регуляторного типа. Общее повышение ро
ли внутренних факторов морфогенеза по сравнению с
внешними и возникновение регуляций в процессе стабили
зирующего отбора и представляет собой ту фундаменталь
ную закономерность эволюции, по отношению к которой
наследственная фиксация модификаций через отбор высту
пает как частный случай. Косвенный отбор Кирпичникова
[1935], совпадающий— Лукина [1942], органический и
совпадающий отбор Л. Моргана [Morgan, 1900], Осборна
[Osborn, 1897], Болдуина [Baldwin, 1896], отбор модифи
каторов Фишера [Fisher, 1928, 1931 ] — это частные про
явления общего принципа стабилизирующего отбора, вы
двинутого И. И. Шмальгаузеном.
Анализ эволюции нормы реакции и взаимодействия
всех уровней организации живого (макромолекулярного,
клеточного, организменного, популяционно-видового и
биоценотического) в процессе исторического преобразова
ния форм — наиболее сильная сторона «Факторов эволю
8 V. Я Берг
225
ции» и она делает книгу непревзойденной до настоящего
времени.
К самой эволюции изменчивости И. И. Шмальгаузен
подошел совершенно своеобразно. Повышение реактивно
сти организма на воздействие среды, увеличение количест
ва связей со средой, усовершенствование физиологических,
в том числе поведенческих реакций на мельчайшие изме
нения в среде, активное овладение жизненными ресурсами,
само ускорение темпов эволюции он рассматривает как ре
зультат прогрессирующего высвобождения из-под зависи
мости от среды, как результат завоевания известного гос
подства над нею, как следствие повышения устойчивости
живых систем.
Книга «Факторы эволюции» не принадлежит к числу
творений, которые, сыграв свою роль на коротком отрезке
времени, превращаются в лучшем случае в достояние исто
рии науки. И сейчас, через четверть века после ее написа
ния, книга И. И. Шмальгаузена содержит еще далеко не
воплощенную в жизнь программу исследований. Можно
утверждать, не боясь впасть в преувеличение, что мысль
Шмальгаузена намного опередила свое время, и «Факторы
эволюции» как раз и являются одним из тех основных тру
дов, которыми открывается новая эра в биологии.
И. И. Шмальгаузен дал научное обоснование идее о за
кономерном характере эволюционного процесса. Законо
мерности эволюции связаны с ограничениями, которые на
лагает на дальнейшие преобразования структура эволюци
онной системы. Эволюция — это движение, осуществляю
щееся по определенным путям, игра с обязательным со
блюдением правил.
Рассматривая эволюцию как регулируемый процесс,
И. И. Шмальгаузен во главу угла ставит не изменяемость
органических форм, а — парадоксальным образом — их
устойчивость. Это не устойчивость неорганической приро
ды, а способность сохранить стационарное состояние при
меняющихся взаимоотношениях со средой, основанная на
регуляциях. Динамический режим, лежащий в основе ус
тойчивости органических форм, сам создается в эволюции
и оказывает направляющее действие на дальнейшее ее те
чение. Предшествующие усовершенствования, обеспечива
ющие надежность существования живой системы в меняю
щихся условиях среды, предопределяют дальнейшие собы
тия. Регулируются не только отклонения от нормального
морфогенеза, вызванные изменением условий существова
ния, но и проявление вновь возникающих мутаций и их
комбинаций, а тем самым и сам эволюционный процесс.
226
В том случае, когда регуляции на клеточном и онтогенетиче
ском уровнях организации материи не сработали, и организм
погиб, вступил в действие отбор — главный фактор эволю
ции, ее контролер и регулятор. То, что на клеточном и онто
генетическом уровнях является смертью, отсутствием регу
ляции, на популяционном уровне выступает как регуляция.
Взаимодействие клеточных, онтогенетических и популяци
онных регуляций и создает закономерный ход эволюции.
И. И. Шмальгаузен впервые в понятие генетической
системы включил кроме клеточного и популяционного
уровней уровень онтогенетический. Но он не ограничился
этими тремя уровнями. В последнее десятилетие своей
жизни он рассмотрел эволюцию популяций как проявление
жизнедеятельности системы высшего по отношению к попу
ляции уровня организации — биоценоза. Эволюция популя
ций — это в конечном счете механизм поддержания устой
чивости биоценоза как целостной системы, и стабилизиру
ющий отбор — главнейшая компонента этого механизма.
СПИСОК ЛИТЕРАТУРЫ
Кирпичников В. С. Роль наследственной изменчивости в процессе естест
венного отбора / / Биол. ж урн.— 1935.— Т. 4, № 5.
Лукин Е. И. Приспособительные ненаследственные изменения организмов
и их эволюционная судьба / / Журн. общ. биол.— 1942.— Т. 3, № 4.
Ш мальгаузен И. И. Регуляция формообразования в индивидуальном раз
витии..— M.: Наука, 1964.
Шмальгаузен И. И. Кибернетические вопросы биологии.— Новосибирск:
Наука. Сиб. отд-ние, 1968.
Baldwin J. М. A new factor in evolution / / Amer. Naturalist.— 1896.— N 30.
F isher R. A. The possible modification of the response of the wild type to re
current mutations / / Amer. Naturalist:— 1928. — Vol. 62, N 115.
F isher R. A. The evolution of dominance / / Biol. Rev.— 1931.— Vol. 6.
Morgan C. L. Animal Behaviour.— L., 1900.
Osborn H. F. The limits of organic selection / / Amer. Naturalist.— 1897. —
N 31.
Waddington С. H. The Strategy of the G ene.— L., 1957.
ВИД КАК ЭВОЛЮЦИОНИРУЮЩАЯ СИСТЕМА*
Р. Л . Берг
1.
Говоря о виде как о системе, многие авторы подразу
мевают, что виду присуща какая-либо одна черта, харак
теризующая собой систему: ее расчлененность (Вавилов,
* Реферат работ учреждений Отделения биологических наук
АН СССР за 1941— 1943 гг.— М.: Изд-во АН СССР, 1944.— С. 184.
227
Синская, Дубинин, Соколов и др.), взаимодействие частей
(Парамонов, Северцов С. А.), коррелятивная связь между
отдельными свойствами (Васнецов, Северцов С. А.) или со
подчинение части и целого (Спенсер). Спенсер рассматри
вает вид, как систему, чья активность направлена на под
держание ее существования. Все эти представления о виде
не вскрывают специфичности системы вида, ее отличий от
системы организма, с одной стороны, от других таксономи
ческих единиц или групп особей, с другой стороны. Неуди
вительно поэтому, что представление о виде, как о систе
ме, подверглось критике (Комаров).
2. В настоящее время из цитологических, генетических,
цитогенетических, экологических и эмбриологических исс
ледований (Дарлингтон, Мёллер, Гекели, Малиновский и
др.) выкристаллизовывается новое представление о виде,
как об эволюционирующей системе.
Теоретический анализ, так же как собственное экспе
риментальное изучение естественных популяций вида
Drosophila melanogaster, позволили автору сформулировать
представление о виде, как о системе, приспособленной в
процессе эволюции претерпевать дальнейшие эволюцион
ные преобразования.
3. Способность изменять свое взаимоотношение со сре
дой путем эволюционных преобразований отличает систе
му вида от системы организма.
То обстоятельство, что эволюционные преобразования,
в частности дифференциация частей (образование всех ви
дов полиморфизма, включая возрастной, половой, социаль
ный, модификационный, наследственный систематический
и несистематический и др.), не только не нарушают цело
стности вида, а напротив, совершенствуют способы поддер
жания его целостности, отличают вид от всех других так
сономических единиц, ставя его в этом отношении в один
ряд с такими системами, как биоценоз и живое вещество
биосферы (как ее понимает Вернадский). Однако степень
дифференциации частей, а отсюда и характер соподчине
ния частей и целого в пределах вида (в частности, особен
ности борьбы за существование) отличают систему вида и
от этих, близких ему в других отношениях, систем. От
биоценоза вид отличается единством его геохимической
функции в биосфере. Независимость эволюционного пути
от путей эволюции ближайших систем (ближайших как в
смысле пространственном, так и в смысле сходства) отли
чает вид от низших таксономических единиц. Независи
мость путей эволюции покоится на обособленности вида от
близких видов (физиологической, географической, эко
228
логической изоляции), обособленности, которой друг в
отношении друга не обладают низшие таксономические
единицы.
4. Удалось показать, что виду присущи все черты,
свойственные системе: 1) ее расчлененность; 2) наличие
взаимодействия между различными частями; 3) соподчине
ние части и целого; 4) закономерное строение, основанное
на наличии корреляций между различными свойствами си
стемы; 5) отграниченность от окружающей среды; 6) цело
стный характер во взаимодействии со средой; 7) способ
ность входить как целое в системы высшего порядка, обра
зуя их дискретную часть; 8) активность в процессе взаимо
действия со средой (свойство, присущее всем живым систе
мам) и, наконец, 9) способность к регуляции.
Регуляция в системе вида отличается от регуляции в
системе организма тем, что она: 1) сказывается не только
в отношении процесса жизнедеятельности, но и в отноше
нии темпов, путей и способов эволюции вида как целого:
2) совершается через отбор. В организме отбор и регуля
ция противополагаются друг другу — на уровне вида регу
ляция совершается с помощью отбора, как это ясно пони
мал еще Спенсер.
5. Регуляция особенностей вида, на которых покоится
его эволюция, совершается через отбор групп в пределах
вида, эволюционирующих наиболее успешно. Работы эко
логов (Дайвера, Элли, Синской, Андерсона и др.) показа
ли, что неполная изоляция отдельных популяций, т. е. та
самая структура вида, которая допускает возможность
межгруппового отбора, присуща огромному количеству ви
дов и представляет собой, по-видимому, всеобщую законо
мерность расселения особей по ареалу вида.
6. В свою очередь межвидовой отбор сохраняет те ви
ды, у которых регуляция темпов, путей и способов эволю
ции развита в наибольшей мере, т. е. те виды, где течение
индивидуального (внутригруппового) отбора контролирует
ся межгрупповым отбором.
7. В процессе эволюции устанавливается оптимум на
следственной и ненаследственной пластичности вида, выра
батывается приспособительный для вида в целом характер
наследования мутаций, создаются оптимальные условия
использования гетерозиготных и гомозиготных мутаций
видом, оптимальное соотношение внутри- и межгрупповой
борьбы в пределах вида. Иными словами, устанавливается
закономерное соотношение внутренних факторов эволюции
вида, предопределяющее в значительной мере способы и
темпы его дальнейшей эволюции.
229
8.
Представление о виде, как о системе, позволило дать
следующее определение вида. Видом называем эволюцио
нирующую систему, слагающие части которой объединены
не только общностью эволюционного пути, но и единством
геохимической функции в биосфере. От всех других видов
данный вид отличается своим только ему присущим путем
эволюции.
СЛУЧАЙНА ИЛИ ЗАКОНОМЕРНА ЭВОЛЮЦИЯ?
Р. Л . Берг
1. ЭВОЛЮ ЦИЯ -
ИЗМЕНЧИВОСТЬ, ИМЕЮЩАЯ ВЕКТОР
Эволюция органического мира — особая разновидность
изменчивости. Это изменчивость, имеющая направление,
изменчивость векторизованная. «Если взять процесс эво
люции в целом, то для него характерно постепенное усо
вершенствование организации, или прогресс» [Берг, 1922,
с. 15]. Слова эти принадлежат Л. С. Бергу — известному
географу и зоологу, автору книги «Номогенез, или эволю
ция на основе закономерностей».
Повышение уровня организации и возрастающее разно
образие частей — тот вектор, который отличает органиче
скую эволюцию от простой изменчивости.
Эволюция случайна, ибо строится на статистических
закономерностях, на закономерностях множеств. Это мно
жество не только организмов, но и ситуаций, причинноследственных рядов, их пересечений. Она случайный по
иск, игра использованных шансов.
Эволюция закономерна, она — номогенез. Развитие ор
ганических форм устремляется по определенным каналам.
Закон — ограничение. Там, где не все позволено, разными
способами образ>ется сходство действий, повадок, струк
тур. И вместе с тем, на разных путях возникает разнооб
разие. Эволюция — неогенез. Эволюционировать — зна
чит изобретать, конструировать, пользуясь осуществлен
ным для изобретения нового. Ход эволюции предопределен
внутренним строением того, что эволюционирует. Строе
ние эволюционирующей системы — род программы, сово
купность запретов и разрешений. Эволюция — согласова
ние со своим окружением и тут уже запреты и разрешения
налагаются и даются средой. Исход эволюции непредсказу
ем и предопределен в одно и то же время. Предопределен
прогресс, и в этом смысле эволюция закономерна.
230
2. МЕРА ПРОГРЕССА
Прогресс, повышение уровня организации — до недав
него времени эти слова воспринимались скептически все
ми, кто требовал строгости определения понятий. В настоя
щее время создалась возможность количественно оценить
сложность системы, а тем самым и высоту уровня ее орга
низации.
Критерий прогресса коренится в самой организации
жизни на нашей планете. Органический мир подчинен
трем законам. Первый из них — зональный принцип раз
мещения комплексов живых существ по поверхности Зем
ли. Ландшафтно-географические зоны — это прежде всего
зоны жизни: тундра, тайга, степь, лесостепь...
Второй закон — прерывистое иерархическое разнообра
зие живых существ, наличие царств, типов, классов, отря
дов... Групповое разнообразие, при котором сходства в пре
делах групп не менее поразительны, чем различия между
группами.
Третий закон — иерархия уровней организации, вели
кое членение живого компонента биосферы на целостные
компоненты — биоценозы, которые и дальше подлежат
членению. Популяция, организм, клетка и, наконец, макромолекулярный комплекс — носитель элементарного
свойства жизни, способный построить свою копию и оттор
гнуть ее от себя,— таковы главные ступени иерархии*.
Каждый представитель каждого главного уровня рас
членен на части, которые обладают известной автономией
в смысле как строения, так и функции. Организм делится
на системы органов, орган состоит из тканей, ткани из
клеток. Популяция включает в себя более тесные объеди
нения особей: куртины у растений, стада, стаи, колонии,
семьи у животных. Наилучшее описание членений систем
высшего ранга на системы подчиненные, как и наиболее
ясное понимание тектологических принципов построения
живого покрова Земли, мы находим у В. Н. Беклемишева
[1964, 1970].
Именно соподчинение систем дает нам в руки способ
количественно оценить высоту организации любого объек
та. Мы вычленяем функционально или морфологически
целостные части, из которых состоит система, а затем про
делываем ту же процедуру в отношении этих частей. Мы
относим число различных частей к числу элементов ниже* В дальнейшем о принципе вычленения уровней организации будет
сказано подробно, здесь же я ограничусь этим перечислением.
231
стоящего уровня организации. Отношение и служит мери
лом сложности. У системы с более высоким уровнем орга
низации разнообразие достигнуто при меньшей затрате
строительного материала. Принцип отношения числа час
тей к числу составляющих их дискретных отдельностей в
качестве мерила высоты организации применим везде и
всюду: в неорганической природе и в органическом мире, в
технике и в социальных системах.
Биоценоз устроен тем более сложно, чем больше разно
видовых популяций он содержит и чем меньшим числом
особей представлена каждая из них. Е. Одум [1968] поль
зуется этим критерием для оценки степени разнообразия
видового состава биоценоза. Чем больше отношение числа
видов к логарифму числа особей, тем больше степень раз
нообразия, тем стабильнее система. Е. Одум подчеркивает
значение видов, насчитывающих минимальное количество
особей. Вклад этих видов в поддержание устойчивости сис
темы может оказаться весьма большим.
Организация цветка тем выше, чем больше разнород
ных частей он содержит и чем меньшим числом представ
лена каждая часть. Большой набор органов разнообразного
назначения — лепестков, чашелистиков, пестиков, тычи
нок, нектарников, выростов, указывающих путь к нектару,
специализированных рычагов, обеспечивающих локализа
цию пыльцы на теле насекомого — переносчика пыль
цы,— показатель высоты организации. Малое число уст
ройств каждого типа указывает на совершенство в выпол
нении функции каждым инструментом. Чем совершеннее
система, тем экономичнее строение. Максимум разнообра
зия достигается при минимуме строительного материала.
О направлении эволюции в сторону снижения числа
гомологичных органов пишет Ч. Дарвин в «Происхожде
нии видов». В. А. Догель [1954] приводит множество при
меров уменьшения числа гомологичных частей в процессе
эволюции
животных.
Олигомеризацию
В. А. Догель
рассматривает как доказательство закономерного хода
эволюции.
Еще один пример, иллюстрирующий всеобщность на
шего критерия,— его применимость к любым системам.
Культура человеческого общества тем выше, чем больше
специальностей и чем меньшим числом лиц представлена
каждая из них. Число родов деятельности по мере разви
тия общества возрастает. По мере рационализации труда
для выполнения каждой функции требуется все меньшее
количество людей.
232
3. ПЕРВИЧНОСТЬ ПРОГРЕССА
Но ведь кроме прогресса, усложнения организации су
ществует и другое направление эволюции — инволю
ция — упрощение, регресс, дегенерация. В иных случаях
вид процветает за счет вырождения своих представителей.
Увеличение его численности, расширение ареала, возник
новение разнообразных форм в его пределах достигнуто
ценой снижения уровня организации. А. Н. Северцов
[1934] предложил различать биологический и морфофизи
ологический прогресс. Увеличение численности особей,
расширение ареала вида и его распадение на низшие по
отношению к виду таксономические единицы А. Н. Север
цов называет биологическим прогрессом. Классификация
типов прогресса усовершенствована И. И. Шмальгаузеном
[1969].
Если каждому представителю вида грозит ежеминут
ная, никакими силами не предотвратимая гибель, преиму
щество получают те, кто поставил на вооружение наиболь
шую плодовитость, наименьшие размеры, наиболее корот
кий цикл развития. Тогда ничтожность повышает шансы
выжить. Множество форм регрессировало этим путем. Но
инволяция не была основным путем истории органического
мира. Прогресс первичен, регресс вторичен.
Чтобы было чему претерпевать упрощение, нужно,
чтобы сперва совершилось усложнение. Эволюция органи
ческого мира в целом идет в направлении повышения
уровня организации, по пути прогресса. Она — векторизо
ванная изменчивость.
4. ЗАПРЕТ -
ОСНОВА ЗАКОНА
Раз есть вектор, путь, канал, направление, значит есть
ограничения, налагающие запрет на движение по всем ос
тальным направлениям.
Запрет — основа закона.
Эволюция — движение по разрешенным путям. Этим
она отличается от броуновского движения молекул, от
флюктуаций атомов в молекуле.
Разнообразие органического мира грандиозно. Оно гип
нотизирует нас, заставляя забыть об ограничениях. О том,
чего нет, труднее думать, чем о том, что есть. Глазу не на
чем остановиться. Но нет-нет и заходит разговор об огра
ничениях.
233
Колесо. Читателю ничего не стоит догадаться, почему
нет колеса. Способ образования многоклеточного организ
ма путем деления клеток, объединение всех частей тела в
процессе нервной регуляции, централизованность питания
и выделения исключают такую самостоятельную часть как
колесо из набора технологических устройств. Колесо есть
перекати-поле, но это колесо катится, а не способствует
передвижению других. Организмы могли бы использовать
друг друга в качестве транспортных средств типа колеса,
но до этого они не додумались*. Мне кажется, дело стало
за втулкой. Об ограничениях говорит К. Уоддингтон [1970,
с. 11]. «Мыслимо ли существование биологической тополо
гии, которая могла бы объяснить, почему существуют
большие классы животных с двумя, тремя и четырьмя па
рами конечностей, или же неопределенным их числом, как
у многоножек, но неизвестно ни одной крупной группы
животных с шестью или семью парами конечностей. Поче
му существует двух-, четырех-, пяти- и шестилучевая сим
метрия, но нет, по-видимому, семилучевой симметрии?»—
спрашивает он.
Уоддингтон дает и решение проблемы запрета в самом
общем виде с помощью следующей модели: «Вы можете
получить треугольник лишь в том случае, если а + Ь>с».
Эволюция и закономерное изменение — синонимы. Са
мо слово «эволюция» употребляется с целью обозначить
нечто, от простой изменчивости отличающееся.
Направленные процессы идут и в неорганической при
роде. В закрытой системе в отсутствии притока энергии в
соответствии со вторым законом термодинамики неуклонно
совершается выравнивание энергетических потенциалов —
свободная энергия стремится к минимуму. Энтропия возра
стает. Направление изменений задано. Все, что может со
вершиться, совершается по линии наименьшего сопротив
ления, все переходит из состояния менее вероятного в со
стояние более вероятное. Но и в открытых системах, подо
бных нашей планете, купающейся в потоке льющейся из
вне энергии солнечных и космических излучений, и в от
крытых системах, ставших на путь самовоспроизведения,
осуществляется то, чему надлежит осуществиться, притом
императив долженствования диктуется максимумом веро
ятности.
В иных случаях вероятнее хаос. Там, где разрыв связей
ведет к более устойчивому состоянию, связи рвутся, ка
мень крошится, вода испаряется. Планеты пенепленизиру* Известны, однако, симбионты, паразиты и т. д.
234
ются, крутизна падения водопадов уменьшается, они сдви
гаются к истоку реки. Но если устойчивость достигается
взаимосвязанностью первоначально разрозненных частей,
происходит усложнение.
Направленность эволюционных изменений прекрасно
понимал А. А. Богданов [1925, с. 103]. Преобразование,
осуществляемое живыми системами, ведет к повышению
энергетического потенциала. А. А. Богданов пишет: «...для
сохранения в изменяющейся, т. е., в конечном счете, во
всякой среде, недостаточно простого обменного равновесия.
Единственно, что может давать относительную гарантию
сохранения, это возрастание суммы активностей, перевес
ассимиляции: тогда новые неблагоприятные воздействия
встречают не прежнее, а увеличенное сопротивление.
Именно этим путем идет природа в деле сохранения жиз
ненных форм, и человек в своем коллективном самосохра
нении: путем роста комплексов, накопления запаса актив
ностей. Каждый шаг в эту сторону увеличивает возмож
ность поддержания жизни при изменяющихся условиях.
Иными словами, динамическим элементом сохранения
комплекса является возрастание его активностей за счет
среды».
В 1963 г. в Вакула-Спрингсе во Флориде на конферен
ции, посвященной возникновению пробиологических сис
тем, одному из участников совещания А. Липману был за
дан вопрос — чем определяется уникальная роль фосфора
в процессе биологического переноса энергии. Устойчивость
фосфоангидридных соединений типа аденозинтрифосфата
(АТФ) в условиях разбавленных нейтральных растворов,
характерных для среды, где идет биосинтез, больше устой
чивости других соединений — был ответ. «Причина устой
чивости фосфорильных производных заключается, по-ви
димому, в том, что богатая энергией связь защищена отри
цательным зарядом фосфорила, отталкивающим молекулы
воды и ионы ОН- . В ацетангидриде, напротив, аналогич
ная связь оказывается незащищенной и легко гидролизует
ся. Таким образом, преимущество АТФ состоит в том, что
связь Р— О— Р экранирована и ее энергия сохраняется до
тех пор, пока не вступит в реакцию соответствующий ка
тализатор. Кроме того, создается впечатление, что ста
бильность макроэрги ческой связи возрастает, если фосфорильный радикал присоединяется к органическому остат
ку»,— сказал А. Липман [1966].
Дж. Уолд [1964] пишет: «В выборе атомов нет ничего
случайного... В общем и целом наиболее легкие элементы
являются наиболее широко распространенными; однако,...
235
отбор элементов, из которых построены организмы, опре-/
делялся какими-то иными критериями, чем их доступ-у
ность» (с. 102). Выбор строительного материала диктуете^
его пригодностью для той роли, которую элементы и их со,1
четания призваны играть в организме, утверждает
Дж. Уолд. Возможности выбора чрезвычайно ограниченны,
так что в большинстве случаев организмы вынуждены бы
ли сделать именно тот выбор, который они сделали. «По
этой причине я полагаю,— пишет Дж. Уолд,— что не
только химические элементы, но и химическая конститу
ция
организмов
одинаковы
во
всей
Вселенной»
(с. 102— 103). Около 99 % тканей организма построено из
четырех элементов: Н, О, N и С. Это те элементы перио
дической системы, которые имеют атомы наименьшего раз
мера и приобретают стабильные электронные конфигура
ции после присоединения соответственно 1, 2, 3 и 4 элект
ронов. Размеры атома — очень существенная черта, от ко
торой зависит, войдет или не войдет элемент в состав жи
вого. Атомы наименьшего размера образуют устойчивые
связи, и только эти атомы, как правило, образуют кратные
связи. Элементы как будто соревнуются друг с другом за
место в живых организмах. В этом соревновании фтор по
терпел поражение, так как атомы водорода имеют мень
ший размер. Кремний встречается в поверхностных геосфе
рах и оболочках Земли, включая атмосферу и гидросферу,
в 135 раз чаще, чем углерод. Но и он не занял почетного
места среди атомов, строящих живое. Углерод вытеснил
его. Дж. Уолд перечисляет три преимущества углерода, ко
торые обеспечили ему победу над кремнием: связи крем
ния с самим собой и с другими атомами менее прочные,
чем С— С-связи. Он не способен образовать двухвалент
ную связь. Соединения кремния инертны. Цепи кремния в
присутствии кислорода, воды, аммиака легко рвутся. Вода
и кислород противопоказаны соединениям кремния. Имен
но поэтому кремний противопоказан живому. Сера и фос
фор, напротив, обладают особыми преимуществами в пере
носе групп атомов и энергии. Типы их связей разнообраз
ны, а сами связи внутренне нестабильны, что облегчает
обмен. И энергетические потенциалы связей и нестабиль
ность связей имеют меру, ту самую, которая обеспечивает
сере и особенно фосфору участие в построении живых сис
тем. Свойства фосфора и серы обусловливают адекватную
дозировку энергии.
Эволюция материи идет от рассеянных в пространстве
элементарных частиц к образованию атомов, микромоле
кул, макромолекул. Число связей возрастает, сила связей
236
обретает меру, части системы делаются сложнее, возраста
ет организованность. И точно так же, как астрономическое
тело — скопление разнородных элементарных частиц, если
это светило, или атомов и молекул, если перед нами
скромная планета, так и минерал — скопление однород
ных молекул — результат эволюции материи. Звезды,
планеты и минералы возникают на определенном этапе
длйтельной истории развития неорганической материи.
На пути возникновения организованной устойчивости
создалась и та принципиально неустойчивая, перманентно
преобразующаяся констелляция элементов, которая вне
преобразования не существует и существовать не может и
которую мы называем живой. Но прежде чем началось на
копление негэнтропии, должна была возрасти энтропия.
5. ОТБОР -
ПРЕОБРАЗОВАТЕЛЬ МИРА
Неравенство вероятностей сохраниться для объектов
или повториться для состояний носит общеизвестное назва
ние. Это отбор. С предельной ясностью это выражено
А. А. Богдановым [1925]. Отбор царит везде в неорганиче
ской природе и среди живых организмов. Отбор в его со
храняющей форме.
Вероятная картина мира — его выражение. Чем устой
чивее явление, тем оно вероятней, тем чаще оно встретит
ся нам. Звездные системы, минералы, органические фор
мы, общественные формации, болезни и мнения подчине
ны этому принципу. Дифференциальное сохранение объек
тов неуклонно меняет облик Вселенной.
И в органическом мире отбор — единственный фактор
эволюции — причина, но как мы увидим, уже не единст
венная, придающая изменчивости направленный характер.
Но это какой-то другой отбор. Различие между сохраняю
щим отбором, царящим везде, и двигателем органической
эволюции — мы назовем его отбором преобразующим —
коренится в самом различии живого и неживого.
6. РЕДУПЛИКАЦИЯ И ОТБОР
Эволюция привела к образованию органических соеди
нений — макромолекул. Жизнь, вероятно, возникла в тот
момент, когда первые полимеры нашли путь авторепродук
ции, стали матрицами, по образу которых могли синтези
роваться новые матрицы, а на тех — снова новые... Вместе
237
с матричным принципом авторепродукции в мир вошел
новый фактор эволюции — отбор в его преобразующей
форме.
I
Первая четкая формулировка принципа преобразующей
го отбора принадлежит Гете. В 1809 г. великий натуралиут
и поэт писал: «Представьте себе природу, которая как оы
стоит у игорного стола и неустанно выкрикивает: a double!
т. е., пользуясь уже выигранным, счастливо, до бесконеч
ности продолжает игру сквозь все области своей деятельно
сти. Камень, животное, растение — все после таких счаст
ливых ходов постоянно снова идет на ставку, и кто знает,
не является ли весь человек, в свою очередь, только став
кой на высшую цель?» Уберите камень, и Вы получите
идеальное описание механизма органической эволюции.
Цель совсем не звучит здесь телеологически. Трудно ска
зать больше, чем сказано в этих немногих словах. Природа
на каждом туре игры удваивает ставку! Афоризм великого
поэта и натуралиста, каким был Гете, приведен Л. С. Бер
гом [1922, с. 42]. И. И. Канаев [1957] приводит эти слова
в вышедших в его переводе и с его комментариями избран
ных сочинениях Гете (с. 390—391).
Жизнь представляет собой открытую систему. Вещество
и энергия аккумулируются живым и используются в про
цессе конструктивных преобразований. Одно из них —
процесс создания своей копии, другое — эволюция.
Если бы способность самокопирования обрели не поли
меры, а микромолекулы, никакой органической эволюции
не было бы. Постоянство состава, жестко заданное в мире
микромолекул,— непреодолимая преграда для эволюции.
Эволюция — закономерное следствие не только удвоения,
но и химического непостоянства тех нитевидных углерод
ных соединений, которые стали на путь построения своих
копий.
Треугольник возникает из отрезков а, b и с, только ес
ли а + Ь > с и а — Ь <с. Но так обстоит дело, если длина
отрезков я, Ъ и с жестко задана. Растущие и укорачиваю
щиеся отрезки, по трое соединяясь концами, беспрепятст
венно соединяются в треугольники. Устойчивость треуголь
ников требует соблюдения некоторых условий. Необходи
мы жесткость строительного материала и дискретный ха
рактер изменений. Рост, как и укорочение, должны чере
доваться с периодами стойкого сохранения размера. Если
эти условия не соблюдены, треугольники будут превра
щаться в кольца, проходя через последовательность при
чудливых фигур, спадаться, скручиваться и претерпевать
всевозможные другие изменения под влиянием внешних
238
условий. Конвариантная редупликация обеспечила как ус
ловия преобразования живых существ, так и стойкость воз
никающих изменений. Она дала возможность, говоря сло
вами Гете, воспользоваться уже выигранным.
\ Материал для построения копий создавался под дейст
вием космических лучей и коротковолновых излучений
Срлнца. Часть соединений, а именно углеводороды, веро
ятно в частично окисленном состоянии, образовались в не
драх Земли. Аминокислоты, пиримидиновые и пуриновые
основания создавались, как утверждает Холдейн [1966,
с. 18], в самой атмосфере.
Синтезировать из неорганических соединений свой
строительный материал, как это делают сейчас все без ис
ключения зеленые растения, первичные шаблоны не мог
ли. Довольно и того, что они творили по своему образу и
подобию, эти не видимые ни простым глазом, ни в свето
вой микроскоп образования. Укладывать камни, извольте,
за милую душу. Заготовка камней должна совершиться в
небесах. Они строители и только, каменщик и поставщик
строительного материала — Солнце. Когда самовоспроизводящих матриц стало много, наступил острый кризис.
Строительного материала стало не хватать. Шаблоны всту
пили в жесточайшую конкуренцию друг с другом. Те, ко
торые быстрее и с наименьшей затратой энергии способны
были перехватывать пригодные органические соединения,
обрекали на бездеятельность всех остальных. Они побеж
дали на арене экономической борьбы за сырье. Своим пре
имуществом они обязаны непостоянству состава макромо
лекул. Только они одни могли слегка изменить свою хими
ческую природу, не теряя способности самовоспроизведе
ния, оставаясь в существенном самими собой. Их самовос
произведение — это не просто редупликация, а редуплика
ция конвариантная.
Конвариантные редуплицирующиеся макромолекулы и
стали объектом отбора в его преобразующей форме.
Энергия и вещество подавались извне, использовать их
оказалось возможным на много ладов. Конкуренция за
строительный материал открыла возможность прогрессиру
ющего усложнения организации. Образование нуклеопротеида, сочетание нуклеиновой кислоты и белка, было ре
шительным шагом по пути повышения уровня организа
ции. Отбор в пользу наименее вероятного — невозможного
в закрытой системе — вот чем стала эволюция с того са
мого момента, как возникли первые зародыши жизни.
Генетическая концепция жизни, которую развивал
Г. Дж. Мёллер еще в 1926 г. [Muller, 1929; Мёллер, 1937]
239
и которая полностью принимается многими выдающимися
умами и в настоящее время [Тимофеев-Ресовский и др.,
1969; Уоддингтон, 1970], включает эволюцию в самоопре
деление жизни. Мёллер в докладе «Ген, как основа жизни»
на Международном ботаническом конгрессе в Нью-Йорке в
1926 г, говорил: «...гены составляли основу первого живог^)
вещества. В силу своего свойства (находимого тишь в жи
вых телах) мутировать без потери способности к росту они
в соответствии с требованиями своей непрерывности ини
циировали эволюцию более сложных образований, чем они
сами,— протоплазмы, организма и т. д. Даже если бы гены
не составляли фундаментальную основу различия между
живым и неживым (а я попытаюсь показать, что они со
ставляют) , большинство биологов согласится с утверждени
ем, что они лежат в основе всех устройств, отличающих
высшие формы от самых примитивных форм жизни. Когда
я говорю “лежат в основе“ я подразумеваю, что изменение
в структуре гена не только необходимо для возникновения
морфологических и физиологических различий, но что эта
измененная структура гена существует и ныне и что без
нее не могли бы сохраняться эти новые свойства, формы и
физико-химические реакции».
Живой системой является устройство, способное пре
терпевать эволюционные преобразования. Неизбежным
следствием конвариантной редупликации является отбор в
его преобразующей форме. Неизбежным следствием отбо
ра — эволюция. Быть может, редупликацию, отбор и эво
люцию не следует рассматривать как причинно-следствен
ный ряд. Все эти свойства, характеризующие один и тот
же объект — живое,— это свойства сопряженные, друг от
друга неотъемлемые, друг другу комплементарные.
7. ХИМИЯ ЗАПРЕТА
С неизбежностью по строгим законам совершилось
важнейшее событие в истории планеты Земля. Строжай
ший детерминизм существует в выборе веществ, способных
осуществить конвариантную редупликацию. Сходные друг
с другом химические соединения — нуклеиновые кислоты:
рибонуклеиновая кислота (РНК) и дезоксирибонуклеино
вая кислота (ДНК) — одни оказались носителями фунда
ментального свойства живого. Нуклеиновая кислота хранит
и передает информацию, необходимую для ее собственного
воспроизведения и для осуществления в процессе индиви
дуального развития всех признаков организма. «Очевидно,
240
накопление информации чрезвычайно эффективно»,— пи
шет Фред Хойль [Hoyle, 1964, с. 32— 33]. Мы в самом де
ле можем быть уверены, что заметно более эффективные
Уимические системы невозможны. Те, которыми обладаем
мы, заведомо делают свое дело наилучшим образом. Могут
быть и другие пути, но наш путь представляет собой гене
ральную линию жизни во Вселенной. У меня есть уверен
ность, что многие планетные системы обладают живыми
формами, очень похожими на земные.
Класс веществ, пригодных для осуществления элемен
тарной жизненной функции самовоспроизведения, оказал
ся чрезвычайно узким. Эволюция оказалась введенной в
заданное русло с самого начала. Возможности усложнения
организации открылись, но они отнюдь не были безгранич
ными.
Усложнение организации наступало там и только там,
где прежняя функция могла быть выполнена быстрее и
лучше сложной структурой или где возникала новая функ
ция. Там, где наиболее успешное выполнение функции
осуществляла наиболее просто устроенная структура, на
эволюцию наложен запрет. Выражение этого запрета —
единообразие структурных основ наследственности во всем
органическом мире, начиная от бактерий и вирусов и кон
чая высшими животными, включая человека.
Сходство касается нс только химии хромосом. Химиче
ские основы процесса перекодировки с языка нуклеотидов
на диалект белков, перевод текста, написанного с помощью
четырехбуквенного алфавита оснований, на язык, пользу
ющийся двадцатизнаковым кодом аминокислот, осуществ
ляются совершенно сходным образом во всем органическом
мире. Одни и те же триплеты оснований подают команду
определенной аминокислоте занять надлежащее место в
цепи пептида у кишечной палочки и у крысы. Сигнал рав
нения для каждой аминокислоты, будь то в клетке уха
слона или в клетке фасеточного глаза стрекозы, один и тот
же. Процесс считывания информации, если смотреть на
него глазами химика, один и тот же во всем органическом
мире.
8. ГЕОМЕТРИЯ ЗАПРЕТА
Геометрическая конфигурация наследственного аппара
та неизменна везде и всюду. Линейный принцип организа
ции не нарушается нигде. Кольцо, ламповая щетка, ворси
ны которой образованы петлями, перпендикулярно отходя
241
щими от стержня и продолжающими этот самый стер
жень,— структуры, встречающиеся то тут, то там, не от
клоняются от принципа линейности ни на йоту. Не то что
кодирующих объемов, плоскостей, но даже разветвленных
хранилищ информации нет нигде в органическом мире. На
усложнение организации структурных основ наследствен
ности наложен строжайший запрет. Видно, без одномерной
организации не обойтись. Генонема, как назвал самовоспроизводящую часть хромосом Н. К. Кольцов [1938], имеет
линейную структуру совсем не случайно. Единообразие
строения структурных основ наследственности отнюдь не
служит доказательством единства происхождения всех жи
вых существ от одного предка, как думают все без исклю
чения биологи, включая И. И. Шмальгаузена [1969,
с. 37— 38 ], а является выражением запрета на все другие
конфигурации, кроме существующей. Вето на разнообра
зие было наложено в момент возникновения жизни, и не
было оснований снять его.
Две функции хранилища наследственной информации
осуществляются наилучшим из возможных способов с по
мощью линейных структур — самовоспроизведение и пода
ча команд к синтезу белка.
Самовоспроизведение — исконная функция живого —
осуществлялось на заре возникновения жизни в условиях
острой конкуренции за строительный материал. Преиму
щества были на стороне тех, у кого синтезирующая повер
хность была максимальной. Наиболее выгодное соотноше
ние между поверхностью и объемом было у мономолекулярных структур. Те макромолекулы, которые объединяли
свои усилия в улавливании строительного материала, тер
пели урон — вся поверхность их соприкосновения выклю
чалась из синтеза. Действовать порознь было куда спод
ручней. Нехватка строительного материала, необходимость
конкурировать за него сделала гены маленькими. Ли
нейные структуры — биополимеры — обладали доста
точной степенью организованности и возможностью менять
свой химический состав, не теряя способности к редуп
ликации, в силу своей большой длины. Толщина, которая
могла быть достигнута только объединением многих
молекул,
была
в этих условиях противопоказана.
Одномерность, существенная одномерность, конечно, со
ответствовала требованиям экономичности процесса ре
дупликации.
Считывание наследственной информации в процессе
синтеза белков также требует линейной организации хра
нилища информации. Вот что пишет по этому поводу
242
М. В. Волькенштейн [1965, с 327]: «Матрица для синтеза
белка одномерна, это линейная макромолекула мРНК
(матричная, или информационная мРНК, это РНК — по
средник между ДНК и рибосомами, переносящий инфор
мацию из конструкторского и планового бюро фабрикиклетки к цехам, выпускающим готовую продукцию — бел
ки). Очевидна биологическая целесообразность одномерно
сти в этом случае. Представим себе трехмерную матрицу.
Это нечто вроде трехмерных шахмат — игры, которую
предложил некий изобретательный ум. Играть в них, ко
нечно, невозможно.Невозможно было бы извлечь из трех
мерной матричной сетки синтезированные в ней биологи
ческие молекулы. Двумерная матрица ... также крайне не
удобна, так как она должна встречать большое сопротивле
ние при перемещении в клетке, и ее конформационная по
движность (ее способность менять взаимное расположение
частей.— Р. Б.) не может быть достаточной для образова
ния функциональных структур типа комплексов мРНК с
рибосомами. Одномерная матрица мРНК находится, конеч
но, в соответствии с линейным строением синтезируемого
биополимера — белка».
Вся информация, которая считывается во времени, за
писана в одну строчку. Нотные знаки и текст книги, маг
нитофонная запись и патефонная пластинка — тому при
меры. Время одномерно. И линейная запись знаков, подле
жащих последовательной перекодировке, представляет со
бой пространственную проекцию времени.
Дж. Свифт описывает, как Гулливер, попав к лилипу
там, обнаружил у них письменность. Лилипуты писали не
так, как европейцы — слева направо, не так, как арабы —
справа налево, не так, как китайцы — сверху вниз, а так,
как английские дамы — из одного угла бумаги в другой.
Замечу в скобках, что, высмеивая все и вся, Дж. Свифт
направлял острие своей сатиры против англичан — народа,
к которому принадлежал он сам. Разнообразие способов
начертания очевидно, но оно не касается линейной по
следовательности знаков. Линейный принцип осуществ
ляется неизменно. Возник он у разных народов конвергентно в соответствии с требованиями дешифровки.
Конвергентно в соответствии с теми же требованиями
возникла и закрепилась линейная организация струк
турных основ наследственности. Пример конвергенции,
выходящей за пределы органического мира. Бионика,
вернее тектология, осуществленная самим ходом истории
человечества!
243
9. АСТРАЛЬНЫ Й АСПЕКТ ПРОБЛЕМЫ
«БЫТЬ ИЛИ НЕ БЫТЬ»
Геофизические и геохимические условия возникнове
ния жизни на Земле продиктовали химический состав жи
вого и его пространственную организацию. Нельзя назвать
случайным то, что входит как неотъемлемая часть в орга
низованность более высокого порядка, в данном случае в
организованность Земли как планеты. Сама возможность
появления и существования жизни на планете теснейшим
образом связана со свойствами планеты. Множество чисто
астрономических параметров решает, быть или не быть.
Расстояние от планеты до центрального светила, размеры
планеты, наличие или отсутствие вращения вокруг оси,
форма и размер орбиты, скорость движения по ней, наклон
оси планеты к плоскости эклиптики определяют темпера
туру планеты и границы температурных колебаний, состав
атмосферы и направление эволюции газовой оболочки пла
неты. Жизнь входит как необходимый компонент в систе
му взаимоотношений между астрономическими, геофизи
ческими, геохимическими свойствами Земли как планеты.
Американский
астроном
Харлоу
Шепли
[1962,
с. 62— 63] пишет: «Планеты, на которых жизнь могла бы
не только возникнуть, но и развиваться, должны быть не
очень близки к своей центральной звезде, но и не слишком
далеки от нее для поддержания необходимой температуры.
Они должны быть достаточно велики, чтобы удержать кис
лородную атмосферу (если, конечно, мы не собираемся ог
раничиться низшими анаэробными организмами...)
Планеты, имеющие благоприятные размеры, темпера
туру и химический состав, должны также двигаться по ор
битам с малым эксцентриситетом. В случае очень вытяну
тых орбит, таких, как у большинства комет, планеты под
ходили бы слишком близко к звезде в периастре и слиш
ком далеко уходили бы от нее в апоастре. В результате
этого колебания температуры были бы слишком велики
для нормального течения жизни, а может быть, и для воз
никновения и поддержания жизни на ранних стадиях ее
развития. Также нежелательны и слишком резкие колеба
ния температуры при смене дня и ночи. Поэтому лучше
всего было бы, если бы планеты быстро вращались, а их
ось вращения была бы не сильно наклонена (как у нашей
планеты) к плоскости их орбиты».
Наша Земля с ее почти круговой орбитой, с 24-часовым
периодом обращения вокруг своей оси, со всеми свойства
ми является лучшим из мйрор, но таких избранников во
244
Вселенной насчитывается, по самым скромным подсчетам
астрономов, сотни миллиардов. Вопрос о закономерностях
эволюции в наш век познания Космоса отнюдь не является
праздным. Если эволюция органического мира — процесс
закономерный, мы по свойствам планеты можем предска
зать вероятные свойства ее обитателей.
10. НА ГОРИЗОНТЕ ПОЯВИЛАСЬ СМЕРТЬ
Преобразующий отбор вступил в действие в момент
возникновения жизни. Перед живыми существами откры
лась возможность прогрессивной эволюции. Структуры,
обеспечивающие процесс редупликации и считывание ин
формации в процессе биосинтеза, не подлежали дальней
шему существенному усложнению. Отбор, создав их, сам
наложил запрет на повышение уровня их организации. Ус
ложнению подлежали аппараты, направленные на добыва
ние веществ и энергии, необходимых для редупликации.
Использование обмена веществ для добывания энергии и
сырья было первым существенным усовершенствованием и
вместе с тем первым грехопадением живых организмов.
Пока они ждали милостей от природы, пока смирно карау
лили падающие с неба стройматериалы, они вкушали веч
ное блаженство. Они были потенциально бессмертны. За
тем, включив метаболические котлы в состав своего орга
низма, перестав уповать на небо, они грандиозно повысили
надежность притока вещества и энергии и в особенности
их количество, они встали на путь прогресса, широчайшие
горизонты открылись перед ними, их нервная деятельность
усложнилась, достигнув уровня, достойного наименования
психической деятельности, а затем и души, но они утеряли
бессмертие. Однако перед тем как встать на путь неогра
ниченного прогресса, живые организмы чуть было не за
крыли для себя эту сладостную и такую трагическую вме
сте с тем возможность усовершенствования.
11. ИСПОЛНЕНИЕ СМЕРТНОГО ПРИГОВОРА
ОТСРОЧЕНО
В борьбе за освобождение от тягостной необходимости
ждать, когда будут доставлены пригодные для редуплика
ции органические соединения, организмы пошли по пути
создания клеток. Редуплицирующие нити окружили себя
245
протоплазмой, где с помощью белков-ферментов осуществ
лялся процесс создания нужных соединений.
Линейная организация уже не удовлетворяла новым
обстоятельствам. Заработали мембраны — сперва клеточ
ная, а затем ядерная оболочка, осуществляя избирательное
поступление нужных веществ в клетку и выделение отра
ботанных продуктов из нее, обеспечивая постоянство внут
ренней среды. В трехслойных молекулярных устройствах
стали осуществляться метаболические процессы. Достигнув
клеточного уровня организации, живые организмы овладе
ли процессами, происходящими на плоскости. Им стала до
ступна существенно двумерная организация пространства.
В корне изменился характер воспроизведения. Мономолекулярные хранилища наследственной информации с
прежним постоянством совершали процесс редупликации.
Клетка в целом приступила к делению. Но тут-то и таи
лась опасность вечного прозябания. Деление несовместимо с
усложнением организма. Делимое не может достичь той сте
пени усложнения, того увеличения размеров, какое затем
было достигнуто живыми организмами на путях многокле
точное™. Делимость обеспечила метаболизм, она не унич
тожила потенциального бессмертия живых систем. Она на
ложила запрет на усложнение организации, на прогресс.
Единообразие строения клеток во всем органическом ми
ре — выражение этого запрета, а совсем не доказательство
единства происхождения всех живых существ от одного од
ноклеточного прародителя, как думают сейчас все без еди
ного исключения биологи-эволюционисты [Шмальгаузен,
1969, с. 37— 38; Вилли, 1964, с. 44]. Клетка — система де
лящаяся, и как таковая никакого другого устройства, кро
ме организации, совместимой с делением, иметь не может.
Вирусы, а среди протистов
инфузории изобрели час
тичную смерть во имя бессмертия. Вирус при проникнове
нии в клетку хозяина сбрасывает белковый футляр и весь
аппарат, обеспечивающий поиск клетки хозяина и проник
новение в нее. В клетку хозяина проникает нуклеиновая
нить, благополучно редуплицируется, строит оболочку и
бронебойное орудие для проникновения в новую клетку.
У инфузорий отбрасывается макронуклеус. Частичная
смерть позволила усложнить организацию, но вирусы оста
лись вирусами, протисты — протистами. Многого на пути
частичного умирания живые существа нс достигают. Отде
ление большой части, подлежащей уничтожению, несовме
стимо с неделимостью, индивидуальностью высших форм.
Глубокие различия между клетками разных тканей
многоклеточного организма начинаются с дифференциации
246
органов и тканей организма. Возникновение различий со
провождается утерей клетками делимости. Делящиеся
клетки всех живых существ сходны.
12. ПРОГРЕСС И БЕССМЕРТИЕ ДВЕ ВЕЩ И НЕСОВМЕСТНЫЕ
Эволюция не остановилась на создании клеток. Новые
невиданные возможности открылись перед живыми сущест
вами, когда они «напали» на идею многоклеточности. На
дежность поступления энергии во много раз возросла, уве
личилась вероятность сохраниться среди бесчисленных
превратностей бытия. За счет включения метаболических
аппаратов в состав организма возросли его размеры. Гран
диозно увеличилась его сложность, повысился уровень ор
ганизации. Прогресс перестал быть табу, каким он был для
обреченных на деление клеток. Одни запреты оказались
снятыми. Их снятие было причиной наложения новых. Те
перь под запретом оказалась делимость и, что обиднее все
го, сопряженное с делимостью бессмертие.
Усложнение всей организации в целом и в особенности
нервной системы привело к возникновению сложной пси
хической деятельности, к созданию духовного мира челове
ка — его души. Душа человека, по определению, потенци
ально бессмертна. Смерть — возникновение трупа — не
избежна для тела. Необходимость для вида устранить от
размножения организмы, сносившиеся в процессе жизнеде
ятельности, приводит к тому, что уничтожение организма
становится под генный контроль, и смерть наступает по
программе. Организмы как бы насильственно устраняются
с жизненной арены. Иногда отбор идет на сокращение
жизни особи во имя сохранения или процветания вида.
Путь эволюции в сторону дегенерации у свободно живу
щих форм — яркий пример такого генотипически обуслов
ленного прогрессирующего укорочения жизненного цикла.
Человек не идет по пути дегенерации. Но и для него
смерть — неизбежный конец его индивидуального разви
тия. Духовный мир человека — система потенциально бес
смертная, способная неограниченно развиваться, усовер
шенствоваться, гармонически усложняться. Никакой про
граммы самоуничтожения душа нормального человека не
содержит и тлену не подвержена. Бессмертную душу
уволакивает в могилу смертное тело. Эта трагедия тра
гедий — плата за прогресс. Смерть — мзда за бессмерт
ную душу.
247
За неделимость, за неповторимую свою индивидуаль
ность тоже заплачено по дорогой цене. Многоклеточные
организмы усложнились, впервые полностью овладели
трехмерным пространством, сперва они еще удерживали в
лице своих низших представителей остатки делимости —
вегетативное размножение. Потом они утратили и его.
Размножение стало осуществляться с помощью отделения
матриц, содержащих в закодированном виде программу
развития будущего существа. От микроскопически малого
комочка материи, лишенного мысли, как всякое однокле
точное существо, находящегося на грани между жизнью и
смертью, подверженного всем превратностям, всем случай
ностям бытия, к неповторимой индивидуальности взрослого
человека ведет долгий и тяжкий путь. Мы эмбрионы, и со
хранение нашего существования на эмбриональной стадии не
гарантировано законом, мы дети, и наше детство — пора
чудовищной зависимости и бесправия, мы вступаем в пору
юношества с его страданиями. Дорогая цена прогресса, на
шей смертной бессмертной души, нашей неделимости.
Но и этого мало. С возникновением полового процесса
индивид перестал воспроизводить себя, свою индивидуаль
ную неповторимость. Воспроизведению подлежит предста
витель своего вида, существо, наделенное в результате ген
ной комбинаторики, осуществляемой при мейозе, новым,
только ему присущим сочетанием свойств. Человек порож
дает человека, отнюдь не копирующего его самого. Отли
чительные черты характера потомка подчас делают его не
приемлемым для родителей. Мы не выбираем подходящих
для себя родственников, как выбираем мужа или жену,
друзей и знакомых. Закон обязывает нас выращивать су
щества, которым мы, быть может, не будь они нашими
собственными детьми, не подали бы руки при встрече.
И то же касается родителей — они не выбирали нас, но
ведь и мы их не выбирали. Еще один источник трагедий,
порожденный матричным принципом нашего способа восп
роизведения. Только популяция воспроизводит себя с по
мощью своих представителей.
Мы смертны еще и в том смысле, что мы неповторимы.
Запрет на бессмертие — выражение одной из фундамен
тальных закономерностей эволюции.
Переход к многоклеточности подготовлен долгим разви
тием на уровне одноклеточных организмов. Знаменатель
но, что высшие формы среди многоклеточных диплоидны
на всем протяжении жизненного цикла, т. с. обладают во
всех клетках, кроме зрелых половых клеток, двойным на
бором хромосом. Почему наличие гигантских клеточных ко
248
лоний с разделением функций между членами колонии не
совместимо с гаплоидным набором хромосом? Дезинтегриру
ющее действие мутационного процесса воспрепятствовало,
на мой взгляд, переходу гаплоидных организмов к многоклеточности. Соматический мутагенез, нарушая дифференциа
цию клеток, внося помехи в нормальное их функционирова
ние, лишь за редчайшими исключениями использован мно
гоклеточными существами в качестве конструктивного нача
ла. Вот пример конструктивного использования мутационно
го процесса. Мутационный процесс и отбор клонов мутант
ных лимфоцитов — продуцентов специфических антител —
служат, согласно клонально-селекционной теории иммуни
тета Бернета [1964], защитным механизмом против пато
генных микроорганизмов. В недифференцированных кле
точных популяциях кроветворных органов позвоночных
животных мутационный процесс, меняя биохимические
свойства клеток, не только не ведет к деструктивным по
следствиям, а напротив, выполняет охранительную роль.
В дифференцированных тканях высокоорганизованных
существ регулярно совершается полиплоидизация клеток.
Гигантские клетки слюнных желез насекомых из отряда
двукрылых, содержащие гигантские многонитчатые хромо
сомы, тому пример. В тканях самцов многих видов насеко
мых из семейства кокцид так же регулярно происходит
гаплоидизация клеток. Следует оговориться, что измене
ния в числе наборов хромосом, наступающие в онтогенезе
всех особей в строго определенных или во всех клетках,
можно лишь условно назвать мутационным процессом.
Смена гапло- и диплофазы — универсальное средство, по
зволяющее сочетать постоянство числа наборов хромосом с
генной комбинаторикой. Регулярно наступающая гаплои
дизация, так же, как увеличение числа наборов хромосом,
примыкают как к этим процессам, так и к процессу воз
никновения мутаций. От мутационного процесса онтогене
тическую гаплоидизацию и полиплоидизацию отличает де
терминизм, указывающий на существование пускового ме
ханизма, в корне отличного от мутагенеза.
Но возвратимся к преградам, вставшим на пути про
гресса гаплоидов. Прогресс требовал объединения. Деструк
ция, вызванная соматическими мутациями, встала на пути
объединения гаплоидных клеток. Стоит только представить
себе гигантский сонм клеток, строящих организм высоко
организованного существа, сложнейшую дифференциацию
и согласованность их функций, чтобы понять, какой беспо
рядок внесет мутационный процесс в эту систему. Устране
ние мутационных клеток могло бы исправить положение
249
вещей, но дифференциация тканей, жесткость конструк
ции многоклеточного существа почти полностью подавили
действие отбора. Диплоидность — превосходное устройст
во, позволяющее до известной степени погасить вредные
последствия мутационного процесса и одновременно ис
пользовать мутации, возникающие в половых клетках. Со
четание многоклеточности с диплоидностью — выражение
закономерного течения эволюционного процесса. Наличие
одного свойства коренится в другом. На вопрос — почему
мы диплоиды, следует ответ — потому что мы многокле
точные. Объединение двух наборов хромосом в одной клет
ке создало то постоянство, ту степень устойчивости отдель
ной клетки, которая была необходима для объединения
клеток в гигантские клеточные колонии. Многоклеточность
и диплоидия — свойства сопряженные, сочетающиеся не
случайно, а во имя одной цели — повышения сопротивляе
мости разрушению.
Мы рассмотрели запрет на усложнение организации.
Подведем итог сказанному.
Эволюция или усложнение организации совершается
там, где функция может быть выполнена более сложной
структурой лучше, чем структурой более простой. В тех
случаях, когда максимум эффективности системы не сов
падает с максимумом сложности, на усложнение налагает
ся запрет. Примером свойств живого, налагающих запрет
на дальнейшее усложнение после того, как определенная
высота организации достигнута, являются конвариантная
редупликация и делимость.
Единообразие структурных основ наследственности во
всем органическом мире, как сказано выше, является сви
детельством запрета на усложнение. Единообразие в дан
ном случае — результат конвергенции. Конвергентная эво
люция макромолекулярных комплексов осуществлялась на
самых ранних этапах биопоэза, а также, по-видимому, в
предбиологический период эволюции материи.
13. ПОПУЛЯЦИЯ -
АРЕНА ОТБОРА
Сама возможность действия преобразующего отбора по
коится на структурных предпосылках. Отбор эффективен
только в системе, имеющей определенное строение. Нару
шение определенных организационных принципов системы
кладет запрет на отбор.
Построим логическую модель объекта, в котором воз
можно протекание процессов преобразующего отбора.
250
1. Иерархическая двухступенчатая структура, или на
личие частей, входящих в состав объекта.
2. Пространственная разобщенность частей, отсутствие
запрета на перемещение частей друг по отношению к дру
гу, или рассеяние частей.
3. Конечный срок существования частей.
4. Способность частей к редупликации.
5. Существенный изоморфизм частей, не предполагаю
щий их тождества.
6. Способность частей меняться, переходя из одного ус
тойчивого состояния в другое.
7. Способность воспроизвести измененное состояние в
нисходящем ряду поколений.
8. Избыток воспроизведенных частей.
Выше мы охарактеризовали целый ряд живых сис
тем — макромолекулярный комплекс, осуществляющий
самовоспроизведение, клетку, многоклеточный организм.
На эволюцию одних наложено табу, другие беспрепятст
венно рвутся к высотам бытия, увенчанным мыслью. Само
строение этих систем не гарантирует действия в них отбо
ра. Ни одна система не состоит из взаимозаменимых частей,
срок существования которых меньше срока существования
системы как целого. Ни генонема, ни клетка, ни организм
рассеянными в пространстве системами не являются. Им не
свойствен изоморфизм частей. Отбор оперирует ими как
внешняя сила. В их пределах он невозможен. Они подпадают
под действие селективных сил как целостные образования.
Единственной системой, удовлетворяющей всем пере
численным условиям, является популяция — одновидовое
население замкнутого местообитания. Отбор идет внутри
нее. Популяция — целостная система, части которой суще
ственно изоморфны друг другу, способны к самовоспроиз
ведению, меняются и воспроизводят измененное состояние.
Части рассеяны в пространстве, обладают конечным сро
ком существования, далеко уступающим сроку существова
ния того целого, которое они слагают. Поддержание устой
чивости системы покоится не на размножении системы как
целого, как это свойственно всем сплошным системам, а на
самообновлении за счет замены старых частей новыми.
Популяция — арена деятельности преобразующего от
бора. Организм — его объект. Но сама популяция в силу
изоморфизма ее частей, ограниченности срока их сущест
вования, в силу их разобщенности в пространстве, пред
ставляет собой систему низкоорганизованную. Взаимозаменимость изоморфных друг другу частей налагает вето на
усложнение организации, вето на дифференциацию час
251
тей, на прогресс. Принцип взаимоисключения прогресса и
потенциального бессмертия в полной мере приложим к по
пуляции. Она бессмертна за счет самообновления, регуляр
ной замены износившихся частей новыми. Взаимозаменимость исключает дифференциацию. Запрет на разнообра
зие исключает прогресс. Популяция — не конечное звено
в иерархии конкурирующих за овладение жизненными ре
сурсами и пространством систем. Население замкнутого,
относительно замкнутого, конечно, местообитания — по
пуляция, каждая из них — только часть системы более
высокого ранга — вида. Биологический вид — система как
нельзя более пригодная, чтобы стать ареной преобразую
щего отбора. Его части — популяции — воспроизводят се
бя, изменчивы, способны удержать измененное состояние,
смертны и способны перемещаться в лице своих представи
телей — особей. Внутренняя дифференциация — разнооб
разие особей в пределах популяции — позволяет ей овла
деть пространством, жить в разных средах — в воде, на
суше, в поднебесье, использовать разнообразные источники
питания, выполнять разнообразные геохимические функ
ции... Преимущества дифференциации неисчислимы. Спо
собность меняться, претерпевать конструктивные преобра
зования и способность сохранять status quo — среди них.
На пути использования преимуществ разнообразия особей
стоит барьер. Барьер этот — взаимозаменимость. Она са
ма, конечно, в лице принципиального равенства всех пред
ставителей вида есть превеликое благо, если обратить мыс
ленно взор на самого себя, на человечество. Как ни рассе
лялся этот вид по планете, как ни лез в воду, в атмосферу,
в Космос, наконец, а принципиальное равенство его пред
ставителей — основа взаимопонимания, столп демокра
тии — сохранялось. А о видах растений, животных, микро
организмов и говорить не приходится. Взаимозаменимость
придает виду постоянство, очерчивает границы вида, дела
ет многообразие органического мира прерывистым.
Преимущественное выживание более дифференциро
ванных популяций все же совершалось. Не могло не совер
шаться, поскольку более дифференцированные имели пре
имущество перед менее дифференцированными. Однако
нашлись способы сочетать разнообразие с взаимозаменимостью. Углубимся в этот предмет.
Есть два типа разнообразия. Первый из них создается
мутационным процессом — поставщиком материала для
естественного отбора. По определению это переходное раз
нообразие, в конфликт с преобразующим отбором не всту
пающее. Другой тип разнообразия относится к структуре,
252
к организованности популяции, составляет неотъемлемую
и постоянную ее характеристику. Это балансированный
полиморфизм [Берг, 1957]. Прежде, чем заняться им, об
ратимся к первому типу. Темп мутационной изменчивости
находится под двойным контролем. Он регулируется инди
видуальным отбором внутри популяции и межпопуляцион
ным отбором на арене вида как целостной системы. На
этой арене сохраняются наиболее пластичные, те, кто спо
собен выработать в кратчайший срок иммунитет по отно
шению к новому губителю или сопротивляемость к новым
условиям температуры и влажности.
Устойчивый, или балансированный полиморфизм —
великая уловка природы, Взаимозаменимость сочетается с
разнообразием. И каким разнообразием! Насекомое с пол
ным превращением — шелкопряд. Сперва он существо од
ноклеточное, потом — эмбриончик в яйце, потом — гусе
ница, пожирающая листья шелковицы, невинное дитя, ни
о чем, кроме питания, не помышляющее. Бьет час, гусени
ца берется за работу, вьет кокон и через положенный срок
выходит из него в виде бабочки — существа ничем, кроме
воспроизведения не занимающегося, крылатого транспор
тера своих половых клеток с заправкой, накопленной
тогда, когда невинное дитя ело. Взрослые шелкопряды в
пище не нуждаются. Сделал свое дело — и на покой, на
вечный покой. Сочетание разнообразия с взаимозаменимостью возможно, если все представители популяции
проделывают один и тот же цикл превращений на протя
жении своего индивидуального бытия. Популяции и состо
ят из особей, претерпевающих ряд превращений в течение
онтогенеза.
Другой способ сочетать дифференциацию с взаимозаменимостью — воспроизведение многообразия каждым
представителем множества. Каждая самка вида, имеющего
и самцов и самок, порождает и тех и других. Различия бы
вают подчас фундаментальными, функции родных братьев
и сестер могут быть весьма различными. Муравьи тому
пример. Одни солдаты, другие добытчики, кормильцы,
третьи родители и ничего больше.
Вы оставлены на племя,
Я назначен на убой,
как сказал генерал-боец, поэт Денис Давыдов в стихотворе
нии, обращенном к штабным генералам. А порождаются и
те, и другие одной матерью. У муравьев во всяком случае.
Различия между членами одной семьи могут быть гене
тическими, как различия между маткой и трутнем, а мо
253
гут зависеть от различий в условиях выращивания, как в
случае матки и рабочей пчелы.
Генераторы разнообразия сами становятся объектами
группового отбора. Образование колоний, где часть чле
нов — евнухи, знаменует тупик эволюции. Невыгодность
для социума иметь много евнухов великолепно выразил в
1727 г. Монтескье [Montesquieu, 1964, с. 183] в своих сюр
реалистических «Персидских письмах». Евнухи потеряны
для грядущего. Оставить за каждой особью возможность
воспроизведения и участия в создании разнообразия —
наилучшее разрешение конфликта между взаимозаменимостью и прогрессом. Именно такова генеральная линия
эволюции человека и нет сомнения, что выработка совер
шенного механизма поддержания разнообразия — одно из
важных условий антропогенеза.
Но и на этом пути имеются свои западни и капканы.
По мере образования надорганизменных систем — семей,
колоний, стад — число объектов отбора уменьшается, а
значит падает его эффективность: в надорганизменной сис
теме отбор, если и не сходит на нет, то ограничивается.
Чем дальше прогрессирует разделение функций, чем
больше развиваются взаимосвязи между слагающими сис
тему представителями популяции — самцами и самками,
родителями и детенышами, вожаками и рядовыми членами
стада, между рабочими, сторожами и солдатами муравей
ника,— тем выше интеграция внутрипопуляционных надо
рганизменных систем, тем меньше взаимозаменимость. Те
перь уже табу на усложнение системы, на отбор! Про
изошло самоустранение отбора, как это великолепно выра
зил в свое время В. И. Кремянский [1939, 1940] примени
тельно к антропогенезу. С. А. Северцов [1951 ] обратил
внимание на взаимные приспособления органов разных
особей одного вида друг к другу — сосательные устройства
новорожденных и сосцы матери млекопитающих, аромати
ческие железы самцов оленей и органы обоняния самок.
С. А. Северцов назвал их конгруэнциями.
14. К ЛАССИФ И КАЦИ Я Ж ИВЫ Х СИСТЕМ
Мы познакомились, таким образом, с четырьмя систе
мами, различно построенными, находящимися в различном
отношении к отбору. Одни из них способны претерпевать
прогресс, другие отлично функционируют, сохраняя status
quo. Каждая из систем, описанных выше, отличается своим
закономерно построенным комплексом свойств, неповтори254
Принцип класси
фикации
Сист емы
Наличие или
отсутствие
самовоспроизведения
самовос-
Размножение
Характер
размножения
Деление
Живые
^ения
Редупли
кация
Редупли
кация
Характер
преобразования
Деление
Косные
Самовозобно
вление
•Замена час
тей, им ею си,и ос конеч
ный срок
сущ ест во
вания
Эволю ция
От деление
мат риц
П роисхож де
ние ж и зн е н
ного ц и кл а
7 Организация
пространства
М икроско
п и ч е ск и й
Субмикро- „
скопическии
1. Размер
больш ой
Гигант ский
\
Овладение
Трехмерt
Одномерность ^Хострансл
пространством ность
2. мерность
Распределе
ние частей
Двумерность,
разм ещ ение
н а плоскост и
J.
4. Морризм
Организация
времени
1. Квант
Сплошное^
Сущ естаенный
Е-
i
М / У n / c u i. —
Очень.малень
кий
2. Наличие
или отсут
ствие смерти
нет смерти
Рассеянное
Сущ ест венны й
изом орф изм
Гетероморфизм
/ \
.. г большой
_Л
Маленький
\
нет
~ Г ~
£етб
Есть
к____
Нет
Геологичесное
врем я
♦
Нет
Щ. Прост ранст
венно-врем енны е
хар акт ер и ст и ки
1. Делимость
нет
2. Обмен
веществ
3. Индивидуаль
ное развитие
4. Отбор частей
внутри системы
*
нет
6. Прогресс
нет
^ет
*
нет
\
I
Генонема
J
нЧет
е/
О граниченная
делим ост ь
v
------------------- >
Есть
Есть
♦
нет
4 Ограниченный
играниченнь/и
т\
тштт.
прогресс
нь'й прогресс
I
плетка
1
Многоклеточныр организм
Нет
Есть
П9г>/уи////^>ииьт
прогресс
~
т
Популяция
вид
Классификация живых систем.
Подчеркнуты те свойства, которыми данная система существенно отличается от всех
остальных.
мым их сочетанием. Комплексный характер различий по
набору до известней степени жестко сопряженных призна
ков указывает кД существование типов. Напрашивается
следующая классификация (см. рисунок).
255
Первую и наиболее существенную демаркационную ли
нию между системами разных типов проводит наличие или
отсутствие
самовоспроизведения.
Самовоспроизведение
есть — перед нами живая система* Самовоспроизведения
нет — и мы имеем дело с системой, которую вслед за
В. И. Вернадским [1960] целесообразно назвать косной.
Живые системы по способу самовоспроизведения делят
ся на два подтипа — размножающиеся и самообновляющи
еся. Размножаются генонемы, клетки, организмы. Самооб
новляются популяции, виды, биоценозы, живой компонент
биосферы. Четыре последние системы осуществляют само
воспроизведение сходным образом, три первые размножа
ются по-разному. Генонема строит свою копию и отторгает
ее от себя, клетка делится, организм отделяет от себя ма
ленькую часть, содержащую программу развития.
Перед нами три принципиально разных способа раз
множения — три класса систем. Четвертый класс — систе
мы самообновляющиеся.
Обладание каждым из способов самовоспроизведения
имеет далеко идущие последствия. Организация простран
ства и времени, течение процессов, или иначе говоря про
странственно-временная характеристика, набор запретов и
разрешений принципиально различен у систем в зависимо
сти от того, размножаются они редупликацией, делением,
или отделяют от себя матрицы. Глубочайшим пространст
венно-временным своеобразием обладают, в отличие от
размножающихся систем, системы самообновляющиеся.
Проследим за этими различиями.
15. О РГАНИЗАЦИЯ СИСТЕМ В ПРОСТРАНСТВЕ
Генонема субмикроскопически мала и существенно од
номерна. Она одномерна потому, что размножается с по
мощью построения копии, которую она создает по своему
образцу и подобию. Она творит, как божество, и только
она одна обладает, при всей ничтожности своих размеров и
простоте в сравнении с высшими формами жизни, этим ат
рибутом божественности. Каков способ воспроизведения,
таковы и размеры, такова и мерность. Генонема неделима.
Ее индивидуальность — это индивидуальность молекулы,
но уже ею-то она обладает в полной мере. Всегда, во все
времена с момента возникновения и по сей день генонема
могла бы сказать «я», умей она говорить.
Клетки в огромном большинстве микроскопически ма
лы, они овладели двумерным пространством, процессами,
256
осуществляющимися на плоскости, но они делимы. Инд и
видуальность они утеряли. «Я, я...» начинает говорить
клетка и вдруг останавливается, и вот ей уже надлежит
сказать «мы». Клетка мала и проста в сравнении с орга
низмом потому что она делима, дивидуальна.
Организм — многоклеточное существо. Он гигант —
кит, слон, секвойя, бизон, акула, человек. Он овладел
трехмерным пространством. Он неделим, он индивидуаль
ность, я, я, я... Способность поддерживать свой род с по
мощью отделения матриц обеспечила ему неделимость,
большие размеры, даровала ему трехмерное пространство.
Увы! Мы знаем, чего нам стоили эти дары циклично-мат
ричного способа самовоспроизведения!
Все три класса систем — сплошные, их части лежат
бок о бок. Все они гетероморфны, если не морфологически,
то функционально.
Системы надиндивидуальные.
Популяция. Она имеет гигантские размеры даже по
сравнению со слоном и секвойей. Она насчитывает порой
миллионы миллионов особей. Ее мерность не ограничена
изнутри, как мерность генонемы. Размещение на поверх
ности большого геоида ограничивает ее и делает сущест
венно двумерной. Она делима, если угодно, но делимость
не является способом поддержания ее устойчивости. Меж
ду нею и клеткой есть различие. Межиндивидуальные вза
имодействия и коллективное воздействие на среду, переме
шивания всех родов от скрещивания и до простого переме
щения спаивают ее в единое целое, и деление может при
нести популяции гибель. Механизма деления у популяции
в отличие от клетки нет. Ее фактическая делимость осно
вана на изоморфизме ее частей. Изоморфизм ее частей от
личает ее от всех сплошных, гетероморфных систем. Ее
части подобны друг другу. Они рассеяны в пространстве.
Разобщенность частей делает популяцию живых организ
мов подобной звездно-планетной системе или галактике.
16. О РГАНИЗАЦИЯ ВО ВРЕМЕНИ
Живая система — система устойчиво преобразующая
ся. Каждому классу систем присущ свой тип преобразова
ния. Тип самовоспроизведения это и есть тип преобразова
ния. Для генонемы преобразование и есть редупликация,
для клетки — деление, для организма — цикл его сущест
вования от образования зиготы до смерти. Для популяции
акт преобразования это уже событие эволюционного плана.
9 Р. Л. Берг
257
Что это — завершение элементарного эволюционного яв
ления — завоевание мутантным аллеломорфом всей попу
ляции — или акт видообразования, когда различия между
представителями данного поколения и представителями
следующих поколений достигли ранга видовых? Не знаю.
Что-нибудь в эгом роде.
Как бы то ни было, при переходе от систем, устроен
ных более примитивно, к системам, устроенным более
сложно, от уровня к уровню длительность времени, отде
ляющего один акт преобразования от другого, возрастает.
Каждая система характеризуется своим квантом времени,
и глубокие различия между системами включают в себя
длительность кванта времени.
Время, отделяющее один акт редупликации от другого,
меньше, чем время от деления до деления. Длительность
жизни многоклеточного организма больше, чем промежу
ток между делениями бластомеров, а элементарное эволю
ционное явление, осуществляясь в процессе смены поколе
ний, требует больше времени, чем жизнь отдельного пред
ставителя вида. В филогенетическом ряду развивающихся
форм высшие формы имеют оптимальный, а не максималь
ный срок жизни.
Конец преобразования может быть разным. Смерть —
одна из разновидностей финиша. Смерть означает возник
новение трупа. Конец преобразования генонемы заканчи
вается созиданием новой нити, конец деления завершается
появлением двух клеток вместо одной, популяция преобра
зуется, оставаясь бессмертной. Организм гибнет, умирает в
буквальном смысле этого слова. Его настигает смерть. Ор
ганизм — система терминально циклическая. Все осталь
ные потенциально бессмертны, персистентны.
17.
ТЕЧЕНИЕ ПРОЦЕССОВ
ИЛИ ПРОСТРАНСТВЕННО-ВРЕМЕННАЯ
ОРГАНИЗАЦИЯ СИСТЕМ
Живая система открыта для вещества и энергии.
В этом сходны все четыре класса систем. Все они находят
ся в вещественно-энергетическом взаимоотношении со сре
дой. Они аккумулируют вещество и энергию и используют
их в процессе конструктивных преобразований. Но не все
они обладают обменом веществ. Обмен веществ — замени
мость атомов, поглощение одних веществ и выделение дру
гих — ток вещества и энергии через саму систему присущ
белку. Он отсутствует у генонемы. Нуклеиновые кислоты
258
хранят наследственную информацию, воспроизводят себя,
подают команды для биосинтеза, они мутируют. Все их
действия ничего общего с окислительно-восстановительны
ми реакциями, составляющими суть обменных процессов,
не имеют. Генонема, сама оставаясь стационарной, лишь
использует притекающие извне вещества и энергию. Квазистационарны белки. Нуклеиновые кислоты — инициато
ры обмена — собственным обменом веществ не обладают.
Задолго до того, как Мессельсон и Сталь для кишечной
палочки Esherichia coli и Тейлор для конских бобов Vicia
faba с помощью меченых атомов азота и водорода показа
ли, что исходная матрица остается немеченой, т. е. не уча
ствует в обмене веществ, Н. К. Кольцов [1938], основыва
ясь на данных генетики, с предельной убедительностью до
казал, что обмена веществ у генонемы нет и что сама воз
можность его в принципе исключена. Устойчивость геноне
мы подобна устойчивости атома. Как атом «равнодушен» к
своему пребыванию в молекуле, так «равнодушен» к свое
му окружению ген. Ни наличие разнообра:ршх наборов ал
лелей в других локусах, ни наличие отличающегося от не
го партнера в гомологичной хромосоме в том самом локусе,
где лежит он сам, не меняют химического состава гена.
Безотносительно к тому, осуществил он свое действие в
биосинтезе или его команды оказались заглушенными сиг
налами, которые подают другие гены, он передается в не
изменном виде из поколения в поколение и вероятность
его передачи ни в какой связи с его участием в биосинтезе
не стоит. Как атом кислорода не хранит воспоминания о
своем пребывании в составе воды, или углекислоты, или
оксида кремния, так не хранит воспоминания о своем про
шлом ген. Его устойчивость — свидетельство молекулярно
го уровня его организации. В театральной феерии обмена,
которая невозможна без его участия, он режиссер. Он спо
собствует перевоплощению других, сам не перевоплощаясь.
Будучи молекулой, или частью большой молекулы, он не
делим продольно. Удьоение генонемы может осуществлять
ся только одним единственным способом — построением
копии и отторжением ее от себя. Устойчивость, молекуляр
ный уровень строения, отсутствие обмена, неделимость,
матричный принцип самовоспроизведения — свойства,
друг другу комплементарные. Таков был ход мысли
Н. К. Кольцова. Обменом веществ хранилище информации
не обладает. Самая примитивная система, которая способ
на активно добывать энергию из среды — клетка. Начиная
с нее, живые системы всех уровней организации обладают
обменом веществ. Организм включает в себя не только за
259
нятые метаболизмом клетки. Целые грандиозные вспомога
тельные конструкции, обеспечивающие надежность подачи
вещества и энергии и рациональное выделение продуктов
обмена: листья, устьица, сосуды растений, пищеваритель
ная система, трахеи и бронхи, сердце и сосуды, почечные
канальцы животных обслуживают метаболизм. Популяция
также обладает обменом веществ. Она черпает ресурсы из
своего окружения, выделяет в него продукты обмена. По
коление за поколением ложится костьми, возвращая взя
тое. Популяция осуществляет обмен со средой, выступая в
основном как сумма особей. Биоценоз и живой компонент
биосферы занимаются прогонкой вещества и энергии по
тому же принципу. Единственной в своем роде является
генонема. Только она одна не подвержена регулярной сме
не состава.
Посмотрим теперь, как обстоит дело с онтогенезом, с
цикличностью превращений в пределах одного полного
цикла преобразования. Под цикличностью следует пони
мать наличие дискретных фаз, регулярно сменяющих друг
друга в жестко заданных пределах. С известным основани
ем можно говорить о цикле редупликации, о цикле клеточ
ного деления. Истинной цикличностью, включающей диск
ретные фазы фундаментальной перестройки организации,
образование неделимого и смерть, обладает многоклеточ
ный организм и только он один. Изменение размеров орга
низма в течение жизни от зиготы до взрослого состояния
беспрецедентно. Организм человека увеличивается в тече
ние жизни в 250 раз. Из одноклеточного он превращается в
представителя другого типа — в существо многоклеточное.
Клетка подготавливается к делению, выходит из фазы де
ления, в промежутке пребывает в метаболической фазе.
Эти превращения не ведут по смыслу вещей к созданию
неделимого. Популяция, проделав цикл превращений, да
же если мы примем за элементарный процесс преобразова
ния возникновение нового вида, отличается от исходного
состояния, как отличаются виды одного рода друг от друга.
Организм же человека превращается из представителя од
ного крайнего типа в представителя другого крайнего типа
животного царства. Циклом развития с его неизбежными
бедствиями, о которых мы имели случай говорить, облада
ет только многоклеточный организм.
С известным правом можно говорить об индивидуаль
ном развитии биоценоза, понимая под ним сукцессию или
закономерную смену форм в пределах сообщества. Биоце
нозы вступают в зрелую фазу. Естественной смерти сооб
щество не знает.
260
Достаточно сказано о запрете на отбор, который нало
жен жестким строением всех живых систем за исключени
ем систем надиндивидуальных.
Последняя характеристика пространственно-временной
организации систем, о которой уже шла речь,— это про
гресс, усложнение организации. Многоклеточный орга
низм — единственная система, перед которой открыт неог
раниченный путь к усложнению организации.
Мы говорили об отдельных свойствах систем и об их
распределении среди систем различного рода. Теперь по
смотрим, как сочетаются эти свойства в пределах каждого
класса систем.
Вся совокупность свойств системы каждого класса жест
ко задана характером ее самовоспроизведения. Причина
проявления каждого свойства лежит в другом. Спрашивая,
почему наш объект обладает тем или иным свойством, мы,
отвечая, указываем на другое свойство, ибо в нем коренит
ся причина первого. Почему ген маленький, почему он не
обладает собственным обменом веществ, почему он неде
лим, бессмертен, почему закрыт перед ним путь усложне
ния? Ответ гласит — все эти свойства комплементарны ре
дупликации.
Почему мала клетка, почему так ограничены ее воз
можности прогрессировать, почему она бессмертна? От
вет — делимость.
Организм — единственная система, сочетающая слож
ность организации с неизбежностью трагического конца.
Только в лице человека природа достигла самопознания, и
только на путях возникновения самопознающих систем
возникла смерть. Циклически — матричный принцип ре
дупликации — причина прогресса, причина гибели.
Самообновление, обеспечивая бессмертие популяции,
обрекает ее на изоморфизм частей, на малую их взаи
мосвязанность, на низкий уровень ее собственной орга
низации.
Популяция человека, населяющая Землю, далеко не
достигла как система того совершенства, которое являют
лучшие ее представители. Она даже не достигла уровня
многоклеточного организма, она устроена менее совершен
но, чем амеба. Она воюет. Сборище смертных убийц!
Создать мыслящее существо, сочетающее высокую ор
ганизацию с бессмертием,— задача человека. Решена она
будет с помощью эмбрионализации соматических клеток.
Место мумии займет живое существо, обреченное на муки
онтогенеза, лишенное родительской опеки, подверженное
всем превратностям бытия.
261
18. УРОВНИ ОРГАНИЗАЦИИ
Что такое уровень? Когда мы говорим уровень, мы под
разумеваем некое множество равных. Уровень воды в ста
кане, в озере — это наличие плоскости, в которой распо
лагаются лежащие выше всех молекулы воды. Их положе
ние по отношению к усредненной поверхности дна равно.
Падающая вода не имеет уровня.
Когда мы говорим о молекулярном, клеточном, ткане
вом уровне организации живого объекта, мы имеем в виду
наличие множества сходных элементов. Каждый элемент
всегда часть системы более высокого порядка. Высший уро
вень включает в себя объект низшего уровня, как свою
часть. В этом смысле он высший. Сложность системы вы
сшего уровня может быть меньше сложности системы низ
шего уровня. Популяция — пример системы, которая как
таковая устроена менее сложно, менее совершенно, чем со
ставляющие ее особи. Ряд живых систем, выделенных по
этому принципу, начинается с клетки. Живой компонент
биосферы состоит из биоценозов, биоценоз — из популя
ций, популяция — из особей, особь — из клеток.
Макромолекулярный комплекс — носитель элементар
ного свойства жизни, самовоспроизведения,— стоит особ
няком. Клетка не состоит из хромосом в том смысле, в ка
ком организм состоит из клеток или популяция — из орга
низмов.
Кажется тем не менее целосообразным выделить в ка
честве главных шесть уровней организации — генонему,
клетку, организм, популяцию, биоценоз и биосферу. Это
участники драмы, разыгрывающейся с соблюдением един
ства времени и места. Выделены эти уровни организации
потому, что им всем без исключения присуща фундамен
тальная черта живого — способность воспроизвести себя
как целое.
Есть в живом й другие целостные образования, тоже
образующие уровни, тоже целостные и пространственно
ограниченные, обладающие всеми признаками систем.
Ткань, орган, система органов — примеры таких комплек
сов, выделенных по функциональному признаку в преде
лах организма. Семья, стая — у животных, куртина — у
растений — в пределах популяции. Ярус, цепь питания —
в пределах биоценоза. Способностью воспроизвести себя
как целое эти комплексы не обладают. Они образуют авто
номные образования в пределах главных систем.
262
19. ВИД КАК ЭВОЛЮЦИОНИРУЮЩАЯ СИСТЕМА
Представление о виде, как об эволюционирующей сис
теме, развито мною в 1944 г. [Берг, 1944, 1957]. Тогда
уже на основе генетических и экологических данных, под
впечатлением исследований, вскрывающих целостность
живых систем надорганизменного ранга и в результате соб
ственных наблюдений над изменчивостью географически
разобщенных
популяций
плодовой мухи
Drosophila
melanogaster у меня сложилось представление, что вид это
надиндивидуальная система особого типа, а именно систе
ма эволюционирующая. Способность вида претерпевать
эволюционные преобразования, специфические для каждо
го вида, поддерживается межгрупповым отбором — заме
щением одних популяций другими, эволюционирующими
более успешно. Отсюда вытекает, что на службе эволюции
самой эволюции находится целая иерархия отборов. Под
их совместным действием устанавливается закономерное
соотношение внутренних факторов эволюции вида, предоп
ределяющее в значительной мере способы и темпы его
дальнейшей эволюции. Видообразование — не только обо
собление форм, но обособление глубоко своеобразных пу
тей эволюции. В каком отношении стоит вид, как система,
к нашей генеральной последовательности живых систем?
Каждая из систем нашего главного ряда обладает свое
образием, ее свойства образуют неповторимое сочетание —
комплекс, закономерно построенный и строго согласую
щийся с характером самовоспроизведения системы.
Совсем особое место в нашей системе систем принадле
жит популяции. Это элементарная единица эволюционных
преобразований, арена отбора, единственная система, сла
гающаяся из существенно изоморфных частей. Она входит
как часть в систему высшего порядка — биоценоз. Но в
отличие от всех других систем популяция является состав
ной частью не одной системы высшего по отношению к ней
порядка, а двух. Популяция — одновидовое население
замкнутого местообитания — входит в состав вида, как си
стемы. Она составляет его целостную часть. Она как целое
является объектом отбора. Но это уже не тот отбор, кото
рый оперирует внутри нее. По отношению к популяции в
целом этот отбор действует как внешняя сила, точно так
же как отбор, происходящий внутри популяции, выступает
как внешняя сила по отношению к особям. Это отбор груп
повой.
Пространственно разобщенные, относительно изолиро
ванные друг от друга популяции, входящие в состав раз263
ных биоценозов, конкурируют друг с другом по способам
существования, по экономичности использования жизнен
ных ресурсов, по способности устоять в критические мо
менты жизни биоценоза.
Но не только жизнь множества меняется в результате
межгруппового соревнования. Меняется сама эволюция.
Групповому отбору подлежат групповые свойства, та
кие как полиморфизм или разнообразие форм внутри по
пуляции, не только сам факт существования различий
между представителями популяций, но и численное соот
ношение форм. Сама частота возникновения наследствен
ных изменений регулируется групповым отбором.
Представим себе две популяции дрозофил, заселяющих
два винных завода. Заводы находятся на таком расстоянии
друг от друга, что обмен особей между ними затруднен, но
не исключен. Численность обеих популяций сдерживается
не только недостатком корма для личинок зимой и весной.
Мухи гибнут от инфекционных болезней и от инсектици
дов. Время от времени вспыхивают эпидемии, или вводит
ся новый инсектицид. Численность мух резко снижается.
Выживают отдельные представители вида, носители редких
мутаций, невосприимчивые к возбудителю болезни или к
действию инсектицида. Их тем больше, чем больше часто
та возникновения мутаций. Теперь представим себе, что
популяции отличаются по частоте возникновения мутаций.
Популяция, у которой мутации возникают чаще, имеет
больше шансов сохраниться, чем та, у которой мутантов
слишком мало, чтобы вся группа в целом могла противо
стоять катастрофическому изменению среды. Менее пла
стичная популяция погибнет, более мутабильная сохранит
ся. А затем опустевший очаг снова заселится резистентны
ми пришельцами извне.
Акт группового отбора совершился. Победителем в
межпопуляционном соревновании оказалась популяция с
более высоким уровнем мутабильности. Ее устойчивость
покоилась на относительно большей неустойчивости ее ге
нов. Мутационный процесс — генератор новшеств — ока
зался элементом защитного механизма, охранившего мно
жество от уничтожения. Устойчивость одного уровня орга
низации гарантирована в данном случае, как впрочем и во
множестве других, неустойчивостью другого [Берг и др.,
1941]. Шмальгаузен [1940, с. 365], размышляя над защи
щенностью организма не только по отношению к вредонос
ному действию среды, но и по отношению к гибельному
эффекту мутаций, высказал мысль, что аппарат наследст
венности — генотип популяции — менее устойчив, чем
264
находящийся под его контролем фенотип популяции,
Шмальгаузен не подозревал, что в генотипе популяции
имеется устройство, меняющее в зависимости от измене
ния требований, предъявляемых популяции средой, часто
ту и характер возникающих мутаций.
Более чем полувековые исследования мутабильности
свободно-живущих дрозофил Европы, Азии и Северной
Америки вскрыли устройство, регулирующее через отбор
наследственную изменчивость. Изучение флюктуаций му
табильности в популяциях человека показало, что анало
гичные механизмы частоты возникновения мутаций опери
руют и у человека [Berg, 1982, 1985, 1986, 1987]. Сорев
нуясь друг с другом по приспособляемости, популяции всех
видов животных, растений и самого человека оказались об
ладателями устройства, предназначенного предвосхищать
возможные катастрофические изменения среды и противо
поставлять им вспышки наследственной изменчивости, по
ставляя мутантов, способных выжить среди повальной ги
бели немутантных собратий по популяции.
Это устройство — набор генов мутаторов, генов, вызы
вающих мутации других генов. Ген мутатор действует не
на все гены. Он заставляет мутировать один из них, не за
трагивая строение других. Он действует слепо, вне зависи
мости от пертурбаций в среде обитания своего носителя.
Множество вызываемых им мутаций вредны при всех усло
виях. Некоторые мутации могут оказаться полезными
только при наступлении катастрофы.
Частота генов мутаторов в генотипе популяции регули
руется индивидуальным естественным отбором в зависимо
сти от пользы или вреда продуцируемых каждым из них
мутаций. Вероятность иметь в своем генотипе помимо му
тации, вызванной мутатором, еще и сам ген мутатор, вы
ше для мутанта, чем для тех, чьи гены не претерпели из
менения. Прямой отрицательный отбор мутантов, возник
ших под действием одного из мутаторов, это косвенный от
рицательный отбор этого мутатора. Число аллелей генов
мутаторов в. генотипе популяции сдерживается отбором на
низком уровне. Но вот наступает катастрофа. Введен но
вый инсектицид.
Если есть в генотипе популяции мутатор, вызывающий
мутации резистентности по отношению к нему, отбор на
правит мутационный процесс в приспособительное русло.
Прямой положительный отбор мутантов это косвенный по
ложительный отбор мутатора. Обладая набором мутаторов,
популяция имеет все шансы выжить и восстановить свою
численность. Генные частоты генов мутаторов и частота
265
вызываемых ими мутаций возрастают не беспредельно.
Когда популяция состоит из одних резистентных особей,
отбор снижает мутабильность до прежнего уровня. Он
вступает в действие, потому что мутаторы продолжают
свою, теперь уже преимущественно губительную, деятель
ность. Гибнущие мутанты увлекают за собой вызвавшие их
мутаторы. Аллельные частоты мутаторов снижаются,
вспышка мутабильности гаснет.
Удалось показать, что в одних популяциях снижение
частоты возникновения мутаций идет за счет снижения ал
лельных частот генов мутаторов, а в других — за счет по
ложительного отбора генов, подавляющих действие генов
мутаторов.
Не подлежит сомнению, что система генов мутаторов и
их супрессоров, регулирующая частоту и характер возни
кающих мутаций, вырабатывается и совершенствуется под
действием группового отбора. Аллельная частота каждого
из мутаторов находится под контролем индивидуального
отбора. Но быть или не быть каждому из них порознь и
той или иной численности всей их совокупности решает
групповой отбор — соревнование между не полностью раз
общенными популяциями и видами одного рода, изолиро
ванными друг от друга барьерами для скрещивания в пре
делах биоценоза.
Групповой отбор — регулятор популяционных свойств.
Не только изменчивость, но и сам индивидуальный отбор
подпадает под его действие.
Индивидуальный отбор становится менее жестким по
мере того, как развивается забота о потомстве, создаются
средства коллективной защиты, появляется совместное до
бывание пищи, совершенствуются межиндивидуальные
связи внутри популяции. Но наиболее парадоксальны, наи
б о л ее интересны явления, когда в эволюции создаются спе
циальные устройства, повышающие интенсивность индиви
дуального отбора, если не в отношении всех предствителей
популяции, то в отношении части их. Мы будем иметь
случай познакомиться с приспособлениями, выработанны
ми со специальной целью повысить интенсивность отбора
самцов, раз уж они оказываются в силу бурного темпера
мента в избытке. Здесь же обратимся к механизму отсева
вредных мутаций с помощью возвращения от диплоидного
состояния к гаплоидному.
Гигантские объединения клеток, образующих много
клеточный организм, могли образоваться только после до
стижения диплоидного состояния. Диплоидия не устранила
мутации, она погасила до известной степени их действие.
266
Мутации получили возможность накапливаться. Половой
процесс обеспечил возможность встречи вредоносных генов
и наряду со счастливыми обладателями удачных сочетаний
генов стали во множестве появляться особи ослабленные,
неполноценные, хилые. Отбор выметал их, и вид процве
тал за счет размножения своих лучших представителей.
Но так обстоит дело, пока одни представители вида бес
препятственно заменяются другими. Процветание в этом
случае — функция изоморфизма. Но популяция эволюци
онирует в сторону усложнения внутренних связей. Одной
заботы о потомстве достаточно, чтобы слабые получили
шанс выжить. Непроизводительная затрата сил со стороны
родителей, выхаживающих неполноценное потомство, ло
жится тяжким бременем на вид. Кто обречен, должен гиб
нуть ради всеобщего блага как можно раньше. Еще лучше,
если он не родится вовсе. И там, где выхаживание каждого
существа требует затраты больших сил со стороны других
членов вида, вырабатываются специальные сита, барьеры,
не дающие слабым прорваться на жизненную арену. Дип
лоидные организмы возвращаются к гаплоидии или выра
батывают различные ее субституции.
Пчелы. Выращивание потомства сопряжено с гигант
скими трудностями. Развивающееся дитя кормят, защища
ют, обеспечивают ему гигиенические условия, кондицио
нируют среду. Целый штат нянь — бесплодных сестер —
на службе каждой личинки. Но дитя должно соответство
вать принятым стандартам. И вредным мутациям не дано
накапливаться под прикрытием нормальных аллеломорфов.
Самцы развиваются из неоплодотворенных яиц, они начи
нают свое развитие как гаплоиды. Одиночный набор их
хромосом удваивается в первой же клетке, дающей начало
самцу. Первая редупликация хромосом не сопровождается
делением клетки. Образуется диплоидный организм, но он
полностью гомозиготен и любая мутация, даже если она у
самки с ее диплоидным набором хромосом была скрыта,
перейдет в явное состояние у самца. Он погибнет. Участво
вать в распространении вредоносного гена в нисходящих
рядах потомков он не будет.
Наездники. Забота о потомстве выражается в обеспече
нии личинки обильной пищей. Самка наездника отклады
вает яйцо в гусеницу бабочки или в личинку мухи. Если
гусеница может выкормить только одного представителя
паразитического вида, самка кладет одно яйцо, если пищи
хватит для многих, она положит много яиц. Она не рас
суждает, конечно. Виды наездников и их хозяев связаны
друг с другом неразрывной связью и самка не имеет свобо
267
ды выбора, пища ее детей ограничена одним видом, и она
откладывает яйца, не считая, откладывает столько, сколь
ко следует, чтобы все вылупившиеся личинки выкормились
[Солт, 1964]. После того как яйца отложены, самка метит
зараженную гусеницу запахом, чтобы другой самке или ей
самой не повадно было снова отложить яйца. Личинкам
предоставлены наилучшие из возможных условия выкарм
ливания. Угроза накопления вредоносных мутаций налицо.
Самцы наездников гаплоидны. Предоставлять заботу
всяким уродам виду нерентабельно. Кто предоставлял —
погиб.
Многие виды клещей, щитовки тоже имеют гаплоидных
самцов. Можно не сомневаться, что технология использо
вания жизненных ресурсов видом требует и у них своевре
менного отсева вредоносных мутантов.
И мы, мы, окружающие наших детей порой растлеваю
щей заботой, выращивающие миллионы умственно отста
лых, отягощенных разнообразными наследственными не
дугами потомков, мы наделены ситом, без которого число
мутантов было бы еще больше. Мы начинаем наше раз
витие, будучи гаплоидными. Спермий привносит в
яйцеклетку набор хромосом, способный включиться в дело
лишь через некоторое время после того, как набор хро
мосом яйцеклетки вступил в действие. Клеточным ле
тальным генам предоставлена возможность проявить свое
действие.
Отбор, способный вырабатывать механизмы, повышаю
щие вероятность гибели, это групповой отбор.
Вид, как эволюционирующая система, расчленен на по
пуляции, конкурирующие друг с другоми по способам,
темпам, направлениям эволюции. Его цельность поддержи
вается групповым отбором. Сам индивидуальный отбор, са
мо течение эволюции предстают перед нами как регулиру
емый процесс. В качестве регулятора выступает взаимозаменимость популяций.
Эволюция не только закономерный процесс, она про
цесс регулируемый.
Самая жестокая схватка, борьба не на жизнь, а на
смерть, конкуренция, не имеющая иного исхода, кроме ги
бели одного из конкурирующих партнеров, совершается
между видами одного рода. Внутривидовая, межгрупповая
борьба нечто вроде маневров, репетиции этой гигантской
битвы.
Никаких других отношений, кроме антагонистических,
между видами одного рода не существует. Их представите
ли не только не вступают друг с другом во взаимоотноше
268
ния симбиоза, им чужды взаимоотношения паразитизма и
хищничества. Симбионтами, паразитами и хозяевами,
хищниками и жертвами становятся виды, относящиеся к
систематическим категориям выше рода, чаше всего пред
ставители разных типов.
Паразитизм и хищничество — примеры взаимоотноше
ний, антагонистических только на уровне отдельных пред
ставителей вида, но отнюдь не на видовом уровне. Вид
хищника и вид жертвы находятся в трогательном альянсе,
жить друг без друга не могут, их эволюции сопряжены и
носят прогрессивный характер.
Лиса бежит за зайцем. Заяц пойман и конец его ясен.
Если добыча ускользнула, заяц поставил лису и ее потом
ков на грань голодной смерти. На войне, как на войне. Но
виду жертвы хищник нужен ничуть не меньше, чем жерт
ва хищнику. Жертвы конкурируют друг с другом за обла
дание высокоорганизованным фактором отбора, способным
осуществить избирательную браковку. Роль вида хищника
в жизни жертвы двоякая. Хищник выполняет санитарную
роль, препятствуя распространению эпизоотий, уничтожая
заболевших. Он губит немногих во имя большинства.
Хищник — могучий фактор прогрессивной эволюции вида
жертвы. Способствуя избирательной смертности, он ведет
жертву по пути прогресса. В результате его деятельности
уничтожаются менее совершенные, сохраняются и получа
ют преимущество в борьбе за жизнь самые зоркие, чуткие,
быстрые, выносливые, склонные к социальным формам
жизни, способные выдвигать из своей среды лидеров и под
чиняться им, имеющие средства общения — сигналы тре
воги, понятные им одним, и приемы коллективной оборо
ны. И точно ту же роль фактора прогрессивной эволюции
играет вид жертвы в отношении вида хищника. Отноше
ния, антагонистические на уровне индивидуальном, на
уровне ансамблей становятся симбиотическими. До поры
до времени. Вид жертвы выходит из игры победителем.
Олени, зубры, бизоны стали неуязвимыми. Хищники от
ступили. Почитайте Моуэта «Не кричи: волки!» [1967].
Большое удовольствие получите.
Виды одного рода не вступают ни на один из путей, ве
дущий к симбиозу. Борьба идет не на жизнь, а на смерть.
Миллионы видов погибли в этой борьбе. Ее результат —
резкие различия между видами. Если каждый вид остался
единственным в своем роде, различия между видами дан
ного семейства достигли ранга родовых. Повышение уровня
организации биоценоза совершается именно в процессе
межвидовой конкуренции. Она результат элиминации всех
269
видов каждого рода, кроме одного. Формирование биоцено
за, его онтогенез сопровождается увеличением отношения
числа родов к числу видов. Это отношение — родовой ко
эффициент — в пределе равно единице. Виды расходятся
по разным биоценозам, но множество видов сгинуло с лица
Земли.
В. Л. Комаров сделал остроумный подсчет. Он подсчи
тал, какое количество родов в семействе сложноцветных
насчитывает один вид, какое количество — два, три и т. д.
Следовало бы ожидать, что распределение числа родов
окажется близким к нормальному: наибольшее число родов
будет состоять из некоторого среднего числа видов; родов с
очень маленьким числом видов и с очень большим будет
тем меньше, чем дальше от средней отстоит число видов.
Ничуть не бывало. Максимальное число родов насчитывает
в своем составе один вид. От максимальной величины идет
крутой спад. В. Л. Комаров [1940] говорит, что семейство
сложноцветных не представляет в этом отношении исклю
чения. Зоологи сделали аналогичные подсчеты с тем же ре
зультатом.
Максимальное количество
родов содержит один
вид. Какой вид имеет шансы сохраниться? Тот, эволюция
которого совершается в направлении повышения устой
чивости всей системы в целом, повышения надежности
добывания вещества и энергии из среды. Наибольшие
шансы выжить имеет вид, сочетающий приспособленность
с приспособляемостью, устойчивый и пластичный одно
временно.
Вид — эволюционирующая система не только в том
смысле, что его строение разрешает отбор. Его устройст
во — наличие не только взаимозаменимых особей, но и
взаимозаменимых групп — позволяет менять само течение
отбора. Случайный поиск осуществляется не только в от
ношении строения организмов, но и в отношении путей
эволюции, и этот случайный поиск создает ситуацию, от
случайного поиска весьма отличающуюся.
Э.
Майр [1968, с. 308] задается вопросом: «почему
существуют виды, какова их роль в системе явлений?»
Ответ на вопрос гласит: «Организация всего разнообразия
органического мира в форме видов создает систему, ко
торая допускает возникновение генетического разнообра
зия и накопления благоприятных генов и генных ком
бинаций без всякой угрозы разрушения основного генного
комплекса».
270
20. ИЕРАРХИЯ ОТБОРОВ
Иерархии живых систем соответствует иерархия отбо
ров. Каждый высший компонент иерархии служит регуля
тором по отношению к низшему. Везде, где есть комплек
сы рассеянных в пространстве изоморфных друг другу,
способных к самовоспроизведению недолговечных частей,
осуществляет отбор.
Внутри клетки возможен отбор пластид, митохондрий.
В организме, в кроветворных органах, в гонадах не исклю
чен отбор клеток.
Точно так же, как внутриклеточный отбор подчинен
отбору клеток, а в конце концов отбору организмов, сопод
чинены индивидуальная и межгрупповая сортировка внут
ри вида. Межгрупповой отбор находится на поводу у меж
видового отбора. То, что вполне может разрешить индиви
дуальный отбор, подчас наталкивается на непреодолимое
препятствие со стороны отбора группового.
Самка наездника отыскала подходящую гусеницу, от
ложила в нее яйцо, пометила ее соответствующим запахом
и отогнала тем самым от забронированной добычи других
претендентов. Она имеет все шансы принять участие в воспроиззедении следующего поколения, влить свои гены в
генный фонд идущих на смену наездников. Но если вид
наездника в результате этих успешных действий самок до
стигнет численности вида, поставляющего ему своих гусе
ниц, кормовая база личинок наездника будет подорвана.
Хозяин не сможет воспроизвести себя. Он погибнет, а вме
сте с ним погибнет его процветавший дотоле паразит.
Групповой отбор вступил в свои права.
В биоценозе беспрестанно идет приспособление техно
логий жизни разных видов друг к другу [Беклемишев,
1970]. Пары видов — хищник и жертва, паразит и хозяин,
цветковое растение и насекомое — переносчик его пыль
цы — вступают на арену жизненной борьбы с парами ви
дов, занимающими сходное с ними положение в биоценозе.
Это уже надвидовой отбор — один из путей эволюции био
ценоза как целого [Lotka, 1925].
Другой пример — насекомые-браконьеры. Цветки рас
тений, имеющих узкий в таксономическом отношении круг
переносчиков пыльцы, не бросают свою пыльцу на ветер.
Они даже не осыпают ею насекомое с ног до головы. Они
экономно локализуют ее на маленьком местечке на теле
насекомого, там, где насекомому труднее всего ее счи
стить,— на мембране, поддерживающей хоботок, на сте
бельке, соединяющем грудь и брюшко, или на верхней сто271
роне головы [Шванвич, 1956]. Но цветок устроен так, что
бы насекомому не только снять с себя уже прилипшую
пыльцу, но и добыть эту драгоценную малость было бы
чрезвычайно трудно. Скаредности растений насекомые
противопоставляют «бандитизм». Насекомые-браконьеры
отказываются от транспортировки, вспарывают челюстями
щели в венчиках цветков и грабят нектар. Индивидуаль
ный отбор поощряет их действия. Ничего преступного в
экономии трудовых затрат, в остроумном использовании
ротовых частей — инструментов, предназначенных для
других операций,— он не находит. Но вид переносчика,
отказавшись доставлять по месту назначения пыльцу свое
го кормильца, лишает его возможности воспроизвести себя.
Гибнут оба. Групповой отбор в его надвидовой форме вы
нес свой приговор. Растениям не надлежит быть уж черес
чур скаредными в использовании пыльцы, им лучше иметь
не один, а несколько способов опыления. Насекомым не
следует скупиться на трудовые затраты, добывая пыльцу.
Чрезмерная эксплуатация и массовое браконьерство запре
щены. И тем и другим нужно иметь запас пластичности,
разнообразие и изменчивость, его создающую. Отборы —
межгрупповой, внутривидной, межвидовой и, наконец,
надвидовой — обеспечивают меру и в этом трудном деле.
Серая мухоловка с поражающим упорством выкармли
вает кукушонка. Что он ей? Он выбросил ее собственных
детей из гнезда, он требует все больше и больше внимания
к своей особе. Мухоловка идет на все. Как может сохра
ниться такой инстинкт? Как может он возникнуть? Ведь
другие мухоловки не дали провести себя, выкинули яйцо
кукушки из гнезда и делу конец. Они — умные — приня
ли посильное участие в воспроизведении следующего поко
ления, а она осталась бездетной. Мимикрия яиц кукушки,
по размерам, форме и цвету похожих на яйца приемных
родителей, неоспоримо доказывает, что птицы различают
подлог [Southern, 1954]. Иначе, как могла бы выработать
ся мимикрия? Индивидуальный отбор на стороне умных:
принимая образ кукушонка, он устраняет глупых. За одно
поколение все без исключения дураки должны вымереть и
в следующем поколении ни одно яйцо кукушки не будет
насижено. Так было бы, буть дети умных умными. Умст
венные способности наследуются, инстинкт охраны гнезда
у птиц — тем более. Но гнездовой паразитизм кукушки
сохраняется в течение тысячелетий. Умные рождают глу
пых. Наследуемость ума мала. Но ведь и сама жестокость
наследственной программы могла бы измениться под дейст
вием отбора.
272
Умные, порождающие глупцов, неспособных отличить
свое яйцо от яйца кукушки, имеют меньше шансов влить
свои гены в генофонд следующего поколения, чем умные,
имеющие только умных детей. Отбор должен неукосни
тельно действовать в пользу самых умных и притом умных
генотипически. Почему же глупцы не перевелись? Очевид
но, они нужны виду серой мухоловки, не отдельным ее
особям, а виду в целом. Выкармливая птенца кукушки,
мухоловка исключает себя из процесса самовоспроизведе
ния. Казалось бы, она действует во вред не только себе, но
и своим собратьям по виду. Ее выкормыш со временем
подбросит яйца в гнезда ее племянников, сократит числен
ность ее собственной популяции. Но так обстоит дело
только на первый взгляд. Глупцы действительно нужны.
Они просто необходимы. Таланты рождают тупиц именно
потому, что виду серой мухоловки нужна кукушка. Ку
кушка уничтожает тех крупных насекомых, вредителей
леса, которых не может истреблять мухоловка. Не будь
кукушки, не было бы леса, не было бы мухоловок.
Хорошо, говорите Вы. Так вот, пусть кукушка и вы
кармливает сама своих детей крупными гусеницами, кото
рых она ест. Охрана леса станет еще надежней.
Как бы не так. Птенцам нужен мелкий корм, много
мелкого корма. И тут кукушка — пас. Проворная птичка,
шныряющая в поисках корма, собирая всякую мелочь, спе
циализируется на добыче этого корма, ест его и детей кор
мит. Это уже не кукушка. Сдерживать численность круп
ных насекомых некому. Серая мухоловка, вынянчивая
птенца кукушки, выполняет не менее великую биологиче
скую миссию, не меньше заботится о сохранении своего
вида в нисходящем ряду поколений, чем мухоловка, ус
пешно выращивающая своих собственных птенцов. Она
способствует поддержанию равновесия в биоценозе. Мис
сия глупцов экологическая.
Виды умников, рождавшие одних умников, сгинули с
лица Земли со своим биоценозом. Надвидовой отбор, отбор
множеств сосуществующих в биоценозе видов — деревьев
и насекомых, кукушек и мухоловок, сочетаясь с индивиду
альным отбором, выработал гнездовой паразитизм и неже
сткий генотипический контроль умственных способностей
обоих видов — участников драмы. Индивидуальный и
групповой отбор, действуя в прямо противоположных на
правлениях, создали оптимальные взаимоотношения между
популяцией мухоловок и популяцией кукушек. Саусерн
[Southern, 1954] показал, что кукушка запоминает биотоп,
где она выросла, находит гнездо птицы не только того ви
273
да, который вырастил ее, но той же самой популяции и
подбрасывает в него £вое яйцо.
Индивидуальный отбор поощряет тех из кукушек, кто
способен находить наиболее глупых мухоловок. Среди му
холовок оставить потомство имеют шанс только умные. За
вершается процесс гибелью кукушек. Они истребили глуп
цов, а умные повыбрасывали их яйца из своих гнезд.
Раз кукушки существуют, значит индивидуальный от
бор натолкнулся на преграду в лице группового отбора.
Групповой отбор поощрял глупцов среди кукушек, тех, кто
забывал место, где его выкормили, когда он был птенцом,
и глупцов среди мухоловок, не способных отличить свои
яйца от чужих. Разнообразие поведения и кукушек и му
холовок приобрело черты балансированного полиморфизма.
Существование вцдов кукушки и мухоловки — условие
стабильности биоценоза.
Но если предметом регуляции служат групповые свой
ства, включая пластичность, если сама мутабильность ме
няется в соответствии с требованиями устойчивости, разве
не мог отбор в его высшей форме наложить печать и на
характер возникающих изменений, приспособить саму из
менчивость к наиболее вероятному ходу событий?
Определенные каналы могли открыться для изменчиво
сти, иные пути могли оказаться закрытыми.
Если условия периодически меняются, если изменения
среды в данное время похожи на изменения в прошлом,
эта! повторность сходного создаст благоприятные условия
для самонастройки. Не только жизнь, не только изменчи
вость, сама эволюция становится благодаря существованию
видов самонастраивающимся процессом. Успешно эволю
ционировать — это своего рода искусство. Нужно иметь
врожденные способности, чтобы заниматься им. Но кое-че
му, технике во всяком случае, можно и подучиться. Мы
говорим обучение. Может быть, лучше сказать игра, где
проигрыш — уничтожение.
Виды — это детские сады, школы, университеты, где
популяции учатся эволюционировать. Педагогика этих
учебных заведений должна составить новую отрасль биоло
гии.
По аналогии с другими ветвями науки о жизни Джули
ан Гекели [Huxley, 1942] предлагает назвать ее сравни
тельной революцией.
274
21. СТРУКТУРА И ПРЕОБРАЗОВАНИЕ
Подведем итог сказанному.
Эволюция совершается по разрешенным путям. В каж
дой ситуации достигается максимум устойчивости системы.
Устойчивость живых систем обеспечивается процессом ак
кумуляции вещества и энергии. Живые системы это систе
мы, устойчиво преобразующиеся. Преобразование — род
изменения, при котором как течение процесса, так и его
результат с известной жестокостью заданы предсуществующей организацией системы. Если устойчивость — цель
преобразования, то отбор во всех его формах — сохраняю
щий и преобразующий — является единственным факто
ром эволюции. Целесообразность в этом понимании и от
бор — понятия сопряженные.
С самого начала, еще в пробиологической фазе эволю
ции материи, усложнение организации шло по разрешен
ным путям. Разрешено то, что на путях повышения веро
ятности сохраниться в наибольшей мере сообразуется с
максимальным количеством физических, химических, гео
физических, географических и астрономических парамет
ров среды. Не случайно углерод лег в основу органических
соединений, и так же не случайно аккумуляция энергии
оказалась связанной с соединениями фосфора, с узким
кругом этих соединений. Не случайно медь и железо вов
лечены в процессы обмена. Сходство химического строения
хлорофилла и гемоглобина тому доказательство. Голубая
кровь моллюсков и красная кровь позвоночных — пример
различий, только подчеркивающих независимое возникно
вение сходств.
Не случайно живыми стали полимеры. Жизнь, согласно
В. И. Вернадскому [19606], возникла в виде биосферы, и
для отбора в его преобразующей форме открылось широ
чайшее, не отнюдь не беспредельное поле деятельности.
Отбор как фактор преобразования сам требует опреде
ленной организации системы, малой жесткости связей
между частями системы, взаимозаменимости частей и
главное — способности частей к саморепродукции. Услож
няя систему, отбор работает против самого себя. По мере
возрастания числа связей, на течение отбора налагается
запрет. Колонии клеток с течением времени утеряли свой
ство взаимозаменимости и многоклеточные организмы
вступают в процесс отбора как целостные образования. От
бор применительно к ним это внешняя сила. Внутри орга
низма возможен отбор, но он ведет за немногими исключе275
ниями (клональный отбор продуцентов специфических ан
тител по Бернету) к деструктивным последствиям.
Есть и другой барьер, помимо жесткости связей, для
деятельности отбора. Это достижение предельных состоя
ний. Если функция в совершенстве выполняется простой
структурой, то на усложнение налагается запрет. Единооб
разие строения и функций хранилища наследственной ин
формации во всем органическом мире, поразительное сход
ство делящихся клеток всех многоклеточных организ
мов — выражения этого запрета.
Жизнь на нашей планете представлена большим иерар
хически построенным разнообразием живых систем. Систе
мы отличаются друг от друга по способу самовоспроизведе
ния. Есть четыре приема воссоздать себя — можно постро
ить свою копию по своему шаблону и отделить ее от себя,
иной способ — это разделиться, еще иной — отделить ма
ленькую матрицу, способную проделать цикл превраще
ний, завершающийся организмом, подобным исходному, и,
наконец, можно возобновить себя с помощью замены изно
сившихся частей новыми. Четырем типам воспроизведения
соответствует четыре типа систем — генонема, клетка,
многоклеточный организм и популяция. Все свойства каж
дого типа систем жестко заданы типом самовоспроизведе
ния, сопряжены с ним, от него неотъемлемы. Тип самовос
произведения налагает категорический, а иногда только
более или менее категорический запрет на одни свойства и
так же категорически предопределяет другие. Строение си
стемы предопределяет возможности ее преобразования.
Иерархия систем сочетается с иерархией отборов. То,
что разрешено на индивидуальном уровне, наталкивается
на запрет на уровне групповом. Межвидовой отбор контро
лируется надвидовым. Кто окажется победителем, решают
активность в овладении жизненными ресурсами, быстрота
миграции атомов, их разнообразие, их количество.
Эволюция биосферы приобретает направленный характер — увеличивается число атомов, вовлеченных в сферу
жизни, срок пребывания атомов в составе живого вещества
биосферы удлиняется. Направленным процессом эволюции
является возрастание связанности частей, жесткости строе
ния живых систем.
В. И. Вернадский [1960а] в докладе, прочитанном на
заседании Ленинградского общества естествоиспытателей
5 февраля 1928 г. говорил: «Взятая в целом, палеонтологи
ческая летопись имеет характер не хаотического измене
ния, идущего то в ту, то в другую сторону, а явления оп
ределенного, развертывающегося все время в одну и ту же
276
сторону — в направлении усиления сознания, мысли, и со
здания форм, все более усиливающих влияние жизни на
окружающую среду... Если бы создание видов происходило
вслепую, случайно, вне зависимости от окружающей сре
ды, т. е. независимо от организованности биосферы, то все
же не всякий вид, случайно создавшийся, мог бы выжить и
войти в сложный биоценоз планеты; выжил бы тот, кото
рый был бы в нем устойчив, т. е. который увеличивал бы
биогенную миграцию атомов биосферы... Едва ли, однако,
возможно в настоящее время так элементарно просто про
тивопоставлять организм среде, т. е. биосфере, как это де
лалось раньше. Мы знаем, что организм в среде — не слу
чайный гость: он часть сложной организованности. И час
тью той же организованности является его эволюция».
Дифференциация кладет на взаимозаменимость частей
строжайший запрет, будь то органеллы клетки или орга
низма. Создавая самовоспроизводящиеся жесткие целост
ности — генонему, клетку, организм — отбор как бы вос
ходит по ступеням. Отбор в популяции клеток привел к
образованию многоклеточных организмов и уже не клетка,
а гигантская колония их подпадает как единое целое под
его действие. Объектом отбора становится организм —
консолидированное множество клеток. По мере увеличения
жесткости связей между представителями популяции она,
как целое, становится объектом отбора. Межгрупповой от
бор ограничивает возможности внутригруппового, способ
ствует дифференциации и интеграции популяций в систе
ме вида и уже виды, как целое, выступают на арену меж
видовой борьбы. И это еще не финал. Конечный этап —
отбор надвидовой. Исключая взаимозаменимость внутри
объекта, отбор становится по отношению к нему внешней
силой. Если объектов много, если они воспроизводят себя,
создается новая игра, где случайный поиск осуществляется
с помощью сопоставления уже не частей объектов внутри
них, а самих объектов друг с другом, и снова консолида
ция, и снова случайный поиск объектов высшего ранга.
Иерархия отборов — движитель закономерного хода эво
люции. Восхождение по ступеням иерархии отборов — за
кон эволюции.
Малиновский [1970] и Геодакян [1970] с предельной
ясностью выразили эту идею эволюции форм отбора. Гео
дакян [1970, с. 62] пишет: «Наблюдаемое нами богатство
форм живых систем — это различные стадии эволюции, в
которых и сейчас идут процессы усиления взаимодействия
между элементами систем. Происходит всеобщее укрепле
ние связей, утрата степеней свободы элементов и появле
277
ние новых программ в системах. Поэтому системы популя
ционного типа постепенно превращаются в системы организменного типа. В свою очередь однотипные организменные системы образуют популяционные системы на новом
уровне организации». Малиновский утверждает, что слож
ность организма создает предпосылку для образования
множества сходных, заменимых, то преходящих, то устой
чиво повторяющихся актов взаимодействия с физико-хи
мическим, биологическим, социальным окружением. Диск
ретные, повторяющиеся, постоянно генерируемые организ
мом акты поведения могут поощряться или тормозиться в
течение жизни индивидуума, и этот процесс в сочетании с
памятью тоже является частным случаем отбора.
Нельзя не согласиться с Бергсоном [1914, с. 20— 21 ],
когда он пишет: «Непрерывность в изменчивости, сохране
ние прошлого в настоящем, истинная длительность — вот,
по-видимому, свойства живого существа, общие со свойст
вами сознания. Нельзя ли пойти дальше и сказать, что
жизнь есть изобретательство, подобное сознательной дея
тельности, что подобно последней она есть непрерывное
творчество?»
Есть глубокое сходство в эволюции органических форм
и в эволюции психики. Как эволюция неорганической ма
терии завершается возникновением хранилищ информации
о прошлом, способных предвосхищать будущее и преумно
жать себя, так эволюция высшей нервной деятельности
животных привела к созданию памяти — к способности
удержать образ.
Великие прозрения мы находим у Тейара де Шардена
[1965, с. 290— 291 ]. Он пишет: «...как в этом знаменатель
ном союзе технического упорядочивания и психического
сосредоточения (centration) не видеть все еще действия из
вечной великой силы, той самой, которая нас произвела?..
Через социализацию человека... продолжает свой ход сама
ось космического вихря интерьеризации».
Память — основа отбора состояний. Сперва она только
атрибут внутреннего устройства нервной системы. Затем
она материализуется вовне. Создаются внешние хранили
ща информации — наскальная живопись, письмена. Спер
ва они единичны и как уникальные творения человеческо
го гения только умножают повторность восприятия — их
видят многие много раз. Потом возникает редупликация.
Печатание — только частное ее проявление.
Отбор в сфере духовного творчества оперирует объекта
ми, срок существования которых конечен. Идеи, теории
имеют свое деление. Они существуют в пространстве —
278
времени Бергсона. Они — «смертной мысли водомет». Так
сказал Тютчев. А Державин перед самой смертью написал
мелом на грифельной доске:
Река времен в своем теченьи
Уносит все дела людей
И топит в пропасти забвенья
Народы, царства и царей.
А что случайно остается
По воле лиры и трубы,
То жерлом вечности пожрется
И общей не уйдет судьбы.
Идеи не только воспроизводятся, они эволюционируют.
Из единичных мыслей, теорий возникает путем консо
лидации, сосредоточения концепций, и она подвергается
испытанию на целесообразность. Внутренняя непротиворе
чивость и соответствие с действительностью обеспечивают
концепции устойчивость. Идет стабилизирующий отбор
концепций, совершенствуются механизмы их образования.
Создается иерархия слагающих психический мир целост
ных образований — идей, теорий, концепций, метаконцеп
ций. И им соответствует иерархия отборов.
Закрыв для себя прежние возможности, отбор на опре
деленной ступени развития сам создает новые.
Отбор, уничтожение, смерть. Сама наша смерть — де
ло его рук. Отбор долговечных сделал нас смертными, пре
вратил нас в слагаемые систем высшего уровня. Как части
этих систем мы бессмертны.
Ленинград , СССР 1972
Сент-Луис, США 1984
Вашингтон , США 1991
Новосибирску СССР 1991
СПИСОК ЛИТЕРАТУРЫ
Беклемишев В. Н. Об общих принципах организации жизни / / Бюл.
МОИГ1. Отдел биол.— 1964.— Т. 69, № 2.— С. 22— 38.
Беклемишев В. Н. Биоценотические основы сравнительной паразитоло
гии.— M.: Наука, 1970.
Берг Л. С. Теории эволюции.— Петербург: Academia, 1922.
Берг Р. Л. Вид как эволюционирующая система / / Рефераты работ уч
реждений Отдел, биол. наук АН СССР за 1941 — 1943 гг.— 1944.—
С. 184— 186.
Берг Р. Л. Типы полиморфизма / / Вести. ЛГУ Сер. биол.— 1957.—
№ 21, вып. 4 .— С. 115— 138.
Берг Р. Л., Бриссенден Э. Б., Александрийская В. Т., Галковская К. Ф.
Генетический анализ двух природных популяций Drosophila
melanogaster / / Журн. общ. биол.— 1941.— Т. 2, № 1.—
С. 143— 158.
279
Бергсон А, Творческая эволюция.— Москва; Санкт-Петербург: Книгоиз
дательство «Русская мысль», 1914.
Вернет Ф . Целостность организма и иммунитет.— М.: Мир, 1964.
Богданов А. А. Общая организационная наука (тектология).— М.: Книга,
1925.
Вернадский В. И* Эволюция видов и живое вещество.— Избранные сочи
нения.— М.: Изд-во АН СССР, 1960а.— Т. 5 .— С. 238—251.
Вернадский В. И. Биосфера.— Избранные сочинения.— М.: Изд-во АН
СССР, 1 9 6 0 6 .- Т. 5.
Вилли К. Биология.— 4-е изд.— М.: Мир, 1964.
Волькенштейн М. В. Молекулы и жизнь.— М.: Наука, 1965.
Геодакян В. А. Организация систем — живых и неживых / / Системные
исследования. Ежегодник.— М.: Наука, 1970.— С. 49— 62.
Догель В. А. Олигомеризация гомологичных органов.— Л.: Изд-во ЛГУ,
1954.
Канаев И. И. Иоганн Вольфганг Гете.— Избранные сочинения по естест
вознанию.— Л.: Изд-во АН СССР, 1957.
Кольцов Н. К. Строение хромосом и обмен веществ в них / / Биол.
ж урн.— 1938.— № 1.
Комаров В, Л* Учение о виде у растений.— М.: Изд-во АН СССР, 1940.
Кремянский В. И. Самоустранение направляющего влияния естественного
отбора / / Сб. студенческих работ. Биология.— М.: Изд-во МГУ,
Кремянский В. И. О значении различных Форм филогенетической преем
ственности / / Журн. общ. биол.— 1940.— Т. 1, № 3.
Липман Ф . Современный этап эволюции биосинтеза и предшествовавшее
ему развитие / / Происхождение предбиологических систем.— М.:
Мир, 1968.
Майр Э. Зоологический вид и эволюция.— М.: Мир, 1963.
Малиновский А. А. Теория структур и ее место в системном подходе / /
Системные исследования. Ежегодник.— М.: Наука, 1970.—
С. 10— 32.
Моуэт Ф . Не кричи: волки!— М.: Мир, 1967.
Одум Е. Экология.— М.: Просвещение, 1968.
Северцов А. Н. Главные направления эволюционного процесса.— М.:
Биомедгиз, 1934.
Северцов А. Н. О конгруэнциях и понятии целостности вида / / Пробле
ма экологии животных. Неопубликованные работы С. А. Северцова.— М.: Изд-во АН СССР, 1951.— Т. 1.— С. 30—57.
Солт Д. Конкуренция между паразитоидными насекомыми / / Механизм
биологической конкуренции.— М.: Мир, 1964.— С. 124— 154.
Тейар д е Ш арден П. Феномен человека.— М.: П ротесе, 1965.
Тимофеев-Ресовский Н. В., Воронцов Н. Н., Яблоков А. В* Краткий
очерк теории эволюции.— М.: Наука, 1969.
Уоддингтон К .-Х . Основные биологические концепции / / На пути к тео
ретической биологии. I. Пролегомены.— М.: Мир, 1970.—
С. 11— 38.
Уолд Дж. Почему живое вещество базируется на элементах второго и
третьего периодов периодической системы? Почему фосфор и сера
способны к образованию макроэргических связей? / / Горизонты
биохимии.— М.: Мир, 1964.— С. 102— 113.
Холдейн Дж. Информация, необходимая для воспроизведения первичного
организма / / Происхождение предбиологических систем.— М.:
Мир, 1966.
Шванвич Б. Н. О работе медоносной пчелы на мохнатой вике и на лядвенце / / Вести. ЛГУ. Сер. биол.— 1956.— № 9 .— С. 55—61.
Ш епли X. Звезды и лю ди.— М.: Изд-во иностр. лит., 1962.
Ш мальгаузен И. И. Возникновение и преобразование системы морфоге
нетических корреляций в процессе эволюции / / Журн. общ. би
ол.— 1940.— Т. 1, № 3 .— С. 349— 370.
Ш мальгаузен И. И. Проблемы дарвинизма.— М.: Наука, 1969.
Berg R. L. Mutability Changes in Drosophila melanogaster Populations of
Europe, Asia and North America and Probable Mutability Changes in
280
Human populations of the USSR / / Jap. J. Genetics.— 1982.—
Vol. 5 7 .— P. 171 — 183.
Berg R. L. Probable Mutability Fluctuation in Human Populations of the
USSR, Paralleled by Mutability fluctuations / / Recent Trends in
Medical Genetics. Proc. Symp. 15th Inrern. Congr. of Genetics,
Madras, India, 1983.— Oxford; New York: Pergamon Press, 1986.—
P. 3 2 1 - 3 5 0 .
Berg R. L. Mutator — suppressor system, a decisive factor in successional
speciation going on in Drosophila melanogaster on a global scale / /
Abstract Genetics, Suppl.— 1987.— Vol. 116, № 1, Pt 2.— P. 18.
H oyle F. Of Men and G alaxies.— Univ. of Washington Press, 1964.
M ontesquieu. Letters Persanes.— Paris: Gamier — Flammarion, 1964.
H uxley
J.
Evolution.
The
Modern
Synthesis.— N. Y.;
L.,
1942.— P. 125— 150.
M uller H. T. The Gene as the Basis of the Life / / Proc. Intern. Congr. Plant
Sci.— 1929.— Vol. 1.— P. 807—921. (Мёллер Г. Ген как основа
жизни / / Избр. работы по генетике.— М.; Л.: Сельхозгиз, 1937.—
С. 148— 177.
Southern Н. Mimicry in Cuckoos Eggs / / Evolution as a Process / Ed.
Huxley J., Hardy A. G., Ford E. B.— L.: Allen and Unwin, 1954.
СОДЕРЖАНИЕ
Предисловие.................................................................................
3
Ра з д е л I
Эволюционная генетика
О путях эволюции генотипа (Берг Р. Л .у ТимофеевРесовский Н. В) ................................................................
Относительная роль стабилизации и передифференцировки гена в эволюции наследственной суб
станции (Берг Р . Л .)
Историческое развитие механизма реализации пола
(Берг Р. Л .) ........................................................................
Гетероцикличность хромосом и их комплексов как
механизм, обеспечивающий последовательность
вступления в действие отдельных элементов
кода наследственной информации (Берг Р. Л
Типы полиморфизма (Берг Р. Л,)
Значение валентности ядра в жизни и эволюции
вида (О некоторых механизмах, обеспечива
ющих оптимальное число наборов хромосом)
{Берг Р. Л .) ........................................................................
6
26
31
56
63
96
Р а з д е л II
Корреляционные плеяды
Экологическая интерпретация корреляционных плеяд
{Берг Р. Л .) ........................................................................
Корреляционные плеяды и стабилизирующий отбор
(Берг Р. Л .) ............................................. ".........................
О закономерностях эволюции на примере комплекс
ных преобразований признаков при дивергенции
видов вероник {Veronica L., Scrophulariaceae)
(Берг Р. У/., Колосова Л. Д .) ......................................
123
137
178
Р а з д е л III
Закономерности эволюции
Межвидовая и внутривидовая изменчивость жилкова
ния крыла в семействе дрозофил ид ( Drosophilidae ) (Берг Р. Л.)
Проблема целостности живых систем в трудах
И. И. Шмальгаузена (Берг Р. Л .) .............................
Место теории стабилизирующего отбора И. И. Шмаль
гаузена в современных эволюционных воззрени
ях (Берг Р. Л .) ..................................................................
Вид как эволюционирующая система (Берг Р. Л . ) .......
Случайна или закономерна эволюция? (Берг Р. Л.)
188
205
224
227
230
Научное издание
Берг Раиса Львовна
ГЕНЕТИКА
И
ЭВОЛЮ ЦИЯ
ИЗБРАННЫЕ ТРУДЫ
Редактор Н. Л. Шварцкопп
Художественный редактор В. А. Реймхе
Художник Н. А. Пискун
Технический редактор Т. Н. Драгун
Корректор Г. Д. Смоляк
ИБ №51036
Сдано в набор 24.06.92. Подписано к печати 16.09.92. Формат 84х 108^/32* Бумага тип. N° 2.
Офсетная печать. Уел. печ. л. 15,1 + вкладка. Уел. кр.-отт. 15,4. Уч.-изд. л. 17,5. Тираж 1000 экз.
Заказ №271. С024.
Ордена Трудового Красного Знамени ВО «Наука», Сибирская издательская фирма.
630099 Новосибирск, ул. Советская, 18.
Новосибирская типография №4 ВО «Наука». 630077 Новосибирск, ул. Станиславского, 25.
В Н И М А Н И Ю Ч И ТАТЕЛ ЕЙ !
Сибирская издательская фирма
ВО «НАУКА»
готовит к выпуску следующие книги:
Генетика кошки / Я. М. Бородину А. С. Графодатскийу А И . Железова и др. 17 л.
В монографии впервые дается исчерпывающее описание
генетики одного вида млекопитающих — домашней кошки.
Содержатся сведения по эволюции и цитогенетике семей
ства кошачьих, полный перечень всех известных мутаций,
систематизированные данные по молекулярной и биохи
мической генетике, генетике поведения, иммуногенетике,
популяционной генетике, биологии размножения и разви
тия кошки. Особый интерес для любителей кошек пред
ставляют сведения об основных породах кошек и принци
пах их разведения.
Книга заинтересует специалистов по генетике и эволю
ции животных, а также любителей кошек.
Жимулев И . Ф. Гетерохроматин и эффект положения
гена. 30 л.
В монографии рассмотрено современное состояние зна
ний об организации и генетических функциях особых рай
онов хромосом: прицентромерного и интеркалярного гете
рохроматина. Систематизированы сведения о таких его
свойствах, как компактность в клеточном цикле, генети
ческая инертность, поздняя репликация, соматическая
конъюгация, образование хромосомных перестроек. Сфор
мулирована современная модель интеркалярного гетеро
хроматина. Предложена гипотеза о необратимой компактизации укладки материала хромосомы в ходе генетической
инактивации при эффекте положения.
Книга адресована генетикам, цитологам, молекулярным
биологам.
Книги высылаются наложенным платежом. Заказы
просим направлять по адресу: 630090 Новосибирску
Морской проспекту 22. Магазин «Наука».
В Н И М А Н И Ю ЗА КА ЗЧИ КО В!
Книги можно предварительно заказать в магазинах
торговой фирмы «Академкнига», в местных магазинах кни
готоргов или потребительской кооперации.
480091 Алма-Ата, Казахстан, ул. Фурманова, 9 1 /9 7
(«Книга — почтой»);
370001 Баку, Азербайджан, ул. Коммунистическая, 51
(«Книга — почтой»);
720001 Бишкек, Кыргызстан, бульвар Дзержинского, 42
(«Книга — почтой»);
232600 Вильнюс, Литва, ул. Университето, 4;
690088 Владивосток, Россия, Океанский проспект, 140
(«Книга — почтой»);
320093 Днепропетровск, Украина, проспект Гагарина, 24
(«Книга — почтой»);
734001 Душанбе, Таджикистан, проспект Ленина, 95
(«Книга — почтой»);
620151 Екатеринбург, Россия, ул. Мамина-Сибиряка, 137
(«Книга — почтой»);
375002 Ереван, Армения, ул. Туманяна, 31;
664033 Иркутск, Россия, ул. Лермонтова, 289 («Книга —
почтой»);
420043 Казань, Татарстан, ул. Достоевского, 53 («Кни
га — почтой»);
252030 Киев, Украина, ул. Ленина, 42;
252142 Киев, Украина, проспект Вернадского, 79;
252025 Киев, Украина, ул. Осипенко, 17;
252208 Киев, Украина, проспект Правды, 80-а («Книга —
почтой»);
277012 Кишинев, Молдова, проспект Штефана Велико^
148;
343900 Краматорск Донецкой обл., Украина, ул. Марату
1 («Книга — почтой»);
660049 Красноярск, Россия, проспект Мира, 84;
220012 Минск, Беларусь, Ленинский проспект, 72 («Кнц
га — почтой»);
103009 Москва, Россия, Тверская ул., 19-а;
117312 Москва, Россия, ул. Вавилова, 5 5 /7 ;
117393 Москва, Россия, ул. Академика Пилюгина, Ц
корп. 2 («Книга — почтой»);
630076 Новосибирск, Россия, Красный проспект, 51;
630090 Новосибирск, Россия, Морской проспект, ^
(«Книга — почтой»);
142284 Протвино Московской обл., Россия, ул. Победы, 8;
142292 Пущино Московской обл., Россия, МР «В», |
(«Книга — почтой»);
443002 Самара, Россия, проспект Ленина, 2 («Книга ^
почтой»);
191104 Санкт-Петербург, Россия, Литейный проспект, 57;
199164 Санкт-Петербург, Россия, Таможенный пер., 2;
194064 Санкт-Петербург, Россия, Тихорецкий проспект, <(
700000 Ташкент, Узбекистан, ул. Ю. Фучика, 1;
700029 Ташкент, Узбекистан, ул. Ленина, 73;
700070 Ташкент, Узбекистан, ул. Шота Руставели,43;
700185 Ташкент, Узбекистан, ул. Дружбы народов, <
(«Книга — почтой»);
634050 Томск, Россия, наб. реки Ушайки, 18;
450059 Уфа, Башкортостан, ул. Р. Зорге, 10 («Книга
почтой»);
450025 Уфа, Башкортостан, ул. Коммунистическая, 49;
310078 Харьков, Украина, ул, Чернышевского, 87.